




































































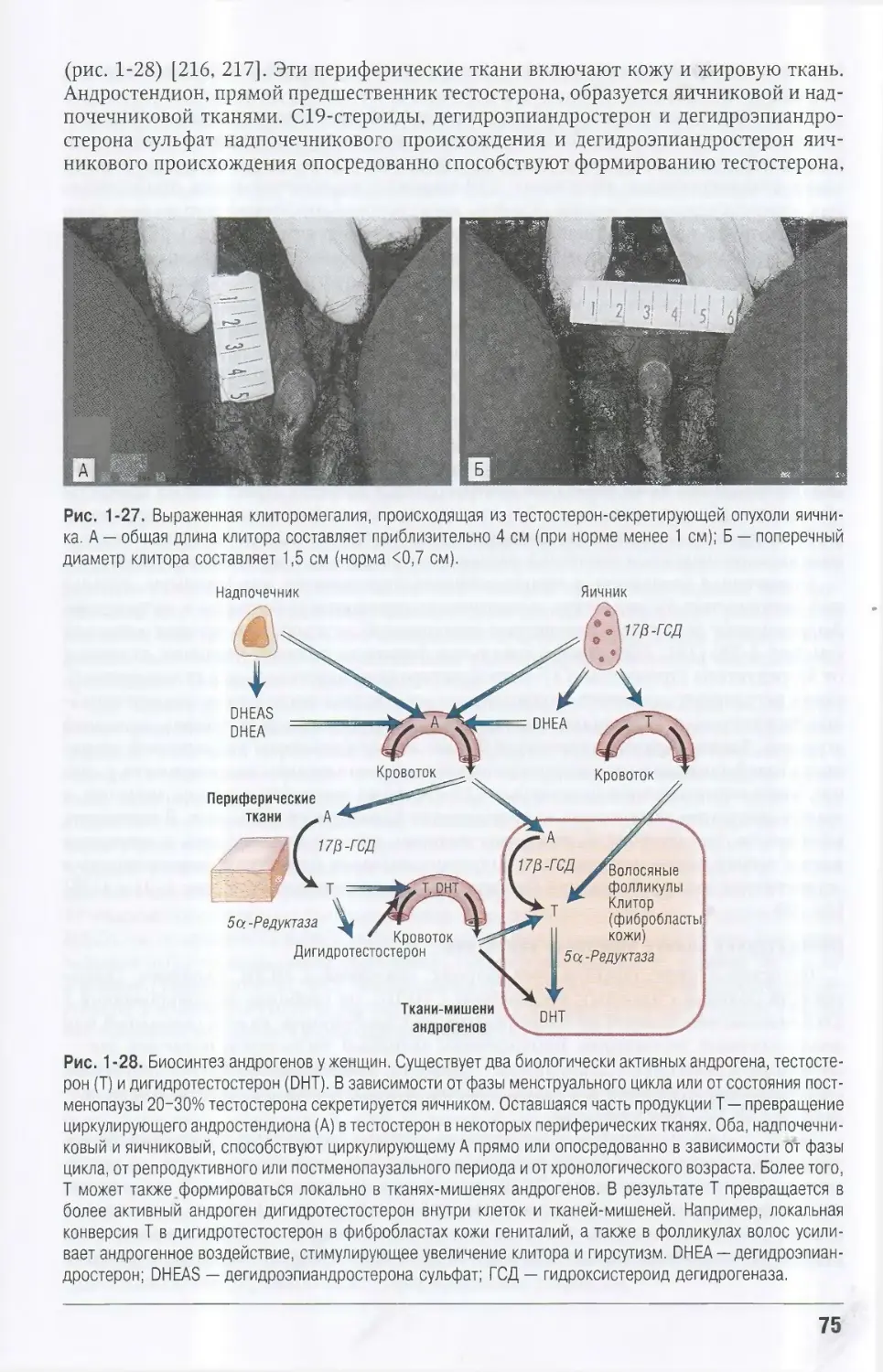





































































































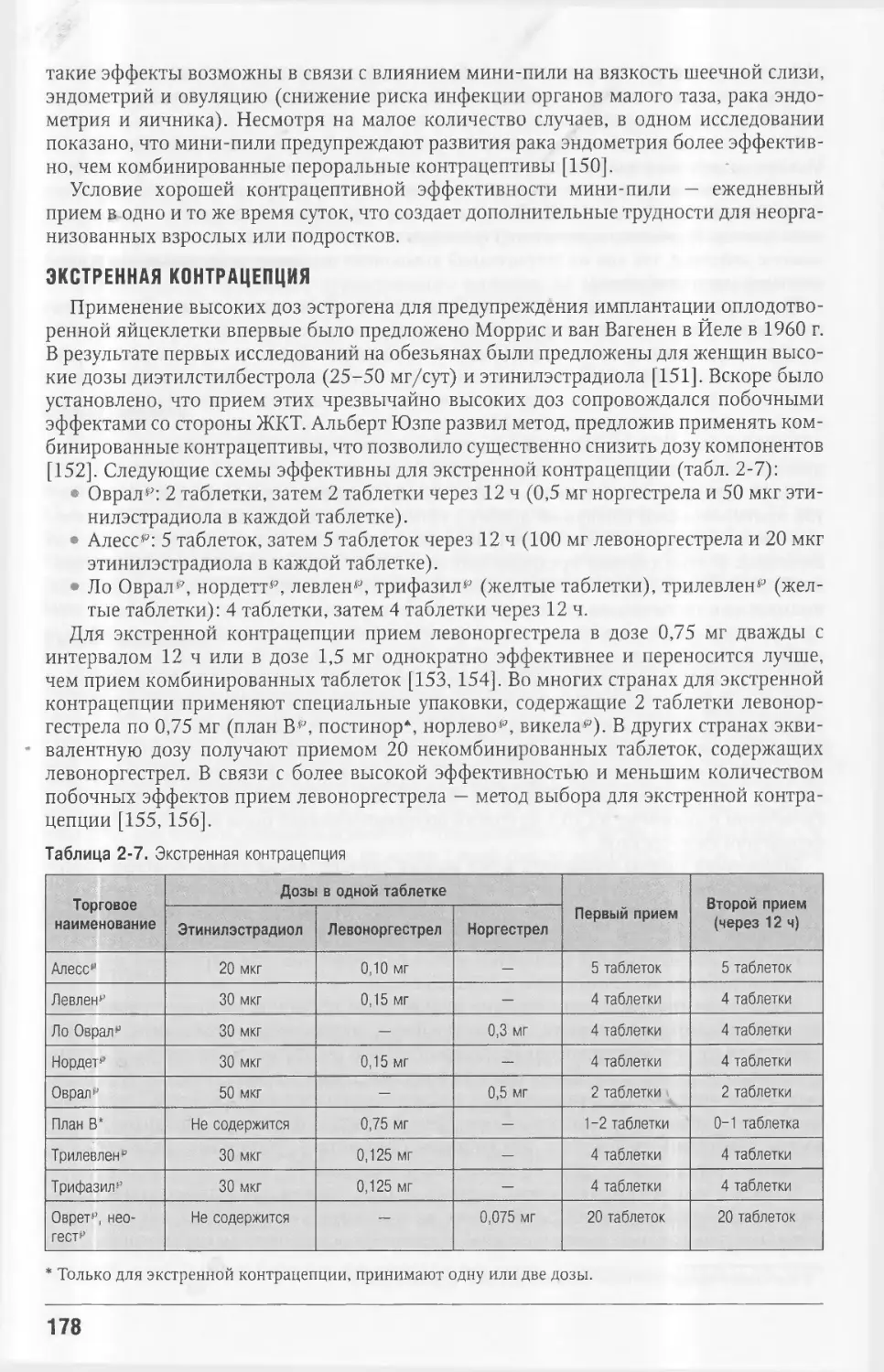



































































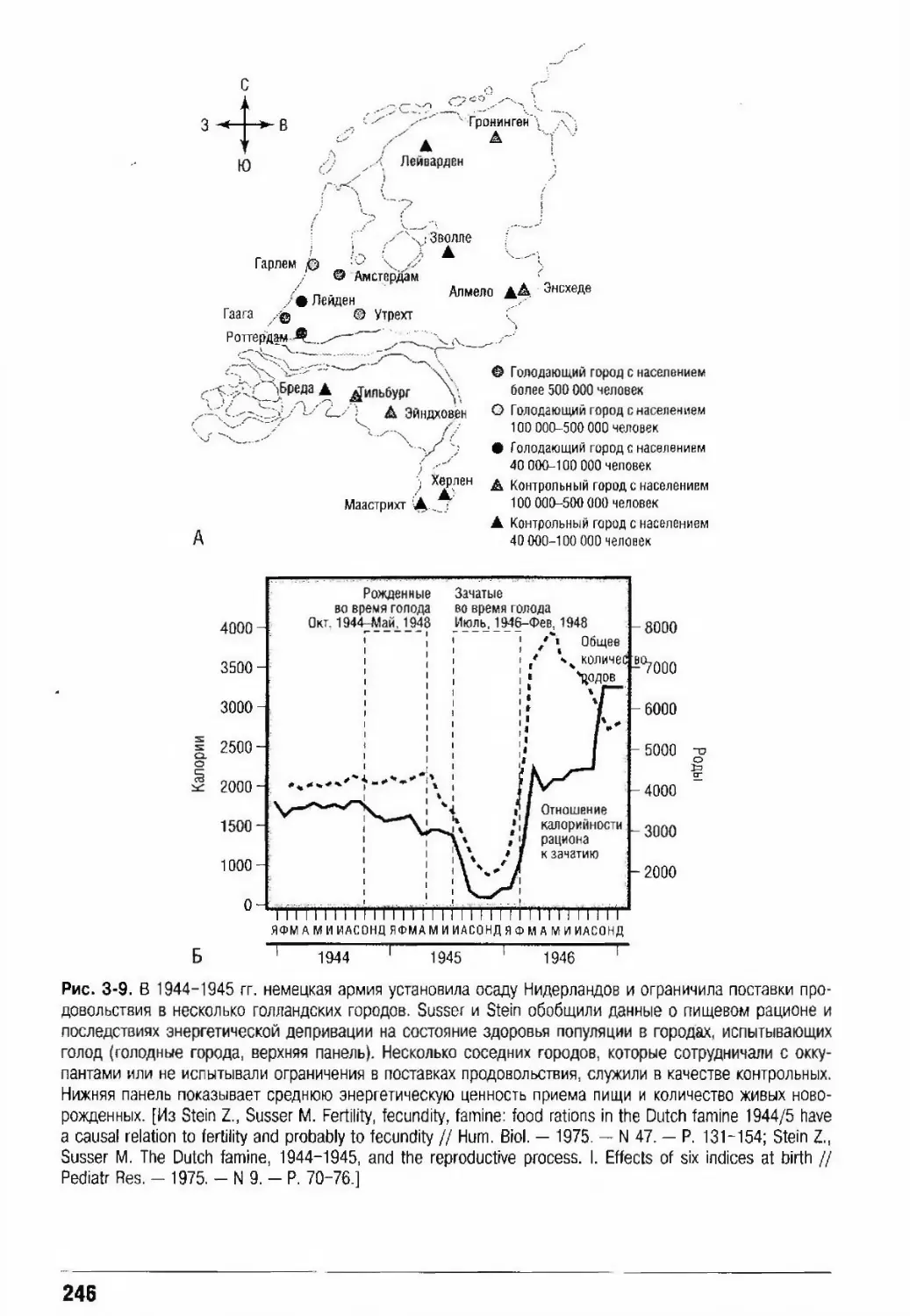


































































































































































Автор: Кроненберг Генри М. Мелмед Шлеме Полонски Кеннет С. Ларсен П. Рид
Теги: гигиена нервной системы гигиена и этика заболевания мочеполовой системы гинекология акушерство антропология урология медицина эндокринология
ISBN: 978-5-91713-029-3
Год: 2011




РЕПРОДУКТИВНАЯ
ЭНДОКРИНОЛОГИЯ
Перевод с английского под редакцией
акад. РАН и РАМН И.И. Дедові
”. КІП ІГАП ОЛМІІ г А Мапк^иііиоиїтА
■
т..
-V ^
чл.-кор. РАМН Г.А. Мельниченко
ч
ИЗДАТЕЛЬСКАЯ ГРУППА
«ГЭОТАР-Медиа»
EDITION
11
Williams Textbook of
ENDOCRINOLOGY
Henry M. Kronenberg, MD
Professor of Medicine
Harvard Medical School
Chief, Endocrine Unit
Massachusetts General Hospital
Boston, Massachusetts
ShJomo Melmed, MD, FRCP
Helene and Philip Hixon Chair
in Investigative Medicine
Senior Vice President for Academic Affairs
Cedars Sinai Medical Center
Los Angeles, California
Kenneth S. Polonsky, MD
Adolphus Busch Professor and Chairman
Department of Medicine
Washington University School of Medicine
St. Louis, Missouri
P. Reed Larsen, MD, FACP, FRCP
Professor of Medicine
Harvard Medical School
Chief, Division of Endocrinology, Diabetes,
and Hypertension
Brigham and Women's Hospital
Boston, Massachusetts
Генри М. Кроненберг, Шлеме Мелмед,
Кеннет С. Полонски, П. Рид Ларсен
ЭНДОКРИНОЛОГИЯ
по ВИЛЬЯМСУ
РЕПРОДУКТИВНАЯ
ЭНДОКРИНОЛОГИЯ
Перевод с английского под редакцией
акад. РАН и РАМН И.И. Дедова,
чл.-кор. РАМН Г.А. Мельниченко
Москва
Рид Элсивер
2011
УДК [612-6+613.888.15+616.69+618.1] (035.3)
ББК 28.707.3я81+56.9я81+.57.0я81+57.1я81
Р41
Издание представляет собой перевод с английского книги
«Williams Textbook of Endocrinology, 11*^ edition«^
авторов Henry M. Kronenberg, Shiomo Melined, Kenneth S. Polonsky и P. Reed Larsen.
Перевод опубликован по контракту с издательством «Elsevier Ltd.«^.
Научное редактирование перевода
Ледов Иван Иваиовиг,
акад. РАН и РАМН, д-р мед. наук, проф., директор Эндокринологического научного центра
Росмедтехнологий, зав. кафедрой эндокринологии Первого МГМУ им. И.М. Сеченова
Мельнигенко Галана Афанасьевна,
чл.-кор. РАМН, д-р мед. наук, проф. кафедры эндокринологии Первого МГМУ им. И.М. Сеченова,
директор Института клинической эндокринологии Эндокринологического научного центра
Росмедтехнологий
Перевод с английского осуществлен
ООО Издательская группа «ГЭОТАР-Медиа^^.
ИЗДАТЕЛЬСКАЯ ГРУППА
«ГЭОТАР-Медиа»
Репродуктивная эндокринология / Генри М. Кроненберг, Шлогио Мелмед, Кеннет С.
Р41 Полонски, П. Рид Ларсен ; пер. с англ. под ред. И. И. Дедова, Г. А. Мельниченко. - М. ;
ООО «Рид Элсивер», 2011. — 416 с. - (Серия «Эндокринология по Вильямсу^»). — Перевод изд.
Williams Textbook of Endocrinology, ll”* edition / Henry M. Kronenberg, Shiomo Melmed, Kenneth
S. Polonsky и P. Reed Larsen.
ISBN 978-5-91713-029-3 (pyc.)
В книге представлены разделы из фундаментального руководства «Эндокринология по
Вильямсу», посвященные заболеваниям репродуктивной системы. Затронуты вопросы особен¬
ностей физиологии, гормональной регуляции, биохимических процессов репродуктивной си¬
стемы мужчин и женщин. Отдельно рассмотрены вопросы контрацепции, .заболевания мужских
половых желез, включая заболевания яичек, бесплодие и снижение фертильности, новообразо¬
вания яичек. Также обсуждается вопрос незаконного использования андрогенов спортсменами.
Отдельная глава посвящена сексуальной дисфункции у мужчин и женщин.
Руководство предназначено для врачей-эндокринологов, гинекологов, урологов и сексопа¬
тологов, а также студентов медицинских вузов, ординаторов, интернов и аспирантов, специали¬
зирующихся на лечении заболеваний щитовидной железы,
УДК [612.6+613.888.154б16.69+618.1](035,3)
ББК 28.707.3я81+56.9я81+57.0я81з 57.1я81
Предупрежденне. Знания и практический опыт в области эндокринологии постояшю развиваются. Новые
исслсдовзЕгия и клинический опыт расвигряют наши знания, поэтому может возникнуть необходимость в изме¬
нении методов диагностики, способов лечения и лекарственной терапии. В любом случае советуем читателям,
помимо этой книги, обращаться к дру]‘им источникам информации. При назначении больным лекарственных
средств необходимо ознакомиться с сонроводительной инструкцией, вложенной в упаковку, чтобы столпить
рекомендуемую дозу, способ и продолжительность применения, а также определить противопоказания. Только
профессионал, полагаясь на собственный опыт и знания о больном, отвечает за постановку диагноза, выбор до¬
зировки и оптимальной схемы лечения для каждого пациента. По закону ни издатель, ни автор не несут какой-
либо ответственности за любой вред и/или ущерб, нанесенный людям или собственности в результате любого
использования материала, содержащегося в этой книге, или в связи с ним.
Все права защищены. Не допускается воспроизведение или распространелше в любой форме или любым спо¬
собом (электронным или механическим, включая фотокопирование, видеозаЕшсь или любую систему хранения
и поиска информации) без письменного разрешения издателя. Разрешение можно запросить непосредственно у
Elsevier’s Health Sciences Rights Department в Филадельфии, Пенсильвания. США. Телефон: <1 (215) 239-38-04;
факс: +1 (215) 239-38-05; e-mail; healthperraissions@elsevier.coTn или on-line на http;//M'v\'M'.elsevier.coni.
© Elsevier Ltd.. 2008, 2003,1998,1992,1985,1981,1974,1968,
1962,1955. Бее права защищены
ISBN 978-5-91713-029-3 (рус.) © ООО «Рид Элсивер», ООО Издательская группа
ISBN 978-1-4160-2911-3 (англ.) «ГЭОТАР-Медиа», перевод на русский язык, 2011
Предисловие к русскому изданию 6
Предисловие к иностранному изданию 7
Авторы 8
Список сокращений и условных обозначений 14
Глава 1. Физиология и патология репродуктивной системы женщины 17
Физиология репродукции 17
Репродуктивные функции гипоталамуса 19
Репродуктивные функции переднего гипофиза 24
Яичники 26
Эндометрий 54
Обследование женщин с репродуктивной дисфункцией 63
Расстройства женской половой системы 66
Дифференциальная диагностика и лечение ановуляторных маточных
кровотечений 107
Гормонозависимые доброкачественные гинекологические заболевания 117
Ведение менопаузы 120
Заместительная гормональная терапия в постменопаузе 126
Глава 2. Гормональная контрацепция 157
Пероральная контрацепция 158
Влагалищное и чрескожное (трансдермальное) применение стероидов
для контрацепции 181
Инъекционная контрацепция 186
Имплантационная контрацепция 194
Глава 3, Заболевания мужских половых желез 221
Физиологическая регуляция функции яичек: продукция и действие половых
стероидных гормонов 221
Заболевания яичек 235
Бесплодие и снижение фертильности у мужчин 276
Новообразования яичек 289
Незаконное использование андрогенов спортсменами и бодибилдерами 292
Глава 4. Сексуальные расстройства у мужчин и женщин 331
Сексуальные расстройства у мужчин 332
Сексуальная дисфункция у женщин 364
Предметный указатель 407
Уважаемые коллеги!
Для нас большая честь представить вашему вниманию первый русский перевод
Итго издания «Эндокринологии по Вильямсу» — настольной книги каждого эндо¬
кринолога мира.
Многие поколения врачей всех специальностей обращаются ежедневно к этому
уникальному изданию как к основной справочной литературе по всем вопросам, свя¬
занным не только с физиологией и патофизиологией, клиническими проявлениями и
методами диагностики первичных эндокринопатий, но и с эндокринными проявле¬
ниями соматических заболеваний, психических нарушений, болезней репродуктив¬
ной системы, с физиологическими и патологическими изменениями в эндокринной
сфере при беременности, родах, старении, тяжелых неэндокринных заболеваниях,
последствиях лечения и со многими-многими другими проблемами, возникающими
во врачебной практике.
Всеобъемлющий и универсальный характер этого поисгине грандиозного учебного
пособия таков, что путеводную нить для решения любой клинической проблемы, даже
в минимальной степени связанной с гормональной регуляцией деятельности организ¬
ма человека, и нужное направление поиска всегда можно найти на его страницах.
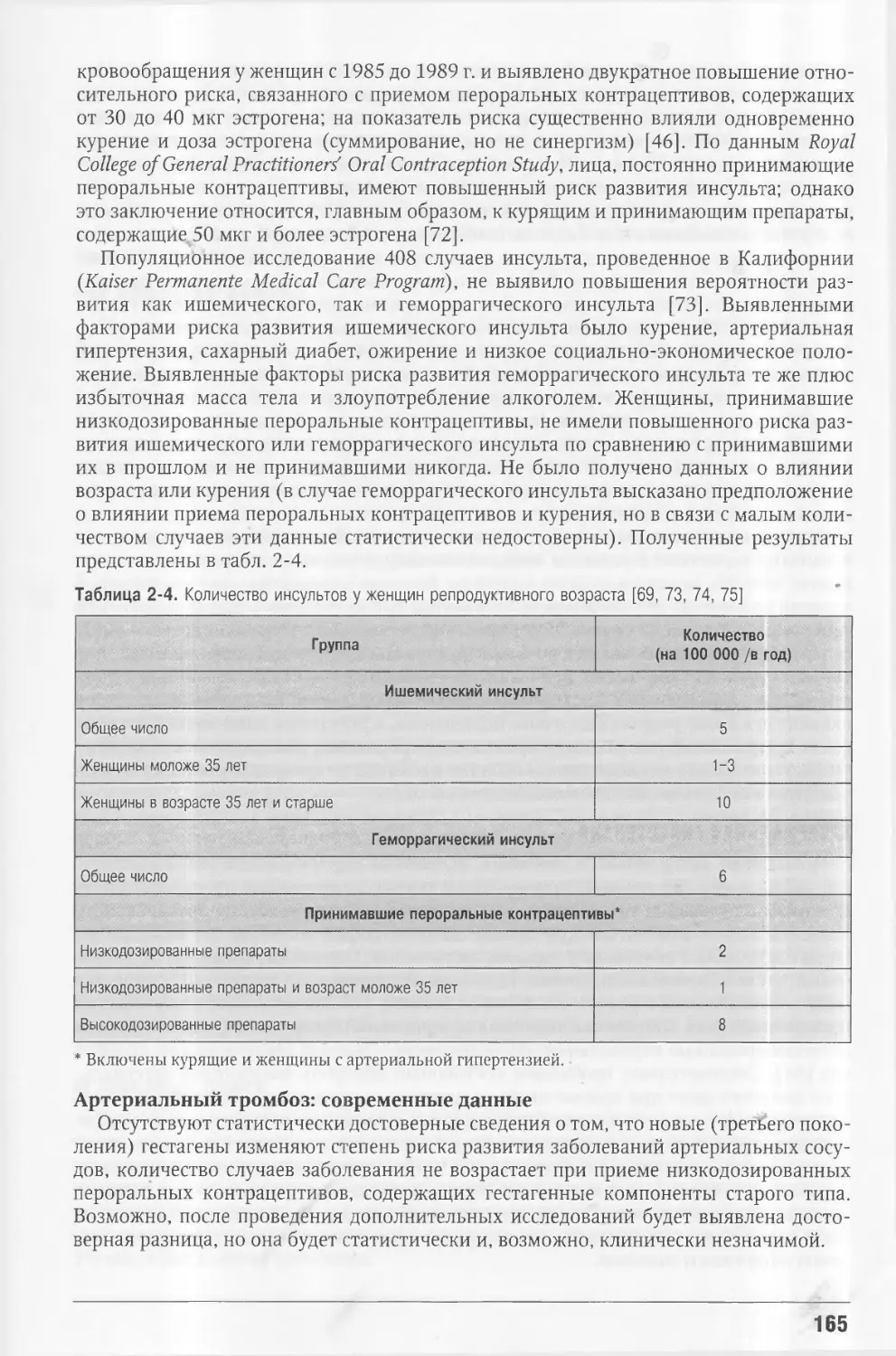
Этот уникальный справочный и обучающий материал непрерывно пополняется и
интегрируется с уже имеющимися сведениями, и каждое издание обновляется в соот¬
ветствии с полученными новыми данными, при этом сохраняется удивительная чет¬
кость изложения и стремление сделать даже самую сложную информацию доступной
для практического врача.
Неизбежное при переводе на русский язык расширение объема издания стало
одной из основных причин решения выпустить его в ином, отличном от оригинала
формате (в виде серии книг), что, по нашему мнению, сделает чтение более удобным.
При переводе столь масштабного и концептуального фактического материала мы
в осторожной, корректной форме старались включать результаты международных
консенсусов, определивших современные международные и отечественные стандар¬
ты и алгоритмы (протоколы) в области диагностики, лечения и профилактики всех
болезней эндокринной системы.
Разумеется, первый перевод такого фундаментального труда не может не иметь
технических погрешностей и неточностей, и участники проекта будут благодарны за
полученные замечания и уточнения. Мы надеемся, что и студент-медик, и врач любой
специальности найдут на страницах книги важную информацию.
Акад. РАН и РАМН, д-р мед. наук, проф. \ ] И.И. Дедов
о ‘
Чл.-кор. РАМН, д-р мед. наук, проф. ГЛ. Мельнигенко
ПРЕДИСЛОВИЕ К ИНОСТРАННОМУ ИЗДАНИЮ
Приглашаем вас ознакомиться с 11-м изданием «Эндокринологии по Вильямсу».
Хотя Роберт Вильямс написал эту книгу более 50 лет назад, ее суть не изменилась.
Вильямс считал своей целью «собрать и обсудить опыт эндокринологов-практиков
и данные лабораторных исследований и обследований пациентов». Естественно, в
данную работу мы должны были включить «генетические и эпидемиологические
исследования и результаты анализа медицинских баз данных» как основные факты
для постановки диагноза в современной медицине. В действительности синтетические
науки — это самый обширный и непрерывный источник новой информации, поэтому
данная книга лучше других поможет врачу-эндокринологу лечить своих пациентов.
Чтобы поддержать идею объединить несколько наук, мы продолжаем привлекать
к работе над книгой наиболее выдающихся врачей и ученых и стараемся показать
читателям наиболее полную картину современной эндокринологии. Задача книги —
представить материал в сжатом виде и обеспечить быстрый и удобный поиск нужной
информации, что очень важно для врача.
Текст обновлен цветными иллюстрациями, что способствует лучшему усвоению
материала. Чтобы выгоднее представить и применить алгоритмы диагностики и лече¬
ния, выбран единый стиль подачи информации в книге.
Мы выражаем глубокую признательность нашим сотрудникам: Ирме Саббаг, Линн
Моултон, Грэйс Лабрадо, Льюис Ишибаши и Шерри Тюнер, стараниями которых эта
книга издана. Благодарим также наших коллег из издательства «Эльзевир» — Долорес
Мелони и Энн Снидер, которые следили за всеми переизданиями книги в других стра¬
нах, стараясь сохранить наши основные идеи и не задерживать выход руководства. Их
усилия гарантировали успех издания и сохранение высокого качества нашей книги.
Генри М. Кроненберг,
Шломо Мелмед,
Кеннет С. Полонски,
П. Рид Ларсен
Джон Ц. Ачерманн (fohn С. Achermann,
MD)
Wellcome Trust Senior Research Fellow
in Clinical Science, Developmental
Endocrinology
Research Group, UCL Institute of Child
Health; Honorary Consultant in Paediatric
Endocrinology, Great Ormond Street Hospital
for Children NHS Trust, London, United
Kingdom
Disorders of Sex Development
Эли Ю. Адаши (Eli Y, Adashi, MD)
Dean of Medicine and Biological Sciences,
Division of Biology and Medicine, Brown
University Medical School, Providence, Rhode
Island
The Physiology and Pathology of the Female
Reproductive Axis
Ллойд П. Аело (Lloyd P. Aiello, MD,
PhD)
Head, Section of Eye Research, Director.
Beetham Eye Institute, Joslin Diabetes Center;
Associate Professor of Ophthalmology,
Harvard Medical School, Boston,
Massachusetts
Complications of Diabetes Mellitus
Эндрю Арнольд (Andrew Arnold, MD)
Murray-Heilig Chair in Molecular Medicine
and Professor of Medicine and Genetics,
University of Connecticut School of Medicine;
Chief. Division of Endocrinology
and Metabolism, and Director, Center
for Molecular Medicine, University
of Connecticut Health
Center, Farmington, Connecticut
Pathogenesis of Endocrine Tumors
Дженифер M. Баркер (|eimifer M.
Barker, MD)
Assistant Professor of Pediatrics,
University of Colorado School of Medicine;
Endocrinologist,
Barbara Davis Center for Childhood Diabetes,
Aurora, Colorado
The Immunoendocrinopathy Syndromes
Розмари Бессон [Rosemary Basson, MD,
FRCP(UK)]
Clinical Professor, University of British
Colombia Faculty of Medicine; Director, UBC
Sexual
Medicine Program, University of British
Columbia; Director, Sexual Medicine Program,
Vancouver Hospital. Vancouver, British
Columbia, Canada
Sexual Dysfunction in Men and Women
Томас П. Берсот (Thomas P. Bersot, MD,
PhD)
Associate Investigator, Gladstone Institute
of Cardiovascular Disease, Professor
of Medicine,
University of California, San Francisco
Disorders of Lipid Metabolism
Шалендер Басин (Shalender Bhasin, MD)
Professor of Medicine, Section
of Endocrinology, Diabetes, and Nutrition,
Boston University
School of Medicine; Chief, Section
of Endocrinology, Diabetes, and Nutrition,
Boston
Medical Center, Boston, Massachusetts
Testicular Disorders
Sexual Dysfunction in Men and Women
Эндрю Дж. М. Боултон (Andrew J. М.
Boulton, MD, FRCP)
Professor of Medicine, University
of Manchester. Consultant Physician,
Manchester Royal
Infirmary, Manchester, United Kingdom;
Division of Endocrinology, Metabolism, and
Diabetes, University of Miami School
of Medicine, Miami, Florida
Complications of Diabetes Mellitus
Глен Д. Браунштейн (Glenn D.
Braunstein, MD)
Professor of Medicine, David Geffen School
of Medicine at UCLA; Chairman, Department
of Medicine, and James R. Klinenberg Chair
in Medicine, Cedars-Sinai Medical Center, Los
Angeles, California
Endocrine Changes in Pregnancy
Грегори A. Брент (Gregory A. Brent, MD)
Professor of Medicine and Physiology, David
Geffen School of Medicine at UCLA; Chief,
Endocrinology and Diabetes Division, VA
Greater Los Angeles Healthcare System, Los
Angeles, California
Hypothyroidism and Thyroiditis
Ф, Ричард Брингхарст (F. Richard
Bringhurst, MD)
Associate Professor of Medicine Harvard
Medical School, Physician, Endocrine Division,
Massachusetts General Hospital, Boston,
8
Massachusetts
Hormones and Disorders of Mineral
Metabolism
Мишель Браунли (Michael Brownlee, MD)
Anita and Jack Saltz Professor of Diabetes
Research, Director. JDRF International
Center for Diabetic Complications Research,
Department of Medicine and Pathology,
Albert Einstein
College of Medicine, Bronx, New York
Complications of Diabetes Mellitus
Сердар И. Булун (Serdar E. Bulun, MD)
George H. Gardner Professor of Clinical
Gynecology and Chief, Division
of Reproductive Biology Research,
Department of Obstetrics and Gynecology,
Northwestern University Feinberg School
of Medicine, Chicago, Illinois
The Physiolog)' and Pathology of the Female
Reproductive Axis
Чарльз Ф. Бурант (Charles F, Burant,
MD, PhD)
Professor of Internal Medicine University
of Michigan Health System
Ann Arbor, Michigan
Type 2 Diabetes Mellitus
Джон Б. Буз 0ohn В. Buse, MD, PhD)
Professor of Medicine, University of North
Carolina at Chapel Hill School of Medicine,
Chapel Hill, North Carolina
■R'pe 2 Diabetes Mellitus
Type 1 Diabetes Mellitus
Дэвид A. Бушиинский (David A.
Bushinsky, MD)
Professor of Medicine, Pharmacology and
Physiology, University of Rochester School
of Medicine and Dentistry: Chief. Division
of Nephrology, University of Rochester Medical
Center, Rochester, New %rk
Kidney Stones
Эрнесто Каналис (Ernesto Canalis, MD)
Professor of Medicine, University
of Connecticut School of Medicine,
Farmington: Director of Research,
St. Francis Hospital Medical Center, Hartford,
Connecticut
Metabolic Bone Disease
Кристин Картер-Су (Christin Carter-Su,
PhD)
Professor of Molecular and Integrative
Physiology, Associate Director and Head of the
Michigan Diabetes Research and Training
Center, University of Michigan Medical
School,
Ann Arbor, Michigan
Mechanism of Action of Hormones That Act
at the Cell Surface
Роджер д. Кон (Roger D. Cone, PhD)
Director, Center for the Study of Weight
Regulation and Associated Disorders, Oregon
Health and Science University, Portland;
Senior Scientist, Vollum Institute, Oregon
Health and Science University, Portland,
Oregon
Neuroendocrine Control of Energy Stores
Марк И. Купер (Mark E. Cooper, MBBS,
PhD, FRACP)
Professor of Medicine and Immunology,
Eastern Clinical School, Monash University;
Professor of Medicine and Physiology,
University of Melbourne, Senior
Endocrinologist,
Alfred Hospital, Melbourne: Head, JDRF
Danielle Alberti Memorial Centre for Diabetic
Complications, Diabetes and Metabolism
Division, Baker Heart Research Institute,
Melbourne, Victoria, Australia
Complications of Diabetes Mellitus
Филипп и. Краер (Philip E. Cryer, MD)
Irene E, and Michael M. Karl Professor
of Endocrinology and Metabolism in Medicine,
Washington University in St. Louis School
of Medicine; Physician, Barnes-Jewish Hospital,
St. Louis, Missouri
Glucose Homeostasis and Hypoglycemia
Филипп Д, Дарней (Philip D. Damey,
MD, MSc)
Professor of obstetrics. Gynecology,
and Reproductive Sciences, University
of California,
San Francisco, School of Medicine; Chief,
Obstetrics and Gynecology, San Francisco
General
Hospital. San Francisco, California
Hormonal Contraception
Терри Ф. Дэвис (Terry F. Davies, MBBS,
MD, FRCP, FACE)
Florence and Theodore Baumritter Professor
of Medicine, Mount Sinai School of Medicine;
Attending Physician, Mount Sinai Hospital;
Director, Division of Endocrinology
and Metabolism, James J. Peters VA Medical
Center, New York, New York
Thyroid Physiology and Diagnostic Evaluation
of Patients with Thyroid Disorders
Thyrotoxicosis
Hypothyroidism and Thyroiditis
Мэри Б. Демай (Marie В, Demay, MD)
Associate Professor of Medicine, Harvard
Medical School. Massachusetts General
Hospital:
Boston, Massachusetts
Hormones and Disorders of Mineral
kjetabolism
Дэниел Дж. Друкер (Daniel J. Dnicker,
MD)
Director, Banting and Best Diabetes Centre,
University of Toronto, Toronto General
Hospital, Toronto, Ontario, Canada
Gastrointestinal Hormones and Gut Endocrine
Tumors
Джорж С. Эйсенбарт (George S,
Eisenbarth, MD, PhD)
Professor, Departments of Pediatrics,
Medicine, and Immunology, University
of Colorado
School of Medicine, Denver: Executive
Director, Barbara Davis Center for Childhood
Diabetes, Aurora; Physician, University
Hospital and the Children’s Hospital, Denver,
Colorado
Type 1 Diabetes Mellitus
The Immunoendocrinopathy Syndromes
Джоэл К. Элмквист (Joel К. Elmquist,
DVM, PhD)
Maclin Family Professor in Medical Science,
in Honor of Dr. Roy A. Brinkley, Departments
of Medicine and Pharmacology; Director,
Division of Hypothalamic Research, University
of Texas Southwestern Medical Center, Dallas,
Texas
Neuroendocrine Control of Energy Stores
Дэниел Д. Федерман (Daniel D.
Federman, MD)
Carl W. Walter Distinguished Professor
of Medicine, Harvard Medical School, Boston,
Massachusetts
The Endocrine Patient
Себастьяно Филетти (Sebastiano Filetti,
MD)
Professor of Internal Medicine, Department
of Clinical Sciences, University of Rome La
Sapienza School of Medicine; Head,
Department of Internal Medicine, Policlinico
Umberto
I, Rome, Italy
Nontoxic Diffuse and Nodular Goiter
and Thyroid Neoplasia
Делберт A. Фишер (Delbert A. Fisher,
MD)
Professor of Pediatrics and Medicine
Emeritus, David Geffen School of Medicine
at UCLA,
Los Angeles; Chairman Emeritus, Department
of Pediatrics, and Serous Scientist, Walter
Martin Research Center, Harbor-UCLA
Medical Center, Torrance, California
Endocrinology of Fetal Development
Роберт Ф. Гагел (Robert F. Gagel, MD)
Professor and Head, Division of Internal
Medicine, the University of Texas MD
Anderson
Cancer Center; Professor of Cell Biology,
Baylor College of Medicine, Houston, Texas
Multiple Endocrine Neoplasia
Эзио Хиго (Ezio Ghigo, MD)
Professor of Endocrinology, Department
of Internal Medicine, University of Turin
Faculty of Medicine; Chief, Division
of Endocrinology and Metabolism, University
Hospital, Turin, Italy
Hormones and Athletic Performance
Питер A. Готлиб (Peter A. Gottlieb, MD)
Associate Professor of Pediatrics and
Medicine, University of Colorado School
of Medicine, Aurora; Physician, University
of Colorado Hospital and the Children’s
Hospital, Denver, Colorado
The Immunoendocrinopathy Syndromes
Стивен К. Гринспун (Steven К.
Grinspoon, MD)
Associate Professor of Medicine, Harvard
Medical School: Director, MGH Program
in Nutritional Metabolism, and Clinical
Director, Neuroendocrine Clinical Center,
Massachusetts General Hospital, Boston,
Massachusetts
Endocrinology of HIV/AIDS
Мелвин M. Грумбач (Melvin M.
Grumbach, MD)
Edward B. Shaw Emeritus Professor
of Pediatrics and Emmtus Chairman,
Department of Pediatrics, University
of California, San Francisco, California
Puberty: Ontogeny, Neuroendocrinology,
Physiology, and Disorders
10
Джоэл Ф. Хабенер (Joel F, Habener, MD)
Professor of Medicine, Harvard Medical
School: Associate Physician, Massachusetts
General Hospital, Boston, Massachusetts
Genetic Control of Peptide Hormone
Formation
Ян Д. Хей [Ian D. Hay, BSc, MBChB, phD,
MRCP(UK), FACE, FACP, FRCP (Edin
Lond), FRCPl (Hon), FRCPS (Glasg)]
Richard F. Emslander Professor in
Endocrinology and Nutrition Research
and Professor of Medicine, Mayo Clinic
College of Medicine Consultant, Division
of Endocrinology, Metabolism, Nutrition &
Internal Medicine, Mayo Clinic, Rochester,
Minnesota Thyroid Physiology and Diagnostic
Evaluation of Patients with Thyroid Disorders
Nontoxic Diffuse and Nodular Goiter
and Thyroid Neoplasia
Иеуан A. Xarxec (leuan A. Hughes, MD,
FRCP, FRCPCh, FMedSci)
Professor of Paediatrics and Head,
Department of Paediatrics, University
of Cambridge;
Addenbrooke’s Hospital, Cambridge, United
Kingdom Disorders of Sex Development
Джорж Дж. 1Слии (George G. Klee, MD,
PhD)
Professor of Laboratory Medicine, Mayo
Medical School; Consultant, Mayo Clinic,
Rochester, Minnesota
Laboratory Techniques for Recognition
of Endocrine Disorders
Сэмюэль Клейн (Samuel Klein, MD)
William H. Danforth Professor of Medicine
and Nutritional Science, Washin^on
University in St. Louis, School of Medicine;
Attending Physician, Barnes-Jewish Hospital.
St. Louis, Missouri Obesity
Дэвид Клейнберг (David Kleinberg, MD)
Professor of Medicine, Director,
Neuroendocrine Unit, New York University
School of Medicine; Chief of Endocrinology.
Veterans Administration Medical Center, New
York, New York
Anterior Pituitary
Генри M. Кроненберг (Henry M.
Kronenberg, MD)
Professor of Medicine, Harvard Medical
School; Chief, Endocrine Unit, Massachusetts
General Hospital. Boston, Massachusetts
Principles of Endocrinology
Hormones and Disorders of Mineral
Metabolism
Стивен В. Дж. Ламберте (Steven W J.
Lamberts, MD, PhD)
Professor of Medicine, Erasmus University
Faculty of Medicine; Endocrinologist, Erasmus
Medical Center. Rotterdam, the Netherlands
Endocrinology and Aging
Фабио Ланфранко (Fabio Lanfranco, MD)
Assistant Professor of Endocrinology,
Department of Internal Medicine, University
of Turin Faculty of Medicine; Clinical Practice
in Endocrinology, University Hospital,
Turin, Italy
Hormones and Athletic Perfonnance
П. Рид Ларсен (P. Reed Larsen, MD,
FACP, FRCP)
Professor of Medicine, Harvard Medical
School: Chief, Division of Endocrinology,
Diabetes, and Hypertension, Brigham
and Women's Hospital, Boston, Massachusetts
Principles of Endocrinology
Thyroid Physiology and Diagnostic Evaluation
of Patients with Thyroid Disorders
Thyrotoxicosis
Hypothyroidism and Thyroiditis
Митчелл A, Лазар (Mitchell A. Lazar,
MD, PhD)
Sylvan H. Eisman Professor of Medicine
and Professor of Genetics; Chief, Division
of Endocrinology, Diabetes, and Metabolism;
and Director, Institute for Diabetes, Obesity,
and Metabolism, University of Pennsylvania
School of Medicine, Philadelphia,
Pennsylvania
Mechanism of Action of Honnones That Act
on Nuclear Receptors
Джозеф A. Лоренсо 0oseph A, Lorenzo,
MD)
Professor of Medicine, University
of Connecticut School of Medicine: Director,
Bone Biology
Research, University of Connecticut Health
Center; Attending Physician, John Dempsey
Hospital, Farmington, Connecticut
Metabolic Bone Disease
Малколм Дж. Лоу (Malcolm J. Low, MD,
PhD)
Senior Scientist and Associate Director, Center
for the Study of Weight Regulation: Professor
of Behavioral Neuroscience, School
of Medicine; Affiliate Scientist, Vollum
Institute, Oregon
11
Health & Science University, Portland, Oregon
Neuroendocrinology
Роберт в. Махлей (Robert W. Mahley,
MD, PhD)
President, the J. David Gladstone Institutes,
Senior Investigator, Gladstone Institute
of Cardiovascular Disease: Professor
of Pathology and Medicine, University
of California, San Francisco, California
Disorders of Lipid Metabolism
Стефен Дж. Маркс (Stephen J. Marx, MD)
Chief, Metabolic Diseases Branch, Genetics
and Endocrinology Section, National Institute
of Diabetes and Digestive and Kidney
Diseases, National Institutes of Health,
Bethesda, Maryland
Multiple Endocrine Neoplasia
Шломо Мелмед (Shiomo Melmed, MD,
FRCP)
Helene and Philip Hlxon Chair in Investigative
Medicine, Senior Vice President for Academic
Affairs, Cedar'S Sinai Medical Center, Los
Angeles, California
Principles of Endocrinology
Anterior Pituitary
Ребека Д. Монк (Rebeca D. Monk, MD)
Associate Professor of Medicine, University
of Rochester School of Medicine and Dentistry,
Rochester, New York
Kidney Stones
Ричард В. Несто (Richard W. Nesto, MD)
Associate Professor of Medicine, Harvard
Medical School, Boston; Chair, Department
of Cardiovascular Medicine, Lahey Clinic
Medical Center, Burlington, Massachusetts
Complications of Diabetes Mellitus
Кджел И. Йоберг (Kjell E. Oberg, MD,
PhD)
Professor, Department of Medical Sciences,
and Dean, Medical Faculty, Uppsala
University:
Attending, Department of Endocrine
Oncology, University Hospital, Uppsala,
Sweden
Carcinoid Tumors, Carcinoid Syndrome, and
Related Disorders
Кеннет С. Полонски (Kenneth S.
Polonsky, MD)
Adolphus Busch Professor and Chairman,
Department of Medicine, Washington
University in St. Louis, School of Medicine,
St, Louis, Missouri
Principles of Endocrinology
Type 2 Diabetes Mellitus
Type 1 Diabetes Mellitus
Лоуренс Г. Райз (Lawrence G. Raisz, MD)
Board of Trustees Distinguished Professor
of Medicine, University of Connecticut
School of Medicine; Physician, John Dempsey
Hospital, Farmington, Connecticut
Metabolic Bone Disease
Эдвард 0. Рейтер (Edward О. Reiter,
MD)
Professor of Pediatrics, Tufts University
School of Medicine, Boston; Chair.
Department of Pediatrics, Baystate Children’s
Hospital, Springfield, Massachusetts
Normal and Aberrant Growth
Алан Г. Робинсон (Alan G. Robinson,
MD)
Professor of Medicine and Executive Associate
Dean, David Geffen School of Medicine
at UCLA; Associate Vice Chancellor, Medical
Sciences, University of California at Los
Angeles,
Los Angeles, California
Posterior Pituitary
Джоанес A. Ромиджн (Johannes A.
Romijn, MD, PhD)
Professor, Department of Endocrinology,
Leiden University Medical School; Physician,
Department of Endocrinology, Leiden
University Medical Center, Leiden, the
Netherlands
Obesity
PoH Г. Посенфилд (Ron G. Rosenfeld, MD)
Professor of Pediatrics, Oregon Health
& Science University School of Medicine,
Portland,
Oregon, and Stanford University School
of Medicine, Stanford, California; Senior Vice-
President for Medical Affairs, Lucile Packard
Foundation for Children’s Health, Palo Alto,
California
Normal and Aberrant Growth
Ричард Сантен (Richard Santen, MD)
Professor of Medicine, University of Virginia
School of Medicine; Associate Director,
Clinical Research, Division of Endocrinology
and Metabolism, University of Virginia Health
System, Charlottesville, Virginia
Endocrine-Responsive Cancer
12
Мартин-Джин Шлумбергер (Martin-
Jeaiï Schlumberger, MD)
Professor of Oncology, University of Paris,
Paris SWD; Chairman, Department of Nuclear
Medicine and Endocrine Oncology, Institute
Gustave Roussy, Villejuif, France
Thyroid Physiology and Diagnostic Evaluation
of Patients with Thyroid Disorders
Nontoxic Diffuse and Nodular Goiter
and Thyroid Neoplasia
Алан Спигел (Allen Spiegel, MD)
Marilyn and Stanley M. Katz Dean, Albert
Einstein College of Medicine, Bronx, New
York
Mechanism of Action of Hormones that Act
at the Cell Surface
Пол M. Стьюард (Paul M. Stewart, MD,
FRCP. FMedSci)
Professor of Medicine, the University
of Birmingham Institute of Biomedical
Research:
Honorary Consultant Physician, Department
of Medicine, University Hospital Birmingham
Foundation, NHS IVust, Birmingham, United
Kingdom
The Adrenal Cortex
Кристиан Дж. Страсбургер (Christian J.
Strasburger, MD)
Professor of Clinical Endocrinology
and Division Chief, Department of Medicine,
Charité
Universitatsmedizin Berlin, Berlin, Germany
Hormones and Athletic Performance
Гордон Дж. Стревлер (Gordon J.
Strewler, MD)
Vice-Chairman, Department of Medicine,
Beth Israel Deaconess Medical Center;
Professor of Medicine; Master, Walter
Bradford Cannon Society, Harvard Medical
School, Boston,
Massachusetts
Humoral Manifestations of Malignancy
Дэннис M, Стайн (Dennis M. Styne, MD)
Rumsey Chair of Pediatric Endocrinology,
Professor of Pediatrics, University
of California, Davis School of Medicine,
Sacramento, California
Puberty; Ontogeny, Neuroendocrinology,
Physiology, and Disorders
Симеон И. Тейлор (Simeon I, Taylor, Md,
PhD)
Vice President, Discovery Biology,
Pharmaceutical Research Institute, Bristol-
Myers Squibb, Hopewell, New Jersey
Mechanism of Action of Hormones that Act
at the Cell Surface
Джозеф Г. Вербалис (Joseph G. Verbalis,
MD)
Professor of Medicine and Physiology
and Interim Chair, Department of Medicine,
Georgetown University School of Medicine;
Physician-in-Chief of Medicine, Georgetown
University Hospital, Washington, DC
Posterior Pituitary
AapoH И. Виник (Aaron I. Vinik, MD,
PhD)
Professor, Internal Medicine, Pathology,
Neurobiology, Eastern Virginia Medical
School;
Director, Strelitz Diabetes Research Institutes,
Norfolk, Virginia
Complications of Diabetes Mellitus
Карл X. Вейсграбер (Karl H, Weisgraber,
PhD)
Professor of Pathology, University
of California, San Francisco, School
of Medicine; Senior Investigator, Gladstone
Institute of Neurological Disease,
San Francisco, California
Disorders of Lipid Metabolism
Вильям Ф. Йонг (William F, Young, Jr.,
MD, MSc)
Professor of Medicine, Mayo Clinic College
of Medicine; Consultant, Division
of Endocrinology, Diabetes, Metabolism,
and Nutrition, Mayo Clinic, Rochester,
Minnesota
Endocrine Hypertension
I щш
* — обозначение торгового наименования лекарственного средства
^ — обозначение не зарегистрированного в России лекарственного средства
® — обозначение препаратов, аннулированных из Государственного реестра ЛС
АКТГ — адренокортикотропный гормон
ВДКН — врожденная дисфункция коры надпочечников
ВИП — вазоактивный интестинальный полипептид
ВИЧ — вирус иммунодефицита человека
ВМС — внутриматочные средства
ГнРГ — гонадотропин-рилизинг-гормон
ДГТ — дигидротестосгерон
ДГЭА — дегидроэпиандростерон
ДГЭАС — дегидроэпиандростерона сульфат
ДМПА — депо-медроксипрогестерона ацетат
ЖКТ — желудочно-кишечный тракт
ЗГТ - заместительная гормональная терапия
ЗППП — заболевания, передающиеся половым путем
ИБС — ишемическая болезнь сердца
ИГГ — идиопатический гипогонадотропный гипогонадизм
ИФР — инсулиноподобные факторы роста
КГРП — кальцитонин-генродственный пептид
КТ — компьютерная томография
КЭ — конъюгарованные эстрогены
ЛГ — лютеинизирующий гормон
ЛПВП — липопротеины высокой плотности
ЛПНП — липопротеины низкой плотности
МПА — медроксипрогестерона ацетат
МРТ — магнитно-резонансная томография
НПВС — нестероидные противовоспалительные средства
ОК — оральные контрацептивы
ПИЯ - преждевременное истощение яичников
ПР — рецепторы к прогестерону
ПСА — простатоспецифический антиген
ПСПС — протеин, связывающий половые стероиды
ПЦР — полимеразная цепная реакция
РКИ — рандомизированные контролируемые исследования
СГЯ — синдром гиперстимуляции яичников
СД - сахарный диабет
СМЭР — селективный модулятор эстрогеновых рецепторов
СПИД — синдром приобретенного иммунодефицита
СПКЯ — синдром поликистозных яичников
ССЗ — сердечно-сосудистые заболевания
Т — тестостерон
ТТГ — тиреотропный гормон
ТЭЛА — тромбоэмболия легочной артерии
УЗИ — ультразвуковое исследование
ФДЭ — циклические нуклеотидфосфодиэстеразы
ФДЭ-5 — фосфодиэстераза-5
ФСГ — фолликулостимулирующий гормон
ХГЧ — хорионический гонадотропин
цАМФ — циклический адснозинмонофосфат
14
ЦНС — центральная нервная система
ЧМГ — человеческий менопаузальный гонадотропин
ЭД — эректильная дисфункция
ЭКО — экстракорпоральное оплодотворение
HERS — сердце и заместительная гормональная терапия эстроген/гестагенами
ICSI — внутрицитоплазматическая инъекция сперматозоидов
LRH-1 — гомологичный рецептор печени 1
SF-1 — стероидогенный фактор-1
StAR — белок острой регуляции стероидогенеза
WHI — Инициатива по охране здоровья женщин
лава
Физиология и патология
репродуктивной системы женщины
ФИЗИОЛОГИЯ РЕПРОДУКЦИИ
Точно скоординированное взаимодействие гипоталамуса, гипофиза, яичников и
эндометрия обеспечивает регулярность менструаций, что, в свою очередь, позволяет
судить о регулярной овуляции. Также необходимое условие регулярной овуляции —
нормальное функционирование щитовидной железы и надпочечников. Например,
пациентки с гипертиреозом или гипотиреозом, синдромом Кушинга или резистентно¬
стью к глюкокортикоидам могут страдать ановуляцией. Именно поэтому для точной
диагностики и выработки правильной стратегии лечения врачам важно иметь четкое
представление как о функционировании гипоталамуса, гипофиза, яичников и матки,
так и о взаимодействии их с другими органами и системами.
Наиболее важная функция гипоталамуса — пульсирующая секреция гонадотропин-
рилизинг-гормона (ГнРГ), Обратная отрицательная связь определяется рядом факто¬
ров, включая гормоны яичников, которые регулируют секрецию гипоталамусом ГнРГ
в портальные сосуды. Допамин, норадреналин, серотонин, опиоиды, продуцируемые
в головном мозге, могут ретушировать секрецию ГйРГ через яичниковые гормоны
или другие сигналы. В ответ на активость ГнРГ клетки передней доли гипофиза
секретируют фолликулостимулирующий гормон (ФСГ) и лютеинизирующий гормон
(ЛГ). Стероиды (эстрадиол и прогестерон) и пептиды (ингибин) яичникового проис¬
хождения, а также активин или фоллистатин гипофизарного происхождения влияют
на секрецию ФСГ и ЛГ. ЛГ стимулирует продукцию андростендиона в тека-клетках
яичника, тогда как ФСГ регулирует продукцию эстрадиола и ингибина В гранулезны¬
ми клетками, а также рост фолликулов. Высвобождение ооцита из зрелого фолликула
зависит от повышения уровня ЛГ в середине цикла. После овуляции фолликул транс¬
формируется в желтое тело, которое секретирует вместе эстрадиол и прогестерон под
контролем ФСГ и ЛГ. ЛГ также стимулирует лютеинизированные гранулезные клетки
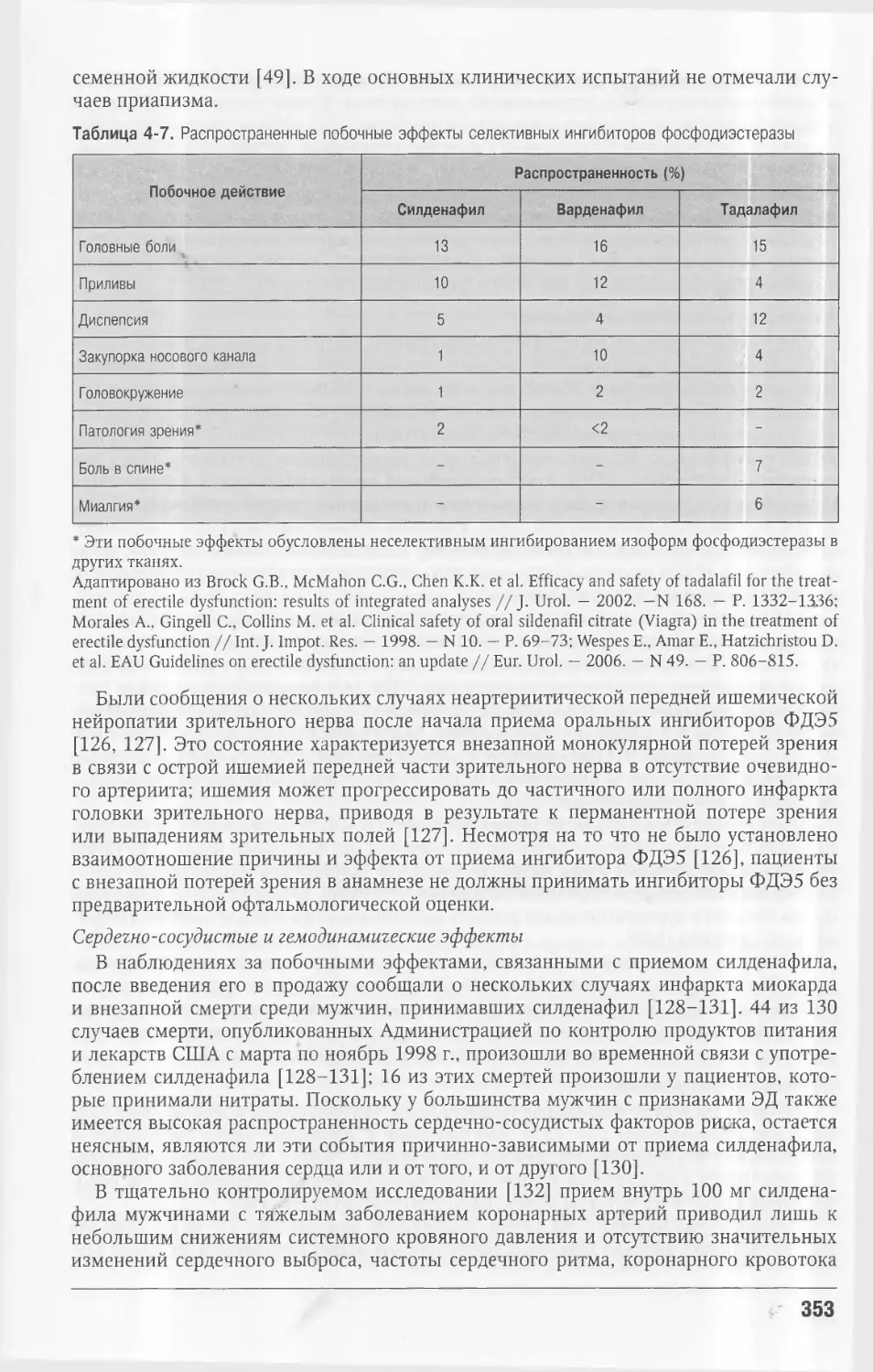
желтого тела секретировать ингибин А (рис. 1-1, А).
Эндокринные эффекты ФСГ, ЛГ, эстрадиола, прогестерона, ингибина А, ингиби¬
на В показаны в связи с изменениями уровня гормонов в сыворотке крови в течение
менструального цикла (см. рис. 1-1, А). Данные эндокринные эффекты позже были
продемонстрированы в клеточных исследованиях и исследованиях in vivo (рис. 1-1,
Б). Активин и фоллистатин синтезируются как в яичнике, так и в гипофизе. Эти
гормоны влияют на деятельность гипофиза через аутокринные и паракринные, но
не через эндокринные механизмы. Активин стимулирует продукцию ФСГ, тогда как
фоллистатин подавляет этот эффект активина.
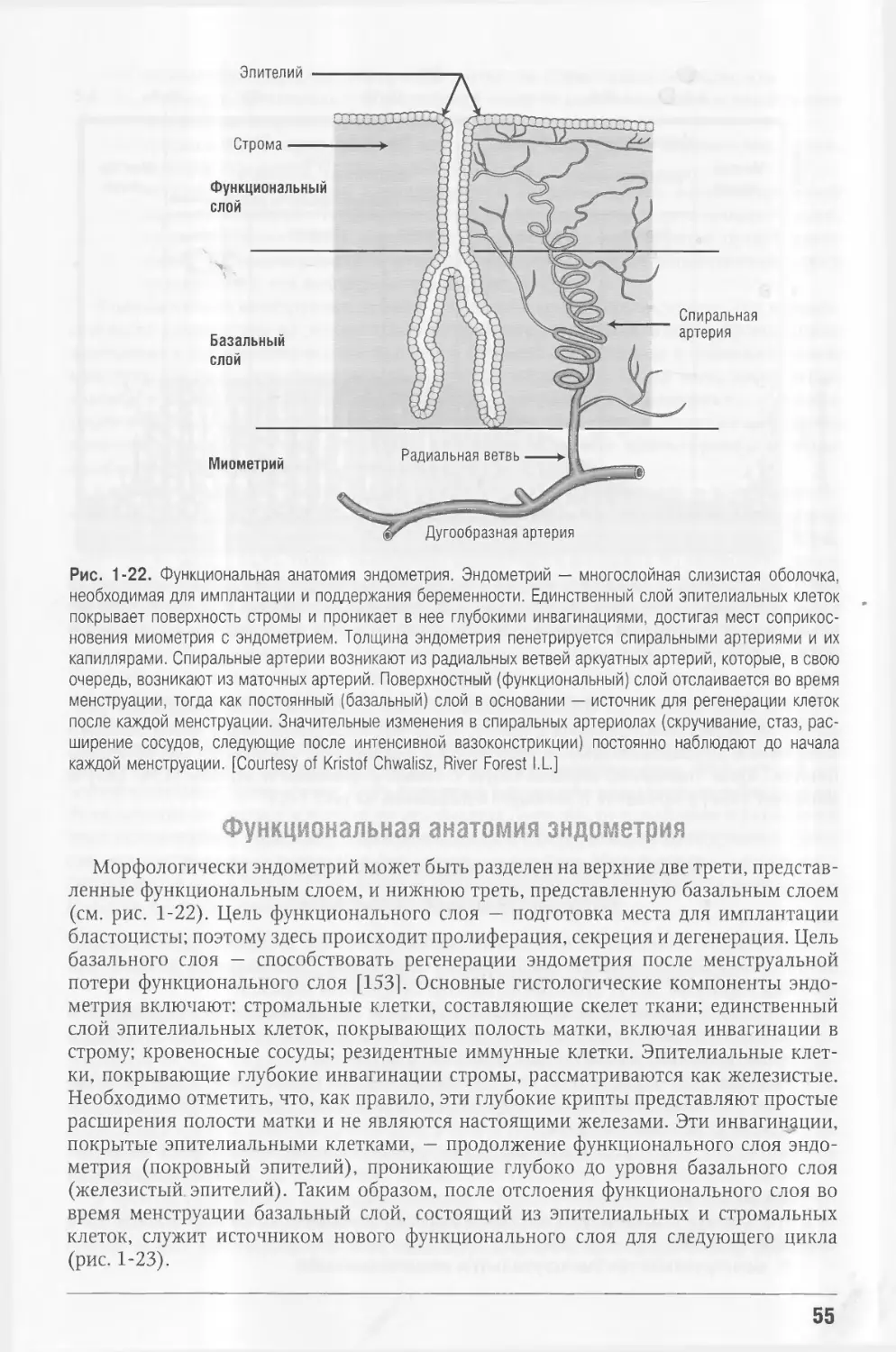
Эндометрий — слизистая оболочка полости матки; содержит вьгсокую концентра¬
цию рецепторов к эстрогенам и прогестерону и является чрезвычайно чувствитель¬
ной к этим гормонам. Биологачески активный гормон эстрогенового ряда эстрадиол
стимулирует рост эндометрия, а прогестерон контролирует этот эффект и усиливает
Фолликулярная фаза
Овуляция
Лютеиновая фаза
Пролиферативная фаза
Секреторная фаза
Менструации
Ингибин В
Лютеини¬
зирующий
гормон
^Фолликул
''стимулирующий
гормон
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28
Дни
Рис. 1-1. А — изменения в фоллику¬
ле яичника, толщине эндометрия, а
также уровене гормонов в сыворотке
в течение 28-днёвного менструально¬
го цикла; Б — эндокринные взаимо¬
действия в женской репродуктивной
системе. На рисунке изображены неко¬
торые хорошо известные эндокринные
взаимодействия между гипоталамусом,
гипофизом, яичниками и эндометрием,
характеризующие регуляцию менстру¬
ального цикла.
18
дифференцировку. Отслоение функционального слоя эндометрия следует за падением
уровня эстрогена или прогестерона. Остающийся базальный слой способен полно¬
стью регенерировать в ответ на новую стимуляцию эстрогеном.
Яичники не функционируют до начала пубертата, поскольку гипоталамус является
незрелым в препубертатном возрасте, и ФСГ и ЛГ не стимулируют яичники. После
менопаузы репродуктивная функция целиком и эндокринная функция в частности
прекращаются, поскольку яичники теряют все ооциты и окружающие клетки, отве¬
чающие за стероидогенез. Таким образом, препубертатный и постменопаузальный
периоды характеризуются отсутствием функции яичников и соответственно отсут¬
ствием менструаций.
Репродуктивную систему женщины от препубертата до менопаузы можно сравнить
с очень точными часами. Нормальное функционирование этого механизма зависит
от скоординированных взаимодействий гипоталамуса, гипофиза, яичников и эндоме¬
трия. Результат данного взаимодействия — регулярные менструации каждые 24-35
дней. Любое заболевание тканей и органов репродуктивной системы может вызвать
ановуляцию и нерегулярные маточные кровотечения.
ГнРГ — пептид, содержащий 10 аминокислот, который синтезируется в специ¬
альных нейрональных тельцах аркуатных ядер медиобазального гипоталамуса [1].
Аксоны от ГйРГ-нейронов достигают срединное возвышение и заканчиваются в
капиллярах, которые впадают в портальные сосуды.
Портальная вена представляет собой медленную транспортную систему, соеди¬
няющую гипоталамус с передней долей гипофиза. Направление тока крови — от
гипоталамуса к гипофизу. Таким образом, ГнРГ, синтезируемый в ядрах срединного
возвышения гипоталамуса, секретируется в портальный кровоток, который доносит
гормон к передней доле гипофиза (рис. 1-2) [2] . Зрелый декапептид ГнРГ получается
из пре-про-ГнРГ, подвергшегося посттрансляционному процессингу (см. рис. 1-2)
[3], Эта молекула-предшественница является продуктом функционирования гена,
локализованного в коротком плече хромосомы 8 [4]. Пре-про-ГнРГ содержит 92
аминокислоты и состоит из четырех частей (от N до С окончания): (1) — домен,
состоящий из 23 аминокислот [5], (2) — декапептид ГнРГ, (3) — аминокислотная
протеолитическая процессинговая часть и (4) — домен, содержащий 56 аминокислот,
называемый ГнРГ-ассоциированным пептидом [2, 6]. Продукты расщепления этого
предшественника ГнРГ и ГнРГ-ассоциированный пептид трнспортируются в нервное
окончание и секретируются в портальный кровоток (см. рис. 1-2) [3, 7, 8]. Роль ГнРГ-
ассоциированного пептида до конца не определена [7].
У человека нейроны ГнРГ раполагаются преимущественно в аркуатных ядрах
медиобазального гипоталамуса и преоптической области переднего гипоталамуса.
Количество нейронов ГнРГ относительно лимитировано и находится в пределах
1000-3000. Нейроны, продуцирующие ГнРГ, происходят из обонятельной области
во время эмбриогенеза. Эти клетки мигрируют вдоль черепных нервов, соединяю¬
щих нос и передний мозг с гипоталамусом. Нейрональный гликопротеин, распола¬
гающийся на поверхности клеток, вероятно медиирующий адгезию клетки к клетке,
является определяющим фактором миграции. Мутация в гене, который кодирует эту
миграционную молекулу, ассоциирована с изолированным дефицитом гонадотропи¬
на, связанного с Х-хромосомой (синдром Калльмана) и проявляющегося в виде поте¬
ри обоняния, дефицитом ГнРГ и гипогонадотропным гипогонадизмом [9].
19
-23 I
Сигнальный пептид
С
1
8 9 10
Глу Гис Три Сер Три Гли Лей Apr Про Гли
Гли
Гонадолиберин (10 аминокислот)
(
Г онадолиберин-ассоциированный
протеин (56 аминокислот)
J[ +92
Рис. 1-2. Продукция гонадотропин-рилизинг-гормона (ГнРГ). А — ген ГнРГ кодирует предшественник
белка (пре-про-ГнРГ) в нейрональных тельцах. Внутри нейронального тельца происходит синтез ГнРГ и
ГнРГ-ассоциированного протеина путем протеолитического процессинга [2]. И ГнРГ, и ассоциирован¬
ный протеин транспортируются по аксону в терминальное окончание, откуда происходит их секреция
в портальный кровоток; Б — пре-про-ГнРГ — 92-компонентный аминопротеин. Биологически активный
декапептид {аминокислоты 1-10 расположены между 23-аминокислотным сигнальным белком и после¬
довательностью Гли-Арг-Лиз). Стрелкой указано место протеолитического процессинга. Для синтеза
ГнРГ-ассоциированного протеина С-окончание 5б-аминокислотного белка расщепляется. {Yen S.S.C.
Endocrine regulation of the reproductive system // Yen S.S.C., Jaffe R.B., Barbieri R.L (eds). Reproductive
Endocrinology. - 4th ed. - Philadelphia: WB Saunders. - 1999. - P. 44.]
20
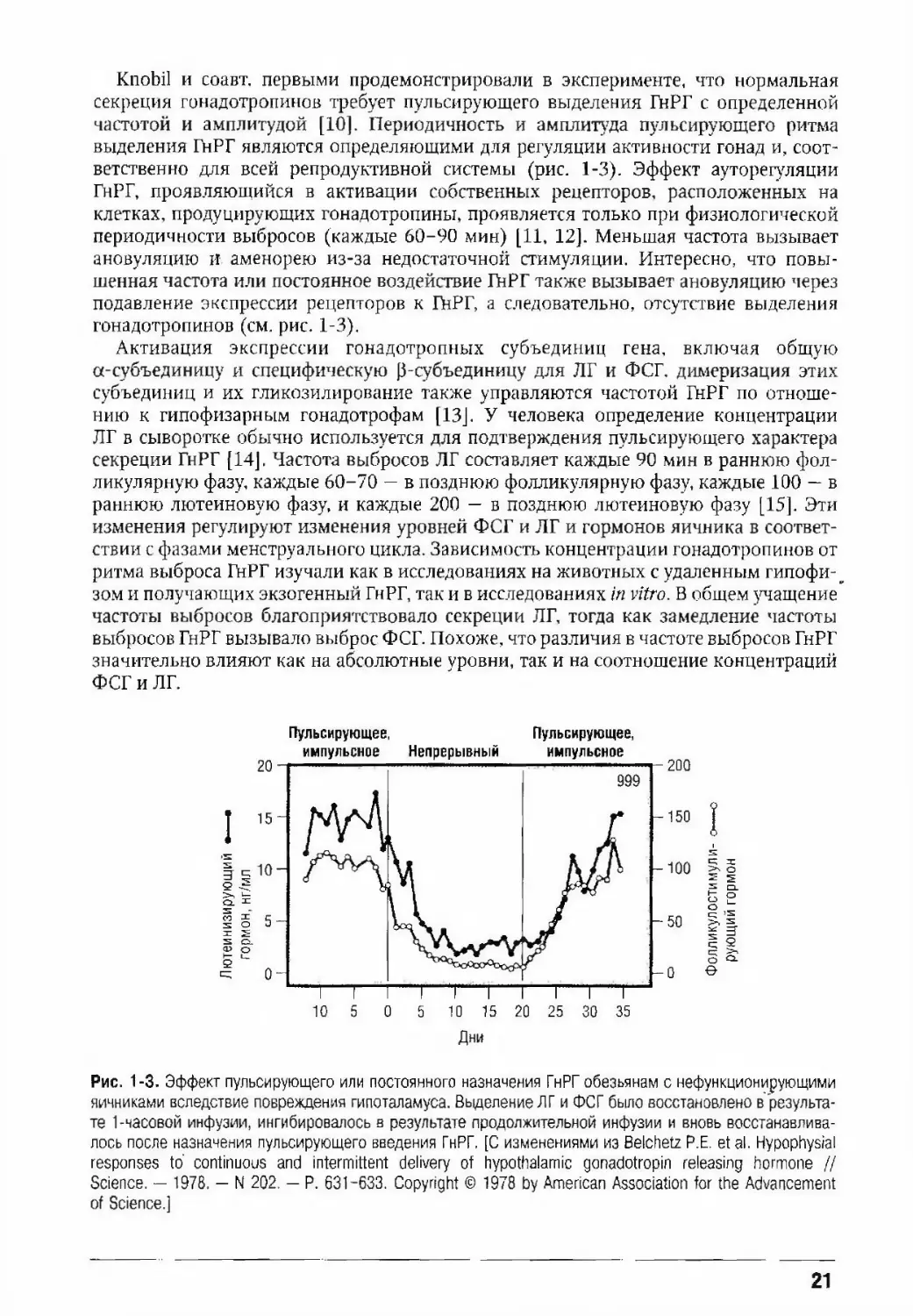
Knobil и соавт. первыми продемонстрировали в эксперименте, что нормальная
секреция гонадотропинов требует пульсирующего выделения ГнРГ с определенной
частотой и амплитудой [10]. Периодичность и амплитуда пульсирующего ритма
выделения ГнРГ являются определяющими для регуляции активности гонад и, соот¬
ветственно для всей репродуктивной системы (рис. 1-3). эффект ауторегуляции
ГнРГ, проявляющийся в активации собственных рецепторов, расположенных на
клетках, продуцирующих гонадотропины, проявляется только при физиологической
периодичности выбросов (каждые 60-90 мин) [11, 12]. Меньшая частота вызывает
ановуляцию и аменорею из-за недостаточной стимуляции. Интересно, что повы¬
шенная частота или постоянное воздействие ГнРГ также вызывает ановуляцию через
подавление экспрессии рецепторов к ГйРГ, а следовательно, отсутствие выделения
гонадотропинов (см. рис. 1-3).
Активация экспрессии гонадотропных субъединиц гена, включая общую
а-субъединицу и специфическую р-субъединицу для л Г и ФСГ, димеризация этих
субъединиц и их гликозилирование также управляются частотой ГнРГ по отноше¬
нию к гипофизарным гонадотрофам [13]. У человека определение концентрации
ЛГ в сыворотке обычно используется для подтверждения пульсирующего характера
секреции ГнРГ [14]. Частота выбросов ЛГ составляет каждые 90 мин в раннюю фол¬
ликулярную фазу, каждые 60-70 — в позднюю фолликулярную фазу, каждые 100 — в
раннюю лютеиновую фазу, и каждые 200 — в позднюю лютеиновую фазу [15]. Эти
изменения регулируют изменения уровней ФСГ и ЛГ и гормонов яичника в соответ¬
ствии с фазами менструального цикла. Зависимость концентрации гонадотропинов от
ритма выброса ГнРГ изучали как в исследованиях на животных с удаленным гипофи-^
зом и получающих экзогенный ГнРГ, так и в исследованиях in vitro. В общем учащение
частоты выбросов благоприятствовало секреции ЛГ, тогда как замедление частоты
выбросов ГнРГ вызывало выброс ФСГ. Похоже, что различия в частоте выбросов ГнРГ
значительно влияют как на абсолютные уровни, так и на соотношение концентраций
ФСГ и ЛГ.
Пульсирующее.
Пульсирующее,
II
ье
=1
si
о ^
е
Дни
Рис. 1-3. Эффект пульсирующего или постоянного назначения ГнРГ обезьянам с нефункционирующими
яичниками вследствие повреждения гипоталамуса. Выделение ЛГ и ФСГ было восстановлено в результа¬
те 1-часовой инфуэии, ингибировалось а результате продолжительной инфузии и вновь восстанавлива¬
лось после назначения пульсирующего введения ГнРГ, [С изменениями из Belchetz Р.Е. et а1. Hypophysial
responses to' continuous and intermittent delivery of hypothalamic gonadotropin releasing hormone //
Science. — 1978. — N 202. — P. 631-633. Copyright © 1978 by American Association for the Advancement
of Science.]
21
Регуляция секреции ГнРГ
Для регулярных менструаций необходима пульсирующая секреция ГнРГ с харак¬
теристиками, описанными выше. Способность к пульсирующим ритмичным выбро¬
сам — специфическое свойство ГнРГ-нейронов, на которое влияют различные гормо¬
ны и нейротрансмиттеры (рис. 1-4).
Вариации частоты выброса ГнРГ достигаются отчасти посредством обратной связи,
осуществляемой гонадными стероидами. Эстрадиол увеличивает частоту выбросов
ГнРГ, тогда как увеличение концентрации прогестерона снижает частоту выбросов
[13]. В связи с этим возможно, что повышенный уровень прогестерона может вызвать
уменьшение частоты выбросов ГнРГ и поэтому приводить к преимущественному син¬
тезу и секреции ФСГ, которые наблюдаются в поздней лютеиновой фазе [13].
Уровень эстрогенов, улавливаемый ГнРГ-нейронами, критичен для подавления как
ФСГ, так и ЛГ, а также для координации преовуляторного выброса ЛГ. ГнРГ-нейроны
регулируются напрямую эстрадиолом преимущественно через (3-рецепторы и опосре¬
дованно через эстрадиолчувствительные афферентные нейроны. Прямая стимуляция
ГнРГ-нейронов подавляет экспрессию ГнРГ, но в организме (ш vivo) эта депрессия в
скором времени перекрывается опосредованными эстрадиолзависимыми сигнала¬
ми от афферентных нейронов. Таким образом, эстрадиол может ингибировать или
стимулировать секрецию ГнРГ и координировать как созревание фолликулов, так и
преовуляторный выброс ГнРГ [16].
На выбросы ГнРГ также влияют местные нейротрансмиттеры. Норадреналин сти¬
мулирует выброс ГнРГ, тогда как допамин демонстрирует супрессирующий эффект (см.
рис. 1-4) [17]. р-Эндорфины и другие опиоиды также могут подавлять выброс гипота¬
ламусом ГнРГ [18,19]. Стероидные гормоны яичников модулируют активность опиои-
дов, поэтому обратное отрицательное влияние стероидных гормонов на гонадотропины
частично реализуется через эндогенные опиоиды [20]. В связи с этим предполагают, что
половые стероиды усиливают активность эндогенных опиоидов, что последовательно
ведет к ингибирующему эффекту на секрецию ГнРГ [21]. Негативный эффект опиоидов
на. секрецию ГнРГ также можно объяснить клинически, так как сниженная секреция
Рис. 1-4. Регуляция секреции
ГнРГ, ФСГ и Л Г. Ритмичную
секрецию ГнРГ в портальный кро¬
воток регулируют локальные и
системные гормоны. В свою оче¬
редь, ГнРГ вместе с некоторыми
стероидными и пептидными гор¬
монами регулирует синтез а- и
|3-субъединиц гонадотропина, а
также формирование и секрецию
ФСГ и ЛГ.
22
ГнРГ, связанная с гипоталамической аменореей, может быть медиирована повышением
до ингабирующего уровня эндогенного опиоида [22].
Кисспептины, кодируемые геном Kissl, и их рецептор GPR54 играют ключевую
роль в регуляции секреции ГнРГ [23]. Мутации или целевое выключение GPR54
вызывают изолированный гипогонадотропный гипогонадизм у людей и мышей, что
подтверждает, что сигнал, передаваемый посредством этого рецептора, необходим
для полового развития и функционирования [24,25]. Кисспептины стимулируют ГнРГ
и гонадотропную секрецию у препубертатных и взрослых животных [26], Более того,
нейроны, экспрессирующие кисспептин и GPR54, являются целями для негативной и
позитивной обратной связи половых гормонов [23].
АНАЛОГИ ГОНАДОТРОЛИН-РИЛИЗИНГ-ГОРМОНА
Время полувыведения ГнРГ незначительное (2-4 мин), поскольку он быстро раз¬
рушается пептидазами в гипоталамусе и гипофизе [27]. Эти пептидазы расщепляют
связи между следующими аминокислотами: 5 и 6, 6 и 7, 9 и 10. Аналоги ГнРГ были
синтезированы путем изменения аминокислот в этих позициях. До сих пор произво¬
дят множество агонистов и антагонистов аналогов ГнРГ с различными биологически¬
ми свойствами.
АГОНИСТЫ ГОНАДОТРОПИН-РИЛИЗИНГ-ГОРМОНА
Несколько агонистов ГнРГ были созданы щт-ем замещения аминокислот в 6-й и
10-й позиции. Высокая биологическая активность агонистов ГнРГ обусловливается
высоким сродством к рецепторам ГнРГ и пониженной восприимчивостью к фермента¬
тивной деградации. Аминокислотное замещение в позиции 6 усиливает метаболиче¬
скую стабильность, а замещение глицинами да в С-окончании на эти л амидную группу
значительно усиливает сродство к рецепторам [27-29]. Агонисты ГйРГ назначают
подкожно, интраназально или внутримышечно. Первичный эффект агонистов (так
называемый эффект вспышки) связан с увеличением концентрации ЛГ и ФСГ.
Наиболее выраженный агонистический ответ наблюдают во время ранней фолли¬
кулярной фазы, когда сочетание эффекта агониста ГнРГ и повышенного уровня эстра¬
диола создает значительный резервный пул гонадотропинов [30]. Десенситизация
и подавление гипофиза вызывают гипогонадотропный гипогонадизм. Назначение
длительно действующей формы депо-агониста ГнРГ приводит к подавлению
гонадотропин-гонадной оси в течение 1-3 нед. Первичный подавляющий эффект
связан с десенситизацией, тогда как продолжительный ответ связан с потерей рецеп¬
торов и их отщеплением от эффекторных систем, к тому же агонисты могут вызывать
биологическое состояние покоя яичников через секрецию биологически неактивных
гонадотропинов.
До сих пор PDA одобряло использование агонистов ГнРГ для лечения зависимых от
ГнРГ преждевременного полового созревания, эндометриоза, а также рака простаты.
Другим показанием было улучшение гематологических показателей у пациенток с
анемией, развившейся в связи с миомой матки. Показаниями для назначения аго¬
нистов ГнРГ, не указанными в инструкции по применению (Off-label), считают пода¬
вление функции гипофиза во время индукции овуляции, атрофию эндометрия перед
хирургической аблацией эндометрия, а также предотвращение кровотечения у паци¬
енток с дефектами гемостаза. Кроме того, агонисты ГнРГ используют для подавления
стероидогенеза в яичниках при гирсутизме [31].
Наиболее выраженный побочный эффект, который вызван длительным примене¬
нием депо-формы агониста ГнРГ, вызван возникновением эстрогенного дефицита.
Использование депо вызывает состояние, напоминающее перименопаузу: приливы,
сухость влагалища, резорбцию кости, остеопению. Остеопения обратима у молодых
женщин, получающих лечение депо-формой не более 6 мес [32. 33], поэтому перед
23
длительным назначением агонистов ГнРГ необходимо взвесить все за и против.
Дополнительное назначение низких доз эстрогенов, гестагенов или обоих совместно с
агонистами ГнРГ возможно для преодоления побочных эффектов [34].
АНТАГОНИСТЫ ГОНАДОТРОПИН-РИЛИЗИНГ-ГОРМОНА
Путем замещения нескольких аминокислот были синтезированы антагонисты
ГйРГ. Для этого модифицировали пироглютаминовые и глициновые окончания в
положениях 1 и 10 или удалили либо заменили гидрофобные аминокислоты в поло¬
жениях 2 и 3 [29]. Антагонисты ГнРГ связываются с рецепторами ГнРГ, тем самым
вызывая их конкурентное ингибирование. Антагонисты имеют преимущество в виде
моментального снижения уровней циркулирующих гонадотропинов и быстрого
обратного восстановления [35]. Первые антагонисты обладали слабым эффектом,
или их применение было связано с нежелательными побочными явлениями в резуль¬
тате выброса гистамина. Синтез новых антагонистов находится на этапах разработки
[36-41], эти новые продукты более эффективны и не обладают побочным эффектом
выброса гистамина. Совсем недавно новые антагонисты потеснили агонисты в пода¬
влении естественного выброса ЛГ при индукции овуляции инъекционным ФСГ [42].
Использование антагонистов ГнРГ стало особенно популярным в протоколах индук¬
ции овуляции при искусственном оплодотворении (ЭКО) [42].
РЕПРОДУКТИВНЫЕ ФУНКЦИИ ПЕРЕДНЕГО ГИПОФИЗА
Гонадотрофы
Гонадотрофы — специализированные клетки переднего гипофиза, которые син¬
тезируют и секретируют ЛГ и ФСГ. Эти клетки состав.ляют от 7 до 15% всех гипо¬
физарных клеток и определяются в этом месте с ранних сроков развития плода [43].
.Большинство этих гонадотрофов способны синтезировать как ФСГ, так и ЛГ [43, 44].
И ЛГ, и ФСГ состоят из двух связанных нековалентной связью белковых субъединиц,
называемых аир (см. рис. 1-4). В гонадотрофах гены субъединиц переписываются
в транспортные РНК, которые последовательно переводятся в предшественников
субъединиц. Гонадотрофы содержат на поверхности ГнРГ-рецепторы, которые опо¬
средуют эффект ГнРГ. Эти рецепторы относят к 7-му трансмембранному домену и
семейству связанных с С-белком рецепторов.
РЕЦЕПТОР К ГОНАДОТРОПИН-РИЛИЗИНГ-ГОРМОНУ
У млекопитающих встречают два вида рецепторов, связанных с С-белком [45].
Рецепторы I типа, расположенные в гонадотрофах, хорошо описаны у человека.
Недавнее клонирование рецептора ГнРГ II типа показало, что он структ>'рно и функ¬
ционально отличается от рецетора I типа. Тем не менее гомолог гена рецептора II типа
у человека имеет фреймшифт ф-атезЫА, сдвиг рамки считывания генетического кода)
и преждевременный стоп-кодон; вероятно, рецептор полной длины не встречается у
человека [45], Именно поэтому последующая информация относится.только к I типу
ГнРГ рецепторов.
У людей ГнРГ гипоталамуса регулирует секрецию гонадотропинов через рецепто¬
ры I типа, активируя Gq/ll [46], Несмотря на то что преимущественное связывание
рецепторов I типа происходит через стимуляцию Сд/11, передача сигнала может про¬
исходить через другие С-белки, и потенциально путем, независимым от С-белка [45,
46]. ГнРГ активирует несколько нисходящих каскадов. Они включают в себя РКС-,
Са^^- и тирозин-киназазависимые пути [45]. Рецепторы ГнРГ, расположенные в гона¬
дотрофах гипофиза мыши, активируют несколько МАРК-каскадов, включая ЕЕК1/2,
24
c-Jun amino-tenninal kinase QNK), p38 МАРК, a также большой МАРК (BMK1/ERK5)
[45]. Возможность перекреста этих путей требует дальнейшего прояснения.
Активация рецепторов, связанных с G-белком, типично происходит после их десен-
ситизации и интернализации, и эти процессы включают быстрое фосфорилирование
рецепторов, вызванное агонистами через вторую мессенджерзависимую протеин-
киназу и киназу рецепторов, связанных с G-белком. Поскольку остатки серина и
треонина, фосфорилированные киназой G-белок связанного рецептора, часто рас¬
полагаются в карбоксильных терминальных хвостах, которые уникально отсутствуют
в ГнРГ рецепторах млекопитающих, несколько исследований показало, что рецепто¬
ры ГнРГ, не обладающие таким хвостом, не подвергаются ни быстрой гомологичной
десенситизации, ни вызванным агонистами фосфорилированием рецепторов. Кроме
того, рецепторы интернализуются медленно через покрытые клатрином пузырьки, и
этот процесс происходит независимо от (1-аррестина и динамина [45].
ЛЮТЕИНИЗИРУЮЩИЙ и ФОЛЛИКУЛОСТИМУЛИРУЮЩИЙ ГОРМОН
Каждый гонадотропин является гетеродимером. Как ЛГ, так и ФСГ содержат
две пептидные субъединицы, называемые а и р (см. рис. 1-4). Субъединицы аир
связаны нековалентными связями. а-Субъединица человеческих ЛГ, ФСГ, тиро-
тропина и человеческого хорионического гонадотропина (ХГЧ) имеет идентичную
полип ептидную структуру, и наоборот, р-субъединица каждого гормона имеет уни¬
кальную последовательность аминокислот и определяет специфическую активность
аР-гетеродимера. Каждая субъединица богата цистеином и содержит множество ди-
сульфидных связей и углеводных половинок, которые играют значительную роль в
биологической активности и метаболизме этих гормонов. Идентичная а-субъединица
содержит 92 аминокислоты, р-Субъединицы человеческих ФСГ, ЛГ и ХГЧ содержат
117, 121 и 145 аминокислот соответственно (табл. 1-1) [47-50]. После связывания
ГнРГ с его рецептором происходит биосинтез гонадотропинов, включающий транс¬
крипцию генов субъединиц, трансляцию мРНК субъединиц, посттрансляционные
модификации предшественников субъединиц, сборку субъединиц, упаковку «зрелых»
гормонов и секрецию гормонов (см. рис. 1-4),
Таблица 1*1. Свойства гонадотропинов
Гонадотретнм
Локализация гена
^-еубъедтицы
Мубьединшуы
Время циркутфоФания в
ФСГ
Хромосома 11р13
117 аминокислот
3-4 ч
ЛГ
Хромосома 19q13.3
121 аминокислота
20 мин
ХГЧ
Хромосома 19ql3.3
145 аминокислот
24 ч
Ген а-субъединицы у человека расположен на коротком плече хромосомы 6.
Кодируемый предшественник-полипептид имеет 24-аминокислотную последователь-
ность,котораярасщепляетсяпослетрансляциидляполучениязрелой92-аминокислотной
субъединицы.
Гены человеческого ЛГ и а-субъединицы ХГЧ локализуются на хромосоме 19qlЗ.З,
содержащей кластер из 7 подобных р-субъединице генов [48]. Пять из этих последо¬
вательностей являются некодируемыми псевдогенами, собранными в группы друг за
другом и образующими инвертированные лары. Только ЛГ и р-субъединица ХГЧ дают
начало двум различным и функциональным видам мРНК. мРНК р-субъединицы ХГЧ
кодирует 145-а минокислотный протеин-предшественник, который позже расщепляет¬
ся на 24-аминокислотный лидер-пептид и 121-аминокислотный биологически актив¬
ный зрелый пептид. мРНК р-субъединицы ХГЧ также кодирует 145-аминокислотный
протеин. Как правило, этот протеин не обрабатывается посттрансляционно и функ¬
25
ционирует как биологически активная р-субъединица ХГЧ. Последовательность
аминокислот человеческого ЛГ и р-субъединица ХГЧ гомологичны в 82%. Эти две
субъединицы обеспечивают схожую биологическую активность, когда связаны с
а-субъединицей [48, 49, 50].
Единственный ген, расположенный на коротком плече хромосомы 11, кодирует
р-субъединицу ФСГ, содержащую последовательность из 117 аминокислот [51].
Дополнительная ДНК, кодирующая человеческие р-ФСГ, р-ЛГ или р-ХГЧ в ком¬
бинации с дополнительной ДНК а-субъединиц, была воспроизведена в культуре
клеток млекопитающих. Эти клетки могли синтезировать вышеуказанные про¬
теины, модифицировать их после трансляции, гликозилировать и комбинировать их
субъединицы, а затем секретировать их интактным как ФСГ, ЛГ так и ХГЧ [52]. Эти
рекомбинантные гонадотропины в настоящее время используются в клинике для сти¬
муляции функции гонад.
РЕГУЛЯЦИЯ КОНЦЕНТРАЦИЙ ФСГ И ЛГ
Молекулярный механизм, отвечающий за формирование и комбинацию а- и
Р-субъединиц ФСГ и Л Г, до конца неизвестен. Уровень продукции обеих субъединиц
аир регулируется, как минимум, обратной отрицательной связью эстрогенами, кото¬
рые контролируют пульсирующий выброс ГнРГ из гипоталамуса [52, 53]. Гипофиз
всегда содержит больше мРНК а-субъединиц, чем р-субъединиц, а в сыворотке крови
определяется уровень свободной а-субъединицы. С другой стороны, уровень свобод¬
ной р-субъединицы определяется в сравнительно низкой концентрации в гипофизе и
практически не определяется в сыворотке крови или моче, в связи с этим специфи¬
ческая р-субъединица может быть фактором, определяющим скорость синтеза этих
гормонов-гликопротеинов.
Ингибин, активин и фоллистатин были сначала идентифицированы как поло¬
вые гормоны, которые могут оказывать влияние на секрецию ФСГ. Несмотря на то
что первичный источник ингибина остается в яичнике, и активин, и фоллистатин
синтезируются в экстрагонадных тканях и могут оказывать влияние на ФСГ через
цутокринные-паракринные механизмы. Ингибин В секретируется гранулезными клет¬
ками во время фолликулярной фазы (под контролем ФСГ), а ингибин А — желтым
телом в лютеиновую фазу цикла под контролем ЛГ. Ингабины действуют совместно
с эстрадиолом для ингибирования секреции ФСГ. Активин может напрямую стиму¬
лировать синтез ФСГ и секрецию гормона гонадотрофами гипофиза. Фоллистатин
может подавлять эти эффекты, связывая активин и предупреждая его взаимодействие
с рецепторами на мембране клеток [54].
Уровни гонадотропинов в плазме крови пропорциональны уровню секреции и вре¬
мени полувыведения, которые, в свою очередь, регулируются количеством углевод¬
ных остатков. Чем более высокое содержание углеводных остатков, особенно остат¬
ков сиаловых кислот, тем ниже зфовень метаболизма и больше время полувыведения.
Содержание сиаловых кислот в гонадотропных гормонах и других гликопротеинах
имеет выраженный эффект на степень клиренса и также влияет на размер молекул.
Более высокая концентрация сиаловой кислоты в ФСГ по сравнению с ЛГ определяет
более медленное исчезновение ФСГ из кровотока. ХГЧ имеет самую высокую концен¬
трацию сиаловых остатков и самое длительное время полувыведения (см. табл. 1-1).
яичники
Яичник — основное место образования яйцеклеток и синтеза половых гормо¬
нов, эстрадиола и прогестерона. Эти события организованы в циклический про¬
цесс созревания фолликула, овуляции и формирования и регрессии желтого тела.
26
Следовательно, яичник выполняет две основные функции: генерация яйцеклеток,
способных к оплодотворению, и подготовка эндометрия к имплантации в процессе
выработки эстрадиола и прогестерона. Овариальный фолликул, содержащий яйце¬
клетку, окруженную клетками гранулезы и теки, составляет фундаментальную функ¬
циональную единицу яичника.
Яичники человека представляют собой овальные тела длиной от 2 до 5 см, шири¬
ной от 1,5 до 3 см и толщиной от 0,5 до 1,5 см. Общая масса нормальных яичников
в репродуктивном периоде от 10 до 20 г (в среднем 14 г). Яичники находятся в про¬
странстве между задней и боковой стенками таза и прикрепляются к задней поверхно¬
сти широкой связкой складки брюшины, называемой брыжейкой яичника (мезовари-
ум). Кровеносные сосуды, нервы и лимфатические сосуды проходят через брыжейку
яичника и входят в него в области хилуса [56].
Яичник состоит из трех структурно различающихся областей:
1) наружный корковый слой, содержащий поверхностный зародышевый эпителий
и фолликулы;
2) мозговое вещество, располагающееся в центре и состоящее из стромы;
3) хилус вокруг места прикрепления яичника к мезоварию.
Функциональная анатомия яичника проиллюстрирована на рис. 1-5. Хилус — место
прикрепления мезовария к яичнику. Он содержит сосуды, нервы и хилусные клетки,
которые имеют возможность активно участвовать в стероидогенезе или формировать
андроген-секретирующие опухоли. Эти клетки схожи с тестостерон-продуцирующими
клетками Лейдига яичка. Наружная поверхность коркового вещества, называемая
Зародышевый
эпителий Атрезированный
Первичные \ Развивающийся фолликул
фолликулы \
Брыжейка \\ \ I Зрелый
фолликул
Фолликулярная фаза
яичника
Ворота
Белое тело
Желтое тело
\
Яйцеклетка
Гранулезные
клетки
Базальная
мембрана
Внутренний
клеточный
слой
Наружный
фиброзный
слой
Спавшиеся ворота яичника
(фибрин и тромб)
Тека-лютеиновые клетки
Гранулезные клетки
Лютеиновая фаза
Рис. 1-5. функциональная анатомия и эволюционные сдвиги в яичнике взрослого во время овариаль¬
ного цикла. [Carr B.R., Wilson J.D. Disorders of the ovary and female reproductive tract // Braunwald E.,
Isselbacher K.J., Petersdorf R.G. et al. (eds). Harrison’s Principles of Internal Medicine. — 11th ed. — New
York: McGraw-Hill, 1987. - P. 1818-1837.]
27
белочной оболочкой, покрыта одним слоем поверхностных кубических клеток
эпителия (герминативный эпителий). Ооциты, собранные в комплексы, называе¬
мые фолликулами, располагаются во внутренней части коркового слоя и окружены
стромой (см. рис. 1-5). Один доминантный фолликул овулирует в течение каждого
цикла. Преовуляторный фолликул трансформируется в желтое тело после овуляции
(см. рис. 1-5). При отсутствии беременности желтое тело регрессирует в белое тело
(см. рис. 1-5).-€тромальные ткани состоят из соединительной ткани и интерстициаль¬
ных клеток, происходящих из мезенхимальных тканей и имеющих способность отве¬
чать на стимуляцию ЛГ или ФСГ продукцией андростендиона. Центральная, мозговая
часть яичника происходит в основном из мезонефрических клеток.
Дифференцировка яичников и фолликулогенез зависят от координированной экс¬
прессии и взаимодействия множества генов [57]. Целенаправленное разрушение или
внедрение генов у мышей позволило получить информацию о функционировании
специфических генов дифференцировки яичников и фолликулогенеза. На рис. 1-6
суммированы биологические эффекты некоторых из этих генов [57]. Генетически
измененные мыши представляют собой первый этап попыток понять ш vivo взаимо¬
действия различных генов, результатом которых является функционирование яични¬
ка. Несомненно, патологические состояния яичников у трансгенных мышей близко
напоминают нарушения, наблюдаемые у мутантных человеческих гомологов, как
проиллюстрировано в случаях с р-субъединицей ФСГ и рецептором к ФСГ. Доступны
многие опытные модели на мышах, демонстрирующие патологические состояния
яичников. Вообще они могут быть разделены на мышей с пренатальным выключе¬
нием функции яичников с нарушенным формированием гонад и уменьшенным коли¬
чеством зародышевых клеток или отсутствием зародышевых клеток и мышей с пост-
натальным нарушением функции яичников как результатом дефектов на различных
стадиях фолликулогенеза (см. рис. 1-6) [.57]. Эти опытные модели должны приводить
к идентификации генетических и молекулярных механизмов, ответственных за раз¬
витие и функционирование яичников человека.
У людей определенные дефекты в генах приводят к определенным нарушениям
в фолликулогенезе. Это было установлено в результате открытия гетерозиготных
мутаций в гене морфогенетического протеина кости-15 (BMPJS), которые вызывают
дисгенезию яичников. ВМР15 является ооцит-слецифическим фактором роста/диф¬
ференцировки, стимулирующим фолликулогенез и рост гранулезных клеток. In vitro
мутантный ВМР15 снижает рост гранулезных клеток и противодействует стимули¬
рующей активности немутировавшего протеина на гранулезные клетки. In vivo эта
мутация ассоциирована с семейной формой дисгенезии яичников, что демонстрирует
необходимость ВМР15 для развития фолликулогенеза у человека [58].
ооцит
Известно, что примордиальные зародышевые клетки рождаются вне эмбриона из
энтодермы желточного мешка. В этом месте они могут быть выявлены уже на 3-й нед
беременности с помощью окрашивания щелочной фосфатазой. Зародышевые клетки
мигрируют на достаточно большое расстояние от желточного мешка до полового
бугорка благодаря амебообразным движениям с помощью псевдоподий [59]. Этот
28
г онадогропиннезависимый рост
Преантральный
►
Гонадотропинзависимый рост
Антральный
Антральный
Предовуляторный
Первичный
примордиальный
П Т
Figla Amh
Kit
KitI
Nobox
Вторичный
Шт
Fshb
^Ат2
Fshr
Inha
ИФР1
Inhba
Ccnd2
Gja4
Smad3
Esri
Taf4b
Esr2
Размер фол¬
ликула. мм <20
20-70
70
4
Овуляция
Оплодотворение
Втр15
Образование
желтого тела
Рис. 1-6. Схема, иллюстрирующая стадии развития, на которых определенные гены мыши влияют
на оогенез. Данные от трансгенных мышей с различными разрушенными генами, показывающие кри¬
тическую роль определенного количества генов, разные фазы развития фолликулов. Преантральный
рост фолликулов гонадотропинзависим, тогда как формирование полости и созревание фоллику¬
лов требует стимуляции срСГ. Acvr2 — активиновый рецептор II типа; Amh — антимюллеров гормон;
б/лр 15 —костный морфогенетический протеин-15; Сспс12 — циклин <12; Esrt — эстрогеновый рецептор а;
Esr2 — эстрогеновый рецептор ß; Figla — фактор в зародышевой линии а; Foxl2 — транскрип¬
ционный фактор «forkhead box L2»; Fshb— ß-субъединица фолликулостимулирующего гормона;
Fshf — рецептор ФСГ; Gdf9 — фактор-9 дифференциации роста; Gja4 — белок коннексин 37 щеле¬
вого контакта: ИФР1 — инсулиноподобный cjiaiaop роста-1; Inha — а-субъединица ингибина:
Inhba - субъединица ßA активина; Kit — kit-рецептор, KitI — kit-лиганд; Nobox — гомеобокс-ген яич¬
ника новорожденного; Smad3 — транскрипционный фактор «Sma mothers against decapentapiegic-3»;
Taf4b — <t)aicTop-4b, ассоциированный с протеином, связывающим ТАТА-бокс. [Модифицировано из
Simpson J.L., Rajkovic A. Ovarian differentiation and gonadal failure // Amer. J. Med. Genet. — 1999. —
N 89. — P. 186-200; Choi Y., Rajkovic A. Genetics of mammalian folliculogenesis // Cell. Mol. Life. Sei, —
2006.-N63 (5).-P. 579-590.]
длинный путь миграции вдоль дорсального мезентерия задней кишки прерывается
только необходимым пересечением целомического угла на уровне полового бугор¬
ка (рис. 1-7). Механизмы, заставляющие клетки перемещаться, остаются неразга¬
данными полностью до сих пор, хотя не исключается некоторая роль хемотаксиса.
Зародышевые (половые) клетки не обладают способностью существовать вне полово¬
го бугорка, который служит единственным местом, поддерживающим развитие гонад.
Это подтверждает, что зародышевые клетки играют ключевую роль в развитии гонад.
В действительности в отс}тствие зародышевых клеток не может происходить форми¬
рование функционирующих гонад.
К моменту завершения перемещения к половому бугорку к 5-й нед беременности
предмейотические половые клетки называются оогониями [60]. В течение последую¬
щих 2 нед внутриутробного развития (от 5 до 7 нед беременности, или на индиффе¬
рентной стадии) примордиальные гонадные структуры представлены не более чем
выпуклостью на медиальном выступе урогенитального бугорка (см. рис. 1-7). Это
возвышение образовано вследствие пролиферации поверхностного герминативного
29
Аоота
Вольфов
проток
Симпатические ганглии
Надпочечниковое
мозговое вещество
Надпочечниковая кора
Начало
мюллерова
протока
Гонадный
гребень
Рис. 1 -7. Поперечный разрез каудальной области 5-недельного эмбриона, показывающий расположе¬
ние половых бугорков, зачатка надпочечников и миграционные пути первичных половых клеток. С 3-й
недели половые клетки, возникающие вокруг желточного мешка, пересекают дорзальный мезентерий
задней кишки и мигрируют к половым гребешкам. Быстрое деление первичных половых клеток, эпите¬
лия гонад и мезенхимы, которое начинается к концу 5-й недели, последовательно приводит к развитию
яичника у плода с генотипом 46,XX. ПЦ — полость целома. [Модифицировано из Moore К. The Developing
Human. — Philadelphia: WB Saunders, 1983.]
эпителия за счет роста подлежащей мезенхимы и размножения оогониев. Количество
оогониев достигает 10 ООО от 6 до 7 нед внутриутробного существования. Поскольку
мейоза и атрезии оогониев не происходит, реальное число зародышевых клеток
определяется числом митозов.
Во время этой индифферентной фазы впервые определяются очертания будущего
коркового и мозгового слоя, в то же время не имеется четких доказательств половой
принадлежности эмбриона. Тем не менее отсутствие признаков развития яичек до
7-й нед беременности расценивается как предположительные признаки развития яич¬
ника. Дополнительными признаками половой принадлежности гонад можно считать
мейоз оогониев приблизительно на 8-й нед развития, поскольку подобные процессы не
наблюдаются в яичке до половой зрелости. Половая принадлежность гистологически
ясна к 16-й нед, когда можно визуализировать первые примордиальные фолликулы.
С 8-й нед внутриутробного развития постоянные митозы увеличивают количество
оогониев до 600 ООО (рис. 1-8). С этого момента оогонии участвуют в трех одновре¬
менно протекающих процессах: митозе, мейозе и атрезии. Начало мейоза и атрезии
клеток совпадает с процессом митоза оогониев. В результате к 20-й нед количество
зародышевых клеток достигает 6-7x10^ (см. рис. 1-8). В это время две трети всех
зародышевых клеток являются первичными ооцитами на стадии митоза, оставшаяся
треть представляет собой оогонии. Пиковое увеличение числа и последующее сниже¬
ние вычисляются по прогрессивному снижению числа митозов оогониев, продолжаю¬
щемуся до 7-го мес внутриутробного развития плода. В равной степени важен уровень
атрезии оогониев, который достигает пика на 5-м мес беременности. В этот период
регуляция развития яичника комплексная и, вероятно, включает различные группы
генов (см. рис. 1-6) [61, 62].
С середины беременности происходит постоянное уменьшение количества половых
клеток в гонадах. В конечном итоге 50 лет спустя яичники окончательно истощаются.
30
Рис. 1 -8. Изменения количества половых клеток, зависящие от возраста яичника человека. Наибольшее
количество ооцитов определяется у плода в середине беременности. Это количество резко снижается
в III триместре. После рождения постоянное снижение количества ооцитов продолжается вплоть до
наступления менопаузы. [Baker T.G. А quantitative and cytological study of germ cells in the human ovaries //
Proc. R. Soc. Biol. Sei. - 1963. - N 158. - P. 417-433.]
Чаще всего этот процесс начинается с 6 мес беременности и продолжается в течение
всей жизни, происходя более в форме фолликулярной атрезии, чем атрезии оогониев
(см. рис. 1-8). И наоборот, атрезия оогониев является основным процессом до 7 мес
внутриутробного развития, тогда как фолликулярная атрезия только начинается.
Фолликулярная атрезия влияет на содержание половых клеток, уменьшая их количе¬
ство до 1-2x10^ к моменту рождения (см. рис. 1-8) [63]. Это драматическое снижение
происходит в течение короткого периода, равного 20 нед внутриутробного развития.
Ничего подобного не происходит ни раньше, ни позже. В результате новорожденные
девочки вступают в жизнь, все еще не достигнув репродуктивного потенциала, теряя
около 80% запасов своих половых клеток. К пубертатному возрасту это количество
составит 300 ООО. И только от 300 до 400 фоликулов (менее 1% общего числа) овули¬
рует в течение последз^ющего репродуктивного периода жизни.
На 8-13-й нед беременности часть оогониев покидает митотический цикл, для того
чтобы войти в профазу первого мейотического деления. Такое изменение определяет
превращение этих клеток в первичные ооциты перед последующим образованием
фолликулов. Мейоз (начинающийся с 8-й нед беременности) обеспечивает временную
защиту оогониев от атрезии, позволяя им вместе с гранулезными клетками сформи¬
ровать примордиальные фолликулы. Соответственно оогонии, которые существуют
31
более 7 нед, не входя в мейоз, подвергаются последующей атрезии. Именно поэтому к
моменту рождения обычно не определяется ни одного оогония.
Однажды образованный первичный ооцит существует в профазе первого мейо¬
тического деления до овуляции, когда мейоз возобновляется и формируется первое
полярное тельце (рис. 1-9). Несмотря на то что клеточный механизм, ответственный
Мейоз I
с количеством ДНК 2п
(длительная
остановка)
Первичный ооцит
с удвоенным набором ДНК
Мейоз II
Рис. 1-9. Мейотическое деление клеток. Мейоз происходит исключительно в половых клетках и обеспе¬
чивает два важных процесса: (1) образование половых клеток, отличающихся от соматических клеток; (2)
образование молодой яйцеклетки с рецуцированным набором хромосом с 46 до 23. Генетическая реком¬
бинация путем скрещивания генов между гомологичными хромосомами и случайным набором материн¬
ских и отцовских хромосом в дочерних клетках во время первого мейотического деления обеспечивает
первую функцию мейоза — обеспечение генетического многообразия. Вторая функция обеспечивается
через редуцирование числа хромосом так, что каждая дочерняя клетка, или яйцеклетка, получает случай¬
ным образом одну из каждой из 23 пар. Во время оплодотворения в результате соединения яйцеклетки
и сперматозоида, содержащих по 23 хромосомы, получается генетически новый индивидуум с набором
из 46 хромосом. Хромосомы, обозначенные белым в оогонии (верхний левый угол), происходят от отца
плода, а голубые хромосомы предоставлены матерью плода. Случайный обмен генами (аллелями) между
гомологичными хромосомами (кроссинговер) происходит прежде прекращения мейоза в профазе 1.
После родов зти ооциты находятся в таком состоянии до периода пубертата. В развивающемся ооците
мейоз возобновляется сразу после преовуляторного выброса ЛГ во время каждого овуляторного цикла.
Мейотическое созревание определяется как период с момента разрушения ядра ооцита (зародышево¬
го пузырька) до того момента, как ооцит достигнет метафазы II (то есть перехода из ооцита в зрелую
яйцеклетку). Второе короткое прекращение мейоза происходит во II метафазе до того момента, как
ооцит будет оплодотворен сперматозоидом. ДНК — дезоксирибонуклеиновая кислота; СУВО — прорыв
зародышевого пузырька; мат — материнская хромосома; п — количество ДНК в гаплоидном наборе хро¬
мосом; отц — отцовская хромосома.
32
за прекращение мейоза, неясен, в настоящий момент признана роль ингибитора
мейоза, выделенного из гранулезных клеток. Эта гипотеза базируется на наблюдени¬
ях за ооцитами, лишенными гранулезных клеток, которые были способны спонтанно
завершить мейотическое созревание in vitro.
Первичный ооцит преобразуется во вторичный ооцит после завершения метафазы
первого мейоза и формирования первого полярного тельца, до момента овуляции, но
после выброса ЛГ. Во время овуляции вторичный ооцит и окружающие гранулезные
клетки {cumulus oophorus) изгоняются и входят в фаллопиеву трубу. Если происходит
проникновение спермы, вторичный ооцит подвергается второму мейотическому деле¬
нию, после чего выбрасывается второе полярное тельце (см. рис. 1-9).
КЛЕТКИ ГРАНУЛЕЗЫ
Базальная пластинка разделяет ооцит и гранулезные клетки от окружающих стро-
мальных клеток [64]. Таким образом, гранулезные клетки не имеют прямого досту¬
па к кровоснабжению (рис. 1-10). Отсутствие сосудов требует от клеток гранулезы
Рис. 1-10. Структурное взаимодействие между
гранулезными клетками и ооцитом. А — микро¬
ворсины ооцита с цитоплазматическими отрост¬
ками гранулезных клеток, пенетрирующих zona
pellucida (ZP). Б — пенетрация zona pellucida цито¬
плазматическими отростками гранулезных клеток.
Маленькие щелевые контакты (тонкие стрелки)
наблюдаются между гранулезными клетками и
мембраной ооцитов. Толстые стрелки показывают
щелевые контакты между гранулезными клетками.
[С разрешения из Erickson G.F. Ап analysis of follicle
development and ovum maturation // Semin. Reprod.
Endocrinol, — New York: Thieme Medical Publishers,
1986. - N 4. - P. 233.]
33
контактов с соседними клетками. Так, гранулезные клетки взаимосвязаны развитой
системой межклеточных щелевых контактов, что в результате образует обильный,
интегрированный и функциональный синцитий (см. рис. 1-10) [65, 66]. Щелевые
контакты образованы протеином, называемым коннексином. Коннексин-37 пред¬
ставлен в щелевых контактах в фолликулах, а недостаток этого протеина вызывает
недостаток граафовых фолликулов, нарушение овуляции и развитие недостаточности
желтого тела [67]. Эти специализированные межклеточные контакты могут быть важ¬
ными при метаболическом обмене, а также при транспорте маленьких молекул между
соседними гранулезными клетками. Более того, гранулезные клетки распространяют
цитоплазматические отростки для пенетрации zona pellucida, чтобы сформировать
контакты с мембраной ооцита (см. рис. 1-10). У мышей с отсутствием контактного
белка коннексин-37 развитие ооцита прекращается до достижения мейотической
компетентности [67]. Таким образом, межклеточные контакты представляют клю¬
чевую коммуникационную систему, которая необходима для тщательного контроля,
приводимого в действие множеством гранулезных клеток для возобновления мейоза
первичным ооцитом.
Гранулезные клетки в полностью созревшем граафовом фолликуле непосред¬
ственно до овуляции структурированы таким образом, что можно различить разные
популяции клеток [68, 69]. Отдельные популяции гранулезных клеток проявляют
специфические, специализированные функции [70-72]. Гранулезные клетки в преде¬
лах наружного слоя содержат высокий уровень рецепторов к гонадотропным гормо¬
нам, а также стероидогенные ферменты, и поэтому более относятся к стероидогенезу
в фолликуле [68, 69] (рис. 1-11). Яйценосный бугорок {cumulus oophorus) содержит
яйцеклетку, а окружающие массы гранулезных клеток формируют множество меж¬
клеточных контактов с яйцеклеткой и поэтому играют ключевую роль в развитии
ооцита [70-72] (см. рис. 1-11).
ВНУТРИФОЛЛИКУЛЯРНОЕ ОКРУЖЕНИЕ: ТЕКА-ИНТЕРСТИЦИАЛЬНЫЕ КЛЕТКИ
Ryan и Petro [73] показали, что тека-интерстициальные клетки продуцируют С19-
стероиды, используемые в качестве предшественников для эстрогенов и андрогенов.
Rice и соавт. [74] отметили возможность клетками theca interna (см. ниже) и интер¬
стициальными клетками совершать синтез С19-стероидов de novo (рис. 1-12). Клетки,
продуцирующие С19-стероиды, происходят из популяции мезенхимальных клеток
в стромальном окружении и располагаются в свободном пространстве коркового и
мозгового вещества.
Клетки, создающие тека-интерстициальное окружение, являются гетерогенны¬
ми по природе. Современный взгляд на динамические изменения этого окружения
в яичнике представлен Erickson и соавт. [75]. Было определено несколько классов
интерстициальных клеток. Среди этих классов тека-интерстициальные клетки пред¬
ставляли постоянную характеристику развивающихся фолликулов (см. рис. 1-11 и
1-12) [11, 12], Эти клетки идентифицируются как theca interna, стромальный слой
клеток, располагающийся на базальной пластинке вокруг гранулезных клеток, и theca
externa, менее развитый слой стромальных клеток, который создает наружную часть
фолликула (см. ниже, рис. 1-11 и 1-12) [11,12]. Тека-интерстициальные клетки пред¬
ставляют в фолликуле основной компонент, продуцирующий С19^стероиды.
РЕЗИДЕНТНЫЕ БЕЛЫЕ КРОВЯНЫЕ КЛЕТКИ ЯИЧНИКА
Б отличие от семявыносящих протоков яичка, яичник не является иммунологи¬
чески привилегированным местом. Именно поэтому мононуклеарные яичниковые
фагоциты (макрофаги), лимфоциты и полиморфноядерные гранулоциты могут быть
определены на разных стадиях развития фолликула. Например, макрофаги, как
известно, составляют большой клеточный компонент интерстициального (так назы-
34
Примордиальный
фолликул
Первичный
фолликул
Вторичный
фолликул
Третичный
фолликул
Базальная мембрана
Гранулезные клетки
Базальная мембрана
Гранулезные клетки
Ооцит
Прозрачная оболочка
Рис. 1-11. Стадии развития фол¬
ликула в яичнике. Примордиальный
фолликул состоит из одного слоя
гранулезных клеток и одного незре¬
лого ооцита в диплотеновой стадии
первого мейотического деления.
Примордиальный фолликул отде¬
лен от окружающей стромы тон¬
ким базальным листком (базальной
мембраной). Ооцит и гранулезные
клетки не имеют прямого контак¬
та с кровоснабжением. Первым
признаком вовлечения фолликула
является кубоидальная дифферен¬
цировка веретенообразных клеток
внутри базальной мембраны, кото¬
рая далее подвергается последую¬
щим митотическим делениям для
формирования многослойной гра-
нулезоклеточной зоны. Ооцит уве¬
личивается и секретирует слизепо¬
добную субстанцию, содержащую
гликопротеид и называемую zona
pellucida, которая окружает ооцит
и отделяет его от гранулезных
клеток. Это структура первичного
фолликула. Вторичный фолликул
формируется путем дальнейшей
пролиферации гранулезных клеток
и во время финальной фазы роста
ооцита, в которой ооцит достига¬
ет диаметра 120 мкм, происходя¬
щего совместно с пролиферацией
клеток, расположенных сразу же
снаружи от базальной пластинки и
формирующих теку. Тека, располо¬
женная на базальной мембране,
называется theca interna. Клетки
теки, соприкасающиеся со стромой, называются theca externa. Вторичный фолликул обзаводится неза¬
висимым источником питания, состоящим из одной или нескольких артериол, которые заканчиваются в
капиллярном русле у базальной пластинки. Капилляры не пенетрируют базальную мембрану, и грануле-
за, и ооцит остаются бессосудистыми. Третичный фолликул характеризуется дальнейшей гипертрофией
теки и возникновением заполненного жидкостью пространства внутри гранулезных клеток, называемого
«ворота яичника» (antrum). Жидкость состоит из транссудата плазмы и секрета гранулезных клеток,
некоторые из них (эстрогены) находятся в очень высокой концентрации в сравнении с периферической
кровью. Фолликул быстро увеличивается в размерах под влиянием гонадотропинов и формирует зре¬
лый, или граафов, фолликул. В граафовом фолликуле гранулезные клетки и ооцит окружены базальной
мембраной и отделены от прямого кровоснабжения. Антральная жидкость увеличивается в объеме, и
ооцит, окруженный накоплением гранулезных клеток (яйценосный бугорок), находится в полярной, экс¬
центричной позиции внутри фолликула. Зрелый граафов фолликул готов к освобождению яйцеклетки бо
время овуляции. [Адаптировано из Erickson G.F., Magoffin D.A., Dyer C.A. The ovarian androgen producing
cells: a review of structure-function relations // Endocr. Rev. — 1985. — N 6. — P. 371-379. Copyright © 1985
by The Endocrine Society.]
Граафов
фолликул
Холмик
из фолликулярных
клеток
Базальная мембрана
Гранулезные клетки
Ооцит
Прозрачная оболочка
- Оболочка
Внутренний
клеточный слой
Наружный фиброзный слой
Базальная мембрана
Прозрачная оболочка
Полностью созревший ооцит
Ворота яичника
Множественные слои
гранулезных клеток
Ворота яичника
(фолликулярная жидкость)
Наружный фиброзный слой
Прозрачная оболочка
Мембрана из гранулезных
клеток
Базальная мембрана
Полностью созревший ооцит
Внутренний
клеточный слой
35
X я г
Фолликулярная
жидкость
Гранулезные
клетки
Б
Внутренний
клеточный слой
Наружный
клеточный слой
Строма
яичника
Рис. 1-12. Гистология граафова фолликула человека. А — 10-12-й день цикла: граафов фолликул при¬
ближается к зрелости (X — холмик из фолликулярных клеток; Г — слой гранулезных клеток; Я — яйцеклет¬
ка; НС — наружный клеточный слой; ВС — внутренний клеточный слой); Б — разрез через стенку зрелого
граафова фолликула. [Из Cunningham F.C. et al. Pregnancy: overview and diagnosis; ovarian function and
ovulation // Williams Obstetrics. — 19th ed. — Stamford, CT: Appleton & Lange, 1993. — P. 11-55.]
36
ваемого интрафолликулярного) окружения в яичнике [76]. Частично эти макрофаги
представлены в пределах стромы яичника возле перифолликулярных капилляров.
Лимфоциты и полиморфноядерные лейкоциты, с другой стороны, также описаны в
фолликуле и желтом теле в разном количестве в процессе развития фолликула, обра¬
зования желтого тела и атрезии фолликулов [77-79].
Значение вышеописанных наблюдений может быть в том, что резидентные пред¬
ставители белых кровяных клеток в яичнике создают на месте систему потенциаль¬
ных модуляторов функции яичников, которая действует через локальную секрецию
регуляторных цитокинов [80]. Поскольку направление движения информации скорее
всего многостороннее, одни и те же клетки могут получать стероидные и пептидерги-
ческие сигналы. Более того, иммунные клетки богаты стероидогенными возможно¬
стями, которые могут, с их стороны, влиять на стероидную структуру [81].
ФОЛЛИКУЛЫ
Фолликул представляет наиболее важную функциональную единицу в яичнике
с учетом развития половых клеток и продукции стероидов. Фолликулы расположе¬
ны в свободной части соединительной ткани коркового вещества яичников и могут
быть разделены на две условные группы: неразвитые (примордиальные) и разви¬
тые. Большая часть фолликулов не развита (90-95%) на протяжении всей жизни.
Бовлечение примордиального фолликула инициирует критические изменения в
развитии, структуре и функции. Развивающиеся фолликулы представлены четырьмя
стадиями: первичные, вторичные, третичные и граафовы (см. рис. 1-11). Первые три
стадии роста могут происходить в отсутствие гипофизарной поддержки и поэтому
регулируются интраовариальными механизмами (рис. 1-13). Фолликул, предполагае¬
мый для овуляции, определяется в первые дни менструального цикла [82].
Ранний рост фолликулов происходит в течение нескольких менструальных циклов,
но овуляторным фолликулом становится только один из группы во время перехода
предшествующей лютеиновой фазы в настоящую фолликулярную (см. рис. 1-13) [83,
84]. Общее время для достижения преовуляторного статуса составляет 85 дней (см.
рис. 1-13) [83, 84]. Большая часть этого периода развития зависит от стимуляции
ФСГ. Б конечном счете эта группа фолликулов вступает в стадию атрезии, за исклю¬
чением единственного, вовлеченного в дальнейшее развитие ФСГ. Таким образом,
группа фолликулов, имеющих размеры от 2 до 5 мм, может отвечать на воздействие
ФСГ. Бозрастание уровня ФСГ в позднюю лютеиновую фазу является критическим в
спасении этой группы фолликулов от атрезии. Кроме того, поддержка роста уровня
ФСГ в течение определенного периода является обязательной [85].
Быбранный примордиальный фолликул либо развивается в преовуляторный с
последующей овуляцией, либо подвергается атрезии [86]. Среднее время разви¬
тия выбранного фолликула до момента овуляции составляет от 10 до 14 дней (см.
рис. 1-13). Если фолликул не рекрутируется, он подвергается процессу, называемому
атрезией, во время которого ооцит и окружающие его в пределах базальной пластин¬
ки гранулезные клетки погибают и замещаются фиброзной тканью. Наоборот, клетки
тека снаружи от базальной пластинки не погибают, а подвергаются редифференци-
ровке и возвращаются в пул клеток, состоящих из яичниковых интерстициальных или
стромальных клеток [56]. Считается, что процесс атрезии происходит при недостатке
гормонов или факторов роста, которые формируют зрелый доминантный фолликул
через яичниковые механизмы. Существует общее мнение, что атрезия фолликулов
происходит через апоптоз [87]. Апоптоз является активным и регулируемым про¬
цессом, вызываемым каскадом протеаз, называемых каспазами, которые приводят к
характерной фрагментации ДНК и «пузырению» (блеббингу) мембран.
37
Инициация
фазы роста
Фаза тонического роста
Фаза роста, регулируемая
гонадотропином
Овуляция М Овуляция М Овуляция М
1-й цикл 2-й цикл 3-й цикл
Рис. 1-13. Полная схема развития фолликула. Фолликул класса I — вторичный фолликул, имеющий
тека-клетки и способный отвечать на стимуляцию гонадотропинами. Поскольку ранняя стадия развития
фолликула (классы I-IV) гонадотропинзависима, финальные стадии развития фолликула (классы V-VIII)
полностью зависят от гонадотропной стимуляции. Согласно этой теории в поздней лютеиновой фазе
фолликулы V класса представлены группой, из которой будет выбран фолликул, предназначенный для
овуляции в текущем цикле. Быстрый гонадотропинзависимый рост (классы V-VIII) происходит во время
фолликулярной фазы сразу после третьей менструации, считая от момента инициации развития. Во
время этой фазы происходит селекция и выбор доминантного фолликула. Общая длительность процес¬
сов от момента, когда фолликул класса I превращается в преовуляторный класса VIII, оценивается в 85
дней и занимает три овуляторных цикла. М — менструация. [Courtesy of А. Gougeon, Clamart, France.]
Овуляция
в середине цикла происходит резкое повышение уровня циркулирующего эстра¬
диола (см. далее). Это увеличение уровня эстрадиола сменяется пиковым выбросом
ЛГ и в меньшей степени ФСГ. Все это вызывает овуляцию в доминантном фолликуле.
В каждом менструальном цикле овулирует один фолликул, из которого образуется
одно желтое тело, у человека для стимуляции разрыва зрелого фолликула необходим
или ЛГ, или ХГЧ. Считают, что локальное увеличение биосинтеза простагландинов
медиирует овуляторный эффект ЛГ [88, 89].
Овуляция представляет собой быстрое увеличение размеров фолликула с после¬
дующим его выпячиванием над поверхностью яичника. В дальнейшем происходит
разрыв фолликула и изгнание комплекса, содержащего яйцеклетку, в брюшную
полость (рис. 1-14). Разрыв фолликула, или овуляция, происходит через 34-36 ч от
38
Надрыв
Рис. 1-14. Овуляция комплекса «яйце-
клетка-яйценосный бугорок» через стигму.
[С разрешения из Erickson G.F. Ап analysis
of follicle development and ovum matura¬
tion // Semin. Reprod. Endocrinol. — New
York: Thieme Medical Publishers, 1986. —
N 4. - P. 233.]
начала выброса ЛГ. Появление конической стигмы на поверхности выпячивающегося
фолликула предшествует разрыву (см. рис. 1-14). Разрыв этой стигмы сопровожда¬
ется аккуратным освобождением яйцеклетки и антральной жидкости. Продукция
гонадотропинзависимых протеаз, действующих локально на протеиновые субстраты
в базальной пластинке, может играть важную роль в формировании стигмы и раз¬
рыве фолликула [90, 91]. В частности, до овуляции в фолликуле возрастает уровень
плазминогена [92]. Таким образом, превращение плазминогена в плазмин с помощью
активатора плазминогена может относиться к протеолитическому разрушению фол¬
ликулярной стенки, предшествующему разрыву фолликула.
ЖЕЛТОЕ ТЕЛО
После овуляции доминантный фолликул превращается в желтое тело (рис. 1-15).
После разыва фолликула капилляры и фибробласты из окружающей стромы раз¬
множаются и проникают через базальную мембрану. Т^ая быстрая васкуляризация
желтого тела может проводиться с помощью ангиогенных факторов, некоторые из
них обнаружены в фолликулярной жидкости [93]. Сосудистый эндотелиальный фак¬
тор роста, выделенный из желтого тела, вместе с фактором роста фибробластов могут
быть потенциальными ангиогенным агентами в желтом теле [94]. Гранулезные клетки
и тека-клетки подвергаются одновременным изменениям, называемым лютеиниза-
цией. Гранулезные клетки становятся большими гранулезолютеиновыми клетками, а
тека-клетки трансформируются в тека-лютеиновые — малые — клетки (см. рис. 1-15,
Б) [95]. Так называемые К-клетки, разбросанные по всему желтому телу, считаются
макрофагами.
Желтое тело — эндокринная железа, которая служит основным источником гормо¬
нов, секретируемых яичником во время лютеиновой фазы цикла. Желтое тело чело¬
века секретирует около 40 мг прогестерона в день в середине лютеиновой фазы мен¬
струального цикла [96]. Несмотря на маленькие размеры желтого тела, это наиболее
активная стероидогенная ткань у человека. Важный момент формирования желтого
тела — пенетрация базальной мембраны кровеносными сосудами, снабжающими гра-
нулезолютеиновые клетки липопротеинами низкой плотности (ЛПНП) [92]. ЛПНП
холестерин служит в желтом теле субстратом для продукции прогестерона.
Ключевой регулятор стероидогенеза в желтом теле — ЛГ. У людей рецепторы к ЛГ
поддерживаются в течение времени существования желтого тела и не удаляются при
39
Рис. 1-15. Желтое тело беременности. А — малое увеличение; Б — Большое увеличение. Г — гранулез¬
ные клетки; К — клетки внутреннего слоя. [Из Cunningham F.G., MacDonald Р.С., Gant N.F. et al. Pregnancy:
overview and diagnosis; ovarian function and ovulation // Williams Obstetrics. — 19th ed. — Stamford, CT:
Appleton & Lange. — 1993. — P. 11-55.]
40
возникновении у матери беременности [97]. Скорость формирования прогестерона
под влиянием ЛГ в лютеинизированных гранулезных клетках определяется поступле¬
нием холестерина в митохондрии, которые регулируются белком острой регуляции
стероидогенеза (StAR; см. ниже) [98]. Таким образом, доступность холестерина и про¬
никновение его через стероидогенный регуляторный протеин в митохондрии — два
критических фактора, определяющих производство больших количеств прогестерона
в желтом теле.
Период функциональной активности желтого тела составляет 14±2 сут.
Впоследствии желтое тело спонтанно регрессирует. Если не возникает беременность,
то желтое тело замещается бессосудистым рубцом, называемым белым телом.
Длительность жизни желтого тела может регулироваться следующими факторами;
гормоном ХГЧ, васкуляризацией желтого тела и иммунными клетками [99]. Имеется
некоторая неопределенность во мнениях о центральной роли ЛГ в поддержании функ¬
ции желтого тела. Отключение стимуляции ЛГ во множестве экспериментов неизменно
приводило к регрессии желтого тела [99]. Во время беременности ЛГ заменяет ХГЧ,
который секретируется трофобластом, поддерживая способность желтого тела выра¬
батывать прогестерон. Эта стимуляция помогает поддерживать беременность вплоть
до смены желтого тела плацентой [100]. Соответственно желтое тело удваивается в
размерах (по сравнению с размером до беременности) в течение первых 6 нед бере¬
менности (см. рис. 1-15). Это происходит благодаря пролиферации соединительной
ткани и кровеносных сосудов вместе с гипертрофией лютеинизированной гранулезы и
тека-клеток. Эта ранняя гипертрофия позже сменяется регрессией. К сроку доношенной
беременности желтое тело составляет половину того, что развивается в течение мен¬
струального цикла.
Такие гормоны, как эстрогены и простагландины, играют важную роль в гибели
желтого тела [101, 102], Иммунные факторы также могут иметь значение, так как
регрессия желтого тела сопровождается массивной инфильтрацией макрофагами и
лимфоцитами [99].
Апоптоз может быть последним механизмом выключения желтого тела. Желтое
тело в ранней лютеиновой фазе и на ранних сроках беременности не имеет фрагмен¬
тированной ДНК, характерной для апоптоза [103]. С другой стороны, фрагментация
ДНК определяется в середине и конце лютеиновой фазы [103]. Таким образом, пола¬
гают, что апоптоз — главный механизм угас^ия желтого тела. ЛГ или ХГЧ ингиби¬
руют апоптоз в желтом теле. При отсутствии'этих трофических факторов происходит
апоптоз. Остатки желтого тела состоят из плотной соединительной ткани и называ¬
ются белым телом.
РЕЦЕПТОРЫ К . И ЛЮТЕИНИЗИРУЮЩЕМУ
ГОРМОНУ В ЯИЧНИКЕ
Рецептор к ФСГ экспрессируется исключительно гранулезными клетками.
Рецепторы к ЛГ/ХГЧ первично экспрессируются тека-интерстициальными клетками
всех фолликулов и гранулезными клетками крупных преовуляторных фолликулов.
Гранулезные клетки первичных или вторичных фолликулов, находящихся на
ранней стадии развития, до формирования полости (преантральные фолликулы) пер¬
вично связываются с ФСГ, но не Л Г. В этих преантральных фолликулах связывание с
ЛГ и ХГЧ ограничено тека-интерстициальными клетками [104]. Гранулезные клетки
у более зрелых третичных фолликулов со сформированной полостью способны свя¬
зывать как ЛГ, так и ФСГ. Таким образом, рецепторы к ФСГ находятся в гранулезных
клетках фолликулов всех размеров и стадий, а рецепторы к ЛГ находятся только в
гранулезных клетках крупных преовуляторных фолликулов [105-107]. Эти наблю¬
дения согласуются с концепцией появления рецепторов к ЛГ на гранулезных клетках
под влиянием ФСГ [108,109].
41
Рецепторы к гормонам-гликопротеинам имеют схожую структуру (рис. 1-16).
Рецепторы относятся у большой семье рецепторов, связанных с 0-белком, мембраны
которых имеют трансмембранный домен, состоящий из семи пересекающих мембра¬
ну а-спиралей, связанных тремя внеклеточными и тремя интрацеллюлярными пет¬
лями (см. рис. 1-16). Гликопротеиновые рецепторы гормона формируют отдельную
подгруппу внутри большой семьи посредством их большого внеклеточного гормон-
связывающего домена на уровне Н-окончания. ФСГ связывается с рецептором к ФСГ,
а ЛГ и ХГЧ — с одним и тем же рецептором к ЛГ. Гены рецепторов к ЛГ и ФСГ локали¬
зованы на хромосоме 2 в области р21 [51]. Отношение гликопротеиновых рецепторов
гормона к другим рецепторам, связанным с С-белком, определяется гомологией их
последовательностьи в С-окончании половины всех рецепторов. Этот домен, коди¬
руемый последним экзоном, содержит семь трансмембранных сегментов и домен,
связанный с С-белком. Необычно большой внеклеточный домен гликопротеиновых
рецепторов гормона кодируется первыми 9 из 10 экзонов (рис. 1-17).
Роль фолликулостимулирующего гормона в функции яичника
Как понятно из названия, ФСГ — основной стимулятор созревания фолликулов. С
учетом того что рецепторы к ФСГ локализуются исключительно на гранулезных клет¬
ках, считают, что ФСГ влияет на функциушальное состояние яичников через гранулез¬
ные клетки. Возможностью ФСГ управлять ростом и дифференцировкой фолликулов
определяется его способность оказывать несколько воздействий одновременно.
Фенотипы женщин, имеющих мутации, которые нарушают функцию р-субъеди¬
ницы ФСГ-гена, подтверждают, что ФСГ необходим для нормального развития фол¬
ликулов, овуляции и фертильности [51]. Более того, пубертатное развитие затруднено
при отсутствии достаточного количества фолликулов поздних стадий, имеющих
гранулезные клетки, необходимые для адекватной продукции эстрогенов. Лечение
таких пациенток экзогенным ФСГ приводит к созреванию фолликулов, овуляции и
нормальной беременности [51]. Демонстрируемый фенотип дефицита р-субъединицы
ФСГ практически идентичен фенотипу, причина которого — мутация, инактиви¬
рующая рецептор к ФСГ [51]. Женщины с мутациями рецепторов к ФСГ клинически
подобны пациенткам с дисгенезией гонад, с отсутствием или плохо развитыми вто¬
ричными половыми признаками и высоким уровнем ФСГ и ЛГ в плазме крови. У них
присутствуют фолликулы в яичнике, что указывает на независимое от ФСГ развитие
NH2
соон
Внеклеточная
часть
Плазматическая
мембрана
Цитоплазматическая
часть
Рис. 1-16. Рецептор к гонадотропину.
Седьмой трансмембранный домен и
связанные с С-белком рецепторы к ЛГ
и ФСГ, локализованные в мембране
гранулезных и тека-клеток яичника,
типично имеют большие внеклеточ¬
ные домены. NHj — аминогруппа;
СООН — карбоксильная группа. [Из
Bulun S.E., Simpson E.R., Mendelson
C.R. The molecular basis of hormone
action // Carr B.R., Blackwell R.E. (eds).
Textbook of Reproductive Medicine. —
2nd ed. — Stamford, CT: Appleton &
Lange. - 1998. - P. 137-156.]
42
Экзон
1 2,3,4,5,6 7,8
10
11
Ген рецептораЛГ
Экзон
3 4 5,6 7.8
9 1Q Ген рецептора ФСГ
Рис. 1-17. Схематическое представление генов рецептора к гонадотропину человека. Структура генов
отображена в верхней части рисунка. Светлые участки отображают экзоны, кодирующие нетранслиро-
ванные участки мРНК; закрашенные учаештпоказывают последовательности, кодирующие протеин. Оба
гена размером по меньшей мере 80 кб. Взаимоотношение интрон/экзон-структуры гена с доменами
протеина обозначено линиями, соединяющими ген и протеин. Горизонтально выделенная часть протеина
показывает сигнальный пептид, поперечно заштрихованный участок обозначает семь сегментов транс¬
мембранного домена. Числа ниже протеина показывают начало и конец сигнального пептида. Сбратите
внимание, что рецепторные гены похожи по структуре, за исключением дополнительных экзонов в гене
рецептора ЛГ. Экзон 1 кодирует сигнальный пептид и малую часть внеклеточного домена, последую¬
щие девять или восемь экзонов кодируют остаток внеклеточного домена, включая богатые лейцином
повторяющиеся фрагменты. У обоих генов последний экзон наибольший и содержит информацию для
трансмембранного сигнального трансдукционного домена. ФСГ — фолликулостимулирующий гормон.
[Из Themmen A.P.N., Huhtaniemi І.Т. Mutations of gonadotropins and gonadotropin receptors: elucidating the
physiology and pathophysiology of pituitary-gonadal function // Endocr. Rev. — 2000. — N 21. — P. 551-583.
Copyright © 2000 by The Endocrine Society.]
примордиальных фолликулов, a также рост фолликулов на ранних стадиях. Напротив,
в исследованиях, в ходе которых не были выявлены мутации рецептора к ФСГ, было
определено полное отсутствие всех фолликулов, включая примордиальные стадии
[51]. Таким образом, фенотип яичников при дефиците рецепторов отличается от
типичных форм дисгенезии гонад, характерных для синдрома Тернера с гонадами-в
виде полосок и отсутствием растущих фолликулов [51].
Исследования на грызунах in vivo свидетельствуют, что ФСГ способен увеличивать
количество собственных рецепторов в гранулезных клетках. В то время как эстрадиол
сам по себе не может влиять на распределение, количество или сродство рецепторов
гранулезных клеток к ФСГ, эстрогены вместе с ФСГ увеличивают общее количество
рецепторов к ФСГ в гранулезных клетках [110]. Следовательно, изменения в про¬
43
дукции эстрадиола преантральными фолликулами может усиливать их ответ на сти¬
муляцию ФСГ через регуляцию рецепторов к ФСГ, расположенных на поверхности
гранулезных клеток. Это взаимодействие ФСГ с эстрадиолом было продемонстриро¬
вано на грызунах. Похоже, что оба вида рецепторов к эстрогену а (ER-a) и ß (ER-ß)
могут медиировать эстрогенный эффект на развитие яичников и фолликулов у мышей
[111]. С другой стороны, в настоящее время неизвестно, имеются ли подобные взаи-
моотйошения у человека. ER-a не определяется в яичнике человека в значительных
количествах. Присутствие ER-ß в яичниках человека, напротив, подтверждает взаи¬
модействие ФСГ с эстрогенами при регуляции нормального развития фолликулов и
овуляции у женщин [112].
Один из важных эффектов ФСГ — индукция ароматазной активности в гранулезных
клетках [113,114]. Таким образом, небольшое количество или отсутствие эстрогенов
может производиться не ФСГ-гранулезными клетками при условии, что они снабжа¬
ются ароматизируемыми андрогенными предшественниками. В то же время лечение
ФСГ усиливает ароматизирующую способность гранулезных клеток — эффект, сопро¬
вождающийся увеличением содержания ароматаз в гранулезных клетках [115,116].
Один из эффектов лечения ФСГ — увеличение рецепторов к ЛГ в гранулезных клет¬
ках. Способность ФСГ индуцировать рецепторы к ЛГ усиливается сопутствующим
присутствием эстрогенов [117]. Более того, гестагены, андрогены и ЛГ сами по себе
могут индуцировать ЛГ-рецепторы. После того как произошла индукция, рецептор к
ЛГ гранулезной клетки требует постоянного присутствия ФСГ для поддержки.
Результаты, которые были получены в ходе исследования женщин с мутациями
генов, кодирующих ФСГ- и ЛГ-рецепторы, а также ароматазу Р450, свидетельствуют
о том, что действие ФСГ, а не эстрогенов или ЛГ, необходимо для роста фолликулов
у человека [51, 118]. У женщин с недостаточной стимуляцией ЛГ или недостаточ¬
ным биосинтезом эстрогенов были зафиксированы рост и развитие фолликулов до
антральной стадии, но без последующей овуляции [51, 118]. С другой стороны, у
женщин с мутациями ß-субъединицы ФСГ или рецептора к ФСГ определяются только
примордиальные фолликулы в яичниках [51]. Эти данные показывают, что эстрогены
или ЛГ не являются критичными для развития фолликулов, по крайней мере до тре¬
тичной стадии (см. рис. 1-11 и 1-13) [11,13]. Необходимо помнить, что, как правило,
ФСГ сам по себе недостаточен для достижения нормального развития фолликулов и
овуляции.
Роль лютеинизирующего гормона в функции яичника
ЛГ необходим для овуляции (разрыва фолликула) и поддержки функции желтого
тела. К тому же ЛГ играет другие важные роли в функционировании фолликула. Прежде
всего, ЛГ стимулирует продукцию андрогенов тека-интерстициальными клетками.
Более того, ЛГ может действовать совместно с ФСГ на более развитых стадиях фол¬
ликулогенеза. И последнее: небольшие и постоянные увеличения циркулирующего ЛГ
необходимы для роста и развития фолликулов до преовуляторных стадий [119,120].
Предполагают, что ЛГ действует на тека-интерстициальные клетки маленьких фол¬
ликулов, которые поддерживают биосинтез С19-стероидов [121]. Последовательное
увеличение продукции эстрогенов относится к росту и развитию фолликулов. Лечение
малыми дозами ЛГ также предположительно приводит к увеличению содержания
рецепторов к ЛГ и к увеличению продукции стероидогенных протеинов, таких как
StAR, расщепление боковых цепей Р450 (P450scc), Sß-rnnpoKCHCTeponn дегидрогеназа
тип II (3ß-rCA-II) и 17-гидроксилаза (Р450с17).
Роль ЛГ в физиологии яичников человека была показана на примере женщин с
мутацией гена рецептора к ЛГ [51]. У пациенток клиническая аменорея, нормально
развитые вторичные половые признаки, увеличены уровни ФСГ и ЛГ, низкий уровень
эстрадиола и прогестерона, они были нечувствительны к лечению ХГЧ [51]. Яичник,
44
содержащий фолликулы, развитые до антральной стадии с хорошо развитым слоем
тека-клеток, не содержит ни преовуляторных фолликулов, ни желтого тела. Эти
наблюдения подтверждают мнение, что ЛГ необходим для овуляции и достаточной
продукции эстрогенов, тогда как развитие фолликулов первично автономно и на позд¬
них стадиях зависит от ФСГ.
Стероидогенез в яичниках
Проводили, сравнение содержания стероидных гормонов в крови, оттекающей по
яичниковой вене, и в периферической венозной крови для определения уровня стеро¬
идов, секретируемых яичником, надпочечником и получаемых в результате перифери¬
ческой конверсии предшественников [122]. Эти исследования показали, что яичники
секретируют прегненолон, прогестерон, 17а-гидросипрогестерон, дегидроэпиандро¬
стерон (ДГЭА), андростендион, тестостерон, эстрон и эстрадиол [123]. Несмотря на
то что эти измерения облегчают понимание исследуемых стероидогенных путей, они
не определили специфически вовлеченные клетки яичника. Исследования микропре-
парированных яичников показали, что эстрон и эстрадиол — основные стероидные
продукты (рис. 1-18). Прогестерон и, с другой стороны, 17а-гидроксипрогестерон —
основные продукты желтого телаЛ<й. рис. 1-18).
Общий путь стероидогенеза для производства эстрогенов и андрогенов показан на
рис. 1-18. Биологически активные стероиды в яичнике — эстрадиол и прогестерон
(см. рис. 1-18). Главный С19-стероидный продукт яичника, андростендион, не явля¬
ется биологически активным. Андростендион действует как двойной предшествен¬
ник и способствует концентрации циркулирующих эстрона и тестостерона путем их-
конверсии в экстрагенитальных тканях (жировая ткань, кожа) [124-126]. Бероятно,
слабый эстроген эстрон далее превращается в эстрадиол, а тестостерон превраща¬
ется в более активный дигидротестостерон локально в органах-мишенях — мозге,
молочных железах, простате и коже гениталий, где проявляет свое биологическое
действие [126]. Присутствие множества протеинов с ферментативной активностью
(17р-гидроксистероиддегидрогеназа и 5а-редуктаза), которые катализируют эти пре¬
образования в большом количестве тканей человека, подтверждают этот факт [127].
Преовуляторный фолликул секретирует эстрадиол в первую половину менструаль¬
ного цикла, а желтое тело секретирует эстрадиол и прогестерон во вторую половину
(рис. 1-19) [128-130]. Продукция этих двух биологически активных стероидов в фол¬
ликуле и желтом теле происходит под контролем ФСГ и ЛГ.
Имеется три основных типа яичниковых стероидов.
С18-СТЕР0ИДЫ
Бозникшие естественным путем С18-стероиды характеризуются ароматическим
А-кольцом, фенольной гидроксильной группой в положении С-3 и присутствием
либо гидроксильной группы (эстрадиол), либо кетоновой группы (эстрон) в положе¬
нии С-17. Ключевой фермент для продукции эстрогенов — ароматаза (см. рис. 1-18).
Протеин ароматаза Р450 (Р450агот) определяет специфическую активность комплек¬
са «фермент-ароматаза». Продукция в яичниковых гранулезных клетках Р450агот
первично регулируется ФСГ [131]. Основной и наиболее активный эстроген, секре-
тируемый яичником, — эстрадиол. Эстрон синтезируется и секретируется как в яич¬
никах, так и путем экстрагенитальной конверсии в периферических тканях [132].
Эстриол (16-гидроксиэстриол) определяется в моче в наибольшей концентрации и
является продуктом метаболизма эстрадиола и эстрона в периферических тканях.
Бее С18-стероиды, включая эстрадиол, эстрон и эстриол, относятся к эстрогенам.
Необходимо отметить, что эстрон и эстриол — слабые эстрогены, которые могут
45
P45ÖC17 ф Q
Р450С17
НО
4
он
зр-гсд-н
Щ-ГСД-1
Дегидроэпиандростерон
О
Андростендион
Р450агот ^ Q
НО
О
Тестостерон
Р450агот ^
Щ-ГСД-1
Эстрон
¥
НО
Эстрадиол
Рис. 1-18. Стероидогенез в яичнике. Биологически активные стероиды прогестерон и эстрадиол обра¬
зуются первично в яичнике женщины репродуктивного возраста. Продукция эстрадиола требует актив¬
ности шести стероидогенных протеинов, включая StAR, и происходит в шесть этапов. Р450с17 (продукт
гена CYP17) катализирует две ферментные реакции. Hejbipe молекулы холестериновых колец и их
производные стероиды обозначаются первыми буквами алфавита, а углероды нумеруются в последова¬
тельности, показанной во вставке. Sß-fCfl-ll — Sß-rnflpoKCHCTeponfl дегидрогеназа А5, четыре изомераза
типа II; 17ß-rCfl-l — 17ß-rHApoKcncTepoMA дегидрогеназа типа I; P450arom — ароматаза; Р450с17 — 17а-
гидроксилаза/17,20-лиаза; StAR — белок острой регуляции стероидогенеза.
превращаться в эстрадиол, обладающий полным эстрогенным эффектом. Имеется по
крайней мере семь ферментов в семье 17р-гидроксистероиддегидрогидрогеназ, обла¬
дающих перекрывающейся активностью, которые конвертируют эстрон в эстрадиол в
яичнике и экстрагенитальных тканях [133]. ^
Катехиновые эстрогены формируются гидроксилированием эстрогенов в позициях
С-2 или С-4. Физиологическая роль катехиновых эстрогенов неизвестна. Низкая масса
тела и гипертиреоидизм связаны с увеличенным формированием катехиновых эстро¬
генов [134]. Эстрона сульфат, получаемый периферическим превращением эстрадиола
и эстрона, имеет наибольшую концентрацию в крови, но не является физиологически
активным. Эстрона сульфат используется как источник для формирования эстрона в
некоторых тканях, включая те, которые служат мишенями для эстрогенов. Эстрадиол
регулирует секрецию гонадотропина и способствует развитию вторичных половых
46
Рис. 1-19. Две клеточные гипотезы яичникового стероидогенеза. А — преовуляторный фолликул
образует эстрадиол через паракринное взаимодействие тека- и гранулезных клеток. В ответ на стиму¬
ляцию гонадотропинами стероидогенный фактор-1 (SF-1, относится к семейству ядерных рецепторов)
действует как мастер-включатель для инициации транскрипции серии стероидогенных генов в тека-
клетках. В гранулезных клетках фолликула другой ядерный рецептор, гомологичный рецептор печени
(LRH-1), первично медиирует снижение эффекта ФСГ на яичники грызунов [128] [129]. Роль SF-1 и
LRH-1 в стероидогенезе у человека в преовуляторных гранулезных клетках до конца не определена.
Поскольку гранулезные клетки не имеют прямого контакта с кровоснабжением, ароматаза (P450arom)
в гранулезных клетках зависит от андростендиона, который необходим в качестве субстрата, диффун¬
дирующего из тека-клеток. Два важных этапа в фомировании эстрадиола — внедрение холестерина в
митохондрии, которому способствует белок острой регуляции стероидогенеза (StAR) в тека-клетках, и
превращение андростендиона в эстрон, катализируемое ароматазой (P450arom) в гранулезных клетках;
47
признаков, росту матки, увеличению толщины слизистой влагалища, истончению
слизистой эндоцервикса и линейному росту протоковой системы груди.
С21-СТЕРОИДЫ
Главные прогестагены — С21-стероиды, включая прегненолон, прогестерон и
ІУ-гидроксипрогестерон (см. рис. 1-18). Прегненолон особенно важен для яичника,
поскольку является предшественником всех стероидных гормонов. Прогестерон —
главный продукт желтого тела, отвечающий за прогестагенный эффект (то есть
дифференцировку клеток, индукцию секреторной активности эндометрия, ранее
получившего воздействие эстрогенами). Прогестерон требуется для имплантации
оплодотворенного яйца и поддержки беременности. Он также индуцирует децидуали-
зацию эндометрия, ингибирует сокращения матки, увеличивает вязкость цервикаль¬
ной слизи, способствует развитию альвеол молочной железы и увеличивает базаль¬
ную температуру. С другой стороны, 17-гидроксипрогестерон, также секретируемый
желтым телом, имеет незначительную биологическую активность.
С19-СТЕРОИДЫ
Яичник — источник различных С19-стероидов, включая дегидроэпиандростерон
(ДГЭА), андростендион и тестостерон (см. рис. 1-18). Они образуются тека-клетками
и клетками овариальной стромы. Главный С19-стероид — андростендион, часть
которого секретируется прямо в плазму, а меньшая часть превращается в эстроген
гранулезными клетками. Андростендион может конвертироваться в эстроген или
тестостерон в яичнике или экстагенитальных тканях. Только тестостерон и дигидро¬
тестостерон, а не андростендион, являются настоящими андрогенами с возможностя¬
ми связывания с андрогеновыми рецепторами (см. ниже).
Стероиды, синтезированные в яичниках и в других стероидпродуцирующих тка¬
нях, происходят из холестерина (см. рис. 1-18). Существует несколько источников
холестерина, которые могут снабжать яичник субстратами для стероидогенеза. Они
включают: холестериновый липопротеин плазмы, холестерин, синтезированный в
яичнике, и холестерин из внутриклеточных запасов холестериновых эфиров внутри
липидных капель. Холестериновый липид низкой плотности — важный источник
холестерина, используемого в стероидогенезе яичника человека [96]. ЛГ стимулирует
активность аденилатциклазы, увеличивая продукцию циклической аденозинмо-
нофосфорной кислоты (цАМФ), которая служит вторым мессенджером, увеличи¬
вающим количество рецепторов мРНК, связывание холестериновых ЛПНП, а также
формирование холестериновых эфиров [95, 96]. Производный ЛПНП-холестерин
чрезвычайно важен для нормального уровня продукции прогестерона в лютеинизи¬
рованных гранулезных клетках желтоію тела (см. рис. 1-19) [96].
Первой и определяющей скорость стадией в синтезе всех яичниковых стероидных
гормонов является процесс проникновения холестерина в митохондрии, регулируе¬
мый StAR (см. рис. 1-18 и 1-19) [18,19,135]. Это проникновение следует за превраще-
Б — в желтом теле гранулезные клетки хорошо васкуляризованы, что является критичным для проник¬
новения холестерина в эти клетки через первичные рецепторы липопротеинов низкой плотности и для
секреции прогестерона в сосудистое русло. Вхождение холестерина в митохондрии (с помощью StAR)
представляет наиболее важный этап для формирования прогестерона в лютеинизированных гранулезных
клетках. Андростендион, вырабатываемый в тека-лютеиновых клетках, используется как субстрат для
продукции эстрадиола в лютеинизированных гранулезных клетках. Предполагают, что у человека LRH-1
может регулировать часть гонадотропинзависимого стероидогенеза в желтом теле [130]. Таким обра¬
зом, гонадотропины SF-1 и, возможно, LRH-1 играют ключевые роли для важных стероидогенных шагов
в яичнике. АТФ — аденозинтрифосфат; цАМФ — циклический аденозинмонофосфат; FSH-R — рецептор
ФСГ; ГСД — гидроксидегидрогеназа; LH-R — рецептор ЛГ.
48
ниєм холестерина в прегненолон, катализируемым митохондриальным ферментным
комплексом, состоящим из P450SCC, адренодоксина и флавопротеина. ЛГ индуцирует
стероидогенез через увеличение внутриклеточного цАМФ, который усиливает кон¬
версию холестерина в прегненолон двумя путями:
1) быстрая регуляция (около 1 мин), происходящая через фосфорилирование
существующего StAR и быстрого синтеза нового StAR-протеина;
2) хроническая стимуляция, требующая от часа до нескольких дней путем индукции
экспрессии P450SCC и, как следствие, увеличения стероидогенеза (см. рис. 1-19). StAR
увеличивает количество холестерина, поступающего в митохондрии, таким образом
регулируя доступность субстрата независимо от количества имеющегося P450scc на
внутренней митохондриальной мембране [135]. В отсутствие StAR только 14% макси¬
мально индуцированного StAR уровня стероидогенеза существует независимо.
Эспрессия StAR в преовуляторном граафовом фолликуле первично ограничена
тека-клетками (см. рис. 1-19) [136]. Наиболее важным продуктом тека-клеток во
время фолликулярной фазы является андростендион — предшественник эстроге¬
нов, продукция которого контролируется первично с помощью StAR (см. рис. 1-19).
Биологически активный продукт яичника во время фолликулярной фазы представ¬
лен эстрадиолом, который синтезируется в гранулезных клетках, расположенных по
соседству с тека-клетками (см. рис. 1-19). Этапом, лимитирующим скорость продук¬
ции эстрадиола гранулезными клетками, служит регуляция активности фермента аро¬
матаза в циклической форме с помощью ФСГ (см. рис. 1-19) [114]. Во время лютеи¬
новой фазы клетки желтого тела, включая лютеинизированные гранулезные клетки,
также показывают интенсивную иммунореактивность StAR с мозаичным распределе¬
нием (см. рис. 1-19) [98,136]. Доставка холестерина в митохондриальную фермент¬
ную систему расщепления боковых цепей в желтом теле — реакция, определяющая
скорость биосинтеза прогестерона и регулируемая StAR (см. рис. 1-19) [98]. Таким
образом, продукция эстрадиола регулируется первично с помощью StAR и P450arom,
тогда как биосинтез прогестерона может быть первичен под контролем StAR.
Яичниковые клетки гранулезы, теки и желтого тела обладают StAR плюс пятью
различными протеинами со специфической ферментной активностью, необходимы¬
ми для образования стероидов. Это стероидогенные ферменты — P450scc, 3ß-HSD-II,
Р450СІ7, P450arom, и 17ß-HSD-l [131]. Данные ферменты отвечают за превращение
холестерина в два больших биологически активных продукта — эстрадиол и проге¬
стерон [137].
Стероидогенез, зависимый от ЛГ и ФСГ, в тека- и гранулезных клетках медиируется
общими сигнальными молекулами, включая цАМФ и специфические транскрипци¬
онные факторы: стероидогенный фактор 1 (SF-1), гомологичный рецептор печени 1
(LRH-1), которые относятся к семейству ядерных рецепторов (см. рис. 1-19) [138,139].
SF-1 и LRH-1 регулируют экспрессию генов, кодирующих StAR, P450scc, 3ß-HSD-II,
Р450СІ7 и P450arom (см. рис. 1-19). Таким образом, SF-1 и, возможно, LRH-1 могут
рассматриваться как основные регуляторы овариального стероидогенеза [139].
Заключение по усовершенствованной двухклеточной теории
стероидогенеза
Классическая двухклеточная теория подтверждается нижеследующими молекуляр¬
ными данными. Стероидогенез в яичнике в преовуляторном фолликуле регулируется
через ЛГ-рецепторы на тека-клетках и ФСГ-рецепторы (плюс, возможно, ЛГ) — на
гранулезных клетках (см. рис. 1-19). Продукция циклической АМФ-кислоты и уве¬
личенное связывание SF-1 с множеством стероидогенных промотеров опосредуют
активность ЛГ в тека-клетках. Б частности, StAR является первичным регулятором
49
продукции андростендиона, который впоследствии диффундирует в гранулезные
клетки и служит, в свою очередь, предшественником эстрогенов, в тека-клетках
преовуляторного фолликула холестерин возникает из циркулирующих липопротеи-
дов путем биосинтеза de novo. ФСГ отвечает за рост фолликулов, а также формиро¬
вание эстрогенов. ФСГ индуцирует формирование цАМФ, увеличенную активность
связывания LRH-1 и/или SF-1, а также экспрессию F450arom в преовуляторных
гранулезных клетках, чтобы впоследствии способствовать первичной ароматизации
андростендиона в эстрадиол (см. рис. 1-19). Голь SF-1 и LRH-1 в клетках яичниковой
гранулезы до овуляции неизвестна. Анализ данных, полученных в ходе исследований
в отношении грызунов, позволяют предположить, что LRH-1 может быть первичным
регулятором [128,129,140].
Большие запасы холестерина в желтом теле создаются из циркулирующих липо¬
протеинов для поддержки продукции сверхвысоких концентраций прогестерона.
Другие ключевые события в формировании желтого тела — разрушение базальной
мембраны между гранулезными и тека-клетками и интенсивная васкуляризация
лютеинизированных гранулезных клеток (см. рис. 1-15). Тека-лютеиновые клетки
имеют рецепторы к ЛГ и образуют андростендион. Циклическая АМФ, SF-1 и StAR,
индуцированные ЛГ, требуются как ключевые регуляторы для биосинтеза андростен¬
диона в тека-клетках, который, в свою очередь, является предшественником синтеза
эстрогенов в соседних лютеинизированных гранулезных клетках (см, рис. 1-19).
Лютеинизированные гранулезные клетки желтого тела и анатомически, и функ¬
ционально отличаются от эквивалента в преовуляторном фолликуле. Бо-первых, эти
клетки лютеинизированы, хорошо васкуляризированы и содержат большое коли¬
чество холестерина. Бо-вторых, лютеинизированные гранулезные клетки содержат
большое количество рецепторов к ЛГ в добавление к ФСГ-рецепторам, Б-третьих,
они образуют большие количества прогестерона и первично регулируются ЛГ и
StAR. Лютеинизированные гранулезные клетки также ароматизируют андростендион
текального происхождения и в итоге способствуют продукции эстрадиола через сти¬
муляцию ФСГ и F450arom. Хорошо известные медиаторы ЛГ и ФСГ в человеческих
лютеинизированных гранулезных клетках — увеличенные уровни цАМФ и LRH-1
[130]. Голи LRH-1 и SF-1 для продукции прогестерона и эстрадиола в лютеинизиро¬
ванных гранулезных клетках до конца не ясны. Специфические функции двух гонадо¬
тропинов (то есть дифференцирцировка, рост и формирование прогестерона в отли¬
чие от формирования эстрадиола), вероятно, определяются пока еще не изученными
модифицирующими факторами (см. рис. 1-19)
Пептидные гормоны, продуцируемые яичником
Яичник образует большое количество пептидов, которые могут действовать интра-
кринно, паракринно и эндокринно. Они включают несколько ростовых факторов
[например, инсулиноподобные факторы роста (ИФГ)] и цитокины (в частности,
интерлейкин-1Ь). ИФГ вместе с ФСГ-зависимым каскадом усиливают эффект ФСГ на
гранулезные клетки.
Группа пептидов, включая ингибин, активин и фоллистатин, образуется под кон¬
тролем ФСГ и ЛГ в гранулезных клетках яичника (рис. 1-20). Продукция ингибина
и активина не ограничивается яичником. Определенное количество других тканей,
включая надпочечник, гипофиз, а также плацента синтезируют эти пептиды. Были
выделены две изоформы ингибина — ингибин А и ингибин Б. Оба содержат схожие
субъединицы а, но отличающиеся субъединицы (3 (РА и РБ). Каждая субъединица
кодируется определенным геном. Гетроспективно гетеродимеры ингибина аРА и аРБ
определены как ингибин-А и ингибин-Б (см. рис. 1-20). Несмотря на то что ингибин
50
а-Субъединица ингибина Рлв-Субъединица ингибина
I Предшественник ^„
Рге Pro а1\1 аС м КТ'Д Предшественник
Рге Pro р
Y?A Ц-у- Pro-aN-aC I ' I р Мономер
aN
Активин
Р
Ингибин \ ^ \ р Активин А, АВ или В
6к 23к 20к
а
43к 15к
105 кДа
Ра,в Фоллистатин
Е 95 кДа 31_44|^
ХЦ 75 кДа
55 кДа
Рис. 1-20. Схематическое представление предшественников субъединиц ингибина и обработанных
форм в плазме, активины и фоллистатины. Точный вклад каждого ингибина определенной молекулярной
массы в биологическую активность до конца не ясен, однако показано, что 55-кДа- и 32-кДа-формы
биологически активны. [Из Burger Н. Inhibin, activin and neoplasia // Yen S.С., Jaffe R.B., Barbieri R.L. (eds).
Reproductive Endocrinology. —4th ed. — Philadelphia: WB Saunders. — 1999. — P. 669-675.]
образуется определенными тканями, наибольшее количество синтезируется в гона¬
дах, В яичнике источник ингибина — гранулезные клетки. Главная роль ингибина
заключается, как следует из названия, в подавлении синтеза продукции ФСГ в яични¬
ке (см. рис. 1-1, Б и 1-4 [1, 4]).
Хотя обе изоформы ингибина, похоже, имеют схожие биологические свойства,
их синтез регулируется по-разному во время фолликулярной и лютеиновой фаз (см.
рис. 1-1, А). Под влиянием ФСГ ингибин В секретируется в основном в раннюю фол¬
ликулярную фазу, далее концентрация снижается в середине фолликулярной фазы
и исчезает после пика ЛГ. Индуцированный ЛГ уровень ингибина А, как правило,
низкий в первую половину фолликулярной фазы, но градиентно возрастает во время
середины фолликулярной фазы с пиком во время лютеиновой фазы. Все три субъе¬
диницы определяются в молодых антральных фолликулах иммуногистохимически
и методом гибридизации in situ [141, 142]. а- и (ЗА-Субъединицы обнаруживают в
доминантных фолликулах и в желтом теле. Все три единицы экспрессируются в ответ
на воздействие гонадотропинов или факторов, которые увеличивают внутриклеточ¬
ную цАМФ [143].
Активин структурно относится к ингибинам, но может проявлять противоположное
действие. Активины содержат две субъединицы, которые идентичны р-субъединицам
ингибинов А и в. Таким образом, активин представлен тремя изоформами: активин
А (рАрА), активин В (рВрВ) и активин АВ (рАрВ). В гипофизе активин стимулиру¬
ет освобождение ФСГ. В фолликуле яичника активин усиливает действие ФСГ (см.
рис. 1-20). Как в случае ингибина, активины также производятся в гранулезных клет¬
ках яичника и в гонадотрофах гипофиза. В противоположность ингибину локально
синтезируемый в гипофизе активин в большей степени регулирует ФСГ, чем яичнико¬
вый активин (см. рис. 1-4).
51
Фоллистатин — пептид, состоящий из одной единицы, который продуцируется в
некоторых тканях человека, включая гипофиз и яичник (см. рис. 1-20). Он связыва¬
ется и нейтрализует биологические функции активина. Предполагают, что локальные
уровни фоллистатина в тканях модулируют эффект активина. Этим объясняется
ингибирующий эффект фоллистатина на секрецию ФСГ гипофизом (см. рис. 1-4).
Гормональные изменения во время овариального цикла
Секреция ФСГ подавляется по механизму отрицательной обратной связи продукци¬
ей овариальных гормонов эстрогена, ингибина, а также прогестерона во время ранней
и середины лютеиновой фазы. Во время регрессии желтого тела в позднюю лютеино¬
вую фазу обычно резкое падение уровней эстрогенов, ингибина и прогестерона анну¬
лирует эту обратную связь (см. рис. 1-1, А). Это, в свою очередь, увеличивает секрецию
ФСГ незадолго до и сразу после менструации. Такое начальное увеличение ФСГ необхо¬
димо для выбора фолликула, его роста и стероидогенеза (см. рис. 1-1, А). С последую¬
щим ростом фолликула аутокринные и паракринные факторы, производимые внутри
фолликула, поддерживают чувствительность фолликула к ФСГ. Продолжительное и
сочетанное воздействие ФСГ и активина приводит к возникновению рецепторов к ЛГ
на гранулезных клетках, предпосылке для овуляции и лютеинизации.
Овуляция инициируется за счет быстрого подъема содержания эстрадиола.
Положительная обратная связь на уровне переднего гипофиза и, возможно, гипотала¬
муса приводит к выбросу ЛГ, который необходим для выхода яйцеклетки в середине
цикла и формирования желтого тела (см. рис. 1-1, А). После овуляции происходит
подъем уровня прогестерона в течение 14 дней лютеиновой фазы, сопровождающийся
вторым подъемом эстрадиола, а также низким уровнем ФСГ и ЛГ. Гибель желтого тела
вместе с падением уровня гормонов (прогестерона, эстрадиола и ингибина А) позволяет
ФСГ вновь увеличить концентрацию до конца лютеиновой фазы, таким образом ини¬
циируя последующий цикл (см. рис. 1-1, А). Если возникает беременность и происходит
имплантация бластоцисты, как правило, структурная целостность и функция (продук¬
ция прогестерона и эстрадиола) желтого тела поддерживается ХГЧ, который секретиру¬
ется из трофобласта. ХГЧ действует по отношению к желтому телу как суррогат ЛГ.
Кроме ФСГ и ЛГ, локальные факторы (активин и ингибин) регулируют развитие
фолликулов и стероидогенез. в ранней фолликулярной фазе активин, продуцируемый
гранулезными клетками в незрелых фолликулах, усиливает эффект ФСГ на арома-
тазную активность и формирование рецепторов к ФСГ и ЛГ, одновременно подавляя
образование С19-стероидов в тека-клетках. В поздней фолликулярной фазе увели¬
ченная продукция ингибина гранулезными клетками и сниженный уровень активина
способствуют синтезу С19-стероидов в слое тека-клеток в ответ на ЛГ и локальные
ростовые факторы, а также цитокины, что приводит к увеличению содержания андро¬
стендиона — предшественника, необходимого для синтеза эстрона и эстрадиола в
гранулезных клетках [144].
Оба процесса и опосредованный ЛГ-синтез андростендиона в тека-клетках и опо¬
средованный ФСГ-синтез эстрадиола потенцируются ИФР [145]. Основной эндоген¬
ный ИФР, продуцируемый в гранулезных и тека-клетках, — ИФР-П (в сравнении с
ИФР-1). Эффекты ИФР-1 и ИФР-П опосредуются рецепторами к ИФР-1 типа в обоих
типах клеток. Рецептор к ИФР типа I структурно похож на рецептор к инсулину.
Получается, что зависящее от гонадотропина действие ИФР в яичнике первично регу¬
лируется рецепторами ИФР-П и ИФР-1 [145].
Таким образом, овуляция находится под контролем субстанций, функционирую¬
щих как классические гормоны (ФСГ, ЛГ, эстрадиол и ингибин), осуществляющих
связь между яичником и гипоталамо-гипофизарной осью, и паракринными и ауто-
52
кринными факторами, такими как ИФР-П, ингибин и активин, которые координи¬
руют свои действия для того, чтобы фолликул овулировал. Обратная отрицательная
связь между продуктами желтого тела (эстрадиол, прогестерон и ингибин) и ФСГ
приводит к критически важному подъему уровня ФСГ непосредственно до и во время
менструации, а обратная положительная связь между эстрадиолом и ЛГ обеспечи¬
вает овуляцию (см. рис. 1-1). в яичнике ИФР-П, ингибин и активин модифицируют
фолликулярные ответы, необходимые для роста и развития. Эти эндокринные, пара¬
кринные и аутокринные факторы, несомненно, представляют только часть общей
картины. Причины ановуляции разнообразны и могут быть обусловлены дефектами
рецепторов на йоверхности клеток, внутриклеточных элементов сигнальной транс-
дукции или межклеточными взаимодействиями [146].
Экстраовариальный стероидогенез
У женщин репродуктивного возраста образование эстрадиола происходит в опреде¬
ленных тканях трех категорий: яичник, периферические ткани, такие как подкожный
жир и кожа, а также физиологические и патологические органы-мишени — гипота¬
ламус, клетки опухоли груди, эндометриоидные клетки (рис. 1-21) [126]. Последние
два источника эстрогенов особенно важны при ановуляции в пре- и постменопаузе.
Поскольку небольшое количество эстрогенов также производится в фибробластах
жировой ткани или кожи, эти клетки вследствие своей распространенности влияют на
Надпочечник
Яичник
Ароматаза
Щ-ГСД
\
Жировая
ткань
Периферические
ткани
Рис. 1-21. Биосинтез эстрогенов у женщин. Биологически активный эстроген эстрадиол (Е^) образуется
в трех основных местах: (1) прямая секреция из яичника у женщин репродуктивного возраста; (2) конвер¬
сия циркулирующего андростендиона (А) надпочечникового и/или овариального происхождения в эстрон
(Е,) в периферических тканях; (3) превращение андростендиона в Е^ в органах-мишенях. В последних
двух субстанциях слабый эстроген Е, превращается далее в Е в тех же самых тканях. Присутствие
ферментов ароматазы и 17р-гидроксистероиддегидрогеназы (17р-ГСД) критично для формирования Е^
в этих тканях. Образование Е^ путем периферической и локальной конверсии особенно важно у пост¬
менопаузальных пациенток и при эстрогензависимых заболеваниях, таких как рак груди, эндометриоз
и рак эндометрия.
53
концентрацию циркулирующего эстрадиола (см. рис. 1-21) [126]. Этот эффект более
выражен у тучных женщин из-за увеличенного количества жировой ткани и площади
кожного покрова [124].
Р450агот в жировой ткани и в фибробластах кожи ответствен за периферическую
ароматизацию андростендиона, который в пременопаузе вырабатывается в надпо¬
чечниках и яичнике и первоначально в надпочечнике в постменопаузе (см. рис. 1-21).
Продукт этой реакции — эстрон — является слабым эстрогеном. Далее эстрон пре¬
вращается в сульфат эстрона, который служит резервуаром для эстрона в крови и
других тканях. Эстрон, образующийся из андростендиона и эстрона сульфата, затем
становится биологически активным эстрадиолом в органах-мишенях, таких как
эндометрий или грудь, с помощью ряда ферментов с перекрывающейся восстанови¬
тельной активностью 17(3-ГСД (см. рис. 1-21) [133, 147, 148]. Считают, что локаль¬
ная экспрессия Р450агот в гипоталамусе критически важна для регуляции секреции
гонадотропинов [149]. В итоге эстрогензависимые патологические ткани, такие как
опухоль молочной железы и эндометриоз, содержат исключительно высокие уровни
Р450агот, которые улучшают рост ткани посредством увеличения локальной концен¬
трации эстрадиола (см. рис. 1-21) [150]. Циркулирующий андростендион — основ¬
ной субстрат для ароматазной активности этих физиологических и патологических
органов-мишеней [147,150].
Значительное количество циркулирующего андростендиона может быть конверти¬
ровано в тестостерон в периферических тканях (см. ниже). Вероятно, это совершается
большим количеством 17р-ГСД с перекрывающейся восстановительной активностью
в периферических тканях [133]. Андрогенная активность тестостерона существенно
увеличивается при его конверсии в дигидротестостерон в периферических тканях и
органах-мишенях (кожа, предстательная железа). По крайней мере два фермента,
кодируемые двумя разными генами, 5а-редуктаза типа 1 и типа 2, катализируют
конверсию тестостерона в дигидротестостерон в коже, простате и печени [127].
Локальная продукция дигидротестостерона фибробластами в коже гениталий важна
для нормальной внутриутробной маскулинизации наружных гениталий у плода муж¬
ского пола [127]. Дигидротестостерон, формирующийся в коже, играет важную роль
в этиопатогенезе гирсутизма (см. ниже) [151].
ЭНДОМЕТРИЙ
Эндометрий — слизистая оболочка матки. Децидуальной оболочкой называется
высокоспециализированный эндометрий во время беременности. С точки зрения эво¬
люции человеческий эндометрий приспособлен для гемохориоэндотелиального типа
плацентации, который требует спиральных артерий (рис. 1-22). Клетки трофобласта
бластоцисты внедряются в спиральные артерии во время имплантации, что приводит
к возникновению маточно-плацентарных сосудов.
Спиральные артерии эндометрия человека также подвергаются другому уникальному
процессу, называемому менструацией. Менструация — отслаивание ткани эндометрия,
сопровождающееся кровотечением, зависящим от изменения уровня половых гормонов
в спиральных артериях. Присутствие спиральных артерий существенно для менструации,
поскольку только человек и несколько других приматов обладают подобным строением
эндометрия. При овуляторном менструальном цикле, не закончившемся оплодотворени¬
ем, менструация вызывает десквамацию эндометрия. Новый рост эндометрия и его раз¬
витие должны быть инициированы при каждом яичниковом цикле, так чтобы созревание
эндометрия соответствовало следующей возможности возникновения беременности.
Существует узкое «окно» между 20-м и 24-м днями 28-дневного менструального цикла,
когда эндометрий обладает возможностью принять бластоцисту [152].
54
Рис. 1-22. Функциональная анатомия эндометрия. Эндометрий — многослойная слизистая оболочка,
необходимая для имплантации и поддержания беременности. Единственный слой эпителиальных клеток
покрывает поверхность стромы и проникает в нее глубокими инвагинациями, достигая мест соприкос¬
новения миометрия с эндометрием. Толщина эндометрия пенетрируется спиральными артериями и их
капиллярами. Спиральные артерии возникают из радиальных ветвей аркуатных артерий, которые, в свою
очередь, возникают из маточных артерий. Поверхностный (функциональный) слой отслаивается во время
менструации, тогда как постоянный (базальный) слой в основании — источник для регенерации клеток
после каждой менструации. Значительные изменения в спиральных артериолах (скручивание, стаз, рас¬
ширение сосудов, следующие после интенсивной вазоконстрикции) постоянно наблюдают до начала
каждой менструации. [Courtesy of Kristof Chwalisz, River Forest I.L.]
Функциональная анатомия эндометрия
Морфологически эндометрий может быть разделен на верхние две трети, представ¬
ленные функциональным слоем, и нижнюю треть, представленную базальным слоем
(см. рис. 1-22). Цель функционального слоя — подготовка места для имплантации
бластоцисты; поэтому здесь происходит пролиферация, секреция и дегенерация. Цель
базального слоя — способствовать регенерации эндометрия после менструальной
потери функционального слоя [153]. Основные гистологические компоненты эндо¬
метрия включают: стромальные клетки, составляющие скелет ткани: единственный
слой эпителиальных клеток, покрывающих полость матки, включая инвагинации в
строму: кровеносные сосуды; резидентные иммунные клетки. Эпителиальные клет¬
ки, покрывающие глубокие инвагинации стромы, рассматриваются как железистые.
Необходимо отметить, что, как правило, эти глубокие крипты представляют простые
расширения полости матки и не являются настоящими железами. Эти инвагинации,
покрытые эпителиальными клетками, — продолжение функционального слоя эндо¬
метрия (покровный эпителий), проникающие глубоко до уровня базального слоя
(железистый эпителий). Таким образом, после отслоения функционального слоя во
время менструации базальный слой, состоящий из эпителиальных и стромальных
клеток, служит источником нового функционального слоя для следующего цикла
(рис. 1-23).
55
3-5
8-9
Дни
14 15-18
25
I
28-1
I
3-5
га ’I
:£ 2
Менстру¬
альная
Менстру¬
альная
Рис. 1-23. Циклические изменения в толщине и морфологии эндометрия и отношение этих изменений
к событиям в овариальном цикле. [Из Cunningham F.G., MacDonald Р.С., Gant N.F. et al. The endometrium
and decidua: menstruation and pregnancy // Williams Obstetrics. — 19th ed. — Stamford, CT: Appleton &
Lange.- 1993.-P. 81-109.]
Клеточный компонент функционального слоя подвергается выраженному раз¬
витию во время менструального цикла, тогда как базальный лишь незначительно
изменяется. Последовательность изменений в эндометрии, связанная с овуляторным
циклом, была тщательно изучена Noyes с соавт. у человека и Магкее [154, 155] и
Bartelmez [156] у приматов и детально изображена на рис. 1-23.
Морфологические изменения эндометрия,
индуцированные гормонами
Циклические изменения в гистологии эндометрия точно воспроизводятся каждый
овуляторный менструальный цикл. Данные изменения, зависящие от воздействия
половых гормонов, могут быть представлены следующим образом.
• В преовуляторной, или фолликулярной, фазе цикла эстрадиол секретируется
(главным образом единственным доминантным фолликулом одного яичника) в
увеличивающихся количествах до момента овуляции.
• Во время постовуляторной, или лютеиновой, фазы цикла прогестерон секрети¬
руется желтым телом в увеличивающихся количествах (от 40 до 50 мг/день) до
середины лютеиновой фазы.
« Начиная с 7-8-го дня после овуляции уровни секреции прогестерона (и эстро¬
гена) желтым телом начинают постепенно снижаться до начала менструации
(см. рис. 1-1).
В ответ на эти циклические изменения в эндометриальном цикле определяют пять
основных стадий:
<7 менструально-постменструальную реэпителизацию;
56
^ эндометриальную пролиферацию в ответ на стимуляцию эстрадиолом:
^ обильную эпителиальную секрецию в ответ на комбинированное воздействие
эстрадиола и прогестерона:
предменструальную ишемию как результат инволюции эндометрия, вызы¬
вающей стаз крови в спиральных артериях;
^ менструацию, которая предшествует и сопровождается вазоконстрикцией
эндометриальных спиральных артерий и коллапсом и десквамацией всех,
кроме глубоких, слоев эндометрия, в результате менструация представляет
собой последовательность отмены факторов, которые поддерживают рост
эндометрия и его дифференцировку (рис. 1-23).
в целом начало менструации связано с падением уровня прогестерона. Эта концеп¬
ция была разработана на основе опыта назначения эстрогенов, а затем прогестерона
женщинам в постменопаузе с последующей отменой прогестерона и возникновением
менструации даже при продолжении введения эстрогенов. Более того, прогестерон
способствует децидуализации эндометрия и поддерживает беременность, а отмена
прогестерона приводит к началу менструации, лактации и родов. Имеется множество
дополнительных факторов (отличных от снижения уровня прогестерона), которые
важны для успешного воплощения каждого из событий.
Как преовуляторная (фолликулярная или пролиферативная), так и постовулятор-
ная (лютеиновая или секреторная) фазы менструальных циклов привычно делятся на
раннюю и позднюю стадии (см. рис. 1-1). Нормальная секреторная фаза менструаль¬
ного цикла может быть точно разделена (практически по дням) по гистологическим
критериям на постовуляторную и фазу начала менструации. В действительности^
Noyes и другие исследователи детально описали гистологические особенности секре¬
торной фазы эндометрия, которая позволяет расписывать во времени лютеиновую
фазу цикла. Гинекологи используют гистологическое датирование эндометриальных
биопсий, полученных во время лютеиновой фазы, для оценки овуляции, продукции
прогестерона или уровня биологического ответа эндометрия на стимуляцию проге¬
стероном. Считают, что эндометрий развивается нормально при совпадении данных
гистологического и хронологического датирования в пределах 2 дней. При отличии
более чем на 2 дня эндометрий считают несоответствующим фазе. Эндометрий, не
соответствующий фазе, может быть причиной нарушения имплантации и бесплодия.
Если лечение бесплодия основано на результатах биопсий, то понимание ограничений
этих тестов важно, поскольку чувствительность и специфичность датирования биоп¬
сии эндометрия при определении бесплодия неизвестны. Например, одно из важных
ограничений этого теста — различие в интерпретации гистологических результатов
биопсии в рамках проводимого исследования.
Эффекты яичниковых стероидов на эндометрий
Эстрадиол или синтетические эстрогены вызывают увеличение толщины эндоме¬
триальной ткани. И стромальные, и эпителиальные клетки эндометрия быстро про¬
лиферируют под влиянием эстрадиола. Эстрогены увеличивают митотическую актив¬
ность и синтез ДНК в обоих типах клеток (рис. 1-24). Наряду с поддержкой роста
эстрогены подготавливают эндометриальную ткань к ответу на воздействие прогесте¬
рона, индуцируя экспрессию рецепторов к прогестерону, поскольку для эффектов про¬
гестерона необходимо предшествующее или одновременное воздействие эстрогенов
на эндометрий [157].
в противоположность пролиферативному эффекту эстрогена действие прогесте¬
рона первоначально способствует дифференцировке эндометрия. Например, проге¬
стерон может ингибировать и даже оказывать действие, противоположное пролифе-
57
Эпителий
/Пролифераций /Пролифераций /ПтролиферацияХ
Ангиогенез
Дифференцировп
Ангиогенез
Дифференцировке
Ангиогенез
Дифференцировф
Эстрадиол
Рис. 1-24. Важные эффекты эстрогенов на эпителий (например, синтез ДНК, пролиферация и экспрес¬
сия генов) первоначально опосредуются рецепторами к эстрогенам в стромальных клетках эндометрия
через паракринные механизмы. Это было продемонстрировано на мышах [158]. Также в исследованиях
на мышах и людях было показано, что антиэстрогенный эффект прогестерона на эпителиальные клетки
(например, снижение пролиферации и улучшенная дифференцировка) первоначально опосредуется
через рецепторы к прогестерону на стромальных клетках [159, 160]. РП — рецептор прогестерона;
РЭ — рецептор эстрогена.
ративному эффекту эстрогенов на функциональный слой (см. рис. 1-24) [158-160].
Более того, прогестерон подготавливает эндометрий к имплантации эмбриона через
процесс дифференцировки эпителиальных и стромальных клеток. Прогестерон инду¬
цирует продукцию и секрецию богатых гликогеном субстанций эпителиальными
клетками, также гормон увеличивает объем цитоплазмы стромальных клеток (псев-
додецидуализация).
ДЕЙСТВИЕ ЭСТРОГЕНА
Эстрадиол, наиболее активный, имеющий естественное происхождение эстроген,
секретируемый гранулезными клетками доминантного фолликула яичника, оказывает
влияние на эндометрий через схожие с другими стероидными гормонами механизмы
действия. Эстрадиол попадает в клетки посредством простой диффузии, но в эстроген-
чувствительных клетках связывается с рецепторами эстрадиола. Эстрогеновые рецеп¬
торы — протеины с высоким сродством к эстрадиолу и другим биологически актив¬
ным эстрогенам, то есть синтетическим эстрогенам. Несмотря на то что в эндометрии
представлены оба типа эстрогеновых рецептора — а и (3, считают, что рецептор а
является первоначальным медиатором эстрогенного воздействия на эндометрий [161,
162]. Комплекс эстрадиол-рецептор после трансформации становится транскрипци¬
онным фактором, который связан с эстроген-отвечающими участками специфических
генов. Это взаимодействие извещает о начале эстроген-специфической транскрипции
генов в организме, которые способствуют синтезу специфических мРНК, а затем
синтезу специфических протеинов [163]. Среди многих протеинов, синтезируемых
в большинстве эстрогенчувствительных клеток, имеются дополнительные рецепто¬
ры к эстрогенам и рецепторы к прогестерону. Таким образом, эстрадиол действует в
эндометрии и в других эстрогенчувствительных тканях с целью поддержки эффекта
эстрогена и обеспечения чувствительности данной ткани к прогестерону.
58
Эндометриальные эпителиальные клетки эстрогенчувствительны, но, вероятно,
они не пролиферируют в результате прямого действия эстрадиола. Пролиферация
эндометриальных эпителиальных клеток в культуре значительно не увеличивается в
ситуациях добавления эстрогенов к среде. Более того, эстрогены действуют на стро¬
мальные клетки, способствуя синтезу ростовых факторов эпителиальных клеток (см.
рис. 1-24) [158]. Эти ростовые факторы используют паракринные механизмы для
того, чтобы увеличить синтез ДНК и размножение соседних клеток эпителия. Данный
тип паракринного взаимодействия может быть общим механизмом, который способ¬
ствует действию эстрогенов в гормонзависимых тканях.
ДЕЙСТВИЕ ПРОГЕСТЕРОНА
Прогестерон также проникает в клетки с помощью диффузии и в прогестерончув-
ствительных клетках связывается с рецепторами, обладающими сродством к прогесте¬
рону. Обе изоформы рецепторов к прогестерону (ПР), ПР-А и ПР-Б представлены в
эндометрии [164]. Поскольку ПР-Б-, а не ПР-А-уровни в эндометрии точно регулиру¬
ются во время менструального цикла человека, считают, что ПР-Б играет более важную
биологическую роль [164]. Обычно количество рецепторов к прогестерону зависит
от предшествующего действия прогестерона. Комплекс «рецептор к прогестерону-
прогестерон» также вызывает транскрипцию генов, но ответ на прогестерон отличен от
ответа, вызваемого комплексом «рецептор к эстрадиолу-эстрадиол».
Эффекты прогестерона включают снижение синтеза молекул рецепторов к эстро¬
генам [165]. Один этот факт свидетельствует о том, что прогестерон (и синтетиче¬
ские гестагены) уменьшает эффекты эстрогенов. Прогестерон также увеличивает
ферментную реакцию инактивации эстрадиола в эстрон за счет увеличения актив¬
ности окислительного типа фермента 17р-ГСД. Прогестеронзависимая транскрипция
специфических генов 2-го типа 17р-ГСД ответственна за эту ферментную активность
[166]. Прогестерон также увеличивает другой способ инактивации эстрогенов —
сульфатирование эстрогенов (эстрогенсульфотрансфераза) [167]. Следовательно,
прогестерон действует как антиэстроген — по крайней мере тремя путями: через
уменьшение синтеза эстрогеновых рецепторов, посредством уменьшения концен¬
трации эстрадиола в тканях путем конверсии в эстрон и благодаря усилению инак¬
тивации эстрогенов путем сульфатации. Как и при действии эстрогенов на матку,
эксперименты с рекомбинацией тканей, с использованием ткани нормальной матки
мыши и с выключенными рецепторами к прогестерону показали, что многие эффекты
гормона на эпителиальные клетки опосредуются паракринными механизмами через
рецепторы к прогестерону в стромальных, а не эпителиальных клетках (см. рис. 1-24)
[159]. Прогестеронзависимая индукция 2-го типа фермента 17р-ГСД и последующая
инактивация эстрадиола первоначально медиируются через стромальные рецепторы
к прогестерону в эндометрии человека [160].
Наиболее значимый результат действия прогестерона в эндометрии — дифферен¬
цировка эндометрия. Гистологическое соотношение дифференцировки, децидуали¬
зации стромы секреции эпителия коррелирует с присутствием ядерных рецепторов
к прогестерону и увеличенным уровнем циркулирующего прогестерона во время
лютеиновой фазы. Корреляция на молекулярном уровне с учетом дифференцировки
включает продукцию лактоферрина и гликоделина в эпителиальных клетках, а также
пролактина и протеина, связывающего инсулиноподобный фактор роста-1, -в стро¬
мальных клетках эндометрия.
Рецептивная фаза эндометрия для имплантации
Если яйцеклетка не оплодотворяется в течение 24 ч после овуляции, она погибает.
Оплодотворение происходит в ампулярной части трубы (дистальная треть). Б течение
59
следующих 2 дней оплодотворенная яйцеклетка остается свободной в просвете трубы,
получая питательные вещества и энергию для ранних этапов деления из секрета
трубы и оставшихся прикрепленными гранулезных клеток яйценосного бугорка. По
завершении этого этапа плотный шарик из клеток (морула) покидает маточную трубу
и попадает в полость матки. К этому времени появившийся под воздействием про¬
гестерона секрет эндометрия заполняет полость матки и окружает эмбрион, снабжая
его питательными веществами. Это первое из многих точно синхронизированных
событий, которые происходят в системе «оплодотворенная яйцеклетка-эндометрий».
