/











































































































































































Текст
ЛЮСЬЕН ЛОБЬЕ
ОАЗИСЫ НА ДНЕ ОКЕАНА
Предисловие Ж. М. Переса
ЛЕНИНГРАД Г И ДРОМЕТ ЕОИЗДАТ
1990
ББК 28.082
Л 68
LUCIEN LAUB1ER
DES OASIS
AU EOND DES MENS
'' A
LE KOCHER, 1986
Перевод с французского M. А. Долголенко Научный редактор д-р биол. наук К- Н. Несис
Рисунки в тексте Виолен Мартен Рисунки в приложении С. В. Галкина Фото на вкладке Ю. А. Володина
© Le Rocher 1986
ISBN 5-286-00329-Х (СССР)
ISBN 2-268-00464-3 (Франция)
© Перевод на русский язык, «К советскому читателю», послесловие, приложения. Гидрометеоиздат, 1990
К СОВЕТСКОМУ ЧИТАТЕЛЮ
Еще подростком я страстно увлекался изучением морской прибрежной фауны Средиземного моря, неудивительно поэтому, что меня заинтересовали тайны животных, затерянных в больших глубинах океана, в ледяных безднах, где царствует вечная тьма... В 1960 году я смог увидеть, благодаря «ныряющему блюдцу» Жака Ива Кусто, на 300-метровой глубине у каталонского берега великолепие зарослей белых кораллов, залитых светом прожекторов.
Спустя несколько лет во Франции появилось новое научно-исследовательское учреждение — Национальный центр исследования океанов (CNEXO). Его создание позволило использовать для исследования больших глубин океанов такие новые стредства, как океанографическое судно «Жан Шарко». Примерно тогда же, в 1966 году, я впервые побывал в СССР, в Москве на Международном океанографическом конгрессе, а затем, на следующий год, во время научной командировки я познакомился с Ленинградом, Одессой, Мурманском. В то время советская школа океанологии, руководимая такими всемирно известными учеными, как Л. А. Зенкевич, Я. А. Бирштейн, А. В. Иванов, Н. Г. Виноградова, 3. А. Филатова, мой друг профессор П. В. Ушаков и др., занимала одно из ведущих мест в изучении абиссальной и ультраабиссальной (или хадальной) фауны. Результаты, полученные в северо-западной части Тихого океана, в частности в Курило-Камчатской впадине, были очень интересны и значительны как в области зоологии, так и в области экологии.
Моя мечта сбылась, когда мне удалось организовать в новой лаборатории в Бресте небольшую исследовательскую группу, вместе с которой в период 1969—1978 годов
я изучал глубоководную фауну и ее связи с окружающей средой. Постепенно мы приходили к мысли, что эти животные, вопреки распространенному мнению, страдают не столько от неблагоприятных физических условий (очень низкая температура, от —0,5 до 2 °C, полная темнота, высокое давление), сколько от крайне малого количества пищи, попадающей сюда из поверхностных слоев после долгого опускания в виде микроскопических частиц — именно они образуют тот самый «морской снег», который часто описывали первые наблюдатели на батискафах.
Вот почему открытие глубоководных оазисов, расположенных в непосредственной близости от горячих гидротермальных источников, сделанное геологами в 1976— 1977 годах, явилось для всех морских биологов знаменательным событием. В 1979 году мне выпала удача изучать очень интересную форму многощетинковых червей, открытую во время франко-американских погружений на Восточно-Тихоокеанском поднятии на 21° с. ш. Вместе с моим учеником Даниэлем Дебрюйером мы назвали этого червя помпейским — Alvinella pompejana, так как, подобно жителям античного города Помпеи, трагически погибшего во время извержения Везувия, он непрерывно посыпается дождем частичек, как пеплом.
Познакомиться с этим фантастическим миром, где сталкиваются силы земли и воды, давая начало жизни, я и приглашаю вас. Советским исследователям уже знакомо это зрелище: начиная с 1986 года с борта подводного обитаемого аппарата они исследуют гидротермальное излияние, открытое канадцами на широте Британской Колумбии. Их сборы позволили описать новые формы животных, тесно связанных с гидротермальными излияниями. Я надеюсь, что будущее позволит нам теснее сотрудничать, чтобы лучше понять функционирование этих подводных оазисов жизни.
Люсьен Лобъе
ПРЕДИСЛОВИЕ
С огромным удовольствием пишу я это предисловие для книги, которую Люсьен Лобье посвятил «оазисам глубин». Большие океанические глубины влекут меня еще с той поры, когда я участвовал в погружениях на французских батискафах, оставивших у меня, пожалуй, одни из самых ярких за всю мою долгую жизнь воспоминаний.
С момента открытия гидротермальных источников на некоторых подводных поднятиях я прочитал множество статей, посвященных ключевым биологическим и экологическим моментам этих явлений, и убедился, что приоткрылась еще одна дверь в изучение больших глубин. И вот сейчас, когда я познакомился с переданной мне автором рукописью, я понял, что дверь-то распахнута гораздо шире, чем я думал. Я нашел на страницах рукописи счастливое сочетание самих наблюдений и их биологической и экологической интерпретации, с одной стороны, и того синтетического подхода — с другой,— который поразил меня, когда я был членом комиссии, присуждавшей Люсьену Лобье ученую степень доктора наук за диссертацию, тема которой весьма далека от этой книги, хотя бы потому, что она посвящена изучению гораздо меньших глубин.
Автор взялся за нелегкую задачу, попытавшись ввести неискушенного читателя в микромир, достаточно непривычный даже для большинства специалистов, весьма сведущих в некоторых проблемах, связанных с науками об океанах, но я думаю, что он справился с этим.
Прочитайте введение, которое открывает книгу. Оно представит вам подводный мир, гораздо более захватывающий, чем тот, что открывается при глубоководных погружениях на батискафах, осуществляемых вот уже 20 лет, когда проплываешь надо дном километры и кило
метры, прежде чем увидишь хоть каких-нибудь представителей фауны, которых, к тому же, часто бывает очень трудно определить.
Последуйте за Люсьеном Лобье в эти оазисы жизни, разбросанные здесь и там по подводным хребтам, где выходы теплых вод рождают удивительные пейзажи, вероятно, недолговечные, обреченные исчезнуть через несколько десятков лет, но могущие возникнуть опять чуть в стороне и вновь достичь расцвета. Разве не замечательно, что виды, обитающие в этих оазисах, по большей части являются видами архаичными, настоящими живыми ископаемыми,— палеонтологи обнаружили в геологических отложениях, часто довольно древних, виды, родственные тем, о биологии которых автор рассказывает столь подробно.
В истоке этого микромира лежат хемотрофные бактерии, способные осуществлять синтез органического вещества из минеральных элементов (вот вам подходящий случай, чтобы воспользоваться словарем терминов в конце книги). Разумеется, животные, с которыми вы знакомитесь, рассматривая рисунки, должны откуда-то получать необходимую им энергию, и весь этот маленький мирок существует только благодаря бактериям, которые изобилуют в гидротермальной сети вблизи поверхности дна. Некоторые из этих беспозвоночных включают бактерии в ткани своего тела, другие покрыты ими, третьи отфильтровывают микроскопические скопления бактерий из толщи омывающей их воды, подобно тому, как обыкновенная мидия отфильтровывает микроводоросли, а равно и бактерии. И конечно, есть здесь и хищники, например крабы, головоногие, даже рыбы. Короче говоря, в этих глубоководных оазисах жизни существует то, что называется трофической сетью, то есть существуют такие отношения между видами — начинай от видов, способных улавливать энергию,— когда одни виды служат добычей для других видов (мы называем их хищниками), в свою очередь являющихся добычей другого вида-хищника. Весьма возможно, что эти маленькие экосистемы, включающие виды, прекрасно приспособленные к существованию в столь необычной среде, идеально соответствуют условиям в гидротермалях, где основную энергию вырабатывают бактерии.
Я уверен, что если вас хоть немного интересует долгая история нашей планеты, возникновение жизни в тех
физико-химических условиях, которые можно назвать по меньшей мере неблагоприятными, то, прочитав эту увлекательную книгу, вы будете просто поражены, насколько невероятно богата и разнообразна жизнь в океане,— такая, какой вы видите ее, листая энциклопедию,— по сравнению с теми малыми, но удивительно интенсивными очагами жизни, каковы эти оазисы.
История подводного гидротермализма и жизни, которая там эволюционировала,— это еще и урок альтруизма и довод в защиту сотрудничества специалистов различных отраслей знаний. Изучение подводных гидротермалей и их биологических и экологических особенностей не развивалось бы столь быстро, если бы один геофизик не увидел чуть больше, чем показание своих приборов, не вспомнил о своих коллегах-биологах и не сообщил им, что заметил каких-то странных животных. Потому что, если уж тебе повезло и ты спустился в глубины океана, ты не имеешь права интересоваться только объектом изучения той узкой области знаний, в которой ты специализируешься, не говоря уже о том, что подобные экспедиции требуют высокой технической оснащенности и значительных финансовых средств.
Средства Люсьен Лобье нашел во Французском Институте исследования морей (IFREMER), но он сумел также заинтересовать специалистов самых разных областей из университетских лабораторий, и каждый из них внес свой вклад в понимание этой экосистемы, которая представляется столь отклоняющейся от нормы. В самом деле, экосистема оазисов, в том что касается движения энергии, кажется в целом похожей на экосистемы, извлекающие необходимую энергию из фотосинтеза: энергия движется от первоначального источника — в данном случае бактериального хемосинтеза — через первичных потребителей к видам, которые получают ее после одного или нескольких переходов через промежуточных потребителей.
Известно, что во всех освещенных экосистемах, где органическая продукция высока, имеет место некоторая потеря первичной энергии. И хотя экосистемы, порожденные гидротермальными выходами, гораздо более продуктивны, чем самые богатые экосистемы поверхности, изоляция оправдывает сравнение их с оазисами в пустыне. Тем не менее некоторая часть энергии, протекающей через экосистему оазисов, не реализуется немедленно на
месте, что позволяет существовать на периферии этих оазисов небольшим сообществам животных, которые пользуются крохами этой «манны небесной».
Не так ли, по сути, и нищие поселения окружают некоторые крупные города и существуют лишь за счет расточительства общества потребления.
Ж. М. Перес, член Академии наук
ВВЕДЕНИЕ
В XViii веке, когда первые гидрографы начали исследовать океан на глубинах, превосходящих несколько десятков метров, был поставлен важный научный вопрос: до каких пределов в этой неблагоприятной среде, где царят полная тьма, огромное давление и очень низкие температуры, может существовать жизнь? Для разрешения этого основополагающего вопроса в течение более ста лет снаряжались многочисленные океанографические экспедиции.
В начале XIX века, использовав средства, обычно употребляемые рыбаками-промысловиками, один натуралист установил, что на глубинах порядка одного километра живут рыбы и ракообразные: вблизи Ниццы рыбацкие донные яруса и тралы позволили аптекарю и натуралисту Риссо описать многочисленных представителей этих глубоководных животных. Однако экологический интерес этих первых наблюдений остался незамеченным.
Несколькими десятилетиями позднее в ходе систематических исследований глубин близ британских берегов англичанин Форбс анализирует распределение фауны рыб и беспозвоночных, обитающих на дне, и констатирует заметное уменьшение плотности поселений фауны с увеличением глубины; продолжая свои наблюдения в Эгейском море (восточная часть Средиземного моря) до глубин около 400 м, Форбс наблюдает еще более быстрое уменьшение плотности поселений фауны. Экстраполируя это утверждение на глубину, он выдвигает гипотезу, что ниже 550-метровой глубины всякая жизнь исчезает. Едва успев возникнуть, эта гипотеза подвергается сомнению.
Спорам положило конец одно решающее наблюдение — речь идет об открытии, сделанном случайно в
1859 году, когда на поверхность был поднят один из первых подводных телеграфных кабелей, проложенных двумя годами раньше между Сардинией и Алжиром на глубине 1800 м: с каким же изумлением инженеры обнаружили прикрепившихся к кабелю животных, которые, очевидно, жили на этой глубине и чувствовали себя там превосходно; в основном это были мадрепорарии и моллюски. Вот так своеобразно была отодвинута граница распространения жизни, установленная Форбсом. Следует подчеркнуть, что этот, казалось бы, сугубо научный вопрос вызвал живой интерес: на борту судна, прибывшего для подъема кабеля, не было ни одного натуралиста, и лишь любознательность инженера Дженкинса, ответственного за проведение операции, позволила французскому зоологу Мильн-Эдвардсу представить результаты исследований образцов фауны, собранной с кабеля, на заседании Академии наук в 1861 году.
В эти же годы неожиданное открытие придает новый смысл изучению больших океанических глубин: в глубоком норвежском фиорде священник и натуралист М. Саре вылавливает с глубины более 600 м несколько экземпляров изящной морской лилии и знакомит с ними ученый мир. До того времени считалось, что эти животные, чьи ископаемые остатки были хорошо известны геологам, навсегда исчезли с лица земли. Еще через несколько лет будет открыт другой образчик живого ископаемого с больших глубин, на этот раз на северо-востоке Атлантического океана; последняя находка подтвердит гипотезу, что в ходе геологического времени абиссальные глубины могли служить убежищем для примитивных форм жизни. Эту гипотезу можно найти среди утверждений, выдвинутых организаторами крупнейшей кругосветной экспедиции на английском судне «Челленджер» для ее обоснования; некоторые даже ожидали — ни больше, ни меньше,— что в ходе экспедиции будет пойман живой белемнит или аммонит... В этот период— 1870-е годы — натуралисты разработали целый арсенал средств сбора животных, который за последующие 60 лет существенно не изменился. Если говорить только о Франции, то здесь, не считая нескольких экспедиций в Бискайский залив и в Средиземное море, организацией больших океанографических экспедиций, материалы которых изучались французскими зоологами, мы обязаны в основном князю Монако Альберту I. В ходе экспедиций Альберта I по исследованию
больших глубин было обнаружено, что кашалоты способны заныривать очень глубоко, более чем на 1000 м, и что на этой глубине они ловят гигантских кальмаров, прежде науке не известных, и охотно их поедают; использование больших тралов с металлической рамой позволяет в 1910 году установить рекорд, который не будет превзойден в течение более чем 60 лет,— поймать живую рыбу с глубины более 6000 м.
В конце первой половины нашего века получен окончательный ответ на жгучий вопрос о границах жизни на глубинах: в июле 1951 года датское судно «Галатея», проводя лов тралом с металлической рамой на глубинах от 9820 до 10 210 м на дне Марианской впадины в Тихом океане, подняло на поверхность несколько беспозвоночных (актинии, голотурии). Эти результаты стали настоящей сенсацией: итак, ничто, ни температура чуть ниже 0 °C, ни давление более 1000 бар, ни вечная ночь на глубинах, не может противостоять колонизирующей активности жизни, которая стремится закрепиться во всех средах планеты.
Тогда морские биологи заинтересовались другими проблемами, и в частности источником пищи для глубоководной фауны. При полном отсутствии света, естественно, не может быть и растительной продукции — результата фотосинтеза, который имеет место повсюду в осыхающей приливной полосе и на мелководьях до глубин 50—100 м. Значит, органические вещества, необходимые для жизни животных, должны поступать из освещенных слоев океана или с прибрежной приливной зоны, в некоторых случаях расположенной совсем недалеко от глубоких океанских впадин. Некоторые технические нововведения позволили довольно быстро установить связь между плотностью поселений фауны и глубиной: плотность поселения фауны (число особей на единицу площади дна) тем меньше, чем больше глубина. В то же время сравнение плотности и особенно биомассы (количества живого вещества на единицу площади дна) фауны, живущей на одной и той же глубине, но в разных районах океана, показывает, что простой и универсальной зависимости между биомассой и глубиной не существует. Следует учитывать третий фактор — величину первичной продукции фотосинтеза, которая образуется в поверхностном освещенном слое воды: именно здесь находится источник органического вещества, используемого глубоководными животными. Речь идет о
маленьких частицах диаметро>м от нескольких десятых до нескольких сотых долей миллиметра, которые медленно опускаются в глубины со скоростью порядка ста метров в день. Чтобы достичь глубины 10 км, этим микрочастичкам требуется более трех месяцев, и все это время их атакуют бактерии, которые потребляют часть погружающегося вещества. Вот так схематически объясняется довольно быстрое уменьшение биомассы фауны по мере увеличения глубины.
Вопреки недавно бывшим в ходу идеям, ни крайне низкая температура, ни огромное давление, ни отсутствие света не ответственны за обеднение на глубинах — причина этого явления гораздо проще: чем глубже, тем меньше количество доступной пищи. С точки зрения эколога, глубоководная фауна представляет собой уникальный пример живой системы, на протяжении нескольких километров от поверхности использующей в качестве .источника энергии органическое вещество, синтезируемое на поверхности. Выявлена также другая особенность глубоководных фаун, которая может интерпретироваться как адаптация к жизни в среде, где катастрофически мало пищи: имеется в виду присутствие на глубинах хищных животных больших размеров — ракообразных и рыб,— чья биомасса почти равна биомассе животных — потребителей мелких частиц. Если вспомнить, что в пищевой пирамиде перенос органического вещества с одного уровня на другой, вышестоящий (например, с уровня растительноядных на уровень хищников, поедающих этих растительноядных) происходит с 10-процентной отдачей веса органического вещества, то здесь налицо экологический парадокс. Объяснение его заключается, очевидно, в том, что трупы крупных животных, обитающих у поверхности (рыб, рептилий и морских млекопитающих), после их гибели быстро опускаются на глубины и становятся добычей упомянутых глубоководных хищников. Таким образом, оказывается, что в одной среде сосуществуют животные,, как говорят экологи, разных функциональных групп, жизнь которых основана на потреблении двух типов частиц разного размера.
Знания постепенно накапливались, и в какой-то момент специалисты сочли возможным утверждать, что основные характеристики глубоководных экосистем теперь известны: все они базируются на фотосинтетической продукции, создаваемой вблизи поверхности, и в основ
ном жестко связаны в своем развитии с количеством наличной пищи.
Но вдруг весной 1977 года несколько погружений американского подводного аппарата «Алвин» на глубину 2500 м на Галапагосском рифте в зоне, обследованной годом ранее по фотографиям, сделанным аппаратом, буксируемым в 10—30 м над дном, опрокидывают сложившиеся представления морских биологов: на значительной глубине, в зоне, где продукция фотосинтеза с поверхности океана крайне мала, наблюдатели с борта подводного аппарата обнаруживают в лучах прожектора изобильную фауну, ярко окрашенную, какое-то невероятное кипение жизни, и описывают все это такими образными выражениями, как «Райский сад», «Розовый сад», «Ми-диевая банка»... Настроенные поначалу скептически, биологи вскоре восторженно принимают это открытие, которое, вероятно, останется наиболее важным результатом биологической океанографии за последнее десятилетие. В 1979 году группа американских биологов получает возможность посетить странные подводные оазисы, в основе существования которых лежит не поток органического вещества с поверхности, но использование бактериями химической энергии, черпаемой из веществ, выносимых гидротермальной циркуляцией.
Начиная со времени этих первых наблюдений, разные страны организуют всё новые экспедиции, оснащенные необходимой техникой (то есть обитаемыми подводными аппаратами, способными погружаться на глубины до 3000 м). Несмотря на географическую удаленность этих зон, — в основном расположенных в восточной части Тихого океана,— Франция, которая около 30 лет назад была первой страной, предпринявшей глубоководные погружения, имея два батискафа «hNRS-iii» и «Архимед», играет, наряду с США и Канадой, важнейшую роль в этих исследованиях. Совсем недавно аналогичными экологическими явлениями заинтересовалась Япония, так что можно предвидеть быстрое развитие знаний в этой области, которая частенько еще считается лишь биологическим курьезом.
1.
ГЕОЛОГИЧЕСКИЕ И ГЕОХИМИЧЕСКИЕ УСЛОВИЯ
Открытие гидротермальных подводных явлений на больших глубинах вдоль активных океанических поднятий — это косвенное следствие теории тектоники плит, и их существование было предсказано лет за двадцать до самого открытия. Сегодня теория тектоники плит широко известна: согласно этой теории, поверхность Земли состоит из шести—восьми основных гигантских плит, постоянно перемещающихся одна по отношению к другой под влиянием движений в мантии Земли. Плиты сложены главным образом океанической молодой корой (более половины поверхности Земли); на них, как на огромных плотах, покоятся континенты, образованные более легкими породами, прочно спаянными с глубинной частью плит. На границе между двумя плитами происходят различные процессы. Когда две плиты сталкиваются друг с другом, более тяжелая океаническая плита постепенно погружается под континентальную плиту и опускается в мантию, где исчезает (вследствие этого явления, называемого суб-дукцией, дно океанов относительно молодо — его возраст не превышает 180 млн. лет). Одновременно погружение плиты приводит к образованию глубокого желоба в месте столкновения; например, в западной части Тихого океана глубина такого желоба свыше 10 км. Наоборот, когда две плиты раздвигаются, на поверхность поднимается вещество земной мантии и возникает новая океаническая кора (явление аккреции). Как правило, зоны аккреции планеты расположены под океанами. Они образуют систему подводных хребтов протяженностью примерно 70 тыс. км, знакомую широкой публике по великолепным цветным картам. Из таких хребтов наиболее известен, пожалуй, Срединно-Атлантический хребет, расположенный примерно на одинаковом расстоянии от американского,
европейского и африканского берегов и вздымающийся более чем на 2000 м над абиссальными равнинами, лежащими по обе стороны от него. Обычно эти хребты состоят из серий линейных сегментов протяженностью в несколько сотен километров, разделенных поперечными разломами длиной в несколько километров. Все вместе образует зигзагообразный рисунок с очень характерными неравновеликими ответвлениями. Эти хребты имеют высокую активность: геофизики установили, что плиты отодвигаются друг от друга и что для различных типов хребтов характерна различная средняя годовая скорость раздвижения. В Атлантическом океане, где хребет представляет собой настоящую горную цепь, годовая скорость раздвижения низка, порядка 1—2 см; наоборот, на относительно невысоком Восточно-Тихоокеанском поднятии среднее годовое раздвижение больше — от бСм в год на севере до 16—16 см на широте острова Пасхи. Долгое время предполагали, что существует связь между скоростью раздвижения и интенсивностью гидротермальных процессов: так, гидротермальные проявления в Атлантическом океане считались недостаточными для возникновения оазисов жизни, какие развиваются на хребтах восточной части Тихого океана. Но совсем недавно стало известно, что в Атлантическом океане тоже существуют гидротермальные проявления с высокой температурой,— в двух местах на Срединно-Атлантическом хребте на 24 и 26" с. ш. с помощью подводного телевидения наблюдались сообщества животных, но мы пока еще не знаем ни их состава, ни структуры*.
Но осям активных хребтов магма, состоящая из пород в вязком состоянии вследствие плавления земной мантии на глубине нескольких сот километров, поднимается к поверхности. 1'емпература ее примерно 1200 "С. Приближаясь к поверхности, магма остывает и постепенно затвердевает, образуя новую океаническую кору; потоки лавы, которые изливаются в результате этого процесса, создают характерные образования: базальтовые пилоны поддерживают настоящие потолки, возникшие вследствие опорожнения лавового озера после частичного остывания, и делают опасным плавание подводных аппаратов на дне долины. Поскольку магма, поднимаясь на поверхность, расширяется, глубина над океаническими хребтами отно-
* См. Приложение 2.— Прим. ред.
Гидротермальное 50°С/ 350°С
Рис. 1. Схематический разрез через срединно-океанический хребет, показывающий подъем магмы при высокой температуре, гидротермальную циркуляцию и металлоносные осадки по обе стороны хребта.
сительно невелика: их вершины расположены на глубинах 2500—3000 м. Чем дальше от осей хребтов, тем сильнее сжимается магма в ходе охлаждения и тем соответственно глубже становится океаническое дно. Это явление заглубления подчиняется простому закону: глубина океанической коры возрастает как корень квадратный от ее возраста.
Поднимаясь и остывая, магма сжимается и глубоко растрескивается. Морская вода проникает вглубь по сети трещин и щелей на многие сотни метров, а иногда и больше. При соприкосновении с породами в ходе их отвердевания морская вода разогревается и преобразуется во флюид, состав которого весьма далек от ее первоначального химического состава. Удалось установить температуру, до которой морская вода нагревается в ходе своего подземного следования: по соотношению изотопов (изотопы — атомы химического элемента, обладающие одинаковыми химическими свойствами, но разнящиеся своими атомными массами) кислорода, содержащегося в минералах, отложившихся в трещинах и добытых бурением, максимальная температура образования этих отложений оценивается в 350—370 °C (действительно, известно, что в химических реакциях, где принимает участие кислород, соотношение двух естественных изотопов кислорода 16О и 18О зависит, в частности, от температуры реакции;
зная эту пропорцию в отложившихся минералах, можно узнать температуру их образования). Другие методы расчетов, базирующиеся на осаждении из морской воды некоторых элементов, таких, как магний, дают большие величины, порядка 450 °C.
Удалось оценить и величину потока морской воды, проходящей в масштабе Мирового океана через подземную сеть под океаническими хребтами. Метод основан на измерении потока тепла, идущего из земных недр и поступающего в океан через дно. Существуют способы оценки потерь тепла на теплопроводность (способность твердых тел передавать тепло) через неподвижную среду, в данном случае через осадки, покрывающие базальтовую кору. Установлено, что вблизи океанических хребтов потери на теплопроводность падают, хотя в отсутствие всякого другого явления следовало бы ожидать обратного: ведь молодая океаническая кора теплее, чем более древняя океаническая кора, и результирующий поток тепла должен был бы быть сильнее. Эта аномалия объясняется гидротермальной циркуляцией. Морская вода, циркулирующая в сети трещин, поглощает в результате конвекции (движение жидкости, вызываемое разницей температур) тепло лавы и высвобождает его в океан. Конвективный перенос тепла за год в результате гидротермальной циркуляции оценивается с помощью этого метода в 5*1019 кал. Принимая максимальную температуру, достигаемую в ходе этой циркуляции, равной 350 °C (а для повышения температуры одного грамма морской воды от 2 до 350 "С требуется приблизительно 350 кал), рассчитали, что весь объем океанов (1,37- 1021 л) проходит через гидротермальные сети приблизительно за 8 млн. лет.
Под конец своего долгого подземного путешествия флюид вновь выходит на поверхность дна океанов в виде настоящих горячих источников, вокруг которых появляется жизнь. Но температура выходящего флюида и его химический состав сильно изменчивы, что предполагает существование более или менее интенсивной вторичной циркуляции, которая приводит к разбавлению первичного гидротермального флюида, перед тем как он поступит в море. В действительности среди известных местонахождений есть такие гидротермальные излияния, в которых температура колеблется от нескольких градусов до приблизительно 40 °C и отложения минералов не наблюдается; есть излияния, в которых температура колеблется
приблизительно от 100 до 250 °C и испускается более или менее непрозрачная белесоватая жидкость с отложениями ангидрита (сульфат кальция); наконец, есть излияния с высокой температурой от 250 до 370 °C, сопровождаемые мощными выбросами полиметаллических сульфидов и известные под названием черных курильщиков. На Восточно-Тихоокеанском поднятии на 13° с. ш. существуют все три типа гидротермальных источников, тогда как на Галапагосском рифте отмечены излияния только первого типа. Каждому типу излияний соответствует свой состав сообществ животных.
Биолог, изучающий подводные оазисы, должен обратить внимание не столько на температуру, сколько на химический состав гидротермальных флюидов: именно в нем кроется источник этой обильной жизни в форме восстановленных соединений (восстановление — удаление из соединения кислорода, иными словами, реакция, вследствие которой атом или ион присоединяет периферические электроны, то есть уменьшает свой положительный электрический заряд), в частности сероводорода (H2S). Чтобы понять происхождение этих соединений, надо знать, в какие реакции вступает морская вода под большим давлением и при высокой температуре в присутствии базальта: магний, содержащийся в морской воде, соединяется с силикатами базальта и образует нерастворимый в воде гидроксисиликат (Mg(OH)SiOa), который выпадает в осадок; ионы водорода Н+, высвобождающиеся в ходе реакции, сильно окисляют морскую воду и замещают кальций и калий в кристаллической решетке базальта. Высвобожденный таким образом кальций в свою очередь вступает в реакцию с сульфатами, содержащимися в морской воде, и образует сульфат кальция, который выпадает в осадок в форме ангидрита, как только достигается соответствующая температура; сульфаты морской воды реагируют также с металлами, содержащимися в базальте, и образуют сернистые соединения, которые затем при охлаждении жидкости выпадают в осадок. Когда кислотность жидкости достаточно высока, базальт высвобождает оксид кремния (SiCh), который переходит в раствор. На основе лабораторных опытов и наблюдений над составом гидротермальных флюидов и минеральных отложений, добытых бурением, геохимики разработали непротиворечивую схему, включающую всю совокупность химических реакций (рис. 2). Выпадение
Рис. 2. Схема химических реакций, которые происходят при циркуляции морской воды в сети трещин и расщелин остывающей лавы с выходами флюида высокой и средней температуры.
в осадок сульфата кальция происходит при проникновении морской воды в подземную сеть. Еще глубже, в зоне реакции, происходит образование сероводорода и СО2 путем восстановления сульфатов и разложения карбонатов, высвобождение ионов Н+, которые подкисляют среду и высвобождают ионы металлов и кремний. Затем горячий гидротермальный флюид, богатый металлами и сульфидами, поднимается к поверхности. Под землей флюид, разбавляясь, охлаждается, и в трещинах осаждаются сульфиды металлов. Когда гидротермальный флюид выходит на поверхность дна неразбавленным, он создает настоящие трубы, высотой иногда до 20 м, образованные выпавшими в осадок полиметаллическими сульфидами и сульфатом кальция. На выходе из трубы находящееся в растворе железо соединяется с сероводородом и дает
черный султан, образованный частичками пирита (FeS2) во взвешенном состоянии, а марганец остается в растворенном состоянии. В некоторых случаях железо и марганец затем окисляются и отлагаются на дне, образуя металлоносные осадки.
Сооружение черного курильщика начинается с фазы роста при промежуточной температуре из отложений сульфата кальция, образующегося в результате смешения гидротермального флюида с холодной морской водой. Когда курильщик вырастает, сульфат кальция начинает подвергаться воздействию неразбавленного гидротермального флюида, соответственно гораздо более горячего и кислого: тогда происходит частичное или полное вторичное растворение отложений ангидрита и замещение его полиметаллическими сульфидами.
Когда гидротермальный флюид несколько разбавляется, перед тем как излиться в океан, отложения полиметаллических сульфидов не происходит, образуются только отложения ангидрита, которые часто напоминают распылители: через малейшие поры они выбрасывают белесоватую более или менее опалесцирующую жидкость. Геохимики, исследуя содержание в гидротермальном флюиде магния и сульфатов, входящих в состав и обычной морской воды, умеют оценивать степень разбавленности флюида: она тем ниже, чем выше температура флюида, и достигает нуля при температуре порядка 350—370 °C, то есть как раз такой, которая измерялась на выходе черных курильщиков. Концентрация растворенного кремния, напротив, с температурой возрастает.
Гидротермальный флюид температурой 370 °C, изливаемый черными курильщиками, содержит повышенное количество различных металлов: 100 млн-1 (частей на миллион) железа, несколько млн-1 цинка, меди и никеля, что в 108 раз больше содержания этих металлов в морской воде, а также более 200 млн-1 сероводорода, тогда как нормальная морская вода не содержит его совсем (за исключением крайне редких случаев, как, например, в глубинах Черного моря или некоторых других аноксических, то есть полностью лишенных кислорода океанических бассейнов). Что касается некоторых токсичных металлов, то если бы такое их содержание было отмечено в прибрежной зоне, где имеют место сбросы промышленных и городских сточных вод, немедленно зафиксировали бы значительный уровень загрязнения! Следова
тельно, животные, которые обитают рядом с источниками, должны обладать специальными биологическими механизмами, позволяющими переносить подобные концентрации.
