/




































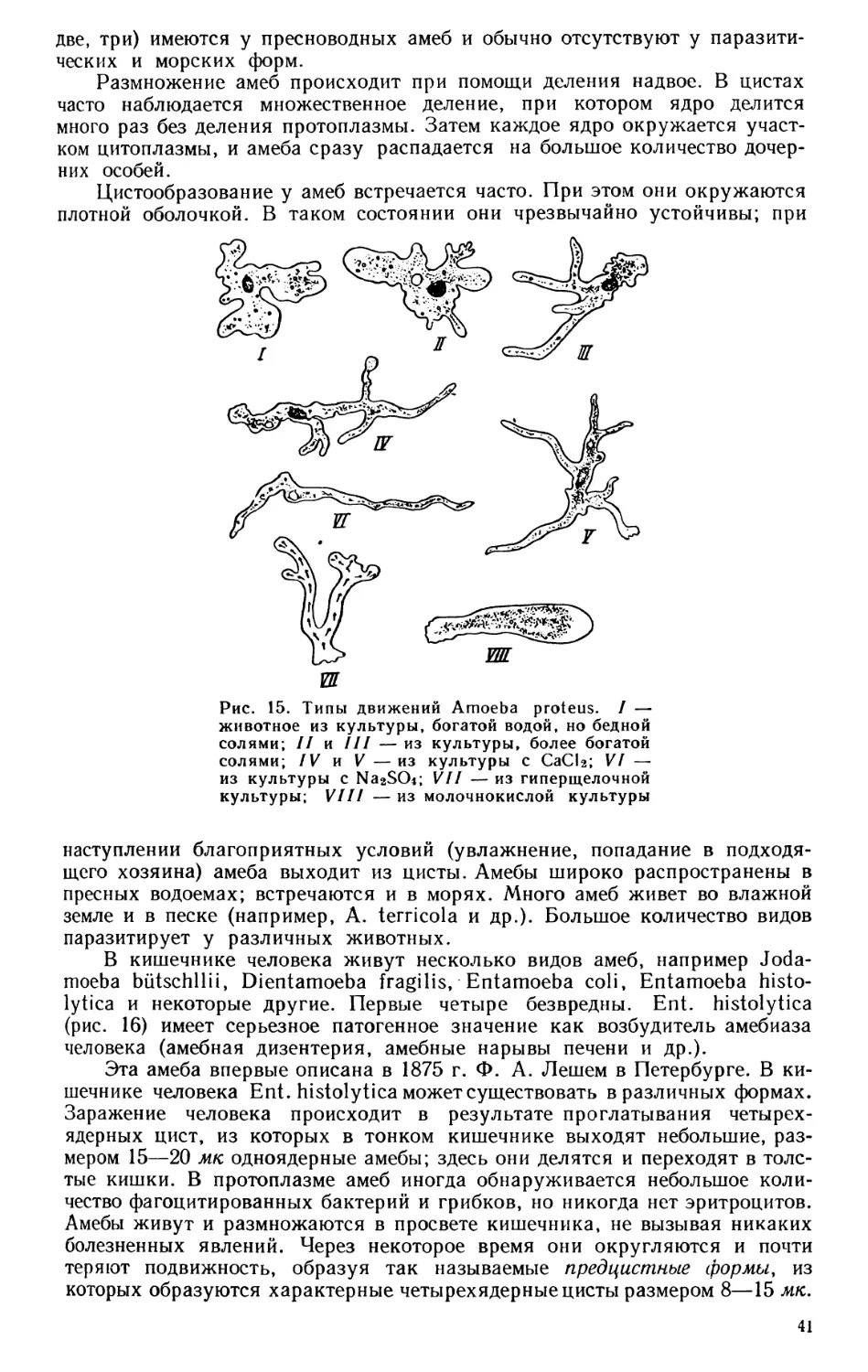
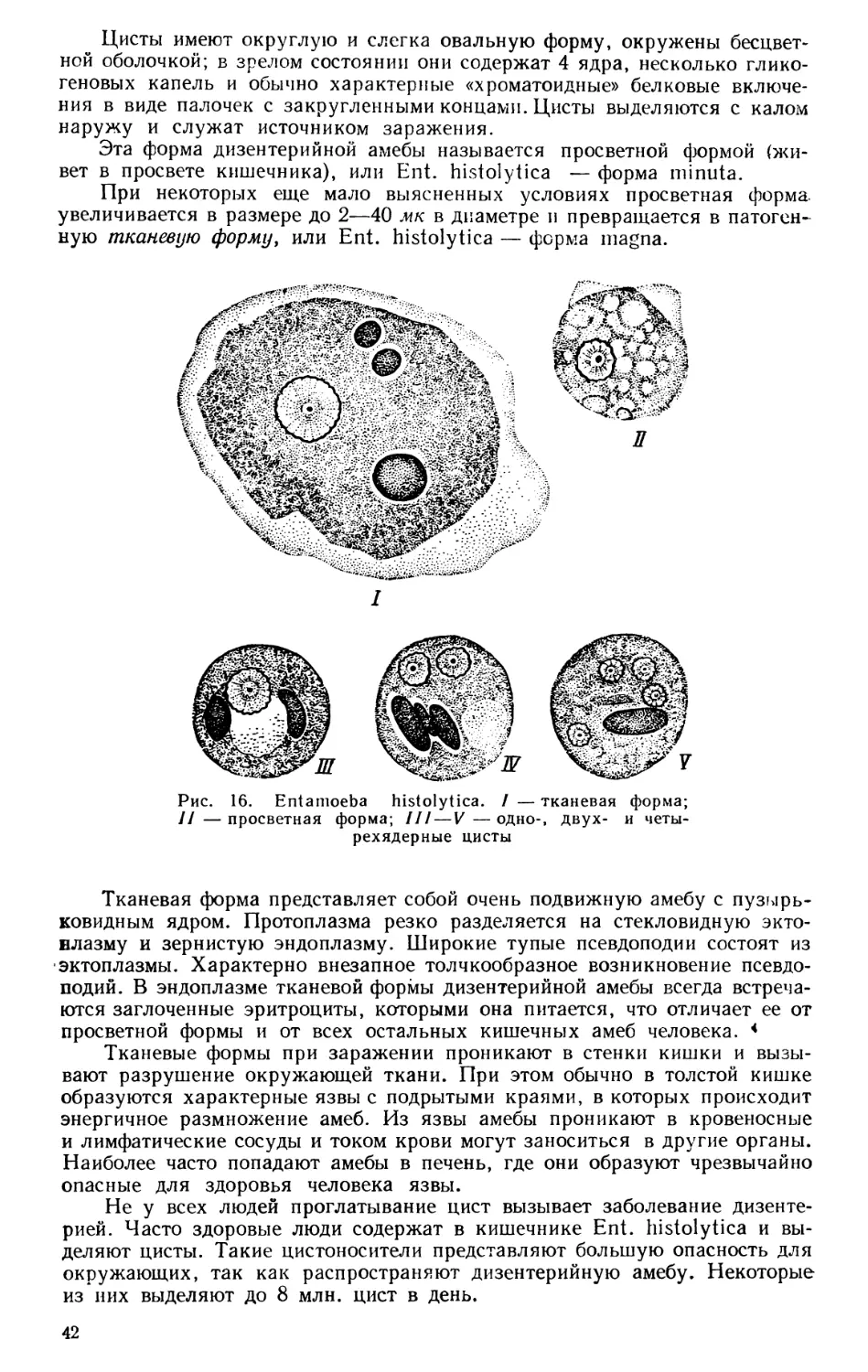


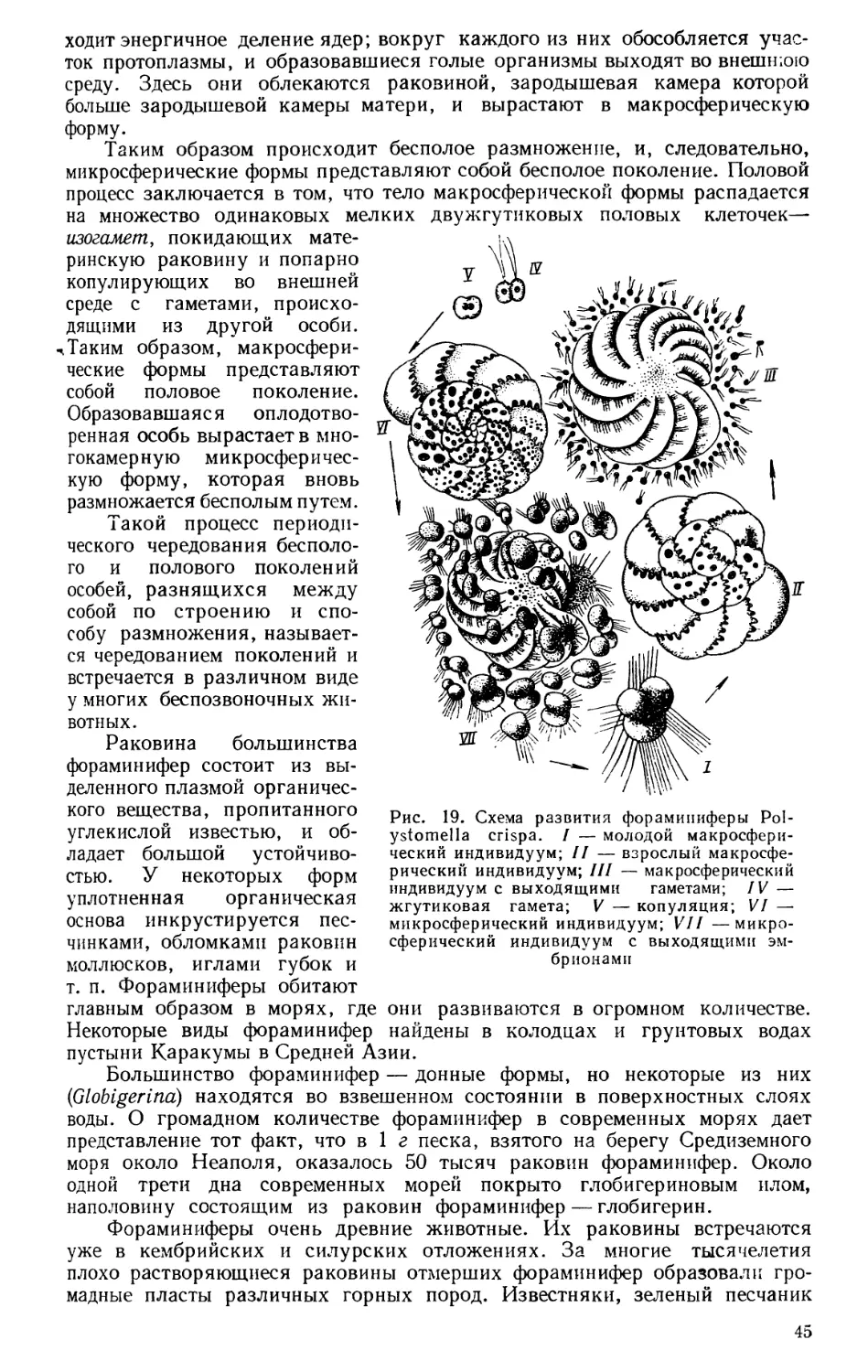





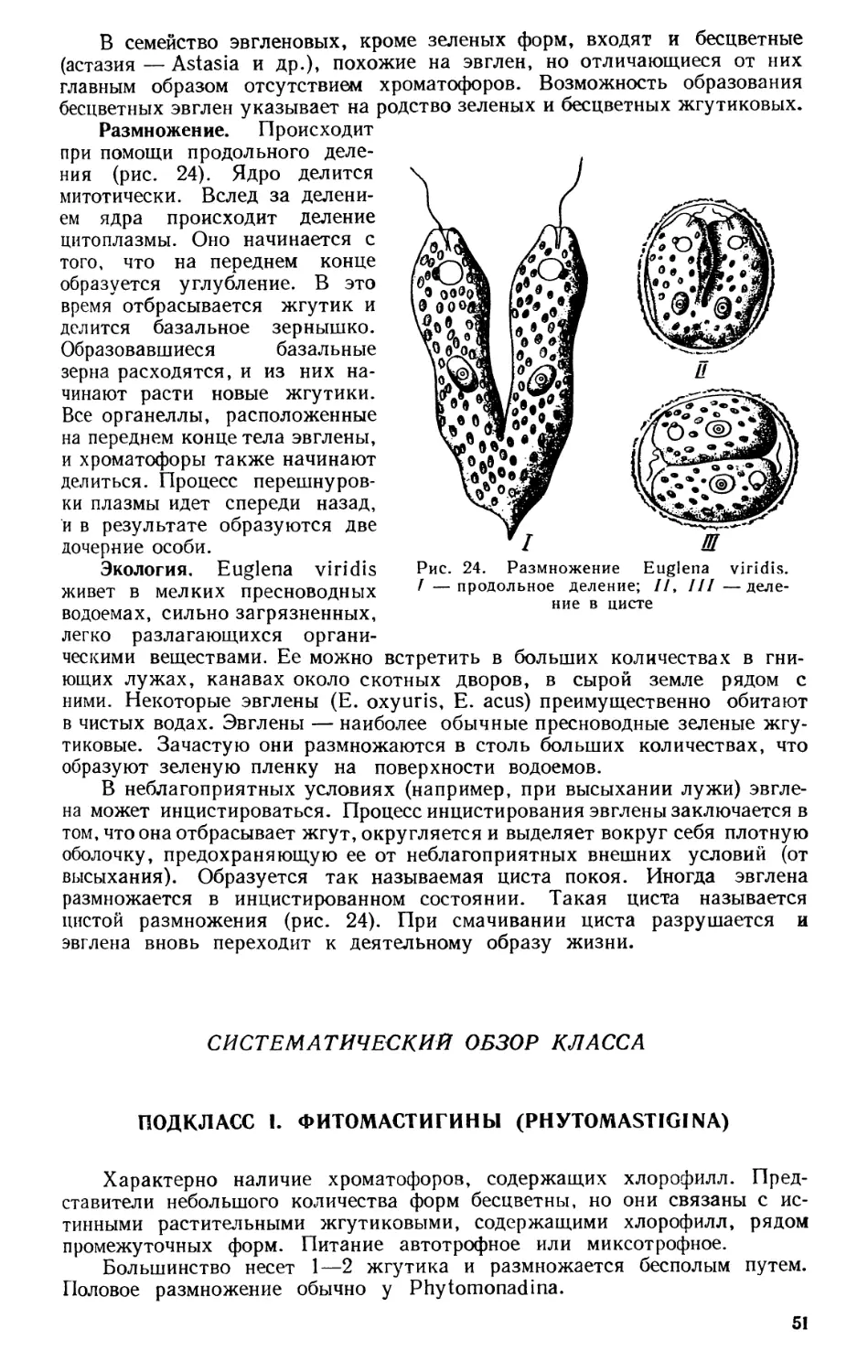

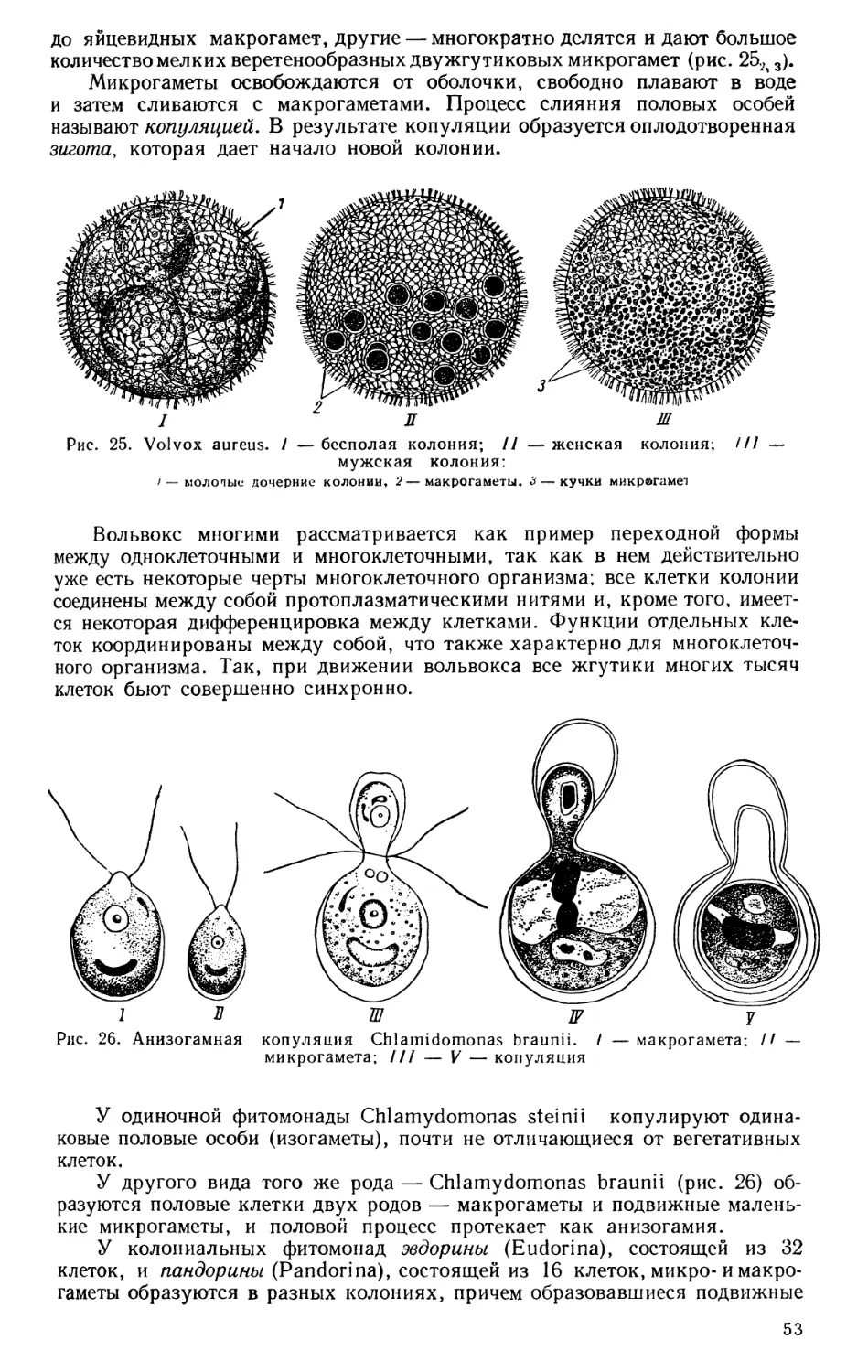


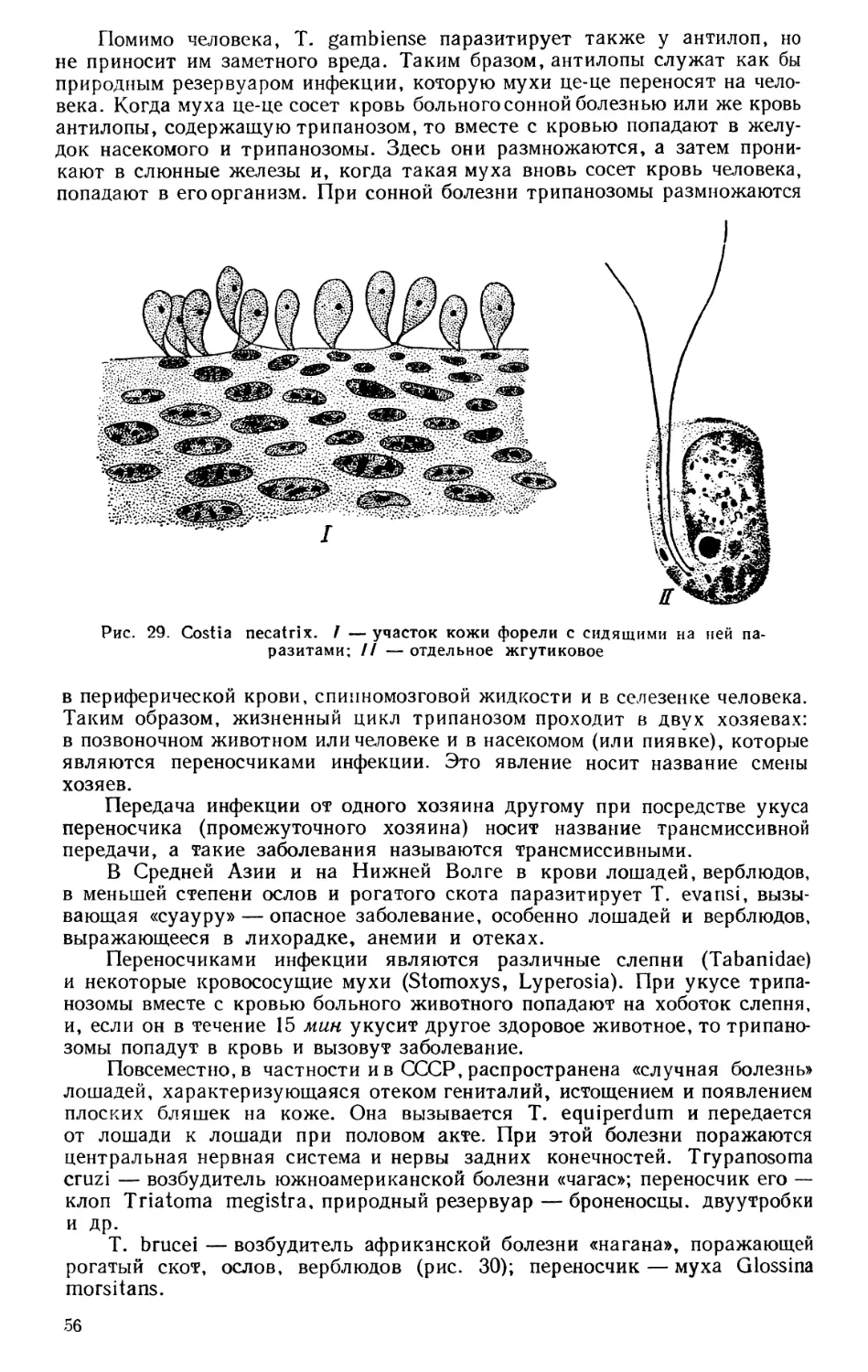
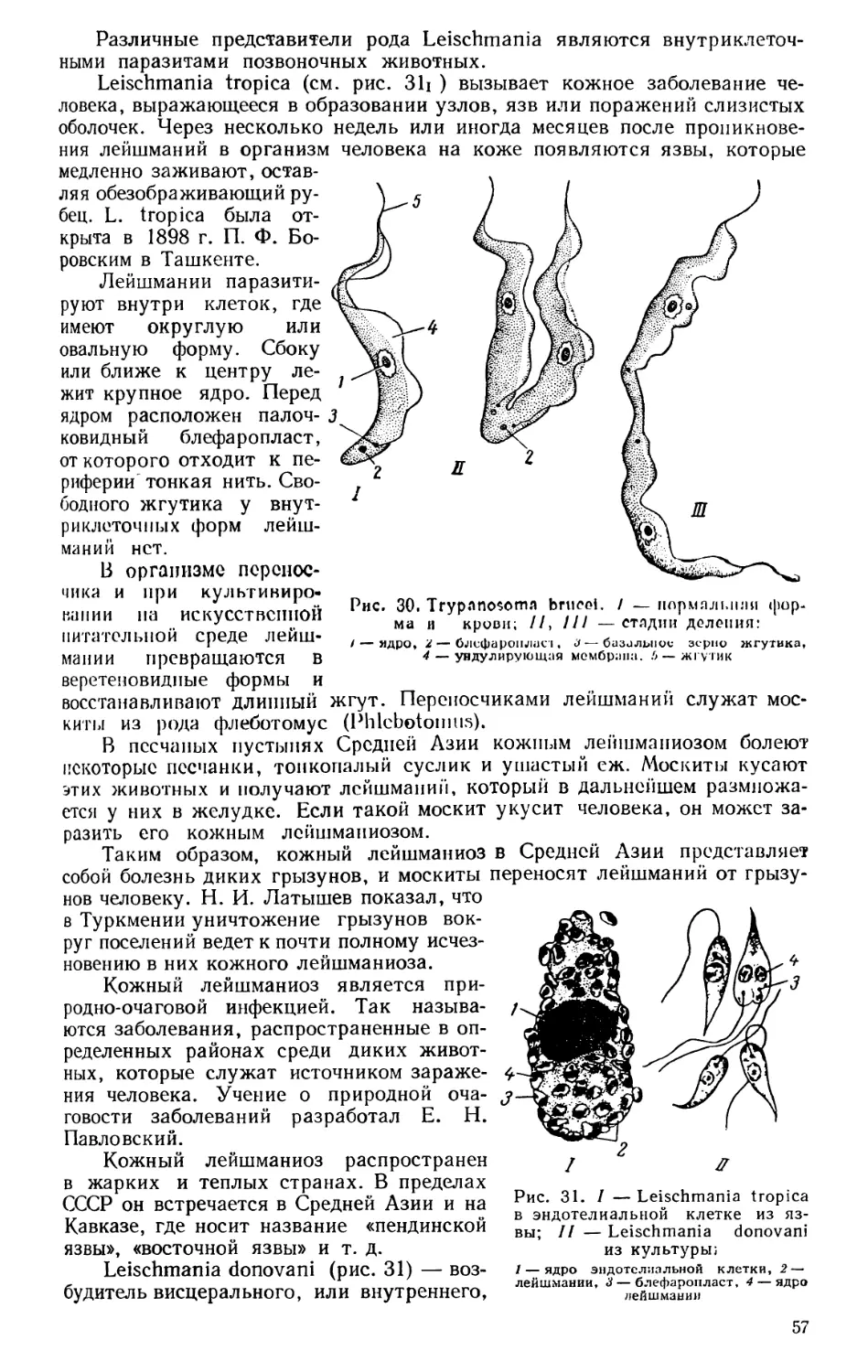
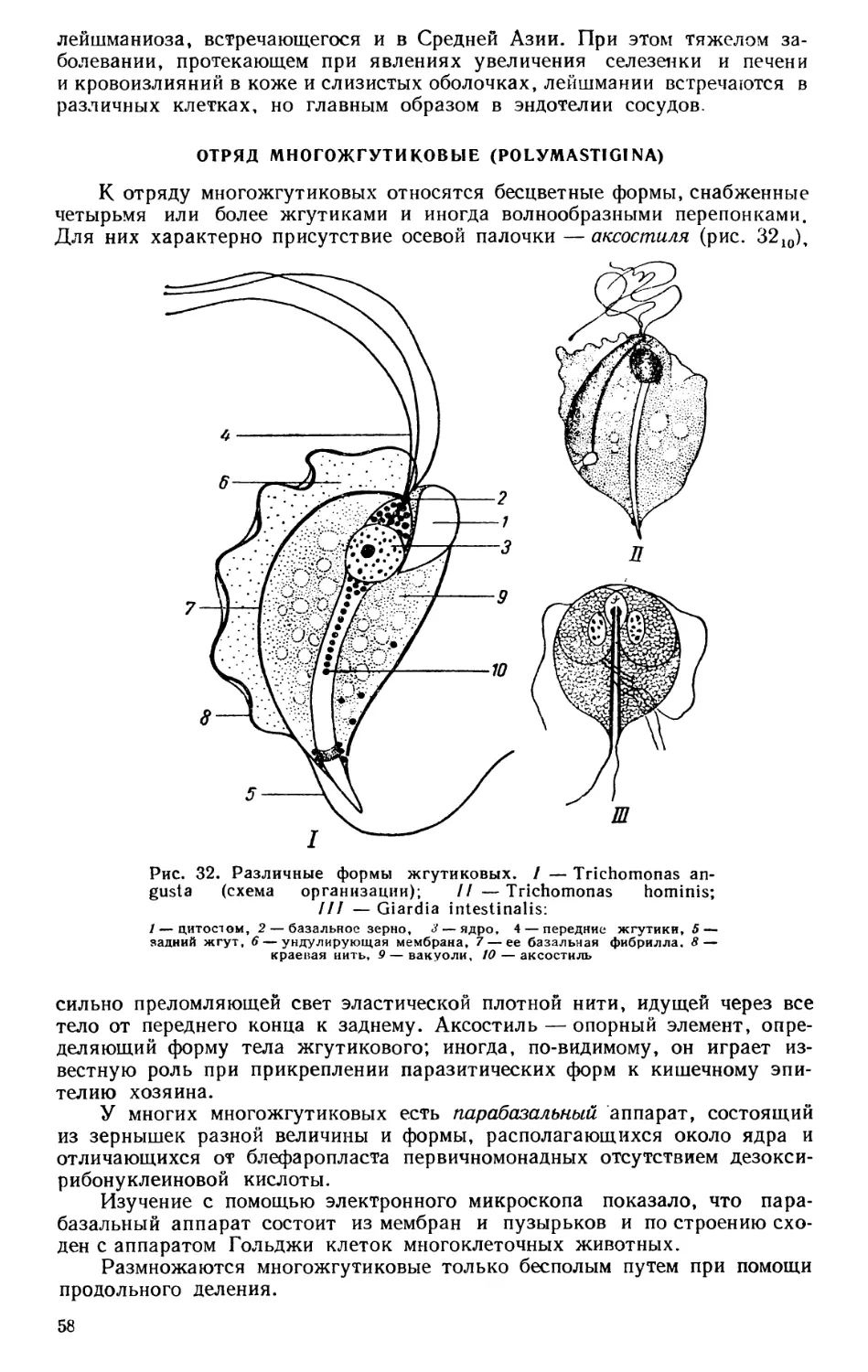
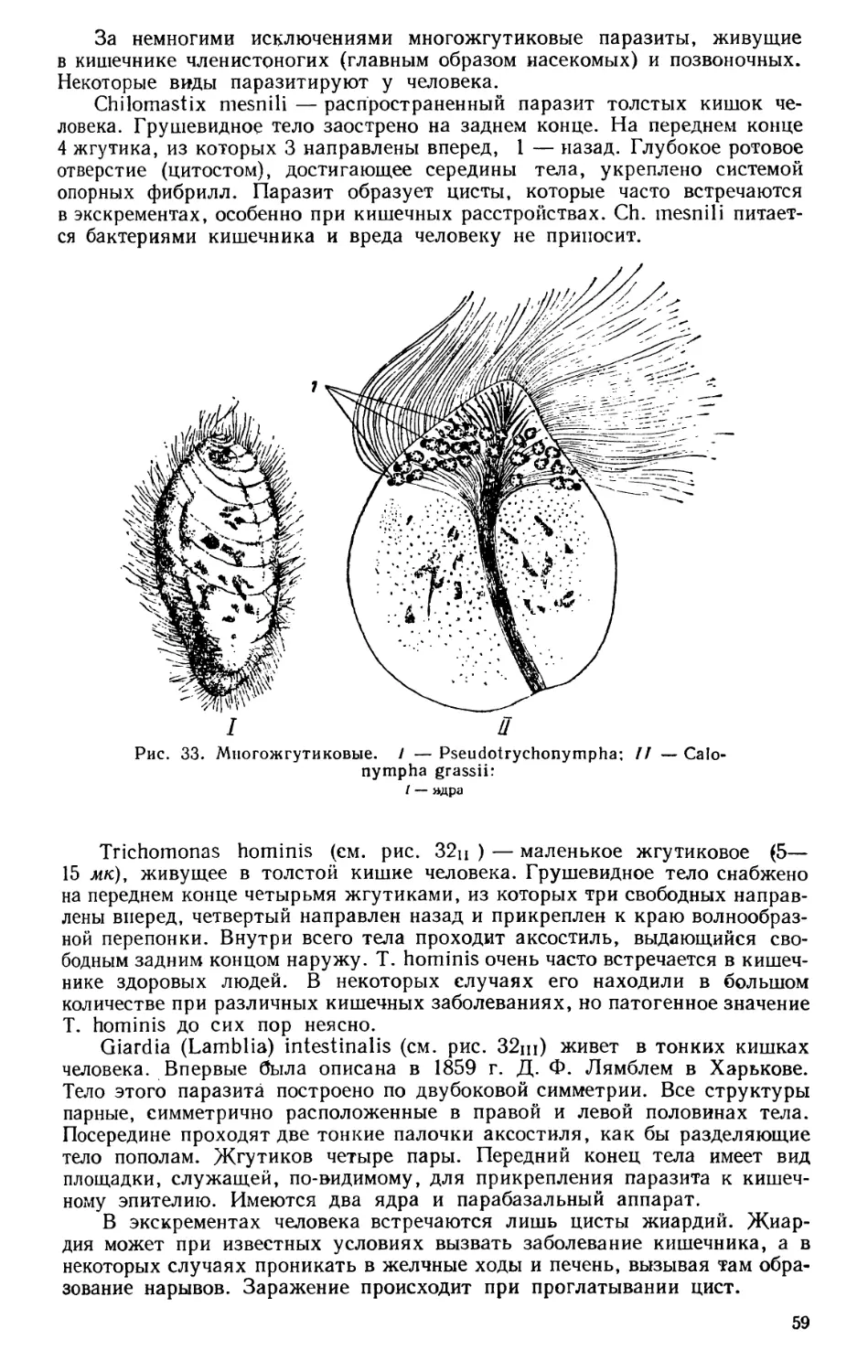


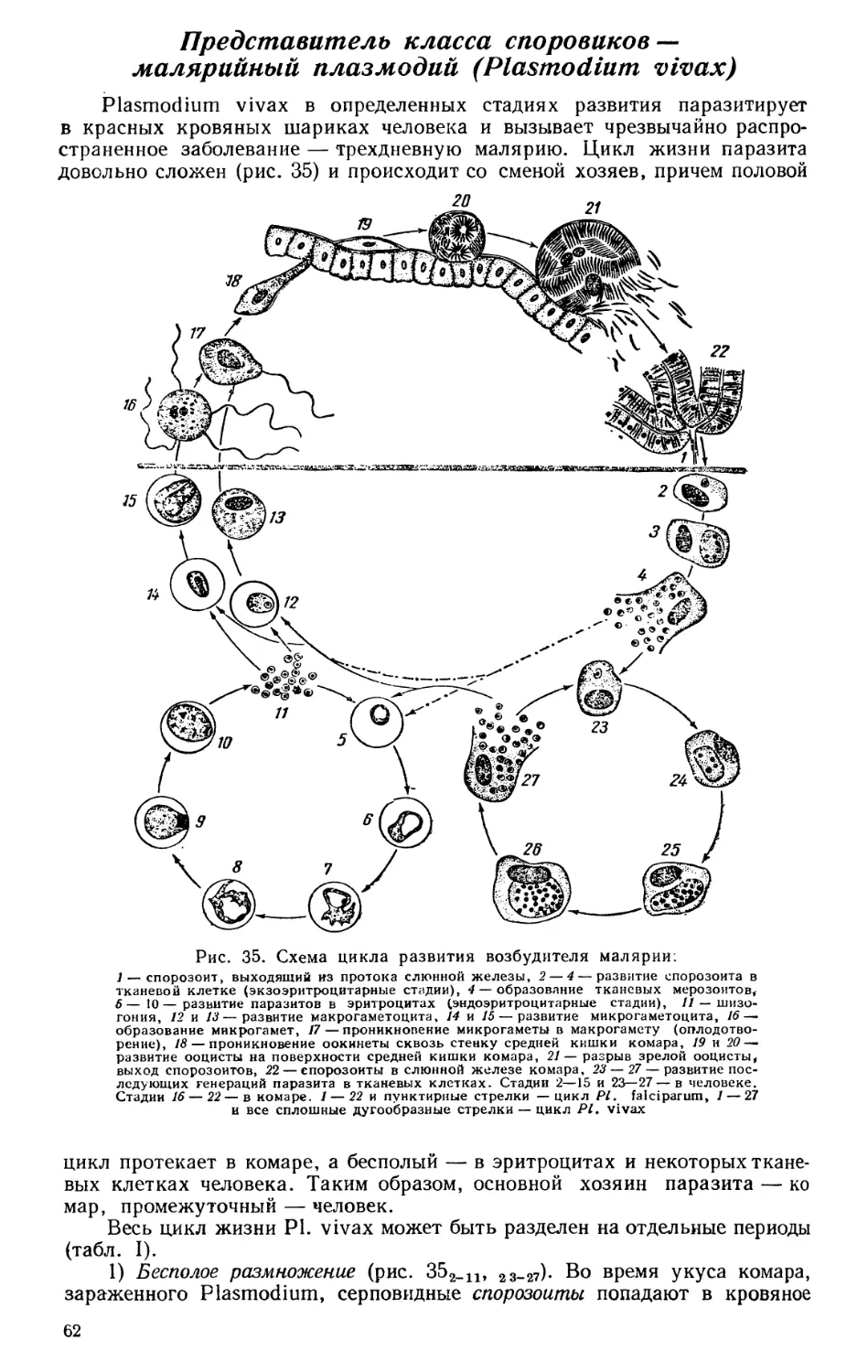


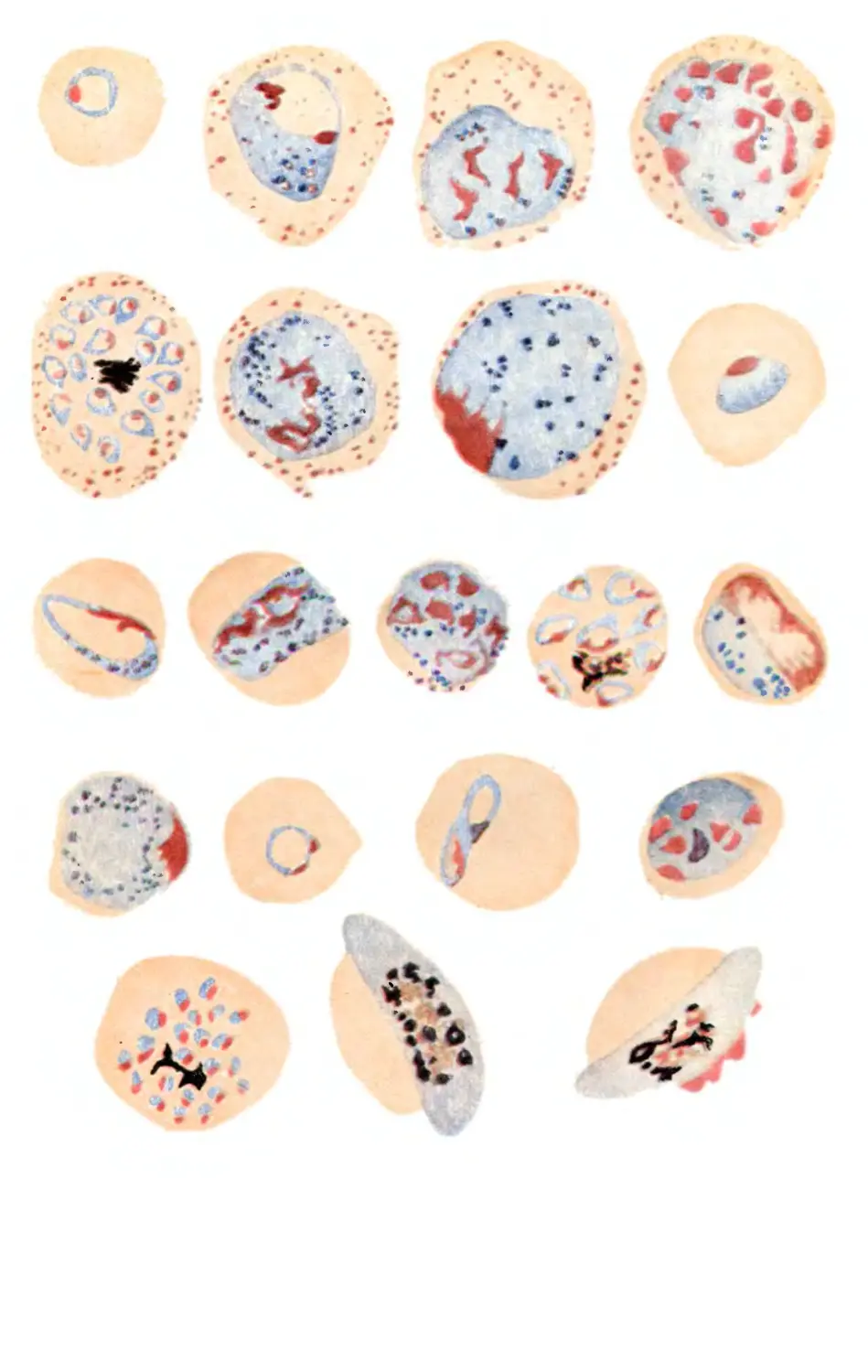



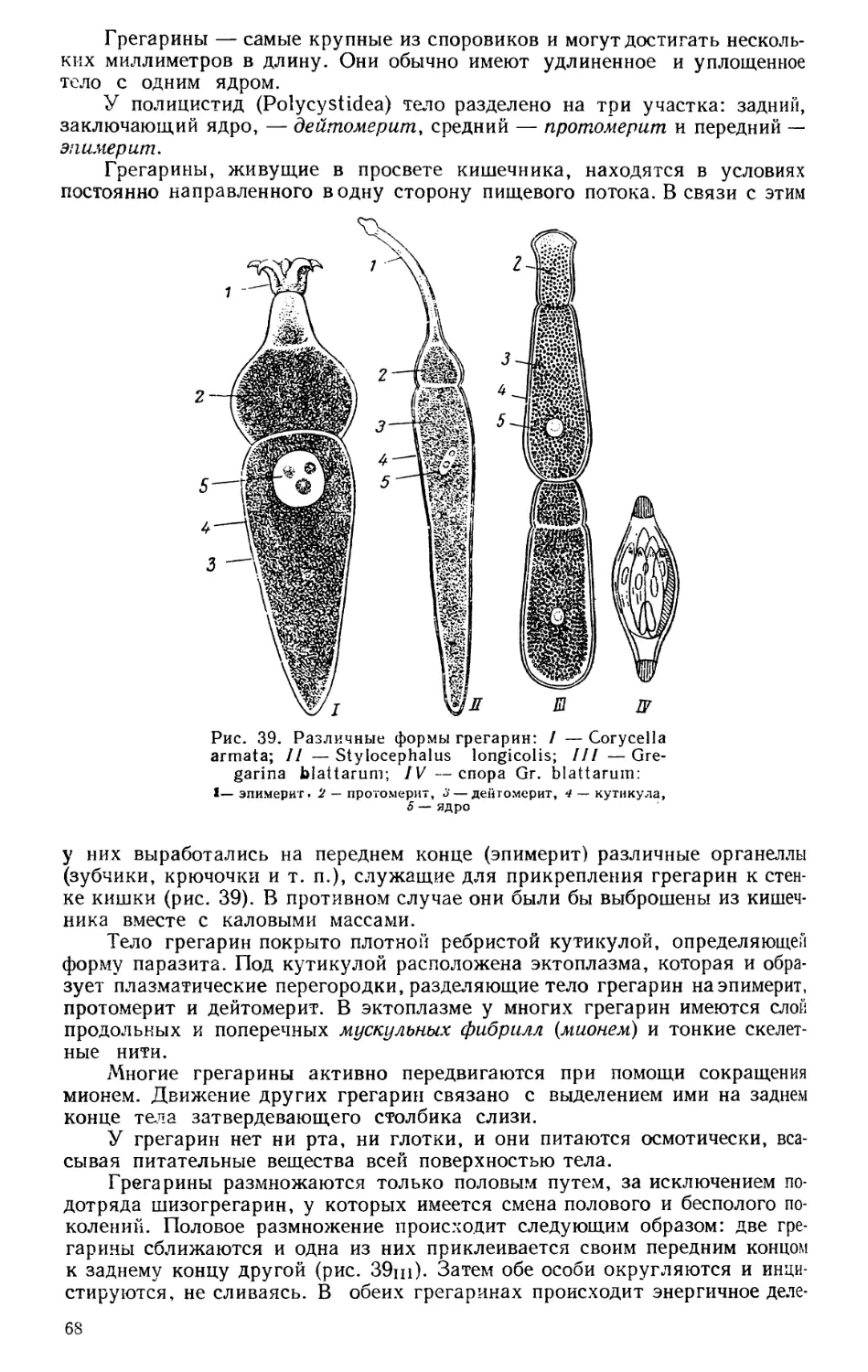
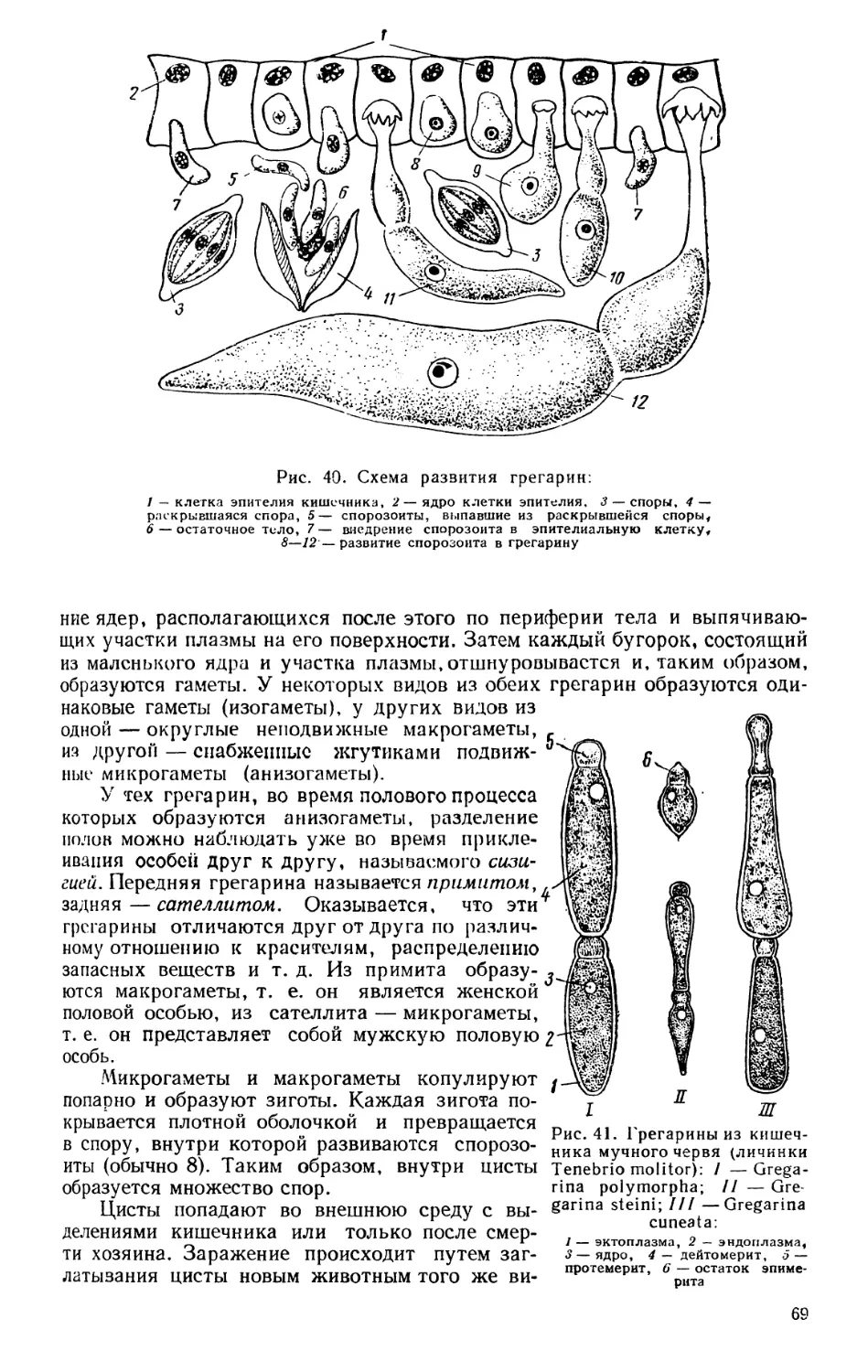
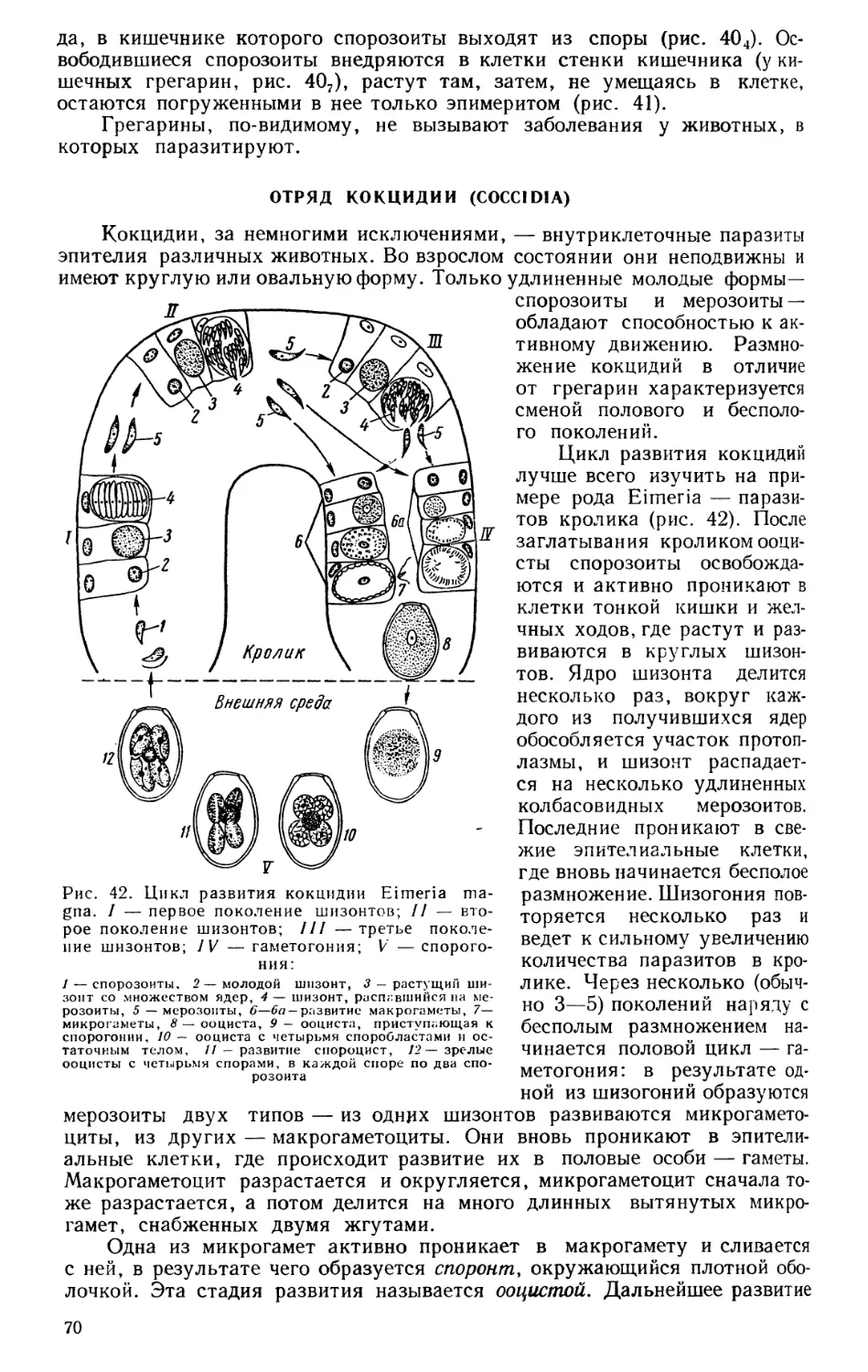
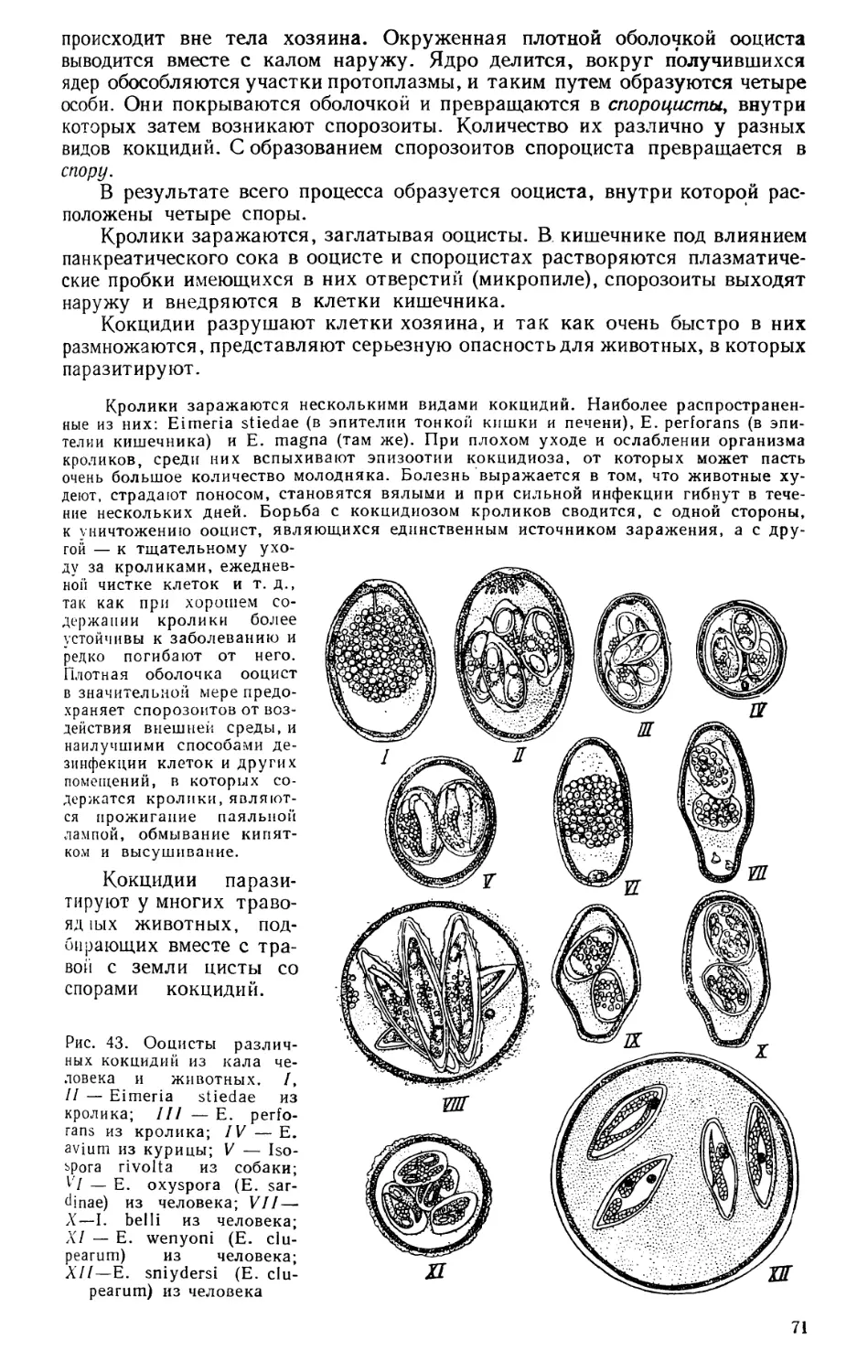


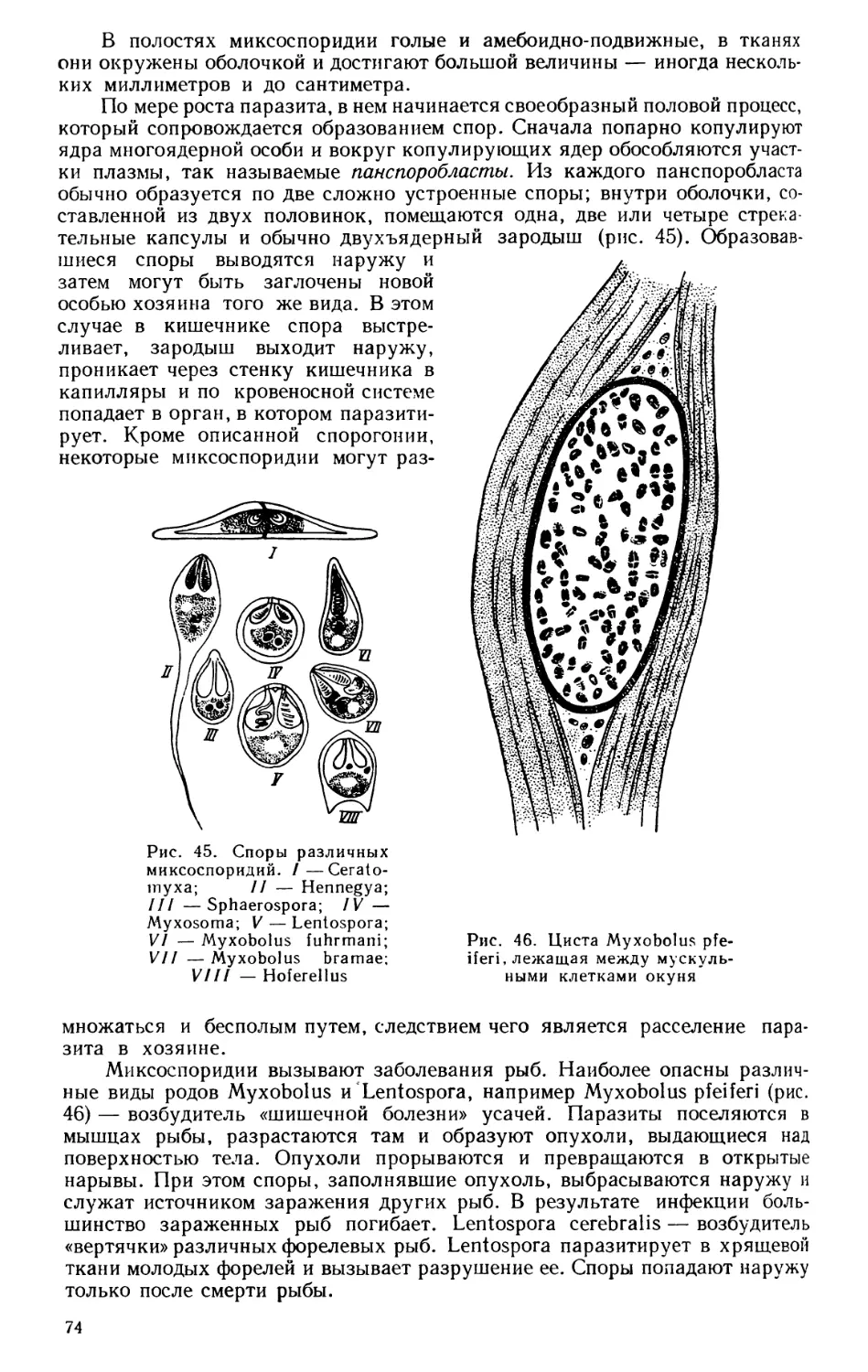



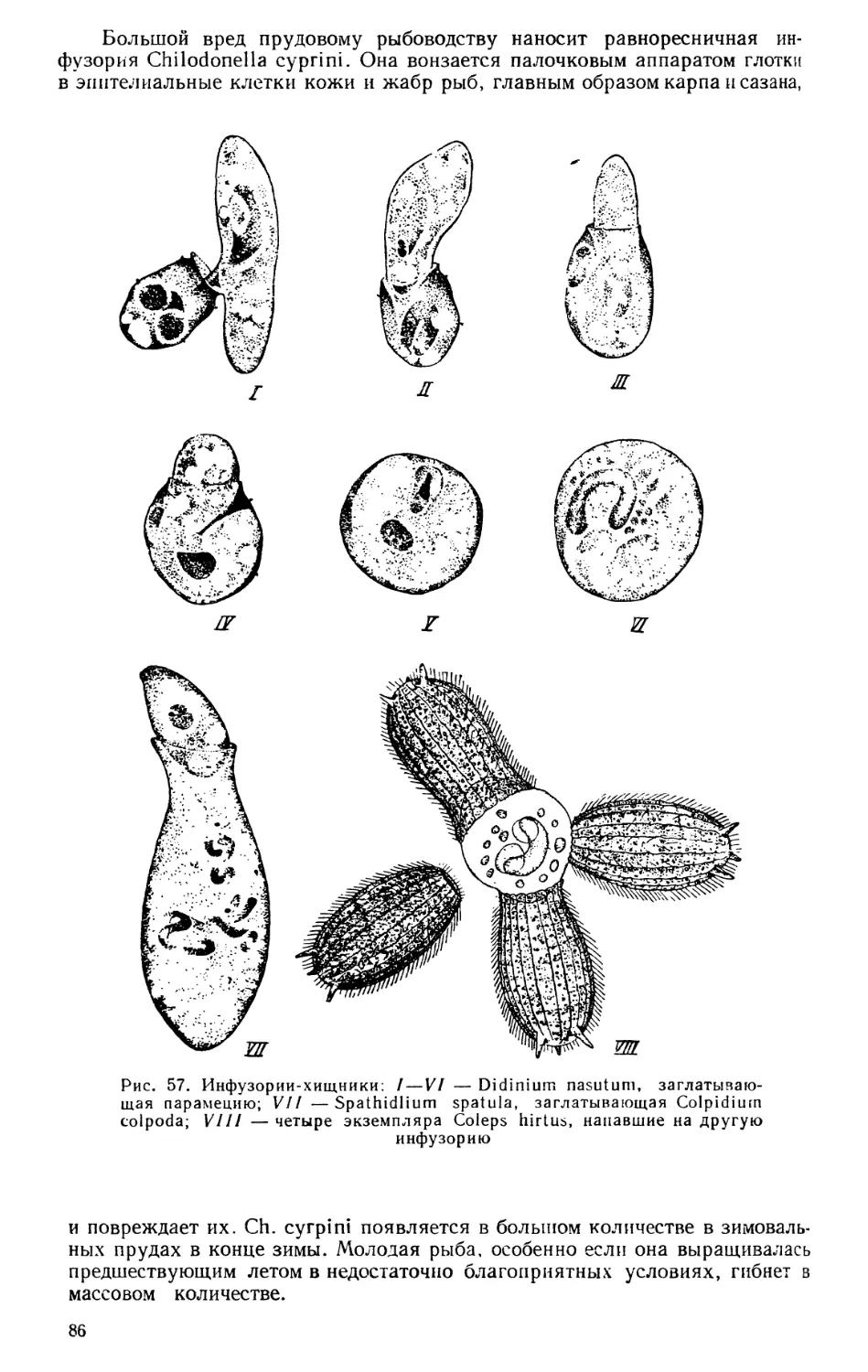
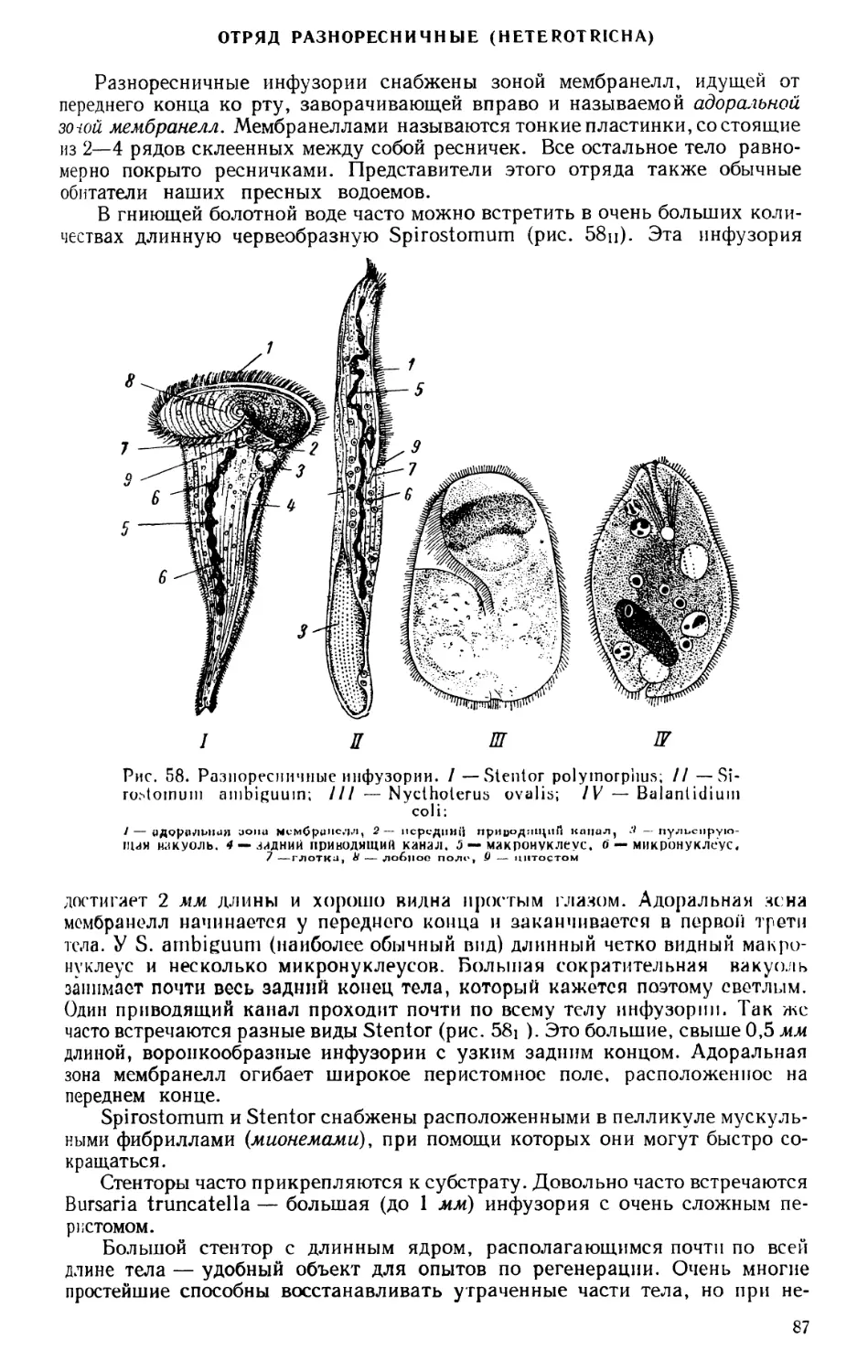

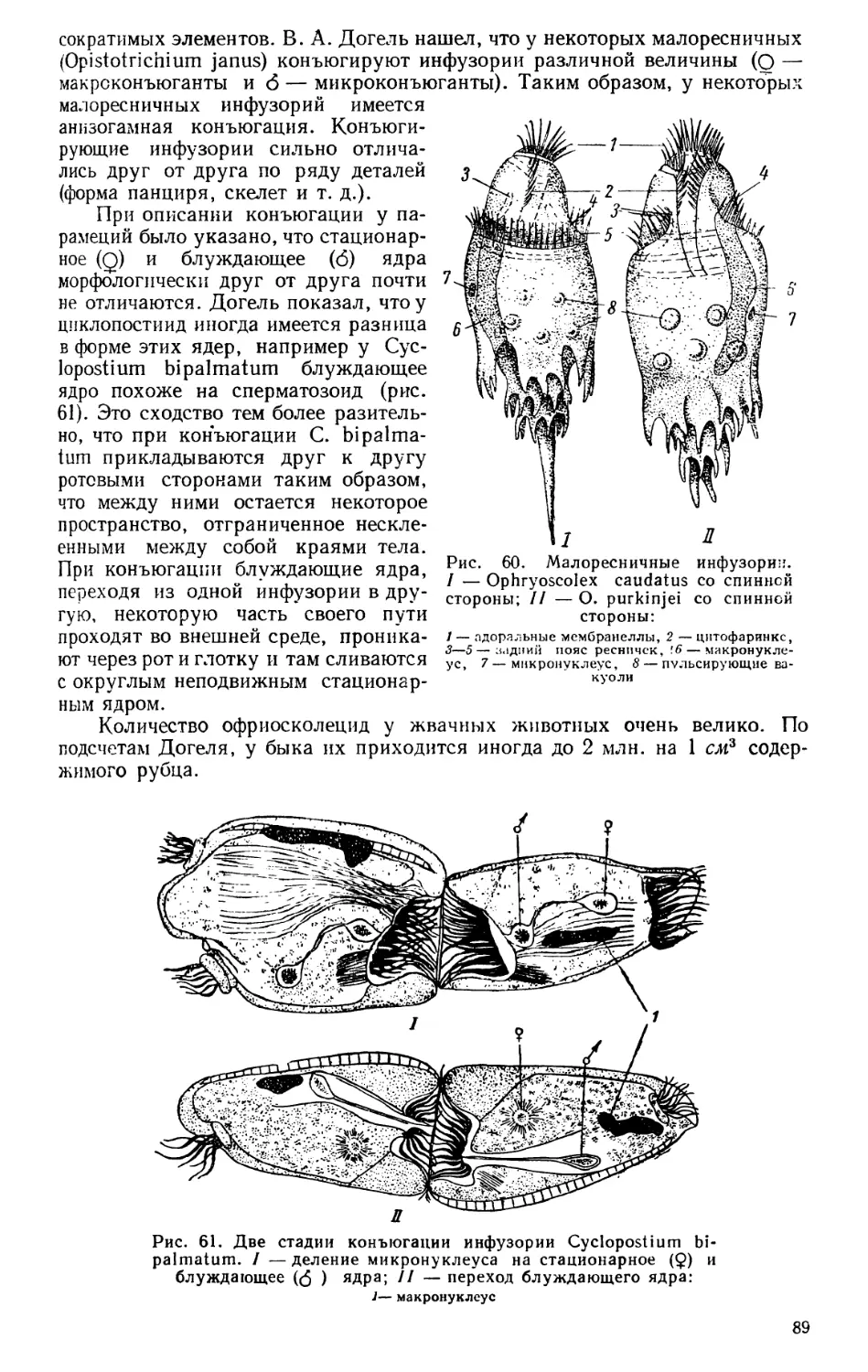
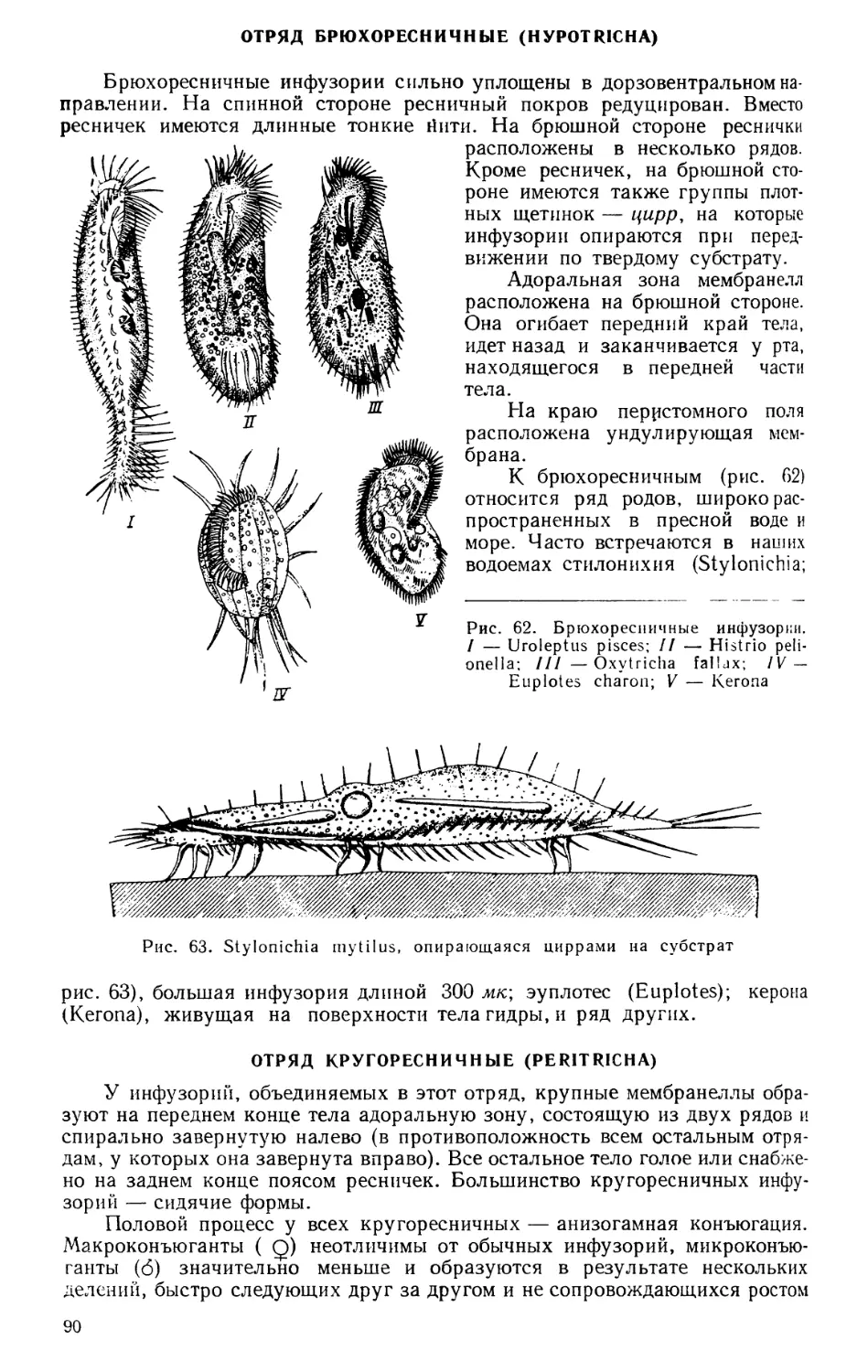

















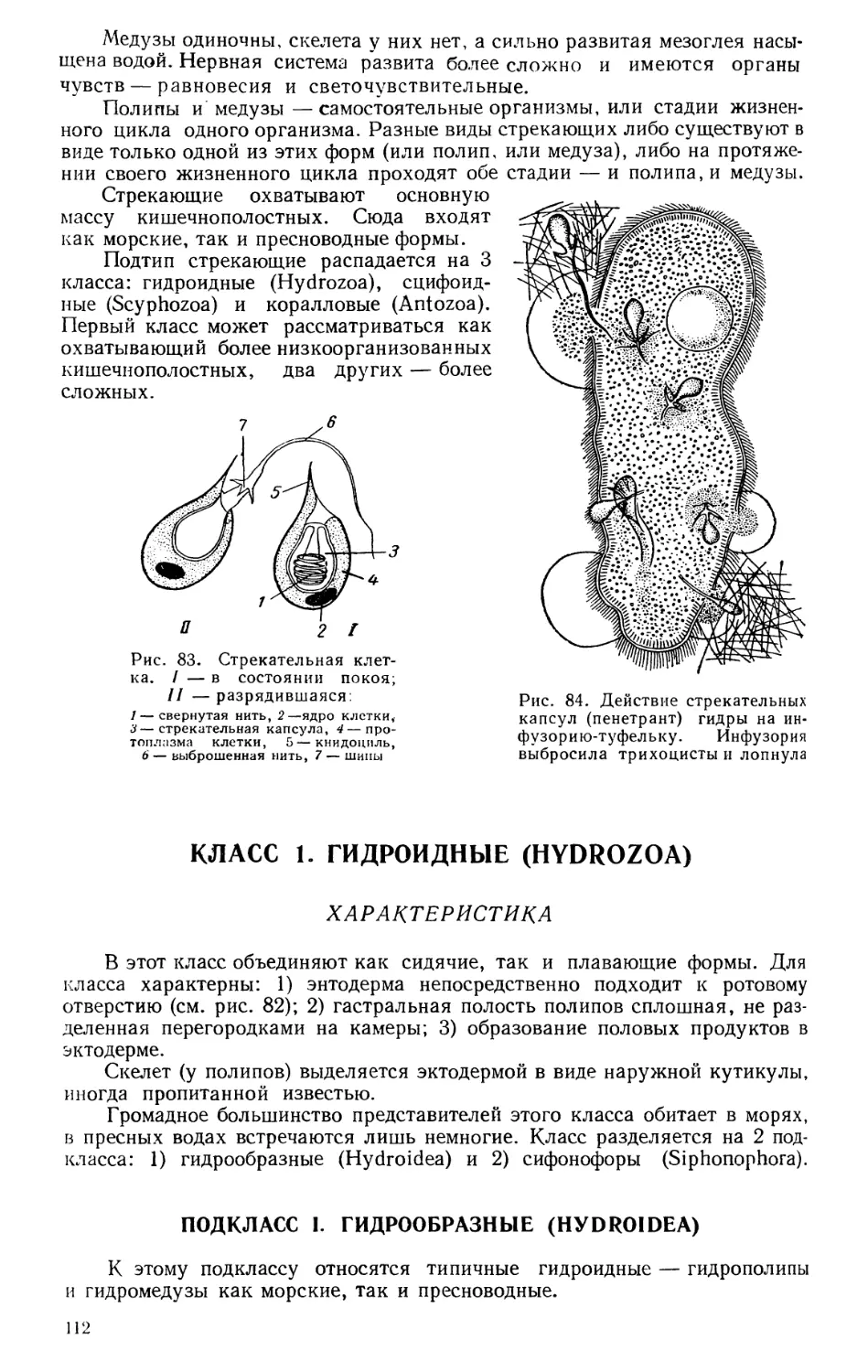
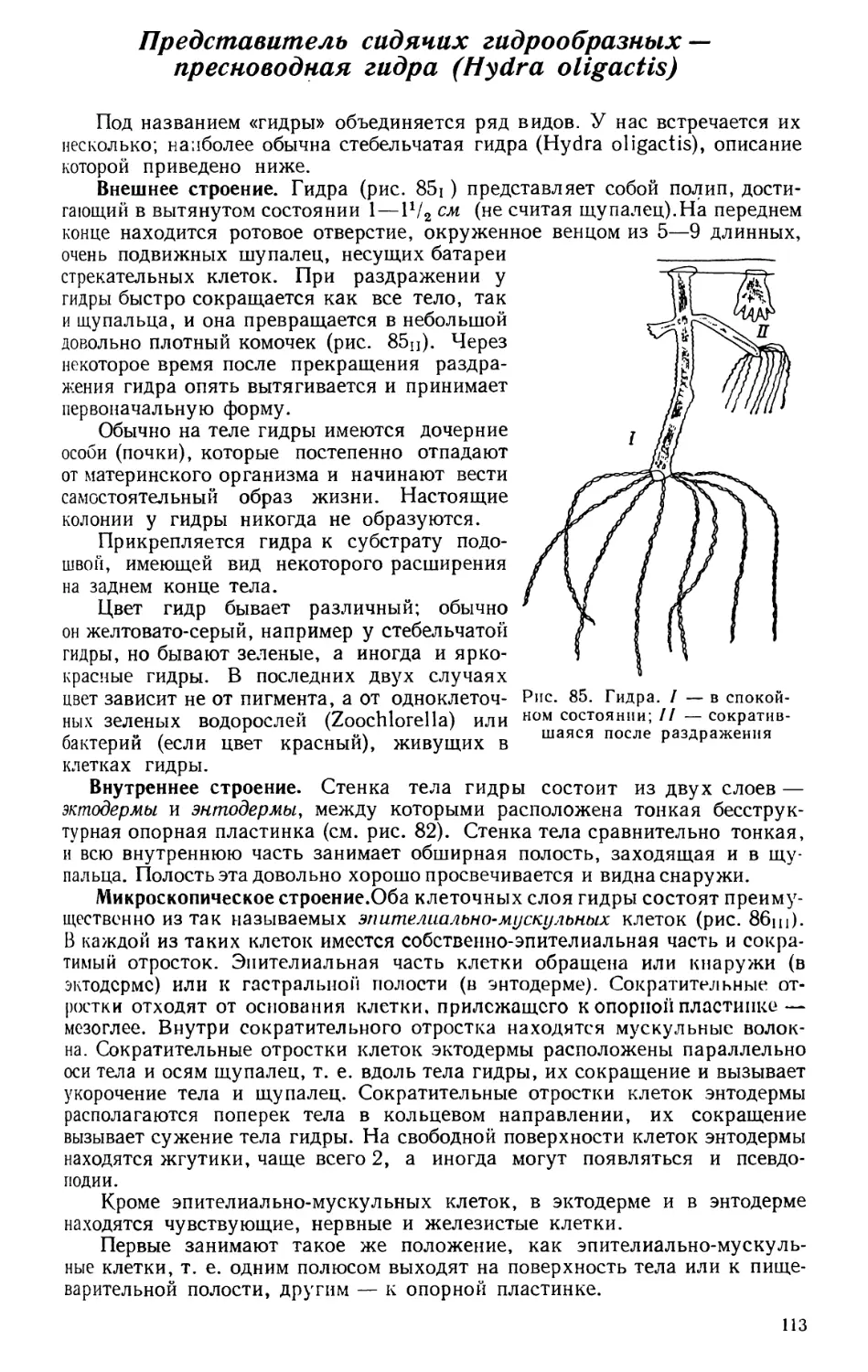
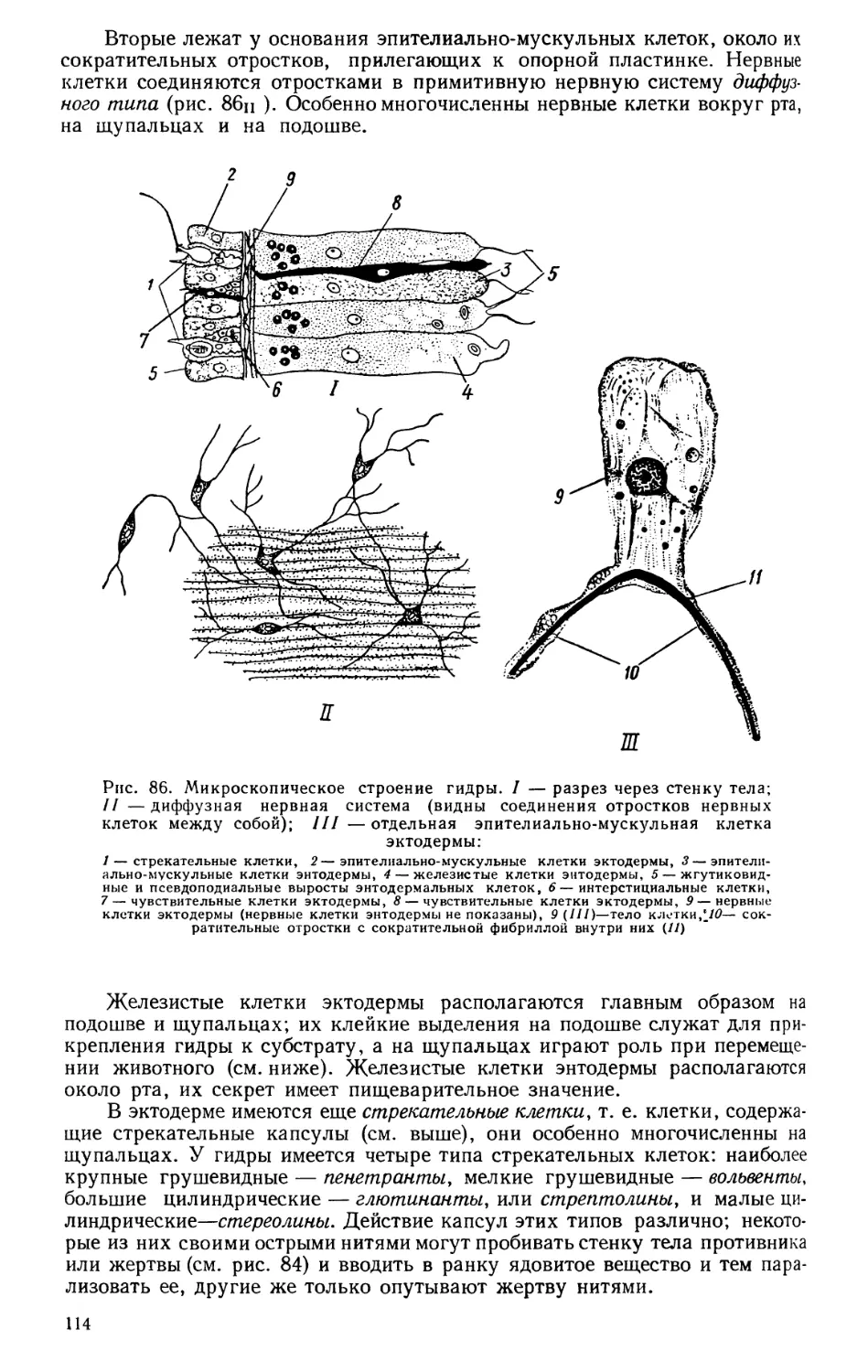
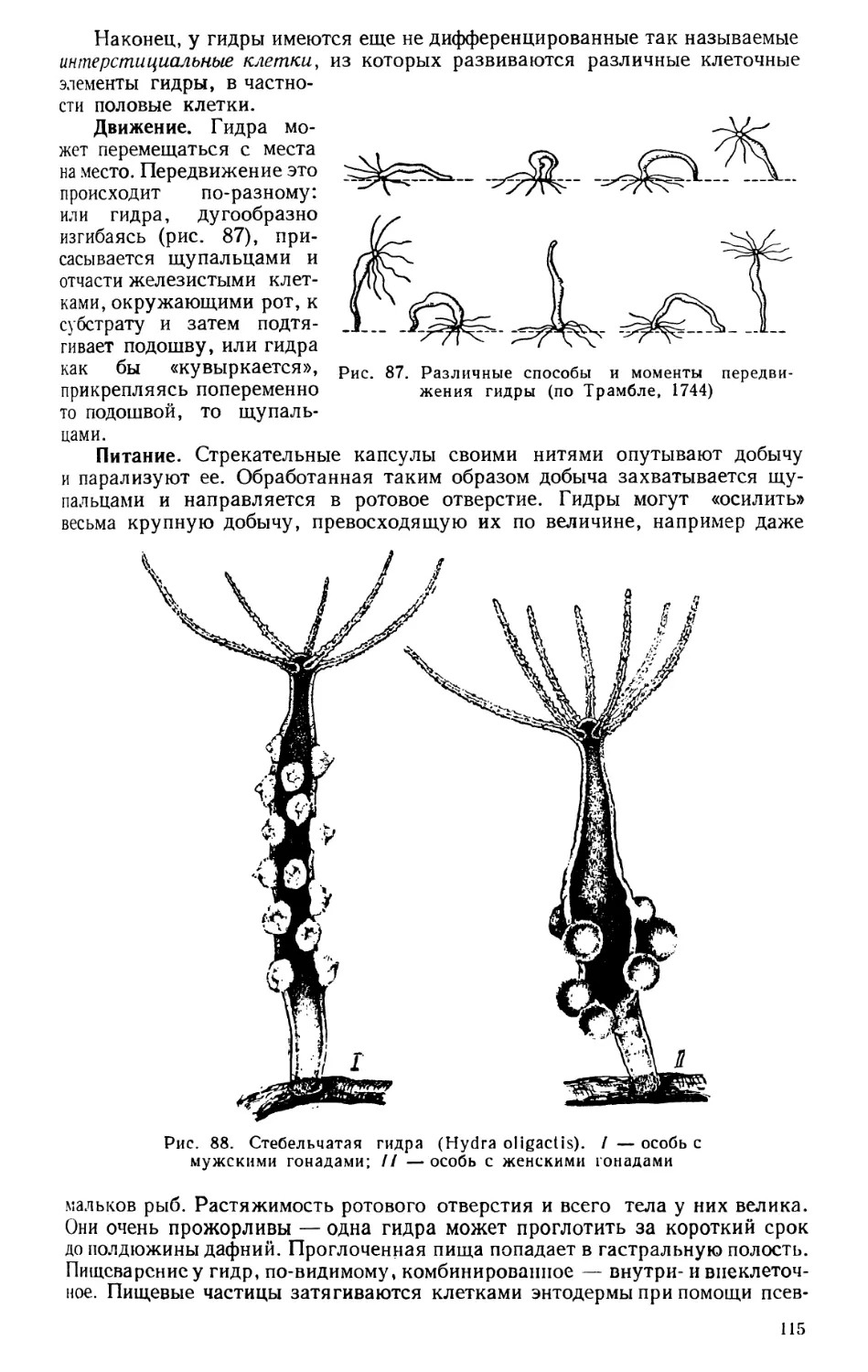


















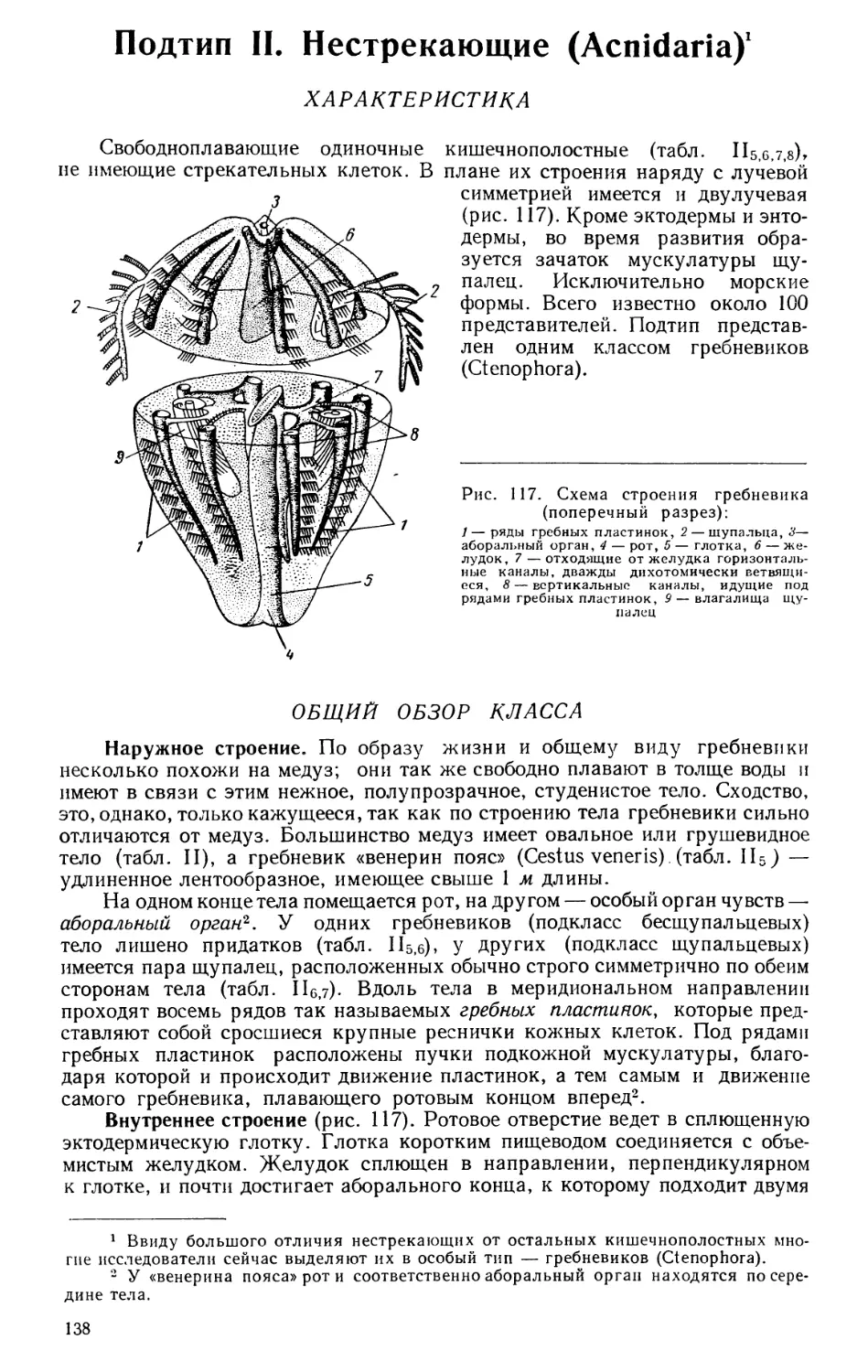










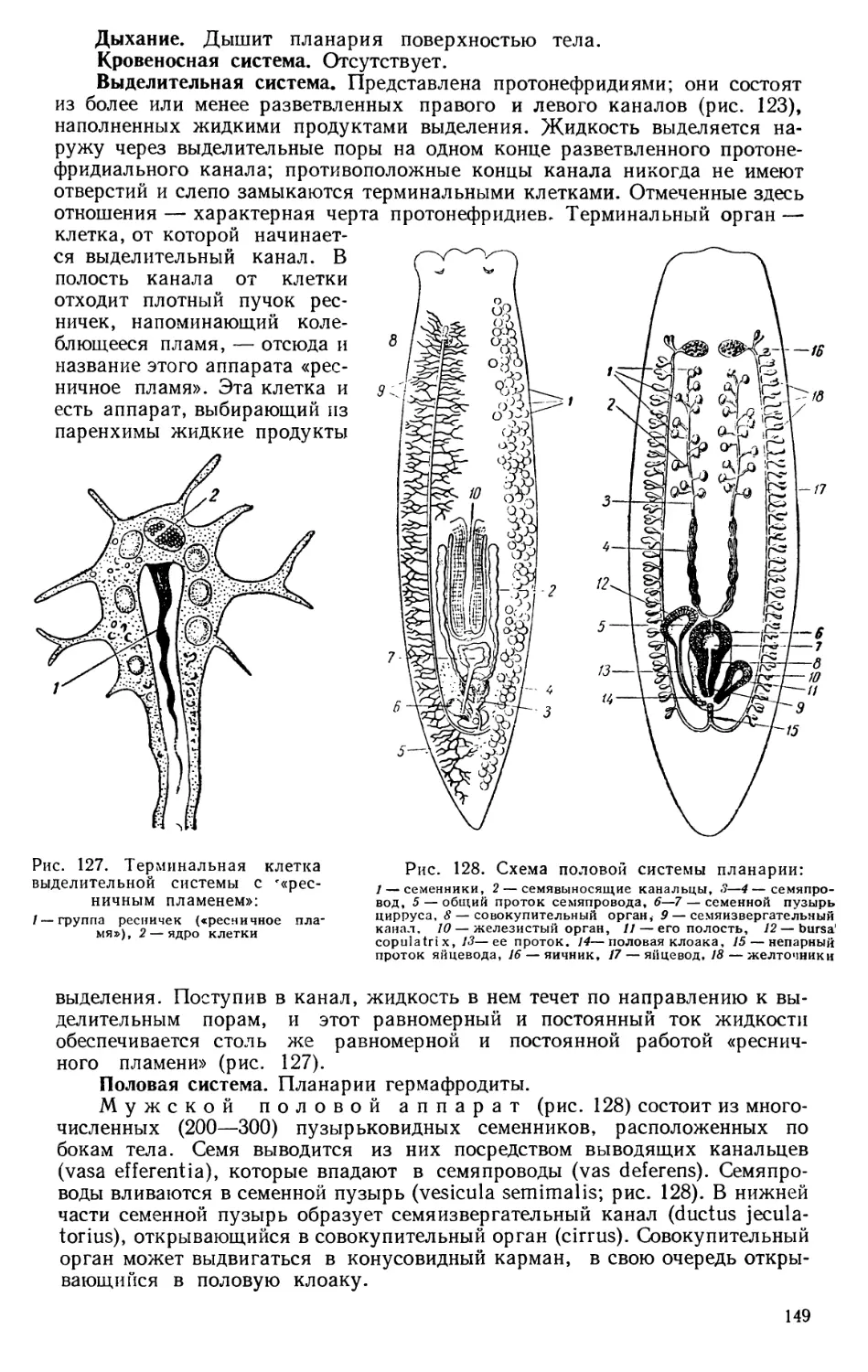


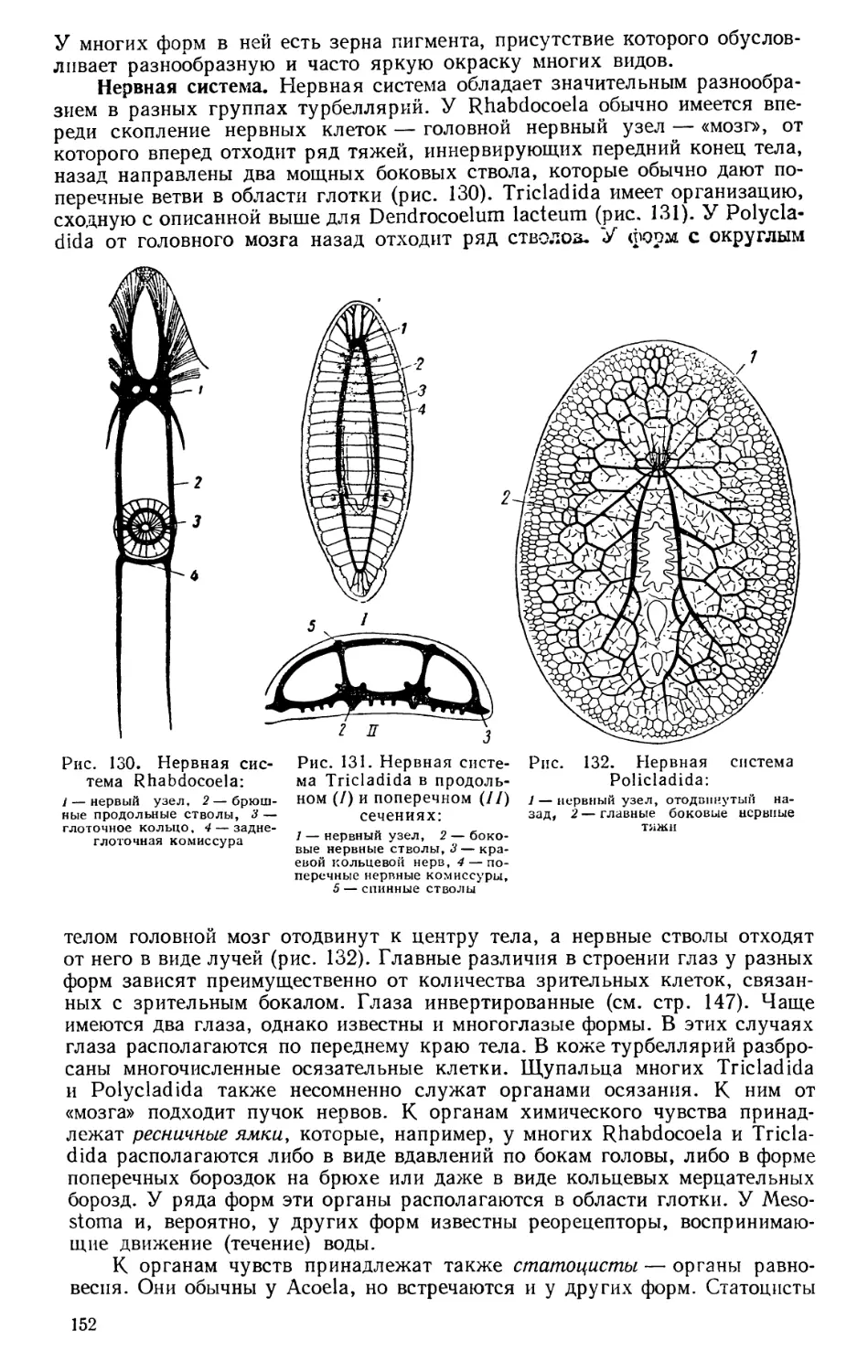



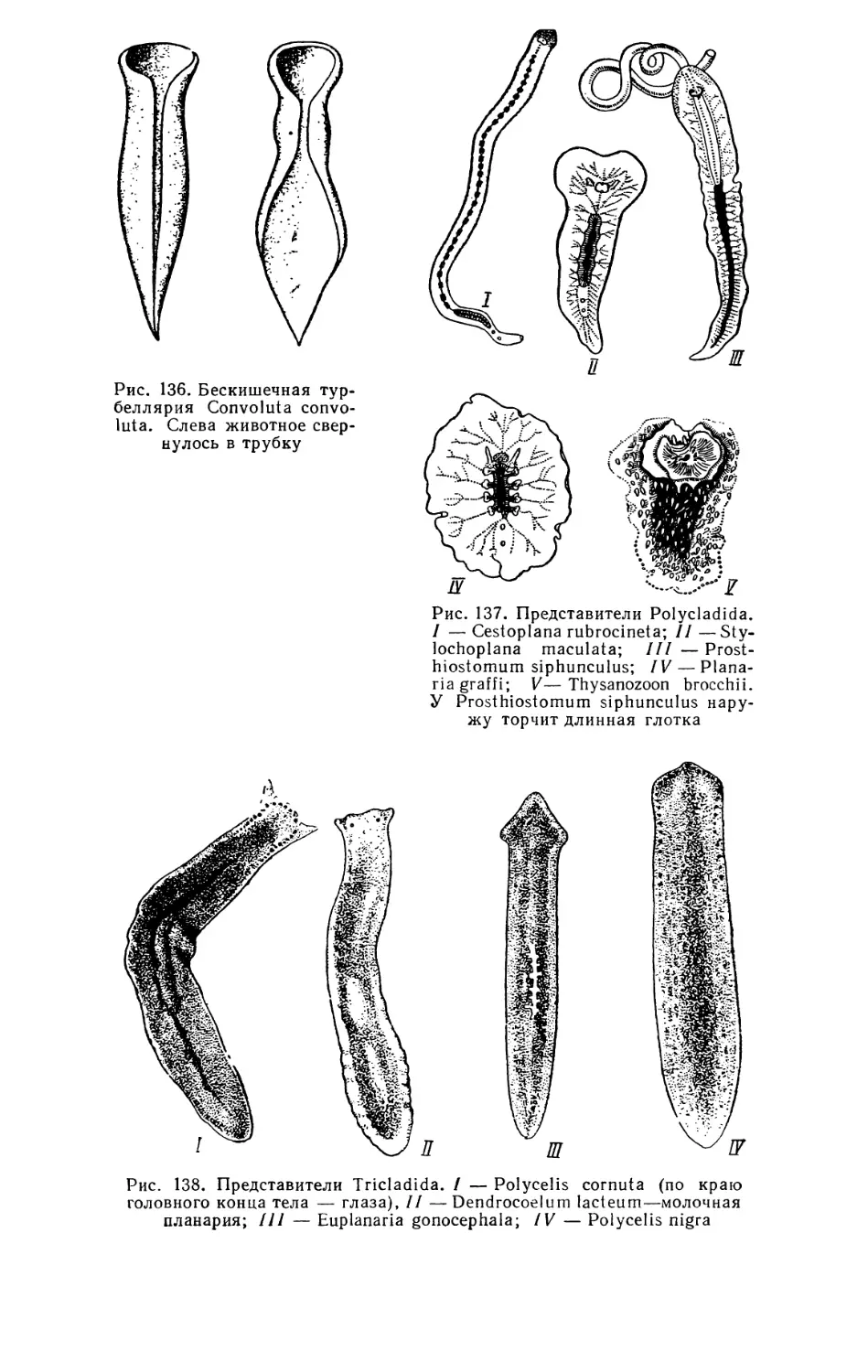
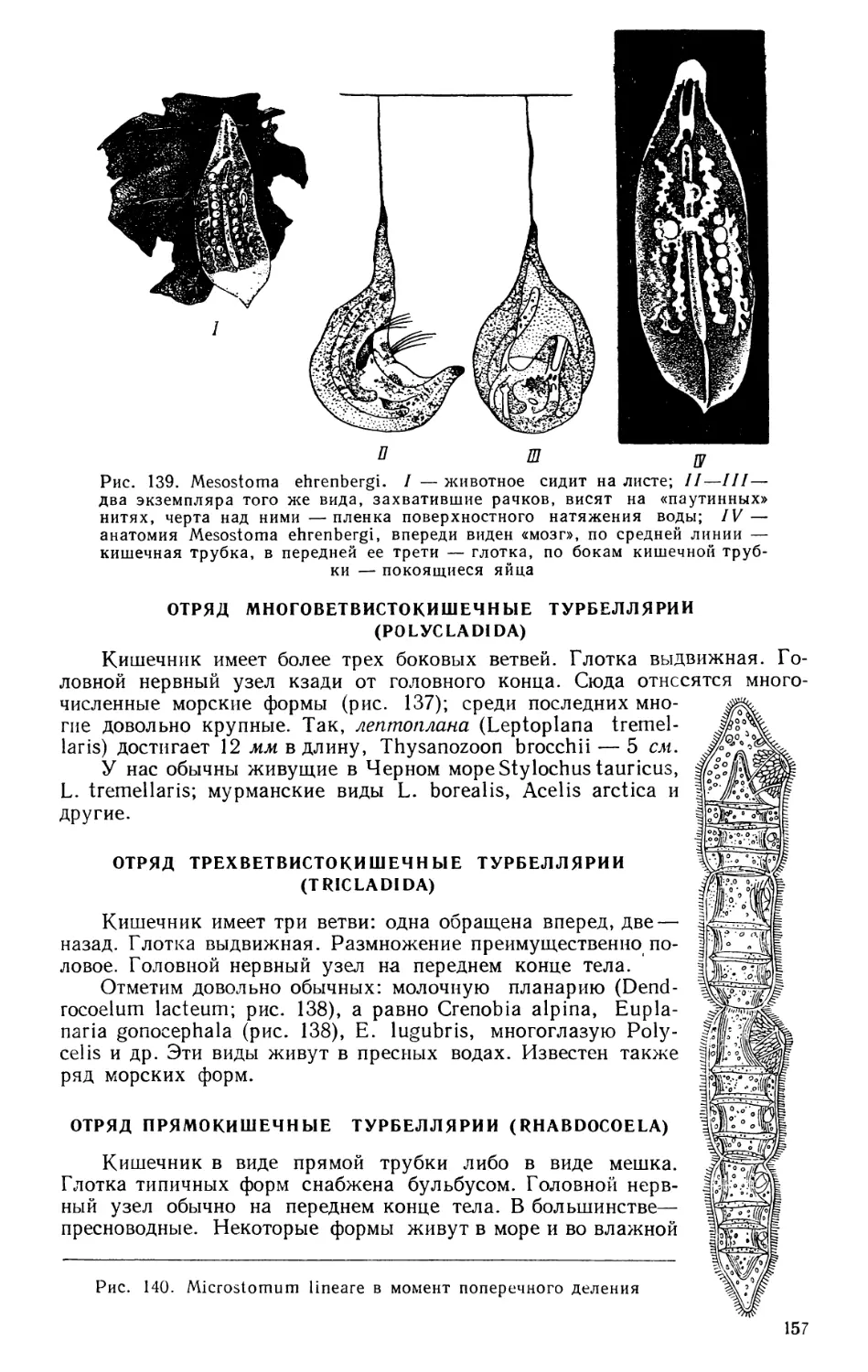