К 6-му дню после овуляции эмбрион на стадии бластоцисты готов к прикреплению
в матке. К этому времени он определил часть эндометрия достаточной глубины,
васкуляризации и богатую питательными веществами для поддержки последующей
плацентации. Сразу под эпителиальным слоем формируется богатая капиллярная
сеть, способная к участию в создании кровяного трофобласт-материнского взаи¬
модействия. Позже окружающая порция функциональной зоны, занимающая все
большую полость эндометрия, образует плотную пластинку, сохраняющую строение
эндометрия, несмотря на активность растущего трофобласта.
Для поддержания беременности важно действие прогестерона. Бластоциста зави¬
сит от прогестерона, продуцируемого в это время желтым телом. Секретируемый
трофобластом ХГЧ предупреждает регрессию желтого тела, действуя вместо ЛГ. Это
способствует снабжению прогестероном, необходимым для поддержания беремен¬
ности до того момента, как плацентарная ткань не станет продуцировать прогестерон
самостоятельно на 6-7-й нед после оплодотворения.
Исследования на экспериментальных и домашних животных показали, что для
нормальной имплантации и развития необходимо синхронное развитие плода и эндо¬
метрия. У лабораторных животных для оплодотворения существует лишь небольшое
временное окно, в некоторых случаях занимающее несколько часов.
Рецептивная фаза эндометрия — временной промежуток созревания эндометрия,
в течение которого трофэктодерма бластоцисты может прикрепиться к эндометри¬
альным эпителиальным клеткам и далее внедриться в строму эндометрия. Ключевой
вопрос исследования восприимчивости человеческого эндометрия — определение
этого временного промежутка имплантации. Только факторы, которые экспрессиру¬
ются во время этого окна, могут считаться маркерами или функциональными медиа¬
торами рецептивной стадии.
Окно имплантации может быть определено исходя из знаний, полученных от пере¬
носа эмбриона в матку женщины, предварительно получившей экзогенные эстроге¬
ны и прогестерон (рис. 1-25). Существует отдельное окно для переноса эмбриона и
имплантации, которое соответствует 16-20-му дню. Предварительно реальное окно
для имплантации следует за окном для переноса, поскольку эмбриону необходимо
определенное время для развития от 4-клеточной стадии до 8-клеточной, затем до
бластоцисты и инициации имплантации и внедрения.
Окно имплантации было определено в 20-24-й день цикла путем многократных
серийных измерений уровня ХГЧ в плазме как маркера первоначального взаимо¬
действия эмбриона и матери [168]. Таким образом, окно имплантации относительно
широкое (около 4 дней). Эти наблюдения подтверждаются ранее опубликованными
морфологическими данными Adam с соавт. [169].
60
Реципиент с яичниковой недостаточностью
Рис. 1-25. Схематичное представление донорства яйцеклетки в цикле экстракорпорального оплодотво¬
рения пациентке-реципиенту с прекращением функции яичников, пролеченной экзогенными эстрогенами
и прогестероном. А — женщина с прекращением функции яичников, получившая лечение эстрогенами
с 1-го по 14-й день цикла. Экзогенный прогестерон был добавлен к эстрогенам с 15-го дня цикла до
28-го дня и был продолжен, если была диагностирована беременность. Семь донорских яйцеклеток
были оплодотворены спермой мужа и пять эмбрионов были перенесены в матку с 16-го до 18-й день;
Б — донор ооцитов был пролечен человеческим менопаузальным гонадотропином до 8-го дня, потом
был введен ХГЧ, и через 32-36 ч были получены ооциты. Половина яйцеклеток была введена реципиенту,
другая половина была оплодотворена спермой мужа; пять оплодотворенных яйцеклеток были перенесе¬
ны в матку донора. Показаны уровни эстрадиола (Е^) и прогестерона (Р^) в плазме у обеих женщин. Для
конвертации эстрадиола в пикомоли на литр значение необходимо умножить на 3,671. Для конвертации
прогестерона в наномоли на литр необходимо умножить на 3,180. [Из Rosenwaks Z. Donor eggs: their
application in modern reproductive technologies // Fertil. Steril. — 1987. — N 47. — P. 895-909.]
Механизм менструации
Если нет беременности, отсутствие ХГЧ приводит к вазомоторным изменениям,
связанным с прекращением эстроген-прогестеронной стимуляции, и к менструальной
десквамации. Инициируется программа восстановления эндометрия: повреждения
во внеклеточном матриксе, инфильтрации лейкоцитами приводят к гипоксйческому
повреждению тканей и отслоению функционального слоя эндометрия с последующей
активацией гемостатического и регенеративного процессов. Основные гистологиче¬
ские критерии предменструальной фазы — деградация ретикулярной стромальной
сети, стромальная инфильтрация полиморфонуклеарными и мононуклеарными лей¬
коцитами, а также секреторное истощение желез эндометрия, чьи эпителиальные клет-
61
ки имеют базальные ядра. Эндометрий уменьшается перед менструацией в результате
уменьшения секреторной активности и катаболизма внеклеточного матрикса.
Наиболее значимый эффект прекращения стимуляции эстрогенами и прогесте¬
роном — менструация (см. рис. 1-23). Классические исследования Магкее [154]
показали, что ишемия, вызанная вазоконстрикцией артериол и спиральных артерий,
предшествует менструации от 4 до 24 ч. Кровотечение, возникающее после рассла¬
бления артериол и артерий, происходит из-за гипоксическо-реперфузионной травмы.
Поверхностные слои эндометрия, разрыхленные образованием гематом и щелей,
приводят к отделению фрагментов тканей. Происходят лизис и фрагментация клеток.
Менструальные выделения состоят из фрагментов эндометрия, крови и фибриноли-
зированных фрагментов крови (см. рис. 1-23). Могут быть сгустки различных разме¬
ров в случае значительного кровотечения.
Контроль эндометриальной функции у получающих
синтетические гормоны
Фертильность женщины определяется биологическим качеством ооцитов — СВОЙ¬
СТВОМ, которое частично отражается способностью оплодотворенной яйцеклетки к
делению и содержанием нормального набора хромосом. Такое качество яйцеклеток
резко снижается после 35 лет. В то же время эндометрий остается неизменным на про¬
тяжении продолжительного времени [170]. Получение ооцитов от женщин-доноров и
последующее искусственное оплодотворение этих яйцеклеток спермой реципиента-
партнера, перемещение в полость матки женщины, имеющей нефункционирующие
яичники (преждевременное прекращение функции яичников), с успехом использо¬
вали в качестве терапевтической стратегии для лечения бесплодия (см. рис. 1-25)
[170, 171]. Это клиническое применение дает уникальные возможности изучения
гормональных требований для созревания эндометрия. Предложено несколько про¬
токолов гормонотерапии, в результате были получены беременности с использовани¬
ем донорского генетического материала у женщин с преждевременным прекращением
функции яичников (см. рис. 1-25). Успех этих процедур достигает 50% (уровень бере¬
менностей) на перенос эмбрионов.
Уровень дифференцировки эндометрия в ответ на стимуляцию экзогенными гормо¬
нами определяется гистологическим анализом образцов эндометрия. Эпителиальные
элементы показывают запоздалое созревание во время назначения прогестерона на
20-22-й день, которое диагностируется до 26-го дня. Несмотря на видимую дисин¬
хронизацию, частота беременностей у данных пациенток с донорскими яйцеклетками
выше, чем у обычного искусственного оплодотворения [171].
Большинство специалистов по бесплодию в настоящее время используют поэтап¬
ное назначение пероральной формы микронизированного эстрадиола в 2,4, 6 мг еже¬
дневных дозах с последующим назначением 4-6 мг эстрадиола, комбинированного
с в/м прогестероном в дозе 50 мг, необходимым для секреторной трансформации.
Эстрадиол у данных пациенток во время заместительной «фолликулярной» фазы
достигает высоких уровней для стимуляции роста эндометрия. Бнутримышечные
инъекции масляного раствора прогестерона 50 мг/сут приводят к уровню прогесте¬
рона более 20 нг/мл. Длительность действия прогестерона, но не абсолютная концен¬
трация прогестерона в плазме крови, достигается после предварительной подготовки
эндометрия эстрогенами и является основным фактором развития чувствительности
матки. Таким образом, экзогенного назначения только эстрадиола и прогестерона
достаточно для подготовки эндометрия к имплантации в отсутствие функциони¬
рующих яичников. Это наблюдение подчеркивает ключевую роль таких стероидов в
физиологии матки.
62
ОБСЛЕДОВАНИЕ ЖЕНЩИН С РЕПРОДУКТИВНОЙ ДИСФУНКЦИЕЙ
Репродуктивная дисфункция у женщин проявляется чаще всего нарушением мен¬
струального цикла. Точное определение причин, лежащих в основе, требует хорошего
понимания физиологии и патологии женской репродуктивной системы, а также акку¬
ратного сбора информации, относящейся к заболеванию, и объективного исследова¬
ния. Без критического анализа клинических случаев, основанного на точном знании
нормы и патологии репродуктивной функции, применение алгоритмов лабораторных
исследований приводит к ненужному определению гормонов и откладыванию време¬
ни постановки диагноза.
Анамнез заболевания
Бажный момент для исследования женщины с репродуктивным заболеванием —
аккуратно записанный анамнез заболевания. Анамнез заболевания, полученный от
пациентки, должен оценивать биологические эффекты каждого из многообразия гор¬
монов. Запись деталей пубертатного развития как ссылки на начало важного симпто¬
ма снабжает критическими данными этиологии определенных репродуктивных забо¬
леваний. Например, ановуляция, проявляющаяся нерегулярными кровотечениями,
ассоциируется с синдромом поликистозных яичников (СПКЯ), часто начинающемся
во время пубертатного периода. Начало градиентно-прогрессирующего во время
пубертата гирсутизма предполагает неклассический вариант врожденной дисфункции
коры надпочечников (ВДКН) или СПКЯ. В этих случаях измерение уровня в плазме
17-гидроксипрогестерона может быть полезно в дифференцировке неклассической
ВДКН от СПКЯ (см. ниже). Возникновение гирсутизма до или несколько лет спустя
с начала пубертата должно просигнализировать врачу о возможности яичниковой
или надпочечниковой неоплазии. Неожиданное (или постепенное) возникновение
гирсутизма в любом возрасте или присутствие вирилизации должно напомнить врачу
о необходимости исключить яичниковые стероид-секретирующие или надпочечни¬
ковые опухоли. Большинство женщин с симптоматическим эндометриозом страдают
от серьезных эпизодов болезненных менструаций (дисменореи), начавшихся в пубер¬
татные годы.
Исследование женской репродуктивной функции включает детальную историю
менструальной функции. Например, СПКЯ маловероятен без длительной истории
нерегулярных с самого пубертата менструаций. Справедливо и то, что история
периода регулярных, предсказуемых менструаций до начала нерегулярных эпизодов
должно обратить внимание на гипоталамические или другие причины ановуляции.
Частота, регулярность, длительность и количество маточных кровотечений должны
быть тщательно зафиксированы в связи с несколькими причинами. Бо-первых, эта
информация отражает тесные взаимосвязи нескольких тканей, включая гипоталамус,
гипофиз, яичники и эндометрий. Бо-вторых, регулярные, предсказуемые менструа¬
ции способствуют овуляции. Б-третьих, определение типа нерегулярных менструаций
помогает с диагнозом лежащей в основе патологии. Например, длительная аменорея
у худой и эстрогендефицитной женщины предполагает ановуляцию гипоталамиче¬
ской этиологии. Нерегулярные периоды различной длительности и различной кро-
вопотери у женщины с нормальным уровнем эстрогенов предполагают первичную
овариальную дисфункцию, например СПКЯ. Необходимо помнить, что ановуляция
у худощавой женщины с нормальным уровнем эстрогенов может также возникнуть
в результате СПКЯ. Регулярные, но обильные и удлиненные менструации с переме¬
жающимися кровянистыми выделениями могут быть вызваны анатомическими нару¬
шениями, такими как аденомиоз или лейомиома. Наконец, неопластические заболе¬
вания эндометрия, включая эндометриальные полипы, гиперплазии или онкологию,
63
могут манифестировать нерегулярными кровянистыми выделениями. Комбинация
влагалищного ультразвукового исследования (УЗИ) и биопсии эндометрия исключи¬
тельно чувствительны в диагностике эндометриальной неоплазии [172].
Расстройство менструального цикла и непредсказуемые менструации — общий
и беспокоящий симптом, который первоначально приводит пациентку к врачу.
После тщательной оценки менструальных сиптомов врач должен обнаружить дру¬
гие заметные симптомы эндокринных расстройств, лежащих в основе меноррагий.
Беременность — наиболее частая причина аменореи (и, возможно, любой иной мен¬
струальной нерегулярности) у женщин репродуктивного возраста. У женщин с аме¬
нореей или иным нерегулярным проявлением менструации первоначально должна
быть исключена беременность (в том числе внематочная). Тщательная оценка репро¬
дуктивного анамнеза, сексуальной активности и контрацептивной практики позво¬
ляет предположить вероятность беременности. Более того, репродуктивный анамнез
позволяет заподозрить синдром Шиена (послеродовый пангипопитуитаризм), если
менструации не возобновились после родов, сопровождавшихся значительной кро-
вопотерей [173]. Б таких случаях также необходимо исключить гипотиреоз и надпо¬
чечниковую недостаточность. Классический пример синдрома Шиена проявляется
отсутствием лактации после родов, обусловленным дефицитом пролактина.
Аменорея традиционно подразделяется на первичную (отсутствие менструации
в анамнезе) и вторичную (задержка менструации после определенного периода).
Причины первичной аменореи различны. Несмотря на то что различия между пер¬
вичной и вторичной аменореей важны для идентификации механизмов, лежащих в
основе заболевания и дифференциальной диагностики, врач должен знать, что неко¬
торые заболевания первоначально представлены либо первичной, либо вторичной
аменореей. Например, большинство женщин с дисгенезией гонад имеют первичную
аменорею, однако у некоторых пациенток присутствуют остаточные фолликулы и ову¬
ляция; у таких женщин с частичной дисгенезией редкие менструации и редкие бере¬
менности могут возникать до нарушения функции яичников [174, 175]. Пациентки с
СПКЯ имеют обычно вторичную аменорею и очень редко первичную.
Бторичная аменорея часто возникает в связи с хронической ановуляцией, кото¬
рая может быть связана с дисфункцией гипоталамуса, галактореей, прекращением
функции яичников, избытком андрогенов, хроническими заболеваниями и первич¬
ной патологией матки (например, внутриматочные синехии, возникшие в резуль¬
тате послеродового кюретажа) [176]. Нахождение любых ассоциаций вторичной
аменореи с событиями в жизни весьма информативно. Тяжелые физические нагруз¬
ки часто ассоциируются с аменореей. Потеря массы тела нередко предшествует
или сопровождает вторичную аменорею, а также может быть доказательством дис¬
функции гипоталамуса. Необычная история питания может предполагать булимию
или нервную анорексию. Быскабливание полости матки в анамнезе, послеродовой
эндометрит или диссеминированный туберкулез и отсутствие или скудные мен¬
струации могут свидетельствовать о внутриматочных спайках [177]. Присутствие
любых симптомов или знаков дефицита эстрогенов, включая болезненный поло¬
вой акт, атрофию влагалища, эмоциональную лабильность, а также вазомоторную
лабильность, может говорить о центральном механизме ановуляции и низкой кон¬
центрации циркулирующих гонадотропинов (гипогонадотропном гипогонадизме)
или прекращении функции яичников при повышенном уровне гонадотропинов
(гипергонадотропном гипогонадизме).
Галакторея при отсутствии в анамнезе недавней беременности зачастую является
проявлением увеличенной секреции пролактина, а также может быть результатом
повышенной чуствительности ткани груди к гормонам, необходимым для продук¬
ции молока. Б анамнезе может быть выявлено употребление различных препаратов,
включая некоторые психотропные агенты, антигипертензивные средства, оральные
64
контрацептивы. Первичный гипотиреоз может быть связан с преждевременным поло¬
вым созреванием у детей, с галактореей, аменореей или и тем, и другим у взрослых
женщин. Чрезмерная стимуляция сосков или заболевания грудной клетки в анамнезе
могут служить причиной галактореи. Пролактиномы, пролактин-секретирующие аде¬
номы гипофиза — частая причина галактореи, проявляющейся аномально высокими
уровнями пролактина,
Физикальное обслбдование
Количество и распределение избыточного роста волос следует оценить с учетом
семейного анамнеза. Гипертрихоз — избыточный рост волос на ногах, шее, тулови¬
ще - должен быть дифференцирован от истинного гирсутизма, проявляющегося
развитием волос на лице, груди, а также мужским типом оволосения лобка в ответ
на увеличенную продукцию биологически активных андрогенов или вследствие
повышенной чувствительности к ним. Некоторый уровень гипертрихоза характе¬
рен для женщин, живущих в районах бассейна Средиземного моря, тогда как любое
проявление гирсутизма на лице у азиатских женщин требует дополнительного
изучения. Гирсутизм в наилучшей степени документируется с помощью фотографий.
Вирилизация характеризуется огрублением голоса, значительными кистозными акне,
потерей волос, увеличением мышечной массы, а также клиторомегалией и требует
значительно большего избытка андрогенов, чем тот, при котором обнаруживается
гирсутизм. Синдром полной андрогенной нечувствительности характеризуется ред¬
ким (или вовсе отсутствующим) лобковым и подмышечным оволосением в связи с
резистентностью к андрогенам.
Тщательное исследование молочных желез — необходимая часть физикально-
го обследования. Классификация степени развития молочных желез Маршалла и
Таннера [178] — удобное и ценное дополнение. И недоразвитие молочных желез (зна¬
чительный избыток андрогенов), и хорошо сформированные и пигментированные
ареолы (как во время беременности), и выделения из сосков должны быть тщательно
исследованы.
Женщины с СПКЯ, никогда не имевшие овуляции или получавшие гестагенсодер¬
жащие медикаменты, могут иметь 4-ю стадию развития молочных желез по Танеру,
что обусловливается адекватной продукцией эстрогенов, тогда как прогрессирование
к 5-й стадии требует воздействия прогестерона либо при овуляции, либо при приеме
гестагенов (например, назначения оральных контрацептивов).
Вульва, влагалище, а также шейка матки представляют собой чувствительные
индикаторы воздействия половых гормонов. Поскольку чувствительность кожи гени¬
талий и слизистой оболочки влагалища уменьшается с момента ранних стадий раз¬
вития плода до репродуктивного возраста, выраженность вирилизации может быть
полезной для определения времени воздействия андрогенов. Наиболее выраженные
андрогенные эффекты — слияние половых губ с образованием губчатой части муж¬
ского мочеиспускательного канала, часто наблюдают у пациенток, подвергавшихся
воздействию андрогенами в I триместре (12 нед) беременности. Подобные случаи
были описаны у пациенток с вирилизирующей формой ВДКН, истинным гермаф¬
родитизмом, а также с лекарственной вирилизацией. Значительная клиторомега-
лия после родов, с другой стороны, требует стимуляции гормонами, и в отсутствие
значительных экзогенных стероидов свидетельствует об андроген-секретирующей
опухоли. По сравнению с измерением длины клитора измерение его основания —
более точный метод определения андрогензависимого увеличения клитора. Индекс
клитора (определяемый как произведение сагиттального и поперечного диаметров в
основании) более 35 мм^ находится за пределами 95% доверительного интервала, то
есть выше нормы.
65
Влагалище и шейка матки — наиболее чувствительные индикаторы для влияния
эстрогенов. Под влиянием эстрогенов слизистая оболочка влагалища развивается в
период полового созревания из ткани с блестящей ярко-красной окраской и редкими
секретами в ткань с толстой серо-розовой складчатой поверхностью с обильными
секретами. Хорошо эстрогенизированная слизистая оболочка влагалища с растя¬
жимым секретом (кристаллизация слизи в шейке матки) может быть индикатором
пролиферативной фазы менструального цикла у женщин с овуляторным циклом или
внеовариального формирования эстрогенов у женщин с ановуляторным менструаль¬
ным циклом при СПКЯ. Биологическая активность эстрогенов также может быть
оценена с помощью цитологического метода.
Нерегулярные менструации — частый повод для обращения за врачебной помо¬
щью. Различные заболевания гипоталамуса, гипофиза, яичников или матки, а также
других тканей могут быть причиной этого беспокоящего симптома. После исключения
беременности необходимо собрать детальный анамнез и провести тщательное физи¬
кальное обследование. Очень важно, чтобы врач обратил внимание на яркие моменты
в анамнезе и биологические индикаторы действия гормонов в органах-мишенях во
время объективного обследования. Анализ этих данных нередко приводит к точному
диагнозу. В последующем диагноз должен быть подтвержден лабораторным исследо¬
ванием.
РАССТРОЙСТВА ЖЕНСКОЙ ПОЛОВОЙ СИСТЕМЫ
Хроническая ановуляция
Хроническая ановуляция — одна из наиболее частых проблем в гинекологической
практике. Эти женщины могут быть с симптомами вторичной аменореи, редкими,
нерегулярными менструациями (олигоменореей) или нерегулярными, но выражен¬
ными метроррагиями. Бесплодие — логичный результат хронической ановуляции.
Одна группа ановуляторных пациенток эстрогендефицитна. Общие симптомы этой
группы включают гипоталамическую ановуляцию, галакторею-гиперпролактинемию
(например, гипотиреоз, пролактинома, нефункционирующая опухоль гипофиза), а
также преждевременное прекращение функции яичников у женщин репродуктивного
возраста. Эти пациентки обычно страдают аменореей и имеют дефицит эстрогенов.
Одно из важных следствий — потеря костной ткани, приводящая к остеопении и осте-
опорозу. По возможности лежащие в основе причины должны быть корригированы.
В случае если овуляция не может быть восстановлена, должна проводиться замести¬
тельная гормональная терапия (ЗГТ).
Женщины с избытком андрогенов составляют вторую группу пациенток с ано¬
вуляцией. Серьезное следствие ановуляции в этой группе — более высокий риск
развития карциномы эндометрия вследствие неуравновешенного воздействия эстро¬
генов, формируемых в экстрагенитальных тканях. Наиболее частое заболевание,
связанное с избытком андрогенов и ановуляцией, — СПКЯ. Инсулин играет важную
роль в данном заболевании и вместе с гиперандрогенией увеличивает риск развития
сердечно-сосудистых расстройств и диабета [146]. Врач должен помнить о долго¬
срочном эффекте СПКЯ и разработать терапевтический план для этих ановуляторных
пациенток, дабы избежать нежелательных последствий заболевания. Врач должен
также разработать вместе с пациенткой план в отношении возможных долгосрочных
осложнений, вызванных неуравновешенными эстрогенами (например, опухоль эндо¬
метрия). Для предупреждения гиперплазии или рака эндометрия могут быть назначе¬
ны оральные контрацептивы или периодически гестагены.
66
Для практических целей представлены следующие пять групп, включающие боль¬
шинство этиологических факторов, приводящих к хронической ановуляциии у жен¬
щин репродуктивного возраста:
• гипоталамическая ановуляция;
• гиперпролактинемия;
• избыток андрогенов;
преждевременное прекращение функции яичников;
• хронические заболевания (например, печеночная или почечная недостаточность,
СПИД).
Существует множество механизмов, приводящих к ановуляции у женщин с хрони¬
ческим заболеваниями. Эффективная терапия первичного заболевания может вос¬
становить нормальную менструацию. С другой стороны, ановуляторное кровотечение
может быть лечено экзогенными гормонами, как будет показано ниже. Последующий
текст детально опишет специфические заболевания, вызывающие хроническую ано¬
вуляцию у женщин репродуктивного возраста.
ГИПОТАЛАМИЧЕСКАЯ АНОВУЛЯЦИЯ
Продукция ГнРГ в нейронах аркуатного ядра гипоталамуса и его секреция в пор¬
тальный кровоток в срединной возвышенности в пульсирующем режиме необходимы
для продукции и секреции ФСГ и ЛГ в гипофизе [10]. Нейроны ГнРГ деполяризуют и
секретируют ГнРГ с частотой от 60 до 200 мин во время специфических фаз менстру¬
ального цикла, для того чтобы увеличить или уменьшить секрецию ФСГ или ЛГ [13,
15]. Вариации в частоте выбросов ГнРГ определяются механизмами обратной связи.
Локальные нейромодуляторы в головном мозге, включая норадреналин, дофамин,
(3-эндорфин, опосредуют эффекты половых стероидов на гипоталамус (см. рис. 1-4)
[21]. Любое расстройство центральной нервной системы (ЦНС), которое пересекается
с этим запутанным процессом, может стать причиной ановуляции. Некоторые из этих
нарушений могут быть представлены как генетические или анатомические заболевания,
такие как изолированный дефицит гонадотропинов (с/без аносмии), инфекции, супра-
селлярные опухоли (аденомы гипофиза, краниофарингиома), а также травма головы
[10]. Эти генетические и анатомические заболевания влияют на гипоталамус, и некото¬
рые из них могут быть исключены на этапах сбора анамнеза, физикального исследова¬
ния, а также посредством применения лучевых методов диагностики (табл. 1-2).
Таблица 1-2. Классификация ановуляций, вызванных гипоталамо-гипофизарными заболеваниями
Функциональная гипоталамическая ановуляция
• Стресс (психогенный или физический).
• Диета.
• Интенсивные физические нагрузки.
• Хронические заболевания (например, хроническая почечная или печеночная недостаточность, ВИЧ)
Психические нарушения
• Нервная анорексия
Прием лекарственных препаратов
• Антагонисты допамина (например, галоперидол).
• Опиаты.
• Старые антигипертензивные препараты (например, метилдопа, резерпин)
Гипотиреоз
Анатомически или генетически определяемая патология гипоталамо-гипофизарного комплекса
• Опухоли гипофиза.
• Пролактинома.
• Клинически нефункционирующая аденома.
67
Окончание табл. 1-2
• СТГ-секретирующая аденома (акромегалия).
' АКТГ-секретирующая аденома (болезнь Кушинга).
' Другие гипофизарные опухоли (например, метастазы, менингиома).
' Повреждение ножки гипофиза.
' Геморрагическая деструкция гипофиза, включая апоплексию гипофиза и синдром Шиена.
' Аневризма гипофиза.
' Инфильтративные заболевания гипофиза (например, лимфоцитарный гипофизит, саркоидоз, гистиоцитоз X,
туберкулез).
’ Синдром пустого турецкого седла.
' Опухоли, влияющие на функцию гипоталамуса (например, метастазы, краниофарингиомы).
' Инфильтративные гранулематозные заболевания гипоталамуса (например, саркоидоз, гистиоцитоз X, туберку¬
лез).
' Травма головы.
' Облучение головы.
' Инфекция ЦНС.
' Изолированный дефицит гонадотропинов (включая синдром Кальмана).
’ Другое.
Наиболее часто наблюдаемая форма гипоталамической ановуляции, как правило,
не сопровождается видимыми нейроанатомическими дефектами [10]. Эта форма, име¬
нуемая функциональной гипоталамической ановуляцией, характеризуется обратимы¬
ми нейроэндокринными нарушениями. Изменения в образе жизни часто приводят к
возвращению нормальных овуляторных циклов. Функциональная гипоталамическая
ановуляция может быть обусловлена избыточными физическими нагрузками, резким
похудением, эмоциональным стрессом. Предполагают, что стресс вызывает ановуля¬
цию через влияние на пульсирующую секрецию ГнРГ в мозге. Другие причины ано¬
вуляции, демонстрируемые нейроанатомическими или генетическими изменениями,
обнаруживаются реже (см. табл. 1-2).
Функциональная гипоталамическая ановуляция
Ановуляция гипоталамического происхождения характеризуется дефицитом
эстрогенов, а также низким уровнем гонадотропинов. У большинства пациенток не
находят ни анатомических, ни генетических нарушений. Концепция функциональной
гипоталамической ановуляции была впервые предложена в 1940-х гг., тогда причиной
считали нарушение гипоталамо-гипофизарных связей, необходимых для секреции
ЛГ из передней доли гипофиза [179]. Позже многие клинические исследования под¬
твердили эту концепцию [14]. Полученные данные до сих пор свидетельствуют о том,
что в основе лежит нарушение импульсного характера секреции ГнГГ. Интересно, что
разные причины, такие как нарушение питания, депрессия или стресс, избыточные
нагрузки или их сочетание, предшествовали развитию функциональной гипотала¬
мической ановуляции. Строгая диета, изнуряющие физические нагрузки, а также
нерациональное снижение массы тела привели к эпидемии этой патологии среди
женщин.
Диагноз фукциональной гипоталамической ановуляции
Пациентки с функциональной гипоталамической ановуляцией чаще всего имеют
вторичную аменорею, характеризующуюся отсутствием менструальных циклов более
6 мес при отсутствии органических признаков заболевания. Необходимо еще раз
отметить, что диагноз гипоталамической ановуляции — диагноз исключения. Имеется
много органических или генетических заболеваний, схожих с функциональной гипо¬
таламической ановуляцией (см. табл. 1-2). Таким образом, тщательное и целостное
исследование необходимо для постановки диагноза.
Женщины с функциональной гипоталамической ановуляцией имеют в анамнезе
период нормального менструального цикла. Затем этот период нормального функци-
68
онирования яичников (по анамнезу) прерывается ановуляцией, которая проявляется
вторичной аменореей. Также необходимо отметить, что изредка женщины с функ¬
циональной гипоталамической аменореей могут быть представлены и первичной
аменореей.
Кроме того, они обычно имеют нормальную или сниженную массу тела; их работа,
как правило, связана с высоким уровнем стресса. Род занятий этих пациенток (напри¬
мер, балерина, спортсменка) может быть чрезвычайно важным фактором. Детальный
опрос способствует выявлению большого количества эмоциональных стрессов (раз¬
вод, смерть близкого человека), предшествовавших аменорее. Во время интервью
могут быть выявлены следующие важные факторы: академическая нагрузка, плохая
социальная адаптация, психосексуальные проблемы. При обследовании пациентки
необходимо отметить режим питания, использование седативных или гипнотических
препаратов, строгость привычек и упражнений пациентки. Несмотря на тщательность
опроса, у некоторых пациенток не удается выявить стрессов, избыточных физических
нагрузок или расстройств питания. Эти женщины обычно не жалуются на приливы
крови к лицу, которые, напротив, зачастую наблюдаются при яичниковой недостаточ¬
ности.
Врач должен исключить возможную гиперпролактинемическую этиологию (про¬
лактинома, гипотиреоз), а также избыток андрогенов (СПКЯ) во время объективно¬
го обследования. Эти женщины имеют нормальные вторичные половые признаки.
Гинекологическое исследование выявляет истонченную слизистую оболочку влага¬
лища, которая сопровождается скудным секретом, нормальную или уменьшенную
матку, а также другие признаки дефицита эстрогенов. Признаки хорошо эстрогенизи-.
рованных влагалища и шейки матки при такой патологии нехарактерны.
Лабораторные тесты проводятся для исключения других причин ановуляции и
вторичной аменореи. Необходимо определить уровни ФСГ и ЛГ. Обычно уровни
гонадотропинов ниже, чем нормальные значения в раннюю фолликулярную фазу.
Уровни тиреотропного гормона (ТТГ) и пролактина определяют для исключения
гипотиреоза и гиперпролактинемии. Тест с прогестагенами (медроксипрогестерона
ацетат по 10 мг в день в течение 10 дней) демонстрирует либо минимальные мажущие
выделения, либо их отсутствие (при наличии данной патологии). Это подтверждает
очень небольшое действие эстрогенов на эндометрий или его отсутствие, поскольку
концентрация эстрогенов в крови равна или ниже, чем в раннюю фолликулярную
фазу. Измерение уровня эстрадиола в крови не является обязательным. Необходимо
также провести магнитно-резонансную томографию (МГТ) головного мозга для
исключения опухоли турецкого седла. Подобное исследование также важно, если
аменорея возникла неожиданно или ассоциируется с возникновением неврологиче¬
ских расстройств.
Патофизиология функциональной гипоталамической ановуляции
При функциональной гипоталамической ановуляции отсутствует повышение уров¬
ня гонадотропинов, несмотря на недостаток ингибирующих факторов яичникового
происхождения, таких как эстрадиол и ингибин. Уровень секреторного ЛГ нормаль¬
ный. Причина развития этого типа ановуляции кроется в снижении частоты выбросов
ГнГГ [14, 180, 181]. С целью непрямого изучения выделения ГнГГ проводили частый
забор крови для изучения эпизодичности секреции ЛГ, выделяющегося под влиянием
ГнГГ [181].
Существует значительная вариабельность в амплитуде и частоте секреции ЛГ
при функциональной гипоталамической ановуляции. При сравнении секреторных
образцов ЛГ с уровнем секреции в нормальную фолликулярную фазу менструального
цикла были получены данные об аномалии по частоте и амплитуде секреции, а также
регрессии до соответствия препубертатному состоянию [14,180,181]. В тяжелых слу¬
чаях частота и амплитуда выбросов ЛГ значительно уменьшались. Эти данные также
69
свидетельствуют о том, что секреция ГнРГ не нарушается в такой же степени. Во время
фазы восстановления при гипоталамической ановуляции часто наблюдается возврат к
секреции ЛГ, соответствующей раннему пубертатному периоду.
Ответ гипофиза на стимуляцию ГнРГ по отношению к продукции и выбросу гона¬
дотропинов не нарушается при функциональной гипоталамической ановуляции.
Внутривенное пульсирующее введение ГнРГ может восстановить нормальные уровни
ЛГиФСГ-,[182,183].
В исследованиях на животных норадреналин, дофамин, а также серотонин, синте¬
зируемые в головном мозге, могли модулировать секрецию ГнРГ или ЛГ. Пациентки,
получавшие лекарства, которые блокировали эти нейротрансмиттеры (седативные,
антидепрессанты, стимуляторы, антипсихотики), имели нарушения менструального
цикла. Таким образом, данные этих исследований подтвердили, что нарушение нерв¬
ных путей приводит к нарушению процессов, регулируемых у людей ГнРГ. Согласно
данным исследованиям активация норадренергических нейронов приводит к акти¬
вации освобождения ГнРГ [184, 185], а дофаминергические и серотонинергические
нейроны могут стимулировать или ингибировать секрецию ГнРГ и ЛГ [185].
Несколько препаратов использовали в качестве проб, для того чтобы определить,
может ли быть нормализована секреция ГнРГ. Например, метоклопрамид блокирует
действие дофамина [186]. Инъекция метоклопрамида приводит к соответствующему
выбросу ЛГ у пациенток с функциональной ановуляцией. Эти исследования показы¬
вают также увеличенную дофаминергическую активность при данном заболевании.
Необходимо отметить, что обычно хроническое назначение антагонистов дофамина
(галоперидол, метоклопрамид) может вызвать ановуляцию.
Другая группа веществ, оказывающих ингибирующее действие на ГнРГ, представ¬
лена эндогенными опиоидными пептидами [187,188]. У женщин с данной патологией
блокады эндогенных опиоидных рецепторов назначение налоксона, опиатного анта¬
гониста, приводила к увеличению частоты и амплитуды выбросов ЛГ [186]. Секреция
гонадотропинов возобновлялась, если активность опиатных рецепторов блокирова¬
лась длительнодействующим налоксоном, в некоторых случаях вплоть до восстанов¬
ления функции яичников [189]. Данные исследования показывают, что имеется общее
увеличение активности эндогенных опиатов, которое может снизить пульсирующие
выбросы ГнРГ при функциональной гипоталамической ановуляции.
Репродуктивная функция может быть нарушена при хроническом воздействии
стресса [179]. В действительности активация гипофизарно-надпочечниковой систе¬
мы — обычный ответ у пациенток с хроническим стрессом [190]. При функциональ¬
ной гипоталамической ановуляции такие стрессорные факторы, как физические
нагрузки или эмоциональный стресс, могут активировать гипоталамо-гипофизарно-
надпочечниковую систему. Наблюдается повышение уровня кортизола днем и приту¬
пление ответа гипофиза на выделение кортикотропина [183, 191]. Стрессовый ответ
связан с усиленной секрецией кортикотропина, адренокортикотропного гормона
(АКТГ), кортизола, пролактина, окситоцина, вазопрессина, адреналина и норадрена-
лина.
Взаимосвязь эмоционального или физического стресса с расстройством репродук¬
тивной функции комплексная и включает несколько механизмов. На моделях живот¬
ных кортикотропин был важным фактором, ингибирующим выбросы ГнГГ [192,193].
Ингибирующий эффект может быть предупрежден одновременным назначением аго¬
ниста кортикотропина или прекращен введением антагониста опиатов налоксона, что
позволяет предположить перекрестное взаимодействие эффектов кортикотропина с
активацией опиоидергической системы. Более того, назначение АКТГ блокировало
ответ гипофиза на ГнГГ [194, 195]. Другой стрессовый гормон, окситоцин, может
ингибировать гипоталамическую секрецию [188]. Итак, повышенная продукция кор¬
тикотропина и других стрессовых гормонов в мозге, а также активация хроническим
70
стрессом гипофизарно-надпочечниковой системы служат причиной возникновения
ингибирования секреции гонадотропинов при функциональной гипоталамической
ановуляции.
Недавно была исследована роль дефицита лептина как причины гипоталамиче¬
ской аменореи. Действительно, назначение лептина лептин-дефицитным пациенткам
улучшало репродуктивную, тиреоидную, соматотропную функции, а также маркеры
формирования кости. Это позволяет предположить, что лептин — периферический
сигнал, отражающий адекватность энергетических запасов, который необходим для
нормального функционирования репродуктивной и нейроэндокринной систем [196].
Гипоталамическая ановуляция и физические нагрузки
Регулярные интенсивные нагрузки могут приводить к нарушению цикла, задержке
менструаций, дисфункции лютеиновой фазы и вторичной аменорее. 30% юных тан¬
цовщиц балета имеют подобную проблему в пубертатном возрасте. Средний возраст
менархе задерживается до 15 лет. Вообще увеличение пубертатного возраста совпадает
со временем отдыха или восстановлением после травмы [197-202]. Интенсивность,
длительность и вид спорта определяют тяжесть заболевания. К занятиям, связанным с
увеличением риска репродуктивной дисфункции и способствующим снижению массы
тела, относят бег на средние и длинные дистанции, плавание, гимнастику, а также балет.
Спортсмены, участвующие в соревнованиях, могут иметь гипоталамо-гипофиза-
рные нарушения, аналогичные таковым при других формах функциональной гипо¬
таламической ановуляции. К ним относят повышение уровней кортиколиберина и
(3-эндорфина.
Ведение данного типа ановуляции зависит от выбора и ожиданий пациентки-.
Побочные эффекты (остеопороз и задержка пубертатного периода) должны быть
обсуждены с пациенткой [203]. Для возвращения овуляции необходимо снижение
уровня нагрузок и изменение поведения. ЗГТ назначают при отсутствии эффекта от
вышеописанных мероприятий. Низкодозированные оральные контрацептивы — воз¬
можный вариант для женщин репродуктивного возраста.
Гипоталамическая ановуляция, связанная с нарушением питания
Существуют два заболевания, связанных с нарушением питания: нервная анорексия и
булимия. При нервной анорексии наблюдают экстремальную потерю массы тела (более
25% первоначальной массы) и искаженное представление о теле, сопровождающееся
навязчивым страхом ожирения. Булимия представлена эпизодами объедания, сменяю¬
щимися эпизодами воздержания от еды либо избыточным использованием диуретиков
и слабительных. От 90 до 95% этих пациентов женщины. Большинство пациенток —
европеоиды из семей со средним и выше среднего достатком. Гаспространенность
классической формы нервной анорексии составляет 1 на 100 ООО случаев в общей
популяции [204]. Среди студенток высших школ и колледжей булимия довольно рас¬
пространена [5]. Гаспространенность нервной анорексии имеет два пика — в возрасте
13 и 17 лет. Булимия возникает позже, в 17-25 лет. Нервная анорексия в 9% случаев
заканчивается летально, поэтому считается неотложной проблемой. Смерть может
наступить вследствие сердечной аритмии, которая возникает в результате уменьшения
мышечной массы органа и электролитных нарушений [205]. Пациентки с анорексией
характеризуются высоким риском суицидальных попыток [206].
Секреция гонадотропинов соответствует препубертатному состоянию и похожа на
другие формы функциональной гипоталамической ановуляции. Переходные образцы
ЛГ регистрируют во время эпизодов восстановления, также наблюдают нормальный
или чрезмерный ответ на стимуляцию ГнГГ, Ановуляция персистирует примерно у
50% таких пациенток даже после восстановления нормальной массы тела. И ано¬
рексичные, и булимические пациентки демонстрируют гиперактивацию гипоталамо-
гипофизарной системы. Несмотря на то что присутствует дневная вариация, имеется
71
постоянная гиперсекреция кортизола в течение дня [207]. Особенности, характерные
для синдрома Кушинга, как правило, не представлены, частично потому, что имеется
умеренная гиперкортизолемия и редукция периферических глюкокортикоидных
рецепторов, в ЦНС увеличены уровни и кортиколиберина, и р-эндорфина [208, 209].
При нервной анорексии основной метаболизм снижен вследствие уменьшения
периферической конверсии тироксина (Т^) в биологически активный трийодтиро-
нин (Тз).-Напротив, конвертируется в реверсивный Т3, в неактивную изоформу.
Такое нарушение наблюдается у тяжелобольных пациенток и во время голода [210].
Страдающие анорексией также имеют частичный несахарный диабет, они неспособны
адекватно концентрировать мочу из-за недостатка секреции вазопрессина [211],
И нервную анорексию, и булимию тяжело лечить. Наиболее распространенный
подход включает индивидуальную и групповую психотерапию и модификацию пове¬
дения. Пациентки с нарушением питания должны получать психиатрическую кон¬
сультацию и в последующем наблюдаться у психиатра. Это помогает и в диагностике,
и в лечении. Пациентки с дефицитом массы тела более 25% идеального должны быть
немедленно госпитализированы с безотлагательным началом лечения. Хронические
нарушения нервной анорексии включают остеопороз, дефицит эстрогенов и общий
результат недостаточного питания [200]. Необходимо назначить комбинированные
оральные контрацептивы до восстановления овуляторной функции.
Лечение функциональной гипоталамической ановуляции
Лечение хронической ановуляции зависит от расстройств системы «ЦНС-
гипоталамус» и должно быть направлено на исключение первопричины (коррекция
стресса и массы тела, снижение физической нагрузки). Успешность лечения этого
болезненного состояния важна и потому, что эти пациентки в будущем могут страдать
остеопорозом. Для значительного количества женщин спонтанное восстановление
менструальной функции происходит после изменения стиля жизни, психологических
установок или приспособления к окружающему стрессу. В связи с этим первоначаль¬
ное лечение должно быть направлено на то, чтобы изменить стиль жизни пациенток
-(при этом важно учитывать потребности каждой). Для тех, у кого сохраняется амено¬
рея, разумно исследование репродуктивного статуса 1 раз в 4-6 мес.
В ситуациях, когда ановуляция длится более 6 мес и невозможно прекратить
действие причины, вызвавшей развитие этого состояния (у профессиональных спор¬
тсменов, балерин), важно понимание долговременных эффектов гипоэстрогении,
особенно в отношении костного обмена. В добавление к эстрогенному дефициту в
развитии потери костной ткани принимают участие дефицит ИФР-1, гиперкорти-
цизм, а также пищевой фактор [212]. К сожалению, эпидемиологические данные по
риску возникновения переломов и о пользе заместительной гормонотерапии скудны
[203, 212]. На основе исследования женщин репродуктивного возраста с удаленными
яичниками или леченных ГнРГ по поводу эндометриоза определено значительное
снижение плотности костной ткани даже в течение 6 мес аменореи. Поскольку эти
пациентки часто сопротивляются назначению терапии, необходимо использовать
результаты исследования плотности костной ткани поясничных позвонков и бедрен¬
ной кости для убеждения начать заместительную гормонотерапию. Если пациентка
не входит в группу риска по тромбоэмболии и не курит, для заместительной гормо¬
нотерапии показано использование низкодозированных оральных контрацептивов.
Альтернативой может служить комбинация конъюгированных эстрогенов (0,625 мг)
и медроксипрогестерон ацетат (2,5 мг) в день. Гестаген (медроксипрогестерон ацетат)
добавляют отдельно для предупреждения гиперплазии эндометрия.
Если пациентка желает восстановить овуляцию, с тем чтобы забеременеть в буду¬
щем, наиболее физиологическим будет назначение индукции овуляции посредством
пульсирующего режима ГнРГ, Это в настоящее время лучший метод, поскольку
72
причина ановуляции — снижение эндогенного уровня ГнРГ. Пульсирующее внутри¬
венное введение 5 мкг ГнРГ каждые 90 мин доказало свою эффективность [213].
Интенсивность мониторирования уровня эстрадиола в плазме или развития фол¬
ликулов можно минимизировать, так как ответ фолликулов яичников и уровень
гонадотропинов подражает нормальному менструальному циклу. У этих пациенток
продолжение пульсирующего режима ГнРГ или введение 4 доз ХГЧ по 1500 ME вну¬
тримышечно каждые 3 дня может поддерживать функцию желтого тела. Результаты
внутривенного введения ГнРГ приводят к восстановлению овуляции в среднем в
90% случаев, к возникновению беременности — в 30%, а уровень гиперстимуляции
составляет менее 1% на каждый цикл лечения [213]. Поскольку внутривенная помпа
с ГнРГ не является практическим выбором для многих женщин, альтернативная стра¬
тегия предполагает использование подкожного введения рекомбинантного ФСГ для
развития 1-3 фолликулов с последующей индукцией овуляции в/м введением ХГЧ и
поддержкой лютеиновой фазы в/м ХГЧ или масляным раствором прогестерона.
ХРОНИЧЕСКАЯ АНОВУЛЯЦИЯ ВСЛЕДСТВИЕ ЗАБОЛЕВАНИЙ ГИПОФИЗА
Наиболее частая причина ановуляции, связанная с гипофизом, — гиперпролакти¬
немия, вызываемая пролактиномами или другими функциональными или анатомиче¬
скими состояниями гипофиза.
ХРОНИЧЕСКАЯ АНОВУЛЯЦИЯ ВСЛЕДСТВИЕ ИЗБЫТКА АНДРОГЕНОВ
Наиболее распространенным заболеванием, связанным с яичником и хронической
ановуляцией, является СПКЯ. Нерегулярные менструации и/или аменорея и выброс
андрогенов — часто наблюдаемые проявления СПКЯ. Другие причины ановуляции —
стероид-секретирующие опухоли яичников и преждевременное прекращение функ¬
ции яичников. Избыток андрогенов, происходящий из экстрагонадных источников
(заболевания надпочечников), также ассоциируется с ановуляцией.
Подход к пациентке с избытком андрогенов
Два натуральных андрогена — тестостерон, транспортируемый к органу-мишени
с кровью, и дигидротестостерон, который первично образуется тканями-мишенями.
Увеличенные уровни этих андрогенов могут приводить к гирсутизму, который про¬
является избыточным ростом волос на теле или вирилизацией (более выраженная
форма избытка андрогенов). Гирсутизм — грубые терминальные волосы в неха¬
рактерных для женщин местах — на лице в области щек, над верхней губой и на
подбородке (рис. 1-26, А и Б). Присутствие волос на срединной линии груди также
важно (рис. 1-26, В). Кроме того, мужской тип оволосения — волосы на внутренней
поверхности бедер, пояснице, продолжающиеся между ягодиц, — признаки избытка
андрогенов. Умеренное количество волос на предплечьях и голенях само по себе не
является ненормальным, даже несмотря на то, что это смущает пациентку и может
быть ошибочно принято за гирсутизм. Предложено несколько систем подсчета для
измерения гирсутизма. Одна из наиболее детализированных шкал была предложена
Ferriman, Gallwey и Lorenzo [214]. Практическая и клиническая польза измерения гир¬
сутизма заключается в детальном описании оволосения с помощью простых рисунков
и фотографий. В частности, фотографии являются незаменимым способом точного
документирования степени гирсутизма.
В противоположность гирсутизму вирилизация является более выраженной фор¬
мой избытка андрогенов и предполагает значительно более высокие уровни продук¬
ции тестостерона. Эти проявления включают височное облысение, понижение тембра
голоса, уменьшение размера груди, увеличение мышечной массы, потерю женских
контуров тела и увеличение клитора (рис. 1-27). Даже если уровни тестостерона уме¬
ренно увеличены (<1,5 нг/мл), височное облысение и клиторомегалия могут наблю-
73
Рис. 1-26. Гирсутизм. А — легкие проявляения на лице; Б — выраженные проявления на лице (под¬
бородок), которые требуют регулярного бритья; В — выраженный гирсутизм на груди. [Б и В из Dunaif
A., Hoffman A.R., Scully R.E. et al. The clinical, biochemical and ovarian morphologic features in women with
acanthosis nigricans and masculinization // Obstet. Gynecol. — 1985. — N 66. — P. 545-552.]
даться после длительного промежутка времени (>1 года) присутствия постоянного
избытка андрогенов. Значительное увеличение в секреции андрогенов (как это про¬
исходит при продукции опухолями) приводит к более яркой картине вирилизации в
течение короткого промежутка времени (менее нескольких месяцев).
Измерение увеличенного клитора может быть использовано для подсчета степени
вирилизации. Длина клитора более 10 мм считается аномальной (см. рис. 1-27). Как
правило, это величина изменяющаяся. Увеличение диаметра — более выраженный
индикатор андрогенного воздействия. Нормальным считают диаметр клитора менее
7 мм в основании головки (см. рис. 1-27). Наиболее точное определение клиторо-
мегалии включает использование клиторального индекса (соотношение ширины и
длины головки клитора). Клиторальный индекс более 35 мм^ считается аномальным
и статистически коррелирует с избытком андрогенов [215].
Происхождение андрогенов
Два природных С19-стероида способны действовать как андрогены на органы
мишени: тестостерон и дигидротестостерон. В этой главе использование термина
«андроген» относится к каждому из этих стероидов. Тестостерон у женщин репродук¬
тивного возраста образуется двумя основными путями: прямой секрецией яичником,
составляющей приблизительно треть от общей продукции тестестерона, и превраще¬
нием предшественника андростендиона в тестостерон в периферических экстраго¬
надных тканях, составляющим приблизительно две трети от продукции тестостерона
74
(рис. 1-28) [216, 217]. Эти периферические ткани включают кожу и жировую ткань.
Андростендион, прямой предшественник тестостерона, образуется яичниковой и над¬
почечниковой тканями. С19-стероиды, дегидроэпиандростерон и дегидроэпиандро¬
стерона сульфат надпочечникового происхождения и дегидроэпиандростерон яич¬
никового происхождения опосредованно способствуют формированию тестостерона,
Рис. 1-27. Выраженная клиторомегалия, происходящая из тестостерон-секретирующей опухоли яични¬
ка. А — общая длина клитора составляет приблизительно 4 см (при норме менее 1 см); Б — поперечный
диаметр клитора составляет 1,5 см (норма <0,7 см).
Надпочечник
17Р-ГСД
5а-Редуктаза
^ Кровоток
Дигидротестостерон
Волосяные
фолликулы
І^итор
(фибробласты
кожи)
5(х-Редуктаза
Ткани-мишени
андрогенов
онт
Рис. 1 -28. Биосинтез андрогенов у женщин. Существует два биологически активных андрогена, тестосте¬
рон (Т) и дигидротестостерон (ОНТ). В зависимости от фазы менструального цикла или от состояния пост¬
менопаузы 20-30% тестостерона секретируется яичником. Оставщаяся часть продукции Т—превращение
циркулирующего андростендиона (А) в тестостерон в некоторых периферических тканях. Оба, надпочечни¬
ковый и яичниковый, способствуют циркулирующему А прямо или опосредованно в зависимости ют фазы
цикла, от репродуктивного или постменопаузального периода и от хронологического возраста. Более того,
Т может также формироваться локально в тканях-мишенях андрогенов. В результате Т превращается в
более активный андроген дигидротестостерон внутри клеток и тканей-мишеней. Например, локальная
конверсия Т в дигидротестостерон в фибробластах кожи гениталий, а также в фолликулах волос усили¬
вает андрогенное воздействие, стимулирующее увеличение клитора и гирсутизм. ОНЕА —дегидроэпиан¬
дростерон; ОНЕАЗ — дегидроэпиандростерона сульфат; ГСД — гидроксистероид дегидрогеназа.
75
будучи сначала конвертированными в андростендион, который далее конвертируется
в тестостерон (см. рис. 1-28).
Тестостерон является андрогеном, а дегидроэпиандростерона сульфат — биологи¬
чески инертным стероидом. До 20 мг дегидроэпиандростерона сульфата синтезирует¬
ся каждый день в сравнении с 3 мг андростендиона и 8 мг дегидроэпиандростерона,
также синтезируемыми ежедневно. С19-стероиды надпочечникового происхожде¬
ния-(дегидроэпиандростерона сульфат, дегидроэпиандростерон) проявляют свои
эффекты после конверсии в активный андроген тестостерон (см. рис. 1-28). Только
андростендион может конвертироваться напрямую в тестостерон. Уровень конверсии
циркулирующего андростендиона в тестостерон в экстрагонадных тканях составляет
около 5% как у мужчин, так и у женщин.
Тестостерон связывает ядерный рецептор к андрогенам и активирует ген-мишень
андрогена. Тестостерон должен быть конвертирован в более активный стероид,
дигидротестостерон, для того чтобы проявить полный андрогенный эффект на
таких тканях-мишенях, как волосяные фолликулы и наружные гениталии [127,151].
Например, для активности фибробластов в коже гениталий необходимо заполне¬
ние рецепторов дигидротестостероном. Эта конверсия катализируется ферментом
5а-редуктаза и происходит в печени внутри андрогенных клеток-мишеней, напри¬
мер фибробластах кожи гениталий (интракринный эффект). Протеиновые продукты
обоих генов (5а-редуктаза 1-го и 2-го типов) проявляют ферментативную активность
[127]. Андрогенная активность дигидротестостерона по отношению к росту волос и
вирилизации наружных гениталий значительно выше, чем эффект тестостерона.
Андрогенная активность в тканях-мишенях определяется, как минимум, локаль¬
ной активностью 5а-редуктазы и частично содержанием рецепторов к андрогенам.
Андрогеновые рецепторы опосредуют андрогенную активность в органах-мишенях
(см. рис. 1-28) [151, 218]. Другие локальные ферменты в тканях-мишенях, отличные
от 5а-редуктазы (ароматазы и 17-гидроксистероиддегидрогеназа или кеторедуктаза),
также регулируют активность андрогенного воздействия посредством конвертирова¬
ния тестостерона в андрогенно неактивный андростендион или эстрадиол, активный
эстроген. Таким образом, существует баланс между усилением андрогенной актив¬
ности при формировании дигидротестостерона и уменьшением андрогенности в слу¬
чае, когда синтезируются неактивные С19-стероиды или когда в тканях-мишенях и
других экстрагонадных тканях из тестостерона формируется эстрадиол. В частности,
метаболизм тестостерона в дигидротестостерон либо андростендиона в эстрадиол
в этих тканях важен при развитии андрогензависимых (гирсутизм, вирилизация) и
эстрогензависимых заболеваний (опухоли груди и эндометрия) (см. рис. 1-21 и 1-28)
[21, 28].
Лабораторная оценка эффектов андрогена
Тестостерон циркулирует в трех формах: связанной с ПСПС (протеин, связы¬
вающий половые стероиды); несвязанной с ПСПС, но свободно ассоциированной с
альбумином: несвязанной ни с глобулином, ни с альбумином, то есть свободный или
диализируемый тестостерон. Биологически активный тестостерон включает две —
свободную и связанную с альбумином — фракции. Таким образом, тестостерон крови,
способный диффундировать в ткани-мишени, называется биодоступным, или не свя¬
занным с ПСПС, тестостероном.
ПСПС — один из первичных регуляторов, которые определяют количества цирку¬
лирующих связанного и биодоступного тестостерона, способного влиять на ткани-
мишени. Условия, снижающие ПСПС-связывание (например, избыток андрогенов,
ожирение, акромегалия, гипотиреоидизм, заболевание печени), также увеличивают
биодоступный тестостерон, тем самым повышая его эффект. ПСПС, кроме того,
регулирует количество циркулирующего биодоступного эстрадиола через связывание
76
значительной фракции циркулирующего эстрадиола. Следовательно, условия, снижа¬
ющие уровни ПСПС, способствуют увеличению уровней биоактивного эстрадиола.
Измерение связанных не с ПСПС (биодоступных) форм тестостерона предполагает,
что в состоянии избытка андрогенов возможно более точное определение субтильных
форм гирсутизма. Несмотря на то что диагостическое значение этого измерения дей¬
ствительно превосходит измерение общего тестостерона, корреляция между общим и
не связанным с ПСПС тестостероном сильная. Биоактивный тестостерон, как прави¬
ло, может быть предсказан исходя из известного общего уровня тестостерона [219].
Цель измерения общего тестостерона в плазме — оценка избытка циркулирующего
андрогена и определение сверхвысоких уровней гормона, которые могут происходить
из андроген-секретирующих опухолей.
Нормальные уровни андрогенов в плазме, особенно свободного тестостерона,
определяемого радиоиммунологическим анализом, отличаются от лаборатории к
лаборатории. По этой причине группа исследователей сравнила уровни свободных
андрогенов в плазме, измеренные методом равновесного диализа, с результатами,
полученными от прямых радиоиммунологических измерений, а также полученны¬
ми при расчете андрогенового индекса, 100 х тестостерон/ПСПС, простого индекса,
коррелирующего с уровнем свободного тестостерона [220]. Рассчитанные с исполь¬
зованием индекса свободного андрогена значения свободного тестостерона хорошо
коррелировали с результатами, полученными путем равновесного диализа. Наоборот,
прямой радиоиммунологический метод имел недопустимо большую систематическую
ошибку и случайную изменчивость и не коррелировал с результатами, полученными
методом равновесного диализа. Более того, нижний предел определения был наи¬
большим для прямого радиоиммунологического метода в сравнении с равновесным
диализом или рассчитанным свободным тестостероном [220]. Таким образом, врач
должен быть осведомлен об ограничениях радиоиммунологического исследования,
проводимого без жесткого контроля качества.
Для практических целей определение уровней всех С19-стероидов не является
клинически необходимым у большинства пациентов, имеющих избыток андрогенов.
Наиболее полезный первичный тест — определение уровня общего тестостерона в
плазме. Аномальный уровень в присутствии гирсутизма или вирилизации может
быть ассоциирован с СПКЯ, гипертекозом, неклассической формой БДКН или
андроген-секретирующей опухолью. Большинство андроген-секретирующих опу¬
холей происходят из яичника. Бероятность опухоли сильно коррелирует с ростом
уровней тестостерона. Нижеперечисленные исследования могут лежать в основе
диагностики соответствующей клинической картины заболевания: уровень в плазме
17-гидроксипрогестерона (неклассическая форма дисфункции коры надпочечников,
БДКН, см. подробности ниже), уровень пролактина и тиротропина в плазме (незначи¬
тельный избыток андрогенов, связанный с гиперпролактинемией), уровень ФСГ и ЛГ
в плазме (увеличение ЛГ/ФСГ соотношения при СПКЯ), уровень дегидроэпиандро¬
стерона в плазме (опухоли коры надпочечников), а также визуализирующие методы
исследования яичников и коры надпочечников (СПКЯ, опухоли).
Источники избытка андрогенов
Различные заболевания приводят к избытку андрогенов. Это могут быть «необыч¬
ные» состояния: ятрогенный, или вызванный лекарствами, избыток андрогенов,
врожденная неопределенность гениталий (например, избыточное внутриутробное
образование андрогенов при женском псевдогермафродитизме), а также состояния,
связанные с беременностью (лютеома беременности, множественные лютеинизиро¬
ванные кисты). Эти «необычные» причины и относительно более распространенные
заболевания, связанные с избытком андрогенов, представлены в табл. 1-3. Термин
«экстраовариальный стероид» используют как синоним экстрагландулярного, экс-
трагонадного или периферического формирования стероидов.
77
Таблица 1-3. Причины избытка андрогенов у женщин репродуктивного возраста
Яичники
. СПКЯ,
• Гипертекоз (тяжелый вариант СПКЯ).
• Опухоли яичников (например, опухоль Сертоли-Лейдига)
Надпочечники
• Неклассическая надпочечниковая гиперплазия.
• Синдром Кушинга.
• Резистентность к глюкокортикоидам.
• Опухоли надпочечников (например, аденома, карцинома)
Особые состояния во время беременности
Лютеома беременности.
Лютеинизация фолликулярных кист яичников.
Дефицит ароматазы у плода
Другое
• Гиперпролактинемия, гипотиреоз.
• Лекарственные препараты (даназол, тестостерон, анаболические средства).
• Идиопатический гирсутизм (нормальный уровень тестостерона у овулирующих женщин).
• Идиопатическая гиперандрогения (пациентки, не попадающие ни в одну из вышеперечисленных категорий)
Вообще распространенность заболеваний, связанных с избытком андрогенов,
составляет: СПКЯ — 72,1% (пациентки без овуляции — 56,6%; умеренно вовлечен¬
ные пациентки с овуляцией — 15,5%); идиопатическая гиперандрогения — 15,8%;
идиопатический гирсутизм — 7,6%; неклассическая дисфункция коры надпочечников
вследствие дефицита 21-гидроксилазы — 4,3%: андроген-секретирующие опухоли —
0,2% [221].
Во всех гиперандрогенных заболеваниях андрогены происходят из более чем
одного источника (см. рис. 1-28). Так, секреция тестостерона увеличена при СПКЯ,
при этом большая часть образуется экстраовариально из существенно увеличенно¬
го уровня андростендиона яичникового происхождения. Более того, у пациенток с
СПКЯ также увеличена продукция надпочечниками дегидроэпиандростерона суль¬
фата, который (после периферической конверсии в дегидроэпиандростерон, а далее
в андростендион) опосредованно способствует экстраовариальной продукции тесто¬
стерона (см. рис. 1-28).
Избыток андрогенов при первичной аменорее требует исключения внутриутроб¬
ной неправильной половой дифференцировки. Более того, до начала серьезного
исследования гирсутизма или вирилизации врачу следует исключить употребление
экзогенных андрогенов. Было бы правильным попросить пациентку перечислить
все употребляемые лекарственные препараты, включая инъекции. Как правило, это
более полезно, чем просто спросить пациентку, употребляет она андрогены или нет.
Препараты, которые могут вызывать вирилизацию или гирсутизм, связаны с тесто¬
стероном. Их перечень включает анаболические стероиды или препараты со схожим
составом.
Наиболее частая определяемая причина избытка андрогенов — СПКЯ. СПКЯ
обсуждается в другом разделе этой главы. Б настоящем разделе сначала опишем
некоторые другие причины, связанные с гирсутизмом или вирилизацией. Затем будет
изложена упрощенная стратегия лечения, которая может быть применена к большин¬
ству гирсутных пациенток внутри группы СПКЯ с неклассической формой БДКН и
идиопатическим гирсутизмом.
78
Идиопатический гирсутизм
Избыточный рост волос в отсутствие заметных признаков избытка андрогенов
относят к идиопатическому или конституциональному; он встречается чаще всего
в определенных этнических группах, особенно у жительниц районов бассейна
Средиземного моря [151]. Он носит название гирсутизма при наличии регулярного
менструального цикла и нормального уровня тестостерона плазмы. Идиопатический
гирсутизм не ассоциируется ни с какими признаками вирилизации. Его причины
окончательно не выяснены. Считается, что женщины с идиопатическим гирсутизмом
имеют значительно повышенный уровень активности 5а-редуктазы в коже [222].
Присутствие или отсутствие подобной ассоциации окончательно не установлено.
Также неясно, какая из изоформ ферментов 5а-редуктазы (тип 1 или 2) доминирует
при развитии идиопатического гирсутизма [151].
Идиопатический гирсутизм диагностируют у женщин с гирсутизмом, нормальной
овуляторной функцией и нормальным уровнем общего или свободного тестостерона,
в целом более 80% женщин с циклическими предсказуемыми менструациями имеют
овуляцию. Овуляторная функция может быть верифицирована по уровню прогесте¬
рона, который должен быть по меньшей мере 5 нг/мл на 7-й день лютеиновой фазы
цикла. 7-й день лютеиновой фазы соответствует 21-му дню 28-дневного менструаль¬
ного цикла или 28-му дню 35-дневного менструального цикла. Присутствие олиго¬
овуляции или ановуляции у гирсутных женщин после исключения определенных
заболеваний (гипотиреоза, гиперпролактинемии или неклассической ВДКН) связы¬
вают с СПКЯ [151]. Дисфункция щитовидной железы и гиперпролактинемия долж¬
ны быть исключены посредством измерения уровня ТТГ и пролактина. Базальный
уровень 17-гидроксипрогестерона в фолликулярную фазу измеряется для исключе¬
ния дефицита 21-гидроксилазы, неклассической ВДКН. Использование экзогенных
андрогенов также должно быть исключено. Итак, в заключение следует отметить, что
диагноз идиопатического гирсутизма — диагноз исключения, при котором овулятор¬
ная дисфункция, повышенный уровень циркулирующего тестостерона, а также другие
причины избытка андрогенов должны быть исключены.
Андроген-секретирующие опухоли яичников и надпочечников
Большинство андроген-секретирующих опухолей происходят из яичника. Эти
яичниковые опухоли секретируют большие количества тестостерона или его предше¬
ственника андростендиона. Список опухолей включает: опухоль из клеток Сертоли-
Лейдига, опухоли из клеток хилуса, опухоли из липоидных клеток, а также редко
опухоли из гранулезных клеток. Стероидогенные яичниковые опухоли, такие как
эпителиальные цистаденомы или цистаденокарциномы, могут продуцировать фак¬
торы, которые стимулируют стероидогенез в прилежащей неопластической строме
и индуцировать продукцию значительных количеств предшественников андрогенов,
таких как андростендион, приводящих к клинически заметному увеличению андро¬
генов. Около 5% андростендиона превращается в тестостерон в экстраовариальных
тканях, опухоли из клеток способствуют в конечном счете избытку андрогенов (см.
рис. 1-28).
Опухоли из клеток Сертоли-Лейдига, которые составляют менее 1% всех солидных
яичниковых опухолей, чаще всего возникают во 2-4-й декаде жизни, тогда как опухо¬
ли из клеток хилуса чаще образуются у женщин в постменопаузе. С течением времени
симптомы избытка андрогенов вызывают необходимость у пациентки обратиться за
медицинской помощью: при этом опухоли из клеток Сертоли-Лейдига увеличивают¬
ся до размера, пальпируемого при бимануальном исследовании, а опухоли из клеток
хилуса остаются небольшими. У женщин с этими типами опухолей наблюдают зна¬
чительное увеличение уровня тестостерона. Гранулезоклеточные опухоли первично
образуют эстрадиол, но иногда могут продуцировать и тестостерон.
79
Быстропрогрессирующие симптомы избытка андрогенов предполагают андроген-
продуцирующую опухоль, если не доказано обратное. Быстрое прогрессирование
характерно как для яичниковых, так и надпочечниковых андроген-секретирующих
опухолей, возникают больше признаки дефеминизации (потеря женских очерта¬
ний тела, уменьшение размеров груди), нежели андрогенизации. С ростом опухоли
образуется больше и больше тестостерона, который усиливает гирсутизм и вирили¬
зацию. Для всех опухолей яичников характерно увеличение уровня тестостерона.
Этот процесс может быть медиирован двумя механизмами: продукцией и секрецией
тестостерона непосредственно опухолью [223] или секрецией больших концентраций
андростендиона, которые конвертируются в тестостерон в экстрагонадных тканях.
Уровни тестостерона, продуцируемые определенными опухолями яичников (напри¬
мер, опухолью из клеток Сертоли-Лейдига), могут быть уменьшены назначением
агониста ГнРГ [224]. Следовательно, использование ГнРГ не может основываться на
дифференциации опухоли от другого функционального состояния.
При интерпретации уровней тестостерона врач должен в первую очередь знать
нормальные значения для данной конкретной лаборатории. Значение, превышающее
нормальное в 3 раза (или >2 нг/мл), предполагает опухоль, особенно если клиниче¬
ские симптомы подтверждают этот диагноз. Необходимо помнить, что более низкий
уровень тестостерона иногда наблюдается при вирилизирующих опухолях яичника.
При подозрении на андроген-секретирующую опухоль измерение уровня андростен¬
диона может быть также полезным. Кроме того, существенно увеличенный уровень
андростендиона может быть проявлением опухоли яичника или надпочечника. При
определении высокого уровня тестостерона и сопутствующих клинических про¬
явлений тщательное ультразвуковое трансвагинальное исследование должно быть
доступным для выявления опухоли яичника. Трансвагинальное УЗИ — наиболее чув¬
ствительный метод для определения опухолей яичника.
Б противоположность андроген-секретирующим опухолям яичников тестостерон-
секретирующие опухоли надпочечников чрезвычайно редки. Клетки надпочечнико¬
вой тестостерон-продуцирующей опухоли похожи на клетки хилуса яичника, кото¬
рые, в свою очередь, подобны клеткам Лейдига. Эти опухолевые клетки образуют
тестостерон и могут быть стимулированы и ЛГ, и ХГЧ. Таким образом, у пациенток
с тестостерон-продуцирующими аденомами надпочечников секреция тестостерона
обычно снижается после подавления ЛГ и повышается после стимуляции ХГЧ.
Бирилизирующие надпочечниковые опухоли обычно секретируют большие коли¬
чества дегидроэпиандростерона сульфата, дегидроэпиандростерона и андростендио¬
на, тогда как тестостерон появляется, как правило, в результате экстраовариальной
конверсии этих предшественников, уровни дегидроэпиандростерона сульфата в плаз¬
ме значительно повышены при большинстве вирилизирующих опухолей надпочечни¬
ков [225]. При превышении дегидроэпиандростероном сульфата 8 мг/мл необходимо
провести сканирование компьютерным или магнитно-резонансным томографом, за
исключением случаев с длительной предшествующей картиной СПКЯ (длительная
клиническая картина симптомов без вирилизации). Б последнем случае, вероятно,
следует искать функциональную аномалию надпочечников: ферментный дефект
(врожденная гиперплазия коры) или необъяснимое гиперфункциональное состояние,
часто ассоциируемое с СПКЯ. При подобных обстоятельствах сканирование может
быть задержано до получения результатов других исследований.
Уровни различных надпочечниковых стероидов, включая кортикостерои¬
ды, могут быть увеличены в различных комбинациях при опухолях надпочечни¬
ков. Таким образом, невозможно описать особую комбинацию гормонов, харак¬
теризующую надпочечниковую опухоль [225]. Б общем, сверхвысокие уровни
дегидроэпиандростерона сульфата (>8 мг/мл) предполагают надпочечниковую опу¬
холь. Тестостерон-секретирующие опухоли надпочечников встречаются чрезвычайно
80
редко. Бирилизирующие яичниковые опухоли, с другой стороны, диагностируют
намного чаще, чем таковые в надпочечниках. Если картина напоминает андроген-
секретирующую опухоль и яичники нормальные по данным трансвагинального УЗИ,
следующими должны быть изучены надпочечники.
Увеличение концентрации тестостерона в 3 раза по сравнению с нормальной
(или >2 нг/мл), а также дегидроэпиандростерона сульфата выше 8 мг/мл — повод для
дальнейшего исследования яичникового или надпочечникового источника избыточ¬
ного андрогена. Необходимо особо отметить, что приведенные цифры указываются
как рекомендации, а не правила. Особое внимание должно быть уделено следующим
исключениям. Бо-первых, поскольку опухоли секретируют андрогены эпизодически,
необходимо более одного исследования для определения значительно повышенного
уровня андрогенов [226]. Бо-вторых, другие стероиды-предшественники часто также
имеют повышенные концентрации (особенно андростендион), что соответственно
диктует необходимость их измерения, и наконец, опухоли могут вызывать умеренное
повышение дегидроэпиандростерон сульфата и тестостерона. Б частности, даже уме¬
ренное повышение уровня андрогенов у женщин в постменопаузе подозрительно на
предмет андроген-секретирующей опухоли. Б подтверждение сказанного значительно
повышенный уровень тестостерона в плазме может быть обнаружен у женщин с тяже¬
лой формой гипертекоза яичников даже при отсутствии опухоли.
Недавнее начало вирилизации и ее небольшая продолжительность — показа¬
ние для дальнейшего исследования даже при незначительном увеличении уровня
тестостерона и дегидроэпиандростерон сульфата. С учетом прогресса в технологиях
сканирования (трансвагинальное УЗИ — для яичниковой патологии, абдоминальное
УЗИ, компьютерная томография и магнитно-резонансная томография — для надпо¬
чечников) возможно обнаружение опухолей даже небольшого размера. Однако если
опухоль не была диагностирована, может быть полезным исследование яичника или
надпочечников после внутривенного введения йодометилнорхолестерина, меченного
радиоактивным изотопом, который определяет стероид-продуцирующие опухоли
[227], Данные диагностические исследования должны быть активно проведены до
хирургического вмешательства по поводу подозрения на опухоль.
Неопухолевые заболевания надпочечников и избыток андрогенов
Определенное количество надпочечниковых заболеваний, таких как классическая
БДКН, синдром Кушинга, резистентность к глюкокортикоидам, способствует избы¬
точной продукции андрогенов в связи с избытком в надпочечниках предшественни¬
ков тестостерона. Эти заболевания рассматриваются в других главах. Б настоящей
главе рассматривается неклассическая форма БДКН.
Несмотря на то что заболевание очевидно существует, продолжаются дискус¬
сии, касающиеся диагноза и распространенности неклассической БДКН. Другие
термины, используемые для описания данной патологии, — поздняя неполная
и скрытая форма БДКН. Эта форма БДКН вызывается частичным дефицитом
активности 21-гидроксилазы. Несмотря на то что недостаток Ир-гидроксилазы
и Зр-гидроксистероид дегидрогеназы может приводить к заболеванию, дефекты в
21-гидроксилазе встречаются в 90% случаев [228].
Клиническая картина практически такая же, как у пациенток с СПКЯ.
Распространенность этого заболевания варьирует в соответствии с этническим фонбм,
данные, сообщаемые различными авторами, также сильно отличаются. Характерные
клинические проявления включают ановуляторные маточные кровотечения и про¬
грессирующий с пубертатного возраста гирсутизм. Эти пациентки рождаются с нор¬
мальными гениталиями, отсутствует потеря солей, а также клинические проявления
до начала пубертата. Пациентки, предки которых вышли из Северной Европы, имеют
меньшую частоту этого заболевания, тогда как евреи ашкенази, испаноязычные
81
американцы, а также пациентки, предки которых были выходцами из Центральной
Европы, в большей степени подвержены этому заболеванию [229]. На основании
этого рекомендуется проводить скрининг групп высокого риска.
Скрининг пациенток с ановуляторным циклом может начинаться с анализа в 8 ч
утра в любой день концентрации 17-гидроксипрогестерона в плазме крови. Несмотря
на то что большинство женщин с неклассической ВДКН ановуляторны, некоторые
могут иметь регулярные менструации и гирсутизм с начала полового созревания либо
только идиопатическое бесплодие [228]. Если у овуляторной пациентки подозревают
неклассическую дисфункцию коры надпочечников на основе клинической картины,
необходимо во время фолликулярной фазы проанализировать в 8 ч утра концентра¬
цию 17-гидроксипрогестерона, поскольку этот гормон имеет более высокую концен¬
трацию в лютеиновую фазу по сравнению с пролиферативной как у больных, так и у
здоровых женщин [228]. Уровень ниже 2 нг/мл помогает достоверно исключить этот
диагноз [228].
Диагноз неклассической ВДКН может быть поставлен, если базальный уровень
17-гидроксипрогестерона выше 8 нг/мл. В этом случае нет необходимости проводить
другие исследования. Значения между 2 и 8 нг/мл расценивают как увеличенные, но
не подтверждающие диагноз неклассической ВДКН. Так, здоровые женщины или
пациентки с СПКЯ также могут иметь базальный уровень 17-гидроксипрогестерона
в этих пределах [228]. При таких обстоятельствах единственным способом диффе¬
ренцировать неклассическую ВДКН от СПКЯ будет стимуляционный тест с АКТГ.
Тест считается положительным в отношении диагноза неклассической ВДКН при
повышении уровня 17-гидроксипрогестерона по меньшей мере до 10 нг/мл через
60 мин после в/в введения АКТГ [230]. Необходимо отметить, что, как правило,
более высокий базальный уровень 17-гидроксипрогестерона в пределах от 2 до 8 нг/мл
ассоциируется с более высокой вероятностью неклассической ВДКН. Например, в
8:00 уровень 17-гидроксипрогестерона выше 4 нг/мл имеет чувствительность 90% по
отношению к диагнозу неклассической ВДКН [228].
У пациенток, принадлежащих к этнической группе, в которой высока распростра¬
ненность заболевания, уровень 17-гидроксипрогестерона должен измеряться в 8:00.
К тому же следующие группы пациенток должны иметь скрининговое исследование
17-гидроксипрогестерона: с преждевременным пубархе, у которых избыток андро¬
генов возник с началом пубертата, женщины с прогрессирующим гирсутизмом или
вирилизацией, а также при семейном анамнезе выраженного избытка андрогенов.
Лабораторные методы в дифференциальной диагностике гиперандрогении
Существует несколько алгоритмов для дифференциальной диагностики ановуля¬
ции, ассоциированной с гирсутизмом и/или вирилизацией. Яркие клинические осо¬
бенности имеют огромное значение для проведения лабораторного исследования: наи¬
более важные из них включают время появления и степень выраженности симптомов,
а также скорость, с которой они прогрессируют. Высокая скорость прогрессирования
выраженного избытка андрогенов предполагает андроген-секретирующую опухоль,
пока не будет доказано обратное. В дальнейшем подчеркивается вероятность опухо¬
ли у женщин в постменопаузе или репродуктивного возраста, имеющих в анамнезе
циклические месячные. Гипертекоз яичников, тяжелый вариант СПКЯ, также при¬
водит к выраженному избытку андрогенов, который может прогрессировать быстро,
особенно во время предполагаемого пубертата. Избыток андрогенов, развивающийся
в пубертатном периоде, может обозначать СПКЯ или неклассическую ВДКН.
Наиболее полезным первоначальным тестом для оценки избытка андрогенов в
плазме крови является общий тестостерон плазмы (табл. 1-4). Уровни тестостерона у
большинства нормально овулирующих женщин определяются ниже 0,6 нг/мл, несмо¬
тря на то что значения могут варьироваться от лаборатории к лаборатории. Женщины
82
с идиопатическим гирсутизмом имеют регулярные менструации и нормальный уро¬
вень тестостерона. В отношении данной группы пациенток дальнейшие исследования
не нужны.
Таблица 1-4. Лабораторные методы в дифференциальной диагностике гиперандрогении
Первичное исследование
Общий тестостерон.
Пролактин.
Тиреотропный гормон
Последующие исследования, базирующиеся на основе клинических проявлений*
• 17-гидроксипрогестерон (анализ выполняют в 8 ч утра).
• 17-гидроксипрогестерон спустя 60 мин после в/в введения АКТГ.
• Кортизол (8 ч утра) после 1 мг дексаметазона в полночь.
• Дегидроэпиандростерона сульфат.
• Андростендион.
• Исследование яичников (трансвагинальное УЗИ).
• Визуализация надпочечников (абдоминальное УЗИ, КТ, МРТ).
• Сцинтиграфия после в/в введения радиоактивного меченого холестерина
* Пояснения в тексте.
При повышенном уровне тестостерона у женщин с ановуляторным циклом для
исключения ановуляции, вызванной гиперпролактинемией, необходимо измерить
концентрацию в плазме ТТГ и пролактина. Для подтверждения или исключения__
опухоли яичников или СПКЯ также полезно УЗИ яичников. Если этнический фон
пациентки (евреи ашкенази, испаноязычные американцы, а также имеющие предков
из стран центральной Европы) или семейный анамнез предполагает неклассическую
ВДКН, следует провести измерение базального уровня 17-гидроксипрогестерона в
плазме в 8 ч утра. Редкие причины избытка андрогенов включают опухоль надпочеч¬
ников, синдром Кушинга, резистентность к глюкокортикоидам. Для исключения или
подтверждения диагноза опухоли надпочечников необходимо провести измерение
концентрации дегидроэпиандростерона сульфата в плазме крови и исследование над¬
почечников с помощью методов лучевой диагностики. В зависимости от оснащенно¬
сти радиологической лаборатории для оценки надпочечников могут быть использова¬
ны КТ, МРТ, абдоминальное УЗИ. Для диагностики редких надпочечниковых причин
избытка андрогенов может быть проведено скрининговое исследование на синдром
Кушинга или резистентности к глюкокортикоидам [231].
Большинство пациенток с хронической ановуляцией и гирсутизмом от слабого до
умеренного, возникшего в пубертате, попадают в группу СПКЯ. Эти женщины имеют
нормальный или повышенный уровень тестостерона и отсутствие других лаборатор¬
ных аномалий. Если в результате лабораторного исследования или клинической кар¬
тины другие диагнозы исключены, может быть выставлен диагноз СПКЯ.
Лечение гирсутизма
Лечение избытка андрогенов должно быть этиотропным и направленным на
подавление аномальной секреции андрогенов. Специфическое лечение гирсутизма
и вирилизации должно проводиться в следующих ситуациях: опухоли яичников и
надпочечников, гипертекоз, синдром Кушинга, дисфункция коры надпочечников.
Опухоли, требующие хирургического лечения, детально не описываются. Подавление
аналогами ГнРГ может быть первично использовано в лечении гипертекоза яичников.
К сожалению, для контроля избытка андрогенов двустороннее удаление яичников
необходимо большинству пациенток (см. ниже). Специфическое лечение проводится
в отношении пациенток с заболеваниями надпочечников. При синдроме Кушинга
83
лечение проводят в соответствии с источником гиперкортицизма. При классической
ВДКН необходимо заместительное введение глюкокортикоидов, как при недостаточ¬
ности надпочеников. При лечении избытка андрогенов, связанного с неклассической
ВДКН, антиандрогенный препарат (например, спиронолактон) может быть исполь¬
зован в комбинации с оральными контрацептивами или глюкокортикоидами. При
длительных курсах лечения дозы глюкокортикоидов, необходимые для подавления
надпочечников, как правило, могут вызывать симптомы и признаки избытка глюко¬
кортикоидов. В связи с этим при лечении избытка андрогенов предпочтение должно
быть отдано комбинации антиандрогена и оральных контрацептивов, особенно если
пациентка отвечает на проводимое лечение снижением гирсутизма.
Общие принципы лечения избытка андрогенов направлены на предупреждение
аномального роста волос и вирилизации. Для практических целей подобный подход
используется при лечении идиопатического гирсутизма, СПКЯ и неклассической
ВДКН. Существующие волосяные фолликулы и проявления вирилизации (огру¬
бление голоса, клиторомегалия, височное облысение) остаются даже после пре¬
кращения избыточной продукции андрогенов. В связи с этим пигментированные
волосы должны быть удалены механическим способом (например, электролизом)
по крайней мере через 3 мес после снижения уровня андрогенов. Пациентки с кли¬
торомегалией могут быть направлены к урологу для хирургического уменьшения
размеров клитора после эффективного устранения источника вирилизации. Ниже
обсуждаются лекарства, которые используются для лечения избытка андрогенов и
гирсутизма.
Оральные контрацептивы
Оральные контрацептивы (ОК) снижают концентрацию циркулирующих пред¬
шественников тестостерона и андрогенов через супрессию ЛГ и стимуляцию ПСПС
и, таким образом, уменьшают проявления гирсутизма у гиперандрогенных пациен¬
ток [151]. ОК снижают уровень циркулирующих андрогенов при СПКЯ и действу¬
ют совместно с антиандрогенами. Также возможно, что ОК могут в дальнейшем
улучшать результаты антиандрогенной терапии при идиопатическом гирсутизме.
'Рекомендуется использовать ОК, содержащие 30 или 35 мкг этинилэстрадиола, для
достижения эффективной супрессии ЛГ [151].
Спиронолактон
Наиболее распространенный блокатор андрогенов, используемый для лечения гир¬
сутизма в США, — спиронолактон, антагонист альдостерона, структурно относящийся
к гестагенам. Спиронолактон эффективен для лечения аномального роста волос, свя¬
занного с СПКЯ или идиопатическим гирсутизмом.
В связи с тем что механизм действия спиронолактона отличается от механиз¬
ма действия оральных контрацептивов, общая эффективность увеличивается при
совместном назначении этих препаратов, в том числе в отношении пациенток с
идиопатическим гирсутизмом. Кроме ингибирования стероидогенеза и действия как
антагониста андрогенов, спиронолактон обладает значительным эффектом ингиби¬
рования активности 5а-редуктазы [151, 232]. Фундаментальные и отдельные клини¬
ческие исследования четко подтвердили эффективность спиронолактона для лечения
гиперандрогении; предполагают, что основной эффект относится к периферическому
блокированию продукции и действия андрогенов [151].
Дозы спиронолактона различаются в различных клинических исследованиях от 50
до 400 мг ежедневно. Несмотря на то что дозы в 100 мг ежедневно эффективны для
лечения гирсутизма, более высокие дозы (от 200 до 300 мг/день) могут быть более
предпочтительными у женщин с экстремальной формой гирсутизма или у пациенток
со значительным ожирением [151, 232]. Таким образом, рекомендуется начинать с
дозы в 100 мг ежедневно и постепенно увеличивать ее на 25 мг в день каждые 3 мес
до достижения 200 мг в день с учетом клинического ответа. Этот подход может быть
84
полезным для минимизации таких побочных эффектов, как гастрит, сухая кожа и
ановуляция.
У пациенток с нормальной функцией почек практически никогда не наблюдает¬
ся гиперкалиемия. Гипотензия также редка — за исключением пожилых женщин.
Мониторинг, как правило, обязателен для контроля уровня электролитов и артери¬
ального давления в течение первых 2 нед каждого изменения дозы. Регулирование
дозы, как и у других антиандрогенов, должно проводиться после 3-6 мес, что объ¬
яснимо скоростью цикла развития волоса. Пациентки обычно первично отмечают
проходящий диуретический эффект. Некоторые женщины, имеющие нормальные
менструальные циклы, жалуются на нерегулярные менструации при применении
спиронолактона. Последнее ликвидируется за счет снижения дозы или добавления
ОК. Механизм аномального кровотечения неясен. У женщин с олигоменореей, напри¬
мер при СПКЯ, могут восстановиться нормальные менструации. Частично это может
быть объяснено изменениями в уровнях циркулирующих андрогенов, несмотря на
то что лишь изредка отмечено снижение уровня ЛГ [233]. Другое важное соображе¬
ние — потенциальный феминизирующий эффект антиандрогенов на гениталии плода
с генотипом 46,ХУ. Таким образом, женщины, принимающие спиронолактон, должны
быть обеспечены эффективной контрацепцией.
Ципротерона ацетат
Ципротерона ацетат — производное 17-гидроксипрогестерона ацетата с выражен¬
ными прогестагенными свойствами. Ципротерона ацетат действует как антиандроген,
конкурирующий с дигидротестостероном и тестостероном за связывание с андроген¬
ными рецепторами. Существуют также некоторые факты, что ципротерона ацетат и
этинилэстрадиол в комбинации могут ингибировать активность 5а-редуктазы в коже
[234]. Ципротерона ацетат в настоящее время недоступен в США, но используется в
других странах. Лекарство в основном назначается в дозах от 50 до 100 мг с 5-го по
15-й день лечебного цикла. В связи с медленным метаболизмом он назначается в начале
лечебного цикла, тогда как этинилэстрадиол при добавлении обычно используется по
50 мг с 5-го по 26-й день. Этот режим необходим для контроля менструации и обычно
называется обратным режимом. Ципротерона ацетат в дозах от 50 до 100 мг/день в
комбинации с этинилэстрадиолом от 30 до 35 мг/день столь же эффективен, как ком¬
бинация спиронолактона и орального контрацептива для лечения гирсутизма [151].
В меньших дозах (2 мг) ципротерона ацетат назначают как ОК в ежедневной ком¬
бинации с 50 или 35 мкг этинилэстрадиола. Этот режим подходит для пациенток с
мягкой формой гиперандрогении [151].
Финастерид
Финастерид ингибирует активность 5а-редуктазы и первоначально использовался
для лечения гиперплазии простаты. Он также может быть применен для лечения
гирсутизма [235, 236]. После 6 мес приема препарата в дозе 5 мг/день наблюдается
значительное уменьшение гирсутизма без проявления побочных эффектов. У женщин
с гирсутизмом падение уровня циркулирующих андрогенов незначительно и не может
быть использовано для мониторирования терапии поскольку эти лечебные режимы
повышают уровень тестостерона, при этом уровень ПСПС остается неизменным
[235].
Финастерид первично ингибирует 5а-редуктазу 2-го типа. Этот препарат имеет
лишь частичную эффективность, так как в развитии гирсутизма играют роль рецеп¬
торы обоих типов. Несмотря на то что длительное использование финастерида не
исследовано, одним из преимуществ этого препарата является слабая выраженность
побочного действия. В ходе одного из исследований показана годичная эффектив¬
ность лечения гирсутизма [237]. Также было отмечено, что финастерид менее эффек¬
тивен, чем спиронолактон, в отношении уменьшения гирсутизма [151]. Тем не менее
финастерид полезен при лечении женщин с гирсутизмом в дозе 5 мг/день в течение
85
длительного промежутка времени с учетом слабовыраженных побочных эффектов и
хорошей переносимостью пациентами. Как и в случае спиронолактона, финастерид
может вызывать нарушение строения гениталий у плода с генотипом 46,Х¥, поэтому
во время применения необходимо обеспечить надежную контрацепцию.
Флутамид
Флутамид — потенциальный антиандроген, используемый в лечении рака проста¬
ты. Он также показал эффективность при лечении гирсутизма [238,239]. Тем не менее
редкая, но тяжелая степень гепатотоксичности делает использование этого препарата
невозможным для лечения гирсутизма [240].
Заключение
Вышеперечисленные препараты могут быть эффективными при индивидуальном
подборе. Пациентки с наиболее распространенными формами гирсутизма (напри¬
мер, СПКЯ) часто первично получают комбинацию двух препаратов: один подавляет
секрецию в яичнике (например, оральный контрацептив), а другой подавляет экстра-
овариальное периферическое действие андрогенов (спиронолактон). Таким образом,
ОК, содержащие 30-35 мкг этинилэстрадиола, комбинируемый со спиронолактоном,
100 мг/день, — препараты первой линии. Даже у женщин с идиопатическим гирсу¬
тизмом добавление ОК к антиандрогену спиронолактону может увеличить эффектив¬
ность и предотвратить аномальное кровотечение. У женщин с легким гирсутизмом
использование только ОК может быть адекватным первым шагом.
Поскольку фаза роста волос на теле длится от 3 до 6 мес, не стоит ожидать эффекта
ранее 6 мес от начала лечения. Для оценки роста волос необходимо применять объек¬
тивные критерии. Системы подсчета и оценки фазы роста волосяного стержня затруд¬
нительны; фотографирование — наиболее простой и объективный метод. До момента
появления объективных признаков пациентки часто не подозревают о происходящих
изменениях. Фотографии лица и определенных участков срединной линии до и после
курса лечения особенно полезны для поддержки пациенток и согласия с лечением.
Супрессия продукции и активности андрогенов ингибирует только рост новых
волос. Именно поэтому существующие грубые волосы необходимо удалять механиче¬
ски. Выдергивание, удаление воском и бритье малоэффективны для удаления волос и
вызывают раздражение, фолликулиты и врастание волос. Электролиз — метод выбо¬
ра. Лазерная эпиляция — относительно новая процедура, требующая дальнейшего
изучения [151].
Большинство пациенток с СПКЯ и идиопатическим гирсутизмом отвечают на
лечение в течение года. Пациенты должны быть готовы продолжить лечение, как
минимум, в течение 2 лет. После этого в зависимости от их пожеланий и клиническо¬
го ответа терапия может быть остановлена и проведена оценка результатов. Многие
пациентки требуют продолжительного лечения для супрессии гирсутизма.
СИНДРОМ поликистозных яичников
СПКЯ — самая частая форма хронической ановуляции, возникающая у 5-10%
женщин репродуктивного возраста и связанная с избытком андрогенов [146]. Диагноз
СПКЯ ставят на основании исключения других причин гиперандрогении (например,
неклассическая ВДКН, андроген-секретирующие опухоли, гиперпролактинемия) у
женщин с хронической ановуляцией и избытком андрогенов,
В репродуктивном возрасте СПКЯ ассоциируется с серьезным нарушением репро¬
дуктивной функции, включая бесплодие, нерегулярные менструации, невынашивание
беременности. Эндометрий у пациенток с СПКЯ должен быть оценен с помощью
биопсии в связи с длительным воздействием эстрогенов, без адекватного воздействия
прогестероном, которое может привести к раку эндометрия. СПКЯ также ассоции¬
руется с увеличением метаболических и сердечно-сосудистых факторов риска [241].
86
Эти риски связаны с инсулиновой резистентностью и ожирением, однако СПКЯ также
обнаруживают у женщин, не страдающих ожирением [146].
СПКЯ определяют как гетерогенное расстройство мультифакторной этиологии.
Риск СПКЯ значительно увеличивается в случае наследственной хронической ано¬
вуляции и избытка андрогенов, и этот комплекс расстройств может наследоваться
полигенно [242, 243].
Исторический экскурс
в своих первых исследованиях Штейн и Левенталь [244] описали связь между
увеличенными с обеих сторон поликистозными яичниками и аменореей, олигоме¬
нореей, гирсутизмом и ожирением (рис. 1-29). В то время данные симптомы рассма¬
тривались сугубо как проявление симптомокомплекса, позже названного синдромом
Штейна-Левенталя. Эти исследователи также сообщили о результатах двусторонней
клиновидной резекции яичников, проведенной по поводу СПКЯ, с удалением по
меньшей мере половины каждого яичника, после которой у пациенток восстанавли-
Рис. 1-29. Поликистоз яичников. А — классически увеличенный поликистозный яичник (материал опе¬
раций). Матка располагается рядом с двумя увеличенными яичниками; Б — разрезанный поликистозный
яичник с несколькими фолликулами; В — гистологический препарат поликистозных яичников с множе¬
ственными субкапсулярными фолликулярными кистами и стромальной гипертрофией (малое увеличение,
слева). При большом увеличении (хЮО) в строме определяются островки лютеинизированных клеток
(справа). Эти морфологические изменения называются стромальным гипертекозом и прямо связаны
с уровнем циркулирующего инсулина. [Из Dunaif А. Insulin resistance and the polycystic ovary syndroime:
mechanism and implications for pathogenesis // Endocr. Rev. — 1997. — N 18. — P. 774-800. Copyright ©
1997 by The Endocrine Society.]
87
вался менструальный цикл и наступали беременности. На основании этого постули¬
ровали, что удаление утолщенной капсулы с поверхности яичника восстанавливает
овуляцию, способствуя достижению доминантным фолликулом поверхности яичника
(см. рис. 1-29). Точный механизм, определяющий терапевтический эффект удаления
части ткани яичника, до конца не понятен.
На основании работы Штейна и Левенталя первичный дефект был определен
в яичнике, а расстройство было названо синдромом поликистозных яичников.
Последующие клинические, морфологические, гормональные и метаболические
исследования выявили множество расстройств, лежащих в основе заболевания
(см. ниже). Термин «поликистозные яичники» был предложен для отражения гетеро¬
генности этого расстройства.
Одно из наиболее важных открытий в патофизиологии СПКЯ — уникальная форма
инсулинорезистентности и гиперинсулинемии [146]. Впервые Burghen с соавт. сооб¬
щили об этой находке в 1980 г. [245]. Инсулинорезистентность при СПКЯ в последую¬
щем была подтверждена исследованиями по всему миру [146].
Диагностика синдрома поликистозных яичников и лабораторные исследования
Одно из наиболее важных проявлений СПКЯ — история овуляторной дисфункции
(аменорея, олигоменорея или другие формы нерегулярных маточных кровотечений),
начинающейся в пубертате. Таким образом, наличие в прошлом циклических пред¬
сказуемых менструаций с самого менархе делает диагноз СПКЯ маловероятным.
Приобретенную инсулинорезистентность связывают либо с избыточной массой тела,
либо с неизвестной причиной, которая, тем не менее, может иногда приводить к кли¬
нической картине СПКЯ у женщин с нормальной овуляторной функцией. Гирсутизм
может возникать в препубертате или в подростковом возрасте или может отсутство¬
вать до 3-й декады жизни. Себорея, акне и алопеция — обычные признаки избытка
андрогенов. При экстремальных случаях гипертекоза яичников (тяжелый вариант
СПКЯ) может наблюдаться клиторомегалия. Тем не менее быстрое нарастание сип¬
томов андрогенизации и вирилизации нехарактерно для СПКЯ. Некоторые женщины
. могут никогда не иметь симптомов избытка андрогенов из-за врожденной разницы в
чувствительности в тканях-мишенях к андрогенам [151]. Бесплодие, связанное с ано¬
вуляцией, может быть единственным симптомом.
Во время объективного исследования необходимо провести поиск и зафиксировать
признаки избытка андрогенов (гирсутизм и вирилизация или и то, и другое), инсу¬
линорезистентности (акантоз, acanthosis nigricans), а также присутствие прикрытого
прогестероном эстрогенового воздействия (влагалище с выраженными складками,
растяжимая прозрачная цервикальная слизь) для подтверждения диагноза СПКЯ
(рис. 1-30). Необходимо отметить, что ни один из вышеперечисленных симптомов не
характерен для СПКЯ и может быть связан с любым из состояний, перечисленных при
дифференциальной диагностике с СПКЯ (табл. 1-5).
Таблица 1-5. Дифференциальный диагноз синдрома поликистозных яичников
' Идиопатический гирсутизм.
I Гиперпролактинемия, гипотиреоз.
' Неклассическая ВДКН.
' Опухоли яичников.
I Опухоли надпочечников.
' Синдром Кушинга.
' Резистентность к глюкокортикоидам.
' Другие редкие причины избытка андрогенов
До недавнего времени СПКЯ определяли согласно рекомендациям экспертов кон¬
ференции, проведенной под патронажем Национального института здоровья (НИЗ,
National Institutes of Health) [246] в 1990 г., согласно которым заболевание должно
88
Рис. 1-30. Акантоз [acanthosis nigricans). А —умеренный акантоз (потемнения и утолщения кожи) по
бокам и ниже складок кожи шеи. Обратите внимание на гирсутизм на лице (бакенбарды); Б — выра¬
женный акантоз у другой пациентки с инсулинорезистентностью. [Б — Courtesy of Dr. R. Ann Word,
Dallas, TX.l
иметь: гиперандрогенизм и/или гиперандрогенемию, олиго/ановуляцию и исключе¬
ние известных заболеваний, связанных с избытком андрогенов и ановуляцией. При
этом другая конференция экспертов в Роттердаме в 2003 г. определила СПКЯ как
заболевание, которое после исключения известных заболеваний удовлетворяет двум
из трех критериев: олиго/ановуляция, клинические и/или биохимические признаки
гиперпандрогенизма и поликистозные яичники (рис. 1-31). Критерии, сформулиро¬
ванные в Роттердаме (2003), расширяют определение Национального института здо¬
ровья (1990) созданием двух новых фенотипов — женщины с овуляторным циклом
с поликистозными яичниками и гиперандрогенизмом, а также пациентки с олиго/
ановуляцией с поликистозными яич¬
никами, но без гиперандрогенизма.
Клиническая польза от включения
этих новых групп по отношению к
повышенному риску бесплодия,
инсулиновой резистентности, дли¬
тельным метаболическим осложне¬
ниям в настоящее время четко не
определена [247].
Исключение гиперпролактине¬
мии, гипотиреоза, неклассической
ВДКН, опухоли требует тщательно¬
го сбора анамнеза и объективного
исследования, а также лабораторно¬
го обследования, как описано ранее
(см. табл. 1-4). Синдром Кушинга и
резистентность к глюкокортикоидам
могут приводить к избытку андро¬
генов и ановуляции после периода
нормальной овуляторной функции
у подростков. Определение уровня
кортизола в 8 ч утра после назна¬
чения дексаметазона в дозе 1 мг в
Рис. 1-31. Ультразвуковой снимок при трансвагиналь¬
ном УЗИ. Обратите внимание на множественные фол¬
ликулы среднего размера по периферии и увеличенные
солидные участки в середине. [Из Franks S. Medical
progress: polycystic ovary syndrome // New Engl. J. Med. —
1995.-N 333. - P. 853-861.1
89
полночь — полезный скрининговый тест для обоих состояний. Синдром Кушинга
можно определить по характерным клиническим признакам, тогда как уровни кор¬
тизола в 8 ч и в 16 ч важны для диагностики резистентности к глюкокортикоидам
[248]. Резистентность к глюкокортикоидам характеризуется фиксированным днев¬
ным ритмом, несмотря на значительно увеличенный кортизол, АКТГ и уровень С19-
надпочечникового стероида, а также отсутствием симптомов синдрома Кушинга [248]
(табл. 1-6) [249, 250].
Таблица 1-6. Диагностические критерии синдрома поликистозных яичников
Все нижеперечисленные симптомы
1. Гиперандрогенизм и/или гиперандрогенемия
2. Олиго/ан(«уляция
3. Исключение схожих (связанных) заболеваний*
ESHRS/ASRM (ÜütteTdænb 20031^50]
Наличие двух из нижеперечисленных и исключение других возможных заболеваний*
1. Олигоовуляция или ановуляция (например, аменорея или нерегулярные маточные кровотечения)
2. Клинические и/или биохимические признаки гиперандрогенизма (гирсутизм и/или увеличенная концентрация
общего или связанного тестостерона в плазме)
3. Поликистоз яичников (по данным УЗИ)
* Б том числе дефицит 21-гидроксилазы неклассической ВДКН, дисфункции щитовидной желе¬
зы, гиперпролактинемии, неопластической андрогенной секреции, лекарственно-индуцированного
избытка андрогенов, синдрома Кушинга или резистентности к глюкокортикоидам. [Модифицировано
из Azziz R. Diagnosis of Polycystic Ovarian Syndrome: The Rotterdam Criteria Are Premature // J. Clin.
Endocrinol. Metab. - 2006. - N 91. - P. 781-785. CopjTight © 2006 by The Endocrine Society.]
Примечание. ASRM — American Society for Reproductive Medicine, ESHRE — European Society for Human
Reproduction and Embryohgy.
Как было отмечено ранее, повышенный уровень тестостерона — наиболее прямое
свидетельство избытка андрогенов. У женщин с СПКЯ уровень тестостерона разли¬
чен, Редко выявленный уровень тестостерона более 2 нг/мл может быть ассоциирован
с наиболее тяжелой формой СПКЯ, гипертекозом яичников. Более того, часто уровень
тестостерона у женщин с СПКЯ не превышает верхнюю границу нормы,
Пролактин и ТТГ должны быть рутинно исследованы для исключения мягкой
формы избытка андрогенов и ановуляции, которые могут быть связаны с гиперпро¬
лактинемией. Базальный уровень Л Г не используется как маркер СПКЯ, поскольку
у значительного количества пациенток нет увеличения уровня ЛГ или повышения
отношения ЛГ/ФСГ. Консенсус под медицинским патронажем НИЗ в 1990 г. пришел
к выводу, что ни определенный уровень ЛГ, ни определенное соотношение ЛГ/ФСГ
не требуются для установления диагноза СПКЯ [249, 251]. Гетерогенность уровней
ЛГ у пациенток с СПКЯ может объясняться пульсирующей природой секреции ЛГ и
негативными эффектами ожирения на уровни ЛГ. Таким образом, увеличенное соот¬
ношение ЛГ /ФСГ поддерживает диагноз СПКЯ и может быть полезным при диффе¬
ренциации мягких форм СПКЯ без ожирения, без выраженного избытка дндрогенов
от гипоталамической ановуляции. Согласно определению, неклассическая дисфунк¬
ция коры надпочечников не проявляется как врожденная вирилизация наружных
гениталий. Гиперандрогенные симптомы наиболее часто возникают в препубертате.
Клиническая оценка и диагноз неклассической ВДКН, основанные на лабораторных
данных, обсуждались выше, в рубрике «Неопухолевые заболевания надпочечников и
избыток андрогенов». Скрининговый тест для синдрома Кушинга или резистентности
к глюкокортикоидам должен быть проведен по клиническим показаниям.
90
Уровень сывороточного ДГЭАС может быть повышен (до 8 мг/мл) у 50% женщин
с ановуляторным циклом с СПКЯ. ДГЭАС вырабатывается практически весь в над¬
почечнике. Причины гиперактивности надпочечников [252] при СПКЯ неизвестны.
Измерение уровня ДГЭАС у женщин с СПКЯ рутинно не рекомендовано, так как это
не изменяет диагноз или план ведения больного. С другой стороны, если подозрева¬
ется опухоль надпочечников, должен быть измерен уровень ДГЭАС. Уровень ДГЭАС
более 8 мг/мл может быть связан со стероидоактивной опухолью надпочечников,
вследствие чего показано проведение лучевой диагностики.
Роттердамские критерии включают использование УЗИ как диагностического
теста. Использование УЗИ в диагностике СПКЯ должно быть ограничено в связи
с большим количеством женщин, имеющих ультразвуковую картину поликистоз¬
ных яичников. УЗ-картина поликистозных яичников — неспецифический признак,
характерный для ановуляции (см. рис. 1-31). Установлен ли диагноз по УЗИ или по
традиционным клиническим и биохимическим критериям, исследование женщин с
ановуляторным циклом, проведенное в этот период, выявляет, что около 75% имеют
поликистозные яичники (что подтверждают результаты УЗИ) [253]. Поскольку суще¬
ствует несколько причин ановуляции, соответственно есть несколько причин возник¬
новения поликистозных яичников. Похожая клиническая картина и состояние яич¬
ников могут отражать любое из дисфункциональных состояний, отмеченных выше.
Другими словами, поликистоз яичников — результат функционального расстройства,
а не специфический признак центрального или локального нарушения.
Биохимические доказательства резистентности к инсулину или нарушения толе¬
рантности к глюкозе также не всегда необходимы для диагноза СПКЯ. Нарушение
толерантности к глюкозе, тем не менее, должно быть исключено. Вследствие этого
уровень глюкозы в плазме должен быть измерен после введения 75 г глюкозы в каче¬
стве теста на толерантность к глюкозе\
Женщины с СПКЯ обычно имеют нерегулярный менструальный цикл (олигоме¬
норея или аменорея). Необязательно документировать ановуляцию с помощью УЗИ,
измерения уровня прогестерона, особенно если менструации нерегулярные с эпизода¬
ми аменореи. Для подтверждения диагноза хронической ановуляции и относительной
гиперэстрогении на фоне дефицита гестагенов обьпшо проводят тест с гестагеном
(после отрицательного теста на беременность). Поскольку эндометрий хронически
подвергается воздействию эстрогенов при СПКЯ, эти женщины отвечают на тест- с
гестагеном [например, медроксипрогестерона ацетат (МПА) 10 мг в день внутрь в
течение 10 дней] маточным кровотечением через несколько дней после последнего
приема гестагена. Причины отсутствия маточного кровотечения после теста с геста-
генами — беременность, недостаточность предшествующего воздействия эстрогенов
на эндометрий либо анатомический дефект. Если маточное кровотечение не следует
за гестагеновым тестом, необходимо вновь исключить беременность, а также другие
причины хронической ановуляции, как это было описано ранее. Анатомические
дефекты, такие как внутриматочные спайки, могут быть исключены с помощью гисте-
росальпингограммы или гистероскопии.
Наконец, при первичном исследовании рекомендовано получить биопсию эндоме¬
трия с использованием пластиковой канюли (например, Пайпеля), Если хроническая
ановуляция сохраняется, биопсия эндометрия должна быть проведена несколько раз.
Необходимо исключать беременность перед каждой биопсией эндометрия. Ответ
на ОК или периодическое назначение гестагенов с предсказуемым кровотечением
отмены — обнадеживающий фактор. Пациентки с предсказуемым кровотечением не
требуют биопсии эндометрия во время этого лечения, у нелеченных, даже молодых
* Наличие СПКЯ рассматривается как риск диабета и следует исключить собственно диабет (под¬
робнее см. том «Эндокринология по Вильямсу. Сахарный диабет и нарушения углеводного обмена»),
(Иримег. ред.)
91
пациенток риск гиперплазии и рака эндометрия значительно увеличен с СПКЯ в связи
с относительной гиперэстрогенией на фоне дефицита гестагенов.
Продукция гонадотропинов при синдроме поликистозных яичников
Женщины с СПКЯ имеют более высокие средние концентрации ЛГ, но низкие или
в нижней границе нормы концентрации ФСГ, сравниваемые с уровнями, найденны¬
ми у здоровых женщин в ранней фолликулярной фазе [254]. Повышенный уровень
ЛГ частично связан с увеличенной чувствительностью гипофиза к стимуляции ГнРГ,
что проявляется увеличением частоты и особенно амплитуды ЛГ [255, 256, 257].
Интересно, что и биоактивность ЛГ также возрастает [256].
Повышенный уровень ЛГ при СПКЯ обусловлен усилением выбросов ГнРГ.
Центральная опиоидная активность оказывается подавленной, так как секреция ЛГ
не изменяется в ответ на налоксон [257]. Действительно, усиленной пульсирующей
секреции ГнРГ приписывается уменьшение ингибирования опиоидов в гипоталаму¬
се, которое возникает при хроническом отсутствии прогестерона [181]. Увеличение
амплитуды и частоты секреции ЛГ также коррелирует с установившимися уровнями
циркулирующих эстрогенов.
у женщин с СПКЯ и ожирением уровень ЛГ не увеличен. Увеличение частоты
выбросов ЛГ характерно для ановуляторного состояния независимо от содержания
жира в теле [259]. Амплитуда выброса ЛГ, однако, сравнительно нормальная у тучных
женщин с СПКЯ и повышена у женщин с СПКЯ, но нормальной массой тела [260].
Общее снижение ЛГ у женщин с ожирением и СПКЯ может быть связано с факторами,
отличными от влияющих на них амплитуд ЛГ. Необходимо отметить вновь, что низ¬
кий уровень Л Г не является основанием для исключения диагноза СПКЯ, тогда как
высокое значение отношения ЛГ/ФСГ подтверждает этот диагноз у ановуляторных
женщин.
Инсулин также считают потенциальным регулятором секреции ЛГ при СПКЯ.
Инсулин усиливает транскрипцию гена р-субъединицы Л Г [262, 263]. С другой сто¬
роны, увеличение секреции ЛГ у женщин с СПКЯ, так же как у нормальных женщин,
было неизменным при длительной индукции инсулином [264]. Более того, уровни
ЛГ, частота и амплитуда выбросов ЛГ, так же как и ответ гонадотропинов на ГнРГ,
не изменялись пиоглитазоном с/без инсулинотерапии [264]. Таким образом, ш vivo
исследования на людях не подтверждают, что инсулинорезистентность или гиперин-
сулинемия ответственны за аномальный выброс гонадотропинов.
Продукция стероидов при синдроме поликистозных яичников
Овуляторные циклы характеризуются циклическими колеблющимися уровнями
гормонов, регулирующих овуляцию и менструацию (см. рис. 1-1, А).
Ановуляция у женщин с СПКЯ сопровождается стабильными уровнями гонадо¬
тропинов и яичниковых стероидов, у пациенток с постоянной ановуляцией средний
уровень продукции эстрогенов и андрогенов в течение дня повышен и зависит от
стимуляции ЛГ (рис. 1-32) [265]. Это отражено в увеличенных циркулирующих уров¬
нях тестостерона, андростендиона, ДГЭА, ДГЭАС, 17-ГСД и эстрона. Тестостерон,
андростендион и ДГЭА секретируются яичником, тогда как ДГЭАС, повышенный
у половины женщин с ановуляцией и СПКЯ, секретируется надпочечниками [252].
Уровень циркулирующего андростендиона, секретируемого поликистозными яични¬
ками. значительно увеличен (см. рис. 1-32),
Эстрон возникает прежде всего в результате периферической ароматизации андро¬
стендиона и частично за счет секреции яичниками (рис. 1-33) [266]. Он не является
активным эстрогеном и может рассматриваться как предшественник, конвертируе¬
мый в эстрадиол, который обладает полной эстрогенной активностью. Присутствует
некоторое количество фермента 1713-ГСДГ с перекрывающейся активностью, который
катализирует конверсию эстрона в эстрадиол в периферических (экстраовариальных)
92
Надпочечник
МФР
ФСГ Инсулин
1
Измененные уровни
Гипоталамус
половых гормонов
Резистентность
к инсулину
Инсулин!
Биодоступный Ег!
Биодоступный Т !
иодоступный ИФР-1!/
Измененная
секреция ГнРГ
Гипофиз
ЛСПСІ
ИФРС^-1 і
Жировая ткань
Е2 онт
Периферические ткани и ткани-мишени
Печень
Рис. 1 -32. Патологические механизмы при СПКЯ. Недостаточный ответ фолликула на физиологические
количества ФСГ, возможно, в связи с ослабленным взаимодействием сигнальных путей ФСГ с инсулино'-
подобным фактором роста (ИФР) или инсулином, что может быть серьезным нарушением при СПКЯ. Это
может быть ключевым событием, ответственным за ановуляцию при СПКЯ. Резистентность к инсулину
связана с увеличенными циркулирующими и тканевыми уровнями инсулина и биодоступного эстрадиола
(Е^), тестостерона (Т), а также ИФР-1, способствующими аномальной продукции гормона в определен¬
ных тканях. Избыточная секреция гипофизом ЛГ, сниженная секреция ФСГ и сниженная выработка про¬
теина, связывающего половые стероиды (ПСПС), а также ИФР-1-связывающего протеина (ИФРСП-1) в
печени, увеличенная секреция в надпочечниках дегидроэпиандростерона сульфата (ДГЭАС) и увеличен¬
ная секреция яичниками андростендиона (А), — все это относится к порочному кругу, который поддер¬
живает ановуляцию и избыток андрогенов при СПКЯ. Избыточные количества Е^ и Т возникают первично
в результате конверсии (А) в периферических тканях и тканях-мишенях. 17Р-ГСД — 17()-гидроксистероид
дегидрогеназа; 5а-ред — 5а-редуктаза.
тканях [132, 266]. Повышение уровня андростендиона приводит к ощутимому уве¬
личению циркулирующего эстрадиола у женщин с СПКЯ в сравнении с уровнями
эстрадиола, измеренными во время первых дней овуляторного менструального цикла.
Этот процесс происходит через активность ароматазы и ІУр-ГСДГ в таких экстраова¬
риальных тканях, как кожа и подкожный жировой слой. Также локальная конверсия
эстрона в эстрадиол является важным физиологическим процессом для опреде¬
ляемых тканей-мишеней, таких как здоровая молочная железа и кожа гениталий, В
результате локальная конверсия также может способствовать росту патологических
эстрогензависимых тканей, таких как рак молочной железы и эндометриоз (см. рис.
1-33) [133, 267-269].
Андростендион яичникового происхождения наиболее заметно повышается при
СПКЯ феди всех стероидов. Андростендион не является биологически активным,
но служит двойным предшественником для андрогенов (тестостерон, который позже
превращается в биологически активный дигидротестостерон) и эстрогенов (эстрон,
позже превращающийся в биологически активный эстрадиол в клетках-мишенях, см.
рис. 1-33) [124]. Эстрадиол — самый активный стероид. Биологически активные уров¬
ни эстрадиола измеряются в пг/мл, или пмоль/л, тогда как биологически активные
93
Рис. 1 -33. Экстраовариальная конверсия андростендиона в андроген и эстроген. Андростендион надпо¬
чечникового или яичникового происхождения действует как двойной предшественник для андрогенов и
эстрогена. 5% циркулирующего андростендиона превращается в циркулирующий тестостерон, тогда как
1,3% циркулирующего андростендиона превращается в циркулирующий эстрон в периферических тка¬
нях. Тестостерон и эстрон в дальнейшем конвертируются 8 биологически активные стероиды дигидро¬
тестостерон и эстрадиол в периферических тканях-мишенях. Биологически активные количества эстра¬
диола в плазме измеряются в пг/мл (пмоль/мл), тогда как биологически активный уровень тестостерона
в сыворотке измеряется в нг/мл (нмоль/мл). Таким образом, 1,3% конверсии нормального количества
андростендиона в эстрон может иметь критическое биологическое влияние в случае постменопаузаль¬
ного рака эндометрия или рака молочной железы. Более того, значительный избыток андрогенов наблю¬
дается в случаях аномально высокой продукции андростендиона (например, при СПКЯ).
уровни тестостерона — в нг/мл, или нмоль/л (что больше в 10-100 раз по сравнению
с уровнем эстрадиола). Таким образом, даже небольшая конверсия андростендиона
в эстрон может иметь значительное биологическое значение, тогда как значительно
возросшая продукция андростендиона требуется для производства значительных
количеств тестостерона и проявления избытка андрогенов (см. рис. 1-33). Поскольку
такая возросшая продукция андростендиона проходит при СПКЯ, экстраовариальная
продукция тестостерона биологически важна при данном заболевании. У женщин в
постменопаузе, которые имеют намного более низкие уровни андростендиона, экс¬
траовариальная продукция тестостерона менее важна. С другой стороны, относитель¬
но маленькие количества эстрона (и эстрадиола). продуцируемые первично при пери¬
ферической ароматизации андростендиона, имеют биологическое значение у мужчин
и женщин в постменопаузе [124].
Продукция протеина, связывающего половые стероиды, при синдроме
поликистозных яичников
ПСПС связывает тестостерон и эстрадиол и таким образом уменьшает биологиче¬
скую активность этих важных стероидов. При СПКЯ увеличена продукция андрогенов
и эстрогенов. Увеличенные эстроген- и андрогенные эффекты при СПКЯ, как прави¬
ло, также связаны со снижением концентрации ПСПС, который способствует увели¬
чению свободного или биологически активного эстрадиола и тестостерона (см. рис.
1-32), Уровень ПСПС контролируется балансом гормонального влияния на их синтез
в печени. Тестостерон и инсулин подавляют, а эстрогены и тироксин стимулируют
образование ПСПС [270]. У женщин с ановуляцией и СПКЯ циркулирующие уровни
ПСПС снижены примерно на 50%; это может быть ответом печени на увеличенные
уровни циркулирующего тестостерона и инсулина (см. рис, 1-33) [270]. Свободный
94
циркулирующий эстрадиол и тестостерон повышены в связи с существенным сниже¬
нием ПСПС у пациентов с СПКЯ.
Таким образом, три механизма обеспечивают присутствие увеличенного количе¬
ства биологически активного эстрадиола при СПКЯ:
« повышенная продукция эстрадиола из эстрона в периферических (экстраовари¬
альных) тканях, поддерживающая увеличенные уровни циркулирующего эстра¬
диола;
* повышенный уровень биологически активного циркулирующего эстрадиола
вследствие сниженного уровня ПСПС;
« локальная конверсия эстрона в эстрадиол в тканях-мишенях [268]. Последний
локальный механизм физиологически значим в таких тканях-мишенях, как
молочная железа, которая пролиферирует в ответ на действие эстрогенов (см.
рис. 1-32).
Помимо повышения уровня биологически активного эстрадиола, сниженная
концентрация в плазме ПСПС вызывает увеличение биологически доступного тесто¬
стерона. В свою очередь, тестостерон снижает уровень ПСПС в плазме, способствуя
возникновению порочного круга, связывающего низкий ПСПС и высокий биодоступ¬
ный тестостерон (см. рис. 1-32). Инсулин напрямую снижает концентрацию ПСПС
у женщин с СПКЯ независимо от любого действия половых стероидов [270J, Таким
образом, инсулин увеличивает свободный тестостерон при СПКЯ через два незави¬
симых механизма: увеличение яичниковой секреции предшественников тестостерона
(например, андростендион) и супрессию ПСПС [270J,
Фолликулы при синдроме поликистозных яичников
Под влиянием относительно низких, но стабильных концентраций ФСГ постоянно
стимулируется рост фолликулов, но не до стадии зрелого фолликула и овуляции [271].
Даже несмотря на то что полный потенциал роста не реализуется, жизнь фолликула
может продолжаться несколько месяцев в форме множественных фолликулярных
кист. Большинство фолликулов имеет размер 2-10 мм в диаметре, некоторые достига¬
ют 15 мм. Гиперпластические тека-клетки, часто лютеинизированные в ответ на высо¬
кий уровень ЛГ, окружают эти фолликулы (см. рис. 1-29). Накопление фолликулов,
прекративших развитие на разных стадиях, способствует увеличенному и относитель¬
но постоянному синтезу стероидов в ответ на стабильные уровни гонадотропинов.
Эти фолликулы также подвергаются атрезии и замещаются новыми фолликулами
с подобным ограниченным потенциалом роста. Обновление стромальных клеток
связано со стромальным компартментом яичника и поддерживается тканями, полу¬
ченными после атрезии фолликулов. Дегенерирующий гранулезный компартмент
способствует атрезии, оставляя тека-клетки пополнять стромальный компартмент
яичника (рис 1-29). Эта функционирующая стромальная ткань секретирует значи¬
тельное количество андростендиона под влиянием увеличенного ЛГ. Андростендион
посредством описанных механизмов приводит к увеличению свободного тестостерона
и эстрадиола и уменьшению ПСПС (см. рис. 1-33). С точки зрения стероидогенеза
и эффекта стероидов СПКЯ сопровождается порочным кругом, который включает
определенное количество положительных и отрицательных механизмов обратной
связи (см. рис. 1-32).
На рис. 1-32 представлена схема недавно установленных механизмов, леждщих в
основе СПКЯ. Поскольку ФСГ и инсулин/ИФР могут действовать совместно, считают,
что эта синергия не возникает в присутствии инсулиновой резистентности и может
приводить к относительной резистентности яичниковых фолликулов по отношению
к ФСГ. Исследования in vitro не подтверждают эти данные. Культивированные гра¬
нулезные клетки, полученные из маленьких фолликулов поликистозных яичников,
образуют незначительное количество эстрадиола, но при добавлении к культуре ФСГ
95
или ИФР-1 количество продуцируемых эстрогенов значительно возрастает. Более
того, когда ФСГ и ИФР-1 добавляли совместно in vitro, они взаимодействовали, уси¬
ливая биосинтез эстрогенов в гранулезных клетках, полученных из поликистозных
яичников [272].
Индукция овуляции при СПКЯ достигается повышением уровня ФСГ, что, вероят¬
но, устраняет предполагаемую резистентность к ФСГ на уровне гранулезных клеток.
Два популярных лекарственных средства — кломифепа цитрат для приема внутрь и
инъекционный рекомбинантный ФСГ — повышают уровень эндогенного и экзоген¬
ного ФСГ, что приводит к овуляции. Некоторым пациенткам с СПКЯ могут потребо¬
ваться большие дозы для достижения овуляции. Парадоксально, но поликистозный
яичник может прореагировать на стимуляцию ФСГ вовлечением большого количе¬
ства развивающихся фолликулов одновременно, изредка приводя к синдрому гипер¬
стимуляции яичников (см. ниже) [273]. Терапевтическое окно между отсутствием
эффекта и гиперстимуляцией, как правило, узкое. До сих пор остаются нерешенными
вопросы, которые не позволяют полностью совместить данные, полученные in vitro и
in vivo, относящиеся к патофизиологии СПКЯ.
Гипертекоз яичников
Гипертекоз яичников — тяжелый вариант СПКЯ. Этот термин относят к увеличе¬
нию стромальной ткани вместе с лютеинизированными тека-клетками, разбросан¬
ными между большими пластами клеток, похожих на фибробласты. И клинические,
и гистологические находки представляют тяжелую форму СПКЯ [274]. Этот диагноз
может быть поставлен на основе клинической картины; яичниковая биопсия необяза¬
тельна, за исключением необходимости исключить опухоль яичника.
Увеличенная продукция андрогенов приводит к клинической картине наиболее
интенсивной андрогенизации. Более высокий уровень тестостерона вызывает сни¬
жение Л Г путем блокирования эффектов эстрогенов на уровне гипоталамус-гипофиз
[261]. Гипертекоз характеризуется наиболее выраженной тяжестью процесса, приво¬
дящего к хронической ановуляции при СПКЯ, Существует корреляция между выра¬
женностью текоза и степенью инсулинорезистентности [261]. Кроме того, поскольку
инсулин и ИФР-1 стимулируют пролиферацию тека-интерстициальных клеток, гипе-
ринсулинемия может быть важным патофизиологическим фактором в этиологии
гипертекоза.
Нередко встречают значительное увеличение уровня тестостерона, более 2 нг/мл,
при гипертекозе яичников; в таких случаях обычно диагностируют вирилизацию.
Овуляция в ответ на назначение кломифена цитрата или рекомбинантного ФСГ у
этих пациенток не возникает. Часто тяжело подавить продукцию тестостерона, даже
используя антагонисты ГнРГ. Двустороннее удаление яичников следует использовать
при неэффективности других методов, но, к сожалению, у некоторых больных, несмо¬
тря на вмешательство может быть необходим дополнительный контроль уровня
тестостерона.
Генетика синдрома поликистозных яичников
СПКЯ часто наблюдают у членов одной семьи, что говорит о наследственной пред¬
расположенности [275, 276], Ранее установлено, что СПКЯ имеет мультифакториаль-
ную природу и его развитие происходит за счет множества разных патофизиологи¬
ческих механизмов. Эти особенности подтверждаются полигенностью исследования
признаков. Значительные усилия прилагают к пониманию генетической основы
СПКЯ. По меньшей мере у одной группы пациенток с СПКЯ заболевание имело доми¬
нантный, Х-сцепленный тип наследования. Показан увеличенный в два раза риск
возникновения гирсутизма и олигоменореи при наследовании по мужской линии, но
со значительной вариабельностью фенотипической экспрессии. С другой стороны,
исследования больших семей выявили наследование по аутосомно-доминантному
96
типу с преждевременным облысением у мужчин [277,278]. Кроме того, сильная связь
между гиперинсулинемией и гиперандрогенией позволяет предположить, что сти¬
мулирующий эффект инсулина на продукцию яичниковых андрогенов регулируется
генетической предрасположенностью. Действительно, женщины с гиперандрогенной
ановуляцией и поликистозными яичниками часто имеют родственниц с гиперинсули¬
немией и родственников по мужской линии с облысением [279]. в результате наслед¬
ственная предрасположенность к увеличенному уровню тестостерона при СПКЯ пред¬
полагает избыток уровня андрогенов сам по себе. Исследования генетической связи
находится на пути определения индивидуальных генов, дефекты которых приводят к
СПКЯ [242, 280, 281].
Инсулинорезистентность при синдроме поликистозных яичников
Инсулинорезистентность — главный фактор в патогенезе сахарного диабета
2-го типа (СД-2). Под термином «инсулинорезистентность» понимают нарушение
усвоения глюкозы тканями всего тела. Диагностируют ее с помощью эугликемиче-
ского клэмп-теста; инсулинорезистентность — нарушение утилизации глюкозы [146].
Клинически заболевание определяют как невозможность эндогенного или экзоген¬
ного инсулина увеличивать потребление и утилизацию глюкозы у индивидуума по
сравнению со здоровыми. Инсулинорезистентность наблюдают как у худощавых, так
и у тучных женщин с СПКЯ. Более выраженные степени инсулинорезистентности или
нарушение толерантности к глюкозе чаще встречают среди тучных женщин с СПКЯ
[146].
Взаимосвязь нарушения обмена углеводов с избытком андрогенов впервые была
описана в 1921 г. Archard и Thiers и названа «диабетом бородатых женщин», С этого
времени связь между СПКЯ и инсулинорезистентностью или нарушением толерант¬
ности к глюкозе хорошо определена [146]. Клиническая взаимосвязь инсулинорези¬
стентности и гиперандрогенной ановуляции обычно выявляют по всему миру и среди
различных этнических групп [282]. К тому же избыток андрогенов и инсулинорези¬
стентность нередко сопровождаются акантодермией. Акантодермия представлена
серо-коричневыми бархатистыми участками кожи, чаще всего шеи, паха, подмышек,
а также под молочными железами, что является проявлениями инсулинорезистентно¬
сти (см. рис. 1-30). Гиперкератоз и папилломатоз — гистологические характеристики
акантодермии. Присутствие акантодермии у гиперандрогенных женщин зависит от
выраженности гиперинсулинемии и инсулинорезистентности [283]. Механизм разви¬
тия акантодермии до конца не ясен. Этот аномальный ответ кожи может быть опосре¬
дован рецепторами к различным ростовым факторам, включая таковые для инсулина
и ИФР-1. Акантодермия неспецифична для инсулинорезистентности, поскольку она
может наблюдаться в отсутствие инсулинорезистентности или избытка андрогенов.
Инсулинорезистентность характеризуется слабым ответом глюкозы на опреде¬
ленное количество инсулина. У многих подобных пациентов нормальные уровни
глюкозы поддерживаются под действием повышенной концентрации инсулина для
устранения дефекта, лежащего в основе. Более выраженные формы инсулинорези¬
стентности при СПКЯ варьируют от нарушения толерантности к глюкозе до СД-2,
Нарушение усвоения глюкозы, стимулированной инсулином, является относительно
общим феноменом в популяции, иногда называемым Х-синдромом, или метаболиче¬
ским синдромом, фундаментальная аномалия, приводящая к проявлению симптомов
метаболического синдрома, — нарушение усвоения глюкозы мышцами и усиление
липолиза, приводящие к увеличению концентрации жирных кислот [284], Такие
пациенты имеют также дислипидемию, гипертензию и увеличенный риск развития
сердечно-сосудистых заболеваний. Неудивительно, что дислипидемия и сердечно¬
сосудистые заболевания значительно распространены среди женщин с СПКЯ [285,
286]. Вероятность возникновения гипертензии резко повышена после менопаузы у
97
женщин с СПКЯ в анамнезе [146]. Таким образом, имеется клиническое и патологи¬
ческое «перекрывание» между метаболическим синдромом и СПКЯ [287].
Клиническая картина пациенток с инсулинорезистентностью зависит от способ¬
ности поджелудочной железы компенсировать тканям-мишеням резистентность к
инсулину. На ранних стадиях развития этого состояния компенсация эффективна,
единственное нарушение — гиперинсулинемия. у многих пациенток р-клетки под¬
желудочной железы в конечном счете не удовлетворяют требованиям, снижение
уровня инсулина приводит к недостаточной толерантности к глюкозе и в итоге к СД.
в действительности дисфункция р-клеток показательна у женщин с СПКЯ до начала
нарушения толерантности к глюкозе [288].
Исследования хорошо известных причин гиперинсулинемии и избытка андроге¬
нов отражают различные механизмы инсулинорезистентности. Такие факторы, как
снижение способности связывать инсулин из-за антител к рецепторам инсулина,
пострецепторные дефекты, а также уменьшение количества инсулиновых рецепторов
в тканях-мишенях, участвуют в развитии инсулинорезистентности [289]. Эти редкие
синдромы обычно определяются у небольшой группы женщин с ановуляцией, избыт¬
ком андрогенов и инсулинорезистентностью, оставляя большую часть пациенток с
СПКЯ без очевидных нарушений количества и качества рецепторов и формирования
антител. Природа инсулинорезистентности у большей части женщин с СПКЯ четко не
определена.
Для понимания молекулярных дефектов, лежащих в основе инсулинорезистент¬
ности при СПКЯ, Dunaif и соавт. [146] исследовали различия между фибробластами
кожи женщин с и без СПКЯ, при этом особое внимание уделяли трансдукции сигнала.
Фибробласты женщин с СПКЯ показали отсутствие изменений связывании инсулина
или сродстве рецепторов, у половины этих женщин, однако, наблюдается пострецеп-
торный дефект [146], который характеризуется повышенным фосфорилированием
серина инсулинового рецептора и снижением инсулинзависимого фосфорилирования
тирозина [146]. Аномальное фосфорилирование специфических остатков инсули¬
новых рецепторов может представлять молекулярный механизм, ответственный
за инсулинорезистентность, ановуляцию и избыток андрогенов при СПКЯ [146].
Причины этих аномальных фосфорилирований и последствия для инсулиновых
эффектов — важные направления для будущих исследований.
В рамках объединенной гипотезы инсулинорезистентность представляется крити¬
ческим механизмом, который описывает большинство дефектов, наблюдаемых при
СПКЯ (см. рис, 1-32). Инсулинорезистентность ассоциирована с аномальным отве¬
том яичникового фолликула на ФСГ, который способствет ановуляции и секреции
андрогенов. Это приводит к нециклическому формированию эстрогенов из андроге¬
нов в периферических тканях. Эстрадиол вместе с увеличенным уровнем андрогенов
служит причиной аномальной секреции гонадотропинов. Как итог — ановуляторное
состояние, содействующее постоянному формированию ЛГ, предшественников сте¬
роидов, андрогенов и эстрогенов (см. рис. 1-32).
Роль ожирения в инсулинорезистентности и ановуляции
Увеличенное соотношение окружности талии к окружности бедер называется
андроидным ожирением, поскольку это тип распределения жировой ткани чаще
наблюдают среди мужчин. Женщины с избыточной массой тела с ановуляторным
избытком андрогенов, как правило, имеют подобный тип распределения жировой
ткани [290]. Андроидное ожирение — результат отложения жира в брюшной стенке
и висцеральных медиастернальных локализациях. Этот жир более чувствителен к
катехоламинам, менее чувствителен к инсулину и более метаболичен. Андроидное
ожирение ассоциируется с инсулинорезистентностью, нарушением толерантности к
глюкозе, СД и увеличением продукции андрогенов, проявляющимися в снижении
98
уровня ПСПС и увеличении уровней тестостерона и эстрадиола [290]. Неудивительно,
что андрогенное ожирение ассоциируется с сердечно-сосудистыми факторами риска,
включая гипертензию и дислипидемию. Важно отметить, что андроидное ожирение
некоторым образом коррелирует с заметным риском рака молочной железы с плохим
прогнозом [291, 292]. Однако не выявлено прямой взаимосвязи СПКЯ с риском рака
молочной железы [293].
Несмотря на то что комбинация инсулинорезистентности и избытка андрогенов
часто наблюдается среди тучных женщин, женщины с андрогенным ожирением
имеют значительно более высокий риск инсулинорезистентности и избыток андро¬
генов. Тем не менее инсулинорезистентность и избыток андрогенов определяют не
только у женщин с ановуляцией и ожирением, но также у пациенток без ожирения
с ановуляцией [259]. Несмотря на то что ожирение само по себе вызывает инсули¬
норезистентность, сочетание инсулинорезистентности и избытка андрогенов харак¬
терно для СПКЯ. Неудивительно, что сочетание ожирения и СПКЯ связано с более
выраженными степенями инсулинорезистентности, нежели определяется у женщин
без ожирения с СПКЯ [259, 294]. Андроидный тип ожирения в противоположность
общему ожирению — более специфичный фактор риска СПКЯ.
Диагноз инсулинорезистентности
в повседневной клинической практике критерии для диагноза инсулинорезистент¬
ности не стандартизированы и представляют собой исключительно сложные вопросы.
Во-первых, четверть здоровой популяции имеет натощаковый глюкозозависимый
уровень инсулина, превосходящий таковой у инсулинорезистентных пациенток
[146], что обусловлено значительной вариабельностью чувствительности к инсулину
у здоровых женщин. Во-вторых, клинически доступные методы оцеипки активности
инсулина, такие как голодание и глюкозостимулированный уровень инсулина, плохо
коррелируют с более точными методами оценки чувствительности инсулина в усло¬
виях лаборатории.
С учетом данных ограничений оправданно считать всех женщин с СПКЯ в группе
риска по инсулинорезистентности и с ассоциированными нарушениями — по мета¬
болическому синдрому, дислипидемии, гипертензии, сердечно-сосудистым расстрой¬
ствам [287]. Липидный профиль должен быть получен во всех случаях СПКЯ. У жен¬
щин с СПКЯ и ожирением необходимо исследовать уровень глюкозы натощак и через
2 ч после введения 75 г глюкозы в качестве скрининга нарушений углеводного обмена.
Врач должен убедить пациентку использовать любые методь[ (например, уменьшение
массы тела и физические нагрузки) для снижения инсулинорезистентности.
Использование антидиабетических лекарств для лечения ановуляции и избытка
андрогенов
Логичный подход к ведению СПКЯ — использование лекарств, улучшающих чув¬
ствительность к инсулину в тканях-мишенях, таким образом способствуя уменьше¬
нию секреции инсулина и стабильности толерантности к глюкозе. Антидиабетические
препараты — метформин, тиазолидиндион^’, пиоглитазон и росиглитазон^' — были
использованы для уменьшения инсулинорезистентности. Хотя метформин допол¬
нительно влияет на яичниковый стероидгенез напрямую, этот эффект не является
первично ответственным за истощение продукции андрогенов яичниками у женщин
с СПКЯ. Скорее, метформин ингибирует производство глюкозы печенью, что Требует
меньшей концентрации инсулина и, таким образом, снижает продукцию андрогенов
тека-клетками [295].
Метформин по 500 мг в день снижает гинеринсулинемию, базальный и стимулиро¬
ванный уровни ЛГ, а также свободный уровень тестостерона у женщин с ожирением
и СПКЯ [296, 297]. У значительного числа ановуляторных женщин вновь возникали
овуляция и беременность [298]. Среди опубликованных исследований метформина и
99
СПКЯ исследуемые популяции, материалы, методы и исходы широко различались.
Метаанализ 13 исследований, в ходе которых метформин назначали 543 участникам,
свидетельствует о том, что у женщин, получавших метформин, отношение шансов
(ОК) начала овуляции составляло 3,88 (95% ДИ, 2,25-6,69) в сравнении с плацебо
и 4,41 (95% ДИ, 2,37-8,22) для комбинации метформина и кломифена в фавнении
с монотерапией кломифеном [299]. Метформин улучшает также уровень инсулина
натощак, артериальное давление и уровень ЛПНП, холестерина. Эти эффекты были
определены как независимые от любых изменений массы, которые были связаны с
метформином. Однако мнения расходятся в отношении того, какой из двух положи¬
тельных эффектов метформина полностью независим от снижения массы тела, обыч¬
но наблюдаемого в начале лечения [295,300].
Тиазолидиндионы - фармакологические лиганды для ядерного гамма-рецептора
(РРАКу), активируемого пролифератором пероксисом, которые увеличивают эффек¬
ты инсулина в печени, скелетной мускулатуре и жировой ткани и обладают скромным
эффектом на продукцию глюкозы печенью. Как и метформин, тиазолидиндионы
влияют напрямую на синтез стероидов в яичнике, однако более убедительны данные,
что снижение уровня инсулина ответственно за сниженную концентрацию циркули¬
рующих андрогенов [295].
Женщины с СПКЯ, получившие троглитазон, имели стойкое снижение выра¬
женности иммунорезистентности, гиперандрогении и улучшение толерантности к
глюкозе [301, 302] К тому же лечение троглитазоном было связано с относитель¬
ным улучшением функции клеток поджелудочной железы [301, 302]. Эти открытия
послужили поводом для проведения двойного слепого рандомизированного плацебо-
контролируемого исследования троглитазона при СПКЯ [303]. Овуляция происходила
значительно чаще у женщин, получавших троглитазон, чем у принимавших плацебо;
уровень свободного тестостерона снижался, а уровень ПСПС повышался в дозозави¬
симой форме. Практически все показатели гликемии выявили дозозависимое сниже¬
ние при лечении троглитазоном. Несмотря на то что троглитазон недоступен в настоя¬
щее время в связи с гепатотоксичностью, последующие исследования, в ходе которых
применяли более современные производные росиглитазон^ и пиоглитазон, имели
схожую эффективность [304-306]. Невозможность использования тиазолидиндионов
во время беременности ограничивает их применение для рутинного лечения СПКЯ.
Успех лечения инсулинорезистентности в рамках коррекции нарушений при СПКЯ
подтверждает центральное значение этого дефекта патогенеза заболевания.
Ведение отдаленных осложнений синдрома поликистозных яичников
Отдаленные последствия при СПКЯ представлены нерегулярными маточными кро¬
вотечениями, ановуляторным бесплодием, избытком андрогенов (вирилизация или
гирсутизм), хронически увеличенным уровнем свободного эстрогена, ассоциирован¬
ного с повышенным риском развития рака эндометрия, а также инсулинорезистентно¬
стью, обусловленной большим риском сердечно-сосудистой патологии и СД. Именно
поэтому лечение должно преследовать следующие цели: помощь в организации здо¬
рового образа ЖИ.ЗНИ и достижении нормальной массы тела, защиту эндометрия от
эффектов гиперэстрогении, а также уменьшение уровня тестостерона.
Если пациентка желает забеременеть, ей необходима медикаментозная индукция
овуляции. Беременные с СПКЯ относятся к группе повышенного риска по спонтан¬
ному прерыванию беременности [307]. Этот повышенный риск может быть связан с
повышенным уровнем Л Г, который способствует развитию неблагоприятного окру¬
жения для ооцита и эндометрия. По этой причине до индукции овуляции Л Г пода¬
вляется ОК. Это подавление может быть достигнуто у любой пациентки с СПКЯ при
использовании ОК от 4 до 6 мес до индукции овуляции с кломифена цитратом или
рекомбинантным ФСГ.
100
Если пациентка не желает забеременеть, лечение направлено на прекращение
гиперэстрогении на эндометрий. Постоянный уровень эстрадиола в отсутствие про-
1'естерона вызывает нерегулярные маточные кровотечения, аменорею и бесплодие
и увеличивает риск развития рака эндометрия. Женщины с отсутствием овуляции
вследствие СПКЯ могут заболеть раком эндометрия в возрасте 20 лет [308], По этой
причине женщинам, страдающим СПКЯ и не получающим лечение, независимо от
возраста должна проводиться биопсия эндометрия. Перед каждой биопсией эндоме¬
трия должна быть исключена беременность. Маточное кровотечение не должно стать
препятствием к проведению биопсии. Аменорея не исключает гиперплазию эндоме¬
трия. Критические факторы, определяющие риск опухоли эндометрия. — длительная
ановуляция и воздействие гиперэстрогении. Длительное лечение гестагенами или ОК
значительно снижает риск развития рака эндометрия.
Один из наиболее простых и эффективных способов длительного назначения
гестагенов — использование ОК, которые обладают двумя дополнительными положи¬
тельными эффектами: уменьшением избытка андрогенов и контрацепцией. Снижение
ОК-уровня циркулирующего ЛГ и стимуляция уровня ПСПС приводит к снижению
гирсутизма у пациенток с гиперандрогенией [151].
Возрос интерес к возможному инсулин-десенсибилизирующему эффекту ОК [146].
Старые препараты вызывали увеличение инсулинорезистентности в связи с высо¬
ким уровнем эстрогенного компонента [309]. Тем не менее продолжительные иссле¬
дования не выявили увеличения частоты СД у женщин, применявших высокодо-
зированные ОК в прошлом или употребляющие их в настоящее время [310, 311].
Действительно, многие исследования показывают, что современные ОК либо не
изменяют, либо существенно снижают инсулинорезистентность у женщин с СПКЯ.
ОК, не вызывающие инсулинорезистентность у женщин с СПКЯ, содержат 30 мг или
менее этинил эстрадиола и дезогестрел, норгестимат или гестоден в качестве геста-
генового компонента [312]. К тому же у употреблявших ОК в прошлом отсутствует
повышение риска сердечно-сосудистых заболеваний [313]. Назначение низкодо¬
зированных ОК женщинам с гестационным или инсулинозависимым диабетом не
сопровождалось развитием побочных эффектов [314, 315, 316]. Низкодозированные
ОК не увеличивали риск ретинопатии или нейропатии у пациенток с диабетом и не
ухудшали показатели биохимических маркеров липидов [314, 315, 316]. Необходимо
повторить, что наиболее важный фактор негликемической природы, способствующий
гиперинсулинемии и инсулиновой резистентности при СПКЯ, — ожирение [250,294].
Лечение ановуляции и гиперинсулинемии ОК у женщин с избытком андрогенов не
увеличивает риск развития сердечно-сосудистых заболеваний.
Для пациенток, которые не жалуются на гирсутизм, но у которых отсутствуют
овуляция и регулярные кровотечения, лечение исключительно гестагенами может
представлять альтернативу ОК. Терапия гестагенами направлена на прерывание
хронического воздействия на эндометрий гиперэстрогении. МПА можно назначать с
перерывами (например, 10 мг ежедневно в течение первых 10 дней каждого месяца)
для обеспечения кровотечения, отмены и избегания гиперплазии эндометрия. Это
лечение не снижает уровень избыточных андрогенов и не является контрацепцией.
Поскольку новые ОК (с содержанием этинилэстрадиола 30 мкг или легких гестаге¬
нов) подавляют избыток андрогенов яичникового происхождения, обеспечивают
контрацепцию, защищают эндометрий и не увеличивают инсулинорезистентность,
новые низкодозированные ОК — лечение выбора для некурящих с СПКЯ. ОК вместе
с антиандрогеном спиронолактоном (100 мг в день) рекомендованы как начальное
лечение в отношении женщин с гирсутизмом при СПКЯ. Доза спиронолактона может
быть постепенно увеличена для подавления роста волос, как это было описано ранее.
Лечение ОК (±спиронолактон) может быть неэффективным для подавления андро¬
генов при тяжелых случаях СПКЯ. Таким пациенткам, резистентным к ОК, может
101
потребоваться подавление яичника антагонистом ГнРГ. Поскольку глюкокортикои-
ды усиливают инсулинорезистентность, они должны с осторожностью применяться
у пациенток с гиперинсулинемией. Спиронолактон не влияет на чувствительность
к инсулину у женщин с ановуляцией, может быть использован без последствий для
углеводного или липидного метаболизма [317].
Врачи, консультирующие женщин с СПКЯ, должны предупредить о высоком риске
развития'тахарного диабета. Возраст развития инсулиннезависимого сахарного диа¬
бета значительно ниже у женщин с СПКЯ, чем в популяции [241]. Пациентки с СПКЯ
чаще заболевают гестационным диабетом [318]. Продолжительные исследования
показали высокий риск развития обычного сахарного диабета у женщин с ановуляци¬
ей и СПКЯ [146]. Именно поэтому важно мониторировать переносимость глюкозы с
периодическими уровнями глюкозы натощак и после нагрузки глюкозой 75 г.
Поскольку инсулинорезистентность приводит к аномальному липидному профилю
и увеличению кардиоваскулярного риска у женщин с СПКЯ, снижение массы тела —
одна из главных целей у пациенток с ожирением [319]. И инсулинорезистентность, и
избыток андрогенов при снижении избыточной массы тела могут быть уменьшены по
крайней мере на 5% [320,321]. Значительное снижение массы тела приводит к восста¬
новлению овуляции и возникновению беременности у некоторых пациенток с СПКЯ
[322], Следовательно, консультирование по изменению питания и стиля жизни —
важный компонент при продолжительном лечении СПКЯ.
Место инсулин-сенсибилизаторов, таких как метформин и тиазолидиндионы*“, при
продолжительном лечении СПКЯ требует дальнейшего уточнения с у^1етом данных из
будущих крупных клинических исследований [301, 302, 323]. При продолжительном
наблюдении за пациентками с СПКЯ андрогенное ожирение и гиперинсулинемия
сохраняются в постменопаузе [324]. Таким образом, женщины постменопаузального
возраста, у которых в прошлом отсутствовала овуляция, но присутствовали гипе¬
рандрогения и гиперинсулинемия, имеют значительный риск развития сердечно¬
сосудистых заболеваний и сахарного диабета, В связи с этим необходимо проведение
превентивных мероприятий по снижению риска сердечно-сосудистых заболеваний и
других неблагоприятных последствий СПКЯ [146].
Врач должен предупредить женщину с СПКЯ, что около половины родственниц
первой линии могут иметь СПКЯ или по крайней мере избыток андрогенов и в при¬
сутствии регулярных менструаций [243]. Эти пациентки могут иметь более высокий
риск сердечно-сосудистых заболеваний и получат пользу от превентивных мероприя¬
тий, снижающих его.
Индукция овуляции при синдроме поликистозных яичников
Кломифена цитрат
Для индукции овуляции при СПКЯ повышение уровня ФСГ достигается исполь¬
зованием, например, кломифена цитрата или инъекцией рекомбинантного ФСГ.
Вероятно, фармакологические уровни ФСГ преодолевают яичниковый дефект, ответ¬
ственный за ановуляцию при СПКЯ.
Кломифена цитрат - нестероидный лиганд эстрогеновых рецепторов, индуци¬
рующий овуляцию, со смешанными агонистически-антагонистическими свойствами
[325]. Считают, что кломифена цитрат, действуя как антиэстроген, вытесняет эстро¬
ген из рецепторов, расположенных в гипоталамусе, снимая отрицательную обрат¬
ную связь, осуществляемую эндогенными эстрогенами. Результирующее изменение
пульсирующего выброса ГнРГ нормализует освобождение гипофизарных ФСГ и ЛГ,
последующий выбор фолликулов, достижение доминантности и в конечном счете
овуляции [325].
Пациентки с ановуляцией и аменореей могут начинать лечение кломифеном в
любое время, предварительно убедившись в отсутствии беременности, в качестве
102
альтернативы маточное кровотечение может быть индуцировано приемом ОК в тече¬
ние 21-го дня или назначением МПА на протяжении 10 дней (10 мг в день). Прием
кломифена цитрата в дозе 50 мг в день начинается на 3-й, 4-й или 5-й день от начала
менструации и далее в течение 5 дней. В последние 20 лет использовали практический
подход, называемый «три семерки», для того чтобы мониторировать индикаторы
овуляции после приема кломифена цитрата [325]. Протокол представлен на рис.
1-34. Этот подход используется для того, чтобы определить преовуляторный выброс
эстрадиола в плазме на 7-й день после приема последней дозы кломифена (указано
стрелкой вверх, см. рис. 1-34), а также уровень прогестерона в плазме на 14-й день
после последней дозы кломифена (стрелка вниз, см. рис. 1-34), что необходимо
для документирования овуляции. Повторные визиты планируют на 7-й день после
определения концентрации прогестерона, что соответствует 21-му дню после приема
последней дозы кломифена (см. рис. 1-34) [325]. Пациентка должна быть проинфор¬
мирована о необходимости коитуса каждый день в течение 10-дневного периода, сле¬
дующего за приемом последней дозы кломифена цитрата. Альтернативное изменение
ЛГ в моче для определения пика ЛГ может быть использовано для расчета времени
коитуса. В этом случае пара должна быть инструктирована о необходимости совер¬
шить половой акт в день пика ЛГ.
Если овуляция не наступает после первого курса терапии кломифена цитратом в
дозе 50 мг в день, может быть назначен второй курс — по 100 мг в день на протяжении
5 дней. Отсутствие ответа на 150-200 мг в день — показание для изменения лечения.
Большинство пациенток, готовящихся стать матерями, беременеют на стартовой дозе
(50 мг в день в течение 5 дней). Большинство оплодотворений, стимулированных
кломифеном, обычно происходит в течение первых шести овуляторных циклов [325].
Кломифена цитрат
Менструации
Рис. 1-34. Мониторирование
гормонов при овуляции, стиму¬
лированной кломифена цитра¬
том: использование режима
«три семерки». [Из Adashi E.Y.
Clomiphene citrate-initiated ovula¬
tion: a clinical update // Semin.
Reprod. Endocrinol. — 1986. —
N 4. - P. 255-276,]
День цикла
103
Частота многоплодной беременности при индукции кломифеном составляет 7,9%,
причем 6,9% — двойни [325].
Ингибиторы ароматазы
Ингибиторы ароматазы — летрозол и анастрозол — назначаются как эксперимен¬
тальные лекарства для индукции овуляции [326, 327]. Механизм действия схож с
кломифеном. Назначение летрозола по 2,5 мг в день или анастрозола по 1 мг в день
внутрь на 3-й и 7-й дни после кровотечения эффективно стимулирует овуляцию при
ановуляторном бесплодии [328]. Предполагают, что индукция овуляции ингибитора¬
ми ароматазы происходит за счет дефицита эстрогенов, который возникает на уровне
гипоталамуса [118, 326]. Применение ингибиторов ароматазы может уменьшить
неблагоприятный эффект на эндометрий, часто наблюдаемый при использовании
антиэстрогенов (например, кломифена цитрат) для индукции овуляции. Клинические
данные этого экспериментального лечения очень скудны. Проведено лишь несколько
контролируемых исследований, а оставшаяся часть — пилотные или предваритель¬
ные сравнения [328]. Несмотря на то что мировой опыт использования ингибиторов
ароматазы для индукции овуляции возрастает, рандомизированных контролируемых
исследований, сравнивающих кломифен и ингибиторы ароматазы, недостаточно.
Стандартная терапия гонадотропинами
Женщинам, у которых не наступает овуляция в ответ на кломифена цитрат, под¬
кожно вводят рекомбинантный ФСГ в стартовой дозе 2 ампулы (эквивалентно
150 МЕ ФСГ), начиная с 3-го дня индуцированного гестагенами маточного кровоте¬
чения. Дозу увеличивают на 75 МЕ через интервалы в 3-7 дней до тех пор, пока кон¬
центрация эстрадиола не начнет повышаться. Далее дозу поддерживают до разрыва
фолликула, который индуцируется внутримышечным введением ХГЧ (10 ООО МЕ).
Рост фолликулов наблюдают с помощью трансвагинального УЗИ и анализа концен¬
трации эстрадиола плазмы крови, который служит биохимическим маркером для
гранулезо-клеточной массы в растущем фолликуле [329]. Основыми осложнениями
лечения гонадотропинами при СПКЯ являются значительное учащение многоплод¬
ной беременности, синдром гиперстимуляции яичников тяжелой степени и спонтан¬
ное прерывание беременности [329].
Синдром гиперстимуляции яичников (СГЯ) — осложнение стимуляции яичников.
Легкие формы достаточно часты и характеризуются набором массы тела, дискомфор¬
том в области живота и увеличением яичников. При этой форме необходимы постель¬
ный режим и достаточный прием жидкостей. Тяжелый СГЯ происходит в 0,1-0,2%
случаев стимуляции и сопровождается выраженным асцитом, гидротораксом, дис¬
балансом электролитов и гиповолемией с олигурией. Наиболее тяжелые осложне¬
ния — тромбоз глубоких вен и эмболизация. Этиология до конца не ясна. Большое
количество фолликулов, выброс эстрадиола более 2000 пг/мл, а также беременность
связаны с высокой верояргностью СГЯ. Профилактика заключается в воздержании от
инъекций больших доз ХГЧ и соблюдении диеты. Лечение тяжелого СГЯ заключает¬
ся в госпитализации, поддержании баланса жидкости и электролитов, профилактике
тромбоэмболии гепарином и дренировании тяжелого асцита или гидроторакса. Часто
поддерживающих мероприятий достаточно для ведения этого состояния.
Низкодозная терапия гонадотропинами
Терапия гонадотропинами в обычных дозах вызывает два серьезных осложнения:
» тревожное увеличение количества многоплодной беременности (в пределах от 14
до 50% на лечебный цикл);
» значительное увеличение риска развития тяжелой формы СГЯ (предел от 1,3 до
9,4% на каждый лечебный цикл) [329].
Низкодозные режимы ФСГ для индукции овуляции у женщин с СПКЯ успеш¬
ны в отношении уменьшения уровня многоплодных беременностей до 6% [273].
Низкодозный режим также практически элиминирует осложнения тяжелого СГЯ
104
[273]. Подобные результаты были получены достижением, но не превышением
порогового уровня ФСГ, начиная с дневной дозы 75 МЕ в течение более 10 дней, и
использованием, при необходимости, малого повышения дозы. Этот режим инду¬
цирует развитие единственного фолликула в 70% циклов. Уровень оплодотворения
сравним с терапией обычными дозами. Уровень невынашивания остается несколько
выше, чем при спонтанном оплодотворении (20 к 25%). Время лечения для режима
низких доз значительно больше, чем для терапии гонадотропинами в обычных дозах,
рекомбинантный ФСГ в большей степени, чем гонадотропины мочи, укорачивает
время лечения [330].
ПРЕЖДЕВРЕМЕННОЕ ИСТОЩЕНИЕ ФУНКЦИИ ЯИЧНИКОВ
Преждевременное угасание функции яичников, которое ранее определяли как
истощение яичниковых фолликулов до достижения 40 лет, является формой гиперго-
надотропного гипогонадизма. У этих пациентов диагностируют аменорею или олиго¬
менорею. Они имеют нормальный пубертатный возраст и различный период цикли¬
ческих менструаций с последующей олигоменореей и аменореей. Следовательно,
преждевременное истощение функции яичников всегда должно быть включено
в дифференциальный диагноз хронической ановуляции. Анамнез и объективное
исследование могут выявить нерегулярность менструаций или вторичную аменорею,
сопровождаемую симптомами дефицита эстрогенов, такими как приливы и урогени¬
тальная атрофия. Повышенные уровни ФСГ (более 95% доверительного интервала
пика гонадотропина в середине нормального менструального цикла, например более
40 МЕ/л), выявленные, по крайней мере, дважды, подтверждают диагноз [331].
В среднем менопауза возникает в возрасте 50 лет, при этом 1% женщин продол¬
жают менструировать до 60 лет, еще у 1% женщин менопауза возникает до 40 лет.
Таким образом, определение преждевременной менопаузы или истощения яични¬
ков как отсутствия менструаций у женщин моложе 40 лет достаточно произвольно
[332]. В большинстве случаев этиология преждевременного истощения яичников
неясна. Пациентку можно информировать, что заболевание имеет генетическую
основу, вследствие чего исчезновение фолликулов происходит быстрее, чем в норме.
Специфические аномалии половых хромосом могут быть обнаружены в подгрзшпе
женщин, имеющих преждевременную менопаузу [333]. Среди них наиболее частые
45,Х и 47,ХХ¥, за которыми следует мозаицизм в разных комбинациях [334].
Лежащий в основе дефект яичников может проявляться в различном возрасте в
зависимости от количества фолликулов, оставшихся в яичнике. Симптомы могут рас¬
сматриваться как фазы в процессе менопаузальных изменений независимо от реаль¬
ного возраста пациентки. Если потеря фолликулов происходит быстро до пубертата, то
возникает первичная аменорея и отсутствие вторичных половых признаков. Уровень,
до которого разовьется фенотип взрослого и когда возникнет вторичная аменорея,
зависит от того, когда произойдет потеря фолликулов: во время или после пубертата.
В случае первичной аменореи, связанной с половым инфантилизмом, зачатки яични¬
ков выглядят как полоски; УЗИ обычно не определяет никаких яичников.
Преждевременное истощение яичников может происходить из-за аутоиммунного
процесса, поскольку это состояние часто определяется одновременно с аутоиммун¬
ными полиэндокринными синдромами [335]. Другие причины преждевременной
менопаузы могут быть обусловлены внезапной гибелью фолликулов под воздействи¬
ем таких факторов, как химиотерапия, радиация или инфекция (оофорит, вызванный
свинкой). Эффект радиации зависит от возраста и дозы рентгеновских лучей [336].
После радиационного облучения яичников уровень стероидов начинает падать, а
уровень гонадотропинов растет в пределах 2 нед. Молодые женщины, подвергшие¬
ся радиационному воздействию, в меньшей степени страдают истощением яични¬
ков в связи с большим количеством ооцитов, существующих в молодом возрасте.
105
в ситуациях, когда поля радиации исключают малый таз или яичники выведены за
пределы малого таза с помощью лапароскопической операции до облучения, риск
преждевременного истощения яичников значительно снижается [337]. Большинство
химиотерапевтических лекарств, используемых для лечения опухолей, токсичны по
отношению к яичникам и вызывают их истощение [338]. После завершения химио-
или лучевой терапии описаны восстановление менструаций и беременность [339]. С
другой стороны, преждевременное истощение яичников может происходить через
годы после химио- или радиотерапии [332].
Наконец, такие генетические дефекты, как мутации рецепторов к ФСГ и ЛГ, хруп¬
кая Х-премутация и галактоземия, могут приводить к истощению яичников [340].
Мутация галактозы-1 фосфатуредилтрансферазы, например, может приводить к
истощению яичников, поскольку аккумуляция галактозы-1-фосфат происходит на
токсических уровнях [5].
Диагностика и ведение преждевременного истощения яичников
Преждевременное истощение яичников (ПИЯ) может быть заподозрено у женщин
моложе 40 лет, которые имеют аменорею, олигоменорею или другую форму нарз^ше-
ний менструального цикла. Менопаузальный уровень ФСГ в сыворотке (40 МЕ/мл
и более), полученный по крайне мере два раза, существен для диагноза ПИЯ. У
молодых женщин можно диагностировать ПИЯ и бесплодие, если уровень гонадо¬
тропинов неоднократно зафиксирован как увеличенный. Получено некоторое коли¬
чество данных о беременностях у заболевших женщин, получавших ЗГТ [341, 342].
Рандомизированное исследование роли ЗГТ в такой ситуации показало, что фолли¬
кулогенез происходит часто, однако редко сопровождается овуляцией и еще реже —
беременностью (до 14%); терапия эстрогенами не увеличивает частоту фолликулоге¬
неза, овуляций или беременности [331]. Следовательно, врач должен информировать
пациенток с диагнозом ПИЯ о том, что имеется небольшая, но реальная вероятность
спонтанной беременности в будущем. Женщины, желающие забеременеть, получают
лучшие результаты от вспомогательных репродуктивных технологий, использую¬
щих донорские яйцеклетки, поскольку вероятность спонтанной беременности очень
низка. Пациентке, которая желает самостоятельно выносить ребенка, предлагают
использование донорских ооцитов с последующим оплодотворением спермой пар¬
тнера и переносом эмбриона в полость матки после синхронизации эндометрия
пациентки-рецепиента с донорским циклом с использованием экзогенных эстрогена
и прогестерона (см. рис. 1-25). Этот подход дает прекрасные шансы на беременность
(более 50% на донорский ооцит-цикл искусственного оплодотворения).
Пациентки с П11Я находятся в группе повышенного риска аномального карио-
типа [333]. Риск увеличивается по мере уменьшения возраста возникновения ПИЯ.
Некоторым из таких пациенток рекомендуют хромосомный анализ, поскольку
1[рисутствие У-хромосомы связано с увеличенным риском опухолей гонад [343].
Возрастная группа для хромосомного анализа включает женщин 30 лет и менее,
поскольку чрезвычайно редко можно встретить опухоль гонад у пациенток с ПИЯ
старше 30 лет [344].
Мозаицизм с включением У-хромосомы ассоциируется с высокой частотой опу¬
холей гонад [343]. Эти злокачественные опухоли возникают из зародышевых клеток
и включают гонадобластому, дисгерминому, опухоль желточного мешка и хори-
карциному. В действительности присутствие вторичной вирилизации у пациенток с
аномальным кариотипом и ПИЯ значительно увеличивает риск дисонтогенетических
опухолей гонад. Точные цифры риска опухоли в различных подгруппах этих пациен¬
ток неизвестны, поскольку значительное количество женщин носит У-хромосому и
не имеет симптомов вирилизации. Частота У-хромосомы, определяемая с помощью
полимеразной цепной реакции (ПЦР), высока при синдроме Тернера (12,2%), но
106
возникновение опухоли гонад среди У-позитивных пациенток встречается в 7-10%
случаев [345].
ПИЯ может также встречаться и как изолированное аутоиммунное заболевание, и
совместно с гипотиреозом, диабетом, надпочечниковой недостаточностью и гипопа-
ратиреозом или системной красной волчанкой [346]. Исследования, перечисленные
в табл. 1-7, должны проводиться каждые несколько лет, поскольку ПИЯ может быть
частью аутоиммунного полиэндокринного синдрома [335]. Тиреоидная или адрена¬
ловая (надпочечниковая) недостаточность и диабет — наиболее частые эндокринные
заболевания, которые связаны с ПИЯ. Необходимо отметить, что, как правило, соче¬
тание эндокринного заболевания с ПИЯ бывает чрезвычайно редко [347].
Таблица 1-7, Лабораторная оценка преждевременного истощения функции яичников
ФСГ (для подтверждения диагноза преждевременного истощения яичников).
Кариотип (возраст моложе 30 лет или сексуальный инфантилизм),
Уровень кортизола после стимуляции АКТГ, антиадреналовые антитела (надпочечниковая недостаточность).
ТТГ, антитиреоидные антитела (гипотиреоз).
Глюкоза (натощак и через 2 ч после введения 75 г глюкозы, сахарный диабет).
Кальций и фосфор (гипопаратиреоз).
' СОЭ, общий анализ крови, антиядерные антитела, ревматоидный фактор (аутоиммунные антитела).
' Прегненолон (для оценки дефицита 17а'Гвдроксилазы у сексуально-инфантильных женщин)
Лечение ПИЯ по возможности должно быть направлено на специфическую причи¬
ну. В большинстве случаев невозможно определить специфическую этиологию, если
отсутствуют аномалии кариотипа. Если пациентка хочет забеременеть, ей должна
быть предложена донация яйцеклетки и искусственное оплодотворение спермой ее
партнера (см, рис. 1-25). Если беременность нежелательна, пациентка должна быть
пролечена ОК- или ЗГТ-эстрогенами и гестагенами. Терапия эстрогенами поддержи¬
вает вторичные половые характеристики и препятствует раннему остеопорозу.
Ациклическая продукция эстрогенов во время ановуляторных циклов приводит
к нерегулярному отслоению эндометрия. Эти кровотечения — проявления анову¬
ляторных циклов при отсутствии внуриматочной патологии или системных забо¬
леваний, в целом называемых дисфункциональными маточными кровотечениями.
Ановуляторное маточное кровотечение — наиболее частая причина хронических
нерегулярных менструаций и служит диагнозом исключения. Беременность, миома
матки, эндомеп'риальный полип и аденомиоз должны быть исключены, как и анато¬
мические причины нерегулярных менструаций. Злокачественная опухоль влагалища,
шейки, эндометрия, миометрия, фаллопиевых труб и яичников должны быть исклю¬
чены до установления диагноза «ановуляторное маточное кровотечение». Кроме того,
должны быть исключены аномалии коагуляции.
Ановуляторные маточные кровотечения могут быть вылечены без хирургического
вмешательства либо через восстановление овуляции, либо путем искусственного соз¬
дания овуляторного профиля назначением экзогенных стероидов. Логичным обосно¬
ванием использования экзогенных стероидов является информация о предсказуемом
ответе эндометрия на эстрогены и прогестерон. Физиологический ответ эндометрия
на естественные стероиды яичника был обнаружен по макро- и микроскопическим
изменениям эндометрия во время тысяч нормальных овуляторных циклов у людей и
приматов [154, 156, 348]. Фармакологическое использование экзогенных эстрогенов
и гестагенов у женщин с ановуляторны ми маточными кровотечениями направлено на
107
коррекцию продукции локальных тканевых факторов, которые опосредуют физио¬
логическое действие стероидов и таким образом прекращают избыточное и пролон¬
гированное кровотечение, типичное для ановуляторных циклов.
Лечение нерегулярных маточных кровотечений с помощью экзогенных гормо¬
нов — проверенный метод, обладающий диагностической ценностью. Невозможность
контролировать кровотечение из влагалища с помощью гормональной терапии,
несмотря «а соответствующее применение и использование, делает диагноз аномаль¬
ного маточного кровотечения менее вероятным. В этом случае внимание направлено
на обнаружение патологического объекта внутри репродуктивной оси как на причину
аномального кровотечения.
Обильные, но регулярные менструации (гиперменорея) могут происходить у жен¬
щин с овуляцией. Это может быть вызвано такими анатомическими причинами, как
миома тела матки, деформирующая полость матки, или диффузное и патологическое
присутствие доброкачественных эндометриальных желез в миометрии (аденомиоз).
В отсутствие специфической патологической причины считается, что гиперменорея
отражает легкие нарушения в механизмах эндометриальной ткани. Во всех случаях
оценка и лечение — идентичные подходы, описанные в этом разделе.
Нормальные менструации происходят через 14 дней после каждого эпизода овуля¬
ции как следствие постовуляторного падения эстрогенов и прогестерона. Такая пред¬
сказуемость приводит к тому, что многие женщины ожидают схожие характеристики
менструального кровотечения. Каждое незначительное отклонение (например, плюс
или минус один день длительности или незначительное отклонение от ожидаемого
количества используемых тампонов) является причиной беспокойства у пациенток.
Большинство женщин репродуктивного возраста могут предсказать время своих мен-
С1 руаций так точно, что даже небольшие изменения могут потребовать оценки врача.
Несмотря на то что вариабельность менструального цикла — обычное свойство в
подростковые годы и перименопаузальном периоде, характеристики менструального
кровотечения не подвергаются ощутимым изменениям у женщин 20-40 лет [349].
Для женщин с овуляторным циклом изменения в длительности менструальных
циклов в течение репродуктивного периода предсказуемы. Между менархе и 20 года¬
ми продолжительность цикла для большинства женщин с овуляцией относительно
увеличена. С 20 до 40 лет происходит увеличение регулярности, но цикл укорачивает¬
ся. После 40 лет цикл снова удлиняется. Наибольшая частота ановуляторных циклов
встречается до 20 и после 40 лет [350]. В этой возрастной группе средняя продол¬
жительность цикла 25-28 дней. У женщин с овуляцией длительность цикла меньше
21 дня и более 35 дней встречается очень редко (менее 2% случаев) [351]. Вообще
большинство женщин имеют циклы от 24 до 35 дней (рис. 1-35) [349]. Между 40 и 50
годами менструальный цикл удлиняется и начинает преобладать ановуляция [352].
Среднее постовуляторное кровотечение длится от 4 до 6 дней. Нормальный объем
кровопотери составляет 30 мл. Кровопотеря более 80 мл считается аномальной.
Большая часть кровопотери происходит в первые 3 дня, так что обильные выделения
могут происходить без пролонгирования менструации.
При овуляторном цикле время от овуляции до менструации относительно посто¬
янно и составляет 14 дней (см. рис. 1-1, А). Большая вариабельность в длительности
пролиферативной фазы создает распределение в длительности менструального цикла.
Менструальное кровотечение, возникающее чаще, чем раз в 24 дня или реже раза в 35
дней, требует оценки [349, 352]. Выделения, длящиеся более 7 дней, также требуют
оценки. Выделения, составляющие более 80 мл в месяц, обычно приводят к анемии
108
Продолжительность менструального цикла в днях
Рис. 1-35. Изменение длительности менструального цикла у женщин с регулярными циклами. [Из
Cunningham F.G., MacDonald Р.С., Gant N.F, et al. The endometrium and deciduas; menstruation and preg¬
nancy II Williams Obstetrics. — 19th ed. — Stamford, CT; Appleton & Lange. — 1993. — P. 81-109.]
и должны быть медикаментозно корректированы [355]. В клинической практике, как
правило, трудно измерить менструальные кровопотери, поскольку оценка и лечение
базируются исключительно на ощущениях пациентки касательно длительности, коли¬
чества и времени менструального кровотечения. Несмотря на сложности в подсчете
менструальной кровопотери, врач должен оценить причину избыточного маточного
кровотечения. Необходимо исключить анемию с помощью общего анализа крови
[356]. Низкое значение гемоглобина, сопровождающееся микроцитозом, гипохро-
мией эритроцитов, предполагает значительное кровотечение во время менструаций.
Эти пациентки должны дополнительно получать препараты железа. Вероятность
присутствия дефектов коагуляции, миомы матки или аденомиоза, лежащих в осно¬
ве продолжительных менструаций, должна быть изучена у пациенток с анемией с
помощью тщательного сбора анамнеза, объективного исследования с последующей
лабораторной оценкой.
Олигоменорея — увеличение интервалов между маточными кровотечениями до 35
дней и более, а термин «полименорея» используют для описания интервалов менее
24 дней. Гиперменорея — регулярные (через 24-35 сут) маточные кровотечения с
увеличением либо длительности, либо объема теряемой крови, либо и того, и другого.
Гкпоменорея — снижение показателей либо длительности, либо объема регулярных
маточных кровотечений, либо обоих показателей.
109
КРОВОТЕЧЕНИЕ ПОСЛЕ ОТМЕНЫ ЭСТРОГЕНОВ
Маточное кровотечение следует за резким прекращением эстрогеновой поддержки
эндометрия. Таким образом, этот тип кровотечения может возникать после двусто¬
роннего удаления яичников, облучения зрелых фолликулов или назначения, а затем
отмены эстрогенов женщинам, у которых отсутствуют яичники, кровотечение, кото¬
рое возникает после стерилизации, может быть задержано назначением эстрогенов.
Кровотечение возникает на отмену экзогенных эстрогенов. Таким образом, отмена
эстрогенов сама по себе (в отсутствие прогестерона) практически всегда вызывает
маточное кровотечение.
ЭСТРОГЕННЫЕ СПОНТАННЫЕ КРОВОТЕЧЕНИЯ
Продолжительное воздействие различных количеств эстрогенов постоянно стиму¬
лирует рост эндометрия в отсутствие прогестерона, например, как при избыточном
экстра гонадном синтезе эстрогенов у лиц с СПКЯ. После определенного времени
количество эстрогенов, продуцируемых в экстраовариальных тканях, становится
недостаточным для оказания поддержки эндометрия. Это приводит к непредска¬
зуемым эпизодам поверхностного отслоения эндометрия. Относительно низкие дозы
эстрогенов способствуют перемежающимся маточным выделениям, которые могут
быть продолжительными, но обычно незначительны по количеству кровопотери. с
другой стороны, высокий уровень эстрогенов и продолжительное воздействие приво¬
дят к продолжительным периодам аменореи, сменяющимся острыми, часто профуз-
ными эпизодами кровотечения с избыточной потерей крови.
КРОВОТЕЧЕНИЯ ПРИ ОТМЕНЕ ПРОГЕСТЕРОНА
Типичное кровотечение при отмене прогестерона происходит после овуляции при
отсутствии беременности. Удаление желтого тела — другая причина, вызывающая
десквамацию эндометрия. Фармакологически подобное событие может быть полу¬
чено назначением и последующей отменой прогестерона или синтетического геста¬
гена. Кровотечение после отмены прогестерона возникает, только если эндометрий
первично подготовлен эндогенными или экзогенными эстрогенами. Если терапия
эстрогенами продолжается, а прогестерон отменен, вскоре наступает кровотечение
вследствие отмены прогестерона. Только в случае высокого уровня эстрогенов кро¬
вотечение, возникающее в ответ на отмену прогестерона, задерживается [357]. Таким
образом, кровотечение после отмены прогестерона хорошо предсказуемо при пред¬
шествующем или одновременном воздействии эстрогенов.
ГЕСТАГЕНОВЫЕ СПОНТАННЫЕ КРОВОТЕЧЕНИЯ
Этот фармакологический феномен происходит в присутствии неблагоприятного
высокого отношения гестагенов к эстрогенам. В отсутствие достаточного количе¬
ства эстрогенов продолжающаяся терапия гестагенами приводит к эпизодаческим
кровотечениям различной длительности, похожим на прорывные кровотечения
при стимуляции низкими дозами эстрогенов, как это было описано ранее. Этот тип
кровотечения ассоциирован с приемом комбинированных ОК, которые содержат
низкодозированные эстрогены или продолжительно действующий гестаген, напри¬
мер норплант®, депо-провера* [358]. Гестагеновые спонтанные кровотечения непред¬
сказуемы и характеризуются значительной вариабельностью.
110
п
ричины нерегулярных маточных кровотечений
Беременность и ее осложнения представляют одну из наиболее частых причин
нерегулярных маточных кровотечений (табл. 1-8). Необходимо исключить беремен¬
ность по анализу мочи у любой женщины репродуктивного возраста, имеющей нере¬
гулярные кровотечения (табл. 1-9).
Таблица 1-8. Причины нерегулярных маточных кровотечений
Осложнения беременности
• Угрожающий аборт.
• Неполный аборт.
• Эктопическая беременность
Ановуляция
• Физиологическая:
- неосложненная беременность (аменорея);
- пубертатная (постменархе) ановуляция;
- предменопаузальная ановуляция.
• Лекарственные средства (оральные контрацептивы, агонисты ГнРГ, даназол).
• Гипоталамические (часто представлены как аменорея):
- функциональные (диета, физические нагрузки, стресс).
• Анатомические (опухоль, гранулематозное заболевание, инфекция).
• Лекарственные препараты.
• Другие.
• Гиперпролактинемия, другие расстройства гипофиза:
- пролактинома;
- другие опухоли гипофиза, гранулематозные заболевания:
- гипотиреоз;
- медикаменты;
- Другое.
• Избыток андрогенов:
- СПКЯ, гипертекоз;
- опухоли яичников (опухоль Сертоли-Лейдига);
- неклассическая дисфункция коры надпочечников;
- синдром Кушинга;
- резистентность к глюкокортикоидам;
- опухоли надпочечников (аденома, карцинома);
- медикаменты (тестостерон, даназол);
- другие.
• Преждевременное прекращение функции яичников (часто представлено аменореей).
• Хронические заболевания:
- печеночная недостаточность;
- почечная недостаточность:
- СПИД.
• Другие
Анатомические дефекты, затрагивающие матку
• Лейомиома матки.
• Полипы эндометрия.
• Аденомиоз (часто представлен гиперменореей).
• Внутриматочные сращения (часто представлены как спайки).
• Эндометриты.
• Гиперплазия эндометрия, рак эндометрия.
• Хроническое воздействие эстрогенов (СПКЯ, лекарственные препараты, печеночная недостаточность).
• Эстроген-секретирующая опухоль яичника (гранулезоклеточная опухоль).
• Распространенный рак шейки матки.
• Другое
111
Окончание табл. 1-8
Дефекты коагуляции (обычно представлены как гиперменорея)
’ Болезнь фон Виллебранда.
' Дефицит XI фактора.
’ Другие
Экстрагенитальные кровотечения (могут напоминать маточные кровотечения)
• Вагиниты.
• Генитальная травма.
• Инородные тела.
• Влагалищные неоплазии.
• Вульварная неоплазия.
• Другое
Таблица 1-9. Диагностические тесты для оценки нерегулярных маточных кровотечений
Часто используемые тесты
• Определение уровня ХГЧ в моче.
• Уровень ХГЧ в плазме крови (неполный аборт, внематочная беременность).
• Трансвагинальное УЗИ (маточная и внематочная беременность, лейомиома матки, полип или неоплазия эндоме¬
трия, опухоль яичника).
• Уровень ФСГ, ЛГ в плазме крови (ановуляция, прекращение функции яичников).
• Уровень пролактина, ТТГ в сыворотке крови (ановуляция, гиперпролактинемия).
• Развернутый общий анализ крови, протромбиновые тесты (дефект коагуляции).
• Почечная и печеночная функции, ВИЧ [почечные и печеночные пробы (ановуляция, хронические заболевания)].
• Биопсия эндометрия (заболевания эндометрия; полип, неоплазия, эндометрит)
Реже используемые тесты
• Оценка СПКЯ, яичниковой или надпочечниковой опухоли, неклассической ВДКН, синдрома Кушинга и резистент¬
ности к глюкокортикоидам (избыток андрогенов).
• КТ головы или МРТ (гипоталамическая ановуляция, гиперпролактинемия).
• МРТ таза (аденомиоз, лейомиома матки).
• Гистеросонография вместе с инстилляцией физиологического раствора (полип эндометрия, лейомиома матки).
• Гистероскопия (полип эндометрия, лейомиома матки).
• Дилатация и кюретаж (заболевание эндометрия, недиагностированное УЗИ и биопсией)
Как было отмечено ранее, ановуляторные маточные кровотечения, возникающие
как ответ эндометрия на неадекватную продукцию яичниковых стероидов, также
называются дисфункциональными маточными кровотечениями, поскольку лечение,
восстанавливающее овуляторную функцию, потенциально прекращает нерегулярные
маточные кровотечения. Распространенные примеры ановуляторных кровотечений
включают ановуляцию, связанную с физическими нагрузками, гиперпролактинемию,
гипотиреоз или СПКЯ [359]. В этих случаях восстановление овуляции путем коррек¬
ции лежащего в основе расстройства либо использование экзогенных гормонов могут
привести к предсказуемым маточным кровотечениям. С другой стороны, разные
патологические состояния гениталий (яичников, матки, влагалища или вульвы) или
дефект коагуляции могут также вызывать отклонения от характеристик нормальной
менструации (см. табл. 1-8).
Ановуляторное маточное кровотечение — диагноз исключения по следующим при¬
чинам. Опухоли вульвы, влагалища и матки вызывают нерегулярные кровотечения.
Более того, эстроген- или андроген-секретирующие опухоли яичников также могут
способствовать появлению аномальных маточных кровотечений (см. табл. 1-8).
Беременность или проблемы, с ней связанные (внематочная беременность или спон¬
танное прерывание беременности), — чрезвычайно распространенные причины ано¬
мального маточного кровотечения. Действительно, наиболее частая причина нару-
112
шения нормальной менструации — беременность или ее осложнения. Другая частая
причина нерегулярных кровотечений у некоторых женщин — прием оральных кон¬
трацептивов. Гестагеновые спонтанные кровотечения распространены при проведе¬
нии ЗГТ больным в постменопаузе (см. ниже). Пациентки могут использовать другие
гормональные препараты, не осознавая влияния на эндометрий. Так, использование
корня женьшеня связано с эстрогенной активностью и аномальным кровотечением
[360]. Поскольку маточное кровотечение — частый побочный эффект продолжи¬
тельных гормональных схем лечения, врач должен быть убежден в отсутствии иной
патологии. Органическая патология, приводящая к патологической менструальной
кровопотере, включает гиперплазию эндометрия, рак, полипы эндометрия, миому
матки, аденомиоз и эндометрит. Нерегулярные тяжелые кровотечения могут быть
связаны с хроническими заболеваниями, такими как печеночная и почечная недоста¬
точность, а также СПИД. Необходимо провести тщательное изучение для исключения
повреждения гениталий или присутствия чужеродного объекта (см. табл. 1-8).
В пубертате наиболее частая причина нерегулярных маточных кровотечений —
ановуляция. Около 20% таких подростков с массивным нерегулярным кровотечением
имеют дефект свертывания крови [361, 362]. Среди всех женщин репродуктивного
возраста с гиперменореей распространенность дефекта коагуляции составляет 17%.
Самый частый дефект — болезнь Виллебранда, а дефицит XI фактора — вторая рас¬
пространенная причина. Кровотечения, которые возникают вторично вследствие
коагуляционного дефекта, представлены регулярными обильными кровотечениями
(гиперменореей), причем подобная картина может также наблюдаться у пациенток,
получавших антикоагулянты [363]. Кровотечения обычно ассоциируются с гиперме¬
нореей с возраста менархе, а также с кровотечениями после травм или хирургического ‘
вмешательства. Гиперменорея может быть и единственным признаком наследствен¬
ного заболевания, проявляющегося кровотечением [364].
При жалобах на нерегулярные кровотечения в репродуктивном периоде в первую
очередь должны быть исключены беременность раннего срока и ее осложнения с
помощью измерения ХГЧ в моче (см. табл. 1-9). Угрожающий или неполный аборт и
внематочная беременность — причины нерегулярных маточных кровотечений. При
первичном клиническом исследовании при необходимости могут быть назначены
другие тесты. Эти исследования предназначены для оценки расстройств различной
этиологии, проявляющихся ановуляцией (см. табл. 1-9). У пациенток с продолжи¬
тельными обильными менструациями (гиперменорея) пубертатного возраста в анам¬
незе необходимо провести исследования состояния коагуляции (например, протром-
биновое время, частичное тромбопластиновое время, время кровотечения), а также
полный анализ крови.
Ультразвуковое исследование влагалищным датчиком исключительно полезно
для оценки нормальной или аномальной беременности, лейомиомы матки, неопла¬
зии эндометрия, а также опухоли яичников (см. табл. 1-9). Другие исследования
могут быть использованы для исключения патологий гипоталамуса, гипофиза или
надпочечников (см. выше). Известна польза МРТ таза для исключения аденомио¬
за, заболеваний матки, характеризующихся аномальным присутствием диффузной
эндометриальной ткани в миометрии (см. табл. 1-9). Распространенный аденомиоз
ассоциируется с диффузным увеличением матки, гиперменореей и анемией.
В отношении пациенток с риском развития гиперплазии или рака эндометрия
(например, СПКЯ, печеночная недостаточность, диабет, ЗГТ) необходимо провести
биопсию эндометрия с последующим гистологическим исследованием биоптата.
Доброкачественный полип эндометрия или миома матки, располагающиеся в поло¬
сти, могут быть диагностированы гистеросонографией с помощью внутриматочного
введения физраствора или гистероскопией. Гистеросонография или гистероскопия
не являются достаточными исследованиями для оценки гиперплазии или рака эндо¬
113
метрия, поскольку эти процедуры могут вызывать диссеминацию злокачественных
клеток. Подозрение на опухоль должно быть исключено амбулаторной биопсией
эндометрия (см. табл. 1-9). Изредка амбулаторная биопсия не может быть проведена
или не является информативной. В этих редких случаях для получения достаточного
количества тканей необходимо проведение кюретажа эндометрия под анестезией.
Вновь тщательно собранный анамнез и объективное исследование уменьшают
необходимость в большинстве этих диагностических исследований. До проведения
диагностической процедуры врачу полезно спросить самого себя, изменит ли резуль¬
тат этого теста план клинического ведения.
Ведение ановуляторного маточного кровотечения
Термины «дисфункциональное маточное кровотечение» и «ановуляторное крово¬
течение» взаимозаменяемы и описывают неадекватную стимуляцию эндометрия во
время дисфункциональных состояний репродуктивной системы. Если овуляторная
функция может быть восстановлена, ановуляторные кровотечения сменяются цикли¬
чески предсказуемыми. Поскольку восстановление овуляторной функции может быть
невозможным у большого количества таких женщин, в различных целях назначают
экзогенные эстрогены и гестагены. Показания для гормонального лечения маточных
кровотечений — необходимость остановки маточного кровотечения, поддержка регу¬
лярности кровотечений или предупреждение гиперплазии эндометрия. Для остановки
ановуляторного маточного кровотечения или индуцирования регулярных эпизодов
кровотечения используют гормональные схемы. Необходимо повторить, что анову¬
ляторные маточные кровотечения — диагноз исключения. До назначения лечебных
схем и режимов следует исключить различные анатомические аномалии генитального
тракта, представленные в табл. 1-8.
ОРАЛЬНЫЕ КОНТРАЦЕПТИВЫ
.Использование ОК в кратковременном и длительном режимах — наиболее часто
применяемое лечение нерегулярных маточных кровотечений. Эстрогеновый компо¬
нент комбинированного препарата стабилизирует эндометриальную ткань, снижает
секрецию яичниками половых стероидов через подавление гонадотропинов в течение
нескольких дней. Гестагеновый компонент препарата напрямую воздействует на эндо¬
метрий, отдаляя отслоение на несколько дней, и поддерживает супрессию яичников
эстрогенами. Гестагены (в присутствии эстрогенов) индуцируют дифференцировку
эндометрия в стабильную псевдодецидуальную форму. Обычно предпочитают моно-
фазный ОК, содержащий 30 или 35 мкг этинилэстрадиола. Трехфазные ОК или иные
с менее чем 30 мкг эстрадиола неприемлемы для лечения выраженного ановуля¬
торного кровотечения. Комбинированные ОК в высоких дозах (2 или 3 таблетки в
день) могут быть использованы для кратковременных интервалов (недели) лечения
эпизодов выраженного маточного кровотечения. Обычная доза (1 таблетка в день)
может назначаться годами для ведения хронических ановуляторных кровотечений,
ассоциированных с СПКЯ или гиперпролактинемией.
Оральные контрацептивы и острые массивные маточные кровотечения,
ассоциированные с анемией
Воздействие гиперэстрогении на женщин с ановуляторными маточными кровоте¬
чениями часто ассоциируется с хронической гиперплазией эндометрия и эпизодами
профузных кровотечений. Назначаемая терапия — 1 таблетка дважды в день в тече¬
ние одной недели. У женщин с ожирением ОК могут назначаться 3 раза в день. Эта
терапия продолжается, несмотря на задержку прекращения выделений в течение 2
дней. Если кровотечение не уменьшается, целесообразно исключить другие причины
114
(полипы, неполные аборты, неоплазию). В случае ановуляторного кровотечения оно
уменьшается в течение 2 дней от начала высокодозной терапии (1 таблетка 2 или 3
раза в день). В течение последующих нескольких дней изучают возможные причины
ановуляции и исключают коагулопатию. В это время врач также определяет необ¬
ходимость использования компонентов крови или железосодержащих препаратов в
лечении. Высокодозные эстроген-гестагеновые комбинации вызывают характерные
структурные изменения (в частности, плотности), свойственные компактной псев-
додецидуальной реакции. Таким образом, прекращаются произвольные разрушения
хрупкой эндометриальной ткани и, как следствие, кровотечение останавливается.
Тем не менее значительное количество тканей остается чувствительной на отмену
эстроген-гестагенов. Пациентка должна быть предупреждена о тошноте, вызываемой
высокими дозами ОК, и о возможности обильных выделений в течение нескольких
дней после прекращения такой терапии.
Спустя неделю от начала лечения высокими дозами ОК проводится временная
отмена ОК, вследствие чего в течение нескольких дней начинаются обильные выде¬
ления. На 3-й день этого кровотечения отмены вновь назначают прием комбиниро¬
ванных ОК в стандартной дозе (1 таблетка в день) в течение 3 нед с перерывом на 1
нед. Ожидаемый результат — уменьшение объема выделений с каждым завершенным
циклом. ОК снижает менструальную кровопотерю более чем в 2 раза у большинства
женщин [365].
Раннее назначение комбинации эстрогенов и гестагенов ограничивает рост и
способствует регрессу избыточной толщины эндометрия до нормального уровня.
Поскольку ОК не воздействуют на причины, лежащие в основе ановуляции, но способ¬
ствуют уменьшению проявлений через прямое воздействие на эндометрий, задержка
в приеме ОК приводит к возобновлению патологического маточного кровотечения.
Невзирая на критерии контрацепции, ОК представляют собой лучший выбор для
гормонального лечения ановуляторных кровотечений и должны быть предложены в
качестве одного из вариантов продолжительного лечения.
Оральные контрацептивы и хронические нерегулярные маточные кровотечения
СПКЯ — распространенная форма ановуляции, связанная с хронически устано¬
вившимися уровнями гиперэстрогении, которые могут приводить к гиперплазии и
раку эндометрия (см. выше). Гипоталамическая ановуляция и гиперпролактинемия, с
другой стороны, связаны с низким уровнем эстрогенов, недостаточным для предупре¬
ждения потери костной ткани. Комбинированные ОК подходят для лечения обеих
форм проявлений ановуляции.
ОК представляют наиболее подходящее продолжительное симптоматическое лече¬
ние для любого варианта ановуляторного маточного кровотечения, включая олиго¬
менорею. До назначения ОК необходимо исключить беременность. ОК назначают
по 1 таблетке в день в течение 3-недельного периода с последующим перерывом на 1
нед. Во время недельного перерыва происходит кровотечение отмены. Гестагеновый
компонент способствует предупреждению гиперплазии эндометрия, которая связана
с воздействием гиперэстрогении при СПКЯ (см. выше), в случаях ановуляций, ассо¬
циированных с гипоэстрогенией (например, гипоталамическая ановуляция, гипер¬
пролактинемия), эстрогенный компонент препарата обеспечивает достаточное заме¬
щение для предупреждения потери костной ткани. Риск тромбоэмболии, инсульта
или инфаркта миокарда, связанный с продолжительным назначением, чрезвычайно
низок у некурящих и при отсутствии эпизодов тромбоэмболии, с учетом того, что ОК
эффективно контролируют аномальные маточные кровотечения, женщины с хрони¬
ческой ановуляцией могут продолжать этот режим до менопаузы.