Когда гидротермальный флюид перед излиянием претерпевает вторичное разбавление, то есть в случае теплых источников с опалесцирующей водой без минеральных отложений и излияний белесоватой жидкости с промежуточной температурой, невозможно определить, на какой именно глубине под базальтовым основанием происходит смешение: в нескольких десятках метров под поверхностью или всего в нескольких десятках сантиметров? Дольчатые и подушковидные вытекания лавы и лавовые озера образуют иногда большие подземные резервуары, открывающиеся на дне в разных местах, часто весьма удаленных друг от друга. Не исключено, что поселения животных и особенно микроорганизмов, которые можно наблюдать на поверхности базальтового основания, представляют собой (по крайней мере для некоторых видов) лишь видимую часть более обширного подземного населения, способствующего, в частности, постоянному обмену генами между разбросанными популяциями, наблюдаемыми на поверхности. Пока еще не было возможности каким-либо образом подтвердить или опровергнуть эту гипотезу, правда, американские биологи наблюдали поселения двустворчатых моллюсков, глубоко погруженных в расщелину в дольчатой лаве. Факт существования в одной и той же зоне трех основных типов гидротермальных излияний на расстоянии порядка сотен метров друг от друга согласуется с идеей о наличии подповерхностных локализованных сетей вторичного разбавления.
' Если на Восточно-Тихоокеанском поднятии, к примеру, есть множество горячих излияний и активных черных курильщиков, то число неактивных курильщиков, где полиметаллические сульфиды покрыты слоем поверхностных оксидов, часто окрашенных в охряные и темно-зеленые цвета, еще больше. В изученной зоне протяженностью несколько десятков километров, расположенной на Восточно-Тихоокеанском поднятии на 12—13° с. ш., насчитывается в четыре раза больше мертвых курильщиков, чем действующих!
Некоторые неактивные постройки из полиметаллических сульфидов, расположенные примерно на 21° с. ш., удалось датировать: оказалось, что они имеют возраст
Рис. 3. Так художник изобразил устье черного курильщика; на заднем плане — бездействующие гидротермальные постройки. На врезке — разрез устья черного курильщика, поясняющий механизмы отложения и вторичного растворения ангидрита.
от 23 лет до 61 года. Продолжительность жизни курильщика оценивается в 50—100 лет. Образцы, взятые на активных постройках на 13° с. ш., имеют возраст примерно 10 лет.
Известны также случаи, когда активный хребет не выходит на поверхность, будучи перекрыт плотным слоем осадков. Таков район гидротермальных излияний в бас-
Рис. 4. Гидротермальные местонахождения и места высачивания минерализованной воды, обследованные биологами: 1, 3 и 4 — гидротермальные излияния на Восточно-Тихоокеанском поднятии на 18° ю. ш., 13 и 21° с. ш.; 5— гидротермальное местонахождение в осадках бассейна Гуаймас; 6—высачивание минерализованных вод у подножия Флоридского эскарпа; 7 — Орегонская зона суб-дукции; 8, 9 и 10 — гидротермальные местонахождения на хребтах Хуан-де-Фука, Эндевор и Эксплорер; 11 и 12 — желоб Нанкай, впадина Дзенису и каньон Тенрю; 13 и 14 — подводные вулканы Эримо и Касима.
сейне I уаймас в Калифорнийском заливе на 27° с. ш. Этот бассейн является зоной аккреции, которая входит в комплекс сегментов срединного хребта и трансформных разломов, связывающих Восточно-Тихоокеанское поднятие с разломом Сан-Андреас. Базальтовое основание покрыто почти 400-метровым слоем современных осадков, обязанных своим происхождением выносам реки Колорадо в Калифорнийский залив, и местной продукции фитопланктона и имеющих четвертичный возраст. Процесс аккреции проявляется в проникновении лавы внутрь рыхлых осадков. Активная гидротермальная циркуляция увлекает к поверхности значительное количество органического вещества, возникшего в результате трансформации органического вещества осадков под действием высокой температуры и давления, которым эти осадки подвержены. Коротко говоря, эти условия значительно ускоряют трансформацию органического вещества в сложные углеводороды — в самих осадках и на их поверхности были выявлены различные алканы, ароматические и нафтеновые соединения, опаны*. В некоторых гидротермальных источниках, которые открываются на поверхности осадков, находят также большое количество сероводорода. Осадки довольно часто покрыты плотным ковром нитчатых бактерий. В некоторых случаях вокруг источников развиваются большие поселения животных, но по видовому составу они значительно отличаются от оазисов Галапагосского рифта и Восточно-Тихоокеанского поднятия.
Заслуживают внимания совсем недавние и весьма важные для понимания биологического функционирования подводных оазисов и его обобщения открытия, хотя речь в них не идет непосредственно о гидротермалях. Хронологически первое открытие датируется мартом 1984 года. Оно было сделано на глубине 3266 м в Мексиканском заливе, в геологически неактивном районе, у подножия огромного известнякового подводного обрыва — Флоридского эскарпа, который вздымается более чем на 2000«м над абиссальными осадками. Процесс глубинной седиментации постепенно погребает под осадками подножие этого
* Опаны— органические молекулы, появившиеся в ходе диагенеза органического вещества, заключенного в осадках; до некоторой степени предвестники нефтяных углеводородов. Опаны хранят след биологического происхождения (микробы, зеленые растения) и могут играть роль хронологической метки в эволюции органического вещества.— Прим. ped.
обрыва. Здесь, на стыке глубинных осадков и подножия склона, развиваются животные и микробные сообщества, очень схожие с сообществами оазисов Восточно-Тихоокеанского поднятия; судя по первым наблюдениям, эти сообщества, вероятно, черпают свою энергию от восстановленных соединений, возможно сульфидов, в пользу чего говорит хотя бы наличие таких восстановленных минералов, как пирит, образовавшихся здесь же в результате выделения кислоты, растворившей близлежащий известняк. В этом месте не отмечено никакого повышения температуры; однако у подножия склона высачивается флюид, насыщенный минеральными солями,— вероятно, это действительно вода, вышедшая на поверхность после циркуляции внутри эскарпа. Об этом явлении, важном с точки зрения демонстрации значения неорганических восстановленных соединений для развития подводных оазисов, известно пока еще слишком мало.
Второе открытие было сделано несколько месяцев спустя, в августе 1984 года, в Тихом океане против берегов штата Орегон на глубине 2036 м, в совершенно иных геологических условиях: теперь дело происходило в зоне субдукции, где океаническая Тихоокеанская плита погружается под континентальную Американскую плиту. Это приводит к образованию складчатости осадков и обильному их накоплению у окраины континента. В этой зоне наблюдали высачивание воды из осадков. Речь идет об интерстициальной воде, содержащейся в древних осадках и выдавливаемой наружу при сжатии под действием субдукции. Установлено, что в Орегонском районе температура изливающейся воды не отличается от температуры окружающей воды, известен также ее состав: она не обогащена сульфидами, в частности сероводородом, зато содержит метан, радон, аммиак, железо и марганец,— наличие этих веществ как раз и свидетельствует о том, что это действительно интерстициальная вода, выжимаемая из осадков силами субдукции. В качестве рабочей гипотезы авторы этого открытия высказывают предположение, что очень богатое поселение животных, весьма близкое к поселениям гидротермальных оазисов, черпает энергию за счет окисления метана бактериями в основании пищевой цепи. Ровно через год, в августе 1985-го, похожие наблюдения были сделаны в другой зоне субдукции, в системе желобов к востоку от Японских островов, на глубинах 3800—4000 м в каньоне Тенрю и в
бассейне Дзенису и на глубине 5800 м в двух районах, расположенных севернее. Животное население, которое здесь наблюдали, менее разнообразно, чем у берегов Орегона, но зато эти сообщества — наиболее глубоководные сообщества такого типа из известных ныне. В этом случае авторы открытия также предполагают, что метан, содержащийся в интерстициальной воде, вытесняемой из осадков под действием огромного давления, которое они испытывают на глубине, является основой этих сообществ.
В настоящее время, когда пишется эта книга, еще не получены неоспоримые доказательства хемосинтетического происхождения этих вновь открытых оазисов. Для этого потребуются опыты на месте и сложные лабораторные исследования, которые еще не было возможности провести. Но при современном состоянии знаний эта гипотеза остается единственно правдоподобной. Единственное, что еще возможно добавить,— это то, что аммиак, другое минеральное восстановленное соединение, также может играть какую-то роль. Во всяком случае, упомянутые открытия показывают, что глубоководные оазисы распространены шире, чем об этом думали еще в 1984 году.
2
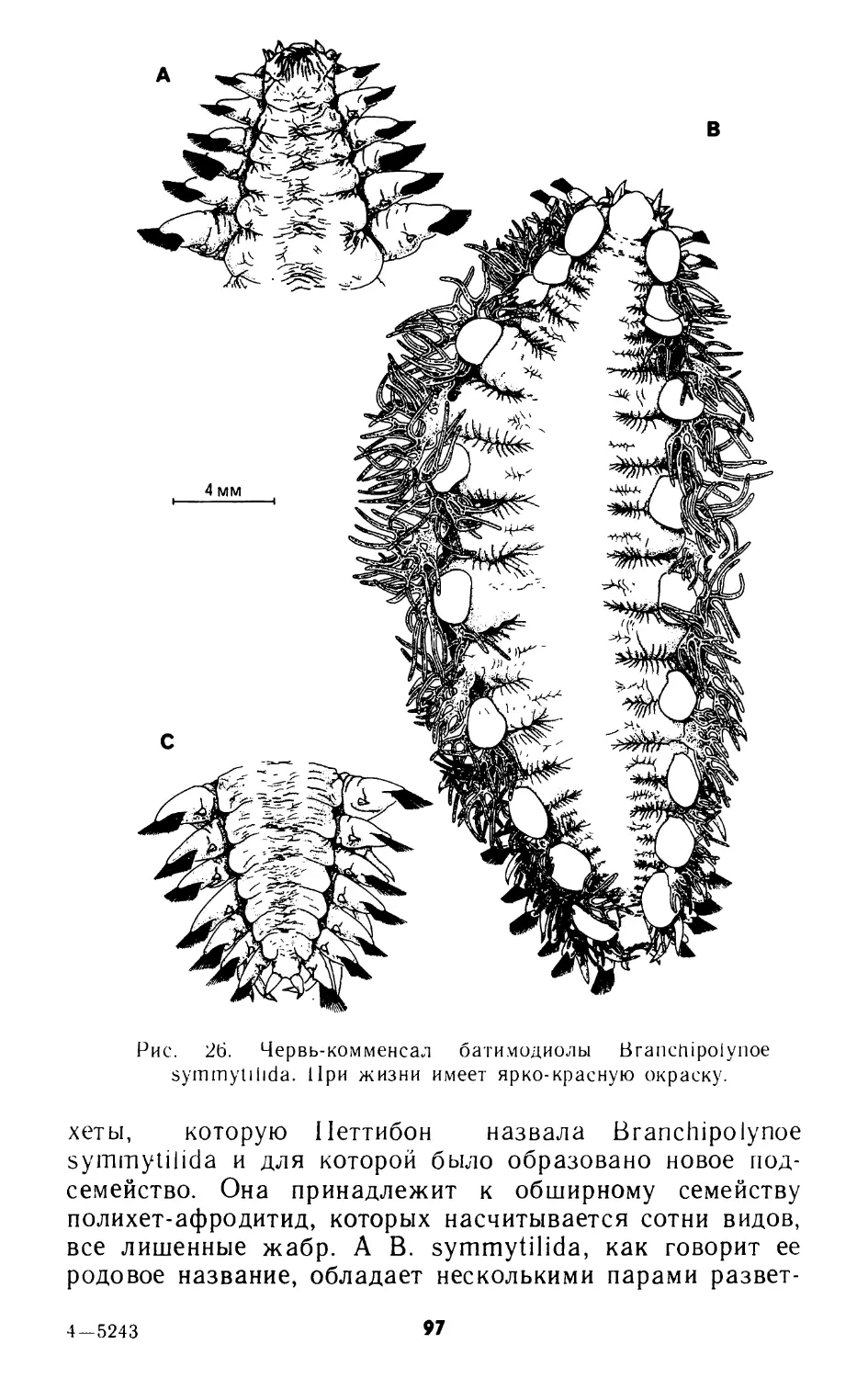
ОТКРЫТИЕ ГЛУБОКОВОДНЫХ ОАЗИСОВ
Общим местом стала мысль о том, что развитие научных знаний в конце XX века в большой степени зависит от технического прогресса, это утверждение особенно верно для океанографии в целом и глубоководной морской экологии в частности. Открытие глубоководных оазисов прямо связано с двумя сопряженными фактами: с одной стороны, с прогрессом морских наук о Земле, позволившим изучить критические зоны, включающие океанические хребты и огромные желоба субдукции, с другой стороны, с использованием систем физико-химических измерений и фотографирования дна на больших глубинах с помощью буксируемых океанографическими судами аппаратов. Такие буксируемые системы — специалисты назвали их «рыбами» — позволяют опознавать и отбирать местонахождения с аномальными значениями некоторых измеренных параметров (аномалии). Так, «рыба», с помощью которой в 1976 году были открыты первые оазисы подводной жизни на Галапагосском рифте и на Восточно-Тихоокеанском поднятии недалеко от экватора, измерила температуру морской воды в нескольких десятках метров над дном океана и сфотографировала дно. Положительные аномалии температуры — ее отклонение на несколько десятых градуса выше температуры окружающей среды — послужили указателями для определения наиболее интересных зон и отбора для анализа фотографий, одновременно сделанных «рыбой».
Однако буксируемые аппараты, снабжаемые все более и более сложными измерительными приборами, не позволяют ни непосредственно наблюдать, ни брать пробы, необходимые для работы биологов, и тем более не допускают проведение эксперимента непосредственно в местах обитания животных. Незаменимые орудия для предва-
рительного обследования в масштабе от нескольких километров до десятков километров, «рыбы» не могут, однако, заменить прямого наблюдения и прицельного отбора проб под визуальным контролем специалиста.
Только благодаря использованию подводных аппаратов для глубоководных исследований было должным образом оценено значение открытия глубоководных оазисов, и не лишним будет напомнить кратко об основных этапах разработки этих обитаемых подводных аппаратов,— той области, где Франция сумела занять особо почетное место.
Если говорить очень сжато и не упоминать о разработках некоторых предшественников, можно обозначить первый период, который длился от конца второй мировой войны до конца 60-х годов, как период технологической концепции батискафов и покорения самых больших глубин. Второй период отмечается глубоким технологическим переворотом, позволившим создать легкие подводные аппараты, более маневренные и менее дорогостоящие, чем батискафы.
Батискаф, изобретенный физиком Пиккаром накануне второй мировой войны,— это транспонированный для океана стратосферный аэростат: гондола, сопротивляющаяся давлению, более тяжелая, чем объем вытесняемой ею жидкости, поддерживается на плаву надлежащим объемом жидкости, более легкой, чем вода. В случае стратосферного аэростата гондола должна противостоять уменьшению атмосферного давления с высотой, и ее «поплавок» представляет собой непроницаемую оболочку, наполненную более легким, чем воздух, газом — гелием. В случае батискафа судно должно противостоять огромному давлению, царящему на больших глубинах, ведь давление в морской воде возрастает более чем на 1 бар (1 атмосферу) на каждые 10 м глубины, что дает на глубине 10 км давление свыше 1000 бар. «Поплавок» представляет собой оболочку из тонкого листового металла, которая заключает в себе серию резервуаров; в целях безопасности резервуары разделены и наполнены веществом, как можно менее сжимаемым и более легким, чем вода: в данном случае это авиационный бензин или керосин, плотность которого близка к 0,65. Подобно тому как в стратосферном аэростате можно выбирать высоту полета, утяжеляя его путем выпуска гелия или облегчая путем выбрасывания балласта, в батискафе можно вытра
вить железную дробь, служащую балластом, или бензин, обеспечивающий его плавучесть.
После первых попыток, мало убедительных с точки зрения управляемости первого батискафа, сконструированного Пиккаром и Козинсом, Франция построила и ввела в действие один за другим два батискафа: «bNRS-iii»— название является аббревиатурой Бельгийского национального фонда научных исследований (bonds National de la Recherche Scienlilique) и повторяет название первого батискафа Пиккара «BNRS-ii» («bNRS-i» был стратосферным аэростатом, сконструированным Пиккаром до войны), модифицированная устойчивая оболочка которого была использована для «HNRS-iil»,— и «Архимед», полностью построенный в нашей стране. Мы еще помним первый рекорд погружения «bNRS-IiI»: в июле 1954 года на широте Дакара с двумя исследователями на борту, Уо и Вильмом, он достиг глубины 4050 м*. Из соображений безопасности, учитывая прочность обитаемой сферы, батискаф «HNRS-111» не мог превзойти этот рубеж глубины в 4000 м. «Архимед», построенный в 1961 году, был сконструирован в расчете на погружение на самые большие глубины Мирового океана, то есть немногим меньше 11000 м. В 1962 году «Архимед» с Уо и Вильмом на борту без труда достиг глубины 9500 м в Курильской впадине, северо-восточнее Японии; на основании батиметрических карт, составленных советскими учеными из Института океанологии Академии наук СССР, экспедиция (в которой я имел счастье участвовать как ответственный за океанографические измерительные приборы, взятые на борт батискафа) рассчитывала погрузиться ниже отметки 10 000 м; после нескольких дней, проведенных в поисках этой глубины, пришлось удовольствоваться тем, что имели. В ту пору ошибки измерения глубины порядка 10%, к сожалению, были обычны для больших глубин океана.
За период с 1955 по 1974 год в ходе многих десятков экспедиций в разные моря Мирового океана «FNRS-i 11» и «Архимед» осуществили в общей сложности 232 погружения, из них 93 — «FNRS-iii» и 139 — «Архимед». Свои последние погружения «Архимед» выполнил во время Франко-Американской экспедиции по исследованию
* См. подробнее: Уо Ж. 20 лет в батискафе. Л., Гидрометеоиздат, 1976.— Прим, перев.
Срединно-Атлантического хребта в 1973—1974 годах (FAMOUS, French-American Mid Ocean Underwater Survey)*, в ходе которой его могли сравнить с новыми подводными аппаратами: «Сианой» (Франция) и «Алвином» (США).
Батискафы, вследствие использования для создания их плавучести соответствующих материалов, тяжелы и малоподвижны. Жилая сфера «Архимеда» диаметром 2,1 м весит на воздухе 22 т. Чтобы поддерживать на плаву этот вес вместе с дополнительным весом — батареями аккумуляторов, хранилищем балластной дроби, всевозможными надстройками легкой оболочки и разнообразными аппаратами для взятия проб и проведения наблюдений, требуется объем бензина 160 000 л, что дает общее водоизмещение порядка 180 т. Такое устройство, полное бензина, практически невозможно поднять на палубу обеспечивающего судна, и приходится буксировать батискаф на малой скорости от порта приписки до места погружения. Кроме того, бензин, используемый как поплавок, имеет два неподходящих физических свойства — он сжимается при понижении температуры с глубиной и при повышении давления. Оба эти фактора действуют в одном направлении: чтобы достичь глубины 9500 м в Курильской впадине, где разница температур между поверхностью и дном составляет 20 градусов, и уравновесить батискаф на дне, пришлось выбросить более 10 т балласта, что значительно повысило стоимость работы.
Разумеется, не появись новые материалы для создания плавучести, батискафы продолжали бы использоваться, и надо сказать, что «Архимед», который легко ввести в строй, если понадобится, является и сегодня единственным аппаратом в мире, способным достигнуть глубины более 10 000 м, а научных задач не убавилось... Но появление новых материалов, в частности композитных, пошатнуло технические основы, на которых построена концепция батискафов. Эти новые легкие материалы, состоящие из микроскопических пустотелых и поэтому очень легких шариков из полимера с высоким сопротивлением, позволяют достичь плотности 0,6, близкой к плотности бензина, используемого в батискафах. В отличие от бензина, эти
* См. подробнее: Риффо К., Ле Пишон К. Экспедиция «FAMOUS». Три тысячи метров в глубь Атлантики. Л., Гидрометеоиздат, 1979.— Прим, перев.
композитные материалы почти несжимаемы ни при понижении давления, ни при понижении температуры, что составляет их первое ценное преимущество. Кроме того, освоение металлургии титана — металла, более прочного, чем лучшие сорта стали, позволило уменьшить толщину стенок обитаемой сферы и, следовательно, ее вес. В настоящее время композитные материалы выдерживают давление вплоть до глубин порядка 6000 м. Для погружения на большие глубины подводного аппарата, вес которого не превышает 20—25 т — предел, позволяющий судну-носителю поднимать и спускать его на воду,— потребуется добиться новых успехов в разработке композитных плавучих материалов в смысле их сопротивления и плотности. Вот критическая точка, на которую сегодня должны быть направлены усилия технологического поиска.
Современные глубоководные аппараты весят от 10 до 20 т, что позволяет спускать их на воду и поднимать с воды судну-носителю средних размеров, водоизмещением порядка 1000 т. В ходе погружения плавучесть этих устройств практически не меняется, и работа их обходится дешевле, чем работа батискафов. Сегодня в мире существует шесть типов глубоководных аппаратов, предназначенных для использования в научных целях: «Пайсисы», сконструированные в Канаде (СССР закупил два «Пайсиса», которые опускаются с океанографического судна), как и «Синкай-2000», построенный японцами, могут достигать глубины 2000 м; французская «Сиана», появившаяся в начале 70-х годов, может погружаться на глубину 3000 м; американский «Алвин» — на глубину 4500 м (благодаря использованию прочной оболочки из титана); наконец, французский «Наутилус» и американский «Си Клиф», оба построенные в 1985 году, могут достигать глубины 6000 м. Самые тяжелые из них весят 22 т на воздухе. По сравнению с батискафами эти современные подводные аппараты обладают значительным преимуществом в управляемости: более легкие, они имеют повышенную маневренность, что весьма облегчает действия на дне, они снабжены одним или двумя манипуляторами, несущими различные системы захвата и управляемыми пилотами; кроме фотографических аппаратов, устанавливавшихся еще на батискафах, они оборудованы телевизионными камерами непрерывного действия, ведущими во время погружения видеозапись, которая затем может быть использована многими специалистами: наконец, они
Рис. 5. Крупномасштабная батиметрическая карта местонахождения Шенетт на 13° с. ш., построенная при помощи фотограмметрии на основе полученных при погружении видеозаписей (по Фюстеку, 1985 ).
1 — разрушенная дольчатая лава; 2— бездействующий курильщик; 3 — белый курильщик; 4 — черный курильщик.
имеют систему точной ориентировки на дне при помощи акустической триангуляции: подводный аппарат перемещается в пределах поля, ограниченного акустическими буями с расстоянием между ними 5—10 км, и, непрерывно посылая сигналы к каждому из буев, определяет свое местонахождение по отношению к ним; в некоторых случаях судно-носитель лоцирует подводный аппарат на поле буев во время погружения и руководит его перемещением, связываясь с ним по подводному телефону, в настоящее время уже надежно функционирующему.
Впоследствии по видеозаписям и навигационным данным, также непрерывно регистрируемым в центре сбора данных, можно восстановить путь подводного аппарата и составить точную карту дна. Точность акустической навигации находится в пределах 10—20 м, что совсем неплохо, но все еще недостаточно для картирования подводных оазисов. Метод анализа изображений, позволяющий установить взаимное расположение различных заметных форм рельефа дна и локализовать затем подводный аппарат в поле таким образом определенных природных ориентиров, был разработан Фюстеком. Расстояния между ориентирами и между подводным аппаратом и ориентирами определяются методом фотограмметрии, приспособленной для фотографирования в наклонной плоскости и использующей в качестве масштаба предметы-реперы известного размера, разбросанные по местности («межевые столбы»). Такой метод, который, к сожалению, применялся пока только на «Сиане» в ходе двух экспедиций, проведенных в 1982 и 1984 годах в излиянии на 13° с. ш., позволяет составлять карты масштаба 1:200, достаточного для представления пространства, занятого различными гидротермальными сообществами.
Биологу необходимо, очевидно, не только получить описания, но и поймать тех животных, что он наблюдает — будь то для изучения еще неизвестных науке видов, которые нередко встречаются в среде такого типа, или же для подтверждения прямым изучением материала первоначальных определений, сделанных визуально при плохой видимости организмов. Манипуляторы глубоководных подводных аппаратов и сачки для сбора проб довольно хорошо приспособлены для обращения с твердыми объектами, такими, например, как обломки лавы. Но когда их используют для сбора животных, они дают неудовлетворительные результаты: захваты, сила которых нерегулируема, раздавливают трубки червей, разбивают раковины двустворчатых моллюсков и панцири крабов; сачки из металлической сетки, лишенные крышек, практически невозможно использовать: мелкие организмы проскакивают сквозь ячею, а более крупные теряются при подъеме на поверхность из-за водоворотов, вызываемых самим сачком. Так что пришлось придумать систему транспортировки материала и образцов, могущую отделяться от подводного аппарата: настоящий /автономный подъемник, она представляет собой емкость в форме
Рис. 6. Крупномасштабная батиметрическая карта местонахождения Париго, 13° с. ш., построенная так же, как и предыдущая (по Фюстеку,' 1985).
1 — активный гидротермальный курильщик; 2 — белый курильщик; 3 — бездействующая постройка; 4 — базальтовый столб.
большого снаряда с полукруглой крышкой, имеющую две крупные закрывающиеся камеры, защищенные от водоворотов съемной металлической юбкой. Подводный аппарат локализует опущенную на дно емкость и с помощью манипулятора достает из нее необходимые инструменты и помещает туда собранные образцы. С таким ценным дополнением подводный аппарат являет собой единственный инструмент, действительно приспособленный для изучения оазисов на глубинах.
Хотя в настоящее время известно более десятка гидротермальных оазисов, ставших предметом подводных исследований, подробно описано население только двух или трех из них. Поскольку рассказать о всем разнообразии типов животного населения оазисов невозмож
но, мы остановимся на двух гидротермальных излияниях, наиболее изученных к настоящему времени,— это район на Галапагосском рифте на глубине примерно 2500 м, расположенный на экваторе около 86° з. д., и излияния на 13° с. ш. на Восточно-Тихоокеанском поднятии, на глубине примерно 2600 м. Галапагосское местонахождение, открытое в 1976 году, дважды изучалось американскими биологическими экспедициями, которые имели в своем распоряжении многочисленные данные, полученные ранее геологами и геохимиками. Что касается местонахождения на 13° с. ш., то оно исследовалось двумя экспедициями, организованными французскими биологами, также использовавшими данные геологов.
Общим правилом любой исследовательской работы является описание открытых объектов и присвоение им названий. Гидротермальные оазисы — не исключение из этого правила, и воображению первых наблюдателей было где разгуляться. На Галапагосском рифте четыре гидротермальных излияния, растянувшиеся в общей сложности километров на двадцать и расположенные в 3—8 км один от другого, получили названия Розовый сад, Мидиевая банка, Райский сад и Мелюзга (по-английски Small Fry). На широте 13° с. ш. топонимы, менее образные, отражают положение или внешний вид местонахождения либо его населения: Погонор, Актинуар, Погосюд, Погомор, Пари-го...* Если галапагосские излияния принадлежат к типу теплых источников (температура на выходе менее 30— 40 °C), лишенных минеральных отложений, то на излияниях на 13° с. ш. встречаются как теплые источники, так и распылители белесоватой жидкости и знаменитые черные курильщики с высокой температурой изливающейся жидкости.
Если список определенных в гидротермальных оазисах неизвестных или известных науке видов уже перевалил за сотню, прямое наблюдение и исследования фотографий и видеозаписей не позволяют насчитать такое количество: мелкие виды, длиной менее 1 см, невозможно, кроме исключительных случаев, с уверенностью определить ви-
* Погомор — от французских слов Pogonophore (погонофора) « niort (мертвый); Актинуар — actif (активный) и noir (черный); Поп-сюд — Pogonophore и sud (юг); Погонор — Pogonophore и nord (север/, Париго — от французского Paris (Париж) и английского to go (идти). — Прим, перев.
Рис. 7. Цепочки раковин крупной двустворки Calyptogena magnifica, тянущиеся вдоль трещин, откуда выходит разбавленный гидротермальный флюид, 21° с. ш.
зуально. На Галапагосском рифте было опознано 22 вида против 26 на Восточно-Тихоокеанском поднятии на 13° с. ш. С количественной точки зрения, таким образом, разница невелика, но качественный анализ выявляет более значительные различия.
Среди четырех излияний у Галапагосских островов наибольшую биомассу имеет Розовый сад. Большая расщелина, откуда выходит опалесцирующий гидротермаль
ный флюид, окружена скоплениями плотных кустов высотой до двух метров — это погонофоры, гигантские черви, живущие в белых, отливающих перламутром трубках, увенчанных султаном ярко-красных щупалец. Здесь также обитают два вида крупных двустворчатых моллюсков — Bathymodiolus (батимодиола), связанный с погонофорами, и Calyptogena (калиптогена), чьи белые раковины располагаются по краям банок батимодиол и тянутся вдоль тонких трещин, удаленных от основных скоплений. Среди погонофор изобилуют крабы и креветки, а также мелкие брюхоногие моллюски в форме китайской соломенной шляпы. Дно здесь сложено плоскими базальтовыми лавами, перерезанными трещинами, вдоль которых селятся калиптогены. Вблизи основной гидротермальной расщелины базальты в форме подушек покрыты белыми трубками кольчатых червей — серпулид в сопровождении актиний (морских анемонов). В этой зоне, образующей кольцо вокруг плотных поселений погонофор, изобильны мелкие ракообразные — белые галатеиды.
Мидиевая банка, расположенная на склоне из подушечных лав, представляет собой настоящую стену, образованную батимодиолами, растущими в несколько слоев; здесь имеются также несколько разбросанных букетов погонофор и незамкнутое кольцо серпулид. Невдалеке видны скопления мертвых раковин батимодиол в процессе растворения, свидетельствующие об иссякшем гидротермальном источнике. Часто встречаются кишечнодышащие животные, прозванные «спагетти» за свой характерный облик, обусловленный множеством переплетенных трубок на фоне базальтовых лав.
Райский сад раскинулся на лавовой базальтовой подушке. Выход гидротермального флюида здесь слабее, чем в двух рассмотренных выше излияниях. Погонофоры, так же как и батимодиолы, встречаются реже. Серпулиды и галатеиды изобилуют вокруг излияния.
Наконец, на излиянии Мелюзга выхода гидротермального флюида не отмечено, и оно отличается от окружающей среды лишь относительным изобилием серпулид и «спагетти».
Были сделаны некоторые оценки биомассы: на Мидие-вой банке моллюсков насчитывается более 300 экз./м2, что равно биомассе 10 кг/м2 (в сыром весе, включая раковину). Плотность поселения гигантских погонофор Riftia (рифтия) составляет 22 экз./м2, что соответствует
Рис. 8. Знаменитая белая галатеида Munidopsis subsquamosa, верный страж подводных оазисов.
биомассе 6—10 кг/м2. Максимальная температура, измеренная на каждом излиянии, варьирует от 15 °C (Розовый сад) до 13 (Райский сад) и 12 °C (Мидиевая банка). На излиянии Мелюзга температура не превышает температуру окружающей среды (2 °C). Значение температуры является прекрасным показателем степени разбавленности гидротермального флюида к тому моменту, когда он изливается в море. Рифтии предпочитают температуры от 4 до 12 '"С, батимодиолы — от 3 до 8 °C. Трудно получить правильные значения температуры в этих микросредах, где уже само введение зонда-измерителя изменяет выход флюида и соответственно установившийся температурный градиент. Серпулиды живут в зонах, где температура изменяется от 2,5 до 3°С. Микрораспространение видов
зависит в значительной степени от физических законов, которые управляют выходом гидротермальных флюидов и их смешением с морской водой: восходящие течения, порожденные высокой температурой, быстро увлекают вверх большую часть излившегося флюида, в соответствии с относительно слабым градиентом разбавления; по сторонам от источника влияние гидротермального флюида, напротив, ощущается на очень небольших расстояниях — от нескольких дециметров до нескольких метров — и соответствующий градиент гораздо сильнее. Это объясняет распределение калиптоген вдоль трещин, а также в целом вытянутую вверх форму кустов рифтии, которые скапливаются вокруг излияния, будто нарочно для того, чтобы сохранить как можно дольше питательный флюид, препятствуя его разбавлению морской водой. Точно так же можно наблюдать рифтий, лежащих на дне таким образом, что султан их щупалец располагается как раз над микротрещиной, откуда выходит флюид.