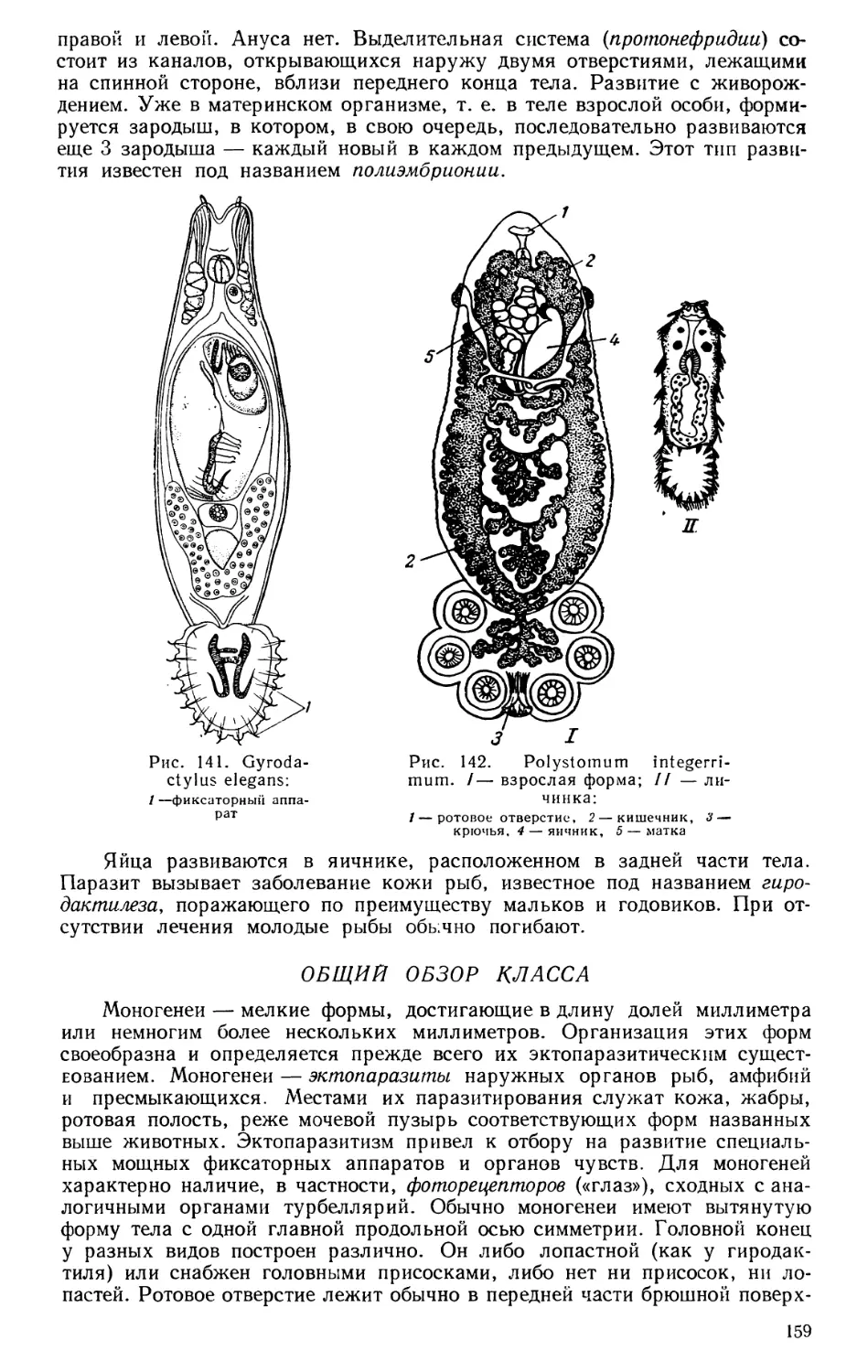


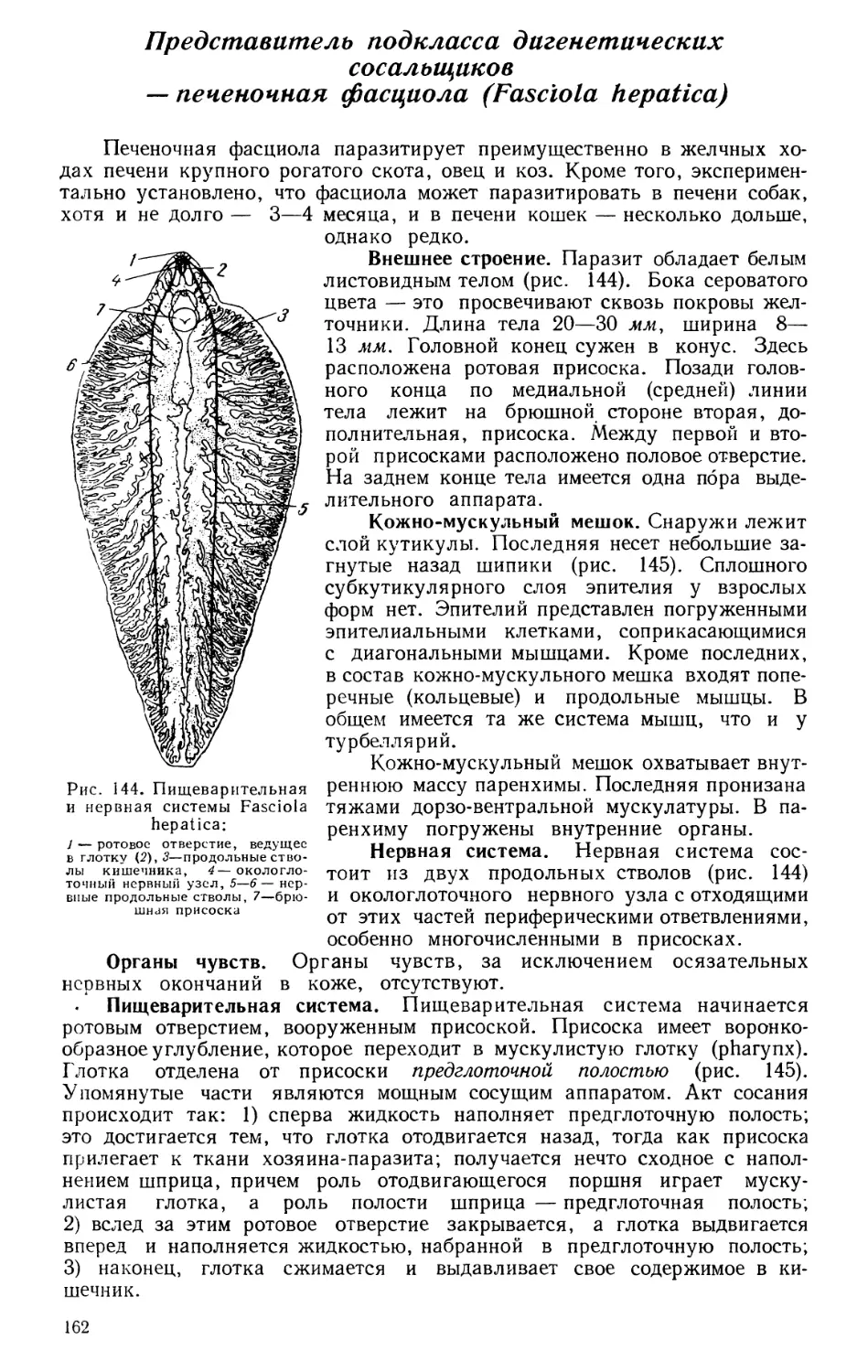
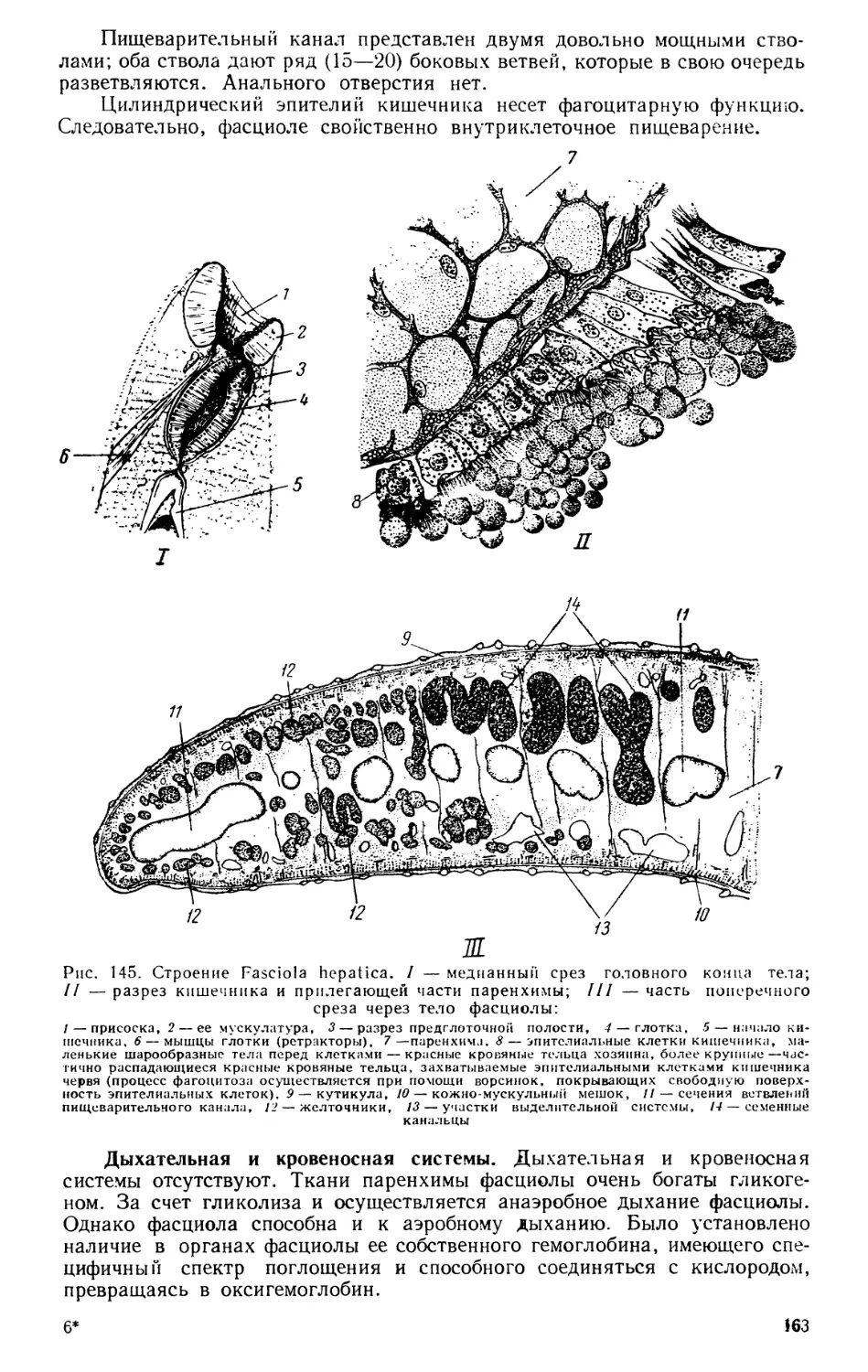















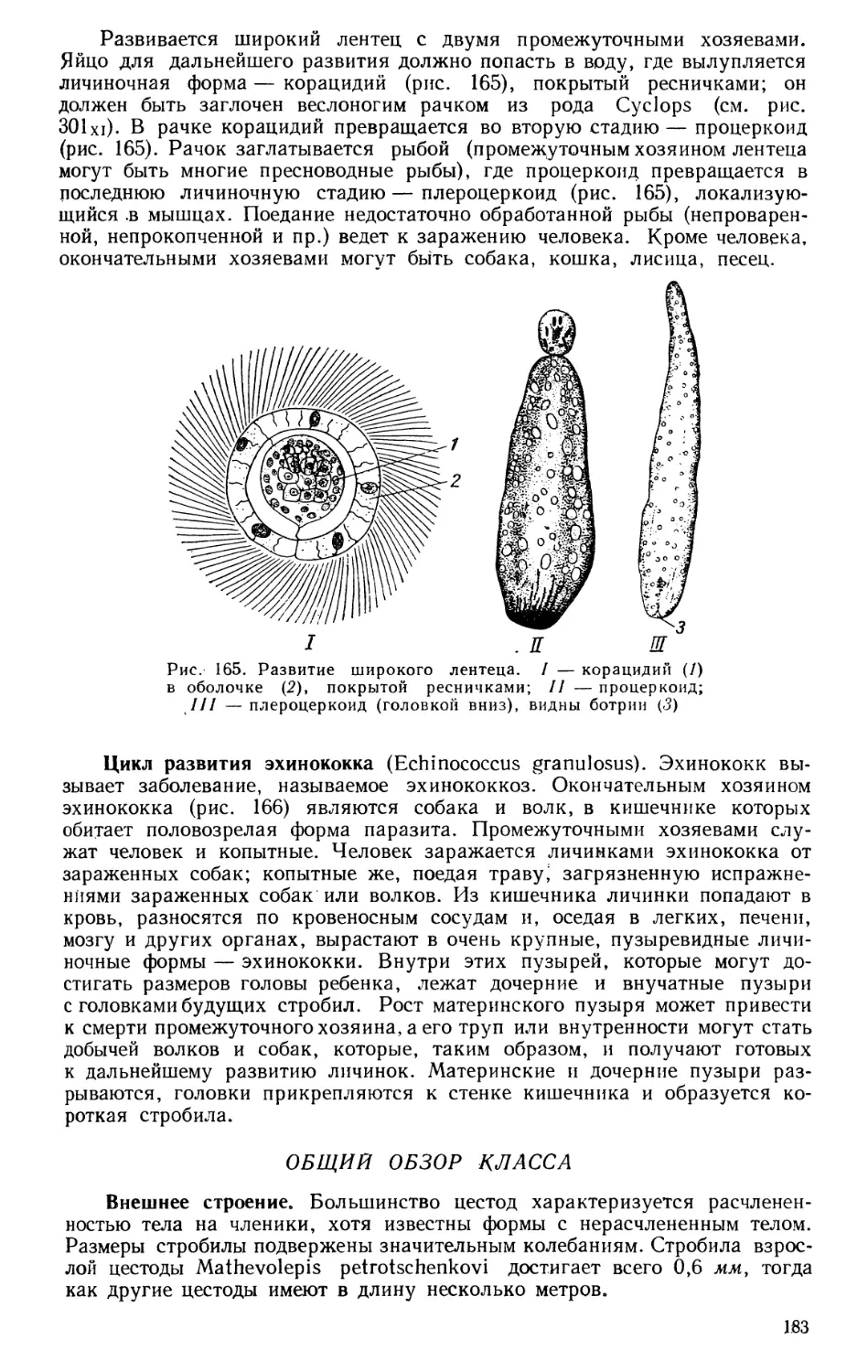




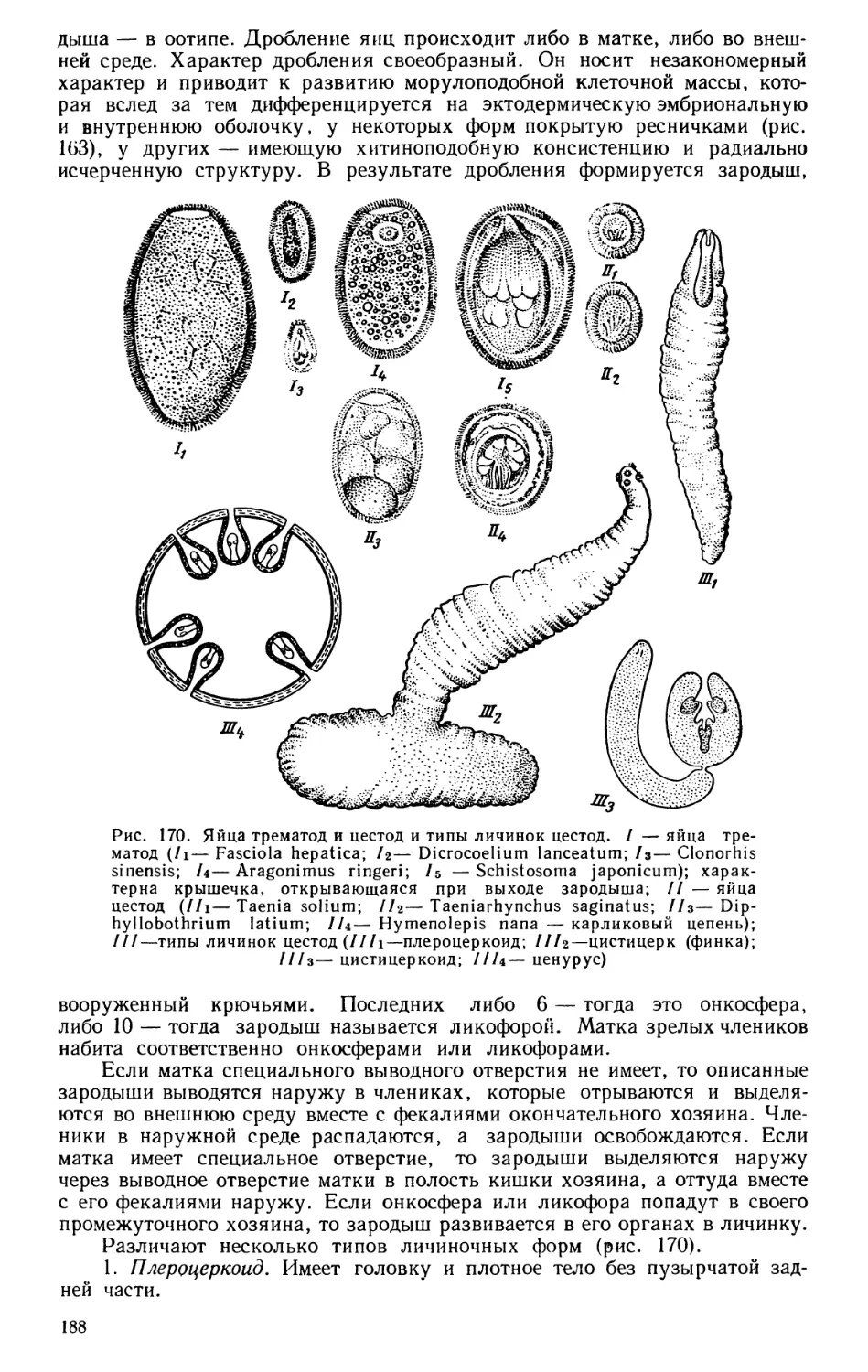


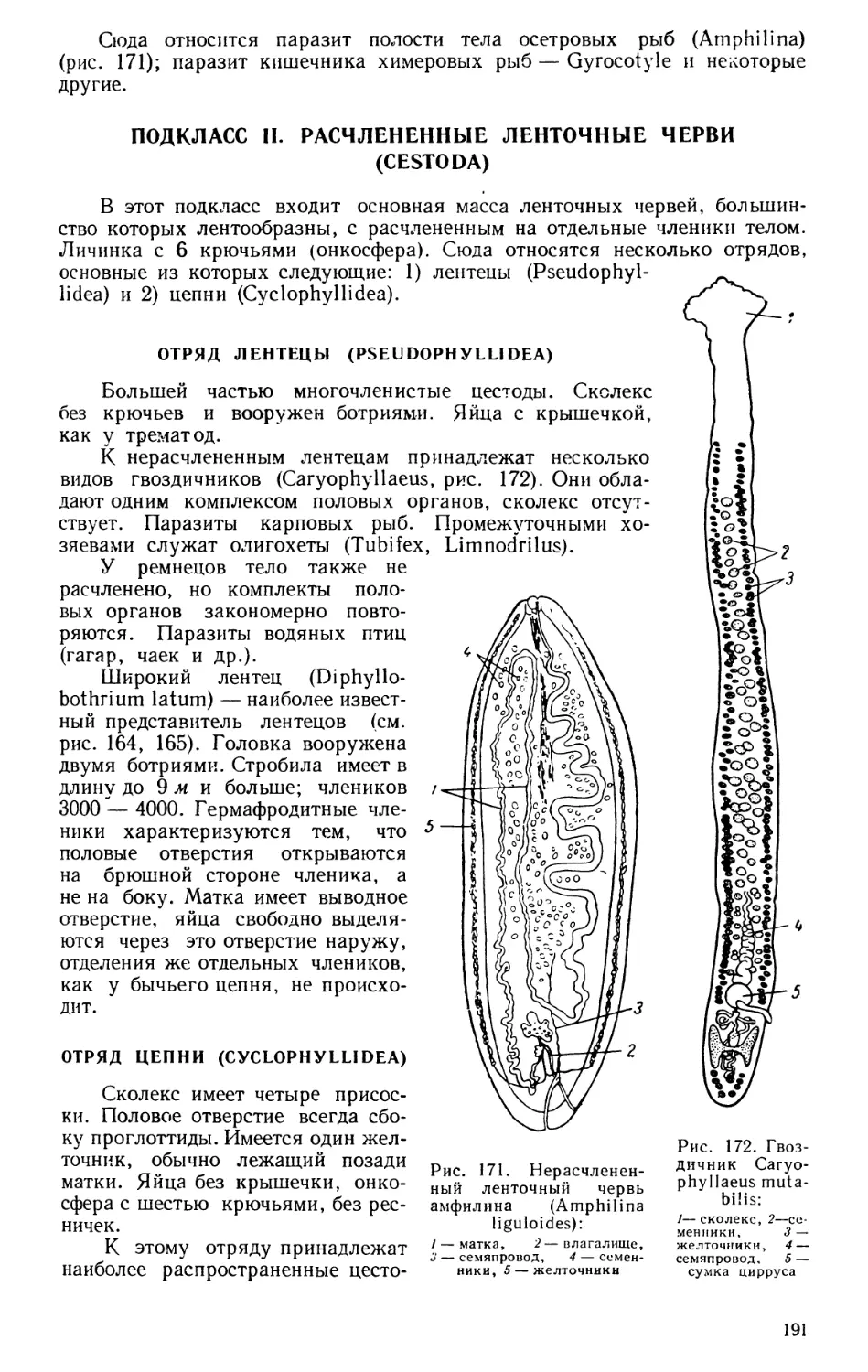


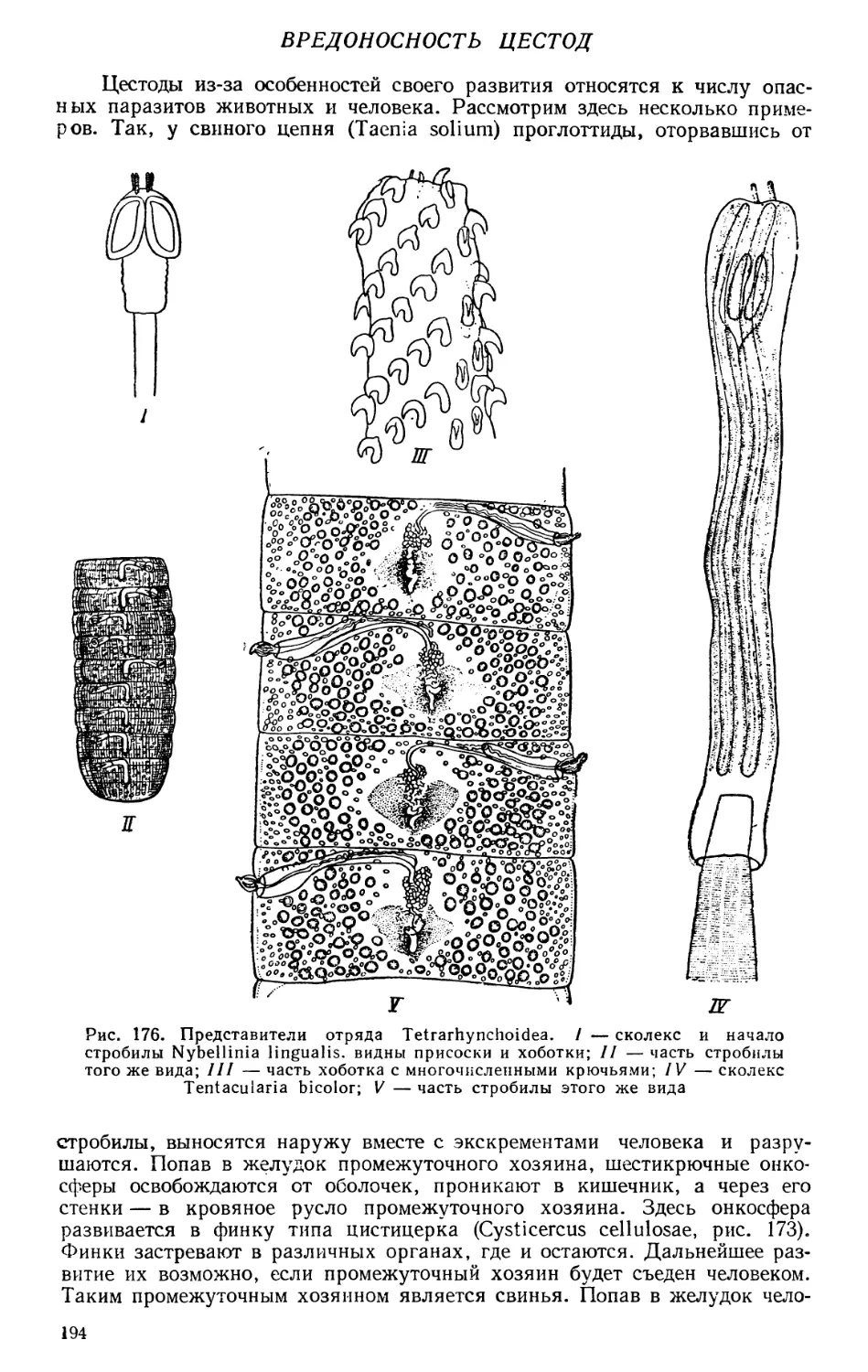




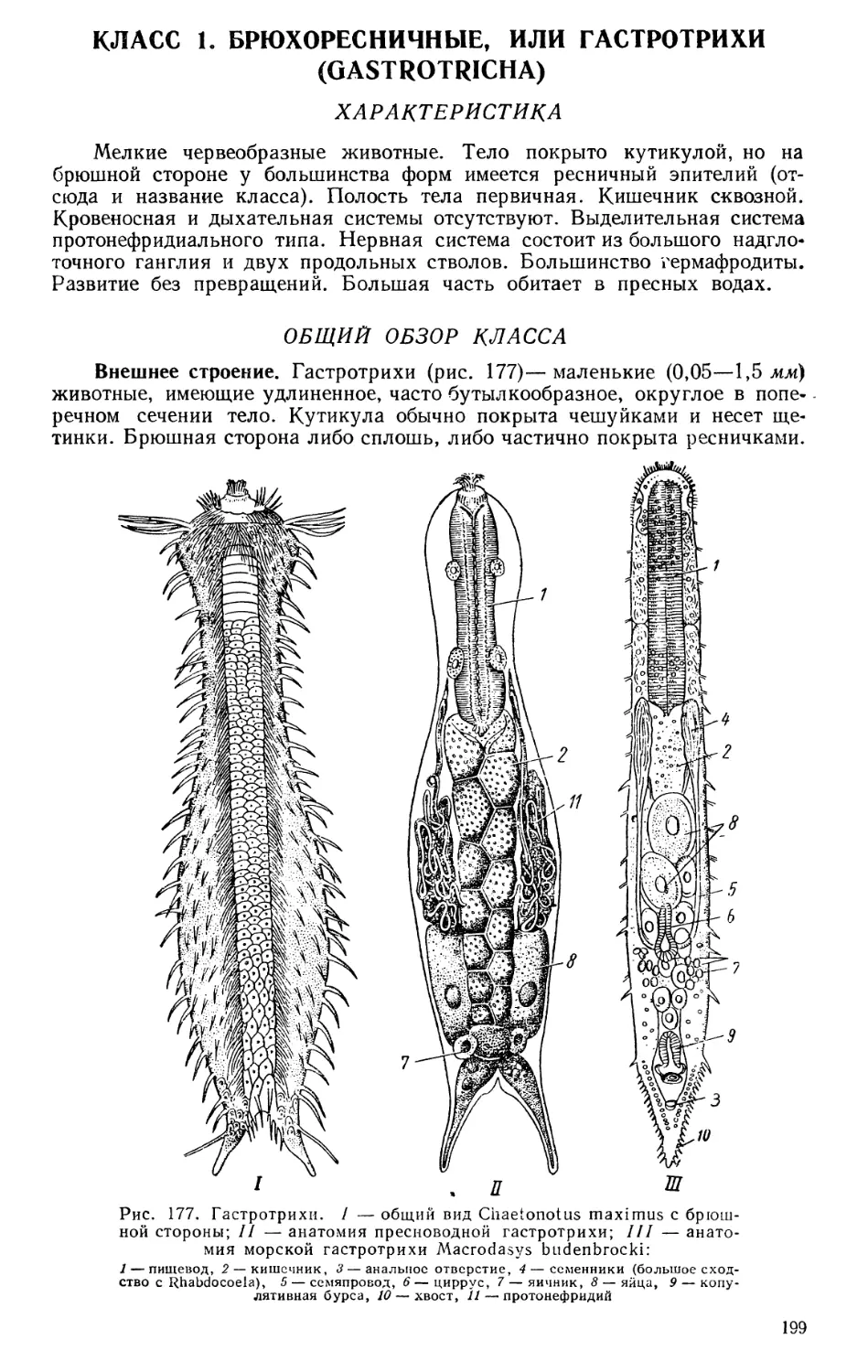


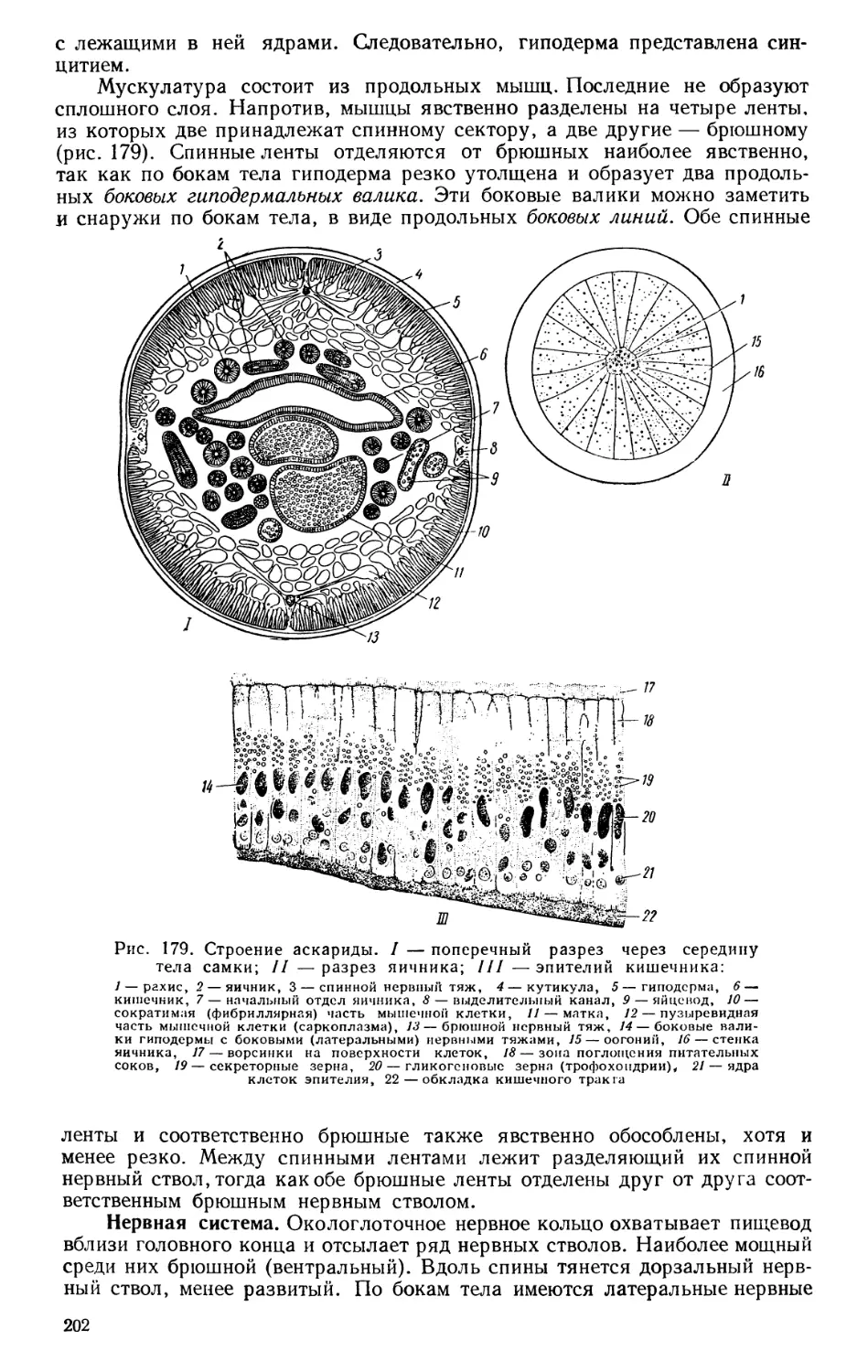


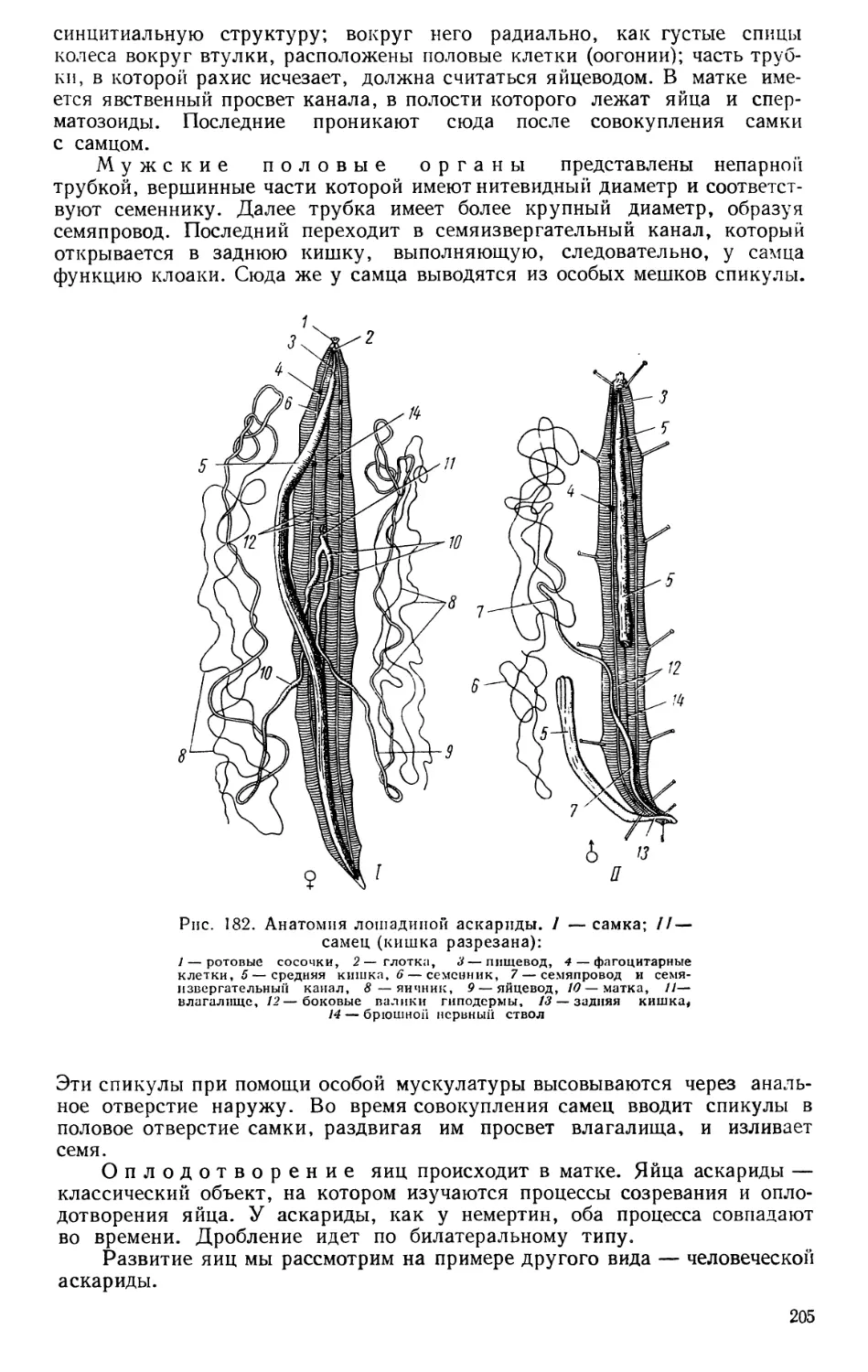

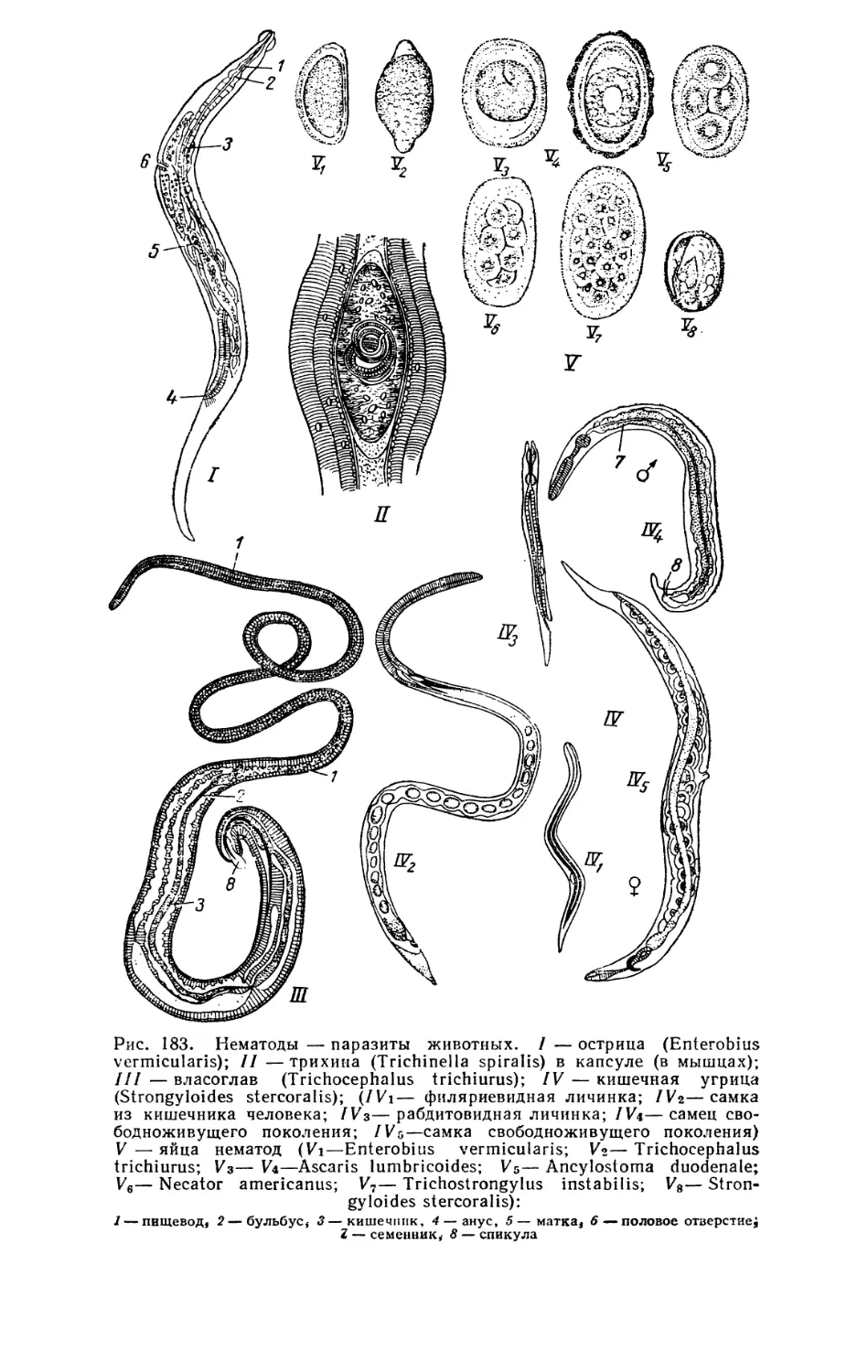















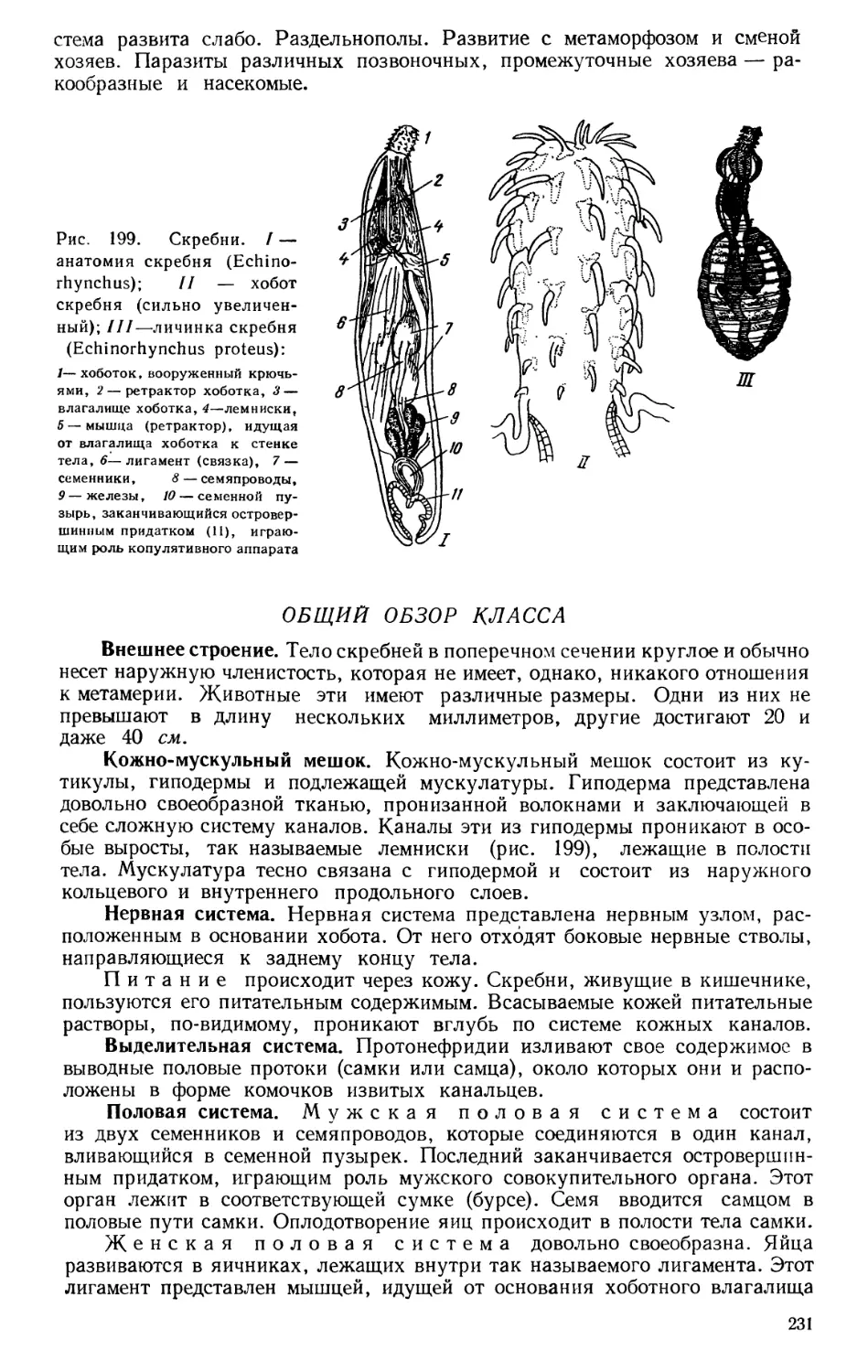


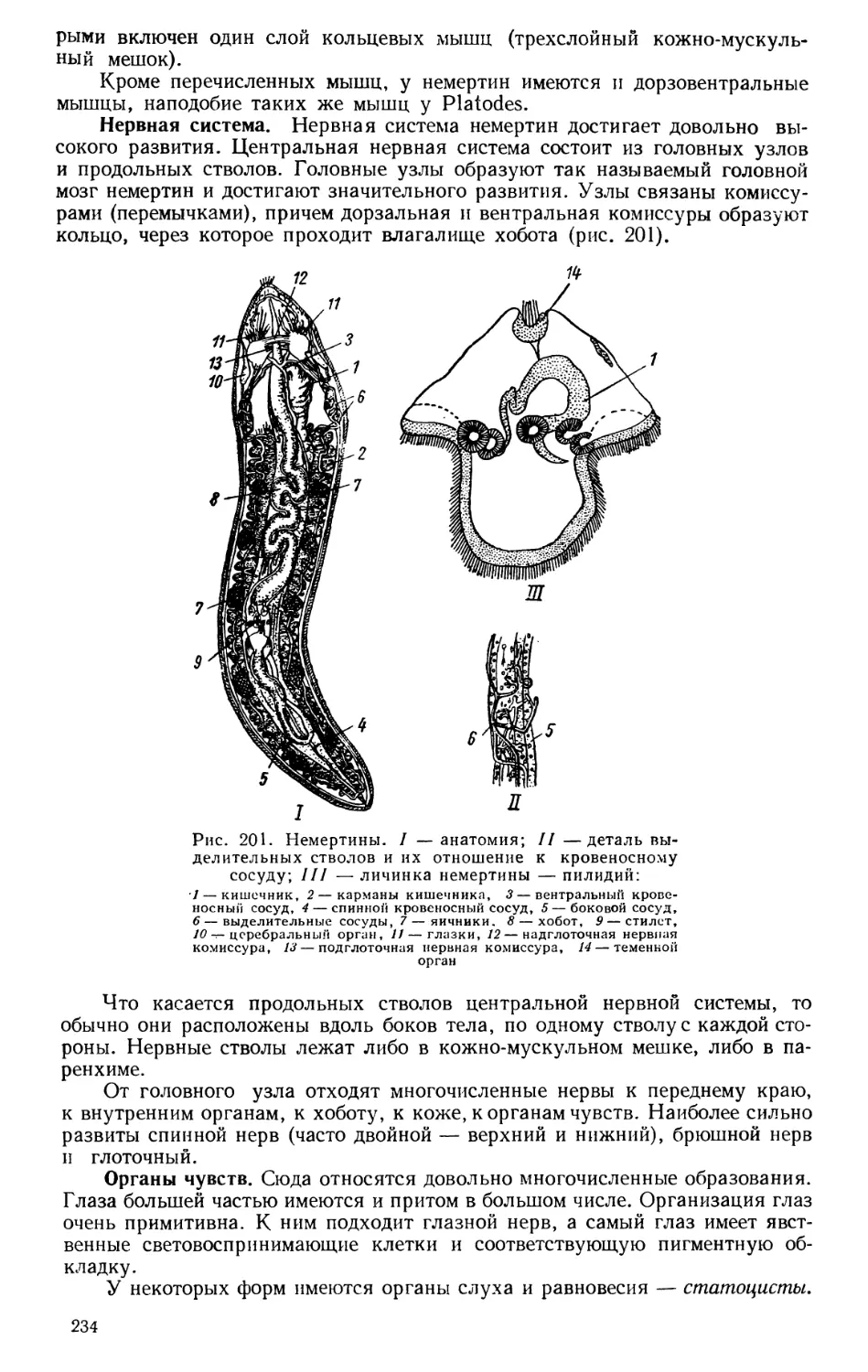




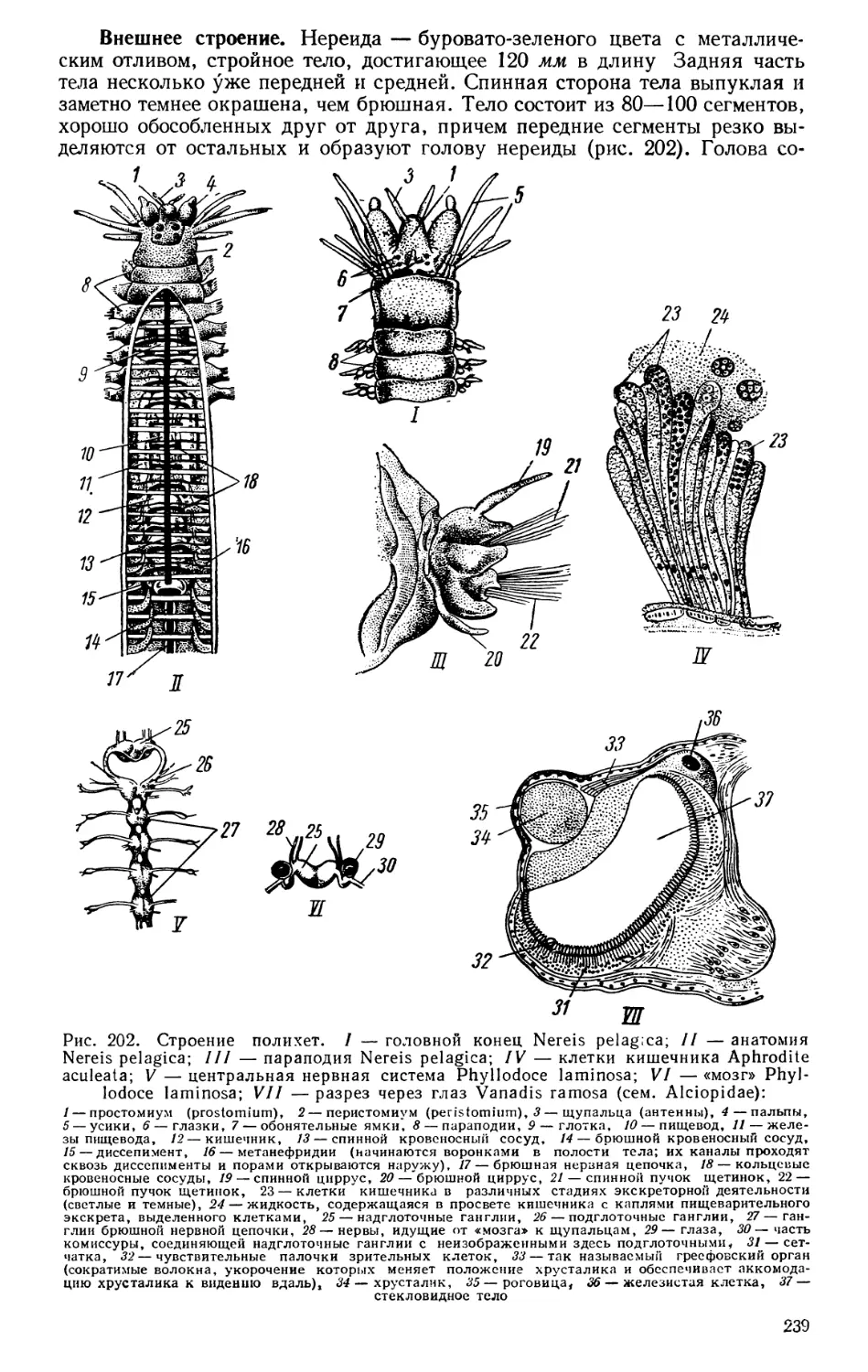






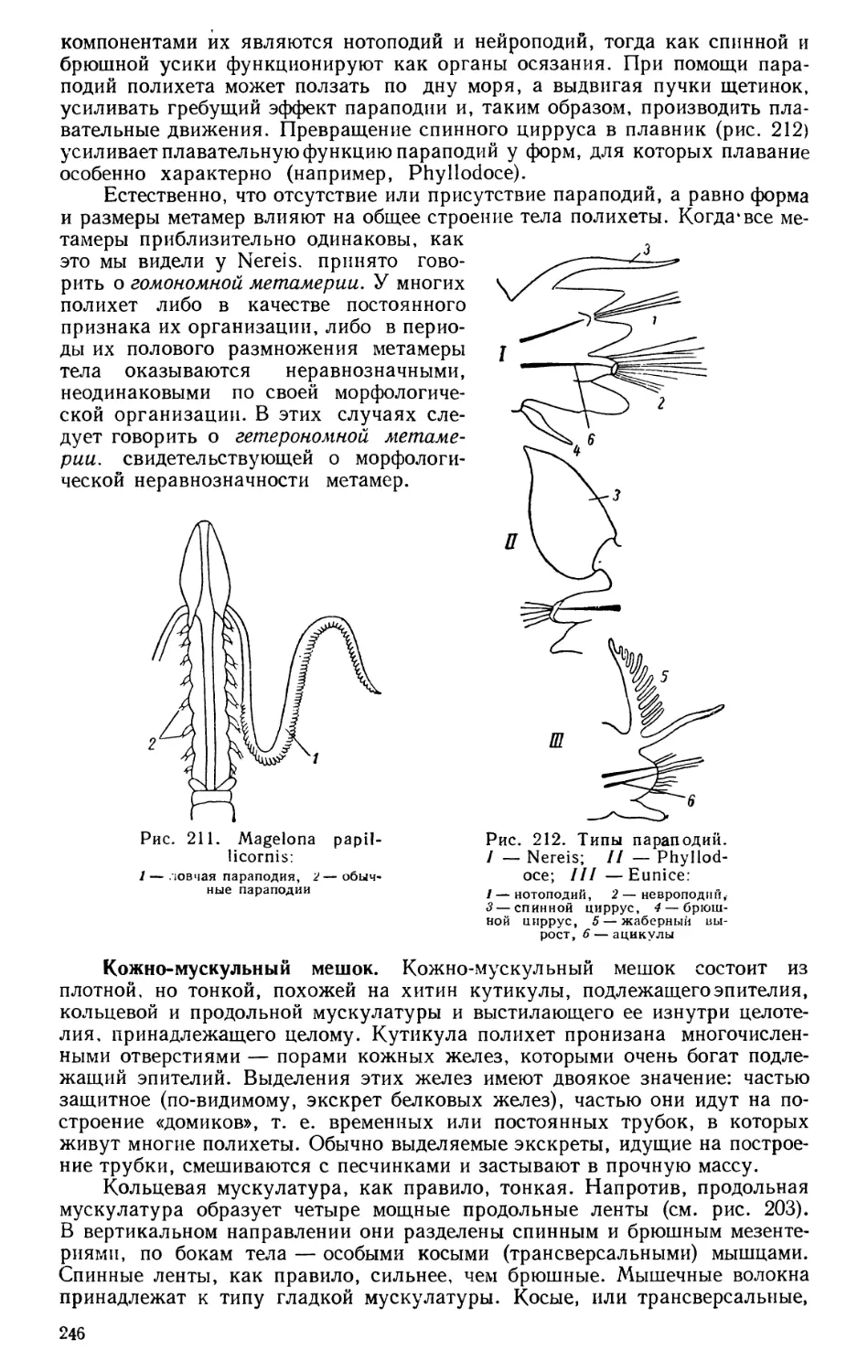


























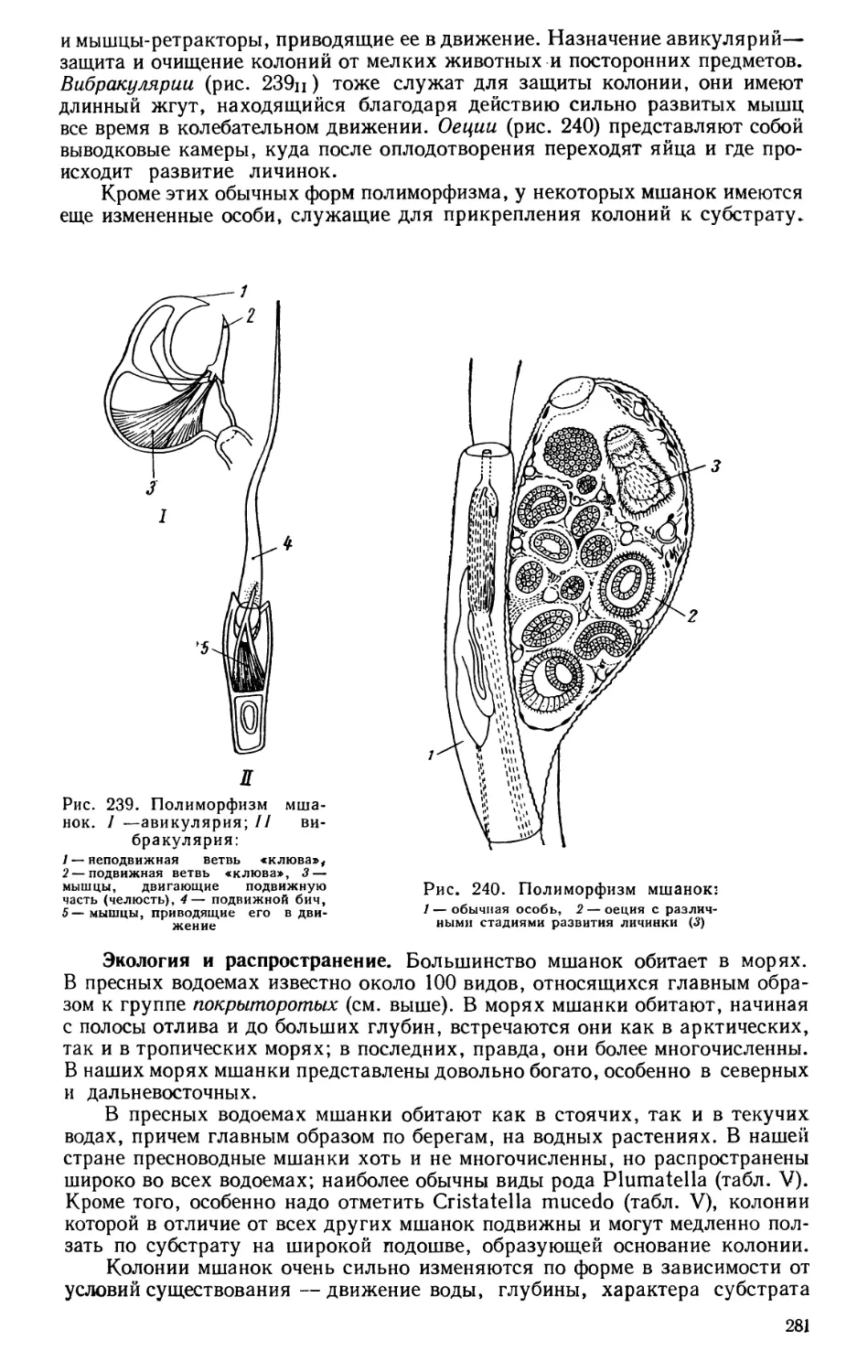


























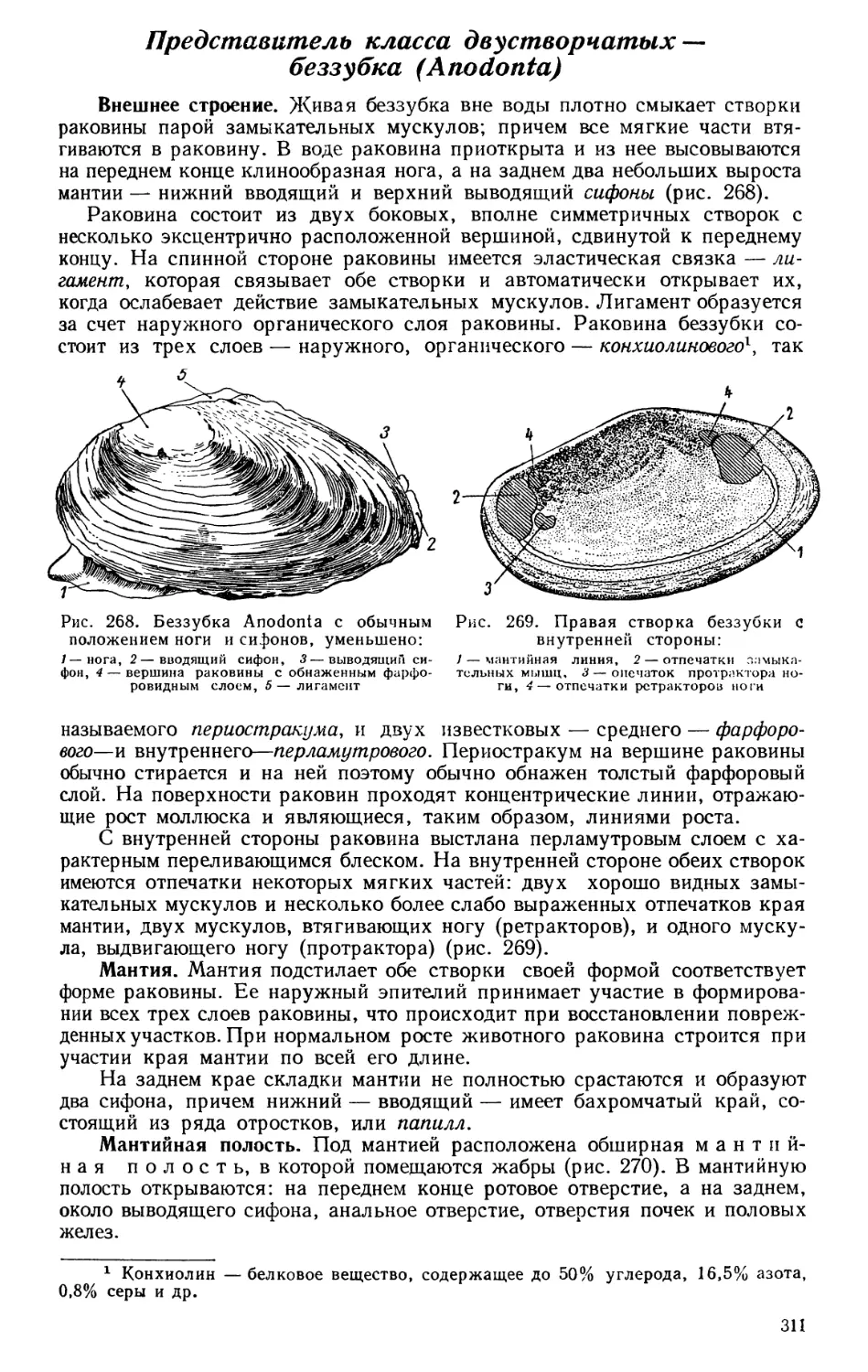
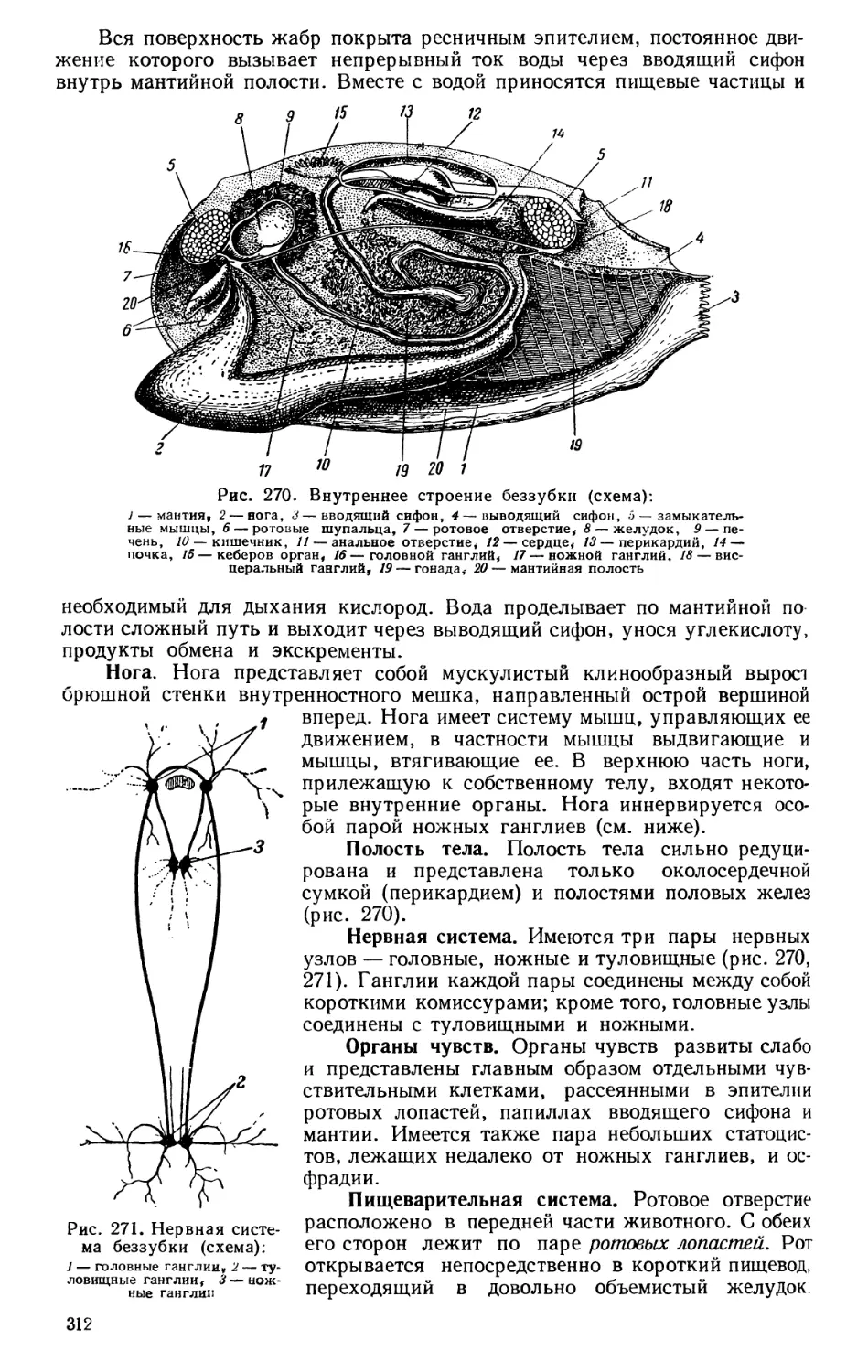



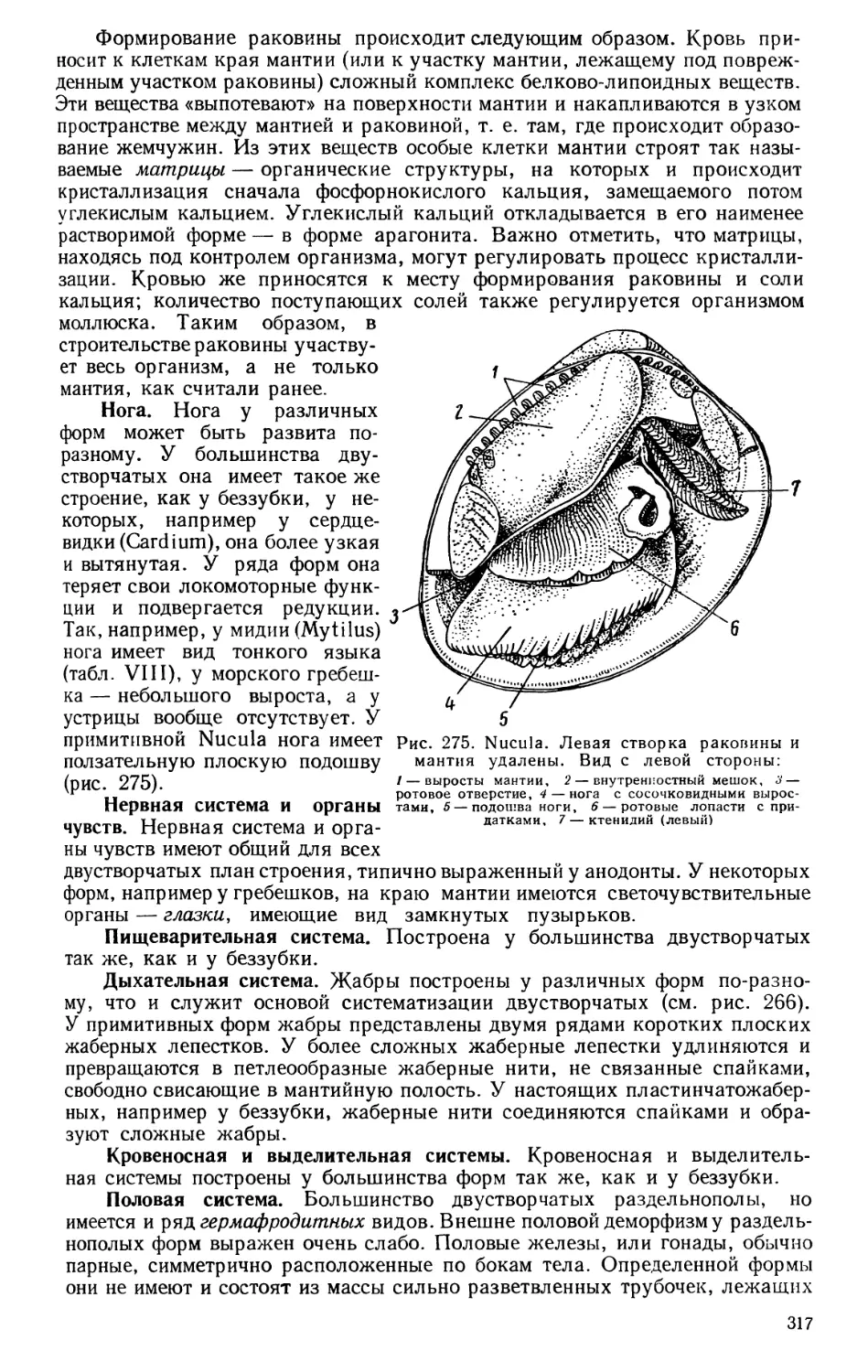






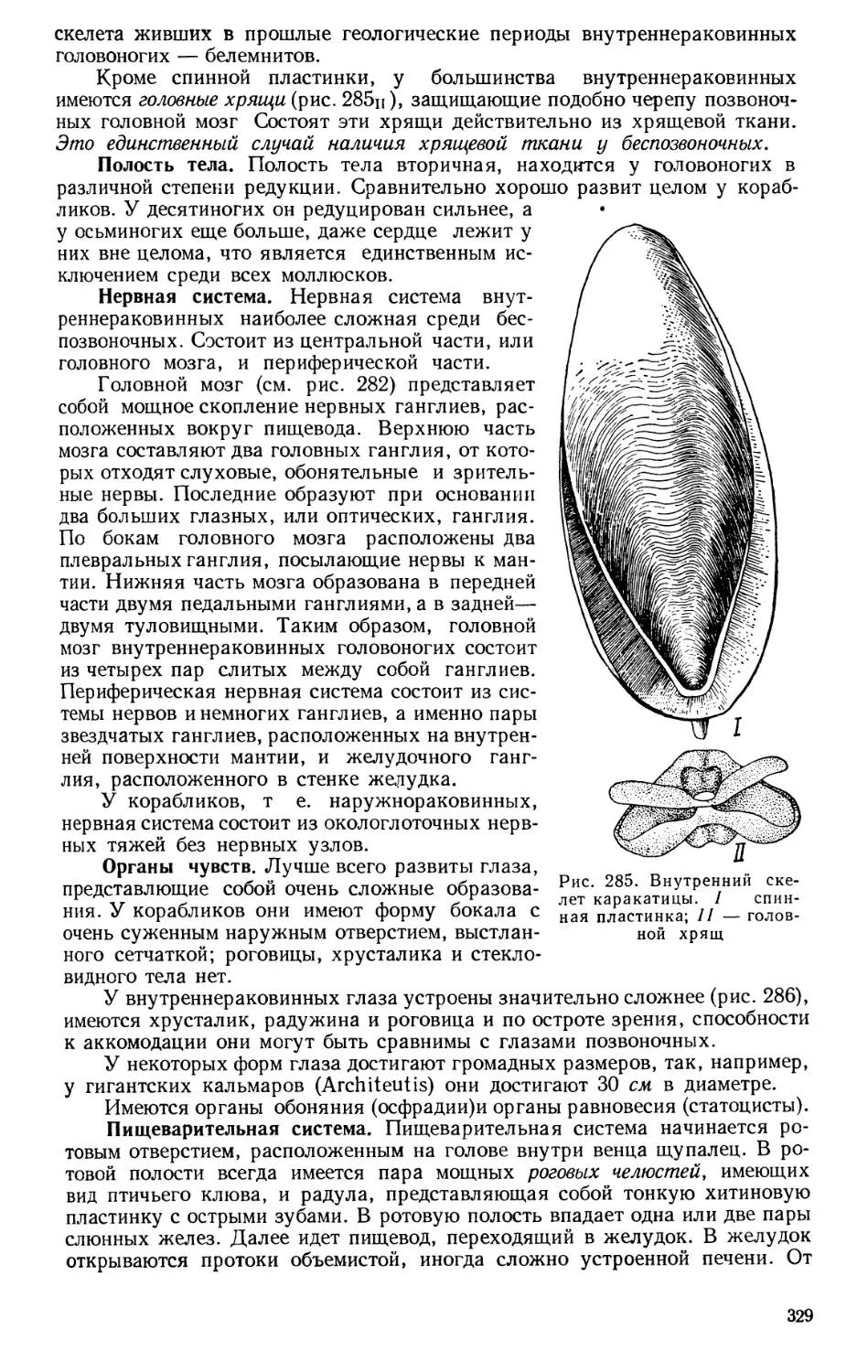
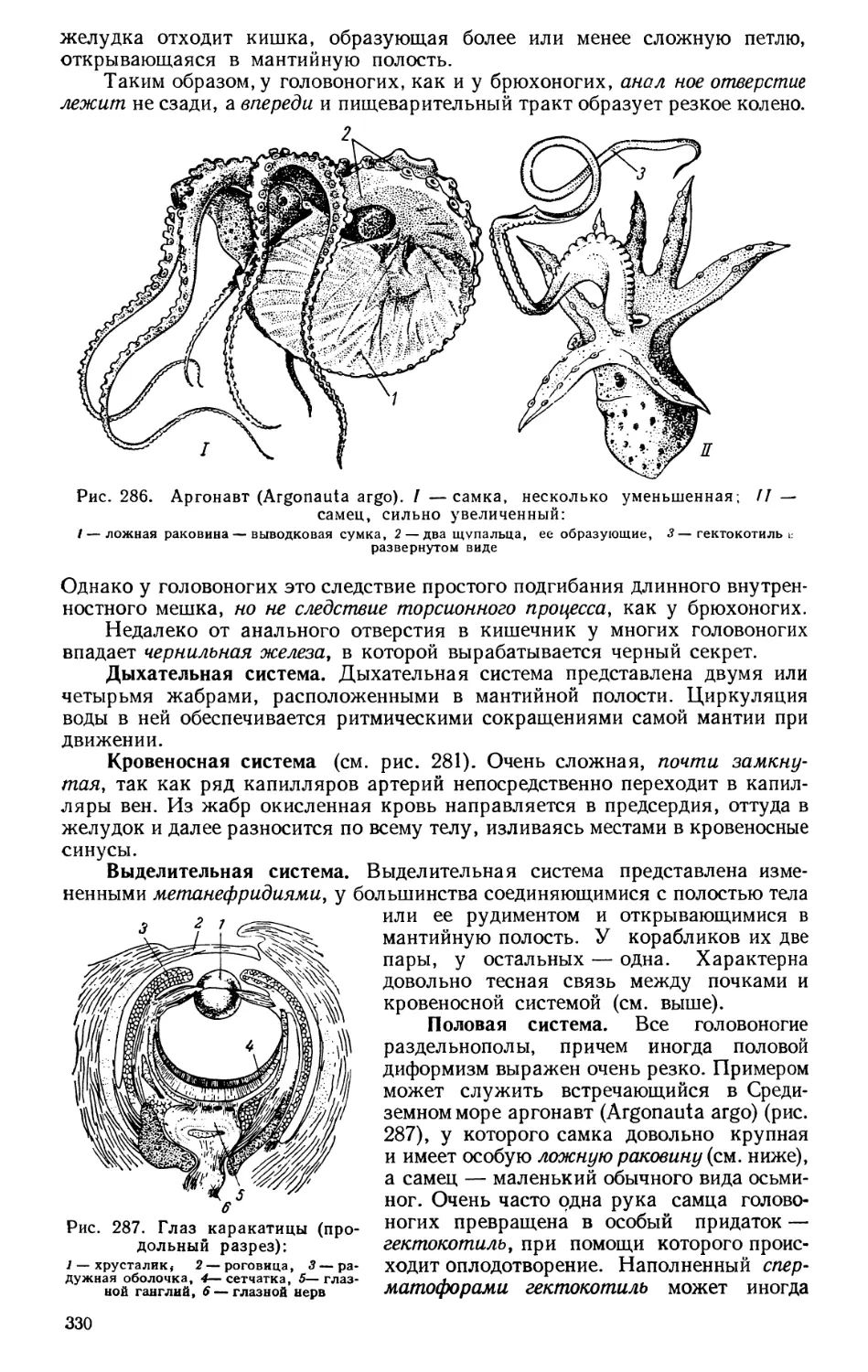
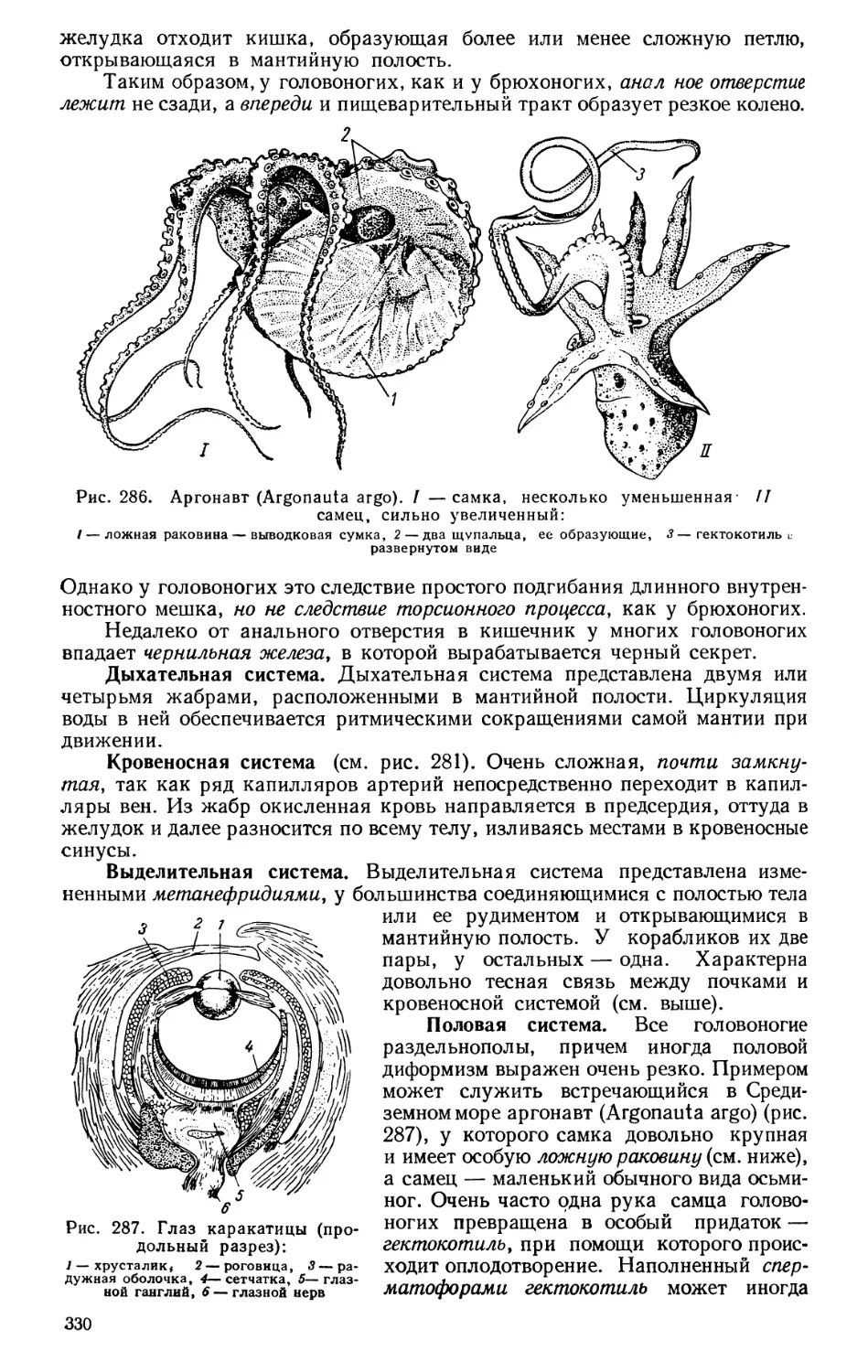










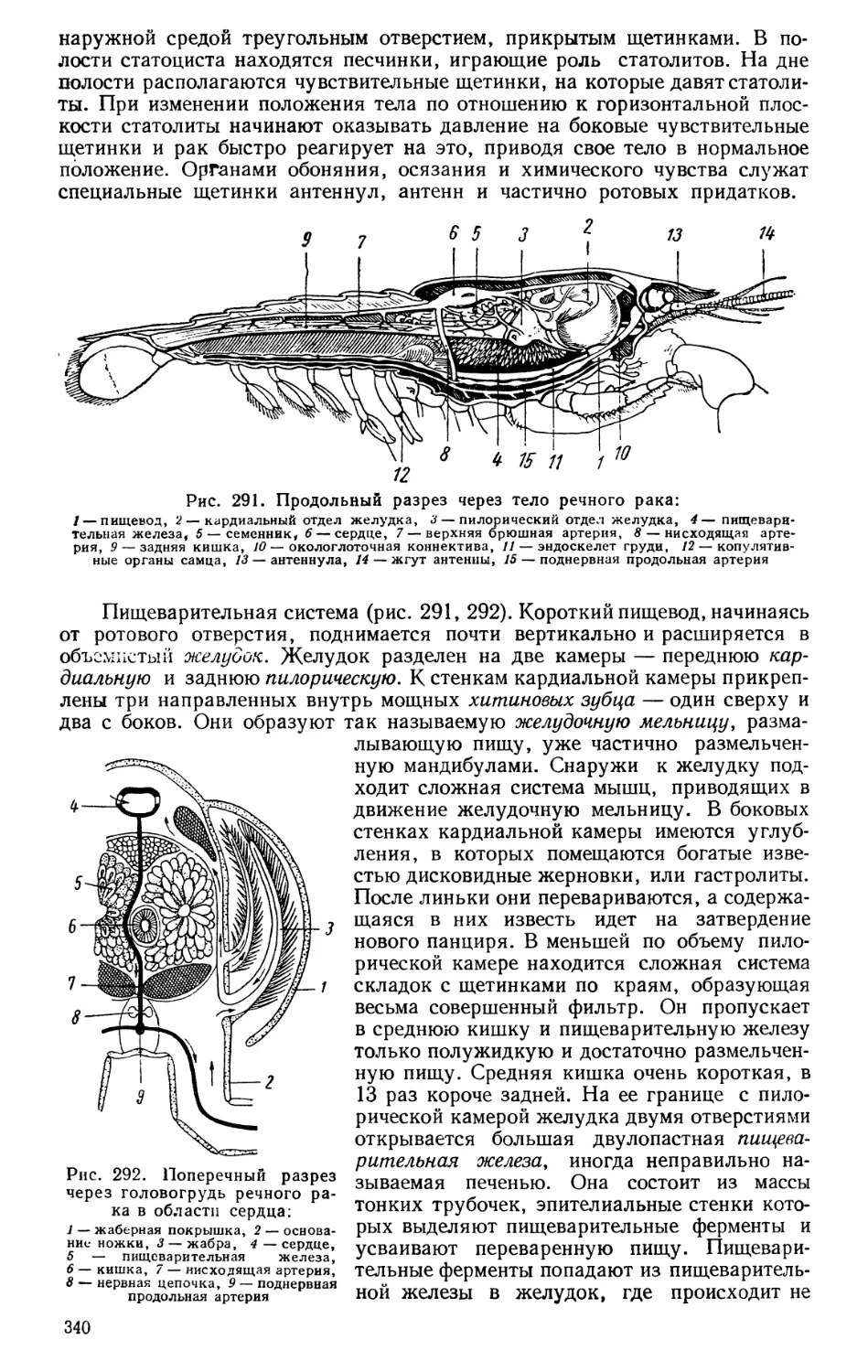












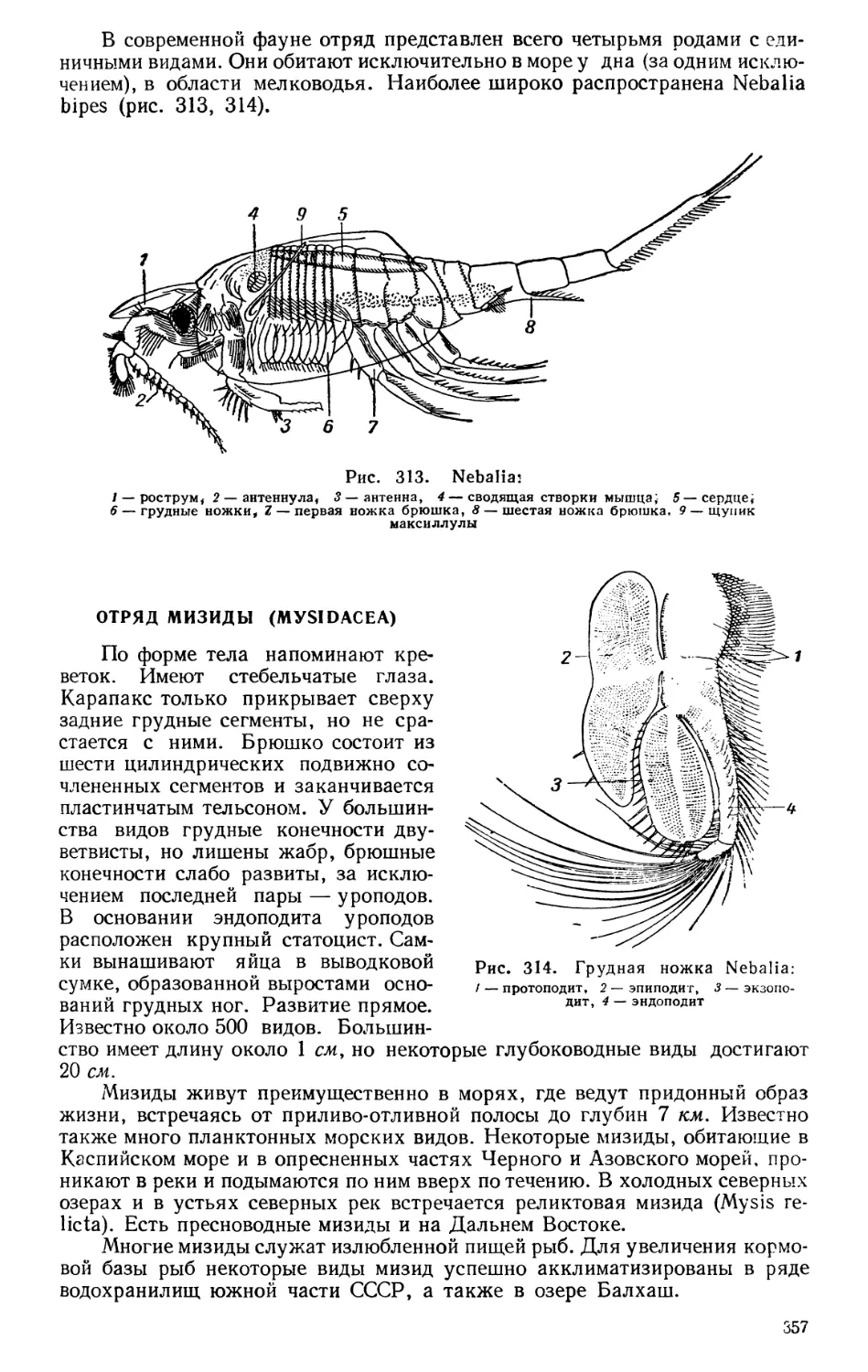



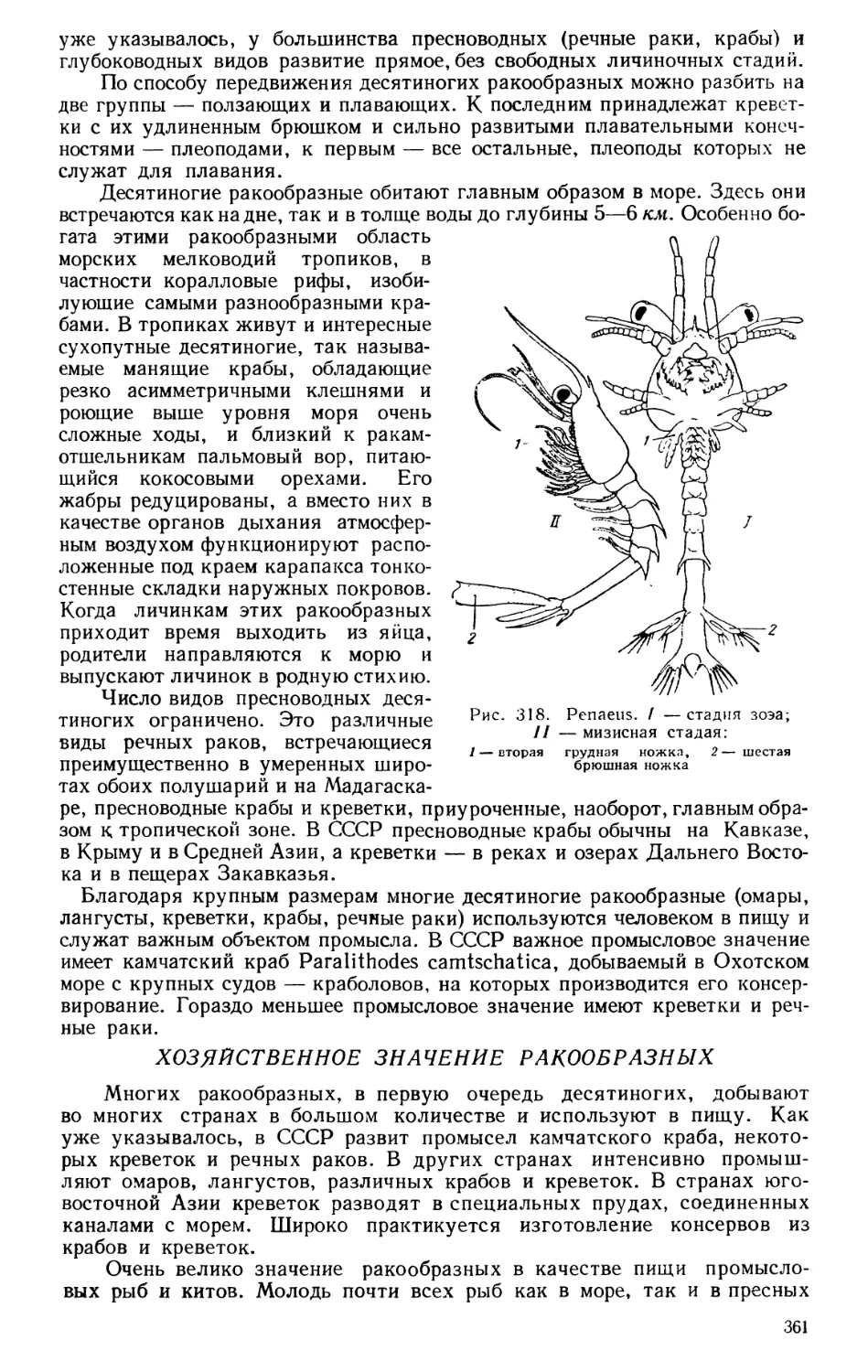

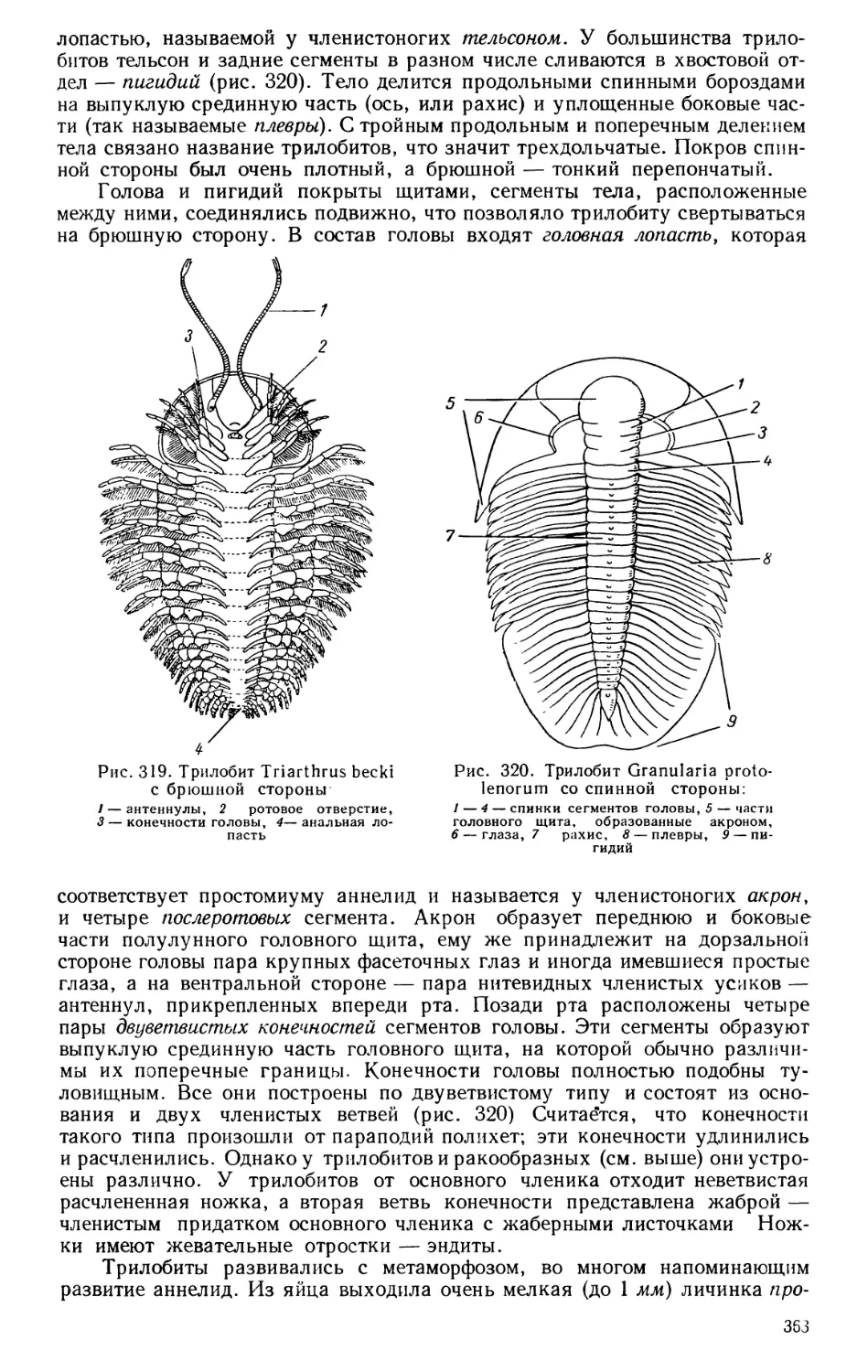

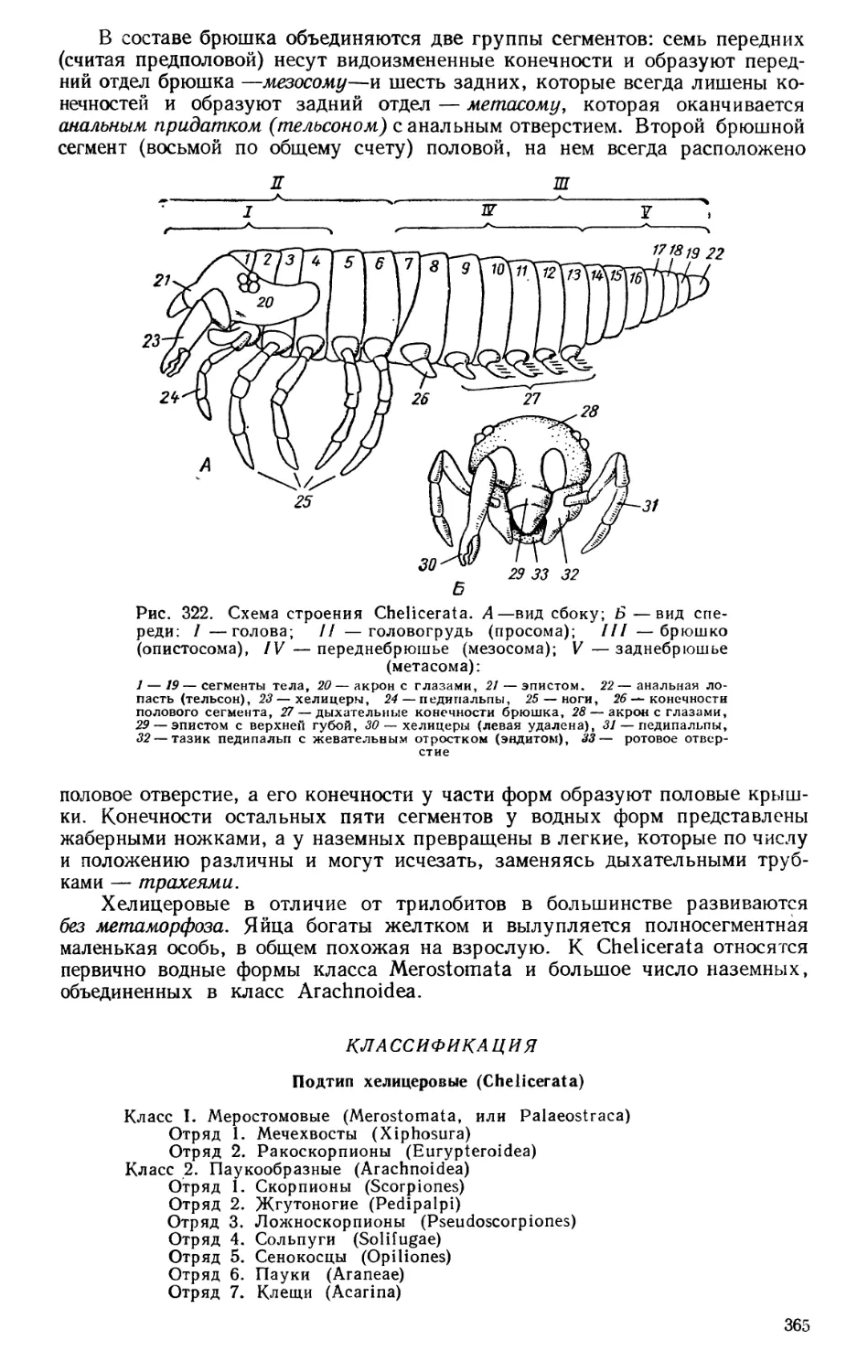
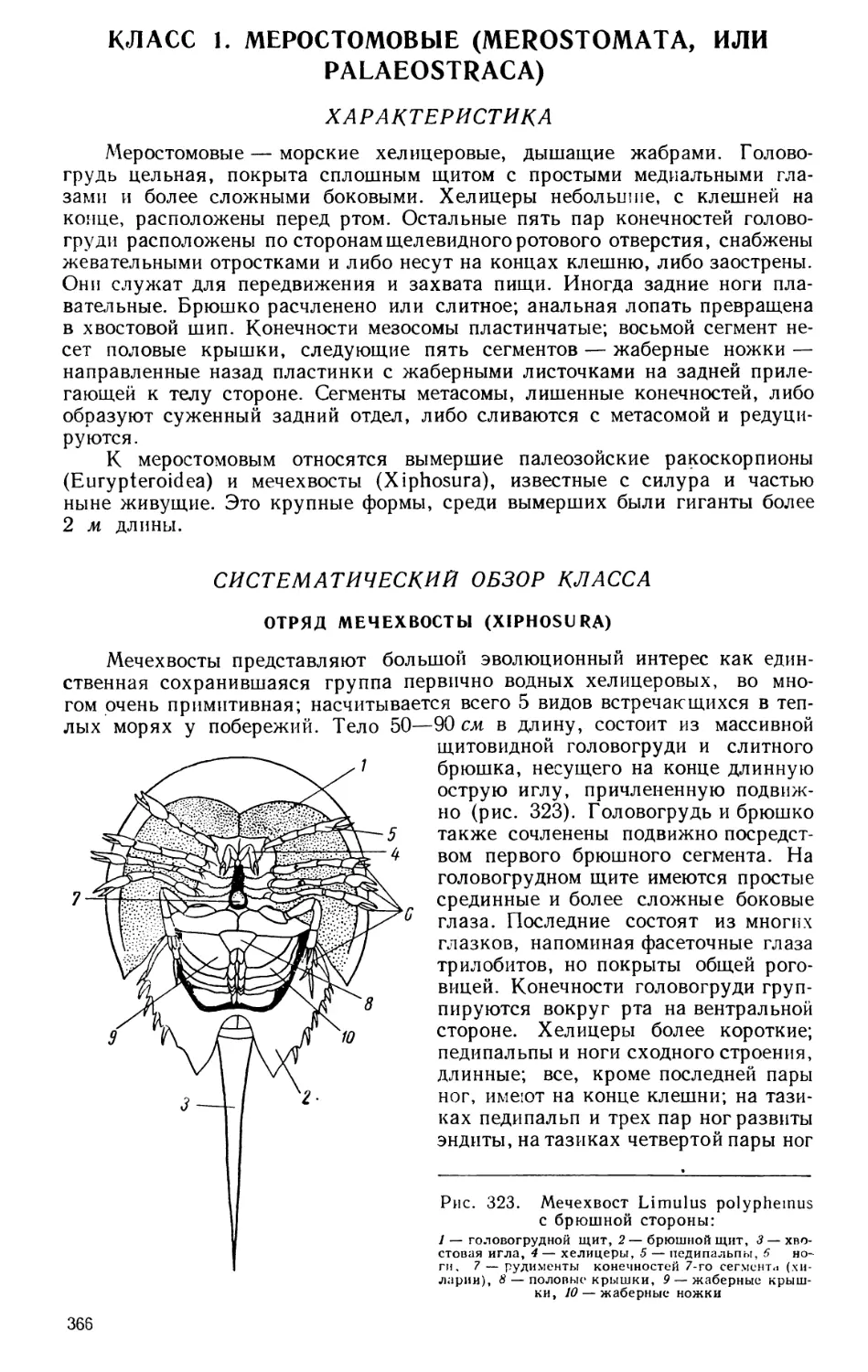