115
СИНТЕТИЧЕСКИЕ ГЕСТАГЕНЫ
Синтетические гестагены способствуют дифференцировке эндометрия и обладают
антагонистическим действием по отношению к пролиферативному эффекту эстроге¬
нов на эндометрий (см. рис. 1-24) [165, 366]. Эффекты натурального прогестерона
или гестагенов включают ограничение индуцированного эстрогенами роста эндоме¬
трия и предупреждение гиперплазии эндометрия. Отсутствие естественно синтезиро¬
ванного прогестерона при ановуляторных состояниях — основание для назначения
гестагенов.
Наиболее частым показанием для длительного циклического назначения геста¬
генов служит предупреждение малигнизации эндометрия у пациенток с СПКЯ и
длительным неоппонированным воздействием эстрогенов на эндометрий. В таких
случаях комбинированные ОК — препараты выбора. Если пациентка не может
использовать ОК по какой-то причине (например, эпизод тромбоэмболии), гестагены
могут быть назначены в циклическом режиме для предупреждения гиперплазии эндо¬
метрия. До назначения гестагенов или ОК необходимо исключить беременность. При
лечении олигоменореи, связанной с СПКЯ, назначение МПА 10 мг в день минимум
10 дней каждые 2 мес приводит к снижению кровотечения отмены. Альтернатива —
норэтиндрон ацетат*“ 5 мг в день или мегэстрол 20 мг в день каждые 2 мес в течение 10
дней. Отсутствие кровотечения отмены требует дальнейшего обследования.
При лечении массивного маточного кровотечения (гиперменореи или полимено¬
реи) эти гестагены в более высоких ежедневных дозах (МПА 20 мг в день, норэтин¬
дрон ацетат*“ 10 мг в день или мегэстрол 40 мг в день) назначают на 2 нед для индуци¬
рования предецидуальных стромальных изменений в эндометрии. В ответ на отмену
препарата в течение 3 дней после приема последней дозы происходит выраженное
кровотечение. Впоследствии для поддержания терапевтического эффекта повторное
лечение гестагенами (медроксипрогестерон 10 мг в день, норэтиндрон ацетат*“ 5 мг в
день, мегэстрол 20 мг в день) назначают циклически по крайней мере первые 10 дней
каждого следующего месяца. Если лечение гестагенами неэффективно, может потре¬
боваться проведение биопсии эндометрия, с другой стороны, предсказуемое крово¬
течение отмены после каждого цикла назначения гестагенов предполагает отсутствие
опухолей эндометрия.
ВЫСОКИЕ ДОЗЫ ЭСТРОГЕНОВ ПРИ ОСТРОМ МАССИВНОМ МАТОЧНОМ КРОВОТЕЧЕНИИ
Как было описано ранее, ОК, назначаемые 2 или 3 раза в день, — препараты выбо¬
ра для остановки сильных ановуляторных кровотечений. Высокодозный режим ОК
должен быть назначен женщинам с выраженным маточным кровотечением и асим-
птоматической анемией после исключения иной патологии генитального тракта (см.
табл. 1-8). С другой стороны, пациентка с острой и тяжелой формой ановуляторного
кровотечения, с сопутствующей симптоматической анемией требует неотложной
терапии. Такие пациентки должны быть немедленно госпитализированы, и им (по
показаниям) должно быть проведено переливание крови. При исключении патологии
гениталий с помощью анамнеза, объективного исследования, а также УЗИ препарат
выбора для остановки кровотечения, угрожающего жизни, — внутривенное назначе¬
ние высокой дозы эстрогенов. Хорошо зарекомендовал себя режим 25 мг конъюги¬
рованных эстрогенов, назначаемых каждые 4 ч до значительного уменьшения кро¬
вотечения или, по крайней мере, в течение 24 ч [367]. Эстрогены преимущественно
оказывают влияние на капиллярном уровне, где индуцируют сгущение крови [368].
Прием ОК по схеме 3 раза в день начинается до того момента, как будет прекращено
внутривенное введение эстрогенов. Лечение ОК продолжают по описанному выше
режиму.
Поскольку высокие дозы эстрогенов — фактор риска для тромбоэмболии, прием
двух или трех доз ОК в течение недели или высокие дозы конъюгированных эстроге¬
116
нов в течение 24 ч рассматривают как фактор значительного риска. В настоящее время
отсутствуют данные для оценки риска, связанного с использованием гормональной
терапии для подобных коротких интервалов. Врач и пациентка должны принять
решение относительно высокодозной гормональной терапии с учетом известных
рисков и преимуществ. Пациенткам с выраженными факторами риска могут быть
предложены альтернативные методы лечения. У женщин с эпизодами идиопатическо¬
го венозного тромбоза в прошлом назначение высоких доз эстрогенов должно быть
отменено. Высокие дозы гормонального лечения не должны применяться женщина¬
ми с тяжелыми формами заболеваний, такими как недостаточность печени и почек.
Единственным альтернативным методом лечения для этих пациенток служит дилата¬
ция и кюретаж с последующим назначением ОК по 1 таблетке в день до тех пор, пока
маточное кровотечение не будет взято под контроль.
АНАЛОГИ ГОНАДОТРОПИН-РИЛИЗИНГ-ГОРМОНА ДЛЯ ЛЕЧЕНИЯ ВЫРАЖЕННОГО
МАТОЧНОГО КРОВОТЕЧЕНИЯ, ВЫЗВАННОГО АНОВУЛЯЦИЕЙ
Аналог ГнРГ может быть назначен женщинам с выраженным ановуляторным кро¬
вотечением или гиперменореей, вызванной тяжелыми хроническими заболеваниями,
такими как печеночная недостаточность или коагулопатия. Необходимо отметить,
что ежемесячные инъекции депо-агониста ГнРГ неэффективны для лечения выра¬
женного маточного кровотечения, они могут даже увеличить маточное кровотечение
первые 2 нед. Антагонист ГнРГ, с другой стороны, снижает ФСГ и ЛГ без задержки
и быстрее приводит к аменорее. Агонист ГнРГ депо лепролид ацетат^ 3,75 мг может
назначаться внутримышечно ежемесячно в течение 6 мес и более для контроля
маточных кровотечений, связанных с хроническим заболеванием на долговремен¬
ной основе. Антагонисты ГнРГ могут быть использованы для остановки острого или
хронического ановуляторного кровотечения; однако в настоящее время не опубли¬
ковано ни одной значимой работы, определяющей эффективную дозу. Отдаленный
побочный эффект лечения аналогами ГнРГ — остеопороз, что делает нежелательным
выбор этого препарата для продолжительного лечения. При выборе длительного
лечения аналогами ГнРГ необходимо дополнительно назначить норэтиндрон ацетат*“
в дозе 2,5 мг ежедневно. Данного назначения обычно достаточно для предупреждения
остеопороза, при этом, как правило, отсутствует усиление маточного кровотечения.
Г0Р1У10Н03А8ИСИ[\^ЫЕ ДОБРОКАЧЕСТВЕННЫЕ
ГИНЕКОЛОГИЧЕСКИЕ ЗАБОЛЕВАНИЯ
Эндометриоз
Эндометриоз определяют как присутствие ткани, напоминающей эндометрий, за
пределами полости матки, наиболее часто на поверхности брюшины и яичников. Это
наиболее частая причина бесплодия и хронических тазовых болей; поражает 1 из 10
женщин репродуктивного возраста [369]. Распространенность увеличивается до 30%
у пациенток с бесплодием и до 45% у пациенток с хронической тазовой болью [370].
Точный диагноз эндометриоза может быть сделан путем визуализации этих пери¬
тонеальных изменений с помощью лапароскопии или лапаротомии. Эндометриоз
наследуется полигенно, как диабет или астма [369]. Родственники женщин с этим
заболеванием имеют в 7 раз более высокую заболеваемость эндометриозом в срав¬
нении с родственниками группы контроля [369]. По мнению Сампсон, наиболее
вероятный механизм развития эндометриоза на перитонеальных поверхностях —
имплантация эндометриальной ткани на брюшину при ретроградной менструации
117
[369]. Поскольку ретроградная менструация происходит у 90% женщин, эндометриоз
может быть вызван генетическими дефектами, предрасполагающими к выживанию
эндометриальной ткани менструального содержимого на брюшине [369].
Эндометриодная ткань, как и нормальная эндометриальная ткань, отвечает на
эстрогены и прогестерон схожими гистологическими изменениями [269]. Эстрогены
поддерживают рост участков эндометриоза, тогда как прогестерон может уменьшить
митогенный эффект эстрогенов [269]. Некоторые эндометриоидные импланты под¬
вергаются атрофии Б ответ на пролонгированную терапию ОК, так же как это проис¬
ходит у нормального эндометрия. Тем не менее эндометриальная ткань не отвечает
на терапию гестагенами или натуральным прогестероном так же предсказуемо, как
это делает нормальный эндометрий. Эндометриальная ткань при эктопической лока¬
лизации на брюшине или яичнике значительно отличается от эутопического эндоме¬
трия внутри полости матки по следующим характеристикам; продукция цитокинов
и простагландинов, стероидный биосинтез или метаболизм, содержание стероидных
рецепторов и клинический ответ на гестагены [148, 269, 370].
Хотя эффективность современной гормональной терапии бесплодия, связанного с
эндометриозом, не является окончательно доказанной, подобное лечение может быть
полезно при тазовых болях, связанных с эндометриозом [369]. Ремиссия, которая
достигается лекарственным лечением, как правило, относительно кратковременна
[372]. Различные препараты, используемые для лечения, сравнимы по эффективно¬
сти. Большинство современных схем лечения были созданы либо с целью снижения
секреции эстрогенов яичниками (например, агонисты ГнРГ, ОК, даназол или геста¬
гены), либо с целью снижения эффектов эстрогенов на эндометриоидные импланты
(например, ОК, даназол и гестагены). Возможные альтернативные механизмы воз¬
действия с использованием андрогенного стероида даназола или гестагенов имеют
прямое антипролиферативное воздействие на эндометриальную ткань.
Многие пациентки и врачи относятся к даназолу настороженно в связи с его
анаболическим и андрогенным побочным эффектом (набор массы тела, судороги
мышц, редкая необратимая вирилизация, клиторомегалия, изменение голоса) [373].
Действительно, почти половина женщин с эндометриозом не завершают 6-месячный
курс лечения даназолом [374]. Оставшиеся гормональные препараты — ОК, гестагены,
а также агонисты ГнРГ — показывают сопоставимую эффективность контроля болей,
связанных с эндометриозом [375-377]. 6-месячный курс применения одного из этих
препаратов приводит к значительному снижению болей у 50% пациенток и более
[375-377]. Лечение болей назначением ОК или гестагенов в непрерывном режиме
занимает больше времени, чем использование агониста ГнРГ. Уровень персистенции
заболевания остается значительным после завершения лекарственной терапии [378].
Спустя полгода после завершения 6-месячного курса лечения гестагенами, ОК или
агонистами ГнРГ половина первичных респондентов отмечают возобновление боли
от умеренной до тяжелой интенсивности [376]. у остальных боли рецидивируют в
5-20% случаев в течение следуюших 5 лет [378]. 6-месячный курс лечения агони¬
стом ГнРГ — в настоящее время самый популярный режим. Наиболее выраженный
побочный эффект лечения агонистами ГнРГ — потеря костной ткани, которая связана
с недостатком эстрогенов. Для его минимизации назначают эстроген-гестагеновые
препараты или бисфосфонаты [34].
Наука далека от излечения эндометриоза, а современные методы лечения не дают
удовлетворительного контроля боли. Радикальное лечение — удаление обоих яични¬
ков, причем даже этот вариант не считается эффективным в ряде случаев постменопа¬
узального эндометриоза [376]. Необходимы новые решения, которые можно было бы
предложить женщинам с эндометриозом, чтобы дать реальный шанс жить без стра¬
даний от хронических тазовых болей в течение десятилетий. Существуют два важных
предупреждения, которые не адресованы лечению агонистами ГнРГ. Во-первых,
118
большие количества эстрогенов могут продуцироваться локально вн>три эндоме-
триоидных клеток, что представляет интракринный механизм эстрогенной актив¬
ности в противоположность секреции яичниками, которые, согласно представлениям
об эндокринной функции, снабжают этими стероидами ткани мишени (см. рис. 1-21)
[269, 376]. Во-вторых, эстрадиол, продуцируемый в периферических тканях, напри¬
мер в жировой ткани или в фибробластах кожи, может приводить к патологически
значимому увеличению концентрации эстрадиола у части женщин. Агонисты ГйРГ не
ингибируют периферическое формирование эстрогенов или локальную продукцию
эстрогенов внутри эстрогенчувствительного образования. Как было отмечено выше,
эндометриоз устойчив к определенным эффектам прогестерона и используемым
сегодня гестагенам [269]. Следовательно, ингибиторы ароматазы и селективные про-
гестероновые модуляторы ответа — кандидаты в терапевтические агенты для лечения
эндометриоза.
Экспрессия ароматазы и локальный эстрогеновый биосинтез в эндометриоид-
ных имплантах подсказывает пилотные исследования для определения ароматаз в
эндометриоидных гетеротопиях с использованием третьего поколения ингибиторов.
Назначение ингибиторов третьего поколения — анастрозола и летрозола — было
успешным при лечении эндометриоза у женщин в пре- и постменопаузальном возрас¬
те [379-383]. Эти исследования позволяют предположить следующее;
» ингибиторы ароматазы эффективно лечат тазовые боли, связанные с эндометри¬
озом, даже такие, которые резистентны к ранее разработанным терапевтическим
способам воздействия;
« ингибиторы ароматазы — лекарства выбора для лечения персистирующего эндо¬
метриоза в постменопаузе;
* использование ингибиторов ароматазы у женщин в пременопаузе с эндометрио¬
зом требует супрессии яичников через добавление аналогов ГйРГ, гестагенов или
комбинированных ОК.
Пилотные исследования подтвердили, что профили побочных эффектов были
крайне благоприятны и не включали потерю костной ткани в режимах, включавших
комбинацию ингибитора ароматазы и ОК или гестаген [381, 383].
Таким образом, ингибиторы ароматазы представляют один из наиболее обещаю¬
щих методов лечения боли, связанной с эндометриозом.
ЛЕЙОМИОМА МАТКИ
Лейомиома матки происходит из миометрия и является наиболее частой солид¬
ной опухолью таза [384]. В связи с лейомиомой ежегодно в США проводится более
200 тыс. гистероэктомий. Подавляющее большинство этих опухолей доброкаче¬
ственные и представляют собой экспансию клона одной клетки. Лейомиомы могут
вызывать различные симптомы, включая нерегулярные профузные маточные кро¬
вотечения, ощущение давления внизу живота, боль во время полового акта, тазо¬
вые боли, невынашивание беременности, бесплодие, а также компрессию близко
расположенных органов таза или могут быть совершенно асимптоматичными.
Распространенность лейомиомы матки составляет от 25 до 30% [384]. Лейомиомы
наиболее часты у афроамериканских женщин, а также имеют полигенную природу
наследований. Диагноз может быть подтвержден с помощью трансвагинального или
абдоминального УЗИ. Трансвагинальное УЗИ — чувствительный метод для определе¬
ния размера, локализации и количества леномиом матки.
Миома матки возникает в репродуктивном возрасте и регрессирует в постмено¬
паузе, что указывает на зависимость от стимуляции яичниковыми стероидами. Роль
стероидов или других факторов роста в инициации и росте этих опухолей четко не
определена. Неопластическая трансформация миометрия в лейомиому, вероятно,
вовлекает соматические мутации нормального миометрия, а также комплексное воз¬
119
действие половых стероидов и ростовых факторов [385]. Традиционно эстрогены
считаются основными промоутерами роста миомы. Современные биохимические,
гистологические, а также клинические данные подтверждают важную роль прогесте¬
рона в росте лейомиомы матки. Биохимические и клинические исследования пред¬
полагают, что прогестерон и гестагены, действуя через прогестероновые рецепторы,
могут стимулировать пролиферативную активность в лейомиомах [385].
Терапевтическая тактика зависит от цели терапии, причем гистерэктомия чаще
всего используется для окончательного лечения, а миомэктомия — при необходимо¬
сти сохранения способности к вынашиванию беременности. Лапароскопическая мио¬
мэктомия в настоящее время технически доступна, но имеется значительный риск раз¬
рыва матки во время беременности. Частота рецидивов после миомэктомии различен
(10-50%). Индуцированный агонистом ГйРГ гипогонадизм, способствующий умень¬
шению общего объема матки с лейомиомами и снижению кровоснабжения опухоли, с
одной стороны, и тяжелые побочные эффекты и быстрые рецидивы, с другой, делают
использование агонистов ГнРГ полезным только для получения кратковременных
эффектов, таких как уменьшение анемии, связанной с маточным кровотечением, или
снижение кровоснабжения опухоли до гистероскопической резекции [386].
ВЕДЕН1^Е МЕНОПАУЗЫ
Штвтшт шттшщы
КЛИМАКТЕРИЙ
Под менопаузой понимается прекращение менструаций как результат необрати¬
мой потери определенного количества яичниковых функций, включая овуляцию и
продукцию эстрогенов. Климактерий — критический период жизни, в котором на
пути к менопаузе происходят волнующие изменения в эндокринной, соматической и
психологической сферах. Климактерий также называется перименопаузой; он вклю¬
чает изменения циклов от овуляторных до задержки менструаций и характеризуется
нерегулярностью менструальных выделений.
Наиболее чувствительный клинический признак климактерия — прогрессивно уве¬
ли чивающиеся задержки менструаций. Менструальный цикл у большинства женщин
с овуляцией длится от 24 до 35 дней, тогда как 20% всех женщин репродуктивного
возраста имеют нерегулярные циклы [349]. Когда возраст женщины достигает 40 лет,
начинает преобладать ановуляция; до ановуляции длительность менструального
цикла возрастает в течение нескольких лет до менопаузы [352]. Медиана возраста
начала климактерических изменений составляет 47,5 года [387], Невзирая на возраст
начала, менопаузе предшествует период удлинения интервалов между менструациями
[388]. Увеличенная концентрация ФСГ определяет изменения менструального цикла
до менопаузы и сопровождается уменьшением уровней ингибинов, нормальным уров¬
нем ЛГ, а также слегка повышенным уровнем эстрадиола [389]. Эти изменения уровня
гормонов в плазме отражают сниженный овариальный резерв фолликулов и могут
быть определены более точно на 2-й или 3-й день менструального цикла.
Уровень эстрадиола в климактерии начинает снижаться за год до менопаузы. В
связи с усиленным ответом фолликулов на повышение ФСГ средний уровень эстра¬
диола выше у женщин в перименопаузе, чем у молодых женщин [390]. Уменьшение
продукции ингибина фолликулом, способствующее повышению ФСГ в позднем
репродуктивном периоде, отражает уменьшение фолликулярного резерва и возмож¬
ностей фолликулов. Снижение выработки ингибина фолликулами яичников начи¬
нается после 30 лет, причем это снижение становится более ощутимым после 40 лет.
120
Гормональные изменения происходят параллельно снижению фертильности, которое
начинается в возрасте 35 лет.
Климактерий — переходный период, во время которого может быть определен уро¬
вень ФСГ, характерный для постменопаузы, несмотря на продолжающиеся менструа¬
ции; в то же время уровень ЛГ остается в пределах нормы. У женщин в перименопаузе
сохраняется способность к беременности, поскольку могут происходить редкие ову¬
ляции и формирование функционирующего желтого тела. Следовательно, для преду¬
преждения нежелательных беременностей необходимо рекомендовать контрацепцию
до тех пор, пока не произойдет полного прекращения менструаций или уровень ФСГ
будет определен выше 40 МЕ/л двумя независимыми исследованиями.
Климактерий представляет оптимальный период для оценки общего состояггая
здоровья зрелой женщины и для разработки мероприятий, цель которых — подго¬
товка женщины к тем физиологическим изменениям, которые придут с менопаузой.
Пациентка и ее врач должны попытаться достичь несколько важных целей во время
климактерия. Отдаленная цель — поддержка оптимального качества физической и
социальной жизни. Другая непосредственная цель — определение любых знататель-
ных хронических заболеваний, которые могут возникнуть с возрастом. В общем, врач
должен рассказать женщине в перименопаузе о симптомах и отдаленных последстви¬
ях менопаузы. Необходимо уделить достаточное внимание риску и пользе замести¬
тельной гормонотерапии.
МЕНОПАУЗА
Средний возраст менопаузы около 51 года [391]. Возраст постменопаузы, вероят¬
но, частично определяется генетическими факторами, поскольку у матерей и дочерей'
менопауза возникает приблизительно в одно время [392, 393, 394]. Определенные
факторы окружения могут изменять возраст менопаузы, например курение свя¬
зано с ранней менопаузой, а употребление алкоголя отдаляет менопаузу [391].
Использование ОК не изменяет возраст менопаузы.
Симптомы, часто наблюдаемые и относящиеся к сниженному синтезу эстрогенов
в менопаузе, включают нерегулярнз^ю частоту менструаций, за которыми следуют
аменорея, вазомоторная нестабильность, проявляющаяся горячими приливами и
потами, урогенитальной атрофией, которая делает болезненными половые контакты,
и разнообразными зоологическими симптомами, также впоследствии появляются
остеопороз и сердечно-сосудистая патология. Комбинация и продолжительность
этих симптомов различна для каждой женщины. У некоторых развивается множество
симптомов, которые приводят к недееспособности, тогда как у других симптомы либо
отсутствуют, либо умеренно выражены.
БИОСИНТЕЗ ЭСТРОГЕНОВ И ДРУГИХ СТЕРОИДОВ У ЖЕНЩИН В ПОСТМЕНОПАУЗЕ
После менопаузы в яичнике невозможно обнаружить гистологи^іеские фоллику¬
лярные единицы [395]. У женщин в репродуктивном возрасте гранулезные клетки
овуляторного фолликула — основной источник ингибина и эстрадиола. Б отсутствие
этих факторов, ингибирующих секрецию гонадотропинов, уровни ФСГ и ЛГ резко
возрастают после менопаузы. Они максимальны в течение нескольких лет после
менопаузы, после чего постепенно и медленно снижаются [396]. Уровень гонадотро¬
пинов в плазме крови в постменопаузе может быть выше 100 МЕ/л. Уровень ФСГ
обычно выше, чем ЛГ, потому что ЛГ исчезает из крови .значительно быстрее, а также,
возможно, потому, что низкий уровень ингибина селективно повышет секрецию ФСГ.
Тем не менее высокий уровень ЛГ — основной фактор, поддерживающий значитель¬
ные количества андростендиона и секрецию тестостерона яичником, так как общий
уровень продукции обоих стероидов снижается после менопаузы.
121
Главные стероидные продукты постменопаузального яичника — андростендион и
тестостерон [397]. Уровень продукции андростендиона в пременопаузе, составляю¬
щий в среднем 3 мг в день, снижается в 2 раза — до 1,5 мг в день [397]. Это снижение
обусловлено значительным снижением овариального андростендиона. У женщин в
постменопаузе основное количество андростендиона секретируется надпочечниками
и только небольшая часть — яичниками [398]. И ДГЭА, и ДГЭАС в постменопаузе
составляют четверть концентрации у молодых женщин [399].
После менопаузы продукция тестостерона сокращается на у, [397]. Общая про¬
дукция тестостерона может быть рассчитана суммированием яичниковой секреции
и периферического формирования из андростендиона (см. рис. 1-28). У женщин в
пременопаузе значительное количество тестостерона образуется из андростендиона
в экстраовариальных тканях. Поскольку секреция андростендиона яичниками зна¬
чительно снижается после менопаузы, снижение продукции тестостерона в постмено¬
паузе определяется снижением относительного вклада из экстраовариальных источ¬
ников [397]. По мере исчезновения фолликулов и снижения эстрогенов повышенный
уровень гонадотропинов стимулирует оставшуюся стромальную ткань яичника для
поддержания секреции тестостерона на уровне пременопаузального возраста. Таким
образом, вклад постменопаузального яичника в общую продукцию тестостерона уве¬
личивается в присутствии внешне не измененной яичниковой секреции.
Наиболее выраженное эндокринное изменение в климактерии включает снижение
концентрации эстрадиола в плазме крови, а также его общего уровня продукции.
Средний уровень циркулирующего эстрадиола в менопаузе составляет менее 20 пг/мл.
Уровень эстрадиола и эстрона у женщин в постменопаузе обычно несколько ниже,
чем у взрослого мужчины, Эстрадиол, определяемый в плазме крови у женщин (и
мужчин) в постменопаузе, получается путем периферической конверсии андростен¬
диона в эстрон, который далее периферически превращается в эстрадиол (см. рис.
1-21) [386, 400]. Средний уровень циркулирующего эстрона (37 пг/мл) у женщин в
постменопаузе выше, чем эстрадиола. Средний постменопаузальный уровень продук¬
ции эстрона приблизительно составляет 42 мг в течение 24 ч. После менопаузы прак¬
тически весь эстрон и эстрадиол получаются путем периферической ароматизации
андростендиона. Таким образом, имеется существенное изменение в соотношении
андрогены-эстрогены вследствие острого снижения уровня эстрадиола и небольшого
снижения тестостерона. Частое появление умеренного гирсутизма после менопаузы
отражает этот сдвиг в гормональном балансе. Во время постменопаузы уровни ДГЭАС
и ДГЭА продолжают снижаться по мере старения, тогда как андростендион, тесто¬
стерон, эстрон и эстрадиол существенно не изменяют своей концентрации в плазме
крови [396].
Ароматизация андростендиона в эстрон в экстраовариальных тканях положительно
коррелирует с массой тела и увеличивающимся возрастом (см. рис. 1-21 и 1-33) [125].
Масса тела находится в положительной корреляции с уровнями циркулирующего
эстрона и эстрадиола. Поскольку ферментная активность ароматазы присутствует в
значительных количествах в жировой ткани, повышенная ароматизация андростен¬
диона у тучных женщин может отражать увеличенное количество ткани, содержащей
этот фермент [124]. Дополнительно определяется от 2- до 4-кратного увеличения
специфической активности ароматазы на клетку с учетом увеличивающегося возрас¬
та [126]. Увеличение общего количества фибробластов жировой ткани и уменьшение
уровней ПСПС вызывает увеличение уровня свободного эстрадиола, а также является
фактором высокого риска развития рака эндометрия у тучных женщин [126]. Уровень
продукции и концентрации эстрадиола после менопаузы недостаточен для поддерж¬
ки урогенитальных тканей и костей. Таким образом, остеопороз и урогенитальная
атрофия — одни из наиболее драматических и нежелательных проявлений дефицита
эстрадиола во время менопаузы.
122
Эстроген синтезируется локально в таких патологических тканях, как рак груди,
с помощью ароматазы и 17р-гидростероид-дегидрогеназы (рис. 1-36) [126, 401]. У
постменопаузальных женщин андростендион надпочечникового происхождения —
наиболее важный субстрат для ароматазы в тканях опзхоли [126,401]. Эстрон образу¬
ется первично ароматазой, которая сохраняется в недифференцированных жировых
фибробластах, окружающих злокачественные эпителиальные клетки (см. рис. 1-36)
[126, 401]. Далее эстрон диффундирует в злокачественные эпителиальные клетки,
которые содержат сниженную по активности 17р-гидростероид-дегидрогеназу, и пре¬
вращается в биологически активный эстрадиол (см. рис. 1-36) [147]. Таким образом,
паракринные взаимоотношения в опухоли молочной железы способствуют про¬
дукции злокачественными эпителиальными клетками эстрадиола, который, в свою
очередь, стимулирует пролиферацию. Клиническое значение этих находок было про¬
демонстрировано успешным использованием ингибиторов ароматазы как в первой,
так и во второй линии лечения постменопаузального рака груди [402].
МАТОЧНОЕ КРОВОТЕЧЕНИЕ В ПОСТМЕНОПАУЗЕ И ЕГО ВЕДЕНИЕ
Перименопаузальное или постменопаузальное кровотечение может быть связа¬
но с назначением гормонов или избыточным экстраовариальным формированием
эстрогенов. Нерегулярное маточное кровотечение часто наблюдается во время
Рис. 1-36. Тканевый источник эстрогенов при раке молочной железы в постменопаузе. Рисунок пока¬
зывает патологическую роль экстраовариального и локального биосинтеза эстрогена при эстрогенза-
висимом заболевании у постменопаузальных женщин. Предшественник эстрогена андростендион (А)
первично происходит из надпочечников постменопаузальных женщин. Экспрессия ароматазы и .актив¬
ность ферментов в экстраовариальных тканях, например жировой, нарастает с возрастом. Активность
ароматазы в коже и подкожно-жировых фибробластах способствует формированию системно-активного
эстрона (Е^) и в меньшей степени — эстрадиола (Е^). Конверсия циркулирующего А в Е, в недифферен¬
цированных фибробластах жировой ткани груди, концентрирующихся вокруг злокачественных эпители¬
альных клеток, и последующая конверсия Е, в ^ 8 злокачественных эпителиальных клетках способствуют
высокой концентрации в тканях Е^, необходимого для роста опухоли. Клиническое значение этих данных
подтверждено успешным использованием ингибиторов ароматазы для лечения рака груди.
123
перименопаузального перехода овуляторных циклов в ановуляторные. Маточное
кровотечение после менопаузы встречается менее часто, если пациентка не при¬
нимает ЗГТ [403]. Пациенты с ожирением чаще имеют постменопаузальное крово¬
течение в связи с увеличенной периферической ароматизацией андростендиона из
надпочечников. У женщин, получающих длительно комбинированный режим ЗГТ,
может развиваться неожиданное маточное кровотечение (см. ниже). При таких
обстоятельствах главной целью является исключение рака эндометрия. Это может
быть легко достигнуто исследованием ткани, полученной с помощью биопсии, про¬
веденной пластиковой канюлей в офисе врача. Для уменьшения числа ненужных
биопсий возможно использование ультразвукового измерения толщины эндоме¬
трия у женщин в постменопаузе [172]. Биопсия требуется, если толщина эндометрия
больше или равна 5 мм.
До использования УЗИ или биопсии эндометрия для изучения этиологии крово¬
течения из полости матки врач должен исключить заболевание вульвы, влагалища и
шейки матки как потенциальных источников кровотечения. Внимательное исследо¬
вание этих органов, а также нормальный ПАП мазок с шейки матки в течение послед¬
него года являются важными для исключения этих органов как источников потен¬
циального кровотечения. С одной стороны, маточное кровотечение в постменопаузе
является наиболее частым первым знаком, предупреждающим пациентку и ее врача
о возможности рака эндометрия; с другой стороны, причинами постменопаузально¬
го маточного кровотечения бывают в основном доброкачественные состояния. Рак
эндометрия среди пациенток с кровотечением составляет от 1 до 2% среди биопсий
эндометрия в постменопаузе [404]. В ходе примерно трех четвертей этих биопсий
выявляют либо отсутствие патологии, либо атрофический эндометрий. Другая гисто¬
логическая находка включает гиперплазию (15%) или полипы эндометрия (3%).
Постоянные внеплановые маточные кровотечения требуют повторного исследова¬
ния, биопсии, гистероскопии, а также выскабливания полости матки.
Неожиданные нерегулярные маточные кровотечения наблюдаются приблизитель¬
но у 20% женщин в постменопаузе, полу^шющих длительную комбинацию эстроген-
гестагеновых препаратов. Они также должны быть исследованы с помощью УЗИ или
биопсии либо совместно (см. ниже) [172].
ПРИЛИВЫ
Наиболее частый и наиболее выраженный симптом климактерия представлен горя¬
чими приливами. Приливы обычно возникают во время перехода от пери менопаузы
к постменопаузе. Приливы являются главными симптомами постменопаузы и могут
продолжаться до 5 лет после менопаузы [405]. Более чем у. всех женщин испытывают
приливы в течение 3 мес после прекращения функции яичников, произошедшего есте¬
ственным образом или хирургическим путем. Из этих женщин более чем три четверти
испытывают приливы 1 год, а примерно половина — более 5 лет [405]. С увеличением
возраста приливы уменьшаются по частоте и интенсивности, в то время как другие
проявления менопаузы прогрессируют с течением времени.
Прилив представляет собой субъективное ощущение интенсивного потепления
верхней половины туловища, которое обычно длится в течение 4 мин, но может быть
в пределах от 30 с до 5 мин. Вслед за приливом возникает продрома в виде сердце¬
биения или головной боли, которая часто сопровождается слабостью, общим недо¬
моганием или головокружением. Эпизод обычно заканчивается профузными потами
или ощущением холода. Частота может варьировать от чрезвычайно редкой до одного
раза каждые несколько минут. Ночами приливы возникают намного чаще и тяжелее,
приводя к пробуждению женщины во время сна. Они также более интенсивны во
время стресса. В прохладном помещении приливы встречаются реже, менее интенсив¬
ны и коротки по сравнению с помещением с высокой температурой [406].
124
приливы возникают вследствие резкого снижения уровня эстрогенов, большем,
чем при развитии гипоэстрогении. Следовательно, несмотря на причину менопаузы
(естественная, хирургическая либо связанная с использованием длительно действую¬
щего агониста ГнРГ), приливы всегда связаны с резким и значительным падением
уровня эстрогенов.
Связь между началом приливов и острой отменой эстрогенов доказывается эффек¬
тивностью терапии эстрогенами и отсутствием приливов при пролонгированных
гипоэстрогенных состояниях, таких как дисгенезия гонад. Женщины с гипогонадиз¬
мом имеют приливы, только если эстроген был назначен, а затем отменен [407]. Не
все приливы связаны с дефицитом эстрогенов. Неожиданные эпизоды потоотделения
и приливов могут быть связаны с катехоламин- или гистаминсекретирующими опу¬
холями (феохромоцитома, карциноид), гипертиреозом или хронической инфекцией
(например, туберкулезом). Приливы могут быть психосоматическими по происхожде¬
нию и не связанными с отменой эстрогенов. При таких сомнительных обстоятельствах
врач должен исследовать уровень ФСГ в плазме для подтверждения климактерия или
менопаузы до начала ЗГТ. Женщины с ожирением редко имеют приливы, женщины
без симптомов имеют ббльшую массу тела в сравнении с женщинами с тяжелыми
симптомами, даже с учетом возраста, состояния яичников и времени после менопаузы
[408]. Низкая частота и интенсивность приливов у женщин с ожирением могут быть
объяснены увеличенным уровнем циркулирующего эстрадиола [409].
УРОГЕНИТАЛЬНАЯ АТРОФИЯ
Урогенитальный синус трансформируется в нижнюю треть влагалища, вульву и
уретру во время развития эмбриона, и эти ткани эстрогензависимы. Снижение уровня-
эстрогенов во время менопаузы вызывает побледнение стенок влагалища в связи со
снижением васкуляризации и уменьшением толщины до 3-4 клеток. Клетки эпителия
влагалища у постменопаузальных женщин содержат меньше гликогена, который до
менопаузы метаболизировался лактобациллами, создающими кислую реакцию среды
и, таким образом, обеспечивающими защиту влагалища от роста бактериальной
флоры. Потеря этого защитного механизма делает ткань чувствительной к инфекции
и изъязвлениям. Влагалище может потерять свои складки, а также стать короче и
неэластичнее. Женщины в постменопаузе могут жаловаться на симптомы, вторичные
по отношению к сухости влагалища, такие как боли во время полового контакта, вла¬
галищные выделения, жжение, зуд или кровотечение, урогенитальная атрофия при¬
водит к различным симптомам, которые влияют на качество жизни.
Уретриты с дизурией, стрессовое недержание мочи, учащение мочеиспуска¬
ния и диспареуния являются следствием истончения слизистой уретры и пузыря.
Внутри влагалищное введение эстрогенов пациенткам в постменопаузе может быть
эффективно использовано при лечении влагалищных симптомов и периодически
повторяющихся инфекций мочевыводящих путей [410]. Прием эстрогенов внутрь
способствует быстрому восстановлению влагалища и уменьшению уретральных сим¬
птомов, вызванных дефицитом эстрогенов.
ПОСТМЕНОПАУЗАЛЬНЫЙ ОСТЕОПОРОЗ
Остеопороз — заболевание, характеризующееся низкой массой кости и микроар-
хитектурным повреждением ее ткани, что в итоге приводит к увеличению хрупкости
и, таким образом, повышает риск развития переломов. Наиболее частые места пере¬
лома — тела позвонков, дистальная часть лучевой кости и шейка бедра. Остеопороз
стал глобальной мировой проблемой. В Соединенных Штатах он находится в ряду
эпидемических заболеваний, затрагивая более 20 млн людей [411], Большинство
пациенток, страдающих остеопорозом, — женщины в постменопаузе.
125
Остеопороз у женщин в постменопаузе является результатом увеличения возраста и
дефицита эстрогенов. 75% и более потери костной ткани у женщин в течение первых
15 лет после менопаузы является проявлением дефицита эстрогенов в большей сте¬
пени, чем старение [412, 413]. В течение первых 20 лет после прекращения секреции
эстрогенов яичниками постменопаузальный остеопороз приводит к 50% уменьшению
трабекулярной кости и 30% потере кортикальной костной ткани [412,413]. Особенно
уязвймы позвонковые кости, поскольку трабекулярные кости тел позвонков мета¬
болически активны и в ответ на дефицит эстрогенов значительно уменьшаются в
количестве. Масса позвонковой кости значительно уменьшается у женщин в пери¬
менопаузе и ранней постменопаузе, когда происходит повышение ФСГ и снижение
уровня эстрогенов, при этом потеря костной ткани лучевой кости не определяется, как
минимум, год после менопаузы [413].
Риск переломов зависит от двух факторов: пика костной массы, достигаемой в зре¬
лости (в возрасте 30 лет), и последующего уровня потери кости. Увеличение скорости
потери костной ткани после менопау.зы гарантированно предсказывает увеличение
риска переломов. Женщины с низкой массой костной ткани в пременопаузе и уско¬
ренной потерей костной ткани в менопаузе имеют самый высокий риск возникнове¬
ния переломов. Ускоренная потеря костной массы во время менопаузы — индикатор
ни.зкого эндогенного уровня эстрогенов; это может быть объяснено тем, что постме¬
нопаузальная потеря костной ткани происходит в меньшей степени у женщин с боль¬
шим количеством жировой ткани и, следовательно, увеличенным периферическим
формированием эстрогенов [267],
Многочисленные исследования показали, что ЗГТ. начатая в климактерии, преду¬
преждает постменопаузальную потерю костной ткани [414]; начатая в любом возрасте
в постменопаузе, она имеет потенциальный положительный эффект, как минимум,
предупреждая дополнительную потерю костной массы. Несмотря на это, необходимо
отметить, что частота переломов или уровень потери массы не снижался в течение 4
лет наблюдения у женщин, которые начали ЗГТ в среднем возрасте 66,7 (±6,7) года.
Недавно результаты исследования WHI подтвердили уменьшение количества пере¬
ломов позвонков и бедренной кости в группе пациенток в постменопаузе, получав¬
ших комбинацию эстроген/гестаген или только эстроген; эти важные сведения были
первыми среди рандомизированных исследований, предполагавших, что эстрогены
предупреждают переломы [415, 416].
Первое, что хотелось бы отметить: женщины, у которых была удалена матка, в
постменопаузе получают ЗГТ только с эстрогенами (ЗГТ-Э). Гестаген добавляют к
эстрогенами (ЗГТ-ЭП) у женщин в постменопаузе с сохраненной маткой, для того
чтобы предупредить гиперплазию Э11дометрия или рак. ЗГТ-ЭП назначают значи¬
тельно чаще, чем ЗГТ-Э. Польза и риски этих режимов различны. Именно поэтому в
данном разделе будет описан каждый режим.
До конца 1990-х гг. наиболее частой практикой лечения женщин, страдающих
от дефицита гормонов (приливов), было назначение эстрогенов и использование
продолжительной гормональной профилактики остеопороза. Предположение, что
ЗГТ обладает кардиопротективным действием, поддерживало убеждение женщин
оставаться на этом режиме. Эстрогены и гестагены, используемые для ЗГТ в пост¬
менопаузе, были среди наиболее часто назначаемых лекарств в США. Национальное
анкетирование показало, что имеется существенная тенденция к увеличению исполь¬
зования любых форм ЗГТ от 1970 до 1990 гг. Многие женщины, начавшие лечение
126
ЗГТ во время менопаузы или позже, продолжали его, 46% женщин с естественной
менопаузой и 71% женщин, в отношении которых было проведена двусторонняя
оофорэктомия, сообщили об использовании постменопаузальной ЗГТ. Средняя дли¬
тельность использования ЗГТ в США в 1992 г. составляла 6,6 года, в то же время лишь
20% продолжали лечение в течение 5 лет.
С 2000 г. после публикации основных результатов двух больших рандомизиро¬
ванных исследований «Сердце и ЗГТ эстроген/гестаген» (HERS) в 1998 г., а также
«Инициатива по охране здоровья женщин» (WHI) серьезно изменили эту тенденцию
[418, 419, 420]. Результаты WHI стали причиной прекращения ЗГТ у 30% женщин в
постменопаузе [420]. Активно обсуждается применимость результатов WHI и HERS
ко всем или отдельным группам женщин в постменопаузе. В то время как использо¬
вание ЗГТ значительно снизилось, эти влиятельные исследования подняли опреде¬
ленное количество важных вопросов, которые требуют дополнительного изучения.
Ожидается, что дальнейшие эпидемиологические экспериментальные исследования
прояснят некоторые из обсужденных вопросов в следующих редакциях книги.
Исследование WHI чрезвычайно важно для представления о поддержке менопаузы.
Большинство ранних когортных ретроспективных и проспективных обсервационных
исследований показало значительное (40-60%) снижение заболеваемости ишеми¬
ческой болезнью сердца (ИБС) у женщин в постменопаузе, получавших ЗГТ-Э и
ЗГТ-ЭП. Эти исследования также подтвердили снижение смертности от всех причин,
уменьшение количества переломов, связанных с остеопорозом, однако одновременно
свидетельствовали об увеличении риска заболеваемости раком груди от 20 до 30%.
Отдельные подобные изучения продемонстрировали снижение заболеваемости болез¬
нью Альцгеймера в группе женщин, использовавших ЗГТ-ЭП, по сравнению с группой
женщин, не получавших ЗГТ. В связи с систематическими ошибками, которые присут¬
ствуют в обсервационных исследованиях для решения вопроса, обеспечивает ли ЗГТ
защиту от сердечно-сосудистых заболеваний (ССЗ) или деменции, необходимо прове¬
дение рандомизированных клинических исследований. Исследование HERS и другие
вторичные профилактические исследования показали отсутствие пользы от примене¬
ния ЗГТ у женщин с ранее поставленным диагнозом ИБС, которые начали лечение в
сроки от 8 до 23 лет после менопаузы. В исследовании WHI у женщин с отсутствием
в анамнезе ИБС ЗГТ-ЭП (конъюгированные эстрогены с МПА) в среднем в течение
12 лет после менопаузы слабо и незначительно повышала риск ИБС, тогда как только
эстрогены (конъюгированные эстрогены без МПА) не увеличивали риск ССЗ, но и
существенно его не снижали. Наблюдали незначительное повышение тромбоэмбо¬
лических заболеваний и инсультов в обоих исследованиях, а также статистически
достоверное пограничное увеличение частоты рака груди при использовании ветви
ЗГТ-ЭП исследования WHI. Оценка когнитивных функций у женщин в WHI показало
повышение заболеваемости деменцией и отсутствие разницы на уровне когнитивных
расстройств. Было сделано заключение, что общее соотношение «риск/польза» для
ЗГТ неблагоприятно. Результаты из WHI показывают, что комбинированные постме¬
нопаузальные гормоны, представленные конъюгированными эстрогенами 0,625 мг в
день+МПА по 2,5 мг в день, не должны назначаться или продолжаться с целью про¬
филактики ССЗ. Более того, польза от назначения ЗГТ, представленная снижением
риска развития переломов, должна быть тщательно сопоставлена с риском развития
ССЗ и рака груди.
127
Историческая перспектива научных исследований, касающихся
использования заместительной гормональной терапии
Начиная с 1950-х гг. среди врачей и пациенток возникло убеждение, что замеще¬
ние исчезаюпщх во время менопаузы эстрогенов может предотвратить старение, ССЗ,
переломы, связанные с остеопорозом, а также снижение когнитивных и сексуальных
функций. Это привело к широкому использованию ЗГТ-Э после менопаузы. Другими
значимыми фактами в поддержку защиты от ССЗ стало следующее: обсервацион¬
ные исследования показали меньшее количество ССЗ среди женщин, принимающих
эстрогены; патофизиологические механизмы, обеспечивающие биологическую досто¬
верность; клинические исследования, которые выявили улучшения в уровнях липи¬
дов крови и других подобных показателях [421].
Наиболее часто используемыми для ЗГТ-Э были конъюгированные эстрогены
(КЭ). В 1980 г. стало ясно, что использование ЗГТ-Э в постменопаузе вызывает рак
эндометрия. Несмотря на невысокую частоту и курабельность, этот тип опухоли
может быть предупрежден с помощью гестагена — антагониста эстрогена, после чего
было разработано несколько комбинаций эстрогенов и гестагенов. С 1990 г., после
того как было продемонстрировано, что комбинация конъюгированных эстрогенов с
МПА значительно улучшает липидный профиль, этот особый режим Э+П (КЗ+МПА)
стал наиболее популярным режимом в США, назначаемым постменопаузальным жен¬
щинам с интактной маткой [421].
В конце 1990-х гг. было доказано, что ЗГТ-ЭП увеличивает частоту рака груди
[422]. Это повышение бьшо минимальным, но достоверным. В связи с тем что ЗГТ
сопровождалось положительными эффектами на ССЗ и оказывало защитный эффект
на костную ткань, многие женщины и их врачи полагают, что общая польза значи¬
тельно превьппает риск [421].
В 1998 г. HERS, первое крупное рандомизированное исследование, неожиданно
обнарз^ило увеличение числа ССЗ в течение первого года, а также отсутствие поль¬
зы по отношению к ССЗ при длительном наблюдении, когда ЗГТ-ЭП сравнивался с
плацебо у 2763 пациенток, страдающих ИБС [423]. Это исследование также обнару¬
жило, что ЗГТ-ЭП увеличивает частоту тромбозов. В 2002 г. исследование WHI было
остановлено в связи с тем, что ветвь ЗГТ-ЭП достоверно причиняла вред пациентке
[415]. Среди 16 608 здоровых женщин постменопаузального возраста с сохраненной
маткой ЗГТ-ЭП повышало риск коронарных осложнений, инсульта, рака груди, тром¬
боэмболии легочной артерии (ТЭЛА). Гяобальный индекс, сформулированный авто¬
рами, показал, что эти негативные эффекты перевешивают пользу от профилактики
переломов костей бедра и рака кишечника (табл. 1-10). Следующий доклад выявляет
повышенное в 2 раза количество деменции среди женщин старше 65 лет, 5^1аств)то-
щих в исследовании WHI, что подтверждает вышесказанное [424]. Результаты HERS
и WHI побудили FDA требовать предупреждения о присутствии потенциального
вреда и рекомендовать информировать о том, что эстогенсодержащие препараты не
должны использоваться для предупреждения ССЗ или быть первой линией препа¬
ратов, используемых для предупреждения остеопороза. Согласно полученным WHI
результатам, ряд профессиональных сообществ изменили свои руководства, ЗГТ не
должна быть использована для предупреждения заболевания, а в ситуациях, когда
используется для лечения симптомов, доза препарата и время приема должны быть
минимально необходимыми.
128
Таблица 1-10. Отношение рисков по данным трех исследований лечения гормонами
Клиническое событие
Отношение рисков (95% доверительный интервал)
HERS
(эстроген + гестаген)
WHI
(эстроген + гестаген)
WHI
(эстроген отдельно)
ИБС
0,99 (0,80-1,22)
1,29 (1,02-1,63)
0,91 (0,75-1,12)
Инсульт
1,23 (0,89-1,70)
1,41 (1,07-1,85)
1,39(1,10-1,77)
ТЭЛА
2,79 (0,89-8,75)
2,13(1,39-3,25)
1,34 (0,87-2,06)
Рак груди
1,30 (0,77-2,19)
1,26 (1,00-1,59)
0,77(0,59-1,01)
Рак толстого кишечника
0,69 (0,32-1,49)
0,63 (0,43-0,92)
1,08 (0,75-1,55)
Перелом бедра
1,10 (0,49-2,50)
0,66 (0,45-0,98)
0,61 (0,41-0,91)
Смерть
1,08 (0,84-1,38)
0,98 (0,82-1,18)
1,04 (0,88-1,22)
Общий индекс*
1,15(1,03-1,28)
1,01 (0,91-1,12)
* Общий индекс составлен из любого упоминания ситуации, представленной выше.
Женщины и их врачи с волнением ожидали результатов ветви ЗГТ-Э исследова¬
ния WHI, проводимого у 10 739 здоровых женщин после гистерэктомии, которые
были рандомизированы на группы, получавшие КЭ и плацебо [416]. Использование
ЗГТ-Э не уменьшало риск развития ИБС, как и в HERS, и WHI ветви ЗГТ-ЭП, в то
же время утяжеление ИБС на ранних этапах приема ЗГТ было выражено слабее, чем
при комбинации с гестагенами [416]. Риск ЗГТ, полученный по результатам трех
рандомизированных исследований (HERS, WHI-Э, WHI-ЭП), может быть суммиро¬
ван нижеследующим образом. Необходимо помнить, что эти риски минимальны, но
статистически достоверны.
РИСКИ и ПРОТИВОПОКАЗАНИЯ К ЗАМЕСТИТЕЛЬНОЙ ГОРМОНАЛЬНОЙ ТЕРАПИИ
Ишемическая болезнь сердца
Учитывая отсутствие доказательств о пользе ЗГТ для предупреждения ССЗ во всех
трех исследованиях, используемый ранее постулат о благоприятном влиянии ЗГТ на
течение ИБС считается ошибочным (см. табл. 1-10). И ЗГТ-ЭП, и ЗГТ-Э не предупре¬
ждают ИБС, как полагали ранее. Наоборот, имеется слабое и достоверное повышение
ИБС у пациенток, получавших ЗГТ в режиме ЗГТ-ЭП [415, 423]. Обе женщины — и с
ИБС, и здоровая — находятся в группе риска. С другой стороны, ЗГТ-Э не увеличивает
риск для здоровых женщин [416].
Инсульт
Другой результат, полученный в результате трех исследований, — возросший риск
развития инсульта у женщин при назначении ЗГТ-Э и ЗГТ-ЭП. Увеличенный риск
инсульта, возможно, связан с эстрогенным компонентом данных гормональных пре¬
паратов, поскольку это единственный статистически достоверный неблагоприятный
побочный эффект ЗГТ-Э [415, 416, 423].
Тромбоэмболия легочной артерии
Увеличенный риск ТЭЛА наблюдали во всех исследованиях, причем в WHI ЗГТ-Э
риск был слабым и статистически недостоверным [415, 416, 423].
129
Рак молочной железы
Результаты WHI ЗГТ-Э значительно отличались от результатов HERS и WHI
ЗГТ-ЭП в отношении рака груди [415, 416, 423J. Исследования комбинации Э
плюс П (HERS и WHI ЗГТ-ЭП) выявляли увеличение риска развития рака груди
на 25%, а исследование WHI-Э — на 23%. Многочисленные доказательства под¬
тверждают увеличение риска рака груди при воздействии эстрогенов: исследования
на культурах клеток и моделях животных, многие обсервационные исследования, а
также факт, что антагонисты эстрогенов снижают риск развития рака груди у моло¬
дых женщин. Таким образом, это противоречие стало определенным сюрпризом.
Вероятно, более высокий риск развития рака груди, определяемый в исследовании
комбинированных эстроген-гестагеновых препаратов, представляет негативный
эффект МПА. Увеличение риска, полученное в ветви ЗГТ-ЭП исследования WHI,
было статистически достоверным. Данные результаты были подтверждены больши¬
ми обсервационными наблюдениями, предполагавшими, что добавление МПА или
другого гестагена к эстрогену может значительно увеличивать риск развития рака
груди (см. табл. 1-10).
Рак яичников
Ретроспективное когортное исследование 44 241 женщины в постменопаузе выяви¬
ло, что ЗГТ-Э, особенно проводимое в течение 10 лет и более, существенно увеличи¬
вает риск развития рака яичников. Относительный риск 10-19 лет и 20 лет и более
был 1,8 (95%, доверительный интервал 1,1-3,0) и 3,2 (95%, доверительный интервал
1,7-5,7) соответственно (значение р для тренда <0,001). Это исследование не пока¬
зало повышенного риска для женщин, использующих кратковременную ЗГТ-ЭП, но
есть предположение, что риск ассоциирован с более длительным применением ЗГТ-
ЭП (это требует дальнейших исследований). Исследование WHI ЗГТ-ЭП выявило
тенденцию к увеличению риска рака яичников [416].
Деменция
у женщин 65 лет или старше ЗГТ-ЭП значительно увеличивало риск и приводи¬
ло к дополнительным 23 случаям деменции на 10 ООО женщин в год [426]. Болезнь
Альцгеймера - наиболее распространенное проявление деменции. Похожая тенден¬
ция, однако без статистической достоверности, наблюдалась в группе ЗГТ-Э [424].
Когда данные были объединены, оказалось, что ЗГТ достоверно увеличивает риск
развития деменции [426].
Гиперлипидемия
Этот редкий побочный эффект наблюдали у пациенток с тяжелой наследственной
гиперлипидемией. Прием эстрогенов перорально может ускорять тяжелую гиперли-
пидемию или панкреатит у женщин с увеличенным уровнем триглицеридов [427],
Именно поэтому повышенный уровень триглицеридов служит относительным проти¬
вопоказанием для назначения эстрогенов с заместительной целью.
Заболевание желчного пузыря
оба WH1 исследования (ЗГТ-Э и ЗГТ-ЭП) показали повышенный риск развития
заболеваний желчного пузыря или операции, связанной с эстрогенами [428]. Оба
исследования выявили высокий риск развития холецистита или холелитиаза. Кроме
того, женщины, получавшие любой из режимов ЗГТ, чаще подвергались холецистэк¬
томии [428]. Эти данные свидетельствуют в пользу увеличения рисков болезней били¬
арного тракта среди женщин в постменопаузе, принимающих лечение эстрогенами.
Заболеваемость и стоимость лечения, связанные с данными исходами, должны быть
обоснованы относительно терапии эстрогенами. Заболевания желчного пузыря —
относительное противопоказание к ЗГТ эстрогенами.
130
ПОЛЬЗА ЗАМЕСТИТЕЛЬНОЙ ГОРМОНАЛЬНОЙ ТЕРАПИИ
Приливы
ЗГТ-Э и ЗГТ-ЭП надежно избавляют от приливов большинство женщин. Б настоя¬
щее время приливы — самое частое показание для короткого курса ЗГТ (<5 лет).
Переломы
ЗГТ-Э и ЗГТ-ЭП значительно уменьшают количество переломов бедренной кости,
тел позвонков,щ также других переломов, связанных с остеопорозом [415, 416]. На
сегодня результаты обсервационных исследований связи эстрогенов с риском пере¬
ломов. а также исследования, использующие суррогатные показатели (минеральная
плотность кости), совпадают с результатами клинических исследований о предупре¬
ждении переломов.
Рак толстой кишки
Рак толстой кишки по непоняп ным причинам встречается достоверно реже при гор¬
мональном лечении в исследовании ЗГТ-ЭП, но не в исследовании ЗГТ-Э (WHI) [415,
416]. Вполне вероятно, что гестаген оказывает протективное действие на кишечник.
Другие результаты
Негативные результаты WHI важны для ранее постулированной пользы ЗГТ. При
WHI ЗГТ-ЭП отсутствуют достоверные эффекты на общее здоровье, жизнеспособ¬
ность, психическое здоровье, депрессивные синдромы или сексуальное удовлетворе¬
ние [429]. Использование комбинации Э+П более года может быть связано со стати¬
стически небольшой, но достоверной и клинически незначимой пользой в отношении
расстройства сна, физической активности или физической боли. Через 3 года не
обнаруживали значительной пользы по какому-либо из параметров качества жизни.
Среди женщин 50-54 лет, имеющих вазомоторные симптомы от слабой до выражен¬
ной степени, ЗГТ-ЭП эффективна в отношении приливов и приводит к незначитель¬
ному улучшению при расстройствах сна, однако при этом отсутствуют улучшения в
других параметрах качества жизни [429]. Группа, которая получала ЗГТ-Э (WHI) в
течение 3 лет, также не подтвердила значимого улучшения качества жизни [430].
ЗГТ-ЭП (WHI), несмотря на ожидание, не улучшает когнитивных функций [424],
Группа, получавшая ЗГТ-Э, показала невыраженную тенденцию к ухудшению когни¬
тивных расстройств слабой степени [426]. Среди женщин в постменопаузе в возрасте
старше 65 лет ЗГТ-ЭП не улучшало когнитивных функций при сравнении с плацебо,
тогда как ЗГТ-Э оказывала неблагоприятный эффект на когнитивную функцию, и он
был более выражен среди женщин с более слабой когнитивной функцией в начале
лечения [431,432].
Средний возраст участников данных рандомизированных исследований состав¬
лял 60 лет; это поддерживает мнение о том, что полученные результаты могут быть
неприменимы к женщинам, находящимся в ранней менопаузе [415, 416, 423]. В этом
отношении в исследовании WHI ЗГТ-Э было показано, что подгруппа более молодых
женщин (50-59 лет) отвечает на лечение эстрогенами более благоприятно, чем пожи¬
лые, по многим показателям, включая глобальный индекс. Тем не менее различия в
коэффициентах риска среди подгрупп были статистически достоверны только для 1
из 23 тестов [416].
Даже если коэффициенты риска схожи у женщин всех возрастов, абсолютные
риски различаются существенно. В целом абсолютный риск для многих заболеваний
131
приблизительно удваивается с каждой декадой жизни, поэтому женщины в возрас¬
те 50 лет имеют половину риска женщин 60 лет и четверть риска женщин в возрасте
70 лет. Это означает, что многие эффекты гормонотерапии этих заболеваний будут
менее выражены у молодых женщин. Возможность более благоприятных эффектов в
возрасте 50-59 лет, а также низкий абсолютный риск побочных эффектов в этой воз¬
растной группе предполагает, что использование ЗГТ-Э для лечения менопаузальных
симптомов в течение ограниченного времени в начале менопаузы обосновано.
Исследования WHI предполагают, что ЗГТ-Э имеет преимущества перед ЗГТ-ЭП
при лечении женщин в постменопаузе: всего один или два неблагоприятных эффек¬
та (увеличение инсультов и, вероятно, ТЭЛА) в сравнении с четырьмя (увеличение
инсультов, ИБС, ТЭЛА, а также рак грзщи). Оба режима способствуют уменьшению
количества переломов. Однако оценка «риск/выгода» неприменима при деменции,
причем при отсутствии деменции проведение ЗГТ-Э не приведет к уменьшению гло¬
бального индекса, к тому же ЗГТ-Э у женщин с интактной маткой увеличивает риск
рака матки и число маточных кровотечений, биопсий и гистерэктомий. Проведение
более взвешенных и подробных анализов результатов этих исследований, скорее
всего, приведет к изменениям в новых рекомендациях и руководствах.
В завершение необходимо указать, что в настоящее время происходят дебаты
относительно типа эстрогенов и вариантов их назначения. Возможно, эстрадиол
имеет преимущество перед КЭ и трансдермальный путь введения позволяет избежать
высокой концентрации в портальной циркуляции, а прямой путь к печени, возможно,
обладает преимуществом над оральным путем [433]. Новые исследования планируют
изучение эффектов трансдермального эстрадиола [434].
Рекомендаций для заместительной гормональной терапии
после исследования ЮНІ
ЗГТ эффективна для лечения приливов, и с этой точки зрения ЗГТ-Э ассоциирует¬
ся с меньшими рисками в сравнении с ЗГТ-ЭП. Тем не менее ЗГТ-Э имеет побочные
эффекты, и это заставляет благоразумно поддерживать дозу на низком уровне, а курс
лечения — максимально коротким. Разумным вариантом может быть непрерывное
лечение низкими дозами эстрадиола, вводимого трансдермально, и периодическим
введением гестагенов через большие интервалы (например, 15 дней лечения геста¬
генами каждые 3-6 мес вместо ежемесячного режима). Долговременный профиль
безопасности для этого режима неизвестен. Большинство женщин в постменопаузе
завершают ЗГТ в течение нескольких лет после начала лечения, поскольку приливы к
этому времени снижаются или исчезают совсем. Большинство врачей убеждены в том,
что, несмотря на приостановку ЗГТ, инициированную в последние годы, уменьшение
приливов либо сохраняется, либо приливы становятся менее суровыми, чем во время
климактерия [435, 436].
В отсутствие доказательств об общей пользе использования в постменопаузе ЗГТ-Э
и при наличии доказательств о побочном действии ЗГТ-ЭП ни одна из схем лечения
не должна использоваться для профилактики ССЗ или для улучшения интеллекту^аль-
ных функций. Несмотря на то что другие формы и дозы гормонов могут быть более
полезными, эти факты должны быть подтверждены в исследованиях до того, как
любой гормональный режим будет рекомендован для предупреждения заболевания.
Целевые гщшы для шшвтшівпьшт гормональной тераом^
у женщин с дисгенезией гонад или хирургической постменопаузой время отсут¬
ствия эстрогенов продолжительно. Этим пациенткам рекомендуется ЗГТ эстрогена¬
132
ми с целью уменьшения приливов и длительной профилактики ССЗ, остеопороза и
атрофии органов-мишеней. Некурящим женщинам в возрасте до 45 лет могут быть
назначены низкодозированные ОК, После этого возраста более подходящими будут
дозы эстрогенов, эквивалентные 0,625 мг в день КЭ, определенные в зависимости
от связанного с возрастом повышения риска тромбоэмболических событий. Врач
должен рекомендовать режим непрерывного приема комбинации Э+П пациенткам с
сохраненной маткой и режим только эстрогенами женщинам с удаленной маткой.
Во время климактерия приливы могут быть подавлены комбинацией эстрогенов
с гестагенами. Дополнительное преимущество заключается в том, что ЗГТ предот¬
вращает наряду с приливами начало остеопороза, которое также имеет эстроген-
дефицитный характер [437]. При необъяснимых маточных кровотечениях у женщин
в климактерии необходима биопсия эндометрия до начала ЗГТ.
Расчет ш штвшт доза зотрогенов
Эффективные в отношении приливов дозы эстрогенов различны. Для подавле¬
ния приливов разумно начать с 0.3 мг КЭ в день (или трансдермальный эстрадиол
0,025 мг в день) и постепенно увеличивать эту дозу до 0,625 мг в день (эквивалентно
дозе трансдермального эстрадиола 0,05 мг в день), а затем 1,25 мг в день (эквивалент¬
но трансдермальному эстрадиолу 0,1 мг в день). Если приливы не исчезли при 1,25 мг
в день КЭ или эквивалентной дозе трансдермального эстрадиола, маловероятно, что
большие дозы будут эффективными, в этом случае необходимо исключить другие
заболевания (туберкулез, депрессия).
Низкие дозы эстрогенов (0,3 мг КЭ в день или 0,025 мг в день трансдермальных
эстрогенов) поддерживают уровень эстрадиола крови между 17 и 32 пг/мл (в среднем
22 нг/мл) и могут быть достаточными для сохранения плотности кости и снижения
симптомов менопаузы [438]. Эффект эстрогенов на артериальный тромбоз дозоза¬
висим [438]. Для примера: ОК с высокими дозами эстрогенов чаще ассоциируются
с повышенным риском инфаркта миокарда и инсульта, особенно у курящих. Таким
образом, при выборе дозы для ЗГТ необходимо достигать и поддерживать минималь¬
ный уровень циркулирующего эстрадиола и избегать более высокого уровня для того,
чтобы минимизировать риск тромбозов.
Добавление к принимаемому эстрогену циклических или непрерывных гестагенов
не только снижает риск развития эстрогензависимой гиперплазии и рака эндометрия,
но и приносит дополнительные проблемы [439]. Эти проблемы включают регулярное
кровотечение отмены у 90% женщин, получающих циклическую терапию, нерегуляр¬
ные кровянистые выделения у 20% получающих непрерывную терапию комбинацией
эстрогенов и гестагенов. Более того, гестагены снижают положительный эффект
эстрогенов на липопротеидь[ низкой плотности, холестерин и повышают риски ТЭЛА,
ССЗ и рака груди [415,416, 423].
Хорошо известный режим представлен назначением 0,625 мг КЭ или эквива¬
лентными дозами доступных препаратов с 1-го по 25-й день каждого месяца (см.
рис. 1-37). Ежедневная доза МПА в 10 мг добавляется с 12-го по 25-й день цикла или
с 16-го по 25-й день каждого месяца. Кровотечение отмены ожидают на 26-й день
каждого месяца. Другой распространенный циклический режим включает непрерыв¬
ный прием 0,625 мг в день КЭ или эквивалентную ежедневную дозу (см. рис. 1-37).
Ежедневная до.за МПА 5-10 мг в день добавляется с 10-го по 14-й день каждого меся¬
ца. Одногодичное рандомизированное исследование показало, что 5 мг доза защища¬
ет эндометрий так же, как и 10 мг доза [440]. Кровотечение отмены возникает у 90%
женщин, получающих непрерывный или циклический режим [441, 442]. Эти режимы
также могут вызывать побочные симптомы, связанные с высокими дозами гестагенов.
133
такими как болезненность молочных желез, задержка жидкости и депрессия. Именно
поэтому рекомендуется использовать минимальную эффективную дозу гестагенов.
Непрерывный комбинированный метод лечения, с другой стороны, имеет потен¬
циальное преимущество в виде снижения кровотечений и возникновения аменореи,
но изредка осложняется прорывными кровотечениями (см. рис. 1-37) [441, 442].
При подобном режиме пациентке назначают перорально комбинацию 0,625 мг КЭ
и 2,5 мг МПА ежедневно. Непрерывный комбинированный режим прост, удобен и
связан с высокой частотой аменореи у 80% пациенток после 6 мес использования. У
оставшейся части пациенток изредка возникают неожиданные кровотечения. Таким
образом, общая переносимость гораздо выше у пациенток, использующих непрерыв¬
ный комбинированный режим. Более того, низкая ежедневная доза МПА связана с
низкой частотой возникновения нагрубания груди. Возможны и другие комбинации
эстрогенов и гестагенов для подобного непрерывного использования.
Помимо непрерывного ежедневного и циклического добавления, гестагены могут
быть использованы еще реже — 1 раз в 3-6 мес. При добавлении к стандартной дозе
эстрогенов 10 мг МПА каждые 3 мес на 14 дней гиперплазия эндометрия определяется
в 1,5% случаев (такой низкий уровень интерпретируется как защита эндометрия), а
длительное использование МПА с 6-месячными интервалами ассоциируется с низ¬
ким уровнем возникновения рака эндометрия. Несмотря на это, врачи не определили
оптимальную дозировку гестагена и график для совместного использования с низ-
День 1 25 1 25 1 25
12/16 25 12/16 25 12/16 25
День 1:^™^ Непрерывно (ежедневно)
- мес '/2 мес —
1 10/14 1 10/14 1 10/14
Б
День 1^™»^Непрерывно (ежедневно)
|Е 1
День 1=™^ Непрерывно (ежедневно)
в
День 1:зв^Непрерывно (ежедневно)
мес
1 14 1 14
Г
Рис. 1-37. Режимы ЗГТ [403]. Эстроген (Е) назначают женщинам в постменопаузе для предупрежде¬
ния урогенитальной атрофии, остеопороза и приливов. У постменопаузальных женщин с сохраненной
маткой к эстрогенам необходимо добавлять гестагены (Г), чтобы предупредить развитие гиперплазии
и рака эндометрия. Е и Г могут быть назначены разными путями. А и Б — женщины в постменопаузе,
получающие ЗГТ, имеют предсказуемые кровотечения после каждого курса гестагенов; В — эти женщины
получают Е и Г непрерывно. После года непрерывной терапии уровень кровяных мажущих выделений
составляет 20%; Г — для минимизации неблагоприятных эффектов гестагенов предложен относительно
новый режим. В настоящее время неизвестны результаты долговременного изучения безопасности отно¬
сительно возникновения гиперплазии эндометрия и рака. Предсказуемые кровотечения каждые 3 мес
после курса гестагенов считают нормальным явлением.
134
кими дозами эстрогенов. Низкие дозы эстрогенов могут потребовать меньшей дозы
гестагенов для защиты эндометрия.
Большинство женщин в постменопаузе могут изменить режим ЗГТ со стандартного
на низкодозированный с добавлением МПА каждые 3 мес или начать ЗГТ с низких доз
[438]. Поскольку отдаленная безопасность по отношению к гиперплазии эндометрия
не подтверждена, следующие режимы представляются разумными компромиссами
для лечения приливов и предупреждения остеопороза при минимизации побочных
эффектов гестагенов и высоких доз эстрогенов [438]. КЭ 0,3 мг в день или трансдер¬
мальный эстрадиол в дозе 0,025 мг назначают непрерывно. Каждые 3 мес к данному
режиму должен быть добавлен 14-дневный курс 5 мг МПА (см. рис. 1-37, Г). Биопсия
эндометрия не требуется при кровотечениях отмены по завершении каждого курса
гестагенов, а также в отсутствие нерегулярных кровотечений. Этот режим может быть
продолжен в течение 5 лет. После прекращения ЗГТ женщинам для защиты костной
ткани в постменопаузе может быть рекомендован бисфосфонат или селективный
модулятор эстрогеновых рецепторов (СМЭР).
ВЕДЕНИЕ КРОВОТЕЧЕНИЙ ОТМЕНЫ ВО ВРЕМЯ ЗАМЕСТИТЕЛЬНОЙ ГОРМОНАЛЬНОЙ
ТЕРАПИИ В ПОСТМЕНОПАУЗЕ
Около 90% женщин, получающих эстрогены плюс циклические гестагены, демон¬
стрируют предсказуемое ежемесячное кровотечение отмены, тогда как непрерывный
прием комбинации Э+П-препаратов приводит к кровотечению ориентировочно у
40% женщин во время первых месяцев (оставшаяся часть пациенток, использующих
этот режим, имеют аменорею). Возникновение влагалищного кровотечения при
непрерывном режиме плохо предсказуемо и вызывает состояние тревожности у боль-'
шинства пациенток. К счастью, частота спонтанных кровотечений при непрерывном
комбинированном режиме снижается на 20% за год лечения [441-443]. Прорывные
кровотечения при непрерывном комбинированном режиме остаются наиболее важ¬
ным фактором для прекращения режима. Большинство пациенток находят его мало¬
приемлемым и предпочитают переключиться на циклический режим приема геста¬
генов или вовсе прекратить ЗГТ. Не существует эффективного фармакологического
метода для остановки прорывных кровотечений, связанных с непрерывным комбини¬
рованным Э+П-режимом. Можно только убедить пациентку, что кровотечение пре¬
кратится в течение года от начала ЗГТ. Если спонтанные кровотечения продолжаются
более года, то режим должен быть заменен на ежедневный прием эстрогена плюс
циклический прием гестагена.
У пациенток с аменореей ЗГТ можно начать в любое время. Пациенткам в пери¬
менопаузе с олигоменореей, приливами и другими связанными симптомами также
может быть назначена ЗГТ. Пациенткам с олигоменореей ЗГТ рекомендуют на 3-й
день одной из нечастых менструаций. Если кандидатка на ЗГТ не имеет нерегулярных
менструаций, нет необходимости в проведении биопсии эндометрия до начала лечения.
Исследованиями показано, что асимптомные женщины в постменопаузе редко имеют
аномалии эндометрия [443-445]. Биопсия до начала лечения с использованием тон¬
кой пластиковой биопсийной канюли может быть ограничена пациентками с высоким
риском гиперплазии эндометрия (непредсказуемые маточные кровотечения, СПКЯ или
хроническая ановуляция в анамнезе, ожирение, заболевания печени и СД).
Назначение женщинам комбинированной эстроген-гестагеновой терапии не пре¬
дотвращает развития рака эндометрия [446]. Имеено поэтому очень важно исключить
злокачественную опухоль эндометрия у женщин, получающих ЗГТ и имеющих нере¬
гулярные маточные кровотечения. Важная задача — дифференцировка прорывных
кровотечений от кровотечений, вызванных гиперплазией или раком. В связи с тем
что прорывные кровотечения достаточно часты, для выявления относительно редких
случаев рака эндометрия необходимо провести количество биопсий. Для того чтобы
135
уменьшить количество биопсий эндометрия, рекомендуется использовать трансваги¬
нальное УЗИ [172]. Толщина эндометрия, измеренная в постменопаузе, коррелирует
с отсутствием или присутствием патологии [172]. Пациентки, получающие любой из
циклического или ежедневного комбинированного режима ЗГТ и имеющие толщину
эндометрия меньше 5 мм, требуют консервативного ведения [447-449]. Толщина
эндометрия, равная или больше 5 мм, требует биопсии. Следуя этому алгоритму, счи¬
тают, что от 50 до 75% пациенток с кровотечениями, которые получают ЗГТ, показана
биопсия [172].
ЗАМЕСТИТЕЛЬНАЯ ГОРМОНАЛЬНАЯ ТЕРАПИЯ ПОСЛЕ ДИАГНОЗА РАКА МОЛОЧНОЙ
ЖЕЛЕЗЫ
От ЗГТ традиционно воздерживаются при диагнозе рак молочной железы, так
как считается, что эстрогены мохут стимулировать рост опухоли. Удивительно, но
несколько сравнительно небольших исследований показали либо отсутствие влияния,
либо уменьшение риска рецидива и смертности у женщин, применявших ЗГТ, в срав¬
нении с теми, кто не использовал ЗГТ после диагноза рак груди [450,451]. ЗГТ в боль¬
шинстве из этих исследований начиналась после по меньшей мере 5-летнего периода
без признаков заболевания [450, 451]. На базе этих недостаточных, но ободряющих
данных решение о проведении ЗГТ может зависеть от индивидуального выбора паци¬
ентки. У подобных пациенток реальную альтернативу замещению эстрогенов с целью
длительной профилактики остеопороза представляют тамоксифен, ралоксифен или
бисфосфанаты.
СЕЛЕКТИВНЫЕ МОДУЛЯТОРЫ ЭСТРОГЕНОВЫХ РЕЦЕПТОРОВ И БИСФОСФОНАТЫ
КАК АЛЬТЕРНАТИВА ЗАМЕСТИТЕЛЬНОЙ ГОРМОНАЛЬНОЙ ТЕРАПИИ
СМЭР созданы таким образом, что, действуя как эстрогены в некоторых тканях-
мишенях, они одновременно оказывают антагонистический эффект в других [452].
Одним из первых СМЭР является тамоксифен, для которого агонистическая эстро-
генподобная активность по отношению к костной ткани была показана с одно¬
временным антагонистическим эффектом по отношению к молочной железе [453].
Нежелательный эффект тамоксифена состоит в эстрогеноподобном действии на
эндометрий. Со временем было разработано второе поколение СМЭР, наиболее
известным представителем которого является ралоксифен. Эти препараты харак¬
теризуются эстрогенподобным действием на костную ткань, липиды и на систему
коагуляции крови и антагонистическим эффектом на молочную железу, при этом
отсутствует заметное влияние на эндометрий [454]. В рандомизированных плацебо-
контролируемых исследованиях женщин в постменопаузе или пациенток с остеопо¬
розом назначение ралоксифена в дозе 60-150 мг в день приводило к возрастанию
минеральной плотности кости в течение 12-36 мес [454]. В ходе другого рандоми¬
зированного исследования ралоксифен уменьшал риск перелома позвонков, но не
влиял на риск других переломов [455]. Ралоксифен идентичен плацебо в эффекте на
эндометрий, но в то же время как эстроген он в 2-3 раза увеличивает риск инсульта и
тромбоэмболий [455,456]. Способность ралоксифена вызывать приливы препятству¬
ет его применению у женщин с вазомоторными симптомами [457].
В ходе одного из рандомизированных исследований сравнивали тамоксифен и
ралоксифен по влиянию на предупреждение рака груди. Ралоксифен был столь же
эффективен, как и тамоксифен, в отношении снижения риска развития инвазивного
рака груди и имел более низкий риск тромбоэмболии и развития катаракты, но более
высокий, статистически недостоверный риск неинвазивного рака груди. Риск по дру¬
гим опухолям (включая рак матки), переломам, ИБС и инсульту был схож в обеих
группах [458]. Таким образом, тамоксифен и ралоксифен могут быть использованы
для снижения частоты переломов позвоночника и инвазивного рака груди. Гтавные
136
помехи для обоих препаратов — приливы и увеличение частоты тромбоэмболических
событий,
Бисфосфонаты являютсй широко назначаемыми антирезорбтивными агентами
и эффективными препаратами для лечения постменопаузального остеопороза. Эти
агенты подавляют резорбцию через ингибирование присоединения остекластов к
матриксу кости и усилению апоптоза остеокластов. Бисфосфонаты первого поколе¬
ния включают этидронат и хлодронат; ни один из препаратов не одобрен для лечения
остеопороза. Алендроновая кислота (от 5 до 10 мг в день или 35-70 мг в неделю) и
ризедронат^* (5 мг в день или 30-35 мг в неделю) представляют второе поколение
бисфосфонатов, которые показали в рандомизированных исследованиях повышение
минеральной плотности костной ткани у женщин в постменопаузе с остеопенией или
остеопорозом; у женщин с остеопорозом было продемонстрировано снижение коли¬
чества переломов бедра, позвонков и других переломов до 50%, особенно на первом
году лечения [459]. И алендроновая кислота, и ризедронат*“ одобрены FDA для лече¬
ния или предупреждения постменопаузального остеопороза [459].
Алендронат может быть назначен по меньшей мере на 7 лет без негативного влия¬
ния на прочность кости [459], более того, прекращение длительного курса алендроно-
вой кислоты приводило к минимальной потере костной ткани в последующие 3-5 лет.
Алендроновая кислота или ризедронат^, назначаемые однократно в неделю, показали
снижение эзофагитов, вызванных ежедневным приемом препаратов [460]. В недавнем
годичном сравнительном исследовании алендронат увеличил плотность позвонков и
бедренной кости чуть больше, чем ризендронат, причем клиническое значение этих
находок неоднозначно [459, 460]. Совсем недавно FDA одобрило к использованию
ибандронат*“ по 2,5 мг в день или 150 мг в месяц для предупреждения и лечения пост¬
менопаузального остеопороза [461], Ежедневный прием ибандроната^ показал сни¬
жение частоты переломов позвонков у женщин с остеопорозом, в том числе 1'яжелым
[461]. Другие внутривенные формы бисфосфонатов используются off label лш лечения
остеопороза. Внутривенный палмидронат^ и золедронат*“ используются для лечения
женщин, которые плохо переносят оральные формы бисфосфонатов [460].
СПИСОК ЛИТЕРАТУРЫ
1. Seeburg Р.Н., Adelman J.P. Characterization of cDNA for precursor of human luteinizing hor¬
mone releasing hormone //Nature. — 1984, — Vol. 311. — N 5987. — P. 666-668.
2. Berger S.L. et al- Genetic isolation of ADA2: a potential transcriptional adaptor required for func¬
tion of certain acidic activation domains // Cell. — 1992. — Vol. 70. — N 2. — P. 251-265.
3. Seeburg P.H. et al. The mammalian GnRH gene and its pivotal role in reproduction // Rec. Prog.
Horm. Res. - 1987. - Vol. 43. - P. 69-98.
4. Hayflick J.S. et al. The complete nucleotide sequence of the human gonadotropin-releasing hor¬
mone gene // Nucleic. Acids. Res. - 1989. — Vol. 17. — N15. — P. 6403-6404.
5. Guerrero N.V. et al. Risk factors for premature ovarian failure in females with galactosemia //
J. Pediatr. - 2000. - Vol. 137. - N 6. - P. 833-841.
6. Nikolics K. et al. A prolactin-inhibiting factor within the precursor for human gonadotropin-
releasing hormone j j Nature. — 1985. — Vol. 316. — N 6028. — P. 511-517.
7. Ackland J.F. et al. Molecular forms of gonadotropin-releasing hormone associated pep¬
tide (GAP): changes within the rat hypothalamus and release from hypothalamic cells in vitro //
NeuroendoCTinology. - 1988. — Vol. 48. - N 4. - P. 376-386.
8. Ronnekleiv O.K. et al. Combined immunohistocheraistry for gonadotropin-releasing hormone
(GnRH) and pro-GnRH, and in situ hybridization for GnRH messenger ribonucleic acid in rat brain //
Mol. Endocrinol. - 1989. - Vol. 3. - N 2. - P. 363-371.
9. Віск D. et al. Brief report: intragenic deletion of the KALIG-1 gene in Kallmann’s syndrome //
N. Engl. J. Med. - 1992. - Vol. 326. - N 26. - P. 1752-1755.
10. Knobil E. The neuroendocrine control of the menstrual cycle // Rec. Prog, Horm. Res. —
1980. - Vol. 36, - P. 53-88.
137
11. Van Vugt D.A. et al. Gonadotropin-releasing hormone pulses in third ventricular cerebro¬
spinal fluid of ovariectomized rhesus monkeys: correlation with luteinizing hormone pulses //
Endocrinology. - 1985. - Vol. 117. - N 4. - P. 1550-1558.
12. Gross K.M. et al. Evidence for decreased luteinizing hormone-releasing hormone pulse frequen¬
cy in men with selective elevations of follicle-stimulating hormone //J. Clin. Endocrinol. Metab. —
1985. - Vol. 60. - N 1. - P. 197-202.
13. Haisenleder DJ. et al. A pulsatile gonadotropin-releasing hormone stimulus is required to
increase transcription of the gonadotropin subunit genes: evidence for differential regulation of tran¬
scription by pulse frequency in vivo // Endocrinology. — 1991. — Vol. 128. — N 1. — P. 509-517.
14. Reame N.E. et al. Pulsatile gonadotropin secretion in women with hypothalamic amenorrhea:
evidence that reduced frequency of gonadotropin-releasing hormone secretion is the mechanism of
persistent anovulation //J. Clin. Endocrinol. Metab. — 1985, — Vol. 61. — N 5. - P. 851-858.
15. Filicori M. et al. Characterization of the physiological pattern of episodic gonadotropin secre¬
tion throughout the human menstrual cycle //J. Clin. Endocrinol. Metab. — 1986. — Vol. 62. —
N6.-P. 1136-1144.
16. Petersen S.L. et al. Direct and indirect regulation of gonadotropin-releasing hormone neurons
by estradiol //Biol. Reprod. — 2003. — Vol. 69. — N 6. — P. 1771-1778.
17. Herbison A.E. Noradrenergic regulation of cyclic GnRH secretion // Rev. Reprod. — 1997. -
Vol. 2. - N 1. - P. 1-6.
18. Gindoff P.R. et al. Endogenous opioid peptides modulate the effect of corticotropin-releasing
factor on gonadotropin release in the primate // Endoainology. — 1987. - Vol. 121. - N 3. -
P. 837-842.
19. Rabinovici J, et al. Endocrine effects and pharmacokinetic characteristics of a potent new
gonadotropin-releasing hormone antagonist (Ganirelix) with minimal histamine-releasing proper¬
ties: studies in postmenopausal women //J. Clin, Endoainol. Metab. — 1992. — Vol. 75. — N 5. -
P. 1220-1225.
20. Shoupe D. et al. The effects of estrogen and progestin on endogenous opioid activity in
oophorectomized women //J. Clin. Endocrinol, Metab. — 1985, - Vol. 60. - N 1. - P. 178-183.
21. Goodman R.L. et al. Endogenous opioid peptides control the amplitude and shape of gonado¬
tropin-releasing hormone pulses in the ewe // Endocrinology. — 1995. — Vol. 136. — N 6. — P. 2412-
2420.
22. Wildt L. et al. TVeatment with naltrexone in hypothalamic ovarian failure: induction of ovulation
a'nd pregnancy // Hum. Reprod. — 1993. - Vol. 8. - N 3. — P. 350 358.
23. Gottsch M.L, et al. Kisspeptin-GPR54 signaling in the neuroendocrine reproductive axis // Mol.
Cell. Endocrinol. - 2006. - Vol. 254-255. - P. 91-96.
24. Seminara S.B. et al. The GPR54 gene as a regulator of puberty // N. Engl. J, Med. - 2003. -
Vol. 349. - N 17. - P. 1614-1627.
25. de Roux N. et al, Hypogonadotropic hypogonadism due to loss of function of the KiSSl-derived
peptide receptor GPR54 // Proc. Natl. Acad. Sci U S A. - 2003, - Vol. 100, - N 19. - P. 10972-
10976.
26. Messager S. et al. Kisspeptin directly stimulates gonadotropin-releasing hormone release via G
protein-coupled receptor 54 // Proc. Natl. Acad. Sci USA. — 2005. — Vol. 102. — N 5. — P. 1761-
1766.
27. Handelsman D.J. et al. Pharmacokinetics of gonadotropin-releasing hormone and its analogs //
Endoa, Rev. - 1986. - Vol. 7. - N 1. - P. 95-105.
28. Galbiati M. et al. Transforming growth factor-beta and astrocytic conditioned medium influ¬
ence luteinizing hormone-releasing hormone gene expression in the hypothalamic cell line GTl //
Endocrinology. - 1996. - Vol. 137. - N 12. - P. 5605-5609.
29. Kartell M.J. et al. Gonadotropin-releasing hormone analog design, Structure-function studies
toward the development of agonists and antagonists: rationale and perspective // Endocr. Rev. -
1986. - Vol. 7. - N 1. - P. 44-66.
30. Lemay A. et al. Reversible hypogonadism induced by a luteinizing horm one-releasing hormone
(LH-RH) agonist (Buserelin) as a new therapeutic approach for endometriosis // Fertil. Steril. —
1984. - Vol. 41. - N 6. - P. 863-871.
31. Carr B.R. et al. Oral contraceptive pills, gonadotropin-releasing hormone agonists, or use in
combination for treatment of hirsutism; a clinical research center study //J. Clin. Endocrinol. Metab. -
1995. - Vol. 80, - N 4. - P. 1169-1178.
138
32. Caiiii C.E. et ai. Decreased spinal mineral content in amenorrheic women //JAMA. — 1984. —
Vol. 251. - N 5. - P. 626-629.
33. Malta W.H. et al. Reversible trabecular bone density loss following induced hypo-oestrogenism
with the GnRH analogue buserelin in premenopausal women j j Clin. Endocrinol (Oxf). — 1988. —
Vol. 29. - N 1. - P. 45- 51.
34. Surrey E.S. Add'back therapy and gonadotropin-releasing hormone agonists in the treatment
of patients with endometriosis: can a consensus be reached.? Add-Back Consensus Working GrouP //
Fertil. Steril. - 1999. - Vol. 71. - N 3. - P. 420-424.
35. Cetel N.S. et al. The d>Tiamics of gonadotropin inhibition in women induced by an antagonistic
analog of gonadotropin-releasing hormone // J. Clin. Endocrinol. Metab. - 198^ — Vol. 57. —
N 1. - P. 62-65.
36. Pavlou S.N. et al. Single subcutaneous doses of a luteinizing hormone-releasing hormone
antagonist suppress serum gonadotropin and testosterone levels in normal men //j. Clin. Endocrinol.
Metab. - 1986. - Vol. 63. - N 2. - P. 303-308.
37. Pavlou S.N. et al. Mode of suppression of pituitary and gonadal function after acute or pro¬
longed administration of a luteinizing hormone-releasing hormone antagonist in normal men //J. Clin.
Endocrinol. Metab. - 1989. - Vol. 68. - N 2. - P. 446-454.
38. Edelstein M.C. et al. Single dose long-term suppression of testosterone secretion by a gonad¬
otropin-releasing hormone antagonist (Antide) in male monkeys // Contraception. - 1990. -
Vol. 42. - N 2. - P. 209-214.
39. Behre H.M. et al. High loading and low maintenance doses of a gonadotropin-releasing hormone
antagonist effectively suppress serum luteinizing fiormone, follicle-stimulating hormone, and testoster¬
one in normal men //J. Clin. Endocrinol. Metab. - 1997. - Vol. 82. - N 5. - P. 1403-1408.
40. Fujimoto V.Y. et al. Dose-related suppression of serum luteinizing hormone in women by a
potent new gonadotropin-releasing hormone antagonist (Ganirelix) administered by intranasal spray //
Fertil. Steril. - 1997. - Vol. 67. - N 3. - P. 469-473.
41. Andreyko J.L. et al. Concordant suppression of serum immunoreactive luteinizing hormpne
(LH), follicle-stimulating hormone, alpha subunit, bioactive LH, and testosterone in postmenopausal
women by a potent gonadotropin releasing hormone antagonist (detirelix) // J. Clin. Endocrinol.
Metab. - 1992. - Vol. 74. - N 2. - P. 399-405.
42. Gonzalez-Barcena D. et al. Treatment of uterine leiomyomas with luteinizing hormone-releasing
hormone antagonist Cetrorelix // Hum. Reprod. — 1997. - Vol. 12. - N 9. — P. 2028-2035.
43. Childs G.V. et al. Heterogeneous luteinizing hormone and follicle-stimulating honnone storage
patterns in subtypes of gonadotropes separated by centrifugal élutriation // Endocrinology. - 1983. -
Vol. 113. - N 6. - P. 2120-2128.
44. Childs G.V. Functional ultrastructure of gonadotropes: a review // Curr. Top. Neuroend ocrinol. —
1986.-P. 49-97.
45. Cheng C.K. et al. Molecular biology of gonadotropin-releasing hormone (GnRH)-l, GnRH-II
and their receptors in humans // Endocr. Rev. — 2005. — Vol. 26. — N 2. — P. 283-306.
46. Millar R.P. et al. Gonadotropin-releasing honnone receptors // Endocr. Rev. — 2004. —
Vol. 25. - N 2. - P. 235-275.
47. Gharib S.D. et al. Molecular biology of the pituitary gonadotropins // Endocr. Rev. - 1990. -
Vol. 11. - N 1. - P. 177-199.
48. Talmadge K. et al. Evolution of the genes for the beta subunits of human chorionic gonadotropin
and luteinizing hormone // Nature. - 1984. - Vol. 307. - (5946). - P. 37-40.
49. Jameson J.L. et al. Human follicle-stimulating hormone beta-subunit gene encodes multiple
messenger ribonucleic acids // Mol. Endocrinol. — 1988. — Vol. 2. — N 9. — P. 806-815.
50. Jameson L. et al. The gene encoding the beta-subunit of rat luteinizing hormone. Analysis of
gene structure and evolution of nucleotide sequence //J. Biol. Chem. — 1984. - Vol. 259. — N 24. —
P. 15474-15480.
51. Themmen A.P.N. et al. Mutations of gonadotropins and gonadotropin receptors: elucidating the
physiology and pathophysiology of pituitary-gonadal function // Endocr. Rev. - 2000. -- Vol. 21. —
N5. - P. 551-583.
52. Shupnik M.A. Gonadotropin gene modulation by steroids and gonadotropin-releasing hor¬
mone // Biol. Reprod. - 1996. - Vol. 54. - N 2. - P. 279-286.
53. Abbud R.A. et al. Chronic hypersecretion of luteinizing hormone in transgenic mice selectively
alters responsiveness of the alpha-subunit gene to gonadotropin-releasing hormone and estrogens //
Mol. Endocrinol. - 1999. - Vol. 13. - N 9. - P. 1449-1459.
139
54. Gregory SJ. et al. Regulation of gonadotropins by inhibin and activin // Semin. Reprod.
Endocrinol. - 2004. - Vol. 22. - P. 253-267.
55. de Leeuw R. et al, Stmcture-functioii relationship of recombinant follicle stimulating hormone
(Puregon) // Mol. Hum. Reprod. - 1996. - Vol. 2. - N 5. - P. 361-369.
56. Mossniaii K.D. Comparative Morphology of the Mammalian Ovary. — Madison, WI: Wisconsin
Press, 1973.
57. Simpson J.L. et al. Ovarian differentiation and gonadal failure // Am. J. Med. Genet. - 1999. -
Vol. 89. -'N 4. - P. 186-200.
58. Di Pasquale E. et al. Hypergonadotropic ovarian failure associated with an inherited mutation of
human bone morphogenetic protein-15 (BMP15) gene // Am. J. Hum. Genet. — 2004. — Vol. 75. —
N1.-P. 106-111.
59. Witschi E. Migration of the germ cells of human embryos from the yolk sac to the primitive
gonadal folds // Carnegie Institute Contributions to Embryology No. 209. — 1948. - Vol. 32. —
P. 67-97.
60. Baker T.G. et al. The fine structure of oogonia and oocytes in human ovaries // J. Cell. Sci. —
1967. - Vol. 2. - N 2. - P. 213-224.
61. Shifren J.L. et al. Human fetal ovaries and uteri: developmental expression of genes encoding
the insulin, insulin-like growth factor I and insulin-like growth factor II receptors j j Fertil. Steril. —
1993, - Vol. 59. - N 5. - P, 1036-1040.
62. Bennett R.A. et al. Immunohistochemical localization of transforming growth factor-alpha,
epidermal growth factor (EGF) and EGF receptor in the human fetal ovary //J. Clin. Endocrinol.
Metab. - 1996. - Vol. 81. - N 8. - P. 3073-3076.
63. Himelstein-Braw R. et al. Follicular atresia in the infant human ovary //J. Reprod. Fertil. —
1976. - Vol. 46. - N 1. - P. 55-59.
64. Weakly J.N. et al. Synaptic drive and postsynaptic inhibition: a physiological and pharmacologi¬
cal investigation // Proc. West. Pharmacol. Soc. - 1966. — Vol. 9. - P. 42-44.
65. Albertini D.F. et al. The appearance and structure of intercellular connections during the ontog¬
eny of the rabbit ovarian follicle with particular reference to gap junctions // J. Cell. Biol. — 1974. —
Vol. 63. - N 1. - P. 234-250.
66. Amsterdam A, et al. Hormonal regulation of cytodifferentiation and intercellular communication
in cultured granulosa cells // Proc. Natl. Acad. Sci. USA. — 1981. - Vol. 78. — N 5. — P. 3000-3004.
67. Simon A.M. et al. Female infertility in mice lacking connexin 37 // Nature. — 1997. —
Vol. 385. -- (6616), - P. 525-529.
68. Zoller L.C. et al. Identification of cytochrome P-450, and its distribution in the membrana
granulosa of the preovulatory follicle, using quantitative cytochemistry // Endocrinology. - 1978. —
Vol. 103. - N 1. - P. 310-311.
69. Zoller L.C. et al. A quantitative cytochemical study of glucose-6-phosphate dehydrogenase and
delta 5-.3 beta-hydroxysteroid dehydrogenase activity in the membrana granulosa of the ovulable type
of follicle of the rat // Histochemistry. — 1979. — Vol. 62. ~ N 2. — P. 125-135.
70. Hillensjo T. et al. Gonadotropin releasing hormone agonists stimulate meiotic maturation of
follicle-enclosed rat oocytes in vitro // Nature. — 1980. - Vol. 287. - (5778). — P. 145-146.
71. Lawrence T.S. et al. Binding of human chorionic gonadotropin by rat cmnuii oophori and granu¬
losa cells: a comparative study j j Endocrinology. — 1980. - Vol. 106. — N 4. - P. 1114-1118.
72. Magnusson C. et al. Comparison between the progestin secretion responsiveness to gonado¬
trophins of rat cumulus and mural granulosa cells in vitro // Acta. Endoainol. (Copenh). - 1982, -
Vol. 101. - N 4. - P. 611-616.
73. Ryan K.J. et al. Steroid biosynthesis by human ovarian granulosa and thecal cells // J. Clin.
Endocrinol. Metab. - 1966. - Vol. 26. - P. 46-52.
74. Rice B. Steroid hormone formation in the human ovary, IV Ovarian stromal compartment: for¬
mation of radioactive steroids //J. Clin. Endocrinol. Metab. - 1966. - Vol. 26. — P. 593-609.
75. Erickson G.F. et al. The ovarian androgen producing cells; a review of structure/function rela¬
tionships // Endocr. Rev. - 1985. - Vol. 6. - N 3. - P. 371-399.
76. Hume D.A. et al. The mononuclear phagocyte system of the mouse defined by immunohis¬
tochemical localisation of antigen F4/80: macrophages associated with epithelia // Anat. Rec. —
1984. - Vol. 210. - N 3. - P. 503-512.
77. Nakamura Y. et al. Increased number of mast cells in the dominant follicle of the cow: relationships
among luteal, stromal, and hilar regions // Biol. Reprod. — 1987. — Vol. 37. - N 3. — P. 546-549.
140
78. Krishna A. et al. Histamine and increased ovarian blood flow mediate LH-induced superovula¬
tion in the cyclic hamster // J. Reprod. Fertil. — 1986. — Vol. 76. — N 1. — P. 23-29.
79. Cavender J.L. et al. Morphological studies of the microcirculatory system of periovulatory ovine
follicles // Biol. Reprod. - 1988. - Vol. 39. - N 4. - P. 989-997.
80. Takemura R. et al. Secretoiy products of macrophages and their physiological functions // Am.
j. Physiol. - 1984. - Vol. 246. - (1 Pt 1). - P. Cl-9.
81. Reynolds H, et al. Release of estradiol from fetal bovine serum by rat thymus, spleen, kidney,
lung and lung macrophage cultures // Endoainology. — 1982. - Vol. 110, - N 6. - P. 2213-2215.
82. Mais V. et al. The dependency of folliculogenesis and corpus luteum function on pulsatile gonad¬
otropin secretion in cycling women using a gonadotropin-releasing hormone antagonist as a probe //
J. Clin. Endocrinol. Metab. - 1986. - Vol. 62. - N 6. - P. 1250-1255.
83. Gougeon A. Regulation of ovarian follicular development in primates: facts and hypotheses //
Endocr. Rev. - 1996. - Vol. 17. - N 2. - P. 121-155.
84. Gougeon A. Dynamics of follicular growth in the human: a model from preliminary results //
Hum. Reprod. - 1986. - Vol. 1. - N 2. - P. 81-87.
85. Schipper I. et al. The follicle-stimulating hormone (FSH) threshold/window concept exam¬
ined by different interventions with exogenous FSH during the follicular phase of the normal men¬
strual cycle; duration, rather than magnitude, of FSH increase affects follicle development //J. Clin.
Endocrinol. Metab. - 1998. - Vol. 83. - N 4. - P. 1292-1298.
86. Peters H. et al. The Ovary. — London, Toronto, Sydney, New York; Granada Publishing,
1980. - P. 12-34.
87. Tilly J.L. et al. Involvement of apoptosis in ovarian follicular atresia and postovulatory regres¬
sion // Endocrinology. - 1991. - Vol. 129. - N 5. - P. 2799-2801.
88. Bauminger A. et al. Periovulatory changes in ovarian prostaglandin formation and their hor¬
monal control in the rat // Prostaglandins. — 1975. — Vol. 9. - N 5. — P. 737-751.
89. Tsafriri A. et al. Physiological role of prostaglandins in the induction of ovulation //
Prostaglandins. - 1972. - Vol. 2. - N 1, — P. 1-10.
90. Espey L.L. Ovarian proteolytic enzymes and ovulation // Biol. Reprod. — 1974. — Vol. 10. —
N 2. - P. 216-235.
91. Bjersing L. et al. Ovulation and the mechanism of follicle rupture. IV. Ultrastructure of ineni-
brana gi'anulosa of rabbit graafian follicles prior to induced ovulation // Cell. Tissue. Res. — 1974. —
Vol. 153.-Nl.-P. 1-14.
92. Beers W.H. et al. Ovarian plasminogen activator; relatiomship to ovulation and hormonal regula¬
tion // Cell. - 1975. - Vol. 6, - N 3. - P. 387-394.
93. Frederick J.L. et al. Initiation of angiogenesis by human follicular fluid // Science. - 1984. —
Vol. 224. - (4647). - P. 389-390.
94. Kamat B.R. et al. Expression of vascular permeability factor/vascular endothelial growth factor
by human granulosa and theca lutein cells. Role in corpus luteum development // Am. J. Pathol. —
1995. - Vol. 146. - N 1. - P. 157-165.
95. Ohara A. et al. Functional differentiation in steroidogenesis of two types of luteal cells iso¬
lated from mature human corpora lutea of menstrual cycle // J. Clin. Endocrinol. Metab. - 1987. - -
Vol. 65. - N 6. - P. 1192-1200.
96. Carr B.R. et al. The role of lipoproteins in the regulation of progesterone seaetion by the human
corpus luteum // Fertil. Steril. - 1982. - Vol. 38. - P. 303-311.
97. Duncan W.C. et al. Luteinizing hormone receptor in the human corpus luteum: lack of down-
regulation during maternal recognition of pregnancy // Hum. Reprod. - 1996. - Vol. 11. - N 10. -
P. 2291-2297.
98. Strauss 3rd J.F. et al. Providing progesterone for pregnancy: control of cholesterol flux to the
side-chain cleavage system //J. Reprod. Fertil. Suppl. — 2000. — Vol. 55. — P. 3-12.
99. Bukovsky A. et al. Is irregular regression of corpora lutea in climacteric women caused by age-
induced alterations in the “tissue control system?” // Am. J. Reprod. Immunol. — 1996. Vol. 36,
N 6. - P. 327-341.
100. Casper R.F. et al. Induction of luteolysis in the human with a long-acting analog of luteinizing
hormone-releasing factor // Science. — 1979. — Vol. 205. — N 4404. — P. 408-410.
101. Schoonmaker et al A receptive period for estradiol-induced luteolysis in the rhesus monkey //
Endocrinology. - 1981. - Vol. 108. - N 5. - P. 1874-1877.
102. O’Gray J. et al. Inhibition of progesterone synthesis in vitro by prostaglandin F2a //]. Reprod.
Fertil. - 1972. - Vol. 30. - N 1. - P. 153-156.
141
103. Shikone T. et al. Apoptosis of human corpora lutea during cyclic luteal regression and early
pregnancy //J, Clin, Endocrinol. Metab. - 1996. - Vol. 81. - N 6. - P. 2376-2380.
104. Bortolussi M. et al. Autoradiographic study of the distribution of LH(HCG) receptors in the
ovarv of untreated and gonadotrophin-primed immature rats // Cell. Tissue. Res. — 1977. — Vol. 183. -
N 3.'- P. 329-342.
105. Nimrod A. et al. A specific FSH receptor in rat granulosa cells: properties of binding in vitro //
Endocrinology. - 1976. - Vol. 98. - N 1. - P. 56-64.
106. Nimrod A. et al. Appearance of LH-receptors and LH-stimulable cyclic AMP accumulation in
granulosa cells during follicular maturation in the rat ovary // Biochem. Biophys. Res. Commun. -
1977. - Vol. 78. - N 3. - P. 977-984.
107. Jaaskelainen K. et al. Internalization of receptor-bound human chorionic gonadotrophin in
preovulatory rat granulosa cells in vivo jf Acta. Endocrinol (Copenh). — 1983. — Vol. 103. — N 3. —
P. 406-412.
108. Uilenbroek J.T. et al. Ovarian follicular development during the rat estrous cycle; gonadotropin
receptors and follicular responsiveness // Biol. Reprod. — 1979. — Vol. 20. — N 5. — P. 1159-1165.
109. Zeleznik A.J. et al. Granulosa cell maturation in the rat: increased binding of human chori¬
onic gonadotropin following treatment with follicle-stimulating hormone in vivo j j Endocrinology. —
1974. - Vol. 95. - N 3, - P. 818-825.
110. Richards J.S. et al. Ovarian follicular development in the rat: hormone receptor regulation by
estradiol, follicle stimulating hormone and luteinizing hormone//Endocrinology. — 1976. - Vol. 99. -
N 6. - P. 1562-1570.
111. Couse J.F. et al. Estrogen receptor null mice: what have we learned and where will they lead us.? //
Endocr. Rev. - 1999. - Vol. 20. - N 3. - P. 358-417.
112. Brandenberger A.W. et al. Estrogen receptor alpha (ER-alpha) and beta (ER-beta) mRNAs
in normal ovary, ovarian serous cystadenocarcinoma and ovarian cancer cell lines: down-regulation
of ER-beta in neoplastic tissues //J. Clin. Endocrinol .Vietab. — 1998. - Vol. 83. - N 3. - P. 1025-
1028.
113. Dorrington J.H. et al. Estradiol-ITbeta biosynthesis in cultured granulosa cells from hypophy-
sectomized immature rats; stimulation by follicle-stimulating hormone // Endocrinology. — 1975. —
Vol. 97. - N 5. - P. 1328-1331.
114. Simpson E.R. et al. Aromatase cytochrome P450, the enzyme responsible for estrogen biosyn¬
thesis // Endocr. Rev. - 1994. - Vol. 15. N 3. - P. 342-355.
115. Moon Y.S. et al. Stimulatory action of follicle-stimulating honnone on estradiol-i? beta
secretion by hvpophysectomized rat ovaries in organ culture // Endocrinology. — 1975. - Vol. 97. —
N 1. - P. 244-247.
116. Armstrong D.T. et al. Stimulation of aromatization of exogenous and endogenous androgens
ill ovaries of hypophysectomized rats in vivo by follicle-stimulating hormone // Endocrinolog}^ -
1976. - Vol. 99. - N 4. - P. 1144-1151.
117. Rani C.S- et al. Follicle-stimulating hormone induction of luteinizing hormone receptor
in cultured rat granulosa cells: an examination of the need for steroids in the induction process //
Endocrmolog}^ - 1981. - Vol. 108. - N 4, - P. 1379-1385.
118. Bulun S.E. Aromatase deficiency and estrogen resistance: from molecular genetics to clinic //
Seminars in Reproductive Medicine. - 2000. - Vol. 18. - N 1. - P. 31-39.
119. Richards J.S. et al. Evidence that changes in tonic luteinizing hormone secretion determine the
growth of preovulatory follicles in the rat // Endocrinology. — 1980. — Vol. 107. — N 3. — P. 641-
648.
120. Richards J.S. et al. Effects of human chorionic gonadotropin and progesterone on follicular
development in the immature rat // Endocrinolog>'. - 1982. - Vol. 111. - N 5. - P. 1429-1438.
121. Bogovich K. et al. Androgen biosynthesis in developing ovarian follicles: evidence that luteiniz¬
ing hormone regulates thecal 17 alpha-hydroxylase and C17-20-lyase activities // Endocrinology. —
1982. - Vol. 111. - N 4. - P. 1201-1208.
122. Barlow JJ. et al. Estradiol production after ovariectomy for carcinoma of the breast // N. Engl.
J. Med. - 1969. - Vol. 280. - N 12. - P. 633-637.
123. Baird D. et al. Concentration of estrone and estradiol-17beta in follicular fluid and ovarian
venous blood of women // Clin. Endocrinol. (Oxf). - 1969. - Vol. 4. - N 3. - P. 259-266.
124. Hcmsell D.L. et al. Plasma precursors of estrogen. II. Correlation of the extent of conversion
of plasma androstenedione to estrone with age // J, Clin. Endocrin. Metab. — 1974, — Vol. 38. —
P. 476-479.
142
125. Bulun S.E. et al. Competitive reverse transcription-polymerase chain reaction analysis indi¬
cates that levels of aromatase cytochrome P450 transcripts in adipose tissue of buttocks, thighs, and
abdomen of women increase with advancing age //J. Clin. Endocrinol. Metab. — 1994. - Vol. 78. -
N 2. - P. 428-432.
126. Bulun S.E. et al. Aromatase in aging women // Semin. Reprod. Endocrinol. - 1999. — Vol.
17.-N 4.-P. 349-358.
127. Mahendroo M.S. et al. Male and female isoenzymes of steroid .5alpha-reductase //
Rev. Reprod. - 1999. - Vol. 4. - N 3. - P. 179-183.
128. Hinshelwood M.M. et al. Expression of LRH-1 and SF-1 in the mouse ovary: localization in
different cell types correlates with differing function // Mol. Cell. Endocrinol. - 2003. - Vo). 207. -
N 1-2. - P. 39-45,
129. Falender A.E. et al. Differential expression of steroidogenic factor-1 and FTF/LRH-1 in the
rodent ovary // Endocrinology. - 2003. - Vol. 144. - N 8. - P. 3598-3610.
130. Peng N. et al. The role of the orphan nuclear receptor, liver receptor homologue-1, in the
regulation of human corpus luteum 3beta-hydroxysteroid dehydrogenase type II //J. Clin. Endocrinol.
Metab. - 2003. - Vol. 88, - N 12. - P. 6020-6028.
131. Simpson E.R. et al. Aromatase cytochrome P450, the enzyme responsible for estrogen biosyn¬
thesis // Endocr. Rev. - 1994. - Vol. 15. - P. 342-355.
132. Siiteri P.K. et al. Role of extraglandular estrogen in human endocrinology // Creep R.O.,
Astwood E.B. (ed.). Handbook of Physiology. — Vol 2. — Washington, DC: American Physiological
Society, 1973. - P. 619-629.
13^ Peltoketo H. et al. 17[i-Hydroxysteroid dehydrogenase (rcH)/17-ketosteroid reductase (KSR)
family; nomenclature and main characteristics of the 17HSD/KSR enzymes // J. Mol, Endocr. -
1999. - Vol. 23. - P. 1-11.
134. Merriam G.R. et al. Catechol estrogens and the control of gonadotropin and prolactin secretion
in man //J. Steroid. Biochem, - 1983. — Vol. 19. - (IB). - P. 619-625.
135. Miller W.L. et al. Molecular pathology and mechanism of action of the steroidogenic acute
regulatory protein, StAR //J. Steroid. Biochem. Mol. Biol. - 1999. - Vol. 69. - P. 131-141.
136. Pollack S.E. et al. Localization of the steroidogenic acute regulatory protein in human tis¬
sues //J. Clin. Endocrinol. Metab. - 1997. - Vol. 82. - N 12. - P. 4243-4251.
137. Ying S.Y. Iiihibins, activins, and follistatins: gonadal proteins modulating the secretion of
follicle-stimulating hormone // Endocr. Rev. — 1988. - Vol. 9. — N 2. - P. 267-293.
138. Leers-Sucheta S. et al. Synergistic activation of the human type II 3beta-hydroxysteroid dehy-
drogenase/delta5-delta4 isomerase promoter by the transcription factor steroidogenic factor-1/adrenal
4-binding protein and phorbol ester //J. Biol. Chem. — 1997. - Vol. 272. - N 12. - P. 7960-7967.
139. Hanley N.A. et al. Steroidogenic factor 1 (SF-1) is essential for ovarian development and func¬
tion // Mol. Cell. Endocrinol. - 2000. - Vol. 163. - N 1-2. - P. 27-32.
140. Week J. et al. Switching of NR5A proteins associated with the inhibin (alpha)-subunit gene
promoter following activation of the gene in granulosa cells // Mol. Endocrinol. — 2006. - Vol. 20. —
N 5. - P. 1090-1103.
141. Jaatinen T.A. et al. Expression of inhibin alpha, beta A and beta B messenger ribonucleic acids
in the normal human ovary and in polycystic ovarian syndrome // J. Endocrinol. — 1994. — Vol. 143. —
Nl.-P. 127-137.
142. Roberts V.J. et al. Expression of inhibin/activin subunits and follistatin messenger ribonucleic
acids and proteins in ovarian follicles and the corpus luteum during the human menstrual cycle //
J. Clin. Endocrinol. Metab. - 1993. - Vol. 77. - N 5. - P. 1402-1410.
143. Eramaa M. et al. Regulation of inhibin alpha- and beta A-subunit messenger ribonucleic add
levels by chorionic gonadotropin and recombinant follicle-stimulating hormone in cultured human
granulosa-luteal cells //J. Clin. Endocrinol. Metab. — 1994. — Vol. 79. — N 6. — P. 1670-1677.
144. Groome N.P. et al. Measurement of dimeric inhibin B throughout the human menstrual
cycle // J. Clin. Endocrinol. Metab, — 1996. — Vol. 81. — N 4. — P. 1401-1405.
145. Adashi E.Y. The IGF family and folliculogenesis //J. Reprod. Immunol. — 1998. — VoL 39. —
N 1-2. - P. 13-19.
146. Dunaif A. Insulin resistance and the polycy'Stic ovary syndrome: mechanism and implications
for pathogenesis // Endocr. Rev. - 1997. - Vol. 18. - N 6. — P. 774-800.
147. Sasano H. et al. Aromatase and 17beta-hydroxysteroid dehydrogenase type 1 in human breast
carcinoma //J. Clin. Endoainol, Metab. — 1996. - Vol. 81. - P. 4042-4046.
143
148. Zeitoun K.M. et al. Deficient ITbeta-hydroxysteroid dehydrogenase type 2 expression in
endometriosis: failure to metabolize estradiol-17alpha // J. Clin. Endocrinol. Metab. — 1998. —
Vol 83. - P. 4474-4480.
149. Bulun S.E. et al. Expression of dioxin related trans-activating factors and target genes in human
endometrium and endometriosis // Am. J. Obstet. Gynecol. — 2000. — Vol. 182. — P. 767-77,5.
150. Bulun S.E. et al. Endocrine disorders associated with inappropriately high aromatase expres¬
sion //J. Steroid. Biochem. Mol. Biol. - 1997. - Vol. 61. - P. 133-1.39.
151. Azziz R. et al. Idiopathic hirsutism // Endocr. Rev. - 2000. - Vol. 21, - N 4. - P. 347-362.
152. Psychoyos A, Uterine receptivity for nidation // Ann. N. Y. Acad. Sci. — 1986. — Vol. 476. —
P. 36-42.
153. Wynn R.M, Histology and ultrastructure of the human endometrium // Wynn R.M. (ed.).
Biology of the Uterus. — New York: Plenum Press, 1977. — P. 341-376.
154. Markee J.E. Menstruation in intraocular endometrial transplants in the rhesus monkey //
Carnegie Institute Coutributions to Embryology, — 1940. — Vol, 28. — P. 219-308.
155. Markee J. Morphological basis for menstrual bleeding: relation of regression to the initiation
of bleeding// Bull. N. Y. Acad. Med. - 1948. - Vol. 24. - P. 253-268.
156. Bartelmez G.W, The phases of the menstrual cycle and their interpretation in terms of the
pregnancy cycle // Am. J. Obstet. Gynecol. - 1957. - Vol. 74. - P. 931-955.
157. Eckert R. et al. Human endometrial cells in primary tissue culture: modulation of the progester¬
one receptor level by natural and synthetic estrogens in vitro //J. Clin, Endocrinol. Metab, — 1981. —
Vol. 52. - P. 699-708.
158. Cooke P.S. et al. Stromal estrogen receptors mediate mitogenic effects of estradiol on uterine
epithelium // PNAS. - 1997. - Vol. 94. - P. 6535-6540.
159. Kurita T. et al. Stromal progesterone receptors mediate the inhibitory effects of progesterone on
estrogen-induced uterine epithelial cell deoxyribonucleic acid synthesis // Endocrinology, — 1998. —
Vol. 139. - P. 4708-4713.
160. Yang S. et al. Stromal progesterone receptors mediate induction of 17beta-hydroxysteroid
dehydrogenase type 2 expression in human endometrial epithelium: a paracrine mechanism for inacti¬
vation of estradiol II Mol. Endocrinol. — 2001. ~ Vol. 15, — P. 2093-2105.
161. Cooke P. et al. Mechanism of estrogen action: lessons from the estrogen receptor-alpha knock¬
out mouse // Biol- Reprod. - 1998. — Vol. 59, — P. 470-475.
162. Matsuzaki S. Oestrogen receptor alpha and beta mRNA expression in human endometrium
throughout the menstrual cycle // Mol. Hum. Reprod. — 1999. — Vol. 5. — P. 556-564.
163. Katzenellenbogen B. Estrogen receptors: Woactivities and interactions with cell signaling path¬
ways // Biol. Reprod. - 1996. - Vol. 54. - P. 287-293.
164. Attia G.R. et al. Progesterone receptor isoform A but not B is expressed in endometriosis //
J. Clin. Endocrinol, Metab. - 2000, - Vol. 85. - P. 2897-2902.
165. Tseng L. et al. Effects of progestins on estradiol receptor levels in human endometrium //
J. Clin. Endocrinol. Metab. - 1975. - Vol. 41. - P. 402-404.
166. Casey M.L. et al. 17p-Hydroxysteroid dehydrogenase type 2: chromosomal assignment
and progestin regulation of gene expression in human endometrium // J, Clin. Invest. — 1994. —
Vol. 94. - P. 2135-2141.
167. Tseng L. et al. Stimulation of acylsulfotransferase activity by progestin in human endometrium
in vitro //J. Clin. Endocrinol. Metab. - 1981. - Vol. 53. - P. 418-421.
168. Bergh P.A. et al. The impact of embryonic development and endometrial maturity on the timing
of implantation // Fertil. Steril. - 1992. - Vol. 58. - N 3. - P. 537-542.
169. Adams E.C. et al. A description of 34 human ova within the first 17 days of development // Am.
J. Anat. - 1956. - Vol. 98. - N 3. - P. 435-493.
170. Sauer M.V. et al. Reversing the natural decline in human fertility. An extended clinical trial of
oocyte donation to women of advanced reproductive age //JAMA. — 1992. — Vol. 268. — N 10. —
P. 1275-1279.
171. Rosenwaks Z. Donor eggs: their application in modem reproductive technologies // Fertil,
Steril. - 1987. - Vol. 47. - N 6. - P. 895-909.
172. Langer R.D, et al. Transvaginal ultrasonography compared with endometrial biopsy for the
detection of endometrial disease. Postmenopausal Estrogen/Progestin Interventions Trial // N. Engl.
J. Med. - 1997. - Vol. 337. - N 25. - P. 1792-1798.
173. Sheehan H.L. The recognition of chronic hypopituitarism resulting from postpartum pituitary
necrosis // Am. J. Obstet. Gynecol. - 1971. - Vol.'111. - N 6. - P. 852-8.54.
144
174. Simpson J.L. et al Gonadal dysgenesis in individuals with apparently normal chromosomal
complements: tabulation of cases and compilation of genetic data // Birth. Defects, Orig. Artie. Ser. -
1971. - Vol. 7. - N 6. - P. 215-228.
175. Hague W.M. et al. 45 X Turner’s syndrome in association with polycystic ovaries. Case
report //Br.J. Obstet. Gynaecol. - 1989. - Vol. 96. - N 5. - P. 613-618.
176. Division of Reproductive Sciences, O.R.P.R.C., Beaverton 97006. Progesterone-dependent
expression of keratinocyte growth factor mRNA in stromal cells of the primate endometrium: kératino¬
cyte growth factor as a progestomedin //J, Cell. Biol. - 1994, - Vol. 125. — N 2. — P. 393-401.
177. Asherman J.G. Amenorrhea traumatica (atretica) //J. Obstet. Gynecol. Br. Emp. — 1948. —
Vol. 55. - P. 23-27.
178. Marshall W. et al. Variations in patterns of pubertal changes in girls // Arch, Dis. Child. -
1969. - Vol. 44. - P. 291-303.
179. Klinefelter H.J. et al. Experience with a quantitative test for normal or decreased amounts of
follicle-stimulating hormone in urine in endocrinological diagnosis // J. Clin. Endocrinol. Metab.
1943. - Vol. 3. - P. 529-544.
180. Khoury S.A. et al. Diurnal patterns of pulsatile luteinizing hormone secretion in hypothalamic
amenorrhea: reproducibility and responses to opiate blockade and an alpha 2-adrenergic agonist //
J. Clin. Endocrinol. Metab. - 1987. - Vol. 64. - N 4. - P. 755-762.
181. Berga S.L. et al. Neuroendocrine aberrations in women with functional hypothalamic amenor¬
rhea //J. Clin. Endocrinol. Metab. - 1989. - Vol. 68. - N 2. - P. 301-308.
182. Yen S.S. et al. Hypothalamic amenorrhea and hypogonadotropinism: responses to synthetic
LRF//J. Clin. Endocrinol. Metab. - 1973. - Vol. 36. - N 5. - P. 811-816.
183. Loucks A.B. et al. Alterations in the hypothalamic-pituitary-ovarian and the hypothalamic-
pituitary-adrenal axes in athletic women //J. Clin. Endocrinol. Metab. ~ 1989. - Vol. 68. - N 2. -
P. 402-411.
184. Wilson R.C. et al. Central electrophysiologic correlates of pulsatile luteinizing hormone secre-^^
tion in the rhesus monkey // Neuroendocrinology. — 1984. — Vol. 39. — N 3. — P. 256-260.
185. Kalra S.P. Catecholamine involvement in preovulatory LH release: reassessment of the role of
epinephrine // Neuroendocrinology. — 1985. — Vol. 40. — N 2. — P. 139-144,
186. Quigley M.E. et al. Evidence for increased dopaminergic inhibition of secretion of thyroid-
stimulating hormone in hyperprolactinemic patients with pituitary microadenoma // Am. J. Obstet,
Gynecol. - 1980. - Vol. 137. - N 6. -- P. 653-655.
187. Ropert J.F. et al. Endogenous opiates modulate pulsatile luteinizing hormone release in
humans //J. Clin. Endocrinol. Metab. - 1981. - Vol, 52. - N 3. - P. 583-585.
188. Gambacciani M. et al. GnRH release from the mediobasal hypothalamus: in vitro inhibition by
corticotropin-releasing factor // Neuroendocrinology. — 1986. — Vol. 43. - N 4. — P. 533-536.
189. Geiiazzani A.D. et al. Naltrexone treatment restores menstrual cycles in patients with weight
loss-related amenorrhea//Fertil. Steril. — 1995. ~ Vol. 64. — N 5. — P. 951-956.
190. Selye H. The stress syndrome // Nature. - 1936. - Vol. 138. — P. 32.
191. Suh B.Y. et al. Hypercortisolism in patients with functional hypothalamic amenorrhea //
J. Clin. Endocrinol. Metab. - 1988. - Vol. 66. - N 4. - P. 733-739.
192. Xiao E. et al. Acute inhibition of gonadotropin seaetioii by corticotropin-releasing hormone
in the primate: are the adrenal glands involved? // Endoainology. — 1989. - Vol. 124. - N 4. -
P. 1632-1637.
193. Rivier C. et al. Influence of corticotropin-releasing factor on reproduaive functions in the rat //
Endocrinology. - 1984. - Vol. 114. - N 3. - P. 914-921.
] 94. Matteri R.L. et al. Adrenocorticotropiii-induced changes in ovine pituitary gonadotropin secre¬
tion in vitro Ii Endocrinology. - 1986. - Vol. 118. — N 5. - P. 2091-2096.
195. Kamel F. et al. Modulation of gonadotropin seaetion by corticosterone: interaction with gonad¬
al steroids and mechanism of action // Endocrinology. - 1987. - Vol. 121. - N 2. - P. 561-568.
196. Welt C.K. et al. Recombinant human leptin in women with hypothalamic amenorrhea //
N. Engl. J. Med. - 2004. - Vol. 351. - N 10. - P. 987-997.
197. Baranowska B. et al. The role of endogenous opiates in the mechanism of inhibited luteiniz¬
ing hormone (LH) secretion in women with anorexia nervosa: the effect of naloxone on LH, follicle-
stimulating hormone, prolactin and beta-endorphin secretion //J. Clin. Endocrinol. Metab. - 1984, —
Vol. 59. - N 3. - P. 412-416.
198. Atkinson R.L. Naloxone decreases food intake in obese humans // J. Clin. Endocrinol.
Metab. - 1982. - Vol. 55. - N 1. - P. 196-198,
145
199. Motley J.E. et al. Stress-induced eating is mediated through endogenous opiates // Science. —
1980. - Vol. 209. - (4462). - P. 1259-1261.
200. Rigotti N.A. et al. The clinical course of osteoporosis in anorexia nervosa. A longitudinal study
of cortical bone mass //JAMA. - 1991. - Vol. 265. - N 9. - P. 113.3-1138.
201. Frisch R.E. et al. Delayed menarche and amenorrhea in ballet dancers // N. Engl. J. Med. —
1980. - Vol. 303. - N 1. - P. 17-19.
202. Frisch R.E. et al. Delayed menarche and amenorrhea of college athletes in relation to age of
onset of training //JAMA. — 1981. — Vol. 246. — N 14. — P. 1559-1563.
203. Drink water B.L. et al. Bone mineral content of amenorrheic and eumenorrheic athletes //
N. Engl. J. Med. - 1984. - Vol, 311. - N 5. - P. 277-281.
204. Willi J. et al. Epidemiology of anorexia nervosa in a defined region of Switzerland //
Am. J. Psychiatry. — 1983. - Vol. 140. - N 5. - P. 564-567.
205. Schwartz D,M. et al. Do anorectics get well? Current research and future needs //
Am. J. Psychiatry. - 1981. - Vol. 138. - N 3. - P. 319-32.3.
206. Swift W.J. The long-term outcome of early onset anorexia nervosa. A critical review //
J. Am. Acad. Child. Psychiatry. - 1982. - Vol. 21. - N 1, - P. 38-46.
207. Bovar R.M. et al. Cortisol secretion and metabolism in anorexia nervosa // N. Engl. J. Med. -
1977. - Vol. 296. - N 4. - P. 190-193.
208. Gold P.W. et al. Abnormal hypothalamic-pituitary-adrenal function in anorexia nervosa.
Pathophysiologic mechanisms in underweight and weight-corrected patients // N. Engl. J. Med. -
1986. - Vol. 314. - N 21. - P. 1335-1342.
209. Kaye W.H. et al. Elevated cerebrospinal fluid levels of immunoreactive corticotropin-releasing
hormone in anorexia nervosa: relation to state of nutrition, adrenal function, and intensity of depres¬
sion//J. Clin. Endocrinol. Metab. - 1987. - Vol. 64. - N 2. — P. 203-208.
210. Moshang TJ. Low triiodothyronine euth>Toidism in anorexia nervosa j j Vigersky R.S. (ed.).
Anorexia Nervosa. — New York: Raven Press, 1977. — P. 263-270.
211. Gold P. et al. Abnormalities in plasma and cerebrospinal fluid arginine vasopressin in patients
with anorexia nervosa j j N. Engl.J. Med. — 1983. — Vol. 308. — P. 1117-1123.
212. Soyka L.A. et al. The effects of anorexia nervosa on bone metabolism in female adolescents j j
J. Clin. Endocrinol. Metab. - 1999. - Vol. 84. - N 12. - P. 4489-4496.
213. Liu J.H. et al. The use of gonadotropin-releasing hormone for the induction of ovulation //
Clin. Obstet. Gynecol. - 1984. - Vol. 27. - N 4. - P. 975-982.
'214. Hatch R. et al. Hirsutism; implications, etiology, and management // Am. J. Obstet. Gynecol. —
1981. - Vol. 140. - N 7. - P. 815-830.
215. Tagatz G.E. et al. The clitoral index: a bioassay of androgenic stimulation // Obstet. Gynecol. —
1979. - Vol. 54. - N 5. - P. 562-564,
216. Bardin C.W. et al. Testosterone and androstenedione blood prodciction rates in normal women
and women with idiopathic hirsutism or polycystic ovaries // J, Clin. Invest. — 1967. — Vol. 46. —
N 5. - P. 891-902.
217. Youngblood G.L. et al. Isolation and characterization of the mouse P45017a-hydroxylase/
cl7-20 lyase gene (CYP17): traiiscTiptional regulation of the gene by cyclic adenosine 3',5’-mono-
phosphate in M A-10 Leydig cells j j Mol. Endocrinol. — 1992. — Vol. 6. — P. 927-934.
218. Mowszowicz 1. et al. Androgen binding capacity and 5 alpha-reductase activity in pubic skin
fibroblasts from hirsute patients //J. Clin. Endocrinol. Metab. - 1983. - Vol. 56. - N 6. - P. 1209-
1213.
219. Schwartz U. et al. The diagnostic value of plasma free testosterone in non-tmnorous and tumor¬
ous hyperandrogenism // Fertil. Steril. — 1983. — Vol. 40. — N 1. — P. 66-72.
220. Miller K.K, et al. Measurement of free testosterone in normal women and women with andro¬
gen deficiency: comparison of methods //J. Clin. Endocrinol. Metab. - 2004. — Vol.. 89. — N 2, -
P. 525-533.
221. Carmina E. et al. Extensive clinical experience: relative prevalence of different androgen excess
disorders in 950 women referred because of clinical hyperandrogenism // J. Clin. Endocrinol. Metab. —
2006. - Vol. 91. - N 1. - P. 2-6.
222. Paulson R.J. et al. Measurements of 3 alpha,17 beta-androstanediol glucuronide in serum and
urine and the correlation with skin 5 alpha-reductase activity // Fertil. Steril. — 1986. — Vol. 46. —
N 2. - P. 222-226.
146
223. Barbieri R.L. et al. Presence of 17 beta-hydroxysteroid dehydrogenase type 3 messenger ribo¬
nucleic acid transcript in an ovarian Sertoli-Leydig cell tumor // Fertil. Steril. - 3997. — Vol. 68. —
N 3, - P. 534-537.
224. Kennedy L. et al. Short terra administration of gonadotropin-releasing hormone analog to a
patient with a testosterone-secreting ovarian tumor //J. Clin. Endocrinol. Metab. — 1987. - Vol. 64. -
N 6. - P. 1320-1322.
225. Derksen J. et al. Identification of virilizing adrenal tumors in hirsute women j j N. Engl.
J. Med. - 1994. - Vol. 331. - N 15. - P. 968-973.
226. Friedman C.I. et al. Serum testosterone concentrations in the evaluation of androgen-produc¬
ing tumors // Am. J. Obstet. G}mecol. — 3985. — Vol. 1.53. — N 1. — P. 44-49.
227. Taylor L. et al. Diagnostic considerations in virilization: iodomethyl-iiorcholesterol scanning in
the localization of androgen secreting tumors // Fertil. Steril. — 1986. — Vol. 46. — N 6. — P. 1005-
1010.
228. Azziz R. et al. Screening for 21-hydroxylase-deficient nonclassic adrenal hyperplasia among
hyperandrogénie women: a prospective study // Fertil. Steril. — 1999. — Vol. 72. - N 5. — P. 915-
925.
229. Speiser P.W. et al. High frequency of nonclassical steroid 21-hydroxylase deficienq^ // Am.
J. Hum. Genet. - 1985. - Vol. 37. - N 4. - P. 650-667.
230. New M.L et al. Genotyping steroid 21-hydroxylase deficiency: hormonal reference data //
J. Clin. Endocrinol. Metab. - 1983. - Vol. 57. - N 2. - P. 320-326.
231. Stratakis C.A. et al. Glucocorticosteroid resistance in humans. Elucidation of the molecular
mechanisms and implications for pathophysiology' // Ann. N. Y. Acad, Sci. — 1994. — Vol. 746. -
P. 362-374. discussion 374-376.
232. Lobo R.A. et al. The effects of two doses of spironolactone on serum androgens and anageii hair
in hirsute women // Fertil. Steril. - 1985. - Vol. 43. - N 2. - P. 200-205.
233. Evron S. et al. Induction of ovulation with spironolactone (Aldactone) in anovulatory oligom-
eiiorrbeic and hyperandrogénie women // Fertil. Steril. — 1981. — Vol. 36. — N 4. — P. 468-471.
234. Mowszowicz I. et al. Androgen metabolism in hirsute patients treated with cyproterone
acetate //J. Steroid. Biochem. - 1984. — Vol. 20. — N 3. - P. 757-761.
235. Rittmaster R.S. Finasteride // N. Engl.J. Med. - 1994. - Vol. 330. - N 2. - P. 120-125.
236. Wong I.L, et al. A prospective randomized trial comparing finasteride to spironolactone in the
treatment of hirsute women //J. Clin. Endocrinol. Metab. — 1995. — Vol. 80. — N 1. — P. 233-238.
237. Castello R. et al. Outcome of long-term treatment with the 5 alpha-reductase inhibitor finas¬
teride in idiopathic hirsutism: clinical and hormonal effects during a 1 -year course of therapy and
l-yearfollow-uP // Fertil. Steril. - 1996. - Vol. 66. - N 5. - P. 734-740.
238. Diamanti-kandarakis E. et al. The effect of a pure antiandrogen receptor blocker, flutamide, on
the lipid profile in the polvcy'stic ovary symdrome //J. Clin, Endocrinol. Metab. - 1998. - Vol. 83. -
N 8. - P. 2699-2705.
239. Moghetti P. et al. Flutamide in the treatment of hirsutism: long-term clinical effects, endocrine
changes, and androgen receptor behavior j j Fertil. Steril, — 1995. — Vol. 64. - N 3, - P. 511-517.
240. Wysowski D.K. et al. Fatal and nonfatal hepatotoxicity associated with flutamide // Ann.
Intern. Med. - 1993. - Vol. 118. - N 11. - P. 860-864.
241. Legro R.S. et al. Prevalence and predictors of risk for type 2 diabetes mellitus and impaired glu¬
cose tolerance in polycystic ovary syndrome: a prospective, controlled study in 254 affected women //
J. Clin. Endocrinol. Metab. - 1999, - Vol. 84. -Nl.-P. 165-169.
242. Urbanek M. et al. Thirty-seven candidate genes for polycystic ovary syndrome: strongest
evidence for linkage is with follistatin // Proc. Natl. Acad. Sci. USA. — 1999. — Vol. 96. — N 15. -
P. 8573-8.578.
243. Legro R.S. et al. Evidence for a genetic basis for hyperandrogenemia in polycystic ovary syn¬
drome // Proc. Natl. Acad. Sci. USA. - 1998. - Vol. 95. - N 25. - P. 14956-14960.
244. Stein I. et al. Amenorrhea associated with bilateral polycystic ovaries // Am. J. Obstet.
Gynecol. - 1935. - Vol. 29. - P. 181,
245. Burghen G.A, et a). Correlation of hyperandrogenism with hyperinsulinism in polycystic ovar¬
ian disease //J. Clin. Endocrinol. Metab. — 1980. — Vol. 50. — N 1. — P. 113-116.
246. Tamura M. et al. Estrogen up-regulates cyclooxygenase-2 via estrogen receptor in human uter¬
ine microvascular endothelial cells // Fertil. Steril, — 2004. — Vol. 81, - - P. 1351-1356.
147
247. Azziz R. Controversy in clinical endocrinology: diagnosis of polycystic ovarian syndrome:
the Rotterdam criteria are premature // J. Clin. Endocrinol, Metab. — 2006. — Vol. 91. - N 3. -
P. 781-785.
248. Stratakis C.A. et al. Glucocorticosteroid resistance in humans. Elucidation of the molecular
mechanisms and implications for pathophysiology // Ann. N. Y. Acad. Sci. — 1994. — Vol. 746. —
P. 362-374.
249. Z'awadzki J.K. et al. Diagnostic criteria for polycystic ovary syndrome: towards a rational
approach // Dunaif A. et al. (ed,). Polycystic Ovary Syndrome. — Boston, MA: Blackwell, 1992. —
P. 377-384.
250. Revised 2003 consensus on diagnostic criteria and long-term health risks related to polycystic
ovary syndrome // Fertil. Steril, - 2004. - Vol. 81. - N 1. - P. 19-25.
251. Dunaif A. Insulin resistance in polycystic ovarian syndrome // Ann. N. Y. Acad. Sci. — 1993. —
Vol. 687. - P. 60-64.
252. Hoffman D. et al. The prevalence and significance of elevated dehydroepiandrosterone sulfate
levels in anovulatory women // Fertil. Steril. - 1980. — Vol. 42. — P. 853-861.
253. Franks S. Polycystic ovary syndrome // N. Engl. J. Med. - 1995. - Vol, 333. - N 13. -
P. 853-861.
254. Kletzky O.A. et a). Clinical categorization of patients with secondary amenorrhea using pro¬
gesterone-induced uterine bleeding and measurement of serum gonadotropin levels j j Am. J. Obstet.
Gynecol. - 1975. - Vol. 121. - N 5. - P. 695-703.
255. Venturoli S. et al. Episodic pulsatile secretion of FSH, LH, prolactin, oestradiol, oestrone, and
LH circadian variations in polycystic ovary syndrome // Clin. Endocrinol. (OxL). — 1988. - Vol, 28. —
N 1. - P. 93-107.
256. Imse V. et al. Comparison of luteinizing honnone pulsatility in the serum of women suffering
from polycystic ovarian disease using a bioassay and five different immunoassays //J. Clin. Endocrinol.
Metab, - 1992. - Vol. 74. - N 5. - P. 1053-1061.
257. Hayes F.J. et al. Use of a gonadotropin-releasing hormone antagonist as a physiologic probe
in polycystic ovary syndrome: assessment of neuroendocrine and androgen dynamics // J. Clin.
Endocrinol. Metab. - 1998. - Vol. 83, - N 7. - P. 2343-2349.
258. Barnes R.B. et al. Central opioid activity in polycystic ovary syndrome with and without dop¬
aminergic modulation //J. Clin. Endocrinol. Metab. — 1985. — Vol. 61. — N4. — P. 779-782.
259. Morales A.J. et al. Insulin, somatotropic, and luteinizing hormone axes in lean and obese
women with polycystic ovary syndrome: common and distinct features //J. Clin. Endocrinol. Metab. —
1996. - Vol. 81. - N 8, - P. 2854-2864.
260. Taylor A.E. et al. Determinants of abnormal gonadotropin seaetion in clinically defined
women with polycystic ovary syndrome //J. Clin. Endoainol. Metab. — 1997. — Vol. 82. — N 7. -
P. 2248-2256.
261. Nagamani M. et al. Hyperinsulinemia in hyperthecosis of the ovaries // Am. J. Obstet.
Gynecol. - 1986. - Vol. 154. - N 2, - P. 384-389.
262. Dorn C. et al. Insulin enhances the transaiption of luteinizing hormone-beta gene // Am.
J. Obstet. Gynecol. - 2004. - Vol. 191. - N 1. - P. 132-137.
263. Adashi E.Y. et al. Insulin enhancement of luteinizing hormone and follicle-stimulating hormone
release by cultured pituitary cells //Endocrinology. - 1981. - Vol. 108. - N 4. — P. 1441-1449.
264. Mehta R.V. et al. Luteinizing hormone secretion is not influenced by insulin infusion in women
with polycystic ovary syndrome despite improved insulin sensitivity during pioglitazone treatment //
J. Clin. Endocrinol. Metab. - 2005. - Vol. 90. - N 4. - P. 2136-2141.
265. Chang RJ. Ovarian steroid secretion in polycystic ovarian disease // Semin. Reprod,
Endocrinol. - 1984. - Vol. 2. - P. 244.
266. Wajchenberg B.L, et al. The source(s) of estrogen production in hirsute women with polycystic
ovarian disease as determined by simultaneous adrenal and ovarian venous catheterization // Fertil.
Steril. - 1988. - Vol. 49. - N 1. - P. 56-61.
267. Bulun S. et al. Aromatase in aging women j j Seminars in Reproductive Endocrinology. —
1999. - Vol. J7. - P. 349-358.
268. Reed M.J. et al. Regulation of estradiol 17beta-hydroxysteroid dehydrogenase in breast tissues;
the role of growth factors //J. Steroid. Biochem. Mol. Biol. - 1991. — Vol. 39. — P. 791-798.
269. Bulun S.E. et al. Regulation of aromatase expression in estrogen-responsive breast and uterine
disease: from bench to treatment // Pharmacol. Rev. - 2005. - Vol, 57. - N 3. - P. 359-383.
148
270. Nestler J.E. Obesity, insulin, sex steroids and ovulation //Int.J. Obes. Relat. Metab. Disord. —
2000. - Vol. 24. - Suppl. 2. - P. 71-73.
271. Fauser B.C. Observations in favor of normal early follicle development and disturbed dominant
follicle selection in polycystic ovary syndrome // Gynecol. Endoainol. — 1994, — Vol. 8, - N 2. —
P. 75-82.
272. Mason H.D. et al. Insulin-like growth factor-I (IGF-I) inhibits production of IGF-binding
protein-1 while stimulating estradiol secretion in granulosa cells from normal and polycystic human
ovaries //J. Clin. Endocrinol. Metab. - 1993. - Vol. 76. - N 5. - P. 1275-1279.
273. Homburg R. et al. Low-dose FSH therapy for anovulatory infertility associated with polycystic
ovary syndrome: rationale, results, reflections and refinements // Hum. Reprod. Update. — 1999. —
Vol. 5. - N 5. - P. 493-499.
274. Judd H.L. et al. Familial hyperthecosis: comparison of eiidocrinologic and histologic findings
with polycystic ovarian disease // Am. J. Obstet. Gynecol. — 1973. — Vol. 117, — N 7, — P. 976-982,
275. Cooper H.E. et al. Hereditary factoi-s in the Stein-Leventhal syndrome // Am. J. Obstet.
Gynecol. - 1968. - Vol. 100. - N 3. - P. 371-387.
276. Ferriman D. et al. The inheritance of polycystic ovarian disease and a possible relationship to
premature balding // Clin. Endocrinol. (Oxf.). — 1979. — Vol. 11. — N 3. — P. 291-300.
277. Carey A.H. et al. Evidence for a single gene effect causing polycystic ovaries and male pattern
baldness // Clin. Endocrinol. (Oxf.). - 1993. - Vol. 38. - N 6. - P. 653-658.
278. Govind A. et al. Polycystic ovaries are inherited as an autosomal dominant trait: analysis
of 29 polycystic ovary syndrome and 10 control families // J. Clin. Endocrinol, Metab, — 1999. —
Vol. 84. - N1. - P. 38-43.
279. Norman R.J. et al. Hyperinsulinemia is common in family members of women with polycystic
ovary syndrome // Fertil. Steril. — 1996. — Vol. 66. — N 6. — P. 942-947.
280. Urbanek M. et al. Candidate gene region for polycystic ovary syndrome on chromosome
19pl3.2 //J. Clin. Endocrinol. Metab. - 2005. - Vol. 90. - N 12. - P. 6623-6629.
281. Ho C.K. et al. Increased transcription and inaeased messenger ribonucleic add (mRNA) stabil¬
ity contribute to inaeased GATA6 mRNA abundance in polycystic ovary syndrome theca cells //J. Clin.
Endocrinol. Metab. - 2005. - Vol. 90. - N 12. - P. 6596-6602.
282. Osei K. et al. Ethnic differences in secretion, sensitivity, and hepatic extraction of insulin in
black and white Americans // Diabet. Med. — 1994. — Vol. 11. — N 8. — P. 755-762.
283. Dunaif A. et al. Acanthosis nigricans, insulin action, and hyperandrogenism: clinical, histological,
and biochemical findings //J. Clin. Endoainol. Metab. — 1991. — Vol. 73. — N 3. — P. 590-595.
284. Reaven G.M, et al. Hypertension and associated metabolic abnormalities — the role of insu¬
lin resistance and the sympathoadrenal svstem // N. Engl. J. Med. — 1996. - Vol. 334. - N 6. -
P. 374-381.
285. Mather K.J. et al. Hyperinsulinemia in polycystic ovary syndrome correlates with increased
cardiovascular risk independent of obesity // Fertil. Steril. — 2000. — Vol. 73. - N 1. — P. 150-156.
286. Talbott E. et al. Coronary heart disease risk factors in women with polycystic ovary syn¬
drome // Arteriosder. Thromb. Vase. Biol. - 1995. - Vol. 15. - N 7. - P. 821-826.
287. Ehrmann D.A. et al. Prevalence and predictors of the metabolic syndrome in women with poly¬
cystic ovary syndrome //J. Clin. Endocrinol. Metab. — 2006. — Vol. 91. — N 1. — P. 48-53.
288. Dunaif A. et al. Beta-cell dysfunction independent of obesity and glucose intolerance in the
polycystic ovary syndrome //J. Clin. Endocrinol. Metab. — 1996. — Vol. 81. — N 3. — P. 942-947.
289. Reddy S.S. et al. Epidermal growth factor receptor deferts in leprechaunism. A multiple growth
factor-resistant syndrome //J. Clin. Invest. — 1989. — Vol. 84. — N 5. — P. 1569-1576.
290. Kirschner M.A. et al. Androgen-estrogen metabolism in women with upper body versus lower
body obesity//J. Clin. Endocrinol. Metab. - 1990. - Vol. 70. - N 2. - P. 473-479.
291. Schapira D.V. et al. Abdominal obesity and breast cancer risk // Ann. Intern. Med. - 1990, -
Vol. 112. - N 3. - P. 182-186.
292. Kumar N.B. et al. Android obesity at diagnosis and breast carcinoma survival; evaluation of
the effects of anthropometric variables at diagnosis, including body composition and bod^ fat distribu¬
tion and weight gain during life span, and survival from breast carcinoma // Cancer. — 2000. — Vol.
88. - N 12. - P. 2751-2757.
293. Anderson K.E. et al. Association of Stein-Leventhal syndrome with the incidence of postmeno¬
pausal breast carcinoma in a large prospective study of women in Iowa // Cancer. — 1997. — Vol. 79. —
N 3. - P. 494-499.
149
294. Campbell P.J. et al. Impact of obesity on insulin action in volunteers with normal glucose toler¬
ance: demonstration of a threshold for the adverse effect of obesity //J, Clin. Endocrinol. Metab. —
1990. - Vol. 70.-N 4. - P. 1114-1118.
295. Ehrmann D.A. Polycystic ovary'syndrome j j N. Engl.J. Med, 2005. — Vol. 352. — N 12. —
P. 1223-1236.
296. Velazquez E.M. et al. Metformin therapy in polycystic ovary syndrome reduces hyperinsu¬
linemia, insulin resistance, hyperandrogenemia, and systolic blood pressure, while facilitating normal
menses-and pregnancy //Metabolism. - 1994. — Vol. 43. — N 5. — P. 647-654.
297. Nestler J.E. et al. Decreases in ovarian cytochrome P450cl7 alpha activity and serum free
testosterone after reduction of insulin secretion in polycystic ovary syndrome // N. Engl, J. Med. —
1996. - Vol. 335. - N 9. - P. 617-623.
298. Velazquez E. et al. Menstrual cyclicity after metformin therapy in polycystic ovary syndrome //
Obstet. Gynecol. - 1997. - Vol. 90. - N 3. - P. 392-395.
299. Lord J.M. et al. Insulin-sensitising drugs (metfonniii, troglitazone, rosiglitazone, pioglita¬
zone. D-chiro-inositol) for polycystic ovary syndrome // Cochrane Database Syst. Rev. — 2003. -
CD003053.
300. Crave J.C. et al. Effects of diet and metformin administration on sex hormone-binding globulin,
androgens, and insulin in hirsute and obese women //J. Clin. Endocrinol. Metab. — 1995. - Vol. 80. —
N 7. - P. 2057-2062.
301. Dunaif A. et al. The insulin-sensitizing agent troglitazone improves metabolic and reproductive
abnormalities in the polycystic ovary' syndrome //J. Clin. Endocrinol. Metab. - 1996. - Vol. 81. -
N 9. - P. 3299-3306.
302. Ehrmann D.A. et al. Troglitazone improves defects in insulin action, insulin secretion, ovar¬
ian steroidogenesis, and fibrinolysis in women with polycystic ovary' syndrome //J. Clin. Endocrinol.
Metab. - 1997. - Vol. 82. - N 7. - P. 2108-2116.
303. Azziz R. et al. Troglitazone improves ovulation and hirsutism in the polycystic ovary syndrome:
a multicenter, double blind, placebo-controlled trial //J. Clin. Endocrinol, Metab. — 2001. - Vol. 86. -
N4.-P. 1626-1632.
304. Ghazeeri G. et al. Effect of rosiglitazone on spontaneous and clomiphene citrate-induced
ovulation in women with polycystic ovary s}'ndrome j j Fertil. Steril. — 2003. — Vol. 79. — N 3. —
P. 562-566.
305. Belli S.H. et al. Effect of rosiglitazone on insulin resistance, growth factors, and reproductive
disturbances in women with polycystic ovary S3mdronie // Fertil. Steril. — 2004. — Vol. 81. — N 3. —
P. 624-629.
306. Romualdi D. et al. Selective effects of pioglitazone on insulin and androgen abnormalities in
normo- and hyperinsulinaeraic obese patients with polycystic ovary syndrome // Hum. Reprod. —
2003. - Vol. 18. - N 6. - P. 1210-1218.
307. Regan L. et al. Hypersecretion of luteinising hormone, infertility, and miscarriage // Lancet. -
1990. - Vol. 336. - (8724). - P. 1141-1144,
308. Gitsch G. et al. Endometrial cancer in premenopausal women 45 years and j'ounger // Obstet.
Gynecol. - 1995. - Vol. 85. - N 4. - P. 504-508.
309. Godsland I.F. et al. Update on the metabolic effects of steroidal contraceptives and their rela¬
tionship to cardiovascular disease risk j j Am. J. Obstet. Gynecol. — 1994. — Vol. 170. — (5 Pt 2). —
P. 1528-1536.
310. Duffy T.J. et al. Oral contraceptive use: prospective follow-up of women with suspected glucose
intolerance // Contraception. - 1984. — Vol. 30. — N 3. — P. 197-208,
311. Hannaford P.C. et al. Oral contraceptives and diabetes mellitus // BMJ. — 1989. - Vol. 299. -
(6711). - P. 1315-1316.
312. Escobar-Morreale H.F. et al. Treatment of hirsutism with ethinyl estradiol-desogestrel contra¬
ceptive pills has beneficial effects on the lipid profile and improves insulin sensitivity // Fertil. Steril. -
2000. - Vol. 74. - N 4. - P. 816-819,
313. Colditz G.A, Oral contraceptive use and mortality during 12 years of follow-up: the Nurses’
Health Study //Ann. Intern. Med. - 1994. - Vol. 120. - N 10. - P. 821-826.
314. Kjos S.L. et al. Contraception and the risk of type 2 diabetes mellitus in Latina women with
prior gestational diabetes mellitus //JAMA. — 1998, — Vol. 280. — N 6. - P. 533-538.
315. Garg S.K. et al. Oral contraceptives and renal and retinal complications in young women with
insulin-dependent diabetes mellitus //JAMA. - 1994. — Vol. 271. - N 14. — P. 1099-1102.
150
316. Petersen K.R. et al. Effects of contraceptive steroids on cardiovascular risk factors in women
with insulin-dependent diabetes mellitus // Am. J. Obstet. Gynecol. — 1994. — Vol. 171. — N 2. - -
P. 400-405.
317. Diamanti-Kandarakis E. et al. Insulin sensitivity and antiandrogenic therapy in women with
polycystic ovary syndrome // Metabolism. — 1995. — Vol. 44. — N 4. — P. 525-531.
318. Lanzone A. et al. Preconceptional and gestational evaluation of insulin secretion in patients
with polyq'stic ovary syndrome // Hum. Reprod. - 1996. - Vol. 11. - N 11. - P. 2382-2386.
319. Wild R.A. et al. Lipid and apolipoprotein abnormalities in hirsute women. I. The association
with insulin resistance // Am. J. Obstet. Gynecol. — 1992. — Vol. 166. — N 4. — P. 1191-1196. discus¬
sion 1196-1197.
320. Kiddÿ D.S. et al. Improvement in endocrine and ovarian function during dietary treatment
of obese women with polycystic ovary s>'udrome // Clin. Endocrinol. (Oxf.). - 1992. — Vol. 36. —
N 1.-P. 105-111.
321. Guzick D.S. et al. Endocrine consequences of weight loss in obese, hyperandrogénie, anovula-
toiy women // Fertil. Steril. — 1994. — Vol. 61. — N 4. — P. 598-604.
322. Clark A.M. et al. Weight loss results in significant improvement in pregnancy and ovulation
rates in anovulatory obese women // Hum. Reprod. - 1995. - Vol. 10. - N 10. - P. 2705-2712.
323. Nestler J.E. et al. Effects of metformin on spontaneous and clomiphene-induced ovulation in
the polycystic ovary syndrome // N. Engl. J. Med. - 1998. — Vol. 338. - N 26. — P. 1876-1880.
324. Dahlgren E. et al. Women with polycystic ovary syndrome wedge resected in 1956 to 1965: a
long-term follow-up focusing on natural history and circulating hormones // Fertil. Steril. - 1992. -
Vol. 57, - N 3. - P. 505-513.
325. Adashi E.Y. Clomiphene citrate-initiated ovulation: a clinical update // Semin. Reprod.
Endocrinol. - 1986. - Vol. 4. - P. 225-276.
326. Mitwally M,F. et al. Use of an aromatase inhibitor for induction of ovulation in patients with an
inadequate response to clomiphene citrate j j Fertil. Steril. - 2001. — Vol. 75. — N 2. — P. 305-309.
327. Al-Oraari W.R. et al. Comparison of two aromatase inhibitors in women with cloiniphene-resis¬
tant polycystic ovary syndrome // Int.J. Gynaecol. Obstet. — 2004. — Vol. 85. — N 3. — P. 289-291.
328. Casper R.F. et al. Review: aromatase inhibitors for ovulation induction //J. Clin. Endocrinol.
Metab. - 2006. - Vol. 91. - N 3. - P. 760-771.
329. Hamilton-Fair ley D. et al. Common problems in induction of ovulation j j Baillieres. Clin.
Obstet. Gynaecol. - 1990. - Vol. 4. - N 3. - P. 609-625.
330. Marci R. et al. A low-dosc stimulation protocol using highly purified follicle-stimulating
hormone can lead to high pregnancy rates in in vitro fertilization patients with polycystic ovaries who
are at risk of a high ovarian response to gonadotropins // Fertil. Steril. — 2001. — Vol. 75. — N 6. -
P. 1131-1135.
331. Taylor A.E. et al. A randomized, controlled trial of estradiol replacement therapy in women
with hypergonadotropic amenorrhea //J. Clin. Endocrinol. Metab. — 1996. — Vol. 81. — N 10. —
P. 3615-3621.
332. Conway G.S. Premature ovarian failure // Br. Med. Bull. — 2000. — Vol. 56. — N 3. —
P. 643-649.
333. Dewald G. et al. Sex chromo.some anomalies associated with premature gonadal failure //
Semin. Reprod. Endocrinol. - 1983. — Vol. 1. - P. 79.
334. Devi A.S, et al. 45,X/46,XX mosaicism in patients with idiopathic premature ovarian failure //
Fertil. Steril. - 1998. - Vol, 70. - N 1. - P. 89-93.
335. Myhre A.G. et al. Autoimmune polyendocrine syndrome type 1 (APS I) in Norway // Clin.
Endocrinol. (Oxf.). - 2001. - Vol. 54. - N 2. - P. 211-217.
336. Wallace W.H. et al. Ovarian failure following abdominal irradiation in childhood: natural his¬
tory and prognosis // Clin. Oncol (R. Coll. Radiol). - 1989. - Vol. 1. — N 2. — P. 75-79.
337. Morice P. et al. Fertility results after ovarian transposition for pelvic malignancies treated by
external irradiation or brachj'therapy // Hum, Reprod. — 1998. — Vol. 13. — N 3. — P. 660-663.
338. Bines J. et al. Ovarian function in premenopausal women treated with adjuvant chemotherapy
for breast cancer //J. Clin. Oncol. - 1996. - Vol. 14. - N 5. - P. 1718-1729.
339. Byrne J. et al. Effects of treatment on fertility in long-term survivors of childhood or adolescent
cancer// N. Engl.J. Med. - 1987. - Vol. 317. - N 21. - P. 1315-1321.
340. Aittomaki K. The genetics of XX gonadal dysgenesis // Am. J. Hum. Genet. - 1994. -
Vol. 54. - N 5. - P. 844-851.
151
341. Rebar R.W. et al. Clinical features of young women with hypergonadotropic amenorrhea //
Fertil. Steril. - 1990. - Vol. 53, - N 5. - P. 804-810.
342. Nelson L.M. et al. Development of luteinized graafian follicles in patients with karyotypically
normal spontaneous premature ovarian failure // J. Clin, Endocrinol. Metab. — 1994. — Vol. 79. —
N5.-P. 1470-1475.
343. Giltay J.C. et al. Short stature as the only presenting feature in a patient with an isodicentric (Y)
(qll.23) and gonadoblastoma. A clinical and molecular cytogenetic study // Eur. J. Pediatr. - 2001. —
Vol. 160. - N 3.' - P. 154-158.
344. Manuel M. et al. The age of occurrence of gonadal tumors in intersex patients with a Y chromo¬
some // Am. J. Obstet. Gynecol. - 1976. - Vol. 124. — N 3. — P. 293-300.
345. Gravholt C.H. et al. Occurrence of gonadoblastoma in females with Turner syndrome and Y
chromosome material: a population study //J. Clin. Endocrinol. Metab, - 2000. — Vol. 85. - N 9. -
P. 3199-3202.
346. Nelson L.M. Autoimmune ovarian failure: comparing the mouse model and the human disea¬
se //J. Soc. Gynecol. Invest. — 2001. — Vol. 8. — (1 Suppl. Proceedings). — P. S55-S57.
347. Wheatcroft NJ. et al. Identification of ovarian antibodies by immunofluorescence, enzyme-
linked immunosorbent assay or immunoblotting in premature ovarian failure // Hum. Reprod, —
1997. - Vol. 12. - N 12. - P. 2617-2622.
348. Markee J. Morphological basis for menstrual bleeding: relation of regression to the initiation
of bleeding // Bull. N. Y. Acad, Med. - 1948. - Vol. 36. - P. 153.
349. Belsey E.M. et al. Menstrual bleeding patterns in untreated women. Task Force on Long-Acting
Systemic Agents for Fertility Regulation // Contraception. — 1997. — Vol. 55. — N 2. — P. 57-65.
350. Chiazze Jr. L, et al. The length and variability of the human menstrual cycle // JAMA. —
1968. - Vol. 203. - N 6. - P. 377-380.
351. Munster K. et al. Length and variation in the menstrual cycle—a cross-sectional study from a
Danish county // Br.J. Obstet. Gynaecol. - 1992. — Vol. 99. — N 5. — P. 422-429.
352. Treloar A.E, et al. Variation of the human menstrual cycle through reproductive life // Int.
J. Fertil. - 1967. - Vol. 12. - (1 Pt 2). - P. 77-126.
353. Rybo G. Menstrual blood )o.ss in relation to parity and menstrual pattern // Acta Obstet.
Gynecol. Scand. — 1966. — Vol. 45. — suppl. 7. - P. 25-45.
354. Haynes P.J. et al. Measurement of menstrual blood loss in patients complaining of menorrha¬
gia // Br.J, Obstet. Gynaecol. - 1977. - Vol. 84. - N 10. - P. 763-768.
355. Higham J.M, et al. Assessment of menstrual blood loss using a pictorial chart // Br. J. Obstet.
Gynaecol. - 1990. - Vol. 97. - N 8. - P. 734-739.
356. Fraser I.S. et al. A preliminary study of factors influencing perception of menstrual blood loss
volume //Am. J. Obstet. Gynecol, - 1984. — Vol. 149. - N 7. - P. 788-793.
357. de Ziegler D. et al. Effects of luteal estradiol on the secretory transformation of human
endometrium and plasma gonadotropins j j J. Clin. Endocrinol. Metab. — 1992. — Vol. 74. — N 2. —
P. 322-331.
358. Belsey E.M. Vaginal bleeding patterns among women using one natural and eight hormonal
methods of contraception // Contraception. - 1988. - Vol. 38. - N 2. - P. 181-206.
359. Wilansky D.L. et al. Early hypothyroidism in patients with menorrhagia // Am. J. Obstet.
Gynecol. - 1989. - Vol. 160. - N 3. - P. 673-677.
360. Hopkins M.P. et al. Ginseng face cream and unexplained vaginal bleeding // Am. J. Obstet,
Gynecol. - 1988. - Vol. 159. - N 5. - P. 1121-1122.
361. Claessens E.A. et al. Acute adolescent menorrhagia // Am. J. Obstet. Gynecol. — 1981. —
Vol. 139. - N 3. - P. 277-280.
362. Smith Y.R. et al. Menorrhagia in adolescents requiring hospitalization //J. Pediatr. Adolesc.
Gynecol. - 1998. - Vol. 11. - N 1. - P. 13-15.
363. van Eijkeren M.A. et al. Measured menstrual blood loss in women with a bleeding disorder or
using oral anticoagulant therapy // Am. J. Obstet, Gynecol. - 1990. — Vol. 162. — N 5. — P. 1261-
1263.
364. Edlund M. et al. On the value of menorrhagia as a predictor for coagulation disorders // Am.
J. Hematol. - 1996. - Vol. 53. - N 4. - P. 234-238.
365. Nilsson L. et al. Treatment of menorrhagia // Am. J, Obstet. Gynecol. — 1971, — Vol. 110. —
N .5. - P. 713-720.
152
366. Kirkland JX. et al. Progesterone inhibits the estrogen-induced expression of c-fos messenger
ribonucleic acid in the uterus // Endocrinology. — 1992. - Vol. 130. - N 6. - P. 3223-3230.
367. DeVore G.R. et al. Use of intravenous Premarin in the treatment of dysfunctional uterine
bleeding—a double-blind randomized control study // Obstet. Gynecol. — 1982. — Vol. 59. - N 3. —
P. 285-291.
368. Livio M. et al. Conjugated estrogens for the management of bleeding associated with renal
failure //N. Engl.J. Med. - 1986. - Vol. 315. - N 12. - P. 731-735.
369. Olive D. et al. Endometriosis // N. Engl.J. Med. - 1993, - Vol. 328. — P. 1759-1769.
370. Vercellini P. et al. Progestins for symptomatic endometriosis: a critical analysis of the evi¬
dence // Fertil. Steril. - 1997. - Vol. 68. - N 3. - P. 393-401.
371. Collaborative Group on Hormonal Factors in Breast Cancer; Breast cancer and hormone
replacement therapy: collaborative reanalysis of data from 51 epidemiological studies of 52,705 women
with breast cancer and 108,411 women without breast cancer // Lancet. - 1997. - Vol. 350. -
P, 1047-1059,
372. Waller K.G. et al. Gonadotropin-releasing hormone analogues for the treatment of endometri¬
osis: long-term foilow-uP j j Fertil. Steril. - 1993. - Vol. 59. - N 3. - P. 511-515.
373. Shaw R.W. An open randomized comparative study of the effect of goserelin depot and danazol
in the treatment of endometriosis. Zoladex Endometriosis Study Team // Fertil. Steril. — 1992. —
Vol. 58. - N 2. - P. 265-272,
374. Menopause. A decision tree for the use of estrogen replacement therapy or hormone replace¬
ment therapy in postmenopausal w'omen: consensus opinion of the North American Menopause
Society. - 2000. - Vol. 7. - P. 76-86.
375. Vercellini P. et al. Progestins for symptomatic endometriosis; a critical analysis of the evi¬
dence // Fertil. Steril. - 1997. - Vol. 68. - P. 393-401.
376. Takayaina K. et al. Treatment of severe postmenopausal endometriosis with an aromatase
inhibitor // Fertil. Steril, - 1998. - Vol. 69. - N 4. - P. 709-713.
377. Vercellini P. et al. A gonadotropin-releasing hormone agonist versus a low-dose oral contra¬
ceptive for pelvic pain associated with endometriosis // Fertil. Steril. — 1993. — Vol. 60, — N 1. —
P. 75-79.
378. Waller K.G. et al. Gonadotropin-releasing hormone analogues for the treatment of endometri¬
osis; long-term follow-uP // Fertil. Steril. - 1993. - Vol. 59. — N 3. - P. 511-515.
379. Takayaina K. et al. Treatment of severe postmenopausal endometriosis with an aromatase
inhibitor // Fertil. Steril. - 1998. - Vol. 69. - P. 709-713.
380. Amsterdam L. et al. Treatment of endometriosis-related pelvic pain with a combination of
an aromatase inhibitor (aiiastrozole) plus a combination oral contraceptive: a novel approach //
Proceedings of the 85th Annual Endocrine Society Meeting. — 2003. — Vol. 1. — P. 360.
381. Ailawadi R. et al. Treatment of endometriosis and chronic pelvic pain with letrozole and nore-
thindrone acetate: a pilot study // Fertil. Steril. - 2004. — Vol. 81. - N 2. - P. 290-296.
382. Soysal S. et al. The effects of post-surgical administration of goserelin plus anastrozole com¬
pared to goserelin alone in patients with severe endometriosis; a prospective randomized trial // Hum.
Reprod. - 2004. - Vol. 19. - P. 160-167.
383. Amsterdam L. et al. Anastrozole and oral contraceptives: a novel treatment for endometriosis //
Fertil. Steril. - 2005. - Vol. 84. - N 2. - P. 300-304.
384. Stewart E.A. Uterine fibroids // Lancet. — 2001. — Vol. 357. — (9252). - P. 293-298.
385. Maruo T. et al. Effects of progesterone on uterine leiomyoma growlh and apoptosis //
Steroids. - 2000. - Vol. 65. - N 10-11. - P. 585-592.
386. Judd H.L. et al. Endocrine function of the postmenopausal ovary: concentration of androgens
and estrogens in ovarian and peripheral vein blood //J. Clin. Endocrinol. Metab. - 1974. — Vol. 39. -
N 6. - P. 1020-1024.
387. McKinlay S.M. et al. The normal menopause transition // Maturitas. — 1992. — Vol. 14, —
N2.-P. 103-115.
388. den Tonkelaar I. et al, Menstrual cycle length preceding menopause in relation to age at meno¬
pause // Maturitas. - 1998. - Vol. 29. - N 2. - P. 115-123.
389. Buckler H.M. et al. Gonadotropin, steroid, and inhibin levels in women with incipient ovarian
failure during anovulatory and ovulatory rebound cycles j j J. Clin. Endocrinol. Metab. — 1991. —
Vol. 72. - N 1. - P. 116-124.
153
390. Santoro N. et a). Characterization of reproductive hormonal dynamics in the perimenopause //
J. Clin. Endocrinol. Metab. - 1996. - Vol. 81. - N 4. - P. 1495-1501.
391. McKinlay S.M. et al. Smoking and age at menopause in women // Ann. Intern. Med. —
1985. - Vol. 103. - N 3. - P. 350-356.
392. Torgerson D.J. et al. Factors associated with onset of menopause in women aged 45-49 //
Maturitas. - 1994. - Vol. 19. - N 2. - P. 83-92.
393. Torgerson DJ. et al. Alcohol consumption and age of maternal menopause are associated with
menopause onset j j Maturitas. - 1997. - Vol. 26. — N I. - P. 21-25.
394. Cramer D.W. et al. Family history as a predictor of early menopause // Fertil. Steril. - 1995. -
Vol. 64. - N 4. - P. 740-745.
395. Gosden et al. Follicular status at the menopause // Hum. Reprod. — 1987. - Vol. 2. —
N 7. - P. 617-621.
396. Jiroutek M.R. et al. Changes in reproductive hormones and sex hormone-binding globulin
in a group of postmenopausal women measured over 10 years // Menopause. — 1998. - Vol. 5. —
N 2. - P. 90-94.
397. Adashi E.Y. The climacteric ovary as a functional gonadotropin-driven androgen-producing
gland // Fertil. Steril. - 1994. - Vol. 62. - N 1. - P. 20-27.
398. Grodin J.M. et al. Source of estrogen production in postmenopausal women // J. Clin,
Endocrinol. Metab. - J973. - Vol. 36. - P. 207-214,
399. Labrie F. et al. Marked decline in serum concentrations of adrenal C19 sex steroid precursors
and conjugated androgen metabolites during aging //J. Clin. Endocrinol. Metab. — 1997. - Vol. 82. —
N 8, - P. 2396- 2402.
400. Judd H.L. et al. Origin of serum estradiol in postmenopausal women // Obstet. Gynecol. -
1982. - Vol. 59. - P. 680-686.
401. Bulun S.E. et al. Endocrine disorders associated with inappropriately high aromatase expres¬
sion //J. Steroid. Biochem. Mol, Biol. - 1997. - Vol. 61. - N 3-6. - P. 133-139.
402. Bonneterre J. et al. Anastrozole is superior to tamoxifen as first-line therapy in hormone recep¬
tor positive advanced breast carcinoma // Cancer. - 2001. - Vol. 92. - N 9. - P. 2247-2258.
403. Kroman N. et al. Time since childbirth and prognosis in primary breast cancer: population
based study // BMJ. - 1998. - Vol. 315. - P. 851-8.55.
404. Feldman S. et al. Two-year follow-up of 263 patients with post/periinenopausal vaginal bleed¬
ing and negative initial biopsy // Gynecol. Oncol. — 1994. — Vol. 55. — Nl.-P. 56-59.
405. Oldenhave A, et al. Impact of climacteric on well-being. A sutv'ey based on 5213 women 39 to
60 years old //Am. J. Obstet. Gynecol. - 1993. - Vol. 168. - (3 Pt 1). - P. 772-780.
406. Kronnenberg F. et al. Modulation of menopausal hot flashes bv ambient temperature //
J. Therm. Biol. - 1992. - Vol. 17. - P. 43.
407. Yen S.S. The biology of menopause //J. Reprod. Med. - 1977. - Vol. 18. - N 6. - P. 287-296.
408. Erlik Y. et al. Estrogen levels in postmenopausal women with hot flashes // Obstet. Gvnecol. —
1982. - Vol. 59. - N 4. - P. 403-407.
409. Davidson B.J. et al. Free estradiol in postmenopausal women with and without endometrial
cancer //J. Clin. Endocrinol. Metab. - 1981. - Vol. 52. - N 3. - P. 404-408.
410. Raz R. et al. A controlled trial of intravaginal estriol in postmenopausal women with recurrent
urinary tract infections // N. Engl.J. Med. - 1993. - Vol. 329. - N 11. - P. 753-756,
411. Dempster D.W. et al. Pathogenesis of osteoporosis // Lancet. - 1993. - Vol. 341, - N
8848. - P. 797-801.
412. Richelson L.S. et al. Relative contributions of aging and estrogen deficiency' to postmenopausal
bone loss // N. Engl. J. Med. - 1984. - Vol. 311. - N 20. - P. 1273-1275.
413. Nilas L. et al. Bone mass and its relationship to age and the menopause //J. Clin. Endocrinol,
Metab. - 1987. - Vol. 65. - N 4, - P. 697-702.
414. Christiansen C. Hormone replacement therapy and osteoporosis // Maturitas, — 1996. —
Vol. 23. - supP. 1. - P. S71-S76.
415. Rossouw J.E, et al. Risks and benefits of estrogen plus progestin in healthy postmenopausal
women: principal results from the Women’s Health Initiative randomized controlled trial //JAMA. —
2002. - Vol. 288. - N 3. - P. 321-333.
416. Anderson G.L. et al. Effects of conjugated equine estrogen in postmenopausal women with
hysterectomy; the Women’s Health Initiative randomized controlled trial // JAMA. — 2004. -
Vol. 291. - N 14. - P. 1701-1712.
154
417. Brett K.M. et al. Use of postmenopausal hormone replacement therapy: estimates from a nation¬
ally representative cohort study //Am. J. Epidemiol. — 1997. — Vol. 145. — N 6. - P. 536-545.
418. Ness J. et al. Use of hormone replacement therapy by postmenopausal women after publication
of the Women's Health Initiative Trial // J. Gerontol. A. Biol. Sci. Med. Sci. - 2005. - Vol. 60.
N 4. - P. 460-462.
419. Hoffmann M. et al. Changes in women’s attitudes towards and use of hormone therapy after
HERS and WHI // Maturitas. - 2005. - Vol. 52. - N 1. - P. 11-17.
420. Thunell L. et al. Scientific evidence changes prescribing practice — a comparison of the man¬
agement of the climacteric and use of hormone replacement therapy among Swedish gynaecologists in
1996 and 2003 // Br.J. Obstet. Gyenaecol. - 2006. - Vol. 113. - N 1, - P. 15-20.
421. Nicholson W.K. et al. Hormone replacement therapy for African American women: missed
opportunities for effective intervention j I Menopause. — 1999. — Vol. 6. — N 2. — P. 147-155.
422. Bergkvi.st L. et al. The risk of breast cancer after estrogen and e.strogen-progestin replace¬
ment // N. Engl.J. Med. - 1989. - Vol. .321. - P. 293-297.
423. Hulley S. et al. Randomized trial of estrogen plus progestin for secondary prevention of
coronary heart disease in postmenopausal women. Heart and Estrogen/progestin Replacement Study
(HERS) Research GrouP //JAMA. - 1998. - Vol. 280. - N 7. - P. 605-613.
424. Shumaker S.A. et al. Estrogen plus progestin and the incidence of dementia and mild cognitive
impairment in postmenopausal women: the Women’s Health Initiative Memory' Study: a randomized
controlled trial //JAMA. - 2003. - Vol. 289. - N 20. - P. 2651-2662.
425. Lacey Jr. J.V. et al. Menopausal hormone replacement therapy and risk of ovarian cancer //
JAMA. - 2002. - Vol. 288. - N 3. - P. 334-341.
426. Shumaker S.A. et al. Conjugated equine estrogens and incidence of probable dementia and
mild cognitive impainiient in postmenopausal women: Women’s Health Initiative Memory Study //
JAMA. - 2004. - Vol. 291. - N 24. - P. 2947-2958.
427. Glueck C.J. el al. Severe hypertriglyceridemia and pancreatitis when estrogen replacement
therapy is given to hypertriglyceridemic women // J. Lab. Clin. Med. — 1994. — Vol. 123. - N 1. —
P. 59-64.
428. Cirillo D.J. et al. Effect of estrogen therapy on gallbladder disease // JAMA. — 2005, -
Vol. 293, - N 3. - P. 330-339.
429. Havs J. et al. Effects of estrogen plus progestin on health-related quality of life // N. Engl.
J. Med. - 2003. - Vol. 348. - N 19. - P. 1839-1854.
430. Brunner R.L. et al. Effects of conjugated equine estrogen on health-related quality of life in
postmenopausal women with hysterectomy: results from the Women’s Health Initiative Randomized
Clinical Trial//Arch. Intern. Med. - 2005. - Vol. 165. - N 17. - P. 1976-1986.
431. Rapp S.R. et al. Effect of estrogen plus progestin on global cognitive function in postmenopaus¬
al women: the Women’s Health Initiative Memory Study: a randomized controlled trial // JAMA. —
2003. - Vol. 289. - N 20. - P. 2663-2672.
432. Espeland M.A. et al. Conjugated equine estrogens and global cognitive function in postmeno¬
pausal women: Women’s Health Initiative Memory Study //JAMA. - 2004. - Vol. 291. - N 24. -
P, 2959-2968.
433. Reed S.D. et al. Indications for hormone therapy: the post-Women’s Health Initiative era //
Endocrinol. Metab. Clin. North. Am. — 2004. — Vol. 33. — N 4, — P. 691-715.
434. Harman S.M. et al. Is the estrogen controversy over.? Deconstructing the Women’s Health
Initiative Study: a critical evaluation of the evidence // Annals of NY Acad. Sci. - 2005. -
Vol. 1052. - P. 43-56.
435. Speroff T. et al. A risk-benefit analysis of elective bilateral oophorectomy; effect of changes
in compliance w'ith estrogen therapy on outcome // Am, J, Obstet. Gynecol. - 1991. - Vol. 164. —
(1 Pt 1). - P. 165-174.
436. Berman R. et al. Compliance of women in taking estrogen replacement therapy //J. Womens.
Health. - 1996. - Vol. 701. - N 2. - P. 213.
4,37. Riis B.J. et al. Low bone mass and fast rate of bone loss at menopause: equal risk factors for
future fracture: a 15-year follow-up study // Bone. - 1996. - Vol. 19. - N1. - P. 9-12.'
438. Ettingcr B. et al. Low-dosage esterified estrogens opposed by progestin at 6-month intervals //
Obstet. Gynecol. - 2001. - Vol. 98. - N 2. - P. 205-211,
155
439. Lindheim S.R. et al. A possible bimodal effect of estrogen on insulin sensitivity in postmeno¬
pausal women and the attenuating effect of added progestin // Fertil. Steril. — 1993. — Vol. 60. —
N 4. - P. 664-647.
440. Woodruff J.D. et al. Incidence of endometrial hyperplasia in postmenopausal women taking
conjugated estrogens (Premarin) with medroxyprogesterone acetate or conjugated estrogens alone. The
Menopause Study GrouP // Am. J. Obstet. Gynecol. - 1994. - Vol. 170, - (5 Pt 1). - P. 1213-1223.
441. Archer D.F. et al. Bleeding patterns in postmenopausal women taking continuous combined
or sequential regimens of conjugated estrogens with medroxyprogesterone acetate. Menopause Study
Group // Obstet. Gynecol. - 1994. - Vol. 83. - (5 Pt 1). - P. 686-692.
442. The Writing Group for the PEPI Trial: (No authors Listed) Effects of estrogen or estrogen/
progestin regimens in heart disease risk factors in postmenopausal women: the Postmenopausal
Estrogen/Progestin Interventions (PEPI) trial // JAMA. — 1995. — Vol. 273. — P. 199-208.
443. Nand S.L. et al. Bleeding pattern and endometrial changes during continuous combined
honnone replacement therapy. The Ogen/Provera Study GrouP // Obstet. Gynecol. — 1998. —
Vol. 91. - (5 Pt 1). - P. 678-684.
444. Archer D.F, et al. Endometrial morphology in asymptomatic postmenopausal women // Am. J.
Obstet. Gynecol. - 1991. - Vol. 165. - N 2. - P. 317-320. discussion 320-322.
445. Korhonen M.O. et al. Histologic classification and pathologic findings for endometrial biopsy
specimens obtained from 2964 perimenopausal and postmenopausal women undergoing screening for
continuous hormones as replacement therapy (CHART 2 Study) // Am. J. Obstet. Gynecol. — 1997, —
Vol. 176. - N 2. - P. 377-380.
446. McGonigle K.F, et al. Development of endometrial cancer in women on estrogen and progestin
hormone replacement therapy // Gynecol. Oncol. — 1994. — Vol. 55. — N 1. — P. 126-132.
447. Karlsson B. et al. TVansvaginal ultrasonography of the endometrium in women with post¬
menopausal bleeding - a Nordic multicenter study // Am. J. Obstet. Gynecol. - 1995. - Vol. 172. -
N 5. - P. 1488-1494.
448. Bakos O. et al. Transvaginal ultrasonography for identifying endometrial pathology in post¬
menopausal women // Maturitas. — 1994, — Vol. 20. — N 2-3. — P. 181-189.
449. Granberg S. et al. Endometrial sonographic and histologic findings in women with and without
hormonal replacement therapv suffering from postmenopausal bleeding // Maturitas. — 1997. —
Vol. 27. - N 1. - P. 35-40.
450. O’Meara E.S, et al. Hormone replacement therapy after a diagnosis of breast cancer in relation
to recurrence and mortality //J. Natl. Cancer. Inst. - 2001. — Vol. 93. — N 10. — P. 754-762.
451. Col N. et al. Hormone replacement therapy after breast cancer: a systematic review and quan¬
titative assessment of risk //J. Clin. Oncol. -• 2001. — Vol. 19. — P. 2357-2363.
452. Dardes R.C. et al. Novel agents to modulate oestrogen action // Br. Med. Bull. — 2000. —
Vol. 56. - N 3. - P. 773-786.
453. DiezJ.L. Skeletal effects of selective oestrogen receptor modulators (SERMs) // Hum, Reprod.
Update. - 2000. - Vol. 6. - N 3. - P. 255-258.
454. Clemett D. et al. Raloxifene: a review' of its use in postmenopausal osteoporosis // Drugs. —
2000. - Vol. 60. - N 2. - P. 379-411.
455. Barrett-Connor E. et al. Effects of raloxifene on cardiovascular events and breast cancer in
postmenopausal women j j N. Engl.J. Med. — 2006, — Vol. 355. — N 2. — P. 125-137.
456. Burger H.G. Selective oestrogen receptor modulators // Horm. Res. — 2000. — Vol. 53, —
Suppl. 3. - P. 25-29.
457. Land S.R. et al. Patient-reported symptoms and quality of life during treatment with tamoxifen
or raloxifene for breast cancer prevention: the NSABP Study of Tamoxifen and Raloxifene (STAR) P-2
trial //JAMA. - 2006. - Vol. 295. - N 23. - P. 2742-2751.
4Sk Vogel V.G. et al. Effects of tamoxifen vs raloxifene on the risk of developing invasive breast
cancer and other disease outcomes: the NSABP Study of Tamoxifen and Raloxifene (STAR) P-2 trial //
JAMA. - 2006. - Vol. 295. - N 23. - P. 2727-2741.
459. Bonnick S. et al Comparison of weekly treatment of postmenopausal osteoporosis with
alendronate versus risedronate over two years // J, Clin. Endocrinol. Metab. - 2006. - Vol. 91, -
N 7. - P. 2631-2637.
460. Epstein S. Update of current therapeutic options for the treat-ment of postmenopausal osteo¬
porosis // Clin. Ther. - 2006. - Vol. 28. - N 2. - P. 151-173.
461. Close P. et al. Developments in the pharmacotherapeutic management of osteoporosis //
Exp. Opin, Phannacother. — 2006. - Vol. 7. - N 12. — P. 1603-1615.
Глава 2
Гормональная контрацепция
Обзор
Гормональные контрацептивы, содержащие синтетические аналоги половых сте¬
роидов, остаются основными способами нехирургического контроля рождаемости во
всем мире. Данная глава отражает, как изменялись пути доставки контрацептивных
доз гормонов, включая обычный пероральный прием, а также новые пути введения —
парентеральный (чрескожный или трансдермальный, подкожный), влагалищный и
внутриматочный. Каждый путь введения имеет свои преимущества и недостатки, но
эффективность контрацепции — предупреждение нежелательной беременности —
зависит в большей степени от самой женщины, чем от особенностей метода. Чем
больше выбор контрацептивных средств, тем больше вероятность, что каждая жен¬
щина или супружеская пара подберет для себя наилучший и наиболее приемлемый
в соответствующих ситуациях вариант. Большое количество новых методов контра¬
цепции, появившееся в последние годы, отражает избирательный подход женщин к
их выбору и направлено на необходимость повышения ее эффективности во многих
странах. Однако в некоторых странах, например в США и большинстве менее разви¬
тых стран, по меньшей мере половина всех случаев беременности относится к неже¬
лательным, следовательно, остаются актуальными разработка, внедрение и успешное
применение современных способов контрацепции.
В табл. 2-1 представлена сравнительная эффективность различных методов кон¬
трацепции (по данным клинических испытаний) — как прогнозируемая (приведен
самый низкий показатель), так и фактическая. На эффективность контрацепции
влияют возраст и материальное положение женщины: количество нежелательных
беременностей больше в молодом возрасте и при низком доходе. Эффективность кон¬
трацепции выше у рожавших женщин, чем у желающих предупредить возникновение
первой беременности.
Таблица 2-1. Сравнительная эффективность различных методов контрацепции, по данным США [1, 2]
Метод контрацепции
Количество беременностей на 100 женщин в год
прогнозируемое
(наименьший показатель)
фактическое
Не применявшие контрацептивы
85
85
Таблетки
Комбинированные
0,1
7,6
Гестагенные
0,5
3,0
157
Окончание табл. 2-1
Внутриматочные средства
Левоноргестрел
0,1
0,1
Соррег Т ЗбОА*“
0,6
0,8
Импланты
6 капсул с левоноргестрелом (норплант*)
0,05
0.2
2 капсулы с левоноргестрелом (яделл»’)
0,06
1 капсула с левоноргестрелом (импланон*')
0,01
Другие методы
Инъекционные
0,3
3.0
Стерилизация женщин
0,05
0,05
Стерилизация мужчин
0,1
0,15
ПЕРОРАЛЬНАЯ КОНТРАЦЕПЦИЯ
Пероральные контрацептивы начали применять в 50-е гг. прошлого столетия; они
содержали дозы эстрогена и гестагена в 10 раз выше, чем современные таблетки. В
последующие годы входящие в их состав синтетические стероидные эстрогены, такие
как местранол и хлормадинон, были заменены на новые аналоги с меньшим числом
побочных эффектов. Значительно позже вместо стандартной схемы применения
(таблетки принимают в течение 3 нед с перерывом на 7 дней) были предложены дру¬
гие режимы приема, более удобные, повышающие эффективность контрацепции и
ослабляющие побочное действие.
С внедрением новых дозировок пероральных контрацептивов, синтетических сте¬
роидов и режимов применения появились сведения о физиологических и эпидемио¬
логических результатах их приема. Ни одно лекарственное средство не изучали так
подробно, как комбинированные пероральные контрацептивы. Улучшение состава
таблеток и их назначения настолько снизило показатели заболеваемости и смертно¬
сти, обусловленные приемом пероральных контрацептивов, что на сегодняшний день
их применение имеет существенное положительное влияние на здоровье общества и
отдельного индивидуума.
Механизм действия
Первыми противозачаточными гормональными средствами, разрешенными для
применения, были комбинированные таблетки, содержащие эстроген и гестаген; их
следовало принимать ежедневно каждые 3 нед с перерывом на 7 дней/: целью ими¬
тации нормальных изменений содержания гормонов в организме во время менстру¬
ального цикла. Значительно позже появились другие схемы применения, например
прием таблеток, содержащих только эстроген, вместо 7-дневного перерыва или еже¬
дневный (без перерывов) прием комбинированных таблеток в течение 3 мес.
Комбинированные таблетки подавляют овуляцию вследствие угнетения секреции
гонадотропных гормонов гипофиза и гипоталамических гормонов, стимулирую¬
щих синтез и высвобождение половых гормонов. Гестагенный компонент таблетки
158
преимущественно подавляет секрецию ЛГ (подавление овуляции); эстрогенный ком¬
понент угнетает выброс ФСГ (предупреждение развития доминантного фолликула).
Следовательно, эстрогенный компонент вносит существенный вклад в развитие кон¬
трацептивного действия. Однако даже если рост и развитие фолликула не подавлены,
гестагенный компонент препарата предупреждает пик высвобождения Л Г, необходи¬
мый для овуляции.
Эстрогенный компонент таблеток определяет два эффекта. Он поддерживает
стабильность функционального слоя эндометрия, препятствует преждевременной
отслойке и таким образом предупреждает возникновение межменструальных кро¬
вотечений, а также потенцирует действие гестагенного компонента, что позволяет
снизить дозу последнего в таблетке. Предположительно этот механизм действия
эстрогенов связан с увеличением концентрации внутриклеточных рецепторов проге¬
стерона. Следовательно, для обеспечения эффективности комбинированных таблеток
достаточно минимальной эффективной дозы эстрогена.
В связи с тем что основная роль в комбинированных таблетках всегда принад¬
лежит гестагенному компоненту (если дозу эстрогена не увеличить в значительной
степени), изменение состояния эндометрия, свойств шеечной слизи, сократимости
маточных труб отражает действие гестагена, В эндометрии происходят изменения,
затрудняющие имплантацию яйцеклетки. Шеечная слизь становится более вяз¬
кой, что затрудняет продвижение сперматозоидов. Возможно, гестагены влияют
на секрецию и перистальтику маточных (фаллопиевых) труб, что также имеет
значение для контрацептивного действия. Именно поэтому даже при сохранении
незначительного роста фолликула (что возможно при приеме низкодозированных
препаратов) вышеперечисленные эффекты обеспечивают высокую степень надеж¬
ности контрацептива [3].
Контрацептивная эффективность новых пероральных препаратов (многофазных и
содержащих очень низкие дозы эстрогенов) такая же, как у ранних монофазных — как
низкодозированных (<50 мкг этинилэстрадиола), так и с более высоким содержанием
гормона [3]. Б тщательно контролируемых исследованиях в отношении мотивиро¬
ванных женщин установлено, что нежелательная беременность возникла в среднем
у 0,1% в течение первого года, однако в популяции этот показатель равен 7,6% [4].
Показатели представлены Национальной службой роста семьи и контроля рождаемо¬
сти (National Survey of Family Growth) за 1995 г. по .зарегистрированнь[м случаям искус¬
ственного прерывания беременности [2, 4, 5].
Препараты обеспечивают надежную контрацепцию в течение первого менстру¬
ального цикла после начала их приема при условии, что женщина начала принимать
таблетки не позже 5-го дня цикла и принимала их ежедневно (без пропусков). Прием
таблеток с 1-го дня цикла гарантирует быстрое развитие их противозачаточного дей¬
ствия. В США большинство женщин начинают прием таблеток в ближайшее со дня
начала менструации воскресенье, так как этот день легче запомнить, а также преду¬
преждаются менструальные кровотечения в конце рабочей недели. Вполне вероятно,
что даже если при начале приема таблеток в воскресенье у женщин развился доми¬
нантный фолликул, у них не повысится секреция ЛГ и не произойдет овуляция [6].
Некоторые врачи предлагают начинать прием таблеток сразу после их приобрете¬
ния, независимо от дня менструального цикла, так как в дни, оставшиеся до начала
159
нового цикла или ближайшего воскресенья, может возникнуть нежелательная бере¬
менность [7]. В течение первой недели приема таблеток сразу после приобретения
следует использовать дополнительные методы контрацепции, предпочтительно пре¬
зервативы. Предлагаемая схема способна предотвратить возникновение нежелатель¬
ной беременности в оставшиеся дни менструального цикла (то есть до начала приема
таблеток по традиционной схеме с 1-го дня следующего цикла). Так как наступление
менструаций в этом случае будет задержано, для исключения беременности при нали¬
чии сомнений разумно провести чувствительный тест на беременность. У женщин,
принимающих таблетки по данной схеме, не происходит усиления прорывных кро¬
вотечений [8],
Начало менструации можно отсрочить, если после 21-го дня приема таблеток не
сделать 7-дневного перерыва.
Рекомендация о необходимости 7-дневного перерыва в приеме таблеток необосно¬
ванна, так как подобная практика слишком часто приводит к возникновению неже¬
лательной беременности, к тому же не предупреждает развитие серьезных побочных
эффектов.
Несмотря на то что последствия непрерывного приема таблеток полностью не
изучены, нет основания полагать, что перерыв в приеме таблеток ослабляет спон¬
танные кровотечения. Кроме того, непрерывный прием удобнее для формирования у
женщины привычки ежедневно принимать контрацептивные таблетки.
Оредрреждение тттрщпьыът щйвйттвтш
Все больше женщин верят в то, что более редкие менструации облегчают восста¬
новление организма после кровотечений и менструальных симптомов. Появилась
схема ежедневного приема таблеток в течение 84 дней, приводящая к уменьшению
числа менструаций до четырех в год [9]. Более того, врачи давно назначали дли¬
тельный ежедневный прием пероральных контрацептивов для лечения, например,
эндометриоза, кровотечений, менструальных судорог, головных болей при мен¬
струации, а также для предупреждения наступления менструации у спортсменов
и деловых женщин. Многие женщины не нуждаются в наступлении менструации
как показателе отсутствия беременности и могут сами выбирать желаемую частоту
менструаций.
Любые пероральные комбинированные контрацептивы можно принимать еже¬
дневно; каждый из них, даже с самым низким содержанием эстрогена, в режиме
длительного применения вызывает нормальное менструальное кровотечение и пере¬
носимые побочные эффекты [10, 11]. Длительный режим приема более удобный и
надежный. Если начинается спонтанное кровотечение, женщину можно успокоить,
что это практически всегда временная проблема. При длительном кровотечении
может быть эффективен 3- или 4-дневный перерыв в приеме таблеток [12].