Для этих четырех излияний характерен в основном один и тот же тип сообществ, в котором доминируют три гигантских вида: рифтия, батимодиола и калиптогена, указывающая на выходы, гидротермального флюида с пониженной температурой. Над самым излиянием часто наблюдают рыбу в вертикальном положении, головой вниз. Поймать ее пока не удалось. Возможно, она питается скоплениями бактерий, выбрасываемыми с гидротермальным флюидом. Эта рыба, как предполагают, принадлежит к роду Dipjacanthopoma из семейства бити-тидовых (Bythitidae).
Излияния на Восточно-Тихоокеанском поднятии на 13" с. ш. известны, пожалуй, лучше, чем галапагосские: их изучали в ходе двух следовавших друг за другом экспедиций одни и те же специалисты. Общая морфология поднятия тоже хорошо известна: рифтовая долина, расположенная по оси хребта, средней глубиной 2650 м и шириной на этой широте 250—300 м, окаймлена двумя эскарпами, наибольшая высота которых достигается на глубине 2600 м, то есть на 50 м выше дна долины. Доминирующий рельеф этой долины обусловлен широкими то сохранившимися в целости, то обрушившимися растеками дольчатой лавы со стекловидной поверхностью. Эти обрушившиеся лавовые озера достигают глубины 10 м. Явления местного переохлаждения в ходе опорожнения озер оставляют внушительные базальтовые контрфорсы,
Рис. 9. Эта гигантская актиния, часто встречающаяся на 13° с. ш. по краям обрушившихся лавовых озер и на лавовых столбах, может достигать длины 1,5 м. Ее длинные щупальца ориентируются по течению. Ее не удалось поймать, так что, эта актиния до сих пор остается не определенной.
тонкослоистый профиль которых свидетельствует о резких переменах уровня лавы в процессе ее излияния из озера. Глубокие многометровые трещины образуются параллельно оси хребта. Здесь, среди величественных форм подводного ландшафта, гораздо более беспорядочных, чем на Галапагосском рифте, следы прошлых гидротермальных
Рис. 10. Карта района Восточно-Тихоокеанского поднятия, в котором работали в 1982 и 1984 годах на борту подводного аппарата «Сиана» французские биологи. Прерывистыми линиями показаны два эскарпа, окаймляющие осевую рифтовую долину (по Фюстеку, 1985).
явлений предстают в форме массивных высоких сооружений из полиметаллических сульфидов, покрытых пест-роокрашенным окисленным слоем. Эти постройки могут достигать в высоту несколько метров, даже более 10 м; их основание занимает площадь в несколько квадратных метров. Неактивные трубы встречаются чаще активных: примерно четыре неактивных излияния на одну действующую трубу. Действующие трубы встречаются здесь гораздо чаще, чем на Галапагосском рифте: на 13° с. ш. на отрезке хребта длиной 7,5 км были обнаружены 24 действующих трубы, то есть в
Рис. II. Белый курильщик на 13 “ с. ш.: по массивной колонии полихет Alvinella ловко передвигается большой краб Cyanagraea; справа — несколько трубок, в которых живут погонофоры.
среднем приблизительно по одной на каждые 300 м.
Выбранные пять точек стали объектами тщательного топографического изучения на основе анализа полученных описательных документов. С севера на юг последовательно располагаются излияния Шенетт («Цепочка»), Погонор, Актинуар, Погосюд и Погомор. Каждое излияние служит иллюстрацией того или иного аспекта гидротермальных
Рис. 12. Распределение поселений серпулид на местонахождении Погонор в 1982 и 1984 годах (батиметрия 1984 года). За два года в этом местонахождении произошло общее снижение численности серпулид: 1982 год— 150 экз./м2,1984 год: а — от 50 экз./м2, б — от 100 экз./м2, в — от 150 до 200 экз./м2. 1 — разрушившаяся дольчатая лава, 2 — бездействующий курильщик, 3 — белый курильщик.
(По Фюстеку, 1985).
поселений. У трех излияний — Погонор, Актинуар и Погосюд — побывала «Сиана» с интервалом в два года, в марте 1982-го и марте 1984-го (именно в ходе этой последней экспедиции мне и довелось с ними познакомиться) .
Излияние Шенетт («Цепочка») было открыто в 1982 году. Оно образовано большим распылителем, расположенным на глубине 2643 м на западном берегу обрушившегося лавового озера, ориентированного вдоль центральной впадины. Распылитель покрыт массивной колонией кольчатых червей Alvinella (альвинелла), живущих в пергаментных трубках и первоначально названных помпейскими червями, так как на них постоянно сыплется дождь вулканических частиц. У подножия главного рас
пылителя имеется несколько небольших черных курильщиков, достигающих всего несколько десятков сантиметров в высоту. На вершине массива альвинелл также наблюдается несколько действующих черных курильщиков. У подножия большого курильщика с помпейскими червями поселилась маленькая колония — около сотни особей — гигантских погонофор. Вокруг этого местонахождения видны прикрепившиеся к дольчатой лаве серпу-лиды, в большинстве мертвые. В нескольких метрах к северу от основного местонахождения есть неактивные трубы. В 1984 году это излияние заново не исследовалось.
Погонор представляет собой поднятый на высоту 5— 6 м надо дном долины массив, выделенный двумя параллельными расщелинами глубиной 2640 м. Центральный массив эллипсовидной формы занимает площадь примерно 8X3 м2. На западной стороне массива вздымается сложная гидротермальная постройка: она состоит из цилиндроконического курильщика высотой 3 м и прилегающего к нему шаровидного распылителя метрового диаметра. В 1982 году поверхность площадью примерно 7 м2 покрывало плотное поселение рифтий, отмечались также несколько иерихонских червей (другой вид погонофор). Зона серпулид развита хорошо, плотность трубок может достигать 150—300 на 1 м2. Активные части системы курильщиков заселены помпейскими червями, а температура, измеренная в глубине массы трубок сантиметрах в двадцати от ее поверхности, достигает 270 °C! Двумя годами позже это же местонахождение разительно изменилось: большая часть рифтий исчезла, и их лишенные поддержки пустые трубки усыпают дно северо-восточной расщелины. Занявшие их место батимодиолы кажутся более многочисленными, чем в 1982 году: тогда часть раковин была скрыта рифтиями. Шаровидный распылитель вырос по крайней мере на полметра в высоту, а флюид, белесоватый в 1982 году, в 1984-м стал прозрачным. Популяция рыб, связанная с этим излиянием и насчитывавшая в 1982 году более 300 экземпляров, сократилась до нескольких десятков, да и серпулиды тоже регрессировали. Что же произошло за два года? Некоторые наблюдения позволяют сказать, что существенно снизилось количество гидротермального флюида на глубине и это повлекло за собой гораздо большее его разбавление: флюид стал менее теплым и менее насыщенным минеральными компонентами. В то же время под действием текто-
Рис. 13. Крупномасштабная батиметрическая карта местонахождения Актинуар на 13° с. ш. (батиметрия 1982— 1984 годов); прерывистой линией показана ось разреза рис. 14, проходящая через большой черный курильщик.
1 — разрушившаяся дольчатая лава, 2 — бездействующий курильщик, 3 — белый курильщик, 4 — черный курильщик.
нических подвижек разрушился язык дольчатой лавы на севере массива альвинелл. Такие явления, конечно, случаются часто и более или менее быстро приводят к истощению гидротермальных излияний, а затем и к полному исчезновению оазисов жизни. Так объясняют целые кладбища раковин калиптоген, наблюдаемые с 1978 года на 21° с. ш., свидетели былого существования исчезнувших оазисов. По оценкам, в местных условиях известковая раковина должна полностью раствориться самое большее за несколько столетий.
Рис. 14. Схематический разрез местонахождения Актинуар; пунктиром обозначен небольшой черный курильщик, располагавшийся в 1982 году на середине высоты большого, а в 1984 году закрывшийся.
Над излиянием Актинуар, расположенным чуть южнее Погонора, возвышается черный курильщик в стадии наибольшей активности; его средний диаметр 2 м, а высота с 1982 по 1984 год возросла с 15 до 17 м. Эта внушительная колонна из сульфидов покрыта на разных уровнях колониями помпейских червей, белесоватая окраска которых выделяется на охристом фоне — результате окисления поверхности сульфидов. В основании курильщика в 1982 году была отмечена популяция актиний — два года спустя она исчезла. Во время одного из подъемов «Сианы» удалось заснять этот черный курильщик во всю высоту: незабываемое зрелище длилось долгих полминуты и завершился подъем у двух черных султанов температурой 300 °C, испускаемых двумя смежными жерлами диаметром в десяток сантиметров. А в каких-нибудь двадцати сантиметрах от них уже селятся помпейские черви! У подножия этой гигантской трубы расположен шаровидный
курильщик диаметром более 2 м, серовато-белый, покрытый хорошо развитой колонией помпейских червей. Белесоватый гидротермальный флюид (содержащий сульфаты кальция и кремния) образует над колонией как бы маленькое облачко. Эти колонии помпейских червей — первые наблюдатели назвали их снежками—часто связаны с черными курильщиками; такая связь объясняется подземной циркуляцией и вторичным разбавлением. Невдалеке от этого массива довольно часто встречаются серпудиды, сопровождаемые непременным конвоем гала-теид. В отличие от излияния Шенетт, здесь нет поселений погонофор. В десятке метров на север от излияния значительный район занят пустыми трубками серпулид. Там же отмечены несколько створок раковин, уже почти полностью растворившихся: это раковины батимодиол и, что самое удивительное, крупных белых калиптоген, которые до сих пор не встречались в живом состоянии в излияниях на 13° с. ш. Следовательно, скорее всего это излияние затухло самое большее лет двадцать назад.
Излияние Погосюд, расположенное еще дальше к югу, характеризуется скоплением больших кустов погонофор, покрывающих в общей сложности площадь около 15 м5, причем каждый куст занимает от половины до целого квадратного метра. Отложения неактивных сульфидов в центре излияния свидетельствуют о былой гидротермальной активности с высокой температурой. В настоящее время температура здесь не превышает 20 °C. Вокруг погонофор на площади примерно в 100 м2 изобилуют серпулиды, плотность их поселений составляет 200— 300 трубок/м2. Среди зарослей рифтий и на базальтовых лавах обитают богатые популяции колпачковидных брюхоногих моллюсков. У основания трубок погонофор селятся батимодиолы. Среди колоний рифтий ползают крабы, галатеиды, белые осьминоги. Целые облака крохотных частиц над излиянием поедаются мелкими планктонными ракообразными; наконец, тут масса рыб, как это наблюдалось на излиянии Погонор в 1982 году. Излияние Погосюд мало изменилось за два года, однако можно отметить, что некоторые заросли рифтий и сопутствующие им серпулиды регрессировали.
Наконец, в нескольких десятках метров к югу от Пого-сюда простирается сложная зона, сформировавшаяся под действием многочисленных тектонических и гидротермальных событий, как о том свидетельствует множество
Рис. 15. Скопление гигантских погонофор Riftia pachyptila с многочисленными батимодиолами; крабы, галатеиды и рыбы изобильны на границе поселений рифтий. Местонахождение Погосюд, 13° с. ш.
неактивных труб, часть которых ранее были заселены помпейскими червями. Оба излияния — Погомор i и II — имеют сходную фауну. Все крупные погонофоры погибли, остались только их трубки, лишенные опоры. Иерихонские черви, более выносливые, еще держатся. Трубки серпулид в основном пустые. Лишь одна популяция батимодиол с плотностью поселения 150 экз./м2, в большинстве живых,
все еще существует. Изобилуют колпачковидные брюхоногие моллюски и любопытный маленький гребешок (двустворчатый моллюск). Выходов гидротермального флюида в двух этих местонахождениях не обнаружено, вероятно, они находятся на довольно далеко зашедшей стадии регрессии.
Приведенное описание нескольких местонахождений на Галапагосском рифте и на 13° с. ш. на Восточно-Тихоокеанском поднятии позволяет сделать несколько замечаний общего характера:
— Прежде всего, гидротермальные сообщества поражают своей невероятно высокой биомассой по сравнению с биомассой окружающей среды на тех же глубинах: в некоторых случаях она в 10 000—100 000 раз больше.
— Во-вторых, эти сообщества сильно структурированы в зависимости от расстояния от источника, природы и количества выбрасываемого флюида. Если говорить схематично, наблюдается концентрическое расположение видов вокруг точки излияния.
— Существуют два типа гидротермальных сообществ: первый характеризуется наличием рифтии, калиптогены и батимодиолы, ищущих невысокие температуры — от 3 до 12 °C, второй характеризуется наличием помпейских червей и поедающих их крабов, которые переносят более высокую температуру и более концентрированный флюид. Оба эти типа сообществ могут сосуществовать, если выбросы гидротермального флюида достаточно горячи. В противном случае представлено лишь низкотемпературное сообщество.
— Физико-химические условия, складывающиеся у гидротермальных источников и способствующие развитию оазисов жизни на глубинах, недолговечны. В случае единичного гидротермального излияния, длительность его функционирования составляет максимум несколько сотен лет, быть может и меньше. Часто наблюдаются сообщества регрессирующие. Именно это основополагающее наблюдение рождает вопрос: каким образом виды, строго связанные с гидротермальными излияниями, расселяются от одного местонахождения к другому.
3
ПРОДУКЦИЯ БАКТЕРИИ И ТЕРМОФИЛЬНЫЕ БАКТЕРИИ
Любопытно, что первыми наблюдениями подводных оазисов, связанных с гидротермализмом, мы обязаны геофизику Лонсдейлу: по фотографиям, полученным при помощи буксируемого устройства, протянутого на расстоянии 10—30 м над дном на излияниях Галапагосского рифта и примерно на 3° с. ш. над Восточно-Тихоокеанским поднятием, он описал в 1977 году изобильные поселения крупных раковин и, потрясенный их невероятной для подобной глубины плотностью, выдвинул две первые гипотезы. Первая из них предложила чисто физическое объяснение: эмиссия гидротермального флюида, температура которого, как предполагали уже тогда, достигает 100—200 °C, вызывает образование конвективных ячеек, и частицы органического вещества из приповерхностного освещенного слоя под влиянием горизонтальных течений, возникших в результате конвекции, концентрируются в непосредственной близости от горячих источников. Вторая же гипотеза, которая тогда казалась менее правдоподобной, предполагала существование бактерий, способных быстро развиваться на некоторых соединениях, несомых гидротермальными флюидами. Уже во время первых погружений американского подводного аппарата «Алвин» на излиянии на Галапагосском рифте в 1977 году именно вторая гипотеза подтвердилась прямыми наблюдениями. Наиболее активные источники испускали опалесцирующую молочного цвета жидкость, что может указывать на наличие в ней серы в коллоидальном состоянии, серы, возникающей либо при спонтанном окислении сульфидов, либо под действием бактерий. Высокое содержание сероводорода и сульфидов разных металлов, установленное в этих флюидах, было первым указанием в пользу такой гипотезы: оно позволило предполагать, что здесь
существует настоящая первичная продукция, очень локализованная и производимая автотрофными хемосинтезирующими бактериями. Если первая гипотеза сегодня не может быть окончательно отброшена, то вторая — как подтверждают многочисленные указания, верная в том, что касается участия в процессе хемосинтезирующих бактерий,— должна быть глубоко переработана в той части, которая касается использования этой продукции беспозвоночными животными. В действительности оказалось, что беспозвоночные не собирают, как думали раньше, бактерий, обитающих во внешней среде, но создают с ними весьма тесную ассоциацию: «полезные» бактерии живут внутри клеток животных, которые ими питаются!
Перед тем, как подойти вплотную к этому вопросу, следует вкратце напомнить, что же представляет собой явление автотрофного хемосинтеза. Автотрофные растения способны синтезировать необходимый им органический углерод из СО2, используя солнечную энергию, улавливаемую в ходе фотосинтеза, тогда как животные, которых называют гетеротрофными, могут лишь преобразовывать произведенные растениями органические вещества для построения своих собственных клеточных компонентов. Кроме этих двух основных типов обмена веществ, фотоавтотрофии и гетеротрофии, существует третий, совсем особый, представленный хемосинтезирующими бактериями, или просто хемотрофами. Эти бактерии, для того чтобы превратить минеральный углерод в углерод органический, используют энергию, полученную при окислении неорганических веществ. Рисунок 16 показывает место, занимаемое хемосинтезом среди разных автотрофных путей обмена веществ.
Первыми фотосинтезирующими организмами были прокариоты (группа микроорганизмов, лишенных мембраны, отделяющей ядро от содержимого клетки; к ней принадлежат бактерии и синезеленые водоросли, или цианобактерии), которые жили в воздушной или жидкой среде, лишенной кислорода, и использовали свет как источник энергии для восстановления СО2, при участии сероводорода или других восстановленных соединений. В ходе эволюции у синезеленых, а позднее и у растений, вода заместила сероводород, а кислород соответственно серу как продукт экскреции, выбрасываемый в окружающую среду. Высвобождаемый таким образом кислород очень быстро захватил почти все биотопы суши и,
Рис. 16. Упрощенная схема положения бактериального хемосинтеза по отношению к фотосинтезу примитивных микроорганизмов и зеленых растений.
как настоящий яд, вытеснил примитивные анаэробные организмы, которые вынуждены были либо искать убежища в немногих защищенных от этого «загрязнения» биотопах, либо полностью исчезнуть.
Именно на основе сероводорода, как это имело место на Земле несколько миллиардов лет назад, и функционируют хемосинтезирующие бактерии.
Первые наблюдения были сделаны на излиянии на Галапагосском рифте, в месте, где температура гидротермальных источников колеблется в пределах 15—40 °C. Прежде всего было установлено, что в пробах опалесцирующего флюида содержится очень много бактерий: 105—106 бактерий в миллилитре воды, а вблизи дна на тех же глубинах насчитывается 102—104 клеток в миллилитре воды. Повышенная изменчивость этих величин объясняется, вероятно, большей или меньшей разбавленностью воды, из которой взяты пробы. Эти цифры говорят о такой плотности бактерий, а следовательно, о таком количестве доступного для животных органического вещества (бактериальная биомасса порядка 0,1 — 1 мг/л), которые значи
тельно превышают норму. Другой метод определения биомассы бактерий состоит в оценке содержания в определенном объеме морской воды аденозин-5-трифосфата (более известного под сокращенным названием АТФ), внутриклеточного компонента, играющего роль высокоэнергетического горючего. Если в нормальной придонной воде концентрация АТФ составляет 1,7 нг/л (1 нг — одна миллиардная часть грамма) при максимуме в 100 нг/л в высокопродуктивных поверхностных водах, то в пробах гидротермального флюида были получены значения 300—500 нг/л. Равным образом и другой биохимический показатель, характеризущий скорость клеточного биосинтеза,— отношение количества гуанозин-5-три-фосфата к количеству АТФ (ГТФ/АТФ), измеренный в тех же пробах гидротермального флюида, оказался в четыре-пять раз выше, чем в продуктивных поверхностных водах, и в сотни раз выше, чем в обычных придонных водах. В тех же самых теплых излияниях с опалесцирующей жидкостью можно невооруженным глазом увидеть частицы, которые представляют собой бактериальные агрегаты. Один из видов рыб, свойственный излияниям Галапагосского рифта (уже упомянутый выше), часто держится прямо над источником головой вниз, будто собирая агрегаты бактерий, что являлось бы уникальным примером столь короткой пищевой цепи.
Для получения более точной оценки продукции органического вещества бактериями, как правило, измеряется количество ассимилированного СО2, меченого радиоактивным изотопом углерода 14С. С помощью манипуляторов аппарата «Алвин» удалось наполнить гидротермальным флюидом шприцы с заранее введенным в них определенным количеством радиоактивного СОг. После инкубации в течение определенного времени достаточно профильтровать воду и отделить бактерии. Величина радиоактивности, зарегистрированной в бактериальных клетках, соответствует доле меченого минерального углерода, преобразованного в органический углерод. Результаты, полученные таким методом, очень интересны: наибольшая величина продукции, полученная при температуре около 3—4 °C, то есть после сильного разбавления гидротермального флюида окружающей морской водой, составляет 3,6—4,8 мг углерода на 1 м3 воды в сутки, что примерно соответствует той величине, какую можно получить в глубоком аноксическом бассейне между насыщенным кислородом
поверхностным слоем и глубинным слоем, например во впадине Карьяко в Карибском море. Эти величины не покажутся очень высокими, если сравнить их с 11,7 мг ассимилированного углерода на 1 м” в сутки, установленными для глубоководных слоев Черного моря, то есть для среды, лишенной кислорода в результате естественных причин. Исходя из повышенной концентрации сероводорода в гидротермальном флюиде, пытались также доказать наличие бактерий, способных окислять сульфиды. Среди сотен штаммов бактерий, изолированных и выделенных в культуры, многие оказались способны окислять сероводород: одни из них — это облигатные хемотрофы, их метаболическая активность приводит к выделению в среду кислых веществ; другие — факультативные хемотрофы, они выделяют основные вещества (политионаты). Удалось определить некоторые штаммы, переносящие повышенное содержание сероводорода при термическом оптимуме 25—35 °C, но при давлении 250 бар (давление, соответствующее глубине излияния на Галапагосском рифте) они сохраняют только 75% своей активности на поверхности (например, бактерии, принадлежащие к роду Thiomicrospira).
Общий вывод из исследований, проведенных на излияниях Галапагосского рифта, ясен: в физико-химических условиях, свойственных этому местонахождению, ассимиляция СО2 (мера хемосинтетической активности) в гидротермальном флюиде в ходе перемешивания его с окружающей морской водой низка по сравнению с биомассой бактерий, обнаруженных в этом флюиде. Следовательно, эта биомасса бактерий, по всей вероятности, создается в подповерхностной сети трещин и разломов, во всяком случае до выхода флюида в морскую воду. Микробиологи даже считают, что метаболическая активность бактерий, присутствующих в гидротермальном флюиде, в общем много меньше той, которая могла бы удовлетворить нужды процветающих сообществ животных.
Не следует ли вернуться к первой гипотезе, согласно которой конвективные ячейки, порожденные гидротермальной эмиссией, способствуют тому, что как раз на уровне излияний концентрируется достаточное количество взвешенного органического вещества, приносимого с поверхности моря? Нет, не следует, ибо простые расчеты показывают, что количество органического вещества, которое подобным образом может постоянно поступать
сюда, намного ниже потребностей плотных сообществ животных. В действительности бактериальная продукция, создающаяся в основном внутри гидротермальной сети, под поверхностью базальтов, вероятно, достаточна, чтобы объяснить большую плотность поселений некоторых беспозвоночных-фильтраторов, способных улавливать взвешенные частицы, и в частности агрегаты бактерий. Это объясняет также наличие мелких планктонных животных над излияниями.
Опять же в пробах, взятых главным образом на Галапагосском рифте, были определены разные типы хемосинтезирующих бактерий. Различают шесть крупных морфологических категорий:
— Одноклеточные бактерии округлой формы, часто заключенные в плотные железисто-марганцевые отложения.
— Бактерии, имеющие очень длинную ножку, типа Hyphomonas-Hyphomicrobium.
— Формы, напоминающие цианобактерии, морфологически очень близкие к Calothrix.
— Формы, содержащие сильно развитые цитоплазматические мембранные системы, определенные как метило-трофные бактерии.
— Нитчатые бактерии, напоминающие роды fhiotrix и Leucolhrix, которые составляют большинство проанализированных бактерий.
— Нитчатые неприкрепленные бактерии, напоминающие Beggiatoa.
На излияниях черных курильщиков, и в частности на местонахождении 21° с. ш. на Восточно-Тихоокеанском поднятии, было проведено меньше исследований. Здесь были получены более оригинальные, но порой спорные результаты. Во-первых, удалось показать существование термофильных бактерий, то есть таких бактерий, у которых оптимальная метаболическая активность наблюдается при температуре выше 50 °C. Для некоторых проб бактерий был установлен температурный оптимум 105 °C; эти экстремально термофильные бактерии при температурах ниже 70 °C становятся неактивными. Метаболическая активность этих бактерий чрезвычайно интересна: в культуральных средах, содержащих минералы (S2O3, Мп и Fe), при температуре 100 °C и давлении 1 атмосфера они способны синтезировать метан (СН4), окись углерода (СО) и водород, а также небольшое количество закиси
азота (N2O); они могут также окислять метан. Если до этого открытия геологи считали, что метан, водород и окись углерода, содержащиеся в гидротермальных флюидах, возникают либо при дегазации мантии Земли, либо в результате химических реакций между базальтом и морской водой, то это открытие объясняет явление с биологической точки зрения. Эти термофильные бактерии быстро размножаются: штамм, принадлежащий к роду Methanococcus, обнаруженный в отложениях минеральных частиц в основании черного курильщика, имеет оптимум роста при 86 °C и время удвоения 26 минут.
Но вернемся к бактериям, выделенным из гидротермального флюида черного курильщика, взятого при температуре 350 °C: в июне 1983 года в авторитетном научном журнале «Нейчер» («Природа») появилась статья, всего на три страницы, озаглавленная «Рост бактерий из черных курильщиков при температуре как минимум 250 °C»! Эта короткая статья знакомит с опытами, проведенными с пробой воды, взятой осенью 1979 года на Восточно-Тихоокеанском поднятии на 21° с. ш. из султана черноватой жидкости температурой 306 °C. Бактериальные культуры были выращены в камере, позволяющей воссоздать температуру и давление, свойственные этому местонахождению (то есть температуру 350 °C и давление 265 атмосфер), на среде, в состав которой входили различные минеральные соли. При подсчете бактерий в эпифлуоресцентном микроскопе (процедура состоит в стимуляции флуоресценции красителем DAPi;::, который избирательно фиксируется на молекулах ДНК хромосомного вещества бактерий) были установлены следующие скорости удвоения бактерий: 8 часов при температуре 150 °C; 1,5 часа при температуре 200 °C и всего лишь 40 минут при температуре 250 °C. При последнем значении температуры масса белка удваивается с той же скоростью, что и число бактерий. При температуре 300 °C после некоторого возрастания количества клеток наблюдается довольно быстрое его уменьшение, что может быть объяснено разрушением клеток во время необходимой для взятия проб декомпрессии или даже во время самого опыта, поскольку, в частности, при высоких давлении и температуре в морской воде понижается показатель кислотности. На некоторых снимках,
* 4,6-диамидино-2-фенилиндол.— Прим. ред.
сделанных в электронном микроскопе, видны организмы, выросшие в культуре при температуре 250 °C и давлении 265 атмосфер.
Чтобы лучше понять значение этой публикации и тот отпор, который ей дали микробиологи, необходимо сделать некоторые пояснения общего характера. Самая высокая экспериментально измеренная температура, при которой возможны жизнь и рост микроорганизмов, в начале века составляла 55 °C, в 1970 году — 95 °C и в 1982-м—105 °C. Последний результат был получен на бактериях, взятых из геотермальной скважины на морской сольфатаре на малой глубине, а также, как говорилось выше, на некоторых штаммах, изолированных из проб черноватого гидротермального флюида. Только микроорганизмы — имеются в виду прокариоты — способны жить при температурах такого порядка. В начале 1970-х годов один микробиолог, специалист по термофильным бактериям, на основе изучения микрофлоры гейзеров Йеллоустонского Национального парка выдвинул идею о том, что наличие воды в жидком состоянии — гораздо более важное условие для существования жизни, чем максимальная температура. Все бактерии, способные переносить температуры выше 80—85 °C, принадлежат к группе архебактерий, которые, как указывает их название, считаются наиболее близкими к предковым формам: много среди них метаногенных бактерий, способных синтезировать метан (роды Thermoplasma, Sulfolobus, Methanococcus). Недавно описанный вид из пробы черноватого гидротермального флюида, Methanococcus jan-naschii, отличается от других известных архебактерий оригинальной биохимической структурой клеточной мембраны: у архебактерий мембранные липиды состоят из изопреноидных углеводородов и алкилглицериновых эфиров, а у вновь открытого вида мембранные липиды на 95% состоят из макроциклического диэфира глицерина — молекулы, доселе не встречавшейся в живом мире.
Палеонтологам давно знакомы ископаемые структуры, являющиеся результатом жизнедеятельности морских микроорганизмов,— строматолиты, наиболее древние из ныне известных свидетельств жизни. Некоторые строматолиты имеют возраст около 3,5 млрд, лет и отложения, в которых они находятся, иногда носят следы гидротермальной деятельности. Эти факты следует рассматривать в общехМ контексте эволюции нашей планеты: миллиард
лет тому назад примитивный океан был совсем лишен кислорода, и его температура, по мнению некоторых специалистов, должна была составлять 50—70 "С. В более отдаленные эпохи температура океана, возможно, была еще выше и достигала 150 °C, оставаться жидкой при такой температуре воде позволяло высокое давление земной атмосферы, связанное с очень большим содержанием СО2. Задолго до образования континентов этот архейский океан, похоже, был совсем неглубоким и являлся ареной интенсивных гидротермальных проявлений.
Из этого следует, что микроорганизмы, которые жили на Земле первые два миллиарда лет после возникновения жизни, были термофильными формами, обитавшими в аноксической среде и использовавшими для своего развития минеральные элементы, происходящие из базальтовой магмы. В той особенной среде, которая представляет собой гидротермальную подповерхностную сеть, до наших дней сохраняются физико-химические условия если не идентичные, то очень близкие к условиям архейского океана. В гидротермальных выходах, относящихся к этой сети, до сих пор находят архебактерий, и можно утверждать, что изучение этой среды представляет особый интерес для познания условий развития первых сообществ микроорганизмов.
Не удивительно поэтому, что сообщение об открытии бактерий, способных размножаться при температуре 250 °C и давлении 265 атмосфер, стимулировало исследование этих проблем и повлекло за собой серию работ, призванных проверить полученные результаты. В самом деле, если бы они подтвердились, была бы поставлена под сомнение распространенная теория происхождения жизни на Земле, согласно которой первые органические молекулы образовались фотохимическим способом из газов, содержащихся в атмосфере, аминокислоты появились в результате конденсации углеводов и формамида, и все это привело к появлению настоящего органического «супа», плавающего на поверхности вод. В этом примитивном «супе» якобы зародились первые молекулы, способные к самовоспроизведению (нуклеиновые кислоты), затем первые прокариотные организмы. Эта теория не учитывает недавние открытия подводных гидротерм, но является в настоящее время официально принятой. Отсюда становится понятным то значение, которое биологи придают этим новым результатам. С момента выхода в 1983 го
ду упомянутой статьи в США развернулась полемика.
Многочисленные последующие работы опровергли реальность полученного результата. Так, один биохимик показал, что максимальная температура, которую могут переносить живые существа,— это такая температура, при которой наступает естественный гидролиз их биохимических компонентов (гидролиз — химическое разложение молекулы путем фиксации воды), а температура гидролиза, по мнению исследователя, ниже 250 °C. Поэтому он приходит к благоразумному заключению: если ультра-термофильные бактерии действительно существуют и их метаболические реакции происходят в водной среде, то они не могут жить при температуре 250 °C, если состоят из биологических молекул, таких, как белки и нуклеиновые кислоты. Можно критиковать и эту работу, основанную на опытах, проведенных при давлении 39,2 атмосферы (давление водяного пара при температуре 250 °C) и на изолированных молекулах, не заключенных в биологическую мембрану. Другие авторы показали, что штамм термофильных архебактерий (Pyrodictium occultum), способных жить при температуре 110 °C при атмосферном давлении в широком диапазоне солености, разрушается гидролизом за 6 часов при температуре 250 °C под давлением 260 атмосфер.
Какой же вывод можно сделать из этих работ, проведенных с таким тщанием? Для многочисленных микробиологов этих доказательств достаточно: микроорганизмов, способных жить при температуре 250 °C и давлении 265 атмосфер, не существует, а результаты, опубликованные в 1983 году, должны быть отнесены к экспериментальным артефактам. Для других вопрос остается открытым, здесь необходимы новые исследования.
Однако доказано, что в гидротермальных флюидах с высокой температурой существуют бактерии, у которых температурный оптимум жизнедеятельности близок к 110 °C. Из недавней работы Деминга, одного из двух авторов первоначального открытия ультратермофильных бактерий, доложенной на совещании в Океанографическом институте (Париж, ноябрь 1985 года), следует, что те же самые бактерии размножаются при температуре 120 °C под давлением 200 атмосфер, при культивировании их на новой твердой среде, которая остается стабильной при этой температуре. Этот результат не противоречит наблюдениям над бактериями, полученными из морских
прибрежных сольфатар в Тирренском море. Откуда появляются бактерии, обнаруживаемые в гидротермальных флюидах? Если они действительно происходят, как на это указывают все имеющиеся данные, из подповерхностной гидротермальной сети, значит, в той или иной форме эти существа живут при очень высоких температурах.