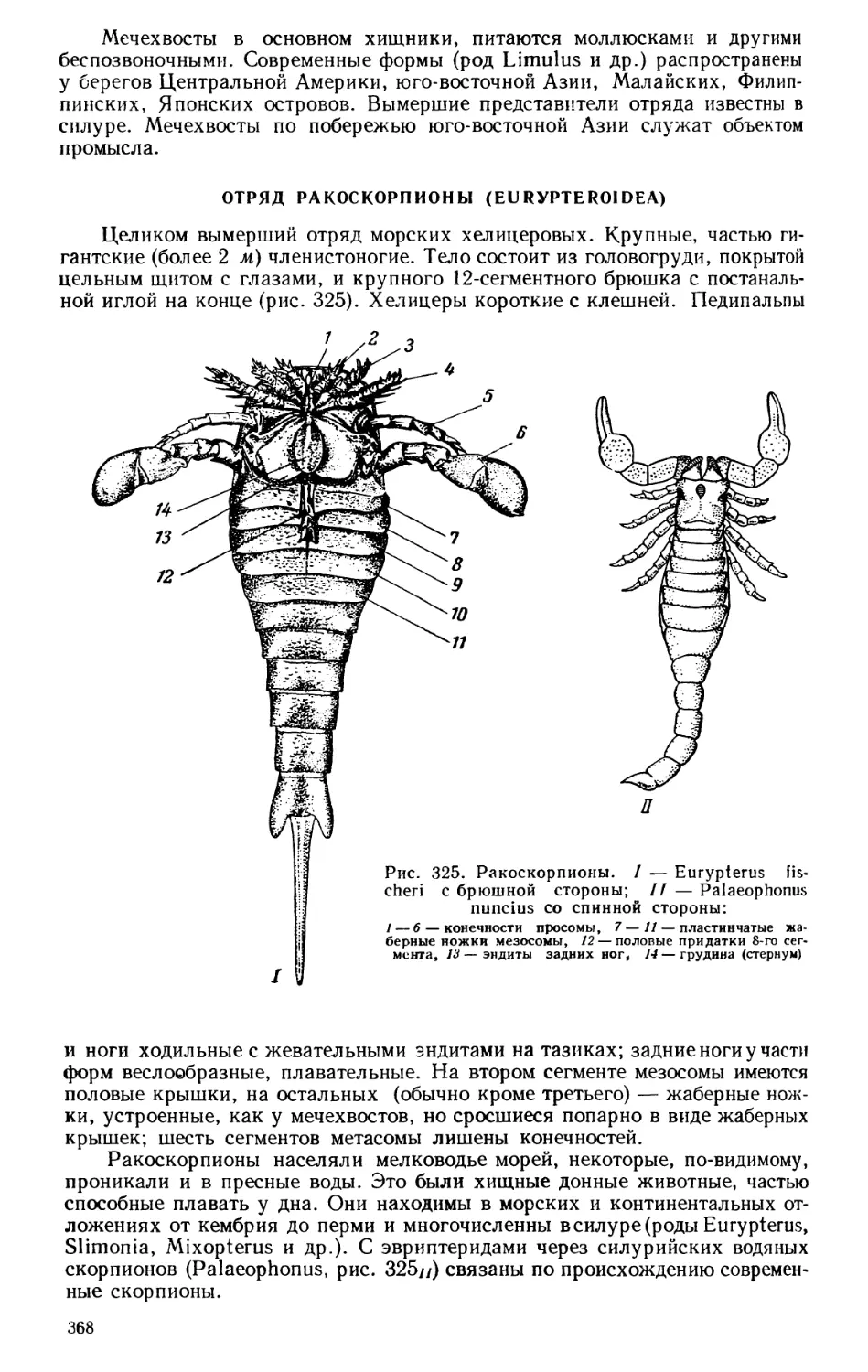
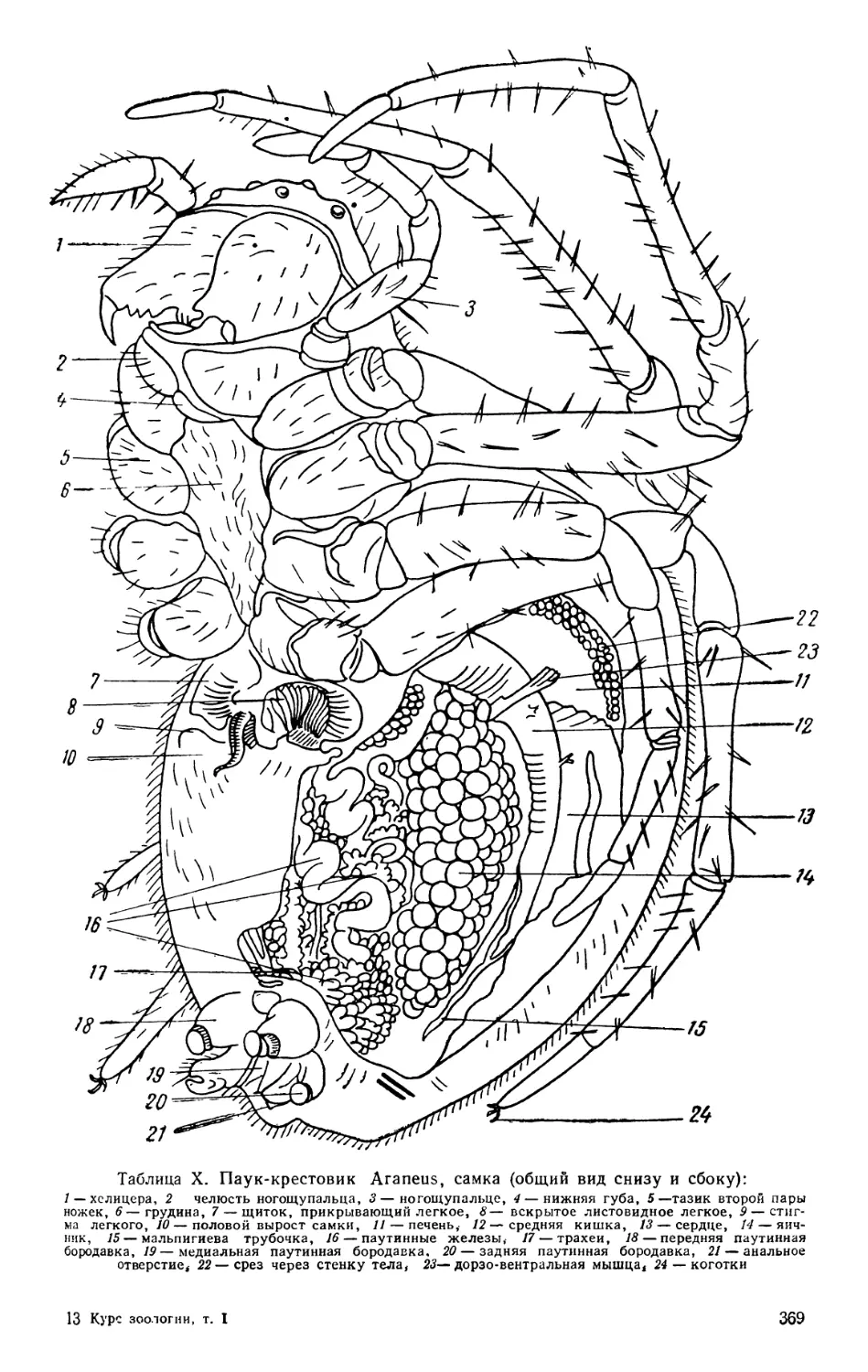
































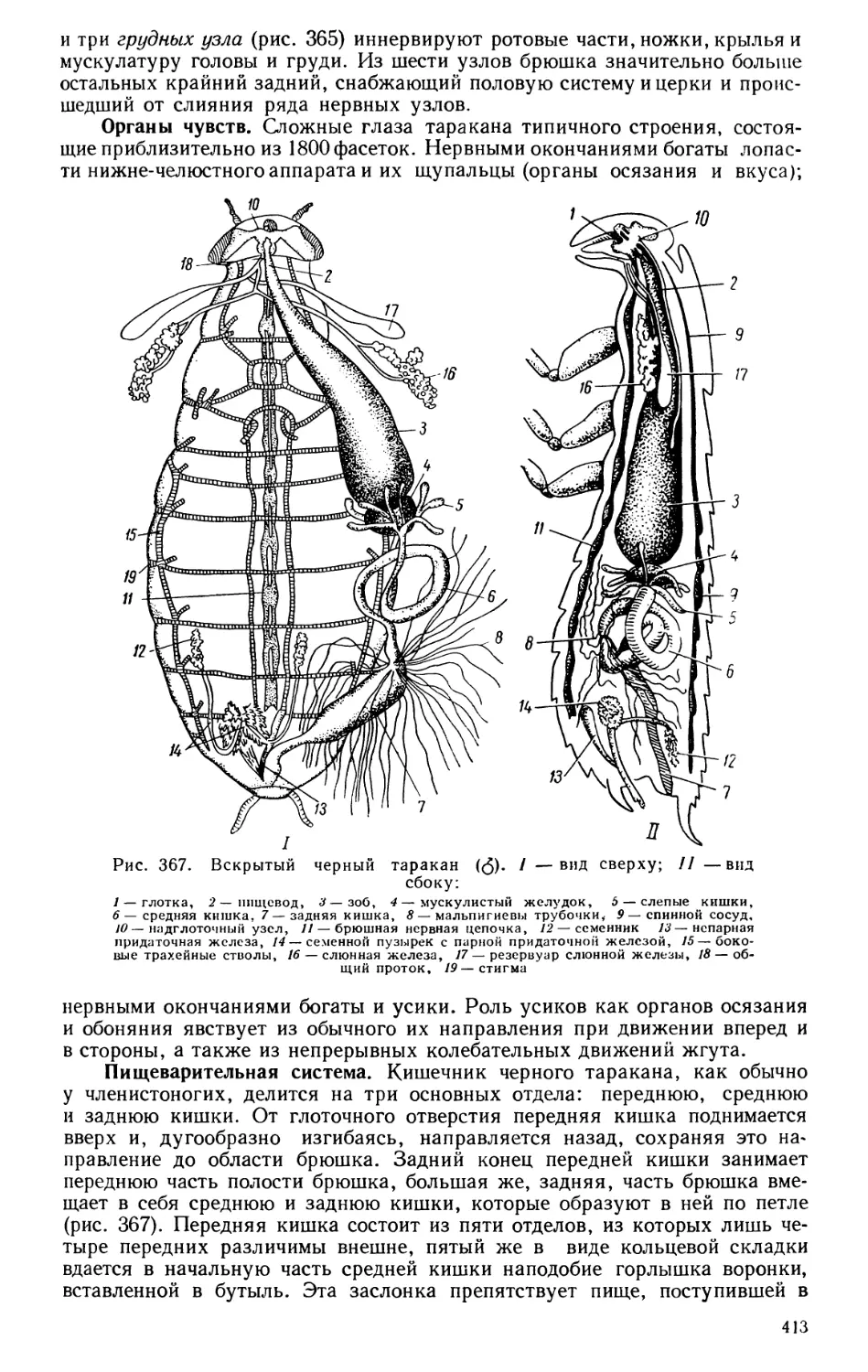


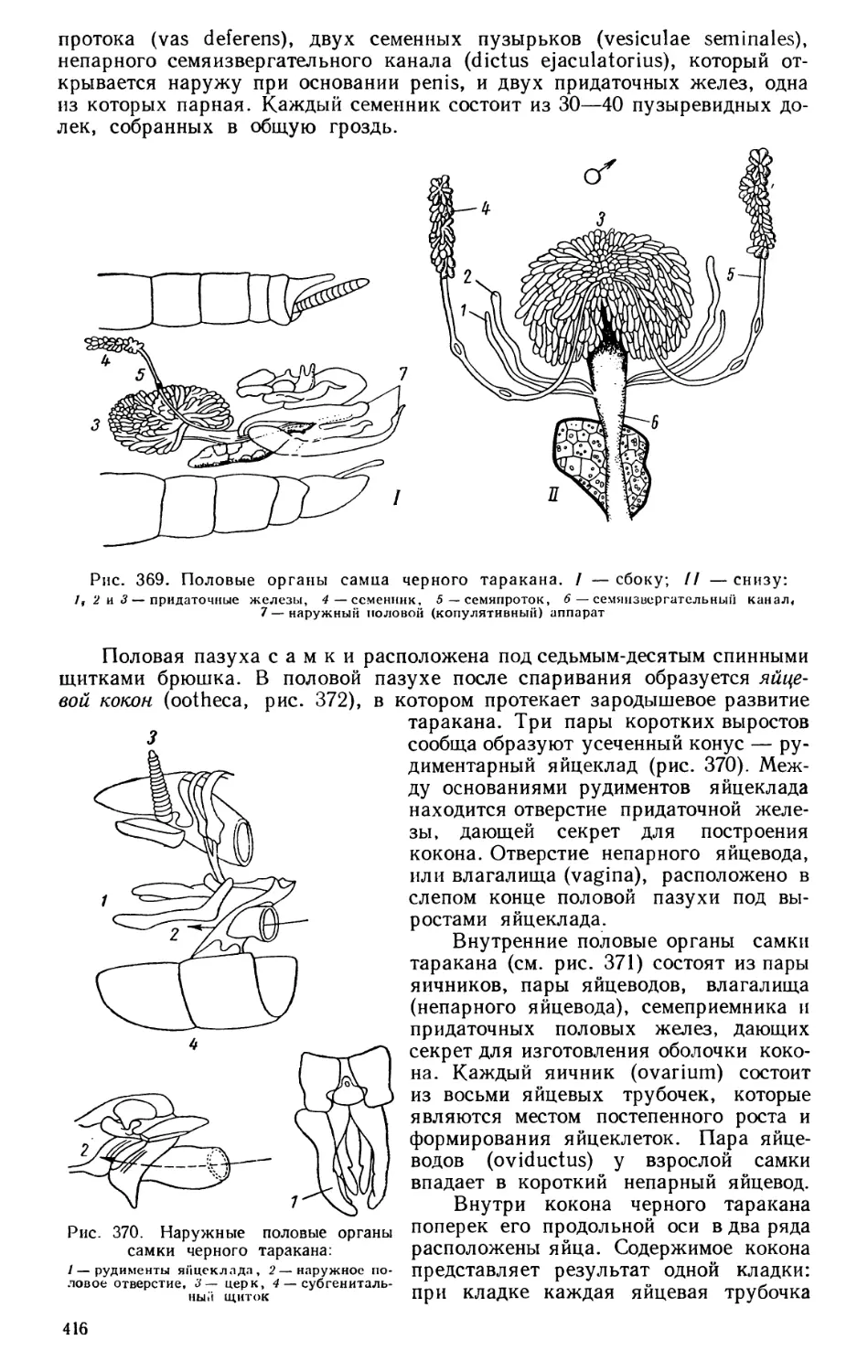
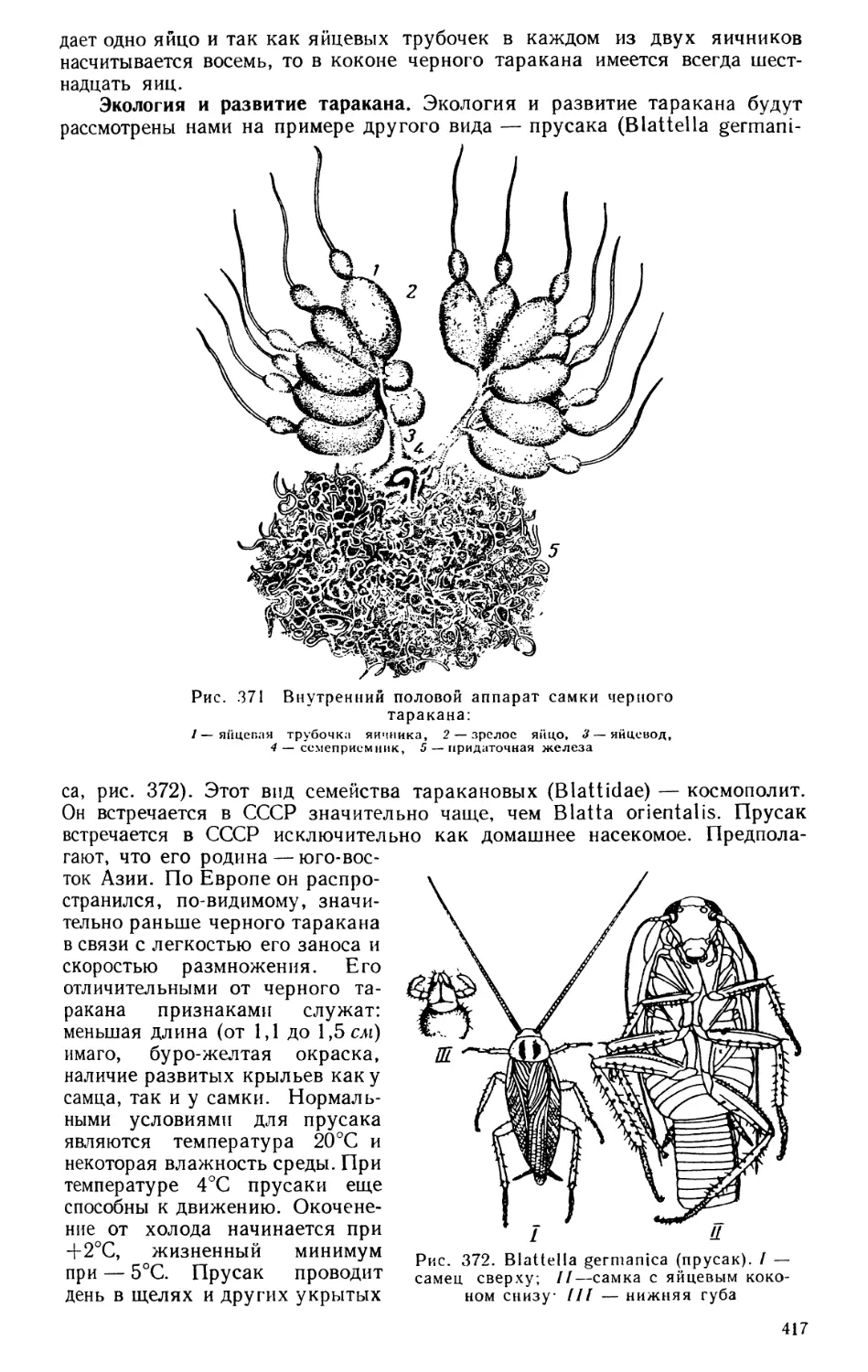


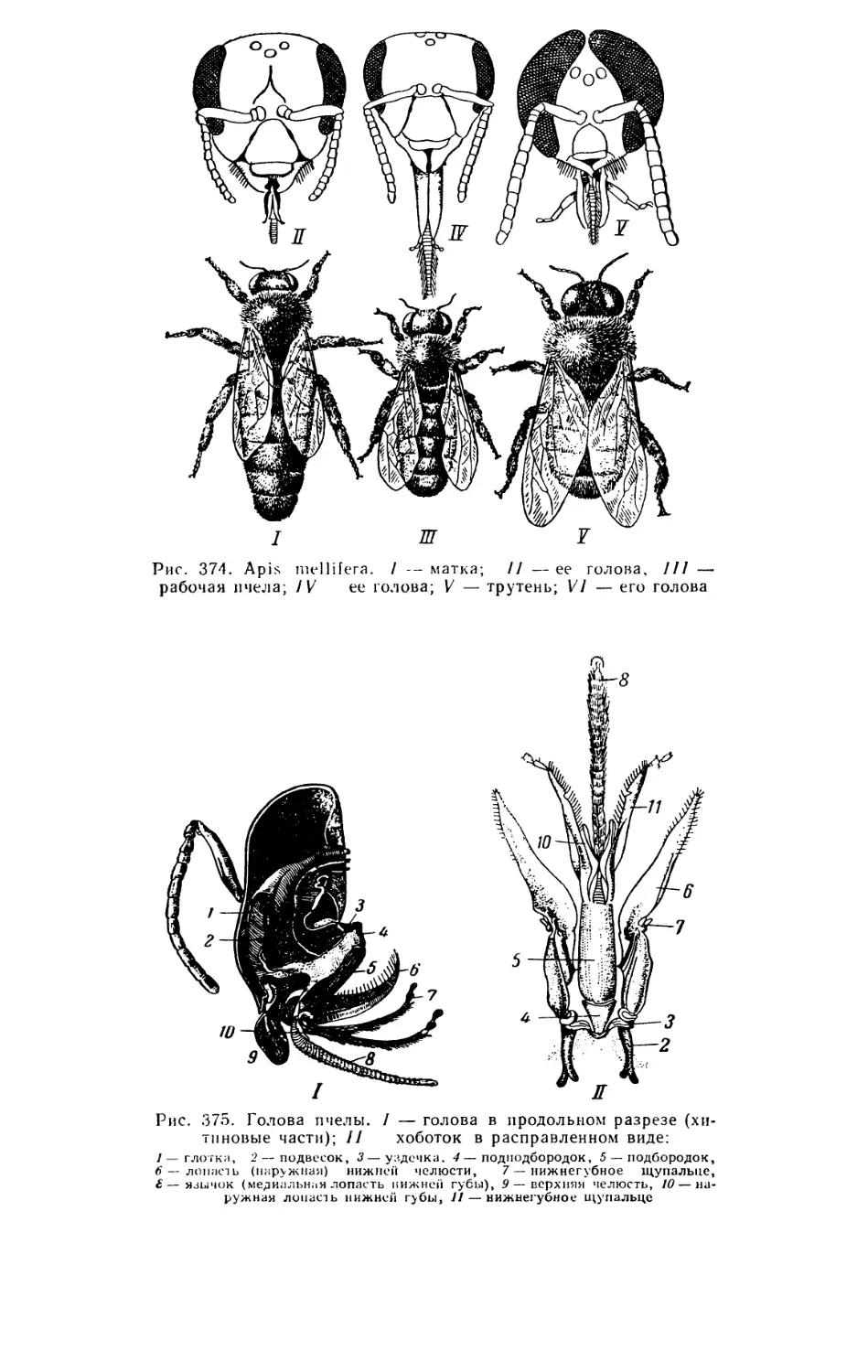
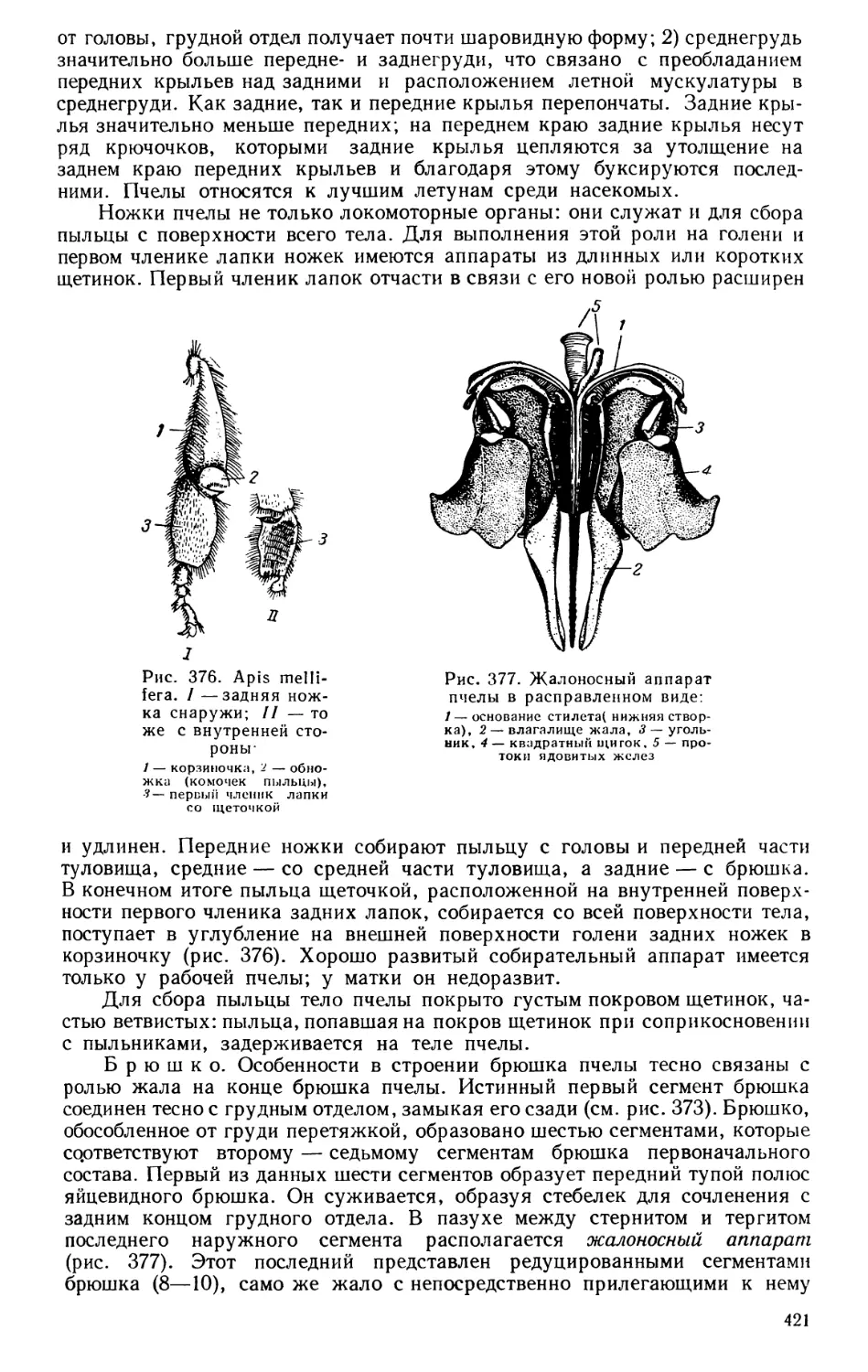


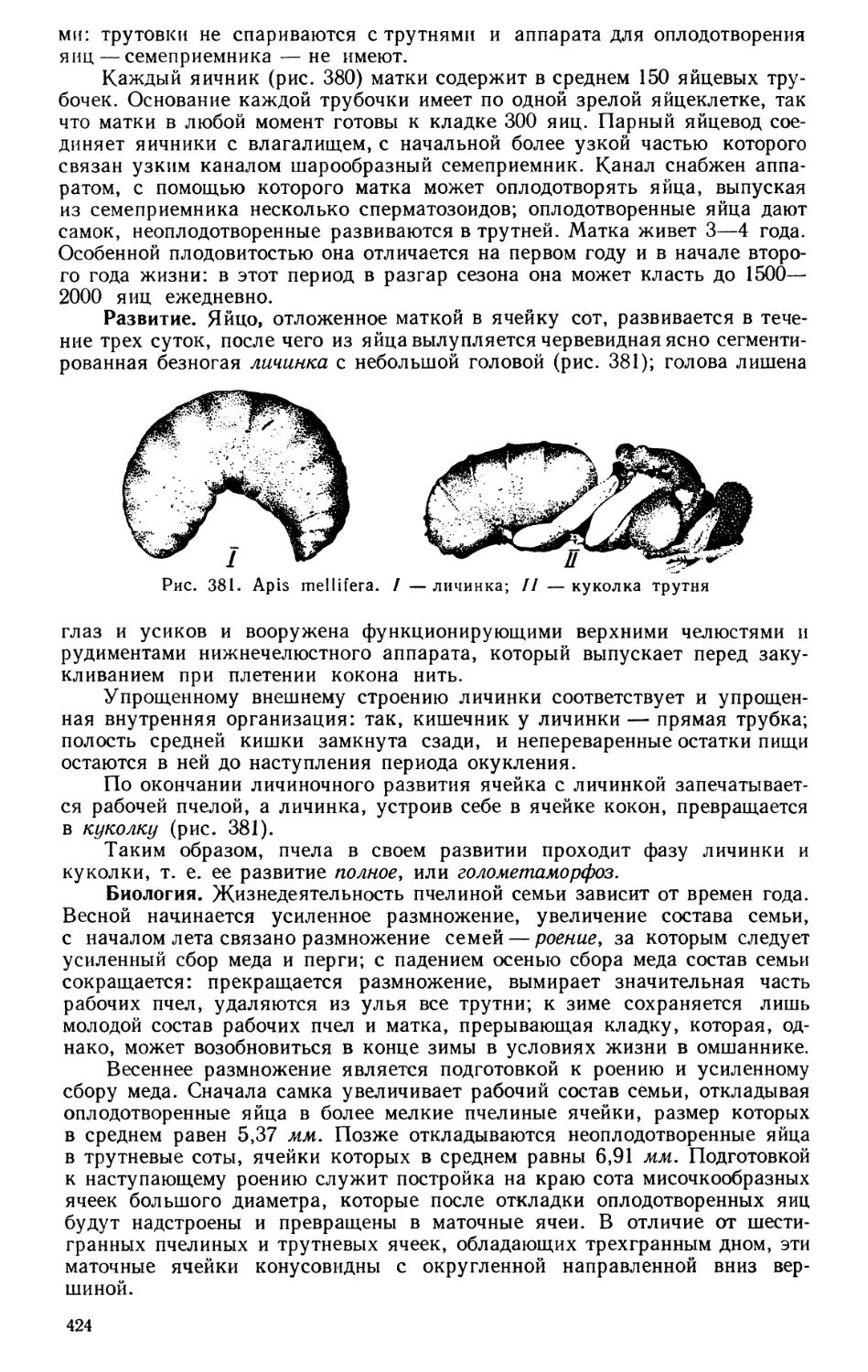

















































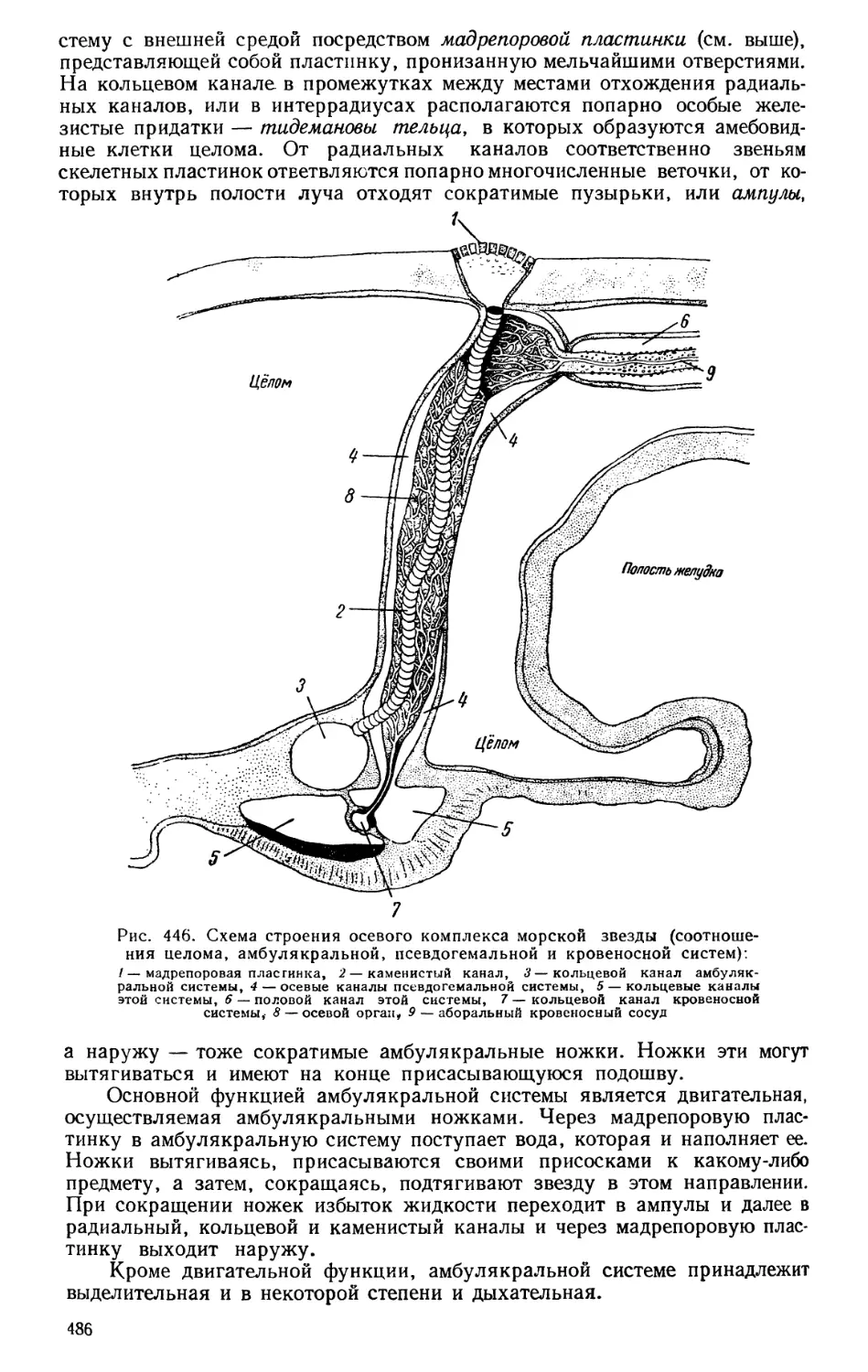

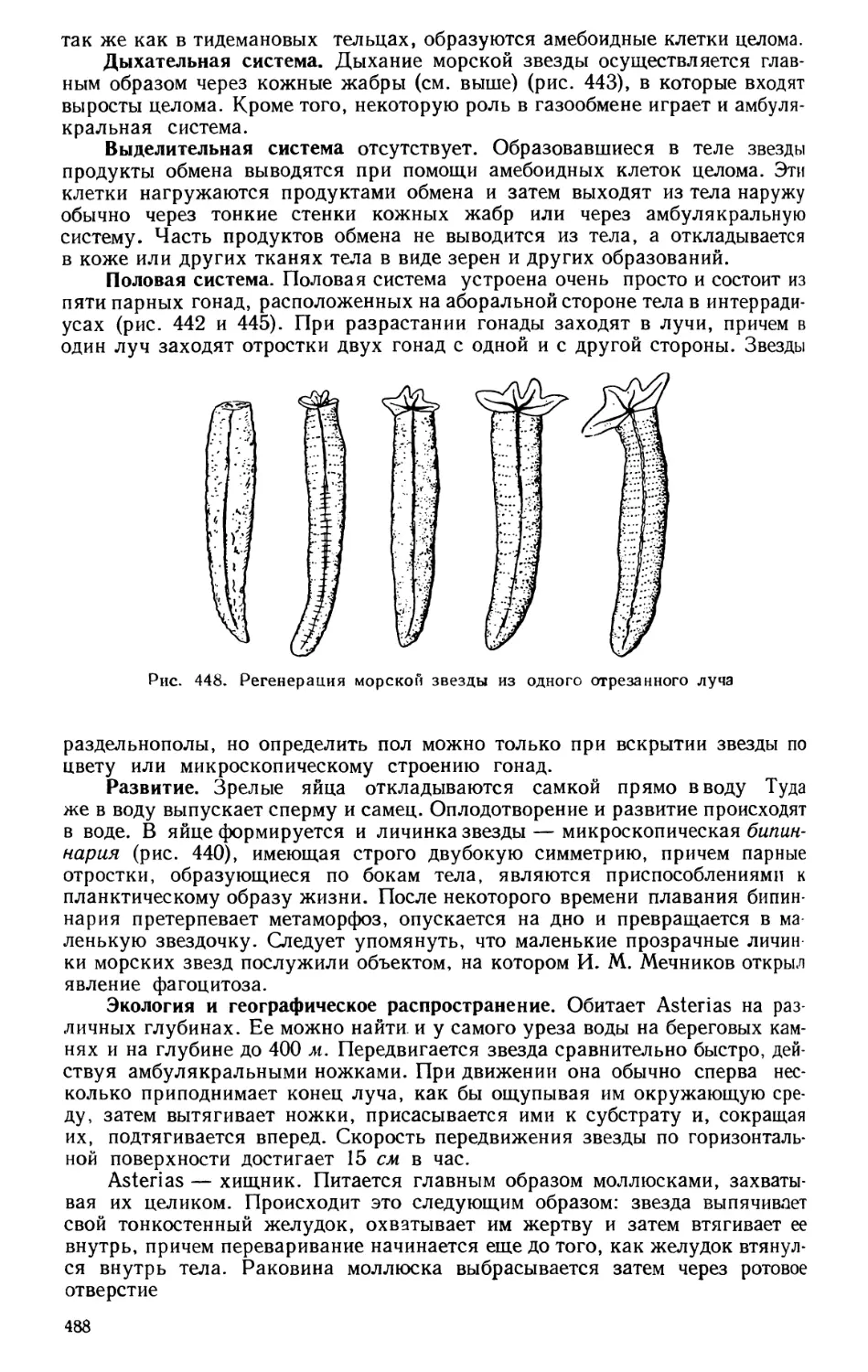
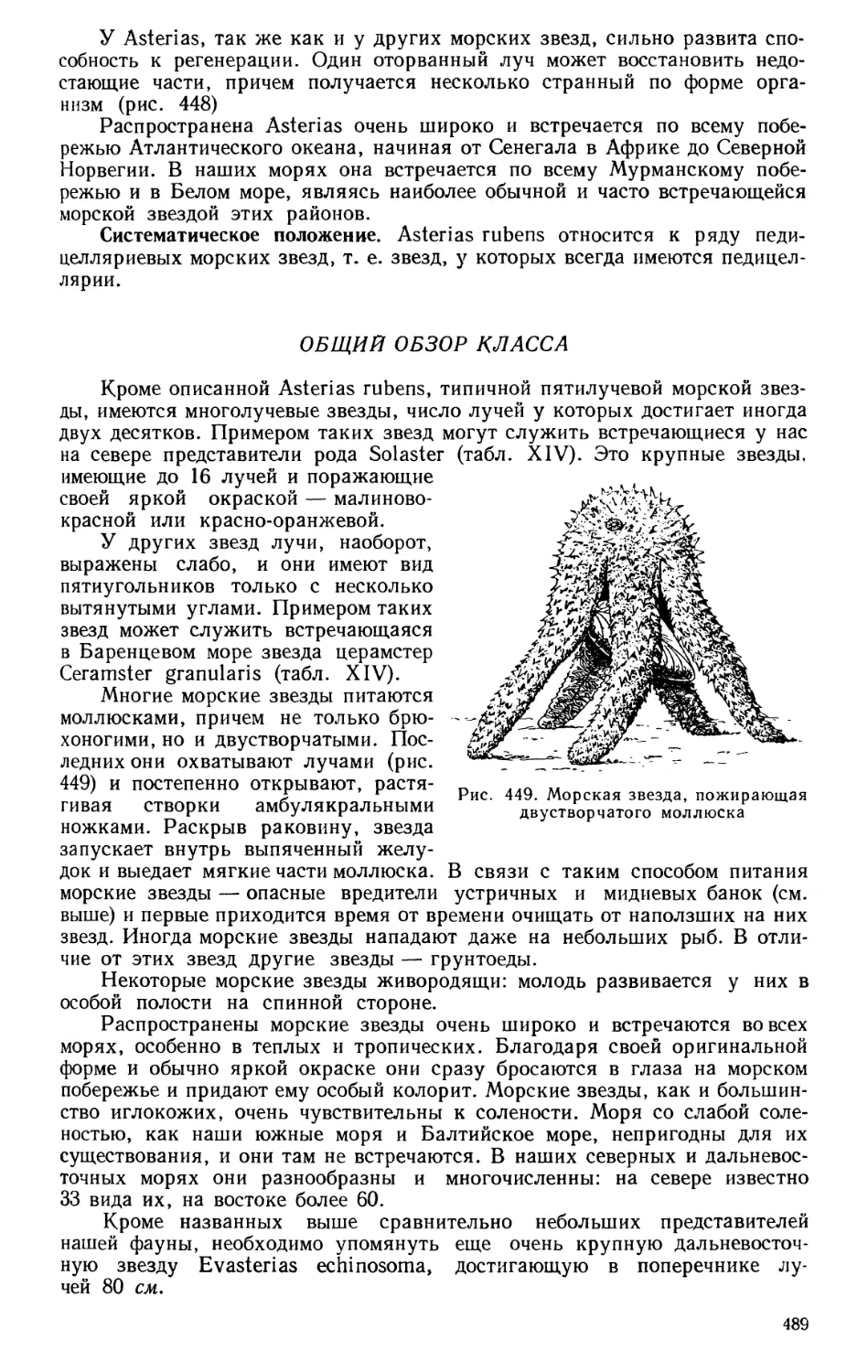

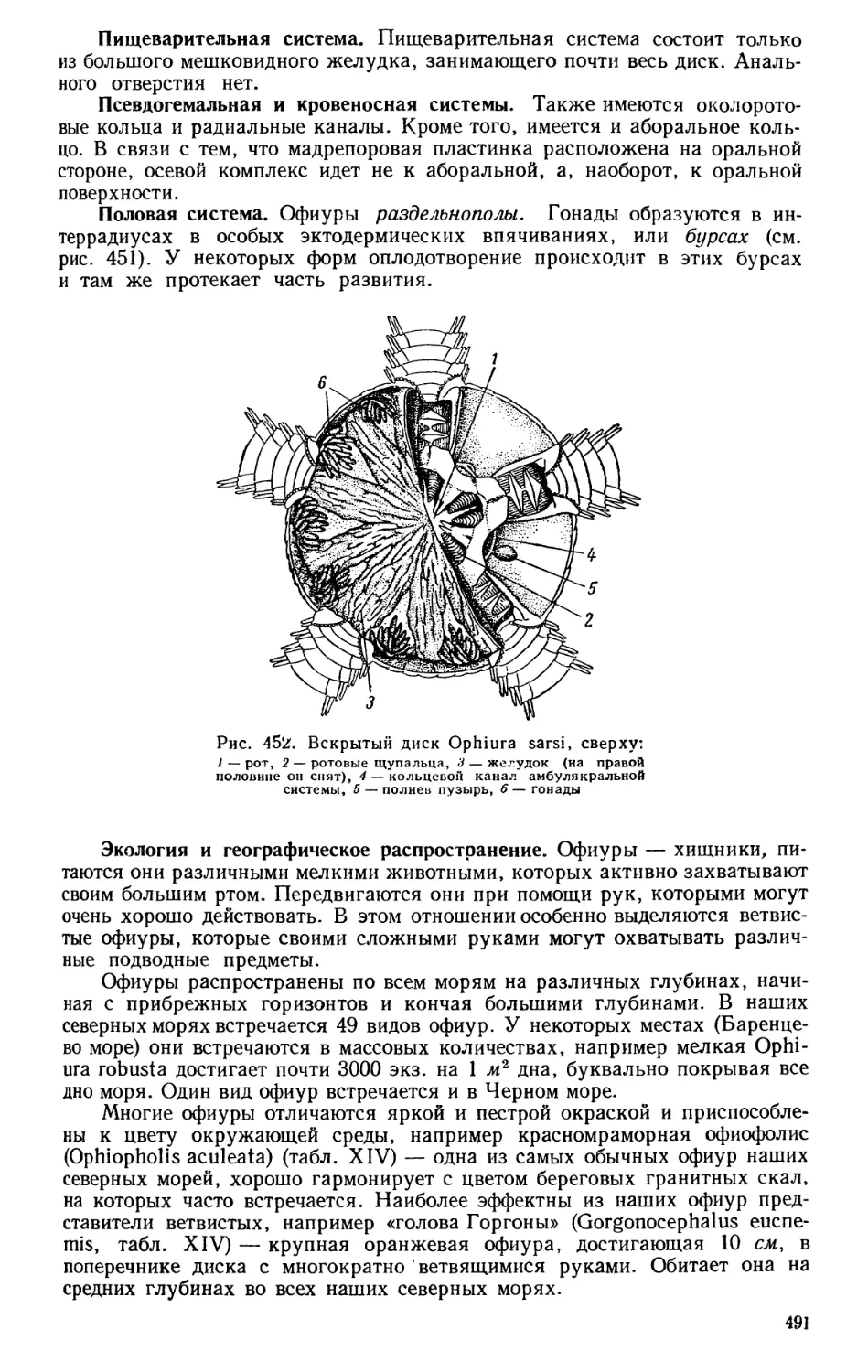

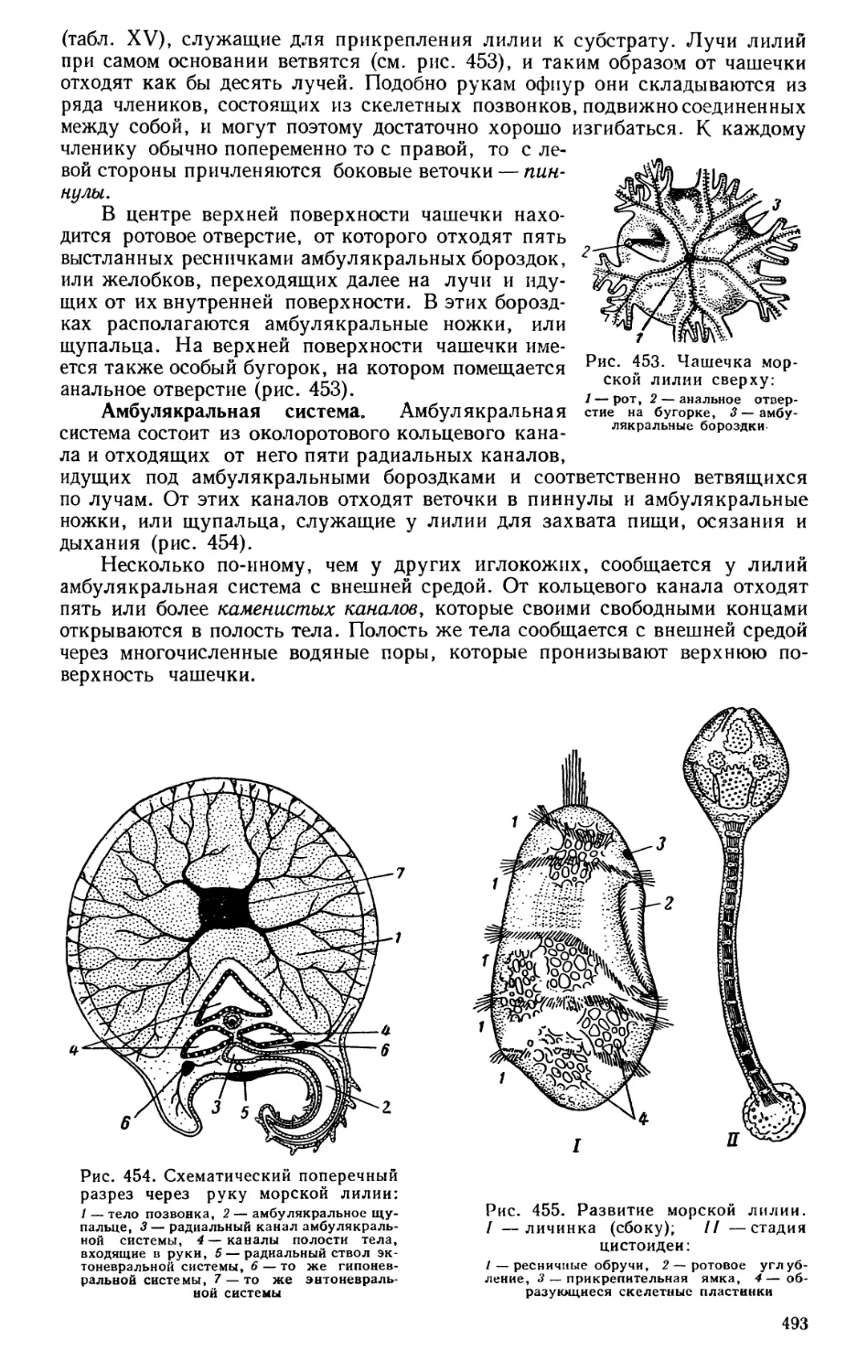


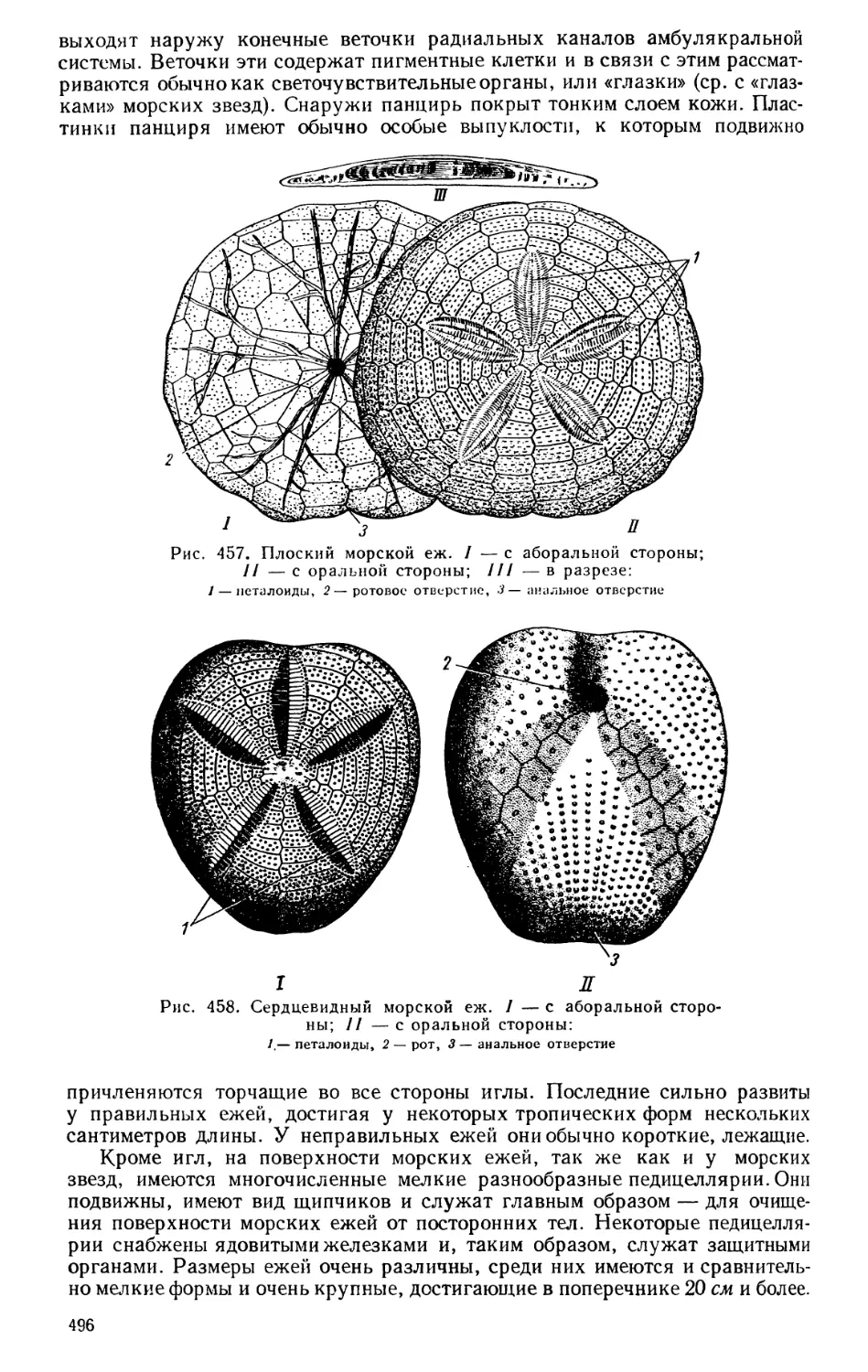




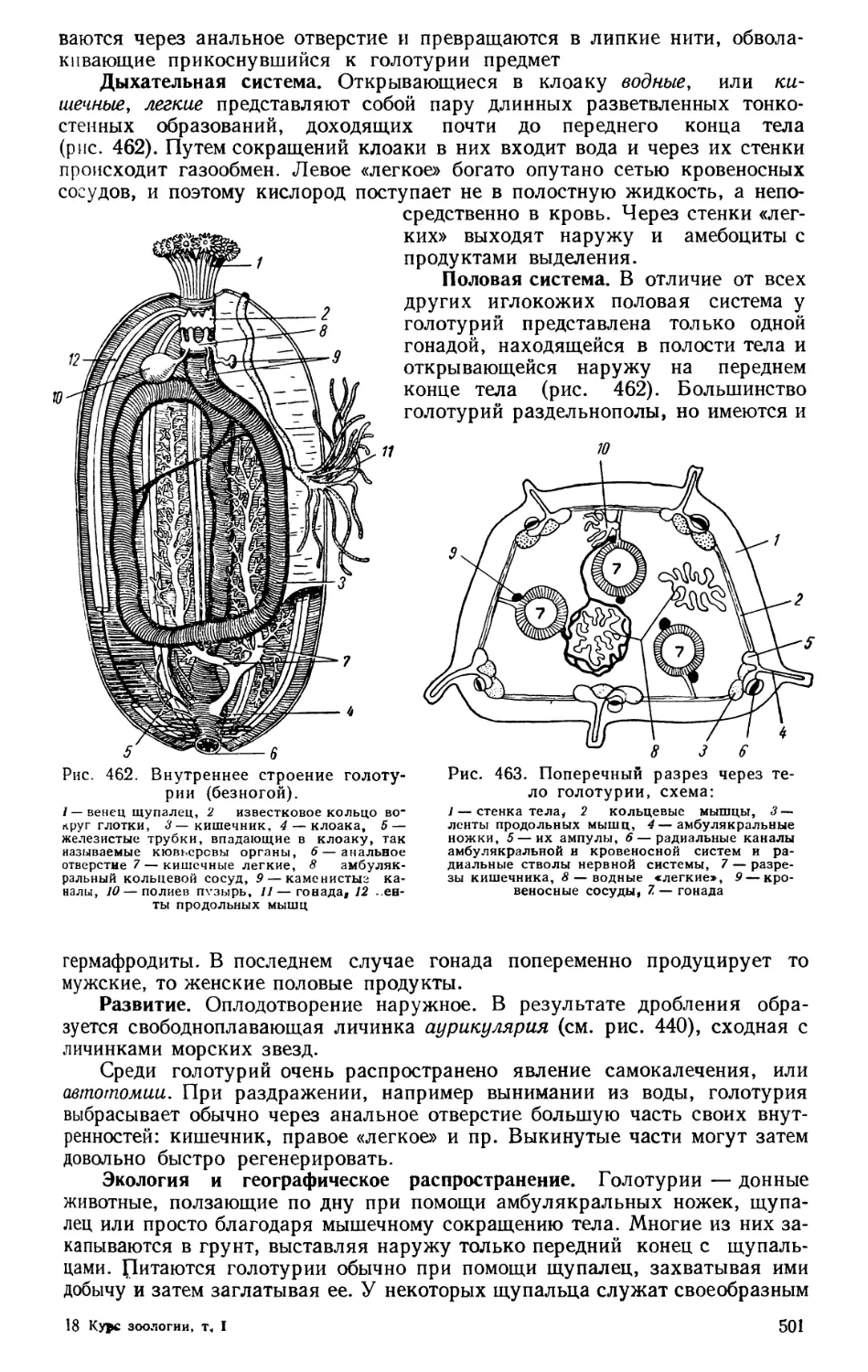
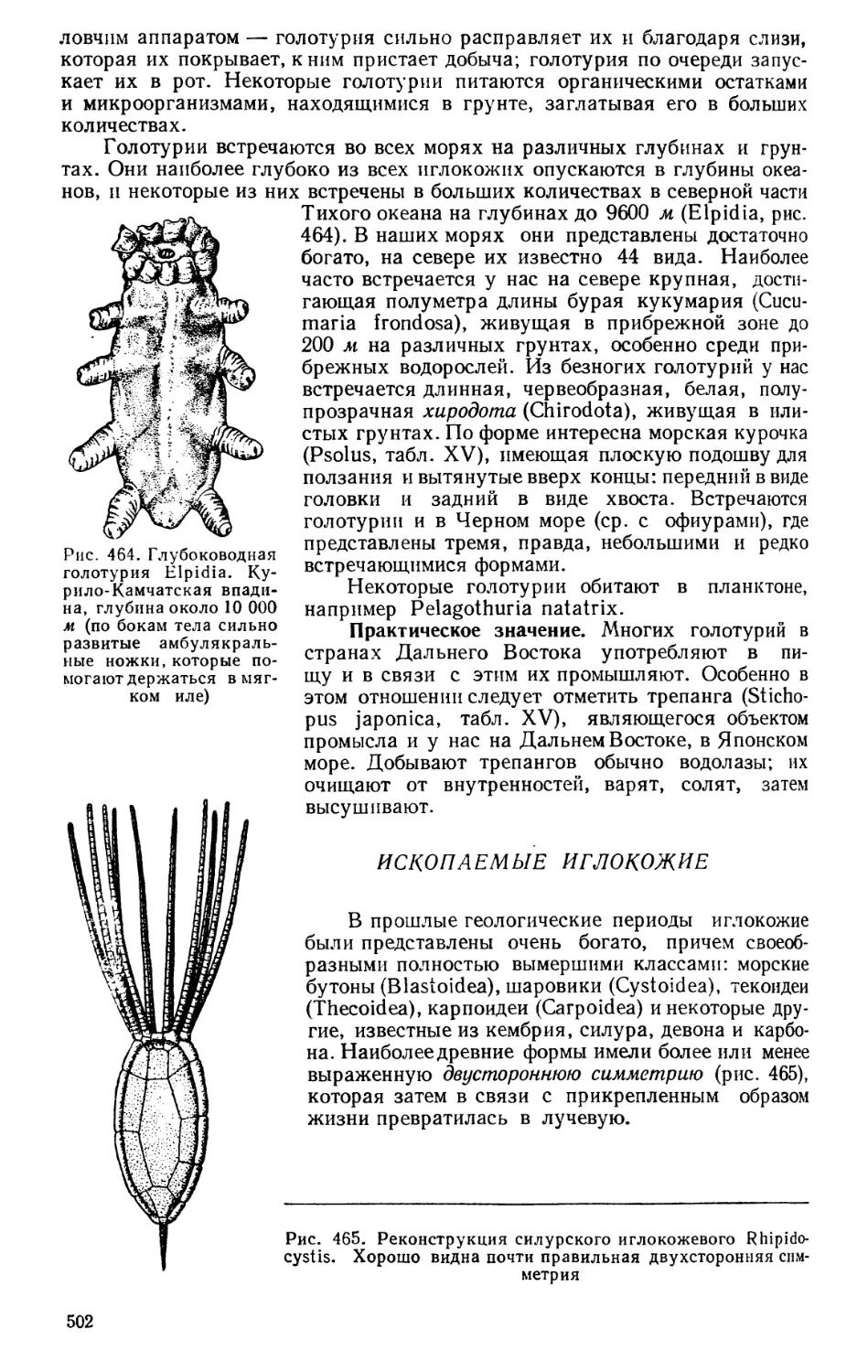



















