Щ&щт в пщешв 1дйтт%
Часто женщина пропускает прием очередной таблетки. Электронный мониторинг
показал, что женщины пропускают прием таблеток чаще, чем признаются в этом;
лишь 33% женщин ни разу не пропустили прием таблеток в течение первого цикла, в
течение третьего цикла приблизительно треть женщин пропустили прием трех и более
таблеток и часто пропускали прием таблеток несколько дней подряд [13]. Эти данные
свидетельствуют о том, что женщины со временем менее внимательно относятся к
приему противозачаточных таблеток, поэтому необходимо повторно напоминать, что
нужно делать, если пропущено время приема очередной таблетки.
160
Проведены исследования, посвященные влиянию пропуска в приеме таблеток на
эффективность контрацепции. Согласно результатам одной работы, пропуск в приеме
четырех таблеток подряд в различные периоды цикла не привел к возникновению ову¬
ляции [6]. У женщин, добровольно увеличивших 7-дневный перерыв в приеме таблеток
до 11 дней, также не выявлено признаков овуляции [14,15]. Следовательно, отсутству¬
ют доказательства, что снижение дозы гормона приводит к уменьшению эффективно¬
сти контрацепции. Несмотря на то что при приеме пероральных контрацептивов, содер¬
жащих очень низкие дозы гормонов, фолликулярная активность выше, овуляция, тем
не менее, эффективно подавляется [16]. Гестагенное действие на эндометрий и шеечную
слизь обеспечивает хорошую эффективность контрацепции [3].
Наиболее частыми установленными причинами неэффективности пероральной
контрацепции служат рвота и диарея [17, 18]. Женщину следует предупредить, что
даже если она не пропускала время приема таблеток, ей необходимо использовать
другие методы предохранения от беременности в течение не менее 7 дней после эпи¬
зода гастроэнтерита.
Кттчвтшв пробле1).1Ь1 связанные с при1^внением пероральных
контрацегтшов
СПОНТАННЫЕ КРОВОТЕЧЕНИЯ
Существует два типа проблем, связанных с кровотечением: нерегулярные кровоте¬
чения в первые месяцы после начала приема пероральных контрацептивов и возник-”
новение внеочередной менструации после нескольких месяцев приема препаратов.
Следует предпринять все усилия для решения этих проблем таким образом, чтобы
женщина продолжала пользоваться низкодозированными препаратами. Начало
кровотечения не служит показателем снижения эффективности контрацептива вне
зависимости от его состава и низких доз. В тщательно проведенном исследовании
показано отсутствие корреляции между возникновением спонтанного кровотечения и
изменением концентрации контрацептивных стероидов в крови [19]. Перед назначе¬
нием пероральных контрацептивов женщину следует информировать о возможности
спонтанного кровотечения.
Наиболее часто спонтанное кровотечение возникает в первые несколько месяцев
приема препарата. Частота возникновения наибольшая в первые 3 мес, она колеблет¬
ся от 10 до 30% в первый месяц и снижается до 10% и менее в третий месяц приема.
Спонтанное кровотечение несколько чаще возникает при применении низкодозиро¬
ванных препаратов [20, 21]. Частота прорывных кровотечений выше у курящих жен¬
щин, а среди них выше у тех, кто принимает препараты, содержащие 20 мкг этинил¬
эстрадиола [22]. Однако разница в этом показателе у женщин, принимающих таблет¬
ки разного состава, имеет минимальное клиническое значение. Основной вывод этой
работы тот же: частота кровотечений наибольшая в первый месяц приема и у куря¬
щих женщин.
Спонтанное кровотечение, развивающееся после приема контрацептивов в течение
нескольких месяцев, является следствием вызванной гестагеном децидуализации.
Эндометрий и пронизывающие его кровеносные сосуды становятся ломкими, что
приводит к возникновению спонтанных и несинхронных кровотечений.
Известны две причины (обе устранимые), повышающие вероятность возникно¬
вения кровотечения. При пропусках в приеме таблеток и курении чаще появляются
кровянистые выделения и кровотечения, причем более существен первый фактор,
сильнее отражающийся в последующие циклы, в то время как курение постоянно
оказывает свое влияние [23]. Возобновление непрерывного режима приема таблеток
161
способно практически устранить возможность развития спонтанного кровотечения.
Другой причиной кровотечения может служить инфекция цервикального канала, осо¬
бенно вызванная хламидиями [24].
ТРОМБОТИЧЕСКИЕ ОСЛОЖНЕНИЯ
Эстрогенный компонент в дозах, содержащихся в высокодозированных комбини¬
рованных пероральных контрацептивах, вызывает повышение образования факторов
свертывания крови, таких как факторы V, VIII, X и фибриноген [25]. Гестагенный
компонент также влияет на факторы свертывания [26]. В ходе отдельных исследо¬
ваний выявлено, что монофазные и многофазные низкодозированные перораль¬
ные контрацептивы не оказывают клинически значимого влияния на эту систему.
Незначительное повышение образования фибрина компенсируется повышением
фибринолитической активности крови [27, 28]. Другие исследования препаратов,
содержащих 30 или 35 мкг этинилэстрадиола, свидетельствуют об увеличении содер¬
жания свертывающих факторов и повышении активности тромбоцитов [29]. Однако
практически все эти изменения находятся в пределах нормальных показателей, и их
клиническая значимость неизвестна [26].
Курение вызывает волнообразное повышение свертываемости крови [30].
Согласно опубликованным сведениям, препараты, содержащие 20 мкг эстрогена, не
оказывают влияния на показатели свертывания крови даже у курящих [30, 31]. Лишь
в ходе одного сравнгггельного исследования препаратов, содержащих 20 или 30 мкг
эстрогена, обнаружено небольшое повышение свертывающей и фибринолитической
активности с тенденцией к большему изменению последней меньшей дозы препарата
[32]. Эти результаты имеют значение для эпидемиологических исследований клини¬
ческих случаев.
Отсутствуют доказательства повышения риска развития сердечно-сосудистых забо¬
леваний у лиц, принимающих пероральные контрацептивы [33-35]. В исследованиях
Nurses' Health Study Королевского колледжа врачей общей практики (The Royal College
of General Practitioner^ Study) и Оксфордского центра планирования семьи (The Oyford
Family Planning Association Study) показано, что длительное применение пероральных
контрацептивов не приводит к повышению показателей общей смертности [36-38].
Некоторое беспокойство о возможных отдаленных последствиях применения перо¬
ральных контрацептивов основано на их предполагаемом неблагоприятном влиянии
на атеросклеротический процесс, которое в сочетании с возрастными изменениями
может проявиться впоследствии. При этом имеется множество данных, что развитие
сердечно-сосудистых заболеваний, вызванных приемом пероральных контрацепти¬
вов, является следствием их острых эффектов, особенно тромбозом — дозозависимым
побочным эффектом эстрогена.
Венозный тромбоз
Тромбоэмболия вен
Более ранние эпидемиологические исследования связи приема пероральных кон¬
трацептивов и развития сосудистых заболеваний свидетельствовали о том, что веноз¬
ный тромбоз может быть вызван эстрогеном; риск развития осложнения исчезал
через 3 мес после прекращения приема препарата [39,40]. Считали, цто тромбоэмбо¬
лические заболевания — следствие применения эстрогена и степень риска зависит от
дозы [41-43]. Подтверждено, что курение служит дополнительным фактором риска
развития тромбоза артерий [44-46], но не венозной тромбоэмболии [47, 48].
В первые годы применения пероральных контрацептивов, когда препараты содер¬
жали 80 и 100 мкг этинилэстрадиола (чрезвычайно высокие дозы), было отмечено
6-кратное повышение риска развития венозного тромбоза, в том числе у беременных
[49]. В связи с повышением вероятности этого осложнения, а также инфаркта мио-
162
карда и инсульта на фармацевтическом рынке стали преобладать низкодозированные
препараты (<50 мкг эстрогена), и врачи стали проявлять большую осторожность в
определении индивидуальных показаний для назначения пероральных контрацепти¬
вов и выбора препаратов. В итоге, согласно исследованиям Puget Sound study, в США
риск развития венозного тромбоза снизился вдвое [50].
Венозная тромбоэмболия и мутация V фактора Лейдена
Риск развития идиопатического венозного тромбоза в группе принимающих низ¬
кодозированные пероральные контрацептивы приблизительно в 3-4 раза выше, чем
в популяции [51-55]. Однако наследственная резистентность к активированному
белку С, мутация V фактора Лейдена, может иметь значение для значительной части
пациентов с венозным тромбозом и принимающих пероральные контрацептивы
(табл. 2-2).
Таблица 2-2. Относительный риск и количество случаев венозной тромбоэмболии [56, 57, 58, 59]
Группы
Относительный
риск
Количество случаев
(на 100 ООО в год)
Молодые женщины (основная группа)
4-5
Беременные
12
48-60
Принимающие вызокодозированные пероральные контрацептивы
6-10
24-50
Принимающие низкодозированные пероральные контрацептивы
3-4
12-20
Носители мутации V фактора Лейдена
5-8
20-40
Носители мутации V фактора Лейдена и принимающие пероральные
контрацептивы
10-30
40-150
Мутация Лейдена, гомозиготы
80
320-400
Врожденная резистентность к активированному белку С, мутация V фактора
Лейдена, — наиболее типичная наследственная патология свертываемости крови, пере¬
даваемая по аутосомально-доминантному типу [60, 61]. У гетерозиготных носителей
риск развития венозной тромбоэмболии повышается в 5-8 раз, а у гомозиготных — в
80. Сообщают, что у лиц с вышеупомянутой мутацией, принимающих пероральные
контрацептивы, риск развития венозного тромбоза повышается в 30 раз [62, 63].
Правда, по мнению других авторов, эта цифра преувеличена и приближается к 10-15
[59,64]. Вероятность возникновения венозного тромбоза наибольшая в первые месяцы
применения препаратов; высказано мнение, что развитие этого осложнения в первый
месяц предположительно свидетельствует о наличии патологии свертывания [65].
Комбинированные пероральные контрацептивы противопоказаны женщинам с
идиопатической тромбоэмболией в анамнезе, а также при наличии этой патологии
у ближайших родственников. У этих женщин могут быть обнаружены врожденная
недостаточность важных показателей системы коагуляции, особенно антитромбина
III, белка С, белка 5, и резистентность к активированному белку С [83]. Женщины, у
которых не выявлена врожденная патология свертываемости крови* могут выбрать
пероральную контрацепцию, но это трудное и рискованное для женщины и для врача
решение, и благоразумнее применять другие методы контрацепции. К другим факторам
риска развития тромбоэмболии, которые следует учитывать врачу, относятся такие
приобретенные заболевания, как волчаночный антикоагулянт или злокачественные
новообразования, длительная иммобилизация или травма. Варикозное расширение вен
не относится к факторам риска, если оно не очень распространенное [49].
При отягощенном семейном анамнезе. (Примег. ред.)
163
Артериальный тромбоз
Острые нарушения мозгового кровообращения (тромботический инсульт или
преходящие ишемические нарушения) у молодых женщин встречаются чаще, чем
венозная тромбоэмболия и инфаркт миокарда, и могут привести к смерти или инва-
лидизации. В связи с более высокой частотой развития и риска для жизни тромбоз
мозговых артерий — наиболее важный возможный побочный эффект пероральных
контрацептивов. Инсульт у молодых женщин возникает очень редко, поэтому показа¬
тель абсолютного риска повышается незначительно. Однако в связи с тем, что тром¬
ботический инсульт чаще развивается у женщин старше 40 лет, им можно принимать
пероральные контрацептивы при условии хорошего здоровья и отсутствия факторов
риска развития сердечно-сосудистых заболеваний (особенно артериальной гипертен¬
зии, мигрени с аурой и курения).
Доказан синергизм курения и приема пероральных контрацептивов как причина
развития инфаркта миокарда (табл. 2-3). Сложно установить связь между развитием
артериального тромбоза и дозой эстрогенного компонента в связи с малым количе¬
ством случаев. Тем не менее доза эстрогена служит важным фактором для риска раз¬
вития инфаркта миокарда или тромботического инсульта [66, 67].
Таблица 2-3. Количество случаев развития инфаркта миокарда у женщин репродуктивного возраста
[68]
Группы
Количество
(на 100 ООО женщин в год)
Все [69]
5
Женщины моложе 35 лет
Некурящие
4
Некурящие, принимающие пероральные контрацептивы
4
Курящие
8
Курящие, принимающие пероральные контрацептивы
43
Женщины в возрасте 35 лет и старше
Некурящие
10
Некурящие, принимающие пероральные контрацептивы
40
Курящие
88
Курящие, принимающие пероральные контрацептивы
485
Примечание. В группы включены женщины, принимающие контрацептивы и имеющие другие фак¬
торы риска, преобладающие в общей популяции. При более тщательном отборе групп количество слу¬
чаев может быть меньше. Повышение риска развития заболевания в группе курящих и принимающих
пероральные контрацептивы отражает наличие необнаруженных факторов риска развития сердечно¬
сосудистых заболеваний, особенно артериальной гипертензии.
Артериальный тромбоз — инсульт
Более ранние исследования, а также когортные исследования выявили повышен¬
ный риск развития тромбоза артерий мозга у лиц, принимающих высокодозирован-
ные пероральные контрацептивы [70-72]. Однако у здоровых некурящих женщин,
принимавших пероральные контрацептивы, содержащие менее 50 мкг этинилэстра¬
диола, количество тромботических инсультов не было повышено [71, 72]. В Дании
был проведен анализ всех 794 случаев острого тромботического нарушения мозгового
164
кровообращения у женщин с 1985 до 1989 г. и выявлено двукратное повышение отно¬
сительного риска, связанного с приемом пероральных контрацептивов, содержащих
от 30 до 40 мкг эстрогена; на показатель риска существенно влияли одновременно
курение и доза эстрогена (суммирование, но не синергизм) [46]. По данным Royaî
Collège of General Practitioner^ Oral Contraception Study, лица, постоянно принимающие
пероральные контрацептивы, имеют повышенный риск развития инсульта; однако
это заключение относится, главным образом, к курящим и принимающим препараты,
содержащие 50 мкг и более эстрогена [72].
Популяционное исследование 408 случаев инсульта, проведенное в Калифорнии
(Kaiser Permanente Médical Саге Program), не выявило повышения вероятности раз¬
вития как ишемического, так и геморрагического инсульта [73]. Выявленными
факторами риска развития ишемического инсульта было курение, артериальная
гипертензия, сахарный диабет, ожирение и низкое социально-экономическое поло¬
жение. Выявленные факторы риска развития геморрагического инсульта те же плюс
избыточная масса тела и злоупотребление алкоголем. Женщины, принимавшие
низкодозированные пероральные контрацептивы, не имели повышенного риска раз¬
вития ишемического или геморрагического инсульта по сравнению с принимавшими
их в прошлом и не принимавшими никогда. Не было получено данных о влиянии
возраста или курения (в случае геморрагического инсульта высказано предположение
о влиянии приема пероральных контрацептивов и курения, но в связи с малым коли¬
чеством случаев эти данные статистически недостоверны). Полученные результаты
представлены в табл. 2-4.
Таблица 2-4. Количество инсультов у женщин репродуктивного возраста [69, 73, 74, 75]
Группа
Количество
(на 100 ООО /в год)
Ишемический инсульт
Общее число
5
Женщины моложе 35 лет
1-3
Женщины в возрасте 35 лет и старше
10
Геморрагический инсульт
Общее число
6
Принимавшие пероральные контрацептивы*
Низкодозированные препараты
2
Низкодозированные препараты и возраст моложе 35 лет
1
Высокодозированные препараты
8
* Включены курящие и женщины с артериальной гипертензией.
Артериальный тромбоз: современные данные
Отсутствуют статистически достоверные сведения о том, что новые (третьего поко¬
ления) гестагены изменяют степень риска развития заболеваний артериальных сосу¬
дов, количество случаев заболевания не возрастает при приеме низкодозированных
пероральных контрацептивов, содержащих гестагенные компоненты старого типа.
Возможно, после проведения дополнительных исследований будет выявлена досто¬
верная разница, но она будет статистически и, возможно, клинически незначимой.
165
Наиболее важно, что новые исследования не выявили риска развития ише¬
мического или геморрагического инсульта при применении низкодозированных
пероральных контрацептивов здоровыми молодыми женщинами. Исследование,
проведенное ВОЗ, выявило отрицательное влияние курения на этот показатель
у женщин моложе 35 лет, в то время как Kaiser study не выявило. Эту разницу
можно объяснить дополнительным влиянием артериальной гипертензии, основ¬
ного доказанного фактора риска. В исследовании ВОЗ наличие артериальной
гипертензии основывалось лишь на сообщениях пациентов об эпизодах подъема
артериального давления в прошлом (исключая период беременности), не под¬
твержденных записями врача. В Kaiser study женщин относили к группе с артери¬
альной гипертензией, если они сообщали о приеме антигипертензивных препа¬
ратов (менее 5% женщин, принимавших пероральные контрацептивы, лечились
от артериальной гипертензии, и среди них не было ни одной, принимавшей
высокодозированные контрацептивы). В исследовании ВОЗ прием пероральных
контрацептивов как фактор риска при наличии артериальной гипертензии был
определен при сравнении разнородных групп: данные по Европе (женщин в груп¬
пы отбирали врачи очень тщательно) сравнивали с данными по развивающимся
странам (менее тщательный отбор), следовательно, большее количество женщин
с сердечно-сосудистыми факторами риска в развивающихся странах принимали
пероральные контрацептивы.
Пероральные контрацептивы, содержащие менее 50 мкг этинилэстрадиола, не
повышают вероятность развития инфаркта миокарда или инсульта у здоровых неку¬
рящих женщин независимо от их возраста. Влияние курения на этот показатель у
женщин моложе 35 лет невозможно установить при отсутствии у них артериальной
гипертензии. У женщин старше 35 лет практически неуловимое наличие артериальной
гипертензии затрудняет анализ, но Kaiser study показывает, что увеличение возраста и
курение сами по себе имеют небольшое влияние на риск возникновения инсульта
у женщин, принимающих низкодозированные пероральные контрацептивы. Отбор
пациентов в Kaiser program был очень тщательным, в результате лишь несколько жен¬
щин с артериальной гипертензией принимали пероральные контрацептивы. Согласно
результатам новых исследований, основная проблема — артериальная гипертензия,
особенно как фактор риска возникновения инсульта.
АРТЕРИАЛЬНАЯ ГИПЕРТЕНЗИЯ
Повышение артериального давления, вызванное пероральными контрацептива¬
ми, наблюдали у 5% женщин, принимавших высокодозированные препараты. Более
поздние исследования показывают, что незначительное повышение артериального
давления может отмечаться при приеме даже монофазных таблеток, содержащих
30 мкг эстрогена в сочетании с новыми гестагенами. Однако нет сведений об увели¬
чении числа клинически значимых подъемов артериального давления [77-80], что
может быть связано с редким их возникновением. The Nurses' Health Study отметил
повышенный риск клинически значимой артериальной гипертензии у принимающих
низкодозированные пероральные контрацептивы — частота случаев 41,5 на 10 ООО в
год [81]. Следовательно, необходим постоянный контроль показателей артериаль¬
ного давления даже при приеме низкодозированных препаратов. По данным Rancho
Bernardo Study у женщин в постменопаузе, ранее принимавших пероральные контра¬
цептивы (возможно, высокодозированные), было слегка повышено (на 2-4 мм рт.ст.)
диастолическое давление [82]. В связи с тем что частота возникновения и показатели,
свидетельствующие о риске развития сердечно-сосудистых заболеваний у женщин,
принимавших пероральные контрацептивы в прошлом и не принимавших никогда,
была одинаковой, сомнительно, что эти различия в показателях артериального дав¬
ления клинически значимы.
166
Низкодозированные пероральные контрацептивы безопасны для здоровых моло¬
дых женщин. Отбор в группы женщин старше 35 лет и с сердечно-сосудистыми фак¬
торами риска, особенно с артериальной гипертензией, позволяет практически исклю¬
чить повышенный риск развития артериальной патологии, связанный с приемом
низкодозированных пероральных контрацептивов. Риск возникновения сердечно¬
сосудистой патологии не возрастает в зависимости от возраста женщины (при дли¬
тельном применении препаратов). В больших когортных исследованиях смертность
в группах женщин, применявших и не применявших пероральные контрацептивы,
была одинаковой [36-38].
РАК МОЛОЧНОЙ ЖЕЛЕЗЫ
Применение в прошлом и в настоящее время пероральных контрацептивов,
согласно исследованиям, может привести к возрастанию на 20% риска возникнове¬
ния раннего (в возрасте до 35 лет) предменопаузального рака груди, обычно локаль¬
ного. Предполагается, что умеренное возрастание относительного риска связано с
незначительным повышением количества случаев заболевания (настолько неболь¬
шим, что не влияет на показатели заболеваемости). Выявление умеренного возрас¬
тания относительного риска у молодых женщин может быть частично или целиком
связано с повышением уровня диагностики и профилактических осмотров, а также
ростом уже существовавших злокачественных новообразований; аналогичные дан¬
ные получены при изучении влияния заместительной гормонотерапии в климак¬
терическом периоде на риск возникновения рака молочной железы. Успокаивает
факт, что рост количества случаев рака молочной железы в период 1973-1994 гг. у
немолодых, не принимавших пероральные контрацептивы американок был больше
[125]. У женщин моложе 50 лет в этот же период отмечено лишь небольшое увеличе¬
ние частоты возникновения заболевания. Большое исследование связи заболеваемо¬
сти раком молочной железы женщин в возрасте 35-65 лет с приемом пероральных
контрацептивов, проведенный в США, дало полностью отрицательный результат и
внушает оптимизм.
Факт применения в прошлом или длительность (вплоть до 15 лет непрерывно)
приема пероральных контрацептивов (том числе высокодозированных) не влияет на
риск возникновения рака молочной железы. С использованием в прошлом перораль¬
ных контрацептивов может быть связано понижение риска возникновения метастати¬
ческого и постменопаузального рака молочной железы.
Прием пероральных контрацептивов не повышает степень риска развития рака
молочной железы у женщин с наследственной предрасположенностью, а также при
доброкачественной опухоли молочной железы. Однако врачам следует обращать
внимание на все факторы, способствующие развитию этого заболевания. Одними из
факторов его профилактики служат кормление грудью и отказ от злоупотребления
алкоголя. Это особенно важный аргумент для убеждения женщины кормить грудью.
Кормление грудью относится к условиям профилактики, главным образом, пред¬
менопаузального рака молочной железы, что особенно важно для женщин молодого
возраста, применяющих пероральные контрацептивы.
УГЛЕВОДНЫЙ ОБМЕН
Высокодозированные пероральные контрацептивы первого поколения изменяли
толерантность к глюкозе у многих женщин, при исследовании этого показателя отме¬
чено повышение концентрации инсулина в плазме крови и глюкозы в крови. В целом
пероральные контрацептивы снижают чувствительность периферических тканей к
инсулину, у большинства женщин это компенсируется повышением секреции инсули¬
на, изменения в тесте толерантности к глюкозе не выявляются, хотя показатели через
1 ч могут быть слегка повышены.
167
На чувствительность к инсулину влияет прежде всего гестагенный компонент
таблеток [84]. К нарушениям углеводного обмена может также приводить влияние
эстрогенного компонента на липидный обмен, ферменты печени и повышение кон¬
центрации свободного кортизола. Изменение толерантности к глюкозе дозозависимо:
меньшее влияние оказывают низкодозированные препараты. Изменения секреции
инсулина и концентрации глюкозы настолько минимальны, что их считают клиниче¬
ски не значимыми [80. 85, 86, 87, 88]. Эти исследования включают изучение показа¬
телей в течение длительного срока с определением гликированного гемоглобина Aie
(HbAlc).
ПЕЧЕНЬ
Половые стероиды влияют на функции печени разнообразнее, чаще и сильнее,
чем на любой другой орган (исключая половые органы). Эстрогены оказывают
влияние на синтез печеночной ДНК и РНК, ферменты гепатоцитов, сывороточные
ферменты, синтезируемые в печени, и белки плазмы крови. Эстрогенные гормоны
также влияют на синтез липидов и липопротеинов в печени, промежуточный обмен
углеводов и активность внутриклеточных ферментов. Тем не менее анализ результа¬
тов проспективных когортных исследований женщин, проведенный в Royal College
of General Practitioners' Oral Contraception Study и the O)ford Family Planning Association
Contraceptive Study, не выявил повышения риска развития серьезной патологии пече¬
ни у женщин, принимающих пероральные контрацептивы [89].
Эстрогены и некоторые гестагены изменяют активный транспорт компонентов
желчи. Механизм этого действия неясен, но при приеме высокодозированных пре¬
паратов могут развиться холестатическая желтуха и зуд. Подобно рецидивирующей
желтухе при беременности этот холестаз доброкачественный и обратимый. Частота
осложнения при приеме низкодозированных препаратов неизвестна, но должна быть
очень низкой.
Единственное абсолютное противопоказание для приема пероральных контрацеп¬
тивов — острое или хроническое холестатическое заболевание печени. Цирроз печени
и перенесенный гепатит не усугубляются приемом препаратов. После выздоровления
от острой фазы заболевания печени женщина может принимать пероральные контра¬
цептивы.
Применение пероральных контрацешшш
и медицинские проблемы
МЕТАБОЛИЧЕСКИЕ НАРУШЕНИЯ
Сахарный диабет
Пероральные контрацептивы можно принимать больным сахарным диабетом,
если они моложе 46 лет, не курят и не имеют сопутствующих заболеваний (особенно
сосудистых осложнений). Установлено, что пероральные контрацептивы не влияют
на развитие ретинопатии или нефропатии у молодых женщин с инсулинзависимым
сахарным диабетом [98]. Исследование в течение одного года женщин с инсулинзави¬
симым сахарным диабетом, принимающих низкодозированные пероральные контра¬
цептивы, не выявило изменений липопротеиновых и гемостатических маркеров риска
сердечно-сосудистых заболеваний [99]. Не обнаружено также влияния пероральных
контрацептивов на показатели смертности от сердечно-сосудистых заболеваний в
группе больных сахарным диабетом [100]. При сахарном диабете с сопутствующи¬
ми сосудистыми заболеваниями или основными факторами риска возникновения
168
сердечно-сосудистой патологии следует избегать приема препаратов, содержащих
фармакологические дозы эстрогена.
Заболевания желчного пузыря
Пероральные контрацептивы способны провоцировать обострение желчекаменной
болезни или других заболеваний желчного пузыря, следовательно, применять эти
препараты следует с осторожностью или не применять совсем.
Гиперлипидемия
Низкодозированные пероральные контрацептивы оказывают незначительное
влияние на содержание липопротеинов, поэтому гиперлипидемия не относится
к абсолютным противопоказаниям, за исключением резкого повышения содер¬
жания триглицеридов (эстрогены могут вызвать еще большее повышение). При
повышении концентрации триглицеридов в крови выше 250 мг/дл эстрогены
следует принимать с большой осторожностью. Если уже имеется сосудистая
патология, пероральные контрацептивы противопоказаны. При наличии других
факторов риска, особенно курения, пероральная контрацепция не рекомендована.
Пациенткам с дислипидемией, начавшим прием пероральных контрацептивов,
следует в течение нескольких месяцев ежемесячно проводить биохимический ана¬
лиз крови, при отсутствии такой возможности следует применять другие способы
контрацепции [108].
Пероральные контрацептивы, содержащие синтетические гестагены с очень низкой
андрогенной активностью (дезогестрел, норгестимат или гестоден), увеличивают в
крови содержание липопротеинов высокой плотности, однако данные о клинической
значимости этих изменений отсутствуют, при наличии только гипертриглицери-
демии следует учитывать, что действие эстрогена на концентрацию триглицеридов
развивается быстро. Концентрация триглицеридов в крови выше 750 мг/дл служит
абсолютным противопоказанием к применению эсгрогенсодержащих препаратов в
связи с риском развития панкреатита.
Заболевания печени
Пероральные контрацептивы можно применять после нормализации функцио¬
нальных проб печени. Пробы следует повторить после 2-3 мес приема пероральных
контрацептивов.
Ожирение
Женщины с ожирением при отсутствии других заболеваний могут применять низ¬
кодозированные пероральные контрацептивы. Однако при ожирении следует учиты¬
вать следующее.
Ожирение является независимым фактором риска развития венозного тромбоза,
этот риск возрастает при приеме пероральных контрацептивов [109-111].
Эффективность гормональных контрацептивов может быть ниже у женщин с
избыточной массой тела [масса более 155 фунтов (около 70 кг)] [112-114]. При
клинических испытаниях в исследуемые группы не включают женщин с большой
массой тела, поэтому влияние массы тела на эффективность контрацепции до конца
не изучено. Назначение женщинам с избыточной массой тела препарата, содер¬
жащего 50 мкг эстрогена, могло бы повысить надежность контрацепцци, но при
этом риски развития венозного тромбоза, вызванного высокой дозой эстрогена и
ожирением, дополняют друг друга. Следует у^штывать, что заключения о снижении
эффективности контрацепции при повышенной массе тела основаны на разнице в
2-4 нежелательных беременности на 100 женщин в год. Тем не менее у женщин с
повышенной массой тела эффективность пероральной контрацепции выше, чем
барьерных методов.
169
Поликистоз яичников и резистентность к инсулину
в связи с тем что более ранние высокодозированные пероральные контрацептивы
снижали чувствительность периферических тканей к инсулину, было высказано пред¬
положение, что их следует избегать женщинам с ановуляторными циклами и повы¬
шенной массой тела. Низкодозированные пероральные контрацептивы практически
не влияют на углеводный обмен, поэтому, возможно, не окажут неблагоприятного
действия на большинство женщин с повьппенной секрецией инсулина и гиперандро-
генным фоном [115].
Изменения концентраций инсулина и глюкозы в крови при применении низкодо¬
зированных пероральных контрацептивов (<50 мкг этинилэстрадиола) настолько
незначительны, что не имеют клинического значения [87]. Долговременные иссле¬
дования не выявили повышения частоты возникновения сахарного диабета или
нарушения толерантности к глюкозе даже у женщин, постоянно принимавших в
прошлом высокодозированные препараты [116, 117]. Отсутствуют свидетельства о
повышении риска развития сердечно-сосудистых заболеваний у женщин, в прошлом
применявших пероральные контрацептивы [35, 36]. Более того, низкодозированные
препараты принимали женщины с диабетом беременных в анамнезе без каких-либо
неблагоприятных последствий; у женщин с инсулинзависимым диабетом низкодози¬
рованные препараты не ухудшали показатели липидных и биохимических маркеров
сердечно-сосудистой патологии или не вызывали учащения развития ретинопатии и
нефропатии [96-99J. Прием низкодозированных препаратов женщинами с высокой
степенью ожирения и очень выраженной инсулиновой резистентностью приводил
лишь к незначительному снижению толерантности к глюкозе [118].
Чрезвычайно интересны и поразительны данные длительного (приблизительно
10-летнего) наблюдения женщин с поликистозом яичников и повышением секреции
инсулина, свидетельствующие о том, что показатели обмена веществ, включая массу
тела, толерантность к глюкозе, содержание инсулина и липопротеинов высокой плот¬
ности в крови, у применявших пероральные контрацептивы не только не ухудшались
по сравнению с не применявшими, но даже улучшались [119]. Эти данные подтверж¬
дают безопасность лечения комбинированными контрацептивами женщин с отсут¬
ствием овуляций, повышенным андрогенным и гиперинсулиновым состоянием.
Нарушение приема пищи (пищевого поведения)
у пациенток с нарушениями пищевого поведения плотность костной ткани корре¬
лирует с массой тела. Гормонотерапия не улучшает показателей плотности костной
ткани, пока масса тела остается патологически измененной [120]. Отсутствие уве¬
личения плотности костной ткани при лечении эстрогенами может быть связано с
побочным действием на костную ткань гиперкортицизма, вызванного стрессовыми
нарушениями. В пубертатном возрасте значительно повышается плотность костной
ткани, и, если по какой-либо причине этого не произошло, гормонотерапия может
быть неэффективна. Снижение менструальной функции по любой причине в раннем
возрасте может вызвать остаточное снижение плотности костной ткани, не корри¬
гируемое полностью ни нормализацией менструального цикла, ни гормонотерапией
[121,122].
СЕРДЕЧНО-СОСУДИСТЫЕ ЗАБОЛЕВАНИЯ
Артериальная гипертензия
Низкодозированные пероральные контрацептивы можно применять женщинам
моложе 35 лет с артериальной гипертензией, хорошо контролируемой фармако¬
терапией, при условии отсутствия сопутствующих заболеваний и некурящим. Мы
рекомендуем применять препараты с наименьшей дозой эстрогенного компонента.
170
Тем не менее имеются данные бразильских исследователей о том, что артериальная
гапертензия хуже контролируется у женщин, принимающих пероральные контрацеп¬
тивы [90]. Несомненно, женщинам с контролируемой артериальной гипертензией,
но имеющим и другие заболевания или курящим, не следует принимать эстроген-
гестагенсодержащие контрацептивы (включая чрескожные и влагалищные пути
введения). Молодым женщинам с контролируемой артериальной гипертензией бе.з
сопутствующих заболеваний необходим частый и тщательный мониторинг показа¬
телей артериального давления. В связи с тем что инфаркт миокарда и инсульт чаще
развиваются после 35 лет, женщинам с контролируемой артериальной гипертензией
в этот период не следует принимать комбинированные эсгроген-гестагенсодержащие
препараты, а остановиться на препаратах, содержащих только гестаген.
Пролапс митрального клапана
при пролапсе митрального клапана пероральные контрацептивы можно назна¬
чать некурящим женщинам при отсутствии клинических симптомов (клинических
признаков митральной регургитации). У небольшого количества пациентов с про¬
лапсом митрального клапана повышен риск развития тромбоэмболии. Пациентам
с фибрилляцией предсердий, мигренозными головными болями или нарушениями
свертывания крови следует предпочесть препараты, содержащие только гестаген, или
внутриматочные средства (при наличии митральной регургитации перед введением
последних необходимо профилактическое назначение антибиотиков).
Курение
Пероральная контрацепция абсолютно противопоказана курящим женщинам
старше 35 лет. Она относительно противопоказана женщинам 35 лет и моложе,
если они выкуривают 15 сигарет и более в день. Относительный риск сердечно¬
сосудистых нарушений при применении контрацептивов возрастает у курящих жен¬
щин любого возраста, но в связи с низкой частотой сердечно-сосудистых нарушений
у молодых показатели абсолютного риска у них тоже низки (с возрастом увеличи¬
ваются). Женщин, не курящих в течение года и более, можно считать некурящими.
Вышеупомянутый риск касается только активных курильщиков. При отс)'тствии
какого-либо другого фактора риска низкодозированные пероральные контрацеп¬
тивы можно применять малокурящим или использующим никотиновые пластыри.
Препараты, содержащие 20 мкг эстрогена, — препараты выбора для курящих женщин
любого возраста, так как эстроген в этой дозе не влияет на факторы свертывания
крови и активацию тромбоцитов [30, 31].
Курение остается серьезной проблемой не только для здравоохранения в целом,
но и затрудняет анализ результатов исследований. В больших американских обзорах
1982 и 1988 гг. количество курящих и некурящих женщин в группах принимающих
и не принимающих пероральные контрацептивы было одинаковым; однако 24,3%
женщин от 35 до 45 лет, пользующихся пероральными контрацептивами, курили
[76]! В этой группе 85,3% женщин выкуривали 15 сигарет или более в день (много
курящие). Несмотря на широкое распространение информации, что курение служит
противопоказанием для приема пероральных контрацептивов для женщин старше
35 лет, многие женщины этого возраста, принимавшие пероральные контрацептивы,
курили, и курили много по сравнению с молодыми. Это служит явным свидетельством
того, что курящие женщины старше 35 лет не совсем искренни во время консультации
с врачом о выборе противозачаточных средств, что, в свою очередь, затрудняет про¬
ведение различных статистических исследований и анализ результатов. Курившая
женщина считается некурящей, если она не выкуривала ни одной сигареты в течение
предшествовавших 12 мес. Женщин, в крови которых имеется никотин, поступивший
из пластырей или жвачек, следует относить к группе курящих.
171
Врожденные заболевания сердца или пороки клапанов сердца
Пероральная контрацепция противопоказана лишь при тяжелой декомпенсации
или состояниях, предрасполагающих к тромбозу.
ЗАБОЛЕВАНИЯ, СВЯЗАННЫЕ С БЕРЕМЕННОСТЬЮ
Артериальная гипертензия, вызванная беременностью
Женщинам, у которых во время беременности развилась артериальная гипер¬
тензия, в послеродовом периоде можно начать прием пероральных контрацептивов
после нормализации показателей артериального давления.
Гестационный сахарный диабет
Низкодозированные препараты не вызывают изменений толерантности к глюкозе
у женщин с гестационным сахарным диабетом в анамнезе, также не выявлено увели¬
чения частоты возникновения клинически выраженного сахарного диабета при при¬
еме комбинированных пероральных препаратов [96, 97]. Женщины с гестационным
сахарным диабетом в анамнезе могут применять пероральные препараты при условии
ежегодного определения содержания глюкозы в крови.
Холестатическая желтуха при беременности
Не у всех пациентов с холестатической желтухой при беременности в анамнезе
разовьется желтуха при приеме пероральных контрацептивов, особенно это малове¬
роятно при назначении низкодозированных препаратов.
ДОБРОКАЧЕСТВЕННЫЕ ОПУХОЛИ
Лейомиома матки
Риск развития заболевания снижается на 31% у женщин, применявших высокодо¬
зированные пероральные контрацептивы в течение 10 лет [91]. Однако исследования
случаев развития заболевания у применявших низкодозированные препараты не
'выявили ни снижения, ни повышения риска, хотя в The Nurses' Health Study сообща¬
ли о небольшом повышении вероятности развития заболевания при начале приема
пероральных контрацептивов в раннем подростковом периоде [92-94]. В одном
исследовании показано снижение риска развития миоматоза матки при увеличении
длительности приема пероральных контрацептивов [95]. Прием низкодозированных
препаратов женщинами с лейомиомой не стимулирует фиброидный рост и сопрово¬
ждается уменьшением менструального кровотечения [95].
Доброкачественная опухоль молочной железы
Доброкачественная опухоль молочной железы не является противопоказанием к
пероральной контрацепции. После 2 лет приема препаратов опухоль иногда умень¬
шается в размерах.
Аденомы гипофиза, секретирующие пролактин
Эстроген стимулирует секрецию пролактина и вызывает рост лактотрофов гипо¬
физа, но пероральные контрацептивы не повышают риск возникновения аденомы
гипофиза и могут применяться при наличии микроаденом, не оказывая влияния на
их количество и рост.
НЕВРОЛОГИЧЕСКИЕ И ПСИХИЧЕСКИЕ НАРУШЕНИЯ
Головные боли по типу мигрени
у некоторых женщин головные боли уменьшаются с началом приема пероральных
контрацептивов. Низкодозированные препараты (содержащие наименьшую дозу
172
эстрогена) можно применять женщинам, страдающим мигренями без ауры, при усло¬
вии тщательного наблюдения- Ежедневный прием препаратов может предупредить
менструальные головные боли. При ауре или дополнительных факторах риска разви¬
тия инсульта (особенно возраст, курение, артериальная гипертензия) лзшше избегать
приема пероральных контрацептивов.
Судорожные расстройства
Пероральные контрацептивы не ухудшают течения эпилепсии, у некоторых жен¬
щин, напротив, даже облегчают [101,102]. Однако противоэпилептические средства,
влияющие на функции печени, могут снизить эффективность пероральной контра¬
цепции. Некоторые врачи выступают за применение высокодозированных (50 мкг
эстрогена) препаратов; тем не менее исследований, подтверждающих необходимость
повышения дозы, не проводили. Другая проблема состоит в том, что переход к более
высокодозированным препаратам приводит к повышению содержания в них эстро¬
генного компонента (и риску развития побочных эффектов), но существенно не изме¬
няет дозу гестагена, то есть компонента, подавляющего овуляцию. Благоразумнее
применять внутриматочный способ контрацепции, пролонгированные лекарственные
формы, содержащие гестаген, или прибегнуть к хирургической стерилизации.
Депрессия
Низкодозированные пероральные контрацептивы практически не оказывают
влияния на настроение.
НАРУШЕНИЯ ГЕМОПОЭЗА
Серповидноклеточная анемия
Женщины, страдающие серповидноклеточной анемией, могут применять перо¬
ральные контрацептивы. Потенцирование риска развития тромбоза при серпо¬
видноклеточной анемии возможно теоретически. Однако, по нашему убеждению,
эффективное предупреждение беременности оправдывает у этих больных прием
низкодозированных пероральных контрацептивов. В единственном сообщении о
длительном (в течение 10 лет) наблюдении за женщиной, страдающей этим заболе¬
ванием и применявшей пероральные контрацептивы (в это время на фармацевтиче¬
ском рынке преобладали высокодозированные препараты), не приводится сведений
о выявленных побочных эффектах [103]. В исследовании деформации эритроцитов
у женщин с серповидноклеточной анемией не выявлено побочных эффектов контра¬
цептивных препаратов на этот показатель [104]. Следует учитывать, что применение
депо медроксипрогестерона, используемого для контрацепции, сопровождается тор¬
можением образования серповидных клеток и улучшением показателей крови у этих
больных [105].
Геморрагические расстройства
Женщины с геморрагическими нарушениями и женщины, принимающие анти¬
коагулянты, могут использовать пероральную контрацепцию. Торможение овуляции
способно предотвратить у них серьезную проблему кровоизлияния в желтое тело.
Дополнительное положительное следствие — уменьшение менструальной кровопо¬
тери.
ВОСПАЛИТЕЛЬНЫЕ И ИММУННЫЕ РАССТРОЙСТВА
Системная красная волчанка
Пероральные контрацептивы способны усугубить течение системной красной
волчанки и сопутствующего поражения сосудов, поэтому эстрогенсодержащие кон¬
173
трацептивы противопоказаны при этом заболевании [106]. Препараты выбора — кон¬
трацептивы, содержащие только гестаген. Однако возможно применение низкодози¬
рованных пероральных контрацептивов, если заболевание протекает без обострений
или в неактивной фазе, отс}аствует поражение почек или нет высокой концентрации
антифосфолипидных антител [107].
Инфекционный мононуклеоз
Пероральные контрацептивы можно применять при нормальных функциональных
пробах печени.
Неспецифический язвенный колит
Связь между течением неспецифического язвенного колита и приемом перораль¬
ных контрацептивов отсутствует. Заболевание не является противопоказанием к
приему пероральных контрацептивов [123]. Эти препараты всасываются в основном
в тонком кишечнике.
Болезнь Крона
В проспективных когортных исследованиях не было выявлено отрицательного
влияния пероральных контрацептивов на клиническое течение заболевания, в част¬
ности влияния на обострение [124],
Плановое хирургическое вмешательство
Рекомендация прекратить прием пероральных контрацептивов за 4 нед до пла¬
нового большого хирургического вмешательства во избежание повышенного риска
развития послеоперационного тромбоза основана на результатах изучения высокодо¬
зированных препаратов. Безопаснее по возможности следовать этой рекомендации,
если в послеоперационном периоде пациентке предстоит длительная иммобилизация.
Женщинам, принимавшим в прошлом или продолжающим принимать пероральные
контрацептивы, при большой операции с последующей иммобилизацией рекомен¬
довано профилактическое введение антикоагулянтов. Лучше использовать другие
методы контрацепции вплоть до женской стерилизации.
ПОЛОЖИТЕЛЬНЫЕ ПОСЛЕДСТВИЯ ПРИМЕНЕНИЯ ПЕРОРАЛЬНЫХ КОНТРАЦЕПТИВОВ
Положительные последствия применения низкодозированных пероральных кон¬
трацептивов можно разделить на две основные группы: польза от их применения с
целью контрацепции (табл. 2-5) и польза от их применения для лечения некоторых
заболеваний и нарушений.
Таблица 2-5. Положительные последствия применения пероральных контрацептивов
Эффективная контрацепция
• Уменьшение случаев искусственного прерывания беременности.
• Уменьшение показаний к проведению женской стерилизации
Более регулярные менструации
Уменьшение кровопотери.
Ослабление болей при менструации.
Уменьшение выраженности анемии
Другие
Снижение риска рака эндометрия.
Снижение риска рака яичников.
Уменьшение вероятности возникновения внематочной беременности.
174
Окончание табл. 2-5
• Уменьшение вероятности развития сальпингита.
• Повышение плотности костной ткани.
• Вероятное уменьшение риска развития эндометриоза.
• Вероятное уменьшение риска развития доброкачественных опухолей молочной железы.
• Вероятное уменьшение риска развития ревматоидного артрита.
• Вероятная профилактика атеросклероза.
• Вероятное уменьшение риска развития кист яичника
Особенности применения пероральных контрацептивов
ТАБЛЕТКИ, СОДЕРЖАЩИЕ ТОЛЬКО ГЕСТАГЕН (МИНИ-ПИЛИ)
Мини-пили содержат низкую дозу гестагена, и их следует принимать ежедневно
[126, 127]. Имеющиеся на фармацевтическом рынке препараты практически не раз¬
личаются клиническими эффектами, однако дезогестрел более надежно подавляет
овуляцию. Практически все препараты, представленные в табл. 2-6, могли быть
использованы в высоких дозах (10-20 таблеток) для экстренной контрацепции, но
доказана эффективность только левоноргестрел а.
Таблица 2-6. Мини-пили, доступные на мировом фармацевтическом рынке
Торговые названия
Состав
Микронор®, нор-ОО, норидэй, нород
0,350 мг норэтистерона
МикровалР, норгестон^, микролют*
0,030 мг левоноргестрела
Оврет*, неогестр
0,075 мг норгестрела (эквивалентно 0,0375 мг левоноргестрела)
Экслютон*
0,500 мг линестренола^
Фемулен*’
0,500 мг этинодиала диацетата*’
Церазет»’
0,075 мг дезогестрела
Механизм действия
После приема некомбинированной таблетки создается небольшая концентрация
гестагена в крови (приблизительно 25% его содержания в комбинированных перо¬
ральных контрацептивах), оказывающая существенное действие только на ткани,
очень чувствительные к прогестерону. Контрацептивное действие в большей степени
зависит от влияния гестагена на состояние эндометрия и свойства цервикальной
слизи, так как секреция гонадотропных гормонов не подавляется в постоянном режи¬
ме. Шеечная слизь становится более густой, что значительно препятствует прохожде¬
нию через нее сперматозоидов, а в эндометрии развиваются изменения, затрудняю¬
щие возможную имплантацию оплодотворенной яйцеклетки. Приблизительно у 40%
женщин овуляция не подавляется [128, 129]. Высказано также предположение, что
препараты влияют и на сократимость маточных труб. Более эффективны мини-пили,
содержащие 0,075 мг дезогестрела, предположительно в связи с тем, что он сильнее
подавляет овуляцию по сравнению с другими гестагенами [130].
В связи с низкой дозой содержащегося гестагена некомбинированные таблетки
(мини-пили) необходимо принимать ежедневно в одно и то же время (строго герез каж¬
дые 24 г). Изменения слизи развиваются через 2-4 ч после приема препарата и, что
важно, уменьшаются через 22 ч, а полностью исчезают через 24 ч.
175
Мини-пили не предупреждают возникновения внематочной беременности столь
же эффективно, как нормальной. Несмотря на то что количество случаев возникно¬
вения внематочной беременности не повышается (оно все равно намного ниже, чем
у женщин, не принимающих контрацептивы), следует учитывать, что если все же
на фоне приема мини-пили беременность наступила, то наиболее вероятно, что она
внематочная. Внематочная беременность в анамнезе не считается противопоказани¬
ем для приема мини-пили, однако женщинам с риском возникновения внематочной
беременности следует пользоваться иными способами контрацепции, например ком¬
бинированными таблетками.
Мини-пили не оказывают существенного влияния на метаболические процессы
(содержание липидов в крови, углеводный обмен, факторы свертывания крови) [84,
131, 132, 133]. Фертильность восстанавливается сразу после прекращения приема
таблеток (в отличие от комбинированных таблеток, при приеме которых ее восста¬
новление отсрочено).
Эффективность
Контрацептивная эффективность мини-пили составляет от 1,1 до 9,6 беременности
на 100 женщин в течение первого года применения [134]. Количество нежелатель¬
ных беременности на фоне применения препаратов выше у более молодых женщин
(ЗД беременности на 100 женщин в год) по сравнению с женщинами старше 40 лет
(0,3 беременности на 100 женщин в год) [135]. У мотивированных женщин кон¬
трацептивная эффективность мини-пили сопоставима с таковой комбинированных
пероральных контрацептивов и составляет менее 1 беременности на 100 женщин в
год 1136,137).
Прием мини-пили
Прием мини-пили следует начинать с 1-го дня менструального цикла; в течение
первой недели использования необходимо применять дополнительный метод кон¬
трацепции, так как овуляция может возникнуть на 7-9-й дни от начала менструации.
Препараты следует принимать ежедневно в одно и то же время (с этой точки зре¬
ния их удобнее принимать перед какой-либо ежедневной привычной процедурой).
Если женщина пропустила время приема таблетки или в связи с нарушением работы
желудочно-кишечного тракта (ЖКТ) она могла не всосаться, необходимо принять
таблетку, как только женщина вспомнит о пропущенной таблетке (а следующую — в
обычное время), и далее в течение не менее 2 дней (после возобновления регулярного
приема) использовать дополнительньЕЙ метод контрацепции. Если женщина пропу¬
стила прием двух и более таблеток подряд и через 4-6 нед не наступила менструация,
следует провести тест на беременность. При задержке времени приема таблетки более
чем на 3 ч в течение 48 ч после ее приема следует использовать дополнительный метод
контрацепции.
Проблемы
в связи с тем что при приеме мини-пили овуляция в некоторых сл>'чаях проис¬
ходит, основная клиническая проблема — нерегулярность менструальных кровоте¬
чений, связанная также с влиянием гестагена на состояние эндометрия. У пациентов
возможны нормальные овуляторные менструальные циклы (у 40-50%), короткие,
нерегулярные циклы (у 40%) или полное их отсутствие (от нерегулярных кровоте¬
чений до кровянистых мажущих выделений и аменореи — у 10%). Нерегулярность
кровянистых выделений из половых путей относят к основной причине, по которой
женщины прекращают использовать этот метод контрацепции [137].
У женщин, применяющих мини-пили, чаще развиваются кисты яичника, чем у при¬
меняющих другие методы, полностью подавляющие секрецию гонадотропных гормо¬
нов [138,139]. В связи с тем что практически все кисты подвергаются регрессии, этот
176
эффект не относят к клиническим проблемам, однако при наличии множественных
кист лучше применять препараты, полностью подавляющие овуляцию (например,
комбинированные пероральные контрацептивы).
Мини-пили, содержащие левоноргестрел, могут привести к появлению угрей.
Механизм действия аналогичен использованию норпланта* (см. ниже). Андрогенная
активность левоноргестрела приводит к снижению концентрации в крови андроген-
связывающего глобулина [140]. Следовательно, содержание свободного тестостерона
повышается. Комбинированные пероральные контрацептивы оказывают противопо¬
ложное действие, так как их эстрогенный компонент вызывает повышение андроген-
связывающего глобулина.
Мини-пили не уступают по контрацептивной эффективности комбинирован¬
ным таблеткам у женщин старше 40 лет и у кормящих грудью. У последних они не
оказывают неблагоприятного действия на количество грудного молока, рост и раз¬
витие младенца [141-143]. Установлено, что у женщин, принимающих мини-пили,
выработка грудного молока прекращается немного позже, чем обычно [144]. В связи
с небольшим положительным влиянием на лактацию можно начинать прием мини-
пили сразу после родов. Не установлено нежелательного влияния раннего начала
приема этих препаратов на лактацию [145].
Мини-пили можно принимать в случаях, когда прием эстрогенсодержащих табле¬
ток противопоказан (сахарный диабет с сосудистыми осложнениями, тяжелое тече¬
ние системной красной волчанки [146], тяжелые заболевания сердечно-сосудистой
системы). В этих сл)гчаях предполагают отсутствие осложнений, связанных с влия¬
нием эстрогена. С другой стороны, есть все основания полагать, что все эффекты,
вызываемые гестагенным компонентом комбинированных таблеток, проявятся при ,
приеме мини-пили, но в связи с дозозависимостью при приеме последних они будут
слабее. Исследование, проведенное ВОЗ {The World Health Organizntion case-control
study), и межнациональное исследование (Transnational case-control study) не выявили
повышения риска развития инсульта, инфаркта миокарда или венозной тромбоэм¬
болии при приеме некомбинированных таблеток, но исследования были посвящены
эффектам комбинированных пероральных контрацептивов и включали лишь неболь¬
шое число случаев приема мини-пили [147,148]. Не выявлено влияния мини-пили на
систему свертывания крови [131, 149]. Вероятно, их можно применять женщинам с
тромбозом в анамнезе; в США из списка противопоказаний была исключена болезнь
периферических сосудов.
Мини-пили можно применять в тех редких случаях, когда прием комбинирован¬
ных пероральных контрацептивов вызвал снижение полового влечения, предполо¬
жительно вследствие антиандрогенного действия. Их также можно рекомендовать
некоторым женщинам, отказавшимся от применения комбинированных таблеток
вследствие развития таких неопасных побочных эффектов, как нарушения функции
ЖКТ, нагрубание молочных желез, головные боли.
Мини-пили содержат относительно низкие дозы гестагена, поэтому при лечении
лекарственными препаратами, активирующими печеночный метаболизм, следует
отказаться от некомбинированных контрацептивов (таблеток, имплантов). К лекар¬
ственным средствам, способным снизить контрацептивную эффективность перораль¬
ных, в том числе некомбинированных, таблеток относятся карбамазепин (тегретол),
фелбамат*“, невирапин, окскарбазепин, фенобарбитал, фенитоин (дилантин), при-
мидон (мисолин), рифабутин, рифампицин (рифампин), топирамат, вигабатрин и,
возможно, этосуксимид, гризеофульвин*“ и троглитазон*“*.
В связи с меньшим количеством женщин, использующих для контрацепции мини-
пили, неизвестно, вызывают ли они такие же положительные эффекты, как комбини¬
рованные пероральные контрацептивы. Некоторые исследователи предполагают, что
* Б настоящее время препарат не используется. {Примег. ред.)
177
такие эффекты возможны в связи с влиянием мини-пили на вязкость шеечной слизи,
эндометрий и овуляцию (снижение риска инфекции органов малого таза, рака эндо¬
метрия и яичника). Несмотря на малое количество случаев, в одном исследовании
показано, что мини-пили предупреждают развития рака эндометрия более эффектив¬
но, чем комбинированные пероральные контрацептивы [150].
Условие хорошей контрацептивной эффективности мини-пили — ежедневный
прием в одно и то же время суток, что создает дополнительные трудности для неорга¬
низованных взрослых или подростков.
ЭКСТРЕННАЯ КОНТРАЦЕПЦИЯ
Применение высоких доз эстрогена для предупреждения имплантации оплодотво¬
ренной яйцеклетки впервые было предложено Моррис и ван Вагенен в Йеле в 1960 г.
В результате первых исследований на обезьянах были предложены для женщин высо¬
кие дозы диэтилстилбестрола (25-50 мг/сут) и этинилэстрадиола [151]. Вскоре было
установлено, что прием этих чрезвычайно высоких доз сопровождался побочными
эффектами со стороны ЖКТ. Альберт Юзпе развил метод, предложив применять ком¬
бинированные контрацептивы, что позволило существенно снизить дозу компонентов
[152]. Следующие схемы эффективны для экстренной контрацепции (табл. 2-7):
• Оврал*“: 2 таблетки, затем 2 таблетки через 12 ч (0,5 мг норгестрела и 50 мкг эти¬
нилэстрадиола в каждой таблетке).
• Алесс^: 5 таблеток, затем 5 таблеток через 12 ч (100 мг левоноргестрела и 20 мкг
этинилэстрадиола в каждой таблетке).
• Ло Оврал*“, нордетт*“, левлен*“, трифазил^ (желтые таблетки), трилевлен*“ (жел¬
тые таблетки): 4 таблетки, затем 4 таблетки через 12 ч.
Для экстренной контрацепции прием левоноргестрела в дозе 0,75 мг дважды с
интервалом 12 ч или в дозе 1,5 мг однократно эффективнее и переносится лучше,
чем прием комбинированных таблеток [153,154]. Во многих странах для экстренной
контрацепции применяют специальные упаковки, содержащие 2 таблетки левонор¬
гестрела по 0,75 мг (план В^, постинор*, норлево*“, викела*“). В других странах экви¬
валентную дозу получают приемом 20 некомбинированных таблеток, содержащих
левоноргестрел. В связи с более высокой эффективностью и меньшим количеством
побочных эффектов прием левоноргестрела — метод выбора для экстренной контра¬
цепции [155,156].
Таблица 2-7. Экстренная контрацепция
Торговое
наименование
Дозы в одной таблетке
Первый прием
Второй прием
(через 12 ч)
Этинилэстрадиол
Левоноргестрел
Норгестрел
Алесс^
20 мкг
0,10 мг
-
5 таблеток
5 таблеток
Левлен*’
30 мкг
0,15 мг
-
4 таблетки
4 таблетки
Ло Оврал*»
30 мкг
-
0,3 мг
4 таблетки
4 таблетки
Нордет*’
30 мкг
0,15 мг
-
4 таблетки
4 таблетки
Оврал*'
50 мкг
-
0,5 мг
2 таблетки
2 таблетки
План В*
Не содержится
0,75 мг
-
1-2 таблетки
0-1 таблетка
ТрилевленР
30 мкг
0,125 мг
-
4 таблетки
4 таблетки
Трифазил*’
30 мкг
0,125 мг
-
4 таблетки
4 таблетки
Оврет»’, нео-
гест»’
Не содержится
-
0,075 мг
20 таблеток
20 таблеток
* Только для экстренной контрацепции, принимают одну или две дозы.
178
Этот метод обычно называют посткоиталъной контрацепцией, но более подходя¬
щее название — экстренная контрацепция, отражающая ее применение лишь шора-
дически, например в случаях нарушения целостности презерватива, изнасилования,
смещения диафрагмы или шеечного колпачка, а также при неправильном исполь¬
зовании другого метода. Анализ случаев искусственного прерывания беременности
показал, что 50-60% женщин могли бы избежать возникновения нежелательной
беременности, если бы захотели или имели возможность использовать экстренную
контрацепцию [157, 158J. Б США установлено, что экстренная контрацепция еже¬
годно могла бы предотвратить 1,7 млн нежелательных беременностей, а количество
искусственного:их прерывания снизить на 40% [159].
Многие женщины не знают о существовании такого метода [158,160], а даже если
знают, то недостаточно подробно [161]. Те, кто пользовался экстренной контрацеп¬
цией, удовлетворены этим методом и, что наиболее важно, не намерены заменить им
регулярную контрацепцию [162]. Информацию для пациентов и врачей, в том числе
о новейших препаратах, можно получить на веб-сайте Office of Population Research at
Princeton University: http://ec.princeton.edu.
Врачам следует рассмотреть вопрос обеспечения каждой женщины специальным
набором для экстренной контрацепции (в простейшем варианте это может быть кон¬
верт с необходимым количеством таблеток и инструкцией по их применению), кото¬
рый она сможет использовать при необходимости. Было установлено, что молодые
женщины в Шотландии и Калифорнии стали чаще прибегать к экстренной контра¬
цепции без нежелательных последствий в виде увеличения количества половых кон¬
тактов без предохранения или заражения заболеваниями, передающимися половым
путем [163-167],
Гестагенсодержащие препараты для экстренной контрацепции в настоящее время
продаются без рецептов во многих странах, в том числе США, где их может купить
любая женщина, достигшая 18 лет, без консультации с врачом. Лицам более моло¬
дого возраста, кому они наиболее необходимы, следует иметь рецепт, кроме редких
исключений, предусмотренных законом [168]. Доступность таблеток для экстренной
контрацепции не приводит к ее использованию в качестве регулярного метода предо¬
хранения [168]. Подростки прибегают к экстренной контрацепции в тех же случаях,
что и взрослые [167].
Механизм действия и эффективность
Механизм действия окончательно не выяснен, справедливо полагают, что он соче¬
тает торможение овуляции с местным изменением эндометрия и предупреждением
оплодотворения [169-173]. Вопрос о том, как действуют препараты после произо¬
шедшего оплодотворения, неясен, но этот механизм не считают основным [172,174].
Эффективность экстренной контрацепции подтверждена в больших клиниче¬
ских исследованиях и обобщена в подробных литературных обзорах [175-177],
Количество возникших беременностей после приема высоких доз эстрогена или
применения левоноргестрела составляет приблизительно 1%, а при применении ком¬
бинированных пероральных контрацептивов — 2-3%. Этот показатель наименьший
при приеме высоких доз этинилэстрадиола в первые 72 ч после полового акта (0,1%),
но развившиеся побочные эффекты ограничивают использование этого метода. Б
целом применение пероральных контрацептивов позволяет снизить риск возникнове¬
ния нежелательной беременности приблизительно на 75% [179-181].
Результаты применения левоноргестрела даже лучше и снижают риск возникнове¬
ния беременности приблизительно на 85%; согласно обширному исследованию, про¬
веденному БОЗ, риск возникновения беременности был на 60% ниже при применении
левоноргестрела по сравнению с приемом комбинированных таблеток, и при этом
практически вдвое реже развивались тошнота и рвота [154].
179
Методика приема препаратов
Прием таблеток необходимо начать как можно скорее после полового акта, но не
позднее 120 ч. Тщательный анализ литературных данных показывает, что эффектив¬
ность экстренной контрацепции одинаковая, если прием таблеток начат в 1-й, 2-й или
3-й день после полового акта, и может сохраняться и через 72 ч [182, 183]. Однако
результаты рандомизированного клинического испытания, проведенного ВОЗ, под¬
тверждают зависимость эффективности применения экстренной контрацепции от
времени, прошедшего после полового контакта, и свидетельствуют, что эффектив¬
ность снижается после 72 ч, а наибольшая — при приеме препарата в первые 24 ч
[184]. Опоздание с приемом препарата на 12 ч повышает вероятность возникновения
беременности на 50%. Именно поэтому прием препарата необходимо начать как
можно скорее после полового контакта, что подтверждает целесообразность всегда
иметь при себе таблетки для экстренной контрацепции.
Если пероральные контрацептивы приняты при наступившей беременности, то
эстроген и гестаген в дозах, содержащихся в них, не окажут на эмбрион тератогенного
действия [185, 186, 187]. Кроме того, экстренная контрацепция неэффективна при
наличии беременности. Задержка менструации после применения препаратов служит
показанием для проведения теста на беременность и оправдывает предположение о
возможности внематочной беременности.
При экстренной контрацепции можно применять и противорвотные средства,
например длительно действующий препарат меклизин^’ (бонин®, драмамин
антиверт^'’) в дозе 25 или 50 мг за 1 ч до первого приема контрацептивов. Побочные
эффекты высоких доз гормонов: тошнота (50%), рвота (20%), нагрубание молочных
желез, головные боли и головокружение. Если рвота развивается в течение первого
часа после приема контрацептива, нужно как можно скорее повторить прием пре¬
парата. Так как прием одного левоноргестрела обычно не вызывает тошноту и рвоту,
принимать противорвотные средства необязательно.
The UK General Practice Research Database не выявил повышения риска развития
венозной тромбоэмболии при применении пероральных контрацептивов для экс¬
тренной контрацепции. Не было выявлено ни одного случая этого осложнения в тече¬
ние 60 дней после экстренной контрацепции среди 100 ООО женщин, ее проводивших
[188,189].
Установлено, что эффективность комбинаций норэтиндрон-этинилэстрадиол и
левоноргестрел-этинилэстрадиол одинаковая, вероятно, любой комбинированный
пероральный контрацептив будет эффективным [190]. Однако это спорная точка зре¬
ния, так как применение только левоноргестрела в настоящее время считают методом
выбора экстренной контрацепции.
Тремя основными проблемами экстренной контрацепции являются высокая
частота развития побочных эффектов, необходимость начать прием таблеток сразу
после полового акта и пусть невысокая, но важная для каждой женщины частота ее
неэффективности. Применение однократно внутрь антагониста прогестерона мифе-
пристона в дозе 600 мг реже вызывает тошноту и рвоту, чем комбинированные кон¬
трацептивы, и эффективно почти в 100% случаев [191,192]. В рандомизированных
испытаниях доза мифепристона, равная 19 мг, была равноэффективна 25, 50 или
600 мг и предупреждала 80-85% возможных беременностей (эффективность такая
же, как при применении левоноргестрела); эффективность незначительно снижает¬
ся, если прием препарата был начат через 5 дней после полового акта [155,193,194].
В связи с тем что мифепристон вызывает задержку начала менструации, сразу после
его приема необходимо использовать методы контрацепции. Мифепристон может
внести существенный вклад в предупреждение нежелательных беременностей и
искусственных абортов.
180
ВЛАГАЛИЩНОЕ И ЧРЕСКОЖНОЕ (ТРАНСДЕРМАЛЬНОЕ)
ПРИМЕНЕНИЕ СТЕРОИДОВ ДЛЯ КОНТРАЦЕПЦИИ
Влагалищное и трансдермальное применение половых стероидов имеет потенци¬
альное преимущество перед пероральным приемом в том, что исключается эффект
первого прохождения препаратов через печень и создаются высокие их концентрации
в крови. Например, можно уменьшить влияние препаратов на свертываемость крови,
так как не стимулируется синтез белков в печени. Более высокая безопасность влага¬
лищного и трансдермального применения половых стероидов требует дальнейшего
эпидемиологического подтверждения, однако теоретически возможна. Следующее
возможное преимущество этих методов контрацепции — большее удобство для жен¬
щины, так как отсутствует необходимость ежедневного приема таблеток.
Влагалищная стероидная контрацепция
Слизистая оболочка влагалища представляет собой превосходное место для вса¬
сывания стероидных препаратов. Бсасывание из ЖКТ может быть непредсказуемым
и зависит от многих факторов (наличие рвоты, лекарственного взаимодействия или
снижения всасывающей способности кишечника). Избежать эффекта первого про¬
хождения через печень особенно важно для препаратов, подвергающихся в значи¬
тельной степени печеночному метаболизму; природные эстрогены, например, при
пероральном приеме на 95% метаболизируются в печени. Пероральный прием вызы¬
вает значительные колебания концентрации стероидов в сыворотке крови, что может
привести к таким побочным эффектам, как нерегулярные маточные кровотечения и
тошнота. Эти суточные колебания концентрации в крови при влагалищном введении
менее выражены, чем при пероральном или трансдермальном, и наименьшие при
имплантационном и внутриматочном путях применения [195,196].
Благалищные контрацептивные кольца изучали в течение 30 лет. Было создано
шесть видов колец, содержащих только гестагены, и семь видов колец, содержащих
комбинации гестаген-эстроген, предназначенные для оказания контрацептивного
действия длительностью от 1 нед до 1 года. Короткодействующие (в течение 1 нед)
кольца содержали прогестерон и медроксипрогестерон, а длительнодействующие (до
1 года) — более сильные левоноргестрел и нестерон.
Лишь влагалищный комбинированный гормональный контрацептив НуваРинг^ раз¬
решен к применению в США (рис. 2-1).
Он представляет собой гибкое мягкое
прозрачное кольцо, изготовленное из
этиленвинилацетатного кополимера
(60%), в котором содержатся кристаллы
этоногестрела (биологически активного
метаболита дезогестрела, ранее извест¬
ного как 3-кетодезогестрел) и этини¬
лэстрадиола. Кольцо покрыто мембра¬
ной из этиленвинилацетата толщиной
2 мм, его толщина 4 мм, диаметр 54 мм
(меньше, чем диафрагма). НуваРинг*“
выделяет 15 мкг этинилэстрадиола и
120 мкг этоногестрела в сутки [197].
Концентрация в крови эстрогена дости¬
гает максимальных значений через 2-3 2-1, Влагалищное кольцо, содержащее комби-
дня, а этоногестрела - через 7 дней, и нацию эстроген-гестаген.
181
эти значения сохраняются в течение 35 дней [196] (рис. 2-2). Кольцо женщина вво¬
дит самостоятельно на 3 нед. Обычно кольцо заменяется на новое через каждые 4 нед
(перерыв на 1 нед на время менструального кровотечения), но можно его применять
и в непрерывном режиме.
Содержание в крови гестагена и эстрогена, вьщеляющихся из кольца, составляет
соответственно лишь 40 и 30% максимальной концентрации гормонов, выделяемых
из пероральных контрацептивов, содержащих 150 мкг дезогестрела и 30 мкг этинил-
эстрадиола [196]. Эти количества вызывают эффективное подавление овуляции,
согласно клиническим исследованиям, нежелательная беременность возникает менее
чем у 1% женщин [197-199]. Содержание гормонов в кольце достаточно для тормо¬
жения овуляции в течение 5 нед [198]. Если влагалищное кольцо удалено более чем
на 3 ч, производитель рекомендует применять барьерные методы контрацепции в
течение последующих 7 дней.
Связывание с белками плазмы крови влияет на биодоступность всосавшихся гормо¬
нов, поэтому системное действие этоногестрела, содержащегося во влагалищном коль¬
це, такое же, как 150 мкг дезогестрела. содержащегося в таблетках. Однако системное
действие этинилэстрадиола, выделяющегося из кольца, составляет 50% такового из
таблеток, содержащих 30 мкг вещества [196]. Эти данные могут объяснить меньшее
количество эстрогензависимых побочных эффектов, таких как тошнота, нагрубание
молочных желез, головные боли, развивающихся при применении влагалищного
кольца [197, 199], Межменструальные кровотечения и кровянистые выделения воз¬
никают реже (приблизительно у 6% женщин) (рис. 2-3), чем при приеме пероральных
контрацептивов, содержащих 30 мкг этинилэстрадиола, и значительно реже, если его
содержание составляет 15 или 20 мкг [199, 200].
Не имеет значения, где и как располагается кольцо во влагалище, необходимо
лишь, чтобы оно соприкасалось со слизистой оболочкой. При анатомических осо¬
бенностях строения влагалища или инфекциях следует применять другие методы
контрацепции. Наиболее частая причина отказа от этого метода (в клинических
исследованиях приблизительно 2-4% женщин) — неприятные ощущения во влагали¬
ще, сомнения в его наличии на месте, проблемы во время полового акта или его выпа¬
дение (в течение года использования возникает приблизительно у 2-3% женщин). По
мнению женщин, кольцо легко помещать и извлекать из влагалища, и хотя около 15%
женщин и 30% половых партнеров сообщают, что они чувствуют наличие кольца во
время полового акта, обычно это не служит причиной отказа от использования такого
Нормальное
использование
Расширенное
использование
Количество дней после введения
Рис. 2-2. Фармакокинетические показатели влагалищного кольца при длительном применении. [Из
Timmer C.J., Mulders Т.М. Pharmacokinetics of etonogestrel and ethinylestradiol released from a combined
contraceptive vaginal ring // Clin. Pharmacokinet. - 2000. — N 39. — P. 233-242.]
182
40-
35-1
30-
25-
20-
15-
10-
54
О
^ ^ ^ ^
■ ■ ■ ».Ж! 1..1.
15ЭЭ/60ГСТ
20 3Э/150ДЗГ
ЗОЭЭ/150ДЗГ
М.М
1 2 3
6 7
Цикл
Т
9 10 11 12 13
Рис. 2-3. Нерегулярные кровянистые выделения из матки при применении влагалищного кольца. ДЗГ —
дезогестрел; ЭЭ — этинилэстрадиол: ГСТ — гестоден. [Из Roumen F.J., Apter D., Mulders T.M., Dieben T.O.
Efficacy, tolerability and acceptability of a novel contraceptive vaginal ring releasing etonogestrel and ethinyl
estradiol // Hum. Reprod. — 2001. — N 15. — P. 469-475.]
метода [199, 201]. He рекомендовано извлечение кольца перед половым актом, хотя
эффективность контрацепции сохраняется, если его поместить обратно в ближайшие
3 ч. Применение кольца не вызывает цитологических и микробиологических измене¬
ний во влагалище [199, 202, 203].
Спермицидное средство ноноксилон-9*“ не оказывает влияния на выделение из
НуваРинга*“ и абсорбцию этинилэстрадиола и этоногестрела, что показано при
изучении концентрации этих веществ в сыворотке крови [204]. Одновременное
использование барьерного метода с применением спермицидного средства (широко
одобренный контрацептивный метод, обеспечивающий также защиту от заболева¬
ний, передающихся половым путем) не влияет на контрацептивную эффективность
кольца. Ни влагалищное применение противогрибковых препаратов (миконазола),
ни использование тампонов не изменяют всасывание контрацептивных компонентов
кольца [204].
Црттжшт (транодермальше) гщшдштътш
контрацептивов
МЕТОД
Трансдермальный контрацептивный пластырь (орто евра*“) представляет собой
трехслойный квадрат площадью 20 см^ (4,5 смх4,5 см). Наружный полиэстерный слой
образует защитную пленку, средний слой содержит гормоны и адгезивные вещества, а
нижний слой - прокладка из полиэстера, которую снимают с липкой стороны перед
накладыванием пластыря. Размер пластыря обеспечивает поступление эффективной
дозы гормонов. Пластырь содержит 750 мкг этинилэстрадиола и 6,0 мг норелгестро-
мйна и после наложения на разные участки кожи нижней части живота, наружной
стороны плеча, ягодичной области и верхней половины туловища (кроме грудной
области) обеспечивает ежедневное поступление в организм 20 мкг этинилэстрадио¬
ла и 150 мкг норелгестромина. Норелгестромин — активный метаболит перорально
примененного норгестимата — ранее был известен как 17-деацетилноргестимат.
Норелгестромин при трансдермальном применении все же подвергается печеночно¬
му метаболизму, однако образующийся метаболит, левоноргестрел, в значительной
степени взаимодействует с андрогенсвязывающим глобулином, что ограничивает его
биологическое действие. Приблизительно 97% норелгестромина связывается с альбу¬
минами плазмы крови [205].
183
Пластырь следует накладывать еженедельно в один и тот же день на разные
участки тела в течение 3 нед, затем необходим недельный перерыв. Время наложе¬
ния пластыря не имеет значения, так как эффективные концентрации препаратов
в крови поддерживаются 9 дней. В течение первой недели применения пластыря
следует использовать дополнительные методы контрацепции (если начало приме¬
нения .не совпало с 1-м днем менструального цикла). Как и в случае с пероральными
контрацептивами, пациент и врач могут выбрать режим непрерывного применения
пластыря, что предупреждает развитие менструальных кровотечений, но приводит к
нерегулярным кровянистым выделениям.
приблизительно 5% пластырей самопроизвольно отлипает от кожи, из них поло¬
вина — в течение первого цикла их применения [206, 207]. В исследованиях, прово¬
димых в течение 1 года и более, установлено, что пациентки повторно накладывали
от 2 до 5% пластырей [208,209], Если пластырь не был в контакте с кожей более 24 ч,
накладывают новый пластырь, и этот день считается 1-м (новым) днем нового цикла
применения (рекомендованы дополнительно барьерные методы контрацепции в тече¬
ние 7 дней). При опоздании с началом нового цикла необходимо начать новый цикл
(с дополнительным способом контрацепции в течение первых 7 дней). Опоздание с
заменой пластыря не более чем на 2 дня в пределах одного цикла не повышает риск
возникновения беременности и не изменяет схемы применения, однако, если опозда¬
ние превышает 2 дня, следует начать новый цикл (с дополнительным способом предо¬
хранения в первую неделю). Однако в сравнительном исследовании возникновения
овуляции при трехдневном перерыве в применении пероральных контрацептивов и
пластырей установлено, что овуляция значительно реже возникала в группе женщин,
использовавших пластыри [210].
ЭФФЕКТЫ
Концентрация гормонов в крови после наложения пластыря возрастает быстро и
достигает в среднем приблизительно 0,7 нг/мл, концентрация норелгестромина нахо¬
дится в пределах 0,6-1,2 нг/мл, в среднем около 50 пг/мл, а этинилэстрадиола — в
пределах 25-75 пг/мл. Это те же концентрации, которые отмечают при приеме табле¬
ток, содержащих 250 мкг норелгестромина и 35 мкг этинилэстрадиола (рис. 2-4) [211,
212]. Однако изменения концентраций гормонов в крови при применении пластырей
и таблеток различны; пластыри не вызывают колебаний концентрации в течение дня,
но площадь под кривой (АОС) в течение недели применения пластыря больше, чем
160 Н
I 140'
І 120
Il 100
ОТ 2
і со
ш о.
80
60-
40
20-
О-
— НуваРинг^
Пластырь
Пилюли
14
21
Дни
Рис. 2-4. Сравнительная характеристика фармакокинетики этинилэстрадиола после его приема в
контрацептивном кольце, пластыре и таблетке. [Из Abrams LS., Skee D.M., Natarajan J. et al. Multiple-
dose pharmacokinetics of a contraceptive patch in healthy women participants // Contraception. — 2001. —
N 64. - P. 287-294,]
184
при ежедневном приеме 35 мкг этинилэстрадиола. Не установлено, приводит ли эта
разница к повышению риска развития тромбоза при использовании пластырей, так
как имеется лишь два исследования с противоречивыми заключениями. Эффективные
концентрации гормонов в крови достигаются даже в том случае, если замена пластыря
задержана на 2 дня (старый пластырь находился на коже 9 дней) [213]. Концентрации
гонадотропных гормонов возвращаются к исходным значениям через 6 нед после
прекращения применения пластырей [210]. Ежедневное ношение пластыря хорошо
изучено; такие факторы, как физические нагрузки, купание, плавание, сауна или
горячая ванна, не вызывают его отлипания или изменений концентрации гормонов
в крови [214].
Трансдермальная контрацепция вызывает такой же спектр фармакологических
эффектов и имеет такой же показатель эффективности в клинических испытаниях,
что и пероральные контрацептивы. Следовательно, все приведенные выше сведения
о пероральных контрацептивах относятся и к трансдермальной контрацепции, в том
числе противопоказания и положительные стороны. При трансдермальном приме¬
нении препаратам не свойствен эффект первого прохождения через печень, что дает
основание предполагать меньшую вероятность лекарственных взаимодействий, одна¬
ко это лишь предположение, поэтому женщинам, принимающим лекарственные пре¬
параты, влияющие на печеночный метаболизм, следует использовать иные методы
контрацепции. Прием тетрациклина не изменяет концентрации стероидных гормонов
в крови при применении трансдермальной контрацепции [215].
При трансдермальном применении происходят те же изменения факторов сверты¬
вания крови и липопротеинов, что и при приеме низкодозированных пероральных
контрацептивов. Установлено, что параметры гемо коагуляции клинически значимо
не меняются, уровень триглицеридов повышается умеренно, соотношение липопро¬
теинов низкой плотности и липопротеинов высокой плотности уменьшается незна¬
чительно [216]. Женщинам с высокой степенью риска развития тромбоза, связанного
с врожденными дефектами, подобно дефициту фактора V Лейдена или белков С или
S, а также при очень высоком содержании триглицеридов в крови следует (как и при
выборе пероральных контрацептивов) предпочесть гормональные препараты, не
содержащие эстрогена.
КЛИНИЧЕСКАЯ КАРТИНА
Степень торможения овуляции при трансдермальном методе сопоставима с тако¬
вой при приеме пероральных контрацептивов, а показатель возникновения нежела¬
тельных беременностей менее 1,0% [113, 206, 208, 209]. Согласно рандомизирован¬
ным испытаниям, частота маточных кровотечений и кровянистых выделений такая
же, как при применении монофазных и двух трехфазных препаратов, за исключением
незначительного учащения кровянистых выделений в первые два цикла [206, 208].
Современные гормональные контрацептивы не вызывают повышения массы
тела. Трансдермальный метод не является исключением; в сравнительных рандо¬
мизированных исследованиях показано, что изменение массы тела при применении
контрацептивных пластырей и идентичных пластырей, содержащих плацебо, было
одинаковым [217]. При клинических испытаниях контрацептивных пластырей выяв¬
лено возникновение нежелательной беременности у 15 женщин, причем у пятерых
масса тела превышала 90 кг [113]. Это совпадает с данными о меньшей эффективности
пероральных контрацептивов у женщин с ожирением [112]. Однако даже у этой кате¬
гории пациенток в целом контрацептивная эффективность высокая, так как только
10-20% изменений концентрации гормонов в крови связано с повышенной массой
тела [113].
Основная причина возникновения нежелательной беременности при приеме перо¬
ральных контрацептивов — нарушение правила необходимости ежедневного их
185
приема (вследствие забывчивости, недисциплинированности и т.д.). Трансдермальная
контрацепция с режимом замены пластыря лишь 1 раз в неделю удобнее и менее
чувствительна к опозданиям с заменой старого пластыря на новый. Согласно данным
рандомизированных исследований, охватывающих 4-13 циклов применения пласты¬
рей, на 10-20% женщин больше сочли применение пластырей более удобным, чем
прием пероральных контрацептивов [206, 208], В клинических исследованиях более
низкий показатель беременностей, возникших на фоне применения пластырей, чем
таблеток, был объяснен большей приверженностью (комплаентностью) женщин.
Самое важное, что женщины молодого возраста, особенно моложе 20 лет, чаще пред¬
почитают применение трансдермальной контрацепции, чем пероральной, по сравне¬
нию с более старшим поколением [218].
Приблизительно у 20% женщин развивалась небольшая кожная реакция на месте
прикрепления пластыря, в связи с которой 2% отказались от этого метода контра¬
цепции [206, 208, 209, 217]. Неприятные ощущения в области молочной железы
появлялись в первые несколько месяцев у 20% женщин, чаще при трансдермальной
контрацепции, чем при пероральной, но обычно были незначительными и привели к
отказу от этого метода лишь 1% пользователей [217].
ИНЪЕКЦИОННАЯ КОНТРАЦЕПЦИЯ
Депо-медроксипрогестерона ацетат
Депо-медроксипрогестерона ацетат (ДМПА, депо-провера^) — наиболее подробно
изученный гестагенсодержащий контрацептив. Он был разрешен к применению в
США в 1992 г., хотя в некоторых странах применялся с середины 60-х гг. Очень боль¬
шая часть сведений о безопасности, эффективности и приемлемости длительно дей¬
ствующей контрацепции поступает из Индонезии, Шри-Ланка, Таиланда и Мексики,
^’де ДМПА применяли на протяжении десятилетий. Значительно более позднее раз¬
решение применения в США было обосновано больше политическими и экономиче¬
скими, чем научными соображениями [219].
ДМПА представляет собой водную суспензию микрокристаллов гестагена. Для
контрацепции препарат показан в дозе 150 мг внутримышечно (в ягодичную или
дельтовидную мышцу) каждые 3 мес. В сравнительном исследовании установлена
существенно меньшая эффективность внутримышечного введения препарата в дозе
100 мг [220], однако в 2005 г, был выпущен новый препарат, введение которого в дозе
104 мг подкожно каждые 3 мес было равноэффективно внутримышечному введению
предыдущего в дозе 150 мг. Эффективная концентрация гормона в крови достигается
через 14 нед после введения. Из существующих контрацептивов ДМПА имеет наи¬
большую эффективность, которая составляет в феднем 1 беременность на 100 жен¬
щин после непрерывного применения в течение 5 лет [220, 221].
ДМПА не является лекарственной формой с замедленным высвобождением гор¬
монов; из него выделяются пиковые концентрации, достаточные для подавления
овуляции и изменения вязкости цервикальной слизи. Минимальные концентрации
гестагенов в крови при применении ДМПА в 10 раз выше, чем при применении под¬
кожных и внутриматочных депо-препаратов с замедленным высвобождением гор¬
монов. К другим широко применяемым инъекционным контрацептивам относятся
норэтиндрона энантат^ по 200 мг каждые 2 мес, люнелл*^ (25 мг медроксипрогестеро-
на ацетата и 5 мг эстрадиола ципионата) и мезигина^ (50 мг норэтиндрона энантата и
5 мг эстрадиола валерата) ежемесячно.
Показания и противопоказания для клинического применения ДМПА приведены
в табл. 2-8.
186
Таблица 2-8. Показами и противопоказания по применению депо-проверы
Показания
Контрацепция у женщин, не желающих иметь детей в течение ближайшего года и более
Высокоэффективная длительная контрацепция у женщин, не желающих заботиться о предотвращении беремен¬
ности перед каждым половым актом
Контрацепция у женщин, по каким-либо причинам скрывающих факт применения контрацептивов
Случаи, когда показаны препараты, содержащие только гестаген
Контрацепция у кормящих матерей
• Контрацепция у женщин с серповидноклеточной анемией
Контрацепция у женщин, страдающих судорожными расстройствами
Противопоказания
Абсолютные
Беременность
• Влагалищные кровотечения неясного генеза
Выраженные нарушения свертывания крови
• Аденома печени, вызванная применением половых гормонов в анамнезе
Относительные
• Заболевания печени
Тяжелые сердечно-сосудистые заболевания
Необходимость в быстром восстановлении фертильности
проблемы с инъекционным введением
Тяжелая депрессия
МЕХАНИЗМ ДЕЙСТВИЯ
Механизм действия ДМПА такой же, как у других контрацептивов, содержащих
только гестаген: повышение вязкости шеечной слизи, изменение состояния эндо¬
метрия и торможение стимулируемой лютропином овуляции [222]. Подавление
секреции ФСГ не столь значительно, как при приеме комбинированных пероральных
контрацептивов; концентрация эстрогена в крови сопоставима с таковой в ранней
фолликулярной фазе нормального менструального цикла [223]. Проявления недо¬
статочности эстрогенных гормонов, такие как атрофические изменения слизистой
оболочки влагалища или уменьшение размеров молочных желез, не развиваются, но
возможно снижение плотности костной ткани.
По мнению отдельных исследователей, случаи возникновения беременности,
наступившие в самом начале применения ДМПА, сопровождаются повышением
частоты смерти плода или новорожденного, предположительно связанные с повыше¬
нием вероятности нарушения внутриутробного развития [224, 225]. Следовательно,
наиболее важно соблюдать меры предосторожности в период после первой инъек¬
ции препарата. Для надежности контрацепции первую инъекцию следует провести
187
в первые 5 дней менструального цикла (до разрыва доминантного фолликула) или в
течение 2 нед после первой инъекции применять дополнительные методы контрацеп¬
ции [226-228J, Несоблюдение правил применения препарата снижает длительность
контрацептивного эффекта. Препарат вводят глубоко в мышцу, меняя место инъекции
(Z-образно); место инъекции нельзя массировать. Инъекции не следует проводить в
места, подвергаемые естественным растираниям во время движений.
ЭФФЕКТИВНОСТЬ
Эффективность этого метода такая же, как после женской стерилизации, и выше,
чем у пероральных методов [2, 229]. В связи с созданием высоких концентраций
гормона в крови эффективность не зависит от массы тела женщины или применения
лекарственных препаратов, повышающих активность ферментов печени. Напротив,
ДМПА служит препаратом выбора для женщин, принимающих противоэпилептиче¬
ские средства, так как высокие концентрации гестагена в крови повышают судорож¬
ный порог [230].
ПРЕИМУЩЕСТВА
Как и при применении контрацептивных препаратов с постепенным высво¬
бождением гормонов, при приеме ДМПА отсутствует необходимость ежедневно
принимать таблетки и заботиться о предотвращении беременности перед каждым
половым актом. По сравнению с пероральными контрацептивами при применении
ДМПА у подростков показатели длительности применения выше, а количество
повторных беременностей меньше: однако эти показатели не различаются, когда
подростки начинают использовать эти методы контрацепции в раннем послеродо¬
вом периоде [231, 232]. Применение ДМПА предпочтительнее для женщин, в силу
разных причин забывающих регулярно принимать таблетки, имеющих беспоря¬
дочные половые связи, а также при нарушениях умственного развития. Однако в
-некоторых случаях могут возникнуть затруднения с плановым посещением врача
для проведения инъекций каждые 3 мес. Тогда хорошей альтернативой может
служить препарат ДМПА для подкожного введения, который можно вводить само¬
стоятельно.
Благодаря отсутствию побочных эффектов эстрогена ДМПА можно применять
при врожденном пороке сердца, серповидноклеточной анемии или тромбоэмболии
в анамнезе, а также у женщин старше 30 лет с такими факторами риска, как курение,
артериальная гипертензия или сахарный диабет. Абсолютная безопасность ДМПА в
отношении развития тромбоза в основном теоретическая, не подтвержденная контро¬
лируемыми исследованиями. Однако повышение риска не выявлено в эпидемиологи¬
ческом исследовании лиц, применявших ДМПА, а исследование, проведенное БОЗ,
не выявило повышения риска развития инсульта, инфаркта миокарда или венозной
тромбоэмболии [147, 221].
В отношении женщин с серповидноклеточной анемией показано, что препарат
угнетает in vivo образование патологических клеток и улучшает гематологические
показатели [105]. Понижается также частота и тяжесть болевых кризов при серповид¬
ноклеточной анемии [233].
Другое преимущество ДМПА - увеличение количества материнского молока у
кормящих грудью, эффект, противоположный действию комбинированных таблеток.
Препарат не проникает в грудное молоко, не выявлено его отрицательного действия
на рост и развитие младенцев [234-236]. Тщательное исследование новорожденных
мальчиков, матери которых применяли ДМПА, не выявило метаболитов в моче, а
также изменений концентрации ФСГ и ЛГ, тестостерона и кортизола в крови [237].
В связи с некоторым стимулирз'ющим влиянием на лактацию ДМПА можно приме¬
188
нять сразу после родов. Раннее применение препарата не оказало побочного действия
на качество и количество грудного молока [145].
Можно применять ДМПА при судорожных расстройствах; облегчение контроля
судорог, возможно, связано с седативными свойствами гестагенов [230].
Применение ДМПА также снижает риск развития рака эндометрия, эффект сопо¬
ставим с аналогичным действием пероральных контрацептивов [238], а также,
подобно гестагенам в пероральных контрацептивах, уменьшает объем менструальной
кровопотери и анемию, частоту возникновения воспалительных заболеваний органов
малого таза, эндометриоза, фиброзных поражений матки [239], внематочной бере¬
менности. Неудачу ВОЗ при попытке доказать снижение частоты возникновения рака
яичников можно предположительно объяснить недостаточностью статистических
данных и большим числом родов у применяющих ДМПА (в прошлом) [240].
Подобно пероральным контрацептивам ДМПА снижает риск возникновения вос¬
палительных заболеваний органов малого таза, однако единственное исследование
было затруднено недостаточностью наблюдений [241]. В связи с подавлением ову¬
ляции исключается вероятность возникновения внематочной беременности и резко
снижается вероятность развития кист яичников.
Чем больше выбор противозачаточных средств, тем легче женщине подобрать себе
оптимальное. Для некоторых женщин основное преимущество ДМПА — секретность
и легкость применения. Никто, кроме самой женщины, не знает о получаемых инъ¬
екциях, а введение препарата каждые 3 мес незатруднительно для тех, кто не боится
инъекций. В некоторых популяциях в силу тех или иных причин к инъекциям лекар¬
ственных препаратов относятся с особым доверием, в этих случаях ДМПА является
самым популярным контрацептивом, несмотря на вызываемые им нарушения мен¬
струального цикла и другие побочные эффекты. Преимущества ДМПА приведены в
табл. 2-9.
Таблица 2-9. Преимущества применения депо-проверы
Простота применения: нет необходимости принимать ежедневно или заботиться о контрацепции перед каждым
половым актом
Безопасность, отсутствие серьезных побочных эффектов
Высокая контрацептивная эффективность, инъекция так же эффективна, как женская стерилизация, внутрима-
точная и имплантационная контрацепция
Отсутствие побочных эффектов, связанных с эстрогенным компонентом
Возможность скрыть от окружающих факт применения
• Стимуляция лактации
Наличие других положительных фармакологических эффектов
ПРОБЛЕМЫ
Основные проблемы, возникающие при приеме ДМПА, — нерегулярные кровя¬
нистые выделения, нагрубание молочных желез, головные боли, повышение массы
тела и депрессия [220, 221]. Наиболее частая проблема — нарушения менструального
цикла. До 25% женщин прекращают применение ДМПА в течение первого года из-за
нерегулярных кровотечений [242]. Кровянистые выделения из половых путей редко
бывают интенсивными, количество гемоглобина крови у применяющих ДМПА обыч¬
но повышается. Нерегулярные кровотечения возникают у 70% женщин в течение
первого года применения и у 10% в более поздние сроки. Вероятность возникновения
кровотечений и мажущих выделений снижается после каждой повторной инъекции,
189
при применении препарата в течение 5 лет у 80% женщин менструации отсутствуют
(при применении норпланта^ — у 10%) [243J. Нерегулярность кровянистых выде¬
лений из половых путей приносит неудобства и у многих женщин снижает половую
активность, поэтому некоторые пациентки испытывают облегчение, когда после дли¬
тельного применения ДМПА развивается аменорея.
При возникновении межменструальных кровотечений для лечения можно при¬
менить препараты, содержащие экзогенный эстроген, 1,25 мг конъюгированных
эстрогенов или 2 мг эстрогена ежедневно в течение 7 дней. Эффективен также прием в
течение 7 дней нестероидных противовоспалительных средств; или можно назначить
пероральные контрацептивы на 1-3 мес. Внеочередная инъекция ДМПА не изме¬
няет характера кровотечений [244]. Большинство женщин, если их заранее инфор¬
мировать, могут просто ожидать, что у них со временем прекратятся менструации.
Трансдермально введенный эстроген не снижал вероятности возникновения нере¬
гулярных кровотечений в такой степени, чтобы улучшить показатели продолжения
применения ДМПА у молодых женщин после абортов [245].
Приблизительно треть женщин прекращают применение ДМПА через год, поло¬
вина — через 2 года и 80% — через 3 года [246]. Длительность применения ДМПА
в течение года в общественных клиниках Техаса составляла лишь 29%, показатель
значительно меньший, чем где-либо [247]. Такой же показатель у женщин, решивших
сделать первую инъекцию сразу после добровольного прерывания беременности
[245]. Согласно большому международному исследованию, наиболее распространен¬
ными медицинскими причинами прекращения применения ДМПА в первые 2 года
были головные боли (2,3%), повышение массы тела (2,1%), головокружение (1,2%),
боли в животе (1,1%) и беспокойство (0,7%) [221].
В странах Запада в этот список также включают депрессию, усталость, снижение
либидо, артериальную гипертензию. Трудно выяснить, связаны ли эти жалобы с
действием медроксипрогестерона ацетата, поскольку они типичны и для женщин, не
принимающих этот контрацептив [248]. При более тщательном исследовании было
установлено, что у женщин, принимающих ДМПА, не усиливались симптомы депрес¬
сии, причем даже у тех, кто ее испытывал до начала приема препарата [249, 250].
Попытки подтвердить способность ДМПА повышать массу тела дали противоречи¬
вые результаты: некоторые исследователи не обнаружили этого влияния, другие выяви¬
ли небольшое повышение массы тела на фоне препарата (например, приблизительно на
4 К1’ после более 5 лет применения в одном исследовании и на 11 кг после 10 лет приме¬
нения в другом) [251-254]. Согласно результатам плацебо-контролируемого исследо¬
вания, ДМПА не влиял на количество принимаемой пищи, энергетические потребности
или массу тела [255]. Так же как при приеме пероральных контрацептивов, повышение
массы тела может быть вызвано не гормоном, а следствием образа жизни и старения.
С другой стороны, отдельные пациенты и этнические группы могут быть предрасполо¬
жены к ожирению, например было сообщение о значительном повышении массы тела
после применения ДМПА у женщин в Навайо [256] и у подростков с уже повышенной
массой тела. У лиц с нормальной массой тела после начала применения ДМПА ее повы¬
шение было таким же, как в контрольной группе [257].
Если появившиеся побочные эффекты связаны с действием гестагена, то после пре¬
кращения применения ДМПА они исчезают лишь после 6-8 мес (в отличие от мини-
пили, имплантов, колец и пластырей). Клиренс препарата замедлен у женщин с повы¬
шенной массой тела. Примерно у 50% женщин после окончания применения ДМПА
нормальный менструальный цикл восстанавливается через 6 мес после последней
инъекции, а у 25% — через год [243].
190
Злокачественные новообразования
Рак молочной железы
При анализе клинических случаев, проведенном ВОЗ в течение 9 лет в трех разви¬
вающихся странах, установлено, что применение ДМПА сопряжено с очень незначи¬
тельным повышением риска возникновения рака молочной железы в первые 4 года,
но этот показатель не возрастает при увеличении длительности приема препарата
[258]. Количество случаев заболевания у лиц, применявших ДМПА в прошлом, было
небольшим. Возможным объяснением такого результата служит сочетание изменения
качества диагностики с ростом имевшейся до начала приема препарата опухоли, как и
в случае, приведенном выше, при применении пероральных контрацептивов.
Злокачественные новообразования другой локализации
Повышение риска возникновения дисплазии шейки матки не установлено даже
при длительном (4 года и более) применении ДМПА [259]. Согласно данным БОЗ, не
происходит увеличения частоты развития аденокарциномы или аденоскваматозной
карциномы [260]. Исследования, проведенные ВОЗ, не выявили повышения риска
возникновения инвазивных скваматозных раковых клеток в шейке матки у при¬
меняющих ДМПА, однако риск возникновения карциномы шейки матки ш situ был
слегка повышен. Неясно, отражают ли полученные результаты реальную ситуацию
или изменение качества диагностики [261,262].
Метаболические эффекты
Влияние ДМПА на содержание липопротеинов неясно. Отдельные исследования
не обнаруживают побочного влияния препарата на этот показатель, что объясняют
тем, что ДМПА не подвергается эффекту первого прохождения через печень; другие
исследования выявили снижение ЛПВП и повышение общего холестерина и ЛПНП
[263, 264]. По данным многоцентрового клинического исследования, проведенного
ВОЗ, преходящее отрицательное влияние препарата на содержание липопротеинов
отмечали лишь в первые несколько недель после инъекции в случаях, когда оно было
исходно повышено [265].
ДМПА не вызывает существенных изменений углеводного обмена и количества
факторов свертывания крови [266, 267]. Данные о влиянии препарата на женщин,
больных сахарным диабетом или с гестационным диабетом беременности в анамнезе,
отсутствуют.
Влияние на плотность костной ткани
Врачи озабочены тем, что применение в качестве контрацептива ДМПА сопряжено
со снижением плотности костной ткани, что оговорено в инструкции к применению
препарата. Обоснование для применения эстрогена для предупреждения снижения
плотности костной ткани — установленный факт, что концентрации в крови эстрогена
при применении ДМПА сравнительно ниже, чем в аналогичный период нормального
менструального цикла [268]. В патологоанатомических исследованиях было установ¬
лено снижение плотности крестцовой и бедренной костей у женщин, применявших
ДМПА [269, 270]. Согласно данным патологоанатомического исследования амери¬
канских ученых, снижение плотности костей усиливается с увеличением длительно¬
сти применения ДМПА, особенно у женщин 18-21 лет [271].
Снижение плотности костей было слабее, чем в раннем климактерическом периоде.
К тому же оно было частично обратимым. Определение плотности костной ткани у
женщин, прекративших применение (даже длительное) ДМПА, установило значи¬
тельное восстановление этого показателя в поясничной области, но не в шейке бедра,
в течение 2 лет; в другом когортном исследовании показано, что в обеих областях этот
показатель частично восстанавливался через 30 мес [272, 273]. Наиболее важно, что в
патологоанатомических исследованиях женщин в постменопаузальном периоде, про¬
191
веденных в Новой Зеландии, и в большом мультицентровом исследовании не выяв¬
лено различия в плотности костной ткани у применявших и не применявших ДМПА,
что свидетельствует о том, что препарат не оказывает выраженного действия на этот
показатель [274, 275].
Плотность костей повышается быстро и значительно в подростковом периоде.
Практически полностью костная масса бедренной кости и тел позвонков сформиро¬
вана у девушек в возрасте 18 лет, причем наиболее важны первые годы после насту¬
пления менструаций [276, 277]. Именно поэтому любое лекарственное средство, тор¬
мозящее повышение плотности кости в этот период, способно увеличить вероятность
развития в дальнейшем остеопороза. Проспективное исследование 48 подростков
выявило, что применение ДМПА (16 человек) сопровождалось снижением плотности
костей поясничной области (приблизительно на 1,5% в течение года), в то время как
у применявших норплант**^ (7 человек) и пероральные контрацептивы (9 человек) она
повысилась в соответствии с возрастными изменениями [278].
Противоречивость результатов исследований, различная степень снижения плот¬
ности костей, а также некоторая обратимость изменений свидетельствуют о том, что
вероятность развития этого побочного эффекта не должна ограничивать применение
ДМПА и дополнительная профилактическая терапия эстрогеном не показана (она
к тому же может отрицательно повлиять на выбор данного метода контрацепции).
Ввиду возможности развития этого побочного эффекта женщины, применявшие
ДМПА в прошлом, должны находиться под соответствующим наблюдением. Однако
обеспокоенность относительно возможного снижения плотности костной ткани не
должна быть причиной отказываться от этого метода контрацепции. Маловероятно,
что этот эффект настолько значителен, чтобы существенно повысить в последующем
риск развития остеопороза.
Влияние на фертильность
Поздние сроки восстановления фертильности после прекращения приема ДМПА —
проблема, характерная только для инъекционной контрацепции, так как при всех
остальных существующих методах контрацепции фертильность восстанавливается
быстро [279]. Однако медроксипрогестерон не окончательно подавляет функцио¬
нальную активность яичников, при этом тот факт, что возможность бесплодия в
сочетании с нарушением менструальной функции может быть обусловлена при¬
менением ДМПА не подтвержден эпидемиологическими данными. Количество воз¬
никших желаемых беременностей у женщин, прекративших введение препарата,
не отличается от нормальных показателей [280]. Через 18 мес после последней
инъекции препарата у 90% развилась беременность; такой же показатель выявлен
у женщин, применявших другие методы контрацепции [281]. Фертильность восста¬
навливается приблизительно через 9 мес после последней инъекции независимо от
длительности применения ДМПА. Следовательно, женщины, желающие забереме¬
неть немедленно после прекращения приема контрацепитива. не должны применять
ДМПА. Подавление менструальной функции в течение более 18 мес после последней
инъекции не связано с действием препарата и служит показанием к дополнительному
обследованию женщины.
Подкожное введение ДМПА
Внутримышечное введение ДМПА имеет ряд недостатков. Максимальные кон¬
центрации препарата в сыворотке крови выше, чем это необходимо для подавления
овуляции, и вызывают уменьшение образования эндогенного эстрадиола, что может
приводить к снижению плотности костной ткани. Пролонгированное поступление
препарата из мьпиечной ткани может привести к восстановлению способности к ову¬
ляции только через несколько месяцев после последней внутримышечной инъекции.
192
Женщины, применяющие ДМПА, должны посещать врача каждые 3 мес для проведе¬
ния повторной инъекции.
Все эти проблемы отсутствуют при подкожной инъекции суспензии, содержащей
104 мг (в большей степени, чем 150 мг) ДМПА в 0,65 мл растворителя, отличающемся
от тех, которые используются в лекарственных формах ДМПА для внутримышеч¬
ных инъекций. Создающиеся максимальные концентрации в крови ДМПА ниже при
сохранении высокой эффективности; согласно первому клиническому испытанию,
нежелательные беременности не возникли у 1783 женщин, из которых 11% страдали
ожирением. Плотность костной ткани не изменялась в течение года применения, а
способность к овуляции восстанавливалась быстрее, чем при применении препарата
для внутримышечных инъекций. Изменения менструального цикла были такими же,
как при внутримышечном введении, за исключением того, что аменорея развивалась
в более поздние сроки; через год у 45% женщин прекращались менструации, а при
внутримышечном приеме — у 54% [282].
Подкожные инъекции ДМПА можно проводить самостоятельно. Часть женщин
в первом клиническом исследовании могли самостоятельно делать инъекции, и они
были эффективны.
Инъекционные контрацептивы короткого действия
Комбинации эстрогена и гестагена для инъекций ежемесячно или один раз в
2 мес не новы, они появлялись в течение нескольких десятков лет [283], Этот метод
контрацепции популярен в Китае, странах Латинской Америки и Восточной Азии.
Лекарственный препарат, широко применяемый в Китае и известный как Китайские
инъекции № 1, содержит 250 мг 17-гидроксипрогестерона капроата и 5 мг эстрадиола
валерата.
ЛЮНЕЛЛ^(ЦИКЛОФЕМ^)
Люнелл^’ содержит 25 мг ДМПА и 5 мг эстрадиола ципионата, его применяют в
виде глубоких внутримышечных инъекций через каждые 28-30 дней (не более чем
через 33 дня). Эффективность препарата такая же, как у депо-провера*, но линелл*’ не
вызывает таких проблем, как нерегулярные кровянистые выделения и сильные кро¬
вотечения, а также аменорею [284-289]. Кроме того, фертильность восстанавливает¬
ся быстро после прекращения применения (в те же сроки, что и при использовании
пероральных контрацептивов) [290].
Одним из недостатков препарата является необходимость ежемесячных инъекций,
другой недостаток — вероятность того, что комбинация эстрогена и гестагена будет
угнетать лактацию. Проблему необходимости ежемесячных введений можно умень¬
шить, если использовать специальные автоматические шприцы для самостоятельного
введения [291], Приблизительно у 80% женщин, у которых развилась аменорея при
применении депо-проверы*, менструальные кровотечения возобновились после
замены его на люнелл^ [292]. Противопоказания, побочные эффекты и предосторож¬
ности, а также возможные лечебные эффекты пероральных контрацептивов могут
быть отнесены и к люнеллу*^, который более не продается в США, но доступен во
многих других странах.
НОРЭТИНДРОНА ЭНАНТАТ^
Норэтиндрона энантат*^ применяют в дозе 200 мг внутримышечно каждые 2 мес.
Этот гестаген действует так же, как депо-провера, и его применение сопряжено с
такими же проблемами. [221]. Препарат мезигина, представляющий собой комбина¬
цию норэтиндрона энантата^’ (50 мг) и эстрадиола валерата (5 мг), при ежемесячном
193
введении обеспечивает надежную контрацепцию с хорошим контролем менструаль¬
ного цикла [293]. По сравнению с люнеллом*“ мезигина*“ реже вызывает проблемы
с возникновением кровотечений [294]. Фертильность восстанавливается быстро (в
течение 1 мес) после прекращения применения [290].
КОМБИНАЦИЯ ДИГИДРОПРОГЕСТЕРОНА АЦЕТОФЕНИДИКА И ЭСТРАДИОЛА ЭТАНТАТА
Комбинация 150 мг дигидропрогестерона ацетофенидина с 10 мг эстрадиола
этантата (выпускается под разными торговыми наименованиями) является наи¬
более широко применяемым инъекционным контрацептивом в странах Латинской
Америки. Как и при приеме мезигины^ и люнелла^, при ежемесячном применении
этого препарата менструальный цикл остается регулярным, и даже ослабляются
менструальные кровотечения [295]. Более низкие дозы (90 мг дигидропрогестерона
ацетофенидина с 6 мг эстрадиола этантата) равноэффективны более высоким дозам с
аналогичным влиянием на менструальный цикл [296].
ИМПЛАНТАЦИОННАЯ КОНТРАЦЕПЦИЯ
Подкожные контрацептивные импланты обеспечивают длительное контролируе¬
мое выделение гестагенов. С 1980 г. эти импланты внедрены более чем в 60 странах
и используются 11 млн женщин. Высокая контрацептивная эффективность и про¬
стота применения делают их препаратами выбора для женщин, которым показаны
контрацептивы, содержащие только гестаген (противопоказаны эстрогенсодержащие
препараты), подросткам, которым трудно заботиться о предотвращении нежелатель¬
ной беременности перед каждым половым актом или ежедневно, а также здоровым
женщинам, нуждающимся в длительном предохранении от беременности. Имплант,
содержащий левоноргестрел, отсутствующий в настоящее время на фармацевтиче¬
ском рынке США, создавал иногда проблемы с введением и удалением, С появлением
высокоэффективного и длительно действующего импланта этоногестрела (импланон
в виде одной вживляемой капсулы) имплантационная контрацепция вновь стала
доступной в США.
Преимущества имплантационной контрацепции
Этот метод имеет множество преимуществ. Он обеспечивает длительную эффек¬
тивную контрацепцию без необходимости ежедневного или еженедельного приме¬
нения (в отличие от таблеток, колец или пластырей), а также регулярных инъекций.
Соотношение цена/эффективность при имплантационной контрацепции лучше, чем у
короткодействующих методов, или при возникновении незапланированной беремен¬
ности [297]. Постепенное высвобождение низких доз гестагена и создание стабиль¬
ных концентраций его в крови обеспечивают высокую эффективность и большую
продолжительность контрацепции, быстрое восстановление фертильности и отсут¬
ствие серьезных сердечно-сосудистых осложнений [298].
Изыскание и разработка подкожных имплантов, содержащих .только гестаген,
были начаты в шестидесятых годах прошлого века, однако первое исследование
имплантов с очень низкой дозой гестагенов выявило их низкую эффективность в
предотвращении возникновения внематочной беременности. Был внедрен имплант,
содержащий активный гестаген левоноргестрел в виде 6 капсул, вживляемых под
кожу, в 1991 г. появился первый имплант, содержащий этоногестрел; 1 млн женщин
в США выбрали для себя в качестве контрацептива имплант, содержащий левонор¬
гестрел, который подтвердил высокую эффективность: в течение 7 лет применения
нежелательная беременность возникла лишь у 1% женщин [299]. Несмотря на высо-
194
кую эффективность и малое количество серьезных побочных эффектов, в связи с
дефицитом поставки силиконовых капсул в 2002 г. имплант, содержащий левой орге-
стрел, был снят с производства, не оставив для американских женщин альтернативы
для имплантационной контрацепции [300].
15-летний опыт применения импланта, содержащего левоноргестрел, стимулировал
разработку и улучшение его дизайна, В 1998 г. был разрешен к применению имплант
левоноргестрела, представляющий собой 2 капсулы для вживления, но его никогда
не продавали в США, хотя используют в странах Европы и Азии. FDA пересмотрело
отношение к контрацептивному импланту, содержащему 68 мг этоногестрела, актив¬
ного метаболита дезогестрела, в виде одной капсулы для вживления, изготовленной
из винилацетата. Этот контрацептив считают наилучшим из существующих гормо¬
нальных методов контроля рождаемости, его используют более 2 млн женщин в стра¬
нах Европы и Азии [301]. В Австралии 25% женщин, использующих контрацептивы,
в 2004 г. предпочли этот имплант.
Левоноргестрел обладает высокой активностью к рецепторам прогестерона,
андрогенов, минерале- и глюкокортикоидов, но не связывается с эстрогеновыми
рецепторами. Этоногестрел не обладает эстрогенной, противовоспалительной или
минералокортикоидной активностью, но установлены его слабое андрогенное и
анаболическое и сильное антиэстрогенное действия. В отличие от левоноргестрела,
взаимодействующего главным образом с андрогенсвязывающимся глобулином, это¬
ногестрел связывается преимущественно с альбуминами, что существенно не зависит
от изменений концентраций эндогенного и экзогенного эстрадиола. Безопасность
этоногестрела была показана в исследованиях с применением комбинированных
пероральных контрацептивов, влагалищных колец, содержащих комбинацию гор¬
монов, а также мини-пили; все эти препараты содержали дезогестрел, ближайший
аналог этоногестрела.
МЕХАНИЗМ ДЕЙСТВИЯ
Импланты, содержащие гестаген, обладают двумя основными механизмами дей¬
ствия: торможение овуляции и изменение свойств шеечной слизи, препятствующее
проникновению сперматозоидов [305]. Имплант левоноргестрела препятствует росту
фолликулов и тормозит овуляцию, воздействуя по принципу отрицательной обрат¬
ной связи на высвобождение стимулирующих гормонов гипоталамуса и гипофиза,
что приводит к разнообразным изменениям (от отсутствия овуляции до недосгаточ-
ност’и функции желтого тела). У большинства женщин, использующих импланты
левоноргестрела, овуляции возобновляются по мере снижения концентрации лево¬
норгестрела в крови [306]. Имплант этоногестрела подавляет овуляцию, влияя на ось
«гипоталамус-гипофиз» и тормозя волнообразное повышение секреции ЛГ, необхо¬
димое для образования, роста и овуляции фолликула [307]. У некоторых женщин,
особенно с высоким содержанием эндогенного эстрогена, при применении импланта
образуются кисты яичника, очень редко вызывающие болевые ощущения и не тре¬
бующие лечения.
Даже при сохранении роста фолликула на фоне применения гестагенсодержащих
имплантов оплодотворения ооцитов не происходит. После разрыва фолликула нару¬
шенный процесс овуляции препятствует образованию жизнеспособного желтого тела.
Антиэстрогенные эффекты гестагенов изменяют свойства шеечной слизи, повышая
ее вязкость, что препятствует проникновению сперматозоидов [308]. Перечисленные
изменения функций яичников и свойств шеечной слизи обеспечивают высокую кон-
195
трацептивнз^ю эффективность. Импланты не влияют на развитие эмбриона и не вызы¬
вают прерывания наступившей беременности.
ИМПЛАНТ, СОДЕРЖАЩИІ ЭТОНОГЕСТРЕЛ (ОДНА ВЖИВЛЯЕМАЯ ПОД КОЖУ КАПСУЛА)
Дизайн
Высокая контрацептивная эффективность этого нового средства обусловлена
способностью низкой дозы этоногестрела подавлять овуляцию и его большой дли¬
тельностью действия. Однократного введения препарата под кожу достаточно для
обеспечения противозачаточного эффекта в течение 3 лет [308]. Вживление всего
одной капсулы значительно удобнее, чем 6 капсул, содержащих левоноргестрел [309,
310]. Исследования импланта, содержавшего этоногестрел, начатые в 1995 г. в США и
европейских странах, показали, что введение подкожно капсулы занимает в среднем
1 мин, а удаление — 3 мин, то есть значительно меньше, чем имплантов, содержащих
левоноргестрел [301].
Имплант, содержащий этоногестрел, имеет и другие преимущества в дизайне.
После введения одна капсула находится строго в определенном месте, в то время
как при введении системы из 6 капсул высока вероятность смещения предыдущей
капсулы после вживления последующей [311]. Кроме того, капсула, содержащая это¬
ногестрел, не силиконовая и не приводит к образованию фиброзной ткани, затруд¬
няющей последующее извлечение капсулы [312]. Эти различия облегчают введение и
извлечение капсулы, что причиняет меньше неудобств и не отпугивает пациента, а для
клиницистов ускоряет и облегчает процедуру ее введения и удаления.
Фармакологические свойства
Имплант, содержащий этоногестрел (импланон^’), состоит из одной не разру¬
шающейся в тканях капсулы размером 40 ммх2 мм, содержащей 40% этинилви-
нилацетата и 60% этоногестрела. Она покрыта оболочкой из этинилвинилацетата
толщиной 0,06 мм, обеспечивающей постепенное высвобождение гормона. Капсула
содержит 68 мг этоногестрела, поступающего в кровоток с первоначальной скоростью
60 мг/сут, которая постепенно снижается до 30 мг/сут после истечения 2 лет. Быстрая
первоначальная абсорбция предположительно связана с большим количеством это¬
ногестрела, выделяющегося из непокрытых оболочкой концов импланта. Пиковые
концентрации (266 пг/мл) этоногестрела в сыворотке крови достигаются в течение
1 сут после введения и приводят к подавлению овуляции [307, 308]. Подобно осталь¬
ным гормональным контрацептивам содержание этоногестрела в сыворотке крови
ниже у женщин, принимающих индукторы микросомальных ферментов печени,
например карбамазепин, гризеофульвин, фенитоин и рифампицин, но не изменяется
при применении антибиотиков.
Этоногестрел постоянно поступает в кровоток, не подвергаясь эффекту первичного
прохождения через печень. Биодоступность этоногестрела составляет приблизитель¬
но 100% в течение 2 лет применения. Период полувыведения этоногестрела равен
25 ч (у левоноргестрела — 42 ч). Этоногестрел полностью выводится из организма в
течение 1 нед после удаления импланта. У 94% женщин способность к овуляции вос¬
станавливается через 3-6 нед после удаления капсулы [307, 308].
Эффективность
Контрацептивная эффективность капсулы, содержащей этоногестрел, в клиниче¬
ских исследованиях на 1100 женщинах (общая длительность применения равна 26 700
циклов) составила более 99% [313]. Из этого количества у 835 женщин капсула была
удалена через 2-3 года, а дополнительно 526 женщин применяли имплант в течение
3 лет и более. В этих исследованиях не зарегистрировано ни одного случая возникнове¬
ния беременности, в том числе у женщин (365) с массой тела 70 кг и более [308].
196
Метаболические эффекты
Влияние импланта, содержащего этоногестрел, на обмен веществ минимальный и
клинически незначимый. Имплант практически не влияет на метаболизм липидов,
углеводный обмен, функции печени, факторы гемостаза, артериальное давление, функ¬
циональную активность щитовидной железы и надпочечников [314,315, 316, 317].
Безопасность
В целом импланты, в том числе содержащие этоногестрел, считаются безопас¬
ными, частота осложнений (внезапная смерть, злокачественные новообразования,
анемия, артериальная гипертензия, изменение плотности костной ткани, сахарный
диабет, заболевания желчного пузыря, тромбоцитопения, воспалительные заболе¬
вания органов малого таза) при их применении сопоставима с таковой у женщин,
не применяющих импланты [318]. Имплант, содержащий этоногестрел, обладает
лечебным действием при болезненных менструациях (эффект выявлен у 88% жен¬
щин с дисменореей); боли усиливались лишь у 2% женщин, применявших имплант с
этоногестрелом [318]. В сравнительном исследовании 42 младенцев, находящихся на
естественном вскармливании, и их матерей, применяющих импланты, содержавшие
этоногестрел, с 38 младенцами и их матерями, использующими внутриматочные сред¬
ства, не обнаружено существенных различий в количестве и составе грудного молока,
времени введения и количества прикорма, а также в показателях роста [304].
Контрацептивы, содержащие низкие дозы гестагена, имеют мало противопоказа¬
ний к применению. Их эффективность может быть снижена у женщин с ожирением,
а также при лечении лекарственными средствами, стимулирующими печеночный
метаболизм стероидов, такими как некоторые антибиотики (например, рифампин) и
противоэпилептические средства (например, фенитоин).
Способ применения
Несмотря на то что система, содержащая этоногестрел, появилась для обеспечения
более быстрого и легкого введения имплантов, для вживления капсулы необходимо
специальное обучение персонала [319]. Введение одной капсулы легче и быстрее (в
среднем за 1-2 мин), чем 6 капсул с левоноргестрелом [320]. Система снабжена спе¬
циальным аппликатором для введения капсулы [321]. У кончика иглы есть два лез¬
вия, каждый с различным наклоном (рис. 2-5). Крайний кончик иглы более острый,
чем остаток, чтобы облегчить начальный прокол кожи. Верхняя часть менее острая,
чтобы уменьшить риск перфорации кожи во время продвижения после начального
проникновения.
Печать аппликатора
Аппликатор
Опора обтуратора
/
3 Обтуратор
Игла
Месторасположение
папочки
(ттттттшт^ ^ ' Г Канюля
Ч
Палочка
гч п >- .1 см
Рис. 2-5. Имплант, содержащим
этоногестрел. I- Колпачок иглы
197
Капсулу вводят под кожу внутренней поверхности левого плеча на расстоянии
6-8 см выше локтевого сустава в стерильных условиях. Место инъекции можно
предварительно обезболить введением небольшого количества (0,1 мл) местного
анестетика, но, как правило, в этом нет необходимости. После введения капсула не
выступает над поверхностью кожи, но пальпируется.
Женщинам, ранее не применявшим гормональные контрацептивы, введение
импланта следует произвести в 1-5 дни менструального цикла. При переходе с ком¬
бинированных таблеток, мини-пили или внутриматочной контрацепции на имплант
с этоногестрелом его следует вводить во время «свободной» от приема препаратов
недели. При замене инъекционной контрацепции на имплантационную имплант вво¬
дят в день очередной плановой инъекции. Контрацептивный эффект импланта прояв¬
ляется быстро, поэтому (если время введения соответствует инструкции) отсутствует
необходимость в применении барьерных методов. При введении импланта с этоноге¬
стрелом сразу после медицинского аборта или родов не нужно пользоваться допол¬
нительными мерами контрацепции. Во всех случаях перед применением импланта
необходимо исключить возможную беременность, несмотря на отсутствие данных о
том, что гормональные контрацептивы вызывают нарушения развития плода.
Имплант с этоногестрелом удаляют в конце третьего года или в любое время по
желанию женщины. Удаление проводят через небольшой (2-3 мм) разрез в дисталь¬
ной части плеча надавливанием на противоположный конец капсулы. Удаление обыч¬
но занимает менее 5 мин (2,6-5,4 мин) [321]. На месте введения и удаления импланта
МОГ)”! развиться отек, покраснение или гематома. При использовании импланта неже¬
лательная беременность может возникнуть в конце срока его действия, поэтому необ¬
ходимо вовремя заменить имплант или применить другие методы контрацепции.
Недостатки
Несмотря на то что контрацептивы, содержащие только гестаген, эффективны и
безопасны, они имеют некоторые недостатки. Хирургический метод введения препа¬
рата требует специально обученного персонала. Соотношение «цена-.эффективность»
зависит от длительности применения, менее выгодно краткосрочное использование
[297]. Подобно остальным методам гормональной контрацепции, импланты не пред¬
упреждают заражения заболеваниями, передающимися половым п^^тем.
Побочные эффекты импланта с этоногестрелом вкаючают нарушение менструаль¬
ного цикла [редкие менструальные кровотечения (26,9%), отсутствие менструаций
(18,6%), длительные менструальные кровотечения (15,1%), частые менструальные
кровотечения (7,4%)], повышение массы тела (20,7%), угри (15,3%), болезненность
в молочных железах (9,1%), головную боль (8,5%), но эти эффекты редко служат
причиной для удаления импланта [301, 318]. При применении любого чистого геста¬
генного метода контрацепции развивается нарушение менструального цикла [322,
323], Сравнительное исследование менструальных циклов у женщин с одной капсулой
с этоногестрелом и шестью капсулами с левоноргестрелом выявило статистически
значимое уменьшение дней кровотечений в первой группе (15,9-19,3 и 19,4-21,6 дня
соответственно; Р=0,0169) [324]. В связи с уменьшением общей потери крови у жен¬
щин, применяющих контрацептивы, содержащие только гестаген (в том числе перо¬
ральные), менее вероятно развитие анемии. Однако в этом исследовании установлено,
что при применении имплантов с этоногестрелом менструальный цикл нарушается в
большей степени, чем в группе с имплантами с левоноргестрелом. [324]. В табл. 2-10
представлены различия в менструальном цикле при применении разных имплантов,
однако какое из этих нарушений может развиться, непредсказуемо. Несмотря на
возможные побочные эффекты и зависимость от навыков медицинского персонала,
большинство женщин удовлетворены имплантационной контрацепцией в связи с ее
удобством, длительностью и высокой эффективностью [325, 326].
198
Таблица 2-10. Сравнительная оценка нарушений менструального цикла при применении имплантов с
этоногестрелом и левоноргестрелом [326, 327]
Характер нарушений*
Имплант с этоноге¬
стрелом, % (п=1б9)
Имплант с левонорге¬
стрелом, % {п=163)
Статистическая достовер¬
ность
Аменорея
21
Да
Редкие менструальные кро¬
вотечения
27
22
Да
Частые менструальные кро¬
вотечения
Нет
Длительные менструальные
кровотечения
12
Нет
* Наблюдения в течение 90 сут.
Досрочное удаление импланта
Частота досрочного удаления импланта с этоногестрелом различается в раз¬
ных странах: от 30,2% в странах Европы и Канаде до 0,9% в странах Южной Азии
[318, 321]. Основная причина досрочного удаления — нерегулярные менструации.
Метаанализ 13 исследований, опубликованных в 1989-1992 гг., установил, что из
1716 женщин, применявших имплант с этоногестрелом, 5,3% удалили его в первые
6 мес применения, 6,4% — через 7-12 мес, 4,1% — через 13-18 мес и 2,8% — через
19-24 мес. Следовательно, 82% женщин применяли имплант в течение 24 мес [301].
Рекомендации
Частоту досрочного удаления импланта можно уменьшить, если заранее предупре¬
дить женщину о вероятности возникновения нерегулярных менструаций. Женщины
должны получить полную информацию о возможных нарушениях менструального
цикла, чтобы соотнести неудобства, связанные с побочными эффектами, и пользу
от высокой контрацептивной активности и принять осознанное решение о примене¬
нии этого метода. Существенным для успешного длительного применения импланта
является предварительная беседа с женщиной с последующим наблюдением после
введения импланта.
У женщин, имеющих множество половых партнеров, повышена вероятность
как возникновения нежелательной беременности, так и заражения заболеваниями,
передающимися половым путем, например СПИДом, гепатитом В, хламидиозом,
сифилисом, гонореей, которые могут иметь очень серьезные, угрожающие жизни
последствия. Имплантационная контрацепция не влияет на степень риска заражения
этими заболеваниями [326]. Женщин этой категории следует предупредить об опас¬
ности заражения заболеваниями, передающимися половым путем, и посоветовать
дополнительно использовать барьерные методы контрацепции, что предохранит и от
нежелательной беременности, и от заражения. Имплант с этоногестрелом, подобно
пероральным контрацептивам и депо-медроксипрогестерона ацетату, снижает риск
возникновения инфекций органов малого таза.
Клинический опыт применения импланта с этоногестрелом свидетельствует о том,
что эффективность и популярность этого метода тесно связаны с информированно¬
стью женщины и навыками медицинского персонала. В первые 18 мес с начала при¬
менения импланта с этоногестрелом в Австралии (май 2001 г.) появились сообщения
о непредвиденно большом количестве развившихся осложнений, а также о 100 слу¬
чаях нежелательных беременностей. Практически все эти осложнения были связаны
с нарушениями техники (из-за неопытности медицинского персонала) или времени
199
введения импланта, неправильным подбором женщин, которым показан имплант,
а также недостаточным их информированием. Royal Australian College of General
Practitioners документировал процесс, процедуру и отбор пациентов и, таким образом,
решил проблему [327].
Внутриматочная гормональная контрацепция
Для контрацепции применяют внутриматочные средства (ВМС), как выделяющие
лекарственные вещества, так и не выделяющие их. Внутриматочные средства, содер¬
жащие медь или синтетические гестагены, эффективнее и имеют меньше побочных
эффектов, чем просто пластмассовые или стальные, и практически вытеснили более
старые образцы. Значительная часть информации о внутриматочных средствах уста¬
рела и неверна, так как основана на исследовании этих старых вариантов контрацеп¬
тивов.
ЛНГ-20 (мирена*), производимая Leiras-Schering AG в Финляндии, выделяет in vitro
20 мг левоноргестрела в сутки [328]. Мирена* имеет Т-образную форму с покрытием,
содержащим 52 мг левоноргестрела в полидиметилсилоксане. Левоноргестрел снача¬
ла выделяется со скоростью 20 мг/сут in vivo, постепенно снижающейся и достигаю¬
щей половины указанного значения через 5 лет. Длительность применения, согласно
инструкции, составляет 5 лет, однако действие продолжается до 10 лет, препарат
ослабляет менструальные кровотечения и частоту возникновения инфекций органов
малого таза [329-331]. Внутриматочное средство, содержащее левоноргестрел, при¬
близительно равноэффективно выскабливанию эндометрия при лечении меноррагий
[332]. Местное действие гестагена на эндометрий можно использовать у женщин,
принимающих тамоксифен [333], при дисменореє [334], лечении эстрогенами в кли¬
мактерическом периоде [335, 336]. В Европе разработаны внутриматочные средства,
выделяющие 5 мг или 10 мг левоноргестрела, для применения в течение не менее 5 лет
у женщин в постменопаузе [336, 337].
. Реакцию матки на присутствие инородного тела дополняет влияние гестагена,
выделяющегося из внутриматочного средства, на состояние эндометрия. Развиваются
изменения слизистой оболочки эндометрия с атрофией желез [338]. Гестаген имеет
два механизма действия: препятствует имплантации оплодотворенной яйцеклетки
и увеличивает вязкость шеечной слизи, что затрудняет продвижение через нее спер¬
матозоидов. Концентрации в сыворотке крови левоноргестрела, выделяющегося из
внутриматочного средства, составляют приблизительно половину таковых при при¬
менении норпланта*“; таким образом, развитие фолликул в яичнике также частично
угнетается. После года применения ВМС овуляция сохраняется у 50-75% женщин
независимо от характера менструаций [339]. В конечном итоге гестагенсодержащие
ВМС изменяют свойства шеечной слизи, что препятствует проникновению сперма¬
тозоидов. При применении гестагенсодержащих ВМС менструации становятся менее
обильными (приблизительно на 40-50%), исчезают или уменьшаются боли перед
и во время менструаций; через год применения ВМС, содержащие левоноргестрел,
могут уменьшить кровопотерю во время менструации на 90% [340], а в среднем у 50%
женщин развивается аменорея [341, 342]. Среднее содержание гемоглобина и железа
в крови повышается по сравнению с исходными значениями [343].
ЭФФЕКТИВНОСТЬ
ВМС, постепенно выделяющее медь (Т Ку-380А или Парагард^), разрешено в США
для непрерывного применения в течение 10 лет. Однако показано, что его эффектив¬
ность сохраняется не менее 12 лет [344]. Т Ку-200*“ нужно заменять каждые 4 года,
а Нову^ — каждые 5 лет. Левоноргестрелсодержашие ВМС могут непрерывно нахо¬
200
диться в полости матки не менее 7 лет, а возможно, 10 лет [345]. Эффективность ВМС,
содержащего левоноргестрел, выделяющийся со скоростью 15-20 мг/сут, такая же,
как новых медьсодержащих ВМС [329, 346, 347, 348].
Длительность нахождения в полости матки ВМС, не выделяющих лекарственные
вещества, не ограничена. ВМС с отложенными на его поверхности солями кальция
оказывает раздражающее действие на эндометрий. Если после установки ВМС, не
выделяющей лекарственного вещества, постепенно усиливается кровотечение, эту
ВМС следует заменить. Некоторые врачи рекомендуют заменить старые ВМС на
новые, содержащие медь, как на более эффективные (что соответствует истине).
Вероятность возникновения внематочной беременности не зависит от длитель¬
ности применения ВМС (Т Ку-380А или выделяющей левоноргестрел) [346, 349]. В
проспективном исследовании при использовании ВМС, содержащей левоноргестрел,
не выявлено ни одного случая возникновения внематочной беременности в течение
7 лет, в другом исследовании — 1 случай за 5 лет [346, 350]. В рандомизированных
мультицентровых испытаниях зарегистрирован 1 случай возникновения внематоч¬
ной беременности при применении Т Ку-380А при пересчете на 8000 женщин/лет
(что в 10 раз меньше, чем при применении инертных ВМС или Т Ку-200) [346].
Следовательно, риск развития внематочной беременности у женщин, использующих
медь- или левоноргестрелсодержащие ВМС, значительно меньше, чем у женщин, не
пользующихся контрацепцией; однако если при применении ВМС беременность воз¬
никла, вероятность, что она внематочная, высока [351].
Низкая степень риска возникновения внематочной беременность при применении
ВМС Т Ку-380А или выделяющих левоноргестрел делает их контрацептивами выбора
для женщин, имевших внематочную беременность в прошлом (табл. 2-11).
Таблица 2-11. Частота возникновения внематочной беременности на 1000 женщин/лет [352, 353]
Метод контрацепции
Частота
Не использовали, все возрастные группы
3,00-4,50
ВМС с левоноргестрелом
0,20
ВМС Т Ку-ЗвОА
0,20
ПОБОЧНЫЕ ЭФФЕКТЫ
При условии правильного выбора показаний к внутриматочной контрацепции
и соблюдения техники введения медьсодержащие и вьщеляющие гестаген ВМС не
повышают вероятности развития бесплодия после их удаления. Даже при досрочном
удалении в связи с побочными эффектами фертильность не изменяется [348, 354,
355].
Симптомами, наиболее часто служащими причиной досрочного удаления ВМС,
являются увеличение кровопотери при менструации и болезненные менструации. В
связи с этими проблемами 5-15% женщин удаляют ВМС в первый год применения.
При применении ВМС с меньшим содержанием меди или левоноргестрела значитель¬
но снижается вероятность возникновения вышеперечисленных осложнений, однако
при консультации женщины при выборе этого способа контрацепции важно обратить
внимание на характер менструации. Женщины с продолжительными, обильными
или болезненными менструациями могут плохо переносить медьсодержащие ВМС,
а гестагенсодержащие ВМС могут оказать благоприятное влияние на характер мен¬
струации [340]. Обильные менструации и боли внизу живота наиболее выражены
в течение первых месяцев после установки ВМС; эти симптомы можно ослабить
приемом нестероидных противовоспалительных средств (НПВС; угнетают синтез
простагландинов) в первые менструальные циклы. НПВС эффективны даже при
201
длительных обильных менструациях [356]. Прием НПВС следует начать в 1-й день
менструации и продолжить в течение 3 дней. В Китае применяют медьсодержащие
ВМС, выделяющие также небольшие количества индометацина, что ослабляет выра¬
женность кровотечений [357].
У женщин с ВМС могут возникать межменструальные кровянистые выделения из
влагалищам, причиняющие неудобства, но не приводящие к существенной кровопо¬
тере. При появлении этих симптомов следует обратиться к врачу для определения
причины кровотечения (исключения патологии шейки матки или эндометрия).
Нерегулярность кровянистых выделений из половых путей может внести непредви¬
денные коррективы в ритм половой жизни.
Современные медьсодержащие ВМС приводят к увеличению кровопотери во время
каждой менструации приблизительно на 55% в течение всего времени применения
[358]. Увеличение кровопотери связано с небольшим (1-2 дня) удлинением мен¬
струации и не приводит в течение года к развитию лабораторных признаков желе¬
зодефицитной анемии (например, изменение концентрации ферритина в сыворотке
крови). Однако при более длительном применении ВМС концентрации ферритина
в сыворотке крови снижаются, что предполагает истощение запасов железа в орга¬
низме [359]. Необходимо своевременное выявление признаков истощения запасов
железа и анемии у женщин, длительно применяющих медьсодержащие ВМС, а также
у предрасположенных к развитию железодефицитной анемии. В регионах с высокой
заболеваемостью анемией перечисленные изменения развиваются быстрее, поэтому
рекомендована заместительная терапия препаратами железа [360].
При применении гестагенсодержащих ВМС постепенно могут развиться децидуа-
лизация, атрофические изменения в эндометрии. У 30-40% женщин, использующих
ВМС, выделяющих левоноргестрел, менструации становятся менее обильными, а
у 30-40% в течение 2 лет развивается аменорея [342, 361]. При применении ВМС,
выделяющих левоноргестрел, более 12 лет у 60% женщин менструации отсутство¬
вали, у 12% были редкие и скудные, а у 28% — регулярные, но необильные [343].
Некоторые женщины так расстраиваются из-за отсутствия менструаций, что просят
удалить ВМС. С другой стороны, следствием отсутствия менстр5'’аций является повы¬
шение содержания гемоглобина в крови [346,362].
ВМС, содержащие левоноргестрел, очень эффективны для лечения меноррагий
[340,363]. Этот лечебный эффект настолько выражен, что сопоставим с результатами
хирургического лечения (выскабливания полости матки или гистерэктомии) [332,
364, 365, 366]. Кровотечения уменьшаются даже при наличии лейомиомы одно¬
временно с уменьшением размеров матки [367]. ВМС, содержащие левоноргестрел,
успешно применяют при лечении эндометриоза, особенно при дисменореє, связанной
с эндометриозом [368].
Гестаген, выделяющийся из левоноргестрелсодержащих ВМС, попадает в систем¬
ный кровоток, поэтому может вызвать побочные эффекты, связанные с андрогенным
действием, такие как угри и гирсутизм, однако в одном исследовании не обнаружено
изменений концентрации андрогенсвязывающих глобулинов, поэтому развитие этих
побочных эффектов маловероятно [140]. Для заключения о влиянии ВМС, содержа¬
щих левоноргестрел, на соотношение липопротеинов в крови необходимы тщатель¬
ные клинические исследования, однако маловероятно, что низкие дозы левонорге¬
стрела могут повлиять на риск развития сердечно-сосудистых заболеваний.
ИНФЕКЦИИ
В настоящее время считают, что развитие воспалительных заболеваний малого
таза, связанных с бактериями, вызвано бактериальным инфицированием полости
матки во время введения ВМС. В ходе классического исследования МізЬеІІ было под¬
тверждено, что эта процедура обычно сопровождается бактериальным инфицирова-
202
ниєм [369]. Инфекции, развившиеся через 3-4 мес после введения ВМС, переданы
половым путем, а не являются результатом введения ВМС. Инфекции, вызванные
ВМС, развиваются в ранние сроки после введения, они полимикробные и связаны с
распространением эндогенной микрофлоры, преимущественно анаэробной, из влага¬
лища и шейки матки.
В обзоре ВОЗ всех клинических испытаний ВМС, проведенных под эгидой этой
организации, сделано заключение, что вероятность воспалительных инфекционных
заболеваний органов малого таза в 6 раз выше в первые 20 дней после введения,
чем в более поздние сроки, но, что наиболее важно, этот показатель в поздние сроки
чрезвычайно низкий [370J. Однако из 23 ООО введений ВМС лишь в 81 случае были
диагностированы воспалительные инфекционные поражения органов малого таза, и
очень их малое количество отмечали в случаях, когда практически исключали веро¬
ятность ЗППП. Достоверной разницы при сравнении медьсодержащих, инертных
или гестагенсодержащих ВМС не выявлено. Эти сведения подтверждают полученные
ранее результаты о том, что наибольший риск развития инфекций малого таза отме¬
чен сразу после введения ВМС, и он не повышается при длительном применении [371,
372]. Проблема инфицирования органов малого таза решается тщательным отбором
женщин, которым можно вводить ВМС, и соблюдением всех правил асептики. Риск
развития инфекций малого таза не повышен даже у женщин, страдающих сахарным
диабетом 1-го типа [373, 374].
Для профилактики возникновения инфекций органов малого таза за 1 ч до введе¬
ния ВМС можно назначить внутрь доксициклин (200 мг) или азитромицин (500 мг),
однако профилактический прием антибиотиков, вероятно, нецелесообразен при низ¬
кой степени риска инфицирования заболеваниями, передающимися половым путем.
Нет оснований предполагать, что не содержащие лекарственных средств или покры¬
тые медью ВМС обеспечат защиту от заболеваний, передающихся половым путем, в
частности воспалительных заболеваний органов малого таза [375]. Тем не менее
существует мнение, что ВМС, выделяющие левоноргестрел, могут оказать защит¬
ное действие в отношении инфекций органов малого таза, а при применении ВМС,
покрытых медью, снижается титр противохламидийных антител [330, 376]. In vitro
медь тормозит рост хламидий в клетках эндометрия [377]. Таким образом, версия, что
использование ВМС сопряжено с риском возникновения инфекций органов малого
таза (и бесплодием), не нашла подтверждения [378]. Женщин, использующих ВМС,
следует предупредить о необходимости дополнительного применения презервативов
при половом контакте с новым партнером для исключения заражения ЗППП. В связи
с тем что вероятность возникновения инфекций малого таза в наибольшей степени
зависит от постоянства полового партнера, при решении вопроса о возможности при¬
менения ВМС врач должен расспросить женщину о количестве половых партнеров,
характере половых контактов, их частоте, вероятности беспорядочных контактов у
партнера, времени начала половой жизни, наличии ЗППП в прошлом [379]. У женщин
с малой степенью риска заражения ЗППП маловероятно развитие инфекций органов
малого таза, связанных с применением ВМС [372].
СПИСОК ЛИТЕРАТУРЫ
1. Trussell J, Contraceptive efficacy of the personal hormone monitoring system Persona // Br. J.
Fam. Plann. - 1999. - Vol. 25. - P. 178-179,
2. Fu H., Darroch J.E., Haas T„ Ranjit N. Contraceptive failure rates: new estimates from the 1995
National Survey of Family Growth // Fain. Plann. Pcrspect. — 1999. — Vol. 31. - P. 58-^3.
3. Rossmanith W.G., Steffens D., Schramm G. A comparative randomized trial on the impact of two
low-dose oral contraceptives on ovarian activity, cervical permeability and endometrial receptivity //
Contraception. - 1997. - Vol. 56. — P. 23-30.
203
4. Trussell J., Vaughan Б. Contraceptive failure, method-related discontinuation and resumption
of use: results from the 1995 National Survey of Family Growth // Fam. Plann. Perspect. - 1999. -
Vol. 31. - P. 64-72.
5. Trussell J. Contraceptive failure in the United States // Contraception. ~ 2004. — Vol. 70. —
P. 89-96.
6.,Xetterie G.S., Chow G.E. Effect of “missed” pills on oral contraceptive effectiveness // Obstet.
Gynecol. - 1992. - Vol. 79. - P. 979- 982.
7. Westhoff C., Kems J„ Morroni C. et al. Quick-start: a novel oral contraceptive initiation method //
Contraception. — 2002. - Vol. 66. — P. 141-145.
8. Westhoff C., Morroni C., Kems J., Murphy P.A. Bleeding patterns after immediate vs. conventional
oral contraceptive initiation: a randomized, controlled trial // Fertil. Steril. - 2003. - Vol. 79. —
P. 322-329.
9. Anderson F.D., Hait H. The Seasonale-301 Study Group. A multicenter, randomized study of an
extended cycle oral contraceptive // Contraception. — 2003. — Vol. 68. — P. 89 -96.
10. Kwiecien M., Edehnan A., Nichols M.D., Jensen J.T. Bleeding patterns and patient acceptability
of standard or continuous dosing regimens of a low-dose oral contraceptive: a randomized trial //
Contraception. — 2003. - Vol. 67. - P. 9-13.
11. Miller L., Hughes J.P. Continuous combination oral contraceptive pills to eliminate withdrawal
bleeding: a randomized trial // Obstet. Gynecol. — 2003. — Vol. 101. — P. 653-661.
12. Sulak P.J., Carl J., Gopalakrishnan I. et al. Outcomes of extended oral contraceptive regiments
with a shortened hormone-free interval to manage breakthrough bleeding // Contraception. —
2004. - Vol. 70. - P. 281-287.
13. Potter L.. Oakley D., de Leon-Wong E., Canamar R. Measuring compliance among oral
contraceptive users // Fam. Plann. Perspect. — 1996. ~ Vol. 28. — P. 154-158.
14. Killick S.R., Bancroft K., Oelbaum S, et al. Extending the duration of the pill-free interval during
combined oral contraception // Adv.Contracept. - 1990. - Vol. 6. - P. 33-40.
15. Elomaa K., Rolland R., Brosens I. et al. Omitting the first oral contraceptive pills of the cycle does
not automatically lead to ovulation // Am. J. Obstet. G5mecol. — 1998. — Vol. 179. — P. 41-46.
16. van Heusden A.M., Fauser B.C. Activity of the pituitary-ovarian axis in the pill-free interval
during use of low-dose combined oral contraceptives // Contraception. — 1999. — Vol. 59. —
P. 237-243.
17. Sparrow M.J. Pill metliod failures // N. Z. Med. J. — 1987. - Vol. 100. — P. 102-105.
18. Hansen Т.Н., Lundvall F. Factors influencing the reliability of oral contraceptives // Acta Obstet.
Gynecol. Scand. - 1997. - Vol. 76. - P. 61-64.
19. Jung-Hoffmann C., Kuhi H. Intra- and interindividual variations in contraceptive steroid levels
during 12 treatment cycles: no relation to irregular bleedings // Contraception. — 1990. — Vol. 42. —
P. 423-438.
20. Endrikat J., Müller U., Düsterberg В. A twelve-month comparative clinical investigation of
two low-dose oral contraceptives containing 20 jLg ethinylestradioI/75 jig gestodene and 30 |ig
ethinylestradiol/75 jig gestodene. with respert to efficacy, cycle control, and tolerance //
Contraception. — 1997. — Vol. 55. — P. 131-137.
21. Rosenberg M.J., Meyers A. Roy V Efficacy, cycle control, and side effects of low- and lower-dose
oral contraceptives, a randomized trial of 20 micrograms and 35 micrograms estrogen preparations j j
Contraception. — 1999. - Vol. 60. - P. 321-329.
22. Rosenberg M.J., Waugh M.S., Stevens C.M. Smoking and cycle control among oral contraceptive
users // Am. J. Obstet. Gynecol. - 1996. - Vol. 174. - P. 628-632.
23. Rosenberg M.J., Waugh M.S., HigginsJ.E. The effect of desogestrel, gestodene, and other factors
on spotting and bleeding // Contraception. — 1996. — Vol. 53. — P. 85-90.
24. Krettek J.E., Arkin S.L, Chaisilwattana P., Monif G.R. Chlamydia trachomatis in patients who
used oral contraceptives and had intermenstrual spotting // Obstet, Gynecol. — 1993. Vol. 81. —
P. 728-731.
25. Meade T.W. Oral contraceptives, dotting factors, and thrombosis // Am. J. Obstet. Gynecol. —
1982. - Vol. 142. - P. 758-761.
26. The Oral Contraceptive and Hemostasis Study Group. The effect of seven monophasic oral con¬
traceptive regimens on hemostatic variables: conclusions from a large randomized multicenter study //
Contraception. — 2003. — Vol. 67. — P. 173-185.
204
27. Jespersen J., Petersen K.R., Skouby S,0. Effects of newer oral contraceptives on the inhibition
of coagulation and fibrinolysis in relation to dosage and type of steroid // Am. J. Obstet. Gynecol. —
1990. - Vol. 163. - P. 396-403.
28. Notelovitz M., Kitchens C.S., Khan F.Y. Changes in coagulation and anticoagulation in women
taking low-dose triphasic oral contraceptives; a controlled comparative 12-month clinical trial // Am.
J. Obstet. Gynecol. - 1992. - Vol. 167. - P. 1255-1261.
29. Schlit A.F., Grandjean P., Domiez J., Lavenne E. Large increase in plasmatic ll-dehydro-TXB2
levels due to oral contraceptives // Contraception. — 1995. — Vol. 51. — P. 53-58.
30. Fruzzetti F., Ricci C., Fioretti P. Haemostasis profile in smoking and nonsmoking women taking
low-dose oral contraceptives j j Contraception. — 1994. - Vol. 49. - P. 579-.592.
31. Basdevant A., Conard J., Pelissier C. et al. Hemostatic and metabolic effects of lowering
the ethinyl-estradiol dose from 30 p.g to 20 pg in oral contraceptives containing desogestrel //
Contraception. — 1993. - Vol. 48. — P. 193-204.
32. Winkler U.H., Schindler A.E., Endrikat J., Dusterberg B. A comparative study of the effects of
the hemostatic system of two monophasic gestodene oral contraceptives containing 20 |U.g and 30 |ig
ethinylestradiol // Contraception. - 1996. - Vol. 53. - P. 75-84.
33. Croft P., Hannaford P.C. Risk factors for acute myocardial infarction in women: evidence from
the Royal College of General Practitioners' oral contraception study j j BMJ. - 1989. — Vol. 298. -
P. 165-168.
34. Rosenberg L., Palmer J.R., Lesko S.M., Shapiro S. Oral contraceptive use and the risk of myocar¬
dial infaraction // Am. J. Epidemiol. — 1990. — Vol. 131. - P. 1009-1016.
35. Stampfer M.J., Willett W.C., Colditz G.A. et al. Past use of oral contraceptives and cardiovas¬
cular disease: a meta-analysis in the context of the Nurses’ Health Study // Am. J. Obstet. Gynecol. —
1990. - Vol. 163. - P. 285-291.
36. Colditz G.A. Oral contraceptive use and mortality during 12 years of follow-up: the Nurses'
Health Study // Ann, Intern. Med. - 1994. - Vol. 120. - P. 821-826.
37. Beral V., Hermon C., Kay C. et al. Mortality associated with oral contraceptive use: 25-year
follow-up of cohort of 46,000 women from Royal College of General Practitioners’ oral contraception
study // BMJ. - 1999. - Vol. 318. - P. 96-100.
38. Vessey M., Painter R., Yeates D. Mortality in relation to oral contraceptive use and cigarette
smoking j j Lancet. - 2003. - Vol. 362. — P. 185-191.
39. Bottiger L.E., Boman G., Eklund G., Westerholm B. Oral contraceptives and thromboembolic
disease: effects of lowering oestrogen content // Lancet. — 1980. — Vol. 1. — P. 1097-1101.
40. Gerstman B.B., Piper J.M., Tomita D.K. et al. Oral contraceptive estrogen dose and the risk of
deep venous thromboembolic disease // Am. J. Epidemiol. — 1991. - Vol. 133. - P. 32-37.
41. Vessey M., Mant D., Smith A., Yeates D. Oral contraceptives and venous thromboembolism: find¬
ings in a large prospective study // BMJ. — 1986. - Vol. 292. - P. 526-533.
42. Helmrich S.P., Rosenberg L., Kaufman D.W. et al. Venous thromboembolism in relation to oral
contraceptive use j j Obstet. Gynecol. - 1987. — Vol. 69. - P. 91-95.
43. Thorogood M.. Mann J., Murphy M., Vessey M, Risk factors for fatal venous thromboembolism
in young women: a case-control study // Int. J. Epidemiol. — 1992. — Vol. 21, — P. 48-52.
44. Rosenberg L., Hennekens C.H., Rosner B. et al. Oral contraceptive use in relation to nonfatal
myocardial infarction// Am. J. Epidemiol. — 1980. — Vol. 11. — P. 59-66.
45. Royal College of General Practitioners’ Oral Contraceptive Study. Incidence of arterial disease
among oral contraceptive users //J. R. Coll. Gen. Pract. - 1983. - Vol. 33, - P. 75-82.
46. Lidegaard O. Oral contraception and risk of a cerebral thromboembolic attack: results of a case-
control study // BMJ. - 1993, - Vol. 306. - P. 956-963.
47. Lawson D.H., Davidson J.F., Jick H. Oral contraceptive use and venous thromboembolism:
absence of an effect of smoking // BMJ. - 1977. - Vol. 2. - P. 729-730.
48. Petitti D.B., Wingerd J., Pellegrin F., Ramcharan S. Oral contraceptives, smoking, and other fac¬
tors in relation to risk of venous thromboembolic disease// Am. J. Epidemiol, — 1978. — Vol. 108. — P.
480-485.
49. Royal College of General Practitioners : Oral contraceptive study: oral contraceptives, venous
thrombosis and varicose veins //J. R, Coll. Gen. Pract. — 1978. — Vol. 28. — P. 393-399.
50. Forter J.B., Hershel J„ Walker A.M. Mortality among oral contraceptive users // Obstet.
Gynecol. - 1987. - Vol. 70. - P. 29-32.
205
51. WHO Collaborative Study of Cardiovascular Disea.se and Steroid Hormone Contraception. Effect
of different progestagens in low oestrogen oral contraceptives on venous thromboembolic disease //
Lancet. - 1995. - Vol. 346. - P. 1582-1588.
52. Spitzer W.O., Lewis M.A., Heinemann L.A. et al. Third generation oral contraceptives and
risk of venous thromboembolic disorders: an international case-control study // BMJ. — 1996. —
Vol. 312. - P. 83-88.
53. Jiek H., Jick S.S., Gurewich V. et al. Risk of idiopathic cardiovascular death and nonfatal venous
thromboembolism in women using oral contraceptives with differing progestagen components //
Lancet. - 1995. - Vol. 348. - P. 1589-1593.
54. Bloemenkamp K.W., Rosendaal F.R., Helmerhorst F.M. et al. Enhancement by factor V Leiden
mutation of risk of deep-vein thrombosis associated with oral contraceptives containing a third-gener¬
ation progestagen // Lancet. — 1995. — Vol. 348. — P. 1593-1596.
55. Farmer R.D.T., Preston T.D. The risk of venous thromboembolism associated with low estrogen
oral contraceptives //J. Obstet. Gynaecol. — 1995. — Vol. 15, - P. 195-200.
56. Vandenbroucke J.P., van der Meer F.J., Helmerhorst F.M., Rosendaal F.R. Factor V Leiden //
BMJ. - 1996, - Vol. 313. - P. 1127-1130.
57. Ridker P.M., Miletich J,P„ Hennekens C.H., Buring J.E. Ethnic distribution of factor V Leiden
in 4047 men and women: implications for venous thromboembolism screening //JAMA. — 1997. -
Vol. 277. - P. 1305-1307.
58. Sidney S., Petitti D.B., Soff G.A. et al. Venous thromboembolic disease in users of low-estrogen
combined estrogen-progestin oral contraceptives // Contraception. — 2004, — Vol, 70. — P. 3-10.
59. Emmerich J„ Rosendaal F.R., Cattaneo M. et al. Combined effect of factor V Leiden and
prothrombin 20210A on the risk of venous thromboembolism-pooled analysis of 8 case-control
studies including 2310 cases and 3204 controls. Study Group for Pooled-Analysis in Venous
Thromboembolism //Thromb. Haemost. — 2001. - Vol. 86. - P. 809-816.
60. Hajjar K.A. Factor V Leiden; an unselfish gene.? // New. Engl.J. Med. - 1994. — Vol. 331. -
P. 1585-1587.
61. Vensson P.J., Dahlback B. Resistance to activated protein С as a basis for venous thrombosis //
New-. Engl.J. Med. - 1994. - Vol. 330. - P. 517-522.
62. Hellgren М., Svensson P.J., Dahlback B. Resistance to acti\’ated protein С as a basis for venous
thromboembolism associated with pregnancy' and oral contraceptives // Am. J. Obstet. Gynecol. —
1995. - Vol. 173. - P. 210-213.
63. Vandenbroucke J.P., Koster Т., Briet E. et al. Increased risk of venous thrombosis in oral-
contraceptive users who are carriers of factor V Leiden mutation // Lancet, — 1994. - Vol. 344. —
P. 1453-1457.
64. Spannagl М., Heinemann A.J., Schramm W. Are factor V Leiden carriers who use oral contra¬
ceptives at extreme risk for venous thromboembolism? // Eur. J. Contracept. Reprod. Health. Care. —
2000. - Vol. 5. - P. 105-112.
65. Bloemenkamp K.W., Rosendaal F.R., Helmerhorst F.M., Vandenbroucke J.P. Higher risk of
venous thrombosis during early use of oral contraceptives in women with inherited clotting defects //
Arch. Intern. Med. — 2000. — Vol. 160. — P. 49-52.
66. Lidegaard O., Kreiner S. Cerebral thrombosis and oral contraceptives. A case-control study //
Contraception, — 1998. - Vol. 57. - P. 303-314.
67. Lidegaard O., Edstrom B, Oral contraceptives and myocardial infarction. A case-control study
(abstract) // Eur. J. Contracept. Reprod. Health, Care. - 1996. - Vol. 1. - Suppl. - P. 72-73.
68. WHO Collaborative Study of Cardiovascular Disease and Steroid Hormone Contraception.
Acute myocardial infarction and combined oral contraceptives: results of an international multicentre
case-control study // Lancet. — 1997, — Vol. 349. - P. 1202-1209.
69. Petitti D.B., Sidney S., Quesenberry C.P. Jr., Bernstein A. Incidence of stroke and myocardial
infarction in women of reproductive age // Stroke. — 1997. — Vol. 28. — P, 280-283.
70. Jick H., Porter J., Rothman K.J. Oral contraceptives and nonfatal stroke in healthy young
women // Ann. Int. Med. - 1978. - Vol. 89. — P. 58-60.
71. Vessey M.P., Lawless М., Yeates D. Oral contraceptives and stroke: findings in a large prospective
study // BMJ. - 1984. - Vol. 289. - P. 530-531.
72. Hannaford P.C., Croft P.R., Kay C.R. Oral contraception and stroke: evidence from the
Royal College of General Practitioners’ Oral Contraception Study // Stroke. - 1994. - Vol. 25. -
P. 935-942.
206
73. Petitti D.B„ Sidney S., Bernstein A. et al. Stroke in users of low-dose oral contraceptives j I New.
Engl. J. Med, - 1996. - Vol. 335. - P. 8-15.
74. WHO Collaborative Study of Cardiovascular Disease and Steroid Hormone Contraception.
Ischaemic stroke and combined oral contraceptives: results of an international, multicentre case-control
study // Lancet. - 1996. - Vol. 348. - P. 498-505.
75. WHO Collaborative Study of Cardiovascular Disease and Steroid Hormone Contraception.
Haemorrhagic stroke, overall stroke risk, and combined oral contraceptives: results of an international,
multicentre, case-control study j j Lancet. — 1996. — Vol. 348. - P. 505-510,
76. Barrett D.H., Anda R.F., Escobedo L.G. et al. Trends in oral contraceptive use and cigarette
smoking j j Arcb, Fam, Med. — 1994. — Vol. 3. — P. 438-443.
77. Kovacs L., Baitfai G„ Apro G. et al. The effect of the contraceptive pill on blood pressure: a
randomized controlled trial of three progestogen-oestrogen combinations in Szeged, Hungary //
Contraception. - 1986. — Vol. 33. - P. 69-77.
78. Nichols M-, Robinson G., Bounds W. et al. Effect of four combined oral contraceptives on blood
pressure in the pill-free interval // Contraception. — 1993, - Vol. 47. - P. 367-376.
79. Shen Q-. Lin D.. Jiang X. et al. Blood pressure changes and hormonal contraceptives //
Contraception. - 1994. - Vol. 50. — P. 131-141.
80. Damey P. Safety and efficacy of a triphasic oral contraceptive containing desogrestrel: results of
three multicenter trials // Contraception. — 1993. — Vol. 48. — P. 323-337.
81. Chasan-Taber L., Willett W.C., Manson J.E. et al. Prospective study of oral contraceptives and
hypertension among women in the United States // Circulation, — 1996. — Vol. 94. — P. 483-489.
82. Brady W.A., Kritz-Silverstein D., Barrett-Connor E., Morales A.J. Prior oral contraceptive use
is associated with higher blood pressure in older women //J. Women’s. Health. - 1998. - Vol. 7. -
P. 221-227.
83. Pabinger L, Schneider B„ the GTH Study Group. Thrombotic risk of women with hereditary
antithrombin III, protein C, and protein S deficiency taking oral contraceptive medication // Thromb.
Haemost. - 1994. - Vol. 5. - P. 548-552.
84. Godsland I.F., Crook D., Simpson R. et al. The effects of different formulations of oral contra¬
ceptive agents on lipid and carbohydrate metabolism // New. Engl. J. Med. - 1990. - Vol. 323. —
P. 1375-1381.
85. van der Vange N., Kloosterboer H.J., Haspels A.A. Effect of seven low-dose combined oral
contraceptive preparations on carbohydrate metabolism // Am. J. Obstet. Gynecol. — 1987. — Vol.
156. - P. 918-922.
86. Bowes W.A. Jr., Katta L.R., Droegemueller W„ Bright T.G. Triphasic randomized clinical trial:
Comparison of effects on carbohydrate metabolism // Am. J. Obstet. Gynecol. — 1989. — Vol. 161. —
P. 1402-1407.
87. Gaspard U.J., Lefebvre PJ. Clinical aspects of the relationship between oral contraceptives,
abnormalities in carbohydrate metabolism, and the development of cardiovascular disease // Am. J.
Obstet. Gynecol. - 1990. - Vol. 163. - P. 334-343.
88. Troisi R.J., Cowie C.C., Harris M.I. Oral contraceptive use and glucose metabolism in a national
sample of women in the United States // Am. J. Obstet. Gynecol. — 2000. — Vol. 183. — P. 389-395.
89. Hannaford P.C., Kay C.R., Vessey M.P. et al. Combined oral contraceptives and liver disease //
Contraception. — 1997. — Vol. 55. — P. 145-151.
90. Lubianca J.N., Faccin C.S., Fuchs F.D. Oral contraceptives: a risk factor for uncontrolled blood
pressure among hypertensive women // Contraception. - 2003. — Vol. 67, — P. 19-24.
91. Ross R.K., Pike M.C., Vessey M.P. et al. Risk factors for uterine fibroids: reduced risk associated
with oral contraceptives // BMJ. - 1986. - Vol. 293. - P. 359-362.
92. Parazzini F., Negri E., La Vecchia C, et al. Oral contraceptive use and risk of uterine fibroids //
Obstet. Gynecol. - 1992. - Vol. 79. - P. 430-433.
93. Samadi A.R., Lee N.C., Flanders W.D. et al. Risk factore for self-reported uterine fibroids: a case-
control study // Am. J. Public. Health. — 1996. — Vol. 86. — P. 858-862.
94. Marshall L.M., Spiegelman D., Goldman M.B. et al. A prospective study of reproductive factors
and oral contraceptive use in relation to the risk of uterine leiomyomata // Fertil. Steril. - 1998. -
Vol. 70. - P. 432-439.
95. Chiaffarino F., Parazzini F., La Vecchia C. et al. Use of oral contraceptives and uterine fibroids:
results from a case-control study // Br. J. Obstet. Gynaecol, - 1999. — Vol. 106. — P. 857-860.
207
96. Kjos S.L., Shoupe D., Douyan S. et al. Effect of low-dose oral contraceptives on carbohydrate
and lipid metabolism in women with recent gestational diabetes: results of a controlled, randomized,
prospective study // Am. J. Obstet. Gynecol, - 1990. — Vol. 163. — P. 1822-1827.
97. Kjos S.L., Peters R.K., Xiang A. et al. Contraception and the risk of type 2 diabetes in Latino
women with prior gestational diabetes //JAMA. — 1998. — Vol. 280. — P. 533-538.
98. Garg S.K., Chase H.P., Marshall G. et al. Oral contraceptives and renal and retinal comphca-
tions in young women with insulin-dependent diabetes mellitus // JAMA, — 1994. — Vol. 271. -
P. 1099-1102.
99. Petersen K.R., Skouby S.O., Sidelmann J. et al. Effects of contraceptive steroids on cardiovascular
risk factors in women with insulin-dependent diabetes mellitus // Am. J. Obstet. Gynecol. — 1994. —
Vol. 171. - P. 400-405.
100. Klein B.E., Klein R„ Moss S.E. Mortality and hormone-related exposures in women with dia¬
betes // Diabetes. Care. - 1999. - Vol. 22. - P. 248-252.
101. Mattson R.H., Cramer J.A., Damey P.D., Naftolin F. Use of oral contraceptives by women with
epilepsy //JAMA. - 1986. - Vol. 256. - P. 238-240.
102. Vessey M., Painter R., Yeates D. Oral contraception and epilepsy: findings in a large cohort
study // Contraception. — 2002. — Vol. 66. — P. 77-79.
103. Lutcher C.L., Milner P.F. Contraceptive-induced vascular occlusive events in sickle cell disor¬
ders — fact or fiction,? // [abstract]. Clin. Res. — 1986. — Vol. 34, - P. 217A.
104. Yoong WC., Tuck S.M., Yardumian A. Red cell deformability in oral contraceptive pill users with
sickle cell anemia // Br.J. Haematol. - 1999. - Vol. 104. - P. 868-870,
105. De Ceulaer K„ Gruber C., Hayes R., Serjeant G.R. Medroxyprogesterone acetate and homozy¬
gous sickle-cell disease // Lancet. - 1982. - Vol. 2. — P. 229-231.
106. Jüngers P., Dougados M.. Pelissier C. et al. Influence of oral contraceptive therapy on the activ¬
ity of systemic lupus erythematosus //Arthritis. Rheum. — 1982, — Vol. 25. - P. 618-623.
107. Petri M., Robinson C. Oral contraceptives and systemic lupus erythematosus // Arthritis.
Rheum. - 1997. - Vol. 40. - P. 797-803.
108. Knopp R.H., LaRosa J.C., Burkman Jr. R.T. Contraception and dyslipidemia // Am. J. Obstet.
Gynecol. - 1993. - Vol. 168. - P. 1994-2005.
109. WHO Collaborative Study of Cardiovascular Disease and Steroid Hormone Contraception.
Venous thromboembolic disease and combined oral contraceptives; results of international multicentre
case^-control study //Lancet. - 1995. — Vol. 346. - P. 1575-1582.
110. Lidegaard O., Edstrom B., Kreiner S. Oral contraceptives and venous thromboembolism: a five-
year national case-control study // Contraception. — 2002. — Vol. 65. — P. 187-196.
111. Abdollahi M., Cushman M., Rosendaal F.R. Obesity: risk of venous thromboembolism and the
interaction with coagulation factor levels and oral contraceptive use // Thromb. Haemost. — 2003. —
Vol. 89. - P. 493-498.
112. Holt V.L., Cushing-Haugen K.L., Daling J.R. Body weight and risk of oral contraceptive
failure // Obstet. Gynecol. — 2002, — Vol. 99. - P. 820-827.
113. Zieman M., Guillebaud J., Weisberg E. et al. Contraceptive efficacy and cycle control with the
Ortho Evra/Evra transdermal system; the analysis of pooled data // Fertil. Steril. - 2002. — Vol. 77. —
Suppl. 2. - P. 13-18.
114. Norris P.M., Kamat A., Estes C. et al. Contraceptive failure in overweight patients taking com¬
bination oral contraceptive pills. Presented at Association of Reproductive Health Professionals Annual
Meeting. — San Diego. Calif.. 2003.
115. Azziz R. The hyperandrogenic-insulin-resistant acanthosis nigricans syndrome; therapeutic
response // Fertil. Steril. - 1994. - Vol. 61. - P. 570-572.
116. Duffy T.J., Ray R. Oral contraceptive use: prospective follow-up of women with suspected glu¬
cose intolerance // Contraception. — 1984. — Vol. 30. — P. 197-208.
117. Hannaford P.C., Kay C.R, Oral contraceptives and diabetes mellitus // BMJ. - 1989. —
Vol. 299. - P. 1315-1316.
118. Nader S., Riad-Gabriel M.G., Saad M. The effect of a desogestrel-containing ora! contracep¬
tive on glucose tolerance and leptin concentrations in hyperandrogénie women //J. Clin. Endocrinol.
Metab. - 1997. - Vol. 82. - P. 3074-3077.
119. Pasquali R., Gambineri A., Anconetani B. et al. The natural history of the metabolic syndrome
in young women with the polycystic ovary syndrome and the effect of long-term oestrogen-progestogen
treatment // Clin. Endocrinol. - 1999. - Vol. 50. - P. 517-527.
208
120. Klibanski A., Biller B.M., Schoenfeld D.A. et al. Tlie effects of estrogen administration on
trabecular bone loss in young women with anorexia nervosa // J. Clin. Endocrinol. Metab. - 1995. -
Vol. 80. - P. 898-904.
121. Drinkwater B.L., Braemner B„ Chesnut 3rd C.H. Menstrual history as a determinant of current
bone density in young atliletes // JAMA, - 1990. — Vol. 263, - P. 545-548.
122. Jonnavithula S., Warren M.P., Fox R.P., Lazaro M.I. Bone density is compromised in amen-
orrheic women despite return of menses; a 2-year study // Obstet. Gynecol. — 1993. — Vol. 81. —
P. 669-674.
123. Lashner B.A., Kane S.V., Hanauer S.B. Lack of association between oral contraceptive use and
ulcerative colitis // Gastroenterology. — 1990. — Vol. 99. — P. 1032-1036.
124. Cosnes J., Carbonnel F., CaiTat F. et al. Oral contraceptive use and the clinical course of Crohn’s
disease; a prospective cohort study j j Gut. — 1999, — Vol. 45, — P. 218-222.
125. American Cancer Society: Breast Cancer Facts and Figures 2003-2004 {accessed December 9.
2006). http://www.cancer.org/docroot/STT/content/STT_lx_Breast_Cancer_Facts_Figures_2003-
2004.asp
126. Chi I. The safety and efficacy Issues of progestin-oniy oral contraceptives ~ an epidemiologic
perspective /7 Contraception. — 1993. — Vol. 47. — P. 1-21.
127. McCann M.F., Potter L.S, Progestin-only oral contraception: a comprehensive review //
Contraception. — 1994. — Vol. 50. — Suppl. 1. — P. S9-S195.
128. Moghissi K.S., Marks C, Effects of microdose progestogens on endogenous gonadotrophic and
steroid hormones, cervical mucus properties, vaginal cytology and endometrium // Fertil, Steril. —
1971. - Vol. 22. - P. 424-434.
129. Moghissi K.S., Syner F.N.. McBride LC. Contraceptive mechanism of microdose norethin-
drone // Obstet. G^ecol. - 1973. - Vol. 4. - P. 585-594.
130. Collaborative Study Group on the Desogestrel-Containing Progestogen-Only Pill: A double-
blind study comparing the contraceptive efficacy, acceptability and safety of two progestogen-only pills
containing desogestrel 75 irg/day or levonorgestrel 30 pg/day // Eur. J. Contracept. Reprod. Health.
Care. - 1998. - Vol. 3. - P. 169-178.
131. Fotherby K. The progestogen-only pill and thrombosis j j Br. J. Fam. Plann. - 1989. -
Vol. 15. - P. 83-85.
1.32. Ball M.J., Ashwell E., Gilhner M.D. Progestagen-only oral contraceptives; comparison of
the metabolic effects of levonorgestrel and norethisterone // Contraception. - 1991. — Vol. 44. -
P. 223-233.
133. Winkler U.H. Blood coagulation and oral contraceptives. A critical review// Contraception. —
1998. ' Vol. 57. - P. 203-209.
134. Trussell J., Kost K. Contraceptive failure in the United States: a critical review of the litera¬
ture II Stud. Fam. Plann. - 1987. - Vol. 18. - P. 237-283.
135. Vessey M.P, et al. Progestogen-only contraception: findings in a large prospective study with
special reference to effectiveness // Br. J. Fam. Plann. — 1985. — Vol. 10. — P. 117-121,
136. Bisset A.M., Dingwall-Fordyce I., Hamilton M.J.K. The efficacy of the progestogen-only pill as
a contraceptive method // Br. J. Fam. Plann. - 1990. - Vol. 16. - P. 84-94.
137. Broome М., Fotherby K. Clinical experience with the progestogen-only pill // Contraception. -
1990. - Vol. 42, - P. 489-495.
138. Tayob Y. et al. Ultrasound demonstration of increased frequency of functional ovarian cysts in
women using progestogen-only ora) contraception // Br. J. Obstet. Gynaecol. — 1985. — Vol. 92. — P.
1003-1009.
139. Vessey М., Metcalfe A., Wells C. et al. Ovarian neoplasms, functional ovarian cysts, and oral
contraceptives // BMJ. - 1987. - Vol. 294. - P. 1518-1520.
140. Pakariiien P., Lahteenmaki P., Rutanen E.M. The effect of intrauterine and oral levonorgestrel
administration on serum concentrations of sex hormone-binding globulin, insulin and insulin-like
growth factor binding protein-1 //Acta Obstet. Gynecol. Scand. — 1999. — Vol. 78. — P. 423-428.
141. Tankeyoon М., Dusitsin N., Chalapati S. et al. Effects of hormonal contraceptives on milk
volume and infant growth, WHO Special Programme of Research, Development and Research Training
in Human Reproduction Task Force on Oral Contraceptives // Contraception. — 1984. — Vol. 30. —
P. 505-522.
209
142. WHO Task Forte for Epidemiological Research on Reproductive Health, Special Programme of
Research, Development and Research Training in Human Reproduction, Progestogen-only contracep¬
tives during lactation, I. Infant growth // Contraception. - 1994. - Vol. 50. — P. 35-53.
143. WHO Task Force for Epidemiological Research on Reproductive Health, Special Programme of
Research, Development and Research Training in Human Reproduction. Progestogen-only contracep¬
tives during lactation. II. Infant development // Contraception. — 1994. - Vol. 50. - P. 55-68.
144. McCann M.F., Moggia A.V., HigginsJ.E. et al. The effects of a progestin-only oral contracep¬
tive (levonorgestrel 0.03 jig) on breast-feeding // Contraception. — 1989. — Vol. 40. — P. 635-648.
145. Halderraan L.D., Nelson A.L. Impact of early postpartum administration of progestin-only
hormonal contraceptives compared with nonhormonal contraceptives on short-term breast-feeding
patterns // Am. J. Obstet. Gynecol. — 2002. — Vol. 186, — P. 1250-1258.
146. Mintz G., Gutierrez G., Deleze M., Rodriguez E. Contraception with progestogens in systemic
lupus erythematosus // Contraception. - 1984. - Vol. 30. - P. 29-38,
147. World Health Organization Collaborative Study of Cardiovascular Disease and Steroid
Hormone Contraception. Cardiovascular disease and use of oral and injectable progestogen-only
contraceptives and combined injectable contraceptives. Results of an international, multicenter, case-
control study // Contraception, — 1998. — Vol. 57. — P. 315-324.
148. Heinemann L.A., Assmann A., DoMinh T., Garbe E. Oral progestogen-only contraceptives
and cardiovascular risk: results from the Transnational Study on Oral Contraceptives and the Health of
Young Women // Eur. J. Contracept. Reprod. Health. Care. — 1999. - Vol. 4. - P. 67-73.
149. Winkler U.H., Howie H., Buhler K. et al. A randomized controlled double-blind study of the
effects on hemostasis of two progestogen-only pills containing 75 pg desogestrel or 30 pg levonorg¬
estrel // Contraception. - 1998. - Vol. 57. - P. 385-392.
150. Weiderpass E., Adami H.O., Baron J.A. et al. Risk of endometrial cancer following estrogen
replacement with and without progestins // J. Natl. Cancer. Inst. — 1999. — Vol. 91. — P. 1131-
1137.
151. Morris J.M., van Wagenen G. Compounds interfering with ovum implantation and develop¬
ment. III. The role of estrogens // Am. J. Obstet. Gynecol. - 1966. - Vol. 96. - P. 804-815.
152. Yuzpe A.A., Smith R.P., Rademaker A.W. A multicenter clinical investigation employing ethinyl
estradiol combined with dl-norgestrel as a postcoital contraceptive agent // Fertil, Steril. — 1982. —
Vol. 37.-P. 508-513.
153. Ho P.C., Kwan M.S.W. A prospective randomized comparison of levonorgestrel with the Yuzpe
regimen in post-coital contraception // Hum. Reprod. — 1993, - Vol. 8. — P. 389-392.
154. Task Force on Postovulatory Methods of Fertility Regulation. Randomised controlled trial of
levonorgestrel versus the Yuzpe regimen of combined oral contraceptives for emergency contracep¬
tion // Lancet. - 1998. - Vol. 352. - P. 428-433.
155. von Hertzeii H., Piaggio G., Ding J. WHO Research Group on Post-ovulatory Methods of
Fertility Regulation. Low dose mifepristone and two regimens of levonorgestrel for emergency contra¬
ception: a WHO multicentre randomised trial // Lancet. — 2002. — Vol. 360. - P. 1803-1810.
156. Arowojoulu A.O., Okewole I. A., Adekunie A.O. Comparative evaluation of the effectiveness and
safety of two regimens of levonorgestrel for emergency contraception in Nigerians // Contraception. —
2002. - Vol. 66. - P. 269-273.
157. Burton R., Savage W., Reader F. The «morning after pill». Is this the wrong name for it.? //
Br.J. Fam. Plann. - 1990. - Vol. 15. - P. 119-121.
158- Young L., McGowan L.M., Roberts H.E., Farquhar C.M. Emergency contraception-why
women don't use it // N. Z. Med. J. - 1995. - Vol. 108. - P. 145-148.
159. Harper C.C., Ellertson C.E. The emergency contraceptive pill: a survey of knowledge and
attitudes among students at Princeton // Am. J. Obstet. Gynecol. - 1995. - Vol. 173. - P. 1438-
1445.
160. Delbanco S.F., Mauldon J., Smith M.D. Little knowledge and limited practice: emergency
contraceptive pills, the public, and the obstetrician-gynecologist // Obstet. Gynecol. — 1997. —
Vol. 89. - P. 1006-1011.
161. Trussell J., Stewart F., Guest P., Hatcher R.A. Emergency contraceptive pills: a simple proposal
to reduce unintended pregnancies // Fam. Plann. Perspect, - 1992. - Vol. 24. - P. 269-273.
162. Harvey S.M., Beckman L.J., Sherman C., Petitti D. Women's experience and satisfaction with
emergency contraception // Fam. Plann. Perspect. — 1999. — Vol. 31. — P. 237-240, 260.
210
163. Glasier A., Baird D. The effects of self-administering emergency contraception // New. Engl.
J. Med. - 1998. - Vol. 339. - P. 1-4.
164. Raine Т., Harper C., Leon K., Damey P. Emergency contraception: advance provision in a
young, high-risk clinic population // Obstet. Gynecol. - 2000. — Vol. 96, - P. 1-7.
165. Jackson R.A., Sv^^arz E.B., Freedman L., Darney P. Advance Supply of emergency contraception:
effect on use and usual contraception — a randomized trial // Obstet. Gynecol. — 2003. — Vol. 102. —
P. 8-16.
166. Raine T.R., Harper C.C., Rocca C.H. et al. Direct access to emergency contraception through
pharmacies and effect on unintended pregnancy and STIs: a randomized controlled trial j j JAMA, —
2005. - Vol. 293. - N 1. - P. 54-62.
167. Harper C.C-, Gheong М., Rocca C.H. et al, The effect of inaeased access to emergency contiaception
among young adolescents // Obstet. Gynecol. - 2005. - Vol. 106. — N 3. - P, 483-491.
168. Gainer E., Blum J., Toverud E.L. et al. Bringing emergency contraception over the counter:
experiences of nonprescription users in France, Norway, Sweden and Portugal // Contraception. —
2003. - Vol. 68. - P. 117-124.
169. Young D.C., Wiehle R.D., Joshi S.G., Poindexter A.N. Emergency contraception alters
progesterone-associated endometrial protein in serum and uterine luminal fluid // Obstet. Gynecol, —
1994. - Vol. 84. - P. 266-271.
170. Swahn M.L., Westlund P.. Johannisson E., Bygdeman M. Effect of postcoital contraceptive
methods on the endometrium and the menstrual cycle // Acta Obstet. Gynecol. Scand. — 1996. —
Vol. 75. - P. 738-744.
171. Trussell J„ Raymond E.G. Statistical evidence about the mechanism of action of the Yuzpe regi¬
men of emergency contraception // Obstet. Gynecol. — 1999. - Vol. 93. - P. 872-876.
172. Marions L., Hultenby K„ Lindell I. et al. Emergency contraception with mifepristone and
levonorgestrel: mechanism of action // Obstet. Gynecol. - 2002. - Vol. 100. - P. 65-71.
173. Croxatto Н.Б., Fuentealba B., Brache V.et al. Effects of the Yuzpe regimen, given during the
follicular phase, on ovarian function // Contraception, — 2002. — Vol. 65. — P. 121-128.
174. Trussell J., Eliertson C., Dortlinger L. Effectiveness of the Yuzpe regimen of emergency contra¬
ception by cycle day of intercourse: implications for mechanism of action // Contraception. - 2003. —
Vol. 67. - P. 167-171.
175. Fasoli М., Parazzini F., Cecchetti G., La Vecchia C. Post-coital contraception: an overview of
published studies // Contraception. - 1989. — Vol. 39. — P. 459-468.
176. Haspels A.A. Emergency contraception: a review // Contraception. - 1994. - Vol. 50, —
P. 101-108.
177. Glasier A. Emergency postcoital contraception // New. Engl. J. Med. — 1997. — Vol. 337. —
P. 1058-1064.
178. Wilcox A.J., Weinberg C.R., Baird D.D. Timing of sexual intercourse in relation to ovulation.
Effects on the probability of conception, survival of the pregnancy and sex of the baby // New. Engl. J.
Med. - 1995. - Vol. 333. - N 23. - P. 1517-1521.
179. Trussell J., Eliertson C., Stewart F, The effectiveness of the Yuzpe regimen of emergency con¬
traception j j Fam. Plann. Perspect. — 1996. — Vol. 28. — P. 58-64.
180. Trussell J., Rodriguez G., Eliertson C. New estimates of the effectiveness of the Yuzpe regimen
of emergency contraception// Contraception. — 1998. - Vol. 57. - P. 363-369.
181. Trussell J., Rodriguez C., Eliertson C. Updated estimates of the effectiveness of the Yuzpe regi¬
men of emergency contraception // Contraception. - 1999. — Vol. 59. — P. 147-151.
182. Trussell J-, Eliertson C., Rodriguez G. The Yuzpe regimen of emergency contraception; how
long after the morning after.^ // Obstet. Gynecol. — 1996. — Vol. 88. — P. 150-154.
183. Rodrigues L, Grou F., Joly J. Effectiveness of emergency contraceptive pills between 72 and
120 hours after unprotected sexual intercourse // Am. J. Obstet. Gynecol. — 2001. — Vol. 184. —
P. 531-537.
184. Piaggio G„ von Hertzen H.. Grimes D.A., Van Look P.F. Timing of emergency contraception
with levonorgestrel or the Yuzpe regimen // Lancet. — 1999. — Vol. 353. — P. 721.
185. Simpson J.L., Phillips O.P. Spermicides, hormonal contraception and congenital malforma¬
tions // Adv. Contraception. — 1990. - Vol. 6. - P. 141-167.
186. Bracken M.B. Oral contraception and congenital malformations in offspring: a review and
meta-analysis of the prospective studies // Obstet. Gynecol. — 1990, - Vol. 76. — P. 552-557.
211
187. Raman-Wflms L., Tseng A.L., Wighardt S. et al. Fetal genital effects of first trimester sex hor¬
mone exposure: a meta-analysis // Obstet. Gynecol. — 1995. — Vol. 85. — P. 141-149.
188. Vasilakis C., Jick S.S., Jick H. The risk of venous thromboembolism in usere of postcoital con¬
traceptive pills // Contraception. — 1999. — Vol. 59. — P. 79-83.
189. Webb A., Tabemer D. Clotting factors after emergency contraception // Adv. Contracept. —
1993. - Vol. 9. - P. 75-82.
190. Ellertson C., Webb A., Blanchard K. et al. Modifying the Yuzpe regimen of emergency con¬
traception: a multicenter randomized controlled trial // Obstet. Gynecol. — 2003. - Vol. 101. —
P. 1160-1167.
191. Webb A.M.C., Russell J., Elstein M. Comparison of Yuzpe regimen, danazol and mifepristone
(RU486) in oral postcoital contraception // BMJ. — 1992. — Vol. 305. — P. 927-931.
192. Glasier A., Thong K.J., Dewar M. et al. Mifepristone (RU 486) compared with high-dose
estrogen and progestogen for emergency postcoital contraception // New. Engl. j. Med. — 1992. —
Vol. 327. - P. 1041-1044.
193. Task Force on Postovulatory Methods of Fertility Regulation. Comparison of three single doses
of mifepristone as emergency contraception: a randomised trial j j Lancet. - 1999. - Vol. 353. -
P. mi-ira.
194. Xiao B.L., von Hertzen H., Zhao H., Piaggio G. A randomized double-blind comparison of
two single doses of mifepristone for emergency contraception // Hum. Reprod. — 2002. — Vol. 17. —
P. 3084-3089.
195. Alexander N.J., Baker E., Kaptein M. et al Why consider vaginal dnig administration? // Fertil.
Steril. - 2004. - Vol. 82. - P. 1-12.
196. Timmer C.J., Mulders T.M. Pharmacokinetics of etonogestrel and ethinylestradiol released from
a combined contraceptive vaginal ring // Clin. Pharmacokinet. — 2000. - Vol. 39. - P. 233-242.
197. Roumen F.J. et al. Efficacy, tolerability and acceptability of a novel contraceptive vaginal ring
releasing etonogestrel and ethinyl estradiol // Hum. Reprod. - 2001. - Vol. 16, - P. 469-475.
198. Mulders T.M., Dieben T.O. Use of the novel combined contraceptive vaginal ring NuvaRingfor
ovulation inhibition // Fertil. Steril. — 2001. — Vol. 75. — P. 865-870.
199. Dieben T.O., Roumen F.J., Apter D. Efficacy, cycle control, and user acceptability of a novel
combined contraceptive vaginal ring// Obstet. Gynecol. — 2002. — Vol. 100. — P. 585-593.
200. Bjamad ttir R.L, Tuppurainen M., Killick S.R. Comparison of cycle control with a combined
vaginal ring and oral levonorgestrel/ethinyl estradiol // Am. J. Obstet. Gynecol. — 2002. — Vol, 186. —
P. 389-395.
201. Szarewski A, High acceptability and satisfaction with NuvaRing use // Eur. J. Contracept.
Reprod. Health. Care. - 2002. - Vol. 7. - P. 31-36.
202. Roumen F.J., Boon M.E., van Velzen D. et al. The cervico-vaginal epithelium during 20 cycles’
use of a combined contraceptive vaginal ring // Hum. Reprod. — 1996. — Vol. 11. — P. 2443-2448.
203. Davies G.C., Feng L.X., Newton J.R. et al. The effects of a combined contraceptive vaginal
ring releasing ethinyloestradiol and 3-keto-desogestrel on vaginal flora // Contraception. — 1992. -
Vol. 45. - P. 511-518.
204. Haring T., Mulders T.M. The combined contraceptive ring NuvaRing and spermicide co¬
medication /7 Contraception. - 2003. - Vol. 67. — P. 271-272.
205. Hammond G.L., Abrams L.S., Creasy G.W. et al. Serum distribution of the major metabolites
of noregestimate in relation to its pharmacological properties jf Contraception. — 2003. - Vol. 67. —
P. 93-99.
206. Dittrich R., Parker L.. Rosen J.B., Ortho Evra/Evra 001 Study Group et ai. Transdermal con¬
traception: evaluation of three transdermal norelgestromin/ethinyl estradiol doses in a randomized
multicenter, dose-response study // Am. J. Obstet. Gynecol. — 2002. — Vol. 186. - P. 15-20.
207. Zacur H.A., Hedon B„ Mansour D. et al. Integrated summaiy of Ortho Ewa/Evra contraceptive
patch adhesion in varied climates and conditions // Fertil. Steril. — 2002. — Vol. 77. — Suppl. 2. —
P. 32-35.
208. Audet M.C., Moreau M., Koltun W.D. et al. Ortho Evra/Evra 004 Study Group et al. Evaluation
of contraceptive efficacy and cycle control of a transdermal contraceptive patch vs an oral contraceptive.
A randomized controlled trial //JAMA. — 2001. — Vol. 285, — P. 2347-2354.
209. Smallwood G.H., Meador M.L., Lenihan J.P. et al. Ortho Evra/Evra 002 Study Group et al.
Efficacy and safety of a transdeniial contraceptive system // Obstet. Gynecol. - 2001. - Vol. 98. -
P. 799-805.
212
210. Pierson R.A., Archer D.F., Moreau M. et al. Ortho Evra/Evra versus oral contraceptives: fol¬
licular development and ovulation in normal cycles and after an intentional dosing error // Fertil.
Steril. - 2003. - Vol. 80. - P. 34-42.
211. Abrams L.S., Skee D.M., Natarajan J. et al. Multiple-dose pharmacokinetics of a contraceptive
patch in healthy women participants // Contraception. - 2001. — Vol. 64. - P. 287-294.
212. Abrams L.S., Skee D.M., Natarajan J. et al. Pharmacokinetics of a contraceptive patch (Evra/
Ortho Evra) containing norelgestromin and ethinyloestradiol at four application sites j j Br. J, Clin.
Pharmacol. - 2002. - Vol. 53. - P. 141-146.
213. Abrams L.S., Skee D.M., Wong F.A. et al. Pharmacokinetics of norelgestromin and ethinyl
estradiol from two consecutive contraceptive patches // J. Clin. Pharmacol. — 2001. — Vol. 41. —
P. 1232-1237.
214. Abrams L.S., Skee D.M., Natarajan J. et al. Pharmacokinetics of norelgestromin and ethinyl
estradiol delivered by a contraceptive patch (Ortho Evra/Evra) under conditions of heat, humidity, and
exercise //J. Clin. Pharmacol. - 2001, - Vol. 41. - P. 1301-1309.
215. Abrams L.S., Abrams L.S., Skee D., Natarajan J., Wong F.A. Pharmacokinetic overview of
Ortho Evra/Evra // Fertil. Steril. — 2002. — Vol. 77. — Suppl. 2. — P. S3-S12.
216. Creasy G.W., Fisher A.C., Hall N., Shangold G.A. Transdermal contraceptive patch deliver¬
ing norelgestromin and ethinyl estradiol. Effects on the lipid profile //J. Reprod. Med. — 2003. —
Vol. 48. - N 3. - P. 179-186.
217. Sibai B.M., Odlind V., Meador M.L. et al. A comparative and pooled analysis of the safety
and tolerability of the contraceptive patch (Ortho Evra/Evra) j j Fertil. Steril. - 2002. - Vol. 77. -
Suppl. 2. - P. S19-S26.
218. Archer D.F., Bigrigg A., Smallwood G.H. et al. Assessment of compliance with a weekly
contraceptive patch (Ortho Evra/Evra) among North American women // Fertil. Steril. — 2002, —
Vol. 77. - Suppl. 2. - P. 27-31.
219. Rosenfield A., Maine D., Rochat R. et al. The Food and Drug Administration and medroxypro¬
gesterone acetate: what are the issues? // JAMA. — 1983. — Vol. 249. — P. 2922-2928.
220. Said S., Omar K., Koetsawang S. et al. A multicentered phase III comparative clinical trial of
depot-medroxyprogesterone acetate given three-monthly at doses of 100 pg or 150 pg: I. Contraceptive
efticacy and side effects. World Health Organization Task Force on Long-Acting Systemic Agents for
Fertility Regulation. Special Programme of Research, Development and Research 'lYaining in Human
Reproduction // Contraception. - 1986. - Vol. 34. — P. 223-235.
221. World Health Organization ; Multinational comparative clinical evaluation of two long-acting
injectable contraceptive steroids: norethisterone enanthate and medroxyprogesterone acetate. Final
report // Contraception. - 1983. - Vol. 28. - P. 1-20.
222. Mishell Jr. D.R- Effect of 6 methyl- 17-hydroxyprogesterone on urinary excretion of luteinizing
hormone // Am. J. Obstet. Gynecol. - 1967. - Vol. 99. - P. 86-90.
223. Fraser I.S., Weisberg E.A. A comprehensive review of injectable contaception with special
emphasis on depot medroxyprogesterone acetate // Med. J, Aust. - 1981. -- Vol. 1. — Suppl, 1. -
P. 3-19.
224. Pardthaisong T., Gray R.H. In utero exposure to steroid contraceptives and outcome of preg¬
nancy // Am. J. Epidemiol. - 1991. - Vol. 134. - P. 795-803.
225. Gray R.H. Pardthaisong. In utero exposure to steroid contraceptives and survival during
infancy // Am. J. Epidemiol. — 1991. — Vol. 134. — P. 804-811.
226. Siriwongse T., Snidvongs W„ Tantayaporn P., Leepipatpaiboon S. Effect of depot-medroxy¬
progesterone acetate on serum progesterone levels, when administered on various cycle days //
Contraception. - 1982. - Vol. 26. - P. 487-493.