Одна из проблем, связанных с этим спором, чисто техническая: каким образом из хорошо уравновешенного подводного аппарата взять металлическим батометром, заканчивающимся управляемой трубкой для забора воды, пробу гидротермального флюида, не разбавленного морской водой? Проблема эта совсем не простая, до настоящего времени она не нашла решения. Прибавим, что число проб черноватого флюида, изученных микробиологами, вряд ли превышает десяток, и тогда станет ясно, что окончательное заключение формулировать рановато. И хотя для выводов время еще не пришло, тем не менее с самой общей точки зрения тот факт, что до наших дней сохранились первоначальные физико-химические условия, существовавшие при появлении жизни на нашей планете три с половиной миллиарда лет назад, заслуживает не мимолетного внимания.
Бактерии, о которых мы вели речь, живут или свободно, или прикрепляясь к разным субстратам, или взвешенными в морской воде. В подводных оазисах есть и другие бактерии, более или менее тесно связанные с различными гидротермальными беспозвоночными. Эти ассоциации, составляющие, пожалуй, наиболее яркую особенность гидротермальных сообществ, будут предметом обсуждения в двух последующих главах.
4
ПОЧТИ СОВЕРШЕННОЕ СООБЩЕСТВО
Из всех новых форм животных, открытых в гидротермальных сообществах, большой трубчатый червь, названный в 1981 году Riftia pachyptila, является сегодня наиболее известным. Его длина может превышать 1,5 м, а ярко-красный, киноварный цвет его султана, контрастирующего с перламутрово-белой окраской трубки, придают подводным фотографиям с его изображением неоспоримую эстетичность. Внутренее его строение также удивительно: этот червь не имеет ни рта, ни пищеварительной системы, ни анального отверстия!* Наконец, по биомассе этот червь во много раз превосходит все другие виды гидротермальных сообществ.
Для специалистов, однако, рифтия не является совершенной незнакомкой. Она входит в тип беспозвоночных, открытый в 1914 году во время экспедиции на голландском океанографическом судне «Сибога» в воды Индонезийского архипелага. Описавший червя французский зоолог Коллери, пораженный характерным видом его трубок, окрестил червя Siboglinum (от Siboga — название судна и linum — нить по-латыни) и не колеблясь отнес его к новому типу беспозвоночных. Коллери отметил полное отсутствие у червя пищеварительной системы и, пытаясь объяснить это, предположил, что Siboglinum — колониальное животное, у которого функции питания и размножения могут выполняться разными индивидуумами, как это часто имеет место у гидрозоев: отсутствие в собранных материалах других индивидуумов, ответственных за питание колонии, было, однако, трудно понять.
* В 1988 году установлено, что личинки и очень ранняя молодь вестиментифер имеют рот, кишку и анальное отверстие, которые исчезают вскоре после установления симбиоза с бактериями.— Прим. ред.
Эти крохотные червеобразные животные живут в хитиновых трубках правильной цилиндрической формы диаметром примерно 1 мм.
Лет через тридцать после их открытия швед Йохансон объединяет Siboglinum с другой, весьма отличной формой из Охотского моря, Lamellisabella в новый класс морских беспозвоночных — погонофоры (от греческого «кто носит бороду», другими словами, бородатые...), быстро возведенный зоологами в ранг подтипа. Открытия множатся, замечено, что представители «бородатых», считавшиеся обитателями исключительно больших глубин, могут жить и на глубине нескольких десятков метров и встречаются гораздо чаще, чем предполагалось ранее. Но все эти погонофоры невелики по размерам и все лишены пищеварительной системы. Первую гипотезу, разрешающую эту загадку, выдвинул советский ученый А. В. Иванов. Согласно его гипотезе эпидермальные клетки передних щупалец погонофор способны вылавливать мелкие органические частицы путем фагоцитоза. Но поскольку наблюдать фагоцитоз не удалось, Иванов позднее выдвинул другую гипотезу: собранные султаном щупалец частицы перевариваются вне тела животного ферментами, выделяемыми в наружную среду, а мелкие молекулы, образующиеся в ходе ферментативного разложения частиц, затем абсорбируются проницаемой клеточной мембраной. Второй гипотезе повезло не больше, чем первой, и она так и не была доказана.
В 1966 году на глубине 1125 м у побережья Калифорнии открыты погонофоры, которые зоолог Вебб назвал Lamellibrachia barhami. Они отличаются от всех других наличием особого вестиментального участка тела, на котором на спинной стороне открываются парные половые отверстия. Этот участок расположен между щупальцевой короной и туловищем и снабжен двумя складками, или крыльями, благодаря которым между вестиментумом и стенкой трубки червя образуется дорсальная камера. Чтобы отметить эту особенность, а также другие, не менее важные черты строения, в типе погонофор создаются новый отряд — вестиментиферы и новый класс — афренуляты. Характерная для типа «бородатых» сегментированная и снабженная хитиновыми щетинками задняя часть тела позволяет сблизить их с кольчатыми червями. Но по-прежнему неизвестно, как и чем питаются эти странные животные.
Рис. 17. Гигантская погонофора-вестиментифера Riitia pachyptila, извлеченная из трубки, и фрагменты трубки рифтии и кольчатой трубки вестиментиферы Tevnia jerichona-па. На врезке — циркуляция крови в султане щупалец и газообмен между морской средой и эндосимбиотическими бактериями в трофосоме.
Вот так в общих чертах обстояло дело, когда в 1977 году на излиянии на Галапагосском рифте был открыт новый вид Riftia pachyptila. Изучением первых собранных образцов занимается американский зоолог Джоунс; менее чем через четыре года после первых находок опубликовано описание вида. Это действительно вестименти-фера, но столь сильно отличающаяся от Lamellibrachia,
что для нее приходится создать особое семейсто — Rif-tiidae. Одновременно Джоунс устанавливает, что туловище, занимающее около 3/4 длины тела, заполнено тканью, образованной дольками, обильно снабженными кровеносными сосудами,— трофосомой; эта ткань буквально насыщена округлыми бактериями диаметром около 3—5 мкм. Трофосома различима даже у самых молодых изученных особей (длиной 1,44 мм) и содержит во множестве кристаллы серы.
Спустя несколько месяцев, в июне 1981 года, в одном из номеров высокоавторитетного журнала «Сайнс» («Наука») — органа Американской ассоциации развития науки — появляется целая серия статей, посвященных рифтиям. В них сообщается множество результатов, подтверждающих, что рифтия получает питание от бактерий, содержащихся в трофосоме, или потребляет их метаболиты. Хотя доказательства еще далеко не исчерпывающие, приведенные данные в целом весьма определенно ориентированы в эту сторону.
Первый комплекс данных о содержащихся в трофосоме бактериях получен с помощью электронного микроскопа и свидетельствует о том, что это сферические грам-отрицательные бактерии, мембрана которых содержит липополисахариды. Полученные изображения весьма сходны с ранее опубликованными фотографиями бактерий из трофосомы Lamellibrachia, первой известной вестиментиферы (тогда Вебб чисто гипотетически отвел трофосоме роль органа-накопителя продуктов, необходимых для сперматогенеза, или органа детоксикации, не имея каких бы то ни было доказательств).
Интереснее и существеннее результаты, полученные биохимиком Фельбеком. Прежде всего, ткани трофосомы имеют повышенную активность многих ферментов, входящих в цикл окисления — восстановления восстановленных соединений серы (например, тиосульфат-редуктазы, или роданезы, и аденозинтрифосфатсульфурилазы). Это доказывает, что сферические бактерии из трофосомы рифтии участвуют в цикле серы (в частности, окислении сероводорода). Другие опыты показали, что бактерии трофосомы содержат также характерные ферменты фиксации СО2 в цикле, называемом Сз-циклом или циклом Кальвина—Бенсона, широко распространенном у высших растений. Первое органическое соединение, образованное из атмосферного СО2, содержит три атома углерода и его
синтез идет с участием фермента рибулозо-1,5-бифос-фаткарбоксилазы (более известного биохимикам под названием RuBP-аза). Эта RuBp-аза содержится в трофосо-ме рифтии, но полностью отсутствует в мышцах червя. То же относится и к другому ферменту, рибулозо-5-фос-фаткиназе (или Ри5Р-киназе). 1аким образом, можно предположить, что рифтия, благодаря заключенным в ее трофосоме бактериям, способна кормиться либо полностью как автотрофный организм, либо частично, за счет энергии окисления сероводорода, способствующего восстановлению СО2 в органические соединения.
Еще один, последний результат касается измерения отношения двух стабильных изотопов углерода МС и 12С в трофосоме и в остальном теле рифтии. Известно, что отношение двух природных изотопов углерода практически не меняется при переходе с одного уровня пищевой пирамиды на другой, так как оно зависит от величины этого отношения в источнике минерального углерода и изначально определяется способом фиксации СО2. Это свойство позволяет использовать изменение отношения двух природных изотопов элемента, обычно присутствующего в цикле органического вещества, в качестве метки для пищевой цепи.
Измерения соотношения изотопов углерода (3С/НС), проведенные Рау в трофосоме и в мышцах рифтии, дали одинаковые значения — примерно — 11%0 (эта величина выражает отклонение между соотношением изотопов в исследуемой пробе и в стандартном образце, например в воздухе, вычисленное по формуле: отношение в исследуемой пробе, поделенное на отношение в стандартном образце, минус 1, выраженное в промилле). Тот факт, что и в трофосоме и в мышцах было получено одно и то же значение, указывает, что эти две ткани непосредственно связаны трофически, иными словами, что бактерии или их метаболиты служат пищей для погонофор. Абсолютное значение отношения интерпретировать труднее. В самом деле, у морских животных, которые питаются органическими частицами, произведенными одноклеточными водорослями фитопланктона, значения отношения колеблются от —15 до —18%0. По сравнению с этими величинами, относительно высокие значения, полученные для тканей рифтии, могут быть объяснены тремя причинами: либо тем, что их пищу составляют остатки растений, функционирующих по циклу С4, что представляется
маловероятным; либо тем, что бактериальный хемосинтез идет в условиях недостатка СО2, вследствие чего ферменты цикла Кальвина оказываются менее селективными по отношению к двум изотопам углерода; либо тем, что для переноса СО2 бактерии, подобно некоторым зеленым растениям, используют четырехуглеродный сахар малат.
Фельбек продемонстрировал правильность последнего объяснения: он вносил радиоактивно меченую СО2 в рифтий, через несколько часов после сбора помещенных в охлажденный до 8 °C аквариум под давлением НО атмосфер, и после нескольких часов инкубации обнаруживал в крови рифтий меченые молекулы малата и сукцината. Таким образом, СО2 поступает до трофосомы, где высвобождается путем декарбоксилирования.
Наконец, изучение крови рифтий показывает, что своим цветом она обязана повышенному содержанию гемоглобина (в крови рифтий нет оформленных элементов, как у большинства беспозвоночных) и что она обладает повышенной способностью переносить кислород (до 11 мл кислорода на 100 мл крови). Подобное свойство может объясняться большой потребностью в кислороде для бактериального хемотрофного метаболизма, которая добавляется к потребности в кислороде для дыхания.
Эти результаты еще не позволяют утверждать, что рифтия питается за счет хемосинтезирующих бактерий, живущих в ее трофосоме: надо еще объяснить, каким образом сероводород, извлекаемый султаном щупалец из морской воды, доносится до бактерий, которые его окисляют, откуда берется азот, необходимый для построения белков, и как осуществляется перенос бактериального органического вещества к тканям погонофор.
Тем временем открытие Riftia pachyptila стимулировало исследования классических погонофор. Шаг за шагом было экспериментально показано, что поглощения растворенных в морской воде органических веществ, таких, как аминокислоты, при их естественных концентрациях недостаточно для удовлетворения метаболических нужд даже такого маленького червя, как Siboglinum fjordicum; затем в 1981 году — что и эти организмы, как и большие вестиментиферы, содержат хемосинтезирующие внутриклеточные бактерии (были исследованы три вида Siboglinum и один вид Oligobrachia). Отношение изотопов 13С/12С очень низкое (—45%О, что позволяет предполагать использование метана. Наиболее важная сторона
этого исследования заключается в том, что оно показало: связь между хемосинтезирующими бактериями и погонофорами отнюдь не ограничивается только вестименти-ферами, классические мелкие погонофоры также получают по меньшей мере часть пищи от хемосинтезирующих бактерий, содержащихся в их клетках.
Все это время не прекращались исследования рифтии, которая становится настоящим лабораторным животным. Перенос сульфидов кровью в принципе сталкивается с многочисленными трудностями: сульфиды образуют с гемоглобином стабильные соединения, которые не могут связывать кислород; более того, в присутствии этого газа они спонтанно окисляются и не могут быть далее использованы бактериями; наконец, они сильно токсичны и подавляют внутриклеточные ферментативные реакции дыхания. В 1983 году две американские исследовательские группы одновременно выявили в крови рифтии белок с повышенной молекулярной массой, способный соединяться с очень большим количеством сульфидов и таким образом защищать гемоглобин крови и внутриклеточные дыхательные ферменты от вредных воздействий. Благодаря этому белку кровь рифтии способна накапливать большие количества сероводорода на уровне султана щупалец и транспортировать его до трофосомы. Более поздние работы показывают, что этот любопытный белок является не чем иным, как гемоглобином, который может, таким образом, обеспечивать перенос кислорода и сульфидов на двух разных участках: кислорода — на геме, а сульфидов — на глобине. Действительно, количество фиксированных сульфидов, как оказалось, прямо зависит от количества гема, и, кроме того, к тому времени было уже известно, что молекулярная масса этого белка весьма близка к молекулярной массе гемоглобина.
Годом позже публикация Боска и Грассе об особо тщательном исследовании трофосомы рифтии в электронном микроскопе приносит целую серию интереснейших результатов. Прежде всего, сферические бактерии трофосомы локализованы внутри клеток ткани трофосомы, что прежде доказано не было. Во-вторых, эти внутриклеточные бактерии присутствуют во всех клетках трофосомы за исключением тех, которые окаймляют кровеносные сосуды, и самых наружных клеток долек. Внимательный анализ клеток, окаймляющих кровеносные сосуды, показывает, что составные элементы крови, представленные
структурированными частицами, приклеиваются к наружной поверхности клеточной мембраны и образуют очень характерную сплошную выстилку. Эти частицы поступают в цитоплазму клеток путем пиноцитоза (пиноцитоз — это формирование вакуолей, которые образуют ножку, затем полностью отделяются от клеточной мембраны, на которой они возникли, и проникают в тело клетки). Полученные снимки показывают, что, вероятно, в клетку поступает белок крови, который доносит сероводород до трофосомы. В более удаленных от сосуда клетках появляются бактерии, которые быстро размножаются. Судя по цитоплазматическим включениям клетки-хозяева имеют пониженную метаболическую активность. Этот процесс продолжается в периферических клетках и ведет к гибели бактерий: измененная бактериальная стенка не препятствует более проникновению воды, диаметр бактерии увеличивается более чем на 20%, затем бактерии подвергаются настоящему внутриклеточному пищеварению — бактериальному лизису. Клетка червя, нагруженная продуктами бактериального распада, завершает процесс дегенерации, начинающийся с момента появления первого бактериального симбионта: смешанные продукты бактериального распада и разрушения клетки, которая их содержит (бактериоцита), поступают в кровь погонофоры,— они-то и являются теми веществами, которые обеспечивают выживание и рост червя.
Чтобы быть вполне доказательным, ультраструкгурный анализ трофосомы рифтии должен быть подтвержден физиологическими опытами. В частности, сам по себе он не позволяет оценить соотношение органических метаболитов, выделяемых бактериями в ходе их жизнедеятельности, и продуктов посмертного распада бактерий. Американские исследователи провели в 1985 году специальную экспедицию, посвященную физиологическому изучению гидротермальных организмов. Они располагали системой, позволяющей поместить животных, взятых .со дна за несколько часов до эксперимента, в замкнутую камеру под давлением, и снабжать их различными необходимыми минеральными соединениями (СО2, сероводород, кислород), непрерывно измеряя определенные физико-химические параметры крови. Нет сомнения, что такие методы помогут нам глубже понять функционирование ассоциации рифтий и связанных с ними бактерий.
Здесь многое еще неясно. Прежде всего, даже если ассоциация с хемосинтезирующими бактериями явно доказана, то значение трансэпидермального питания органическими молекулами, растворенными в морской воде, остается неизвестным: комбинируется ли оно с первым способом питания или же его следует решительно отвергнуть? Во-вторых, цикл азота, необходимого для синтеза белков, еще мало изучен; установлено лишь наличие в трофосоме фермента, способного извлекать азот из молекул нитрата путем денитрификации,— нитратредуктазы. В-третьих, в гидротермальных флюидах присутствуют другие восстановленные соединения: метан и некоторые азотистые вещества. Используются ли они ассоциацией бактерии-рифтия или эта ассоциация специфически тяготеет к сероводороду, что, возможно, и является результатом адаптации червя к гидротермальной среде?
Последний вопрос заставляет задуматься о связях, объединяющих бактерий и погонофор: настоящий ли это симбиоз, как в случае лишайников, которые являются примером постоянного и обязательного симбиоза гриба и водоросли определенных видов, всегда одних и тех же, не способных развиваться один без другого, или же это ассоциация, которая не предполагает такой жесткой специфической связи между двумя организмами? Сейчас еще слишком рано отвечать на этот вопрос. Бактерии, которые содержатся внутри клеток трофосомы, еще не определены, и самые последние наблюдения, похоже, свидетельствуют, что существует, возможно, не меньше двух категорий клеток-хозяев, что предполагает изменчивость на уровне самих штаммов бактерий.
Наконец, пока еще неизвестен механизм передачи бактерий взрослыми червями своим потомкам. Созревающие яйца извлекали из яичников самки рифтии и изучали под электронным микроскопом — оказалось, что они лишены симбиотических бактерий. Однако у трех очень молодых экземпляров рифтии длиной 2 мм Джоунс установил наличие ресничного отверстия в основании султана щупалец, связанного с тонким ресничным канальцем, который проходит через мозг и достигает полости тела. Эта структура исчезает на ранней стадии. Возможно, она представляет собой путь для проникновения бактерий. Но каким образом молодые черви выбирают в окружающей их среде те бактериальные штаммы, с которыми
они затем образуют длительную и обязательную ассоциацию, настоящий симбиоз? Или они способны включать в свои клетки бактерии, принадлежащие к разным штаммам? Сейчас невозможно ответить на эти вопросы. Можно лишь подчеркнуть, что у рифтии столько биологических и физиологических адаптаций, что гипотеза настоящего симбиоза с вестиментиферами кажется наиболее вероятной.
Наконец, вестиментиферы представляют особенно показательный пример эволюционной модели, которая известна нам пока лишь частично. В самом деле, вновь открытые гидротермальные излияния, например те, что расположены в зоне субдукции или в местах выхода интерстициальных вод, принесли Джоунсу большое количество новых для науки форм. Судите сами: в 1982 году было известно всего три вида вестиментифер, сгруппированных в два семейства, каждое с одним родом: Lamellibrachiidae с видами Lamellibrachia barhami, первой известной вестиментиферой, и L. luymesi, Riftiidae с видом Riftia pachyptila.
В ноябре 1985 года на международном совещании по гидротермальной фауне, состоявшемся в Океанографическом институте в Париже, Джоунс сообщил, что число известных видов вестиментифер утроилось, что они объединяются в пять семейств, не считая некоторых видов, которые пока еще не определены. Шесть новых видов, чьи названия говорят о зонах, где они были собраны, распределены в четыре новых семейства:
— Ridgeiidae (от английского ridge — хребет), с родом Ridgeia, содержащим два вида, собранных на гидротермальных излияниях хребтов Хуан-де-Фука и Эксплорер: Ridgeia piscesae и R. phaeophiale.
— Escarpiidae, с родом Escarpia (от английского escarp—обрыв, склон, эскарп), включающим два вида, собранных в оазисах у подножия Флоридского эскарпа, Escarpia spicata и Е. laminata.
— Tevniidae, с двумя родами: Tevnia (поскольку название Ventia, производное от английского слова vent — выход, источник, уже использовалось в номенклатуре животных, то, чтобы сохранить упоминание о гидротермальной среде, Джоунс прибег к анаграмме) и Oasisia. Род Tevnia с единственным видом Т. jerichonana был собран «Сианой» в 1982 и 1984 годах на 13° с. ш. на Восточно-Тихоокеанском поднятии. Род Oasisia, также
включающий единственный вид О. alvinae, был собран «Алвином» на 21° с. ш. на том же поднятии.
В обрабатываемых коллекциях есть еще второй иерихонский червь, добытый на 18° ю. ш. на Восточно-Тихоокеанском поднятии на расстоянии более 3500 км от местонахождения, где был найден Т. jerichonana, и три вида рода Ridgeia.
Создается впечатление, будто эти крупные животные, еще двадцать лет назад не известные науке, способны эволюционировать с такой скоростью, что образуют четко различающиеся виды, роды и семейства, оставаясь строго приуроченными к определенным физико-химическим условиям среды, которые изменяются в узких пределах и, кроме того, значительно различаются в разных гидротермальных излияниях. Следует ожидать, что у всех этих видов будут открыты физиологические механизмы, схожие с теми, которые были обнаружены у Riftia pachyptila, или даже идентичные им.
Наконец, Джоунс предлагает повысить старый подтип погонофор Obturata (иначе говоря, вестиментифер) до ранга типа и числить их под названием Vestimentifera, принимая во внимание следующие их анатомические отличия от собственно погонофор: различия в числе и размере внутренних полостей тела (целомических полостей), конфигурацию вентральной нервной цепочки и наличие особого тканевого листка, мезентерия, в терминальном районе тела. Итак, отныне погонофоры и вестиментиферы составляют два различных типа морских беспозвоночных.
5
БОЛЕЕ СКРОМНЫЕ СООБЩЕСТВА
С замечательной вестиментиферой рифтией и ее симбиотическими бактериями соседствуют некоторые беспозвоночные, часто больших размеров, которые также научились извлекать пользу из наличия в гидротермальной среде разнообразных типов бактерий. Таковы, например, два крупных двустворчатых моллюска Calyptogena magniiica и Bathymodiolus thermophilus и помпейские черви, полихеты из рода Alvinella. Эти три формы, довольно широко распространенные в гидротермальных излияниях на Восточно-Тихоокеанском поднятии и на Галапагосском рифте, установили с бактериями связи, по-видимому, не такие тесные, как рифтия, и в настоящее время гораздо хуже изученные.
Calyptogena magniiica, вид, описанный в 1980 году, принадлежит к известному с миоцена роду, виды которого в большинстве живут на глубинах от 200 до 1500 м. Матово-белые раковины калиптоген располагаются более или менее равномерно вдоль трещин, откуда выходит гидротермальный флюид. Длина раковин может достигать 25 см. У живых моллюсков между приоткрытыми створками видны ткани тела, окрашенные в ярко-красный цвет, и нога — напоминание о роющем образе жизни других видов рода. Пищеварительная система уменьшена, в частности, лабиальные пальпы, служащие для направления ко рту собранных на жабрах пищевых частиц, рудиментарны. У всех исследованных экземпляров кишечник пуст.
Хотя двустворчатые моллюски обычно питаются взвешенными в воде частицами, наличие красного пигмента в крови, обусловливающей красный цвет тканей, и высокая биомасса популяций калиптоген, как и сама локализация животных, привели биологов к мысли, что и здесь
Рис. 18. Вверху живая Calyptogena magnifica, достигающая в длину 25 см. Внизу — раковины в процессе растворения в зоне, ставшей неактивной.
возможен оригинальный способ питания с участием хемосинтезирующих бактерий.
Предварительные результаты были получены путем определения отношения изотопов углерода /3С/12С (см. гл. 4, стр. 65): отношение изотопов для калиптоген с Восточно-Тихоокеанского поднятия на 21° с. ш. и с Галапагосского рифта оказалось равным —32%О. В то же время органический углерод частиц, образовавшихся из при
поверхностного планктона, характеризуется более высокими значениями: от —15 до —25%о- Чтобы объяснить аномально низкое значение изотопного отношения, Рау предположил, что моллюски питаются в основном хемосинтезирующими бактериями. Известно, кстати, что некоторые хемотрофные бактерии предпочтительно фиксируют 12СОг, что понижает отношение изотопов на величину, которая может достигать —30%о по отношению к первоначальному источнику минерального углерода. Если принять, что хемосинтезирующие бактерии фиксируют только СОг, содержащийся в гидротермальном флюиде, изотопное отношение которого равно —4%о> то, учитывая, что перенос бактериальной биомассы в ткани двустворок не меняет этого отношения, можно заключить, что значение —32%О, обнаруженное у калипто-гены, есть результат питания преимущественно хемосинтезирующими бактериями. Аналогичные результаты дает изучение отношения изотопов азота (15N/l2rN): его значение у калиптогены колеблется между +2 и +5%о-В планктоне это отношение колеблется между +5 и + 10%о и слегка повышается (до +13%о) в органической фракции глубоководных осадков. Причем отношение изотопов азота зависит от положения организма в пищевой цепи и оно тем выше, чем выше трофический уровень. Значение, измеренное у калиптогены, не позволяет заключить, что эта двустворка кормится органическими частицами, образовавшимися в поверхностном слое. Напротив, морские микроорганизмы, способные непосредственно фиксировать газообразный азот, характеризуются низкими значениями изотопного отношения — между 0 и +2%о, что сравнимо со значением, установленным для калиптогены.
Следовательно, весьма возможно, что эти двустворки питаются хемосинтезирующими бактериями, использующими локальный источник СО2, связанный с наличием гидротермального флюида, и способными прямо фиксировать азот. Остается только определить, где живут эти бактерии.
Как раз в этот момент американским биохимикам, работавшим с рифтией, удалось показать, что калиптогена тоже содержит фермент RuBP-азу, необходимую для фиксации СО2 по циклу Сз (циклу Кальвина — Бенсона). Этот фермент встречается исключительно в тканях жабр, что, впрочем, характерно и для других двустворчатых
моллюсков, живущих в прибрежной зоне с повышенным содержанием сероводорода (мангровые болота и прибрежные лагуны). Жабры калиптогены, сильно развитые и окрашенные в темно-красный цвет, резко отличаются от мелких бесцветных жабр двустворок, лишенных подобных ферментов.
Тогда еще можно было сомневаться: те хемосинтезирующие бактерии, которые ответственны за необычные значения отношения изотопов углерода и азота и активность ферментов цикла Кальвина,— сконцентрировались ли они на жабрах в ходе активного фильтрационного питания двустворки или же они связаны с двустворчатыми моллюсками значительно теснее? Два исследования ультраструктуры жабр, проведенные независимо в США и во Франции, дают однозначные ответы: высокие концентрации сопутствующих бактерий отмечены в клетках наружного края жаберных пластинок, пребывающих в непосредственном контакте с окружающей средой. Эти бактерии заключены в карманы, ограниченные двойной мембраной, вероятно, выделяемой клеткой-хозяином. Эти сферические или палочковидные бактерии меньше, чем бактерии из рифтии, и достигают в диаметре примерно 0,6 мкм. В основании карманов с бактериями наблюдаются очень плотные гранулы, возможно, металлические конкреции. Таким образом, внутри клеток жабр калиптогены действительно имеются бактерии. Однако в том, что касается функционирования ассоциации бактерии — двустворка, еще очень много неясного. Было установлено также повышенное содержание в крови калиптогены сероводорода, фиксированного на белке, который, подобно гемоглобину рифтии, возможно, осуществляет перенос сульфидов из окружающей среды к клеткам жаберного эпителия, содержащим хемосинтезирующие бактерии. К тому же, теперь уже в отличие от рифтии, калиптогена имеет пищеварительную систему, которая хотя отчасти и редуцирована, может быть, функционирует. Значит, калиптогена, возможно, питается органическими частицами, осевшими на жаберные фильтры, в дополнение к пище, получаемой от внутриклеточных бактерий в жабрах в результате еще не окончательно выясненного процесса. Другими словами, нельзя полностью отбросить гипотезу смешанного питания, которое в экологии называется миксо-трофией.
Самая последняя работа, посвященная калиптогене,— это ультраструктурный анализ животных, собранных в 1982 году на 21° с. ш. американской экспедицией «Оазис» на подводном аппарате «Алвин». Животные были зафиксированы на борту в отличных условиях, и их изучение в большой степени подтверждает важную роль, которую играют включенные в ткань моллюска бактерии.
Жабры имеют нормальное ресничное покрытие. Вентральная борозда, предназначенная у других двустворчатых моллюсков для проталкивания собранных на жаберных нитях частиц в рот в форме слизистого шнура, значительно редуцирована и не функционирует: из 24 собранных и проанализированных экземпляров ни у одного не было отмечено слизистых шнуров. Значит, обильное выделение слизи связано с очисткой жабр, а реснички, должно быть, способствуют притоку морской воды, омывающей жабры. За исключением небольшой поверхности, занятой ресничками, подавляющее большинство жаберных клеток преобразованы в бактериоциты, заключающие палочковидные внутриклеточные бактерии. Наблюдалось деление этих бактерий, что служит прямым доказательством их метаболической активности. Бактериоциты имеют разный вид — свидетельство того, что, как и у рифтии, здесь имеет место циклический процесс: после фазы роста и деления бактерий, возможно, наступает их распад — лизис и клетки двустворок так или иначе переваривают их. Наблюдавшиеся светлые зоны, более или менее гранулированные, объяснены как следствие распада бактерий. Незараженные клетки среди бактериоцитов составляют, быть может, резерв бактериоцитов. Наконец, обилие складок на наружной стенке бактериоцитов, увеличивающих поверхность обмена с внешней средой, с очевидностью свидетельствует о возможности прямого проникновения сульфидов, а может быть и мелких растворенных органических молекул.
Как видно, многое еще остается сделать, чтобы понять точное назначение внутриклеточных хемосинтезирующих бактерий в жабрах Calyptogena magnifica с точки зрения физиологических функций, не говоря уже о других видах того же рода, живущих в аналогичных условиях: С. pacifica на пронизанных гидротермальными флюидами осадках бассейна Гуаймас в Калифорнийском заливе, пока
Рис. 19. На этой раковине Bathymodiolus thermophilus видны основания биссусных нитей других особей.
не описанных трех видов Calyptogena*, обнаруженных на глубинах 3800 и 5800 м в зоне субдукции близ Японии, и еще одного вида, живущего у подножия Флоридского эскарпа в Мексиканском заливе».
Второй вид крупных двустворчатых моллюсков, открытый в подводных оазисах, принадлежит к хорошо известному семейству митилид, к которому относится съедобная мидия, обычная для побережья Франции. Правда, «мидии» из подводных оазисов, собранные в 1977 году на Галапагосском рифте, принадлежат к группе менее известных митилид—к модиолам, отличающимся от настоящих мидий положением замка раковины и закругленной формой ее переднего края. Модиолы известны палеонтологам в отложениях, относящихся еще к девону, то есть это довольно древняя группа моллюсков. Найденные в оазисах батимодиолы достигают в длину 18 см, иначе говоря, они намного крупнее других видов модиол. Раковины, от желтоватых до темно-коричневых, часто имеют следы прикрепления фиксирующих нитей других особей (так называемого биссуса, имеющегося и у прибрежных мидий), обнаружены на них и мелкие колпачковидные брюхоногие моллюски. Батимо-
* Их оказалось не три, а четыре вида: Calyptogena nautilei, С. kaikoi и С. laubieri описаны с глубин около 3800 м юго-восточнее полуострова Кии, С. phaseoliformis — с глубины около 5650 м восточнее Токио.— Прим. ред.
диолы способны образовывать плотные банки с многорядным расположением особей; часто они прикрепляются к трубкам рифтий на различной высоте. Измерение температур на разных местонахождениях показало, что батимо-диолы выносят температуру от 3 до 8—9 °C и удаляются от гидротермального источника не более чем на десяток метров. Митилиды с Галапагосского рифта после нескольких лет изучения наконец-то описаны и названы Bathymodiolus thermophilus. Батимодиолы, найденные на Восточно-Тихоокеанском поднятии на 13° с. ш. принадлежат, вероятно, к тому же виду. «Мидии» отмечены также у подножия Флоридского эскарпа и на хребте Эксплорер, вблизи Британской Колумбии, но они пока еще плохо изучены.
