/




























































































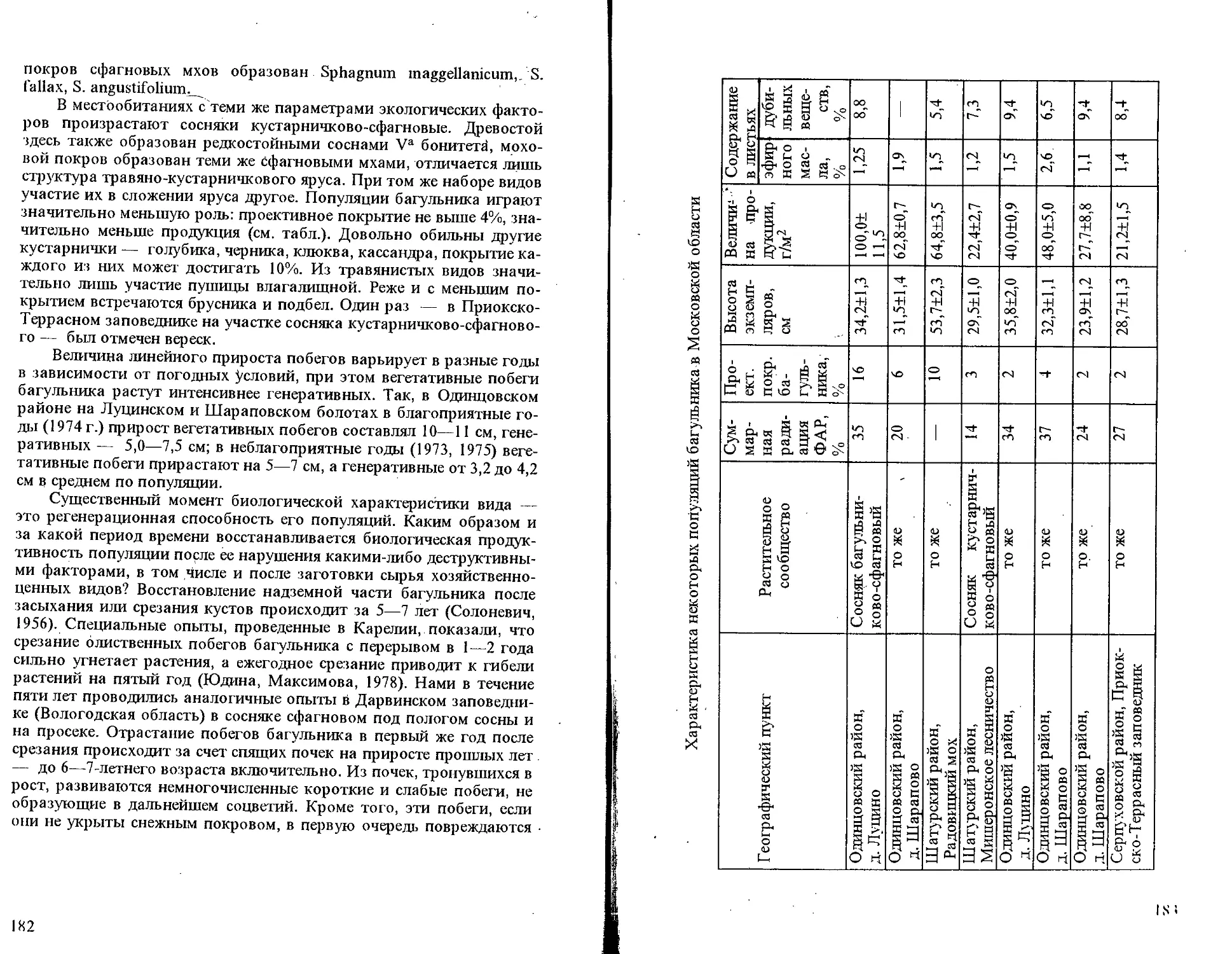













Автор: Тихомиров В.Н. Павлов В.Н.
Теги: география растений фитогеография флора географическое распространение растений фауна
ISBN: 5-211-02853
Год: 1995
Текст
БИОЛОГИЧЕСКАЯ ФЛОРА
МОСКОВСКОЙ ОБЛАСТИ
Москва
1995
БИОЛОГИЧЕСКАЯ ФЛОРА
МОСКОВСКОЙ
ОБЛАСТИ
Под редакцией
профессора В.Н. Павлова,
члена-корреспондента РАН В.Н. Тихомирова
Выпуск десятый
Издательство Московского университета
Издательство «Аргус»
1995
УДК 581.9 (471.311)
Биологическая флора Московской области. Вып. 10. Под
ред. В.Н.Павлова, В. Н. Тихомирова. — М.: Изд-во МГУ; Изд-
во «Аргус», 1995. — 208 с.
ISBN 5-211-02853
В книге обобщены итоги оригинальных наблюдений, экспери-
ментальных работ и литературные данные по систематике, географи-
ческому распространению, морфологии, онтогенезу, экологии, фито-
ценологии, консортивным связям, хозяйственному значению и охране
видов растений, произрастающих в Московской области. Охаракте-
ризованы как широко распространенные (вейник наземный, чистяк
весенний, бересклет бородавчатый, виды родов Манжетка и Герань),
в том числе водные (сем. рясковых) и лекарственные (багульник бо-
лотный) растения, так и редкие виды растений (виды тайника и ят-
рышник шлемоносный).
Для студентов, аспирантов, преподавателей высших учебных за-
ведений биологического профиля, широкого круга специалисгов-био-
логов, работников сельского и лесного хозяйства, деятелей охраны
природы.
Редакторы выпуска М.Г. Вахрамеева, Н.Г. Уланова
Рецензенты: доктор биологических наук А.П. Меликян,
кандидат биологических наук В.В. Экзерцева
Печатается по постановлению
Редакционно-издательского Совета
Московского университета
Biological Flora of the Moscow Region. Part 10.
Chief editors - Prof. V.'N. Pavlov, Prof. V.N. Tikhomirov
Edited by M.G. Vakhrameeva, N.G. Ulanova
Издание осуществлено при поддержке Российского фонда
фундаментальных исследований по проекту № 95-04-286506.
077(02) - 95 — заказное
1995
ISBN 5-211-02853
© Коллектив авторов,
ПРЕДИСЛОВИЕ
Начиная с 1974 г., в 9 выпусках сборника «Биологическая флора
Московской области», было охарактеризовано свыше 130 видов рас-
тений. В настоящем выпуске, как и в прошлых, принимали участие
сотрудники ряда вузов и научных учреждений: Московского
государственного университета, Московского государственного
педагогического университета, Тверского государственного универ-
ситета, Института лекарственных растений и других. Растения опи-
саны в градационном для настоящего издания плане: морфология,
географическое распространение, онтогенез, сезонное развитие, спо-
собы размножения и распространения, экология, фитоценология, хо-
зяйственное значение. Во всех статьях раздел «Номенклатура» напи-
сан чл.-корр. РАН В.Н. Тихомировым.
Экологическая характеристика видов так же, как и в предыду-
щих выпусках, дана с использованием экологических шкал Л.Г.
Раменского (Раменский Л.Г. др. Экологическая оценка кормовых
угодий по растительному покрову. М... 1956), Г. Элленберга (Н.
Ellenberg. Zeigerwerte der Gefasspflanzen Mitteleuropas. In: Scripta
Geobotanica. 1974. Bd. 9) и Э. Ландольта (E. Landolt. Okologische
Zeigerwerte zur Schweizer Flora. In: VerofT. Geobot. Inst. ETH., 1977. H.
64). Краткий перевод последних двух шкал опубликован в 6-м выпус-
ке «Биологической флоры Московской области» (1980). В тексте при-
ведены только фамилии авторов экологических шкал без ссылки на
источники.
Основную работу по комплектованию выпуска и его редактиро-
ванию осуществили сотрудники кафедры геоботаники МГУ проф.
В.Н. Павлов, ст.н.с. М.Г. Вахрамеева, доц. Ю.Е. Алексеев, а также
ст.н.с. А.А. Маслов. Существенную помощь в подготовке рукописи к
печати оказали научные сотрудники кафедры Н.Г. Уланова, Т.В. Ба-
гдасарова и Т.П. Баландина.
3
Н.Г. Уланова
ВЕЙНИК НАЗЕМНЫЙ
Номенклатура, систематическое положение, внутривидовая
систематика
Calamagrostis epigeios (L.) Roth., 1788, Tent. Fl. Germ. 1:34; Кауф-
ман, 1866, Моск, фл., изд. 1: 556; Цингер, 1886, Сборн. свед.: 487;
Шмальгаузен, 1897, Фл. Сред, и Южн. Росс. 2: 609; Сырейщиков,
1906, Илл. фл. Моск. губ. 1: 123; он же, 1914, пит. сОч. 4: 14; он же,
1927, Опред. раст. Моск, губ.: 61; Рожевиц, 1934, Фл. СССР, 2: 194;
он же, 1940, в Маевский, Фл., изд. 7: 114; Цвелев, 1954, в Маевский,
Фл., изд. 8: 833; он же, цит. соч., 1964, изд. 9: 795; Скворцов, 1966, Оп-
ред. расг. Моск, обл.: 75; Цвелев, 1974, Фл. Евр. части СССР, 1: 222;
Скворцов, 1966, Опр. раст. Моск, обл.: 75; Цвелев, 1974, Фл. Европ.
части СССР, 1: 222; Clarke, 1980, Fl. Europ. 5: 237; Черепанов, 1981,
Сосуд, раст. СССР: 340; Е. Алексеев, 1986, Опред. раст. Мещеры, 1:
83. — Arundo epigeios L. 1753, Sp. Pl.: 81 — Вейник наземный (сем.
Poaceae — злаковые).'
Вейник наземный — полиморфный вид. Н.Н. Цвелев (1976) де-
лит видна 5 подвидов, из них 4 распространены в европейской части
России. ,
1. С. epigeios subsp. glomerata (Boiss. et Buhse) Tzvel. 1965, l.c.: 41
— В. скученноколосковый. Отличается очень густыми метелками с
тесно скученными колосками. В европейской части встречается на
Южном Урале, в Средне-Днепровском, Волжско-Донском, Заволж-
ском районах и далее во всех более южных районах. Обитает на су-
хих лугах, в степях, среди кустарников, на лесных полянах, у дорог,
на приречных Песках и галечниках, в горах не поднимается выше
среднего пояса.2п = 28.
2. С. epigeios subs, meinshausenii Tzvel. 1965, l.c.: 41 — В. Мейн-
схаузена. Отличается отсутствием шероховатости на стволе под ме-
телкой и на листьях. В европейской части распространен в Карелии,
Прибалтике, в Ладожско-Ильменском районе и по Сев. Двине, Печо-
ре, Вычегде. 2п = 28; по данным А.П. Соколовской и Н.С. Пробато-
вой (1977) 2п = 56.
3. С. epigeios subsp. epigeios — Arundo epigeios L. 1753, l.c.: 81 —
Calamagrostis epigeios (L.) Roth., 1788, l.c. — 1 . наземный. Отличается
шероховатостью стебля под метелкой, метел си довольно густые, но
колоски менее скучены, чем у других подвидов. 2п = 28, 42, 56. В ев-
ропейской части встречает, >, зсеместно, т роме Северного Урала,
обычен в лесной и лесосге "ах.
4. С. epigeios subs, п acidlepis ( tzvel. 1974, l.c.: 154 — В.
крупночешуйный.жОтлич ’ ся до вольв крупными колосками (7,5—9
4
мм длиной) и длинными, узкими метелками. В европейской части
распространен на приречных и приморских песках, каменистых
склонах и осыпях в Причерноморье, на юге Нижне-Волжского рай-
она. 2п = ?
Внутри вида выделяются разновидности (Маевский, 1954):
1. Var. macra Trin. — соцветие густое, продолговатое, колоски 5 мм
длиной, зеленые, нижняя цветковая чешуя 2,5 мм длиной, волоски
при основании цветка 3—3,5 мм длиной, ость выходит из середины
чешуи или чуть выше ее; 2. Var. variegata Schur. — то же, но колоски
с узкой черно-фиолетовой каймой у колосковых чешуй, реже сплошь
фиолетовые, 6—7 мм длиной. 3. Var. glauca Blytt. —влагалища ниж-
них листьев покрыты вниз направленными волосками, нижняя цвет-
ковая чешуя с глубоким доходящим до основания ости надрезом,
ость сидит около середины чешуи. 4. Var. tenuiflora Trin. — соцветие
густое, узкое, почти линейное, колоски бледные, 5 мм длиной, ость
прямая, выходит около середины нижней цветковой чешуи, несколь-
ко превышая ее. Влагалища нижних листьев голые.
В роде Calamagrostis особенно широко представлены межвидо-
вые, часто межсекционные, гибриды, и большинство видов этого ро-
да могут скрещиваться со всеми другими растущими в этой же мест-
ности видами, независимо от степени родства между ними (Цвелев,
1965, 1976). В то же время благодаря интенсивному вегетативному
размножению даже стерильные гибриды могут образовывать боль-
шие заросли, долго существовать и производить впечатление само-
стоятельных видов.
Наиболее часто встречаются гибриды:
1. С. arundinacea s.l. х epigeios s.l. = С. х acutiflora (Schrad.) Reichb.
1830, Fl. Germ. Excurs.: 26; Griseb. 1852, l.c.: 427; Рожев., 1934, цит.
соч.: 223. — Arundo acutiflora Schrad. 1806, l.c.: 217. — Calamagrostis
trimi Rupr. 1845, l.c.: 36. — Exc. Kneuck. Gram. Exs. n 629; H.F.R. n
2198, a, b — описан из Средн. Европы. Один из наиболее часто
встречающихся гибридов. — 2п = ?
2. С. neglecta s.l. х epigeios s.l. = С. strigosa (Wahl.) Hartm., 1820, l.c.:
46; Griseb. 1852, l.c.: 428, p.p.; Цвел., 1965, цит. соч.: 49. — Arundo
strigosa Wahl., 1812, l.c.: 29 — Описан из Швеции, возможный изотип
в Санкт-Петербурге — 2п = ?
3. С. canescens х epigeios s.l. = С. х rigens Lindgr. 1843, Bot. Not.
(Lund.). 1843: 4; Цвел. 1965, цит. соч.: 48. — С. х neumaniana Torges,
1902, Mitt. Tharing Bot. Ver., N.F. 17: 93. — Описан из Швеции. — 2n
- у
Географическое распространение
Вейник наземный распространен почти по всей внетропической
Евразии, кроме Арктики и северо-восточной Сибири. Вид — евроа-
5
зиатский бореальный. Как заносное растение отмечен во многих
других странах (Флора СССР, 1934).
На территории бывшего СССР ареал вида охватывает почти
всю лесную, лесостепную, полупустынную и пустынную зоны. В
Большеземельской тундре обнаружены отдельные реликтовые место-
нахождения. В горах Кавказа и Средней Азии вейник не поднимает-
ся выше среднего горного пояса (Цвелев, 1965, 1976).
Морфологическое описание
Многолетнее травянистое поликарпическое длиннокорневищное
растение. Взрослое растение состоит из многих разновозрастных ро-
зеточных, полициклических интравагинальных и экстравагинальных
побегов. Вейник наземный имеет ди- и полициклические побеги с хо-
рошо выраженной многоузловой розеткой и малоузловой соломиной
с резко выраженной акротонией. Удлиненных вегетативных побегов
не бывает (Серебрякова, 1971). Побеги — дициклические, четвертого
года зацветания, первый и второй год существуют в виде почки, на
третий год — в виде розетки, а на четвер тый год — в виде цветонос-
ного побега. Наблюдается изменчивость циклов развития монокар-
пических побегов в пределах одного растения.
Корневище длинное, прочное, не ветвится, белого, соломенного
цвета, не имеет на своей поверхности почек и чешуй, толщиной 1—2
мм. Молодые корневища — сочные и большего диаметра (2,0—2,5
мм). Новые кусты образуются только из конечных почек, загибаю-
щихся вверх на конце корневища. В месте изгиба образуется новый
узел кущения, при этом часть новых почек развивается в надземные
побеги, часть — в новые корневища. Корневища растут в горизон-
тальном направлении, обычно на глубине 10—20 см.
Придаточные корни располагаются исключительно в зонах ку-
щения. Глубина проникновения корневой системы зависит от почвы
и режима увлажнения. Так, на подзолистых почвах она проникает не
глубже 15—40 см (Печникова, 1931; Тольский, 1922). На бедных пес-
чаных почвах развивается мощная вертикальная корневая система,
проникающая на глубину до 2,5 м в горизонте В (Глухов, Федякова,
1938; Патраболова, 1953; Саутин, 1957), часто достигая уровня грун-
товых вод.
Все растение сизое или серо-зеленое. Генеративные побеги име-
ют высоту 80—150 см, прямые, крепкие, с двумя-пятью узлами. Сте-
бель под соцветием шероховатый от густо покрывающих его мелких
щетинок. В кусте чаще всего 2—4 вегетативных розетки, редко их
число достигает 10. При наличии генеративных побегов в кусте, их
бывает 1—3, и несколько вегетативных розеток.
Листья серо-зеленые 2—12 мм шириной, плоские или свернутые,
сверху шероховатые, жесткие, с отчетливо выраженной ребристо-
стью. Влагалища листьев длинные, остро-шероховатые. Обычно 3—
5 длинных влагалища листьев у вегетативных розеток образуют как
6
бы ложный стебель высотой до 50 см. Листовая пластинка суживает-
ся с середины к основанию. Язычок листьев до 12 мм длиной, тупой,
часто разорванный. Клетки эпидермиса листьев с утолщенными на-
ружными стенками, несут толстостенные, вздутые в основаниях ту-
пые шипики. Особый интерес представляет опушенность влагалища,
характерная для var. glauca. Вероятнее всего, это признак клонально-
го полиморфизма. П.А. Смирнов (1958) и К.В. Зворыкина (1983) от-
мечают приуроченность этой разновидности вида к более влажным
лесным экотопам.
Метелки до 20 (30) см длиной, 1,5—6 см шириной, довольно гус-
тые, сжатые, с толстыми вверх торчащими ветвями, на вершине за-
остренные. Нижние ветви могут быть длинными, до 10 см. Ветви ме-
телки усажены мелкими шипиками. ' .
Колоски линейно-ланцетные, собраны густыми пучками (часто
однобокими), 4,5—7 мм длиной, состоят из 1 цветка, зеленые, иногда
е фиолетовым или пурпурным оттенком. На освещенных местах ко-
лоски более темные. Зачаток второго цветка имеет вид бугорка или
плохо выраженного отростка, не покрытого волосками, и по длине
не превышает основания развит ого цветка.
Колосковые чешуи шиловидно-ланцетные, заостренные наподо-
бие короткой ости, верхняя чешуя не более чем на 1 мм короче ниж-
ней, на спинке острошероховатая. Нижняя цветковая чешуя 2,7—3,5
мм длиной, пленчатая, часто на верхушке с двумя зубцами, с тремя
жилками, центральная жилка переходит в тонкую ость, отходящую
от середины или немного выше, при этом ость длиннее чешуи. Верх-
няя цветковая чешуя вдвое короче нижней, чаще без ости. На каллусе
расположен венчик из длинных белых простых шелковистых волос-
ков, длиннее или равных колосковой чешуе. Цветки — обоеполые во
всех колосках. В цветке 3 тычинки, 1 пестик с двумя перистыми
рыльцами.
Зерновка — продолговатая, светло-коричневая, часто полупро-
зрачная, снизу по жилке слегка выпуклая или почти ровная. Отпада-
ет вместе с цветковыми чешуями. Ребра на верхней стороне закруг-
ленные или с утолщенной вершинкой; киль полукруглый, располо-
жен не точно посредине плода, иногда киля нет. Зерновка вместе с
цветковыми чешуями и волосками па каллусе очень легкая, хорошо
разносится ве'п>ом на большие расстояния. Волоски в сухую погоду
раздвигаются в стороны, образуя «параппотик».
Вид довольно полиморфен по числу хромосом. Это, возможно,
связано с гибридизацией с другими видами этого рода. Наиболее
часто указывается 2п - 28 (Gadella, Kliphuis, 1967), 2n = 56 (Polya,
1949; Skalinska et.al., 1957; Сорокин, 1990), 2n = 28, 42, 56 (Цвелев,
1976; Nygrcn, 1946; Jones, 1957), реже 2n = 70 (Абдулов, 1931;
Rohwedcr, 1937).
7
Онтогенез
Для прорастания зерновок вейника требуются довольно высокие
температуры. В лабораторных условиях оптимальная температура
прорастания — 17—30°С. Время прорастания при 5—7° составляет
39 дней, при 7—15° — 24 дня, а при 17—30° — 8 дней (Бельков, 1957).
Последний срок подтверждают и наши данные. Всхожесть семян вей-
ника за 8 дней составляет 20—50% (Печникова, 1931), при этом в ла-
бораторных условиях добиться прорастания удалось только в песке
и почве при переменной температуре. После зимнего проморажива-
ния энергичность прорастания возрастает при t=16—18°; особенно
энергичное прорастание наблюдалось в мае в природных условиях
при достаточной влажности почвы (Бельков, 1957). Всхожесть семян
в лабораторных условиях составляет 73% (Бельков, 1957). По нашим
данным (север Московской области) всхожесть семян составляет 50—
80% и в среднем близка к 70%. Хранение семян в течение 3 лет не
оказало существенного влияния на их всхожесть (Печникова, 1931).
В природных условиях зерновки способны прорастать сразу после
созревания, Что и наблюдалось на вырубках в Ленинградской облас-
ти (Бельков, 1957).
Развитие проростков на фильтровальной бумаге при t=18—22°,
по нашим данным, идет очень быстро. На второй день у проростков
уже сформирован первичный корешок и влагалищный лист, который
пробивает цветковые чешуи. На третий день — заметен вертикально
стоящий линейный первый лист. На 5—7-й день развития (после по-
садки в землю) начинается формирование горизонтальных прида-
точных корней и узла кущения, расположенного чуть выше зернов-
ки. Первые 1—2 боковых побега образуются экстравагинально в те-
чение 1 месяца. И.Г. Патраболова (1953) указывает на интраваги-
нальное происхождение боковых побегов на кусте первого года. Да-
лее из нижних почек в зоне! кущения начинают формироваться пла-
гиотропные корневища. Достигнув некоторой длины, они загибают-
ся вверх и выходят на дневную поверхность. Перед этим на корневи-
ще образуются корни, закрепляя в почве растение; одновременно об-
разуется новая зона кущения. Период образования корней и загиба-
ния корневища кверху является началом формирования побега у до-
чернего куста (Юнаш, 1953).
Надземный побег имеет несколько низовых листьев и 2—3 сре-
динных влагалищных листа. Листья довольно жесткие и закрывают,
как футляр, более глубоко расположенные растущие части побега и
служат молодому побегу защитой от высыхания и перегрева (Патра-
болова, 1953). На второй месяц развития у таких дочерних кустов
первого поколения в нижней части зоны кущения бывают уже сфор-
мированы новые открытые почки, которые начинают плагиотропно
расти, образуя корневища. В течение первого вегетационного сезона
может быть сформирована особь, состоящая из 1 материнского кус-
8
та (1—3 побега), 5—10 кустов первого поколения (1—4 побега) и 5—
10 кустов второго поколения. Куст второго поколения представляет
собой едтничный побег с несколькими открытыми почками в зоне
кущения или уже начавшими расти корневищами. Таким образом, к
осени первого года существования особь может занять участок до 1
м2.
Все почки по внутреннему строению одинаковы и определить за-
ранее, из какой почки в дальнейшем разовьется корневище, а из ка-
кой — надземный побег, не представляется возможным (Патраболо-
ва, 1953). Почки сохраняют способность к развитию в побег до пол-
ного отмирания материнского побега (3—5 лет). Однако обычно они
существуют 2 года, постепенно развиваясь в открытую почку (Сереб-
рякова, 1971). Почки способны трогаться в рост без заметного пе-
риода покоя (Патраболова, 1953; Серебрякова, 1971). Поэтому ин-
тенсивное развитие корневищ и побегов начинается сразу после от-
таивания почвы весной в мае и оканчивается в октябре при пониже-
нии температуры почвы. Обычно за вегетационный периоде сомкну-
тых травостоях вейник наземный образует одно поколение корне-
вищ (Патраболова, 1953; Любарский, 1964). Однако из-за того, что
приросты корневищ могут быть и не ежегодные, определить относи-
тельный возраст куста не представляется возможным.
Большой жизненный цикл вейника наземного протекает с усиле-
нием побегов последовательных порядков (Серебрякова, 1971); глав-
ная ось и побеги развиваются, но не проходят полного цикла разви-
тия, они меньше и слабее побегов последующего порядка. Только по-
беги в кустах третьего порядка могут пройти полный цикл развития.
Все побеги — ди- и полициклические, четвертого года зацветания
(Серебрякова, 1971). В благоприятных условиях вейник наземный, по
нашим данным, может цвести на второй год, хотя обычно цветение
наблюдается на третий год жизни особи (системы парциальных кус-
тов), причем цветут кусты 3—4 поколения.
После цветения у вейника отмирают все длинные междоузлия
цветоносного стебля, а из числа коротких — только более высоко
расположенные. Нижние же междоузлия остаются живыми и, про-
должая существовать в составе куста, являются резервом почек во-
зобновления. На вырубках в Южном Предуралье главная ось куста
зацветает на 3-й год его жизни, после чего куст может существовать
еще 1—2 года за счет развития боковых побегов и затем полностью
отмирает (Патраболова, 1953). На вырубках в Тверской и Москов-
ской областях нами обнаружены кусты, существующие 7—8 лет, ко-
гда боковые побеги даже 2-го порядка проходят полный цикл разви-
тия. Обычно куст существует 3—4 года. В среднем число побегов со-
ставляет в одном кусте 2—3, но может доходить и до 20. Число кор-
невищ, отходящих от куста в среднем 2, но может варьировать от 1
до 10. В различных типах сосняков Татарии среднее число побегов в
кусте составляет от 1,2 до 3,0, а число живых кустов особи — 2—9
9
(Любарский, 1964). Среднее число стеблей вейника на 1 м2 на пусты-
рях в Южном Предуралье составляет 16—71 (Юнаш, 1953).
Как показал наш анализ динамики популяций на вейниковых
вырубках Тверской области, численность кустов достигает макси-
мальной величины на третий год (180 на м2). В этот период происхо-
дит смыкание особей, и образуется сплошной покров травостоя вы-
сотой до 1,5 метров. Одновременно наблюдается массовый переход
кустов в генеративное состояние. Интенсивность вегетативного раз-
множения падает, сокращается число почек возобновления. На чет-
вертый год число генеративных кустов равно числу виргинильных
или в 2 раза меньше. Численность кустов уменьшается до 110 на 1 м2,
новых кустов появляется мало. На вырубках старше 5 лет вейник ос-
тается только на участках, свободных от конкурентов — подроста
деревьев и малины. Численность генеративных кустов вновь стано-
вится в 2—6 раз меньше, чем виргинильных. При этом увеличивается
доля отмирающих кустов. На старых вырубках 11—13 лет на освет-
ленных участках наблюдается увеличение численности генеративных
кустов, популяция как бы «выцветает»: происходит более быстрое
прохождение побегами малого жизненного цикла. Так, побег зацве-
тает уже на первый год жизни, при этом резко сокращается число
молодых вегетативных розеток, преобладают субсенильные и се-
нильные кусты.
В березняках, ельниках и сосняках на месте вырубок популяции
вейника наземного сохраняются в угнетенном состоянии. Велика
численность виргинильных, субсенильных и сенильных кустов при
незначительном участии генеративных.
На лугах лесостепи отмечена динамичность возрастного состоя-
ния популяции вейника наземного (Куркин, 1976). В периоды, благо-
приятные для вегетативного размножения, она резко омолаживается,
а в многолетние периоды засухи — катастрофически стареет. Изме-
нение доли генеративных побегов по годам меняется весьма сильно.
Это связано с изменением возраста кустов в популяциях вейника на-
земного. Повышение доли генеративных побегов, по мнению К.А.
Куркина (1976), отражает преобладание в популяциях двух-трехлет-
них кустов В среднем за 10 лет один генеративный побег приходится
на 2,9 тыс. вегетативных.
Общая длительность жизни клона составляет, возможно, не ме-
нее 450 лет для популяций, выросших после пожара в сосняках Юж-
ной Финляндии (Oinonen, 1969), при этом средний размер колонии
достигает диаметра 50 метров.
Сезонное развитие
Всходы вейника наземного появляются в мае—июне, хотя могут
появиться и в сентябре сразу после опадения семян. Наиболее интен-
сивный отпад всходов наблюдается в течение первого летнего месяца
10
их жизни, в дальнейшем число всходов меняется мало (Бельков',
1957).
Новые кусты вегетативного происхождения могут появляться у
вейника наземного в течение всего вегетационного периода, но в ле-
состепной зоне Южного Предуралья наиболее интенсивно этот про-
цесс идет в июле—августе (Патраболова, 1953). По нашим данным, в
Тверской области на вырубках новые кусты вегетативного происхо-
ждения появляются с июня по август. При этом в год образования
побеги чаще всего не выходят на поверхность почвы, а перезимовы-
вают в ней, иногда в подстилке, а в июне появляются над почвой и
начинают развиваться (Бельков, 1957).
Рост открытых почек в узлах кущения (под землей) начинается в
начале мая после полного схода снега и оттаивания почвы. Побеги
появляются над землей во второй декаде мая. В мае, июне идет раз-
витие многоузловой розетки у вегетативных побегов. Одновременно
происходи'!' формирование генеративных побегов и зачатков соцве-
тий. К началу июля цветоносный стебель бывает уже сформирован.
Начало цветения в значительной степени зависит от погодных
условий. Так, после холодного мая 1990 г. на вырубках в Тверской
области цветение началось только в начале августа и продолжалось
практически до конца лета. В другие годы цветение начиналось в се-
редине июля и продолжалось до середины августа. В лесу цветение
начинается на одну—две недели позже и наблюдается только в са-
мые жаркие дни. По данным И.Г. Серебрякова (1947), на лесных по-
лянах Лосиного острова (Московская область) цветение вейника на-
земного продолжается от начала и до конца июля. В августе, сентяб-
ре формируются плоды. Обсеменение наблюдается в течение октяб-
ря—ноября.
По нашим наблюдениям, сначала готовы к опылению пестики (у
всех цветков в метелке одновременно), а через неделю полностью со-
зревают тычинки. Синхронность начала созревания пестиков харак-
терна также для всех растений одной колонии. Для лесостепной зоны
Зауралья, Западного Предуралья А.Н. Пономарев (1966) и для степей
Забайкалья М.Б. Русакова (1969) отмечают только утреннее время
открывания цветков (6—9 ч.). На Украине вид также относят к ут-
ренним злакам (Злаки Украины, 1977). По нашим наблюдениям в
Московской и Тверской областях, цветение более растянуто по вре-
мени (от 6 до 12 ч. утра). Вейник наземный цветет при температуре
10—22° и относительной влажности воздуха 60—98%. Цветение на-
растает и убывает постепенно.
Отмирание генеративных побегов и надземной части вегетатив-
ных розеток наблюдается в октябре, ноябре. Подземные междоузлия
стеблей, укоренившиеся в узлах, остаются живыми и сохраняются до
следующего года. При этом решающим моментом в определении чис-
ла сохранившихся междоузлий является положение верхней почки
кущения: все находящиеся над нею междоузлия погибают, находя-
щиеся под нею — сохраняются (Рожевиц, 1937). В ветошь переходят
11
убитые морозом растения, не закончившие цикл своего развития и
сохраняющие в значительной степени свой химизм. Сухие части рас-
тения могут сохраняться над снегом до весны. В мае весь запас вето-
ши прошлого года, выщелоченный осадками и вымоченный талыми
водами, спрессовывается в подстилку. В июле ветошь уже состоит из
отмерших растений вегетации данного сезона, прошедших полно-
стью цикл развития и трансформировавших свой химический состав
(Куркин, 1976).
Динамика надземной фитомассы вейника наземного на вейнико-
вом лугу в Барабинской лесостепи (Западная Сибирь) проанализиро-
вана А.А. Титляновой (1979). Сухой вес возрастает с 275 кг/га в июне
до 483 кг/га в июле и уменьшается к август) до 405 кг/га. При этом
наблюдается такой же пик в июле по содержанию азота (5,8 кг/га),
кремния (15,6 кг/га), калия (6,6 кг/га), кальция (11,4 кг/га), хлора (3,1
кг/га)! Содержание магния в течение сезона уменьшается от 1,5 кг/га
до 0,7 кг/га; содержание натрия и фосфора не изменяется по месяцам.
На вейниковой вырубке в Московской области нами исследова-
ны изменения среднего веса надземного побега в течение вегетацион-
ного сезона.
Тип побега Вес побега, г
май июнь ИЮЛЬ август сентябрь
Вегетативный 0,6±0,03 0,22±0,02 0,33±0,03 0,45±0,09 0,25±0,03
Генеративный — 0,96±0,06 1,44±0,10 1,32±0,10 0,55±0,06
Способы размножения и распространения
Вейник наземный — строго ветроопыдяемое растение. Широко
распространена межвидовая и межродовая гибридизация (Цвелев,
1965, 1976).
В кусте обычно бывает 1 генеративный побег, часто — 2, редко
— 3—4. Число цветущих побегов на вырубках составляет 236 на 100
м2 (Печникова, 1931). Такая низкая «генеративность» свойственна
видам, интенсивно размножающимся вегетативным путем (Любар-
ский, 1967). «Генеративность» по годам изменяется весьма резко. Это
связано с динамикой возрастов парциальных кустов в популяциях
вейника. В метелке образуется в среднем 1,5 тыс. семян (Бельков,
1956) и даже 1—6 тыс. (Быков, 1962). На 1 га вырубки падает до 50
млн. семян, что в 50 раз больше, чем их дает, например, сосна; одна-
ко семена могут и не выпадать из соцветия (Бельков, 1956). Даже в
прошлогодних метелках можно видеть невыпавшие семена. Зерновки
вместе с цветковыми чешуями распространяются исключительно вст-
12
ром. Парашютик из волосков на каллусе увеличивает парусность се-
мени, и позволяет им улетать на большие расстояния.
Во всех природных сообществах численность проростков и мо-
лодых ювенильных растений мала. Существует даже представление
об исключительно вегетативном размножении вейника наземного
(Смирнов, 1958; Работнов, 1984). На лугах Барабинской лесостепи
всходов вейника практически не имеется (Куркин, 1976). По нашим
данным, вейник наземный в массе появляется из семян только на
сильно* нарушенных почвах однолетних вырубок, достаточно увлаж-
ненных, Но без застоя воды (Тверская и Московская области). На не-
больших участках численность молодых семенных растений может
достигать 30 на 1 м2. Анализ 2—3-летних колоний вейника на таких
вырубках показал, что 80% колоний имеет семенное происхождение,
в 20% возникли из особей вегетативного происхождения, оставшихся
от исходного лесного ценоза. По данным В.П. Белькова (1956), всхо-
ды вейника появляются на вырубках только на 3-й год после рубки,
если запас семян на вырубке вначале отсутствовал и первые семена
образовались только у растений вегетативного происхождения. На
пожарищах вейник наземный также появляется из семян на нарушен-
ной почве обычно в течение нескольких лет (Oinonen, 1969).
Вегетативная подвижность обеспечивается приростом плагио-
тропных побегов, функционирующих как органы вегетативного во-
зобновления. Общая длина живых корневищ, связывающих между
собой надземные побеги кустов, может достигать при благоприят-
ных условиях 20 м (Любарский, 1967). В среднем же в вейниковых со-
сняках (Волжско-Камский заповедник) общая длина корневищ одно-
го куста составляет от 120 до 437 см при числе побегов в кусте 3,3—
7,5 (Любарский, 1964). Средний годичный прирост корневищ в этих
условиях — 18—31 см в год, при максимуме 64 см в год. В сосново-
березовых лесах Тюменской области прирост корневищ составляет
15—40 см в год (Чижов, 1981). В сосняках и ельниках лишайниковых
и брусничных Финляндии средняя скорость роста колонии по диа-
метру составляет 12,5 см в год, при максимальном приросте 13,5 см в
год (Oinonen, 1969).
Старческой партикуляции колонии не происходит, т.к. корневи-
ща с возрастом не перегнивают, и сохраняется связь всех кустов в ко-
лонии. Интересно отметить, что часто на корневищах находятся
много засохших кустов, однако корневище продолжает функциони-
ровать как проводящая система. Выкопанные корневища быстро от-
мирают, гак как они плохо защищены от высыхания.
Экология
Вейник наземный растет в самых разных экотопах и зонах, от
тундр до пустынь. Г. Эллепбсрг (Ellenberg, 1974) относит вид к числу
индифферентных к влаге. Хотя целый ряд авторов считают данный
вид мезоксерофитом (Цыганов, 1983; Каразия, 1977), и даже ксеро-
13
фитом (Каменецкая, Михайленко, 1963), на самом деле вейник назем-
ный весьма требователен к влаге, и его обилие всегда указывает на
хорошее водоснабжение (Смирнов, 1958). Даже на почвах с избыточ-
ным и застойным увлажнением, в березняках черничных и долго-
мошных, вид этот весьма устойчив, что указывает на отсутствие у
него резко выраженной отрицательной реакции на недостаток аэра-
ции почвы (Зворыкина, 1983). По нашим данным, вейник наземный
успешно растет на вырубках на месте сведенного сосняка сфагново-
го. Однако затопление в разгар вегетации, особенно при высоких
температурах, (даже если слой воды очень небольшой) ведет к гибели
вейника (Куркин, 1976).
Многолетнее иссушение почвы препятствует вегетативному раз-
множению вида, вызывая резкое уменьшение численности молодых и
гибель появившихся корневищ (Куркин, 1976). Прекращение корне-
образования связано с тем, что для формирования корневищ требу-
ется больше пластических веществ, чем для развития ортотропных
побегов (Патраболова, 1953). Для укоренения молодых корневищ
при их переходе к формированию молодого парциального куста тре-
буется хорошее увлажнение почвы на глубине, где закладываются уз-
лы кущения (Смелов, 1947; Патраболова, 1953). При высыхании пес-
чаной почвы в сосняках в лесостепи наблюдалась гибель не только
корневищ, но и парциальных кустов. При глубине залегания узлов
кущения менее 5 см кусты засыхали, а при глубине 8 см и более —
нет.
Вейник наземный растет преимущественно на открытых местах:
на лугах, вырубках, залежах, на обочине дорог, на полянах, в освет-
ленных лесах. Г. Эппенберг (Ellenberg, 1974) относит его к группе по-
лусветовых растений (балл 7). Любое затенение резко подавляет «ге-
неративность» вейника, так как не только ослабляет жизнедеятель-
ность, но и направляет ее в сторону вегетативного роста и размно-
жения (Рысин, Рысина, 1966; Любарский, 1967). По нашим данным,
при зарастании вырубок мелколиственными породами обилие вей-
ника резко снижается именно из-за уменьшения освещенности. В ель-
никах вейник наземный чаще имеет вид угнетенных одиночных веге-
тативных побегов и только в окнах образует кусты, которые цветут.
Вейник наземный произрастает на разнообразных почвах: пес-
чаных, суглинистых, известняковых, черноземных, солонцеватых, ал-
лювиальных. Так, несмотря на репутацию псаммофита, в березняках
Ярославской области вейник наземный распространен на суглини-
стых почвах шире, чем на супесчаных и даже песчаных почвах (Зво-
рыкина, 1983). На супесях и песках он встречается только при усло-
вии близкого залегания суглинистых прослоек или валунной море-
ны. В Приокско-Террасном заповеднике вейник наземный произра-
стает как на песчаных, так и на суглинистых и даже торфяных поч-
вах (Смирнов, 1958).
По экологическим шкалам Элленберга (Ellenberg, 1974) вейник
наземный индифферентен к реакции почвы; по содержанию доступ-
14
ного аммонийного и нитратного азота в минеральных горизонтах
предпочитает почвы между средними и богатыми (балл 6). Е. Лан-
дольт (Landolt, 1977) относит вейник к видам, растущим на почвах с
небольшим гумусовым горизонтом (балл 2).
Фитоценология
Вейник наземный широко распространен на лугах, в лесах, на
вырубках и гарях, вдоль дорог, по нарушенным пескам. На лугах
часто бывает доминантом. Выделяется даже формация Calamag-
roslideta epigeii (Быков, 1962), широко распространенная в степной,
лесостепной и полупустынной зонах. Наиболее часто в этой форма-
ции встречаю тся Phragmites communis и Agropyron repens. Сообщест-
ва могут быть пионерными. Всего флора формации (34 ассоциации)
содержит около 150 видов (Быков, 1962).
11аземповсйниковые луга в лесной зоне распространены исклю-
чительно в поймах рек, на прирусловых гривах, на повышенных ма-
ло заливаемых участках (Методические рекомендации..., 1987). В
классе Molinio-Arrhenatheretea (порядок Arrhenatheretalia, союз
Festucion pratensis (Sipaylova et al. 1985) выделяется, в частности, ас-
социация Calamagrostietum epigeii Sapegin 1986, описанная в Белорус-
ском Полесье (Сапегин, 1986).
В луговых степях наземновейниковые луга тяготеют к склонам
балок, к пониженным частях пойм малых рек. В степной зоне вейни-
ковые луга распространены на разбитых и плохо заросших песках,
повышенных и выровненных участках пойм крупных и средних рек,
в пониженных частях пойм малых рек, в неглубоких западинах водо-
разделов. В полупустынной зоне вейник растет по лиманам и подам.
В лесной зоне вейник наземный наиболее характерен для гарей и
вырубок союза Epilobion angustifolii Тх 37 em. 50 класса Epilobietea
angustifolii (Vlieger 37) Tx et Preising in Tx 50. Наземновейниковые вы-
рубки распространены в европейской части и в Западной Сибири.
Они возникают на месте вырубленных ельников, сосняков и березня-
ков кисличной и 4qjHH4Hott групп.
В условиях вырубок вейник наземный — типичный эксплерент.
В течение первого года вид быстро захватывает участки, освободив-
шиеся после рубки леса. Интенсивное вегетативное размножение, по-
явление семенных экземпляров, позволяют вейнику наземному доми-
нировать уже на 3-летних вырубках после ельников в Тверской и
Московской областях. На вырубках после сосняков в условиях дюн-
ных песков во Владимирской, Нижегородской областях вейник ста-
новится доминантом только с 7—8-го года после рубки (Любимепко,
1902), а в Татарии — с 5-го года (Печникова, 1931). В сообществах
(па отдельных лугах, вырубках), где вейник наземный прочно удер-
живает терриз оршо, он выступает как сильный эдификатор. Под его
15
влиянием происходит значительное изменение условий внешней сре-
ды как в надземной, так и в подземной части.
Листья и стебли вейника — жесткие, малопитательные, плохо
поедаются животными. Отмирая осенью, они образуют слой войло-
ка. Остатки листьев медленно разлагаются из-за большой механиче-
ской прочности (Смирнов, 1958). Накапливаясь каждый год, войлок
формирует подстилку толщиной до 2—3 см (Саутин, 1957). Эта под-
стилка плотным слоем прикрывает минеральную часть почвы и пре-
пятствует появлению всходов любых других растений. Однако под-
стилка и войлок не препятствуют нормальному развитию самого
вейника, у которого весенние надземные побеги имеют острые, жест-
кие верхушки, свободно проходящие через слой почвы в 12—15 см и
через подстилку.
В подземной части вейник образует очень густую сеть корневищ
на глубине от 2 до 40 см в зависимости от почвенных условий. Раз-
ные авторы отмечают различную глубину размещения корневищ:
2—15 см (Печникова, 1931), 12—20 см (Hilf, 1928), 10—40 см (Быков,
1962), 3—14 см (Любарский, 1964), 5—8 см (Чижов, 1981), 4—7 см
(Рысин, Рысина, 1987). Насыщенность почвы корневищами составля-
ет до 100—110 м на 1 м2 , причем они приурочены к нижней части гу-
мусового горизонта (Любарский, 1964). Встречаются корневища и в
горизонтах AjA2 (Чижов, 1981), А2 (по нашим данным). Таким обра-
зом, вейник наземный является сильным конкурентом за воду и пита-
тельные вещества, и в его зарослях могут произрастать очень немно-
гие виды, в основном, корнеподстилочные.
На вырубках под вейником формируется более мощный дерно-
вый горизонт с высоким содержанием гумуса (Уланова, Тощева,
1989). Возможно также повьппение плотности почвы в гумусовом го-
ризонте и подкисление почвы (Глухов, Федякова, 1938).
В условиях чистого вейникового травостоя продукты опада и
выделения вейника, по мнению Е.Л. Любарского (1964), создают в
почве неблагоприятную, токсичную для него самого среду, что при-
водит к общему самоугнетению. Лабораторные исследования под-
тверждают, что энергия прорастания семян и рост проростков сосны
и ели тормозится вытяжками из корней и сена вейника наземного
(Миронов, Шильников, 1972), но в полевых условиях влияние вытя-
жек на всходы и 1—2-летние растения отсутствует.
По нашим данным, наземновейниковые сообщества сохраняют-
ся на вырубках до тех пор, пока вейник не будет вытеснен березой,
ивой, елью. Вытеснение малиной как более сильным конкурентом на-
блюдается на четвертый год существования вырубки. Чаще всего на
5—7-й год формируются березово-вейниковые группировки, в кото-
рых вейник только вегетирует, и группировки вейниковые на освет-
ленных участках вдоль посадок елей. Последние группировки встре-
чаются и на 12—15 летних вырубках, на опушках березняков. В виде
угнетенных экземпляров вейник сохраняется в старых ельниках и со-
сняках. Вейниковые вырубки, возникшие на месте сосняков могут
16
просуществовать очень долго, так как сосна не может сама возоб-
новляться в этих условиях (Смирнов, 1958).
В лесах вейник наземный широко распространен в осветленных
сосняках и березняках, значительно реже он встречается в осветлен-
ных ельниках. Эколого-ценотический оптимум вейника наземного
совпадает с эдафическим ареалом кисличной и черничной групп ти-
пов л?са в Финляндии (Oksanen, 1983), в Ленинградской, Ярослав-
ской и Московской областях (Зворыкина, 1983; Маслов, 1990). По
данным А.А. Маслова (1990), в лесах Московской области наивыс-
шая встречаемость вейника наземного наблюдается в сосняке-чер-
ничнике. Однако на юге Московской области, на Украине и в Бело-
руссии, в зоне лесостепи вейник наземный часто встречается и в ряде
случаев достигает большого обилия в остепненных сосняках редко-
травно-лишайниковых (Рысин, 1975).
При изучении мозаичности лесных сообществ было показано,
что вейник наземный произрастает, как правило, вместе с Vaccinium
vitis-idaea, Melampyrum pratense, Luzula pilosa, Hylocomiurn splendens,
Aulacomnium palustre. He обнаружено тяготения вейника к стволам
берез, сосен, елей (Маслов, 1990). Однако внутри сообществ распре-
деление вейника сильно зависит от фитоценотической обстановки;
при этом нередко он образует особую группу, как в сосняке-зелено-
мошнике или на семилетней вырубке (Уланова, Маслов, 1989)
По мере увеличения возраста сосняков (с возрастанием освещен-
ности на уровне нижних ярусов) покрытие, численность надземных
побегов, а также общий вес вейника наземного могут возрастать
(Любарский, 1964). Такая же связь вида с освещенностью наблюдает-
ся в ельниках.
Консортивные связи
Вейник наземный — микосимбиотроф. На корнях образуются
микоризы. Степень микотрофности варьирует. Вид относится к сред-
немикотрофным. Наблюдается только эндотрофная микориза, отно-
сящаяся к фикомицетной или тамнискофаговой и образуемая гриба-
ми из сем. Endogonaceae (Селиванов и др., 1961). Эндофитный гриб
сосредоточен преимущественно в эндодерме, причем и гифы гриба, и
продукты его переваривания находятся в клетках коры, а не в меж-
клетниках. Наличие микоризы характерно только для самых тонких
ответвлений питающих корней, имеющих диаметр от 80—120 до
200—300 мкм. Корни большего диаметра, как правило, без мико-
ризы.
Вейник наземный поражается ржавчинными грибами (Ростов-
цев, 1908; Ульянищев, 1978), являясь промежуточным хозяином для
в торой и третьей стадии развития гриба. Основные грибы-паразиты:
Puccinia calamagrostidis Sydow., Р. coronata Corda, P. coronifera
Klebhn., P. gluinarum Schmidt., P. graminis Pers., P. Rubigo Vera DC.,
P. pygmaea, P. epigeios Ito. Кроме этого вейник поражается спорынь-
17
ей Claviceps microcephala Wallr. Другие грибы также обитают на ли-
стьях и стеблях: Ascochyta calamagrostidis Brun., Epichloe typhina
Pers., Fusoma triseptatum Sacc., Septoria calamagrostidis Sacc., Tilletia
calamagrostidis Fuck., T. striformes Westend., Uromyces calamagrostidis
Uljanish., Ustilago hypodyles Schlecht.
Насекомые плохо поедают грубые листья вейника. Однако на
лугах Барабинской лесостепи саранчовые поедают начинающие от-
растать молодые побеги (Куркин, 1976). В результате постоянного
объедания молодых побегов запас почек в зоне кущения постепенно
истощается, что ведет к гибели зарослей вейника.
Особую роль на этих лугах играют земляные муравьи Lasius
niger L., L. flavus Fabr., Mynnica scabrinodis Nyl. (Куркин, 1976).
Своими земляными выбросами они засыпают куртины вейника на-
земного, что ведет к постоянному перемещению узлов кущения
вверх. Это, в свою очередь, стимулирует вегетативное размножение.
На корневищах вейника наземного паразитирует Melampyrum
pratense (Логинова, 1985), что подтверждается при анализе межвидо-
вых сопряженностей (Маслов, 1990).
Биологическая продуктивность
Фитомасса вейника наземного в 25 летнем смешанном лесу на
месте вырубки в Московской области составляет 23 г/м2 (Холопова,
Солнцева, 1987). В березовой парцелле его фитомасса составила 3,2
г/м2, а в елово-мелколиственной только 2,2 г/м2. В вейниковых сосня-
ках Волжско-Камского заповедника средний вес вегетативного побе-
га 364—633 мг, генеративного побега — 1350—2650 мг при общей
надземной биомассе 37—86 г/м2 (Любарский, 1964). Общая фитомас-
са вейниковой вырубки (смешанные леса Загорского района Москов-
ской области) составляет 50 ц/га, при этом на вейник наземный при-
ходится 80—90% (Миронов, Шильников, 1972).
Хозяйственное значение
Вейник наземный дает грубое жесткое сено, плохо поедаемое
всеми видами животных. Такое сено плохо переваривается. Р.Ю. Ро-
жевиц (1937) считал этот вид не имеющим хозяйственного значения,
однако в зоне Нечерноземья вейниковые луга скашиваются и исполь-
зуются под выпас.
В зависимости от условий произрастания вейник наземный по-
разному относится к выпасу. На песчаных легких почвах он не выно-
сит даже слабого выпаса и быстро сокращает численность па выпа-
саемых лесосеках, сохраняясь только внутри зарослей кустарников, в
которые не заходит скот (Смирнов, 1958). Т.А. Работнов (1984) так-
же относит вейник наземный к видам, чувствительным к вытаптыва-
18
нию, однако Л.П. Рысин и Г.П. Рысина (1987), напротив, считают
вейник довольно устойчивым.
На суглинистых почвах вид может быть устойчив к выпасу и се-
нокошению (Кормовые растения..., 1950). Однако косить его можно
не позднее фазы колошения (хотя при этом и получается малый сбор
сена), так как ко времени цветения сено грубеет. Урожайность вейни-
ковых лугов в Центре России составляет 20—80 ц/га (Быков, 1962), а
в пойме р. Енисей — 22—53 ц/га (Номоконов, 1959).
Особо активно выпасаются наземновейниковые луга на солон-
цеватых И солончаковых почвах в лесостепной, степной и полупус-
тынной зонах. Эти пастбища расположены в поймах рек, на склонах
балок, в неглубоких западинах водоразделов (Методические реко-
мендации..., 1987).
Вейник наземный имеет большое значение в лесоводстве. Вейник
— антагонист молодых поколений леса на начальных этапах разви-
тия. Он угнетает сеянцы и культуры сосны в питомниках и культу-
рах. Так, В.П. Бельков (1960) отмечает ухудшение роста и увеличение
отпада саженцев сосны в питомниках при заселении вейника. В Бе-
лоруссии на легких супесчаных и песчаных почвах вейник наземный
образует сплошной покров на вырубках, при этом 85% посадок со-
сны гибнет за 7 лет существования (Саутин, 1957). Семенные культу-
ры сосны (Нижегородская область) менее устойчивы против вейни-
ка, чем посадки (Набатов, 1964). На наиболее плодородных почвах в
сосняке кислично-черничном и ельнике травяно-липовом посевные
культуры сосны просто обречены на гибель.
Для борьбы с вейником рекомендуется до посадки культур про-
водить ряд специальных мероприятий, направленных на уничтоже-
ние его корневищ. Основной способ — вспашка на глубину 12—20
см всего участка, через 15—20 дней — дискование бороной; одновре-
менно проводят боронование (Саутин, 1957). В течение лета еще 2—3
раза ведут культивацию почвы. Весеннюю посадку саженцев прово-
дят после осенней глубокой вспашки на глубину 22—25 см. Более
простой способ предлагал П.А. Смирнов (1958) для уничтожения
вейника на легких почвах при высоком положении почек возобнов-
ления. При сенокошении полян в течение 3—4 лет вейник исчезает
из—за постоянного удаления оснований стеблей с почками. После
этого можно сажать саженцы сосны. Однако при достаточно редком
стоянии стеблей вейника процесс лесовозобновления может идти до-
вольно успешно (Мелехов, 1980).
19
П.Ю. Жмылев, И.С. Кривохарченко, А.В. Щербаков
СЕМЕЙСТВО РЯСКОВЫЕ
Номенклатура и систематика
Lemnaceae S. Gray, 1821, Nat. Arr. Brit. Pl. 2: 729, (nom
abortivum), correxit: Dumort., 1827, Fl. Belg.: 147 — Рясковые.
Тип семейства — Leinna L. Название типового рода возможно
произошло от греческого "limne" — болото (Каден, Терентьева,
1975).' На местном говоре некоторых областей России слово "ряска"
(уменьшительное от,"ряса") означает ряд, низка, нитка ожерелья, се-
точка и т.п. (Даль, 1882).
Рясковые — небольшое семейство класса однодольных растений.
По данным разных авторов насчитывает от 4 до 6 родов и от 28 до
43 видов, распространенных почти по всему земному шару, исключая
полярные области (Daubs, 1965; Иванова, 1973; Hendrych, 1985;
Landolt, 1986; Тахтаджян, 1987; Kandeler, 1988 и др.).
Все рясковые по строению соцветия, наличгпо корней, количест-
ву и расположению кармашков объединяются в два подсемейства,
взаимоотношения между которыми до конца не выяснены. Подсе-
мейство Lemnoideae включает роды Leinna и Spirodela Schleid., а под-
семейство Wolffioideae Engler — роды Wolffiella Hegelm. и Wolffia
Hork. Во флоре России семейство представлено тремя родами и пя-
тью видами, из которых только Wolffia arrhiza (L.) Horkel ex Wimmer
отсутствует в Московской области. Северная граница ее естествен-
ного распространения в Восточной Европе проходит по Украине,
Липецкой, Воронежской и Самарской областям (Флора европ. час-
ти..., 1979; Цвелев, 1988). Нахождение вольфии в заводи р. Осетр в
Зарайском районе, куда она, по-видимому, была занесена аквариу-
мистами (Игнатов и ДР-, 1990), пока не подтверждается гербарными
Материалами.
Рясковые издавна привлекали к себе внимание ботаников, и в
настоящее время библиография по этому семейству, обобщенная Е.
Landolt и R. Kandeler (Landolt, 1986; Landolt, Kandeler, 1987), насчи-
тывает более 4000 работ. Впрочем, многие стороны их жизни по-
прежнему остаются недостаточно изученными. Новые литературные
данные во многом противоречивы, и, как следствие, нарастающий
информативный поток не столько сужает, сколько расширяет-круг
нерешенных вопросов по биологии, морфологии и систематике этих
воистину растений-загадок.
Морфологическая характеристика
Рясковые — свободноплавающие вегетативные однолетники. В
результате перехода к водному существованию их строение претср-
20
пело настолько значительное упрощение, что среди современных
цветковых растений они занимают первое место как по миниатюр-
ности, так и по простоте своей организации. Внешним обликом веге-
тативное тело рясковых напоминает слоевище низших растений. Оно
не дифференцировано на стебель и лист и обозначается по-разному:
побег, лист, филлокладий, вайя, щиток, пластинка, листоветвь,
фронд, листец. Последнее название кажется наиболее удачным. Зару-
бежные ботаники уже более столетия в аналогичном смысле исполъ-
зуется'термин "frond" (Hillman, 1961), а в германоязычной литерату-
ре наряду с ним используют название "Glied".
Листецы рясковых представляют собой симметричные или слег-
ка асимметричные, округлые или эллиптические, уплощенные или
вздутые с нижней стороны зеленые пластинки не более 1,5 см длиной.
От нижней (вентральной) поверхности листеца отходит один корень
(у Lemna) или пучок из нескольких корней (у Spirodela). Место их от-
хождения обозначается как "узел", который разграничивает листец
на две зоны: проксимальную (базальную) и дистальную (рис. 1).
Проксимальная зона в основании сужена и переходит в так на-
зываемую "ножку" или "стипу" листеца. В дистальной зоне распола-
гаются жилки, которые отходят от "узла". Кроме того, в области "уз-
ла" находятся два углубления, именуемые почечными (выводковыми)
кармашками или просто кармашками. В этих углублениях формиру-
ются дочерние листецы или соцветие. Кармашки расположены лате-
рально, по обе стороны от дистально-проксимальной оси, которую
рассматривают как главную морфологическую ось растения (Каси-
нов, 1973). Дочерние листецы развиваются в кармашках и отделяют-
ся от материнской особи поочередно, что обусловливает своеобраз-
ную, лево-правую изомерию строения вегетативного тела рясковых.
Для различения кармашков принято располагать материнский лис-
тец проксимальным концом к наблюдателю и устьичной стороной
вверх. Тогда дочерний листец, формирующийся в правом кармашке,
обозначается как "правый" или знаком а дочерний листец проти-
воположного кармашка — как "левый" или знаком "+" (рис. 2).
Относительно морфологической природы листецов рясковых
высказаны различные точки зрения (Hegelmaier, 1868; Caldwell, 1899;
Ростовцев, 1905; Blodgett, 1915; Arber, 1919; Goebel, 1921; Lawalree,
1943, 1945, 1952; Ashby et al., 1949; Иванова, 1973; Landolt, 1986 и
др.). Значительная часть из них — модернизированные варианты од-
ной и той же гипо тезы, рассма тривающей листец как листоветвь осо-
бой природы: дистальная се часть образована листом, а проксималь-
ная имеет стеблевое происхождение (Engler, 1889). Различия между
вариантами касаются интерпретации стенок кармашка.
Корни рясковых не ветвятся и лишены корневых волосков. Раз-
виваются они из клеток, расположенных около "узла", в направле-
нии к дистальному концу листеца и параллельно его главной морфо-
логической осн (Hegelmaier, 1868; Касимов, 1973; Landolt, 1986). В
дальнейшем растущий корень изгибается и располагается перпендп-
21
Рис. 1. Строение особи (/) и лнстеца (Я) многоеоренника обыкновенного (по: Hegelmaier, 1868).
—• листец материнский. Л। — листец дочерний первой генерации, — листец дочерний второй генерации; 1 — кармашек. 2 — ряды вы-
тянутых клеток от узла к ножке, 3 — ножка, 4 — корень, 5 — узел, 6 — жилки, 7 — базальная часть, 8 — дистальная часть, 9 — часть профиллю-
ма, переходящая с дорсальной на вентральную сторону, 10— профиллюм с дорсальной стороны.
22
Рис. 2. Строение особи ряски трехдольной.
JIQ — материнский листец; 1, 2, 3. ... 5 — дочерние листецы первой, второй, треть-
ей, ... пятой генерации; а — первый дочерний листец; б — второй дочерний листец; "+"
— "плюс" сторона листеца; "-" — "минус" сторона листеца.
кулярно плоскости листеца. Вода и минеральные соли абсорбируют-
ся всей или только нижней поверхностью листеца, и корни не имею т
существенного значения в этом процессе (Ice et al., 1987). Это обще-
признанное суждение было принято еще в прошлом веке на основа-
нии экспериментальных работ F. Hegelmaier (1868) и с тех пор изред-
ка подвергается сомнению (Музафаров и др., 1968; Король, 1988).
Рясковые размножаются преимущественно вегета тивно. Дочер-
ние особи формируются поочередно в каждом кармашке и остаются
некоторое время соединенными друг с другом посредством "ножки",
образуя одно- или многоспиральные труппы листецов. При этом ха-
рактер изомерии материнского раст ения сохраняется в потомстве ве-
гетативного происхождения. Однако левые и правые формы могут
быть представлены в равных соотношениях в потомстве искусствен-
но самоопыленттых растений (Hillman, 1961; Касинов, 1971, 1973).
У цвет ущих особей в одном из кармашков, реже в двух, фор-
мируются т оперативные органы: две, редко три тычинки и один
пест ик, которые окружены одно- или двуслойной пленкой. Элемен ты
околоцветник:! отсутствуют. Традицпонпо пленка ("спада") рассмат-
23
ривается как покрывало, а совокупность тычинок и пестика — как
сильно редуцированный початок ароидных, с которыми рясковые
имеют общее происхождение (Engler, 1889; Ростовцев, 1905; Daubs,
1965; Иванова, 1970, 1973; Куприянова, Тарасевич, 1984; Landolt,
1986; Тахтаджян, 1987 и др.). Реже совокупность генеративных орга-
нов принимается за обоеполый цветок (Lawalree, 1943, 1945;
Maheshwari, Kapil, 1963; Лукина, 1988 и'др.).
Цветение, опыление и диссеминация
В биологии генеративного размножения рясковых до сих пор
много неясного. Прежде всего, это связано с ограниченностью на-
блюдения цветущих растений в природе. Так в Польше с 1679 г. бы-
ло отмечено всего 2 случая цветения рясковых (Czopek, 1960), а в
Финляндии с 1895 по 1947 г. — 33 (Luther, 1947-1948). На территории
бывшего СССР с 1814 г. в литературе имеются упоминания о 108 на-
блюдениях этого явления. Предполагается, что из-за миниатюрности
растений их цветение трудно обнаружить (Шпор, 1915; Кузнецов,
Зайченко, 1979; Печенюк, 1985 и др.). Впрочем, в культуре пока выде-
лено всего несколько клонов, у особей которых при определенных ус-
ловиях можно спровоцировать образование цветков (Hillman, 1961;
Nakashima, 1969; Oda, 1969; Posner, 1969; Miyata, 1970; Касинов,
1971; Landolt, Kandeler, 1987 и др.).
Экологические условия, благоприятствующие цветению, точно
не установлены. Вероятно, разные виды или даже клоны зацветают
при разных условиях. Правда известны случаи одновременного цве-
тения в одном водоеме нескольких видов рясковых. Это позволяет
либо утверждать обратное, либо допустить существование механиз-
ма биохимической индукции (например, этиленом) цветения одного
вида (клона) другим (Landolt, 1986).
Весьма противоречивые суждения высказываются разными ис-
следователями и относительно механизма опыления цветков. Одни
относят рясковые к анемофильным растениям (Ростовцев, 1905), дру-
гие — к автофильным (Hegelmaier, 1868; Engler, 1889), третьи — к
зоо- или энтомофильным. В качестве доказательства последнего
предположения приводятся следующие доводы: скульптурирован-
ность поверхности и небольшое количество пыльцевых зерен, кото-
рые могут склеиваться в комочки; наличие сахаров в жидкости, вы-
деляющейся на поверхности рыльца; нахождения пыльцы рясковых
на брюшках мелких насекомых (Иванова, 1970; Куприянова, Тарасе-
вич, 1984 и др.). Однако вполне возможно, что рясковые не имеют
какого-либо жестко детерминированного механизма опыления, и в
зависимости от стадии цветения, могут опыляться как ветром, так и
насекомыми (см. Иванова, 1970). В некоторых случаях, например, ко-
гда цветущие листецы разобщены вегетирующими, плоды завязыва-
ются, вероятно, и в результате гейтоногамии.
24
Данные по диссеминации рясковых немногочисленны. Неясно,
насколько эффективно распространение листецов при помощи раз-
личных животных, жизнь которых тесно связана с водной средой.
1^значительная устойчивость растений к высыханию вероятно'огра-
ничивает дальность их переноса таким способом. Так, по данным
D.L. Jacobs (1947), на поверхности оперения птиц во время полета
создаются неблагоприятные условия, и в силу этого листецы быстро
погибают. Возможно, в виде семян рясковые могут распространяться
на большие расстояния. Однако семена, например, ряски маленькой
мало устойчивы к высыханию (Ростовцев, 1901, 1905). Влажная пого-
да или попадание листецов и семян внутрь оперения птиц, несомнен-
но, может благоприятствовать эпиорнитохорному расселению ряско-
вых, но максимальная дальность такого переноса не установлена.
(Заметим, что пока ничего не известно и о всхожести семян после их
прохождения через желудочно-кишечный тракт птиц). Вероятнее все-
го расселение рясковых происходило и происходит постепенно в ре-
зультате гидрохории и отчасти зоохории. В некоторых случаях скач-
кообразно-дальнюю миграцию можно объяснить исключительно ан-
тропохорией. Например, появление ряски маленькой на Галапагос-
ских островах (Ridley, 1930; Landolt, 1986).
Фитоценология
Рясковые, покрывая сплошным зеленым ковром всю поверхность
водоема или ее значительную часть, формируют одно- или малови-
довые группировки, а также нередко выступают субдоминантами в
сообществах многих прибрежно-водных растений. Часто они растут
вместе с другими свободноплавающими гидрофитами. Для террито-
рии бывшего СССР было описано более десятка таких ассоциаций
(Араратян, Агаджанян, 1948; Марков и др., 1955; Алиев, 1958; Барсе-
гяп, 1958; Быков, 1960; Экзерцев, I960; Абдиев, 19716; Таубаев, Аб-
днев, 1973; Кузнецов, Зайченко, 1979; Дубына, 1986; Соломещ и др.,
1988). Однако плавающие "ковры" рясковых, названные Б.А. Быко-
вым (1957) "кочующими ступенями", могут свободно перемещаться
по водоему под действием ведра или течения из одного сообщества в
другое. Поэтому формируемую ими синузию (ярус) предлагается вы-
делять как самостоятельную синтаксономическую единицу (Корча-
I пи, 1976; Landolt. 1986).
О взаимоотношениях рясковых с другими водными растениями
известно немного. Несомненно, что такие крупные воздушно-водные
рисз епня, как тростник, рогоз ияи камыш, перехватывая часть свето-
вого потока, в определенной степени могут подавлять развитие ряс-
ковых. Но в то же время заросли этих растений представляют собой
Ш1СТОЯЩПС "убежища" для свободноплавающих гидрофитов от воз-
действия ветра, волнения и течения. Вероятно только некоторые во-
доросли являю тся для рясковых сильными конкурентами. Например,
оиразовапие сплошного покрова спирогиры или водяной сеточки <н-
25
рицательно сказывается па развитии рясковых, а интенсивное обрас-
тание их корней или нижней поверхности листецов эдогониумом или
сценедесмусом ускоряег отмирание растений (Эргашев, Абдиев,
1971).