227. Petta C.A., Faundes A., Dunson T.R. et al. Timing of onset of contraceptive effectiveness in
Depo-Provera users. I. Changes in cervical mucus // Fertil, Steril. — 1998. — Vol. 69. — P. 252-257.
228. Petta C.A., Faundes A., Dunson T.R. et al. Timing of onset of contraceptive effectiveness in Depo-
Provera users, n. Effects on ovarian function // Fertil. Steril. — 1998. — Vol. 70. — P. 817-82Q.
229. Harlap S., Kost K., Forrest J.D. Preventing Pregnancy, Protecting Health: A New Look at Birth
Control Choices in the United States, New York, The Alan Guttmacher Institute, 1991.
230. Mattson R.H., Cramer J.A., Caldwell B.V., Siconolfi B.C. Treatment of seizures with medroxy¬
progesterone acetate: preliminary report // Neurology. - 1984, - Vol. 34. — P. 1255-1258.
231. O’Dell C.M., Forke C.M., Polaneczky M.M. et al. Depot medroxyprogesterone acetate or oral
contraception in postpartum adolescents // Obstet. Gynecol. — 1998. — Vol. 91. — P. 609-614.
213
232. Polaneczky M., Guarnacda M., Alon J., Wiley J. Early experience wth the contraceptive use
of depo-medroxyprogesterone acetate in an inner-city population // Fam. Plann. Perspect. - 1996. -
Vol. 28. - P. 174-178.
233. de Abood M., de Castillo Z., Guerrero F. et al. Effect of Depo-Provera or Microgynon on the
painful crises of sickle-cell anemia patients // Contraception. - 1997. — Vol. 56. — P. 313-316.
234. Jimenez J„ Ochoa M., Soler M.P., Portales P. Long-term follow-up of children breast-fed
bv mothers receiving depot-medroxyprogesterone acetate // Contraception. — 1984. — Vol. 30. —
R .523-533.
235. Zacharias S., Aguilera E., Assenzo J.R., Zanartu J. Effects of hoimonal and non-honuonal
contracepters on lactation and incidence of pregnancy // Contraception. — 1986. — Vol. 33. —
P. 203-213.
236. Pardthaisong T., Yenchit C., Gray R, The long-term growth and development of children
exposed to Depo-Provera during pregnancy or lactation // Contraception. — 1992. — Vol. 45. -
P. 313-324.
237. Virutamasen P., Leepipatpaiboon S., Kriengsinyot R. et al. Pharmacodynamic effects of depot-
medroxyprogesterone acetate (DMPA) administered to lactating women on their male infants j j
Contraception. - 1996. - Vol. 54. - P. 153-157.
238. Depot-medroxyprogesterone acetate (DMPA) and risk of endometrial cancer. The WHO
Collaborative Study of Neoplasia and Steroid Contraceptives j j Int. J. Cancer. - 1991. - Vol. 49. -
P. 186-190.
239. Lumbiganon P., Rugpao S., Phandhu-fung S. et al. Protective effect of depot-medroxypro¬
gesterone acetate on surgically treated uterine leiomyomas: a multicentre case-control study // Br. J.
Obstet. Gynaecol. — 1996. — Vol. 103. — P. 909-914.
240. Depot-medroxyprogesterone acetate (DMPA) and risk of epithelial ovarian cancer. The WHO
Collaborative Study of Neoplasia and Steroid Contraceptives // Int.J. Cancer. - 1991. - Vol. 49. -
P. 191-195.
241. Gray R.H. Reduced risk of pelvic inflammatory disease with injectable contraceptives //
Lancet. - 1985. - Vol. 1. - P. 1046-1049.
242. Cromer B.A., Smith R.D., Blair J.M. et al. A prospective study of adolescents who choose
among levonorgestrel implant (Norplant), medrox^'progesterone acetate (Depo-Provera), or the coin-
^ bined oral contraceptive pill as contraception // Pediatrics. - 1994. — Vol, 94. - P. 687-694.
243. Gardner J.M., Mishell D.R. Jr. Analysis of bleeding patterns and resumption of fertility fol¬
lowing discontinuation of a long-acting injeaable contraceptive // Fertil. Steril. — 1970. — Vol. 21. —
P. 286-291.
244. Harel Z., Biro P.M., Kollar L.M. Depo-Provera in adolescents: effects of early second injection
or prior oral contraception //J, Adolesc. Health. - 1995. - Vol. 16. - P. 379-384.
245. Goldberg A.B., Cardenas L.H,, Hubbard A.E., Darney P.D. Post-aboition depot medroxypro¬
gesterone acetate continuation rates: a randomized trial of cyclic estradiol //Contraception. — 2002. -
Vol. 66. - P. 215-220.
246. Smith R.D., Cromer B.A., Hayes J.R. Medroxyprogesterone acetate (Depo-Provera) use in
adolescents: uterine bleeding and blood pressure patterns, patient satisfaction, and continuation
rates //Adolesc. Pediatr. Gynecol. — 1995. — Vol. 8. - P. 24-28.
247. Saiigi-Haghpeykar H. et al. Experiences of injectable contraceptive users in an urban setting //
Obstet. Gynecol. - 1996. - Vol. 85. - P. 227-233.
248. Westhoff C., Wieland D., Tiezzi L. Depression in users of depo-medroxyprogesterone acetate //
Contraception. - 1995. - Vol. 51. - P. 351-354.
249. Westhoff C., Truman C., Kalmuss D. et al. Depressive symptoms and Depo-Provera //
Contraception. - 1998, - Vol. 57. - P. 237-240.
250. Gupta N.. O’Brien R., Jacobsen L.J. et al. Mood changes in adolescents using depot-me¬
droxyprogesterone acetate for contra-ception: a prospective study // J. Pediatr, Adolesc. Gynecol, -
2001. - Vol. 14, - P. 71-76.
251. Moore L.L., Valuck R., McDougall C„ Fink W. A comparative study of one-year weight gain
among users of medroxj’progesterone acetate, levonorgestrel implants, and ora) contraceptives //
Contraception. - 1995. - Vol. 52. - P. 215-219.
252. iMainwaring R„ Hales H.A., Stevenson K. et al. Metabolic parameters, bleeding, and weight
changes in U.S. women using progestin only contraceptives j j Contraception. — 1995, — Vol. 51. —
P. 149-153.
214
253. Taneepanichskul S., Reinprayoon D., Khaosaad P. Comparative study of weight change between
long-term DMPA and lUD acceptors // Contraception. — 1998. — Vol. 58. — P. 149-151.
254. Bahamondes L., Del Castillo S., Tabares G. et al. Comparison of weight increase in users
of depot medroxyprogesterone acetate and copper lUD up to 5 years // Contraception. — 2001. —
Vol. 64. - P. 223-225.
255. Pelkman C.L. et al. Short-term effects of a progestational contraceptive drug on food intake,
resting energy expenditure, and body weight in young women j j Am. J. Clin. Nutr. — 2001. -
Vol. 73. - P. 19-26.
256. Espey E. et al. Depo-provera associated with weight gain in Navajo women // Contraception. —
2000. - Vol. 62. - P. 55-58.
257. Croatner B.A., Lazebnik R., Rome E. et al. Double-blinded randomized controlled trial of estro¬
gen Supplementation in adolescent girls who receive depot medroxyprogesterone acetate for contracep¬
tion // Am. J. Obstet. Gynecol. - 2005. - Vol. 192. - № 1. - P. 42-47.
258. WHO Collaborative Study of Neoplasia and Steroid Contraceptives: Breast cancer and depot-
medroxyprogesterone acetate: a multinational study // Lancet. - 1991. - Vol. 338. - P. 833-838.
259. The New Zealand Contraception and Health Study Group. History of long-term use of depot-
medrox}'progesterone acetate in patients with cervical dysplasia; case-control analysis nested in a
cohort study // Contraception. — 1994. — Vol. 50. — P. 443-449.
260. Thomas D.B., Ray R.M. Depot-medroxyprogesterone acetate (DMPA) and risk of invasive
adenocarcinomas and adenosquamous carcinomas of the uterine cervix. WHO Collaborative Study of
Neoplasia and Steroid Contraceptives // Contraception. — 1995. — Vol. 52. — P. 307-312.
261. WHO Collaborative Study ofNeoplasia and Steroid Contraception. Depot-medroxy'progesterone
acetate (DMPA) and risk of invasive squamous cell cervical cancer // Contraception. — 1992. —
Vol. 45. - P. 299-312.
262. Thomas D.B., Ye Z., Ray R.M. Cervical carcinoma in situ and use of depot-medroxypro¬
gesterone acetate (DMPA). WHO Collaborative Study of Neoplasia and Steroid Contraceptives I I
Contraception. — 1995. — Vol. 51. — P. 25-31.
263. Garza-Flores J., De la Cruz D.L., Valles de Bourges V, et al. Long-term effects of depot-
medroxyprogesterone acetate on lipoprotein metabolism // Contraception. — 1991. — Vol. 44. —
P. 61-71.
264. Enk L., Landgren B.M., Lindberg U.B. et al. A prospective, one-year study on the effects of two
long acting injectable contraceptives (depot-medroxyprogesterone acetate and norethisterone enan¬
thate) on senim and lipoprotein lipids // Horm. Metab. Res. - 1992. - Vol. 24. - P. 85-89.
265. Kongsayreepong R., Chutivongse S., George P. et al. A multicentre comparative study of
serum lipids and apolipoproteins in long-term users of DMPA and a control group of lUD users //
Contraception. - 1993. - Vol. 47. - P. 177-191.
266. Fahmy K., Khairy M., Allam G. et al. Effect of depo-medroxyprogesterone acetate on coagulation
factors and serum lipids in Egyptian women j j Contraception. — 1991. — Vol. 44. — P. 431-444.
267. Fahmy K„ Abdel-Razik M„ Shaaraway M. et al. Effect of long-acting progestagen-only inject¬
able contraceptives on carbohydrate metabolism and its hormonal profile // Contraception. — 1991. —
Vol. 44. - P. 419-430.
268. Cundy T., Ames R., Horne A. et al. A randomized controlled trial of estrogen replacement
therapy in long-term users of depot medroxyprogesterone acetate // J. Clin. Endocrinol. Metab. -
2003. - Vol. 88. - P. 78-81.
269. Cundy T., Evans M., Roberts H. et al. Bone density in women receiving depot medroxyproges¬
terone acetate for contraception //BMJ. — 1991. ~ Vol. 303. — P. 13-16.
270. Cundy T., Cornish J., Roberts H. et al. Spinal bone density in women using depot medroxypro¬
gesterone contraception // Obstet. Gynecol. - 1998. -- Vol. 92. - P. 569-573.
271. Scholes D„ Lacroix A.Z., Ott S.M. et al. Bone mineral density in women using depot medroxy¬
progesterone acetate for contraception // Obstet. Gynecol. - 1999. - Vol. 93. - P. 233-238.
272. Cundy T., Cornish J., Evans M.C. et al. Recoveiy of bone density in women who stop using
medroxyprogesterone acetate // BMJ. — 1994. — Vol. 308. — P. 247-248.
273. Scholes D., LaCroix A.Z., Ichikawa L.E. et al. Injectable hormone contraception and bone den¬
sity: results from a prospective study // Epidemiology. — 2002. — Vol. 13. — P. 581-587.
274. Orr-Walker B.J., Evans M.C., Ames R.W. et al. The effect of past use of the injectable contracep¬
tive depot medroxyprogesterone acetate on bone mineral density in normal post-menopausal women //
Clin. Endocrinol. - 1998. - Vol. 49. - P. 615-618.
215
275. Petitti D.B., Piaggio G., Mehta S. et al. Steroid hormone contraception and bone mineral den¬
sity: a cross-sectional study in an international population // Obstet. Gynecol. — 2000. — Vol. 95. —
P. 736-744.
276. Theintz G., Buchs B., Rizzoli R. et al. Longitudinal monitoring of bone mass accumulation
in healthy adolescents: evidence for a marked reduction after 16 years of age at the levels of lumbar
spine and femoral neck in female subjects // J. Clin. Endocrinol. Metab. — 1992, — Vol. 75, —
P. 1060-1065.
277. Matkovic V., Jelic T„ Wardlaw G.M. et al. Timing of peak bone mass in Caucasian females and
its implication for the prevention of osteoporosis: inference from a cross-sectional model // J. Clin.
Invest. - 1994. - Vol. 93. - P. 799-808.
278. Cromer B.A., Blair J.M., Mahan J.D. et al. A prospedive comparison of bone density in adoles¬
cent girls receiving depot medroxyprogesterone acetate (Depo-Provera), levonorgestrel (Norplant), or
oral contraceptives //J. Pediatr. - 1996, - Vol. 129. - P. 671-676.
279. Garea-Flores J., Cardenas S., Rodriguez V.et al. Return to ovulation following the use of
long-acting injectable contraceptives: a comparative study // Contraception. — 1985, — Vol. 31. —
P. 361-366.
280. Pardthaisong T. Return of fertility after use of the injectable contraceptive Depo Provera: up¬
dated analysis //J. Biosoc. Sci. — 1984. - Vol. 16. — P. 23-34.
281. Schwallie P., Assenzo J. The effect of depo-medroxyprogesterone acetate on pituitary and
ovarian fundion, and the return of fertility following its discontinuation: a review // Contraception. —
1974. - Vol. 10. - P. 181-202.
282. Darney P. Bleeding patterns shift to amenorrhea with continued use of subcutaneous or intra¬
muscular depot medroxyprogesterone acetate // Obstet. Gynecol. — 2005. — Vol. 105. — N 4. — P. 54.
283. Newton J.R., d’Arcangues C., Hall P.E. A review of «once-a-month» combined injectable con¬
traceptives //J. Obstet. Gynaecol. - 1994. - Vol. 14. - Suppl, 1. — P. 1-34.
284. World Health Organization Task Force on Long-Acting Systemic Agents for Fertility
Regulation. A multicentered phase III comparative study of two hormonal contraceptive preparations
given once-a-month by intramuscular injection // Contraception. — 1989. — Vol, 40. — P. 531-551.
285. Cuong D.T., Huong M. Comparative phase III clinical trial of two injedable contraceptive prep¬
arations, depot-medroxyprogesterone acetate and Cyclofem in Vietnamese women // Contraception. —
1996. - Vol. 54. - P. 169-179.
286. Hall P. et al. Introdudory study of the once-a-month, injedable contrac'eptive Cyclofem in
Brazil, Chile, Columbia, and Peru // Contraception. - 1997. - Vol. 56. — P, 353-359.
287. Garza-Flores J.. Moraks del Olmo A., Fuziwara J.L. et al. Introdudion of Cyclofem once-a-
month injectable contraceptive in Mexico j j Contraception. - 1998. — Vol. 58. - P. 7-12.
288. Kaunitz A.M.. Garceau R.J., Cromie M.A. Comparative safety, efficacy, and cycle control of
Lunelle monthly contraceptive injedion (medroxyproge-sterone acetate and estradiol cypionate inject¬
able suspension) and Ortho-Novum 7/7/7 oral contraceptive (Norethindrone/ethinyl estradiol tripha¬
sic) // Contraception. - 1999. - Vol. 60. - P. 179-187.
289. Garceau R.J., Wajszczuk C.J., Kaunitz A.M.; Lunelle Study Group. Bleeding patterns of women
using Lunelle monthly contrace-ptive injedions (medroxyprogesterone aceate and estradiol cypionate
injedable suspension) compared with those of women using Oitho-Novum 7/7/7 (norethindrone/
ethinyl estradiol triphasic) or other oral contraceptives // Contraception. — 2000. — Vol. 62. —
P. 289-295.
290. Bahamondes L„ Lavin P., Ojeda G. et al. Return of fertility after discontinuation of the once-a-
month injedable contraceptive Cyclofem // Contraception. — 1997. — Vol. 55. — P. 307-310.
291. Bahamondes L., Marchi N.M., Nakagava H.M. et al. Self-administration with Unijed
of the once-a-month injedable contraceptive Cyclofem // Contraception. - 1997, - Vol, 56. —
P. 301-304.
292. Piya-Anant M., Koetsawang S., Patrasupapong N. et al. Effediveness of Cyclofem in the treat¬
ment of depot medroxyprogesterone acetate induced amenorrhea // Contraception. — 1998, — Vol.
57. - P. 23-28.
293. von Kesseru E., Aydinlik S., Etchepareborda J.J. Multicentered, pliase HI clinical trial of nore-
thisteone enanthate 50 pg plus estradiol valerate 5 pg as a monthly injectable contraceptive: final three-
year report II Contraception. — 1994. — Vol. 50. — P. 329-337.
294. Sang G.W., Shao Q.X., Ge R.S. et al. A inulticentred phase III comparative clinical trial of
Mesigyna, Cyclofem, and injectable no, 1 given monthly by intramuscular injection to Chinese women.
I- Contraceptive efficacy and side effeds // Contraception, - 1995. - Vol. 51. - P. 167-183.
216
295. Martinez G.H., Castaneda A., Correa J.E. Vaginal bleeding patterns in users of Perlutal, a
once-a-month injectable contraceptive consisting of 10 pg estradiol enanthate combined with 150 pg
dihydroxyprogesterone acetophenide. A trial of 5462 woman-months j j Contraception. — 1998. —
Vol. 58. - P. 21-27.
296. Coutinho E.M., Spinola P., Barbosa I. et al. Multicenter, double-blind, comparative clinical
study on the efficacy and acceptability of a monthly injectable contraceptive combination of 150 pg
dihydroxyprogesterone acetophenide and 10 pg estradiol enanthate compared to a monthly injectable
contraceptive combination of 90 pg dihydroxyprogesterone acetophenide and 6 pg estradiol enantha-
le//Contraception. - 1997. - Vol. 55. - P. 175-181.
297. Trussell J., Leveque J.A., Koenig J.D. et al. The economic value of contraception: a comparison
of 15 methods (j Am. J. Public. Health. - 1995. - Vol. 85. - P. 494-503.
298. Diaz S. et al. Recoveiy of fertility and outcome of planned pregnancies after the removal of
Norplant subdermal implants or Copper-T lUDs // Contraception. - 1987. — Vol. 35. — P. 569-579.
299. Sivin I., Mishell Jr. D.R., Darney P. et al. Levonorgestrel capsule implants in the United States:
a 5-year study // Obstet. Gynecol. — 1998. — Vol. 92. — P. 337-.344.
300. Kuiper H., Miller S., Martinez E. et al. Urban adolescent females’ views on the implant and
contraceptive decision-making: a double paradox j j Fam. Plann. Perspect. — 1997. — Vol. 29. —
N 4, - P. 167-172.
301. Croxatto H.B., Urbancsek J.. Massai R. et al. A multicentre efficacy and safety study of the
single contraceptive implant Implanon. Implanon Study Group // Hum. Reprod. - 1999. - Vol. 14. -
P. 976-981.
302. Meirik O., Farley T.M., Sivin I. Safety and efficacy of levonorgestrel implant, intrauterine
device and sterilization // Am. J. Obstet. Gynecol. - 2001. - Vol. 97. — P. 539-547.
303. Diaz S.. Herreros C., Juez G. et al. Fertility regulation in nursing women: influence of Norplant
levonorgestrel implants upon lactation and infant growth // Contraception. — 1985, — Vol. 32. —
P. 53-74.
304. Reinprayoon D.. Taneepanichskul S., Bunyavejchevin S. et al. Effects of the etonogestrel-
releasing contraceptive implant (Implanon) on parameters of breastfeeding compared to those of an
intrauterine device // Contraception. - 2000. — Vol. 62. — P. 239-246.
305. Shaaban M.M., Salem H.T., Abdullah K.A. Influence of levonorgestrel contraceptive implants,
Norplant, initiated early postpartum, upon lactation and infant growth // Contraception. — 1985. —
Vol. 32. - P. 623-635.
306. Brache V.et al. Anovulation, inadequate luteal phase, and poor sperm penetration in cervi¬
cal mucus during prolonged use of Norplant implants // Contraception. — 1985. — Vol. 31, —
P. 261-273.
307. Makarainen L., van Beek A., Tuomivaara L. et al. Ovarian function during the use of a single
contraceptive implant: Implanon compared with Norplant // Fertil. Steril. — 1998. — Vol. 69. —
P. 714-721.
308. Croxatto H.B., M к г inen L. The pharmacodynamics and efficacy of Implanon //
Contraception. — 1998. — Vol. 58. — P. 91S-97S.
309. Sivin I., Mishell Jr. D.R., Diaz S. et al. Prolonged effectiveness of Norplant capsule implants; a
7-year study j j Contraception. — 2000. — Vol. 61. — №3. — P. 187-194.
310. Dunson T.R., Amatya R.N., Krueger S.L. Complications and risk factors associated with the
removal of Norplant implants // Obstet. Gynecol. — 1995. — Vol. 85. — P. .543-548.
311. Zieman М.. Klaisle C, Walker D. et al. Fingers versas instruments for removing levonorgestrel
contraceptive implants (Norplant) //J. Gynecol. Tech. — 1997. — Vol. 3. — P. 213-217.
312. Meckstroth K.R., Darney P.D. Implant contraception // Semin. Reprod. Med. — 2001. —
Vol. 19. - N 4. - P. 339-354.
313. Meirik 0„ d’Arcangues C. For the WHO Consultation on Implantable Contraceptives for
Women. Implantable contraceptives for women j j Hum. Reprod. Update. — 2003. — Vol. 9. —
P. 49-59.
314- Mascarenhas L. et al. TVenty-four month comparison of apolipoproteins A-1, A-II and
В in contraceptive implant users (Norplant and Implanon) in Birmingham, United Kitigdom //
Contraception. - 1998, - Vol. 58. - P. 215-219.
315. Biswas A., Viegas O.A., Bennink H.J. et al. Effect of Implanon use on selected parameters of
thyroid and adrenal function // Contraception. - 2000. — Vol. 62. — P. 247-251.
217
316. Bisv\'as A., Viegas O.A., Coding Bennink HJ. et al. Implanon contraceptive implants: effects on
carbohydrate metabolism j j Contraception. — 2001. — Vol. 63. — P. 137-141.
317. Biswas A.. Viegas O.A., Roy A.C. Effect of Implanon and Norplant subdermal contraceptive
implants on serum lipids — a randomized comparative study // Contraception. — 2003. — Vol. 68. —
P. 189-193.
318. Groxatto H.B., Urbanesek J., Massai R. et al. A multicentren efficacy and safety study of the
single contraceptive implant Implanon j j Hum. Repro. - 1999. — Vol. 14. — P. 976-981.
319. Bromham D.R. et ai. Materials, methods and results of the Norplant training program j j Br. J.
Fam. Plann. - 1995. - Vol. 10. - P. 256-262.
320. Mascarenhas L. Insertion and removal of Implanon: practical considerations // Eur. J.
Contracept. Reprod. Health. Care. — 2000. — Vol. 5. — Suppl. 2. — P. 29-34.
321. Smith A., Reuter S. An assessment of the use of Implanon in three community sei-vices //
J. Fam. Plann. Reprod. Health. Care. - 2002. - Vol. 28. - P. 193-196.
322. Darney P.D., Taylor R.N., Klaisle C. et al. Serum concentrations of estradiol, progesterone,
and levonorgestrel are not determinants of endometrial histology or abnormal bleeding in long-term
Norplant implant users // Contraception. - 1996. -- Vol. 53. - P. 97-100.
323. Alvarez-Sanchez F., Brache V., Thevenin F. et al. Hormonal treatment for bleeding irregulari¬
ties in Norplant implant users // Am. J. Obstet. Gynecol. - 1996. - Vol. 174. - P. 919-922.
324. Zheng S.R., Zheng H.M., Qian S.Z. et al. A randomized multicenter study comparing the effi¬
cacy and bleeding pattern of a sing!e-rod (Implanon) and a six-capsule (Norplant) hormonal contra¬
ceptive implant // Contraception. — 1999. — Vol. 60. — P. 1-8.
325. Darney P.D., Atkinson E., Tanner S. et al. Acceptance and perceptions of Norplant among users
in San Francisco, USA // Stud. Fam. Plann. - 1990. - Vol. 21. - P. 152-160.
326. Damey P.D., Callegari L.S., Swift A. et al. Condom practices of urban teens using Norplant
contraceptive implants, oral contraceptives, and condoms for contraception // Am. J. Obstet.
Gynecol. - 1999. - Vol. 180. - P. 929-937.
327. Harrison-Woolrych M„ Hill R. Unintended pregnancies with the etonogestrel implant
(Implanon): a case .series from postmarketing experience in Australia // Contraception. - 2005. -
Vol. 71. - N 4. - P. 306-308.
328. Luukkainen T., Allonen H.. Haukkamaa M. et al. Five years’ experience with levonorgestrel-
releasing lUDs // Contraception. - 1986. — Vol. 33. - P. 139-148.
329. Sivin I., Stern J., Coutinho E. et al. Prolonged intrauterine contraception: a seven-year
randomized study of the levonorgestrel 20 p-cg/day (LNg 20) and the copper T380 Ag lUDs //
Contraception. - 1991. - Vol. 44. - P. 473-480.
330. Toivonen J., Luukkainen T., Allonen H. Protective effect of intrauterine release of levonorgestrel
on pelvic infection: three years' comparative experience of levonorgestrel and copper-releasing
iiilrauterine devices // Obstet. Gynecol. — 1991. — Vol. 77. — P. 261-264.
331. Xiao B.L., Zhou L.Y., Zhang X.L. et al. Pharmacokinetic and pharmacodynamic studies of
levonorgestrel-releasing intrauterine device // Contraception. - 1990. - Vol. 41. — P. 353-362.
332. Crosignani P.G., Vercellini P., Mosconi P. et al. Levonorgestrel-releasing intrauterine device
versus hysteroscopic endometrial resection in the treatment of dysfunctional uterine bleeding //
Obstet. Gynecol. - 1997. - Vol. 90. - P. 257-263.
333. Gardner F.J.E. Endometrial protection from tamoxifen-stimulated changes by a levonorg¬
estrel-releasing intrauterine system: a randomised controlled trial // Lancet. - 2000. - Vol. 356. -
P. 1711-1717.
334. Vercellini P., Aimi G.. Panazza S. et al. A levonorgestrel-releasing intrauterine system for the
treatment of dysmenorrhea associated with endometriosis: a pilot study // Fertil. Steril. — 1999. —
Vol. 72. - P. 505-508.
335. Varila E., Wahlstrom T,, Rauramo I. A 5-year follow-up study on the use of a levonorgestrel
intrauterine system in women receiving hormone replacement therapy // Fertil. Steril. - 2001. —
Vol. 76. - P. 969-973.
336. Raudaskoski T., Tapanainen J., Tomas E. et al. Intrauterine 10 microg and 20 microg levonorg¬
estrel systems in postmenopausal women receiving oral oestrogen replacement therapy: clinical, endo¬
metrial and metabolic responses // BJOG. — 2002. — Vol. 109. — P. 136-144.
337. Wollter-Svensson L.O., Stadberg E., Andersson K. et al. Intrauterine administration of
levonorgestrel 5 and 10 |ig/24 hours in perimenopausal hormone replacement therapy. A randomized
clinical study during one year // Acta Obstet. Gynecol. Scand. — 1997. — Vol. 76. — P. 449-454.
218
338. Critchley H.O., Wang H„ Jones R.L. et al. Morphological and functional features of endo¬
metrial decidualization following long-term intrauterine levonorgestrel delivery // Hum. Reprod, —
1998. - Vol. 13. - P. 1218-1224.
339. Barbosa 1., Olsson S.E., Odlind V.et al. Ovarian function after seven years' use of levonorgestrel
lUD // Adv. Contraception. - 1995. - Vol. 11. - P. 85-95.
340. Andersson J., Rybo G. Levonorgestrel-releasing intrauterine device in the treatment of menor¬
rhagia II Br. J. Obstet. Gynaecol. - 1990. - Vol. 97. - P. 690-694.
341. Baldszti E., Wimmer-Puchinger B., Loschke K. Acceptability of the long-term contraceptive
levonorgestrel-releasing intrauterine system (Mirena): a 3-year follow-up study // Contraception. -
2003.-Vol. 67.-P. 87-91.
342. Hidalgo М., Bahamondes L„ Perrotti M. et al. Bleeding patterns and clinical performance
of the levonorgestrel-releasing intrauterine system (Mirena) up to two years // Contraception. —
2002. - Vol. 65. - P. 129-132.
343. Ronnerdag М., Odlind V. Health effects of long-term use of the intrauterine levonorgestrel-
releasing system. A follow-up study over 12 years of continuous use // Acta Obstet. Gynecol. Scand. -
1999. - Vol. 78. - P. 716-721.
344. United Nations Development Programme/United Nations Population Fund/World Health
Organization/World Bank, Special Programme of Research, and Development and Research Training
in Human Reproduction. Long-term reversible contraception. Twelve years of experience with the
ТС11З8ОА and TCu220C // Contraception. - 1997. - Vol. 56. - P. 341 352.
345. Chi I-C: The TCu-380A (AG), MLCu375, and Nova-T lUDs and the lUD daily releasing 20 p.g
levonorgestrel — four pillars of lUD contraception for the nineties and beyond.? // Contraception. —
1993. - Vol. 47. - P. 325-347.
346. Shin I., Stern J. Health during prolonged use of levonorgestrel 20 fig/d and the copper TCu 380
Ag intrauterine contraceptive devices: a multicenter study. International Committee for Contraception
Research // Fertil. Steril. — 1994. - Vol. 61. - P. 70-77.
347. Sivin L, Stern J., Diaz J. et al. Two years of intrauterine contraception with levonorgestrel and
with copper: a randomized comparison of the TCu 380Ag and levonorgestrel 20 peg/day devices' //
Contraception. — 1987. - Vol. 35. - P. 245-255.
348. Sivin I., Stern J., Diaz S. et al. Rates and outcomes of planned pregnancy after use of Norplant
capsules, Norplant II rods, or levonorgestrel-releasing or copper TCu 380Ag intrauterine contraceptive
devices //Am. J. Obstet. Gynecol. - 1992. - Vol. 166. - P. 1208-1213.
349. WHO Special Programme of Research, Development and Research Training in Human
Reproduction and Task Force on the Safety and Efficacy of Fertility Regulating Methods. The TCu
380A, TCu 220C, Multiload 250 and Nova T lUDs at 3, 5, and 7 years of use // Contraception. -
1990. - Vol. 42. - P. 141-158.
350. Andersson K., Odlind V., Rybo G. Levonorgestrel-releasing and copper-releasing (Nova T)
lUDs during five years of use: a randomized comparative trial // Contraception. — 1994. - Vol. 49. —
№1.-P.56 72.
351. Backman T. Pregnancy during the use of levonorgestrel intrauterine system / T, Backman,
I. Rauramo, S. Huhtala, M. Koskenvuo // Am. J, Obstet. Gynecol. — 2004. — Vol. 190. - P. 50-54.
352. Sivin I, Dose- and age-dependent ectopic pregnancy risks with intrauterine contraception //
Obstet. Gynecol. - 1991. - Vol. 78. - P. 291-298.
353. Franks A.L. et al. Contraception and ectopic pregnancy risk // Am. J. Obstet. Gynecol. —
1990. - Vol. 163. - (4 Pt 1). - P. 1120-1123.
354. Wilson J.C. A prospective New Zealand study of fertility after removal of copper intrauterine
devices for conception and because of complications: a four-year study // Am. J. Obstet. Gynecol. —
1989. - Vol. 160. - P. 391-396.
355. Skjeldestad F.E., Bratt H. Fertility after complicated and non-complicated use of lUDs, A con¬
trolled prospective study //AdV.Contracept. - 1988. - Vol. 4. — P. 179.
356. Cameron I.T., Haining R., Lumsden M.A. et al. The effects of mefenauiic acid and norethister¬
one on measured menstrual blood loss // Obstet. Gymecol. — 1990. — Vol. 76. - P. 85-88.
357. Zhao G., Li М., Zhu P. et al. A preliminary morphometric study on the endothetrium from
patients treated with indomethadn-releasing copper intrauterine device // Hum. Reprod. — 1997. —
Vol. 12. - P. 1563-1566.
358. Milsom I., Andersson K., Jonasson K. et al. The influence of the Gyne-T 380S lUD on men¬
strual blood loss and iron status // Contraception. - 1995. — Vol. 52. — P. 175-179.
219
359. Task Force for Epidemiological Research on Reproductive Health, United Nations Development
Programme/United Nations Population Fund/Worid Health Organization/World Bank Special
Programme of Research, and Develpment and Research Training in Human Reproduction. Effects of
contraceptives on tiemoglobin and ferritin // Contraception. — 1998. - Vol. 58. — P. 261-273.
360. Hassan E.O., El-Husseini М., El-Nahal N. The effect of 1-year use of the CuT 380A and
oral contraceptive pills on hemoglobin and ferritin levels // Contraception. - 1999. - Vol. 60. -
P. 101-105.
361. Backman T., Huhtala S., Blom T. et al. Length of use and s}rmptoms associated with premature
removal of the levonorgestrel intrauterine system: a nation-wide study of 17,360 users j j Br.J. Obstet.
Gynaecol. - 2000. - Vol. 107. - P. 335-339.
362. Sivin L, Schmidt F. Effectiveness of lUDs; a review // Contraception. — 1987. — Vol. 36. —
P. 55-84.
363. Tang G.W., Lo S.S. Levonorgestrel intrauterine device in the treatment of menorrhagia in
Chinese women: efficacy versus acceptability // Contraception. — 1995. — Vol. 51. — P. 231 -235.
364. Crosignani P.G., Vercellini P., Apolone G. et al. Endometrial resection versus vaginal hyster¬
ectomy for menorrhagia: long-term clinical and quality-of-life outcomes // Am. J. Obstet. Gynecol. —
1997. - Vol. 177. - P. 95-101.
365. Römer T. Prospective comparison study of levonorgestrel lUD versus Roller-Ball endometrial
ablation in the management of refractory recurrent hypermenorrhea // Eur. J. Obstet. Gynecol. Reprod.
Biol. - 2000. - Vol. 90. - P. 27-29.
366. Istre 0., Trolle В. Treatment of menorrhagia with the levonorgestrel intrauterine system versus
endometrial resection // Fertil. Steril. — 2001. — Vol. 76. — P. 304-309.
367. Fong Y.F., Singh K. Effect of the levonorgestrel-releasing intrauterine system on uterine myo¬
mas in a renal transplant patient // Contraception. — 1999. — Vol. 60. — P. 51-53.
368. Lockhat F.B., Emembolu J.O., Konje J.C. The evaluation of the effectiveness of an intrauterine-
administered progestogen (levonorgestrel) in the symptomatic treatment of endometriosis and in the
staging of the disease // Hum. Reprod. — 2004. — Vol. 19. — P. 179-184.
369. Mishell D.R. Jr. et al. The intrauterine device: a bactériologie study of the endometrial cavity //
Am. J. Obstet. Gynecol. - 1966. - Vol. 96. - P. 119-126.
370. Farley T.M., Rosenberg M.J., Rowe P.J. et al. Intrauterine devices and pelvic inflammatory
disease: an international perspective // Lancet. — 1992. - Vol. 339. — P. 785-788.
371. Lee N.C. et al. Type of intrauterine device and the risk of pelvic inflammatory disease // Obstet.
Gynecol. - 1983. - Vol. 62. - P. 1-6.
372. Lee N.C., Rubin G.L., Bomcki R. The intrauterine device and pelvic inflammator}' disease revis¬
ited: new results from the Womens’ Health Study // Obstet. Gynecol. — 1988. — Vol. 72. — P. 1-6.
373. Skouby S.O., Molsted-Pedersen L., Kosonen A. Consequences of intrauterine contraception in
diabetic women // Fertil. Steril. — 1984. — Vol. 42. — P. 568-572.
374. Kimmerle R. et al. Effectiveness, safety, and acceptability of a copper intrauterine deivce (Cu
Safe 300) in type T diabetic women // Diabetes. Care. - 1993. — Vol. 16. — P. 1227-1230.
375. Buchan H., Villard-Mackintosh L., Vessey M. et al. Epidemiology of pelvic inflammator)'
disease in parous women with special reference to intrauterine device use // Br.J. Obstet. Gynaecol. —
1990. - Vol. 97. - P. 780-788.
376. Mehanna M.T. et al. Chlamydial serologic characteristics among intrauterine contraceptive
device users: does copper inhibit chlamydial infection in the female genital tract.? // Am. J. Obstet.
Gynecol. - 1994. - Vol. 171. - P. 691-693.
377. Kleinman D., Insler V., Sarov I. Inhibition of Chlamydia trachomatis growtli in endometrial
cells by copper: possible relevance for the use of copper lUDs // Contraception. — 1989. — Vol. 39. —
P. 665-676.
378. Mohllajee A., Curtis K., Peterson H. Does insertion and use of an intrauterine device increase
the risk of pelvic inflammatory disease among women with sexually transmitted infection.? A systematic
review // Contraception. — 2006. — Vol. 73. — P. 145-153.
379. Lee N.C., Rubin G.L., Grimes D.A. Measures of sexual behavior and the risk of pelvic inflam¬
matory disease // Obstet. Gynecol. — 1991. — Vol. 77. — P. 425-430.
Хотя яички не являются обязательными для выживания организма, эти органы
играют важную роль в воспроизводстве вида посредством продукции сперматозои¬
дов и половых стероидных гормонов. Развитие половых клеток в яичках ограничено
семенными канальцами, а продукция тестостерона (Т) происходит в интерстициаль¬
ном отделе клетками Лейдига. Размер яичек, обусловленный в большой мере отделом,
содержащим половые клетки, значительно варьирует среди млекопитающих даже с
учетом размеров тела и никак не связан с локализацией яичек (абдоминальная или
мошоночная), формой тела и локомоцией [1]. В целом млекопитающие с моногамной
системой размножения имеют меньшие яички по сравнению с полигамными видами,
когда несколько самцов могут спариться с самкой во время одной течки [1]. Более
крупный размер яичек у полигамных млекопитающих приписывают высокой часто¬
те спаривания, необходимости высокой скорости продукции семени и конкуренции
семени различных самцов в плане оплодотворения яйцеклетки [1].
Тестостерон — 19-углеродный стероидный гормон, синтезируемый яичками,
который преобладает у большинства видов млекопитающих. Однако существенное
количество Д^-андростендиона секретируется у самок некоторых видов, включая
кротов и пятнистых гиен iCrocuta crocutä) [2, 3]. Самки пятнистых гиен имеют более
высокие уровни андростендиона, тяжелее и агрессивнее, чем самцы, и имеют крупный
псевдопенис, способный эрегироваться [2-5], У некоторых видов 5а-андростан-За,
17|3-диол — основной андроген, секретируемый яичками в течение периода раз¬
вития простаты и полового члена [6, 7]. У этих видов во время дифференциации
дигидротестостерон (ДГТ) образуется альтернативным путем из 5а-андростана-За,
17р-диола (5-андростандиол), который из 17а-гидроксипрогестерона превращается в
5а-восстановленные прогестероны, далее конвертирующиеся в 5-андростандиол.
Важность тестостерона в регуляции вторичных половых признаков, росте мышц и
сексуальном и несексуальном поведении у самцов млекопитающих была установле¬
на давно. В своих трансплантологических экспериментах Адольф Бертольд (Adolph
Berthöld) наблюдал, что цыплята, кастрированные во время своего развития, стано¬
вились послушными каплунами, а не обычными петухами [8]. Когда яички от других
цыплят были имплантированы в абдоминальные полости этих каплунов, животные
221
развивались в петухов, становились агрессивными и показывали нормальное пове¬
дение самцов. Эти ранние трансплантологические эксперименты показали, что яички
влияют на развитие самцов и их поведение посредством секреции сильного гормона
прямо в кровоток.
Андрогены прямо или опосредованно влияют почти на все системы организма во
время фетального и пубертатного развития, а также и во взрослой жизни. Половая
дифференцировка плода млекопитающего требует не только генетической инфор¬
мации, заложенной в половых хромосомах, но и гормональных секретов феталь¬
ных яичек, называемых тестостероном и мюллеровым ингибирующим гормоном.
Тестостерон регулирует дифференцировку вольфовых протоков в придаток яичка,
семявыносящий проток, а у некоторых видов и в семенные пузырьки [9]. у людей
превращение Т в 5а-дигидротестостерон требуется для дифференциации урогени¬
тального бугорка и урогенитального синуса в половой член, мужскую уретру и пред¬
стательную железу [10]. Кроме того, увеличивающиеся уровни Т во время полового
созревания обеспечивают соматический рост и вирилизацию мальчиков. Т играет
критическую роль в репродукции млекопитающих; он необходим для поддержания
сексуальной функции, развития половых клеток и дополнительных половых органов,
У взрослого животного Т оказывает дополнительное воздействие на мышцы, жиро¬
вую ткань, кости, гемопоэз, свертывание крови; липидный, белковый и углеводный
обмен; психосексуальное и когнитивное поведение.
ПРОДУКЦИЯ АНДРОГЕНОВ У МУЖЧИН
у самцов большинства видов млекопитающих 95% циркулирующего Т секрети¬
руется яичками. У мужчин ежедневно синтезируется яичками 3-10 мг Т; прямая
секреция Т надпочечниками и периферическое превращение андростендиона, секре¬
тируемого надпочечниками, в общей сложности составляют еще 500 мкг Т ежеднев¬
но. При этом только малое количество ДГТ (около 70 мкг ежедневно) секретируется
непосредственно яичками; большая часть циркулирующего ДГТ образуется путем
периферической конверсии т.
Т образуется в яичках гетерогенной группой клеток, которая включает взрос¬
лые клетки Лейдига, предшественники клеток Лейдига и незрелые клетки Лейдига.
Исследования на гипогонадотропных мышах показывают, что фетальное развитие
клеток Сертоли и клеток Лейдига происходит независимо от гонадотропинов; однако
нормальная дифференцировка и пролиферация популяции взрослых клеток Лейдига
требует присутствия гонадотропинов. Ряд клеток Сертоли после рождения также
регулируется гонадотропинами. 46,ХУ-Мужчины с мутациями рецептора ЛГ харак¬
теризуются агенезией или гипоплазией клеток Лейдига, указывающими на важную
роль ЛГ в регуляции развития клеток Лейдига у людей. Клетки Лейдига происходят из
пока недостаточно охарактеризованных мезенхимальных клеток-предшественников
под влиянием Л Г, инсулиноподобного фактора роста-1, трансформирующего фактора
роста-а, трансформирующего фактора роста-р, интерлейкина-1 и основного фактора
роста фибробластов. Клетки Лейдига у высших млекопитающих существуют в двух
различных генерациях, фетальной и взрослой, которые разделены препубертатным
периодом, во время которого яички лишены клеток Лейдига.
Секреция тестостерона клетками Лейдига находится под контролем Л Г (гликопро-
теиновый гормон гипофиза). ЛГ связывается со специфическими рецепторами, ассо¬
циированными с С-белком, на клетках Лейдига и активирует каскад циклического
аденозинмонофосфата [11,12]. Хотя ЛГ также активирует путь фосфолипазы С, пока
неясно, является ли этот путь обязательным для опосредованной ЛГ-стимуляции
продукции т. Основная цель ЛГ — боковая ветвь расщепляющего фермента С¥Р11А1,
Рецептор пролактина не экспрессируется на клетках Лейдига в яичках человека, и нет
данных, что пролактин регулирует продукцию Т у мужчин [13].
222
Продукция Т клетками Лейдига модулируется рядом паракринных ростовых фак¬
торов, регуляторных пептидов и цитокинов в пределах семявыносящих канальцев и
интерстиция яичек, включая инсулиноподобный фактор роста-1, белки, связываю¬
щие инсулиноподобный фактор роста-1, ингибины, активины, трансформирующий
фактор роста-а, эпидермальный фактор роста, интерлейкин-1, фактор некроза
опухоли-а, основной фактор роста фибробластов, гонадотропин-высвобождающий
гормон и вазопрессин [14].
Холестерин, необходимый для биосинтеза Т, может быть импортирован в клетку
из циркулирующего холестерина, или он может быть синтезирован в клетке либо
de novo из ацетата, либо из эфиров холестерина [15]. Стадией биосинтеза Т, лимити¬
рующей его скорость, служит доставка холестерина к внутренней митохондриальной
мембране, где расположен комплекс отщепления боковой цепи холестерина, который
превращает холестерин в прегненолон (рис. 3-1). StAR делает холестерин доступным
комплексу холестериновой боковой цепи, что регулирует скорость биосинтеза т.
Периферический рецептор бензодиазипинов (митохондриальный холестеринсвязы-
Холестерин плазмы крови
ф —
Первое
отщепление _
боковой цепи
Второе
отщепление
боковой
цепи
Восстановление
17-кетогруппьГ
Ацетат
Холестерин ^
4
StAR
CYP11A1
Эфиры холестерина
Зр-ГСДИ
Прегненолон > Прогестерон
CYP17
t
CYP17
17-Оксигидропрегненолон - - 17-Оксигидропрогестерон
CYP17 CYP17
г
Дегидроэпиандростерон ► Андростендион
17Ц-ГСДЗ
f
Андростендиол
зв-гсдн
17В-ГСДЗ
Тестостерон
Окисление кольца А
Рис. 3-1. Пути биосинтеза тестотерона (Т) в яичках человека. Имеется два пути, которыми прегненолон,
происходящий от холестерина, может быть превращен в Т. А®-Путь, который проходит через прегнено¬
лон, 17-гидроксипрегненолон, дигидроэпиандростерон и андростендиол, включает отщепление боковой
цепи и восстановление 17-кетогруппы до окисления кольца А. A''-Путь включает окисление кольца А пре-
гненолона перед отщеплением боковой цепи и восстановления 17-кетогруппы. В яичках человека преоб¬
ладает А®-путь. Доставка холестерина к внутренней митохондриальной мембране для отщепления боковой
цепи — этап, лимитирующий скорость биосинтеза Т, который находится под контролем ЛГ. Как показано
на рисунке, холестерин для биосинтеза Т может быть синтезирован de novo в клетке, или он может быть
получен из циркулирующего холестерина или из эфиров холестерина. StAR — белок острой регуляции сте-
роидогенеза; CYP11A1 — фермент отщепления боковой цепи; зр-ГСДП — Зр-гидроксистероид дегидроге¬
наза; CYPM — 17а-гидроксилаза; 17Р-ГСДЗ — 17р-гидроксистероид дегидрогеназа. [Из Griffin J.E., Wilson
J.D. Disorders of the testis and the male reproductive tract // Larsen P.R., Kronenberg H.M., Melmed S., Polonsky
K.S. (eds). Williams Textbook of Endocrinology. — 10th ed. — Philadelphia: Saunders, 2003. — P. 709-763.]
223
вающий протеин), как известно, вовлечен в процесс транспорта холестерина и при¬
сутствует в высокой концентрации во внешней митохондриальной мембране; также
было предложено рассматривать его как сильный регулятор стероидогенеза в клетках
Лейдига. Кроме того, в контроль стероидогенеза в клетках Лейдига была вовлечена
другая группа недостаточно пока изученных митохондриальных протеинов.
Превращение холестерина в Т может происходить посредством двух биосинтети¬
ческих путей [16]: А^- и А’*-путь. В яичках человека А^-путь доминирует и включает
отщепление боковой цепи прегненолона, восстановление 17-кетогруппы и окисления
кольца А. При А’*-пути окисление кольца А предшествует отщеплению боковой цели и
восстановлению 17-кетогруппы.
Тестостерон — главный циркулирующий андроген у мужчин. Секреция Т регули¬
руется по механизмам прямой и обратной связи, которые действуют в пределах оси
«гипоталамус-гипофиз-гонады». Гипоталамический выброс гонадолиберина стимули¬
рует передний гипофиз к вьщелению ЛГ, который, в свою очередь, стимулирует клетки
Лейдига в яичках к синтезу Т. Секреция Т имеет пульсирующий, суточный и годичный
ритмы; наиболее высокие концентрации наблюдают в часы после пробуждения, наиболее
низкие — после полудня и вечером. Концентрации Т могут сильно варьировать у отдель¬
но взятого пациента из-за суточного ритма и пульсирующего характера секреции.
ДГЭА и А'’-андростендион — слабые андрогены [17]; А'*-андростендион может
быть конвертирован в эстроген в некоторых неполовых тканях. В настоящее
время не описана биологическая роль прегнолона, 17-гидроксипрегненолона и
17-гидроксипрогестерона.
ТРАНСПОРТ АНДРОГЕНОВ В ОРГАНИЗМЕ
Большая часть циркулирующего Т связана с двумя белками плазмы крови, с глобу¬
лином, связывающим половые гормоны, и альбумином; лишь от 0,5 до 3% Т находит¬
ся в свободном состоянии. Циркулирующий Т прочно с высокой аффинностью связан
с глобулином, связывающим половые гормоны, свободно — с альбумином, и только
0,5-3% гормона содержится в несвязанной форме (рис. 3-2). Аффинность связывания
Т и ДГТ с глобулином, связывающим половый гормоны, по меньшей мере в 4 раза
выше аналогичного показателя для альбумина [18]. У мужчин Т главным образом
связан с альбумином (50-68%), меньшая его часть — с глобулином, связывающим
половые гормоны (30-45%). У женщин около 70% циркулирующего гормона связано
с глобулином, связывающим половые гормоны, а 25% — с альбумином. Биодоступным
Т называют свободный или несвязанный Т плюс фракция, связанная с альбумином.
Гипотеза свободного гормона предполагает, что только несвязанная фракция био¬
логически активна [19] и что несвязанный Т, будучи липофильным, входит в клетку
посредством свободной диффузии; однако существенный массив данных указывает на
то, что гормон, связанный с альбумином, может в различной степени диссоциировать
в капиллярах и становиться биодоступным [20]. ЗаИуата и соавт. [20] показали, что
андрогены, связанные с альбумином и глобулином, связывающим половые гормо¬
ны, представляют собой главный циркулирующий пул биодоступного гормона для
яичек и предстательной железы. Более того, некоторые утверждают, что комплекс
«половой гормон-глобулин, связывающий половые гормоны» способен проходить
через гематотестикулярный барьер или мембрану «простата-плазма крови» [21]; это
мнение разделяется не всеми. Было доказано, что мегалин, представитель семейства
рецепторов липопротеинов низкой плотности, облегчает эндоцитоз Т, связанного с
глобулином, связывающим половые гормоны, посредством эндоцитозных ямок [22,
23]. Животные с дефицитом мегалина показывают резистентность к некоторым, но
не всем, эффектам андрогенов [22]. Эти данные вызывают сомнения по поводу обо¬
снованности гипотезы свободного гормона.
Глобулин, связывающий половые гормоны, — гликопротеин, синтезируемый в
печени, который обладает высокой аффинностью связывания Т и эстрадиола [18].
224
Несвязанный,
Мужчины
Женщины
Свободный тестостерон = Несвязанный тестостерон
Биодоступный = несвязанный + связанный с альбумином
Рис. 3-2. Связывание циркулирующего тестостерона (Т) с белками плазмы крови у мужчин и женщин.
Большая часть циркулирующего гормона связана с высокой аффинностью с глобулином, связывающим
половые гормоны, и с низкой аффинностью — с альбумином. Только 0,5-3,0% циркулирующего Т не
связано или свободно и измеряется равновесным диализным методом. Термин «биодоступный Т» обо¬
значает фракцию циркулирующего Т, которая является несвязанной или связана с альбумином и может
быть измерена с помощью метода преципитации сульфатом аммония.
Инсулин, тиреоидные гормоны, диетические факторы, андрогены и эстрогены регу¬
лируют печеночную продукцию глобулина, связывающего половые гормоны. Этот
белок служит в качестве главного транспортного протеина половых гормонов в плазме
крови, его концентрация в значительной степени определяет их распределение между
протеинсвязанным и несвязанным состояниями, хотя его функциональная роль в
доставке Т к органам-мишеням вызывает значительные дебаты [18]. Назначение
андрогенов, ожирение, гиперинсулинемические состояния и нефротический син¬
дром ассоциированы с низкими концентрациями глобулина, связывающего половые
гормоны [18]. И наоборот, уровни этого белка увеличены у пациентов с гипертирео-
идным состоянием, различными типами хронических воспалительных состояний и
при старении. Локус, ассоциированный с концентрациями глобулина, связывающего
половые гормоны, у афроамериканцев и представителей белой расы определен в 1ц44
[24]. Кроме того, несколько других локусов у представителей афроамериканской расы
показывают связь с концентрациями глобулина, связывающего половые гормоны.
Это позволяет предположить, что многие гены, вероятно, регулируют уровни этого
белка, в отличие от человека и крыс, мыши не имеют глобулина, связывающего поло¬
вые гормоны, и экспрессируют очень низкие уровни андрогенсвязывающего протеина
в яичках, но не проявляют нарушения фертильности или поведения самцов [25].
МЕТАБОЛИЗМ ТЕСТОСТЕРОНА
Тестостерон метаболизируется главным образом в печени (50-70%), хотя некоторая
часть распада также происходит в периферических тканях, особенно в предстатель¬
ной железе и коже. Печень забирает Т из крови и путем серии химических реакций,
которые включают 5а- и 5р-редуктазы, зр-, За-гидроксистероиддегидрогеназы и 17-
гидроксистероиддегидрогеназу, превращает его в андростерон, этиохоланолон (оба —
интактные метаболиты), ДГТ и 3-андростандиол (рис. 3-3). Эти соединения подвер¬
гаются глюкуронидации или сульфатации перед их экскрецией почками. Свободный
225
Продукция
тестостерона
(5 мг/сут)
i
I
X...
Эстрадиол
плазмы
(0,02 мг/сут)
I
Ароматаза
г
5а-Редуктаза
17Р-ГСД,
0,3%
6-8%
5р-редуктаза
i
40%
1
Гидроксилаза;
конъюгирующие
ферменты
50%
Дигидро¬
тестостерон
плазмы
(0,3 мг/сут)
17-кетостероиды-
андростерон +
этиохоланолон
(2,0 мг/сут)
Полярные
метаболиты —
диолы, триолы
и конъюгаты
(2,5 мг/сут)
Активные метаболиты
Выводимые метаболиты
Рис. 3-3. Тестостерон метаболизируется путем серии ферментативных реакций в активные и неактив¬
ные метаболиты. Ароматизация кольца А, или 5а-восстановление, превращает его в активные метабо¬
литы эстрадиол и дигидротестостерон. Дополнительно посредством действия ряда 5а- и 5|3-редуктаз
17р-гидроксистероиддегидрогеназа и гидроксилазтестостерон превращается в ряд неактивных метабо¬
литов, которые конъюгируются и выводятся с мочой. 17р-ГСД — 17р гидроксистероиддегидрогеназа.
[M3GriffinJ.E., Wilson J.D. Disorders of the testis and the male reproductive tract//Larsen P.R., Kronenberg H.M.,
Melmed S., Polonsky K.S. (eds). Williams Textbook of Endocrinology. — 10th ed. — Philadelphia: Saunders,
2003. - P. 709-763.]
и конъюгированный андростерон и этиохоланолон — основные метаболиты Т в моче.
Несмотря на то что большинство из этих метаболитов неактивны, некоторые могут
быть прямо или опосредованно активными. Например, 5-андростандиол потенциаль¬
но может быть конвертирован в 5-дигидротестостерон (5а-ДГТ). Также доказано, что
метаболиты 5р-андрогена стимулируют гемопоэз [26].
ТЕСТОСТЕРОН В КАЧЕСТВЕ ПРОГОРМОНА: РОЛЬ ЭСТРАДИОЛА Щ-
И 5а-ДИГИДР0ТЕСТ0СТЕР0НА
Тестостерон превращается во многих периферических тканях в активные мета¬
болиты, 17р-эстрадиол и 5а-ДГТ (рис. 3-4). Ароматизация кольца А превращает Т
в 17[3-эстрадиол. Дополнительно восстановление двойной связи 4,5 превращает Т в
ДГТ. Действие Т во многих тканях осуществляется посредством этих метаболитов.
Эффекты Т на трабекулярную резорбцию кости, липиды плазмы крови, про¬
грессирование атеросклероза и половую дифференциацию головного мозга требуют
ароматизации эстрадиола. Значительные сведения о роли эстрогенов в физиоло¬
гии млекопитающих были получены из исследований естественных мутаций генов
рецептора-а эстрогена и CYP19 ароматазы у человека и мышей с мутациями генов
рецепторов-а [27] и рецепторов-р эстрогенов и CYP19 ароматазы, делающих эти гены
недействующими. Эти модели эстрогеновой недостаточности демонстрируют зна¬
чительные нарушения сперматогенеза и фертильности, повышенные уровни Т и ЛГ,
задержку созревания эпифизов, сниженную массу костной ткани и нарастание массы
тела, свидетельствующие о важной роли эстрогенов в регуляции костной массы и
гонадотропной функции, структуре тела и сперматогенеза. Описано очень мало людей
226
А^-Андростендион
17р -гидроксистероид-
дегидрогеназа 3
Тестостерон
17р-Эстрадиол
4
Дигидротестостерон
• Резорбция кости
> Эпифизиальное слияние
> Половая дифференциация
головного мозга
- Поведение
• Липиды плазмы крови
- Прогрессирование атеросклероза
■ Рост предстательной железы
■ Кожа
■ Волосяные фолликулы в андро-
генчувствительных областях
Рис. 3-4. Тестостерон в качестве прогормона. Тестостерон может быть превращен действием
5а-редуктазы в 5а-дигидротестостерон или действием СУР19 ароматазы в эстрадиол. А'*-Андростендион
также может быть ароматизирован в эстрон. Кроме того, эстриол может быть образован путем аро¬
матизации 16а-гидроксипрогестерона (не показано на данном рисунке). Действие тестостерона на
предстательную железу, кожу и волосяные фолликулы в андрогенчувствительных областях требует его
облигатного 5а-восстановления в 5а-дигидротестостерон. Ароматизация тестостерона в 17р-эстрадиол
требуется для опосредования его эффектов на костную резорбцию, эпифизиальное слияние, половую
дифференциацию головного мозга, некоторые типы поведения, липиды плазмы крови и прогрессиро¬
вание атеросклероза.
с инактивирующими мутациями СУР19 ароматазы [28-31]. Женщины с мутациями
гена СУР19 ароматазы маскулинизированы, у них не происходит полового развития,
они имеют повышенные уровни андрогенов, ЛГ и ФСГ, поликистозные яичники и
высокий рост [28-31]. Мужчины с мутациями гена СУР19 ароматазы характери¬
зуются увеличенными уровнями Т, низкими уровнями эстрадиола, остеопорозом,
ускорением костной резорбции, задержкой созревания эпифизов и высоким ростом
[28-31].
Выделено и охарактеризовано по меньшей мере две изоформы стероид-5а-
редуктазы [32]. Тип 1 этого фермента экспрессируется во многих экстрагенитальных
тканях, картирован в хромосомальной области 5р15 и имеет оптимальный рН=8.
227
Тип 2 изофермента стероид-5а-редуктазы экспрессируется в предстательной железе
и других половых органах, был картирован в 2р23 и имеет оптимальный рН=5,0
[32], Биологическая роль типа 1 5а-редуктазы полностью не была установлена.
Генонаправленные эксперименты предполагают, что первый тип фермента играет
роль в метаболизме прогестерона в конце беременности [33]. Мыши с инактивиро¬
ванным типом 1 фермента испытывают трудности в созревании шейки матки и родах
[33].
Эффекты Т на предстательную железу и железы кожного покрова требуют его
5а“Восстановления в ДГТ с помощью 2-го типа 5а-редуктазы. Этот фермент доми¬
нирует в предстательной железе, была установлена его роль в патофизиологии
доброкачественной гипертрофии предстательной железы, гирсутизма и облысения
по мужскому типу. Во время эмбрионального развития Т контролирует дифферен¬
циацию вольфовых протоков в придаток яичка, семявыносящие протоки и семен¬
ные пузырьки. Развитие структур из урогенитального синуса и полового бугорка,
таких как мошонка, половой член и мужская уретра, требует действия ДГТ. Роль
5а-восстановления Т в опосредовании его эффектов на мышцы и сексуальную функ¬
цию остается неясной. Хотя Т и ДГТ могут оказывать анаболические эффекты на
мышцы, активность стероид 5а-редуктазы в скелетной мускулатуре очень низкая или
отсутствует [34], и неизвестно, является ли 5а-восстановление Т облигатным для опо¬
средования андрогенных эффектов на мышцы. Опубликованные данные также раз¬
норечивы в том, осуществляются ли андрогенные эффекты на сексуальную функцию
через Т или его 5а-восстановленный метаболит, ДГТ.
МЕХАНИЗМ ДЕЙСТВИЯ АНДРОГЕНОВ
Большинство эффектов андрогенов опосредованы через их связывание с внутри¬
клеточным андрогеновым рецептором, который действует как лигандзависимый
фактор транскрипции [35-37]. Рецептор андрогенов гомологичен другим ядерным
рецепторным белкам, включая рецепторы к глюкокортикоидам, прогестерону и
мийералокортикоидам [38-40]. Преобладающий белок андрогенового рецептора
из 919 аминокислот массой 110 кДа имеет три консервативных функциональных
домена: стероидсвязывагощий домен, ДНК-связывающий домен и транскрипцион¬
ный активирующий домен (см. рис. 3-5); из них центральный ДНК-связывающий
домен, богатый цистеином. наиболее консервативен [38]. Единственная копия гена
андрогенового рецептора охватывает область 90 кЬ в регионе Xqll-12. При отсут¬
ствии лиганда белок рецептора андрогенов распределен и в ядре, и в цитоплазме.
Однако связывание андрогена с рецептором вызывает перенос последнего в ядро;
аминокислотная последовательность между 617 и 633 положениями аминокислот
рецептора андрогена важна для его миграции в ядро и функции трансактивации
[35]. Взаимодействие связанного комплекса «андроген-рецептор» с соответствую¬
щей последовательностью в промоутерной области гена-мишени изменяет скорость
транскрипции гена-мишени.
Связывание андрогенов с рецептором андрогенов приводит к конформационным
изменениям .этого белка; связывание антагониста андрогенов с рецептором андроге¬
нов может индуцировать другой набор конформационных изменений, чем агонистов
андрогенов. Рецептор андрогенов может использовать два трансактивационных доме¬
на — АЕ1 и АЕ2. Домен трансактивации АЕ1 (включающий регионы 1 и 5) локализо¬
ван в М-терминальной части белка-рецептора, а АЕ2 локализован в С-терминальном
лигандзависимом домене. В интактном рецепторе и АР1, и АЕ2 являются лигандза-
висимыми, на них влияют коактиваторы ядерного рецептора. В противоположность
этому в «урезанном» рецепторе андрогенов, в котором отсутствует лигандсвязываю-
щий домен, АЕ1 становится активным. Связывание гормона с рецептором андрогенов
приводит к образованию одного комплекса с тканеспецифичными коактиваторами
228
Регуляторный ДНК-связыва-
ющий домен
Андрогенсвязывающий
домен
т1
т5
NH2
AF1
617-634
HR
Спираль,
^локализованная
в ядре
AF2
HBD
-СООН
Активация
транскрипции
101 ?71
Гормоннезависимая
трансактивация
361
528
ДНК-связывающий
домен
537 621
Связывание лиганда
I 1
671
919
Рис. 3-5. А ~ схематическое представление гена рецептора андрогенов. Каждый бокс представляет
экзон. Андроген-связывающий домен включает около 250 аминокислот в С-терминальной области проте¬
ина, которые кодируются экзонами 4-8. Регуляторная область кодируется экзоном 1 и частью экзона 2. а
ДНК-связывающий домен — экзоном 3 и частью экзона 2; Б — функциональные домены белка рецептора
андрогенов. N-терминальная область протеина рецептора андрогена включает домен активации 1 {AF1)
с двумя функциями трансактивации Т1 (101-371 аминокислоты) и Т5 (361-528 аминокислоты). ДНК-
связывающий домен (DBD) расположен в середине, и за ним следуют сигнал ядерной локализации (NLS)
и шарнирная область (HR). Гормонсвязывающий домен (HBD) локализован у С-терминального конца
белка и связывает лиганды и белок теплового шока. [Из Klocker Н., Gromoll J., Cato A.C.В. Androgen
receptor: molecular biology// Nieschlag E., Behre H.M. (eds). Testosterone: Action. Deficiency, Substitution. —
Cambridge, UK: Cambridge University Press, 2004. — P. 39-92.]
и корепрессорами, которые определяют специфичность и тканевую селективность
действия гормона.
В тканях-мишенях андрогенов либо Т, либо его 5а-восстановленный метаболит
ДГТ связываются с андрогеновым рецептором и регулируют экспрессию генов.
Тестостерон связывается с рецептором андрогенов с аффинностью в 2 раза ниже, чем
данный показатель у ДГТ, хотя максимальная связывающая емкость сопоставима
у обоих андрогенов. Комплекс «ДГТ-рецептор андрогенов» более термостабилен и
имеет более низкую скорость диссоциации, чем комплекс «Т-рецептор андрогенов».
Это может обеспечивать более высокую эффективность ДГТ в опосредовании андро¬
генных эффектов в некоторых андрогенчувствительных органах, таких как предста¬
тельная железа. Однако почему 5а-восстановление необходимо для опосредования
андрогенных эффектов в одних тканях и не требуется в других, неясно. Неизвестно
также, почему этот этап метаболического превращения происходит для Т и не проис¬
ходит в отношении других стероидных гормонов.
Имеются неубедительные свидетельства, что некоторые эффекты андрогенов могут
быть опосредованы через негеномные рецепторы на клеточной мембране.
229
Б ходе первых исследований Kochakian и соавт. подтвердили, что андрогены при¬
водят к задержке азота в организме, стимулируют синтез мышечного белка и соответ¬
ственно повышают мышечную массу. В ходе недавних исследований выяснилось, что
андрогены увеличивают скелетную мышечную массу путем способствования диффе-
ренцировке мезенхимальных стволовых клеток по миогенной линии и ингибирова¬
нию их адиггоцитарной дифференциации [41, 42]. Кроме того, андрогены могут вли¬
ять на вступление клеток-сателлитов в клеточный цикл, а также стимулируют синтез
мышечных белков [43-48]. Тестостерон ингибирует дифференциацию преадипоцитов
в адипоциты, Singh и соавт. [41] показали, что андрогены способствуют ассоциации
андрогенового рецептора с р-катенином, стабилизации р-катенина и облегчению его
переноса в ядро, где комплекс «андрогеновый рецептор-(З-катенин» связывается с
TCF-4 и регулирует некоторое количество Wnt-направленных генов, таким образом
определяя клеточную линию, способствуя миогенной дифференцировке и ингибируя
адипогенную дифференцацию.
РАЗВИТИЕ ПЕРВИЧНЫХ ПОЛОВЫХ КЛЕТОК В ЯИЧКАХ
Сперматогенез, процесс, посредством которого сперматозоиды формируются из
сперматогенных стволовых клеток в семенных канальцах яичек, происходит в три
стадии (рис. 3-6): сперматогенная репликация, мейоз и спермиогенез. Во время пер¬
вой стадии сперматогенные стволовые клетки митотически делятся несколько раз,
давая начало последующим поколениям сперматогоний, из которых в канальцах у
человека имеется по меньшей мере три основных типа: темный тип А, светлый тип А
и тип Б. Сперматогонии типа Б пролиферируют, давая начало первичным спермато-
цитам на стадии прелептотены мейоза, в которой активно синтезируется ДНК.
Во время второй стадии первичные диплоидные сперматоциты входят в мейоз,
который состоит из двух последовательных делений первичного сперматоцита,
сопровождающихся только одним удвоением хромосом. По завершении мейоза обра¬
зуются 4 сперматиды, каждая из которых содержит единичный, или гаплоидный,
набЪр хромосом.
Спермиогенез — финальная стадия сперматогенеза — включает структурную транс¬
формацию и дифференциацию сперматид. Во время спермиогенеза хроматин сперма-
тид конденсируется в компактную массу, ядро становится облаченным мембранным
производным аппарата Гольджи (акросома), которое содержит ферменты, лизи-
рующие оболочки яйцеклетки и позволяющие сперматозоиду проникнуть в ооцит в
процессе оплодотворения, а цитоплазма сперматиды удлиняется и окружает жгутик,
образованный из центриоли. В процессе спермиогенеза большая часть цитоплазмы
отбрасывается в форме резидуального тельца.
Сперматиды заканчивают свое превращение в сперматозоид путем образования
сложного хвоста с помощью аксонемального комплекса из двух внутренних единич¬
ных и девяти наружных двойных микротрубочек, у человека общая продолжитель¬
ность сперматогенеза составляет 74 сут; сперма проводит еще 21 сут в придатке яичка,
где подвергается дальнейшему созреванию.
Гормональная регуляция развития половых клеток
Нормальный сперматогенез требует комплексных взаимодействий половых клеток,
клеток Сертоли и Лейдига, а также синергического влияния гипофизарных гонадотро¬
пинов ЛГ и ФСГ. ЛГ после его связывания с рецептором G-белка стимулирует продук¬
цию Т клетками Лейдига; высокие внутрияичковые концентрации Т — необходимое
условие инициации и поддержания сперматогенеза в гонадах. ФСГ воздействует на
клетки Сертоли и стимулирует секрецию ряда белков и ростовых факторов, таких как
андрогенсвязывающий белок, ингибин, активин, фактор стволовых клеток, активатор
плазминогена, трансферрин, сульфатированные гликопротеины и лактат. Во время
полового развития увеличивающиеся концентрации ФСГ повышают чувствитель-
230
Рис. З-б. Схематическое изображение развития зародышевых клеток во время сперматогенеза у чело¬
века. Предшественники зародышевых клеток способны самообновляться путем митотического деления.
После полового созревания некоторые сперматогонии проходят дифференциацию в первичные сперма-
тоциты, которые претерпевают два последовательных меойтических деления для образования четырех
гаплоидных сперматид. Круглые сперматиды путем прохождения нескольких этапов дифференциации
становятся удлиненными, у них развивается хвост, они трансформируются в сперматозоиды. Процесс
сперматогенеза занимает 73 дня в яичках человека. Сперматогенные стволовые клетки, сперматогонии
и первичные сперматоциты являются диплощными (то есть они содержат 46 хромосом), в то время как
сперматиды и сперматозоиды гаплоидны (23 хромосомы).
ность клеток Лейдига к стимуляции ЛГ. После того как сперматогенез установился во
взрослых яичках, клетки Сертоли становятся менее восприимчивыми к ФСГ.
Точная роль ФСГ в регуляции сперматогенеза у человека полностью не изучена.
У половозрелых людей, которые имеют спонтанный или индуцированный гипого¬
надотропный гипогонадизм, Л Г или ХГЧ, могут реинициировать сперматогенез. У
мужчин, которые стали гипогонадотропными в результате назначения Т-энантата,
сперматогенез может быть инициирован повторно путем изолированного назначе¬
ния ХГЧ или ФСГ; комбинированное применение ФСГ и ХГЧ ассоциировано с более
высокой концентрацией спермы, чем при назначении только ХГЧ. Таким образом,
у взрослых добавление ФСГ не является обязательным для реинициации или под¬
держания сперматогенеза, но он улучшает сперматогенный ответ на ЛГ или ХГЧ,
В противоположность этому у мужчин, гонадотропная недостаточность у которых
231
появилась до пубертатного периода, только Л Г или ХГЧ обычно недостаточно для
начала сперматогенеза; у этих пациентов для инициации сперматогенеза необходимо
добавление ФСГ. Таким образом, ФСГ, по всей видимости, обязателен для программи¬
рования сперматогенного механизма во время полового созревания. После того как
это программирование произошло и сперматогенез был инициирован, Л Г в одиночку
может поддержать и реинициировать сперматогенез.
Большой ряд Y-специфичных, а также аутосомных генов вовлечен в регуляцию
развития половых клеток в яичках. Хотя количество генов, про которые известно,
что они вовлечены в патофизиологию бесплодия у человека, небольшое, значительно
более крупные базы данных существуют для мышей и Drosophila. Так, показано, что
локусы 2400 Drosophila причастны к развитию бесплодия у мужчин. Дополнительные
аутосомные и X- и Y-специфичные гены-кандидаты, которые ассоциированы с дефек¬
тами репликации зародышевых клеток, мейоза или сперматогенеза, вероятно, еще
будут выявлены у бесплодных мужчин.
Роль Y-хромосоиы в сперматогенезе человека
Человеческая Y-хромосома состоит из маленькой псевдоаутосомной области, реком¬
бинирующей с гомологичной областью Х-хромосомы; оставшиеся 95% Y-хромосомы
(приблизительно 65 Mb), которые не рекомбинируют с Х-хромосомой, называют специ¬
фическим мужским регионом Y-хромосомы. Page и соавт. установили, что мужеспе¬
цифическая область Y-хромосомы содержит 156 транскрипционных блоков, которые
кодируют 27 белков [49]. Эти транскрипционные блоки сгруппированы в три класса
эзгхроматиновых последовательностей; Х-транспозированная, Х-дегенерированная
и ампликоновая {ampliconic) (рис. 3-7) [49]. Ампликоновая область организована в
крупные палиндромы, которые содержат несколько семейств мультикопированных
Y-специфичных генов, все из которых экспрессируются в яичках [49]. Высокая степень
аналогичности последовательностей между плечами палиндромов — результат конвер-
Специфический
мужской регион
■ Х-транспозированный Ш Ампликоновый В Псевдоаутосомный
□ Х-дегенерированный ^ Гетерохроматиновый □ Другой
Рис. 3-7. Схематическое представление Y-хромосомы человека. Она состоит из маленького псевдоау-
тосомного региона, который рекомбинирует с гомологичными областями Х-хромосомы. Оставшиеся
95% Y-хромосомы (приблизительно 65 Mb), которые не рекомбинируют с Х-хромосомой, называют
специфическим мужским регионом Y-хромосомы. Раде и соавт. установили, что мужеспецифический
регион Y-хромосомы содержит многие транскрипционные блоки, которые сгруппированы в три класса:
Х-транспозированный, Х-дегенерированный и ампликоновый [49]. Ампликоновые области содержат
девять семейств мультикопийных Y-специфичных генов, все из которых экспрессируются в яичках
[35]. Х-транспозированные последовательности имеют высокую степень гомологии с Х-хромосомой,
Х-дегенерированные последовательности представляют собой то, что осталось от древних ауто-
сом, которые были транспозированы для формирования X- и Y-хросомосом. Yp — короткое плечо
Y-хромосомы. Yq — длинное плечо Y-хромосомы. [Из Skaletsky Н., Kuroda-Kawaguchi Т., Minx P. et al. The
male-specific region of the Y-chromosome is a mosaic of discrete sequence classes // Nature. — 2003. — N
423. - P. 825-837.]
232
сий интра-У-генов [50]. Page и соавт. [49] предполагают, что конверсии интра-У-генов
не только возможны, но и происходят достаточно часто.
Х-транспозированные последовательности имеют высокую степень гомологии
с Х-хромосомой, а Х-дегенерированные последовательности представляют то, что
осталось от древних аутосом, которые были транспозированы для формирования
X- и У-хромосом [49]. Таким образом, выходит, что У-хромосома содержит большое
количество яичкоспецифических генов, необходимых для сперматогенеза [49. 50], и
является далеко не «отстойной свалкой» для «старых» аутосомных генов [49].
ЭНЕРГЕТИЧЕСКИЙ БАЛАНС И РЕПРОДУКТИВНАЯ ФУНКЦИЯ
Нормальная репродуктивная функция требует оптимального режима питания;
недостаточная энергетическая ценность и последующее снижение массы тела, так же
как избыточный прием пищи и ожирение, ассоциируются с нарушением репродуктив¬
ной функции. Состав тела, особенно количество жировой ткани в организме, влияет
на начало полового созревания, продолжительность репродуктивного периода,
количество потомков и возраст менопаузы [51-53]. Снижение массы тела, вызван¬
ное голоданием, расстройствами питания и физической нагрузкой, может нарушить
репродуктивную функцию [54-60]. У женщин потеря массы тела ассоциируется с
задержкой полового созревания, прекращением менструаций и сниженной секрецией
гонадотропинов [52, 61-69]; снижение уровней ФСГ и ЛГ коррелирует со степенью
потери массы тела.
ПУТИ, КОТОРЫЕ СВЯЗЫВАЮТ ЭНЕРГОЧУВСТВИТЕЛЬНЫЕ МЕХАНИЗМЫ
И РЕПРОДУКТИВНУЮ ОСЬ
На рубеже XXI в. стало очевидным значительное продвижение нашего понимания
биохимических путей, которые связывают репродуктивную систему с энергетическим
балансом (рис. 3-8). Метаболические сигналы, регулирующие секрецию гонадолибе¬
рина гипоталамусом, опосредованы через лептин и нейропептид Y [58,70]. Лептин -
гормон, секретируемый жировыми клетками, — регулирует активность эффекторной
системы ЦНС, которая поддерживает энергетический баланс [70-73]. Энергетическая
нехватка обусловлена пониженными уровнями лептина, так же как и Л Г [74, 75].
Назначение лептина людям и мышам с нехваткой энергетической ценности рациона
устраняет ингибирование секреции гонадотропинов, связанной с пищевым ограниче¬
нием [76,77]. Аналогично генетические ob/ob мыши с недостаточностью лептина [78]
и люди с врожденной лептиновой недостаточностью страдают гипогонадотропным
гипогонадизмом и бесплодием [79]; лечение пациентов и мышей с дефицитом лепти¬
на восстанавливает секрецию гонадотропинов и фертильность [80]. Таким образом,
нарушенная секреция гонадолиберина, ассоциированная с энергетическим дефици¬
том и потерей массы тела, — по меньшей мере частично результат сниженной секре¬
ции лептина. Несомненно, лептин — важная связь между энергетическим балансом и
репродуктивной осью, хотя неясно, служит ли он основным активатором для начала
продукции гонадолиберина в начале полового созревания. Появляющиеся данные
позволяют предполагать, что лептин обязателен, но недостаточен для инициации
полового созревания.
Лептин стимулирует секрецию ЛГ путем активации синтазы оксида азота в клетках,
образующих гонадотропины [81]. Лептин также ингибирует секрецию нейропептида
Y [82]. Нейропептид Y оказывает тонический ингибирующий эффект на секрецию
лептина и гонадолиберина [83, 84]. Лептин также стимулируех^продукцию оксида
азота в медиобазальном гипоталамусе [81]; оксид азота стимулирует секрецию гона¬
долиберина гипоталамическими гонадолиберинсекретирующими нейронами [81].
Именно поэтому итоговым эффектом действия лептина посредством стимуляции
оксида азота и ингибирования нейропептида Y служит стимуляция гипоталамической
секреции гонадолиберина и гипофизарной секреции ЛГ.
233
Рис. 3-8.*Биохимические пути, которые связывают энергетический гомеостаз и центральный контроль
репродукции. Биохимические сигналы, свидетельствующие о состоянии энергетических запасов, проис¬
ходят из адипоцитов и опосредованы через лептин и нейропептид У. Лептин (продукт оЬ-гена) секре¬
тируется адипоцитами и регулирует секрецию гонадолиберина путем снижения нейропептида V (NPY),
который, в свою очередь, уменьшает секрецию гонадолиберина. Конечный эффект лептина заключается
в стимуляции выброса гонадолиберина. Кроме того, лептин также стимулирует секрецию ЛГ через меха¬
низм, опосредованный оксидом азота. Было показано, что грелин и пептид YY, которые образуются в
ЖКТ, также регулируют секрецию гонадотропина. Кисспептин, естественный лиганд рецептора С-белка
54, объединяет сигналы от поступления питательных веществ и энергетических запасов и передает их
нейронам, секретирующим гонадолиберин, и клеткам, образующим гонадотропины. Таким образом,
сложный комплекс сигналов, исходящих из энергетических запасов, ЖКТ и других неизвестных мест,
объединяются в ЦНС для регуляции секреции гонадотропина и поступления и расхода энергетических
запасов. КРГ — кортикотропин-рилизинг-гормон; срСГ — фолликулостимулирующий гормон; СРВ54 —
рецептор, связанный с С-белком; ПОМК — пропиомеланокортин.
Грелин — естественный лиганд рецептора, усиливающего секрецию гормона роста
[85], — был предложен в качестве дополнительного звена между энергетическим
дефицитом и репродуктивной функцией [86-88]. Грелин, высвобождающийся из
эндокринных клеток в ЖКТ [89, 90], ингибирует секрецию ЛГ, ответ ЛГ на гонадо¬
либерин и ответ Т на ЛГ [86, 87] независимо от его эффектов на аппетит и секрецию
гормона роста [89-91]. Полипептид ¥¥3-36 является другим гормоном желудочно-
кишечного происхождения; он связывается с подтипами ¥2 и ¥5 рецептора нейропеп¬
тида [92], угнетает прием пищи, стимулирует секрецию ЛГ и ФСГ и усиливает ответ
ЛГ на гонадолиберин.
234
Недавно клонированный рецептор 54, связанный с С-белком, играет важную роль
в интеграции сигналов от путей энергетического гомеостаза для регуляции секреции
гонадотропина. Пищевая депривация ассоциирована со снижением кисспептина
(лиганда к рецептору 54, связанному с С-белком) и увеличением экспрессии матрич¬
ной РНК этого рецептора у крыс [93]. В модели пониженного питания постоянное
назначение кисспептина вызывает секрецию гонадотропинов и восстановление
открытия влагалища [93]. В совокупности эти данные позволяют предположить, что
гормоны, происходящие из ЖКТ и энергетических запасов, действуют совместно с
сигналами от рецептора 54, связанного с С-белком, в плане обеспечения важных сиг¬
налов для центральной регуляции репродуктивной функции.
ЗАБОЛЕВАНИЯ ЯИЧЕК
Заболевания яичек можно классифицировать на расстройства продукции или
действия половых гормонов, нарушения отдела половых клеток и новообразования
яичек. Андрогенная недостаточность и гинекомастия — наиболее частые расстрой¬
ства продукции половых гормонов. Дефекты действия андрогенов включают мута¬
ции андрогенового рецептора, стероид-5а-редуктазы, CYP19 ароматазы или генов
а-рецептора эстрогенов. Дефекты сперматогенеза и функции спермы проявляются в
виде бесплодия и пониженной фертильности. Описаны также клинические послед¬
ствия злоупотребления андрогенами спортсменами и бодибилдерами.
Синдром андрогенной недостаточности у мужчин
Гипогонадизм — синдром, ассоциированный с нарушенной продукцией или дей¬
ствием андрогенов. Андрогенная недостаточность может быть результатом аномалий
тестикулярной функции (первичный гипогонадизм), гипоталамической или гипофи¬
зарной регуляции тестикулярной функции (вторичный гипогонадизм) [94] или нару¬
шения действия андрогенов в тканях-мишенях (нечувствительность к андрогенам).
Первичный гипогонадизм характеризуется низким уровнем Т, нарушением спер-
маї'огенеза и повышенной концентрацией ЛГ и ФСГ [94\. Вторичный гипогонадизм
происходит в результате патологии гипоталамуса или гипофиза и ассоциирован с
низким содержанием Т и низкими или неадекватно нормальными концентрация¬
ми ЛГ и ФСГ [94]. Классификация мужчин с дефицитом андрогенов на пациентов
с первичной и вторичной тестикулярной недостаточностью клинически полезна,
поскольку фертильность у мужчин с вторичным гипогонадизмом можно индуциро¬
вать путем назначения гонадотропинов или пульсирующей гонадолибериновой тера¬
пии [94]. Мужчины с первичной тестикулярной недостаточностью, напротив, вряд
ли достигнут спонтанной фертильности, а варианты решения проблемы включают
усыновление, искусственную инсеминацию донорской спермой или в редких случаях
внутрицитоплазматическую инъекцию сперматозоида (ICSI) с использованием тести¬
кулярной спермы (ТЕСА, ПЕСА).
ПРИЧИНЫ АНДРОГЕННОЙ НЕДОСТАТОЧНОСТИ У МУЖЧИН
Состояния, ассоциированные с низкими уровнями Т, перечислены в табл. 3-1.
Причины первичного гипогонадизма
Типичными причинами первичной тестикулярной недостаточности служат син¬
дром Клайнфелтера, неоткорригированный крипторхизм, химиотерапия и лучевая
терапия при злокачественных опухолях, травма и орхидэктомия.
235
Таблица 3-1. Состояния, ассоциированные с низкими уровнями тестостерона у мужчин
Типичные
Реже встречающиеся
Редкие
Первичный гипогонадизм
Синдром Клайнфелтера.
Некорректированный крипторхизм
Радиация
Химиотерапия
Орхит
Травма
Анорхия, обусловленная двусторон¬
ним перекручиванием или синдро¬
мом исчезающих яичек
Лекарственные средства
Синдром Нунана.
Миотоническая дистрофия.
Мутации рецептора Л Г или его
субъединиц.
Тип 1 аутоиммунного полигландуляр-
ного синдрома
Вторичный гипогонадизм
Хронические заболевания
(ВИЧ, хроническая обструктивная
болезнь легких, терминальная
почечная недостаточность).
Критическое заболевание.
Г иперпролактинемия.
Лекарственные средства (например,
опиаты, анаболические гормоны)
Опухоли гипоталамо-гипофизарной
области
Использование наркотиков, таких
как алкоголь и марихуана
Повреждение головы
Хирургическое вмешательство
Инфильтративные процессы (напри¬
мер, гемохроматоз, саркоидоз,
гистиоцитоз)
Тяжелое ожирение
Избыточная физическая нагрузка
Расстройства питания
Идиопатический гипогонадотропный
гипогонадизм, синдром Калльмана.
Инфекции (например, туберкулез).
Апоплексия гипофиза.
Синдром Прадера-Вилли
Смешанные формы
Возраст
Алкоголизм
Гемохроматоз
Серповидноклеточная анемия
Врожденная гипоплазия надпочечни¬
ков (мутации ОАХ-7)
Синдром Клайнфелтера
Синдром Клайнфелтера — наиболее частое хромосомное заболевание, обуслов¬
ленное первичной тестикулярной недостаточностью и мужским бесплодием, которое
выявляют у 1:500-1:1000 живорожденных мальчиков [95-97]; оно является наи¬
более узнаваемой причиной первичной тестикулярной недостаточности. Наиболее
распространенный кариотип у мужчин с синдромом Клайнфелтера — 47,ХХ¥ (93%),
однако были сообщения о кариотипах 46,ХУ/47,ХХУ, 48,ХХХУ, 48,ХХУУ и 49,ХХХХУ
[98]. ХХУ-хромосомное строение было описано у других млекопитающих, включая
мышей, китайских хомяков, кошек, собак, овец, быков и свиней, оно было связано с
бесплодием [99]. Семенники 47,ХХУ-млекопитающих лишены половых клеток [99].
Азооспермия, как правило, характерна для мужчин с синдромом Клайнфелтера
(47,ХХУ) [98, 100]. Мужчины с мозаицизмом могут иметь половые клетки в яичках,
особенно в молодом возрасте.
При гистологическом исследовании гонад мужчин с синдромом Клайнфелтера
отмечают гиалинизацию семенных канальцев и отсутствие сперматогенеза [98, 100].
Пациенты с мозаицизмом могут иметь яички нормальных размеров и сперматогенез в
пубертатном периоде. Однако вскоре после полового созревания происходит прогрес¬
сивная дегенерация и гиалинизация семенных канальцев [100]. У некоторых мужчин
дисгенез канальцев неоднороден; дегенерированные канальцы перемежаются с явно
нормальными. Клетки Лейдига выглядят увеличенными, хотя их функция нарушена
[100].
Кариотип 47,ХХУ у пациентов с синдромом Клайнфелтера образуется в результате
нерасхождения хромосом во время первого мейотического деления у одного из роди¬
236
телей. Отсутствие расхождения материнских хромосом — причина 47,XXY кариотипа
у двух третей больных мужчин. Увеличение возраста матери является фактором риска
заболевания [97.101]. Механизм нерасхождения Х-хромосомы, которое делает паци¬
ентов бесплодными, неизвестен. В мужских половых клетках инактивация единичной
Х-хромосомы в первичных сперматоцитах гетерогаметных мужчин необходима для
прохождения клеток через мейоз. Неизвестно, почему инактивация Х-хромосомы
служит обязательным фактором для дифференциации мужских половых клеток у
гетерогаметных видов; возможно, что экспрессия некоторых Х-связанных генов
вредна для сперматогенеза. Также инактивация единичной Х-хромосомы может тре¬
боваться для нормального образования пары половых хромосом.
Большинство мужчин с синдромом Клайнфелтера живут без постановки диагноза
[102]. У мальчиков с синдромом Клайнфелтера диагноз, скорее всего, установят при
обследовании на предмет задержки развития и проблем поведения [102]. Мужчины
с синдромом Клайнфелтера обычно попадают в поле зрения во время обследования
на предмет гипогонадизма и бесплодия [98, 102]. Дети с синдромом Клайнфелтера
имеют нормальное половое развитие в пубертате [103]. Биопсии гонад у мальчиков
с данным диагнозом на различных стадиях полового развития показывают, что на
ранних этапах в яичках имеются диплоидные половые клетки, но они утрачиваются
во время полового созревания [104,105].
По сравнению с общей популяцией мужчины с синдромом Клайнфелтера имеют
более высокую смертность от рака легкого, грудной железы и неходжкинской лимфо-
мы [106] и более низкую — от рака предстательной железы [107,108]. Особенно высок
относительный риск рака грудной железы и неходжкинской лимфомы [107,108].
Гонадная токсичность химиотерапии при злокачественных опухолях
Дисфункция половых желез возникла как важное позднее осложнение химиотера¬
пии при раке [109]. Это особенно актуально для гематологических и тестикулярных
новообразований, при которых комбинированная химиотерапия значительно увели¬
чила показатели выживаемости. Поскольку эти пациенты обычно молоды и прогноз
исхода заболевания после лечения благоприятен, бесплодие является значимым
результатом для пациентов, выживших после перенесенного рака. Кроме того, сни¬
женные уровни Т могут вызвать повышенную утомляемость, сексуальную дисфунк¬
цию, измененное настроение, потерю мышечной массы и пониженную минеральную
плотность костной ткани [110, 111]. Хотя сообщалось о повышенной частоте анеу-
плодии и хромосомных аномалий у мужчин, получавших химиотерапию по поводу
злокачественного новообразования [112,113], в настоящее время не выявлено сз^^ще-
ственного повышения частоты дефектов у новорожденных [114].
Компартмент половых клеток показывает более высокую чувствительность к
повреждению противоопухолевыми химиотерапевтическими средствами; в то же
время сообщали о дисфункции клеток Лейдига [109, 115]. Эффекты химиотерапии
на тестикулярную функцию зависят от типа химиотерапевтического агента, куму¬
лятивной дозы и природы злокачественного новообразования [109, 1006, 117].
Алкилирующие агенты циклофосфамид и прокарбазин особенно токсичны для яичек.
У 90% пациентов, леченных комбинацией химиотерапевтических средств (мустин,
винбластин, прокарбазин и преднизолон или мустин, винкристин, прокарбазин
и преднизолон), содержащих прокарбазин, развивается азооспермия [118, 119],
Намного меньшая часть содержит низкие концентрации Т и приблизительно 20-25%
имеют нормальные уровни Т с повышенными концентрациями ЛГ, предполагающими
умеренную дисфункцию клеток Лейдига. Новейшие химиотерапевтические режимы,
такие как ABVD, которые не включают прокарбазин, ассоциированы с более низкой
частотой азооспермии, чем режимы, содержащие прокарбазин, такие как MOPP
[120]. Высокодозовая химиотерапия, используемая перед и во время трансплантации
костного мозга, также ассоциирована с необратимым повреждением герминативного
237
эпителия, олигоспермией или азооспермией и повышенными уровнями ФСГ у боль¬
шинства леченых мужчин 1121J.
Прогноз для фертильности и нормальной функции гонад лучше в отношении муж¬
чин с тестикулярным раком, которые получали комбинации, содержащие платину;
хотя у многих из этих пациентов после получения химиотерапии отмечают снижение
концентрации спермы, 80% мужчин, выживших в течение 5 лет, имеют нормальную
концентрацию сперматозоидов в эякуляте [122]. Частота низкой концентрации Т
и повышенного содержания ЛГ у пациентов с длительной выживаемостью после
тестикулярного рака также низкая [122]. У пациентов, леченных высокими дозами
интерлейкина-2 по поводу метастазов рака, могут быть транзиторные снижения
уровня Т. Частота тестикулярной дисфункции после химиотерапии у мальчиков в пре-
пубертатном периоде и взрослых значительно не различается.
Изучали несколько стратегий предупреждения повреждения половых клеток во
время химиотерапии по поводу рака, но ни одна из них не эффективна у человека.
Именно поэтому криоконсервация эякулята для последующего использования в
циклах ЭКО стала стандартной практикой для мужчин, подвергающихся химиотера¬
пии по поводу злокачественного новообразования, и должна быть предложена паци¬
ентам. Терапевтические подходы, основанные на супрессии тестикулярной функции
путем введения аналогов гонадолиберина или энантата Т, не были эффективными в
предупреждении повреждения половых клеток у мужчин, получающих MOPP- или
MVPP-режимы [123,124]. Аутологическую имплантацию стволовых клеток в яички
для репопуляции отделения половых клеток и восстановления сперматогенеза после
завершения химиотерапии рассматривают и считают обещающей.
Травма яичек
Из-за локализации за пределами живота яички особенно уязвимы для травмы.
Тестикулярная атрофия может развиться почти у 50% мужчин, перенесших тупую
травму мошонки [125].
Инфекционный орхит
Инфекционное поражение яичек эпидемическим паротитом, ЕСНО-вирусами,
группой В арбовирусов и лимфоцитарным хориоменингитом может привести к тести¬
кулярной атрофии. В последние годы заболеваемость эпидемическим паротитом сни¬
зилась, орхит осложняет течение этого заболевания приблизительно у 25% мужчин,
обычно на 4-8-е сут после начала паротита [126]. У трети пациентов с орхитом при
эпидемическом паротите постепенно развивается тестикулярная атрофия в течение
нескольких месяцев после разрешения острого периода заболевания [126]. Орхит
может поражать только одно яичко более чем у 50% пациентов; у мужчин с односто¬
ронним орхитом концентрация эякулята обычно возвращается к норме в течение 1-2
лет после инфекции [126].
ВИЧ-инфекция
В ранний период эпидемии ВИЧ до внедрения антиретровирусной лекарственной
терапии у 40-50% мужчин, пораженных ВИЧ, снижались уровни Т [127]. Недавние
обзоры свидетельствуют, что даже среди ВИЧ-инфицированных мужчин, получаю¬
щих высокоактивную антиретровирусную терапию, 20-30% имеют низкий уровень
общего или свободного Т [128,129]. Уровень глобулина, связывающего половые гор¬
моны, обычно повышен у ВИЧ-инфицированных мужчин. В то же время у 80% ВИЧ-
инфицированных мужчин с низким содержанием Т уровни ЛГ и ФСГ нормальные,
20% — повышенные [128]. Таким образом, патофизиология низкого содержания Т у
ВИЧ-инфицированных мужчин комплексная, и за низкое содержание этого гормона
ответственны дефекты на всех уровнях гипоталамо-гипофизарно-тестикулярной
оси. При тестикулярных биопсиях ВИЧ-инфицированных мужчин выявили утрату
архитект>’ры семенных канальцев, гиалинизацию, потерю половых клеток и инфиль¬
трацию мононуклеарными клетками [130]. Низкие уровни Т у ВИЧ-инфицированных
238
мужчин ассоциированы с потерей массы тела, ускоренным прогрессированием болез¬
ни, снижением мышечной ткани и возможностью совершать физические нагрузки,
а также с увеличенным счетом по шкале депрессии [131]. Снижение биодоступных
уровней Т предшествует началу истощения [132].
Анорхия (синдром исчезающих яичек)
Анорхией называют отсутствие тестикулярной ткани у мужчины с фенотипом
46,XY. Эти пациенты имеют нормальные внешние мужские половые органы, нормаль¬
ные структуры вольфова протока и полное отсутствие структур мюллерова протока
[133,134]. Это свидетельствует о том, что у этих лиц во время критических периодов
половой дифференцировки должны были присутствовать яички, Т и Мюллеров инги¬
бирующий гормон. Патогенетические факторы, которые служат причиной атрофии и
исчезновения яичек, неизвестны. Семейные случаи и связь анорхии с 46,XY гонадной
дисгенезией показывают, что в процесс могут быть вовлечены генетические факторы,
ассоциированные с детерминацией или опущением яичек [135,136]. Однако мутаций
SRY, INSL3, и LGR8 (рецептор к INSL3) не было выявлено при анализе небольших
групп пациентов с анорхией [137]. Сообщали, что распространенность двусторонней
анорхии составляет среди мужчин 1:20 ООО и 1:5000 — односторонней анорхии [134],
но возможно, что показатель распространенности выше (из-за того, что некоторые
случаи анорхии могут быть ошибочно расценены как крипторхизм).
У мужчин с кариотипом 46,XY и отсутствием при пальпации яичек необходимо
провести системную работу по локализации яичек. Определение содержания мюл¬
лерова ингибирующего гормона, тест на стимуляцию ХГЧ с измерением концентра¬
ции Т, МРТ живота помогут определить локализацию внутрибрюшных яичек. Если
результаты теста со стимуляцией ХГЧ и МРТ не подтверждают присутствие внутри¬
брюшных яичек, может потребоваться лапароскопическое исследование или в неко¬
торых случаях диагностическая лапаротомия для локализации яичек, их репозиции и
закрепления в мошонке.
Инактивирующие мутации гена рецептора ЛГ
Инактивирующие мутации гена рецептора ЛГ ассоциируются с гипогонадизмом и
гипоплазией клеток Лейдига. Имеются сообщения о ряде семей с резистентностью к
действию ЛГ, обусловленной инактивирующими мутациями рецептора ЛГ [138-148],
Мужчины с мутациями рецептора ЛГ обращаются с целым спектром фенотипических
аномалий, варьирующих от феминизации внешних половых органов у 46,XY-мужчин
до гипоплазии клеток Лейдига, первичного гипогонадизма и задержки полового раз¬
вития [138-148]. При гистологическом исследовании яичек фенотипических мужчин
с мутациями рецептора ЛГ обычно выявляют отсутствие зрелых клеток Лейдига и
остановку сперматогенеза на стадии удлиненных сперматид, 46,XX-Женщины с мута¬
цией рецептора ЛГ обычно проходят нормальное развитие вторичных половых при¬
знаков, но имеют повышенные уровни ЛГ и аменорею.
Инактивирующие мутации гена рецептора ФСГ
Мутация гена-рецептора ФСГ (2р21) ассоциирована с гипергонадотропной дисге¬
незией яичников [149-153]. Обширное исследование многочисленных больных фин¬
ских семей выявило С566Т перемещение, обусловливающее Alal89Val-замену в гене
рецептора ФСГ, которое было ассоциировано с первичной аменореей, остановкой
фолликулярного развития и бесплодием [149-153]. Экспрессия гена в трансфектных
клетках показало снижение проведения сигнала или отсутствие проведения. По срав¬
нению с пациентами с дисгенезией яичников, которые не имеют мутации, пациенты
с мутацией рецептора ФСГ ниже ростом и имеют больше яичниковых фолликулов. В
отличие от пациенток больные мужского пола с этой мутацией были фертильными,
но имели сниженное количество сперматозоидов. Популяционные исследования
обнаружили эту мутацию у 1% финнов с увеличением больных в различных геогра¬
фических областях, предполагающим влияние первородителей популяции [151].
239
другие тестикулярные заболевания, ассоциированные с первичной тестику¬
лярной недостаточностью
Гипогонадизм может развиться как часть 1-го, 2-го или 4-го типа аутоиммунного
полиэндокринного синдрома [154]. Эти пациенты типично имеют низкие концентра¬
ции Т и повышенные уровни ЛГ и ФСГ. Сообщали об антителах к 17а-гидроксилазе
и ферменту, отщепляющему CYP боковую цепь, у мужчин с аутоиммунным полиэндо-
кринным синдромом и низкими уровнями Т [154].
Мужчины с синдромом Дауна (трисомия 21) могут иметь повышенные уровни ЛГ
и ФСГ и сниженный сперматогенез.
Причины вторичного гипогонадизма
Гиперпролактинемия, хронические заболевания, старение, использование опиои-
дов и других наркотических средств, расстройства питания и опухоли гипофиза —
наиболее частые причины вторичного гипогонадизма.
Гиперпролактинемия может индуцировать андрогенную недостаточность раз¬
личными механизмами. Большинство исследований механизмов пролактин-ассо-
циированного гипогонадизма проведено у женщин, и данные были экстраполированы
на мужчин. Частота импульсов выделения ЛГ ниже у пациентов с гиперпро л актине-
мией, она нормализуется при терапии бромокриптином [155,156]; также было пока¬
зано, что пролактин ингибирует выделение гонадолиберина в линии нейрональных
клеток [157]. Эти сведения привели к гипотезе, что основным механизмом действия
пролактина служит супрессия гипоталамической секреции гонадолиберина [157].
Амплитуда импульсов ЛГ не снижена у пациентов с гиперпролактинемией [155], хотя
у крыс с удаленными яичниками высокие концентрации пролактина ингибируют
частоту импульсов ЛГ, а также их амплит}^ду. Появилось предположение, что пролак¬
тин может также влиять на точку отсчета для гонадостата (механизма, ответственного
за стабильность гонадотропной функции). Имеются некоторые данные, хотя и не
окончательные, что пролактин может также угнетать ответ гонад на гонадотропины.
У пациентов с пролактинсинтезирующими гипофизарными макроаденомами также
может развиться гипогонадотропный гипогонадизм из-за эффекта «опухолевой
массы» на гонадотропные клетки.
Общий, свободный и биодоступный Т и дегидроэпиандростерон в сыворотке
крови ниже у пожилых, нежели у молодых мужчин, даже после учета потенциальных
неблагоприятных факторов, таких как время получения образца, сопутствующие
заболевания, лекарственные средства и технические особенности, связанные с мето¬
дом определения гормонов [158-164]. Старение ассоциировано с аномалиями на
всех уровнях гипоталамо-гипофизарно-тестикулярной оси; ответ Т на ЛГ и ответ ЛГ
на гонадолиберин ослаблены у пожилых лиц по сравнению с молодыми мужчинами.
Кроме того, взаимоотношения по типу обратной и прямой связи между гипотала¬
мусом, гипофизом и яичками искажены [165-168]. Повышения концентрации ЛГ у
пожилых мужчин ниже тех значений, которые предсказаны из степени снижения кон¬
центраций Т в сыворотке крови, что отражает как ослабление гипофизарной функции,
так и переустановку гонадостата [169,170]. Таким образом, у многих пожилых муж¬
чин с возрастным снижением концентраций Т сывороточные уровни ЛГ либо в норме,
либо слегка повышены.
Применение опиоидов пациентами с болью злокачественного или неканцероге-
незного характера или героиновыми наркоманами и пациентами на метадоновой
поддержке ассоциировано с центральным гипогонадизмом, характеризующимся
низкими уровнями Т и низкими концентрациями ЛГ и ФСГ [171-175]. Степень
тестостероновой супрессии связана с дозой опиоидов и типом применяемого опиоида
[171]. Сообщали, что бупренорфин вызывает супрессию гонад в меньшей степени,
чем метадон [176]. Механизмы, посредством которых опиоиды угнетают функцию
240
гонад, полностью не изучены, однако известно, что они включают супрессию секре¬
ции гонадолиберина [177J. Veldhuis предположил, что опиоиды изменяют чувстви¬
тельность центральных нейронов, секретиругощих гонадолиберин, по типу обратной
связи эндогенным Т, «переустанавливая», таким образом, гонадостат на более низкие
концентрации Т [178-180]. Супрессия Т опиоидами имеет клинические последствия,
поскольку это ассоциировано с нарушением сексуальной функции, остеопорозом,
повышенной утомляемостью, беспокойством и депрессией, потерей мышечной массы
и силы, сниженным качеством жизни [171,174].
Генетические синдромы, ассоциированные с гипоталамическим гипогонадиз¬
мом >
У мужчин, страдающих гипогонадотропным гипогонадизмом, в отношении кото¬
рых обнаруживаемые причины гонадотропной недостаточности были исключены,
специфические гипоталамические синдромы могут быть распознаны с помощью ассо¬
циированных соматических стигм [181]. Ряд гипоталамических синдромов харак¬
теризуются гипогонадотропным гипогонадизмом в сочетании с дисм орфическими
соматическими признаками, такими как подковообразная почка, нейросенсорная
тугоухость, выраженное ожирение, гиперфагия, полидактилия, пигментозный рети¬
нит и задержка умственного развития [181,182]. Диагноз данных гипоталамических
синдромов устанавливают путем распознавания картины синдрома.
Мужчины с гипогонадотропным гипогонадизмом при отс>тствии обнаруживаемой
причины диагностируют идиопатический гипогонадотропный гипогонадизм (ИГГ),
представляют собой гетерогенную группу заболеваний, характери.зующихся селек¬
тивной гонадотропной недостаточностью, возникающей в результате изолированного
дефекта секреции гонадолиберина [182-184]. Первичный патогенетический дефект у
этих пациентов является гипоталамическим, а нарушенная секреция гонадотропинов
вторична по отношению к гипоталамической аномалии в секреции гонадолиберина.
Хотя аносмия и гипосмия — первые и наиболее известные симптомы, описанные
приданном синдроме, были отмечены другие соматические аномалии [181]. Наиболее
типичные сочетанные нарушения включают цветовую слепоту, расщепленные губу и
нёбо, дефекты черепных нервов (включая ГЛ3ГХ0Т}» вследствие поражения 8 нерва),
подковообразные почки, крипторхизм и атрофию зрительного нерва.
Гетерогенность пульсирующей секреции гонадотропина у пациентов с идиопатигеским
гипогонадотропным гипогонадизмом
Существует значительная гетерогенность клинического проявления ИГГ [182,184].
Лица с наиболее тяжелым дефицитом могут жаловаться на полное отсутствие полово¬
го развития, половой инфантилизм и в некоторых случаях различные степени гипо-
спадии и неопущения яичек. Пациенты мужского пола могут иметь полное отсутствие
вторичных половых признаков, инфантильные гонады и азооспермию. Пациенты
женского пола могут обращаться с первичной аменореей. Пациенты с частичным
дефицитом гонадолиберина могут иметь различные степени задержки полового раз¬
вития пропорционально тяжести гонадотропной недостаточности.
Пациенты с ИГГ полностью гетерогенны по их профилю секреции ЛГ [184],
Наибольшую подгруппу составляют пациенты, у которых совсем нет пульсирующей
секреции ЛГ; эти лица имеют наиболее тяжелую гонадолибериновую недостаточ¬
ность. Меньшая подгруппа имеет низкоамплитудные импульсы секреции. Другая
часть пациентов имеет импульсы секреции Л Г с выраженно сниженной частотой.
Оставшаяся, четвертая подгруппа характеризуется преимущественно импульсами во
сне {sleep-entrained), напоминающими картину ранних стадий полового созревания:
эту ситуацию можно рассматривать как форму «задержки развития».
Два варианта ИГГ особенно интересны. Термин «синдром фертильного евнуха»
был использован при описании пациентов, которые имели пропорции евнухов и
задержку полового развития, но яички нормальных размеров. Такие мужчины, веро¬
241
ятно, имеют достаточно гонадотропинов для стимулирования высоких интратести-
кулярных уровней Т и для инициации сперматогенеза, но недостаточную секрецию
Т в кровь для адекватной вирилизации периферических тканей; они фактически
страдают частичной гонадотропной недостаточностью [185]. Другой вариант с преоб¬
ладанием фолликулостимулирующей недостаточности также был описан, хотя такие
случаи довольно редки [186].
Генетика идиопатигеского гипогонадотропного гипогонадизма
Треть пациентов с ИГГ имеют отягощенный семейный анамнез [183,187]. Из этих лиц
в среднем 20% имеют Х-сцепленный характер наследования, порядка 30% — аутосомно-
рецессивный и 50% — аутосомно-доминантный тип наследования [183,187].
Локус для Х-сцепленной формы синдрома Калльмана был определен в хромосомной
области Хр22.3 [188,189]. Две группы независимо клонировали из этой области проте¬
ин, кодируемый геном KALIG-l (Kallmann’s syndrome interval-l — интервал-1 синдрома
Калльмана) [188,190]. Белковый продукт гена KALIG-1 предположительно регулирует
миграцию гонадолибериновых и обонятельных нейронов и их морфогенез. Мутации в
гене KALIG-1 были обнаружены у небольшой группы пациентов с Х-сцепленной фор¬
мой ИГГ [191]. Кроме того, вероятно, в патогенез других подгрупп синдрома Калльмана
вовлечены пока еще не идентифицированные Х-сцепленные гены.
Гонадолибериновые нейроны впервые появляются в эпителии обонятельных
плакод у эмбриона мыши и далее мигрируют в передний мозг и затем окончательно
в их конечную гипоталамическую локализацию [192, 193]. Подобные наблюдения
позволяют предполагать, что ИГГ может быть дефектом развития, возникающим в
результате нарушенной миграции нейронов, содержащих гормон, высвобождающий
ЛГ. МРГ-исследования показали: обонятельные луковицы борозды плохо развиты у
пациентов с ИГГ, которые страдают аносмией и гипосмией [194].
Были также сообщения о связи ИГГ и аносмии с мутациями гена рецептора 1 фак¬
тора роста фибробластов (FGFR1) [195]. Родственники с FGPRI-мутациями демон¬
стрируют аутосомно-доминантное наследование [195].
Мутации генов DAX-1 и SF1, ассоциированные с гипогонадотрофигеским гипогонадиз¬
мом и врожденной гипоплазией надпогегников
Мутации по меньшей мере двух локусов (NR0B1, который кодирует ген DAX-1,
и NR5A1, который кодирует ген стероидогенного фактора-1) были ассоциированы
с врожденной гипоплазией надпочечников и гипогонадотропным гипогонадиз¬
мом [196], Продуктом гена DAX-1 служит ядерный рецептор [197, 198]. Мутации
С-терминального конца гена DAX-1 были ассоциированы с Х-сцепленным гипого¬
надотропным гипогонадизмом и недостаточностью надпочечников (врожденной
гипоплазией надпочечников). Однако мутации гена DAX-1 — необычная причина
ИГГ (менее 1% случаев). Эти пациенты обычно имеют нормальный ответ Т на ХГЧ,
свидетельствуя о нормальной функции клеток Лейдига, хотя у некоторых пациентов
был описан первичный гипогонадизм.
Мутации гена стероидогенного фактора-1 у 46,XY-пациентов также ассоциированы
с врожденной гипоплазией надпочечников, половым изменением по женскому фено¬
типу, гонадами-стреками и гипогонадотропным гипогонадизмом [196]. Мыши с целе¬
направленным разрушением мышиного гена SF-1 характеризуются агенезией гонад,
женскими внутренними половыми органами и первичной надпочечниковой недоста¬
точностью [199]. Продуктом SF-1 гена служит рецептор, который взаимодействует с
ВАХ-1 для регуляции половой дифференцировки и развития надпочечников [200].
Мутации гена рецептора гонадолиберина, ассоциированные с гипогонадотропным гипо¬
гонадизмом
Были сообщения о нескольких семьях с гипогонадотропным гипогонадизмом,
обусловленным мутациями рецептора гонадолиберина [200]. Гонадолибериновый
242
рецептор — рецептор, связанный с С-белком, имеющий внеклеточный Ы-конец, семь
трансмембранных регионов и внутриклеточный С-конец [202]. Пациенты с мутация¬
ми рецептора гонадолиберина имеют нормальное обоняние, В этих семьях характер
наследования ИГГ аутосомно-рецессивный, часто имеется кровосмешение. Мужчины
и женщины среди пораженных лиц встречаются в равных количествах. Мутации
рецептора гонадолиберина составляют почти 40% случаев в семьях с аутосомно-
рецессивным типом наследования и 10% спорадических случаев. Мутации гена рецеп¬
тора гонадолиберина могут изменить связывание гонадолиберина или лигандиндуци-
рованное проведение сигнала [202].
Мутации рецептора 54, связанного с С-6елком
Мутации рецептора 54, связанного с С-белком (СРК54), ассоциированы с задерж¬
кой полового созревания и гипогонадотропным гипогонадизмом у людей и мышей
[203]. СРК54 — член родопсинового семейства рецепторов, связанных с С-белком,
который вместе с лигандом метастином играет важную роль в регуляции пульсирую¬
щей секреции гонадолиберина и начала полового созревания [204]. Метастины обра¬
зуются путем процессинга одного протеина-предшественника, кисспептина 1 [205];
метастин 1-54, более короткий С-терминальный фрагмент, метастин 45-54 служат
мощными стимуляторами выброса гонадолиберина и менее сильными стимулятора¬
ми выброса гипофизом ЛГ. Метастин-индуцированная стимуляция гонадолиберина,
вероятно, независима от лептина. Совпадая с началом полового созревания, экспрес¬
сия мРНК Кисс 1 и СРЕ54 усиливается в гипоталамусе [206].
Крупные делеции и перестановки гена гонадолиберина не были обнаружены у
пациентов с ИГГ посредством саузерн-блоттинга или секвенирования продуктов ПЦР,
Однако более тонкие дефекты в гене гонадолиберина не были полностью исключены.
Мутации назального эктодермального фактора гормона, высвобождающего ЛГ
Мутации назального эктодермального фактора гормона, высвобождающего ЛГ,
продукт гена которого вовлечен в миграцию нейронов, были отмечены у небольшо¬
го количества пациентов с ИГГ [207]. Аналогично задержка полового созревания и
гипогонадотропный гипогонадизм были ассоциированы с мутациями лептина или его
рецепторов [208, 209].
Известные мутации генов КАЫ0-1, рецептора гонадолиберина, СРЕ54, рецептора 1
фактора роста фибробластов и ВАХ-1 могут составлять только 30-40% случаев гипо-
гонадотропного гипогонадизма [208]. Именно поэтому, вероятно, будет установлена
роль в патогенезе других случаев ИГГ дополнительных аутосомных и X-сцепленных
генов.
Мутации гена ^-субъединицы ФСГ
Наследственные мутации генов р-субъедининицы ФСГ нетипичны, есть сообще¬
ния, что они вызывают гипогонадизм у представителей мужского пола и задержку
полового созревания у мальчиков [210]. Самцы мышей с дефицитом ФСГ фертиль-
ны, хотя имеют маленькие тестикулы и субнормальный сперматогенез [211]. Самки
мышей с дефицитом р-субъединицы ФСГ, полученные с помощью технологии эмбри¬
ональных стволовых клеток, бесплодны и имеют блок фолликулогенеза до образова¬
ния антрального фолликула [211].
Пример. 46,ХХ-Пациентка, гомозиготная по точечной мутации р-субъединицы
ФСГ, обратилась с первичной аменореей, бесплодием и низкими уровнями ФСГ в
крови [210]. Анализ гена р-субъединицы ФСГ выявил двунуклеотидную делецию,
которая привела к структурному сдвигу последующих кодонов и преждевременной
терминации [210]. Родственница указанной пациентки была в постменопаузальном
периоде и имела субнормальные уровни ФСГ. В этой семье наблюдали две точечные
мутации гена р-субъединицы ФСГ, обе локализованные в экзоне 3 в кодонах 51 и 61
[210].
243
Гипогонадизм, ассоциированный с инактивирующими мутациями гена ^-субъединицы ЛГ
Был описан единственный пациент с мутацией гена р-субъединицы ЛГ [212]: муж¬
чина обратился с задержкой полового развития. Он имел повышенные уровни сыво¬
роточного иммунореактивного ЛГ, но пониженные биоактивные концентрации этого
гормона. Его измененный ЛГ имел пониженную активность связывания с рецептором
из-за гомо.зйготного замещения глицина в положении 54 аргинином (G54R) [212],
Члены семьи мужского рода, которые были гетерозиготны по данной мутации, имели
сниженные уровни т.
Полиморфный вариант ЛГ был описан в Финляндии и Японии [213, 214], Вариант
гормона имел замещение двух аминокислот, W8R и I15T, которые ассоциированы
с повышенной биоактивностью и уменьшенным периодом полувыведения [214].
Клиническое значение этого полиморфизма неи.звестно.
Активирующие мутации генов рецепторов ЛГ и ФСГ
Активирующие мутации гена рецептора ЛГ ассоциированы с гонадотропиннеза-
висимым ранним половым развитием. Активирующие или ул)гчшающие функцию
мутации рецептора ЛГ связаны с гонадотропин независимым ранним половым раз¬
витием у мальчиков, но они не вызывают видимых изменений фенотипа у девочек
[215]. Анализ рецептора ЛГ у такого пациента показал замену С на Т в экзоне 11,
который приводит к замене аланина на валин. Когда C0S-7 клетки были транзиент-
но трансфектированы цДНК «дикого» типа или А1а373Val-мутанта рецептора ЛГ,
образование с мутантной цДНК имело более высокую базальную и стимулированную
ХГЧ-аккумуляцию цАМФ.
Известен только один случай активирующей мутации рецептора ФСГ : этот паци¬
ент был фертилен даже после хирургической гипофизэктомии, которая снизила его
иммуннореактивность ФСГ до недетектируемых уровней [216].
Синдром Прадера-Вилли
Пациенты с синдромом Прадера-Вилли обычно имеют ожирение с детства, неона¬
тальную гипотонию, задержку развития, умственную отсталость, гипогонадизм,
короткое телосложение и маленькие ладони и ступни [217]. Типичны гипогонадо-
тропный гипогонадизм, крипторхизм и микропенис [218.219]. Ответ Л Г на одиночное
болюсное введение гонадолиберина снижено по сравнению с контрольными лицами,
страдающими ожирением [218, 219]. Степень гонадотропной недостаточности у этих
пациентов вариабельна. Также были описаны немногочисленные пациенты с гипер-
1’онадотропным гипогонадизмом.
Синдром Прадера-Вилли — генное импринтинговое заболевание, которое обычно
возникает в результате сниженной экспрессии отцовских генов в PWS критической
области хромосомы 15qll. Делеции проксимальной части отцовской хромосомы 15q
служат наиболее частой причиной сниженной экспрессии отцовских генов в этой
области [220]. Материнские копии генов, ответственных за синдром Прадера-Вилли,
в проксимальной 15q в норме «молчат» [220]. Именно поэтому делеция отцовской
копии активных генов приводит к болезни. Синдром Прадера-Вилли также может
развиться, если обе копии гена произойдут от матери, поскольку материнские копии
инактивируются предположительно путем метилирования ДНК [217]; это состояние
известно как однородительская дисомия. Структурные аномалии импринтингового
центра также могут вызвать синдром Прадера-Вилли.
Гены, ответственные за синдром Прадера-Вилли, не были идентифицированы.
Генетический диагноз можно поставить, используя комбинацию цитогенетического
и молекулярного подходов, включая флуоресцентную гибридизацию in situ (FISH) и
анализ различий аллельспецифического метилирования ДНК в 15qll-ql3 [220].
244
Нарушения развития гипофиза, обусловленные мутациями гомеодоменных факторов
транскрипции
Ряд гомеодоменных факторов транскрипции вовлечен в развитие и дифференциа¬
цию различных гормонпродуцирующих клеток в пределах гипофиза. Мутации этих
факторов транскрипции были ассоциированы с недостаточностью гипофизарных
гормонов [221-226J.
Мутации Pit-1 гомеодоменного фактора транскрипции были обусловлены нару¬
шением развития нескольких типов дифференцированных клеток в пределах гипо¬
физа и с дефицитом гормона роста, пролактина и тиреотропина [223, 226]. Эти
пациенты имеют маленькие или нормальных размеров гипофизы. Были описаны
аутосомно-доминантные и рецессивные формы наследования в зависимости от ДНК-
связывающих способностей измененного протеина.
Пациенты с мутациями Propl имеют, наряду с дефицитом гормона роста, про¬
лактина и тиреотропина, недостаточность ЛГ и ФСГ [222, 224]. Секреция АКТГ нор¬
мальная при рождении, но клетки, секретирующие этот гормон, могут деградировать
вторично. У этих пациентов объем гипофиза вариабелен и может быть нормальным,
уменьшенным или увеличенным.
Gsx-J, «сиротский» ген гомеобокса, требуется для нормального развития гипофиза.
Гомозиготные мутации этого гена ассоциированы с экстремальной карликовостью,
половым инфантилизмом и повышенной перинатальной смертностью [222]. Гипофиз
больных маленький, содержит мало клеток и имеет уменьшенное количество клеток,
продуцирующих гормон роста и пролактин.
Другой ген гомеобокса Hesxl кодирует гипофизарный фактор транскрипции, мута¬
ции которого ассоциированы с септооптической дисплазией [225].
Синдром Ригера
Гомеодоменный ген, который транскрибируется как две мРНК, проходящие
альтернативный сплайсинг, кодирующие два отдельных протеина — Ptx2a и Ptx2b,
служит геном-кандидатом синдрома Ригера, аутосомно-доминантного заболевания с
различными черепно-лицевыми, зубными, офтальмологическими и гипофизарными
аномалиями [227].
Lhx3
Была установлена связь комбинированной недостаточности гипофизарных гормо¬
нов с мутациями, приводящими к замене аминокислоты, гена Lhx3, который кодирует
один из членов ЫМ-класса гомеодоменных протеинов [222,228]. Гомозиготные мута¬
ции в Lhx3 у членов двух неродственных единокровных семей были ассоциированы с
дефицитом множества гипофизарных гормонов, за исключением адренокортикотроп¬
ного гормона, и с ригидностью шейного отдела позвоночника [228].
Дисфункция гонад при расстройствах питания
Голландская голодная зима
Stein и Susser описали эффекты недостатка еды во время немецкой оккупации
Нидерландов между октябрем 1944 г. и маем 1945 г. [54, 55]. Во время немецкой
осады поставки продовольствия в определенные голландские города (рис. 3-9) были
в значительной степени урезаны. В некоторых соседних городах поставки продоволь¬
ствия не были уменьшены немцами; Susser и Stein использовали их как контроль [54,
55]. В голодающих местностях 50% женщин отмечали аменорею, а частота зачатия
снизилась до значения 53% показателя, наблюдавшегося в контрольных городах [54,
55]. Также отмечали увеличение частоты перинатальной смертности/ врожденных
аномалий, шизофрении и ожирения [54, 55].
Изменение массы тела и фертильность в !Kung San из Ботсваны
Антропологическое исследование в ÎKung San из Ботсваны, племени охотников-
собиральщиков до 1970-х гг., ярко иллюстрирует взаимоотношения массы тела с
245
ю
с..’:'?
‘ Гронинген \
А
Лейварден
Гарлем ^
/
• Лейден
;о
® Амстердам
Зволле
А
Алмело
Энсхеде
Гаага
Роттердам
© Утрехт
'^■'V Л/'7
^ильбург
' i А Эйндховен
-у5
)
■j Херлен
/ JL
Маастрихт А-ГТ
® Голодающий город с населением
более 500 ООО человек
О Голодающий город с населением
100 000-500 ООО человек
• Голодающий город с населением
40 000-100 ООО человек
А Контрольный город с населением
100 000-500 ООО человек
А Контрольный город с населением
40 000-100 ООО человек
4000-
3500-
3000-
s
1
2500-
I
2000-
1500-
1000-
0-
Рожденные
BO время голода
Окт, 194<ММай^1_948
1
1
1
i
1
1
1
1
i
1
1
1
1
! Зачатые
[ во время голода
- Июль_М^6-Фев, 1948
[ [ Общее
1 КОЛИЧеС
I 1 • 'чодов
1 1; \ Г”
1 (] Ч
\ !; р
' '! 1
;
• |>*\
1 V
1
1
1
1
1
1
.........
г
^ г 1 Отношение
,* 1 / калорийности
, [/ рациона
!\\ * 1 к зачатию
1 \ /|
1 V 1
........ ... .Г ......
M M M M I! Ml II M M 1 И M I M M Ml M M
ЯФМ АМИИАСОНДЯФМАМИИАСОНД ЯФМ A МИИАСОНД
-8000
-°7000
-6000
- 5000 ^
-4000
-3000
-2000
1944
1945
1946
Рис. 3-9. В 1944-1945 гг. немецкая армия установила осаду Нидерландов и ограничила поставки про¬
довольствия в несколько голландских городов. Susser и Stein обобщили данные о пищевом рационе и
последствиях энергетической депривации на состояние здоровья популяции в городах, испытывающих
голод (голодные города, верхняя панель). Несколько соседних городов, которые сотрудничали с окку¬
пантами или не испытывали ограничения в поставках продовольствия, служили в качестве контрольных.
Нижняя панель показывает среднюю энергетическую ценность приема пищи и количество живых ново¬
рожденных. [Из Stein Z., Susser М. Fertility, fecundity, famine: food rations in the Dutch famine 1944/5 have
a causal relation to ferhlity and probably to fecundity // Hum. Biol. — 1975. — N 47. — P. 13Ы54; Stein Z.,
Susser M. The Dutch famine, 1944-1945, and the reproductive process. I. Effects of six indices at birth //
Pediatr Res. - 1975. - N 9. - P. 70-76.]
246
Месяцы
Рис. 3-10. Регуляция фертильности и масса тела по отношению к сезонным изменениям в энер¬
гетической ценности приема пищи в IKung San из Ботсваны. Члены племени San были охотникам и-
собирателями еще 30 лет назад, когда проводились эти исследования. Во время весны и лета пищевое
обеспечение было изобильнее, чем зимой. Соответственно масса тела достигала пика поздним летом.
Количество рождений в племени было ассоциировано с массой тела после поправки на 9-месячный
гестационный период. [Из: van der Walt LA., Wilmsen E.N., Jenkins T. Unusual sex hormone patterns among
desert-dwelling hunter-gatherers // J. Clin. Endocrinol. Metab. — 1978. — N 46. — P. 658-663; and van der
Watt LA., Wilmsen E.N., Levin J., Jenkins T. Endocrine sürdies on the San {«bushmen») of Botswana // S. Afr.
Med. J. - 1977. - N 52. - P. 230-232.]
фертильностью [56, 57]. Масса тела мужчин и женщин в племени в значительной
степени варьирует на протяжении года в зависимости от доступности пищи. В летние
месяцы пищевое обеспечение более обильное и масса тела увеличивается; количество
рождений в племени достигает пика приблизительно через 9 мес после пика массы
тела (рис. 3-10) [56, 57], иллюстрируя, как доступность пищи влияет на характер
фертильности в природе.
Отсрогенное наголо менструаций у балерин
На начало и прогрессирование полового созревания у девочек заметно влияют
физические тренировки и расход энергии. Менархе бывает более поздним у бале¬
рин по сравнению с контрольной группой (начало менархе в 15,4 года у балерин
против 12,6 года в контрольной группе) [229-231]. Периоды отдыха или снижения
интенсивности физической нагрузки, обусловленные травмой, связаны с быстрым
половым развитием и возобновлением менструаций [229-231]. Хотя у балерин име¬
ется задержка развития молочных желез и менархе, развитие лобкового оволосения
не изменяется, доказывая, что различные механизмы регулируют начало адренархе
и менархе. Хотя балерины имеют более низкую массу тела и количество жира в
теле, чем сопоставимые по возрасту контрольные лица, неизвестно, обусловлена ли
переустановка генератора импульсов секреции гипоталамического гонадолиберина
расходом энергии, низкой массой тела или низким содержанием жира вд^ле [231].
Frisch [52, 62] предположил, что для запуска менархе необходимо достижение
минимального соотношения «жир-масса тела» (примерно 17% жира на массу тела).
Однако некоторые исследователи задаются вопросом, является ли жир тела крити¬
ческой детерминантой начала полового созревания [232], поскольку спортсмены с
нормальными менструациями и с аменореей не различаются по составу тела [233],
247
Репродуктивная дисфункция и гипогонадотропный гипогонадизм у спортсменов
У взрослых спортсменов женского пола, особенно у бегунов на длинные дистанции,
танцоров и пловцов, отмечают высокую частоту аменореи, ановуляторных циклов
и других нарушений менструации [230J. У спортсменов меньшая масса тела и более
низкий процент жировой ткани тела, нежели у здоровых лиц сопоставимого возраста
[63]. У бегунов на длинные дистанции количество миль, пробегаемых каждую неделю,
коррелирует со степенью менструальной дисфункции, у женщин-спортсменок обыч¬
но отмечают гипогонадотропный гипогонадизм, предположительно обусловленный
приобретенной недостаточностью гонадолиберина [234, 235]. Некоторые, но не все,
спортсменки с гипоталамической аменореей отвечают на пульсирующее назначение
гонадолиберина, подтверждая, что дополнительные патофизиологические механиз¬
мы могут быть действующими.
Frisch выдвинул гипотезу, что поддержание нормальной репродуктивной функции
у женщин требует определенного минимального отношения «жир-масса тела» [52].
Жировая ткань тела играет важную роль в продукции и метаболизме эстрогена. Т
и андростендион превращаются в эстрогены с помощью CYP19 ароматазы в адипо-
цитах. У чрезмерно худощавых женщин, таких как марафонские бегуньи или лица,
страдающие анорексией, образуется большее количество катехоловых эстрогенов
по сравнению с женщинами, имеющими больший процент жировой ткани в теле.
Катехоловые эстрогены имеют меньшую эстрогеновую активность, чем некатехоло-
вые эстрогены, в частности эстрадиол. Существует также обратная взаимосвязь про¬
цента жира в организме с концентрациями глобулина, связывающего половые гор¬
моны. Именно поэтому изменения концентраций глобулина, связывающего половые
гормоны, может влиять на метаболизм и клиренс эстрогенов у изящных женщин.
Спортсменки имеют более низкую частоту рака молочной железы и других эстро¬
гензависимых репродуктивных новообразований, чем контрольные лица — не атле¬
ты, по-видимому, из-за уменьшенного воздействия эстрогенов [236-241].
Несмотря на то что аномалии секреции гонадолиберина и менструальной функции
хорошо задокументированы у спортсменок, схожие репродуктивные нарушения не
были широко известны у спортсменов мужского пола. Клинически значимый гипого¬
надизм необычен для выносливых спортсменов мужского пола. Концентрации Т и ЛГ
в сыворотке крови выносливых спортсменов мужского пола обычно нормальные или
находятся у нижней границы нормы [242]. В одном из исследований частота и ампли¬
туда импульсов ЛГ были значительно ниже у бегунов по сравнению с контрольными
лицами соответствующего возраста [243], несмотря на то что средние концентрации
ЛГ, ФСГ и Т в плазме крови незначительно различались между группами. При всех
дозах гонадолиберина ответ ЛГ на гонадолиберин был ниже у бегунов, чем у кон¬
трольных лиц [243]. Эти данные свидетельствуют о том, что марафонские бегуны
мужского пола также имеют расстройства гипоталамического импульсового генера¬
тора гонадолиберина, хотя клинически явная андрогенная недостаточность менее
типична, нежели у бегунов женского пола [243].
Гипогонадотропный гипогонадизм у пациентов с расстройствами питания
Нервная анорексия — расстройство, характеризующееся искажением образа тела и
самоиндуцированной потерей массы тела, с препубертатной секрецией ЛГ и ФСГ, при
котором содержание обоих гормонов низкое и они плохо отвечают на стимуляцию
гонадолиберином [60,67,244]. Восстановление питания и массы тела ассоциировано
с нормализацией пульсирующей секреции ЛГ и ФСГ [60], что подтверждает корре¬
ляцию между энергетическим балансом, массой тела и репродуктивной функцией.
Варианты расстройств питания, такие как нервная булимия и легкие формы нерацио¬
нальной диеты, обычны среди молодых женщин и могут быть пропущены, если врач
не проявляет высокую степень настороженности.
248
Эффекты экспериментального ограниге-
ния питания на репродуктивную функцию
у молодых людей: миннесотский экспери¬
мент
В конце 1940-х гг. Ancel Keys и соавт.
изучали голодание человека экспери¬
ментально: 32 человека добровольно
согласились жить в кампусе университета
Миннесоты и соблюдать диету с энерге¬
тической ценностью около 1600 ккал в
сутки, то есть примерно две трети от их
нормальной энергетической потребно¬
сти (рис. 3-11) [245, 246]. Добровольцы
потеряли в среднем 23% их начальной
массы тела, более чем 70% жира тела и
24% мышечной ткани. Группа Keys обна¬
ружила, что снижение поступления энер¬
гии и последующая потеря массы тела
вначале вызывают утрату либидо [246].
Результатом продолжающейся потери
массы тела служит уменьшение простати¬
ческого секрета и снижение подвижности
спермы, продукция спермы снижалась,
когда потеря массы тела достигала при¬
близительно 25% [246]. Прирост массы
тела восстанавливал репродуктивную
функцию у этих лиц [246].
Репродуктивная дисфункция и эндокрин¬
ные аномалии при ожирении
Ожирение связано со спектром эндо¬
кринных изменений, включающих изме¬
нения плазмы крови, характеров секреции и скорости клиренса циркулирующих
половых стероидных гормонов, у большинства мужчин с ожирением от легкой до
средней степени сдвиг уровня общего Т обусловлен изменениями в циркулирующих
концентрациях глобулина, связывающего половые гормоны [247-250]. Поскольку
концентрация глобулина, связывающего половые гормоны, снижается обратно про¬
порционально степени ожирения, содержание общего Т в сыворотке крови уменьша¬
ется по мере того, как растет масса тела [249]. Сывороточные уровни свободного Т,
измеряемые путем равновесного диализа, при этом остаются в пределах нормальных
значений у подавляющего большинства мужчин с ожирением от легкой до средней
степени [249, 251].
Более низкие уровни глобулина, связывающего половые гормоны, у тучных муж¬
чин были приписаны частично гиперинсулинемии, ассоциированной с ожирением
[252-254]. Ожирение и прибавка массы тела ассоциированы с более высокими уров¬
нями инсулина, который подавляет продукцию глобулина, связывающего половые
гормоны, в печени [255]. В противоположность этому старение и состояние хрониче¬
ского воспаления связывают с более высокими уровнями глобулина, связывающего
половые гормоны [158, 160, 256, 257]. Таким образом, у тучных мужчин среднего и
пожилого возраста эффекты ожирения ослабляются признаками старения и заболе¬
вания.
Мужчины с выраженным ожирением могут иметь дефект гипоталамо-гипофизар¬
ной оси, как это предполагают на основании низкого содержания свободного Т при
Рис. 3-11. Ancel Keys с одним из добровольцев
в его исследовании. В конце 1940-х гг. Ancel Keys
и его коллеги изучали эффекты энергетического
ограничения в историческом эксперименте, в
котором 32 мужчины добровольно согласились
жить в кампусе университета Миннесоты и соблю¬
дать диету, обеспечивающую приблизительно две
трети их нормальной энергетической потребности.
Добровольцы в среднем потеряли 23% начальной
массы тела, более 70% жира тела и 24% мышеч¬
ной ткани.
249
отсутствии повышенного уровня гонадотропинов [258]. У тучных мужчин по сравне¬
нию со здоровыми исследуемыми лицами без ожирения могут быть более высокие
уровни сывороточного эстрадиола из-за ароматизации Т в эстрадиол в жировых клет¬
ках [259-260]. Исследователи предположили, что очень высокие уровни эстрогенов
у мужчин с массивным ожирением могут угнетать секрецию гонадолиберина и гона¬
дотропинов [258]. Ответ Т на стимуляцию ХГЧ нормальный у большинства тучных
мужчин, что свидетельствует о нормальных тестикулярном резерве и функции [261].
Снижение массы тела ассоциированно связано с обратным развитием многих из этих
отклонений, включая увеличение сывороточных уровней общего и свободного Т, а
также снижение уровней эстрадиола.
Ряд врожденных гипоталамических синдромов, таких как синдром множествен¬
ных веснушек, синдромы Лоренса-Муна и Барде-Бидля, синдром Коэна, синдром
Берьесона-Форссмана-Леманна, врожденный ихтиоз, синдром Руд, мозжечковая
атаксия, оптико-септальная дисплазия и синдром Мебиуса, обусловлены ожирением
и гипогонадотропным гипогонадизмом. Патофизиология гипогонадотропного гипо¬
гонадизма при этих расстройствах неизвестна, и диагноз ставят путем распознавания
специфических соматических аномалий, присущих этим синдромам [181].
ДИАГНОСТИКА АНДРОГЕННОЙ НЕДОСТАТОЧНОСТИ
Диагностика андрогенной недостаточности проходит в три этапа (рис. 3-12),
Первый включает общее обследование состояния здоровья для выявления признаков
и симптомов андрогенной недостаточности и исключения системного заболевания,
расстройств питания и нарушений образа жизни, таких как чрезмерная физическая
нагрузка или злоупотребление наркотическими средствами (этанол, марихуана и
опиаты). Второй этап — определение уровня общего Т, предпочтительнее в образ¬
це крови, полученном рано утром, с использованием надежной методики анализа.
Третий этап — измерение уровня ЛГ у лиц с подозрением на андрогенный дефицит
для определения, расположен ли дефект на тестикулярном уровне или в гипоталамо-
гипофизарной области.
Клиническое обследование мужчин с подозрением на андрогенную
недостаточность
Комиссия экспертов Эндокринолошческого общества рекомендовала, что диагноз
андрогенной недостаточности следует выставлять только мужчинам с характерными
симптомами и признаками и недвусмысленно низкими уровнями Т [94]. Диагноз
андрогенной недостаточности также не следует ставить во время острой или под-
острой стадии болезни [94].
Симптомы и признаки андрогенной недостаточности неспецифичны и изменя¬
ются в зависимости от возраста, начала, тяжести и продолжительности андрогенной
недостаточности, сопутствующих заболеваний, чувствительности к андрогенам и
предыдущей терапии Т [94]. Если андрогенная недостаточность появилась до завер¬
шения полового созревания пациента, она часто маскируется под отсроченное или
неполное половое развитие и евнухоидальные пропорции (размах рук больше роста
на 2 см и более). Пациенты с препубертатным началом андрогенной недостаточности
также сохраняют высокочастотный голос, и у них не происходит исчезновения волос
в височной области с возрастом. У мужчин, андрогенная недостаточность у которых
развивается после завершения полового созревания, отмечают сниженное половое
влечение и активность, уменьшение спонтанных эрекций, утрату волос на теле и
сниженную частоту бритья, бесплодие, сниженную мышечную массу и силу, приливы
тепла и потливости, снижение роста из-за атравматического перелома, маленькие или
сморщенные яички, увеличение или болезненность молочных желез. Менее специфи¬
ческие симптомы включают слабость, снижение мотивации и инициативности, чув¬
ства печали или подавленности, сниженное настроение, дистимию, плохую концен-
250
Общая оценка состояния здоровья; исключить системные
заболевания, расстройства питания, злоупотребление рас¬
слабляющими веществами, такими как марихуана, алкоголь, кокаин
Общий тестостерон
ниже нижней границы
нормы для данной
методики исследо¬
вания у здоровых
молодых мужчин
Подтвердить путем
повторного иссле¬
дования общего
тестостерона
Общий тестостерон
275-400 нг/дл
Измерить
свободный Т
путем диализа
или калькуляции
Общий
тестостерон
более 400 нг/дл
Андрогенная
недостаточность
маловероятна
Свободный Т меньше
нижней границы нормы
для здоровых молодых
мужчин при данной
методике анализа
Подтвердить
путем повторного
анализа
Рис. 3-12. Диагностический алгоритм у мужчин с подозрением на андрогенную недостаточность.
Референсные значения Т варьируют в зависимости от типа методики и лаборатории. Именно поэто¬
му следует использовать методические и лабораторно-специфические показатели. Т — тестостерон.
[С изменениями из: Bhasin S., Cunningham G.R., Hayes F.J. et al. Testosterone therapy in adult men with
androgen deficiency syndromes: an endocrine society clinical practice guideline // J. Clin. Endocrinol. Metab. —
2006.-N 91.-P. 1995-2010.]
трацию и память, расстройства сна и повышенную сонливость, повышение жировой
составляющей тела и сниженную способность к физическим нагрузкам и работе. У
пожилых возможно наличие этих признаков на фоне неспецифических симптомов,
обусловленных возрастом.
Пороговый уровень Т, ниже которого развиваются симптомы и признаки, неизве¬
стен. В European Male Aging Study (Европейское исследование старения мужчин), попу¬
ляционном проспективном исследовании 3369 мужчин из 8 стран Европы, сниженные
утренние эрекции, сниженное половое влечение, эректильная дисфункция и низкая
физическая активность имели четкую связь с низкими уровнями Т [262]. Физические
и сексуальные симптомы, по-видимому, обладают большей специфичностью, нежели
психологические симптомы [262].
Лабораторные исследования
Измерение уровня общего Т в крови рано утром с использованием надежного мето¬
да — критический этап в диагностике андрогенной недостаточности (см. рис. 3-12).
На общие концентрации Т, представляющие сумму связанного и свободного Т, влия¬
ют пульсирующий, циркадный и годичный ритмы секреции Т, вариабельность анали¬
за и превалирующие концентрации глобулина, связывающего половые гормоны [94,
263]. Уровни Т наиболее высоки в утренние часы после прогулки и ниже в вечерние
часы [257, 264]. Именно поэтому концентрации Т следует измерять утром.
251
На уровень общего Т влияют изменения концентраций глобулина, связывающего
половые гормоны, которые обусловлены старением, системным заболеванием, ожи¬
рением, тиреоидными гормонами, а также многие лекарственные средства, включая
Т [18, 265]. Состояния, связанные с пониженными уровнями глобулина, связываю¬
щего половые гормоны, включают ожирение, нефротический синдром, гипотиреои¬
дизм, исйъльзование глюкокортикоидов, гестагенов и андрогенных стероидов [265].
И наоборот, старение, цирроз печени, гипертиреоидизм, эстрогены, ВИЧ-инфекция
и многие системные заболевания бывают вызваны увеличенными концентрациями
глобулина, связывающего половые гормоны.
Измерение концентраций общего и свободного тестостерона
Концентрации общего Т обычно измеряют с помощью радиоиммупного анализа,
им му неметрических методик или жидкостной хроматографии в тандеме с масспек-
троскопией (ГС-М5/М5). Исторически измерение концентраций Т в сыворотке крови
включало экстракцию органическими растворителями, отделение Т от перекрестно-
реагирующих стероидов посредством хроматографии и далее анализ концентраций
Т. Однако в последние годы для достижения высокого качества анализов и автомати¬
зации большинство коммерческих и многие исследовательские лаборатории минуют
этапы экстракции и хроматографического разделения. Эти прямые, автоматизирован¬
ные радиоиммунные и иммунометрические методики недостаточно точны при низких
диапазонах концентраций Т, которые превалируют у женщин и некоторых мужчин с
андрогенной недостаточностью [266]. Нормативные диапазоны различных методик
варьируют, ограничивая сравнения данных, полученных различными лаборатория¬
ми. Однако с использованием адекватных методикоспецифических нормативных диа¬
пазонов большинство аналитических методов могут различить мужчин с эугонадным
и гипогонадным состоянием [266], Методика ЬС-М8/М8, которую считают «золотым
стандартом» измерения половых гормонов, в настоящее время становится все более
доступной, ее предполагают в качестве стандартного метода измерения концентраций
половых гормонов в плазме крови.
Свободный, или несвязанный, Т, составляющий 0,5-3% циркулирующего Т,
обычно измеряется с помощью равновесного диализа, рассматриваемого в качестве
«золотого стандарта». Значительный массив данных поддерживает точку зрения, что
альбуминсвязанный Т может диссоциировать на тканевом уровне и становится био-
доступным в некоторых органах [267]. Биодостзшные уровни Т измеряются посред¬
ством метода осаждения сульфатом аммония, который осаждает глобулин, связываю¬
щий половые гормоны, и Т, связанный с этим протеином, оставляя в супернатанте
несвязанный и альбуминсвязанный Т (см, рис. 3-2). Однако методы равновесного
диализа и сульфата аммония технически сложны и не всегда доступны в лаборато¬
риях больниц. Концентрации свободного и биодоступного Т могут быть рассчитаны
из концентраций Т и глобулина, связывающего половые гормоны, с использованием
утвержденных уравнений действующих масс [268].
Алгоритмы для расчета концентраций свободного и биодоступного Т на основании
концентраций общего Т, альбумина и глобулина, связывающего половые гормоны,
можно найти Б Интернете {1У\тЛ5ат.ск и wwwMm-Unk.com), Поскольку концентрации
глобулина, связывающего половые гормоны, и общего Т легче измеримы в лаборатор¬
ных условиях, нежели уровни свободного и биодоступного Т, использование рассчи¬
танных концентраций свободного Т служит удобной и благоразумной альтернативой.
Методики с заменой меченым аналогом, которые доступны во многих больничных
лабораториях, неточны, их использование не рекомендовано [269].
Дальнейшее обследование мужчин с андрогенной недостаточностью
У мужчин с андрогенной недостаточностью, которая диагностирована на осно¬
вании признаков, симптомов и низкого уровня Т, определение концентраций ЛГ и
252
ФСГ может помочь определить, на каком уровне есть дефект — на тестикулярном
или гипоталамо-гипофизарном (рис. 3-13). Мужчины с первичной тестикулярной
недостаточностью, о чем свидетельствуют повышенные уровни ЛГ и ФСГ и низкие
уровни Т, должны быть подвергшты кариотипированию для исключения синдрома
Югайнфелтера, типичной причины первичной тестикулярной недостаточности.
В отношении мужчин с вторичным гипогонадизмом требуется дальнейшее инди¬
видуальное обследование путем измерения уровня пролактина в сыворотке крови,
других гипофизарных гормонов, сывороточного железа и насыщения трансферрина
и МРТ для исключения гиперпролактинемии, других дефицитов гипофизарных гор¬
монов, гемохроматоза и объемных патологических процессов гипоталамуса и гипо¬
физа, Экономическая эффективность такого диагностического подхода неизвестна,
поскольку у мужчин среднего и пожилого возраста с вторичным гипогонадизмом
процент лиц, которые имеют обнаруживаемые объемные патологические процессы
гипоталамо-гипофизарной области, низок. Диагностический выход можно улучшить
путем ограничения лиц, у которых проводят поиск патологии гипофиза, мужчинами
с уровнем Т в сыворотке крови ниже 150 нг/дл, пангипопитуитаризмом, гиперпро¬
лактинемией и эффектами массы опухоли, о чем свидетельствуют головные боли или
дефекты поля зрения [94]. В исследованиях мужчин, обращающихся с сексуальной
дисфункцией и низкими уровнями Т, средняя распространенность гипоталамо-
гипофизарных объемных поражений низкая; кроме того, в этих исследованиях муж¬
чины с объемными повреждениями гипоталамо-гипофизарного места имели уровень
Т в сыворотке крови ниже 150 нг/дл [270, 271J.
Скрининг
Скринингом называют идентификацию нераспознанной болезни или дефекта с
помощью тестов или процедур, которые могут быть быстро применены на популя-
Измерить уровни ЛГ и ФСГ
ПовышенныеуровниЛГ и/или ФСГ: |
первичный гипогонадизм
Кариотипирование для исключения
синдрома Клайнфелтера
Низкий или неадекватно
нормальный уровень ЛГ и ФСГ:
вторичный гипогонадизм
Исключить системные заболевания,
расстройства питания, использование
расслабляющих средств
и чрезмерную физическую нагрузку;
измерить уровень пролактина, других
гипофизарных гормонов, сыворо¬
точного железа, насыщения транс¬
феррина и провести магнитно-
резонансную томографию гипо¬
таламо-гипофизарной области
Если исключены другие причины гипогонадотропного гипогонадизма, -
провести поиск ассоциированных дисморфологических признаков
и установить гипоталамические синдромы путем дифференциальной
диагностики
Рис. 3-13. Дальнейшее обследование мужчин, считающихся андрогендефицитными. ФСГ — фолликуло¬
стимулирующий гормон; Л Г — лютеинизирующий гормон.
253
ционном уровне, в то же время при случайной находке используют процедуры для
идентификации существующей, но ранее не подозреваемой болезни, которая может
быть не связана с главными жалобами пациента, обратившегося за медицинской
помощью. Комиссия экспертов Эндокринологического общества сделала заключение,
что недостаточно свидетельств, оправдывающих проведение скрининга на андроген¬
ную недостаточность в общей популяции [94]. Андрогенная недостаточность не имеет
необходимого критерия для популяционного скрининга из-за недостатка данных
о вреде андрогенной недостаточности на здоровье населения. Более того, характе¬
ристики проведения доступных скрининговых инструментов — андрогенная недо¬
статочность у пожилых мужчин (ADAM) [272], массачз'^сетская шкала исследования
старения мужчины {Massachusetts Male Aging Study, MMAS) [273] и шкала симптомов
пожилых мужчин {Aging Males'Symptom Scale, AMS) [274]; предсказательная ценность
и экономическая эффективность скрининговой стратегаи в общей популяции неиз¬
вестны. Также ограничена информация о долгосрочных последствиях андрогенной
недостаточности для здоровья.
Комиссия экспертов Эндокринологического общества рекомендует «врачам рас¬
смотреть возможность выявления случаев болезни путем измерения уровней общего
Т у мужчин с определенными клиническим расстройствами, при которых имеется
высокая частота низких концентраций Т, или у тех, кому рекомендована терапия Т»
[94]. Высокая распространенность низких уровней Т была описана у мужчин, обра¬
щающихся с эректильной дисфункцией, переломом при небольшой травме [275,276],
ВИЧ-инфекцией, снижением массы тела [127-129,132, 277-279], терминальной ста¬
дией болезни почек [280J, хронической обсгруктивной болезнью легких [281-283]
и сахарным диабетом [284], а также у пациентов, принимающих глюкокортикоиды
[285] или опиоиды [171,172,176].
ТЕСТОСТЕРОНОВАЯ ТЕРАПИЯ У МУЖЧИН С ДЕФИЦИТОМ АНДРОГЕНОВ
В отношении молодых андрогендефицитных мужчин терапия Т приносит много
'пользы и ассоциирована с низким риском серьезных неблагоприятных эффектов.
В систематическом обзоре по большей части открытых исследований среди здоро¬
вых андрогендефицитных мужчин терапия Т была ассоциирована со значительным
положительным эффектом на мышечную ткань, максимальную произвольную силу
и костную минеральную плотность позвонков, а также со значительным снижением
массы жировой ткани всего тела [131].
Терапия Т улучшает показатели половой активности и полового влечения [286-
289], Назначение Т влияет на многие сферы половой функции: оно увеличивает
частоту спонтанных сексуальных мыслей и фантазий [288], улучшает отзывчивость
на эротические стимулы [290] и увеличивает частоту и продолжительность ночных
эрекций полового члена [291]. Терапия Т не изменяет эректильный ответ на визуаль¬
ный эротический стимул [288] или частоту оргазмов у гипогонадных мужчин [94],
хотя увеличивает объем эякулята. В ходе открытых исследований было выявлено, что
терапия Т улучшает положительные и снижает отрицательные аспекты настроения
[292]; данные рандомизированных клинических исследований о влиянии терапии на
настроение ограничены и не показывают значительно большего улучшения настрое¬
ния по сравнению с плацебо.
Однако андрогендефицитные мужчины сообщают об улучшении чувства здоровья
и энергии после начала терапии Т. Некоторые исследования описывают небольшие
и непостоянные эффекты на зрительно-пространственное распознавание, слуховую
память и беглость речи [293, 294]. Б исследованиях эффектов терапии Т на чувстви¬
тельность к инсулину получены противоречивые результаты [295, 298]. Большинство
исследований были открытыми, отбор пациентов проводили на основании низких
концентраций Т.
254
Препараты тестостерона и их клиническая фармакология
Заместительная терапия Т может быть назначена с использованием одной из
нескольких доступных лекарственных форм, при этом особое внимание должно быть
уделено ее фармакокинетике (табл. 3-2 и 3-3) [2, 3].
Эфиры тестостерона, вводимые посредством инъекции
Эстерификация Т в 17-р-гидроксильном положении делает молекулу гидрофоб¬
ной и увеличивает продолжительность ее действия. Медленное высвобождение
эфира Т из масляного депо в мышце объясняет его увеличенную продолжительность
действия. Больше боковая цепь, больше гидрофобность эфира и больше продолжи¬
тельность действия. Таким образом, Т-энантат и ципионат с более длинными боко¬
выми цепями имеют более долгую продолжительность действия, чем Т-пропионат.
Деэсгерификация эфиров Т быстро происходит в плазме, по скорости не ограничена
и не может объяснить большую продолжительность действия.
в пределах 24 ч после внутримышечного введения 200 мг Т-энантата или ципиона-
та сывороточные уровни Т растут до верхней границы нормы или до супрафизиологи-
ческого диапазона и далее постепенно снижаются до гипогонадного диапазона через
следующие 2 нед [289,299]. Режим введения Т-энантата или ципионата 2 раза в месяц
приводит к пикам и спадам уровней сывороточного Т, что может ассоциироваться
с изменениями настроения пациента, сексуального желания, уровня активности и
бодрости [289, 299]. Кинетика Т-энантата и ципионата схожа [300]. Хотя концен¬
трации эстрадиола и ДГТ увеличиваются во время назначения эфира Т, соотношения
эстрадиол:Т и ДГТ:Т остаются нормальными [289,299]. При соответствующем мони¬
торинге можно подобрать адекватные переносимые дозы инъецируемых эфиров Т
(они являются наименее дорогими, доступными препаратами Т).
Чрескожный тестостерон
Тестостероновый гель
В продаже доступны два тестостероновых геля [287,301-306]. Фармакокинетичес¬
кие исследования подтвердили, что 5-10-граммовые дозы, наносимые ежедневно на
кожу, могут повысить и поддержать концентрации общего и свободного Т в сыво¬
ротке крови в средненормальном диапазоне у гипогонадных мужчин [287, 303], хотя
у отдельных пациентов возможны значительные вариации на протяжении дня. В
настоящее время рекомендовано начинать с 5 г и подбирать дозу на основании сыво¬
роточных уровней Т.
Преимуществами тестостеронового геля служат легкость применения, его невиди¬
мость после нанесения и гибкость дозирования. Главной особенностью использова¬
ния геля является потенциальная возможность его переноса сексуальному партнеру
или ребенку, которые вступают в близкий контакт с пациентом. Сывороточное соот¬
ношение концентраций ДГТ к Т выше у гипогонадных мужчин, леченных тестосте¬
роновым гелем, чем у эугонадных мужчин. Кожная переносимость хорошая, кожные
раздражения достаточно редки.
Трансдермальный тестостероновый пластырь
Один или два 5 мг внеполовых тестостероновых пластыря могут быть приклеены
на кожу вне области мошонки [286, 307-309]. Уровни Т и эстрадиола находятся в
средненормальном диапазоне на протяжении 4-12 ч после нанесения пластыря [308-
309]. Внеполовые пластыри обеспечивают физиологические уровни сывороточного
ДГТ, Один 5 мг пластырь может быть недостаточен для увеличения сывороточных
концентраций Т до средненормального диапазона у всех гипогонадных мужчин; неко¬
торые пациенты могут нуждаться в ежедневном назначении двух 5 мг пластырей для
достижения целевого уровня [280]. У некоторых пациентов использование внеполо¬
вых пластырей сопровождается кожным раздражением.
255
го
сл
о>
Таблица 3-2. Клиническая фармакология некоторых лекарственных форм тестостерона
Лекарственная
форма
Режим терапии
Фармакокинетический
профиль
Дигидротестостерон
и эстрадиол
Преимущества
Недостатки
Т-энантат или
ципионат
100 мг/нед или 200 мг в
2 нед внутримышечно
После одной внутримы¬
шечной инъекции уровни
сывороточного Т становятся
выше физиологического диа¬
пазона и далее постепенно
снижаются до гипогонадного
диапазона к концу интервала
между введением доз
Уровни дигидротестостерона
и эстрадиола растут про¬
порционально увеличению
концентраций Т;
отношения Т;ДГТ и
Т:эстрадиол (кЕ^) не изме¬
няются
Корригирует симптомы
андрогенной недостаточ¬
ности;
относительно недорогой,
если вводить самостоя¬
тельно;
гибкость дозирования
Требует внутримышечной инъ¬
екции;
пики и спады сывороточных
уровней Т
Внеполовая чре¬
скожная система
Один или два пластыря,
номинально обеспечивающие
поступление 5-10 мг Т за
24-часовой промежуток вре¬
мени, наложенные ежедневно
на области, не подвергаю¬
щиеся давлению
Восстанавливает сыво¬
роточные уровни Т, ДГТ
и эстрадиола до уровней
физиологического диапазона
у мужчин
Т:ДГТ- и Т:Е2-уровни находят¬
ся в физиологическом диа¬
пазоне для мужчин
Легкость применения, кор¬
ригирует симптомы андро¬
генной недостаточности и
подражает нормальному
суточному ритму секреции Т;
меньше увеличивает гемо¬
глобин, чем эфиры, вводи¬
мые с помощью инъекции
Сывороточные уровни Т у
некоторых мужчин с дефици¬
том андрогенов могут быть у
нижней границы нормы; этим
мужчинам может понадобиться
применение двух пластырей
ежедневно;
раздражение кожи в месте при¬
крепления может быть пробле¬
мой для некоторых пациентов
Тестостероновый
гель
5-10 г геля, содержащего
50-100 мг Т, должен быть
применен ежедневно
Восстанавливает сывороточ¬
ные уровни Т и эстрадиола
до уровней физиологическо¬
го диапазона у мужчин
Сывороточные уровни ДГТ
выше, а отношение Т:ДГТ
ниже у мужчин с гипогона¬
дизмом, леченных гелем,
чем у здоровых мужчин с
нормальной функцией поло¬
вых желез
Корригирует симптомы
андрогенной недостаточно¬
сти, обеспечивает гибкость
дозирования, легкость при¬
менения, хорошо переносит¬
ся кожей
Потенциальная возможность
переноса на партнера женского
пола или ребенка при прямом
контакте кожа-кожа;
умеренно высокие уровни ДГТ
17а-
метилтестостерон
Это 17а-алкилированное
производное не должно
использоваться из-за воз¬
можности печеночной ток¬
сичности
Активно при приеме внутрь
Клинические ответы вариабель¬
ны; потенциально токсичен для
печени;
не должен быть использован
для лечения андрогенной недо¬
статочности
Окончание табл. 3-2
Щечные биоадге-
зивные тестостеро¬
новые таблетки
30 мг биоадгезивные таблет¬
ки с контролируемым высво¬
бождением, используемые 2
раза в сутки
Абсорбируется через слизи¬
стую оболочку щеки
Нормализует сывороточные
концентрации Т и ДГТ у муж¬
чин с гипогонадизмом
Корригирует симптомы
андрогенной недостаточ¬
ности у здоровых мужчин с
гипогонадизмом
У 16% пациентов неблагопри¬
ятные эффекты, связанные с
деснами
Т-ундеканоат для
приема внутрь’
40-80 мг внутрь 2-3 раза в
сутки с приемом пищи
Когда назначается в олеи¬
новой кислоте, Т-ундеканоат
абсорбируется через лимфа¬
тические сосуды портальной
системы;
значительная вариабельность
у одного и того же пациента
в различные дни и среди
различных пациентов
Высокое отношение ДГТ:Т
Удобство приема внутрь
Не разрешено в США;
вариабельные клинические
ответы, вариабельные уровни
Т в сыворотке крови, высокое
отношение ДГТ:Т
Вводимый с помо¬
щью инъекции,
долго действующий
Т-ундеканоат в
масле*
1000 мг, вводимые внутри¬
мышечно с последующим
введением 1000 мг через
6 нед и далее по 1000 мг
каждые 12 нед
Когда назначают в дозе
1000 мг внутримышечно,
сывороточные уровни Т под¬
держиваются в нормальном
диапазоне у большинства
мужчин
Уровни ДГТ и эстрадиола
растут пропорционально уве¬
личению концентраций Т;
отношения Т:ДГТ и Т:Е2 не
изменяются
Корригирует симптомы
андрогенной недостаточ¬
ности;
требует нечастого назна¬
чения
Требует внутримышечной инъ¬
екции большого объема (4 мл)
Импланты тестосте¬
рона
4-6 имплантов по 200 мг
подкожно
Сывороточный Т достигает
пика в 1 мес и далее под¬
держивается в нормальном
диапазоне 4-6 мес
Отношения Т:ДГТ и не
изменяются
Корригирует симптомы
андрогенной недостаточ¬
ности
Требует хирургического раз¬
реза для введения; спонтанные
экструзии
* Эта лекарственная форма доступна за пределами США, в настоящее время не одобрена FDA.
Воспроизведено из Bhasin S., Cunningham G.R., Hayes FJ. et al. Testosterone therapy in adult men with androgen deficiency syndromes: an Endocrine Society clinical
practice guideline //J. Clin. Endocrinol. Metab. — 2006. — N 91. — P. 1995-2010.
Таблица 3-3. Некоторые рекомендованные режимы тестостеронзаместительной терапии
Форма
Дозирование
В США
Т-энантат.ил1и ципионат
75-100 мг/нед или 150-200 мг каждые 2 нед, назначенные внутри¬
мышечно
Внеполовые тестостероновые пластыри
Один или два 5 мг-пластыря, наложенные ночью на кожу спины,
бедра или верхней части плеча, вдали от областей, подвергающихся
давлению
Тестостероновый гель
5-10 г, нанесенные на укрываемые участки кожи
Биоадгезивные щечные тестостероновые
таблетки
Таблетки 30 мг, наложенные на слизистую оболочку щеки 2 раза
сутки
За пределами США*
Т-ундеканоат для приема внутрь
Обычно 40-80 мг внутрь 2 или 3 раза в сутки вместе с приемом
пищи
Т-ундеканоат для инъекций
Обычно 1000 мг внутримышечно вначале и через 6 нед и далее по
1000 мг каждые 12 нед
Импланты тестостерона
4-6 имплантов по 200 мг подкожно каждые 4-6 мес
* Эти формы доступны для клинического использования во многих странах: врачи, желающие исполь¬
зовать эти лекарственные формы, должны следовать режимам назначения препаратов, одобренным
в их странах.
Примечание. Эти режимы дозирования должны рассматриваться как предполагаемые для начала
Т-заместительной терапии; дозу и режим следует откорригировать на основании измерения уровней
сывороточного Т.
Адаптировано из Bhasin S., Cunningham G.R., Hayes F.J. et al. Testosterone therapy in adult men with
■androgen deficiency syndromes: an endocrine society clinical practice guideline // J. Clin. Endocrinol.
Metab. - 2006. - N 91. - P. 1995-2010.
Биоадгезивные щегные тестостероновые таблетки
Биоадгезивные 30-мг тестостероновые таблетки с контролируемым высвобождени¬
ем, наложенные каждые 12 ч на слизистую оболочку щеки, нормализуют сывороточные
уровни Т, ДГТ и эстрадиола у гипогонадных мужчин [310, 312]. У 16% пролеченных
мужчин использование щечных таблеток ассоциировано с проблемами десен.
Лекарственные формы тестостерона, недоступные в США
Т-ундеканоат при назначении внутрь в олеиновой кислоте абсорбируется преимуще¬
ственно через лимфатические сосуды в системный кровоток и избегает разрушения пер¬
вого прохождения через печень. Обычно назначают 40-80 мг 2-3 раза в сутки. Однако
клинические ответы бывают вариабельными и субоптимальными [313-321]. Отношения
ДГТ к Т в сыворотке крови выше у гипогонадных пациентов, леченных путем приема
внутрь Т-ундеканоата, по сравнению с эугонадными мужчинами [313-321].
Импланты кристаллического Т вводят в подкожную ткань с помощью троакара
через маленький кожный разрез. Т высвобождается за счет поверхностной эрозии
импланта и абсорбируется в системный кровоток. Четыре-шесть 200-мг имплантов
могут поддержать сывороточные концентрации Т в средненормальном и верхненор¬
мальном диапазоне в течение 6 мес [322-326]. Необходимость кожного разреза для
введения и удаления, спонтанные экструзии и фиброз в месте введения импланта —
потенциальные недостатки этой лекарственной формы.
Внутримышечная форма Т-ундеканоата в виде масляного раствора может под¬
держивать уровни Т в сыворотке крови в нормальном диапазоне на протяжении
258
10-12 нед после внутримышечной инъекции [327, 328]. Обычный режим назначения
Т-ундеканоата включает начальную инъекцию 1000 мг препарата внутримышечно,
вторую инъекцию 1000 мг через 6 нед и далее внутримышечные инъекции каждые
12 нед по 1000 мг. Относительным преимуществом этой инъецируемой формы слу¬
жит ее продолжительное действие и более слабые степени колебаний сывороточных
уровней Т, нежели те, что отмечают при терапии Т-энантатом или ципионатом.
Однако Т-ундеканоат назначают в большом объеме масла, что вызывает дискомфорт
у пациента.
17а-алкилированные производные тестостерона, назначаемые внутрь
Т хорошо всасывается после приема внутрь, но при этом быстро подвергается
пресистемному метаболизму. Именно поэтому невозможно добиться устойчивых
уровней Т в крови после приема внутрь кристаллического Т. 17а-Алкилированные
производные Т относительно резистентны к печеночной деградации и могут быть
назначены внутрь, однако из-за потенциальной гепатотоксичности и доступности
альтернативных и более безопасных лекарственных форм 17а-алкилированные про¬
изводные Т для заместительной терапии не рекомендованы [329, 330].
Новейшие андрогеновые лекарственные формы
Ряд новейших лекарственных форм андрогенов с лучшей фармакинетикой или бо¬
лее селективным профил ем активности находятся в стадии разработки. Лекарственная
форма биодеградивных тестостероновых микросфер обеспечивает физиологические
уровни Т на протяжении 10-11 нед [331, 332]. Длительно действующие эфиры Т,
Т-буциклат [333, 334] и Т-ундеканоат [335-340] при внутримышечном введении
могут поддержать циркулирующие концентрации Т в мужском диапазоне на про¬
тяжении 7-12 нед. Начальные клинические испытания показали возможносаь
назначения Т сублингвальным или щечным путем. 7а-Метил-19-нортестостерон —
андроген, который не может быть 5а-восстановлен; поэтому по сравнению с Т он
проявляет относительно более сильную агонистическую активность на мышцы и
супрессию гонадотропинов, но меньшую активность на простату [341-343].
Ряд нест'ероидных селективных модуляторов андрогеновых рецепторов (8АЯМ)
с тканевой селективностью в настоящее время находятся в стадии разработки. Эти
агенты могут селективно оказывать андрогеновую активность на мышцы, костную
ткань и сексуальную функцию с минимальным действием на другие ткани, такие как
простата и сердечно-сосудистая система [131, 344].
Противопоказания к терапии тестостероном
Тестостероновая терапия не должна применяться в отношении мужчин с метаста¬
тическим раком простаты или раком грудной железы, поскольку она может способ¬
ствовать росту этих опухолей. Терапия может ухудшить существующий эритроцитоз,
нелеченное тяжелое обструктивное ночное апноэ и тяжелую застойную сердечную
недостаточность. Поскольку узел в простате, уплотнение или повышенные уровни
простатоспецифического антигена (ПСА) могут быть индикаторами нераспознанного
рака простаты, мужчины с этими состояниями должны пройти урологическое обсле¬
дование до рассмотрения возможности терапии Т. Противопоказания перечислены в
табл.3-4.
Неизвестно, насколько безопасна Т-терапия в отношении мужчин, которые под¬
верглись радикальной простатэктомии на предмет локализованного рака простаты и
кажутся излеченными; отдельные специалисты считают, что терапия"! может быть
назначена в индивидуальном порядке таким мужчинам, в случае если у них в течение
более 2 лет ПСА не обнаруживают [345-347]. Однако данных рандомизированных
контролируемых исследований на эту тему нет, а опубликованные исследования
включают очень немногочисленных пациентов. Соответственно недостаточно дан¬
ных, чтобы рекомендовать такой подход.
259
Таблица 3-4. Противопоказания к назначению тестостерона
Очень большой риск серьезных неблагоприятных исходов
Метастатический рак простаты.
Рак грудной железы
Умеренный и высокий риск неблагоприятных исходов
Недиагностированный узел или уплотнение в простате.
Неуточненное увеличение уровня ПСА*.
Эритроцитов (гематокрит более 50%).
Выраженные симптомы доброкачественной гиперплазии предстательной железы, о чем свидетельствует значение
Американской урологической ассоциации/международной шкалы простатических симптомов более 19.
Нестабильная тяжелая застойная сердечная недостаточность (III или IV класса)
* Базовое значение уровня ПСА, выше которого Т-терапия не должна назначаться без зфологического
обследования, было объектом значительной дискуссии. Комиссия экспертов Эндокринологического
общества указала, что Т-терапия без урологического обследования для исключения рака простаты не
должна назначаться мужчинам с уровнем ПСА более 3 нг/мл.
Из Bhasin S., Cunningham G.R., Hayes F.J. et al. Testosterone therapy in adult men with androgen defi¬
ciency syndromes: an Endocrine Society clinical practice guideline //J. Clin. Endocrinol. Metab. — 2006. —
N 91. - P. 1995-2010.
Неблагоприятные эффекты, ассоциированные с тестостероновой терапией
Частота неблагоприятных эффектов, связанных с Т в открытых исследованиях
с участием молодых гипогонадных мужчин, была низкой [348]. Наиболее частыми
Т-связанными неблагоприятными эффектами служат эритроцитоз, акне, жирная
кожа и болезненность грудных желез. Хотя есть сообщения о гинекомастии и индук¬
ции или ухудшении обструктивного ночного апноэ, частота этих неблагоприятных
эффектов во время терапии низкая. Побочные эффекты перечислены в табл. 3-5.
Широко распространенное мнение, будто Т-терапия увеличивает риск атеросклеро¬
тического поражения сердца, не подкрепляется имеющимися данными. Долгосрочные
последствия добавления Т для риска заболевания сердца остаются неизвестными.
Хотя супрафизиологические дозы неароматизируемых андрогенов, обычно приме¬
няемые бодибилдерами и спортсменами, снижают уровни ЛПВП в плазме крови [349,
350], физиологическое замещение Т у пожилых мужчин было ассоциировано только
с умеренным снижением ЛПВП в плазме крови или вообще отсутствием такового
[94, 351, 352]. Профильные исследования мужчин среднего возраста выявили скорее
прямую, нежели обратную взаимосвязь сывороточных уровней Т с концентрациями
ЛПВП в плазме крови и обратную корреляцию между уровнями Т и объемом висце¬
рального жира [356, 357]. Более низкие уровни Т у мужчин ассоциированы с более
высокими уровнями частичек ЛПНП и протромбиновых факторов [354, 355].
Открытые исследования применения эфиров Т в основном у молодых гипогонад¬
ных мужчин показали небольшое снижение общего холестерина, холестерина ЛПВП
и ЛПНП в плазме крови [358]. Плацебо-контролируемые исследования у пожилых
мужчин не выявили значительных изменений в уровнях ЛПВП в плазме крови при
долгосрочном назначении Т [351, 352].
Спонтанная [359] и экспериментально индуцированная [360] андрогенная недо¬
статочность связана с повышенной массой жировой ткани. Терапия Т снижает массу
жировой ткани у пожилых мужчин, которые имеют низкие уровни Т [131]. Были
сообщения о снижении объема висцерального жира, сывороточной концентрации
глюкозы и артериального давления и повышении чувствительности к инсулину при
добавлении Т мужчинам среднего возраста [295, 296], хотя повышение чувствитель¬
ности к инсулину при назначении Т не было подтверждено в других исследованиях.
Хирургическая кастрация крыс понижает чувствительность к инсулину; физиоло¬
гическое замещение Т приводит к обратному развитию этого нарушения [361]. Тем
260
Таблица 3-5. Потенциальные неблагоприятные эффекты заместительной терапии тестостероном
Явно связанные
• Эритроцитоз.
• Акне и жирная кожа.
• Выявление субклинического рака простаты.
• Рост метастатического рака простаты.
• Сниженные продукция сперматозоидов и бесплодие
Возможно связанные
• Гинекомастия.
• Облысение по мужскому типу (семейное).
• Ухудшение симптомов доброкачественной гиперплазии предстательной железы.
• Рост рака грудной железы.
• Индукция или ухудшение обструктивного ночного апноэ
Специфичные для лекарственной формы
Таблетки для приема внутрь
• Нарушения функции печени (метилтестостерон).
• Снижение холестерина высокой плотности (метилтестостерон)
Имплантируемые пилюли
• Инфекция и рубцовое изгнание пеллеты
Внутримышечная инъекция тестостерона энантата или ципионата
• Изменения настроения и либидо.
• Боль в месте инъекции.
• Эритроцитоз (особенно у пожилых пациентов)
Чрескожные пластыри
• Кожные реакции в месте применения
Трансдермальный гель
• Потенциальный риск переноса партнеру (необходимо напоминать пациенту покрывать места нанесения одеждой
и мыть кожу и руки с мылом перед прямым контактом с другим человеком)
Щечные таблетки
• Изменения вкуса.
• Раздражение десен
H3 Bhasin S., Cunningham G.R., Hayes F.J. et al. Testosterone therapy in adult men with androgen deficiency
syndromes: an endocrine society clinical practice guideline //J. Clin. Endocrinol. Metab. — 2006. — N 91. —
P. 1995-2010.
не менее высокие дозы Т нарушают чувствительность к инсулину у кастрированных
крыс. Андрогены увеличивают инсулиннезависимый захват глюкозы [362] и модули¬
руют активность липопротеин липазы тканеспецифическим образом [363].
Эффекты тестостерона на риск атеросклеротического поражения сердца
Из 30 профильных исследований, обзор которых провел А1ехапбег5еп [364], в 18
сообщали о более низких уровнях Т у мужчин с ИБС, в 11 обнаружили схожие уровни
у контрольных лиц и мужчин с ИБС, в одном выявили более высокие уровни сульфа¬
та дегидроэпиандростерона. В проспективных исследованиях не удалось обнаружить
ассоциацию между уровнями общего Т и началом ИБС [365, 366].
В одном из исследований по приему Т-ундеканоата внутрь сообщали об улуч¬
шении стенокардии напряжения у мужчин с ИБС [367]. Инфузия Т резко улучшает
коронарный кровоток в модели у лошади [166-169]. Вазодилатирующие эффекты
Т опосредованы через кальциевые каналы Т-типа [368, 369]. Мужчины с дефицитом
андрогенов имеют нарушенную реактивность сосудов [370], но эффекты терапии Т
на сосудистую реактивность у андрогендефицитных мужчин были противоречивы,
в ходе некоторых исследований показано ухудшение сосудистой реактивности после
лечения [370-372]. В модели инсулиновой резистентности у грызунов фруктозное
261
питание приводит к развитию инсулиновой резистентности и гипертензии [373]. У
этих крыс, питающихся фруктозой, гонадэктомия предупреждает ухудшение артери¬
ального давления; терапия Т была необходимым условием развития гипертензии у
крыс, питающихся фруктозой [373].
В модели атеросклероза у мышей с недостаточностью рецептора к ЛПНП [173]
хирургй'ческая кастрация ускоряет, а терапия Т замедляет прогрессирование атеро¬
склероза. Величина эффекта Т на прогрессирование атеросклероза схожа с таковой
при применении эстрогенов. Благоприятным эффектам Т на атеросклероз в этой
модели на мышах противодействовало сопутствующее назначение ингибитора С¥Р19
ароматазы; предполагается, что эффекты Т опосредованы через его превращение в
эстрогены в сосудистой стенке [374], Эффекты Т на задержку прогрессирования ате¬
росклероза являются независимыми от липидов плазмы крови [374]. Такие положи¬
тельные эффекты Т у мышей с недостаточностью рецептора к ЛПНП не наблюдались
постоянно в ходе других экспериментальных моделей [375].
Эти данные дают возможность предположить, что и супрафизиологические, и суб-
физиологические, циркулирующие концентрации Т связаны с повышенным сердечно¬
сосудистым риском, хотя очевидность этого заключения далеко не окончательна
[366]. Эффекты терапии Т на прогрессирование атеросклероза и частоту сердечно¬
сосудистых событий не изучали в отношении мужчин в ходе рандомизированных
клинических исследований, однако они важны, поскольку даже небольшие изменения
в показателях заболеваемости сердечно-сосудистыми заболеваниями могут оказать
существенное влияние на здоровье популяции.
Тестостерон и риск рака простаты
Нет доказанных подтверждений того, что терапия Т служит причиной рака про¬
статы [376]. Также не существует устойчивой взаимосвязи эндогенных сывороточных
уровней Т с риском рака простаты. Однако имеется ряд опасений.
У многих пожилых мужчин в их предстательной железе «затаились» микроскопи¬
ческие очаги рака. Неизвестно, будет ли терапия Т приводить к росту этих субклини-
ческих локусов рака и делать их клинически очевидными.
Рак простаты - типичная андрогензависимая опухоль, и назначение андрогенной
терапии может способствовать ее рост}» [377,378]. Именно поэтому терапия Т проти¬
вопоказана мужчинам с метастатическим раком простаты.
Кроме того, Morgentaler и соавт. [379] сообщили о высокой частоте рака простаты,
который можно выявить посредством биопсии у мужчин с низкими уровнями общего
и свободного Т, несмотря на нормальное содержание ПСА и результаты пальцевого
ректального обследования. Однако это исследование не включало контрольную груп¬
пу, и неясно, будет ли такая же высокая частота рака, обнаруживаемого с помощью
биопсии, при секстантной биопсии у контрольных лиц сопоставимого возраста с нор¬
мальными уровнями Т.
Сывороточные уровни ПСА ниже у мужчин с дефицитом Т и восстанавливаются
до нормальных значений после терапии Т [380, 381]. Однако сывороточные уровни
ПСА не увеличиваются прогрессивно у здоровых гипогонадных мужчин при назна¬
чении замещающих доз Т. В двух плацебо-контролируемых исследованиях по поводу
назначения Т пожилым мужчинам изменение в уровнях ПСА в сывфотке крови на
протяжении 3 лет не отличалось в значительной степени у мужчин, пролеченных
плацебо и Т [382,383].
Увеличение уровней ПСА во время Т-терапии может послужить триггером для
обследования и биопсии у некоторых пациентов [376]. Более интенсивный скрининг
ПСА и наблюдение за мужчинами, получающими Т-терапию, может вести к повы¬
шенному количеству биопсий простаты и обнаружений субклинического рака про¬
статы, который в другом случае остался бы незамеченным. Мужчины, получающие Т,
имеют повышенный риск биопсии простаты и обнаружения повышенного количества
малигнизации в простате [348].
262
Сывороточные уровни ПСА колеблется при повторном измерении у одного и того
же пациента с течением времени [376], а также из-за вариабельности методик анали¬
за, последствий возраста и других биологических факторов. Именно поэтому, когда
сывороточные уровни ПСА у мужчин с недостаточностью андрогенов, получающих
Т-терапию, отличаются от ранее полеченных значений, врач должен решить, требует
ли это изменение детального обследования пациента на предмет рака простаты или
оно обусловлено лишь вариабельностью между тестами по измерению ПСА.
Заместительная терапия Т может быть безопасно назначаема мужчинам с добро¬
качественной гиперплазией предстательной железы, которые имеют сумму по шкале
симптомов от легкой до умеренной. Тяжесть симптомов, ассоциированных с доброкаче¬
ственной гиперплазией предстательной железы, может быть оценена с помощью опро¬
сника Международной шкалы простатических симптомов/Симптомов Американской
урологической ассоциации. Андрогенная недостаточность ассоциирована с умень¬
шенным объемом простаты, и замещение андрогенов увеличивает объем простаты до
значения у контрольных лиц соответствующего возраста [380,381]. У пациентов с уже
существующими тяжелыми симптомами доброкачественной гиперплазии предстатель¬
ной железы даже малое увеличение объема простаты во время назначения Т может уси¬
лить симптомы обструкции, в отношении этих мужчин Т либо не должен применяться,
либо назначается при тщательном мониторинге симптомов обструкции.
Терапия тестостероном и эритроцитоз
Препараты Т дозозависимо увеличивают эритроцитарную массу [348], по-видимо-
му, через воздействие Т на эритропоэтин и пролиферацию стволовых клеток [384,
385]. Именно поэтому замещение Т не показано в отношении мужчин с базовым
уровнем гематокрита 50% и выше без соответствующего обследования и лечения
эритроцитоза. Назначение Т молодым мужчинам с дефицитом андрогенов обычно
обусловлено небольшим увеличением уровня гематокрита. Клинически значимый
эритроцитоз необычен у молодых гипогонадных мужчин во время Т-терапии, но он
может появиться у мужчин с синдромом ночного апноэ при выраженном курении или
хронической обсгруктивной болезни легких.
Повышение гемоглобина во время Т-терапии выше у пожилых лиц, нежели у
молодых мужчин [386]. Эритроцитоз — наиболее частый неблагоприятный эффект,
вызванный лекарственным препаратом, у мужчин среднего и пожилого возраста, а
также наиболее частая причина прекращения терапии [348]. Было выявлено, что
Т-терапия посредством трансдермальных систем вызывает меньшее увеличение
уровней гемоглобина, чем то, что ассоциировано с эфирами Т [309]; по-видимому,
обычные режимы трансдермальных Т-систем способствуют поступлению меньших
доз Т, чем эфиры т.
Т-терапия должна быть прекращена при превышении уровня гематокрита до 54%,
и следует воздерживаться от терапии до тех пор, пока гематокрит не снизится до
безопасного уровня (менее 50%), после чего Т-терапия может быть начата повторно
в меньшей дозе.
Тестостерон превращается путем ароматизации в эстрадиол во многих перифериче¬
ских тканях и может обострить рак молочной железы, поэтому его не следует назна¬
чать мужчинам с раком грудной железы.
Тестостерон может индуцировать синдром ночного апноэ или усилить существую¬
щий синдром ночного апноэ из-за его нейромышечных эффектов на верхние дыха¬
тельные пути, и он не должен назначаться мужчинам с тяжелым синдромом обструк¬
тивного ночного апноэ без соответствующего обследования и лечения-[387, 388].
Мужчины с синдромом ночного апноэ могут иметь низкие уровни Т [389].
Назначение Т может транзиторно индуцировать задержку натрия и воды, следователь¬
но, его не следует давать (^или назначать с большой осторожностью) пациентам с тяжелой
застойной сердечной недостаточностью и симптомами IV функционального класса.
263
Мониторинг тестостероновой терапии
Согласно рекомендациям Эндокринологического общества (Endocrine Society)
мужчины, получающие Т-терапию, должны быть осмотрены через 3 мес после начала
терапии и далее ежегодно с использованием стандартного плана мониторинга эффек¬
тивной коррекции симптомов и признаков, ассоциированных с недостаточностью Т,
и для содействия раннему обнаружению неблагоприятных эффектов [94]. Т-терапия
должна быть нацелена на повышение сывороточных уровней Т до средненормального
диапазона. Даже у мужчин с первичной тестикулярной недостаточностью сывороточ¬
ные уровни ЛГ не нормализуются под действием тех доз Т, которые восстанавлива¬
ют сексуальную функцию и индуцируют клиническое улучшение. Именно поэтому
сывороточные уровни ЛГ не используют для мониторинга адекватности Т-терапии у
мужчин с дефицитом андрогенов. Определение гемоглобина, гематокрита, концентра¬
ции ПСА и пальцевые исследования простаты должны проводиться через регулярные
интервалы (табл. 3-6) в добавление к общему обследованию состояния здоровья, как
рекомендовано местными стандартами.
Таблица 3-6. Наблюдение мужчин, получающих Т-терапию
Обследование пациента через 3 мес после начала лечения и далее ежегодно для определения, ответили ли
симптомы на терапию и страдает ли пациент от какого-либо неблагоприятного эффекта терапии.
Мониторинг уровней Т 2 или 3 мес после начала тестостероновой терапии
• Терапия должна быть нацелена на повышение уровня Т в сыворотке крови до средненормапьного диапазона,
• Т-энантат и ципионат для инъекций; измерение сывороточных уровней Т в середине интервала времени
между инъекциями. Если концентрация Т более 700 нг/дл или 350 нг/дл. следует подобрать дозу или частоту.
• Трансдермальный пластырь; измерение уровней Т через 3 и 12 ч после нанесения пластыря; подобрать дозу
для достижения уровней Т в средненормальном диапазоне.
• Биоадгезивная щечная тестостероновая таблетка; измерить уровни непосредственно перед и после примене¬
ния новой системы.
• Трансдермальный крем; измерить уровень Т в любое время, после того как пациент находится на лечении, по
меньшей мере 1 нед; подобрать дозу для достижения уровней Т в средненормапьном диапазоне.
• Т-ундеканоат для приема внуфь*: мониторинг сывороточных уровней Т 3-5 ч после приема внутрь.
• Т-ундеканоат для инъекций*; измерить сыаорочный уровень Т перед каждой последующей инъещией и подо¬
брать междозовый интервал для поддержания сывороточного Т в средненормапьном диапазоне.
Проверить гематокрит в начале лечения, через 3 мес и далее ежегодно. Если гематокрит более 54%, остано¬
вить терапию до тех пор, пока гематокрит не снизится до безопасного уровня; обследовать пациента на гипок¬
сию и синдром ночного апноэ; начать повторно терапию в меньшей дозе.
Измерить минеральную плотчостъ костъюй ткани поясничного отдела позвоночника и/или шейки бедренной
кости после 1-2 лет Т-терапии у гипогонадных мужчин с остеопорозом или переломом кости, возникшим при
малой травме, согласно местному стандарту медицинской помощи.
Выполнить палытевое ректальное обследование и проверить уровень ПСА до начала лечения через 3 мес и
далее в соответствии с руководствами по скринингу pai«i простаты в зависимости от возраста и расы пациента.
Оценить при каждом визите неблагоприятные эффекты, специфичные для лекарственной формы
• Щечные тестостероновые таблетки; спросить об изменениях вкуса и обследовать десну и слизистую оболочку
ротовой полости на предмет раздражения.
• Эфиры Т для инъекции: спросить об изменениях настроения и либидо,
• Тестостероновые пластыри; осмотреть места нанесения на предмет кожной реакции.
• Тестостероновые гели: посоветовать пациентам покрывать место нанесения сорочкой и мыть с мылом кожу
перед контактом кожа-кожа, поскольку при использовании геля тестостерон может остматься на коже.
Сывороточные ^фовни Т поддерживаются, когда место нанесения моют через 4-6 ч после применения тесто¬
стеронового геля
* Не о/1обрены для клинического применения в США.
Модифицировано из Bhasin S., Cunningham G.R., Hayes F.J, et al. Testosterone therapy in adult men
with androgen deficiency syndromes: an endocrine society clinical practice guideline //J. Clin. Endocrinol.
Metab. - 2006. - N 91. - P. 1995-2010.
264
Наиболее трудный момент наблюдения гипогонадных мужчин, получающих
заместительную Т-терапию, связан с критериями, которые необходимо использо¬
вать при принятии решения о проведении биопсии простаты (табл. 3-7). Существует
значительная вариабельность в измерении ПСА от теста к тесту. В некоторой части
эта вариабельность обусловлена неотъемлемой вариабельностью методики анализа,
в некоторой степени — неизвестными факторами [390-400]. Вариабельность может
быть еще выше, если исследования выполняют в лабораториях, которые использу¬
ют различные методики анализа [390-400]. 90% доверительный лимит изменения
уровней ПСА между двумя тестами, выполненными порознь за 3-6 мес у принимав¬
ших плацебо мужчин, в исследовании финастерида у мужчин с доброкачественной
гиперплазией предстательной железы был 1,4 нг/мл [401]. Именно поэтому изме¬
нение более чем на 1,4 нг/мл в образцах, взятых за 3-6 мес, был использован как
критерий для урологического обследования в некоторых клинических исследованиях
финастерида [402]. По предварительным оценкам, коэффициент вариации ПСА
между исследованиями в среднем составляет 15% [403, 404]. Коэффициент вариа¬
ции исследования ПСА следует принимать во внимание при оценке изменения ПСА
между образцами, забранными в различное время, или при интерпретации скорости
изменения ПСА.
Таблица 3-7. Показания для консультации уролога у мужчин, получающих Т-терапию
• Уровень ПСА более 4 нг/мл.
• Увеличение сывороточной концентрации ПСА более чем на 1,4 нг/мл в течение 12 мес Т-тврап»»1,
• Скорость на{»стания ПСА более 0,4 нг/мл в год, при этом в качестве референсного значения используют уро¬
вень ПСА через 6 мес после нгвначения Т (гфименимо, только ест« данные о содержшжи ПСА доступны за
период более 2 лет).
• Выявлшие аномалии простаты при пальцевом решлыюм исследовании,
• Значение по шкале простатических симптомов Американсю^ урологической аосодиации или Международной
шкалы простатических симптомов более 19
Из Bhasin S., Cunningham G.R., Hayes FJ. et al. Testosterone therapy in adult men with androgen deficiency
syndromes: an endocrine society clinical practice guideline //J. Clin. Endocrinol. Metab. - 2006, - N 91. -
P. 1995-2010.
Назначение замещающих доз T мужчинам с дефицитом андрогенов увеличивает
сывороточные уровни ПСА. В системном обзоре среднее увеличение ПСА после
инициации Т-терапии у молодых гипогонадных мужчин было 0,3 нг/мл, а у пожи¬
лых мужчин с низким или на нижней границе нормы уровнями Т этот показатель
равнялся 0,44 нг/мл [376]. Повышение уровней ПСА после замещения Т у мужчин с
дефицитом андрогенов обычно не превышает 0,5 нг/мл, а рост на 1 нг/мл за период
3-6 мес встречается крайне редко. У некоторых мужчин назначение Т при базовом
содержании ПСА от 2,5 до 4 нг/мл вызывает повышение уровней ПСА выше 4 нг/мл.
Повышение уровней ПСА выше 4 нг/мл будет требовать консультации уролога, мно¬
гим из этих мужчин проведут биопсию простаты, что увеличит вероятность обнару¬
жения заболевания простаты.
В отношении пациентов, у которых доступны сведения о последовательных изме¬
рениях ПСА на протяжении более 2 лет, Carter и соавт. [391,397,405,406] предложи¬
ли использовать критерий скорости нарастания ПСА для идентификации мужчин с
высоким риском рака простаты. В Балтиморском продольном исследовании старения,
когда базовое содержание ПСА было от 4 до 10 нг/мл, скорость изменения уров¬
ня ПСА более 0,75 нг/мл в год было необычным для мужчин с доброкачественной
гиперплазией предстательной железы [391, 397, 405, 406]. Поскольку большинство
мужчин, получающих Т-терапию, имели базовые уровни ПСА ниже 4 нг/мл, кри¬
терий, установленный для базовых значений ПСА между 4 и 10 нг/мл, может быть
неприемлем для использования в исследованиях Т или во время Т-терапии.
265
Недавний анализ данных балтиморского продольного исследования старения Fang
и соавт. [407] показал скорость изменения уровня ПСА у мужчин с базовыми уров¬
нями ПСА менее 4 нг/мл. У этих мужчин с базовым содержанием ПСА менее 4 нг/мл
использование пороговой скорости изменения ПСА в 0,2 нг/мл в год давало чувстви¬
тельность 52,4% и специфичность 79,4% в плане выявления рака простаты (эти сведе¬
ния требуют дальнейшего подтверждения). Однако следует принять во внимание зна¬
чительную вариабельность уровней ПСА в период между тестом и повторным тестом,
когда оценивают скорость изменения уровней ПСА, поскольку пороговые уровни для
назначения биопсии простаты значительно ниже межтестовой вариабельности будут,
вероятно, приводить к большому количеству ошибочных направлений на биопсию.