Текст
курс зоологии
Г. Г. АБРИКОСОВ, А. Г. БАННИКОВ,
| 3. Г, БЕККЕР, I Я. А. БИРШТЕЙН,
| Н. Г. БОБРИНСКИЙ, | А. Б. ЛАНГЕ,
Б. С. МАТВЕЕВ, П. В. МАТЕКИН,
А. А. МАХОТИН, А. А. ПАРАМОНОВ
ПРИ УЧАСТИИ В. Д. ЛЕБЕДЕВА,
А. Н. ФОРМОЗОВА и Н. Г. ДЕМЕНТЬЕВА
курс зоологии
В ДВУХ ТОМАХ
под редакцией
профессора Б. С. Матвеева
ИЗДАТЕЛЬСТВО «ВЫСШАЯ ШКОЛА»
МОСКВА — 1966
Г. Г. АБРИКОСОВ, 3. Г. БЕККЕР,
Я. А. БИРШТЕЙН, А. Б. ЛАНГЕ,
Л. Б. ЛЕВИНСОН, Б. С. МАТВЕЕВ,
П. В. МАТЕКИН, А. А. МАХОТИН,
А. А. ПАРАМОНОВ
курс зоологии
ЗООЛОГИЯ БЕСПОЗВОНОЧНЫХ
Под редакцией
Б. С. Матвеева и П. В. Матекина
Издание 7
ДОПУЩЕНО
МИНИСТЕРСТВОМ ВЫСШЕГО
И СРЕДНЕГО СПЕЦИАЛЬНОГО
ОБРАЗОВАНИЯ СССР
В КАЧЕСТВЕ УЧЕБНИКА
ДЛЯ СТУДЕНТОВ УНИВЕРСИТЕТОВ
Георгий Георгиевич Абрикосов,
|Эрнест Георгиевич Беккер,|
Яков Авидьевич Бирштейн,
Александр Борисович Ланге,
Леон Бенцианович Левинсон,
Борис Степанович Матвеев,
Петр Владимирович Матекин,
Анатолий Анатольевич Махотин,
Александр Александрович Парамонов
КУРС ЗООЛОГИИ
ТОМ I
ЗООЛОГИЯ БЕСПОЗВОНОЧНЫХ
Редактор В. С. Капышева
Художественный редактор Э. А. Марков
Технический редактор С. С. Горохова
Корректор М. А. Якушева
Т-09543. Сдано в набор 4/XI 1965 г.
Подп. к печати 4/VII 1966 г. Формат
70xl08,/ie- Объем 34,3 п. л.+6 цв. вкле-
ек (0,375 п. л.), 48,3 усл. печ. л.,
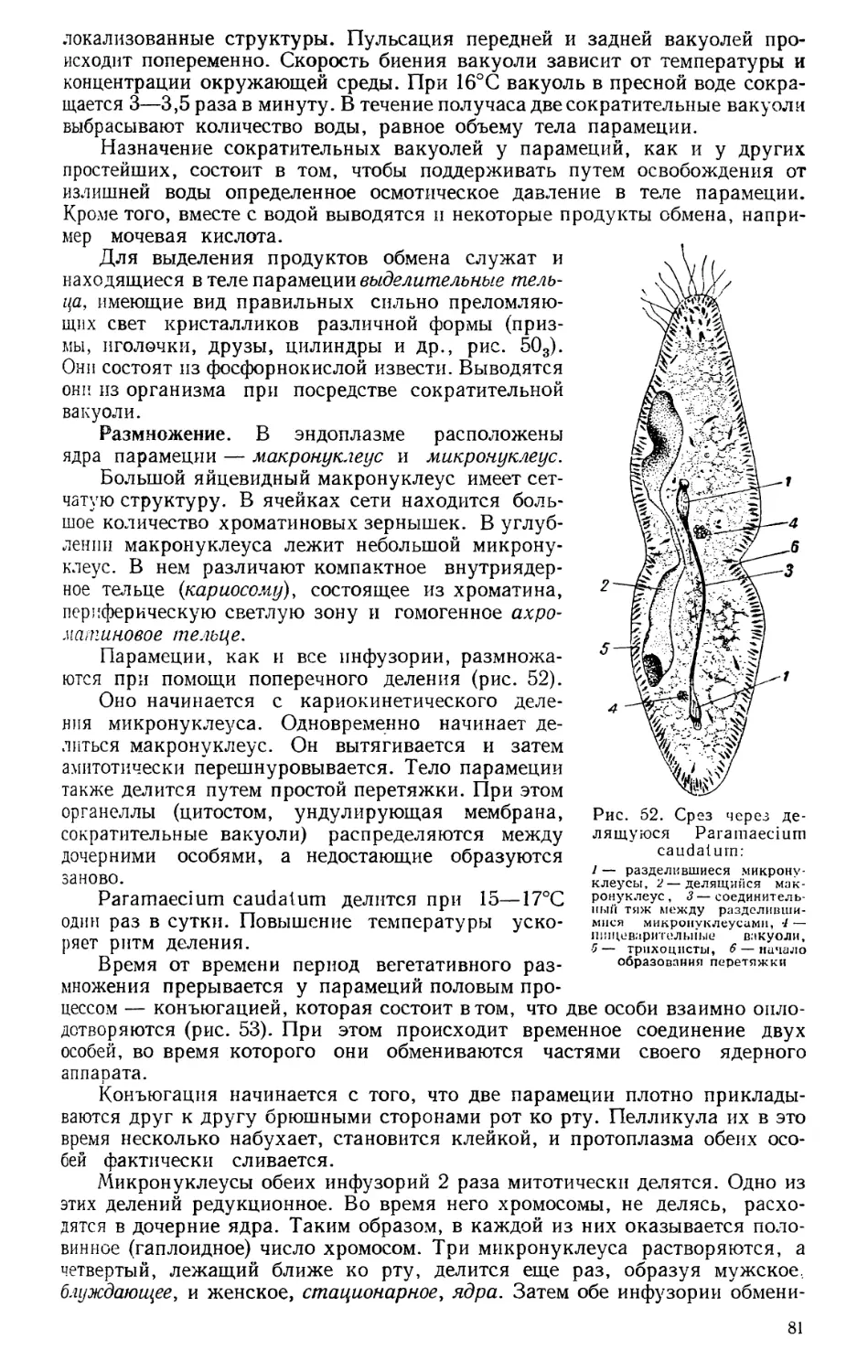
45,09 уч.-изд. л. Изд. № Е—14/64.
Тираж 40000 экз. Зак. 857. Цена 1 р. 75
Тематический план издательства «Выс-
шая школа» (вузы и техникумы) на
1966 г. Позиция № 169.
Москва, И-51, Неглинная ул., д. 29/14,
издательство «Высшая школа».
Ярославский полиграфкомбинат Главпо-
лиграфпрома Комитета по печати при
Совете Министров СССР. Ярославль,
ул. Свободы, 97.
2—10—6
169—66
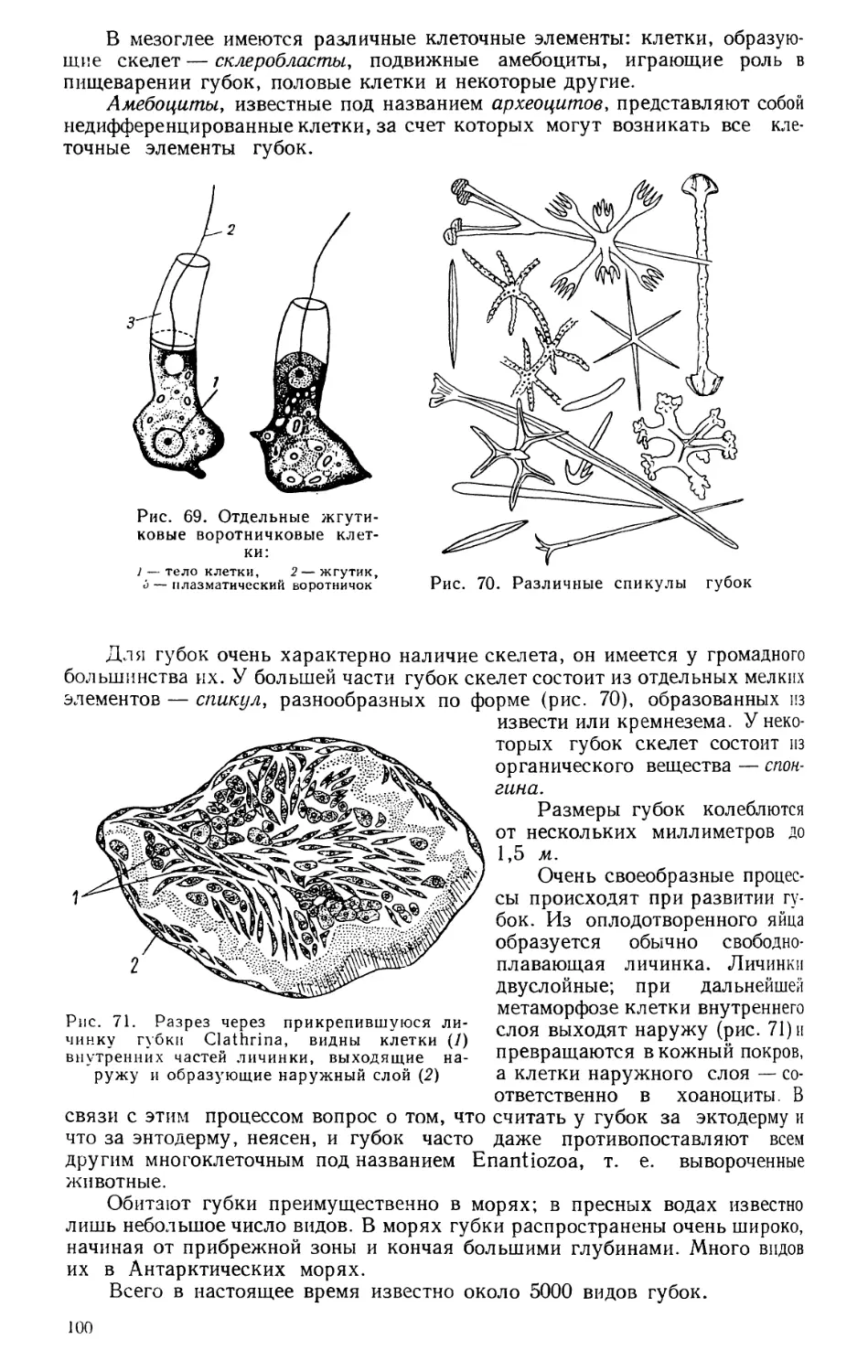
ПРЕДИСЛОВИЕ К СЕДЬМОМУ ИЗДАНИЮ
Два тома «Курса зоологии» — том I «Зоология бес-
позвоночных» и том II «Зоология позвоночных» — за
тридцать лет своего существования получили широкое
распространение не только в республиках Советского Со-
юза, но и во многих зарубежных демократических странах.
В настоящем, седьмом издании текст и рисунки полностью
пересмотрены, внесены дополнения, изменения, сделаны
замены соответственно новым научным данным.
При переработке I тома «Курса зоологии» внесены
следующие изменения: «Введение» сильно сокращено и в
значительной части заново написано проф. Б. С. Матвее-
вым. «Простейшие» пересмотрены проф. Л. Б. Левин-
соном; в этот раздел внесены новые данные по изучению
строения клетки с применением электронной микроскопии
и биохимии. Разделы, посвященные губкам, кишечнополост-
ным, мшанкам, плеченогим, иглокожим и погонофорам,
пересмотрены Г. Г Абрикосовым при участии Ф. А. Лав-
рехина; внесены дополнения взамен старого текста. Раз-
делы, посвященные типам плоских червей, круглых чер-
вей, немертинам и кольчатым червям, пересмотрены проф.
А. А. Парамоновым. Соответственно современным научным
данным по систематике и классификации «беспозвоноч-
ных животных» общее наименование для всех них «чер-
ви» изъято, так как в настоящее время принято не счи-
тать их единой генетической группой; они развивались
самостоятельными путями от примитивных двусторонне-
симметричных животных. Соответственно этому общий
раздел поделен на два самостоятельных: «Плоские, круг-
лые черви и немертины» и «Кольчатые черви». В текст
внесены дополнения путем замены старых текстов новыми.
Тип моллюски заново переработан доктором биологиче-
ских наук П. В. Матекиным, внесены новые рисунки.
Тип членистоногие вследствие смерти Э. Г. Беккера
переработан рядом новых авторов.
5
Подтип жабернодышащие, включающий класс рако-
образных, заменен новым текстом, написанным проф.
Я. А. Бирштейном, заменены некоторые рисунки. Подти-
пы трилобитов и подтип хелицероносных, включающий
паукообразных и клещей, класс меростомовых также за-
ново составлены доц. А. Б. Ланге. Подтип трахейнодыша-
щих, включающий классы первичнотрахейных, многоножек
и насекомых, пересмотрен заново А. А. Махотиным. За-
менены общие характеристики, сделаны дополнения путем
замены старых текстов и перестроен порядок раздела «Си-
стематический обзор».
Эволюционное развитие беспозвоночных пересмотрено
Г. Г. Абрикосовым и Б. С. Матвеевым, внесены допол-
нения соответственно новому тексту книги.
Редакторы Б. С. Матвеев, П. В. Матекин
ВВЕДЕНИЕ
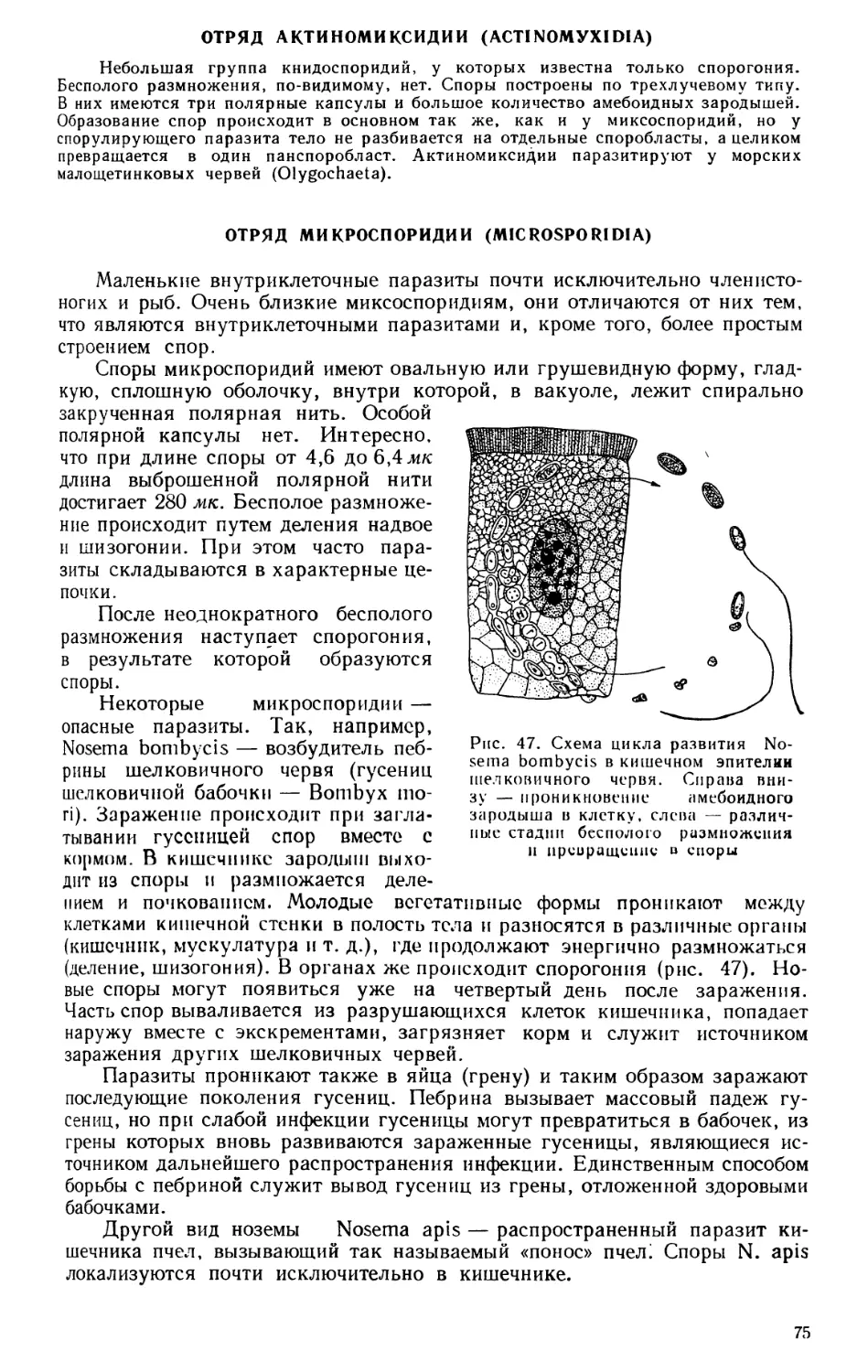
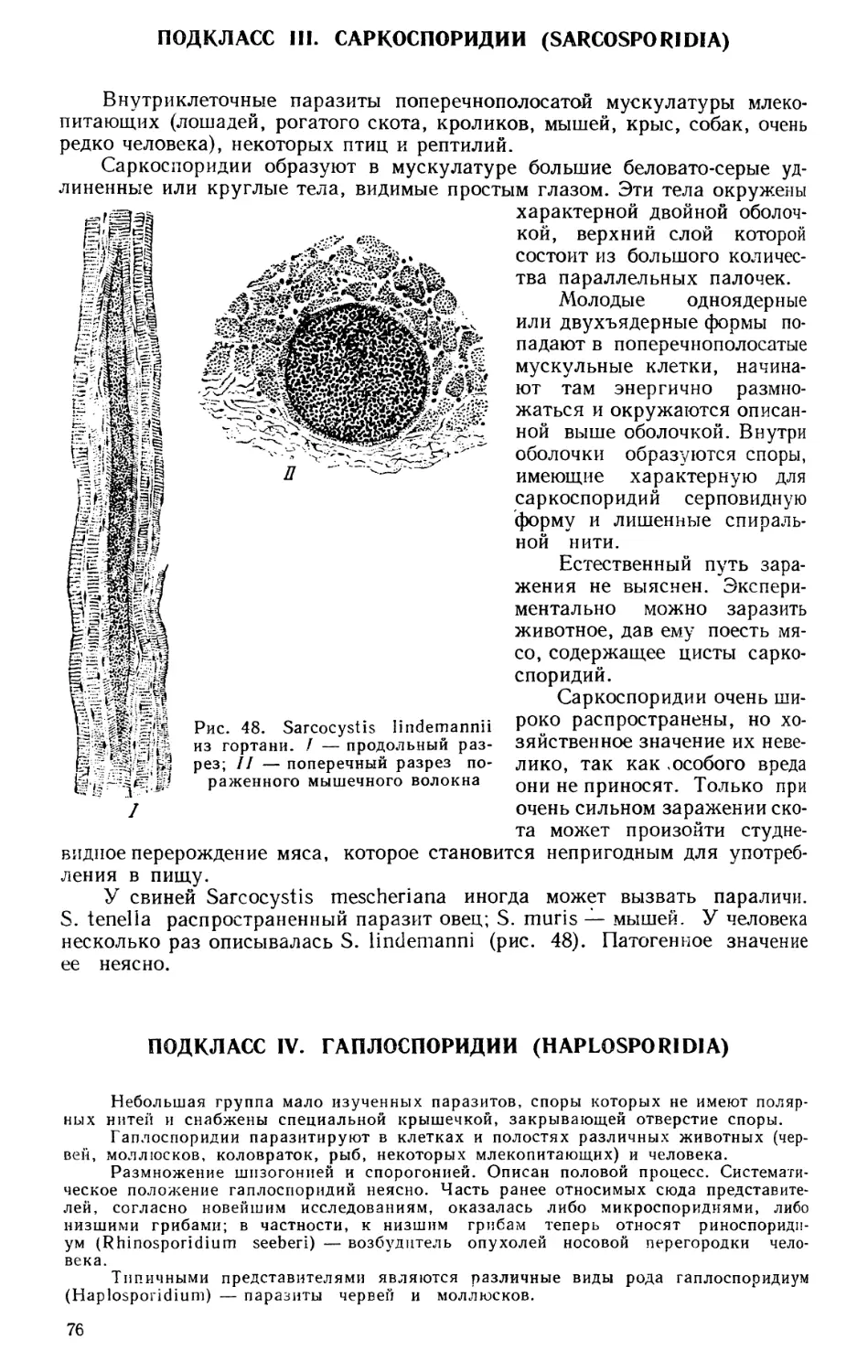
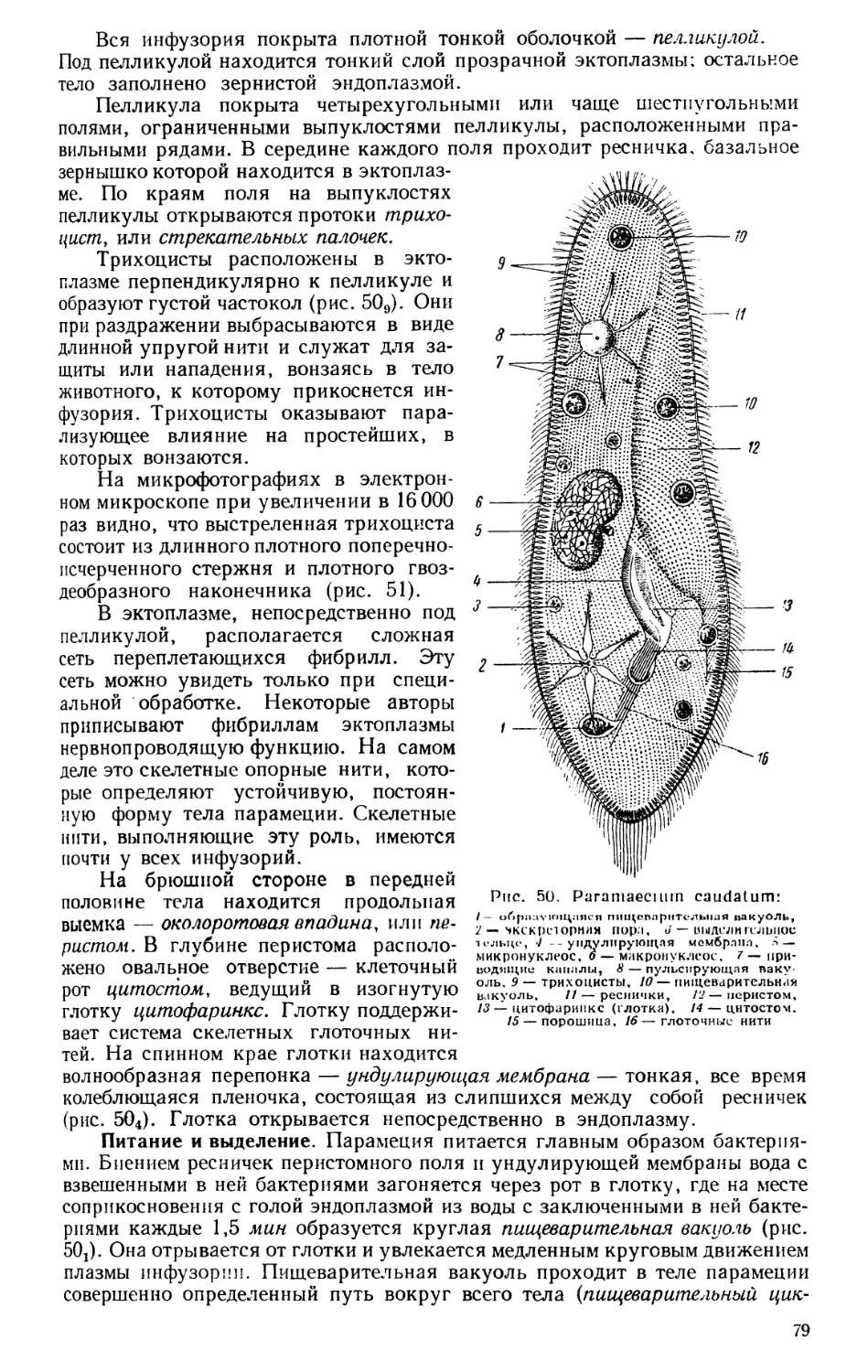
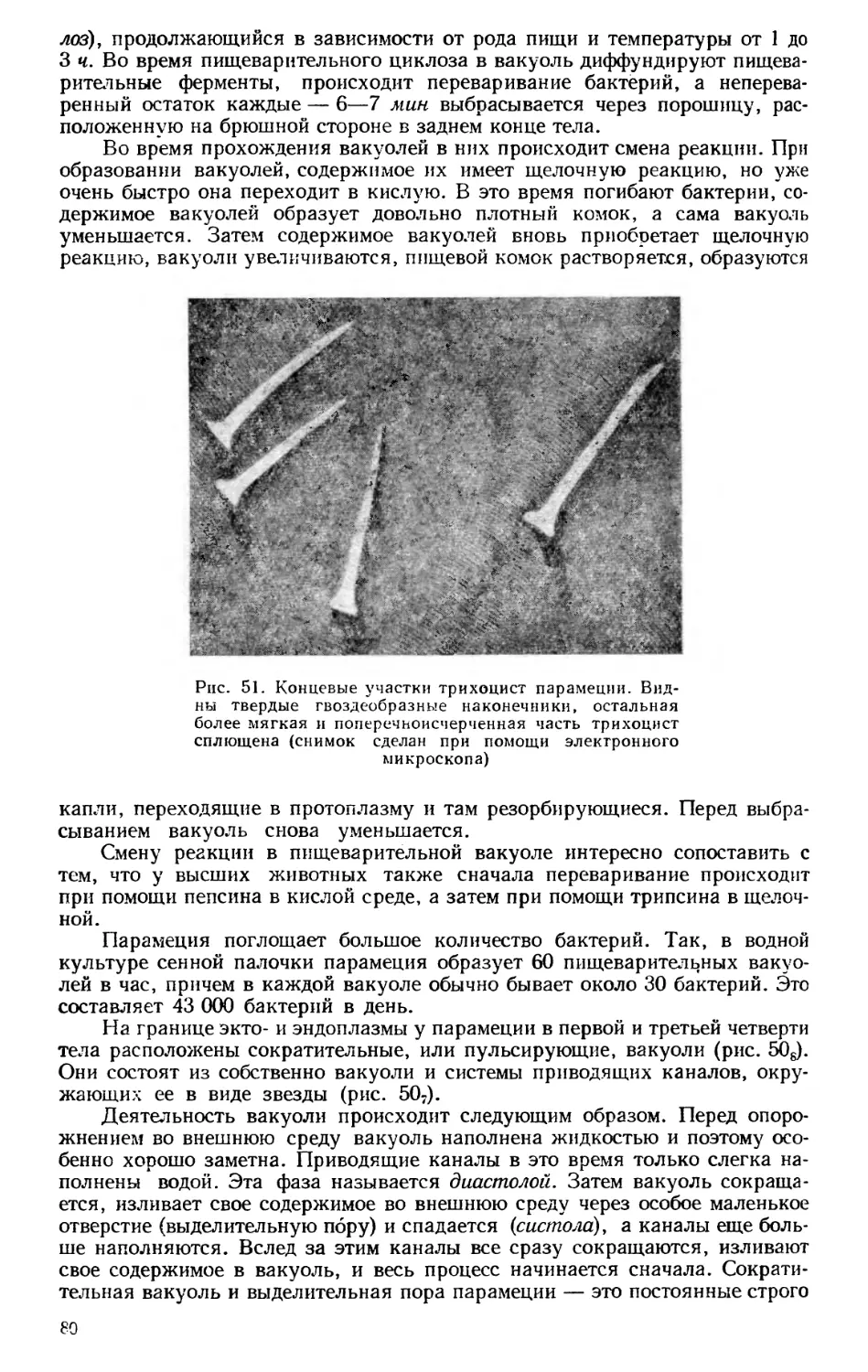
ЗООЛОГИЯ КАК СИСТЕМА НАУК
Зоология — наука о животных — в настоящее время представляет
собой не единую науку, а систему наук, изучающих животный мир с самых
различных точек зрения. Изучаются строение и жизненные отправления
животных как во взрослом состоянии, так и в разные периоды развития,
состав фауны того или иного района и особенности географического рас-
пространения животных, связь животных организмов с условиями сущест-
вования и, наконец, закономерности эволюционного развития животных.
Кроме того, изучаются виды животных, полезные человеку в хозяйст-
венной деятельности, с целью увеличения добычи диких и повышения
продуктивности домашних, а также и вредные животные с целью, наоборот,
выработки рациональных мер борьбы.
В зоологии выделяются, с одной стороны, науки, изучающие отдель-
ные стороны жизни животных — их строение, развитие, жизнедеятель-
ность, распространение, связь с внешней средой и т. д.; с другой стороны, —
науки, изучающие отдельные, наиболее крупные и практически важные
группы животных (частная зоология). К первой группе относятся следую-
щие зоологические науки.
Морфология — наука о преобразовании формы, т. е. о строении
животных в зависимости от изменения функций и условий существования
организмов в индивидуальном и историческом развитии. В морфологию
входит ряд более частных дисциплин: анатомия — изучающая строение
и соотношение органов; сравнительная анатомия — изучающая изменения
и превращения формы и функции органов различных животных путем
сравнения сходства и различий в организации как современных, так и
ископаемых животных; гистология — изучающая микроскопическое строе-
ние животных.
Эмбриология — наука об индивидуальном развитии животных
и закономерностях этого развития.
Физиология — наука о жизненных отправлениях животных,
т. е. о процессах, совершающихся в организме (пищеварение, дыхание,
выделение, кровообращение, передача нервного возбуждения, размножение
и т. д.), и об обмене веществ между организмом и внешней средой. Особой
наукой, изучающей химический состав и химические процессы, которые
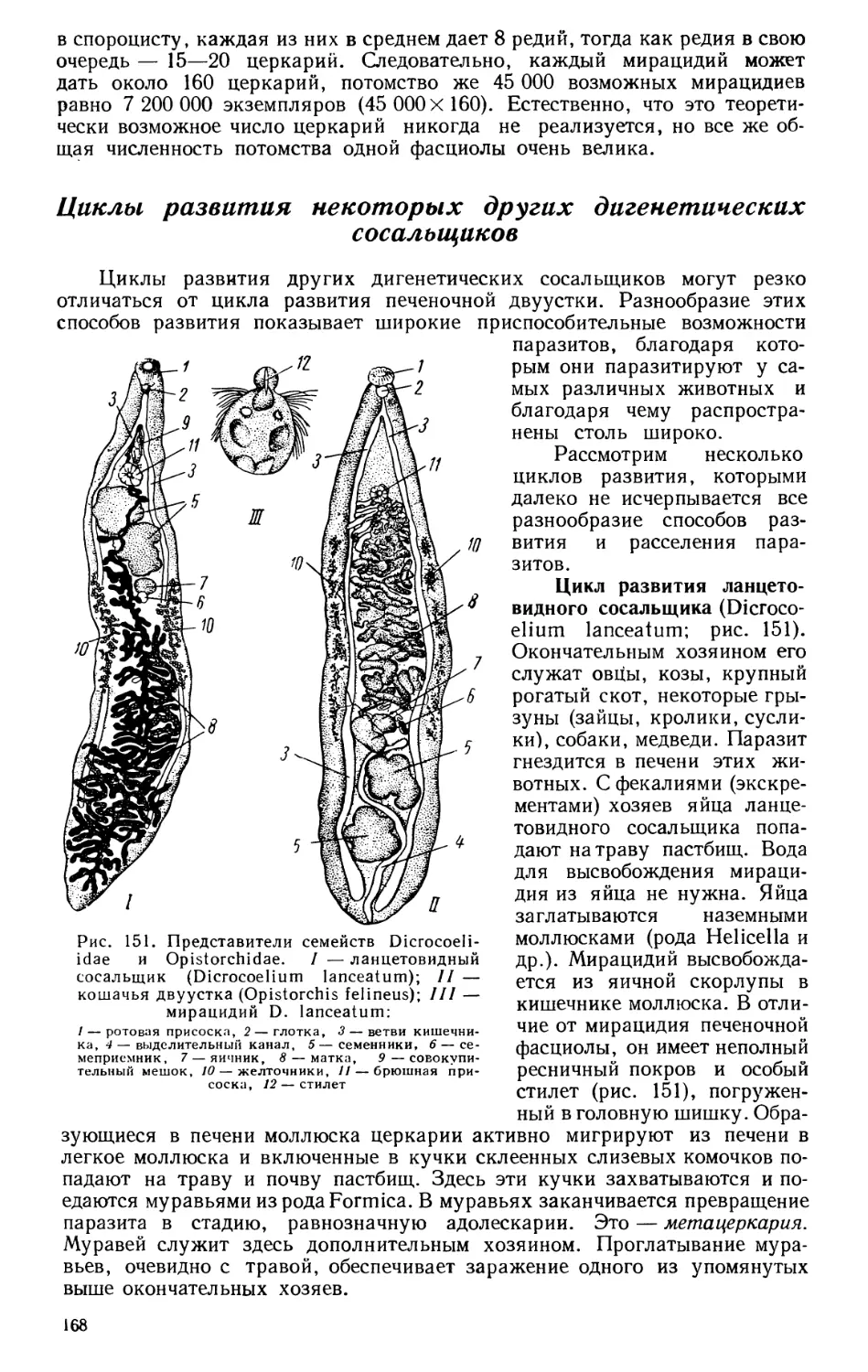
протекают в животном организме, является биохимия животных.
Экология — наука о взаимодействии животных с внешней средой,
как неорганической, так и органической, и о закономерностях зависимости
жизни совокупностей особей (популяций), составляющих виды, и динамики
их численности при различных условиях существования.
7
Тесно связаны с экологией две комплексные биологические дисципли-
ны, имеющие большое практическое значение: паразитология и гидро-
биология.
Паразитология — наука, изучающая явление паразитизма (см.
ниже) — строение паразитов, их приспособление к специфическим ус-
ловиям существования, их жизненные циклы и разрабатывающая био-
логические основы борьбы с паразитами, т. е. с паразитарными болез-
нями человека, домашних животных и культурных растений.
Гидробиология — наука, изучающая закономерности жизни
в водоемах, приспособления к условиям существования водных животных
и растений, их распределение в водоемах и разрабатывающая меры повы-
шения продуктивности водоемов, а также вопросы санитарного состояния
их.
Зоогеография — наука о географическом распространении жи-
вотных и о закономерностях как современных, так и исторических, опреде-
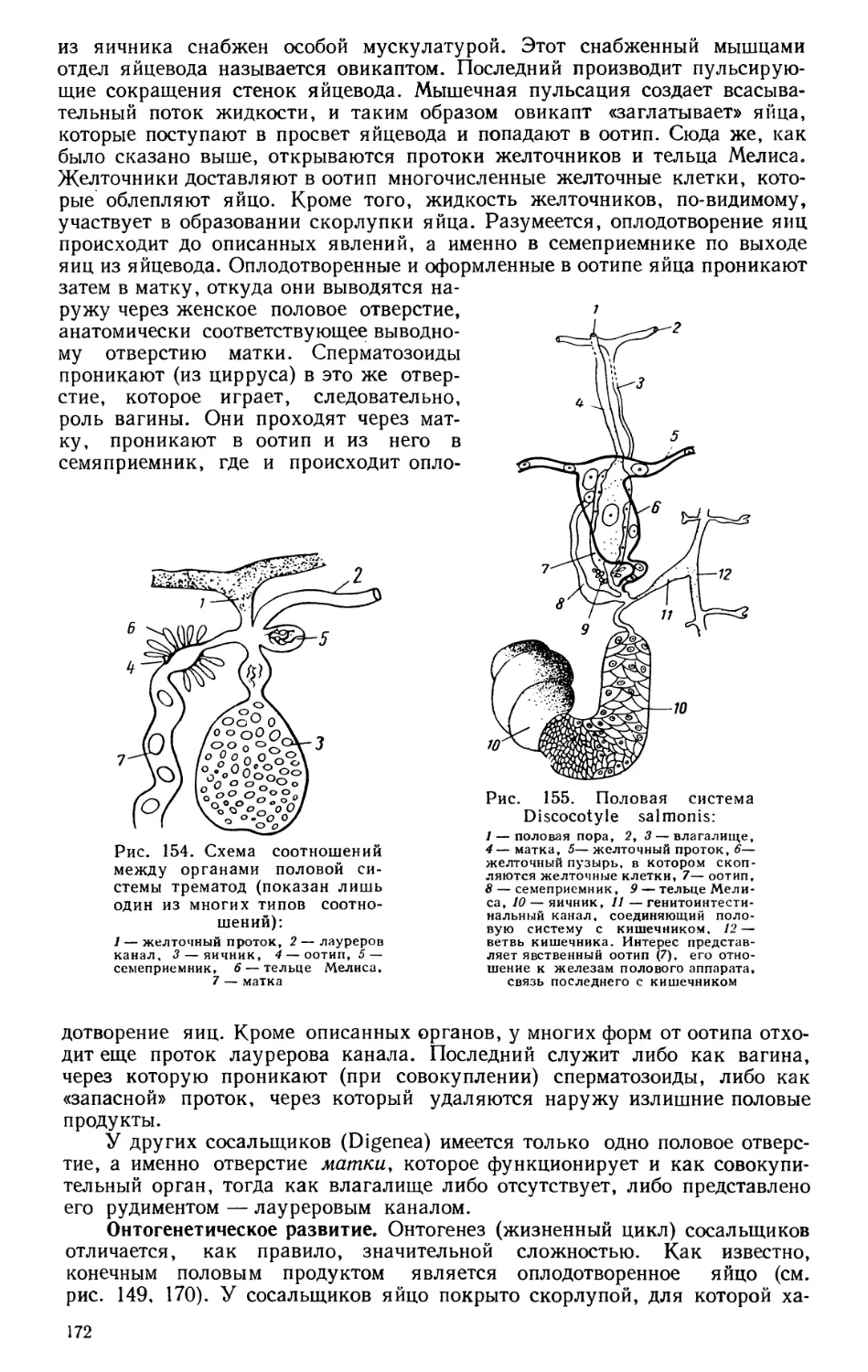
ляющих это распространение.
Палеозоология — наука о вымерших животных, их строении,
геологическом распределении, историческом развитии, происхождении,
а также взаимоотношении с современными организмами.
Генетика — наука о закономерностях наследственности и ее
изменчивости.
Систематика — наука о многообразии животных, их классифи-
кации и ее закономерностях. Систематика использует данные всех зоологи-
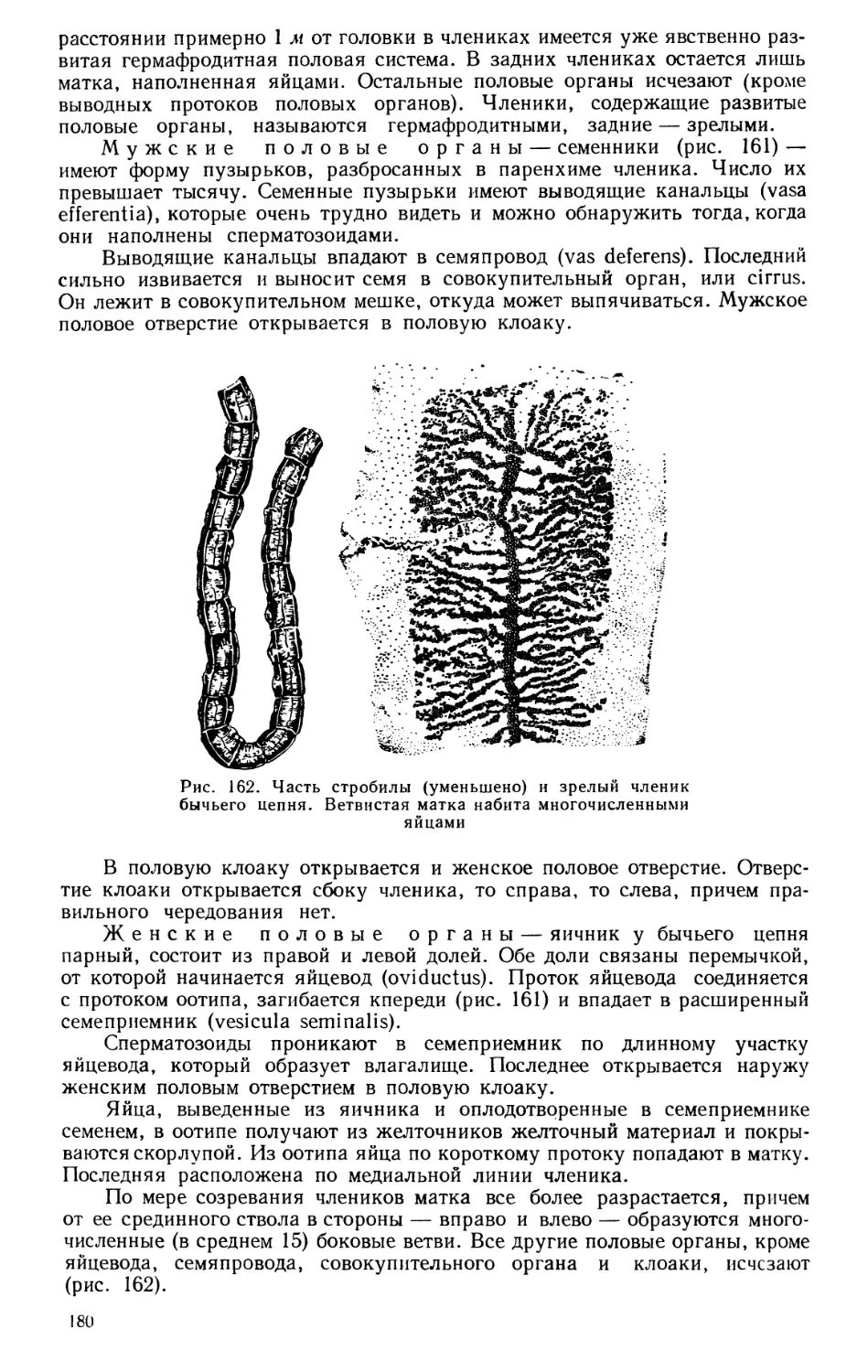
ческих наук и строит естественную, эволюционную систему животного
мира.
Во всех науках большое значение имеет сравнительный метод изучения,
позволяющий применять исторический метод в биологии при изучении
мира животных. При помощи исторического метода рассматривают живот-
ных не только в их настоящем состоянии, но и их развитие в прошлом,
а также стремятся управлять их жизненными процессами путем выяснения
закономерностей их развития. Такое направление в биологических науках
получило название эволюционной морфологии, эволюционной физиологии,
эволюционной гистологии, эволюционной биохимии и т. д.
Ко второй группе зоологических наук относятся:
Протозоология — наука об одноклеточных животных — изу-
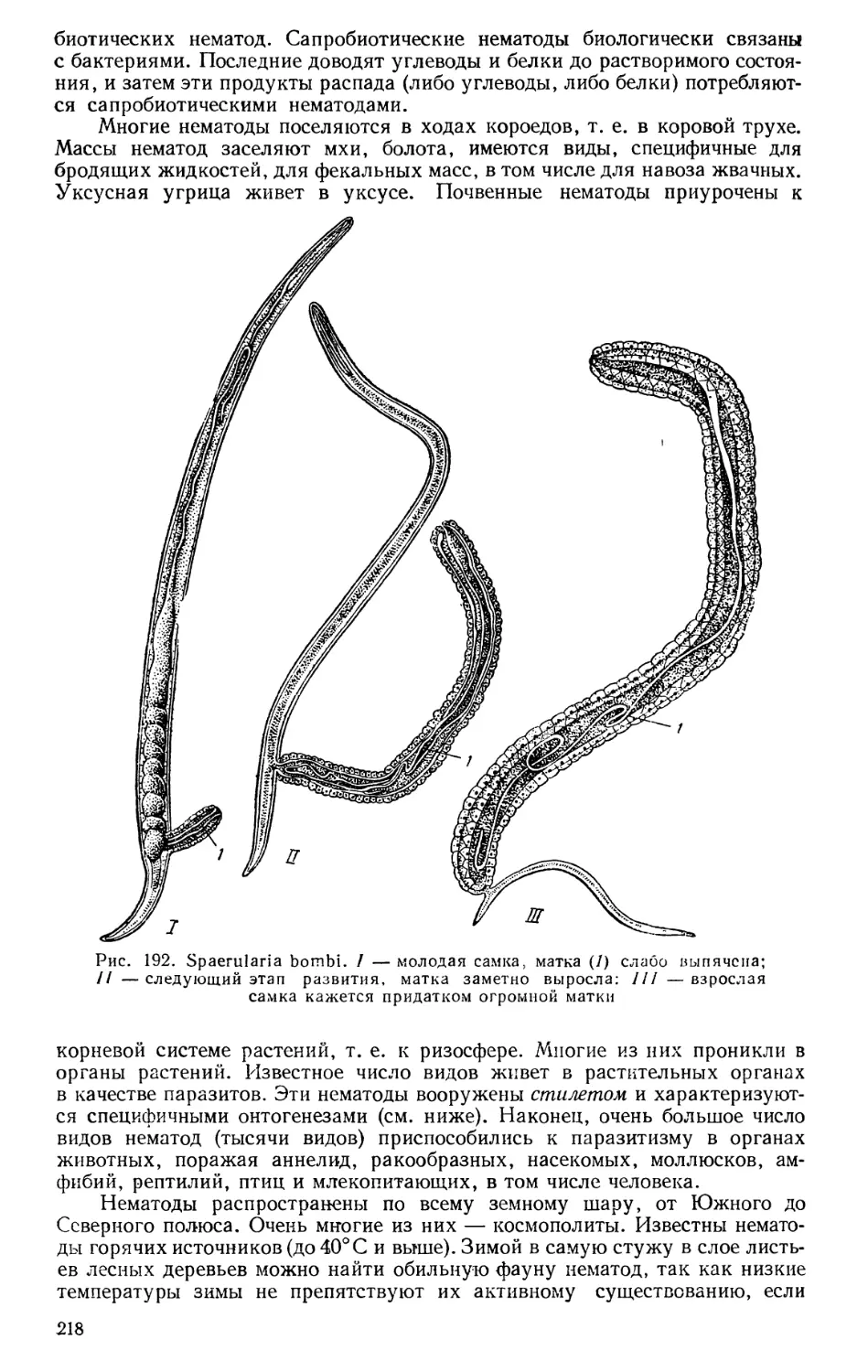
чает строение, жизнедеятельность и жизненные циклы одноклеточных как
свободноживущих, так и паразитических.
Малакология — наука о моллюсках — изучает систематику и
биологию моллюсков.
Гельминтология — наука о паразитических червях — изу-
чает строение, систематику и жизненные циклы этих червей и разрабаты-
вает биологические основы борьбы с ними.
Энтомология — наука о насекомых, самом многочисленном
и многообразном классе животных, — изучает их строение, жизнедеятель-
ность и значение в сельском хозяйстве и здравоохранении.
Ихтиология — наука о рыбах — занимается изучением всех
сторон жизнедеятельности рыб, их строения, систематики, экологии и
разрабатывает биологические основы рыболовства и рыбоводства.
Герпетология — наука о земноводных и пресмыкающихся —
изучает систематику и биологию этих групп позвоночных.
Орнитология — наука о птицах, изучает их систематику, биоло-
гию, особенности распространения, охотничье и промысловое значение и по-
лезную и вредную деятельность в сельском и особенно лесном хозяйствах.
Териология, или маммология — наука о млекопитаю-
щих — изучает систематику, анатомию и биологию представителей этого
высокоорганизованного класса животных, а также их значение в охотничь-
ем и пушном хозяйстве, и их роль как вредителей в сельском хозяйстве
и носителей инфекции.
8
В связи с выявлением практического значения той или иной группы жи-
вотных выделяются и приобретают большое значение новые отрасли зоо-
логии; например, за последние годы выделилась и приобретает все большее
и большее значение с практической точки зрения в медицине и ветерина-
рии акарология — наука о клещах.
В тесной связи с зоологией стоит и животноводство — наука
о домашних животных, которая, в свою очередь, распадается на ряд част-
ных отраслей — скотоводство, овцеводство, птицеводство, пчеловодство
и пр.
ИСТОРИЯ зоологии
Развитие зоологии в древности наиболее известно по тому состоянию,
в котором она находилась у древних греков. Крупнейший греческий фило-
соф и естествоиспытатель Аристотель (384—322 гг. до нашего летоисчисле-
ния) оставил весьма солидное литературное наследие и в области зоологии.
Он не только вполне овладел знаниями своей эпохи, но и чрезвычайно
расширил их своими личными исследованиями, особенно в области системы
животных, их морфологии и эмбриологии. Животный мир был разделен
Аристотелем на две группы: имеющих красную кровь и «бескровных».
Таким образом, были отделены друг от друга позвоночные и беспозвоноч-
ные. К первым Аристотель относил пять более мелких групп, приблизи-
тельно соответствующих современным классам позвоночных: 1) «живородя-
щие четвероногие», соответствующие современным млекопитающим, но без
китообразных; 2) птицы; 3) «яйцекладущие четвероногие» и «безногие с
легочным дыханием», соответствующие пресмыкающимся и земноводным
современной классификации; 4) «живородящие безногие с легочным ды-
ханием» — китообразные и 5) «покрытые чешуей безногие, дышащие
жабрами» — рыбы.
Соединение земноводных и пресмыкающихся в одну группу весьма
долго сохранялось в зоологии, и группу «гадов» (земноводные и пресмыкаю-
щиеся) можно встретить в книгах по зоологии даже начала нашего века.
Это объединение — характерный пример искусственной системы, основан-
ной на сходстве случайно выбранных признаков; по совокупности
известных в настоящее время признаков пресмыкающиеся ближе
к птицам, чем к земноводным. Что касается огромной группы беспозво-
ночных, то подразделение ее Аристотелем на более мелкие группы было
уже менее удачным, что вполне понятно, если принять во внимание огром-
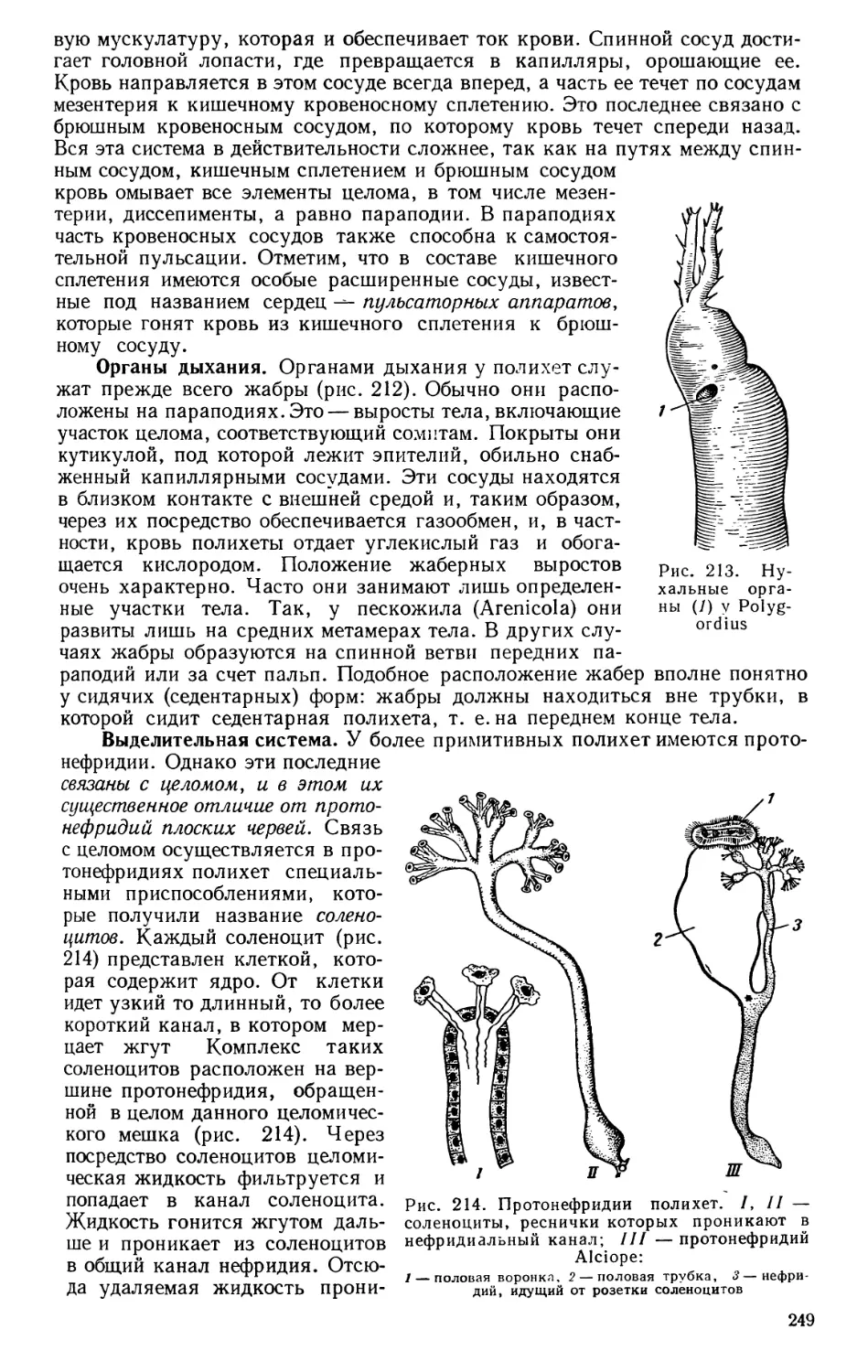
ное число и разнообразие относящихся сюда форм. Аристотель считал бес-
позвоночных «бескровными». Всего у Аристотеля описано 520 форм живот-
ных. В наше время количество форм животных исчисляется сотнями ты-
сяч; впрочем, Аристотель не ставил, по-видимому, себе задачи описать все
известные ему формы.
Мы можем не останавливаться на истории зоологии в средние века.
Религиозная нетерпимость не только препятствовала непосредственному,
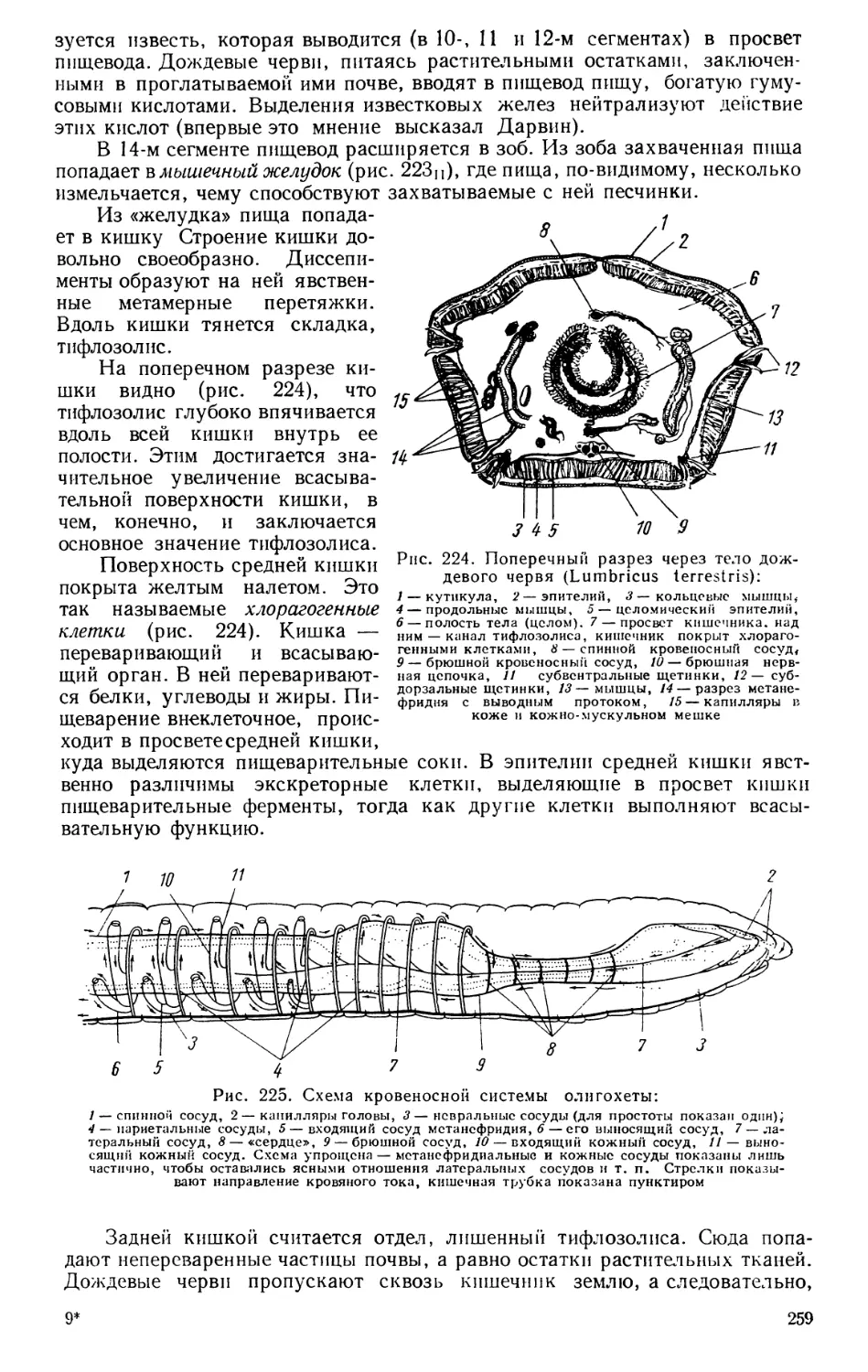
свободному изучению природы, но даже и написанное древними, дохристи-
анскими, авторами находилось под запретом. Написанное в эту эпоху
стоит, как правило, неизмеримо ниже произведений античных авторов.
Лишь в эпоху Возрождения наступает оживление в области зооло-
гических исследований. Наряду с описаниями животных появился усилен-
ный интерес к анатомии человека в связи с запросами медицины; Леонардо-
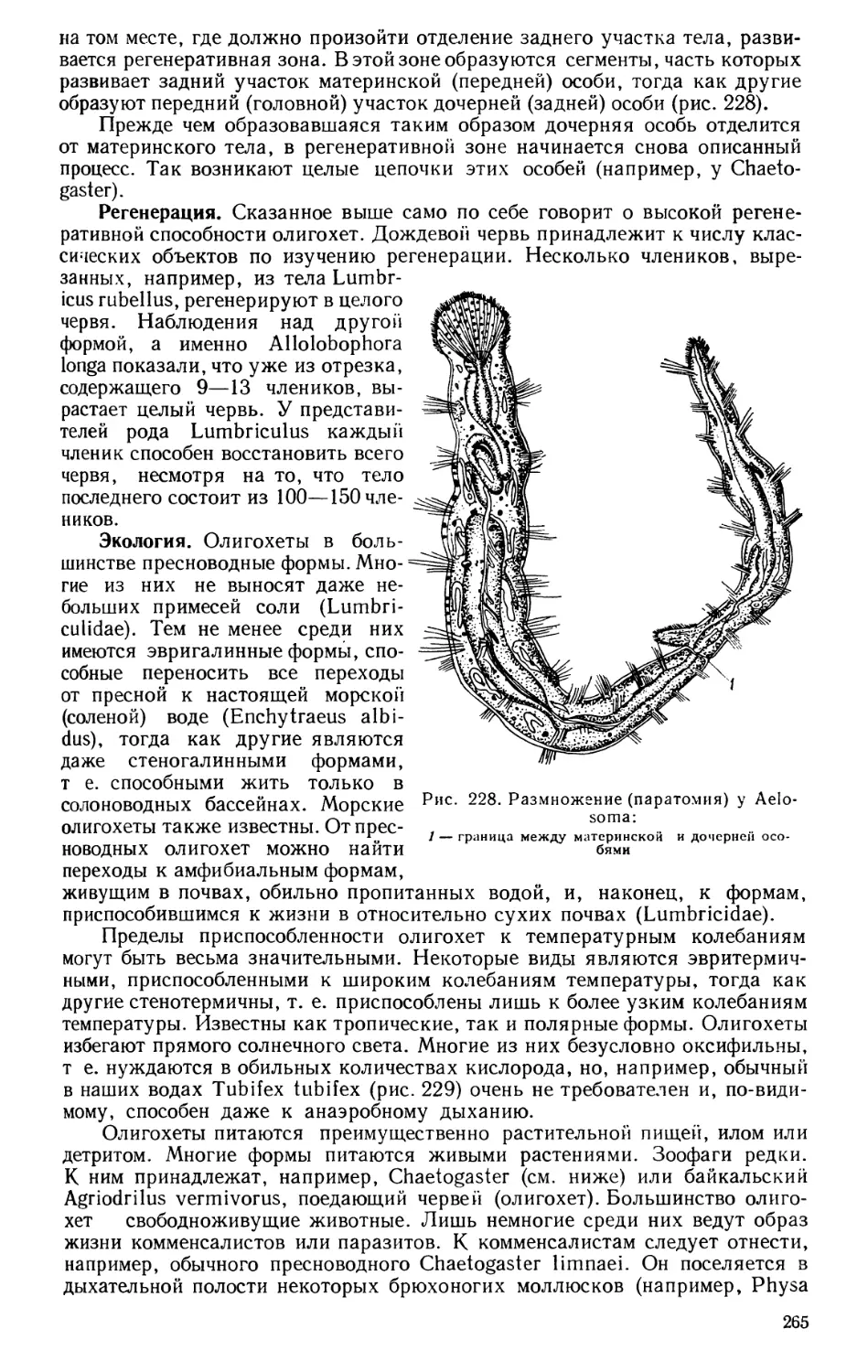
да-Винчи (1452—1519), Везалий (1514—1564), Евстахий, Варолий, Бота-
лий, Фаллопий и другие работали в этой области, причем впервые стали
обращаться к изучению анатомии на человеческих трупах. Крупнейший
анатом древности Гален (131—201) изучал анатомию на животных и пере-
носил свои выводы на человека, а в средние века занятия на трупах людей
были еще менее возможны. Изучение анатомии животных продолжалось
9
еще некоторое время с целью нахождения ошибок Галена, труды которого
принялись изучать так же энергично, как и труды Аристотеля. Впослед-
ствии же стали изучать анатомию животных ради нее самой. Так возникла
описательная анатомия животных, накопившая материал для позднейшей
сравнительной анатомии. В те же времена более интенсивно начала разви-
ваться и физиология. Из физиологии этого периода особенно известен
Вилиам Гарвей (1578—1657) своими исследованиями кровообращения;
эмбриологические наблюдения привели его к выводу, что всякое животное
развивается из яйца.
В XVI в. впервые стали употреблять очки и увеличительные стекла.
Примитивные комбинации последних привели к созданию микроскопов,
дававших вначале лишь весьма слабые увеличения и пригодных для рас-
сматривания довольно крупных объектов вроде мелких насекомых. Усо-
вершенствование микроскопа зависело от искусства шлифования стекол,
которым и владел один из первых микроскопистов — Антони Левенгук
(1632—1723), открывший инфузорий, кровяные тельца и нашедший сперма-
тозоиды в семенной жидкости (совместно с Гаммом). Марчелло А\альпиги
(1628—1694) изучал при помощи линз развитие цыпленка в яйце и много
сделал в области анатомии насекомых, изучая тутового шелкопряда.
Таким образом, из зоологии и ботаники, благодаря применению ми-
кроскопа, постепенно выделилась отдельная дисциплина — микроскопи-
ческая анатомия, изучающая микроскопическое строение животных и
растений. Однако самостоятельная наука — гистология — выделилась лишь
в первые годы XIX в., когда выступил со своим учением о строении тканей
французский анатом К. Биша (1801). Огромную роль в развитии гистологии
сыграла клеточная теория, сформулированная Шванном в 1839 г.
Применение микроскопа при изучении развития животных дало тол-
чок к выделению самостоятельной науки — эмбриологии. Начиная с
XVII в. ряд исследователей: И. Фабриций (1537—1614), М. Мальпиги
(1628—1694), А. Левенгук (1632—1723) и другие занимались изучением
зародыша, строения яйца и мужских половых элементов.
Открытие микроскопа — лишь один из частных примеров развития
естествознания, начавшегося в Европе с XVII в. В ряде стран Европы
этот период связан с переходом от феодального строя общества к капитали-
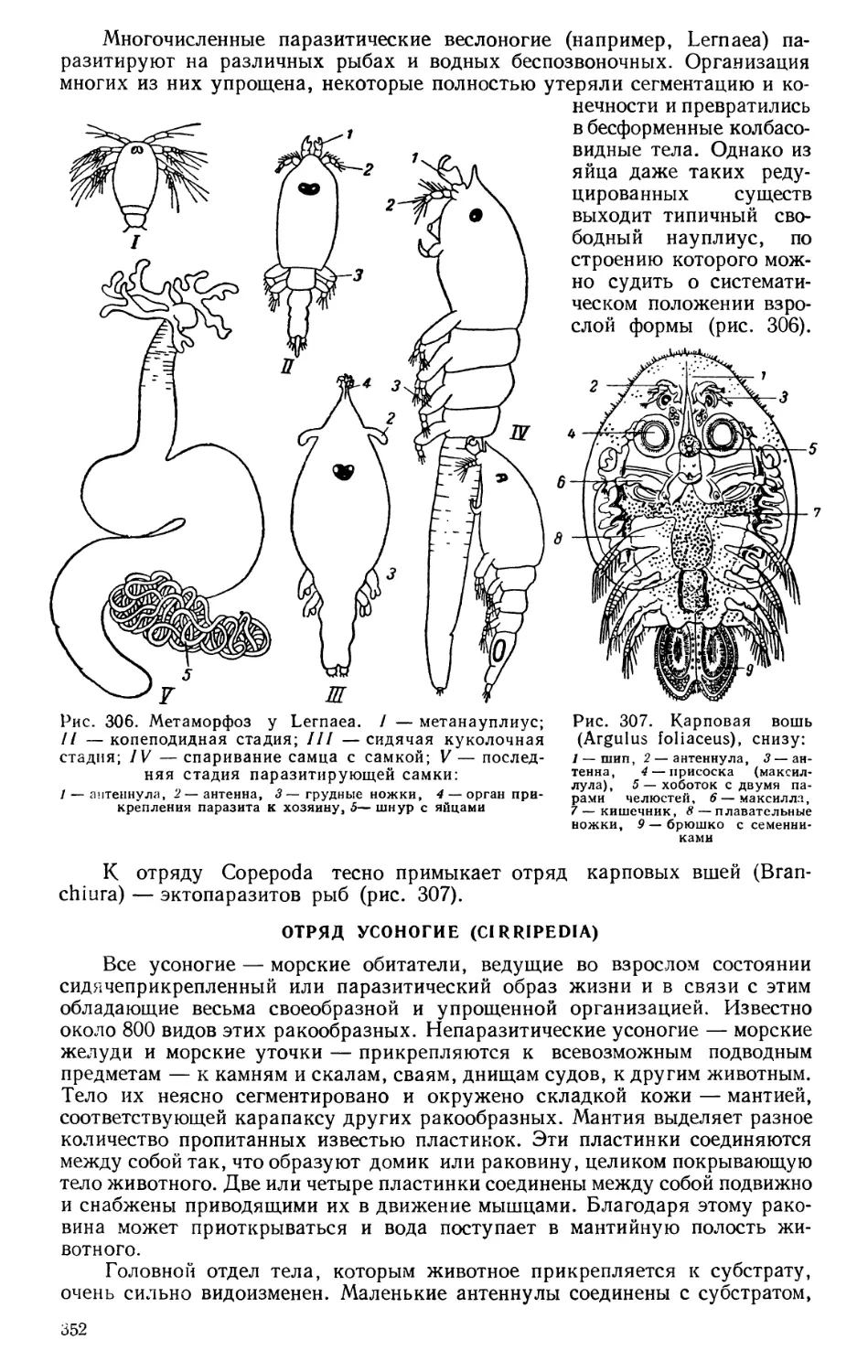
стическому. ' Это была эпоха интенсивного развития производительных
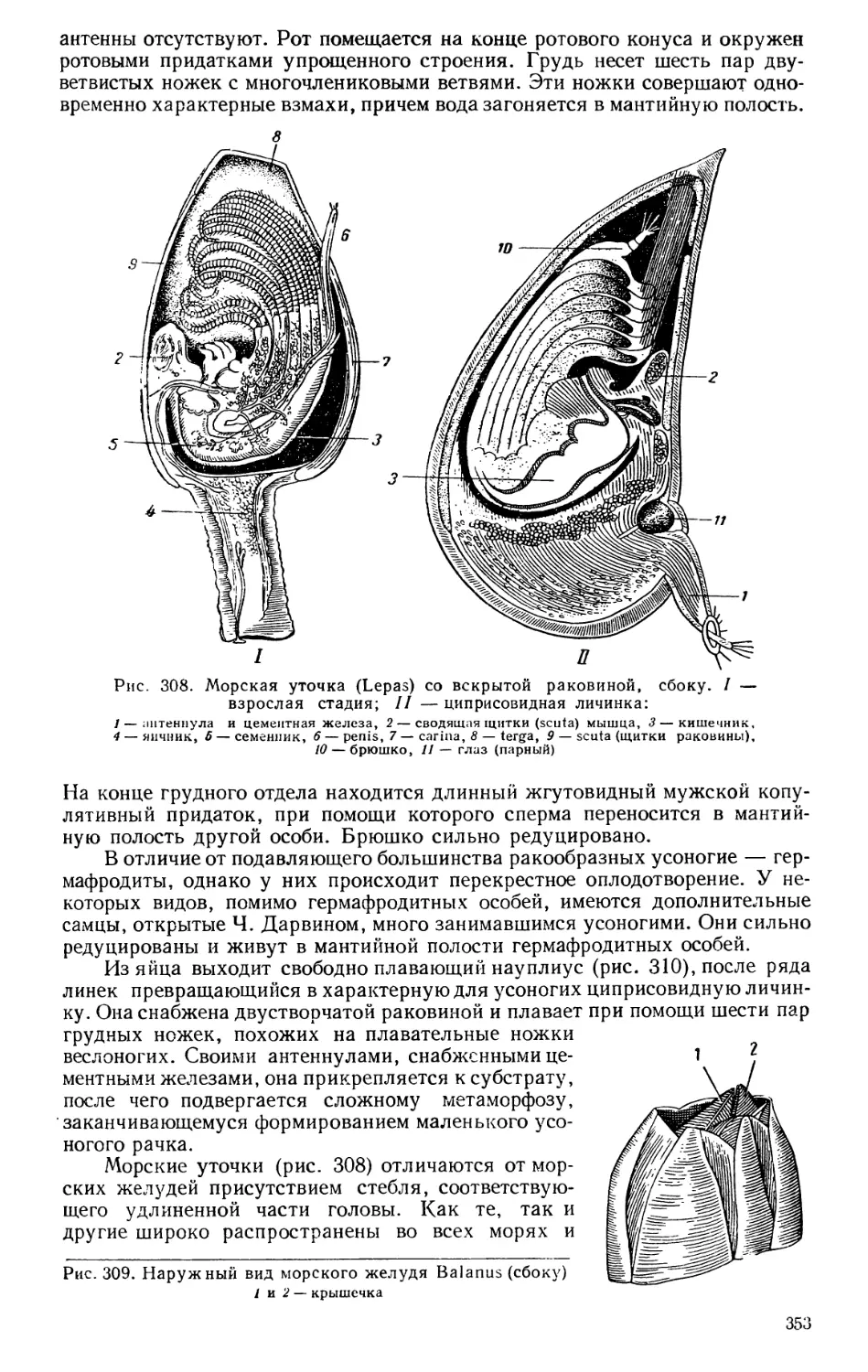
сил. Необычайно быстро начинают расти различные отрасли промышлен-
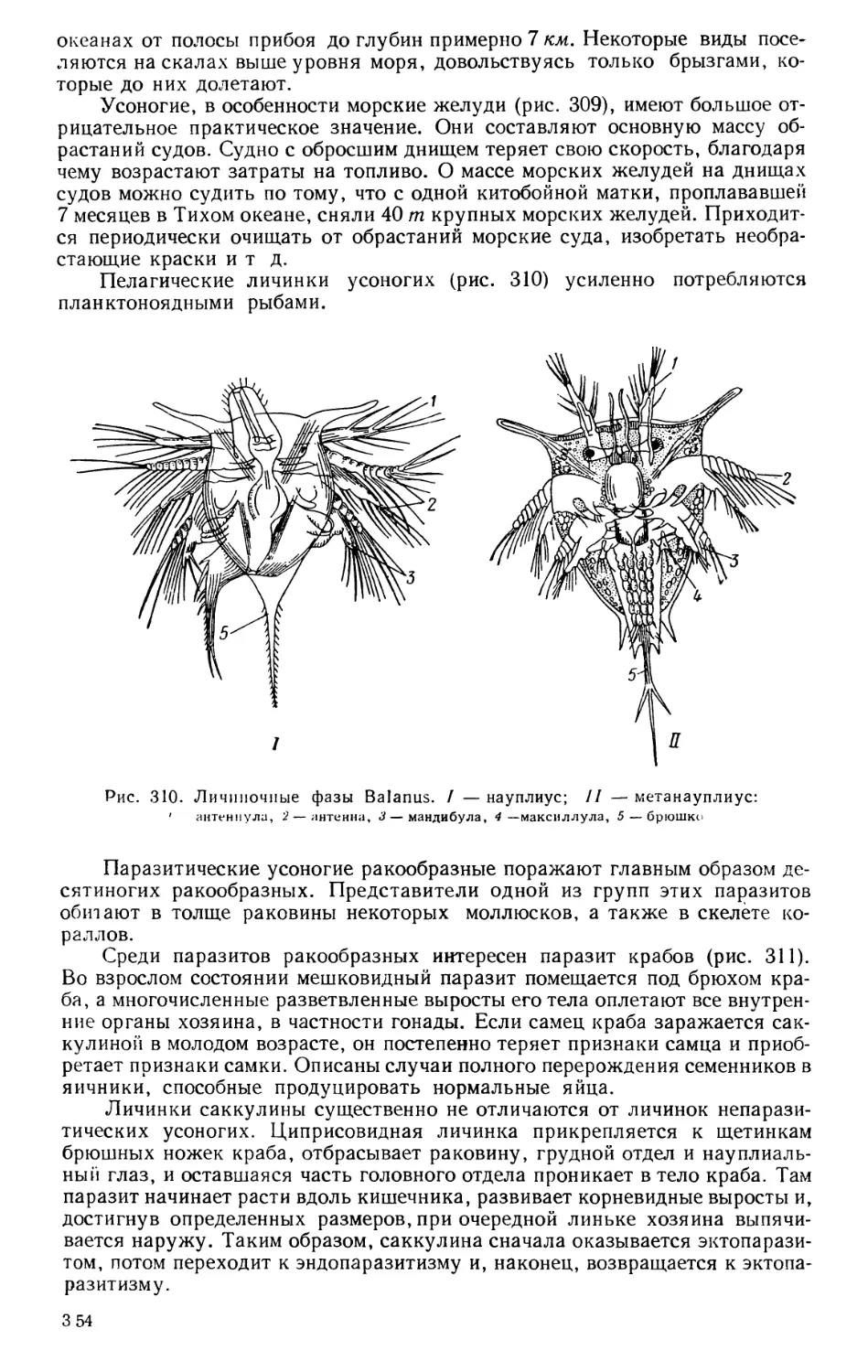
ности (горная, металлургическая, ткацкая, кожевенная, судостроительная
и др.). Растет потребность в новых рынках, что стимулирует многочислен-
ные путешествия, приводящие к великим географическим открытиям.
Во время этих путешествий происходит громадное накопление новых
знаний о животных и растениях, обитающих в других странах. В Англии,
Франции учреждаются академии наук, зоологические музеи и сады, где
концентрируются коллекции, привозимые путешественниками. Выде-
ляется как самостоятельная наука зоология, включающая систематику
и биологию животных; анатомия и физиология человека приобретают
значение самостоятельных дисциплин.
Параллельно с развитием морфологических дисциплин (анатомии,
эмбриологии) шло развитие учения о системе животных — систе-
матики. Джон Рей (1627—1705) впервые ввел понятие о виде как о совокуп-
ности особей, отличающихся друг от друга не более, чем дети от родителей.
Крупнейшим систематиком XVIII в. был Карл Линней (1707—1778),
сочинение которого «Система природы» вышло в свет в 1735 г. Линней ввел
единые общеобязательные латинские наименования животных и этим по-
кончил с величайшей путаницей в зоологических сочинениях, проистекав-
шей из различных наименований одного и того же животного авторами
различных национальностей. Линней ввел также двойное наименование —
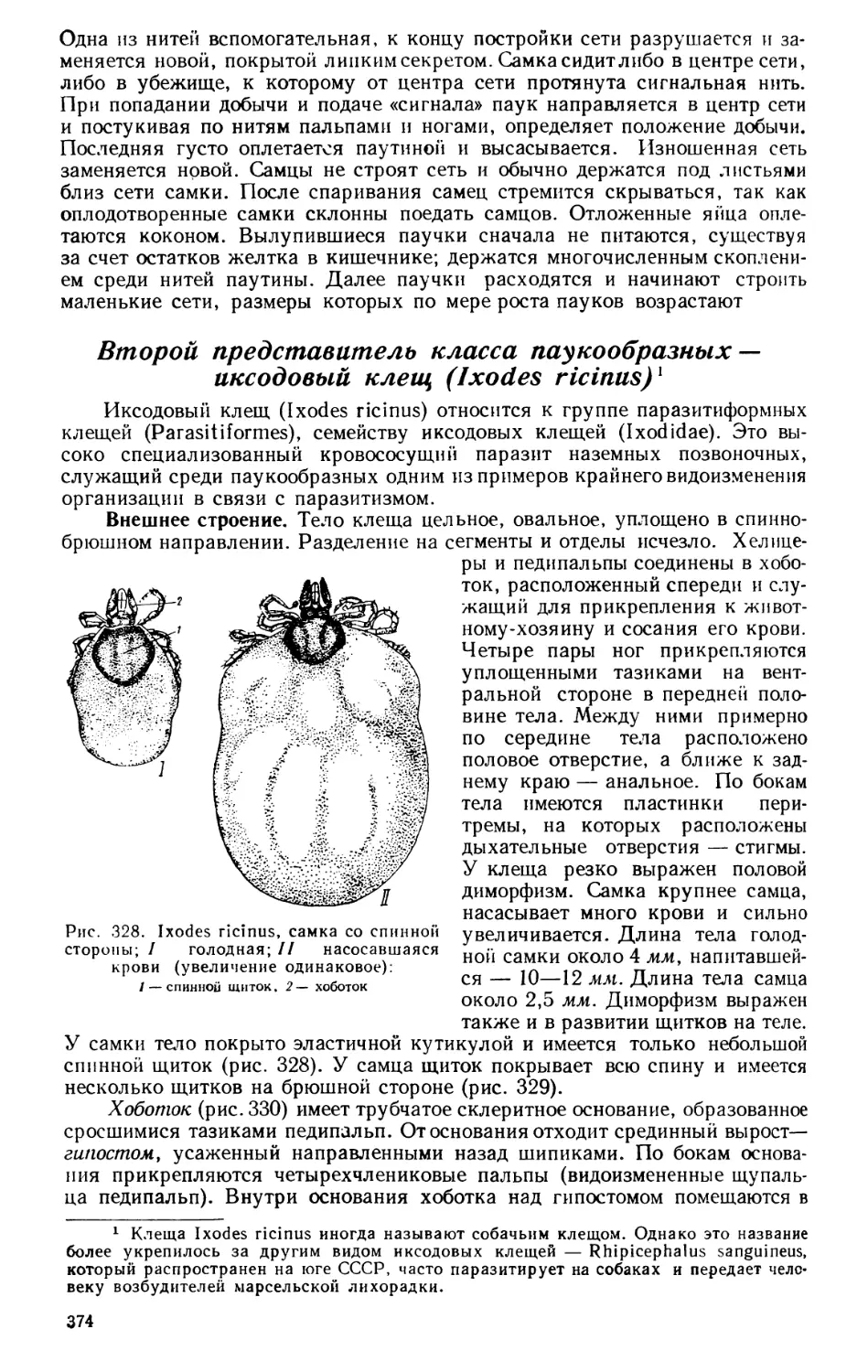
бинарную номенклатуру — для видов (см. ниже). Эта интернациональная
систематическая терминология Линнея касалась не только названий видов,
10
но и наименований других систематических групп. Последних Линней
ввел три категории, иерархически подчиненные друг другу: сходные виды
им объединены в роды, сходные роды — в отряды, отряды — в классы*, по-
следние были самыми крупными систематическими категориями, на кото-
рые распадалось «царство животных». Практиковавшиеся ранее подробные
описания животных Линней заменил короткими диагнозами, в которые
включались лишь важные с точки зрения классификации признаки данной
категории; диагноз более крупной группы не повторяется в диагнозах
всех, входящих в нее более мелких групп. В своей системе животного мира
Линней удачно объединил членистоногих в один класс («насекомые»), но
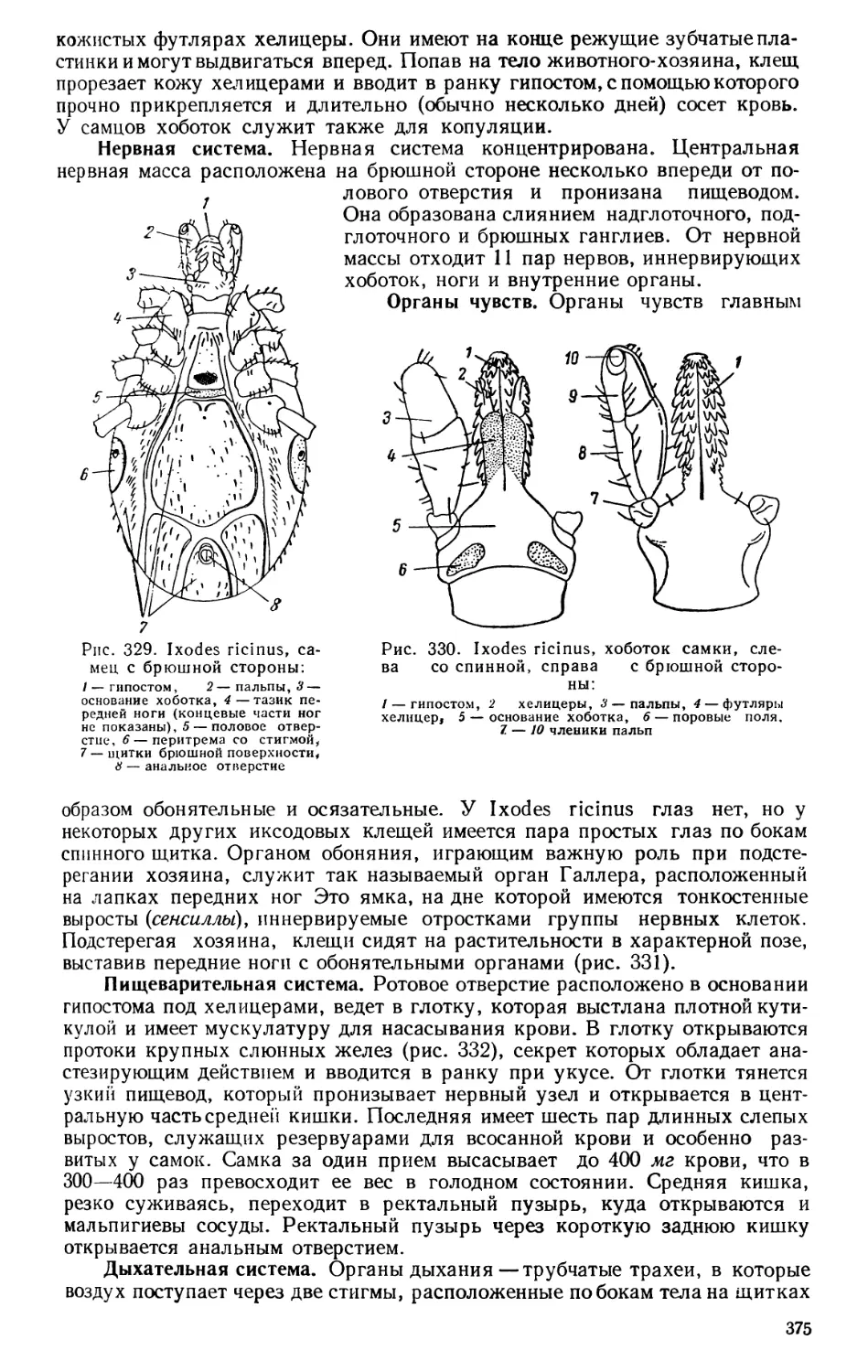
в класс «червей» включил всех остальных беспозвоночных. Шагом назад
по сравнению с Аристотелем было отсутствие объединения млекопитающих,
птиц, «гадов» и рыб в более крупную группу, которая соответствовала бы
современным классам позвоночных.
К началу XIX в. изучение анатомии животных сильно продвинулось
вперед. В частности, гораздо лучше, чем во времена Линнея, стал известен
огромный мир так называемых «беспозвоночных» животных, разделенный
Жаном Б. Ламарком (1744—1828) на целых десять классов. Ламарк вос-
становил также разделение животного мира на беспозвоночных и позвоноч-
ных, видя в этом одно из основных проявлений «градации» в усложнении
организации. Описательная анатомия переросла в сравнительную и сдела-
лась основой для построения системы животных главным образом благодаря
исследованиям по сравнительной анатомии современных и ископаемых
позвоночных животных знаменитого французского зоолога Жоржа Кювье
(1769—1832). Кювье разделил (1812) царство животных на четыре крупней-
шие группы, впоследствии названные типами (теория типов): позвоночных,
членистых, моллюсков и лучистых. Ко второй группе — к членистым —
были отнесены, кроме членистоногих современной системы, еще и кольча-
тые черви; к моллюскам были ошибочно отнесены, кроме настоящих мол-
люсков, усоногие ракообразные, а лучистые представляли собой искусствен-
ную группу — собрание недостаточно изученных в то время животных —
инфузорий, кишечнополостных, иглокожих, паразитических червей. Между
типами Кювье не находил никаких переходных форм, считал их совершенно
обособленными в настоящем и. в прошлом и такими же независимыми друг
от друга по происхождению, постоянными и неизменными, какими Линней
считал свои виды.
Теория типов Кювье вступила в резкое противоречие с другим на-
правлением в зоологии, развившимся из идеи единого плана строения
животных. Это второе направление было широко распространено еще в
XVII—XVIII вв. в форме учения о «лестнице организмов», о том, что расте-
ния и животные располагаются по степени своей сложности и совершенства
в один непрерывный ряд. Наиболее ярким представителем теории лестницы
организмов является Ш. Боннэ (1720—1793), согласно теории которого
самые нижние ступени лестницы занимают огонь, воздух, вода и земля,
а самые верхние — четвероногие животные, человек, ангелы, бог. Возник-
шее в последнем десятилетии XVIII в. и широко распространенное в первой
четверти XIX в. также натурфилософское направление в зоологии воспри-
няло идею лестницы существ и распространило ее на зародышевое развитие.
Зародыш высшего животного проходит в течение своего развития через
состояния низших животных, стоящих ниже его на «лестнице организмов».
Эволюционисты начала XIX в. также поддерживали идею лестницы, по-
скольку единство и непрерывность животного мира хорошо согласовались
с эволюционными взглядами. Напомним учение Ламарка (1809) о града-
ции животных, о постепенном усложнении их организации. Э. Жоффруа
Сент-Илер (1772—1844)—крупнейший представитель французской натур-
философии и эволюционист, развивал мысль о единстве плана строения
во всем животном мире, резко противоречившую учению Кювье о четырех
постоянных типах.
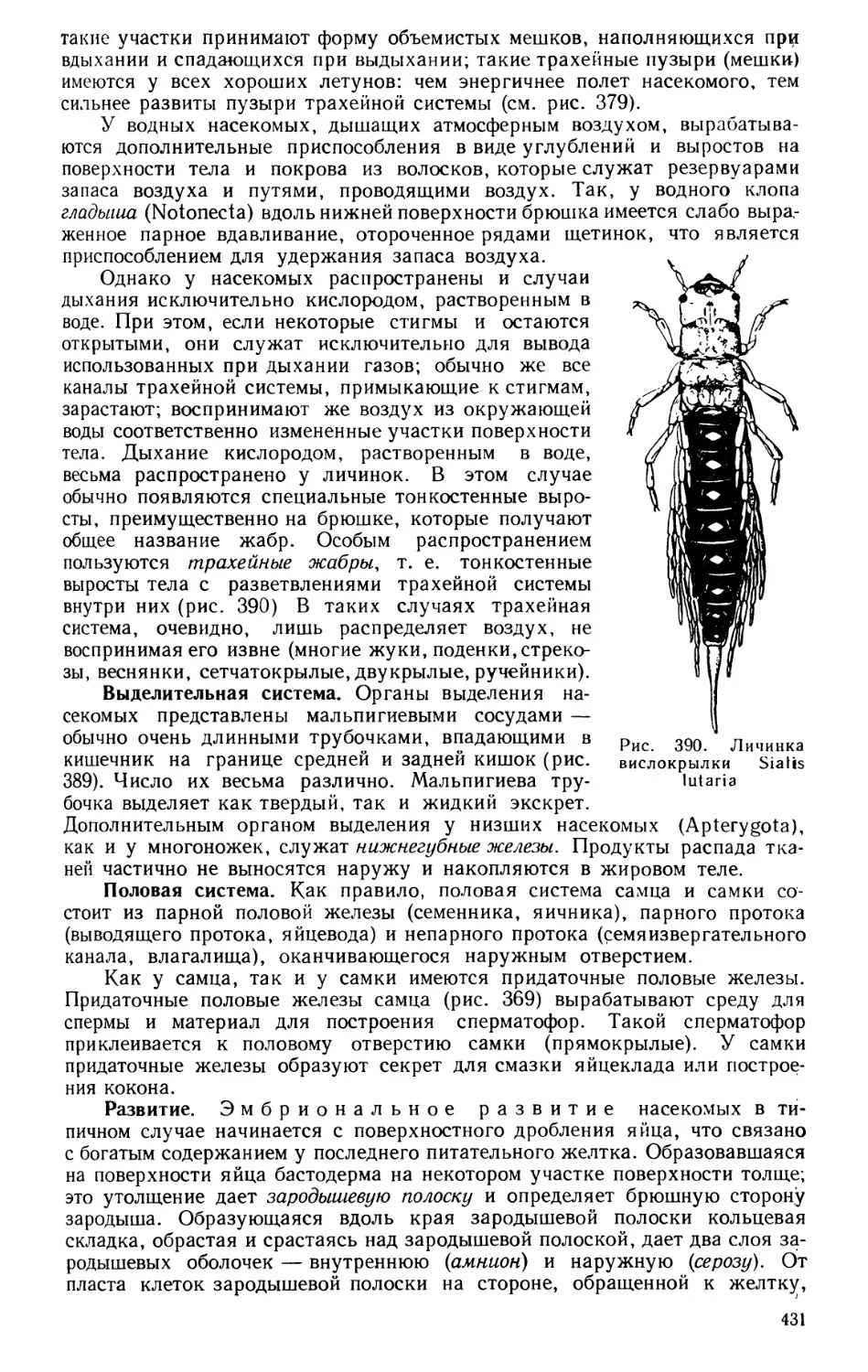
11
После открытия в 1725 г. Академии наук, а в 1755 г. Московского
университета в России начинается быстрое формирование русской науки.
Еще в 1720 г. Петр Великий отправил в Сибирь ученого врача Даниила
Мессершмидта, который семь лет путешествовал по Сибири и привез оттуда
богатые коллекции зверей и птиц. Еще более богатые сборы и открытия
сделали участники второй Камчатской северной экспедиции 1733—1742 гг.:
С. П. Крашенинников, Гмелин Старший, Стеллер. Громадное влияние на
развитие русского естествознания оказали исследования гениального сына
русского народа М. В. Ломоносова, своими блестящими открытиями опе-
редившего западную науку. Почти всю жизнь прожил в России академик
П. С. Паллас. Он и его современники И. Лепехин, Гмелин Младший и
А. Гюльденштедт обследовали восток и юг Европейской части России,
Западную Сибирь, Алтай, Байкал и Забайкалье. В XIX в. зоологическое
обследование России продолжалось экспедициями Академии наук (К. Бэр,
А. Ф. Миддендорф), Московскими обществами испытателей природы и лю-
бителей естествознания (С. Карелин, Н. А. Северцов, А. П. Богданов,
А. П. Федченко) и Русским географическим обществом (Н. АГ Пржеваль-
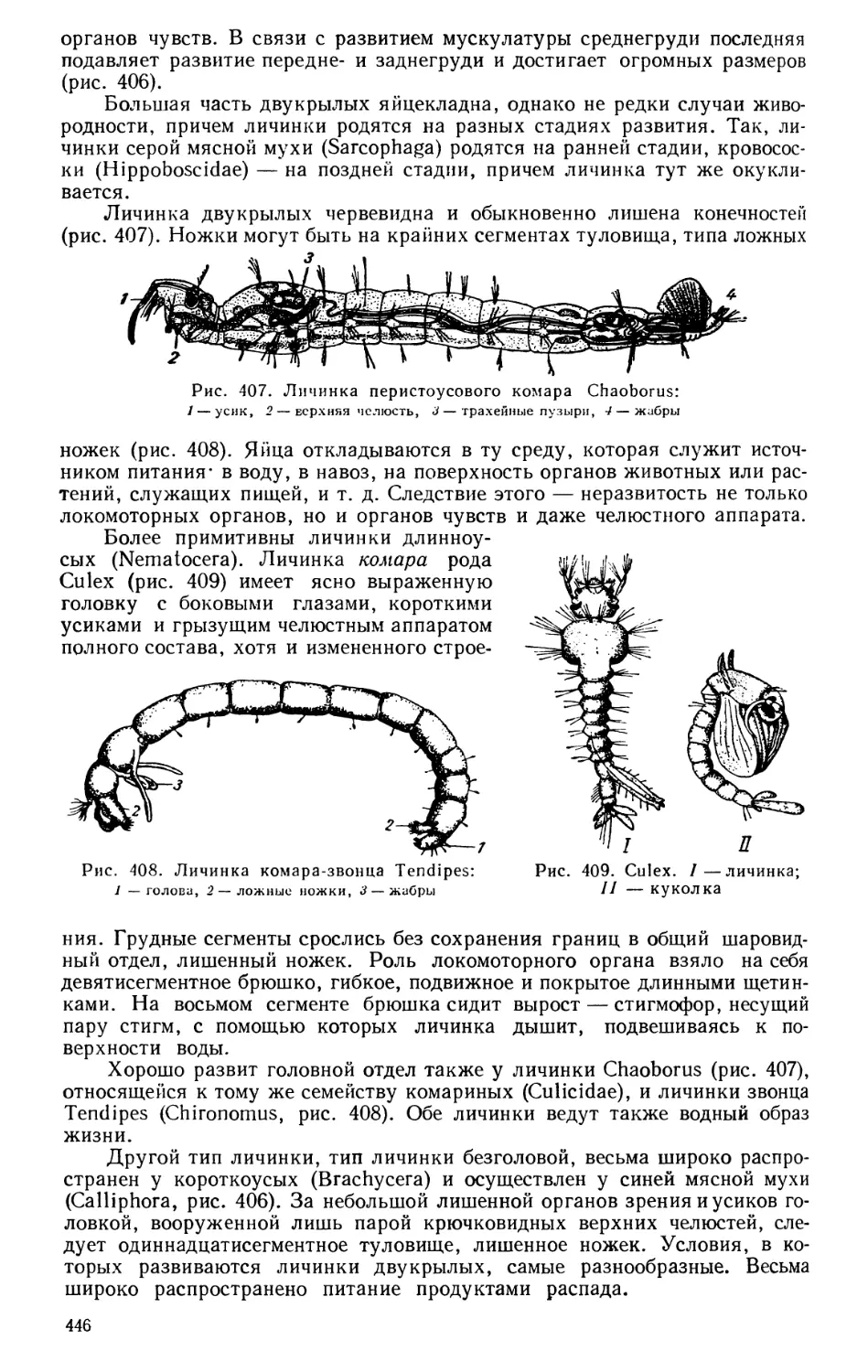
ский, И. Потанин, П. К. Козлов, М. Н. Богданов, П. П. Семенов-Тян-
Шанский).
Громадное значение для изучения фауны морей сыграло открытие
биологических станций: Севастопольской биологической станции (основан-
ной А. О. Ковалевским в 1871 г.), Неаполитанской зоологической станции
(А. Дорн, 1872), пресноводной станции на Глубоком озере под Москвой
(Н. Ю. Зограф, 1891), Мурманской станции (К. М. Дерюгин, 1896), Бай-
кальской лимнологической станции и др.
Выход в свет в 1859 г. «Происхождения видов» Ч. Дарвина яви-
лось крупным поворотным этапом в истории зоологии.
После появления учения Дарвина (1859) установившиеся понятия и
представления во всех областях биологии подверглись коренному пере-
смотру. Вид перестали считать неизменным. Разновидность стали пони-
мать как образующий вид, систему животных — как соотношение между
группами, возникающее в результате эволюционного процесса. Известные
уже до Дарвина явления сходства в развитии и основном плане строения
органов (гомологи, см. ниже) получили естественное объяснение. Стало
понятным единство многоклеточных животных в отношении их клеточного
строения. В эмбриологии начало бурно развиваться учение о зародышевых
пластах (см. ниже); явление сходства зародышей у животных, сильно раз-
личающихся во взрослом состоянии, стало понятно и послужило исходным
пунктом учения о повторении зародышевым развитием развития эволюцион-
ного. Стали понятны многочисленные интереснейшие факты географиче-
ского распространения животных и их геологической истории и т. д.
В России учение Дарвина, несмотря на противодействие царского пра-
вительства и духовенства, нашло благодарную почву для дальнейшего
развития.
Передовые слои русского общества были подготовлены к восприятию
материалистического учения Дарвина блестящей проповедью революцио-
неров-материалистов 40-х и 60-х годов XIX в.: Н. Г. Чернышевского
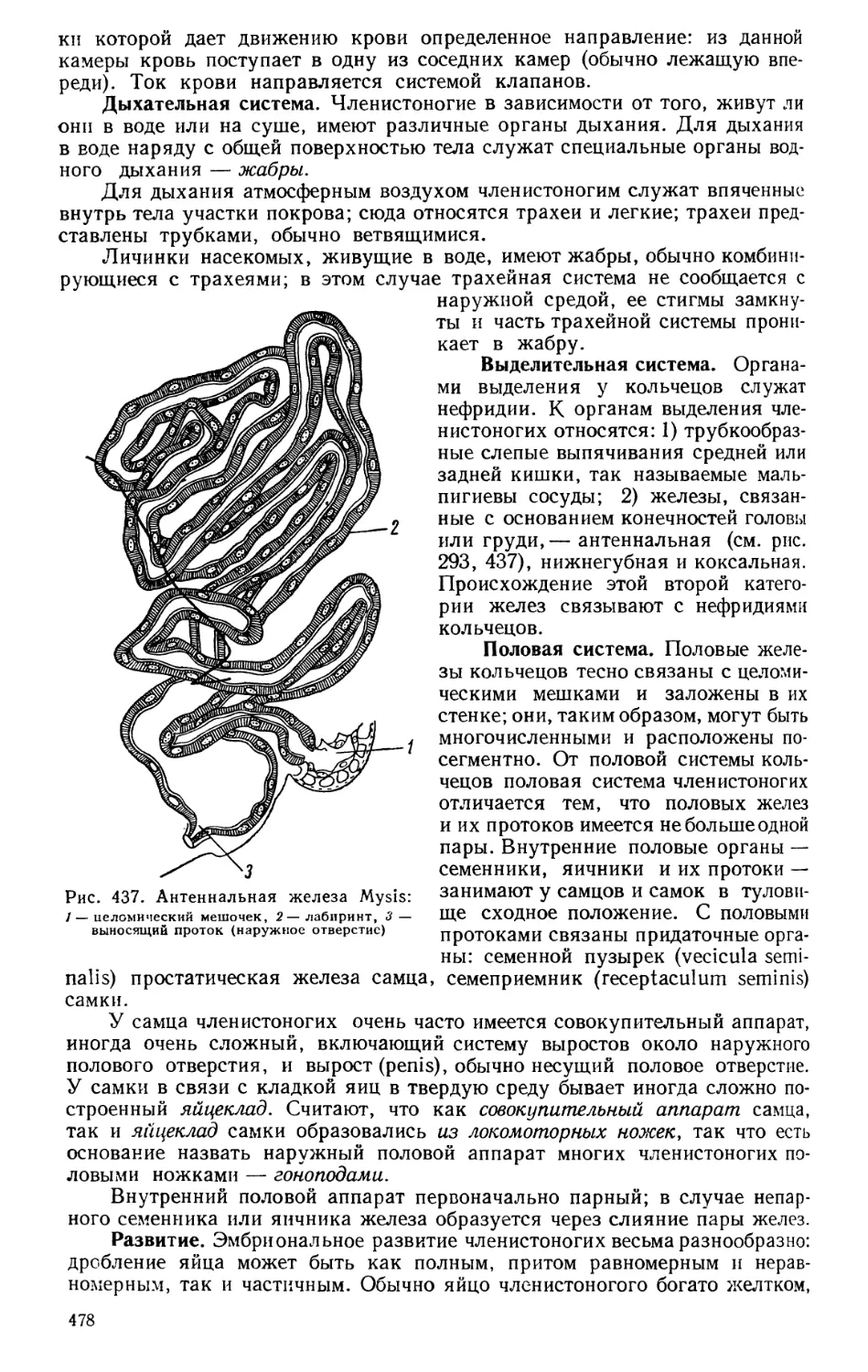
(1828—1889), Д. И. Писарева (1840—1868), Н. П. Огарева (1813—1877),
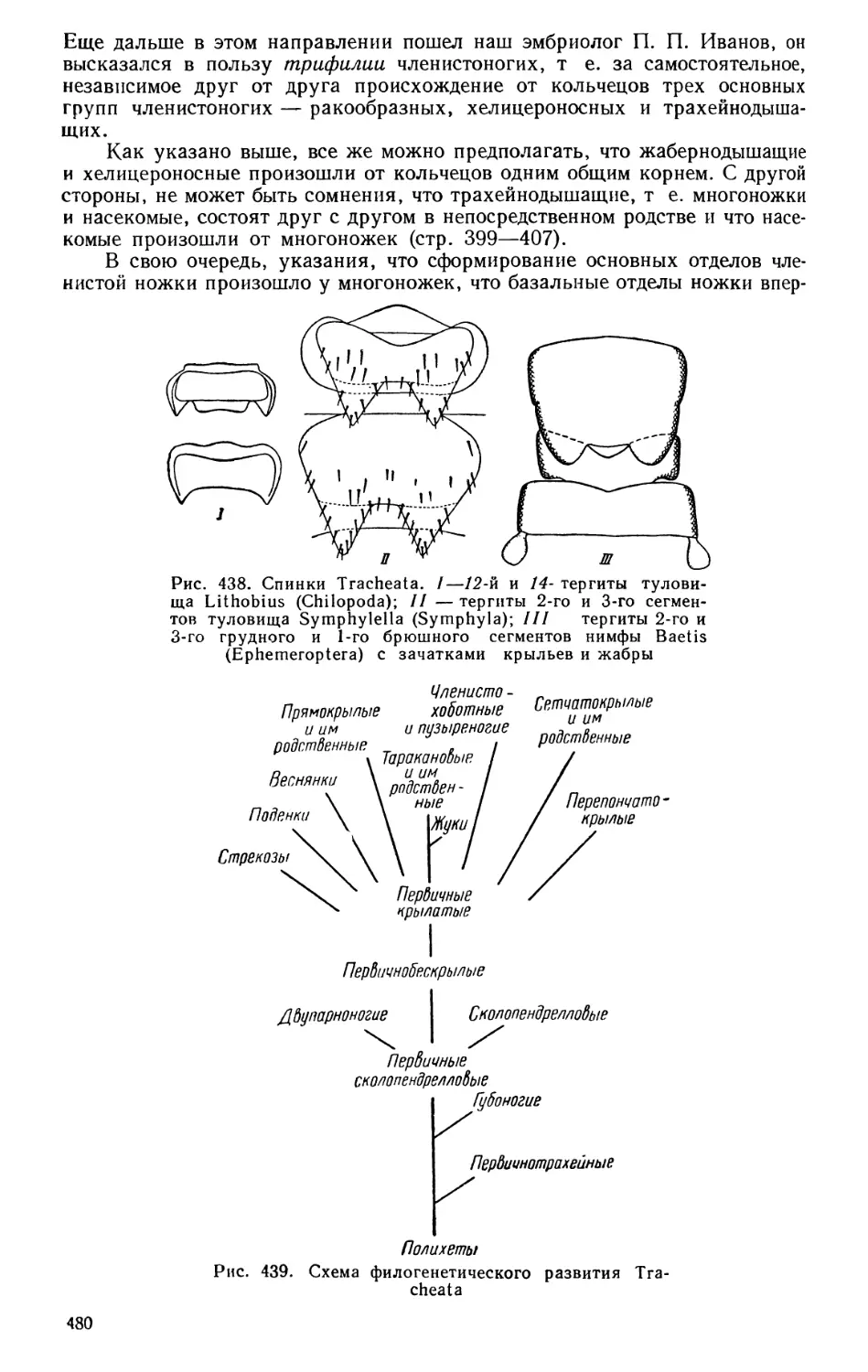
А. Н. Герцена (1812—1870) и других гуманистов этойэпохи. Плеяда моло-
дых русских зоологов — А. О. Ковалевский (1840—1901), И. И. Мечни-
ков (1845—1915), В. В. Заленский (1846—1900) — своими замечательными
исследованиями по морфологии и эмбриологии различных беспозвоночных
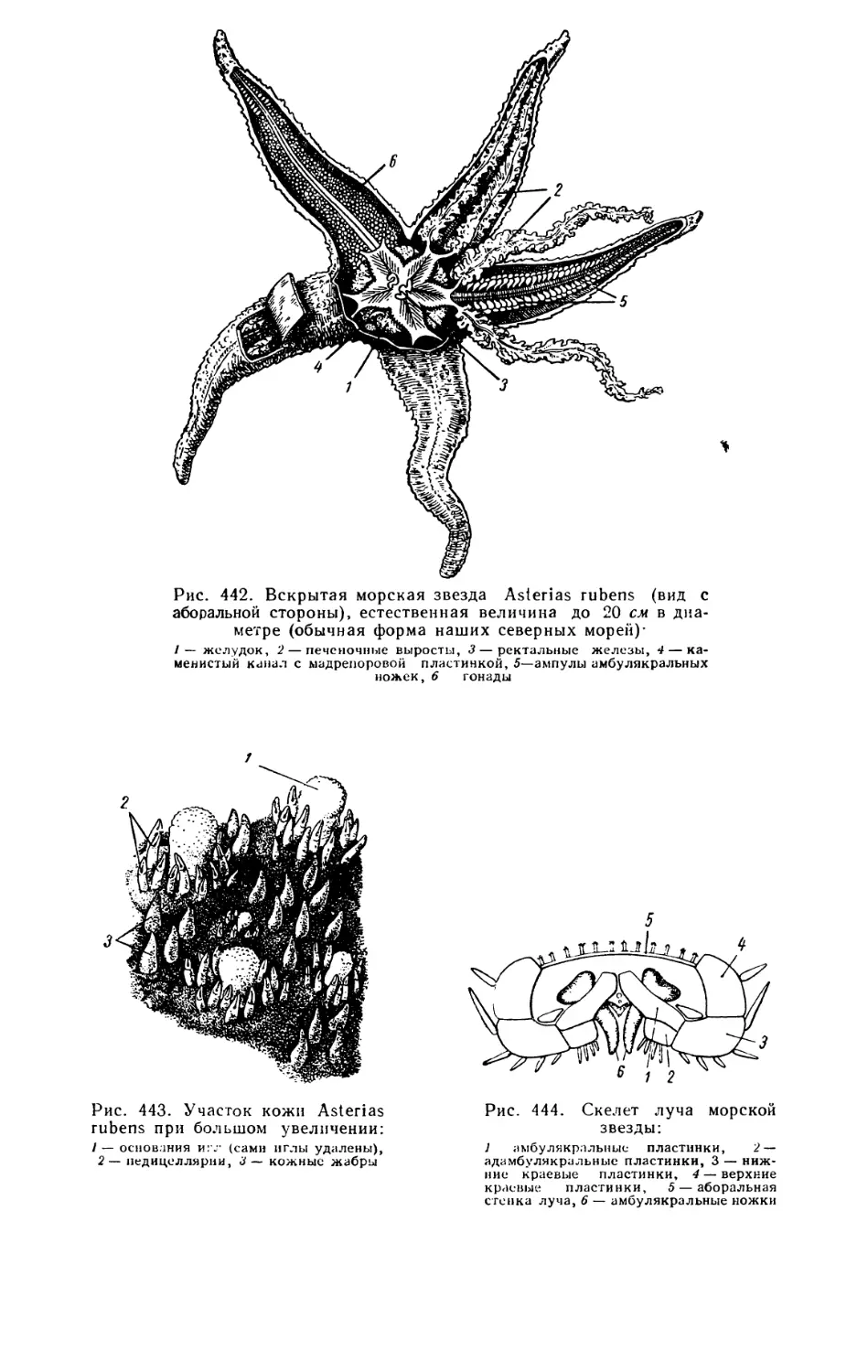
животных, а также низших хордовых дали неопровержимые доказатель-
ства родства позвоночных и беспозвоночных животных и общего происхож-
дения типов животного мира из единого корня, как того требовало учение
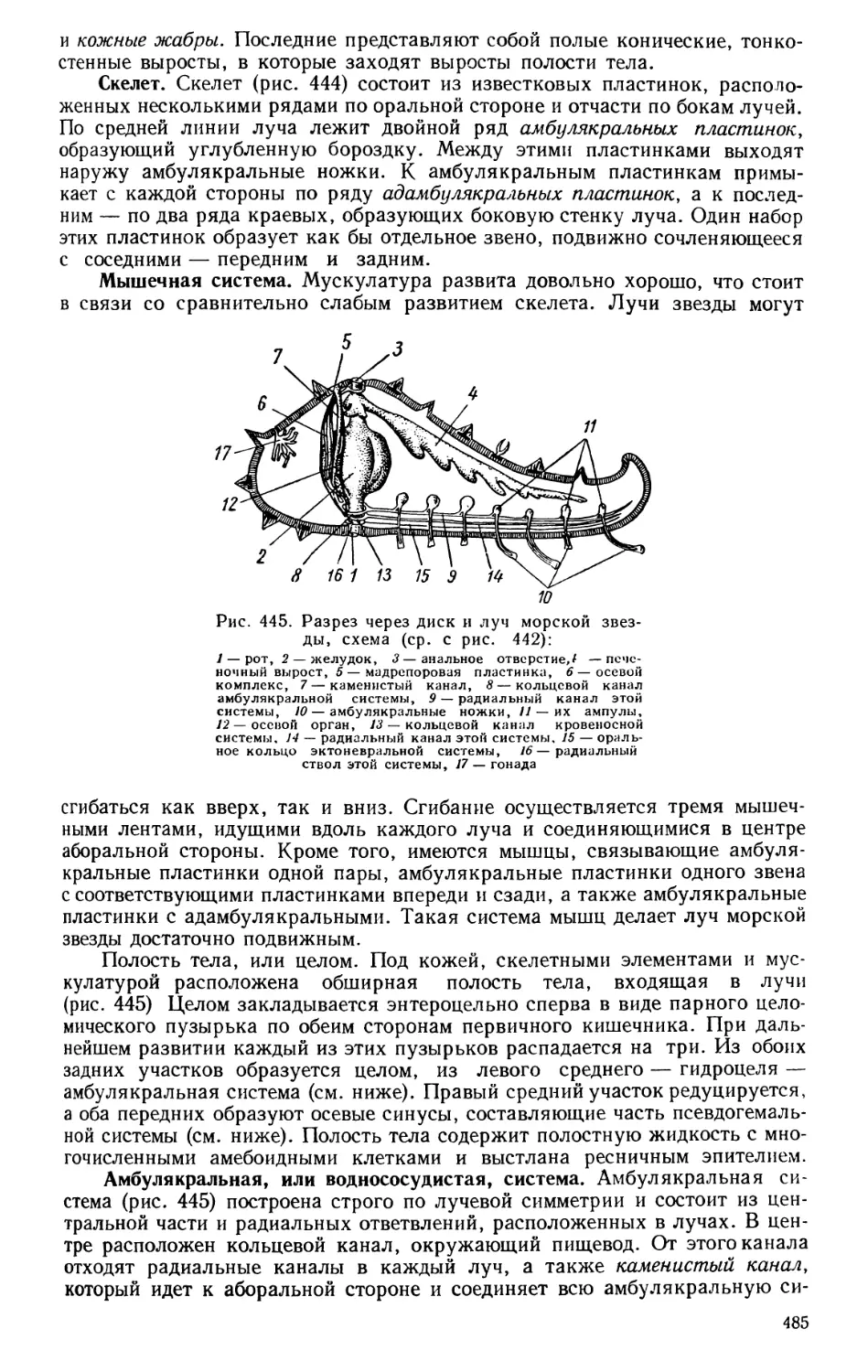
Дарвина.
Это эволюционное направление в морфологии в дальнейшем плодотвор-
но развивалось В. М. Шимкевичем, М. А. Мензбиром и его учениками,
12
из которых особенно большую роль сыграл А. Н. Северцов, создавший
русское направление эволюционной морфологии и воспитавший большую
школу учеников.
Громадную роль в развитии идей Дарвина сыграли другие направле-
ния эволюционной биологии.
В. О. Ковалевский (1842—1883) заложил основу эволюционной палеон-
тологии, а И. М. Сеченов (1829—1905) — русской школы физиологии жи-
вотных. Наконец, необходимо отметить громадную роль в развитии эволю-
ционных идей в зоологии, блестящей пропаганды исторического метода
в биологии К. А. Тимирязева (1843—1920) — яркого борца за идеи Дар-
вина.
К этому же славному поколению первых зоологов-эволюционистов
относятся основатели русской орнитологии (наука о птицах) и зоогеогра-
фии — Н. А. Северцов, известный путешественник по Средней Азии и его
ученик М. А. Мензбир, а позднее — П. П. Сушкин. Много дали также в
изучении зверей и птиц России замечательные путешествия в Центральной
Азии и Монголии Н. М. Пржевальского, П. К. Козлова и других русских
фаунистов.
ОРГАНИЗМ КАК ЦЕЛОЕ
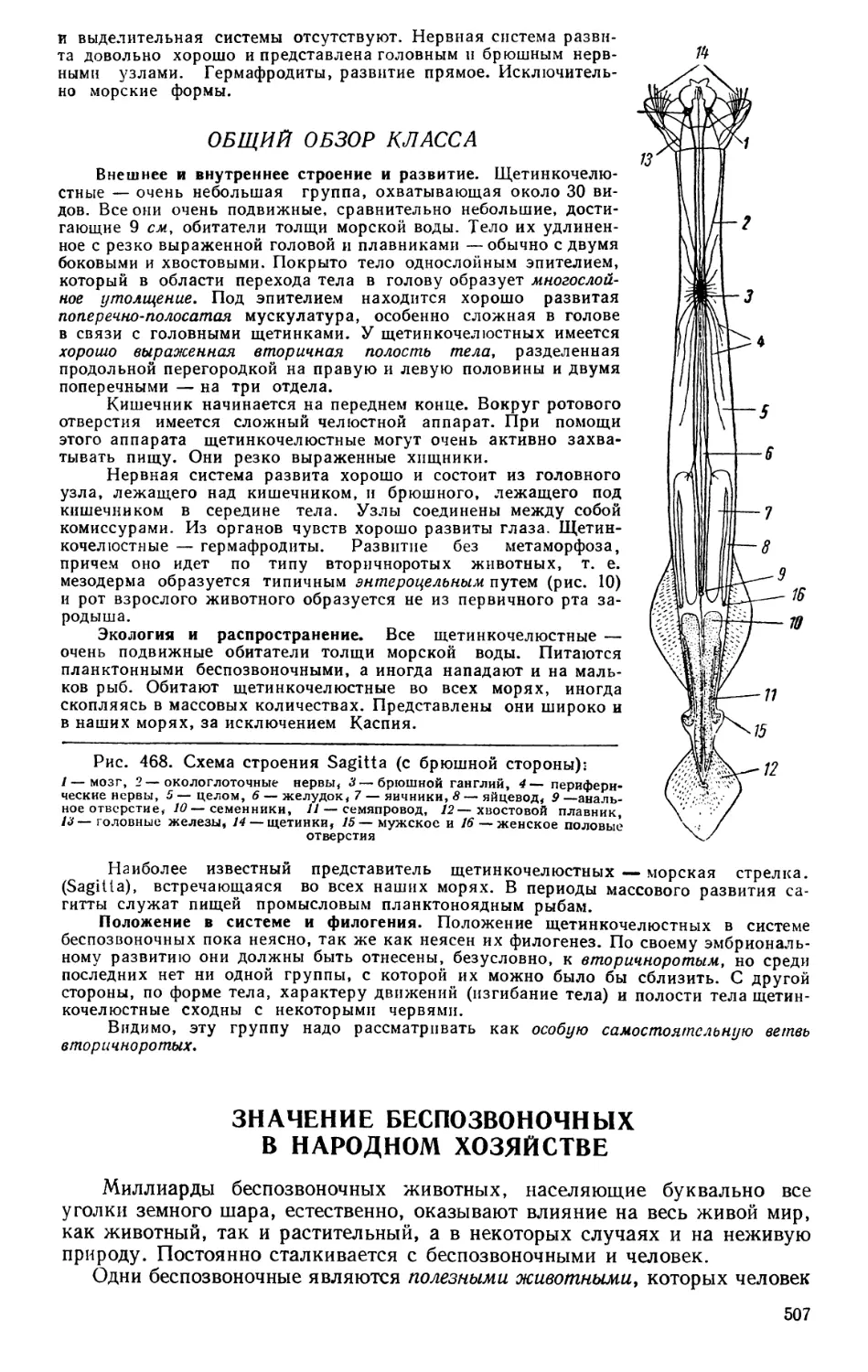
Обмен веществ. Организм — живое существо — является единст-
венным реальным носителем жизни, характеризующимся всеми свойствами
живого. Основной признак жизни — обмен веществ. Проявление жизни
всякого организма выражается во взаимодействии организма с окружаю-
щей средой. Это взаимодействие заключается в том, что организм воспри-
нимает из окружающей среды некоторые вещества, ассимилирует их и в
свою очередь отдает в нее другие вещества. В этих непрерывно идущих
процессах созидания и разрушения выражается обмен веществ. «...Всякое
органическое существо, — писал Энгельс, — в каждое данное мгновение
является тем же самым и не тем же самым; в каждое мгновение оно пере-
рабатывает получаемые им извне вещества и выделяет из себя другие ве-
щества, в каждое мгновение одни клетки его организма отмирают, другие
образуются; по истечении более или менее длительного периода време-
ни вещество данного организма полностью обновляется...».1
Эти две стороны обмена веществ носят название ассимиляции и дис-
симиляции.
Ассимиляцией называется преобразование веществ, поступающих в
организм из окружающей среды, и уподобление их составным частям
самого организма. Воспринимаемые организмом вещества претерпевают
различные превращения и входят в состав протоплазмы клетки или от-
кладываются про запас.
Диссимиляцией называется преобразование воспринятых организмом
веществ в более простые соединения, распадение веществ в организме,
причем происходит освобождение энергии, необходимой для осуществле-
ния различных функций организма.
Оба эти процесса неразрывно связаны друг с другом. Создание внутри
организма составляющих его веществ (ассимиляция) не может происходить
без затраты энергии, освобождающейся при протекающих одновременно
процессах распада. Процессы распада (диссимиляция) в свою очередь
нуждаются в принятии ассимилируемых веществ и в накоплении энергии.
В этом единстве ассимиляции и диссимиляции, единстве создания и распада
и заключается способность организма сохранять свою индивидуальность
ив то же время, непрерывно разрушаясь, освобождать энергию для
нового усвоения.
Круговорот веществ в природе. Различные организмы по-разному
воспринимают из окружающей их среды необходимые для них пищевые
1 Ф. Энгельс. Анти-Дюринг. В кн.: К. Маркс и Ф. Энгельс. Соч., изд. 2-е,
т. 20 , стр. 21—22.
13
вещества. Особое значение в круговороте веществ в природе имеет способ-
ность зеленых растений под влиянием солнечных лучей синтезировать
органические вещества из неорганических солей, получаемых с водой из .
земли и из углекислоты воздуха. Это явление фотосинтеза, открытое
К. А. Тимирязевым, устанавливает единство законов превращения энер-
гии в неорганическом и органическом мире. От этого свойства зеленых
растений зависит жизнь животных, не обладающих этой способностью
и благодаря этому связанных в своей жизнедеятельности с растительными
организмами или другими животными.
Некоторые бактерии синтезируют из углекислоты воздуха и воды
органические вещества без участия солнечных лучей. Они используют для
этой цели энергию, которая получается вследствие окисления бактериями
неорганических соединений. Зеленые растения, обладающие способностью
синтеза органических веществ на свету, т. е. фотосинтезом, и бактерии,
способные к синтезу за счет энергии химических реакций окисления (хе-
мосинтез), объединяются вместе в группу организмов автотрофных. Жи-
вотные и растения, питающиеся готовыми органическими веществами, на-
зываются гетеротрофными организмами.
Разложение органических веществ в организме в процессе его жизне-
деятельности происходит двумя путями: окислением, или дыханием и рас-
щеплением, или брожением. Процесс окисления состоит в том, что органи-
ческое вещество под влиянием различных ферментов распадается на более
простые составные части путем присоединения кислорода, поэтому такой
процесс аналогичен горению. Брожение заключается в разложении орга-
нического вещества под влиянием ферментов без участия кислорода. При
брожении освобождается во много раз меньше энергии, чем при дыхании,
и продукты, получающиеся в результате брожения, содержат неиспользо-
ванную потенциальную энергию.
Путь, проделываемый химическими веществами, которые поступают
в организм из окружающей природы, перерабатываются в организме и
вновь выделяются в окружающую среду, носит название круговорота ве-
ществ в природе. Все вещества, входящие в состав живой материи, прохо-
дят этот путь, но наиболее полно он изучен для двух важнейших элементов,
входящих в состав живого вещества: для углерода и азота. В качестве
примера разберем круговорот углерода.
Углерод попадает в воздух из почвы в виде углекислоты вместе с
почвенными водами, которые бывают насыщены углекислотой в результате
дыхания животных и растений, при горении и т.д. Углекислота поглоща-
ется зелеными растениями и под воздействием солнечных лучей распадается
на кислород и углерод. Углерод входит в состав углеводов (сахара и крах-
мала), образующихся в листьях при фотосинтезе. В самом растении обра-
зуется малый круг круговорота углерода: растение потребляет образую-
щиеся при фотосинтезе углеводы, расщепляет их в процессе диссимиляции
до углекислоты, которая снова возвращается в воздух. Однако большая
часть углерода идет на построение углеводов, белков и жиров растительного
организма, которые служат объектами питания животных. В организме жи-
вотного сложные органические вещества, благодаря процессу диссимиля-
ции, расходуются как источник энергии при дыхании и брожении и рас-
щепляются до углекислоты, которая выделяется в процессе дыхания.
При сжигании органических веществ также получается углекислота, пос-
тупающая в воздух. Ткани животных и растений при гниении, благодаря
бактериальному разрушению, также освобождают углекислоту, и таким
путем замыкается большой круг круговорота углерода в природе.
Организм. Организм представляет собой сложно организованную
систему органических веществ, среди которых основное значение имеют
белковые тела. Всякий организм находится в постоянном взаимодействии
с внешней средой и сохраняет свою целостность и жизнеспособность бла-
годаря постоянному процессу обмена веществ. Обмен веществ — основное,
14
свойство живых существ, все остальные свойства являются его производ-
ными. Важнейшая черта организма как носителя жизни — способность
к воспроизведению, т. е. к явлениям размножения, роста и развития.
Не менее характерна для живых существ способность к реагированию на
внешние воздействия сложными проявлениями. Реагирование организма,
его чувствительность к внешним воздействиям, проявляется либо в виде
возбудимости или раздражимости, либо в виде постепенного изменения его
свойств в сторону их большего соответствия условиям среды, что носит
название приспособляемости. Способность приспособляться к меняющимся
условиям среды представляет одно из наиболее ярких отличий живого от
неживого.
Организм как целостная система может быть представлен одной един-
ственной клеткой, которая выполняет все функции, как то имеет место
у простейших одноклеточных животных и растений. У всех остальных
животных организм состоит из большого количества клеток, разнородных
по своему составу и функции. Процесс преобразования комплекса однород-
ных клеток в разнородные благодаря разделению функций носит название
процесса дифференциации. При процессе дифференциации строение орга-
низма усложняется, отдельные части организма (органы, системы органов)
несут различные функции (пищеварение, дыхание, восприятие ощущений
и т. д.).
Понятие отдельного организма как особи, или индивидуума (неделимо-
го), для животного, вполне ясно: каждая особь вполне обособлена от окру-
жающей среды, она имеет определенное строение и обладает способностью
выполнять главнейшие жизненные отправления. В некоторых случаях
при размножении особи остаются связанными друг с другом — образуется
колония особей.
СТРОЕНИЕ ОРГАНИЗМА ЖИВОТНЫХ И ФУНКЦИИ
ЕГО ОРГАНОВ
В основе строения и развития животных и растительных организмов
лежит клетка — основная структурная форма организации живого веще-
ства. Учение о клетке как об основном стуктурном элементе, из которого
построен и развивается организм (животный и растительный), получило
название клеточной теории. Сформулированная в первой половине прош-
лого века клеточная теория показала единство многообразия основных
структур животных и растений и установила, что, кроме частных законо-
мерностей, характерных для тех или иных групп животного мира, сущест-
вуют общебиологические закономерности, общие законы развития для
всех животных и растений. Этим клеточная теория дала доказательства
общности их происхождения и подготовила почву для победы учения Дар-
вина об эволюции органического мира.
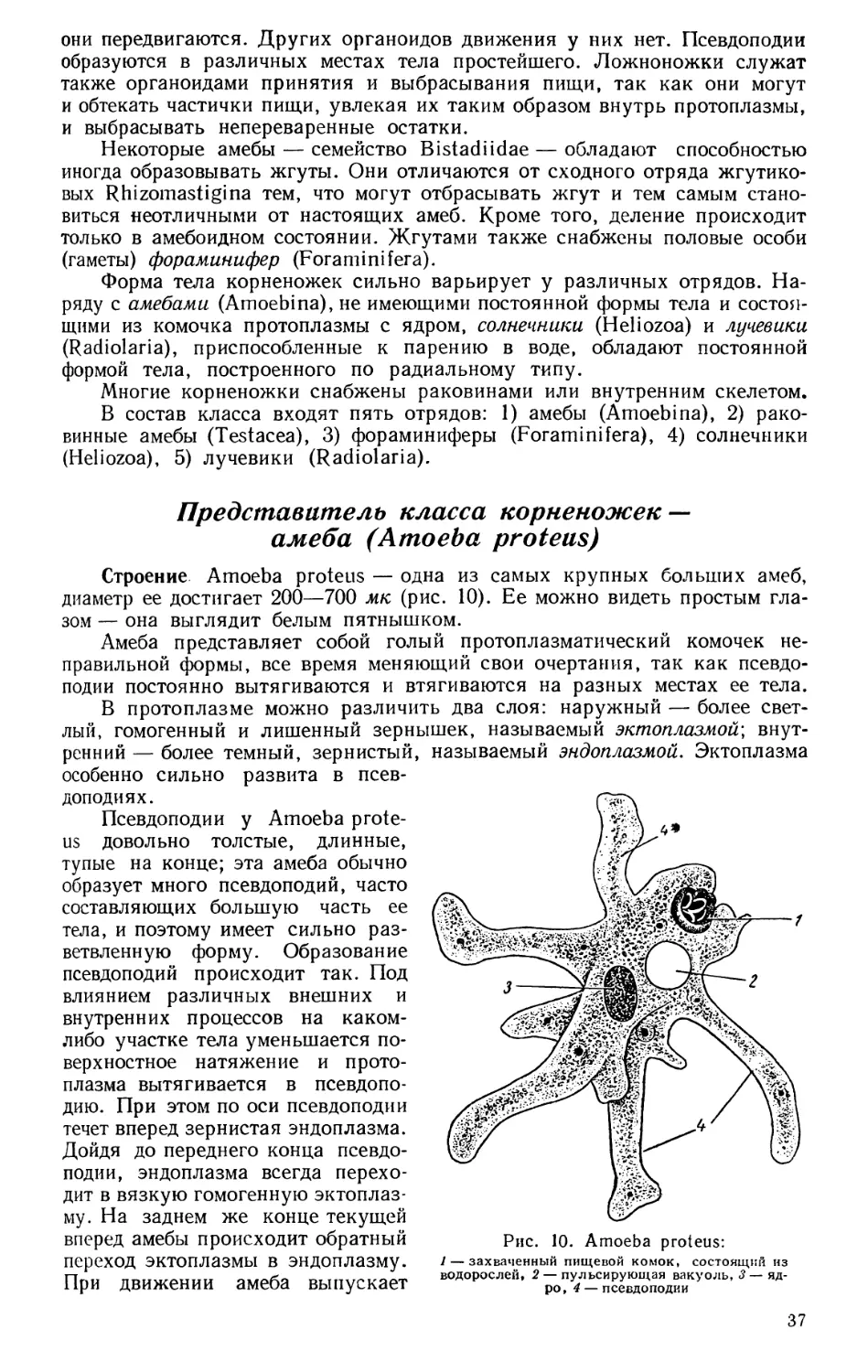
Строение клетки. Форма клеток весьма разнообразна (рис. 1.). Наи-
более обычны шаровидные, овальные, призматические, веретеновидные
клетки, встречаются также звездчатые, нитчатые, дисковидные и др. Коли-
чество клеток, входящих в состав организма, весьма велико, например
в коре больших полушарий головного мозга человека имеется 14—15 млрд,
нервных клеток, вместе с тем в состав некоторых коловраток и круглых
червей входит всего 400—900 клеток. Во всякой клетке различают прото-
плазму, ядро, оболочку, органоиды и включения.
Протоплазма состоит из сложных органических соединений. Главные
из них — белки, жиры, углеводы и нуклеиновые кислоты. В протоплазме
содержатся также различные минеральные соли и большое количество
воды; тело медузы состоит из воды на 95%, тело шелковичного червя —
на 72—82%.
15
Веществами, которые определяют основную структуру и жизнедея-
тельность протоплазмы и ядра, служат белки. В них происходят основные
процессы обмена веществ.
Во всех клетках имеется ядро, отграниченное от протоплазмы. В не-
которых клетках бывают и два, и три ядра. В ядре различают оболочку,
внутреннее полужидкое содержимое, состоящее из различных белков и
нуклеиновых кислот и одного или нескольких небольших округлых плот-
ных ядрышек.
Определенные методы исследования ядра позволяют установить в нем
наличие сложного строения их структуры —хроматиновых нитей. Они пред-
ставляют собой скопление глыбок хроматина, состоящего из нуклеопро-
теида. В период деления клетки эти хроматиновые нити реорганизуются
в более плотные образования — хромозомы. Количество и форма хромозом
считаются постоянными для каждого вида животных.
Ядро, являющееся необходимой составной частью клетки, и все вхо-
дящие в него структуры находятся в непрерывном взаимодействии с окру-
Рис. 1. Различные животные клетки. / — нервная
клетка; // — IV — эпителиальные клетки; V — сое-
динительнотканая клетка; VI — яйцеклетка; VII —
мышечная клетка
16
жающей ядро протоплазмой клетки. Ядро принимает участие во всех жиз-
ненных процессах, в развитии и размножении.
Органоиды (хондриозомы, аппарат Гольджи, клеточный центр) рас-
положены в протоплазме и служат постоянными и необходимыми состав-
ными частями клетки. В протоплазме имеется также обычно ряд непостоян-
ных включений, питательных веществ и продуктов обмена веществ. Хонд-
риозомы имеют вид нитей, палочек, зернышек. Они играют важную роль
в окислительных и других ферментативных процессах, происходящих в
клетке. Аппарат Гольджи в клетках позвоночных обычно имеет вид сеточ-
ки, расположенной около ядра, у беспозвоночных чаще состоит из отдель-
ных нитей и чешуек, разбросанных по всей клетке. Этот органоид принима-
ет участие в процессах концентрации веществ в клетке и выделения раз-
личных секретов. Клеточный центр, или центрозома. расположен обычно
около ядра и состоит из 1—2 зернышек, окруженных участками более
светлой плазмы. Центрозома принимает участие в процессах, происходящих
при кариокинетическом делении клетки.
Существует два основных типа деления клеток: митоз (кариокинез),
при котором оболочка ядра исчезает и образующиеся хромозомы оказывают-
ся лежащими непосредственно в протоплазме, и амитоз, при котором
происходит перешнуровка ядра и протоплазмы пополам.
В одних случаях митоз сопровождается удвоением числа хромозом
благодаря тому, что каждая из них синтезирует себе подобную. Деление
клетки надвое снова приводит к прежнему числу хромозом. В другом слу-
чае — при формировании половых клеток — митоз протекает без удвоения
числа хромозом. В силу этого каждая из двух вновь возникших при деле-
нии клеток получает вдвое меньшее количество хромозом. Такой тип митоза
при делении созревающих половых клеток носит название мейоза. или редук-
ционного деления созревания. Обычное для данного вида животного коли-
чество хромозом восстанавливается при половом процессе, т. е. при слия-
нии мужских и женских половых клеток и их ядер.
Ткани. У громадного большинства многоклеточных клетки отли-
чаются как по форме, так и по функции. Комплексы клеток, сходные
по структуре, функции и происхождению, образуют ткани. Обычно разли-
чают эпителиальную, соединительную, кровь, мышечную и нервную ткани.
Каждая ткань в известной степени обособлена и имеет свои характерные
черты, но вместе с тем в организме они тесно связаны и переплетаются
одна с другой. Названные ткани ясно выражены только у высокоорганизо-
ванных животных и во взрослом состоянии. У низших животных (губки
и кишечнополостные) имеются не все ткани, и одни и те же клетки могут
выполнять функции, свойственные различным тканям, например эпители-
ально-мышечные клетки кишечнополостных. На ранних стадиях развития
все клетки зародыша более или менее сходны.
Эпителиальная ткань, с одной стороны, защищает
организм от вредных воздействий внешней среды, с другой, через нее про-
исходит весь обмен веществ между организмом и внешней средой. Эпите-
лиальная ткань покрывает всю наружную поверхность организма, высти-
лает пищеварительную трубку, мочевые пути и вообще все полости тела.
Клетки эпителиальной ткани обычно тесно прилегают одна к другой, обра-
зуя сплошные пласты без межклеточного вещества. Пласты эти могут
состоять из одного слоя или ряда клеток, образуя однослойный эпителий,
или из многих слоев, образуя многослойный эпителий. Особым видом эпите-
лия является ресничный, или мерцательный, эпителий. Клетки его на сво-
бодной поверхности несут реснички, а в некоторых случаях жгутики.
Этот вид эпителия очень распространен, встречается во всех группах живот-
ных и играет большую роль в жизненных процессах — движении, питании,
дыхании и выделении.
Специальной дифференцировкой эпителиальных клеток служат же-
лезистые клетки, в протоплазме которых вырабатывается тот или иной
17
секрет. Эти клетки образуют железы, которые могут быть одноклеточными
или многоклеточными, в том случае, если большое число железистых клеток
собрано вместе.
Соединительная ткань объединяет на первый взгляд
очень разнообразные виды тканей — рыхлую соединительную ткань, связки,
сухожилия, хрящ и кость. Все эти образования, однако, имеют ряд общих
черт. Во-первых, все виды соединительной ткани имеют большое количество
промежуточного (межклеточного) вещества: или мягкого (рыхлая соедини-
тельная ткань), или более или менее плотного (хрящ), пропитанного в
некоторых случаях неорганическими веществами (кость). Во-вторых, ос-
новная функция всех видов соединительной ткани опорная; ткань дает
прослойки между внутренними органами, связывает их, а у иглокожих
и хордовых образует скелет. В-третьих, все виды соединительной ткани
происходят из одной и той же эмбриональной ткани —мезенхимы. Помимо
основной функции — опорной, соединительная ткань (главным образом
рыхлая) несет и другие функции: питательную, так как через нее идет
весь ток питательных веществ, ассимилируемых кишечным эпителием;
защитную, так как некоторые клетки ее способны заглатывать и перевари-
вать попавшие в организм чужеродные образования, в частности болезне-
творных бактерий. Этот процесс фагоцитоза впервые был описан великим
русским ученым И. И. Мечниковым.
Кровь — жидкая ткань, состоит из неклеточной жидкой плазмы,
в которой плавают кровяные клетки: красные и белые кровяные тельца,
называемые форменными элементами крови. Кровь нередко относят к соеди-
нительным тканям.
Мышечная ткань выполняет сократительную функцию.
Сокращение мышечных клеток происходит благодаря наличию в их про-
топлазме особых сократительных нитей, или миофибрилл. Мышечная ткань
бывает двух видов: гладкая и поперечнополосатая. Гладкая состоит из длин-
ных веретеновидных клеток. Характерна гладкая мышечная ткань для
большинства беспозвоночных, у позвоночных из гладкой мышечной ткани
построена вся мускулатура внутренних органов, кроме сердца.
Поперечнополосатая мышечная ткань образована мышечными волок-
нами, которые могут достигать 10 см длины. Они представляют собой уже
не клеточные образования, а симпласты, так как в их протоплазме находит-
ся много ядер. В поперечнополосатых мышечных волокнах имеются также
мышечные фибриллы, состоящие из двоякого рода участков — дисков,
правильно чередующихся друг с другом. Одни из них обладают способ-
ностью к двойному лучепреломлению — они анизотропны, другие этой
способностью не обладают — они изотропны. Благодаря правильному че-
редованию дисков все волокна и кажутся поперечноисчерченными. Попе-
речнополосатая мускулатура характерна для позвоночных, где образует
всю скелетную мускулатуру. Из беспозвоночных эта мускулатура имеется
только у членистоногих, некоторых кольчатых червей и частично моллюс-
ков (например, замыкательные мышцы раковин у двустворчатых).
Нервная ткань. Основная функция нервной ткани — регу-
ляция всех жизненных процессов в организме. Нервные клетки способны
к восприятию и переработке раздражений, получаемых организмом из
внешней среды, и передаче возбуждения. С этими функциями связана и
форма нервных клеток — для них характерно наличие отростков (см. рис. 1),
которые бывают двух родов: короткие и длинные. Короткие, обычно сильно
ветвящиеся, носят название дендритов', их может быть один или несколько.
Они несут функцию восприятия раздражения и передают его в тело клетки.
Длинные отростки обычно ветвятся на конце и отходят только по одному
от каждой клетки. Они называются невритами и передают возбуждение
по направлению от тела клетки. Нервная клетка со всеми отростками на-
зывается невроном. Отдельные нервные клетки соединяются в той или иной
форме вместе и образуют нервы, нервные стволы и узлы. Нервная ткань
18
той или иной степени сложности имеется у всех животных, за исключени-
ем губок, хотя и у них по исследованиям последних лет обнаружены
нервные клетки.
ОРГАНЫ И СИСТЕМЫ ОРГАНОВ ЖИВОТНЫХ
Помимо гистологической дифференциации тела животных на ткани,
тело сложных многоклеточных расчленено на органы и системы органов.
Органами называются части тела, выполняющие отдельные жизненные
функции. Органы обыкновенно состоят из разных тканей. Отдельные жиз-
ненные функции могут выполняться одним органом, как это имеет место
у низших животных, а у высших отдельные жизненные отправления выпол-
няются целым комплексом органов, которые называются системами орга-
нов. Так, например, у человека различают 11 систем органов: 1) покровная
система покрывает снаружи все тело, защищает его и воспринимает воз-
действия окружающей среды; 2) скелетная система служит опорой тела
и обеспечивает возможность движения; 3) мышечная система вместе со
скелетной осуществляет движение; 4) нервная система воспринимает раз-
дражения от органов чувств, проводит к мозгу и по всему телу, вызывая
различные реакции организма; 5) органы чувств принимают раздражения
из внешнего мира и от различных участков тела; 6) пищеварительная си-
стема воспринимает пищу и перерабатывает ее в простые химические ве-
щества, всасываемые в кровь; 7) система кровообращения переносит веще-
ства внутри организма; 8) дыхательная система обеспечивает поступление
в кровь кислорода и освобождение крови от углекислоты; 9) выделительная
система удаляет из организма ненужные продукты обмена веществ; 10) эн-
докринная система обеспечивает организм особыми веществами — гормо-
нами, координирующими функции организма; И) система органов раз-
множения готовит новое поколение, продолжающее дальнейшее существо-
вание вида.
ПЛАН СТРОЕНИЯ И СИММЕТРИЯ
Рассматривая общий план строения какого-либо животного, мы за-
мечаем обычно известную правильность в расположении частей тела —
они повторяются в определенной закономерности вокруг некоторой централь-
ной его оси. Такое правильное положение частей называют симметрией.
Симметрия может быть разной; определение характера симметрии важно
для общего анализа организации животных. Так, низшим животным боль-
шей частью свойственна бесконечная радиальная симметрия, при которой
бесконечным количеством плоскостей, проходящих через центр тела, оно
рассекается на совершенно одинаковые участки. В процессе эволюции
бесконечная радиальная симметрия сменяется ограниченной радиаль-
ной симметрией, при которой через центр тела можно провести лишь
определенное количество плоскостей, рассекающих его на одинаковые
участки. При ограниченной радиальной симметрии органы обычно боль-
ше располагаются в радиальных направлениях от центра и у жи-
вотного можно различить лишь верхнюю и нижнюю стороны тела.
Радиальная симметрия свойственна пассивно плавающим и прикреплен-
ным к субстрату животным.
У подвижных животных, двигающихся в одном направлении вперед
или назад, развивается двусторонняя, или билатеральная, симметрия,
при которой только одна плоскость может разделить тело на две равные
половины — правую и левую. У высших двусторонне-симметричных
животных, кроме того, обособляются передний (головной) и задний (хвосто-
вой) отделы тела. В связи с разным положением тела в пространстве перед-
ний отдел тела называют краниальным, задний — каудальным, спинную
сторону — дорзальной, брюшную — вентральной, плоскость, проходящую
19
по средней линии тела, называют медиальной, параллельные ей плоскости —
сагиттальными, а плоскость, делящую тело на брюшную и спинную сто-
роны, — фронтальной. Органы, располагающиеся на боках тела, зани-
мают латеральное положение. Наконец, плоскость, делящая тело перпен-
дикулярно к продольной оси его, называют поперечной.
Сравнительно очень немногие животные не имеют определенно выра-
женной симметрии и являются таким образом асимметричными. Обычно
асимметрия — явление вторичное и вызвано тоже определенными условия-
ми существования, при которых одна из частей тела животного попадает
в условия, неодинаковые с другой.
У ряда высокоорганизованных двусторонне-симметричных животных
(кольчатые черви, членистоногие, бесчерепные, позвоночные) различают
по длине тела ряд участков — сегментов, или метамер. Такая сегментация,
или метамеризация, обычно охватывает тело не только снаружи, но и
распространяется на внутренние органы: мышцы, нервную систему, органы
кровообращения и выделения, отдельные элементы половой системы. У ряда
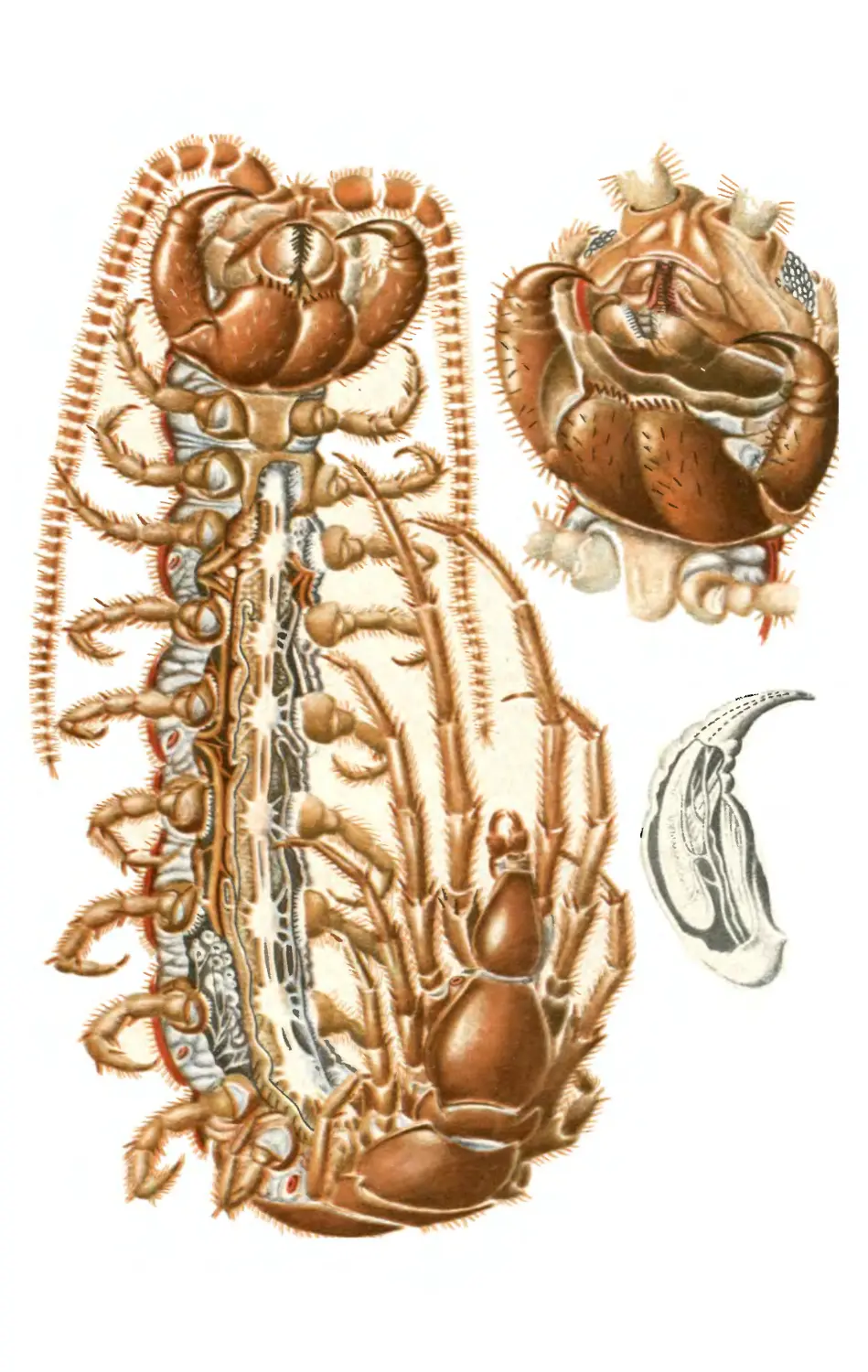
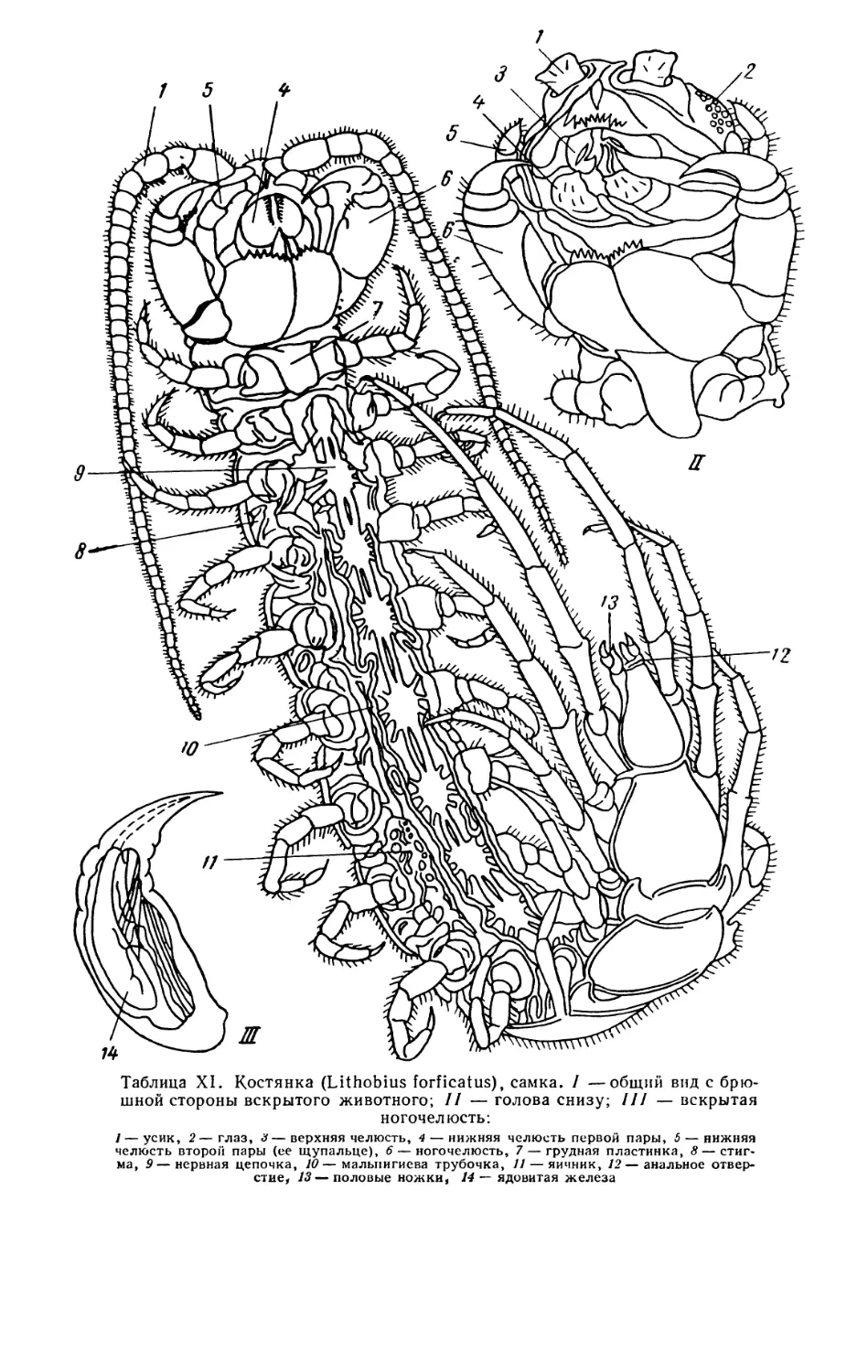
сегментированных животных сегментация хорошо видна снаружи (табл. XI),
у других выражена только во внутреннем строении (бесчерепные, позвоноч-
ные). Иногда сегментация может вторично исчезать, например у клещей
(рис. 341). Образование сегментации — результат активного образа жизни
животного и было очень прогрессивным явлением в эволюционном разви-
тии животных.
РАЗМНОЖЕНИЕ И РАЗВИТИЕ
Размножение. Размножение — основное свойство живых существ, поз-
воляющее существовать каждому виду животных в течение многих поколе-
ний. Существует два основных типа размножения: бесполое и половое.
В бесполом размножении участвует только одна родительская особь, кото-
рая делится или почкуется для образования двух или большего числа новых
особей. При этом дочерние организмы по наследственным свойствам сход-
ны с родительской особью. Соответственно этому различают формы беспо-
лого размножения: 1) деление, если животное делится на две более или
менее одинаковые части, 2) почкование, когда небольшая часть родитель-
ского организма отделяется и превращается во взрослую особь, и, наконец,
3) спорообразование, когда родительская особь распадается на большое
число спор — особых клеток с защитными оболочками, способных выдер-
живать неблагоприятные условия среды.
В половом размножении участвуют две особи, каждая из которых
образует специализированную клетку — гамету. У самок гамета пред-
ставляет собой яйцеклетку. Обычно это крупная неподвижная клетка
Рис. 2. Яйцеклетки. 1 — морской звезды; 11 — кости-
стой рыбы:
1 — ядро
20
(«яйцо»), содержащая запас желтка, служащего источником питания заро-
дыша, и называется яшрж. Гаметы самцов представлены сперматозои-
дами— очень мелкими и подвижными клетками, обычно снабженными
хвостом, который позволяет им активно двигаться, приближаясь к яйцу.
Процесс слияния двух половых клеток — гамет носит название оплодо-
творения, а оплодотворенная яйцеклетка называется зиготой.
Для водных животных характерно наружное оплодотворение’, яйца
и сперма выделяются животными в воду, где происходит слияние половых
клеток. У наземных и некоторых водных животных наблюдается внутрен-
нее оплодотворение: при спаривании сперма вносится самцом в половые
пути самки. У некоторых животных встречается девственное размножение,
называемое партеногенезом, когда происходит развитие новой особи из
неоплодотворенного яйца без участия сперматозоида.
Индивидуальное развитие. Индивидуальное развитие живых существ
продолжается от момента оплодо-
творения яйцеклетки до конца
жизни организма и называется
онтогенезом. Он осуществляется
сложным преобразованием зиготы,
которая даже у высших животных
организмов и человека состоит из
единственной клетки. Зигота за-
ключает в себе в потенции все
видовые особенности родителей,
так как она образовалась путем
слияния яйцеклетки и спермато-
зоида.
Первым этапом процесса раз-
вития является деление оплодотво-
ренного яйца и превращение зи-
готы в многоклеточный организм:
этот этап носит название дробле-
ния яйца, а клетки дробящегося
яйца называются бластомерами.
Дробление отличается от деления
клетки как способа бесполого раз-
множения тем, что при дроблении
яйца не происходит роста дочер-
них клеток до размеров клетки
перед делением, а наоборот, клет-
ки становятся все мельче — до
размеров клеток тела данного
вида. Однако темпы деления кле-
ток при дроблении неодинаковы
и зависят от количества питатель-
ного материала — желтка, заклю-
ченного в яйцеклетку. Мелкие
клетки дробления называются ми-
кромерами, а крупные макроме-
рами. Соответственно порядку де-
ления клеток различают несколь-
ко типов дробления — полное и
неполное. Полное дробление может
быть равномерным и неравномер-
ным. Порядок дробления разли-
чают радиальный или спиральный.
Неполное дробление может быть
дискоидальным или поверхностным
Рис. 3. Сперматозоиды. / — ската;
II — чайки; III — улитки; IV — меду-
зы; V — щуки; VI — жука; VII —
двоякодышащей рыбы; VIII — рака;
IX — круглого червя; X, XI — мелких
рачков
21
Рис. 4. Схема радиального дробления. I— стадия восьми бластомеров;
// — стадия шестнадцати бластомеров
Рис. 5. Схема спирального дробления. 1 — стадия восьми бласто-
меров; II — стадия шестнадцати бластомеров
Рис. 6. Неполное дробление. / — схема
дискоидального дробления яйца головоно-
гого моллюска; // — схема поверхностно-
го дробления яйца насекомого:
1 — поверхностный слой образовательной плазмы,
2—желток, 3—ядра, получившиеся в результате
дробления яйцевого ядра, окруженные участками
образовательной плазмы
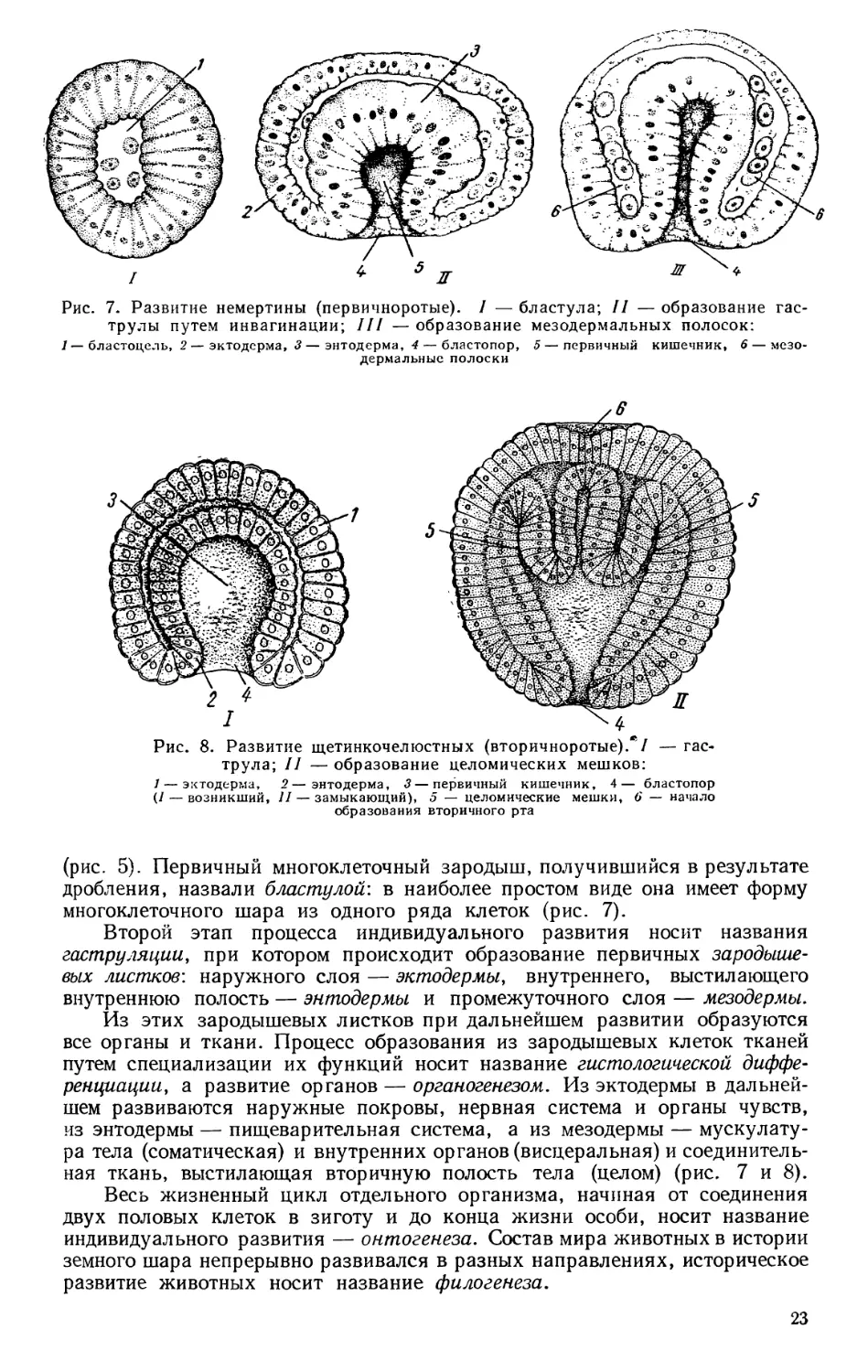
Рис. 7. Развитие немертины (первичноротые). / —бластула; II —образование гас-
трулы путем инвагинации; III —образование мезодермальных полосок:
1— бластоцель, 2 — эктодерма, 3—энтодерма, 4 — бластопор, 5 — первичный кишечник, 6 — мезо-
дермальные полоски
Рис. 8. Развитие щетинкочелюстных (вторичноротые)// — гас-
трула; II — образование целомических мешков:
1 — эктодерма, 2—энтодерма, 3— первичный кишечник, 4— бластопор
(/— возникший, // — замыкающий), 5 — целомические мешки, 6 — начало
образования вторичного рта
(рис. 5). Первичный многоклеточный зародыш, получившийся в результате
дробления, назвали бластулой’, в наиболее простом виде она имеет форму
многоклеточного шара из одного ряда клеток (рис. 7).
Второй этап процесса индивидуального развития носит названия
гаструляции, при котором происходит образование первичных зародыше-
вых листков', наружного слоя — эктодермы, внутреннего, выстилающего
внутреннюю полость — энтодермы и промежуточного слоя — мезодермы.
Из этих зародышевых листков при дальнейшем развитии образуются
все органы и ткани. Процесс образования из зародышевых клеток тканей
путем специализации их функций носит название гистологической диффе-
ренциации, а развитие органов — органогенезом. Из эктодермы в дальней-
шем развиваются наружные покровы, нервная система и органы чувств,
из энтодермы — пищеварительная система, а из мезодермы — мускулату-
ра тела (соматическая) и внутренних органов (висцеральная) и соединитель-
ная ткань, выстилающая вторичную полость тела (целом) (рис. 7 и 8).
Весь жизненный цикл отдельного организма, начиная от соединения
двух половых клеток в зиготу и до конца жизни особи, носит название
индивидуального развития — онтогенеза. Состав мира животных в истории
земного шара непрерывно развивался в разных направлениях, историческое
развитие животных носит название филогенеза.
23
СИСТЕМА МИРА ЖИВОТНЫХ
Мир животных в прошлом и настоящем населяет земной шар не как
множество отдельных организмов. Организмы связаны друг с другом в
естественные группировки, носящие название вида, а виды составляют
высшие группировки, носящие название систематических категорий, ко-
торые соподчинены друг с другом в стройную систему — классификацию
животных. Отдел зоологии, занимающийся классификацией мира живот-
ных, носит название систематики.
Систематика и распределяет животные организмы по группам в зависи-
мости от степени сходства или различия между ними и строит, таким обра-
зом, систему животного мира. При построении системы современная систе-
матика учитывает не отдельные признаки, а их комплекс, не только внеш-
нюю форму, но и внутреннее строение, физиологию, развитие, а иногда
экологию и распространение животных. Таким образом, систематика
представляет собой синтетическую науку, опирающуюся на достижения
других зоологических дисциплин.
Значение систематики велико как теоретически, так и практически.
Теоретически она важна тем, что строит естественную систему, отражаю-
щую существующие в природе связи и историческое развитие (эволюцию)
животного мира. Практически она важна тем, что без объединения живот-
ных в систему невозможно разобраться в их разнообразии.
Единицей систематики является вид (species). К одному виду относят
совокупность особей, сходных между собой, имеющих общее происхождение,
свободно скрещивающихся и дающих плодовитое потомство.
Обычно особи, принадлежащие к различным видам, не скрещиваются
между собой или дают неплодовитое потомство, но из этого правила извест-
ны многочисленные исключения. Межвидовое скрещивание применяют как
в животноводстве, так и в растениеводстве для улучшения пород и сортов.
Близкие виды объединяются в роды (genus), и каждый известный вид
имеет двойное наименование', первое —родовое и второе — видовое, при-
чем наименования эти даются по общепринятому международному правилу
по-латыни. Волк, например, имеет следующее наименование — Canis lupus.
Такой способ обозначения животных (это в полной мере относится и к
растениям) получил название двойной, или бинарной, номенклатуры.
Близкие роды объединяются в семейства (familid), семейства — в
отряды (ordo), отряды — в классы (classis) и классы — в типы (typus).
Иногда система осложняется тем, что приходится применять промежуточ-
ные группы — подтипы, подклассы, подотряды и т. д. Род кошек (Felis)
вместе с родами рысей (Lynx), леопардов (Pardus) и некоторыми другими
объединяются в семейство кошачьих (Felidae). Это семейство вместе с семей-
ствами псовых (Canidae), куньих (Mustelidae), медведей (Ursidae) и некото-
рыми другими объединяется в отряд хищных (Carnivora). Этот отряд вместе
с другими отрядами — копытными, ластоногими, грызунами, приматами
и другими образуют класс млекопитающих (Mammalia). Этот класс вместе
с классами птиц, пресмыкающихся, амфибий, рыб и круглоротых образуют
подтип позвоночных (Уertebrata), входящий вместе с подтипами бесчереп-
ных и личиночнохордовых в тип хордовых (Chordata).
Таким образом, соотношение главнейших систематических категорий
представляется в следующем виде:
Класс 1 Отряд 1 Семейство 1 Род 1 Вид 1
Класс 2 Отряд 2 Семейство 2 Род 2 Вид 2
Тип Класс 3 Отряд 3 Семейство 3 Род 3 Вид 3
и т. д. и т. д. и т. д. и т. д. и т. д.
Кроме систематических категорий выше вида (род, семейство и т. д.),
различают более мелкие внутри вида, подвиды для диких и породы для до-
машних животных.
24
В данном учебнике принята следующая система. Таблица 1
Типы Количество классов, входящих в тип Примерное количе- ство видов Среда обитания Пара- зиты
моря прес- ные воды почвы суша
I. Одноклеточные — Protozoa II. Губки— Spongia III. Кишечнополост- ные — Coelente- rata IV. Плоские, или бес- полостные чер- ви — Plathelmint- hes V. Круглые, или пер- вичнополостные , черви — Nemathel- minthes VI. Немертины— Ne- mertini .... VII. Кольчатые, или вторичнополост- ные, черви — Ап- nelides . . . VIII. Членистоно- гие— Arth- ropoda IX. Моллюски — Mol- lusca X. Мшанки — Bryozoa XI. Плеченогие—Bra- ch iopoda XII. Иглокожие — Echinodermata XIII. Погонофоры — Pogonophora . XIV. Хордовые—Chor- data .... -f- Основные среды оби' Примечание. В чи которых в системе неясно. Относительное рирует следующая 5 1 4 7 8 1 5 9 7 2 2 5 1 12 Вес гания; (-f-) Hi ело классов количеств схема: 20 000 5 000 9 000 6 000 10 000 750 7000 900 000 100 000 3 000 250 4500 50 50 000 то пример зибольшее ч не вошли к. о видов +* + + + + 4- 4- 4- 4- + 4- 4- 4- IHO 1 1 [ИСЛО ви лассы разли (+) 4~ 4~ + (+) 4~ + 4“ (+) 4- 00 000 ДОВ. Phorono чных 4- 4- 4- 4- idea и групп (+) (+) + 4- Ghaeto живс Е 1 III 11+ + + ±+ + + । + X “ Более 800 000 насеко- мых Несколь- ко тысяч ископае- мых положение иллюст-
25
Само собой разумеется, что никакая система не является чем-то окон-
чательным и незыблемым. Новые данные по строению и развитию отдель-
ных форм и групп могут изменить наши представления о той или иной
группе животных.
ВЗАИМООТНОШЕНИЕ ЖИВОТНЫХ С ОКРУЖАЮЩЕЙ
СРЕДОЙ И МЕЖДУ СОБОЙ
У животных обоеполых состав вида однороден, т. е. особи отличаются
только индивидуальными признаками. У всех же раздельнополых живот-
ных особи, составляющие вид, разделяются приблизительно в равных
соотношениях на самцов и самок, отличающихся строением органов размно-
жения. Среди животных есть много видов, где самцы и самки отличаются
не только строением органов размножения, но и целым рядом наружных
признаков. При этом различия выражаются в органах, не имеющих прямо-
го отношения к размножению. Такие различия называются вторичнополо-
выми признаками, а наличие двух форм (половых) в популяции носит на-
звание полового диморфизма. Так, у многих бабочек самцы отличаются
более ярким и пестрым рисунком крыльев. У многих насекомых (например,
у светляков) самцы крылаты, а самки бескрылы, у птиц самцы значительно
более ярко окрашены, чем самки (например, у куриных — петухи, фазаны,
павлины и др.), и обладают гребнями и другими кожными выростами на
голове, а на ногах — шпорами. У млекопитающих половой диморфизм
выражается также в более яркой окраске, в наличии рогов (у самцов оле-
ней), гривы (у самцов львов) и т. д.
Помимо полового диморфизма, у бабочек встречается еще сезонный
диморфизм, когда поколения, выведшиеся весной, резко отличаются по
окраске от осенних поколений, что связано с развитием в различных тем-
пературных условиях и др. В тех случаях, когда животные живут сооб-
ществами, как это имеет место у общественных насекомых (пчел, муравьев
и термитов), а также у колониальных животных, в составе такого сообщест-
ва встречается несколько форм, отличающихся рядом особенностей строе-
ния и функций, причем разные полиморфные особи несут в сообществе
различные функции. Такое явление получило название полиморфизма.
Состав вида, особи которого заселяют свойственную ему территорию,
неоднороден. Вся совокупность вида разделяется на отдельные группы
особей, отличающиеся друг от друга рядом особенностей соответственно
условиям их жизни. Такие группы организмов отдельных видов носят
название популяций. Соответственно условиям существования, свойствен-
ным данной популяции, каждая из них отличается рядом особенностей.
К таким особенностям относятся: плотность населения популяции, рож-
даемость, смертность, возрастной состав, скорость расселения и др.
Условия существования животных на земной поверхности и в водое-
мах очень различны; это различие определяется разными факторами как
неорганического порядка (абиотические факторы — температура, влаж-
ность, соленость, глубина и пр.), так и органического (биотические факто-
ры — почвенные условия, растительность, окружающее животное населе-
ние и пр.). Каждый участок суши или водоема имеет определенный
комплекс этих факторов и образует так называемый биотоп.
Тот или иной биотоп населяют животные, приспособленные как мор-
фологически, так и физиологически к данным условиям, в чем и заключает-
ся единство организмов с условиями существования. Животные населяют
самые разнообразные биотопы, иногда как будто бы совершенно непригод-
ные для жизни, например водоемы, перенасыщенные солями, в которых
содержание солей достигает иногда ЗОО°/оо (нормальная соленость океанов
36—37°/оо)1, горячие источники с температурой до 53°.
°/00—обозначает промилле, т. е. число единиц на 1000.
26
Некоторые группы животных или отдельные виды могут существовать
при очень больших колебаниях того или иного фактора; такие группы
или виды носят название широкосолевых, широкотепловых и т. д., или
эвригалинных, эвршпермных и т. д. Другие, наоборот, могут существовать
только в пределах небольших колебаний данного фактора и носят названия
узкосолевых, узкотепловых и т. д., или стеногалинных, стенотермных
и т. д. Иглокожие, например, стеногалинный тип, так как все иглокожие
могут существовать только в морской воде и не переносят опреснения.
Моллюски в целом, наоборот, эвригалинны, так как они хорошо сущест-
вуют и в морях и в пресных водах, т. е. выдерживают колебания солености
от (Хб0/^ до 37°/00. Вместе с тем головоногие моллюски — стеногалинная
группа, так как они обитают только в морях. Примером стенотермной
группы могут служить рифообразующие кораллы (Madreporaria), не встре-
чающиеся при температурах ниже 20—22° С. Примером узкоглубинной,
или стенобитной группы могут служить стебельчатые морские лилии
(см. табл. XV), обитающие только на больших глубинах.
Взаимоотношения животных с другими организмами определяются
прежде всего пищевыми связями. По характеру питания животных можно
поделить на всеядных, растительноядных и плотоядных, или хищных.
Низшие животные не имеют обычно избирательной способности к пище
и питаются всем, что попадается, например животные-фильтраторы (поли-
пы, двустворчатые моллюски, мшанки, усоногие рачки и Др.), которые
при помощи различных приспособлений отфильтровывают из воды как
различных микроскопических животных и растений, так и различную
органическую взвесь (детрит). У более высокоорганизованных животных
избирательная способность к составу пищи, наоборот, имеется и выражена
иногда очень резко, так, например, некоторые насекомые могут питаться
только определенными растениями.
В природе между животными существует разная степень взаимосвя-
зей. Иногда одни животные вступают с другими в более тесные взаимоотно-
шения. Можно различить следующие виды взаимоотношений.
Симбиозом называется такая взаимосвязь, когда обе стороны взаим-
но полезны друг другу. Например, пчелы, питаясь нектаром цветков, одно-
временно опыляют их. В некоторых случаях симбиоза животные вступают
в более тесный контакт, так, например, актиния поселяется на раковине,
в которой обитает рак-отшельник (см. рис. 114) и своими стрекательными
клетками защищает его; в свою очередь рак-отшельник, передвигаясь
по дну, переносит вместе с собой актинию, что дает ей возможность более
интенсивно питаться.
Комменсализмом называется такая взаимосвязь, когда одна
сторона служит местообитанием или защитой для другой, не получая для
себя ничего взамен. Так, например, довольно часто обыкновенные воробьи
устраивают свои гнезда в громадном гнезде орлана-белохвоста, пользуясь
им для защиты от врагов; ряд сидячих животных поселяется на других —
подвижных, не причиняя им никакого вреда, но используя их как субстрат
и передвигаясь при их помощи, например, некоторые усоногие рачки (см.
рис. 309) поселяются на коже китов, ряд сидячих инфузорий (см. рис. 65) —
на рачках и т. д.
Паразитизмом называется такая взаимосвязь организмов,
когда один организм (паразит) живет на теле или в теле другого (хозяина),
питаясь содержимым его кишечника или его тканями и принося таким
образом ему вред (заболевание) или приводя даже к смерти.
Паразитический образ жизни очень сильно отражается на строении
(как внешнем, так и внутреннем) организмов—паразитов. У них, с одной
стороны, происходит в той или иной степени редукция ряда органов (орга-
нов движения, органов чувств, пищеварительной системы), с другой —
развиваются специальные органы, связанные с паразитизмом, например
органы прикрепления к хозяину (см. рис. 159). Внешний вид животного
27
в связи с паразитизмом настолько изменяется, что определить его система-
тическое положение без глубокого анализа становится невозможным.
Ярким примером в этом отношении могут служить паразитические брюхо-
ногие моллюски (см. рис. 264) и паразитический усоногий рачок Sacculina
(см. рис. 312), которые совершенно утратили облик представителей исход-
ных групп.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ ЖИВОТНЫХ
Распространение животных по земному шару зависит как от свойств
животных, позволяющих им жить лишь в определенных условиях среды,
так и от различного рода преград, препятствующих их расселению. По-
скольку и свойства животных и лик Земли изменялись во времени, распро-
странение животных зависит не только от современных условий, но и от
предшествующей истории развития.
Некоторые виды распространены очень широко в самых разнообразных
условиях почти по всему земному шару; они носят название космополитов.
Таких сравнительно немного, большинство имеет свою определенную об-
ласть распространения, или ареал. Ареал каждого вида определяется дву-
мя факторами. Во-первых, требованиями представителей вида к условиям
окружающей среды как абиотическим, так и биотическим; эти требования
дают возможность обитать только в тех местах, где существуют для вида
благоприятные условия. Во-вторых, геологическим прошлым, определяю-
щим распределение условий существования в современный период.
Некоторые виды имеют достаточно большой ареал и широко распро-
странены, другие, наоборот, очень небольшой — обитают только в опреде-
ленных участках суши или в определенных обычно изолированных водое-
мах, такие животные носят название эндемиков данного участка суши или
данного водоема. Примером водоема, где очень резко выражен эндемизм
фауны, может служить озеро Байкал, в котором около 3/4 населяющих
его животных — эндемики.
Обычно ареал того или иного вида сплошной, но ряд видов имеет ра-
зорванные ареалы. В таких случаях имеются основная часть ареала и ряд
изолированных участков, оторванных от нее. Такие ареалы объясняются
тем, что в прошлые геологические периоды распространение данного вида
было более широким, а затем при изменении условий существования ареал
сократился, и животные остались только в тех местах, где сохранились
благоприятные для них условия. Так, например, в ледниковый период
ряд холодолюбивых видов был распространен далеко на юг, т. е. имел
более широкий ареал. При потеплении климата, после отступления ледни-
ка, ареал этих форм, естественно, сократился, но в некоторых местах ста-
рого ареала сохранились благоприятные для них условия, и они продол-
жают существовать. Такие обитатели, оторванные от основного ареала
вида, носят название реликтов, в данном случае это ледниковые реликты.
Различные районы суши и океанов имеют свое животное население,
свою фауну, представляющую собой сложный комплекс, сложившийся
в результате определенных условий, имеющихся в данном районе, а также
геологической истории его.
Изучение географического распространения животных приводит к
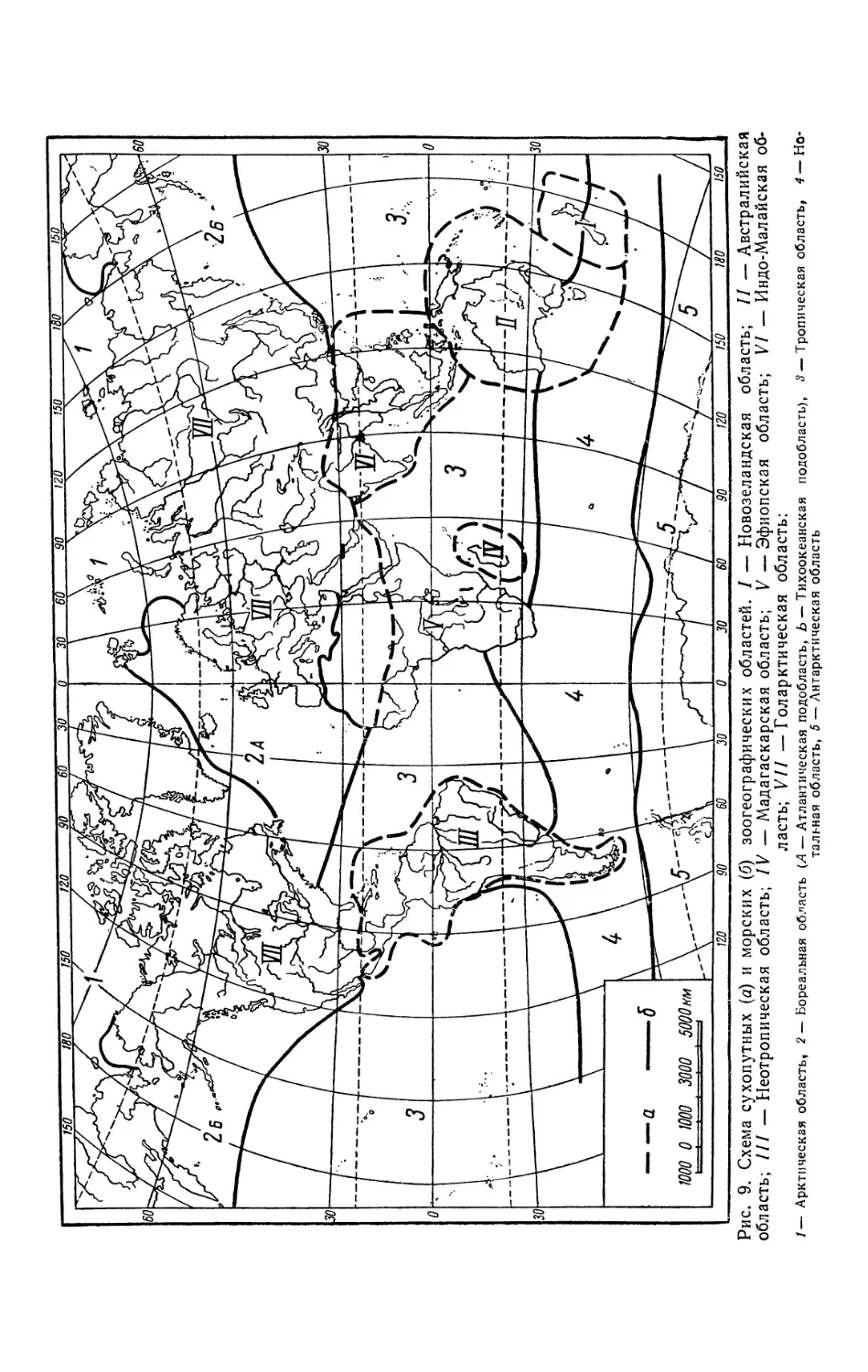
установлению зоогеографических областей суши и океанов (рис. 9), ко-
торые в большой степени не совпадают с очертаниями современных матери-
ков и океанов, что объясняется также историческими причинами.
Так, например, фауны Европы, Северной Азии, Северной Америки и
Северной Африки настолько сходны между собой, что их объединяют в
единую Голарктическую область. Это сходство объясняется тем, что Берин-
гов пролив образовался сравнительно недавно, и до того времени сущест-
вовал свободный обмен животными между Евразией и Северной Америкой.
28
Рис. 9. Схема сухопутных (а) и морских (б) зоогеографических областей. / — Новозеландская область; // — Австралийская
область; /// — Неотропическая область; IV — Мадагаскарская область; V —Эфиопская область; VI — Индо-Малайская об-
ласть; V// — Голарктическая область:
1— Арктическая область, 2 — Бореальная область (Л — Атлантическая подобласть, Ь — Тихоокеанская подобласть), <? —Тропическая область, 4 — Но-
тальная область, 5 — Антарктическая область
Наоборот, фауны Северной и Южной Америк, несмотря на имеющееся
в настоящее время между этими материками соединение, резко отличаются,
и Южная Америка выделяется в особую, Неотропическую, область.
ГЕОЛОГИЧЕСКАЯ ИСТОРИЯ ЖИВОТНОГО МИРА
Обычно палеозоология объединяется с палеоботаникой, т. е. с наукой
об ископаемых растениях, в единую науку — палеонтологию.
Основной задачей палеонтологии является изучение исторического
развития органического мира, т. е. появления, строения, развития и выми-
рания видов и групп организмов в течение истории Земли.
Земная кора в значительной своей части имеет слоистое строение,
обусловленное отложением остатков организмов и минеральных частиц
на дне морей, а также и на суше в течение огромных промежутков времени.
Прерывистость слоев, смена одних слоев другими вызывалась поднятиями
и опусканиями земной коры, причем море могло отступать, сменяясь сушей,
и наоборот. Чем глубже залегает слой в толще земной коры, тем, как пра-
вило, он древнее (ранее образовался) и тем больше отличаются от совре-
менных форм находимые в нем остатки организмов. Геологи классифици-
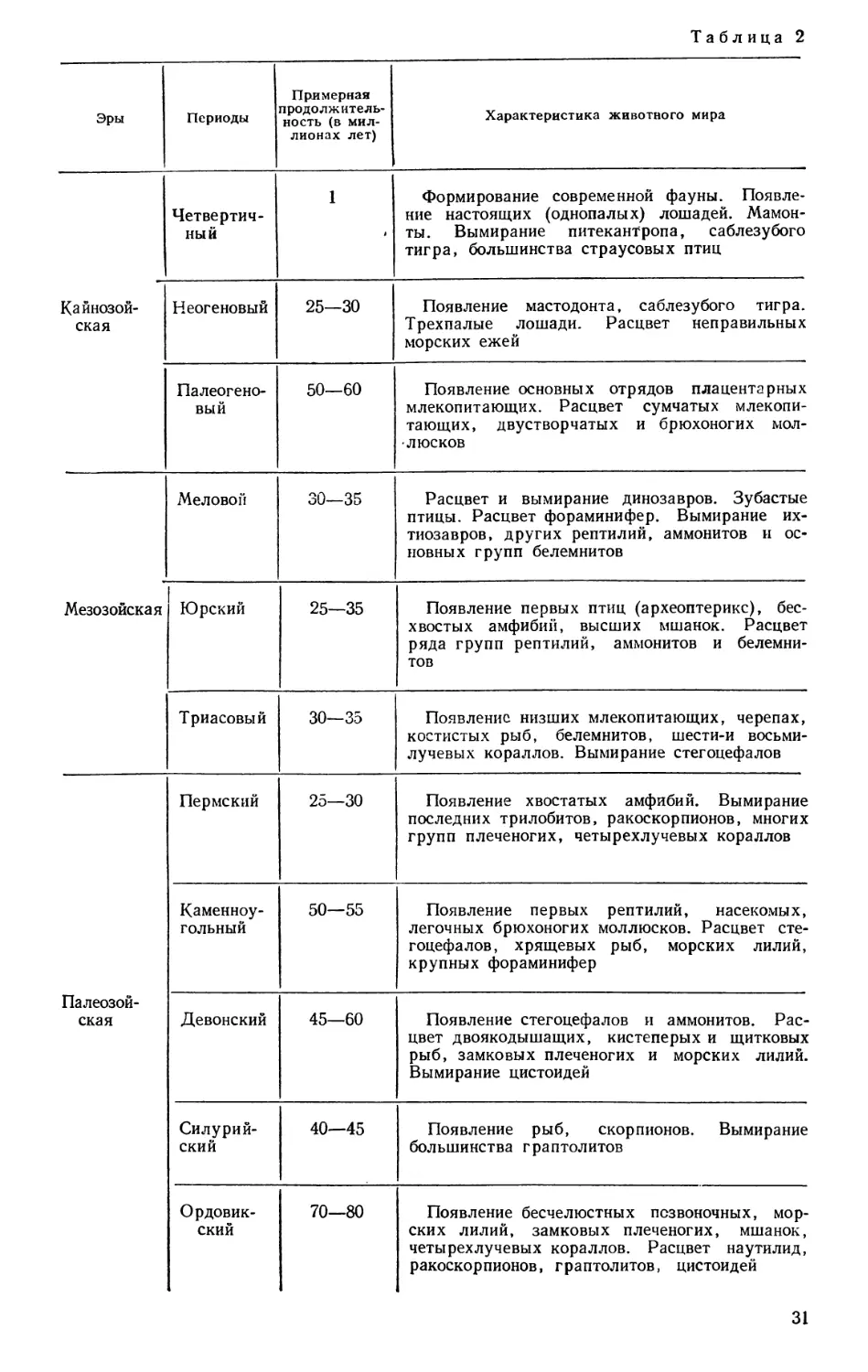
руют слои, подразделяя их на крупные группы, а группы делят затем на
системы слоев. Время, в течение которого образовалась группа слоев,
называют эрой', время, соответствующее отложению системы, — периодом.
В таблице 2 приведены названия групп и систем и некоторые крупнейшие
факты истории животного мира.
Из таблицы видно, что эры и периоды характеризуются своими фауна-
ми. Отдельные представители фаун настолько характерны для определенных
слоев, что по их нахождению можно судить, к какой системе относятся
те или иные слои; это имеет большое значение для геологии при изучении
местонахождений полезных ископаемых. Такие характерные представители
называются руководящими ископаемыми.
Изучая ископаемые остатки отдельных представителей на протяжении
ряда геологических периодов, можно проследить последовательное преоб-
разование этих форм. Очень хорошим примером этого является эволюция
современной лошади. Древнейший предок ее относится к началу третичного
периода, затем можно проследить постепенную смену форм, и в четвертич-
ном периоде появляется уже настоящая лошадь. Процесс этот идет в тесной
связи с теми условиями существования, в которых обитал тот или иной
предок лошади (жизнь в лесу или, наоборот, на открытых пространствах).
УЧЕНИЕ ОБ ЭВОЛЮЦИИ
Животный и растительный мир на земном шаре стал таким, какой
он есть в настоящее время вследствие длительного процесса эволюции,
которая непрерывно продолжается от возникновения жизни на Земле до
настоящего времени. Как показывает распределение остатков вымерших
животных, живших в предшествующие геологические эпохи, преобразова-
ние организации живых существ шло от простых форм к сложным в тесной
связи с изменением условий существования. Высшие формы растительного
и животного мира являются, таким образом, исторически более молодыми,
а низшие группы — более древними. Однако низшие группы не останови-
лись в своем развитии, а наряду с древними примитивными чертами орга-
низации приобрели новые черты, соответствующие современным условиям
их существования.
Соответственно этому общие черты организации в ряде животных
являются свидетельством их родства, т. е. происхождения от исходной
родоначальной формы, а черты различия — это новые приспособления,
приобретенные соответственно новым условиям существования. Однако
в ряде случаев имеет место и другой процесс, когда сходные черты органи-
30
Таблица 2
Эры Периоды Примерная продолжитель- ность (в мил- лионах лет) Характеристика животного мира
Кайнозой- ская Четвертич- ный 1 Формирование современной фауны. Появле- ние настоящих (однопалых) лошадей. Мамон- ты. Вымирание питекантропа, саблезубого тигра, большинства страусовых птиц
Неогеновый 25—30 Появление мастодонта, саблезубого тигра. Трехпалые лошади. Расцвет неправильных морских ежей
Палеогено- вый 50—60 Появление основных отрядов плацентарных млекопитающих. Расцвет сумчатых млекопи- тающих, двустворчатых и брюхоногих мол- люсков
Мезозойская Меловой 30—35 Расцвет и вымирание динозавров. Зубастые птицы. Расцвет фораминифер. Вымирание их- тиозавров, других рептилий, аммонитов и ос- новных групп белемнитов
Юрский 25—35 Появление первых птиц (археоптерикс), бес- хвостых амфибий, высших мшанок. Расцвет ряда групп рептилий, аммонитов и белемни- тов
Триасовый 30—35 Появление низших млекопитающих, черепах, костистых рыб, белемнитов, шести-и восьми- лучевых кораллов. Вымирание стегоцефалов
Палеозой- ская Пермский 25—30 Появление хвостатых амфибий. Вымирание последних трилобитов, ракоскорпионов, многих групп плеченогих, четырехлучевых кораллов
Каменноу- гольный 50—55 Появление первых рептилий, насекомых, легочных брюхоногих моллюсков. Расцвет сте- гоцефалов, хрящевых рыб, морских лилий, крупных фораминифер
Девонский 45—60 Появление стегоцефалов и аммонитов. Рас- цвет двоякодышащих, кистеперых и щитковых рыб, замковых плеченогих и морских лилий. Вымирание цистоидей
Силурий- ский 40—45 Появление рыб, скорпионов. Вымирание большинства граптолитов
Ордовик- ский 70—80 Появление бесчелюстных позвоночных, мор- ских лилий, замковых плеченогих, мшанок, четырехлучевых кораллов. Расцвет наутилид, ракоскорпионов, граптолитов, цистоидей
31
Эры Периоды Примерная продолжи- тельность (в миллионах лет) Характеристика животного мира
Кембрий- ский 70—90 Появление кишечнополостных, листоногих раков, примитивных двустворчатых и наути- лоидей. Расцвет трилобитов и беззамковых плеченогих
Протерозой- ская — 600—700 Редкие находки радиолярий и губок
Архейская — >800 Остатки организмов неизвестны
зации возникают у различных, не родственных друг другу форм совершенно
самостоятельно как приспособление к одинаковому образу жизни. Так,
например, в разных типах животных мы встречаемся с самостоятельным
развитием органов зрения. Наоборот, у глубоководных форм и у форм,
живущих в пещерах, т. е. в полной темноте, и относящихся к различным
систематическим группам, происходит редукция органов зрения.
Первым крупным эволюционистом был Ж. Б. Ламарк (см. выше),
который в своей книге «Философия зоологии» (1809) отбросил идею о по-
стоянстве видов и пытался обосновать теорию превращения видов. Ламарк
указывал, что условия существования на Земле за огромный период ее
истории неуклонно изменялись. Соответственно этим изменениям среды
изменялись в ряде поколений и организмы, отличаясь по своим признакам
от предков. Ламарк считал, что изменение растений и низших животных
происходит путем прямого воздействия на организм окружающих условий
среды, а высших животных — под воздействием упражнения органов при
прогрессивных изменениях и неупражнения их при регрессивных измене-
ниях. Передача по наследству приобретенных изменений в следующие
поколения является, по Ламарку, мощным творческим фактором, повышаю-
щим организацию животных и растений в эволюционном развитии. Ламарку
принадлежит большая заслуга в установлении связи изменений организма
с изменениями внешней среды, но он не нашел еще материалистического
объяснения приспособительного характера этих изменений.
Только Ч. Дарвину (1809—1882) удалось окончательно обосновать
эволюционное учение и дать ему материалистическое толкование. В 1859 г.
вышла его книга «Происхождение видов путем естественного отбора или
сохранение благоприятствуемых пород в борьбе за жизнь», где он дал
исчерпывающие доказательства эволюционной теории.
Дарвин доказал, что изменчивость является основным свойством жи-
вых существ. Наследуемые изменения являются в следующих поколениях
источником развития приспособлений к новым условиям существования,
благодаря чему близкородственные виды отличаются друг от друга.
Человек при разведении домашних животных и культурных растений
использует их способность к изменчивости и закреплению полученных
отклонений наследственностью. Отбирая лучшие образцы сортов и пород
для дальнейшего размножения, человек накапливает в ряде поколений
их полезные особенности и путем такого искусственного отбора совершен-
ствует качество сортов и пород и создает новые. Дарвин установил, что в
природе также происходит отбор особей с полезными индивидуальными
различиями и гибель особей с неблагоприятными признаками и назвал
этот процесс естественным отбором. Естественный отбор является основ-
32
ним фактором закрепления приобретенных изменений в потомстве
вида.
Изменчивость, наследственность и естественный отбор — основные
факторы, объясняющие возникновение многообразия живых существ на
Земле. Систематика, объединяя животных и растения в последовательные
ряды групп от низших к высшим, отражает процесс длительной эволюции
от простейших к высшим, наиболее сложным организмам. Таким образом,
мир животных связан общим происхождением от некогда существовавших
родоначальных предков. Сродство организмов друг с другом в процессе
исторического развития — важнейшее теоретическое обобщение Дарвина.
Приспособленность всех черт организации каждого организма и целе-
сообразность его поведения соответственно окружающим условиям сущест-
вования не является следствием изначальной целесообразности, свойствен-
ной живым существам, как утверждали философы телеологического направ-
ления. Целесообразность взаимосвязи организма со средой, по учению
Дарвина, есть следствие выживания в ряде поколений наилучше приспо-
собленных к данным условиям жизни и гибели в борьбе за существование
неприспособленных особей. В этом и заключается сущность творческой роли
естественного отбора как основного фактора прогрессивной эволюции живых
существ в их историческом развитии. Однако эволюционное развитие не
имеет предустановленной направленности к совершенствованию, как учит
философия автогенеза1. Биологический прогресс вида в некоторых случаях
достигается и регрессивным развитием — редукцией органов, потерявших
в изменившихся условиях свое значение. Это особенно хорошо видно у
форм, перешедших от свободного образа жизни к паразитическому.
Следовательно, целесообразность взаимоотношений организма со средой
относительна, так как любое животное и растение приспособлено лишь к
определенной среде и образу жизни. Так, например, глубоководные рыбы
приспособлены к жизни в глубинах морей, но быстро гибнут при выносе
их в поверхностные слои воды.
Таким образом, Ч. Дарвин еще 100 лет назад дал материалистическое
объяснение происхождению органического мира на Земле и доказал несо-
стоятельность господствующего до него представления о неизменности
видов со времени «сотворения мира живых существ божественным творцом».
Учение Ч. Дарвина среди биологов в России нашло широкую под-
держку, так как они были подготовлены к восприятию идей эволюции
блестящими лекциями К. Ф. Рулье еще до Дарвина, проповедывающего
идеи изменяемости природы. Получившие мировое признание исследо-
вания А. О. Ковалевского и И. И. Мечникова по эмбриологии и сравни-
тельной анатомии беспозвоночных животных и В. О. Ковалевского по
палеонтологии позвоночных утвердили учение Дарвина о родстве разных
типов животного мира. Основатель русской школы физиологов И. М. Се-
ченов и ботаник П. А. Рачинский были первыми переводчиками собра-
ния сочинений Дарвина на русский язык, а физиолог растений К. А. Ти-
мирязев был горячим проповедником исторического метода в биологии.
Среди борцов за учение Дарвина в России надо назвать зоологов
Н. А. Северцова, М. А. Мензбира, сравнительных анатомов В. М. Шим-
кевича, П. П. Сушкина, А. Н. Северцова. П. П. Сушкин и А. А. Борисяк,
А. П. Карпинский в советский период возглавили направление эволю-
ционной палеонтологии, А. Н. Северцов, И. И. Шмальгаузен — эволю-
ционной морфологии, Н. К. Кольцов и Ю. А. Филипченко были основа-
телями русской школы генетиков. Направление эволюционной физиоло-
гии возглавлял И. П. Павлов и Л. А. Орбели, а эволюционной
гистологии А.. А. Заварзин и Н. Г. Хлопин. 1 2
1 Автогенезом называется учение, признающее существование в организме внут-
ренней силы — способности к совершенствованию независимо от внешней среды.
2 Курс зоологии, т. I
33
После Дарвина возникли различные теоретические толкования, объяс-
няющие вопросы о возникновении изменчивости и наследственности.
В настоящее время широким распространением пользуются теории
наследственности и изменчивости, разрабатываемые генетикой. Согласно
воззрениям генетиков, жизнь обладает тремя первичными свойствами, яв-
ляющимися главными факторами эволюции. Первое — способность к са-
мовоспроизведению. Второе — способность к возникновению новых на-
следственных изменений, называемых мутациями — мутациогенез. Третье-
наследственность, т. е. закрепление в последующих поколениях изменений,
вызванных мутациями. Изучением способности к мутированию, т. е. из-
менению наследственности, современная генетика занимается путем экспе-
риментов с носителями наследственности, заключенными в хромозомах
половых клеток и называемых генами. Новейшие достижения биохимии,
физики и генетики открыли новую область биологии — молекулярную
биологию. Новейшими исследованиями уточнено молекулярное строение
носителей наследственности — генов — и химический состав веществ, со-
держащихся в половых клетках и передающих наследственные свойства.
Таким веществом является дезоксирибонуклеиновая кислота (ДНК), за-
ключенная в ядерном веществе всех клеток.
ТИП I. ПРОСТЕЙШИЕ (PROTOZOA)
ХАРАКТЕРИСТИКА
К типу простейших относятся микроскопические животные, тело
которых состоит из одной единственной клетки. Этим Protozoa резко от-
личаются от всех прочих животных, состоящих из очень большого количе-
ства клеток и объединяемых под названием многоклеточных — Metazoa.
Простейшие — одноклеточные животные, и морфологически тело их
соответствует одной клетке многоклеточного организма. Физиологически
же каждая особь простейших представляет самостоятельный организм
в отличие от клетки, являющейся частью многоклеточного организма.
Каждому простейшему животному присущи все основные жизненные
функции, характерные для целого организма: обмен веществ с ассимиляцией