Перед биологами очень быстро встал вопрос о питании батимодиол. Как и у калиптогены, было измерено отношение изотопов углерода с целью установить источник органического углерода, используемого батимодио-лами. Значения от —32 до —34%о Для батимодиол из подводных оазисов значительно отличаются от более высоких значений (от —16 до —18%0) для прибрежных мидий и, наоборот, близки к значениям, полученным для калиптоген. Поэтому были предприняты попытки узнать, имеются ли в жабрах батимодиол, как в жабрах калиптоген, ферменты, характерные для цикла фиксации СОг. Хотя их активность у батимодиол не столь высока, как у калиптоген, удалось выявить наличие в жабрах батимодиол с Галапагосского рифта RuBP-азы, характерной для цикла Кальвина — Бенсона. Жабры этого моллюска, как показало исследование в электронном микроскопе, также содержат бактерии. Эти бактерии мельче, чем бактерии из калиптоген, не больше 0,4 мкм в диаметре, и тоже внутриклеточные. Они помещаются в эпителиальных клетках жабр в карманах, которые могут занимать большую часть клетки-хозяина, так что ее ядро оказывается сдвинутым к базальной части. Кроме то’го, на поверхности жаберных клеток обнаружены нити длиной 10—20 мкм и диаметром 0,5 мкм, которые, видимо, являются нитчатыми бактериями, и другие нити, более тонкие и с менее правильной морфологией, возможно, актиномицеты.
В отличие от калиптогены, у батимодиолы есть функционирующая пищеварительная система и развитые лабиальные пальпы. Жабры, как и у прибрежных мидий,
имеют ресничную оболочку и систему бороздок, окруженных слизистыми клетками. Отсюда можно заключить, что ток воды, вызванный биением ресничек, обеспечивает перенос окутанных слизью частиц до лабиальных пальп, которые препровождают их в рот. Микроскопический анализ содержимого кишечника батимодиолы подтверждает это заключение: там обнаружены раковины фора-минифер, фрустулы (створки) диатомовых водорослей, опустившихся с поверхности вод, разные остатки и многочисленные бактериальные клетки.
Экспериментальное доказательство роли хемосинтезирующих внутриклеточных бактерий было получено в 1984 году на батимодиолах с излияния на 13° с. ш. на Восточно-Тихоокеанском поднятии. В отличие от американских исследователей, выполнявших работы на рифтиях, французские специалисты Фиала-Медиони, Алэз и Каэ выбрали способ in situ: батимодиол помещали манипулятором «Сианы» в закрытую камеру объемом 25 л. Под водой производили и инъекции веществ, помеченных радиоактивной меткой (карбонат, помеченный ИС, и различные аминокислоты, помеченные 3Н), и, после инкубации в течение 20 часов, животных поднимали на поверхность (практически это промежуток времени между двумя последовательными погружениями «Сйаны»). Результаты этих опытов показали, что радиоактивный углерод концентрируется в жабрах (75—90% общей радиоактивности), в зонах, богатых внутриклеточными бактериями. Добавка антибиотиков перед инъекцией меченого карбоната значительно уменьшает фиксацию СО2, подтверждая ведущую роль бактерий в этом процессе. Биохимические анализы показывают, что радиоактивный углерод включается в мелкие молекулы (сахара, аминокислоты), а также обнаруживается в макромолекулах белков и липидов. Наконец, меченые аминокислоты тоже абсорбируются батимодиолами, особенно в жабрах. Таким образом, батимодиола, в зависимости от местных условий, вероятно, может использовать три способа питания: симбиоз с хемосинтезирующими бактериями, абсорбцию мелких органических молекул, таких, как аминокислоты, которые могут встречаться в измеримых количествах вблизи гидротермальных источников, бактерий, связанных с гидротермалью, или органических остатков, опускающихся из поверхностных вод.
Как видно, ситуация с батимодиолами посложнее,
Рис. 20. Помпейский червь Alvinella pompejana с немногочисленными сегментами и длинными дорсальными папиллами.
чем с калиптогеной, что, возможно, объясняет, почему батимодиолы могут жить на большем удалении от гидротермальных источников.
Третья форма, обнаруженная в непосредственной близости от устьев гидротермальных излияний, тоже живет в тесном контакте с бактериями,— это два вида
кольчатых червей (полихет): помпейские черви, названные так потому, что они постоянно посыпаются настоящим дождем частиц серы, словно вулканическим пеплом, и описанные над названием Alvinella pompejana и А. caudata. Помпейские черви впервые наблюдались у излияния на 21° с. ш. на Восточно-Тихоокеанском поднятии. Они достигают 20 см в длину и имеют 80—190 одинаковых сегментов тела. Появляются эти трубчатые черви, когда температура выбрасываемого гидротермального флюида превосходит 50 °C. Появившись, они образуют колонии, ковром выстилающие стенки труб полиметаллических сульфидов высокой температуры (черных курильщиков) и покрывающие распылители с белесоватой жидкостью промежуточной температуры — так называемые снежки, открытые геологами в 1976 году на 21° с. ш. и тогда же ими описанные. Альвинеллы — самые термофильные формы из всех известных в настоящее время беспозвоночных, связанных с гидротермальными излияниями. Судя по наблюдениям «Сианы» на 13° с. ш., они переносят температуру от 20 до 40 °C. На распылителях с белесоватой жидкостью измерения температуры дают невероятно высокий градиент: непосредственно на поверхности колонии температура равна 30 °C, а при погружении зонда на глубину каких-нибудь 10 см она достигает 100 °C, на глубину 20 см, в толщу червей — уже 200 °C! Конечно, помпейские черви не всегда вынуждены жить при такой температуре, которую они, вероятно, и не вынесли бы, но это наблюдение показывает, что гидротермальный флюид температурой 250 °C, испускаемый распылителем, постепенно смешивается с морской водой в системе пор, образованных пустыми трубками помпейских червей, более или менее срощенными и уплощенными. В этой лакунарной массе обильно размножаются бактерии, результаты деятельности которых проявляются в повышенном содержании органического вещества бактериального происхождения над колониями альвинелл.
Исследуя первые экземпляры помпейских червей, которые я описал вместе с Даниэлем Дебрюйером в 1980 году, я был удивлен тем, что небольшая коллекция десятка в полтора животных, добытых «Алвином», содержит две формы червей: два экземпляра более крупных животных с 90 сегментами тела и 15 червей меньшего диаметра, но имеющих 120—190 сегментов. У всех кольчатых червей, известных на сегодня, соматический рост идет путем
возрастания числа сегментов тела, образующихся как раз перед последним сегментом, так называемым пигидием. Поэтому я ожидал увидеть крупных червей с большим количеством сегментов и мелких червей с малым количеством сегментов, но столкнулся с абсолютно противоположной ситуацией. Кроме того, черви были одеты обильной ресничной оболочкой, по-разному расположенной у этих двух форм: у крупных особей ресничный покров образует настоящий войлок из ворсинок, различимых невооруженным глазом и правильно расположенных на спинной части среднего отдела тела червя; у мелких особей с большим количеством сегментов обильно покрыта очень тонкими нитями лишь задняя часть, сильно уплощенная, с ножками (параподиями), развитыми гораздо сильнее, чем параподии переднего отдела телц. Но поскольку все характеристики, используемые обычно для различения видов этого отряда полихет, у обоих форм идентичны, мы выдвинули гипотезу о двух последовательных стадиях развития одного и того же вида, предположив, что крупные особи — результат автотомии (рефлекторное самокалечение, наиболее известный пример — отбрасывание клешни у крабов) особей с многими сегментами. К тому же эта гипотеза поддерживалась существованием подобного явления у других полихет, явления, связанного с размножением.
Описывая как можно полнее обе эти формы червей, мы все же пришли тогда к заключению, что существует единственный вид помпейского червя. По мере того как наши работы по изучению этой формы продвигались, мы все более и более скептически относились к этому первоначальному заключению. В частности, несмотря на новые, весьма обильные сборы, нам не удалось найти экземпляра на промежуточной между двумя описанными формами стадии развития. Четыре года спустя с помощью генетиков проблема была решена, к великому смущению специалистов, на основе анализа ферментного полимор> физма. Генетики называют полиморфизмом одновременное существование в популяции многочисленных вариантов одного и того же признака. Измерение ферментного полиморфизма сотни помпейских червей, принадлежащих к двум описанным формам, показало, что на самом деле это два хорошо различающихся вида, не связанных близким родством. Мы назвали форму с большим количеством сегментов Alvinella caudata, оставив первоначальное
Рис. 21. Помпейский червь Alvinella caudata с многочисленными сегментами и измененной задней частью тела.
название A. pompejana за крупными особями с меньшим количеством сегментов, что соответствует Международному кодексу зоологической номенклатуры.
Как бы то ни было, оба вида Alvinella имеют одну общую черту—плотную оболочку из тонких или более толстых нитей, совершенно необычную для кольчатых червей. Очень скоро было показано, что эти реснички и нити в действительности — нитевидные бактерии, обитающие изолированно или группами на прозрачной органической основе. Эти нитевидные бактерии имеют диаметр 4—5 мкм и могут образовывать цепочки длиной до нескольких миллиметров. На уровне прикрепления этих бактерий к стенке тела червя замечены изменения стенки, или кутикулы. У A.caudata различаются три основные зоны: в передней части сферические и нитевиднче бактерии локализованы в межсегментных спинных участках, образующих глубокие борозды на кутикуле; в 10— 15 сегментах переходной зоны кутикула видоизменена появлением маленьких пиннул, к которым прикрепляются бактерии; наконец, в заднем отделе пучки бактериальных нитей оседают на всех выростах полностью измененных параподий, длина которых превосходит диаметр тела. Расположение бактерий у A. pompejana еще примечательнее: они представляют собой очень длинные нити, собранные в пучки прозрачной органической основой, выделяемой червем. Кажется, что эти структуры принадлежат червю: в самом деле, в основании этих папилл наблюдаются отверстия диаметром примерно 100 мкм, открывающиеся на короткой тегументальной ножке. Отверстия расположены вдоль межсегментных границ по обе стороны от них на спинной стороне среднего отдела тела. Папиллы достигают длины 3—4 мм. Микроскопическое исследование показывает, что они образованы нитчатыми бактериями, расположенными продольно вокруг прозрачной центральной оси.
На внутренней стороне трубки помпейских червей также имеется множество различных бактерий. А вот внутриклеточные бактерии у них, в отличие от описанных выше беспозвоночных, никогда не наблюдались.
Этот исключительный бактериальный эпибиоз существует у обоих видов помпейских червей у всех особей, насчитывающих более 50 сегментов. Имеет ли такая ассоциация функциональное значение, сравнимое со значением внутриклеточных бактерий? Большое морфологи
ческое разнообразие бактерий-эпибионтов заставляет предположить, что ответ на этот вопрос не будет однозначным.
Изучение отношения изотопов углерода в тканях червей (значения от —10 до —11%0) показало, что источник используемого неорганического углерода местный — его производят хемосинтезирующие бактерии. Измерение отношения изотопов азота, наоборот указывает на то, что помпейские черви сильнее удалены от источника минерального азота, чем погонофоры и двустворчатые моллюски. Следовательно, сообщество, если оно существует, должно быть более открытым, чем в случае бати-модиол.
Тогда попытались определить, действительно ли бактерии, ассоциированные с покровами червей, имеют автотрофный метаболизм и синтезируют органическое вещество из минерального углерода. Первое указание в пользу этого дало обнаружение в гомогенате покровов тела весьма активной RuBP-азы, уже известной у погонофор и двустворчатых моллюсков. В гомогенате трубок присутствует другой фермент — РЕР-аза. Следовательно, по крайней мере часть бактерий, связанных с червями или их трубками, имеет хемосинтетический обмен веществ. Лабораторные культуры бактерий, полученных из покровов червей, позволили изолировать 36 разных штаммов, из них лишь три являются в условиях культуры строгими автотрофами. Правда, метаболизм бактерий цикла серы значительно варьирует в зависимости от условий среды; Баросе и Деминг показали, что в условиях, приближенных к тем, которые наблюдаются в излияниях, и при полном отсутствии кислорода бактерии с трансформированных параподий A. caudata осаждают сульфиды из гидротермального флюида.
Это повлекло за собой проведение экспериментов in situ с системой, представленной червем, его трубкой и сопутствующими бактериями.
Пищеварительный тракт альвинеллы, похоже, функционирует: буккальные щупальца могут улавливать разнообразные частицы, а изучение пищевого комка показывает, что черви заглатывают значительное количество минералов (барит, металлическая сера, ангидрит...), осажденных на бактериях и некоторых частицах, опустившихся с поверхности. Некоторые из этих бактерий являются спорами термофилов.
В 1982 году сначала на Галапагосском рифте, а затем на 13° с. ш. была обнаружена новая форма альвинеллид. Названная Paralvinella grasslei в честь Грассля, руководителя первой американской биологической экспедиции на Галапагосский рифт, она представляет новый род, более древний, чем род Alvinella, если судить по модификации самых передних сегментов, несущих жабры. В самом деле, у наиболее примитивных кольчатых червей все сегменты одинаковы, тогда как у более продвинутых форм имеет место специализация некоторых передних сегментов и, главное, более или менее полное слияние сегментов друг с другом. У Paralvinella четыре отдельных передних сегмента, каждый из которых несет по паре жабр, тогда как у Alvinella только один сегмент, образовавшийся в результате слияния четырех сегментов, и он несет все четыре пары жабр. Паральвинелла полностью лишена бактерий-эпибионтов. Червь живет среди массы трубок рифтий, на которых он строит свои собственные прозрачные слизистые трубки, но встречается также среди колоний альвинелл, расположившихся на беловатых распылителях.
В ходе экспедиции 1984 года на излиянии на 13° с. ш. с борта «Сианы» удалось провести много новых опытов. Животных собирали с помощью манипулятора и помещали в закрытую камеру — инкубатор, такую же, как использованная для батимодиол. Первый опыт показал, что меченый радиоактивной меткой углерод, предложенный альвинеллам, включается в органические молекулы примерно за 20 часов инкубации, причем в отсутствие антибиотиков, следовательно, по крайней мере часть бактерий-эпибионтов способна фиксировать СОг; а некоторые продукты их обмена затем передаются червям. Метод авторадиографии позволяет определить, что метку содержат только бактерии, собранные в пучки в межсегментных пространствах, и те, что прикрепляются к внутренней поверхности трубки. Добавка антибиотиков не подавляет, как ожидалось, включения меченого углерода в ткани альвинелл. Может быть, примененные антибиотики с широким спектром действия не влияют на имеющиеся здесь штаммы, или имеет место прямое включение меченого СО2 червями? Во втором опыте использовались две радиоактивные метки: 14С в минеральной форме и 3Н в органической (тимидин), а паральвинелла — полихета, начисто лишенная эпибионтных бактерий,— служила
контролем. Абсорбция углерода происходит у обоих родов на одних и тех же участках тела, в частности в жабрах и нежелезистом эпидермисе. 3Н обнаруживается в основном в жабрах, буккальных щупальцах и железистом эпидермисе.Нитчатые бактерии крупных спинных папилл альвинелл не включают углерод в экспериментальных условиях, но биологически активны, что видно по включению трития. Хемосинтезирующими бактериями являются как раз бактерии, собранные в пучки в межсегментных участках у альвинелл, а также бактерии внутри трубок. Топографические отношения между бактериями-эпибион-тами и путями проникновения органических меченых метаболитов не установлены.
Из всего набора экспериментальных фактов можно сделать вывод, что хемосинтезирующие бактерии фиксируют и включают в обмен веществ СО2, затем высвобождают органические метаболиты, которые червь поглощает через жабры и нежелезистый эпидермис. Однако экспериментальные условия весьма далеки от естественных, в которых функционирует ассоциация бактерии—альвинеллы. В самом деле, когда комок трубок, взятых манипулятором, помещается в камеру-инкубатор, многое меняется. Прежде всего, останавливается приток гидротермального флюида, который в нормальных условиях просачивается из глубины колонии к периферическим районам, постепенно смешиваясь с морской водой. В результате количество сероводорода значительно уменьшается, тем самым некоторые бактериальные процессы меняются и даже начинают идти в противоположном направлении. Далее, можно предположить, что большая часть органических метаболитов, высвобожденных хемосинтезирующими бактериями, происходит из глубинных слоев колоний, где температура порядка 100 °C и выше, что соответствует потребностям термофильных бактерий,— там, вероятно, и расположен очаг наиболее интенсивного хемосинтеза. Кроме того, возможно, что в экспериментальных условиях концентрация органических метаболитов гораздо ниже, чем в естественных условиях, а ведь многочисленные опыты, вот уже лет пятнадцать проводимые в лаборатории над прибрежными животными, выявили, что абсорбция мелких органических молекул и их перенос через эпидермис в большой мере зависит от их концентрации. И наконец, на трубах курильщиков или распылителей, заселенных альвинеллами, очень трудно найти горизон
тальные поверхности достаточного размера, чтобы расположить там в приемлемых условиях стабильности камеру-инкубатор; это значит, что температура воды в камере во время инкубации не превышает десятка градусов, тогда как ассоциация бактерии—альвинеллы функционирует обычно при температурах 20—40 °C.
Оригинальность ассоциации бактерий и помпейских червей, может быть, заключается в использовании не только сложных физиологических механизмов, но и физико-химических свойств разбавленных флюидов, которыми омываются микроорганизмы и помпейские черви. Эта оригинальность подтверждается биогеохимическим исследованием, проведенным независимо от описанных выше опытов, которое заключалось в анализе углеводородов и жирных кислот, присутствующих в пробах воды, взятых в десятке сантиметров над тремя основными зонами поселений: распылителями с альвинеллами, ореолами с рифтиями и с серпулидами. Результаты анализов показывают, что концентрации углеводородов и жирных кислот, растворенных или взвешенных, над тремя зонами поселений больше, чем в придонной воде, взятой на той же глубине в стороне от гидротермальных излияний, и, главное, что самые высокие концентрации жирных кислот обнаруживаются над колониями альвинелл. Если вспомнить, что в зоне рифтий биомасса гораздо выше, чем в зоне альвинелл, и что ткани тела помпейских червей содержат мало углеводородов, но много жирных кислот, то мы получаем первое интересное указание. Жирные кислоты разделяются на две основные химические группы — насыщенные и ненасыщенные; ненасыщенные жирные кислоты являются прямым продуктом метаболизма живых организмов и используются ими как источник пищи; они также гораздо менее устойчивы, чем насыщенные жирные кислоты. В отдельную группу выделяются раз-ветйленные жирные кислоты с 15—17 атомами углерода — они встречаются только в мембранах бактерий и, следовга-тельно, могут использоваться как показатель наличия бактерий в среде. Изучение отношения между насыщенными и ненасыщенными жирными кислотами подтверждает предыдущее наблюдение: содержание ненасыщенных жирных кислот максимально над альвинеллами. Содержание разветвленных жирных кислот над колониями альвинелл также намного выше, чем где бы то ни было, затем идут воды, омывающие поселения рифтий, а зона,
заселенная серпулидами, дает самые низкие показатели. Это свидетельствует о важности бактерий-эпибионтов альвинелл и микроорганизмов, живущих в массе трубок червей: значительные количества разветвленных жирных кислот высвобождаются в результате жизнедеятельности этих бактерий и после их гибели. Наоборот, рифтии после гибели хемосинтезирующих внутриклеточных бактерий используют их и вовлекают их в обмен веществ непосредственно внутри тела, и поставлять эти вещества в воду, омывающую поселения больших погонофор, могут лишь свободноживущие бактерии, обитающие на трубках рифтий или под поверхностью, в протоках, где циркулирует разбавленный гидротермальный флюид. Над серпулидами концентрации минимальны, что объясняется как малой биомассой в этой зоне, так и удаленностью ее от гидротермальных излияний: действительно, серпулиды составляют последний круг жизни гидротермального сообщества, который сменяют почти пустынные глубины океана.
Эти биогеохимические данные равным образом доказывают существование мелкомасштабной конвективной циркуляции частиц, выходящих в морскую среду как раз над колониями альвинелл, циркуляции, возникающей в результате излияния гидротермального флюида. Часть органических частиц, образованных в колониях альвинелл путем бактериального хемосинтеза, таким образом переносится в другие зоны сообщества и используется организмами, способными собирать взвешенные в морской воде частицы,— батимодиолами и серпулидами.
6
ХИЩНИКИ, КОММЕНСАЛЫ, ПАРАЗИТЫ И ОППОРТУНИСТЫ
Невероятно высокая биомасса, свойственная основным видам гидротермалей (Riftia, Calyptogena, Bathymodiolus, Alvinella), и ощутимое количество органического вещества, выделяемого в морскую воду во взвешенном и растворенном виде, используются животными некоторых экологических групп, более или менее известных в настоящее время: хищниками, комменсалами, паразитами и в основном так называемыми оппортунистами (приспособленцами) — видами, которые кормятся чаще всего взвешенными частицами и способны жить в глубине океана также и вдали от влияния гидротермальных источников. Естественно, мы здесь не собираемся приводить обзор 90 видов, описанных и определенных в разных гидротермальных сообществах, изученных в настоящее время, а только покажем на ряде примеров разнообразие биологических и экологических адаптаций этой фауны, которая в действительности оказалась гораздо более разнообразной, чем предполагалось на заре этих открытий.
Среди хищных форм представлены три четко различающиеся группы: моллюски, ракообразные и рыбы. Из моллюсков по крайней мере на двух излияниях наблюдали хищного брюхоногого моллюска-охотника (среди прибрежных гастропод к таким можно отнести, например, моллюсков-сверлильщиков, которые опустб-шают плантации мидий, выращиваемых на плотах) из рода Phymorhynchus, семейство Turridae. Этот брюхоногий моллюск размером с хорошую виноградную улитку давно известен как обитатель глубин океана, а в гидротермальных оазисах он находит для себя особенно благоприятные условия. На 13° с. ш. он был встречен, в частности, в умирающем оазисе Погомор, где аномалии температуры не обнаруживается. Является ли он обычным
Рис. 22. Маленький белый осьминог быстро продвигается по базальтовой лаве среди серпулид, распустивших султаны щупалец.
обитателем всех оазисов, где прячется, например, в массе трубок рифтий, или же его привлекают ослабевшие гидротермальные животные — этот вопрос остается открытым.
Второй пример хищных моллюсков относится к головоногим. На 13° с. ш. на Восточно-Тихоокеанском поднятии довольно часто наблюдают осьминога с совершенно белым телом и шероховатыми руками, снабженными одним рядом присосок, который активно перемещается по базальтовым лавам, покрытым трубками серпулид. Он может достигать 50 см в размахе рук. Это еще не определенный до вида представитель семейства Octopodidae, к которому принадлежит и обычный осьминог французских прибрежных вод. В ходе экспедиции 1984 года один такой осьминог проник в ловушку с наживкой, поставленную для поимки крабов и рыб, и с борта «Сианы» наблюдали, что он сидел внутри нее во время ее поднятия со дна и помещения в один из отсеков аппарата, предназначенного для подъема собранного материала на поверхность. В про-
Рис. 23. Большой «ушастый осьминог» достаточно часто встречался на местонахождении 13° с. ш. Он не боится прожекторов «Сианы».
цессе подъема осьминог умудрился сбежать, а рыбам и крабам, также бывшим в ловушке, не удалось этого сделать! Глубоководные осьминоги столь же умны, как и их прибрежные родичи. О питании этого белого осьминога, конечно, ничего неизвестно, но сам факт, что он был привлечен приманкой, показывает, что он — хищник и его присутствие в гидротермальных оазисах не случайно.
Во время той же экспедиции «Сианы» на некотором расстоянии от гидротермальных источников много раз видели представителей другого вида крупных головоногих, достигающих 1,5 м в длину. Это также были осьминоги (в отличие от головоногих, у которых десять рук,— кальмаров и каракатиц), но принадлежащие к группе плавниковых (Cirrata), называемых в просторечии ушастыми осьминогами. Для плавниковых характерно наличие одного ряда присосок, окаймленных с обеих сторон рядом усиков, или циррей, которые и дали им латинское название. «Уши» на самом деле — это плавники, подобные имеющимся у прибрежных кальмаров и каракатиц. Осьминог, наблюдавшийся с «Сианы», возможно, относится
к роду Grimpoteuthis, характеризующемуся, в частности, широкой мембраной, соединяющей руки и расположенной несимметрично на их оконечностях. Похожие формы были отмечены в Атлантическом океане вне гидротермального влияния с советского подводного аппарата «Пайсис». Пользуются ли эти животные, безусловно хищные, биомассой гидротермальных сообществ или оказались там случайно? Пока еще слишком рано давать ответ на этот вопрос.
Из ракообразных известны по меньшей мере три вида плотоядных крабов, строго привязанных к гидротермальным оазисам. С 1979 года ловушки, поставленные «Алвином» на Галапагосском рифте, принесли более 150 экземпляров белого краба Bythograea thermydron, для которого Вильямс создал новое надсемейство. Этот краб живет среди зарослей рифтий, но отдельные особи часто встречаются и на некотором расстоянии от источников. Родственные связи этого надсемейства еще плохо известны. Происходит ли оно от прибрежных крабов, которые, подобно Neolepas (см. стр. НО), опустились на глубины вдоль тектонически активных зон, или же от малочисленных крабов, обитающих на глубинах 1000— 3000 м? Как бы то ни было, Bythograea thermydron — активный хищник, его наблюдали пожирающим красные фрагменты султана щупалец рифтии и запечатлели это на пленку. Подобно молодым камбалам, которые обкусывают концы сифонов у закапывающихся в песок моллюсков, Bythograea не убивает свою добычу, а лишь откусывает куски ее тела, которыми и питается. Поврежденные ткани погонофор, по-видимому, быстро регенерируют. С точки зрения экологической такое равновесие между двумя популяциями более устойчиво во времени, чем при убийстве хищником своей жертвы. В содержимом желудка краба находят и остатки мелких ракообразных. Битогрея имеет интересные физиологические особенности: этот краб способен регулировать обмен веществ и несколько часов выживать при полном отсутствии кислорода. Кроме того, дыхательный пигмент крови бито-греи работает в широком диапазоне температур и кислотности внутренней среды. Личинок битогреи ловили в планктоне над гидротермальными источниками, а молодые крабы разных размеров попадались в ловушках и в пробах среди трубок рифтий. Их изучение позволило понять в подробностях процесс, который приводит к тому,
Рис. 24. Два характерных краба гидротермальных источников; вверху — крупный Cyanagraea praedator, живущий на колониях помпейских червей; внизу— мелкий Bythograea thermyd- . гоп, живущий вблизи колоний погонофор.
что взрослые особи почти полностью слепнут: рудиментарный глазной стебелек, доходящий до уровня роговицы глаза, частично скрыт лобной складкой карапакса.
Три года спустя, в 1982 году, новые сборы, проведенные «Сианой» на 13° с. ш., позволили обнаружить наряду с многочисленными Bythograea thermydron две другие
Рис. 25. Рыба из семейства бельдюговых из гидротермального оазиса на 13° с. ш., по мнению французских специалистов идентичная с Thermarces cerberus, обнаруженной на местонахождении 21° с. ш.
формы: второй вид этого же рода, гораздо меньших размеров и с развитым волосяным покровом у самок, которого де Сен-Лоран назвала В. microps, и краба гораздо больших размеров с нормально развитыми, хорошо заметными глазными стебельками, которого наблюдатели окрестили крабом курильщиков, так как он живет в неровностях на поверхности черных курильщиков и беловатых распылителей, заселенных помпейскими червями. Этот краб, названный Cyanagraea praedator, имеет карапакс шириной не меньше сантиметров десяти, так что из-за своих размеров не может пролезть в ловушки для битогрей. Он живет исключительно на колониях помпейских червей, и плотность его поселений может достигать 2—3 экз./м2. Анализ содержимого желудка двух самок, пойманных в 1982 году, показал присутствие многочисленных фрагментов альвинелл, хорошо заметных по характерным щетинкам, а также остатки молодых битогрей. Если судить по фотографиям, полученным «Алвином», С. praedator встречается и на излиянии на 21° с. ш., где тоже приурочен к колониям помпейских червей. Как и последние, он отсутствует на Галапагосском рифте, что свидетельствует о специфичности его питания и об адаптированности к особым физико-химическим условиям, которые свойственны поверхности колонии помпейских червей. В отличие от битогреи, он никогда не встречался вне гидротермальных излияний. Вполне здоровый краб, пойманный в 1984 году, погиб при подъеме «Сианы», тогда как битогреи прекрасно переживают и декомпрессию, и физико-химический шок. До настоящего момента не было возможности уточнить отношения между этими двумя родами: Cyanagraea отпочковалась от Bythograea или наоборот?
Из рыб на излияниях Галапагосского рифта и Восточно-Тихоокеанского поднятия отмечено несколько видов.
На 13 и 21° с. ш. в изобилии встречается похожая на толстого угря рыба длиной не более 30 см с беловатой кожей, покрытой плотной слизью,— это рыба семейства бельдюговых (Zoarcidae), но ее видовая принадлежность не уточнена. Американские зоологи создали для нее новый род Thermarces с двумя видами: Т. anderssoni с 13° с. ш. и Т. cerberus с 21° с. ш. Французские же специалисты полагают, что это один и тот же вид, который по правилам приоритетного названия должен называться Т. cerberus.
Эта рыба — хищник, который питается мелкой добычей — мелкими блюдечковидными брюхоногими моллюсками, альвинеллами, мелкими ракообразными... Два наблюдения, проведенные на 13° с. ш. на излиянии Погонор, показывают важность пищевой связи между рыбой и гидротермальным оазисом: за период между 1982 и 1984 годом гидротермальное население сильно уменьшилось и численность популяции рыб упала с 350 до нескольких десятков экземпляров. Thermarces способны жить, по крайней мере некоторое время, на больших расстояниях от гидротермальных источников: на 13° с. ш. рыбы отмечались почти в километре от осевой долины, что подразумевает способность этих животных находить оазисы по градиентам концентрации определенных веществ.
Эксплуатируются ли глубоководные оазисы только специфическими хищниками, тесно с ними связанными, или же время от времени на них нападают шайки проходимцев, постоянно живущих на больших глубинах,— бокоплавов и рыб, иногда очень крупных, о которых говорилось в начале этой главы? Пока никто не может утверждать обратного, иначе пришлось бы допустить, что даже очень разбавленные гидротермальные флюиды играют роль защитного барьера против этих опасных животных. Чтобы это узнать, надо наблюдать за жизнью гидротермального оазиса в течение недель и месяцев, что требует использования автономных систем видео,-записи. С точки зрения экологии такой процесс был бы очень важен, поскольку он обеспечивал бы перенос значительного количества биомассы гидротермального происхождения за весьма короткие промежутки времени в абиссальные океанические пустыни.
В настоящее время известен лишь один пример комменсализма в гидротермальных оазисах: многие батимодиолы служат хозяином для одной или двух особей поли-
Рис. 26. Червь-комменсал батимодиолы Branchipolynoe syminyiilida. При жизни имеет ярко-красную окраску.
хеты, которую Петтибон назвала Branchipolynoe symmylilida и для которой было образовано новое подсемейство. Она принадлежит к обширному семейству полихет-афродитид, которых насчитывается сотни видов, все лишенные жабр. А В. symmytilida, как говорит ее родовое название, обладает несколькими парами развет
вленных жабр. Окраска живых червей ярко-красная. Их кишечник содержит частицы, склеенные слизью в комочки, псевдофекалии, и фрагменты жабр двустворчатого моллюска. Эта ассоциация напоминает сожительство съедобных мидий с настоящим комменсалом — маленьким крабом-пиннотерой, который получает свою долю от собранных хозяином пищевых частиц.
Копеподы, группа мелких ракообразных, изобилующих в планктоне, которая также включает многочисленных паразитов беспозвоночных и рыб, дает нам прекрасный пример паразитизма с двумя животными из гидротермальных оазисов. Две наружные паразитические копеподы с двух видов первого описанного рода погонофор-вестиментифер Lamellibrachia уже были известны, хотя и сравнительно недавно. Эти две формы составляли особое семейство Dirivultidae. Два новых вида паразитических копепод, найденных в подводных оазисах, также относятся к этому семейству, но отличаются от первых столь четко, что для них созданы два новых рода: Ceuthoecetes aliger, паразитирующий на Riftia pachyptila с Галапагосского рифта, и Benthoxynus spiculifer,. паразитирующий на Ridgeia piscesae с хребта Хуан-де-Фука. Диривультиды принадлежат к группе сифоностомных копепод, у которых три модифицированных буккальных сегмента образуют настоящий стилет, способный проткнуть покровы хозяина, чтобы сосать его кровь, как это делают комары. На 13° с. ш. был найден еще один эктопаразит, пока еще не определенный. Судя по паразитарной специфичности других диривультид, это может быть связанный с рифтией С. aliger.