Судя по экспериментальным данным, в оптимальных условиях
произрастания разные виды ига» даже клопы рясковых могут всту-
пать в конкурентные взаимоотношения друг с другом за свет и эле-
менты минерального питания (Clatworthy, Harper, 1962; Rejmankova,
1975а, 19756; Keddy, 1976; Mcliraith el al, 1989 и др.). При этом отме-
чается, что конкурентная мощность растений не связана со скоро-
стью их роста, а зависит, прежде всего, от особенностей морфологи-
ческого строения листецов. Так, вздутые .листецы L. gibba затеняют
плоские листецы L. minor и S. polyrrhi/.a. В свою очередь наличие
большого количества корней позволяет' S.polyrrhiza использовать
больший объем воды в конкуренции за элементы минерального пи-
тания по сравнению с однокорешковыми рисками. Виды рода Leinna
адсорбируют минеральные cojui нижней поверхность листеца и их
конкурентная мощность зависит уже от размера формируемого ими
"ковра". Исключение составляет L. Irisulca, которая может использо-
вать фосфор, поступающий с атмосферными осадками, и в связи с
этим нередко доминирует в водоемах с постоянно низкой концентра-
цией этого элемента. Однако не ясно, в какой степени результаты
экспериментальных исследований отражают реальные взаимоотно-
шения видов рясковых в естественных условиях. По-видимому, про-
веденные в этой области эксперименты можно пока рассматривать
как модели, реализация которых в природе ограничена, по крайней
мерс, двумя обстоятельствами. Во-первых, клоны одного и того же
вида нередко проявляют разную конкурентную мощность (Wollek,
1984: цит. по: Landolt, 1986). Во-вторых, конкурентная мощность
рясковых изменяется в зависимости от климатических и погодных ус-
ловий. Например, на территории университета штата Манитоба
(Канада) L. Irisulca и L.minor формируют совместные группировки в
эуТрофиых заболоченных прудиках. При этом первый вид доминиру-
ет весной, а второй — летом (Mcliraith et al, 1989). По данным Е.
Landolt (1986), L. gibba доминирует только в оптимальных для нее
климатических и гидрологических условиях, а при недостатке эле-
ментов минерального питания вытесняется в областях с теплым ле-
том S. polyrrhiza, а в более холодных областях L. minor.
Ключ для определения видов, встречающихся в
Московской области
1. Растения очень мелкие (не более 1,5 мм), без корней. Wolffia
arriza (L.) Horkel ex Wimmer. — Вольфия бескорпсвая.
26
— Растения более крупные (обычно более 2 мм), с одним или не-
сколькими маленькими корнями ............................ 2
2. Листец с пучком корней, с нижней стороны и по краю верхней
фиолетово-пурпурный. 4. Spirodela polyrrhiza (L.) Schleid. —
Многокоренпик обыкновенный.
— Листец с одним корнем, с нижней стороны зеленый, белый или
розоватый ............................................... 3
3. Растения, погруженные в воду, и только во время цветения пла-
вающие на поверхности; листецы продолговато-ланцетные, тон-
кие, полупрозрачные, светло-зеленые или (редко) красноватые, в
верхней части выемчато-зубчатые, в нижней сужены в ножку,
почти равную половине их длины. 3. Lcmna trisulca L. — Ряска
трехдольная.
— Растения, плавающие на поверхности воды; листецы непрозрач-
ные, эллиптические, обрагпояйцевидныс или округлые, цельно-
крайние.................................................. 4
4. Листецы округлые или обратнояйцевидные, сверху плоские, зе-
леные, нередко красноватые или буроватые, снизу выпуклые, бе-
лые или розоватые; самые крупные воздушные полости разме-
ром более 0,3 мм. 2. Lemna gibba L. — Ряска горбатая.
— Листецы эллиптические, обратнояйцевидные или почти округ-
лые, сверху и снизу плоские, зеленые, иногда в начале и конце
вегетационного сезона сверху буроватые; самые крупные воз-
душные полости не превышают 0,25 мм. 1. Lcmna minor L. —
Ряска маленькая.
1. Ряска маленькая
Номенклатура, систематическое положение,
внутривидовая систематика
Lemna minor L. 1753, Sp. Pl.: 970; Кауфман, 1866, Моск. фл„ изд.
1: 601; Цингер, 1885, Сборник свед.; 406; Шмаш.гаузен, 1897, Фл.
Средн, и Южн. Росс. 2: 526; Сырейщиков, 1906, Илл. фл. Моск. губ. к
219; он же, 1927, Опред. раст. Моск, губ.; 84; Кузенева, 1935, Фл.
СССР, 3: 493; Назаров, 1940, в Маевск., Фл., изд. 7: 209; Е.Бобров,
1954, цит. соч., изд. 8: 704; он же, 1964, цит. соч., изд. 9: 691; В. Тихо-
миров, 1966, Опред. раст. Моск, обл.: 108; Иконников, 1979, Фл. ев-
роп. части СССР, 4: 320; Lawalree, 1980, Fl. Europ. 5: 273; Черепанов,
1981, Сосуд, паст. СССР: 284; Новиков, 1986, Опред. раст. Мещеры,
1:122.
Тип: "Europae aquis quietis" (ВМ). Типовой вид секции Lcmna,
объединяет 13 видов. Ряска маленькая - полиморфный вид, из со-
става которого Е. Landolt (1975, 1986) предложил выделить L,
turioniiera. Растения згой расы образуют специализированные зп-
27
мующие "почки" (турионы) и отличаются от L. minor s. str. следую-
щими образом:
1. Растения не образуют зимующих почек; верхняя половина листе-
ца ровная; отношение дайны листеца к его ширине не менее чем
1,6. L. minor L. s.str.
— Растения образуют мелкие 1,5 2 мм, бескорневые зимующие
почки (турионы); верхняя поверхность листеца с небольшим вы-
ступом; отношение дайны листеца к его ширине не более чем 1,5.
L. turionifera Landolt.
Географическое распространение
Ряска маленькая — гемикосмополитный вид, распространенный
по всей внетропической Евразии и заходящий на север до 68°30' с.ш.
в Европе (Постовалова, 1969) и до 63° с.ш. в Азии (Флора Сибири,
1987). Встречается в Средиземноморье, горах Йемена, Абиссинии,
Эритреи, в районе озер Виктория и Танганьика, в ЮАР и в умерен-
ных областях Северной Америки. На юг Австралии и в Новую Зе-
ландию, вероятно, занесен человеком (Landolt, 1986). В Московской
области — одно из обычнейших водных растений, и по данным А.В.
Щербакова, отмечена более чем в 97% от числа всех обследованных
водоемов.
По мнению Е. Landolt (1982, 1986), типовая раса ряски малень-
кой не встречается к востоку от Уральского хребта. В Сибири и на
Дальнем Востоке она замещается соответственно L. turionifera и L.
japonica Landolt.
Морфологическое описание
Ряска маленькая — вегетативный однолетник с эллиптическими,
обратнояйцевидными или почти округлыми, цельнокрайними, упло-
щенными (не более 1 мм толщины) листецами, 1-8 (10) мм дайной и
0,6-5 (7) мм шириной, соединенными в группы по 2-6. Ножка белесая,
очень короткая и не выдается за край материнского растения. Верх-
няя поверхность листеца зеленая или немного буровато-красноватая,
блестящая, слегка выпуклая или плоская, с немногочисленными мел-
кими бугорками, расположенными вдоль средней жилки, и .иногда, с
более крупными бугорками выше "узла" и на конце дистальной час-
ти листеца. Нижняя поверхность бледно-зеленая или слегка красно-
ватая, плоская. Жилок 3, редко 4—5. Корешок один, 0,15—0,2 мм в
диаметре и до 15 см дайной; корневой колпачок притупленный. Воз-
душные полости в 2—3 рядах, менее 0,25 мм в диаметре. Зимующие
листецы меньшего размера и с менее развитой аэренхимой.
Генеративные растения морфологически сходны с вегетирующи-
ми. Соцветие из двух мужских и одного женского цветка, окружен-
ных пленчатой спатой. Завязь с одной амфитропной семяпочкой
28
(Daubs, 1965; Sculthorpe, 1967; Landolt, 1986). Стилодий 0,1-0,15 мм
длиной. Пыльцевые зерна шаровидные, уплощенные с проксималь-
ной стороны, (15,4) 21,6—25,2 мкм в диаметре (Куприянова, Тарасе-
вич, 1984). Плоды односемянные, овальные, часто Краснова тыс,
0,8 -1 мм длиной, 0,8-1,1 мм шириной, с крыловидным выступом
0,05—0,1 мм шириной. Семена 0,7—1 мм длиной, 0,4 -0,6 мм толщи-
ной, обратнояйцевидиые, несколько сплюснутые, белесые, с 10 16
продольными ребрами (Лодкина, 1985; Landolt, 1986).
2п = 20, 30, 40, 42, 50, 63, 80, 126 (Blackbum, 1933; Urbanska-
Worytkiewicz, 1975; Landolt, 1986).
Онтогенез и сезонное развитие
Прорастание семян ряски маленькой описано в работах F.
Hegelmaier (1868), O.W. Caldwell (1899), С. Ростовцевой (1901, 1905),
К. Goebel (1921), М.М. Лодкиной (1985) и др. При прорастании за-
родышевая крышечка открывается по кольцевой линии и отодвига-
ется в сторону разрастающимся влагалищем (выростом основания
семядоли) и суспензор-гипокотилем. Семядоля, которую иногда обо-
значают как первый листец, остается в семени и выполняет гаустори-
алъную функцию. Суспензор-гипокотиль и влагалище по мере своего
развития изгибаются и занимают почти горизонтальное положение.
Питание проростка обеспечивается эндоспермом и фотосинтезирую-
щим влагалищем, на верхней поверхности которого расположено
7—8 устьиц. Зародышевый главный корень не развивается. Первый
настоящий листец закладывается в углублении, образованном влага-
лищем и суспензор-гипокотилем, еще во время внутрисемейного раз-
вития зародыша. Его корень пробивает гипокотиль, а листоподоб-
ная часть выдвигается из-под влагалища и несет только один карма-
шек на выпуклой стороне. Морфологический облик последующего
листеца уже ничем не отличается от взрослого растения.
Особи живут недолго. В зависимости от условий культивирова-
ния максимальный возраст отдельного листеца колеблется от 5 до 10
недель (Ashby et al., 1949; Wangennann, 1952; Wangennann, Lacey,
1955; Boszonnenyi, Boszormenyi, 1957 и др.). При этом наблюдается
следующая закономерность: с увеличением скорости вегетативного
размножения длительность жизни рамет уменьшается (Landolt,
1986). В культуре одно растение продуцирует нс более 15-21 дочер-
них особей (Pirson, Gdllner, 1953; Касинов, 1981 и др.). Причем по ме-
ре старения материнского растения величина ее рамет постепенно
уменьшается. Однако вырождения клона не происходит. С опреде-
ленного момента уменьшенные в размерах раметы продуцируют бо-
лее крупные особи и в течение четырех генераций восстанавливается
изначальная величина листецов (Ashby, Wangennann, 1949; Ashby et
al., 1949; Wangennann, Ashby, 1950 и др.). В результате общие разме-
ры клона не уменьшаются из-за отмирания стареющих особей, а в ес-
тественных условиях разрастающийся "ковер" может быстро запол-
29
нить всю поверхность водоема. Факторы, обусловливающие такой
процесс циклического восстановления исходного размера и сто про-
должителыгость в природе, пока неизвестны.
В онтогенезе от дельного листеца можно выделить пят ь периодов
или этапов: развитие внутри кармашка материнской особи, вытяги-
вание, появление корня, образование дочерних особей, отмирание.
Полная дифференциация листеца закапчивается на первом этапе. В
результате меристематической деятельности тканей "узла" первые
боковые лисгецы закладываются, когда ширина материнского расте-
ния превышает 0,08 мм (Кауфман, 1886). В дальнейшем увеличение
количества клеток происходит одновременно и в материнском зача-
точном растении, и в зачатках его двух будущих дочерних особей.
Однако последующее развитие одного из них задерживается. В опти-
мальных условиях, как правило, преимущественное развитие получа-
ет зачаток, обращенный к дистальному концу материнского расте-
ния (Касинов, 1973). Формирование его воздушных полостей и жи-
лок начинается, когда он достигает длины около 0,2 мм (Hegehnaier,
1868). В конце первого этапа ширина листеца колеблется от 0,8 до I
мм.
Выдвижение полностью сформировавшегося .листеца из кармаш-
ка материнского растения осуществляется за счет вытягивания кле-
ток и меристематической активности в зоне его "узла". В результате
в течение второго этапа онтогенеза форма листеца изменяется от уп-
лощенной до почти округлой. Корень пробивает нижний эпидермис,
когда дайна листеца примерно равгга его ширине, и растет до дости-
жения окончательных размеров особи. Скорость его роста изменяет-
ся в зависимости от условий и может достигать 0,5 мм/час. В культу-
ре максимальная длина корней отмечается при высокой интенсивно-
сти света или низком содержании азота (White, 1937; White,
Tcmplcman, 1937; Pirson, Gollner, 1953; Hillman, 1961; Landolt, 1986 и
ДР)-
Увеличение размеров растения, главным образом в длину, про-
должается до появления его первого, а иногда и второго бокового
листеца. Причем'последний выдвигается из кармашка, когда длина и
ширина первого бокового листеца примерно равны. Продолжитель-
ность связи с материнской особью зависит от многих факторов: ус-
ловий среды, интенсивности роста, возрастное™ и др. Иногда, в ос-
новном в культуре, формируется длинная ножка (до 2 мм), которая
вытягивается в сторону материнского растения и раздвигает плотно
расположенные листецы. Чаще ножка нс превышает в длину 0,5 мм и
достигает своего максимального размера непосредственно перед раз-
делением группы in 6 7 листецов.
В оптимальных условиях культивирования новый дочерний лис-
тец появляется через двое суток (Музафаров и др., 1968; Таубасв, Аб-
диев, 1971 и др.). Однако последовательное формирование листецов
из одного кармашка происходит нерегулярно даже при постоянных
условиях культивирования, и экспоненциальный характер роста об-
30
паруживается только при подсчете листецов для всего клона
(Landolt, 1986). В естественных условиях скорость вегетативного раз-
множения проявляет сезонную изменчивость. Максимальные значе-
ния отмечаются в середине летнего периода, а минимальные - вес-
ной и осенью. По данным В.С. Степановой (1933), в прудах Ленин-
градской области скорость размножения ряски маленькой с мая по
июль увеличилась почти в 3 раза, а в сентябре не превышала значе-
ний весеннего периода роста. Наибольший прирост численности, со
160 до 417 листецов на 0,25 кв. м, отмечен с 1 по 9 июля.
В конце лета или в начале осени, при понижении температуры
воды, формируются небольшие, округлые, слегка утолщенные листе-
цы, которые из-за большого количества крахмальных зерен и реду-
цированной аэренхимы погружаются в толщу воды (Hcgelniaicr,
1868; Landolt, 1986; Лукина, 1988 и др.). Нередко развитие таких по-
коящихся листецов останавливается на первом этапе их онтогенеза,
и в таком состоянии они опускаются на дно после заполнения водой
воздушных полостей отмирающего мазеринского растения.
Перезимовавшие листецы начинают всплывать ранней весной
по мере освобождения водоема ото льда. Значительная часть их по-
гибает из-за повторяющихся ночных заморозков. Другие образу-
ют нормальные листецы. Например, в середине апреля 1991 г. в не-
большом водоеме в Москве на площади 0,1 кв. м отмечено 240 лнете-
цов, из которых 38% вскоре отмерли.
По данным Е. Landolt (1986), ряска маленькая относится к груп-
пе редко цветущих и плодоносящих растений. Из просмотренных им
1500 гербарных листов лишь 86 образцов были с цветками и 11 с
плодами. На территории бывшего СССР, начиная с 1814 г„ цветение
этого вида было документально зарегистрировано всего 60 раз
(Goldbach, 1817; Горожанкин, 1888; Ростовцев, 1901, 1905; Сидоров,
1910; Шпор, 1915; Жуковский, 1923, 1948; Булавкина, 1925; Доброхо-
това, 1949; Иванова, 1970; Матвеев, 1977; Кузнецов, Зайченко, 1979;
Чорна, 1979; Печенюк, 1984, 1985; Голубинский, Гапои, Секач, 1987;
Еремеева, 1988 и др.). При э том цветущие растения чаще всего обна-
руживались в середине лета (koiда вода в водоемах прогревается в
наибольшей степени) и в течение немногим более одного месяца. Ре-
же цветение наблюдалось на протяжении поч ти всего лета. Так, в не-
большом водоеме в Хоперском заповеднике в 1983 г. ряска зацвела
16 мая, когда температура на поверхности воды достигала 23.6°. На
100 кв. см насчитывалось от 3 до 53 цветущих листецов. Заметное
цветение продолжалось до 20 июня, а 11 августа отмечались лишь
единичные цветущие растения (Печенюк, 1985).
Во время цветения первым из-под спаты выдвигается рыльце, а
затем поочередно тычинки (Lawahee, 1961; Sculthorpe, 1967). При
этом не исключается возможность гейтоногамии (Hillman, 1961;
Landolt, 1986). По мнению Л.А. Кузнецова и О.Б. Зайченко (1979),
ветер нс имеет существенного значения в опылении растения. Даже
при сильном ветре авторы нс обнаружили пыльцевых зерен на пред
И
мсгных стеклах, смазанных глицерином и экспонируемых от 60 до
120 мин. на разных расстояниях от цветущих листецов.
Семена ряски маленькой прорастают на следующий год после
цветения (Ростовцев, 1901, 1905). Они сохраняют всхожесть в течение
нескольких педель при воздействии небольшой отрицательной тем-
пературы и могут тронуться в рост при температуре воды около 5°
(Guppy, 1894; Таубаев, Абдиев, 1973 и др.). При высыхании семена
быстро погибают (Ростовцев, 1905; Аржанов, 1926). Однако, по дан-
ным Л.А. Кузнецова и О.Б. Зайченко (1979), при выдерживании пло-
дов ряски маленькой без воды is течение четырех месяцев всхожесть
семян составляла 80° о.
Экология
Ряска маленькая — свободноплавающий на поверхности воды
гидрофит, обитающий в самых разнообразных пресных водоемах.
Особенно хорошо она развивается в небольших эутрофных прудиках
и озерках. В крупных водоемах и реках приурочена к защищенным
от ветра и слабо проточным участкам. По некоторым сведениям, ус-
тойчива к колебаниям уровня воды в прибрежной зоне (Булавкина,
1925; Sculthorpe, 1967) и на сырой илистой почве долго сохраняет
способность к вегетативному возобновлению (Guppy, 1894). Однако
ио данным Р.А. Keddy (1976), вынутые из воды листецы высыхают
через два часа при температуре 25° и влажности воздуха 70%.
Ряска маленькая — гелиофит, но хорошо развивается и при сла-
бом затенении (7-я ступень шкалы Элленберга; 4-я ступень шкалы
Ландольта). С увеличением сигал света с 50 до 300 cd отмечается ли-
нейное возрастание ее фотосинтеза (White, Temoleman, 1937). В сте-
рильной культуре световое насыщение фотосинтетического выделе-
ния О2 при температуре 15-35° набгаодается в интервале 300-600
мкэишгейн/м-с. При этом оптимальным условием культивирования
является постоянное освещение (Лукина, Смирнова, 1988; Лукина,
Микрякова, 1990).
Температурный интервал устойчивости ряски маленькой, по-ви-
димому, составляет около сорока градусов. Растение прекращает ве-
гетативное возобновление при 2-5° (Таубаев и др., 1971; Landolt,
1986), но листецы не отмирают подо льдом в течение нескольких
дней при температуре воды ниже 0°. В водоемах Ташкентской облас-
ти интенсивная вегетация не прекращается и зимой при 5-12° (Муза-
фаров и др., 1968). Экспериментально установленный порог теплоус-
тойчивости листецов достигает 41,5° (Лукина, Смирнова, 1988). Во
флоре водохранилищ-охладителей ряска маленькая вполне обычное
растение (Казанская, 1979, 1981). Известны се местонахождения и в
термальных источниках с температурой воды 32-35° (Воронихин,
1953; Библь, 1965). Однако в экспериментальных условиях при такой
icMiiepaiype наблюдается остановка роста (Landolt, 1957, 1982,
32
\
\
1986). Оптимальная температура культивирования ряски при осве-
щенности 7 тыс. лк составляет 25-26° (Лукина, Микрякова, 1990). Ве-
роятно, это значение ненамного превышает оптимальную темпера-
туру для роста растения в естественных условиях (Hillman, 1961;
Landolt, 1986).
Область возможного распространения ряски маленькой характе-
ризуется, по Е. Landolt (1982, 1986), следующими климатическими
показателями. Число дней со средней температурой выше 10° — не
менее 50. Средняя минимальная температура трех наиболее холод-
ных месяцев — минус 15°. Абсолютный минимум — минус 30°. Сред-
няя температура трех наиболее теплых месяцев — не ниже 10° и не
выше 24°.
Другим важным фактором, опзаничивающим распространение и
развитие ряски маленькой, является содержание в воде азота, фосфо-
ра, калия и других элементов минерального питания (Hillman, 1961;
Landolt, Wildi, 1977; Liiond, 1980, 1990; Zimmermann, 1981 и др.). Из-
менения скорости вегетативного размножения от концентрации не-
которых их этих элементов и соединений отражены на рис. 2 и 3.
Минимальная потребность ряски маленькой в кальции, магнии, ка-
лии, азоте и фосфоре соответственно составляет (в мг/л): 10, 1,0, 1,0,
0,04 и 0,006 (Landolt, 1986). В слабо эфтрофицированных водоемах
отмечается низкая урожайность растения. Например, в некоторых
озерах Волжского бассейна при незначительном содержании в воде
биогенов фитомасса (сырой вес) не превышает 0,5 кг/кв. м (Лукина,
Микрякова, 1990). В условиях культуры наибольшая продуктивность
наблюдается на органических средах. При этом ряска одинаково хо-
рошо реагирует на увеличение концентрации аммония и нитратов
(Landolt, 1986), но вегетативное размножение интенсивнее протекает
на средах с солями аммония (Liiond, 1990).
Активная кислотность воды, вероятно, опосредованно влияет на
распространение и развитие ряски маленькой. В одних случаях опти-
мальной для роста является нейтральная или слабощелочная среда
(Sculthorpe, 1967), в других — кислая или слабокислая (Aschy, 1929;
Музафаров и др., 1968). В естественных условиях ряска вполне ус-
пешно развивается в озерах с pH 5 и меньше (Hick, 1932), а в культу-
ре устойчива к pH 3,2 (Landolt, 1986). Однако сильнокислая и кислая
вода, как правило, бедна элементами минерального питания, и в
природе ряска маленькая обычно встречается в водоемах с pH 5-7,5
(3-я ступень шкалы Ландольта).
Биологическая продуктивность
В водоемах Ленинградской области урожайность ряски малень-
кой достигает 1,73 кг/кв. м, в Рязанской — 0,94 кр/кв. м, в других
районах не превышает 3 — 4 кг/кв. м в сыром весе (Таубаев, Абдисв,
1973; Кузнецов, Зайченко, 1979; Евдушенко, 1988; Мельник, 1988 и
33
др.). Наибольшие значения фитомассы отмечаются в чистых зарос-
лях. В Ташкентской области, в озерах бассейна р. Чирчик, продук-
ция сырой массы колеблется от 2,8 до 3,8 кг/кв. м, сухой — 0,19—0,32
кг/кв. м. При многократном изъятии суммарное значение фйтомассы
возрастает в несколько раз и может превышать 20 кг/кв. м сырого ве-
са (Музафаров и др., 1968; Пашкевич, Юдин, 1978). В условиях куль-
туры продуктивность значительно выше. Так в бетонированных бас-
сейнах при внесении органических удобрений и периодическом сборе
урожая суммарная фитомасса за апрель-ноябрь составляет 279 т/га
(Таубаев и др., 1971).
Консортивные связи
С ряской маленькой топически или трофически связано большое
число организмов. Поверхность растения заселяют различные бакте-
рии, сине-зеленые, зеленые и диатомовые водоросли. Микроорганиз-
мы прочно прикрепляются к корням благодаря наличию на их по-
верхности слизистого слоя или приникают в межклеточные полости
паренхимы, как, например, виды родов Gleotrichia, Anabaena,
Calotrix, Cylindrospennum (Zuberer, 1984; Landolt, 1986). С некоторы-
ми сине-зелеными водорослями, возможно, формируются симбиоти-
ческие отношения (Landolt, 1986; Лукина, Микрякова, 1990). Число
бактерий на верхней поверхности листецов достигает 0,1х106 кле-
ток/кв. см, на нижней — 9,4x10б клеток/кв. см, а на корнях — 4,4x106
клеток/кв. см (Hossel, Baker, 1979, цит. по: Landolt, 1986). Различные
микроскопические простейшие, питающиеся бактериями и водорос-
лями, находят в зарослях ряски благоприятные для себя условия (на-
пример, Vorticella convallaria, V. microstoma, Pyxidium invaginatum,
Chilodonella uncinata). Высокая степень ассоциированности с зарос-
лями ряски маленькой отмечается для многих планктонных живот-
ных (например, Cyclopodia, Calanoida, Cladocera, Bosmina, Ostracoda,
Ainphipoda).
Ряд грибов (например, Hyphochytriomycetes) участвует в минера-
лизации отмирающих растений. Другие (Rhizoctonia solani, Pythium
myriotylum) поражают живые листецы и при определенных условиях
могут привести к гибели всей популяции (Rejmankova et al., 1986).
По данным Н.С. Гаевской (1966), ряска маленькая входит в пи-
щевой рацион 24 видов фитофагов. Среди них моллюски (Calba
palustris, Limnaea stagnalis, Radix sp., Coretus comeus), ракообразные
(Asellus aquaticus, Gammarus pulex, Leander modestus, Palaemon sp.),
ногохвостки (Sminthurius aquaticus), равнокрылые хоботные (Aphis
sambuci, Rhopalosiphuna nympheae), жуки (Hydrothassa marginella),
двукрылые, бабочки (Nymphula obliteralis), рыбы, птицы и млекопи-
тающие (Ondatra zibetica, Myocastor coypus). Суточное потребление
ряски некоторыми из них в несколько раз превышает вес их тела.
Возможно, некоторые моллюски и ракообразные используют расте-
34
ние, прежде всего, как дополнительный источник кальция (Гаевская,
1966; Landolt, 1986). Ряд насекомых (например, Scirtes tibialis,
Tanysphyrus lemnae) являются строгими монофагами, другие обнару-
живают более сложные отношения (Natanson-Grodzinska, 1932;
Scotland, 1934, 1940; Кокин, 1982). Например, жизненный цикл ряско-
вой мушки и бабочки Cataclysta lemnata полностью связан с ряской
маленькой.
Хозяйственное значение
Ряска маленькая известна как ценное кормовое растение. Насе-
ление некоторых приобских поселков издавна использует ее для под-
кормки домашней птицы, собирая растение на вырытых в болоте ма-
леньких водоемчиках (Пашкевич, Юдин, 1978). В смеси с отрубями,
зерном или другими компонентами, ряска хорошо поедается всеми
сельскохозяйственными животными (Гаевская, 1966; Музафаров и
др., 1968, Таубаев, Абдиев, 1971 и др.). В сухой массе растения содер-
жится 30-32% белков, 4-5% жиров, 30-35% крахмала. Протеина в ряс-
ке содержится на 12-14% больше, чем в пшенице, и на 18-19% боль-
ше, чем в кукурузе. По содержанию общего и белкового азота расте-
ние нисколько не уступает бобовым, а в некоторых условиях накап-
ливает его даже больше (Рахимов, 1966). Значительное содержание
витаминов (Е — 35,5-36,8 мг/кг, В} — 2,0-2,1 мг/кг, В6 — 2,1-2,6
мг/кг, Вг, РР и др.), каротина (85-103 мг/кг) и микроэлементов (ко-
бальт — 0,48 мг/кг, бром — 0,18 мг/кг, медь — 0,32 мг/кг, никель —
0,7 мг/кг, титан — 4,8 мг/кг, йод, цинк, ванадий, золото и др.) позво-
ляет использовать это растение как ценную витаминную и биостиму-
ляторную добавку в кормовых рационах домашних животных (Му-
зафаров и др., 1968, 1971 и др.).
Ряска маленькая активно абсорбирует'различные биогенные эле-
менты и может быть использована как активный биоаккумулятор на
специальных очистных сооружениях (Смиренский, 1952; Мережко,
1973; Clandon, McNabb, 1978; Reddy, Debnsk, 1985; Таубаев, Джаман-
гариева, 1988; Краснова, Кузьмичев, 1990).
Как и другие виды семейства, ряска маленькая представляет цен-
ный экспериментальный объект для морфогенетических, биохимиче-
ских, физиологических и радиобиологических исследований
(Hillman, 1961; Касипов, 1973; Landolt, 1986; Landolt, Kandeler, 1987).
В опытах с некоторыми флавоноидами, производными кумарина,
бензойной п коричной кислот показана возможность ее использова-
ния как бпотеста в аллелопатии (Einhelling et al., 1985).
35
2. Ряска горбатая
Номенклатура, систематическое положение, внутривидовая
систематика
Leinna gibba L. 1753, Sp. PL: 970; Lawalree, 1980, Fl. Europ. 5: 273;
Черепанов, 1981, Сосуд, раст. СССР: 284; Щербаков, 1987, Бюл. МО-
ИП, отд. биол., 92. 3: 90 — Telmatophace gibba (L.) Schleid. 1839,
Linnaea, 13: 391; Циигер, 1885, Сборник свед.: 407.
Тип: "in Europae aquis segnibus". Местонахождение типового об-
разца неизвестно (Daubs, 1965; Landolt, 1986).
Таксономический статус L. gibba до конца не ясен. Вид входит в
состав самой крупной секции Lemna и отличается от L. minor, преж-
де всего, выпуклостью нижней стороны листеца. Однако этот при-
знак подвержен сезонной и экологической изменчивости (Kandeler,
1975; De Lange, 1975; Pieterse, 1975; Юнусов, 1979; Печенюк, 1984; De
Lange et al., 1984 и др.), что представляет практически непреодоли-
мые трудности в определении таксономической принадлежности рас-
тений с плоскими листецами. Из других признаков наибольшую ди-
агностическую значимость приписывают размерам воздушных по-
лостей (Landolt, 1986), строению плодов (Kandeler, 1975) и некото-
рым биохимическим особенностям (McClure, 1975). Однако, по дан-
ным U. Starfinger (1985), между типичными формами L. gibba и L.
minor существует плавный переход по форме и симметричности лис-
тецов, отношению их длины к ширине, окраске, числу жилок, форме
корешка, величине воздушных полостей и морфологическим особен-
ностям плодов.
Географическое распространение
Ряска горбатая — гемикосмополитный вид, встречающийся поч-
ти на всех обитаемых континентах. В Австралии и в Новой Зеландии
замещается близким и морфологически плохо отграниченным видом
— L. disperma Hegelm. В Старом Свете ряска горбатая распростране-
на в Европе (проникает на север до 60° с.ш.), Средиземноморье, в
горных районах Афганистана, Пакистана, Индии и юго-запада Ара-
вийского полуострова, а также в Восточной и Южной Африке. В Но-
вом Свете отмечена в горных системах Кордильер и Анд от Орегона
до Огненной Земли (Landolt, 1982, 1986).
Для территории бывшего СССР ряска горбатая приводится для
южных и западных областей европейской части (Флора европейской
части..., 1979), Северного Прикаспия (Иванов, 1969), Армении (Бар-
сегян, 1964), Прибалхашья (Доброхотова и др., 1982) и Средней Азии
(Таубаев, 1970; Мадалиева, 1975). В Средней полосе европейской
России до последнего времени считалась редким видом, характерным
36
лишь для ее южной части (Матвеев, 1961; Маевский, 1964; Камышев,
1978; Лисицына, Экзерцев, 1984).
На территории современной Московской области ряска горба-
тая отмечалась еще в начале прошлого века Ф. Стефаном, Г. Мар-
циусом и М. Максимовичем (Цингер, 1885). Однако Н. Кауфман
(1866, 1889) критически отнесся к этим сведениям и после безуспеш-
ных поисков не включил ее в список "Московской флоры". Почти че-
рез сто пет ряска горбатая была обнаружена Ю.В. Рычиным (1948) "в
истоках речки Апрелевки и при впадении ее в Десну", но из-за отсут-
ствия гербарных сборов вид не был включен в "Определитель расте-
ний Московской области" (Ворошилов и др., 1966). В 1986 г. ряска
горбатая была снова обнаружена в Московской области (Щербаков,
1987), а в последующие года отмечалась в каждом седьмом обследо-
ванном А.В. Щербаковым водоеме. В эти же годы было зарегистри-
ровано ее произрастание в Тверской, Ярославской, Ульяновской
(Лисицына, 1990) и Владимирской областях (Шилов, Богачев, 1988).
В 1988 1989 гг. А.В. Щербаков наблюдал повсеместное развитие
растения в Оке на участке от Мценского района Орловской обл. до
Каширского района Московской, а также в Дону (Кимовский район
Тульской обл.). Летом 1994 г. отмечалась А.В. Щербаковым и В. Лу-
киным в нескольких водоемах Сухиничского района Калужской об-
ласти. Возможно, 'ли многочисленные находки связаны со следую-
щим один за другим теплыми годами или с увеличением загрязнен-
ности водоемов. Если так, то они отражают современное изменение
северной границы распространения вида в европейской России.
Впрочем, не менее вероятно обратное: ряска горбатая и ранее произ-
растала во многих водоемах Московской области (исключая Ист-
ринское водохранилище, Косинские и Шатурские озера). Но, нахо-
дясь на северном пределе своего распространения, образовывала
преимущественно плоские листецы, которые принимались за L.
minor. Очевидно, что и в том, и в другом случае регистрация ряски
горбатой как адвентивного растения Московской области (Игнатов
и др., 1990) не совсем оправдана.
Морфологическое описание
Вегетативный однолетник с округлыми или обратнояйцевидны-
ми, цельнокрайними, вздутыми (до 4 мм толщиной) или уплощенны-
ми листецами 1-8 мм длиной, 0,8-6 мм шириной, соединенными в
группы по 2-4. Ножка белесая, очень короткая и не выдается за край
материнской особи. Верхняя поверхность листеца желтовато-зеле-
ная, иногда блестящая, с ясными красноватыми точками, плоская
или немного выпуклая, нередко буровато-красноватая, особенно по
краю. Жилок 4-5, редко 3-7. Корешок до 12 см длиной; корневой кол-
пачок 0,6-1,8 мм дайной. Воздушные полости в несколько рядов, бо-
лее 0,3 мм в диаметре. Зимующие листецы темно-зеленые, уплощен-
ные и меньшего размера.
37
Цветущие листецы морфологически не отличаются от вегети-
рующих. Соцветие состоит из двух мужских и одного женского цвет-
ков, которые окружены пленчатой спатой. Завязь с 1-7 анатропными
или одной амфитропной семяпочкой (Sculthorpe, 1967; Landolt, 1986).
Стилодий 0,05-1 мм длиной. Пыльцевые зерна почти шаровидные,
(20,0) 21,6-22,1 мкм в диаметре (Куприянова, Тарасевич, 1984). Пло-
ды с 1-6 семенами, шаровидные, 0,6-1,4 мм длиной, 0,8-1,2 мм шири-
ной, с крыловидным выступом 0,1-0,2 мм шириной. Семена 0,7-0,9 мм
длиной, 0,4-0,6 мм толщиной, продолговато-эллиптические, белесые,
с 8-16 продольными ребрами (Лодкина, 1985; Landolt, 1986).
211 = 40, 42, 44, 50, 60, 64, 70, 80, 84 (Urbanska-Worytkiewicz, 1975;
Landolt, 1986).
Онтогенез и сезонное развитие
Проростки ряски горбатой по своему строению сходны с проро-
стками ряски маленькой (Hegelmaier, 1868; Landolt, 1986; Witztum,
1987). Семена засухоустойчивы и выдерживают в течение нескольких
недель небольшие отрицательные температуры (Landolt, 1957; Ива-
нова, 1971; Rejmankova, 1975; Кузнецов, Зайченко, 1979). Прораста-
ют на следующий год после цветения (Янишевский, 1921). При не-
благоприятных условиях, вероятно, сохраняют всхожесть в течение
нескольких лет. Например, семена из поверхностных соскобов со дна
пересохшего водоема в Израиле сохраняли всхожесть в течение пяти
лет (Witztum, 1987).
Отдельные листецы живут недолго, в оптимальных условиях
культивирования —- до 29 дней (Landolt, 1986). За это время они об-
разуют 10—12 дочерних особей (Kang, Cleland, 1985, цит. по:
Landolt, 1986). Онтогенетическое развитие листеца, по-видимому,
полностью совпадает со схемой, приведенной для ряски маленькой.
Покоящиеся листецы темно-зеленые, плоские, с редуцированной
аэренхимой и небольшим корнем (Guppy, 1894; Sculthorpe, 1967). Они
поднимаются со дна на поверхность водоема ранней весной и обра-
зуют вначале плоские дочерние листецы, 'грудноотличимые от листе-
цов ряски маленькой. Растения последующих генераций буровато-
красные и с хорошо развитой аэренхимой, отчего они явно вздуты с
нижней стороны.
По данным Е. Landolt (1986), ряска горбатая о тносится к группе
часто цветущих и плодоносящих растений. Из просмотренных им
1100 гербарных образцов 154 были с цветками и 102 с плодами. На
территории бывшего СССР с 1905 г. зарегистрировано всего 9 случа-
ев цветения этого вида (Ростовцев, 1905; Янитггсвский, 1921; Ма т веев,
1963; Иванова, 1970; Кузнецов, Зайченко, 1979; Чорна, 1979; Печс-
нюк, 1985). Цветение чаще всего наблюдалось в середине лета (когда
вода в водоемах прогревается в наибольшей степени) и в течение
около 6 недель. Реже оно растягивается почти на 3 месяца (Янишев-
ский, 1921). В Саратовской области в разгар цветения на 0,1 м2 па-
38
считывалось до 167 генеративных особей (Матвеев, 1963). Судя по
литературным данным, одинаково хорошо цветут как крупные, так и
мелкие растения, как с хорошо развитой, так и с почти редуцирован-
ной аэренхимой. Элементы соцветия закладываются, когда длина
листеца не превышает 0,07 мм (Landolt, 1986). По наблюдениям И.Е.
Ивановой (1970), цветущий листец развивается трое суток. Первым
из кармашка выдвигается столбик, рыльце которого способно вос-
принимать пыльцу в течение 24 часов. У одних растений он распола-
гается вертикально, и рыльце направлено вверх, у других — изгиба-
ется таким образом, что рыльце обращено к поверхности листеца.
Позднее, но до увядания рыльца, выдвигается терминальная, а через
10—12 часов и базальная тычинка.
Экология
Ряска горбатая — гидрофит, свободноплавающий на поверхно-
сти различных пресных водоемов. В Московской области встречает-
ся в естественных и искусственных водоемах: озерах, водохранили-
щах, прудах, мелиоративных канавах и заводях рек (Щербаков, 1987,
1990). В Ташкентской области особенно хорошо развивайся в за-
грязненных и заболоченных прудиках (Абдиев, 1971).
Ряска горбатая — гелиофит, но хорошо развивается и при не-
большом затенении (7-я ступень шкалы Элленберга; 4-я ступень шка-
лы Ландольта). Листецы не отмирают подо льдом при температуре
воды немногим ниже 0° в течение нескольких дней, хотя вегетатив-
ное возобновление прекращается при 4°. В водоемах Узбекистана
интенсивное возобновление наблюдалось при 28 — 35° (Абдиев,
1971а), а в культуре при 35° скорость роста была минимальна
(Landolt, 1986).
Область возможного распространения ряски горбатой характе-
ризуется, по Е. Landolt (1982, 1986), следующими климатическими
показателями. Число дней со средней температурой выше 10° не ме-
нее 50. Средняя минимальная температура трех наиболее холодных
месяцев минус 1°. Абсолютная минимальная температура мйнус 20°.
Средняя температура трех наиболее теплых месяцев не ниже +10° и
не выше +26°.
Содержание в воде азота, фосфора, магния, кальция и других
элементов минерального питания является важным лимитирующим
фактором. Изменение скорости вегетативного размножения в зави-
симости оз концензрации некоторых веществ о гра жены на рис.З и
рис. 4.
Е. Landolt (1986) приводит следующую характеристику мини-
мальных потребностей в кальции, магнии, калии, азоте и фосфоре:
18; 4,5; 4,5; 0,2 и 0,03 мг/л соответственно. При культивировании
наибольшая продуктивность отмечалась иа органических средах с
добавлением солей магния и железа (Абдиев, 1971а). В естественных
39
Рис. 3. Изменение скорости вегетативного размножения в зависимости от концентрации
азота (по: Ltlond, 1980).
w — Lemna minor, s — Spirodela polyrrhiza, g — Lemna gibba.
условиях ряска горбатая наиболее чувствительна к недостатку азота
(8-я ступень шкалы Ландольта) и одинаково реагирует на его содер-
жание как в нитратной, так и в аммонийной форме (Landolt, 1986).
Однако, по данным A. Liiond (1990), вегетативное размножение про-
текает несколько интенсивнее на средах с солями аммония.
Ряска горбатая предпочитает водоемы с нейтральной или слабо-
щелочной водой (4-я ступень шкалы Ландольта) и не отмирает в
сильнощелочных водах (Landolt, Wild!, 1977). Однако в культуре гиб-
нет при рН=-Х (Landolt, 1986).
Биологическая продуктивность
Фитомасса ряски горбатой в озерах Рязанской области -колеб-
лется от 113 до 150,4 г/кв. м сырого веса; в воздушно-сухом состоя-
нии — от 3,1 до 20,0 г/кв. м (Кузнецов, Зайченко,. 1979). В условиях
культуры интенсивный рост наблюдается в теплые месяцы года. Вес-
ной и осенью прирост не превышает 20-25 г на 300 г маточной куль-
туры в сыром весе. При внесении органических удобрений и неодно-
кратном снятии урожая суммарная фитомасса за шесть месяцев куль-
40
Рис. 4. Изменение скорости вегетативного размножения в зависимости от концентрации
кальция и магния (/), фосфора (//) (по: Zimmermann, 1981; Liiond. 1980).
Обозначения как на рис. 3.
41
тивирования может достигать 185 — 190 т/га сырого или 12 — 14
т/га сухого веса (Абдаев, 1971 а).
Консортивные связи
По данным Н.С. Гаевской (1966), известны только два фитофага
(Rhopalosiphum numpheae, Desmana moschata), в пищевой рацион
которых входит ряска горбатая. В штате Луизиана (США) было от-
мечено массовое поражение листецов пероноспоровым грибом
Pythium myriotyluin (Rejmankova el al., 1986). Первые симптомы забо-
левания обнаруживались в марте, а через несколько дней более 90%
растений погибло. По характеру развития все пероноспоровые qjn-
бы — эфемеры (Новотельнова, 1976) Однако, по мнению авторов,
исследовавших подверженность рясковых заболеванию питиумом,
быстрая гибель популяции растения связана не с ритмом развития
гриба, а с выделяемым им фитотоксином. В условиях культуры неко-
торые плесневые грибы способствуют усилению роста листецов.
Предполагается, что это связано с выделением грибами этилена
(Wetsteynet al., 1984).
Хозяйственное значение
Ряску горбатую можно включать в кормовые смеси почти для
всех сельскохозяйственных животных (Абдиев, 1971а).
3. Ряска трехдольная
Номенклатура, систематическое положение, внутривидовая
систематика
Lemna trisulca L. 1753, Sp. Pl.: 970; Кауфман, 1866, Моск, фл.,
изд.1: 601; Цингер, 1885, Сборник свед.: 406; Шмальгаузен, 1897, Фл.
Средн, и Южн. Росс. 2: 526; Сырейщиков, 1906, Илл. фл. Моск. губ. 1:
218; он же, 1927, Опред. раст. Моск, губ.: 84; Кузспева, 1935, Фл.
СССР, 3: 493; Назаров, 1940, в Маевск., Фл., изд. 7: 208; Е. Бобров,
1954, цит. соч., изд. 8: 704; он же, 1964, цит. соч., изд. 9: 690; В.Тихо-
миров, 1966, Опред. раст. Моск, обл.: 108; Иконников, 1979, Фл. ев-
роп. части СССР, 4: 321; Lawalree, 1980, Fl. Europ. 5: 273; Черепанов,
1981, Сосуд, раст. СССР: 284; Новиков, 1986, Опред. раст. Мещеры,
1: 122.
Тип: "in Europae sub aquis pigris puris" (BM). Ряска грсхдольпая
занимает- обособленное положение в роде Lein па и выделяется в мо-
нотипную секцию Hydrophilla Dumort. (Landoil, 1986) пли в само-
стоятельный род Staurogeton Reichenb. (1830, Fl. Genn. Excurs.: 10).
42
Географическое распространение
Ряска трехдольная — гемикосмополитный вид, отсутствующий в
Южной Америке. В Южном полушарии вид редок и пока отмечен
только в водоемах горных систем Центральной Африки, Новой Гви-
неи и на крайнем юге Австралии (Landolt, 1982, 1986). В Голарктике
распространен повсеместно в умеренных и умеренно холодных об-
ластях; заходя на юг по горным системам до Мексики, Южного Сре-
диземноморья, Ирана, Кашмира и Ньянмы. Как наиболее холодо-
устойчивый вид семейства, достигает на северном пределе своего
распространения полосы южных тундр: до 71° с.ш. (Ребристая, Тока-
ревских, 1967; Landolt, 1982, 1986; Флора Сибири, 1987).
В Московской области ряска трехдольная встречается нередко: в
среднем в 41% от числа всех обследованных А.В. Щербаковым водо-
емов. Причем с севера на юг частота ее встречаемости уменьшается с
46% до 36%. Еще реже встречается эта ряска на территории Средне-
русской возвышенности, где количество сравнительно крупных стоя-
чих или слабо проточных водоемов уменьшается. Например, в Туль-
ской области встречаемость L. trisulca не превышает 20%.
Морфологическое описание
Вегетативный однолетник с продолговатыми, тонкими, светло-
зелеными, иногда прозрачными, в верхней части выемчато-зубчаты-
ми листецами. Листецы 3-15 мм длиной (без ножки) и 1-5 мм шири-
ной, соединенные в группы по 5-30. У вегетирующих особей ножка
длинная, выступающая за край материнского листеца, зеленая, 2-10
мм длиной. Верхняя поверхность листецов без бугорков и устьиц.
Жилок 3, редко 1. Воздушные полости формируются только в дис-
тальной части. Корень один, очень редко два, до 2,5 см длиной.
Цветущие листецы меньшего размера, овальные, темпо-зеленые,
с укороченной ножкой и загнутой вниз верхушкой. Эпидермис верх-
ней поверхности с устьицами. Корень маленький. Соцветие из двух
мужских и одного женского цветка, окруженных пленчатой спатой.
Завязь с одной амфитропной семяпочкой (Daubs, 1965; Landolt.
1986). Стилодий около 0,15 мм длиной. Пыльцевые зерна шаровид-
ные, уплощенные с проксимальной стороны, (19,3) 21,0-25,2 мкм (Ку-
приянова, Тарасевич, 1984). Плоды односемянные, уплощенно-шаро-
видные, часто красноватые, 0,6-0,9 мм длиной, 0,7-1,2 мм шириной, с
крыловидным выступом 0,15 мм шириной. Семена 0,6-1,1 мм длиной,
0,5-0,8 мм толщиной, продолговато- >ллиптнческие, несколько сплюс-
нутые, с 12-18 продольными ребрами (Лодкина, 1985).
43
Онтогенез и сезонное развитие
О прорастании семян ряски трехдольной известно немного. Про-
росток мало чем отличается от такового ряски маленькой
(Hegelmaier, 1868; Sculthorpe, 1967). Влагалище не имеет устьиц
(Landolt; 1986), и семена, вероятно, прорастают на дне.
Листецы живут недолго, в естественных условиях не более 6-7 не-
дель. Ножка отмирает поздно, и в результате растение образует за
один вегетационный период небольшое число дочерних особей (не
более 4-5).
В онтогенезе листеца можно выделить пять или шесть периодов
(этапов): развитие внутри кармашка материнской особи, вытягива-
ние, появление корня, образование дочерних листецов и ножки, от-
мирание. Боковые листецы закладываются, когда материнский дос-
тигает 0,07-0,08 мм в ширину (Кауфман, 1886). В конце первого эта-
па ширина листеца колеблется от 0,6 до 0,8 мм. Выдвижение полно-
стью сформировавшегося листеца из кармашка осуществляется за
счет вытягивания клеток и меристематической активности в зоне его
узла. В результате, в течение второго этапа онтогенеза форма листе-
ца изменяется от уплощенной до почти овальной (более быстрый
рост в длину, чем в ширину).
Корень пробивает нижний эпидермис, когда длина листеца не-
много больше его ширины, и обычно растет до становления оконча-
тельных размеров особи. Увеличение размеров листеца, главным об-
разом в длину, продолжается до появления его первого, а иногда и
второго бокового листеца. В это же время или несколько позже из
кармашка материнской особи начинает выдвигаться ножка, деление
клеток в которой направлено в сторону дочернего листеца (Landolt,
1986). Причем по мере развития угол отхождения ножки уменьшает-
ся с 90° до 45°. Возможно, в результате такой переориентации дости-
гается наиболее оптимальное расположение в пространстве связан-
ных в цепочку листецов.
В конце вегетационного периода при понижении температуры
воды формируются относительно более широкие и менее длинные
листецы, которые устойчивы к небольшим отрицательным темпера-
турам воды. Такие покоящиеся, по Е. Landolt (1986), особи, в отли-
чие от других видов семейства, могут вегетировать в течение всей зи-
мы. Не исключено, что именно благодаря этой способности ряска
трехдольная может проникать, столь далеко на север.
Lemna trisulca относится к ipynne иногда цветущих и спорадиче-
ски плодоносящих раст ений. Из просмотренных Е. Landolt 2500 гер-
барных листов 37 образцов были с цветками и лишь 10 с плодами
(Landolt, 1986). В литературе отмечено только 30 случаев цветения
этого вида на территории бывшего СССР (Кауфман, 1889; Флеров,
1898; Ростовцев, 1901, 1905; Шпор, 1915; Булавкина, 1925; Иванова,
1970, 1972; Матвеев, 1977; Чориа, 1979; Печенюк, 1985 и др.). Цвете-
ние растения чаще всего наблюдалось в середине лета и в течение
44
около одного месяца. Так в Харьковской области ряска трехдольная
цвела в самое жаркое время года, когда водоемы начинали пересы-
хать и вода в них хорошо прогревалась (Флора европейской части...,
1979). Цветущие листецы мельче вегетативных и изогнуты таким об-
разом, что их верхушка и основание остаются погруженными в воду,
а средняя часть располагается на поверхности воды. Ножка укороче-
на или вообще не развивается (Ростовцев, 1901, 1905; Иванова, 1970).
Наличие воздушных полостей обусловливает всплывание генератив-
ных растений. Цветущие листецы одиночные или соединены с не-
большой группой вегетирующих. Первая форма образуется, если "це-
почка" Глубоко погружена в толщу воды, вторая — при ее поверхно-
стном расположении (Лукина, 1988).
По наблюдениям И.Е. Ивановой (1970), генеративный листец
развивается в течение четырех дней и цветет около семи суток. Пер-
вым из кармашка выдвигается столбик, который изгибается так, что
воспринимающая поверхность рыльца обращена к плоскости листе-
ца. Рыльце не увядает около 50 ч., вплоть до появления или пыления
тычинок. Тычинки выдвигаются поочередно. В момент высыпания
пыльцы столбик выпрямляется, и воспринимающая поверхность
рыльца оказывается повернутой в сторону нависающего пыльника.
Вторая тычинка выдвигается из кармашка через 24 ч. после первой,
и ко времени ее пыления женский и первый мужской цветок увядают.
Экология
Ряска трехдольная — свободно плавающий в толще вода (ис-
ключая цветущие листецы) гидрофит разнообразных пресных водо-
емов. В Северном Казахстане встречается и в солоноватых озерках
(соленость 8%о — Шенников, 1950). Особенно хорошо развивается в
местах впадения дренажных вод с удобряемых полей (Мяэметс, 1980).
Неустойчива к колебаниям уровня воды в прибрежной зоне и быстро
отмирает на влажной почве (Sculthorpe, 1967).
Ряска грехдольная - гелиофит (8-я ступень шкалы Элленберга;
3-я ступень шкалы Ландольта). На глубине 4,5 м баланс фотосинтеза
отрицателен даже при хорошем освещении (541 кал/кв. см в сутки) и
прозрачности воды 3,8 м. Оптимальные условия освещения отмеча-
ются в этом случае на глубине менее 3 м, но и здесь фотосинтез ха-
рактеризуется высоким положением компенсационной точки. В сред-
них широтах ряска фотосинтезирует и зимой под прозрачным льдом.
Однако в этих условиях фотосинтез малоэффективен и практически
не приводит к увеличению первичной продукции (Покровская, 1976;
Лукина, Смирнова, 1988).
Область возможного распространения ряски грехдольной харак-
теризуется, по Е. Landolt (1986), следующими климатическими пока-
нтгелями. Число дней со средней температурой выше 10° не менее
50. Средняя минимальная темпера тура грех наиболее холодных меся-
45
цев выше минус 40°. Абсолютная минимальная температура — ниже
минус 40°. Средняя температура трех наиболее теплых месяцев — не
ниже +10° и не выше +22°. Минимальная температура, при которой
возможен рост, колеблется от +4° до +6°, а максимальная немногим
выше +30°.
Скорость вегетативного возобновления ряски трехдольной зави-
сит не только от температуры воды, но и от концентрации в ней азо-
та, фосфора, магния, кальция, карбоната калия и других элементов
минерального питания. Е. Landolt (1986) приводит следующие харак-
теристики минимальных потребностей растения в кальции, магнии,
калии, азоте и фосфоре: 40; 4,5; 4,5; 0,04; 0,01 мг/л, соответственно.
При этом отмечается очень низкая активность нитратредуктазы и
растение восполняет расходуемый азот лишь за счет аммонийных со-
единений.
Ряска трехдольная встречается в водоемах со слабокислой, ней-
тральной или слабощелочной водой (3-я ступень шкалы Ландольта).
Биологическая продуктивность
Фитомасса ряски трехдольной в Валдайском озере изменяется в
зависимости от условий от 20 до 124 г/кв. м и в зимний период года
не превышает 100 г/кв. м (Федорова, 1976). В пойменных водоемах
Средней Оби она составляет 0,6 кг/кв. м, а в озерах лесостепной по-
лосы достигает 6 — 7 кг/кв. м сырого веса (Пашкевич, Юдин, 1978).
Консортивные связи
Листецы ряски трехдольной обильно заселяются различными
микроорганизмами, включая бактерии, сине-зеленые, зеленые и диа-
томовые водоросли. Трудности культивирования растения на средах,
свободных от водорослей, и наличие в нижнем эпидермисе листецов
водорослей из родов Nostoc, Gleotrichia, Anabaena, Calotrix и
Cylindrospennum позволяют предполагать, что ряска трехдольная
вступает в симбиотические отношения с некоторыми сине-зелеными
водорослями (Sculthorpe, 1967; Landolt, 1986). Многие микроскопиче-
ские животные, питающиеся водорослями и бактериями, находят для
себя благоприятные условия в зарослях L. trisulca. Количество мик-
рофауны достигает здесь 1674 особей на 1 кв. м поверхности
(Kobusziewska, 1973, цит. по: Landolt, 1986).
Ряска трехдольная входит в пищевой рацион 11 видов животных
(Гаевская, 1966). Среди них моллюски (Limnaea stagnalis, Radix
ovata, Coretus comeus), ракообразные (Asellus aquaticus, Leander
modestus, Palaemon superbus, Palaemonetes sinensis), бабочки
(Cataclysta lemnata), рыбы (Ctenopharyngodon idella), птицы (Anas
penelope, A. strepera) и млекопитающие (Myocastor ceypus). Суточное
потребление растения рядом фитофагов в несколько раз превышает
вес их тела. Например, доля листецов этой ряски от всего объема со-
держимого кишечника свиязи составляет 43% (Гаевская, 1966).
46
Хозяйственное значение
Ряска трехдольная — ценное кормовое растение, содержащее до
30% протеина, 2,7% жиров, 20,8% клетчатки и 24% безазотистых экс-
трактивных веществ на единицу сухого веса (Смиренский, 1952; Во-
ронихин, 1953; Таубаев, Абдиев, 1971; Пашкевич, Юдин, 1978). Вме-
сте с роголистником ряска грехдольная за 15 дней снижала содержа-
ние соединений азота и фосфора в сточных водах животноводческих
комплексов в 1,5 — 2 раза (Шаларь и др., 1983).
4. Многокоренник обыкновенный
Номенклатура, систематическое положение, внутривидовая
систематика
Spirodela polyrrhiza (L.) Schleiden, 1839, Linnaea, 13: 392; Кауф-
ман, 1866, Моск, фл., изд. 1: 600; Цингер, 1885, Сборник свед.: 407;
Сырейщиков, 1906, Илл. фл. Моск. губ. к 219; он же, 1927, Опред.
раст. Моск, губ.: 84; Кузенева, 1935, Фл. СССР, 3: 492; Назаров, 1940,
в Маевск., Фл., изд. 7: 208; Е.Бобров, 1954, цит. соч., изд. 8: 703; он
же, 1964, цит. соч., изд. 9: 690; В.Тихомиров, 1966, Опред. раст. Моск,
обл.: 108; Иконников, 1979, Фл. европ. части СССР, 4: 318; Lawalree,
1980, Fl. Europ. 5: 273; Черепанов, 1981, Сосуд, раст. СССР: 284; Но-
виков, 1986, Опред. раст. Мещеры, 1: 122. Lemna polyrrhiza L., 1753,
Sp. Pl.: 970; Шмальгаузен, 1897, Фл. Средн, и Южн. Росс. 2: 527.
Тип: "in Europae paludibus fossis”. Местонахождение типового
образца неизвестно (Daubs, 1965; Landolt, 1986).
S. polyrrhiza — один из трех видов секции Spirodela.
$ Географическое распространение
Многокоренник обыкновенный — космополитный вид, встре-
чающийся па всех обитаемых континентах. Распространен практиче-
ски по всей Европе, в Средиземноморье, на Кавказе, в Сибири, Сред-
ней Азии и Казахстане, на Дальнем Востоке, Японских островах, в
Китае, Индии, Индокитае, на островах Зондского архипелага, в гор-
ных районах Северной, Восточной и Юго-Восточной Австралии,
Центральной, Восточной и Южной Африки, на большей части Се-
верной и Центраш.ной Америки. В Южную Америку проникает
только па крайнем северо-западе, замещаясь далее к югу и востоку S.
intermedia W. Koch и S. punctata (G.F.W. Meyer) Thompson (Landolt,
. 1982, 1986).
В Московской области, как и на сопредельных территориях,
многокоренник обыкновенный — одно из наиболее обычных водных
47
растений. Обнаружен А.В.Щербаковым в 81,4% от числа всех обсле-
дованных им водоемов Московской области.
Морфологическое описание
Многокоренник обыкновенный — вегетативный однолетник с
округлыми или обратнояйцевидными, цельнокрайними, уплощенны-
ми листецами (3)5-8(10) мм длиной и 2,5-8(10) мм шириной, соединен-
ными в группы по 2-5. Ножка до 2 мм длиной, иногда выступает за
край материнской особи. Верхняя поверхность листеца желто-зеле-
ная, иногда с красным пятнышком выше "узла", плоская, с выдаю-
щимися дугообразными жилками. Нижняя поверхность — фиолето-
во-пурпурная, плоская юш слегка вздутая. Жилок 5-15. Корней 7-21
до 3 см длиной. Воздушные полости в 2-4 рядах, почти достигают
края листеца. У молодых листецов в базальной части находится
пленчатое образование — профиллюм, который прикрепляется лате-
рально ниже "узла" и свободными краями покрывает с одной сторо-
ны основание верхней поверхности листеца, с другой — зону отхож-
дения корней от его нижней поверхности. Турионы округлые или
почти почковидные, 0,5-3,5 мм в диаметре, темно-зеленые или пур-
пурные, бескорневые или с недоразвитыми корнями.
Цветущие листецы морфологически сходны с вегетирующими.
Соцветие состоит из двух (редко трех) мужских и одного женского
цветка, окруженных пленчатой спатой. Завязь с одной амфитропной
или двумя анатропными семяпочками (Daubs, 1965). Стилодий около
0,3 мм длиной. Пыльцевые зерна почти шаровидные, (10) 17-18 (20)
мкм в диаметре (Куприянова, Тарасевич, 1984). Плоды с одним, ред-
ко с двумя семенами, 1-1,5 мм длиной, 1 -1,5 мм шириной, с крыловид-
ным выступом 0,15 мм шириной, часто красноватые. Семена 0,7-1 мм
длиной и около, 0,7 мм толщиной, с 12-20 продольными ребрами.
2п=30, 32, 38, 40, 50, 80 (Landolt, 1986).
Онтогенез и сезонное развитие
Условия прорастания семян неизвестны. Строение проростка,
вероятно, очень сходно с таковым ряски маленькой. В культуре от-
дельный листец живет 33 дня (Boss et al., 1963, цит. по: Landolt, 1986)
и образует от 8 до 10 дочерних особей. В его развитии можно выде-
лить пять периодов или этапов: развитие внутри кармашка материн-
ского растения, вытягивание, появление первых корней, образование
дочерних листецов, отмирание. Первый боковой листец закладыва-
ется, когда ширина материнского листеца составляет 0,09 мм. Про-
цесс дифференциации этого зачатка наступает после увеличения его
ширины до 0,12 мм (Кауфман, 1886). В конце первого этапа ширина
полностью сформировавшегося листеца колеблется от 0,6 до 0,8 мм,
и его выдвижение из кармашка осуществляется за счет вытягивания
те
клеток и меристематической активности в зоне "узла". Причем до по-
явления первого корня рост в ширину протекает интенсивнее, чем в
длину. Первый корень пробивает профиллюм, когда листец достига-
ет в ширину около 4 мм, и растет до становления окончательных раз-
меров растения. Последующие корпи появляются поочередно с ин-
тервалами от нескольких часов до двух суток. Первый дочерний лис-
тец первой генерации выдвигается из кармашка, когда растение име-
ет почти максимальное число корней, а его диипа почти равна или
немного превышает ширину. Ножка разрывается в двух местах
(Hegelmaier, 1868) после появления второю лигзеца второй генера-
ции.
По данным В.С. Степановой (1933), в прудах Jlciiiiin радекой об-
ласти скорость размножения многокоренника обыкновенною с сере
дины апреля до начала августа увеличивалась почти в 4 раза, л в
сентябре не превышала весенних значений. Наибольший нриршт
численности (со 120 до 242 листецов на 0,25 кв. м) был отмечен в не
риод с 28 июля по 1 августа.
В конце лета или осенью при понижении температуры воды вме-
сто боковых листецов развиваются так называемые турионы, кото-
рые более устойчивы к действию низких температур. Воздушные по-
лости у них почти не развиты, а клетки заполнены крахмальными
зернами. Турионы погружаются на дно водоема и до весны не про-
растают (Hegelmaier, 1868; Guppy, 1894; Czopek, 1964). Однако в ус-
ловиях культуры покой турионов можно прервать повышением тем-
пературы воды в течение нескольких дней (Jacobs, 1947; Henssen,
1954; Sculthorpe, 1967).
По данным Е. Landolt (1986), многокоренник обыкновенный от-
носится к группе очень редко цветущих растений. Из просмотренных
им 3500 гербарных листов только 12 образцов имели цветки. На тер-
ритории бывшего СССР с 1884 г. документаш.но зарегистрировано
всего 9 случаев цветения (Раевский, 1884; Ростовцев, 1905; Булавки-
на, 1925; Матвеев, 1977; Кузнецов, Зайченко, 1979; Псчешок, 1985).
Цветущие растения ни по форме, ни по окраске, пи по величине не
отличались от вегетирующих. Сначала раскрывается женский цве-
ток, а затем поочередно — мужские (Ростовцев, 1905; Landolt, 1986 и
ДР-)-
Экология
Многокоренник обыкновенный — гидрофит пресных водоемов,
свободноплавающий на поверхности воды. Хорошо развивается в
небольших эутрофных прудиках и озерках, а в крупных водоемах и
реках приурочен к защищенным от ветра и слабо проточным участ-
кам. По некоторым данным, устойчив к колебаниям уровня воды в
прибрежной зоне (Guppy, 1894; Sculthorpe, 1967). По другим — нс
только листецы, но и турионы, если они не покрыты влажным грун-
том, высыхают очень быстро (Jacobs, 1947).
49
Многокоренник обыкновенный — гелиофит (7-я ступень шкалы
Элленберга; 4-я ступень шкалы Ландольта). Соотношение хлорофил-
ла а и хлорофилла b составляет- 2,7 , что характерно для светолюби-
вых растений. У листецов, выросших при освещении 1400 лк, 12-ча-
совая экспозиция светом в 10, 25 и 50 тыс. лк вызывает повышение
дыхания, ассимиляционных чисел и скорости фотосинтеза до 60-85%.
При этом максимальное увеличение активности фотосинтетического
аппарата совпадает с наибольшей интенсивностью света (Гапонен-
ко, Стражецкий, 1969; Лукина, Смирнова, 1988).
Температурный режим устойчивости многокоренника весьма
широк. Известны его местонахождения в термальных источниках с
температурой воды 35° (Библь, 1965). В культуре остановка роста от-
мечается при 38° (Docauer, 1983, цит. по: Landolt, 1986). Вегетатив-
ное возобновление прекращается при 9-13°, но турионы не отмирают
и при значительно более низкой температуре. При минус 4° они со-
храняют всхожесть в течение трех недель, а при минус 24° — в тече-
ние 24 часов (Jacobs, 1947; Landolt, 1986).
Область возможного распространения многокоренника обыкно-
венного характеризуется, по Е. Landolt (1982, 1986), следующими
климатическими показателями. Число дней со средней температурой
выше 10° — не менее 110. Средняя минимальная температура трех
наиболее холодных месяцев — выше минус 40°. Абсолютный мини-
мум — ниже минус 40°. Средняя температура грех наиболее теплых
месяцев — более +28°, но не ниже +16°.
Важным фактором, лимитирующим распространение и развитие
многокоренника, является содержание в воде азота, фосфора, маг-
ния, кальция, карбоната калия и других элементов минерального пи-
тания (Landolt, 1957, 1982, 1986; Hillman, 1961; Landolt, Wildi, 1977; L
uond, 1980, 1990; Zimmermann, 1981 и др.). Изменение скорости веге-
тативного размножения от концентрации некоторых из этих элемен-
тов отражены на рис. 3, 4. Е. Landolt (1986) приводит следующие
данные по минймальной потребности многокоренника в кальции,
магнии, калии, азоте и фосфоре: 18; 2,5; 1,5; 0,1; 0,01 мг/л соответст-
венно. При этом отмечается, что восполнение ресурсов азота может
происходить за счет поглощения как аммония, так и нитратов. Од-
нако на средах с солями аммония вегетативное размножение проте-
кает интенсивнее (Luond, 1990).
Многокоренник обыкновенный встречается в водоемах со слабо-
кислой, нейтральной или слабощелочной водой (3-я ступень шкалы
Ландольта). В естественных условиях он устойчив в сильнощелочных
водах, а в культуре отмирает- при pH 4 (Landolt, Wildi, 1977; Landolt,
1986).
50
Консортивные связи
Листецы многокоренника обыкновенного активно заселяются
бактериями и водорослями. Некоторые сине-зеленые водоросли
(Nostoc, Gleotrichia, Anabaena, Calotrix, Cylindrospennum) проника-
ют в нижний эпидермис и, возможно, вступают в симбиотические от-
ношения с растением. При э том многокоренник, вероятно, получает
дополнительно соединения азота, а водоросли — защиту от прямого
солнечного света, стимуляторы роста и некоторые органические ве-
щества (Landolt, 1986). Число бактерий на 1 г сырого веса растения
достигает 105 клеток (Antipchuck, 1974, цит. по: Landolt, 1986). Мно-
гие питающиеся бактериями микроскопические животные находят
благоприятные для себя условия в зарослях многокоренника. Чис-
ленность микрофауны достигает' 155 особей на 1 кв. м поверхности
(Kobusziewska, 1973, цит. по: Landolt, 1986). Некоторые грибы засе-
ляют только отмирающие листецы (Hyphochytrioinycctcs). Другие
(Tracya lemnae, Pythium myriotylum) поражают живые растения и при
определенных условиях могут вызывать гибель всей популяции
(Rejmankova et al., 1986).