и диссимиляцией, раздражимость, движение, размножение, приспособление
к среде и т. д. В теле простейших дифференцируются отдельные клеточные
участки, выполняющие различные жизненные функции и в этом смысле
аналогичные органам многоклеточных животных. Но так как эти приспо-
собления не имеют клеточного строения и не состоят из тканей, они назы-
ваются не органами, а органеллами, или органоидами.
Органеллами движения служат либо ложноножки (псевдоподии), пред-
ставляющие собой временные выступы протоплазмы, возникающие по мере
необходимости в различных местах тела, либо постоянно существующие
образования в виде длинных и немногочисленных жгутов или же коротких
многочисленных ресничек. У некоторых форм под оболочкой расположены
сократительные тяжи (мионемы), у ряда простейших имеется сложно уст-
роенная система скелетных образований.
Органеллами питания служат у некоторых простейших клеточный
рот, различно устроенная глотка, пищеварительные вакуоли. У многих
форм имеются сократительные (пульсирующие) вакуоли, служащие для
экскреции и регулирования осмотического давления внутри организма.
Организация простейших может в связи с наличием тех или других
органелл достигать значительной сложности (особенно у некоторых ин-
фузорий).
Простейшие — обычно очень мелкие организмы. Наименьшие из них
равны 3—4 мк\ в среднем они достигают 50—150 м/с. Но есть сравнительно
крупные формы. Например, инфузория стентор (Stentor) достигает 1,5 мм,
спиростомум (Spirostomum) до 3 мм (см. рис. 58), раковины ископаемых
корненожек нуммулитов имеют до 2,5—3 см в диаметре; размер раковины
фораминиферы (Cornuspira violacea), живущей в полярных морях у берегов
СССР, достигает 2—3 см.
В зависимости от строения тех или иных органелл движения тип Pro-
tozoa делится на два подтипа, включающих пять классов.
2*
35
КЛАССИФИКАЦИЯ
Тип простейшие (Protozoa)
Подтип I. Плазматические (Plasmodroma)
Класс. 1. Корненожки (Rhizopoda)
Отряд 1. Амебы (Amoebina)
Отряд 2. Раковинные амебы (Testacea)
Отряд 3. Фораминиферы (Foraminifera)
Отряд 4. Солнечники (Heliozoa)
Отряд 5. Лучевики (Radiolaria)
Класс 2. Жгутиковые (Flagellata)
Подкласс I. Фитомастигины (Phytomastigina)
Отряд 1. Хризомонады (Chrysomonadina)
Отряд 2. Гетерохлоридиновые (Heterochloridina)
Отряд 3. Криптомонады (Cryptomonadina)
Отряд 4. Панцирные (Dinoflagellata)
Отряд 5. Эвгленовые (Euglenoidina)
Отряд 6. Хлоромонады (Chloromonadina)
Отряд 7. Фитомонады (Phytomonadina)
Подкласс I. Зоомастигины (Zoomastigina)
Отряд 1. Первичномонадные (Protomonadina)
Отряд 2. Многожгутиковые (Polymastigina)
Отряд 3. Корнежгутиковые (Rhisomastigina)
Класс 3. Споровики (Sporozoa)
Подкласс I. Телоспоридии (Telosporidia)
Отряд 1. Грегарины (Gregarina)
Отряд 2. Кокцидии (Coccidia)
Отряд 3. Гемоспоридии (Haemosporidia)
Подкласс II. Книдоспоридии (Cnidosporidia)
Отряд 1. Слизистые споровики (Myxosporidia)
Отряд 2. Актнномиксидии (Actinomyxidia)
Отряд 3. Микроспоридии (Microsporidia).
Подкласс III. Саркоспоридии (Sarcosporidia)
Подкласс IV. Гаплоспоридии (Haplosporidia)
Подтип II. Ресничные, или инфузории (Ciliophora)
Класс 1. Ресничные инфузории (Ciliata)
Подкласс I. Протоцилиаты (Protociliata)
Подкласс II. Собственно инфузории (Euciliata)
Отряд 1. Равноресничные (Holotricha)
Отряд 2. Разноресничные (Heterotricha)
Отряд 3. Малоресничные (Oligotricha)
Отряд 4. Брюхоресничные (Hypotricha)
Отряд 5. Круглоресничные (Peritricha)
Класс 2. Сосущие инфузории (Suctoria)
Подтип I. Плазматические (Plasmodroma)
Передвигаются при помощи псевдоподий (Rhizopoda) или жгутиков
(Flagellata) и никогда не имеют многочисленных мелких ресничек. Веге-
тативные особи споровиков вообще не имеют органелл движения, и только
половые особи их могут передвигаться либо амебоидно, либо при помощи
жгутиков. К этому подтипу относятся 3 класса: 1) корненожки (Rhizopoda),
2) жгутиковые (Flagellata) и 3) споровики (Sporozoa).
КЛАСС 1. КОРНЕНОЖКИ (RHIZOPODA)
ХАРАКТЕРИСТИКА
Характерным признаком, объединяющим в один класс всех разнород-
ных по форме и строению корненожек, является способность их голого,
не покрытого плотной оболочкой тела образовывать временные протоплаз-
матические выросты — ложноножки (псевдоподии), при помощи которых
36
они передвигаются. Других органоидов движения у них нет. Псевдоподии
образуются в различных местах тела простейшего. Ложноножки служат
также органоидами принятия и выбрасывания пищи, так как они могут
и обтекать частички пищи, увлекая их таким образом внутрь протоплазмы,
и выбрасывать непереваренные остатки.
Некоторые амебы — семейство Bistadiidae — обладают способностью
иногда образовывать жгуты. Они отличаются от сходного отряда жгутико-
вых Rhizomastigina тем, что могут отбрасывать жгут и тем самым стано-
виться неотличными от настоящих амеб. Кроме того, деление происходит
только в амебоидном состоянии. Жгутами также снабжены половые особи
(гаметы) фораминифер (Foraminifera).
Форма тела корненожек сильно варьирует у различных отрядов. На-
ряду с амебами (Amoebina), не имеющими постоянной формы тела и состоя-
щими из комочка протоплазмы с ядром, солнечники (Heliozoa) и лучевики
(Radiolaria), приспособленные к парению в воде, обладают постоянной
формой тела, построенного по радиальному типу.
Многие корненожки снабжены раковинами или внутренним скелетом.
В состав класса входят пять отрядов: 1) амебы (Amoebina), 2) рако-
винные амебы (Testacea), 3) фораминиферы (Foraminifera), 4) солнечники
(Heliozoa), 5) лучевики (Radiolaria).
Представитель класса корненожек —
амеба (Amoeba proteus)
Строение Amoeba proteus — одна из самых крупных больших амеб,
диаметр ее достигает 200—700 мк (рис. 10). Ее можно видеть простым гла-
зом — она выглядит белым пятнышком.
Амеба представляет собой голый протоплазматический комочек не-
правильной формы, все время меняющий свои очертания, так как псевдо-
подии постоянно вытягиваются и втягиваются на разных местах ее тела.
В протоплазме можно различить два слоя: наружный — более свет-
лый, гомогенный и лишенный зернышек, называемый эктоплазмой; внут-
ренний — более темный, зернистый,
особенно сильно развита в псев-
доподиях.
Псевдоподии у Amoeba prote-
us довольно толстые, длинные,
тупые на конце; эта амеба обычно
образует много псевдоподий, часто
составляющих большую часть ее
тела, и поэтому имеет сильно раз-
ветвленную форму. Образование
псевдоподий происходит так. Под
влиянием различных внешних и
внутренних процессов на каком-
либо участке тела уменьшается по-
верхностное натяжение и прото-
плазма вытягивается в псевдопо-
дию. При этом по оси псевдоподии
течет вперед зернистая эндоплазма.
Дойдя до переднего конца псевдо-
подии, эндоплазма всегда перехо-
дит в вязкую гомогенную эктоплаз-
му. На заднем же конце текущей
вперед амебы происходит обратный
переход эктоплазмы в эндоплазму.
При движении амеба выпускает
называемый эндоплазмой. Эктоплазма
Рис. 10. Amoeba proteus:
1 — захваченный пищевой комок, состоящий из
водорослей, 2— пульсирующая вакуоль, 3 — яд-
ро, 4 — псевдоподии
37
псевдоподии в определенном направлении, и постоянно все тело ее как
бы переливается в направлении выпущенных ложноножек. Постоянных
полюсов тела у амебы нет.
Движение при помощи псевдоподий, состоящее как бы в медленном
перетекании тела с места на место, получило название амебоидного. Оно
встречается не только у простейших, но и у некоторых клеток многоклеточ-
ных (лейкоцитов, макрофагов и т. п.). Различные амебы движутся со ско-
ростью от 0,5 до 3 мк в минуту. Псевдоподии обладают довольно значитель-
ной силой. Так, они могут подтягивать по отвесной стенке аквариума тя-
желую раковину фораминиферы Orbitolites.
Протоплазма амебы находится в постоянном движении, хорошо за-
метном благодаря наличию в эндоплазме большого количества светлых
и темных зернышек.
Благодаря движению протоплазмы все органеллы передвигаются в
плазме животного. Овальное, слегка уплощенное ядро заметно в виде свет-
лого пятна в эндоплазме; пульсирующая, или сократительная, вакуоль
также имеет вид светлого пятнышка несколько меньшей величины, чем
ядро. Кроме того, в эндоплазме расположены пищеварительные вакуоли.
Пульсирующая, или сократительная, вакуоль представляет собой
пузырек водяной жидкости, который ритмически сокращается раз в 5—
8 мин, выбрасывая свое содержимое в окружающую среду. Концентрация
солей и других веществ в протоплазме амебы значительно больше, чем в
окружающей ее пресной воде. Вследствие этого вода осмотически прони-
кает внутрь тела амебы. Кроме того, вода образуется при дыхании, захва-
тывается вместе с пищей и т. д. Пульсирующая вакуоль выводит наружу
излишнюю воду и не допускает таким образом разжижения протоплазмы;
кроме того, через вакуоль выводятся продукты обмена. У морских и пара-
зитических простейших, живущих в среде с более высокой концентрацией
солей, пульсирующей вакуоли обычно нет или же она сокращается значи-
тельно реже, чем у пресноводных форм.
Если.поместить пресноводную амебу в среду с более высокой концент-
рацией «солей, скорость пульсирования сократительной вакуоли умень-
шится.
Питание. Amoeba proteus питается бактериями, водорослями и различ-
ными, часто довольно крупными, простейшими. Стоит большой инфузории,
например стентору, остановиться около амебы, как вытянувшаяся псевдо-
подия прикрепляется к инфузории, обволакивает и начинает втягивать
внутрь тела.
После тогб как стентор целиком втянется в эндоплазму, он окружается
капелькой жидкости, в которую из протоплазмы амебы выделяются, по-
видимому, пищеварительные ферменты. Это образование называется пи-
щеварительной вакуолью. Для некоторых амеб доказано присутствие в
пищеварительной вакуоли фермента пепсина и какой-то кислоты, nj-ви-
димому, соляной.
Пищеварительные вакуоли довольно долго остаются в эндоплазме.
За это время происходит переваривание и всасывание их содержимого в
протоплазму амебы.
После того как переваривание закончилось, пищеварительные вакуоли
подходят к стенке тела амебы и непереваренный остаток выбрасывается
наружу (рис. 11).
Амеба может также захватывать пищевую частицу, окружая ее псев-
доподиями таким образом, что в конце концов она оказывается включенной
непосредственно в протоплазму амебы (рис. 12). Длинные водоросли амеба
медленно втягивает в протоплазму, постепенно переваривая их (рис. 12).
Описанный процесс внутриклеточного пищеварения И. И. Мечников
назвал фагоцитозом, но амебы захватывают не только оформленные пище-
вые частички (бактерий и т. п.), но и капельки жидкости, этот процесс
называется пиноцитозом. Капелька жидкости втягивается внутрь прото-
38
плазмы животного. При этом в амебу поступают вода, соли и, по-видимому,
некоторые белки. Пиноцитоз имеет большое значение в физиологии амебы.
Достаточно сказать, что в течение 3 ч в амебу проникает такое количество
жидкости, которое составляет 30% объема ее тела.
Рис. 11. Amoeba verrucosa. Выбрасывание непереваренных остатков пищи
Рис. 12. Захватывание пищи амебами. / — последовательное положение псевдопо-
дий амебы, захватывающей жгутиковое Chilomonas; // — обтекание цисты эвглены;
Ш—втягивание нт?ги водоросли Oscillaria; /V —одновременное захватывание
инфузории и жгутикового (сверху)
Выбрасывание непереваренных остатков, так же как и втягивание
различных организмов, может совершаться в любом месте тела амебы.
Размножение. Amoeba proteus размножается только бесполым путем
при помощи деления надвое (рис. 13). При этом она сначала округляется,
а затем вытягивается; на экваторе намечается перетяжка, постепенно уг-
лубляющаяся и, наконец, разделяющая простейшее на две приблизительно
равные части. Ядро делится кариокинетически. Дочерние организмы сразу
переходят к самостоятельной жизни. Весь процесс продолжается меньше
часа.
Экология. При наступлении неблагоприятных условий, например при
высыхании водоема или при недостатке пищи, амеба инцистируется. При
этом она округляется и выделяет двуслойную плотную оболочку.
В природе Amoeba proteus часто встречается на дне стоячих водоемов,
в особенности в гниющих прудах и болотах, в которых есть много бактерий.
Культивировать ее хорошо в небольших стеклянных чашках с болот-
39
ной водой, в которую посажены, кроме того, небольшие водоросли или зе-
леные жгутиковые (например, эвглены). Воду нужно каждые 2—3 дня
менять. Один раз в 2 дня амеб нужно кормить инфузориями-парамециями.
Рис. 13. Наружные морфологические изменения во время деления Amoeba proteus
(увелич. в 20 раз). I и 11 —самое начало деления; ///—стадия, предшествующая
вытягиванию тела; IV — постепенное вытягивание; V—деление почти закончено
СИСТЕМАТИЧЕСКИЙ ОБЗОР КЛАССА
ОТРЯД АМЕБЫ (AMOEBINA)
К амебам относятся одноядерные, реже многоядерные простейшие,
не обладающие постоянной формой тела, передвигающиеся при помощи
временно образующихся ложноножек и не имеющие ни раковины, ни внут-
реннего скелета. Ложноножки могут быть у разных амеб различной формы,
чаще лопастные или пальцевидные (A. proteus), стрельчатые (A. radiosa)
(рис. 14).
Рис. 14. Псевдоподии различных амеб: I —Amoeba limax; 11—Pelo-
myxa binucleata; /// — Amoeba proteus; IV — Amoeba radiosa; V — Am-
oeba verrucosa; VI — Amoeba polypodia
Форма псевдоподий и тип движения могут сильно изменяться в зави-
симости от воздействия внешней среды. Если помещать A. proteus в среды
с различными химическими свойствами, общий вид амебы сильно изменится.
Это же относится и к другим амебам.
Некоторые амебы могут временно образовывать жгутики (семейство
Bistadi i dae).
Протоплазма обычно резко разделяется на зернистую эндоплазму
и гомогенную эктоплазму. Пульсирующие вакуоли (обычно одна, реже
40
две, три) имеются у пресноводных амеб и обычно отсутствуют у паразити-
ческих и морских форм.
Размножение амеб происходит при помощи деления надвое. В цистах
часто наблюдается множественное деление, при котором ядро делится
много раз без деления протоплазмы. Затем каждое ядро окружается участ-
ком цитоплазмы, и амеба сразу распадается на большое количество дочер-
них особей.
Цистообразование у амеб встречается часто. При этом они окружаются
плотной оболочкой. В таком состоянии они чрезвычайно устойчивы; при
Рис. 15. Типы движений Amoeba proteus. / —
животное из культуры, богатой водой, но бедной
солями; // и III — из культуры, более богатой
солями; IV и V — из культуры с CaCh; VI —
из культуры с Na2SOi; VII — из гиперщелочной
культуры; VIII — из молочнокислой культуры
наступлении благоприятных условий (увлажнение, попадание в подходя-
щего хозяина) амеба выходит из цисты. Амебы широко распространены в
пресных водоемах; встречаются и в морях. Много амеб живет во влажной
земле и в песке (например, A. terricola и др.). Большое количество видов
паразитирует у различных животных.
В кишечнике человека живут несколько видов амеб, например Joda-
moeba biitschllii, Dientamoeba fragilis, Entamoeba coli, Entamoeba histo-
lytica и некоторые другие. Первые четыре безвредны. Ent. histolytica
(рис. 16) имеет серьезное патогенное значение как возбудитель амебиаза
человека (амебная дизентерия, амебные нарывы печени и др.).
Эта амеба впервые описана в 1875 г. Ф. А. Лешем в Петербурге. В ки-
шечнике человека Ent. histolytica может существовать в различных формах.
Заражение человека происходит в результате проглатывания четырех-
ядерных цист, из которых в тонком кишечнике выходят небольшие, раз-
мером 15—20 мк одноядерные амебы; здесь они делятся и переходят в толс-
тые кишки. В протоплазме амеб иногда обнаруживается небольшое коли-
чество фагоцитированных бактерий и грибков, но никогда нет эритроцитов.
Амебы живут и размножаются в просвете кишечника, не вызывая никаких
болезненных явлений. Через некоторое время они округляются и почти
теряют подвижность, образуя так называемые предцистные формы, из
которых образуются характерные четырехядерные цисты размером 8—15 мк,.
41
Цисты имеют округлую и слегка овальную форму, окружены бесцвет-
ной оболочкой; в зрелом состоянии они содержат 4 ядра, несколько глико-
геновых капель и обычно характерные «хроматоидные» белковые включе-
ния в виде палочек с закругленными концами. Цисты выделяются с калом
наружу и служат источником заражения.
Эта форма дизентерийной амебы называется просветной формой (жи-
вет в просвете кишечника), или Ent. histolytica — форма niinuta.
При некоторых еще мало выясненных условиях просветная форма,
увеличивается в размере до 2—40 мк в диаметре и превращается в патоген-
ную тканевую форму, или Ent. histolytica — форма magna.
Рис. 16. Entamoeba histolytica. / — тканевая форма;
11 — просветная форма; 111 — V — одно-, двух- и четы-
рехядерные цисты
Тканевая форма представляет собой очень подвижную амебу с пузырь-
ковидным ядром. Протоплазма резко разделяется на стекловидную экто-
плазму и зернистую эндоплазму. Широкие тупые псевдоподии состоят из
эктоплазмы. Характерно внезапное толчкообразное возникновение псевдо-
подий. В эндоплазме тканевой формы дизентерийной амебы всегда встреча-
ются заглоченные эритроциты, которыми она питается, что отличает ее от
просветной формы и от всех остальных кишечных амеб человека. 4
Тканевые формы при заражении проникают в стенки кишки и вызы-
вают разрушение окружающей ткани. При этом обычно в толстой кишке
образуются характерные язвы с подрытыми краями, в которых происходит
энергичное размножение амеб. Из язвы амебы проникают в кровеносные
и лимфатические сосуды и током крови могут заноситься в другие органы.
Наиболее часто попадают амебы в печень, где они образуют чрезвычайно
опасные для здоровья человека язвы.
Не у всех людей проглатывание цист вызывает заболевание дизенте-
рией. Часто здоровые люди содержат в кишечнике Ent. histolytica и вы-
деляют цисты. Такие цистоносители представляют большую опасность для
окружающих, так как распространяют дизентерийную амебу. Некоторые
из них выделяют до 8 млн. цист в день.
42
Цисты в воде месяцами сохраняют жизнеспособность и вместе с сырой
водой могут попадать в организм человека. Немытые овощи с огородов,
удобряемых фекалиями, пищевые продукты, загрязненные руками челове-
ка, и т. п. также могут быть источником заражения.
Заболевание обычно начинается кровавым поносом и в острых случаях
приводит к смерти через 4—5 дней. Часто первый приступ проходит благо-
получно, но через некоторое время болезнь может повториться. Смертность
от амебной дизентерии, если заболевание не лечить, колеблется от 18 до
40%.
Амебная дизентерия распространена главным образом в тропических
и субтропических странах. В Европе имеются очаги амебной дизентерии
на Балканах и в южной Испании. В СССР она встречается главным образом
в Средней Азии и Закавказье.
ОТРЯД РАКОВИННЫЕ АМЕБЫ (TESTACEA)
Раковинные амебы представляют собой группу, близкую к амебам,
'м являются одноядерными корненожками, которые тоже передвигаются
и воспринимают пищу при помощи псевдоподий. Но их голое протоплазма-
тическое тело заключено в однокамерную раковину, снабженную отверстием
(устьем), через которое выходят псевдоподии. Раковины состоят из хитино-
подобного или студневидного вещества и могут иметь различную форму
(мешковидную, блюдцевидную и т. п.). У некоторых родов раковина чисто
органическая (арцелла — Arcella, рис. 17; хламидофрис — Chlamydophris
Рис. 17. Раковинные амебы. / — Difflugia pyriformis; II — Arcella vulgaris; III —
Arcella dentata; IV — Lesquereusia modesta; V —Centropyxis; VI —Difflugia coro-
na; VII — Euglypha alveolata
и др.), но у большинства она состоит из выделенного амебой основного
вещества, инкрустированного песчинками, панцирями диатомовых водо-
рослей и т. п. (центропиксис — Centropyxis, диффлюгия — Difflugia и
др., см. рис. 17). Животное сначала втягивает внутрь тела твердые частич-
ки, а затем выделяет их на поверхность основного вещества раковины.
Псевдоподии обычно нитевидны, но могут быть и лопастными, как у амеб.
В теле раковинных амеб часто можно различить три зоны. В задней,
прилегающей к вершине раковины, расположены ядро и различные вклю-
чения, затем идет средняя, сильно вакуолизированная зона с пищевари-
тельными и сократительными вакуолями и, наконец, ближе к отверстию
43
раковины расположена зона протоплазмы, обычно свободной от включений,
из которой образуются псевдоподии.
У форм, снабженных мягкой раковиной, размножение происходит при
помощи поперечного деления, сопровождаемого перешнуровкой раковины.
В случае же наличия твердой раковины половина плазмы выходит через
отверстие наружу и образует новую раковину. Ядро делится митотически.
Встречается также и множественное деление, которое происходит
внутри раковины, после чего голые амебы вылезают из раковины, снова
делятся и затем каждая облекается новой раковиной.
При неблагоприятных условиях все раковинные амебы образуют
цисты. Живут они только в пресной воде на водорослях и на дне водоемов
в иле. В болотах, влажных мхах они особенно многочисленны.
В нашей фауне обычны различные виды родов Arcella, Difflugia, Еug-
lypha, Centropyxis (см. рис. 17) и др.
ОТРЯД ФОРАМИНИФЕРЫ (FORAMINIFERA)
Фораминиферы — в большинстве морские корненожки, обладающие
раковиной, которая обычно состоит из большого числа камер.
Раковины фораминифер бывают самой различной формы. У однокамер-
ных (Monothalamia) это обычно более или менее разветвленные трубоч-
ки или мешочки. У молодых многокамерных фораминифер (Polythalam-
Рис. 18. Различные представители Foraminife-
ra. I — Nodomorphina compressiuscula; II —
Rhabdammina abyssorum; /// — Spiroloculina
depressa; IV — Textullaria sagittula; V — Lage-
na stricta; VI — Ammodiscus incertus;
VII —Globotruncana area; VIII— Peneroplis
planatus
ia) раковина вначале бывает
однокамерной. Затем время от
времени из отверстия раковины
выходит часть плазмы, которая
облекается раковиной, и таким
образом образуется вторая ка-
мера. Этот процесс повторяется
много раз, и в результате ино-
гда образуется раковина, содер-
жащая несколько сот камер
(рис. 18). Первая камера назы-
вается зародышевой. Отдельные
камеры связаны между собой
соединительными отверстиями,
через которые сообщается нахо-
дящаяся в них плазма.
Псевдоподии, при помощи
которых фораминиферы пере-
двигаются и захватывают пищу,
выходят у непрободенных (Inp-
erforata) только через устье
раковины, а у прободенных
(Perforata), кроме того, через
многочисленные отверстия, про-
низывающие стенки раковины.
Размножение фораминифер
происходит при помощи смены
полового и бесполого поколе-
ний, отличающихся друг от дру-
га размерами зародышевой ка-
меры (рис. 19). Микросфериче-
ские формы обладают маленькой
зародышевой камерой, макросфе-
рические — большой. В теле
микросферической формы проис-
44
ходит энергичное деление ядер; вокруг каждого из них обособляется учас-
ток протоплазмы, и образовавшиеся голые организмы выходят во внешнюю
среду. Здесь они облекаются раковиной, зародышевая камера которой
больше зародышевой камеры матери, и вырастают в макросферическую
форму.
Таким образом происходит бесполое размножение, и, следовательно,
микросферические формы представляют собой бесполое поколение. Половой
процесс заключается в том, что тело макросферической формы распадается
на множество одинаковых мелких двужгутиковых половых клеточек—
изогамет, покидающих мате-
ринскую раковину и попарно
копулирующих во внешней
среде с гаметами, происхо-
дящими из другой особи.
Таким образом, макросфери-
ческие формы представляют
собой половое поколение.
Образовавшаяся оплодотво-
ренная особь вырастаете мно-
гокамерную микросферичес-
кую форму, которая вновь
размножается бесполым путем.
Такой процесс периоди-
ческого чередования бесполо-
го и полового поколений
особей, разнящихся между
собой по строению и спо-
собу размножения, называет-
ся чередованием поколений и
встречается в различном виде
у многих беспозвоночных жи-
вотных.
Раковина большинства
фораминифер состоит из вы-
деленного плазмой органичес-
Рис. 19. Схема развития фораминиферы Ро1-
ystomella crispa. I — молодой макросфери-
ческий индивидуум; II — взрослый макросфе-
рический индивидуум; III — макросферический
индивидуум с выходящими гаметами; IV —
жгутиковая гамета; V — копуляция; VI —
микросферический индивидуум; VII —микро-
сферический индивидуум с выходящими эм-
брионами
кого вещества, пропитанного
углекислой известью, и об-
ладает большой устойчиво-
стью. У некоторых форм
уплотненная органическая
основа инкрустируется пес-
чинками, обломками раковин
моллюсков, иглами губок и
т. п. Фораминиферы обитают
главным образом в морях, где они развиваются в огромном количестве.
Некоторые виды фораминифер найдены в колодцах и грунтовых водах
пустыни Каракумы в Средней Азии.
Большинство фораминифер — донные формы, но некоторые из них
(Globigerina) находятся во взвешенном состоянии в поверхностных слоях
воды. О громадном количестве фораминифер в современных морях дает
представление тот факт, что в 1 г песка, взятого на берегу Средиземного
моря около Неаполя, оказалось 50 тысяч раковин фораминифер. Около
одной трети дна современных морей покрыто глобигериновым илом,
наполовину состоящим из раковин фораминифер — глобигерин.
Фораминиферы очень древние животные. Их раковины встречаются
уже в кембрийских и силурских отложениях. За многие тысячелетия
плохо растворяющиеся раковины отмерших фораминифер образовали гро-
мадные пласты различных горных пород. Известняки, зеленый песчаник
45
состоят главным образом из раковин фораминифер, иногда достигающих
большой величины (размер дисковидных нуммулитов равен нескольким
сантиметрам). Из мелких раковин фораминифер состоит писчий мел.
ОТРЯД СОЛНЕЧНИКИ (HELIOZOA)
Солнечники — шаровидные корненожки с отходящими во все стороны
подобно лучам псевдоподиями (рис. 20). Псевдоподии имеют постоянную
игольчатую форму благодаря присутствию в них осевой плотной скелетной
нити; такие псевдоподии называются аксоподиями. У некоторых солнечни-
ков имеется скелет, составленный из радиально расположенных кремне-
земных палочек.
Иногда солнечники сливаются своими псевдоподиями и образуют не-
постоянные колонии. Actinophrys sol довольно часто образует колонии из
10—20 индивидуумов.
6
Рис. 20. Различные формы солнечников. / —Actinosphaerium eichhornii:
// — Actinophrys sol:
/ — эктоплазма, 2 — эндоплазма, 3 — псевдоподии, 4 — ядра, 5 — пищеварительные ва-
куоли, 6— пульсирующие вакуоли, 7 — заглоченны • жгутиковые
Протоплазма делится на более плотную мелкозернистую эндоплазму,
в которой расположены ядра, и вакуолизированную эктоплазму, в которой
находятся пульсирующие вакуоли.
Питаются солнечники мелкими организмами, которые они захватывают
псевдоподиями. Они ловят не только мелкие водоросли, простейших жи-
вотных и т. д., но нападают даже на мелких многоклеточных — коловра-
ток, маленьких рачков.
Размножение обычно бесполое при помощи деления надвое или реже
почкования. У широко распространенных в наших пресных водоемах
представителей родов Actinosphaerium и Actinophrys наблюдается поло-
вое размножение. У Actinophrys sol оно протекает следующим образом:
солнечник делится, облекается студневидной оболочкой и в ядрах дочер-
них клеток происходят два деления созревания. Затем две дочерние клетки
вновь сливаются в зиготу, которая после некоторого периода покоя разви-
вается во взрослого солнечника. Эта форма полового процесса, при которой
одна особь делится надвое и две образовавшиеся дочерние клетки снова
сливаются, образуя зиготу, называется педогамией.
Солнечники живут главным образом в пресных водоемах. Большинство
из них свободно носится в воде, но есть и сидячие формы, прикрепляющиеся
к субстрату при помощи особого стебелька.
Солнечники предпочитают чистую воду, богатую кислородом, но не-
которые из них (Actinophrys и Actinosphaerium, рис. 20) встречаются и в
загрязненной воде болот. В большом количестве можно найти солнечников
в торфяных болотах.
46
ОТРЯД ЛУЧЕВИКИ (RADIOLARIA)
Лучевики обладают радиально расположенными псевдоподиями, часто
имеющими плотную осевую нить (рис. 21). Они во многом похожи на сол-
нечников, но отличаются от них присутствием плотной псевдохитиновой
центральной капсулы — особой оболочки, которая окружает центральную
часть тела и делит протоплазму лучевиков на внутрикапсулярную и вне-
капсулярную части, сообщающиеся через имеющиеся в капсуле от-
верстия.
Во внутрикапсулярной более плотной плазме расположены одно или
много ядер и обычно жировые капли. Внекапсулярная плазма имеет круп-
ноячеистое строение и содержит всевозможные включения (пищевые час-
тички, продукты обмена и часто симбиотические водоросли). Вакуоли,
Рис. 21. Различные формы лучевиков. / — Acantometra elastica; // — Lithop-
tera elegans:
/ — центральная капсула, 2—внекапсулярная плазма, 3— ядро, *— игвлки скелета, 5 — псевдо
подии
расположенные в большом количестве во внекапсулярной плазме, по-ви-
димому, имеют гидростатическую функцию, т. е. поддерживают тело лу-
чевиков в воде во взвешенном состоянии. Большинство лучевиков снабжено
очень красивым скелетом, построенным обычно по радиальному типу и
состоящим из кремнезема или сернокислого стронция. Радиолярии размно-
жаются главным образом бесполым путем, причем делятся надвое и цент-
ральная капсула, и минеральный скелет. Другая половина этих образова-
ний создается в дочерних особях заново. У многих лучевиков описан половой
процесс, при котором из внутрикапсулярной плазмы образуются жгути-
ковые гаметы, затем копулирующие.
Лучевики — исключительно морские организмы, которые в громад-
ных количествах живут главным образом в поверхностных слоях воды.
После смерти радиолярий их скелеты падают на дно моря в виде как бы
непрерывного «дождя». В тропических частях Индийского и Тихого океа-
нов на больших глубинах от 4 до 8 тыс. м часть ила морского дна состоит
из раковинок лучевиков. На этих глубинах известковые раковинки
фораминифер растворяются и остаются только кремнеземные — лу-
чевиков.
47
КЛАСС 2. ЖГУТИКОВЫЕ (FLAGELLATA)
ХАРАКТЕРИСТИКА
имеют гораздо оолее сложное
Рис. 22. Электронная микрофото-
графия поперечного разреза жгу-
тиков Pseudotrychonympha. Хоро-
шо видны оболочка, девять пери-
ферических двойных фибрилл и
две центральные фибриллы в каж-
дом жгутике. Увеличение 90 000
Основным признаком, объединяющим в один класс все разнообразные
формы жгутиковых, служит наличие у них одного или нескольких (иногда
очень много) жгутиков, необходимых для передвижения этих организмов.
Жгутики отходят от переднего конца тела. Обычно их бывает 1 или 2, реже
4, 8 и более. Жгутик представляет собой тонкий нитевидный или лентовид-
ный вырост протоплазмы, способный к быстрому вибрационному движению.
Жгутики отходят от базальных зерен — особых маленьких телец, располо-
женных в протоплазме.
При помощи электронного микроскопа было установлено, что жгутики
строение (рис. 22), чем считали до сих пор.
На поперечном разрезе хорошо видно,
что жгутик покрыт оболочкой, представ-
ляющей непосредственное продолжение
оболочки тела жгутикового. В цитоплаз-
ме жгутика расположено 11 продольных
фибрилл, 9 из них двойные и образуют
кольцо по периферии цитоплазмы жгути-
ка под оболочкой; 2 самостоятельные
фибриллы лежат в центре. Центральные
фибриллы заканчиваются у поверхности
простейшего, а 9 периферических прони-
кают в эктоплазму, сливаются здесь и
образуют плотное тельце, которое в све-
товом микроскопе называют базальным
зерном.
Электронно-микроскопическая струк-
тура жгутиков у Flagellata и ресничек
инфузорий весьма сходна. Жгутики и рес-
ничники многоклеточных животных почти
неотличимы по структуре от одноименных
органоидов у простейших.
Некоторые формы (отряд Rhizoma-
stigina) обладают способностью наряду
со жгутиками образовывать псевдоподии.
Это указывает на близость жгутиковых к корненожкам. Почти все жгути-
ковые покрыты довольно плотной эластичной оболочкой, носящей назва-
ние пелликулы, которая определяет постоянную обычно удлиненную фор-
му тела. Кроме того, некоторые формы могут образовывать наружную
защитную оболочку или панцирь, составленный из клетчатковых пла-
стинок.
Протоплазма разделяется на тонкий наружный слой прозрачной эк-
топлазмы и зернистую глубжележащую эндоплазму.
По способу питания жгутиковых делят на 3 группы. г
Автотрофные организмы синтезируют органические вещества (угле-
воды) из углекислоты и воды при помощи хлорофилла на солнечном свету,
служащем источником энергии. Эти жгутиковые питаются как настоящие
растения и не нуждаются в органической пище, поэтому этот способ пита-
ния называется также голофитным, или растительным. Хлорофилл пропи-
тывает особые органоиды — хроматофоры, которые бывают зеленого, бу-
рого, красного или желтого цвета. Хлорофилл есть во всех хроматофорах,
но в незеленых он зетемнен желтым, красным или коричневым пигментами.
К автотрофным организмам принадлежат все окрашенные жгутиковые.
Гетеротрофные организмы не обладают хлорофиллом и не могут по-
этому синтезировать углеводы из неорганических веществ. Они питаются
48
КЛАСС 2. ЖГУТИКОВЫЕ (FLAGELLATA)
ХАРАКТЕРИСТИКА
имеют гораздо оолее сложное
Рис. 22. Электронная микрофото-
графия поперечного разреза жгу-
тиков Pseudotrychonympha. Хоро-
шо видны оболочка, девять пери-
ферических двойных фибрилл и
две центральные фибриллы в каж-
дом жгутике. Увеличение 90 000
Основным признаком, объединяющим в один класс все разнообразные
формы жгутиковых, служит наличие у них одного или нескольких (иногда
очень много) жгутиков, необходимых для передвижения этих организмов.
Жгутики отходят от переднего конца тела. Обычно их бывает 1 или 2, реже
4, 8 и более. Жгутик представляет собой тонкий нитевидный или лентовид-
ный вырост протоплазмы, способный к быстрому вибрационному движению.
Жгутики отходят от базальных зерен — особых маленьких телец, располо-
женных в протоплазме.
При помощи электронного микроскопа было установлено, что жгутики
строение (рис. 22), чем считали до сих пор.
На поперечном разрезе хорошо видно,
что жгутик покрыт оболочкой, представ-
ляющей непосредственное продолжение
оболочки тела жгутикового. В цитоплаз-
ме жгутика расположено 11 продольных
фибрилл, 9 из них двойные и образуют
кольцо по периферии цитоплазмы жгути-
ка под оболочкой; 2 самостоятельные
фибриллы лежат в центре. Центральные
фибриллы заканчиваются у поверхности
простейшего, а 9 периферических прони-
кают в эктоплазму, сливаются здесь и
образуют плотное тельце, которое в све-
товом микроскопе называют базальным
зерном.
Электронно-микроскопическая струк-
тура жгутиков у Flagellata и ресничек
инфузорий весьма сходна. Жгутики и рес-
ничники многоклеточных животных почти
неотличимы по структуре от одноименных
органоидов у простейших.
Некоторые формы (отряд Rhizoma-
stigina) обладают способностью наряду
со жгутиками образовывать псевдоподии.
Это указывает на близость жгутиковых к корненожкам. Почти все жгути-
ковые покрыты довольно плотной эластичной оболочкой, носящей назва-
ние пелликулы, которая определяет постоянную обычно удлиненную фор-
му тела. Кроме того, некоторые формы могут образовывать наружную
защитную оболочку или панцирь, составленный из клетчатковых пла-
стинок.
Протоплазма разделяется на тонкий наружный слой прозрачной эк-
топлазмы и зернистую глубжележащую эндоплазму.
По способу питания жгутиковых делят на 3 группы. г
Автотрофные организмы синтезируют органические вещества (угле-
воды) из углекислоты и воды при помощи хлорофилла на солнечном свету,
служащем источником энергии. Эти жгутиковые питаются как настоящие
растения и не нуждаются в органической пище, поэтому этот способ пита-
ния называется также голофитным, или растительным. Хлорофилл пропи-
тывает особые органоиды — хроматофоры, которые бывают зеленого, бу-
рого, красного или желтого цвета. Хлорофилл есть во всех хроматофорах,
но в незеленых он зетемнен желтым, красным или коричневым пигментами.
К автотрофным организмам принадлежат все окрашенные жгутиковые.
Гетеротрофные организмы не обладают хлорофиллом и не могут по-
этому синтезировать углеводы из неорганических веществ. Они питаются
48
движутся по
Рис. 23. Euglena
v iridis:
1 — жгут, 2—стигма,
3 — резервуар, 4 —
пульсирующая ваку-
оль, 5 — приводящие
вакуоли, 6—зерна
парамила, 7 —хрома-
тофоры, 8 —ядро
нескольких жировых капель, пропитанных красным пигментом — гема-
тохромом.
Обычно эвглены обладают положительным фототаксисом, т. е. они
авлению к источнику света. Если осветить стеклянную
банку с водой, в которой плавают эвглены, с одной
стороны, то все они соберутся на стороне, обращенной
к источнику света. Если же экспериментально разру-
шить стигму, то эвглена перестает реагировать на свет.
Этот опыт показывает, что стигма действительно слу-
жит примитивным светочувствительным органом.
Около резервуара помещается пульсирующая ваку-
оль, несущая выделительную и осморегуляторную функ-
ции. Пульсирующая вакуоль —образование, встречаю-
щееся у многих простейших (жгутиковых, корнено-
жек, инфузорий). Она представляет собой пузырек,
который работает ритмически, попеременно то расши-
ряется и медленно наполняется жидкостью (стадия
диастолы), то быстро сокращается (стадия систолы),
выталкивая свое содержимое наружу. Пульсирующая
вакуоль у эвглены окружена несколькими маленьки-
ми приводящими вакуолями, которые время от вре-
мени в нее изливаются. Пульсирующая вакуоль сокра-
щается 3 раза в минуту при температуре 30° С. На-
полняющая ее жидкость при этом попадает в резерву-
ар и через глотку выводится наружу. В задней части
тела эвглены расположено пузырьковидное ядро (см.
рис. 23).
В плазме находится большое количество зеленых,
пропитанных хлорофиллом хроматофоров (см. рис. 23),
образующих одну или две характерные звездчатые фи-
гуры.
Питание. Эвглена принадлежит к миксотрофным ор-
ганизмам. На свету она питается подобно настоящему
зеленому растению, т. е. синтезирует углеводы за счет
углекислоты и воды. Продуктом ассимиляции является парамил — угле-
вод, близкий к крахмалу, но не дающий под действием иода фиолето-
вого окрашивания. Не только в хроматофорах, но и в протоплазме имеется
большое количество зерен парамила.
Минеральные соли и азотные соединения поступают в организм эвгле-
ны осмотически.
При помещении в темноту эвглена теряет зеленую окраску, потому
что хлорофилл исчезает из хроматофоров. Последние сильно уменьшаются
в размерах. Стигма также редуцируется.
Бесцветная эвглена не имеет, конечно, возможности синтезировать
углеводы и питается исключительно органическими веществами, которые
она либо воспринимает осмотически, либо захватывает в виде оформлен-
ных частичек, попадающих в резервуар через глотку с током воды. При этом
из стенки резервуара выступают маленькие нитевидные псевдоподии,
увлекающие частицы пищи внутрь тела.
При перенесении бесцветных форм на свет, в них восстанавливаются
хлорофилл и стигма, они вновь переходят к растительному питанию.
Бесцветные эвглены могут образовываться и другим путем. Единст-
венными носителями хлорофилла в протоплазме являются хроматофоры,
обладающие способностью самостоятельно делиться. Иногда темп деления
особи быстрее темпа деления хроматофоров. Описаны случаи, когда в ре-
зультате повторных делений эвглены одна дочерняя особь оставалась ли-
шенной хроматофоров, бесцветной. Таким образом могут возникнуть стой-
кие бесцветные эвглены.
50
В семейство эвгленовых, кроме зеленых форм, входят и бесцветные
(астазия — Astasia и др.), похожие на эвглен, но отличающиеся от них
главным образом отсутствием хроматофоров. Возможность образования
бесцветных эвглен указывает на родство зеленых и бесцветных жгутиковых.
Размножение. Происходит
при помощи продольного деле-
ния (рис. 24). Ядро делится
митотически. Вслед за делени-
ем ядра происходит деление
цитоплазмы. Оно начинается с
того, что на переднем конце
образуется углубление. В это
время отбрасывается жгутик и
делится базальное зернышко.
Образовавшиеся базальные
зерна расходятся, и из них на-
чинают расти новые жгутики.
Все органеллы, расположенные
на переднем конце тела эвглены,
и хроматофоры также начинают
делиться. Процесс перешнуров-
ки плазмы идет спереди назад,
и в результате образуются две
дочерние особи.
Экология. Euglena viridis
живет в мелких пресноводных
водоемах, сильно загрязненных,
Рис. 24. Размножение Euglena viridis.
I — продольное деление; //, III —деле-
ние в цисте
легко разлагающихся органи-
ческими веществами. Ее можно встретить в больших количествах в гни-
ющих лужах, канавах около скотных дворов, в сырой земле рядом с
ними. Некоторые эвглены (Е. oxyuris, Е. acus) преимущественно обитают
в чистых водах. Эвглены — наиболее обычные пресноводные зеленые жгу-
тиковые. Зачастую они размножаются в столь больших количествах, что
образуют зеленую пленку на поверхности водоемов.
В неблагоприятных условиях (например, при высыхании лужи) эвгле-
на может инцистироваться. Процесс инцистирования эвглены заключается в
том, что она отбрасывает жгут, округляется и выделяет вокруг себя плотную
оболочку, предохраняющую ее от неблагоприятных внешних условий (от
высыхания). Образуется так называемая циста покоя. Иногда эвглена
размножается в инцистированном состоянии. Такая циста называется
цистой размножения (рис. 24). При смачивании циста разрушается и
эвглена вновь переходит к деятельному образу жизни.
СИСТЕМАТИЧЕСКИЙ ОБЗОР КЛАССА
ПОДКЛАСС 1. ФИТОМАСТИГИНЫ (PHYTOMASTIGINA)
Характерно наличие хроматофоров, содержащих хлорофилл. Пред-
ставители небольшого количества форм бесцветны, но они связаны с ис-
тинными растительными жгутиковыми, содержащими хлорофилл, рядом
промежуточных форм. Питание автотрофное или миксотрофное.
Большинство несет 1—2 жгутика и размножается бесполым путем.
Половое размножение обычно у Phytomonadina.
51
ОТРЯД ЭВГЛЕНОВЫЕ (EUGLENOIDINA)
Сравнительно большие веретеновидные или вытянутые жгутиковые,
обычно покрытые плотной пелликулой. На переднем конце находится
глубокая ямка, со дна которой отходят жгуты, чаще всего 1, иногда 2 и
больше. У гетеротрофных форм на переднем конце имеется ротовое отвер-
стие.
Характерно для отряда наличие одной или нескольких сократитель-
ных вакуолей, изливающихся в несократимый резервуар (см. описание
эвглены). На переднем конце многих эвгленовых находится красная стигма.
Хроматофоры зеленого цвета. Часто содержат пиреноиды — овальные тела,
лежащие в центре хроматофоров. Функция их неизвестна. Продуктом асси-
миляции служит парамил.
Бесцветные формы питаются бактериями, мелкими простейшими и
водорослями, которых переваривают в пищеварительных вакуолях. Раз-
множаются бесполым путем при помощи продольного деления.
Эвгленовые очень распространены в небольших загрязненных водое-
мах. Встречаются также в почве. Многие (например, астазия) паразитируют
в кишечнике различных животных.
ОТРЯД ФИТОМОНАДЫ (PHYTOMONADINA)
Тело фитомонад покрыто оболочкой, состоящей из клетчатки, благо-
даря чему они имеют постоянную форму. На переднем конце в оболочке
обычно имеется пора, сквозь которую проходят жгутики (обычно 2 или
4, реже 1 или 8).
У большинства видов есть зеленые хроматофоры и пиреноиды. Многие
формы окрашены в красный цвет благодаря присутствию в хроматофорах
пигмента — гематохрома.
Фитомонады — автотрофные или миксотрофные формы. Только неко-
торые бесцветные виды гетеротрофны. На переднем конце обычно располо-
жены пульсирующие вакуоли.
Некоторые фитомонады, объединяемые в семейство вольвоксовых
(Volvocidae), включают колониальные формы. Наиболее просто устроен-
ные из вольвоксовых —Gonium — состоят из 4—16 совершенно одинако-
вых клеток, соединенных вместе студневидной оболочкой, образованной
из оболочки особей, составляющих колонию. Все клетки расположены в
один ряд, и колония имеет вид пластинки. Каждая клетка, выйдя из коло-
нии, может размножаться обычным путем, образуя новую колонию. У Р1е-
odorina (128 клеток) наблюдается уже дифференцировка между особями,
составляющими колонию. Последняя содержит два рода клеток: одни —
маленькие, снабженные стигмой, — это соматические особи, не способ-
ные к размножению; другие — значительно большей величины, — пред-
ставляют собой генеративные клетки, из которых образуются макро- и
микрогаметы.
Дифференцировка особей одной и той же колонии наиболее резко
выражена у видов рода вольвокс (Volvox,) образующих колонии в виде
наполненного жидкостью пузыря из сотен и тысяч двужгутиковых особей
(иногда до 20 000; рис. 25), громадное большинство которых сомати-
ческие.
Дифференцировка различных клеток у вольвокса состоит не только
в наличии соматических и генеративных особей, но, кроме того, и в том,
что те соматические особи, которые при движении организма направлены
вперед, обладают значительно большими стигмами.
Генеративные особи вольвокса весной начинают делиться и образуют
дочерние колонии. Они падают внутрь материнского пузыря, который по-
степенно отмирает, освобождая дочерние колонии (рис. 25j ). Осенью из
генеративных особей образуются половые клетки. Часть из них вырастает
52
до яйцевидных макрогамет, другие — многократно делятся и дают большое
количество мелких веретенообразных двужгутиковых микрогамет (рис. 25,А 3).
Микрогаметы освобождаются от оболочки, свободно плавают в воде
и затем сливаются с макрогаметами. Процесс слияния половых особей
называют копуляцией. В результате копуляции образуется оплодотворенная
зигота, которая дает начало новой колонии.
Рис. 25. Volvox aureus. 1 — бесполая колония; // — женская колония; III —
мужская колония:
/— ыолочыс дочерние колонии, 2—макрогаметы. Ь — кучки микрогамет
Вольвокс многими рассматривается как пример переходной формы
между одноклеточными и многоклеточными, так как в нем действительно
уже есть некоторые черты многоклеточного организма; все клетки колонии
соединены между собой протоплазматическими нитями и, кроме того, имеет-
ся некоторая дифференцировка между клетками. Функции отдельных кле-
ток координированы между собой, что также характерно для многоклеточ-
ного организма. Так, при движении вольвокса все жгутики многих тысяч
клеток бьют совершенно синхронно.
1 27 ТП W У
Рис. 26. Анизогамная копуляция Chlamidomonas braunii. I —макрогамета: И —
микрогамета; /// — V — копуляция
У одиночной фитомонады Chlamydomonas steinii копулируют одина-
ковые половые особи (изогаметы), почти не отличающиеся от вегетативных
клеток.
У другого вида того же рода — Chlamydomonas braunii (рис. 26) об-
разуются половые клетки двух родов — макрогаметы и подвижные малень-
кие микрогаметы, и половой процесс протекает как анизогамия.
У колониальных фитомонад эвдорины (Eudorina), состоящей из 32
клеток, и пандорины (Pandoriпа), состоящей из 16 клеток, микро-и макро-
гаметы образуются в разных колониях, причем образовавшиеся подвижные
53
микрогаметы плывут к макрогаметам и копулируют с ними. Таким образом,
среди этих жгутиковых имеются мужские и женские особи. У вольвокса
мужские и женские половые особи также часто образуются в разных коло-
ниях. Но есть виды, у которых они имеются в одной и той же колонии.
Самооплодотворение исключается в этих случаях тем. что микро- и макро-
гаметы созревают в разное время.
Фитомонады широко распространены в наших пресных водоемах.
ПОДКЛАСС II. 300МАСТИГИНЫ
(ZOOMAST1GINA)
В отличие от фитомастигин жгутиковые никогда не содержат хромато-
форов. Питание гетеротрофное. Многие зоомастигины паразиты и питаются
сапрофитно. Размножение бесполое, главным образом путем продольного
деления. Количество жгутиков может быть различно: от 1—2 до многих
десятков.
ОТРЯД ПЕРВИЧНОМОНАДНЫЕ (PROTOMONADINA)
Маленькие бесцветные формы, несущие обычно 1—2 жгутика.
Кроме жгутиков, у некоторых первичномонадных, например трипано-
зом (Trypanosoma), есть волнообразная перепонка (ундулирующая мембрана),
являющаяся выростом оболочки тела. По свободному краю ее расположен
жгут. Для семейства трипанозомид (Trypanosomidae) характерно наличие
блефаропласта (см. рис. 302).
Блефаропластом называется круглое или овальное тельце, содержа-
щее дезоксирибонуклеиновую кислоту (которая входит, кроме того, в
состав хроматина клеточных ядер) и расположенное около базального
зерна жгутика.
Электронно-микроскопические исследования последних лет показали,
что у трипанозом блефаропласт, или, как часто его называют, кинетопласт,
тесно связан с митохондрией, и фактически обе органеллы представляют
собой одно целое (рис. 27). Поскольку митохондрии содержат большое
количество ферментов и принимают участие в дыхании и выделении энер-
Рис. 27. Электронная микро-
фотография участка тела Try-
panosoma cruzi. Хорошо видно,
что блефаропласт (В/) соединен
с большой митохондрией (7И):
BZ — базальное зерно; вертикальная
; стрелка (справа) указывает место со-
единения поверхностных мембран
блефаропласта и митохондрии. Го-
ризонтальные стрелки (слева) указы-
вают на двойную мембрану фибрилл
блефаропласта. Увеличение 59200
54
гии, можно предполагать, что блефаропласт имеет большое значение в об-
мене веществ протозойной клетки.
Бесполое размножение происходит у первичномонадных обычно при
помощи продольного деления.
Все первичномонадные по способу питания относятся к гетеротрофным
формам; питательные вещества либо воспринимаются ими осмотически,
Рис. 28. Воротничковые жгутиковые. / — Codonisiga alliodes, колония
на разветвленной ножке; II — Codonisiga botrytis (при большом увеличе-
нии)
либо захватываются в виде оформленных кусочков. Свободноживущие
формы, например Monas vulgaris, обитают в большинстве случаев в сильно
загрязненной воде.
Особый интерес представляет небольшое семейство воротничковых
жгутиковых (Craspedomonadinae). В большинстве это колониальные сво-
бодноживущие формы. Основание жгутика у них окружено тонким прото-
плазматическим воротником (рис. 28). Это семейство интересно тем, что
его представители имеют большое сходство в строении с энтодер-мальными
воротничковыми клетками губок — хоаноцитами. В связи с этим губок
ставят в генетическую связь с воротничковыми жгутиковыми.
Многие первичномонадные паразитируют у различных животных и
человека и являются возбудителями серьезных болезней, например, раз-
личные представители родов костия (Costia), трипанозома (Trypanosoma)
и лейшмания (Leischmania). Грушевидная четырехжгутиковая Costia пе-
catrix паразитирует на коже рыб (форели, карпа и др.), к которой прикреп-
ляется двумя жгутами (рис. 29). При сильной инфекции она сплошь по-
крывает кожу и жабры. У молодых рыб инфекция Costia necatrix иногда
вызывает массовую смертность.
Большое хозяйственное и медицинское значение имеют различные
трипанозомы (см. рис. 30). Это небольшие (20—30 мк) продолговатые жгу-
тиковые, заостренные на обоих концах. Блефаропласт лежит у них позади
ядра; от него начинается жгут, который идет по краю волнообразной пере-
понки, огибая все тело трипанозомы, и свободно кончается на переднем
конце. Трипанозомы паразитируют в крови и тканях различных позвоноч-
ных животных и человека.
Важнейшие представители. Trypanosoma gambiense — возбудитель
африканской «сонной болезни» человека. Переносчиком трипанозом слу-
жит муха це-це (Glossina palpalis).
55
Помимо человека, Т. gambiense паразитирует также у антилоп, но
не приносит им заметного вреда. Таким бразом, антилопы служат как бы
природным резервуаром инфекции, которую мухи це-це переносят на чело-
века. Когда муха це-це сосет кровь больного сонной болезнью или же кровь
антилопы, содержащую трипанозом, то вместе с кровью попадают в желу-
док насекомого и трипанозомы. Здесь они размножаются, а затем прони-
кают в слюнные железы и, когда такая муха вновь сосет кровь человека,
попадают в его организм. При сонной болезни трипанозомы размножаются
Рис. 29. Costia necatrix. I — участок кожи форели с сидящими на ней па-
разитами; // — отдельное жгутиковое
в периферической крови, спинномозговой жидкости и в селезенке человека.
Таким образом, жизненный цикл трипанозом проходит в двух хозяевах:
в позвоночном животном или человеке и в насекомом (или пиявке), которые
являются переносчиками инфекции. Это явление носит название смены
хозяев.
Передача инфекции от одного хозяина другому при посредстве укуса
переносчика (промежуточного хозяина) носит название трансмиссивной
передачи, а такие заболевания называются трансмиссивными.
В Средней Азии и на Нижней Волге в крови лошадей, верблюдов,
в меньшей степени ослов и рогатого скота паразитирует Т. evansi, вызы-
вающая «суауру» — опасное заболевание, особенно лошадей и верблюдов,
выражающееся в лихорадке, анемии и отеках.
Переносчиками инфекции являются различные слепни (Tabanidae)
и некоторые кровососущие мухи (Stomoxys, Lyperosia). При укусе трипа-
нозомы вместе с кровью больного животного попадают на хоботок слепня,
и, если он в течение 15 мин укусит другое здоровое животное, то трипано-
зомы попадут в кровь и вызовут заболевание.
Повсеместно, в частности и в СССР, распространена «случная болезнь»
лошадей, характеризующаяся отеком гениталий, истощением и появлением
плоских бляшек на коже. Она вызывается Т. equiperdum и передается
от лошади к лошади при половом акте. При этой болезни поражаются
центральная нервная система и нервы задних конечностей. Trypanosoma
cruzi — возбудитель южноамериканской болезни «чагас»; переносчик его —
клоп Triatoma megistra, природный резервуар — броненосцы, двуутробки
и др.
Т. brucei — возбудитель африканской болезни «нагана», поражающей
рогатый скот, ослов, верблюдов (рис. 30); переносчик — муха Glossina
morsitans.
56
Различные представители рода Leischmania являются внутриклеточ-
ными паразитами позвоночных животных.
Leischmania tropica (см. рис. 3L ) вызывает кожное заболевание че-
ловека, выражающееся в образовании узлов, язв или поражений слизистых
оболочек. Через несколько недель или иногда месяцев после проникнове-
ния лейшманий в организм человека на коже появляются язвы, которые
медленно заживают, остав-
ляя обезображивающий ру-
бец. L. tropica была от-
крыта в 1898 г. П. Ф. Бо-
ровским в Ташкенте.
Лейшмании паразити-
руют внутри клеток, где
имеют округлую или
овальную форму. Сбоку
или ближе к центру ле-
жит крупное ядро. Перед
ядром расположен палоч-
ковидный блефа ропласт,
от которого отходит к пе-
риферии тонкая нить. Сво-
бодного жгутика у внут-
риклеточных форм лейш-
маний нет.
В организме перонос-
I
Рис. 30. Trypanosoma brtiooi. / — нормальная фор-
ма и крови; //, /// —стадии деления!
/ — ядро, 2—блсфароплас 1, базальное зерно жгутика,
4 — ундулирующая мембрана. Ь — жгу гик
чика и при культивиро-
вании на искусственной
питательной среде лейш-
мании превращаются в
веретеновидные формы и
восстанавливают длинный жгут. Переносчиками лейшманий служат мос-
киты из рода флеботомус (Phlcbotoinus).
В песчаных пустынях Средней Азии кожным лейшманиозом болеют
некоторые песчанки, тонкопалый суслик и ушастый еж. Москиты кусают
этих животных и получают лейшманий, который в дальнейшем размножа-
ется у них в желудке. Если такой москит укусит человека, он может за-
разить его кожным лейшманиозом.
Таким образом, кожный лейшманиоз в Средней Азии представляет
собой болезнь диких грызунов, и москиты переносят лейшманий от грызу-
нов человеку. Н. И. Латышев показал, что
в Туркмении уничтожение грызунов вок-
руг поселений ведет к почти полному исчез-
новению в них кожного лейшманиоза.
Кожный лейшманиоз является при-
родно-очаговой инфекцией. Так называ-
ются заболевания, распространенные в оп-
ределенных районах среди диких живот-
ных, которые служат источником зараже-
ния человека. Учение о природной оча-
говости заболеваний разработал Е. Н.
Павловский.
Кожный лейшманиоз распространен
в жарких и теплых странах. В пределах
СССР он встречается в Средней Азии и на
Кавказе, где носит название «пендинской
язвы», «восточной язвы» и т. д.
Leischmania donovani (рис. 31) — воз-
будитель висцерального, или внутреннего,
Рис. 31. / — Leischmania tropica
в эндотелиальной клетке из яз-
вы; II — Leischmania donovani
из культуры;
1 — ядро эндотелиальной клетки, 2 —
лейшмании, 3 — блефаропласт, 4 — ядро
лейшмании
57
лейшманиоза, встречающегося и в Средней Азии. При этом тяжелом за-
болевании, протекающем при явлениях увеличения селезенки и печени
и кровоизлияний в коже и слизистых оболочках, лейшмании встречаются в
различных клетках, но главным образом в эндотелии сосудов.
ОТРЯД МНОГОЖГУТИКОВЫЕ (POLYMASTIGINA)
К отряду многожгутиковых относятся бесцветные формы, снабженные
четырьмя или более жгутиками и иногда волнообразными перепонками.
Для них характерно присутствие осевой палочки — аксостиля (рис. 3210),
Рис. 32. Различные формы жгутиковых. / — Trichomonas ап-
gusta (схема организации); // —Trichomonas hominis;
/// — Giardia intestinalis:
1 — цитостом, 2 — базальное зерно, 3 — ядро, 4 — передние жгутики, 5 —
задний жгут, 6 — ундулирующая мембрана, 7 — ее базальная фибрилла. 8—
краевая нить, 9 — вакуоли, 10 — аксостиль
сильно преломляющей свет эластической плотной нити, идущей через все
тело от переднего конца к заднему. Аксостиль — опорный элемент, опре-
деляющий форму тела жгутикового; иногда, по-видимому, он играет из-
вестную роль при прикреплении паразитических форм к кишечному эпи-
телию хозяина.
У многих многожгутиковых есть парабазальный аппарат, состоящий
из зернышек разной величины и формы, располагающихся около ядра и
отличающихся от блефаропласта первичномонадных отсутствием дезокси-
рибонуклеиновой кислоты.
Изучение с помощью электронного микроскопа показало, что пара-
базальный аппарат состоит из мембран и пузырьков и по строению схо-
ден с аппаратом Гольджи клеток многоклеточных животных.
Размножаются многожгутиковые только бесполым путем при помощи
продольного деления.
58
За немногими исключениями многожгутиковые паразиты, живущие
в кишечнике членистоногих (главным образом насекомых) и позвоночных.
Некоторые виды паразитируют у человека.
Chilomastix mesnili — распространенный паразит толстых кишок че-
ловека. Грушевидное тело заострено на заднем конце. На переднем конце
4 жгутика, из которых 3 направлены вперед, 1 — назад. Глубокое ротовое
отверстие (цитостом), достигающее середины тела, укреплено системой
опорных фибрилл. Паразит образует цисты, которые часто встречаются
в экскрементах, особенно при кишечных расстройствах. Ch. mesnili питает-
ся бактериями кишечника и вреда человеку не приносит.
Рис. 33. Многожгутиковые. 1 — Pseudotrychonympha; II — Calo-
nympha grassii:
i — ядра
Trichomonas hominis (см. рис. 32ц ) — маленькое жгутиковое (5—
15 jhk), живущее в толстой кишке человека. Грушевидное тело снабжено
на переднем конце четырьмя жгутиками, из которых три свободных направ-
лены вперед, четвертый направлен назад и прикреплен к краю волнообраз-
ной перепонки. Внутри всего тела проходит аксостиль, выдающийся сво-
бодным задним концом наружу. Т. hominis очень часто встречается в кишеч-
нике здоровых людей. В некоторых случаях его находили в большом
количестве при различных кишечных заболеваниях, но патогенное значение
Т. hominis до сих пор неясно.
Giardia (Lamblia) intestinalis (см. рис. 32щ) живет в тонких кишках
человека. Впервые была описана в 1859 г. Д. Ф. Лямблем в Харькове.
Тело этого паразита построено по двубоковой симметрии. Все структуры
парные, симметрично расположенные в правой и левой половинах тела.
Посередине проходят две тонкие палочки аксостиля, как бы разделяющие
тело пополам. Жгутиков четыре пары. Передний конец тела имеет вид
площадки, служащей, по-видимому, для прикрепления паразита к кишеч-
ному эпителию. Имеются два ядра и парабазальный аппарат.
В экскрементах человека встречаются лишь цисты жиардий. Жиар-
дия может при известных условиях вызвать заболевание кишечника, а в
некоторых случаях проникать в желчные ходы и печень, вызывая там обра-
зование нарывов. Заражение происходит при проглатывании цист.
59
В кишечнике термитов паразитируют представители семейства Calo-
nymphidae (о симбиозе между термитами и жгутиковыми см. стр. 96). У
этих форм (например, Calonympha grassii, рис. 33ц) много ядер, базаль-
ных зерен, аксостилей и парабазальных аппаратов. От каждого базального
зерна отходят три-четыре жгутика, образуя целые жгутиковые зоны. Пи-
таются С. grassii деревом, находящимся в кишечнике термитов.
У этих же животных паразитируют представители семейства Hype-
rmast iginae (рис. 33i ), которые также обладают большим количеством
жгутиков (иногда покрывающих почти все тело), парабазальных аппаратов
и базальных зерен, но имеют только одно ядро. Аксостили есть не у всех
видов.
ОТРЯД КОРНЕЖГУТИКОВЫЕ (RHIZOMASTIGINA)
Это группа своеобразных организмов, которые в своем строении сое-
диняют признаки жгутиковых и корненожек и, таким образом, являются
как бы переходными формами между двумя классами. У них имеется один
или несколько жгутиков, соединенных тонкой нитью (ризопластом) с ядром.
Рис. 34. Корнежгутиковые. / — Mastigamoeba aspera;
II —Actinomonas mirabilis:
/ — жгуты, 2 — псевдоподии
Наряду со жгутиками они образуют ложноножки (псевдоподии) и об-
ладают амебоидной подвижностью, но никогда не отбрасывают жгута.
Обычно одноядерны. Питаются бактериями, водорослями, простейшими
и т. п.
Корнежгутиковые размножаются только бесполым путем при помощи
деления в свободноплавающем состоянии. Примеры: Mastigamoeba aspera,
довольно часто встречающаяся в пресных водоемах (рис. 34i ), Actinomonas
mirabilis (рис. 34ц ) и др.
КЛАСС 3. СПОРОВИКИ (SPOROZOA)
ХАРАКТЕРИСТИКА
Класс споровиков включает паразитических простейших, которые во
время своего цикла развития образуют особые стадии размножения, со-
стоящие из зародыша, в большинстве случаев окруженного плотной обо-
60
лочкой. Эти образования называются спорами. В споре может быть и один
и несколько зародышей.
По сравнению с другими классами простейших споровики сильно из-
менены под влиянием паразитического образа жизни. У них нет органелл
движения, рта, пищеварительных вакуолей. Питаются они только осмоти-
чески.
Споровики размножаются либо только половым путем (грегарины —
Gregarina), либо происходит смена полового и бесполого размножения (кок-
цидии—Coccidia, гемоспоридии — Haemosporidia и др.). Развитие может
протекать внутри одного хозяина, и тогда заражение другого животного
происходит вследствие проглатывания выделенных во внешнюю сре-
ду спор (грегарины, кокцидии, слизистые споровики — Myxosporidia
и др.).
У других споровиков происходит смена хозяев (например, у гемоспори-
дий: комар и человек, птица или другое позвоночное животное). В последнем
случае зародыш не покрыт твердой оболочкой, так как передача происхо-
дит непосредственно от одного животного к другому и зародыш не попадает
во внешнюю среду.
Споровики очень широко распространены и паразитируют у предста-
вителей почти всех классов многоклеточных, особенно у кольчатых червей,
моллюсков, членистоногих и позвоночных.
Споровики, являясь главным образом внутриклеточными паразитами,
в процессе своей жизнедеятельности разрушают клетки хозяина и во многих
случаях служат причиной тяжелых заболеваний животных и человека
(кокцидиоз кроликов, малярия человека, пироплазмоз крупного рогатого
скота и овец, слизистые споровики у рыб и т. Д.).
Ввиду относительной сложности цикла развития споровиков и громозд-
кости терминологии, применяемой в его описании, ниже приводится объяс-
нение встречающихся терминов.
В жизни простейших различают три стадии: 1) вегетативное суще-
ствование; 2) бесполое размножение; 3) половое размножение. Трофозои-
ты — вегетативные формы, типичные для вида. Шизонт — стадии подго-
товки к множественному бесполому делению, называемому шизогонией.
в результате которой образуются дочерние особи, называемые мерозои-
тами. Они служат главным образом для распространения инфекции в
том же самом хозяине.
При половом размножении — гаметогонии — сначала образуются не-
зрелые половые клетки — гаметоциты, которые могут быть двух родов:
мужские — микрогаметоциты и женские — макрогаметоциты.
При дальнейшем развитии гаметоциты превращаются в зрелые поло-
вые особи — гаметы, соответственно — в микрогаметы и макрогаметы.
В результате слияния (копуляции) гамет образуется оплодотворенная
зигота.
Зиготу и продукты ее развития, окруженные плотной оболочкой, на-
зывают ооцистой.
После копуляции в зиготе начинается спорогония, т. е. процесс деления,
в результате которого вначале образуются споробласты, из которых путем
дальнейшего деления образуются мелкие зародыши — спорозоиты — ко-
нечный продукт спорогонии. Спорозоиты служат для перехода паразита
от одного животного к другому, т. е. распространяют инфекцию. Споро-
зоиты, происшедшие из одного споробласта и окруженные общей плотной
оболочкой, образуют спору.
В состав класса споровиков входят 4 подкласса: 1) телоспоридии
(Telosporidia), 2) книдоспоридии (Cnidosporidia), 3) саркоспоридии (Sar-
cosporidia), 4) гаплоспоридии (Haplosporidia).
6i
Представитель класса споровиков —
малярийный плазмодий (Plasmodium vivax)
Plasmodium vivax в определенных стадиях развития паразитирует
в красных кровяных шариках человека и вызывает чрезвычайно распро-
страненное заболевание — трехдневную малярию. Цикл жизни паразита
довольно сложен (рис. 35) и происходит со сменой хозяев, причем половой
Рис. 35. Схема цикла развития возбудителя малярии:
1— спорозоит, выходящий из протока слюнной железы, 2 — 4 — развитие спорозоита в
тканевой клетке (экзоэритроцитарные стадии), 4 — образование тканевых мерозоитов,
5— 10—развитие паразитов в эритроцитах (эндоэритроцитарные стадии), /1 — шизо-
гония, 12 и 13—развитие макрогаметоцита, 14 и 15 — развитие микрогаметоцита, 16—
образование микрогамет, 17 — проникновение микрогаметы в макрогамету (оплодотво-
рение), 18 — проникновение оокинеты сквозь стенку средней кишки комара, 19 и 20 —
развитие ооцисты на поверхности средней кишки комара, 21— разрыв зрелой ооцисты,
выход спорозоитов, 22— спорозоиты в слюнной железе комара, 23— 27 — развитие пос-
ледующих генераций паразита в тканевых клетках. Стадии 2—15 и 23—27—в человеке.
Стадии 16— 22—в комаре. 1 — 22 и пунктирные стрелки — цикл Pl. falciparum, 1 — 27
и все сплошные дугообразные стрелки — цикл Pl. vivax
цикл протекает в комаре, а бесполый — в эритроцитах и некоторых ткане-
вых клетках человека. Таким образом, основной хозяин паразита — ко
мар, промежуточный — человек.
Весь цикл жизни Pl. vivax может быть разделен на отдельные периоды
(табл. I).
1) Бесполое размножение (рис. 352_п, 23_27). Во время укуса комара,
зараженного Plasmodium, серповидные спорозоиты попадают в кровяное
62
русло человека и проникают в клетки эндотелия сосудов печени и других
органов.
В тканевых клетках спорозоиты округляются, растут и в результате
образуются большие формы — шизонты. Через некоторое время начинает-
ся процесс шизогонии', ядро делится, вокруг дочерних ядер обособляются
участки протоплазмы и шизонт распадается на столько мерозоитов, сколь-
ко образовалось ядер. Тканевые клетки затем распадаются, мерозоиты
попадают в ток крови и большая часть их активно проникает в красные
кровяные клетки — эритроциты.
У Pl. vivax часть мерозоитов вновь проникает в тканевые клетки, и
повторяется вышеописанный процесс.
Проникшие в эритроциты мерозоиты, или молодые шизонты, округ-
ляются и растут. Через 2—3 ч в них появляется вакуоль, придающая им
характерную форму кольца. Вакуоль вскоре исчезает. Через 6 ч в теле ши-
зонта появляется бурый пигмент, представляющий собой продукт расщепле-
ния гемоглобина, использованного паразитом в процессе ассимиляции.
В это время паразит, представляющий собой взрослую вегетативную
форму плазмодия, оживленно амебоидно двигается. Через 36 ч после начала
развития в эритроците период роста паразита заканчивается, он округля-
ется, перестает двигаться, начинается деление его ядра на 10—20 частей
(обычно 16); дочерние ядра располагаются по периферии шизонта, вокруг
каждого из них обособляется участок плазмы, и через 48 ч после проникно-
вения спорозоита в эритроцит наступает множественное деление — шизо-
гония, в результате которой образуются мерозоиты.
Пигмент и часть протоплазмы остаются в виде остаточного тела и по-
гибают. К моменту образования мерозоитов эритроцит разрывается, меро-
зоиты выходят в кровь, проникают в новые эритроциты, и процесс начи-
нается сначала.
Такое многократное повторение шизогонии приводит к увеличению
числа паразитов в крови человека.
Стадии цикла малярийного паразита, размножающиеся в тканевых
клетках, называются тканевыми стадиями, или экзоэритроцитарными,
развивающиеся в эритроцитах — эндоэритроцитарными.
2. Образование половых форм (гамет) — макрогамет и микрогамет —
и оплодотворение (рис. 3512_17). После ряда следующих друг за другом
шизогоний обычно на 10—11-й день после заражения часть мерозоитов
начинает образовывать половые формы: макро- и микрогаметоциты.
Период роста макро- и микрогаметоцитов вдвое больше, чем у шизон-
тов (94—96 ч), и они не проходят стадии кольца. Зерен пигмента образуется
больше, и они крупнее. На окрашенных препаратах микро- и макрогаме-
тоциты хорошо различимы. Макрогаметоциты обладают густой, хорошо
красящейся протоплазмой, сравнительно небольшим периферически распо-
ложенным ядром (оно все же больше ядер шизонтов) и большим количест-
вом грубозернистого пигмента, имеющего форму палочек. Макрогамето-
циты приблизительно вдвое больше шизонта. Протоплазма микрогамето-
цитов значительно бледнее, ядро большое, компактное, богатое хроматином.
Пигмента в протоплазме еще больше, чем у макрогаметоцита. Микрогаме-
тоцит также несколько больше шизонта.
По сравнению с шизонтами гаметоциты представляют следующие
биологические особенности: они не способны к размножению в организме
человека и развиваются только в организме комара; поэтому они не пред-
ставляют опасности для человека, в котором живут, но их присутствие
делает человека носителем малярии, опасным для окружающих. Если
комар из рода анофелес (Anopheles) насосется крови человека, содержащей
гаметоцитов, то в нем произойдет дальнейшее развитие паразитов, и комар
получит способность заражать других людей.
Созревание гаметоцитов и оплодотворение происходит в средней кишке
анофелеса. Ядро микрогаметоцита делится на 6—8 ядер, которые распола-
63
Таблица I. Развитие Plasmodium vivax (1—7), Pl. malariae (5—14), Pl.
falciparum (15—20) (окрашено по Романовскому):
1— молодая кольцевидная форма, 2—шизонт (ядро разделилось на два ядра), 3—взро-
слый шизонт (ядро разделилось на 4 ядра, в протоплазме расположен пигмент),- 4 — под-
готовка к шизогонии (ядро разделилось на 14 дочерних ядер), 5 — шизогония (образо-
вание мерсзоитов), 6—микрогаметоцит, 7—макрогаметоцит, 8— трофозоит, 9—раз-
вивающийся шизонт, принимающий характерную форму кольца, 10—развивающийся
шизонт, имеющий характерную форму пояска, 11 — взрослый шизонт (в протоплазме
имеются 8 ядер, но мерозоиты еще не образовались), 12 — шизогония (образование ме-
розоитов), 13—микрогаметоцит, 14 — макрогаметоцит, 15—молодая кольцевидная
форма, 16—растущий трофозоит, 17 — деление ядер во взрослом шизонте, 18— шизо-
гония (образование мерозоитов). 19—микрогаметоцит (хроматин и пигмент расположе-
ны более рыхло, чем у макрогаметоцита (20)
Рис. 36. Температурная кривая трехдневной малярии. Внизу — стадии развития
Pl. vivax, соответствующие той или иной температуре больного
гаются по периферии, вокруг них обособляются участки плазмы; затем в
теле микрогаметоцита возникают сильные протоплазматические токи, в
результате чего каждый ядросодержащий участок плазмы вытягивается
в длинное жгутоподобное образование — микрогамету, отрывающуюся от
сравнительно небольшого центрального участка тела микрогаметоцита —
остаточного тела. После образования микрогамет остаточное тело погибает.
Микрогаметы обладают активной подвижностью.
Подвижная микрогамета проникает в макрогамету, образовавшуюся
из макрогаметоцита, и оплодотворяет ее. Вслед за этим вокруг зиготы
образуется слизеподобный слой, препятствующий проникновению других
микрогамет.
3. Спорогония (рис. 3518_22). Оплодотворенная макрогамета, пробыв-
шая в кишечнике комара несколько часов, принимает веретеновидную
форму. Эта стадия называется оокинетой (рис. 3518). Активно двигаясь,
оокинета проталкивается между эпителиальными клетками, и дальнейшее
развитие происходит в эластическомышечной оболочке кишечника комара.
Здесь оокинета округляется и одевается эластичной капсулой — превра-
щается в ооцисту. Ооциста быстро растет, в ней происходит деление ядер,
число которых может дойти до нескольких тысяч. Протоплазма разбива-
ется на отдельные тяжи, по их периферии располагаются ядра. Эти тяжи —
споробласты — распадаются на отдельные одноядерные тельца — споро-
зоиты. После образования спорозоитов ооциста лопается, серповидные
подвижные спорозоиты выходят в полость тела комара и собираются в
слюнных железах.
При укусе комаром человека в его кровь вместе со слюной попадают
и спорозоиты. Весь процесс развития Pl. vivax в теле комара протекает
10—14 дней.
Приступы малярии у человека связаны, по-видимому, с тем , что при
разрушении эритроцитов во время выхода мерозоитов в кровь попадают
накопившиеся в зараженных красных кровяных тельцах ядовитые продук-
ты жизнедеятельности паразитов, которые и отравляют организм.
Повышение температуры при малярии (приступ) совпадает с моментом
шизогонии (рис. 36). Так как шизогония у Pl. vivax длится 48 ч, то приступы
повторяются один раз в три дня, в связи с чем малярия, вызванная этим
плазмодием, называется трехдневной лихорадкой.
Приступ может наступить только при условии, что количество парази-
тов в крови не ниже определенной нормы. Поэтому первый приступ наступа-
ет после инкубационного периода , т. е. после того, как прошло несколько
шизогоний и плазмодии достаточно размножились в организме человека.
3 Курс зоологии, т. I
65
(возбудитель тропической лихорадки; рис. 37, III) и
Рис. 37. Сравнительная таблица развития малярий-
ных плазмодиев. / — Pl. vivax; // — PI. malariae;
III — Pl. falciparum
Существуют две разновидности Pl. vivax — северная и южная. При
заражении северной разновидностью инкубационный период длится 11—
16 и более дней. При заражении южной разновидностью инкубационный
период длится 8—10 и более месяцев; человек, заразившийся в летне-осен-
ние месяцы, заболевает в весенне-летние месяцы следующего года.
Во многих случаях после окончания первых приступов трехдневной
малярии наступает рецидив, т. е. возврат приступов. Это связано с тем,
что паразиты, полностью исчезнувшие из крови после первых приступов
и сохраняющиеся только в тканевых клетках, вновь выходят в кровь в
виде мерозоитов и начинают размножаться в эритроцитах.
В России до Великой Октябрьской социалистической революции каждый год
заболевало малярией свыше 3 млн. человек.
Для борьбы с малярией в Советском Союзе выработан ряд мер: развернута боль-
шая сеть противомалярийных учреждений, проведено массовое обследование населе-
ния для выявления и лечения больных малярией, разработаны новые методы лечения,
систематически в широких размерах ведется уничтожение малярийных комаров и т. д.
В результате заболеваемость малярией в основном ликвидирована.
При малярии, кроме лихорадочных приступов, наблюдается прогрессирующая
анемия (уменьшение в крови количества красных кровяных шариков). В нормальной
крови содержится 5 млн. эритроцитов на 1 мм3. Во время заболевания число их может
снизиться до 4, 3, 2 млн., а иногда в тяжелых случаях—до 1 млн. на 1 мм3. Вследствие
этого сильно уменьшается и количество гемоглобина в крови. Эритроциты разрушают-
ся частью непосредственно самими паразитами, частью же под влиянием выделяемых
последними токсинов. Кроме того, токсины вредно действуют и на другие органы
человека.
Малярия распространена почти по всей тропической и умеренной полосами зем-
ного шара и является опасным заболеванием; например в Индии ежегодная смерт-
ность от разных видов малярии превышает 1 млн. человек (при заболеваемости в
100 млн.).
Кроме Pl. vivax, возбудителями малярии являются Pl. malariae (возбудитель
четырехдневной лихорадки, впервые открытый Н. А. Сахаровым), PI. falciparum
PI. ovale.
Шизогония у PL malariae
продолжается 72 ч (в связи с
чем приступы повторяются
один раз в четыре дня), у Р1.
falciparum — 48 ч, PL ovale—
48 ч. Развитие этих видов про-
исходит примерно так же,как п
развитие Pl.vivax.Имеются раз-
личия в морфологии паразитов;
этими различиями пользуют-
ся для дифференциального ди*
агноза.
Для Pl. malariae харак-
терны форма пол у взрослых
шизонтов в виде ленты, ле-
жащей в средней части эрит-
роцита, и меньшее, чем у Р1.
vivax число мерозоитов (6—
14). Pl. falciparum хорошо от-
личим уже в стадии молодого
гадия характеризуется большой
вакуолью и малым количеством протоплазмы. Наиболее характерны для Pl. falcipa-
rum полулунная форма гаметоцитов, темно-коричневый цвет пигмента, специфическая
красная пятнистость (после окраски по Романовскому) эритроцитов.
Шизогония Pl. falciparum происходит не в периферической крови, а в капиллярах
мозга, селезенки и костного мозга. Число мерозоитов от 8 до 25.
Pl. ovale был впервые описан в 1922 г. Вызывает лихорадку типа трехдневйой.
В СССР встречается редко. Кольца и шизонты похожи на те же стадии Pl. malariae,
но содержат более крупное ядро. При шизогонии образуется В 12. чаще 8 мерозои-
тов. Паразит вызывает увеличение и сильное обесцвечивание пораженных эритроци-
тов. часть из них становится овальными.
Малярия переносится от человека к человеку только комарами различных ви-
дов рода анофелес. В качестве переносчиков малярии известно около 36 видов этого
рода. В СССР наиболее распространен Anopheles maculipennis. Этим определяется
эпидемиология малярии: заболевания возникают только там, где распространены
комары анофелес, причем в этой же местности должны находиться люди, в крови
которых имеются гаметоциты малярийных плазмодиев. Отсюда вытекают рациональ-
шизонта, благодаря малой величине кольцевидная
66
Рйс. 38. Различия между Си lex (слева) и Anophel (справа)
ные меры борьбы с малярией: необходимо, во-первых, уничтожить комаров анофелес
и, во-вторых, тщательно лечить больных малярией для того, чтобы они не стано-
вились источниками новой инфекции.
Практически важно отличать обычных у нас и совершенно безвредных в смыс-
ле малярии комаров рода кулекс (Culex) от комаров рода анофелес (рис. 38, см. также
стр. 445—449).
СИСТЕМАТИЧЕСКИЕ ОБЗОР КЛАССА
ПОДКЛАСС I. ТЕЛОСПОРИДИИ
(TELOSPORIDIA)
Сюда относятся споровики, которые во взрослом состоянии обычно
неподвижны и одноядерны. Бесполое размножение происходит при помощи
множественного деления (шизогония). Характерно наличие жгутов у мик-
рогамет, что, по-видимому, указывает на происхождение телоспоридий
от жгутиковых. При половом процессе происходит анизогамная копуля-
ция, после которой начинается образование спор, обычно заключенных
в цисту. Споры образуются только в определенные моменты цикла разви-
тия организма.
В состав этого подкласса входят 3 отряда: 1) грегарины (Gregarina),
2) кокцидии (Coccidia) и 3) гемоспоридии (Haemosporidia).
ОТРЯД ГРЕГАРИНЫ (GREGARINA)
Грегарины — паразиты беспозвоночных животных. Во взрослом со-
стоянии в противоположность остальным телоспоридиям, паразитирую-
щим внутриклеточно, грегарины живут в полостях тела хозяина (кишеч-
ник, реже полость тела, семенные пузырьки и т. д.). Внутриклеточными
паразитами бывают только некоторые из них на ранних стадиях развития.
3* 67
Грегарины — самые крупные из споровиков и могут достигать несколь-
ких миллиметров в длину. Они обычно имеют удлиненное и уплощенное
тело с одним ядром.
У полицистид (Polycystidea) тело разделено на три участка: задний,
заключающий ядро, — дейтомерит, средний — протомерит и передний —
эпимерит.
Грегарины, живущие в просвете кишечника, находятся в условиях
постоянно направленного в одну сторону пищевого потока. В связи с этим
Рис. 39. Различные формы грегарин: / — Corycella
armata; II —Stylocephalus longicolis; III —Gre-
garina blattarum; IV —спора Gr. blattarum:
1— эпимерит. 2 — протомерит, 3 — денгомерит, 4 — кутикула,
5 — ядро
у них выработались на переднем конце (эпимерит) различные органеллы
(зубчики, крючочки и т. п.), служащие для прикрепления грегарин к стен-
ке кишки (рис. 39). В противном случае они были бы выброшены из кишеч-
ника вместе с каловыми массами.
Тело грегарин покрыто плотной ребристой кутикулой, определяющей
форму паразита. Под кутикулой расположена эктоплазма, которая и обра-
зует плазматические перегородки, разделяющие тело грегарин наэпимерит,
протомерит и дейтомерит. В эктоплазме у многих грегарин имеются слой
продольных и поперечных мускульных фибрилл (мионем) и тонкие скелет-
ные нити.
Многие грегарины активно передвигаются при помощи сокращения
мионем. Движение других грегарин связано с выделением ими на заднем
конце тела затвердевающего столбика слизи.
У грегарин нет ни рта, ни глотки, и они питаются осмотически, вса-
сывая питательные вещества всей поверхностью тела.
Грегарины размножаются только половым путем, за исключением по-
дотряда шизогрегарин, у которых имеется смена полового и бесполого по-
колений. Половое размножение происходит следующим образом: две гре-
гарины сближаются и одна из них приклеивается своим передним концом
к заднему концу другой (рис. 39ш). Затем обе особи округляются и инщь
стируются, не сливаясь. В обеих грегаринах происходит энергичное деле-
68
Рис. 40. Схема развития грегарин:
/ — клетка эпителия кишечника, 2— ядро клетки эпителия. 3 — споры, 4 —
раскрывшаяся спора, 5— спорозоиты, выпавшие из раскрывшейся споры,
6 — остаточное тело, 7 — внедрение спорозоита в эпителиальную клетку,
5—12— развитие спорозоита в грегарину
ниеядер, располагающихся после этого по периферии тела и выпячиваю-
щих участки плазмы на его поверхности. Затем каждый бугорок, состоящий
из маленького ядра и участка плазмы, отшнуровывастся и, таким образом,
образуются гаметы. У некоторых видов из обеих грегарин образуются оди-
наковые гаметы (изогаметы), у других видов из
одной — округлые неподвижные макрогаметы,
из другой — снабженные жгутиками подвиж-
ные микрогаметы (анизогаметы).
У тех грегарин, во время полового процесса
которых образуются анизогаметы, разделение
полов можно наблюдать уже во время прикле-
ивания особей друг к другу, называемого сизи-
гией. Передняя грегарина называется приматом,
задняя—сателлитом. Оказывается, что эти
грегарины отличаются друг от друга по различ-
ному отношению к красителям, распределению
запасных веществ и т. д. Из примита образу-
ются макрогаметы, т. е. он является женской
половой особью, из сателлита — микрогаметы,
т. е. он представляет собой мужскую половую
особь.
Микрогаметы и макрогаметы копулируют
попарно и образуют зиготы. Каждая зигота по-
крывается плотной оболочкой и превращается
в спору, внутри которой развиваются спорозо-
иты (обычно 8). Таким образом, внутри цисты
образуется множество спор.
Цисты попадают во внешнюю среду с вы-
делениями кишечника или только после смер-
ти хозяина. Заражение происходит путем заг-
латывания цисты новым животным того же ви-
Рис. 41. Грегарины из кишеч-
ника мучного червя (личинки
Tenebrio molitor): / —Grega-
rina polymorpha; 11 — Gre-
garina steini; III —Gregarina
cuneata:
1 — эктоплазма, 2 — эндоплазма,
3—ядро, 4— дейтомерит, 5 —
протемерит, 6 — остаток эпиме-
рнта
69
да, в кишечнике которого спорозоиты выходят из споры (рис. 40 4). Ос-
вободившиеся спорозоиты внедряются в клетки стенки кишечника (у ки-
шечных грегарин, рис. 407), растут там, затем, не умещаясь в клетке,
остаются погруженными в нее только эпимеритом (рис. 41).
Грегарины, по-видимому, не вызывают заболевания у животных, в
которых паразитируют.
ОТРЯД КОКЦИДИИ (COCC1DIA)
Кокцидии, за немногими исключениями, — внутриклеточные паразиты
эпителия различных животных. Во взрослом состоянии они неподвижны и
имеют круглую или овальную форму. Только удлиненные молодые формы-
Рис. 42. Цикл развития кокцидии Eimeria ша-
gna. I — первое поколение шизонтов; II — вто-
рое поколение шизонтов; 111 —третье поколе-
ние шизонтов; IV — гаметогония; V — спорого-
ния:
1 — спорозоиты, 2— молодой шизонт, 3 — растущий ши-
зонт со множеством ядер, 4 — шизонт, распавшийся на ме-
розоиты, 5 — мерозоиты, 6—6а — развитие макрогаметы, 7—
микрогаметы, 8—ооциста, 9— ооциста, приступающая к
спорогонии, 10 — ооциста с четырьмя споробластами и ос-
таточным телом, // — развитие спороцист, 12— зрелые
ооцисты с четырьмя спорами, в каждой споре по два спо-
розоита
спорозоиты и мерозоиты —
обладают способностью к ак-
тивному движению. Размно-
жение кокцидий в отличие
от грегарин характеризуется
сменой полового и бесполо-
го поколений.
Цикл развития кокцидий
лучше всего изучить на при-
мере рода Eimeria — парази-
тов кролика (рис. 42). После
заглатывания кроликом ооци-
сты спорозоиты освобожда-
ются и активно проникают в
клетки тонкой кишки и жел-
чных ходов, где растут и раз-
виваются в круглых шизон-
тов. Ядро шизонта делится
несколько раз, вокруг каж-
дого из получившихся ядер
обособляется участок протоп-
лазмы, и шизонт распадает-
ся на несколько удлиненных
колбасовидных мерозоитов.
Последние проникают в све-
жие эпителиальные клетки,
где вновь начинается бесполое
размножение. Шизогония пов-
торяется несколько раз и
ведет к сильному увеличению
количества паразитов в кро-
лике. Через несколько (обыч-
но 3—5) поколений наряду с
бесполым размножением на-
чинается половой цикл — га-
метогония: в результате од-
ной из шизогоний образуются
мерозоиты двух типов — из одних шизонтов развиваются микрогамето-
циты, из других — макрогаметоциты. Они вновь проникают в эпители-
альные клетки, где происходит развитие их в половые особи — гаметы.
Макрогаметоцит разрастается и округляется, микрогаметоцит сначала то-
же разрастается, а потом делится на много длинных вытянутых микро-
гамет, снабженных двумя жгутами.
Одна из микрогамет активно проникает в макрогамету и сливается
с ней, в результате чего образуется споронт, окружающийся плотной обо-
лочкой. Эта стадия развития называется ооцистой. Дальнейшее развитие
70
происходит вне тела хозяина. Окруженная плотной оболочкой ооциста
выводится вместе с калом наружу. Ядро делится, вокруг получившихся
ядер обособляются участки протоплазмы, и таким путем образуются четыре
особи. Они покрываются оболочкой и превращаются в спороцисты, внутри
которых затем возникают спорозоиты. Количество их различно у разных
видов кокцидий. С образованием спорозоитов спороциста превращается в
спору.
В результате всего процесса образуется ооциста, внутри которой рас-
положены четыре споры.
Кролики заражаются, заглатывая ооцисты. В кишечнике под влиянием
панкреатического сока в ооцисте и спороцистах растворяются плазматиче-
ские пробки имеющихся в них отверстий (микропиле), спорозоиты выходят
наружу и внедряются в клетки кишечника.
Кокцидии разрушают клетки хозяина, и так как очень быстро в них
размножаются, представляют серьезную опасность для животных, в которых
паразитируют.
Кролики заражаются несколькими видами кокцидий. Наиболее распространен-
ные из них: Eimeria stiedae (в эпителии тонкой кишки и печени), Е. perforans (в эпи-