Использование критерия скорости нарастания ПСА, возможно, будет более полез¬
ным в отношении мужчин, у которых данные о ПСА доступны на протяжении многих
лет, с тем чтобы повышение ПСА над базовым уровнем превышало ожвдаемуго меж-
тестовую вариабельность. Carter [391,397.405,406] обращает внимание, что скорость
изменения ПСА не следует использовать при периоде наблюдения менее 2 лет. Также
установление скорости изменения ПСА требует многочисленных определений ПСА
на протяжении времени. Скорость изменения ПСА рассчитывают как общее измене¬
ние ПСА от базового уровня, разделенное на время в годах. Скорость изменения ПСА
более 0,2 нг/мл в год не означает, что пациент имеет рак простаты и что ее следует
использовать в качестве показания для немедленной биопсии простаты; скорее она
означает риск развития рака простаты выше среднего во время последующих деся¬
тилетий. Именно поэтому скорость изменения ПСА может быть полезна во время
долгосрочного наблюдения за пациентами на протяжении многих лет. В отношении
периодов менее 3 лет предполагают, что скорость изменения ПСА более 0,4 нг/мл в
год должна слуэкить показанием для урологического обследования и более интенсив¬
ного наблюдения в плане рака простаты в будущем [376], Для периодов наблюдения
более 3 лет скорость изменения ПСА более 0,2 нг/мл в год, вероятно, ассоциирована
с более высоким риском рака простаты, и она должна быть причиной более тщатель¬
ного наблюдения за пациентом. Специфичность или чувствительность критерия ско¬
рости изменения ПСА не была исследована в ходе проспективных исследований Т.
ТЕСТОСТЕРОН-ТЕРАПИЯ У ПОЖИЛЫХ МУЖЧИН С ВОЗРАСТНЫМ СНИЖЕНИЕМ
КОНЦЕНТРАЦИИ ТЕСТОСТЕРОНА
Ряд поперечных (срез популяции в определенное время) эпидемиологических
исследований подтверждает, что уровни общего, свободного и биодоступного Т в
сыворотке крови ниже у пожилых мужчин, нежели у молодых, даже после учета
потенциальных мешающих факторов — время забора образца, сопутствующие забо¬
левания, лекарственные препараты и технические моменты, связанные с анализом
гормонов [159, 160, 162-164, 408]. Несколько продольных (во времени) исследо¬
ваний [158, 160, 161] подтвердили постепенное, но прогрессирующее снижение
концентраций сывороточного Т в возрасте от 20 до 80 лет. Суточный ритм секреции
Т угнетен у пожилых мужчин [388]. Поскольку уровни глобулина, связывающего
половые гормоны, выше у пожилых мужчин по сравнению с молодыми^[409], воз¬
растное снижение содержания свободного и биодоступного Т по величине больше,
чем снижение уровней общего т. На возрастные изменения уровней Т влияют комор-
бидные состояния, ожирение, физическая активность и факторы образа жизни, вклю¬
чая курение. Мужчины, которые набирают массу тела и становятся тучными, имеют
более высокую скорость снижения общего уровня Т [410]; более высокая скорость
снижения содержания общего, но не свободного Т у мужчин, которые приобретают
избыточную массу тела, обусловлена главным образом более низким возрастным
увеличением глобулина, связывающего половые гормоны [410].
266
Взаимосвязь низких уровней тестостерона с отдаленными осложнениями
Уровни Т ассоциированы с рядом клинических последствий у пожилых мужчин
в популяционных эпидемиологических исследованиях, хотя эти связи слабые. Так,
ни.зкие концентрации биодоступного Т связаны со сниженной массой скелетных
мышц конечностей [276, 411-413] и сниженным мышечным напряжением [414].
Уровни общего и свободного Т также ассоциированы с самостоятельно описываемой
физической функцией, оцениваемой с помощью показателя физического компонента
и физической работоспособности [415]. В обзоре здоровых пожилых мужчин [416]
уровни биодоступного Т положительно коррелировали с мышечным напряжением
и минеральной плотностью кости и отрицательно коррелировали с массой жировой
ткани. Наилучшим предиктором минеральной плотности кости служил эстрадиол, не
связанный с глобулином, связывающим половые гормоны. В ходе других исследова¬
ний уровни общего Т отрицательно коррелировали с отношением «талия-бедро» и
массой висцерального жира [357], хотя коэффициенты корреляции невысокие. Это
дает основания предполагать, что существуют другие важные детерминанты состава
тела, помимо Т. После учета возраста и других сопутствующих факторов уровни Т
слабо связаны с половым желанием и эректильной дисфункцией у мужчин среднего и
пожилого возраста [262,417].
В двух [418, 419] из трех эпидемиологических исследований [418-420], в ходе
которых изучали взаимосвязь уровней Т с симптомами депрессии, выявили отсут¬
ствие значительной корреляции между этими двумя показателями; одно из иссле¬
дований [420] указывало на обратную связь между уровнями биодоступного Т и
показателем опросника депрессии Веек (Beck's Depression Inventory). При дальнейшем
анализе данных массачусетского исследования старения мужчин Seidman и соавт.
[421] выявили значительную взаимосвязь уровней Т, длины CAG повторов в гене
рецептора андрогенов с показателями депрессии.
Результаты отдельных эпидемиологических исследований также свидетельствуют
об обратной ассоциации между уровнями свободного Т в сыворотке крови и висце¬
ральным ожирением [356, 357], инсулиновой резистентностью [422, 423], метаболи¬
ческим синдромом [424] и риском сахарного диабета 2-го типа [^55,422, 425]. В ходе
некоторых исследований концентрации свободного Т были измерены посредством
метода меченого аналога. На измерение свободного Т с помощью данного метода
влияют превалирующие концентрации глобулина, связывающего половые гормоны
[269]: этот факт заставляет отдельных экспертов сомневаться в обоснованности и
точности данного метода. Из-за зависимости уровней общего и свободного Т при
методе меченого аналога от концентраций глобулина, связывающего половые гормо¬
ны, нельзя исключить вероятность того, что взаимосвязь измеренных концентраций
Т, висцерального жира с атеросклерозом отражает взаимосвязь глобулина, связы¬
вающего половые г ормоны, с этими последствиями. Поскольку известно, что инсулин
ингибирует глобулин, связывающий половые гормоны, следует ожидать, что тучные и
инсулинрезистентные мужчины с высокими уровнями инсулина имеют более низкие
уровни глобулина, связывающие половые гормоны, и, следовательно, более низкие
концентрации Т,
Эффекты тестостероновой терапии
в ходе нескольких рандомизированных плацебо-контролируемых исследований
продолжительностью до 3 лет изучали эффекты добавления Т у пожилых мужчин
с концентрациями Т от низких нормальных до низких [382, 383, 426]. В целом
исследования Т у пожилых мужчин характеризовались малым размером выборки,
включением здоровых пожилых мужчин с низкими или низкими нормальными уров¬
нями т. которые не имели симптомов, и использованием суррогатных исходов. Эти
исследования не имели достаточной силы для обнаружения либо многозначительных
положительных преимуществ в исходах, важных для пациентов, либо изменений в
частоте событий, связанных с простатой или сердечно-сосудистой системой. Именно
267
поэтому риски и клинические выгоды долгосрочной Т-терапии у пожилых мужчин с
возрастным снижением уровней Т остаются неизвестными.
Состав тела
В системном обзоре плацебо-контролируемых рандомизированных исследований
у мужчин среднего и пожилого возраста [44, 287, 351, 383, 426-431] замещение Т
было ассоциировано со значительно более весомым увеличением мышечной массы
и большим снижением массы жировой ткани (-2,0 кг; 95% доверительный интервал
от -3,1 до +0,8), чем плацебо. Изменение массы тела значительно не различалось
между группами (-0,6 кг; 95% доверительный интервал от -2,0 до +0.8). Т-терапия
была ассоциирована с более значительным улучшением мышечной силы, чем плацебо
(3,3 кг; 95% доверительный интервал от 0,7 до 5,8) [351, 428, 429, 432]. Изменения
измерений физической активности, основанных на производительности, упомянуты
только в немногочисленных исследованиях и были непостоянными. Анализ одного из
исследований свидетельствует об отсутствии изменений физической функции [383],
два других — об улучшении [428].
Т-терапия не влияет в значительной степени на средние показатели качества
жизни, оцененные с помощью опросника 8Р-36 в исследовании медицинских исходов,
однако по сравнению с плацебо Т-терапия была связана со значительно более замет¬
ным улучшением показателей физической функции [94]. При системном обзоре не
было выявлено значительного улучшения на фоне Т-терапии показателей депрессии
или познавательной функции у пожилых мужчин,
В исследованиях, которые включали мужчин с уровнями Т ниже 300 нг/дл,
Т-терапия была ассоциирована со значительным улучшением либидо и непостоян¬
ным улучшением эректильной функции [94]. Однако Т-терапия не улучшала либидо
или эректильную функцию у мужчин с базовым уровнем Т более 300 нг/дл.
Неблагоприятные эффекты
В недавнем метаанализе оценивали неблагоприятные эффекты, ассоциированные
с Т-терапией в рандомизированных исследованиях у мужчин среднего и пожилого
возраста [348]. Б 19 исследованиях, которые соответствовали приемлемым критери¬
ям, 651 мужчина получал терапию Т, 433 лечили плацебо. Комбинированная частота
всех событий, связанных с простатой, была значительно выше у мужчин, леченных Т,
чем у мужчин, принимавших плацебо (1,78; 95% доверительный интервал от 1,07 до
2,95). Частота рака простаты, концентрации ПСА выше 4 нг/мл и биопсий простаты
была в числовом соотношении выше в группе Т, чем в группе плацебо, хотя различия
между группами отдельно не были статистически достоверными. У мужчин, получав¬
ших Т, вероятность гематокрита более 50% была почти в 4 раза выше по сравнению с
пациентами, принимавшими плацебо (3,69; 95% доверительный интервал 1,82-7,51).
Увеличение гематокрита было наиболее частым неблагоприятным событием, связан¬
ным с замещением Т. Частота сердечно-сосудистых событий, синдрома ночного апноэ
или смертельного исхода значительно не различалась между двумя группами.
Таким образом, существует сильное предубеждение в отношении обнаружения
большего количества событий, связанных с простатой, в исследованиях Т. Лечение
Т повышает уровни ПСА у мужчин с дефицитом андрогенов. Поскольку биопсии
простаты обычно назначаются в ответ на увеличение ПСА, а повышение ПСА более
вероятно у мужчин, леченных Т, нежели плацебо, мужчины, получающие гормон,
имеют более высокий риск подвергнуться биопсии простаты, чем мужчины, полу¬
чающие плацебо. В действительности в метаанализе мужчины в группе Т перенесли
биопсию более чем в 20 раз чаще по сравнению с лицами из группы плацебо. Из-за
более высокого риска подвергнуться биопсии простаты мужчины, получавшие Т,
будут иметь диагноз субклинического заболевания простаты с большей долей вероят¬
ности, чем пациент из группы плацебо, В клинических исследованиях, так же как и в
клинической практике, требуются стратегии минимизации риска с использованием
268
стандартизированного плана мониторинга. Эти рассуждения еще раз подтверждают
необходимость мониторинга гематокрита, ПСА и пальцевого исследования простаты
во время Т-терапии у пожилых мужчин на основании тщательного и стандартизиро¬
ванного плана мониторинга.
Оценка -«риск-польза»
Доступные сведения препятствуют оценке клинической пользы и долгосрочных
рисков Т-терапии в отношении пожилых мужчин с возрастным снижением уров¬
ней т. Комиссия экспертов Эндокринологического общества выступает «против
политики назначения Т-терапии всем пожилым мужчинам с низкими уровнями Т».
«...Врачи рассматривают предложение Т-терапии на индивидуальной основе пожи¬
лым мужчинам с постоянно низкими уровнями Т, установленными при более чем
одном исследовании, и выраженными симптомами андрогенной недостаточности
после соответствующего обсуждения неопределенностей риска и пользы Т-терапии
у пожилых мужчин». Отдельная комиссия экспертов Института медицины также
сделала заключение, что требуются рандомизированные исследования адекватной
мощности по эффективности Т-терапии у пожилых мужчин с симптоматическими
состояниями, ассоциированными с недостаточностью андрогенов и недвусмыслен¬
но низкими уровнями Т. Однако комиссия Института медицины предполагает, что
крупные исследования для определения рисков Т-терапии следует отложить до тех
пор, пока не будет установлена эффективность этого лечения. Именно поэтому споры
вокруг Т-терапии у пожилых мужчин вряд ли будут разрешены в ближайшее время.
Гинекомастия
Гинекомастией называют увеличение мужской грудной железы, обусловленное
доброкачественной пролиферацией железистой ткани [433,434], Увеличение желези¬
стой ткани следует отличать от чрезмерного накопления жировой ткани; железистая
ткань плотная и содержит фиброзоподобные тяжи [433,434].
Гинекомастия обычно вызвана нарушением баланса между действием эстрогенов и
андрогенов или повышенным соотношением эстрогены/андрогены, обусловленным
увеличенной продукцией эстрогенов, сниженным образованием андрогенов или неко¬
торой комбинацией этих процессов [433, 434]. Даже у мужчин с тяжелой гинекома¬
стией галакторея является необычной, так как полное развитие молочной ацинарной
ткани по женскому типу требует участия прогестагенов [435].
ПРОДУКЦИЯ ЭСТРОГЕНОВ у МУЖЧИН
в отличие от овулирующих женщин, у которых большая часть циркулирующих
эстрогенов происходит из яичников. 85% циркулирующего эстрадиола и более чем
95% циркулирующего эстрона у мужчин образуется путем экстрагонадного превра¬
щения Т и Д^'-андростендиона большей частью в жировой ткани и коже [436, 437].
Ежедневная скорость образования эстрадиола и эстрона установлена на уровне 45
и 65 мкг соответственно [438]. Приблизительно 7 мкг эстрадиола секретируется
непосредственно яичками, 17 мкг образуется путем периферического превращения
Т и 22 мкг из эстрона путем восстановления 17-кетогруппы под действием 17-р-
гидроксистероид дегидрогеназы [438].
Превращение Т и Д'*-андростендиона в эстрадиол и эстрон соответственно требу¬
ет координированного действия белкового продукта гена ароматазы, именуемого
СУР19, и флавопротеина НАДФ-Н-цитохром Р450 редуктазы, которая переносит вос¬
станавливающие эквиваленты от НАДФ-Н к цитохрому Р450 [436]. Человеческий ген
СУР19 транскрибируется в гонадах и многих экстрагонадных тканях под контролем
тканеспецифичных промоутеров, которые локализованы в пределах крупной 90 кЬ
регуляторной области в 5'-направлении [436, 439]. Под влиянием различных гормо¬
269
нов и ростовых факторов эти промоутеры регулируют экспрессию гена СУР! 9 арома¬
тазы тканеспецифическим образом, управляя транскрипцией мРНК, которые имеют
идентичную кодирующую последовательность, но различные 5'-нетранслируемые
области [439]. Продукция эстрогена в гранулезных клетках яичников у менструирую¬
щих женщин находится под контролем проксимально расположенного промоутера II,
который регулируется ФСГ [436]. В жировой ткани и коже экспрессия С¥Р19 регули¬
руется промоутерами 1,3 и 1,4, которые локализованы на 0,2 и 73 кЬ в направлении 5'
от кодирующей области [345, 440].
ПРИЧИНЫ и ПАТОФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ
Высокая частота гинекомастии наблюдается в неонатальный период, во время
полового созревания и при старении [433, 434, 441]. Причины гинекомастии пере¬
числены в табл. 3-8.
Таблица 3-8. Причины гинекомастии, классифицированные по механизмам
Повышенная продукция предшественников эстрогенов
• Опухоли надпочечников.
• Заболевания печени.
• Недостаточность 17Р-гидроксистероидцегидрогеназы
Повышенная активность ароматазы
Повышенная активность в нормальной ткани
• Ожирение.
• Старение
Нарушение регуляции ароматазы
• Семейный синдром избытка ароматазы.
• Новообразования, характеризующиеся эктопической продукцией эстрогенов: опухоли из клеток Сертоли (изоли¬
рованные, синдром Пейтца-Егерса, комплекс Карни), трофобластические опухоли, продуцирующие ХГЧ.
•|1овообразования, характеризующиеся эктопической продукцией эстрогенов: феминизирующая адренокорти-
'кальная карцинома, гепатоцеллюлярная карцинома
Измененное соотношение Т-эстрогены, обусловленное сниженной продукцией или действием андрогенов
• Синдром нечувствительности к андрогенам.
• Недостаточность 5а-редуктазы.
• Тиреотоксикоз.
• Синдром Клайнфелтера
Модифицировано из Braunstein G.D. Aromatase and gynecomastia // Endocr. Relat. Cancer. — 1999. —
N 6. - P. 315-324.
Кратковременное увеличение молочных желез отмечают у 20-30% новорожден¬
ных. Оно обычно служит результатом трансплацентарного переноса материнских и
плацентарных эстрогенов.
Гинекомастия также обычна для мальчиков в период полового созревания [442,
443]; увеличение грудных желез появляется сначала в раннем пубертате, цо-видимому,
отражая высокое отношение эстрогенов к Т на ранних этапах полового созревания до
тех пор, пока продукция тестикулами Т не становится полностью установившейся.
Уровни эстрадиола растут более быстро, чем уровни Т во время раннего пубертата,
что приводит к высокому соотношению «эстрогены-андрогены» [444]. У большин¬
ства мальчиков увеличение грудных желез регрессирует с прогрессированием поло¬
вого развития и подъемом уровней Т, так что к 21 году только малая часть мальчиков
имеют персистирующую гинекомастию.
Распространенность гинекомастии растет с возрастом и индексом массы тела [445]
из-за увеличенной активности ароматазы в жировой ткани. Индекс массы тела кор-
270
релирует с риском гинекомастии [445] и диаметром ткани молочной железы [441].
Масса тела служит хорошим предиктором скорости продукции эстрогенов у тучных
мужчин и женщин в постменопаузе [259]. По мере увеличения возраста уровни Т
падают, уровень глобулина, связывающего половые гормоны, растет, а перифериче¬
ская скорость ароматизации становится выше, чем у молодых лиц, приводя к повы¬
шенному отношению «эстрогены-андрогены». Более высокая скорость продукции
эстрогенов у пожилых мужчин связана с возрастным увеличением активности С¥Р19
в жировой ткани [446] и увеличением в среднем массы жировой ткани.
Мужчины с синдромами андрогенной недостаточности часто имеют гинекомастию;
непрерывный синтез эстрадиола путем ароматизации остаточных надпочечных и
гонадных андрогенов ведет к повышенному отношению «эстрогены-андрогены».
Гинекомастия — характерная черта синдрома Клайнфелтера. Расстройства с нечув¬
ствительностью к андрогенам также служат причиной гинекомастии из-за отсутствия
противодействия эффектам эстрогенов.
Избыток продукции эстрогенов может быть вызван опухолями клеток Лейдига или
Сертоли, которые секретируют эстрогены, или опухолями половых клеток, которые
секретируют ХГЧ (447). Эстроген-продуцирующие опухоли из клеток Сертоли могут
случаться спорадически, при синдроме Пейтца-Егерса [448] или комплексе Карни.
ХГЧ также стимулирует синтез эстрогена клетками Лейдига.
У пациентов с феминизирующими опухолями надпочечников или врожденной
ВДКН повышенные уровни эстрогенов — результат увеличенной скорости продукции
Д'*-андростендиона, который может быть превращен в эстрон, У пациентов с болезнью
печени катаболизм андростендиона в печени снижен; в результате более значительное
количество этого предшественника становится доступным для ароматизации в пери¬
ферических тканях [449].
Семейная форма препубертатной гинекомастии, наследуемая как аутосомно-
доминантное или Х-связанное расстройство, развивается из-за повышенной про¬
дукции эстрогенов, обусловленной чрезмерной ароматазной ативностью в пери¬
ферических тканях [450, 451], Пациенты мужского пола с синдромом избыточной
периферической ароматазной активности характеризуются высокими уровнями
эстрадиола, препубертатной гинекомастией, ускоренным костным возрастом в дет¬
стве и уменьшением окончательного роста во взрослом состоянии из-за преждев¬
ременного эпифизиального слияния [452]. Пациентки с этим синдромом обраща¬
ются с жалобами на преждевременное развитие грудных желез [451]. У некоторых
[іациентов с этим синдромом была описана гетерозиготная инверсия в хромосоме
15д21.2-3; инверсия служит причиной того, что ген С¥Р19 чрезмерно транскриби¬
руется с помощью конституционально активного промоутера других прилегающих
генов, что приводит к повышенной продукции эстрогенов в зкстрагонадных тканях
[450, 453].
Назначение некоторых терапевтических препаратов ассоциировано с гинекомасти¬
ей. Так, например, оральные контрацептивы, фитоэстрогены и препараты наперстян¬
ки, имеют прямую эстрогенную активность; другие, такие как изониазид, могут инду¬
цировать повьЕшенную активность эстрогенов. Немногочисленные препараты инги¬
бируют биосинтез андрогенов (например, кетоконазол) или их действие (в частности,
спиронолактон, флутамид, ципротерона ацетат и бикалутамид). Описаны отдельные
случаи гинекомастии, обусловленной нечаянным воздействием эстрогенных произ¬
водных, содержащихся в промышленных продуктах и косметике. Так. гинекомастия
была выявлена у сотрудников предприятий, которые производят диэтилстилбестрол
или другие эстрогены, и у детей матерей, использующих эстрогеновые кремы. Многие
косметические кремы и лосьоны содержат различное количество эстрогенов. Кроме
того, мясо, молоко и другие молочные продукты, полученные от животных, леченных
эстрогенами, могут служить случайными источниками эстрогенов [454].
271
ДИАГНОСТИКА
Клиническое обследование
Поскольку гинекомастию в некоторой степени можно обнаружить у половины
госпитализированных мужчин и у подавляющего большинства пациентов гине¬
комастия доброкачественная, детализированное диагностическое обследование
не показано всем мужчинам, обращающимся по поводу гинекомастии. Обширное
обследование рекомендовано в отношении худощавых мужчин и тех, кто имеет
недавнее начало, быстрый рост, болезненную или очень большую ткань грудной
железы. Диагностическое обследование должно включать тщательный сбор анам¬
неза, обследование гонад, оценку вторичных половых признаков и обследование
функции печени.
Лабораторные исследования
Следует измерить активность печеночных ферментов, концентрации сывороточно¬
го Т, эстрадиола, андростендиона, ЛГ и ХГЧ (рис. 3-14). Если гонады маленькие, для
исключения синдрома Клайнфелтера нужно провести кариотипирование. Диагноз
устанавливают менее чем у половины пациентов, вероятно, из-за того, что изменения
Увеличение молочных желез
т
Верифицировать, что увеличение молочной железы
обусловлено разрастанием действительно железистой ткани
Начало в неонатальном или
перипубертатном периоде
Воздействие известного
причинного агента
Болезнь печени
Размер железистой ткани менее
4 см, не было недавнего роста
і
Периодическое повторное \
наблюдение
Низкий уровень Т,
высокое отношение Е/т
Обследовать на синдром
андрогенной недостаточности
Болезненность молочной
железы
Железистая ткань более 4 см
в диаметре
Отсутствие причинных
лекарственных средств
или болезни печени
Симптомы и признаки
андрогенной недостаточности
Определить сывороточные
концентрации тестостерона,
ЛГ и ФСГ, Е, и ХГЧр
і
Обследовать на опухоли,
секретирующие ХГЧр
Повышенный уровень ХГЧр
Твердое образование,
фиксированное к подлежащим
тканям, быстрый рост, увеличенные
подмышечные лимфатические узлы
Повышенный уровень ХГЧ(3
Повышенный уровень
нормальное содержаниеТ,
высокое отношение Е/Г
Повышенная ароматизация
андрогенов в эстрогены
(ожирение, феминизирующие
опухоли надпочечников,
опухоли из клеток Сертоли
или Лейдига, врожденное
нарушение регуляции
ароматазы)
Рис. 3-14. Обследование пациента с гинекомастией. Е2 — эстрадиол; ХГЧ-р — (3-субъединица челове¬
ческого хорионического гонадотропина; Т — тестостерон.
272
соотношения «эстрогены-андрогены» часто бывают неуловимыми. Большинство
коммерческих методик определения эстрогенов имеют недостаточную чувствитель¬
ность при низких мужских диапазонах концентрации гормона, что затрудняет обна¬
ружение малых изменений в уровнях эстрогенов и отношениях эстроген-Т.
ГИНЕКОМАСТИЯ И РИСК РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ
Риск рака молочной железы заметно повышен у мужчин с синдромом Клайнфелте¬
ра [107,108,455-461]. Хотя относительный риск рака молочной железы повышен у
мужчин с первичной тестикулярной дисфункцией и гиперэстрогенизмом, абсолютное
увеличение риска небольшое [462]. Ожирение — также важный фактор риска молоч¬
ной железы [462]. Неясно, повышен ли риск рака грудной железы у всех мужчин с
гинекомастией [462].
ЛЕЧЕНИЕ
Когда первичную причину удается выявить и откорригировать, увеличение грудной
железы обычно исчезает спустя несколько месяцев. Однако если гинекомастия сохра¬
няется более года, ее полное обратное развитие бывает невозможным, и наиболее
эффективно хирургическое вмешательство. Показания для хирургического лечения
включают тяжелые психологические и косметические проблемы, продолжающийся
рост или болезненность ткани грудной железы либо подозрение на озлокачествле-
ние.
Отдельные сообщения содержат ссылку на положительные эффекты ингибиторов
ароматазы при лечении пациентов с чрезмерной ароматазной активностью [463,464].
Однако опыт использования ингибиторов ароматазы у пациентов с гинекомастией в
значительной мере не оправдан. Хотя уменьшение размеров молочных желез было
описано при назначении тестолактона у мальчиков с пубертатной гинекомастией
[465] и в ходе открытого исследования у взрослых с идиопатической гинекомастией
[434], полное исчезновение гинекомастии является необычным при лекарственной
терапии. Количество исследуемых в этих открытых исследованиях было мини¬
мальным [434, 465]. В рандомизированном клиническом исследовании мальчиков
с пубертатной гинекомастией [466] количество пациентов с 50% и более снижением
общего объема молочных желез, измеренным посредством ультрасонографии, не раз¬
личалось значительно в группах анастрозола и плацебо.
У пациентов, которые имеют болезненную гинекомастию и не претендуют на
хирургическое лечение, терапия антиэстрогенами, такими как тамоксифен, снижала
боль и размер ткани молочной железы приблизительно у двух третей [467-471].
Также было показано, что тамоксифен предупреждает появление боли в молочных
железах и их увеличение у пациентов с раком простаты, которых лечили антиандро¬
генной терапией [467]. Ингибиторы ароматазы, по-видимому, бывают эффективны¬
ми на ранней пролиферативной стадии болезни; на поздних стадиях, когда фиброз
закрепляется, не следует ожидать, что лекарственная терапия приведет к уменьшению
размеров молочной железы. Необходимы плацебо-контролируемые исследования с
более сильными ингибиторами ароматазы. в частности анастрозолом, фадрозолом,
летрозолом или фроместаном.
Нарушения действия половых гормонов
Т служит не только как гормон, но и превращается во многих периферических
тканях в два активных метаболита — эстрадиол 17(3- и 5-ДГТ. Эффекты Т во многих
тканях опосредованы через эти метаболиты. Именно поэтому действие андрогенов
потенциально может быть нарушено в результате мутаций генов рецептора андроге¬
нов, стероид-5а-редуктазы, СУР19-ароматазы или рецептора эстрогенов.
273
МУТАЦИИ ГЕНА РЕЦЕПТОРА АНДРОГЕНОВ
Многие эффекты андрогенов опосредованы через связывание их с внутрикле¬
точным рецептором андрогенов, который действует как лигандзависимый фактор
транскрипции. Мутации гена рецептора андрогена были ассоциированы с широким
спектром фенотипических аномалий [40, 472J. Пациенты с полной нечувствитель¬
ностью к андрогенам имеют мужской псевдогермафродитизм, характеризуются жен¬
ской внешностью с хорошо развитыми молочными железами, женскими внешними
половыми органами, слепым вагинальным мешком и отсутствием волос в андроген¬
чувствительных областях [473J. Внутренние структуры, которые происходят из мюл-
леровых протоков, отсутствуют; таким образом, у этих пациентов нет матки, шейки
матки и маточных труб. Структуры вольфова протока также обычно отсутствуют. Эти
пациенты часто обращаются за медицинской помощью по поводу первичной амено¬
реи [473]. Иногда нео пустившиеся яички в паховом канале могут быть ошибочно рас¬
ценены как паховая грыжа, что требует дальнейшего обследования. Синдром полной
тестикулярной феминизации наследуется как Х-сцепленное рецессивное заболевание,
хотя треть пациентов не имеют семейной истории болезни и, вероятно, имеют мута¬
цию de novo [473].
Лица с синдромом частичной андрогенной нечувствительности могут иметь раз¬
личный фенотип [474]. Некоторые пациенты с частичной андрогенной нечувстви¬
тельностью имеют мужской фенотип и легкие аномалии, такие как гипоспадия,
гинекомастия и бесплодие; эта форма частичной андрогенной нечувствительности
также известна как синдром Рейфенштейна [475, 476]. Другие пациенты с частичной
андрогенной нечувствительностью обращаются по поводу неопределенных половых
органов или женской внешности и женских наружных половых органов с различны¬
ми степенями слияния и клитеромегалии [476]. Мутации рецептора андрогенов были
отмечены также у полностью маскулинизированных беплодных мужчин, имевших
нормальные гормональные профили [475].
У диц с кариотипом 46,XY и мутациями рецепторов андрогенов обычно повышен
уровень Т, ЛГ и эстрадиола, а сывороточные отношения Т к ДГТ и Т к эстрадиолу —
нормальные. Нормальные значения отношения Т к ДГТ отличают пациентов с мута¬
циями рецептора к андрогенам от тех, которые имеют мутации гена 5а-редуктазы;
последние имеют высокие соотношения Т к ДГТ.
Полные или крупные делеции гена рецептора к андрогенам, как правило, нети¬
пичны для лиц с синдромом полной нечувствительности к андрогенам; большинство
пациентов с формами частичной и полной андрогенной нечувствительности имеют
точечные мутации в нескольких различных местах в экзонах 2-8, кодирующих ДНК-
связывающий и андрогенсвязывающие домены, в гене рецептора к андрогенам, при¬
водящие к заменам аминокислот или преждевременному стоп-кодону [40, 472, 477].
Кроме того, сообщали о нуклеотидных вставках, ведущих к сдвигам рамки считыва¬
ния (фрейма) или преждевременной терминации, делециях и мутациях интронов в
донорских или акцепторных местах сплайсинга [477].
Мутации в ДНК-связывающем домене приводят к измененному протеину рецептора
андрогенов, который имеет нормальное связывание с гормоном, но является дефект¬
ным в димеризации или связывании ДНК, результатом чего служит уменьшенная
транскрипционная активность [472]. ДНК-связывающий домен содержит два цинко¬
вых пальца (штифта): первый цинковый штифт важен для связывания ДНК, а второй
вовлечен в протеин-протеин-взаимодействие и димеризацию [35]. Были описаны
мутации обоих цинковых пальцев (они происходят с равной частотой) [478,479].
Мутации лигандсвязывающего домена приводят либо к отсутствию, либо к умень¬
шенному связыванию гормона. Относительно большое количество мутаций было
описано в экзонах 5 и 7, но мутации в экзоне 1 и шарнирной области экзоны 4 встре¬
чаются редко.
274
Adachi и соавт. [480] описали пациента с фенотипическими признаками, характер¬
ными для синдрома андрогенной нечувствительности, но у которого не было обнару¬
жено мутации рецептора андрогенов. Сигнал активации от области AF-1 андрогеново¬
го рецептора был прерван, что свидетельствовало о дефекте в протеине коактивации,
взаимодействующем с этой областью [480].
Длины CAG и GCG тринуклеодитных повторов, которые кодируют полиглута-
минный и полиглицинный тракты в экзоне 1 гена рецептора андрогенов, обратно
коррелируют с транскрипционной активностью протеина рецептора андрогенов.
Аномальная длина полиглутаминного тракта (более 40 глутаминных остатков) ассо-
цирована со спинномозговой и бульбарной мышечной атрофией, так же известной
как болезнь Кеннеди [481,482]. Протеин рецептора андрогенов с увеличенным поли-
глутаминным трактом цитотоксичен для клеток, и его протеолитическое расщепление
вызывает смерть нейрональных клеток [483]. Мужчины с болезнью Кеннеди часто
имеют гинекомастию, повышенные концентрации Т и ЛГ и признаки сниженного
действия андрогенов. Хотя в некоторых исследованиях сообщали о связи продолжи¬
тельности полиглутаминного и полиглицинного трактов с мужским бесплодием (см.
раздел по бесплодию в этой главе) и риском рака простаты, другие специалисты не
подтвердили эти данные [484].
МУТАЦИИ ГЕНА СТЕР0ИД-5а-РЕДУКТАЗЫ
Пациенты с аутосомно-рецессивным типом 2 недостаточности стероид-5а-
редуктазы обычно являются мужчинами с 46,XY кариотипом, имеющими нормальные
мужские внутренние органы, включая яички, но у которых при рождении отмечают
неопределенные или женские внешние половые органы [10]. в пубертатном периоде у
этих детей начинается частичная вирилизация и некоторые идентифицируют себя как
мужчин, даже если они были воспитаны в раннем детстве как девочки. Это развитие
предполагает, что сам по себе Т может стимулировать психосексуальное поведение,
либидо, развитие эмбрионального вольфова протока, мышечное развитие, огрубение
голоса, сперматогенез и рост волос в подмышечной впадине и на лобке. В противопо¬
ложность этому ДГТ требуется для развития и роста простаты и для развития наруж¬
ных половых органов и роста волос на лице и теле по мужскому типу или облысения
по мужскому типу.
Все дети с дефицитом 5а-редуктазы, которые были обследованы к настоящему вре¬
мени, имели мутации 2-го типа стероид-5а-редуктазы, доминирующей формы в пред¬
стательной железе. У многих пациентов с врожденной недостаточностью 2-го типа
5а-редуктазы низкие или у нижней границы нормы концентрации ДГТ в пубертатном
периоде, что обусловлено либо активностью изофермента 1-го типа, либо резидуаль-
но низкой активностью изофермента 2-го типа. Неясно, обусловлено ли нормальное
развитие мышц и мужское психосексуальное поведение, наблюдаемое у этих пациен¬
тов после полового созревания, низкими уровнями ДГТ или самим Т.
Мутации 1-го типа стероид-5а-редуктазы не были описаны у человека.
МУТАЦИИ ГЕНА CYP19 АРОМАТАЗЫ
Продукт гена ароматазы катализирует финальную стадию превращения С19 поло¬
вых гормонов в С18 эстрогены п>тем конвертации кольца А молекулы стероида в
ароматическое кольцо. Эффекты Т на резорбцию трабекулярной кости, половую
дифференциацию мозга, липиды плазмы крови, прогрессирование атеросклероза,
эпифизиальное слияние и некоторые типы поведения опосредованы через превра¬
щение Т в эстроген. Пациентки с мутациями гена ароматазы маскулинизированы,
они не подвергаются женскому половому созреванию, имеют повышенные уровни
андрогенов, ЛГ и ФСГ, поликистозные яичники и высокий рост [31]. Пациенты
мужского пола с мутациями СУР19 имеют отсроченное эпифизиальное слияние, про¬
275
должающийся линейный рост, высокий рост, евнухоидные пропорции, остеопороз,
повышенный оборот костной ткани, нарушение сперматогенеза и бесплодие [30]. У
этих мужчин нормальные или повышенные уровни Т, повышенные концентрации ЛГ
и ФСГ и заметно сниженные уровни эстрадиола. Пациенты и мужского, и женского
пола с мутациями гена ароматазы также имеют нарушенную толерантность к глюкозе
и гиперинсулинемию [29,31].
Smith и соавт. [27] сообщили о единичном случае резистентности к эстрогенам, обу¬
словленной инактивирующей мутацией гена а-рецептора эстрогенов. Этот пациент
мужского пола отличался высокой фигурой, продолжающимся после пубертатного
периода во взрослом состоянии линейным ростом, неслившимися эпифизами, остео¬
порозом, нормальными наружными мужскими половыми органами, нормальными
концентрациями Т и повышенными концентрациями эстрадиола, ЛГ и ФСГ [27].
Лечение высокими дозами эстрогена, которое увеличило уровни эстрадиола до супра-
физиологического диапазона, было неэффективно в индуцировании созревания кости
или эпифизарного слияния.
БЕСПЛОДИЕ и СНИЖЕНИЕ ФЕРТИЛЬНОСТИ У МУЖЧИН
Определение и эпидемиология
ВОЗ определила бесплодие как неспособность сексуально активной пары достичь
беременности, несмотря на незащищенный половой акт во время фертильной фазы
менструального цикла, в течение периода более 12 мес [485]. Установлено, что коли¬
чество пар, обращающихся за медицинской помощью для лечения бесплодия, состав¬
ляет от 4 до 17% [486]. 3-4% всех пар остаются ненамеренно бездетными до конца их
репродуктивного периода.
Установление показателей распространенности бесплодия и снижения фертиль¬
ности решающим образом зависит от метода определения этих состояний [487,488].
Определение ВОЗ было основано на исследованиях, исходивших из времени воз¬
никновения беременности; обнаружили, что вероятность оплодотворения составляет
20% за цикл, или 85-90% за год [485, 489-492]. Даже среди пар, которые не зачали
в течение 12 мес, 55% имеют живого новорожденного в течение следующих 36 мес
[493]. Когда продолжительность бесплодия превышает 4 года, частота оплодотворе¬
ния за месяц падает до 1,5% [494].
Результаты двух исследований [495, 496] показали: частота спонтанного зачатия
выше, чем было установлено ранее. Например, в ходе проспективного исследования
с участием китайских женщин - работниц текстильной промышленности в возрасте
от 20 до 34 лет [495] 50% забеременели в первые два цикла и 90% - в первые шесть
циклов, давая месячную плодовитость около 30-35%. В проспективном исследова¬
нии немецких пар [496] установленная кумулятивная вероятность оплодотворения
составляла 38, 68,81 и 92% за 1,3, 6 и 12 циклов соответственно,
У 20% бесплодных пар основная проблема - у партнера мужского пола; в других
26% проблемы существуют у обоих партнеров; таким образом, партнер мужского
пола «вносит вклад» в бесплодность примерно в половине случаев. Распространение
бесплодия значительно влияет на взаимоотношения пары, качество жизни и расходы
на медицинские услуги.
Причины и патофизиология
Частота этиологических факторов варьирует в обзорах из разных центров из-за
различий в порядке направления пациентов, степени диагностического обследования
276
и географии: однако эти обзоры едины в том, что снижение фертильности и бесплодие
у мужчин — гетерогенная группа нарушений [497] (табл. 3-9). От 15 до 20% бесплодных
мужчин имеют азооспермию [498]. У других 10% выявлена тяжелая олигозооспермия,
когда концентрация сперматозоидов в эякуляте менее 1 млн/мл. Специфическую при¬
чину бесплодия невозможно определить у 40-60% мужчин [494]. Большинство бес¬
плодных мужчин имеют идиопатическую олигозооспермию [498, 499, 500].
Таблица 3-9. Типичные диагнозы у мужчин, обследованных на предмет бесплодия
Диагностическая категория
Частота, %
Идиопатическое бесплодие
50-60
Первичный гипогонадизм (хромосомные расстройства, включая синдром Клайнфелтера, микро-
делеции У-хромосомы, неопустившиеся яички, иррадиация, орхит, лекарственные средства)
10-20
Обструкция полового тракта (врожденное отсутствие семявыносящих протоков, вазэктомия,
обструкция придатка яичка)
Коитальные нарушения
<1
Гипогонадотропный гипогонадизм (аденомы гипофиза, пангипопитуитаризм, идиопатический
гипогонадотропный гипогонадизм, гиперпролактинемия)
3-4
Варикоцеле*
15-35
Другие (антиспермальные антитела, лекарственные средства, токсины, системное заболевание)
* Хотя варикоцеле наблюдают у бесплодных мужчин и при снижении фертильности с более высокой
частотой, чем у фертильных мужчин, его роль и вклад в развитие бесплодия остается неясным.
Адаптировано и модифицировано из: Вакег е! а1. [4] с разрешения [707].
Корректируемые и излечимые причины бесплодия, такие как недостаточность
гонадотропинов и обструкция, имеются только у малого количества мужчин, но
важно распознавать их, поскольку доступны эффективные методики лечения [500].
Варикоцеле наблюдают у 10-30% мужчин с бесплодием; его роль, какая бы она ни
была, в патофизиологии мужского бесплодия остается неясной. Увеличивается коли¬
чество генетических расстройств, вовлеченных в специфические аномалии развития
половых клеток; кроме того, ряд системных поражений влияет на сперматогенез. Из
них синдром Клайнфелтера и микроделеции ¥-хромосомы — наиболее распростра¬
ненные расстройства, поражающие вместе 10-20% пациентов (см. табл. 3-9). Хотя
распространенность антиспермальных антител у бесплодных мужчин выше, чем
у фертильных [501], механизмы, с помощью которых антиспермальные антитела
вызывают бесплодие, неизвестны.
ГЕНЕТИЧЕСКИЕ РАССТРОЙСТВА, АССОЦИИРОВАННЫЕ С БЕСПЛОДИЕМ
Генетические расстройства, ассоциированные с нарушенной секрецией
гонадотропина или их действия
Генетические расстройства, ассоциированные с нарушенной секрецией гонадотро¬
пина или их действия, подробно описаны в главе «Гипогонадизм». Этот раздел будет
сфокусирован только на генетических нарушениях развития половых клеток.
Первичные дефекты сперматогенеза
Нарушения половых хромосом
Около 5% бесплодных мужчин имеют хромосомные аномалии: большинство из
них включают половые хромосомы (в среднем 4%) и 1% — аутосомы [499, 502].
Распространенность аномалий половых хромосом и аутосом у бесплодных муж¬
277
чин соответственно в 15 и 6 раз выше, чем в общей популяции [95, 502]. Синдром
Клайнфелтера — наиболее частое хромосомное заболевание, ассоциированное с муж¬
ским бесплодием.
Синдром Нунан, синдром Тернера у мужгин
Такие пациенты с кариотипом 46,XY имеют мужские наружные половые органы и
клинические стигмы синдрома Тернера [503]. Размер яичек уменьшен, а уровни Т в
целом низкие. У этих мужчин обычно отмечают крипторхизм и бесплодие.
Синдром XYY
Кариотип 47,XYY часто встречается у высоких мужчин с узловыми кистозными
акне среди заключенных и пациентов психиатрических клиник [504]. Для 47,XYY-
мужчин харктерны низкий интеллект и более низкий уровень образования по сравне¬
нию со здоровыми лицами.
Смешанная дисгенезия гонад
Пациенты со смешанной гонадной дисгенезией обычно имеют кариотип 45,X/46,XY,
яички на одной стороне тела и гонаду-стрек на другой [505,506]. Наружные половые
органы часто имеют неопределенное строение [505]. У фенотипических мужчин со
смешанной гонадной дисгенезией часто наблюдают абдоминальные яички с нормаль¬
ными клетками Лейдига, но со сниженным количеством или отсутствием половых
клеток, в дисгенетичных гонадах высок риск неопластической дегенерации.
ХХ~мужгины
46,ХХ-мужчины имеют мужскую внешность и нормально выглядящие яички, но
эти пациенты обычно страдают азооспермией, а уровни ЛГ и ФСГ у них повышены
[507], У некоторых ХХ-мужчин часть Y-хромосомы, содержащая Sry ген, может быть
транслоцирована на Х-хромосому или аутосому; у немногих пациентов может отме¬
чаться мозаицизм и некоторые 46.XY-KneTKn. Эти люди обычно стерильны, посколь¬
ку у них нет других Y-специфичных генов, требуемых для сперматогенеза.
Микроделеции Y-xpoMOCOMbi
Во время тщательного исследования кариотипов Tiepolo и Zuffardi обнаружили,
что мужчины с большими промежуточными делециями длинного плеча У-хромосомы
были бесплодны: ученые предположили, что локус «фактора азооспермии» располага¬
ется на длинном плече Y-хромосомы. Крупные делеции длинного плеча Y-хромосомы,
которые видимы под световым микроскопом в позднюю профазу, необычны для бес¬
плодных мужчин. Однако мелкие делеции Y-хромосомы, невидимые под микроско¬
пом в позднюю профазу и поэтому называемые микроделециями, выявляют у 5-10%
бесплодных мужчин с азооспермией или тяжелой олигозооспермией [508-519]. Эти
микроделеции можно обнаружить посредством основанного на ПЦР картирования с
использованием меченых последовательностей [519].
Большинство бесплодных мужчин с Y-делециями имеют тяжелые дефекты сперма¬
тогенеза — они страдают либо азооспермией, либо тяжелой олигозооспермией [514,
516]. Значительная часть мужчин с Y-микроделециями имеют слегка уменьшенные
объемы яичек (менее 15 мл) и повышенные уровни ФСГ, однако у других пациентов
нормальные объемы яичек и уровни этого гормона [498, 508, 520]. Гистологические
исследования у мужчин с Y-делециями выявили присутствие либо только клеток
Сертоли, либо только половых клеток. Существует плохая корреляция между тести¬
кулярной гистологией и локализацией и размером микроделеций. Однако Vogt и кол¬
леги [514] сообщили, что в Yq можно идентифицировать три локуса, названные AZFa,
AZFb и AZFc, в которых делеции ассоциированы со специфичной гистопатологией
яичек.
Несколько семейств Y-специфичных генов, клонированные путем картирования
делений у бесплодных мужчин с Yq-делециями, были предложены в качестве канди¬
278
датов для предполагаемого AZF локуса; они включают семейство генов RBM {RNA
binding motif) [521,522] и DAZ {deleted in azoospermia) [510,523, 524]. Оба семейства —
многокопийные гены. Семейство генов RBM имеет более 30 копий, разбросанных по
всей Y-хромосоме, а большинство копий локализовано в интервале делеций 6А и 6В
[525]. По меньшей мере два члена семейства RBM генов - RBM-1 и RBM-2 — экспрес¬
сируются в яичках [522]. Гены играют роль в процессинге РНК. Тем не менее точная
роль RBM белков в ра.звитии половых клеток до конца не изучена.
Семейство генов DAZ — гены с многочисленными копиями [526]. Гомологи семей¬
ства DAZу мышей и Drosophila были картированы в хромосоме 17 и 3 соответственно
[527, 528]. Аутосомные гомологи семейства DAZ также были идентифицированы
у человека и картированы в хромосоме 3 [524, 523]. Гомологи аутосомного DAZ-
похожего гена, DAZL1, имеются у всех видов млекопитающих; гомологи DAZ при¬
сутствуют на Y-хромосоме только у высших обезьян и людей. Мутации гена DAZL1 у
Drosophila связаны с остановкой мейоза и а.зооспермией [527]. Схожим образом мыши
с отсутствием DAZL1 стерильны, что свидетельствует о важной роли белкового про¬
дукта DAZL1 в развитии половых клеток [529]. У бесплодных мужчин с делениями
DAZ были описаны картины остановки мейоза и поражения клеток Сертоли; возмож¬
но, дегенерация половых клеток происходит вторично.
Точная фи.зиологическая функция и роль семейств генов RBM и DAZ в сперма¬
тогенезе человека остается неясной. Молекулы РНК, которые служат целями РНК-
связывающих протеинов, не были полностью охарактеризованы. Используя DAZ в
качестве приманки в двугибридной системе, группа Tsui [530] идентифицировала
два новых протеина, DAZ-ассоциированные белки {DAZAP) 1 и 2, которые взаимо¬
действуют с DAZ и DAZL1. Гены DAZAP были картированы в хромосомных областях
19р13.3 и 2q33-q34.
Хотя делеции, включающие DAZ-гены, являются наиболее частыми у бесплод¬
ных мужчин с Yq-делециями, большая порция Y-делеций располагается вне области
DAZ; некоторые из них включают ЙВЛ4-гены. Дополнительные семейства генов-
кандидатов, включающие BPY2, CDYÏ, PRY и TTY2, были идентифицированы в AZFc
регионе Y-хромосомы. Роль этих дополнительных семейств Y-специфичных генов в
развитии половых клеток и бесплодия не была выяснена. Неясно также, как делеции
одной или двух копий RBM или DAZ могут объяснить бесплодие, когда имеются
многочисленные копии этих генов в других местах Y-хромосомы. Значительная часть
бесплодных мужчин с DAZ-делециями страдают олигозооспермией, а не азоосперми¬
ей. Кроме того, только 5-10% бесплодных мужчин имеют Y-делеции. Эти сведения
позволяют предположить, что дополнительные Y-специфичные и аутосомные гены
могут быть вовлечены в другие фенотипы бесплодия.
Хсугя Y-микроделеции были описаны у фертильных мужчин, частота Y-микроделеций
очень низкая у таких мужчин и пациентов с концентрацией спермы выше 1 млн/млн.
Длина полиглутаминного тракта белка рецептора андрогенов и бесплодие
Экзон одного человеческого рецептора андрогенов содержит два полиморфных
'факта: полиглутаминный и полиглицинный. Длина CAG тринуклеотидных повторов в
экзоне одного гена рецептора андрогенов, которые кодируют полиглутаминный тракт,
имеет обратную корреляцию с трансактивационной активностью протеина рецептора
андрогена [531, 532]. Некоторые сообщения свидетельствуют о том, что мужчины с
идиопатической олигозооспермией имеют большую вероятность более длинного Г[ОЛИ-
глутаминного тракта, нежели фертильные мужчины [531, 532]. Dadze и соавт. [533], с
другой стороны, не обнаружили взаимосвязи длины повтора CAG с нарушенным спер¬
матогенезом у исследуемой группы бесплодных мужчин германского происхождения.
Именно поэтому истинность этой гипотезы еще предстоит верифицировать.
Экспрессия гена модулятора звена цАМФ-ответа и остановка сперматогенеза
Ген модулятора звена цАМФ-ответа {CREM — cAMP-response element modulator)
кодирует фактор транскрипции, который экспрессируется в постмейотических поло¬
279
вых клетках и регулирует баланс между дифференциацией и апоптозом половых
клеток во время сперматогенеза [534]. В процессе сперматогенеза в постмейотиче-
ских половых клетках происходит переключение с CREM-репрессорных на CREM-
активаторные изоформы [534]. Биопсии яичек, взятые от бесплодных мужчин с
остановкой половых клеток, показывают отсутствие активаторных изоформ CREM в
постмейотических половых клетках и повышенный апоптоз [535, 536]. Эти данные
наводят на мысль, что остановка половых клеток может быть результатом нарушения
этого нормального перехода от репрессорных к активаторным изоформам CREM в
семенных канальцах. Мыши с мутациями СЯБМ-протеина бесплодны, у них обна¬
руживают блок сперматогенеза на поздних сперматидах и повышенное количество
апоптозных половых клеток в семенных канальцах [537].
ДРУГИЕ РАССТРОЙСТВА, АССОЦИИРОВАННЫЕ С БЕСПЛОДИЕМ
Двустороннее врожденное отсутствие семявыносящего протока и мутации СРТЯ
Большинство пациентов с классической легочной формой кистозного фиброза
страдают от обструктивной азооспермии, обусловленной врожденным отсутствием
семявыносящего протока. Дополнительно мутации области, кодирующей ген регу¬
лятора проводимости кистозного фиброза {(ystic fibrosis conductance regulator gene,
CFTR), могут привести к врожденному отсутствию протока без развития классической
легочной формы болезни [538]. Почти у 70% мужчин с врожденным отсутствием
семявыносящего протока наблюдают мутации гена CFTR. Около 50% гомозиготны по
типичным аномалиям гена кистозного фиброза, таким как F508, а некоторые имеют
составную гетерозиготность.
Дисфункция гонад, ассоциированная с сорловидноклвточной анемией
и ß-талассвмией
Значительная часть мужчин с серповидноклеточной анемией имеют низкие уров¬
ни Т. Большинство мужчин с серповидноклеточной анемией, которые имеют низкие
уровни Т, страдают первичной тестикулярной дисфункцией [539]. Считают, что
тестикулярная дисфункция обусловлена микроинфарктами в яичках, происходящи¬
ми в результате вазоокклюзивной болезни. Однако сообщали о гипогонадотропном
гипогонадизме, обусловленном гипоталамо-гипофизарной дисфункцией у мужчин с
серповидноклеточной анемией.
Гипофизарная и гонадная дисфункции случаются при талассемии из-за отложения
железа в этих тканях [540]. Гипогонадотропный гипогонадизм — доминирующая
форма синдрома андрогенной недостаточности у мужчин с талассемией и может
быть эффективно пролечена с помощью гонадотропной заместительной терапии.
Гйпофизарная и тестикулярная перегрузка и возникающий гипогонадизм могут быть
предотвращены с помощью профилактической железохелатирующей терапии.
Гипогонадизм при миотоничаской дистрофии
Распространение СГС-повторов в гене DMPK вызывает миотоническую дистро¬
фию, аутосомно-доминантное заболевание, характеризующееся миотонией, лобным
облысением, прогрессирующей мышечной слабостью и тестикулярной атрофией
[541]. 75% пациентов с миотонической дистрофией имеют тестикулярную атрофию,
главным образом обусловленную дегенерацией семенных канальцев. Хотя клетки
Лейдига сохранены, сывороточные уровни Т низкие у многих пациентов [542].
Снижение фертильности, связанное с диабетом
Мужчины с сахарным диабетом могут страдать бесплодием по многим причинам,
например из-за ретроградной эякуляции, обусловленной автономной нейропатией.
Эректильная дисфункция также широко распространена у мужчин, которые страдают
диабетом более 10 лет. У мужчин с сахарным диабетом 2-го типа чаще обнаруживают
более низкий уровень Т, чем у контрольных лиц соответствующего возраста [284].
280
КЛИНИЧЕСКОЕ ОБСЛЕДОВАНИЕ
Обследование бесплодной пары должно включать одновременно партнеров и
женского, и мужского пола (рис. 3-15). Начальное обследование должно быть сфо¬
кусировано на общем состоянии здоровья, сопутствующих медицинских проблемах
(эректильная дисфункция, сахарный диабет и автономная нейропатия), которые
могут быть ассоциированы с ретроградной эякуляцией, а также факторах образа
жизни (расстройства питания, чрезмерная физическая нагрузка) и злоупотребле¬
нии расслабляющими веществами (марихуаной, кокаином, опиатами и алкоголем).
Продолжительность бесплодия, раннее имевшиеся свидетельства фертильности муж¬
чины или женщины, использование контрацептивов, половая функция, частота и
время полового акта по отношению к менструальному циклу и половая практика
должны быть известны. Время пубертатного развития, частота бритья, потеря волос
и распределение волосяного покрова должны быть учтены. Дополнительно выясняют
историю травмы мошонки, мочеполовой инфекции, болезней, передающихся поло¬
вым путем, и хирургических вмешательств на мошонке или паховой области, включая
герниопластику и вазэктомию, рака, прежнего лечения рака химиотерапевтическими
препаратами и лучевого воздействия на паховую и мошоночную области.
Обследуют пациента в плане андрогенной недостаточности. Пропорции тела (отно¬
шения роста к размаху рук и верхнего сегмента к нижнему), голос (высокий или нет),
распределение волосяного покрова, включая низ живота, мышечная масса и тело¬
сложение, отсутствие или наличие гинекомастии могут указывать на гипогонадцзм.
Тестикулярный объем следует измерять с помощью орхидометра Прадера. Очень
маленькие яички могут указывать на синдром Клайнфелтера или тяжелую гонадо-
I Обследование лары
Мужчина I
Анамнез
и физикальное
обследование
Лабораторное
обследование
і Нормальное количество
сперматозоидов,
их подвижность
и морфология
Нормальное количество I
сперматозоидов,
аномальная подвижность:
или морфология |:
Общий анализ крови,
биохимический анализ
крови, ЛГ и ФСГ,
тестостерон,
анализ спермы
Аномальное
количество
сперматозоидов
I Обследовать партнера женского пола;
I если норма, провести специальные
! исследования функции сперматозоидов
Рис. 3-15. Обследование бесплодной пары. ФСГ — фолликулостимулирующий гормон; ЛГ — лютеини¬
зирующий гормон.
281
тропную недостаточность. Пациента осматривают для исключения крипторхизма,
варикоцеле или изменений семявыносящего протока. Для оценки размеров простаты
выполняют пальцевое ректальное обследование.
ЛАБОРАТОРНЫЕ ИССЛЕДОВАНИЯ
Лабораторные (рис. 3-15—3-17 [15-17]) исследования должны включать оценку
общего состояния здоровья путем проведения полного анализа крови, биохимическо¬
го анализа крови и анализа мочи [500,543]. Три и более образцов семени, полученных
путем мастурбации после по меньшей мере 48-часового периода воздержания следует
оценить с точки зрения обьема, концентрации семени и количества сперматозоидов,
их подвижности и морфологии. Согласно ВОЗ нормальный образец семени имеет
объем более 2 мл, концентрацию более 20 миллионов/мл и общее количество сперма¬
тозоидов более 40 млн в эякуляте. Более 50% сперматозоидов должны проявлять под¬
вижность вперед, а более 30% клеток должны иметь нормальную морфологию [544,
545]. Значимость лейкоцитов в сперме неясна. Лейкоциты в семени необязательно
свидетельствуют об инфекции желез дополнительной секреции.
Измерение уровней общего Т, ЛГ и ФСГ поможет диагностировать гипогонадизм
и определить, является ли он первичным или вторичным. Высокие уровни ЛГ и ФСГ
при наличии низких концентраций Т характерны для первичной тестикулярной недо¬
статочности. Мужчинам с первичной тестикулярной недостаточностью следует про¬
вести анализ кариотипа для исключения синдрома Клайнфелтера (47,ХХ¥) или его
вариантов. Повышенные уровни Т и ЛГ у мужчин, которые выглядят гипогонадными,
предполагают нечувствительность к андрогенам. Мужчинам с синдромом нечувстви¬
тельности к андрогенам следует провести анализ кожных фибробластов на связывание
андрогенов или ДНК периферических лимфоцитов для генетического тестирования
мутаций в генах рецептора андрогенов или 5а-редуктазы. Изолированное повышение
сывороточного содержания ФСГ с нормальными уровнями ЛГ и Т часто предполагает
селективное нарушение отделения половых клеток. Мужчины с гипогонадотропным
Аномальное количество слерматозоидов
или их морфология
Низкое
Нормальное
1 Низкое 1
содержание Т,
содержание Т
1 содержание Т, |
высокое ЛГ и ФСГ
и высокий ФСГ
[ ЛГиФСГ 1
,,
Нормальное
содержание Т,
ЛГ и ФСГ
Первичный
гипогонадизм
Кариотипирование
для исключения
синдрома Клайнфелтера
Поражение
яичек
Скрининг на наличие
микроделеций
У-хромосомы
См. рис. 3-13
Г
Рис. 3-16. Обследование бесплодных мужчин с аномальным количеством сперматозоидов и их мор¬
фологией.
282
Если подтверждено отсутствие
семявыносящего протока,
показано исследование
на наличие мутации СРТП
Рис. 3-17. Обследование мужчин с азоспермией с нормальными уровнями ФСГ, Л Г и Т. CFTR — транс¬
мембранный регулятор кистофиброза.
гипогонадизмом нуждаются в дополнительном обследовании для определения'при¬
чины гонадотропной недостаточности и исключения гиперпролактинемии, гипофи¬
зарных объемных поражений и гемохроматоза.
У мужчин с азоспермией и нормальными уровнями ЛГ и Т следует исключить
обструкцию мочеполового тракта, обусловленную врожденным отсутствием протока
или придатка яичка или приобретенным обструктивным нарушением (см. рис. 3-17).
Следует проверить постэякуляторную мочу для исключения ретроградной эякуляции.
Очень низкие концентрации фруктозы в семенной жидкости предполагают отсутствие
семенных пузырьков или их обструкцию. Таких пациентов направляют к урологу для
исследования и биопсии яичек для исключения обструкции или недостатка половых
клеток. Мужчин с врожденным отсутствием семявыносящего протока или семенных
пузырьков целесообразно протестировать на мутации в гене муковисцидоза.
СПЕЦИАЛИЗИРОВАННЫЕ ИССЛЕДОВАНИЯ ФУНКЦИИ СПЕРМЫ
Полезность таких исследований спермы ограничена, и эти тесты следует проводить
строго в специализированных лабораториях и только у мужчин, которые имеют нор¬
мальные уровни гормонов и низкое или нормальное количество сперматозоидов [485,
544, 545]. Исследования функции спермы включают тест на пенетрацию шеечной
слизи, тесты на акросомальную реакцию, пенетрацию свободной зоны яйцеклетки
хомячка и тест на связывание с zona pellucida человека. Клиническая польза теста на
акросомальную реакцию неясна. Положительный тест пенетрации ооцита хомячка
свидетельствует о том, что сперматозоиды способны подвергаться капацитации и
акросомной реакции и могут пенетрировать и сливаться с яйцеклеткой хомячка [544].
Хотя этот тест имеет хорошую конкордантность с оплодотворением in vitro, наблюда¬
ют высокую частоту ложноотрицательных результатов.
Компьютерные системы анализа семени удобны, но они не предоставляют реаль¬
ных преимуществ перед ручными методами в плане оценки морфологии сперматозои¬
дов, кроме того, они допускают ошибки в определении концентрации спермы [544].
283
БИОПСИЯ ЯИЧЕК
Тестикулярная биопсия может быть полезна у мужчин с азооспермией и нормаль¬
ными концентрациями Т и ЛГ, у которых следует исключить ретроградную эякуля¬
цию путем анализа постэякулярной мочи на наличие спермы и семенной жидкости
на наличие фруктозы. Если в семенной жидкости имеется фруктоза, следует провести
тестикулярную биопсию и исследование для исключения обструкции и установления
сперматогенеза в яичках.
Тестикулярная биопсия также полезна для забора сперматозоидов или сперматид
для внутрицитоплазматической инъекции сперматозоидов у мужчин с азооспермией,
у которых сперматозоиды не могут быть получены из эякулята. Была достигнута фер¬
тильность с помощью внутриплазматической инъекции сперматозоидов, забранных
из тестикулярного биоптата, у мужчин с азооспермией и синдромом Клайнфелтера, а
также другими заболеваниями.
ГЕНЕТИЧЕСКОЕ ТЕСТИРОВАНИЕ
Поскольку генетические расстройства составляют значительную часть причин
бесплодия у мужчин, генетическое тестирование бесплодного мужчины и потомков
будет иметь все большее значение, особенно у пар, у которых рассматривается вопрос
об внутриплазматической инъекции сперматозоидов (ICSI). Бесплодные мужчины,
которые страдают врожденным отсутствием семявыносящего протока или семен¬
ных пузырьков, должны быть обследованы на предмет мутаций гена муковисцидоза
[538].
Проводят скрининг бесплодных мужчин с азооспермией или тяжелой олиго¬
зооспермией, у которых причина бесплодия не может быть выяснена, на предмет
микроделеций У-хромосомы [520]. Обдумывают определение кариотипа у бес¬
плодного мужчины с необструктивной азооспермией, особенно если рассматривают
вопрос о проведении ICSI, Это обусловлено тем, что потомки, рожденные после
ICSI, имеют более высокую частоту анеуплодии половых хромосом, чем в общей
популяции [546]; кроме того, распространенность нарушений половых хромосом
существенно выше у бесплодных мужчин, особенно среди тех, которые страдают
азооспермией [502, 547].
Обсуждают мутации гена СШМ у мужчин с постмейотическим блоком половых
клеток. Многие системные заболевания (сахарный диабет, гемоглобинопатии и мио-
тоническая дистрофия) имеют генетическую основу; эти пациенты также нуждаются
в генетическом консультировании.
Эндокринолог может играть основную координирующую роль в лечении бес¬
плодной пары, начав рациональное диагностическое обследование для выяснения
специфической излечимой причины бесплодия и направив на обследование тех, кому
требуется более специализированная помощь. Особенно важно для эндокринолога
идентифицировать и лечить гонадотропную недостаточность, поскольку заместитель¬
ная гормональная терапия при этом расстройстве высокоэффективна. Эндокринолог
должен быстро выяснить, страдает ли пациент неизлечимой стерильностью, в слу¬
чае которой пара должна быть соответствующим образом проконсультирована на
предмет усыновления или искусственной инсеминации донорской спермой. Во всех
слу^іаях эндокринолог должен предоставлять паре реалистичный прогноз, «за» и
«против» различных вариантов лечения, давать оценку расходов и уводить пару от
вмешательств, заведомо неэффективных.
284
ИНДУКЦИЯ ФЕРТИЛЬНОСТИ МУЖЧИН с ГИПОГОНАДОТРОПНЫМ ГИПОГОНАДИЗМОМ
у пациентов с идиопатическим гипогонадотропным гипогонадизмом сперматогенез
может быть индуцирован путем назначения либо гонадотропинов, либо импульсных
инфузий гонадолиберина. И пульсирующая терапия гонадолиберином, и гонадотроп¬
ная заместительная терапия одинаково эффективны в индуцировании сперматог'енеза
у мужчин с идиопатическим гипогонадотропным гипогонадизмом [548-552].
Liu и соавт. [551] сравнили режим назначения ХГЧ/ЧМГ (хорионический гона¬
дотропин человека/человеческий менопаузальный гонадотропин) с пульсирующей
терапией гонадолиберином в плане их эффективности индуцирования спермато¬
генеза. После 2-летнего лечения 40% мужчин, леченных гонадолиберином, и 80%,
леченных ХГЧ/ЧМГ, продуцировали сперму. Концентрация спермы бьша сравнима
в обеих группах. Buchter и соавт. [553, 554] выяснили, что назначение ХГЧ/ЧМГ и
пульсирующая терапия гонадолиберином одинаково эффективны в индуцировании
сперматогенеза. Сперматогенез был индуцирован у 54 из 57 курсов терапии, беремен¬
ности развились у 26 из 36 пар.
Два вида терапии значительно не различалась ни во времени первого появления
семени, ни в частоте беременностей. Успешный терапевтический ответ на гонадоли¬
берин требует интактного гонадотропного механизма, поэтому эта форма лечения не
служит вариантом терапии у пациентов с пангипопитуитаризмом.
Препараты гонадотропина
ХГЧ выделяют из мочи беременных, он имеет главным образом активность ЛГ;
стимулирует клетки Лейдига продуцировать Т путем взаимодействия с рецепторами
ЛГ/ХГЧ. ХГЧ, произведенный из мочи постменопаузальных женщин, обладает актив - -
ностями ЛГ и ФСГ в равных пропорциях.
Высокоочищенные препараты человеческих ЛГ и ФСГ, выделенные из гипо¬
физов трупов человека, которые стали доступны для исследовательских целей от
Национального гипофизарного агентства NIDDK, более не рекомендованы. Развитие
болезни Крейцфельда-Якоба у небольшого количества пациентов, леченных ранни¬
ми препаратами человеческого гормона роста [555], привело к отзыву и прекращению
использования всех человеческих гипофизарных гормонов, полученных из трупов
для терапевтических целей. Высокоочищенные препараты человеческого ФСГ ком¬
мерчески доступны для использования в гинекологической сфере и были применены
для лечения мужчин с гонадотропным гипогонадизмом [549].
Рекомбинантный человеческий ФСГ, экспрессируемый линией яичниковых клеток
китайского хомяка, был одобрен Управлением по пищевым продуктам и лекарствам
США для индукции сперматогенеза у мужчин с идиопатическим гипогонадотропным
гипогонадизмом [556, 557]. Готовая {5-субъединица рекомбинантного человеческого
ФСГ имеет на семь аминокислот меньше, чем сообщается в опубликованных данных
[60], но она неотличима от очищенного человеческого ФСГ, полученного из мочи,
по биологической активности in vitro и in vivo. Фармакокинетика человеческого ФСГ,
полученного из мочи, и рекомбинантного ФСГ схожа [556].
Гонадотропиновая терапия
Лечение мужчин с идиопатическим гипогонадотропным гипогонадизмом начина¬
ют с 1000 Ед ХГЧ, назначаемого внутримышечно 3 раза в неделю. Измеряют сыво¬
роточные уровни Т через 6-8 нед после начала терапии ХГЧ и через 48-72 ч после
инъекции препарата. Подбирают дозу для достижения сывороточных уровней Т
средненормального диапазона. Во время терапии гонадотропином проводят монито¬
ринг количества сперматозоидов на ежемесячной основе.
Если после 6 мес монотерапии ХГЧ сывороточные уровни Т находятся в средненор¬
мальном диапазоне, но концентрации се.мени низкие, добавляют ФСГ. Это может быть
285
осуществлено с использованием ЧМГ, высокоочищенного ФСГ или рекомбинантного
ФСГ. Выбор дозы ФСГ является эмпирическим. Начинают с 75 МЕ препарата 3 раза
в неделю в сочетании с инъекциями ХГЧ.
Если после 3 мес комбинированной терапии концентрация спермы все еще низ¬
кая, увеличивают дозу ЧМГ или рекомбинантного ФСГ до 150 Ед 3 раза в неделю.
Предупреждают пациентов вначале о потенциальной продолжительности и стоимости
лечения и предоставляют консервативные оценки частоты успеха лечения, поскольку
оно может занять 18-24 мес или более, чтобы сперматогенез восстановился [548,549,
553, 557]. Пары часто проявляют нетерпение и преждевременно отказываются про¬
должать лечение.
Осложнения
Гонадотропная терапия обычно хорошо переносится и имеет низкую частоту
неблагоприятных эффектов, но она дорогостоящая. Неудача лечения, обусловлен¬
ная появлением антител к человеческому гонадотропному гонадотропину, является
необычным событием [558, 559], развивающимся менее чем у 1% леченых мужчин.
Гинекомастия — необычное осложнение терапии. Аллергические реакции чрезвычай¬
но редки.
Исходы
Двумя благоприятными прогностическими факторами гонадотропной терапии
у гипогонадотропных мужчин служат объем яичек при обращении к врачу и время
начала гипогонадизма (в препубертатный или постпубертатный период) [548, 551],
У мужчин с постпубертатным началом гипогонадизма сперматогенез обычно можно
повторно инициировать монотерапией ХГЧ; частота успеха при этом достаточно
высока [548]. С другой стороньг, мужчины, у которых гипогонадотропный гипогона¬
дизм развился до того, как период полового созревания завершился, обычно не отве¬
чают на монотерапию ХГЧ, а требуют комбинированной терапии ХГЧ и ФСГ более
продолжительное время, и средняя частота успеха ниже.
Прежнее лечение андрогенами, похоже, не влияет на последующий ответ на гона¬
дотропную терапию [560].
Степень гонадотропной недостаточности (что отражает объем яичек до лечения)
является важной детерминантой ответа на гонадотропную терапию. В целом мужчи¬
ны с объемом яичек более 8 мл имеют более высокую частоту ответов, чем те, у кого
этот показатель был менее 4 мл [551]. Наличие сопутствующей или ассоциированной
первичной тестикулярной аномалии будет ослаблять тестикулярный ответ на гона¬
дотропную терапию. Некоторые пациенты с идиопатическим гипогонадотропным
гипогонадизмом также страдают крипторхизмом; эти пациенты могут не отвечать
хорошо на терапию ХГЧ.
Пульсирующая терапия гонадолиберином
Пульсирующее назначение гонадолиберина требуется для поддержания нормально¬
го выделения ЛГ и ФСГ гипофизом [561]. Непрерывная инфузия гонадолиберина или
назначение агонистов гонадолиберина нарушает регуляцию секреции Л Г и ФСГ [561,
562]. Успех гонадолибериновой терапии предполагает нормальную функцию гипофи¬
за и гонад. Пульсирующее назначение гонадолиберина следует начинать с начальной
дозы 25 нг/кг на пульс, вводимой подкожно каждые 2 ч переносным насосом для инфу¬
зии [552, 563]. Следует проводить мониторинг уровней сывороточного Т, ЛГ и ФСГ.
Подбирают дозу гонадолиберина до тех пор, пока сывороточные уровни Т не войдут
в федненормальный диапазон. Существует значительная вариабельность требуемой
дозы гонадолиберина среди различных субъектов, и для индуцирования вирилизации
может потребоваться доза, варьирующая от 25 до 200 нг/кг массы тела [563].
После успешной индукции вторичных половых признаков доза гонадолиберина
может быть снижена без любого неблагоприятного эффекта на сывороточные уровни
Т, ЛГ и ФСГ. Секреция гонадотропина и функции гонад могут сохраняться в течение
286
протяженных периодов времени (от месяцев до нескольких лет) у большинства тща¬
тельно отобранных пациентов с идиопатическим гипогонадотропным гипогонадиз¬
мом с помощью пульсирующей терапии гонадолиберином [552, 563].
Осложнения
Появление антител к гонадолиберину служит редкой причиной неудачи лечения
[552, 563]. Краснота, уплотнение и подкожные инфекции в месте инфузии могут
развиться, но являются необычными, когда соблюдаются меры предосторожно¬
сти. Ношение портативного устройства для инфузий достаточно обременительно.
Последующее ведение этих пациентов часто требует значительного врачебного над¬
зора и лабораторного мониторинга.
Исходы
Об увеличении количества сперматозоидов и объема яичек сообщают у более чем
70% леченых мужчин, а улучшения половой функции и вирилизации могут быть
индуцированы у более чем 90% субъектов [552, 563]. Некоторые пациенты с идио¬
патическим гипогонадотропным гипогонадизмом имеют ассоциированный криптор¬
хизм, и эти мужчины, вероятно, не будут отвечать или будут отвечать субоптимально
на пульсирующую терапию гонадолиберином. Хотя индуцирование вирилизации с
помощью пульсирующей терапии гонадолиберином у пациентов с идиопатическим
гипогонадотропным гипогонадизмом обеспечило важное понимание механизмов
полового созревания и регуляции секреции гонадотропинов под действием гонадоли¬
берина, этот лечебный подход не имеет заметного преимущества перед гонадотроп¬
ной терапией [551, 554],
ВАРИАНТЫ ЛЕЧЕНИЯ ПАЦИЕНТОВ С АЗООСПЕРМИЕЙ
Прогноз у мужчин с азооспермией, тотальной тератозооспермией и первичной
тестикулярной недостаточностью с азооспермией плохой [564]. Для этих мужчин
усыновление и искусственная инсеминация с использованием донорской спер¬
мы служат благора.зумными вариантами. С появлением ICSI прогноз у мужчин с
азооспермией улучшился. Существуют сообщения о нескольких случаях успешных
беременностей у партнерш мужчин с азооспермией или синдромом Клайнфелтера,
достигнутых с помощью интрацитоплазматической инъекции спермы, полученной
при тестикулярной биопсии, в ооцит [565,566]. Palermo и соавт. сообщили о высокой
частоте успеха при внутрицитоплазматической инъекции спермы при использовании
сперматозоидов, хирургически забранных у мужчин с азооспермией [565]. Частота
успеха значительно варьирует среди центров в зависимости от выбора пациентов и
опыта проведения процедуры.
Пары, которые подвергаются процедуре ICS1 с использованием тестикулярных
сперматозоидов, должны быть проконсультированы на предмет риска передачи
анеуплодии половой хромосомы и других генетических нарушений потомкам [567].
Вспомогательные репродуктивньге технологии, включая внутриплазматическую инъ¬
екцию спермы, дорогие варианты лечения. Следует делать реалистичный прогноз и
уменьшать ожидания в начале лечения.
ВНУТРИЦИТОПЛАЗМАТИЧЕСКАЯ ИНЪЕКЦИЯ СПЕРМАТОЗОИДА
Внутрицитоплазматическая инъекция сперматозоида (ICSI) впервые была успешно
использована в 1992 г. для лечения бесплодия, сегодня она стала широко и.сщользуе-
мым вариантом лечения при идиопатическом мужском бесплодии во всем мире [565,
568-573]. В 2001 г. более 114 ООО лечебных циклов внутриплазматической инъекции
сперматозоида было проведено в 539 центрах по всему миру [574].
Обзор, проведенный Европейским обществом по репродукции человека и эмбрио¬
логии, показал, что клиническая частота беременностей для каждой аспирации и
переноса при ICSI составила в 2002 г. 27 и 29% соответственно [575, 576]. Частота
287
многоплодной беременности составляет 25-35% [577]. Показатели фертильности
лучше при использовании эякулированной и эпидидимальной спермы, чем тести¬
кулярных сперматозоидов [570]. Частота беременностей при обструктивной и необ¬
структивной азооспермии одинакова. Частота ооцитов, поврежденных при процедуре,
остается низкой (ниже 10%). Пока.затель частоты беременностей на перенос выше
для свежей спермы, нежели для криоконсервированного семени, и выше для свежих
эмбрионов, чем для замороженных-размороженных эмбрионов. На результаты вну¬
трицитоплазматической инъекции спермы влияют возраст партнера женского пола и
качество ооцита. Частота успеха внутриплазматической инъекции спермы значитель¬
но ниже у мужчин, сперма которых была забрана из яичек путем биопсии, и у мужчин
с некро- или глобозооспермией.
Широкое распространение методики внутрицитоплазматической инъекции спер¬
мы привело к появлению озабоченности в плане безопасности этого новейшего
метода. Европейское общество репродукции человека и эмбриологии распорядилось
ежегодно собирать сведения о клинических результатах, исходах беременностей и
последующем наблюдении за детьми, родившимися в результате использования дан¬
ной методики, для изучения этих вопросов.
Многоплодная беременность с рисками низкой массы тела при рождении и пре¬
ждевременных родов является частым осложнением циклов ЭКО/ICSI [569]. Риск
гинекологических и перинатальных осложнений выше при беременностях, возник¬
ших в результате внутрицитоплазматической инъекции семени, чем при беременно¬
стях, появившихся естественным путем [578]. Частота хромосомных аномалий выше
у детей, родившихся в результате внутрицитоплазматической инъекции спермы, чем
у контрольных лиц; существует также небольшое, но значимое увеличение частоты
хромосомной анеуплодии, особенно анеуплодии половой хромосомы, среди потомков
при внутрицитоплазматической инъекции спермы [568, 570, 579-581].
Частота врожденных мальформаций схожа при ICSI и ЭКО (оплодотворение
in vitro) [580]. Небольшое увеличение частоты врожденных мальформаций по сравне¬
нию с естественным оплодотворением было объяснено более высокой частотой мно¬
гоплодных родов. Когда принимают во внимание многоплодность, частота крупных и
малых мальформаций не повышена. Однако даже среди одиночных новорожденных,
родившихся в результате вспомогательных репродуктивных технологий, риск низкой
массы при рождении, преждевременных родов и неблагоприятных перинатальных
исходов повышен [569, 572, 573,578,579].
Исходы развития детей, рожденных после внутрицитоплазматической инъекции
спермы, в возрасте 2 и 5 лет значительно не отличаются от тех, которые рождены
после ЭКО [571, 582]. Также некоторые сообщения позволяют предполагать повы¬
шенный риск болезней импринтинга, гипоспадии и некоторых типов детского рака
[583, 584]; эти моменты требуют дальнейшего исследования. Долгосрочные данные
об умственном и физическом самочувствии детей, рожденных путем использования
внутрицитоплазматической инъекции спермы, недоступны.
Самопроизвольные беременности у бесплодных женщин ассоциированы с более
высоким риском акушерских осложнений и перинатальной смертности, чем спон¬
танные беременности у фертильных женщин. Именно поэтому неясно, являются ли
осложнения, наблюдаемые при беременностях и родах после внутрицитоплазматиче¬
ской инъекции спермы, результатом самой процедуры или бесплодия родителей.
Внутрицитоплазматическая инъекция спермы может быть эффективна во многих
случаях мужского бесплодия. Однако эта методика является дорогой и сложной про¬
цедурой, долгосрочная безопасность которой все еще не ясна. Пары должны быть
проконсультированы перед их направлением на ICSI [578]; усыновление или искус¬
ственная инсеминация донорской спермы всегда служат благоразумными альтерна¬
тивами. Пары также должны быть подвергнуты генетическому консультированию
288
и тестированию. У мужчин с идиопатической необструктивной олигозооспермией
или азооспермией следует провести кариотипирование и тестирование на наличие
микроделеции Y-хромосомы перед процедурой ЭКО/ICSI; некоторым пациентам
может быть показано тестирование на дополнительные гены-кандидаты. Пациенты
с обсгруктивной азооспермией должны быть обследованы с точки зрения мутации
в CTRF гене. Дополнительно пренатальное тестирование и забор образцов ворсинок
хориона могут быть показаны парам, подвергающимся процедуре ICSI.
ОБСТРУКТИВИЫЕ ПОВРЕЖДЕНИЯ
Микрохирургические техники для лечения обструктивных нарушений значительно
улучшились. Хотя процент успеха восстановления проходимости высокий (70-90%),
частота беременностей остается значительно ниже (40-50%).
ВАРИКОЦЕЛЕ У БЕСПЛОДНЫХ МУЖЧИН
Варикоцеле имеется у 10-30% бесплодных мужчин; однако оно также встречается
у фертильных мужчин. Именно поэтому роль варикоцеле в патофизиологии мужского
бесплодия остается неясной [585-589].
Хотя многие неконтролируемые исследования сообщают об улучшении концентра¬
ции спермы и подвижности сперматозоидов после резекции варикоцеле, эти данные
сложны для интерпретации из-за недостатка соответствующей контрольной группы.
Многие исследования варикоцеле имеют недостаточную на)щную строгость и неинтер-
претируемы. Было проведено немного контролируемых клинических исследований, в
ходе которых не удалось показать значительного более значительного улучшения фер¬
тильности после хирургической резекции варикоцеле, чем после только консультирова¬
ния [585,586]. Именно поэтому недостаточно данных для поддержания рекомендации
хирургической резекции варикоцеле у большинства бесплодных мужчин. Однако при
отсутствии других альтернатив врачи часто не могут преодолеть искушения рекомендо¬
вать резекцию варикоцеле или инъекцию склерозирующего агента.
Эксперты-урологи отдают предпочтение хирургической коррекции варикоцеле у
мальчиков подросткового возраста; рациональная гипотеза заключается в том, что
после варикоцелэктомии существует возможность наверстать рост яичек, что может
не случиться без хирургической коррекции. Однако проспективных данных клиниче¬
ских исследований недостаточно.
НОВООБРАЗОВАНИЯ ЯИЧЕК
Эпидемиология
Опухоли яичек составляют 1-2% всех злокачественных опухолей у мужчин, хотя
у лиц молодого и среднего возраста они составляют более высокий процент всех
видов рака [590, 591]. Распространенность этих заболеваний выше у представителей
белой расы (по сравнению с негроидной), а наиболее высокие показатели частоты
рака яичек отмечают в Дании [590, 591]. Показатели заболеваемости тестикуляр¬
ными герминомными опухолями устойчиво повышаются у белых мужчин, особенно
в Северной Европе, и они удвоились по сравнению с 1960-ми гг. [592]. Риск тести¬
кулярной неоплазии повышен в 50 раз у мужчин с крипторхизмом или‘ранее кор¬
ригированным крипторхизмом [593]. Атрофия яичек, паховая грыжа и бесплодие
служат дополнительными факторами риска тестикулярных опухолей из половых
клеток [594]. Среди членов семьи мужчин с тестикулярными опухолями из половых
клеток отмечают более высокую распространенность паховых грыж, крипторхизма
и гидроцеле, предполагая, что эти состояния могут быть причинно связаны с общим
289
дефектом в урологическом развитии [595]. Опухоли надпочечников развиваются
преимущественно у мужчин с плохо контролируемой врожденной ВДКН.
Опухоли яичек из половых клеток возникают во время фетального развития из
клеток линии половых клеток, которые останавливаются в период их созревания
[596]. Эти опухоли, как правило, прогрессируют по завершении неинвазивной стадии,
называемой неклассифицированной интратубулярной герминальной неоплазией или
карциномой in situ [596,597]. Примордиальные половые клетки во время фетального
развития подвергаются значительному импритингу; многие из этих моделей импри-
тинга отсутствуют в тестикулярных герминомных опухолях [598]. Таким образом,
часто наблюдалось гипометилирование многих генов, включая 5' конец гена XÎST при
семиномных и несеминомных опухолях [599].
Тестикулярные герминомные опухоли обычно проявляют анеуплодию, а неклас¬
сифицированные интратубулярные герминальные неоплазии часто триплоидны [596,
597, 600]. Усиление функции нескольких генов, локализованных на хромосомном
плече 12р, либо из-за изохромосомы, либо как результат амплификации, гена являет¬
ся обычной находкой при тестикулярных герминомных опухолях [601, 602]. Анализ
других исследований показал измененную экспрессию нескольких генов на 12р,
включая CCND2, регулятор клеточного цикла; NANOG и STELLAR, которые вовле¬
чены в поддержание мультипотентности половых клеток [603, 604]; гены в других
областях хромосом, включая KIT, STAT и RAS, которые регулируют пролиферацию и
ми1’рацию половых клеток [605, 606]. Молекулярная основа высокой чувствительно¬
сти опухолей из половых клеток к терапии цисплатином полностью непонятна; повы¬
шенная экспрессия р53 и недостаточность механизмов восстановления нуклеодитных
эксцизий были предложены в качестве возможных путей [607, 608].
Большинство тестикулярных опухолей имеют герминомное происхождение; негер-
миномные опухоли яичек относительно редки [596]. обычные типы гермином¬
ных опухолей включают семиному, эмбрионально клеточную карциному, тератому,
хорионкарциному и смешанные опухоли, которые состоят более чем из одного типа
клеток. Негерминомные опухоли яичек включают неоплазмы из клеток Лейдига и
Сертоли, опухоли добавочного надпочечника, стромальные опухоли и смешаннокле¬
точные опухоли.
Карцинома in situ яичек характеризуется наличием анеуплоидных злокачествен¬
ных половых клеток с ясной цитоплазмой, локализованных в пределах семенных
канальцев, вдоль базальной мембраны трубочек или в просвете канальцев [596,
597]. Приблизительно 50% неклассифицированных интратубулярных герминальных
неоплазий могут развиться в злокачественные герминомные опухоли в течение 5 лет,
а большинство развиваются в злокачественные опухоли в пределах 7 лет. Эти данные
указывают на злокачественную природу и инвазивный потенциал неклассифициро¬
ванных интратубулярных герминальных неоплазий и позволяют предположить, что
большинство герминомных опухолей яичек развивается из этого типичного предше¬
ствующего повреждения.
Опухоли яичек обычно обнаруживают посредством пальпации твердой тестикуляр¬
ной массы или метастазов [596]. От 5 до 10% опухолей из половых клеток развивают¬
290
ся в экстрагонадных местах, главным образом ретроперитонеально или в средостении
[609J. Внутричерепные опухоли из половых клеток редки и в целом локализованы в
шишковидной железе или надседловидной области [610].
Многие опухоли из половых клеток секретируют рХГЧ или а-фетопротеин, а
немногочисленные опухоли секретируют карционэмбриональный антиген, которые
могут служить в качестве полезных маркеров опухолей для диагностики, лечения и
последующего наблюдения этих опухолей после химиотерапии.
Опухоли из клеток Лейдига или Сертоли могут выделять андрогены или эстрогены;
пациенты с этими опухолями могут обращаться по поводу изосексуального ранне¬
го развития иди гинекомастии. Герминомные опухоли, секретирующие ХГЧ, также
могут манифестировать изосексуальным ранним половым созреванием у мальчиков
или гинекомастией у мужчин.
Диагностика основана на гистологической оценке биопсии яичек, хотя было опи¬
сано применение тонкоигольной аспирации для обнаружения карциномы in situ. У
мальчиков, обращающихся с преждевременным половым созреванием или гинеко¬
мастией, следует определить сывороточные уровни Т, эстрадиола и ХГЧ. Опухоли из
клеток Лейдига секретируют и Т, и эстрогены, а из клеток Сертоли обычно только
эстрогены. У пациентов с опухолями из половых клеток следует измерить содержа¬
ние ХГЧ, а-фетопротеина и карционэмбрионального антигена, поскольку, если они
имеются, эти опухолевые маркеры мог>т быть полезны в последующем наблюдении
ответа на терапию и для ранней диагностики рецидивов.
Опухоли яичек из половых клеток хорошо отвечают на лечение, даже когда паци¬
ент обращается с запущенной болезнью [590, 591]. Орхидэктомия или лучевое лече¬
ние — эффективное лечение для пациентов с односторонней неклассифицированной
интратубулярной герминальной неоплазией, а низкодозовое облучение рекомендова¬
но пациентам с двусторонней неклассифицированной интратубулярной герминаль¬
ной неоплазией [590].
Лечением тестикулярных герминомных опухолей служит хирургическое удаление
первичного очаг а путем орхидэктомии с последующей адъювантной терапией [590].
Выживаемость при семиноме I стадии составляет 98% и выше [596]. Радиотерапия на
околоаортальные и паракавальные лимфатические узлы ассоциирована с частотой
излечения 95-98%. Аналогично монотерапия карбоплатином была ассоциирована
с очень низкой частотой рецидивов [611]. На стадии семином ПА/В орхидэктомия
с последующей лучевой терапией обеспечивает частоту излечения 90-95% [612].
Стандартное лечение запущенного процесса состоит из химиотерапии первой линии
цисплатином, этопозидом и блеомицином с последующим хирургическим вмешатель¬
ством в случаях резидуальной опухоли [590, 596]. Из тех больных, кто не отвечает
или имеет остаточный патологический процесс после химиотерапии первой линии,
химиотерапия второй линии может спасти значительную часть. Почти 80% мужчин
с заболеванием в запущенной стадии могут достичь излечения при соответствующем
лечении [590,596].
У пациентов с несеминомными опухолями из половых клеток 5-летняя выживае¬
мость составляет 94, 83 и 71% соответственно для опухолей с хорошим, промежуточ¬
ным и плохим прогнозом [613]. Даже у пациентов, считающихся рефрактерными
к препаратам платины, было отмечено, что частота ответов на комбинированную
терапию, включавшую гемцитабин и паклитаксель или оксалиплатин-гемцитабин,
составляла до 45% [613].
Краткосрочные токсические эффекты системной химиотерапии опухолей яичка
из половых клеток — миелосупрессия, тошнота и рвота, легочная токсичность,
291
нефротоксичность и нейропатия. Долгосрочные токсические эффекты включают
ототоксичность, нарушение фертильности, нейропатии, сердечно-сосудистые заболе¬
вания и вторичные новообразования. Из-за молодого возраста этих пациентов и их
относительно хорошего прогноза в плане долгосрочной выживаемости фокус следует
сместить на минимизацию токсичности лекарственных средств.
Точную распространенность использования андрогенных стероидов спортсменами
трудно определить, поскольку данные основаны на самостоятельном сообщении,
и многие лица могут не признаваться в использовании этих препаратов из боязни
обвинения. Среди профессиональных спортсменов и олимпийцев в обзорах выяв¬
лена высокая частота использования андрогенных стероидов [614, 615]. От 4 до 6%
старшеклассников и 1-2% старшеклассниц сообщают, что по меньшей мере один раз
использовали андрогенные стероиды [616-619]. Аналоптчно высокие показатели
применения андрогенных стероидов были выявлены в исследованиях, проведенных
в других странах [620,621].
Распространенность использования андрогенных стероидов у девочек увеличилась
с 1980-х гг, [615,616]. В исследовании, опубликованном в 1993 г., УезаИз установил, что
примерно 1 млн американцев в некоторые периоды их жизни назначали андрогенные
стероиды [622]. Наиболее вопиющий пример спонсированного государством допинга
андрогенными стероидами был раскрыт в прежней Германской Демократической
Республике после падения коммунистического режима [623].
Хотя известно, что злоупотребления были с большим количеством андрогенных
стероидов, пять наиболее часто используемых с этой целью андрогенных стероидов
включают Т, нандролон, станозолол, метандиенон и метенолол. Обзоры добровольно
признанного использования препаратов спортсменами и бодибилдерами свидетель¬
ствуют о нескольких типичных моделях [624-625].
» Внутримышечные лекарственные формы андрогенных стероидов применяют
намного чаще, чем формы для приема внутрь [624]: приблизительно 80% спор¬
тсменов, сообщающие о применении андрогенных стероидов, делали самостоя¬
тельные внутримышечные инъекции.
« Комбинации андрогенных стероидов применялись более часто, чем одиночные
препараты [624,626]. Обычно один или более андрогенных стероидов назначают
в прогрессивно увеличивающихся дозах в течение нескольких недель, что в прак¬
тике известно как «нагружение».
* Спортсмены и бодибилдеры используют супрафизиологические дозы Т или
других андрогенных стероидов. В одном из исследований [624] 50% лиц, при¬
меняющих андрогенные стероиды, сообщали об использовании по меньшей
мере 500 мг Т в неделю или эквивалентной дозы другого андрогенного стероида:
в другом исследовании [626] почти четверть лиц, применяющих андрогенные
стероиды, использовали 1000 мг Т еженедельно или эквивалентную дозу другого
андрогенного стероида.
• Многие лица, применяющие андрогенные стероиды, злоупотребляют другими
препаратами, которые воспринимаются в спортивном сообществе как улучшаю¬
292
щие наращение, форму мышц или работоспособность [624]. Эти дополнительные
препараты включают стимуляторы, такие как амфетамин, кленбутерол, эфедрин
и тироксин, другие анаболические вещества, в частности гормон роста, ИФР-1
и инсулин, а также препараты, считающиеся уменьшающими неблагоприятные
эффекты, такие как ХГЧ, ингибиторы ароматазы или антагонисты эстрогенов
[624]. Потенциальные неблагоприятные последствия некоторых дополнитель¬
ных препаратов могут быть более серьезными, чем у андрогенных стероидов.
Андрогенные етерощы ш тпщшвныв результаты
Андрогенные стероиды увеличивают мышечнзто массу, произвольное максималь¬
ное напряжение и силу [350, 627, 628]; выигрыш в мышечной массе и напряжении
зависит от дозы [627]. Именно поэтому можно считать, что эти препараты улучшают
производительность в видах, таких как подъем тяжестей (пауэрлифтинг), в которых
производительность зависит от мышечного напряжения. Неудивительно, что сообща¬
ли о высокой распространенности использования андрогенных стероидов среди лиц,
занимающихся пауэрлифтингом. Бодибилдеры используют андрогенные стероиды
для увеличения массы скелетных мышц и снижения массы жировой ткани, которое
обеспечивает более выраженную рельефность мышц.
Применение андрогенных стероидов спортсменами, участвующими в видах спор¬
та, требующих выносливости, таких как бег на длинные дистанции и велогонки,
малообоснованно, поскольку не было показано, что андрогены улучшают пре¬
делы выносливости, такие как пороговый уровень лактата и [629, 630].
Андрогенные стероиды увеличивают уровни гемоглобина, который теоретически
улучшает кислород-переносящую емкость крови, но даже это не было показано в
контролируемых исследованиях [631]. Специалисты выдвигают гипотезу о том, что
андрогенные стероиды могут позволить спортсменам тренироваться усерднее благо¬
даря улучшению регенеративного ответа скелетных мышц на повреждение и их моти¬
вационным эффектам (эти гипотезы не были точно протестированы).
Широко распространенное использование андрогенных стероидов бейсбольными
игроками и спринтерами также нельзя легко объяснить на основании имеющихся
данных об эффектах андрогенных стероидов. Способность ударить по мячу, чтобы
послать его со скоростью 100 миль в час за пределы поля, требует экстраординарной
степени координации «руки-глаза», — способность локализовать мяч в определенной
точке пространства и направить биту в точно определенное место со значительным
напряжением и силой. Имеются отдельные подтверждения того, что андрогенные
стероиды уменьшают время реакции путем улучшения нейромышечной передачи
[632, 633]. Улучшенное время реакции в сочетании с повышенным напряжением и
силой могут потенциально объяснить ощущаемое улучшение спортивной произво¬
дительности игроками в бейсбол, хотя свидетельств для поддержания этой гипотезы
недостаточно.
Аналогично у спринтеров, у^іаствующих в соревнованиях по бегу на 100 метров,
увеличение массы тела, индуцированное андрогенами, может быть рассмотрено
как контрпродуктивная мера, поскольку повышенная масса тела будет увеличивать
количество совершенной работы при переносе этой массы тела против силы тяже¬
сти и сопротивления вдоль беговой дорожки. Тогда почему легендарные спринтеры,
наподобие Бена Джонса, применяли андрогены? И снова улучшенное время реакции,
повышенный психологический предел из-за мотивационных эффектов андрогенов и
способность усерднее тренироваться были предложены в качестве возможных объ¬
яснений без поддающихся проверке свидетельств.
293
йвтещшвлъмът и&бтгвщштиьт вффшы штвпьвтшшш^
андрогенных стероадов
Систематические исследования неблагоприятных эффектов андрогенных стерои¬
дов у .спортсменов и бодибилдеров являются редкими. Этим исследованиям препят¬
ствуют огромная вариабельность типов используемых препаратов, дозы, частоты и
продолжительности, возраста начала применения и конкурирующее использование
дополнительных препаратов. Более того, правдивость добровольного признания
использования препаратов всегда подозрительна.
Удивительно, что частота серьезных неблагоприятных эффектов, ассоциирован¬
ных с использованием андрогенных стероидов, была такой же низкой, как об этом
и сообщали. Был описан ряд летальных исходов, обусловленных неожиданными
сердечно-сосудистыми и цереброваскулярными осложнениями среди лиц, применяв¬
ших андрогенные стероиды [634, 635]; но эти сообщения по большей степени мало¬
вероятны и не установлена причинно-следственная в.заимосвязь. Неблагоприятные
эффекты, ассоциированные с использованием андрогенных стероидов, включают
вредные изменения факторов сердечно-сосудистой системы, включая выраженное
снижение уровня липопротеидов высокой плотности [636], факторов свертывания
крови [637], угнетение сперматогенеза, приводящее к бесплодию, и увеличение актив¬
ности ферментов печени [638-640].
Изменения липидов плазмы крови варьируют в зависимости от дозы, способа
введения (оральный или парентеральный), может ли андроген быть ароматизирован
или нет. Таким образом, 17-а-алкилированный неароматизируемый андроген, при¬
нятый внутрь, приводит к более выраженному снижению уровней холестерина высо¬
кой плотности в плазме крови, чем парентерально назначенный Т. Схожим образом
подъемы активности печеночных ферментов, новообразования печени и печеночная
пурпура были описаны при приеме внутрь 17-а-алкилированных андрогенных сте¬
роидов [638, 639], а не при парентеральном введении Т или его эфиров [348].
Опубликованы сообщения о свя.зи андрогенных стероидов с «реакциями гнева».
Однако плацебо-контролируемые исследования Т не продемонстрировали статисти¬
чески достоверного увеличения показателей гнева или агрессивного поведения [641-
646]. Для измерения агрессивного поведения применяли различные инструменты, и
возможно, что самостоятельно заполняемые опросники были недостаточно чувстви¬
тельны для обнаружения небольших, но значимых изменений агрессии. Замечено, что
небольшое количество участников контролируемых исследований продемонстриро¬
вали выраженное увеличение показателей агрессии при назначении супрафизиологи-
ческих доз Т, но большинство участников показали небольшое изменение или отсут¬
ствие такового. Возможно, что высокие дозы андрогенных стероидов провоцируют
реакции гнева в подгруппе пациентов с предсуществующей психопатологией.
Коип и соавт. [645] сообщили о том, что назначение супрафизиологических доз
(600 мг в неделю) Т-энантата здоровым молодым мужчинам связано со значитель¬
ным увеличением агрессивности по сравнению с плацебо. Дозы Т. которые в среднем
равны замещающим или были слегка выше, не вызвали значительных изменений в
агрессивном ответе. В этом исследовании участников попросили сы!рать в игру про¬
тив вымышленного оппонента (участники не знали, что оппонент был вымышлен),
в которой можно было нажать клавишу А для получения финансовой награды или
клавишу В, которая забирала бы деньги у вымышленного оппонента (агрессивный
ответ). Даже хотя целью игры было достижение наибольшего денежного выигрыша
и наилучшей стратегией для этого было нажатие клавиши А, участники, получавшие
супрафизиологические дозы (600 мг в неделю) Т-энантата, выбирали клавишу В с
более высокой частотой (для наказания вымышленного оппонента) и, таким образом,
имели более высокие показатели агрессивного ответа, чем те, кто получал меньшие
дозы Т или не получал его [645].
294
Назначенные внутрь 17а-алкилированные андрогены также были ассоцииро¬
ваны с резистентностью к инсулину и нарушением толерантности к глюкозе [647].
Существуют беспокойства по поводу потенциальных долгосрочных эффектов на
риск заболеваний простаты и сердечно-сосудистой системы, однако долгосрочные
эффекты супрафизиологических доз андрогенных стероидов неизвестны. Сообщали
об увеличениях массы левого желудочка среди лиц, использующих андрогенные сте¬
роиды [648]; неизвестно, является ли увеличение массы левого желудочка полезным
или вредным.
Болезненность и увеличение молочной железы («волчий сосок» на уличном жар¬
гоне) часто связаны с использованием ароматизируемых андрогенных стероидов. Для
спортсменов не является чем-то необычным использование ингибиторов ароматазы
или антагонистов эстрогенов в комбинации с андрогенными стероидами для преду¬
преждения увеличения молочных желез.
Назначение андрогенных стероидов угнетает продукцию эндогенного Т и семени
путем супрессии гипоталамо-гипофизарно-тестикулярной оси [649, 650], Мужчины,
применяющие андрогенные стероиды, могут страдать пониженной фертильностью
или бесплодием [651]. После прекращения экзогенного введения андрогенов вос¬
становление гипоталамо-гипофизарно-тестикулярной оси может занять промежуток
времени от недель до месяцев в зависимости от дозы и продолжительности приме¬
нения андрогена [652-655]. После прекращения приема андрогенов циркулирующие
уровни Т очень низкие, эти пациенты могут испытывать беспокоящие симптомы
андрогенной недостаточности, включая потерю сексуального желания и функции,
пониженное настроение и горячие приливы. Некоторые мужчины, которые находят
эти симптомы отмены непереносимыми, снова начинают применять андрогенные
стероиды, укрепляя порочный круг злоупотребления, симптомов отмены и зависи¬
мости [653-655]. Другая надежда на безрецептурное применение ингибиторов аро¬
матазы или ХГЧ, приобретенных незаконно, основана на народном понятии, широко
распространенном в учебных учреждениях, что эти агенты могут ускорить восста¬
новление гипоталамо-гипофизарно-тестикулярной оси, хотя нет свидетельств, под¬
тверждающих эту предпосылку. Долгосрочная супрессия гипоталамо-гипофизарно-
тестикулярной оси и сопровождающий ее риск зависимости - серьезное осложнение
применения андрогенных стероидов, которое не было широко принято во внимание.
Самостоятельное проведение внутримышечных инъекций увеличивает риск инфек¬
ции, мышечного абсцесса и даже сепсиса [625]. Сообщали о передаче ВИЧ-инфекции
среди лиц, использующих андрогенные стероиды, по-видимому, из-за общей иглы
или использования неадекватно простерилизованных игл и шприцев.
Чрезмерная гипертрофия мышц без соразмерной адаптации прилегающих сухо¬
жилий и соединительной ткани может привести к риску повреждения и разрыва
сухожилия, а также необычного напряжения суставов у спортсменов, применяющих
андрогенные стероиды [656].
Кроме того, 90% лиц, применяющих андрогенные стероиды, злоупотребляют
дополнительными препаратами [624]. Некоторые из этих препаратов, такие как
кокаин, амфетамин и эфедра, могут иметь потенциально серьезные медицинские
осложнения.
Вьшттт незаконных андрогенных отарощов
Применение синтетических андрогенных стероидов было запрещено на
Олимпийских играх в 1974 г. Хотя вначале для обнаружения андрогенных стероидов
в образцах мочи были использованы радиоиммунные методики анализа, начиная с
1981 г. аккредитованные лаборатории используют либо газовую хроматографию с
295
масс-спектроскопией, либо — в некоторых случаях — жидкостную хроматографию
с масс-спектроскопией для выявления тех андрогенных стероидов, которые невоз¬
можно обнаружить при газовой хроматографии или которые термолабильны [657],
Для улучшения чувствительности газовой хроматографии образцы мог}»т быть допол¬
нительно обработаны [658], Таким образом, введение силильной группы обычно
используется для дериватизации образцов при анализе на андрогенные стероиды; эта
реакция превращает полярные группы (гидрокси- и кетогруппы) в менее полярные
триметилсилильные эфиры, что улучшает отношение сигнала к шуму [658]. Также
с конца 1990-х гг. внедрение масс-спектроскопии высокого разрешения и танде¬
ма «масс-спектроскопия-хроматография» еще больше улучшили чувствительность
методик обнаружения андрогенных стероидов.
Для обнаружения использования Т применяют анализ отношения Т к эпи¬
тестостерону в сочетании с отношением изотопов с помощью окислительной
масс-спектроскопии [659-666]. Отношение Т к эпитестостерону в моче обычно
составляет менее 6:1 и является постоянным у каждого индивидуума. Существуют
генетические различия отношения Т к эпитестостерону. Назначение экзогенного Т
увеличивает экскрецию с мочой тестостерон-глюкоронида и увеличивает отноше¬
ние Т к эпитестостерону. Соотношение Т к эпитестостерону более 6:1 подозритель¬
но. Большие соотношения требуют исследования прежних или дополнительных
образцов мочи, взятых через временной интервал. Если высокое соотношение
обусловлено генетической вариацией, тогда все образцы, полученные от субъекта,
будут иметь высокое отношение. Повышенное соотношение Т к эпитестостерону в
настоящих образцах по сравнению с предыдущими значениями расценивают как
положительный тест.
Если результаты теста на отношение Т к эпитестостерону ненормальные и пред¬
полагают использование экзогенного Т, требуется дополнительное подтверждение
путем использования газовой хроматографии с определением отношения изотопов
при окислительной масс-спектроскопии [658, 661]. Этот метод основан на измере¬
нии отношения изотопов углерода в Т. В природе 1,1% углерода существует
в виде С’\ Однако во время химического синтеза Т атомы углерода вступают
в реакцию медленнее, чем атомы Именно поэтому синтетический Т схожим
образом имеет более низкое отношение чем справочный газовый стандарт.
Во время процесса газовой хроматографии с определением отношения изотопов
при окислительной масс-спектроскопии стероиды отделяются посредством газовой
хроматографии и окисляются в оксид углерода в камере для сжигания. Отношение
СО2 с (молекулярная масса 45) и СО2 с (молекулярная масса 44) наблюдают
с помощью определяющего отношения изотопов масс-спектрометра и рассчитыва¬
ют значение (1 (показателем с1 называют снижение относительно справочного
газа со стандартизированным отношением [667]. Отрицательное значение
(1 наряду с высоким отношением Т к эпитестостерону предполагает введение экзо¬
генного Т.
Процедура сбора и перевозки образцов на допинг-тесты выполняется по строгим
правилам, которые были установлены спортивными организациями [658]. Обычно
каждый образец мочи, собранный под непосредственным контролем аккредитован¬
ного наблюдателя, разделяется на две части (А и В образцы) и транспортируется в
тестирующую лабораторию под охраной. Если образец А окажется положительным,
тогда анализируют образец В в присутствии спортсмена или авторизованного пред¬
ставителя спортсмена. Если проба В также положительна, допинг с андрогенными
стероидами подтверждают, и спортивная организация может наложить карательные
санкции [658].
296
1^ог1011ьзование оредшаотвенникое андрогенов
и специально разработанных стероидов
А^-АНДРОСТЕНДИОН
A“*-Андростендион, предшественник Т в каскаде биосинтеза Т, превращается в орга¬
низме с помощью 17-гидроксил стероид-дегидрогеназы в Т, Признание известного
бейсболиста Марка Магвайра в использовании андростендиона вызвало настоящий
ажиотаж и быстрый рост розничных продаж пищевых добавок, таких как андростен¬
дион и дегидроэпиандростерон.
Андростендион продавался до недавнего времени как пищевая добавка согласно
Закону о пищевых добавках для здоровья и образования [668, 669]. Доступность
андростендиона в пищевых магазинах сделала его популярным средством для злоу¬
потребления среди бодибилдеров [617, 670], В отличие от других андрогенных
стероидов, продажа которых регулировалась Законом о контроле андрогенных
стероидов, андростендион продавали в розницу, и его продажи не контролировали
Администрация по пищевым продуктам и лекарствам и Агентство по ограниченным
препаратам. Однако недавно конгресс США добавил андростендион к запрещенному
списку анаболических стероидов.
В ранних исследованиях назначение 100 мг андростендиона внутрь ежедневно на
7 дней не вызывало значительного увеличения циркулирующих концентраций Т, а
назначение 300 мг приводило только к умеренному возрастанию площади Т под кри¬
вой, но намного более выраженному увеличению концентраций эстрадиола и эстрона
[671-675]. В более поздних исследованиях Jasuja и соавт. [676] продемонстрировали,
что 500 мг андростендиона, назначаемые трижды в сутки в течение 12 нед гипогонад-
ным мужчинам, увеличили сывороточные уровни Т и свободного Т до эугонадного
диапазона, а также увеличили мышечную массу и напряжение. Эти исследования
показали, что андростендион связывается с андрогеновым рецептором, хотя и со
значительно меньшей связывающей аффинностью, чем Т, и что он способствует мио¬
генной дифференциации в мезенхимальной мультипотентной линии клеток [676].
Схожим образом у женщин назначение 100 мг андростендиона значительно увеличи¬
вает сывороточные уровни Т выше физиологического диапазона [671]. Как обсужда¬
ли ранее, у самок гиен и некоторых других видов млекопитающих циркулируют более
высокие концентрации андростендиона, чем у человека, что приводит к вирилизации
[671]. Таким образом, андростендион отвечает всем критериям анаболического стеро¬
ида: он имеет структурное сходство с Т, связывает рецептор андрогенов, способствует
миогенной дифференцировке in vitro и увеличивает мышечную массу, когда назнача¬
ется в достаточно высоких дозах [676]. Основываясь на этих данных, конгресс США
недавно классифицировал андростендион как анаболический стероид и запретил его
розничную продажу.
После назначения андростендиона внутрь большая часть дозы инактивируется во
время его пресистемного метаболизма, о чем свидетельствует выраженное увеличение
метаболитов андростендиона в моче, включая тестостерон глюкуронид, с небольшим
повышением сывороточной концентрации Т [276]. Контаминация диетических доба¬
вок другими андрогенными стероидами из-за плохого контроля качества может при¬
вести к тому, что допинг-тесты станут положительными [677],
Проблемы контроля качества в розничных препаратах андростендиона^
Поскольку производство и продажи андростендиона не регулируются Управлением
по пищевым продуктам и лекарственным средствам, существуют серьезные проблемы
контроля качества розничных препаратов андростендиона [678, 679]. В одном из
исследований ученые выявили значительную вариабельность содержания андростен-
297
диона в препаратах и даже среди разных партий одного и того же производителя;
контаминация другими андрогенами достаточно частая [677,680].
Потенциальные неблагоприятные эффекты андростендиона
Отдаленные побочные эффекты использования андростендиона неизвестны.
Краткосрочное назначение препарата ассоциировано со значительным повышением
уровней эстрадиола. У мужчин повышение концентраций эстрогенов в сыворотке
крови может влиять на качество семени, увеличивать содержание воспалительных
маркеров, повышающих риск сердечно-сосудистых заболеваний, приводить к гинеко¬
мастии и индуцировать эпигенетические и цитогенетические эффекты на сперму.
Прием андростендиона снижает уровни ЛПВП и увеличивает отношение
ЛПНП:ЛПВП. Другие неблагоприятные эффекты андростендиона обусловлены потен¬
циальным увеличением уровня т. Это неблагоприятные эффекты на липиды плазмы
крови, эритроцитоз, акне, синдром ночного апноэ и повышенный риск обнаружения
патологии простаты.
Из-за недостатка данных об эффективности и тотального отсутствия данных о
долгосрочной безопасности использование андростендиона клинически не реко¬
мендовано при любом показании, включая лечение андрогенной недостаточности у
мужчин и женщин.
ДЕГИДРОЭПИАНДРОСТЕРОН
ДГЭА в норме образуется в ретикулярной зоне надпочечников. Как слабый андро¬
ген, он превращается в периферических тканях в Т и эстрадиол. Циркулирующие
уровни ДГЭА выраженно снижаются с увеличением возраста, так что сывороточные
концентрации ДГЭА на 7-м десятилетии жизни составляют приблизительно 20%
пиковых уровней, наблюдавшихся ранее во взрослой жизни [681, 682]. Кроме дей¬
ствия в качестве предшественника и слабого андрогена, ДГЭА может выступать в
качестве нейростероида [683].
ДГЭА связывает рецептор андрогенов со значительно более низкой аффинностью,
нежели ДГТ. Было высказано предположение об отдельном мембранном рецеп¬
торе ДГЭА, связанном с С-белком [684]; однако существование подобного ДГЭА-
специфичного мембранного рецептора требует дальнейшего подтверждения. Также
было показано, что ДГЭА модулирует активность рецептора М-метил-О-аспартата и
гамма-аминобутановой кислоты [685].
Исследования эффективности ДГЭА
Опубликованные сведения о ДГЭА трудно интерпретировать по нескольким при¬
чинам.
» Многие исследования ДГЭА. сообщающие о положительном нейротропном и
антиканцерогенном эффекте и иммунном улучшении, были проведены на грызу¬
нах, которые имеют очень низкие уровни циркулирующего ДГЭА. Следовательно,
данные исследований на грызунах мало применимы у человека.
* В исследованиях на людях изучались разные дозы, лекарственные формы и были
различные исследуемые популяции. Были использованы дозы очень высокие
(1500 мг/сут) и очень низкие (25 мг/сут).
® Большая часть исследований на людях включала малое количество испытуемых
и имела относительно короткую продолжительность.
Исследования ДГЭА были проведены у пациентов с надпочечникововой недоста¬
точностью [686-690], у пожилых мужчин и женщин [691-696], пери- и постменопау¬
зальных женщин [697,698] и пациентов с аутоиммунными заболеваниями [699-701].
Большинство интервенционных исследований использовали 50 мг ДГЭА ежедневно
на протяжении 3-6 мес. Кокрейновский обзор ДГЭА позволил сделать заключение
о недостаточности свидетельств положительного влияния ДГЭА на познавательную
298
способность у пожилых мужчин и женщин [695]. Не было обнаружено, что даже более
высокие дозы ДГЭА, назначенные пожилым мужчинам, приводят к значительному
улучшению когнитивной функции [696]. в рандомизированных контролируемых
исследованиях сообщали о более значительном улучшении минеральной плотности
костной ткани у пожилых мужчин и женщин, получавших 50 мг ДГЭА ежедневно, по
сравнению с плацебо [702].
Ряд исследований добавления ДГЭА женщинам с надпочечниковой недостаточ¬
ностью был обусловлен предположением, что, несмотря на адекватную замести¬
тельную глюкокортикоидную терапию, пациенты с надпочечниковой недостаточно¬
стью сообщают о субоптимальном качестве жизни. Результаты этих исследований
были противоречивыми. Агк и соавт. [686] использовали двойной слепой гшацебо-
контролируемый перекрестный дизайн исследования у женщин с первичной или вто¬
ричной надпочечниковой недостаточностью, которые получали либо плацебо, либо
50 мг ДГЭА в сутки в течение 16 нед. Назначение ДГЭА было ассоциировано с улучше¬
ниями показателей депрессии и тревожности, половой функции и циркулирующими
уровнями остеокальцина, но не было отмечено значительных изменений состава тела
[686]. Другие исследования добавления ДГЭА женщинам с надпочечниковой недо¬
статочностью не подтвердили положительные эффекты ДГЭА на настроение, само¬
чувствие или половую функцию, наблюдавшиеся в ходе исследования Агк [687-689].
Аналогично не было найдено стойких эффектов на состав тела, физическую функцию
или чувствительность к инсулину. Несмотря на необоснованные заявления в сред¬
ствах массовой информации и в Интернете, не было проведено рандомизированных
контролируемых исследований адекватной мощности и достаточной продолжитель¬
ности для определения эффектов назначения ДГЭА на частоту сердечно-сосудистых
заболеваний или рака.
Сообщали, что фармакологические дозы ДГЭА (200 мг в сутки) связаны с уме¬
ренным улучшением исходов системной красной волчанки и более выраженным
снижением обострений заболевания и активности болезни, чем плацебо [699-701].
Эффекты ДГЭА на минеральную плотность костной ткани у пациентов с системной
красной волчшнкой были противоречивы.
Таким образом, рандомизированные контролируемые исследования ДГЭА не про¬
демонстрировали стойкой эффективности при любом болезненном состоянии и не
оправдали использования ДГЭА по любому клиническому показанию в настоящее
время.
другие предшественники андрогенов и новейшие стероиды, специально
разработанные как спортивные допинги
Ряд дополнительных стероидов, предшественников Т (4-андростендиол и
5-андросгандиол в добавлении к 4-андростендиону и ДГЭА), ДГТ (5а-андростан-
Зр-17р-диол, 5а-андростан-3(х,178-диол, 5а-андростан-3,17-дион, 5а-андрост-1-
ен-Зб17-дион, 17(5-гидрокси-5а-андрост-1-ен-3-он, 5ос-андрост,1-енД7р-диол) или
нортестостерона (4-норандростендион, 4-норандростендиол и 5-норандростендиол),
появился на черном рынке [658]; эти соединения слабые, но в организме превращаются
в мощные андрогены. Даже предшественник (андросга-1,4-диен-3,17-дион) больдено-
на (17[5-гидроксиандроста-1,4-диен-3-он) был рекомендован в Интернете [658].
До недавнего времени андрогенными стероидами, которые потребляли спортсме¬
ны, служили препараты, синтезируемые вначале для медицинских и ветеринарных
целей. Недавнее появление на рынке специально разработанных стероидов, таких
как тетрагидрогестринон [703, 704] и мадол [705], которые были созданы для упо¬
требления спортсменами и чей синтез не был продиктован каким-либо медицинским
показанием, представляет собой неблагоприятный вариант развития данной области
медицины [706]. Выявление злоупотребления новейшими андрогенными стероидами,
299
для которых не были стандартизированы методы обнаружения, является серьезным
вызовом антидопинговым лабораториям. Поскольку специально разработанные
вещества ранее никогда не были тестированы на животных или людях, их использо¬
вание спортсменами потенциально опасно для здоровья. Поскольку нет опубликован¬
ных данных или предшествующего опыта использования этих новейших стероидов и
поскольку требуется значительное время для получения данных об их андрогенной
и анаболической эффективности, которые позволили бы воздержаться от научного
и правового испытания, службы правительства были поставлены в тупик в попытках
регуляции подпольного рынка.
СПИСОК ЛИТЕРАТУРЫ
1. Kegaiiy G.J., Trombulak S.G, Size and function of mammalian testes in relation to body size //
J. Mammology. - 1986. - Vol. 67. - P. 1-22.
2. Glickman S.E., Coscia E,M., Frank L.G. et al. Androgens and masculinization of genitalia in the
spotted hyaena (Crocuta crocuta). 3. Effects of juvenile gonadectomy //J. Reprod. Fertil. — 199S. —
Vol. 113. - P. 129-135.
3. Glickman S.E., Frank L.G., Pavgi S., Licht P. Hormonal correlates of “masculinization” in female
spotted hyaenas (Crocuta crocuta). 1. Infancy to sexual maturity //J. Reprod. Fertil. - 1992. - Vol.
95. - P. 451-462.
4. Glickman S.E., Frank L.G., Davidson J.M. et al. Androstenedione may organize or activate
sex-reversed traits in female spotted hyenas // Proc. Natl. Acad. Sci. USA. — 1987. — Vol, 84. —
P. 3444-3447.
5. Goymann W., East M.L., Hofer H. Androgens and the role of female “hyperaggressiveness” in
spotted hyenas (Crocuta crocuta) //Horm. Behav. — 2001. — Vol. 39. — P. 83-92.
6. Wilson J.D., Auchus R.J., Leihy M.W. et al. 5a-androstane-3a,17b-diol is formed in tammar ^val-
laby pouch young testes by a pathway involving 5a-pregnane-3a,17a-diol-20-one as a key intermedi¬
ate// Endocrinology. - 2003. - Vol. 144. - P. 575-580.
7. Wilson J.D., Shaw G., Renfree M.B. et al. Ontogeny and pathway of formation of 5a-androstaiie-
3a,17p-diol in the testes of the immature brushtail possum Trichosurus vulpecmla // Reprod. Fertil.
Dev. - 2005. - Vol. 17. - P. 603-609.
8. Stechell B.P. The testis and tissue transplantation; historical aspects // J. Reprod. Immunol. —
1990. - Vol. IS. - P. 1-8.
9. George F.W., Wilson J.D. Sex determination and differentiation // Knobil E., Wilson J.D. (ed.).
The Physiologs' of Reproduction. - 2nd edition. - New York: Raven Press, 1994. - P. 3-26.
10. Wilson J.D., Griffin J.E., Russel D.W. Steroid 5a reductase-2 deficiency // Endocr. Rev. —
1993. - Vol. 14. - P. 557-593.
11. Meiiderlson C.R., Dufau M.L., Catt K.J. Gonadotropin stimulation of cyclic adenosine 3'-5'
monophosphate and testosterone production in isolated Leydig cells //J. Biol. Chem. — 1975. — Vol.
250. - P. 8818-8823.
12. Payne A.H., Quinii P.G., Rani C.S. Regulation of microsome cytochrome P-450 enzymes and tes¬
tosterone production in Leydig cells // Recent. Prog. Horm. Res. — 1985. — Vol. 41. — P. 153-197.
13. Wahlstrom J.T., Huhtaniemi I., Hovatta O. et al. Localization of luteinizing hormone, follicle-
stimulating hormone, prolactin, and their receptors in human and rat testis using immunohistochemis-
try and radioreceptor assays //J. Clin. Endocrinol. Metab. - 1983. - Vol. 57. — P. 825-830.
14. Saez J.M. Leydig cells: endocrine, paracrine, and autocrine regulation // Endocr, Rev. — 1994. —
Vol. 16. - P. 574-626.
15. Carr B.R., Parker Jr. C.R., Ohashi M. et al. Regulation of human fetal testicular secretion of
testosterone: low-density lipoprotein-cholesterol and choleterol synthesized de novo as steroid precur¬
sor // Am. J. Obstet. Gynecol. - 1983. - Vol. 146. - P. 241-247.
16. Miller W.L. Molecular biology of the steroid hormone synthesis // Endocrine. Rev. — 1988. —
Vol. 9. - P. 295-318.
17. Jasuja R., Ramaraj T„ Mac R.P. et al. Androstenedione binds androgen receptor, promotes
myogenesis in vitro, and increases fat-free mass and muscle strength in hypogonadal men j j J. Clin.
Endocrinol. Metab. - 2005. - Vol. 90. - P. 855-863.
18. Rosner W. Plasma steroid-binding proteins // Endocrinol. Metab. Clin, North. Am. — 1991. —
Vol. 20. - P. 697-720.
300
19. Mendel С.М. The free hormone liypothesis: a physiologically based mathematical model j j
Endocr. Rev. - 1989. - Vol. 10. - P. 232-274.
20. Sakiyama R., Pardridge W.M., Musto N.A. Influx of testosterone-binding globulin (TeBG) and
TeBG-bound sex steroid honnones into rat testis and prostate //J. Clin. Endocrinol. Metab. 1988. —
Vol. 67. - P. 98-103.
21. Nakhla A.M., Rosner W. Stimulation of prostate cancer growth by androgens and estrogens
through the intermediacy of sex hormone-binding globulin j j Endocrinology. — 1996. — Vol. 137. —
P. 4126-4129.
22. Hammes A., Andreassen Т.К., Spoelgen R. et al. Role of endocytosis in cellular uptake of sex
steroids // Cell. - 200.‘>. - Vol. 122. - P. 751-762.
23. Andreassen Т.К. The role of plasma-binding proteins in the cellular uptake of lipophilic vitamins
and steroids // Horm. Metab. Res. — 2006. — Vol. 38. — P. 279-290.
24. Ukkoia O., Rankinen Т., Gagnon J. et al. A genome-wide linkage scan for steroids and SHBG
levels in black and white families: the НЭР-ITAGE Family Study // J. Clin. Endocrinol. Metab. —
2002. - Vol. 87. - P. 3708-3720.
25. Joseph D.R. Structure, function and regulation of androgen-binding protein/sex hormone-
binding globulin // Vitam. Horm. - 1994. - Vol. 49. - P. 197-280.
26. Besa E.C., Bullock L.P. The role of the androgen receptor in erythropoiesis // Endocrinology. —
1981. - Vol. 109. - P. 1983-1989.
27. Smith E.P., Boyd J., Frank G.R. et al. Estrogen resistance caused by a mutation in the estrogen
receptor gene in a man // N. Engl. J. Med. — 1994. — Vol. 331. - P. 1056-1061.
28. Carani C., Rochira V., Faustini-Fustini M. et al. Role of oestrogen in male sexual behaviour:
insights from the natural model of aromatase doficiencv // Clin. Endocrinol. (Oxf.). — 1999. —
Vol. 51. - P. 517-524.
29. Deladoey J., Fluck C., Bex M. et al. Aromatase deficiency caused by a novel P450arom gene
mutation: impact of absent estrogen production on serum gonadotropin concentration in a boy //J.
Clin. Endocrinol. Metab. - 1999. - Vol. 84. - P. 4050-4054.
30. Morishima A., Grumbach M.M., Simpson E.R. et al. Aromatase deficiency in male and female
siblings caused by a novel mutation and the physiological role of estrogens // J. Clin. Endocrinol.
Metab. - 1995. - Vol. 80. - P. 3689-3698.
31. Mullis P.E., Yoshimura N.. Kuhlinann B. et al. Aromatase deficiency in a female who is com¬
pound heterozygote for two new point mutations in the P450arom gene: impact of estrogens on
hypergonadotropic hypogonadism, multicystic ovaries, and bone densitometry in childhood //J. Clin.
Endocrinol. Metab. - 1997. - Vol. 82. - P. 1739-1745.
32. Russell D.W., Wilson J.D. Steroid 5a reductase: two gene/two enzymes j j Ann. Rev. Biochem. —
1994. Vol. 63. - P. 25-61.
33. Mahcndroo M.S., Porter A., Russell D.W., Word R.A. The parturition defect in steroid
5a-reductase type 1 knockout mice is due to impaired cervical ripening // Mol. Endocrinol. — 1999. —
Vol. 13. - P. 981-992.
34. BartschW., KriegM., Voigt K.D. Quantification of endogenoustestosterone,5a-dihydrotestosterone
and 5a-androstaiie-3a,17p-diol in subcellular fractions of the prostate, bulbocavernosus/levator ani
muscle, skeletal muscle and heart muscle of the rat // J. Steroid. Biochem. — 1980. — Vol. 13. —
P. 259-264.
35. Simental J.A., Sar М., Wilson E.M. Domain functions of the androgen receptor //J. Steroid.
Biochem. Mol. Biol. - 1992. - Vol. 43. - P. 37-41.
36. McPhatil MJ. Molecular defects of the androgen receptor // J. Steroid. Biochem. Mol. Biol. —
1999. - Vol. 69. - P. 315-322.
37. McPhaul M.J., Marcelli М., Zoppi S. et al. Genetic basis of endocrine disease. 4. The spectrum
of mutations in the androgen receptor gene that causes androgen resistance // J. Clin. Endocrinol.
Metab. - 1993. - Vol. 76. - P. 17 -23.
38. Lubahn D.B., Brown T.R., Simental J.A. et al. Sequence of the intron/exon junctions of the cod¬
ing region of the human androgen receptor gene and identification of a point mutation in a family with
complete androgen insensitivity // Proc. Natl. Acad. Sci. USA. — 1989. — Vol. 86. — P. 9534-9538.
39. Slagsvold Т., Kraus I., Bentzen T. et al. Mutational analysis of the androgen receptor AF-2 (acti¬
vation function 2) core domain reveals functional and mechanistic differences of conserved residues
compared with other nuclear receptors // Mol. Endocrinol. - 2000. — Vol. 14. — P. 1603-1617.
40. Quigley C.A., De Beilis A., Marschke K.B. et al. Androgen receptor defects: historical, clinical,
and molecular perspectives // Endocr, Rev. — 1995. — Vol. 16. — P. 271-321.
301
41. Singh R., Artaza J.N., Taylor W.E. et al. Testosterone inhibits adipogénie differentiation in 3T3-
L1 cells: nuclear translocation of androgen receptor complex with p-catenin and T-cell factor 4 may
bypass canonical Wnt signaling to down-rcgulate adipogénie transcription factors j j Endocrinology. -
2006. - - Vol. 147. - P. 141-154.
42. Singh R., Artaza J.N., Taylor W.E. et al. Androgens stimulate myogenic differentiation and
inhibit adrpogenesis in C3H lOTl/2 pluripotent cells through an androgen receptor-mediated path¬
way // Endocrinology. - 2003. - Vol. 144. - P. 5081-5088.
43. Ferrando A.A., Raj D., Wolfe R.R. Amino acid control of muscle protein turnover in renal
disease //J. Ren. Nutr. - 2005. - Vol. 15. - P. 34-38.
44. Ferrando A.A., Sheffield-Moore М., Paddon-Jones D. et al. Differential anabolic effects of
testosterone and amino acid feeding in older men //J. Clin. Endocrinol. Metab. - 2003. - Vol, 88. -
P. 358-362.
45. Ferrando A.A., Sheffield-Moore М., Wolf S.E. et al. Testosterone administration in severe burns
ameliorates muscle catabolism // Crit. Care. Med. - 2001. - Vol. 29, ■ P. 1936-1942.
46. Ferrando A.A., Sheffield-Moore М., Yeckel C.W. et al. Testosterone administration to older men
improves muscle function: molecular and physiological mechanisms // Am, J. Physiol. EndocTinol.
Metab. - 2002. - Vol. 282. - P. 601-607.
47. Sheffield-Moore М., Urban R.J., Wolf S.E. ct al. Short-term oxandrolone administration
stimulates net muscle protein synthesis in young men //J. Clin. Endocrinol. Metab. — 1999. —
Vol. 84. - P. 2705-2711.
48. Wolfe R„ Ferrando A., Sheffield-Moore М., Urban R. Testosterone and muscle protein
metabolism // Mayo Clin. Proc. — 2000. - Vol. 75. - Suppl. - P. 55- 59; discussion S59-S60.
49. Skaletsky H., Kuroda-Kawaguchi T., Minx P. et al. The male-specific region of the Y chromosome
is a mosaic of discrete sequence classes // Nature. — 2003. - Vol. 423. — P. 825-837.
.50. Hawley R.S. The human Y chromosome; the rumors of its death have been greatly exagge¬
rated // Cell. - 2003. - Vol. 113. - P. 825-828.
51. Knuth U.A., Hull M.G., Jacobs H.S. Amenorrhoea and lo.ss of weight // Br. J. Obstet,
Gynaecol. - 1977. - Vol. 84. - P. 801-807.
52. Frisch R.E, Body fat, menarche, fitness and fertility // Hum. Reprod. — 1987, - Vol. 2. —
P. 521-533.
53. Penny R., Goldstein I.P., Frasier S.D. Gonadotropin excretion and body composition //
'pediatrics. - 1978. - Vol. 61. - P. 294-300.
54. Stein Z., Susser M. Fertility, fecundity, famine: food rations in the Dutch famine 1944/5 have a
causal relation to fertility and probably to fecundity // Hum. Biol. — 1975. — Vol, 47. — P. 131-154.
55. Stein Z,, Susser M. The Dutch famine, 1944-1945, and the reproductive process. I. Effects of six
indices at birth j j Pediatr, Res. - 1975. - Vol. 9. - P. 70-76.
56. van der Walt L.A., Wilmsen E.N., Jenkins T. Unusual sex hormone patterns among desert-
dwelling hunter-gatherers //J. Clin. Endocrinol. Metab. - 1978. — Vol. 46. — P. 658-663.
57. van der Walt L.A., Wilmsen E.N., Levin J., Jenkins T. Endocrine studies on the San (“bushmen”)
of Botswana // S. Afr. Med. J. - 1977. - Vol. 52. - P. 230-232.
58. Foster D.L., Nagatani S. Physiological perspectives on leptin as a regulator of reproduction; role
in timing puberty // Biol. Reprod. — 1999. — Vol. 60. — P. 205-215.
59. Halmi K.A., Sherman B.M. Gonadotropin response to LH-RH in anorexia nervosa // Arch. Gen.
Psychiatry. - 1975. - Vol. 32. - P. 875-878.
60. Sherman B.M., Halmi K.A,, Zamudio R. LH and FSH response to gonadotropin-releasing
hormone in anorexia nervosa: Effect of nutritional rehabilitation // J. Clin. EndocTinol, Metab. —
1975. - Vol. 41. - P. 135-142.
61. Baker E.R. Body weight and the initiation of puberty j j Clin. Obstet. Gynecol - 1985. - Vol.
28. - P. 573-579.
62. Frisch R.E. Fatness, menarche, and female fertility // Perspect. Biol. Med. - 1985. - Vol 28. --
P. 611-633.
63. Frisch R.E., Gotz-Welbergen A.V., McArthur J.W. et al. Delayed menarche and amenorrhea of
college athletes in relation to age of onset of training //JAMA. - 1981. — Vol. 246. - P. 1559-1563.
64. Van Der Spuy Z.M., Jacobs H.S. Weight reduction, fertility and contraception // IPPF. Med.
Bull. - 1983. - Vol 17. - P. 2-4.
65. Laughlin G.A., Dominguez C.E., Yen S.S. Nutritional and endocrine-metabolic aberrations in
women with functional hypothalamic amenorrhea //J. Clin. Endocrinol. Metab. — 1998. — Vol. 83. -
P. 25-32.
302
66. Laughlin G.A., Yen S.S. Nutritional and endoaine-metabolic aberrations in amenorrheic
athletes //J. Clin. Endocrinol. Metab. - 1996, — Vol. 81. - P. 4301-4309.
67. Misra M., Aggarwal A., Miller K.K. et al. Effects of anorexia nervosa on clinical, hematologic,
biochemical, and bone density parameters in community-dwelling adolescent girls // Pediatrics. -
2004, - Vol. 114. - P. 1574-1583.
68. Bates G.W., Bates S.R., Whitworth N.S. Reproductive failure in women who practice weight
control // Fertil. Steril. - 1982. - Vol. 37. - P. 373-378.
69. Rock C.L., Gorenflo D.W., Drewnowski A., Demitrack M.A. Nutritional characteristics,
eating pathology, and hormonal status in young women // Am. J. Clin. Nutr. - 1996. - Vol. 64. -
P. 566-571.
70. Cunningham M.J., Clifton D.K., Steiner R.A. Leptin’s actions on the reproductive axis; perspec¬
tives and mechanisms // Biol. Reprod. — 1999. — Vol. 60. - P. 216-222.
71. Zhang Y., Proenca R., Maffei M. et al. Positional cloning of the mouse obese gene and its human
homologue // Nature. - 1994. - Vol. 372. - P. 425-432.
72. Campfield L.A., Smith F.J., Burn P. The OB protein (leptin) pathway — a link between adipose
tissue mass and central neural networks // Horm. Metab. Res. - 1996. — Vol. 28. — P. 619-632.
73. Campfield L.A., Smith F.J., Guisez Y. et al. Recombinant mouse OB protein: evidence for a
peripheral signal linking adiposity and central neural networks // Science. — 1995. - Vol. 269. -
P. 546-549.
74. Weigle D.S., Duell P.B., Connor W.E. et al. Effect of fasting, refeeding, and dietary fat restriction
on plasma leptin levels //J. Clin. Endocrinol Metab. — 1997. — Vol. 82. — P. 561-565.
75. Keim N.L., Stem J.S., Havel P.J. Relation between circulating leptin concentrations and appetite dming
a prolonged, moderate energy deficit in women // Am. J. Clin. Nutr. — 1998. - Vol. 68, - P. 794-801.
76. Chan J.L., Matarese G., Shetty G.K. et al. Differential regulation of metabolic, neuroendocrine,
and immune function by leptin in humans // Proc. Natl. Acad, Sci. USA. — 2006. — Vol. 103. —
P. 8481-8486.
77. Ahima R.S., Prabakaran D., Mantzoros C. et al. Role of leptin in the neuroendocrine resjronse to
fasting // Nature. - 1996. - Vol. 382. - P. 250-252.
78. Swerdloff R.S., Batt R.A., Bray G.A. Reproductive hormonal function in the genetically obese
(ob/ob) mouse /7 Endocrinology. - 1976. - Vol. 98. - P. 1359-1364.
79. Farooqi I.S., Matarese G., Lord G.M. et al. Beneficial effects of leptin on obesity, T cell hypo-
respoiisiveness, and neuroendocrine/metabolic dysfunction of human congenital leptin deficiency j j
J. Clin. Invest. - 2002. - Vol. 110. - P. 1093-1103.
80. Chehab F.F., Lim M.E., Lu R. Correction of the sterility defect in homozygous obese female mice
by treatment vdth the human recombinant leptin // Nat. Genet. - 1996. - Vol. 12. - P. 318-320.
81. Yu W.H., Walczewska A., Karanth S., McCann S.M. Nitric oxide mediates leptin-induced
luteinizing honnone-releasing hormone (LHRH) and LHRH and leptin-induced LH release from the
pituitary gland // Endocrinology. — 1997. — Vol. 138. — P. 5055-5058.
82. Barb C.R., Kraeling R.R., Rampacek G.B., Hausman G.J. The role of neuropeptide Y and interac¬
tion with leptin in regulating feed intake and luteinizing hormone and growth hormone secretion in the
pig // Reproduction. - 2006. - Vol. 131. - P. 1127-1135.
83. Kaynard A.H., Pau K.Y., Hess D.L., Spies H.G. Third-ventricular infusion of neuropeptide Y
suppresses luteinizing hormone secretion in ovariectomized rhesus macaques // Endocrinology, —
1990. - Vol. 127. - P. 2437-2444.
84. Catzeflis C„ Pierroz D.D., Rohner-Jeanrenaud F. et al. Neuropeptide Y administered chronically
into the lateral ventricle profoundly inhibits both the gonadotropic and the somatotropic axis in intact
adult female rats j j Endocrinology. — 1993, - Vol. 132. — P. 224-234.
85. Zizzari P., Halem H.. Taylor J, et al. Endogenous ghrelin regulates episodic growth hormone
(GH) secretion by amplifying GH pulse amplitude: evidence from antagonism of the GH secretagogue-
Rla receptor // Endocrinology. — 2005. — Vol. 146. — P. 3836-3842.
86. Fernandez-Fernandez R„ Martini A.C., Navarro V.M. et al. Novel signals for the integration of
energy balance and reproduction // Mol. Cell. Endocrinol. - 2006. — Vol. 254-255. — P. 127-132.
87. Fernandez-Fernandez R., Tena-Sempere M., Navarro V.M. et al. Effects of ghrelin upon gonad¬
otropin-releasing hormone and gonadotropin secretion in adult female rats: in vivo and in vitro stu¬
dies II Neuroendocrinology. - 2005. - Vol. 82. - N 5-6. - P. 245-255, Epub, 2006 Apr. 20.
88. Fernandez-Fernandez R., Tena-Sempere M., Navarro V.M. et al. Effects of ghrelin upon gonad¬
otropin-releasing hormone and gonadotropin secretion in adult female rats: in vivo and in vitro stu¬
dies iI Neuroendocrinology. — 2006. - Vol. 82. — N 5-6. — P. 245-255 [Epub. ahead of print].
303
89. Date Y„ Kojima M., Hosoda H. et al. Ghrelin, a novel growth hormone-releasing acylated pep¬
tide, is synthesized in a distinct endocrine cell type in the gastrointestinal tracts of rats and humans //
Endocrinology. - 2000. - Vol, 141. - P. 4255-4261.
90. Kojima M., Hosoda H„ Date Y. et al. Ghrelin is a growth-hormone-releasing acylated peptide
from stomach // Nature. — 1999. — Vol. 402. — P. 656-660.
91. Nakazato M., Murakami N.. Date Y. et al. A role for ghrelin in the central regulation of
feeding //Nature. - 2001. - Vol. 409. - P. 194-198.
92. Fernandez-Femandez R,, Aguilar E., Tena-Sempere M„ Pinilla L. Effects of polypeptide YY(3-
36) upon luteinizing honnone-releasing hormone and gonadotropin secretion in prepubertal rats: in
vivo and in vitro studies // Endocrinology. - 2005. - Vol. 146. - P. 1403-1410.
93. Navarro V.M., Castellano J.M., Fernandez-Fernandez R. et al. Effects of KiSS-1 peptide, the
natural ligand of GPR54, on follicle-stimulating hormone secretion in the rat // Endocrinology. -
2005. - Vol. 146. - P. 1689-1697.
94. Bhasin S., Cunningham G.R., Hayes FJ. et al, Testosterone therapy in adult men with androgen
deficiency syndromes: an endocrine society clinical practice guideline //J. Clin. Endocrinol. Metab. —
2006. - Vol. 91. - P. 1995-2010.
95. Jacobs P.A., Melville M., Ratcliffe S. et al. A cytogenetic survey of 11,680 newborn infants //
Ann. Hum. Genet. - 1974. - Vol. 37. - P. 359-376.'
96. Hamerton J.L., Canning N., Ray M., Smith S. A cytogenetic survey of 14,069 newborn infants. I,
Incidence of chromosome abnormalities j j Clin. Genet. — 1975. - Vol. 8. — P. 223-243.
97. Bojesen A., Juul S., Gravholt C.H, Prenatal and postnatal prevalence of Klinefelter syndrome: a
national registry study //J. Clin. Endocrinol. Metab, - 2003. - Vol. 88. - P. 622-626.
98. Paulsen C.A., Gordon D.L., Carpenter R.W, et al. Klinefelter’s syndrome and its variants: a hor¬
monal and chromosomal study // Recent. Prog. Horm. Res, - 1968. - Vol. 24. - P. 321-363.
99. Huckins C., Bullock L.P., Long J.L, Morphological profiles of cryptorchid XXY mouse testes j j
Anat. Rec. - 1981. - Vol. 199. - P. 507-518.
100. Bandmann H.J., Breit R., Perwein E. Klinefelter’s Syndrome. — Berlin, Springer-Verlag, 1984.
101. Hook E.B. Rates of chromosome abnormalities at different maternal ages // Obstet. Gynecol. —
1981. - Vol. 58. - P. 282-285.
102. Abramsky L„ Chappie J. 47,XXY (Klinefelter syndrome) and 47,XYY: estimated rates of and
indication for postnatal diagnosis with implications for prenatal counselling // Prenat. Diagn. —
1997. - Vol. 17. - P. 363-368.
103. Wikstrom A.M., Dunkel L., Wickman S. et al. Are adolescent boys with Klinefelter syndrome
androgen deficient.? A longitudinal study of Finnish 47,XXY boys // Pediatr. Res. — 2006. — Vol.
59. - P. 854-859.
104. Aksglaede L„ Wikstrom A.M., Rajpert-De Meyts E. et al. Natural history of seminiferous tubule
degeneration in Klinefelter syndrome // Hum. Reprod. Update. - 2006. — Vol. 12. - P. 39-48.
105. Wikstrom A.M., Raivio T., Hadziselimovic F. et al. Klinefelter syndrome in adolescence: onset
of puberty is associated with accelerated germ cell depletion //J, Clin. Endocrinol. Metab. — 2004. —
Vol. 89. - P. 2263-2270.
106. Bojesen A., Juul S., Birkebaek N., Gravholt C.H. Increased mortality in Klinefelter syndrome //
J. Clin, Endocrinol. Metab. — 2004. — Vol. 89. - P. 3830-3834,
107. Swerdlow A.J., Hermon C., Jacobs P.A. et al. Mortality and cancer incidence in persons with
numerical sex chromosome abnormalities: a cohort study // Ann. Hum. Genet. - 2001, - Vol. 65. -
P. 177-188.
108. Swerdlow A.J., Schoemaker M.J., Higgins C.D. et al. Cancer incidence and mortality in men with
Klinefelter syndrome: a cohort study //J. Natl. Cancer. Inst. - 2005. - Vol. 97. - P. 1204-1210.
109. Howell S.J., Shalet S.M. Spermatogenesis after cancer treatment: damage and recovery //J.
Natl. Cancer. Inst. Monogr. - 2005. - Vol. 34. - P. 12-17.
110. Howell S.J., Radford J.A., Smets E.M., Shalet S.M. Fatigue, sexual function and mood following
treatment for haematological malignancy: the impact of mild Leydig cell dysfunction // Br. J. Cancer. —
2000. - Vol. 82. - P. 789-793.
111. Holmes S.J., Whitehouse R.W., Clark S.T. et al. Reduced bone mineral density in men following
chemotherapy for Hodgkin’s disease // Br.J. Cancer, — 1994. — Vol. 70. - P. 371-375.
112. Monteil M„ Rousseaux S., Chevret E. et al. Increased aneuploid frequency in spermatozoa from
a Hodgkin’s disease patient after chemotherapy and radiotherapy // Cytogenet. Cell. Genet. — 1997. -
Vol. 76. - P. 134-138.
304
113. Robbins W.A., Meistrich M.L., Moore D. et al. Chemotherapy induces transient sex chromo¬
somal and autosomal aneuploidy in human sperm // Nat. Genet. — 1997. - Vol. 16. - P. 74-78.
114. Robbins W.A. Cytogenetic damage measured in human sperm following cancer chemothera¬
py // Mutat. Res. - 1996. - Vol. 355. - P. 235-252.
115. Howell S.J., Shalet S.M. Effect of cancer therapy on pituitary-testicular axis // Int. J. Androl. —
2002. - Vol. 25. - P. 269-276.
116. Meistrich M.L., Chawla S.P., Da Cunha M.F. et al. Recovery of sperm production after chemo¬
therapy for osteosarcoma // Cancer. — 1989. — Vol. 63. — P. 2115-2123,
117. da Cunha M.F., Meistrich M.L., Fuller L.M. et al. Recovery of spermatogenesis after treatment
for Hodgkin’s disease: limiting dose of MOPP chemotherapy //J. Clin, Oncol. - 1984. — Vol. 2. —
P. 571-577.
118. Whitehead E., Shalet S.M., Blackledge G. et al. The effects of Hodgkin’s disease and com¬
bination chemotherapy on gonadal function in the adult male // Cancer. - 1982. - Vol. 49. —
P. 418-422.
119. Whitehead E., Shalet S.M., Jones P.H. et al. Gonadal function after combination chemotherapy
for Hodgkin’s disease in childliood // Arch. Dis. Child. — 1982. — Vol. 57. - P. 287-291,
120. Viviani S., Santoro A., Ragni G. et al. Gonadal toxicity after combination chemotherapy for
Hodgkin’s disease. Comparative results of MOPP vs ABVD // Eur. J. Cancer. Clin. Oncol. — 1985. —
Vol. 21. - P. 601-605.
121. Chatterjee R., Mills W., Katz M. et al. Germ cell failure and Leydig cell insufficiency in post-
pubertal males after autologous bone marrow transplantation with BEAM for lymphoma // Bone
Marrow. Transplant. — 1994. — Vol. 13. — P. 519-522.
122. Hansen S.W.. Berthelsen J.G., von der Maase H. Long-term fertility and Leydig cell function
in patients treated for germ cell cancer with cisplatin, vinblastine, and bleomycin versus surveillance //
J. Clin. Oncol. - 1990. - Vol. 8. - P. 1695-1698.
123. Johnson D.H., Linde R., Hainsworth J.D, et al. Effect of a luteinizing hormone releasing hor¬
mone agonist given during combination chemotherapy on posttherapy fertility in male patients with
lymphoma: preliminary observations //Blood. — 1985. — ’Yol. 65. — P. 832-836.
124. Waxman J.H., Ahmed R., Smith D. et al. Failure to preserve fertility in patients with Hodgkin’s
disease // Cancer. Chemother. Pharmacol. — 1987. - Vol. 19. - P. 159-162.
125. Cross J.J., Berman L.H., Elliott P.G., Irving S. Scrotal trauma: a cause of testicular atrophy //
Clin. Radiol. - 1999. - Vol. 54. - P. 317-320.
126. Manson A.L. Mumps orchitis // Urology. — 1990. — Vol. 36. — P. 335-338.
127. Dobs A.S., Dempsey M.A., Ladenson P.W., Polk B.F. Endocrine disorders in men infected with
human immunodeficiency virus // Am. J. Med. - 1988. — Vol. 84. - P. 611 616.
128. Arver S., Sinha-Hikim L, Beall G. et al. Serum dihydrotestosterone and testosterone concentra¬
tions in human immunodeficiency vinis-infected men with and without weight loss // J. Androl. —
1999. - Vol. 20. - P. 611-618.
129. Rietschel P., Corcoran C., Stanley T, et al. Prevalence of hypogonadism among men with weight
loss related to human immunodeficiency virus infection who were receding highly active antiretroviral
therapy j j Clin. Infect. Dis. - 2000. - Vol. 31. - P. 1240-1244.
130. Salehian B„ Jacobson D., Swerdloff R.S. et al. Testicular pathologic changes and the pituitary-
testicular axis during human immunodeficiency vims infection j j Endocr. Pract. - 1999. — Vol. 5. -
P. 1-9.
131. Bhasin S., Calof O., Storer T.W. et al. Drug insights: anabolic applications of testosterone and
selective androgen receptor modulators in aging and chronic illness // Nature. CPEM. — 2006. — Vol.
2. - P. 133-140.
132. Dobs A.S., Few 3rd W.L., Blackman M.R. et al. Serum hormones in men with human
immunodeficiency virus-associated wasting j/ J. Clin. Endoainol. Metab. — 1996. — Vol. 81. — P.
4108-4112.
133. Edman D.C., Winter A.J., Porter J.C. Embryonic testicular regression: a clinical spectrum of XY
agonadal individuals // Obstet. Gynecol. — 1977. — Vol. 49. — P. 208-217.
134. Borrow M., Gough M. Bilateral absence of the testes // Lancet, - 1970: - Vol. 1. - P. 366.
135. Hall J.. Morgan A., Blizzard R.M. Familial congenital anorchia // Birth. Defects. Orig. Art.
Ser. - 1975. - Vol. 11. - P. 115-119.
136. Marcantonio S.M,, Fechner P.Y., Migeon CJ. et al. Embryonic testicular regression sequence:
a part of the clinical spectmm of 46, XY gonadal dysgenesis j j Am. J. Med. Genet. - 1994. - Vol.
49. - P. 1-5.
305
137. Vinci G., Anjot M., Trivin C, et al. An analysis of the genetic factors involved in testicular
de.scent in a cohort of 14 male patients with anorchia //J. Clin. Endocrinol. Metab. - 2004. — Vol.
89. - P. 6282-6285.
138. de Roux N., Milgrom E, Inherited disorders of GnRH and gonadotropin receptors // Mol. Cell.
Endocrinol. - 2001. - Vol. 179. - P. 83-87.
139. Gromoll J., Eiholzer U„ Nieschlag E., Simoni M. Male hypogonadism caused by homozygous
deletion of exon 10 of the luteinizing hormone (LH) receptor; differential action of human chorionic
gonadotropin and LH //J. Clin. Endocrinol. Metab. - 2000. - Vol. 85. - P. 2281-2286.
140. Latronico A.C., Arnhold I.J. Inactivating mutations of LH and FSH receptors — from genotype
to phenori'pe // Pediatr. Endocrinol. Rev. — 2006. - Vol. 4. - P. 28-31.
141. McDonough P.G. Inactivating mutations in the LH receptor - as rare a “a hen with teeth” or as
frequent as polycystic ovary syndrome.? // Fertil. Steril. — 2000. — Vol. 73. — P. 655-656.
142. Richter-Unruh A., Martens J.W., Verhoef-Post M. et al. Leydig cell hypoplasia: cases with new
mutations, new polymorphisms and cases without mutations in the luteinizing hormone receptor gene //
Clin. Endocrinol. (Oxf.). - 2002. - Vol. 56. - P. 103-112.
143. Salameh W,, Choucair M., Guo T.B. et al. Leydig cell hypoplasia due to inactivation of
luteinizing hormone receptor by a novel homozygous nonsense truncation mutation in the seventh
transmembrane domain // Mol. Cell. Endocrinol. - 2005. - Vol. 229. - P. 57-64.
144. Themmen A.P., Brunner H.G. Luteinizing hormone receptor mutations and sex differentia¬
tion // Eur. J. Endocrinol. - 1996. - Vol. 134. - P. 533-540.
145. Toledo S.P., Brunner H.G., Kraaij JR. et al. An inactivating mutation of the luteinizing hormone
receptor causes amenorrhea in a 46, XX female // J. Clin. Endocrinol. Metab. — 1996. — Vol. 81. -
P. 3850-3854.
146. Tsigos C., Latronico C., Chrousos G.P. Luteinizing hormone resistance syndromes // Ann. N.
Y. Acad. Sci. - 1997. - Vol 816. ~ P. 263-273.
147. Wu S.M., Hallermeier K.M., Laue L. et al. Inactivation of the luteinizing hormone/chorionic
gonadotropin receptor bv an insertional mutation in Le3»dig cell hypoplasia // Mol. Endoainol. —
1998. - Vol. 12, - P. 1651-1660.
148. Wu S.M., Leschek E.W., Rennert O.M., Chan W.Y. Luteinizing hormone receptor mutations in
disorders of sexual development and cancer // Front. Biosci. — 2000. — Vol. 5. — P. 343-352.
149. Aittomaki K., Herva R., Stenman U.H. et al. Clinical features of primary ovarian failure caused
bv'a point mutation in the follicle-stimulating hormone receptor gene //J. Clin. Endocrinol. Metab. —
1996. - Vol. 81. - P, 3722-3726.
150. Allen L.A., Achermann J.C., Pakarinen P. et al. A novel loss of function mutation in exon 10 of
the FSH receptor gene causing hypergonadotrophic hypogonadism: clinical and molecular characteris¬
tics // Hum. Reprod. - 2003. - Vol. IS. - P. 251-256.
151. Jiang M., Aittomaki K., Nilsson C. et al. The frequency of an inartivating point mutation
(566C-^T) of the human follicle-stimulating hormone receptor gene in four populations using allele-
specific hybridization and time-resolved fluorometry //J. Clin, Endocrinol. Metab, - 1998. — Vol.
83. - P. 4338-4343.
152. Layman L.C., Amde S., Cohen D.P. et al. The Finnish follicle-stimulating hormone receptor
gene mutation is rare in North American women wth 46, XX ovarian failure // Fertil. Steril. — 1998. —
Vol. 69. - P. 300-302.
153. Loutradis D., Patsoula E., Stefanidis K. et al. Follicle-stimulating hormone receptor gene muta¬
tions are not evident in Greek women with premature ovarian failure and poor responders // Gynecol.
Obstet. Invest. - 2006. - Vol. 61. - P. 56-60.
154. Betterle C., Pra C,D,, Mantero P., Zanchetta R. Autoimmune adrenal insufficiency and autoim¬
mune polyendocrine syndromes: aiitoantibodies, autoantigens, and their applicability in diagnosis and
disease prediction // Endocr. Rev. - 2002. - Vol. 23. - P. 327-364.
155. Sauder S.E., Frager M., Case G.D. et al. Abnormal patterns of pulsatile luteinizing hormone
secretion in women with byperprolaainemia: responses to bromocriptine // J, Clin. Endocrinol,
Metab. - 1984. - Vol. 59. - P. 941-948.
156. Tay C.C., Glasier A.F., Illingworth P.J.. Baird D.T. Abnormal 24-hour pattern of pulsatile LH
seaetion and response to naloxone in women with hyperprolartinemic amenorrhea // Clin. Endocrinol.
(Oxf.). - 1993. - Vol. 39. - P. 599-606.
157. Miienkovic L. et ai. Inhibition of gonadotropin hormone-releasing hormone release by prolac¬
tin from GTl neuronal cell lines through prolactin receptors // Proc, Natl. Acad. Sci USA. - 1994. -
Vol. 91. - P. 1244-1247,
306
158. Feldman H.A., Longcope С.. Derby C.A. et al. Age trends in the level of serum testosterone and
other hormones in middle-aged men: longitudinal results from the Massachusetts Male Aging Study //
J. Clin. Endocrinol. Metab. - 2002. - Vol. 87. - P. 589-598.
159. Gray A. An examination of research design effects on the association of testosterone and
male aging: results of a meta-analysis / A. Gray, J.A. Berlin, J.B. McKinlay, C. Longcope //J. Clin.
Epidemiol. - 1991. - Vol. 44. - P. 671-684.
160. Harman S.M., Metter EJ.. Tobin J.D. et al. Longitudinal effects of aging on serum total and
free testosterone levels in healthy men. Baltimore Longitudinal Study of Aging //J. Clin. Endocrinol.
Metab. - 2001. - Vol. 86. - P. 724-731.
161- Morley J.E., Kaiser F.E., Perry 3rd H.M. et al. Longitudinal changes in testosterone, luteiniz¬
ing hormone, and follicle-stimulating hormone in healthy older men // Metabolism. — 1997. - Vol.
46. - P. 410-413.
162. Simon D., Preziosi P., Barrett-Connor E. et al. The influence of aging on plasma sex hormones
in men: the Telecom Study // Am. J. Epidemiol. - 1992. - Vol. 135. - P. 783-791.
163. Zmuda J.M., Cauley J.A., Kriska A. et al. Longitudinal relation between endogenous testoster¬
one and cardiovascular disease risk factors in middle-aged men. A 13-year follow-up of former Multiple
Risk Factor Intervention Trial participants j j Am. J. Epidemiol. — 1997. — Vol. 146. — P. 609-617.
164. Ferrini R.L., Barrett-Connor E. Sex hormones and age: a cross-sectional study of testosterone
and estradiol and their bioavail-able fractions in community-dwelling men j j Am. J. Epidemiol. —
1998. - Vol. 147. - P. 750-754.
165. Harman S.M., Tsitouras P.D. Reproductive hormones in aging men. i. Measurement of sex
steroids, basal luteinizing hormone, and Leydig cell response to human chorionic gonadotropin // J.
Clin. Endocrinol. Metab. - 1980. - Vol. 51. - P. 35-40.
166. Veldhuis J.D., Veldhuis N.J., Keenan D.M, Iranmanesh A. Age diminishes the testicular ste¬
roidogenic response to repeated intravenous pulses of recombinant human LH during acute GnRH-
receptor blockade in healthy men // Am. J. Physiol. Endocrinol. Metab. — 2005. - Vol. 288. —
P. 775-781.
167. Takahashi P.Y., Liu P.Y., Roebuck P.D. et al. Graded inhibition of pulsatile luteinizing hormone
secretion by a selective gonadotropin-releasing hormone (GiiRH)-receptor antagonist in healthy men:
evidence that age attenuates hypothalamic GnRH outflow // J. Clin. Endocrinol. Metab. — 2005. —
Vol. 90. - P. 2768-2774.
168. Keenan D.M., Takahashi P.Y., Liu P.Y. et ai. An ensemble model of the male gonadal axis: illus¬
trative application in aging men // Endocrinology. — 2006. — Vol. 147. - P. 2817-2828.
169. Mulligan Т., Iranmanesh A., Johnson M.L. et al. Aging alters feed-foi-ward and feedback
linkages between LH and testosterone in healthy men // Am. J. Physiol. — 1997. — Vol. 273. —
P. 1407-1413.
170. Veldhuis J.D., Zwart A., Mulligan T„ Iranmanesh A. Muting of androgen negative feedback
unveils impoverished gonadotropin-releasing hormone/luteinizing hormone secretory reactivity in
healthy older men //J. Clin. Endocrinol. Metab. - 2001. - Vol. 86. - P. 529-535.
171. Daniel! H.W. Hypogonadism in men consuming sustained-action oral opioids // J. Pain. —
2002. - Vol. 3. - P. 377-384.
172. Rajagopal A.. Vassilopoulou-Sellin R., Palmer J.L. et al. Symptomatic hypogonadism in male
survivors of cancer with chronic exposure to opioids // Cancer. — 2004. — Vol. 100. — P. 851-858.
173. Abs R.. Verhelst J., Maeyaert J. et al Endocrine consequences of long-term intrathecal adminis¬
tration of opioids //J. Clin. EndoCTinol. Metab. - 2000. - Vol. 85. - P. 2215-2222.
174. Daniel] H.W., Lentz R., Mazer N.A. Open-label pilot study of testosterone patch therapy in men
with opioid-induced androgen deficiency //J. Pain. - 2006. - Vol. 7. - P. 200-210.
175. de la Rosa R.E., Hennessey J.V. Hypogonadism and methadone: Hypothalamic hypogonadism
after long-term use of high-dose methadone // Endocr. Pract. - 1996. - Vol. 2. - P. 4-7.
176. Bliesener N.. Albrecht S., Schwager A. et al. Plasma testosterone and sexual function in men
receiving buprenorphine maintenance for opioid dependence //J. Clin. Endocrin&l. Metab. — 2005. —
Vol. 90. - P. 203-206.
177. Rasmussen D.D,, Liu J.H., Wolf P.L., Yen S.S. Endogenous opioid regulation of gonadotropin-
releasing hormone release from the human fetal hypothalamus in vitro //J. Clin. Endocrinol. Metab. —
1983. - Vol. 57. - P. 881-884.
178. Ellingboe J., Veldhuis J.D., Mendelson J.H. et al. Effect of endogenous opioid blockade on
the amplitude and fi-equency of pulsatile luteinizing hormone secretion in normal men // J. Clin.
Endocrinol. Metab. - 1982. - Vol. 54. - P. 854-857.
307
179. Evans W.S., WeltmanJ.Y., Johnson M.L. et al, Effects of opioid receptor blockade on luteinizing
hormone (LH) pulses and interpulse LH concentrations in normal women during the early phase of the
menstrual cycle //J. Endocrinol. Invest. — 1992. — Vol. 1.5. — P. 525-531.
180. Veldhuis J.D., Rogol A.D., Johnson M.L. Endogenous opiates modulate the pulsatile secretion of
biologically active luteinizing hormone in man //J. Clin. Invest. - 1983. - Vol. 72. - P. 2031-2040.
181. Rimoin D.L., Schimke R.N. Mechanisms of gene action in disorders of the endocrine glands //
Birth. Defects, Orig. Art. Ser. — 1971, — Vol. 7. — P. 5-11.
182. Crowley Jr. W.F., Whitcomb R.W., Jameson J.L. et al. Neuroendocrine control of human repro¬
duction in the male // Recent. Prog. Horm. Res. — 1991. — Vol. 47. — P. 27-62.
183. Waldstreicher J., Seminara S.B., Jameson J.L. et al. The genetic and clinical heterogeneity of
gonadotropin-releasing hormone deficiency in the human //J. Clin. Endocrinol. Metab. — 1996. —
Vol. 81. - P. 4388-4395.
184. Spratt D.I., Carr D.B., Merriam G.R. et al. The .spectrum of abnormal patterns of gonadotropin-
releasing hormone secretion in men with idiopathic hypogonadotropic hypogonadism: clinical and
laboratory correlations //J. Clin. Endocrinol. Metab. — 1987. — Vol. 64. — P. 283-291.
185. Smals A.G., Kloppenborg P.W., van Haelst Lf.J. et al. Fertile eunuch syndrome versus classic
hypogonadotrophic hypogonadism // Arta Endocrinol. (Copenh). - 1978. - Vol. 87. - P. 389- 399.
186. Mozaffarian G.A., Higley M., Paulsen C.A. Clinical studies in an adult male patient with “iso¬
lated follicle stimulating hormone (FSH) deficiency” //J. Androl. — 1983. - Vol. 4. - P. 393 -398.
187. Seminara S.B., Oliveira L.M., Beranova M. ct al. Genetics of hypogonadotropic hypogona¬
dism //J. Endocrinol. Invest. — 2000. — Vol. 23. — P. 560-565,
188. Franco B„ Guioli S., Pragliola A. et al. A gene deleted in Kallmann’s syndrome shares homol¬
ogy with neural cell adhesion and axonal path-finding molecules // Nature. - 1991. - Vol. 353. ■-
P. 529-536.
189. Ballabio A„ Bardoni B., Carrozzo R. et al. Contiguous gene syndromes due to deletions in the
distal short arm of the human X chromosome j j Proc. Natl. Acad. Sci USA. - 1989. - Vol, 86. —
P. 10001- 10005.
190. Legouis R-, Hardelin J.P., Levilliers J. et al. The candidate gene for the X-linked Kallmann syn¬
drome encodes a protein related to adhesion molecules // Cell. — 1991. — Vol. 67. — P. 423-435.
191. Georgopoulos N.A., Pralong F.P., Seidman C.E. et al. Genetic heterogeneity evidenced by low
incidence of KAL-1 gene mutations in sporadic cases of gonadotropin-releasing hormone deficiency //
J. Clin': Endocrinol. Metab. - 1997. - Vol. 82. - P. 213-217.
192. Schwanzel-Fukuda M., Pfaff D.W. Origin of luteinizing hormone-releasing hormone
neurons j j Nature. — 1989. — Vol. 338. — P. 161-164.
193. Schwanzel-Fukuda M., Pfaff D.W. The migration of luteinizing hormone-releasing hormone
(LHRH) neurons from the medial olfactoiy' placode into the medial basal forebrain j j Experientia. —
1990. - Vol. 46. - P. 956-962.
194. Klingmuller D., Dewes W., Krahe T. et al. Magnetic resonance imaging of the brain in patients
with anosmia and hypothalamic hypogonadism (Kallmann’s syndrome) j j J, Clin. Endocrinol.
Metab. - 1987. - Vol. 65. - P. 581-584.
195. Sato N.,Katsumata N.,Kagami M. etal. Clinical assessment and mutation analysis of Kallmann
syndrome 1 (KALI) and fibroblast growth factor receptor 1 (FGFRl, or KAL2) in five families and 18
sporadic patients //J. Clin. Endocrinol. Metab. - 2004. - Vol. 89. — P. 1079-1088.
196. Phelan J.K., McCabe E.R.B. Mutations in NROBl (DAXl) and NR5A1 (SFl) responsible for
adrenal hypoplasia congenita // Human. Mutations. - 2001. - Vol. 18. — P. 472-487,
197. Burris T.P., Guo W., McCabe E.R. The gene responsible for adrenal hypoplasia congenita,
DAX-1, encodes a nuclear hormone receptor that defines a new class within the superfamily // Recent.
Prog. Horm. Res. - 1996. — Vol. 51. - P. 241-259. discussion 259-260.
198. Habiby R.L., Boepple P., Nachtigall L. et al. Adrenal hypoplasia congenita with hypogonado¬
tropic hypogonadism: evidence that DAX-1 mutations lead to combined hypothalmic and pituitary
defects in gonadotropin production //J. Clin. Invest. — 1996. — Vol. 98. — P. 1055-1062.
199. Luo X., Ikeda Y., Parker K.L. A cell-specific receptor is essential for adrenal and gonadal devel¬
opment and sexual differentiation // Cell. — 1994. — Vol. 77. - P. 481-490.
200. Crawford P.A., Dorn C., Sadovsky Y., Milbrandt J. Nuclear receptor DAX-1 recruits nuclear
receptor corepressor N-CoR to SF-1 // Mol. Cell. Biol. - 1998. - Vol. 18. - P. 2949-2956.
201. de Roux N., Young J-, Misrahi M. el al. A family with hypogonadotropic hypogonadism and
mutations in the gonadotropin-releasing hormone receptor // N. Engl. J. Med. - 1997. - Vol. 337. -
P. 1597-1602,
308
202. Caron P., Chauvin S., Christin-Maitre S. et al. Resistance of hypogonadic patients wilh mutated
GnRH receptor genes to pulsatile GnRH administration //J. Clin. Endocrinol. Metab. — 1999. — Vol.
84. - P. 990-996.
203. Seminara S.B., Messager S., Chatzidaki E.E. et al. The GPR54 gene as a regulator of puberty //
N. Engl. J. Med. - 2003. - Vol. .349. - P. 1614-1627.
204. Seminara S.B. The first kiss—a crucial role for kisspeptin-l and its receptor, G-protein-coupled
receptor .54, in puberty and reproduction // Nature. CPEM. — 2006. — Vol. 2. — P. 328-334.
205. Ohtaki T., Shintani Y., Honda S. et al. Metastasis suppressor gene kiss-1 encodes peptide ligand
of a G protein coupled receptor // Nature. — 2001. - Vol. 411, — P. 613-617.
206. Navarro V.M,, Castellano J.M., Fernandez-Fernandez R. et al. Developmental and hormon¬
ally regulated mRNA expression of kiss-1 and its putative receptor, GPR54, in the rat hypothalamus
and potent LH releasing activity of KISS-1 peptide // Endocrinology. — 2004. - Vol. 145. —
P. 4565-4574.
207. Miura K., Aderiio Jr. J.S., Seminara S.B. Characterization of the human nasal embryonic LHRH
factor gene, NELF, and a mutation screening among 65 patients with idiopathic hypogonadotropic
hypogonadism (IHH) //J. Hum. Genet. - 2004. - Vol. 49. - P. 265-268.
208. Bbagavath B., Podolsky R.H., Ozata M. et al. Clinical and molecular characterization of a
large sample of patients with hypogonadotropic hypogonadism // Fertil, Steril. — 2006. - Vol. 85. —
P. 706-713.
209. Layman L.C. The molecular basis of human hypogonadotropic hypogonadism // Mol. Genet.
Metab. - 1999. - Vol. 68. - P. 191-199.
210. Layman L.C., Lee E.J., Peak D.6. et al. Delayed puberty and hypogonadism caused by muta¬
tions in the follicle-stimulating hormone fi-subunit gene // N. Engl. J. Med. - 1997. — Vol. 337. —
P. 607-611.
211. Kumar T.R. Follicle stimulating hormone is required for ovarian follicle maturation but not
male fertility / T.R, Kumar, Y. Wang, N. Lu, M.M. Matzuk j j Nat. Genet. — 1997. — Vol. 15. -
P. 201-204.
212. Weiss J.. Axelrod L., Whitcomb R.W. et al. Hypogonadism caused by a single amino acid
substitution in the p-subunit of luteinizing hormone // N. Engl. J. Med. - 1992. - Vol. 326. —
P. 179-183.
213. Nilsson C., Pettersson K., Millar R.P. et al. Worldwide frequency of a common genetic variant
of luteinizing hormone: an international collaborative research. International Collaborative Research
Group // Fertil. Steril. - 1997. - Vol. 67, - P. 998-1004,
214. Raivio T., Huhtaniemi L, Anttila R. et al. The role of luteinizing hormone-p gene polymorphism
in the onset and progression of puberty in healthy boys // J. Clin. Endocrinol. Metab. -- 1996. —
Vol. 81. - P. 3278-3282.
215. Gromoll J., Partsch C.J., Simoni M.N. et al. A mutation in the first transmembrane domain
of the lutropin receptor causes male precocious puberty // J, Clin. Endocrinol, Metab. — 1998. —
Vol. 83. - P. 476-480.
216. Gromoll J., Simoni M., Nordhoff V. et al. Functional and clinical consequences of mutations in
the FSH receptor // Mol. Cell. Endocrinol. - 1996. - Vol. 125. - P. 177-182.
217. Gunay-Aygun M., Schwartz S., Heeger S. et al. The changing purpose of Prader-Willi syndrome
clinical diagnostic criteria and proposed revised criteria // Pediatrics. — 2001. — Vol. 108. — P. 92.
218. Gunay-Aygun M„ Cassidy S.B., Nicholls R.D. Prader-Willi and other .syndromes associated
with obesity and mental retardation // Behav. Genet. - 1997. — Vol. 27. — P. 307-324,
219. Saitoh S., Buiting K., Cassidy S.B. et al. Clinical spectrum and molecular diagnosis of Angelman
and Prader-Willi syndrome patients with an imprinting mutation // Am. j. Med. Genet. — 1997. —
Vol. 68. - P. 195-206.
220. LaSalle J.M. et al. Clonal heterogeneity at allelic méthylation sites diagnostic for Prader-Willi
and Angelman syndromes // Proc. Natl. Acad. Sci. USA. - 1998. — Vol. 95. - P. 1675-1680.
221. Amhold I.J., Nery M., Brown M.R et al. Clinical and molecular characterization of a Brazilian
patient with Pit-1 deficiency //J. Pediatr. Endocrinol. Metab. - 1998. — Vol. 11. — P. 623-630.
222. Parks J.S., Brown M.R. Transcription factors regulating pituitary development // Growth.
Horm. IGF Res. - 1999. - Vol. 9. - Suppl. B. - P. 2-8. discussion 8-11.
223. Pfaffle R.W. et al. Pit-1 and pituitary function //J. Pediatr. Endocrinol. — 1993. — Vol. 6. ~
P. 229-233.
224. Wu W„ Cogan J.D., Pfaffle R.W. et al. Mutations in PROPl cause familial combined pituitary
hormone deficiency // Nat. Genet. - 1998. — Vol. 18. — P. 147-149.
309
225. Dattani M.T., Martinez-Barbera J.P,, Thomas P.Q, et al. Mutations in the homeobox gene
HESXl/Hesxl associated with septo-optic dysplasia in human and mouse j j Nat. Genet. - 1998. -
Vol. 19. - P. 125-133.
226. Radovick S., Nations M., Du Y. et al. A mutation in the POU-homeodomain of Pit-1 responsible
for combined pituitary hormone deficiency // Science. — 1992. — Vol. 257, - P. 1115-1118.
227. Semina E.V., Reiter R., Leysens N.J. et al. Cloning and characterization of a novel bicoid-related
homeobox transcription factor gene. RTEG, involved in Rieger syndrome // Nat. Genet. — 1996. — Vol.
14. - P, 392-399.
228. Netchine 1., Sobrier M.L., Krude H. et al. Mutations in LHX3 result In a new syndrome revealed
by combined pituitary hormone deficiency j j Nat. Genet. — 2000. - Vol. 25. - P. 182-186.
229. Brooks-Gunn J., Warren M.P., Hamilton L.H. The relation of eating problems and amenorrhea
in ballet dancers // Med. Sd. Sports. Exerc. — 1987. — Vol. 19. — P. 41-44.
230. Warren M.P., Perlroth N.E. The effects of intense exercise on the female reproductive sys¬
tem //J. Endocrinol. - 2001. - Vol. 170. - P. 3-11,
231. Frisch R.E., Wyshak G., Vincent L. Delayed menarche and amenorrhea in ballet dancers j j
N. Engl. J, Med. - 1980. - Vol, 303. - P. 17-19.
232. Glass A.R., Harrison R„ Swerdloff R.S. Effect of undeniutrition and amino acid deficiency on
the timing of puberty in rats // Pediatr. Res. — 1976. — Vol. 10. - P. 951-955.
233. Loucks A.B., Horvath S.M. Exercise-induced stress responses of amenorrheic and eumenor¬
rheic runners//J. Clin. Endocrinol. Metab. - 1984. — Vol. 59. - P. 1109-1120.
234. Veldhuis J.D., Evans W.S., Demers L.M. et al. Altered neuroendocrine regulation of gonado¬
tropin secretion in women distance runners //J. Clin. Endocrinol. Metab. — 1985. — Vol. 61. —
P. 557-563.
235. Loucks A.B, et al. Alterations in the hypothalamic-pituitary-ovarian and the hypothalamic-
pituitary-adrenal axes in athletic women // J. Clin. Endocrinol. Metab. — 1989. - Vol. 68. —
P. 402-411.
236. Frisch R.E., Wyshak G., Albright N.L. et al. Lower prevalence of non-reproductive system
cancers among female former college athletes // Med. Sci. Sports. Exerc. — 1989. — Vol. 21. —
P. 250-253.
237. Frisch R.E., Wyshak G., Albright N.L, et al. Lower prevalence of breast cancer and cancers of
the reproductive system among former college athletes compared to non-athletes // Br. J, Cancer. -
1985. - Vol. 52. - P. 885-891.
238. Frisch R.E., Wyshak G., Albright N.L. et al. Lower lifetime occurrence of breast cancer and
cancers of the reproductive system among former college athletes // Am. J. Clin. Nutr. - 1987. — Vol.
45. - P. 328-335.
239. Frisch R.E., Wyshak G„ Witschi J. et al. Lower lifetime occurrence of breast cancer and can¬
cers of the reproductive system among former college athletes // Int.J. Fertil. -■ 1987. — Vol. 32. —
P. 217-225.
240. Wyshak G., Frisch R.E., Albright N.L. et al. Lower prevalence of benign diseases of the breast
and benign tumours of the reproductive system among former college athletes compared to non¬
athletes // Br.J. Cancer. - 1986. - Vol. .54. - P. 841-845.
241. Malin A., Matthews C.E,, Shu X.O. et al. Energy balance and breast cancer risk // Cancer.
Epidemiol. Biomarkers. Prev. — 2005. — Vol. 14. - P. 1496-1501.
242. Rogol A.D. et al. Pulsatile secretion of gonadotropins and prolactin in male marathon runners.
Relation to the endogenous opiate system //J. Androl. — 1984. - Vol. 5. — P. 21-27.
243. MacConiiie S.E. et al. Decreased hypothalamic gonadotropin-releasing hormone secretion in
male marathon runners j j N. Engl. J, Med. — 1986. — Vol. 315. — P. 411-417.
244. Comerci G.D. Medical complications of anorexia nerv'osa and bulimia nervosa // Med. Clin.
North. Am, - 1990. - Vol. 74. - P. 1293-1310.
245. Kalm L.M., Semba R.D. They starved so that others be better fed: remembering Ancel Keys and
the Minnesota experiment //J. Nutr. — 2005. — Vol. 135. — P. 1347-1352.
246. Keys A„ Brazek J., Henschel A., Mickelsen 0., Taylor H.L. The Biolog>' of Human Starva¬
tion. — Minneapolis: University of Minnesota Press, 1950.
247. Glass A.R., Swerdloff R.S.. Bray G.A. et al. Low serum testosterone and sex-hormone-binding-
globnlin in massively obese men //J. Clin. Endocrinol. Metab. — 1977. - Vol. 45. - P. 1211-1219.
248. Amatruda J.M. et al. Depressed plasma testosterone and fractional binding of testo.sterone in
obese males //J. Clin. Endocrinol. Metab. - 1978. - Vol. 47. - P. 268-271.
310
249. Vermeulen A. Decreased androgen levels and obesity in men // Ann. Med. — 1996. — Vol.
28. - P. 13-15.
250. Vermeulen A. et al. Attenuated luteinizing hormone (LH) pulse amplitude but normal LH pulse
frequency and its relation to plasma androgens in hypogonadism of obese men //J. Clin. Endocrinol.
Metab. - 1993. - Vol. 76. - P. 1140 -1146.
251. Zumoff B„ Strain G.W.. Miller L.K. et al. Plasma free and non-sex-hormone-binding-globulin-
bound testosterone are decreased in obese men in proportion to their degree of obesity // J, Clin.
Endocrinol. Metab. - 1990. - Vol. 7L - P. 929-931.
252. Birkeland K.I. et al. Level of sex hormone-binding globulin is positively correlated with
insulin sensitivity in men with type 2 diabetes //J. Clin. Endocrinol. Metab. — 1993. — Vol. 76. —
P. 275-278.
253. Laing I. et al. Serum sex-hormone-binding globulin is related to hepatic and peripheral insulin
sensitivity but not to beta-cell function in men and women with type 2 diabetes // Diabet. Med. —
1998. - Vol. 15, - P. 473-479.
254. Giagulli V.A.. Kaufman J.M., Vermeulen A. Pathogenesis of the decreased androgen levels in
obese men // J. Clin. Endocrinol. Metab. — 1994. - Vol. 79. - P. 997-1000.
255. Plymate S.R. et al. Inhibition of sex honnone-binding globulin production in the human hepatoma
(Hep G2) cell line by insulin and prolactin //J. Clin. Endocrinol. Metab. - 1988. - Vol. 67. — P. 460-464.
256. Crave J.C., Lejeune H., Brebant C. et al. Differential effects of insulin and insulin-like growth
factor I on the production of plasma steroid-binding globulins by human hepatoblastoma-derived (Hep
G2) cells //J. Clin. Endocrinol. Metab. - 1995. - Vol. 80. ^ P. 1283-1289.
257. Plymate S.R., Tenover J.S., Bremner W.J, Circadian variation in testo.sterone, sex hormone-
binding globulin, and calculated non-sex hormone-binding globulin bound testosterone in healthy
young and elderly men //J. Androl. — 1989. — Vol. 10. — P. 366-371.
258. Strain G.W., Zumoff B., Kream J. et al. Mild hypogonadotropic hypogonadism in obese men //
Metabolism. - 1982. - Vol. 31. - P. 871-875.
259. Schneider G, et al. increased estrogen production in obese men // J. Clin. Endocrinol.
Metab. - 1979. - Vol. 48. - P. 633-638.
260. Zumoff B., Miller L.K., Strain G.W. Reversal of the hypogonadotropic hypogonadism of obese
men by administration of the aromatase inhibitor testolactone // Metabolism. - 2003. - Vol. 52, -
P. 1126-1128.
261. Amatruda J.M. et al. Hypothalamic and pituitary dysfunction in obese males // Int. J. Obes. -
1982. - Vol. 6. - P. 183-189.
262. Arnott J.M., Wu F.C.W., Pye S.et al. A clinically relevant approach to defining the prevalence of
late onset hypogonadism in men from the general population. Presented at the 88th Endocrine Society
Meeting, Boston, June 20-24,2006.
263. Diver M.J. Analytical and physiological factors affecting the interpretation of serum testoster¬
one concentration in men //Ann. Clin. Biochem. — 2006. — Vol. 43. — P. 3-12.
264. Cooke R.R., McIntosh J.E., McIntosh R.P, Circadian variation in scrum free and non-SHBG-
bound testosterone in normal men: measurements and simulation using a mass action model // Clin.
Endocrinol. (Oxf,). - 1993. - Vol. 39, - P. 163-171.
265. Rosner W. Sex steroids and the free hormone hypothesis // Cell. — 2006. — Vol. 124. —
P. 455-456. author reply 456-457.
266. Wang C-, Catlin D.H., Demers L.M. et al. Measurement of total serum testosterone in adult
men: comparison of current laboratoty^ methods versus liquid chromatography-tandem mass spectrom¬
etry /7 J. Clin. Endocrinol. Metab. - 2004. — Vol. 89. - P. 534-543.
267. Manni A„ Pardridge W.M., Cefalu W. et al. Bioavailability of albumin-bound testosterone //
J. Clin. Endocrinol. Metab. - 1985. - Vol. 61. - P. 705-710.
268. Vermeulen A., Verdonck L.. Kaufman J.M. A critical evaluation of simple methods for
the estimation of free testosterone in serum // J. Clin. Endocrinol. Metab. — 1999. — Vol. 84. —
P. 3666-3672.
269. Winters S.J., Kelley D.E., Goodpaster B. The analog free testosterone assay: are the results in
men clinically useful? // Clin. Chem. — 1998. — Vol. 44. — P. 2178-2182.
270. Buvat J.. Lemaire A. Endocrine screening in 1,022 men with erectile dysfunction: clinical sig¬
nificance and cost-effective strategy //J. Urol. - 1997. - Vol. 158. — P. 1764-1767.
271. Citron J.T., Ettinger B„ Rubiiioff H. et al. Prevalence of hypothalamic-pituitan^ imaging
abnormalities in impotent men with secondary hypogonadism // J. Urol. — 1996. — Vol. 155. —
P. 529-533.
311
272. Morley J.E., Chariton E., Patrick P. et al. Validation of a screening questionnaire for androgen
deficiency in aging males // Metabolism. — 2000. — Vol. 49. — P. 1239-1242.
273. Smith K.W., Feldman H.A., McKinlay J.B. Construction and field validation of a self-adminis-
tered screener for testosterone deficiency (hypogonadism) in ageing men //Clin. Endocrinol. (Oxf.). —
2000. - Vol. 53. - P. 703-711.
274. Moore C., Huebler D., Zimmermann T. et al. The Aging Males’ Symptoms scale (AMS) as out¬
come measure for treatment of androgen deficiency // Eur, Urol. — 2004. — Vol. 46. — P. 80-87.
275. Stanley H.L. et al. Does hypogonadism contribute to the occurrence of a minimal trauma hip
fracture in elderly men? //J. Am. Geriatr, Soc. - 1991. - Vol. 39. - P. 766-771.
276. Riggs B.L., Khosla S., Melton 3rd L.J. Sex steroids and the constniction and conservation of the
adult skeleton // Endocr. Rev. - 2002. - Vol. 23. - P. 279-302.
277. Wahlstrom J.T., Dobs A.S. Acute and long-term effects of AIDS and injection drug use on
gonadal function //J. Acquir. Immune Defic. Syndr. — 2000. — Vol. 25. — Suppl. 1. — P. 27-36.
278. Wahlstrom J.T.. Tang A., Cofrancesco J. et al. Gonadal hormone levels in injection drug
users // Drug. Alcohol. Depend. - 2000. — Vol. 60. — P. 311-313.
279. Grinspoon S., Corcoran C., Lee K, et ai. Loss of lean body and muscle mass correlates with
androgen levels in hypogonadal men with acquired immunodeficiency syndrome and wasting //J. Clin.
Endocrinol. Metab. — 1996. - Vol. 81. - P. 4051-4058.
280. Singh A.B., Norris K., Modi N. et al. Pharmacokinetics of a transdermal testosterone system in
men with end stage renal disease receiving maintenance hemodialysis and healthy hypogonadal men //
J. Clin. Endocrinol. Metab. - 2001. - Vol. 86. - P. 2437-2445.
281. Casaburi R. Rationale for anabolic therapy to facilitate rehabilitation in chronic obstructive
pulmonary disease // Baillieres. Clin. Endocrinol. Metab. — 1998. - Vol. 12. - P. 407-418.
282. Casaburi R. Skeletal muscle dysfunction in chronic obstructive pulmonary disease // Med. Sci.
Sports. Exerc. - 2001. ~ Vol. 33. - P. 662-670.
283. Creutzberg E.C., Casaburi R. Endocrinological disturbances in chronic obstructive pulmonary
disease // Eur. Respir. J. Suppl. — 2003. - Vol. 46. — P. 76-80.
284. Dhindsa S., Prabhakar S., Sethi M. et al. Frequent occurrence of hypogonadotropic hypogo¬
nadism in type 2 diabetes //J. Clin. Endocrinol. Metab. — 2004. — Vol. 89. — P. 5462-5468.
285. Reid I.R. et al. Plasma testosterone concentrations in asthmatic men treated with glucocorti¬
coids // Br. Med. J. (Clin. Res. Ed.). - 1985. - Vol. 291. - P. 574.
"286. Arver S.. Dobs A.S., Meikle A.W. et al. Improvement of sexual function in testosterone deficient
men treated for 1 year with a permeation enlianced testosterone transdermal system // J. Urol. —
1996. - Vol. 155. - P. 1604-1608.
287. Steidle C., Schwartz S., Jacoby K. et al. AA2500 testosterone gel normalizes androgen levels
in aging males with improvements in body composition and sexual function // J. Clin. Endocrinol.
Metab. - 2003. - Vol. 88. - P. 2673-2681.
288. Kwan M., Greenleaf W.J., Mann J. et al. The nature of androgen action on male sexuality: a
combined laboratory-self-report study on hypogonadal men //J. Clin. Endocrinol, Metab. — 1983. —
Vol. 57. - P. 557-562.
289. Snyder P.J., Lawrence D.A. Treatment of male hypogonadism with testosterone enanthate //
J. Clin. Endocrinol. Metab. — 1980. — Vol. 51. — P. 1335-1339.
290. Alexander G.M., Sherwin B.B. Tlie association between testosterone, sexual arousal and selec¬
tive attention for erotic stimuli in men // Horm, Behav. - 1991. - Vol. 25. - P. 367-381.
291. Cuiuiingham G.R. et al. Testosterone replacement therapy and sleep-related erections in
hypogonadal men //J. Clin. Endocrinol. Metab. — 1990. — Vol. 70. — P. 792-797.
292. Wang C., Alexander G., Berman N. et al. Testosterone replacement therapy improves mood
in h}'pogonadal men — a clinical research center study // J. Clin. Endocrinol. Metab. — 1996. — Vol.
81. - P. 3578-3583.
293. Janowsky J.S.. Oviatt S.K., Orwoll E.S. Testosterone influences spatial cognition in older
men // Behav. Neurosci. — 1994. — Vol. 108. — P. 325-332.
294. Cherrier M.M., Asthana S., Plymate S. et al. Testosterone Supplementation improves spatial
and verbal memory in healthy older men //Neurology. - 2001. - Vol. 57, - P. 80-88.
295. Marin P., Holmang S., Jonsson L. et al. The effects of testosterone treatment on body composi¬
tion and metabolism in middle-aged obese men j j Int. J. Obes. Relat. Metab. Disord. - 1992. — Vol.
16. - P. 991-997.
296. Marin P., Krotkiewski M„ Bjomtorp P. Androgen treatment of middle-aged, obese men: effects
on metabolism, muscle and adipose tissues // Eur. J. Med. — 1992. — Vol. 1. — P. 329-336.
312
297. Singli A.B., Hsia S., Alaupovic P. et al. The effects of varying doses of T on insulin sensitivity,
plasma lipids, apolipoproteins, and C-reactive protein in healthy young men //J. Clin. Endocrinol.
Metab. - 2002, - Vol. 87. - P. 136-143.
298. Kapoor D. et al. Testosterone replacement therapy improves insulin resistance, glycaemic con¬
trol, visceral adiposity and hypercholesterolaemia in hypogonadal men with 1}фе 2 diabetes // Eur. J,
Endocrinol. - 2006. - Vol. 154. - P. 899-906.
299. Sokol R.Z. et al. Comparison of the kinetics of injectable testosterone in eugonadal and hj^pogo-
nadal men j j Fertil. Steril. - 1982. — Vol. 37. — P. 425-430.
300. Schulte-Beerbuhl M„ Nieschlag E. Comparison of testosterone, dihydrotestosterone, luteiniz¬
ing hormone and follicle-stimulating hormone in serum after injection of testosterone enanthate of
testosterone cypionate // Fertil. Steril. — 1980. — Vol. 33. — P. 201-203.
301. Wang Z.M., Pierson Jr. R.N.,. Heymsfield S.B. The five-level model; a new approach to organiz¬
ing body-composition research // Am. J. Clin. Nutr. — 1992. — Vol. 56. — P. 19-28.
302. Wang C., Swedloff R.S., Iranmanesh A. et al. Transdermal testosterone gel improves sexual
function, mood, muscle strength and body composition parameters in hypogonadal men. Testosterone
Gel Study Group /7 J. Clin. Endocrinol. Metab. - 2000. - Vol. 85. - P. 2839-2853.
303. Wang C-, Berman N„ Longstreth J.A. et al. Pharmacokinetics of transdermal testosterone gel
in hypogonadal men: application of gel at one site versus four sites: a General Clinical Research Center
Study//J. Clin. Endocrinol. Metab. - 2000. - Vol. 85. - P. 964-969.
304. Wang C., Swerdloff R.S., Iranmanesh A. et al. Effects of transdermal testosterone gel on bone
turnover markers and bone mineral density in hypogonadal men // Clin, Endocrinol, (Oxf.). — 2001. —
Vol. 54. - P. 739- 750.