По данным Н.С. Гаевской (1966), многокоренник обыкновенный
входит в пищевой рацион 6 фитофагов. Среди них моллюски (Galba
palustris, Physa acuta, Coretus comeus), бабочки (Cataclysta lemnata,
Nymphula nymphaeata) и рыбы (Ctenophaiyngodon idella). Суточное
потребление листецов некоторыми фитофагами может в несколько
раз превосходить вес их тела. В Индии были получены положитель-
ные результаты использования прудовика и жука-слоника в ограни-
чении разрастания многокоренника в водоемах (Sahai, Roy, 1977).
Хозяйственное значение
Многокоренник обыкновенный — ценное кормовое растение,
содержащее до 20,9% протеина, 2,7% жира, 14,6% зольных веществ
26,6% клетчатки и 35,2% безазотистых экстрактивных веществ на
единицу сухого веса (Смиренский, 1952; Таубаев, Абдиев, 1971; Паш-
кевич, Юдин, 1978). Способность растения накапливать токсичные
соединения позволяет использовать его для очистки сточных вод
(Лукина, Смирнова, 1988). По наблюдениям противомалярийных
станций, большое скопление многокоренника отрицательно сказы-
вается на размножении малярийного комара (Воронихин, 1953).
51
Т.И. Варлыгина
РОД ТАЙНИК
Род Тайник (Listera) относится к подсемейству Orchidoideae се-
мейства Orchidaceae. Свое название он получил в честь Мартина
Листера — ботаника и лейбмедика английской королевы Анны. Род
содержит около 30 видов, распространенных в холодных и умерен-
ных областях Евразии и Северной Америки. В Европе встречаются
два вида, оба они растут в Московской области.
Ключ для определения видов
1 Кисть многоцветковая, 10—15 см длиной. Цветки на сильно
скрученных цветоножках. Листья эллиптические, яйцевидные,
крупные, 6—14,5 см длиной и 2,8—9 см шириной. Listera ovata
(L.) R. Br. — Тайник яйцевидный.
— Кисть из 4—12 цветков, более короткая, 2—3,5(5) см длиной.
Цветки на прямых нескрученных или очень слабо скрученных
цветоножках. Листья сердцевидные или сердцевидно-почковид-
ные 1—2,5 см длиной и при основании такой же ширины. Listera
cordata (L.) R. Br. — Тайник сердцелистный.
Тайник сердцелистный
Номенклатура
Listera cordata (L.) R. Br. 1813, in Aiton, Hort. Kew. ed. 2, 5: 201;
Кауфман, 1866, Моск, фл., изд. 1: 477; Цингер, 1885, Сборник свед.:
422; Шмальгаузен, 1897, Фл. Средн, и Южн. Росс. 2: 449; Сырейщи-
ков, 1906, Илл. фл. Моск. губ. к 267; он же, 1927, Опред. раст. Моск,
губ.: 97; Невский, 1935, Фл. СССР, 4: 613; Назаров, 1940, в Маевский,
Фл., изд. 7: 243; Голицын, 1954, цит. соч., изд. 8: 693; он же, 1964, цит.
соч.: 680; Скворцов, 1966, Опред. раст. Моск, обл.: 124; Смольянино-
ва, 1979, Фл. европ. части СССР, 2: 21; Moore, 1980, Fl. Europ. 5: 329;
Черепанов, 1981, Сосуд, раст. СССР: 310; Новиков, 1986, Опред.
раст. Мещеры, к 133. Ophrys cordata L. 1753, Sp. PL: 946. — тайник
сердцелистный (семейство Orchidaceae — орхидные).
Географическое распространение
Голарктический циркумбореаяьный вид, ареал которого вклю-
чает всю Европу (в северных районах он встречается чаще, в южных
— спорадически, в горах — до Средиземноморья, Крыма и Кавказа),
52
значительную часть Азии (Малую Азию, Западную и Восточную Си-
бирь, Дальний Восток, Японию), а также Северную Америку.
На территории России распространен в лесной зоне, почти во
всех районах. В европейской части северная граница ареала доходит
до Полярного круга, отдельные местонахождения известны на Со-
ловках, близ Архангельска, на реках Сотке и Няндоме, в нижнем те-
чении-Мезени. В северной подзоне тайги и в лесотундре, на Урале и
в его отрогах встречается довольно часто; для тундровой зоны извес-
тен из Воркуты и Малоземельской тундры. К югу спускается до
Тульской области, среднего течения Дона и Волги. За пределами лес-
ной зоны — в горах Кавказа.
Тайник сердцелистный — вид монтанно-субальпийский с очень
разорванным ареалом. В горах встречается на высоте до 1500 м (Еф-
ремов, Антосяк, 1990).
В Сибири вид на востоке доходит до линии: р. Селенга — вос-
точное побережье оз. Байкал — р. Витим, а на севере не поднимается
выше 64° с.ш., причем в южных районах он встречается чаще. На
Дальнем Востоке отмечен на островах и на Камчатке (Флора СССР,
1935; Флора европ. части СССР, 1976; Флора Сибири, 1987 и др.).
В Московской области тайник сердцелистный встречается очень
редко, в сырых мшистых хвойных лесах. В настоящее время досто-
верно известно два местонахождения вида: на Клинско-Дмитровской
гряде и в Шатурском районе.
Морфологическое описание
Травянистый многолетник с тонким ползучим столоновидным
корневищем. Придаточные корни (в небольшом количестве) распо-
ложены близко к поверхности почвы. Стебель (4,5)6—15(22) см высо-
той, тонкий, слабый, при основании с двумя буроватыми влагалища-
ми, выше листьев слегка железисто-опушенный, более тонкий, без
маленьких редуцированных листочков. Ближе к середине стебля рас-
положены 2 супротивных сидячих листа. Листья имеют сердцевидно-
яйцевидную или почти треугольную форму с сердцевидным основа-
нием, на конце заострены в коротенькое остроконечие, тонкие, снизу
более бледные, 1—2,5(3) см длиной и шириной.
По форме побегов Е.С. Смирнова (1990) относит вид к катего-
рии VII — «корневищных растений с побегами, междоузлия которых
разновелики». Ветвление побеговой системы монохазиальное. Эле-
ментарная единица системы однопорядковая, малометамерная,
двулистная. Все количественные показатели у данного вида в 3—5
раз меньше, чем у тайника яйцевидного. Отличительной чертой тай-
ника сердцелистиого также является то, что междоузлия нижних ме-
тамеров длинные, как это видно из формулы листовой серии (Смир-
нова, 1990). Междоузлие под нижним листом длинное, тогда как под
в торым листом оно укорочено.
53
Соцветие — редкая малоцветковая кисть (4—12 цветков) дайной
2—3,5(5) см. У мощных особей этого вида на многоцветковых соцве-
тиях имеются 2 срединные брактеи, у малоцветковых — их нет
(Смирнова, 1990). Все 6 листочков околоцветника почти одинако-
вые, до 2,5 мм дайной, продолговато эллиптические, тупые; губа
вдвое длиннее листочков (3,5—4 мм), обратно-клиновидная, при ос-
новании с двумя ланцетными зубчиками. Наружные листочки около-
цветника зеленые, внутренние и губа — грязновато-пурпурного цве-
та (Флора СССР, 1935; Флора европ. части СССР, 1976; Флора Си-
бири, 1987).
Колонка очень короткая, прямая, с прямым, немного выемчатым
носиком; поллинии 2-раздельные. Завязь веретенообразная, голая.
Плод — продолговато-овальная коробочка грязно-зеленого цвета.
Семена мелкие темно-коричневые (Собко, 1989).
Растение незаметное и легко просматривается. Диплоидное чис-
ло хромосом на территории республики Коми 2п=39—42, макси-
мальное — 2п=44 (Аверьянов и др., 1982). На побережье оз. Байкал
2п=40 (Иванова, 1987).
Онтогенез и сезонное развитие
По онтогенезу два представленных вида мало отличаются от он-
тогенеза .других наземных орхидей, подробно описанного в прошлом
столетии Т. Ирмишем (1853). От прорастания семени до появления
цветков проходит 13—15 лет. Для прорастания необходимо наличие
влажной среды и присутствие гифов гриба. Гифы проникают в семя
при его набухании или еще раньше. Первые годы растение ведет под-
земный образ жизни, растет медленно, питаясь при помощи гриба.
Первый зеленый лист развивается на 4-ый год (Summerhayes, 1951).
Корневище тайника сердцелистного, в отличие от яйцевидного,
забирает гораздо меньше питательных веществ и сравнительно ко-
роткоживущее. Корни длинные, почти постоянно сильно инфициро-
ваны грибом.
Цветет с середины июня до середины августа, но иногда — ив
сентябре (Summerhayes, 1951).
Способен к медленному росту и в осенне-зимний период (Тата-
ренко, 1991).
Способы размножения и распространения
Тайник сердцелистный размножается семенным и вегетативным
путем. Цветки опыляются мелкими насекомыми (двукрылыми и пере-
пончатокрылыми), которых привлекает неприятный запах и нектар.
Изредка наблюдается самоопыление. Плоды образуют 61—78% (до
80 %) цветков (Вахрамеева, 1988, 1991). На открытых местах зрелые
плоды находят чаще.
54
Для вида более характерно вегетативное размножение побегами,
образующимися из почек на придаточных корнях (Ziegenspeck, 1936;
Summerhayes, 1951; Nilsson, 1979; и др.). X. Расмуссен (Rassmussen,
1986) отмечал способность апикальной меристемы корня орхидных
преобразовываться в стеблевую и формировать у некоторых.видов
придаточные почки на корнях. Это явление было обнаружено и у ви-
дов рода Listera. Особенно часто побеги из таких почек образуются у
совсем, молодых особей. Растения, возникшие таким путем, зацвета-
ют на*3-й год и после этого тоже обретают способность формиро-
вать новые вегетативные особи из почек на корнях. Корневые почки
играют большую роль в жизни тайника сердцелистного, так как по-
зволяют в благоприятных условиях за короткое время образовать
большое количество особей. У данного вида такое размножение про-
исходит интенсивнее, чем у тайника яйцевидного (Summerhayes,
1951).
Экология
Тайник сердцелистный — оксиломезофит, предпочитает сырые и
очень влажные почвы (4-я ступень шкалы Ландольта). Встречается
как на верховых болотах, так и в заболоченных лесах с моховым по-
кровом на очень кислых почвах (pH 3—4,5) и никогда не растет на
почвах нейтральных или щелочных (1-я ступень шкалы Ландольта,
2-я — Элленберга). Растение нетребовательно к богатству почвы,
растет и на почвах, очень бедных минеральным азотом (1-я ступень
шкалы Ландольта). Является индикатором очень кислых и очень
бедных почв. Вид произрастает почти исключительно на почвах бо-
гатых гумусом, избегает минеральные почвы (Landolt, 1977). Часто
встречается на грубых болотных почвах в глубоком торфе среди мха
и хвойных иголок (Mijitzer, 1956). В.Н. Павлов собирал в Теберде у
Северного приюта (ущ. Гоначхир) тайник сердцелистпый на пова-
ленных сгнивших стволах елей и пихт, в моховом покрове, и в таких
же условиях в ущелье Муруджу (личное сообщение).
Г. Элленберг (Ellenberg, 1974) и Э. Ландольт (Landolt, 1977) от-
носят тайник cq?3uejmcTHbni к теневым растениям, которые чаще
всего встречаются в условиях 10% освещенности и практически не
растут при 3%. На освещенных местах встречается редко и в незна-
чительном количестве.
Тайник сердцелистный — геофит, почки его зимуют в почве, но
все же он нс выносит сильных морозов и поздних заморозков. На
территориях с кон тинен тальным климатом встречается только в бла-
гоприятных местообитаниях (Landolt, 1977; Ellenberg, 1974).
55
Фитоценология
На территории России тайник сердцелистный распространен в
тенистых заболоченных или сырых хвойных и смешанных лесах, пре-
имущественно мшистых, и на моховых болотах. Обычно встречается
небольшими группами, но иногда образует скопления до нескольких
тысяч надземных побегов. Растения часто скрыты опадом и поэтому
могут просматриваться.
В хвойных заболоченных моховых лесах Карпатского заповед-
ника на высоте 600—1500 м по берегам ручьев отмечались группы из
нескольких десятков особей (Ефремов и др., 1990). На Украине, Т.Л.
Андриенко (1983) связывает ареал тайника сердцелистного с распро-
странением хвойных лесов. Так, в сосновом лесу между селами Усово
и Червонка ею найдена популяция из 11 особей: 6 из них были гене-
ративными, молодые полностью отсутствовали. Позже в окрестно-
стях с. Усово в заболоченном сосновом лесу с примесью березы, дуба
и ольхи найдена популяция этого вида, занимающая площадь около
15 га. Местами встречается по 5—10 экземпляров на 1 кв. м. Покры-
тие кустарников и кустарничков доходит до 70%, травостой изре-
женный (покрытие около 20%), значительная площадь покрыта
сфагновыми и зелеными мхами (Смык, Бортняк, 1984). Многочислен-
ные популяции тайника сердцелистного, до 104 особей на 1 кв. м,
найдены в Закарпатье (Загульский, 1994). Здесь преобладали взрос-
лые вегетативные особи (28—49%), генеративные составляли от 2,3
до 48%.
В Западной Европе вид обитает как на совсем открытых участ-
ках — в заболоченных вересковых зарослях, так и в лесах, чаще —
сосновых, в тени или на склонах северной экспозиции. Лесные расте-
ния — более крупные и мощные, чем болотные, возможно,они пред-
ставляют две разные расы. На болотах тайник может встречаться
вместе с клюквой, росянкой, болотным миртом и т.д. В лесах приуро-
чен скорее к открытым участкам, где растет среди вереска
(Summerhayes, 1951). В горах и предгорьях Европы и Урала растет в
пихтовых, еловых и елово-пихтовых лесах (Militzer, 1956; Флора Се-
веро-востока..., 1976).
В северных областях европейской части России тайник сердцели-
стный произрастает на облесенных переходных и низинных болотах
в ельниках зеленомошных, черничных, голубичных, хвощево-сфагно-
вых; в сосняках сфагново-черничных и долгомошных; в смешанных
лесах — на кочках, изредка в ерниковой тундре (Флора северо-восто-
ка..., 1976).
В восточной части Московской области обнаружена значитель-
ная по численности популяция в сосняке чернично-сфагновом (Алек-
сеев и др., 1986); найдены также несколько экземпляров на большом
верховом болоте на севере Московской области. Средообразующая
роль вида очень невелика.
56
Консортивные связи, хозяйственное значение и охрана вида
Оба вида тайников имеют эумицетную толипофаговую эндоми-
коризу. Корни к началу инфицирования достигают в длину 5—10 см.
Грибная инфекция распространена по корню очень неравномерно и
се интенсивность невелика (7 -45%). Гриб в корнях часто отсутству-
ет (Татаренко, 1991). Опыление осуществляется двукрылыми и пере-
пончатокрылыми насекомыми (Zicgcnspcck, 1936).
Сведений о каком-либо утилитарном использовании данного
вида нет.
Как вид с многолетним!! подземными органами тайник доволь-
но устойчив к сенокошению, слабому выпасу, слабой рекреации и
беглым пожарам (Татаренко, 1991). Чувствит елен к осушению участ-
ков и другим изменениям гидрологического режима, в том числе и
подтоплению. Так. в Полесской низменности этот вид исчез из-за
сильного заболачивания (Королева, 1990). Растение — мало замет-
ное и пол ому не страдает- от сбора в букеты.
Тайник сер днелистный занесен в Красные книги Белоруссии
(1981), Украины (1980) и Карелии (1985), включен в список редких
видов Московской области.
В культуре данный вид не разводили, так как создать условия
для его выращивания очень сложно (Собко, 1989). Единственный
возможный путь его сохранения — охрана местообитаний, т.е. соз-
дание заказников и заповедников в местах произрастания вида. В
Московской области следует взять под охрану все немногочисленные
популяции тайника сердцелистного. В настоящее время лишь одно
его местообит ание охраняется в заказнике.
Тайник яйцевидный
Номенклатура
Listera ovata (L.) R. Вт. 1813, in Alton, Hort. Kew. cd. 2, 5: 201; Ка-
уфман, 1866, Моск, фл., пзд. 1: 477; [Дингер, 1885 .Сборник свод.: 421;
Шмальгаузен. 1897, Фл. Средн, и Южп. Росс. 2; 448: Сырешциков,
1906, Ил.ч. <]>. !. Моск. губ. 1; 266; он же. 1927, Опред. раст. Моск, губ.:
97; Невский, 1936, Фл. СССР, 4: 612; Назаров, 1940, в Маевск., Фл.
ищ. 2; 243; Голицын. 1954, цит. соч., изд. 8: 693; он же, 1964, цит. соч.
|нд. 9: 681); Скворцов, 1966. Опред. раст. Моск, об.'!.: 124; Смольяни-
нова, 19 '6. ф.ч. свроп. части СССР, 2; 21; Moore. 1980, Fl. Europ. 5:
329; Черепанов, 1981, Сосуд, раст. СССР: 310; Новиков, 1986, Опред.
раст. Мещеры. I: 133. Ophrys ovata I... 1753, Sp. Pl.: 946. тайник
яйцевидный (еемейетво Orchidaceae орхидные).
S7
Географическое распространение
Евроазиатский вид, ареал которого охватывает всю Европу,
Кавказ, Крым, Средиземноморье, Малую Азию, Среднюю Азию
(Тянь-Шань, Джунгарию, Тарбагатай), Западную Сибирь и юго-за-
падную часть Восточной Сибири (Флора СССР, 1935; Флора свроп.
части СССР, 1976; Флора Сибири, 1987 и др.).
Вид равнинно-субальпийский (Ефремов, Антосяк, 1990). В горах
встречается до высоты 2000 м (Ziegenspeck, 1936).).
На территории России довольно широко распространен в лес-
ной зоне. В европейской части северная граница ареала проходит не-
много южнее Полярного круга по территории Соловецких островов
и Архангельской области (Флора северо-востока..., 1976); южная —
совпадает с границей лесной зоны, спускается до среднего течения
Волги и Дона. Восточная граница в Сибири доходит почти до Анга-
ры и оз. Байкал, северная — за Уралом не поднимается выше 59°
с.ш. (Флора Сибири, 1987).
В Московской области тайник яйцевидный встречается по свет-
лым лесам, кустарникам и полянам, спорадически во всех лесных
районах.
Морфологическое описание
Травянистый многолетник с коротким толстоватым корневи-
щем, несущим многочисленные длинные шнуровидные корни. Сте-
бель (20)25—60(70) см высотой, ниже листьев — голый и более тол-
стый, с буроватыми влагалищами, выше листьев — коротко-желези-
сто опушенный, с 1—3 маленькими редуцированными листочками.
Листья широкояйцевидные, суженные к основанию, сидячие, со
стеблеобъемлющим основанием, на верхушке тупЫе или с коротким
заострением, (4)6—10(15) см длиной и 3—5 см шириной. Листьев, ча-
ще всего, два (иногда 3—4). Они — сближенные, почти супротивные,
расположены в средней части стебля. Нижний лист голый и более
толстый, с буроватым влагалищем; верхний — коротко железисто
опушенный.
По форме роста побегов Е.С. Смирнова (1990) относит этот вид,
к той же категории, что и тайник сердцелистный. Формула листовой
серии показывает, что в отличие от тайника сердцелистного, междо-
узлия корневищного участка и поворота к вертикальному росту
(междоузлия нижних метамеров) — короткие. Как и у предыдущего
вида междоузлие под нижним листом длинное, а под вторым листом
оно предельно укорочено.
Соцветие - узкая многоцветковая кисть 10—20(25) см длиной.
Цветки мелкие, желтовато-зеленоватые. Число цветков в соцветии
может доходить до 40 и более (no Summerhayes, 1951, — до 100). Е.С.
Смирнова (1990) обращает внимание на то, что у всех образцов из
58
Московской области, имеющих многоцветковое соцветие, развиты
две срединные брактеи, а у малоцветковых — они отсутствуют.
Прицветники 3,5—5 мм длиной, яйцейидно-ланцетные, заострен-
ные; цветоножки длинные (4—6 мм), железисто-волосистые. Около-
цветник зеленый. Три наружных листочка околоцветника — яйце-
видные, два внутренних — более узкие, линейно-продолговатые, все
— одинаковой длины (4 4,5 мм). Губа желтовато-зеленая, в 2-3
раза Длиннее (0,7—1,1 см), обратно-клиновидная, почти до середины
надрезанная на две линейно-продолговатые лопасти (Флора СССР,
1935; Флора европ. части СССР, 1976; Флора Сибири, 1987). У от-
дельных особей все 3 внутренних листочка околоцветника имею т вид
губы, или, наоборот, губа имеет вид простого лепестка. Некоторые
растения имеют цветки, где присутствуют две губы нормального
строения. В большинстве случаев губа направлена вверх или гори-
зонтально (Suminerhayes, 1951).
Колонка, как и у предыдущего вида, — очень короткая, прямая,
с прямым немного выемчатым носиком, образующим свод над рыль-
цевой ямкой. Поллинии 2-раздельные, мягкие, не имеют каудикулы,
но могут быть снабжены прилипальцами. Пыльники акротопные,
пыльцевые гнезда сближены между собой, так как отсутствует выра-
женный связник (Жизнь растений, 1982). Завязь веретенообразная
или почти шаровидная, нескрученнная, опушенная. Плод — оваль-
ная коробочка зеленого цвета, раскрывающаяся 6-ю щелями. Семена
очень мелкие, светло-коричневые (Собко, 1989).
Диплоидный набор хромосом на территории республики Комн
— от 34 до 36 (Аверьянов, 1982). Число хромосом по «Флоре Сиби-
ри» (1987) 211-32.
Онтогенез и сезонное развитие
Жизненный цикл этого вида, как и предыдущего, в общих чер-
тах описан выше. Надземный побег с одним листом появляется на 4-
й т од после прорас т ания семени, в т орой лис т на 5-й год. Цветение
наступает на 11 15-й год. В условиях культуры развитие может' ид-
ти быстрее. В Ботаническом саду МГУ нами были обнаружены в
1992 т. растения с двумя маленькими листочками (до 15 мм в ..диамет-
ре), обра ювавшнсея не ранее 1988 г. самосевом из семян урожая.
Кроме т ого, в условиях куль туры у одной геттера пинтой особи в тече-
ние двух лег наблюдалось состояние вторичного покоя. После выхо-
да из нет о растение зацвело через год.
Ост, проростка может сохраняться до 15 лег. В начале жизни
корнтт появляются осенью, а йотом весной, парами до четырех
в т од. В первые годы жизни прибавляется по одном)' междоузлию в
год. Длина междоузлии зависти' от глубины их залегания. Корин
продолжают' расти 10 лет. В литературе описано корпевшие с остат-
ками 4-х соцветий и содержащее 60 корней, половина которых не
59
могла проводить воду, но все корни содержали крахмал
(Summerhayes, 1951; Elliot, 1975).
Каждый год корневище дает надземный цветоносный стебель, а
подземный рост продолжается за счет боковой почки, которая рас-
положена у основания цветоноса, так же как у Cypripcdium calceohis.
Растение живет довольно долго: на одном корневище можно насчи-
тать до 24 старых цветоносов.
Семена тайника яйцевидного в отсутствие гифов симбиотиче-
ского гриба не прорастают ни в воде, ни в питательном растворе
(Dowme, 1949). Микоризные грибы необходимы не только для про-
растания семян, но и для развития растений в первые годы жизни.
Однако во взрослом состоянии корни полностью свободны от гри-
бов. С этого времени растения переходят на самостоятельное пита-
ние, чем этот вид существенно отличается от тайника сердцелистно-
го (Summerhayes, 1951). Связь внутреннего мицелия с почвой почти
отсутствует, грибы выходят лишь в нескольких местах под ризодер-
мой. Микориза обнаруживается в более толстых корнях. Заражение
происходит, в основном, осенью. Летом наблюдается активное пере-
варивание триба (Zigcnspeckt, 1936).
Тайник яйцевидный довольно длительно цветущее растение. За-
цветает в середине лета — в шоле. Цветение одной особи обычно
продолжается более месяца. Как исключение, может цвести в сентяб-
ре (Summerhayes, 1951).
Способы размножения и распространения
Как и предыдущий вид, тайник яйцевидный размножается семе-
нами, но чаще — вегетативно. Для него характерна строгая энтомо-
филия. Цветки имеют сладкий запах, привлекающий насекомых.
Специальные анализы показали наличие смеси моно- и сесвкитерпе-
нов. В их составе в значительном количестве обнаружен линалоол,
присутствие которого и определяет характерный запах (Nilsson,
1981). По данным Г. Эллиота (1975), этот запах исходит от всего рас-
тения. Окраска цветков не является привлекающим фактором.
Попав на цветок, насекомое ползет по губе к центральному же-
лобку, чтобы добраться до нектара, и касается хоботком короткой
колонки. Носик колонки подобен рычагу. Даже при легком прикос-
новении из него с силой выталкивается липкая жидкость, которая
прикрепляет к опылителю свободно лежащие поллинии. Затем насе-
комое несет поллинии на другой цветок (Summerhayes, 1951; Жизнь
растений, 1982). Контакт подлиннее с клювиком осуществляется че-
рез их удлиненную верхушку (Жизнь растений, 1982),
Семенная продуктивность у этого вида не связана с видом опы-
лителя. Однако завязывание плодов сильно зависит от погодных ус-
ловий: при цветении в дождливую погоду оно может снижаться до
10—15%, в теплую и сухую — возрастает до 100 % (Собко, 1989). По
данным других авторов, плодов завязывается много; от 37—66%
60
(Вахрамеева и др., 1991) до 85- -89% (Загульский, 1989). У. Пернула
(Perttula, 1941) указывает на то, что опыляется 42 75% цветков, из
них плоды образуют 81...100%, причем только 75 90"-о плодов
нормальные. Урожай семян зависит типа сообщества и от гкологичс-
ских факторов; он может составлять и 310, и 1620 семян на одну ко-
робочку (Загульский, 1989). В Крыму потенциальная семенная про-
дуктивность (ПСП) одного цветка составила 1102 ссмезачатка, а ре-
альная-семенная продуктивность (РСП) плода достигала лишь 825.
Для одного генеративного побега показатели были следующими:
ПСП — 35715, а РСП — 19967 (Назаров. 1995). Некоторые исследо-
ватели проводили успешные эксперименты по самоопылению, пере-
нося поллинии с одного цветка на другой на одном растении. Само-
опыление в природе наблюдается довольно часто (Prochazka, Velisek,
1983).
Жизнеспособность семян, которую определяли путем окрашива-
ния эмбрионов, была доволг.но низкой (20,6%) и полностью утрачи-
валась через 4 недели хранения семян при комнатной температуре. В
условиях холодильника семена теряли жизнеспособность к 8-й неделе
(Rouse, 1989). В природных условиях число всходов па 1 кв. м. со-
ставляло от 1 до 14 (Perttula, 1941).
Новые растения чаще образуются не из семян, а вегетативным
путем. Как и у тайника сердцелистного, они возникают из почек, ко-
торые закладываются вблизи или на конце корня и дают начало воз-
душным побегам, а затем и маленьким растениям, которые зацвета-
ют через 3- --4 года. Почки могут- закладываться и па корневище, да-
вая начало новым особям после его распада. Таким способом проис-
ходит- размножение растений в раннем возрасте, задолго до начала
их цветения. Этому медленно растущему виду данный способ раз-
множения помогает быстрее расселяться в более благоприятных ус-
ловиях. Кроме того, он является единственным шансом на размноже-
ние в сильно затененных местах, где цветки не опыляются насекомы-
ми или вовсе не образуются (Summerhayes, 1951).
Экология
Для тайника яйцевидного характерна широкая экологическая
амплитуда. Эго мезофит', который встречается па почвах от средне
сухих до влажных (3-я ступень шкалы Ландольта) гиги на свежих
почвах (6-я ступень шкалы Г. Элленберга). К богатству почвы вид
нетребователен, растет как на средне богатых, так и гга средне бед-
ных минеральным азотом почвах (2-я ступень шкалы Ландольта, 5-я
— Элленберга). Предпочитает- хороню проницаемые, богатые гуму-
сом почвы из песка и гравия, частично достигая корнями минераль-
ного горизонта (Ellenberg, 1974; Landolt, 1977). Обычно растет на
слабокислых почвах (р/7 5,5 7,5), но может расти на нейтральных и
слабощелочных, па очень кислых почвах никогда не встречается. М.
Mililzer (1956) указывает, что тайник яйцевидный часто встречается
61
пн известняках, па богатых и свежих почвах, в каменистых гпнромо-
ждениях.
Растения комфортно чувствуют себя в условиях полутени, при
освещенности выше 10°•<>, иногда встречаются на открытых местах
(3-я ступень шкалы Ландольта), по светлым лесам, кустарникам и
полянам. Лесные растения обычно более мощные и высокие, чем рас-
тения, росшие на откры том месте.
Тайник яйцевидный — геофит, почки его зимуют в почве и не
страдают от заморозков (Ellenberg, 1974).
Фитоценология
Тайник яйцевидный это, в основном, равнинное растение, но
может подниматься в горы до субальпийского пояса, где встречается
на лугах и по берегам ручьев.
В северных областях растет на осоковых, сфагновых, пушице-
вых, гипповых болотах, на заболоченных лугах, в кустарниках, осо-
ковых березняках, высокотравных осинниках, травяно-сфагновых
сосняках, сырых приручейных смешанных лесах (Флора северо-вос-
тока..., 1976).
В Западной и Средней Европе он довольно обычен и встречается
в разнообразных местообитаниях. Часто произрастает в широколи-
ственных и хвойно-широколиственных лесах, в связи с чем некото-
рые исследователи относят этот вид к неморальным элементам. Это
одна из немногих орхидей, найденных в Англии в молодых посад-
ках хвойных и широколиственных пород деревьев, в разреженных ле-
сах и на опушках. В этих условиях встречается много вегетативных
растений, видимо, еще не достигших генеративного состояния. На
открытых местах тайник яйцевидный был обнаружен в злаковых и
злаково-вересковых сообществах и даже на песчаных шопах
(Suinmerhayes, 1951). Может встречаться па нарушенных местооби-
таниях, по обочинам шоссе и железных дорог (Вахрамеева и др.,
1991), натурализовался в Северной Америке (Goltz, 1982). На Украи-
не в урочище Билс найдены популяции на краю букового 'леса и на
лугово-степных участках (Загульский, 1990). В лесу преобладали вс-
етативныс особи, а на от крытых местах генера тивные.
Столь большое разнообразие сообществ с участием этого вида
свидетельствует о его большой выносливости. Нахождение тайника
яйцевидного в искусственных и нарушенных фитоценозах указывает
на его слабую конкурент ную способность.
В Московской области вид встречается в сырых лиственных п
смешанных лесах, на заболоченных опушках, среди куст арников и па
полянах, па травяных боло тах.
Довольно большую 'по численности популяцию тайника яйце-
вишюго мы наблюдали в Клинском районе, в смешанном слово-бере-
зовом лесу. Плотность особей в популяции была 1,58. Преобладали
вс1етативныс растения 97,5".., i оперативных было очень мало
62
2,5%. Среди вегетативных было немного ювенильных (1,3%) и имма-
турпых (5,1%). Более заметным оказалось участие молодых вегета-
тивных особей (13,9%). Преобладали в популяции взрослые вегета-
тивные растения (11,2%). Значительную их часть, вероятно, состав-
ляли генератнвные особи в состоянии перерыва в цветении, так как
по данным за 1987 г. генера тивных растений в данной популяции
было около 24%. Очевидно сказалось отрицательное воздействие за-
сухи в год наблюдения (1992 г.), а также значительное затенение, соз-
данное подростом древесных пород. Недостаток влаги в почве отри-
цательно повлиял на развитие растений: отмечалось их полегание и
недоразвитие генера тивных побегов.
Консортивные связи
Для данного вида, как уже отмечалось, характерна эумицетная
толипофаговая эндомикориза. Она формируется на корнях растений
с двумя видами грибов из рода Rizoctonia (Dowme, 1949).
Спектр опылителей очень широк (около 100 видов). Согласно
исследованиям, проведенным в Швеции в 1974—1980 it. (Nilsson,
1981), основными опылителями являются наездники (Hymenoptera,
Parasitica, и, особенно, сем. Ichneumonidae), пилильщики (Hymeno-
ptera, Symphyla) и жуки (Coleopters). Наездники переносят 28—56%
поллиниев, пилильщики -- 28—54%, жуки -- 10—15%. Наиболее
приспособлены цветки к опылителям средней длины (8 — 12 мм), с ко-
роткой и довольно маленькой ротовой частью. Вероятно, оптималь-
ное взаимодействие происходит с самцами наездников (Nilsson,
1981). В указанной работе приводится наиболее полный список вы-
явленных опылителей.
Хозяйственное значение и охрана вида
В Верхнем Поволжье тайник яйцевидный использовали для при-
дания лицу яркого румянца, который долго сохраняется, если на те-
реть щеки массой, приготовленной из его корней. В народной меди-
цине это растение применяли дм лечения сыпей у детей (Флора
СССР, 1935). Используют его в медицине и как ви таминное растение
(Травянистые растения СССР, 1971; Кондратюк, Остапко, 1990).
Как и предыдущий вид, тайник яйцевидный устойчив к сеноко-
шению, слабому выпасу, слабой рекреации и к беглым пожарам.
Вид занесен в Красные книги Украины (1980), Карелии (1985) и
в сводку редких видов Мурманской области (1990), а также некото-
рых других областей. В Московской области растение охраняется на
территории четырех заказников.
По нашим наблюдениям и по литературным данным (Arnold,
1927; Собко, 1989;), в первичной куль туре растение хорошо размно-
жается как семенами, так и вегета тивно, о тмечается самосев.
63
М.Г. Вахрамеева, М.Н. Загульский, Т.М. Быченко
ЯТРЫШНИК ШЛЕМОНОСНЫЙ
Номенклатура и внутривидовая систематика
Orchis militaris L. 1753, Sp. PI.: 941; Кауфман, 1866, Моск, фл.,
изд. 1: 467; Цингер, 1885, Сборник свел,: 418; Шмальгаузен, 1897, Фл.
Средн, и Южн. Росс., 2: 454; Сырейщиков, 1906, Илл. фл. Моск. губ.
1: 253; он же, 1927, Опред. раст. Моск, губ.: 94; Назаров, 1940, в Ма-
евский, Фл., изд. 7: 248; Голицын, 1954, цпт. соч., изд. 8: 699; он же,
1964, цит. соч., изд. 9: 680; Скворцов, 1964, Опред. раст. Моск, обл.:
127; Смольянинова, 1976, Фл. свроп. части СССР, 2: 47; Soo, 1980, FL
Europ. 5: 340; Черепанов, 1981. Сосуд, раст. СССР: 312; — ятрышник
шлемоносный (семейство Orchidaceae — орхидные).
Вид довольно однороден, однако иногда выделяют формы, раз-
личающиеся главным образом соотношением размеров отдельных
частей цветка или общими размерами растения (Prochazka. Velisek,
1983), например, Г. longibracteata Schur — растение высотой 50 см с
прицветниками в 3- 4 раза короче завязи; f. sibirica (Schlechter) Soo
низкорослые растения с небольшим числом цветков и пр. О.М.
Шмейдт и Ы.А. Яагомяэ (1987) указывают на белоцветковую форму
О. militaris в Эстонии. С.А. Невский (Флора СССР, 1935) полагает,
что описанный Рейхенбахом Orchis steveni Rchb., встречающийся в
Крыму и на Кавказе, па самом деле — О. militaris L., но имеющий бо-
лее длинные лопасти средней доли губы и более длинное соцветие,
чем типичная форма. Он же считает, что Ю.Н. Воронов и А.А. Гросс-
гейм ошибочно принимали часть кавказских растений О. militaris за
О. steveni. Нельзя исключить, по его мнению, также, что О. steveni
представляет собой гибрид между О. militaris и О. simia, 1ем более
что оба вида в Крыму и на Кавка te нередко раст ут it непосредствен-
ной близости друг от друга. О. militaris вообще склонен к образова-
нию гибридов с другими,видами рода О. purpurea, О. simia, О.
punctulata, О. mono, О. tridentata и др. Особенно часто с первыми
двумя видами. Встречаются и межродовые I пбриды с Dactylorhiza
maculata. Aceras anlhropophorum (Camus, 1885; Reimers, 1958;
Dupperex, 1961,; Billy, Allci/.ctte, 1964; Hunt, 1975: Флора европейской
части СССР, 1976; Heinrich, 1980; Farrcl, 1985 п др.). В хсловиях Мос-
ковской области । ибридныс формы не отмечены.
Географическое распространение
1 ,г,ра шатскин палеарктический вид. В Европе ареал ыо nxi’.aii.t
вает 1 ер pi 11 оршо от Вс ли кобр и i а пип и южной I Мнении ,то I (ей i pan.
пой И i;i Ilin. JO; о: ыг.Ш!. |>о г i-niiom !"ц кой mu 1 л ) '. ринп и jo
64
лее через всю Центральную и Восточную Европу, включая Прибал-
тику, Белоруссию, Украину, европейскую Россию. На азиатском кон-
тиненте ячрышиик шлемоносный встречается в Малой Азии, Иране,
на Кавказе, в Сибири, где доходит до Забайкалья, найден также на
Алтае, в Северной Монголии, северо-западном Китае (Ziegenspeck,
1936; Meusel et al., 1965; Flora Europaea, 1980; Флора европейской
части СССР, 1976; Heinrich, 1980; Farrel, 1985; Собко, 1989).
В. России распространен в пределах лесной зоны (преимущест-
венно-в подзоне широколиственных лесов) от Калиниградской об-
ласти до-Забайкалья, где в пос. Ямкун Читинской области отмечают
крайнюю восточную точку его ареала в России. Вне сплошного ареа-
ла имеет ряд островных местонахождений в Архангельской, Воло-
годской областях, на южном Урале, юге Якутии, кроме того, изоли-
рованная часть ареала охватывает Дагестан, Ставропольский и
Краснодарский края (Снятков и др., 1922; Флора СССР, 1935; Флора
европейской части СССР, 1976; Флора Сибири, 1987; Красная книга
РСФСР, 1988; Орлова, 1993).
В Московской области изредка встречается почти по всей терри-
тории (реже па севере и востоке), но нигде не образует крупных скоп-
лений.
Морфологическое описание
Многолетнее травянистое растение. По И.Г. Серебрякову (1962,
1964), его следует относить к классу наземных травянистых поликар-
пических растений с ассимилирующими побегами нссуккулентного
типа, подклассу клубнеобразующих многолетников. Корневая систе-
ма ятрышника шлемоносного по В.Н. Голубеву (1984), — кистекор-
невая, образована 5—8 (реже более) придаточными корнями, отходя-
щими от основания побега. Клубни ятрышника довольно крупные
(до 2,5 см длиной и 1,2 см шириной), овальные. Одновременно можно
видеть два клубня — старый, слегка сморщенный, функционирую-
щий в текущем году, и новый, молодой, плотный, беловатый, распо-
ложенный несколько выше старого. Иногда на влажной почве в пер-
вой половине лета хорошо заметен короткий столон, несущий моло-
дой клубень. Во многих как отечественных, гак и зарубежных рабо-
тах отмечают, что клубень наземных орхидных корневого проис-
хождения. Однако исследование ряда видов из родов Platanthera,
Orchis, Habenaria и др. показало, что в образовании клубня многих
наземных орхидей принимает участие не только корень, но, частич-
но, и стебель (Shannan, 1939; Ogura, 1953; Kumazava, 1956, 1958). В
связи с этим Р. Дресслер (Dressier, 1981) предложил называть э тот ор-
ган стеблекорпевым тубероидом. Однако э тот термин еще мало упот-
ребим, и мы позволили себе сохрани ть термин «клубень».
С тебель яярышника шлемоносного, 20 50(60) см высотой с дву-
мя низовыми (Серебряков, 1952; Смирнова, 1990) влагалищными ли-
65
стьями при основании, выше них с 3—4(5) сближенными в розетку
глянцевитыми светло-зелеными эллиптическими или яйцевидными,
суженными к основанию срединными листьями, 8—18 см длиной и
2,5—5 см шириной, выше — с двумя стеблеобъемлющими верховыми
влагалищными листьями (низовыми брактеями в понимании Е.С.
Смирновой, 1990). Е.С. Смирнова (1990) относит ятрышник шлемо-
носный к клубневым розетконосным растениям. При сравнении эк-
земпляров из Московской области и с Кавказа она отмечает у кав-
казских образцов более крупные размеры и большее число листовых
элементов, в частности, число розеточных листьев соответственно 3
и 5, а также большее число брактей. Соцветие — густой колос, со-
стоящий из 7—42(66) цветков, в начале цветения пирамидальное, за-
тем цилиндрическое, 4—10(15) см длиной и 3,5—5 см в диаметре.
Прицветники розовато-фиолетовые, яйцевидные, заостренные, 2—3
см длиной, в два раза короче завязи, с одной жилкой или без нее.
Цветки со сладковатым запахом кумарина, неправильные, с простым
венчиковидным околоцветником, состоящим из двух трехчленных
кругов. Окраска их сильно варьирует от почти белой до розовато-
фиолетовой. Все листочки околоцветника (кроме губы) сложены в
шлем (отсюда и название шлемоносный). Листочки наружного круга
околоцветника яйцевидно-ланцетные, заостренные, 0,9—1,3 мм дли-
ной и 0,3 мм шириной, снаружи беловато-розовые, изнутри — с тре-
мя фиолетово-пурпурными жилками. Два внутренних листочка око-
лоцветника розоватые, линейные, заостренные, с одной жилкой, не-
много короче и втрое уже наружных. Губа 10—14 мм длиной, равна
шлему, при основании розовато-беловатая с пурпурными крапинка-
ми и мельчайшими сосочками, трехлопастная, боковые лопасти ее
фиолетово-розовые, до 8 мм длиной, линейные, тупые; средняя ло-
пасть — обратнояйцевидная, короткодвураздельная, с короткими
(до 4 мм) продолговатыми расходящимися цельнокрайними или зуб-
чатыми долями и с маленьким шиловидным зубчиком в выемке меж:
ду ними. Ширина губы между концами боковых долей 7—10 мм.
Шпорец беловатый или бледнорозовый узкоцилиндрический, слабо
согнутый, 5—6 мм длиной, вдвое короче завязи. Тычинка одна,
пыльник ее с параллельно расположенными гнездами, между основа-
ниями которых выдается отросток клювика, пыльца склеена в була-
вовидные поллинии. Завязь нижняя, сидячая, скрученная. Плод —
коробочка, 15—20 мм дайной, из трех плодолистиков, раскрывается
шестью щелями. Семена мелкие (400x150 Мкм), масса одного семени
7,0 7,1 г (Флора СССР, 1935; Маевский, 1954, 1964; Флора европей-
ской части СССР, 1976; Heinrich, 1980; Farrel, 1985). Диплоидный на-
бор хромосом равен 42 (Lid, 1963; Wcinarck, 1963; Majovski et al.,
1974; Farrel, 1985).
66
Онтогенез
О первых этапах развития ятрышника шлемоносного в природ-
ных условиях известно крайне мало. Лишь в работе К. Сипкес
(Sipkes, 1979) сообщается о выращивании растений из семян на лугах
в Голландии. Попытки вырастить растения из семян в природных ус-
ловиях, в Великобритании оказались безуспешными (Farrel, 1985).
Известно, что в природе семена могут прорасти только в присутст-
вии гриба-. В лабораторных условиях на специально подготовленной
среде семена прорастали и без участия грибов на 12-й день (Linden,
1980). Особенно успешно это происходит под белыми флюоресцент-
ными лампами при температуре 12—17°. Отмечено, что лучше про-
растают недозрелые семена.
При прорастании семени формируется маленькое яйцевидное те-
ло 0,2—0,6 мм в диаметре, чаще, по предложению М. Treub (1879),
называемое протокормом. Первые этапы развития ятрышника шле-
моносного были прослежены В. Linden (1980). По его наблюдениям,
через некоторое время после появления протокорма происходит его
поляризация с образованием конического побегового апекса. Через
3—6 месяцев развиваются поглощающие волоски эпидермального
происхождения. За период 6—12 месяцев формируется побег 1 см
длиной (часто с зеленоватой верхушкой) и придаточные корни. Пер-
вый клубень формируется через 1—2 года после прорастания семени.
Однако Н. Ziegenspeck (1936) считает, что первый клубень образуется
на 3-й год, а первый надземный побег с зеленым листом и новый клу-
бень появляются весной 4-го года. Таким образом, после трехлетнего
подземного развития растение переходит в ювенильное состояние,
характеризующееся формированием укороченного побега и неболь-
шого (длиной до 6 см и шириной до 0,5 см) зеленого листа с 2—4
жилками, 1—2 придаточных корней и клубня до 0,5 см длиной и 0,2
см толщиной (см. рис.). В имматурном состоянии ятрышник имеет
также 1 зеленый лист длиной до 8—9 см и шириной до 1,5 см, с
6—8(10) жилками, 3—4 придаточных корня, клубень до 1 см длиной
и 0,5 см шириной. Во взрослом виргинильном состоянии число зеле-
ных листьев увеличивается до 2—3, длина их — до 11 см, ширина —
до 2,5 см, жилок 10—14, придаточных корней 6—7, клубень до 2,5 см
длиной и 1,5 см шириной. В генеративном состоянии — 3—5 разви-
тых зеленых листьев, длиной до 12,5 см и шириной до 4 см, число жи-
лок 16—22, клубень по размеру остается почти таким же. Число цвет-
ков и длина цветоноса значительно варьируют в зависимости от ус-
ловий местообитания и возраста особи (в молодом и старом генера-
тивном состоянии по сравнению со средневозрастным эти параметры
значительно меньше). Генеративного состояния ятрышник шлемо-
носный достигает не ранее, чем через 7—8 лет после прорастания се-
мени (Farrel, 1985; Summerhayes, 1951). Следовательно, в благоприят-
ных условиях каждая стадия онтогенеза (ювенильная, нмматурная,
67
взрослая виргинильная) длится 1—2 года. Несомненно, что в мало-
благоприятных условиях эти сроки могут значительно увеличиться:
Для сравнения мы приводим биоморфологическую характери-
стику возрастных состояний ятрышника шлемоносного, произра-
стающего на территории Украины (Львовская область) — см.
таблицу.
Изменение биоморфологических признаков ятрышника шлемоносно-
го в зависимости от возрастного состояния.
Признак Возрастная группа
Ювениль- ная Имматур- ная Взрослая вирги- нильная Генера- тивная
Число листьев 1 1 2—3 3 и более
Длина листа, см 6,33+0,68 8,87+0,67 10,43±0,70 11,64±0,90
Ширина листа, см 0,48±0,06 1,46+0,16 2,55+0,17 3,56+0,33
Число жилок 3,36+0,52 10,00+0,88 14,50+0,50 22,25±1,88
Длина клубня, см 0,42±0,04 0,80+0,05 2,23±0,23 22,20+0,12
Ширина клуб- ня, см 0,24+0,02 0,50+0,04 1,46±0,11 1,22+0.08
Число корней 2,36±0,27 4,00+0,72 6,72+0,33 8,50+0,92
Длина цветоно- са, см 37,95+1,25
Длина соцве- тия, см 11,50+0,69
Число цветков 31,14+1,88
Примечание. Для взрослых виргинильных и i енера гивных растений
даны размеры нижнего ассимилирующего листа.
В условиях культуры язрышник шлемоносиый может зацвести
значительно раньше на 4 5-й и даже па 3-й год после прораст а-
ния семени (Witncr, 1974; Farrel, 1985).
68
()нтогенсти'1ескпе состояния ятрышника шлемопоеного: j ювенильное.
bn нммагурное. » взрослое виргиинльное. генеративное.
69
Сезонное развитие
На значительной части ареала в условиях континентального
климата с суровой зимой ятрышник шлемоносный — летнезеленое
растение, листья его появляются только весной после таяния снега.
Однако В.Н. Голубев (1984) утверждает, что в Крыму рост листьев
происходит зимой, в связи с чем он относит этот вид к эфемероидам,
отрастающим зимой. Это же явление отмечено и в условиях Англии,
где верхушка новых побегов появляется над поверхностью почвы в
конце декабря — начале января, а в марте начинается рост листьев.
Причем раньше, как правило, начинают раскрываться почки генера-
тивных растений (Farrel, 1985).
В Московской области ятрышник шлемоносный — типичное
летнезеленое растение, его почка появляется над почвой не ранее се-
редины апреля, а листья начинают развертываться в конце апреля —
начале мая. Цветение на большей части ареала (в том числе и в Мос-
ковской области) происходит с середины мая до середины июня, в хо-
лодные годы задерживается недели на две, и тогда растение цветет
до конца июня. На севере ареала и в горах цветет еще позже — в ию-
не—июле. Цветение длится около трех недель (до 25 дней). Раскрыва-
ются цветки в соцветии акропетально. После опыления цветок увяда-
ет через день. Желтеют листья в конце июля—августе, а полностью
отмирают к началу сентября. Плоды созревают и семена высыпаются
в конце августа — начале сентября.
Способы размножения и распространения
Ятрышник шлемоносный размножается преимущественно семен-
ным путем. Семенная продуктивность довольно высока — от 510 до
4600 семян в среднем в одном плоде (Назаров, 1987). Однако отмече-
но, что далеко не в<:е семена жизнеспособны. Доля завязавшихся пло-
дов сильно варьирует; в Московской области 10—60%, на Волыно-
Подолии этот процент более высок — до 65^75% (Загульский, 1990),
в Великобритании значительно ниже — около 28% (Farrel, 1985).
Причем, просмотрев гербарные материалы, этот исследователь уста-
новил, что слабое завязывание плодов наблюдалось в Великобрита-
нии в течение последних 150 лет. Низкий процент завязывания пло-
дов отмечают в Германии, что С. Godfery et al. (1933) и Н.
Ziegenspeck (1936) связывают с относительно низкой численностью
необходимых этому виду опылителей.
По данным различных авторов, число цветков в соцветии колеб-
лется от 12 до 70, число плодов — от 5 до 36 (Godfery et al., 1933;
Farrel, 1985; Назаров, 1987 и др.). М.Н. Загульский (1989) подсчитал,
что для условий Волыно-Подолии число семян в одном плоде в сред-
нем — 3232 (от 2950 до 3520), что составляет на одну особь около
46000, а на кв. м — от 71672 до 8067077 семян.
70
Периодичность цветения у разных растений весьма различна.
Некоторые особи могут цвести в течение 10 лет без перерыва, другие
цветут с интервалами в 1—3 года и более (Summerhayes, 1951; Farrel,
1985). Отмечают наличие вторичного покоя (когда растение пребы-
вает под землей), которое особенно часто наступает после первого
цветения (Farrel, 1985). Клубни ятрышника шлемоносного, как и у
большинства других ятрышников, замещающего типа, т.е. образуют-
ся по одному в год и служат лишь для продолжения жизни особи, а
не даН вегетативного размножения. Однако в исключительных случа-
ях происходит образование одновременно двух клубней, из которых
на следующий год вырастают две самостоятельные особи (Farrel,
1985). Как правило, возникновение второго клубня связано с каки-
ми-либо механическими повреждениями подземной части растения.
В.Г. Собко (1989) указывает на возможность у этого вида ризорести-
туционного размножения — при отделении молодого клубня и нане-
сении порезов при основании старого клубня возникают один или
несколько новых.
Экология
Ятрышник шлемоносный — мезофит (Вахрамеева и др., 1994),
растущий на почвах от сухих до влажных (2—3-я ступени шкалы
Ландольта, 3-я ступень шкалы Элленберга), избегает как очень влаж-
ных, так и сухих местообитаний, обитает преимущественно на хоро-
шо дренированных склонах (2-я ступень шкалы Ландольта). Отмеча-
ют его отсутствие как в странах с очень сухим и жарким летом,.так и
в районах с очень влажным океаническим климатом (Summerhayes,
1951). Вид предпочитает теплое и влажное лето, удовлетворительно
переносит холодные зимы (Farrel, 1985).
Относится к светолюбивым видам, однако способен выдержи-
вать затенение (4-я степень шкалы Ландольта, 7-я ступень шкалы
Элленберга).
На значительной части ареала встречается на почвах с высоким
содержанием карбонатов, иногда на известняках (например, в Эсто-
нии, Швеции), на почвах щелочных и нейтральных с pH от 5,5 до 9,
кислых почв избегает (4-я ступень шкалы Ландольта, 9-я ступень
шкалы Элленберга). Близ восточной границы ареала в Прибайкалье
(Тункинская долина) растет на почвах с pH 6,5—8,25 и содержанием
гумуса от 1,4 до 5,6%. Может встречаться н на небогатых почвах (2-я
ступень шкал Элленберга и Ландольта).
Растет как па равнине, так и в горах, поднимаясь до 1800 м
(Farrel, 1985), чаще па склонах северной экспозиции, чем южной
(Summerhayes, 1951); может рас гп на очень крутых склонах до 40''.
Чешские исследователи указываю г на наличие двух экологиче-
ских опшмумов у этого вида: один па в лажных (даже сырых) лу i ах.
второй — на солнечных каменистых склонах (Prochazka, Velisek,
1983).
Фитоценология
Ятрышник шлемоносный растет в самых разнообразных фито-
ценозах — на сыроватых, сухих остепненных и альварных лугах, сре-
ди кустарников, в светлых лесах, на лесных полянах, дюнных песках
(Vollman, 1914; Soo, Javorka, 1951; Oostrom, 1956; Nilsson, 1979;
Heinrich, 1980; Загульский, 1986 и др.). В Московской области растет
по светлым лесам, лесным полянам, опушкам, кустарникам, на лугах,
обычно не образуя больших скоплений. Предпочитает участки с раз-
реженным травостоем. Часто встречается на пограничных участках
между фитоценозами. Более или менее успешно конкурирует только с
низкорослыми видами, а при высоком проективном покрытии высо-
корослых растений или при мощном развитии мохового покрова его
жизненность и численность снижаются.
Иногда ятрышник шлемоносный можно встретить на нарушен-
ных местообитаниях или во вторичных фитоценозах — по обочинам
шоссе, на железнодорожных насыпях, по краям полей, в искусствен-
ных лесных насаждениях. Вид способен довольно быстро заселять ос-
вободившиеся от хозяйственного использования территории, напри-
мер, заброшенные поля, пастбища, дороги (Hylander, 1966; Nillson,
1979). В Швеции вид найден на лугу, сформировавшемся на месте от-
стойников бумажной фабрики (50-летней давности) (Nilsson, 1988). В
Прибайкалье найдено несколько цветущих экземпляров на железно-
дорожной насыпи. В Прикарпатье часто встречается на залежах, где
образует популяции с высокой плотностью (до 18 особей на 1 м2).
Появление ятрышника шлемоносного на нарушенных местообитани-
ях, характеризующихся низким проективным покрытием травяного
покрова, в значительной мере следует связывать со стремлением из-
бежать конкуренции со стороны более сильных растений.
. В Подмосковье нами были изучены две популяции ятрышника
на юге области: одна — на опушке липово-березового леса, нормаль-
ная, полночленная из 116 особей (соотношение возрастных групп:
ювенильные 6%, имматурные 28%, взрослые вегетативные 34%, гене-
ративные 32%); вторая — на лесном лугу, состоящая всего из 23 осо-
бей (ювенильные 15%, имматурные 65%, взрослые вегетативные 15%,
генеративные 5%). Средняя плотность этих популяций 2,3 особи на
м2, максимальная — 12 особей на м2. На Украине (Львовская об-
ласть) популяция ятрышника шлемоносного в ненарушенном фито-
ценозе имела примерно такой же возрастной спектр: ювенильные - -
12%, имматурные — 27%, взрослые вегетативные -- 55%, генератив-
ные — 6% , средняя плотность популяции 13,0+3,1 особей на м2. Но
во вторичном местообитании (там же) картина совсем иная: юве-
нильные — 49%, имматурные — 22%, взрослые виргинильные
72
15%, генеративные — 14%, средняя плотность популяции 19,25±5,5
особей на м2 (последняя популяция имеет инвазионный характер). В
благоприятных условиях плотность (и численность) популяций мо-
жет быть значительно выше. Например, в Туапсинском районе Крас-
нодарского края — до 15 особей (среди зарослей можжевельника), в
Эстонии — до 18 (на лютиково-разнотравном лугу), во Львовской
области (в посадках сосны) — до 30 особей на м2.
Консортивные связи
Ятрышник — микоризообразующее растение. Особенно важную
роль играют грибы в первые годы жизни растения. В природных ус-
ловиях прорастание семян происходит только в присутствии гриба.
Однако и во взрослом состоянии (в отличие от некоторых других на-
земных орхидных) гифы гриба находят в корнях. Указывалось на
присутствие в корнях ятрышника шлемоносного несовершенного
гриба Rhizoctonia repens (Witner, 1974), позже был обнаружен гриб
Tulasnella sp., относящийся к одной из примитивных групп базиди-
альных грибов (Farrel, 1985).
Из клубней ятрышника шлемоносного был выделен орхинол (ди-
гидроксифенантрен), который, возможно, играет роль фитоалексина
(Gaumann, 1956; Fisch et al., 1973 и др.), правда истинная его роль
еще не совсем ясна (Stroessel, Arditti., 1984). ’
Ятрышник шлемоносный — насекомоопыляемое растение, посе-
щаемое опылителями из различных групп насекомых (Diptera,
Hymenoptera, Lepidoptera и др.) (Ziegenspeck, 1936; Summerhayes,
1951; Farrel, 1985). В Австрии в течение 5 лет подробно изучал опы-
ление ятрышника W. Voth (1987). Он считает наиболее реальными
опылителями шмелей и пчел, самоопыление отсутствует.
Разные части растения нередко служат пищей различным живот-
ным — клубни поедают мелкие млекопитающие и гусеницы некото-
рых бабочек, личинки других насекомых, листья и соцветия — олени,
кролики, улитки (Ziegenspeck, 1936; Summerhayes, 1951; Farrel, 1985).
Соцветия часто повреждают слизни, особенно в сухую погоду. По-
вреждение соцветий не мешает растениям зацвести на следующий год
(Farrel, 1985).
Хозяйственное значение и охрана вида
Клубни ятрышника шлемоносного содержат крахмал (до 30%),
слизь (50%), декстрин (13%), а также сахарозу, пентозу и другие ве-
щества, используемые при различных желудочно-кишечных заболе-
ваниях, язвах, катарах, а также при болезнях верхних дыхательных
путей и как противоядие (Телятьев и др., 1987). Более широко они
применяются в народной, в том числе и тибетской, медицине при
камнях в почках, туберкулезе легких, в качестве общеукрепляющего
73
и пр. (Асеева и др., 1985). Но в связи с тем, что численность и ареал
этого вида в последние годы неуклонно сокращаются, вид отнесен к
охраняемым растениям, и его использование практически прекраще-
но.
Во многих странах видЛанесен в Красные книги И списки охра-
няемых видов, в том числе и в Красные книги России, Украины, Бе-
лоруссии, стран Балтии, Молдавии, Казахстана. Редок вид и во мно-
гих странах Западной Европы, например, в Великобритании сохра-
нились всего четыре местонахождения (Farrel. 1985). Исключение со-
ставляют лишь Швеция и Эстония, где до сих пор еще находят круп-
ные скопления этого вида. Одной из главных причин исчезновения
ятрышника шлемоносного служит усиление хозяйственного воздей-
ствия на занимаемые им территории (распашка полей, выпас скота,
внесение удобрений, мелиорация и пр.). В малоснежные зимы ятрыш-
ник нередко страдает от кабанов (Цепурите, Страздинып, 1983).
На территории Московской области ятрышник шлемоносный
стал очень редок — сохранилось лишь несколько местонахождений,
большинство исчезло. Ряд прежних местонахождений (например, в
Раменках и Хорошево) вошли в черту Москвы или заняты под строи-
тельство мест отдыха (Сальково) и т.д. Ятрышник шлемоносный вне-
сен в список растений, охраняемых на территории Московской об-
ласти.
Ятрышник шлемоносный культивируется в ряде ботанических
садов (Киева, Москвы, Санкт-Петербурга и др.), но самосева не дает
(Кукк, 1980; Вавриш, Собко, 1984; Собко, 1989).
Р.П. Барыкина
ЧИСТЯК ВЕСЕННИЙ
Номенклатура, систематическое положение, внутривидовая
систематика
Ficaria vema Huds. 1762, Fl. Angl. ed. 1: 214; Овчинникова, 1937,
Фл. СССР, 7: 333; Голицын, 1954, в Маевский, Фл. изд. 8: 85; Мацен-
ко, 1964, цит. соч., изд. 9: 86; Ворошилов, 1966, Опред. раст. Моск,
обл.: 167; Черепанов, 1981, Сосуд, раст. СССР: 421; Новиков, 1986,
Опред. раст. Мещеры, к 179. — F. ranunculoides Moench, 1794, Meth.:
215; Кауфман, 1866, Моск, фл., изд. 7: 372. — Ranunculus ficaria L.,
Sp. Pl.: 550; Цингер, 1885, Сборник свед.: 38; Шмальгаузен, Фл.
Средн, и Южн. Росс, к 16; Сырейщиков, 1907, Илл. фл. Моск. губ. 2:
163; он же, 1927, Опред. раст. Моск, губ.: 134. — R. ficaria L. subs,
bulbifer Lawalree, 1955, in Rohyns, Fl. Gen. Belg. (Spermat.) 2: 60; Tutin,
1964, Fl. Europ. 1: 234. — Чистяк весенний (сем. лютиковые —
Ranunculaceae).
Вопрос о систематическом положении рода Ficaria Guett. до сих
пор остается спорным. Некоторые авторы не признают родовой са-
мостоятельности Ficaria и рассматривают его в составе рода
Ranunculus. Недостаточно ясна также систематическая структура ви-
да F. vema Huds. s. 1. Некоторыми исследователями он понимается
широко; многочисленным родственным таксонам, объединяемым под
этим названием, приписывается ранг подвида или разновидности. В
нашей флоре, во всяком случае, встречаются две четко различающие-
ся морфологически и экологически формы. Одна из них, более широ-
ко распространенная, образует надземные клубни, служащие целям
вегетативного размножения, в то время как семенное размножение
подавлено. Другая форма, свойственная окским заливным лугам, не
образует надземных клубней, но прекрасно размножается семенами.
Последняя форма была описана П.А. Смирновым под названием F.
stepporum Р. Smim. 1958, Тр. Приокско-Террасн. заповеди. 2: 142, но,
вероятно, описывалась неоднократно и до этого. Возможно, что и
настоящий типовой F. vema — это как раз «бесклубеньковая» раса,
но это еще требуется установить.
В настоящем очерке речь идет о «F. vema Huds.» в традицион-
ном, привычном для нашей флористики понимании, т.е. только о ра-
се, способной к образованию надземных аксиллярных клубней.
75
Географическое распространение
Чистяк весенний — широко распространенный европейско-за-
падносибирский вид. Ареал его охватывает почти всю территорию
Европы, за исключением районов Крайнего Севера; наиболее север-
ные местонахождения известны близ устьев Печоры и Северной Дви-
ны. Произрастает на Северном Кавказе и в Западном Закавказье, в
Сибири восточная граница проходит по р. Оби; изолированно встре-
чается в Казахстане (Флора СССР, 1937; Atlas Florae Europaeae,
1989). В Московской области широко распространен во всех районах
(Ворошилов и др., 1966).
Морфологическое описание
Чистяк весенний — кистекорневой травянистый эфемероидный
поликарпик, 15—30 см высотой с восходящим, слабым, простым или
ветвистым полурозеточным малоцветковым побегом. Характерено
наличие пучка клубневидно утолщенных яйцевидных или яйцевидно-
продолговатых придаточных корней, приуроченных к основаниям
пазушных почек низовых и розеточных листьев, а также слабо раз-
ветвленных поглощающих корней. Нижняя, несколько утолщенная
часть побега с укороченными междоузлиями и сближенными узлами
несет 2—3 чешуевидных листа и до 3 длинночерешковых очередных
ассимилирующих розеточных листьев. Пластинки их цельные, округ-
ло-серцевидные, до 2—5 см в поперечнике, по краю крупно- или вы-
емчато-угловато-городчатые. «Стеблевые» листья в числе 4—6, час-
то сближенные по 2—3, короткочерешковые (самый верхний лист
может быть сидячим), разной формы — от угловато-яйцевидной до
лопастно-выемчатых. Близ терминального цветка нередко развива-
ется маленький узкоовальный цельнокрайний лист. Пластинки го-
лые, сверху блестяще- или глянцевато-зеленые, мясистые. В пазухах
всех листьев часто формируются маленькие клубни. Мезотонное
ветвление главного побега может достигать трех порядков. Боковые
побеги развиваются, минуя фазу розетки, их удлиненные междоуз-
лия, в числе 1—2 (до 3), достигают 5—6 см. У некоторых более мощ-
ных особей наблюдается кущение в результате пробуждения пазуш-
ных почек 1—2 розеточных листьев. Побеги кущения, подобно глав-
ному побегу, — полурозеточные.
Цветки одиночные терминальные, 2,5 -3,5 см диаметром, на
длинных цветоножках, актиноморфные, гемициклические, с двойным
околоцветником. Чашечка из 3-х желтовато-зеленых чашелистиков.
Листочки чашечки значительно короче лепестков венчика, продол-
говатые, опадающие вскоре после распускания цветка. Лепестки в
числе 8 14, золотисто-желтые, глянцевитые, удлиненно-эллиптиче-
ские или продолговато-обратпояйцевидные, 10 18 мм длиной и
4—7 мм шириной, в основании с медовой железкой, прикрытой ма-
76
пенькой, двухлопастной чешуйкой (Arber, 1936). Тычинки многочис-
ленные, с длинными нитями и неподвижными продолговатыми пыль-
никами. Пестиков много, они расположены по спирали на кониче-
ском цветоложе. Завязь верхняя, опушенная, с сидячим рыльцем. В
пасмурную погоду и на ночь цветки закрываются, не наклоняясь.
Плод — многоорешек; плодики округло-обратнояйцевидные с редки-
ми волосками, вверху вздутые, к основанию сужено-оттянутые, с
очень коротким, слегка заостренным или туповатым носиком (Фло-
ра СССР, 1937) и мясистой белой тканью (элайосомом) при основа-
нии (РысИНа, 1973). Орешки опадают с еще недифференцированным
зародышем.
В подземной части растение имеет очень короткое малолетнее
корневище с мочкой придаточных корней, среди которых различа-
ются: тонкие — питающие, и утолщенные в виде клубней — запа-
сающие; ниже расположенные перезимовавшие клубни более темные,
выше формируются новые, интенсивно накапливающие продукты
ассимиляции на следующую зиму.
Понимаемый широко вид F. vema кариологически дифференци-
рован на ди-, три-, тетра- и гексаплоиды с основным числом хромо-
сом п=8, имеющие в диплоидном наборе соответственно 16, 24, 32 и
48 хромосом (Langlet, 1932; Marsden-Jones, Turrill, 1952; Heywood,
Walker, 1961; Хромосомные числа..., 1969; Gill et al., 1972: Marchant,
Brighton, 1974). Тетраплоидный цитотип указывается в качестве наи-
более распространенного и считается типовым. Установлена корре-
ляция между числом хромосом и наличием аксиллярных клубеньков.
Последние образуются у всех тетраплоидов, иногда у триплоидов и,
как правило, — отсутствуют у диплоидов.