телии кишечника) и Е. magna (там же). При плохом уходе и ослаблении организма
кроликов, среди них вспыхивают эпизоотии кокцидиоза, от которых может пасть
очень большое количество молодняка. Болезнь выражается в том, что животные ху-
деют, страдают поносом, становятся вялыми и при сильной инфекции гибнут в тече-
ние нескольких дней. Борьба с кокцидиозом кроликов сводится, с одной стороны,
к уничтожению ооцист, являющихся единственным источником заражения, а с дру-
гой — к тщательному ухо-
ду за кроликами, ежеднев-
ной чистке клеток и т. д.,
так как при хорошем со-
держании кролики более
устойчивы к заболеванию и
редко погибают от него.
Плотная оболочка ооцист
в значительной мере предо-
храняет спорозоитов от воз-
действия внешней среды, и
наилучшими способами де-
зинфекции клеток и других
помещений, в которых со-
держатся кролики, являют-
ся прожигание паяльной
лампой, обмывание кипят-
ком и высушивание.
Кокцидии парази-
тируют у многих траво-
ядных животных, под-
бирающих вместе с тра-
вой с земли цисты со
спорами кокцидий.
Рис. 43. Ооцисты различ-
ных кокцидий из кала че-
ловека и животных. I,
// — Eimeria stiedae из
кролика; /// — Е. perfo-
rans из кролика; IV — Е.
avium из курицы; V — Iso-
spora rivolta из собаки;
v7 — Е. oxyspora (Е. sar-
dinae) из человека; VII —
X—I. belli из человека;
XI — Е. wenyoni (Е. clu-
pearum) из человека;
XII—Е. sniydersi (Е. clu-
pearum) из человека
71
Вследствие способности к бесполому размножению внутри зараженного
животного количество кокцидий в одной особи быстро увеличивается, а
следовательно, увеличивается количество уничтожающихся паразитами
кишечных и других клеток и усиливается тяжесть заболевания.
Кокцидии — весьма опасные паразиты многих домашних животных:
крупного рогатого скота, овец, кур, гусей и т. д. Особенно опасно зараже-
ние кокцидиями молодняка; иногда гибнет до 50% заболевших животных.
Массовые заболевания, вызываемые кокцидиями, наблюдаются и у диких
животных: зайцев, куропаток и др.
У человека также описывались кокцидии (Isospora belli, I. hominis и
др.) (рис. 43). По-видимому, они причиняют кишечные расстройства.
ОТРЯД ГЕМОСПОРИДИИ (HAEMOSPORIDIA)
Гемоспоридии — внутриклеточные паразиты красных кровяных ша-
риков (эритроцитов) или эндотелиальных клеток млекопитающих, птиц1
и рептилий, в которых размножаются бесполым путем (шизогония) и обра-
зуют гаметоциты. Оплодотворение и спорогония происходят в организме
различных комаров семейства Culicidae и некоторых других насекомых.
Типичными формами служат описанные выше виды рода Plasmodium1.
Цикл развития гемоспоридий очень сходен с таковым у кокцидий,
поэтому некоторые авторы соединяют их в один отряд кокцидиоморфных
(Coccidiomorpha). На близкое родство кокцидий и возбудителей малярии
впервые в 1887 г. указал И. И. Мечников. Основные отличия гемоспоридий
от кокцидий состоят в наличии у них амебообразно подвижных шизонтов,
в образовании у большинства гемоспоридий характерного коричневого
пигмента, являющегося продуктом превращения воспринятого паразитом
гемоглобина эритроцитов, и в отсутствии спор.
Важную в хозяйственном отношении группу кровепаразитов представ-
ляет подотряд Piroplasmoidea, включающий в себя семейства бабезиид (Ва-
Besiidae) и тейлериид (Theileriidae) — возбудителей тяжелых заболеваний
млекопитающих, в частности домашнего скота, заболеваний, известных под
названием пироплазмозов.
В организме позвоночных животных грушевидные бабезииды размно-
жаются бесполым путем в красных кровяных клетках (эритроцитах) (рис. 44).
Передача пироплазм от животного к животному происходит через иксодовых
клещей, которые служат переносчиками инвазии. Когда клещи Boophilus
сосут кровь больных животных, то вместе с эритроцитами в их организм
попадают грушевидные трофозоиты бабезий. Здесь они размножаются
Рис. 44. Babesia canis (схематично). 1 — V —
последовательные стадии размножения пара-
зита в эритроцитах собаки
бесполым путем в просвете ки-
шечника и внутриклеточно и
распространяются по всем внут-
ренним органам. Затем спорозо-
иты бабезиид проникают в яйце-
вые клетки клещей, продолжая
и там бесполое размножение.
Таким образом зараженные кле-
щи передают инфекцию своему
потомству. Вследствие этого за-
ражать позвоночных животных
могут на протяжении несколь-
ких поколений клещи, сами до
этого не сосавшие кровь зара-
женных животных.
1 Малярийных паразитов птиц впервые описал в 1884 году В. Я. Данилевский.
Это открытие имело большое значение для доказательства того, что паразит, впослед-
ствии описанный в эритроцитах человека, является возбудителем малярии.
72
У тейлериид множественное деление происходит в организме крупного
рогатого скота, главным образом в лимфатических узлах. Часть продуктов
деления вновь проникает в клетки селезенки, другая часть попадает в эри-
троциты. Переносчиками тейлериид служат иксодовые клещи Ripicephalus,
Hyalomma и др. По-видимому, в организме клещей происходит образование
гамет, которые копулируют и образуют зиготы. В результате размножения
в слюнных железах образуются спорозоиты, которые и служат источником
заражения крупного рогатого скота.
Все представители бабезиид и тейлериид паразитируют в эритроцитах
крупного рогатого скота, лошадей, свиней, собак и других млекопитающих
и вызывают тяжелые заболевания, приносящие большой вред животновод-
ству.
Babesia bigemina — возбудитель «техасской лихорадки» крупного ро-
гатого скота. В СССР это заболевание распространено на Северном Кавказе,
в Закавказье и Средней Азии. Заболевшие животные худеют, становятся
малокровными, моча их окрашивается в красный или бурый цвет. На Кав-
казе переносчиком служит клещ Margaropus annulatus calcaratus. Nutallia
equi паразитирует в эритроцитах лошади и является возбудителем опасного
заболевания, часто кончающегося смертью животного; распространен в
южных районах СССР. Переносчики — несколько видов клещей рода Ripi-
cephalus. Piroplasma ovis — возбудитель пироплазмоза овец, Babesiella
bovis вызывает тяжелый пироплазмоз коров, переносится обыкновенным
собачьим клещом Ixodes ricinus.
Так как инфекцию переносят клещи, то профилактика пироплазмозов
состоит в уничтожении клещей путем купания зараженных животных в
дезинфицирующих жидкостях. Другим способом профилактики являются
перегонка стад на свежие пастбища и уничтожение клещей на зараженных
площадях, а также запрещение ввоза в благополучные районы больного
пироплазмозом скота и т. п.
ПОДКЛАСС II. КНИДОСПОРИДИИ (CNIDOSPORIDIA)
К этому подклассу относятся споровики. в спорах которых, кроме за-
родыша» находятся полярные капсулы. похожие на стрекательные капсулы
кишечнополостных и состоящие из грушевидного тельца, внутри которого
находится спирально закрученная нить. Под влиянием кишечных соков пить
выстреливает и прикрепляет спору к стойко кишечника. Вслед за этим из
споры выходит зародыш.
Книдоспоридии паразитируют в полостях и в клетках. На ранних
стадиях развития они движутся при помощи псевдоподий, что, по-нпди-
мому, указывает на происхождение их от корненожек. В противополож-
ность телосноридиям книдоспоридии производят споры в течение всей
жизни.
В состав этого подкласса входят 3 отряда: 1) слизистые споровики (Му-
xosporidia), 2) актиномиксидии (Actinomyxidia), 3) микроспоридии (Micro-
sporidia).
ОТРЯД СЛИЗИСТЫЕ СПОРОВИКИ (MYXOSPORIDIA)
Миксоспоридии — чрезвычайно распространенные паразиты рыб, реже
амфибий и рептилий. Они поражают все органы и ткани хозяина. Наиболее
часто встречаются в мускулатуре, подкожной соединительной ткани,
жабрах, почках, селезенке и печени, в желчном пузыре и мочевом пузыре.
На ранних стадиях развития миксоспоридии одноядерны, затем выра-
стают до сравнительно больших протоплазматических масс со многими
ядрами.
73
В полостях миксоспоридии голые и амебоидно-подвижные, в тканях
они окружены оболочкой и достигают большой величины — иногда несколь-
ких миллиметров и до сантиметра.
По мере роста паразита, в нем начинается своеобразный половой процесс,
который сопровождается образованием спор. Сначала попарно копулируют
ядра многоядерной особи и вокруг копулирующих ядер обособляются участ-
ки плазмы, так называемые панспоробласты. Из каждого панспоробласта
обычно образуется по две сложно устроенные споры; внутри оболочки, со-
ставленной из двух половинок, помещаются одна, две или четыре стрека-
тельные капсулы и обычно двухъядерный зародыш (рис. 45). Образовав-
шиеся споры выводятся наружу и
затем могут быть заглочены новой
особью хозяина того же вида. В этом
случае в кишечнике спора выстре-
ливает, зародыш выходит наружу,
проникает через стенку кишечника в
капилляры и по кровеносной системе
попадает в орган, в котором паразити-
рует. Кроме описанной спорогонии,
некоторые миксоспоридии могут раз-
Рис. 45. Споры различных
миксоспоридий. /—Cerato-
myxa; // — Hennegya;
III —Sphaerospora; IV —
Myxosoma; V — Lentospora;
VI — Myxobolus fuhrmani;
VII — Myxobolus bramae;
VIII — Hoferellus
Рис. 46. Циста Myxobolus pfe-
iferi, лежащая между мускуль-
ными клетками окуня
множаться и бесполым путем, следствием чего является расселение пара-
зита в хозяине.
Миксоспоридии вызывают заболевания рыб. Наиболее опасны различ-
ные виды родов Myxobolus и Lentospora, например Myxobolus pfeiferi (рис.
46) — возбудитель «шишечной болезни» усачей. Паразиты поселяются в
мышцах рыбы, разрастаются там и образуют опухоли, выдающиеся над
поверхностью тела. Опухоли прорываются и превращаются в открытые
нарывы. При этом споры, заполнявшие опухоль, выбрасываются наружу и
служат источником заражения других рыб. В результате инфекции боль-
шинство зараженных рыб погибает. Lentospora cerebralis — возбудитель
«вертячки» различных форелевых рыб. Lentospora паразитирует в хрящевой
ткани молодых форелей и вызывает разрушение ее. Споры попадают наружу
только после смерти рыбы.
74
ОТРЯД АКТИНОМИКСИДИИ (ACTINOMYXIDIA)
Небольшая группа книдоспоридий, у которых известна только спорогония.
Бесполого размножения, по-видимому, нет. Споры построены по трехлучевому типу.
В них имеются три полярные капсулы и большое количество амебоидных зародышей.
Образование спор происходит в основном так же, как и у миксоспоридий, но у
спорулирующего паразита тело не разбивается на отдельные споробласты, а целиком
превращается в один панспоробласт. АктиномиксиДии паразитируют у морских
малощетинковых червей (Olygochaeta).
ОТРЯД МИКРОСПОРИДИИ (MICROSPORIDIA)
Рис. 47. Схема цикла развития No-
sema bombycis в кишечном эпителии
шелковичного червя. Справа вни-
зу — проникновение амебоидного
зародыша в клетку, слева — различ-
ные стадии бесполого размножения
и превращение в спори
Маленькие внутриклеточные паразиты почти исключительно членисто-
ногих и рыб. Очень близкие миксоспоридиям, они отличаются от них тем,
что являются внутриклеточными паразитами и, кроме того, более простым
строением спор.
Споры микроспоридий имеют овальную или грушевидную форму, глад-
кую, сплошную оболочку, внутри которой, в вакуоле, лежит спирально
закрученная полярная нить. Особой
полярной капсулы нет. Интересно,
что при длине споры от 4,6 до 6,4 мк,
длина выброшенной полярной нити
достигает 280 мк. Бесполое размноже-
ние происходит путем деления надвое
и шизогонии. При этом часто пара-
зиты складываются в характерные це-
почки.
После неоднократного бесполого
размножения наступает спорогония,
в результате которой образуются
споры.
Некоторые микроспоридии —
опасные паразиты. Так, например,
Nosema bombycis — возбудитель пеб-
рины шелковичного червя (гусениц
шелковичной бабочки — Bombyx пто-
ri). Заражение происходит при загла-
тывании гусеницей спор вместе с
кормом. В кишечнике зародыш выхо-
дит из споры и размножается деле-
нием и почкованием. Молодые вегетативные формы проникают между
клетками кишечной стенки в полость тела и разносятся в различные органы
(кишечник, мускулатура и т. д.), где продолжают энергично размножаться
(деление, шизогония). В органах же происходит спорогония (рис. 47). Но-
вые споры могут появиться уже на четвертый день после заражения.
Часть спор вываливается из разрушающихся клеток кишечника, попадает
наружу вместе с экскрементами, загрязняет корм и служит источником
заражения других шелковичных червей.
Паразиты проникают также в яйца (грену) и таким образом заражают
последующие поколения гусениц. Пебрина вызывает массовый падеж гу-
сениц, но при слабой инфекции гусеницы могут превратиться в бабочек, из
грены которых вновь развиваются зараженные гусеницы, являющиеся ис-
точником дальнейшего распространения инфекции. Единственным способом
борьбы с пебриной служит вывод гусениц из грены, отложенной здоровыми
бабочками.
Другой вид ноземы Nosema apis — распространенный паразит ки-
шечника пчел, вызывающий так называемый «понос» пчел; Споры N. apis
локализуются почти исключительно в кишечнике.
75
ПОДКЛАСС 111. САРКОСПОРИДИИ (SARCOSPOR1DIA)
Внутриклеточные паразиты поперечнополосатой мускулатуры млеко-
питающих (лошадей, рогатого скота, кроликов, мышей, крыс, собак, очень
редко человека), некоторых птиц и рептилий.
Саркоспоридии образуют в мускулатуре большие беловато-серые уд-
линенные или круглые тела, видимые простым глазом. Эти тела окружены
Рис. 48. Sarcocystis lindemannii
из гортани. / — продольный раз-
рез; II — поперечный разрез по-
раженного мышечного волокна
характерной двойной оболоч-
кой, верхний слой которой
состоит из большого количес-
тва параллельных палочек.
Молодые одноядерные
или двухъядерные формы по-
падают в поперечнополосатые
мускульные клетки, начина-
ют там энергично размно-
жаться и окружаются описан-
ной выше оболочкой. Внутри
оболочки образуются споры,
имеющие характерную для
саркоспоридий серповидную
форму и лишенные спираль-
ной нити.
Естественный путь зара-
жения не выяснен. Экспери-
ментально можно заразить
животное, дав ему поесть мя-
со, содержащее цисты сарко-
споридий.
Саркоспоридии очень ши-
роко распространены, но хо-
зяйственное значение их неве-
лико, так как «особого вреда
они не приносят. Только при
/ очень сильном заражении ско-
та может произойти студне-
видное перерождение мяса, которое становится непригодным для употреб-
ления в пищу.
У свиней Sarcocystis mescheriana иногда может вызвать параличи.
S. tenella распространенный паразит овец; S. muris — мышей. У человека
несколько раз описывалась S. lindemanni (рис. 48). Патогенное значение
ее неясно.
ПОДКЛАСС IV. ГАПЛОСПОРИДИИ (HAPLOSPORID1A)
Небольшая группа мало изученных паразитов, споры которых не имеют поляр-
ных нитей и снабжены специальной крышечкой, закрывающей отверстие споры.
Гаплоспоридии паразитируют в клетках и полостях различных животных (чер-
вей, моллюсков, коловраток, рыб, некоторых млекопитающих) и человека.
Размножение шизогонией и спорогонией. Описан половой процесс. Системати-
ческое положение гаплоспоридий неясно. Часть ранее относимых сюда представите-
лей, согласно новейшим исследованиям, оказалась либо микроспоридиями, либо
низшими грибами; в частности, к низшим грибам теперь относят риноспоридп-
ум (Rhinosporidium seeberi) — возбудитель опухолей носовой перегородки чело-
века.
Типичными представителями являются различные виды рода гаплоспоридиум
(Haplosporidium) — паразиты червей и моллюсков.
76
Подтип II. Ресничные или инфузории
(Ciliophora)
ХАРАКТЕРИСТИКА
К подтипу Ciliophora относятся простейшие, снабженные многочислен-
ными мелкими ресничками или более крупными — мембранеллами, состоя-
щими из склеенных между собой ресничек.
Реснички представляют собой многочисленные очень тонкие и корот-
кие выросты наружного слоя протоплазмы. Оболочка стенки тела (пелли-
кула) непосредственно переходит в оболочку реснички. Реснички покрывают
тело инфузории либо сплошь, либо собраны в отдельные пучки, ленты и т. п.
Рис. 49. Электронная микрофотография срезов ресничек инфузории Ichtiophti-
rius inulIifili>. / — поперечный срез реснички (видны оболочка реснички,
кольцо из дспятн двойных периферических фибрилл и две центральные фибрил-
лы. справа от каждой двойной фибриллы отходят отростки) увеличение
20 000; // — продольный срез стейки тела (видны реснички с центральными
и периферическими фибриллами, под стопкой тела реснички переходят в базаль-
ные зерна):
1 — стенка тела инфузории, 2 — росшгпкп, 3 — периферические фибриллы, * —центральные
фибриллы, J — базальное зерно
Как правило, реснички значительно короче и часто тоньше жгутиков. Элек-
тронно-микроскопическое изучение показало, что структура ресничек очень
сходна со строением жгутиков.
Непосредственно под оболочкой в цитоплазме расположено кольцо
из девяти двойных фибрилл, в центре реснички проходят по всей длине 2
центральные фибриллы. Они оканчиваются на уровне стенки тела, а пери-
ферические фибриллы проникают в эктоплазму и входят в состав базального
зерна (рис. 49).
Инфузории, объединяемые в класс Ciliata, покрыты* ресничками в тече-
ние всей жизни. Только в цистах они сбрасывают реснички. Сосущие же
инфузории (Suctoria) покрыты ресничками только на ранних стадиях раз-
вития в свободноплавающем состоянии. Затем они переходят к сидячему
образу жизни и теряют реснички.
Форма тела инфузорий благодаря присутствию плотной пелликулы, а
часто и скелетных фибрилл всегда определенная.
77
Организация инфузорий часто достигает большой сложности благодаря
тому, что у многих из них имеются мускульные и скелетные элементы, сложно
устроенный перистом (предротовой аппарат) и дифференцированные орга-
неллы, служащие для передвижения (реснички, щетинки, мембранеллы
и т. д.).
Второй, кроме присутствия ресничек, признак, который отличает почти
всех инфузорий (кроме протоцилиат — Protociliata) от остальных простей-
ших,— строение ядерпого аппарата. У них имеются два типа ядер: большое—
макронуклеус и меньшее — микронуклеус. У многих видов имеется по не-
скольку макронуклеусов и микронуклеусов.
Инфузории размножаются делением или почкованием, причем микро-
нуклеус делится митотически, макронуклеус — амитотически.
Оплодотворение в виде пзогамной или анизогамной конъюгации*, только
для протоцилиат характерна копуляция (см. стр. 83).
В состав подтипа входят 2 класса: 1) ресничные инфузории (Ciliata) и
2) сосущие инфузории (Suctoria).
КЛАСС 1. РЕСНИЧНЫЕ ИНФУЗОРИИ (CILIATA)
ХАРАКТЕРИСТИКА
Ресничные инфузории покрыты ресничками всю жизнь. Питаются они
через цитостом (клеточный рот) или осмотически. Размножение происходит
при помощи поперечного деления или почкования. Время от времени бес-
полое размножение прерывается половым процессом — конъюгацией или
копуляцией.
Инфузории распространены как в пресной воде, так и в морях, но боль-
шинство видов обитает в пресной воде. Особенно многочисленны они в мел-
ких водоемах. Имеются и паразитические формы.
В состав класса Ciliata входят 2 подкласса: 1) протоцилиаты (Protoci-
liata) и 2) собственно инфузории (Euciliata).
Представитель класса ресничных инфузорий
— парамеция (Paramaecium caudatum)
Строение. Парамеция1 — обычно встречающаяся в водоемах сво-
бодноживущая инфузория (рис. 50). Удлиненноовальное тело парамеции
равномерно покрыто рядами одинаковых ресничек, поэтому она и относится
к отряду равноресничных (Hoiotricha). Только на заднем конце реснички
несколько длиннее. Реснички морфологически сходны со жгутиками, но
отличаются от них тем, что они короче, тоньше и обычно их гораздо больше,
чем жгутиков; так, у парамеции около 10 000 — 14 000 ресничек. Каждая
ресничка начинается от базального зерна— небольшого зернышка, распо-
ложенного в эктоплазме. Движение реснички состоит в коротком резком
ударе вперед и плавно медленном — назад. Реснички бьют ритмично и
последовательно одна за другой, начиная от переднего конца тела к зад-
нему. Согласованная работа ресничек и определяет поступательное движе-
ние парамеции. Реснички расположены продольными рядами, несколько
спирально изогнутыми по отношению к длинной оси тела. Вследствие этого
парамеция плывет вперед, кружась вокруг продольной оси тела, и движение
идет по винтовой линии.
Парамеция движется со скоростью 2647 мк в секунду. Это одна из на-
иболее быстро движущихся инфузорий. Вообще скорость передвижения
различных инфузорий варьирует в пределах от 310 до 2647 мк в сек.
1 Парамецию часто называют «туфелькой».
78
Рнс. 50. Paramaecium caudatum:
/— образующаяся пищеварительная вакуоль,
2 — чкскрс1орю1я пор.), и — ИЫДСЛНrCJlbllOC
тельце, '/ --ундулирующая мембрана, л —
микронуклеос, 3 — микронуклеос, 7 — при-
водящие каналы, 8 — пульсирующая ваку
оль. 9 — трихоцисты, 10— пищеварительная
вакуоль, 11— реснички, Г2—перистом,
13 — цитофаринкс (глотка), 14 — цитостом.
15 — порошица, 16—глоточные нити
Вся инфузория покрыта плотной тонкой оболочкой — пелликулой.
Под пелликулой находится тонкий слой прозрачной эктоплазмы; остальное
тело заполнено зернистой эндоплазмой.
Пелликула покрыта четырехугольными или чаще шестиугольными
полями, ограниченными выпуклостями пелликулы, расположенными пра-
вильными рядами. В середине каждого поля проходит ресничка, базальное
зернышко которой находится в эктоплаз-
ме. По краям поля на выпуклостях
пелликулы открываются протоки трихо-
цист, или стрекательных палочек.
Трихоцисты расположены в экто-
плазме перпендикулярно к пелликуле и
образуют густой частокол (рис. 509). Они
при раздражении выбрасываются в виде
длинной упругой нити и служат для за-
щиты или нападения, вонзаясь в тело
животного, к которому прикоснется ин-
фузория. Трихоцисты оказывают пара-
лизующее влияние на простейших, в
которых вонзаются.
На микрофотографиях в электрон-
ном микроскопе при увеличении в 16000
раз видно, что выстреленная трихоциста
состоит из длинного плотного поперечно-
исчерченного стержня и плотного гвоз-
деобразного наконечника (рис. 51).
В эктоплазме, непосредственно под
пелликулой, располагается сложная
сеть переплетающихся фибрилл. Эту
сеть можно увидеть только при специ-
альной обработке. Некоторые авторы
приписывают фибриллам эктоплазмы
нервнопроводящую функцию. На самом
деле это скелетные опорные нити, кото-
рые определяют устойчивую, постоян-
ную форму тела парамеции. Скелетные
нити, выполняющие эту роль, имеются
почти у всех инфузорий.
На брюшной стороне в передней
половине тела находится продольная
выемка — околоротовая впадина, или пе-
ристом. В глубине перистома располо-
жено овальное отверстие — клеточный
рот цитостом, ведущий в изогнутую
глотку цитофаринкс. Глотку поддержи-
вает система скелетных глоточных ни-
тей. На спинном крае глотки находится
волнообразная перепонка — ундулирующая мембрана — тонкая, все время
колеблющаяся пленочка, состоящая из слипшихся между собой ресничек
(рис. 504). Глотка открывается непосредственно в эндоплазму.
Питание и выделение. Парамеция питается главным образом бактерия-
ми. Биением ресничек перистомного поля и ундулирующей мембраны вода с
взвешенными в ней бактериями загоняется через рот в глотку, где на месте
соприкосновения с голой эндоплазмой из воды с заключенными в ней бакте-
риями каждые 1,5 мин образуется круглая пищеварительная вакуоль (рис.
50J. Она отрывается от глотки и увлекается медленным круговым движением
плазмы инфузории. Пищеварительная вакуоль проходит в теле парамеции
совершенно определенный путь вокруг всего тела (пищеварительный цик-
79
лоз), продолжающийся в зависимости от рода пищи и температуры от 1 до
3 ч. Во время пищеварительного циклоза в вакуоль диффундируют пищева-
рительные ферменты, происходит переваривание бактерий, а неперева-
ренный остаток каждые — 6—7 мин выбрасывается через порошицу, рас-
положенную на брюшной стороне в заднем конце тела.
Во время прохождения вакуолей в них происходит смена реакции. При
образовании вакуолей, содержимое их имеет щелочную реакцию, но уже
очень быстро она переходит в кислую. В это время погибают бактерии, со-
держимое вакуолей образует довольно плотный комок, а сама вакуоль
уменьшается. Затем содержимое вакуолей вновь приобретает щелочную
реакцию, вакуоли увеличиваются, пищевой комок растворяется, образуются
Рис. 51. Концевые участки трихоцист парамеции. Вид-
ны твердые гвоздеобразные наконечники, остальная
более мягкая и поперечноисчерченная часть трихоцист
сплющена (снимок сделан при помощи электронного
микроскопа)
капли, переходящие в протоплазму и там резорбирующиеся. Перед выбра-
сыванием вакуоль снова уменьшается.
Смену реакции в пищеварительной вакуоле интересно сопоставить с
тем, что у высших животных также сначала переваривание происходит
при помощи пепсина в кислой среде, а затем при помощи трипсина в щелоч-
ной.
Парамеция поглощает большое количество бактерий. Так, в водной
культуре сенной палочки парамеция образует 60 пищеварительных вакуо-
лей в час, причем в каждой вакуоле обычно бывает около 30 бактерий. Это
составляет 43 000 бактерий в день.
На границе экто- и эндоплазмы у парамеции в первой и третьей четверти
тела расположены сократительные, или пульсирующие, вакуоли (рис. 508).
Они состоят из собственно вакуоли и системы приводящих каналов, окру-
жающих ее в виде звезды (рис. 507).
Деятельность вакуоли происходит следующим образом. Перед опоро-
жнением во внешнюю среду вакуоль наполнена жидкостью и поэтому осо-
бенно хорошо заметна. Приводящие каналы в это время только слегка на-
полнены водой. Эта фаза называется диастолой. Затем вакуоль сокраща-
ется, изливает свое содержимое во внешнюю среду через особое маленькое
отверстие (выделительную пору) и спадается (систола), а каналы еще боль-
ше наполняются. Вслед за этим каналы все сразу сокращаются, изливают
свое содержимое в вакуоль, и весь процесс начинается сначала. Сократи-
тельная вакуоль и выделительная пора парамеции — это постоянные строго
80
локализованные структуры. Пульсация передней и задней вакуолей про-
исходит попеременно. Скорость биения вакуоли зависит от температуры и
концентрации окружающей среды. При 16°С вакуоль в пресной воде сокра-
щается 3—3,5 раза в минуту. В течение получаса две сократительные вакуоли
выбрасывают количество воды, равное объему тела парамеции.
Назначение сократительных вакуолей у парамеций, как и у других
простейших, состоит в том, чтобы поддерживать путем освобождения от
излишней воды определенное осмотическое давление в теле парамеции.
Кроме того, вместе с водой выводятся и некоторые продукты обмена, напри-
мер мочевая кислота.
Для выделения продуктов обмена служат и
находящиеся в теле парамеции выделительные тель-
ца, имеющие вид правильных сильно преломляю-
щих свет кристалликов различной формы (приз-
мы, иголочки, друзы, цилиндры и др., рис. 503).
Они состоят из фосфорнокислой извести. Выводятся
они из организма при посредстве сократительной
вакуоли.
Размножение. В эндоплазме расположены
ядра парамеции — макронуклеус и микронуклеус.
Большой яйцевидный макронуклеус имеет сет-
чатую структуру. В ячейках сети находится боль-
шое количество хроматиновых зернышек. В углуб-
лении макронуклеуса лежит небольшой микрону-
клеус. В нем различают компактное внутриядер-
ное тельце (кариосому), состоящее из хроматина,
периферическую светлую зону и гомогенное ахро-
матиновое тельце.
Парамеции, как и все инфузории, размножа-
ются при помощи поперечного деления (рис. 52).
Оно начинается с кариокинетического деле-
ния микронуклеуса. Одновременно начинает де-
литься макронуклеус. Он вытягивается и затем
амитотически перешнуровывается. Тело парамеции
также делится путем простой перетяжки. При этом
органеллы (цитостом, ундулирующая мембрана,
сократительные вакуоли) распределяются между
дочерними особями, а недостающие образуются
заново.
Paramaecium caudatum делится при 15—17°С
один раз в сутки. Повышение температуры уско-
ряет ритм деления.
Время от времени период вегетативного раз-
множения прерывается у парамеций половым про-
цессом — конъюгацией, которая состоит в том, что две особи взаимно опло-
дотворяются (рис. 53). При этом происходит временное соединение двух
особей, во время которого они обмениваются частями своего ядерного
аппарата.
Конъюгация начинается с того, что две парамеции плотно приклады-
ваются друг к другу брюшными сторонами рот ко рту. Пелликула их в это
время несколько набухает, становится клейкой, и протоплазма обеих осо-
бей фактически сливается.
Амикронуклеусы обеих инфузорий 2 раза митотически делятся. Одно из
этих делений редукционное. Во время него хромосомы, не делясь, расхо-
дятся в дочерние ядра. Таким образом, в каждой из них оказывается поло-
винное (гаплоидное) число хромосом. Три микронуклеуса растворяются, а
четвертый, лежащий ближе ко рту, делится еще раз, образуя мужское.
блуждаюицее, и женское, стационарное, ядра. Затем обе инфузории обмени-
Рис. 52. Срез через де-
лящуюся Paramaecium
caudatum:
1 — разделившиеся микрону-
клеусы, 2—делящийся мак-
ронуклеус , 3 — соединитель-
ный тяж между разделивши-
мися микронуклеусами, •/ —
пищеварительные вакуоли,
б— трихоцисты, 6 — начало
образования перетяжки
81
ваются блуждающими ядрами, которые сливаются со стационарными ядрами
другой особи. В результате в ядре восстанавливается диплоидное число
хромосом. Этот процесс называется кариогамией. Вслед за образованием
ядра, получившегося в результате кариогамии и называемого синкарион,
наступает расхождение конъюгантов.
Конъюгация у Paramaecium caudatum длится 12 ч (от момента спарива-
ния двух особей до момента их расхождения).
у 5 б
7 2 3
Рис. 53. Конъюгация у Paramaecium caudatum (схема):
1 — 4 — первое деление мнкронуклеуса, 5—6—второе и третье деление
микронуклеуса, 7—8 — обмен ядрами и кариогамия, 9—13 — три деления
синкариона, 14—16 — два последующих деления эксконъюгантов
В разошедшихся особях, называемых эксконъюгантами, начинается
распад микронуклеуса, и он постепенно растворяется и исчезает. Синкарион
каждой из парамеций делится вновь 3 раза. Из получившихся 8 ядер 3 рас-
творяются, 1 является новым микронуклеусом, а 4 — закладками новых
макронуклеусов. При двух последующих делениях микронуклеус делится
обычным образом, а макронуклеусы, не делясь, расходятся в дочерние особи.
Только после этого в каждом эксконъюганте оказывается нормальный ядер-
ный аппарат.
Прежде предполагали, что если инфузории не имеют возможности
время от времени конъюгировать, то после большего или меньшего числа
обычных делений они обнаруживают признаки старения, дегенерируют и
погибают. Считали, что конъюгация оказывает омолаживающее действие
на культуру парамеций и стимулирует ее жизнедеятельность.
Теперь выяснено, что гибель парамеций, размножающихся только
бесполым путем, вызывалась не внутренними причинами, а условиями куль-
тивирования, главным образом накоплением в культурах ядовитых про-
дуктов обмена инфузорий. Если применять хорошие методы культивирова-
82
ния, то можно не давать парамециям возможности конъюгировать в течение
многих тысяч поколений, и при этом у них не будет никаких признаков
депрессии.
У Paramaecium caudatum через каждые 50—60 дней наступает перио-
дическая реорганизация ядерного аппарата без конъюгации. При этом не
происходит соединения инфузорий в парочки, а в каждой отдельной инфу-
зории макронуклеус распадается и затем его части растворяются; микро-
нуклеус делится 2 раза. Из образовавшихся 4 ядер 3 растворяются, а чет-
вертое делится еще 1 раз. Образовавшиеся 2 ядра сливаются, в результате
чего получается синкарион. Этот процесс называется аутогамией. Он отли-
чается от конъюгации тем, что вместо перекрестного оплодотворения про-
исходит самооплодотворение. Синкарион делится, из одной части образует-
ся микронуклеус, из другой макронуклеус.
Аутогамию удобнее всего наблюдать при ежедневном отсаживании
парамеций поодиночке, вследствие чего не может наступить конъюгация.
Парамеции живут в пресной воде. В небольших, особенно в загниваю-
щих водоемах (лужах, небольших прудах и т. д.) они встречаются в больших
количествах, так как здесь много бактерий, которые служат парамециям
пищей.
При наступлении неблагоприятных условий — высыхании, накоплении
продуктов обмена во внешней среде — парамеции могут инцистироваться.
Род Paramaecium включает несколько сходных видов. Из них особый
интерес представляет Р. bursaria. Она меньше, но относительно шире и
площе Р. caudatum и кажется окрашенной в зеленый цвет. При более
близком рассмотрении оказывается, что вся плазма Р. bursaria заполнена
небольшими зелеными тельцами. Эти тельца — микроскопически малые
водоросли — зоохлореллы, содержащие зеленый хроматофор. Но это не
заглоченная инфузорией пища — здесь мы имеем дело с внутриклеточным
симбиозом, т. е. с таким сожительством простейшего и водоросли, которое
выгодно обоим сожителям. Зоохлореллы получают все необходимые вещества
из плазмы инфузории и отдают ей кислород, вырабатываемый ими в процессе
фотосинтеза. Кроме того, в плазму инфузории поступает избыток синтези-
рованного водорослями углевода. Р. bursaria, в плазме которой имеются зо-
охлореллы, может жить и размножаться в чисто неорганической среде, не
получая извне никакой органической пищи. Зоохлореллы встречаются в
плазме некоторых амеб, солнечников и инфузорий,
СИСТСМАТНЧБСКНП ОБЗОР КЛАССА
ПОДКЛАСС I. ПРОТОЦИЛИАТЫ (PROTOC1LIATA)1
В состав этого подкласса, включающего одно семейство опал ин ид
(Opalinidae), входят инфузории, паразитирующие главным образом в ки-
шечнике бесхвостых амфибий. Встречаются они также у некоторых змей
и ящериц.
Опалиниды сходны с остальными инфузориями в том, что покрыты
ресничками, но отличаются от них целым рядом существенных признаков.
У них нет ядерного диморфизма. Опалиниды имеют два или много оди-
наковых ядер и в этом отношении примитивнее остальных инфузорий.
У них отсутствует характерная для остальных инфузорий конъюгация,
а имеется копуляция. Цикл развития опалины (Opalina ranarum) — очень
1 В последнее время ряд исследователей, основываясь на том, что в противопо-
ложность другим ресничным инфузориям у протоцилиат нет разделения ядерного ап-
парата на макропуклеус и микронуклеус и половой процесс происходит по типу копу-
ляции, а не конъюгации, считает, что опалины представляют особый отряд жгутико-
вых (Opalinina)
83
распространенного, большого, многоядерного паразита полости прямой
кишки лягушки (рис. 54), происходит следующим образом. В течение всего
года, кроме весны, происходит бесполое размножение опалин при помощи
Рис. 54. Opalina гапагшп. / — целое животное с
многочисленными пузырьковидными ядрами;
// —деление опалины на две одинаковые особи
деления надвое. Весной, ко
времени откладки лягушкой
икры, часть опалин начинает
быстро делиться, и в резуль-
тате нескольких следующих
одно за другим делений обра-
зуются маленькие особи, со-
держащие от 3 до 12 ядер.
Эти формы инцистируются, и
цисты с калом выбрасываются
в воду. Здесь их заглатывают
молодые головастики, в киш-
ках которых опалины осво-
бождаются от цист. Часть
опалин вырастает до обыкно-
венных форм, другая же часть
делится и образует одноядер-
ные большие широкие макро-
гаметы и маленькие узкие
микрогаметы; происходитiо
Рис. 55. Цикл Opalina ranarum:
/ — деление, 2 — циста, 3 — гаметы. 4 — копуляция, 5 — инцистиро-
ванная зигота
пуляция. Образовавшаяся зигота вновь инцистируется, выбрасывается в
воду и заглатывается другим головастиком, в кишке которого освобожда-
ется от цисты и вырастает в обычную опалину (рис. 55).
ПОДКЛАСС II. СОБСТВЕННО ИНФУЗОРИИ (EUCILIATA)
В этот подкласс объединены типичные инфузории с ясно выраженнььм
ядерным диморфизмом. Половой процесс — конъюгация. В состав подкласса
входит пять отрядов: 1) равноресничные (Holotricha), 2) разноресничные
(Heterotricha), 3) малоресничные (Oligotricha), 4) брюхоресничные (Нуро-
tricha) и 5) кругоресничные (Peritricha).
84
ОТРЯД РАВНОРЕСНИЧНЫЕ (HOLOTR1CHA)
Равноресничные инфузории равномерно покрыты сравнительно ко-
роткими ресничками, только около рта могут иметься несколько более длин-
ные реснички, никогда не образующие спирали (рис. 56).
У некоторых форм вместо равномерного ресничного покрова могут
быть один или несколько поясов одинаковых ресничек.
К этому отряду принадлежит много обычных родов нашей пресновод-
ной фауны: трахелиус (Trachelitis) с характерным палочковым аппаратом,
Рис. 56. Равноресничные инфузории. / — Lacrimaria sp.; // —
Ciclidium glaucoma; III — Lionotus fasciola; IV — Holophrya dis-
color; V — Trachelius ovum; VI — Ichtyphthirius multifilis
поддерживающим глотку; дидиниум (Didinium) — небольшие хищные ин-
фузории, покрытые рядами ресничек; колепс (Colcps), покрытый панцирем,
и многие другие.
Как и большинство других инфузорий, равпорссничные питаются бак-
териями, но среди них встречаются паразиты, а также хищники, питающиеся
главным образом инфузориями. Так, представители рода Didinium нападают
преимущественно на парамеций, представители рода Coleps, обладающие
растягивающимся ротовым отверстием, могут заглатывать сравнительно
больших инфузорий (рис. 57).
К этому же отряду относится ихтиофтириус — Ichtyophthirius multi-
filis, довольно опасный паразит жабр и кожи различных рыб (рис. 56vi).
Это большие инфузории, достигающие 0,5—0,8 мм длины с большим под-
кововидным макронуклеусом и многочисленными мелкими сократительными
вакуолями. Ихтиофтириусы залегают в кожном эпителии рыб, образуя не-
большие ранки, которые у сильно зараженных рыб покрывают все тело в
виде маленьких беловатых пятнышек. Для размножения инфузория выходит
из кожи рыбы в воду. Здесь она образует нежную студенистую цисту и в ней
делится много раз. В цисте может быть 256 и больше особей. Покинув
цисту, молодые инфузории вбуравливаются в кожу других рыб. Заражение
I. multifilis особенно опасно для молоди рыб (карпов, форели), которая
гибнет от них в больших количествах.
85
Большой вред прудовому рыбоводству наносит равноресничная ин-
фузория Chilodonella cyprini. Она вонзается палочковым аппаратом глотки
в эпителиальные клетки кожи и жабр рыб, главным образом карпа и сазана,
Рис. 57. Инфузории-хищники: / — VI — Didinium nasutum, заглатываю-
щая парамецию; VII —Spathidlium spatula, заглатывающая Colpidiurn
colpoda; VIII — четыре экземпляра Coleps hirtus, напавшие на другую
инфузорию
и повреждает их. Ch. cyrpini появляется в большом количестве в зимоваль-
ных прудах в конце зимы. Молодая рыба, особенно если она выращивалась
предшествующим летом в недостаточно благоприятных условиях, гибнет в
массовом количестве.
86
ОТРЯД РАЗНОРЕСНИЧНЫЕ (НЕТЕ ROTRICH А)
Разноресничные инфузории снабжены зоной мембранелл, идущей от
переднего конца ко рту, заворачивающей вправо и называемой адоральной
зо-юй мембранелл. Мембранеллами называются тонкие пластинки, со стоящие
из 2—4 рядов склеенных между собой ресничек. Все остальное тело равно-
мерно покрыто ресничками. Представители этого отряда также обычные
обитатели наших пресных водоемов.
В гниющей болотной воде часто можно встретить в очень больших коли-
чествах длинную червеобразную Spirostomum (рис. 58ц). Эта инфузория
Рис. 58. Разпоресничные инфузории. / —Stentor polymorphic; // —Si-
roMomum ambiguum; /// — Nycthoterus ovalis; IV — Balantidium
coli:
1— UAQpcWiPHdfl иона мембране.').'*, 2— передний приводящий капал, — пульсирую-
щая нзкуоль. 4 — задний приводящий канал. 5 — макронуклеус, б — микронуклеус.
7 —глотка, М — лобное поле, б — цитостом
достигает 2 мм длины и хорошо видна простым глазом. Адоральная чсна
мембранелл начинается у переднего конца и заканчивается в первой трети
тела. У S. arnbiguum (наиболее обычный вид) длинный четко видный макро-
нуклеус и несколько микронуклеусов. Большая сократительная вакуоль
занимает почти весь задний конец тела, который кажется поэтому светлым.
Один приводящий канал проходит почти по всему телу инфузории. Так же
часто встречаются разные виды Stentor (рис. 58] ). Это большие, свыше 0,5 мм
длиной, воронкообразные инфузории с узким задним концом. Адоральная
зона мембранелл огибает широкое перистомное поле, расположенное на
переднем конце.
Spirostomum и Stentor снабжены расположенными в пелликуле мускуль-
ными фибриллами (мионемами), при помощи которых они могут быстро со-
кращаться.
Стенторы часто прикрепляются к субстрату. Довольно часто встречаются
Bursaria truncatella — большая (до 1 мм) инфузория с очень сложным пе-
ристомом.
Большой стентор с длинным ядром, располагающимся почти по всей
длине тела — удобный объект для опытов по регенерации. Очень многие
простейшие способны восстанавливать утраченные части тела, но при не-
87
прехменном условии, чтобы в отрезанном кусочке, кроме протоплазмы, был
и участок ядра. Отрезанный участок протоплазмы без ядра погибает. Стен-
тора довольно легко разрезать на несколько частей при помощи маленького
скальпеля, который можно изготовить, наточив обыкновенную препаро-
вальную иглу. Уже через 2—3 ч заметно, что бесформенные вначале кусочки
начинают «залечивать раны», регенерировать недостающие органы и быстро
принимать форму нормального стентора только меньшей величины (рис. 59).
Отрезанный задний участок восстанавливает передний конец с адоральной
зоной мембранелл и перистомом, средний участок восстанавливает передний
и задний концы тела и т. д. Обычно весь процесс формирования нормального
животного из отрезанного кусочка протоплазмы с ядром продолжается
около 24 ч.
К этому отряду относится и ряд паразитических форм. В первую оче-
редь балантидий (Balantidium coli; рис. 58iv) — опасный паразит кишечника
человека, вызывающий хронические
поносы. Овальная инфузория 50—
80 мк в длину и 40—60 мк в ширину.
На переднем конце находится перис-
том, по спинной стенке которого рас-
положена адоральная зона мембра-
нелл. В центре инфузории расположен
овальный или бобовидный макронук-
леус, в выемке которого лежит ми-
кронуклеус. Имеются две сократи-
тельные вакуоли. Балантидий пара-
зитирует в толстых кишках, иногда
проникает в стенку кишки, разруша-
ет окружающие ткани и вызывает
образование язв.
В. coli инцистируется в кишечни-
ке. Инфузория теряет реснички и
окружается плотной двуслойной обо-
лочкой. Овальные или шаровидные
цисты имеют по сравнению с цистами
других простейших очень большой размер; они достигают 50—60 мк в диа-
метре. Заражение происходит через заглатывание цист. Некоторые авторы
считают, что В. coli идентичен с В. suis свиней и что свинья служит источ-
ником заражения человека.
Помимо балантидия в кишечнике человека изредка встречается Nyctho-
terus faba — бобовидная инфузория, передний конец которой загнут на
брюшную сторону. Щелевидный перистом достигает середины тела. Длина
26—28 мк, ширина 16—18 мк, Другой представитель этого рода N. ovalis
(рис. 58ш ) живет в кишечнике черного таракана, N. cordiformis — у лягу-
шек и жаб.
ОТРЯД МАЛОРЕСНИЧНЫЕ (OLIGOTRICHA)
Ресничный покров этих инфузорий сильно редуцирован, так что у не-
которых видов сохранилась только завернутая вправо адоральная зона мемб-
ранелл. У других видов, кроме адоральной зоны, имеются еще другие пояса
мембранелл. В состав этого отряда, кроме некоторых пресноводных форм и
почти исключительно морских планктических тинтиннид (Tintinnidae),
имеющих студневидные или псевдохитиновые домики, входят своебразные
инфузории, обитающие в желудке млекопитающих — офриосколециды (Oph-
ryoscolecidae) и в толстых кишках непарнокопытных — циклопостииды
(Cyclopostiidae). Они широко распространены у жвачных животных, у кото-
рых живут в рубце и сетке. Форма этих инфузорий часто весьма сложна
(рис. 60). У них обычно встречаются ряд сложно устроенных скелетных и
88
сократимых элементов. В. А. Догель нашел, что у некоторых малоресничных
(Opistotrichium janus) конъюгируют инфузории различной величины (Q —
макрсконъюганты ид — микроконъюганты). Таким образом, у некоторых
малоресничных инфузорий имеется
анизогамная конъюгация. Конъюги-
рующие инфузории сильно отлича-
лись друг от друга по ряду деталей
(форма панциря, скелет и т. д.).
При описании конъюгации у па-
рамеций было указано, что стационар-
ное (Q) и блуждающее (д) ядра
морфологически друг от друга почти
не отличаются. Догель показал, что у
цпклопостиид иногда имеется разница
в форме этих ядер, например у Cyc-
lopostium bipalmatum блуждающее
ядро похоже на сперматозоид (рис.
61). Это сходство тем более разитель-
но, что при конъюгации С. bipalma-
tum прикладываются друг к другу
ротовыми сторонами таким образом,
что между ними остается некоторое
пространство, отграниченное нескле-
енными между собой краями тела.
При конъюгации блуждающие ядра,
переходя из одной инфузории в дру-
гую, некоторую часть своего пути
проходят во внешней среде, проника-
ют через рот и глотку и там сливаются
I л
Рис. 60. Малоресничные инфузории.
/ — Ophryoscolex caudatus со спинкой
стороны; II — О. purkinjei со спинной
стороны:
1 — адоральные мембранеллы, 2 — цитофаринкс,
3—5 — задний пояс ресничек, !б — макронукле-
ус, 7—мнкронуклеус, 8 — пульсирующие ва-
с округлым неподвижным стационар-
куоли
НЕМ ЯДрОМ.
Количество офриосколецид у жвачных животных очень велико. По
подсчетам Догеля, у быка их приходится иногда до 2 млн. на 1 см3 содер-
жимого рубца.
Рис. 61. Две стадии конъюгации инфузории Cyclopostium bi-
palmatum. I —деление микронуклеуса на стационарное ($) и
блуждающее (<5 ) ядра; // — переход блуждающего ядра:
1— макронуклеус
89
ОТРЯД БРЮХОРЕСНИЧНЫЕ (НУРОТЦ1СНА)
Брюхоресничные инфузории сильно уплощены в дорзовентральном на-
правлении. На спинной стороне ресничный покров редуцирован. Вместо
ресничек имеются длинные тонкие Нити. На брюшной стороне реснички
расположены в несколько рядов.
Кроме ресничек, на брюшной сто-
роне имеются также группы плот-
ных щетинок — цирр, на которые
инфузории опираются при перед-
вижении по твердому субстрату.
Адоральная зона мембранелл
расположена на брюшной стороне.
Она огибает передний край тела,
идет назад и заканчивается у рта,
находящегося в передней части
тела.
На краю перрстомного поля
расположена ундулирующая мем-
брана.
К брюхоресничным (рис. 62)
относится ряд родов, широко рас-
пространенных в пресной воде и
море. Часто встречаются в наших
водоемах стилонихия (Stylonichia;
Рис. 62. Брюхоресничные инфузории.
/ — Uroleptus pisces; // — Histrio peli-
onella; /// —Oxytricha fallax; IV —
Euplotes charon; V — Kerona
Рис. 63. Stylonichia mytilus, опирающаяся циррами на субстрат
рис. 63), большая инфузория длиной 300 мк; эуплотес (Euplotes); керона
(Kerona), живущая на поверхности тела гидры, и ряд других.
ОТРЯД КРУГОРЕСНИЧНЫЕ (PERITRICHA)
У инфузорий, объединяемых в этот отряд, крупные мембранеллы обра-
зуют на переднем конце тела адоральную зону, состоящую из двух рядов и
спирально завернутую налево (в противоположность всем остальным отря-
дам, у которых она завернута вправо). Все остальное тело голое или снабже-
но на заднем конце поясом ресничек. Большинство кругоресничных инфу-
зорий — сидячие формы.
Половой процесс у всех кругоресничных — анизогамная конъюгация.
Макроконъюганты ( Q) неотличимы от обычных инфузорий, микроконъю-
ганты (6) значительно меньше и образуются в результате нескольких
делений, быстро следующих друг за другом и не сопровождающихся ростом
90
дочерних особей. Микроконъюгант отрывается от стебелька, образует на
заднем конце второй пояс ресничек и начинает свободно плавать.
Через некоторое время он отыскивает макроконъюганта и приклады-
вается к нему боковой стороной, после чего весь ядерный аппарат и часть
протоплазмы переливаются из микроконъюганта в макроконъюгант, а ос-
тавшаяся небольшая часть гибнет. Таким образом, здесь имеется не взаим-
ное оплодотворение, что характерно для инфузорий, а оплодотворение боль-
шего конъюганта меньшим.
Кругоресничные снабжены сократимым или несократимым стебельком.
Часто образуют большие колонии. В наших пресных водоемах обычны су-
войка (Vorticella) — одиночные формы на сократимом стебельке (рис. 64 и
65); кархезиум (Carchesium polypinum), образующий большие колонии на
Рис. 64. Сувойка Vorticella nebtili-
f era;
1 — M.'Kponviwicyc. 2— клеточная глотка
(цптофлрнпке), клеточный poi (цито-
стом). 4 — микронуклсус, 5 — адоральная
зона мембранелл, 6 — край перистома, 7—
ундулнрующая мембрана, <9—резервуар,
9 - пульсирующая вакуоль. 10 — пищен рн-
тильные иакуолн, 11— мионемы, Г2— MIIO-
нема в стебельке
Рис. 65. Сувоикн:
1— сократившаяся еупойкл, 2 — мнкрпкопъ-
югаит, J—i — деление, й — конъюгация
сократимых стебельках, прикрепляющихся к водным растениям, обломкам
дерева и т. д., кампанелла (Campanella umbcllaria), образующая древовид-
ные колонии на несократимых стебельках, и ряд других.
КЛАСС 2, СОСУЩИЕ ИНФУЗОРИИ (SUCTORIA)
ХА РАКТЕРИСТИКА
Сосущие инфузории покрыты ресничками только на ранних стадиях
развития. Во взрослом состоянии они не имеют никаких органелл движения,
и большинство видов при помощи стебелька прикрепляется к мертвому суб-
страту или к водным растениям и животным.
Сосущие инфузории снабжены сосательными палочками (рис. 66i ), ко-
торые представляют собой тонкие сократимые трубочки, полость которых
с одной стороны открывается во внешнюю среду, а с другой — сообщается
91
с эндоплазмой. Сосущие инфузории питаются главным образом ресничными
инфузориями. Если какая-либо ресничная инфузория соприкоснется с со-
сательной палочкой, то последняя немедленно прикрепляется к ней. К до-
быче наклоняются несколько соседних палочек, пелликула ресничной ин-
фузории в местах прикрепления сосательных палочек растворяется, и вся
Рис. 66. Сосущие инфузории. / — Ephelota gemmipara с вытянутыми со-
сательными и хватательными палочками; II —Sphaerophrya magna, схватив-
шая и высасывающая пять инфузорий (Colpoda parviformis и Cyclidium):
1 — сосательные палочки, 2— хватательные палочки
протоплазма переливается в протоплазму сосущей инфузории. У некоторых
форм, кроме сосательных, имеются хватательные палочки, которые толь-
ко захватывают и убивают добычу, но не высасывают ее. Имеются
макро- и микронуклеус и сократительные
Рис. 67. Образование бродяжки сосущей инфу-
зории Tocophrya: /—IV — последовательные
стадии, V — бродяжка:
/ — пульсирующая вакуоль, 2 — бродяжка, 3—макронук-
леус, 4— стебелек, 5—сосательные палочки
вакуоли.
Конъюгация в основных
чертах протекает так же, как
и у остальных инфузорий.
Суктории размножаются
при помощи деления и наруж-
ного или внутреннего почко-
вания.
При наружном почкова-
нии, например у Paracineta,
ядра инфузории делятся,
верхняя часть животного оде-
вается ресничками, и в нее
врастают макро- и микронук-
леус; затем почка отделяется
от материнского оранизма и
уплывает. Образовавшийся
таким путем организм обыч-
но называют бродяжкой.
При внутреннем почко-
вании в теле инфузории об-
разуется кольцевая впадина,
которая постепенно углубля-
ется и как бы вырезает учас-
ток протоплазмы. Затем в
этом участке образуются со-
кратительная вакуоль и ряды
ресничек, а из материнского
организма вырастает ядро
(рис. 67).
Б родяжка, образовавша-
яся таким образом, выходит
92
из материнского организма, некоторое время свободно плавает, затем при-
крепляется к субстрату, теряет реснички, образует стебелек, сосательные
палочки и превращается во взрослую инфузорию. У некоторых сосущих
инфузорий наблюдается множественное почкование.
Suctoria — главным образом сидячие формы, снабженные стебельками.
Живут в пресных водоемах и морях. Некоторые формы прикрепляются к
определенным животным, например Tocophrya cyclopum — к рачкам цик-
лопам. Некоторые являются паразитами других инфузорий, например Spha-
erophrya stentoris. Свободноплавающие почки этой суктории прикрепляются
к инфузориям, проникают внутрь тела, размножаются там делением, затем
в свою очередь начинают образовывать покрытые ресничками почки, которые
выходят и внедряются в других инфузорий.
Наличие ресничек у молодых форм Suctoria указывает на происхожде-
ние их от инфузорий, покрытых ресничками всю жизнь. Потеря ресничек
взрослыми формами, по-видимому, связана с переходом сосущих инфузорий
к сидячему образу жизни.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
И экология ПРОСТЕЙШИХ
Простейшие живут всюду, где есть влажная среда. В морях, реках,
озерах, болотах, в лужах, оставшихся после дождя, во влажной земле и
мху — всюду можно найти простейших; они чаще встречаются в стоячей
воде — болотах, прудах, озерах, — чем в проточной. Большая группа про-
стейших паразитирует у различных одноклеточных и в клетках, кровяной
плазме, содержимом кишечника многоклеточных.
При неблагоприятных условиях существования простейшие обычно
инцистируются. Инцистирование наступает при неблагоприятной темпера-
туре, высыхании, недостатке кислорода в среде обитания, накопления в
ней вредных продуктов обмена и главным образом при недостатке пищи.
Они отбрасывают жгутики, реснички, втягивают псевдоподии, принимают
шарообразную форму и выделяют на своей поверхности одну или две плот-
ные оболочки. Цисты могут длительно оставаться живыми в сухом состоя-
нии, при низких температурах и т. п. Например, цисты инфузории Colpoda
остаются живыми после того, как они пролежали в сухом состоянии 14—16
месяцев, цисты инфузории Gastrostyla переносят двухлетнее пребывание в
сухом месте, цисты жгутикового Eicomonas termo — 5—5 х/2 лет, а цисты
Peridinium cinctum — даже 161/алет. Когда цисты попадают во влажное
место, они раскрываются, и простейшие переходят к деятельному образу
жизни.
Процесс инцистировання представляет собой явление адаптации —
приспособления к неблагоприятным условиям жизни —• и служит для про-
стейших мощным средством борьбы за существование. Инцистируясь, жи-
вотное защищается оболочкой цисты от вредных для него воздействий внеш-
ней среды.
Ветер подхватывает эти микроскопические цисты и может отнести их
на довольно большое расстояние. Кроме того, они прикрепляются к водо-
плавающим птицам, к водным насекомым, которые также переносят их с
места на место.
Если положить в банку сена, соломы или травы, налить туда воды и
оставить ее открытой, то через несколько часов или дней там будет много
различных простейших. Часть их выйдет из цист, которые были прикреплены
к сену, а часть цист попадает туда из воздуха.
Большинство пресноводных простейших имеют очень широкое геогра-
фическое распространение. Много среди них и космополитов.
Одинаковых простейших можно встретить под Москвой, в Сиби-
ри, в Индии, в Африке и в Америке. Даже большинство простейших,
93
найденных на островах Атлантического океана, лежащих далеко от мате-
риков, те же самые, какие живут повсюду. Такое широкое распространение
простейших объясняется тем, что их цисты очень легко переносятся на до-
вольно большие расстояния, и тем, что они питаются очень разнообразной
пищей, которую можно всюду найти — бактериями, одноклеточными водо-
рослями, другими простейшими, детритом и т. д.
Но, с другой стороны, фауна простейших в пресных водоемах, лежащих
недалеко друг от друга, может быть различной. Это зависит от того, какие
соли и в каком количестве содержатся в воде, каково в ней содержание орга-
нических веществ, кислорода, какова активная кислотность или щелочность
воды, наконец, как освещается данный водоем. Отличия в содержании солей,
количестве органических веществ, содержании кислорода в водоемах носят
местный, локальный, характер. Эти условия могут быть различными в рядом
лежащих водоемах и одинаковыми в водоемах различных частей света.
Иначе обстоит дело с морскими простейшими: между разными морями
существуют постоянные различия в температуре, иногда и в солености,
поэтому и фауна простейших там в основном разная. Морские лучевики,
например, являются планктическими животными, т. е. они свободно плавают
в верхних слоях воды; множество отростков увеличивает поверхность их
тела и этим повышает способность к парению. Это же относится и к ряду
панцирных жгутиковых. Оказывается, что у видов, живущих в теплых морях,
где вода менее плотная, отростки, а тем самым и поверхность относительно
объема тела больше, чем у видов, обитающих в холодных морях, в которых
вода плотней.
Некоторые отряды простейших живут или только в морях, или только
в пресной воде: фораминиферы, за немногими исключениями, населяют
только моря с достаточной соленостью. В Аральском и Каспийском морях
встречаются только единичные виды; несколько больше их в Черном море.
Обильная фауна фораминифер имеется в наших северных и дальневосточ-
ных морях. Лучевики также только морские животные, причем они обитают
в морских водоемах с большой соленостью. В Аральском, Каспийском и
Черном морях их нет совсем. Кроме того, они избегают холодной воды;
поэтому в наших северных морях их очень немного, а в южных дальнево-
сточных встречается довольно большое количество видов. Почти все солнеч-
ники, наоборот, обитатели пресных водоемов.
Географическое распространение паразитических простейших зависит
от распространения хозяина. Виды, которые для своего развития нуждаются
в переносчике или промежуточном хозяине, встречаются только там, где
есть и хозяин и переносчик. Некоторые паразитические простейшие распро-
странены по всему миру, например кишечные жгутиковые и кишечные амебы
человека (включая Entamoeba histolytica). Но распространение некоторых
видов в высокой степени зависит от климатических, в первую очередь темпе-
ратурных, условий. Так, например, малярийные паразиты человека могут
развиваться в теле комара только при условии, если температура окружаю-
щего воздуха будет не ниже + 15°С.
Условия, в которых живут простейшие в пресных водоемах, чрезвычайно
разнообразны. Как уже было указано, особенно много простейших в мел-
ких водоемах — прудах, болотах, лужах и т. д. В быстро текущих реках,
очень чистых прудах их мало. То, что мы называем «пресной водой», в мелких
водоемах часто представляет собой растворы, сильно разнящиеся друг от
друга. Почва определяет собой содержание различных солей в водоемах
(сульфатов, карбонатов, хлоридов и т.п.). Содержание нитратов, фосфатов,
углекислоты, органических соединений в общем зависит от разложения
органических веществ в водоеме. Различно также содержание кислорода.
Большое значение для простейших имеет активная кислотность или щелоч-
ность воды (концентрация водородных ионов). Большое влияние оказывает
также свет. Понятно, что для простейших, содержащих хлорофилл, послед-
ний фактор имеет первенствующее значение. Температура воды, которая так
94
сильно меняется в течение года в пресных водоемах, — также один из важ-
нейших факторов для жизни простейших и приспособления их к условиям
окружающей среды. Простейшие весьма чувствительны к этим разнообраз-
ным условиям. При-описании жгутиковых мы уже упоминали о различных
типах питания у простейших. Различна потребность в кислороде у разных
видов. Инфузория Spirostomum живет обычно в слегка щелочной среде,
жгутиковые Polytoma, Astasia — в кислой. Есть также холодо- и теплолю-
бивые виды.
Большое влияние на динамику фауны водоема оказывают взаимоотно-
шения организмов между собой. Например, инфузории Didinium и Coleps
питаются парамециями и обычно очень сильно снижают численность их в
водоеме.
Все эти факторы среды, с одной стороны, и приспособления различных
простейших к определенным условиям среды, с другой, определяют состав
фауны простейших в отдельных водоемах. Этим пользуются для так называ-
емой биологической характеристики воды в санитарных целях. Чистота
воды и пригодность ее для питья зависят в основном от содержания органи-
ческих соединений. Так как различные простейшие приспособлены к жизни
в воде с определенным содержанием этих веществ, то были выделены группы
видов, нахождение которых может характеризовать ту или другую степень
загрязнения воды. Так, например, в сильно загрязненной, гниющей воде
живут Euglena viridis, Colpidium colpoda, Paramaecium putrinum, Vorti-
cella microstoma и многие другие. В более чистой воде, в которой наряду с
расщеплением органических веществ идет энергичное окисление, живут
Cryptomonas, Chlamydomonas, Actinophrys sol, Actinosphaerium eichhor-
nii, Paramaecium caudatum, Stentor, Spirostomum и др.; для воды с малым
содержанием органических веществ и большим количеством минеральных
характерны: Volvox, Gonium, Eudorina, Amoeba proteus, Lacrymaria и др.
В пресной воде, так же как и в море, имеются донные (бентические) и
планктические, т. е. свободноплавающие в толще воды, формы.
Принадлежность к той или иной экологической группе можно часто
различить уже по строению животных, так как у них развиваются соответ-
ствующие приспособления.
Планктические виды, к которым принадлежат главным образом мор-
ские панцирные жгутиковые, хризомонады, лучевики, некоторые форамини-
феры, пресноводные лучевики и многие жгутиковые, обладают обычно боль-
шой поверхностью тела, различными плавательными отростками, и скелет
их не компактен (например, решетчатый скелет лучевиков). Донные же
формы, к которым принадлежат, например, большинство морских фора-
мпнифср и пресноводные раковинные амебы, обладают плотной раковиной.
Большое количество простейших живет в почве либо в инцистироваи-
ном состоянии, либо в свободножнвущем (там, где почва более влажная).
Паразитизм широко распространен среди простейших. Нет ни одного
класса, среди представителей которого не было бы паразитов. Из тридцати
отрядов простейших десять включают в себя только паразитов, в семнадцати
есть различное количество паразитов и только в трех отрядах (Chrysomo-
nadina, Foraminifera, Radiolaria) до сих пор не найдены паразитические
представители.
В одном хозяине может жить много различных видов паразитических
простейших. Так, например, в человеке паразитирует 25 видов.
По расположению на хозяине различают эктопаразитов, живущих на
поверхности тела хозяина, и эндопаразитов, живущих внутри него. При-
мером первых может служить Costia necatrix, паразитирующая на коже
рыб, ко вторым относятся трипанозомы, миксоспоридии, кокцидии и др.
Эндопаразитов по образу жизни можно разделить на три группы: 1) живущие
в полостях тела, наполненных жидкостью (кишечник, выделительная си-
стема, полость тела и кровеносная система); к ним относятся уже знакомые
нам кишечные простейшие трипанозомы и многие другие; 2) межклеточные,
95
или тканевые, паразиты, как, например, дизентерийная амеба, инфузория
Balantidium coli, проникающие через стенку кишечника и живущие в тка-
нях, затем саркоспоридии, миксоспоридии; 3) внутриклеточные паразиты,
к которым относятся малярийный плазмодий, кокцидии, отчасти грегарины
и др.
По взаимоотношению с хозяином среди паразитических простейших
следует различать комменсалистов, симбионтов и собственно паразитов.
Комменсалистами называются такие формы, которые хотя и живут в каком-
либо хозяине, но не причиняют ему вреда (непатогенны) и питаются только
остатками пищи или другими организмами (например, бактериями), живу-
щими в том же хозяине. К ним относится большинство жгутиковых и амеб,
живущих в кишечнике высших животных и человека.
При симбиозе и простейшее, и хозяин, в котором оно живет, получают
выгоды от совместного жительства. Например, симбиоз жгутиковых и
термитов. Многие виды термитов питаются древесиной. В кишечнике этих
видов всегда живет большое количество жгутиковых, которые питаются
этой древесиной. Оказывается, что термиты сами переварить древесину не
могут и если удалить из них жгутиковых и кормить термитов только древе-
синой, то они погибнут от голода. Таким образом, жгутиковые, переваривая
древесину, переводят ее в усвояемое для своего хозяина состояние и, по-
гибая, доставляют ему обильную пищу.
Настоящие паразиты живут за счет своего хозяина и питаются соками
его тела, клетками или переваренной им пищей и приносят ему этим часто
большой вред, вызывая различные заболевания, а иногда и смерть хозяина.
На паразитических простейших образ жизни наложил определенный
отпечаток. Они эволюционировали в сторону приспособления к специфиче-
ским, своеобразным условиям среды, в которой они обитают.
Приспособительные изменения привели у различных видов, с одной
стороны, к потере, а с другой, — к приобретению ряда признаков, и в
этом смысле изменения простейших аналогичны тем изменениям, которые
мы находим у паразитических многоклеточных.
У тех форм, которые живут в вязкой среде полостей тела (кровь, со-
держимое кишечника и т. п.), сильно развивается двигательный аппарат.
У паразитических жгутиковых имеется такая мощная органелла движе-
ния, как ундулирующая мембрана. Среди свободноживущих до сих пор
известен только один вид морских жгутиковых, снабженных ундулирующими
мембранами.
Интересно, что когда трипанозомы из вязкой плазмы крови позвоноч-
ных животных попадают в значительно менее плотную среду в кишечнике
переносчика или в культуре, то они теряют свою ундулирующую мембрану.
Можно сказать, что в общем у паразитических жгутиковых жгутов
больше, чем у близких свободноживущих видов. Процесс увеличения коли-
чества жгутов достиг высшей фазы у отряда многожгутиковых, паразити-
рующих в кишечнике термитов (см. рис. 21). Увеличение количества жгутов
также служит целям усиления двигательного аппарата.
Паразитический образ жизни может в соответствующих условиях вести
и к потере двигательного аппарата: лейшмании, живущие внутриклеточно и,
следовательно, не имеющие возможности свободно передвигаться, не имеют
жгута. Но стоит им попасть в организм насекомого-переносчика или в куль-
туру, как они восстанавливают жгут и начинают двигаться так же, как и
остальные жгутиковые.
Трипанозома Schizotrypanum cruzi часть своего цикла проводит внутри-
клеточно и в это время не имеет ни жгута, ни ундулирующей мембраны.
Когда же одна из клеток переходит в ток крови того же хозяина, то она
вновь восстанавливает свой двигательный аппарат. Некоторые панцирные
жгутиковые, паразитирующие в морских ракообразных, во взрослом со-
стоянии также не имеют жгутов.
96
Пульсирующая вакуоль, столь распространенная у свободноживущих
простейших, у паразитических в большинстве случаев отсутствует. Она
служит главным образом для осморегуляции; в морской воде и в среде,
окружающей паразитов, концентрация солей значительно выше, чем в
пресной воде, и приближается к концентрации в плазме простейшего или
даже выше нее. Поэтому надобность в пульсирующей вакуоле отпа-
дает.
Паразиты воспринимают пищу в растворенной форме всей поверхно-
стью тела. В связи с этим у многих из них отсутствует рот. Отсутствие рта
характерно для всего класса споровиков. Среди инфузорий есть целое се-
мейство безротых инфузорий (Astomata), паразитирующих главным обра-
зом у кольчатых червей.
У некоторых свободноживущих инфузорий также нет рта, а именно у
тех из них, у которых в плазме живут симбиотические зооксантеллы, спо-
собствующие питанию инфузорий. Так, например, целиком редуцируется
рот у Mesodinium rubrum.
Приспособление грегарин к паразитическому образу жизни выражается,
в частности, в том, что у них сильйо дифференцировался передний конец
тела (см. рис. 39), при помощи которого они либо проникают в клетку (на
ранних стадиях развития) — в этом случае он заострен и покрыт плотной
пелликулой, — либо удерживаются в кишечнике, и тогда на эпимерите
развиваются разнообразные прикрепительные органеллы. У паразитических
безротых инфузорий (например, Hoplytophrya) также образовались плотные
скелетные образования, служащие для прикрепления к тканям хо-
зяина.
Приспособительным изменением является и повышенная плодови-
тость паразитических простейших. Цисты или споры паразитов выделяются
во внешнюю среду и для дальнейшего развития должны вновь попасть в
организм соответствующего животного. В других случаях, перед тем как
попасть в окончательного хозяина, они должны пройти организм переносчи-
ка. Понятно, что у них значительно меньше возможности попасть в условия,
необходимые для их развития, и не погибнуть, чем у свободноживущих
форм. Поэтому у паразитических простейших так распространено множест-
венное деление (трипанозомы, споровики и т. д.), при котором один мате-
ринский организм быстро образует большое количество дочерних особей.
Большинство паразитических простейших размножается чрезвычайно
быстро. Хорошим примером может служить дизентерийная амеба. За одни
сутки человск-цнстоиоснтсль может выделить до 8 000 000 цист амсб. Бы-
строта размножения, повышенная плодовитость обеспечивают сохранение и
расселение вида.
В процессе паразитизма изменяется нс только паразит. Изменяется и
хозяин. В нем вырабатываются защитные приспособления. У многоклеточ-
ных животных в ответ на проникновение паразита вырабатываются анти-
тела, специальные клетки-фагоциты захватывают и пожирают паразитов
и т. д.
Эволюция взаимоотношения паразита и хозяина, эволюция парази-
тизма идет в направлении утраты паразитом болезнетворных свойств,
патогенности и выработки защитных свойств у хозяина. Действительно,
острое заболевание и смерть хозяина служат для паразита фактором небла-
гоприятным, так как в случае смерти хозяина он погибает вместе с
ним.
Например, при амебной дизентерии амебы удаляются из кишечника
хозяина в вегетативном состоянии до того, как они инцистировались, и все
погибают. Из организма же «носителя», который от амеб не страдает, они
выделяются в виде цист и могут заражать других людей.
Ясно, что когда те или иные простейшие, например трипанозомы,
кокцидии, малярийный плазмодий, вызывают смерть хозяина, они погибают
вместе с ним.
4 Курс зоологии, т. 1
97
ФИЛОГЕНИЯ ПРОСТЕЙШИХ
Наиболее просто морфологически устроенными простейшими являются
амебы. Для них характерно отсутствие постоянной формы тела, скелетных
и сократимых образований, специализированных органелл, непостоянство
формы и числа псевдоподий. Все эти признаки указывают на примитивность
строения амеб. Считают, что от предков саркодовых и произошли все осталь-
ные группы простейших.
Ранее было высказано предположение о том, что самыми древними фор-
мами следует считать тех простейших, которые могут питаться за счет не-
органической среды. Среди Protozoa только жгутиковые питаются как расте-
ния, синтезируя органические вещества из неорганических. Однако уста-
новлено, что аутотрофный способ питания не может считаться первичным,
так как для него необходим более сложный комплекс органелл. В связи с
этим трудно согласиться с представлениями о происхождении современных
простейших с гетеротрофным питанием от аутотрофных жгутиковых.
Раковинные корненожки, фораминиферы и все другие более сложно
устроенные корненожки произошли, по-видимому, от амеб путем их даль-
нейшего усложнения.
Споровики представляют собой сборную группу, в которой различные
подклассы имеют разное происхождение. Подкласс Telosporidia, по-види-
мому, произошел от жгутиковых, потому что половые особи грегарин кок-
цидий весьма с ними сходны (наличие жгутов, постоянная форма тела).
Гаметы гемоспоридий, хотя и не имеют жгутиков, но родство гемоспоридий
и кокцидий не вызывает сомнения.
Представители других подклассов споровиков, например Myxosporidia
из подкласса Cnidosporidia, обладают амебоидным движением, снабжены
псевдоподиями и ни в одной стадии не имеют жгутов. Это сближает их с
корненожками. Можно думать, что класс споровиков имеет двойственное
происхождение — от жгутиковых и от корненожек. Многие авторы поэтому
рассматривают Telosporidia и Cnidosporidia как самостоятельные классы,
не объединяя их в класс Sporozoa.
Характер органов движения сближает инфузорий и жгутиковых. Ди-
морфизм ядерного аппарата и наличие полового процесса в виде конъюгации
делают вопрос о происхождении инфузорий еще неясным, но в последнее
время описана свободноживущая инфузория Stephanopogon, у которой
имеется несколько одинаковых ядер, т. е. нет разделения на макро- и микро-
нуклеус. Это заставляет думать, что инфузории произошли от жгутиковых,
тем более, что резкого различия между жгутиками и ресничками нет, как
это видно на примере различных представителей Hypermastigidae.
Наличие ресничек у свободноплавающих «эмбрионов» сосущих инфу-
зорий ясно указывает на происхождение Suctoria от Ciliata.
ТИП II. ГУБКИ (SPONGIA, ИЛИ PORIFERA)
ХАРАКТЕРИСТИКА
Губки — наиболее примитивные многоклеточные. Примитивность их
выражается в том, что клеточные элементы не образуют у них настоящих
тканей и обособленных органов. Тело губок (рис. 68) состоит из двух кле-
точных слоев — наружного, образованного уплощенными эпителиальными
Рис. 68. Туалетная губка Euspongia officinalis (общий вид). Кусок
губки вырезан, чтобы показать внутреннее строение — систему
каналов. Стрелки показывают направление токов воды:
1 — поры, 2—спонгоцель, 3—оскулу.мы
клетками, и внутреннего, состоящего из воротничковых клеток — хоано-
цитов. Между ними находится студенистое вещество — мезоглея с клеточ-
ными элементами разного рода.
У губок отсутствуют мышечные и, по-видимому, настоящие нервные
клетки1. В связи с этим губки — животные неподвижные, почти не реаги-
рующие на внешние раздражения. Они прогоняют через систему каналов,
пронизывающих их тело, воду. Движение воды производится воротничко-
выми клетками — хоаноцитами (рис. 69). Вместе с водой в тело губок по-
падают пищевые частицы и кислород; с выходящей водой из их тела выво-
дятся продукты обмена.
1 В литературе сообщается о наличии у губок клеток, внешне похожих на нервные.
4* 99
В мезоглее имеются различные клеточные элементы: клетки, образую-
щие скелет — склеробласты, подвижные амебоциты, играющие роль в
пищеварении губок, половые клетки и некоторые другие.
Амебоциты, известные под названием археоцитов, представляют собой
недифференцированные клетки, за счет которых могут возникать все кле-
точные элементы губок.
Рис. 69. Отдельные жгути-
ковые воротничковые клет-
ки:
1 — тело клетки, 2 — жгутик,
«3 — плазматический воротничок
Рис. 70. Различные спикулы губок
Рис. 71. Разрез через прикрепившуюся ли-
чинку губки Clathrina, видны клетки (/)
внутренних частей личинки, выходящие на-
ружу и образующие наружный слой (2)
Для губок очень характерно наличие скелета, он имеется у громадного
большинства их. У большей части губок скелет состоит из отдельных мелких
элементов — спикул, разнообразных по форме (рис. 70), образованных из
извести или кремнезема. У неко-
торых губок скелет состоит из
органического вещества — спон-
гина.
Размеры губок колеблются
от нескольких миллиметров до
1,5 м.
Очень своеобразные процес-
сы происходят при развитии гу-
бок. Из оплодотворенного яйца
образуется обычно свободно-
плавающая личинка. Личинки
двуслойные; при дальнейшей
метаморфозе клетки внутреннего
слоя выходят наружу (рис. 71) и
превращаются в кожный покров,
а клетки наружного слоя — со-
ответственно в хоаноциты. В
связи с этим процессом вопрос о том, что считать у губок за эктодерму и
что за энтодерму, неясен, и губок часто даже противопоставляют всем
другим многоклеточным под названием Enantiozoa, т. е. вывороченные
животные.
Обитают губки преимущественно в морях; в пресных водах известно
лишь небольшое число видов. В морях губки распространены очень широко,
начиная от прибрежной зоны и кончая большими глубинами. Много видов
их в Антарктических морях.
Всего в настоящее время известно около 5000 видов губок.
100
КЛА ССИФИКА ЦИЯ1
Тип губки (Spongia, или Porifera)
Отряд 1. Известковые губки (Calcarea)
Отряд 2. Трехосные губки (Triaxonida)
Отряд 3. Четырехосные губки (Tetraxonida)
Отряд 4. Кремнероговые губки (Cornacuspongia)
Представитель типа губок —сикон (Sycon)
Рио. 72. Sycon rnphaniK (общий
вил). Кусок губки вырезан, что-
бы показать внутреннее строение
/ — устье, окруженное спикулами, 7 —
СПОИГОЦ0Л1., ./— Я4ГУТПКОЫ.1Л КпрМ МП4.
V — пучки уиикул uvp
Внешнее строение. Sycon (рис. 72) имеет вид веретеновидного мешка,
несколько суживающегося книзу, достигающего 2 см высоты. Основанием
своим губка либо непосредственно прикрепляется к субстрату, либо в слу-
чае колониальности сращена с другими членами колонии. На конце, проти-
воположном основанию, у губки находит-
ся большое отверстие — устье (osculum),
окруженное воротничком из длинных ске-
летных игл. Через устье вода выходит из
тела губки, втягивается она через много-
численные поры, покрывающие все тело
губки. Поверхность обычно шероховатая от
торчащих из нее мелких скелетных игл.
Цвет губки белый, иногда с несколько
желтова тым оттенк ом.
Внутреннее строение. Внутри тела на-
ходится обширная полость, называемая
некоторыми авторами спонгоцелем, от ко-
торой в радиальных направлениях отхо-
дят многочисленные цилиндрические кар-
маны. Карманы эти выстланы жгутико-во-
ротничковыми клетками (хоаноцитами).
Короткими каналами и порами они сообща-
ются с внешней средой, действием жгути-
ков создается ток воды через поры внутрь
тела губки.
Между жгутиковыми карманами рас-
положена меюелея, в которой залегают
у Sycon, как и у Других известковых гу-
бок, мшу г быть трех видов: одноосные,
трехосные и четырехосные. Крупные одно-
осные спикулы окружают устье, более мелкие тесно облегают наружные
концы жгутиковых карманов и целыми пучками торчат наружу (рис. 72).
У наружной поверхности тела расположены главным образом трехосные
спикулы, причем все три оси их лежат в одной плоскости, параллельной
наружной поверхности тела. Трехосные спикулы облегают и каждый
жгутиковый карман. У концов этих карманов, обращенных к срединной
полости, наряду с трехосными лежат и четырехосные спикулы, причем их
четвертая ветвь, перпендикулярная к трем остальным, направлена в сре-
динную полость. Таким образом, несмотря на то что скелет Sycon состоит
из отдельных спикул, их строгое закономерное расположение приводит к
образованию как бы общего каркаса тела.
Спикулы образуются в мезоглее особыми клетками — склеробластами,
происходящими из археоцитов.
Спикулу строят две клетки: клетка-образовательница, постепенно
образующая иглу, удлиняя ее с одного конца, и клетка-усилительница.
известковые иглы, или спикулы. Спикулы
Даны наиболее обычные отряды.
101
накладывающая новые массы извести на образованную первой клеткой
иглу. Образование трехосной спикулы происходит путем срастания трех
игл в одной точке. У четырехосных спикул образование четвертого луча
происходит позднее первых трех.
Жизнедеятельность. Как и все губки, Sycon неподвижен. Питание осу-
ществляется пассивным способом. Воротничковыми клетками создаются
токи воды, вовлекающие ее внутрь тела губки.
Вместе с водой в тело губок засасываются и взвешенные в ней частицы,
достаточно мелкие, чтобы пройти через поры. Проносясь вместе с током воды
мимо жгутиково-воротничковых клеток, эти частицы попадают в водоворо-
ты, производимые жгутиками, и засасываются в воротнички, к которым и
прилипают. Затем движением плазмы они передвигаются по воротничку к
самой клетке и втягиваются внутрь нее. Из этих клеток частицы переходят в
мезоглею, где захватываются подвижными амебоцитами, внутри которых и
происходит переваривание. Таким образом, пищеварение у губок внутри-
клеточное. Питается Sycon бактериями, мелкими планктическими водо-
рослями и органическими остатками (детритом).
Развитие. Sycon — гермафродит: мужские и женские половые клетки
образуются в мезоглее одной особи. Яйцеклетки подвижные и могут пере-
двигаться при помощи псевдоподий. Сперматозоиды вместе с током воды
выносятся наружу, проникают в другие особи и там оплодотворяют их
яйцеклетки.
Развитие начинается в теле материнского организма. В результате дроб-
ления получается бластула, состоящая из клеток двух видов — мелких
жгутиковых и более крупных зернистых, лишенных жгутиков. Затем про-
исходит образование двуслойного зародыша путем инвагинации, причем
Рис. 73. Развитие Sycon raphanus. I — инвагинация
в мезоглее материнского организма; II — плавающая
личинка — амфибластула; III — впячивание жгути-
ковых клеток; IV — прикрепление и начало метамор-
фоза
впячиваются внутрь крупные клетки (рис. 73j ). На этом развитие в мате-
ринском организме заканчивается, и зародыш выходит наружу. Здесь про-
исходит обратное выпячивание крупных клеток, в результате чего образуется
однослойная, состоящая как бы из двух половинок, свободноплавающая
личинка — амфибластула.
Через некоторое время амфибластула превращается в двуслойную ли-
чинку, но на этот раз впячиваются мелкие жгутиковые клетки. Впоследствии
личинка прикрепляется к субстрату тем полюсом, на котором находятся
мелкие жгутиковые клетки; на противоположном полюсе образуется устье—
оскулум. Дальнейшее развитие состоит в образовании пор, устья и других
структур.
Особенности развития Sycon и других губок многие авторы рассматри-
вают как своеобразную смену зародышевых листиков: крупные клетки,
которые сначала находились внутри зародыша и являлись как бы энтодер-
102
мой, впоследствии образуют наружный слой, а мелкие клетки, находившие-
ся снаружи, уходят внутрь, где превращаются в хоаноциты.
Экология и географическое распространение. Виды рода Sycon очень
часто встречаются во всех морях на сравнительно небольших глубинах.
Поселяются они обычно на водорослях, гидроидах или мшанках. В наших
северных морях, например в Белом море, очень обычен Sycon raphanus.
Положение в системе. Сикон относится к отряду известковых губок.
Второй представитель типа губок — бодяга
Под названием бадяги известен ряд видов, относящихся к двум близким
родам Spongilla и Ephydatia.
Внешний вид. Бадяги не имеют определенной формы. Они могут быть
или в виде подушкообразных наростов, или в виде довольно сложных вет-
вистых образований (рис. 74i ). Цвет бадяг желтовато-бурый или зеленый.
Рис. 74 Баля га (Spongilla). / — общий вид колонии; // — схема внутреннего стро-
ения; /// — часть скелета:
/—поры, 2 — субдермальные полости, 3—приводящие каналы, 4— жгутиковые камеры, 5—выво-
дящие каналы, 6 — спонгоцель, 7 — устье
Последний зависит от присутствия в тканях губки симбиотических зеленых
водорослей. Бадяги имеют обычно очень специфический неприятный запах.
Один экземпляр бадяги представляет собой в большинстве случаев коло-
ниальное образование. О количестве особей, входящих в него, можно су-
дить по числу устьев (рекулумов). Устья расположены у бадяг обычно на
небольшом возвышении, так называемой оскулярной трубочке (рис. 75),
которая может несколько вытягиваться и втягиваться. Движение оскуляр-
ных трубочек — единственная форма движения, которую можно наблюдать
у бадяг.
Поверхность бадяг покрыта кожной, или дермальной, мембраной, про-
низанной порами.
Внутреннее строение. Под дермальной мембраной расположены обыч-
но обширные подкожные, или субдермальные, полости (рис. 74). На дне
103
этих полостей начинается сложная система каналов, идущих к особым рас-
ширениям в мезоглее. Расширения эти выстланы жгутико-воротничковыми
клетками и носят название жгутиковых камер. От камер идут каналы к
спонгоцелю. Между каналами и камерами в мезоглее находятся скелетные
Рис. 75. Часть поверхности
бадяги. Видна вытянутая
оскулярная трубка
Рис. 77. Геммулы бадяг. / — геммулы Spongilla на
субстрате (ест. величина); // — разрез геммулы Ephy-
dathia (сильно увелич.):
/ — внутренняя масса клеток, 2— внутренняя оболочка, 3 — на'
ружная оболочка, 4— амфиднски
Рис. 76. Личинка бадя-
ги
образования. Последние
состоят из кремневых од-
ноосных спикул, различ-
ных по величине: боль-
шие спикулы, или MQK-
росклеры, и мелкие —
микросклеры. Макро-
склеры расположены
обычно определенными
рядами и склеены орга-
ническим веществом
(рис. 74).
Размножение. Раз-
множаются бадяги по-
ловым и бесполым пу-
тем.
Половое раз-
множение. Все ви-
ды бадяг раздельнопо-
лы. Половые клетки об-
разуются в мезоглее.
Оплодотворение проис-
ходит в женской особи,
куда с током воды при-
носятся сперматозоиды.
В результате дробления
развивается личинка,
состоящая из внутрен-
ней массы клеток и на-
ружного слоя, несущего
на поверхности реснич-
ки. Такая личинка
(рис« 76) выходит из ма-
теринского организма и
короткое время (около
12 ч) свободно плавает,
затем прикрепляется к
субстрату и превраща-
ется в губку. Это прев-
ращение сопровождает-
ся погружением клеток наружного слоя в глубину тела, где они и
становятся жгутиковыми воротничковыми клетками.
Бесполое размножение. Бесполое размножение про-
исходит путем почкования, причем последнее может быть двоякое: наруж-
ное, ведущее к образованию массивных колоний, и внутреннее, очень харак-
терное для бадяг, в результате которого образуются так называемые гем-
мулы. Геммулы (рис. 77) образуются в мезоглее в конце лета и представляют
собой комочек клеток с запасом питательного материала, окруженного
двумя плотными оболочками, между которыми располагаются воздушные
камеры и микросклериты (у рода Spongilla) или особые скелетные образо-
вания, так называемые амфидиски (у рода Ephydatia).
Биологическое значение геммул очень большое; они представляют собой
приспособление для переживания неблагоприятного времени года, т. е. в
наших широтах зимы. К началу зимы сама бадяга обычно погибает и разру-
104
шается, геммулы же благодаря двум оболочкам и находящемуся между ними
воздушному слою хорошо перезимовывают, и весной из них развиваются
молодые губки. Следует отметить, что наличие геммул характерно именно
для пресноводных губок, подвергающихся резким периодическим измене-
ниям окружающей среды по временам года. Подобные образования встре-
чаются и у других групп пресноводных животных: коловраток, мшанок, не-
которых рачков и др. (см. в соответствующих главах).
Регенерация. Бадяги, как и губки вообще, обладают большой способ-
ностью к регенерации. Если, например, потереть кусочек бадяги через
мелкоячеистую ткань, получается эмульсия из отдельных изолированных
клеток. Эти клетки слипаются друг с другом, образуя клеточные скопления.
Скопления эти растут и через несколько дней при благоприятных условиях
могут превратиться в маленькую губку.
Такая большая способность к регенерации подтверждает примитивность
губок и еще очень незначительную специализацию у них отдельных клеток.
Экология и географическое распространение. Бадяги распространены
главным образом в тропиках и субтропиках. В умеренных широтах они хотя
и довольно обычны, но менее разнообразны. В нашей стране известно де-
сять видов бадяг, принадлежащих к четырем родам. Особое место занимают
губки озера Байкал, где встречается эндемичное (т. е. только там обнару-
женное) семейство Lubomirskiidae с несколькими видами, отличающееся от
других пресноводных форм рядом признаков, например отсутствием геммул.
Бадяги встречаются в различных водоемах как в стоячих, так и в те-
кучих, хотя в последних они обитают у берегов, где нет особо сильного тече-
ния. Субстратом для бадяг служат самые разнообразные подводные пред-
меты — сваи, каряги, подводная растительность, береговые камни и пр.
Довольно часто поселяются они и на других животных, например на мол-
люсках. Поселяясь в различных гидротехнических и водопроводных соору-
жениях, губки могут сильно засорить их, принося тем самым довольно зна-
чительный вред.
Благодаря пористому строению бадяги служат хорошим убежищем
для ряда личинок различных насекомых, червей, некоторых ракообразных,
образуя вместе с ними характерный биоценоз.
Загрязнение воды органическими веществами обычно благоприятно
для роста бадяг. Поэтому они часто сильно развиваются в реках ниже
больших городов н играют важную роль в естественном очищении водоемов.