305. Wang C., Cunningham G., Dobs A. et al. Long-term testosterone gel (AndroGel) treatment
maintains beneficial effects on sexual function and mood, lean and fat mass, and bone mineral density
in hypogonadal men //J. Clin, Endocrinol. Metab. - 2004. - Vol. 89. - P. 2085-2098.
306. McNicholas T.A., Dean J.D., Mulder H. et al. A novel testosterone gel formulation normalizes
androgen levels in hypogonadal men, with improvements in body composition and sexual function //
BJU Int. - 2003. - Vol. 91. - P. 69-74.
307. Arver S., Dobs A.S., Meikle A.W. et al. Long-term efficacy and safety of a permeation-enhanced
testosterone transdermal system in hypogonadal men // Clin. Endocrinol. (Oxf.). — 1997. — Vol, 47. —
P. 727-737.
308. Dobs A.S. et al. Pharmacokinetic characteristics, efficacy and safety of buccal testosterone in
hypogonadal males: a pilot study //J. Clin. Endocrinol. Metab. — 1998. — Vol. 83. - P. 33-39.
309. Dobs A.S., Meikle A.W., Ai-ver S. et al. Pharmacokinetics, efficacy, and safety of a permeation-
enhanced testosterone transdermal system in comparison with bi-weekly injections of testosterone
enanthate for the treatment of hypogonadal men //J. Clin. Endocrinol. Metab. — 1999. — Vol. 84. —
P. 3469-3478.
310. Wang C., Swerdloff R., Kipnes M. et al. New testosterone buccal system (Striant) delivers
physiological testosterone levels: pharmacokinetics study in hypogonadal men // J. Clin. Endocrinol.
Metab. - 2004. - Vol. 89. - P. 3821 -3829.
311. Ross R.J., Jabbar A., Jones Т.Н. et al. Pharmacokinetics and tolerability of a bioadhesive buccal
testosterone tablet in hypogonadal men // Eur. J. Endocrinol. — 2004. - Vol. 150. — P. 57-63.
312. Dobs A.S. et al. Short-term pharmacokinetic comparison of a novel testosterone buccal system
and a testosterone gel in testosterone deficient men // Curr. Med. Res. Opin. — 2004. — Vol. 20. —
P. 729-738.
313. Hirschhauser C. et al. Testosterone undecanoate; a new orally active androgen // Acta
Endocrinol. (Copenh.). - 1975. - Vol. 80. - P. 179-187.
314. Franchimont P. Effects of oral testosterone undecanoate in hypogonadal male patients // Clin.
Endocrinol. (Oxf.). - 1978. - Vol. 9. - P. 313-320.
315. Skakkebaek N.E. et al. Androgen replacement with oral testosterone undecanoate in
hypogonadal men; a double blind controlled study // Clin. Endocrinol. (Oxf.). — 1981. — Vol. 14. —
P. 49-61.
316. Maisey N.M., Bingham J., Marks V. et al. Clinical efficacy of testosterone un'decanoate in male
hypogonadism // Clin. Endocrinol. (Oxf.). - 1981. — Vol. 14. — P. 625-629.
317. O’Carroll R., Shapiro C., Bancroft J. Androgens, behaviour and nocturnal erection in hypogo¬
nadal men: the effects of varying the replacement dose // Clin. Endocrinol. (Oxf.). — 1985. - Vol.
23. - P. 527-538.
313
318. Conway A.J., Boylan L.M., Howe C. et al. Randomized clinical trial of testosterone replacement
therapy in h>'pogonadal men // Int.J. Androl. — 198S. — Vol. 11. — P. 247-264.
319. Gooren L.J. A ten-year safety study of the oral androgen testosterone undecanoate //
J. Androl. - 1994. - Vol. 15. - P. 212-215.
320. Jockenhovel F. et al. Effects of various modes of androgen substitution therapv on erythropoi-
esis /7 Eur. J, Med, Res. -- 1997. - Vol. 2. - P. 293-298.
-321. Zitzmann M., Nieschlag E. Hormone substitution in male hypogonadism // Mol. Cell.
Endocrinol. - 2000. - Vol. 161. - P. 73-88.
322. Cantrill J.A., Dewis P., Large D.M. et al. Which testosterone replacement therapv? // Clin.
Endocrinol. (Oxf.), - 1984. - Vol. 21. - P. 97-107.
323. Handelsman D J., Conway A.J., Boylan L.M. Pharmacokinetics and pharmacodynamics of tes¬
tosterone pellets in man //J. Clin. Endocrinol. Metab. - 1990. - Vol. 71. — P. 216-222.
324. Jockenhovel F., Vogel E., Kreutzer M. et al. Pharmacokinetics and pharmacodynamics of
subcutaneous testosterone implants in hypogonadal men // Clin. Endocrinol. (Oxf.). — 1996. — Vol.
45. - P. 61-71.
.325. Handelsman D.J., Mackey M.A., Howe C. et al. An analysis of testosterone implants for andro¬
gen replacement therapy // Clin. Endocrinol. (Oxf.). - 1997, - Vol. 47. - P. 311-316.
326. Kelleher S., Turner L., Howe C. et al. Extrusion of testosterone pellets: a randomized controlled
clinical study // Clin. Endocrinol, (Oxf.). — 1999. - Vol. 51. — P. 469-471.
327. Kamischke A., Venherm S., Ploger D. et al. Intramuscular testosterone undecanoate and nore-
thisterone enanthate in a clinical trial for male contraception //J. Clin. Endocrinol. Metab. — 2001. —
Vol. 86. - P, 303-309.
328. von Eckardstein S., Nieschlag E. Treatment of male hypogonadism with testosterone unde¬
canoate injected at extended intervals of 12 weeks; a phase II study //J. Androl. — 2002. - Vol. 23. -
P. 419-425.
329. Loughlin K.R., O’Leary M.P. Re; Oral androgens in the treatment of hypogonadal impotent
men //J. Urol. - 1995. - Vol. 153. - P. 1645.
330. Kelleher S., Conway A.J,, Handelsman D.J. Blood testosterone threshold for androgen defi¬
ciency symiptoms // J. Clin, Endocrinol. Metab. — 2004. — Vol. 89. — P. 3813-3817.
331. Bhasin S., Swerdloff R.S., Steiner B. et al. A biodegradable testosterone microcapsule formula¬
tion provides uniform eugonadal levels of testosterone for 10-11 weeks in hj'pogonadal men //J. Clin.
Endocrinol. Metab. - 1992. — Vol. 74. - P. 75-83.
332. Amory J.K., Anawalt B.D., Blaskovich P.D. et al. Testosterone release from a subcutaneous,
biodegradable microcapsule fonnulation (Viatrel) in hc'pogonadal men //J. Androl. — 2002. — Vol.
23.-P. 84-91.
333. Behre H.M,, Nieschlag E. Testosterone buciclate (20 Aet-1) in h>'pogonadal men; pharmacoki¬
netics and pharmacodynamics of the new long-acting androgen ester //J. Clin. Endocrinol. Metab, -
1992. - Vol. 75. - P. 1204-1210.
334. Kinger S., Pal P.C., Rajalakshmi M. et al. Effects of testosterone buciclate on testicular and
epididymal sperm functions in bonnet monkevs (Macaca radiata) // Contraception. — 1995. — Vol.
52. - P. 121-127.
335. Schubert M„ Minnemann T., Hubler D. et al. Intramuscular testosterone undecanoate: phar¬
macokinetic aspects of a novel testosterone formulation during long-term treatment of men with
hypogonadism //J. Clin. Endocrinol. Metab. — 2004. — Vol. 89. — P. 5429-5434.
336. Partsch C.J. et al. Injectable testosterone undecanoate has more favourable pharmacokinetics
and pharmacodynamics than testosterone enanthate // Eur. J. Endocrinol. — 1995. — Vol. 132. —
P. 514-519.
337. Zhang G.Y., Gu Y.Q., Wang X.H. et al. A pharmacokinetic study of injectable testosterone
undecanoate in hypogonadal men //J, Androl. - 1998. - Vol. 19. — P. 761-768.
338. Behre H.M., Abshagen K., Oettel M. et al. Intramuscular injection of testosterone undecanoate
for the treatment of male hypogonadism: phase I studies // Eur. J. Endocrinol. - 1999. - Vol. 140. -
P. 414-419.
339. Zhang G.Y., Gu Y.Q., Wang X.H. et al. A clinical trial of injectable testosterone undecanoate as
a potential male contraceptive in normal Chinese men // J. Clin. Endocrinol. Metab. — 1999. - Vol.
84. - P. 3642-3647.
340. Nieschlag E., Buchter D., Von Eckardstein S. et al. Repeated intramuscular injections of tes¬
tosterone undecanoate for substitution therapy in li)'pogonadal men // Clin. Endocrinol. (Oxf.). —
1999. - Vol. 51. - P. 757-763.
314
341. Anderson R.A., Wallace A.M., Sattar N. et al. Evidence for tissue selectivity of the s}nithetic
androgen 7«-methvl-19-nortestosterone in hypogonadal men //J. Clin. Endocrinol. Metab. - 2003. -
Vol. 88. - P. 2784-2793.
342. Anderson R.A., Martin C.W., Kung A.W. et al. 7a-Methyl-19-nortestosterone maintains
sexual behavior and mood in hypogonadal men //J, Clin. Endocrinol. Metah. — 1999, - Vol. 84. —
P. 3556-3562.
343. Suvisaari J., Sundaram K., Noe G. et al. Pharmacokinetics and pharmacodynamics of
7a-methvl-19-iiortestosterone after intramuscular administration in healthy men // Hum. Reprod. -
1997. - Vol. 12. - P. 967- 973.
344. Negro-Vilar A, Selective androgen receptor modulators (SARMs): a novel approach to androgen
therapy for the new millennium //J. Clin. Endocrinol. Metab. - 1999. — Vol. 84. — P. 3459-3462.
345. Agarwal P.K., Oefelein M.G. Testosterone replacement therapy after primary treatment for
prostate cancer//J. Urol. — 2005. — Vol. 173. — P. 533-.536.
346. Kaufman J. A rational approach to androgen therapy for hjqjogonadal men with prostate can¬
cer // Int. J. Impôt. Res. - 2006. - Vol. 18. - P. 26-31.
347. Kaufman J.M., Graydon R.J. Androgen replacement after curative radical prostatectomy for
prostate cancer in hypogonadal men //J. Urol. - 2004. - Vol. 172. - P, 920-922.
348. Calof 0., Singh A.B., Lee M.L. et al. Adverse events associated with testosterone replacement
in middle-aged and older men: a meta-analysis of randomized, placebo-controlled trials //J. Gerontol.
A. Biol. Sci. Med. Sci. - 2005. - Vol. 60. - P. 1451-1457.
349. Thompson P.D., Cullinane E.M., Sady S.P. et al. Contrasting effects of testosterone and
stanozolol on serum lipoprotein levels //JAMA. - 1989. - Vol. 261. — P. 1165-1168.
350. Bhasin S., Storer T.W., Bennan N. et al. The effects of supraphysiologic doses of testo.sterone on
muscle size and strength in normal men j j N. Engl. J. Med. - 1996. — Vol, 335. — P. 1-7.
351. Sih R., Morley J.E., Kaiser F.E, et al. Testosterone replacement in older hypogonadal men:
a 12-month randomized controlled trial // J. Clin. Endocrinol. Metab. — 1997. — Vol. 82. —
P. 3661-1667.
352. Snyder PJ., Peachey H., Berlin J.A. et al. Effect of transdennal testosterone treatment.on serum
lipid and apolipoprotein levels in men more than 65 years of age // Am. J. Med. -- 2001. — Vol. 111. —
P. 255-260.
353. Barrett-Connor E., Khaw K.T. Endogenous sex hormones and cardiovascular disease in men.
A prospective population-based study // Circulation. — 1988. — Vol. 78. - P. 539-545.
354. De Pergola G., De Mitrio V, Sciaraffia M. et al. Lower androgenicity is associated with higher
plasma levels of prothrombotic factors irrespective of age, obesity, body fat distribution and related
metabolic parameters in men // Metabolism. - 1997. - Vol. 46. - P. 1287-1293.
355. Haffner S.M., Laakso M„ Miettinen H. et al. Low levels of sex hormone-binding globulin
and testosterone are associated with smaller, denser low density lipoprotein in normoglycemic men //
J. Clin. Endocrinol. Metab. - 1996. - Vol. 81. - P. 3697-3701.
356. Khaw K.T., Barrett-Connor E. Lower endogenous androgens predict central adiposity in men //
Ann. Epidemiol. - 1992. - Vol. 2. - P. 675-682.
357. Seidell J.C,, Bjorntorp P., Sjostrom L. et al. Visceral fat accumulation in men is positively
associated with insulin, glucose, and C-peptide levels, but negatively with testosterone levels //
Metabolism. - 1990. - Vol. 39. - P. 897-901.
358. Whitsel E.A., Boyko E.J., Matsumoto A.M. et al. Intramuscular testosterone esters and plasma
lipids in hypogonadal men: a meta-analysis // Am. J. Med. — 2001. — Vol. 111. — P. 261-269.
359. Katznelsori L., Rosenthal D.L, Rosol M.S. et al. U.sing quantitative CT to assess adipose distri¬
bution in adult men with acquired hypogonadism // AJR. Am. J. Roentgenol. — 1998. — Vol. 170, —
P. 423 427.
360. Mauras N., Hayes V., Welch S. et al. Testosterone deficiency in young men: marked alterations
in whole body protein kinetics, strength and adiposity //J. Clin. Endocrinol. Metab. - 1998. - Vol.
83.--P. 1886-1892.
361. Holmang A., Bjorntorp P. The eff ects of testosterone on insulin sensitivity in male rats // Acta.
Physiol. Scand. - 1992. - Vol. 146. - P. 505-510.
362. Hobbs C.J., Jones R.E., Plymate S.R. Nandrolone, a 19-nortestosterone, enhances insulin-
independent glucose uptake in normal men // J. Clin. Endocrinol. Metab. — 1996. — Vol. 81. —
P. 1582-1585.
363. Ramirez M.E., McMurry M.P., Wiebke G.A. et al. Evidence for sex steroid inhibition of lipopro¬
tein lipase in men: comparison of abdominal and femoral adipose tissue // Metabolism. — 1997. — Vol.
46. - P. 179-185.
315
364. Alexandersen P., Haarbo J., Christiansen C. The relationship of natural androgens to coronary
heart disease in males: a review // Atherosclerosis. — 1996. — Vol. 125. — P. 1-13.
365. Bhasin S.. Herbst К. Testosterone and atherosclerosis progression in men // Diabetes. Care. -
2003. - Vol, 26. - P. 1929-1931.
366. Liu P.Y., Death A.K., Handelsman D.J. Androgens and cardiovascular disease // Endocr.
Rev. - 2003. - Vol. 24. - P. 313-340.
367.' English K.M., Steeds R.P., Jones Т.Н. et al. Low-dose transdermal testosterone therapy
improves angina threshold in men with chronic stable angina: A randomized, double-blind, placebo-
controlled study // Circulation. - 2000. — Vol. 102. - P. 1906-1911.
368. Hall J., Jones R.D., Jones Т.Н. et al. Selective inhibition of L-type Ca2+ channels in A7r5 cells
by physiological levels of testosterone // Endocrinology. - 2006. — Vol. 147. — P. 2675-2680,
369. Scragg J.L., Jones R.D., Channer K.S. et al. Testosterone is a potent inhibitor of L-type Ca2+
channels // Biochem. Biophys. Res. Commun. — 2004. - Vol. 318. — P. 503-506.
370. Ong P.J., Patrizi G., Chong W.C. et al. Testosterone enhances flow-mediated brachial arteiy'
reactivity in men with coronary artery disease // Am. J. Cardiol, — 2000. — Vol. 85. - P. 269-272.
371. Bernini G.P,, Sgro M., Moretti A. et al. Endogenous androgens and carotid intimal-medial
thickness in women //J. Clin. Endocrinol. Metab. — 1999. — Vol. 84. — P. 2008-2012.
372. Zitzmann M., Bmne M., Nieschlag E. Vascular reactivity in hypogonadal men is reduced by
androgen substitution //J. Clin. Endocrinol. Metab. - 2002. - Vol. 87, — P. 5030-5037.
373. Vasudevan H„ Nagareddy P.R., McNeill J.H. Gonadectomy prevents endothelial dysfunction
in fructose-fed male rats, a factor contributing to the development of hypertension // Am. J. Physiol.
Heart. Circ. Physiol. - 2006. - Vol, 291. - N 6. - P. 3058 -3064.
374. Nathan L., Shi W., Dinh H, et al. Testosterone inhibits early atherogenesis by conversion to estra¬
diol: critical role of aromatase // Proc. Natl. Acad. Sci. USA. — 2001, - Vol. 98. — P. 3589-3593.
375. Eckardstein A., Wu F.C. Testosterone and atherosclerosis // Growth. Horm. IGF Res. —
2003. -- Vol. 13. - Suppl. - P. 72-84.
376. Bhasin S., Singh A.B., Mac R.P. et al. Managing the risks of prostate disease during testos¬
terone replacement therapy in older men: recommendations for a standardized monitoring plan //
J. Androl. - 2003. - Vol. 24. - P. 299-311.
377. Huggins C., Hodges C.V. Studies on prostatic cancer: I. The effect of castration, of estrogen
and of androgen injection on serum phosphatases in metastatic carcinoma of the prostate. 1941 //
J. Urol. - 2002. - Vol. 168. - P. 9-12.
378. Fowler Jr. J.E,, Whitmore Jr. W.F. The response of metastatic adenocarcinoma of the prostate
to exogenous testosterone //J. Urol. — 1981. - Vol. 126. — P. 372-375.
379. Morgentaler A,, Bruning 3rd C.O., DeWolf W.C. Occult prostate cancer in men with Ionv serum
testosterone levels //JAMA, - 1996. - Vol. 276. - P. 1904-1906.
380. Behre H.M., Bohmeycr J., Nieschlag E. Prostate volume in testosterone-treated and untreated
hypogonadal men in comparison to age-matched normal controls // Clin. EndocTinol. (Oxf.). —
1994. - Vol. 40. - P. 341-349.
381. Meikle A.W., Arver S., Dobs A.S. et al. Prostate size in hypogonadal men treated with a
nonscrotal permeation-enlianced testosterone transdermal system // Urology. — 1997. — Vol. 49. —
P. 191-196.
382. Amory J.K., Watts N.B., Easley K.A. et al. Exogenous testosterone or testosterone with finas¬
teride increases bone mineral density in older men with low serum testosterone //J. Clin. Endocrinol.
Metab. - 2004. - Vol. 89. - P. 503-510.
383. Snyder P.J., Peachey H., Haiinoush P. et al. Effect of testosterone treatment on body composi¬
tion and muscle strength in men over 65 years of age //J. Clin. Endocrinol. Metab. - 1999. - Vol.
84. - P. 2647-2653.
384. Mirand E.A., Gordon A.S., Wenig J. Mechanism of testosterone action in erythropoiesis //
Nature. - 1965. - Vol. 206. - P. 270-272.
385. Gordon A.S. et al. Stimulation of mammalian erythropoiesis by 5p-H steroid metabolites //
Proc. Natl. Acad. Sci. USA. - 1970. - Vol. 65. - P. 919-924.
386. Bhasin S., Woodhouse L., Casaburi R. et al. Older men are as responsive as young men to the
anabolic effects of graded doses of testosterone on the skeletal muscle //J. Clin. Endocrinol. Metab. —
2005. - Vol. 90. - P. 678-688.
387. Matsumoto A.M., Sandblom R.E., Schoene R.B. et al. Testosterone replacement in hypogonadal
men: effects on obstructive sleep apnoea, respiratory drives and sleep // Clin. Endocrinol. (Oxf,). —
1985,-Vol. 22.-P. 713-721.
316
388. Sandblom R.E., Matsumoto A.M., Schoene R.B. et al. Obstructive sleep apnea syndrome
induced by testosterone administration // N. Engl. J. Med. - 1983. — Vol. 308. - P. 508-510.
389. Gruiistein R.R., Handelsman D.J., Lawrence S.J. et al. Neuroendocrine dysfunction in sleep
apnea: reversal by continuous positive airways pressure therapy // J. Clin. Endocrinol. Metab. —
1989. - Vol. 68. - P. 352-358.
390. Riehmann M., Rhodes P.R., Cook T.D. et al. Analysis of variation in prostate-specific antigen
values // Urology. - 1993. - Vol. 42. - P. 390-397.
391. Carter H.B., Pearson J.D., Waclawiw Z. et al. Pro.state-specific antigen variability in men
without prostate cancer: effect of sampling interval on prostate-specific antigen velocity // Urology. —
1995. - Vol. 4.5. - P. 591-596.
392. Brawer M.K. et al. Assay variability in serum prostate-specific antigen determination //
Prostate. - 1995. - Vol. 27. - P. 1-6.
393. Wener M.H., Daum P.R., Brawer M.K. Variation in measurement of prostate-specific antigen:
importance of method and lot variability // Clin. Chem. — 1995. — Vol. 41. — P. 1730-1737.
394. Roehrbom C.G., Pickens G.J., Carmody 3rd T. Variability of repeated serum prostate-spe¬
cific antigen (PSA) measurements within less than 90 days in a well-defined patient population //
Urology. - 1996. - Vol. 47. - P, 59-66.
395. Kadmon D., Weinberg A.D., Williams R.H. et al. Pitfalls in interpreting prostate spedfic antigen
velocity //J. Urol. - 1996. - Vol. 155. - P. 1655-1657.
396. Prestigiacomo A.F.. Stamey T.A. Physiological variation of serum prostate specific antigen in
the 4.0 to 10.0 ng/mL range in male volunteers //J. Urol. - 1996. — Vol. 155. - P. 1977-1980.
397. Carter H.B. PSA variability versus velocity // Urology. — 1997. — Vol. 49. — P. 305.
398. Oesterling J.E., Roy J., Agha A. et al. Biologic variability of prostate-specific antigen and its
usefulness as a marker for prostate cancer: effects of finasteride. The Finasteride PSA Study Group //
Urology. - 1997. - Vol. 50. - P. 13-18.
399. Oesterling J.E., Roy J., Agha A. et al. Biologic variability of prostate-specific antigen and its
usefulness as a marker for prostate cancer: effects of finasteride. Finasteride PSA Study Group //
Urology. - 1998. - Vol, 51. - P. 58-63.
400. Lujan M., Paez A., Sanchez E. et al. Prostate specific antigen variation in patients without clini¬
cally evident prostate cancer //J. Urol. — 1999. — Vol. 162, — P. 1311-1313.
401. Gormley G.J., Stoner E., Bruskewtz R.C. et al The effect of finasteride in men with benign
prostatic hyperplasia. The Finasteride Study Group // N. Engl. J. Med. — 1992. — Vol. 327. —
P. 1185-1191.
402. McConnell J.D., Bruskewitz R., Walsh P. et al. The effect of finasteride on the risk of acute
urinary retention and the need for surgical treatment among men with benign prostatic Ьурефіазіа,
Finasteride Long-Term Efficacy and Safety Study Group // N. Engl. J. Med. — 1998, - Vol. 338. —
P. 557-563.
403. Omstein D.K., Smith D.S., Rao G.S. et al. Biological variation of total, free and percent free
serum prostate specific antigen levels in screening volunteers // J. Urol. — 1997. — Vol. 157. —
P. 2179-2182.
404. Arcangeli C.G. et al. Prostate-specific antigen as a screening test for prostate cancer. The United
States experience j j Urol. Clin. North. Am. — 1997. — Vol. 24. — P. 299-306.
405. Carter H.B., Pearson J.D. PSA velocity for the diagnosis of early prostate cancer. A new con¬
cept // Urol. Clin. North. Am. — 1993. — Vol. 20. — P. 665-670.
406. Carter H.B., Pearson J.D., Metter EJ. et al. Longitudinal evaluation of serum androgen levels
in men with and without prostate cancer // Prostate. - 1995. - Vol. 27. — P. 25-31.
407. Fang J, et al. PSA velocity for assessing prostate cancer risk in men with PSA levels between 2.0
and 4.0 g/mL // Urology. - 2002. - Vol. 59. - P. 889-893. discussion 893-884,
408. Dai W.S., Kuller L.H., LaPorte R.E. et al. The epidemiology of plasma testosterone levels in
middle-aged men //Am. J. Epidemiol. — 1981. — Vol. 114. — P. 804-816.
409. Gray A. et al. Age, disease and changing sex hormone levels in middle-aged men: results of the
Massachusetts Male Aging Study //J. Clin. Endocrinol. Metab. — 1991. — Vot. 73. — P. 1016-1025.
410. Mohr B.A., Bhasin S., Link C.L. et al. The effect of changes in adiposity on testosterone levels
in older men: longitudinal results from the Massachusetts Male Aging Study // Eur. J. Endocrinol, -
2006. - Vol. 155. - P. 443-452.
411. Roy T.A., Blackman M.R., Harman S.M, et al. Interrelationships of serum testosterone and
free testosterone index with FFM and strength in aging men // Am. J. Physiol. Endocrinol. Metab. —
2002. - Vol. 283. - P. 284-294.
317
412. BauMrartner R.N., Waters D.L., Gallagher D. et al. Predictors of skeletal muscle mass in elderly
men and women // Mech. Ageing. Dev. - 1999, - Vol. 107. — P. 123-136.
413. Lau E.M., Lynn H.S., Woo J.W. et al. Prevalence of and risk factors for sarcopenia in elderly
Chinese men and women //J. Gerontol. A. Biol. Sci. Med. Sci. — 2005. — Vol. 60. - P. 213-216.
414. Morley J.E., BauMrartner R.N„ Roubenoff R. et al. Sarcopenia //J. Lab. Clin. Med. — 2001. —
Vol. 137. - P. 231-243.
415. O’Donnell A.B., Travison T.G., Harris S.S. et al. Testosterone, dehydroepiandrosterone, and
physical performance in older men: results from the Massachusetts Male Aging Study j j J, Clin.
Endocrinol. Metab. - 2006. - Vol. 91. - P. 425-431.
416. van den Beld A.W., Bots M.L., Janssen J.A, et al. Endogenous hormones and carotid atheroscle¬
rosis in elderly men // Am. J. Epidemiol. - 2003, - Vol. 157. - P. 25-31.
417. Travison T.G., Morley J.E., Araujo A.B. et al. The relationship between libido and testosterone
levels in aging men //J. Clin. Endocrinol. Metab. — 2006. — Vol. 91. - P. 2509-2513.
418. Araujo A.B., Durante R., Feldman H.A. et al. The relationship between depre.ssive symptoms
and male erectile dysfunction: cross-sectional results from the Massachusetts Male Aging Study //
Psychosom. Med. - 1998. — Vol. 60. - P. 458-465.
419. Booth A., Johnson D.R., Granger D.A. Testosterone and men’s health //J. Behav. Med. —
1999. - Vol. 22. - P. 1-19.
420. Barrett-Connor E., Von Muhlen D,G., Kritz-Silverstein D. Bioavailable testosterone and
depressed mood in older men: the Rancho Bernardo Study // J. Clin. Endocrinol, Metab. — 1999. -
Vol. 84. - P. 573-577.
421. Seidman S.N. et al. Testosterone level, androgen receptor polymorphism and depressive s\mip-
tnms in middle-aged men // Biol. Psychiatry. — 2001. — Vol. 50. - P. 371-376.
422. Haffner S,M. et al. The relationship of sex hormones to hyperinsulinemia and hyperglyce¬
mia// Metabolism. - 1988. - Vol. 37. - P. 683-688.
423. Pitteloud N., Mootha V.K., Dwj'er A.A. et al. Relationship between testosterone levels, insulin
sensitivity and mitochondrial function in men // Diabetes. Care. — 2005. — Vol. 28. — P. 1636-
1642.
424. Blouin K., Despres J.P., Couillard C. et al. Contribution of age and declining androgen levels to
features of the metabolic 5)и1Ьготе in men // Metabolism. — 2005. - Vol. 54. — P. 1034-1040.
425. Haffner S.M. Sex hormones, obesity, fat distribution, type 2 diabetes and insulin resistance:
epidemiological and clinical correlation // Int.J. Obes. Relat. Metab. Disord. - 2000. - Vol, 24. -
Suppl. 2. - P. 56-58.
426. Kenny A.M., Prestwood K.M., Gruman C.A. et al. Effects of tran.sdermal testosterone on bone
and muscle in older men with low bioavailable testosterone levels // J. Gerontol. A. Biol. Sci. Med.
Sci. - 2001. - Vol. 56. - P. 266-272.
427. Ten over J.S. Effects of testosterone Supplementation in the aging male //J. Clin. Endocrinol.
Metab. - 1992. - Vol. 75. - P. 1092-1098.
428. Page S.T., Amory J.K., Bowman F.D. et al. Exogenous testosterone (T) alone or with finasteride
increases physical performance, grip strength, and lean body mass in older men with low serum T //
J. Clin. Endocrinol. Metab. - 2005. - Vol. 90. - P. 1502-1510.
429. Wittert G.A., Chapman I.M., Haren M.T. et al. Oral testosterone Supplementation increases
muscle and decreases fat mass in healthy elderly males with low-normal gonadal status //J. Gerontol.
A. Biol. Sci. Med. Sci. - 2003. - Vol. 58. - P. 618-625.
430. Blackman M.R., Sorkin J.D., Munzer T. et al. Growth hormone and sex steroid administration
in healthy aged women and men: a randomized controlled trial // JAMA. - 2002. - Vol. 288. -
P. 2282-2292.
431. Harman S.M., Blackman M.R. The effects of growth hormone and sex sterojd on lean body
mass, fat mass, muscle strength, cardiovascular endurance and adverse events in healthy elderly women
and men // Horm. Res. - 2003. - Vol. 60. — P. 121-124.
432. Morley J.E., Perry 3rd H.M., Kaiser F.E. et al. Effects of testosterone replacement therapy in old
h^ogoiiadal males: a preliminary study //J. Am. Geriatr. Soc. — 1993. — Vol. 41. — P. 149-152,
433. Braunstein G.D. Gynecomastia //N. Engl.J. Med, — 1993. — Vol. 328. — P. 490-495.
434. Braunstein G.D. Aromatase and gynecomastia // Endocr. Relat. Cancer. — 1999. — Vol. 6. —
P. 313-324.
435. Kanhai R.C., Hage J.J., van Diest P.J. et al. Short-term and long-term histologic effects of
castration and estrogen treatment on breast tissue of 14 male-to-female transsexuals in comparison
with two chemically castrated men // Am. J. Surg. Pathol. - 2000. - Vol. 24. - P. 74-80.
318
436. Simpson E.R., Mahendroo M.S., Means G.D. et al. Aromatase cytochrome P450, the enzyme
responsible for estrogen biosynthesis // Endocr. Rev. - 1994. — Vol. 15. — P. 342-355.
437. Hemsell D.L., Grodin J.M., Brenner P.F. et al. Plasma precursors of estrogen. II. Correlation
of the extent of conversion of plasma androstenedione to estrone with age // J. Clin. Endocrinol.
Metab. - 1974. - Vol. 38. - P. 476-479.
438. McDonald P.C., Maldden J.D., Brenner P.F. et al. Origin of estrogen in normal men and in
women with testicular feminization //J, Clin. Endocrinol. Metab. - 1979. — Vol. 49. — P. 905-916.
439. Sebastian S., Bulun S.E. A highly complex organization of the regulatory region of the human
CYP19 (aromatase) gene revealed by the Human Genome Project //J. Clin. Endocrinol. Metab. -
2001. - Vol. 86. - P. 4600-4602.
440. Mahendroo M.S., Mendelson C.R., Simpson E.R. Tissue-specific and hormonally controlled
alternative promoters regulate aromatase cytochrome P450 gene expression in human adipose tissue //
J, Biol. Chem. - 1993. - Vol. 268. - P. 19463-19470.
441. Georgiadis E„ Papandreou L., Evangelopoulou C. et al. Incidence of gynaecomastia in 954
young males and its relationship to somatomctric parameters // Ann. Hum. Biol. - 1994. — Vol.
21. - P. 579-587.
442. Ersoz H., Onde M.E., Terekeci H. et al. Causes of gynaecomastia in young adult males and fac¬
tors associated with idiopathic gynaecomastia // Int. J, Androl. - 2002. - Vol. 25. — P. 312-316.
443. Einav-Bachar R. et al. Prepubertal gynaecomastia: aetiology, course and outcome // Clin.
Endocrinol. (Oxf.). - 2004. - Vol. 61. - P. 55-60.
444. Lee P.A. The relationship of concentrations of serum hormones to pubertal gynecomastia //
J. Pediatr. - 1975. - Vol. 86. - P. 212-215.
445. Niewoehner C.B., Nuttall F.Q. Gyneomastia in a hospitalized male population // J. Clin.
Endocrinol. Metab. - 1984. - Vol. 77. - P. 633-638.
446. Cleland W.H., Menderlson C.R., Simpson E.R. Effects of aging and obesity on aromatase activ¬
ity of human adipose cells //J. Clin. Endocrinol. Metab, - 1985. - Vol. 60. — P. 174-177.
447. Young S.. Gooneratne S., Straus 2nd F.H. et al. Feminizing Sertoli cell tumors in beys with
Peutz-Jeghers s>mdrome // Am. J. Surg, Pathol. — 1995. — Vol. 19. -- P. 50-58.
448. Lefevre H., Bouvattier C., Lahlou N. et al. Prepubertal gynecomastia in Peutz-Jeghers syn¬
drome: incomplete penetrance in a familial case and management with an aromatase inhibitor // Eur.
J. Endocrinol. - 2006. - Vol. 154. - P. 221-227.
449. Gordon G.G., Olivo J., Rafi F. et al. Conversion of androgens to estrogens in cirrohosis of the
liver //J. Clin. Endocrinol. Metab. — 1975. - Vol. 40. — P. 1018-1026,
450. Shozu M., Sebastian S., Takayaina K. et al. Estrogen excess associated with novel gain-of-
function mutations affecting the aromatase gene // N. Engl. J. Med. - 2003. — Vol. 348. - P.
1855-1865.
451. Stratakis G.D., Vottero A., Brodie A. et al. The aromatase excess syndrome is associated with
feminization of both sexes and autosomal dominant transmission of aberrant P450 aromatase gene
transcription //J. Clin. Endocrinol. Metab. — 1998. — Vol. 83. — P. 1348-1357,
452. Hemsell D.L., Edman C.D., Marks G.G. et al. Massive extraglandular aromatization of plasma
androstenedione resulting in feminization of a prepubertal boy//J. Clin. Invest. — 1977. - Vol. 60. —
P. 455-464.
453. Tiulpakov A., Kalintchenko N.. Semitcheva T. et al. A potential rearrangement between CYP19
and TRPM7 genes on chromosome 15q21.2 as a cause of aromatase excess syndrome j j J. Clin.
Endocrinol. Metab. - 2005. - Vol. 90. - P. 4184-4190.
454. Hendricks D,M„ Gray S.L., Hoover J. Residue levels of endogenous estrogens in beef tissues //
J. Anim. Sci. - 1983. - Vol. 57. - P. 247-255.
455. Beuers U., Richter W.O., Ritter M.M. et al. Klinef'elter’s syndrome and liver adenoma //J. Clin.
Gastroenterol. - 1991. - Vol, 13. - P. 214-216.
456. D’Avanzo B., La Vecchia C. Risk factors for male breast cancer // Br. J. Cancer. — 1995. - Vol.
71. - P. 1359-1362.
457. Hultborn R.. Hanson C., Kopf I. et al. Prevalence of Klinefelter's s),Tidrome.iii male breast can¬
cer patients j I Anticancer. Res. - 1997. — Vol. 17. - P. 4293-4297.
458. Meguerditchian A.N., Falardeau M., Martin G. Male breast carcinoma // Can. J. Surg. -
2002. - Vol. 45. - P. 296-302.
459. Sasco A.J., Lowenfels A.B., Pasker-de Jong P. Review article: epidemiology of male breast can¬
cer. A meta-analysis of published case-control studies and discussion of selected aetioiogical factors //
Int.J. Cancer. - 1993. - Vol. 53. - P. 538-549.
319
460. Wolman S.E. et al. Breast cancer in males: DNA content and sex chromosome constitution j j
Mod. Pathol. - 1995. - Vol. 8. - P. 239-243.
461. Swerdlow A.J., Higgins C.D., Schoemaker M.J. et al. Mortality in patients with Klinefelter syn¬
drome in Britain: a cohort study //J. Clin. Endocrinol. Metab. — 2005. - Vol, 90. - P. 6516-6522.
462. Weiss J.R., Moysidi K.k, Swede H. Epidemiolog)' of male breast cancer // Cancer. Epidemiol,
Biomarkers. Prev. — 2005. - Vol. 14. — P. 20-26.
•463. Coen P., Kulin H., Ballantine T. et al. An aromatase-producing sex-cord tumor resulting in
prepubertal gynecomastia // N. Engl. J. Med. — 1991. - Vol. 324. - P. 317-322.
464. Leiberman E., Zachmann M. Familial adrenal feminization probably due to increased steroid
aromatization // Horm. Res. — 1992. — Vol. 37. — P. 96-102.
465. Zachmann M„ Eiholzer U., Muritano M. et al. Treatment of pubertal gynaecomastia with tes-
tolactone // Arta. Endocrinol. Suppl, (Copenh.). — 1986. — Vol. 279. - P. 218-226.
466. Plourde P.V., Reiter E.O., Jou H.C. et al. Safety and efficacy of anastrozole for the treatment
of pubertal gynecomastia: a randomized, double-blind, placebo-controlled trial //J. Clin. Endocrinol.
Metab. - 2004. - Vol. 89. - P. 4428-4433.
467. Perdoiia S„ Autorino R., De Placido S. et al. Efficacy of tamoxifen and radiotherapy for pre¬
vention and treatment of gynaecomastia and breast pain caused by bicalutamide in prostate cancer: a
randomised controlled trial // Lancet. Oncol. - 200k - Vol. 6. - P. 295-300.
468. Derman O., Kanbur N.O., Kutluk T, Tamoxifen treatment for pubertal gynecomastia j j Int. J.
Adolesc, Med. Health. - 2003. - Vol. 15. - P. 359-363.
469. Lawrence S.E. et al. Beneficial effects of raloxifene and tamoxifen in the treatment of pubertal
gjmecomastia //J. Pediatr, - 2004. - Vol. 145. - P. 71-76.
470. Hanavadi S. et al. The role of tamoxifen in the management of gynaecomastia // Breast. —
2006. - Vol. 15. - P. 276-280.
471. Khan H.N., Rampaul R., Blarney R.W. Management of physiological gynaecomastia with
tamoxifen // Breast. — 2004, — Vol. 13. - P. 61-65.
472. Brinkmann A.O. Molecular basis of androgen insensitivity // Mol, Cell. Endocrinol. -
2001. - Vol. 179. - P. 105-109.
473. Griffin J.E., Leshin M., Wilson J.D. Androgen resistance syndromes // Am. J. Physiol.
1982.-Vol. 243.-P. 81-87.
474. Wilson J.D., Harrod M.J., Goldstein J.L. et al. Familial incomplete male pseudohermaphrodit¬
ism, type 1. Evidence for androgen resistance and variable clinical manifestations in a family with the
Reifenstein syndrome // N. Engl.J. Med. - 1974. - Vol. 290. - P. 1097-1103.
475. AimanJ., Griffin J.E., GazakJ.M. et al. Androgen insensitivity as a cause of infertility in other¬
wise normal men // N. Engl.J. Med. — 1979. — Vol. 300. — P. 223-227.
476. Madden J.D. et al. Clinical and eiidocrinologic characterization of a patients with the syn¬
drome of incomplete testicular feminization // J. Clin. Endocrinol, Metab. — 1975. — Vol. 41. —
P. 751-760.
477. Gottlieb B., Beitel L.K., Lumbroso R. et al. Update of the androgen receptor gene mutations
database j j Human. Mutations. — 1999. — Vol. 14. — P. 103-114.
478. Marcelli M., Zoppi S., Grino P.B. et al. A mutation in the DNA-binding domain of the andro¬
gen receptor gene causes complete testicular feminization in a patient with receptor-positive androgen
resistance //J. Clin. Invest. - 1991. - Vol. 87. - P. 1123-1126.
479. Zoppi S., Marcelli M., Deslypere J.P. et al. Amino acid substitutions in the DNA-binding
domain of the human androgen receptor are a frequent cause of receptor-binding positive androgen
resistance // Mol. Endocrinol. — 1992, - Vol, 6. — P. 409-415.
480. Adachi M., Takayanagi R., Tomura A. et al. Androgen-insensitivity syndrome as a possible
coactivator disease // N. Engl. J. Med. — 2000. — Vol. 343. - P. 856-862.
481. La Spada A.R., Wilson E.M., Lubahn D.B. et al. Androgen receptor gene mutations in X-linked
spinal and bulbar muscular atrophy // Nature. — 1991. — Vol. 352. — P. 77-79.
482. Nance M.A, Clinical aspects of CAG repeat diseases j j Brain. Pathol. - 1997. - Vol. 7. —
P. 881-900.
483. LaFevre-Bernt M.A., Ellerby L.M. Kennedy’s disease. Phosphorylation of the polyglutamine-
expanded form of androgen receptor regulates its cleavage by caspase-3 and enhances cell death //J.
Biol. Chem. - 2003. - Vol. 278. - P. 34918-34924.
484. Yong E.L., Loy C.J., Sim K.S, Androgen receptor gene and male infertility // Hum. Reprod.
Update. - 2003. - Vol. 9. - P. 1-7.
320
485. World Health Organization : WHO manual for the standardised investigation and diagnosis of
infertile couples. - Cambridge, UK: Cambridge University Press, 2000,
486. Templeton A. Infertility and the establishment of pregnancy — overview j j Br. Med. Bull. —
2000. - Vol. .56. - P. 577-587,
487. Giioth C., Godehardt E., Frank-Herrmann P. et al. Definition and prevalence of subfertility and
infertility// Hum. Reprod. - 2005. — Vol. 20. - P. 1144-1147.
488. van der Steeg J.W., Steures P., Hompes P.G. et al. Investigation of the infertile couple: a basic
fertility work-up performed within 12 months of trying to conceive generates costs and complications
for no partiailar benefit // Hum. Reprod. - 2005. - Vol. 20. - P. 2672-2674.
489. Juul S., Karmaus W., Olsen J. Regional differences in waiting time to pregnancy: pregnancy-
based suiYeys from Denmark, France, Germany, Italy and Sweden. The European Infertility and
Subfecundity Study Group // Hum. Reprod. — 1999. - Vol. 14, — P. 1250-1254.
490. Jensen Т.К., Slama R„ Ducot B. et al. Regional differences in waiting time to pregnancy among
fertile couples from four European cities // Hum. Reprod. - 2001. - Vol. 16. - P. 2697-2704.
491. Hull M.G., Glazener C.M., Kelly N.J. et al. Population study of causes, treatment and outcome
of infertility // Br. Med. J. (Clin. Res. Ed.). - 1985. - Vol. 291. - P. 1693-1697.
492. Snick H.K. et al. The spontaneous pregnancy prognosis in untreated subferiile couples: the
Walcheren primary care study // Hum. Reprod. — 1997. - Vol. 12. — P. 1582-1588.
493. Habbema J.D.. Collins J., Leridon H. et al. Towards less confusing terminology in reproductive
medicine: a proposal // Fertil. Steril. - 2004. - Vol. 82. - P. 36-40.
494. Dohle G.R., Colpi G.M., Hargreave T.B. et al. EAU guidelines on male infertility // Eur. Urol. -
2005. - Vol. 48. - P. 703-711.
495. Wang X., Chen C„ Wang L. et al. Conception, early pregnancy loss and time to clinical preg¬
nancy: a population-based prospective study // Fertil. Steril. — 2003. - Vol. 79. — P. 577-584.
496. Gnoth C„ Godehardt D., Godehardt E. et al. Time to pregnancy: results of the German pro¬
spective study and impact on the management of infertility // Hum, Reprod. - 2003. - Vol. 18. —
P. 1959-1966.
497. Wong W.Y., Thomas C.M., Merkus J.M. et al. Male factor subfertility: possible causes and the
impact of nutritional factors j j Fertil. Steril. - 2000. — Vol. 73. - P. 435-442.
498. Bhasin S., de Kretser D,M„ Baker H.W. Clinical review 64: Pathophysiology and natural history
of male infertility //J. Clin. Endocrinol. Metab. — 1994. - Vol. 79. — P. 1525-1529.
499. De Kretser D.M,, Burger H.G., Fortune D. et al. Hormonal, histological and chromosomal
studies in adult males with testicular disorders // J. Clin. Endocrinol. Metab. — 1972. — Vol. 35. -
P. 392-401.
500. Swerdloff R.S., Boyers S.P. Evaluation of the male partner of an infertile couple. An algoritlimic
approach //JAMA. - 1982. - Vol. 247. - P. 2418-2422.
501. Mazumdar S., Levine A.S. Antisperm antibodies: etiology, pathogenesis, diagnosis, and treat¬
ment // Fertil. Steril. - 1998. - Vol. 70. - P. 799-810.
502. Chaiidley A.C. The chromosomal basis of human infertility // Br, Med. Bull. — 1979. — Vol.
35. - P. 181-186.
503. Sharland M. et al. A clinical study of Noonan syndrome // Arch. Dis. Child. - 1992. - Vol.
67. - P. 178-183.
504. Santen R.J. et al. Gonadotrophins and testosterone in the XYY syndrome // Lancet. — 1970. -
Vol. 2. - P. 371.
505. Davidoff F., Federman D.D. Mixed gonadal dysgenesis // Pediatrics. - 1973. - Vol. 52. —
P. 725-742.
506. Federman D.D., Davidoff F.M., Ouellette E. Presumptive Y/D translocation in mixed gonadal
dysgenesis //J. Med. Genet. - 1967. - Vol. 4, - P. 36-40.
507. Page D.C., Brown L,G., de la Chapelle A. Exchange of terminal portions of X- and
Y-chromosomal short arms in human XX males // Nature. — 1987. — Vol. 328. — P. 437-440.
508. Najmabadi H., Huang V., Yen P. et al. Substantial prevalence of microdeletions of the
Y-chromosome in infertile men with idiopathic azoospermia and oligozoospermia detected using a
sequence-tagged site-based mapping strategy // J. Clin. Endocrinol. Metab. — 1996. - Vol. 81. -
P. 1347-1352.
509. Reijo R. et al. Severe oligozoospermia resulting from deletions of azoospermia factor gene on
Y-chroinosome // Lancet. — 1996. — Vol. 347. — P. 1290-1293.
510. Reijo R., Lee T.Y., Salo P. et al. Diverse spermatogenic defects in humans caused by
Y-chromosome deletions encompassing a novel RNA-binding protein gene // Nat. Genet, - 1995. —
Vol. 10. - P. 383-393.
321
511. Kent-First M.. Muallem A., Shultz J. et al. Defining regions of the Y-chromosome responsible
for male infertility and identification of a fourth AZF region (AZFd) by Y-chromosome microdeletion
detection // Mol. Reprod. Dev. — 1999. — Vol. 53. — P. 27-41.
512. Piryor J.L., Kent-First M„ Muallem A. et al. Microdeletions in the Y-chromosome of infertile
men // N. Engl. J. Med. - 1997. - Vol. 336. - P. 534-539.
513. Ma K., Sharkey A., Kirsch S. et al. Towards the molecular localisation of the AZF locus: map¬
ping 6f microdeletions in azoospermie men within 14 subintervals of interval 6 of the human Y chromo¬
some // Hum. Mol. Genet. — 1992. — Vol. 1. — P. 29-33.
514. Vogt P., Chandley A.C., Hargreave T.B. et al. Microdeietions in interval 6 of the Y-chromosome
of males with idiopathic sterility point to disruption of AZF, a human spermatogenesis gene // Hum.
Genet. - 1992. - Vol. 89. - P. 491-496.
515. Ferlin A. et al. Human male infertility and Y-chromosome deletions: role of the AZF-candidate
genes DAZ, RBM and DFFRY // Hum. Reprod. - 1999. - Vol. 14. - P. 1710-1716.
516. Foresta C., Мого E., Ferlin A. Y-chromosome microdeietions and alterations of spermatogen¬
esis // Endocr. Rev. - 2001. - Vol. 22. - P. 226-239.
517. Maurer B. et al. Prevalence of Y-chromosome microdeietions in infertile men who consulted a
tertiary care medical centre: the Munster experience // Andrologia. — 2001. — Vol. 33. — P. 27-33.
518. Simoni M., Gromoll J., Dworniczak B. et al. Screening for deletions of the Y-chromosome
involving the DAZ (Deleted in AZoospermia) gene in azoospermia and severe oligozoospermia j j
Fertil. Steril. - 1997. - Vol. 67. - P. 542-547.
519. Simoni M., Kamischke A., Nieschlag E. Current status of the molecular diagnosis of
Y-chromosomal miaodeletions in the work-up of male iufeitility. Initiative for international quality
control // Hum. Reprod. - 1998. - Vol. 13. - P. 1764-1768.
520. Bhasin S., Ma K., de Kretser D.M. Y-chromosome microdeietions and male infertility // Ann.
Med. - 1997. - Vol. 29. - P. 261-263.
521. Ma K., înglis J.D., Sharkey A. et al. A Y-chromosome gene family with RNA-binding protein
homology: candidates for the azoospermia factor AZF controlling human spermatogenesis // Cell. —
1993. - Vol. 75. - P. 1287-1295.
522. Najmabadi H.. Chai N.. Kapali A. et al. Genomic structure of a Y-specific ribonucleic add bind¬
ing motif-containing gene: a putative candidate for a subset of male infertility //J. Clin. Endocrinol.
Metab. - 1996, - Vol. 81. - P. 2159-2164.
523. Chai N.N. et al. A putative human male infertility gene DAZLA: genomic structure and méthy¬
lation status // Mol. Hum. Reprod. - 1997. - Vol. 3. - P. 705-708.
524. Yen P.H., Chai N.N., Salido E.C. The human autosomal gene DAZLA: testis specifidty and a
candidate for male infertility // Hum. Mol. Genet. — 1996. - Vol. 5. — P. 2013-2017.
525. Chai N.N., Zhou H., Hernandez J. et al. Structure and organization of the RBMY genes on the
human Y-chromosome: transposition and amplification of an ancestral autosomal hnRNPG gene //
Genomics. - 1998. - Vol. 49. - P. 283-289.
526. Saxena R., Brown L.G., Hawkins T. et ai. The DAZ gene duster on the human Y-chromosome
arose from an autosomal gene that was transposed, repeatedly amplified and pruned // Nat. Genet. -
1996. - Vol. 14. - P. 292-299.
527. Eberhart C.G., Maines J.Z., Wasserman S.A. Meiotic cell cycle requirement for a fly homologue
of human DAZ // Nature. - 1996. - Vol. 381. - P. 783-785.
528. Cooke H. et al. A murine homologue of the human DAZ gene is autosomal and expressed in
male and female gonads // Human. Mol. Genet. — 1996. — Vol. 5. — P. 513-516.
529. Ruggiu M.. Speed R., Taggart M. et al. The mouse Dazla gene encodes a cytoplasmic protein
essential for gametogenesis // Nature. - 1997. - Vol. 389. - P. 73-77.
530. Tsui S., Dai T., Roettger S. et al. Identification of two novel proteins that interact with germ-
cell-spedfic RNA-binding proteins DAZ and DAZLl // Genomics. — 2000. — Vol. 65. — P. 266-273.
531. Lim H.N., Chen H., McBride S. et al. Longer polyglutamine tracts in the androgen receptor are
assodated with moderate to severe underraasculinized genitalia in XY males // Hum. Mol. Genet. —
2000. - Vol. 9. - P. 829-834.
532. Tut T.G., Ghadessy F.J., Trifiro M.A. et al. Long polyglutamine tracts in the androgen receptor
are associated with reduced trans-activation, impaired sperm produdion and male infertility // J. Clin.
Endocrinol. Metab. - 1997. - Vol. 82. - P. 3777-3782.
533. Dadze S., Wieland C., Jakubiczka S. et al. The size of the CAG repeat in exon 1 of the androgen
receptor gene shows no significant relationship to impaired spermatogenesis in an infertile caucasoid
sample of German origin // Mol. Hum. Reprod. — 2000. — Vol. 6. — P. 207-214.
322
534. Tamai K.T., Monaco L., Nantel F. et al. Coupling signalling pathways to transcriptional control:
nuclear factors responsive to cAMP // Recent. Prog. Horm. Res, - 1997. - Vol. 52. - P. 121-139.
discussion 139-140.
535. Weinbauer G.F. et al. Testicular cAMP responsive element modulator (CREM) protein is
expressed in round spennatids but is absent or reduced in men with round spermatid maturation
arrest // Mol. Hum, Reprod. - 1998. - Vol. 4. - P. 9 -15.
536. Peri A., Krausz C., Cioppi F. et al. Cyclic adenosine 3'„5'-monophosphate-responsive element
modulator gene expression in germ cells of normo- and oligoazoospermic men //J. Clin. Endocrinol.
Metab. - 1998. - Vol. 83. - P. 3722-3726.
537. Blendy J.A., Kaestner K.H., Weinbauer G.F. et al. Severe impairment of spermatogenesis in
mice lacking the CREM gene // Nature. - 1996. - Vol. 380. - P. 162-165.
538. Anguiano A., Oates R.D., Amos J.A. ct al. Congenital bilateral absence of the vas deferens.
A primarily genital form of cystic fibrosis //JAMA. — 1992. — Vol. 267. — P. 1794-1797.
539. Abbasi A.A., Prasad A.S., Ortega J. et al. Gonadal function abnormalities in sickle cell anemia.
Studies in adult male patients // Ann. Intern. Med. - 1976. - Vol. 85. — P. 601-605,
540. Kletzky O.A., Costin G„ Marrs R.P. et al. Gonadotropin insufficiency in patients with thalas¬
semia major // J. Clin. Endocrinol. Metab. - 1979. - Vol. 48. - P. 901-905.
541. Marchini C., Lonigro R., Verriello L. et al Correlations between individual clinical manifes¬
tations and CTG repeat amplification in myotonic dystrophy j j Clin. Genet. — 2000. — Vol. 57. —
P. 74-82.
542. Takeda R., Ueda M. Pituitary-gonadal function in male patients with myotonic dystrophy -
serum luteinizing hormone, follicle stimulating hormone and testosterone levels and histological
dmaage of the testis // Acta Endoainol. (Copenh.). — 1977, - Vol. 84. - P. 382-389.
543. Skakkebaek N.E., Giwercman A„ de Kretser D. Pathogenesis and management of male infertil¬
ity //Lancet. - 1994. - Vol. 343. - P. 1473-1479.
544. Wang C., Chan S.Y., Ng M. et al. Diagnostic value of sperm function tests and routine semen
analyses in fertile and infertile men //J. Androl. - 1988. — Vol. 9. — P. 384-389.
545. World Health Organization: Laboratory Manual for the Examination of Human Semen and
Semen-Cervical Mucus Interaction. - Cambridge, UK: Cambridge University Press, 1987.
546. Simpson J,L„ Lamb DJ. Genetic effects of intracytoplasmic sperm injection // Semin. Reprod,
Med. - 2001. - Vol. 19. - P. 239-249.
547. Colombero L.T., Hariprashad J.J., Tsai M.C. et al. Incidence of sperm aneuploidy in relation to
semen diaracteristics and assisted reproductive outcome // Fertil. Steril. — 1999. — Vol. 72. — P. 90-96.
548. Finkel D.M., Phillips J.L., Snyder P.J. Stimulation of spermatogenesis by gonadotropins in men
with hypogonadotropic hypogonadism // N. Engl. J. Med. - 1985. - Vol. 313. - P. 651 -655.
549. Burgues S., Calderon M.D. Subcutaneous self-administration of highly purified follicle stimu¬
lating hormone and human chorionic gonadotrophin for the treatment of male hypogonadotrophic
hypogonadism. Spanish Collaborative Group on Male Hypogonadotropic Hypogonadism // Hum.
Reprod. - 1997. - Vol. 12. - P. 980-986.
550. Santen R.J., Paulsen C.A. Hypogonadotropic eunuchoidism, II. Gonadal responsiveness to
exogenous gonadotropins //J. Clin. Endocrinol. Metab. — 1973. — Vol. 36. — P. 55-63.
551. Liu L. et al. Two-year comparison of testicular responses to pulsatile gonadotropin-releasing
hormone and exogenous gonadotropins from the inception of therapy in men with isolated hypogo¬
nadotropic hypogonadism //J. Clin, Endoainol. Metab. — 1988. — Vol. 67. — P. 1140-1145.
552. Spratt D.I., Finkelstein J.S., O'Dea L.S. et al. Long-term administration of gonadotropin-
releasing hormone in men with idiopathic hypogonadotropic hypogonadism. A model for studies of the
hormone’s physiologic effects // Ann. Intern. Med. — 1986. — Vol. 105. - P. 848-855.
553. Buchter D. et al. Pulsatile GnRH or human chorionic gonadotropin/human menopausal
gonadotropin as effective treatment for men with hypogonadotropic hypogonadism; a review of 42
cases // Eur. J. Endoainol. — 1998. — Vol. 139. — P. 298-303.
554. Kliesch S., Behre H.M., Nieschlag E. High efficacy of gonadotropin or pulsatile gonadotropin-
releasing hormone treatment in hypogonadotropic hypogonadal men // Eur. J. Endocrinol. — 1994. -
Vol. 131. - P. 347-354.
555. Fradkin J.E., Schonberger L.B., Mills J.L. et al. Creutzfeldt-Jakob disease in pituitary growth
hormone recipients in the United States //JAMA. - 1991. - Vol, 265. - P. 880-884.
556. Recombinant Human FSH Product Development Group, Recombinant follicle stimulating hor¬
mone: development of the first biotechnology product for the treatment of infertility // Hum. Reprod,
Update. - 1998. - Vol. 4. - P. 862 -881.
323
557. Liu P.Y., Turner L., Rushford D. et al. Efficacy and safety of recombinant human follicle
stimulating honnone (Gonal-F) with urinary human chorionic gonadotrophin for induction of sper¬
matogenesis and fertility in gonadotrophin-deficient men // Hum. Reprod. — 1999. — Vol. 14. —
P. 1540-1545.
558. Sokol R.Z. et al. Gonadotropin therapy failure secondary to human chorionic gonadotropin-
induced antibodies //J, Clin. Endocrinol. Metab. — 1981. - Vol. 52. — P. 929-932.
559. Braunstein G.D. et al, Characterization of antihuman chorionic gonadotropin serum anti¬
body appearing after ovulation induction // J. Clin. Endocrinol. Metab. - 1983. - Vol. 57. •
P. 1164-1172.
560. Burger H.G. et al. Effects of preceding androgen therapy on testicular response to human
pituitary gonadotropin in hypogonadotropic hypogonadism: a study of three patients // Fertil. Steril. —
1981. - Vol. 35. - P. 64-68.
561. Belchetz P.E., Plant T.M., Nakai Y. et al. H5»pophysial responses to continuous and intermit¬
tent delivery of hypopthalamic gonadotropin-releasing hormone // Science, - 1978, - Vol. 202. -
P. 631-633.
562. Knobil E. The neuroendocrine control of the menstrual cycle // Recent. Prog. Horm. Res. —
1980. - Vol. 36. - P. 53-88.
563. Whitcomb R.W., Crowley W.F. Jr. Hypogonadotropic hypogonadism: gonadotropin-releasing
hormone therapy j j Curr. Ther. Endocrinol. Metab. — 1997. — Vol. 6. — P. 353-355.
564. Burger H.G., Baker H.W. The treatment of infertility // Aimu. Rev. Med. — 1987. — Vol.
38. - P. 29-40.
565. Palermo G.D., Schlegel P.N., Hariprashad J.J. et al. Fertilization and pregnancy outcome
with intracytoplasmic sperm injection for azoospermic men // Hum. Reprod. - 1999, ■' Vol. 14. -
P. 741 -748.
566. Bourne H-, Stern K., Clarke G. et al. Delivery of normal twins following the intracytoplasmic
injection of spermatozoa from a patient with 47, XXY Klinefelter's syndrome // Hum. Reprod. —
1997. - Vol. 12. - P. 2447-2450.
567. Cram D.S., Ma K„ Bhasin S. et al. Y-chromosome analysis of infertile men and their sons con¬
ceived through intracytoplasmic sperm injection: vertical transmission of deletions and rarity of de novo
deletions // Fertil. Steril. - 2000. - Vol. 74. - P. 909-915.
568. Bonduelle M., Camus M., De Vos A. et al. Seven years of intracytoplasmic sperm injection
and follow-up of 1987 subsequent children // Hum. Reprod. - 1999. - Vol. 14. - Suppl. 1. -
P. 243-264.
569. Wright V.C., Schieve L.A., Reynolds M.A. et al. Assisted reproductive technology surveil¬
lance - United States, 2001 // MMWR Surveill. Summ. - 2004. - Vol. 53. - P. 1-20.
.570. Tarlatzis B.C., Bili H, Intracytoplasmic sperm injection. Survey of world results // Ann. N. Y.
Acad. Sci. - 2000. - Vol. 900. - P. 336-344.
571. Ponjaert-Kristoffersen L, Bonduelle M., Barnes J. et al. International collaborative study of
intracytoplasmic sperm injection-conceived, in vitro fertilization-conceived and naturally conceived
5-year-old child outcomes: cognitive and motor assessments // Pediatrics. — 2005. — Vol. 115. —
P. 283-289.
572. Schieve L.A., Ferre C., Peterson H.B. et al. Perinatal outcome among singleton infants con¬
ceived through assisted reproductive technology in the United States // Obstet. Gynecol. — 2004. —
Vol. 103. - P. 1144-1153.
573. Van Steirteghem A. et al. Follow-up of children born after ICSI // Hum. Reprod. Update. -
2002. - Vol. 8. - P. 111-116.
574. Nyboe Andersen A., Erb K, Register data on assisted reproductive technology (ART) in
Europe including a detailed description of ART in Denmark // Int. J. Androl. — 2006. — Vol. 29, -
P. 12-16.
575. Andersen A.N., Gianaroli L,, Felberbaum R. et al. Assisted reproductive technology in Europe,
2002. Results generated from European registers by ESHRE // Hum. Reprod. - 2006. - Vol. 21. -
P. 1680-1697.
576. Nygren K.G., Andersen A.N. Assisted reproductive technology in Europe, 1999. Results gener¬
ated from European registers by ESHRE // Hum. Reprod. - 2002. - Vol. 17. - P. 3260-3274.
577. Feenstra J. et al. Acute myocardial infarction associated with sildenafil // Lancet. - 1998. —
Vol. 352. - P. 957-958.
578. Allen V.M., Wilson R.D., Cheung A. Pregnancy outcomes after assisted reproductive technol¬
ogy //J- Obstet. Gynaecol. Can. - 2006. - Vol. 28. - P. 220-250.
324
579. Schicve L.A., Rasmussen S.A., Buck G.M. et al. Are children born after assisted reproductive
technology at increased risk for adverse health outcomes.? // Obstet. Gynecol. — 2004. - Vol. 103. —
P. 1154-1163.
580. Wennerholm U.B., Bergh C„ Hamberger L. et al. Incidence of congenital malformations in
children born after ICSI // Hum. Reprod. - 2000. - Vol. 15. - P. 944-948.
581. Tarlatzis B.C., Bili H. Survey on intracytoplasmic sperm injection: report from the ESHRE ICSI
Task Force. European Society of Human Reproduction and Embryology // Hum. Reprod. — 1998. —
Vol. 13. - Suppl. 1. - P. 165-177.
582. Leslie G.I., Gibson F.L., McMahon C. et al. Children conceived using ICSI do not have an
increased risk of delayed mental development at 5 years of age // Hum. Reprod. - 2003. — Vol. IS. —
P. 2067-207Z
583. Maher E.R. Imprinting and assisted reproductive technology // Hum. Mol. Genet. — 2005. —
Vol. 14. - Spec. N 1. - P. 133-138.
.584. Maher E.R., Afnan M., Banatt C.L. Epigenetic risks related to assisted reproductive tech¬
nologies: epigenetics, imprinting, ART and icebergs? // Hum. Reprod. — 2003. — Vol. IS. —
P. 2508-2511.
585. Nieschlag E„ Hertle L., Fischedick A. et al. Update on treatment of varicocele: counselling as
effective as occlusion of the vena spermatica // Hum, Reprod. - 1998. — Vol. 13. - P. 2147-2150.
586. Nieschlag E. et al. Treatment of varicocele: counselling as effective as occlusion of the vena
spermatica // Hum. Reprod. — 1995, — Vol. 10. — P. 347-353.
587. Ismail M.T., Sedor J., Hirsch I.H. Are sperm motion parameters influenced by varicocele liga¬
tion? // Fertil. Steril. - 1999. - Vol. 71, - P. 886-890.
588. Asci R., Sarikaya S., Buyukalpelli R. et al. The outcome of varicocelectomy in subfertile men
with an absent or atrophic right testis // Br. J. Urol. - 1998. - Vol. 81. - P. 750-752.
589. Segenreich E., Israiluv S., Shmuele J. et al. Evaluation of the relationship between semen
parameters, pregnancy rate of wives of infertile men with varicocele, and gonadotropin-releasing hor¬
mone test before and after varicocelectomy // Urology. - 1998. -• Vol. 52. — P. 853-857,
590. Bosl G.J., Motzer R.J. Testicular germ-cell cancer // N. Engl. J. Med. - 1997. - Vol. 337» ~
P. 242-253.
591. Hartmann J.T., Kanz L., Bokemeyer C. Diagnosis and treatment of patients wih testicular germ
cell cancer // Drugs. - 1999. - Vol. 58. - P. 257-291.
592. Richiardi L., Bellocco R., Adami H.O. et al. Testicular cancer incidence in eight northern
European countries: secular and recent trends j j Cancer. Epidemiol. Biomarkers. Prev. — 2004. — Vol.
13, - P. 2157-2166.
593. Moller H., Skakkebaek N.E. Testicular cancer and cryptorchidism in relation to prenatal fac¬
tors: case control study in Denmark // Cancer. Causes. Control. — 1997. - Vol. 8. — P. 904-912.
594. Jacobsen R., Bostofte E., Enghoim G. et al. Risk of testicular cancer in men with abnormal
.semen characteristics; cohort study // BMJ. — 2000. — Vol. 321. — P. 789-792.
595. Tollerud D., Blattner W.. Fraser M. et al. Familial testicular cancer and urogenital developmen¬
tal anomalies // Cancer. - 1985. - Vol. 55. - P. 1849-1854.
596. Norwich A.. Shipley J., Huddart R. Testicular germ-cell cancer // Lancet. — 2006. — Vol.
367. - P. 754-765.
597. Giwercman A., Muller J.E., Skakkebaek N.E. Prevalence of carcinoma m situ and other histo-
pathological abnormalities in testes from 399 men who died suddenly and unexpectedly j j J. Urol. —
1991. - Vol. 145. - P. 77-80.
598. Smiraglia D.J., Szymanska J., Kraggerud S.M. et al. Distinct epigenetic phenot3'pes in semi-
nomatous and nonseminomatous testknilar germ cell tumors // Oncogene. — 2002. — Vol. 21. —
P. 3909- 3916.
599. Kawakami T. et al. XIST unmethylated DNA fragments in male-derived plasma as a tumor
marker for testicular cancer // Lancet. - 2004. - Vol. 363. - P. 40-42.
600. OosterhuisJ., Castedo S,, De Jong B. Cytogenetics, ploidy, and differentiation of human testicu¬
lar, ovarian and extragonadal germ cell tumors // Cancer. Surveys. - 1990. — Vol. 9. — P. 320-332.
601. Ottesen A.M., Skakkebaek N.E., Lundsteen C. et al. High-resolution comparative genomic
hybridization detects extra chromosome arm 12p material in most cases of carcinoma ih situ adjacent
to overt germ cell tumors, but not before the invasive tumor development // Genes. Chromosomes.
Cancer. - 2003. - Vol, 38. - P. 117-125.
602. Heiiegariu O., Vance G.H., Heiber D. et al. Triple color FISH analysis of 12p amplification in
testicular germ-cell tumors using 12p-band specific paiting probes // J. Mol. Med. - 1998. - Vol.
76. - P. 648 655.
325
603. Rodriguez S., Jafer O., Goker H. et al. Expression profiling of genes from 12p in testicular
germ cell tumors of adolescents and adults associated with i(12p) and amplification of 12pll.2 //
Oncogene. - 2003. - Vol. 22. - P. 1880-1891.
604. Clark A.T., Rodriguez R.T., Bodnar M.S. et al Human STELLAR, NANOG and GDF3 genes are
expressed in pluripotent cells and map to chromosome 12pl.3, a hotspot for teratocarcinoma // Stem.
Cells. - 2004. - Vol. 22. - P. 169-179.
605. Keminer K., Corless C.L., Fletcher J.A. et al. Kit mutations are common in testicular seminoma //
Am. J. Pathol. - 2004. - Vol. 164. - P. 305-313.
606. Li J„ Xia F., Li W.X. Coactivation of STAT and Ras is required for germ cell proliferation and
invasive migi-ation in Drosophila // Dev. Cell. - 2003. - Vol. 5. - P. 787-798.
607. Kcrsemaekers A.M., Mayer F., Molier M. et al. Role of p53 and MDM2 in treatment response
of human germ cell tumors //J. Clin. Oncol. - 2002. - Vol. 20. - P. 1551-1561.
608. Koberle B. et a!. Defective repair of dsplatin-induced DNA damage caused by reduced XPA
protein in testicular germ cell tumors // Curr. Biol. — 1999. — Vol. 9. — P. 273-276.
609. Bokemeyer C., Nichols C., Droz J-P. et al. Extragonadal germ cell tumors of the mediastinum
and retroperitonemn: results from an international analysis // J. Clin. Oncol. — 2002. — Vol. 20. —
P. 1864-1873.
610. Jennings M.T., Gelman R., Hochberg F. Intracranial genii-cell tumors: natural history and
pathogenesis //J. Neurosurgery. - 1985. - Vol. 63. - P. 155-167.
611. Oliver R.T., Mason M.D., Mead G.M. et al. Radiotherapy versus single-dose carboplatin
in adjuvant treatment of stage I seminomas: a randomised trial // Lancet. — 2005. — Vol. 366. —
P. 293-300.
612. Classen J., Schmidberger H., Meisner C. et al. Radiotherapy for stages llA/B testicular semi¬
nomas: final report of a prospective multicenter clinical trial //J. Clin. Oncol. — 2003. — Vol. 21. —
P. 1101-1106.
613. van Dijk M.R., Steyerberg J., Habbema D.F. Survival of noii-seminomatous germ cell cancer
patients according to the IGCC classification: an update based on meta-analysis // Eur. J, Cancer. -
2005. - Vol. 42. - P. 820-826.
614. Yesalis C.E. 3rd, Bahrke M.S., Kopstein A.N., Baruskicwicz C.K. Incidence anabolic steroid
use: a discussion of methodological issues // Yesalis C.E. 3rd. (ed.). Anabolic Steroids in Sports and
Exercise. — 2"‘* ed. — Champaign, IL: Human Kinetics, 2000 — P. 73-115.
615. Bahrke M.S., Yesalis C.E. Abuse of anabolic androgenic steroids and related substances in sport
and exercise // Curr. Opin. Pharmacol. — 2004. — Vol. 4. — P. 614-620.
616. Batirke M.S. et al. Risk factors assodated with anabolic-androgenic steroid use among adoles¬
cents // Sports. Med. - 2000. - Vol. 29. - P. 397-405.
617. Yesalis C.E. et al. Trends in anabolic-androgenic steroid use among adolescents // Arch.
Pediatr. Adolesc. Med. - 1997. - Vol. 151. - P. 1197-1206.
618. Buckley W.E., Yesalis 3rd C.E., Friedl K.E. et al. Estimated prevalence of anabolic steroid use
among male high school seniors //JAMA. — 1988. — Vol. 260. — P. 3441-3445.
619. Irving L.M., Wall M.. Neumark-Sztainer D. et al. Steroid use among adolescents: findings from
project EAT //J. Adolescent. Health. — 2002. — Vol. 30. — P. 243-252.
620. Melia P., Pipe A., Greenberg L. The use of anabolic-androgenic steroids by Canadian stu¬
dents // Clin. J. Sport. Med. - 1996. - Vol. 6. - P. 9-14.
621. Handelsman DJ., Gupta L. Prevalence and risk factors for anabolic-androgenic steroid abuse
in Australian high school students // Int.J. Androl. - 1997. - Vol. 20. - P. 159-164.
622. Yesalis C.E., Kennedy N.J., Kopstein A.N., Bahrke M.S. Anabolic-androgenic steroid use in the
United States //JAMA. - 1993. - Vol. 270. - P. 1217-1221.
623. Franke W.W., Berendonk B. Hormonal doping and androgenization of athletes: a secret
program of the German Democratic Republic government // Clin. Chem. - 1997. — Vol. 43. -
P. 1262-1279.
624. Evans N.A. Gym and tonic: a profile of 100 male steroid users // Br. J. Sports. Med. — 1997. —
Vol. 31. - P. 54-58.
625. Evans N.A. Local complications of self administered anabolic steroid injections // Br.J. Sports.
Med. - 1997. - Vol. 31. - P. 349-350.
626. Pope Jr. H.G., Katz D.L, Psychiatric and medical effects of anabolic-androgenic steroid use.
A controlled study of 160 athletes // Arch. Gen. Psychiatry, — 1994. — Vol. 51. — P. 375-382.
627. Bhasin S., Woodhouse L., Casaburi R. et al. Testosterone dose-response relationships in healthy
young men // Am. J. Physiol. Endocrinol. Metab. - 2001. - Vol, 281. - P. 1172-1181.
326
628. Storer T.W., Magliano L„ Woodhouse L. et al. Testosterone dose-dependently inaeases
maximal voluntary strength and leg power, but does not affect fatigability or specific tension //J. Clin.
Endocrinol. Metab. - 2003. - Vol. 88. - P. 1478-1485.
629. Baume N., Schumacher Y.O., Sottas P.E. et al. Effect of multiple oral doses of androgenic ana¬
bolic steroids on endurance performance and serum indices of physical stress in healthy male subjects //
Eur. J. Appl. Physiol. - 2006. - Vol. 98. - P. 329-340.
630. Georgieva K.N., Boyadjiev N.P. Effects of nandrolone decanoate on Vo2max, running economy
and endurance in rats // Med. Sci. Sports. Exerc. — 2004. - Vol. 36. — P. 1336-1341.
631. Hendler E.D., Solomon L.R. Prospective controlled study of androgen effects on red cell oxy¬
gen transport and work capacity in chronic hemodialysis patients // Acta Haematol. — 1990. - Vol.
83. - P. 1-8.
632. Blanco C.E., Popper P., Micevych P. Anabolic-androgenic steroid induced alterations in choline
acetyltransferase messenger RNA levels of spinal cord motoneurons in the male rat // Neuroscience. —
1997. - Vol. 78. - P. 873-882.
633. Blanco C.E. Exogenous testosterone treatment decreases diaphragm neuromuscular transmis¬
sion failure in male rats / C.E. Blanco, W.Z. Zhan, Y.H. Fang, G.C. Sieck//J. Appl. Physiol. — 2001. —
Vol. 90. - P. 850-856.
634. Wight Jr. J.N., Salem D. Sudden cardiac death and the “athlete’s heart” // Arch. Intern. Med. -
1995. - Vol. 155. - P. 1473-1480.
635. Melchert R.B., Welder A.A. Cardiovascular effects of androgenic-anabolic steroids // Med. Sci.
Sports, Exerc. - 1995. - Vol. 27. - P. 1252-1262.
636. Glazer G. Atherogenic effects of anabolic steroids on serum lipid levels. A literature review j j
Arch. Intern. Med. - 1991. - Vol. 151. - P. 1925-1933.
637. Ansell J.E., Tiarks C„ Fairchild V.K. Coagulation abnormalities associated with the use of ana¬
bolic steroids II Am. Heart. J. - 1993. — Vol. 125. — P. 367-371.
638. Soe K.L., Soe M., Gluud C. Liver pathology associated with the use of anabolic-androgenic
steroids // Liver. - 1992. - Vol. 12. - P. 73-79.
639. Dickerman R.D., Pertusi R.M., Zachariah N.Y. et al. Anabolic steroid-induced hepatotoxicity:
is it overstated? // Clin. J. Sport. Med. - 1999. - Vol. 9. - P. 34-39.
640. Pertusi R., Dickennan R.D., McConathy W.J. Evaluation of aminotransferase elevations in
a bodybuilder using anabolic steroids: hepatitis or rhabdomyolysis? // J. Am. Osteopath. Assoc. —
2001. - Vol. 101. - P. 391-394.
641. Tricker R., Casaburi R., Storer T.W. et al. The effects of supraphysiological doses of testosterone
on angry behavior in healthy eugonadal men — a clinical research center study // J. Chn. Endocrinol.
Metab. - 1996. - Vol. 81. - P. 3754-3758.
642. Daly R.C., Su T.P., Schmidt P.J. et al. Neuroendocrine and behavioral effects of high-dose ana¬
bolic steroid administration in male normal volunteers // Psychoiieuroendocrinology. — 2003, — Vol.
28.-P. 317-331.
643. Su T.P., Pagliaro M., Schmidt P.J. et al. Neuropsychiatric effects of anabolic steroids in male
normal volunteers //JAMA. — 1993. — Vol. 269. — P. 2760-2764,
644. Yates W.R., Perry P.J., Maclndoe J. et al. Psychosexual effects of three doses of testosterone
c>'c!ing in normal men // Biol. Psychiatry. — 1999. — Vol. 45. — P. 254-260.
645. Kouri E.M., Lukas S.E., Pope H.G. Jr., Oliva P.S. Increased aggressive responding in male vol¬
unteers following the administration of gradually increasing doses of testosterone cypionate // Drug,
Alcohol. Depend. - 1995. - Vol. 40. - P. 73-79.
646. Pope Jr. H.G., Kouri E.M., Hudson J.L Effects of supraphysiologic doses of testosterone
on mood and aggression in normal men: a randomized controlled trial // Arch. Gen. Psychiatry. —
2000. - Vol. 57. - P. 133-140. discussion 155-156.
647. Cohen J.C., Hickman R. Insulin resistance and diminished glucose tolerance in powerlifters
ingesting anabolic steroids //J. Clin. Endocrinol. Metab. - 1987. — Vol. 64. — P. 960-963.
648. Karila T.A., Karjalainen J.E., Mantysaari M.J. et al. Anabolic androgenic steroids produce
dose-dependant increase in left ventricular mass in power atheletes and this effect is potentiated by
concomitant use of growth hormone // Int. J. Sports. Med. - 2003. - Vol, 24. - P. 337-343.
649. Gill G.V. Anabolic steroid induced hypogonadism treated with human chorionic gonadotro¬
pin // Postgrad. Med. J. — 1998. — Vol. 74. — P. 45-46.
650. Maclndoe J.H., Perry P.J., Yates W.R. et al. Testosterone suppression of the HPT axis j j
J. Investig. Med. - 1997. - Vol. 45. - P. 441-447.
327
651. Lloyd F.H., Powell P., Murdoch A.P. Anabolic steroid abuse by body buildere and male
subfertility // BMJ, - 1996. - Vol. 313. - P. 100-101.
652. Jarow J.P., Lipshultz L.L Anabolic steroid-induced hypogonadotropic hypogonadism // Am. I.
Sports. Med. - 1990. - Vol. IS. - P. 429-431.
653. Brower K.J. Anabolic steroid abuse and dependence // Curr. Psychiatry. Rep. — 2002. — Vol.
4. - P. 377-387.
654. Brower K.J,, Blow F.C., Young J.P., Hill E.M. Symptoms and correlates of anabolic-androgenic
steroid dependence // Br. J. Addict. - 1991. - Vol. 86. - P. 759-768.
655. Brower K.J., Eiiopulos G.A., Blow F.C. et al. Evidence for physical and psychological
dependence on anabolic androgenic steroids in eight weight lifters // Am. J. Psychiatry. — 1990, — Vol.
147. - P. 510-512.
656. Evans N.A., Bowrey D.J., Newman G.R. Ultrastructural analysis of ruptured tendon from
anabolic steroid users // Injuiy, — 1998. — Vol. 29. — P. 769-773,
657. Schanzer W. Metabolism of anabolic androgenic steroids // Clin. Chem, - 1996. — Vol. 42. —
P. 1001-1020.
658. Schanzer W. Abuse of androgens and detection of illegal use j j Nieschlag E., Behre H.M.
(ed.). Testosterone: Action, Deficiency, Substitution. - ed. - Cambridge, UK: Cambridge University
Press, 2004. - P. 715-736.
659. Aguilera R„ Becchi M., Grenot C. et al. Detection of testosterone misuse: comparison of two
chromatographic sample preparation methods for gas chromatographic-combustion/isotope ratio mass
spectrometric analysis //J. Chromatogr. B. Biomed. Appl. — 1996, — Vol 687. - P. 43-53.
660. Aguilera R., Chapman T.E., Starcevic B, et al. Performance characteristics of a carbon isotope
ratio method for detecting doping with testosterone based on urine diols; controls and athletes with
elevated testosterone/epitestosterone ratios // Clin, Chem. - 2001. - Vol. 47. - P. 292-300.
661. Catlin D.H., Hatton C.K., Starcevic S.H. Issues in detecting abuse of xenobiotic anabolic steroids
and testosterone by analysis of athletes’ urine // Clin. Chem. — 1997. — Vol. 43. — P. 1280-1288.
662. Garle M. et al. Increased urinary testosterone/epitestosterone ratios found in Swedish athletes
in connection with a national control program. Evaluation of 28 cases // J. Chromatogr. B. Biomed.
Appl. - 1996, - Vol. 687. - P. 55-59.
663. Gonzalo-Luinbreras R., Pimentel-TVapero D„ Izquierdo-Hornillos R. Development and method
validation for testosterone and epitestosterone in human urine samples by liquid chromatography
applications //J. Chromatogr. Sci. - 2003. - Vol. 41. - P. 261-265.
664. Hebestreit M„ Flenker U„ Buissoii C. et al. Application of stable carbon isotope analysis to
the detection of testosterone administration to cattle //J. Agric. Food. Chem. - 2006. — Vol. 54. -
P. 2850-2858.
665. Saudan C., Baume N., Robinson N. et al Testosterone and doping control // Br. J. Sports.
Med. - 2006. - Vol. 40. - Suppl. 1. - P. 21-24.
666. Sottas P.E., Baume N., Saudan C., Schweizer C., Kamber M., Saugy M. Bayesian detection of
abnormal values in longitudinal biomarkers with an application to T/E ratio // Biostatistics. - June
19,2006 (accessed January» 5,2007). Available at http://biostatistics.oxfordjournals.org/cgi/content/
abstract/l«1009vl
667. Shackleton C.H. et al. Confirming testosterone administration by isotope ratio mass spectro¬
metric analysis of urinary androstanediols // Steroids. - 1997. - Vol. 62. - P. 379-387.
668. Kottke M.K. Scientific and regulatory aspects of nutraceutical products in the United States //
Drug, Dev. Ind. Pharm. - 1998. - Vol. 24. - P. 1177-1195.
669. Stevens K. Dietary Supplement Health and Education Act: quiet but far-reaching consequences //
Alteni. Ther. Health. Med. — 1995. — Vol. 1. — P. 17-18.
670. Yesalis 3rd C.E. Medical, legal, and societal implications of androstenedione use [editorial;
comment] //JAMA. - 1999. - Vol. 281. - P. 2043-2044.
671. Leder B.Z., Leblanc K.M., Longcope C. et al. Effects of oral androstenedione administration
on serum testosterone and estradiol levels in postmenopausal women //J. Clin. Endoainol. Metab. -
2002. - Vol. 87. - P. .5449-5454.
672. Leder B.Z., Longcope C., Catlin D.H. et al. Oral androstenedione administration and serum
testosterone concentrations in young men //JAMA. - 2000. — Vol. 283. - P. 779-782.
673. Brown G.A., Vukovich M.D., Martini E.R. et al. Endocrine responses to chronic andros¬
tenedione intake in 30- to 56-year-old men // J. Clin. Endocrinol. Metab. - 2000. - Vol. 85. -
P. 4074-4080.
328
674. Brown G.A., Vukovich M.D., Martini E.R. et al. Effects of androstenedione-herbal
Supplementation on serum sex hormone concentrations in 30- to 59-year-old men // Int. J. Vitam.
Nutr. Res. - 2001. - Vol. 71. - P. 293-301.
675. King D.S., Sharp R.L., Vukovich M.D. et al. Effect of oral androstenedione on serum testos¬
terone and adaptations to resistance training in young men: a randomized controlled trial //JAMA. —
1999. - Vol. 281. - P. 2020-2028.
676. Jasuja R., Ramaraj P., Mac R.P. et al. D4-Androstene-3,17-dione binds androgen receptor, pro¬
motes myogenesis in vitro and increases serum testosterone levels, fat-free mass and muscle strength in
hypogonadal men //J. Clin. Endocrinol. Metab. - 2005. - Vol. 90. - P. 855-863.
677. Catlin D.H., Leder B.Z., Ahrens B. et al. Trace contamination of over-the-counter androstene¬
dione and positive urine test results for a nandrolone metabolite // JAMA. — 2000. — Vol. 284. —
P. 2618-2621.
678. Delbeke F.T. et al. Prohormones and sport //J. Steroid. Biochem. Mol. Biol. — 2002. — Vol.
83. - P. 245-251.
679. van der Merwe P.J., Grobbelaar E. Unintentional doping through the use of contaminated
nutritional Supplements // S. Air. Med. J. — 2005. — Vol. 95. — P. 510-511.
680. Geyer H., Part M.K., Mareck U. et al. Analysis of non-hormonal nutritional Supplements for
anabolic-androgenic steroids — results of an international study // Int.J. Sports. Med. — 2004. — Vol.
25. - P. 124-129.
681. Laughlin G.A., Barrett-Connor E. Sexual dimorphism in the influence of advanced aging on
adrenal hormone levels: the Rancho Bernardo Study // J. Clin. Endocrinol. Metab, — 2000. — Vol.
85. - P. 3561-3568.
682. Orentreich N., BrindJ.L., Vogelman J.H. et al. Long-term longitudinal measurements of plasma
dehydroepiandrosterone sulfate in normal men // J. Clin. Endocrinol. Metab. — 1992. — Vol. 75. —
P. 1002-1004.
683. Baulieu E.E., Robel P. Dehydroepiandrosterone (DHEA) and dehydroepiandrosterone sul¬
fate (DHEAS) as neuroactive neurosteroids // Proc, Natl. Acad. Sci. USA.- 1998. - Vol. 95. -
P. 4089-4091.
684. Liu D., Dillon J.S. Dehydroepiandrosterone activates endothelial cell nitric-oxide synthase by
a specific plasma membrane receptor coupled to Ga(i2,3) // J. Biol. Chem. — 2002. — Vol. 277. —
P. 21379-21388.
685. Demirgoren S. et al. Receptor binding and electrophysiological effects of dehydroepi¬
androsterone sulfate, an antagonist of the GABAA receptor // Neuroscience. ~ 1991. — Vol. 45. —
P. 127-135.
686. Arlt W., Callies F., van Vlijmen J.C. et al. Dehydroepiandrosterone replacement in women with
adrenal insufficiency// N. Engl. J. Med. - 1999. - Vol. 341. - P. 1013-1020.
687. Hunt P.J., Gurnell E.M., Huppert F.A. et al. Improvement in mood and fatigue after dehy¬
droepiandrosterone replacement in Addison’s disease in a randomized, double blind trial // J. Clin.
Endocrinol. Metab. - 2000. - Vol. 85. - P. 4650-4656.
688. Lovas K., Gebre-Medhin G., Trovik T.S. et al. Replacement of dehydroepiandrosterone in
adrenal failure: no benefit for subjective health status and sexuality in a 9-month, randomized, parallel
group clinical trial //J. Clin. Endocrinol. Metab. — 2003, — Vol. 88. — P. 1112-1118.
689. Johannsson G.. Burman P., Wiren L et al. Low dose dehydroepiandrosterone affects behavior
in hypopituitary androgen-defident women: a placebo-controlled trial //J. Clin. Endocrinol. Metab. -
2002. - Vol. 87. - P. 2046-2052.
690. Gebre-Medhin G., Husebye E.S., Mallmin H. et al. Oral dehydroepiandrosterone (DHEA)
replacement therapy in women with Addison’s disease // Clin. Endocrinol. (Oxf.). — 2000. — Vol.
52. - P. 775-780.
691. Morales A.J., Haubrich R.H., Hwang J.Y. et al. The effect of six months treatment with a 100
pg daily dose of dehydroepiandrosterone (DHEA) on circulating sex steroids, body composition and
muscle strength in age-advanced men and women // Clin. Endocrinol. (Oxf.). — 1998. — Vol. 49. —
P. 421-432.
692. Arlt W., Callies F., Koehler I. et al. Dehydroepiandrosterone Supplementation in healthy men
with an age-related decline of dehydroepiandrosterone secretion // J. Clin. Endocrinol. Metab. -
2001. - Vol. 86. - P. 4686-4692.
693. Callies R, Arlt W., Siekmann L. et al. Influence of oral dehydroepiandrosterone (DHEA) on
urinary steroid metabolites in males and females // Steroids. — 2000. — Vol. 65. — P. 98-102.
329
694. Wolkowitz O.M., Reus V.I., Keebler A. et al. Double-blind treatment of major depression with
dehydroepiandrosterone //Am.J. Psychiatry. - 1999, - Vol. 156. - P. 646-649.
695. Huppert F.A., van Niekerk J.K. Dehdroepiandrosterone (DHEA) Supplementation for cogni¬
tive function // Cochrane Database Syst. Rev. — 2001. — CD000304.
696. Flynn M.A., Weaver-Osterholtz D., Sharpe-Timms K.L. et al. Dehydroepiandrosterone replace¬
ment in aging humans //J. Clin. Endocrinol. Metab. - 1999. - Vol. 84. - P, 1527-1533.
697. Genazzani A.D., Stomati M., Bernardi F. et al. Long-term low-dose dehydroepiandrosterone
oral Supplementation in early and late postmenopausal women modulates endocrine parameters and
synthesis of neuroactive steroids // Fertil. Steril. — 2003. — Vol. 80. — P. 1495-1501.
698. Barnhart K.T., Freeman E„ Grisso J.A. et al. The effect of dehydroepiandrosterone
Supplementation to symptomatic perimenopausal women on serum endocrine profiles, lipid parameters
and health-related quality of life //J. Clin. Endocrinol. Metab. - 1999. - Vol. 84. - P. 3896-3902.
699. Chang D.M. et al. Dehydroepiandrosterone treatment of women with mild-to-moderate
systemic lupus erythematosus: a multicenter randomized, double-blind, placebo-controlled trial //
Arthritis. Rheum. - 2002. - Vol. 46, - P. 2924-2927.
700. Hartkamp A., Geenen R., Godaert G.L. et al. The effect of dehydroepiandrosterone on lum¬
bar spine bone mineral density in patients with quiescent s3»stemic lupus erythematosus // Arthritis.
Rheum. - 2004. - Vol. 50. - P. 3591-3595.
701. van VoUenhoven R.F., Park J.L., Genovese M.C, et al. A double-blind, placebo-controlled, clini¬
cal trial of dehydroepiandrosterone in severe systemic lupus erythematosus // Lupus. — 1999. — Vol.
8. - P. 181-187.
702. Jankowski C.M., Gozansky W.S., Schwartz R.S. et al. Effects of dehydroepiandrosterone
replacement therapy on bone mineral density in older adults: a randomized, controlled trial //J. Clin.
Endocrinol. Metab. - 2006. - Vol. 91. - P. 2986-2993,
703. Catlin D.H., Sekera M.H., Ahrens B.D, et al. Tetrahydrogestrinone: discovery, synthesis and
detection in urine // Rapid. Commun. Mass. Spectrom, - 2004. - Vol. 18. - P. 1245.1049.
704. Jasuja R., Catlin D.H., Miller A. et al. Tetrahydrogestrinone is an androgenic steroid that stimu¬
lates androgen receptor-mediated, myogenic differentiation in C3H10T1/2 multipotent mesenchymal
cells and promotes muscle accretion in orchidectomized male rats // Endocrinology. — 2005. — Vol.
146. - P. 4472-4478.
705. Sekera M.H., Ahrens B.D., Chang Y.C. et al. Another designer steroid: discovery, s5»nthesis, and
detection of “madol” in urine // Rapid. Commun. Mass. Spectrom. — 2005. — Vol. 19. — P. 781-784.
706. Handelsman D.J. Designer androgens in sport; when too much is never enough // Sci. STKE. —
2004. - Vol. 244. - P. 41.
707. Baker H.W.G., Burger H.G., de Kretser D.M., Hudson B. Relative incidence of etiologic dis¬
orders in male infertility // Santen R.J., Swerdloff R.S. (ed.). Male Reproducture Dysfunction: diag¬
nosis and management of hypogonadism, infectitity and improtence. — New York: Marcel Dekker. -
1986. - P. 342-372.
На протяжении всей истории человечества религиозные догматы и мифология фор¬
мировали наши взгляды на человеческую сексуальность; большинство из этих взгля¬
дов были чрезвычайно ошибочными. В историческом контексте создание Камасутры,
трактата, посвященного человеческой сексологии, практически два тысячелетия
назад стало выдающимся достижением [1]. Сексуальность человека практически не
изучали до середины XX в., когда пионерские эпидемиологические исследования,
проведенные Michael Kinsey, предоставили первые доказательные данные о значи¬
тельной изменчивости сексуального поведения американских мужчин и женщин [2].
Впоследствии на основе изысканий, проведенных William Masters и Virginia E. Johnson
[3], было установлено, что как у мужчин, так и у женщин проявляются предсказуемые
физиологические реакции после сексуальной стимуляции. Эти ставшие поворотным
моментом описания цикла человеческих сексуальных реакций, сделанные Masters и
Johnson, стали основой для рациональной классификации человеческих сексуальных
нарушений [3].
Зигмунд Фрейд приписывал сексуальные проблемы у взрослых предшествующим
сложностям при формировании сексуальности в детском возрасте и взаимоотношени¬
ях родителей и детей. Психоаналитические теории Фрейда были подвергнуты значи¬
тельной критике в связи с недавними данными о физиологических и биохимических
механизмах формирования эрекции полового члена и развития терапии эректильной
дисфункции (ЭД).
В 1980-е и 1990-е гг. говорили о стабильном прогрессе в нашем понимании физико¬
химических механизмов, приводящих к набуханию и упругости полового члена. Было
установлено, что эрекция полового члена — результат релаксации кавернозной гладко¬
мышечной мускулатуры и усиления кровотока в половом члене [4]. Признание оксида
азота в качестве ключевого вазодилататора сосудистой гладкой мускулатуры стало
кардинальным открытием, позднее отмеченным присуждением Нобелевской премии
в области физиологии и медицины Robert F, Furchgott, Louis J. Ignarro и Ferid Murad.
Установление факта, что оксид азота вызывал релаксацию гладкой мускулатуры опо¬
средованно через стимуляцию гуанилатциклазы, составило теоретическую основу для
открытия высокоэффективных видов терапии ЭД.
Великолепные эпидемиологические опросы, такие как Male Agï'rzg Study (MMAS,
исследование взрослых мужчин), проведенное John McKinlay [5J, и National Health
and Social Life Survey (NHSLS, обзор национального здоровья и социальной жизни),
выполненный Laumann и Rosen [6, 7] с применением современных выборочных
методик, выявили высокие уровни распространенности сексуальной дисфункции
среди проживающих в сообществе мужчин среднего и старшего возраста. Установив
важность сексуальной функции как детерминанты качества жизни, Всемирная орга¬
низация здравоохранения декларировала сексуальное здоровье как фундаментальное
331
право для мужчин и женщин. Слияние этих движущих сил в середине 1990-х гг, обу¬
словила разработку эффективной пероральной терапии ЭД у мужчин.
Исторически классификация и номенклатура сексуальных нарушений основы¬
валась на руководстве по диагностике и статистике умственных нарушений (ОСМ),
которое является первоосновой психиатрической номенклатуры, отражающей веру в
то, что сексуальные расстройства у мужчин и женщин психогенны по своей природе
[В]. В 1990-х гг. несколько экспертных групп обновили клинические определения сек¬
суальных расстройств у мужчин [9,10]. Нарастание понимания того, что ЭД обычно
служит проявлением системного заболевания и доступность удобных в применении
методов лечения, в том числе оральных и интрауретральных лекарственных пре¬
паратов, ввели сексуальные расстройства у мужчин в область оказания первичной
помощи, к которой они принадлежат до настоящего времени. В отличие от этого
патофизиология сексуальных расстройств у женщин остается недостаточно понятой;
не было одобрено никаких видов фармакологической терапии по этому показанию,
за исключением эстрогенной терапии диспареунии, сопровождающейся вульво¬
вагинальной атрофией.
СЕКСУАЛЬНЫЕ РАССТРОЙСТВА У МУЖЧИН
Сексуальная дисфункция — более общий термин, который включает дисфунк¬
цию либидо, оргазмическую и эякуляторную дисфункцию в дополнение к ЭД [11].
Эректильная дисфункция, ранее называемая импотенцией, является наиболее специ¬
фическим термином, который свидетельствует о неспособности лица мужского пола
достигать или поддерживать эрекцию, достаточную для удовлетворительного поло¬
вого сношения [9,10],
Регуляция сексуальной функции
И физиология эрекции полового члена
СЕКСУАЛЬНАЯ РЕАКЦИЯ У ЧЕЛОВЕКА
William Masters и Virginia E. Johnson [3] установили, что как у мужчин, так и у жен¬
щин проявляются прогнозируемые физиологические реакции после сексуальной сти¬
муляции, которые могут быть классифицированы в четыре фазы; фаза возбуждения,
плато, оргазмическая и фаза разрешения (рис. 4-1). В фазе возбуждения возрастают
сердечный и дыхательный ритмы, происходит подъем артериального давления, соски
становятся эрегированными, половой член проходит различные степени эрекции,
яички подтягиваются, также происходит прилив крови к коже [3]. Далее следует фаза
плато, при которой сердечный ритм продолжает увеличиваться, сексуальное желание
усиливается, закрывается сфинктер мочевого пузыря, мышцы у основания полового
члена сокращаются ритмически, и у мужчин начинается секреция небольшого коли¬
чества семенной жидкости [3].
Развитие оргазма сопровождается сокращениями тазовых мышц и анального сфин¬
ктера, эякуляцией семенной жидкости, испытанием интенсивного чувства наслажде¬
ния и сбросом напряжения. В первоначальной выделительной фазе семенная жид¬
кость собирается в уретральной луковице [12], для последующей эякуляции семени
из полового члена требуются сокращения периуретральных и других тазовых мышц
[12]. В процессе фазы разрешения мышцы релаксируют, сердечный ритм и артериаль¬
ное давление снижаются, и эрекция полового члена спадает. Оргазм и эякуляция сме¬
няются рефрактерным периодом различной продолжительности, во время которого
мужчины неспособны достичь оргазма вновь [3].
332
Рис. 4-1. Четыре фазы цикла человеческой сексуальной реакции по предположению Masters и Johnson,
Фаза разрешения значительно пролонгирована у мужчин, так что они могут испытывать рефрактерность
к дальнейшей стимуляции длительное время перед тем, как достичь оргазма. Как обсуждается в тексте,
взгляды на цикл женской реакции в настоящее время значительно расширились.
МЕХАНИЗМЫ ЭРЕКЦИИ ПОЛОВОГО ЧЛЕНА
Анатомия полового члена и особенности кровотока
Эректильная ткань полового члена состоит из двух дорсально расположенных
пещеристых тел и вентрально расположенного губчатого тела [13]. Эректильная
ткань пещеристых тел и губчатого тела состоит из множества пещеристых про¬
странств. разделенных трабекулами [14]. Эти трабекулы состоят преимущественно
из гладкомышечных клеток, которые окружены синцитием. Эндотелиальные клетки
покрывают поверхности трабекул.
Артериальное кровоснабжение полового члена осуществляется из половых арте¬
рий, которые являются ветвями внутренних подвздошных артерий (рис. 4-2). Половая
артерия, ветвь подчревной артерии, подразделяется на пещеристую, спинки полового
члена и бульбоуретральные артерии. Пещеристые артерии и их ветви, за витковые
артерии, обеспечивают приток крови к пещеристым телам [14]. Дилатация Завитко¬
вых артерий увеличивает приток крови и давление в кавернозных синусах [15].
Иннервация полового члена
Иннервация полового члена представлена симпатическими (Т11-Ь2), парасимпа¬
тическими (52-54) и соматическими нервами (табл. 4-1). Симпатические и парасим¬
патические волокна сливаются в нижнее подчревное сплетение, где происходит инте¬
грация автономной иннервации к половому члену и ее сообщение с половым членом
через пещеристые нервы, у человека нижнее подчревное ганглионарное сплетение
локализовано ретроперитонеально вблизи прямой кишки [13].
Различные отделы головного мозга, в том числе миндалина, медиальное предзри-
тельное поле, паравентрикулярное ядро гипоталамуса и околоводопроводное серое
вещество, действуют скоординированно для осуществления эрекции полового члена.
Медиальное предзрительное поле гипоталамуса служит в качестве интегративной
области для контроля эрекций со стороны центральной нервной системы; она при¬
нимает чувствительные волокна из миндалины и посылает импульсы к паравентри-
кулярным ядрам гипоталамуса и околоводопроводному серому веществу. Нейроны
в паравентрикулярных ядрах отдают волокна к пояснично-грудным и крестцовым
ядрам, связанным с эрекциями.
333
Таблица 4-1. Иннервация полового члена
ТиШВОЯбКОН
неіЦкмюв
Н€К:р|^8вЛ0)Ш8
фушарв)
Симпатические
Т10-12
Превертебральные исходящие волок¬
на через подчревные и пещеристые
нервы; дополнительно параверте-
бральные исходящие волокна через
парасимпатичесше уалы и половой
или тазовый и пещеристый нервы
В общем антиэрекгильная функ¬
ция; симпатическая иннервация
играет важную роль в {жгулящги
выделения семенной жидкости
Парасимпатические
82-84
Пещеристые и тазовые нервы
Проэректильная функция
Соматические
82-84
Половой нерв
Чувствительность полового члена,
сокращение поперечнополосатых
мышц в процессе эякуляции
Парасимпатическая иннервация полового члена проэректильна, а симпатическая
иннервация в основном ингибирующая. Стимулы от промежности и нижнего отдела
мочевыделительного тракта «доносятся» до полового члена посредством крестцовой
рефлекторной дуги.
Гемодинамические изменения в процессе эрекции полового члена
Эрекция полового органа — результат серий биохимических и гемодинамических
событий, которые связаны с активацией регионов центральной нервной системы,
принимающих участие в регуляции эрекций, релаксации пещеристых гладких мышц,
увеличении притока крови в пещеристых синусах и венозной окклюзии, приводящей
к накоплению крови в половом члене и его твердости [14]. Нормальная эрекция
полового члена требует скоординированного участия непораженной центральной и
периферической нервной системы, пещеристых и губчатых тел, нормального артери¬
ального кровоснабжения и венозного оттока [14].
Пещеристая гладкая мускулатура релаксирует, и приток крови к половому члену
возрастает, накопление крови в пещеристых пространствах приводит к увеличению
полового члена (см. рис. 4-2) [4]. Расширяющиеся пещеристые тела прижимают вену-
лы к напряженной белочной оболочке, ограничивая венозный отток из пещеристых
пространств [4]. Это способствует захвату крови в пещеристых синусах, обеспечивая
твердость полового члена.
Биохимическая регуляция тонуса пещеристых гладких мышц
Тонус гладкомышечных клеток тел определяет эрегированное состояние полового
члена [4, 5]. Когда пещеристые гладкомышечные клетки расслаблены, половой член
переполняется кровью и эрегирует, в то время как кавернозные гладкомышечные
клетки сокращаются, происходит преобладание активности симпатических нервов и
половой член становится мягким.
Тонус гладкой мускулатуры в пещеристых телах поддерживается за счет выделе¬
ния из резервов внутриклеточного кальция в цитоплазму и его проникновения через
мембранные каналы [16]. Трансмембранное поступление кальция в кавернозные
гладкомышечные клетки опосредовано преимущественно потенциалзависимыми
кальциевыми каналами Ь-типа [16], несмотря на то ^гго кальциевые каналы Т-типа
также экспрессируются в кавернозных гладкомышечных клетках [17]. Увеличение
уровня внутриклеточного кальция активизирует киназу легких цепей миозина, при¬
водя к их фосфорилированию, взаимодействиям актина и миозина и сокращению
гладкой мускулатуры [14,18].
Трансмембранный и внутриклеточный кальциевый ток в пещеристых гладко-
мышечных клетках регулируется рядом клеточных процессов, которые включа-
334
Простата
Дорсальная вена
Дорсальная артерия
Кавернозный нерв
(автономный, вегетативный)
Дорсальный нерв
(соматический)
Дорсальный нерв
(соматический)
Синусоидные
пространства
Пещеристые ^
Кавернозная
артерия
Эрегированное состояние
Подоболочечное
венозное сплетение
Расслабленное состояние
Рис. 4-2. Анатомия и механизм эрекции полового члена. Пещеристые тела образованы трабекулярными
пространствами, которые окружены пещеристыми гладкими мышцами. Завитковые артерии обеспечива¬
ют артериальный приток к пещеристым пространствам. Дорсальный нерв обеспечивает чувствительную
иннервацию полового члена. В процессе эрекции релаксация трабекулярных гладких мышц и увели¬
ченный кровоток приводят к увеличению синусоидальных пространств в пещеристых телах. Синусоиды,
расширяясь, прижимают сосуды, осуществляющие венозный возврат, к белочной оболочке, приводя
к накоплению крови. Это придает твердость разбухшему половому члену. [Адаптировано из Lue T.F.
Erectile dysfunction // New Engl. J. Med. - 2000. - N 342. - P. 1802-1813.]
ЮТ течение через калиевые каналы, щелевидные соединения, опосредованные
коннексином-43, и ряд холинергических, адренергических и норадренергических
нехолинергических медиаторов (рис. 4-3-4-6) [3-6, 14, 16]. Норадренергические
нехолинергические медиаторы включают вазоактивный интестинальный пептид,
кальцитонин-ген, родственный пептид и оксид азота (N0) [14].
Адренергические проводящие пути действуют через рецепторы норадреналина
и al-адренергические рецепторы, активизируя фосфолипазу С, образующую диа-
цилглицерол и инозитолтрифосфат. Диацилглицерол активизирует протеинкиназу
С, которая ингибирует К* каналы и активизирует трансмембранный приток кальция
путем активизирования кальциевых каналов L-типа (см. рис. 4-4) [19]. Инозитол
1,4,5-трифосфат (ИФ3) повышает внутриклеточный уровень кальция посредством
обеспечения его выделения из внутриклеточных кальциевых резервов [20, 21].
335
ПГЕі
цАМФ
I
ПКА
О
Са2+
Рис. 4-3. Регуляция сократимости пещеристой глад¬
кой мускулатуры под влиянием ПГЕ,. Релаксация
пещеристой гладкой мускулатуры регулируется вну¬
триклеточными цАМФ и цГМФ. Путем активации
специфических протеинкиназ эти внутриклеточные
вторичные мессенджеры вызывают секвестрацию
внутриклеточного кальция, закрытие кальциевых кана¬
лов и открытие К+-каналов. Это приводит к общему
снижению уровня внутриклеточного кальция, вызы¬
вающему расслабление гладкой мускулатуры. ПГЕ,
посредством связывания с рецепторами увеличива¬
ет внутриклеточную концентрацию цАМФ, который
активизирует ПКА. ПКА способствует секвестрации
внутриклеточного кальция, ингибирует приток кальция
и стимулирует К*-каналы. Общий результат заключа¬
ется в снижении внутрицитоплазматического уровня
кальция и релаксации гладкой мускулатуры. цАМФ —
циклический аденозинмонофосфат; цГМФ — цикличе¬
ский гуанозинмонофосфат; ПГЕ, — простагландин Е,;
ПКА — протеинкиназа А. [Адаптировано из Bhasin S.,
Benson G.S. Male sexual function // De Kretser D. (ed.).
Knobil and Neill’s Physiology ot Reproduction. — 3th ed. —
Boston; Academic Press. — 2006. — P. 1173-1194; Lue
T.F. Erectile dysfunction // New Engl. J. Med. — 2000. —
N 342.-P. 1802-1813.1
НЭ
/ \
ДАГ ИФЗ
ПКС
K+
\
/
Ca2*
I
Ca2+
Рис. 4-4. Регуляция сократимости пещеристой
гладкой мускулатуры под влиянием норадреналина.
Норадреналин передает адренергические сигналы,
связывается с адренергическими рецепторами и сти¬
мулирует диацилглицерол (ДАГ) и инозитол-З-фосфат
(ИФЗ). ДАГ стимулирует протеинкиназу С (ПКС), кото¬
рая совместно с ИФЗ вызывает возрастание уровня
внутрицитоплазматического кальция и ингибирование
К*-каналов. Увеличенный уровень внутриклеточного
кальция приводит к сокращению пещеристой гладкой
мускулатуры и прекращению эрекции полового орга¬
на. НЭ — норэпинефрин. [Адаптировано из Bhasin S.,
Benson G.S. Male sexual function // De Kretser D. (ed.)
Knobil and Neill’s Physiology of Reproduction. — 3th ed. —
Boston: Academic Press, 2006. — P. 1173-1194; Lue
T.F. Erectile dysfunction // New Engl. J. Med. — 2000. —
N 342.-P. 1802-1813.1
Общее повышение внутриклеточного уровня кальция способствует взаимодействию
актин-миозин, что приводит к сокращению гладкой мускулатуры и смягчению поло¬
вого члена.
Простагландин (ПГЕ^) провоцирует формирование цАМФ, которая активизи¬
рует протеинкиназу А. Активизированная протеинкиназа А стимулирует К* каналы,
приводя в результате к оттоку К* из клеток (см. рис. 4-3). Процессы, опосредованные
протеинкиназой А, также завершаются общим снижением внутриклеточного кальция,
способствуя релаксации гладкомышечных клеток.
336
Са2*
Са2^
Рис. 4-5. Взаимосвязи пещеристых гладкомышеч¬
ных клеток в половом члене. Щелевидные соеди¬
нения, опосредованные коннексином-43, объеди¬
няют прилежащие гладкомышечные клетки тела и
дают возможность для тока ионов среди взаимос¬
вязанных гладкомышечных клеток. Таким образом,
колебания потенциала действия и активности
К*-каналов в каждом из миоцитов оказывают влия¬
ние на соседние миоциты. [Адаптировано из Meiman
А., Christ G.J. Integrative erectile biology. The effects
of age and disease on gap junctions and ion channels
and their potential value to the treatment of erectile
dysfunction // Urol. Clin. N. Am. — 2001. — N 28. —
P. 217-230.1
Ca2"
4
1
Ca2+
Ca2+
4
Ca2+
Ингибиторы фосфодиэстеразы
5 типа циклических
нуклеодидов
(силденафил, тадалафил, варденафил)
©
5'-ГМФ
Протеины
Фосфопротеины
(фосфорилированные белки)
Изоляция
внутриклеточного кальци
4
Расслабление
гладкой мышцы
Рис. 4-6. Регуляция расслабления гладкой мускулатуры под влиянием оксида азота. Циклический гуано-
зинмонофосфат (цГМФ) регулирует расслабление пещеристой гладкой мускулатуры путем обеспечения
секвестрации цитоплазматического кальция. Оксид азота (N0) выделяется из норадренергических нор-
холинергических нервных окончаний и, вероятно, из эндотелия. N0 активизирует гуанилатциклазу, кото¬
рая образует цГМФ, которая, в свою очередь, активизирует цГМФ-зависимые киназы^.что в результате
приводит к секвестрации внутриклеточного кальция и релаксации гладкой мускулатуры. цГМФ дегра¬
дирует под влиянием циклических нуклеотид-фосфодиэстераз. Силденафил, варденафил и тадалафил
представляют собой селективные ингибиторы изоформы 5 ПГЕ, который присутствует в пещеристой
гладкой мускулатуре. [Адаптировано из Bhasin S., Benson G.S. Male sexual function // De Kretser D. (ed.).
Knobil and Neill's Physiology of Reproduction. — 3th ed. — Boston: Academic Press, 2006. — P. 1173-1194;
Lue T.F. Erectile dysfunction // N. Engl. J. Med. - 2000. - N 342. - P. 1802-1813.]
337
к*-каналы
Как минимум, три типа калиевых каналов, Kß, и кальцийчувствительные
каналы, относящиеся к или макси-К-каналам, экспрессируются на пещеристых
гладкомышечных клетках. Среди них ВК^-каналы самые важные, поскольку они
несут ответственность за 90% оттока К' из пещеристых гладкомышечных клеток.
Была продемонстрировано, что факторы, открывающие ВК^^,-каналы, вызывают
релаксацию гладкомышечных клеток w vitro [22]. Таким образом, неудивительно,
что стратегии, которые увеличивают экспрессию каналов ВК^ in vivo, улучшают эрек¬
тильную способность у старых грызунов и грызунов с диабетом [23] и исследуются на
предмет использования в качестве терапии ЭД [24].
Коннексин-43. Щелевидные соединения
Гпадкомышечные клетки в пещеристых телах соединяются между собой при помощи
коннексин-43-щелевидных соединений, которые позволяют ионам и некоторым сиг¬
нальным молекулам, таким как инозитолтрифосфат, свободно диффундировать через
гладкомышечные клетки [25]. Об ионных изменениях, индуцируемых стимулом из
одной гладкомышечной клетки, быстро «оповещаются» другие гладкомышечные клет¬
ки, что приводит к скоординированной регуляции всего пещеристого тела [16]. Таким
образом, пещеристые тела могут рассматриваться с функциональной точки зрения как
синцитий взаимодействующих гладкомышечных клеток (см. рис. 4-5) [16].
Оксид азота
Оксид азота, выделяемый нервными окончаниями, иннервирующими пещеристые
тела, эндотелиальной выстилкой артерий полового члена и кавернозными синусами, —
важный биохимический регулятор релаксации пещеристой гладкой мускулатуры. Оксид
азота также индуцирует артериальную дилатацию [26], Воздействия N0 на кавернозные
гладкие мышцы и артериальный кровоток опосредуются через активацию гуанилатци-
клазы, продукцию циклического гуанозинмонофосфата (цГМФ), а также активацию
цГМФ-зависимой протеинкиназы (см, рис. 4-6). цГМФ вызывает расслабление гладкой
мускулатуры путем снижения уровня внутриклеточного кальция. Имеются экспери¬
ментально доказанные сведения о том, что N0 ингибирует Rho-киназаиндуцируемую
чувствительность пещеристой гладкой мускулатуры к кальцию [27].
Циклические нуклеотидфосфодиэстеразы
Циклические нуклеотидфосфодиэстеразы (ФДЭ) гидролизируют циклический
аденозинмонофосфат (цАМФ) и цГМФ, таким образом снижая их концентрации вну¬
три пещеристых гладкомышечных клеток. Среди 13 или более изоформ циклических
нуклеотидфосфодиэстераз, которые были выделены, в половом члене экспрессиру¬
ются изоформы 2, 3, 4 и 5 [14, 28]. Только фосфодиэстераза 5 (ФДЭ5) специфична
для пути NO/цГМФ в пещеристых телах [14,28]. Гидролиз цГМФ под влиянием этого
фермента приводит к обратимости релаксации гладкой мускулатуры и устранению
эрекции полового члена. Силденафил, варденафил и тадалафил — эффективные и
селективные ингибиторы активности ФДЭ5, которые предотвращают распад цГМФ и
приводят к увеличению эрекции полового члена [29].
Регуляция чувствительности к внутриклеточному кальцию под влиянием
сигнализирования Rho A/Rho-киназы
Значительные усилия были сконцентрированы на изу^чении роли Rho-киназы в
модулировании чувствительности пещеристой гладкой мускулатуры к внутриклеточ¬
ному кальцию (см. ссылку 18). На основе увеличивающегося объема доказательных
данных предполагают, что сенситизация к внутриклеточному кальцию регулирует¬
ся путем создания баланса между фосфорилированием регуляторной легкой цепи
миозина II под влиянием киназы легких цепей миозина и его дефосфорилирования
под влиянием фосфатазы легких цепей миозина (рис. 4-7) [18]. Фосфорилирование
регуляторных легких цепей миозина II необходимо для активизации миозин II
аденозинтрифосфатаз (АТФаз) под действием актина, а его дефосфорилирование
338
Миозин II РГ\%
Rho-киназа
4
КЛЦМ I I Фосфатаза ▼ Неактивная
Са Сам I I ЛЦМ Р фосфатаза
ЛЦМ
Миозин II РЛЩ.-Р
Сокращение кавернозной гладкой
мускулатуры и чувствительность
к внутриклеточному Са2+
Рис. 4-7. Роль Rho A-Rho-киназы в регуляции чувствительности к внутриклеточному Са^^. Чувствительность
к кальцию и сократительная способность гладкой мускулатуры регулируются системой Rho A-Rho-
киназы. Баланс между фосфорилированием киназы регуляторного миозина легких цепей (миозин II
РЛЦ^д) и ее дефосфорилированием под влиянием фосфатазы миозина легких цепей представляет собой
основной определяющий фактор для сенситизации гладкой мускулатуры к Са^*. Rho-киназа, нисходяищй
эффектор Rho А, может регулировать способность реагирования гладкой мускулатуры на кальций путем
ингибирования активности фосфатазы миозина легких цепей (МЛЦ). [Адаптировано из Somiyo А.Р.,
Somiyo A.V. Са^' sensitivity of smooth muscle and nonmuscle myosin II: modulated by G proteins, kinases, and
myosin phosphatase // Physiol. Rev. — 2003. — N 83. — P. 1325-1358.]
предотвращает активацию миозин II АТФаз [18]. Соотношение активностей киназ и
фосфатаз — важнейший фактор, определяющий сократительную чувствительность
пещеристых гладкомышечных клеток к внутриклеточному кальцию [18].
Rho А представляет собой гуанозинтрифосфатазу длиной приблизительно 20 кД,
которая изменяет активность Rho-киназы, фосфорилирование миозина легких цепей
и чувствительность к кальцию в гладкомышечных клетках [30]. Комплекс Rho А-ГДФ
(гуанозиндифосфат) обусловливает диссоциацию ингибитора ГДП (НІіоГДИ) до его
неактивного состояния [31]. Ряд внутриклеточных сигналов может обусловливать
обмен ГДП на ГТП в Rho А через регуляцию факторов обмена нуклеотида гуанина.
Rho А-ГТФ взаимодействует со своим нисходящим эффектором Rho-киназой [32],
увеличивая чувствительность сосудистых гладких мышц к внутриклеточному каль¬
цию посредством ингибирования фосфатаз миозина легких цепей [33]. Несмотря
на то что экспрессия Rho A/Rho-киназы не имеет значительных отличий у молодых
и старых крыс, активность Rho-киназы выше в пещеристой гладкой мускулатуре у
старых, нежели у молодых крыс [34]; связанное с возрастом увеличение активности
Rho-киназы было предложено как один из возможных механизмов для объяснения
снижения эректильной способности у крыс в связи с возрастом [34]. Ингибирование
активности Rho-киназы у экспериментальных животных приводит к увеличению
релаксации пещеристой гладкой мускулатуры и совершенствует внутрипещеристое
давление и эрекцию полового члена [34, 35]. Таким образом, ингибиторы Rho А/
Rho-киназы указывают на привлекательные перспективы развития методов терапии
ЭД[18].
Роль тестостерона в регуляции сексуальной активности у мужчин
Тестостерон регулирует многие аспекты сексуальной активности у мужчин и
женщин. Несмотря на то что у мужчин с андрогенной недостаточностью возможно
достижение эрекций полового органа, в целом их сексуальная активность снижена [5,
25]. Тестостерон способствует формированию сексуальных мыслей и желания [36] и
увеличивает сексуальное возбуждение и внимание к эротическим звуковым и другим
стимулам [37]. Исследования визуализации головного мозга позволяют предпола¬
339
гать, что обработка секо^альных стимулов может изменяться у мужчин с дефицитом
андрогенов, при этом отмечают сниженную активацию в тех зонах головного мозга,
которые обычно активизируются у эугонадальных мужчин и у мужчин с андрогенной
недостаточностью после заместительной терапии тестостероном [38].
Воздействуя на допаминергические рецепторы в медиальном предзрительном поле
гипоталамуса, тестостерон устанавливает модель поведения поиска подкрепления
у млекопитающих мужского пола [39]. Это может служить основанием для мотива¬
ционных воздействий тестостерона на сексуальное поведение млекопитающих [39].
Роли систем ароматазы С¥Р19 и стероидной 5а-редуктазы в регуляции эффектов
андрогенов на сексуальную активность остаются неясными. Анализ исследований
позволяет предположить, что 5а-редукция тестостерона не обязательна для регуля¬
ции воздействий тестостерона на желание, однако неясно, необходима ли ароматиза¬
ция эстрадиола [40].
Самопроизвольные, но не связанные со стимулами эрекции являются тесгостерон-
зависимыми (табл. 4-2) [36]. Мужчины с андрогенной недостаточностью способны
достигать эрекции в качестве реакции на эротический фильм [36]. Ночные эрекции,
по времени соотносящиеся с пиками секреции тестостерона в ночное время, имеют
более низкую амплитуду и продолжительность у мужчин с андрогенной недостаточ¬
ностью, а терапия тестостероном увеличивает частоту, полноту и продолжительность
ночного набухания полового члена [41]. Для максимального отвердения может
потребоваться пороговый уровень андрогенной активности [42]: тестостерон регули¬
рует синтез оксида азота в пещеристой гладкой мускулатуре [43], осуществляет трой¬
ные воздействия на пещеристую гладкую мускулатуру [44], подвздошно-пещеристые
и луковично-губчатые мышцы, а также он необходим для веноокклюзионной реак¬
ции. Мужчины с андрогенной недостаточностью демонстрируют задержку оргазма и
низкий эякуляторный объем.
Таблица 4-2. Области сексуальной активности, регулируемые тестостероном
Области сексуальной а1стивности, в которых происходит улучшение после терапии тестост^ном
у мужчин с аидаогмной недостаточностыо
• Сексуальное влечение.
• Самопроизвольные сексуальные мысли.
• Восприимчивость к эротическим слуховым стимулам.
• Частота ночных и дневных эрекций.
• Продолжительность, выраженность и частота ночных эрекций полового члена.
• Обище ]фовни сексуальной активности,
• Объем эякулята
Области сексуальной активности, в копфых не происходит улучшения под влиянием терапии тестостеро¬
ном или относительно которых имеются неудовлетворительные или недостаточно дохаэателы1ые данные
• Эректильная реакция на ю«уальные эротические стимулы.
• Эректильная активность у мужчин с эректильной дисфунктщей, у которхях имеются нормальные уровни тесто¬
стерона.
• Терапевтический ответ на селективные ингибиторы фосфодиэстеразы.
• Оргазмы
Востфоизведено с разрешения из Bhasin S.. Enzlin P., Covieüo A., Basson R. Sexual dysfunction in men and
women with endocerine disorders j j Lancet. — 2007. — N 369. — P. 597-611.
Терапия тестостероном у мужчин с андрогенной недостаточностью оказывает
положительное воздействие на некоторые — не все — области сексуальной активно¬
сти и положительно влияет на сексуальное влечение, самопроизвольные сексуальные
мысли, восприимчивость к эротическим слуховым стимулам, частоту ночных и днев¬
ных эрекций, продолжительность, выраженность и частоту ночных эрекций полового
органа, общие уровни сексуальной активности и объем эякулята [36, 45, 46]. Однако
тестостерон не улучшает эректильную реакцию на визуальные эротические стимулы
340
[36], эректильную активность у мужчин с ЭД, у которых имеются нормальные уровни
тестостерона [47]. Получены доказательства, подтверждающие предположение о том,
что тестостерон улучшает терапевтическую реакцию на селективные ингибиторы фос-
фодиэстеразы [45,48,49].
Андрогенная недостаточность и ЭД — два независимых распространенных наруше¬
ния, которые встречаются совместно у 6-10% лиц среднего и старшего возраста [50].
Клинические испытания тестостерона в случаях неэффективности силденафила были
неубедительными.
КЛАССИФИКАЦИЯ СЕКСУАЛЬНЫХ РАССТРОЙСТВ НА ОСНОВЕ СЕКСУАЛЬНОЙ РЕАКЦИИ
Сексуальные расстройства были классифицированы по четырем категориям в зави¬
симости от фазы цикла сексуальной реакции, в которой эта патология происходит.
« Расстройство, связанное со сниженным сексуальным влечением.
» Эректильная дисфункция.
* Эякуляторные и оргазмические расстройства.
* Болевые расстройства.
Классификация нарушения у пациента по этим категориям очень важна, поскольку
этиологические факторы, диагностические тесты и терапевтические стратегии разли¬
чаются у разных классов сексуальных расстройств.
Расетройотво, связанное со сниженным
сексуальным впттшем
Расстройство, связанное со сниженным сексуальным влечением, представляет
собой персистирующую или периодическую недостаточность (или отсутствие) сексу¬
альных фантазий и влечения в отношении сексуальной активности, которая вызывает
выраженные страдания и сложности при взаимоотношениях и не может быть объяс¬
нена никаким другим заболеванием, непосредственными физиологическими эффек¬
тами определенных веществ (лекарственных препаратов) или общим клиническим
состоянием [51-53].
Диагноз сексуальной дисфункции обоснован, только если пациент сообщает о
душевном страдании или сложностях взаимоотношений, обусловленных низким сек¬
суальным влечением [51-53]. Низкое сексуальное влечение не обязательно является
патологическим, оно может быть адекватным приспособлением при взаимоотноше¬
ниях и вопросах, связанных со здоровьем [51-53].
Расстройство, связанное со снижением сексуального влечения, — мультифак-
торное нарушение, которое может быть следствием андрогенной недостаточности,
применения лекарственных препаратов [селективный ингибитор обратного захвата
серотонина (СИОЗСТ), антиандрогены, аналоги ГнРГ, антигипертензивные, проти¬
воопухолевые препараты, антиконвульсанты], системных заболеваний, депрессии и
других психологических проблем, а также иных причин сексуальной дисфункции или
проблем, обусловленных взаимоотношениями или дифференцировкой. Андрогенная
недостаточность — важная, корригируемая причина расстройства, связанного с недо¬
статочностью сексуального влечения, и она должна быть исключена посредством
измерения общих зфовней тестостерона в плазме.
Истинные уровни распространенности и частоты расстройства, связанного со сни¬
женным сексуальным влечением, в общей популяции неизвестны, в исследованиях
соответствующих популяций пациентов уровень распространенности оценивался
порядка 5% у мужчин и 22% у женщин [7, 53]. Уровни распространенности увели¬
чиваются с возрастом [53, 54]. Расстройство, связанное со сниженным сексуальным
влечением, часто сочетается с другими сексуальными нарушениями, такими как ЭД,
341
и может развиваться как последствие других предшествующих сексуальных рас¬
стройств [53],
Правильная оценка и лечение сниженного сексуального влечения необходимы,
потому что оценка может привести к выявлению андрогенной недостаточности.
Сниженное сексуальное влечение у одного из партнеров может вызывать напряже¬
ние в их взаимоотношениях [55] и провоцировать ЭД. Низкое сексуальное влечение
может препятствовать или снижать эффективность терапии других сексуальных дис¬
функций.
Существует четыре важных предположения относительно диагностики мужчин с
расстройствами, связанными со сниженным сексуальным влечением. Необходимый
первый шаг в оценке — беседа с парой для выяснения, является ли основной пробле¬
мой пациента ЭД или недостаточное сексуальное влечение. Во-вторых, необходимо
понять, есть ли у пары проблемы во взаимоотношениях. Необходимо установить, про¬
должается ли самостимуляция, несмотря на отсутствие влечения к сексу с партнером.
Так, при наличии доступа к Интернету «секс в одиночку» возможен на постоянной
основе, существует возможность для сексуального выражения в случае сложностей во
взаимоотношениях. В-третьих, показано общее исследование состояния здоровья для
исключения системного заболевания, депрессии и приема лекарственных препаратов.
Следует измерить уровни тестостерона для исключения андрогенной недостаточно¬
сти, поскольку это заболевание является важной, поддающейся коррекции причиной
расстройства, связанного со сниженным сексуальным влечением.
Зректшьвая диофунщш
ЭД, ранее называемая импотенцией, представляет собой неспособность достиже¬
ния и/или удержания эрекции, достаточной для удовлетворительного полового акта
[9,10,14]. Сексуальная дисфункция — более общий термин, который также включает
дисфункцию либидо, оргазмическую и эякуляторную дисфункции в дополнение к
невозможности достижения или поддержания эрекции полового члена.
ММА5 и NHSLS [5, 6] выявили неожиданно высокую распространенность ЭД у
мужчин; например, в ММА5 [5], общинной выборке мужчин в Бостоне, 52% пациен¬
тов в возрасте от 40 до 70 лет страдали ЭД в той или иной степени.
ЭД в значительной степени затрагивает качество жизни как страдающего ею муж¬
чины, так и его партнера. В одном исследовании ЭД оказывала негативное влияние
на сексуальную жизнь партнерш, в особенности на их сексуальное удовлетворение и
половое влечение [56].
РАСПРОСТРАНЕННОСТЬ И ЧАСТОТА
Наилучшие данные относительно распространенности ЭД у мужчин были получе¬
ны по результатам двух перекрестных исследований, в ходе которых использовали
популяционные методики формирования выборки. ММА5 [5, 57] и МН5Ь5 [6, 7].
ММА5: перекрестное и одновременно продольное, популяционное эпидемиологиче¬
ское исследование, в котором между 1987 и 1989 гг. исследовали 17р9 мужчин в воз¬
расте от 40 до 70 лет, проживающих в обширной зоне Бостона [5,57]; кроме того, 847
мужчин были обследованы в 1995-1997 гг. [57], Анализ данного исследования пока¬
зал, что у 52% мужчин 40-70 лет была ЭД до определенной степени; у 17,2% мужчин,
принявших участие в исследовании, выявляли минимальную ЭД, у 25,2% — умерен¬
ную ЭД и у 9,6% — полную ЭД [5]. КН5Б5 был Национальным вероятностным иссле¬
дованием англоговорящих американцев в возрасте от 18 до 59 лет, проживающих в
США [6,7]. В ходе этого исследования обнаружили высокую распространенность ЭД
среди мужчин, которая возрастала по мере увеличения возраста [6, 7].
342
Эти два ключевых исследования и некоторые другие данные подтвердили, что ЭД —
распространенная проблема [6, 58, 59], поражающая от 20 до 30 млн мужчин только
в США. Распространенность ЭД увеличивается с возрастом; она поражает менее 10%
мужчин моложе 45 лет и 75% мужчин старше 80 лет [5]. У пациентов, страдающих от
других медицинских проблем, таких как гипертензия, диабет, сердечно-сосудистые
заболевания и последняя стадия почечных заболеваний, отмечали значительно более
высокую распространенность ЭД по фавнению со здоровыми мужчинами [5].
Существует недостаточное количество продольных данных относительно уров¬
ней частоты ЭД у мужчин [57]. В MMAS было установлено, что приблизительный
уровень частоты ЭД феди белых мужчин в районе Бостона составил 25,9 случая на
1000 мужчино-лет [57]. Частота возрастала от 12,4 случая на 1000 мужчино-лет для
мужчин 40-49 лет до 29,8 случая на 1000 мужчино-лет для мужчин в возрасте от 50
до 59 лет и 46,4 на 1000 мужчино-лет для мужчин 60-69 лет [60]. В ходе другого
исследования по данным опроса мужчин, наблюдавшихся в клинике профилактиче¬
ской медицины, было обнаружено, что частота ЭД составляет менее 3 слунаев на 1000
мужчино-лет среди мужчин моложе 45 лет и 52 случая на 1000 мужчино-лет среди
мужчин 65 лет или старше. Эти данные позволяют предположить, что в мире отмече¬
но 152 млн случаев ЭД в 1995 г. и что от 600 ООО до 700 ООО мужчин в Соединенных
Штатах заболевают ЭД каждый год [61].
ФАКТОРЫ РИСКА ДЛЯ ЭРЕКТИЛЬНОЙ ДИСФУНКЦИИ
В эпидемиологических исследованиях указывается на то, что самые достоверные
прогностические факторы риска ЭД — возраст, сахарный диабет, гипертензия, прием
лекарственных препаратов и сердечно-сосудистые заболевания [5, 58]. Увеличение
возраста — важный фактор риска ЭД среди мужчин [5, 58]: менее 10% мужчин в воз¬
расте моложе 40 лет и более 50% мужчин старше 70 лет страдают ЭД. В обоих иссле¬
дованиях - MMAS и NHSLS - распространенность ЭД возрастает с каждой декадой
жизни [5, 6].
Среди всех хронических заболеваний, связанных с ЭД, сахарный диабет - наи¬
более важный фактор риска. В MMAS стандартизированный по возрасту риск полной
ЭД был в 3 раза выше у мужчин с леченым сахарным диабетом [5, 62]. 50% мужчин
с сахарным диабетом будут страдать ЭД через некоторое время по мере прогрессиро¬
вания их заболевания.
В MMAS у лиц, получающих лечение по поводу заболеваний сердца, гипертензии
и гиперлипидемии, значительно увеличивался риск ЭД. Среди мужчин с заболева¬
нием сердца и гипертензией вероятность развития ЭД была более чем в 2 раза выше
для курильщиков, чем для некурящих. Курение также увеличивает риск ЭД у муж¬
чин, принимающих лекарственные препараты от сердечно-сосудистых заболева¬
ний. Сердечно-сосудистые нарушения, включая гипертензию, инсульт, заболевание
коронарных артерий и заболевание периферических сосудов, связаны с возрастани¬
ем риска ЭД. Физическая активность сопровождается снижением риска ЭД [63].
В различных обзорах уделено особое внимание взаимосвязи выписываемых
лекарственных средств и распространенности ЭД. В MMAS использование антиги-
пертензивных препаратов, препаратов для сердечной терапии и оральных гипогли-
кемических средств сопровождалось увеличением риска ЭД. Тиазидные диуретики и
психотропные вещества, используемые для лечения депрессии, могут быть наиболее
распространенными препаратами, связанными с ЭД. просто в связи с высокой рас¬
пространенностью их применения. Однако разнообразные лекарства, включая прак¬
тически все антигипертензивные препараты, дигоксин, антагонисты Нз-рецепторов,
антихолинергетики, цитотоксические агенты и антагонисты андрогенов, участвуют в
патофизиологии ЭД.
343
симптомы НАРУШЕНИЙ НИЖНЕГО ОТДЕЛА МОЧЕВЫДЕЛИТЕЛЬНОГО ТРАКТА
И ЭРЕКТИЛЬНАЯ ДИСФУНКЦИЯ
В недавних исследованиях была выявлена связь симптомов нарушений нижнего
отдела мочевыделительного тракта и ЭД [64-69J даже после стандартизации по воз¬
расту и другим факторам риска. Выраженность симптомов нарушений нижнего отдела
мочевыделительного тракта — независимый прогностический фактор для ЭД [64-70].
Поскольку симптомы нарушений нижнего отдела мочевыделительного тракта и ЭД —
два распространенных состояния у мужчин среднего и старшего возраста, возможно,
что эта связь отражает сочетание двух широко распространенных состояний.
Однако накапливаются данные, "гго эти два состояния могут быть механически
связаны, поскольку биохимические пути регулирования мышц, выталкивающих мочу
из мочевого пузыря, и функции пещеристых гладких мышц имеют много общего [68,
72]. К*-каналы, в особенности кальцийчувствительные К*-каналы (ВК^ каналы),
сигнальная система Rho A/Rho, кальциевые каналы L-типа и щелевидные соедине¬
ния — важные медиаторы сокращения и релаксации мышцы, выталкивающей мочу,
и пещеристых гладких мышц. Возрастание сократительной способности миоцитов,
которое характеризует дисфункцию мышцы, выталкивающей мочу, и ЭД, может быть
механически соотнесено с увеличением активности Rho-киназы или нарушениями
функционирования К“-каналов [71]. Дополнительные гипотезы включают увеличе¬
ние симпатической активности или автономную дисфункцию и нарушения дегенера¬
ции оксида азота или активность PKG в мышце, выталкивающей мочу, и в пещеристой
гладкой мускулатуре [68].
ДИАГНОСТИКА
Анамнез
Диагноз ЭД основывается преимущественно на рассказе пациента и, следователь¬
но, никакой из лабораторных тестов не может быть заменен хорошим сбором анам¬
неза. Диагностическая проработка пациента с ЭД должна начинаться с оценки общего
состояния здоровья (табл. 4-3) [14,72].
Общий медицинский анамнез должен быть направлен на выявление этиологи¬
ческих факторов, а также причин, которые могут повлиять на отбор пациентов и
реакцию на терапию. Сахарный диабет, заболевания коронарных артерий, перифери¬
ческих сосудов и гипертензия у пациента предполагают сосудистую причину. Анамнез
инсульта, травмы спинного мозга или спины, рассеянного склероза или деменции
могут указывать на неврологическое нарушение. Также важна информация о трав¬
мах таза, хирургическом вмешательстве на предстательной железе или приапизме.
Социальный анамнез должен включать установление факта употребления табака,
кокаина, марихуаны, опиоидов и алкоголя. Информация относительно лекарствен¬
ных препаратов, в особенности антигипертензивных, антиандрогенных, антидепрес¬
сантов и антипсихотических, важна, поскольку практически в четверти всех случаев
ЭД может быть обусловлена лекарственными средствами. Психиатрические заболева¬
ния, такие как депрессия или психоз, или препараты, используемые для лечения этих
расстройств, могут быть связаны с сексуальной дисфункцией.
Необходим детальный сексуальный анамнез, включающий выявление ситуаци¬
онных эректильных неудач, беспокойства при половом акте, природы взаимоотно¬
шений, ожиданий со стороны партнеров и супружеских разногласий (см. табл. 4-3)
[14, 72]. Важно выделить среди различных видов неспособности достижения эрекции
изменения в сексуальном влечении, неспособность достичь оргазма и эякуляции,
неудовлетворенность сексуальными взаимоотношениями. Врач должен добиться
сведений относительно начала, количества и продолжительности эрекций и наличия
ночных и утренних эрекций.
344
Таблица 4-3. Диагностическая оценка эректильной дисфункции
Анамнез
Установите психосоциальный анамнез;
• прочности супружеских взаимоотношений и супружеских разногласий;
• депрессии;
• стресса;
• беспокойства при сексуальных отношениях;
• осведомленности и мнения относительно сексуальности
Установите факторы риска:
• сахарный диабет, гипертензия, заболевание коронарных артерий, терминальная стадия почечного заболевания
и болезнь периферических сосудов;
• анамнез травмы спинного мозга, инсульта, болезни Альцгеймера;
• оперативное вмешательство на предстательной железе или тазе;
• травма таза;
• лекарственные препараты, такие как антигипертензивные, антидепрессанты, антипсихотические, антиандроге¬
ны и ингибиторы образования андрогенов;
• употребление алкоголя, кокаина, опиатов и табака
Установите факторы, которые могут повлиять на выбор терапии и реакцию пациента на нее:
• сопутствующее заболевание коронарных артерий и его симптомы и тяжесть;
• толерантность к физической нагрузке;
• использование нитратов или доноров нитратов;
• использование а-адренергических блокаторов;
• использование вазодилататоров для контроля гипертензии или застойной сердечной недостаточности;
• употребление некоторых продуктов (клюквенный сок) или лекарственных препаратов (эритромицин, ингибито¬
ры протеаз, кетоконазол и итраконазол), которые могут повлиять на метаболизм ингибиторов ФДЭ5
Физикальное обследование
• Установите признаки андрогенной недостаточности, такие как отсутствие вторичных половых признаков, евну-
хоидные пропорции, небольшой объем яичек, увеличение молочных желез.
• Исследуйте чувствительность гениталий и промежности для оценки неврологического дефицита вследствие
повреждения спинного мозга, предшествующего инсульта или периферической нейропатии.
• Оцените пульсацию на бедрах и стопах и признаки ишемии нижних конечностей.
• Обследуйте половой член для исключения болезни Пейрони
Основная лабораторная оценка, которая должна быть проведена у всех мужчин с ЭД
• Уровень глюкозы натощак.
• Уровень липидов в плазме.
• Уровень тестостерона в сыворотке
Примечание. ЭД - эректильная дисфункция; ФДЭ - фосфодиэстераза.
Опросники для самостоятельного заполнения пациентами
За последнее десятилетие произошел общий сдвиг от экстенсивных и инвазивных
техник (например, дуплексное допплеровское УЗИ полового члена, исследования
RigiScan) к использованию простых, неинвазивных опросников самосообщений. Эти
опросники обладают ценностью, поскольку многие мужчины с ЭД неохотно обраща¬
ются к своим терапевтам и сообщают о своих сексуальных жалобах. Многие мужчины
с ЭД чувствуют себя смущенными, а другие считают ЭД неизбежным сопутствующим
обстоятельством процесса старения. Некоторые врачи также чувствуют себя неловко
при обсуждении моментов, имеющих такую интимную природу: это создает атмосфе¬
ру, непродуктивную для эффективного общения. Опросники могут помочь «растопить
лед» и облегчить общение. Эти инструменты доступны и легки для выполнения, и они
могут дополнить или усилить процесс исследования сексуальной дисфункции.
Международный индекс эректильной функции (МИЭФ) представляет собой мно¬
гоуровневую шкалу, состоящую из 15 вопросов, которые обращаются к значимым
факторам сексуальной функции у мужчин [73]. Он был утвержден на нескольких язы-
345
ках, применялся во многих многонациональных клинических испытаниях, было уста¬
новлено, что он имеет адекватную чувствительность и специфичность для выявления
изменений, связанных с терапией, включая реакцию на оральные эстрогенные препа¬
раты у мужчин с ЭД. Была разработана «мужская шкала» для оценки ряда факторов
сексуальной функции, и она может быть использована как в качестве инструмента для
первоначального скрининга, так и для оценки реакции на терапию [74].
Были приложены значительные усилия для разработки сокращенных опросников,
таких как реестр сексуального здоровья мужчин, краткий реестр мужской сексуаль¬
ной функции [75] и сокращенная версия МИЭФ [76]. Работа с этими опросниками
занимает меньше времени по сравнению с МИЭФ и более лаконично выражает ана¬
логичные аспекты мужской сексуальности.
Физикальное обследование
Направленное физиологическое обследование должно оценивать вторичные сек¬
суальные характеристики, увеличение молочных желез и объем яичек. Исследование
полового члена направлено на оценку любых нетипичных искривлений, пальпи¬
руемых бляшек или поверхностных повреждений. Оценка бедренного и стопного
пульса может предоставить информацию о заболевании периферических сосудов.
Неврологическое исследование фокусируется на двигательной слабости, чувстви¬
тельности в промежности, тонусе анального сфинктера и луковично-пещеристого
рефлекса.
Лабораторные тесты, рекомендуемые всем мужчинам с эректильной дисфункцией
Диагностическое обследование мужчин с ЭД обычно включает подсчет лейкоци¬
тов, измерения гемоглобина, глюкозы крови, азота мочевины в крови (АМК) и креа-
тинина, липидов плазмы и уровня тестостерона.
Измерения общих концентраций тестостерона в плазме могут оказать помощь при
выявлении андрогенной недостаточности. Исключение андрогенной недостаточности
у мужчин с проявлениями ЭД необходимо, поскольку она может быть проявлением
тяжелого основного заболевания, такого как опухоль гипофиза или инфицирование
вирусом иммунодефицита человека (ВИЧ). Кроме того, заместительная терапия
тестостероном у мужчин с андрогенной недостаточностью восстанавливает сексуаль¬
ную функцию помогает в поддержании минеральной плотности костей, мышечной
массы и благополучия.
Если анамнез, физическое обследование и опросники ЭД не выявляют каких-
либо очевидных медицинских проблем, требующих дальнейшей разработки, в таком
случае подход на основании цены-эффективности заключается в назначении испы¬
тательного курса орального ингибитора ФДЭ5 при условии, что противопоказания
отсутствуют фапример, прием нитратов) (табл. 4-4, см. табл. 4-3) [3, 4].
Таблица 4-4. Пошаговый подход к лечению эректильной дисфункции
• У всех пациентов, а также их сексуальных партнеров может наступить улучшение после психосексуальной кон¬
сультации, и они должны получить ее
Методы терапии первой линии
• Оральные селективные ингибиторы фосфодиэстеразы (силденафил, варденафил или тадалафил)
Методы терапии второй линии
• Внешние вакуумные устройства.
• Внутриуретрапьный альпростадил»’.
• Внутрипещеристая инъекция альпростадила*’
Методы терапии третьей линии
• Протезирование полового органа.
• Сосудистая хирургия
346
Дальнейшая оценка
Оценка кровеносной системы полового члена и кровотока в нем
Исследования, в которых оценивают сохранность сосудистой сети и кровотока в
половом органе [14, 72], не требуются большинству пациентов с ЭД. Они остаются
в резерве для пациентов, у которых результаты этих тестов повлияют на стратегию
ведения или прогноз, и должны проводиться только опытными специалистами в этой
сфере. Индекс кровяного давления в половом члене и в плече прост и специфичен,
однако он нечувствителен при сосудистой недостаточности [77, 78], гфедставляет
только истормеский интерес и редко применяется в настоящее время.
Внутрипещеристая инъекция вазоактивного агента, в частности ПГЕ^, может быть
полезной в качестве диагностической, а также потенциальной терапевтической мето¬
дики [14,72]. Эта процедура может продемонстрировать, будет ли у пациента реакция
на терапевтические методы и увеличит ли осведомленность пациента относительно
процедуры и ее потенциальных побочных эффектов. Отсутствие реакции на внутри-
пещеристую инъекцию подозрительно на сосудистую недостаточность или венозную
утечку, которая требует дополнительной оценки и лечения.
Большинству мужчин с ЭД не требуются дуплексная цветная соиография, ка верно-
зография или ангиография тазовой области [14, 72]. К примеру, ангиография может
оказаться полезной в отношении молодых людей с артериальной недостаточностью,
связанной с травмой таза. Аналогично подозрение на врожденную венозную утечку у
молодых людей, у которых выявляют ЭД, делает обоснованной кавернозографию. В
каждом случае подтверждение сосудистого повреждения может привести к вопросу о
хирургическом вмешательстве. Дуплексное УЗИ может выступать в качестве неинвэ;
зивного метода оценки сосудистой функции [14].
Ночное набухание полового члена
Исследование ночного набухания полового члена не требуется большинству
пациентов, которые проходят обследование на предмет ЭД, и рекомендуется толь¬
ко в случаях подозрения на психогенную ЭД, при ситуационных проблемах, при
документированной предоперационной низкой ригидности полового органа или
по медико-юридическим причинам. Несмотря на то что регистрация формального
ночного набухания полового члена в лаборатории сна в удачные ночи может помочь
дифференцировать органическую и психогенную импотенцию, этот тест имеет высо¬
кую стоимость и трудозатратен. В большинстве случаев тщательный сбор анамнеза
с установлением эрекций в ночное время или рано утром предоставляет удовлетво¬
рительную корреляцию между формальным ночным набуханием и исследованиями
RigiScan [79].
Внедрение портативных приспособлений RigiScan в 1985 г. предоставило врачам
достоверный способ продолжительного мониторирования набухания полового члена
и ригидности в домашних условиях [79]. Это многокомпонентное приспособление,
которое пациент прикре!1ляет к телу на протяжении 2-3 ночей. Б нем имеются две
измерительные проволочные петли, которые помещаются вокруг основания и кон¬
чика полового члена и регистрируют изменения окружности полового органа и его
ригидности. Данные накапливаются и затем загружаются в программу, которая позво¬
ляет проводить интерпретацию.
Диагностические тесты для исключения андрогенной недостаточности и
гипоталамо-гипофизарньЕЕ повреждений
Существуют значительные разногласия относительно полезности и соотношения
«цена-эффективность гормональной оценки» и той протяженности, до которой
андрогенная недостаточность должна исследоваться у мужчин с проявлениями
ЭД. От 8 до 10% мужчин с ЭД имеют низкие уровни тестостерона; распространен¬
ность андрогенной недостаточности возрастает с увеличением возраста [50, 80, 81].
Распространенность низких уровней тестостерона не имеет значительных различий
347
среди мужчин, у которых есть проявления ЭД, и в популяции, распределенной по
возрастным группам [50]. Эти данные согласуются с предположением о том, что ЭД и
андрогенная недостаточность — два распространенных, но независимо представлен¬
ных нарушения [50],
Тем не менее важно исключить андрогенную недостаточность в этой категории
пациентов. Андрогенная недостаточность — корригируемая причина сексуальной
дисфункции, и у некоторых мужчин с ЭД и низкими уровнями тестостерона будет
ответ на заместительную терапию тестостероном. Андрогенная недостаточность
может оказывать дополнительные негативные воздействия на состояние здоровья
пациента — усиливать остеопороз, а также потерю мышечной массы и функциональ¬
ной способности.
В обширных исследованиях [50, 80, 81] было установлено, что лишь у небольшой
части мужчин с ЭД и низкими уровнями тестостерона имеются объемные обра¬
зования гипоталамо-гипофизарной зоны. В одном большом исследовании было
установлено, что все гипоталамо-гипофизарные очаги были у мужчин с уровнями
сывороточного тестостерона менее 150 нг/дл [81]. Таким образом, соотношение
«цена-эффективность диагностических процедур» для исключения основного пора¬
жения гипоталамо-гипофизарной области может быть усилено посредством ограни¬
чения применения его только у мужчин с уровнями тестостерона в сыворотке менее
150 нг/дл.
ЛЕЧЕНИЕ
В современной практике используют пошаговый подход, при котором первоначально
применяют минимально инвазивные методы терапии — простые и имеющие меньшее
количество побочных эффектов; затем спектр расширяется до более инвазивных мето¬
дов, которые могут потребовать инъекций или хирургического вмешательства после
того, как методы первой линии исчерпали свои возможности (рис. 4-8, см. табл. 4-4).
Врач должен обсудить с парой риски, преимущества и альтернативы всех видов тера¬
пии. Выбор терапевтической методики должен быть основан на исходной этиологии,
предпочтениях пациента, природе и силе взаимоотношений пациента и его сексуаль¬
ного партнера, а также на сопутствующих заболеваниях сердечно-сосудистой системы
и других коморбидных состояниях [14, 72]. У всех пациентов с ЭД может произойти
улучшение после психосексуальной консультации [14, 72].
Для достижения надлежащей медицинской практики должно быть оптимизиро¬
вано лечение всех сопутствующих медицинских расстройств, в отношении мужчин
с сахарным диабетом должны быть предприняты попытки оптимизации контроля
гликемии, несмотря на то что это может не привести к улучшению сексуальной функ¬
ции. у мужчин с гипертензией должен быть оптимизирован контроль артериального
давления, и, если возможно, терапевтический режим может быть модифицирован в
направлении исключения тех антигипертензивных препаратов, которые негативно
влияют на сексуальную функцию. Эта стратегия не всегда осуществима, поскольку
практически все антигипертензивные препараты связаны с сексуальной дисфункцией;
распространенность побочных эффектов меньше при использовании ингибиторов
ангиотензин-превращающего фермента и блокаторов рецепторов ангиотензина по
сравнению с другими препаратами.
Методы терапии первой линии
Психосексуальное консультирование
Как подчеркивал Rosen, основные задачи психосексуальной терапии — снижение
тревожности, связанной с выполнением полового акта, развитие сексуальных навы¬
ков и знаний пациента, модификация сексуальных отношений и улучшение общения
между партнерами [82]. Консультирование может оказывать позитивное воздей¬
ствие как на психические, так и на физические причины сексуальной дисфункции
(табл. 4-5) [10].
348
Психосексуальное
консультирование пары
Оптимизировать медикаментозное
лечение сопутствующих состояний
Оценить сердечно-сосудистый риск и потенциальные
противопоказания к применению ИФДЭ5
jL.
Нет противопоказаний
ИФДЭ5
к использованию ИФДЭ5
противопоказаны
I Начать терапию ИФДЭ5 I
Удовлетворительный
клинический
ответ
Неудовлетворительный
клинический
ответ
т
Рекомендуют терапию второй линии:
вакуумный констриктор, интрауретральное
введение альпростадила или внутрикавернозное
введение альпростадила
Оценить: правильность использования
комплаентность, отношения в паре,
побочные эффекты или страх
побочных эффектов
Увеличить дозу до максимально
разрешенной или максимально
переносимой
Неудовлетворительный ответ
Удовлетворительный
клинический
ответ
Неудовлетворительный
клинический
ответ
.
Периодическое дальнейшее М"
медицинское наблюдение I
Имплант
в половой член
Рис. 4-8. Алгоритм подхода к лечению эректильной дисфункции у мужчин. ИФДЭ5 — ингибитор фос¬
фодиэстеразы 5-го типа.
Таблица 4-5. Цели психосексуальной терапии у мужчин с сексуальной дисфункцией
Снижение тревожности, связанной с выполнением полового акта; тренировка пары в направлении избегания
позиции «наблюдения» и «сконцентрированности на ощущениях».
Выявление проблем взаимоотношений и улучшение общения между партнерами и близости.
Модификация сексуальных отношений и убеждений.
Улучшение сексуальных навыков пары.
Адаптировано из Rosen R.C. Psychogenic erectile dysfunction. Classification and management // Urol. Clin.
North Am. - 2001. - N 28. - P. 269-278.
Masters и Johnson постулировали, что концентрация внимания пациента на сексу¬
альных проявлениях, а не на эротической стимуляции, — основной фактор патофи¬
зиологии психогенной ЭД [82, 83]; они называют это поведение «наблюдательным».
Многие эксперты рекомендовали подход, направленный на коррекцию, сконцентри¬
рованную на ощущениях, при котором пара избегает полового акта и практикует неге¬
нитальные, нетребовательные, нацеленные на получение удовольствия упражнения
для того, чтобы снизить тревожность, связанную с выполнением полового акта [84].
Вовлечение пациента в процесс консультирования способствует рассеиванию
неверного понимания проблемы, снижает уровень стресса, усиливает близость и спо¬
собность разговаривать о сексе и увеличивает шансы на достижение успешного исхода
[82, 84]. Сеансы консультирования также способствуют разрешению конфликтов
349
во взаимоотношениях, психиатрических проблем, при злоупотреблении алкоголем
и наркотиками и при значительных неадекватных установках относительно секса.
Поскольку многие мужчины и женщины утаивают свои неадекватные установки и
нереалистичные ожидания относительно выполнения полового акта и связанных с
возрастом изменений сексуальной функции, когнитивные реструктурирующие мето¬
дики могут оказаться полезными для коррекции сексуальных мифов и убеждений
[82]. Несмотря на то что психоповеденческая терапия была нацелена на облегчение
депрессии и тревожности, имеется небольшое количество данных об эффективности
этого терапевтического подхода.
Селективные ингибиторы фосфодиэстеразы
Селективные ингибиторы ФДЭ5 безопасны и эффективны и стали широко рас¬
пространенными для терапии первой линии у пациентов с ЭД, за исключением тех
мужчин, которым эти препараты противопоказаны [85].
Механизмы воздействия
Три класса ферментов — аденилатциклаза, гуанилатциклаза и ФДЭ — игра¬
ют важную роль в регуляции внутрипещеристых концентраций цАМФ и цГМФ.
Фосфодиэстеразы гидролизируют цАМФ и цГМФ, таким образом снижая их концен¬
трации внутри пещеристых гладких мышц [14, 86, 87]. Несмотря на то что в половом
члене экспрессируются изоформы ФДЭ2,3,4 и 5, только ФДЭ5 специфична для мета¬
болизма ЫО/цГМФ в пещеристых телах [88].
Ингибиторы фосфодиэстеразы силденафил, варденафил, и тадалафил — селек¬
тивные ингибиторы ФДЭ5 (табл. 4-6) [89-94]. Эти препараты блокируют гидролиз
цГМФ, индуцируемый N0 [95, 96], обеспечивая, таким образом, релаксацию пещери¬
стой гладкой мускулатуры. Действие этих препаратов требует ненарушенной реакции
N0, равно как и основополагающего синтеза цГМФ гладкомышечными клетками
пещеристых тел. Путем селективного ингибирования катаболизма цГМФ в пеще¬
ристых гладкомышечных клетках ингибиторы ФДЭ5 восстанавливают природную
эректильную реакцию на сексуальную стимуляцию, однако они не вызывают эрекцию
в отсутствие сексуальной стимуляции.
Клинигеская фармакология
Несмотря на то что все три доступных на сегодняшний день ингибитора фосфо¬
диэстеразы имеют некоторое структурное сходство, они различаются по своей селек¬
тивности и фармакокинетике (см. табл. 4-6). Общие побочные эффекты доступных
ингибиторов ФДЭ5 — головные боли, проблемы со зрением, приливы, гиперемия —
обусловлены неселективным ингибированием изоформ фосфодиэстеразы 6 и 11 в
других органах [94]. Селективность ингибитора ФДЭ5 — баланс между его ингиби¬
рующей способностью по отношению к изоформам фосфодиэстеразы, отличным от
типа 5, и его ингибирующей способностью для изоформ фосфодиэстеразы [94]. Для
ФДЭ6 тадалафил наиболее селективен, а силденафил наименее; в отношении ФДЭ11
варденафил наиболее селективен, а тадалафил наименее [94]. Негативные влияния
силденафила на сетчатку связаны с ингибированием ФДЭб в ней. Мышечные боли,
испытываемые небольшим количеством мужчин, принимающих тадалафил, могут
быть обусловлены ингибированием ФДЭ11 в скелетной мускулатуре [97, 98].
Фармакокинетика
После приема внутрь силденафила пиковые плазменные концентрации достига¬
ются в период между 30 и 120 мин, после этого они снижаются с периодом полувы¬
ведения, составляющим 4 ч (см. табл. 4-6) [99,100]. Варденафил достигает пиковых
концентраций в пределах 0,7-0,9 ч и имеет период полувыведения от 4 до 5 ч. В отли¬
чие от него пиковые концентрации тадалафила достигаются через 2 ч, и его период
полувыведения, составляющий у молодых мужчин 16,9 ч, более продолжителен, чем
у силденафила и варденафила. при этом он более продолжителен у мужчин старшего
350
Таблица 4-6. Клиническая фармакология селективных ингибиторов ФДЭ5
Характеристика
Силденафил
Варденафил
Тадалафил
Коммерческое название
Виагра
Левитра
Сиалис
0,5-2,0 ч
0,7-0,9 ч
2ч
3-4 ч
4-5 ч
16,9 ч (у молодых)
21,6 ч (у пожилых)
ФДЭ6 селективность*
25
187
ФДЭ11 селективность*
780
1160
Наступление эрекции (мин)
От 30 до 60
От 15 до 45
От 20 до 30
Влияние пищи и алкоголя
Снижается С_
Минимальные изменения
Изменения отсутствуют
Связывание с белками
96%
94%
Биодоступность
41%
НД
15%
* Селективность обозначается как соотношение IC50 изоформ ФДЭ, отличных от 5, и 1С50 ФДЭ5.
Более высокие значения указывают на более высокую селективность. Таким образом, силденафил
более эффективен, чем тадалафил, для ФДЭ5, схож с ФДЭ11, однако менее селективен по сравнению
с тадалафилом для ФДЭ6, схож с ФДЭ5.
Примечание. — максимальная плазменная концентрация: НД — недоступен: ФДЭ — фосфодиэ¬
стераза: Tj^2 ~ период полувыведения.
Адаптировано из Montague D.K., Jarow J.P., Broderick G.A. et al. Chapter 1: The management of erectile
dysfunction: an AUA update //J. Urol. — 2005. — Vol. 174. — P. 230-239; Yu G., Mason H., Wu X. et al.
Substituted pyrazolopyridopyridazines as orally bioavailable potent and selective PDE5 inhibitors; potential
agents for treatment of erectile dysfunction //J. Med. Chem. — 2003. — Vol. 46. — P. 457-460; Grossman
E.B., Swan S.K., Muirhead GJ. et al. The pharmacokinetics and hemodynamics of sildenafil citrate in male
hemodialysis patients // Kidney Int. — 2004. — N 66. — P. 367-374; Conti C.R., Pepine C.J., Sweeney M.
Efficacy and safety of sildenafil citrate in the treatment of erectile dysfunction in patients with ischemic heart
disease // Am.J. Cardiol. - 1999. - N 83. - P. 29C-34C.
возраста (21,6 ч) по сравнению с молодыми (16,9 ч). В связи с относительно корот¬
кими периодами полувыведения варденафила и силденафила эти препараты должны
приниматься за 2-4 ч перед планируемым сексуальным контактом. В отличие от них
тадалафил в связи со своим более продолжительным периодом полувыведения может,
но не обязательно должен, приниматься по мере необходимости.
Пища, в особенности с высоким содержанием жиров, и алкоголь могут приводить
к задержке и снижению абсорбции силденафила. Однако в ранних фармакокинетиче¬
ских исследованиях не сообщается об изменениях в максимальной плазменной кон¬
центрации или уровнях абсорбции варденафила или тадалафила в связи с приемом
пищи или умеренным количеством алкоголя [101].
Эффективность
В ходе рандомизированных клинических испытаний было показано, что все три
активных при приеме внутрь селективных ингибитора ФДЭ5 — силденафил, вардена¬
фил и тадалафил — безопасны для тщательно отобранных мужчин с ЭД [102-106]. У
мужчин, получающих терапию оральными селективными ингибиторами ФДЭ5, уров¬
ни успешных половых актов варьируют от 50 до 65%, а уровни улучшения эрекции
колеблются от 70 до 75% [102-107]. Селективные ингибиторы ФДЭ5 эффективны у
мужчин всех этнических групп и любого возраста [102-106], несмотря на то что уров¬
ни реакций колеблются у различных подгрупп пациентов.
351
Введенный в продажу в США в марте 1998 г. силденафил (Виагра, Рйгег, Нью-Йорк)
стал первым эффективным оральным лекарственным препаратом для лечения ЭД [29,
96, 108, 109]. Эффективность силденафила была продемонстрирована в рандомизи¬
рованных контролируемых испытаниях [110], в которых 532 мужчин с органической,
психогенной или смешанной ЭД были рандомизированы по группам и получали пла¬
цебо или 25, 50 или 100 мг силденафила на протяжении 24 нед. В этом исследовании
пациенты, принимающие силденафил, испытывали значительное возрастание ригид¬
ности полового органа, частоты проникновения во влагалище и удержания эрекции
по сравнению с теми, кто получал плацебо. Возрастающие дозы силденафила, по дан¬
ным опросника, сопровождались более высокими средними показателями частоты
проникновения и удержания эрекций после сексуального проникновения.
В последующем исследовании увеличения дозы [110] 329 мужчинам в рандоми¬
зированном порядке был назначен прием плацебо или 50 мг силденафила на протя¬
жении 12 нед. При каждой последующей оценке дозу силденафила увеличивали или
снижали на 50% в зависимости от терапевтической реакции или побочных эффектов.
64% попыток полового сношения были успешными у мужчин, которые принимали
силденафил, по сравнению с 22% у мужчин, получавших плацебо. Среднее число
удачных попыток за месяц составило 5,9 у мужчин, получавших силденафил, и 1,5
у тех, кто получал плацебо. Средние показатели оргазмов, удовлетворения половым
сношением и уровень общего удовлетворения были также значительно выше в группе
пациентов, получавших силденафил, по сравнению с группой плацебо [110].
Силденафил также эффективен для терапии ЭД у пациентов, страдающих сахарным
диабетом [96]. В процессе рандомизированного испытания 268 мужчин с сахарным
диабетом и ЭД [96] у 56%, получавших силденафил на протяжении 12 нед, отмечали
улучшение эрекций по сравнению с 10%, принимающими плацебо (Р <0,001).
В исследованиях эффективности варденафила 5, 10, 20 мг препарата показали
свое преимущество перед плацебо в отношении улучшения показателей эректильной
функции, которые оказались дозозависимыми [111-117]. Варденафил улучшает уров¬
ни проникновения во влагалище, ригидности полового члена, успешности полового
сношения и удовлетворения сексуальным опытом у мужчин с ЭД различной природы
[111-117].
Аналогично в рандомизированных клинических испытаниях каждая из 2,5, 5,10 и
20 мг доза тадалафила показала превосходство перед плацебо в отношении улучшения
показателей эректильной функции [118-123]. Положительные эффекты тадалафила
были дозозависимыми [123].
Ингибиторы ФДЭ5 эффективны у мужчин с ЭД, обусловленной различными при¬
чинами, включая повреждение спинного мозга и радикальную простатэктомию [108,
124]. В общем, базовая сексуальная функция имеет положительную корреляцию с
реакцией на ингибиторы ФДЭ5, и у пациентов, страдающих сахарным диабетом или
перенесших хирургическое вмешательство на предстательной железе, реакция менее
выраженная по сравнению с пациентами, страдающими психогенной или васкулоген-
ной ЭД [108]. Поскольку отсутствует базовая характеристика, которая позволяет рас¬
считать вероятность отсутствия реакции на терапию силденафилом, терапевтические
испытания ингибиторов ФДЭ5 должны проводиться у всех пациентов, за исключени¬
ем тех, которым они противопоказаны [108].
Побогные эффекты
В клинических испытаниях частота побочных эффектов — головные боли, при¬
ливы, ринит, диспепсия, нарушения зрения — выше у мужчин, получавших терапию
ингибиторами ФДЭ5, по сравнению с теми, кто получал терапию плацебо (табл. 4-7)
[125]. Распространенность головных болей, приливов и ринита — прямое следствие
неселективного ингибирования ФДЭ5 в других органных системах и находится в
зависимости от принимаемой дозы. Эти препараты не влияют на характеристики
352
семенной жидкости [49]. В ходе основных клинических испытаний не отмечали слу¬
чаев приапизма.
Таблица 4-7. Распространенные побочные эффекты селективных ингибиторов фосфодиэстеразы
Побочное действие
Распространенность (%)
Силденафил
Варденафил
Тадалафил
Головные боли
13
16
15
Приливы
10
12
Диспепсия
12
Закупорка носового канала
10
Головокружение
Патология зрения*
<2
Боль в спине*
Миалгия*
* Эти побочные эффекты обусловлены неселективным ингибированием изоформ фосфодиэстеразы в
других тканях.
Адаптировано из Brock G.B., McMahon C.G., Chen K.K. et al. Efficacy and safety of tadalafil for the treat¬
ment of erectile dysfunction: results of integrated analyses //J. Urol. — 2002. —N 168. — P. 1332-13.36;
Morales A., Gingell C., Collins M. et al. Clinical safety of oral sildenafil citrate (Viagra) in the treatment of
erectile dysfunction //Int.J. Impôt. Res. — 1998. — N 10. — P. 69-73; Wespes E., Amar E., Hatzichristou D.
et al. EAU Guidelines on erectile dysfunction: an update // Eur. Urol. — 2006. — N 49. — P. 806-815.
Были сообщения о нескольких случаях неартериитической передней ишемической
нейропатии зрительного нерва после начала приема оральных ингибиторов ФДЭ5
[126, 127]. Это состояние характеризуется внезапной монокулярной потерей зрения
в связи с острой ишемией передней части зрительного нерва в отсутствие очевидно¬
го артериита; ишемия может прогрессировать до частичного или полного инфаркта
головки зрительного нерва, приводя в результате к перманентной потере зрения
или выпадениям зрительных полей [127]. Несмотря на то что не было установлено
взаимоотношение причины и эффекта от приема ингибитора ФДЭ5 [126], пациенты
с внезапной потерей зрения в анамнезе не должны принимать ингибиторы ФДЭ5 без
предварительной офтальмологической оценки.
Сердегно-сосудистые и гемодинамигеские эффекты
В наблюдениях за побочными эффектами, связанными с приемом силденафила,
после введения его в продажу сообщали о нескольких случаях инфаркта миокарда
и внезапной смерти среди мужчин, принимавших силденафил [128-131]. 44 из 130
случаев смерти, опубликованных Администрацией по контролю продуктов питания
и лекарств США с марта по ноябрь 1998 г., произошли во временной связи с употре¬
блением силденафила [128-131]; 16 из этих смертей произошли у пациентов, кото¬
рые принимали нитраты. Поскольку у большинства мужчин с признаками ЭД также
имеется высокая распространенность сердечно-сосудистых факторов риска, остается
неясным, являются ли эти события причинно-зависимыми от приема силденафила,
основного заболевания сердца или и от того, и от другого [130].
В тщательно контролируемом исследовании [132] прием внутрь 100 мг силдена¬
фила мужчинами с тяжелым заболеванием коронарных артерий приводил лишь к
небольшим снижениям системного кровяного давления и отсутствию значительных
изменений сердечного выброса, частоты сердечного ритма, коронарного кровотока
353
и диаметра коронарных артерий. В раздельном анализе пяти рандомизированных
плацебо-контролируемых исследований варденафила Kloner с соавт. объединили
данные безопасности сердечно-сосудистого профиля [133]. Общая частота сердечно¬
сосудистых осложнений была схожей у мужчин, получавших терапию варденафилом,
и пациентов, принимавших плацебо. Однако терапия варденафилом сопровождалась
умеренным снижением артериального давления (снижение систолического арте¬
риального давления на 4,6 мм рт.ст.) и небольшим увеличением сердечного ритма
(2 уд./мин). Это привело к тому, что Американская кардиологическая ассоциация
сделала вывод: предшествующее заболевание коронарных артерий само по себе не
является противопоказанием для использования ингибиторов ФДЭ5 [130].
Межлекарственные взаимодействия
Силденафил метаболизируется преимущественно по путям Р450 2С9 и Р450 ЗА4
[130]. Циметидин и эритромицин, ингибиторы Р450 ЗА4, увеличивают плазменные
концентрации силденафила. Ингибиторы протеазы ВИЧ могут также изменять актив¬
ность пути Р450 ЗА4 и оказывать влияние на клиренс силденафила [130]. И наобо¬
рот, силденафил является ингибитором метаболического пути Р450 2С9, и его прием
может потенциально повлиять на метаболизм лекарственных препаратов, таких как
варфарин и толбутамид [130]. Комбинированный прием силденафила и ритонавира
приводит к более выраженному повышению плазменных уровней силденафила, чем
прием только лишь силденафила [134]. Имеются схожие реакции с другими лекар¬
ственными препаратами, включая саквинавир и итраконазол. Таким образом, дозы
ингибиторов ФДЭ5 должны быть адекватно снижены у пациентов, принимающих
ингибиторы протеаз или эритромицин.
Грейпфрутовый сок может изменять фармакокинетику оральных лекарственных
препаратов посредством различных механизмов. Сок, принимаемый однократно в
нормальном количестве (200-300 мл) или в виде сегментов целого свежего фрукта,
может необратимо инактивировать кишечные Р450 ЗА4, таким образом снижая пре-
системный метаболизм и увеличивая биодоступность орального препарата ингибито¬
ров ФДЭ5 [135]. Несмотря на то что значение этой проблемы в клинической практике
неизвестно, очевидно, что предусмотрительным будет предупредить мужчин, которые
намереваются принимать ингибиторы ФДЭ5, о том, что не стоит употреблять боль¬
шое количество грейпфрутового сока.
Наиболее серьезные взаимодействия ингибиторов ФДЭ5 происходят с нитратами.
Вазодилатирующие эффекты нитратов усиливаются ингибиторами ФДЭ5; это также
касается ингаляционных форм нитратов, таких как амилнитрат, или нитритов, кото¬
рые продаются под просторечным названием «кнопки» (poppers). Одновременный
прием этих двух препаратов может вызвать потенциально фатальное падение артери¬
ального давления [130].
Тадалафил и варденафил противопоказаны мужчинам, принимающим а-блока-
торы. В отношении пациентов с застойной сердечной недостаточностью, которые
принимают вазодилатирующие препараты или выполняют сложные режимы приема
антигипертензивных препаратов, необходим мониторинг артериального давления
после первоначального назначения ингибиторов ФДЭ5 [130,133,136].
Терапевтигеские режимы
Группами экспертов нескольких обществ были опубликованы великолепные кли¬
нические рекомендации [74, 137]. Для сведения к минимуму риска гипотензии и
неблагоприятных сердечно-сосудистых осложнений в связи с использованием инги¬
биторов ФДЭ5 Американская кардиологическая ассоциация составила список реко¬
мендаций (табл. 4-8), которые необходимо строго выполнять [130].
У большинства мужчин с ЭД прием силденафила начинается с 25-50 мг. Если эта
доза не вызывает никаких побочных эффектов, она может титроваться до 100 мг [29,
354
Таблица 4-8. Рекомендации по применению селективных ингибиторов фосфодиэстеразы
> Не назначать селективные ингибиторы ФДЭб у мужчин, принимающих препараты нитратов длительного дей¬
ствия и короткого действия на регулярной основе.
' Контролировать применение селективных ингибиторов ФДЭб путем тщательного взвешивания рисков, если
пациент страдает стабильным заболеванием коронарных артерий, не принимает нитраты длительного дей¬
ствия и использует нитраты короткого действия лишь нерегулярно.
• Не вводить селективные ингибиторы ФДЭб в течение 24 ч после приема любых форм нитратов.
' Дать рекомендации мужчинам относительно рисков потенциального взаимодействия селективных ингибиторов
ФДЭб и нитратов, доноров нитратов и а-адренергических блокаторов. Совместное использование нитратов,
доноров нитратов или а-адренергических блокаторов может приводить к гипотензии, которая может быть
серьезной или даже фатальной.
' У мужчин с предшествующим заболеванием коронарных артерий оценить риск ишемии сердца во время поло¬
вого акта перед тем, как выписывать ингибиторы ФДЭб. Эта оценка может включать стресс-тест.
' У мужчин, которые принимают вазодилататоры и диуретики для коррекции гипертензии или застойной сер¬
дечной недостаточности, необходимо рассмотреть потенциальный риск гипотензии в связи с потенциальным
взаимодействием ингибиторов ФДЭб и вазодилататоров, в особенности у пациентов с низким объемом крови.
' У мужчин, инфицированных ВИЧ, необходимо рассмотреть межлекарственные взаимодействия селективных
ингибиторов ФДЭб, антиретровирусных и антимикробных препаратов.
Примечание. ВИЧ — вирус иммунодефицита человека; ФДЭ — фосфодиэстераза [4,14,130].
130]. Дальнейшее ее регулирование должно контролироваться на основании терапев¬
тической реакции и проявления побочных эффектов. В типичных случаях не реко¬
мендуют однократные приемы выше 100 мг. Прием варденафила должен начинаться
с 10 мг; доза должна быть повышена до 20 мг или снижена до 5 мг в зависимости от
первоначальной реакции и проявления побочных эффектов. Однократный .прием
выше 20 мг не рекомендуют. Употребление тадалафила начинается с первоначаль¬
ной дозы 10 мг с дальнейшим регулированием на основе эффективности и побочных
эффектов. Тадалафил нельзя принимать чаще, чем 1 раз каждые 48 ч.
У мужчин, принимающих ингибиторы протеазы (в особенности ритонафира или
индинавира), эритромицин, кетоконазол, итраконазол или большие количества
грейпфрута, дозы ингибиторов ФДЭ5 должны быть снижены, а применение выше
25 мг силденафила, 5 мг варденафила или 10 мг тадалафила не рекомендуют.
Силденафил и варденафил принимают, как минимум, за час до полового акта и
не более 1 раза за 24-часовой период; в связи с более продолжительным периодом
полувыведения тадалафил не должен приниматься непосредственно перед половым
актом.
Недостаточно информации для определения, является ли дневное назначение
селективных ингибиторов ФДЭ5 более эффективным, чем их применение по потреб¬
ности [138].
Рекомендации по применению ингибиторов ФДЭ5 у мужгин с заболеваниями коронар¬
ных артерий
Перед тем как назначать ингибиторы ФДЭ5, необходимо оценить сердечно¬
сосудистые факторы риска (см. табл. 4-8) [130]. Если пациент страдает гипертензией
или симптомным заболеванием коронарных артерий, следует вначале провести кор¬
рекцию этих клинических нарушений [14,130]. Нужно убедиться в приеме нитратов,
поскольку ингибиторы ФДЭ5 противопоказаны пациентам, регулярно применяющим
любые формы нитратов. Ингибиторы ФДЭ5 не должны приниматься в течение 24 ч
после приема нитратов или доноров нитратов [130].
Половой акт может спровоцировать коронарную ишемию у мужчин с предшеству¬
ющим заболеванием коронарных артерий [131]; следовательно, мужчины, в отноше¬
нии которых планируется терапия ЭД, должны пройти исследование толерантности
к физической нагрузке. Один из практических путей по исследованию толерантности
к физической нагрузке заключается в том, чтобы заставить пациента подняться на
два лестничных пролета. Если он может свободно пройти один или два лестничных
355
пролета без стенокардии или избыточной экспираторной одышки, он, вероятно,
может принимать участие в половом акте без схожих симптомов. Исследование с
физической нагрузкой перед назначением ингибиторов ФДЭ5 может быть показано
некоторым мужчинам с тяжелым заболеванием сердца для оценки риска сердечной
ишемии во время полового акта [130]. Было выявлено, что селективные ингибиторы
фосфодиэстеразы не ухудшают способность пациентов с хроническим заболеванием
коронарных артерий к физической активности на уровнях, эквивалентных полово¬
му акту [139-142]. Также было показано, что каждый из трех ингибиторов ФДЭ5
не обладает значительными побочными действиями на гемодинамику и сердечную
деятельность у тщательно отобранных мужчин с ЭД, у которых не имеется каких-
либо противопоказаний для использования ингибиторов ФДЭ5 [139-142]. Ни один
из ингибиторов ФДЭ5 не оказывает побочного действия на общее время физической
нагрузки или время ишемии в процессе исследования с физической нагрузкой у муж¬
чин со стабильной стенокардией.
Ведение пациентов, у которых отсутствует реакция на ингибиторы ФДЭ5
Несмотря на то что оральная терапия ингибиторами ФДЭ5 стала революцией в
ведении ЭД, не у всех мужчин происходит реакция на это лечение. Совокупная веро¬
ятность успеха полового акта при приеме силденафила цитрата увеличивает число
попыток, достигающих максимума после восьми попыток [143]. На основании этих
данных [143] отсутствие ответа на терапию ингибиторами фосфодиэстеразы опреде¬
ляется как неудача достижения удовлетворительной реакции даже после восьми
попыток или максимально разрешенной дозы (например, 100 мг силденафила).
Многие факторы вносят вклад в неудачу лечения, включая несоблюдение приема
лекарственных препаратов, субоптимальные дозы, побочные реакции, психологиче¬
ские проблемы, на которые не было обращено внимание, оставшиеся без должного
рассмотрения проблемы партнера и взаимоотношений с ним, неверный диагноз и
пациент-специфические патофизиологические факторы [144]. В клинических испы¬
таниях ингибиторов ФДЭ5 о неудачах лечения сообщали преимущественно в отноше¬
нии мужчин, которые страдали сахарным диабетом, перенесших не щадящую нервов
радикальную простатэктомию и со значительной тяжестью заболевания [145]. При
оценке биоптата пещеристой гладкой мускулатуры у пациентов, не реагирующих на
силденафил, Wespes с соавт. [146] установили, что выраженные сосудистые поврежде¬
ния, а также атрофия и фиброз пещеристой гладкой мускулатуры - патологические
изменения, которые лежат в основе неудач.
Пациенты могут принимать препарат неправильно в связи с неадекватными
инструкциями, отсутствием понимания инструкций, побочными эффектами или
страхом перед ними [144]. Наиболее оптимальный прием оральных ингибиторов
ФДЭ5 — за 1-2 ч перед планируемым половым актом [74,137]. Маловероятно, что
лекарственные препараты будут эффективны, если они принимаются перед половым
актом; пища и алкоголь могут еще больше повлиять на максимальные сывороточные
концентрации силденафила цитрата. Аналогично пациенты могутнє принимать адек¬
ватные дозы в связи с побочными эффектами или страхом перед ними.
Мужчины, у которых сексуальная дисфункция — следствие неверно диагностиро¬
ванной ЭД и у которых основное сексуальное расстройство невосприимчиво к инги¬
биторам ФДЭ5, могут ошибочно рассматриваться как пациенты с неэффективностью
терапии. К примеру, нет оснований ожидать реакции на ингибиторы ФДЭ5 у мужчин
с расстройством, связанным со сниженным половым влечением, болезнью Пейрони
или оргазмическим/эякуляторным расстройством. Тревожность, сопровождающая
возобновление сексуальной активности, равно как и неразрешенные проблемы отно¬
шений с партнером, могут ослабить реакцию на лечение. Партнер пациента может не
стараться или не иметь способности к участию в сексуальной деятельности в связи с
проблемами взаимоотношений, сексуальными расстройствами или реальными или
ощ}тимыми проблемами со здоровьем.
356
Пациентов, которые сообщают о недостаточности удовлетворительной реакции на
первоначальный прием ингибиторов ФДЭ5, необходимо опросить по поводу времени
приема препарата, принимаемой дозы и испытываемых побочных реакций. Следует
оценить психологические и межпартнерские проблемы. Доза ингибитора ФДЭ5
должна увеличиваться постепенно в зависимости от переносимости. Если пациент не
реагирует на максимальные переносимые дозы ингибиторов ФДЭ5, их можно комби¬
нировать с вакуумными приспособлениями или интрауретральной терапией. Должна
быть рассмотрена терапия второй линии, например внутрипещеристые инъекции.
Мужчины, которые не реагируют на ингибиторы ФДЭ5 и методики терапии второй
линии, могут начать считать имплантацию полового члена приемлемой альтернати¬
вой [144].
Цена-эффективность
В ходе нескольких исследований оценивали экономическую стоимость лечения ЭД
у мужчин в контролируемых планах охраны здоровья [147-150]. Смоделированная
на компьютере стоимость силденафила цитрата составила приблизительно $ 11 ООО
на один год «качественной жизни» [149]. Эта сумма меньше, чем в отношении других
одобренных методов терапии медицинских расстройств, которые стоят менее $50 000-
100 ООО за год; таким образом, соотношение «цена-эффективность терапии ингиби¬
торами ФДЭ5» благоприятно в сравнении с одобренными методами лекарственной
терапии. На основе других методов анализа было сделано заключение, что ингиби¬
торы ФДЭ5 и приспособления для вакуумного сокращения наиболее оптимальны по
«цене-эффективности» по сравнению с доступными методами терапии [147-149].
На отдельных последних анализах стоимости, связанных с заботой о пациентах,
страдающих ЭД, было продемонстрировано, что финансовые нагрузки, накладывае¬
мые на пациентов, в контролируемом плане лечения удивительно низки [149-151]. В
ходе одного из таких исследований пользы и стоимости ежемесячная стоимость обе¬
спечения сервисов, связанных с эректильной дисфункцией в плане здравоохранения
с участием 100 ООО членов, составила менее $0,10 на члена [150]. Таким образом, в
анализах стоимости и пользы нет информации об отказе многих страховых компаний
на покрытие расходов терапии ингибиторами ФДЭ5.
Методы терапии второй линии
Вакуумные устройства для индуцирования эрекции
Вакуумное приспособление состоит из пластикового цилиндра, вакуумной помпы
и эластичной стягивающей ленты [14, 72]. Пластиковый цилиндр прилаживается
вокруг полового органа и подсоединяется к вакуумной помпе. Отрицательное давле¬
ние, создаваемое вакуумом внутри цилиндра, загоняет кровь в половой член, приводя
к эрекции. Эластичная лента, соскальзывающая по основанию полового члена, при¬
водит к накоплению в нем крови, поддерживая эрекцию до тех пор, пока резиновая
лента удерживается вокруг основания. Стягивающая лента не должна оставаться на
месте более 30 мин. Только вакуумные приспособления с механизмом ограничения
давления должны рекомендоваться для предотвращения повреждения вследствие
высокого вакуума [85].
Ограниченные данные открытых исследований относительно эффективности ваку¬
умных приспособлений указывают на то, что приборы безопасны, относительно недо¬
роги и умеренно эффективны [152,153]. Они могут ухудшить эякуляцию, приводя к
задержке семенной жидкости. Некоторые пары не любят отсутствиё спонтанности,
порождаемое этими приспособлениями. Для успешного применения этих приборов
важно взаимодействие партнеров [154].
Интрауретральные методы терапии
Интрауретральная система для поставки алпростадила^, называемая MUSE (лекар¬
ственная уретральная система для восстановления эрекции; VIVUS, Menlo Park, СА),
была выпущена в 1997 г. Алпростадил<^ — стабильная синтетическая форма ПГЕ^,
357
которая повышает уровни цАМФ и снижает концентрацию внутриклеточного каль¬
ция, таким образом обеспечивая релаксацию пещеристых гладких мышц и эрекцию
полового члена.
Алпростадил*“, затем вводимый в уретру, абсорбируется через слизистую оболочку
уретры и вентральный отдел белочной оболочки в пещеристое тело, в сравнении с
внутрйпещеристой инъекцией TTFEj^ внутриуретральный ПГЕ^ легче для применения и
обладает меньшей распространенностью побочных эффектов, в особенности фиброза
полового члена.
Алпростадил^ доступен в 125,250,500, и 1000 мкг дозировках. В типичных случаях
первоначальная доза алпростадила*“, составляющая 250 мг, вводится у терапевта для
наблюдения за изменениями артериального давления или уретрального кровотече¬
ния, вторичного по отношению к неправильному введению приспособления в мочеи¬
спускательный канал.
В первоначальных рандомизированных плацебо-контролируемых испытаниях
сообщается о 40-60% уровнях успешности, определяемых как совершение одного
полового акта и более в течение 3-месячного изучаемого периода [155-157]. В кли¬
нической практике приблизительно у трети мужчин, принимающих алпростадил*“
внутриуретрально, будет получен ответ [158].
К распространенным побочным эффектам интрауретрального алпростадила*’ отно¬
сятся боль в половом органе и жжение в мочеиспускательном канале у 30% пациентов
[155-157]; его применение может также вызывать головокружение, гипотензию и
обмороки у небольшого числа принимающих его. Внутриуретральный алпростадил*“
может вызывать умеренное жжение или зуд во влагалище у сексуального партнера.
Внутриуретральный алпростадил*“ не должен применяться мужчинами, чьи партнер¬
ши беременны, или теми, кто планирует беременность.
Интракавернозные инъекции вазоактивных агентов
Использование интракавернозных инъекций вазоактивных препаратов стало кра¬
еугольным камнем медицинской коррекции ЭД с начала 1980-х гг. Пациенты могут
быть обучены вводить вазоактивный препарат в собственные пещеристые тела при
помощи иглы 27- или 30-го калибра перед планируемым половым актом. Эрекции
наступают обычно спустя 15 мин после инъекции и длятся от 45 до 90 мин. Несмотря
на то что терапия интракавернозными инъекциями в высокой степени эффективна,
она сопровождается значительно более высокими уровнями осложнений по сравне¬
нию с оральной терапией и должна применяться только специалистом, имеющим
опыт в использовании этой терапии, и теми, кто может обеспечить неотложную меди¬
цинскую помощь своим пациентам в случае серьезных побочных эффектов, таких как
приапизм. Рекомендации перечислены в табл. 4-9.
Несмотря на то что различные препараты (ПГЕ^, папаверин, фентоламин) приме¬
няли поодиночке или в комбинации [155,159-165], только внутрипещеристый ПГЕ^
был одобрен для клинического использования. Долгосрочные данные относительно
эффективности интракавернозной терапии различаются.
Несколько препаратов алпростадила*“ (ПГЕ,) коммерчески доступны (Caverject,
Pharmacia; Prostin VR, Pharmacia; Edex, Schwarz Pharma). ПГЕ^ связывается с рецепто¬
рами ПГЕ^ на пещеристых гладкомышечных клетках, стимулирует аденилатциклазу,
повышает концентрации цАМФ и служит мощным релаксантом гладкой мускулатуры.
Обычная доза составляет от 5 до 20 мг, реакция на терапию дозозависима и должна
титроваться [162].
В ходе одного из плацебо-контролируемых исследований интракавернозная инъек¬
ция алпростадила*’ привела к удовлетворительному исполнению полового акта в 90%
введений, и в среднем 85% мужчин и их партнерш сообщают об удовлетворительной
сексуальной активности [162]. Интракавернозный алпростадил*“ обладает большей
эффективностью по сравнению с внутриуретральным алпростадилом*“ [166].
358
Таблица 4-9. Рекомендации по интракавернозной терапии
• Не назначайте интракавернозную терапию мужчинам, которые страдают психическими расстройствами, гипер-
коагуляционными состояниями или серпоаиднокпеточной анемией.
• Не выписывайте интракавернозную терапию мужчинам, которые получают антикоагуляктную терапию или не
способны осмыслить риски или принять адекватные действия в случае развития осложнений.
• Назначьте терапевта или уролога, который будет доступен для коррекции неотложных состояний, связанных с
осложнениями интракавернозных инъекций, таких как пролонгированная эрекция и приапизм.
• Проинструктируйте пациента о технике инъекции, рисках интракавернозной терапии и шагах, которые должны
быть осуществлены в случае пролонгированной эрекции или приапизма.
• Введите первую инъекцию в своем кабинете и понаблюдайте за изменениями артериального давления и сер¬
дечного ритма. Это обш1йчивает великолепную возможность обучения пагщента, наблюдения за побочными
реакциями и определения, будет ли у пациента реакция на внутрипещеристую терапию.
• Начните с низких доз алпростадила^ и титруйте дозу на основе эректильной реакции и продолжительности
эрекции.
• Корригируйте дозу алпростадила»’ для достижения эрекции, которая достаточна для полового акта, одаако не
длится более 30 мин.
• Если эрекция не ослабляется ч^зез 30 мин. пациент должен быть проинструктирован в отношении приема
таблетки псевдозфедрина или бретина*’ либо интракавернозной инъекции (^нилэфрика. Если это оказывает¬
ся неэффективным, пациент должен вызвать назначенного терапевта или уролога и обратиться в отделение
неотложной помощи.
Распространенные побочные эффекты интракавернозной терапии включают боль
в половом органе, гематому, узлы в пещеристых телах, фиброз полового члена и
вероятность пролонгированных эрекций (приапизм, если дольше 4 ч) [162, 163,
165, 167]. В процессе открытого 6-месячного исследования мужчин с ЭД, которые
получали терапию внутрипещеристым алпростадилом^’ [168], боль в половом органе
встречалась в 44%, пролонгированная эрекция — в 8%, приапизм — в 0,9% и фиброз
полового члена — в 4% случаев. Несмотря на эффективность этого подхода в отноше¬
нии ригидных эрекций, многих пациентов не привлекает введение иглы в собствен¬
ный половой член; соответственно не вызывает удивления высокий уровень отказа в
долгосрочном периоде.
Папаверин, получаемый изначально из семян мака, — неспецифический ингибитор
фосфодиэстеразы, которая увеличивает внутриклеточные уровни цАМФ и цГМФ.
Он обладает выраженной способностью провоцировать приапизм и фиброз при дли¬
тельном применении, а данные об эффективности и долгосрочной безопасности на
основании рандомизированных, плацебо-контролируемых испытаний недостаточ¬
ны. Таким образом, имеется недостаточное количество информации для оценки его
эффективности и безопасности.