Среди видов-оппортунистов, то есть таких видов, которые живут в непосредственной близости к гидротермальным оазисам и благодаря этому получают достаточно пищи (взвешенные органические частицы, самые разные остатки...), есть виды, не известные, по крайней мере пока, вне гидротермальных оазисов, и виды, живущие в глубинах океана вне оазисов, но имеющие там меньшую плотность поселения.
Среди последних наиболее известный пример представляют галатеиды — десятиногие ракообразные из группы Anomura (характеризующейся умеренно редуцированным брюшком), принадлежащие к роду Munidopsis, широко распространенному в глубинах Мирового океана. На излияниях Галапагосского рифта и Вос^гочно-Тихо-
Рис. 27. На лавовом столбе возникшие из небытия ночи гигантская актиния и стеклянные губки в форме грибов, окруженные галатеидами.
океанского поднятия они представлены видом М. subsquamosa, описанным лет сто назад (сейчас классификация рода Munidopsis подвергается ревизии, и, возможно, его номенклатура изменится), а на канадских излияниях, в частности на хребте Хуан-де-Фука,— видом М. beringiana. На 13° с. ш. эти белые галатеиды образуют популяции большой плотности, иногда более 10 экз./м2. По мере удаления от гидротермального источ
ника плотность поселений очень быстро уменьшается. Пилоты подводных аппаратов хорошо знают эту особенность и судят по ней о близости активных оазисов. Часто эти галатеиды взбираются на лавовые столбы, где они соседствуют с крупными актиниями и стеклянными губками, по форме напоминающими большие белые грибы,— наблюдатели описывают незабываемое впечатление, которое производят эти необыкновенные формы, внезапно возникающие из тьмы и замирающие в ярком свете прожекторов... Munidopsis с 13° с. ш. переносит температуру 12 °C, что позволяет ему проникать в гущу трубок рифтий. Из местонахождения на 21° с. ш. недавно был описан еще один вид мунидопсиса, М. lentigo, который, по-видимому, тесно связан с гидротермальными излияниями.
Другая интересная форма, довольно многочисленная вокруг глубоководных оазисов и известная также из глубин океана,— сифонофора (сифонофоры — группа, близкая к медузам) Thermopalia taraxaca, которую американские геологи назвали одуванчиком (по-латыни Taraxacum). Это прозрачное шаровидное животное диаметром 5—6 см имеет множество массивных коротких щупалец и живет, прикрепившись ими к неровностям базальтовых лав. Вероятнее всего, оно питается планктонными животными, как и его сородичи, плавающие у поверхности.
Среди видов, свойственных гидротермальным источникам и играющих важную роль в составе и общем облике сообществ, необходимо особо упомянуть серпулид. Эти полихеты, строящие белые известковые трубки с перламутровым отливом и питающиеся взвешенными частицами, которые они извлекают из воды с помощью своих развитых жаберных султанов, принадлежат к двум разным родам и видам: Laminatubus alvini с шаровидной крышечкой, замыкающей известковую трубку при втягивании животного, и Protis hydrothermica, лишенный крышечки, как и другие виды этого рода (крышечка серпулид возникает вследствие преобразования одной из двух самых вентральных жабр, левой или правой). Эти две формы встречаются на всех трех местонахождениях: на Галапагосском рифте, на 13 и 21° с. ш. на Восточно-Тихоокеанском поднятии. Оба эти вида намного крупнее других серпулид, живущих на таких же глубинах. Еще удивительнее их очень высокая плотность, превосходящая 300 экз./м" и объяснимая лишь высокой концентрацией
Рис. 28. Червь-серпулида Laminatubus alvini, извлеченный из известковой трубки, справа — крышечка, представляющая собой видоизмененную жаберную нить.
1 мм
взвешенных органических частиц бактериального либо животного происхождения над подводными оазисами.
Другие полихеты, кроме серпулид, широко представлены среди видов-оппортунистов, свойственных гидротермальным оазисам,— на сегодня их насчитывается около 25 видов, принадлежащих к десятку разных семейств. Но надо признать, что за исключением морфологии и положения в системе, мы почти ничего не знаем об этих кольчатых червях.
Среди ракообразных описаны два вида креветок, один вид из местонахождения на Галапагосском рифте (Alvinocaris lusca), второй — на 13° с. ш. (Lebbeus carinatus). Известны также изопода, найденная на Галапагосском рифте, пиявка, клещ... Мы знаем, что существуют по крайней мере две офиуры (родственницы морских звезд, с шершавыми руками и хорошо обособленным от рук диском), которые еще не собраны, различные актинии... «Спагетти» с Галапагосского рифта принадлежат к новому семейству кишечнодышащих животных, к которым относятся также баланоглоссы с французских атлантических пляжей.
Можно утверждать без риска ошибиться, что все, известное ныне, померкнет перед тем, что станет известно через несколько лет. Между сбором проб и публикацией работ специалистов-систематиков неизбежно проходит какое-то время. В отличие от прибрежной фауны, население глубин плохо изучено, хотя бы потому, что число собранных на больших глубинах животных еще чрезвычайно мало. То, что уже опубликовано по фауне подводных оазисов, представляет едва ли половину того, что специалисты знают и изучают. Поэтому было бы преждевременно представлять сегодня окончательный состав гидротермального сообщества в разных географических местонахождениях, тем более что открытие новых оазисов, связанных уже не с гидротермальными излияниями, а с высачиваниями интерстициальных вод, несомненно, будет способствовать расширению списка животных из разных групп.
7
РОСТ, РАЗМНОЖЕНИЕ И ЭКОЛОГИЧЕСКАЯ СТРАТЕГИЯ
С точки зрения экологии подводные оазисы представляют собой совершенно оригинальное явление, не имеющее эквивалента на нашей планете:
— Гидротермальная среда имеет столь специфические физико-химические характеристики, что те из обитающих в ней видов, которые действительно к ней приспособились и количественно составляют основную часть сообщества, не могут существовать ни в каких других условиях.
— Гидротермальная среда принципиально нестабильна: в масштабе отдельного излияния продолжительность жизни гидротермального источника исчисляется десятками лет, самое большее столетием; в региональном масштабе (от нескольких десятков до нескольких сотен километров вдоль хребта) продолжительность жизни гораздо больше, но все же ограниченна.
Каким же образом гидротермальное население приспособилось к этой среде? Другими словами, какова та адаптивная стратегия, которая позволяет этим сообществам выживать на уровне особи (то есть каковы адаптации, имеющие отношение к поведению и физиологии) и на уровне популяции (выживание популяции, если не говорить об индивидуальных возможностях, зависит от того, как происходит воспроизводство особей рассматриваемой популяции)?
В экологической теории, построенной на наземных моделях, но полностью приложимой к морской среде, различаются две противоположные адаптивные стратегии:
— Стратегия, приспособленная к жизни в нестабильных средах, где физико-химические условия меняются в широких пределах; соответствующие вйды очень толе-рантны по отношению к физико-химическим условиям,
имеют высокую скорость роста, раннее половое созревание и большую способность к расселению. То есть, в антропоморфных терминах, эта стратегия заключается в том, чтобы как можно быстрее извлекать пользу из возникших в определенной среде на короткое время благоприятных условий.
— Стратегия, приспособленная к жизни в стабильных средах, где физико-химические условия постоянны или меняются незначительно с течением времени; соответствующие виды узко приспособлены к особенностям своей среды, имеют малую скорость роста, позднее половое созревание и слабую способность к расселению.
В случае гидротермальных сообществ можно было ожидать преобладание видов первой группы, которых экологи называют г-стратегистами, в отличие от видов второй группы — К-стратегистов.
Хотя современные знания еще очень малы, можно отчасти ответить на поставленный вопрос в отношении крупных видов, лучше всего приспособившихся к гидротермальной среде.
Биологи довольно давно занялись исследованием скорости индивидуального роста двух крупных двустворчатых моллюсков, пытаясь выяснить, соответствует ли их высокая биомасса столь же высокой продукции. С большим или меньшим успехом были использованы различные методы. Для батимодиолы с Галапагосского рифта в 1979 году удалось использовать прямой метод, который состоит в мечении особей путем нанесения меток на раковины с помощью зубоврачебной фрезы, управляемой манипулятором подводного аппарата. Через известный промежуток времени меченых животных достают (так называемый метод пометки — поимки). Помеченные батимодиолы были выловлены через 294 дня. Скорость роста моллюска длиной 18 см в среднем равна 1 см в год; она высока у молодых особей и убывает с возрастом. У калиптоген первоначальный расчет по скорости распада естественных радиоизотопов (серии 26UU и 2j2Th) элементов, входящих в состав известковой раковины — измеряли соотношения радиоизотопов в разных точках по оси максимального роста раковины,— дал следующий результат: скорость роста равна 5 см/год для первых 6 см длины раковины и 2—3 см/год на .каждые последующие 2 см, средний прирост за всю жизнь 4 см/год, что является удивительно высоким результатом. Другие методы, осно
ванные на изучении микроструктуры раковины и использовании отношений изотопов *8О/ГбО и 13С/12С в карбонате кальция раковин калиптоген, дали более низкие значения скорости роста и, соответственно, более значительный возраст. В то же время эти методы позволили выявить определенную ритмичность роста, которая, вероятно, зависит от изменений активности гидротермальных излияний. Самые последние подсчеты вводят поправку на скорость растворения раковин: действительно, на такой глубине кислотность разбавленного гидротермального флюида достаточна, чтобы за несколько десятков лет полностью растворить раковины двустворок, и растворение начинается еще при жизни моллюска. При изучении калиптогены этими методами результаты оказались ближе к тем, что были получены на батимодиолах, чем очень высокие значения, которые дал метод измерения распада радиоизотопов, входящих в состав раковины: 1,2 см/год для особей длиной 12,5 см, то есть моллюсков среднего размера. Даже если между американскими и французскими специалистами имеются некоторые разногласия, можно ориентироваться на следующие средние значения скорости роста: 1 см/год для батимодиолы и 0,8—1,1 см/год для калиптогены, причем максимальный возраст у калиптогены выше, чем у батимодиолы, и составляет 20—30 лет.
Таким образом, гидротермальные двустворчатые моллюски растут в два-три раза быстрее, чем живущие на сравнимых глубинах другие виды двустворок, размер которых не превосходит 1 —1,5 см. В случае батимодиолы наблюдаемая скорость роста сравнима со скоростью роста прибрежных мидий. Другими словами, когда количество пищи не ограничено, митилиды растут со скоростью, которая не зависит от особенностей гидротермальных источников.
Достоверных данных, позволяющих оценить скорость роста каких-либо беспозвоночных с мягким телом, помимо двустворок, сейчас практически нет. Некоторые наблюдения, сделанные на 13° с. ш. с интервалом в два года, похоже, свидетельствуют о высокой скорости роста трубок рифтий (но, разумеется, рост самих животных остается неизвестным, поэтому наблюдения следует повторить и дополнить).
Вторая важная адаптивная особенность касается демографической стратегии: размножения видов и су-
шествования расселительных стадий. Имеющиеся немногие результаты касаются, во-первых, батимодиол из излияний на Галапагосском рифте и на 13° с. ш. и, во-вторых, некоторых видов, для которых есть отрывочные данные. В целом результаты несколько противоречивы.
Яйца батимодиолы довольно мелкие (60 мкм в диаметре), и она выметывает их по одному (у прибрежных мидий яйца выметываются пакетами по нескольку десятков). У батимодиол есть планктонная личинка, обеспечивающая распространение вида. Это, правда, требует комментария. Дело в том, что пока никто не видел личинок батимодиол. Но изучение макушек раковин (то есть, самой молодой части раковины) взрослых особей в сканирующем электронном микроскопе позволяет установить существование двух последовательных личиночных стадий, хорошо известных специалистам под названием про-диссоконхи I и II. Продиссоконха I имеет в длину 95 мкм, продиссоконха II — более 400 мкм. Специалисты по двустворчатым моллюскам считают, что маленькие про-диссоконхи I соответствуют пелагическим личинкам, которые долгое время живут и питаются в планктоне (планктотрофные личинки), а большие продиссоконхи I (130—500 мкм) соответствуют личинкам с укороченной (или отсутствующей) пелагической стадией (лецитотроф-ные личинки). Маленьким продиссоконхам I соответствуют гладкие (без скульптуры) и крупные продиссоконхи II, и наоборот. Следовательно, у батимодиолы есть пелагическая планктотрофная личинка, способная жить в планктоне много недель, а может быть даже несколько месяцев, при температуре 2 °C. Тогда при скорости течения 10—20 см/с возможно расселение таких личинок на расстояния во многие тысячи километров; эта особенность свойственна видам с г-стратегией, что, однако, противоречит малому количеству откладываемых яиц. Но ведь личинку батимодиолы еще никто не видел!
Примененный к калиптогене, тот же метод дал противоположный результат: продиссоконха II полностью отсутствует, что свидетельствует об очень короткой или вовсе отсутствующей пелагической стадии и, вследствие этого, крайне низких расселительных способностях. Точно так же нет пелагической личиночной стадии у большинства колпачковидных брюхоногих моллюсков (у брюхоногих говорят о протоконхе I и II, но соотношения те же самые: протоконха I — это первичная раковина,
Рис. 29. Очень молодые особи Paralvinella grasslei, которые перемещаются, ползая по субстрату. Способ распространения альвинеллид и, в частности, помпейских червей с одного местонахождения на другое пока совершенно не ясен.
формирующаяся во время эмбрионального развития, протоконха II — раковина, строящаяся во время пелагической жизни). У недавно описанного маленького гребешка, ставшего одним из примеров живого ископаемого (см. гл. 8), наоборот, есть продиссоконха I маленького размера и планктотрофная личинка.
Яйца рифтии имеют размер 70—100 мкм, а самые маленькие из собранных особи, уже имеющие облик взрослых, не превышают 2 мм. Можно предполагать, исходя из того что мы знаем о развитии погонофор, что между этими двумя стадиями существует личинка, покрытая редкими ресничками, которая ползает по субстрату и имеет весьма ограниченные способности к распространению. Точно так же самая молодая стадия, известная для помпейских червей, имеет длину менее 1 мм и насчитывает примерно 12 сегментов. По аналогии с близкими видами можно полагать, что у этих животных нет рассели-тельной личиночной стадии. Однако геологи наблюдали роение червей, похожих на помпейских, которые плавали в воде в нескольких метрах надо дном, и по крайней мере одну особь Alvinella caudata, плававшую в воде, наблюдал и точно определил биолог. Может быть, это и есть механизм расселения вида на взрослых стадиях, хорошо, кстати, известный экологам для других видов? Наконец, в жаберной полости Bythograea нашли взрослых червей другого вида, Amphisamytha galapagensis: можно полагать, что таким образом краб переносит их из одного местонахождения в другое.
Эти несколько наблюдений зоолога не позволяют установить связи между разными гидротермальными излияниями одного района, идет ли речь о возрасте населения сообщества (то есть о дате начала заселения) или о происхождении первых особей каждого вида, которые постепенно заселяли излияние. Методы экологической генетики, которые, в частности, позволяют установить существование одной или двух генетически различных популяций в одном сборе, могут оказаться чрезвычайно полезными для развития этих направлений. Пока лишь батимодиолы из двух местонахождений на Галапагосском рифте стали объектами генетического исследования американских ученых, еще не опубликованного.
Вопрос о распространении и выживании гидротермальных сообществ реально стоит в малых масштабах времени и пространства, порядка сотни лет и десятка
километров. Проблема биогеографического распространения гидротермальных сообществ и изменений видового состава, которые отмечаются при сравнении двух крупных совокупностей излияний, расположенных на севере (хребты Хуан-де-Фука и Эксплорер) и на юге (21 и 13° с. ш. на Восточно-Тихоокеанском поднятии и Галапагосском рифте) от Орегонской зоны субдукции, ставит другие вопросы, которые касаются уже эволюции. Разрыв между этими двумя большими совокупностями произошел примерно 25 млн. лет назад. Канадская исследовательница Танниклиф считает, что степень дифференциации гидротермальной фауны этих двух областей недостаточно велика для столь большого промежутка времени. Следует ли из этого, что гидротермальные излияния — очень консервативная среда, скорость эволюции в которой мала, или же что существует еще не известная географическая непрерывность распространения, а может быть, просто наши зоологические познания еще слишком несовершенны? И здесь тоже метод эволюционной генетики, позволяющий восстановить степень родства между близкими видами, поможет лучше понять механизм преобразования видов и механизмы организации разных типов сообществ.
Самый предварительный вывод, возможно, состоит в том, что если в локальном масштабе пространственно-временная изменчивость гидротермальной среды очень велика, то в масштабе региональном, и даже мировом, и в ходе геологического времени гидротермальная среда очень постоянна. Эта двойственность, вероятно, объясняет, почему адаптивные стратегии сложнее, чем это может показаться априори, исходя из чисто локальных оценок.
8
ПРОИСХОЖДЕНИЕ ГИДРОТЕРМАЛЬНОЙ фауны И ЖИВЫЕ ИСКОПАЕМЫЕ
Одним из основных вопросов, касающихся фауны, связанной с гидротермальными явлениями, представляется вопрос о ее происхождении. В самом деле, эти явления существуют уже миллиарды лет, они возникли задолго до возникновения первых многоклеточных животных. Наиболее характерные виды этой фауны — строгие эндемики гидротермальных источников, то есть не обнаруживаются ни в каких других средах. Обычно считается, что высокая степень эндемизма — результат долгой эволюции или очень большой скорости эволюции. Чтобы решить, какая из этих двух гипотез верна, следует восстановить палеонтологическую историю современных форм, что не всегда возможно, в частности в отсутствие ископаемых остатков. Хотя сейчас у нас и нет достаточного количества неоспоримых данных о нескольких сотнях новых видов, описанных со времени открытия гидротермальных сообществ, тем не менее можно дать кое-какие элементы ответа на вопрос о происхождении гидротермальных животных.
Хронологически первые полученные результаты касались стебельчатого усоногого рака из группы, которая включает морских уточек, часто поселяющихся на плавающей древесине, и морских желудей, прикрепляющихся к подводным скалам и употребляемых в пищу в Португалии и Бретани. Этот вид, названный Neolepas zevinae*, был добыт на глубине 2600 м на 21° с. ш. на Восточно-Тихоокеанском поднятии вблизи активного гидротермального источника. Как и все стебельчатые усоногие раки, этот вид во взрослом состоянии прикреп
* Назван в честь советского зоолога и гидробиолога, специалиста по усоногим ракам Г. Б. Зевиной.— Прим. ред.
ляется к субстрату ножкой. Сравнив N. zevinae с разными видами, известными в ископаемом состоянии, а также с последовательными стадиями развития современного рода Pollicipes, наиболее продвинутого из усоногих раков, Ньюман показал, что N. zevinae — настоящее живое ископаемое, то есть живой представитель группы, ранее известной только в ископаемом состоянии и до того момента считавшейся исчезнувшей из современной фауны. Основным признаком для доказательства служило число известковых пластинок домика, укрывающего тело рачка: род Neolepas имеет восемь пластинок (ростр, карину, две дорсальные пластинки, два латеральных щитка и две латеральные пластинки), что ставит его между двумя ископаемыми формами — родом Eolepas, который жил с верхнего триаса до нижнего мела (230—130 млн. лет назад) и имел только шесть основных пластинок, и родом Calantica, известным из верхней юры (160—150 млн. лет назад), который имел 13 пластинок (к восьми пластинкам Neolepas добавляются две пары латеральных пластинок и непарная дополнительная под кариной). Следовательно, хотя сегодня неизвестны ископаемые формы, имеющие восемь пластинок, Neolepas происходит от какого-то предка, появившегося в юрский период и бывшего потомком Eolepas. Эти палеонтологические аргументы дополняет изучение онтогенеза Pollicipes: биологический закон, согласно которому в онтогенезе каждого вида сохраняется память о прошлой эволюции, то есть о филогенезе, в этом случае блестяще подтверждается. В ходе развития Pollicipes проходит несколько стадий, которые начинаются циприсовидной личинкой с двумя пластинками в форме створок; сначала свободноживущая, эта личинка затем закрепляется на дне при помощи трансформированного придатка — антеннулы. Следующая стадия имеет пять хитиноидных пластинок (карину, две дорсальных и два щитка). Затем наступает стадия Eolepas, которая имеет шесть обызвествленных пластинок. Далее следует стадия с восемью пластинками, сравнимая с Neolepas, а затем стадия Calantica с 13 пластинками. В продолжение онтогенеза появляется ряд маленьких пластинок в основании стебелька — известны похожие ископаемые формы, датируемые нижним эоценом (50 млн. лет назад). Таким образом, Neolepas zevinae является прекрасной иллюстрацией понятия живого ископаемого, чьи предки появились в юрский период. Как же Neolepas
Рис. 30. Три примера живых ископаемых, открытых в подводных оазисах. Вверху слева — две створки маленького гребешка Bathypecten vulkani, достигающего 15 мм в диаметре. Справа — усоногий рак Neolepas zevinae, замечательный пример сохранения в онтогенезе памяти о филогенезе. Внизу — небольшой колпачковидный брюхоногий моллюск Neomphalus fretterae.
или его предки приспособились к жизни вблизи гидротермальных источников? Можно выдвинуть две гипотезы. Возникнув в прибрежной зоне, эта форма укрылась от возрастающего пресса хищников в зонах активной тектоники на тропических мелководьях, впоследствии
это открыло ей доступ к более глубоким водам вдоль активных хребтов, где имеют место гидротермальные явления. Согласно второй гипотезе, эта форма произошла от предков, уже обосновавшихся в глубинах, а затем приспособившихся к жизни в гидротермальной среде. Эта гипотеза маловероятна, поскольку в этом случае животным пришлось бы преодолевать более резкий физико-химический градиент, к тому же на малых глубинах формы жизни более разнообразны. Интересно, что гипотеза возрастающего пресса хищников в мезозое уже была привлечена для объяснения эволюции современного рода Lithothrya, усоногого рака той же группы, имеющего, как и Neolepas, восемь пластинок, сильно видоизмененных (крайне редуцированный ростр и две латеральные пластинки) из-за специфического местообитания: эта форма живет в приливно-отливной зоне, в норах, просверленных ею в известковых скалах, где она нашла убежище от хищников. Похоже, что Neolepas укрылся от хищников вблизи гидротермальных источников в тот же момент, когда Lithothrya начала сверлить коралловые рифы. Наконец, и в размножении Neolepas имеются интересные особенности: яйца этого вида довольно крупные (500Х Х300 мкм), что может рассматриваться как адаптация к распространению вида с помощью свободных циприсо-видных личинок на большие расстояния. То, что размеры шести первых пойманных особей колеблются от 3,5 до 58 мм, доказывает, что вымет личинок происходит в течение всего года. Крупные яйца, значительная способность к расселению, непрерывное размножение—все это биологические адаптации, свойственные видам, приспособленным к неустойчивым условиям обитания.
Neolepas zevinae был найден и на 13° с. ш. на Восточно-Тихоокеанском поднятии, также в кольце серпулид, то есть на границе влияния гидротермального флюида.
Другой пример живого ископаемого, возможно не столь впечатляющий,— мелкий брюхоногий моллюск, напоминающий морское блюдечко или китайскую шляпу. Он едва достигает в диаметре 1 см и встречается иногда в больших количествах на базальтовых лавах и отложениях сульфидов. Этот вид, Neomphalus fretterae*,—
* Вид назван в честь крупного английского малалколога, специалиста по брюхоногим моллюскам, Веры Фреттер (Vera Fretter).— Прим. ред.
потомок Euomphalacea, группы, исчезнувшей в конце юрского периода. Эти брюхоногие, известные с палеозоя, претерпевали интенсивную эволюционную радиацию, которая продолжалась более 200 млн. лет. Так что и здесь речь идет о живом ископаемом. Вид был найден на излиянии Галапагосского рифта и на многих других излияниях Восточно-Тихоокеанского поднятия (13 и 21° с. ш.). По Маклину, Neomphalus или, вероятнее, его мезозойский предок нашел, как и Neolepas, в зонах тектонической активности в тропических мелководных морях убежище от пресса хищников, усиление которого в морях было характерно для этой геологической эры, что и позволило ему достичь впоследствии глубоководных гидротермальных источников.
После публикации работ о Neomphalus было открыто более трех десятков новых видов мелких колпачковидных брюхоногих моллюсков, напоминающих морские блюдечки. Все они относятся к обширной группе археогастро-под, которые, кроме других особенностей, отличаются отсутствием плавающей личинки, велигера (велигер — «несущий велум», или «парус», большой дольчатый вырост с реснитчатым краем, выполняющий одновременно двигательную и пищедобывающую функцию). Некоторые из них живут на базальтовых лавах, другие перемещаются по белым трубкам рифтий, где они бывают весьма многочисленны. У одних раковина гладкая, полупрозрачная, хрупкая; у других раковина потолще, с разнообразной скульптурой (кили, выемки, радиальные или концентрические штрихи и т. д.) Изучение их буккального аппарата позволяет выделить четыре главных направления эволюционной радиации, которые восходят, как Neomphalus, к мезозойской эре. Эти брюхоногие имеют буккальную терку, или радулу, и ею они обскребывают поверхность, по которой перемещаются. Радула представляет собой как бы ленту, снабженную поперечными, следующими один за другим рядами крошечных зубов. Строение радулы — важный элемент классификации брюхоногих моллюсков. Один из вопросов, который ставит невероятное изобилие новых форм колпачковидных архео-гастропод,— почему они столь разнообразны в гидротермальной среде? В самом деле, дифференциация видов подразумевает относительную изоляцию популяций в течение достаточно долгого времени. В случае с гидротермальными источниками эта изоляция не может быть
географической, потому что все или почти все формы встречаются совместно на разных излияниях. Тогда приходится предположить, что существуют барьеры экологического характера, которые изолируют популяции, позволяя им эволюционировать независимо и приводят к появлению новых, отличных от предыдущих, видов. Эта проблема может быть разрешена только на основе лучшего знания физико-химических условий и пространственной локализации в масштабе нескольких метров разных видов колпачковидных брюхоногих моллюсков.
Замечательно, что в общем, за исключением одной формы, принадлежащей к роду Provanna из мезогастропод, все гастроподы, тесно связанные с гидротермальными источниками,— это колпачковидные брюхоногие моллюски, сравнимые лишь с ископаемыми формами мезозойского возраста.
Последний пример живого ископаемого, открытого в термальных зонах,— бесспорно, самый интересный: это маленький двустворчатый моллюск из семейства Pectini-dae, куда относится обыкновенный гребешок. Предварительное описание этого вида было опубликовано в 1985 году. Названный Bathypecten vulcani, этот небольших размеров моллюск едва достигает диаметра 15 мм; он был обнаружен на Восточно-Тихоокеанском поднятии на 13° с. ш., в местонахождении, где заметны лишь признаки былой гидротермальной активности, при довольно высокой плотности поселения (30—50 экз./м2). Как и у всех современных гребешков, у него имеются два неравных «ушка» и гладкая раковина, лишенная характерных для обыкновенного гребешка радиальных ребер. Вместо них у него ярко выраженная концентрическая волнистость створок. По некоторым признакам структуры кальцита раковины этот моллюск близок к другому семейству этой группы — Propeamussiidae, включающему, в частности, современные глубоководные формы. Два эти семейства, Pectinidae и Propeamussiidae, разделились еще в триасе (приблизительно 230 млн. лет назад), и можно предположить, что Bathypecten — промежуточное звено между двумя этими семействами — является потомком еще более древнего общего предка, существовавшего в палеозое. По этой гипотезе новое живое ископаемое гораздо древнее обоих вышеупомянутых. Точнее, он демонстрирует сильную морфологическую конвергенцию с ископаемым родом Pterinopecten из девонских отложений США.
Кроме этих трех примеров настоящих живых ископаемых, мало что известно о происхождении разных видов гидротермальной фауны. Крупный белый двустворчатый моллюск Calyptogena magnifica, описанный в 1980 году как новый вид хорошо известного рода, интересен в том смысле, что некоторые виды этого рода, живущие в зонах субдукции, кажется, используют высачивающуюся из осадков под действием давления интерстициальную глубинную воду. Сегодня известно около 20 видов Calyptogena, из них десять — только в ископаемом состоянии из миоценовых и плиоценовых отложений (не более 25 млн. лет назад). Значит, это современная группа, батиметрическое распространение которой охватывает, по крайней мере для ныне живущих видов, глубины от 55 до 5900 м, с максимумом разнообразия на глубинах 500—1500 м. Не только С. magnifica, но и другие виды этого рода имеют гемоглобин в крови, и некоторые из них были найдены в биотопах не гидротермальных, но богатых восстановленными соединениями, произошедшими от разложения органического вещества осадков.
В случаях когда более древние ископаемые формы не известны, почти невозможно установить степень древности гидротермальной фауны только на основе изучения организмов, как, например, в случае альвинеллид или рифтиид, столь обособленных, что для них пришлось создать два новых для науки семейства.
Однако есть некоторые основания думать, что даже если бактерии очень рано поселились в примитивных гидротермальных источниках, различные беспозвоночные также смогли извлечь пользу из этой своеобразной среды. Так, изучение массивных полиметаллических сульфидов, отложившихся в офиолитах (скалах, представляющих остатки древней океанической коры) в районе Самаил в Омане на юго-востоке Аравийского полуострова, обнаружило присутствие трубок диаметром 1—5 мм, располагающихся в беспорядке в массе сульфидов; это — окаменевшие трубки червей, связанных с гидротермальными выходами, отложившими полиметаллические сульфиды около 95 млн. лет назад. Возраст этой зоны соответствует срединному хребту обширного мелового океана Тетис. Отсутствие остатков животных, построивших эти трубки, не позволяет утверждать, что речь идет о предках тех червей, что живут сегодня в горячих источниках
Восточно-Тихоокеанского поднятия. Следует, однако, подчеркнуть совершенное сходство современных черных курильщиков, заселенных помпейскими червями, и массивных полиметаллических сульфидов мелового периода: у червей, построивших ископаемые трубки, были те же, либо очень близкие замечательные адаптации, что и у помпейских червей. Более того, диаметр трубок сходен с диаметром трубок современных альвинелл, который колеблется приблизительно от 5 мм до 1 см. Маловероятно, что это трубки вестиментифер, подобных крупным рифтиям: трубки, которые строят рифтии, прямолинейные и правильного диаметра.
Как бы то ни было, если это открытие окаменевших трубок поставить рядом с существованием в современных гидротермальных излияниях многих живых ископаемых, реликтов мезозойской или еще более древних фаун, то можно думать, что вокруг гидротермальных источников с высокой температурой в морях мелового периода существовали богатые сообщества животных, подобных тем, которых мы знаем сегодня. В то же время это подчеркивает интерес дальнейшего изучения ископаемых гидротермальных отложений.
Недавнее открытие подкрепляет последнее замечание. В отложениях свинцовых, цинковых, медных, серебряных минералов и сульфата бария в центре Ирландии (отложение Тинаг) обнаружены ископаемые трубки диаметром 1,5 мм и длиной примерно 1 см. Эти трубки расположены на стенках ископаемых курильщиков, сложенных главным образом пиритом. В некоторых случаях внутри трубок наблюдали пиритовый контур червеобразного животного, на вид сегментированного, чьи ткани были замещены баритом в процессе фоссилизации. Эти организмы имели в диаметре не более 0,8 мм. Отложения в Тинаге сформировались в нижнем карбоне, около 350 млн. лет назад. Ни размер, ни форма трубок не соответствуют ни современным помпейским червям, ни ископаемым трубкам из сульфидных отложений района Са-маил. Тем не менее это подтверждает, что заселение гидротермальных курильщиков началось очень рано, может быть, со времени появления первых водных беспозвоночных.