Онтогенез
Невскрывающиеся мпогоорешки можно обнаружить в конце
мая. Вес 1000 орешков — 1,9 г (Корсмо, 1933). Плодики односемен-
ные с обильным эндоспермом и расположенным в базальной части
маленьким недифференцированным округлым зародышем. Зародыш
погружен в полость, образованную в результате лизиса части приле-
гающих к нему клеток эндосперма. Дальнейшее развитие зародыша
происходит после дисссмпнации (Stercke, 1900; Кузнецов, 1936). Ран-
ние фазы онтогенеза описаны R. Stercke (1900), C.R. Metcalfe (1936) и
Г.П. Рысиной (1973). Свежесобранпые плодики, посеянные Г.П. Ры-
сииой в чашки Петри при комка гной темпера туре, проросли через 12
мес., а при темпера туре 2 5° уже через 7 мес. после посева. Сфор-
мированный зародыш к этому времени достигает в длину около 2
мм; единственная семядоля, сложенная йдол1убредней жилки, приоб-
ретает светло-зеленую окраску, почечка остается несформированноп.
Детальные морфологические п ана томические исследования проро-
стков п поздних стадии развития зародыша, проведенные C.R.
Metcalfe (1936), выявили природу псевдомопокотплии у чистяка. Как
77
оказалось, это — результат недоразвития одного из двух исходных
зачатков семядолей, а не синкотилии, как это полагал R. Stercke
(1900).
В природе орешки прорастают весной, в середине апреля. Про-
растание надземное, котиледонарное. Семядоля первоначально вы-
полняет гаусториальную функцию. Разрастаясь и всасывая пита-
тельные вещества эндосперма, она постепенно занимает весь объем
семени. Затем, в начале мая, благодаря удлинению (до 50 мм) ее че-
решка, дву-, реже трехлопастная пластинка освобождается из покро-
вов семени (орешки выносятся на поверхность почвы) и становится
единственным фотосинтезирующим органом растения первого года
жизни, так как почечка еще не образует ни одного листа. Очень ко-
роткий бесцветный гипокотиль проростка переходит в слабо раз-
ветвленный тонкий главный корень. В июне семядоля желтеет и от-
мирает. К этому времени, как показали наши исследования, прове-
денные в дубраве близ пос. Шихово Одинцовского р-на Московской
области, пазушная почка семядоли, имея в основании утолщающий-
ся за счет притока ассимилянтов придаточный корень, метаморфизи-
руется в первый подземной запасающий клубень. Большая часть это-
го клубня образована корнем.
Весной следующего года из перезимовавшей терминальной поч-
ки развивается длинночерешковый лист с яйцевидной пластинкой.
Форма пластинки у основания — слегка сердцевидная, на верхушке
— тупая. В пазухе листа формируется новый почко-корневый клу-
бень, который отличается от семядольного более светлой окраской.
Таким образом, у 2-летних особей в подземной сфере имеется 2 клуб-
ня. Главный корень отмирает и замещается придаточными стебле-
родными узловыми поглощающими корнями.
Трех- и четырехлетние растения развивают по одному более
крупному листу. Осевая часть укороченного побега преобразуется в
эпигеогенное вертикальное корневище.
В природе чистяк весенний зацветает, видимо, не ранее, чем на
5-й год. Цветоносный побег — полурозеточный. В структуре его дос-
таточно четко выражены две морфо-функциональные зоны (Troll,
1964): базальная зона возобновления и зона обогащения, несущая
боковые генеративные побеги и аксиллярные клубни. Выпадает зона
торможения. После плодоношения большая часть полегающего по-
бега отмирает; оставшиеся живыми базальные метамеры с укорочен-
ными междоузлиями составляют очередной годичный прирост кор-
невища, в области сближенных узлов которого регулярно образуют-
ся клубневидные почко-корни. Монокарпические побеги, кроме
главного, — моноциклические.
Средняя высота взрослых генеративных растений 15—19 см. Са-
мые мощные из них (до 32 см), растущие, как правило, по сырым те-
нистым оврагам, развиваются в результате удлинения междоузлий
при сохранении более или менее постоянного числа метамеров. На
осветленных местах формируются относительно низкорослые расте-
78
ния высотой 8—10 см, слабо ветвящиеся (до второго-третьего поряд-
ка). В подземной сфере имеется очень короткое малолетнее корневи-
ще. От него пучком отходят клубневидно утолщенные запасающие
корни, приуроченные к основаниям пазушных почек, и тонкие узло-
вые — поглощающие корни, проникающие на глубину 8—10 см. У
крупных экземпляров может быть до 20—22 клубней, как старых
морщинистых 2—3-летних, так и молодых 1-летних.
Длительность жизни чистяка весеннего благодаря интенсивному
вегетативному размножению, видимо, весьма продолжительна.
Сезонное развитие
Чистяк весенний — ранневесеиний лесной эфемероид. Его веге-
тация проходит в условиях менее напряженной конкуренции с позже
развивающимися растениями при обилии влаги, солнечного света,
наличии в почве продуктов разложения прошлогоднего растительно-
го опада. Начало вегетации приходится на февраль — март. Почка
возобновления, перезимовав на глубине 1,5—2 см, выходит на по-
верхность в середине февраля (Лавриченко, 1970). Нередко рост по-
бега начинается еще под снегом (Горышина, 1969). В середине марта
развертываются первые листья. Одновременно с ними становятся за-
метными и первые клубни, прорывающие розеточные листья в их ос-
новании. Полного развития разветвленный полурозеточный фото-
фильный побег достигает ко второй половине апреля.
Цветение, часто очень обильное (с середины апреля до конца
мая), приурочено к первым вылетам насекомых (пчел, шмелей), осу-
ществляющих перекрестное опыление. Плодики созревают в конце
мая — начале июня. Ко времени отцветания в пазухах стеблевых ли-
стьев (заметно вытянувшихся в течение лета), полегающих генера-
тивных побегов формируются клубеньки. В одном аксиллярном ком-
плексе их может быть от 1 до 4 (у крупных экземпляров). Надземная
масса отмирает до полного облиствления деревьев (т.е. до того, как
их листья примут горизонтальное положение). Живыми сохраняю гея
лишь основания побегов, входящих в состав малолетнего эпигеоген-
ного корневища. В почке возобновления корневища формируется по-
бег возобновления будущего года, и к осени закладываются элемен-
ты цветка.
В течение лета за счет накопления весной запасов ассимилягов
заметно разрастаются заглубленные в почв) клубни, точнее око-
рененные почки, образовавшиеся в пазухах .чнегьев базальных мета-
меров побега. Оптимальная темпера i ура пх развития 20
(Courduroux, 1966). Поглощающие корни, напротив, развиваются в
основном с осени и быстро прекращают рост весной (Серебряков.
1952). Таким образом, свосоора шып ритм ceioinioii вегетации чистя-
ка весеннего обусловлен наличием питающих органов (короткого
корневища и клупией), ранним iaзо'кенпем ночек во (Обновления. в
которых е осени полное то сформирован нооег о\ тхщего го та.
79
Способы размножения и распространения
Чистяк весенний размножается как семенами, так и вегетатив-
ным путем. Однако семена, способные к прорастанию, образуются
лишь спорадически. Преобладает вегетативное размножение и рассе-
ление. Оно осуществляется с помощью диаспор сложной морфологи-
ческой природы (Metcalfe, 1938). Это — почко-корневые клубни (Ба-
рыкина, Туленкова, 1990), регулярно формирующиеся в пазухах как
розеточных, так и «стеблевых» листьев. Они легко отделяются от
побега и часто с потоком дождевой воды разносятся на значитель-
ные расстояния. Прорастают диаспоры лишь в следующем вегетаци-
онном сезоне, давая начало новым, сильно омоложенным растениям,
начинающим цикл развития с ювенильной фазы. Небезынтересно от-
метить, что почко-корневых клубней как в подземной, так и в над-
земной части растения появляется значительно больше числа ежегод-
но развивающихся листьев. Это обусловлено внутрипочечным ветв-
лением, приводящим к формированию в пазухе кроющего листа мно-
жественных почек. Основная почка аксиллярного комплекса часто
образует боковой генеративный побег обогащения, а остальные поч-
ки преобразуются в клубни, выступающие в качестве специализиро-
ванных органов вегетативного размножения и расселения. В лесах, в
условиях затенения, чистяк весенний размножается исключительно
вегетативным способом (Perttula, 1941; Кожевников, 1950; Работнов,
1951; Irmisch, 1868, и др.). Интенсивное образование клубней подав-
ляет семенное размножение.
Цветет чистяк весенний часто очень обильно, однако плодиков
завязывается обычно немного. Отмечается массовое опадение недо-
развитых орешков даже при большом числе завязей, в результате че-
го конечная семенная продуктивность растений очень мала. Это свя-
зано, отчасти, с недостатком опылителей. В литературе отмечается,
что цветки чистяка посещаются насекомыми лишь на солнечных ме-
стообитаниях, где, проростки и всходы могут появляться в значи-
тельном количестве (Рысина, 1973). В тени же под пологом леса,
орешки, как правило, не завязываются (Жизнь растений, 1980). Ис-
кусственное опыление дает хорошие результаты (Горышина, 1969).
Не исключены и внутренние факторы стерильности, что было под-
тверждено R. Dostal (1963), получившим заметное увеличение зрелых
плодиков при обработке гиббереллиновой кислотой. Недоразвитие
части орешков в цветке обуславливает отток питательных веществ к
формирующимся надземным клубням (Marchant, Brighton, 1974). Это
еще раз подтверждает, что побег выступает как целостная саморегу-
лирующая система, в которой взаимодействие между отдельными ме-
тамерами направлено на оптимизацию его функций и поддержание
жизнеспособности растения в целом (Барыкина, Туленкова, 1983).
80
Фитоценология и экология
В Московской области чистяк весенний особенно часто встреча-
ется в лиственных лесах на участках с разреженным травяным покро-
вом. Это — приствольные круги деревьев, сырые лесные зропинки,
крутые склоны оврагов, берега, а иногда даже русла лесных ручьев,
прогадины. Распространен также среди кустарников, на сыроватых
лугах,-реже —на полях (Флора СССР, 1937; Ядовитые растения...,
1950). Плотность куртин часто бывает очень высокой — до 3—5 эк-
земпляров на 1 дм2, что составляет 100%-ное проективное покрытие.
Такая большая плотность обусловлена очень интенсивным вегета-
тивным размножением при помощи подземных и надземных клуб-
ней.
Чистяк весенний — световой мезофит или гигромезофит, по ти-
пу перезимовывания — геофит. Согласно Л.Г. Раменскому и др.
(1956), массовая встречаемость его наблюдается в условиях от луго-
востепного до сыролугового увлажнения (51—80-е ступени шкалы).
Предпочитает свежие и влажньге, но не пересыхающие почвы. Чис-
тяк растет при различной освещенности. Характеризуется относи-
тельным светолюбием, оптимального развития достигает при сомк-
нутости крон деревьев 0,4—0,5. Г. Элленберг (Ellenberg, 1974) распо-
лагает его между теневыми и полутеневыми растениями (4-я ступень
шкалы).
Распространен на почвах разного гранулометрического состава,
лесных, луговых, слабо аллювиальных, богатых минеральным азо-
том. Предпочитает нейтральные почвы, но также хорошо развивает-
ся на кислых и основных.
Температурный оптимум фотосинтеза 8—10° (Mudrack, 1935). В
ясную погоду цветки раскрьгваются около 10 часов и закрываются в
16—17 часов, т.е. они открыты только в те немногие часы, когда тем-
пература воздуха достаточно высока для работы опылителей. В пас-
мурную и дождливую холодную погоду цветки закрыты (Горышина,
1969). Активная, достаточно высокая физиологическая деятельность
растения ограничена околополуденными часами солнечных весенних
дней и в сумме короче, чем длительность надземной вегетации.
Биологическая продуктивность
Изучение продуктивности чистяка весеннего поводилось в раз-
ных местообитаниях в Филевском парке и на Ленинских горах (г.
Москва). Результаты этой работы приведеньг в таблице.
В зависимости от условий местообитаний биомасса надземных
органов колеблется от 1800 до 1000 г, подземных — от 650 до 340 г
сырого веса на 1 м2.
81
Биологическая продуктивность чистяка весеннего
в разных фитоценозах г. Москвы (г/м2)
Фитоценоз Биомасса
сырая воздушно-сухая
надземная подземная надземная подземная
ЛП 1800 650 200 170
ПК 1000 . 340 100 95
Обозначения: ЛП — липняк волосисто-осоковый (Филевский парк,
3 мая 1993 г.); ПК — посадки клена средневозрастные (Ленинские го-
ры, 18 мая 1993 г.).
Консортивные связи и хозяйственное значение
Сведений о консортивных связях чистяка весеннего очень мало.
Энтомофил, насекомых привлекает нектар и пыльца; цветки его по-
сещают пыльцеядные жуки, мухи, а также пчелы, шмели. Относится
к растениям мирмекохорам. Элайосомы — базальные участки тканей
околоплодника, богатые маслом, привлекают муравьев и поедаются
ими. Растение микотрофное. Микориза везикулярно-арбускулярного
типа. Цветущие особи ядовиты. Действующим началом является
протоанемонин, служащий защитой от поедания их животными
(Кречетович, 1931; Гусынин, 1947). После отцветания он разрушает-
ся, и отдельные органы растения становятся пригодными в пищу
(Полянский, 1968). Употребляются богатые крахмалом клубни (в ва-
реном виде), листья (в салатах), бутоны (маринованные). Клубни, ли-
стья и стебли издавна использовались в народной медицине как ле-
чебное средство от грудных болезней, геморроя и цинги (Флора
СССР, 1937).
82
В.Н. Тихомиров, А.А. Нотов, Л.В. Петухова,
К.П. Глазунова
РОД МАНЖЕТКА
Номенклатура, систематическое положение
Род Alchemilla L. s.str. относится к подсемейству Rosoideae Focke
семейства Rosaceae Juss. Монограф рода В. Ротмалер рассматривает
его в пределах трибы Potentilleae Spreng, (R.othmaler, 1937). Ряд авто-
ров относит этот род к трибе Sanguisorbeae Spreng. (Focke, 1894; Тах-
таджан, 1987), иногда выделяют самостоятельную трибу Alchemilleae
Rydb. (Rydberg, 1963; Колчанов, 1970).
Род Alchemilla объединяет более 300 видов, распространенных в
Европе, Азии, Африке, Северной Америке. Все манжетки Средней
полосы европейской части России принадлежат к подсекции Helio-
drosium Rothm. секции Alchemilla (sect. Brevicaulon Rothm.). Эта
группа представляет весьма сложный в таксономическом отношении
комплекс. Широкое распространение апомиксиса у манжеток послу-
жило основанием для описания множества апогамных видов
(agamospecies).
Вопрос о том, правомерно ли приравнивать подобные aga-
mospecies к видам амфимиктов, остается открытым. Либо сборный
вид A. vulgaris L. s. ampliss., либо (по крайней мере) отдельные его
группы, предложено рассматривать в качестве обширных агамно-по-
ловых комплексов (Глазунова, 1977). Обнаружение нормальной по-
ловой репродукции и двойного оплодотворения наряду с преобла-
дающим апомиксисом (Глазунова, 1983) подтверждает последнюю
точку зрения. Однако в практической систематике agamospecies ман-
жеток, следуя установившейся традиции, с определенной долей ус-
ловности приравнивают к «нормальным» амфимиктическим видам.
В Европе уже отмечено более 265 апогамных видов (Walters,
Pawlowski, 1968). В Центральной России встречаются 38 апогамных
видов (Маевский, 1964; Тихомиров, 1972, 1980; Tichomirov, 1985), из
них 27 произрастают в Московской области.
Наиболее детализированная система этой группы принадлежит
С.В. Юзепчуку (Флора СССР, 1941). К сожалению, он использовал
таксономические категории, не узаконенные «Международным ко-
дексом ботанической номенклатуры» («группа» X. Линдберга, «под-
группа», «цикл») и не снабдил нх названия необходимыми латин-
скими диагнозами. Поэтому на практике приходится йено й, ювап.
83
менее детальную и совершенную схему В. Ротмалера. По этой схеме
наши виды* распределяются следующим образом:
Sect. Akhemilla (sect. Brevicaulon Rothm.)
Subsecl. Hcliodrosium Rothm.
Ser. Pubescentes Buser
A. glaucescens Wallr. 1840, Linnaea, 14: 134. (A. minor
auct.; A. pubescens auct., non Lam.).
A. hirsuticaulis H. Lindb. 1904, Meddel. Soc. Fauna Fl.
Fenn. 30: 143.
A. plicata Buser, 1843, Bull. Herb. Boiss. 1, App. 2: 20.
A. sergii V. Tichom. 1972, Bull. Soc. Natur. Mose., ser. Biol.
77, 4: 130. Ser. Vulgares Buser
Subser. Hirsutae H. Lindb.
A. hebescens Juz. 1931, Acta Horti Petrop. 43: 537.
A. gibberulosa H. Lindb. 1909, Acta Soc. Sci. Fenn.
37(10): 4.
A. propinqua H. Lindb. ex Juz. 1923, Not. Syst. (Petropolis)
4: 184. (A. orbiculata Alechin; A. basilii P. Smirnov; A.
alechinii Zamelis).
A. conglobata H.Lindb. 1909, Acta Soc. Sci. Fenn. 37
(10): 36
A. monticola Opiz, 1838, in Berchtold et Opiz, Okon.-
Techn. Fl. Bohm. 2, 1:13. (A. pastoralis Buser).
A. schistophylla Juz. 1936, Acta Inst. Bot. Acad. Sci. URSS,
1: 121.
A. sarmatica Juz. 1936, Acta Inst. Bot. Acad. Sci. URSS, 3:
202. (A. strigosula auct. fl. Ross. Med. non Buser, p.p.).
A. breviloba H. Lindb. 1909, Acta Soc. Sci. Fenn. 37 (10): 4.
A. substrigosa Juz. 1940, in Majevski, Fl. Sredn. Ross. ed.
7: 446.
A. cymatophylla Juz. 1922, Not. Syst. (Petropolis) 3: 41.
A. semilunaris Alechin, 1922, Not. Syst. (Pctropolis)3: 132.
A. litwinowii Juz. 1933, Acta Inst. Bot. Acad. Sci. URSS,
1: 122.
A. subcrenata Buser, 1893, apud Magnier, Serin. Fl. Select.
12: 285.
A. heptagona Juz. 1922, op. cit.: 45.
A. stellaris Juz. 1933, Acta Inst. Bot. Acad. Sci. URSS,
1:126.
A. acutiloba Opiz, 1838, in Berchtold et Opiz, Okon.-Tcchn.
Fl. Bohm. 2, 1: 15. (A. acutangula Buser).
A. gracilis Opiz, 1838, op. cit.: 14. (A. micans Buser).
A. lindbergiana Juz. 1923, Not. Syst. (Pctropolis) 4: 181.
A. lilicaulis Buser, 1893, Bull. Herb. Boiss. 1, App. 2: 22.
Здесь и далее в тексте словом «вид» мы обозначаем agamospecies.
84
A. leiophylla Juz. 1933, Acta Inst. Bot. Acad. Sci. URSS,
1: 127.
Subser. Subglabrae H.Lindb.
A. baltica G. Sam. ex Juz. 1940, Fl. Sredn. Ross. ed. 7: 449.
(A. nedulosa G. Sam.).
A. psiloneura Juz. 1933, Acta Inst. Bot. Acad. Sci. URSS,
1: 129.
Subser. Glabrae Rothm.
A. glabricaulis H. Lindb. 1909, Acta Soc. Sci. Fenn. 37:10:3.
Исходя из особенностей распространения некоторых видов, из-
вестных из сопредельных с Московской областей, можно ожидать,
что на нашей территории будут обнаружены и другие виды, прежде
всего, A. nemoralis Alechin, A. devestiens Juz., A. sukaczevii V. Tichom.
- одним словом, «инвентаризацию» манжеток области еще нельзя
считать законченной.
Важнейшие биологические особенности оказались общими для
всех изученных agamospecies в пределах комплекса. Что же касается
экологической приуроченности, фитоценологии, географического
распространения наших манжеток, то они нуждаются в специальном
изучении.
Географическое распространение
Виды комплекса A. vulgaris L. s. ampliss. широко распростране-
ны в Европе и Азии (Rothmaler, 1937; Walters, Pawlowski, 1968).
Шесть видов отмечены на северо-востоке Северной Америки
(Bradshaw et al., 1964). В качестве заносного растения для Австралии
приводится Alchemilla xanthochlora Rothm. (Rothmaler, 1955), в на-
шем распоряжении имеется образец A. gracilis Opiz, любезно при-
сланный для определения д-ром S.M. Walters (Cambridge). Наиболь-
шее число видов сосредоточено в Альпах, на Кавказе, Урале, в Тянь-
Шане, Саянах (Флора СССР, 1941; Rothmaler, 1941, 1962; Юзепчук,
1951, 1954).
Большинство отмеченных в Московской области манжеток при-
надлежит к i-руппе бореальных европейских и бореальных восточно-
европейских видов. Бореальными евросибирскими видами являются
Alchemilla hirsulicaulis Н. Lindb., A. monticola Opiz (Тихомиров, 1976).
Некоторые предварительные данные ботанико-гсшрафического
анализа манжеток приведены в систематическом обзоре видов рода
Alchemilla Московской области (Тихомиров, 1969). Из 27 видов Al-
chemilla, произрастающих в Московской области, 6 обнаруживают
явную связь с Западной и Центральной Европой. Эго A. glaucescens
Wallr., A. plicata Buser, A. monticola Opiz, A. gracilis Opiz, A. filicaulis
Buser, A. acutiloba Opiz, A. subcrenata Buser. Два из них - A. glauces-
cens и A. plicata представляют собой типичные европейские расте-
ния, в Цен тральной России проходи т вос точная граница их распро-
85
странсния (Тихомиров, 1969). Видимо, нужно признать европейское
происхождение A. monticola, A. gracilis, A. acutiloba, A. subcrenata,
имеющих очень обширные ареалы. В настоящее время данные виды
интенсивно расселяются по нарушенным местообитаниям (Тихоми-
ров, 1969).
Десять видов в различной степени тяготеют к Уралу. Три из них
— A. glabricaulis Н. Lindb., A. leiophylla Juz., A. conglobata Н. Lindb.
— помимо европейской части России и Урала найдены на Кавказе.
A. lindbergiana Juz., A. litwinowii Juz., A. substrigosa Juz. известны по-
ка только из Центральной России и с Урала. У нас они находятся
близ западной границы своих ареалов. Значительно шире распро-
странены A. heptagona Juz. (доходящая на запад до Прибалтики), А.
hirsuticaulis Н. Lindb. (достигающая Прибалтики и Финляндии), А.
propinqua Н. Lindb. ex Juz. (проникающая до Средней Европы). А.
hebescens Juz., кроме Урала, встречается на Алтае и в Монголии.
Десять видов пока приходится считать сформировавшимися в
европейской части России. Ареалы A. sannatica Juz. (от Предуралья
до Скандинавии), A. baltica G. Sam. ex Juz. (от Поволжья до Финлян-
дии, как заносное — в Швеции и Западной Сибири), A. semilunaris
Alech. и A. cymatophylla Juz. (европейская часть России — до При-
балтики на западе) очень сходны с той картиной, которую обнару-
живают ареалы A. hirsuticaulis или A. heptagona, хотя последние не
найдены на Урале (Тихомиров, 1969).
A. gibberulosa Н. Lindb. и A. breviloba Н. Lindb. свойственны
центру Европейской равнины, но не найдены в Прибалтике. Четыре
вида — A. stellaris Juz., A. psiloneura Juz., A. sergii V. Tichom., A.
schistophylla Juz. — возможно, являются эндемичными для Централь-
ной России (Тихомиров, 1969, 1972, 1986).
Морфологическое описание
Жизненная форма, структура многолетних органов и цветоно-
сов у всех апогамных видов комплекса A. vulgaris L. s. ampliss. одина-
кова. Манжетка — моноподиально-розеточный, короткокорневищ-
ный травянистый поликарпик с плагиотропным эпигеогенным кор-
невищем, гемикриптофит (Петухова, 1977, 1980).
Во взрослом состоянии функционирует только система прида-
точных корней, сравнительно тонких, от 1 до 2 мм в диаметре. При-
даточные корни подузловые, расположены более или менее равно-
мерно по всей длине корневища по I—2 под узлом, образуя бахром-
чатую корневую систему. Придаточные корни нередко имеют раз-
ный диаметр. Наблюдается слабо выраженная анизоризия. Продол-
жительность функционирования корней зависит от возрастного со-
стояния растения.
Побыи манжетки дифференцированы на два типа (как и у дру-
। их мононодиальпо-розеточных раст ений). Многолетние вегетат ив-
ные побы и розеточные, они длительно нарастают моноподиально
86
за счет верхушечной почки и представлены эпигеогенным корневи-
щем. Цветоносные побеги, развивающиеся на корневище, пазушные,
моноциклические, удлиненные (рис. 1).
Рис. 1. Средневозрастное генеративное растение манжетки горной (A. monticola) (боко-
вой побег второго порядка. 10.09.1974).
лк —придаточные корни; ц —отмершие цветоносы: л - листовые рубцы.
87
Корневище горизонтально распростертое, располагается в гори-
зонте А на глубине 0,5 — 1 см, редко — немного глубже. Его терми-
нальная часть, несущая листья, приподнята над землей. По мере от-
мирания листьев благодаря контрактильной способности корней,
надземная часть побега слегка углубляется в почву.
На корневище хороню заметны перехваты, соответствующие
границам годичных приростов. Вследствие незначизельной длины
междоузлий (в среднем 1,2 мм) величина приростов невелика, макси-
мально —- до 2 см. С базального конца корневище постепенно отми-
рает. Число сохранившихся годичных приростов зависит от возрас-
тного состояния. Максимально отмечали до 13 годичных побегов, из
них 6 8 с деятельными придаточными корнями.
На корневище формируются только листья срединной форма-
ции, благодаря чему почки — открытые (по терминологии И.Г. Се-
ребрякова, 1947). Листья располагаются спирально, по формуле 2/5.
Они лишены отделительного слоя, и после их отмирания на корневи-
ще долго сохраняются остатки черешков и прилистников. Узел за-
крытого типа, благодаря чему листовые рубцы имеют вид кольца,
что облегчает подсчет листьев в пределах годичного прироста.
Розеточные листья длинночерешковые, с вытянутым (до 3 см
длиной), замкнутым в виде влагалища основанием, охватывающим
верхушечную почку. Апекс побега достаточно хорошо защищен вла-
галищами листьев. Влагалище разрывается лишь у листьев, закон-
чивших свой рост, в результате увеличения диаметра стебля.
Листовые пластинки розеточных листьев в очертании почковид-
ные, округло-почковидные или округлые, пальчато-лопастные, с 7—
9—11 лопастями. Лопасти по краю зубчатые. Жилкование пальчато-
краевое.
Из пазушных почек, формирующихся на корневище, образуются
боковые побеги различных типов — вегетативные, повторяющие в
своем развитии главный побег, и монокарпические генеративные по-
беги.. Первый лист пазушных почек (и вегетативных, и генератив-
ных) представлен только влагалищной частью. В начале формирова-
ния почки он значительно обгоняет в росте последующие зачатки.
Таким образом, пазушные почки манжеток мы можем считать за-
крытыми. Следует отмстить, что большая часть пазушных почек не
реализуется. Вегетативные почки остаются жизнеспособными в тече-
ние нескольких лег, переходя в разряд спящих и постепенно отмирая
вместе с корневищем. Боковое вегетативное ветвление происходит
редко. Как правило, трогаются в рост верхние почки в пределах веге-
тативной зоны (акротония).
Пазушные генеративные побеги удлиненные, олиствеипые, пред-
ставлены фрондулезным закрытым тирсом. Длина междоузлий изме-
няется, подчиняясь правилу одновершинной кривой. Гипоподий от-
четливо выражен, максимальной длиной отличается зпиподий.
Нижний лист цветоносного побега представлен только влага-
лищной частью. Остальные листья короткочерешковые, с крупными
88
зелеными прилистниками и слабо развитым влагалищем, с 3—5-ло-
пастной пластинкой. Верхние листья имеют вид крупнозубчатых по
краю прицветников (с небольшой пластинкой или без нее).
В пазухах первых одного-двух листьев почки не закладываются,
в пазухах следующих листьев формируются генеративные почки,
причем нижние из них обычно не трогаются в рост.
Строение цветков одинаково у разных видов. Гипантий коло-
кольчатый или обратноконический, с железистым диском в верхней
части.- Цветки четырехчленные. Чашечка с подчашием. Листочки
подчашия могут иметь разную длину. Соотношение длины чашели-
стиков и листочков подчашия в некоторых группах рода Alchemilla
имеет таксономическое значение. Венчик не развит. Тычинок 4. Они
располагаются против листочков подчашия. Пыльники интрорзшДе,
вскрываются поперечной щелью (Федоров, Артюшенко, 1975). Очень
часто встречаются недоразвитые тычинки с укороченной тычиноч-
ной нитью и сморщенным черным или серым пыльником.
Нормальные пыльцевые зерна манжетки двухклеточные, мери-
дионально-трехбороздно-оровые, продолговато-сфероидальные.
Контур пыльцевого зерна гладкий, текстурный рисунок сетчатый
(Демченко, 1967, 1974; Глазунова, Пермяков, 1980). В пыльниках есть
много деформированных пыльцевых зерен.
Гинецей обычно мономерный, но изредка можно встретить от-
дельные цветки с апокарпным гинецеем из двух плодолистиков (пес-
тиков). О. Пенциг отмечал до 4 пестиков в цветке (Penzig, 1921).
Плодолистик занимает на цветоложе центральное положение.
Он имеет короткую ножку, завязь яйцевидной формы, длинный ги-
нобазический стилодий с шаровидным рыльцем. В нижней части
плодолистика заметен короткий брюшной шов (Глазунова, 1986).
Хотя стилодий закладывается как верхушечный, по своему положе-
нию в сформировавшемся плодолистике он оказывается гинобазиче-
ским. Данные о морфогенезе и структуре васкулярной системы пло-
долистика позволяют считать его кондупликатным. В полости завя-
зи расположена одна крупная почти ортотропная семяпочка со сла-
бо дифференцированным фуникулусом. Семяпочка унитегмальная,
крассинуцеллятная. Археспорий многоклеточный.
Плод — орешек, представляющий собой разросшийся плодоли-
стик с опавшим стилодием. Гипантий при орешке сохраняется
(амальтея одночленная, по Кадену, 1968).
Разные виды манжетки различаются, прежде всего, характером и
степенью опушения листьев и цветков, генеративных побегов, Фор-
мой листовой пластинки, лопастей и их зубцов. Структура изменчи-
вости манжеток нуждается в специальном изучении. Проведен ана-
лиз корреляционной структуры и изменчивости признаков у A. mon-
ticola (Глазунова, Мятлев, 1990). Показано наличие двух групп скор-
релированных признаков — вегетативной и генеративной плеяд, вы-
явлены границы изменчивости изученных признаков. Подобный
анализ других видов манжетки представляется интересным.
89
У наших манжеток довольно часто встречаются различные тера-
тологические изменения в структуре побегов и цветков.
Как отмечалось выше, манжетка характеризуется наличием по-
бегов двух типов. Одни из них вегетативные, розеточные, открытые,
полициклические. Другие — удлиненные, генеративные, моноцикли-
ческие. Однако нередко встречаются побеги «переходного типа», по-
разному сочетающие признаки этих побегов (Черноброва, Петухова,
1991). Сравнительно часто можно наблюдать боковые вегетативные
побеги с удлиненными междоузлиями, предшествующими розетке
(рис. 2). Побеги переходного типа могут быть и генеративными, но с
длинно черешковыми листьями и придаточными корнями, заклады-
вающимися под узлами нижних листьев. Очень редко можно наблю-
дать на цветоносе развитие бокового розеточного побега (см. рис. 2).
Побеги переходного типа формируются на границе вегетативной и
генеративной зон.
Наблюдается большой полиморфизм вариантов ветвления на
верхушке цветоносного побега и боковых осях тирса (рис. 3). Часто-
та встречаемости различных вариантов может быть достаточно вы-
сокой, и не всегда удается провести четкую границу между аномаль-
ными структурами и вариантами в пределах нормального полимор-
физма.
Среди уродств в структуре листа известно образование асцидий
и формирование многочисленных придаточных почек, расположен-
ных по краю листа. Эти явления наблюдались и нами. Аномалии
цветка у наших манжеток также довольно обычны. Сравнительно
часто наблюдается срастание цветков, изменение числа частей цвет-
ка, образование пятичленных и трехчленных цветков. Листочки под-
чашия могут быть крупнее, чем обычно, и иметь на верхушке 2 или 3
зубца, реже листочки подчашия не развиваются (рис. 4).
Отмечены случаи прирастания тычиночных нитей к чашелисти-
кам, а также развитие стаминодиев или листочков околоцветника
внутреннего круга. У A. breviloba мы наблюдали ярусную пролифе-
рацию в цветке. При этом чашелистики были очень крупными и име-
ли листовую пластинку (см. рис. 4).
Сравнительно часто встречаются цветки с двумя плодолистика-
ми. В строении плодолистиков были отмечены аномалии. Встреча-
ются плодолистики со слабым срастанием вдоль брюшного шва,
плодолистики с двумя семяпочками в завязи. Единственный раз у А.
baltica была отмечена семяпочка, расположенная не внутри полости
завязи, а снаружи (Глазунова, 1986), а у A. breviloba наблюдалось
развитие нескольких плодолистиков на внутренней поверхности
стенки незамкнутой завязи (см. рис. 4).
Для оценки примерной частоты отклонений в структуре плетка у
4 видов Московской области К.П. Глазуновой в 1977—78 гг. были
проанализированы 1896 цветков на двенадцати цветоносах.
90
[с. 2. Некоторые варианты структуры боковых побегов манжеток.
91
Рис. 3. Некоторые варианты ветвления в области верхушки цветоносного побега
и на боковых осях тирса у манжеток:
7 — терминальный цветок; 2 — монохазий; 3 - лист с листовой пластинкой; 4 — брактея; 5 — норма.
Рис. 4. Некоторые варианты строения аномальных цветков у манжеток.
Обозначения: □ — норма; • — тычинка.
Число аномальных цветков (в %) и число вариантов строения
цветка (т.е. структурных вариантов) в пределах одного цветоноса
для каждого вида приведены в табл. 1. К.П. Глазунова и М.В. Нило-
ва (1994) провели сравнительное изучение популяций A. gracilis из
разных географических пунктов по частотам встречаемости ано-
мальных цветков (общий объем материала — 7356 цветков). Класси-
фикация множества вариантов аномального строения цветков была
проведена в работе А.А. Нотова и К.П. Глазуновой (1994).
Таблица 1.
Таблица встречаемости аномальных цветков
у различных видов манжетки
Вид Цве- те ние Число цветков в соцветии % ано- мальных цветков Число структур- ных вариан- тов
всего аномаль- ных
A. acutiloba I 82 14 17,0 8
A. acutiloba I 84 . 16 19,0 9
A. filicaulis I 165 40 24,2 7
A. gracilis I 145 10 7,4 5
A. gracilis II 60 13 21,6 . 8
A. heptogona I 307 28 9,1 16
A. heptogona I 118 25 21,2 15
A. heptogona I 338 29 7,9 14
A. heptogona I 284 69 24,3 18
A. heptogona II 80 19 23,7 7
A. heptogona II 180 60 33,0 20
A. heptogona II 215 62 28,8 19
Хромосомные числа
В связи с отсутствием в литературе данных по хромосомным
числах манжеток собственно Московской области в табл. 2 мы при-
водим сведения по хромосомным числам наших видов, полученные
на средне- и западноевропейском материале (Wegener, 1967). Воз-
можно, что зафиксированная разница в числе хромосом вида полу-
94
чается за счет наличия в природных популяциях особей разной пло-
идности и анеуплоидов (Глазунова, 1977).
Таблица 2.
Хромосомные числа манжеток (по: Wegener, 1967)
Вид Сред- нее число по Wegener, 1967 По другим источникам
A. gracilis 128 127—132 —
A. hirsuticaulis 105 103—108 —
A. acutiloba 105 102—106 100 (Ehrenberg, 1945) 64 (Murbeck, 1901) 105—107 (Turesson, 1957)
A. cymato- phylla 104 102—106 106—107 (Turesson, 1957)
A. heptagona 104 102—106 —
A. monticola 104 101—110 101—108 103—109 (Turesson, 1957) 101 (Ehrenberg, 1945)
A. sarmatica 104 102—107 —
A. subcrenata 107 103—110 96 (Love, 1958) 104—110 (Turesson, 1957)
A. baltica 103 99—106 —
A. glabricaulis 103 100—106 —
A. filicaulis 103 101—106 96 (Love, 1958) 103—110 (Turesson, 1957) 103 (Bradshaw, 1963)
A. lindbergiana 101 96—109 —
Сезонный ритм развития
Манжетка — летне-зимнезеленое растение. Взрослые растения
выходят из-под снега с почти полностью развернувшимися одним-
двумя листьями и с несколькими прошлогодними, сохранившимися в
отмершем состоянии. Развертывание следующих листьев идет плавно
в течение всего вегетационного сезона с начала апреля до конца ав-
густа.
95
При отсутствии сенокошения рост листьев прекращается в сере-
дине сентября, в конце октября — начале ноября они завядают. Рас-
тения идут под зиму либо без зеленых листьев, либо сохраняются 1 —
2 листа, частично побуревших, отмирающих зимой. Однако наруж-
ный листовой зачаток верхушечной почки достигает значительных
размеров (до 1,5 см длиной).
Листья различаются временем заложения, развертывания, харак-
тером почек в их пазухах и продолжительностью жизни (время от
начала развертывания диета до полного побурения). Весенние ли-
стья, заложенные на конусе нарастания с осени, развертываются с
начала апреля до середины мая. Их отмирание начинается с середи-
ны мая и продолжается до середины июня, продолжительность жиз-
ни 60—70 дней. В пазухах этих листьев закладываются генеративные
почки. Следующие листья развертываются с начала мая, достигают
окончательных размеров в середине — конце июня, отмирают в кон-
це июля. На конусе нарастания они также закладываются с осени,
составляя самые внутренние листовые примордии. В их пазухах фор-
мируются вегетативные почки. Поздне-летние листья закладываются
весной и постепенно развиваются с начала июня. Максимальных
размеров они достигают в конце июля — начале августа, отмирают
в конце сентября — начале октября, отличаясь наибольшей продол-
жительностью жизни — до 80—85 дней. В их пазухах формируются
генеративные почки. Осенние листья на конусе нарастания заклады-
ваются в начале лета, развертываются в конце августа и сохраняют-
ся до зимы. Продолжительность их жизни сокращается до 70—75
дней. В пазухах этих листьев образуются генеративные почки.
Таким образом, при отсутствии сенокошения развертывание но-
вых листьев прекращается в конце августа. Сенокошение удлиняет
продолжительность листообразования. Формирование новых листь-
ев продолжается до октября, в результате чего увеличивается их об-
щее число на побеге. Кривая роста листьев становится двухвершин-
ной (рис. 5). В условиях сенокошения последние по времени образо-
вания листья уходят под зиму без признаков увядания.
С ритмом листообразования связано и состояние верхушечной
почки. Ее емкость (термин Т.П. Серебряковой, 1959, 1961) колеблется
в течение года незначительно. Развертывание листовых зачатков и
их заложение идет одинаковыми темпами, поэтому с середины апре-
ля до сентября емкость почки равна 7 зачаткам. В сентябре — октяб-
ре в связи с прекращением развертывания новых листьев идет накоп-
ление в почке листовых зачатков до 8—9, и в таком состоянии верху-
шечная почка зимует.
Цветет манжетка в конце мая — июне. За счет неравномерного
развертывания цветков цветение одного генеративного побега рас-
тянуто до 15—20 дней: Общая продолжительность жизни цветоноса
от начала роста до полного увядания составляет 85—90 дней. Весен-
ние цветоносы образуются в пазухах последних осенних листьев, к
этому времени уже отмерших, и одного-трех первых весенних.
96
узлы
Рис. 5. Изменение длины листьев в пределах годичного и элементарного побегов А.
monticola. (а) — без сенокошения, (0 — в условиях сенокошения.
В пазухах последующих листьев (май — июнь) формируются ве-
гетативные почки. В пазухах июльских и следующих за ними листьев
(весенне-летних по времени заложения на конусе нарастания) закла-
дываются генеративные почки. В первых генеративных почках со-
цветия полностью формируются уже в начале июля. Часть генера-
тивных почек может трогаться в рост без периода покоя, чем и обу-
словлено цветение растения с конца июля — начала августа до ок-
тября. Это цветение известно в литературе как вторичное, или осен-
нее (Илличевский, 1925; Мальцев, 1938; Игнатьева, 1965). Однако у
манжетки оно скорее должно быть оценено как первичное (за счет
заложившихся в этом же году почек); весеннее цветение в следующем
году служит его продолжением и завершением. Перерыв в цвегенпп
обусловлен исключительно зимним периодом.
Таким образом, на основе краткого анализа ритма развития
можно говорить о таких структурных особенностях моноподпальпо-
го скелетного побега манжетки: на нем наблюдается закономерное
чередование вегетативной и генеративной зон, которому соответет-
вует изменение длины междоузлий, диаметра корневища, размера ли-
стьев (рис. 6). В начале вегетативной зоны междоузлия — короткие,
корневище имеет характерный изгиб, несколько меньший диаметр,
листья сравнительно небольшие. Дальше идет постепенное увеличе-
ние этих показателей до максимума в начале генеративной зоны. К
концу генеративной зоны размеры листьев, длина междоузлий и диа-
97
98
метр корневища снова уменьшаются. В отдельных случаях плавный
ход одновершинной кривой, характеризующей эти показатели, мо-
жет нарушаться, однако общая тенденция изменений выдерживается
довольно четко.
Участки побега, включающие вегетативные и генеративные поч-
ки, отражают ритм его развития и являются структурными единица-
ми этого побега. К ним можно применить термин «элементарный
побег» (Петухова, 1977). Интересно отметить, что у манжетки каж-
дый элементарный побег заканчивает свое развитие уже во второй
период вегетации. Неблагоприятный зимний период мешает нор-
мальному развертыванию сформированных генеративных почек и,
по-видимому, не обязателен для развития растений, о чем можно су-
дить по плавному ходу одновершинной кривой, характеризующей
различные количественные показатели. Так как каждый элементар-
ный побег заканчивает развитие в следующем году, то участок кор-
невища, соответствующий годичному побегу, включает часть преды-
дущего элементарного побега и часть элементарного побега, разви-
тие которого начинается в этом сезоне. Таким образом, у манжетки
наблюдается несоответствие между годичным и элементарным побе-
гом.
* * *
Особый интерес представляет изучение сезонной морфологиче-
ской изменчивости различных видов манжетки. Большинство авто-
ров отмечают, что ранней весной и во второй половине лета и осе-
нью многие виды манжеток становятся трудно узнаваемыми (Флора
СССР, 1941; Станков, Талиев, 1949; Юзепчук, 1954; Тихомиров, 1964,
1969; Frohner, 1975).
Сезонная изменчивость может достигать значительного разма-
ха. 3. Френер, наблюдая за изменениями признаков в течение сезона
у A. straminea Buser и A. sokolowskii Pawl., отмечал, что весной при-
знаки опушения соответствуют тому, что наблюдается у видов серии
Glabrae (Buser) Pawl., но поздним летом эти виды имеют черешки,
опушенные прижатыми волосками, что характерно для ряда Sub-
glabrae (Н. Lindb.) Pawl.
Попытка анализа характера сезонной морфологической измен-
чивости 12 видов манжеток показала, что размах ее у разных видов
различен (Андреев, 1983). Тем не менее, можно отметить некоторые
общие тенденции изменения признаков. Сезонная изменчивость пра-
ктически не касается признаков цветка. Не меняется размер чашели-
стиков, листочков подчашия, сравнительно невелики изменения ха-
рактера опушения цветоножек гипантия.
Наиболее изменчивы признаки, характеризующие форму и раз- •
меры листа и цветоносов. У весенних и осенних листьев уменьшают-
ся общие размеры, становятся более короткими черешки, увеличива-
ются относительные размеры прилистников. Наиболее часто у видов
99
с округлыми в очертании пластинками раннелетних листьев форма
пластинки меняется на почковидную, уменьшается длина лопастей,
они становятся дуговидными или полукруглыми. Увеличиваются от-
носительные размеры зубцов, появляются более глубокие надрезы
между лопастями. Изменяется также характер и густота опушения. У
видов, характеризующихся оголенными в основании главными жил-
ками, в осеннем и весеннем состоянии этот признак не выражен; Цве-
тоносы, развивающиеся осенью, обычно короче, образуют более
компактные парциальные соцветия.
Менее подвержены сезонным изменениям число зубцов на лопа-
стях, характер и густота опушения цветоносов.
Онтогенез
Особенности онтоморфогенеза манжеток были изучены на при-
мере A. monticola на мелкозлаково-разнотравном лугу со средними
условиями увлажнения (Петухова, 1974, 1977, 1980). В большом жиз-
ненном цикле манжетки можно выделить 4 периода и 8 возрастных
состояний.
Латентный период
По способу диссеминации манжетку следует отнести к автобаро-
хорам (Левина, 1957). Орешки опадают заключенными в гипантий.
Далеко не все цветки завязывают плоды. По данным А.О. Джалило-
вой (1965), у A. monticola в зависимости от условий только 30—35%
цветков образуют плоды.
Виргинилъный период
Проростки появляются в середине мая. В условиях ненарушенно-
го травяного покрова значительная часть проростков погибает.
Прорастание надземное, семядоли выносятся наружу гипокотилем,
достигающим 5—10 мм в длину и 0,5 мм в диаметре. Семядоли оваль-
ные, 2—2,5 мм длиной и 1,5—2 мм шириной. Главный корень до 2—3
см длиной, слабо ветвится (рис. 7). К середине — концу июня семядо-
ли засыхают, и растение переходит в ювенильное возрастное состоя-
ние. К этому времени под узлом одного из семядольных листьев за-
кладывается придаточный корень, гипокотиль полегает.
В течение первого вегетационного периода образуются 4—5 зе-
леных листьев. Листовые пластинки их цельные, округло-треуголь-
ные по форме, со слабо сердцевидным основанием, по краю неравно-
зубчатые. В пазухах листьев (за исключением семядолей) закладыва-
ются почки 0,5—0,9 мм длиной, которые, однако, никогда не трога-
ются в рост.
Побег первого года достигает 2,5—3 мм в длину и имеет диаметр
1,5--2 мм. Междоузлия его укорочены — 0,5—0,8 мм длиной. В тече-
100
гип
Рис. 7. Возрастные состояния A. monticola.
а — проросток; б — ювенильное растение; в — имматурное; г — молодое вегетативное; д — молодое генера-
тивное; ГК — главней корень; ПК — придаточные корни; ГИП — гипокотиль; ГП — генеративные побеги.
101
03
Корни прида- точные*, лет 1 1 мп 00 еч
Корневище, мм Д «с сП мп so о 1-Н 20 мп г-
1 тт । Диаметр (max.), мм еч ч- ; 5,5 Г- СП 00
Листья Высота пластин- ки, см сП о о н 00 Tf о тГ О о 44 о 2,1 ±0,06 2,8±0,07 4,1 ±0,08 гч о 4-1 о сП 00 о о 44 so еч
Ширина, см о ф 44 еч со 1,4± 0,06 00 о 44 so СП СП о 44 so тГ Tf О 44 мп 6,1 ±0,56 00 о 44 мп
। Длина, см мп со 44 00 со 2,3+0,15 мп о 44 тГ оо о 44 so 6±‘0Т0‘Я J о 44 о тГ so еч о 44 so .
Число на ЭП so 00 Г—Ц еч О
Возрастное состояние Ювенильные Имматурные Молодые вегетативные Молодые генеративные Средневозрастные генеративные Стареющие генеративные Сенильные
Обозначения: ЭП — элементарный побег, * — продолжительность функционирования.
102
ние июля закладываются придаточные корни под узлами первых
двух листьев. Придаточный и главный корни обладают контрак-
тильной способностью, благодаря чему к осени базальная часть по-
бега слегка втягивается в почву и образуется первое колено корневи-
ща. Поскольку главный корень сокращается в меньшей степени, ги-
покотиль петлевидаю изгибается.
На следующий год трогается в рост верхушечная почка, образуя
элементарный побег второго года. Боковое ветвление отсутствует. В
последующие годы продолжается моноподиальное нарастание глав-
ной оси (табл. 3). Величина элементарных побегов постепенно воз-
растает, достигая на 5—7-й год 5—6 мм. Увеличение диаметра кор-
невища манжетки осуществляется за счет изменения размеров апекса
путем первичного роста усиления (Петухова, 1974). Одновременно
увеличивается число и размеры листьев, изменяется их форма. У им-
матурных особей листья пяти-, у молодых вегетативных — семилопа-
стные. Увеличивается общее число придаточных корней и число их
на каждом последующем элементарном побеге. Каждый элементар-
ный побег подтягивается и слегка углубляется в почву, однако кон-
трактильная способность корней проявляется лишь в год их образо-
вания, и последующего заглубления корневища не происходит. Ветв-
ление отсутствует до наступления генеративного периода.
На 4—5-й год начинают отмирать система главного корня и ги-
покотиль, однако их остатки обычно сохраняются довольно долго,
что позволяет определить абсолютный возраст до 8—10 лет. Расте-
ния с признаками отмирания системы главного корня можно отнести
к молодым вегетативным. Процессы отмирания распространяются и
на первые по времени образования элементарные побеги, благодаря
чему базальная часть корневища легко обламывается.
Генеративный период
По нашим наблюдениям, манжетка переходит к цветению на 7—
9-й год (Петухова, 1977), по данным Линкола (Lincola, 1937) — не
раньше, чем на 9-й год жизни. В первый год цветения, как правило,
развиваются 1—2 соцветия, хотя генеративных почек формируется
больше. Вегетативные почки по-прежнему остаются в нераскрытом
состоянии.
Средневозрастные генеративные растения отличаются наиболь-
шей мощностью. Величина элементарных побегов достигает макси-
мума (до 20 мм в длину и 10—12 в диаметре) и несколько лет сохра-
няется примерно на одном уровне. Ежегодно формируется 15—17 ли-
стьев, листовые пластинки имеют 10—И лопастей. Соответственно
числу листьев увеличивается число придаточных корней. Они закла-
дываются под узлами почти всех листьев, иногда по 3—4 под одним
узлом. Увеличивается и продолжительность функционирования кор-
ней (до 5—7 лет), поэтому растения этой группы отличаются наибо-
лее мощной корневой системой. Изменяется соотношение вегетатив-
103
ных и генеративных почек в пределах элементарного побега. Если у
молодых генеративных растений это соотношение составляет 1,5:1
или 2:1, то на 6—8-й год цветения — 1:3—1:4. Однако реализуется,
как правило, только часть генеративных почек.
После многократного формирования цветоносных боковых по-
бегов верхушечная почка корневища отмирает. Отмиранию ее пред-
шествует ослабление корне-листовой функциональной корреляции
(Казарян, 1969), которое выражается в сокращении продолжительно-
сти функционирования придаточных корней до 2—4 лет, благодаря
чему уменьшается общее число функционирующих корней, хотя ос-
тальные показатели (число и размер листьев, диаметр оси) остаются
почти на прежнем уровне. Все пластические вещества расходуются
на образование цветоносов уже сформированного элементарного по-
бега, у следующего же развертываются только листья, в пазухах ко-
торых формируются вегетативные почки. Продолжительность жизни
верхушечной почки составляет, по-видимому, 20—25 лет. Отмирание
терминальной почки стимулирует развитие одного, реже нескольких
боковых вегетативных побегов, замещающих материнский, который
еще некоторое время (2—3 года) остается в живом состоянии. Реже
боковые вегетативные побеги образуются до отмирания верхушеч-
ной почки. Во всех случаях бокового ветвления трогаются в рост
почки лишь последнего по времени формирования элементарного
побега, причем наблюдается определенная акротония, так как разви-
ваются лишь самые верхние почки в пределах вегетативной зоны.
Обычно формируется не больше 1—3 боковых побегов.
Вегетативные боковые побеги, как и материнский, розеточные,
нарастатрт моноподиально и представляют собой скелетные оси вто-
рого порядка. Зацветают они на 2—3-й год жизни, постепенно уси-
ливаются, достигая мощности побега первого порядка, однако про-
должительность их жизни меньше, примерно 15 лет. На смену моно-
подиальным побегам второго порядка образуются побега третьего и
последующего порядков, причем наблюдается постепенное ослабле-
ние побегов более высоких порядков и сокращение длительности их
жизни.
Сенильный период
Продолжительность жизни скелетных побегов у сенильных осо-
бей уменьшается до 1—3 лет. Генеративные почки у таких растений
могут закладываться, но не развертываться. Уменьшается число при-
даточных корней. Листья становятся мелкими, напоминают листья
имматурных и молодых вегетативных растений (рис. 8).
Определить общую продолжительность большого жизненного
цикла манжетки не представляется возможным, так как боковые ве-
гетативные побеги при разрушении материнского побега приобрета-
ют самостоятельность. По этой же причине невозможно точно опре-
делить, на каком порядке ветвления прекращается жизнь растения;
104
Рис. 8. Сенильные растения A. inonlieola (10.09.1974).
105
по нашим предположениям — на шестом—восьмом. Общая продол-
жительность большого жизненного цикла составляет, по-видимому,
60—70 лет.
Соотношение этапов онтогенеза и фаз морфогенеза показано на
рис. 9. Выделенные фазы различны по своей деятельности и значе-
нию в жизни растения. Переход от одной фазы к другой всегда свя-
зан с важными структурно-биологическими изменениями. В боль-
шом жизненном цикле один моноподиальный побег последовательно
сменяется другим, при этом все биоморфологические признаки побе-
гов каждого порядка проявляют тенденцию к ослаблению, хотя и не
резко выраженную.
прорк-
Ш
юЛиаиме
и иматцриое
utmost
инератИти
срсМозростюе
генеративное
il
*1
odmfeiotoe
растение
система скелетного
моноподиального побега
разбомбленная
скелетная
система
S
tezcmamrfnt
средневозрастное
и стареющее генератибкое
стареющее генеративное
сенильное
гладкий симподий
парциальные скелетные симподии
симподиальная система скелетных осей.
□/ Ш ^4 ij р V yd yS
Рис. 9. Соотношение этапов онтогенеза и фаз морфогенеза в большом жизненном цикле
A. monticola.
1 — главный скелетный побег; 2 — отмирающие части скелетного побега; 3 — ске-
летный побег второго порядка; 4 — третьего и последующих порядков; 5 — функциони-
рующие придаточные корни; 6 — главный корень; 7 — отмерший, но сохранившийся
главный корень; 8— цветоносные побеги последнего года; 9 — отмершие цветоносные
побеги прошлых лет.
106
Наиболее крупная единица в морфогенезе манжетки — монопо-
диальный скелетный побег, обладающий ограниченным циклом. Он
состоит из более мелких единиц — элементарных побегов, отражаю-
щих ритм роста моноподиального побега.
Поскольку вегетативное боковое ветвление скелетных побегов
выражено слабо, а побеги замещения, появляющиеся последователь-
но при отмирании терминальной почки предыдущих осей, имеют
тенденцию к ослаблению, можно сказать, что вегетативное размно-
жение манжетки в луговом ценозе выражено слабо и связано со ста-
рением.
Следует также отметить, что многие признаки, используемые в
качестве таксономических в пределах комплекса A. vulgaris L. s.
ampliss., сильно меняются в онтогенезе. Особенно это касается числа
и формы лопастей листьев, характера опущения. Старые генератив-
ные растения становятся практически неузнаваемыми. При боковом
вегетативном ветвлении листья на первых элементарных побегах
также сильно отличаются от листьев на материнском растении.
Способы размножения и распространения
Манжезжа относится к растениям с устойчивым апомиксисом,
который первоначально был обнаружен у среднеевропейских пред-
ставителей рода (Murbeck, 1901, 1902; Strasburger, 1904). Долгое вре-
мя манжетки группы A. vulgaris считали типичными облигатными
апомиктами (Флора СССР, 1941; Поддубная-Арнольда, 1964; Grant,
1971). Было распространено мнение об отсутствии у них нормальной
пыльцы (Флора СССР, 1941).
В 1973—1983 гг. проведено детальное исследование эмбриоло-
гии восточноевропейских представителей рода Alchemilla (Глазуно-
ва, Пермяков, 1980; Глазунова, 1981, 1983, 1987). Показано, что регу-
лярный апомиксис у восточноевропейских манжеток нельзя считать
облигатным.
У многих апогамных видов отмечено наличие определенного
процента нормальной пыльцы. Подтверждена замеченная Э. Страс-
бургером (Strasburger, 1904) закономерность: пыльники раскрывают-
ся у манжеток только в том случае, если в них преобладают нормаль-
но сформированные пыльцевые зерна. Среда изученных видов лишь
у A. glaucescens и A. hirsuticaulis пыльники не вскрываются, а их со-
держимое представляет бесструктурную массу (стерильность 100%)*.
У остальных видов в раскрывшихся пыльниках содержатся нормаль-
ные и деформированные пыльцевые зерна. Минимальной стерильно-
стью обладает, по нашим наблюдениям, A. filicaulis Buser (до 14%)
(Глазунова, 1986).
Может быть, по этой причине A. glaucescens и A. hirsuticaulis устойчивы в своих
признаках и не представляют трудностей при определении,
107
Строение и развитие генеративной сферы обнаруживает ряд
приспособлений для нормального осуществления полового процесса
(развитие в пыльнике фиброзного слоя, железистый эпидермис рыль-
ца, тяж стигматоидной ткани к семяпочке и др.) (Глазунова, 1986).
В опытных условиях удалось наблюдать прорастание пыльцы А.
schistophylla и A. filicaulis в посевах на 5%-м агар-агаре (при 10—20%
сахарозы) с дождевой водой. Прорастало около 4% пыльцевых зерен.
Без дождевой воды пыльца не прорастала (Глазунова, 1977). По
наблюдениям в природе, капля воды полностью окружает цветок, ох-
ватывает пыльники, рыльце и нектарный диск, и, вероятно, факт
прорастания пыльцы только при наличии дождевой воды не случаен.
В зародышевых мешках раскрывшихся опыленных цветков А.
baltica, A. glaucescens и A. heptagona изредка наблюдали разные ста-
дии двойного оплодотворения, которое идет по премитотическому
типу (рис. 10) (Глазунова, 1983). В данном случае на рыльце А.
glaucescens прорастала пыльца другого апогамного вида.
Отмечено наличие высокой эмбриологической изменчивости, со-
четание признаков апомиктического и нормального полового спосо-
бов образования семян не только у разных особей одного и того же
вида, но и в пределах одного соцветия. В одном пыльнике можно от-
метить пыльцевые зерна разного размера. В разных цветках в одном
и том же соцветии осуществляются разные способы образования за-
родышевых мешков (анэуспория, апоспория, эуспория), а также раз-
ные способы образования зародыша (половым путем, партеногенс-
тически путем псевдогамии). Отмечены также случаи генеративной и
соматической апоспории, апогаметии, ложной полиэмбрионии. По
результатам многолетних наблюдений показано изменение в разные
годы процента стерильных пыльцевых зерен в образцах пыльцы кон-
кретных растений (Глазунова, 1983).
Таким образом, по крайней мере, для многих наших манжеток
характерно сочетание полового и неполового способов образования
семян. Факуль тативный апомиксис восточноевропейских манжеток
является устойчивым способом семенного размножения, которому,
вероятно, свойственно изменение соотношения различных типов об-
разования семян в разные годы.
К.П. Глазуновой в 1978 г. проведен анализ семенной продуктив-
ности различных видов манжеток на опытном участке в Ботаниче-
ском саду МГУ. Полученные данные приведены в табл. 4. Реальная
семенная продуктивность различных видов составляет в среднем от
30,8 до 60,3% от потенциальной. Интересно отмстить, что этот пока
затель у A. alpina L. может достигать 75% (Похилько, 1985).
Интенсивность семенного и вегетативного размножения А.
monticola изучена А.О. Джалиловой (1965). Исследования проводили
в Ленинградской области на мелкозлаково-разнотравиом лугу, па
участках, скашиваемых в обычный, ранний и поздний сроки и на
участке, оставленном без сенокошения. Показано, что A. monticola в
этом луговом ценозе размножается как вегетативно, гак и семенами.
108
Рис. Id. Последовательные стадии двойного оплодотворения у.разных агамных видов
манжетки: I./{ Л. glaiic’eseens: /> \. hallica; /’,// A. heptacona.
109
о
Таблица 4.
Семенная продуктивность различных видов манжетки
Вид NG* Семенная продуктивность одного генеративного побега
потенциальная реальная реальная в % от потенциальной
min х ± шх max. min х ± mx max. min X ± тх max.
A. heptagona 5 131 202,4±24,9 268 62 87,4±11,3 115 32 43,6±2,2 61
A. schistophylla 6 276 348,5±1.9,4 409 143 211,8±19,3 258 51 60,3±2,9 67
A. glaucescens 5 189 250,0±22,4 328 115 152,0±13,5 198 55 59,8±1,5 64
A. sarmatica 4 557 632,0±26,7 684 113 172,8±26,4 226 27 30,8±1,7 35
A. baltica 3 409 440,3± 17,3 ' 469 201 223,7±11,7 240 49 51,0±1,0 52
A. gracilis 11 61 97,1±7,5 116 18 34,6±3,7 59 17 37,5±4,0 57
* NG — число генеративных побегов за сезон.
, Таблица 5.
Запас жизнеспособных и число проросших семян A. monticola (в слое почвы 0-2 см на 1 м2)
на мелкозлаково-разнотравном лугу (по Джалиловой, 1965, с изменениями)
Режим использования луга Число жизнеспособных семян Число пророс- ших семян
до обсе- менения в 1959 г. в % от всех видов в % от разно- травья после обсеме- нения в 1959 г. в % от общего запаса
с сенокошением в обычный срок 11200 78,0 90,8 308 2,7
с сенокошением в ранний срок 6575 53,4 74,5 64 0,9
с сенокошением в поздний срок 17550 61,1 . 79,5 160 0,9
без сенокошения 11400 67,0 82,9 4 0,03
Семенное размножение осуществляется за счет семян, накопив-
шихся в почве в течение длительного времени, а также за счет семян,
осыпающихся ежегодно при существующих сроках сенокошения. По-
верхностный слой почвы насыщен жизнеспособными семенами ман-
жетки. Они составляют более 50% от общего числа семян и более
70% от общего числа семян разнотравья (табл. 5). Из этого запаса
прорастают лишь 0,03 -2,7% жизнеспособных семян.
Всходы появляются в конце мая — начале июля. Их дальнейшая
судьба зависит от погодных факторов (особенно недостатка влаги,
который приводит к гибели 82—85% всходов) и режима использова-
ния луга. На участках с ранними сроками сенокошения и па участке,
оставленном без сенокошения, наблюдается меньшее число всходов
на 1 м2, что связано с замоховелостыо и запущенностью травостоя.
При отсутствии сенокошения высокий травостой и толстый слой
мертвого покрова препятствует прорастанию и дальнейшему разви-
тию ювенильных растений (табл. 6). Особи семенного происхожде-
ния в этих условиях погибают ня ранних стадиях развития.
Вегетативное размножение манжетки в луговом ценозе выраже-
но слабо и связано со старением особи. Позднее сенокошение в тече-
ние длительного периода способствует вегетативному разрастанию
манжетки. При других режимах использования процесс вегетативно-
го разрастания или приостановлен (при раннем сроке скашивания),
или идет очень ограниченно (при обычном сроке сенокошения и при
отсутствии сенокошения).
Экология
Экология различных видов манжетки нуждается в специальном
изучении. Пока можно привести лишь немногочисленные предвари-
тельные наблюдения.,
Многие виды рода Alcheinilla обнаруживают довольно строгую
связь с определенными экологическими условиями. Особенно это
прослеживается в горах, но и в условиях Среднерусской равнины
экологическая приуроченность манжеток в ряде случаев выражена
достаточно отчетливо. Однако наряду с видами, встречающимися в
узком экологическом диапазоне, существуют виды с очень широкой
экологической амплитудой, способные расти на разных почвах и в
разных условиях увлажнения и освещения, на лугах, пашнях, в лесах
и по дорогам, особенно в нарушенных местообитаниях (Тихомиров,
1969). Такие виды особенно легко становятся синантропными.
Среди видов с узкой экологической амплитудой можно отмстить
A. gibberulosa. Это одна из наиболее ксерофилытЫх наших манжеток,
растущая большей частью по сухим склонам, вершинам холмов, на
солнечных полянах и опушках, заметно предпочитающая супесча-
ную почву (Тихомиров, 1966, 1969). Довольно строго приурочены к
112
сухим солнечным местам также A. glaucescens, A. hebescens, в мень-
шей степени A. hirsuticaulis, которая помимо сухих солнечных скло-
нов может расти на влажной почве и, реже, под пологом леса, обра-
зуя теневые формы.
Таблица 6.
Число всходов и возможность дальнейшего развития ювенильных
растений A. monticola (по Джалиловой, 1965).
Г оды наблюдений Число всходов на 1 м2 Из них выжили в течение
1 года 2 лет 3 лет 4 лет
При обычном сенокошении
1959 308 43 32 24 8
1960 540 232 116 52 —
1961 276 140 120 — —
1962 300 244 — —
При раннем сроке сенокошения
1959 64 12 12 8 8
1960 208 64 28 8 —
1961 88 44 36 — —
1962 68 32 — — —
При позднем сроке сенокошения
1959 160 28 1 1 1
1960 536 112 16 16 —
1961 356 80 56 56 —
1962 264 180 — — —
При отсутствии сенокошения
1959 4 — — . — —
1960 36 — — — —
1961 — — — — —
1962 36 — — — —
ИЗ
Преимущественно к сыроватым, тенистым местам приурочены
A. semilunaris, A. cymatophylla, A. heptagona. Затененные места — за-
росли кустарников, рощи, ольшаники, опушки — предпочитает так-
же A. glabricaulis, значительно реже она растет по открытым травя-
нистым местам.
Весьма широкой экологической амплитудой обладают А.
gracilis, A. monticola, A. acutiloba, A. subcrenata, A. baltica. Среди на-
ших манжеток, вероятно, наиболее широка экологическая амплитуда
A. gracilis. Одинаково часто ее можно встретить по сухим лугам, на
склонах холмов (вместе с A. glaucescens, A. hirsuticaubs), в тенистых
лесах (рядом с A. acutiloba, A. semilunaris, A. heptagofia), по берегам
водоемов. A. gracilis вместе с A. acutiloba нередко встречается на за-
болоченных лугах, по окраинам болот. Видимо, эти два вида лучше
других переносят подкисленные почвы (Тихомиров, 1969).
Экологическая изменчивость различных видов также нуждается
в специальном изучении. Широта экологической амплитуды, безус-
ловно, сказывается на внешнем облике растения. Целесообразность
таксономического отражения этой изменчивости можно установить
путем постановки длительных опытов по параллельной культуре
растений разных популяций. Высказано предположение, что впечат-
ление о высокой пластичности возникает в связи с большой генети-
ческой гетерогенностью популяций, присутствием особей разной
плоиднбсти и анэуплоидов (Глазунова, 1977).
Фитоценология
Манжетки в пределах Центральной России приурочены к луго-
вым и лугово-опушечным ценозам. В большом количестве они встре-
чаются на различного рода нарушенных местообитаниях — обочи-
нах грунтовых и шоссейных дорог, по олуговевшим склонам ж.-д.
насыпей, окраинам полей, вырубкам, пустошам, гарям. Манжетки
являются одним из важных компонентов природных сенокосов и па-
стбищ, на лугах они нередко входят в группу доминантов и субдоми-
нантов.
Сообщества, в составе которых имеются манжетки, включают
обычно несколько видов. Для разных условий характерен различный
видовой состав. Предварительные наблюдения показывают, что ви-
довой состав манжеток зависит не только от условий произрастания,
но и от возраста формирующегося сообщества, степени его нарушен-
ности. При сильной антропогенной нагрузке число видов мало, пре-
обладают преимущественно A. gracilis, A. monticola, A. acutiloba, А.
baltica, По более сухим участкам A. hirsuticaulis. Перечисленные выше
виды в настоящее время быстро распространяются по различным на-
рушенным местообитаниям, расширяя свое присутствие на данной
территории. Такое расселение, вероятно, облегчается свойственной
114
им широкой экологической амплитудой, а устойчивость в такого ро-
да растительных группировках может быть связана с наличием фа-
культативного апомиксиса и возможностью сочетания преимуществ
полового и апомиктического размножения, что оказывается весьма
существенным в меняющихся условиях среды.