В нашей народной медицине «сушеная бадяга» употребляется как сред-
ство против ревматизма, а также как косметическое средство, вызывающее
искусственный румянец; по-видимому, имеет значение раздражающее
действие спикул на кожные покровы при втирании сухой губки.
Положение в системе. Бадяги относятся к отряду кремнероговых губок,
в котором образуют особое семейство пресноводных губок — Spongillidae.
ОБЩИЙ ОБЗОР ТИПА
Как указывалось, важнейшим признаком губок является наличие у
них жгутико-воротнпчковых клеток и системы связанных с ними каналов и
полостей, так называемой ирригационной системы. Система эта может быть
различной степени сложности: у сикона она занимает среднее положение,
а у бадяги — ирригационная система наиболее сложная. В более простых
случаях (тип Ascon, рис. 78) поровые каналы открываются непосредственно
в спонгоцель, которая выстлана жгутико-воротничковыми клетками.
Таким образом, у губок можно проследить постепенное усложнение ирри-
гационной системы: тип Ascon— тип Sycon и тип Leucon (к последнему типу
принадлежит бадяга). Это не систематическое деление, а типы морфологи-
ческого усложнения строения.
Процесс фильтрации воды происходит у губок довольно интенсивно:
так, например, губка Leucandra (близка к Sycon) 7 см высоты и 1 см в попе-
105
Рис. 78. Схема стро-
ения губок типа
Ascon:
/ — поры, 2 — спонгоцель
речнике прогоняет через свое тело в течение суток 22,5 л. Выбрасывание
воды через оскулум происходит также энергично — та же Leucandra выки-
дывает твердые частички на расстояние 25—50 см. Объясняется это очень
большим количеством хоаноцитов у губок.
Кроме известковых, все остальные губки имеют кремневый, кремнеро-
говой или чисто роговой скелет. В большинстве случаев скелет, как и у из-
вестковых губок, состоит из разнообразных спикул.
В некоторых случаях спикулы соединяются вместе и
образуют сплошной скелет в виде ажурного каркаса
(стеклянные губки). Степень включения рогового ве-
щества в скелет губок различна. В некоторых случа-
ях оно, как у бадяг, только соединяет спикулы, в
других — спикулы совершенно выпадают, и скелет
делается чисто роговым — сплошным и эластичным.
Строение скелета губок тесно связано с их обра-
зом жизни. Губки с роговым и кремнероговым скеле-
том обитают обычно на сравнительно небольших глу-
бинах, где они могут без вреда выдерживать удары
волн и движение воды; стеклянные губки с их край-
не ломким скелетом, наоборот, обитают только на
больших глубинах, где царствует постоянно полный
покой. Они встречаются на глубине до 7000 м.
Губки редко используются в пищу другими
животными, причиной этому служит, с одной сторо-
ны, скелет, с другой, — специфические свойства губок — они, по-видимому,
ядовиты для многих животных. Эта специфика выражается и в том остром
и неприятном запахе, которым обладает большинство губок. Благодаря
этим свойствам губки занимают особое положение в общей экономике мо-
рей, в которых они широко распространены. Громадные скопления ор-
ганического вещества, которые они образуют, строя иногда очень боль-
шие колонии, не используются непосредственно. Только после гибели
губок, их разрушения и превращения в детрит (взвешенные в воде мелкие
органические частицы) органическое вещество губок используется в пищу
другими животными.
СИСТЕМАТИЧЕСКИЙ ОБЗОР ТИПА
ОТРЯД ИЗВЕСТКОВЫЕ ГУБКИ (CALCAREA)
Известковые губки представляют собой наиболее примитивный отряд
губок. Ирригационная система у них различной сложности, в этом отряде
имеются все три типа ее. Известковые губки обычно небольших размеров.
Все они обитатели морей, причем живут от прибрежных районов до глубины
1000 м.
ОТРЯД ТРЕХОСНЫЕ ГУБКИ (TRIAXONIDA)
В этот отряд входят губки, имеющие кремневый скелет, состоящий из
разнообразных спикул, все вариации которых могут быть, однако, сведены
к основному типу —три равные оси, пересекающиеся по середине своей
длины под прямым углом. У некоторых форм спикулы соединяются вместе
в единый скелет — тонкий и хрупкий каркас.
Жгутиковые камеры располагаются одним слоем.
Все относящиеся к этому отряду губки — обитатели морей, где они
достаточно разнообразны и широко распространены. Особый интерес в
этом отряде представляют так называемые стеклянные губки, имеющие
цельный, иногда очень причудливый по форме, ажурный скелет (рис. 79).
106
Стеклянные губки обитают только на больших глубинах (до 7000м) и явля-
ются характерными представителями глубоководной фауны.
Представитель стеклянных губок — корзинка Венеры (Euplectella) —
с вытянутым несколько изогнутым ажурным скелетом, достигающим
30 см в высоту (рис. 79и). На верхнем конце помещается широкий оскулум,
закрытый скелетной сеточкой, на нижнем — густой пучок одноосных игл,
Рис. 79. Стеклянные губки. / — Pheronema; II — Euplectella
которыми губка укрепляется в илистом дне. В Euplectella живут обычно,
как комменсалы, различные беспозвоночные, например почти в каждой
особи японского вида Е. oweni живет самец и самка креветки Spongicola.
Интересно отметить, что все жители этой губки забираются в нее еще
в стадии личинок, и вырастая, оказываются вечными пленниками, так как
из-за наличия сеточки, закрывающей оскулум, не могут выйти из нее. Дру-
гим представителем стеклянных губок может служить Pheronema (рис. 79i ).
Из-за красивого скелета ряд стеклянных губок в странах Тихого океана
промышляется и используется как украшения.
107
ОТРЯД ЧЕТЫРЕХОСНЫЕ ГУБКИ (TETRAXONIDA)
В этот отряд входят губки, имеющие кремневый скелет. Спикулы
разнообразные по форме; кроме основных, четырехосных, имеются одноос-
ные с утолщением на конце в виде булавки, а также звездчатые. Хоаноциты
образуют камеры. Кожный слой у ряда форм сильно утолщается и обрастает
Рис. 80. Кубок Нептуна
(Poterion neptuni), ест. ве-
личина около 1 м
губку в виде плотной коры, которая иногда при-
нимает вид панциря благодаря скоплению в ней
спикул. Все представители этого отряда оби-
тают в морях, некоторые из них очень массив-
ны и достигают крупных размеров.
Четырехосная губка Geodia обычна для
Баренцева моря, образует местами громадные
скопления, настолько большие, что трал может
вытащить сразу несколько тонн этой губки.
Кубок Нептуна (Poterion neptuni) — губка,
имеющая вид громадного кубка (рис. 80), дости-
гающего 1,5 м высоты и 0,75 м в поперечнике;
распространена в Тихом океане.
Сверлящая губка (Cliona) обитает в створ-
ках двустворчатых моллюсков. Личинки этой
губки садятся на поверхность раковины и углуб-
ляются в нее, разъедая известь своими содержа-
щими кислоту секретами. Затем губка протачи-
вает в толще створок ходы, открывающиеся на
поверхность округлыми отверстиями, из кото-
рых высовываются выросты губок, снабженные
порами и устьями. Ходы постепенно разраста-
ются, створки разрушаются, и моллюск гибнет.
Сверлящие губки наносят большой вред уст-
ричным банкам и устричному хозяйству.
ОТРЯД КРЕМНЕРОГОВЫЕ ГУБКИ (CORNACUSPONGIA)
Наиболее многочисленный отряд, охватывающий как морские, так и
пресноводные формы. Скелет кремневый или органический, состоящий из
спонгина1. В некоторых случаях скелет только кремневый, в других —
только спонгиновый и, наконец, у ряда форм он смешанный, состоящий из
простых одноосных спикул, склеенных органическим веществом (бадяга).
Имеются жгутиковые камеры, а
также обширные подкожные поло-
сти без жгутиковых воротничко-
вых клеток.
К отряду кремнероговых гу-
бок относится так называемая ту-
алетная, или греческая, губка
(Euspongia officinalis) (см. рис.68),
имеющая чисто спонгиновый, эла-
стичный скелет, состоящий из силь-
но переплетенных друг с другом
волокон спонгина (рис. 81). Губка
эта распространена на небольших
глубинах в теплых морях, в част-
Рис. 81. Часть скелета туалетной губки
1 Спонгин по химическому составу может быть причислен к хитиноидным ве-
ществам, т. е. к веществам, близким к хитину. Характерно для спонгина содержание
в нем йода (до 14%).
108
ности в Средиземном море, и с древних времен служила объектом промыс-
ла; употреблялась она в хирургии (вместо ваты) и как туалетная принад-
лежность. В настоящее время в связи с тем, что использование этой губки
сильно сократилось, промысел ее развит слабо и дает от 5 до 15 000 ц губки
в год. Промышляются губки главным образом в Средиземном море (Италия,
Тунис и Греция), а также в южно-атлантических штатах США.
ФИЛОГЕНИЯ ГУБОК
Особенности строения и развития губок заставляют отделять их от
всех других многоклеточных и рассматривать как особую ветвь животного
мира (Parazoa), отходящую от одноклеточных независимо от всех остальных.
Губки — очень древняя группа животных, известная с кембрия.
Можно предположить, что губки сформировались еще в докембрийский
период. Следует отметить, что за свою долгую историю губки сравнительно
мало изменились и многие ископаемые формы почти не отличаются от совре-
менных.
Благодаря скелету губки хорошо сохранялись в ископаемом состоянии.
В кембрии обнаружены стеклянные губки (Triaxonida), четырехосные
(Tetraxonida), кремнероговые (Cornacuspongia). Известковые губки появи-
лись с девона.
С одной стороны среди одноклеточных группой, которая, возможно,
могла быть исходной для губок, являются воротничковые жгутиковые
(Craspedomonadina) (см. рис. 16). Они снабжены жгутиками и плазматиче-
скими воротничками, подобными тем, которые имеются у хоаноцитов губок.
Таких образований нет нигде в животном мире, кроме губок и воротничко-
вых жгутиковых.
С другой стороны, сходство ранних стадий онтогенеза губок с остальными
многоклеточными служит доводом в пользу взгляда о происхождении
губок от общего ствола Metazoa, от которого они, по-видимому, отделились
на очень раннем этапе развития многоклеточных. Органический (спонги-
новый) скелет, видимо, — уже более позднее образование. У кремнероговых
губок можно проследить постепенное вытеснение кремневых спикул и замену
их чисто спонгиновым скелетом.
ТИП. III. КИШЕЧНОПОЛОСТНЫЕ
(COELENTERATA)
ХАРАКТЕРИСТИКА
Кишечнополостные — двуслойные животные, в большинстве своем
построенные по лучевой симметрии. Стенки их тела состоят из двух слоев—
эктодермы и энтодермы (рис. 82). Эктодерма покрывает тело снаружи,
энтодерма выстилает гастральную полость. Эта полость является пищева-
Рис. 82. Схематический продольный
разрез через гидру:
I — эктодерма, 2 — энтодерма, 3— опорная
пластинка, 4 — внутренняя полость, 5—рото-
вое отверстие, 6 — почка, 7 — яйцеклетка
рительнои, так как в нее попадает
пища через единственное отверстие,
соединяющее эту полость с наруж-
ной средой.
В эктодерме большинства ки-
шечнополостных имеются особые
клетки книдобласты, служащие ору-
диями нападения и защиты.
Между эктодермой и энтодермой
имеется или тонкая опорная пластин-
ка, или толстый студенистый слой —
мезоглея, выделяемая клетками экто-
дермы и энтодермы.
У многих кишечнополостных
имеется скелет — либо наружный,
либо расположенный в мезоглее. В
большинстве случаев он известковый,
реже роговой.
Кишечнополостные очень разно-
образны. Среди них имеются как
свободноплавающие формы (табл. II
и III), так и прикрепленные (табл.IV).
В отличие от губок у кишечнопо-
лостных имеются настоящие нервные
элементы. Они образуют у кишечнополостных или очень примитивную
диффузную нервную систему, состоящую из сети разбросанных нервных
клеток, соединенных между собой своими отростками (см. рис. 86ц), или у
свободноплавающих форм более сложную нервную систему, состоящую из
скоплений нервных клеток. Интересной особенностью нервной системы
кишечнополостных, указывающей на их примитивность, является то,
что нервные клетки лежат у них не только в эктодерме, как у большинства
других многоклеточных, но и в энтодерме.
Размножаются кишечнополостные как половым, так и бесполым путем,
причем в результате последнего и образуются колонии, характерные для
ряда кишечнополостных.
ио
Кишечнополостные в громадном большинстве — морские животные,
в пресных водах встречаются лишь немногие представители их. Всего в на-
стоящее время известно около 9000 представителей этого типа.
Многие кишечнополостные благодаря наличию скелета хорошо сохра-
нились в ископаемом состоянии.
КЛАССИФИКАЦИЯ
Тип кишечнополостные (Coelenterata)
Подтип I. Стрекающие (Cnidaria)
Класс 1. Гидроидные (Hydrozoa)
Подкласс I. Гидрообразные (Hydroidea)
Отряд 1. Гидры (Hydrida)
Отряд 2. Гидроиды (Leptolida)
Отряд 3. Трахилиды (Trachylida)
Подкласс II. Сифонофоры (Siphonophora)
Класс 2. Сцифоидные (Scyphozoa)
Класс 3. Коралловые (Actinozoa, или Anthozoa)
Подкласс 1. Восьмилучевые кораллы (Octocorallia)
Подкласс II. Шестилучевые кораллы (HexacoralHa)
Подтип II. Нестрекающие (Acnidaria)
Класс I. Гребневики (Ctenophora)
Подкласс 1. Щупальцевые (Tentaculata)
Подкласс II. Бесщупальцевые (Atentaculata)
систематический обзор типа
Подтип I. Стрекающие (Cnidaria)
ХАРАКТЕРИСТИКА
Стрекающие кишечнополостные характеризуются наличием в клетках
эктодермы так называемых стрекающих, или крапивных, клеток, которые
служат органами защиты и нападения. В стрекательных клетках (рис. 83)
находятся капсулы, имеющие вид пузырьков со свернутой в форме спирали
полой нитью. Внутри такого пузырька имеется длинная полая нить, кото-
рая при спокойном состоянии капсулы лежит в ней в виде свернутой спи-
рали. На наружной поверхности клетки, содержащей капсулу, находится
чувствительный отросток в виде шипика — книдоциль. Книдоциль воспри-
нимает раздражение из наружной среды. В ответ на раздражение стрека-
тельная капсула выбрасывает содержащуюся в ней нить, которая при
этом выворачивается, как палец перчатки. Вместе с нитью из капсулы
выбрызгивается и жидкое содержимое, имеющее едкие, а иногда и ядовитые
свойства. Такой «выстрел» стрекательной капсулы может дать разные
результаты. В некоторых случаях нить прободает тонкие покровы врага
(рис. 84) и анестезирует его, в других — враг или жертва могут быть опу-
таны стрекательными нитями, как тенетами. Некоторые стрекающие ки-
шечнополостные, например крупные медузы из сцифоидных, могут наносить
стрекательными нитями чувствительные ожоги и человеку. После выбра-
сывания нити стрекательная капсула больше не восстанавливается, и со-
державшая ее клетка дегенерирует. Новые клетки со стрекательными кап-
сулами образуются из соответствующих клеток эктодермы (см. ниже).
У стрекающих имеются две жизненные формы: сидячая — полип и
свободноплавающая — медуза.
Полипы в большинстве случаев колониальны, имеют скелет, иногда
развитый очень мощно, нервная система развита у них слабо и органов
чувств обычно нет.
111
Медузы одиночны, скелета у них нет, а сильно развитая мезоглея насы-
щена водой. Нервная система развита более сложно и имеются органы
чувств — равновесия и светочувствительные.
Полипы и медузы—самостоятельные организмы, или стадии жизнен-
ного цикла одного организма. Разные виды стрекающих либо существуют в
виде только одной из этих форм (или полип, или медуза), либо на протяже-
нии своего жизненного цикла проходят обе стадии — и полипа, и медузы.
Стрекающие охватывают основную
массу кишечнополостных. Сюда входят
как морские, так и пресноводные формы.
Подтип стрекающие распадается на 3
класса: гидроидные (Hydrozoa), сцифоид-
ные (Scyphozoa) и коралловые (Antozoa).
Первый класс может рассматриваться как
охватывающий более низкоорганизованных
кишечнополостных, два других — более
сложных.
Рис. 84. Действие стрекательных
капсул (пенетрант) гидры на ин-
фузорию-туфельку. Инфузория
выбросила трихоцисты и лопнула
Рис. 83. Стрекательная клет-
ка. / — в состоянии покоя;
II — разрядившаяся:
1 — свернутая нить, 2—ядро клетки,
3—стрекательная капсула, 4 — про-
топлазма клетки, 5—книдоциль,
6 — выброшенная нить, 7 — шипы
КЛАСС 1. ГИДРОИДНЫЕ (HYDROZOA)
ХАРАКТЕРИСТИКА
В этот класс объединяют как сидячие, так и плавающие формы. Для
класса характерны: 1) энтодерма непосредственно подходит к ротовому
отверстию (см. рис. 82); 2) гастральная полость полипов сплошная, не раз-
деленная перегородками на камеры; 3) образование половых продуктов в
эктодерме.
Скелет (у полипов) выделяется эктодермой в виде наружной кутикулы,
иногда пропитанной известью.
Громадное большинство представителей этого класса обитает в морях,
в пресных водах встречаются лишь немногие. Класс разделяется на 2 под-
класса: 1) гидрообразные (Hydroidea) и 2) сифонофоры (Siphonophora).
ПОДКЛАСС I. ГИДРООБРАЗНЫЕ (HYDROIDEA)
К этому подклассу относятся типичные гидроидные — гидрополипы
и гидромедузы как морские, так и пресноводные.
112
Представитель сидячих гидрообразных —
пресноводная гидра (Hydra oligactis)
Рис. 85. Гидра. / — в спокой-
ном состоянии; // — сократив-
шаяся после раздражения
Под названием «гидры» объединяется ряд видов. У нас встречается их
несколько; наиболее обычна стебельчатая гидра (Hydra oligactis), описание
которой приведено ниже.
Внешнее строение. Гидра (рис. 85i ) представляет собой полип, дости-
гающий в вытянутом состоянии 1—Р/2 см (не считая щупалец).На переднем
конце находится ротовое отверстие, окруженное венцом из 5—9 длинных,
очень подвижных шупалец, несущих батареи
стрекательных клеток. При раздражении у
гидры быстро сокращается как все тело, так
и щупальца, и она превращается в небольшой
довольно плотный комочек (рис. 85и). Через
некоторое время после прекращения раздра-
жения гидра опять вытягивается и принимает
первоначальную форму.
Обычно на теле гидры имеются дочерние
особи (почки), которые постепенно отпадают
от материнского организма и начинают вести
самостоятельный образ жизни. Настоящие
колонии у гидры никогда не образуются.
Прикрепляется гидра к субстрату подо-
швой, имеющей вид некоторого расширения
на заднем конце тела.
Цвет гидр бывает различный; обычно
он желтовато-серый, например у стебельчатой
гидры, но бывают зеленые, а иногда и ярко-
красные гидры. В последних двух случаях
цвет зависит не от пигмента, а от одноклеточ-
ных зеленых водорослей (Zoochlorella) или
бактерий (если цвет красный), живущих в
клетках гидры.
Внутреннее строение. Стенка тела гидры состоит из двух слоев —
эктодермы и энтодермы, между которыми расположена тонкая бесструк-
турная опорная пластинка (см. рис. 82). Стенка тела сравнительно тонкая,
и всю внутреннюю часть занимает обширная полость, заходящая и в щу-
пальца. Полость эта довольно хорошо просвечивается и видна снаружи.
Микроскопическое строение.Оба клеточных слоя гидры состоят преиму-
щественно из так называемых эпителиально-мускульных клеток (рис. 86щ).
В каждой из таких клеток имеется собственно-эпителиальная часть и сокра-
тимый отросток. Эпителиальная часть клетки обращена или кнаружи (в
эктодерме) или к гастральной полости (в энтодерме). Сократительные от-
ростки отходят от основания клетки, прилежащего к опорной пластинке —
мезоглее. Внутри сократительного отростка находятся мускульные волок-
на. Сократительные отростки клеток эктодермы расположены параллельно
оси тела и осям щупалец, т. е. вдоль тела гидры, их сокращение и вызывает
укорочение тела и щупалец. Сократительные отростки клеток энтодермы
располагаются поперек тела в кольцевом направлении, их сокращение
вызывает сужение тела гидры. На свободной поверхности клеток энтодермы
находятся жгутики, чаще всего 2, а иногда могут появляться и псевдо-
подии.
Кроме эпителиально-мускульных клеток, в эктодерме и в энтодерме
находятся чувствующие, нервные и железистые клетки.
Первые занимают такое же положение, как эпителиально-мускуль-
ные клетки, т. е. одним полюсом выходят на поверхность тела или к пище-
варительной полости, другим — к опорной пластинке.
113
Вторые лежат у основания эпителиально-мускульных клеток, около их
сократительных отростков, прилегающих к опорной пластинке. Нервные
клетки соединяются отростками в примитивную нервную систему диффуз-
ного типа (рис. 86ц ). Особенно многочисленны нервные клетки вокруг рта,
на щупальцах и на подошве.
Рис. 86. Микроскопическое строение гидры. / — разрез через стенку тела;
// —диффузная нервная система (видны соединения отростков нервных
клеток между собой); /// —отдельная эпителиально-мускульная клетка
эктодермы:
1—стрекательные клетки, 2—эпителиально-мускульные клетки эктодермы, 3 — эпители-
ально-мускульные клетки энтодермы, 4—железистые клетки энтодермы, 5—жгутиковид-
ные и псевдоподиальные выросты эктодермальных клеток, 6—интерстициальные клетки,
7—чувствительные клетки эктодермы, 8— чувствительные клетки эктодермы, 9—нервные
клетки эктодермы (нервные клетки энтодермы не показаны), 9 (III)—тело клетки,40— сок-
ратительные отростки с сократительной фибриллой внутри них (II)
Железистые клетки эктодермы располагаются главным образом на
подошве и щупальцах; их клейкие выделения на подошве служат для при-
крепления гидры к субстрату, а на щупальцах играют роль при перемеще-
нии животного (см. ниже). Железистые клетки энтодермы располагаются
около рта, их секрет имеет пищеварительное значение.
В эктодерме имеются еще стрекательные клетки, т. е. клетки, содержа-
щие стрекательные капсулы (см. выше), они особенно многочисленны на
щупальцах. У гидры имеется четыре типа стрекательных клеток: наиболее
крупные грушевидные — пенетранты, мелкие грушевидные — вольвенты,
большие цилиндрические — глютинанты, или стрептолины, и малые ци-
линдрические—стереолины. Действие капсул этих типов различно; некото-
рые из них своими острыми нитями могут пробивать стенку тела противника
или жертвы (см. рис. 84) и вводить в ранку ядовитое вещество и тем пара-
лизовать ее, другие же только опутывают жертву нитями.
114
Наконец, у гидры имеются еще не дифференцированные так называемые
интерстициальные клетки, из которых развиваются различные клеточные
элементы гидры, в частно-
сти половые клетки.
Движение. Гидра мо-
жет перемещаться с места
на место. Передвижение это
происходит по-разному:
или гидра, дугообразно
изгибаясь (рис. 87), при-
сасывается щупальцами и
отчасти железистыми клет-
ками, окружающими рот, к
субстрату и затем подтя-
гивает подошву, или гидра
как бы «кувыркается»,
прикрепляясь попеременно
то подошвой, то щупаль-
Рис. 87. Различные способы и моменты передви-
жения гидры (по Трамбле, 1744)
нами.
Питание. Стрекательные капсулы своими нитями опутывают добычу
и парализуют ее. Обработанная таким образом добыча захватывается щу-
пальцами и направляется в ротовое отверстие. Гидры могут «осилить»
весьма крупную добычу, превосходящую их по величине, например даже
Рис. 88. Стебельчатая гидра (Hydra oligactis). / — особь с
мужскими гонадами; // —особь с женскими гонадами
мальков рыб. Растяжимость ротового отверстия и всего тела у них велика.
Они очень прожорливы — одна гидра может проглотить за короткий срок
до полдюжины дафний. Проглоченная пища попадает в гастральную полость.
Пищсварсниеу гидр, по-видимому, комбинированное — внутри- и внеклеточ-
ное. Пищевые частицы затягиваются клетками энтодермы при помощи псев-
115
доподий внутрь и там перевариваются. В результате пищеварения в клет
ках энтодермы накапливаются питательные вещества, там же появляются
зернышки продуктов выделения, выбрасываемые время от времени неболь
шими порциями в гастральную полость. Продукты выделения, а также не
переваренные части пищи выбрасываются наружу через ротовое отверстие
Размножение. Размножаются гидры бесполым и половым путем. Пр:
бесполом размножении на гидрах образуются почки (см. рис. 85), постепенн*
отрывающиеся от материнского организма. Почкование гидр при благопри
ятных условиях питания может проходить очень интенсивно; наблюдени:
показывают, что за 12 дней число гидр может увеличиться в 8 раз. В те
чение летнего периода гидры размножаются обычно почкованием, но с на
ступлением осени начинается половое размножение, причем гидры могу
быть и гермафродитные и раздельнополые (стебельчатая гидра).
Половые продукты образуются в эктодерме из интерстициальных кле
ток. В этих местах эктодерма вздувается в виде бугорков (рис. 88), в которы:
образуются или многочисленные сперматозоиды, или одна амебовидна:
яйцеклетка. После оплодотворения, происходящего на теле гидры, яйце
клетка покрывается оболочкой. Такое покрытое оболочкой яйцо перезимо
вывает, и весной из него выходит молодая гидра. Личиночная стадия ;
гидр отсутствует.
Регенерация. На значительную способность гидр к регенерации об
ратили внимание уже первые исследователи в XVIII в. В дальнейшем гидрь
стали одним из важнейших объектов при изучении явления регенерации
Небольшие частицы тела гидр, менее 1 мм величиной, могут восстанавливать
ся в целое животное.
Экология и географическое распространение. Гидры широко распро
странены по пресноводным водоемам всего мира; в настоящее время их из
вестно 29 видов.
В СССР гидры встречаются повсеместно и представлены 8 видами
Обитают они и в водоемах равнин, и в высокогорных озерах, наприме]
Кавказа, где нередко встречаются ярко-красные формы. Наиболее част<
встречаются гидры в медленно текущих водоемах на водной растительности-
лучшем для них субстрате. Они держатся обычно близко к поверхности
так как нуждаются в кислороде; вместе с тем они избегают сильно освещен
ных мест. В связи с этим очень хорошее место обитания для них — нижня:
поверхность листьев кувшинок и кубышек.
Положение в системе. Гидры относятся к отряду гидр (Hydrida)
Представитель свободноплавающих гидрообразных
— медуза обелия (Obelia)
Медуза эта не является самостоятельным организмом, а лишь звено:
жизненного цикла гидроидного полипа.
Строение. По внешнему виду медуза резко отличается от полипа, п
общему же плану строения она, наоборот, вполне сходна с ним; она отли
чается от полипа тем, что ведет активный образ жизни в толще воды.
Медуза обелия представляет собой небольшой, около 2—3 мм в попе
речнике, зонтикообразный организм (рис. 89). По окружности она несе
большое число небольших щупалец, а от центра нижней поверхности отходи
хоботообразный вырост, на конце которого располагается рот.
Первая важная и вместе с тем характерная черта строения медуз -
зонтикообразная форма тела и сильное развитие мезоглеи. Вместо опорно
пластинки у них имеется толстый слой мезоглеи, студенистой консистенции
почти совершенно прозрачной, сильно насыщенной водой. У некоторы
медуз содержание воды доходит до 98% от их общего веса. Благодаря этом
медузы имеют очень небольшой удельный вес. что помогает им парить в толш
воды. Разрастание мезоглеи влечет за собой сильное сокращение гастрал!
116
ной полости, которая превращается у медуз в систему каналов, или гастро-
васкулярную систему.
Гастральная полость, выстланная энтодермой, начинается в хоботе и
идет в глубь тела медузы. От этой, центральной, части отходят четыре ра-
диальных канала, которые соединяются вместе кольцевым каналом, идущим
по окружности зонтика медузы.
Вторая характерная черта строения медуз — значительно более силь-
ное, чем у полипа, развитие нервной системы, образующей два нервных
скопления по обеим сторонам кольцевого канала. С этим кольцом связаны
специальные органы равновесия, расположенные по краю зонтика, так
называемые статоцисты в числе 8, по 2 в каждом квадрате, образованном
двумя соседними радиальными каналами. Статоцист (рис. 90) представляет
Рис. 89. Медуза Obelia. Один квад-
рант вырезан, чтобы показать внут-
реннее строение:
/ — рот, 2—ротовой хоботок. 3—радиаль-
ный капал, 4 — кольцевой канал (в разрезе),
5 — щупальца, 6 — статоцисты, 7 — гонада
Рис. 90. Строение статоциста
Obelia:
/ — щупальце, 2 — эктодерма,.? — ста-
тоцист, 4 — полость статоциста, 5—ста-
толит, 6 — чувствительные клетки
собой пузырек, образованный клетками эктодермы, в котором помещается
сферический известковый статолит. Изменение положения тела медузы
в толще воды сопровождается перемещениями статолита, которые и
улавливаются чувствительными клетками, расположенными в стенке пу-
зырька.
Плавают медузы очень активно, сокращая зонтик, выталкивая из-под
него воду и получая таким образом толчок для движения вперед.
Размножение и развитие. Размножаются медузы оболии исключитель-
но половым путем. Они раздельнополы. Половые железы, или гонады,
развиваются у них из эктодермы внутренней поверхности зонтика в числе 4.
Расположены они симметрично под каждым из радиальных каналов (см.
рис. 89). Половые продукты выпадают в воду, где происходит оплодотво-
рение. В результате дробления оплодотворенного яйца формируется одно-
слойная бластула. Путем миграции клеток однослойная бластула превра-
щается в двуслойную, покрытую ресничками личинку плапулу (рис. 92).
Плапула некоторое время плавает, а затем передним концом прикрепляется
к субстрату. На противоположном конце ее тела прорывается рот, вокруг
него образуются длинные выросты — щупальца, и плапула превращается
в одиночного полипа. Такой полип начинает размножаться почкованием и
образует на различных подводных предметах небольшие, в несколько сан-
тиметров высоты, древовидные колонии — гидроиды (рис. 911 )♦ состоящие
обычно из нескольких сотен и даже тысяч особей.
Отдельная особь колонии, или так называемый гидрант, по своему
строению походит на гидру (рис. 91ц). Стенка его тела, так же как и гидры
состоит из двух слоев. Основание чашечки гидранта переходит в стебелек,
который связывает каждого гидранта с общим для всей колонии стволом.
В результате этого и получается единая колония, в которой все особи свя-
1 17
заны друг с другом. Эктодерма по всей колонии выделяет плотную оболоч-
ку, которая образует наружный скелет полипа обелии. Около каждого
гидранта эта оболочка образует расширение в виде бокальчика. В расши-
рение может втягиваться при раздражении венчик щупалец. Этот скелетный
бокальчик вокруг гидранта получил название гидротэки, а скелетная обо-
лочка вокруг ветвей колонии — перисарка. Ротовое отверстие каждого
гидранта находится на выросте,
щупалец. Щупальца у обелии,
как и у всех гидроидов, в отли-
чие от гидры плотные, так как
гастральная полость в них не
заходит.
Размножается колония обе-
лии бесполым путем — почкова-
нием. Новые особи образуются
вокруг которого располагается венчик
Рис. 91. Гидроид Obelia. / — колония (немного увеличена); // — отдельная
ветка колонии (при большом увеличении), часть колонии — в разрезе, чтобы
показать внутреннее строение:
а — гидранты в расправленном состоянии, б — гидрант сократившийся,- в — почки, г—гонан-
гии: 1— ротовое отверстие, 2—эктодерма, 3 — опорная пластинка, 4 — энтодерма, 5— по-
лость гидрантов; 6 — гастральная полость ствола, 7 — бластостиль с почкующимися медузами^
8 — перисарк, 9—гидротэка, 10— гонотэка, 11— крышечка
обычно на верхней части колонии, благодаря чему колония и растет.
У обелии можно наблюдать полиморфизм среди особей одной колонии,
а именно некоторые особи отличаются от обычных как по форме, так и по
функциям. По форме это вытянутые в длину палочковидные образования с
расширением наверху (рис. 91ц); ротового отверстия нет, и поэтому, естест-
венно, такая особь не может питаться самостоятельно, она получает пищу
от гидрантов через гастральную систему колоний. Называется такое обра-
зование бластостилем. Скелетная оболочка дает вокруг бластостиля бутыл-
118
кообразное расширение — гонотэку. Все это образова-
ние в целом носит название гонангия (рис. 91ц).
В гонангии, на бластостиле, путем почкования
образуются медузы. Они отрываются от бластостиля,
выходят из гонангия и начинают вести свободный об-
раз жизни.
Биологическое значение подвижных медуз состоит в
том, что благодаря им происходит расселение гидроидов.
Экология и географическое распространение. Род
Obelia принадлежит к одному из самых распространен-
ных родов гидроидов. Различные виды его встречаются
во всех морях как в теплых, так и холодных на различ-
ных глубинах, начиная с прибрежных камней и водо-
рослей. В наших морях очень обычен вид Obelia geni-
culata, образующий небольшие кустики на различных
подводных предметах. Рис 92 Личинка
Медузы отпочковываются от гидроида в определен- планула в разрезе:
ное время года. В отличие от гидроида, который суще- /—эктодерма,
ствует довольно долго, все время увеличивая размеры г—энтодерма
своей колонии почкованием, медузы очень недолговечны, они образуют
половые продукты и отмирают.
Положение в системе. Обелия относится к отряду гидроидов (Leptolida).
СИСТЕМАТИЧЕСКИЙ ОБЗОР ПОДКЛАССА
Подкласс гидрообразных разделяется на 3 отряда: 1) гидр (Hydrida),
2) гидроидов (Leptolida) и 3) трахилид (Trachilida).
ОТРЯД ГИДРЫ (HYDRIDA)
К этому отряду, кроме гидр,
относится еще очень небольшое
число форм, например своеобраз-
ный пресноводный полип полипо-
диум (Polypodium hydriforme),
найденный в Волге, Дону, Сулаке
и в Аральском море, который на
ранних стадиях развития парази-
тирует в икринках осетровых рыб
(рис. 93).
ОТРЯД ГИДРОИДЫ (LEPTOLIDA)
Рис. 93. Polypodiuiii liydriforme. / — яичник
перлидп с несколькими аарижсппымц
икринками; 11 — икринка сторллдп С. ПП'1-
KUMU полпподпумя; /// — епободпожииу-
щпп формч
К этому отряду относятся фор-
мы, имеющие, как Obelia, сложный
жизненный цикл, состоящий па
бесполого — полипного поколения
н полового—мебуэного. Полипное
поколение в большинстве случаев
колониальное, образующее более
или менее сложные колонии, известные под названием гидроидов.
Обитают гидроиды, за редкими исключениями, в морях, где очень ши-
роко распространены; они распадаются на два подотряда: Thecaphora и
Athecata.
Obelia относится к подотряду Thecaphora, характеризующемуся на-
личием гкдротэки. К этому подотряду относится большое число очень разно-
образных форм. У ряда форм гидротэка развивается еще более сильно, чем
у Obelia, и образует ряд защитных приспособлений — крышечки и т. п.
119
У некоторых форм перисарк утолщается, и колония приобретает грубый,
жесткий вид. У ряда форм полиморфизм колонии выражен сильнее, чем у
обелии; встречаются опорные особи, защитные особи и некоторые
другие.
Медузы этого подотряда, как и медуза обелия, имеют плоский зонтик.
Они носят название лептомедуз, например Staurophora (табл. III).
Другой подотряд Athecata охва-
Рис. 94. Колония Tubularia larynx
в ест. величину (обычная форма на-
ших северных морей)
тывает гидроидов, у которых гидро-
тэка не развивается и перисарк дохо-
дит только до основания гидранта.
Примером этого подотряда может слу-
жить род Tubularia (рис. 94). Гид-
ранты многих представителей этого
подотряда отличаются также строе-
нием щупалец. У них имеются два
типа щупалец — длинные и короткие
Рис. 95. Разрез глазка медузы
Sarsia:
1 — стекловидное тело, 2 — светочувст-
вительные клетки, 3—пигментные
клетки
(булавовидные); последние на расширенной вершине содержат большое
количество стрекательных клеток.
Медузы этого подотряда обычно колоколообразные, имеют глазки,
лишены органов равновесия. Они носят название антомедуз. Примером
может служить медуза Sarsia (табл. III) (соответствующий полип носит на-
звание Syncoryne1), высота колокола который достигает 1 см. Эта медуза в
отличие от Obelia имеет четыре светочувствительных органа (глазка) в виде
темных пятнышек, расположенных по окружности колокола на местах впа-
Рис. 96. Схема строения медузы и форма ее редукции в по-
дотряде Athecata. I — типичная медуза; // — медузоид;
/// —гонофор; IV —споросак
1 Разные родовые названия многих гидроидов и медуз объясняются тем, что они
были сначала описаны независимо друг от друга, а связь между ними была установле-
на позднее.
120
дения радиальных каналов в кольцевой. Глазки эти (рис. 95) устроены срав-
нительно просто в виде эктодермических ямок, причем часть их клеток со-
держит пигмент.
Половые продукты образуются у антомедуз в эктодерме ротового хо-
ботка.
В этом подотряде можно наблюдать редукцию медузного поколения
(рис. 96). У некоторых
форм медуза развивается
как бы нормально, но не
отрывается от полипа, а
остается связанной с ним.
Такая форма медузного
поколения получила наз-
вание медузоида.
Следующий шаг в ре-
дукции медуз представляет
так называемый гонофор.
В его строении можно еще
довольно хорошо видеть
черты сходства с настоя-
щей медузой.
Наконец, поел едня я
форма редукции — споро-
сак — представляет собой
просто мешковидное обра-
зование, набитое половыми
продуктами.
У некоторых гидрои-
дов эктодерма выделяет
Рис. 97. Колония гидрокоралла Allopora stej-
negeri
известковые
скелетные
образования, которые заполняют про-
межутки между отдельными верти-
кальными ветвями колонии, в резуль-
тате чего получаются довольно мас-
сивные колонии, внешне несколько
ианоминиющие кораллы. Такие формы
получили название гпдрокораллов.
Медуз у них пет. Примером гидроко-
раллов могут служить встречающиеся
в наших дальневосточных морях пред-
ставители рода ДПорога ножно-розо-
вого цвета (рис. 97).
Экология и географическое рас-
пространение. Гидроиды распростра-
нены во всех морях и занимают п жи-
вотном мире довольно заметное место.
Они образуют более или менее густые
заросли, внешне похожие на расти-
тельные организмы, от прибрежной
полосы до больших глубин. Больше
всего встречаются гидроиды на сред-
них глубинах. Заросли гидроидов
обычно бывают сложные: один гидро-
ид селится на другом, на них часто
Рис. 98. Гидроид Cordylophora caspia:
/ — планулы, выходящие из гонофора
поселяются другие сидячие живот-
ные — губки, мшанки, а также и
водоросли. Среди этого миниатюрного «леса» поселяется и ряд подвижных
видов, часто приспособленных по форме и окраске к окружающим зарос-
121
лям. Здесь можно встретить некоторых полихет, голожаберных моллюсков,
«морских козочек» из разноногих раков, «морских пауков» (пантопод) и ряд
других организмов. Все они так или иначе используют заросли гидрои-
дов. Для некоторых они только субстрат, другие питаются ими, объедая
гидранты. Все это, вместе взятое, образует характерное морское сообщест-
во, или биоценоз, зарослей гидроидов.
Во всех наших морях, кроме Каспийского, гидроиды многочисленны и
разнообразны, так, например, в Белом море число их видов достигает 78.
Интересно распространение солоноватоводного гидроида Cordylophora
caspia (рис. 98). Он в массовых количествах обитает в Каспийском море, в
опресненных частях Азовского и Черного морей и в лиманах рек, впадающих
в эти моря, а также в Балтийском море. Медузная стадия у него выпала,
имеются гонофоры, в которых развиваются непосредственно планулы.
Медузы гидроидов также многочисленны в планктоне, особенно в опре-
деленное время года, когда они отпочковываются. Несмотря на небольшие
размеры, многие из них хорошо заметны благодаря яркой окраске, напри-
мер Sarsia (табл. Ш4).
Гидрокораллы обитают главным образом в тропических и теплых морях,
входя в состав коралловых рифов.
ОТРЯД ТРАХИЛИДЫ (TRACHYLIDA)
статоциста
краевые
энтодер-
К этому отряду относятся медузы, в большинстве не имеющие полип-
ного поколения; если оно и имеется, то рудиментарное, очень слабо развитое.
Характерной чертой медуз, относя-
щихся к этому отряду, является стро-
ение статоцистов (рис. 99). В отличие
от лептомедуз у трахилид они представ-
ляют собой видоизмененные
щупальца со статолитами в
мальных клетках.
К этому отряду относится несколь-
ко десятков форм, из которых некоторые
достигают 10 см в диаметре. Громадное
большинство этих медуз —морские оби-
татели, но некоторые встречаются и в
пресных водах. Среди последних следу-
ет особо отметить маленькую, до 2 см в
диаметре, медузку Craspedacusta (рис.
100i ). Она имеет рудиментарное полипо-
идное поколение. Соответствующий по-
лип, известный под названием Microhy-
3 —статолит
Р1.с. 99. Схема строения
трахимедузы;
1 — эктодерма, 2— энтодерма,
I
Рис. 100. Пресноводная медуза Craspedacusta sowerbii: I — молодой эк-
земпляр; II — полип Microhydra ryderi
122
dra (рис. 100ц), не имеет щупалец и достигает всего 2 мм. Встречается
эта медуза редко, причем в большинстве случаев в аквариумах и искусст-
венных бассейнах. В естественных условиях она найдена у нас в водое-
мах Тульской обл., в Дону, в водоемах Бухары и в Учинском водохрани-
лище под Москвой.
Небольшая медуза из этого отряда «крестовичок» (Gonionemusvertens),
встречающаяся в Японском море и около Южного Сахалина, действует
своими стрекательными клетками и на людей, вызывая у них не только ожоги,
но и общее отравление.
Трахилидыделятся на два подотряда: 1) трахимедузы, представитель—
Aglantha (табл. Ш5) и 2) наркомедузы, представитель — Aeginopsis (табл.
П13).
ПОДКЛАСС II. СИФОНОФОРЫ (SIPHONOPHORA)
ХАРАКТЕРИСТИКА
Свободноплавающие морские гидроидные, образующие в отличие от
медуз сложные колонии с очень резко выраженным полиморфизмом.
ОБЩИЙ ОБЗОР ПОДКЛАССА
Строение колоний. Основа колонии сифонофор — ствол, стенка тела
которого, как и у всех кишечнополостных, состоит из наружного слоя —
выстилающей гастральную по-
эктодермы, внутреннего — энтодермы,
лость и располагающейся между
эктодермой и энтодермой бесструк-
турной студнеобразной пластинки —
мезоглеи (рис. 101). Полость эта со-
общается со всеми особями колонии,
и, таким образом, они связаны в еди-
ное целое. У большинства сифонофор
ствол имеет вид стержня, на котором
располагаются отдельные особи коло-
нии, например физофора (Physophofa,
табл, JU). У некоторых сифонофор
ствол сильно укорочен, например у
португальского кораблика (Physa Ня,
табл. 11или даже превращен в пла-
стинку, ня нижней поверхности кото-
рой располагаются особи, например
у парусника (Vclella, табл. Н4).
Обычно в верхней части колонии
имеются особи, при помощи которых
вся колония держится в толще воды н
движется, ниже — питающие особи,
половые особи и пр.
Размеры сифонофор различны:
имеются и мелкие формы, например
Dimophyes (табл. II3), у которых вся
колония достигает лишь 1 см длины,
и очень крупные, например Apolemia,
достигающая 2—3 см длины.
Многие сифонофоры ярко окра-
шены; так, большой пневматофор пор-
тугальского кораблика голубой с крас-
Рис. 101. Схема строения сифонофоры.
/ — общий вид колонии; // — отдель-
ная кормидия:
/ — ствол, 2 — пневматофор, 3 — плавательные
колокола, 4 — кормидии, 5 — ствол колонии,
6 — гастрозоид, 7 — арканчик с батареями
стрекательных клеток, 8 — цистозоид, 9 — муж-
ские гонозоиды, 10 — женские гонозоиды, 11 —
крышечка
123
Таблица II. Сифонофоры и гребневики:
'Г^ГаЛЬСКИЙК°Раб\и/ <physalia) —теплые моря, 2 — Physophora hydrostatica — теплые
р м и Ю1да встречается у Мурманского берега, 3—Dimophyes — северные мооя СССР 4 пчп\г
Ч^Уе1е^а)7ТеГ1^,Ь1С; М°РЯ’ 5-ве»сРИ” пояс (Cestus vened?) - ?еплы Уморя^ - PleLrob7achiPZ
черное морс, 7 - Mertensia - северные моря СССР, 8 - Beroe cucmuis - северные моря СССР ~
новатым гребнем, а свешивающиеся от него вниз особи ярко-красные.
Парусник — тоже ярко-синего цвета.
Строение отдельных особей и их полиморфизм. На самой вершине
колонии обычно располагается содержащий газ пузырь — пневматофор
(рис. 1011 ). Газ пневматофора вырабатывается особыми, «газовыми»,желез-
ками и состоит главным образом из азота с примесью кислорода и аргона.
У некоторых сифонофор, например у португальского кораблика, он может
достигать 20—30 см в длину. Такой пневматофор поддерживает всю колонию
на поверхности воды и, выступая над нею, служит вместе с тем как бы пару-
сом, при помощи которого колония переносится ветром.
Под пневматофором располагаются плавательные колокола, или некто-
форы. Они напоминают по своему строению медуз, и благодаря их сокраще-
ниям вся колония может плыть верхним концом вперед.
В нижней части колонии располагаются другие особи, часто собранные
в небольшие группы, так называемые кормидии (рис. 101). В состав этой
части колонии обычно входят следующие особи.
1. Питающие особи, или гастрозоиды, имеющие вид полипов. Они снаб-
жены длинным подвижным придатком, или арканчиком, несущим батареи
стрекательных капсул. Пища (мелкие животные) захватывается арканчи-
ками, проглатывается и усваивается гастрозоидами, а затем поступает в
полость ствола, распределяясь по всей колонии.
2. Половые особи, или гонозоиды, имеющие вид медузоидов или гонофор.
В их стенках развиваются половые продукты мужские или женские. Колонии
могут быть раздельнополы, или гермафродитны.
3. Цистозоиды, напоминающие по своему строению гастрозоиды, но
имеющие вместо рта маленькую пору. Функция их еще не вполне выяснена;
по-видимому, они служат для экскреции, и поры их, таким образом, яв-
ляются выделительными отверстиями.
Размножение. Размножаются сифонофоры бесполым и половым путем.
Образовавшиеся в гонозоидах половые продукты выбрасываются в воду.
Из оплодотворенного яйца развивается планула, которая некоторое время
плавает, а затем начинает выпочковывать отдельные особи и превращается
в колонию; развитие последней идет, таким образом, бесполым путем.
Экология и географическое распространение. Сифонофоры — исклю-
чительно морские животные, встречающиеся главным образом в тропиче-
ских и теплых морях. Обитают они обычно в верхних слоях моря, часто вы-
ставляя свой обычно яркоокрашенный плавательный пузырь из воды. Си-
фонофоры — хищники; благодаря своему мощному стрекательному воору-
жению они могут захватывать довольно крупную добычу, даже небольших
рыб.
Иногда сифонофоры встречаются в массовых количествах (например,
«португальский кораблик», образуя медленно плывущие стаи, хорошо вы-
деляющиеся на поверхности моря яркоокрашенными плавательными пу-
зырями).
В наших морях сифонофоры представлены слабо. В южных морях их
нет совсем,в северных и дальневосточных встречается маленькая Dimophyes.
В Баренцево море иногда струями Гольфстрима заносится нежноокрашен-
ная Physophora (табл. И2).
КЛАСС 2. СЦИФОИДНЫЕ (SCYPHOZOA)
ХАРАКТЕРИСТИКА
Класс этот объединяет преимущественно крупных медуз (табл. III 1,2,6)
с сильно разветвленной системой каналов и хорошо развитыми органами
чувств и с очень слабовыраженным полипным поколением. Половые продукты
образуются у них в энтодерме.
125
Сцифоидные — исключительно морские животные, некоторые из них
очень крупных размеров. Так, например, довольно обычная в наших север-
ных морях медуза цианеа (Cyanea arctica; табл. III2) может достигать более
2 м в поперечнике зонтика и до 30 м по длине щупалец.
Представитель класса сцифоидных —
медуза аурелия (Aurelia aurita)
Внешний вид. Медуза аурелия (рис. 102, 103) имеет сравнительно
плоский зонтик, достигающий 30—40 см в диаметре. По краям диска много-
численные короткие щупальца, а также 8 краевых телец(ропалий), несущих
комплекс органов чувств.
На нижней поверхности зонтика расположено ротовое отверстие, от
которого свисают вниз четыре большие ротовые лопасти. Около ротового
отверстия помещаются четыре слепых углубления — ямки, открывающиеся
Рис. 102. Строение медузы Aurelia aurita, вид
сбоку (один квадрант вырезан, чтобы показать
внутреннее строение):
1 — рот, 2 — ротовые щупальца, 3—ропалии, 4— просве-
чивающие гонады, 5 — желудочная полость, 6 — желудоч-
ные нити, 7 — радиальный канал, 8— гонады, 9 — ямка
под гонадой
Рис. 103. Медуза Aurelia aurita,
вид снизу:
/ — рот, 2— ротовые щупальца, 3 — ро-
палии, 4 — желудочная полость, 5 — же-
лудочные нити, 6—радиальные каналы
наружу отверстиями. Аурелия имеет обычно лпловатую окраску, причем
хорошо просвечивают внутренние органы (система каналов и особенно под-
ковообразные фиолетово-оранжевые гонады).
Внутреннее строение (рис. 102, 103). Ротовое отверстие ведет в доволь-
но объемистую желудочную полость, образующую четыре неглубоких карма-
нообразных углубления. На дне их помещаются многочисленные желудоч-
ные нити железистого характера. От желудочной полости берут начало
многочисленные радиальные каналы гастро-васкулярной системы (рис. 1036),
часть которых ветвится, часть нет. Все они впадают в кольцевой канал,
идущий по краю зонтика. Радиальные каналы выстланы внутри мерцатель-
ным эпителием, благодаря которому обеспечивается движение пищи по
каналам. Аурелия — хищник, захватывающий различные планктонные
организмы.
Половые железы, или гонады, образуются из энтодермы нижней по-
верхности желудочной полости в виде подковообразных валиков позади
желудочных нитей. В одной особи развиваются или мужские, или женские
гонады, т. е. аурелия раздельнопола.
Нервная система несколько более концентрирована, чем у гидроидных
медуз: кроме нервного кольца имеются у оснований ропалий скопления
нервных клеток типа примитивных ганглиев.
126
Краевые тельца, или ропалии (рис. 104), располагаются в особых
выемках края зонтика. Сверху каждая ропалия покрыта лопастью, с верх-
ней стороны которой имеется «обонятельная» ямка. Сама ропалия удлинен-
ной, несколько изогнутой формы, в ее вершине помещается орган равнове-
сия — статоцист. На верхней и нижней поверхностях ропалии находится
по глазку, причем они по своему строению сильно друг от друга отличаются.
Верхний глазок представляет собой утолщение эктодермы, состоящее из
Рис. 104. Продольный разрез через ропалию Aurelia aurita:
1 — покровная лопасть (часть не изображена), 2—«обонятельная» ямка,
3— статоцист, 4 — верхний глазок, 5 — нижний бокаловидный глазок
многочисленных клеток, между которыми располагаются светочувствитель-
ные клетки. Нижний глазок помещается под слоем эктодермы, имеет вид
бокала и состоит из двух слоев — наружного эктодермического и внутрен-
него энтодермического.
Размножение и развитие (рис. 105). Образовавшиеся в гонадах половые
продукты выходят через ротовое отверстие. В воде происходит оплодотво-
рение и образуется свободноплавающая покрытая ресничками личинка —
планула. В некоторых случаях оплодотворение и развитие планулы происхо-
дит в материнском организме, и медуза «родит» готовых планул (например,
беломорская аурелия). Планула некоторое время плавает, затем опускается
на дно, прикрепляется, и из нее образуется маленький полип, так называемая
сцифистома, имеющая вид бокала. Сцифистома растет и затем начинает
размножаться путем своеобразного почкования. Почкование это состоит в
том, что сцифистома перешнуровывается поперечно на ряд дисков, которые
постепенно отрываются от материнского организма и превращаются в ма-
леньких медуз, так называемых Эфир. Процесс этот носит название стро-
биляции. Образовавшиеся в результате стробиляции эфиры растут и посте-
пенно превращаются во взрослых медуз. Развитие аурелии от эфиры до
половозрелой особи протекает обычно в течение нескольких месяцев
127
(в Средиземном море около четырех). Таким образом, у аурелии имеет место
чередование поколений, но в отличие от гидроидных, где основное поколе-
ние полипное, у сцифоидных, наоборот, основное поколение медузы, а по-
липное поколение очень слабо выражено.
Экология и географическое распространение. Аурелия — одна из
самых распространенных сцифомедуз. Она найдена почти во всех морях,
в частности, во всех морях СССР, кроме Каспийского и Аральского. В опре-
деленные времена года аурелии появляются массами как в открытом море,
так и у берегов. Обычно после откладки яиц медузы отмирают и иногда в
массе выбрасываются волнами на берег в виде студенистых полупрозрач-
ных дисков.
Положение в системе. Аурелия относится к отряду Discomedusae —
самому многочисленному отряду класса сцифоидных.
ОБЩИЙ ОБЗОР КЛАССА
По внешнему виду сцифомедузы различны. Колокол их может быть или
плоским, как у аурелии, такие медузы носят название дискомедуз, или,
наоборот, высоким у так называемых кубомедуз. Щупальца могут быть
Рис. 106. Схематический продольный разрез
через корнерота:
/ — каналы, 2— сосательные отверстия
короткими, как у аурелии,
или очень длинными, как у
Суапеа (табл. 1П2).
Интерес представляет
особая группа сцифомедуз,
так называемые корнероты
(рис. 106). У этих медуз ро-
товое отверстие зарастает и
вместо него образуются мел-
кие многочисленные отвер-
стия, расположенные на силь-
но развитых ротовых щупаль-
цах и связанные с гастро-
васкулярной системой сетью
каналов. Через эти отверстия
происходит питание медуз.
Пр едста вител ем кор неротов
служит Rhizostoma pulmo
128
(табл. 1116)— довольно крупная медуза, летом встречающаяся у нас в Чер-
ном море.
Очень своеобразны сидячие медузы (отряд Lucernariida) (рис. 107).
Внешне они мало похожи на настоящих медуз. Верхняя сторона их зонтика
вытянута в довольно длинную ножку, при помощи которой они прикрепля-
ются к субстрату, главным образом к водорослям. Обычно от субстрата эти
медузы свешиваются в виде полупрозрачных мягких колокольчиков. В глу-
бине такого колокольчика, на стебельке, помещается ротовое отверстие,
а по краям 8 пучков булавовидных щупалец. Между щупальцами в углуб-
ленных краях колокола у некоторых форм помещаются ропалоиды, пред-
ставляющие собой упрощенные и измененные ропалии плавающих форм.
Рис. 107. Сидячая медуза Lucernaria;
/ — ротовое отверстие, 2 — щупальца
Они не несут органов чувств, за исключением небольшого пигментного пятна
у основания, но имеют многочисленные железистые клетки. Ропалоиды —
своеобразные органы передвижения медузы по субстрату.
Примером сидячих медуз могут служить довольно обычные в наших
северных морях коричневые небольшие достигающие 2—Зсм представители
рода Haliclystus и нежно-розовые, более крупные, до 10—12 см высоты,
представители рода Lucernaria. Последний род представлен и в Черном море.
В Китае некоторые сцифоидные медузы, например Rhopilema esculenta,
являются объектами промысла, так как идут в пищу. Некоторые медузы
ядовиты: они могут сильно «обжигать» стрекательными клетками.
КЛАСС 3. КОРАЛЛОВЫЕ (ACTINOZOA)
ХАРАКТЕРИСТИКА
В этот класс входят одиночные (рис. 108) и колониальные полипы
(табл. IV), имеющие эктодермическую глотку, гастральную полость, раз-
деленную на камеры, и энтодермические поло-
вые продукты.
Ротовое отверстие коралловых полипов
в большинстве случаев не округлое, а оваль-
ное или щелевидное. В связи с этим глоточ-
ная трубка также сплющена с боков. По
узким сторонам глотки проходят две рес-
Рис. 108. Схема строения одиночного кораллового
полипа:
/ — эктодермическая глотка,- 2 — перегородка внутренней по-
лости, 3— скелет
5 Курс зоологии, т. I
129
Таблица III. Медузы морей CCCPj
1 — Aurelia aurita (сцифомедуза) — встречается во всех морях, 2 — Суапеа arctica
(сцифомедуза) — северные моря, 3—Aeginopsis (наркомедуза) — северные моря,
4 — Sarsia (антомедуза) — северные моря, 5 — Aglantlia (трахимедуза) — северные
моря, 6 — Rhizostoma pulmo (корнерот) — Черное море, 7 — Slaurophora (лептоме-
дуза) — северные моря
ничные бороздки, так называемые сифоногливьг, и по ним идет ток воды во
внутреннюю полость полипа. Гастральная полость разделена перегород-
ками (см. рис. ИЗ), отходящими от внутренней поверхности стенки тела и
выстланными энтодермой. Внутри перегородок имеются прослойка мезо-
глеи, а также мышечные валики. В верхней части полипа перегородки сра-
стаются с глоточной трубкой, в нижней — свободно вдаются в полость. На
свободном конце их имеются извилистые утолщения, содержащие желези-
стые клетки, это так называемые мезентериальные нити. У большинства
форм в той или иной степени развит обычно известковый скелет эктодерми-
ческого происхождения, наружный или лежащий в мезоглее.
Коралловые полипы — исключительно морские обитатели, встречаю-
щиеся в большинстве своем в тропических и теплых морях, требующие для
своего существования температуру не ниже 22°С. Многие из них благодаря
очень сильному развитию скелета принимают участие в образовании рельефа
земной поверхности, строя коралловые рифы и острова. Класс этот
объединяет довольно много разнообразных представителей и делится на 2
подкласса: 1) восьмилучевые (Octocorallia) и 2) шестилучевые (Нехасо-
rallia).
ПОДКЛАСС I. ВОСЬМИЛУЧЕВЫЕ КОРАЛЛЫ
(OCTOCORALLIA)
ХАРАКТЕРИСТИКА
Колониальные коралловые полипы с восемью щупальцами и восемью
мезентериальными перегородками в гастральной полости. Скелет в основе
состоит из отдельных элементов — спикул
расположенных в мезоглее. В ряде слу-
чаев спикулы сливаются вместе и обра-
зуют сплошной скелет различной фор-
мы. Восьмилучевые кораллы обитают
главным образом в тропических и теп-
лых морях на различных глубинах.
Некоторые из них очень эффектны как
по форме, например морские перья (Реп-
natularia) (табл. IV\), так и по окраске,
например красный, или благородный, ко-
ралл (Corallium rubrum) (рис. ПО).
Важно отметить, что у восьмилу-
чевых коралловых полипов глотка лишь
с одним сифоногливом. Благодаря это-
му происходит нарушение столь харак-
терной для сцифоидных медуз и гидро-
идных радиальной симметрии. Действи-
тельно, наличие лишь одного сифоногли-
ва позволяет провести только одну
плоскость симметрии, по обеим сторонам
различных по форме (рис. 109),
Рис. 109. Различные спикулы вось-
которой будут лежать одинаковые части милучевых кораллов
тела восьмилучевого коралла. Эта един-
ственная плоскость симметрии параллельна вертикальной оси тела и рассе-
кает глотку вдоль, деля тем самым пополам и единственный сифоноглив.
Проведение второй плоскости симметрии перпендикулярно к первой нево-
зможно, так как по одну ее сторону окажется часть тела с сифоногливом, а
по другую — часть тела без сифоноглива. Таким образом, в организации
5*
131
1
Рис. ПО. Кусок колонии красного коралла (Corallium rubrum), часть
дана в разрезе:
/ — полипы, два правых в разрезе, 2—ценосарк. <« — скелетный стержень
восьмилучевых коралловых полипов, кроме восьмилучевой радиальной сим-
метрии (восемь щупалец, восемь мезентериальных нитей в гастральной
полости), видна и двусторонняя — билатеральная — симметрия.
ОБЩИЙ ОБЗОР ПОДКЛАССА
Восьмилучевые коралловые полипы очень разнообразны по форме.
Среди них имеются и древовидные формы с твердым осевым скелетом на-
пример красный, или благородный, коралл, и массивные формы с трубчатым
Рис. 111. Часть колонии органного коралла
(Tubipora musica)
скелетом вокруг отдельных осо-
бей, например органный коралл
(Tubipora musica) (рис. Ill), и,
наконец, полумягкие формы со
скелетом из отдельных спикул,
например кожистые кораллы
(Alcyonium) (табл. IV).
У благородного коралла
(рис. ПО) снаружи колония по-
крыта мягкой ярко-красной ко-
рой, так называемым ценосар-
ком, в котором расположены
снежно-белые полипы. В мезо-
глее коры имеются многочислен-
ные спикулы. Внутри колонии
проходит сплошной цилиндри-
ческий известковый стержень
тоже красного, реже розового
цвета. Образуется этот стержень путем слияния отдельных спикул, закла-
дывающихся еще у личинки.
У органного коралла (рис. 111) вокруг каждого полипа образуется
скелетная трубка; такие трубки соединяются вместе горизонтальными пере-
кладинами. При росте колоний постепенно образуются новые «этажи» и
колония получается сложного многоярусного строения. У морских перьев
132
(табл. IV) колония имеет основной стержень, на верхней части которого
располагаются боковые ветви. На последних помещаются отдельные полипы.
Нижним концом колония как бы воткнута в грунт. Многие морские перья—
характерные формы глубоководной фауны, например, большое своеобразное
перо умбелулла (Umbelulla encrinus) (табл. IV3), достигающее 1 м высоты,
встречается на больших глубинах северной Атлантики и Полярного бассейна.
Некоторые восьмилучевые кораллы, в частности благородный коралл,
служат объектами промысла. Скелет благородного коралла идет на изго-
товление различных украшений — брошей, бус и пр. Основной район про-
мысла благородного коралла — Средиземное море: берега Италии, южной
Франции и т. п.
ПОДКЛАСС II. ШЕСТИЛУЧЕВЫЕ КОРАЛЛЫ (HEXACORALLIA)
ХАРАКТЕРИСТИКА
Одиночные и колониальные полипы с большим числом щупалец и пере-
городок, причем число их всегда кратно 6. Скелет или отсутствует или он
массивный. В последнем случае скелет формируется в эктодерме и облекает
тело полипов снаружи. Одиночные шестилучевые полипы, или актинии,
распространены по всем морям, колониальные — главным образом в тро-
пических.
Следует отметить, что наличие двух сифоногливов в глотке шестилу-
чевых коралловых полипов значительно ограничивает их радиальную
симметрию. Через тело этих кораллов можно провести две плоскости сим-
метрии. Таким образом в их организации прослеживается двулучевая сим-
метрия на фоне шестилучевой радиальной симметрии.
Представитель подкласса шестилучевых кораллов —
обыкновенная, или конская, актиния
(Actinia equina)
Внешнее строение (рис. 112). Актиния имеет вид плотного, но мягкого
цилиндра, одним концом прикрепленного к субстрату. На противополож-
ном несколько выпуклом конце расположено удлиненное ротовое отверстие,
окруженное многочисленными сравнительно короткими полыми щупаль-
цами. Цвет актиний очень изменчив:
встречаются, например, экземпляры
ярко-красные с голубой каймой (Чер-
ное море) и светло-зеленые (северные
моря).
Внутреннее строение (рис. 113).
Ротовое отверстие ведет в глотку,
несколько сдавленную с боков, с дву-
мя сифоногливами на обоих концах.
Внутренняя полость с многочислен-
ными перегородками (септами). В
верхней части полости перегородки
доходят до глотки и прирастают к ней,
образуя слепые карманы, переходя-
щие кверху в полость щупалец. Септы
выстланы с внутренней стороны энто-
дермой, а в центральной части состо-
ят из мезоглеи, врастающей внутрь
септы. Каждая перегородка имеет мы-
Рис. 112. Обыкновенная, или кон-
ская, актиния (Actinia equina) —
обычная форма всех наших морей (в
ест. величину)
133
шечный валик, состоящий из продольной мышцы. На концах септ, направ-
ленных в полость, располагаются мезентериальные нити. Нервная система
актиний построена несколько более сложно, чем у гидроидных полипов; у
них имеется довольно большая концентрация более крупных нервных
клеток вокруг ротового отверстия.
Размножение. Обыкновенные актинии раздельнополы. Половые про-
дукты образуются в энтодерме перегородок. Оплодотворение и развитие
Рис. 113. Поперечные разрезы Actinia equina. / — на уровне глотки;
II — в нижней части тела:
/ — эктодерма. 2—энтодерма, 3—глотка, 4 — сифоногливы, 5— камеры внут-
ренней полости, 6 — перегородки (септы) между ними, 7—мышечные валики, 3—
мезентериальные нити
происходят в гастральной полости, и вполне сформированные маленькие
актинии выходят из тела матери через ротовое отверстие. Таким образом,
обыкновенную актинию можно считать живородящей.
Рис. 114. Группа актиний на раковине
с раком-отшельником. Две актинии в
расправленном состоянии, две сократи-
вшиеся
Экология и географическое
распространение. Обыкновенная
актиния распространена очень ши-
роко, она встречается во всех мо-
рях в прибрежной зоне. Актинии
поселяются обычно на нижней сто-
роне камней. Иногда актинии
встречаются в очень больших ко-
личествах. Актинии — хищники;
при помощи стрекательных капсул,
в большом количестве располо-
женных на щупальцах, они могут
оглушать даже мальков рыб и по-
том затягивать их в ротовое от-
верстие. Актинии часто поселяют-
ся на других животных. Наиболее
известен случай симбиоза актиний
и рака-отшельника (рис. 114). По-
следние живут обычно в пустых
раковинах брюхоногих моллюсков,
пряча в них задний мягкий отдел
своего тела. Подрастая, рак-от-
134
шельник меняет раковину, переносит на нее клешнями и актинию. Сим-
биоз рака-отшельника и актинии заключается в том, что актиния защищает
рака своими стрекательными капсулами, а он переносит ее с места на место
и дает возможность ей питаться остатками и отбросами своей пищи.
Положение в системе. Актиния относится к отряду Actiniaria, объе-
диняющему одиночных лишенных скелета коралловых полипов.
ОБЩИИ ОБЗОР ПОДКЛАССА
Основная масса шестилучевых кораллов — колониальные формы, имею-
щие хорошо развитый скелет. Среди них наиболее обширная группа — мад-
репоровые, или каменистые, кораллы. Они очень разнообразны по форме и
имеют вид или ветвистых деревьев, иногда достигающих нескольких метров
Рис. 115. Часть Большого австралийского кораллового рифа при
отливе. Видны колонии кораллов, различных по форме
высоты, или округлых массивных образований (табл. IV). Скелет мадрепоро-
вых кораллов известковый, сплошной для всей колонии. Каждая особь
помещается в собственной ячейке. Образование скелета начинается с выде-
лением эктодермой подошвы отдельной особи известковой пластинки (см.
рис. 108). От этой пластинки скелет затем как бы врастает в полипа, образуя
известковые выступы, соответствующие его перегородкам, а также кольцо
вокруг него. При дальнейшем размножении колонии путем почкования
скелеты отдельных особей сливаются и образуется единый мощный скелет
всей колонии. Таким образом, скелет как бы внутренний, покрытый
сверху мягкими частями.
Мадрепоровые кораллы имеют обычно разнообразную яркую окраску.
В тропических морях, где температура не опускается ниже 20°С, они раз-
135
растаются особенно сильно и образуют рифы и острова. Особенно многочис-
ленны они в Тихом океане.
Рифы могут быть различные — береговые и барьерные. Первые распо-
ложены близ берегов и обнажаются при отливе (рис. 115). Они поражают
яркостью своих красок. Обычно на кораллах и среди них поселяется боль-
шое количество различных других животных (иглокожие, моллюски, черви,
рыбы и Др.). Все они имеют также яркую окраску, причем иногда пятнистую
или полосатую, помогающую им делаться невидимыми среди ярких ко-
раллов.
Барьерные рифы располагаются на некотором расстоянии от берега.
Громадный барьерный риф, находящийся около восточного берега Австралии,
тянется на протяжении 1400 км.
Коралловые острова обычно имеют вид атолла, т. е. выдающегося из
моря на небольшую высоту кольца, внутри которого находится лагуна.
Рис. 116. Коралловый остров —атолл
Возникновение атолла объясняется следующим образом. Он образуется
на подводных возвышенностях, которые ранее выдавались над поверхно-
стью океана, азатем—или вследствие опускания морского дна, или вследст-
вие разрушающего действия атмосферы оказались под водой. Внутренняя
часть коралловой постройки постепенно разрушается в результате раство-
ряющего действия морской воды, обогащенной углекислотой, которая обра-
зуется в результате жизнедеятельности полипов. Снаружи, наоборот, все
время продолжается созидательная работа кораллов, и надстраиваются
все новые и новые слои извести.
На атолл постепенно наносятся ветром почва и семена растений,
и он превращается в цветущий островок (рис. 116). В Тихом океане
таких коралловых островов очень много.
Необходимо отметить, что для правильного понимания различных ко-
ралловых построек многое сделано Ч. Дарвином, собравшим обширные
материалы по этому вопросу во время путешествия на корабле
«Бигль».
Рост кораллов происходит довольно быстро. Так, например, корабль,
затонувший в Персидском заливе, через 20 месяцев оброс коркой кораллов
в 60 см толщиной.
Коралловые рифы известны и в ископаемом состоянии с древнейших
геологических эпох, причем самые древние рифы построены ныне вымер-
шими четырехлучевыми кораллами (Tetracorallia).
136
Таблица IV. Коралльи
1 — морское перо Pennatnla (восьмилучевые) — теплые моря, 2— Gorgonaria (восьмилучевыс)
— теплые моря (а— часть колонии при большом увеличении), 3— C’mbelulla cncrinus (вось-
милучевые)— северная часть Карского моря, северная Атлантика, 4—Alcyoninm (восьми-
лучевые) — теплые моря, 5 — Astrea (шестилучевые) — тропические моря
Подтип П. Нестрекающие (Acnidaria)* 1 1
ХА РА ХТЕРИСТИХА
Свободноплавающие одиночные
не имеющие стрекательных клеток. В
3
кишечнополостные (табл. 115,6,7,в),
плане их строения наряду с лучевой
симметрией имеется и двулучевая
(рис. 117). Кроме эктодермы и энто-
дермы, во время развития обра-
зуется зачаток мускулатуры щу-
палец. Исключительно морские
формы. Всего известно около 100
представителей. Подтип представ-
лен одним классом гребневиков
(Ctenophora).
Рис. 117. Схема строения гребневика
(поперечный разрез):
1— ряды гребных пластинок, 2 —щупальца, 3—
аборальный орган, 4 — рот, 5 — глотка, 6 — же-
лудок, 7 — отходящие от желудка горизонталь-
ные каналы, дважды дихотомически ветвящи-
еся, 8— вертикальные каналы, идущие под
рядами гребных пластинок, 9 — влагалища щу-
палец
ОБЩИЙ ОБЗОР КЛАССА
Наружное строение. По образу жизни и общему виду гребневики
несколько похожи на медуз; они так же свободно плавают в толще воды и
имеют в связи с этим нежное, полупрозрачное, студенистое тело. Сходство,
это, однако, только кажущееся, так как по строению тела гребневики сильно
отличаются от медуз. Большинство медуз имеет овальное или грушевидное
тело (табл. II), а гребневик «венерин пояс» (Cestus veneris), (табл. II5) —
удлиненное лентообразное, имеющее свыше 1 м длины.
На одном конце тела помещается рот, на другом — особый орган чувств —
аборальный орган2, У одних гребневиков (подкласс беещупальцевых)
тело лишено придатков (табл. 115,б)» У других (подкласс щупальцевых)
имеется пара щупалец, расположенных обычно строго симметрично по обеим
сторонам тела (табл. IIgj)- Вдоль тела в меридиональном направлении
проходят восемь рядов так называемых гребных пластинок, которые пред-
ставляют собой сросшиеся крупные реснички кожных клеток. Под рядами
гребных пластинок расположены пучки подкожной мускулатуры, благо-
даря которой и происходит движение пластинок, а тем самым и движение
самого гребневика, плавающего ротовым концом вперед2.
Внутреннее строение (рис. 117). Ротовое отверстие ведет в сплющенную
эктодермическую глотку. Глотка коротким пищеводом соединяется с объе-
мистым желудком. Желудок сплющен в направлении, перпендикулярном
к глотке, и почти достигает аборального конца, к которому подходит двумя
1 Ввиду большого отличия нестрекающих от остальных кишечнополостных мно-
гие исследователи сейчас выделяют их в особый тип — гребневиков (Ctenophora).
2 У «венерина пояса» рот и соответственно аборальный орган находятся посере-
дине тела.
138
1
Рис. 119. Развитие гребневиков:
I — эктодерма, 2 — энтодерма, 3— зачаток мускулатуры
щупалец
отростками. От основания желудка отходят каналы, аналогичные радиаль-
ным каналам медуз, но расположенные не по радиальной, а по двубокой сим-
метрии. В горизонтальном направлении от желудка отходят два канала,
которые затем дважды дихотомически ветвятся. В результате этого к по-
верхности тела в местах расположения рядов гребных пластинок подходят
8 каналов. Здесь от этих каналов отходят в меридиональном направлении
2 канала: один — вниз к ротовому концу, другой — вверх к аборальному.
Таким образом все тело гребневика прони-
зано системой каналов, подобной гастро-
васкулярной системе медуз.
Между каналами и поверхностью те-
ла лежит мезоглея, которая по происхо-
ждению (см. ниже), а также по многочис-
ленным мышечным элементам, находящимся
в ней, может рассматриваться как прими-
тивная мезодерма, или средний зародыше-
вый листок.
Нервная система гребневиков состоит
из субэпителиального сплетения нервных
клеток, вокруг рта и под рядами гребных
пластинок сплетения эти более уплотнен-
ные и образуют околоротовое кольцо и 8
продольных тяжей, сходящихся к абораль-
ному органу.
Аборальный орган (рис. 118) является органом равновесия. Основа
его — статолит, который поддерживается четырьмя пучками S-образно
изогнутых ресничек. Сверху все это образование прикрыто в виде колпачка
пучком сросшихся ресничек. Статолит находится все время в колебательном
движении вследствие сгибания и разгибания поддерживающих его ресничек.
Эти колебания передаются к гребным пластинкам и координируют их дви-
жение. Удаление абораль-
ного органа ведет к нару-
шению согласованной ра-
боты гребных пластинок.
Размножение и раз-
витие. Размножаются
гребневики исключительно
половым путем, причем
все они гермафродиты. Го-
нады развиваются из энто-
дермы в каналах, которые
расположены под рядами
гребных пластинок. Поло-
вые продукты выводятся
через ротовое отверстие.
Оплодотворение происхо-
дит в воде. Развитие простое, без личиночных стадий и чередования
поколений. Дробление неравномерное (рис. 119); на одном полюсе зародыша
образуются мелкие бластомеры — микромеры, на другом — крупные макро-
меры. Микромеры обрастают макромеры и образуют эктодерму. Из макро-
мер формируется энтодерма. Часть клеток энтодермы уходит в промежуток
между эктодермой и энтодермой и образует особую группу клеток, из кото-
рой затем развиваются клеточные элементы мезоглеи и мускулатура щу-
палец.
Экология и географическое распространение. Гребневики встречаются
во всех морях, хотя большинство их приурочено к умеренным и теплым.
Населяют они преимущественно верхние горизонты. Вследствие того, что
у них очень нежное тело, они не переносят сильного движения воды и при
139
наступлении бурной погоды опускаются на большие глубины. Многие греб-
невики светятся, испуская довольно яркий голубоватый свет. В наших мо-
рях гребневики представлены хорошо: встречаются как в северных морях,
например довольно крупный, достигающий 20 см Вегбе cucumis (табл. П8),
так и в Черном море, например небольшой, имеющий пару щупалец Pleu-
robrachia rhododactyla (табл. II6).
Особый отряд среди гребневиков составляют так называемые ползаю-
щие, или плоские, гребневики, куда входит небольшое число форм, отно-
сящихся главным образом к двум родам Coeloplana (рис. 120) и Ctenoplana.
Организмы эти представляют большой интерес, так как они внешне напо-
минают ресничных червей (см. ниже).
Рис. 120. Coeloplana gonoctena, вид сверху;
/ — аборальный орган, 2—щупальца
Впервые представитель этой группы (Coeloplana metschnikowi) был
найден в Красном море нашим выдающимся зоологом и эмбриологом
А. О.Ковалевским. Второй представитель (Ctenoplana kowalevski) был
найден в Малайском архипелаге тоже русским ученым А. А. Коротневым.
Плоские гребневики овальны по форме с сильно сплющенным в направ-
лении главной оси, т. е. сверху вниз, телом. На нижней поверхности имеется
ползательная подошва, на верхней — аборальный орган, пара щупалец и у
Ctenoplana еще и восемь рядов гребных пластинок. Благодаря наличию
последних Ctenoplana может ползать и плавать. Coeloplana, не имея таких
гребных пластинок, не может плавать, а способна лишь ползать по суб-
страту и подвешиваться снизу к пленке поверхностного натяжения.
Личинка Coeloplana имеет гребные пластинки, которые в процессе раз-
вития редуцируются. Таким образом Coeloplana может рассматриваться,
как форма, измененная больше, чем Ctenoplana, и дальше отошедшая от
исходных форм.
ФИЛОГЕНИЯ КИШЕЧНОПОЛОСТНЫХ
Кишечнополостные — безусловно примитивные многоклеточные оста-
новившиеся в своем филогенетическом развитии на стадии двуслойного
организма без настоящих органов. Это позволяет ставить кишечнополост-
ных у основания филогенетического древа всех остальных многоклеточных.
Можно думать, что исходной формой кишечнополостных были свободно-
плавающие организмы типа паренхимеллы Мечникова. Наиболее примитив-
ной группой из всех кишечнополостных, стоящей ближе всего к исходным
формам, является класс Hydrozoa, представители которого не имеют экто-
дермической глотки, обладают очень простой мезоглеей и эктодермическими
половыми продуктами.
Остальные классы кишечнополостных представляют дальнейшее услож-
нение исходного типа, оно выражается в образовании эктодермической
глотки, сложной гастральной системы, в формировании более сложной ме-
140
зоглеи и, наконец, в наличии энтодермических половых продуктов. Вместе
с тем необходимо отметить, что филогенетическое развитие двух основных
классов высших кишечнополостных — сцифоидных и коралловых — пошло
по разным путям. У первых почти совершенно выпадает полипная стадия,
и основную роль играет стадия медузная. У вторых, наоборот, медузная
стадия совершенно выпала и остается только полипная, достигающая здесь
максимальной сложности строения.
Несколько особняком стоят гребневики; они относятся безусловно к
высшим кишечнополостным. Обладая наряду с элементами радиальной
симметрии двулучевой симметрией, мускулатурой щупалец, более сложно
устроенной нервной системой и аборальным органом, детерминированным
дроблением, они представляют собой особую ветвь кишечнополостных (см.
табл. XVI, стр. 518).
ПАРЕНХИМА ТОЗНЫЕ ЧЕРВИ,
ПЕРВИЧНОПОЛОСТНЫЕ ЧЕРВИ
И НЕМЕРТИНЫ
До недавнего времени три группы червей: паренхиматозные, или плос-
кие, черви (Plathelminthes), первичнополостные, или круглые, черви (Ne-
mathelminthes) и немертины (Nemertini) вместе с кольчатыми червями (см.
далее) — относили к одному типу — к типу червей (Vermes). В настоящее
время доказано, что плоские черви, первичнополостные черви, немертины,
кольчатые черви — это четко разграниченные отдельные типы. Вместе с
этим в организации животных, принадлежащих к этим типам, есть общие
черты.
Эти важнейшие общие черты организации следующие: а) наличие перед-
него конца тела с тем или иным комплексом органов чувств; б) билатераль-
ная симметрия', в) наличие кожно-мускульного мешка, т. е. стенки тела, со-
стоящей из обособленных друг от друга кожного эпителия и мышц; г) на-
личие в эмбриональном развитии третьего зародышевого листка — мезо-
дермы, за счет которой формируются важнейшие структуры и органы.
Охарактеризуем вкратце основное значение этих новых в сравнении с
кишечнополостными черт организации рассматриваемых далее групп.
Значение выработки переднего конца тела. Передний конец — место
сосредоточения органов чувств', органов зрения, обоняния, осязания, т. е.
системы рецепторов, с помощью которых животные ориентируются в окру-
жающей среде. Естественно, что наличие этих органов связано с сосредото-
чением на переднем конце нервных клеток, т. е. связано с образованием го-
ловных ганглиев или других форм концентрации нейронов. Все это, взятое
вместе, усиливая возможности ориентации, позволяет совершенствовать
поступательное движение; оно фактически становится единственной формой
движения животных.
Значение билатеральной симметрии. В прямой связи с развитием по-
ступательного движения, т. е. движения в одном определенном направлении—
передним концом вперед, развивается и билатеральная симметрия, сменяю-
щая собой радиальную симметрию кишечнополостных. Билатеральная,
или двусторонняя, симметрия характеризуется тем, что тело можно условно
разделить только одной плоскостью, по обеим сторонам которой будут ле-
жать части тела с одинаковыми структурами и органами.
Билатеральная симметрия усугубляет дифференциацию организма,
так как одновременно с правой и левой сторонами тела возникают брюшная
и спинная стороны. В соответствии с возникшими сторонами тела распола-
гаются и внутренние органы, что ведет к усилению структурированности
142
организма. Возникновение брюшной стороны позволяет животным нетолько
плавать в толще воды, но и ползать по субстрату. Это важнейшее приобрете-
ние, позволяющее освоить новые условия обитания.
Значение развития кожно-мускульного мешка. Напомним, что у кишеч-
нополостных защитная и сократительно-двигательная функции совмеща-
лись в одной эпителиально-мышечной клетке. Разделение этих функций и
образование самостоятельной мускулатуры, подстилающей покровный
эпителий, сильно увеличивает подвижность животных, расширяет возмож-
ность проникновения в новые условия жизни и освоение их.
Значение развития мезодермы. Весьма вероятно, что новая функция —
ползание по субстрату — оказалась ведущей причиной развития другого
важного признака — формирования третьего зародышевого листка, или
мезодермы. Выше было указано, что мезодерма, точнее ее зачатки, возникли
уже у ктенофор. Развитие мезодермы у червей протекает несравненно более
интенсивно, и это явление несомненно связано с усилением двигательной
функции, в свою очередь обусловившей дальнейшее развитие мезодерми-
ческих элементов, за счет которых, в частности, развилась мышечная си-
стема. Если двуслойные радиальные животные способны к относительно
медленным и, как правило, плавным движениям, то уже у примитивных
свободноживущих билатеральных животных — ресничных червей — дви-
гательная функция выражена значительно интенсивнее. Интенсификация
двигательной функции увеличила быстроту перемещения в пространстве,
а это новое качество имело разнообразные и важные последствия.
Прежде всего оно имело своим следствием более быструю смену жизнен-
ной обстановки, а следовательно, и усовершенствование рецепторов, по-
вышение их чувствительности и скорости их реакций на внешние факторы.
Кроме того, интенсификация движения и развитие совершенных рецепторов
привели к возможности значительно более успешного захвата пищи и, сле-
довательно, к интенсификации питания. Отсюда общее повышение интенсив-
ности жизненных процессов и прежде всего обмена веществ. Поэтому уже у
плоских червей мы видим значительное развитие именно перечисленных
выше систем: нервной и двигательной, пищеварительной и выдели-
тельной.
Однако развитие мезодермы имело и другие жизненно важные послед-
ствия. Так, за счет мезодермы у плоских червей формируется паренхима —
резервуар питательных веществ. Паренхима приобрела весьма важное зна-
чение как орган, значительно повышающий сопротивление животного голо-
данию, отчасти замедлила процессы высыхания (при выходе на сушу) и т. п.
Эти факторы в их связях с преимуществами билатеральной симметрии от-
крыли перед животным миром значительно более широкие возможности
эволюции и в частности имели своим следствием несравненно более интен-
сивные преобразования организации в самых различных приспособи-
тельных направлениях. Поэтому, если кишечнополостные остаются только
в водной среде, то уже свободноживущие плоские черви — турбеллярии —
частично проникают во влажную наземную среду. Если первые обитают
преимущественно в морях, то вторые перешли и в пресные воды. Если ки-
шечнополостные почти не дали паразитических форм, то плоские черви ши-
роко перешли к паразитизму, и два класса плоских червей — сосальщи-
ки и ленточные черви — представлены только паразитическими фор-
мами.
Итак, становление переднего конца тела, билатеральная симметрия,
развитие мезодермы и кожно-мускульного мешка получили очень большое
значение как фундаментальные признаки повышения организации, более
многостороннего приспособления к различным условиям существования и
многообразной эволюции в самых различных направлениях, к пышному
развитию жизни в условиях более сложной наземной среды. Можно сказать,
что с возникновением плоских червей открывается новая страница в исто-
рическом развитии животного мира.
143
ТИП IV. ПЛОСКИЕ, или
ПАРЕНХИМАТОЗНЫЕ,
ЧЕРВИ (PLATHELMINTHES)
ХАРАКТЕРИСТИКА
Тело у большинства листовидное или лентообразное, обычно сплющен-
ное в спинно-брюшном направлении. Размеры тела колеблются от долей
миллиметра до нескольких метров. У некоторых форм явственно выражена
закономерная повторяемость комплексов органов, соответствующая столь
же закономерному внешнему расчленению (сегментация, или метамерия).
Снаружи тело одето кожно-мускульным мешком, состоящим из эпите-
лия и мускулатуры, как правило многослойной и сложной. У паразитиче-
ских форм тело покрыто плотной кутикулой. Полости тела нет, внутренние
органы всегда погружены в паренхиму, имеющую преимущественно мезо-
дермическое происхождение. Пищеварительная система у большинства
форм в виде замкнутого пищеварительного мешка иногда сложно развет-
вленного. У некоторых видов пищеварительная система представлена только
скоплением пищеварительных клеток, погруженных в паренхиму. У неко-
торых паразитических форм пищеварительная система вторично отсутст-
вует и питание происходит осмотическим путем через кожу. Кровеносная
система и органы дыхания у плоских червей отсутствуют. Процессы дыхания
происходят через кожные покровы, а у кишечных паразитов имеет место
наряду с аэробным—анаэробное дыхание (брожение), состоящее в расщепле-
нии в отсутствие кислорода органических веществ (главным образом гли-
когена), находящихся в теле этих червей. Выделительная система представ-
лена протонефридиями. Нервная система состоит из головных нервных
узлов (ганглиев) и различного числа продольных нервных стволов, в боль-
шинстве случаев связанных поперечными комиссурами; наибольшего раз-
вития обычно достигают два боковых ствола. Половая система, кроме еди-
ничных случаев, гермафродитная с рядом добавочных желез, имеющих
значение в процессе формирования яиц и их оболочек.
Развитие у большинства форм сложное, сопровождающееся сменой
поколений (полового и бесполого) и сменой хозяев.
В настоящее время известно более 5500 видов плоских червей, часть из
них — свободноживущие формы, обитающие в морях, пресных водах и на
суше во влажных условиях, но большинство — паразиты животных. Не-
которые формы паразитируют и у человека.
КЛАССИФИКАЦИЯ
Тип плоские, или паренхиматозные, черви (Plathelminthes)
Класс 1. Ресничные черви, или турбеллярии (Turbellaria)
Отряд 1. Бескишечные турбеллярии (Acoela)
Отряд 2. Многоветвистокишечные (Policladida)
Отряд 3. Трехветвистокишечные (Tricladida)
Отряд 4. Прямокишечные турбеллярии (Rhabdocoela)
Класс 2. Моногенетические сосальщики (Monogenea)
Класс 3. Трематоды (Trematoda)
Подкласс I. Дигенетические трематоды (Prosostomadidea)
Подкласс II. Брюхоротые трематоды (Bucephalididea)
Подкласс III. Аспидогастроиды (Aspidogastrodidea)
Класс 4. Ленточные черви (Cestoidea)
Подкласс I. Нерасчлененные цестоды (Cestodaria)
Подкласс II. Расчлененные цестоды (Cestoda)
Отряд 1. Лентецы (Pseudophyllidea)
Отряд 2. Цепни (Cyclophillidea)
144
КЛАСС 1. РЕСНИЧНЫЕ ЧЕРВИ, ИЛИ ТУРБЕЛЛЯРИИ
(TURBELLARIA)
ХАРАКТЕРИСТИКА '
Турбеллярии — основной класс плоских червей; в основном они сво-
бодноживущие формы, населяющие моря, пресноводные бассейны и влажную
почву. У турбеллярий значительного развития достигает нервная система,
хорошо развиты органы чувств, они быстро реагируют на внешние факторы
и в частности на свет и присутствие пищи. Все они — хищники, и этот факт
несомненно способствовал более интенсивному развитию органов чувств,
ловчих и защитных приспособлений, интенсификации двигательных реак-
ций. Как правило, все они — мелкие формы. Кожа несет ресничный покров.
Рот всегда на брюшной стороне. Ануса нет. Кожно-мускульный мешок
состоит из эпителия и сложной системы мышц. Центральная нервная си-
стема представлена продольными стволами и головными ганглиями. Ки-
шечник либо отсутствует, либо имеет вид прямой или ветвящейся трубки.
Пищеварение, как правило, внутриклеточное. Дыхание кожное. Кровенос-
ная система отсутствует. Функция выделения принадлежит кишечнику и
специальным органам выделения — протонефридиям. Как правило, герма-
фродиты. Половое отверстие на брюшной стороне. Развитие прямое или с
превращением.
Представитель класса ресничных червей
— молочная планария (Dendrocoelum lacteum)
Форма тела и внешняя организация. Молочная планария хорошо отли-
чается от других видов молочно-белым цветом тела. У наевшихся особей
на белом фоне тела ясно просвечивает темный ветвистый кишечник. Эта
относительно крупная форма достигает 15—26 мм в длину при поперечнике
тела примерно 6 мм. Головной конец тела поперечно срезан, задний — за-
остренно округлен. Позади переднего края головного конца расположены
явственные черные глаза, а под основанием его развита присасывательная
бороздка. Все тело молочной планарии покрыто равномерным ресничным
покровом. Работа ресничек обеспечивает плавное скольжение по субстрату,
которое столь характерно для этого червя, как и для других мелких видов.
На брюшной стороне, несколько кзади от середины тела, расположено
ротовое отверстие, а примерно посередине между ним и задним концом
тела — половое. Мельчайшие боковые выделительные отверстия в коли-
честве 8 пар расположены дорзально. Обитающая в наших пресных водах