Что же следует из этих немногих фактов? Прежде всего то, что подводные гидротермальные источники были очень рано заселены разными беспозвоночными,
4екоторые из них находили в этих неблагоприятных /словиях убежища от хищников. В некоторых случаях можно даже задаться вопросом, а не была ли гидротер-маль той первоначальной средой, в которой возникли, а затем эволюционировали на месте целые группы беспозвоночных? Но сегодня мы не располагаем ни одним доказательством, способным подтвердить такую гипотезу. Далее, учитывая несовершенство наших знаний, можно высказать мысль, что доля древних форм среди гидротермальных обитателей гораздо выше, чем в среднем на глубинах океана, иными словами, что гидротермальная фауна имеет древнее происхождение и издавна связана с гидротермальными источниками.
И последнее замечание, чтобы подкрепить это заключение. Речь идет о нескольких фаунистических аномалиях, выявленных в серии ископаемых, в частности в мезозойских отложениях западных Альп. В сериях юрского возраста Черных Земель, в районе массивов Диуа и Барон-ни (Франция) обнаружены карбонатные стяжения в форме узлов, могущих включать углеводороды и различные минеральные отложения. В некоторых местах эти образования содержат весьма многочисленные раковины двустворчатых моллюсков длиной до 15 см, сопровождаемые более скромной, но разнообразной донной фауной. В свете открытий, сделанных в бассейне Гуаймас (гидротермальные излияния в обстановке усиленного осадконакопления), а также в зонах субдукции (побережье штата Орегон, Японский желоб, Флоридский эскарп), можно интерпретировать эти образования как отложения, вызванные идентичными явлениями, а точнее, учитывая отсутствие полиметаллических сульфидов, просачиванием интерстициальной воды сквозь осадки. Таким образом, можно надеяться на полную реконструкцию фауны этих подводных мезозойских оазисов, что позволило бы лучше понять происхождение современных гидротермальных сообществ.
ЗАКЛЮЧЕНИЕ
Заключение — это обычно завершение рассказа. История же сообществ, связанных с гидротермальными явлениями, не только далека от завершения, но, напротив, перед ней открываются все новые перспективы, придающие ей особое значение. Еще слишком рано предлагать общую интерпретацию открытых менее двух лет назад фактов, которые приводят к неожиданному обобщению процессов бактериального хемосинтеза на больших глубинах океана. Однако в заключении следует кратко напомнить о том вкладе, который внесли в науку эти открытия.
Хронологически первым было открытие сообществ на глубине 3266 м к западу от Флориды, у подножия величественного эскарпа, датируемого нижним мелом; у его основания склон покрыт глинистыми осадками. Здесь, в десятке метров от основания подводного обрыва обнаружено пышное поселение мидий, калиптоген и крупных вестиментифер из рода Escarpia в сопровождении целой свиты видов-оппортунистов. На этом уровне высачивается интерстициальная вода, прошедшая сквозь толщу известняка. Она содержит, в частности, аммиак (до 3—4 ммоль/л). Осадки, расположенные неподалеку от обрыва, черного цвета и издают сильный запах сероводорода. Температурных аномалий не отмечено, а кислотность воды, высачивающейся из-под обрыва, обусловлена ее взаимодействием с известняком мелового возраста. Первые результаты, полученные на основе измерений отношения изотопов углерода в мидиях, позволили построить очень сложную модель. Согласно этой модели, свободные хемосинтезирующие бактерии, используя аммиак или сероводород, синтезируют органическое вещество, потребляемое затем в бескислородной среде бактериями, которые выделяют метан органического проис-
хождения, способный служить энергетической базой для бактерий, связанных с мидиями...
Как бы то ни было, биомасса в этом местонахождении по сравнению с биомассой на соседних глубинах настолько велика, что это подразумевает существование независимого от фотосинтетической первичной продукции источника органического вещества хемосинтетического происхождения.
Через несколько месяцев «Алвин» делает новое открытие у берегов Орегона на глубинах немногим больше 2000 м. И здесь обильное животное сообщество, состоящее в основном из вестиментифер, определенных как Lamellibrachia barhami, и крупных двустворчатых, моллюсков: Calyptogena sp. и одного вида из рода Solemya. На этот раз речь идет не о геологически пассивном районе, где вода просачивается сквозь гигантский известковый массив и истекает на уровне мелкозернистого абиссального осадка, представляющего собой настоящий водонепроницаемый слой; наоборот, здесь на поверхность осадков выходит интерстициальная вода, выдавливаемая из осадков под действием огромного давления, которое они испытывают. Обогащенная метаном и аммиаком (минеральные восстановленные молекулы, сравнимые с сероводородом как источник энергии для организмов), эта вода определенно является первоисточником огромной биомассы, открытой «Алвином». Как и у подножия Флоридского эскарпа, вокруг видов, которые, как предполагают, связаны с высачиванием воды, обитает многочисленная фауна, использующая соседство с процветающим сообществом.
Третье открытие, самое последнее по времени, сделано летом 1985 года «Наутилусом» в Японском желобе и в каньоне Тенрю, к востоку от Японских островов. Были обнаружены два типа поселений. В каньоне Тенрю, а также у подножия впадины Дзенису имеются небольшие пятна более насыщенного водой черного осадка диаметром 1—2 м, в который погружены раковины Calyptogena двух разных видов: один крупный, около 20 см в длину, принадлежащий к тому же подроду, что и С. magnifica из гидротермальных источников Восточно-Тихоокеанского поднятия, другой — меньшего размера и принадлежащий к другому подроду. У обоих видов имеется внутриклеточный гемоглобин. Вокруг этого скопления биомассы сосредоточиваются различные животные: хищные брюхоногие
моллюски, голотурии, привлеченные очень высоким, по-видимому, содержанием органического вещества в черных осадках, галатеиды и т. д. Это сообщество развивается на глубине примерно 3500 м. Глубже, на 5800 м на стыке старых подводных вулканов (вулканы Зримо и Касима) и шельфа Японии «Наутилус» обнаружил другие сообщества двустворчатых моллюсков. Это тоже представители рода Calyptogena, изучение которых ведется в настоящее время. Раковины, как правило более вытянутые и узкие, чем у двух предыдущих видов, погружены вертикально в черный насыщенный водой осадок. Сопутствующая фауна менее богата, просто потому что на таких глубинах многие группы животных представлены хуже и плотность поселений очень низка. Как и в двух предшествующих случаях, эти локальные аномалии биомассы объясняются механизмом хемосинтеза. Интерстициальная вода, выходящая под давлением, вызванным субдукцией, содержит метан в значительном количестве; аммиак и сероводород химическим анализом не обнаруживаются. Предварительные измерения, сделанные на самых глубоководных двустворках, взятых с глубины 5800 м, и в пробах интерстициальной воды, показывают равное отношение изотопов 1JC/‘~C в метане интерстициальной воды и в тканях двустворчатых моллюсков. Первые наблюдения с электронным микроскопом выявили присутствие многочисленных бактерий. А исследователи уже отмечали обилие «красной крови», брызжущей из этих моллюсков, если их случайно раздавит механическая рука «Наутилуса». Так что большинство элементов хемосинтеза налицо—остается описать функционирование сообщества в физиологических терминах, что, как минимум, потребует получения новых живых образцов...
Значение этих открытий не следует недооценивать: протяженность зон субдукции в масштабе всего земного шара — 45 000 км и в любом их месте могут произойти подобные явления! Так что путь, который нам остается пройти, гораздо дольше того, что мы уже прошли.
Пусть последние замечания и данная книга в целом убедят морских биологов, что глубоководные хемосинтезирующие поселения представляют собой не просто курьез, но широко распространенное экологическое явление, не имеющее аналогов в наземной среде, и что оно в большой степени способствует снабжению абиссали органическим веществом.
ПОСЛЕСЛОВИЕ
С декабря 1985 года, когда я вручил рукопись этой книги французскому издательству, множество новых открытий быстро последовали одно за другим: минерализованные «рассольные» источники, высачивающиеся у основания Флоридского эскарпа; просачивание обогащенных метаном холодных вод в зонах субдукции близ берегов штата Орегон в восточной части Тихого океана на глубине примерно 2000 м, а затем и сообщество крупных двустворок рода Calyptogena, открытых летом 1985 года в суб-дукционных впадинах юго-восточнее Японских островов на глубинах 3000—6000 м во время экспедиции «Кайко» (от японского «впадины»),— все это я еще смог упомянуть в заключении своей книги. Но сейчас накопились новые наблюдения, способные подтвердить значимость и широкое распространение этих явлений. Прежде всего, это открытие сообщества крупных двустворок и вестиментифер на глубине 600—700 м на континентальном склоне у берегов Луизианы в Мексиканском заливе — сообществ, связанных с естественными выходами углеводородов, содержащих повышенное количество легких углеводородов с одним или двумя атомами углерода (метан, этан). Первые образцы фауны и сопутствующих осадков были получены тралением с поверхности моря в 1984 году. Еще через два года были проведены первые непосредственные наблюдения за этим местонахождением с борта американского подводного аппарата «Алвин».
Летом 1985 года, вопреки ожиданиям геологов и геофизиков, которые в ту пору считали, что между средней скоростью раздвижения срединно-океанических хребтов и гидротермальной активностью существует прямая связь и что, следовательно, гидротермальные проявления на Срединно-Атлантическом хребте, отличающемся крайне
малой скоростью раздвижения, также должны быть слабы, американский геофизик П. Рона открывает на 26° с. ш. на этом хребте, в зоне, которую он тщательно изучал в течение нескольких лет (эту зону назвали TAG*), настоящие черные курильщики с высокой температурой и обычными для Восточно-Тихоокеанского поднятия минерализованными постройками. Однако фауна, обнаруженная вблизи этих гидротермальных построек, сильно отличается от той, которая была известна в то время в Тихом океане: здесь полностью отсутствуют гигантские вестиментиферы, есть двустворчатые моллюски, но большинство сообщества составляют, похоже, креветки семейства Bresiliidae (к которому принадлежит род Alvinocaris, часто встречающийся на гидротермальных излияниях в восточной части Тихого океана), представленные новым родом Rimicaris, включающим по меньшей мере два хорошо различающихся вида. Рыбы и десятиногие раки рода Munidopsis дополняют сообщество, о котором, по правде говоря, известно еще очень мало, зато существование черных курильщиков в Атлантическом океане на поднятии, скорость раздвижения которого мала — в среднем 1—3 см в год, сегодня подтверждено новыми открытиями на 12 и 15° с. ш. метановых, гелиевых и марганцевых аномалий, служащих превосходными индикаторами гидротермальной активности.
В 1986 году канадские геологи, исследовавшие глубоководный Лаврентийский шлейф в северо-западной части Атлантического океана южнее Ньюфаундленда, к своему удивлению обнаружили весьма обширные пространства, покрытые раковинами калиптоген, в большинстве мертвыми, среди которых активно перемещались крупные хищные брюхоногие моллюски. В этой зоне лет пятьдесят назад произошло заметное землетрясение, спровоцировавшее, вероятно, мощное оползание донных осадков, которое повлекло за собой настоящее срезание поверхностной их части и высвободило в некоторых местах интерстициальные флюиды, обогащенные метаном. Эти внезапные изменения способствовали тому, что здесь поселилось, а затем и развилось богатое сообщество двустворчатых моллюсков, включающих в ткани своего тела симбиотические бактерии и сумевших таким образом воспользоваться этим неожиданным поступлением орга
* Трансатлантический геотраверз.— Прим. ред.
ники. Явление тем более интересно, что землетрясение, ответственное за первоначальный оползень, достоверно известно: оно вызвало разрыв многочисленных телефонных кабелей между Америкой и Европой, которые после этого события пришлось восстанавливать во многих местах. В этом районе тектонические явления практически отсутствуют — это так называемая стабильная окраина в терминах геологов. Верхние слои осадка были срезаны, и в результате, вероятно, обнажились глубинные слои, обогащенные углеводородами, возникшими вследствие разложения органического вещества. Ясно, однако, что излияния долго не просуществуют, и обнаруженные многочисленные пустые раковины двустворок об этом, похоже, и свидетельствуют. Систематическое изучение этой интересной зоны еще не проведено.
В том же 1986 году, а затем и в течение 1987 года японские исследователи на борту подводного аппарата «Синкай-2000», способного, как указывает его название, погружаться на глубину 2000 м, открыли в заливе Ха-цусима, километрах в пятидесяти к юго-востоку от Токио, на глубине примерно 700 м сообщество крупных двустворок Calyptogena soyoae в сопровождении разбросанных скоплений вестиментифер из рода Lamellibrachia. Мы с моим молодым японским коллегой особенно заинтересовались некоторыми полихетами-комменсалами, живущими в паллиальной полости различных двустворок, в том числе и у этого вида калиптоген. Как и в предыдущем случае, речь идет не о гидротермальных явлениях, но о высачивании обогащенного метаном холодного флюида, выдавливаемого на поверхность силами субдук-ции, которые проявляются в глубоководной аккреционной призме, образующейся на стыке континентальной плиты (край которой отмечен Японским архипелагом) и океанической плиты.
В то же время французские геологи и биологи обнаружили по фотографиям, полученным «рыбой»,, буксируемой судном, на аккреционной призме у Барбадоса любопытные скопления животных вокруг настоящих грязевых вулканов. Впервые в подобных условиях среди представленных животных присутствовали крупные гидроиды.
В течение 1987 года внимание ученых снова привлекают гидротермальные явления, но на этот раз в западной части Тихого океана. Первое открытие было сделано
«Алвином» на глубине 3600—3700 м близ двух вулканических построек в Марианской впадине на 18° с. ш. Руководитель подводных работ американский биолог Р. Хесслер открыл поблизости от источников с относительно низкой температурой (примерно 15 °C) сообщество животных ни на что не похожего типа: крупные вестимен-тиферы отсутствуют, с функциональной точки зрения их, по-видимому, замещают спирально завитые брюхоногие моллюски Alvinoconcha hessleri из группы Mesogastro-poda, относящиеся к новому семейству, близкому к семейству Trichotropidae или надсемейству Hipponicacea. Эта «мохнатая ракушка», как ее немедленно окрестили американские и японские ученые, участвовавшие в экспедиции, образует настоящие «поленницы» из многих слоев раковин. В ходе подводных работ были обнаружены и собраны крупные коричнево-желтые батимодиолы, множество усоногих ракообразных, десятиногие раки рода Munidopsis и креветки Bresiliidae, наконец, новый представитель полихет из семейства Alvinellidae, которого мы назвали в честь начальника экспедиции Paralvinella hessleri. Геологические условия, в которых было сделано это открытие, совершенно не похожи на те, что характеризуют океанические поднятия.
Наконец, чтобы завершить перечисление последних открытий, следует упомянуть наблюденное по фотографиям и видеозаписям гидротермальное сообщество, очень похожее на сообщество из Марианской впадины. Фотографии и видеозаписи были сделаны в ходе Франко-Японской экспедиции в задуговом бассейне спрединга северо-восточнее островов Фиджи. Это впадина, образовавшаяся самое большее 10 млн. лет назад. Утончение океанической коры, вызванное силами раздвижения, создало сеть глубинных трещин, в которых установилась активная гидротермальная циркуляция. По фотографиям мохнатые глубоководные моллюски и скопления коричнево-желтых батимодиол наблюдались в разных местах, подчеркивая широту распространения явления. Летом 1989 года здесь были проведены работы французского подводного аппарата «Наутилус» и получены чрезвычайно интересные результаты.
Таким образом, всего за несколько лет экологи и биологи получили подтверждение важной роли хемосинтетической продукции на больших глубинах океана в самых разных геологических условиях. Будь то настоящие гидро
термальные проявления или выходящие на поверхность холодные флюиды, обогащенные легкими углеводородами и восстановленными минеральными соединениями, основной механизм биологического продуцирования один и тот же: это хемосинтез, осуществляемый симбиотическими бактериями, включенными в ткани некоторых беспозвоночных. Только химическое «горючее» бывает разным: в случае гидротермальных сообществ — это сероводород, а в холодных высачиваниях Луизианы и Флориды — метан или, может быть, другие легкие углеводороды. Что касается гигантских двустворок из зон субдукции у Орегона, Японии, Ньюфаундленда, то здесь происхождение используемой бактериями энергии остается спорным. Большинство геохимиков, обращающих внимание на повышенное содержание метана и почти полное отсутствие сероводорода в воде непосредственно над колониями животных, считают, что посредником является метан, как и в местонахождении у Луизианы. Биологи, основываясь на том, что структура бактерий, обнаруженных в тканях моллюсков, резко отлична от структуры настоящих метанотрофных бактерий, а в жабрах отмечено большое количество кристаллов серы, предполагают, что двустворчатые используют высвобождаемый в аноксическом слое осадка сероводород, который они улавливают, благодаря своей длинной ноге, прежде чем он достигнет самых верхних, толщиной в несколько сантиметров, слоев осадка, где свободные сероокисляющие бактерии быстро его уничтожают. Как бы то ни было, по одному важному признаку хемосинтетические проявления делятся на две большие категории: одни, по-настоящему гидротермальные, не используют ранее существовавший источник органического вещества, другие, наоборот, жестко зависят от разложения уже существующего органического вещества, скрытого в осадках, под влиянием высоких температур и возрастающего давления.
Гидротермальная фауна предстает все более и болеё как фауна древняя, содержащая высокий процент живых ископаемых, происхождение которых, в зависимости от группы, датируется временем от 150 до 250 млн. лет назад. В изобилии обнаруживаются новые формы: к концу 1985 года были описаны 15 новых семейств, более 70 новых видов. Сегодня следует говорить о более чем 160 описанных новых видах, не считая еще не описанные формы, которых много в американских, французских, канадских,
японских и советских коллекциях фауны. Перечень микроорганизмов, настоящих бактерий и архебактерий приносит не меньше новинок, а спор по поводу существования ультратермофильных бактерий все еще не разрешен. Выявленные в восточной части Тихого океана два крупных региона севернее и южнее зоны субдукции у берегов Орегона, которая на протяжении сотни километров прерывает цепь гидротермальных излияний, широко подтверждены в плане биогеографии. Для некоторых групп уже необходимо предложить филогенетические схемы, например для нового типа вестиментифер, включающего 12 видов, относящихся к пяти семействам, группируемым в два отряда; равным образом, правда в другом масштабе, это относится к полихетам альвинеллидам, насчитывающим в настоящее время восемь видов и два рода. Но наиболее интересным примером являются колпачковидные брюхоногие моллюски, которые насчитывают уже 40 описанных новых видов и представляют несколько разных эволюционных путей.
Биологические механизмы расселения гидротермальных видов от одного местонахождения к другому в нескольких случаях изучались на основе анализа морфологии и биологии личиночных стадий (планктотрофные или лецитотрофные личинки). Для многочисленных форм, у которых личинка имеет очень короткий срок жизни или живет внутри родительской трубки, в настоящее время не существует удовлетворительного объяснения процесса расселения на большие расстояния; однако перенос в местном масштабе, вероятно, могут объяснить локальные гидродинамические явления, связанные с выходами гидротермального флюида.
Точное описание сообществ гидротермальных излияний и холодных высачиваний сильно продвинулось вперед благодаря использованию методов фотограмметрии в анализе видеозаписей. Были отмечены биомассы более 50 кг/м2 в сыром весе. Наблюдения одного и того же местоположения с промежутком в несколько лет подтвердили, иногда самым драматическим образом, крайне малую продолжительность жизни гидротермальных сообществ — порядка нескольких десятков лет. Нарушения равновесия между разными трофическими уровнями, к которым приводит внезапное исчезновение источника химической энергии, наблюдались недавно на Восточно-Тихоокеанском поднятии на 13° с. ш. Было установлено
существование двух фаунистических сообществ, приспособившихся к контрастным температурным и физико-химическим условиям (холодный и горячий полюса гидротермальной экосистемы), и выявлена разница в их экологическом КПД.
Физиологические и биохимические механизмы, лежащие в основе симбиоза хемосинтезирующих бактерий с гигантскими вестиментиферами, частично выяснены, хотя еще неизвестны многочисленные промежуточные звенья. Хуже изучено функционирование симбиоза бактерий с крупными двустворчатыми моллюсками рода Calyptogena, многочисленные новые виды которого были открыты в желобах Японской зоны субдукции. Загадочный случай представляют и большие батимодиолы: по всей видимости, симбиоз у них является лишь второстепенным способом питания, однако для долгосрочного существования двустворчатых моллюсков он необходим. А вот процесс заражения и поддержания ассоциации некоторых бактериальных штаммов с разными видами беспозвоночных все еще остается непонятным как у вестиментифер, так и у двустворок.
Особенно сложными представляются трофические отношения между полихетами-альвинеллидами из рода Alvinella и различными метаболическими группами бактерий, как эпибионтных, так и прикрепленных в массе возведенных червями трубок. Похоже, что черви улавливают часть веществ, высвобожденных в результате деятельности микроорганизмов, усваивающих растворенное и взвешенное органическое вещество.
Значительны достижения ученых в изучении биохимической структуры и механизмов действия металлопро-теинов — переносчиков кислорода и сероводорода (одна и та же молекула у рифтий, две различные молекулы у калиптогены). Обнаружение металлотионеинов у многочисленных групп беспозвоночных подтверждает важную роль процессов детоксикации у животных, наиболее подверженных высоким концентрациям сульфидов, и даже у связанных с ними бактерий.
Концентрация биогеохимических меток в воде, окружающей колонии животных, подтверждает важность потока биогеохимических частиц, образующегося в ходе биологических процессов, потока частиц, которыми питаются и пока еще плохо известные планктонные организмы, и животные-фильтраторы, ореолом расположен-
• Северная стенка кальдеры горы Осевой хребта Хуан-де-Фука. Морские звезды Brisingidae в характерных ловчих позах, мелкие °фиуры, другая фоновая фауна.
2. Гора Осевая. Красные жа- 3. Гора Осевая. Измерение тем-берные участки тела много- пературы в месте сочения
щетинковых червей Paralvi- флюида.
nella и скопления Lepeto-drilus.
4. Основание черного курильщика в бассейне Гуаймас. Упавшие с его склонов Riftia pachyptila и Munidopsis.
5. Буш Riftia с красными многощетинковыми червями Polynoidae на их белых трубках, достигающих в длину 3 м (там же).
6. Непотревоженные вестиментиферы Riftia с высовывающимися из трубок длинными обтуракулами (там же).
7. Крабоиды Neolithodes с размахом лап около 1 м на предвершинных склонах курильщика котловины Гуай-мас.
8. Самая безжизненная часть гидротермальной постройки — вершина с «каминной трубой».
9. Измерение температуры в зоне разгрузки курильщика, белые бактериальные маты.
10. Черные дымы на участке склона рифтовой долины (26° с. ш. Срединно-Атлантического хребта). Многочисленные креветки Вге-siliidae—Rimicaris exoculata и R. chacei. Крабы Segonzacia mesatlantica.
ные на периферии гидротермальных сообществ. Местная конвективная ячейка масштабом в десятки метров, порожденная вытеканием гидротермального флюида, не способна вызвать скопление животных вследствие приноса частиц из поверхностных слоев, хотя некоторые организмы, например большие батимодиолы, умеют собирать частицы планктонного происхождения, которые обнаруживаются в содержимом их пищеварительного тракта.
Присутствие вокруг колоний крупных двустворок, связанных с холодными высачиваниями в желобах Японской зоны субдукции, многих видов хищников и оппортунистов доказывает возможность выноса органических веществ из высокопродуктивных центров, какими являются эти колонии. Но в настоящее время нет доказательств возможности прямого использования этих невероятных концентраций биомассы гидротермальных сообществ обычными глубоководными океанскими хищниками и трупоедами.
Существование симбиотических хемосинтезирующих бактерий, способных окислять метан и, вероятно, другие легкие углеводороды, было неоспоримо показано в единственном случае — в гигантских мидиях, живущих вокруг нефтеносного истечения в Мексиканском заливе: физиологические опыты подтвердили гипотезу, высказанную на основе изучения изотопов углерода и примечательной ультраструктуры этих бактерий (характерная мембрана в виде стопки тарелок). В случае колоний калиптоген из зон субдукции у Японии или Орегона или же с Лаврен-тийского шлейфа общая гипотеза использования метана хемосинтезирующими бактериями сталкивается с многочисленными трудностями и никак не объясняет наличие кристаллов серы внутри жаберной ткани двустворчатых моллюсков.
Таким образом, с каждым годом и с каждым открытием традиционная концепция практически пустынных океанских глубин, в которых жизнь в сильнейшей степени зависит от вертикального потока опускающихся с поверхности частиц, постепенно уступает место гораздо более контрастной системе, где хемосинтез бактерий-симбионтов в той или иной форме играет, похоже, роль, существенно превосходящую пространственные пределы распространения соответствующих сообществ беспозвоночных. СССР включился на полном ходу в работу по
исследованию глубоководных океанических оазисов, поставив на службу делу два подводных аппарата, способных, как и французский «Наутилус», достигать глубины 6000 м. Япония, со своей стороны, заканчивает постройку нового подводного аппарата «Синкай-6600», который сможет погружаться, как указывает его название, на глубину 6600 м. Идея широко скоординированной в международном масштабе программы становится более чем когда-либо важной для изучения больших глубин океана — последних более или менее девственных пространств нашей планеты. В этих заключительных строках я хочу пожелать, чтобы инициативы исследователей ведущих государств, способных проникнуть в эту необычную среду, позволили объединить усилия и средства для изучения функционирования и значения красочных оазисов, скрытых под километрами океанских вод.
БИБЛИОГРАФИЯ
Открытие подводных оазисов, связанных с гидротермальными источниками, произошло в 1976 году, а биологи начали интересоваться этой областью приблизительно с 1979 года. Поэтому пусть читатель не удивляется отсутствию научно-популярного обзора этого явления. Около 250 научных публикаций, посвященных этому предмету, естественно, не могут быть процитированы в приведенном списке работ, который содержит лишь материалы, более или менее доступные неподготовленному читателю. Henri Bougault. Circulation hydrothermale oceanique et metallogenese, Annales des Mines, novembre — decembre 1982, pp. 1—32.
Daniel Desbruyeres, Daniel Prieur, Lucien Laubier, Marcel Le Pennec, Philippe Bouchet, Michele de Saint-Laurent, Patrick Ceistdoerfer, Michel .Van Praet. Les infraterrestres, L’Univers du vivant, n° 1, juin 1985, pp. 16—39.
Jean Francheteau, Lucien Laubier. Naissance des oceans. L’hydrothermalisme profond et la faune associee,
Revue du Palais de la Decouverte, vol. 10, n° 97, 16 avril 1982, pp. 16—40.
Alan Fustec. Microdistribution et variations temporelles de la faune hydrothermale des sites de la zone 13° N sur la ride du Pacifique Est, These de Docteur-Ingenieur en Sciences agronomiques, specialite Halieutique, n° d’ordre 85/6, serie H, n° 2, soutenue le 5 juin 1985 a I’Ecole Nationale Superieure d’Agriculture de Rennes.
Robert R. Hessler. Quand le soufre donne la vie. La Recherche, vol. 13, n° 131, mars 1982, pp. 405—406.
Lucien Laubier. D’etranges animaux, La Recherche, vol. 13, n° 131, mars 1982, pp. 405—406.
Lucien Laubier, Daniel Desbruyeres. Les oasis au fond des oceans, La Recherche, vol. 15, n° 161, decembre 1984, pp. 1506—1517.
Lucien Laubier, Daniel Desbruyeres. Oases at the bottom of the sea, Endeavour, N. S., vol. 9, fas. 2, 1985, pp. 67—76.
Richard A. Lutz, Robert R. Hessler. Life without sunlight. Biological communities of deep-sea hydrothermal vents, The Science Teacher, vol. 50, n° 3, march 1983, pp. 22—29.
Peter A. Rona, Kurt Bostrom, Lucien Laubier, Kenneth L. Smith, Jr. (edits). Hydrothermal processes at seafloor spreading centers, NATO Conference series IV, Marine sciences, Plenum Press, 1983, pp. I—XIV, 1—796.
СЛОВАРЬ ТЕРМИНОВ
Автотрофия — способность живого существа синтезировать органическое вещество из минеральных элементов, имеющихся в среде. Зеленые растения являются растениями-автотрофами.
Анаэробы — микроорганизмы, способные развиваться при полном отсутствии кислорода. Термин «анаэробия» употребляется для определения обмена веществ, который происходит без потребления кислорода.
Аноксический — аноксическая среда — среда, полностью лишенная кислорода. Свойственна некоторым океаническим впадинам, а также большой части Черного моря.
Биомасса — количество живого вещества на единицу площади. Биомасса может быть выражена в сыром весе, но, когда это возможно, предпочтительнее использовать сухой обеззоленный вес.
Восстановление — химическая реакция, выражающаяся в отнятии кислорода от химического вещества, которое его содержит.
Гетеротрофы — живые существа, которые питаются органическими веществами. Животные и некоторые растения без ассимилирующего пигмента, например грибы, являются гетеротрофами.
Живое ископаемое — так образно называют современный вид, открытый недавно, предки которого во множестве известны в ископаемом состоянии, так что вся группа считалась исчезнувшей с лица земли. Наиболее известным примером живого ископаемого является ла-тимерия.
Изотопы — атомы одного химического элемента, различающиеся только массой ядра, а точнее числом содержащихся в нем нейтронов. Так, водород представлен тремя изотопами: легким водородом (1 протон,
1 электрон), дейтерием, или тяжелым водородом (1 протон, 1 нейтрон, 1 электрон) и тритием (1 протон, 2 нейтрона, 1 электрон). Существует множество естественных изотопов, а также искусственных радиоизотопов.
Интерстициальная вода — вода, заполняющая пространства между песчинками и другими твердыми частицами на пляжах и на дне водоемов.
Окисление — присоединение кислорода к химическому элементу или веществу.
Пелагический — относящийся к толще вод; применяется к живым существам, растениям и животным, которые живут в толще вод океана без контакта с дном; например, тунцы — пелагические рыбы.
Пиноцитоз — приспособление, позволяющее некоторым животным клеткам захватывать мелкие частицы, заключая их в пузырьки, образованные на поверхности клетки, которые затем полностью закрываются, после чего частицы перевариваются внутри клеток.
Трофический — имеющий отношение к питанию (от греческого trophe — питание).
Трофосома — слово, созданное для обозначения отдела тела больших погонофор (вестиментифер), которые содержат хемосинтезирующие бактерии; буквально значит «часть тела (сома — совокупность неполовых клеток живого существа), служащая для питания».
Фагоцитоз — приспособление, позволяющее некоторым животным клеткам захватывать более мелкие частицы, обволакивая их псевдоподиями, а затем их переваривать. Белые кровяные тельца способны фагоцитировать микробы.
Фермент — растворимое органическое вещество, вырабатываемое живыми клетками, которое вызывает или ускоряет какую-либо реакцию. Некоторые ферменты секретируются вне клеток, другие являются внутриклеточными.
Фотосинтез — синтез органических веществ (сахаров) с помощью солнечной энергии растениями, имеющими пигмент хлорофилл.
Хемосинтез — свойство некоторых микроорганизмов использовать химическую энергию, высвобожденную в ходе некоторых реакций, для синтеза органического вещества из минеральных элементов. Отличается от фо
тосинтеза тем, что в последнем используется солнечная энергия.
Хемотрофия — способ питания автотрофных хемосинтезирующих организмов (см. хемосинтез).
Эпибионт — организм, который живет на другом живом существе, как правило постоянно.
Эскарп — то же, что уступ, крутой элемент рельефа, разделяющий поверхности, лежащие на разной высоте.
ПРИЛОЖЕНИЕ 1
О БИОЛОГИЧЕСКИХ ГЛУБОКОВОДНЫХ ИССЛЕДОВАНИЯХ С ПОМОЩЬЮ подводных ОБИТАЕМЫХ АППАРАТОВ
Целый ряд исследований и открытий в морской биологии шел вслед за геологическими открытиями и явился в определенной степени их результатом. Начало такому научному тандему было положено в 1872—1876 годах на корвете «Челленджер», и с тех пор уже более ста лет оба направления уживаются в одной экспедиции. Наиболее ярко это проявилось в изучении абиссали и ультраабиссали Мирового океана, то есть глубин от 3000 до 6000 м и от 6000 м до максимальных глубин, (около 11 000 м) в глубоководных желобах. Картирование районов с такими глубинами, всестороннее изучение физико-химических характеристик заполняющих их вод, геологического строения и фауны были одними из самых романтических страниц современной океанологии. Эти исследования уже в самом своем начале опровергли предположение Э. Форбса, бытовавшее в середине прошлого века, об азойности (безжизненности) моря на глубинах более 550 м, где животные якобы так же не способны обитать, как «в пустоте или огне», и позволили В. И. Вернадскому в 1926 году характернейшей чертой биосферы назвать «всюдность жизни». Вместе с тем изучение фауны глубин Мирового океана сформировало мнение, что первичным источником пищи для поддержания жизни в них является органическое вещество, образованное при участии энергии света, то есть продукт фотосинтеза. Правда, в 1887 году С. Н. Виноградским был открыт хемосинтез — процесс образования бактериями органических веществ из двуокиси углерода за счет энергии, полученной при окислении неорганических соединений, но значение его для жизни на абиссальных глубинах не обсуждалось даже в фантастических романах. Таким образом, к середине нашего века начало складываться мнение, что все крупнейшие
открытия в океанологии уже сделаны и этот раздел наук о Земле входит в стадию стабилизации. Человечество привыкло считать, что давно изучило свою планету, и автоматически переносило это представление с суши на всю поверхность Земли, в том числе и прикрытую толщей вод Мирового океана. Научные открытия второй половины XX века не только опровергли это мнение, но показали, что изучение океана отстало от изучения суши на несколько сот лет...