Обилие манжетки в луговом ценозе может изменяться. Система-
тическое раннее сенокошение (в первой-г-второй декаде июня) отри-
цательно оказывается на росте и развитии растений, приводит их к
постепенному ослаблению и отмиранию' По наблюдениям А.О. Джа-
лиловой (1965), на 8—10-й год в условиях раннего скашивания на
участке 1 м2 насчитывалось около 30 взрослых растений. На кон-
трольном участке с обычными сроками сенокошения обилие вида ос-
тавалось более или менее постоянным, на 1 м2 насчитывалось 40—43
плодоносящих растения.
Консортивные связи
Листья манжетки поедаются скотом (Матвеева, Знаменская,
1960; Кормовые растения..., 1951; Кормовые растения европейской
части СССР, 1981). Однако в литературе можно встретить указания,
будто этому препятствует высокое содержание дубильных веществ в
листьях (Кернер, 1903; Нейштадг, 1963). Вероятно, содержание ду-
бильных веществ зависит, прежде всего, от фенофазы, в которой на-
ходятся растения. На Кавказе отмечено поедание листьев А. саи-
casica Buser кустарниковой полевкой (Формозов, Просвирина, 1935);
возможно, и наши виды могут поедаться грызунами. Довольно часто
на листьях манжетки можно обнаружить погрызы листоедов.
Очень часто манжетки поражаются мучнистой росой, что осо-
бенно проявляется во второй половине лета и осенью. Почти ежегод-
но эпифитотии вызывает мучнисторосяной гриб Sphaerotheca al-
chemillae (Grev.) L. Junell., появляющийся в конце мая — начале ию-
ня. При этом на листьях образуется паутинисто-мучнистый налет.
Вскоре после появления конидиеносной стадии происходит образо-
вание клейстотециев (Ростовцев, 1908; Горлейко, 1983). В Армении
отмечено поражение манжеток грибом Sphaerotheca aphanis (Walk.)
V. Braun (Симонян и др., 1988). Нередко наблюдаются пероноспоро-
зы, вызываемые Peronospora potentillae de Вагу. При этом на листьях
развиваются сдювато-дымчатые пятна. Образование пятен на листь-
ях происходит также при развитии сферейного гриба Coleroa alche-
millae (Grev.) Winter (Ростовцев, 1908).
Нередко манжетки поражаются ржавчинными грибами. При
развитии гриба Uromyces alchemillac Pers, на листьях наблюдается
образование черноватых или бурых пятен. При этом увеличивается
длина черешка, уменьшаются размеры листовой пластинки и длина
115
лопастей (Кернер, 1903; Ростовцев, 1908). Кроме этого на манжетках
отмечен ржавчинный гриб Trachispora alchemillae (Pers.) Fckl. (Са-
винцева, 1988).
Имеются сведения о поражении манжеток некоторыми дейтеро-
мицетами. На ослабленных, подвядающих растениях нередко разви-
вается полусапротрофный гриб Cladospora herbarium (Pers.) Link. На
соцветиях манжеток можно наблюдать развитие несовершенного
гриба Botrytis cinerea Pers. (А.Н. Лихачев, устное сообщение). Кав-
казские виды манжеток могут поражаться грибом Ovularia schroeteri
(Kuhn) Saccardo (Осипян, 1987).
У кавказских видов Alchemilla отмечено образование экто-, эн-
дотрофной микоризы (Микеладзе, 1960). A. retinervis Buser может
быть хозяином Pedicularis condensata Bieb.
Имеются сведения о посещении цветков манжетки мухами. Вы-
сказано предположение, что с помощью мух может осуществляться
перекрестное опыление (Нейштадт, 1963).
Биологическая продуктивность
Заведомо очевидно, что разные виды манжетки должны обла-
дать разной продуктивностью. По средним размерам и общей био-
массе A. acutiloba или A. baltica, например, намного превосходят А.
hirsuticaulis, A. glaucescens или A. glabricaulis. Однако этот вопрос ну-
ждается в специальном изучении.
Биологическая продуктивность вида зависит от возрастного со-
стояния и условий произрастания. Максимальной продуктивностью
обладают средневозрастные генеративные растения. В табл. 6 приве-
дены некот орые биометрические показатели, характеризующие раз-
личные возрастные состояния A. monticola. Наблюдения проведены
па мелко злаково-разнотравном лугу со средними условиями увлаж-
нения в. ассоциациях, гребепниково-манжстковой и душистоколоско-
вб-манжстковой (Петухова, 1980).
А.О. Джалилова (1965) определяла вес сухой надземной массы А.
monticola (листьев и генеративных побегов, срезанных на высоте 4—
5 см от поверхности почвы) на мелкозлаково-разнотравном лугу в
Ленитрадской области. Вес сухой надземной массы у растений, раз-
вивающихся вне травостоя, был в 6—7 раз выше (табл. 7). Показано,
что манжетка сильно реагирует на засуху. Так, в засушливый 1.963
год сухой массы было получено в 2 раза меньше, чем в дождливый.
Отсутствие сенокошения снижает обилие манжетки, но растение
при этом становится сверх нормы мощным (Джалилова, 1965).
116
Хозяйственное значение
Подземные части манжетки содержат дубильные вещества (7,2—
11,3% у A. xanthochlora Rothm.), катехины (Теслов, Макарова, 1969;
Tuka, Popescu, 1979). В надземных частях имеются стероиды
(Muhlemann, 1939), витамин С, фенолкарбоновые кислоты и их про-
изводные, дубильные вещества (7,5—9,35%), липиды (Теслов, Мака-
рова, 1969; Schneider, 1957). В листьях отмечены также кумарины,
флавоноиды (Karrer, 1958).
Манжетки находят применение в народной медицине (Романо-
вич, 1969). На Украине их применяют при лечении кожных болезней,
отвар — при гипертонической болезни (Шиманская, 1966), а надзем-
ные части в виде отвара и настоя используют как вяжущее, гемоста-
тическое, отхаркивающее, наружно как ранозаживляющее и для мы-
тья пораженных мест при чесотке (Лекарственные растения, 1968).
Таблица 7.
Сухой вес (г) надземных органов одного растения A. monticola
(среднее из 25 экземпляров) (по Джалиловой, 1965)
Год Общий вес В том числе
листьев генератив- ных побе- гов
В травостое
1962 1,2 1,0 0,2
1963 0,5 0,2 0,3
Травостой нарушен
1962 7,4 5,5 1,9
1963 3,8 1,2 2,6
Подземные и надземные части A. xanthochlora (и, вероятно, дру-
гих видов) в странах Западной Европы в виде настоя, отвара, по-
рошка используют как вяжущее, гемостатическое, диуретическое, ра-
нозаживляющее, при эпилепсии, ожирении, энтеритах, диабете, дис-
менорее, кожных заболеваниях (Растительные ресурсы, 1987). Сок и
пастой листьев применяют наружно при опухолях, ранах, заболева-
ниях глаз. Молодые листья и стебли сьедобны, могут быть использо-
ваны в виде сала га пли приправы к блюдам (Кощеев, 1981).
117
Изучение лекарственных свойств манжетки проводится в Мос-
ковской медицинской академии имени И.М. Сеченова (В.М. Баева).
Небезынтересны манжетки как кормовые растения (Раститель-
ное сырье СССР, 1957), так как содержат большое количество про-
теина. По содержанию питательных веществ, количеству протеина и
белка манжетка не уступает клеверу красному (МатвееЬа, Знамен-
ская, 1960; Кормовые растения..., 1951; Кормовые растения европей-
ской части СССР, 1981).
Н.М. Григорьева
ЗЕМЛЯНИКА ЗЕЛЕНАЯ
Номенклатура, систематическое положение, внутривидовая
систематика
Fragaria viridis Duch. 1766, Hist. Nat. Frais.: 135; Сырейщиков,
1907, Илл. фл. Моск. губ. 2: 250; он же, 1927, Опред. раст. Моск, губ.:
162; Юзепчук и Пояркова, 1940, в Маевский, Фл. изд. 7: 430; Юзеп-
чук, 1941, Фл. СССР, 10: 61; он же, 1954, в Маевский, Фл., изд. 8: 110;
Иванина, 1964, цит. соч., изд. 9: ПО; Ворошилов, 1966, Опред. раст.
Моск, обл.: 202; Tutin, 1968, Fl. Europ. 2: 48; Черепанов, 1981, Сосуд,
раст. СССР: 436; Киселева, 1986, Опред. раст. Мещеры, к 209.— F.
collina Ehrh. 1792, Beitr. 7: 26; Кауфман, 1866, Моск. фл. изд. 1: 155;
Цингер, 1885, Сборник свед.: 165; Шмальгаузен, 1895, Фл. Средн, и
Южн. Росс, к 324. — Земляника зеленая, полуница (семейство розо-
цветных — Rosaceae)
Д.П. Сырейщиков (1914) указывает, что кроме обычной формы в
Московской области встречается f. neglecta Lindm., у которой цвето-
ножки густо покрыты горизонтально отстоящими волосками или
только часть волосков — отстоящая под углом, а остальные — рас-
положены горизонтально.
Asherson et Graebner, Beck и другие флористы (цит. по Сырейщи-
ков, 1914) приводят гибриды всех видов и считают форму neglecta
Lindm. за гибрид F. viridis х moschata. Л.И. Иванова (Маевский,
1964) понимает F. viridis Duch. var. neglecta Lind, как синоним F.
campestris Stev. (F. neglecta Lind., F. praecox Losinsk.).
В.Ф. Васильев (Флора Крыма, 1960) указывает, что F. campestris
Stev. (in Bull. Soc. Mose. XXIX, III, 1856, 176) — вид очень близкий к
Fragaria viridis. Он отмечает, что Э. Буассь и Агеенко полагали, что
нет оснований выделять F. campestris Stev., так как основной отличи-
тельный признак — погруженность семянок в цветоложе встречается
и у Fragaria viridis. В видовой самостоятельности F. campestris сомне-
вался также С.В. Юзепчук (Флора СССР, 1941). Скорее всего, F.
campestris представляет лишь одну из форм Fragaria viridis. Следует
указать, что погруженность семянок в цветоложе встречается и у F.
vesca.
С.К. Черепанов (1981) понимает F. viridis Duch. subsp. campestris
(Stev.) Pawl = F. campestris.
119
Географическое распространение
Ареал вида ограничен северным полушарием и относится, по
А.А. Гроссгейму (1952), к палеарктическому географическому типу
или к европейско-западносибирско-североказахстанскому. Л.М. Но-
сова (1973) называет тип ареала F. viridis среднеземноморско-евро-
пейско-западносибирско-североказахстанско-горно-южносибирско-
среднеазиатским. В европейской части встречается: в Ладожско-Иль-
менском, Верхневолжском, Заволжском, Верхнеднепровском, Средне-
Днепровском, Причерноморском, Волжско-Донском, Волжско-Кам-
ском районах; в Крыму; во всех районах Кавказа. В Западной Сиби-
ри — в Ленско-Колымском, Даурском; в'Средней Азии — в Джунгар-
ско-Тарбагатайском и Тян-Шанском райбнах (Флора СССР, 1941).
За пределами территории бывшего СССР земляника найдена
почти во всей Европе, кроме Крайнего Севера и южного Средизем-
номорья.
В Московской области встречается в Южном, Юго-восточном,
Приокском, Заокском, редко в Западном и Клинско-Дмитровском
ботанико-географическом районах (Ворошилов и др., 1966).
Морфологическое описание
Земляника зеленая — травянистый, короткокорневищный, кис-
текорневой столонообразующий летнезеленый поликарпик с дицик-
лическйми монокарпическими побегами.
Подземная многолетняя побеговая сфера представлена коротким
корневищем, до первого цветения нарастающим моноподиально.
После отмирания генеративного побега нарастание сменяется на
симподиалЬное (Серебряков, 1952; Troll, 1957). Корневище темно-бу-
рое, в почве располагается горизонтально, вертикально, или под уг-
лом к поверхности, почвы. Верхушечная часть корневища с побегом
текущего года часто бывает' отделена перетяжкой и направлена поч-
ти вертикально вверх. На всем остальном протяжении корневище
имеет неодинаковую толщину, густо одето остатками прилистников
и черешков (Флора СССР, 1941).
Корневая система на первых этапах развития представлена сис-
темой главного и придаточных корней. После отмирания главного
корня у растений развивается система придаточных корней. Прида-
точные корни образуются в течение всего лета. По В.Н. Голубеву
(1962), землянику можно отнести к мелкокорневым, кистекорневым
растениям, имеющим поверхностные корни, расположенные в гуму-
совом горизонте. В надземной сфере растение образует два типа по-
бегов: розеточные вегетативные и полурозеточные генеративные.
Монокарпические побеги при прорастании открытых почек разви-
вают розетку зеленых ассимилирующих листьев и только на второй
год после перезимовки завершают свое развитие, образуя цветки и
120
плоды. Таким образом, генеративные побеги дициклические. Весной,
при образовании цветоносного побега, земляника в пазухе верхнего
розеточного листа цветоносного побега образует дочерний боковой
побег. Как правило, в течение первого лета он развивается как веге-
тативный ассимилирующий, образуя 3—4 крупных зеленых листа
(отмирающих к октябрю) и 1—2 короткочерешковых листа (сохра-
няющихся в зеленом состоянии весь зимний период). В пазухе ниж-
них листьев вегетативного побега развиваются надземные столоны,
или усы, служащие для вегетативного размножения растений. В тер-
минальной точке роста в течение августа и сентября формируется со-
цветие будущего года. Весной из терминальной почки перезимовав-
шего побега развивается цветоносный побег с 1—2 листьями и соцве-
тие, а из пазухи верхнего розеточного листа — новый вегетативный
побег, развивающийся по описанному типу (Серебряков, 1952). От-
клонения от этого типа развития проявляются в том, что вместо ве-
гетативного побега в пазухе верхнего розеточного листа весной ино-
гда повторно развивается цветоносный побег, что обычно наблюда-
ется на вырубках и других сильно освещенных местах. Вегетативный
побег тогда образуется в пазухе розеточного листа дочернего цвето-
носного побега. В условиях влажной и теплой осени терминальные
почки вегетативного побега иногда трогаются в рост, образуя побе-
ги второго цветения.
По В.Н. Голубеву (1965), в условиях луговой степи земляника зе-
леная — длительно вегетирующее растение, относящееся к группе
ксероморфных мелкокорневых растений, к подгруппе мезоморфных
розеточных растений нижнего яруса травостоя. В течение вегетаци-
онного сезона земляника образует две генерации листьев, причем
вторая генерация — неполная, из 1—2 листьев, не достигающих нор-
мальных размеров и уходящих под снег. Это типично летне-зимнезе-
леное растение, зимующее с зелеными листьями. После схода снега
эти листья оказываются вполне жизнеспособными и ассимилируют
более или менее продолжительное время весной, а иногда и летом.
Если зима теплая и почва долгое время не замерзает, у земляники мо-
жет происходить интенсивный зимний рост почек возобновления и
зачаточных листьев. Таким образом, земляника характеризуется вы-
нужденным покоем.
Листья вегетативного розеточного побега — тройчатосложные,
на длинных черешках, мохнатых от густо одевающих их оттопырен-
ных волосков. Средний листочек на коротком черешочке, яйцевид-
ный или обратнояйцевидный, по краю зубчатый, причем конечный
зубец маленький, значительно короче соседних. Листья сверху — зе-
леные, блестящие, прижатоволосистые, снизу — сероватые от густо
прилегающих шелковисто-блестящих волосков, особенно густых на
выдающихся жилках. Листья на цветоносном побеге — сидячие или
на очень коротких черешках, косо яйцевидные, с мелкими, туповаты-
ми (нередко почти закругленными) зубцами с каждой стороны.
121
Розеточные зимующие побеги находятся или на уровне поверх-
ности почвы, или приподняты над поверхностью почвы примерно на
0,5 см.
Цветет земляника со второй половины мая по июнь включитель-
но, т. е., по В.Н. Голубеву (1965), это — средневесеннее-раннелетнее
цветущее растение. Соцветие дихазиальное (Troll, 1957). Цветки
крупные, до 2,5 см в диаметре, обычно обоеполые, хотя могут быть
признаки раздельнополости; чашечки треугольные, ланцетные, не
длиннее венчика, прижатоволосистые, при плодах прижатые к ним.
Наружные листочки подчашия одинаковой длины с чашелистиками
или несколько длиннее их, линейно-ланцетные, на верхушке иногда
расщепленные. Лепестки 5—10 мм длиной, округлые или обратнояй-
цевидные, заходящие друг за друга краями, коротконоготковые, жел-
товато-белые. Тычинки в цветке вдвое длиннее рылец пестика (у не-
плодоносящих цветков) или одинаковой с ними длины (у плодущих).
Цветоложе — волосистое, длиной около 1 см (Флора СССР, 1941).
Микроспоры почти сфероидальные, с овальными вытянутыми
по экватору порами; пленка борозд и пор гладкая, поверхность экзи-
ны — мелкозернистая (Флора Армении, 1958). Пыльца сравнительно
мелкая: продольный диаметр — 15,3—17,8 мкм, экваториальный
диаметр — 15,0—17,6 мкм. Завязь одаогнездная с одним семязачат-
ком, состоящим из нуцеллуса и простого интегумента из-за недораз-
вития внутреннего покрова. Семязачаток крассинуцеллятный с хоро-
шо развитой париетальной тканью (Беридзе, 1965). Б цветках рань-
ше развиваются рыльца пестиков, затем тычинки. Насекомые произ-
водят перекрестное опыление. Если насекомых не бывает, то проис-
ходит самоопыление благодаря тому, что собственная пыльца еще
успевает попасть на рыльце своего же пестика. Цветки на ночь и в
плохую погоду поникают, что защищает их от дождя и росы. После
опыления остаются все время поникшими, и плоды вследствие этого
получаются также поникшими, висячими.
Плод фрага — многоорешек на разросшемся цветоложе, или, по
В.Н. Тихомирову (1989), — «антокарпий». Плоды — орешки могут
как исключение выступать в качестве отдельных диаспор, но расте-
ние выработало приспособление для их совокупного распростране-
ния в виде разрастающегося мясисто-сочного тора (Тихомиров, 1989:
64). Плоды шаровидные или обратнояйцевидные, при основании су-
женные, большей частью желтовато-белые, лишь на верхушке крас-
новатые, реже целиком розоватые или бледно-красные, весьма аро-
матные. От чашечки плод отделяется довольно трудно. Орешки слег-
ка погружены в мякоть плода. Семена продолговатые, заостренные с
одной стороны и округлые — с другой, желго-зеленые с красными
пятнышками, без эндосперма. 2п=14 (Хромосомные числа..., 1969).
122
Онтогенез
В природе особи семенного происхождения найдены не были.
Семена, собранные в июле, в октябре были высеяны в чашки Петри.
В лабораторных условиях на 15—18-й день всхожесть достигла 80—
95%. Прорастание надземное. Семядоли — обратнояйцевидные, с ок-
руглой верхушкой, с суживающимся основанием, с почти незамет-
ным черешком, покрыты короткими железистыми волосками. Гипо-
котиль около 5 мм, корень до 1 см длиной, покрыт большим количе-
ством корневых волосков. Из верхушечной почки развивается розе-
точный побег. Проросток с 1—2 почти округлыми листьями, дайной
0,4 и шириной 0,3 см с 2—3 зубцами. Примерно через 1—2 месяца се-
мядольные листья отмирают, и растения переходят в ювенильное со-
стояние.
Дальнейшее описание онтогенеза основывается на описании
особей вегетативного происхождения, так как они несут черты очень
глубокого омоложения, вплоть до ювенильного состояния. Описание
развития особи на столоне основывается на постоянных наблюдени-
ях.
Такие особи возникают из верхушечной почки надземного сто-
лона, которая образует розеточный побег (боковая продолжает раз-
витие столона в длину). У розеточного побега за чешуевидным лис-
том формируется первый лист. Он появляется свернутым, высотой
0,3—0,5 см, черешок его сочный, опушенный, с хорошо развитыми
прилистниками, длительность жизни листа — 35 дней. Придаточные
корни отсутствуют, намечаются только бугорки. Через 3 дня чере-
шок листа удлиняется до 2 см, и пластинка первого листа разворачи-
вается. Срединный листочек первого листа — округлой формы, дли-
ной 0,6 см, шириной — 0,5 см. Зубцы по краю листочка начинаются
выше середины, причем, конечный зубец длиннее трех остальных (у
всех особей данные по числу зубцов срединного листочка приводятся
только с одной стороны, без конечного зубца). На 4—5-й день разви-
вается один придаточный корень розового цвета, закрепляющий
растение в почве. Вслед за первым листом через 10 дней появляется
второй лист. К этому времени длина черешка первого листа состав-
ляет 3—4 см, дайна листовой пластинки увеличивается до 0,91 см,
ширина — до 0,64 см, зубцов 3—4, конечный зубец равен остальным.
Корень — 5 см, еще не ветвится. К концу ювенильного состояния у
растения развито корневище длиной 0,85 см, которое с помощью
корней втягивается в почву. Развитие особи протекает очень быстро,
и примерно за месяц растение проходит путь от ювенильного до им-
матурного состояния.
Имматурные растения имеют розеточный побег с 1—2 отмерши-
ми листьями и с 1—2 хорошо развитыми живыми листьями. Посто-
янные наблюдения показали, что за вегетационный период растение
образует 4—5 листьев. Длительность жизни листьев — около 30
дней, поэтому в любой момент особь имеет 2 листа живых, 1 отмер-
123
ший и 1 формирующийся. Весной и летом особь развивает длинноче-
решковые листья с длиной черешка до 10 см, срединного листочка —
2 см, шириной 1,34 см, с 5—7 зубцами. У некоторых листочков зубцы
начинаются ниже середины, а у других — выше. Встречаются особи,
у которых на одной стороне листочка 5 зубцов начинаются' выше се-
редины, а с другой стороны этого же листочка — 7 зубцов, и начина-
ются они ниже середины. Конечный зубец или почти равен, или не-
много длиннее остальных.
Листья второй генерации — короткочерешковые, с длиной че-
решка 2—3 см, срединный листочек — длиной 1,5 см и 5—7 зубцами,
развивается в конце августа—сентября. Розеточный побег уже проч-
но закреплен в почве, корневище длиной 2,5 см. Придаточных кор-
ней 7—10, они ветвятся. В имматурное состояние особи переходят
или в тот же сезон, или — на следующий; длительность жизни в им-
матурном состоянии — от 1 до 3 лет. В ювенильном и имматурном
состоянии растение земляники представляет собой моноцентриче-
скую особь, где вегетативный розеточный побег нарастает монопо-
диально.
Виргинильные растения имеют розеточный побег, который за
год образует 6—7 листьев, причем в момент наблюдений (один раз в
месяц) в особи насчитывается 2 листа отмерших, 2—3 — живых и 1
— развивающийся. Весенне-летние листья — длинночерешковые, с
длиной черешка — 15 см, срединным листочком — 5,3 см, шириной
— 3,2 см, с 9—13 зубцами. Конечный зубец по длине или равен ос-
тальным, или немного меньше. Длительность жизни этих листьев со-
। ставляет около 23 дней,. Листья второй генерации — короткочереш-
ковые, с длиной черешка в среднем около 5 см, со срединным листоч-
ком 2,2 см длиной, с 8—10 зубцами. Длительность жизни короткоче-
решковых листьев, появляющихся в конце августа—начале октября
— около 8 месяцев. Корневище становится более мощным, длиной до
3,5 см, число ветвящихся придаточных корней увеличивается. В вир-
гинильном состоянии растение образует надземный столон, разви-
вающийся из пазушной почки первого листа, появившегося весной.
Столоны начинают развиваться в последней декаде июня и к концу
июля — началу августа уже полностью развиты. Особь дает 1, реже
2 столрна, на нем развиваются от 2 до 5 дочерних розеток, причем в
это время на столоне наблюдаются как ювенильные, так и имматур-
ные растения.
Итак, в виргинильном состоянии растение в самом начале веге-
тационного сезона представляет собой моноцентрическую особь; во
время образования столона и укоренения дочерних розеток это —
явно полицентрическая особь с ранней специализированной дезинте-
грацией, а после отмирания столонов особь вновь становится моно-
центрической. Длительность жизни особи в виргинильном состоянии
— до 3—4 лет.
Продолжительность дегенеративного периода земляники зеле-
ной в естественных условиях составляет 5—7 лет, что превышает
124
длительность дегенеративного периода земляники лесной на 2—4 го-
да. По данным И.Г. Серебрякова (1952), земляника лесная зацветает
на 3-й год.
Молодые генеративные особи — это полурозеточные растения с
удлиненными генеративными олиственными побегами, развивающи-
мися из верхушечных почек. По В.Н. Голубеву (1965), в Стрелецкой
луговой степи земляника зеленая — розеточное растение, у которой
в основании генеративного побега не образуется розеточных листь-
ев. По нашим наблюдениям (1982—1985 гт.), на остепненном лугу
высокого уровня р. Угры Калужской области в первых числах мая у
генеративных особей уже имеется небольшой 5—10 см цветоносный
бутонизирующий побег, в основании которого развито от 1 до 3 ли-
стьев с укороченными междоузлиями и 1—2 листа на удлиненном по-
беге. Розеточные листья генеративного побега имеют длину 7—10 см
и в июне полностью отмирают. Первый цветоносный побег — поли-
циклический, прямостоячий, около 28 см длиной, густо покрыт отто-
пыренными волосками. Земляника зеленая зацветает в конце мая, за-
канчивает цветение в июне. Число цветков на особь — 3—5. Плоды
созревают в конце июля, число их, в среднем, — 4 при числе плоди-
ков — 63.
Одновременно с появлением генеративного побега начинает раз-
виваться боковая почка верхнего розеточного листа, которая даст
начало новому вегетативному побегу. К концу июня этот побег име-
ет уже 4 листа. Первая генерация листьев вегетативного побега име-
ет длину черешков — 21 см, срединных листочков — 5,2 см, ширину
— 3,2 см, число зубцов — 9—11. Конечный зубец короче соседних.
Этот признак будет характерен и для следующих состояний. В ре-
зультате интенсивного фотосинтеза новый вегетативный побег нака-
пливает большое количество питательных веществ в стебле, оттесня-
ет в сторону тонкий цветоносный побег и занимает верхушечное по-
ложение. В период цветения создается впечатление, что генератив-
ный побег — боковой. Продолжающий свое развитие вегетативный
побег в августе—сентябре образует листья второй генерации. Длина
черешков листьев, в среднем, — 5,7 см, дайна срединного листочка —
2—3 см, ширина — 1,5, число зубцов — 7—10. За год особь образует
до 8 листьев. Длительность жизни листа около 18 дней.
Из пазушных почек первых листьев нового вегетативного побе-
га образуется 1—2 столона. В конце июля они достигают дайны око-
ло 70 см и формируют до 5 дочерних розеток. Корневище (до 4—5 см
длиной) имеет довольно много ветвящихся придаточных корней. На-
растание сменяется на симподиальное.
Средневозрастные генеративные растения развивают 2—3 розе-
точных побега из 5—8 листьев. Листья первой генерации имеют дли-
ну черешка 23 см, срединный листочек — длиной 4,7 см, шириной —
2,9 см и число зубцов 8—11. Высота генеративного побега — до 27
см, число цветков на особь — 4—8. Число семян на одну особь — 123
при среднем числе плодов на особь — 6. Длина черешков листьев
125
второй генерации вегетативного побега — 3—4 см, домна срединных
листочков — 1,5—2 см, ширина — 1—1,5 см, число зубцов — 8—10.
Растение образует 2—3 столона до 80 см длиной и до 8 дочерних ро-
зеток. Длина корневища достигает 6 см, корневище ветвится, образу-
ются 1 или 2 побега II порядка, с базальной части корневище начи-
нает отмирать, и длина корневища I порядка до ветвления составля-
ет около 1,5 см.
Старые генеративные растения образуют розеточный побег, от 3
до 8 листьев с длиной черешков листьев в первой генерации до 20 см,
срединного листочка — 4,9 см, шириной 3,2 см, с 9—11 зубцами.
Длина черешков листьев второй генерации — 4—5 см, длина средин-
ного листочка — 2—2,5 см, ширина — 1,5 см, число зубцов — 8—10.
Высота генеративного побега уменьшается до 24 см, число цвет-
ков до — 2—4, среднее число плодов на особь — до 2, число семян —
до 36. Старые генеративные особи образуют 1—2 столона длиной до
52 см, на которых развивается от 1 до 6 дочерних розеток. В подзем-
ной сфере происходит разделение (партикуляция) корневищ, длина
корневища достигает 6,2—6,6 см. Уменьшается интенсивность обра-
зования придаточных корней на нарастающем участке корневища,
из-за чего оно перестает втягиваться в почву.
Как показали наблюдения за маркированными особями, у зем-
ляники возможны перерывы в цветении: часть особей цветет ежегод-
но, другая — нет. Часть особей, не цветущая весной, зацветает в ав-
густе. Перерывы в цветении связаны с расходованием при цветении
больших количеств запасных веществ и необходимостью накопления
их для следующего цветения (Молиш, 1933). Длительность жизни
особей в генеративном состоянии — 4—6 лет.
У субсенильных растений отсутствуют надземные столоны и не
образуются генеративные побеги. Вегетативный розеточный побег
— с 2—3 листьями. Высота Черешков — около 11 см, длина средин-
ного листочка — 4,4 см, ширина — 2,5 см, число зубцов — 9. Корне-
вище рыхлое, трухлявое, около 5 см длиной, с базальной части раз-
рушающееся, а с апикальной — покрытое остатками черешков ли-
стьев. Точка роста в результате того, что не образуются новые при-
даточные корни, оказывается приподнятой над поверхностью почвы
на 0,5—0,8 см.
Сенильные растения по размерам и по форме листьев (их 1—2)
напоминают ювенильные. Высота черешка листа — 7 см, длина сре-
динного листочка — 0,7 см, ширина — 0,5 см, число зубцов — 5.
Корневище длинное — 4 см, старое, разрушающееся. Точка роста
приподнята над поверхностью на 1,5—2 см и зимой отмирает.
В процессе онтогенеза земляника зеленая проходит следующие
фазы онтогенеза. Проростки, ювенильные, имматурные растения
представлены первичным розеточным побегом — моноцентрической
структурой. Начиная с виргинильного состояния, когда формируют-
ся надземные столоны, структура особей становится явно полицен-
трической, образуется куртина, но виргинильные растения — розе-
126
точные, а генеративные — полурозеточные, явно полицентрические.
Фаза полицентрической особи короткая — около двух месяцев. Се-
нильные и субсенильные — это розеточные парциальные моноцен-
трические побеги.
Таким образом, взрослые особи земляники относятся к явно по-
лицентрической биоморфе, однако структура взрослой особи ла-
бильна и изменяется в течение вегетационного сезона: весной — она
моноцентрическая, летом — явно полицентрическая, осенью — мо-
ноцентрическая. Ритм сезонного развития зависит от географиче-
ского пункта наблюдений и от погодных условий (табл. 1).
Таблица 1.
Ритм сезонного развития земляники зеленой
Признаки Москва, ГБС АН СССР о Калужская обл. Ъ Курская обл. 3> Сев. Казах- стан «>
Начало вегетации апрель апрель апрель апрель
Окончательное отмирание перезимовавших листьев — конец мая — конец июня начало мая — конец июня конец апреля
Выдвижение цветоносов 7 мая ±6 дней 1—9 мая — —
Начало цветения 2 июня ±6 дней 20—25 мая 16—18 мая 10—12 мая
Массовое цветение 4 июня ±5 дней 31 мая— 2 июня 27 мая 15—20 мая
Окончание цветения 29 июня ±7 дней 20—25 июня 10—12 июня 5—7 июня
Начало плодоношения 7 июля ±9 дней — 4—7 июля —
Массовое плодоношение 21 июля ±11 дней — 11—19 июля июнь
Обсеменение — — — июль
Образование столонов 22 мая конец июня 20 июня —
•>Волкова, 1988; 2)наши наблюдения; Борисова, 1954;\Голубев, 1965;4) Борисова, 1961.
127
Способы размножения и распространения
Земляника зеленая размножается как семенами, так и вегетатив-
ным путем. Последний способ в природных сообществах преоблада-
ет. На побеге образуется от 3 до 9 сборных плодов, каждый содержит
от 20 до 138 семян (Калужская обл., с. Дворцы) и даже — до 480 (г.
Таруса). Семенная продуктивность в расчете на особь — невысокая.
Плоды поедают птицы, что обеспечивает расселение на значитель-
ные расстояния. Семена имеют хорошую всхожесть: на 15-й день
проросли все из 25 посаженных семян. Сбор плодов резко ограничи-
вает возможность семенного размножения.
Вегетативное размножение осуществляется с помощью столонов
(усов). Они развиваются в пазухах нижних листьев боковых розеточ-
ных побегов, появляющихся весной. В начале они растут наклонно
вверх, затем их верхушка активно опускается к поверхности почвы,
укореняясь и образуя дочерние розеточные побеги. От материнского
побега до дочерней розетки формируется всего лишь два междоузлия
— гипоподий и мезоподий. Дочерняя розетка развивается или терми-
нально — после 2-го предлиста столона, или из пазушной почки 2-го
предлиста. И в том, и в другом случае формируется новый столон, на
котором развивается новая розетка. Столоны образуются на расте-
ниях земляники виргинильного, молодого, средневозрастного и ста-
рого генеративного возрастных состояний. Жизнь особи вегетатив-
ного происхождения начинается с ювенильного состояния. В 1981 г.
на лугу р. Угры Калужской области был собран материал по интен-
сивности вегетативного размножения (табл. 2).
Наибольшей интенсивностью вегетативного размножения обла-
дают средневозрастные особи. Они образуют больше всего столонов,
больше розеток и на большее расстояние распространяют вегетатив-
ные зачатки. Число столонов и дочерних розеток, формирующихся
старыми генеративными и молодыми растениями, приблизительно
одинаково. Но молодые особи, обладая более интенсивным ростом
по сравнению со старыми, распространяют вегетативные зачатки на
большее расстояние, чем старые генеративные. Наименьшей интен-
сивностью вегетативного размножения обладают виргинильные осо-
би. Число дочерних розеток и длина столонов зависит от погодных
условий вегетационного сезона. Так, во влажном 1982 году столоны
были длиннее, чем в более засушливом году примерно в 2 раза, а чис-
ло розеток уменьшилось в 1,5—2 раза. Наибольшей жизнеспособно-
стью обладают дочерние розетки, ближе всего расположенные к ма-
теринской особи. Обычно сохраняются две первые розетки, осталь-
ные отмирают в первые два года. На 3-й год отмирание практически
прекращается, наступает период стабилизации численности. Как по-
казали наши исследования, в популяции ежегодно появляются юве-
нильные особи вегетативного происхождения, и их выживание со-
128
ставляет 44—50%. Прегенеративный период при вегетативном раз-
множении продолжается от 2 до 5 лет (редко — 7 лет). Длительность
его сокращается в условиях хорошей освещенности и невысокой чис-
ленности популяции. Общая продолжительность онтогенеза варьи-
рует от 5—7 до 13—15 лет (в условиях сильно сомкнутого траво-
стоя).
Таблица 2.
Средняя длина столонов, число столонов и число дочерних розегок
. на одну особь (с. Дворцы, Калужская область).
Показатели Возрастные состояния
V gl «2
Длина столонов 44,2 63,5 64,7 51,9
Число столонов 1,1 1,4 2,5 1,5
Число дочерних 1 розеток 2 3 4 5 6 7 1,1 1,5 2,5 1,5
0,82 1,5 1,9 1,3
0,45 1,2 1,6 0,96
0,09 0,6 1,3 0,54
0,09. 0,2 0,5 0,3
0,09 0,1 0,2 0,2
— — 0,08 —
Всего: 3,0 5,1 8,08 4,7
Экология
Земляника зеленая — вид экологически пластичный. В неболь-
ших количествах растет и на бедных, и на богатых почвах: от черно-
земных до луговых (дерновых) и подзолистых и даже слабосолонча-
ковых, при pH от слабокислой через нейтральную к слабощелочной
и от влажно-лугового до среднестепного увлажнения.
При высоком обилии характерна для богатых луговых степей,
остепненных сухих лугов, а также сухих лесов — сосновых и дубовых
(зона лесостепи) с влажностепным или луговостепным увлажнением
(Раменский и др., 1956). По нашим данным, земляника сочетает от-
носительную ксероморфность (содержанйе воды в листьях невысокое
— 58—-59%) с теневыносливостью (имеет очень тонкую листовую
129
пластинку 108—139 мкм). Водоудерживающая способность листь-
ев у земляники довольно низкая (потеря воды высокая — 80%) и как
считает Л.Г. Добренькова (1979), она — незасухоустойчива, но зимо-
стойка (Колкова, 1988). По типу водного питания в условиях степей
о тносится к омброфитам (Бейдеман, 1954).
Фитоценология
Земляника зеленая — типичный лесостепной вид. По А.Н. Поно-
мареву (1948, 1949, 1952), входит в географо-генетический евроазиат-
ский элемент лесостепного комплекса, одинаково характерный для
лугово-степных сообществ (лугово-степной эколог о-фитоценотиче-
ский тип, по Борисовой и др., 1961), так и для светлых разреженных
лесов, их полян и опушек. За пределами лесостепи встречается в незо-
нальных условиях: в горах наиболее обыкновенна в горно-луговых
степях (Носова, 1973). В.Н. Голубев (1962, 1965) отмечал, что земля-
ника — один из доминирующих видов степных содб-Тцеств, относится
к субдоминантам и имеет средний показатель обилия сор2. Встреча-
ется среди мезофильной, ксеромезофильиой и мезоксерофильной (лу-
говой и остепненной) растительности. Она является типичным видом
травянистого покрова дубовых лесов из пушистого дуба, для красно-
ковыльных разнотравных луговых степей на выщелоченных чернозе-
мах, для змеевково-тырсовых, вострецово-тырсовых, крупнополын-
но-тырсовых степей (Лавренко, 1956), а также типчаково-вейниково-
разнотравных и разнотравно-вейниковых ассоциаций на глубоко-
столбчатых солонцах (Антонова, Главатских, 1985).
В Московской области П.А. Смирнов (1958) отмечал, что в При-
окско-Террасном заповеднике земляника — пойменное растение.
Часто и обильно растет на степных участках, особенно — ра иго-
травно-типчаковых. Тонкий и богатый гумусом наилок благоприят-
ствует ее развитию. При вытеснении боровой флоры можно устано-
вить особую, длительную фазу клубники (земляники зеленой) с от-
меткой «sociales» и «copiosac», которая может существовать 5 10
лет. В пойме, вне степных участков, земляника есть везде по сухим
местам. На правобережье обычна по открытым местам и склонам, на
ровном водоразделе.
В Калужской области в пойме р. Угры характерна для поймен-
ных остепненных участков высокого уровня.
Особенности вегетативного размножения и возможность дли-
тельного существования особей в вегетативном состоянии определя-
ют характер возрастного спектра в цепопопуляциях: локальный мак-
симум образуют старые растения, абсолютный максимум чаще при-
ходится на прегенеративные растения. Появление новых особей про-
исходит ежегодно и неравномерно в зависимости от условий года
(так, в 1982 г. отмечен резкий подъем численности ювенильных осо-
бей). Резкая количественная диспропорция между виргииилытыми и
молодыми генеративными особями, с одной стороны, и зрелыми гс-
130
неративными, с другой, свидетельствует о том, что большая часть
выживших особей развивается с пропуском зрелого генеративного
состояния, пополняя группу старых растений. Сопоставляя возрас-
тные спектры во времени (Калужская область, 1982 и 1984 гг.) можно
предположить, что отмирание старых растений идет медленно: в те-
чение грех лет численность старых генеративных, субсенильных и се-
нильных растений практически оставалась стабильной. Высокая
плотнрсть ценопопуляции (214—233 особей/м2) с преобладанием
взросших растений или высокая сомкнутость травостоя в условиях
луговой степи способствуют накоплению старых растений; в этих ус-
ловиях особенно выражена эпизодичность возобновления (Былова,
Григорьева, 1987).
Изучение пространственной структуры земляники зеленой пока-
зало, что особи размещены диффузно, не образуя скоплений, что
также, видимо, объясняется вегетативным размножением. Наблюде-
ния за маркированными особями земляники в течение четырех лет
выявили, что усы с дочерними розетками, отходящими от материн-
ских растений, никак нс ориентированы по отношению к сторонам
света.
Земляника зеленая проявляет как свойства реактивности (быст-
рый захват территории, энергичное вегетативное размножение, вы-
сокие показатели численности, преобладание молодых растений в
ценопопуляниях), так и свойства толерантности (развитие с пропус-
ком возрастных состояний в ценопопуляниях). Ценотическая и эко-
логическая (к недостатку света) толерантность земляники проявляет-
ся в том, что она способна длительное время существовать в вегета-
тивном состоянии. В связи с мелкими размерами растений земляника
обладает низкой конкурентной мощностью. Благодаря своим цено-
популяционным свойствам она быстро занимает свободное про-
странство, но одновременно способна длительное время существо-
вать под пологом высокорослых трав, создавая высокое обилие.
Консортивные связи
Для земляники характерно перекрестное опыление. Так же, как и
у земляники лесной, цветки земляники зеленой посещаются мухами,
жуками, шмелями и пчелами (Иванова, Токарева, 1966).
На молодых развивающихся листьях между сложенными листоч-
ками обнаружен земляничный клещик (Tarsonemus fragariae Lim.). В
бутонах выявлен малиновый долгоносик (Anthonomus rubi Herbst.).
Часто листья проедены земляничным листогрызом (Galerucella tenella
L.). Паутинный клещик (Tetrahychus telarius L.) затягивает снизу
jhict паутиной, высасывая соки, вызывает- пожелтение сначала вдо:п>
жилок, а затем всего листа. Плоды часто поражаются серой гнилью
(Sclerotinia fusceliana Fuck.). По результатам исследований в ГБС АН
СССР, земляника сильно поражается мучнистой росой (Sphaerotheca
131
inacularis Fr. Mag. f. fragaria Jacz.), но довольно устойчива к бурой,
угловатой и белой пятнистости (Волкова, 1988).
Биологическая продуктивность
Определение био- и фитомассы особей в разных условиях суще-
ствования свидетельствует о чрезвычайной изменчивости этого по-
казателя; он также меняется в одном местообитании по годам в зави-
симости от увлажнения. По данным В.И. Данилова (1981), продук-
тивность земляники менялась с 1975 по 1980 г. следующим образом: 5
— 8,5 — 9,1 — 6,5 — 1,7 — 10,9 г/м2. Биомасса резко снижалась в за-
сушливые годы и увеличивалась во влажные.
В 1982 влажный год в Калужской области был определен сезон-
ный прирост фитомассы по возрастным состояниям (табл. 3).
Таблица 3.
Биометрические показатели и сезонный прирост фитомассы
(Калужская обл., 1982 г.)
Показатели Возрастные состояния
j im V «1 ё2 Из SS S
Высота расте- 5,7 8,9 16,9 21,6 23,3 20,3 10,9 5,8 НИЯ*, см Число живых 1,9 2,0 2,9 3,6 4,3 2,7 1,8 1,9 листьев Число отмерших 0,1 1,6 2,0 1,4 2,0 1,8 1,0 0,1 листьев Масса многолет- 2 120 130 330 390 430 140 50 ней части, мг Масса вегета- Ь 40 210 440 520 340 70 20 тйвной части, мг Масса особи, мг 3 160 340 770 910 770 210 70 Число особей 119 19 37 9 5 9 23 12 на 0,1 м2 Фитомасса, 3,57 30,0 127. 69,3 45,5 .69,3 48,3 8,52 г/1 м2
* — измеренная по длине листьев.
Урожайность плодов земляники зеленой в степных сообществах
Зауралья зависит от почвенных условий, режима использования уча-
стка (заповедный или сенокосный) и от проективного покрытия в
разных ассоциациях. Наибольшая урожайность была зафиксирована
при заповедном режиме в разнотравной ассоциации на выщелочен-
132
ном суглинистом черноземе: 2448 экз. (1297 г) на 100 м2 при проек-
тивном покрытии 20,5%. При сенокосном режиме на таких же почвах
при проективном покрытии 8,6% урожайность была значительно ни-
же 112 экз. (37 г) на 100 м2 (Антонова, Главатских, 1985).
Хозяйственное значение
Земляника зеленая — кормовое растение среднего качества, но
ценное лекарственное растение. Плоды содержат: глюкозы — 4,98%,
сахарозы — 6,33%, свободных кислот — 0,55% (в том числе фолиевой
кислоты — (В9) — 0,24%, аскорбиновой кислоты — 50—70%), Р-ак-
тивных веществ — 0,25—0,75%, азотсодержащих веществ — 1,49%,
золы — 1,22%, воды — 82,3%. Кроме того, плоды содержат каротин,
витамин В,, а также дубильные, флавоновые и пектиновые вещества,
антоциановые соединения, эфирное масло, фитонциды, микроэле-
менты. В листьях найдены дубильные вещества, от 300 до 438 мг ас-
корбиновой кислоты, эфирное масло (Ларин и др., 1951; Вигоров,
1972), Земляника обладает самым разнообразным действием: проти-
вовоспалительным, рано заживляющим, потогонным, мочегонным,
кровоостанавливающим и вяжущим. Регулирует обмен веществ, ока-
зывает противосклеротическое действие, улучшает состав крови.
Плоды ценятся как средство, растворяющее камни печени и почек и
не допускающее их возникновение, а также как средство при подагре
(Свиридонов, 1984). Настой травы употребляется при заболеваниях
легких (бронхиальной астме, туберкулезе легких), при катаре тол-
стых и тонких кишок, воспалении печени, селезенки, желчного пузы-
ря и почек (Землинский, 1949).
Плоды упо требляют в пищу в свежем, сушеном виде и для приго-
товления варенья. В то же время, земляника содержатся аллергены,
вызывающие у некоторых людей крапивницу, тошноту, рвоту, боли
в желудке (Вигоров, 1972).
Н.С. Сугоркина
РОД ГЕРАНЬ
Род Geranium L. — наиболее крупный в семействе гераниевых,
является почти космополитным, включает около 400 видов, распро-
страненных преимущественно в умеренных широтах (Жизнь расте-
ний, 1981), в тропических областях — только в горных странах. На
территории бывшего СССР встречается 55 видов (Флора СССР,
1949), по другим данным — 68 видов (Черепанов, 1981).
Герани — многолетние или однолетние травы, имеющие полуро-
зеточные или удлиненные репродуктивные побеги. Листья — череш-
ковые, чаще всего пальчато рассеченные или раздельные с нечетным
числом долей; края долей зубчатые, реже городчатые, иногда доли
листа линейные. Соцветия цимозные. Цветки актиноморфные с
двойным околоцветником, обычно обоеполые, энто.мофильные. Око-
лоцветник пятичленный, пять медовых железок чередуются с лепест-
ками, окраска венчика от бело-розового до синего, лилового, мали-
нового. Андроцей образован десятью тычинками, расположенными
в два круга. Для некоторых видов характерна гинодиэция (женская
двудомность), когда у части экземпляров популяции в цветках недо-
развивается андроцей. Гинецей образован пятью плодолистиками,
завязь — верхняя пятигнездная. Плод гераней — стеригма (Каден,
Лановая, 1963; Каден, 1964), в описании этого типа плода отражены
особенности, свойственные плодам типа коробочка: многосемян-
ность и вскрываемость (Цыренова, 1986). В каждом гнезде, как пра-
вило, развивается по одному семени. Для гераней характерна автохо-
рия, возможность ее определяется строением плода.
В Московской области встречается 9 видов этого рода.
Ключ для определения видов, встречающихся в Московской
области*
1. Доли всех листьев в очертании ромбические, общее очертание
листовой пластинки угловатое .......................... 2
— Пластинки, по крайней мере, розеточных листьев, в очертании
округлые. Доли их обратноклиновидные, на вершине округлые
или усеченные.......................................... 7
2. Пластинка листа до основания рассечена на 3—5 сегментов,
средний сегмент на удлиненном черешочке. Чашелистики у рас-
пустившихся цветков отходящие. Ноготок лепестка почти равен
отгибу. G. robertianum L. — Г. Роберта.
* За основу взят кшоч, составленный А.К. Скворцовым (Ворошилов и др., 1966).
134
— Пластинка листа надрезана не до основания, доли ее лопастные
или крупнозубчатые. Чашелистики у раскрывшегося цветка рас-
ходящиеся, ноготок много короче отгиба .............. 3
3. Растения без железистого опушения .................. 4
— Растения с железистым опушением (по крайней мере, на цвето-
ножках) ............................................. 5
4. Цветоносы двуцветковые. Лепестки на верхушке округлые, 12—
18;мм длиной, вдвое длиннее чашелистиков. G. palustre L. — Г.
болотная.
— Цветоносы одноцветковые. Лепестки на верхушке усеченные или
выемчатые, 5—7 мм длиной, чуть длиннее чашелистиков. G.
sibiricum L. — Г. сибирская. ।
5. Верхняя часть стебля, черешки верхних листьев, цветоножки, ча-
шелистики опушены как короткими железистыми и нежелези-
стыми волосками, так и длинными, оттопыренными, тонкими и
осзрыми волосками. Двулетник. G. bohemicum L. — Г. чешская.
— Растения без длинных оттопыренных волосков. Многолетники с
развитыми корневищами.................................... 6
6. Цветоножки до и после цветения вниз отклоненные. Лепестки
голубовато-лиловые. Нити тычинок линейные с яйцевидно-тре-
угольным расширением в основании. G. pratense L. — Г. луговая.
— Цветоносы до и после цветения вверх торчащие. Лепестки лило-
вые, розовые или белые, нити тычинок узколанцетные. G.
syhaticum L. — Г. лесная.
7. Листья глубоко раздельные на продолговато-линейные дольки.
Стебли, черешки и цветоносы покрыты жесткими оттопыренны-
ми волосками. Цветоносы одноцветковые. G- sangnineum L. — Г.
кровяно-красная.
— Доли пластинок округлые или усеченные, крупнозубчатые. Дву-
цветковые цветоносы покрыты только мягкими короткими во-
лосками. Чашелистики 4—5 мм длиной, лепестки ие более
8 мм .................................................... 8
8. Лепестки 6 8 мм длиной, в 1,5—2 раза длиннее чашелистиков.
Колонка плода длиной 10 12 мм. G. pyrcnaicum Bunn. til. — Г.
пиренейская.
— Лепестки 3 4 мм длиной, едва превышают чашечку или короче
ее. Колонка плода 5 8 мм длиной. G. pnsillum L. — Г. мелкая.
Герань луговая
Номенклатура и систематическое положение
Geranium pralense L. 1753, Sp. Pl.: 681; Кауфман, 1866, Моск. ф.'1..
изд. 1: 1 1 1; 1 U и пер. 1885, Сборник свод.: 119; Шмальгау зсн. 1895. Фл.
Средн, и 1<)жп. Росс. 1: 196; Сырейщпков. 1907, Илл. фл. Моск. губ. 2:
3Z23: он же. 1927, Опред. расе. Моск, губ.: 178; Федченко, 1940, в Ма-
135
евский, Фл., изд. 7: 486; Е. Бобров, 1949, Фл. СССР, 14: 31: он же,
1954, в Маевский, Фл., изд. 8: 283; он же, 1964, цит. соч., изд. 9: 278;
Скворцов, 1966, Опред. раст. Моск, обл.: 223; Webb a. Ferguson, 1968,
Fl. Europ. 2: 195; Черепанов, 1981, Сосуд, раст. СССР: 257; Октябре-
ва, 1986, Опред. раст. Мещеры, 1: 237 — Герань луговая (семейство
Geraniaceae — гераниевые).
Географическое распространение
Герань луговая имеет обширный евроазиатский ареал (Перву-
хин, 1963; Валеева, 1977). Вид бореальный, в субконтинентальной
Европе поднимается в горные области (Rothmaler, 1972). На терри-
тории бывшего СССР встречаются две расы: типичная европейская с
широко ромбическими и неглубоко надрезанными сегментами листь-
ев и восточная раса с сегментами, глубоко рассеченными на узкие,
почти линейные доли, с более голыми чашелистиками и более корот-
кими цветоножками (Флора СССР, 1949). Этой расе свойственны бо-
лее густо и почти до низу железистые стебли и черешки листьев, ли-
стья снизу опушены только по жилкам (Опред. высш. раст. Якутии,
1974). Типичная раса вида распространена более широко. На терри-
тории бывшего СССР В Арктической Европе она встречается в Ка-
рело-лапландском и Двино-Печерском районах, распространена в
странах Балтии, в Центральных районах европейской части. В степ-
ной зоне вид известен для ее северной части (Причерноморский,
Нижне-Донской, Нижне-Волжский районы), в Крыму не встречается
(Флора СССР, 1949; Flora Europaea, 1968). G. pratense распростране-
на в Предкавказье, редко в северных предгорьях поднимается до
1220—2590 м (Сорные растения СССР, 1934). В высокогорьях Урала,
в подгольцовом поясе, где производится сенокошение, она входит в
состав преобладающих растений высокотравья (Горчаковский,
1966). В Западной Сибири вид встречается во всех районах, в Обском
не севернее 61° (Флора СССР, 1949). В Восточной Сибири типичная
европейская раса растет только на Енисее: Минусинский район. Вос-
точная раса — G. transbaicalicum Serg. — растет от р. Бирюсы в
Предбайкалье до Амура (Попов, 1957). Она встречается на северо-за-
падном и северо-восточном побережье оз. Байкал (Попов, Бусик,
1966). Типичная форма, видимо, отсутствует на территории Якутии,
обычно наблюдается переход к G. transbaicalicum (Караваев, 1958).
А.К. Скворцов в «Определителе высших растений Якутии» (1974)
указывает, что G. pratense встречается в Алданском, Центрально-
Якутском и Восточно-Ленском районах. Многие из якутских расте-
ний, приближаются по своим признакам к G. transbaicalicum Serg.,
весьма нерезко обособленной расе, которая может быть признана не
более как за подвид. В Саянах герань луговая является обычным рас-
тением лесного пояса и лесостепи. В районах с резко сухим климатом
136
поднимается до субальпийского пояса, достигая высоты 2100 м
(Красноборов, 1976).
За пределами России вид распространен на Британских остро-
вах, в Скандинавии, атлантической и континентальной Европе, на
побережье и некоторых островах Средиземного моря, в северо-запад-
ном Китае, Монголии (Флора СССР, 1949; Flora Europaea, 1968). В
Московской области — обычное растение (Ворошилов и др., 1966).
Морфологическое описание
Герань луговая, многолетнее травянистое поликарпическое рас-
тение, может быть отнесена к группе короткокорневищных (Сереб-
рякова, 1953, 1956). В.Н. Голубев (1956а, 1958, 1960, 1962) на основа-
нии структуры корневой системы относит особи вида к кистекорне-
вым растениям, главным признаком которых он считает отложение
запасающих веществ в придаточных .корнях. По классификации Ра-
ункиера (Raunkiaer, 1934) этот вид является гемикриптофитом.
Растения начинают развитие как стержнекорневые, корневая
система виргинильных особей становится кистекорневой, хотя глав-
ный корень сохраняется до 10(12) лет. Генеративные растения имеют
многочисленные равномерно утолщенные придаточные корни; кор-
невую систему их можно назвать «бахромчатой» (Михайловская,
1981). Корни слабо ветвятся, длина их 8—12(15) см, окраска — ко-
ричневая.
Развитие побеговой системы идет по симподиальиой полурозе-
точной модели побегообразования (Серебрякова, 1977, 1979, 1981).
До перехода к цветению особи имеют главный розеточный побег, ко-
торый развивается как полициклический монокарпический. В год
цветения побег становится полурозеточным. После этого происходит
переход к симподиальному возобновлению. Симподнальная система
корневищ формируется из 1—2 верхних почек возобновления, зало-
женных в пазухах розеточных листьев репродуктивного побега, кор-
невища развиваются как эпигеогенные (Серебряков, Серебрякова,
1965).
Система корневищ взрослых растений ветвится до VI- - VII 1(Х)
порядка, число побегов — 2—10, иногда — более. Побеги развива-
ются как розеточные вегетативные и полурозеточные репродуктив-
ные. Высота репродуктивных побегов 60—90 см, а в некоторые годы
или в особых условиях (например, на местах бывших стогов) до
100—120 см. Розеточные листья длинночерешковые с розоватыми
или бледно-зелеными пленчатыми прилистниками, буреющими после
отмирания листьев. Длина черешков 10 25(40) см, они покрыты от-
стоящими жесткими волосками. Листовые пластинки 6 12 см в диа-
метре, округло-почковидные, пальчато-семираздсльныс, иногда при-
ближающиеся к пальчато-девятираздельным, доли их ромбические с
крупнозубчатым краем, листорасположение очередное. Нижние стеб"
левые листья черешковые, пальчато-пятираздсльпыс. верхние
I.V?
трехраздельные, сидячие. Листорасположение становится супротив-
ным.
У моноподиального полурозеточного репродуктивного побега
можно выделить три зоны: возобновления, торможения и обогаще-
ния. В зоне возобновления он имеет укороченные междоузлия.и несет
4—6 длинночерешковых листьев. Из пазушных почек только 1—2
верхние, наиболее крупные, участвуют в формировании побегов во-
зобновления. Таким образом, побеги возобновления образуются по
типу монохазия или дихазия (Смирнова, 1969, 1974). Зона торможе-
ния (неразветвленный участок побега) представлена одним междоуз-
лием, предшествующим соцветию. Зона обогащения содержит цимоз-
ное соцветие. Соцветие монотелическое, поскольку главная и боко-
вые оси обладают ограниченным ростом, а по особенностям измене-
ния листьев оно является фрондулезно-брактеозным (Федоров, Ар-
тюшенко, 1979). У герани луговой можно встретить объединенные
соцветия, несущие дихазиальные парциальные соцветия. При недо-
развитии цветоносов в конечных частях соцветия появляются парци-
альные соцветия типа плейохазия. Цветки образуются на осях II—
XII порядков. Побеги второй генерации — более низкие (до 35—50
см) и просто устроенные, цветки образуются на осях I—VI порядка.
Ветви соцветия и цветоножки покрыты железистыми волосками.
Цветоножки во время цветения стоящие вверх, до начала цветения и
в момент плодоношения поникающие. Цветки актиноморфные 2—
2,5 см в диаметре. Околоцветник двойной, пятичленный. Свободные
до основания продолговатояйцевидные чашелистики, имеющие три
жилки и заканчивающиеся нитевидным заострением, покрыты желе-
зистыми волосками. Лепестки лилово-синие, обратнояйцевидные,
цельные, в основании волосистые, длиннее чашелистиков почти
вдвое. Тычинок 10. Они расположены в 2 круга, нити тычинок в ос-
новании волосистые и широкие. Гинецей ценокарпный из пяти пло-
долистиков, завязь верхняя. Плод — клювоподобная пятигнездная
коробочка, в каждом гнезде, как правило, формируегся по одному се-
мени.
Семена — овальные длиной 3—3,5 см, шириной около 2 мм, име-
ют темно-коричневую окраску, хорошо выступающий шов и коре-
шок. Семена содержат хорошо сформированный зародыш, эндос-
перм не выражен (Рысина, 1973). По данным Н.Н. Кадена(1964), семя
гераней содержит эндосперм в виде тонкого слоя, зародыш согну-
тый, корешок прилегает к спинке одной из семядолей. Масса 1 тыся-
чи семян — 3,5 г, в 1 кг — до 300 тыс. семян (Доброхотов, 1961)
Число хромосом 2п=28 (Хромосомные числа..., 1969, Bate-Smith,
1973).
Онтогенез
Семена герани луговой имеют период покоя. Запас семян в почве
не образуется (Работнов, 1951, 1958). Семена хорошо прорастаю!' без
138
какой-либо обработки (Николаев и др., 1985). По нашим наблюдени-
ям, при посеве семян в чашки Петри в год обсеменения период про-
растания может растянуться до 2—3 месяцев. Лабораторная всхо-
жесть семян, собранных на пойменном лугу, составила 22—66%. По-
беги второй генерации, возникшие после покоса, дали жизнеспособ-
ные семена, всхожесть их достаточно высока — 46%.
Семена прорастают по надземному типу. Над поверхностью
почвы поднимается тонкий, длинный, округлый гипокотиль, окра-
шенный в интенсивный малиновый цвет (Рысина, 1973).Он выносит
две плотно свернутые зеленые семядоли, на которых иногда сохраня-
ется перикарп семени. Семядоли почковидные, неравнобокие, с серд-
цевидным основанием, реснитчатым краем, более мясистые, чем на-
стоящие листья, имеют ширину 0,6—0,8 (1,2) см. Черешки их густо
опушены и достигают длины 3—5(7) см. К концу мая — началу июня
гипокотиль втягивается в почву, формируется розеточный побег.
Проростки в это время имеют главный розет очный побег с 2 семядо-
лями и 1—2 настоящими листьями. Корневая система представлена
главным корнем и тонкими боковыми корнями.
Ювенильные растения формируют главный розеточный побег с
1—3 листьями. Листовая пластинка пальчато-семираздельная, сред-
няя доля заканчивается тремя зубцами (ювенильный тип листа). Диа-
метр листовой пластинки 1,5—2.2 см, длина черешков увеличивается
до 9—10 см. Главный корень равномерно утолщен, светло-коричне-
вого цвета, боковые корни тонкие.
Имматурные особи продолжают нарастать моноподиально,
имеют главный розеточный побег с 2—3 листьями. Диаметр листо-
вой пластинки возрастает до 2,5—3,5 см, средняя доля ее несет 5—10
краевых зубцов (имматурный тип листа), длина черешков достигает
10—17 см. Формируется ортотропное корневище, длина которого
0,8—1,5 см. Корневая, система становится стержне-кистекорневой,
появляются первые придаточные корни.
Виргинильные растения продолжают формирование главного
розеточного побега, имеют 2—5 листьев. Сильно увеличивается дли-
на черешков (до 20—35 см), благодаря чему листовые пластинки вы-
носятся в более светлую часть травостоя. Увеличивается диаметр
листовых пластинок (3,5—8 см). Они пальчато-семираздельные, ино-
гда приближаются к пальчато-девятираздельным, средняя доля име-
ет 11—18 и более краевых зубцов (взрослый тип листа). Корневая
система становится кистекорневой. Главный корень сохраняется, но
скрывается под массой более длинных придаточных корней.
Молодою генеративные растения имеют главный полурозеточ-
ный репродуктивный побег, высота его 40—70 см, цветки образуют-
ся на осях II—VII(X) порядка. Корневая система, как и у виргиниль-
ных растений, представлена системой главного корня, скрытой под
кистью многочисленных придаточных корней, отходящих от орто-
тропного корневища.
139
Средневозрастные (зрелые) генеративные растения — наиболее
мощные в ценопопуляциях. Их подземная система симподиальных
корневищ ветвится до V1II(X) порядка. Они формируют 2—5(10) ре-
продуктивных полурозеточных побегов, высота которых в отдель-
ные годы может достигать 100—120 см. Цветки образуются ра осях
II—VIII(XII) порядков. Корневая система бахромчатого типа обра-
зована многочисленными придаточными корнями.
Старые генеративные растения представляют собой отдельные
генеративные партикулы или клоны из 2—3(8) партикул. Они имеют
1—3 репродуктивных побега высотой 60—100(120) см. Цветки обра-
зуются на осях II—1Х(ХП) порядков. PasMqjbi клонов могут быть
близки к размерам зрелых генеративных растений. Симподиальная
система корневищ ветвится до n+V(VII) порядка. Корневая система
образована придаточными корнями. Старые участки корневищ ли-
шены придаточных корней.
Субсенильные растения также представлены клонами из 2—3(7)
партикул, а чаще — отдельными вегетативными партикулами. Ре-
продуктивных побегов нет, партикулы имеют 1—3 вегетативных ро-
зеточных побега с 2—5 листьями. Листовые пластинки — пальчато-
семираздельные, по сложности края сходные с листьями имматурных
или виргинильных растений. По облику подземной части субсениль-
ные растения похожи на старые генеративные растения.
Сенильные растения очень малочисленны. Обычно это отдель-
ные вегетативные партикулы с 1—2 вегетативными побегами. Листь-
ев образуется 1—2, ювенильного или имматурпого типа. В подзем-
ной части растения имеют участок старого разрушенного корневи-
ща, почти лишенный придаточных корней.
Продолжительность онтогенеза герани луговой составляет 15—
30 и более лет. Растения в течение всей жизни представляют собой
моноцентрические образования. Первый вариант сокращенного он-
тогенеза выражается в неспособности части растений сформировать
разветвленную систему корневищ, а после партикуляции — клон.
Второй вариант сокращенного онтогенеза отмечается у особей еще
более низкой жизненности. Он выражается в том, что растения не
способны перейти к цветению. По мере старения подземной части
они пополняют группу субсенильных растений.
Сезонное развитие
В природе массовое прорастание семян отмечается после перези-
мовки, в начале мая следующего года, хотя единичные проростки
можно встретить и осенью. Начало вегетации взрослых растений
также приходится на первую декаду мая.
Растение формирует побеги разных типов. Главный побег разви-
вается как полициклический монокарпический. Он нарастает моно-
подиально в течение нескольких лег, все его листья отмирают осе-
нью. Весной следующего года розеточный побег возобновляется из
140
перезимовавшей верхушечной почки. По мере развития главного по-
бега (и хода онтогенеза растения) возрастает емкость его верхушеч-
ной почки:j — 3, itn — 5, v — 7 (наблюдения 15.07.85 в Калужской
области*).
У всех растений емкость верхушечной почки главного побега
оказалась выше емкости почек возобновления, заложившихся в пазу-
хах его розеточных листьев. Возможно, что такое соотношение емко-
сти почек определяет моноподиальный характер нарастания главно-
го побега. В ходе онтогенеза увеличивается число листовых зачат-
ков, которые можно одновременно наблюдать на конусе нарастания.
Так, у имматурной особи на конусе главного побега оформлен пол-
ностью один зачаток. На апексе главного побега виргинильного
растения в момент вычленения листовой пластинки одного листово-
го примордия появляется валик очередного листа. Еще быстрее про-
исходит формирование листовых зачатков в почках возобновления
генеративных растений: в листовом зачатке только намечается рас-
членение листовой пластинки, а на апексе уже появляется валик сле-
дующего листа. В ходе онтогенеза интенсивность образования лис-
товых зачатков возрастает, увеличивается емкость верхушечной поч-
ки главного побега, растет число листьев розеточного побега.
В Подмосковье репродуктивные побеги второго и последующих
порядков формируются по моноциклическому, дициклическому, три-
циклическому типу (Серебряков, 1959). Наблюдения подтверждают
эти особенности, можно лишь добавить, что большинство побегов
развивается по дициклическому типу. Кроме того, у части побеюв
отмечается неполный цикл развития. В ходе онтогенеза побег прохо-
дит внутрипочечную, внепочсчную фазы и фазу вторичной деятель-
ности (Серебрякова, 1961, 1971), последняя определяется продолжи-
тельностью существования резида (термин Е.Л. Нухимовского, 1971)
в многолетней побеговой системе взрослого растения.
Характерные для взрослых растений дицикличсские полурозе-
гочиые репродуктивные побеги начинают свое развитие из почек во-
зобновления, заложившихся в пазухах розеточных листьев материн-
ского побега во время его плодоношения или сразу после покоса. Ро-
зеточные побеги с 2—3 листьями формируются в течение июля -ав-
густа из 1 - 2 почек, обычно самых верхних и наиболее крупных
(0,7—1,5 см). Листья засыхают к концу сентября. Герань луговая
летнезеленое растение, зимует с закрытыми почками. В этих почках
заложены 3—5 чешуевидных листьев и до 10 зачатков настоящих ли-
стьев. Соцветие к октябрю сформировано до внешнего круга около-
цветника в отдельных цветках (Серебрякова, 1956).