Фентоламин - конкурирующий а^- и аз-адренергический антагонист, который
участвует в релаксации гладкой мускулатуры. В качестве монотерапии он облада¬
ет минимальной эффективностью, однако применялся в комбинации с другими
средствами для потенцирования эффектов [169], при этом данных рандомизиро¬
ванных клинических испытаний об эффективности и безопасности недостаточно.
Следовательно, имеется неудовлетворительное количество информации для оценки
эффективности и безопасности.
Основная проблема интракавернозных инъекций — приапизм. Пациентам, у кото¬
рых развивается продолжительная или болезненная эрекция под влиянием ПГЕ^,
могут помочь бретин^^ в дозе 5 мг и псевдоэфедрин^^ в дозе 60 мг. Если приапизм
продолжается дольше 4 ч, пациент должен обратиться за медицинской помощью; для
прекращения эрекции будет проведена аспирация сама по себе либо в комбинации
с инъекцией а-адренергического препарата. Если это закончилось неудачей, может
потребоваться хирургическое лечение для прекращения пролонгированной эрекции;
в противном случае могут произойти аксональное повреждение пещеристых гладко¬
мышечных клеток и фиброз.
359
Методы терапии третьей линии
Протезирование полового члена
Протезирование полового члена — инвазивный и дорогостоящий метод, который
может быть эффективен для восстановления эректильной функции у пациентов с
далеко зашедшими органическими заболеваниями (нет реакции на другие методы
терапии, структурные нарушения полового члена, значительная потеря объема пеще¬
ристых тел вследствие рака или травматического повреждения) [170,171].
Имплантаты полового члена — парные поддерживающие приспособления, кото¬
рые устанавливаются в каждом из двух эректильных тел. Существует два основных
типа имплантатов полового члена: гидравлический, или заполняемый жидкостью,
действующий как надуваемые протезы; мягкий, полуригидный — стержень, который
поддается сгибанию, однако всегда остается твердым в половом члене [170, 171].
Выпускаются протезы полового члена разнообразных длин и объемов. Хирургическая
имплантация обычно занимает менее часа и в большинстве случаев может быть
выполнена как амбулаторная процедура под общей или региональной анестезией.
Инфицирование и механическое нарушение функционирования — самые распро¬
страненные проблемы, связанные с протезами полового члена. В связи с последними
усовершенствованиями материалов и дизайна вероятность механического нарушения
функционирования снизилась до 5-10% за первые 10 лет. Инфекции встречаются в
1-3% сл^шаях, однако уровни инфицирования могут быть выше при ревизионном
хирургическом вмешательстве, в особенности у мужчин с сахарным диабетом.
Общая стоимость имплантации протеза полового члена варьирует от $3000 до
$20 ООО в зависимости от типа используемого приспособления и клиники, в которой
выполняется процедура. Рандомизированные исследования, посвященные эффектив¬
ности, отсутствуют, однако в ретроспективных анализах сообщается, что более 80%
пациентов и 70% партнеров довольны протезами и тем объединяющим началом,
которое они привносят в их взаимоотношения [172,173].
. В последнем сообщении прогнозируется возрастание числа процедур установления
имплантатов полового члена, проводимых к 2010 г. в связи со старением популяции,
увеличением осведомленности о ЭД и возрастающим числом мужчин с тяжелыми
формами ЭД, которые будут устойчивы к оральной лекарственной терапии [172].
Заместительная терапия тестостероном
Терапия тестостероном не улучшает сексуальную функцию у мужчин, страдающих
импотенцией, у которых имеются нормальные уровни тестостерона [45, 47,174], Нет
сведений о том, улучшает ли заместительная терапия тестостероном сексуальную
функцию у мужчин, страдающих импотенцией, с пограничными сывороточными
уровнями тестостерона. Многие мужчины (однако не все), страдающие импотенци¬
ей, с низкими уровнями тестостерона ощущают улучшение своего либидо и общей
сексуальной активности при заместительной терапии андрогенами [47, 175]. Такая
реакция на дополнение тестостероном даже в этой группе у мужчин вариабельна [41,
50, 80, 81] в связи с сопутствующими другими заболеваниями, такими как сахарный
диабет, гипертензия, сердечно-сосудистые заболевания и психогенные факторы.
В метаанализе [47] эффективности заместительной терапии андрогенами делается
вывод о том, что введение тестостерона сопровождается более значительными улуч¬
шениями сексуальной функции по сравнению с плацебо у мужчин с ЭД и низкими
уровнями тестостерона.
ЭД у мужчин среднего и старшего возраста — часто мультифакторное расстройство.
Общие причины ЭД включают сахарный диабет, гипертензию, лекарственные препа¬
раты, заболевания периферических сосудов, психогенные факторы и терминальную
стадию заболевания почек. Многие из этих факторов часто сочетаются у одного и того
же пациента. Следовательно, неудивительно, что терапия одним тестостероном не
может улучшить сексуальную функцию у мужчин с андрогенной недостаточностью.
360
Тестостерон индуцирует деятельность СИНТЕЗЫ оксида азота, обладает трофическим
воздействием на пещеристые гладкие седалищно-пещеристые и луковично-губчатые
мышцы и, по всей вероятности, является необходимым фактором для достижения
оптимальной венозной окклюзии в моделях на животных [44,48,176J. Соответственно
существует вероятность того, что тестостерон может улучшать реакцию на ингибито¬
ры ФДЭ5; однако эта гипотеза не была тщательно исследована [48].
Методы терапии с недоказанной или ограниченной эффективностью
Существующие данные об эффективности тразодона [177] или йохимбина [178]
недостаточны для обоснования их использования у мужчин с ЭД. Опубликованные
данные об эффективности фитотерапии сложно интерпретировать в связи с отсутстви¬
ем стандартов производства препаратов, загрязнением фитопродуктов ингибиторами
ФДЭ5 и недостаточностью данных рандомизированных клинических испытаний [85,
179]. Анализ одного из этих испытаний — в отношении корейского красного женьше¬
ня — показал, что продукт эффективен для лечения ЭД [180]; эти данные нуждаются
в дополнительном подтверждении. Таким образом, использование этого или других
фитотерапевтических препаратов не рекомендуется [85].
Перспективы терапии
Пероральные терапевтические препараты, находящиеся в стадии разработки
а-Меланоцитстимулирующий гормон (a-MSH; мелонтан II), неселективный аго¬
нист рецепторов меланокортина, служит центральным инициатором эрекции. Он осу¬
ществляет допаминергическое агонистическое воздействие и положительно влияет
на либидо. В двойном слепом плацебо-контролируемом перекрестном исследовании
отмечали значительное улучшение событий RigiScan, ригидности полового о^ргана и
сексуального влечения у 10 пациентов с документированными факторами риска ЭД.
Тошнота — наиболее распространенный побочный эффект [181].
РТ-141 — другой синтетический аналог a-MSH, агонист рецепторов меланокор¬
тина, включая МКЗР и МК4Р [182, 183, 184]. Введение РТ-141 крысам и приматам,
кроме человека, приводило к эрекциям полового члена. Введение РТ-141 здоровым
мужчинам и пациентам с ЭД приводило к стремительному дозозависимому возрас¬
танию эректильной активности. Исследования эффективности РТ-141 находятся на
ранних стадиях.
Апоморфин, ингибитор ФДЭ5, также функционирует в качестве агониста допамина
и действует центрально для инициации эрекции; его основной побочный эффект —
тошнота.
Генная терапия и эректильная дисфункция
Цель генной терапии ЭД заключается во введении нового генетического материала
в пещеристые гладкомышечные клетки в попытке восстановления нормальной кле¬
точной и физиологической функции для осуществления терапевтического эффекта
[185]. Генная терапия включает введение требуемого гена в тело, доставку гена в
целевую клетку и экспрессию терапевтического продукта.
Генная терапия была предложена в качестве перспективного метода лечения забо¬
леваний, которые имеют сосудистое происхождение, таких как атеросклероз, застой¬
ная сердечная недостаточность и легочная гипертензия [185,186]. Предполагают, что
генная терапия может быть также применена для лечения сосудистых заболеваний
полового члена, поскольку ЭД в большинстве случаев — проявление сосудистого
заболевания.
Одно из преимуществ применения генной терапии для лечения ЭД — легкодо¬
ступное наружное расположение полового органа [187, 188]. Следовательно, вокруг
основания полового члена может быть наложен жгут и требуемый ген может быть
введен непофедственно в пещеристые тела без проникновения в системный кровоток.
Это особое преимущество по сравнению с другими методиками, при которых вектор,
361
кодирующий требуемый ген, вводят в системный кровоток. Системный подход может
спровоцировать системные побочные эффекты, включая введение гена в неверный
орган или сосудистое ложе. Однако потенциальные побочные эффекты минимизи¬
руются, когда векторы для генной терапии применяют для лечения ЭД, поскольку
половой член имеет свою собственную наружную систему циркуляции, позволяющую
перенести и локализовать ген в одном органе, тем самым снижая риск системного
переизбытка.
Кроме того, число клеток, которые могут быть трансфектированы для терапевти¬
ческого воздействия, часто бывает трудно вычислить. Однако в половом члене только
небольшое число клеток нуждается в том, чтобы быть трансфектированными, потому
что гладкомышечные клетки пещеристых тел взаимосвязаны посредством щелевид¬
ных соединений, которые переносят молекулы вторичных мессенджеров и ионов к
ряду взаимодействующих гладкомышечных клеток [188]. Кроме того, низкий уровень
кругооборота сосудистых гладкомышечных клеток в половом органе позволяет тре¬
буемому гену экспрессироваться на протяжении длительного времени.
Современные стратегии генной терапии для лечения ЭД сфокусированы на моле¬
кулах, которые регулируют релаксацию гладкой мускулатуры тел или увеличивают
неоваскуляризацию (табл. 4-10) [189, 190]. Было исследовано несколько генов-
кандидатов, в том числе ген индуцируемой половым членом синтезы оксида азота,
ген СИНТЕЗЫ эндотелиального оксида азота (eNOS), вазоактивный интестинальный
полипептид (VIP), кальцитонин-ген родственный пептид (CGRP), ген макси-К* кана¬
ла, VEGF и ген нейротрофического фактора головного мозга.
Таблица 4-10. Физиологические мишени для генной терапии
Вектор и меха>ц|зм
Изоформы оксида
азота
Увеличение активности еЛЮ5, пЫ08 и 1Ы05 в пещеристых гладких мышцах
Макси-К*-канал
Перенос макси-К*-каналов с использованием плазмидного вектора, который несет Ь51о/
кальций-акгавируемый канал-ген, кодирующий а-су6ъединицу макси*К*-канала
VEGF
Перенос цЩЧК ]/ЕвР в крысиные пещеристые тела для обеспечения неоваскуляризации
VEGF с AAV-BDNF
Перенос ИЕСР и нейротропного фактора, производимого головным мозгом, с использо¬
ванием аденосатеппитного вируса
Ген нейротрофина 3
Перенос гена нейротрофина 3 с использованием вектора ВПГ
VIP
Трансфекиия/трансформация пещеристых тел крыс с сахарным диабетом, получающих
лечение стрелтозотоцином с применением рс01^АЗ/реком6инантной плазмиды pcDNAЗ,
несущих цДНК 14Р
CGRP
Аденовирусный перенос СбЯР у )фыс старшего возраста
Примечание. CGRP — кальцитонин-ген родственный пептид; eNOS — синтаза эндотелиального
оксида азота; ВПГ — вирус простого герпеса; iNOS — индуцируемая синтаза оксида азота: nNOS —
нейрональная синтаза оксида азота; VEGF - сосудистый эндотелиальный фактор роста; VIP - вазо¬
активный интестинальный полипептид.
Воспроизведено с разрешения из Christ GJ., Hodges S. Molecular mechanisms of detrusor and corporal
myocyte contraction: identifying targets for pharmacotherapy of bladder and erectile dysfunction // Br. J.
Pharmacol. - 2006. - N 147. - Suppl 2. - S41-S55.
Группа Garban [191] впервые продемонстрировала, что генная терапия может быть
проведена в половом члене путем использования депротеинизированной ДНК, коди¬
рующей ген СИНТЕЗЫ индуцируемого половым членом оксида азота, приводя к физио¬
логическому улучшению у стареющих крыс. Christ с соавт. позднее показали, что инъ¬
362
екция hSlo цДНК, которая кодирует макси-К“-каналы человеческих гладких мышц, в
крысиные пещеристые тела может усилить формирование щелевидных соединений и
облегчить эректильные реакции на стимуляцию нервов у стареющих крыс [192].
Было продемонстрировано, что аденовирусные конструкции, кодирующие eNOS и
CGRP, возвращают вспять связанную со старением ЭД у крыс [193,194]. В ходе этих
исследований экспрессия обоих eNOS и CGRP поддерживается на протяжении, как
минимум, 1 мес в пещеристых телах крысиного полового члена. Через 5 дней после
трансфекции вирусами AdCMVeNOS или AdRSVeNOS у стареющих крыс отмечается
увеличение эректильной функции, которое определяется стимуляцией пещеристых
нервов и фармакологической инъекцией эндотелий-зависимого вазодилататора аце-
тилхолина и ингибиторов ФДЭ5 запринаста и силденафила [193,194].
В процессе одного из исследований внутривенная инъекция аденосателлитного
вируса, несущего ген производимого головным мозгом нейротрофического факто¬
ра, улучшила эректильную функцию после повреждения пещеристого нерва [195].
Этот нейротрофический фактор предположительно восстановил нейрональную NOS
в большом тазовом ганглии, таким образом облегчая восстановление эректильной
функции после билатерального повреждения пещеристого нерва [195].
Анализ других исследований свидетельствует о том, что внутрипещеристая инъек¬
ция сосудистого эндотелиального фактора роста и опосредованная аденосателлитным
вирусом терапия i'chom сосудистого эндотелиального фактора роста обращают вспять
вазогенную ЭД у крыс [196]. Дополнительные мишени для генной терапии включают
CGRP, супероксиддисмутазу и киназу Rho A/Rho [190]. Эти ранние, однако иннова¬
ционные исследования предоставляют экспериментально доказанные данные о том,
что перенос гена in vivo технически может быть выполнен и оказывает благотворные
физиологические эффекты на эрекцию полового члена. Была инициирована фаза I
клинического исследования терапии генами калиевых каналов у пациентов с ЭД.
Потенциал терапии стволовыми клетками
Человеческие мезенхимальные стволовые клетки (чМСК) — производные костного
мозга, которые обладают потенциалом к дифференцировке в скелетные и гладкомы¬
шечные клетки, равно как и адипоциты, остеоциты и хондроциты. чМСК — привлека¬
тельные переносчики генов для многих целей [190], Эти клетки могут реплицировать¬
ся in vitro, а также in vivo, обеспечивая обширный пул клеток. Аутологические чМСК
неиммуногенны. в конце концов, будучи транспортированными в соответствующую
тканевую среду, они обладают потенциалом к дифференцировке в требуемый клеточ¬
ный тип.
в ходе первоначальных исследований было продемонстрировано, что крысиные
МСК, культивированные ех vivo и трансформированные [190, 197, 198], а затем
имплантированные в пещеристые тела, способны к экспрессии интересующего генно¬
го продукта [197,198]. Неизвестно, могут ли трансплантированные чМСК дифферен¬
цироваться в функциональные пещеристые гладкомышечные клетки и восстанавли¬
вать эректильную способность.
Ъ%щтшщым расетройства
Эякуляторные расстройства включают анэякуляцию, аноргазмию, задержку эяку¬
ляции, ретроградную эякуляцию, преждевременную эякуляцию и болезненную эяку¬
ляцию [11, 12]. в ходе недавнего опроса на передний план были выдвинуты распро¬
страненность и клиническая важность эякуляторных проблем [11]. Преждевременная
эякуляция — наиболее распространенное сексуальное расстройство среди мужчин в
возрасте от 18 до 59 лет. Ретроградная эякуляция, связанная с диабет-ассоциированной
автономной нейропатией, — второе наиболее распространенное эякуляторное рас¬
стройство, Эякуляторные расстройства также могут приводить к бесплодию.
363
МЕХАНИЗМЫ ЭЯКУЛЯЦИИ
Эякуляторные механизмы имеют три стадии: эмиссию, извержение и оргазм [12,
199]. Несмотря на то что оргазм и извержение семенной жидкости часто происходят
одновременно, два этих процесса регулируются отдельными механизмами. Эякуляция
связана с извержением семенной жидкости, содержащей сперму и секреты из семен¬
ных пузырьков, предстательной железы и бульбоуретральных желез: она регулирует¬
ся преимущественно активацией под влиянием симпатической нервной системы [200,
201]. Эмиссия, депонирование семенной жидкости в заднем отделе мочеиспускатель¬
ного канала, зависит от целостности семявыносящих протоков, семенных пузырьков,
предстательной железы и шейки мочевого пузыря. Эта эмиссия эякулирует из моче¬
испускательного канала посредством сокращений луковично-пещеристой мышцы и
мышцы, поднимающей задний проход. Проникновение семенной жидкости в мочевой
пузырь предотвращается путем закрытия сфинктера мочевого пузыря, обусловленно¬
го симпатической активацией. Ощущение, сопровождающее ритмические сокращения
мышц тазового дна, называется оргазмом [200, 201].
Медиальное предзрительное поле и парагигантоклеточное ядро осуществляют
интеграцию эмиссии семенной жидкости и извержения с копулятивным поведением.
Парагигантоклеточное ядро пофедством серотонинэргических проводящих путей
ингибирует пояснично-крестцовые моторные ядра, которые принимают у^1астие
в эякуляции [202, 203]. Когда приток импульсов из медиального предзрительного
поля к парагигантоклеточному ядру приводит к исчезновению такого ингибирова¬
ния, это приводит к эякуляции [199, 203]. Нейрональные проводящие пути, которые
задействуют рецепторы серотонина и 5-ГТ, регулируют скорость эякуляции. Таким
образом, введение ингибиторов обратного захвата серотонина исследуется для лече¬
ния преждевременной эякуляции [204, 205]. Более глубокое понимание нейрохими¬
ческих механизмов, которые регулируют эякуляцию, может обеспечить механизм-
специфические мишени для коррекции эякуляторных расстройств.
Ретроградная эякуляция может быть результатом автономной дисфункции в
связи с нейропатией, обусловленной сахарным диабетом, симпатэктомией, терапией
адренергическими антагонистами, некоторыми типами антигипертензивных препа¬
ратов, антипсихотическими препаратами, или антидепрессантами, функциональной
недостаточностью шейки мочевого пузыря или обструкцией мочеиспускательного
канала.
Вследствие трансуретральной резекции предстательной железы механизм закрытия
шейки мочевого пузыря может быть нарушен. У пациентов сохраняется способность
регуляции экскреторных функций в связи с вторичным, более дистальным механиз¬
мом регуляции экскреторных функций, который имеется в зоне мембранозной части
мочеиспускательного канала; тем не менее многие пациенты, которые перенесли
трансуретральную резекцию предстательной железы, могут страдать ретроградной
эякуляцией.
Исследование сексуальной дисфункции у женщин часто требуется в эндокриноло¬
гической практике. Не только лишь эндокринные расстройства приводят к симпто¬
мам сексуальных нарушений, но также и их коррекция может негативно повлиять
на сексуальную функцию и удовлетворение. Более того, женщины часто обращаются
к эндокринологам для «исключения гормональной основы» своих сексуальных
проблем. Предположение основывается на том, что если эндокринное заболевание
исключено (и при отсутствии других медицинских нарушений), это психологические
проблемы.
364
Тем не менее психологические и биологические системы не отделены друг от друга.
Расстроенные ощущения, эмоции, настроение, а также внутренний и внешний стресс
глубоко изменяют иммунологические, неврологические и эндокринные системы.
Сексуальная функция и дисфункция — примеры обязательного соединения разума и
тела.
Женщина со стресс-индуцированной гипоталамической аменореей — пример того,
каким образом эмоциональный стресс изменяет психологию, которая, в свою очередь,
снижает сексуальную реакцию как в форме гормональных изменений, так и психоло¬
гической реакции на эти изменения. Отсутствие менструаций может сопровождаться
снижением ощущения собственной женственности и сексуальной привлекательности
и, таким образом, сексуального влечения: отсутствие пиков андрогенов в середине
цикла и перед менструацией может еще более снизить ее влечение и реактивность.
Оценка и лечение сексуальной дисфункции у женщин, следовательно, биопсихосоци-
альны.
Цшт ттутътш реакции жшщшны
у женщин и мужчин имеются различные фазы сексуальной реакции. У мужчин
первоначальное сексуальное влечение (сексуальные «желание, голод, влечение»)
часто предшествует осмысленному вниманию к сексуальным стимулам с после¬
дующим половым возбуждением. Женщины, особенно в установленных взаимо¬
отношениях, в основном соглашаются принять участие в сексуальных отношениях
или инициировать секс со своими партнерами по причинам, отличным от влечения
[206, 207]. Причины участия в сексуальных отношениях, выявленные посредством
количественного исследования, включают содействие эмоциональной близости с
партнером, реакцию на романтическую обстановку и в особенности сексуальные
намеки [208]. Эмпирические, количественные и клинические доказательные данные
делают очевидным тот факт, что сексуальное влечение с типичными проявлениями
в виде сексуальных фантазий, положительных предвкушаемых сексуальных опытов
и спонтанных мыслей о сексе в положительном свете широко распространено среди
женщин и что открытое влечение нечасто встречается у многих сексуально активных
и удовлетворенных женщин [207].
Мотивированная одной или несколькими причинами (некоторые из которых не
являются сексуальными) женщина произвольно проявляет внимание к сексуальным
стимулам, и в результате ее последующее субъективное возбуждение и наслаждение
потенцируют ощущения сексуального влечения [209, 210]. Влечение и половое воз¬
буждение затем сопутствуют и стимулируют друг друга, как показано на рис. 4-9.
При условии, что она может оставаться сфокусированной и продолжать испытывать
наслаждение и что продолжительность стимуляции достаточно длительная и отсут¬
ствует негативный исход (боль или дисфункция партнера), сексуальное удовлетво¬
рение с одним или несколькими оргазмами удовлетворяет ее вновь приобретенное
сексуальное влечение. Ее первоначальная цель или мотивация быть сексуальной
также будет достигнута. Реакция циклична с перекрывающимися фазами различной
последовательности. К примеру, влечение может следовать за возбуждением и высо¬
кое возбуждение может следовать за первым оргазмом. На рис. 4-9 показано, что
сексуальное влечение, единожды запущенное, увеличивает мотивацию к проявлению
внимания к сексуальным стимулам и к приему или требованию более интенсивных
эротических форм стимуляции. Любое первоначальное или спонтанное влечение
будет аналогичным образом потенцировать реакцию.
Модель демонстрирует важность как сексуальных стимулов, так и сексуального
контекста; она показывает, что реакция может начинаться с ощущений, которые не
365
Если возбуждение приятно
и стимулирование продол¬
жается, женщина остается
сфокусированной
(сосредоточенной)
Рис. 4-9. Цикл сексуальной реакции, отражающий многие причины, по которым происходит мотивация
женщин быть сексуальными, субъективное половое возбуждение и возбуждение в ответ на стимулы,
так чтобы (ответное) половое влечение могло быть запущено. Спонтанное или первоначальное половое
влечение может усиливать этот цикл.
обязательно являются сексуальными. В этой модели содержится концепция способ¬
ности формирования сексуального возбуждения, обозначающая легкость, с которой
женщина возбуждается под действием сексуальных стимулов. Эта концепция важна
при имеющихся доказательствах влияния на возбуждение многих факторов, в пер¬
вую очередь ощущение себя объектом влечения (а не используемой) и желанной для
партнера, положительное мнение о поведении партнера, а также обладание положи¬
тельным телесным образом и положительным настроением [208, 211], равно как и
биологические факторы, включающие активность тестостерона [7, 212-215].
В последнее время в концепции женской сексуальной реакции большое внимание
уделяют субъективному возбуждению, а не гиперемии гениталий самой по себе [207].
Традиционно женское половое возбуждение приравнивали к появлению влагалищ¬
ной смазки и набуханию наружных половых органов (как в В5М-1У-ТК). Однако
женщины рассматривают появление смазки в большей степени как побочное явление
[208]. Если внутривлагалищная стимуляция — часть взаимодействия пары, некоторое
количество смазки необходимо, однако ни она, ни набухание гениталий и лежащая в
основе гиперемия не имеют положительной корреляции с субъективным возбуждени¬
ем при эмпирическом исследовании. В отличие от эрекции полового члена у мужчин
женская гиперемия гениталий не приводит к прочному укреплению субъективного
возбуждения [216, 217].
Эти факты вызвали множество вопросов к В5М-1У-ТК. Определения В5М-1У
обычно используются для контроля вопросов при эпидемиологических исследова¬
ниях и методиках оценки в терапевтических исследованиях. Таким образом, уровни
распространенности расстройств женского полового влечения и полового возбужде¬
ния — предмет данного рассмотрения.
366
Определение женской сексуальной дисфункции
или расстройства
Пересмотры В5М-1У определений женских сексуальных расстройств были реко¬
мендованы в последнее время Международным консенсусным комитетом, органи¬
зованным фондом Американской урологической ассоциации, ранее известной как
Американский фонд урологических заболеваний [207, 218]. Последние диагностиче¬
ские объекты показаны на рис. 4-10 и 4-11 [10,11].
РАССТРОЙСТВО СЕКСУАЛЬНОГО ВЛЕЧЕНИЯ ИЛИ ИНТЕРЕСА
Расстройство полового влечения или интереса определяют как «отсутствующие
или сниженные ощущения сексуального влечения или интереса, отсутствие сексуаль-
Детальное сексуальное исследование
т
т
Персистирующий
недостаток полового
интереса, неспособность
чувствовать желание
8 любой момент
полового переживания?
Персистирующая <
неспособность 1
возбудиться? ;
Персистирующий
недостаток, |
задержка или ;
снижение интен- 1
сивности оргазма, f
несмотря на сильное \-
возбуждение?
Повторная боль
при попытке
полового акта
или его совер¬
шения?
Расстройство полового
интереса/желания
"“ЗД...; - и
Т
Трудности
с введением во
влагалище,
переменный страх,
избегание, сжатие
мускулатуры?
Расстройство поло-г
вого возбуждения 1
Расстройство
оргазма
1
Диспареуния
Вагинизм
Рис. 4-10. Пересмотренные определения женской сексуальной дисфункции.
Есть негенитальное половое возбуждение... |
фильмы, книги, стимуляция молочных желез, поцелуи?!
Нет
Генитальное
расстройство
полового
возбуждения
Существует осознание некоторой
степени набухания половых
органов, половых ощущений от
прикосновения к половым органам
или выделения влагалищной смазки,
несмотря на отсутствие возбуждения?
Да
\^ет
Субъективное
расстройство
полового возбуждения
Комбинированное
расстройство
полового возбуждения
■
Рис. 4-11. Диагноз «расстройство сексуального возбуждения». Расстройство субъективного полового
возбуждения характеризуется отсутствием или значительно сниженными ощущениями сексуального
возбуждения или сексуального наслаждения под действием любого типа сексуальной стимуляции.
Присутствует смазывание влагалища и/или другие признаки физиологической реакции. Расстройство
генитального сексуального возбуждения характеризуется отсутствием набухания наружных половых
органов, смазывания влагалища и генитального полового ощущения, несмотря на субъективные ощуще¬
ния сексуального возбуждения.
367
ных мыслей или фантазий и недостаточность ответного влечения. Мотивации (здесь
определяемые как причины/стимулы) для попытки становления сексуального воз¬
буждения недостаточны или отсутствуют. Отсутствие или недостаток интереса оцени¬
вается как находящийся ниже нормативного снижения в соответствии с жизненным
циклом и длительностью отношений» [218].
Такое пересмотренное определение отражает тот факт, что недостаточность любого
первоначального или спонтанного полового влечения находится в пределах нормаль¬
ного жизненного опыта. А дополнительное снижение любого инициируемого или
ответного полового влечения, которое составляет расстройство, обычно связано с
недостаточностью или отсутствием субъективного полового возбуждения и наслаж¬
дения при }'частии в сексуальных отношениях и реакции на другие эротические
стим}шы. Есть научно доказанные данные в пользу того, что первоначальное половое
влечение — реальность только для некоторых женщин, оно наиболее заметно с новым
партнером [219, 220].
КОМБИНИРОВАННЫЕ РАССТРОЙСТВА ПОЛОВОГО ВОЗБУЖДЕНИЯ
Диагностические подтипы расстройств полового возбуждения зависят от женского
восприятия изменений в гениталиях и распознавания субъективного возбуждения
или волнения (см. рис. 4-11), Когда ни один из этих факторов не ощущается в ответ на
любой тип сексуальной стимуляции, ставят диагноз «комбинированное расстройство
полового возбуждения» [218]. Это расстройство определяется как «отсутствующие
или значительно сниженные ощущения сексуального возбуждения (сексуальное вол¬
нение или сексуальное наслаждение) в результате любого типа сексуальной стиму¬
ляции, а также жалобы на отсутствующее или нарушенное генитальное сексуальное
возбуждение, набухание внешних половых органов, появление смазки» [218].
РАССТРОЙСТВО СУБЪЕКТИВНОГО ПОЛОВОГО ВОЗБУЖДЕНИЯ
Женщины, которые отрицают субъективное возбуждение и волнение, однако
признают некоторое рефлекторное появление смазки или набухание гениталий,
или и то, и другое; это расценивают как расстройство субъективного полового воз¬
буждения (см. рис. 4-11). Это расстройство определяется как «отсутствующие или
значительно сниженные ощущения от сексуального возбуждения (сексуальное
волнение или сексуальное наслаждение) при любом типе сексуальной стимуляции.
Появление смазки влагалища или других признаков физиологической реакции все
еще происходит» [218].
Доказательства заключаются в том, что по 05М-1У-ТЕ у женщин с диагнозом
расстройства полового возбуждения («недостаточность или отсутствие реакции
появления смазки/набухания») рефлекторная генитальная реакция заполнения
сосудов происходит, даже если женщина не сообщает о том, что она находит эроти¬
ческий стимул субъективно возбуждающим и не отдает себе отчета о генитальных
изменениях [216]. Эти исследования были проведены с использованием приспо¬
собления, похожего на тампон, называемого вагинальным фотоплетгїзмографом,
который записывает увеличение притока крови к влагалищу в то время, когда жен¬
щина регистрирует свое сексуальное возбуждение или волнение. Несмотря на то,
что мужчины при просмотре сексуальных сцен спаривания животных отмечают, что
находят этот фильм сексуальным, но не возбуждающим, и не проявляют никакой
эректильной реакции, женщины, просматривающие тот же самый фильм, аналогич¬
но отмечают, что этот фильм является сексуальным, но не возбуждающим, но при
этом мониторинг их вагинальной реакции демонстрирует быструю рефлекторную
вазоконстрикцию [221]. Аналогичное увеличение клиторального артериального
кровотока у здоровых женщин в пременопаузе сопровождало как эротические, так
и гуморальные стимулы.
368
РАССТРОЙСТВО ГЕНИТАЛЬНОГО СЕКСУАЛЬНОГО ВОЗБУЖДЕНИЯ
Клиническая подгруппа женщин обычно среднего возраста или старше сообщают
о «безжизненности половых органов», даже если негенитальные стимулы, такие
как стимуляция молочных желез, поцелуи, стимуляция своего партнера или чте¬
ние эротических текстов, все еще возбуждающие для них (см. рис. 4-11). Этот тип
дисфункции называется расстройством генитального сексуального возбуждения и
обозначается как «жалобы на отсутствующее или ослабленное генитальное сексу¬
альное возбуждение. Самоотчет должен включать данные о минимальном набу¬
хании наружных половых органов или смазки влагалища вследствие любого типа
сексуальной стимуляции и сниженных сексуальных ощущений от ласк гениталий.
Субъективное сексуальное возбуждение от негенитальных сексуальных стимулов
все еще присутствует» [218].
В очередной раз уделяется особое внимание тому, что это клинический диагноз и
небольшое количество доказательных данных предполагает, что некоторые из этих
женщин демонстрируют сниженный ответ на негенитальные сексуальные возрастаю¬
щие стимулы. Получается, что женщины как бы утратили чувствительность напол¬
ненных кровью тканей.
ЖЕНСКОЕ ОРГАЗМИЧЕСКОЕ РАССТРОЙСТВО
Пациентка страдает женским оргазмическим расстройством, если, «несмотря на
отмечаемое в самоотчете высокое сексуальное возбуждение/волнение, имеется как
отсутствие оргазма, значительно сниженная интенсивность оргазмических ощу¬
щений, так и значительная отсрочка оргазма при любом типе стимуляции» [218].
Женщины с расстройствами возбуждения также редко или никогда не испытывают
оргазм. В пересмотренном определении устанавливается, что это расстройство воз¬
никает, только когда имеются сложности с любым из типов стимуляции. Женщина,
которая может испытывать оргазмы посредством самостимуляции, не страдает оргаз¬
мическим расстройством.
ВАГИНИЗМ
Вагинизм определяют как «персистирующие или повторяющиеся у женщины
сложности проникновения во влагалище полового члена, пальца или любого другого
объекта, несмотря на выражаемое женщиной желание осуществить это». Часто име¬
ется (фобическое) избегание и предчувствие/страх/опыт боли, сопровождающееся
разнообразными и непроизвольными сокращениями тазовых мышц. Должны быть
исключены/исследованы структурные или другие физические аномалии» [218].
Подтверждение этого диагноза невозможно до тех пор, пока нет терапии, достаточной
для того, чтобы позволить тщательное проникающее и вагинальное исследование. До
этого диагноз предположителен.
ДИСПАРЕУНИЯ
Диспареуния определяется как «персистирующая или повторяющаяся боль при
попытке или завершенном влагалищном проникновении и/или при половом сноше¬
нии посредством полового члена и влагалища» [218]. Существует множество причин,
включающих — наиболее часто — синдром вестибулита наружных половых органов
(недостаточно изученный синдром неизвестной этиологии, характеризующийся
диспареунией, болезненностью, локализованной внутри преддверия вульвы, пред-
дверной эритемой и дискомфортом на входе во влагалище) и атрофию влагалища и
вульвы. Часто наблюдают напряжение возвратных мышц вследствие перенесенных
эпизодов боли, однако диагноз «вагинизм» не должен устанавливаться, если имеются
другие симптомы, кроме напряжения мышц.
369
ДОПОЛНИТЕЛЬНЫЕ ФАКТОРЫ
Женская сексуальность в значительной степени зависит от сопутствующих факто¬
ров. Недостаточная стимуляция, отсутствие привлекательности партнера, отсутствие
безопасности могут приводить к симптомам сексуальной дисфункции, даже если нет
никаких внутренних нарушений в женской системе сексуальной реакции [223].
Кроме того, существуют дополнительные факторы в анамнезе, которые обу¬
словливают сексуальные реакции. Международный комитет по консенсусу (АНАР)
настоятельно пропагандирует добавление следующих описаний в любую диагностику
дисфункции:
^ факторы анамнеза, затрагивающие психосоциальное развитие;
» истинные факторы — межличностные, факторы окружающей среды, социальные,
культурные;
ф медицинские факторы.
Фышйття жвшшт вшщшъшт щшщш
ФИЗИОЛОГИЯ полового ВЛЕЧЕНИЯ и СУБЪЕКТИВНОГО ВОЗБУЖДЕНИЯ
Сексуальное влечение — одна из мотиваций быть сексуальной. Ощущения вле¬
чения могут быть потенцированы внутренними сигналами, в частности фантазии
или воспоминания, и внешними сигналами, такими как привлекательный партнер,
и зависят от адекватной нейроэндокринной функции. Многочисленные нейротранс¬
миттеры, пептиды и гормоны регулируют половое влечение и субъективное возбуж¬
дение. Следующие факторы, берущие свое начало преимущественно из положитель¬
ных и отрицательных сексуальных воздействий фармакологических агентов путем
известных или частично известных механизмов воздействия, очевидно, являются
просексуальньши: норадреналин, допамин, окситоцин и серотонин, действующие
на некоторые рецепторы. Пролактин, серотонин, воздействующие на З-ГТ.^ и 5-ГТ^
рецепторы, а также ГАМК, вазопрессин и глутамат ингибируют сексуальную реак¬
цию. Эти нейротрансмиттеры и пептиды сами по себе регулируются сексуальными
гормонами.
Биологические факторы не действуют независимо от факторов окружающей среды.
Даже в моделях на животных было продемонстрировано, что как допамин, так и про¬
гестерон, воздействующие на рецепторы в гипоталамусе, могут вызывать усиление
сексуального поведения у эстрогенизированных крыс женского пола после овариэк-
томии. Однако особь мужского пола неподалеку от клетки может вызывать идентич¬
ные изменения сексуального поведения без введения как прогестерона, так и допами¬
на [224]. Аналогично у женщин возбудимость и интенсивность реакции может быть
усилена под влиянием повышенных уровней сывороточного тестостерона [212-215,
225] путем введения допаминергического препарата (бупропиона) [226] или путем
смены партнера [220]. Даже у грызунов существуют сложные сети, посредством кото¬
рых особь женского пола оценивает контекст потенциальной сексуальной активности
и соотносит его с прошлым опытом и, следовательно, ожиданием награды. У женщин
такие факторы, как отношение к сексу, ощущения от партнера, прошлые сексуальные
опыты, продолжительность взаимоотношений, а также ментальное и эмоциональное
здоровье в большей степени регулируют половое влечение и возбудимость, чем био¬
логические факторы [223, 227-230].
Эти данные получены из исследований, проведенных на репрезентативных обще¬
национальных выборках женщин. Тем не менее хронические заболевания сами по
себе, их лечение, психологическое воздействие и упомянутые межличностные, лич¬
ностные и контекстуальные проблемы влияют на сексуальную реакцию [231].
370
ФИЗИЧЕСКОЕ СЕКСУАЛЬНОЕ ВОЗБУЖДЕНИЕ
Ряд физических изменений сопровождает субъективное физиологическое или воз¬
буждение. к ним относят набухание гениталий, увеличение количества влагалищной
смазки, прилив крови к молочным железам, эрекцию сосков и увеличение кожной
чувствительности к сексуальной стимуляции; изменения сердечного ритма, артери¬
ального давления, мышечного тон)^са, дыхания и температуры; а также мозаичную
окраску кожи, сексуальный прилив вазодилатации на груди и лице. Эти изменения
происходят от рефлекторного воздействия автономной нервной системы, в течение
нескольких секунд усиливается приток крови к влагалищу: вазодилатация артериол в
подслизистом сплетении увеличивает транссудацию интерстициальной жидкости из
капилляров, через эпителий и в просвет влагалища. Одновременно идет релаксация
гладкомышечных клеток вокруг синусоид ветвей, тела и головки клитора, равно как
и в отростках клиторальной ткани, известной как луковицы преддверия. Влагалище
увеличивается и расширяется, приподнимая матку. Губы становятся более набухши¬
ми, темно-красного цвета, нижняя треть влагалища набухает. По мере того как клитор
все больше набухает, он поднимается и располагается возле лобкового симфиза.
Когда накопление крови в ответ на визуальные эротические стимулы фиксируется
при помощи СВВ, отмечается высоковариабельная корреляция с субъективным воз¬
буждением. Это истинно как для сексуально здоровых, так и для женщин, жалую¬
щихся на отсутствие или недостаток полового влечения, сексуального возбужде¬
ния, а также на боль во время полового акта [216, 232]. У женщин с хроническим
отсутствием или недостаточностью отмечается стремительное накопление крови во
влагалище, сравнимое с изменениями у контрольной группы женщин. Тем не менее
они сообщают об отсутствии субъективного сексуального возбуждения в ответ на
эротическую стимуляцию. Некоторые из этих женщин говорят о негативных эмо¬
циях, таких как тревожность, смущение или грусть. Фактически эти последние нега¬
тивные эмоции коррелируют с увеличением кровенаполнения. Функциональная
МРТ демонстрирует активацию ряда зон головного мозга во время визуальной
эротической стимуляции, в отличие от этих экспериментов на мужчинах, активация
зон, организующих накопление крови в генитальных сосудах, не коррелирует с жен¬
ским субъективным возбуждением, когда она просматривает эротический фильм,
находясь в аппарате МРТ [233].
Лежащая в основе нейробиология генитальной вазоконстриктивной реакции недо¬
статочно понятна. Она, очевидно, в высокой степени автоматизирована и включает
выброс оксида азота (N0) из парасимпатических нервов совместно с вазоактивным
интестинальным пептидом (ВИП). Ацетилхолин, который блокирует норадренер¬
гические вазоконстрикторные механизмы и обеспечивает выброс N0 из эндотелия,
также выделяется. Соматические, симпатические и парасимпатические нервные
проводящие пути гораздо менее разделены, чем предполагалось ранее. Существует
взаимосвязь ЫО-содержащего пещеристого нерва, идущего к клитору, с дистальным
отделом соматического дорзального нерва клитора из полового нерва. Тазовые симпа¬
тические нервы выделяют преимущественно вазоконстрикторы норадреналин, адре¬
налин и АТФ, однако некоторые выделяют ацетилхолин, N0 и ВИП. В соответствии
с этими данными было продемонстрировано, что спровоцированная тревожность в
лабораторных условиях может у сексуально здоровых женщин увеличить реакцию
накопления крови в сосудах гениталий в ответ на эротическую стимуляцию [234]. У
женщин проводилось намного меньше исследований, посвященных нейротрансмис¬
сии скопления крови в гениталиях, чем у мужчин, даже если предполагается, что N0
является основным нейротрансмиттером, регулирующим кровенаполнение вульвы.
Другие факторы, регулирующие кровенаполнение влагалища, включают ВИП, N0 и
другие, вероятно, более важные невыявленные нейротрансмиттеры [235].
371
Это может быть «моторный рефлекс шейки», который заключается в том, что при¬
косновение к шейке снижает давление в верхнем отделе влагалища, увеличивая его в
среднем и нижнем отделах, которое сопровождается увеличенной электромиографиче-
ской активностью мышцы, поднимающей задний проход, и лобково-прямокишечной
мышцы. Было высказано предположение, что во время полового акта толчки половым
членом в шейку могут вызывать рефлекторное сокращение мышц таза, таким образом
облегчая раздувание верхнего отдела влагалища, в то время как это же мышечное
сокращение приводит к сокращению нижнего отдела влагалища. Сниженный тонус
влагалища может возникать в ответ на механическую или электрическую стимуляцию
клитора. Стимуляция клитора снимает базовое сокращение мышц матки, так что дав¬
ление в матке снижается. Возможно, этот рефлекс вносит вклад в известное увеличе¬
ние в размере и элевацию матки при сексуальном возбуждении [236].
Клитор — наиболее сексуально чувствительная зона тела. Стимуляция клитора
обычно доставляет только наслаждение, если произошла нефизическая и неге¬
нитальная физическая стимуляция. Без предварительного возбуждения прямая
стимуляция может быть неприятной, слишком интенсивной и даже болезненной.
Было проведено небольшое количество исследований, посвященных сексуальному
восприятию, однако последние иммуногистохимические исследования выявили
нейротрансмиттеры, которые, как предполагают, связаны с ощущениями (субстан¬
ция Р и КГРП), которые концентрируются непосредственно под эпителием головки
клитора. Недавние МРТ-исследования подтвердили наличие протяженной клито-
ралыюй ткани за пределами видимой части, когда головка клитора втянута [237].
Клитор состоит из головки, тела, ветвей, которые распространяются вдоль лобковой
дуги, периуретральной ткани спереди от передней стенки влагалища, а также буль¬
барной ткани, окружающей передний дистальный отдел влагалища и прилегающей
к периуретральной ткани [237].
Проводили минимум исследований по части физиологии, лежащей в основе неге¬
нитальных физиологических изменений, и их корреляции с субъективным возбужде¬
нием.
Большинство эпидемиологических исследований основано на самостоятельных
жалобах пациентов, в некоторых используются утвержденные опросники, то есть диа¬
гностика дисфункции и расстройств не основана на всестороннем опросе пациентов.
Ни один из утвержденных диагностических опросников не был до настоящего време¬
ни модифицирован с целью отражения наиболее поздних определений женской сексу¬
альной дисфункции, смоделированной на женских, а не мужских циклах сексуальной
реакции [218, 238]. Доступные опросники все еще отражают установки о том, чч'о
первоначальное половое влечение необходимо для нормальной реакции, и внимание
сфокусировано на генитальных событиях в большей степени, чем на субъективном
опыте [238]. Исследование женщин моложе 44 лет в Британии выявило, что хотя 41%
женщин сообщают о сниженном сексуальном интересе на протяжении, как минимум,
месяца за последний год, только 10% говорят о снижении интереса, длящегося более
6 мес [241].
Распространенность расстройства, связанного с низким половым влечением, у
женщин неизвестна. От 30 до 35% женщин, сообщающих о сложностях или пробле¬
мах с низким половым влечением [7,219,239], могут в своем большинстве отражать
тех женщин, которые, сравнивая себя со стандартом, осознают, что имеют проблемы
с половым влечением. Стандарт может быть завышен за счет полового влечения
более ранних взаимоотношений или может быть стандартом, более типичным для
372
мужской сексуальности, представляемой в кино и книгах как норма для женщин.
Понимание того, что спонтанное или первоначальное половое влечение часто
встречается лишь у некоторых женщин и наиболее отчетливо при новом партнере
[219, 220, 228], уменьшит число женщин, воспринимающих собственный опыт как
патологический.
Диаграммы распространенности низкого субъективного возбуждения также недо¬
статочны; внимание эпидемиологии обычно было приковано только к одному
компоненту возбуждения, а именно к увлажнению влагалища. В исследовании 979
женщин в национальной репрезентативной Британской когорте в возрасте 18-70 лет
было установлено, что в 17% случаев проблемы с возбуждением связаны с вагиналь¬
ной сухостью [240], тогда как в исследовании SWAN эта цифра составила 5% [219],
Сложности образования смазки имеют распространенность от 10 до 30% у женщин
в национальных репрезентативных выборках [6, 239]. Эти диаграммы имеют огра¬
ниченную пользу, поскольку неизвестно, сопровождается ли формирование слизи
снижением или отсутствием субъективного возбуждения.
Распространенность оргазмического расстройства аналогично неоднозначна,
поскольку анализ некоторых исследований свидетельствует о том, что женщины с
диагностированными оргазмическими расстройствами имеют сопутствующее рас¬
стройство возбуждения. Как AUAF-, так и DSM-IV-TR-определения устанавливают,
что диагноз расстройства возбуждения препятствует установлению диагноза оргаз¬
мического расстройства. Диапазон составляет от 3,7 до 24% [6, 24]. Более низкая
распространенность была в британском исследовании, в котором 3,7% были не
способны испытать оргазм за период более 6 мес, а 14,4% сообщали о сложностях,
как минимум, на протяжении 1 мес в предыдущем году. Распространенность хрони¬
ческой диспареунии варьирует от 8 до 22%, этот показатель снижается с возрастом в
большинстве исследований. Высокая частота СВВ у женщин молодого возраста, оче¬
видно, находится в равновесии с увеличенной распространенностью диспареунии
вследствие вульвовагинальной атрофии у женщин старшего возраста. Вестибулит,
как предполагается, поражает приблизительно 9% женщин, при этом около 5% всех
женщин испытывают это состояние до 25 лет [242]. Диаграммы распространен¬
ности вагинизма еще менее заслуживают доверия. Когда это состояние полностью
препятствует совершению полового акта, оно может быть выявлено, только если
планируется зачатие.
Сочетание дисфркции с друпши заболеваниями
в связи с перекрытием фаз сексуальной реакции неудивительно, что выявлены
высокие уровни перекрытия среди различных типов сексуальной дисфункции у жен¬
щин (рис. 4-12) [243,244]. Становится все более очевидным, что вместо дискретности
расстройства полового влечения или интереса у женщин более распространено посте¬
пенное притупление сексуальной реакции, так что женщины, предъявляющие жалобы
на отсутствие полового влечения, обычно имеют низкую вероятность интенсивных
ощущений возбуждения и нечастые оргазмы [243]. Последние рандомизированные
контролируемые исследования (РКИ), в ходе которых изунали преимущества добавок
трансдермального тестостерона, хотя в них принимали участие женщины с диагно¬
стированным «расстройством, связанным с гипоактивным сексуальным влечением»,
показали улучшение возбуждения, наслаждения и оргазма. В действительности пер¬
вое из таких исследований продемонстрировало лишь улучшение этих параметров, но
не параметра «полового влечения» [212]. Проблематичная реакция была отчетливо
установлена, несмотря на то что исследование было сфокусировано на расстройстве,
связанном с гипоактивным сексуальным влечением.
373
Рис. 4-12. Перекрест и сложность женских сексуальных дисфункций, у женщин с сексуальной дис¬
функцией может встречаться более одного сексуального расстройства. Также нарушение более чем
одной сексуальной функции может затронуть другие аспекты вторично, приводя к сочетанию нескольких
заболеваний.
Не все женщины с сексуальными проблемами переживают по поводу них. В ходе
многих обширных исследований не удалось продемонстрировать увеличения распро¬
страненности проблем, связанных с половым влечением, в зависимости от возраста
[229, 244]; анализ менее крупных исследований подтвердил, что сексуальное влече¬
ние действительно снижается с возрастом, однако переживание по поводу низкого
полового влечения также уменьшается [245]. Большинство женщин с диспареунией
сообщают об умеренном или значительном страдании [246]. Наиболее поздний кон¬
сенсус по поводу пересмотренных определений дисфункции при постановке диагноза
рекомендует придерживаться оценок степеней страдания: отсутствие, легкая, умерен¬
ная, тяжелая.
Как физиологические факторы, так и медицинские состояния влияют на биологию
сексуальной реакции.
МЕДИЦИНСКИЕ СОСТОЯНИЯ
Было продемонстрировано, что депрессия — основной фактор женской сексуальной
дисфункции. Это истинно даже перед началом приема антидепрессантов с потенциаль¬
ными негативными побочными воздействиями на сексуальную функцию [247]. Из 79
женщин с тяжелой депрессией перед приемом лекарственных препаратов 50% сообщали
о снижении полового влечения и аналогичное количество — о значительном снижении
сексуального возбуждения при занятии сексом [247]. Из 914 женщин среднего возраста
в исследовании SWAN те, у которых в анамнезе было депрессивное заболевание, гово¬
рили о меньшей частоте возбуждения, меньшем физическом наслаждении и меньшем
эмоциональном возбуждении в их настоящих взаимоотношениях [219]. Эти результаты
374
сохраняют свое значение после контроля текущих депрессивных симптомов, семейного
положения и назначения психотропной терапии [248].
Антидепрессанты, в особенности серотонинергические, могут снижать половое
влечение и возбуждение и вызывать задержку оргазма. Когда женщину спрашивают
относительно сексуальных побочных эффектов антидепрессантов, о наличии таковых
сообщается вплоть до 70% случаев [249]. Факторы, предрасполагающие к развитию
антидепрессант-ассоциированной сексуальной дисфункции, включают возраст, заму¬
жество в противоположность одиночеству или разведенному состоянию, отсутствие
среднего или высшего образования, отсутствие работы на полный рабочий день, полу¬
чение меньшего количества сексуального наслаждения в период до наступления депрес¬
сии, сопутствующий прием лекарственных препаратов любых типов, страдание сопут¬
ствующим заболеванием, которое может затрагивать сексуальную активность, наличие
в анамнезе антидепрессант-ассоциированного сексуального расстройства [250],
СНИЖЕННАЯ АНДРОГЕННАЯ АКТИВНОСТЬ
Несмотря на то что, согласно предположениям, у молодых женщин яичники про¬
изводят приблизительно 50% андрогенов, у женщин среднего и старшего возраста
ситуация куда более вариабельна. Когда яичники сохраняются, их вклад в общую
андрогенную активность в значительной степени варьирует. Предполагают, что вну¬
триклеточное производство тестостерона в головном мозге и в других частях тела,
включая гениталии, из надпочечниковых прогормонов (в том числе ДГЭА, ДГЭАС,
андро-5-ене-Зр, 17р-диол и Д"*-андростендион) ответственно за основную часть
активности тестостерона у женщин старшего возраста в физиологической менопаузе,
и это значение приближается к 100% у женщин с хирургической менопаузой [251].
Начиная с третьей декады и далее продукция надпочечниковых прогормонов про¬
грессивно снижается, так что в группе лиц в возрасте от 50 до 60 лет плазменный
уровень ДГЭА уже снижен вплоть до 70% по сравнению с пиковыми значениями у
20-30-летних женщин [251].
Клеточную депривацию андрогенов сложно измерить, при условии, что менее
10% тестостерона, производимого внутри клеток (за исключением гонадных и над¬
почечниковых клеток), возвращается в кровоток [252]. Таким образом, плазменные
уровни тестостерона отражают в основном гонадную продукцию. Кроме того, доступ¬
ные методы анализа плазменных уровней тестостерона (свободный, биодоступный,
общий) не разработаны для женских параметров, и они очень недостоверны [45,253].
Методы жидкостной хроматографии и тандемной масс-спектрометрии для изме¬
рения уровней тестостерона должны в скором времени стать широко доступными.
Метаболиты, образующиеся в результате распада тестостерона, где бы он ни был про¬
изведен, — андростерона глюкуронид (АДТ-Г), андростан-За, 17[5-диол глюкуронид
(За-диол-Г) и андростан Зр, 17р-диол глюкуронид (Зр-диол-Г) — могут быть измере¬
ны [252]. Тем не менее методики оценки этих метаболитов в настоящее время доступ¬
ны только лишь на исследовательском уровне, и пока еще не установлены связанные
с возрастом значения у женщин с сексуальными дисфункциями.
Известно, что внезапное прекращение продукции андрогенов может приводить
к сексуальному синдрому, при котором все ранее эффективные сексуальные стиму¬
лы — ментальный, визуальный, прикосновения к частям тела, отличным от гени¬
талий, прикосновения к гениталиям, половой акт — не вызывают., возбуждения.
Аналогично любое предыдущее врожденное или спонтанное половое влечение теря¬
ется. Индуцированная химиотерапией или хирургическая менопауза, в особенности
у молодых женщин, может быть связана с этим синдромом [254]. Однако другие
женщины с индуцированной менопаузой сообщают об отсутствии сексуальных изме¬
нений. В недавнем проспективном нерандомизированном испытании было продемон¬
стрировано, что 106 женщин в перименопаузе, которым требовалась гистерэктомия
375
по причине доброкачественной патологии и которые выбирали гистерэктомию с
билатеральной сальпинго-оофоректомией вместо одной гистерэктомии, отмечали
отсутствие ухудшения всех сексуальных функций через год после хирургического вме¬
шательства по сравнению с предоперационной оценкой [255]. Различные психологи¬
ческие контексты нежелательного или избыточного медицинского вмешательства по
сравнению с информированным выбором могут быть критическими.
Имеется ограниченное количество доказательных данных по корреляции сексу¬
альной функции с плазменными уровнями андрогенов у женщин. В изучении SWAN
у 2900 женщин, находящихся в пре- и пери менопаузе, проживающих в Северной
Америке, смотрели концентрации общего тестостерона и индекса свободных андро¬
генов. Была показана минимальная корреляция уровня гормона с сексуальной
функцией [256]. Аналогично измерения свободного и общего тестостерона не имели
корреляции с сексуальной функцией в исследовании 1021 австралийской женщины в
возрасте от 18 до 75 лет [257]. Низкий показатель сексуальной реакции для женщин
старше 45 лет был связан с более высоким перевесом в сторону плазменного уровня
ДГЭАС ниже 10-й процентили для этой возрастной группы [257]. Тем не менее преоб¬
ладающее большинство женщин с низкими уровнями ДГЭАС не страдали снижением
сексуальной функции [257].
Очевидно, что существует множество неразрешенных вопросов, включая раз¬
личные количества остающихся надпочечниковых предшественников тестостерона,
активность стероидогенических ферментов, изменения числа и чувствительности
андрогеновых рецепторов и корегуляторов и взаимодействие эстрогенов с андроге¬
нами. Предполагают (но не доказано), что улучшение сексуальной функции проис¬
ходит под влиянием воздействия на андрогеновые рецепторы. Однако ароматизация
эстрогена внутри клеток может оказывать по крайней мере частичное влияние. Б
одном недавнем исследовании показан положительный эффект путем воздействия на
андрогеновые рецепторы, поскольку добавление ингибитора ароматазы не отменяет
положительного эффекта от назначения дополнительного тестостерона женщинам
в постменопаузе, уже получающим трансдермальную эстрогеновую терапию [258].
Также неясно, оказывает ли тестостерон положительное воздействие, просто снижая
SHBG, то есть позволяя эстрогену становиться более биоактивным.
СОСТОЯНИЯ с низким УРОВНЕМ ЭСТРОГЕНОВ
Могут возникать сексуальные последствия низких уровней эстрогена после мено¬
паузы или рождения ребенка; в связи с терапией агонистами ГнРГ, индуцированной
химиотерапией яичниковой недостаточностью, идиопатической яичниковой недоста¬
точностью или хирургической менопаузой; и иногда в связи с приемом комбиниро¬
ванных контрацептивов с низким содержанием эстрогенов или с депопрогестероном.
При отсутствии продукции эстрогенов яичниками внутриклеточное производство
эстрогена из прогестерона, ДГЭА, ДГЭАС и андростендиона зависит от надпочечни¬
ковой и яичниковой продукции этих прогормонов, числа жировых клеток (важное
место ароматизации тестостерона в эстрадиол) и соответствующих стероидогениче¬
ских ферментов для синтеза эстрогена или тестостерона и ДГТ из предшественников.
Последние работы разъясняют структуру множества ткань-специфических генов,
которые кодируют стероидогенные ферменты, управляющие трансформацией пред¬
шественников в андрогены и/или эстрогены [251,252]. В особенности внимание сфо¬
кусировано на Зр-гидроксистероид дегидрогеназе (Зр-ГСД), 17р-ГСД, 5а-редуктазе и
ароматазе [252]. Таким образом, у женщин старшего возраста в локальном биосинтезе
половых гормонов участвуют как эстрогены, так и андрогены.
Генитальные эффекты низкого уровня эстрогена
Состояния, связанные с низкими уровнями эстрогенов, обычно сопровождаются
снижением выработки смазки в ответ на сексуальную стимуляцию. Тем не менее эта
376
проблема комплексная. Визуальные признаки атрофии и низких плазменных уровней
эстрогена коррелируют с базальными измерениями гиперемии влагалища, однако
процент возрастания притока крови в ответ на эротические стимулы аналогичен
при состояниях с недостатком и насыщением эстрогенами [216]. Ранние результаты
исследований МРТ демонстрируют аналогичные изменения влагалищной стенки и
клиторального объема при сексуальной стимуляции у женщин в пре- и постмено¬
паузе [259]. Эстрогеновая недостаточность необязательно препятствует адекватному
выделению смазки, если сексуальная стимуляция достаточна. Тем не менее приблизи¬
тельно .30-50% женщин в постменопаузе сообщают о сексуальных симптомах в связи
с влагалищной атрофией [260].
Низкий уровень эстрогена увеличивает pH влагалищного просвета, предрасполагая
к инфицированию, которое, в свою очередь, подрывает женскую сексуальную уверен¬
ность и вносит вклад в развитие диспареунии. Изучение механизма окисления эстро¬
гена во влагалище изменил традиционный взгляд, согласно которому это объяснялось
пероксидом водорода и протонами, секретируемыми эстроген-зависимыми лакто¬
бациллами Дедерлайна [261]. Теперь имеются некоторые доказательные данные о
том, что вагинальные эктоцервикальные клетки закисляют просвет влагалища путем
секреции протонов через апикальную плазматическую мембрану. Предполагается,
что эта активная секреция протонов осуществляется на протяжении всей жизни, одна¬
ко она регулируется эстрогеном [261].
Симптомы сухости во влагалище, диспареунии и сужения влагалища имеют слабую
корреляцию с визуальными атрофическими изменениями, уровнями эстрогена и про¬
должительностью дефицита эстрогена. Симптомы могут развиваться в перименопау¬
зе, однако в некоторых случаях только через десятилетия после менопаузы. Уровни
эстрогена действительно коррелируют с соотношениями между парабазальными,
промежуточными и поверхностными влагалищными клетками. Роль проницаемости
вагинальных эпителиальных клеток остается неясной. Нервные окончания, содержа¬
щие КГРП, присутствуют на вагинальных эпителиальных клетках и могут регулиро¬
вать их проницаемость [262].
Эстроген и сексуальная чувствительность
Снижению сексуальной чувствительности гениталий и кожи вне гениталий посвя¬
щено мало научных исследований. В небольшом количестве исследований с исполь¬
зованием порогов давления было продемонстрировано снижение чувствительности
внешних половых органов при менопаузе и возрастание чувствительности после
использования местных эстрогенов [263]. Как возраст, так и состояние постменопау¬
зы связаны со снижением вибрационной чувствительности в генитальном тракте, а
возраст сам по себе затрагивает периферическое вибрационное чувство вне гениталий
[264].
«Безжизненность» гениталий, связанная с расстройством генитального сексуаль¬
ного возбуждения, которое сохраняется, несмотря на терапию эстрогеном, плохо
изучена [218]. Сниженная генитальная вазоконстрикция, несмотря на эффектив¬
ную эротическую стимуляцию, которая вызывает возбуждение в мыслях, очевидно,
лежит в основе патологии лишь у одной подгруппы женщин с этим расстройством.
Следовательно, при условии большого количества вульварных андрогеновых рецеп¬
торов эта сниженная андрогенная активность может участвовать в исчезновении сек¬
суальной чувствительности гениталий [222].
Низкий уровень эстрогена и субъективное возбуждение и половое влечение
Роль низкого уровня эстрогена в исчезновении полового влечения и возбудимости
остается неясной. В недавнем исследовании показано, что сексуальная реактивность
может быть улучшена при терапии эстрогенами до уровней 650-750 пмоль/л, что
приблизительно в 2 раза выше уровня, требуемого для устранения местных симпто-
377
мов влагалищной сухости [265]. Улучшение показателей тестостерона, как минимум
частично, может происходить вследствие увеличенной продукции эстрогенов из-за
ароматизации внутри клеток или увеличения биодоступности эстрогена.
РАК МОЛОЧНОЙ ЖЕЛЕЗЫ
Сексуальные проблемы после установления диагноза и лечения рака молочной
железы с наибольшей вероятностью возникают из-за стресса, сохраняющегося на
протяжении более 1 года после постановки диагноза [254]. Химиотерапия играет
основную роль в снижении полового влечения, субъективного возбуждения, в разви¬
тии влагалищной сухости и диспареунии [254]. Прогностическая модель сексуального
интереса, активности и удовлетворения после перенесенной терапии рака молочной
железы была разработана на двух независимых группах лиц, выживших после рака
молочной железы [254]. Наиболее важными прогностическими факторами сексуаль¬
ного удовлетворения были отсутствие влагалищной сухости, эмоциональное благо¬
получие, положительный «Я-образ», хорошее качество взаимоотношений, отсутствие
сексуальных проблем у партнера [254].
Временная медикаментозная менопауза после адъювантной терапии агониста¬
ми ГнРГ, очевидно, сопровождается обратимой сексуальной дисфункцией [266].
Тамоксифен не оказывает непосредственного влияния на сексуальную активность
[266]. Однако ингибиторы ароматазы, приводящие к развитию состояния, близкого к
общему отсутствию любой эстрогенной активности, могут быть связаны с тяжелыми
симптомами вульвовагинальной атрофии [266].
ДИАБЕТ
Исследования прошлых лет привели к спорным результатам, так как они были
выполнены на небольшом количестве субъектов без контрольных групп, часто при
смешении пациентов с диабетом 1-го и 2-го типа, с редко выявляемым эстрогенным
статусом.
Недавнее исследование подтверждает биопсихосоциальную природу женской
сексуальности. В пяти контролируемых изучениях была подтверждена связь между
диабет-ассоциированной сексуальной дисфункцией и депрессией, которая была
установлена в предыдущем метаанализе [267]. Была также пока.зана взаимосвязь
разнообразных психологических факторов, в том числе принятие женщиной ее
заболевания. Сексуальные сложности, по всей видимости, имеют корреляцию с
возрастом, длительностью заболевания, ИМТ, уровнем гликемического контроля,
гормональной терапией, осложнениями заболевания или состоянием менопаузы
[268-272].
Снижение выделения смазки — одно из сексуальных расстройств, которое встре¬
чается более часто у женщин с диабетом, чем в контрольной группе [268-271]. Было
показано, что генитальное вибрационное чувство снижается у женщин с диабетом
[270]; тем не менее такое снижение не было связано с сексуальной дисфункцией.
Эмпирические данные не поддерживают непосредственно клиническое впечатление
о том, что у женщин, в особенности при диабете 1-го типа, потеряна оргазмическая
функция. В одном исследовании пациенток с диабетом и почечной недостаточностью
было продемонстрировано значительное ухудшение оргазма и клиторального сексу¬
ального ощущения по сравнению с контрольной группой [273].
Психофизиологических исследований очень мало. В одном небольшом исследова¬
нии по сравнению с контрольной группой у женщин с диабетом отмечали снижение
набухания гениталий и при этом сопоставимое субъективное сексуальное возбуж¬
дение в ответ на эротический стимул [274]. Данные по диспареунии ограниченные.
Прогнозируют высокую распространенность диспареунии при снижении выработки
смазки, а также (несмотря на доказательные данные положительного эффекта) [275]
378
ограничения назначения эстрогеновой терапии после менопаузы при сахарном диа¬
бете [275]. Кроме того, ожидаемая связь с хроническим кандидозом будет увеличи¬
вать распространенность диспареунии, возможно, за счет провоцирования синдрома
вульварного вестибулита у тех, кто имеет генетическую предрасположенность. Тем не
менее распространенность диспареунии высока у молодых и женщин среднего воз¬
раста как с диабетом, так и без. Сообщают об уровнях распространенности от 32 до
62%, от 42 до 46% и от 26 до 39% у женщин с диабетом 1-го, 2-го типа и отсутствием
диабета соответственно [268, 272].
ПОЧЕЧНАЯ НЕДОСТАТОЧНОСТЬ
Было проведено очень мало исследований относительно сексуальной функции
у женщин с заболеваниями почек. Сопутствующая ановуляция (следовательно,
отсутствие пика тестостерона), высокая секреция пролактина, анемия, коморбидная
депрессия и состояние с низким уровнем эстрогена могут нарушить сексуальную
функцию. Была отмечена сниженная генитальная сексуальная функция у женщин, у
которых почечная недостаточность сопровождалась гипертриглицеридемией [273].
Субъективное возбуждение может улучшиться после трансплантации почки, даже
если наблюдаемый вагинальный гилеремический ответ на эротические стимулы ана¬
логичен тому, который отмечается у женщин на гемодиализе, перитонеальном диа¬
лизе или у пациенток с трансплантатом [276].
СЕРДЕЧНО-СОСУДИСТЫЕ ЗАБОЛЕВАНИЯ
Выполнено ограниченное количество исследований сексуальной функции у жен¬
щин с сердечно-сосудистыми заболеваниями. В одном из них атеросклеротические
поражения подчревной и половой артерий сопровождались снижением образования
смазки и снижением секс}»альной функции. Сексуальные проблемы становились более
выраженными после внутрисосудистой интервенции даже несмотря на общее улучше¬
ние здоровья [277]. У женщин в пременопаузе с умеренной гипертензией, полу^шю-
щих или не получающих терапию, было задокументировано снижение образования
смазки совместно с диспареунией и слабыми оргазмами по сравнению с контрольной
группой соответствующего возраста [278].
ЛЕКАРСТВЕННЫЕ ПРЕПАРАТЫ
Ряд часто назначаемых лекарственных препаратов может угнетать сексуальную
функцию и удовлетворение (см. табл. 4-11).
СИНДРОМ поликистозных яичников
До настоящего времени отсутствуют доказательные данные того, что более высокие
уровни андрогенов, связанные с СПКЯ, дают защиту от жалоб на низкое сексуальное
влечение или низкую сексуальную возбудимость. В выборке из 30 женщин с СПКЯ,
обратившихся для лечения гирсутизма, сексуальное влечение было значительно ниже
по сравнению с контрольной группой [279].
ГИПОТАЛАМО-ГИЛОФИЗАРНОЕ ЗАБОЛЕВАНИЕ
Получены отдельные данные о сексуальной дисфункции у женщин с гипопитуи-
таризмом. Редко дополняемый тестостероном фокус обычно направлен на эстроген
(и прогестерон, если матка расположена на месте). В одном из недавних рандомизи¬
рованных плацебо-контролируемых исследований эффектов введения тестостерона
женщинам с гипопитуитаризмом было продемонстрировано улучшение сексуальной
функции от ежедневных приемов 25-50 мг ДГЭА, назначаемых пациенткам с вто¬
ричной надпочечниковой недостаточностью и женщинам с гипопитуитаризмом [281,
282].
379
Таблица 4-11. Лекарственные факторы риска сексуальной дисфункции у женщин
Лекарственный
препарат
Распространенность
Эффект
Лечение
Антипсихотические пре¬
параты
Сексуальная дисфункция
почти у 73% пациенток с
шизофренией
Традиционные антипси¬
хотические препараты и
рисперидон: сниженный
уровень допамина и увели¬
ченный уровень пролактина
вызывают а-блокаду и
мускариновую блокаду
Используйте антипсихоти¬
ческие препараты II поколе¬
ния, которые не увеличива¬
ют уровень пролактина.
Другие механизмы все еще
могут вызывать дисфункцию
Р-Блокаторы
Снижение полового вле¬
чения до уровня ниже
переносимого ранее под
действием антагонистов
ангиотензина II
Селективная или несе¬
лективная блокада может
сопровождаться сексуаль¬
ной дисфункцией
Используйте антагонист
рецепторов АПФ, блока-
тор кальциевых каналов,
а-блокатор с перифериче¬
ским действием
СИОЗСТ
Сексуальные побочные
эффекты почти у 70%
пациенток
Стимуляция серотониновых
рецепторов, в том числе
рецепторов 5-rTj
Добавление бупропиона»’
может оказать положитель¬
ный эффект; улучшение
отмечали в одном из двух
РКИ
Антиандрогены: антаго¬
нисты ГнРГ, ципротерона
ацетат, спиронолактон в
высоких дозах
Подавление ГнРГ или Л Г и/
или антагонизм с андроге-
новыми рецепторами
Наркотики
Подавление ГнРГ
Отсутствуют специфические
данные относительно добав¬
ления половых гормонов
Противоэпилептические
препараты
Индукция Р450 печеночных
ферментов увеличивает
ПСПС и снижает уровень
свободного тестостерона
Некоторые противоэпи¬
лептические препараты
являются нейтральными по
отношению к ферментам,
например окскарбазапин,
ламотриджин, леветирацета,
однако исследования огра¬
ничены
Комбинированные
оральные контрацептивы
Распространенность сопут¬
ствующей дисфункции
неизвестна, однако около
50% женщин прекраща¬
ют прием ОК в течение
12 мес, многие в связи с
сексуальными побочными
эффектами
Увеличение ПСПС, сни¬
жающего биодоступность
тестостерона; подавление
эндогенного тестостерона:
использование неандроген¬
ного гестагена
Недоступны исследования
относительно дополнения
тестостероном или сравне¬
ния различных рецептур.
Рассмотрите альтернатив¬
ный способ контроля дето¬
рождения, например ВВП,
барьерный метод, необра¬
тимый метод
Примечание. АПФ — ангиотензин-превращающий фермент; ПЭП — противоэпилептический препа¬
рат; ГйРГ — гонадотропин-рилизинг-гормон; 5-ГТ — серотонин; ВВП — интравагинальное приспосо¬
бление; ЛГ — лютеинизирующий гормон; ОК — оральный контрацептив; РКИ — рандомизированное
контролируемое испытание; ПСПС — протеин, связывающий половые стероиды; СИОЗСТ — селек¬
тивный ингибитор обратного захвата серотонина.
Адаптировано с разрешения из Basson R. Sexual dysfunction in women // New Engl. J. Med. — 2006. —
N 354 (14). — P. 1497-1506. Copyright 2006. Massachusetts Medical Society. Bee права защищены.
380
НАДПОЧЕЧНИКОВАЯ НЕДОСТАТОЧНОСТЬ
Отсутствуют данные относительно распространенности сексуальной дисфункции у
женщин с надпочечниковой недостаточностью. Результаты добавления ДГЭА непо¬
стоянны [282. 283].
ПРОСТАЯ ГИСТЕРЭКТОМИЯ
В отношении женщин, перенесших гистерэктомию, не подтверждаются различия
сексуального исхода при вагинальной, субтотальной абдоминальной или тотальной
вагинальной гистерэктомии [284, 285, 286]. В целом было отмечено, что сексуальное
удовлетворение повышается у большинства женщин вне зависимости от типа гистер¬
эктомии.
РАДИКАЛЬНАЯ ГИСТЕРЭКТОМИЯ
Автономные нервные окончания, особенно те, которые проходят в маточно-крестцо¬
вой и кардинальной связках, могут быть нарушены при радикальной гастерэктомии,
если не применяются сберегающие методики. Отсутствует информация о последующей
генитальной сексуальной функции, кроме данных, характеризующих влагалищную
реакцию гиперемии на эротические стимулы. В ходе фотоплетизмографической оценки
наполнения кровью в процессе субъективно возбуждающей сексуальной стимуляции у
12 женщин с перенесенной нервосберегающей радикальной гистерэктомией выявился
сниженный максимальный ответ по сравнению с 17 лицами из контрольной группы,
подобранными по возрасту. Предполагают, что проблемы со смазкой после радикального
хирургического вмешательства в области таза могут возникать вследствие повреждения
нервов [287]. Были внедрены нервосберегающие радикальные гистерэктомии при раке
шейки матки, однако до сих пор нет исследований сексуального исхода.
ГИПЕРПРОЛАКТИНЕМИЯ
У женщин с гиперпролактинемией обычно снижены возбуждение и оргазм и уве¬
личена диспареуния при отсутствии доказанных изменений других гормональных
параметров, кроме пролактина [288].
СТАРЕНИЕ
Степень, до которой возраст является фактором риска сексуальной дисфункции,
неясна. Наиболее частая причина прекращения сексуальной активности, приводимая
женщинами, — отс5»тствие партнера. Многие исследования включают только «сек¬
суально активных» женщин (некоторые только «сексуально активны при половом
акте»), и поэтому доступные данные ограничены. Обширные национальные репре¬
зентативные исследования продемонстрировали небольшое увеличение числа сексу¬
альных проблем с возрастом, однако в исследования часто не включают лиц старше
60 лет [229]. В других исследованиях приблизительно 40% женщин старшего возрас¬
та отмечали снижение сексуальной реактивности, сопровождающееся возрастанием
влечения к негенитальным сексуальным проявлениям [220, 289]. В ходе недавнего
анализа была показана более значительная распространенность сниженного полово¬
го влечения как функции менопаузального статуса и возраста, увеличивающаяся от
22% в пременопаузальной группе до 32% в постменопаузальной [244]. Как и в других
данных, низкое половое влечение имело тесную связь со сложностями возбуждения
и оргазма. В одном обширном продольном исследовании женщин с 10-летним сро¬
ком менопаузы и более было продемонстрировано снижение полового влечения и
реактивности под влиянием как возраста, так и менопаузы [220]: высокие значения
менопаузальных симптомов влияли на благополучность состояния, которое действо¬
вало на сексуальную реактивность и, в свою очередь, сексуальное влечение и интерес.
В более крупном исследовании 14 ООО женщин из 29 стран было установлено, что
Э81
уменьшение количества смазки является лишь одним фактором, связанным с возрас¬
том. К сожалению, уровень реактивности в этом исследовании составил лишь 15% и
были включены только те женщины, которые принимали участие в половом акте в
прошедшем году [299].
Анализ многих исследований подтверждает, что женщины старшего возраста сооб¬
щают о меньшем переживании по поводу снижения/отсутствия полового влечения по
сравнению с женщинами молодого возраста [223]. В последних исследованиях был
продемонстрирован сдвиг от генитального секса к интимной близости по мере ста¬
рения людей. Таким образом, любая деятельность с задействованием привязанности,
романтики, близости и дружеского общения может способствовать удовлетворению
сексуальных потребностей взрослых лиц старшего возраста. Возможно, с разработкой
эффективных методов терапии эректильной дисфункции эти модели могут изменять¬
ся. Выраженные предикты сохранения сексуального интереса — сексуальное поведе¬
ние и наслаждение в более раннем возрасте,
ПСИХОЛОГИЧЕСКИЕ ФАКТОРЫ
Психологические факторы в значительной степени влияют на женскую сексуаль¬
ность, в частности партнер, его общее и сексуальное здоровье [239,290], а также сами
отношения [229]. Эти факторы, в свою очередь, определяют, насколько хорошо пара
может адаптироваться к изменениям в сексуальной жизни под действием внешних
факторов и собственного старения. Личностные факторы, влияющие на возбуди¬
мость, содержат несексуальные возбуждающие факторы, сексуальную уверенность в
себе, ожидание награды, чувства к партнеру как в конкретное время, так и в общем,
и факторы безопасности, включающие адекватный контроль рождаемости, безопас¬
ность относительно заболеваний, передаваемых половым путем, незапланированных
беременностей и вопросов конфиденциальности.
Мысли из детства и подросткового возраста могут отрицательно воздействовать на
возбудимость женщины, они могут играть более важную роль в дальнейшей жизни.
Выраженная потребность ощущения контроля или потребность в подавлении эмоций
могли быть установками, которые помогли пережить детство и подростковый возраст,
однако те же самые защитные механизмы могут препятствовать здоровой сексуаль¬
ной реакции во взрослом состоянии. Иногда неразрешенный гнев и чувство обиды
в отношении родителей переживаются в контексте более поздней жизни, к примеру,
необходимость заботиться о родителях, ранее злоупотреблявших какими-либо веще¬
ствами. Женщины редко способны подавлять только негативные или «нежелатель¬
ные» эмоции; обычно подавляются все, в том числе сексуальные, эмоции.
Личностные факторы могут также оказывать влияние на женскую сексуальную
функцию. В исследованиях было показано, что, по сравнению с женщинами с нор¬
мальной сексуальной функцией, те, которые жаловались на низкое половое влечение
и низкую степень возбуждения, имели уязвимую самооценку, более высокие уровни
тревожности и чувства вины, отрицательный телесный образ, а также были более
интровертны и соматизированы [291]. В большинстве исследований не стремятся
выявлять действительную психопатологию у женщин, жалующихся на низкое поло¬
вое влечение [291], у пациенток с оргазмическим расстройством клиническая картина
состоит в том, что многие из них чувствуют чрезвычайный дискомфорт, если они не
контролируют собственные обстоятельства и телесные функции.
у женщин с вагинизмом, очевидно, имеется фобическая составляющая страха
перед проникновением во влагалище. Эти пациентки обычно не имеют других фобий,
кроме неспособности к рождению ребенка (полагая, что это событие нанесет нево¬
образимую физическую травму), В значительной подгруппе женщин с вестибулитом
отмечают выраженный страх негативной оценки другими, повышенную добросовест¬
ность и самокритику [299], а также возрастание соматизации [293].
382
КАЧЕСТВО В ВОЗРАСТ ПАРТНЕРСКИХ ОТНОШЕНИЙ
Подавляющее большинство женщин, жалующихся на низкое половое влечение и воз¬
будимость, утверждают, что их партнерские отношения стабильны и удовлетворитель¬
ны — «у нас только сексуальные проблемы». Отсутствие конфликтов, пренебрежения
или жестокого обращения и уверенность в преданности, тем не менее, сами по себе недо¬
статочны для удовлетворения женского сексуального влечения и содействия возбужде¬
нию. общие темы включают неудовлетворенную потребность в эротизме, разнообразии
сексуальндй стимуляции и большей эмоциональной близости со своим партнером.
Отсутствие ссор не приравнивается к ощ)»щению близости — чувству безопасности в
состоянии уязвимости и при открытии ощущений, страхов и надежд друг другу.
Отношения «очень хороших друзей» — недостаточно сексуальный контекст для того,
чтобы потенцировать женское половое влечение. Б исследованиях демонстрируется
нормальное снижение внутреннего полового влечения по мере продолжения взаимо¬
отношений [294]. Недавно, по данным национальной выборки, было установлено, что
женские чувства к партнеру — одна из основных детерминант свободы от переживаний
по поводу секса [223]. Чувства женщины к партнеру или недавняя смена партнера — две
из трех основных детерминант женского полового влечения и реактивности, по данным
продольного исследования женщин, переходящих в менопаузу [200].
Сексуальная дисфункция у партнера мужского пола также может оказывать влия¬
ние на женскую сексуальную функцию [239]. Лечение ЭД у пациента мужского пола
может привести к исчезновению сексуальных жалоб женщины, в том числе проблема¬
тичного возбуждения, выделения смазки, оргазма и боли [290].
БЕСПЛОДИЕ
Как оценка собственного бесплодия, так и потребность в привлечении репродук¬
тивных технологий могут негативно повлиять на женский сексуальный «Я-образ».
Сфокусированная на половом акте сексуальная активность при попытках зачатия
может оказаться сексуально неподкрепленной и, следовательно, снизить мотивацию.
К несчастью, успешная беременность необязательно приводит к исчезновению сек¬
суальных проблем; неразрешенное чувство вины по поводу личной ответственности
за бесплодие и возмущения в связи с множеством процедур у женщин, по сравнению
только лишь с анализом спермы у мужчин, могут затягивать решение проблем [295].
Оценка женщины с вдтщпьныш щттщтшвш
Необходим детальный опрос относительно текущих сексуальных проблем и их
контекста как обоих партнеров совместно, так и индивидуально каждого. В табл. 4-12
показано общее представление о той информации, которую необходимо оценить при
диагностике сексуальной дисфункции.
Физикальное обследование, включающее обследование таза и половых органов, —
часть рутинного оказания помощи (табл. 4-13). Если не вовлечена диспареуния, то
при физикальном обследовании редко выявляются причины сексуальной дисфунк¬
ции. Обследование может быть терапевтическим для подтверждения нормальной
анатомии и здоровья тканей. У некоторых женщин с опытом сексуального насилия в
анамнезе или жестокости при половом акте такое обследование может вызывать чрез¬
мерное беспокойство. Нужно перед исследованием назвать причину, п'6 которой это
исследование необходимо, и дать объяснение того, что будет и что не будет сделано.
Если проблема в вагинизме, обследование следует отложить до тех пор, пока терапия
не позволит его осуществить с максимально положительным терапевтическим воз¬
действием на женщину (см. табл. 4-13).
383
Таблица 4-12. Оценка сексуальной дисфункции
Цель анализа
Метод анализа
Сексуальная проблема и причина для обра¬
щения в настоящее время
Попросите пациентку описать проблему своими словами.
Уточните проблему при помощи прямых вопросов, предлагайте паци¬
енту варианты ответа, поддерживайте его, объясняйте ему, что сексу¬
альные проблемы довольно распространены
Продолжительность, устойчивость и прио¬
ритет, если более одной проблемы
Проявляются ли проблемы во всех ситуациях? Какая проблема трево¬
жит ее больше всего?
Контекст сексуальных проблем
Эмоциональная близость с партнером.
Активность/поведение непосредственно перед сексуальной активно¬
стью, уединение, сексуальное общение, время суток/уровень утомле¬
ния.
Контроль рождаемости: адекватность, тип, риск ЗППП.
Эффективность сексуальной стимуляции, сексуальных знаний
Другие сексуальные реакции каждого из
партнеров, кроме данной проблемной зоны
Оцените это в настоящее время и перед началом сексуальных проблем
Реакция каждого из партнеров
Как каждый из них реагирует: эмоционально, сексуально, поведенче¬
ски?
Предыдущая помощь
Соответствие с рекомендациями и эффективностью
Вопросы, которые нужно задать каждому из партнеров при индивидуальной консультации
Собственная оценка пациентом ситуации
Иногда проще обнаружить тяжесть симптомов, например общее сни¬
жение полового влечения, в отсутствие партнера
Сексуальная реакция при самостимуляции
Также выясните сексуальные мысли и фантазии
Предыдущие сексуальные опыты*
Положительные, отрицательные аспекты
История развития*
Взаимоотношения с другими домашними в процессе взросления.
Потери, травмы, с кем (если кто-то был); были ли они близки?
Была ли им показана физическая привязанность, любовь, уважение?
Прошлое или настоящее сексуальное,
эмоциональное или жестокое физическое
обращение*
Объясните, что вопросы 0 жестоком обращении являются рутинными
и необязательно устанавливают причинно-следственную связь с про¬
блемами.
Полезно спросить, подвергались ли они когда-нибудь телесным нака¬
заниям или запугиванию и, если да, желают ли они предоставить
больше информации
Физическое здоровье, в особенности
состояния, приводящие к слабости и утом¬
ляемости: сложности с мобилизацией как
при ласкании партнером, так и при само¬
стимуляции; сложности с «Я-образом»,
например вследствие стом, уродующего
хирургического вмешательства, проблем с
удержанием мочи
Прицельно спросите 0 лекарственных препаратах с известными сексу¬
альными побочными эффектами, в том числе СИОЗСТ, р-блокаторах,
антиандрогенах, агонистах ГнРГ, оральных контрацептивах
Оценка настроения
Значительная корреляция сексуальной функции и настроения (в том
числе тревожности и депрессии) направляет рутинный скрининг на
поиск расстройства настроения с применением как опросников (напри¬
мер, опросник Веек), так и полуструктурированных серий вопросов
* Пункт иногда может быть исключен (например, для недавних проблем после десятилетий здоровой
сексуальной активности).
Примечание. ГйРГ — гонадотропин-рилизинг-гормон; СИОЗСТ — селективные ингибиторы обрат¬
ного захвата серотонина.
Адаптировано с разрешения из: Basson R. Sexual dysfunction in women //New Engl.J. Med. — 2006. —
N 354 (14). — P. 1497-1506. Copyright 2006. Massachusetts Medical Society. Bee права защищены.
384
Таблица 4-13. Физикальное обследование женщины с сексуальной дисфункцией
Обследование
На что нацелен поиск
Общее
Признаки системного заболевания, приводящие к низкой энергии, низкому поло¬
вому влечению, низкой возбудимости; анемия, брадикардия, медленно релакси-
рующие рефлексы при гипотиреоидизме.
Признаки заболевания соединительной ткани, такого как склеродермия или син¬
дром Шегрена, которые сопровождаются вагинальной сухостью.
Нетрудоспособность, которая может препятствовать движениям, задействуемым
при ласкании партнера, самостимуляции, половом сношении.
Уродства, стомы, катетеры, которые могут снижать уверенность в собственной
сексуальности, приводя к низкому половому влечению, низкой возбудимости
Наружные половые органы
Редкие лобковые волосы, предполагающие низкое содержание надпочечниковых
андрогенов.
Заболевания кожи вокруг наружных половых органов, в том числе склерозирую-
щий лишай, который может вызывать болезненность при сексуальной стимуляции.
Трещины или надрывы межгубных складок, предполагающие хронический канди-
доз.
Аномалии половых губ, которые могут вызывать стеснение или сексуальную неуве¬
ренность, например чрезмерно длинные половые губы или их асимметрия
В просвете
Заболевания наружных половых органов с вовлечением просвета, например блед¬
ность, рыхлость, снижение эластичности и влажности, вульварная атрофия, скле-
розирующий лишай.
Повторное расщепление задней уздечки половых губ, проявляется едва заметны¬
ми белыми линиями, перпендикулярными краю уздечки половых губ.
Аномалии девственной плевы.
Сращение малых половых губ.
Опухание зоны больших преддверных желез.
Аллодиния (болезненное ощущение от стимула прикосновения) в складке между
внешним краем девственной плевы и внутренним краем малых половых губ, кото¬
рое типично для вульварного вестибулита.
Цистоцеле, ректоцеле или пролапс, мешающие сексуальному «Я-образу» женщины.
Невозможность напрячь и расслабить перивагинальную мускулатуру, часто сопро¬
вождается гипертонусом тазовой мускулатуры и средневагинальной диспареунией.
Патологические выделения из влагалища, сопровохсдающие жгучую диспареунию
Внутреннее
Тонус тазовой мускулатуры.
Болезненность.
Триггерные точки при пальпации глубокой мышцы, поднимающей задний проход,
в связи с основным гипертонусом
Полное бимануальное
Узлы и/или болезненность в замкнутом пространстве или своде влагалища, вдоль
маточно-крестцовых связок.
Матка, фиксированная в положении ретроверсии, как причина глубокой диспареу¬
нии.
Болезненность при пальпации задней стенки мочевого пузыря с передней стенки
влагалища, предполагающая патологию мочевого пузыря
Адаптировано с разрешения из Basson R. Sexual dysfunction in women // New Engl. J. Med. — 2006. — N
354 (14). - P. 1497-1506. Copyright 2006. Massachusetts Medical Society. Bee права защищены.
Лабораторное исследование играет незначительную роль в оценке сексуаль¬
ной функции. Активность эстрогенов лучше всего определяется при исследовании.
Биохимическая оценка ограничена у женщин старшего возраста, поскольку доступ-
385
ные методы оценки направлены на эстрадиол, а не на эстрон, преобладающий
постменопаузальный эстроген. Плазменные уровни тестостерона не коррелируют с
сексуальной функцией, и существующие в настоящее время доступные методы оценки
не имеют требуемой чувствительности при низких уровнях, выявляемых у женщин.
Пролактин или тиротропин могут измеряться, если имеются другие симптомы, пред¬
полагающие аномалию.
Стратегии лечения, основанные на доказательных данных
Терапевтические стратегии для лечения женской сексуальной дисфункции, осно¬
ванные на доказательных данных, только еще начинают появляться. Одна из слож¬
ностей заключается в том, что этиология обычно мультифакторная, приводящая к
тому, что лечение состоит из множества компонентов. Однако на сегодняшний день
в небольшом количестве РКИ обычно применяется одна стратегия лечения. Перед
назначением терапии необходимо точно сформулировать проблему. С использовани¬
ем структуры цикла женской сексуальной реакции оценивают следующие аспекты.
• Разнообразные мотивации или стимулы быть сексуальной.
• Степень эмоциональной близости, доверия, уважения и привлекательности в
отношениях.
• Сексуальный контекст и соответствие стимулов при попытке сексуального кон¬
такта.
• Что происходит в женском разуме, если она пытается возбудиться.
• Любые биологические факторы, влияющие на ее возбудимость.
• Детали исхода в терминах как физического, так и эмоционального удовлетворе¬
ния и свободы от боли.
• Детальное описание любой боли, ее мысли и ощущения, когда появляется боль,
уровень ее возбудимости и физикальные находки.
ПОВЕДЕНЧЕСКИЕ, КОГНИТИВНЫЕ И СЕКСУАЛЬНЫЕ МЕТОДЫ ТЕРАПИИ
Поведенческая, когнитивная и сексуальная терапия продолжает оставаться опорой
терапии сексуальной дисфункции. Исследуемые виды поведения остаются сексуаль¬
ными или несексуальными, включающими взаимоотношения партнеров за предела¬
ми спальни. Могут привлекаться улучшение сексуального окружения, стимулы, их
распределение по времени и, возможно, сексуальные техники. Мнение женщины о
самой себе, ее отношения, ее сексуальность и ее партнер также подвергаются исследо¬
ванию. Сексуальная терапия концентрируется в большей степени на межличностных
моментах, а также на сексуальных деталях, и может включать техники фокусирования
ощущений [296,297,299-303]. К ним относят обмен физическими прикосновениями,
перемещение от несексуальных к сексуальным зонам тела по аналогии с системной
десенситизацией при других методах поведенческой терапии, которые снижают тре¬
вожность. Когда имеют значение более отдаленные факторы, к примеру предполага¬
ется, что сексуальные симптомы происходят из прошлых (несексуальных) событий в
детстве и/или из «низкого» сексуального образа, может потребоваться краткосрочная
психотерапия.
К сожалению, данные исходов всех этих вмешательств значительно ограничены
(табл. 4-14) [304] в связи с различной продолжительностью терапии (от 2 до 20 нед),
различиями в контрольных исследованиях (от конца терапии до 3 лет), разными
путями оценки исхода (клинически определяемый, определяемый участниками,
детальный исход в отличие от «успешного или неуспешного»). Кроме того, диагноз
дисфункции и терапия основываются на более старой модели сексуального функцио¬
нирования. Только начинают появляться исследования, в которых измеряют измене¬
ния потенцируемого полового влечения, а не спонтанное влечение [209].
386
Таблица 4-14. Исходы психологической терапии
Публикации
Лечение
Уровень эффективности
Delehanty (0)
нм и тренинг уверенности в себе
против контрольного списка ожидаю¬
щих
88% начали испытывать оргазм
Hawton с соавт. (В, В)
Модифицированная фокусировка на
ощущении
57% значительного улучшения
Heiman и LoPiccoio (0)
КБТ, НМ, фокусировка на ощущении
против списка ожидающих
Усиление оргазмической реакции и инициация
сексуальной активности
Hurlbert (В, В)
Супружеская и сексуальная терапия ±
тренировка устойчивости оргазма*
Значительное улучшение возбуждения после
сексуальной терапии плюс устойчивость оргаз¬
ма против одной сексуальной терапии
Hurlbert и Apt (0)
НМ и регулирование полового акта
37% пациентов, которые использовали технику
регуляции полового акта и НМ, против 18%,
использовавших только НА, отмечали значи¬
тельное улучшение оргазмов, ощущаемых во
время полового акта
McCabe (В. В)
КБТ
<50% значительного улучшения
McMullen и Rosen
НМ с библиотерапией против НМ с
видеозаписями против контрольного
списка ожидающих
65 и 55% начали испытывать оргазм
Sarwer и Durlak (В, В)
Поведенческая и сексуальная терапия
64% значительного улучшения
Trudel с соавт. (В, В)
КБТ
74% улучшения сексуальной активности и каче¬
ства супружеской жизни
* тренировка устойчивости оргазма включает поощрение самостимуляции, терапию с фокусировкой
на ощущениях с партнером и коитальные техники для облегчения клиторальной стимуляции.
Примечание. КБТ — когнитивная поведенческая терапия; В, В — дисфункция полового влечения и
возбуждения; НМ — направленная мастурбация; О — оргазмическая дисфункция.
Delehanty R. Changes in assertiveness and changes in orgasmic response occurring in sexual therapy of pre-
orgasmic women //J. Sex Marital. Then — 1982. — N 8. — P. 198-208.
Hawton K., Catalan J., Flagg J. Low sexual desire: sex therapy results and prognostic factors // Behav. Res.
Then - 1991. - N 29. - P. 217-224.
Heiman J., LoPiccolo J. Clinical outcome of sex therapy. Effect of daily vs weekly treatment // Arch. Gen.
Psychiatry. 1983. - N 40. - P. 443-449.
Hurlbert D.F. A comparative study using orgasm consistency training in the treatment of women reporting
hypoactive sexual desire //J. Sex. Marital. Then — 1993. — N 19. — P. 41-55.
Hurlbert D.F., Apt C. Coital alignment technique and directed masturbation; a comparative study on female
masturbation //J. Sex. and Marital. Therapy. — 1995. — N 21. — P. 21-29.
McCabe M.P. Evaluation of a cognitive behavior therapy program for people with sexual dysfunction // J.
Sex. Marital. Then - 2001. - N 27. - P. 259-271.
McMullen S., Rosen R.C. Self-administered masturbation training in the treatment of primary orgasmic dys¬
function //J. Consult. Clin. Psychol. - 1979. - N 47 (5). - P. 912-918.
Sarwer D.B., Durlak J.A. A field trial of the effectiveness of behavioral treatment for sexual dysfunctions //J.
Sex. Marital. Then - 1997. - N 23. - P. 7-97.
Trudel G., Marchand A., Ravart M. et al. The effect of a cognitive behavior treatment program on hypoactive
sexual desire disorder in women // Sex. Rel. Then 2001. — N 16. — P. 145-164.
ФАРМАКОЛОГИЧЕСКИЕ ВМЕШАТЕЛЬСТВА
Отсутствуют одобренные FDA лекарственные препараты для лечения сексуальной
дисфункции у женщин, а эстрогеновая терапия остается резервом для диспареунии,
связанной с вагинальной атрофией. В табл. 4-15 показаны лекарственные препараты.
387
которые использовали не по одобренным показаниям; данные об эффективности
немногочисленны [213-215, 222, 225, 226, 280, 281, 283, 305-313]. Отсутствуют
исследования, включающие психологические вмешательства параллельно с назна¬
чением лекарственных препаратов. В клинической же практике после обследования
обоих партнеров [314, 315] обычно назначают комбинации когнитивной поведенче¬
ской и сексуальной терапии для усиления эффекта любой исследовательской медика¬
ментозной терапии.
Таблица 4-15. Применение препаратов для исследовательского лечения женской сексуальной дис¬
функции
Использование препаратов
не по одобренным показаниям
или исследовательские
лекарственные препараты
Комментарии
Расстройство сексуального интереса и влечения, расстройства субъективного
и комбинированного возбуждения
Тестостерон (тестроген)
Б пяти мультицентрических параллельных группах РКИ женщин в хирур¬
гической (четыре исследования [213-215, 225]) и естественной (одно
исследование [306]) менопаузе с диагностированным расстройством,
связанным с гипоактивностью полового влечения (05М-1У-ТР) было отме¬
чено улучшение «общей удовлетворяющей сексуальной активности» и
показателей опросников относительно полового влечения и реактивности.
Отсутствуют данные о долговременной безопасности; отсутствуют данные
у женщин со снижением эстрогена.
Одно РКИ женщин с гипопитуитарным состоянием показало улучшение
сексуальной функции [280].
Данные исследований, включающих женщин с надпочечниковой недоста¬
точностью, не согласуются между собой [281-283]
ДГЭА (предшественник эстрадиола
и тестостерона)
Исследование женщин в перименопаузе со сниженными ощущениями
благополучия и низким уровнем полового влечения не показало улучше¬
ния [307].
Б двух небольших исследованиях был продемонстрирован положительный
эффект у женщин с гипопитуитаризмом и у женщин с вторичным гипокор-
тицизмом [281, 282]
Эфедрин (а- и р-адренергические
агонисты)
Одно перекрестное РКИ 20 женщин [310]: 50 мг эфедрина усилили набу¬
хание влагалища, однако возникло не субъективное возбуждение в ответ
на эротический фильм
Силденафил (ингибитор ФДЭ5)
Одно исследование на 1-м уровне [305] с участием 781 женщины в пре-
и постменопаузе с расстройствами возбуждения и полового влечения
(за исключением специфических расстройств сексуального возбуждения
гениталий) показало отсутствие улучшения показателей сексуального вле¬
чения, ощущений, смазки или удовлетворения
Бупропион*’ (агонист доперимена
норадреналина)
Одно исследование на 1-м уровне [226] женщин с расстройством, связан¬
ным с гипоактивным половым влечением (08М-1\/-ТР), показало улучше¬
ние возбудимости и реактивности, однако не полового влечения
Тиболон (эстрогенный, прогестаген¬
ный андрогенный стероид)
По европейским данным, терапия в постменопаузе сопровождалась сексу¬
альным улучшением по сравнению с плацебо [308], а также по сравнению
с терапией в менопаузе, включающей 17р-эстрадиол (1 мг)+норэтиндрон
(1 мг).
Отсутствуют исследования у женщин с диагностированной сексуальной
дисфункцией.
Безопасность в отношении рака молочных желез под вопросом
388
Окончание табл. 4-15
Йохимбина гидрохлорид (норадре-
нергический препарат центрального
действия) плюс аргинин (предше¬
ственник N0)
В одном перекрестном РКИ 24 женщин: 6 мг йохимбина плюс 6 г аргини¬
на увеличили набухание влагалища, однако не субъективное возбуждение
в ответ на эротический фильм [309]
Расстройство генитального возбуждения, несмотря на возмещение нехватки эстрогенов
Силденафил (ингибитор ФДЭб)
Лабораторные доказательства улучшения [222]; клинически сложно выде¬
лить подгруппу с РГСВ в связи со снижением кровенаполнения сосудов.
Одно небольшое исследование показало положительный эффект [311].
Отсутствуют крупные исследования, сфокусированные на РГСВ
Оргазмическое расстройство
Силденафил (ингибитор ФДЭб)
В одном небольшом РКИ продемонстрирован положительный эффект на
оргазмическое расстройство [312]
Argin Мах*’ (травяная добавка)
В небольшом РКИ продемонстрирован положительный эффект через
4 нед [313]
* Включены только те РКИ, в которых тестостерон использовали для поднятия плазменного уровня
до физиологических значений у молодых женщин.
обратите внимание: перечислены только те препараты, о которых опубликовано хотя бы одно ран¬
домизированное испытание.
Примечание. ДГЭА — дегидроэпиандростерон; РГСВ — расстройство генитального полового воз¬
буждения; ГТ — гормональная терапия; N0 — оксид азота; ФДЭ — фосфодиэстераза; РКИ — рандо¬
мизированное клиническое испытание.
Адаптировано с разрешения из Basson R. Sexual dysfunction in women // New Engl. J. Med. 2006. — N
354 (14). — P. 1497-1506. Copyright 2006. Massachusetts Medical Society. Bee права защищены.
МЕТОДЫ ГОРМОНАЛЬНОЙ ТЕРАПИИ
Эстрогены
Для диспареунии, связанной с вульвовагинальной атрофией, рекомендуется мест¬
ная терапия. Низкие дозы могут быть доставлены при помощи вагинального кольца
из силастика или в форме слизисто-адгезивных вагинальных таблеток с аналогич¬
ным положительным эффектом и очень низкой системной абсорбцией. Доступные
селективные модуляторы эстрогеновых рецепторов (СМЭР) не являются агонистами
эстрогена во влагалище. К счастью, СМЭР оказывают множество индивидуальных
и классовых эффектов, и было показано, что лазофоксифен«^ (в настоящее время
находится на испытании) восстанавливает вагинальную атрофию. Тиболон, синте¬
тический стероид с тканеспецифическим действием, восстанавливает вагинальную
атрофию. Ожидаются дальнейшие испытания у женщин с сексуальными симптомами,
а тиболон пока остается недоступным в Северной Америке.
Когда требуется системная эстрогеновая терапия, необходимо назначить допол¬
нительный эстроген местно, который можно применять неопределенно долгий срок
[316]. Когда эстрогеновая добавка оказала положительное воздействие на инсом-
нию или диспареунию, можно ожидать возрастание мотивации быть сексуальной,
однако это не изучено досконально. Не было установлено значительных различий
между группами эстрогена и плацебо у тех, кто сообщал о сексуальном удовлетво¬
рении в исследовании группы инициативы женского здоровья. Однако сексуальная
дисфункция не была основным фокусом: в частности, были исключены женщины с
выраженными менопаузальными синдромами, да и исследовательский опросник был
неадекватным [316].
389
Тестостерон
Терапия тестостероном для положительного воздействия на жалобы женщин по
поводу низкого сексуального влечения в настоящее время находится в стадии изуче¬
ния. Женщины привлекаются на основе симптомов, а не уровней тестостерона, и пять
из шести последних исследований были проведены на женщинах в хирургической
менопаузе [212-215, 225]. Пациентки получали терапию эстрогеном: оральную — в
пяти исследованиях, трансдермальную - в одном и смесь — в одном. Все исследова¬
ния были спонсированы производителями, использовали (утвержденные) опросники
для оценки изменений при лечении. Первое исследование было перекрестным по
дизайну [212], во всех остальных были параллельные группы.
В первом исследовании при активной медикаментозной терапии было продемон¬
стрировано улучшение сексуальной реакции, однако не в аспектах, касающихся поло¬
вого влечения (сексуальные мысли, фантазии, первоначальное половое влечение,
предшествующее сексуальным опытам), в оставшихся выявили улучшение в отдель¬
ных аспектах реакции и полового влечения. В терминах ежедневно регистрируемых
«сексуально удовлетворительных эпизодов» глобальные отличия были незначи¬
тельными. При общем анализе данных было показано, что женщины, получавшие
тестостерон, имели на 1,9 больше эпизодов удовлетворительной половой активности
в месяц по сравнению с базовыми показателями, в то время как получавшие плаце¬
бо сообщали об увеличении частоты на 0,9 по сравнению с базовыми значениями.
Улучшение после плацебо наблюдали в отдельных группах, которые номинально при¬
нимали 300 мг ежедневно, но не в тех, где получали 150 и 450 мг ежедневно.
Исследования длились 24 нед. Краткосрочный прием не сопровождался значитель¬
ным увеличением гирсутизма, акне или вирилизирующими дефектами. Также отсут¬
ствовали изменения липидных параметров.
Несмотря на документированное положительное влияние трансдермального тесто¬
стерона у женщин с нормальными концентрациями эстрогена, фактически имеются
три основных фактора, которые ограничивают широкую рекомендацию терапии
[314, 315,317]. Первый заключается в том, что женщины обычно желают оставаться
сексуально активными бесконечно, а долгосрочные данные по безопасности терапии
тестостероном недостаточны. Во-вторых, отсутствует информация о положительном
сексуальном влиянии тестостерона у женщин с дефицитом эстрогена; в связи с широ¬
ко распространенными ограничениями долгосрочной терапии эстрогеном долго¬
срочная терапия тестостероном не допускает сопутствующую эстрогеновую терапию.
Третья основная причина — отсутствие четкого определения целевого расстройства.
Последние РКИ основываются на старой модели женской сексуальной реакции, кото¬
рая предполагает, что женщина инициирует или принимает сексуальные действия
преимущественно по причинам, связанным с половым влечением. Требуется прове¬
дение исследований у женщин, которые мотивируются быть сексуально активными
со своими партнерами (по причинам, не касающимся полового влечения), однако
обнаруживают, что их мысли и тело не могут быть возбуждены ранее эффективными
сексуальными стимулами. Многие из таких женщин были исключены из последних
РКИ, так как они, вероятно, могли обладать редкой сексуальной активностью, что
само по себе мешало их включению в исследование.
ЛЕЧЕНИЕ ВАГИНИЗМА
Научные исследования лечения вагинизма очень редки. То, что оба партнера уча¬
ствуют в поддержании незавершенной природы взаимоотношений, было многократ¬
но изучено, однако детали оптимального лечения партнера мужского пола не были
опубликованы.
Стандартная механическая терапия требует серии визитов к врачу в течение дли¬
тельного периода (от нескольких недель до многих месяцев). Она включает помощь
390
женщине, направленную на улучшение информированности о своей анатомии и обу¬
чение произвольному открытию и закрытию просвета влагалища с использованием
упражнений Кегеля, предпочтительно с возможностью контроля в просвете зеркала.
Полезно изменение концепции о том, что «в нее проникают», на ту, согласно которой
она способна активно охватывать объект своим влагалищем. Поощряются ежеднев¬
ные прикосновения к зоне в непосредственной близости от входа во влагалище. На
этой стадии пациентке желательно открывать половые губы собственными пальцами
так, что вход может обозреваться как женщиной при помощи зеркала, так и врачом.
Для максимальной информативности и терапевтического эффекта физикальное
исследование выполняется, когда женщина готова к нему. Диагноз подтверждают
путем исключения анатомической аномалии или аллодинии при вестибулите либо
другой патологии. Обычно первое исследование неполное и может не включать при¬
касаний врача. Позднее могут быть проведены более полные исследования. Обычно
рекомендуют серии приспособлений для проникновения во влагалище возрастающего
диаметра. Некоторые женщины выбирают первоначально самостимуляцию, которая
способствует генитальному возбуждению для облегчения установления приспособле¬
ния. Строго рекомендуется избегать попыток полового сношения до тех пор, пока не
будет возможным вводить более крупное приспособление. Установление его в присут¬
ствии партнера позволяет женщине обу^шться разделению контроля. На протяжении
всей терапии пара поощряется к сексуальному поведению, которое исключает про¬
никновение во влагалище.
Современные принципы лечения вагинизма.
* Механическая терапия.
« Поощрение секса без проникновения (без вагинальной стимуляции).
» Исследование женских мыслей о чем-то, проникающем в ее влагалище, и помощь
в смене этих мыслей.
« Изучение общей пассивности мужчины в отношениях, если предполагается, что
это приносит вред.
« Помощь паре в преодолении тревожности, возникающей под влиянием измене¬
ний, которые они совершают: их ситуация была переносима и безопасна для них,
часто на протяжении многих лет перед тем, как был начат поиск помощи.
Если преследуемая цель пары — беременность, а не половые сношения, после того
как женщина смогла вынести установление приспособления меньшего размера и спо¬
собна перенести полное генитальное обследование, она может избрать самооплодот¬
ворение семенной жидкостью партнера — как в домашних условиях, так и с помощью
врача в клинике.
Исследования исхода механического подхода ограничены, однако предполага¬
ются высокие ^фовни успешности. Клинический опыт, тем не менее, говорит о том,
что установление приспособления большего диаметра или введение полового члена
партнера может оказаться возможным, однако совсем не обязательно следует пере¬
ход обоих партнеров к совместному наслаждению половым актом. Пропагандируется
лучшее понимание и коррекция психологических проблем у обоих партнеров.
ДИСПАРЕУНИЯ
Из двух наиболее распространенных причин хронической диспареунии, СВВ и
вульвовагинальная атрофия, основанная на доказательных данных терапия суще¬
ствует только для последней. Ряд медицинских, психологических и хирургических
подходов ведения СВВ рассматриваются в сообщении №Н [318]. Назначение лекар¬
ственной терапии хронической боли и поощрение непроникающей сексуальной сти¬
муляции — основные направления консервативной терапии. Улучшение понимания
различных этиологических факторов, вероятно, основанных на генетике [263], может
позволить научно обосновать возможности терапии.
391
БУДУЩИЕ ОБЛАСТИ ИССЛЕДОВАНИЯ
Роль внутриклеточной продукции гормонов и их интракринные эффекты требуют
дальнейшего разъяснения. Измерения общей андрогенной активности, отражаю¬
щиеся концентрациями циркулирующих андрогенных метаболитов у женщин с сек¬
суальной дисфункцией, должны еще исследоваться. Разработка более селективных
эстрогейных и андрогенных модуляторов может способствовать улучшению сексуаль¬
ных проблем без нежелательных побочных воздействий любого из гормонов. Могут
потребоваться рандомизированные испытания комбинированных методик, которые
будут включать как медикаментозный, так и психологический подход.
СПИСОК ЛИТЕРАТУРЫ
]. Bhasin S., Benson G.S. Male sexual function // De Kretser D. (ed.). Knobil and Neill’s Physiology
of Reproduction. — 3rd ed. — Boston: Academic Press, 2006. — P. 1173-1194.
2. Kinsey A.C., Pomeroy W.B., Martin C.E. Sexual Behavior in the Human Male. - Philadelphia:
Saimders, 1948.
3. Masters E.H., Johnson V. Human Sexual Response. - Boston: Little, Brown, 1966.
4. Lue T.R, Tanagho E.A. Hemodynamics of erection // Tanagho E.A., Lue T.F., McClure R.D.
(ed.), Contemporary Management of Impotence and Infertility. - Baltimore: Williams and Wilkins,
1988. - P. 28-38.
5. Feldman H.A., Goldstein I., Hatzichristou D.G. et al. Impotence and its medical and psycho¬
social correlates: results of the Massachusetts Male Aging Study //J. Urol. — 1994. — Vol. 151. —
P. 54-61.
6. Laumann E.O., Paik A., Rosen R.C. The epidemiology of erectile dysfunction: results from
the National Health and Social Life Survey // Int.J. Impôt. Res. — 1999. - Vol. 11. — Suppl. 1. —
P. S60-S64.
7. Laumann E.O., Paik A., Rosen R.C. Sexual dysfunction in the United States: prevalence and pre¬
dictors //JAMA. - 1999. - Vol. 281. - P. 537-544.
8. Segraves R., Woodards T. Female hypoactive sexual desire disorder: history and current status j j
J. Sex. Med. - 2006. - Vol, 3. - P. 408-418.
9. NIH Consensus Development Panel on Impotence :NIH Consensus Conference. Impotence //
JAMA. - 1993. - Vol. 270. - P. 83-90.
10. The Process of Care Consensus Panel: The process of care model for evaluation and treatment of
erectile dysfunction//Int.J. Impôt. Res. — 1999. — Vol. 11, — P. 59-70. discussion 70-74.
11. Benet A.E., Melman A. The epidemiology of erectile dysfunction // Urol. Clin. North. Am. -
1995. - Vol. 22. - P. 699-709.
12. Lipshultz L.L, McConnell J., Benson G.S. Current concepts of the mechanisms of ejaculation.
Normal and abnormal states //J. Reprod. Med. — 1981. - Vol. 26. - P. 499-507.
13. Benson G.S., McConnell J., Lipshultz L.L et al. Neuromorphology and iieuropharmacology of
the human penis: an in vitro study //J. Clin. Invest. — 1980. - Vol. 65. - P. 506-513.
14. Lue T.F. Erectile dysfunction //N. Engl.J. Med. — 2000. — Vol. 342. — P. 1802-1813.
15. Christ GJ. The penis as a vascular organ. The importance of corporal smooth muscle tone in the
control of erection // Urol. Clin. North. Am. - 1995. - Vol. 22. - P. 727-745.
16. Christ GJ. Gap junctions and ion channels: relevance to erectile dysfunction // Int. J. Impôt.
Res. - 2000. - Vol. 12. - Suppl. 4. - P. S15-S25.
17. Zeng X., Keyser B., Li М., Sikka S.C. T-type (alG) low voltage-activated calcium channel
interactions with nitric oxide-cyclic giianosine monophosphate pathway and regulation of calcium
homeostasis in human cavernosa! cells j j J. Sex. Med. - 2005. — Vol. 2. - P. 620-630. Discussion
630-633.
18. Somlyo A.P., Somlyo A.V. Ca^* sensitivity of smooth muscle and nonmuscle myosin II: modulated
by G proteins, kinases, and myosin phosphatase // Physiol. Rev. — 2003. — Vol. 83. — P. 1325-1358.
19. 0-Uchi J., Komukai K., Kusakari Y. et al. al-Adrenoceptor stimulation potentiates L-type Ca^'
current through Ca^*/calmodulin-dependent PKII (CaMKII) activation in rat ventricular myocytes //
Proc. Natl. Acad. Sci USA. - 2005. - Vol. 102. - P. 9400-9405,
20. Krall J.F., Fittingoff М., Rajfer J. Characterization of cyclic nucleotide and inositol 1,4,5-trispho-
sphate-sensitive calcium-exchange activity of smooth muscle cells cultured from the human corpora
cavernosa //Biol. Reprod. — 1988. — Vol. 39. — P. 913-922.
392
21. Fittingoff M., Krall J.F. Changes in inositol polyphosphate-sensitive calcium exchange in aortic
smooth muscle cells in vitro //J. Cell. Physiol. - 1988. — Vol. 134. — P. 297-301.
22. Hewawasam P., Fan W., Ding M. et al, 4-Aryl-3-(hydroxyalkyl)quinolin-2-ones: novel maxi-K
channel opening relaxants of corporal smooth muscle targeted for erectile dysfunction // J. Med,
Chem. - 2003. - Vol. 46. - P. 2819-2822.
23. Christ G.J., Day N., Santizo C. et al. Intracorporal injection of hSlo cDNA restores erectile
capacity in STZ-diabetic F-344 rats in vivo // Am, J. Physiol. Heart. Circ. Physiol. — 2004. — Vol.
287. - P. H1544-H1553.
24. Meiman A., Bar-Chama N., McCullough A. et al. The first human trial for gene transfer
therapy for the treatment of erectile dysfunction: preliminary results // Eur. Urol. — 2005. — Vol.
48. - P. 314-318.
25. Christ G.J., Moreno A.P., Meiman A., Spray D.C. Gap junction-mediated intercellular diffusion
of Ca^' in cultured human corporal smooth muscle cells // Am. J. Physiol, - 1992. — Vol. 263. -
P. C373-C383.
26. Ignarro L.J., Bush P.A., Buga G.M. et al. Nitric oxide and cyclic GMP formation upon electri¬
cal field stimulation cause relaxation of corpus cavemosum smooth muscle // Biochem. Biophys. Res.
Commun. - 1990. - Vol. 170. - P. 843-850.
27. Mills T.M., Chitaley K., Lewis R.W., Webb R.C, Nitric oxide inhibits RhoA/Rho-kinase signaling
to cause penile erection // Eur. J. Pharmacol. - 2002. — Vol. 439. - P. 173-174.
28. Naylor A.M. Endogenous neurotransmitters mediating penile erection // Br. J. Urol. — 1998. —
Vol. 81. - P. 424-431.
29. Goldstein I., Lue T.F., Padma-Nathan H. et al. Oral sildenafil in the treatment of erectile dysfunc¬
tion. Sildenafil Study Group [see comments, published erratum appears // N. Engl. J. Med, - 1998. —
Vol. 339 (1). - P. 59] // N. Engl. J. Med. - 1998. - Vol. 338. - P. 1397-1404,
30. Gong M.C., lizuka K., Nixon G. et al. Role of guanine nucleotide-binding proteins - ras-family
or trimeric proteins or both — in Ca2+ sensitization of smooth muscle // Proc. Natl. Acad. Sci. USA. -
1996. - Vol. 93. - P. 1340-1345.
31. Chikumi H., Fukuhara S., Gutkind J.S. Regulation of G protein-linked guanine nucleotide
exchange factors for Rho, PDZ-RhoGEF and LARG by tyrosine phosphorylation: evidence of a role for
focal adhesion kinase //J. Biol Chem. - 2002. - Vol. 277. - P. 12463-12473.
32. Gong M.C., Fujihara H., Somiyo A.V., Somiyo A.P. Translocation of RhoA associated with Ca^*
sensitization of smooth muscle //J. Biol. Chem. — 1997. - Vol. 272. - P, 10704-10709,
33. Wang H., Eto M., Steers W.D. et al. RhoA-mediated Ca2+ sensitization in erectile function j j
J. Biol, Chem. - 2002. - Vol. 277. - P. 30614-30621.
34. Jin L„ Liu T., Lagoda G.A. et al. Elevated RhoA/Rho-kinase activity in the aged rat penis; mecha¬
nism for age-associated erectile dysfunction // Faseb. J. - 2006. - Vol, 20. — P. 536-538.
35. Mills T.M., Chitaley K., Wingard C.J. et al. Effect of Rho-kinase inhibition on vasoconstriction
in the penüe circulation //J. Appl. Physiol. - 2001. - Vol. 91. - P. 1269-1273.
36. Kwan M., Greenleaf W.J., Mann J. et al. The nature of androgen action on male sexuality: a
combined laboratory-self-report study on hypogonadal men //J. Clin, Endocrinol. Metab. — 1983. —
Vol. 57. - P. 557-562.
37. Alexander G.M., Sherwin B.B. The association between testosterone, sexual arousal and selec¬
tive attention for erotic stimuli in men // Horm. Behav, — 1991. — Vol. 25. — P. 367-381.
38. Redoute J.. Stoleru S., Pugeat M. et al. Brain processing of visual sexual stimuli in treated and
untreated hypogonadal patients // Psychoneuroendocrinology, — 2005. — Vol. 30. — P. 461-482.
39. King B.E., Packard M.G., Alexander G.M. Affective properties of intra-medial preoptic area
injections of testosterone in male rats // Neurosci Lett. - 1999. — Vol. 269. - P. 149-152.
40. Bagatell C.J., Heiman J.R., Rivier J.E., Bremner W.J. Effects of endogenous testosterone and
estradiol on sexual behavior in normal young men [published erratum appears // J. Clin. Endocrinol.
Metab. - 1994. - Vol. 78 (6). - P. 1520] // J. Clin. Endocrinol. Metab. - 1994. - Vol. 78. -
P. 711-716.
41. Carani C., Bancroft J., Granata A. et al. Testosterone and erectile function, nocturnal penile
tumescence and rigidity, and erectile response to visual erotic stimuli in hypogonadal and eugonadal
men // Psychoneuroendocrinology. - 1992. - Vol. 17. - P. 647-654.
42. Buena F.. Swerdloff R,S., Steiner B.S. et al. Sexual function does not change when serum testos¬
terone levels are pharmacologically varied within the normal male range j j Fertil. Steril. — 1993. —
Vol. 59. - P. 1118-1123.
393
43. Lugg J.A., Rajfer J., Gonzalez-Cadavid N.F. Diliydrotestosterone is the active androgen in the
maintenance of nitric oxide-mediated penile erection in the rat j j Endocrinology. — 1995. — Vol.
136. - P. 1495-1501.
44. Shabsigh R. The effects of testosterone on the cavernous tissue and erectile function // World.
J. Urol. - 1997. - Vol. 15. - P. 21-26.
45. Bhasin S„ Cunningham G.R., Hayes F.J. et al. Testosterone therapy in adult men with androgen
deficiency syndromes; an endocrine society clinical practice guideline //J. Clin. Endocrinol. Metab. —
2006. - Vol. 91. - P. 1995-2010.
46. Arver S., Dobs A.S., Meikle A.W. et al. Improvement of sexual function in testosterone deficient
men treated for 1 year with a permeation enhanced testosterone transdermal system // J. Urol. —
1996. - Vol. 155. - P. 1604-160S.
47. Jain P., Rademaker A.W., McVary K.T. Testosterone Supplementation for erectile dysfunction:
results of a meta-analysis //J. Urol. — 2000. - Vol. 164. — P. 371-375.
48. Shabsigh R., Kaufman J.M., Steidle C., Padma-Nathan H. Randomized study of testosterone gel
as adjunctive therapy to sildenafil in hypogonadal men with erectile dysfunction who do not respond to
sildenafil alone //J. Urol. - 2004. - Vol. 172. - P. 658-663.
49. Aversa A.. Mazzilli R, Rossi T, et al. Effects of sildenafil (Viagra) administration on seminal
arameters and post-ejaculatory refractory time in normal males // Hum. Reprod. — 2000. — Vol.
15. - P. 131-134.
50. Korenman S.G., Morley J.E., Mooradian A.D, et al. Secondary hypogonadism in older men: its
relation to impotence //J. Clin, Endocrinol. Metab. — 1990. - Vol. 71. — P. 963-969.
51. Beck J.G. Hypoactive sexual desire disorder: an overview //J. Consult. Clin. Psychol. - 1995. -
Vol. 63. - P. 919-927.
52. Rosen R.C., Leiblum S.R. Hypoactive sexual desire // Psychiatr. Clin, North. Am. - 1995. -
Vol. 18. - P. 107-121.
53. Segraves K.B., Segraves R.T. Hypoartive sexual desire disorder; prevalence and comorbidity in
906 subjects //J. Sex. Marital. Ther. - 1991. - Vol, 17. - P. 55-58.
54. Panser L.A., Rhodes T., Girman C.J. et al. Sexual function of men ages 40 to 79 years; the
Olmsted County Study of Urinary Symptoms and Health Status Among Men //J. Am. Geriatr. Soc. -
1995. - Vol. 43. - P. 1107-1111.
55. LoPiccolo J. Diagnosis and treatment of male sexual dysfunction // J. Sex. Marital. Ther. —
1985,-Vol. 11.-P. 215-232.
56. Chevret M., Jaudinot E., Sullivan K. et al. Impact of erectile dysfunction (ED) on sexual life
of female partners: assessment with the Index of Sexual Life (ISL) questionnaire //J. Sex. Marital.
Ther. - 2004. - Vol. 30. - P. 157-172.
57. Johannes C.B., Araujo A.B., Feldman H.A. et al. Incidence of erectile dysfunction in men 40 to
69 years old: longitudinal results from the Massachusetts Male Aging Study //J. Urol. — 2000. — Vol.
163. - P. 460-463.
58. Braun M-, Wassmer G., Klotz T. et al. Epidemiology of erectile dysfunction: results of the
“Cologne Male Survey” // Int.J. Impôt. Res, - 2000. - Vol. 12. — P. 305-311.
59. McKinlay J.B., Digruttolo L., Classer D. et al. International differences in the epidemiology of
male erectile dysfunction // Int. J. Clin. Pract. Suppl. — 1999. — Vol. 102. — P. 35.
60. McKinlay J.B. The worldwide prevalence and epidemiology of erectile dysfunction j j Int. J.
Impôt. Res. - 2000. - Vol. 12. - Suppl. 4. - P. S6-S11,
61. Ayta LA., McKinlayJ.B., Krane RJ, The likely worldwide increase in erectile dysfunction between
1995 and 2025 and some possible policy consequences // BJU Int. - 1999. — Vol. 84. - P. 50-56.
62. Feldman H.A., Johannes C.B., Derby C.A. et al. Erectile dysfunction and coronary risk factors:
prospective results from the Massachusetts Male Aging Study // Prev. Med. — 2000. — Vol. 30. —
P. 328-338.
63. Derby C.A., Mohr B.A., Goldstein I. et al. Modifiable risk factors and erectile dysfunction: can
lifestyle changes modify risk? // Urology. — 2000. — Vol. 56. — P. 302-306.
64. Rosen R., Altwein J., Boyle P. et al. Lower urinary tract symptoms and male sexual dysfunction:
the multinational survey of the aging male (MSAM-7) // Eur, Urol. - 2003. — Vol. 44. — P. 637-
649.
65. Braun M.H., Sommer R, Haupt G. et al. Lower urinary tract symptoms and erectile dysfunction:
co-morbidity or typical “Aging Male” symptoms? Results of the «Cologne Male Survey» j j Eur, Urol. -
2003. - Vol. 44. - P. 588-594.
394
66. Barqawi A., O’Donnell C., Kumar R. et al. Correlation between LUTS (AUA-SS) and erectile
dysfunction (SHIM) in an age-matched racially diverse male population: data from the Prostate Cancer
Awareness Week (PCAW) // Int. J. Impôt. Res. - 2005. - Vol, 17. - P. 370-374.
67. Glina S., Santana A.W., Azank F. et al. Lower urinary tract symptoms and erectile dysfunction
are highly prevalent in ageing men // BJU Int. — 2006. — Vol. 97. — P. 763-765.
68. McVary K. Lower urinarynract symptoms and sexual dysfunction: epidemiolog}^ and pathophysi¬
ology // BJU Int. - 2006. ' Vol. 97. - Suppl. 2. - P. 23-28; discussion 44-45.
69. Paick S.H., Meehan A., Lee M., Penson D.R, Wessells H. The relationship among lower uri¬
nary tract symptoms, prostate specific antigen and erectile dysfunction in men with benign prostatic
hyperplasia: results from the proscar long-term efficacy and safety study //J. Urol. — 2005. — Vol.
173. - P. 903-907.
70. McVary K.T. Interrelation of erectile dysfunction and lower urinary tract symptoms // Drugs.
Today (Bare.). - 2005. - Vol. 41. - P. 527-536.
71. Christ G.J., Hodges S. Molecular mechanisms of detrusor and corporal myocyte contraction;
identifying targets for phannacotherapy of bladder and erectile dysfunction // Br. J. Pharmacol. -
2006. - Vol. 147. - Suppl, 2. - P. S41-S55.
72. NIH Consensus Development Panel on Impotence: HIH Consensus Conference //JAMA. —
1993. - Vol. 270. - P. 83-90.
73. Cappelleri J.C,, Rosen R.C., Smith M.D. et al. Diagnostic evaluation of the erectile function
domain of the International Index of Erectile Function j j Urology, — 1999. - Vol. 54. — P. 346-351.
74. Lue T.R, Giuliano R, Montorsi R et al. Summary of the recommendations on sexual dysfunc¬
tions in men //J. Sex. Med. - 2004. - Vol. 1, - P. 6-23.
75. O’Leary M.P., Fowler F.J., Lenderking W.R. et al, A brief male sexual function inventory //
Urology. - 1995. - Vol. 46. - P. 697-706.
76. Rosen R.C., Cappelleri J.C., Smith M.D. et al. Development and evaluation of an abridged,
5-item version of the International Index of Erectile Function (IIEF-5) as a diagnostic toql for erectile
dysfunction // Int.J. Impôt. Res. — 1999. — Vol. 11. — P. 319-326.
77. Aitchison M., Aitchison J., Carter R. Ls the penile brachial index a reproducible and useful mea¬
surement.? // Br. J. Urol. - 1990. - Vol. 66. - P. 202-204.
78. Mueller S.C., Wallenberg-Pachaly H„ Voges G.E., Schild H.H. Comparison of selective inter¬
nal iliac pharmaco-angiography, penile brachial index and duplex sonography with pulsed Doppler
analysis for the evaluation of vasculogenic (arteriogenic) impotence //J. Urol. — 1990. -- Vol, 143. —
P. 928-932.
79. Brock G. Tumescence monitoring devices; past and present j j Hellstrom W.J.G. (ed.). The
Handbook of Sexual Dysfunction. — San Francisco: American Society of Andrology, Allen Press,
Lawrence, Kansas, 1999. - P. 65-69.
80. Buvat J., Lemaire A. Endocrine screening in 1,022 men with erectile dysfunction: clinical signifi¬
cance and cost-effective strategy [see commentsj //J. Urol. — 1997. — Vol. 158. — P. 1764-1767.
81. Citron J.T„ Ettinger B., Rubinoff H. et al. Prevalence of hypothalamic-pituitary imaging
abnormalities in impotent men with secondary hypogonadism // J. Urol. — 1996. — Vol. 155. —
P. 529-533.
82. Rosen R.C. Psychogenic erectile dysfunction. Classification and management // Urol. Clin.
North. Am. - 2001. - Vol. 28. - P. 269-278.
83. Abrahamson D.J., Barlow D.H,, BeckJ.G. et al. The effects of attentional focus and partner
responsiveness on sexual responding: replication and extension // Arch. Sex. Behav. — 1985. - Vol.
14. - P. 361-371.
84. Kilmann P.R., Boland J.P., Norton S.P., Davidson E., Caid C. Perspectives of sex therapy out¬
come: a survey of AASECT providers // J. Sex, Marital. Ther. — 1986. — Vol. 12. — P. 116-138.
85. Montague D.K., Jarow J.P., Broderick G.A. et al. Chapter 1: The management of erectile dys¬
function; an AUA update //J. Urol. - 2005, - Vol. 174, - P. 230-239.
86. Haning H., Niewohner U., Bisthoff E. Phosphodiesterase type 5 (PDE5) inhibitors j j Prog,
Med. Chem. - 2003. - Vol. 41. - P. 249-306.
87. Bischoff E. Potency, selectivity and consequences of nonselectivity of PDE inhibition // Int. J.
Impôt. Res. - 2004. - Vol. 16. - Suppl. 1. - P. S11-S14.
88. Boolell M., Allen MJ., Ballard S.A. et al. Sildenafil: an orally active type 5 cyclic GMP-specific
phosphodiesterase inhibitor for the treatment of penile erectile dysfunction j j Int. J. Impôt. Res. —
1996. - Vol. 8. - P. 47-52.
395
89. Taher A., Meyer M., Stief C.G. et al. Cj'clic nucleotide phosphodiesterase in human caveimous
smooth muscle // World. J. Urol. - 1997. - Vol. 15. - P. 32-35.
90. Jeremy J.Y., Ballard S.A., Naylor A.M. et al. Effects of sildenafil, a type-5 cGMP phosphodi¬
esterase inhibitor and papaverine on cyclic GMP and cyclic AMP levels in the rabbit corpus cavernosum
in vitro // Br.J, Urol. - 1997. - Vol. 79. - P. 9.58-963.
91. Stief C.G., Uckert S., Becker A.J. et al. The effect of the specific phosphodiesterase (PDE)
inhibitors on human and rabbit cavernous tissue in vitro and in vivo //J. Urol, — 1998. — Vol. 159, —
P. 1390-1393.
92. Carter A.J., Ballard S.A., Naylor A.M. Effect of the selective phosphodiesterase tj'pe 5 inhibitor
sildenafil on erectile dysfunction in the anesthetized dog //J. Urol. - 1998. — Vol. 160. — P. 242-246.
93. Wallis R.M., Corbin J.D., Francis S.H., Ellis P. Tissue distribution of phosphodiesterase families
and the effects of sildenafil on tissue cyclic nucleotides, platelet function, and the contractile responses
of trabeculae carneae and aortic rings in vitro //Am.J. Cardiol. — 1999. — Vol. 83. — P. 3C-12C.
94. Saenz de Tejada I., Angulo J., Cuevas P. et al. The phosphodiesterase inhibitory selectivity and
the in vitro and in vivo potency of the new PDE5 inhibitor vardenafil // Int. J. Impôt. Res. — 2001. —
Vol. 13. - P. 282-290.
95. Goldstein I. A 36-week, open label, non-comparative study to assess the long-term safety of
sildenafil citrate (Viagra) in patients with erectile dysfunction // Int.J. Clin. Pract. Suppl. - 1999. -
Vol. 102. - P. 8-9.
96. Rendell M.S., Rajfer J., Wicker P.A., Smith M.D. Sildenafil for treatment of erectile dysfunction
in men with diabetes: a randomized controlled trial. Sildenafil Diabetes Study Group [see commentsj //
JAMA. - 1999. - Vol. 281. - P. 421-426.
97. Yu G., Mason H„ Wu X. et al. Substituted pyrazolopyridopyridazines as orally bioavailable
potent and selective PDE5 inhibitors: potential agents tor treatment of erectile dysfunction j j J. Med.
Chem. - 2003. - Vol. 46. - P, 457-460.
98. Seftel A.D. Phosphodiesterase type 5 inhibitor differentiation based on selectivity, pharmacoki¬
netic and efficacy profiles // Clin. Cardiol. — 2004. — Vol. 27. — P. 114-119.
99. Grossman E.B,, Swan S.K., Muirhead GJ. et al. The pharmacokinetics and hemodynamics of
sildenafil citrate in male hemodialysis patients // Kidney Int. — 2004. — Vol. 66. — P. 367-374.
100. Sussman D.O. Pharmacokinetics, pharmacodynamics and efficacy of phosphodiesterase type 5
inhibitors //J. Am. Osteopath. Assoc. - 2004. — Vol. 104. — P. S11-S15.
101. Rajagopalan P., Mazzu A., Xia C. et al. Effect of high-fat breakfast and moderate-fat evening
meal on the phamiacokinetics of vardenafil, an oral phosphodiesterase-5 inhibitor for the treatment of
erectile dysfunction // J. Clin. Pharmacol. - 2003. - Vol. 43. - P. 260-267.
102. Blanker M.H., Thomas S., Bohiien A.M. Systematic review of Viagra RCTs // Br. J. Gen.
Pract. ~ 2002. - Vol. 52. - P. 329.
103. Burls A., Gold L.. Clark W. Systematic review of randomised controlled trials of sildenafil
(Viagra) in the treatment of male erectile dysfunction j j Br. J. Gen. Pract. — 2001. — Vol. 51. —
P. 1004-1012.
104. Fink H.A., Mac Donald R., Rutks I.R. et al. Sildenafil for male erectile dysfunction: a systematic
review and meta-analysis // Arch. Intern. Med. - 2002. - Vol. 162. - P. 1349-1360,
105. Markou S., Perimenis P., Gyftopoulos K. et al. Vardenafil (Levitra) for erectile dysfunc¬
tion; a systematic review and meta-analysis of clinical trial reports // Int. J. Impôt. Res. — 2004. —
Vol. 16. - P. 470-478.
106. Montorsi F., McCullough A. Efficacy of sildenafil citrate in men with erectile dysfunction
following radical prostatectomy: a systematic review of clinical data j j J. Sex. Med. - 2005. -
Vol. 2. - P. 658-667.
107. Moore R.A., Derry S., McQuay H.J. Indirect comparison of interventions using published ran¬
domised trials: systematic review of PDE-5 inhibitors for erectile dysfunction // BMC Urol. — 2005. —
Vol.5.-P. 18.
108. Jarow J.P., Burnett A.L., Geringer A.M. Clinical efficacy of sildenafil citrate based on etiology
and response to prior treatment [see comments] //J. Urol. - 1999. - Vol. 162. - P. 722-725.
109. Dinsmore W.W., Hodges M„ Hargreaves C. et al. Sildenafil citrate (Viagra) in erectile dysfunc¬
tion: near normalization in men with broad-spectrum erectile dysfunction compared with age-matched
healthy control subjects [published erratum appears in Urology. - 1999. - Vol. S3 (5). — P. 1072] //
Urology. - 1999. - Vol. 53. - P. 800-805.
110. Goldenberg M.M. Safety and efficacy of sildenafil citrate in the treatment of male erectile dys¬
function // Clin. Ther. - 1998. - Vol. 20. - P. 1033-1048.
396
ni. Brock G., Nehra A., Lipshultz L.I. et al. Safety and efficacy of vardenafil for the treatment of
men with erectile dysfunction after radical retropubic prostatectomy //J. Urol. — 200.3. — Vol. 170. —
P. 1278-1283.
112. Donatucci C., Eardley I., Biivat J. et al. Vardenafil improves erectile function in men with erec¬
tile dysfunction irrespective of disease severity and disease classification j j J. Sex. Med. — 2004. — Vol.
1. - P. 301-309.
113. Donatucci C., Taylor T., Thibonnier M. et al. Vardenafil improves patient satisfaction with erec¬
tion hardness, orgasmic function and overall sexual experience, while improving quality of life in men
with erectile dysfunction //J. Sex. Med. — 2004. — Vol. 1. — P. 185-192.
114. Hatzichristou D., Montorsi R, Buvat J. et al. The efficacy and safety of flexible-dose vardenafil
(Levitra) in a broad population of European men j j Eur. Urol, — 2004. — Vol. 45. — P. 634-641.
115. Hellstrom W.J., Gittelman М., Karlin G. et al. Vardenafil for treatment of men with erectile
dysfunction: efficacy and safety in a randomized, double-blind, placebo-controlled trial //J, Androl, —
2002. - Vol. 23. - P. 763-771.
116. Nehra A., Grantmyre J., Nadel A. et al. Vardenafil improved patient satisfaction with erectile
hardness, orgasmic function and sexual experience in men with erectile dysfunction following nerve
sparing radical prostatectomy //J. Urol. - 2005. - Vol. 173. - P. 2067-2071,
117. Rosen R., Shabsigh R., Berber M, et al. Efficacy and tolerability of vardenafil in men with mild
depression and erectile dysfunction: the depress!on-related improvement with vardenafil for erectile
response study //Am.J. Psychiatry. - 2006. - Vol. 163. - P, 79-87.
118. Brock G.B., McMahon C.G., Chen K.K. et al. Efficacy and safety of tadalafil for the treatment of
erectile dysfunction: results of integrated analyses //J. Urol. - 2002. — Vol. 168. — P. 1332-1336.
119. Carson C., Shabsigh R., Segal S. et al. Efficacy, safety, and treatment satisfaction of tadalafil
versus placebo in patients with erectile dysfunction evaluated at tertiary-care academic centers //
Urology. - 2005, - Vol. 65. - P. 353-359.
120. Padma-Nathan H., McMurray J.G., Pullman W.E. et al. On-demand IC351 (Cialis)'enhances
erectile function in patients with erectile dysfunction j j Int. J. Impôt. Res. — 2001. - Vol. 13. -
P. 2 -9.
121. Porst H., Padma-Nathan H., Giuliano R et al. Efficacy of tadalafil for the treatment of erectile
dysfunction at 24 and 36 hours after dosing: a randomized controlled trial // Urology. — 2003. — Vol.
62. - P. 121-125. discussion 125-126.
122. Saenz de Tejada I., Anglin G., Knight J.R., Emmick J.T. Effects of tadalafil on erectile dysfunc¬
tion in men with diabetes // Diabetes. Care. — 2002. — Vol. 25. — P. 2159-2164,
123. Staab A„ Tillmann C.. Forgue S.T. et al. Population dose-response model for tadalafil in the
treatment of male erectile dysfunction j j Pharm. Res. - 2004. — Vol. 21. - P. 1463-1470.
124. Giuliano R, Hultling C., el Masry W.S. et al. Randomized trial of sildenafil for the treatment
of erectile dysfunction in spinal cord injury. Sildenafil Study Group // Ann. Neurol. — 1999. — Vol,
46. - P. 15-21.
125. Morales A., Gingell C., Collins M. et al. Clinical safety of oral sildenafil citrate (Viagra) in the
treatment of erectile dysfunction // Int.J. Impôt. Res. - 1998. — Vol. 10. — P. 69-73.
126. Hatzichristou D.G. Phosphodiesterase 5 inhibitors and nonarteritic anterior ischemic optic
neuropathy (NAION): coincidence or causality //J. Sex. Med. — 2004. — Vol. 2. — P. 751-758.
127. Buoiio L., Foroozan R., Sergott R.C., Savino P.J, Nonarteritic anterior ischemic optic neuropa¬
thy // Curr. Opin. Ophthamol. - 2002. - Vol. 13. - P. 357-361.
128. Feenstra J., Drie-Pierik R,J„ Lade C.R, Strieker B.H, Acute myocardial infarction assodated
with sildenafil [letter, see comments] // Lancet. - 1998. - Vol. 352. - P. 957-958.
129. Zusman R.M., Morales A., Glasser D.B., Osterloh LH. Overall cardiovascular profile of sildena¬
fil citrate //Am.J. Cardiol. — 1999. - Vol. 83. — P. 35C-44C.
130. Cheitliii M.D., Huttcr Jr. A.M., Brindis R.G. et al. Use of sildenafil (Viagra) in patients with
cardiovascular disease. Technology and Practice Executive Committee [published erratum appears in
Circulation. -■ 1999. — Vol. 100(23). — P. 2389, see comments] // Circulation. ^ 1999, — Vol. 99. -
P. 168-177.
131. MullerJ.E., Mittleman A„ Madure M. et al. Triggering myocardial infarction by sexual activity.
Low absolute risk and prevention by regular physical exertion. Determinants of Myocardial Infarction
Onset Study Investigators [see comments] //JAMA. — 1996. - Vol. 275. — P. 1405-1409.
132. Herrmann H.C., Chang G., Klugherz B.D., Mahoney P.D. Hemodynamic effects of sildenafil in
men w'ith severe coronary artcr>' disease // N. Engl.J. Med. - 2000. - Vol. 342. ~ P. 1622-1626.
397
133. Kloner R.A. Cardiovascular risk and sildenafil 11 Am. J. Cardiol. - 2000. - Vol. 86. -
P. 57F-61F.
134. Highleyman L. Protease inhibitors and sildenafil (Viagra) should not be combined j j BETA. —
1999. - Vol. 12. - P. 3.
135. Bailey D.G., Dresser G.K. Interactions between grapefruit juice and cardiovascular drugs //
Am. J. Cardiovasc. Drugs. - 2004. - Vol. 4. - P. 281-297.
136. Conti C.R., Pepine C.J-, Sweeney M. Efficacy and safety of sildenafil citrate in the treatment of
erectile dysfunction in patients with ischemic heart disease // Am. J, Cardiol. — 1999, — Vol. 83. —
P. 29C-34C.
137. Wespes E., Amar E., Hatzichristou D. et al. EAU Guidelines on erectile dysfunction: an update //
Eur. Urol. - 2006. - Vol. 49. - P. 806-815.
138. McMahon C. Comparison of efficacy, safety, and tolerability of on-demand tadalafil and daily
dosed tadalafil for the treatment of erectile dysfunction //J. Sex. Med. — 2005. - Vol. 2. - P. 415-
425. Discussion 425-417.
139. Carson 3rd C.C. Cardiac safety in clinical trials of phosphodiesterase 5 inhibitors // Am. J.
Cardiol. - 2005. - Vol. 96. - P. 37M-41M.
140. Jackson G. Hemodynamic and exercise effects of phosphodiesterase 5 inhibitors // Am. J.
Cardiol. - 2005. - Vol. 96. - P. 32M-36M.
141. Kloner R.A. Novel phosphodiesterase type 5 inhibitors: assessing hemodynamic effects and
safety parameters // Clin. Cardiol. — 2004. — Vol. 27, - P. 120-125.
142. Thadani U., Smith W., Nash S. et al. The effect of vardenafil, a potent and highly selective
pliosphodiesterase-5 inhibitor for the treatment of erectile dysfunction, on the cardiovascular response
to exercise in patients with coronary artery disease // J. Am. Coll. Cardiol. - 2002. - Vol. 40. -
P. 2006-2012.
143. McCullough A.R., Barada J.H., Fawzy A. et al. Achieving treatment optimization with sildenafil
citrate (Viagra) in patients with erectile dysfunction j j Urology. - 2002. — Vol. 60. — P. 28-38.
144. Lau D.H., Kommu S., Mumtaz F.H. et al. The management of phosphodiesterase inhibitor
failure // Curr. Vase. Pharmacol. - 2006. - Vol. 4. - P. 89-93.
145. Martinez J.M. Prognostic factors for response to sildenafil in patients with erectile dysfunc¬
tion // Eur. Urol. - Vol. 40. - P. 641-646.
146. Wespes E., Rammal A., Garbar C. Sildenafil no-respoiiders: hemodynamic and тофЬотеГгк
studies // Eur. Urol. - 2005. - Vol. 48. - P. 136-139.
147. McGarvey M.R. Tough choices: the cost-effectiveness of sildenafil [editorial; comment] // Ann.
Intern. Med. - 2000. - Vol. 132. - P. 994-995.
148. Smith K.J., Roberts M.S. The cost-effectiveness of sildenafil [see comments] // Ann. Intern.
Med. - 2000. - Vol. 132. - P. 933-937.
149. Tan H.L, Economic cost of male erectile dysfunction using a decision analytic model: for a
h^othetical managed-care plan of 100,000 members // Pharmacoeconomics. — 2000. — Vol. 17. —
P. 77-107.
150. Sun P., Seftel A., Swindle R. et al. The costs of caring for erectile dysfunction in a managed
care setting: evidence from a large national claims database //J. Urol. — 2005. — Vol, 174. — P. 1948-
1952.
151. Plumb J.M., Guest J.F. Annual cost of erectile dysfunction to UK society //Pharmacoeconomics. -
1999. - Vol. 16. - P. 699-709.
152. Vrijhof H.J., Delaere K.P, Vacuum constriction devices in erectile dysfunction: acceptance and
effectiveness in patients with impotence of organic or mixed aetiology // Br. J. Urol. — 1994. ■- Vol.
74. - P. 102-105.
153. Cookson M.S., Nadig P.W. Long-term results with vacuum constriction device j j J. Urol. —
1993. - Vol. 149. - P. 290-294.
154. Lewis J.H,, Sidi A.A., Reddy P.K. A way to help your patients who use vacuum devices j j
Contemp. Urol. - 1991. - Vol. 3. - P. 15-21.
155. Engelhardt P.P., Plas E., Hubner W.A., Pfluger H. Comparison of intraurethral liposomal and
intracavernosal prostaglandin-El in the management of erectile dysfunction j j Br. J. Urol. — 1998. —
Vol. 81.-P. 441-444.
156. Kim E.D.. McVary K.T, Topical prostaglandin-El for the treatment of erectile dysfunction [see
comments] //J. Urol. - 1995. - Vol. 153. - P. 1828-1830.
398
157. Peterson C.A., Bennett A.H., Hellstrom WJ, et al. Erectile response to transurethral alprostadil,
prazosin and alprostadil-prazosin combinations //J. Urol. — 1998. — Vol. 159. — P. 1523-1527.
158. Fulgham P.P., Cochran J.S., Denman J.L. et al. Disappointing initial results with transurethral
alprostadil for erectile dysfunction in a urology practice setting // J. Urol. — 1998. — Vol. 160. —
P. 2041-2046.
159. Buvat J., Lemaire A., RatajczykJ. Acceptance, efficacy and preference of sildenafil in patients
on long term auto-intracavemosal therapy: a study with follow-up at one year // Int. J. Impôt. Res. —
2002. - Vol. 14. - P. 483-486.
160. El-Sakka A.I. Intracavernosal prostaglandin El self vs office injection therapy in patients with
erectile dysfunction // Int.J. Impôt. Res. — 2006. - Vol. 18. — P. 180-185.
161. Heaton J.P., Lording D., Liu S.N. et al. Intracavernosal alprostadil is effective for the treatment
of erectile dysfunction in diabetic men // Int. J. Impôt. Res. - 2001. — Vol. 13. - P. 317-321.
162. Linet O.L, Ogrinc RG. Efficacy and safety of intracavernosal alprostadil in men with erectile
dysfunction. The Alprostadil Study Group //N. Engl.J. Med. - 1996. - Vol. 334. - P. 873-877.
163. Mulhall J.P., Daller M., Traish A.M. et al. Intracavernosal forskolin: role in management of
vasculogenic impotence resistant to standard 3-agent pharmacotherapy // J. Urol. — 1997. — Vol,
158. - P. 1752-1758. disaission 1758-1759.
164. Nehra A., Blute M.L, Barrett D.M., Moreland R.B. Rationale for combination therapy of
intraurethral prostaglandin El and sildenafil in the salvage of erectile dysfunction patients desiring
noninvasive therapy// Int.J. Impôt. Res. - 2002. - Vol. 14. - Suppl. 1. — P. S38-S42.
165. Tsai Y.S., Lin J.S., Lin Y.M. Safety and efficacy of alprostadil sterile powder (S. Po„ CAVDP-
JECT) in diabetic patients with erectile dysfunction // Eur. Urol. - 2000. — Vol. 38. - P. 177-183.
166. Shabsigh R., Padma-Nathan H., Gittleman M. et al. Intracavemous alprostadil alfadex is
more efficacious, better tolerated, and preferred over intraurethral alprostadil plus optional actis: a
comparative, randomized, crossover, multicenter study // Urology. — 2000. — Vol. 55. — P. 109-113,
167. Chew K.K. Intracavernosal injection therapy. Does it still have a role in erectile dysfunction? //
Aust, Fam. Physician. — 2001. — Vol. 30. — P. 43-46,
168. European Alprostadil Study Group: The long-term safety of alprostadil (prostaglandin-El) in
patients with erectile dysfunction // Br. J. Urol. — 1998. — Vol, 82. — P. 538-543.
169. Dinsmore W.W., Gingell C., Hackett G. et al. Treating men with predominantly nonpsychogenic
erectile dysfunction with intracavernosal vasoactive intestinal polypeptide and phentolamine mesylate
in a novel auto-injector system: a multicentre double-blind placebo-controlled study // BJU Int. —
1999. - Vol. 83. - P. 274-279.
170. Hellstrom W.J., Usta M.R Surgical approaches for advanced Peyronie’s disease patients // Int.
J. Impôt. Res. - 2003. - Vol. 15. - Suppl. 5. - P. S121-S124.
171. Usta M.R, Bivalacqua T.J., Sanabria J. et al. Patient and partner satisfaction and long-term
results after surgical treatment for Peyronie’s disease // Urology, - 2003. — Vol. 62. - P. 105-109.
172. Careon C.C., MulcahyJ.J., Govier RE, Efficacy, safety and patient satisfaction outcomes of the
AMS 700CX inflatable penile prosthesis: results of a long-term multicenter study, AMS 700CX Study
Group //J. Urol. - 2000. - Vol. 164. - P. 376-380.
173. Wilson S.K., Cleves M.A„ Delk 2nd J.R. Comparison of mechanical reliability of original and
enhanced Mentor Alpha I penile prosthesis //J. Urol. — 1999. — Vol. 162. - P. 715-718.
174. Caraiii C., Zini D., Baldini A, et al. Effects of androgen treatment in impotent men with normal
and low levels of free testosterone // Arch. Sex. Behav. — 1990. — Vol, 19, — P. 223-234,
175. Hajjar R.R., Kaiser F.E., Morley J.E. Outcomes of long-term testosterone replacement in older
hypogonadal males: a retrospective analysis // J. Clin. Endocrinol. Metab. - 1997. — Vol. 82. —
P. 3793-3796.
176. Mills T.M., Lewis R.W., Stopper V.S. Androgenic maintenance of inflow and veno-occlusion
during erection in the rat // Biol. Reprod. — 1998. — Vol. 59. — P. 1413-1418,
177. Fink H.A,, MacDonald R., Rutks I.R., Wilt T.J, Trazodone for erectile dysfunction: a systematic
review and meta-analysis // BJU Int. — 2003. — Vol. 92. — P. 441-446.
178. Lebret T., Herve J.M., Gorny P. et al. Efficacy and safety of a novel combination of 1-arginine
glutamate and yohimbine hydrochloride: a new oral therapy for erectile dysfunction // Eur. Urol. —
2002. - Vol. 41. - P. 608-613. discussion 613.
179. Fleshner N., Harvey M., Adomat H. et al. Evidence for pharmacological contamination of
herbal erectile function products with type 5 phosphodiesterase inhibitors [abstrart] // J. Urol. —
2004. - Vol. 171. - P. 314.
399
180. Hong B., Ji Y.H., Hong J.H. et al. A double-blind crossover study evaluating the efficacy of
Korean red ginseng in patients with erectile dysfunction: a preliminary report // J. Urol. - 2002. -
Vol. 168. - P. 2070-2073.
181. Wessells H., Gralnek D., Dorr R. et al. Effect of an a-melanocyte stimulating hormone analog
on penile erection and sexual desire in men with organic erectile dysfunction // Urology. — 2000. —
Vol. 56. - P. 641-646.
182. Pfaus J.G., Shadiack A., Van Soest T. et al. Selective facilitation of sexual solicitation in the
female rat by a melanocortin receptor agonist // Proc. Natl. Acad. Sci. USA. — 2004. — Vol. 101. —
P. 10201-10204.
183. Rosen R.C., Diamond L.E., Earle D.C. et al. Evaluation of the safety, pharmacokinetics and
pharmacodynamic effects of subcutaneously administered PT-141, a melanocortin receptor agonist,
in healthy male subjects and in patients with an inadequate response to Viagra j j Int. J. Impôt. Res. —
2004. - Vol. 16. - P. 135-142.
184. Diamond L.E., Earle D.C., Rosen R.C. et al. Double-blind, placebo-controlled evaluation of the
safety, pharmacokinetic properties and phannacodynamic effects of intranasal PT-141, a melanocortin
receptor agonist, in healthy males and patients with mild-to-moderate erectile dysfunction // Int. J.
Impôt. Res. - 2004. - Vol. 16. - P. 51-59.
185. Nabel E.G., Pompil V.J., Plantz G.E., Nabel G.J. Gene transfer and vascular disease j j
Cardiovasc. Res. - 1994. - Vol. 28, - P. 445-455.
186. Heistad D.D., Farad P.M. Gene therapy for cerebral vascular disease j j Stroke. — 1996. —
Vol. 27. - P. 1688-1693.
187. Bivalacqua T.J., Hellstrom W.J. Potential application of gene therapy for the treatment of erec¬
tile dysfunction //J. Androl. — 2001. — Vol. 22, — P. 183-190.
188. Christ G.J., Melnian A. The application of gene therapy to the treatment of erectile dysfunc¬
tion// Int. J. Impôt. Res. - 1998. - Vol. 10. - P. 111-112.
189. Chancellor M.B., Yoshimura N., Pruchnic R., Huard J. Gene therapy strategies for urological
dysfunction //Trends. Mol. Med. - 2001. - Vol. 7. - P. 301-306.
190. Deng W., Bivalacqua T.J., Hellstrom W.J., Kadowitz P.J. Gene and stem cell therapy for erectile
dysfunction // Int. J. Impôt. Res. — 2005. — Vol. 17. — Suppl. 1. - P. S57-S63.
191. Garban H., Marquez D., Magee T. et al. Cloning of rat and human indudble penile nitric oxide
synthase. Application for gene therapy of erectile dysfunction // Biol. Reprod. — 1997. — Vol. 56. —
P. 954-963.
192. Christ G.J., Rehman J„ Day N. et al. Intracorporal injection of hSlo cDNA in rats produces
physiologically relevant alterations in penile function // Am. J. Physiol. — 1998. - Vol. 275. -
P. H600-H608.
193. Bivalacqua T.J., Champion H.C., Mehta Y.S. et al. Adenoviral gene transfer of endothelial nitric
oxide synthase (eNOS) to the penis improves age-related erectile dysfunction in the rat j j Int.J. Impôt.
Res. - 2000. - Vol. 12. - Suppl. 3. - P. S8-S17.
194. Champion H.C., Bivalacqua T.J., Hyman A.L. et al. Gene transfer of endothelial nitric oxide
synthase to the penis augments erectile responses in the aged rat // Proc. Natl, Acad. Sci. USA. -
1999. - Vol. 96. - P. 11648-11652.
195. Burchardt M., Burchardt T., Anastasiadis A.G. et al. Application of angiogenic factors for
therapy of erectile dysfunction: protein and DNA transfer of VEGF165 into the rat penis // Urology. -
2005. - Vol. 66. - P. 665-670.
196. Rogers R.S., Graziottin T.M., Lin G.S. et al. Intracavernosal vascular endothelial growth factor
(VEGF) injection and adeno-associated virus-mediated VEGF gene therapy prevent and reverse veno-
genic erectile dysfunction in rats // Int. J. Impôt. Res. — 2003. — Vol. 15. - P. 26-37.
197. Deng W., Bivalacqua T.J., Chattergooii N.N. et al. Adenoviral gene transfer of eNOS: high-level
expression in ex vivo expanded marrow stromal cells // Am. J. Physiol. Cell. Physiol. - 2003, - Vol.
285. - P. C1322-C1329.
198. Deng W., Bivalacqua TJ„ Chattergoon N.N. et al. Engineering ex vivo-expanded marrow
stromal cells to secrete calcitonin gene-related peptide using adenoviral vector // Stem. Cells. —
2004. - Vol. 22. - P. 1279-1291.
199. McMahon C.G., Abdo C., Incrocci L. et al. Disorders of orgasm and ejaculation in men // J.
Sex. Med. - 2004. - Vol. 1. - P. 58-65.
200. Yeates W.K. Ejaculation and its disorders j j Arch. Ital. Urol. Nefrol. Androl. — 1990. —
Vol. 62. - P. 137-148.
400
201. Gil-Vemet J.M. Jr. Ejaculation in men; a dynamic endorectal ultrasonographical study j j Br.J.
Urol. - 1994. - Vol. 73. - P. 442-448.
202. Olivier B„ Van Oorschot R., Waldinger M, Serotonin, serotonergic receptors, selective serotonin
reuptake inhibitors and sexual behavior // Int. Clin. Psvchopharaiacol. — 1998. — Vol. 13. - Suppl.
6. - P. S9-S14.
203. Waldinger M. The neurobiological approach to early ejaculation //J. Urol. - 2002. - Vol.
168. - P. 2359-2366.
204. Waldinger M.D., Olivier B. Utility of selective serotonin reuptake inhibitors in premature ejacu¬
lation //Curr. Opin. Investig. Drugs. — 2004. — Vol. 5. - P. 743-747.
205. Waldinger M.D., Zwinderman A.H., Olivier B. Antidepressants and ejaculation: a double¬
blind, randomized, fixed-dose study with mirtazapine and paroxetine //J. Clin. Psychopharmacol. —
2003. - Vol. 23. - P. 467-470.
206. Cain VS., Johannes C.B., Avis N.E. et ai. Sexual functioning and practices in a multi-ethnic
study of midlife women; baseline results from SWAN // J. Sex. Res. — 2003. — Vol. 40, — P. 266-
276.
207. Basson R. Clinical practice. Sexual desire and arousal disorders in women // N. Engl.J. Med, -
2006. - Vol. 354. - P. 1497-1506.
208. Graham C.A. Turning on and turning off; a focus group study of the factors that affect women’s
sexual arousal / C.A. Graliam, S.A. Sanders, R.R. Milhausen, K.R. McBride // Arch. Sex. Behav. -
2004. - Vol. 33, - P. 527-538.
209. Basson R. Using a different model for female sexual response to address women’s problematic
low sexual desire //J. Sex. Marital. Ther. — 2001. — Vol. 27. — P. 395-403.
210. Basson R. Sexual desire and arousal disorders in women // N. Engl. J. Med. — 2006. — Vol.
354. - P. 1497-1506.
211. Galyer K.T., Conaglen H.M., Hare A., Conaglen J.V. The effect of gynecological surgery on
sexual desire //J. Sex. Marital. Ther. - 1999. - Vol. 25. - P. 81-88.
212. Shifren J.L., Braunstein G.D., Simon J.A. et al. TYansdermal testosterone treatment in women whth
impaired sexual function after oophorertomy // N. Engl.J. Med. — 2000. - Vol. 343. — P. 682-688.
213. Braunstein G.D., Sundwall D.A., Katz M. et al. Safety and efficacy of a testosterone patch for
the treatment of hypoactive sexual desire disorder in surgically menopausal women; a randomized,
placebo-controlled trial j j Arch. Intern, Med, — 2005. — Vol. 165. — P. 1582-1589.
214. Buster J.E., Kingsberg S.A., Aguirre O. et al. Testosterone patch for low sexual desire in surgi¬
cally menopausal women: a randomized trial //Obstet. Gynecol. — 2005. — Vol. 105. — P. 944-952.
215. Simon J., Braunstein G., Nachtigall L. et al. Testosterone patch increases sexual activity and
desire in surgically menopausal women with hypoactive sexual desire disorder // J. Clin. Endocrinol.
Metab. - 2005. - Vol. 90. - P. 5226-5233.
216. van Lunsen R.H., Laan E. Genital vascular responsiveness and sexual feelings in midlife
women: psychophysiologic, brain and genital imaging studies // Menopause. — 2004. — Vol. 11. —
P. 741-748.
217. Cranston-Cuebas M.A., Barlow D.H. Cognitive and affective contributions to sexual function¬
ing // Ann. Rev. Sex. Res. — 1990. — Vol. 1. — P. 119-161.
218. Basson R., Leiblum S., Brotto L et al. Definitions of women’s sexual dysfunction reconsidered:
advocating expansion and revision //J, Psychosom. Obstet. Gynaecol. — 2003. — Vol. 24. — P. 221-
229.
219. Avis N.. Zhao X., Johannes C.B. et al. Correlates of sexual function among multiethnic middle-
aged women: results from the Study of Women’s Health Across the Nation (SWAN) // Menopause. —
2005. - Vol. 12. - P. 385-398.
220. Dennerstein L., Lehert P. Modeling mid-aged women’s sexual functioning: a pro.spective,
population-based study //). Sex. Marital. Ther. - 2004. - Vol. 30. - P. 173-183.
221. Chivers M.L., Bailey J.M. A sex difference in features that elicit genital response j j Biol.
Psychol. - 2005. - Vol. 70. - P. 115-120.
222. Basson R., Brotto L.A. Sexual psychophysiology and effects of sildenafil citrate in oestrogenised
women with acquired genital arousal disorder and impaired orgasm: a randomised controlled trial //
Bjog. - 2003. - Vol. 110. - P. 1014-1024.
223. Bancroft J., Loftus J., Long J.S. Distress about sex: a national survey of women in heterosexual
relationships // Arch. Sex. Behav, — 2003. — Vol. 32. — P. 193-208.
401
224. Blausteiii J.D. Progestin receptors; neuronal integrators of hormonal and environmental stimu¬
lation//Ann. N. Y, Acad. Sci. - 2003. - Vol. 1007. - P. 238-250.
225. Davis S.R., van der Mooren MJ., van Lunsen R.H. et al. The efficacy and safety of a testoster¬
one patch for the treatment of hypoactive sexual desire disorder in surgically menopausal w’omen: A
randomized, placebo controlled-trial // Menopause. - 2006. - Vol. 13. - P. 387-396.
226. Segraves R.T., Clayton A., Croft H. et al. Bupropion sustained release for the treatment of
hypoactive sexual desire disorder in premenopausal women //J. Clin. Psychopharmacol. — 2004. —
Vol. 24. - P. 339-342.
227. Tiefer L., Hall М., Tavris C. Beyond dysfunction: a new view of women’s sexual problems //J.
Sex. Marital. Ther. - 2002. - Vol. 28. - Suppl. ], - P. 225-232.
228. Avis N.E., Stellate R., Crawford S., Johannes C., Longcope C. Is there an association between
menopause status and sexual functioning.? // Menopause. — 2000. — Vol. 7. — P. 297-309.
229. Laumann E.O., Nicolosi A., Classer D.B. et al. Sexual problem.s among women and men aged
40-80 years: prevalence and correlates identified in the Global Study of Sexual Attitudes and Beha¬
viors // Int. J. Impôt. Res. - 2005. - Vol. 17. - P. 39-57.
230. DeLamater J.D., Sill M. Sexual desire in later life //J. Sex. Res. — 2005. — Vol. 42. — P. 138-149.
231. Barsk}'J.L., Friedman М., Rosen R. Sexual dysfunction and chronic illness: the role of flexibility
in coping//J. Sex. Marital. Ther. - 2006. - Vol. 32. - P. 235-253.
232. Meston C.M., Gorzalka B.B. Differential effects of sympathetic activation on sexual arousal in
sexually dysfunctional and functional women //J. Abnorm. Psychol. — 1996. — Vol. 105. — P. 582-
591.
233. Karama S., Lecours A.R., Leroux J.M. et al. Areas of brain activation in males and females dur¬
ing viewing of erotic film excerpts // Hum. Brain. Mapp. — 2002. — Vol. 16. — P. 1 -13.
234. Palace E.M., Gorzalka B.B. The enhancing effects of anxiety on arousal in sexually dysfunc¬
tional and functional women //J. Abnorm. Psychol. - 1990. - Vol, 99. - P. 403-411.
235. Creighton S,M„ Crouch N.S., Foxwel! N.A., Cellek S. Functional evidence for nitrergic neu¬
rotransmission in a human clitoral corpus cavemosum: a case study // Int. J. Impôt. Res. - 2004. -
Vol. 16. - P. 319-324.
236. Shafik A., El-Sibai 0., Mostafa R. et al. Response of the internal reproductive organs to clitoral
stimulation: the clitorouterine reflex // Int. J. Impôt. Res. - 2005. - Vol. 17. — P. 121-126.
237. O’Connell H.E., DeLancey J.O. Clitoral anatomy in nnlliparous, healthy, premenopausal vol¬
unteers using unenhanced magnetic resonance imaging // J. Urol. — 2005. — Vol. 173. — P. 2060-
2063.
238. Althof S.E., Dean J., Derogatis L.R. et al. Current perspectives on the clinical assessment and diag¬
nosis of female sexual dysfunction and clinical studies of potential therapies: a statement of concern //
J. Sex. Med. - 2005. - Vol. 2. - Suppl. 3. - P. 146-153.
239. Fugl-Meyer K., Fugl-Meyer A.R. Sexual disabilities are not singularities // Int.J. Impôt. Res. —
2002. - Vol. 14. - P. 487-493.
240. Dunn K.M., Croft P.R., Hackett G.I. Sexual problems: a study of the prevalence and need for
health care in the general population j j Fam. Pract. — 1998. — Vol. 15. — P. 519-524.
241. Mercer C.H., Fenton K.A., Johnson A.M. et al. Sexual function problems and help seeking
behaviour in Britain: national probability sample survey j j BMJ. — 2003. — Vol. 327. — P. 426-427.
242. Harlow B.L., Stewart E.G. A population-based assessment of chronic unexplained vulvar pain:
have we underestimated the prevalence of vulvodynia? //J. Am, Med. Womens. Assoc. — 2003. — Vol.
.58. - P. 82-88.
243. Hartmann U., Philippsohn S., Heiser K., Ruffer-Hesse C. Low sexual desire in midlife and older
women: personality factors, psychosocial development, present sexuality // Menopause. — 2004. -
Vol. 11. - P. 726-740.
244. Leiblum S.R., Koochaki P.E., Rodenberg C.A. et al. Hypoactive sexual desire disorder in
postmenopausal women: US results from the Women’s International Study of Health and Sexuality
(WISHeS) // Menopause. - 2006. - Vol. 13. - P. 46-56.
245. Hayes R„ Dennerstein L. The impact of aging on sexual function and sexual dysfunction in
women: a review of population-based studies //J. Sex. Med. - 2005. - Vol. 2. — P. 317-330.
246. Fugl-Meyer A.R., Fugl-Meyer K. Sexual disabilities, problems and satisfaction in 18-74 year
old Swedes // Scand. J. Sexology. - 1999. - Vol. 2. — P. 79-105.
247. Kennedy S.H., Dickens S.E., Eisfeld B.S., Bagby R.M. Sexual dysfunction before anti depressant
therapy in major depression //J. Affect. Disord. — 1999, — Vol. 56. — P. 201-208.
402
248. Cyranowski J.M., Broinberger J., Youk A. et al. Lifetime depression history and sexual function
in women at midlife // Arch. Sex. Behav. — 2004. — Vol. 33. — P. 539-548.
249. Montejo-Gonzalez A.L., Llorca G., Izquierdo J.A. et al. SSRI-induced sexual dysfunction; flu¬
oxetine, paroxetine, sertraline, and fluvoxamine in a prospective, multicenter and descriptive clinical
study of 344 patients //J. Sex. Marital. Ther. — 1997. — Vol. 23. — P. 176-194.
250. Clayton A.H., Pradko J.F., Croft H.A, et al. Prevalence of sexual dysfunction among newer
antidepressants //J. Clin. Psychiatry. - 2002. - Vol. 63. - P. 357-366.
251. Labrie F.. Belanger A., Cusan L. et al. Marked decline in senim concentrations of adrenal
C19 sex steroid precursors and conjugated androgen metabolites during aging //J. Clin. Endocrinol.
Metab. - 1997. - Vol. 82. - P. 2396-2402.
252. Labrie F., Luu-The V.. Labrie C. et al. Endocrine and intracrine sources of androgens in women;
inhibition of breast cancer and other roles of androgens and their precursor dehydroepiandrosterone //
Endocr. Rev. - 2003. - Vol. 24. - P. 152-182.
253. Padero M.C., Bhasin S., Friedman T.C. Androgen Supplementation in older women; too much
hype, not enough data //J. Am. Geriatr. Soc. — 2002. - Vol. 50. - P. 1131-1140.
254. Ganz P.A., Desmond K.A., Belin T.R. et al. Predictors of sexual health in women after a breast
cancer diagnosis // J. Clin. Oncol. - 1999. - Vol. 17. - P. 2371-2380.
255. Aziz A., Brannstrom M., Bergquist C„ Silfverstolpe G. Perimenopausal androgen decline after
oophorectomy does not influence sexuality or psychological well-being // Fertil. Steril. — 2005. — Vol.
83. - P, 1021-1028.
256. Santoro N., Torrens J., Crawford S. et al. Correlates of circulating androgens in mid-life women:
the study of women’s health across the nation //J. Clin. Endocrinol. Metab. - 2005. — Vol. 90. -
P. 4836-4845.
257. Davis S.R., Davison S.L., Donath S., Bell R.J. Circulating androgen levels and self-reported
se.xual function in women // JAMA. — 2005. — Vol. 294. — P. 91-96.
258. Davis S.R., Goldstat R., Papalia M.A. et al. Effects of aromatase inhibition on sexual function
and w'ell-being in postmenopausal women treated with testosterone: a randomized, placebo-controlled
trial /7 Menopause, - 2006, - Vol. 13. - P. 37-45.
259. Maravilla K.R., Cao Y., Heiman J.R. et al. Serial MR imaging with MS-325 for evaluating
female sexual arousal response: determination of intrasubject reproducibility // J. Magn. Reson.
Imaging. - 2003. - Vol. 18. - P. 216-224.
260. van Geelen J.M., van de Weijer P.H., Arnolds H.T. Urogenital symptoms and resulting dis¬
comfort in non-institutionalized Dutch women aged 50- 75 years // Int. Urogynecol. J. Pelvic. Floor.
Dysfunct. - 2000. - Vol. 11. - P. 9-14.
261. Gorodeski G.L, Hopfer U,, Lhi C.C., Margies E, Estrogen acidifies vaginal pH by up-regulation
of proton secretion via the apical membrane of vaginal-ectocervical epithelial cells // Endocrinology. —
2005. - Vol. 146. - P. 816-824.
262. Hoyle C.H., Stones R.W., Robson T. et al. Innervation of vasculature and miCTovasculatiire of
the human vagina by NOS and neuropeptide-containing nerves //J, Anat. — 1996. — Vol. 188. - Pt.
3. - P. 633-644.
263. Foster D.C., Palmer M., Marks J. Effect of vulvovaginal estrogen on sensorimotor response
of the lower genital tract: a randomized controlled trial j j Obstet. Gynecol. — 1999. - Vol. 94. -
P. 232-237.
264. Connell K., Guess M.K., Bleustein C.B. et al. Effects of age, menopause, and comorbidities on
neurological function of the female genitalia //Int.J. Impôt. Res. — 2005. — Vol. 17. — P. 63-70.
265. Dennerstein L., Lehert P., Burger H. The relative effects of hormones and relationship factors
on sexual function of women through the natural menopausal transition // Fertil. Steril. - 2005. -
Vol. 84. - P. 174-180.
266. Berglund G., Nystedt M., Bolund C. et al. Effect of endocrine treatment on sexuality in
premenopausal breast cancer patients; a prospective randomized study //J. Clin. Oncol. — 2001. —
Vol. 19. - P. 2788-2796.
267. de Groot M., Anderson R., Freedland K.E. et al. Association of depression and diabetes
complications: a meta-analysis // Psychosom. Med. — 2001. - Vol. 63. — P. 619-630.
268. Basson R.J., Rucker B.M., Laird P.G., Conry R. Sexuality of women with diabetes //J. Sex.
Reprod. Med. - 2001. - Vol. 1. - P. 11-20.
269. Erol B„ Tefekli A., Ozbey I. et al. Sexual dysfunction in type II diabetic females: a comparative
study //J. Sex. Marital. Ther. - 2002. - Vol. 28. - Suppl. 1. - P. 55-62.
403
270. Его! В., Tefekli А., Sanli О. et al. Does sexual dysfunction correlate with deterioration of
somatic sensory system in diabetic women.? // Int. J. Impôt. Res. — 2003. — Vol. 15, — P. 198-202.
271. Enzlin P., Mathieu С., Van den Bruel A. et al. Sexual dysfunction in women with type 1 diabe¬
tes: a controlled study // Diabetes. Care. - 2002. - Vol. 25. - P. 672-677.
272. Doruk H„ Akbay E., Cayan S. et al. Effect of diabetes mellitus on female sexual function and
risk factors // Arch. Androl. — 2005. - Vol. 51. - P. 1-6.
273. Peng Y.S., Chiang C.K., Kao T.W. et al. Sexual dysfunction in female hemodialysis patients: a
multiceiiter study // Kidney Int. - 2005. — Vol. 68. - P. 760-765.
274. Wincze J.P., Albert A., Bansal S. Sexual arousal in diabetic females: physiological and self-
report measures //Arch. Sex. Behav. — 1993. — Vol. 22. — P. 587-601.
275. Crespo C.J., Smit E., Snelling A. et al. Hormone replacement therapy and its relationship to
lipid and glucose metabolism in diabetic and nondiabetic postmenopausal women: results from the
Third National Health and Nutrition Examination Survey (NHANES III) j j Diabetes. Care. — 2002. —
Vol. 25. - P. 1675-1680.
276. Toorians A.W., Janssen E., Laan E, et al. Chronic renal failure and sexual functioning: clinical
status versus objectively assessed sexual response // Nephrol. Dial. Transplant. — 1997. — Vol. 12. —
P. 2654-2663.
277. Hultgren R., Sjogren Б., Soderberg M. et al. Sexual function in women suifering from aortoiliac
occlusive disease // Eur. J. Vase. Endovasc. Surg. — 1999. — Vol. 17. — P. 306-312.
278. Duncan L.E., Lewis C„ Jenkins P., Pearson T.A. Does hypertension and its pharmacotherapy
affect the quality of sexual function in women? // Am. J. Hypeitens. — 2000. — Vol. 13. — P. 640-
647.
279. Conaglen H.M., Conaglen J.V. Sexual desire in women presenting for antiandrogen therapy //
J. Sex. Marital. Ther. - 2003. - Vol. 29. - P. 255-267.
280. Miller K.K., Biller B.M., Beauregard C. et al. Effects of testosterone replacement in androgen-
deficient women with hypopituitarism: a randomized, double-blind, placebo-controlled studv //J. Clin.
Endocrinol. Metab. - 2006. - Vol. 91. - P. 1683-1690.
281. Arlt W., Callies F„ van Vlijmen J.C. et al. Dehydroepiandrosterone replacement in women with
adrenal insufficiency // N. Engl. J. Med. - 1999. - Vol. 341. - P. 1013-1020.
282. van Thiel S.W., Romijn J.A., Pereira A.M. et al. Effects of dehydroepiandrostenedione, super¬
imposed on growth hormone substitution, on quality of life and insulin-like growth factor I in patients
with secondary adrenal insufficiency: a randomized, placebo-controlled, cross-over trial // J. Clin.
Endocrinol. Metab. - 2005. - Vol. 90. - P. 3295-3303.
283. Lovas K., Gebre-Medhin G., Trovik T.S. et al. Replacement of dehydroepiandrosterone in
adrenal failure: no benefit for subjective health status and sexuality in a 9-month, randomized, parallel
group clinical trial //J. Clin. Endocrinol. Metab. — 2003. - Vol. 88. - P. 1112-1118.
284. Gimbel H„ Zobbe V., Andersen B.M. et al. Randomised controlled trial of total compared with
subtotal hysterectomy with one-year follow up results // BJOG. - 2003. - Vol. ПО. — P. 1088-
1098.
285. Kim D.H., Lee Y.S., Lee E.S. Alteration of sexual function after classic intrafascial supracervi¬
cal hysterectomy and total hysterectomy //J. Am. Assoc. Gynecol. Laparosc. — 2003. — Vol. 10. —
P. 60-64.
286. Roovers J.P., van der Bom J.G., van der Vaart C.H., Heintz A.P. Hysterectomy and sexual well¬
being: prospective observational study of vaginal hysterectomy, subtotal abdominal hysterectomy and
total abdominal hysterectomy // BMJ. - 2003. - Vol. 327. - P. 774-778.
287. Maas C.P,, ter Kuile M.M., Laan E. et al. Objective assessment of sexual arousal in women with
a history of hysterectomy j j BJOG. - 2004. — Vol. Ill, - P. 456-462.
288. Kadioglu P., Yalin A.S., Tiryakioglu O. et al. Sexual dysfunction in women with hyperpro¬
lactinemia; a pilot study report //J. Urol. — 2005. — Vol. 174. — P. 1921-1925.
289. Mansfield P.K., Koch P.B. Qualities midlife women desire in their sexual relationship and their
chaning sexual response // Psychol. Women. Q. - 1998. - Vol. 22. - P. 282-303.
290. Cayan S., Bozlu М., Canpolat B.. Akbay E. The assessment of sexual functions in women with
male partners complaining of erectile dysfunction: does treatment of male sexual dysfunction improve
female partner’s sexual functions.? //J. Sex, Marital, Ther. — 2004, — Vol. 30. — P. 333-341.
291. Hartmann U., Heiser K., Ruffer-Hesse C., Kloth G.Femalc sexual desire disorders: subtypes,
classification, personality factors and new directions for treatment // World. J. Urol. — 2002. —
Vol. 20. - P. 79-88.
404
292. Brotto L.A., Basson R„ Gehriiig D. Psychological profiles among women with vnlvar vestibuli¬
tis syndrome: a chart review //J. Psychosom. Obstet. Gynaecol. - 2003. - Vol. 24. — P. 195-203.
293. Van Lankveld J.J., Weijenborg P.T., ter Kuile M.M. Psychologic profiles of and sexual func¬
tion in women with vulvar vestibulitis and their partners // Obstet. Gynecol. - 1996. - Vol. 8S. —
P. 65-70,
294. Klusmann D. Sexual motivation and the duration of partnership // Arch. Sex. Behav. —
2002. - Vol. 31. - P. 275-287.
295. Verhaak- C.M,, Sineenk J.M., van Minnen A, et al. A longitudinal, prospective study on emo¬
tional adjustment before, during and after consecutive fertility treatment cycles // Hum. Reprod. —
2005. - Vol. 20. - P. 2253-2260.
296. Hawton K., Catalan J., Flagg J. Low sexual desire: sex therapy results and prognostic factors //
Behav. Res. Ther. - 1991. - Vol. 29. - P. 217-224.
297. Hurlbert D.F. A comparative study using orgasm consistency training in the treatment of
women reporting hypoactive sexual desire //J. Sex. Marital. Ther. - 1993. - Vol. 19. - P. 41-55.
298. Sarwer D.B., Durlak J.A. A field trial of the effectiveness of behavioral treatment for sexual
dysfunctions //J. Sex. Marital. Ther. - 1997. - Vol. 23. - P. 87-97.
299. McCabe M.P. Evaluation of a cognitive behavior therapy program for people with sexual dys¬
function //J. Sex. Marital. Ther. - 2001. - Vol. 27. - P. 259-271.
300. Heiman J., LoPiccolo J. Clinical outcome of sex therapy. Effect of daily vs weekly treatment //
Arch. Gen. Psychiatry. - 1983. - Vol. 40. ~ P. 443-449.
301. Trudel G., Marchand A., Ravart M. et al. The effect of a cognitive behavior treatment program
on hypoactive sexual desire disorder in women // Sex. Rel. Ther. — 2001. — Vol. 16. — P. 145-164.
302. Delehanty R. Changes in assertiveness and changes in orgasmic response occurring in sexual
therapy of preorgasmic women //J. Sex. Marital, Ther. — 1982. — Vol. 8. — P. 198-208.
303. Hurlbert D.F., Apt C. Coital alignment technique and directed masturbation: a comparative
study on female masturbation //J. Sex. Marital Ther. - 1995, - Vol. 21. - P. 21-29.
304. Heiman J.R. Psychologic treatments for female sexual dysfunction: are they effective and do we
need them? // Arch. Sex. Behav. - 2002. - Vol. 31. - P. 445-450.
305. Basson B.R., McKinnes R,, Smith M.D. et al. Efficacy and safety of sildenafil citrate in women
with sexual dysfunction associated with female sexual arousal // Gend. Based. Med, - 2002. -
Vol. 11. - P. 367-377.
306. Shifren J.L., Davis S.R., Moreau M. et al. Testosterone patch for the treatment of hypoactive
sexual desire disorder in naturally menopausal women: results from the INTIMATE NMl study //
Menopause. - 2006. - Vol. 13. - P. 770-779.
307. Barnhart K., Freeman E., Grisso J.A. etal. The effect of dehydroepiandrosterone Supplementation
to symptomatic perimenopausal women on serum endocrine profiles, lipid parameters and health relat¬
ed quality of life //J. Clin. Endocrinol. Metab. — 1999. — Vol. 84. — P. 3896-3902.
308. Laan E., van Lunsen R.H., Everaerd W. The effects of tibolone on vaginal blood flow, sexual
desire and arousability in postmenopausal women // Climacteric. - 2001. - Vol. 4. - P. 28-41.
309. Meston C.M., Worcel M. The effects of yohimbine plus 1-arginine glutamate on sexual arousal
in postmenopausal women with sexual arousal disorder // Arch. Sex. Behav. — 2002. — Vol. 31, —
P. 323-332.
310. Meston C.M., Heiman J.R. Ephedrine-activated physiological sexual arousal in women /7 Arch.
Gen. Psychiatry. - 1998. - Vol. 55. - P. 652-656.
311. Caruso S., Intelisano G., Lupo L. et al. Premenopausal women affected by sexual arousal
disorder treated with sildenafil: a double-blind cross-over placebo-controlled study // Brit. J. Obstet.
Gynecol. - 2001. - Vol. 108. - P. 623-628.
312. Caruso S., intelisano G„ Farina M. et al. The function of seldenafil on female sexual pathways:
A double-blind, cross-over placebo controlled study j j Euro J. Obstet. Gynecol. - 2003. - Vol. 110. -
P. 201-206.
313. Ito T.Y., Trant A.S., Polan M.L. A double-blind placebo-controlled study of Argin Max, a
nutritional Supplement for enhancement of female sexual function //J. Sex. Marital. Ther. — 2001. —
Vol. 27. - P. 541-549.
314. Blake j., Belisle S., Basson R. SOGC Clinical Practice Guideline: Canadian Consensus on
Menopause and Osteoporosis //J. Obstet. Gynecol. Can. - 2006. — Vol. 171. - P. S7-S112.
315. The North American Menopause Society: The role of testosterone therapy in postmenopausal
women: position statement of The North American Menopause Society // Menopause, - 2005. —
Vol. 12. - P. 496-511.
405
316. Hays J., Ockene J.K., Brunner R.L. et al. Effects of estrogen plus progestin on health-related
quality of life // N. Engl. J. Med. - 2003. - Vol. 348. - P. 1839-1854.
317. Wierman M., Basson R., Davis S. et al. Androgen therapy in women: an Endocrine Society
Clinical Practice Guideline // J. Clin. Endocrinol. Metab. — 2006. — Vol. 91. — P. 3697-3710.
Андрогены 74
- незаконное использование 292
Андростендион 297
Ановуляция хроническая 66
- вследствие избытка андрогенов 73
- гипоталамическая 67
- заболевания гипофиза 73
Анорхия 239
Атрофия урогенитальная 125
Бесплодие 383
Бесплодие у мужчин 276
- биопсия яичек 284
-диагностика 281
Болезнь
- Виллебранда 113
- Кушинга 68
- Крейтцфельда-Якоба 285
Вагинизм 369
-лечение 390
Гестагены синтетические 116
Гинекомастия 269
Гипертскоз яичников 96
Гипогонадизм 235
- вторичный 240
Гирсутизм 88
- идиопатический 79
Гистерэктомия 381
Гонадотрофы 24
Гормон
- гонадотропин-рилизинг-фактор 17
- аналоги, агонисты, антагонисты 24
- регуляция секреции 22
- лютеинизирующий 17,25
- пептидный 50
- фолликулостимулирующий 17, 25
Дегидроэпиандростерон 298
Депо-медроксипрогестерона ацетат 186
Детерминанты дифференцировки яичника и фолликулогенеза 28
Диабет 378
Диспареуния 369
- лечение 391
Дисфункция коры надпочечников врожденная 81
Заболевания мужских половых желез 221
Изменения гормональные во время овариального цикла 52
Истощение функции яичников преждевременное 105
407
Климактерий 120
Контрацептивы
— оральные 84,114
— пероральные 158
Контрацепция
-гормональная 157
-.-влагалищное и чрескожное применение стероидов 181
— внутриматочная 200
— имплантационная 194
— инъекционная 186
— мини-пили 175
— осложнения 161
— пероральная 158
Лейомиома матки 119
Маточное кровотечение
— ановуляторное 112
— воздействие стероидных гормонов 110
— в постменопаузе 123
— нерегулярное 111
Менопауза 121
Менструация 61,108
Овуляция 38
Олигоменорея 109
Ооцит 28
Опухоли
— яичек 289
— яичников и надпочечников андрогенсекретирующие 79
Орхит инфекционный 238
Остеопороз постменопаузальный 125
Приливы 124
Прогестерон 59
Рак молочной железы 378
Расстройства сексуальные
— у женщин 364
— вагинизм 369
— диспареуния 369
— методы гормональной терапии 389
— расстройство
оргазмическое 369
полового возбуждения 368,369
сексуального влечения 367
— у мужчин
— механизмы эрекции 333
— сексуальная дисфункция 332
— снижение сексуального влечения 341
— эректильная дисфункция 342
— эякуляторные расстройства 363
Репродукция
— дисфункция
408
- обследование женщин 63
- физиология 17
- функции переднего гипофиза 24
- функция гипоталамуса 19
Рецептор
- к гонадотропин-рилизинг-гормону 24
-кФСГиЛГ41
Сертоли-Лейдига клетки 79
Синдром
- XYY 278
- андрогенной недостаточности у мужчин 235
- Барде-Бидля 250
- Берьесона-Форссмана-Леманна 250
- исчезающих яичек 239
- Калльмана 68
- Клайнфелтера 236,278
- Коэна 250
-Кушинга 83
- Лоренса-Муна 250
- Нунан 278
- поликистозных яичников 86,379
- генетика 96
- диагностика 88
- индукция овуляции 102
- инсулинорезистетітность 97
- продукция
гонадотропинов 92
протеина, связывающего половые стероиды 94
стероидов 92
- Прадера-Вилли 244
- Рейфенштейна 274
- Ригера 245
-Тернера 106,278
- Шиена 68
- Штейна-Левенталя 87
Спиронолактон 84
Стероидогенез
- в яичниках 45
- экстраовариалышш 53
Тело желтое 39
Терапия гормональная заместительная 126
- историческая перспектива 128
- польза 131
-рекомендации 132
- риски и противопоказания 129
Терапия тестостероном 254
Тестостерон 221
Травма яичек 238
Физиология женской сексуальной реакции 370
Финастерид 85
409
Флутамид 86
Фолликул 37
Функция яичек
- метаболизм тестостерона 225
- механизм действия андрогенов 228
- продукция андрогенов 222
Ципротерона ацетат 85
Эндометрий 54
- влияние яичниковых стероидов 57
-имплантация 59
- контроль эндометриальной функции 62
- морфологические изменения 56
Эндометриоз 117
Эстрадиол 58
Эстрогены 58
Яичник 26
- онтогенез 28
Cumulus oophorus 33
Theca interna 34
Zona pellucida 34
РЕПРОДУКТИВНАЯ
ЭНДОКРИНОЛОГИЯ
по ВИЛЬЯМСУ
САМОЕ ИЗВЕСТНОЕ И НЕЗАМЕНИМОЕ
РУКОВОДСТВО ПО ЭНДОКРИНОЛОГИИ
•
Уже многие десятилетия «Эндокринология по Вильямсу» остается извест¬
нейшим в мире руководством по заболеваниям желез внутренней секреции.
Издательская группа «ГЭОТАР-Медиа» рада представить вам перевод пос¬
леднего, 11 -го издания ЭТОГО руководства на русский язык.
Изложение материала в «Эндокринологии по Вильямсу» всегда было
построено на тесной связи патогенеза и клинической медицины. Глубокое
понимание механизмов развития патологических состояний - отличительная
черта квалифицированного современного специалиста-клинициста.
Особенности книг серии «Эндокринология по Вильямсу»
• Облегчают принятие клинических решений благодаря подробной конк¬
ретной теоретической и практической информации.
• Представляют собой максимально полное и современное освещение
эндокринологических заболеваний и синдромов в изложении ведущих
мировых экспертов.
• Наглядность изложения, структурирование материала значительно
облегчают поиск нужной информации.
• Содержат цветные фотографии, схемы патогенеза, графики и т.д.
На русском языке серия «Эндокринология по Вильямсу» выходит под об¬
щей редакцией академика Р/УН и РАМН, профессора И.Й. Дедова и члена-у у
корреспондента РАМН, профессора Г.А. Мельниченко. . : :У:
Эта кНигау несомненно, сбудет интересна как начинающим врачам, тайЩ'"
и опытным практикующим специалистам. У У У;:||
Издание на русском языке включает несколько книг:
• Заболевания коры надпочечников й эндокринная артериальная гипертензия
• Сахарный диабет и нарушения углеводного обмена
• Заболевания щитовидной железы
• Ожирение и нарушения липидного обмена
• Нейроэндокринология
• Репродуктивная эндокринология
ISBN 978-5-91713-029-3
Данное издание предстадпяет собой перевод
с английского оригинального издания «Williams
Textbook of Endocrinology». Перевод опубликован
по контракту с издательством «Эльзевир» (Elsevier).
Информация о новых изданиях и наименованиях
на русском языке доступна на сайте издательства:
www.elsevier.ru.
ELSEVIER
9 785917 130293
Перевод с английского осуществлен:
ИЗДАТЕЛЬСКАЯ ГРУППА
«ГЭОТАР-Медиа»
www.geotar.ru
www.medknigaservis.ru