В 1952—1953 годах Р. Дитц и Г. Менард открыли и закартировали в Тихом океане новые крупные структуры океанического дна, среди которых особое значение имели протяженные линейные зоны разломов. В 1956 году М. Юинг и Б. Хизен описали систему срединно-океанических хребтов, возвышающуюся часто более чем на 3000 м над уровнем ложа океана и опоясывающую весь земной шар. Общая протяженность этой горной системы около 80 000 км при ширине более 1000 км — она превосходит по масштабам Альпы и Гималаи! Они же открыли на гребне срединных хребтов узкий трог, или рифтовую долину. Эти конкретные открытия привели в 60-е годы к разработке теорий спрединга и тектоники плит, которые называют вегенеровской революцией — в честь немецкого метеоролога А. Вегенера, первым собравшего многочисленные свидетельства в пользу идеи дрейфа континентов (1913). Все это легло в основу теории новой глобальной тектоники, суть которой сводится к следующему: литосфера разбита на крупные плиты, которые перемещаются по астеносфере в горизонтальном направлении; близ срединно-океанических хребтов литосферные плиты наращиваются за счет вещества, поднимающегося из недр, и расходятся в стороны (спрединг), в глубоководных желобах одна плита поддвигается под другую и поглощается мантией (субдукция). Таким образом, Земля рассматривается как активная планета, динамичное лабильное космическое тело. Эта перестройка взглядов представляет собой настоящую научную революцию, сравнимую с коперниковской в астрономии и дарвиновской в биологии.
Вскоре после появления теории тектоники плит, объяснившей роль срединно-океанических хребтов, Дж. Элдер из Манчестерского университета сделал предположение о существовании на дне океана горячих источников и тесной связи их с подводными хребтами. Вслед за этим
Подводный обитаемый аппарат «Алвин»
с надводных кораблей обнаружили зоны повышенного теплового потока, аномалии гелия-3, метана, марганца (при помощи буксируемого аппарата «Ангус» массой в 1,5 т). Этим аппаратом, «стреляющим», как пулемет, фотокадрами в 20 м от дна, в 1976 году получены фотографии необычной фауны. Для следующего этапа исследований, проводившегося учеными США, были использованы высокоточные глубоководные системы навигации и подводный обитаемый аппарат (поа) «Алвин» с экипажем из трех человек (пилот и два наблюдателя разных специальностей) на борту. «Алвин» построен в 1964 году, его масса 16,5 т, максимальная глубина погружения— 4500 м. В 1968 году из-за аварии при спуске «Алвин» затонул на глубине 1540 м в НО милях к югу от Вудс-Хола (штат Массачусетс), затем был поднят со дна, где пролежал 11 месяцев. К моменту описываемых исследований на нем совершили 712 научных погружений. Первые 15 погружений, ориентиром для которых служили подводные фотографии «Ангуса», «Алвин» провел в районе Галапагосского рифта в феврале—марте 1977 года на глубинах порядка 2500 м. Результаты превзошли все
ожидания. Участники одного из первых погружений этой серии Дж. Корлисс и Дж. Эдмонд так описывают свои впечатления: «...мы оказались в оазисе. Рифы из мидий и целые поля гигантских двустворок, крабы, актинии и крупные розовые рыбы, казалось, купались в мерцающей воде. Оставшиеся пять часов времени пребывания на дне мы провели в состоянии, близком к помешательству». И далее: «Внутри круга диаметром около 100 м теплая вода струилась из каждой расселины, каждого отверстия в морском дне. Температура воды была различной, но максимальная составляла примерно 17 °C. Организмы оказались довольно разборчивыми — они забивались в отверстия с наиболее теплыми струями. Мы проработали до полного истощения запаса энергии, отведенного на „научные цели“». После подъема аппарата на борт судна-носителя «Лулу» водоотборники с пробами воды были переданы на сопровождающее судно «Норр». Анализы проводились всю ночь, и к утру стало ясно, что воды всех теплых источников характеризуются высоким содержанием сероводорода (H2S) и в них широко представлены хемосинтезирующие бактерии, для жизнедеятельности которых этот сероводород необходим. Было выяснено также, что ряд обитающих здесь животных питаются не экзогенно (органическим веществом внешнего происхождения), а эндогенно — органическим веществом внутреннего происхождения, получаемым ими от населяющих их тело хемосинтетических бактерий, и таким образом являются симбиотрофами.
Работы на Галапагосском рифте были продолжены в январе—марте 1978 года, а в апреле—мае и ноябре аналогичные исследования проводились в районе 21° с. ш. Восточно-Тихоокеанского поднятия. Здесь удалось обнаружить то, о чем ученые не смели и мечтать. Гигантские струи черной воды били из труб, образованных сульфидными минералами, высота этих труб достигала нескольких метров, температура воды на глубине 2600 м превышала 300 °C, кроме того, в растворах содержалось 0,02% сероводорода, тогда как в обычной морской воде он вообще отсутствует. В изобилии встречались представители фауны, аналогичной фауне Галапагосского рифта. Подобная картина наблюдалась и в бассейне Гуаймас Калифорнийского залива, где работы велись в январе 1982 года.
Французские исследователи «вступили в бой» в 1982 го
ду, имея в своем распоряжении поа «Сиана» и опыт работ ученых США, принимавших участие в их первых погружениях на 13° с. ш. Восточно-Тихоокеанского поднятия. До 1984 года этот район становится основным во французских исследованиях, и результаты его изучения изложены как оригинальный материал в этой книге, наравне с американскими данными по Галапагосскому рифту. Следует упомянуть, однако, что к 1989 году газофильные сообщества были обнаружены не менее чем на 20 участках Мирового океана, как в зонах спрединга и субдукции, так и на пассивных окраинах континентального склона. При этом среда их обитания не всегда характеризуется повышенными температурами, но в ней обязательно присутствует или сероводород, или метан.
Осенью 1985 года член-корреспондент АН СССР А. П. Лисицын организовал семинар по проблемам гидро-термали и металлогении, куда кроме геологов Института океанологии АН СССР были приглашены представители других профессий и институтов, в том числе зоологи и микробиологи. Первая советская экспедиция (11-й А рейс научно-исследовательского судна «Академик Мстислав Келдыш» под руководством Л. П. Зоненшайна) состоялась в июне—июле 1986 года в районе между островами Парамушир и Атласова, относящимися к Курильской островной дуге, где уже много лет самописцы эхолотов прорисовывали темный столб высотой около 500 м при 800-метровой глубине. Для визуальных наблюдений и отбора проб наравне с традиционными методами использовались два поа «Пайсис», находящиеся в эксплуатации у Института океанологии с 1977 года; их максимальная глубина погружения — 2000 м; экипаж — три человека (пилот, бортмеханик и наблюдатель). «Пайсисы» погружались на дно 11 раз и позволили ученым экспедиции убедиться, что факел на записи эхолота возникает над газовым, а не гидротермальным источником. Газ выходит из толщи осадков на низком (высота 2—3 м) возвышении размером всего 50X50 м2. Анализы показали, что в составе газов преобладает метан. С судна удалось геологическими трубками взять пробы грунта прямо из района факела. В колонках ила на глубине примерно 1,8 м от поверхности дна четко вырисовывались белые слои сантиметровой толщины, которые быстро с шипением разлагались на воздухе. Это были газогидраты, — твердые соединения газов с водой, сущест-
Подводный обитаемый аппарат «Пайсис»
вующие при низких температурах и высоких давлениях. К району газового источника приурочено специфическое сообщество организмов, включающее бактериальные маты, несколько видов многочисленных двустворчатых моллюсков, многощетинковых червей и других животных.
В августе—декабре 1986 года под руководством А. П. Лисицына состоялся 12-й рейс НИС «Академик Мстислав Келдыш» в северо-восточную часть Тихого океана. Были обследованы две зоны гидротермальных проявлений — гора Осевая хребта Хуан-де-Фука и бассейн Гуаймас Калифорнийского залива. Эти районы ранее визуально обследовались учеными Канады и США, но были выбраны для исследования отчасти из-за своей доступности поа «Пайсис» по глубине: дно кальдеры горы Осевой расположено на глубине около 1580 м, а бассейна Гуаймас — на 2000 м. В экспедиции проведено 42 погружения подводных аппаратов, из них 24 непосредственно в зоны гидротермальной активности. Собрана представительная коллекция фауны, проведены ланд-шафтно-биоценотические исследования, подготовлены препараты для анализа стабильных изотопов, для определения микроэлементов и др. Осуществлена широкая программа микробиологических исследований.
В декабре 1987 года приемочная комиссия АН СССР подписала акт об окончании испытаний и приемке комплекса двух поа «Мир» с глубиной погружения до 6000 м (руководитель работ И. Е. Михал ьцев, заместитель А. М. Сагалевич). Срок создания аппаратов был беспрецедентно коротким — менее двух с половиной лет от начала проектирования до завершения испытаний. Они рассчитаны на работу экипажа из трех человек (один или два пилота и один наблюдатель), максимальная скорость подводного хода 5 узлов, аппараты Оснащены всем комплексом океанологических измерительных средств (кроме гравиметрических), имеют два манипулятора с семью степенями свободы и т. д. До сих пор в мире существовало только два аппарата, способных погружаться на глубину 6000 м — это «Си Клиф», принадлежащий ВМС США (построен в 1984 году), и французский «Наутилус», построенный в 1985 году и в том же году проводивший геологические и биологические исследования на пределе абиссальных глубин в районе Японского желоба в совместной франко-японской экспедиции. Расчетная глубина 6000 м для подводного обитаемого аппарата выбирается как разумный желаемый предел, потому что недоступным для исследований остается лишь около 1% площади дна Мирового океана.
Через два месяца после приемки поа «Мир», в феврале— мае 1988 года состоялся 15-й рейс НИС «Академик Мстислав Келдыш» под руководством А. П. Лисицына. Это была первая экспедиция, в которой поа «Мир» эксплуатировались в научных целях. Районами работ избраны 26 и 23° с. ш. Срединно-Атлантического хребта, что отчасти продиктовано желанием первую же экспедицию с «Мирами» провести в зоне гидротермальной деятельности и вместе с тем минимальным переходом от порта приписки судна-носителя — Калининграда (по этой же причине глубинные океанические испытания «Мира» проводились в Центральной Атлантике). Избранные гидротермальные зоны находятся на глубине 3700 и 3500 м соответственно и в мае 1986 года визуально наблюдались учеными США. В рейсе было проведено 16 погружений подводных аппаратов, из них только одно в зону гидротермальной деятельности ,26° с. ш. Полученные биологические материалы и данные менее обильны, чем в 12-м рейсе НИС «Академик Мстислав Келдыш».
Число научных публикаций в мире по проблеме гидро-
Подводный обитаемый аппарат «Наутилус»
термалей и сходных явлений к концу 1987 года (то есть за 10 лет существования проблемы) было не менее 1600. Ознакомление с этой литературой и оригинальные данные убеждают в том, что фауна Мирового океана по происхождению потребляемого ею органического вещества может быть разделена на фотобиос и хемобиос, последний и рассматривается преимущественно в книге Л. Лобье. Складывается впечатление, что хемобиос обладает не менее чем тремя трофическими стратегиями животных: стратегия тиобиоса — симбиоз беспозвоночных с тионовыми бактериями (возможно, правильнее было бы назвать эту стратегию тионобиос, поскольку именно тионовые бактерии являются «посредниками» между сероводородом и многоклеточными животными); стратегия метанобиоса — симбиоз беспозвоночных с метанотроф-ными бактериями; стратегия метанотиобиоса — симбиоз беспозвоночных с метанотрофными и тионовыми бакте
риями одновременно (в определенной степени универсальная стратегия). Вполне вероятно, что три перечисленные стратегии не исчерпывают всех стратегий хемобиоса.
Начиная с 1985 года,ведутся интрмгивные исследования водной экосистемы в кальдере Кратерной (остров Янкича, острова Ушишир, Курильские острова), которая характеризуется проявлением современного вулканизма в виде береговой гидротермальной деятельности; здесь имеют место подводные выходы термальных вод и газов. У подножия источников и на дне найдены многослойные бактериальные маты, в состав которых входят тионовые бактерии. В 1987 году на острове построена небольшая биологическая станция для стационарных наблюдений, работы проводятся под руководством сотрудников Института биологии моря ДВО АН СССР В. Г. Тарасова и М. В. Проппа с привлечением ученых ряда других институтов АН СССР.
Исследования фауны зон спрединга и субдукции, других газофильных сообществ, симбиотрофии как способа эндогенного питания, распределения хемобиоса в Мировом океане и целого ряда смежных вопросов становятся ведущими в современной биоокеанологии. Книга Люсьена Лобье — первая книга (а не сборник статей или книга с главами, написанными разными авторами) на эту тему. Количество публикаций по этому вопросу растет как снежный ком, и в одной книге даже с рядом послесловий и приложений к ней невозможно сейчас успеть обобщить все новые открытия, ежемесячно появляющиеся в печати. В начале века книги К. Куна (1900) «Из глубин океана» и Д. Меррея и Й. Йорта (1912) «Глубины океана» были откровением для океанологов, но на смену им в наше время пришли книги Г. М. Беляева (1989) и Дж. П. Кеннета (русский перевод— 1987), во многом изменившие прежние представления. Возможно, такая судьба к середине XXI века ждет и эту книгу, но сейчас она очень актуальна и своевременна.
Л. И. Москалев, канд. биол. наук, ст. научный сотрудник Института океанологии АН СССР, участник трех экспедиций с поа.
ПРИЛОЖЕНИЕ 2
ЛАНДШАФТНЫЕ ИССЛЕДОВАНИЯ ГИДРОТЕРМАЛЕН
Подводные обитаемые аппараты открыли новую эпоху в биоокеанологии. Дело тут не только в новых методах сбора материала и постановке витальных экспериментов. Наиболее ценным для биоценологии является возможность проведения непосредственных визуальных наблюдений над глубоководными животными. Именно это позволяет исследовать донные биогеоценозы как ландшафтные комплексы, и в этом смысле можно говорить о рождении нового научного направления — глубоководного ландшафтоведения. По своим характерным признакам донные биогеоценозы, подобно наземным, объединяются в крупные ландшафтные системы (биомы). А состав и пространственная структура гидротермальных оазисов настолько характерны, что уже сейчас можно говорить о существовании особого «гидротермального» типа глубоководного донного ландшафта.
В основе ландшафтных исследований лежат визуальные наблюдения над животными, подсчеты их численности, приуроченности их друг к другу и к определенным формам рельефа’ На ранних этапах исследования производится фотографирование участков дна, сплошная или выборочная видеозапись, отбор проб для определения видов организмов, оценка абиотических факторов, на завершающем этапе — комплексное картирование того или иного участка дна. Работы такого направления проводились французскими учеными в 1982 году на Восточно-Тихоокеанском поднятии. Эти же методы лежат и в основе биогеоценотических исследований гидротерм али, начатых в нашей стране (12-й, 15-й рейсы НИС «Академик Мстислав Келдыш»).
В отличие от французских работ, в нашем случае картина ландшафта получается более обобщенной и
строится в трех-, а не в двухмерном изображении; при этом всегда исследуется не только само гидротермальное сообщество, но и так называемый фоновый ландшафт и населяющая его фоновая фауна.
Сложность ландшафтных исследований, проводимых с помощью поа, состоит в том, что исследователь может видеть одновременно лишь небольшой участок дна, а площадь, охватываемая при фотосъемке и видеозаписи и того меньше (см. цветную вкладку). Для получения целостной картины нередко приходится использовать сотни фотографий и многие часы видеозаписи, обобщать результаты целого ряда погружений. Лишь после этого производится реконструкция, отражающая наиболее характерные черты пространственной структуры того или иного сообщества. Примеры таких реконструкций, выполненных для зон гидротермальной активности, изученных в наших экспедициях, приведены на рис. 1—6. Следует иметь в виду, что некоторые животные, наблюдавшиеся из аппарата, так и не были пойманы, поэтому точное их определение невозможно. Этим объясняется, что некоторые представители фауны определены до вида, а другие — лишь до семейства, отряда и даже более высоких таксономических категорий.
Зона гидротермальной активности горы Осевой хребта Хуан-де-Фука расположена на дне впадины (кальдеры), стенка которой, высотой около 100 м, сложена крупными базальтовыми блоками, а дно залито покровными лавами разных типов (рис. 1). Фауна небогата и представлена главным образом сестонофагами и детритофагами твердого субстрата (губки, гидроиды, офиуры, морские звезды и т. п.) (фото 1 цветной вкладки). Встречаются здесь и крупные крабы — майиды. В зоне гидротермальной активности ландшафт резко меняется (рис. 2). Наиболее заметные здесь организмы — мелкие вестиментиферы из рода Ridgeia, образующие кусты вокруг трещин, сквозь которые сочится флюид. При этом на гидротермальном поле довольно четко выделяются две фаунистические ассоциации (сообщества), в одной из которых явно доминируют вестиментиферы, в другой — двустворчатые моллюски калиптогены. Фекалии (пеллеты) последних образуют на камнях легкий наилок, служащий своеобразной микросредой для обитающей здесь фауны — мелких рачков-копепод, полихет-нереид, нематод. Поселения калиптоген, как правило, были расположены на большем удалении
от источника, чем поселения вестиментифер (рис. 3)
Основная особенность гидротермальных проявлений во впадине Гуаймас Калифорнийского залива заключается в том, что это единственный из ныне известных районов, где термальные флюиды перед выходом на поверхность проходят огромный (до 500 м) слой осадка. Над поверхностью осадка в местах истечений возвышаются сульфидные постройки причудливой формы, высотой от нескольких сантиметров до сотни метров (рис. 4). Все зоны, откуда сочится флюид, густо заселены гигантскими вестиментиферами Riftia pachyptila. В их зарослях, как в джунглях, обитает множество видов сопутствующей фауны (фото 5, 6 цветной вкладки). Там, где теплый раствор сочится непосредственно через осадок, образуются плотные толстые бактериальные маты ярчайших цветов. У подножия построек, там, куда падают отмершие рифтии, часты белые крабоиды Munidopsis и крупные Neolithodes (фото 4, 7 цветной вкладки). В котловине Гуаймас также обитают калиптогены, но другого, чем на горе Осевой, вида. Так же как и там, они менее строго приурочены к местам истечения флюида, чем вестиментиферы.
Таким образом, несмотря на значительные различия в характере гидротермальных проявлений на горе Осевой и в котловине Гуаймас, в пространственной структуре сообщества наблюдается определенная закономерность. В обоих случаях можно выделить две фаунистические ассоциации, в одной из которых доминируют вестиментиферы, в другой — калиптогены. Можно предположить, что на распределении фауны сказываются кацие-то общие экологические причины. Исследования, проведенные на нашем материале учеными Института микробиологии АН СССР, подтвердили, что вестиментиферы (вернее, их симбионты-бактерии) в своем питании ориентируются почти исключительно на сульфид (сероводород), а калиптогены могут использовать метан и частично окисленные соединения серы (тиосульфат). Известно, что сероводород в морской воде быстро спонтанно окисляется, в то время как метан имеет менее локальное распространение. Очевидно, этим объясняется тот факт, что все вестиментиферы резко ограничены в своем распределении зоной непосредственного влияния флюида (не далее нескольких метров от источника), калиптогены же могут жить и на большем удалении, используя метан и тиосульфат (то есть уже частично окисленное соединение серы). В со
ответствии с распределением этих видов находится и распространение ассоциированной с ними фауны. Следует при этом учесть, что общих видов для горы Осевой и впадины Гуаймас почти нет. Сообщества, имеющие при различном видовом составе сходную экологическую структуру, называются параллельными, а составляющие их виды — видами-аналогами. Выявление таких видов и сообществ позволяет подойти к вопросу об общей типологии гидротермальных экосистем — проблеме, пока еще практически не разработанной.
Гидротермальное сообщество Срединно-Атлантического хребта в районе 26° с. ш. весьма своеобразно. Активные гидротермальные источники сосредоточены на вершине холма диаметром около 200 м и высотой порядка 50 м (рис. 5). Фоновая фауна довольно бедна как качественно, так и количественно. Здесь представлены главным образом сидячие сестонофаги твердого субстрата и немногочисленные роющие или плавающие организмы. На самом холме можно проследить довольно четкую фаунистическую зональность (рис. 6). Вершину холма, сложенную свежими неокисленными сульфидами, где наблюдается активная гидротермальная деятельность (муар, теплые черные дымы, курильщики), занимает сообщество с резким доминированием ракообразных (прежде всего креветок Rimicaris). Ниже по склону, где нет действующих источников и преобладают рыжие окисленные сульфиды и охры, наиболее обильны крупные белые актинии и крабоиды Munidopsis. Отмечены здесь и некоторые другие животные; к сожалению, ограниченность материала не позволила провести их точную идентификацию. Можно лишь констатировать, что кольцо актиний и мунидопси-сов в определенном смысле соответствует кольцу плотоядных и сестонофагов (крабы, рыбы, морские звезды и т. п.) вокруг гидротермальных полей горы Осевой и котловины Гуаймас. Наличие такого кольца на периферии зон гидротермальной активности можно, по-видимому, считать третьей общей закономерностью пространственной структуры сообществ гидротермали.
Таким образом, исследования пространственной структуры гидротермали, проводимые методом ландшафтной реконструкции, могут дать интересные результаты и выявить некоторые принципиальные закономерности, которые было бы трудно уловить другим способом. Разумеется, эта методика, как и все методы, связанные
с применением подводных аппаратов, находится пока в стадии разработки и не унифицирована, что затрудняет интерпретацию результатов, полученных в разных экспедициях. Однако не подлежит сомнению, что это направление сыграет важную роль в биогеоценотических исследованиях глубоководной гидротермали.
С. В. Галкин,
канд. биол. наук, научный сотрудник Института океанологии АН СССР, участник трех экспедиций с поа
Рис. 1. Северная стенка кальдеры горы Осевая хребта Хуан-де-Фука. Фоновая фауна.
1 — Hyalospongia: Rhabdocalyptus heteraster (non Bathydorus sp.); 2 — Hyalospongia: Staurocalyptus iuca Tabachnik, 1989; 3 — Coelenterata: Hydrozoa; 4 — Polychaeta: Polynoidae; 5 — Decapoda: Brachyura: Majidae: Macroregonia macrochira; 6 — Asteroidea: Astropectinidae; 7 — Asteroidea; fam. sp.; 8 — Asteroidea: Brisingidae; 9 — Ophiuroidea: Ophiura sp.; 10 — Holothurioidea: Stichopodidae; 11 — Pisces: Macrouridae.
Рис. 2. Дно кальдеры горы Осевая хребта Хуан-де-Фука (глубина 1570 м). Распределение фауны на гидротермальном поле (/—5: ассоциация вестиментифер, 6—9: ассоциация калиптоген).
/ — Gastropoda: Lepetodrilacea: Lepetodrilus fucensis; 2 — Polychaeta: Polynoidae: Lepidonotopodiuin piscesae Pettibone, 1987 (= L. petlibonae Detinova, 1988); 3 — Gastropoda: Pe Itospiridae: Depressigyra globulus; 4— Pantopoda: Aminotheidae: Ammothea verenae Child, 1987
(= Scipiolus thermophilus Turpaeva, 1988); 5 — Polychaeta: Alvinellidae: Paralvinella pairniiormis; 6 — Copepoda: Cyclopoida; 7 — Bivalvia: Vesicomyidae: Calyptogena sp.; 8 — Polychaeta: Nereidae: Nereis sp. nov.; 9 — Nematoda.
Рис. 3. Гора Осевая хребта Хуан-де-Фука. Фаунистические ассоциации зоны гидротермальной активности. Слева — ассоциация калиптоген, справа — вестиментифер. Стрелки — место истечения флюида. В дополнение к набору фауны на рис. 2 в ассоциации калиптоген изображена креветка (точное определение не проведено), а в ассоциации вестиментифер — полихета из семейства Ampharetidae — Amphisa-mytha galapagensis (на трубках вестиментифер и вдоль щелей в базальте). Трубки вестиментифер и край трещины в базальте покрыты бактериальным обрастанием.
Рис. 4. Дно котловины Гуаймас Калифорнийского залива (глубина 2000 м). Распределение фауны в зоне гидротермальных построек.
/— Vestimentifera: Riftiidae: Riftia pachyptila; 2— Decapoda: Anomura: Galatheidae: Mumdopsis alvisca; 3 — Coelenterata: Anthozoa. Actiinaria, 4— Polychaeta: Alvinellidae: Paralvinella grasslei; 5 — Decapoda: Anomura: Lithodidae: Neolithodes diomedeae; 6— Polychaela: Sedeiitaria. lain sp (в илистых трубках); 7 — Gastropoda: Littorinoidea: Provanna goniata; 8—Asteroidea: Brisingidae; 9 — Bivalvia: Vesicomyidae: Calxptogena sp , Ю дым черного курильщика; // — сочение флюида; 12—поступление флюида; 13 — диффузное просачивание флюида; 14 — iидрогсрмальные отложения; /5 — бактериальные маты; 16 — алевритово-пелитовый ил
чганртпчпг
Рис. 5. Восточный склон и рифтовая долина в районе Трансатлантического геотраверза, 26° с. ш. Срединно-Атлантического хребта. Стрелкой показано местоположение гидротермального холма. Основные представители фоновой фауны: эпифауна базальтов (/—//), обитатели мягкого осадка (12—18), активно подвижные формы (19—21).
/. 2 — Hyaiospongia: Pheronematidae: Poliopogon sp.; 3 — Hyaiospongia: Hyalonematidae: Hyalonema sp.; 4 — Coelenterata: Hydrozoa; 5 — Coelenterata: Anthozoa: Actiniaria (белые); 6 — Coelenterata: Anthozoa: Gorgonaria (бичевидная); 7 — Coelenterata: Anthozoa: Gorgonaria (канделябровидная); 8 — Polychaeta: Spiomdae; 9 — Crinoidea; 10 — Asteroidea: Brisingidae; // — Ophiu-roidea; 12 — Coelenterata: Anthozoa: Ac-liniaria (фиолетовые); 13 — Decapoda: Macrura: Natantia; 14 — Hololhunoidea; 15—18 — следы жизнедеятельности животных; 19 — Decapoda: Anomura: Galat-heidae: Munidopsis sp.; 20—Holothurioi-dea (плавающая): Pelagothuriidae; 21 — Pisces: Macrouridae: Coryphaenoides ar-matns.
I la EZ2s EZd,? UiE> E
Рис. 6. Склон активного гидротермального холма в районе Трансатлантического геотраверза (глубина 3660— 3700 м).
I — внешний склон; II — промежуточная площадка; III — внутренний склон; IV — верхняя площадка (активные источники сосредоточены в зонах III и IV); 1—7: основные представители фауны периферии (/—5) и вершины холма (6—7); 8—12: типы субстратов, 13—15: гидротермальные проявления;
I — Decapoda: Anomura: Galatheidae: Mumdop-sis sp.; 2 — Coelenterata: Anthozoa- Actima-ria; 3 — Polychaeta: Ampharetidae; 4 — яйцевая капсула неизвестного беспозвоночного животного; 5 — Copepoda: Calanoida; 6 — Decapoda: Brachyura: Bythograeidae: Segonzacia mesatlan-tica; 7 — Decapoda: Macrura: Natantia: Bresil-udae: Rimicaris exoculata; 8 — алевритово-пелитовый ил; 9 — рыхлые гидротермальные отложения (охры); 10—обломки и блоки окисленных сульфидов; 11— кварцевый песок; 12 — блоки неокисленных сульфидов и активные постройки; 13— холодные истечения (муар); 14 — теплые «черные дымы»; 15 — высокотемпературный черный курильщик.
ОГЛАВЛЕНИЕ
К советскому читателю.......................... 3
Предисловие. Ж. М. Перес....................... 5
Введение....................................... 9
1. Геологические и геохимические условия .... 14
2. Открытие глубоководных оазисов.................27
3. Продукция бактерий и термофильные бактерии . 50
4. Почти совершенное сообщество...................61
5. Более скромные сообщества......................72
6. Хищники, комменсалы, паразиты и оппортунисты . 90
7. Рост, размножение и экологическая стратегия . 103
8. Происхождение гидротермальной фауны и живые ископаемые......................................НО
Заключение. ..................................119
Послесловие...................................122
Библиография................................ 131
Словарь терминов ............................ 133
Приложение 1. О биологических глубоководных исследованиях с помощью подводных обитаемых аппаратов. Л. И. Москалев.....................136
Приложение 2. Ландшафтные исследования гид-ротермалей. С. В. Галкин......................145
Л 68 Лобье Л. Оазисы на дне океана. Перевод с франц, под ред. д-ра биол. наук К- Н. Несиса. Л., Гидрометеоиздат, 1990.—156 стр.
Книга известного французского гидробиолога Люсьена Лобье рассказывает об одном из наиболее значительных открытий XX в. в области биологии — о богатейшей фауне горячих и холодных минерализованных источников на дне Тихого и Атлантического океанов. Обилие животных в подобных сообществах базируется на бактериальном хемосинтезе, в отличие от всех других сообществ, где в начале пищевой цепи лежит фотосинтез, а следовательно, и солнечная энергия. Представители этого «мира без солнца» тем не менее поражают как разнообразием форм, так и очень высокими биомассами. Автор подробно рассматривает все аспекты этой уникальной экосистемы нашей планеты.
Книга рассчитана на широкий круг читателей, интересующихся проблемами биологии.
1903040100-037
Л 069(02)—90 55 90
ББК 28.082
Научно-популярное издание
ЛЮСЬЕН ЛОБЬЕ
ОАЗИСЫ НА ДНЕ ОКЕАНА
Редактор М. Г. Тараканова. Художник И. Г. Архипов. Художественный редактор Б. А. Бураков. Технический редактор Е. А. Ступненкова. Корректор Т. В. Алексеева ИБ № 1972
Сдано в набор 9.10.89. Подписано в печать 3.04.90. Формат 84Х 1О8’/з2- Бумага тип. № 2 и офсет. Печать офсетная. Гарнитура литературная. Усл. печ. л. 8.82 с вкл. Усл. кр.-отт. 10,5. Уч.-изд. л. 8,85. Тираж 118 000 экз. Индекс ПЛ-88. Заказ № 5243. Цена 65 коп. Гидрометеоиздат. 199226. Ленинград, ул. Беринга, 38.
443086, г. Куйбышев, проспект Карла Маркса, 201. Ордена Трудового Красного Знамени тип. издательства Куйбышевского обкома КПСС.
65 коп.
ЛЮСЬЕН ЛОБЬЕ
ОАЗИСЫ НА ДНЕ ОКЕАНА
Люсьен Лобье — профессор Океанографического института в Париже, ведущий научный сотрудник Французского Института исследования морей (IFREAAERL— по образованию биолог. Ему 54 года, 33 из которых он работает как биоокеанолог в морских исследовательских учреждениях Франции. Он счастливо сочетает высокие профессиональные качества зоолога (он — известный специалист по многощетинковым червям), подводника (работал в подводных обитаемых аппаратах) и администратора. Во Франции исследование морской фауны в подводных обитаемых аппаратах было начато и развивалось Жаном-Мари Пересом — Л. Лобье стал его преемником- В последние 15 лет исследования с помощью подводных обитаемых аппаратов приобретают широкий размах. Появилась плеяда ученых, обладающих рядом личностных качеств, позволивших им стать лидерами в столь сложных исследованиях. Люсьен Лобье занимает достойное место среди них.
ISBN 5-286-00329-Х (СССР)
ISBN 2-268-00464-3 (Франция)
в
ГИДРОМЕТЕОИЗДАТ