Весной из перезимовавших верхушечных почек формируются ро-
зеточные побеги с 4 6 длнпночерешковымн листьями. В начале се-
редине июня no6ei и становятся полурозеточиымп, растения зацвета-
ют. Цветки герани луговой протерапдрпчны.
j. ini, v индексы возрастных состояний Уралова (1975).
141
Продолжительность функционирования побегов первой генера-
ции — 1—1,5 месяца. После плодоношения или покоса репродуктив-
ный побег отмирает до зоны возобновления, которая входит в состав
многолетней системы симподиальных корневищ в виде резида одно-
го порядка. С этого начинается фаза вторичной деятельности побе-
га. Длительность существования резида в многолетней системе кор-
невищ растения может достигать 8—10 лет. Репродуктивные побеги
второй генерации появляются особенно массово после раннего поко-
са. Они продлевают цветение герани луговой до августа—начала
сентября.
Придаточные корни формируются в момент развития розеточ-
ной части моноподиального побега, как правило, весной. Корневи-
ща характеризуются высокой окорененностыо, хотя количество кор-
ней может быть различным. На резиде одного порядка развивается
от 1—2 до 14—17 придаточных корней.
Способы размножения и распространения
Основным способом самоподдержания популяции в естествен-
ных условиях является семенное размножение. Специализированных
органов вегетативного размножения у растений данного вида нет.
Для вида характерна автохория, семена отлетают от материн-
ского растения на расстояние 170—220 см. Потенциальная семенная
продуктивность отдельных побегов составляет от 150 до 500 семеза-
чатков, средняя потенциальная продуктивность особи — 150—1050.
Реальная семенная продуктивность побега 40 150 семян, на особи
образуется в разных условиях 40—480 полноценных семян. Урожай
семян на отдельных участках пойменного луга р. Угры в Калужской
области составил 330—4880 шт./м2.
Расчеты показывают, что в ценопопуляциях герани луговой
ювенильные особи составляют 3,4—5,7% от исходного урожая семян.
В среднем из тысячи полноценных семян до зрелого генеративного
состояния доходит одно.
Вегетативное размножение герани луговой осуществляется в
форме старческой партикуляции растений высокой жизненности. В
результате формируются компактные клоны из 2—8 партикул. Омо-
ложение дочерних особей, проходящее как частичное освобождение
от отмерших частей, следует считать слабым. Образовавшиеся пар-
тикулы продолжают ход онтогенеза материнского растения. Моло-
дые клоны имеют четкие границы, развитие их может привести к по-
степенному обособлению партикул.
Учет длительности зрелого генеративного состояния,, способ-
ность растений к вегетативному размножению, приводящая к обра-
зованию такого количества старых генеративных растений, которое
в 6—10 раз превышает исходную численность плодоносящих расте-
ний, говорит об устойчивости природных ценопопуляций вида и
способности их к успешному самоподдержанию.
142
Экология
Герань луговая — мезофит, единично встречается в условиях лу-
гово-степного и сыролугового увлажнения. Вид наиболее обилен при
влажнолуговом увлажнении (64—74-е ступени шкалы Раменского,
3-я ступень шкалы Элленберга). Растения герани луговой чувстви-
тельны к переменности увлажнения, обильно распространены в пре-
делах 4.1—12-й ступеней шкалы переменности увлажнения (Рамен-
ский и др., 1956), что соответствует умеренно переменному увлажне-
нию (Работнов, 1974, 1978).
Герань луговая — светолюбивое растение, выносит очень не-
большую затененность (4-я ступень шкалы Элленберга). Отношение
к свету меняется в ходе онтогенеза — на ранних этапах онтогенеза
особи вида более теневыносливы. Площадь листовой поверхности
постепенно увеличивается: ювенильные особи 1—2 см2, взрослые ве-
гетативные растения —243 см2, генеративные растения — 912 см2
(Работнов, 1974). Длина черешков розеточных листьев взрослых рас-
тений коррелирует с высотой травостоя. Благодаря этому листовые
пластинки выносятся на уровень второго, как правило, господствую-
щего яруса травостоя. Этим объясняется формирование в условиях
гераниевого луга двух максимумов в распределении массы надзем-
ных органов (Работнов, 1974, 1978).
Вид является мезотрофом, обильно встречается на довольно бо-
гатых почвах (10—17-е ступени шкалы Раменского, 4-я ступень шка-
лы Элленберга). Растения данного вида часто растут на почвах бога-
тых свободным фосфором (Работнов, 1974).
Фитоценология
Основные формации, в которых встречается герань луговая на
большей части своего ареала, — формации луговой растительности.
Вид характерен для пойменных лугов, участков среднего уровня
(Прозоровский, 1956; Серебрякова, 1956; Работнов, 1974). Так, на За-
лидовских лугах в пойме р. Угры Калужской области герань луговая
широко распространена в центральной пойме на участках среднего
уровня, единично встречается в прирусловой части поймы. В услови-
ях длительного отсутствия половодья она становится обильной в не-
больших депрессиях рельефа, во многих межгривных понижениях
центральной поймы входит в состав доминантов, а в момент массо-
вого цветения образует аспект. Вид встречается в ассоциациях с до-
минированием лисохвоста лугового, овсяницы луговой в составе бо-
бово разнотравных ассоциаций (Алехин, 1925), местами герань луго-
вая может доминировать (Работнов, 1958, 1974). Положительно реа-
гируя на различные нарушения, разрыхление почвы, нарушение дер-
нины, уменьшение замкнутости сообщества после установки стогов,
герань способна занимать большие площади, улучшая при этом жиз-
143
ценность особей. Условия синэкологического оптимума организма и
ценопопуляций для данного вида совпадают и оптимум соответству-
ет начальным этапам демутационной сукцессии на местах бывших
стогов (Сугоркина, 1989, 1991).
Изучение ценопопуляций при разных, способах хозяйственного
использования показало, что средняя численность ценопопуляций
изменяется от 1,4 до 25,2 особей на 0,25 м2. Минимальная числен-
ность отмечена на сильно сбитом пастбище. Базовым спектром явля-
егся двухвершинный спектр с максимумами на ювенильных и старых
генеративных растениях при минимальном участии молодых генера-
тивных.
В степных районах герань луговая иногда может быть эдифика-
тором (Куркин, 1968), встречается по днищам и склонам логов (Голу-
бев, 1965). В высокогорьях она входит в состав травянистых расте-
ний подгольцового пояса, распространена в лесном поясе и лесосте-
пи, а в наиболее сухих местах поднимается в субальпийские луга
(Горчаковский, 1966; Красноборов, 1976).
Биологическая продуктивность
Биологическая продуктивность герани луговой зависит от кли-
матических условий года и особенностей хозяйственного использо-
вания угодий. Урожай сухой надземной массы в разные годы на пой-
менном лугу в Калужской области составлял от 160,7 до 880,2 г/м2.
Величина побеговой фитомассы растений зрелого генеративного
возрастного состояния на разных участках поймы изменялась от 1,7
г до 80,2 г, при этом минимальной она была на пастбище, макси-
мальная величина отмечена на месте бывшего стога.
Хозяйственное значение
Данные о кормовой ценности вида противоречивы. По одним
данным, герань луговая кормовой ценности не имеет (Сорные расте-
ния СССР, 1934), по другим — удовлетворительно поедается в сене,
но плохо сохнет, раструшивается при сушке (Справ, по сенок. и
пастб., 1966). На пастбище скотом поедается плохо, в сене лучше;
удовлетворительно поедается на Алтае маралами, зайцами, сурками,
пищухой; рябчики охотно едят семена (Ларин, 1957).
Герань луговая является дубильным растением за счет содержа-
ния таннидов, состоящих из галлотапнида со следами катехинтанни-
да. В цветках содержится 16,3%, в траве — 3,2% таннидов (Вороши-
лов, 1941); корни содержат 15—17% (Первухин, 1963). По данным Г.
Шлыкова (1932), растения имеют в составе до 20—50% дубильных
веществ, сахаров и геранипа.
Вид относится к важным медоносам. Растения охотно посеща-
ются пчелами, характеризуются продолжительным периодом цвете-
144
ния (около двух месяцев). Герань луговая дает много нектара и явля-
ется перганосом, так как производит значительное количество пыль-
цы (Глухов, 1935; Долгошов, 1948; Иванова, 1956). В народной меди-
цине находит применение как вяжущее средство, используется от по-
носов, кровотечений, женских болезней (Землинский, 1951).
Растения обладают красильными свойствами, пригодны для ок-
рашивания в зеленый цвет (Сорные растения СССР, 1934), и очень
декоративны.
Герань луговая может быть сорным растением в посевах озимых
и яровых пшениц, ржи, проса. На Кавказе на высоте 1200—2500 м ге-
рань — обременительная сорная трава, заглушающая другие расте-
ния и быстро занимающая большие площади (Гроссгейм, 1952). На
пойменных лугах может занимать значительные площади, положи-
тельно реагируя на любые нарушения (разрушение дернины, уста-
новка стогов, умеренный выпас).
Вид отрицательно реагирует на выпас (Смелов, 1947; Работнов,
1974), что объясняется снижением жизненности особей и ценопопуля-
ций, резким падением численности. Мерами борьбы является предот-
вращение обсеменения, опрыскивание гербицидами 2,4Д (Справ, по
сенок. и пастб., 1966).
Герань лесная
Номенклатура и систематическое положение
Geranium sylvaticum L. 1753, Sp. pl.: 681; Кауфман, 1866, Моск,
фл., изд. 1: 111; Цингер, 1885, Сборник свед.: 119; Шмальгаузен,
1895, Фл. Средн, и Южн. Росс. Г 196; Сырейщиков, 1907, Илл. фл.
Моск. губ. 2: 324; он же, 1927, Опред. раст. Моск, губ.: 178; Федченко,
1940, в Маевский, Фл., изд. 7: 487; Е. Бобров, 1949, Фл. СССР, 14: 24;
он же, 1954, в Маевский, Фл., изд. 8: 283; он же, 1964, цит. соч., изд. 9:
278; Скворцов, 1966, Опред. раст. Моск, обл.: 223; Webb a. Ferguson,
1968, Fl. Europ. 2: 195; Черепанов, 1981, Сосуд, раст. СССР: 257; Ок-
тябрева, 1986, Опред. раст. Мещеры, 1: 238. — герань лесная.
Географическое распространение
Ареал герани лесной — евро-западносибирский. Вид встречается
в субокеанической Европе, заходит в арктическую Европу и запад-
ную часть арктической Сибири. Распространен во всех районах ев-
ропейской части, кроме Нижневолжского, Крыма и большей части
Причерноморского, на Кавказе обычен во всех районах, за исключе-
нием Талышского, в Южном Закавказье очень редок, известен в
Прибалхашье. В Западной Сибири герань лесная распространена во
всех районах, за исключением южной части Верхне-Тобольского
145
(Флора СССР, 1949). В Восточной Сибири вид растет в западной
части Ангаро-Саянского и Енисейского районов, на Среднесибир-
ском плато он обычен по р. Бирюсе (Попов, 1957). Общее распро-
странение охватывает Скандинавию, Среднюю и Атлантическую Ев-
ропу, Балканы и Малую Азию.
Морфологическое описание
Герань лесная — многолетнее, травянистое, короткокорневищ-
ное поликарпическое растение с симподиальной системой полурозе-
точных репродуктивных и розеточных вегетативных побегов. По
жизненной форме и строению особи она сходна с геранью луговой.
Корневая система молодых растений представлена системой главно-
го корня. Взрослые растения имеют придаточную корневую систему,
которую можно считать бахромчатой. Длина придаточных корней
— 10—15 см, диаметр — 1,5—2 мм, окраска их коричневая. Корни
равномерно утолщены по всей длине. Симподиальная система эпиге-
огенных корневищ ветвится до IV—VII порядка.
Развитие побеговой системы идет по симподиальной полурозе-
точной модели побегообразования (Серебрякова, 1977, 1979, 1981).
Главный побег — полициклический монокарпический, 6—7(10) лет
развивается как вегетативный розеточный, в год цветения становит-
ся полурозеточным. Розеточные листья — длинночерешковые, диа-
метр листовых пластинок у взрослых растений достигает 8—12(15)
см, длина черешков в 3—4 раза превышает диаметр листовых пла-
стинок, листорасположение очередное. Листовые пластинки пальча-
то-семираздельные, край их крупнозубчатый, число зубцов возраста-
ет в онтогенезе, если ювенильные растения имеют три краевых зубца
в средней доле листовой пластинки, то взрослые растения 11—24 и
более. Прилистники ланцетные, заостренные, 1,5—2 см длиной, свет-
ло-бурые.
Взрослые растения имеют 2—8 побегов, из них 1—3 репродук-
тивных полурозеточных, высота их достигает 30—80 см. Репродук-
тивные побеги имеют сходный план строения с побегами герани лу-
говой, в них также можно выделить три зоны. Розеточных листьев
образуется 3—4, зона торможения представлена одним междоузлием,
зона обогащения — цимозным соцветием. Парциальные соцветия
образованы в основном дихазиями и монохазиями, иногда в конеч-
ных частях соцветия плейохазиями. Удлиненные части побегов более
гладкие, чем у герани луговой, так как покрыты прижатыми волос-
ками, верхние части их, а также цветоносы и чашелистики имеют же-
лезистое опушение. Цветоносы до и после цветения направлены
вверх. Листорасположение в удлиненной части генеративного побе-
га супротивное. Соцветие фрундулезно-брактеозное, монотелическое
(Федоров, Артюшенко, 1979).
146
Цветки актиноморфные с двойным околоцветником, пятичлен-
ные. Лепестки (10)12—16(18) см длиной, розовые, лиловые, малино-
вые, иногда белые, нити тычинок в нижней части узколанцетные.
Плод — пятигнездная коробочка, по мнению других авторов
(Каден, Лановая, 1963; Каден, 1964), — стеригма, вскрывается пятью
створками и выбрасываег семена. Семена овальные (длина 4 мм, ши-
рина 2 мм) с хорошо выраженным швом и корешком. Поверхность
кожуры не блестящая, мелко ячеистая, темно-коричневого цвета (Ры-
сина, 1973).
Для вида характерна гинодиэция (женская двудомность), когда у
части растений в ценопопуляции тычинки недоразвиваются. Такие
цветки, как правило, меньших размеров, лепестки их не превышают
по длине чашечку, а венчик не раскрывается широко (Демьянова и
др., 1979; Демьянова, 1987).
Диплоидный набор хромосом равен 28 (Хромосомные числа...,
1969; Bate-Smith, 1974), указывается еще 2п=24 (Хромосомные чис-
ла..., 1969).
Онтогенез
В лаборатории свежесобранные семена прорастают очень не-
дружно, всхожесть их при комнатной температуре составляет 8—
17%. Низкая лабораторная всхожесть семян может быть связана с
твердосемянностью, так как физиологический покой у них неглубо-
кий. Семена герани лесной хорошо прорастают только после обра-
ботки концентрированной серной кислотой, промывки водой и об-
работки гиббереллинами (Николаева и др., 1985).
Прорастание семян надземное. Гипокотиль выносит на поверх-
ность две семядоли с черешками до 1,5 мм длиной. Семядоли почко-
видные, неравнобокие,, с сердцевидным основанием, длина их —
0,5—0,7 мм, ширина 10—12 мм. Затем гипокотиль втягивается в поч-
ву, проростки образуют главный розеточный побег с двумя семядо-
лями и одним—двумя пальчато-семираздельнымй листьями; длина
черешков составляет 3,5—6,5 см, диаметр листовой пластинки — до
2,3 см. Стержневой корень — белый или желтоватый, имеет боковые
корни II порядка.
У ювенильных растений главный розеточный побег имеет 1—3
листа по размерам сходные с листьями проростков. В отличие от ге-
рани луговой, средняя доля листа заканчивается 3—5 зубцами. Глав-
ный корень равномерно утолщается, становится светло-коричневым.
Имматурные особи, как и у герани луговой, являются стержне-
кистекорневыми растениями. Они имеют систему главного корня и
немногочисленные придаточные корни, формируют ортотропное
корневище, достигающее 0,8—1,8 см. Главный розеточный побег с
1—3 листьями, длина черешков их — 12—16 см, диаметр листовой
пластинки — 3—5 см, ее средняя доля заканчивается 7—13 зубцами.
147
Главный розеточный побег виргинильных растений с 2—5 ли-
стьями, диаметр листовых пластинок — 8—12 см, средняя их доля за-
канчивается 15—25 зубцами. Корневая система становится кистекор-
невой. Главный корень сохраняется, но скрывается под многочислен-
ными придаточными корнями.
Молодое генеративное растение формирует полурозето'чный ре-
продуктивный побег I порядка. Корневая система кистекорневая,
главный корень сохраняется. Иногда главный побег бывает скрыто
генеративным.
Средневозрастные генеративные растения — наиболее мощные.
Главный признак — наличие симподиальной системы эпигеогенных
корневищ, ветвящейся до V—VIII порядка. Все участки корневищ
имеют многочисленные придаточные корни. Генеративных побегов
обычно немного — 1—3, высота их 60—80 см, цветки образуются на
осях II—IX порядка.
Старые генеративные растения чаще всего представлены клона-
ми из 2—4(6) партикул; генеративных побегов 1—2, высота их —
35—65 см, цветки образуются на осях II—VII порядка. Симподиаль-
ная система корневищ ветвится до IV—VI порядка, не все их участки
имеют живые придаточные корни.
Субсенильные растения представлены клонами из 2—4 вегета-
тивных партикул. Симподиальная система корневищ ветвится до
III—V порядка. Придаточные корни сохраняются только на живых
участках корневищ.
Сенильные растения обычно представлены однопобеговыми не-
ветвящимися вегетативными партикулами. Розеточный побег имеет
1—2 листа ювенильного типа, полуразрушенное корневище почти
лишено придаточных корней.
Продолжительность онтогенеза более 20 лет.
Сезонное развитие
Главный побег — полициклический, монокарпический, зацвета-
ет на 6—7(10)-й год. В Подмосковье побеги взрослых растений раз-
виваются по моно-, ди-, или трициклическому типу (Серебряков,
1959). Большинство побегов формируется как дициклические (Кома-
рова, 1988; Сугоркина, 1989). Онтогенез дициклического побега от
возникновения конуса нарастания до завершения деятельности вер-
хушечной меристемы продолжается 3 вегетационных периода (Кома-
рова, 1988). Сначала идет формирование почек, затем развивается
розеточный вегетативный побег, а в последний год формируется по-
лурозеточный репродуктивный побег.
В почках возобновления розеточных побегов к концу лега уже
сформировано соцветие, включая отдельные цветки (Серебряков,
1952). Эти зимующие почки укороченных побегов имеют емкость 6—
7 узлов плюс соцветие (Комарова, 1988).
148
Герань лесная является длительно вегетирующим растением (Го-
лубев, ] 965). За вегетационный сезон образуются две генерации ли-
стьев, что связано с формированием после цветения розеточных по-
бегов возобновления.
Вегетация растений начинается в первой декаде мая, когда фор-
мируется розеточная часть побегов. В конце мая начинает разви-
ваться удлиненная часть репродуктивных побегов. Репродуктивные
побеги зацветают в начале июня, цветение их заканчивается к нача-
лу июля. Цветки цветут 2—3 дня, после чего лепестки венчика опада-
ют, и образуется плод.
Придаточные корни появляются на стадии развития розетки с
конца мая (Серебряков, 1952). Вегетация растений длится иногда до
начала октября, в зависимости от климатических условий. Герань
лесная — летнезеленое растение, зимует с закрытыми почками. По
положению зимующих почек возобновления является гемикриптофи-
том.
Способы размножения и распространения
Герань лесная размножается преимущественно семенным путем.
Виду свойственна гинодиэция (женская двудомность), популяции
представлены обоеполыми и женскими (андростерильными) особями
(Пономарев, Демьянова, 1975). Доля женских особей в них — 8—9%
(Демьянова, 1987). Допустимо предположение, что в пределах ареала
виду свойственна примерно одинаковая и постоянная половая струк-
тура популяций. На Урале и в Мещерской низменности женская
форма составляет 11,1% и 9,4%, соответственно. Близки по участию
женских растений популяции из разных регионов Финляндии —
12,7%, Польши (Татры) — 15,1% и Германии — 20% (Демьянова, По-
номарева, 1979). Исследованные ценопопуляции в Московской и Ка-
лужской областях (Таруба, Павловская Слобода) обычно представле-
ны двуполыми растениями, и только одна ценопопуляция на опушке
липового леса имела 12% женских особей.
В Московской области урожай семян колеблется от 294 до 1050
семян на м2. Максимальный урожай семян отмечен в ценопопуляции,
занимающей дно пересыхающего ручья у края лиственного леса.
Вегетативное размножение происходит в форме старческой пар-
тикуляции, в старом генеративном состоянии образуются клоны из
2—4 партикул.
Экология
Герань лесная — мезофит, растет в условиях от сухо- до сыролу-
гового увлажнения (53—87-я ступени шкалы Раменского, 3-я ступень
шкалы Элленберга). По нашим наблюдениям, ценопопуляции вида
имеют наиболее высокую жизненность в условиях умеренно перемен-
149
ного увлажнения. Вид встречается на бедных и богатых почвах (5—
17-я ступени шкалы Раменского, 4-я ступень шкалы Элленберга). Он
наиболее обилен на небогатых (7—9-я ступень Раменского), слабо-
кислых (3-я ступень Элленберга) почвах.
Герань лесная теневынослива (2-я ступень шкалы Элденберга),
но развивается как под пологом леса, так и на открытых местах (Ни-
китин, 1965; Бельков и др., 1974). Она часто обильна на вырубках,
характерна для опушек. Кроме того, семенное самоподдержание це-
нопопуляций осуществляется наиболее успешно в условиях хорошего
освещения.
Фитоценология
Герань лесную считают горно-лесным эколого-фитоценотиче-
ским элементом (Rothmaler, 1972; Вальтер, 1982). Обычными место-
обитаниями вида являются лиственные и смешанные леса (Гроздов,
1963; Рысин, 1964а, б; Рысин, Рысина, 1966; Ермолова, 1969), негус-
тые сосновые леса (Первухин, 1963).
Исследованные в Московской области ценопопуляций под поло-
грм леса имели численность 1,9—4,7 особей на 0,25 м2. Возрастные
спектры их полночленные, чаще — правосторонние с максимумом на
зрелых или старых генеративных растениях. Проростки в отдельные
годы не развиваются, ювенильные растения составляют 2,2—16,4%.
В луговых сообществах по краю леса (нередко используемых как од-
ноукосные сенокосные угодья) численность ценопопуляций составля-
> ла 4,1—17,1 особей на 0,25 м2. Возрастные спектры — бимодальные с
максимумами на ювенильных и зрелых или старых генеративных
растениях. Участие ювенильных особей составляло 13,9—59,1%.
Консортивные связи
Герань лесная — растение энтомофильное, насекомых привлека-
ет нектар и пыльца. На корнях растений формируются клубеньки,
сходные по строению с клубеньками бобовых, клетки коровой парен-
химы содержат бактерии (Лащинский и др., 1967; Клевенская, 1969).
Есть косвенные доказательства фиксации азота клубеньками небо-
бовых растений.
Биологическая продуктивность
Вследствие небольшого обилия герани лесной в лесных ценозах
биологическая продуктивность ее мала. Так, в лесных ценозах Мос-
ковской области максимальная биологическая продуктивность со-
ставляла 0,04—2,05 г/м2, а полная биологическая продуктивность —
0,13—1,22 г/м2. Максимальные величины отмечены в условиях
сплошной вырубки (Ермолова, 1969). Побеговая фитомасса генера-
150
тивной особи в Московской области в разных условиях составляла
1,8—47,9 г.
Хозяйственное значение
Герань лесная в зеленом виде поедается плохо, в сене — удовле-
творительно (Гроздов, 1963). Элленберг (Ellenberg, 1952) считает ге-
рань удовлетворительным кормом и в зеленом, и в сухом виде. Вид
считается хорошим дубильным растением (Гроссгейм, 1952). Исполь-
зуемую в народной медицине в качестве вяжущего средства и для ос-
тановки кровотечений герань лесную (Землинский , 1951), заготавли-
вают в период цветения обычными методами, оставляя 10—12% рас-
тений нетронутыми для семенного размножения (Саутин, 1978). С
..помощью цветков герани лесной окрашивают ткани в синий цвет
(Гроссгейм, 1952).
Герань болотная
Номенклатура
Geranium palustre L. 1756, Cent. pl. 2: 25; Кауфман, 1866, Моск,
фл., изд. 1: 112; Цингер, 1825, Сборник свед.: 119; Шмальгаузен,
1895, Фл. Средн, и Южн. Росс, к 196; Сырейщиков, 1907, Илл. фл.
Моск. губ. 2: 325; он же, 1927, Опред. раст. Моск, губ.: 178; Федченко,
1940, в Маевский, Фл., изд. 7: 487; Е. Бобров, 1949, Фл. СССР, 14: 40;
он же, 1954, в Маевск., Фл., изд. 8: 283; он же, 1964, цит. соч., изд. 9:
278; Скворцов, 1966, Опред. раст. Моск, обл.: 222; Webb a. Ferguson,
1968, Fl. Europ. 2: 197; Черепанов, 1981, Сосуд, раст. СССР: 257; Ок-
тябрева, 1986, Опред. раст. Мещеры, 1: 237 — герань болотная.
Географическое распространение
Герань болотная имеет европейский ареал, встречается в умерен-
ной и субокеанической зонах (Абеле, 1970; Rothmaler, 1972). Вид рас-
пространен во всех районах европейской части нашей страны и во
всех районах Главного Кавказского хребта, не известен в арктиче-
ской Европе, отсутствует в западном Закавказье и Талыше, является
редким в южном Закавказье (Флора СССР, 1949). В центральных об-
ластях герань болотная относится к группе часто встречаемых расте-
ний во всех районах (Губанов и др„ 1981). Общее распространение
охватывает Скандинавию, среднюю и южную Европу, Балканы
(Флора, СССР, 1949; Flora Europaea, 1968).
,151
Морфологическое описание
Герань болотная — многолетнее травянистое поликарпическое
короткорневищное растение, имеющее симподиальную систему полу-
розеточных репродуктивных и розеточных вегетативных побегов.
Корневища эпигеогенные, симподиальная система корневищ взрос-
лых растений ветвится до VI—VII порядка, корневая система их
близка к бахромчатому типу, образована многочисленными сильно
утолщенными придаточными корнями. По степени выраженности
запасающей функции корней В.Н. Голубев (1956а, б) сближает дан-
ный вид с корнеклубневыми растениями.
Развитие побеговой системы проходит по симподиальной полу-
розеточной модели (Серебрякова, 1977, 1979, 1981). Главный побег
развивается как розеточный до года первого цветения, в год цвете-
ния становится полурозеточным. В начале онтогенеза он образует
1—2 листа, а перед цветением — 3—4(6). Стеблевые листья и листья
в прикорневой розетке — простые, на черешках. Черешки листьев
покрыты оттопыренными волосками, имеют длину 7—20(30) см.
Прилистники пленчатые, светло-коричневые. Листовые пластинки
пальчато-семираздельные, покрыты с обеих сторон прижатыми во-
лосками; доли пластинки ромбические, край их крупнозубчатый.
Репродуктивные побеги — полурозеточные, имеют 3—4 розеточ-
ных листа, нередко при большой высоте полегающие или требующие
опоры, утолщенные в узлах, достигают высоты 30—70(120) см. Побе-
ги имеют сходный план строения с репродуктивными побегами опи-
санных ранее видов, в них Можно выделить те же зоны. Соцветие ци-
мозное, монотелическое, главная и боковые оси заканчиваются, как
правило, двуцветковым цветоносом. Парциальные соцветия — диха-
зий или монохазий. Все части соцветия покрыты простыми волоска-
ми и не имеют железистого опушения. Важным видовым признаком
является то, что цветоносы до цветения и по отцветании отогнуты
вниз. .
Цветки крупные, до 3 см в диаметре, имеют двойной околоцвет-
ник. Чашелистики яйцевидные с 5—7 жилками, прижато-волосистые,
пленчатые по краю, с мягкой остью. Лепестки пурпурные, ярко-ма-
линовые, обратнояйцевидные, вверху цельные, около 1,5 см длиной,
внутри волосистые. Тычинки по длине равны чашелистикам, нити их
расширяются книзу. Створки и столбик плода рассеянно-волосистые,
каждая створка плода скрывает 1 семя (редко — 2). Семена имеют
длину менее 3 мм, ширина их — около 2 мм. Шов выражен хорошо,
корешок — едва заметен, окраска семян серовато-коричневая (Рыси-
на, 1973).
Диплоидный набор хромосом 2п=28 (Хромосомные числа...,
1969; Bate-Smith, 1973).
152
Онтогенез
Лабораторная всхожесть семян оказалась низкой (7—16%). В ес-
тественных условиях для прорастания семян, видимо, необходим не-
который период покоя. Прорастание надземное.
Проростки имеют главный розеточный побег с двумя почковид-
ными, неравнобокими семядолями и 1—3 листьями. Черешки их дос-
тигают длины 3,5(8) см, листовая пластинка пальчато-семираздель-
ная, диаметр ее 1,2—1,8 см, центральная доля имеет 3 краевых зубца.
Главный корень белого цвета с тонкими боковыми корнями.
Главный побег ювенильных растений несет 1—3 пальчато-семи-
раздельных листа, длина черешков — 5—8(10) см, диаметр листовой
пластинки — 1,5—2,3 см, срединная доля заканчивается тремя зубца-
ми (ювенильный тип листа). Главный корень равномерно утолщен,
светло-коричневого цвета, боковые корни остаются тонкими.
Имматурные растения имеют главный розеточный побег с 2—3
пальчато-семираздельными листьями. Длина черешков возрастает до
12—16 см, диаметр листовой пластинки — 2,5—4,5 см, средняя доля
заканчивается 3—6 краевыми зубцами (имматурный тип листа).
Формируется ортотропное эпигеогенное корневище длиной 0,5—0,7
см, появляются придаточные корни, равные по длине главному так-
же коротко утолщенные (иногда клубневидно утолщенные). Расте-
ния данного возрастного состояния имеют кистекорневую жизнен-
ную форму.
Виргинильные особи, также однопобеговые, имеют розеточный
побег с 2—5 листьями. Длина черешков возрастает до 17—30 см, диа-
метр листовой пластинки — до 5—8,5 см. Растения имеют кистекор-
невую жизненную форму, хотя сохраняют главный корень. Отличить
от особей предыдущего возрастного состояния их можно по строе-
нию листовой пластинки (средняя доля имеет 8—13 краевых зубцов
— взрослый тип листа) и по мощности ортотропного корневища,
дайна которого достигает 0,8—1,5 и более.
Молодое генеративное растение формирует полурозеточный ре-
продуктивный побег I порядка, высота которого 45—55 см, розеточ-
ных листьев 2—4. К концу плодоношения в пазухах двух верхних по-
бегов образуются розеточные побеги возобновления. Корневая сис-
тема (как и у виргинильных) представлена системой главного корня,
скрытого под кистью утолщенных придаточных корней. Цветки об-
разуются на осях II—VI(IX) порядка.
Средневозрастные генеративные растения имеют 1 —4 полурозе-
точных репродуктивных побега, высота которых 60 -90(115) см,
нт хи образуются на осях 11 XI(XIII) порядка. Подземная система
вищ ветвится до V - 1Х(Х1П) порядка. Корневая система
... wa, гная, от кистекорневого до бахромчатого типа.
Г. ' ппа старых генеративных растений представлена клонами из
2—4(7) партикул или отдельными паргикулами. Репродуктивных по-
бегов 1—2(4), высота их 60 80(100) см. Система корневищ ветвится
155
до IV—-VII порядка, ранние порядки их лишены придаточных кор-
ней или имеют отмершие придаточные корни.
Субсенильные растения представляют собой клоны из 2—3 веге-
тативных партикул, они имеют 1—4 розеточных побега с 2—5 листь-
ями. Строение подземных частей подобно старым генеративным рас-
тениям.
Сенильные растения — отдельные вегетативные партикулы с
1(2) розеточными побегами, несущими листья ювенильного типа. Ро-
зеточный побет- отходит от участка старого разрушенного корневи-
ща, лишенного придаточных корней.
Продолжительность онтогенеза более 20 лет. Пути сокращенно-
го онтогенеза сходны с описанными у герани луговой.
Сезонное развитие
В природе проростки появляются в первой декаде мая. В это же
время начинается вегетация взрослых растений. Главный побег раз-
вивается как полициклический монокарпический, нарастает монопо-
диально в течение 7—9 лет и зацветает. В начале онтогенеза он об-
разует 1—2 розеточных листа, число их возрастает до 4—6 к момен-
ту цветения.
В Подмосковье у репродуктивных побегов последующих поряд-
ков наблюдается широкая амплитуда колебания цикличности, от
озимых до тетрациклических (Серебряков, 1959), большинство побе-
гов развивается по дициклическому типу. Дициклические полурозе-
। точные репродуктивные побеги начинают развитие из 1—2 верхних
наиболее крупных почек возобновления, заложившихся в пазухах ро-
зеточных листьев материнского побега. На косимых участках розе-
точные побеги возобновления формируются сразу после покоса. Час-
то при отсутствии сенокошения продолжительность функционирова-
ния побегов первой генерации продлевается до конца августа—сере-
дины сентября, побеги возобновления становятся озимыми.
Герань болотная — летнезеленое растение, зимует с закрытыми
почками, по положению почек возобновления — гемикриптофит.
Листья розеточных побегов отмирают к концу сентября. На ранних
этапах онтогенеза растения способны зимовать с открытыми почка-
ми (Рысина, 1973). К октябрю емкость верхушечных почек розеточ-
ных побегов возобновления достигает 9—14 зачатков; сформирова-
ны зачаток соцветия и отдельные цветки (Серебряков, 1952).
Весной из перезимовавших верхушечных почек развиваются ро-
зеточные побеги с 4—6 листьями, к середине июня побеги становятся
полурозеточными. Растения зацветают в конце июня — начале июля.
Массовое плодообразование на низинном лугу в Калужской области
наблюдалось в конце толя — начале августа. Герань болотная явля-
ется длительно цветущим видом, цветение закапчивается в конце ав-
густа — середине сентября. Возможно, это связано с ветвлением со-
154
цветня, цветки образуются на осях XII—XIII порядка (Серебряков,
1948).
Придаточные корни формируются в конце мая — начале июня, а
летом значительно утолщаются (сильно выражена запасающая
функция). На рсзиде одного порядка образуется от I—2 до 5—7 при-
даточных корней.
Способы размножения и распространения
Герань болотная размножается преимущественно семенным пу-
тем. Виду свойственна автохория, семена при вскрывании плода от-
брасываются от материнского растения на расстояние 2,5 м (Кернер,
1902).
Реальная семенная продуктивность растений оказалась низкой,
16—94 семени, и составила 22—40% от потенциальной. Урожай се-
мян колебался значительно (96—1053 семени на 1 м2). Наиболее вы-
сокий урожай семян образовался в ценопопуляции, занявшей дно пе-
ресыхающего ручья, в условиях хорошего освещения, наиболее низ-
кий — на низинном материковом лугу в условйях избыточного ув-
лажнения.
Наибольшие потери при семенном возобновлении наблюдаются
на начальных этапах развития от семени до ювенильного возрастно-
го состояния. Численность ювенильных растений составила 0,6—
9,3% от урожая семян. В исследованных ценопопуляциях до молодо-
го генеративного возрастного состояния семенное самоподдержание
либо стабилизировано, либо осуществляется с большим запасом чис-
ленности.
Вегетативное размножение происходит как партикуляция в ста-
ром генеративном возрастном состоянии, образуются клоны из 2—7
партикул. Отмечается накопление старых генеративных растений,
что продлевает период репродукции.
Экология
Герань болотная особенно требовательна к увлажнению. Она
массово (более 8%) встречается в условиях сыролугового увлажнения
(86—88-я ступени) на довольно богатых почвах (10—11-я ступени
шкал Раменского). В то же время для вида характерна довольно ши-
рокая экологическая амплитуда, так как единичные особи встреча-
ются в условиях сухолугового и болотного увлажнения, на бедных
почвах (Раменский и др., 1956).
Герань болотная, по мнению одних авторов, относится к группе
мезогшрофитов (Рысина, 1973), по мнению других — к гигромезофи-
там (Абеле, 1970). Она является одним из видов-индикаторов умерен-
но избыточного увлажнения (Горышина, 1979).
155
Фитоценология
Герань болотная является горно-лесным фитоценотичрским эле-
ментом (Абеле, 1970; Rothmaler, 1972) и обладает широкой фитоце-
нотической амплитудой. Основные местообитания — влажные и сы-
рые луга, опушки и поляны смешанных, хвойных и мелколиственных
лесов, заросли кустарников (Опред. высш. раст. Северо-запада...,
1981). Она растет по сырым оврагам, вдоль ручьев, по окраинам бо-
лот (Рысина, 1973; Алексеев и др., 1980). В Стрелецкой степи герань
болотная встречается по дну и северным склонам логов (Голубев,
1965).
Исследованные ценопопуляций в Московской и Калужской об-
ластях имели численность 1,3—10,6 особей на 0,25 м2. Возрастные
спектры, как правило, полночленные. В отдельные годы в некоторых
ценопопуляниях наблюдалось отсутствие ювенильных и молодых ге-
неративных растений. Базовым для данного вида является бимодаль-
ный возрастной спектр с максимумами на ювенильных и старых ге-
неративных растениях.
Биологическая продуктивность
Продуктивность герани болотной в разных ценозах Калужской
области колебалась значительно. Так, на низинном лугу воздушно-
сухая надземная биомасса составила 4,9 г/м2, в зарослях ольхи чер-
ной — 62,4 г/м2. Наиболее высокой оказалась биологическая продук-
тивность ценопопуляций, расположенной по дну пересыхающего ру-
чья — 144 г/м2. Величина продуктивности зависит от жизненного со-
стояния растений, интегрально его можно охарактеризовать величи-
ной побеговой фитомассы. Для генеративных растений величина ле-
беговой фитомассы в разных местообитаниях составляла 1,8—47,9 г,
наиболее высокая жизненность особей отмечена в ценопопуляций,
занимающей дно пересыхающего ручья.
Хозяйственное значение
Герань болотная обладает дубильными свойствами, содержит до
12% таннидов, может использоваться для окрашивания тканей (Чер-
нышев, 1934). Вид очень декоративен, имеет яркие цветки, харак те-
ризуется длительным периодом цветения. Он может быть использо-
ван как биоиндикатор избыточного увлажнения.
156
Герань кровяно-красная
Номенклатура
Geranium sanguineum L. 1753, Sp. pl.: 683; Кауфман, 1866, Моск,
фл., изд. 1: 112; Цингер, 1895, Сборник свед.: 118; Шмальгаузен,
1895. Фл. Средн, и Южн. Росс. 193; Сырейщиков, 1907, Илл. фл.
Моск. губ. 2: 326; он же, 1927, Опред. раст. Моск, губ.: 178; Федченко,
1940, в’Маевский, Фл., изд. 7: 486; Е. Бобров, 1949, Фл. СССР, 14: 10;
он же, 1954, в Маевский, Фл., изд. 8: 283; он же, 1964, цит. соч., изд. 9:
277; Скворцов, 1966, Опред. раст. Моск, обл.: 223; Webb a. Ferguson,
1968, Fl. Europ. 2: 195; Черепанов, 1981, Сосуд, раст. СССР: 257; Ок-
тябрева, 1986, Опред. раст. Мещеры, 1: 237 — Герань кровяно-крас-
ная.
Географическое распространение
Вид имеет европейский ареал (Носова, 1973), который охватыва-
ет западное Средиземноморье, Балканский полуостров, южную,
среднюю и атлантическую Европу, южную Скандинавию (Флора
СССР, 1949). Герань кровяно-красная заходит в Прибалтику, рас-
пространена здесь до окрестностей Нарвы, где встречается на извест-
няках. В южной и юго-западной части Литвы она не обнаружена
(Снаркис, 1973). В Эстонии вид встречается на дерново-карбонатных
почвах, в Ленинградской, Псковской, Новгородской областях изред-
ка растет по выходам известняков, склонам холмов на богатых кар-
бонатных почвах и подлежит здесь охране (Опр. высш. раст. Северо-
запада..., 1981). На юге герань кровяно-красная распространена в
Волжско-донском, Бессарабском, Нижне-Донском районах и в Кры-
му. На Кавказе растет.во всех районах, кроме Талышского и боль-
шей части Западно-Закавказского.
В Московской области вид встречается по солнечным склонам в
юго-восточной, восточной, изредка в южной части (Ворошилов и
др., 1966).
Морфологическое описание
Герань кровяно-красная — многолетнее травянистое поликарпи-
ческос растение с симподиадьной системой гипогеогенных корпе-
вши.
В отличие от описанных ранее видов, развитие побеговой систе-
мы герани кровяно-красной идет по симподиальной длиннопобего-
вой модели побегообразования (Серебрякова, 1977, 1979, 1981), что
определяет возможность образования разных жизненных форм, в за-
висимости от условий обитания (Сугоркина, 1989). В хорошо осве-
щенных лугово-степных сообществах в окрестностях Поленова
157
(Тульская обл.) растения герани кровяно-красной можно считать
дииннокорневищными, так как годичные приросты гипогеогенных
корневищ достигают 13—16 см. Взрослые растения являются поли-
центрическими. В условиях затенения, задернения, на каменистых
субстратах величина годичных приростов уменьшается до 0,3—2,2
см, растения становятся короткокорневищными, а взрослые особи
вида можно рассматривать как моноцентрические.
Главный побег развивается как удлиненный, поэтому особенно-
стью растений герани кровяно-красной является переход к симподи-
альному нарастанию уже на второй год после развития из семени.
Последующие побеги, вегетативные и репродуктивные, формируют-
ся как удлиненные или полурозеточные.
Симподиальная система корневищ ветвится до X—XXII поряд-
ка. Корневая система придаточная, придаточные корни формируют-
ся в основании ортотропных побегов.
Репродуктивные побеги достигают высоты 20—70 см. У них вы-
деляются те же три зоны, что и у выше описанных видов. Зона возоб-
новления содержит только укороченные междоузлия. Листья могут
быть черешковыми, но чаще они — чешуевидные, только один зеле-
ный черешковый лист формируется на уровне почвы. Зона торможе-
ния включает 5—7 удлиненных междоузлий с супротивными череш-
ковыми листьями. Зона обогащения содержит цимозное соцветие, все
листья черешковые, несколько уменьшающиеся по величине. Соцве-
тие можноусчитать фрондулезным (Федоров, Артюшенко, 1979).
Парциальные соцветия различно сочетаются в составе объединенно-
го. Начальные оси часто формируются по типу дихазия или плейоха-
' зия, ветвление конечных частей всегда идет по типу монохазия. Пол-
ноценные цветки образуются на осях II—VIII порядка. Листовые
пластинки — округлые или почковидные, глубоко рассеченные на
5—7 долей, средняя доля всегда заканчивается тремя зубцами.
Цветоносы одноцветковые, цветки имеют двойной околоцвет-
ник. Чашелистики с 3—5 жилками, по краю буровато-пленчатые,
снаружи дпинноволосистые. Лепестки кровяно-красные, обратнояй-
цевидные, имеют клиновидное основание и выемку на верхушке, дли-
на их — 1,5—2 см. Плод — клювовидная коробочка, колонка его во-
лосистая, створки слабо пушистые. Семена темно-коричневые, тонко
линейно-точечные, вес 1000 семян — 10,2—14 г (Меньшова, 1985).
Диплоидный набор хромосом равен 56 и 84 (Хромосомные чис-
ла..., 1969; Bate-Smith ,1968)
Онтогенез
Семена имеют сходное строение с другими видами гераней. Ла-
бораторная всхожесть колеблется значительно. В конце второго ме-
сяца прорастает 43—72% семян, что объясняется толстой оболочкой
семени. Лабораторная всхожесть возрастает до 93—97% после обра-
ботки семян в течение 24 часов в 0,0025% p..-.воре янтарной кисло-
158
ты. Рекомендуется проводить посевы осенью, что дает возможность
получить весной дружные всходы (Закиров и др., 1973). По другим
данным, семена не нуждаются в стратификации и способны прорас-
тать сразу после созревания. Лабораторная всхожесть составляла
90%, период прорастания семян — 1—3 месяца. Хорошая всхожесть
семян не утрачивается даже при хранении в течение 3 лет. (Меньшо-
ва, 1985). В весеннем посеве на опытном участке в Московской об-
ласти грунтовая всхожесть составляла 5%.
В условиях хорошо освещенных лугово-степных участков в окре-
стностях Поленова длиннокорневищные растения проходят полный
онтогенез.
Проростки имеют розеточный побег с двумя почковидными се-
мядолями и одним настоящим листом с пальчато-пятирассеченной
листовой пластинкой. Затем главный побег удлиняется и образует
2—3 стеблевых листа. Корневая система представлена системой глав-
ного корня.
Ювенильные растения формируют 1 удлиненный побег I—IV по-
рядка с очередным листорасположением и систему главного корня.
Придаточные корни появляются на 3—4-й год жизни.
Имматурные особи также имеют 1 побег IV—VII порядка, лис-
торасположение становится супротивным. Корневая система сме-
шанная: наряду с системой главного корня есть придаточные корни.
Виргинильные растения формируют разветвленную систему кор-
невищ, образуют 2—5 побегов. Появляются побеги обогащения.
Корневая система представлена системой главного корня и прида-
точными корнями.
В природе растения впервые зацветают в возрасте 10—12 лет.
По данным В.А. Меньшовой (1985), в культуре сеянцы зацветают на
втором году, а растения, полученные из черенков — на первом.
Молодые генеративные растения по строению сходны с вирги-
нильными растениями, но способны к формированию репродуктив-
ных побегов. Появляется способность образовывать удлиненные го-
дичные приросты корневищ длиной до 2,5—4(6) см.
Зрелые генеративные растения представляют собой особь, имею-
щую несколько центров закрепления (парциальный побег с прида-
точными корнями). Величина годичных приростов корневищ дости-
гает 13—16 см, корневая система придаточная, условный возраст
18—22(25) лет.
Старые генеративные растения представлены диффузными кло-
нами, содержащими до 25 партикул или отдельными ветвящимися ге-
неративными партикулами.
Субсенильные растения — отдельные ветвящиеся вегетативные
партикулы, имеющие 2—3(5) побегов, иногда — клоны, состоящие
из подобных партикул, близкие по размерам к старым генератив-
ным.
Сенильные растения представляют собой о тдельные неветвящие-
ся пар тикулы с 1—2 побегами, прида точные корни сохраняю тся в ос-
159
новании побега. В этих же условиях на местах пороев или наруше-
ний на отрезках придаточных корней образуются корневые отпры-
ски. Развитие идет с омоложением до имматурного возрастного со-
стояния. Возрастные группы описаны по сходству с семенными рас-
тениями (Сугоркина, 1987).
Под пологом липы, в условиях затенения формируются растения
короткокорневищной жизненной формы (прирост корневища не пре-
вышает 0,5—2,5 см). Ценопопуляции представлены нецветущими
растениями вегетативного происхождения. Встречаются имматур-
ные, виргинильные, субсенильные и средневозрастные вегетативные
растения, названные так по сходству строения и условному возрасту
с соответствующими растениями семенного ряда (Сугоркина, 1987,
1989).
Сезонное развитие
Проростки в природе появляются в начале—середине мая, в на-
чале мая начинается отрастание взрослых растений.
Моноподиальные побеги (репродуктивные и вегетативные) фор-
мируются, в основном, как моноциклические (Борисова, 1954; Голу-
бев, 1965; Данилов, 1983). Почки возобновления закладываются в па-
зухе каждого листа (чешуевидного и зеленого). Побеги возобновле-
ния развиваются из почек, расположенных в месте перехода плагио-
тропной части побега в ортотропную.
В момент плодоношения материнского побега почки возобнов-
ления трогаются в рост. Формируется плагиотропная часть побега
будущего года с 7—10 чешуевидными листьями. Верхушечная почка,
содержащая 6—8 зачатков, располагается у поверхности почвы, пе-
реходит в состояние покоя и зимует. Весной из перезимовавшей вер-
хушечной почки развивается надземная часть побега возобновления.
В условиях лугово-степных участков на ранних этапах онтогене-
за побеги возобновления образуются из 1 почки, у взрослых расте-
ний из 1—3(7) почек. В условиях затенения побеги следующего по-
рядка формируются из 1—2 почек возобновления, способность к
ветвлению системы корневищ снижается.
Герань кровяно-красная — летнезеленое растение, имеет одну ге-
нерацию побегов (Борисова, 1954; Борисова-Гуленкова, 1960; Голу-
бев, 1965). В годы покоса образуются побеги второй генерации. При
раннем покосе побеги возобновления развиваются из почек, распо-
ложенных в верхних узлах зоны возобновления, на ортотропной час-
ти материнского побега. Формирование побегов возобновления сле-
дующего года идет по описанному выше типу. При позднем покосе,
когда уже начато образование плагиотропных участков к о 'егов во-
зобновления будущего года, розеточные побеги второй генерации
развиваются из верхушечных почек этих побегов возобновления. Ли-
стья их к зиме отмирают, на следующий год побеги продолжают
рост. Наблюдается переход от однолетних побегов незимующего ти-
160
па к однолетним зимующим побегам (Борисова, 1954; Борисова-Гу-
ленкова, 1960).
Некоторые авторы относят герань кровяно-красную к корне-
вищным геофитам (Смык и др., 1983). М.А. Борисова-Гуленкова
(1959) считает ее гемикриптофитом. Наблюдения выявили зависи-
мость способа перезимовки от особенностей использования угодий.
В годы без покоса или при раннем покосе особи вида ведут себя как
геофиты, при позднем покосе — как гемикриптофиты.
Наблюдения показали, что цветение растений отмечается в на-
чале июня.и длится до конца июля. Массовое цветение в окрестно-
стях Поленова (Тульская обл.) можно наблюдать в конце июня. По
данным В.А. Меньшовой (1985), продолжительность жизни одного
цветка составляет от 7 до 12 дней.
Способы размножения и распространения
Герань кровяно-красная размножается семенами и вегетативно.
Семенная продуктивность низкая, что объясняется малым числом
цветков, образующихся на растении (Меньшова, 1985).
Реальная семенная продуктивность побегов в условиях вейнико-
во-разнотравной ассоциации составляет 4,6—10,3 семян. Урожай се-
мян в разных локусах ценопопуляции колебался значительно (25—
719 шт./м2), максимальная величина отмечена внутри клонов. Общий
урожай семян в ценопопуляции, вследствие пространственной неод-
нородности, относительно высок. Ювенильные растения составляют
1,5—6,2% от урожая семян, создающегося вне клонов. Число проро-
стков не превышало 1—8 на 1 м2, что говорит о низкой всхожести се-
мян в природе. Прорастание затрудняется толстой семенной кожу-
рой (Чеврениди и др., 1977). Способность к вегетативному размноже-
нию значительно выше, чем у других описанных видов. Герань кро-
вяно-красную можно размножать корневищными черенками (Смык
и др., 1983; Меньшова, 1985). Она является факультативно корнеот-
прысковым растением и может размножаться участками придаточ-
ных корней (Сугоркина, 1987). Особям данного вида свойственна
парпвкуляция.при которой образуются мощные генеративные пар-
тикулы. Размеры клонов достигают 0,8—1,5 м2.
Экология
Герат, кровяно-красная — наиболее ксерофильный из описы-
ваемых видов. Она массово встречается (с проективным покрытием
2,5 -8% и более) в условиях лугово-степного-сухолугового увлажне-
ния (51- -59-я ступени увлажнения шкалы Раменского), на мезогроф-
иых или довольно богатых почвах — 7—11-я ступени по шкале бо-
гатства почвы Раменского. Единичные особи вида можно всгрегигь
1М
в условиях среднестепного и влажнолугового увлажнения (ступени
45—67), на бедных почвах (5-я ступень шкалы Раменского).
Герань кровяно-красная распространена на слабокислых и сла-
бощелочных почвах, является нейтрофилом — 7—11-й балл по шка-
ле кислотности почвы (Цыганов, 1983). Она часто растет по,выходам
известняков. Г. Элленберг поместил данный вид в кальциефильную
группу видов, растущих на сухих почвах (цит. по: Г. Вальтер, 1982).
Растения герани кровяно-красной особенно хорошо развивают-
ся на открытых пространствах, хотя их можно встретить и под поло-
гом леса (1—8-й балл по шкале освещенности—затенения Цыганова
(1983). Ухудшение освещенности, прежде всего, вызывает уменьше-
ние величины годичных приростов корневищ; при сильном затене-
нии растения не цветут.
Фитоценология
Герань кровяно-красная является лесо-луговым-лугово-степным
эколого-фитоценотическим элементом (Носова, 1973). В разных час-
тях ареала она встречается в составе различных растительных сооб-
ществ. В Литве произрастает в сухих термофильных сосновых или
сосново-еловых лесах, кое-где — в дубовых и грабовых лесах. На Ев-
ропейской равнине герань кровяно-красная растет в светлых лесах,
по сухим лесным опушкам, на сухих лугах. Она встречается в остеп-
ненных борах по левобережью р. Оки, в «келериевых борах» в соста-
ве разнотравья, совместно с Coronilla varia (Алехин, 1947). В Смолен-
ской, Калужской, Брянской областях в небольшом обилии особи ви-
да распространены в сосняках разного типа (Гроздов, 1963). На Ук-
раине герань кровяно-красная растет во всех лесных и лесостепных
районах, часто входит в состав доминантов (Смык и др., 1983). В
Стрелецкой степи она распространена на участках луговой степи
(Борисова-Гуленкова, 1960; Голубев, 1965). На Кавказе и в Крыму
вид растет' в дубняках, невысокой сомкнутости (не более 0,5—0,6) со
слабо выраженным подлеском и густым травостоем.
Изучение возрастной структуры ценопопуляций в лугово-лесной
опушечном комплексе в Тульской области выявило пространствен-
ную неоднородность ценопопуляций, что связано с особенностями
вегетативного размножения (Сугоркина, 1987, 1989). Численность
ценопопуляций в отдельных локусах вейниково-разнотравной ассо-
циации составляла 8,6—20,8 экз./м2. Под пологом липы ценопопуля-
ция была представлена нецветущими растениями короткокорневищ-
ной жизненной формы, ее численность составляла 4,8 особей на 1 м2.
Консортивные связи
В условиях культуры герань кровяно-красная поражается мучни-
стой росой, пятнистостью листьев, ложной мучнистой росой, корне-
162
выми гнилями, скручиванием листьев. Возбудителями являются гри-
бы родов Ramularia, Septoria, Fusarium, Sphaerotheca и др. (Кестен и
др., 1980).
Вредителями герани кровяно-красной являются и щетинистые
долгоносики. Одни из них повреждают бутоны, другие обгрызают
лепестки с краев, иногда объедают цветки. В период максимальной
численности уничтожается 12,8—15,8% цветков (Кестен, 1982), осо-
бенно страдают молодые посадки. Корни повреждают личинки хру-
щей, жуков-щелкунов, гусеницы совок и, особенно, гусеницы орехо-
вой огневки, весной выедающие ходы в корнях (Кестен и др., 1980).
Биологическая продуктивность
Продуктивность герани кровяно-красной изучали в Тульской
области в период наиболее мощного развития растений. Побеговая
фитомасса взрослой особи была минимальной в условиях затенения
— 2,8 г. В вейниково-разнотравной ассоциации побеговая фитомас-
са генеративной особи колебалась от 9,8 до 43,7 г. Наименьшая вели-
чина воздушно-сухой биомассы герани кровяно-красной наблюда-
лась в условиях затенения — 5,6 г/м2. Максимальная величина побе-
говой фитомассы формируется в клонах на открытых участках —
133,2 г/м2, величина биомассы в этих условиях составляла 50,8 г/м2.
Хозяйственное использование
Герань кровяно-красная является удовлетворительным кормом.
В корнях растений содержатся большие концентрации дубильных ве-
ществ из класса энталовой кислоты (Гроссгейм, 1952). Наибольшее
количество дубильных веществ содержится в листьях и корнях в фазе
бутонизации и начала цветения. Во все годы жизни растения это —
лучшее время для заготовки сырья (Чеврениди и др., 1979).
Герань кровяно-красная имеет заметное значение в хозяйстве и
широко вводится в культуру в Узбекистане (Закиров и др., 1973; Чев-
рениди и др., 1979). Сырье, получаемое в культуре для производства
дубильных веществ, имеет низкую себестоимость (Чеврениди, 1979;
Варганов и др., 1985).
В народной медицине Белоруссии отвар корневища рекоменду-
ется пить при раке (Балицкий и др., 1982), растения обладают вяжу-
щими свойствами, могут использоваться от поносов и кровотечений
(Землинский, 1951).
Герань кровяно-красная является перспективным красильным
растением, дающим разные цвета (Каримов и др., 1977). Ее использу-
ют как декоративное растение для озеленения городов (Чеврениди и
др., 1976).
163
Т.В. Багдасарова
БЕРЕСКЛЕТ БОРОДАВЧАТЫЙ
Номенклатура и внутривидовая систематика
Euonymus verrucosus Scop.* 1772, Fl. Cum. ed. 2, k 166; Кауфман,
1866, Моск. фл. изд. 1: 118; Цингер, 1885, Сборник свед.: 124;
Шмальгаузен, 1895, Фл. Средн, и Южн. Росс, к 202; Сырейщиков,
1907, Илл. фл. Моск. губ. 2: 346; он же, 1927, Опред. раст. Моск, губ.:
182; Федченко, 1940, в Маевский, Фл. изд. 7: 503; Проханов, 1949,
Фл. СССР, 14: 559; он же, 1954, в Маевский, Фл., изд. 8: 305; он же,
1964, цит. соч., изд. 9: 298; Ворошилов, 1966, Опред. раст. Моск, обл.:
228; Tutin, 1968, Fl. Europ. 2: 242; Черепанов, 1981, Сосуд, раст.
СССР: 175; Киселева, 1987, Опред. раст. Мещеры, 2: 9.
Имеется много разновидностей бересклета бородавчатого, из
которых следует отметить формы с овальными (Е. verrucosa var.
ovalifolium Gr.), ланцетными (E. verrucosa var. lanceolata Gr.), округ-
лыми (E. verrucosa var. rotundifolium Nest.) и другими листьями (Фл.
СССР, 1949).
Географическое распространение
, Бересклет бородавчатый — европейско-малоазиатский бореаль-
ный вид, распространенный в Европе от юга Скандинавии до Бал-
канского полуострова и Крыма, а в пределах Азии — в Малой
Азии и на Кавказе. В России растет к югу от линии Нарва — Псков
— Старая Русса — Тверь — Ярославль — Вятка — Пермь —
Уфа до Нижнего Дона. Вне сплошного ареала — в Краснодарском,
Ставропольском краях и в Дагестане (Флора СССР, 1949; Юркевич,
1949; Стратонович, 1955; Гроссгейм, 1962).
Северная граница распространения в России изучена особенно
подробно (Юркевич, 1949; Стратонович, 1952). Она извилиста, зна-
чительно отклоняется к северу по долинам рек. По р. Костроме, на-
пример, он доходит почти до р. Буй, по р. Унже — до г. Макарьева,
по р. Вятке — до г. Котельнича, по р. Каме — до г. Оханска и г.
Перми. По мнению И.Д. Юркевича, это связано с расширением ареа-
ла бересклета бородавчатого и передвижением его на север благода-
ря распространению семян малиновками и другими певчими птица-
ми. Другие исследователи объясняют это явление тесной связью вида
с широколиственными лесами, которые в прошлом были распростра-
нены на север значительно дальше, чем в настоящее время, а его при-
уроченность к речным системам связывают с более благоприятными
Sacpe «verrucosa».
164
почвенно-грунтовыми условиями, в которых бересклет при сокраще-
нии ареала еще смог сохраниться.
В Московской области во всех районах обыкновенен, по лесам и
кустарникам (Ворошилов и др., 1966). В культуре известен с 1763 го-
да. Культивируется за пределами ареала в Архангельской, Пермской,
Свердловской, Челябинской областях, на Алтае, в Красноярском
крае, в Приморском крае, Казахстане и Киргизии (Шиманович,
1987).
Морфологическое описание
Бересклет бородавчатый — кустарник, достигающий 1—3 м в
высоту, реже — деревце до 4—6 м. Кора ствола и старых ветвей чер-
ная, морщинистая. Молодые ветви вальковатые или цилиндриче-
ские, зеленые или коричневато-зеленые, голые, густо покрытые чер-
но-бурыми или красноватыми чечевичками (бородавками). Листья
супротивные, яйцевидные или эллиптические, 1,5—6 (10) см длиной и
0,7—4,5 (5,5) см шириной, голые или густо опушенные снизу по глав-
ной и боковым жилкам, сверху — только по главной жилке; на вер-
хушке острые, по краю мелкопильчатые. Выделяют крупнолистные,
поздно распускающиеся формы, и мелколистные рано распускаю-
щиеся.
Соцветия 3—7 цветковые, расположенные по всей длине побега
(кроме верхней пары листьев). Цветок обоеполый, правильный, 4-х
членный с двойным околоцветником. Цветоносы тонкие: цветки ма-
ленькие 6—11 мм в диаметре (цветоложе в диаметре — 2 мм), имеют
спайнолистную чашечку, венчик разнолепестный. Лепестки почти
округлые, желтоватые, с густыми бордовыми точками и пятнами и
густой сетью пальчатых зкилок. Тычинок четыре (реже 5—6), пестик
— один с сидячим рыльцем, из 4-х плодолистиков с 4-мя гнездами
(синкарпный). Плод — коробочка, неглубоко четырехлопастная, с
закругленными по спинке лопастями, 8—12 мм в диаметре, сплюсну-
то-шаровидная, в которой находятся от 3 до 5 (7) семян.
Бересклету бородавчатому свойственна поверхностная корневая
система, однако, в зависимости от сочетания тех или иных окружаю-
щих условий, наблюдаются значительные колебания в глубине про-
никновения корней. Выделяются два основных типа корневых систем
(Габай, 1952): а) горизонтальный тип строения корневой системы,
когда основная масса корней расположена в верхнем 10-сантиметро-
вом слое почвы и имеются единичные вертикальные корни «держатс-
ли», служащие для устойчивости растения; б) наклонно-вертикаль-
ный тип строения корневой системы, с наличием трети и больше кор-
ней, идущих вглубь наклонно или вертикально до 30—40 см. Первый
тип имеет преобладающее распространение и встречается во всех
районах произрастания бересклета. Второй тип встречается на глу-
боких суглинистых почвах в западных районах лесостепи и степи.
165
Имеется и многоярусная корневая система (которая наблюдается
в пойменных дубравах), при которой от основного корня, имеющего
вид «редьки» (старый стебель занесенный песчано-иловатыми нано-
сами уходит вглубь иногда более чем на 50 см), отходят в 2—3 яруса
более мелкие горизонтальные ответвления корней (Габай, 19,52). Ко-
ра корней желтоватого цвета, усеяна пробковым слоем в виде зерен
различной величины. При разрыве коры корней по торцевому на-
правлению обнаруживаются нити гутты (Юркевич. 1949).
Онтогенез и особенности размножения
Семена бересклета бородавчатого прорастают в конце мая —
начале июня. Жизнеспособность семян по сроку не превышает двух
лет. Средний вес одной тысячи семян 20—23 грамма. В 1 кг насчиты-
вается около 45 тысяч семян.
Семенное размножение затруднено, преобладает вегетативный
тип размножения. В природе семена не способны прорастать сразу
после созревания даже в самых благоприятных условиях, а прораста-
ют только на вторую весну. Покой семян является важным
свойством, выработавшимся в процессе эволюции и дающим семени
возможность пережить неблагоприятный период года. Поэтому бо-
лее или менее длительное воздействие низких температур является
непременным условием для преодоления периода покоя и успешного
прорастания семян (Шиманович, 1987). Семена бересклета нуждают-
ся в двухэтапной стратификации: первый — при +10—20° в течение
2—3 месяцев, второй при 0—5°в течение 4—5 месяцев. Обработка се-
мян гиббереллином стимулирует пробуждение семян на холоде, что
позволяет исключить первый этап стратификации (Шиманович,
1987).
Прорастание семян надземное. Гипокотиль утолщенно цилинд-
рический, высокий, 20—25 мм длины и 1,25—1,5 мм ширины, темно-
вато-зеленый. Семядоли,крупные, коротко и широко овальные, на
верхушке широко закругленные, 20—25 мм длины, 12—15 мм шири-
ны, на коротких черешках. Жилкование семядолей хорошо заметно,
петлисто-сетчатое, сверху со слегка выпуклыми жилками. Вслед за
семядолями появляются первые продолговатые листья, по краю зуб-
чатые, на коротких черешках. Жилкование листьев представлено
средней жилкой и очередными восходящими боковыми веточками,
которые соединяются друг с другом с образованием удлиненных пе-
тель; ответвления боковых жилок входят в углубления между зубчи-
ками пластинки. Третий и четвертый листья — продолговатые, по
краю зубчатые. Эпикотиль — около 5 мм (Васильченко, 1960).
На ранних стадиях развития четко выражен главный корень, а с
появлением первых листьев образуются придаточные корни. На вто-
рой—третий год, после отмирания семядолей, молодые особи береск-
лета переходят в ювенильное состояние. Главная ось не ветвится, на-
растает моноподиально или в случае гибели верхушечной почки, —
166
неустойчиво-моноподиально (Истомина, 1994). Под пологом леса в
течение нескольких лет у ювенильного растения боковые пазушные
почки не раскрываются, оставаясь спящими. Время пребывания в
ювенильном состоянии — 3—5 (до 8) лет. Затем надземный побег на-
чинает ветвиться, и растение переходит в имматурное состояние.
По скорости роста бересклет бородавчатый относится к медлен-
но растущим кустарникам (к 10 годам он вырастает до 1,5 м, к 20 го-
дам —до 2,5 м, к 40—50 годам — до 5 м). Первое цветение береск-
лета семенного происхождения начинается с 4—5-летнего возраста,
вегетативного происхождения — с 3-летнего возраста. В условиях
значительного затенения часто встречаются временно нецветущие
генеративные растения, которые трудно отличить от виргинильных.
Длительность жизни около 50 лет.
Бересклет бородавчатый — листопадное растение. Однако у се-
янцев, молодых растений, корнеотпрысковых побегов проявляется
тенденция к вечнозелености. Растения, растущие под пологом леса,
могут сохранять способность к вечнозелености до 7—8-легнего воз-
раста (Иорданская, Серебряков, 1954).
В естественных условиях бересклет бородавчатый размножается
вегетативно. В искусственных условиях он легко размножается от-
водками, корневыми отпрысками, сеянцами, зелеными стеблевыми
черенками (Чистяков, 1935). Особи вегетативного происхождения
возникают из спящих почек эпигеогенных корневищ. Осмотр не-
скольких тысяч экземпляров не дал возможности И.И. Истоминой
(1994) обнаружить особи корнеотпрыскового происхождения, отме-
ченные И.Г. Серебряковым (1962).
Бересклет бородавчатый способен образовывать ряд биоморф
(Истомина, Богомолова, 1991). Детальное исследование онтогенеза
показало, что виду свойственен определенный набор жизненных
форм. Морфогенез каждой из форм является результатом соответст-
вующего пути онтогенеза в определенных фитоценотических услови-
ях. Вне зависимости от происхождения (семенного или вегетативно-
го) бересклет может существовать в лесных ценозах в нескольких
жизненных формах: 1) древовидной; б) аэроксильного вегетативно
неподвижного кустарника; в) геоксильного эпигенно-корневищного
кустарника; г) стланика. Варианты такого онтогенеза (по И.И.
Истоминой, 1^94) показаны на рисунке.
В условиях длительного сильного затенения (0,5—1,2% от пол-
ного света) и достаточной влажности онтогенез бересклета остается
незавершенным. В таких условиях имматурные особи после несколь-
ких лет роста полегают вследствие слабого развития механических
тканей и укореняются, а из спящих почек, расположенных на плагио-
гропной части побега, возникаютювенилоподобные оси следующего
порядка. Таким образом возникают жизненные формы «стланик» и
«полу стланик».
167
Варианты онтогенеза бересклета бородавчатого (по II.И. Иегомлной. 1994).
I - семенной ряд, .4 — онтогенез древовидной формы. А’ - - онтогенез
аэроксильного вегетативно неподвижного кусга. II - вегетативный ряд. .4 онтогенез
эплгеогелпо-геоксильного куста. Г> — формирование стланика.
168
Сезонное развитие
В природе пробуждение бересклета наступает ранней весной.
Полное облиствение происходит в конце апреля — начале мая. Цве-
тение начинается после облиствения с середины мая и продолжается
в июне. Плоды созревают к середине сентября, опадают в середине
октября.
Отмечают неодновременность распускания цветков в пределах
одной особи. Соцветие бересклета относится к симподиальному ти-
пу, формы дихазий. Оно не имеет главной оси, а состоит из началь-
ной оси, выходящей из пазухи листа. Эта ось ниже своей верхушки
Дает две супротивные оси I порядка, которые разветвляясь, образуют
оси II порядка. Начальная и последующие оси оканчиваются одним
цветком. По размеру и развитию бутоны соцветий неодинаковы, и
поэтому их цветение происходит в разное время. Первым развивает-
ся и достигает сравнительно большого размера бутон, которым
оканчивается начальная ось. С этого бутона начинается цветение со-
цветия; он расцветает всегда первым, за ним расцветают бутоны, ко-
торыми заканчиваются оси I порядка, а потом расцветают бутоны,
которыми оканчиваются оси II порядка. Часто цветение следующих
бутонов не наступает до окончания цветения цветов, находящихся
на нижних осях соцветия. Цветение на ветке начинается с соцветия,
расположенного у ее вершины. Неодновременное распускание цвет-
ков сильно растягивает период цветения бересклета, исчисляющийся
у одного куста 23—26 днями. Наблюдение за цветением отдельных
цветков показывает, что тычинки с начала распускания цветка до 4-
х дней сохраняют желтую окраску, Потом, теряя пыльцу, становятся
белыми и опадают. Лепестки могут не опадать 7—10 дней. Одновре-
менно с раскрыванием цветка на цветоложе образуется слизистая
масса, которая сохраняется примерно столько же времени, сколько и
тычинки. Эта масса издает запах, особенно заметный вечером в
тихую погоду (Шиманович, 1987).
Ф.А. Щепотьевым (1941) отмечена интересная особенность рас-
крывания цветков бересклета в течение суток. Наибольшая интен-
сивность приходится на вторую половину дня (в светлый период су-
ток 60%, в темный — 40%). Цветение часто бывает очень обильное,
но плодов образуется немного. Число плодов составляет всего 0,5—
1% от числа цветков. По мнению Ф.А. Щепотьева, такое массовое
опадение цветков и завязей объясняется обычно недостаточностью
опыления и недолговечностью пыльников и самих цветков берескле-
та.
Перекрестное опыление осуществляется благодаря более ранне-
му созреванию пыльников (Шиманович, 1987). Созревание рылец
происходит лишь через несколько дней. Пыльца созревает в закры-
том бутоне. С раскрытием лепестков бутона пыльники лопаются и
из них выступает пыльца. Часто можно наблюдать стерильные ты-
чинки, они меньше размером и имеют белую окраску. Нормальные
169
тычинки окрашены в желтый цвет. Развитые пыльцевые зерна имеют
продолговатую форму, длиной в 2 раза больше ширины. Длина
пыльцевого зерна 27,7 мкм, ширина 13,7 мкм (Сгратопович, 1952;
Шиманович, 1987).
От полного цветения до созревания семян у бересклета прохо-
дит, в среднем, 92 дня. Рост коробочек продолжается чуть дольше од-
ного месяца. После достижения нормальных размеров они начинаю т
белеть, затем желтеют, к началу созревания розовеют, а к моменту
раскрытия в конце августа начале сентября приобретают яркую
розовую, малиновую или красную окраску. Вследствие растянутости
сроков цветения созревание плодов также происходит неравномерно.
Зрелые коробочки раскрываются и семена, одетые в ярко-оранжевый
или красный мясистый присемяиничек, свешиваются на тонких ни-
тях. Через 7-10 дней семена отваливаются, если не склевываются
птицами, которые разносят их на большие расстояния (Шиманович,
1987).
Экология и фитоценология
Бересклет бородавчатый произрастает в подлеске дубовых, ли-
повых, березовых, хвойных и смешанных лесов, на опушках и на
прогалинах, среди кустарников. В горах он поднимается до 2100 м
над уровнем моря.
Бересклет бородавчатый отличается требовательностью к богат-
ству почвы и к ее аэрации. В Московской области растет преимуще-
ственно на свежих плодородных почвах, богатых известью, дерново-
подзолистых крупнопылеватых (зернистых) супесях или суглинках,
на бурых лесных почвах с pH от 4,9 до 6,34 и влажност ью от 9 до
35%.’
Вид засухоустойчив, легко переносит атмосферную засуху, по
хуже — почвенную. Он типичный компонент подлеска и отли-
чается значительной теневыносливостью (до 0,8—-0,9 сомкнутости
крон первого яруса). Интенсивность освещения сказывается на мор-
фологии этого кустарника, в связи с чем выделяются две экологиче-
ские формы (Гроздов, 1960). У особен, растущих на открытых мес-
тах, наблюдается образование большого числа толстых стволиков,
густо покрытых бородавками. Крона компактная, листья плотные,
кожистые. В затененных мест ах образуются кусты с т опкими зелено-
ватыми побегами, почти без бородавок и с тонкими листьями; крона
зонтиковидная, кущение идет слабо. Интересная особенность роста
бересклета в условиях низкой освещенности отмечена Н.П. Иордан-
ской и И.Г. Серебряковым (1954). Под пологом леса у бересклета к
15—20 годам жизни полегает главный стволик, который покрывает-
ся лесной подстилкой и образует придаточные корни, а из спящих
почек развиваются новые побеги, интенсивно раст ущие первые 5 7
лет. Эти побеги сильно ветвятся, к 8 10 годам приобретают типич-
ную для кустарника утолщенную форму,-и живут 30 35, а иногда
170
до 40 лег. Зачем они также могут полегать, укореня ться, из спящих
почек в свою очередь возникают скелетные побеги нового поколе-
ния. В ч еченце жизни растения э го может пов торя т ься неоднократно.
По их мнению, в условиях хорошего освещения бересклет' бородавча-
тый по жизненной форме существенно не отличается от таких де-
ревьев как ель и дуб, но в условиях сильною за тенения верхушечная
почка растения быстро отмирает, развиваются боковые побеги, и бе-
ресклет приобретает форму куста.
В’.Подмосковье бересклет' бородавчатый встречается обычно в
елово-широко.тиственных лесах вместе с лещиной при сомкнутости
крои 0,3- 0,8. Хорошо плодоносит только в условиях достаточной
освещенности (более 20" » от полной), близ опушек, «окон» и дорог.
На осветленных участках до 27% растений образует плоды. Число
плодов на кусте при хорошей освещенности по нашим данным от
50 до 150, при низкой до 10—15 и менее. Число семян в плодах
колеблется от 1 до 7: по 2 семени у 29% плодов, по 3 — у 21% пло-
дов, по 4 -. у 20% плодов, по 7 — у 6% плодов. В Одинцовском
районе, по нашим данным, встречается в лесах разного типа — ши-
роколиственных, мелколиственных, смешанных, в сложных ельниках
и сосняках. Предпочитает фитоценозы со значительным участием
широколиственных элементов: сложный ельник медуницево-кислич-
ный, березняк зеленчуково-волосистоосоковый, липняк с елью кис-
лично-зеленчуковый, сложный ельник зеленчуковый, березняк сныте-
во-волосистоосоковый, березняк с липой снытево-разнозравный.
Консортивные связи
Бересклет' бородавчатый — энтомофил. На это указывает нали-
чие слизи на цветах, запах, чрезвычайно малое количество пыльцы,
шероховатая поверхность пыльцевых зерен, окраска цветка. Слизь
на цветах сохраняется до 4-х дней. Она выделяет неприятный запах,
привлекающий насекомых опылителей. Опылители питаются сли-
зью, а также легко доступным нектаром, выделяемым нектарным
диском. Диск является важной частью цветка, выполняющей разные
функции. С одной стороны, его мясистые, богатые сахаром и летучи-
ми веществами ткани, привлекая насекомых, отвлекают их от поеда-
ния завязей и тычинок, т.е. выполняет защитную функцию. Одновре-
менно к диску, который располагается в центре цветка над венчиком
и кольцеообразпо окружает гинецей, часто прикрепляются основа-
ния тычинок, а иногда и лепестки. На цветках обнаружены следую-
щие насекомые: Tlachiptera Macq. (злаковые мушки), Oxystoma
pomanae F. (долгоножки), Silis nitidula F. (мягкотелки) и муравьи. У
бересклета бородавчатого энтомофилия не облигатна. При недос-
татке пли отсутствии агентов опыления опытным путем доказана не
только возможность самоопыления, но и его преобладание (Щепоть-
ев, 1941).
171
Бересклет бородавчатый больше всего повреждается бересклет-
ной паутиновой молью (Hyponomeuta evonumella Sc.), гусеницы ко-
торой опутывают паутиной ветки и сильно объедают под ней ли-
стья. Вредителем бересклета является также бересклетовая пяденица
(Abraxas adustata Schiff), гусеницы которой объедают листья. Боль-
шой вред бересклету приносит бересклетовая огневка . (Alispa
angustella Hb.), гусеницы которой питаются плодами. Значительные
повреждения могут наносить бересклету почвенные вредители (ли-
чинки хрущей, щелкуна и медведка).
В естественных условиях стволы бересклета очень часто повреж-
даются грибом (Fomes ribis Lili. var. evonymii Bet., etc), поселяющимся
в нижней части стволика и вызывающим буровато-желтую гниль
древесины. В питомниках значительные повреждения сеянцам нано-
сят почвенные грибки из родов Fusarium и Altemaria, вызывающие
болезнь, известную под названием «полегание сеянцев».
Из диких животных, наносящих вред бересклету следует отме-
тить оленей, сильно объедающих побеги, мышей, обгрызающих кор-
ни, а иногда уничтожающих и семена (Леонова, 1974).
Хозяйственное значение
Человек издавна использовал бересклет бородавчатый. О суще-
ствовании у него клеток, выделяющих вещество, похожее на каучук
было известно в прошлом столетии. Однако химическая природа
этого вещества была изучена только в 30-е годы нашего века рус-
ским ученым Г.Г. Боссэ, показавшим, что секреторные клетки запол-
‘ йены гуттой. А.И. Шатерникова в 1938 г. (цит. по Шимановичу,
1987) описала строение гуттовместилищ у бересклета бородавчатого.
Они представляют собой отдельные прямые, сильно удлиненные
клетки с заостренными концами длиной 0,51—0,82 мм и диаметром
5—15 мкм, не сообщающиеся между собой. Гуттовместилища можно
обнаружить в листьях, плодах, наружном слое колленхимы стебля,
даже в цветках, но наибольшее их число образуется во вторичной ко-
ре корня и стебля. Эта кора может служить источником добычи гут-
ты. В среднем, кора корней бересклета содержит 8—16% гулы (от
массы сухой коры), но этот показатель может меняться от 0 до 30%.
Гутта в корнях накапливается из года в год, откладываясь в ежегод-
ных слоях коры рядами.
При эксплуатации бересклета бородавчатого, по данным Стра-
тоновича (1987), кондиционных корней с 1 га получали в 4—5-лет-
нем возрасте— 0,4 тонны, в 7—8-летнем— 1,2 тонны, в 10—I Плет-
нем — 5—6 тонн. При культуре вес корней одного растения в 10-
летнем возрасте достигал 700—800 г, что давало при 10 тыс. расте-
ний на 1 га плантации 7—7,5 тонн корней, из них 70—80% составля-
ли кондиционные корни. При среднем выходе у куста 50—70 г сухой
коры, с 1 га плантации ее получали 0,5—0,7 тонн. При среднем выхо-
де гутты, равном 10—15% веса сухой коры, с 1 га плантаций получа-
172
ли 70—100 кг гутты (или средний годовой прирост гутты от 7 до 10
кг) (Стратонович, 1952).
До недавнего времени значение бересклета в нашей стране опре-
делялось потребностями промышленности в гуттаперче. До появле-
ния синтетических пластмасс гуттаперча находила широкое приме-
нение как электрический изолятор, в том числе — для изоляции мор-
ских кабелей, т.к. она выдерживает длительное пребывание в соле-
ной воде. Ее использовали в качестве очень стойкого материала для
изготовления кислотоупорной посуды, изделий и аппаратуры для хи-
мических лабораторий, в гидротехнических сооружениях, для приго-
товления различных мастик, пластмасс, специальных клеев для обув-
ной промышленности, для протезирования в медицине и некоторых
других областях.
Плотная, твердая, стойкая к гниению, очень прочная и в то же
время легкая (средняя плотность 0,68 г/см3) древесина красивого жел-
товатого цвета нашла применение в токарном деле.
В народной медицине западных областей известно применение
водного отвара цветущих веток бересклета бородавчатого при нерв-
ных расстройствах и сильных головных болях (Юркевич, 1949).
В официальной медицинской практике было опробовано и в ря-
де случаев рекомендовано применение настойки коры бересклета в
70% спирте при лечении гипертонической болезни I и II степени.
Кроме того, известно, что глюкозид эвонилин, содержащийся в коре
ветвей, плодах и корнях бересклета, обладает сердечным действием
наподобие наперстянки. Из листьев бересклета выделен гликозид,
идентичный кемпферол-37-дирампозиду. Бересклет бородавчатый
может служить источником многих химических соединений. В коре
корней и плодах были обнаружены сахара, горькие вещества и смесь
кислот: аспарагиновой, винно-каменной, лимонной, яблочной, азот-
ной и бересклетовой. Сахара, содержащиеся в коре корней берескле-
та в количестве 12% от массы абсолютно сухой коры, по мнению не-
которых специалистов, могут быть использованы для получения
спирта. В семенах бересклета содержится до 54% невысыхающего
масла (Шиманович, 1987).
Бересклет бородавчатый рекомендован для создания лесозащит-
ных полос на полях степных и лесостепных районов России.
И.Л. Крылова, Л.И. Прокошева
БАГУЛЬНИК БОЛОТНЫЙ
Номенклатура и систематическое положение
Ledum palustre L. 1753, Sp. PL: 391; Кауфман, 1866, Моск, фл.,
изд. 1: 318; Цингер, 1885, Сборн. свод.: 289; Шмальгаузен, 1897, Фл.
Средн, и Южн. Росс. 2: 186; Сырейщиков, 1910, Илл. фл. Моск, губ 3:
13; он же, 1927, Опред. раст. Моск, губ.: 204; Федченко, 1940, в Маев-
ский, Фл., изд. 7: 562; Е. Буш, 1952, Фл. СССР, 18: 28; Пояркова,
1954, в Маевский, Фл., изд. 8: 409; она же, 1964, цит. соч., изд. 9: 396;
Скворцов, 1966, Опред. раст. Моск, обл.: 258; Webb, 1972, Fl. Europ.
3: 9; Виноградова, 1981, Фл. европ. части СССР, 5: 42; Черепанов,
1981, Сосуд, раст. СССР: 212; Киселева, 1987, Опред. раст. Мещеры,
2: 40 — Багульник болотный (сем. вересковые — Ericaceae)
Внутривидовая систематика
В последней обработке сем. Ericaceae дая европейской части
CCCt’ (Флора европ. части..., 1981) у вида Ledum palustre L. выделе-
но два подвида — 1) subs, palustre — высокий кустарник с доволь-
но крупными (2—4 см длиной) листьями и слабо завернутыми края-
ми пластинки и 2) subsp. decumbens (Ait.) Hult. — стелющийся кус-
> тарник с мелкими (0,8—2,5 см дайной) листьями и сильно завернуты-
ми краями пластинки.
Однако правомерность подобного выделения очень сомнитель-
на, поскольку форма роста багульника зависит от характера суб-
страта, а размеры листовой пластинки и степень ее завернутости —
от ряда экологических факторов, в первую очередь, — от степени
освещенности местообитания (Крылова, Прокошева, 1980).
Географическое распространение
Багульник болотный — бореальный голарктический вид с цир-
кумполярным ареалом, охватывающим Скандинавию, Среднюю и
Восточную Европу, Балканы, Гренландию, Северную Америку, Мон-
голию, Северо-восточный Китай, Японию.
В России багульник болотный распространен в тундровой и лес-
ной зонах, в европейской части он доходит до берега Баренцева мо-
ря, на юге граница ареала проходит по лесостепи. За Уралом ареал
багульника охватывает всю территорию Сибири и Дальнего Восто-
ка, несколько отступая па севере от побережья, на юге - доходя до
лесостепи Западной Сибири, а в Средней и Восточной-Сибири — до
174
государственной границы, проникая далее в Монголию и Северо-
восточный Китай (Копанева, Шретер, 1976).
Багульник встречается почти по всей территории Московской
области за исключением ее южной и юго-восточной части, т.е. Се-
ребряно-Прудского, Зарайского, Каширского, Озерского, Коломен-
ского, Воскресенского районов (Ворошилов и др., 1966). В Серпухов-
ском районе Отмечены островные местонахождения Приокско-Тер-
расиого заповедника и по левобережью Оки в припойменной части
второй террасы (Смирнов, 1958).
Морфологическое описание
Зимпезеленый кустарник или, чаще, кустарничек 15—120 см вы-
сотой. Жизненная форма варьирует в зависимости от географиче-
ской зоны п характера субстрата. На минеральных грунтах аэро-
ксильньгй кустарник и кустарничек с сохраняющимся всю жизнь
главным корнем (Солопевич, 1956). На сфагновом покрове - гео-
ксилыгый кустарничек или даже стлаиггк без главного корня с укоре-
няющимися нижними ветвями, погребенными нарастающим сфагну-
мом, и отходящими от них парциальными кустами, поднимающими-
ся над поверхностью сфагнового ковра. Нарастание вегетативных
побегов моноподиальное, генеративных — симподиальное. Моло-
дые побеги густо рыжеватоопушенпые, более старые - покрыты
темно-серой корой. Почки защищены почечными чешуями. Пластин-
ки листьев линейно-продолговатые, кожистые, сверху темно-зеленые
с вдавленной срединной жилкой, рассеянными железками и волоска-
ми, снизу - густо ржаво-опушенные простыми и железистыми во-
лосками. Длина пластинок 1,4—7,8 см; ширина 0,14—0,68 см; толщи-
на 10,5— -260,0 мкм. Края пластинок завернуты гга нижнюю сторону,
степень завериутости краев изменяется от 20 до 70%. Морфологиче-
ские показатели размеров листьев у багульника, растущего в евро-
пейской части России, меняются в зависимости от характера место-
обитания в первую очередь от степени освещенности. Наиболее
ксерофитпые листья, т.е. мелкие, с толстым мощным слоем палисад-
ной т кани у багульника из лесотундры и с олиготрофных болот лес-
ной зоны. Самые мезотрофиые листья, с крупной топкой .листовой
пластинкой, — у багульника из среднетаежных сфагновых сосняков
(Крылова, Прокошева, 1980). В Московской области средняя длина
листьев багульника в разных сообществах составляет от 2,47±0,09 до
3,42±0,14 см, средняя ширина листа от 0,84±0,03 до 1,36±0,02 см,
степень завериутости колеблется от 26 до 39?о.
Соцветия щитковидные верхушечные. Цветоножки тонкие, 2 3
см длиной, с опадающими прицветными чешуями при основании. И
цвет оножки, и прицветники, опушены железистыми волосками.
Цветки 1 1,5 см в диаметре, с двойным околоцветником. Ча-
шечка с пятью маленькими округленными долями. Венчик из пяти
175
белых или слегка розоватых лепестков, с расширенными при основа-
нии и реснитчатыми нитями. Завязь верхняя пятигнездная, столбик
прямой с неясно пятилопастным рыльцем. Коробочки поникающие,
продолговато-овальные 4-—7 см длиной, многосеменные, покрытые
железистыми волосками, раскрывающиеся снизу вверх пятыб створ-
ками. Семена мелкие, продолговатые, с перепончатыми крыловидны-
ми выростами. Все растение обладает сильным запахом.
Онтогенез
Прорастание семян надземное. Проростки имеют две овальные
светло-зеленые семядоли, затем появляются 2—3 светло-зеленых ме-
зоморфных листа. В первую зиму верхушка побега войлочно опуше-
на, а почка защищена зелеными листьями, позже развиваются на-
стоящие почечные чешуи; на второй год на гипокотиле и при осно-
вании эпикотиля образуются придаточные корни (Grevilius, Kirchner,
1923). Развитие в первые годы идет очень медленно, ежегодный при-
рост у 2—3-летних растений составляет 3—5 мм, у 5—7-летних 10—
15 мм (Солоневич, 1956).
Первые листья живут 2—3 года, затем опадают. Вновь образую-
щиеся листья более ксероморфны (Мазуренко, 1981). Нарастание мо-
ноподиальное. У имматурных растений в базальной части первично-
го побега развиваются боковые побеги (Гришин и др., 1985). Позже
развитие идет по-разному, в зависимости от характера субстрата, на
котором растет багульник. На минеральном субстрате он существует
в виде кустарника или даже «миниатюрного дерева» (Grevilius,
Kirchner, 1923), всю жизнь сохраняя главный корень. Последующие
фазы развития определяются увеличением размера и числа органов,
интенсивным ветвлением или кущением. Соцветие закладывается в
верхушечной почке ортотропного побега, зачатки листьев при этом
не закладываются. Под соцветием, на приросте предыдущего года,
развиваются пазушные побеги, образуя ложную мутовку. В дальней-
шем эти побеги продолжают нарастать, но дают очень маленькие
приросты. Таким образом, переход в генеративное состояние сопро-
вождается переходом от моноподиального к симподиальному нарас-
танию.
Время наступления цветения и продолжительность отдельных
этапов развития существенно варьируют в разных местообитаниях.
Цветение может быть ежегодным, повторяться через 2—4 года
(Grevilius, Kirchner, 1923) или даже через 5—6 лет (Мазуренко, 1981).
На сфагновом покрове главный корень багульника отмирает
очень быстро: корневая система состоит из многочисленных прида-
точных корней. Последние вырастают на нижней части побегов, ко-
торые погружаются в нарастающий сфагнум, иногда на глубину до 1
метра (Grevilius, Kirchner, 1923). Возраст таких погребенных побегов
доходит до 40 лет. При этом определение возраста возможно по гра-
ницам годичных приростов, а не по годичным кольцам, так как кси-
176
лема нарастает у багульника только в надземной части. Поэтому по-
гребенные побеги имеют не более 6—7 годичных колец (Серебряков,
1962).
Специализированные подземные побеги у багульника не образу-
ются. Нижние части скелетных осей укореняются по всей длине. На
плагиотропных частях из спящих почек образуются новые ортотроп-
ные побеги обычно с крупными листьями. Они интенсивно растут и
на 2—3-й год зацветают (Мазуренко, 1981). В дальнейшем эти побе-
ги образуют новые парциальные кусты, через некоторое время утра-
чивающие связь с материнским растением. Таким образом, создается
клон, отдельные парциали которого продолжают нарастать верхуш-
ками побегов, поднимающимися над поверхностью сфагнового ков-
ра. Именно такая форма роста встречается на сфагновых болотах
Московской области. Интенсивно ветвящихся кустарников с сохра-
няющимся главным корнем здесь обнаружено не было.
По наблюдениям в Томской области, в возрасте 10—17 лет ба-
гульник утрачивает способность к цветению и переходит сначала в
субсенильное, а затем в сенильное состояние (Гришин и др., 1985).
Однако другие исследователи признаков старения у багульника не
обнаруживали (Мазуренко, 1981).
Сезонное развитие
Зачатки соцветия багульника сформированы в почках с осени
(Серебряков, Галицкая, 1951). Рост побегов начинается в мае. Зацве-
тает багульник в тундровых редколесьях в конце июня — начале
июля (Полозова, Боч, 1970); в Карелии — в июне (Кучко, 1975); в
Белоруссии средняя дата зацветания 20 мая (Юркевич, Смеляк, 1963).
По средним многолетним данным, в Подмосковье цветение ба-
гульника начинается 2 июня (Долгошов, 1949). В зависимости от по-
годных условий даты наступления фенофаз и их продолжительность
в разные годы существенно различаются. Так, в 1974 и 1976 г. с ран-
ней, но прохладной весной и частым выпадением осадков, в окрест-
ностях Звенигородской биостанции на Луцинском и Шараповском
болотах бутонизация багульника была отмечена 4 и 1 июня, соответ-
ственно, а цветение — 12 и 16 июня. В годы с теплой весной — 1975
и 1977 — бутонизация багульника наступила 6 и 13 мая, соответст-
венно, и очень быстро — через неделю — 13 и 21 мая — цветение.
Фенологическим индикатором цветения багульника можно считать
зацветание сирени. Обычно в Подмосковье багульник цветет.весь
июнь, лишь в 1975 г. с очень теплой весной и теплым и сухим летом
багульник к концу мая почти полностью отцвел. Во время цветения
первыми распускаются краевые цветки. В это же время под соцвети-
ем начинается рост пазушных побегов, образующих ложную, мутов-
ку.
В июле и августе созревают семена, в сентябре багульник обсе-
меняется. В конце июля начале августа иногда наблюдается вто-
177
ричное цветение. В августе начинают опадать листья на приросте
позапрошлого года. Листья на приросте последних двух лет- осенью с
наступлением холодов краснеют за счет появления в клеточном соке
антоциана и отклоняются вниз, прижимаясь к стеблю.
В соответствии с фазами вегетации в листьях и побегах багуль-
ника изменяется содержание биологически активных веществ. Так,
по наблюдениям в Подмосковье, за период вегетации процент содер-
жания эфирного масла изменяется в 2,4 раза, дубильных веществ —
в 1,9 раза.
Обычно считается, что больше всего эфирного масла растения
содержат во время цветения, однако у багульника в годы с холодным
летом этот пик обычно не выражен. Но независимо от погоды, мак-
симальное количество эфирного масла ежегодно отмечается во время
созревания семян, приблизительно через 40 дней после отцветания —
в конце июля — первых числах августа. После этого содержание
эфирного масла начинает снижаться; минимальное его количество
отмечено в конце и в начале вегетации (Прокошева, 1980).
В содержании дубильных веществ отмечено два максимума — в
начале вегетации до набухания почек (9,2%) и в конце вегетации —
после обсеменения (9,9%). Минимальное же количество дубильных
веществ наблюдается во время созревания семян, т.е. тогда, когда ли-
стья содержат больше всего эфирного масла (Прокошева, 1980).
Способы размножения и распространения
Багульник болотный способен как к семенному, так и к вегета-
тивному размножению. Растения, обитающие в Арктике, образуют
‘ хорошо развитые семена, у багульника из более южных областей от
50 до 80% семян пустые (Grevilius, Kirchner, 1923). Н.Г. Солоневич
(1956) отмечает большую семенную продуктивность багульника в ле-
сотундре. Всхожесть семян высокая; несмотря на то, что багульник
микотроф, присутствие гифов гриба для прорастания не обязатель-
но, однако прорастают семена только на свету (Панкова, 1961). В
природе проростки и всходы багульника можно обнаружить на об-
наженном минеральном грунте без мохового покрова, на галечни-
ках, дорожных насыпях, вдоль тропинок (Мазуренко, 1981).
На сфагновом ковре проростков багульника обнаружить не уда-
ется. Здесь основную роль играет вегетативное размножение за счет
укоренения и нарастания плагиотропных побегов, образующих в
дальнейшем парциальные кусты, которые утрачивают связь с мате-
ринским растением и создают клон.
Экология
Багульник болотный — олиготроф или (в северной тайге и ле-
сотундре) — олигомезотроф (Богдановская-Гиенэф, 1946; Солоне-
178
г
вич, 1970; Хмелев, 1974; Сокол, 1978). Его обычно характеризуют как
умеренно гигрофильный вид или даже гигрофит, распространенный
на болотах средней и умеренной влажности (Богдановская-Гиенэф,
1946; Кац, 1975). Однако на севере лесной зоны и в лесотундре он
встречается и на незаболоченной минеральной, а в горах — даже на
каменистой и щебнистой почве (Солоневич, 1956).
В европейской части России и в Белоруссии местообитания ба-
гульника характеризуются влажнолуговым и болотнолуговым ув-
лажнением (68—92-я ступени шкалы Раменского) и особо бедным и
бедным субстратом (1—6-я ступени шкалы Раменского).
Кислотность субстрата колеблется в широких пределах: от pH
2,2—4,1 в Коми (Сокол, 1978) до pH 4,7—5,1 на южной границе ареа-
ла (Хмелев, 1974).
По нашим данным, в лесной полосе европейской части России
сообщества, где багульник доминирует и содоминирует, занимают
местообитания с pH субстрата 2,8—6,5, увлажнением (от влажнолу-
гового до болотнолугового (69—90-я ступень шкалы Раменского) и
бедными почвами (2—5-я ступени шкалы Раменского). Освещенность
местообитаний колеблется от полной до 3% суммарной радиации в
пределах физиологически активной (ФАР).
В Московской области багульник растет в сосняках с незначи-
тельной сомкнутостью крон, где освещенность под пологом колеб-
лется от 15 до 40% суммарной радиации, на торфянистых, кислых
(pH 4) очень бедных почвах (2—3-я ступени шкалы Раменского) с сы-
ролуговым увлажнением (81—89-я ступени шкалы Раменского).
Таким образом, в Московской области этот вид ведет себя как
теневыносливый олиготроф и оксилофит, умеренный гигрофит.
Фитоценология и биологическая продуктивность
Багульник болотный — доминант и субдоминант третьего яру-
са (Быков, 1965). В европейской части России он встречается в кус-
тарничковых тундрах, в березовых редколесьях лесотундры, образо-
ванных низкорослой Betula tortuosa и очень угнетенной Pinus
silvestris. В север о-таежных лесах багульник распространен в ельни-
ках сфагновых, зеленомошных, долгомошных и черничных. Однако
наиболее характерен он для облесенных олиготрофных болот и со-
сняков багульниково-сфагновых. Эти сообщества различаются лишь
степенью участия сосны и ее высотой, поэтому зачастую трудно про-
вести границу между облесенным болотом и сосняком сфагновым. В
обоих случаях в сложении кустарникового яруса принимают участие
одни и те же виды вересковых, а в моховом покрове — одни и те же
виды сфагнов. Багульник доминирует или содоминирует в травяно-
кустарничковом ярусе. Проективное покрытие его достигает 30%,
высота побегов — 60 см, их годичный прирост колеблется от 4,1+0,2
до 11,9±0,5 см. Это — наиболее продуктивные популяции багульни-
179
ка болотного. Величина фитомассы их годичных побегов составляет
в среднем 158,9+2,3 г/м2 в сосняке багульниково-сфагновом и 115,4±
2,4 г/м2 на облесенном олиготрофном болоте (Крылова, Прокошева,
1982).
Несколько меньше роль багульника в сосняках кустарничково-
сфагновых, произрастающих в тех же местообитаниях, что и сосняки
багульниковые. Здесь содоминантами багульника становятся
Chamaedaphne calyculate (покрытие до 20%), Vaccinium uliginosum и
V. myrtillus (покрытие до 15%). Ту же картину мы видим в сосняках
пушицево-сфагновых, занимающих более увлажненные местообита-
ния. В этих последних, наряду с кустарничками господствует
Eriophorum vaginatum (покрытие до 40%), однако покрытие багуль-
ника, как и в сосняке кустарничковом, достигает 20% и степень раз-
вития его особей не уступает багульнику из сосняков сфагновых.
На менее увлажненных и более богатых почвах произрастают
сосняки зеленомошные и долгомошные, в травяно-кустарничковом
ярусе которых тоже доминируют вересковые. Степень развития мо-
хового покрова может быть различна — от 10 до 953о и преоблада-
ют в нем в соответствии с типом леса зеленые мхи или виды
Polytrichum. Участие популяций багульника в этих лесах сильно
варьирует, так же, как и степень развития особей багульника. Так,
проективное покрытие багульника на отдельных участках изменяет-
ся от 2 до 30%, в/лсота растений в среднем — от 30 до 66 см.
Средняя величина продукции (сырой фитомассы годичных побе-
гов) во всех типах сосняков, кроме сосняка багульниково-сфагново-
' го, колеблется от 82,2±1,5 до 97,9+2,1 г/м2.
Довольно часто багульник встречается на облесенных мез^Ьли-
готрофных болотах, покрытых единичными экземплярами сосны, бе-
резы, изредка ели; в травяно-кустарничковом ярусе обильна'Carex
nigra, но основной состав этого яруса тот же, что и на олиготрофном
болоте. В сложении мохового покрова участвуют не только олиго-
трофные, но и мезотрофные сфагны. Проективное покрытие багуль-
ника колеблется от 3 до 10%, высота растений от 30 до 60 см; величи-
на продукции в разных популяциях от 12,3+1,9 до 192,8±14,8 г/м2.
Анализ данных По величине продукции в пределах европейской
части России показал, что фитоценотический фактор достоверно
влияет на величин)' продукции, однако гораздо сильнее влияние гео-
графического фактора (13 и 35% соответственно). Таким образом,
продуктивность популяции в значительно большей мере зависит не
от типа растительного сообщества, а от географической зоны, в ко-
торой обитает данная популяция. Наиболее продуктивные популя-
ции (в среднем 116,8+6,2 г/м2) произрастают в зоне широколиствен-
но-хвойных лесов, а минимальная продукция (в среднем 85,5+3,9
г/м2), как и следовало ожидать, в популяциях лесотундры (Кры-
лова, Прокошева, 1982).
180
На величину продукции, кроме географического и фитоценоти-
ческого факторов, влияют также отдельные экологические факторы.
Так, во всех географических зонах европейской России продуктив-
ность багульника выше в слабо затененных местообитаниях — при
освещенности 30—60% суммарной радиации в пределах ФАР и на
очень кислом субстрате (pH менее 4). В то же время отношение ба-
гульника к увлажненшо и богатству почвы на протяжении ареала
меняется: в среднетаежных лесах продуктивность багульника выше
на не очень увлажненном (ступени ниже 85-ой по штате Раменского)
и не очень бедном субстрате (ступени выше 3-ей по шкале Раменско-
го), а в более южных зонах наблюдается обратная зависимость.
Интересно сравнить эти данные с влиянием тех же факторов на
линейный прирост побегов. Величина прироста также, в первую оче-
редь, зависит от географического фактора (сила влияния 33%). Сред-
ний годичный прирост побегов в лесотундре, по данным М.С. Боч
(1979), составляет 2,6—2,9 см, а по нашим данным, — 2,4+0,05 см, в
то время как в южнотаежных и широколиственных лесах 7,1 ±0,05 см.
Фитоценотический фактор на величину прироста влияет слабее гео-
графического — сила его влияния составляет лишь 22%: максималь-
ный прирост отмечен в сосняках багульниково-сфагновых — 7,5 см.
Во всех изученных нами сосняках багульниково-сфагновых европей-
ской части России — это средняя величина годичного прироста по-
бегов (Крылова, Прокошева, 1982).
По всей лесной зоне линейный прирост побегов багульника
больше в затененных местообитаниях на не очень увлажненном
(меньше 85-й ступени по шкале Раменского) и не очень бедном суб-
страте (3-я ступень и выше шкалы Раменского).
Если величину продукции признать за критцэий фитоценотнче-
ского оптимума, а по ведичине линейного прироста судить о близо-
сти к экологическому оптимуму (Крылова, 1985), можно сделать вы-
вод, что в лесотундре и в северных лесах экологический и фитоцено-
тический оптимумы этого вида очень близки, а в южной части ареа-
ла они расходятся, поскольку конкурентно мощные виды вытесняют
багульник из самых благоприятных местообитаний.
В Московской области багульник встречается в сосняках багуль-
никово-сфагновых и кустарниково-сфагновых.
Сосняки багульниково-сфагновые произрастают- на бедном,
сильно увлажненном субстрате (2,5—3,5 и 83 - 89 ступени шкалы Ра-
менского соответственно). Древостой образован исключительно со-
сной Va бонитета, диаметром 12- 15 см и высотой 7 9 м. Сомкну-
тость крон невелика освещенность 20 35"» (см. табл.). В подрое-
те встречаются единичные экземпляры Betuia pubesccns. В травяно-
кустарничковом ярусе' господствует багульник с проективным по-
крытием 6 16%. Эго наиболее продуктивные его популяции (см,
табл.). Присутствуют Vaccinium uliginosum (покрытие I 5" и),
Chatnaeilaphnc calyculata, Eriophorum vaginaluin. Почти сплошной
181
покров сфагновых мхов образован Sphagnum maggellanicum,. S.
fallax, S. angustifolium.
В местообитаниях с теми же параметрами экологических факто-
ров произрастают сосняки кустарничково-сфагновые. Древостой
здесь также образован редкостойными соснами Va бонитетй, мохо-
вой покров образован теми же сфагновыми мхами, отличается лишь
структура травяно-кустарничкового яруса. При том же наборе видов
участие их в сложении яруса другое. Популяции багульника играют
значительно меньшую роль: проективное покрытие не выше 4%, зна-
чительно меньше продукция (см. табл.). Довольно обильны другие
кустарнички — голубика, черника, клюква, Кассандра, покрытие ка-
ждого из них может достигать 10%. Из травянистых видов значи-
тельно лишь участие пушицы влагалищной. Реже и с меньшим по-
крытием встречаются брусника и подбел. Один раз — в Приокско-
Террасном заповеднике на участке сосняка кустарничково-сфагново-
го — был отмечен вереск.
Величина линейного прироста побегов варьирует в разные годы
в зависимости от погодных условий, при этом вегетативные побеги
багульника растут интенсивнее генеративных. Так, в Одинцовском
районе на Луцинском и Шараповском болотах в благоприятные го-
ды (1974 г.) прирост вегетативных побегов составлял 10—11 см, гене-
ративных — 5,0—7,5 см; в неблагоприятные годы (1973, 1975) веге-
тативные побеги прирастают на 5—7 см, а генеративные от 3,2 до 4,2
см в среднем по популяции.
Существенный момент биологической характеристики вида —
это регенерационная способность его популяций. Каким образом и
за какой период времени восстанавливается биологическая продук-
тивность популяции после ее нарушения какими-либо деструктивны-
ми факторами, в том числе и после заготовки сырья хозяйственно-
ценных видов? Восстановление надземной части багульника после
засыхания или срезания кустов происходит за 5—7 лет (Солоневич,
1956). Специальные опыты, проведенные в Карелии, показали, что
срезание блиственных побегов багульника с перерывом в 1—2 года
сильно угнетает растения, а ежегодное срезание приводит к гибели
растений на пятый год (Юдина, Максимова, 1978). Нами в течение
пяти лет проводились аналогичные опыты в Дарвинском заповедни-
ке (Вологодская область) в сосняке сфагновом под пологом сосны и
на просеке. Отрастание побегов багульника в первый же год после
срезания происходит за счет спящих почек на приросте прошлых лет
— до 6—7-летнего возраста включительно. Из почек, тронувшихся в
рост, развиваются немногочисленные короткие и слабые побеги, не
образующие в дальнейшем соцветий. Кроме того, эти побеги, если
они не укрыты снежным покровом, в первую очередь повреждаются •
182
Содержание в листьях дуби- льных веще- ств, % оо оо *П сП О\ 1П ч© оо
& ° о _ -§ ® 1,25 j 1П~ СЧ~ 1П~ Ч? СЧ г-Н
Величи- ’ на про- дукции, 1 г/м2 100,0+ 11,5 62,8+0,7 1П +1 оо 3 22,4+2,7 40,0+0,9 48,0+5,0 27,7+8,8 21,2+1,5
Высота экземп- ляров, см +i 31,5+1,4 53,7+2,3 О +1 П 35,8+2,0 +[ СП сч *Н СЧ +i оо сч
о с & , й § Со К Ю u, X о4 ЧО \© О сП СЧ -г С') сч
Saw § § r>>«s «1 з s чо О S X рь га tr о'- сП о СЧ 1 СП СП сч СЧ
Растительное сообщество Сосняк багульни- | ково-сфагновый то же v то же Сосняк кустарнич- ково-сфагновый то же то же т о же то же |
Географический пункт Одинцовский район, д. Луцино Одинцовский район, д. Шарапове Шатурский район, Радовицкий мох Шатурский район, Мишеронское лесничество Одинцовский район, д. Луцино Одинцовский район, д. Шарапове Одинцовский район, 1 д. Шарапове ' Серпуховской район, Приок- ско-Террасный заповедник
при неблагоприятных условиях погоды. За два года, прошедших по-
сле срезания побегов, фитомасса их восстановилась лишь на 40—
60%. Причем существенных различий в скорости восстановления фи-
томассы в хорошо освещенном местообитании (на просеке) и в слабо
затененном (под пологом сосны) не наблюдалось. А следующий, тре-
тий год, привел к резкому уменьшению фитомассы, поскольку в су-
ровую зиму 1977 г. многие замещающие побеги были повреждены.
Таким образом, регендзационная способность багульника очень низ-
ка — полное восстановление исходной величины продукции проис-
ходит не раньше, чем на шестой год и то при условии отсутствия су-
ровых зим.
Консортивные связи
Багульник — микогроф. В клетках эпидермиса молодых корней
находятся гифы грибов (Солоневич, 1956). Микориза эндотрофная
эрикоидного типа (Катенин, 1972). У семян, прорастающих на почве,
гифы грибов обнаруживаются на шестой день, однако присутствие
гриба не является необходимым условием прорастания (Панкова,
1961).
Хозяйственное значение
Листья и молодые побеги багульника содержат эфирное масло,
арбутин, кумарины, дубильные вещества, флавоноиды (Лебедев и
др., 1986). Этот богатый состав биологически активных веществ оп-
( ределяет возможность применения багульника в медицине.
Первое упоминание о лечебных свойствах багульника относится
к XII веку: в Дании, а затем в Швеции, отвар цветков и молодых по-
бегов багульника использовали для лечения пoдaqзы, бронхита, ди-
зентерии и кожных болезней. В России применение его было извест-
но с XVII века (Болотов, 1788). До сих пор применение отваров и на-
стоев багульника в народной медицине чрезвычайно разнообразно:
как отхаркивающее, противовоспалительное, бактерицидное и анти-
гельминтное средство. Кроме того, наружно применяют его при экзе-
ме, диатезах, укусах насекомых, ушибах, обморожениях. В гомеопа-
тии наружно — при ревматизме, подагре, артритах, опухолях,
внутрь — при коклюше, астме и как болеутоляющее.
В ветеринарии багульник используется при колитах, желудочно-
кишечных заболеваниях, чесотке и артритах.
Существуют и другие аспекты применения багульника. Так, от-
вар его добавляют к пиву вместо хмеля для усиления опьяняющего
действия (Лебедев и др., 1986). Отвар, настой и порошок применяют',
как инсектицид и ратицит. Его также можно использовать для вы-
делки и дубления кож. Багульник — медонос, однако есть данные,
184
что багульниковый мед ядовит. Известны случаи отравления овец и
коз (Ядовитые растения, 1950).
В настоящее время настои молодых побегов так называемой
«травы багульника» применяют в медицине как отхаркивающее и
противокашлевое средство (Машковский, 1988). Для аптек ежегодно
заготавливают около 100 т побегов багульника. Из багульника так-
же получают противокашлевой препарат ледии.
Фармакологические исследования показали, что отвар багуль-
ника расширяет бронхи, поэтому его в первую очередь рекомендуют
при бронхиальной астме, коклюше, остром бронхите.
Лечебное действие побегов багульника зависит, главным обра-
зом, от содержания в них эфирного масла, количество которого в ли-
стьях колеблется от 0,1 до 7,5% (Лебедев и др., 1986). Таким образом,
чем богаче эфирным маслом сырье, гем выше лечебный эффект. На
накопление эфирного масла влияют географические и экологические
факторы, существенную роль играет фаза вегетации (см. раздел се-
зонное развитие) и даже характер погоды.
Влияние географического фактора сказывается очень сильно
(сила влияния 27%). Беднее всего эфирным маслом багульник из ле-
сотундры, богаче — из среднетаежных лесов. Характер раститель-
ного сообщества на накопление эфирного масла не сказывается. Воз-
действие остальных факторов в разных частях ареала различно. Так,
в лесотундре, северо- и среднетаежных лесах больше всего эфирного
масла у багульника, растущего в слабо затененных местообитаниях
(освещенность меньше 30% суммарной радиации) на очень кислом
субстрате (pH меньше 4). В южно-таежных и подтаежных лесах на-
оборот — богаче всего эфирным маслом багульник, растущий при
хорошей освещенности и на не очень кислом субстрате.
Интересно, что на накопление дубильных веществ в листьях ба-
гульника изученные факторы влияют совсем по-другому: их содержа-
ние не зависит от географической зоны, в которой произрастает ба-
гульник. Наоборот, характер сообщества, увлажнение и богатство
субстрата сказываются сильно: дубильных веществ больше у багуль-
ника, растущего на сильно увлажненном и очень бедном субстрате
(Крылова, Прокошева, 1979).
В Московской области содержание эфирного масла в листьях
багульника, собранных в разных местообитаниях в фазу созревания
плодов, колеблется от 1 до 1,7%. Максимальное количество эфирно-
го масла было обнаружено у багульника, росшего в сосняке багуль-
никовом на Шараповском болоте. Больше всего дубильных веществ
было в багульнике из сосняка кустарничкового с Лупинского бо-
лота.
Погода существенно влияет на накопление эфирного масла: в го-
ды с теплым сухим летом в листьях багульника содержится больше
эфирного масла, чем в годы с холодным летом. Так, в Подмосковье
среднее содержание эфирного масла в годы с сухим летом (1974, 1975
г.) составляло 1,74+0,12%, а в годы с дождливым летом (1976, 1977)
185
— 1,47±0,04%. Погода влияет и на накопление дубильных вещества:
в жаркое лето их накопление меньше, чем в холодное, в то же время
дождливая погода влияет отрицательно. В сухое лето 1974 и 1975 г,
листья багульника, росшего на Лупинском и Шараповском болотах,
содержали в среднем 9,1 ±0,01° о дубильных веществ, а в дождливое
лето 1976 и 1977 г. - 8,7+0,01%.
ЛИТЕРАТУРА
Л б дне в М. Ряска горбатая (Lemna gibba L.) в культуре // Культивирование во-
дорослей и высших растений в Узбекистане. Ташкент, 1971. С. 131 -138.
А беле Г. Т. Флористический состав разнотравных типов леса (Heteroherbosa) в
Чемгальской равнине // Уч. зап. Латв. гос. ун-та, 1970. Т. 127. С. 4—30.
Лвдуло в II. П. Кариосистематические исследования семейства злаков // Труды
по прикл. бот., генет. и сел. Приложение. 1931.44 с.
Аверьянов Л. В., Аверьянова Е. Л., Л а в р е и к о А. Н. Карио-система-
тичсекос-исследованне орхидных (< Irchidaceae) на территории Коми АССР // Бот. жури.
1982. Т. 67. № 11. С. 1491- -1499.
•Алексеев Е. В., Киселева К. В.. Новиков В. С. и др. Определитель
растений Мещеры. Часть 1. М., 1986. 199 с.
Алексеев К). Е.. Губанов И. А. Флора окрестностей Пущино-на-Оке. М.,
1980.104 с.
Алексеенко Л. Н. Водный режим луговых растений в связи с условиями сре-
ды. Л., 1976. 199 с.
Алехин В. В. Наши поемные луга. М., 1925. 122 с.
Алехин В. В. Растительность и геоботаническое районирование Московской и
сопредельных областей. М., 1947. 69 с.
Алиев Д. А. Растительность заболоченных, водных и влажных местообитаний
Алшерона//Уч. зап. Азерб. ун-та. Сер. биол. 1958. № 2.
Алиев 3. А. Бересклетовая моль и ее энтомофаги // .Защита растений. 1983. №11.
С. 28—29.
А и д р е е в А. А. Сезонная морфологическая изменчивость некоторых подмос-
ковных видов рода Alchemilla 1,. // Дипл. раб. М. Каф. высш. раст. МГУ. 1983.
Андриенко Т. Л. Listera cordata (L.) R. Вг. на Украине // Охрана и культиви-
рование орхидей: Тез. докл. П Всесоюзн. совещ. Киев, 1983. С. 20—21.
Антонова Л. А., Главатских Л. К. Проективное покрытие и урожай-
ность Fragaria viridis Duch. в степных сообществах Зауралья // Растит, рес. 1985. Т. 21,
вып. 2. С. 183—187.
Араратян А. Г., Агаджанян Г. К. Сорные растения рисовых полей Арме-
нии //Изв, АН Арм. ССР. 1948. № I.
Аржанов С. П. Среди води болот. Л., 1926.
Артюшенко 3. Т., Ф е д о р о в Ал. А. Атлас по описательной морфологии
высших растений. Плод. Л., 1986.
Асеева Т. А., Б л и н о в а К. Ф., Яковлев Р. П. Лекарственные растения ти-
бетской медицины. Новосибирск, 1985. 160 с.
Балицкий К. П., Воронцова А. А. Лекарственные растения и рак. Киев,
1982.375 с.
Барсегян А. М. Характерные черты водно-болотной флоры Араратской рав-
нины //Тр. Бот. ин-та Арм. ССР, Ереван, 1964, вып. 4. С. 79—90.
Барыкина Р. П., Туленкова М.А. Элементарный метамер побега цветко-
вого растения // Бюл. МОИП. Отд. биол, 1983. Т. 88, вып. 4. С 114—124.
Барыкина Р. П., Туленкова М.А. Метаморфоз и его значение в жизни
растений // Бюл. МОИП, Отд. биол. 1990. Т. 95, вып, 5, С. 103—111.
Б е й д е м а и И. Н. Методика фенологических наблюдений при геоботанических
наследованиях. М.-Л., 1954. С. 129.
Бельков В. П. Развитие живого покрова ла вырубках в ельниках-кисличниках.
Л., 1956. И с.
Бельков В.П. Особенности главнейших видов травяного покрова вырубок в
кисличниках и черничниках. Л., 1957. 35 с.
Бельков 13.111. Значение густоты посева семян хвойных пород в связи с влияни-
ем травянистых растений // Бот. журн. I960. № 4. С. 608—612. '
Бельков В. П., Мартынов А. Н.. Омельченко А, Я. Регулирование
травяного покровов лесу. М., 1974. 112 с.
1Н7
Бересклет//Сборник трудов. Л.. 1938.
Б с р и д з е Р. К. Род Fragaria. Цитоэмбриологическос и генетическое исследова-
ние // Автореф. дне.., докт. бпол. наук. Тбилиси. 1965. 52 с.
Б и б л ь Р. Цитологические основы экологии растений. М.. 1965. 463 с.
Б о гд ан о в ска я-Г и с и э ф И. Д. О происхождении флоры бореальных лесов
Евразии И Маг-лы по истории флоры и растит. СССР. 1946. вып. 1. М.-Л. С. 425—465.
Б о л о т о в А. Г. (') полезности порста и вонючего вереска И Эконом.«магазин.
1788. ч. 35. №63. С. 174—176.
Борисова И. В. Биология и основные жизненные формы двудольных много-
летних эравянистых растений степных фитоценозов северного Казахстана // Тр. БИН
АН СССР. Сер, 3. Геоботаника. 1961, вып. 13. С. 54—132.
Борисова М. А. Побегообразование и ритм сезонного развития северостеп-
ных растений (на примере растений Стрелецкой степи Курской области) // Автореф.
дпс... канд биол. наук. М., 1954. 15 с.
Борисова-Туленкова М. А. Ритм сезонного развития растений луговой
степи // Бюл. МОИП. Отд. биол. 1960. Т. 64., вып. 6. С. 78—81.
Б о ч М. С. О годичном приросте кустарничков в лесу и на болоте // Тр. Дарйин.
гос. зап. Вологда, 1979, вып. 15. С. 44—51.
Брайтон А. б.. Колесниченко А. И., Бышовец В. В. Вечнозеленые
почвопокровные бересклеты и сезонные изменения пигментного комплекса их листьев //
()храна, изучение и обогащение раститительного мира. 1988, вып. 15.
Булавкина А. Дополнение к статье Э. А. Шпора: некоторые данные о цвете-
нии рясок // Изв. Гл. бот. сада РСФСР. 1925. Т. 24. С. 127—129.
Б ы к о в Б. А. Доминанты растительного покрова Советского Союза. Алма-Ата.
Т. 1-3. 1960 —65.
Былова А. М.. Григорьева Н. М. Биологические особенности, возрастная
и пространственная структура ценопопуляции земляники зеленой (Fragaria viridis Duch.)
и щавеля густого (Rumex confertus Willd.) И Популяционная экология растений. М..
1987. С. 59—65.
Валеева 3. Б. Агрофитоценологическая характеристика посевов ведущих сель-
скохозяйственных культур Удмуртской АССР // Автореф. дне... канд. бпол. наук. Ка-
зань, 1977. 14 с.
Вальтер Г. < )бщая геоботаника. М., 1982. 261 с.
1 .Варганов Л. А., П ау з н е р Л. Е.. Ч с в р е н и д и С. X. Перспективы введе-
ния в культуру глициризинсодержащих и дубильных растений // Пути повышения про-
дуктивности лекарственных и кормовых растений в культуре. Ташкент. 1985. С. 3--9.
Васильченко И. Т. Всходы деревьев и кустарников: Определитель. М.-Л.,
1960.217 с.
В а х р а м е е в а. М. Г. Орхидные // Лесные травянистые растения. М.. 1988. С.
168169.
Вахрамеева М. Г., Денисова Л. В. и др. Орхидеи нашей страны. М..
1991.2'22 с.-
Вах рам сева М. Г., Татар ей к о И. В., Бы чей к о Т. М. Экологические
характеристики некоторых видов евразиатских орхидных // Бюл. МОИН. Отд. биол.
1994. Т. 99, вып. 4. С. 75—82.
Вигоров Л. И. Биоактивные вещества ягод земляники // Культура земляники в
СССР. М., 1972. С. 11 18.
Волкова Т. И. Интродукция видов земляники в Главном ботаническом саду
АН ('ССР//Бюл. Гл. бот. сада. 1988, вып. 148. С. 3 8.
Воронихин Н.П. Растительный мир континентальных водоемов. М.-Л. 1953.
410 с.
Вороши л о в В. II., С к в о р ц о в А. К.. Т и х о м и р о в В. 11. < >ирсделител1.
растений Московской области. М.. 1966. 366 с.
В о р о ш и л о в В. I I. Поиски нового лекарственно! о сырья. М.. 1941. 255 г.
Гавр и л о в а Е. I I. К фармакогпостпческому изучению бересклета бородавчато-
го И Автореф. дне... канд. биол. наук. Молотов., 1956.
Гаевская 11. С. Роль высших растений в питании животных пресных водо-
емов. М„ 1966.327 с.
188
Г а и опенко В. Н., С т р ажецкийВ. изменение интенсивности фотосинтеза
и содержания .хлорофилла у ряски в связи с возрастом и условиями освещения // Физиол.
раст. 1969.Т. 16.№6.('.993-1001,
Г л а з у нова К. II. О возможности применения теории агамно-полового ком-
плекса к систематике покрытосеменных растений (на примере рода Alchemilla L.) I/ Бюл.
МОНИ. Отд. биол. 1977. Т. 82, вып. LC. 129—139.
Г л а з у н о в а К. II. Апомиксис у восточноевропейских представителей рода
Alchemilla L. // Автореф. дне... канд. биол. наук. М.» 1983. 17 с.
Глазунова К. П. Гинецеи манжетки (Alchemilla L.) // Вест. МГУ. Сер биол.
1986. №4. С. 15—19.
Г л а*з у н о в а К. П. < )бразование зародышевых мешков у агамных видов ман-
жетки ( Alchemilla L.) // Бюл. МОЦП. Отд. биол. 1987. 'Г. 92. вып. 5. С. 96—109-
Глазунова К. П., Мят.лев В.Д. Корреляционная структура и изменчивость
признаков при регулярном апомиксисе ( на примере агамного вида Alchemilla monticola
< )piz.) И Бюл. М< )ИП. ()тд. биол. 1990. Т. 95. вып. 6. С. 96 - 110.
Гл а з у н о в а К. П., Нилова М. В. Изменчивость и редукция частей цветка у
факультативно-апомиктической манжетки // Апомиксис у растений: состояние проблемы
и перспективы исследований. Саратов, 1994. С 36—38.
Глазунова К. Г!., Пермяков А. И. Об особенностях пыльцы некоторых
видов рода Alchemilla L. в связи с апомиксисом // Биол. пауки. 1980. № 5. С. 54—59.
Глухов М. А.. Ф е д я к о в а А. А. Влияние зарослей иван-чая и вейников на
физико-химические свойства лесных почв // Леей, хоз-во. 1938. № 3. С. 50—53.
Голубев В.Н. К биоморфологии природных растений Подмосковья с запасаю-
щими органами побегового происхождения // Учен. зап. МО11И. Тр. каф. бот. 1956а. Т.
41. № 1. С'. 41 —104.
Г о л v б с в В. И. К онтогенезу корневищ кистекорнсвых растений // Бот. жури.
19566. Т. 41'. № 2. С. 248 -253.
Г о л у б е в В. Н, О короткокорневищных растениях // Бюл. МОПП. Отд. биол.
1958. Вып. 3. С. 97—103.
Голубев В. Н. К вопросу о классификации жизненных форм // Труды Центр.
Черноз. зап. 1960, вып. 6. С. 117—158.
Голубев В. Н. ()сповы биоморфологии травянистых растений центральной ле-
состепи. Ч. 1. Биоморфология подземных органов .// Труды Центр. Чернот. зап. 1962.
вып. 7. 511 с.
Г о л у б е в В. II. ' Эколого-биологические особениостнт травянистых растений и
растительных сообществ лесостепи. М., 1965. 286 с.
Г о л у б с в В. И. Биологическая флора Крыма. Ялта. 1984. Ден. ВИНИТИ. №
5770 В 84.’
Горленко М. В. Мучнн'сто-росяныс грибы сем. Brisiphaceae. М.. 1983.71 с.
Г о р ч а к и в с к и й П. JI. Флора и растительность высокогории Урала И Тр. I ln-
та биол. 1966. Вып. 48. 269 и.
Г о р ы ш и и а 'Г. К. Раннсвесепние эфемероиды лесостепных дубрав (исследова-
ния но экологии, физиологии, фитоценологии). Л.. 1969.
Г о р ы ш и и а Т. К. 'Экология растений. М., 1979. 368 г.
Г р и hi и и А. В.. И о л о ж и й А. В. и др. Морфологическая характеристика и
сырьевая продуктивность Ledum palustre (Томская область) // Растит, рее. 1985. Т. 21.
вып. I.
Гроздов Б. В. Дендрология. M.-JL. 1952.
Г р о з д о в Б. В. Лесные травы, их индикаторное, кормовое и лекарственное зна-
чение. М.. I 963. 69 с.
Гро г с ) е й м A. A. i )прсделитель растений Кавказа. М.. 1949,
Гро с с । е и м А. А. Растительные богатства Кавказа. М.. 1952. 608 г.
Гросе! е й м А. А- Флора Кавказа. Т. 5. М. -Л.. 1952.
Губанов II. А.. Новиков В Тихомиров В.Н. Определитель высших
растений средней полосы европейской России. М.. 1981.286 с.
Гу г ы и и и II. А. Токеиколо! ня ядовитых растений. М.. 1947.
Дать В.Н. TouhOHbiit словарь живого великорусски! о языка. Г. 4. 1882.
189
Д а и 11 л о в В. И. Степные фитоценозы долины Оки в Московской области и их
происхождение//.Автореф. дис... канд. биол. наук. М.. 1983. 18с.
Демченко Н.П. Палинологическое обоснование самостоятельности подгрибы
Alchemillineac семейства Rosaceae // Сб. научи, тр. каф. бот, Одесск. с.-х. ин-та. Одесса.
1974. Вып 2. О. 75—76.
Демьянова Б. И. Половая структура популяции гвоздичных и двудомных
растений И Популяционная экология растений. М.. 1987. С. 6—10.
Демьянова Е. И., Пономарев А. II. Половая структура природных попу-
ляций гинодиэцичпых и двудомных растений лесостепи Зауралья И Бот. журн. 1979. 'Г.
64,№7.('. 1017-1024.
Джалилова А. Вегетативное л семенное размножение Alchemilla pastoralis
Buser в луговом ценозе // Бот. журн. 1965. Т. 50. № 11. С. 1645—1647.
Добренькова Л. Г. Физиологическая оценка засухоустойчивости сортов и
видов земляники // Бюл. BMP. 1979. Выи. 91. ('. 25—26.
Доброхотов В. Н. Семена сорных растений. М., 1961.
Доброхотова К. В. Цветение ряски в водоемах дельты р. Волги // Природа.
1988. № 10, 192 198.
Доброхотова К. В.. Ролдугин II. И.. Доброхотова О. В. Водные
растения. Алма-Ата. 1982. 192 с.
Д о л г о ш о в В. I I. Календарь цветения главных медоносных растений Подмос-
ковья //Календарь русской природы. Кн. 1. 1948. С. 136—199.
Д о л г о ш о в В. II. Материалы по фенологии древесно-кустарничковых пород
Подмосковья сравнительно с другими районами СССР И Календарь природы СССР.
Кн. 2. 1949. С. 26- 27.
Д у б ы н а Д. В. Классификация свободноплавающей растительности водоемов
Украины // Укр. бот. жури. 1986. Т. 43, № 5. С. 1—15.
Е в д у шенко А. В. Рясковые пойменных водоемов р. Самары Днепропетров-
ской обл. И Вторая Всесоюзн. конф, по высш. водн. и прибрежно-водн. раст. Тез. докл.
Борок. 1988, С. 14.
Ермолова Л.С. Определение максимальной и полной продуктивости надзем-
ной части травяного покрова в лесах Подмосковья с разной степенью вырубки // Земле-
делие и растениеводство. М., 1969. С. 180—188.
Ефремов А; П., Антосяк В. М. Редкие растения Карпатского заповедника.
Деп. ВИНИТИ., № 2791—В.—9. 1990 г. 49 с.
Жизнь растений. М.,Т. 4. 1978: Т. 5(a). 1980; Т. 5(6). 1981;Т. 6. 1982.
Жуковский А, О цветении Lemna в окрестностях г. Ярославля // Север. 1923.
№ 3—4. С. 189—193,
3 а г у л ь с к и й М. Н. Ятрышники секции Orchis флоры Западных областей Ук-
раинской ССР И Тр. Первой конф. мол. ботаников г. Ленинграда. Ленинград, апрель
1986. Ч, 2. Рук. деп. ВИНИТИ. 25.09.86, № 6847—В. С. 179—180.
Загульский М. И. .Некоторые особенности репродуктивной биологии орхид-
ных в западных регионах Украины //Теорет. и прикл. кариология. Тез. докл. Всес. конф.
Кишинев, 1989. С. 179—180.
3 а г у л ь с ь к i й М. Н. Мюцезростання орхщних у Вороняках (Волино-Подшля.
УРСР // Укр. бот. жури. 1990. Т. 47, № 3. С. 60—63.
3 а г у л ь с к и и М. Н. Хоролопя, структура популяцШ та охорона орхщних
(Orchidaceac Joss.) захщних регионов Украши. Автореф. дис... канд. бюл. наук. Knie,
1994.26 с.
Закиров К. 3., Ч е в р е н иди С. X., X а й д м у х а м е д о в Л. П. Особенно-
сти развития герани кровяно-красной на орошаемых землях под Ташкентом И Опыт
культуры сырьевых растений. Ташкент, 1973. С. 48—50.
Зворыкина К. В. Эколого-ценотическая характеристика некоторых предста-
вителен р. ('alamagrostis Adans, (на примере южнотаежных березняков) И Эколого-цено-
тические и географические особенности растительности. М., 1983. С. 85—98.
3 е м л к и с к и й С. Е. Лекарственные растения СССР. М., 1951.508 с.
Злаки Украины. Киев, 1977.518 с.
190
И в а и о в В. В. Определитель некоторых водных высших растений Северного
Прикаспия // Материалы по флоре и растительности Северного Прикаспия. Вып. 4, ч. 1.
Л.. 1969.0.2—55.
И в а и о в а И. Е. Некоторые особенности цветения и опыления ряеок (Lemnaceae
S. Gray) // Бот. журн. 1970. Т. 55, № 5. С. 649—659.
II в а и о в а II. Е. К систематике семейства Lemnaceae S. Gray // Вот. жури. 1973.
№ 10.0. 1413 1423.
II в а и о в а М. А. Медоносные растения в районе реки Гаретовки // Учен. зап.
М< )1 IH. Тр. каф. ботаники. 1956. Т. 41. вып. 1. С'. 105—126.
11 в а и о в а Т. В.. Токарева К). А. К экологии цветения, нектаровыделения и
плодоношения земляники лесной // Взаимоотношение компонентов биогеоценозов в
южной тайге. Калинин, 1966.0. 38—41.
I I г н а т о в М. (Макаров В. В.. Ч и ч е в А. В. Конспект флоры адвентив-
ных растений Московской области // Флористические исследования в Московской об-
ласти. М., 1990. С. 5— 105.
И гнать ев а И .П. Образование побегов и вторичное цветение у стержнекорневых
и кистекорневых травянистых поликарпнков И Бот. журн. 1965. Т. 50, № 1. С. 16 — 28.
• 11 л л и ч е в е к и й С. О. Вторичное цветение, его механизмы и причины // Журн.
русск. бот. общ-ва. 1925. Т. 10. № 1 —2. (168.
И ст о м и н а И. И. Эколого-демографическая характеристика синузии кустарни-
ков//Восточноевропейские широколиственные леса. М, 1994. С. 144—164.
II с т о мина И. И., Богомолова Н. Н. Поливарнантность онтогенеза и
жизненные формы лесных кустарников // Бюлл. МОИП. Отд. биол. 1991. Т. 96, вып. 4. С.
68—78.
Каден Н. Н. Морфология плодов гераниевых И Биол. науки. 1964. № 2. 97--
102.
Каден Н. Н. Эволюция плодов розоцветных И Бюл. МОИП. Отд. биол. 1968. Т.
73. вып. 2. С. 127 — 135.
Каден Н. Н., Лановая В. П. Морфология гинецея и плодов герани // Биол.
науки. 1963. №4. С. 104—109.
Каден Н. Н.. Терентьева Н. Н. Этимологический словарь латинских на-
званий растений, встречающихся в окрестностях агробиостанцни МГУ «Чашниково».
М.. 1975.202 с.
Казарян В. О. Старение у высших растений. М., 1969. 280 с.
Каменецкая II. В., Михайленко М. М.. Савин Е. Н. Сосновые леса
юго-западного Приангарья //Тины лесов Сибири. М.. 1963.
Камышев Н. О. Флора-Центрального Черноземья н ее анализ. Воронеж. 1978.
115 с.
Караваев М. Н. Конспект флоры Якутии. М.-Л.» 1958. 189 с.
К а р а з и я С. Фптоцсно-экологические группы растений лесов Литовской ССР //
Тр. Литовок. НИИ леей. хоз. 1977.Т. 17. ('. 3—10.'
К а р и м о в Г.. Хаидмухамсдов Л. П. Герань кровяно-красная (Geranium san-
guineum I..) как красильное растение И Докл. АН Уз. ССР. 1977. № 5. С. 59 60.
К а с и и о в В. Б. О симметрии в биологии. Л.. 1971.48 е.
К а с и и о в В. Б. Биологическая изомерия. Л.. 1973. 257 с.
К а с и н о в В. Б. Влияние преждевременного удаления дочерних особей на про-
должительность жизни материнской особи Lemna minor (Lemnaceae) // Бот. журн. 1981.
Т.65.№ 11.с. 1603 1613.
К а т а и е к а я В. М. Растительность водохранилищ охладителей тепловых
электростанций Cobctckoi о Союза. Л.. 1979. 278 с.
К а т a i; с к а я В. М. Высшая водная растительность континентальных водоемов
СССРЛ.. 1981.
К аге и и и А. !•’. Микориза растений северо-востока европейской части СССР.
Л.. 1972. (' 73 74.
К а у ф мап Н. Н. Московская флора или описание высших растений и ботппико-
1сографнческнй обзор Московской |убернии. Изд. 1. 2. 1886. 1889.
Кац II. Я. об олиготрофных выпуклых сосново-сфагновых торфяника?;, их
просипи и pacniTcJibnornt И Бюл. М< )l II I. < >гд. биол. 1975. Т. 80. вып. 5. С. 84 9|.
191
Кернер А. Жизнь растений. СПб., 1902. Т. 2. 834 с.
Кестен Л. А. Биологические особенности щетинистого долгоносика (Polydrosus
pilifer Hochuth) — вредителя герани кровяно-красной // Узб. биол. журн. 1982. № 2. ('.
50—52.
Кестен Л. А., Кугми Н. П. Болезни и вредители гераней // Узб. бпол. журн.
1980. №3. С’. 53—55.
К л с в е н с к а я И. Л. Строение корневых клубеньков травянистых нсбобовых
растений Сибири // Пзв. С() АН СССР. 1969. № 10. выл. 2. С. 38—43.
Кожевников А. В. Весна и осень в жизни растений. М.-Л., 1950.
Коки н К. А. Экология высших водных растений. М., 1982. 157 с.
Колчанов Ф. Система розовых (подсемейство Rosoideae) // Авторсф. дне...
канд. биол. наук. Орджоникидзе. 1970. 18 е.
Комарова Т. А. Внутрипочечное и впепочечное развитие генеративного побе-
га герани лесной // Ботан. исслед. на Урале. Свердловск, 1988. С. 49.
Копанова Г. А.. Шрегер А, И. Карта ареала багульника болотного.//Ат-
лас ареалов и ресурсов лекарственных растений. М.. 1987. С. 64- -65.
Коро л ь В. М. Биологические особенности Elodea canadensis Rich, и Lemna
minor L. // Биол. науки. 1988. № 5. С. 67 -71.
К о р с м о Сорные растения современного земледелия. М.-Л., 1933.
Корчагин А. А. Строение растительного сообщества // Полевая геоботаника.
Т. 5. Л.. 1976.
Кощеев А. К. Дикорастущие съедобные растения в нашем питании. М.. 1981.
255 с.
К р а с и о б о р о в II. М. Высокогорная флора Западного Саяна. Новосибирск.
1976. 379 с.
Краснова А. Н.. Кузьмиче в А. II. Флора озер Северо-Двинской водной
системы И Флора и продуктивность пелагических и литоральных фитоценозов водоемов
бассейна Волги. Л.. 1990, С. 95—108.
К р е ч е т о в и ч Л. М. Ядовитые растения, их польза и вред. М.-Л,. 1931.
Крылова П. Л, Ресурсная характеристика лекарственных растений и научная
основа их рациональной эксплуатации // Автореф. дне... докт. биол. наук. М.« 1985.
Крылова II. Л.. Прокошева Л. И. Влияние экологических факторов на со-
держание эфирного масла и дубильных веществ в листьях Ledum palustre L. // Растит,
pec. 1979. Т. 15. выл. 4. С. 575—583.
Крылова И.Л.. Прокошева Л.II. Влияние географического и экологическо-
го факторов на анатомо-морфологические признаки багульника болотного и связь этих
признаков с химическим составом листьев И Растит, рес. 1982. Т. 17, вып. 1.С.З—13.
Кузнецов Л. В., Зайченко О. В. Три вопроса о ряске малой (Lemna minor
L.) // Систематика, анатомия и экология растений европейской части СССР. Л.. 1979. С.
18-34.
К ук к К). Некоторые данные о культуре природных орхидных//Охрана и куль-
тивирование орхидей. Таллинн. 1980. С. 75—78.
Куприянова Л. А.. Тарасевич В. Ф. Ультраструктура поверхности обо-
лочки пыльцевых зерен некоторых родов семейства Lemnaceae и сближенных с ними ро-
дов семейства Агасеае И Бот. журн. 1984. Т. 68, № 12. С. 1656—1662.
Куркин К. А. Эдификаторы и ассектаторы настоящих лугов Барабинской лесо-
степи // Бюлл. МОИП. Отд. биол. 1968. Т. 73, вып. 1. С. 29—41.
Куркин К. А. Системные исследования динамики лугов. М., 1976.284 с.
К уте ев Ф. С. Бересклетовая щитовка//Защита растений. 1969. № 12. С. 45—46.
К у ч к о А. А. Лекарственные травы и кустарнички березняков южной Карелии И
Ресурсы ягодн. и лекарств, раст. и методы их изучения. Петрозаводск, 1975. С. 113—122.
Кормовые растения европейской части СССР. Справочник. Л., 1981.
Кормовые растения сенокосов и пастбищ (’ССР, М.-Л., 1950. Т. 1.. 1951.1'. 2.
Красная книга РСФСР. М., 1985.
Лавренко Е. М. ('тени и сельскохозяйственные земли на месте степей И Растит,
покров СССР. Пояснит, текст к геобот. карте СССР. М.-Л., 1952. Т. 2.
Л а в р и ч е и к о Е, В. Малый жизненный цикл чистяка весеннего //Докл. ТСХА.
1970. Вып. 159. С. 162—166.
192
Ларин И. В. и др. Кормовые растения сенокосов и пастбищ СССР. Т. 2. М.-Л.,
1951.
Л ащинскпй Н. Н., Ронгинская А. В. О корневых клубеньках небобовых
растении Сибири // Микроорганизмы и зеленые растения. Новосибирск, 1967. С. 44—51.
Лебедев А. М., Мазна'я Е. А., Тюлин С. Я. Сем. Ericaceae//Раст. рес.
СССР. Л.. 1986. С. 136—159.
Левина Р. Е. Способы распространения плодов и семян в природе. М., 1957.
328 с.
Лекарственные растения. 4-е изд., Минск, 1968. 390 с.
Л й с и ц и н а Л. II. Флора волжских водохранилищ // Флора и продуктивность
пелагических и литоральных фитоценозов водоемов бас. Волги. Л., 1990. С. 3—49.
Л и с и ц и н а Л. И.. Э кзер цев К А. О редких видах водных и прйбрежно-
водных растений водоемов Волжского бассейна // Состояние и перспективы исслед. фло-
ры средней полосы европ. части СССР. М., 1984. С. 42—43.
Логинова В. Г. Многохозяйственность луговых и лесных попупаразитов //
Бот. исслед. на Урале. Свердловск, 1985. 29 с.
Лодкина М.М. Семейство Lemnaceae // Сравнительная анатомия семян. Т. 1.
Однодольные. Л., 1976.
Лукина Г. А. Жизненный цикл рясковых // Биология внутренних вод. Информ,
бюл. №79. Л., С. 25—27.
Лукина Г. А., М и к р я к о в а Т. Ф. Оптимальные условия культивирования
ряски маленькой (Lemna minor L.) И Флора и растительность пелагических и литораль-
ных фитоценозов водоемов бассейна Волги. Л., 1990. С. 153—158.
Любарский Е. Л. К изучению экологии и взаимоотношений некоторых длин-
нокорневищных растений сосняков // Взаимоотношения растений в растит, сообщест-
вах. Казань. 1964. С. 290—312.
Любарский Е. Л. Экология вегетативного размножения высших растений.
Казань. 1967. 181 с.
• Л ю б и м е н к о В. Н. К вопросу о сорной растительности сплошных вырубок И
Сельское хозяйство и лесоводство. 1902. № 5. С. 290—341.
М а д а л и е в а Г. К. Высшая водная растительность Восточного Арнасоя // Узб.
биол. журн. 1975. № 4. С. 33—35.
Маевский П. Ф. Флора средней полосы европейской части СССР. Изд. 7, 8, 9.
М., 1951. 1954. 1964.
Маевский П.Ф, Флора Средней России. Изд. 1, 2, 3. М., 1892,1895, 1912.
Мазуренко М. Т. Онтоморфогенез Ledum palustre (Ericaceae) в Магаданской
области И Бот. журн. 1981. Т. 66, № 2.
Мальцев А. И. Об осеннем цветении калужницы болотной и некоторых других
растений // Труды бюро по прикл. бот., ген. и сел. Л., 1933. Т. 115, № 5.
Марков М.В., Беляев В. И., Попова Н. К. Растительность водоемов
пойм рек Волги и Камы в пределах Татарской АССР // Учен. зап. Казанск. ун-та. 1955.
Т. 115, № 5.
. Маслов А. А. Количественный анализ горизонтальной структуры лесных сооб-
ществ. М.. 1990. 160 с.
Матвеев В. И. Гидрофиты Куйбышевской области // Учен. зап. Куйбышев,
пед. ин-та. 1961. Вып. 35. С'. 41—45.
М атвеев В.И. О цветении ряски Lemna gibba // Бот. журн. 1963. Т. 48, № 2. С. 272.
Матвеева Е.П., Знаменская Л. А. Химический состав и кормовая харак-
теристика растений, преобладающих на сенокосах и пастбищах в Ленинградской облас-
ти//Тр. БИН АН СССР. Л., 1960. Вып. 12. С. 128—137.
Машковский М. Д. Лекарственные средства. Ч. 1. М., 1988.
Мелехов И.С. Лесоведение. М., 1980. 408 с.
Мельник С. П. Продуктивность растительных сообществ Шацких озер Волын-
ской области // Вторая Всесоюзн. конф, по высшим водным и прибрежноводным расте-
ниям. Борок, 1988. С. 102—104.
Меньшова В. А. Особенности цветения и плодоношения герани кровяно-крас-
ной (Geranium sanguineum L.) и буквицы лекарственной (Betonica officinalis L.) в культу-
ре // Охрана, изучение и обогащение растит, мира. 1985. Вып. 12. С. 87—89.
193
Мережко А.. II. Роль высших водных растении в самоочищении водоемов II
Гидробиол. журн. 1973. Вып. 9,№ 4. С. 118—125.
Методические рекомендации по классификации сенокосов и пастбищ равнинной
территории европейской части СССР. М.. 149 с.
М и к е л а д з е Р. М. К познанию альпийских ковров Юго-Осетии И Проблемы
ботаники. Т. 5. I960.С. ПО—Ш.
Миронов В. В., Шил ь н и к о в Н. Г. () воздействии вытяжек из сена и кор-
ней вейника наземного (Calamagrostis epigeios (L.) Roth.) на сосну и ель в нолевых усло-
виях И Бнол. науки. 1972. № 8. С. 81—84.
Молпш Г. Физиология растений как теория садоводства. М.-Л.. 1933. 343 с.
М у з а ф а р о в А. М., А б д и е в М., Т а у б а е в Т. Т. Биолого-экологические
особенности ряски малой (Lemna minor) в водоемах Узбекистана И Узбекск. биол. журн.
1968. №6. С. 38—41.
- М узафаров А. М., Т а у б а е в Т. Т.. А б дне в М. Об использовании ряски
малой в корме домашней птицы //Узбекск. биол. журн. 1968. № 3. С. 44—46.
М я э м е т с А. А. Изменение высшей водной растительности И Антропоген. возд.
на малые озера-Л., 1980. С. 77—85.
Набатов Н. М. Динамика живого напочвенного покрова и его влияние на
рост культур сосны И Бот. журн. 1964. Т. 49. № 5. С. 669—677.
Н аз ар о в В. В. Семенная продуктивность некоторых орхидей, произрастающих
в Крыму И Охрана и культивирование орхидей. М.. 1987. С. 38—39.
Назаров В. В. Репродуктивная биология орхидных Крыма. Автореф. дне...
канд. биол' наук. СПб. 1995. 26 с.
Нейштадт М. II. Определитель растений средней полосы европейской части
СССР. М.. 1963. 640 с.
Н и к и ти н С. А. Некоторые особенности биологии произрастания лесных расге-
ннй в лесопарковых условиях Серебряноборского лесничества И Леса Подмосковья. М..
1965. С. 169—201.
Николаева М. Г.. Разумова М. В.. Гладкова В. Н. Справочник по
проращиванию покоящихся семян. Л., 1985. 347 с.
Н о в о т е л ь н о в а Н. С. Порядок пероноспоровые (Peronosporales) // Жизнь
растений. Т. 2. М.. 1973. С. 186.
Н о т о в А. А., Глазунова К. П. ()пыт разработай классификации аномаль-
ный вариантов цветка и цветоноса среднерусских манжеток И Флора Тверской области.
Тверь, 1994. С 45—63.
Н у х и м о в с к и Й Е. Л. Осевая и побеговая системы семенных растений И Пзв.
ТОХА. 1971. № I. С. 54—66-
Определитель высших растений Северо-запада европейской части РСФСР. Л..
1981. 371 с.
Определитель высших растений Якутии. Новосибирск. 1974. 543 с.
Определитель растений Мещёры. М., 1986. Т. 1. 240 с.
О сипя и Л. Л. Особенности и динамика развития перонослоровых и паразит-
ных гифальных грибов в высокогорьях Армянской OOP // Вопросы биологии. Ереван,
1987. Вып. 4. С. 170—174.
Панкова И. А. Багульник болотный (Ledum palustre I..) // Тр. БИН. ('ср. 5.
1961. Вып. 9. Л., С. 175—215.
П атраб о л о ва И. Г. Биология вейника наземного в связи с возобновлением
сосны в Бузулукском бору //Автореф. дне... канд. биол. наук. М.. 1953. 16 с.
Пашкевич В.К)., Юдин Б.С. Водные расгсиия и жизнь животных. Новоси-
бирск, 1978- 1 27 с.
Петухова Л. В. Большой жизненный цикл манжетки пастушьей (Alchcmilla
pastoralis Buser.) //Экол. и физиоп. раст. Калинин, 1974. С. 3 13.
Петухова Л. В. Онтогенез н структура систем нобы он манжетки пастушьей //
Бюл. МОИП. Отд. бнол. 1977. Т. 82, вып. 3. С. 120 129.
Петухова Л. В. Сравнительно морфологическое исследование жизненных
форм некоторых моноподиально-розеточных растений сем. Rosaceae // Автореф. дне...
канд. бнол. наук. М., 1980. 12 с.
194
Печенюк Е. В. Lemna gibba (Lemnaceae) в Хоперском государственном запо-
веднике // Бот. журн. 1984. 7’,„69, № 8. С. 1101 —1102.
Печенюк Е. В. О:Дветении рясковых (Lemnaceae) в Хоперском государствен-
ном заповеднике // Бот. журн. 1 985. Т. 70, № 8, С'. 1066—1070,
П е ч я и к о в а ( '. С. Сорная растительность лесосек типа Pinetum hylocomiosum И
11зв. Казанок. лес.-тех. ин-та. 1931. № 2—3, С. 70—122.
П о к р о в с к а я 'Г. Н. Экологические условия фотосинтеза литоральных гидро-
фитов // Антропогенное евзрофирование озер. М., 1976. С*. 17—44..
Полозова 'Г. Г.. Б о ч М. С. Основные черты фенологии растений // Экол. и
биол. растений восточноевроп. лесотундры. Л.. 1970. Ч. 1. С. 307—336.
Полянский И. И. Ботанические экскурсии. М.. 1968.
11 о н о м а р с в А. Н. Лесостепной комплекс северной окраины Кунгурской лесо-
степи // Изв. ест. -научи, ин-та при Пермском гос. ун-те. 1948. Т. 12. вып. 6. С. 225—232.
Пономарев А. Н. О лесостепном флористическом комплексе северного и сред-
него Урала U Бот. журн, 1949. 'Г. 34, № 4, СП 381—388.
П оно марев А.Н. О лесостепном комплексе и сибирских влияниях во флоре Се-
вера Европейской части СССР // Из вест, научн. ин-та при Пермском ун-те. 1952. Т. 13,
вып. 4—5. С. 315—326.
Пономарев А. Н. Некоторые приспособления ’злаков к опылению ветром И
Бот. журн. 1966. Т. 51, № 1. С. 28—39.
Пономарев А. Н., Демьянова Е. И. К изучению гинодиэции у растений
//Бот. журн. 1975. Т. 60, № L С. 3—15.
Попов М. Г. ФЯора Средней Сибири. М.-Л.. 1957. Т. 1.555 с.
Попов М. Г.. Б у с и к В. В. Конспект флоры побережий озера Байкал. М.-Л..
1966. 216е.
П о х и л ь к о А. А. Материалы к биологии Alchemilla alpina L. (манжетки аль-
пийской) // Бот. исслед. за Полярным кругом. Апатиты, 1985. С. 21—37.
Практические указания по производству стратификации семян бересклета. В. Ус-
тюг, 1943. 13 с.
Прозоровский Н. А. Ботаническая география с основами общей ботаники.
М.,1956. 482 с.
Прокошева Л. И. Выявление оптимальных районов заготовок багульника бо-
лотного по урожайности и содержанию действующих веществ // Автореф. дис... дисс.
канд. биол. наук. М., 1980.
Работнов Т. А. Живые семена в почвах лугов Окской поймы (Предваритель-
ное сообщение) // Вопросы кормодобывания. 1951. Вып. 3. С. 111—119.
Ра ботнов Т. А. К методике изучения всхожих семян в почвах лугов // Бот.
журн. 1958. Т. 43, № И. С. 1572—1581.
Работнов Т. А. Луговедение. М., 1-е изд, 1974. 384 с., 2-е изд. 1984. 324 с.
Работнов Т. А. Фитоценология. М., 1978. 383 с.
Раменский Л.Г., Цаценкин И.А., Чижиков О.Н., Антипин Н.А.Эко-
логическая оценка кормовых угодий по растительному покрову. М., 1956. 419 с.
Растительные ресурсы СССР. Семейства Hydrangaceae — Haloragaceae. Л., 1987.
326 с.
Рахимов А. Р. Химический состав ряски малой (Lemna minor), выращенной
при различных условиях питания //Дикорастущие и вводимые в культуру растения в Уз-
бекистане. Ташкент, 1966.
Ребристая О. В., Т о к а р е в с к и х С. А. Новые данные о распространении
водных растений на Крайнем Севере европейской части СССР И Бот. журн. 1967. Т. 52,
№ 1.0.98—103.
Р о ж е в и ц Р. Ю. Злаки. Введение в изучение кормовых и хлебных злаков. М.-Л.,
1937.638 с.
Романович Е. А. Лекарственные растения Северной Осетии и содержание в
них витамина С. Орджоникидзе. 1969. 279 с.
Ростовцев С. И. О цветении ряски под Москвой // Изв. Моск, сельхоз. ин-та.
1901.№7. С. 63.
Росте в ц е в С. 1-1. Биолого-морфологический очерк рясок. М., 1905.108 с.
195
Ростовцев С. И. Пособие по определению паразитных грибов по растениям—
хозяевам. М., 1908. 2-е изд. 252 с.
Русакова М. Б. Антэкология степных злаков Забайкалья // Вопросы антэкол'о-
гии.Л., 1969. С. 46—48.
Р ы с и н Л. П. Растительность некоторых типов леса в Серебряноборском опыт-
ном лесничестве // Стационарные биогеоценотические исслед. в южн. подзоне тайги. М.,
1964. С. 5—12.
Рысин Л. П. Световой режим некоторых хвойных и лиственных типов леса //
Стационарные биогеоценотические исслед. в южн. подзоне тайги. М., 1964. С. 74—89.
Рысин Л.П. Сосновые леса европейской части СССР. М., 1975.212 с.
Рысин Л. П., Р ы с и н а Г. П. Опыт популяционного анализа лесных сооб-
ществ // Бюлл. МОИП, отд. бнол. 1966. Т. 71, вып. 1. С.84—94.
Рысин Л. П., Рысин а Г. П. Опыт стационарного анализа лесных сообществ
// Бюл. МОИП. Отд. биол. 1986, Т. 71, вып. 1. С. 84—94.
Рысин Л. П-, Р ы с и н а Г. П. Морфоструктура подземных органов лесных
травянистых растений. М., 1987. 208 с.
Р ы с н н а Г. П. Ранние этапы онтогенеза лесных травянистых растений Подмос-
ковья. М., 1987. 208 с.
Рычин Ю.В. Флора гигрофитов. М.« 1948. 448 с.
С а п а н к е в и ч П. В. Биология цветения и плодоношения у бересклета бородав-
чатого и бересклета европейского И Автореф. дис... канд. биол. наук. Брянск, 1949. 16 с.
С а п е г и н Л. М. Синтаксономия растительности пойменных лугов Белорусского
Полесья. 1. Пор. Arrhenatheratalia Paul. // 1986. Деп. ВИНИТИ, № 1947—В 86. 30 с.
С а у т и н В. И. Биологические особенности вейника наземного (Calamagrostis
epigeios (L.) Roth.) и меры борьбы с ним при лесоразведении И Бот. журн. 1957. № 6. С.
917—921.
Свиридинов Г. И. Лесной огород. М., 1984. 223 с.
Селиванов И. А., Верещагина В. А. и др. Материалы по изучению ми-
кориз некоторых злаков, произрастающих в степной полосе И Уч. зап. Пермск. гос. ун-
та. 1961. Т. 18, вып. 3. С. 3—10.
Серебряков И. Г. О ритмах сезонного развития растений подмосковных ле-
сов // Вести. МГУ, 1947. N 6.
Серебряков И. Г. Структура и ритм в жизни цветковых растений И Бюл. МО-
141*1. Отд. биол. 1948. Т. 53, № 2. С. 49—63.
Серебряков И. Г. Морфология вегетативных органов высших растений. М.,
1952. 391 с.'
Серебряков И.Г. Типы развития побегов у травянистых многолетников и фак-
торы их формирования И Уч. зап. МГПИ им. Потемкина. 1959. Т. 100. вып. 5. С. 3—37.
Серебряков И. Г. Экологическая морфология растений. М., 1962.
Серебряков И. Г. Жизненные формы высших растений // Полевая геоботани-
ка. Т. 3. М.-Л., 1964. С. 146—205. 1
С е р е р я к о в И. Г., Галицкая Т. М. К биологии сезонного развития бо-
лотных растений Подмосковья в связи с условиями их жизни и происхождением И Уч.
зап. МГПИ. 1951. Т. 19, вып. 11. С. 1947.
Серебряков И. Г., Серебрякова Т. И. О двух типах формирования кор-
невищ у травянистых многолетников И Бюл. МОИП. Отд. биол. 1965. Т. 70, вып. 2. С.
67—81.
Серебрякова Т. И. Побегооборазование и ритм сезонного развития растений
заливных лугов средней Оки//Автореф. дис... канд. биол. наук. М., 1953.
С е р е б р я ко в а Т. И. Побегообразование и ритм сезонного развития растений
заливных лугов средней Оки И Уч. зап. МГПИ им. Ленина. 1956. Т. 9, вып. 3. С. 43—120.
Серебрякова Т. И. О некоторых морфологических закономерностях разви-
тия побегов у овсяницы луговой Festuca pratensis Huds. // Бюл. МОИП. Отд. биол. 1959.
Т. 64, вып. 3. С. 89—98.
Серебрякова Т. И. Некоторые закономерности формирования почек и побе-
гов у луговых злаков И Бюл. МОИП. Отд. биол. 1961. Т. 66, вып. 4. С. 5—12.
Серебрякова Т. И. Морфогенез побегов и эволюция жизненных форм злаков
М., 1971.360 с.
196
Серебрякова Т. И. Об основных «архитектурных моделях» травянистых
многолетников и модусах их побегообразования // Бюл. МОИП. Отд. биол. 1977. Т. 85,
вып. 5. С. 112—128.
Серебрякова Т. И. Модели побегообразования и некоторые пути эволюции
в роде Gentiana // Бюл. МОИП. Отд. биол. 1979. Т. 84, вып. 6. С. 97—109.
Серебрякова Т. И. Жизненные формы и модели побегообразования назем- .
ноползучих многолетних трав // Жизненные формы: структура, спектры и эволюция. М.,
1981.С. 161 — 179.
Симонян С. А., Ульянцев В. И., М е л и я М. С. Конспект рода
Sphaerotheca Lev. (Erisiphaceae) Закавказья // Флора, растительность и растит, ресурсы
Арм. ССР. Ереван, 1988. № 11. С. 9—17.
Смелов С.П. Биологические основы луговодства М., 1947. 230 с.
Смелов С. П. Основные этапы жизни побегов лугового злака при вегетативном
возобновлении и динамика связи между их смежными поколениями // Бот. журн. 1947.
Т. 32, №2. С. 79—89.
С м иренски н А. А. Водные кормовые и защитные растения в охотничье-про-
мысловых хозяйствах, вып. 2. М.. 1952.
Смирнов П. А. Флора Приокско-террасного государственного заповедника//
Труды ПТЗ, вып. 2. М., 1958. 247 с.
Смирнова Е. С. Типы строения вегетативной сферы растений // Бюл. Гл. бот.
сада АН СССР. 1969. Вып. 73. С. 33—40.
Смирнова Е. С. Морфологические типы и формирование облика растений //
Бюл. Гл. бот. сада АН СССР. 1974. Вып. 93. С. 49—57.
Смирнова Е. С. Морфология побеговых систем орхидных. М., 1990. 208 с.
С м ы к Г. К., Б о р т н я к Н. Н. Флористические находки на Словачанско-Ов-
ручском кряже (Центральное Полесье УРСР) // Бот. жур. 1984. Т. 68, № 8.
Смык Г. К., Меньшова В. А., М ы р з а М. В. Восстановление и увеличение
природных запасов лапчатки белой (Potentilla alba L.) и герани кровяно-красной
(Geranium sanguineum L.) на Украине // Экология и физиология растений води, и на-
земн. фитоценозов. Кишинев, 1983. С. 56—64.
С нар скис П. Распространение кровяно-красной герани (Geranium sanguineum
L.) и горного горичника (Peucedanum oreoselinum Moench.) в Литовской ССР // Научи,
тр. высш. уч. зав. Литовск. ССР. Биология. 1973.
Собко В. Г. Opxiaei Украши. Киев, 1989. 192 с.
Сокол А.П. Растительность болот как индикатор химического состава верхних
слоев торфа // Автореф. дис... канд. биол. наук. Л., 1978.
Солоневич Н.Г. Материалы к эколого-биологической характеристике болот-
ных трав и кустарников // Растительность Крайнего Севера СССР и се освоение, вып. 2.
М.-Л., 1956. С. 318—388.
Сорные растения СССР. Л., 1934. Т. 3. 447 с.
Справочник по сенокосам и пастбищам. М., 1966. 504 с.
С т а н к о в С. С., Т а л и е в В. И. Определитель высших растений европейской
части СССР. М.. 1949. Изд. 2. 740 с.
Степанова В. С. Скорость вегетативного размножения ряски в водоемах //
Бот. журн. 1933. Т. 17, №5—6.
С т р а т о н о в и ч А. И. Новый гуптаперченос бересклет бородавчатый
(Evonymus verrucosa Scop.), его природа, свойства и методы разведения. Л., 1936. 79 с.
С тр ат о н о в и ч А. И. Разведение бересклета в СССР. М.-Л., 1952. 120 с.
Стр атоно в и ч А.И. Распространение бересклета бородавчатого на северной гра-
нице ареала и некоторые теоретические выводы И Геогр. сб. М.-Л., 1955. Т. 5. С. 5—15.
Сугоркина Н. С. Особенности популяционной биологии герани кровяно-
красной И Структура и функционирование луговых и степных фитоценозов. Новоси-
бирск, 1987. С. 89—97.
С у г о р к и н а Н. С. Онтогенез и особенности популяционной биологии видов
рода герань//Автореф. дис... канд. биол. наук. дис. М., 1989. 17с.
Сугоркина Н. С. Оценка состояния герани луговой на пойменных лугах //
Популяции раст.: принципы и пробл. охр. природы. Йошкар-Ола, 1991.
197
С ы р е й щ и к о в Д. П. Иллюстрированная флора Московской губернии. М.,
1906, 1907, 1910, 1914. Т. 1—4.
С ырс й щ и к о в Д. Г1. Определитель растений Московской губ. М.» 1927.. '
Т а т а р с и к о I I. В. Орхидные Приморского края: биология, экология, вопросы
охраны // Автореф. дис... канд. биол. наук. дис. М.. 1991.24 с.
Таубаев 'Г. Флора и растительность водоемов Средней Азии и их использова-
ние в народном хозйстве. Ташкент. 1970. 491 с. ,
Т а у б а е в Т. Ф.. А б д и е в М. П. Об изученности рясок в природе и в культуре
// Культивирование водорослей и высш. води. раст. в Узбекистане. Ташкент., 1971. С.
92—98.
Таубаев Т., Абдиев М. Ряски водоемов Узбекистана и их использование в
народном хозяйстве. Ташкент., 1973.
Таубаев Т. Ф., Абдиев М.. Келди беков С. О биологической продук-
тивности ряски малой (Lemna minor L.) в естественных водоемах и культуре // Культиви-
рование водорослей и высш. раст. в Узбекистане. Ташкент, 1971. С. 98—101.
Таубаев Т., Джам ап гарасва А. К. (.) флоре некоторых евтрофных. водо-
емов Казахстана •// Вторая Всесою.зн. конф, но высш. воды, и прибрежноводн. раст. Бо-
рок, 1988. С. 51—52.
Т ахтаджян А.Л. Система н филогения цветковых растений. М.-Л., 1966.
Тахтаджян А.Л. Система магнолиофитов. Л., 1987. 439 с.
Телятьев В. В. и др. Полезные растения Центральной Сибири. Иркутск, 1987.
399 с.
Т е с л о в С'. В., Макарова Л. С. О катехинах некоторых розоцветных в
Пермской области // Научн. тр. Пермск. фармацевт, ин-та. 1969. Вып. 3. С. 217—223.
Т и т л я н о в а А. А. Биологический круговорот азота и зольных элементов в тра-
вяных биогеоценозах. Новосибирск, 1979. 150 с.
Тихомиров В. Н. Alchemilla L. — Манжетка // П. Ф. Маевский. Флора сред-
ней полосы Европ. части СССР. изд. 9, М.-Л., 1964. С. 121—132.
Тихомиров В. Н. Alchemilla L. — Манжетка // Ворошилов В. Н. Скворцов А.
К., Тихомиров В. Н. Определитель растений Московской области. М., 1966.
Тихомиров В. Н. Манжетки Московской области. Систематический обзор //
Растит, и почвы нечериоз. центра европ. части СССР. М.. 1969. С. 98—151.
Тихомиров В. Н. Новый вид рода Alchemilla Ц. из Средней России // Бюл.
(МОИП. Отд. биол. 1972. Т. 77, вып. 4. С. 130—132.
Тихомиров В. Н. Род Alchemilla И Флора северо-востока европейской части
СССР. Т. 3. 1976. Л., 134—145.
Тихомиров В.Н. Alchemilla sukaczevii V. Tichom. — новый вид из Средней
России // Бюл..МОИП. Отд. биол. 1980. Т. 85, вып. 3. С. 98—99.
Тихомиров В. Н. Alchemilla L. — Манжетка И Определитель флоры Мещеры,
ч. 1. М., 1986.
Тихомиров В. Н. О плодах и соплодиях // Бюл. МОИП. Отд. биол. 1989. Вып.
3. с. 59—65.
Т о л ь с к и й А. П. Сорная растительность в лесном хозяйстве и меры борьбы с
ней. М., 1922.
Уланова Н. Г., Маслов А. А. Многомерный анализ горизонтальной струк-
туры растительности вырубки// Бот. журн. 1989. Т. 74, № 9. С. 1316—1323.
Уланова Н. Г., Тоще в а Г.П. Связь растительности микрогруппировок вей-
никовой вырубки с почвами// Бюл. МОИП. Отд. биол. 1989. Т. 94, вып. 4. С. 73—84.
Ульянищев В. И. Определитель ржавчинных грибов СССР. М.» 1987. ч. 2.
У р а н о в А. А. Возрастной спектр фитоценополуляции как функция времени и
энергетических волновых процессов И Биол. науки. 1975. № 2. С. 7—34.
Федоров Ал. А., Артюшенко 3. Т. Атлас по описательной морфологии
высших растений. Л., 1975. Цветок 352 с. 1979. Соцветие. 290 с.
Флора Армении. Ереван, 1958. Т. 3.
Флора европейской части СССР. Л., 1974—1989. Т. 1—8.
Флора Крыма. М., 1960. Т. 2, вып. 2.
Флора’северо-востока европейской части СССР. Л., 1974—1977. Т. 1—4.
Флора Сибири. Новосибирск, 1987, 1988.
198
Флора СССР. М.-Л., 1935. Т. 4; 1936. Т. 5; 1937. Т. 7; 1941. Т. 10.
Флора юго-востока европейской части СССР. М.-Л., 1931.839 с.
Формозов А. Н.. Просвирина II. Б. Деятельность грызунов на пастби-
щах н сенкоспых угодьях. 4. Некоторые данные о грызунах альпийской зоны Кавказа//
Бюл. МОИП. 1935. Т. 42, вып. 1—2. С. 82—89.
Хмелев К. Ф. Ботанико-географическое районирование болот центрального
Черноземья // Биол. пауки. 1974. № 6. С. 65—69.
Холопова Л. Б.. Солнцева О. Н. Растительность и почвенный покров че-
рез 25 лет после сплошной рубки древостоя // Динамика естественных и искусственных
лесных биогеоценозов Подмосковья. М., 1987. С. 52—62.
Хромосомные числа цветковых растений. Л., 1969.
Цвелев Н, Н. Злаки СССР. Л.: Наука., 1976. 788 с.
Цвелев Н. Н. К систематике рода вейник (Calamagrostis Adans.) в СССР // Но-
вости систематики высших растений. Л.. 1 976. 788 с.
Цвелев Н. Н. Флора Хоперского госуд. заповедника. Л., 1988. 191 с.
Цену р п те Б., Страздиньш К). Орхидные Латвии, включенные в «Крас-
ную книгу СССР» // Охрана и культ, орхидей. Киев, 1983. С. 49—51.
Цингер В. Я. Сборник сведений о флоре (вредней России. М., 1886- 520с.
Ц ы г а и о в Д. Н. Фитоиндикация зкологических режимов в подзоне хвойно-ши-
роколиственных лесов. М.,.1983. 196 с.
Ц ы ренова Д. Ю. Род Geranium L. (Geraniaceae Juss.) на Дальнем Востоке
СССР И Автореф. дис... канд. биол. наук. Л.. 1986. 21 с.
Ч с в р е и и д и С. X. Перспективные листовые дубители и опыт их культуры в Уз-
бекистане // Состояние и перспективы изуч. и использ. природных раст. рес. СССР. Таш-
кент, 1 979. С. 66—67.
Чевреннди С.Х.. Хапдмухамедов Л .П. Динамика накопления дубильных
веществ и их распространение в различных органах герани кровяно-красной // Перспек-
тивные сырьевые растения Узбекистана и их культура. Ташкент. 1979. С. 170—173.
Чевреннди С. X., X а и д м у х а м е д о в Л. П., Гнатенко Е. В. Биологи-
ческие особенности прорастания семян герани кровяно-красной — Geranium sanguineum
L. //Опыт культуры новых сырьевых растений. Ташкент. 1977. С. 35—37.
Чевреннди С. X.. Хапдмухамедов Л.П., X у с н у т д и и о в К. X. Ге-
рань кровяно-красная (Geranium sanguineum L.) — новое декоративное растение для озе-
ленения городов Узбекистана//Строительство и архитектура Узбекистана. 1976. № 1.С.
40—41.
Черепанов О. К. (’осудистыс растения СССР. Л.. 1981.509 с.
Черноброва О. Б., Петухова Л. В. Особые случаи бокового ветвления
моноподиально-рсиеточных растений If Флора и растит. южной тайги. Тверь. 1991. С.
96—99.
.Чернышев П. Я. Дубильные расгения нашей страны. М.-Л. 1934.96 с.
Чижов Б. Е. Биоморфологпческая характеристика травянистых растений выру-
бок подзоны сосново-березовых лесов Тюменской области // Научи, тр. Тюмеи. ун-та.
1981. Т. 82. С. 7(1- -91.
Чистяк о в /\.,Р. Биологические особенности бересклета бородавчатого // ('ов.
бот.. 1935. №4.1’. 12-18.
Ill а л а р ь Г. А. и др. Выращивание высших водных растений и водорослей па
средах со сточной водой животноводческих комплексов II Изв. АН МССР. Сер. биол. и
хим. 1983. №6. С. 9 13.
111 с и и и к о в А. 11. 'Экология растений. М.. 1960.
Шилов М.Н.. Богачев В. В. Lcmna gihba во Владимирской облает//Вто-
рая Всссоюзн. конф, ио высш. води, и нрибрежио-водн. раст. Борок. 1988.
Шиманович Ш11. Бересклет. М..1987.65 с.
Шима и с к а я В. 1’. Лекарственные растения народной медицины северо-запад-
ных областей Украинской (ЧТ // Автореф. дис... канд. биол. наук. дис. Львов. 1966. 20 с.
Шлыков Г. Дубильные растения СССР. М.-Л.. 1932. 190с.
Ill м а л ь г а у з с и II. II. Флора Средней н Южной России. Крыма и Северного
Кавказа. 1885. Г. 1.468 с.
111 м а л и ( цузен ll.il. Флора Юго-Западной России. Киев. 1886.783с.
199
ULI м e й д т О. М.< Я а го м я э Ы. А. Некоторые интересные формы орхидных в
Эстонии//Охрана и культ, орхидей. Москва, 1987. С. 18—19.
Шпор Э Некоторые данные о цветении рясок И Тр. Костром, научн. об-ва по
изуч. мести, края. 1915. Т. 4.
Щербаков А. В. Lenina gibba L. в Московской области И Бюл. МОИП. Отд.
биол. 1987. Т. 92, вып. З.С. 90—92.
Щербаков А. В. Конспект флоры водоемов Московской области // Флрристи-
. ческие исследования в Московской области. М., 1990. С. 106—120.
Э к з е р ц е в В. А. Растительность зоны временного затопления южной части
Куйбышевского водохранилища //Тр. ин-та биол. водохр., 1960. № 4.
Э р г а ш е в А. Э.. Абдиев М. К изучению водорослей открытых рясковых
водных бассейнов // Культивирование водорослей и высших водных растений в Узбеки-
стане. Ташкент, 1971. С'. 82—86.
Ю.д и и а В. Ф., Максимова Т. А. О рациональном использовании некото-
рых лекарственных растений Карельской АССР // Тез. докл. 6 дел. съезда ВБО. Киши-
нев. Л.. 1978. С. 67—68.
Ю з е п ч у к С. В. Новые манжетки востока европейской части СССР // Бот. мат.
Гербария БИН АН СССР. 1951. Т. 14. С. 144—185.
Ю з е п ч у к С. В. Новые манжетки европейской части СССР, Сибири и Средней
Азии// Бот. мат. Гербария БИН АН СССР. Т. 16. С. 133—183.
Ю н а ш Г. Г. Меры борьбы с вейником и зубровкой при лесоразведении И Леси,
хоз. 1953. №4. С. 22—25.
Юнусов И. И. Флора и растительность биологических прудов и полей испаре-
ния сточных вод в Узбекистане и их значение // Автореф. дис... канд. бисл. наук. дис.
Ташкент, 1979. 24 с.
IO р к е в и ч И. Д. Закономерности в распространении Evonymus verrucosa Scop,
на территории СССР и некоторые исторические причины, их обуславливающие // Бот.
журн. 1949. Т. 34, № 4. С. 395—409.
Юркевич И. Д., С м е л я к Л. П. Сезонное развитие некоторых видов расте-
ний на болотах // Выращивание продукт, лесов. Минск, 1963. С'. 83—122.
Ядовитые растения лугов и пастбищ. М.-Л., 1950.
Я н и ш е в с к и й Д. Е. Несколько данных о редких растениях водной флоры юго-
» восточного края Европейской России // Работы Волжск, биол. станции. 1921. Т. 6, № 2.
С. 26.
А г b е г A. Studies in flower structure. 2. On the vascular supply to the nectar in
Ranunculus II fain. Bot. 1936. Vol. 50. N 198. P. 305—319.
A г b e r A. Vegetative morphology of Pistia and Letnnaceae // Proc. Roy. Soc. 1919.
Vol.91,N 1.P.96—103.
Arnold R. E. Listera ovata // Orchid. Rev. 1927. N 35. P. 249.
Ashby E. The interaction of factors in the growth of Lenina. IV. The influence of
minute quantities of organic matter upon growth dhd reproduction // Ami. Bot. 1. 1929. Vol.
43.N7.P. 805—816.
Ashby E., Wangermann E., Winter E. J. Studies in the morphologenesis of
leaves. III. Preliminary observations on vegetative growth in Lenina minor // New Phytol.
1949. Vol. 46, N 3. P. 374—384.
Atlas Florae Europaea. Helsinki, 1988. Vol. 8.
Bat e-S m i t h E. C. Chemotaxonomy of Geranium // Bot. Joum. Linn. Soc. 1973. Vol.
67, N 4. P. 347—359.
Billy A., A 11 e i z e 11 e C. Orchis militaris et ses hybrides en Bassin Auvergne // Bui.
Soc. Bot. France 1964. Vol. 3,N 1—2. P. 101 — 102.
Blackburn K.B. Notes on chromosomes on duckweeds (Lemnaceae) introducing the
question of chromosomes size // Proc. Univ. Durham. Phil. Soc. 1933. Vol. 9, N 1. P. 84—90.
Blodgett F. H. Morphology of Lenina frond // Bot. Gaz. 1915. Vol. 60. P. 383.
Boszormenyi E., Boszormenyi E. N and P nutrition and the physiological
age of Lenina minor L. // Aeta Bot. (Acad. Sci. H ungar.) 1957. Vol. 3, N 1. P. 1 —7.
Caldwell O. W. On the life history of Lenina minor // Bot. Gaz. 1899. Vol. 27. P.
37—66.
200
C' a in u s E. G. Note sur les Orchismilitaris L.. 0. purpurea Huds., 0. simia Lam., leurs
varietes et, leurs hybrides dans la flore parisienne // Bui. Soc. Bot. France. 1885. Vol. 32. P.
213- 218, 273—275. 329 -330.
Cheeseman T. F. Manual of the New Zealand Flora. Wellington, 1925.
Clandon R. P., McNabb C.D. The uptake of boron by Lenina minor П Aquat. bot.
1978. Vol. 4.N 1. P. 53—64.
Clalworthy J. N.. Harper J. L. The comparative biology of closely related species
living in the same area. V. Inter- and intraspecific interference within cultures Lenina spp. and
Salviiua natans //J. Exp. Bot. 1962. Vol. 38. N 13. P. 307-324.
(’•ourduroux M. Exigence thermoperiodiques coinparecs de la fieaire et de crosne du
Japan /ABull. Soc. Franc, physiol, vegel. 1966. Vol. 3, N 3. P. 211 —212.
Czopek M. Ekologieznofisiologicznc badania nad zakwitaniem gatunkow z rodziny
Lemnaceae // Wiadow. Bot. 1960. Vol. 4, N 3—4.
Czopek M. Prezegled wynikow badan nad powstawaniem tunonow Spirodela
polyrhiza // Wiadow. Bot. 1964. Vol. 8, N 1 —2. P. 59—78.
Daubs E. N. A monograph of Lemnaceae. Urbana, 1965. 11 Ip.
Postal R. Generative Vehrmehrung bei Ficaria verna und Gibberehnsaure // Natur-
wiss. 1963. Bd. 50, N 7.
D о w m e D. The germination of Listera ovata (L.) R. Br. // Trans. Proc. Bot. Soc.
Edinburgh, 1949. N 2. P- 126—130.
Dressier R. L. The orchids. Natural history and classification. Cambridge, 1981.
332p.
Dupperex A. Orchids of Europe. London, 1961.
De L a n g e L. Gibbo sity in the complex Lemna gibba. Lenina gibba: literature survey
and ecological aspects//Aquatie Bot. 1975. Vol. I,N4. P. 327—332.
DeLange L., Pie terse A.H., Wetsteyn L.P. On the occurrence of the flat
form of Lemna gibba L. in nature 7/ Aeta Bot. neeri. 1984. Vol. 33, N 4. P. 469—474.
E i n h e 11 i g F. A., Leather G. R., H о r r s L. L. Lise of Lenina minor L. as a
bioassay in allelopathy // J. C'hem. Eeol., 1985. V. I l,N 1. P, 65—72.
Ellenberg H. Wiesen und Wieden und ihre standortliche Bewertung. Stuttgart.
1952. Bd. 2. 143 p.
Ellenberg H- Zeigerwerte der Gefasspflanzen Mitteleuropas И Scripta geobotanica.
1974. Bd. 9.
Elliot H. V., G a r 1 у 1 e A. R. The native orchids of the United States and Canada
excluding Florida//New York Bot. Garden, 1975.
Farrel L. Orchis militaris L. (O. galatea Poir., O. rivini Gonau., 0. tephrosanthes
WiHd- el Sw.). The Biol. Flora of the British Isles. N 160 II Joum. Eeol. 1985. Vol. 73, N 3. P.
1041 — 1053.
Fisch V. N., Flick B. N., Arditti J. Structure and fungal activity of hircinol,
loroglossol and orchinol // Phytochemistry. 1973. Vol. 12. P. 437—441.
F о с к e W. (). Rosaceae. In: Engler A., K. Prantl // Die natUrlichen Pflanzen-familicn.
1894 Bd. 3, Abt. 2. P. 1—5.
Fr c li n c r S. Kritik an der e^ropaische Allehemilla-Taxonomic H Fcddes Rep. 1975.
Bd. 86. Heft 3. P. 119—169.
Flora Europaea. Cambridge. 1968, 1980. Vol. 2, 5.
G a d e 11 a T.W.I., Kliphuis Ё. Chromosome number of flowering plants in the Ne-
thrlands. HI // Proc. Roy. Netherlands Acird. Sci. 1967. Ser. (Vol. 70.
G adella Th.. К 1 i p h i u s E. Chromosome numbers of flowering plants in the
Netherlands. IV.//Proc. Roy. Nether!., Acad. Sci. Ser.C. 1968. Vol. 71. P. 168—183.
G a u m a n ft E. Abwehrreaktionen bei Pflanzenkrankheitcn // Eperientia. N 12. P.
411 418.
G i 1 I J. et al. The distribution of chromosome races of Ranunculus ficaria 1.. in the
British Isles//Ann. Bot. (Gr. Brit.) 1972. Vol. 36, N 144. P. 31—47.
Godfcry M.J.. Godfcry H. M. Monograph and Iconography of Native British
Orchidaceae. Cambridge. 1933.
Goebel K. Zur (Jrganographie der Lemnaceae // Flora. 1921. N 14. P. 278 -305.
Grevillius A.. Kirchner O. /iNcrzrcr/tWZ Lebcnsgcschichte der Bliilenpfianzen
Mitteleuropas Stuttgart, 1923. Bd. 4., Abt. 3.
201
G u p p у H. B. On the habits of Lemna minor, L. gihha und L. polvrrhiza //J. Linn.
Soc. Bot. 1894. Vol. 30. P. 323—330.
Hegel in aier F'. Die Lemnaceae — einc monographishe Untcrsuchungcn. Leipzig.
1868. 169p.
H e i n r i e h W. Zur Vcrbrcilung von Orchis militaris L. und Orchis purpurea I luds. ini
Gcbiet utn Jena (Thiiringen)'ll Wiss. 7.. V’. Schiller, Univ. Jena. Math. -Naturwiss. 1980. Vol.
29.N 1.Р. 71—77. '
Hend rveh R. Qualitative I ’bersieht rezenter C ’onnobionten // Preslia. 1985. Bd. 57.
N 4. P. 359—370.
H cnsse n A. Die daucrorgane von Spirodela polyrrhiza (1..) Schleid, in physiolo-
gischcr Belrachlung // Hora. } 954- N 141 - P. 523—566,
II с у w о о d V. H.. Walker S. Morphological separation of cytological races in
Ranunculus ficaria L. // Natur. 1961. Vol. 189.
Hick L. E. Ranges of pH tolerance to the Lemnaceae II Chio J. Sci. 1932. Vol. 32. N 1.
p. 237 - 244.
Hilf H. H. Die Ausbreituiig der «Seggc» (Calamagrostis epigeios) II Forstarchiv. 1928.
N7.P. 111 — 121.
Hill ni a n W. S. The Lemnaceae or duckweeds. A review of descriptive and experimen-
tal literature // Bot. Rev. 1961. Vol. 27. N 2. P, 221—287.
H u n t P.F. Orchis. Orchiductyla and Orchiaceras hybridization and the Hora of the
British Isles. London. 1975. P. 491—494.
H у I a n d c r N. Nordisk K^rlvaxfiora. Stockholm. 1966. Vol. 2.
Ice J., ('ouch R. Nutrient absorption bv duckweed //J. Aquat. Plant. Monag. 1987.
Vol. 25. N l.P. 30—31.
1 r m i s c h T. Bemerkungen uber Ranunculus ficaria und Gagea arxensis // Bot. Zeitung.
Jg. 1868. Bd. 26.
Jacobs D. I. An ecological life-history of Spirodela polyrrhiza (greater duckweed)
with emphasis on the turion phase // Ecol. Monogr. 1947. Vol. 17. N 3. P. 437—469.
К andeler R. Species delimination in the genus Lemna II Aquat. Bot. 1975. Vol. 1. N
4. P. 365—376.
К a r r e r W. Constitution und Vorkoninicn der organischen Pflanzenstoffc. Basel.
Stuttgart, 1958. 1207 p.
К e d d у P. A. Lakes as islands: the distribution of two aquatic plants Lemna minor and
L. trisulca // Ecol. 1976. Vol\57, N 2. P. 353 -359.
К umazava M. /Morphology and development of the sinker in Pecteilis radiala
(Orchidaceae) // Bot. mag. (Tokyo). 1956. Vol. 69, N 10 - -11. P. 455—461.
Kumazava M. The sinker of Plaianlhera and Perularia •- its morphology and
development//Phytomorphology. 1958. Vol.8.N I -2. P. 137 -145.
Landolt E. Distribution pattern and ccophysiological characteristics of European
species Lemnaceae // Ber. Geob. Inst. ETH. 1982. H. 49. S. 127 145.
Landolt E. Okologische* Zeigcnvcrte zur Schweizer Flora II Vcroff. Geob. Inst. ETH.
1977. H. 64. S. 1—208.
Landolt’ E. Physiologische und okologische Gnlcrsuchungen an Letnnaccen H Ber.
Schweiz. Bot. Ges. 1957. Bd. 67, N 2. S. 271 410.
Landolt E. The family of Lemnaceae a monographic study. 1. Biosystcniatic
investigations in the family of duckweeds (Lemnaceae). 2. // Vcroff. Geohot. Inst. ITII. 1986.
IL71. S. 1—556.
Landolt E;, Kandcler R. The family of Lemnaceae a monographic study.
Biosystematic investigations in the family of duckweeds (Lemnaceae). I. Phytochemistrs.
physiology, application, bibliography 2. // Vcroff. (icobot. Inst. ETH. 1987. H. 95. S. 1 638.
L andolt E., Wildi (). Okologische Fckiuntersuchungen bei Wasserlinsen
(Lemnaceae) in den sudwestlichcn Stanton der U.S.A. II Ber. Geobot. Inst. ETH. 1977. II. 44.
S.l 04 146
I.anglet (). fiber (’.hromosomenverhaltnissc und Systematik der Катшсн/штпг //
Svensk Bot. Tidskr. 1932. Bd. 26, N I 2.P. 281 400.
Lawalree A. L’ embryologic des Lemnaceae observations sur Lemna minor I.. //
Cellule. 1952. Vol. 54, N3. P. 303 326:
202
. Lawalree A. La multiplication vegetative des Lemnaceae И Cellule. 1943. Vol. 49, N
3.P, 335- 382.
Lawalree A. La polhnisation de Lemna minor // Natur. Belg. 1961. Vol 42. P.
164 165.
J. a w alree A. La position systematique des Lemnaceae el leur classification // Bui.
Soc. Roy. Bot. Belg. 1946. Vol. 77, N 1. P. 27—38.
Lid .1. Norsk og Svensk Hora. Oslo, 1963.
Linden B. Aseptic germination of seeds of northern terrestrial orchids // Ann. Bot.
Soc. ZooL-Bot. I’enn. Vanamo. 1980. N 17. P. 1 - 23.
L.u о n d A. Effects of nitrogen and phosphorus upon the growth of some Lemnaceae //
VcrotTAieobot. Inst. ETH. 1980. H-70. P. 111 — 141.
L u o n d /\. The development of some Lemnaceae under different nutrient conditions //
Folia Gcobot. cl Phytotax. 1990. Vol. 25, N 3.
M a h c s h w a r i S., Kapil R. Morphological and embryological studies of the
Lemnaceae. II // Amer. J. Bot. 1963. Vol. 50. N 9.
Majovsk у J. et al. Index of chromosome numbers of Slovakian flora. P. 4 // Acta
Fac. rerum natur. Univers. Comentarr. Bratislava. Ser. Bot. Vol. 23. P. 1—23.
Marchant C. J., В r i g h t о n C. A cytological diversity and triploid frequency in a
complex population of Ranunculus ficaria 1.. // Ann. Bot. 1974. Vol. 38, N 154. P. 7-—1 5.
M a r s d c n-J one s E. M., T u r r i 11 W. B. Studies on Ranunculus ficaria // Joum,
Genet. 1952. Vol. 50. P..-7—15, 552—534.
Me ('lure J. N. The applicability of polyphenolic dute to systematic problems in the
Lemnaceae //Aquat. Bot. 1975. Vol. I, N 4. P. 395—405.
Me 11 r a i t h A.L., Robinson G.G., Shay J. M. A field study of completion and
interaction between Lemna minor and Lemna trisulca // Can. J. Bot. 1989. Vol. 67. N 10. P.
2904—2911.
Metcalfe CL R. An interpretation of the morphologies of the single cotyledon of
Ranunculus ficaria based on embryology and seedling anatomy // Ann. Bot. 1936. Vol. 50, N
197.P. 103—120.
Metcalfe C. R. The morphology and mode of development of the axillary tubercules
and root tubers of Ranunculus ficaria // Ann, Bot. 1938. Vol. 2, N 5. P. 145—157 ,
M e u s e I Ha Jager E., Weinert B. Vergleichende Chorologie der Zen-
traleuropaischen Hora, Jena. 1965.
Militzer M. Gesehutzeheimische Pflanzen. 1956. Vol. 65. P. 66.
M i у a t a H. Endogenous light on rhythm in respiration of long-day duckweeds.
Lemna gibba // Plant. Cel. Physiol. 1970. Vol. 11, N 2. P, 293—301.
M u d г a e к F. Ober die Assimilationstatigkeit und das Waehstum von Ficaria vema if
Planta. 1935. T. 23.
M u h 1 c m a n n H. Zur Kenntnis der Inhaltstoffe von Herba Alchemilla vulgaris L. //
Cheni. Zbl. 1939. Jg. 110. Vol. 2, N 7.
M u r b e с к S. Partenogenetische Embryobildung in der Gattung Alchemilla fl Lunds.
Univ. Arsskr. 1901. Bd. 36. Vol. 2. N 7.
Murbeck S. Ober Anomalien in Baue des Nucellus und Embryosaekes bei parteno-
genetischen Arten der Gattung Alchemilla fl Lunds. Univ. Arsskr. 1902. Bd. 38. Vol. 2, N 2.
Nakashima H. Effects of inhibitors of nucleic acid and protein synthesis on the
reappearance of «light interruption rhythm» in a long-day duckweed. Lemna gibba // P|&nt
Cell. Physiol. 1969. Vol. 10, N 2. P. 259—270.
N a i a n s о n-G rodzinska N. Sur la plasticity des instincts chez les larves
aqatiques de C'ataclyste // Bui. Acad. Polon. sci. 1932. Ser. B. Vol. 11, N 5—6.
Nilsson L. A. The pollination ecology of Listera ovata (Orchidaceae) // Nord. J. Bot.
1981. N 1 (4). P. 461—480.
Nilsson O. lohannesnycker vid lohannnistors И Sv. Bot. Tiddskr. 1988, Vol. 82. N
4. P. 257 - 265.
Nilsson S. Orchids of Northern Europe. Middesex. 1979.;
Oda Y. Lhe action of skeleton photopcriods of flowering in Lemna perpusilla // Plant
I'd- Physiol. 1969. Vol. 10. N 2. P. 399—409.
Ogura J. Anatomy and morphology of the subterranean organs in sofoe Orchidaceae
/ Joum. Fac. Sci. Univ. Tokyo. 1953. Vol. 3. N 6 (4). P. 135—157.
203
О i п о n e n E. The time table of vegetation spreading of lily of valley (Convallaria
majalis L.) and the wood sniallreed (Calamagroslis epigeios (L.) Roth.) in southern Finland //
Acta Forest. Fenn. 1969. Vol. 97. 35 p.
Oksancn J. Vegetation of forested inland dunes in Nonh Karelia, eastern Finland H
Aim. Bot. Fenn. 1983. Vol. 20. P. 281—295.
О о s t г о о m S. .1. Flora om van Nederland. Groningen, 1956.
Pcnzig <). PBanzcn-Tcralologie. Berlin, 1921. Bd. 2. P. 213—313. *
Perttula V. Untersuehungen uber die generative und vegetative Vcrmehrung der
Blutenpflanzen//Ann. Acad. Sei. Fenn. Ser. A. 1941. N 158.
P i e t e r s e Л. H. Physiological, morphological and anatomical aspects of gibbersity in
Lemna gibba //Aquat. Bot. 1975. Vol. 1. N 4. P. 333—344.
Pirson A., G о e I n e r E. Beobachlungen zur Entwieklungsphysiologic der Lemna
minor L. //Flora. Jena. 1953. N 140. P. 485—498.
Posner H. B. Inhibitory effect of carbohydrate on flowering in Lemna perpuxilla II
Plant Physiol. 1969. Vol. 44, N 4. P. 562—566. ’ '
Prochazka F.. V с I i s e к V. < Jrchidejc nasi parody. Praha, 1983.
Raunkiaer C. The life form of plants and statistical plant geography. (Ixford. 1943.
632 p.
Redd у К. R.. D e b n a к W. Growth characteristics oi aquatic macrophytes cul-
tured in nutrient-enriched water: Azolla, Duckweed and Sahinia II Econ. Bot. 1985. Vol. 39. N
2. P. 200—208. ‘
Reimers H. Orchis mffitaris L. mit singularis Heidenrcich in Brandenburg // Wilde-
nowia. 1958. Vol. 2,N 1. P. 1—6.
Rcjmankova E. Biology of duckweeds in Pannonian fishponds // Svmp. Biol.
Hung. 1975. N 15. P. 125—131.
R e j m a n к о v a E. Comparison of Lemna gibba and Lemna minor from the produc-
tion ecological viewpoint // Aquat. Bot. 1975. Vol. I, N 4. P. 432—427.
Rej manko v a E.. Blackwell M., Culley D. D. Dynamics of fungal infec-
tion in duckweeds (Lemnaceae] II Vcroff. Geobot. Inst. ETH. 1986. H. 87. S. 178 189.
Ridley H. H. The dispersal of plants throughout the world. England. 1930.
R о n s e A. In vitro propagation of orchids and nature conservation: possibilities and
limitations // Mem. Soe. Roy. Bot. Belg. 1989. N 11. P. 107—114.
R о t h m a 1 e r W. Exkursionsflora fur die Gebiete dor DDR und der BRD. Berlin.
1972.613 s.
R о t h m a I e r W. Systematische Vorarbeiten zu einer Monographic der Gattung
Alchemilla (L.) Scop. V. IX. X. // Feddes Repert. 1937. Bd. 42- P. 111 — 119: Bd. 50. P. 78.80:
Bd. 66. P. 194—234.
Rydberg P. A. Rosaceae Juss. // The North American flora. 1963. Vol. 22. Pt. 3/5. P.
239--560.
S a h a i R., Roy P. S. Report of the natural enemies of Spiradela palyrrhizu (!..}
Schleid. //Gurr. Sci. (India). 1977. Vol. 46, N 23.
Sculthorpe C.l). The biology of aquatic vascular plants. London. 1967. 61 Op
Sharman В. C. The development of the sinker of Ore his mascula II Linn. Soc. J. Hot.
1939. Vol. 59. N 342. P. 145-158.
S i p к e s European orchids from seed II Quart. Bui. Aloine Garden Soc. 1979. Vol.
47. P. 27—-30.
Soo R., J a v о г к a S. A Magyar Novenvvilag Kczikonyve // Akad. Kiado. Buda-
pest, 1957. .
S t a r 1 i n g с r I •. Die flache form von Lemna gihha II Verh. Berlin. Bot. Ver. 1985. N
4. P. 67-77.
S t с г к e R. Rccherches anatomique sur I'embryon et les planlules dans la Camille
Ranunculacees // Arch. inst. Bot. I hiiv. Liege.. 1900. Vol. 2. P. I 120.
St.rasb urgcr IL Die Apogamic dor Eualchcmillcn and allgemcinc Gcsehictpunkte.
der sich aus ihr ergeben II Jahrb. Wiss. Bot. 1904. Bd. 41.
S t г о e s s e 1 A., Arditti J. < Irchids phytoale.xins // ( )rchid Biology: Reviews and
Perspectives. New-York, 1984. P. 151 157.
Summerhayes V.S. Wild orchids of Britain. London. Ed. 1. 1951. Ed. 2. 1981.
204
'Г i с h о m i г о v V. N. •'Alchemilla ventiana — species nova e Rossia Media // Feddes
Report. 1985. Bd. 96, H. 1—2. P.15—16.
'J’ rcub'M. Notes surГ embryogenie de quelques ()rchidees. Amsterdam. 1879.
T roll W. Praktische EihQihrung in die Pflanzenmorphologie. Jena, 1957. 420 p.
Troll W. Die Inflorcszenzen. Jena, 1964. Bd. 1. 1969. Bd. 2.
T u к a L., Po pcsc u H. Determination of tannins in the plants of Alchemilla mollis
(Buser) Rothm. and A. vulgaris L. //Clujul. med. 1979. Vol. 52, N 1. P. 78—83; Chem. Abstr.
1979. Vol. 79. N 198835.
Urbansk a -W о rvt kic wi<?z A. Cytological variation within Lenina L. // Aquat.
Bol. 1975. Vol. LN 4. P. 377 -394.
V^llman F. Flora von Bayern. Stuttgart, 1914.
Vftth W. Bestaubungsbiologische Beobachtungen an Orchis militaris L. Il Orehidcc.
1987. Vol. 38, N 2. P. 77—84,
Walters S. M.. Pawl о w s к IB. Alchemilla L. // Flora Europaca. Cambridge.
1968. Vol. 2. P. 48—64.
W a ngcrmann E. Studies in the morphogenesis of leaves. VIII. A note on the effects
of length of dav and of removing daughter fronds on aging of Lenina minor // New Phytol.
1952. Vol. 51. P. 355 - 358.
Wangcr m a n n E., Ashy E. Morphogenesis in Lenina minor // Proceed. Linn. Soc.
London. 1950. Bd. 162. P. 10—13.
W anger m a n n E.,‘ Lacey H. Studies in the morphogenesis of leaves. IX. Experi-
ments on Lenina minor with adenine, tridobenzoic acid and ultra-violet radiation // New Phy-
tol. 1953. Vol. 52. P. 298 -311.
W e g с n e г К. -A. ('hromosornzahlen aus Wurzelspitzen von Alcheniilla-\rlcn def
Sektionen Pentaphyllon Rothm. und Brevicaulon Rothm. // Biol. Zentralblatt. 1967. H. 6. s.
771-792-
Weinar ck H. Skancs Flora. BokfQrtlaget ('orona. Lund, 1.963.
Wettsicin L. P.. Lange A. 11.. Pictcrse A. H. The influence of modification
of the nutrient medium on the dimensions of Lenina gibba // Acta bot. necrl. 1984. Vol. 33, N 4.
P. 559—560.
White H., Tcmoleman W. The interaction of factors in the growth of Lenina. X.
The interaction of nitrogen and light intensity in relation to respiration // Ann. Bot. 1937. Vol.
l.P. 191—204.
W i t h n с r C. L, The orchids — scientific studies. New-York. 1959. 1974.
W i l s t u m A. Seed viability in Lenina gibba L. // Isr. J. Bol. 1987- Vol. 35. N 3—4.
Zi egensp cck H. Orchidaceae II l.ebensgeschichte der Bliitenpflanzen Mit-
tek-uropas. 1936. Stuttgart Bd. 1.1X.L
Z i m m e r m a n n M. A.^Einfluss von Calcium und Magnesium aufdas Wachslum von
Mitteleuropaischcn Lemnaceeh-.Artcn // Ber. (icob- Inst. Р.ТП 1981.11. 98. P. 120 160.
Zu bercr D. A. Microbial colonization of some duckweeds (Leiiinaceae) examination
bv scanning and transmission electron and light microscopy II Aquat. Bot. 1984. Vol. IS. N 3.
!>'. 274- 285.
СОДЕРЖАНИЕ
Предисловие ......................................................................................... 3
Вейник наземный. Н.Г Уланова..........................................................................4 .
Семейство Рясковые. П.К5. Жмылев. И.С. Кривохарченко. А.В. Щербаков .................................20
Ряска маленькая...................................................................................27
Ряска горбатая ...................................................................................36
Ряска трехдольная ............................................................................... 42
Многокоренник обыкновенный .......................................................................47
Род Тайник. Т.Н. Варлыгина...........................................................................52
Тайник сердцелистный..............................................................................52
Тайник яйцевидный ................................................................................57
Ятрышник шлемоносный. М.Г. Вахрамеева. М.Н. Загулъский, Т.М. Быченко.................................64
Чистяк весенний. Р.П. Барыкина......................................................................... 15
Род Манжетка. В.Н. Тихомиров. А-А. Нотов. Л.В. Петухова. К.П. Глазунова..............................83
Земляника зеленая. Н.М. Григорьева ................................................................................ 119
Род Герань. Н.С. Сугоркина ............................................................................. .......... ... 134
Герань луговая...................................................................................135
Герань лесная..............................................................;.................... 145
Герань болотная .................................................................................151
Герань кровяно-красная...........................................................................157
Бересклет бородавчатый. Т.В. Багдасарова...................................................................... 164
Багульник болотный. И.Л. Крылова, Л.И. Прокошева....................................................174
Литература ........................................................................................ 187
206
CONTENTS
Introduction ....................................................................................... 3
Calamagroslis epigeios (L.) Roth. N.O'. I Hanova.....................................................4
Family Lemnaceae. P. Yu. Zhmylev. I.S. Krivokharchenko. A. V. Shcherbakov ..........................20
Lemna minor 1...................................................................................27
Lemna gibba L...................................................................................36
Lemna trisulca L................................................................................42
Spirodela polyrrhiza (L.) Schleid.............................................................. 47
Genus Listera. 7’./. I'arlygina.....................................................................52
Listera cordata (L.) R. Br......................................................................52
I .istera ovata (L.) R. Br. ....................................................................57
Orchis militaris L. hi.G. Vakhrameeva. M.N. Zagul'sky, T.M. Bychcnka ...............................64
Ficaria vcma lluds. K.P. Barykina .............................................................. /*»
Genus Alchemilla. V.N. Tikhomirov, A.A. Notov, L. V. Petukhova. K.P. (ilazunavu ...................HI
Fragaria viridis Duch. N.M. Grigor'eva ............................................................ 1Ю
Genus Geranium. N.S. Sugorkinu ................................................................... 134
Geranium pratense L.......................................................................... 135
Geranium sylvaticum L......................................................................... 145
Geranium palustre L............................................................................151
Geranium sanguineum L............................................................... 157
Luonymus verrucosus Scop. 7’. К Bagdasarova ................................................................... 164
Ledum palustre L. l.L. Krylova. L.I. Prokosheva .................................................. 174
Literature cited............................................................................ ............................. 187
2.07
Научное издание
БИОЛОГИЧЕСКАЯ ФЛОРА
МОСКОВСКОЙ ОБЛАСТИ
вып. 10
Зав. редакциями И.И. Щехура,
к.б.н. А.В. Щербаков
Редактор О.В. Апентьева
Подписано в печать 27.03.1992 г. ЛР № 040414. Формат 60x90 /)е.
Бумага тип. № 1. Ротапринт. Уч.-изд. л. 13,0. Тираж 500 зкз. 'Заказ
№ 150. Заказное. Изд. № 5901.
Отпечатано с готового оригинал-макета в типографии ПИИСУ.