молочная планария обычно держится под различными подводными предме-
тами, где или неподвижно сидит на месте, или плавно ползет по субстрату
в поисках пищи. Тело молочной планарии отличается присущей всем тур-
белляриям гибкостью и эластичностью, способно удлиняться и сокращаться.
Животное относительно быстро и плавно скользит по субстрату, по подвод-
ным листьям, по затененной поверхности погруженных камней, по нижней
стороне пленки поверхностного натяжения воды.
Кожно-мускульный мешок состоит из однослойного наружного эпи-
телия и подлежащей мускулатуры. Эпителий состоит из тесно поставленных
удлиненных клеток, включающих особые палочковидные образования —
рабдиты, которые могут выбрасываться наружу, расплываясь при этом в
слизь. Местами, пронзая эпителиальный слой, проходят открывающиеся
наружу протоки специальных желез, имеющих отчасти значение приспо-
соблений для прикрепления к субстрату, отчасти же ядовитой природы
(защитные железы). Под эпителием расположена бесструктурная опорная
мембрана, а под ней — мускулатура. Последняя слагается из нескольких
145
ного мешка и кишечником. Она
слоев гладких мышц. Непосредственно под базальной перепонкой находится
слой поперечных кольцевых мышечных волокон (рис. 121). Под этими пос-
ледними расположены более мощный слой продольных мышечных волокон
и тонкая прослойка косых волокон; кроме того, пронизывая внутреннюю
паренхиматозную толщу тела, со спины на брюхо тянутся дорзо-вентраль-
ные мышечные волокна.
Мерцательный эпителий кожи, а равно все три прилегающих к нему
мышечных слоя (продольные, кольцевые и косые мышцы) образуют кожно-
мускульный мешок, облекающий всю внутреннюю массу тела, т. е. парен-
химу с заключенными в ней внутренними органами. Что касается дорзо-
вентральных мышц, то они составляют мускулатуру паренхимы.
Паренхима заполняет все пространство между стенкой кожно-мускуль-
состоит из нежной соединительной ткани,
образующей губчатый остов, в котором
расположены многочисленные и доволь-
но разнородные по функции клетки.
Паренхиматозные клетки имеют много-
угольную форму. Обычно от тела клетки
отходят несколько отростков, которые
переплетаются с отростками соседних
клеток. Однако клетки могут быть и без
отростков или, например, лишь с двумя
отростками и т. д. Паренхима в целом —
важный орган червя. В ней накаплива-
ются запасы питательных веществ и в
то же время за счет этой ткани идут,
например, регенеративные процессы, в
ней образуются различные специализо-
ванные клеточные элементы и т. д. Так,
в паренхиме (рис. 121) находятся осо-
бые клетки, образующие упомянутые
рабдиты. Когда рабдиты достигают нор-
мальной величины, они выходят из обра-
зовательных клеток и затем постепенно
проталкиваются в эпителий кожи. По-
видимому, рабдиты должны рассматри-
ваться как особый внутриклеточный
секрет, который застывает в форме
плотных светопреломляющих телец палочковидной формы. Рабдиты могут
образовываться и в эпителии кожи.
Нервная система слагается из головного нервного узла и нервных ство-
лов. Головной нервный узел, или «мозг», состоит из скопления нервных
клеток и явственно слагается из двух частей (правой и левой). Следователь-
но, мозг молочной планарии — парное образование (рис. 122). От мозга,
расположенного на головном конце тела, отходят вперед короткие голов-
ные нервы, направляющиеся к органам чувств. К заднему концу тела от
головного мозга отходят довольно мощные продольные брюшные нервные
стволы и значительно более слабые спинные.
От продольных брюшных стволов вбок отходят многочисленные боко-
вые нервные тяжи. Нервные стволы состоят из нервных волокон и нервных
клеток, рассеянных вдоль стволов, в особенности в местах отхождений бо-
ковых ветвлений.
Органы чувств. К ним относятся глаза и осязательные головные лопасти.
Молочная планария имеет два глаза (рис. 123). Глаз состоит из бокала
(рис. 124), имеющего пигментную выстилку. В бокал погружены зритель-
ные колбы, несущие на своих концах чувствующие «штифтики». Зри-
тельные колбы служат продолжением лежащих вне бокала многочисленных
удлиненных зрительных клеток, как бы вставленных в бокал. От зритель-
Рис. 121. Часть поперечного среза
через тело молочной планарии Den-
drocoelum lacteum, схема:
1 — эпителий, 2—поперечные (кольцевые)
мышечные волокна, 3—косые мышечные
волокна, под ними кружки поперечных се-
чений продольных мышечных волокон, 4 —
дорзо-вентральные мышечные волокна, 5 —
клетки паренхимы, 6 — клетки, образующие
рабдиты, 7 — рабдиты в эпителиальных
клетках, 8 — одноклеточная кожная железа,
10 — реснички эпителиальных клеток
146
них клеток идут нервные волокна, связанные с головным мозгом посредст-
вом зрительного нерва.
Зрительный бокал обращен вогнутой стороной к свету, следовательно,
к поверхности кожи. Лучи света сперва падают на зрительные клетки, а
затем уже на зрительные колбы. Это обращенные (инвертированные) глаза.
Рис. 122. Передняя часть нервной системы
Crenobia alpina, головной нервный узел
(«мозг») обозначен пунктиром:
/—щупальца, 2—8—мозговые нервы, 9 — зрительный
нерв, 10 — краевой нерв, 11— боковые нервы, 12—
брюшные продольные нервы, 13 —спинные продольные
нервы
Рис. 123. Организация молочной
планарпп Dendrocoelum lacleuni:
/ — ротовое отверстие, 2 — глотка. 3 —
передняя ветвь кишечника, •/ — правая
ветвь кишечника, 5 — задние ветви
кишечника, 6 — головной нервный узел,
7 — боковой нервный ствол, 8 —глаза,
9 — канал выделительной системы, 10 —
выделительные поры
Головные лопасти имеют осязательную функцию. Кроме того, в ко-
же рассеяны многочисленные осязательные клетки, причем к ним под-
ходят разветвленные нервные окончания периферической нервной системы
(рис. 125).
Рис. 124. Глаза планарип. / — Dendrocoelum lacteum; // — Euplanaria gonocephala:
/ — зрительные колбы, 2— зрительные клетки, расположенные вне пигментного бокала, 3—клетки
зрительного бокала, 4—ядро клетки зрительного бокала, 5—эпителий кожи
147
Пищеварительная система. Пищеварительная система молочной пла-
нарии (рис. 123) начинается расположенным на брюшной стороне тела ро-
товым отверстием, снабженным запирательными мышцами. Ротовое отвер-
стие ведет в особую полость, в которой, как в кармане, помещается муску-
листая глотка. Глотка может наподобие языка выдвигаться наружу и тогда
превращается в хоботообразный орган, при помощи которого планария
ощупывает добычу и затем крепко к ней присасывается.
Просвет глотки открывается в полость ки-
шечника, который имеет у молочной планарии
трехветвистую форму: одна ветвь направлена
вперед, а две — назад. Все три ветви имеют
многочисленные (от 20 до 30) вторичные развет-
вления, которые в свою очередь ветвятся на еще
более мелкие веточки. Заднепроходного отвер-
стия нет.
Полость кишечника выстлана однослойным
эпителием (рис. 126). Свободные концы клеток
Рис. 125. Нервные окон-
чания в коже молочной
планарии
имеют округленно-расширенную колбовидную форму. Клетки способны за-
хватывать частицы пищи. Вокруг захваченных пищевых частиц в плазме
клеток образуются пищеварительные вакуоли. Неперевариваемые части
пищи выбрасываются снова в полость кишечника*
Рис. 126. Строение стенки кишечной трубки и паренхимы молоч-
ной планарии:
1 — пищеварительные клетки кишечника (среди них одна железистая—5),
2—3—мышечные волокна паренхимы, 4—6—клеточные элементы паренхи-
мы, 7 — желточные клетки
Из сказанного следует, что эпителиальные клетки кишечника планарии
обладают фагоцитарной функцией и что молочной планарии свойственно
внутриклеточное пищеварение.
Кроме фагоцитарных клеток, в стенке кишечника местами имеются
особые зернистые клетки, выделяющие в просвет кишки клейкий секрет,
обволакивающий частицы пищи.
Непереваренные остатки пищи скопляются в кишечнике и затем вы-
брасываются наружу через ротовое отверстие сильными сократительными
движениями всего тела.
148
Дыхание. Дышит планария поверхностью тела.
Кровеносная система. Отсутствует.
Выделительная система. Представлена протонефридиями; они состоят
из более или менее разветвленных правого и левого каналов (рис. 123),
наполненных жидкими продуктами выделения. Жидкость выделяется на-
ружу через выделительные поры на одном конце разветвленного протоне-
фридиального канала; противоположные концы канала никогда не имеют
отверстий и слепо замыкаются терминальными клетками. Отмеченные здесь
отношения — характерная черта протонефридиев. Терминальный орган —
клетка, от которой начинает-
ся выделительный канал. В
полость канала от клетки
отходит плотный пучок рес-
ничек, напоминающий коле-
блющееся пламя, — отсюда и
название этого аппарата «рес-
ничное пламя». Эта клетка и
есть аппарат, выбирающий из
паренхимы жидкие продукты
Рис. 127. Терминальная клетка
выделительной системы с '«рес-
ничным пламенем»:
/ — группа ресничек («ресничное пла-
мя»), 2— ядро клетки
Рис. 128. Схема половой системы планарии:
1— семенники, 2— семявыносящие канальцы, <?—4—семяпро-
вод, 5 — общий проток семяпровода, 6—7 — семенной пузырь
цирруса, 8— совокупительный орган, 9 — семяизвергательный
канал, 10 — железистый орган, 11— его полость, 12 — bursa1
copulatrix, 13—ее проток. 14—половая клоака, 15 — непарный
проток яйцевода, 16 — яичник, 17 — яйцевод, 18 — желточники
выделения. Поступив в канал, жидкость в нем течет по направлению к вы-
делительным порам, и этот равномерный и постоянный ток жидкости
обеспечивается столь же равномерной и постоянной работой «реснич-
ного пламени» (рис. 127).
Половая система. Планарии гермафродиты.
Мужской половой аппарат (рис. 128) состоит из много-
численных (200—300) пузырьковидных семенников, расположенных по
бокам тела. Семя выводится из них посредством выводящих канальцев
(vasa efferentia), которые впадают в семяпроводы (vas deferens). Семяпро-
воды вливаются в семенной пузырь (vesicula semimalis; рис. 128). В нижней
части семенной пузырь образует семяизвергательный канал (ductus jecula-
torius), открывающийся в совокупительный орган (cirrus). Совокупительный
орган может выдвигаться в конусовидный карман, в свою очередь откры-
вающийся в половую клоаку.
149
Женский половой аппарат состоит из двух яичников,
выводящих протоков и придаточных желез. Яичники расположены вблизи
головного конца тела. Яйца выводятся в длинные боковые яйцеводы. По-
следние соединяются в общее влагалище, которое заканчивается отверстием,
расположенным на границе между карманом совокупительного органа и
половой клоакой (рис. 128ц). Яйцевод молочной планарии сопровождается
желточниками. Желточные клетки, поступающие из этих органов в яйцевод,
идут на питание яйцевых клеток и вместе с яйцами выводятся по яйцеводу
в половую клоаку. Оплодотворенные в половой клоаке яйца облеплены
многочисленными желточными клетками.
К половому аппарату принадлежит также копулятивная (сово :упи-
тельная) сумка (bursa copulatrix). Во время свойственного молочной плана-
рии перекрестного оплодотворения семя партне-
>ра попадает сперва в копулятивную сумку, от-
куда оно по выводному каналу стекает в поло-
вую клоаку, оплодотворяя здесь яйца.
В половую же клоаку у молочной планарии
открывается проток железистого мешка (рис.
128i ). Его функция у D. lacteum остается не-
ясной, у других планарий железистый мешок
может выдвигаться наружу. Вовремя совокупле-
ния он вводится в клоаку партнера и наполня-
ется его семенем. Затем железистый мешок
втягивается обратно и изливает содержимое в
канал копулятивной сумки.
Размножение. Как говорилось выше, опло-
дотворение у молочной планарии перекрестное.
Рис. 129. Кокон планарии Каждый партнер вводит свой cirrus в копуля-
тивную сумку другого партнера. Яйца оплодо-
творяются семенем в половой клоаке.
Комплект таких яиц, облепленных и перемешанных с желточными
клетками, покрывается затем оболочкой, образующейся за счет веществ,
выделяемых желточными клетками. Эта оболочка быстро затвердевает, и в
результате образуется кокон, который и откладывает червь на подводные
предметы. Кокон молочной планарии имеет округлую форму и содержит
от 5 до 42 эмбрионов (рис. 129).
Экология. Dendrocoelum lacteum обитает как в стоячих, так и в теку-
чих водоемах, но все же предпочитает спокойную воду, где обычно прячется
под различными подводными предметами, встречаясь j<aK в чистой, так и в
не слишком сильно загрязненной воде. В то время как многие другие тур-
беллярии весьма оксифильны (кислородолюбивы)-, молочная планария в
этом отношении менее требовательна. Кроме того, она способна жить в соло-
новатой воде лиманов. Эти свойства позволяют определить молочную пла-
нарию как животное эвритопное, т. е. способное приспособляться к
различным физико-химическим условиям существования. Молочная пла-
нария, однако, избегает прямого солнечного света, предпочитая затененные
места. Резкие изменения температуры воды влияют на животное губи-
тельно.
Наевшиеся особи неподвижно сидят на месте, плотно прижавшись к
субстрату. Голод стимулирует движения (см. ниже). Ведя хищный образ
жизни, Dendrocoelum lacteum хватает различных мелких животных, на-
пример водяных блох, водяных осликов, мелких личинок насекомых и т. д.
Описаны случаи, когда D. lacteum овладевала личинками мотыля (Chiro-
nomus). Движения жертвы, по-видимому, связываются слизью, образовав-
шейся за счет распавшихся в воде рабдит.
Мелкие животные просто прилипают к поверхности тела планарии.
В овладении добычей большую роль играет упоминавшаяся выше присасы-
вательная бороздка, а также выдвигающаяся наружу глотка, которая
150
выделяет специфические вещества, разрушающие, по-видимому, ткани
жертвы.
Рабдиты и кожные железы также служат приспособлениями к защите.
Плотно прижавшаяся к грунту планария прочно удерживается на месте
благодаря действию присасывательной ямки и клейких желез брюшной сто-
роны. Эти особенности имеют значение защитных приспособлений.
ОБЩИЙ ОБЗОР КЛАССА
Молочная планария может служить представителем класса; однако ее
организация вовсе не исчерпывает огромного многообразия строения раз-
личных форм, принадлежащих к классу ресничных червей.
Прежде всего значительной видовой изменчивостью отличается форма
тела этих животных. Если плоская форма очень распространена, то наряду
с ней встречаются веретеновидная и лентовидная формы тела. У веретено-
видных форм тело в поперечном сечении, как правило, круглое. Размеры
колеблются от 0,2 до 35 см.
Кожно-мускульный мешок. Кожно-мускульный мешок в общем имеет
указанное выше строение. Однако не всегда эпителий лежит в
виде правильного ряда на подстилающей базальной мембране, и
у многих форм он приобретает характер погруженного эпителия,
граничащего в этих случаях непосредственно с паренхиматозными клетка-
ми. Не всегда тело покрыто сплошным ресничным покровом. Напротив, у
многих форм он редуцирован, и спинная сторона тела частично или полно-
стью лишена его. У некоторых паразитических турбеллярий его вообще
нет. Эпителий богат различными железами и специальными образованиями
защитного значения. Сюда относятся прежде всего рабдиты, обычно более
многочисленные на спинной стороне тела (защитная функция и средство
для ловли добычи, обволакиваемой слизью, образовавшейся за счет расплы-
вающейся массы рабдит). Отметим, что реакция на ранение обычно выра-
жается в выделении множества рабдит вокруг раны. Рана при этом покры-
вается образовавшейся слизью.
Кожные железы погружены в паренхиму, и в эпителии можно
видеть только их протоки. Сюда относятся клейкие железы, расположенные
обычно по краям тела и образующие как бы железистое краевое кольцо.
Ползущая планария оставляет поэтому позади себя слизистый след. Слизь
облегчает движение по субстрату. Указанные железы одноклеточны. Много-
клеточные грушевидные железы белкового состава обладают ядовитыми
свойствами и расположены обычно на брюшной стороне, имея значение
приспособлений для нападания на добычу.
Прядильные железы у Mesostoma herenbergi (см. рис. 139)
выделяют слизистое вещество, в воде тотчас застывающее в нить. При
помощи этой нити животные прикрепляются к поверхностной пленке во-
ды; таким образом нить служит своеобразным якорем, обращенным не
вниз, а к поверхности воды. Железы эти расположены на брюхе, по его
срединной линии.
У представителей Rhabdocoela и Polycladida (см. ниже) в коже наб-
людаются крапивные клетки, вполне соответствующие таким же клеткам
кишечнополостных. В действительности и было показано, что эти крапив-
ные клетки принадлежат кишечнополостным — жертвам хищных турбел-
лярий. У последних крапивные клетки подобно рабдитам попадают в кожу.
Мышцы образуют 3 упоминавшихся слоя мускулатуры: кольцевой,
косой и продольной.
Паренхима. Паренхима имеет описанное выше строение. Толща ее
всегда пронизана дорзо-вентральной мускулатурой. Кроме этой последней,
в паренхиму почвенных Tricladida включены продольные и поперечные
мышечные волокна. В паренхиму включены также различные железы.
151
У многих форм в ней есть зерна пигмента, присутствие которого обуслов-
ливает разнообразную и часто яркую окраску многих видов.
Нервная система. Нервная система обладает значительным разнообра-
зием в разных группах турбеллярий. У Rhabdocoela обычно имеется впе-
реди скопление нервных клеток — головной нервный узел — «мозг», от
которого вперед отходит ряд тяжей, иннервирующих передний конец тела,
назад направлены два мощных боковых ствола, которые обычно дают по-
перечные ветви в области глотки (рис. 130). Tricladida имеет организацию,
сходную с описанной выше для Dendrocoelum lacteum (рис. 131). У Polycla-
dida от головного мозга назад отходит ряд стволоа. У форм с округлым
Рис. 130. Нервная сис-
тема Rhabdocoela:
/ — первый узел, 2 — брюш-
ные продольные стволы, 3 —
глоточное кольцо, 4—задне-
глоточная комиссура
Рис. 131. Нервная систе-
ма Tricladida в продоль-
ном (/) и поперечном (II)
сечениях:
1 — нервный узел, 2 — боко-
вые нервные стволы, 3 — кра-
евой кольцевой нерв, 4 — по-
перечные нервные комиссуры,
5 — спинные стволы
Рис. 132. Нервная система
Policladida:
J — нервный узел, отодвинутый на-
зад, 2 — главные боковые нервные
тяжи
телом головной мозг отодвинут к центру тела, а нервные стволы отходят
от него в виде лучей (рис. 132). Главные различия в строении глаз у разных
форм зависят преимущественно от количества зрительных клеток, связан-
ных с зрительным бокалом. Глаза инвертированные (см. стр. 147). Чаще
имеются два глаза, однако известны и многоглазые формы. В этих случаях
глаза располагаются по переднему краю тела. В коже турбеллярий разбро-
саны многочисленные осязательные клетки. Щупальца многих Tricladida
и Polycladida также несомненно служат органами осязания. К ним от
«мозга» подходит пучок нервов. К органам химического чувства принад-
лежат ресничные ямки, которые, например, у многих Rhabdocoela и Tricla-
dida располагаются либо в виде вдавлений по бокам головы, либо в форме
поперечных бороздок на брюхе или даже в виде кольцевых мерцательных
борозд. У ряда форм эти органы располагаются в области глотки. У Meso-
stoma и, вероятно, у других форм известны реорецепторы, воспринимаю-
щие движение (течение) воды.
К органам чувств принадлежат также статоцисты — органы равно-
весия. Они обычны у Acoela, но встречаются и у других форм. Статоцисты
152
Рис. 133. Выделительная
система Mesostoma ehe-
renbergi:
1 — начальные отделы с тер-
минальными клетками, 2—вы-
делительные каналы, 3—вы-
делительные поры, 4—ки-
шечник
имеют вид замкнутых пузырьков с заключенным внутри гомогенным из-
вестковым отолитом; расположены в области мозга.
Пищеварительный тракт. Пищеварительный тракт у ряда форм пол-
ностью отсутствует (Acoela). В этих случаях ротовое отверстие, лежащее
на брюхе, ведет в глотку, которая переходит в группу пищеварительных
клеток, охваченную паренхиматозными клетками. Эта группа клеток пред-
ставлена плазматической массой с ядрами, следовательно, синцитием.
Жертва, захваченная червем, попадает в синцитий, как в большую амебу.
Здесь пища переваривается, а остатки выбрасыва-
ются через рот.
У всех других турбеллярий, относящихся к
отрядам Rhabdocoela, Tricladida и Polycladida,
кишечник имеется. Ротовое отверстие всегда ле-
жит на брюхе либо ближе к переднему, либо к задне-
му концу тела. Рот ведет в глотку (pharynx), кото-
рая, как у молочной планарии, может выдвигаться
из своего кармана. У многих форм из отряда Tricla-
dida глотка может достигать значительной длины,
высовываясь наружу. Глотка снабжена специаль-
ной мускулатурой — кольцевой и продольной —
и ведет в кишку. Строение последней весьма
различно. У Rhabdocoela кишка имеет либо форму
прямой трубки (отсюда и название отряда), либо
форму объемистого мешка. У Tricladida кишечник
устроен по типу кишечника Dendrocoelum lacteum,
т. е. состоит из трех ветвящихся стволов. У Poly-
cladida он состоит из нескольких стволов, отходя-
щих во все стороны от глотки. Пищеварение идет
по типу, описанному для D. lacteum. Анального
отверстия не бывает.
Кровеносная и дыхательная системы отсут-
ствуют. Дыхание осуществляется через кожные
покровы.
Выделительная система. Выделительная систе-
ма отсутствует только у бескишечных форм, у ко-
торых выделительные функции несет паренхима.
В сущности паренхима остальных турбеллярий,
являясь многофункциональным органом, тоже участвует в процессах вы-
деления. Но образовавшиеся здесь жидкие продукты попадают в русло
специальных выделительных каналов описанных выше протонефридиев.
У Rhabdocoela эти каналы начинаются уже описанными терминальными
(конечными) клетками, то одиночными, то в виде целых пакетов. У неко-
торых форм все мелкие каналы впадают в один срединный выделитель-
ный канал, заканчивающийся задней выделительной порой. У других име-
ются два главных выделительных ствола и одна задняя пора или два
ствола с соответствующими двумя порами, которые открываются либо в
глотку, либо в область полового отверстия (рис. 133). Во всех случаях
протонефридиальные каналы имеют лишь наружные отверстия (выдели-
тельные поры) и никогда не бывают сквозными, что составляет отличите-
льный признак этого органа выделения.
У многих форм функция выделения принадлежит кишечнику, тогда
как протонефридии играют, по-видимому, роль органов регуляции осмо-
тического давления. Снижение последнего у всех форм, живущих в гипото-
нических растворах (пресной воде), достигается путем выведения воды
из тела посредством системы протонефридиев. По-видимому, собственно
выделительная функция протонефридиев является вторичной, и, следова-
тельно, в данном случае мы имеем дело с примером расширения функций
органа.
153
ренная часть последней наполнена
Рис. 134. Половая система Mesostoma
eherenberg-i:
/ — половое отверстие, 2—3 — желточники,
4 — желточный проток, 5 — яичник. 6—7—мат-
ка, 8— семеприемник, 9 и 13 — семяпроводы,
10 — яйцевод. // — совокупительная сумка,
12 — место впадения желточного протока в
матку, 14 — пенис
Половая система. Семенники либо имеют описанное выше строение
семенных пузырьков или долек, либо удлиненных мешковидных тел, от
которых справа и слева отходит соответствующий семяпровод. Яичники
также либо шарообразны, либо представлены удлиненными образованиями,
в которых можно видеть последовательные фазы созревания половых клеток.
Придаточные органы половой системы могут достигать большой слож-
ности. На рис. 134 изображен в качестве примера половой аппарат пред-
ставителя Rhabdocoela — пресноводной турбеллярии мезостомы (Meso-
stoma ehrenbergi). По бокам тела у мезостомы имеются два семенника,
от которых отходят семяпроводы. Семяпроводы изливают семя в cirrus.
Последний имеет форму опрокинутой колбы или, вернее, реторты. Расши-
*менем. Семя по семяизвергательному
каналу (узкая часть реторты) посту-
пает в половую клоаку, которая пере-
ходит в длинный общий проток (duc-
tus communis), ведущий в семеприем-
ник, связанный с непарным яичником
(рис. 134). Наконец, от половой кло-
аки отходит еще двураздельная мат-
ка. Протоки желточников также
изливаются в половую клоаку. Сюда
же открывается проток совокупитель-
ной сумки. При перекрестном оплодо-
творении семя партнера попадает в
совокупительную сумку и отсюда в
ductus communis и семяприемник.
В последнем яйца оплодотворяются и
затем по общему протоку достигают
половой клоаки. Здесь они формиру-
ются, т. е. облепляются желточными
клетками и покрываются скорлупой.
Сформировавшиеся яйца затем
проталкиваются в оба ствола матки,
которая сильно разрастается.
Немногие формы турбеллярий
раздельнополы. Дробление яиц в раз-
ных группах турбеллярий протекает
различно. У представителей отряда
бескишечных (Acoela) процесс дроб-
ления яиц протекает по спиральному типу. Этот тип дробления харак-
терен и для отряда Polycladida.
Иной характер носит дробление яиц у части прямокишечных и трех-
ветвистых турбеллярий. У последних, как сказано при описании молочной
планарии, яйца обильно облеплены желточными клетками. По-видимому,
этот факт влияет на дробление, которое приобретает неправильный харак-
тер (так называемое «анархическое» дробление), причем первоначально
бластомеры дробящегося яйца даже разобщены друг от друга желточным
питательным материалом, в который они погружены. Только позднее на-
чинается дифференцировка эмбриональных органов за счет скопления
части бластомеров. Можно считать, что эти уклонения в форме дробления
вторичного характера и что спиральное дробление яиц все же типично
для класса турбеллярий.
Развитие турбеллярий либо прямое, либо (у Polycladida) с превраще-
нием. В последнем случае развивается так называемая мюллеровская ли-
чинка (рис. 135). Она имеет яйцевидное тело, снабженное восемью лопас-
тями, края которых усажены ресничками. На брюшной стороне личинки
имеется ротовое отверстие, которое ведет в глотку и кишечник, погружен-
ный в паренхиму. Личинка некоторое время плавает, затем ее лопасти
154
редуцируются, она теряет способность плавать и опускаться на дно моря,
превращаясь в молодую турбеллярию.
Некоторым формам свойственно бесполое размножение посредством
деления (см. рис. 140), например Microstomum lineare.
Экология. Турбеллярий широко распространены по всему земному
шару. В морях и океанах они ведут донный образ жизни, в большинстве
в литоральной зоне, заселяя здесь песок, илистые грунты и водоросли.
Море — основная среда обитания этой группы животных; именно в море
они, по-видимому, и возникли. Поэтому морские бассейны наиболее бога-
ты по числу видов турбеллярий.
Турбеллярий, проникшие в пресные воды, принадлежат к более спе-
циализированным группам из числа Rhabdocoela и Tricladida.
Известны также формы, населяющие пещерные водоемы, виды, обита-
ющие во временных водоемах, в озерах, прудах и т. п.
Большое число видов живет во влажном прибре-
жном мху и, наконец, в почве, разумеется, в усло-
виях высокой влажности почвы и воздуха. Почвенные
формы нуждаются также в относительно высокой
температуре, поэтому они приурочены преимущест-
венно к тропическим лесам.
Турбеллярий ведут преимущественно хищный
образ жизни. Круг их жертв довольно обширен.
Так, они поедают многих простейших (инфузорий,
корненожек, жгутиконосцев), нематод, мелких оли-
гохет, мелких ракообразных, личинок мотыля, личи-
нок комаров. Некоторые формы турбеллярий ' напада-
ют на своих собратьев. Гидра с ее защитными крапив-
Рис. 135. Мюллеров-
ская личинка Jungia
aurantiaca, поздняя
стадия. Изображена с
брюшной стороны.
Видны усаженные
ресничками лопасти
(/) и прикрывающая
ротовое отверстие
«верхняя губа» (2).
над ней расположены
глазки (3}
ными клетками также подвергается их нападениям.
Морские поликлады нападают даже на моллюсков и
асцидий. Довольно значительное число видов пере-
шло к паразитическому существованию. При пе-
реходе к паразитной фазе существования у них
редуцируются глаза, исчезают рабдиты и, напротив,
мощно развиваются половые органы. Эти факты, как
увидим ниже, делают вероятным предположение, что
турбеллярий сыграли роль исходных форм, за счет
которых развились классы сосальщиков и цестод.
В биологии турбеллярий значительную роль
играет высокая регенеративная способность. Так х/219 часть тела способна
регенерировать в целое животное. Впрочем, у разных видов регенеративная
способность выражена в разной степени. У некоторых форм она очень вели-
ка, тогда как, например, у Dendrocoelum lacteum она значительно слабее.
Турбеллярий в связи с широким распространением и глубокой эколо-
гической дифференцировкой достигли значительного морфологического и
физиологического разнообразия. В связи с этим они характеризуются
большим систематическим разнообразием.
СИСТЕМАТИЧЕСКИЕ ОБЗОР КЛАССА
ОТРЯД БЕСКИШЕЧНЫЕ ТУРБЕЛЛЯРИЙ (ACOLELA)
Маленькие бескишечные формы. Глотка не выдвижная (рис. 136).
Развитие прямое, без превращения. Большей частью живут в море, преи-
мущественно у берегов (литоральные формы). Наиболее известны предста-
вители родов Proporus и Convoluta, в частности интересна С. rosscoffensis.
В паренхиме этого червя живут зеленые водоросли — зоохлореллы, свя-
занные с червем отношениями симбиоза. Червь способен подгибать боковые
края тела, принимая форму трубки.
155
Рис. 136. Бескишечная тур-
беллярия Convoluta convo-
luta. Слева животное свер-
нулось в трубку
Рис. 137. Представители Polycladida.
/ — Cestoplana rubrocineta; II —Sty-
lochoplana maculata; III — Prost-
hiostomum siphunculus; IV — Plana-
ria graffi; V— Thysanozoon brocchii.
У Prosthiostomum siphunculus нару-
жу торчит длинная глотка
Рис. 138. Представители Tricladida. / — Polycelis cornuta (по краю
головного конца тела — глаза), II —Dendrocoelum lacteum—молочная
планария; III — Euplanaria gonocephala; IV — Polycelis nigra
п Ш дг
Рис. 139. Mesostoma ehrenbergi. I —животное сидит на листе; //—III—
два экземпляра того же вида, захватившие рачков, висят на «паутинных»
нитях, черта над ними — пленка поверхностного натяжения воды; IV —
анатомия Mesostoma ehrenbergi, впереди виден «мозг», по средней линии —
кишечная трубка, в передней ее трети — глотка, по бокам кишечной труб-
ки — покоящиеся яйца
ОТРЯД МНОГОВЕТВИСТОКИШЕЧНЫЕ ТУРБЕЛЛЯРИЙ
(POLYCLADIDA)
Кишечник имеет более трех боковых ветвей. Глотка выдвижная. Го-
ловной нервный узел кзади от головного конца. Сюда отнссятся много-
численные морские формы (рис. 137); среди последних мно-
гие довольно крупные. Так, лептоплана (Leptoplana tremel-
laris) достигает 12 лш в длину, Thysanozoon brocchii — 5 см.
У нас обычны живущие в Черном море Sty lochus tauricus,
L. tremellaris; мурманские виды L. borealis, Acelis arctica и
другие.
ОТРЯД ТРЕХВЕТВИСТОКИШЕЧНЫЕ ТУРБЕЛЛЯРИЙ
(TRICLADIDA)
Кишечник имеет три ветви: одна обращена вперед, две —
назад. Глотка выдвижная. Размножение преимущественно по-
ловое. Головной нервный узел на переднем конце тела.
Отметим довольно обычных: молочную планарию (Dend-
rocoelum lacteum; рис. 138), а равно Crenobia alpina, Eupla-
naria gonocephala (рис. 138), E. lugubris, многоглазую Poly-
cel is и др. Эти виды живут в пресных водах. Известен также
ряд морских форм.
ОТРЯД ПРЯМОКИШЕЧНЫЕ ТУРБЕЛЛЯРИЙ (RHABDOCOELA)
Кишечник в виде прямой трубки либо в виде мешка.
Глотка типичных форм снабжена бульбусом. Головной нерв-
ный узел обычно на переднем конце тела. В большинстве—
пресноводные. Некоторые формы живут в море и во влажной
Рис. 140. Microstomum lineare в момент поперечного деления
157
почве. В пресных водах обычны Microstomum lineare, Mesostomum eheren-
bergi (рис. 139). Некоторым рабдоцелидам, кроме полового, свойственно
бесполое размножение посредством поперечного деления (рис. 140).
ФИЛОГЕНИЯ ТУРБЕЛЛЯРИИ
Вопрос о происхождении турбеллярпй представляет большой теорети-
ческий интерес, так как другие классы плоских червей, а также ряд более
высокоорганизованных групп первичнополостных несомненно связан в своем
происхождении с ними (см. ниже). В связи с этим еще в конце прошлого
века по этому вопросу было высказано несколько теорий.
Заслуживают наибольшего внимания две из них, а именно: первая,
выводящая турбеллярпй из плоских гребневиков (А. Ланг), и вторая —
из планулообразных предков кишечнополостных (Л. Графф).
В настоящее время более обоснованной может считаться вторая теория,
разработанная и обоснованная В. Н. Беклемишевым.
Основана эта теория на сравнительном анализе турбеллярпй и кишечно-
полостных. Анализ показал, что главная ось турбеллярпй, идущая от
переднего конца к заднему, соответствует главной осп кишечнополост-
ных. Несмотря на сдвиг рта на брюшную сторону, повлекший за собой
превращение радиальной симметрии в двустороннюю, в организации тур-
беллярпй (особенно многоветвистокишечных) сохранились следы радиаль-
ной симметрии. Прототипом современных турбеллярии можно считать
первичнопланктонных кишечнополостных. Эти формы являлись равным
образом и предками современных гребневиков.
КЛАСС 2. МОНОГЕНЕТИЧЕСКИЕ СОСАЛЬЩИКИ
(MONOGENEA)
К этому классу принадлежат плоские черви (Platoda), существующие
как эктопаразиты. Название класса определяется характерной биологи-
ческой чертой его — развитием без превращения и без смены хозяев.
Важнейшие дифференциальные признаки моногеней следующие:
1) паразитируют в качестве эктопаразитов рыб, амфибий и рептилий;
2) ротовой мышечной присоски нет, либо имеются одна или две;
3) главный фиксаторный аппарат лежит на заднем конце тела и снаб-
жен, как правило, крючьями;
4) глаза у взрослых форм есть;
5) экскреторная система открывается наружу двумя отверстиями,
расположенными обычно на переднем конце тела;
6) развитие без метаморфоза и без смены хозяев.
Представитель класса моногеней —
гиродактиль (Gyrodactylus elegans)
Мелкие формы, около 0,5 мм в длину. Эктопаразит жабер рыб семей-
ства карповых. Тело удлиненное, суженное к противоположным концам
(рис. 141). На переднем конце тела развиты две головные лопасти с отвер-
стиями головных желез. Кзади от лопастей, на медиальной продольной
линии, расположено ротовое отверстие. Задняя часть тела несет большой
и сложный фиксаторный аппарат (диск) с двумя большими крючка-
ми, скрепленными склеротизированной перемычкой. По краям диска раз-
виты 16 дополнительных крючьев. Глаза (фоторецепторы) отсутствуют.
Ротовое отверстие ведет в мышечную глотку, переходящую в пищевод,
который вливается в среднюю кишку, представленную двумя ветвями —
158
правой и левой. Ануса нет. Выделительная система (протонефридии) со-
стоит из каналов, открывающихся наружу двумя отверстиями, лежащими
на спинной стороне, вблизи переднего конца тела. Развитие с живорож-
дением. Уже в материнском организме, т. е. в теле взрослой особи, форми-
руется зародыш, в котором, в свою очередь,
еще 3 зародыша — каждый новый в каждом
тия известен под названием
последовательно развиваются
предыдущем. Этот тип разви-
полиэмбрионии.
int egerri-
Рис. 142. Polystomum
mum. I— взрослая форма; II — ли-
чи нка:
1 — ротовое отверстие, 2—кишечник, 3 —
крючья, 4 — яичник, 5 — матка
Рис. 141. Gyroda-
ctylus elegans:
l —фиксаторный аппа-
рат
Яйца развиваются в яичнике, расположенном в задней части тела.
Паразит вызывает заболевание кожи рыб, известное под названием гиро-
дактилеза, поражающего по преимуществу мальков и годовиков. При от-
сутствии лечения молодые рыбы обычно погибают.
ОБЩИИ ОБЗОР КЛАССА
Моногенеи — мелкие формы, достигающие в длину долей миллиметра
или немногим более нескольких миллиметров. Организация этих форм
своеобразна и определяется прежде всего их эктопаразитическим сущест-
вованием. Моногенеи — эктопаразиты наружных органов рыб, амфибий
и пресмыкающихся. Местами их паразитирования служат кожа, жабры,
ротовая полость, реже мочевой пузырь соответствующих форм названных
выше животных. Эктопаразитизм привел к отбору на развитие специаль-
ных мощных фиксаторных аппаратов и органов чувств. Для моногеней
характерно наличие, в частности, фоторецепторов («глаз»), сходных с ана-
логичными органами турбеллярий. Обычно моногенеи имеют вытянутую
форму тела с одной главной продольной осью симметрии. Головной конец
у разных видов построен различно. Он либо лопастной (как у гиродак-
тиля) или снабжен головными присосками, либо нет ни присосок, ни ло-
пастей. Ротовое отверстие лежит обычно в передней части брюшной поверх-
159
ности тела. На заднем конце тела расположена либо присоска примерно
того же строения, что у гиродактиля, либо одна присоска без крючьев,
либо целая система фиксаторных аппаратов, как у Polystomum integerri-
mum (рис. 142).
Нервная система. Имеет вид продольных стволов, из них одна
пара дорзальная, другая — латеральная или брюшная. Последнее, по-
видимому, гомологичны нервным стволам турбеллярий. К задним фикса-
торным дискам подходят специальные нервы — иннервирующие присоски,
как это имеет место у Diplozoon paradoxum.
Пищеварительная система. Ротовое отверстие, как правило, распо-
ложено тотчас кзади от переднего конца тела, медиально, на брюшной
Рис. 143. Diplozoon paradoxum. / — внешний вид спаривающихся особей:
// — половая система спаривающихся особей
стороне. Ротовое отверстие ведет в мышечную глотку, впадающую в пище-
вод. Последний переходит в среднюю кишку, чаще двураздельную, в виде
двух длинных стволов. У Dactylogyrus оба ствола смыкаются и образуют
арку.
Дыхательная и кровеносная системы. Моногенеи дышат через кожу.
Кровеносной системы нет.
Выделительная система. Состоит из двух протонефридиальных ка-
налов, всегда раздельных и открывающихся наружу соответственно дву-
мя отверстиями, лежащими обычно в задней части тела.
Развитие. Развитие всегда прямое, без метаморфоза, без смены хозяев.
У Polystomum integerrimum (рис. 142) свободноплавающая личинка силь-
но напоминает взрослую форму, отличаясь меньшими размерами, чем у
взрослых форм и наличием 5 поясов ресничек, при помощи которых она
плавает в воде. В дальнейшем личинка проникает в жаберную полость
головастика; здесь у нее развиваются присоски, а ресничный покров исче-
зает. Обычно дефинитивное развитие паразита (достижение половозрелого
состояния) наступает после превращения головастика в лягушку. К этому
времени личинка паразита по пищеварительному тракту проникает в клоаку
лягушки и в ее мочевой пузырь — место конечной локализации паразита.
Здесь и достигается половая зрелость. Яйца из мочевого пузыря лягушки
попадают в воду, проделывают указанный выше путь развития и служат
источником формирования новых поколений.
Diplozoon paradoxum паразитирует на жабрах рыб. При наступлении
половозрелого состояния две особи попарно и притом крест-накрест спа-
риваются, женские и мужские трубки срастаются, чем обеспечивается
скрепление этих эктопаразитов и возможность перекрестного оплодотворе-
ния, типичного для этого вида (рис. 143).
160
КЛАСС 3. ТРЕМАТОДЫ, ИЛИ ДИГЕНЕТИЧЕСКИЕ
СОСАЛЬЩИКИ (TREMATODA)
ХАРАКТЕРИСТИКА
К классу трематод принадлежат плоские черви (платоды), ведущие
эндопаразитическое существование, локализуясь в органах различных жи-
вотных и человека. У трематод развитие протекает с превращением (мета-
морфозом) и со сменой хозяев — промежуточных и окончательного (дефи-
нитивного). В этой связи у трематод отсутствует и типичный для моноге-
ней (эктоларазитических сосальщиков) фиксаторный аппарат с крючьями.
Органы чувств редуцированы, глаза у взрослых форм всегда отсутствуют.
Ротовое отверстие обычно снабжено ротовой мышечной присоской. Пи-
щеварительный аппарат состоит из переднего глоточного отдела и следую-
щего за ним короткого пищевода, переходящего в среднюю кишку; послед-
няя представлена двумя продольными стволами — правым и левым, иногда
ветвящимися вторично. У некоторых форм оба ствола смыкаются в кольцо.
Ануса нет. На брюшной стороне тела имеется вторая присоска, не сообщаю-
щаяся, однако, с полостью кишечника. Половой аппарат гермафродитный
(за немногими исключениями), сильно развит, продуцирует огромное ко-
личество яиц.
Личинкам трематод присущ свободный образ жизни во внешней
среде. Этим объясняется тот факт, что личинки несут ресничный покров
и способны к свободному плаванию. В ходе развития личинки претерпева-
ют метаморфоз, теряя все приспособления к свободному существованию,
в том числе личиночные глазки, имеющиеся у части видов, и ресничный
покров. В дальнейшем личинки внедряются в органы соответствующих
хозяев. Превращение (метаморфоз) осуществляется со сменой хозяев.
Известно свыше 2000 видов.
Важнейшие дифференциальные признаки класса трематод следующие:
1) паразитируют во внутренних органах позвоночных и человека;
личинки — обычно в органах различных беспозвоночных, в особенности
моллюсков;
2) ротовая присоска образована мышцами; кроме нее, на брюшной
поверхности тела расположена еще одна присоска — брюшная, лежащая
по медиальной линии длинной оси тела кпереди или кзади от его середины
и не имеющая связи с пищеварительной системой;
3) в. задней части тела нет снабженного крючьями фиксаторного ап-
парата;
4) глаза у взрослых форм отсутствуют, только у личинок у части видов
наблюдаются рудиментированные фоторецепторы;
5) экскреторная система открывается наружу одним отверстием;
6) развитие, как правило, протекает с метаморфозом и сменой хозяев
(исключения см. ниже).
В составе класса трематод описаны 3 подкласса: Prosostomadidea, Bu-
cephallididea и Aspidogastrodidea. Значение этих трех подклассов далеко
не однозначно. Наибольший интерес представляет подкласс прозостомади-
дей, или дигенетических сосальщиков (трематод). Показанные выше диффе-
ренциальные признаки класса трематод полно выражены именно в пределах
первого подкласса.
ПОДКЛАСС I. ДИГЕНЕТИЧЕСКИЕ СОСАЛЬЩИКИ
(PROSOSTOMADIDEA)
Типичные трематоды с указанными выше характерными признаками
класса.
6 Курс зоологии, т. I
161
Представитель подкласса дигенетических
сосальщиков
— печеночная фасциола (Fasciola hepatica)
Печеночная фасциола паразитирует преимущественно в желчных хо-
дах печени крупного рогатого скота, овец и коз. Кроме того, эксперимен-
тально установлено, что фасциола может паразитировать в печени собак,
хотя и не долго — 3—4 месяца, и в печени кошек — несколько дольше,
Рис. 144. Пищеварительная
и нервная системы Fasciola
hepatica:
1 — ротовое отверстие, ведущее
в глотку (2), 3—продольные ство-
лы кишечника, 4— окологло-
точный нервный узел, 5—6— нер-
вные продольные стволы, 7—брю-
шная присоска
однако редко.
Внешнее строение. Паразит обладает белым
листовидным телом (рис. 144). Бока сероватого
цвета — это просвечивают сквозь покровы жел-
точники. Длина тела 20—30 мм, ширина 8—
13 мм. Головной конец сужен в конус. Здесь
расположена ротовая присоска. Позади голов-
ного конца по медиальной (средней) линии
тела лежит на брюшной стороне вторая, до-
полнительная, присоска. Между первой и вто-
рой присосками расположено половое отверстие.
На заднем конце тела имеется одна пора выде-
лительного аппарата.
Кожно-мускульный мешок. Снаружи лежит
слой кутикулы. Последняя несет небольшие за-
гнутые назад шипики (рис. 145). Сплошного
субкутикулярного слоя эпителия у взрослых
форм нет. Эпителий представлен погруженными
эпителиальными клетками, соприкасающимися
с диагональными мышцами. Кроме последних,
в состав кожно-мускульного мешка входят попе-
речные (кольцевые) и продольные мышцы. В
общем имеется та же система мышц, что и у
турбеллярий.
Кожно-мускульный мешок охватывает внут-
реннюю массу паренхимы. Последняя пронизана
тяжами дорзо-вентральной мускулатуры. В па-
ренхиму погружены внутренние органы.
Нервная система. Нервная система сос-
тоит из двух продольных стволов (рис. 144)
и окологлоточного нервного узла с отходящими
от этих частей периферическими ответвлениями,
особенно многочисленными в присосках.
Органы чувств. Органы чувств, за исключением осязательных
нервных окончаний в коже, отсутствуют.
Пищеварительная система. Пищеварительная система начинается
ротовым отверстием, вооруженным присоской. Присоска имеет воронко-
образное углубление, которое переходит в мускулистую глотку (pharynx).
Глотка отделена от присоски предглоточной полостью (рис. 145).
Упомянутые части являются мощным сосущим аппаратом. Акт сосания
происходит так: 1) сперва жидкость наполняет предглоточную полость;
это достигается тем, что глотка отодвигается назад, тогда как присоска
прилегает к ткани хозяина-паразита; получается нечто сходное с напол-
нением шприца, причем роль отодвигающегося поршня играет муску-
листая глотка, а роль полости шприца — предглоточная полость;
2) вслед за этим ротовое отверстие закрывается, а глотка выдвигается
вперед и наполняется жидкостью, набранной в предглоточную полость;
3) наконец, глотка сжимается и выдавливает свое содержимое в ки-
шечник.
162
Пищеварительный канал представлен двумя довольно мощными ство-
лами; оба ствола дают ряд (15—20) боковых ветвей, которые в свою очередь
разветвляются. Анального отверстия нет.
Цилиндрический эпителий кишечника несет фагоцитарную функцию.
Следовательно, фасциоле свойственно внутриклеточное пищеварение.
Рис. 145. Строение Fasciola hepatica. / — медианный срез головного конца тела;
// —разрез кишечника и прилегающей части паренхимы; III —часть поперечного
среза через тело фасциолы:
/ — присоска, 2 — ее мускулатура, 3— разрез предглоточной полости, 4 — глотка, 5 — начало ки-
шечника, 6 — мышцы глотки (ретракторы), 7 —паренхима, 8— эпителиальные клетки кишечника, ма-
ленькие шарообразные тела перед клетками — красные кровяные тельца хозяина, более крупные—час-
тично распадающиеся красные кровяные тельца, захватываемые эпителиальными клетками кишечника
червя (процесс фагоцитоза осуществляется при помощи ворсинок, покрывающих свободную поверх-
ность эпителиальных клеток), 9 — кутикула, 10—кожно-мускульный мешок, // — сечения ветвлений
пищеварительного канала, 12— желточники, 13— участки выделительной системы, 14— семенные
канальцы
Дыхательная и кровеносная системы. Дыхательная и кровеносная
системы отсутствуют. Ткани паренхимы фасциолы очень богаты гликоге-
ном. За счет гликолиза и осуществляется анаэробное дыхание фасциолы.
Однако фасциола способна и к аэробному дыханию. Было установлено
наличие в органах фасциолы ее собственного гемоглобина, имеющего спе-
цифичный спектр поглощения и способного соединяться с кислородом,
превращаясь в оксигемоглобин.
6*
163
Выделительная система. Выделительная система хорошо развита и
представлена воднососудистой системой. Последняя состоит из срединного
собирательного канала, который открывается наружу задней выделитель-
ной порой (рис. 146). В этот канал впадает множество боковых каналов,
которые берут свое начало от тон-
чайших разветвлений, снабженных
клетками с ресничным пламенем.
Следовательно, у фасциолы имеется
типичная система протонефридиев.
Половая система. Половая си-
стема достигает огромного разви-
тия. Она гермафродитная.
Мужская половая
система состоит из двух силь-
но ветвящихся семенников — перед-
него и заднего (рис. 147), от кото-
Рис. 147. Половая система Fasciola
hepatica
1— ротовая присоска, 2— глотка, «У—пи-
щевод, 4 — место расположения мужского и
женского половых отверстий, 5 — кишеч-
ник, '6—циррус, 7 — брюшная присоска,
8— матка, 9 — яичник, 10— семяпроток
заднего семенника, 11 — продольные каналы
желточников, 12 — задний семенник, 13 —
левый желточник, 14 — передний семенник,
15— семяпроток переднего семенника, 16 —
тельце Мелиса, 17 — поперечный желточный
Рис. 146. Выделительная
система F. hepatica:
1 — брюшная присоска; все
тело пронизано многочислен-
ными канальцами, впадающи-
ми в главный выделительный
ствол (2), открывающийся
наружу выделительной по-
рой (3)
проток
рых идут два семяпротока (vasa efferentia), впадающие в общий семя-
провод (vas deferens). Последний переходит в семенной пузырь (vesicula
seminal is), который изливается в семяизвергательный канал (ductus jecu-
latorius) совокупительного органа (cirrus). Cirrus (рис. 147) находится в
особом совокупительном мешке и может выпячиваться через половую по-
ру наружу, причем принимает изогнутую форму (рис. 148ц). Семя,
образующееся в семенниках, попадает по vasa efferentia в vas deferens,
оттуда — в vesicula seminalis. Из vesicula seminalis оно проникает в cir-
rus, а из него — наружу.
164
Женская половая система (рис. 147) состоит из яич-
ника, желточников, тельца Мелиса и матки.
Функции этих органов таковы. В яичнике образуются яйца. Отсюда
зрелые яйца попадают в яйцевод. В желточниках вырабатываются питатель-
ные желточные клетки. Желточники у фасциолы представлены множеством
мелких пузырьков (фолликулов) по бокам тела. По тонким каналам жел-
точные клетки, погруженные в облекающую их жидкость, попадают в
продольные желточные каналы, из них — в поперечные правый и левый
желточные протоки. Последние соединяются в непарный желточный про-
ток, посредством которого желточные клетки, как и яйца, попадают в
конечный отдел яйцевода. Тельце Мелиса выделяет жидкий секрет, который
также изливается сюда, здесь же
происходят оплодотворение и фор-
мирование яиц. У фасциолы спер-
матозоиды проходят через вывод-
ное отверстие матки в полость
последней и по длинному извитому
каналу матки проникают в яйцевод.
Когда яйца окажутся оплодотво-
ренными, начинается их формиро-
вание. Каждое яйцо оказывается
как бы облепленным желточными
клетками, и затем на его поверхно-
сти, по-видимому, из жидких
веществ, проникших в яйцевод из
желточников, образуется скорлупа.
Готовые яйца поступают в матку и
в ее выводной проток, а отсюда —
наружу.
На рис. 148 показано, что вы-
водной проток матки открывается
наружу вблизи места, где лежит
cirrus.
У печеночного сосальщика от
конца яйцевода берет начало еще
Рис. 148. Деталь полового аппарата Fasci-
ola hepatica. / — циррус втянут в полость
циррусного мешка; //—циррус выступил
наружу:
/—оба семя выводящих канала, соединяющиеся в
короткий vas deferens, расширяющийся в семенной
пузырь, 2—брюшная присоска, 3 — семенной пу-
зырь. 4 — выводящий проток матки, 5—семяизвер-
гательный канал (ductus jeculatorius) и его продол-
жение (6), в циррусе (7), 8—конец протока матки
и женское половое отверстие (9). открывающееся в
области половой поры
один короткий канал, открываю-
щийся наружу на спинной стороне. Это лауреров канал, рудимент влага-
лища. У фасциолы лауреров канал служит для отвода избытка желточ-
ных продуктов, тогда как сперматозоиды сюда не попадают.
Условия существования и образ жизни фасциолы. Фасциола поселяется
в желчных протоках печени упомянутых выше животных. Забираясь в
глубь печени, червь питается желчью и продуктами распада разрушаемых
им клеток печеночных протоков. Фасциола — типичный паразит, питаю-
щийся соками своих хозяев.
С одной стороны, упрощение организации, столь обычное у паразитов,
проявляется у фасциолы в отношении органов чувств. У нее отсутствуют
глаза, статоцисты, какие-либо органы химического чувства и т. п. С другой
стороны, у фасциолы резко заметны характерные черты специализации:
1) ресничный покров у взрослого червя исчез; 2) эпителиальный слой
кожно-мускульного мешка покрыт кутикулой (ср. Turbellaria, у которых
ее нет); 3) кутикула несет загнутые назад крючочки, приспособленные для
прикрепления (фиксации) к тканям хозяина; 4) развиты присоски — органы
фиксации; 5) половые органы, в особенности матка, достигают огромного
развития, несравненно большего, чем у турбеллярий; в связи с этим
фасциола несравненно более плодовита, чем свободные плоские черви
(турбеллярий); 6) своеобразна и физиология: фасциола способна
к дыханию анаэробного типа. Эти факты указывают на ряд при-
способлений к паразитическому существованию. Эти приспособления,
165
как мы видели, идут по двум направлениям: а) редукции органов, не су-
щественных для
органов, важных
Рис. 149. Яйцо
Fasciola hepatica:
1 — крышечка, 2 — яй-
цевая клетка, 3—жел-
точные клетки
паразитического существования, и б) специализации
для паразита. Сильно развиваются органы фиксации и
половые органы. В то же время, как было сказано,
приобретается способность к анаэробному дыханию.
Таковы морфо-физиологические признаки фасциолы
как паразита, и с этими чертами мы неоднократно
встретимся в дальнейшем при описании других па-
разитических червей.
Здесь же важно познакомиться с факторами, ко-
торые обусловливают одну из типичных черт фасци-
олы — ее огромную плодовитость. С этой целью
остановимся на ее онтогенезе.
Пройдя эмбриональное развитие, яйцо фасциолы
(рис. 149) для дальнейшего развития должно попасть
в воду. Здесь верхняя часть его (крышечка) отгибает-
ся, и из яйцевой оболочки выходит свободноживущая
личинка — мирацидий — первая стадия жизненного
Рис. 150. Развитие Fasciola hepatica. / — мирацидий; //, /// — спо-
роцисты, в которых развиваются редии; IV — редия, в которой повторно
развиваются редии; V — редия, в которой развиваются церкарии; VI —
церкария; VII —адолескария:
1 — кишечник, 2—протонефридий, 3—их выводные каналы, 4 — зародышевые клетки,
5 — рот, 6 — брюшная присоска, 7 — железы, 8— хвост, 9 — передняя присоска
166
цикла печеночной двуустки (рис. 150i ). Мирацидий имеет глазок, его
эпителий состоит из явственно различимых клеток и покрыт ресничками.
Из внутренних органов у него имеются зачаток кишечника, протонефридии
и многочисленные зародышевые клетки. Мирацидий свободно плавает, но
способен к существованию лишь очень ограниченное время —всего 30—40 ч.
К концу этого срока запасные вещества, имеющиеся в его теле, исчер-
пываются, и мирацидий погибает, если не внедрится в промежуточного
хозяина. Таким хозяином для фасциолы являются пресноводные брюхо-
ногие моллюски, обычно малый прудовик (Galba truncatula). Мираци-
дий активно вбуравливается в моллюска, выделяя при этом особый сек-
рет, расплавляющий (лизирующий) ткани, благодаря чему он глубоко про-
никает в тело моллюска и локализуется обычно в печени. Здесь начинается
его регрессивный метаморфоз — он теряет ресничный покров, глазок и все
внутренние органы, за исключением протонефридий и зародышевых кле-
ток, и превращается во вторую стадию — мешкообразную спороцисту
(рис. 150ц, ш). В спороцисте из зародышевых клеток без оплодотво-
рения, т. е. партеногенетически, развиваются многочисленные редии —
третья стадия жизненного цикла (рис. 150iv, v)-
Следует отметить, что партеногенез, или девственное размножение, —
это половой процесс, а не бесполое размножение. Зародышевые клетки,
из которых развиваются редии, представляют собой клетки так называемо-
го герминативного, т. е. полового, эпителия.
Редии по строению отличаются от спороцисты, они имеют ротовое
отверстие и зачаток кишечника, а также многочисленные зародышевые
клетки. Из этих последних также партеногенетически образуются или опять
редии или следующая (четвертая) стадия — церкарии. Церкарии устроены
еще более сложно, чем редии, и имеют уже ряд признаков взрослой фасцио-
лы (рис. 150Vi) — присоски, разветвленный кишечник, нервную систему,
а также особый орган движения — хвостообразный придаток. Церкарии
через особое отверстие выходят из редий и активно покидают моллюска. Во
внешней среде они активно, но очень короткое время плавают, затем при-
крепляются к растительности, теряют хвост, выделяют вокруг себя плотную
оболочку и превращаются в адолескарию (рис. 150Vn) — пятую и послед-
нюю стадию жизненного цикла фасциолы. Весь путь развития от мираци-
дия до церкарии протекает примерно в 50—60 дней.
Адолескария с травой или водой попадает в желудок основного хозяи-
на, чаще всего овцы, где оболочка растворяется, паразит освобождается,
активно проникает к кровеносным сосудам и по ним достигает печени, в
которой локализуется и вырастает во взрослую форму.
Как видим, развитие фасциолы протекает со сменой хозяев: один из
них — моллюск — служит хозяином личиночных стадий развития фасцио-
лы, другой — овца — взрослой половозрелой формы. Первый — это про-
межуточный, второй — окончательный хозяин.
Как видно из приведенного описания цикла развития фасциолы, про-
никновение в хозяев сопровождается существованием двух личиночных
стадий (мирацидий, церкария) во внешней среде. Если встреча с промежуточ-
ным или окончательным хозяином не состоится, то мирацидий и церкария
в конце концов погибают. Естественно, что на сложном пути смены хозяев,
в особенности в условиях внешней среды, гибнут массы личинок фасциолы.
Можно утверждать, что подавляющее большинство ее потомства не достига-
ет полного развития. В этой связи становится понятной огромная плодо-
витость фасциолы. Она компенсирует потери потомства и становится основ-
ным фактором, обеспечивающим, несмотря на массовую гибель мирацидиев
и церкарий, продолжение развития и существование вида.
Добавим к сказанному, что, помимо огромной плодовитости, повыше-
ние численности потомства одной особи фасциолы достигается также и в
процессе жизненного цикла. Каждая особь фасциолы дает примерно 45 000
яиц, каждое из них способно дать мирацидия. Мирацидий превращается
167
в спороцисту, каждая из них в среднем дает 8 редий, тогда как редия в свою
очередь — 15—20 церкарий. Следовательно, каждый мирацидий может
дать около 160 церкарий, потомство же 45 000 возможных мирацидиев
равно 7 200 000 экземпляров (45 000x160). Естественно, что это теорети-
чески возможное число церкарий никогда не реализуется, но все же об-
щая численность потомства одной фасциолы очень велика.
Циклы развития некоторых других дигенетических
сосальщиков
Циклы развития других дигенетических сосальщиков могут резко
отличаться от цикла развития печеночной двуустки. Разнообразие этих
способов развития показывает широкие приспособительные возможности
Рис. 151. Представители семейств Dicrocoeli-
idae и Opistorchidae. / — ланцетовидный
сосальщик (Dicrocoelium lanceatum); II —
кошачья двуустка (Opistorchis felineus); III —
мирацидий D. lanceatum:
/ — ротовая присоска, 2 — глотка, 3 — ветви кишечни-
ка, 4 — выделительный канал, 5— семенники, 6 — се-
меприемник, 7 — яичник, 8 — матка, 9 — совокупи-
тельный мешок, 10—желточники, 11— брюшная при-
соска, 12—стилет
паразитов, благодаря кото-
рым они паразитируют у са-
мых различных животных и
благодаря чему распростра-
нены столь широко.
Рассмотрим несколько
циклов развития, которыми
далеко не исчерпывается все
разнообразие способов раз-
вития и расселения пара-
зитов.
Цикл развития ланцето-
видного сосальщика (Dicroco-
elium lanceatum; рис. 151).
Окончательным хозяином его
служат овцы, козы, крупный
рогатый скот, некоторые гры-
зуны (зайцы, кролики, сусли-
ки), собаки, медведи. Паразит
гнездится в печени этих жи-
вотных. С фекалиями (экскре-
ментами) хозяев яйца ланце-
товидного сосальщика попа-
дают на траву пастбищ. Вода
для высвобождения мираци-
дия из яйца не нужна. Яйца
заглатываются наземными
моллюсками (рода Helicella и
др.). Мирацидий высвобожда-
ется из яичной скорлупы в
кишечнике моллюска. В отли-
чие от мирацидия печеночной
фасциолы, он имеет неполный
ресничный покров и особый
стилет (рис. 151), погружен-
ный в головную шишку. Обра-
зующиеся в печени моллюска церкарии активно мигрируют из печени в
легкое моллюска и включенные в кучки склеенных слизевых комочков по-
падают на траву и почву пастбищ. Здесь эти кучки захватываются и по-
едаются муравьями из рода Formica. В муравьях заканчивается превращение
паразита в стадию, равнозначную адолескарии. Это — метацеркария.
Муравей служит здесь дополнительным хозяином. Проглатывание мура-
вьев, очевидно с травой, обеспечивает заражение одного из упомянутых
выше окончательных хозяев.
168
Цикл развития кошачьей, или сибирской, двуустки (Opistorchis fe-
lineus; рис. 151). Развивается с участием промежуточного хозяина (мол-
люск Bithynia leachi), из которого высвобождаются церкарии и проникают
в дополнительного хозяина — представителя карповых рыб, где превращают-
ся в метацеркарию, и уже при поедании рыб паразитом заражаются окон-
чательные хозяева — кошка, лисица, собака, а также человек.
Цикл развития алярии (Alaria alata). Примером другого усложнения
развития сосальщиков служит онтогенез трематоды, паразитирующей в
тонком кишечнике лисиц и собак — двуустки алярии. Из яиц этого
сосальщика высвобождаются мирацидии, которые плавают в воде, а затем
активно проникают в некоторых пресноводных моллюсков. В моллюсках
в результате спорогонии и партеногонии развиваются церкарии с длинным
вильчатым на конце хвостом. Дальнейшее развитие протекает в дополни-
тельном хозяине, которым служит головастик и лягушка. В тканях голо-
вастиков и лягушек развиваются метацеркарии. Если лягушка будет
съедена лисицей или собакой, то эти окончательные хозяева заражаются,
или, как принято говорить в отношении животных-паразитов, инва-
зируются метацеркариями, и в их органах развиваются взрослые алярии.
Однако опыты показали, что если скормить метацеркарий алярии мышам,
то в их межреберных мышцах снова обнаруживаются метацеркарии.
Дальше они в органах мышей не развиваются. Хозяева, которые способны
инвазироваться метацеркариями, не становятся средой, годной для даль-
нейшего развития метацеркарий, а служат накопителями, носят название
резервуарных (запасных). Мышь — своего рода запасной хозяин, который
сам по себе не определяет дальнейшего развития и не необходим для него.
Однако он биологически оправдан, так как мышь значительно более обычная
пища, например, для лисицы, чем головастики и лягушки, и, следователь-
но, увеличивает возможность развития алярии.
Цикл развития кровяных двуусток (различные виды рода Schistosoma;
см. рис. 156). Половозрелые паразиты обитают в крупных венозных сосу-
дах человека. Промежуточный хозяин один — различные пресноводные
моллюски. Выходящие из них в воду церкарии обладают стилетом, при
помощи которого они активно вбуравливаются через кожу в тело человека.
Онтогенетическое развитие трематод, протекающее по одной из опи-
санных схем, известно под названием развития с чередованием поколений.
ОБЩИЙ ОБЗОР КЛАССА
Внешнее строение. Сосальщики имеют либо сплющенное, либо цилин-
дрическое тело, обычно удлиненное, иногда округлое. Размеры тела колеб-
лются, как это наблюдается и в других группах животных, в очень широких
пределах. В ротовой полости акулы Sciaenia aquila обитает локализующий-
ся в крупных цистах гигантский сосальщик Nematobothrium filarina.
Этот паразит образует клубок; если его распутать, то окажется, что трема-
тода достигает свыше 1 м в длину, при ширине 1,0—2,0 мм. Известны также
сосальщики, не превышающие в длину 0,1 мм.
Кожа всегда покрыта кутикулой, которая у многих форм несет загну-
тые назад крючочки. Головной конец большей частью вооружен присоской,
позади которой обычно, как у печеночной фасциолы, расположена вторая,
брюшная, присоска. Такой тип расположения присосок известен под назва-
нием дистомидного типа (см. рис. 151, 155). В других случаях может раз-
виваться лишь одна присоска — это моностомидный тип. У некоторых
форм брюшная присоска отодвигается далеко назад по сравнению с ее поло-
жением у фасциолы; такой тип расположения присосок обозначают как
парамфистомидный (по имени сосальщиков из рода Paramphistomum). Ин-
тересно отметить, что у трематод, живущих в замкнутых полостях, напри-
мер в кровеносном русле, присоски могут отсутствовать. Обычно наиболее
мощные присоски развиты у тех форм, которые обитают в полостях органов,
169
непосредственно сообщающихся с внешней средой. У некоторых трематод
на брюшной стороне тела развит комплекс присосок, которые объединены
в границах обособленного диска и образуют особый фиксаторный (прикрепи-
тельный) орган, получивший название органа Бэра.
Кожно-мускульный мешок. Кутикула — всегда производное эпите-
лиального покрова животного. Следует подчеркнуть, что для сосальщика
характерен погруженный эпителий, который состоит из явственно обособ-
ленных эпителиальных клеток, погруженных в паренхиму, обычно бо-
гатую гликогеном. У многих сосальщиков в коже развиты кожные железы,
располагающиеся, например, по сторонам ротовой присоски, на вентраль-
ной стороне тела или на заднем конце его. Мышечная система слагается
из наружного кольцевого и внутреннего продольного слоев. Типичны так-
же диагональные мышечные волокна и система спинно-брюшных мышц.
У различных видов развитие мышечной системы очень варьирует. Многие
формы отличаются очень мощной мускулатурой, в особенности виды рода
Paramphistomum, у других она развита слабо. Всегда наиболее сильно раз-
вита мускулатура присосок.
Паренхима. Кожно-мышечный мешок облекает массу внутренней
ткани — паренхимы, подобной паренхиме турбеллярий. Паренхима со-
сальщиков всегда мощно развита, в ней происходит энергичный обмен
веществ и накапливается гликоген — важный продукт питания и объект
гликолиза.
Нервная система. Нервная система сосальщиков состоит из окологло-
точного нервного кольца, включающего два ганглия, и продольных нерв-
ных тяжей. Кпереди направлены нервные тяжи, иннервирующие присоску.
Сзади от нервного кольца тянутся мощные продольные нервные стволы,
обычно три пары. Наиболее мощно развиты боковые нервные стволы. От
этих стволов идут многочисленные нервы, в том числе к брюшной присоске,
если она есть, и к поверхности тела.
Органы чувств. Органы чувств развиты слабо. Глазки, как правило,
наблюдаются у личинок. Иногда развиваются особые придатки по бокам
головного конца тела в форме «ушей», которые рассматриваются как органы
чувств, видимо, химических или осязательных.
Пищеварительная система. Как правило, ротовая присоска располо-
жена на переднем конце тела, несколько сдвинувшись к брюшной поверх-
ности его. Однако у части форм эта присоска переместилась на брюхо.
Ротовое отверстие в таких случаях расположено на брюшной стороне тела
примерно в центре его. Глотка имеет строение, сходное с таковым у печеноч-
ной фасциолы. Однако глотка может отсутствовать; у части форм нет пред-
глотки. Глотка обычно переходит в короткий пищевод. Глотка, предглотка
и пищевод снабжены мускулатурой. Иногда пищевод также отсутствует.
Кишечник (средняя кишка) различен по строению. Иногда его вообще
нет, хотя глотка развита (Nematobothrium). У некоторых форм кишечник
имеет форму широкого мешка. У большинства он, как у Dicrocoelium 1ап-
ceatum, представлен двумя продольными стволами. Эти стволы тянутся
кзади и оканчиваются слепо. Иногда оба ствола сзади соединяются вместе,
и тогда кишечник приобретает форму «арки». Как правило, кишечные ство-
лы заканчиваются слепо; у некоторых форм, однако, они открываются на-
ружу позади анальными отверстиями: либо одним общим, либо особым
для каждой кишечной трубки. Не всегда кишечные стволы ветвятся, как
это мы видели у фасциолы. Чаще они гладкие неразветвленные.
Кровеносная и дыхательная системы. Кровеносная и дыхательная
системы отсутствуют. Однако, как указывалось, в мышцах трематод был
обнаружен гемоглобин, который способен активно поглощать кислород,
отдавая его в дальнейшем медленнее, чем это установлено для гемоглобина
позвоночных. Наличие гемоглобина показывает, что сосальщики вовсе
не должны рассматриваться как формы, способные только к анаэробному
дыханию. В среде кишечника хозяев сосальщиков имеется некоторое ко-
170
личество кислорода, достаточное для дыхания сосальщиков. Именно с
потреблением этого кислорода и связано у них наличие мышечного гемогло-
бина, способного присоединить к себе свободный кислород.
Выделительная система. Выделительная система мощно развита. Она
состоит из многочисленных терминальных клеток, которые впадают в мелкие
экскреторные сосуды, а эти последние в свою очередь выносят свое содер-
жимое — жидкие продукты обмена веществ — в более крупные сосуды.
В конечном счете жидкие продукты обмена изливаются в большие боковые
сосуды, которые либо впада-
ют в «мочевой пузырь» (рис.
152), либо открываются нару-
жу экскреторным отверстием,
расположенным наподобие
того, как это мы выдели у
фасциолы. Иногда в мочевой
пузырь открывается кишеч-
ник. Следует отметить, что
экскреторная система обеспе-
чивает не только выделение
жидких отбросов обмена, но
и имеет также большое значе-
ние как орган, регулирую-
щий осмотическое давление.
Половая система. Поло-
вая система в большинстве
случаев гермафродитная. Она
занимает большую часть па-
ренхиматозной массы тела.
Нередко буквально вся па-
ренхима заполнена огромной
маткой, набитой яйцами (рис.
153).
Мужская половая систе-
ма отличается значительным
разнообразием в частных де-
талях. Число семенников
обычно равно двум, но наблю-
даются формы с одним се-
Рис. 152. Выделитель-
ная система трематод
(на примере АПосге-
adium isoporum):
1 — терминальная клетка
с «ресничным пламенем»,
2 — выводящие стволы, 3—
«мочевой пузырь»
Рис. 153. Dicrocoelium
voluptarium:
/ — матка, заполняющая все
тело и набитая яйцами, 2—
семенники, 3— яичники, 4—
кишечник
менником или с несколькими,
которые образовались за счет фрагментации двух первоначальных семен-
ников. Форма семенников различная. Чаще они цельнокрайние округ-
лые, иногда лопастные (см. рис. 151), реже разветвленные, как у фасци-
олы. Расположение семенников весьма различно — кпереди или кзади от
брюшной присоски, кпереди или кзади от яичника, симметрично или
асимметрично и т. п. Положение семенников имеет большое значение в
систематике. Семяпротоки и семяпроводы впадают в семяизвергательный
канал, который связан с предстательными железами и с семенным пузырь-
ком. Конечным отделом всегда служит совокупительный орган, или циррус.
Обычно конечные отделы системы (семяизвергательный канал и циррус)
лежат в половой бурсе. Иногда бурса отсутствует.
Женская половая система сосальщиков имеет очень сложное строение.
Центральным органом ее нужно считать оотип (от греческих слов oos —
яйцо и typos — печать, формирование), в котором происходит оформление
оплодотворенных яиц. В оотип (рис. 154, 155) открываются протоки сле-
дующих органов: 1) яйцевода, идущего из яичника, 2) семеприемника,
3) протоков из желточников, 4) протоков, идущих из тельца Мелиса.
Впрочем, такая система имеется далеко не у всех сосальщиков. Яйца,
формирующиеся в яичнике, попадают в яйцевод, который в месте выхода
171
из яичника снабжен особой мускулатурой. Этот снабженный мышцами
отдел яйцевода называется овикаптом. Последний производит пульсирую-
щие сокращения стенок яйцевода. Мышечная пульсация создает всасыва-
тельный поток жидкости, и таким образом овикапт «заглатывает» яйца,
которые поступают в просвет яйцевода и попадают в оотип. Сюда же, как
было сказано выше, открываются протоки желточников и тельца Мелиса.
Желточники доставляют в оотип многочисленные желточные клетки, кото-
рые облепляют яйцо. Кроме того, жидкость желточников, по-видимому,
участвует в образовании скорлупки яйца. Разумеется, оплодотворение яиц
происходит до описанных явлений, а именно в семеприемнике по выходе
яиц из яйцевода. Оплодотворенные и оформленные в оотипе яйца проникают
затем в матку, откуда они выводятся на-
ружу через женское половое отверстие,
анатомически соответствующее выводно-
му отверстию матки. Сперматозоиды
проникают (из цирруса) в это же отвер-
стие, которое играет, следовательно,
роль вагины. Они проходят через мат-
ку, проникают в оотип и из него в
семяприемник, где и происходит опло-
Рис. 154. Схема соотношений
между органами половой си-
стемы трематод (показан лишь
один из многих типов соотно-
шений):
1 — желточный проток, 2 — лауреров
канал, 3 — яичник, 4 — оотип, 5 —
семеприемник, 6 — тельце Мелиса,
7 — матка
1
Рис. 155. Половая система
Discocotyle salmonis:
1— половая пора, 2, 3 — влагалище,
4— матка, 5— желточный проток, 6—
желточный пузырь, в котором скоп-
ляются желточные клетки, 7— оотип,
8— семеприемник, 9— тельце Мели-
са, 10 — яичник, 11 — генитоинтести-
нальный канал, соединяющий поло-
вую систему с кишечником, 12 —
ветвь кишечника. Интерес представ-
ляет явственный оотип (7), его отно-
шение к железам полового аппарата,
связь последнего с кишечником
дотворение яиц. Кроме описанных органов, у многих форм от оотипа отхо-
дит еще проток лаурерова канала. Последний служит либо как вагина,
через которую проникают (при совокуплении) сперматозоиды, либо как
«запасной» проток, через который удаляются наружу излишние половые
продукты.
У других сосальщиков (Digenea) имеется только одно половое отверс-
тие, а именно отверстие матки, которое функционирует и как совокупи-
тельный орган, тогда как влагалище либо отсутствует, либо представлено
его рудиментом — лауреровым каналом.
Онтогенетическое развитие. Онтогенез (жизненный цикл) сосальщиков
отличается, как правило, значительной сложностью. Как известно,
конечным половым продуктом является оплодотворенное яйцо (см.
рис. 149, 170). У сосальщиков яйцо покрыто скорлупой, для которой ха-
172
рактерно наличие крышечки, расположенной на одном из полюсов яйца.
Эмбриональные стадии развития у многих форм протекают в матке, где
можно наблюдать процесс дробления — образование бластулы и гаструлы,
а также закладку зародышевых листков. Дробление яиц полное — равно-
мерное или неравномерное. В результате развития яйца в яйцевой скорлу-
пе формируется личинка — мирацидий, наподобие уже известного нам
мирацидия фасциолы. У некоторых форм мирацидий вылупляется не в
наружной среде, а в матке. Однако, как правило, он освобождается от
покрывающей его эмбриональной оболочки и скорлупы яйца уже во внеш-
ней среде или в органах промежуточного хозяина.
Период онтогенеза, завершающийся образованием мирацидия, на-
зывается эмбриогонией. За этим периодом обычно наступает период
партеногонии — размножения за счет неоплодотворенных зародышевых
клеток. Это постэмбриональный период, который протекает в теле проме-
жуточного хозяина; роль последнего у сосальщиков играют различные
моллюски. В теле моллюска, в процессе протекающей в его органах парте-
ногонии, образуется ряд поколений личинок, причем в ходе развития
они подвергаются сложным превращениям, или метаморфозу (спороцисты
и редии, см. фасциолу). Период партеногонии заканчивается развитием
церкарий, которые выходят во внешнюю среду.
С завершением партеногонии онтогенез сосальщиков переходит в
период цистогонии, характеризующийся инцистированием церкарий. При
этом хвост утрачивается, а внутреннее строение церкарий приближается
к организации взрослой формы. Такая инцистированная форма, составляю-
щая новый этап в развитии, называется, как мы уже знаем, адолескарией.
Однако не всегда развитие идет по этой схеме. Во многих случаях
церкария, не превращаясь в наружной среде в адолескарию, активно
проникает в какого-либо дополнительного хозяина; в его органах она пре-
вращается в метацеркарию — стадию, морфологически и по фазе развития
равнозначную адолескарии.
Если адолескария (непосредственно) или метацеркария (через соответст-
венного дополнительного хозяина) попадает в организм окончательного хо-
зяина, то начинается и здесь же и завершается последний этап онтогенеза—
маритогония. В ходе маритогонии в результате формообразовательных
процессов развивается взрослый сосальщик, или марита. Марита развивает
половую систему, продуцирует яйца, и описанный выше путь онтогенети-
ческого развития воспроизводится вновь.
Экология и распространение. На примерах фасциолы и некоторых
других мы познакомились с некоторыми специфичными приспособлениями
трематод к паразитарному образу жизни.
Трематоды поселяются во внутренних органах своих хозяев, в част-
ности в кишечнике, в печени, в поджелудочной железе, в фабрициевой сум-
ке птиц, в легких различных животных, в том числе птиц, в их воздушных
легочных мешках, в мочевом пузыре, в почках; в кровеносной системе
рыб, черепах, птиц и млекопитающих, в евстахиевой трубе, в лобной пазухе,
в полости тела и т. д. Взрослые эндопаразитические сосальщики паразити-
руют в органах позвоночных, реже в органах беспозвоночных, например
в мышцах речных раков (трематода Astacotrema cirrigerum) или в перикарде
и почках многих моллюсков. Личиночные формы эндопаразитических со-
сальщиков поселяются в органах моллюсков — своих типичных промежу-
точных хозяев. В органах этих животных личиночные стадии трематод
нашли очень благоприятную среду для размножения путем спорогонии,
(см. выше). Спороцисты могут размножаться в органах моллюсков даже
путем деления, чаще же уже известным нам партеногенетическим путем,
причем партеногонии может привести к формированию в моллюске сотен
спороцист. Редии, развивающиеся из воспроизводительных клеток споро-
цист, могут давать дочерние поколения редий же. В этих случаях в органах
моллюска развивается огромное количество редий, а затем церкарий.
173
Было установлено, что из прудовика (Limnaea stagnalis) в течение четырех
дней выделились соответственно следующие количества церкарий: 8 000,
5 400, 13 300 и на четвертый день 15 900 экземпляров.
Распространение сосальщиков в большой степени зависит от распро-
странения их хозяев — промежуточных, дополнительных и окончательных.
Поэтому условия существования сосальщиков неразрывно связаны с усло-
виями существования их хозяев. В онтогенезе сосальщиков огромную
роль играет наличие водоемов, пойменных лугов и других угодий, благо-
приятствующих заражению соответствующих хозяев — окончательных и
промежуточных. Онтогенез сосальщиков обеспечивается и другим факто-
ром, а именно наличием контакта между окончательными и промежуточны-
Рис. 156. Кровяная двуустка Schi-
stosoma haematobium.
1 — самка, сидящая в гинскофорном ка-
нале самца
Рис. 157. Bucephalus ро-
lymorphus:
1 — головной конец с мышеч-
ными пальцевидными вырос-
тами. 2 — присоска, 3 — ро-
товая щель, 4 — мешковид-
ная средняя кишка, 5—яич-
ник, 6— матка, 7 — желточ-
ники, 8 — сумка цнрруса,
9 — семенники
ми хозяевами. Поэтому и площади распространения сосальщиков опреде-
ляются распространением окончательных и промежуточных хозяев, а так-
же зонами жизни, в пределах которых имеется контакт между хозяевами,
обеспечивающий все звенья онтогенеза трематод, развивающихся с чередо-
ванием поколений. Следует, однако, отметить, что площади распростране-
ния сосальщиков все же не полностью совпадают с площадями распростра-
нения хозяев и обычно уже их, так как для паразита имеет значение не
только наличие соответствующих хозяев, но и других факторов.
Сюда относятся, например, различные культурные мероприятия че-
ловека — осушение болот, возделывание почвы, развитие и расширение
площадей под культурными посевами и т. п. Наряду с этим распростране-
174
ние и размножение сосальщиков уменьшается и различными естественными
факторами.
Главные представители. Сюда относятся: описанные выше печеночная
и ланцетовидная двуустки, паразиты печени человека, крупного рогатого
скота, овец, коз, кроликов, зайцев и других животных; гигантская двууст-
ка (Fasciola gigantica), достигающая 75 мм длины, паразит печени,
очень опасный для овец и коз; упомянутая выше кошачья, или сибирская,
двуустка (Opistorchis felineus) — паразит желчного пузыря и желчных
протоков печени кошки, собаки, лисицы и человека; легочная двуустка
(Paragonimus westermani) — паразит легких кошки, собаки, свиньи, тигра
и человека, и ряд других.
К этому же подклассу относятся своеобразные кровяные двуустки
(род Schistosoma), отличающиеся от других двуусток рядом признаков.
Особенно следует отметить, что они раздельнополы. Обычно у самца на
брюшной стороне имеется глубокая продольная складка, так называемый
гинекофорный канал (canalis gynaecophorus). В этом канале помещается
самка (рис. 156). Своеобразно у этих двуусток и строение кишечника — он
сперва делится на два боковых ствола, но затем оба ствола образуют круг
и соединяются в один срединный канал, который заканчивается в задней
части тела. Паразитируют кровяные двуустки в крови различных живот-
ных и человека (подробнее о вредоносности сосальщиков см. ниже в специ-
альном разделе).
ПОДКЛАСС II. БУЦЕФАЛИДИДЕИ (BUCEPHALIDIDEA)
Фиксаторный мышечный орган расположен впереди и не связан с
ротовым отверстием, лежащим в середине длины тела или кзади от нее
на брюшной стороне. Ротовой присоски нет.
Глотка и пищевод ведут в мешковидный, ре-
же трубчатый кишечник. Семенники и яич-
ники в середине тела или в его задней части.
Желточники в передней части тела. Половое
отверстие на брюшной стороне тела, около
его заднего конца. Паразиты рыб, изредка
амфибий. Представителем буцефал идидеи,
или брюхоротых трематод, могут служить
Bucephalus polymorphus, относящиеся к се-
мейству Bucephalidae (рис. 157).
Рис. 158. Aspidogaster limacoides, внешний вид с
брюшной стороны
1 — ротовое отверстие, 2—прикрепительный диск
ПОДКЛАСС III. АСПИДОГАСТРОДИДЕИ
(ASPIDOGAST RODIDEA)
Представители этого подкласса трематод характеризуются оригиналь-
ными чертами организации, ставящими их в положение «сборной группы»,
отчасти близкой к моногенетическим сосальщикам, отчасти к типичным
дигенетическим трематодам. Взрослые аспидогастродидеи паразитируют
во внутренних органах (почках и перикардиуме) брюхоногих и пластинча-
тожаберных моллюсков или в кишечном тракте рыб и рептилий. Развитие,
как у моногеней, прямое , т. е. без превращения (метаморфоза) с выхожде-
нием из яиц реснитчатых личинок. Взрослые формы характеризуются сла-
боразвитой ротовой присоской, которая может даже отсутствовать. Кишеч-
175
ник большей частью в форме примитивного мешка. На брюшной стороне
тела развит особый присасывательный диск, состоящий из группы присо-
сок, объединенных в сложный комплекс. Это — орган Бэра (рис. 158).
Представители: Aspidogaster conchicola, паразит моллюсков; в их органах
проходят все стадии развития; хозяевами служат моллюски Anodonta,
Unio, виды рода Vivipara и др. Macraspis elegans паразитирует у химер
(Chimera monstrosa) и др.
ВРЕДОНОСНОСТЬ СОСАЛЬЩИКОВ
Сосальщики, в особенности эндопаразитические, вызывают у позвоноч-
ных животных и человека различные заболевания, известные под общим
наименованием трематодозов. В каждом отдельном случае трематодозы по-
лучают специальные названия, производимые по названию рода паразита.
Так, заболевания, вызываемые фасциолой, называются фасциолезом (от
родового названия Fasciola), болезни, обусловленные присутствием лан-
цетовидного сосальщика, именуются дикроцелиозом (от Dicrocoelium) и
т. д. Трематоды нередко весьма патогенны, они вызывают развитие специ-
фического болезненного состояния животного. Так, при фасциолезе живот-
ное, несмотря на хорошее кормление, худеет, постепенно развивается по-
бледнение слизистых оболочек, глаза становятся впалыми, появляются
боли в области печени и т. д. От фасциолеза особенно тяжело страдают овцы
и телята. Этот трематодоз наносит огромный ущерб животноводству. Многие
другие сосальщики не менее опасны, так как разрушают органы или во
всяком случае нарушают их функции и, кроме того, выделяют в организм
хозяина различные токсические вещества.
Некоторые сосальщики опасны и для человека, причем трематодозы
человека распространены главным образом в тропических и субтропиче-
ских странах. Кроме обычной широко распространенной печеночной двууст-
ки (Fasciola hepatica), которая может встречаться и у человека, упомянем
следующих трематод. Китайская двуустка (Clonorchis sinensis) — окон-
чательными хозяевами ее могут быть человек, кошка, собака, пушные
звери, свинья; промежуточным — пресноводные моллюски из рода Bithynia.
Паразит, проникнув в печень человека, вызывает нарушения ее функций,
склеротические изменения в ней, которые в далеко зашедших случаях
могут перейти в рак печени.
Легочная двуустка (Paragonimus westermani) паразитирует в легких,
где образует опухолевидные цисты, дающие воспалительные процессы,
плевриты и пр. Первым промежуточным хозяином являются пресноводные
моллюски главным образом из рода Melania, вторым — пресноводные раки
и крабы. Распространен паразит в Японии, на п-ове Корея, на о. Тайвань,
в СССР известны завозные паразиты.
Кровяные двуустки (различные виды рода Schistosoma (см. рис. 156),
обитающие в крупных венозных сосудах, вызывают тяжелые заболевания.
Поражается весь организм человека.
Кровяные двуустки встречаются в тропических странах (Африка,
Южная Америка, юго-восточная Азия). В Японии, Китае и прилежащих
странах широко распространен вид Schistosoma japonicum. Борьба с этим
сосальщиком заключается в защите человека, входящего в воду, от нападе-
ния церкарий путем применения различных отпугивающих мазей и пропи-
тывания одежды различными веществами.
БОРЬБА С СОСАЛЬЩИКАМИ
Борьба с сосальщиками ведется методом комплексных мероприятий,
направленных к тому, чтобы разорвать цепь последовательных стадий
развития паразита, преградив для него возможности достигнуть стадии
мариты, т. е. дать новые запасы яиц. Эта общая цель достигается в отноше-
176
нии различных сосалыциков разными путями в зависимости от специфи-
ческих особенностей развития данного вида паразита, круга его промежу-
точных хозяев, условий заражения окончательного хозяина и т. д. В каче-
стве примера рассмотрим здесь методы борьбы с фасциолой — одним из
актуальнейших вредителей животноводства. Существенную роль играет
прежде всего терапевтическое вмешательство в ход заболевания, когда
употреблением специальных лекарств — противоглистных средств, на-
зываемых обычно антигельминтиками (гельминт — паразитический червь),
паразит умерщвляется и затем изгоняется из организма окончательного
хозяина. Этот процесс называется дегельминтизацией. Чтобы дегельмин-
тизация принесла успех, она должна проводиться в соответствии с основны-
ми данными о сроках развития сосальщиков. В отношении фасциолы де-
гельминтизация проводится 2—3 раза в год в соответствии со сроками
развития фасциол в промежуточных и окончательных хозяевах. Это де-
лают, например, непосредственно после постановки скота на стойловое
содержание и за 20—25 дней до выгона на пастбище. В других условиях
приходится проводить дегельминтизацию чаще. Что она дает, видно из
следующего примера. В предгорных районах Армении своевременная де-
гельминтизация привела к резкому снижению фасциолеза с 50—100%
больных животных до нуля. Результаты стойко держались 20 месяцев,
пока не был завезен новый скот, больной фасциолезом, заразивший паст-
бища новыми запасами яиц, почему потребовалась новая дегельминтиза-
ция.
Наряду с борьбой со взрослыми фасциолами планомерно должны вес-
тись работы по уничтожению промежуточного хозяина фасциол, т. е.
малого прудовика. Для уничтожения моллюсков применяются следующие
меры: 1) осушение территории, 2) химические методы борьбы с ними.
Осушение меняет условия существования моллюсков, нуждающихся в из-
вестных запасах влаги для своего развития. Это мероприятие снижает раз-
множение моллюсков. Обрабатывая занятые ими площади растворами
медного купороса или свежегашеной извести, можно добиться существен-
ного снижения численности моллюсков. Наконец, важное значение имеет
режим водопоя, когда принимают меры к ограждению пойм от возможности
занесения в них адолескарий; кроме того, водопои устраиваются там, где
меньше шансов на контакт между окончательными хозяевами фасциолы
(овцы, крупный рогатый скот) и промежуточными (моллюсками). Своевремен-
ным принятием описанных и других мер, основанных на изучении биологии
паразита и промежуточного хозяина, удается снизить заражение (инвазию)
и оздоровить домашних животных.
КЛАСС 4. ЛЕНТОЧНЫЕ ЧЕРВИ, ИЛИ ЦЕСТОДЫ
(CESTOIDEA)
ХАРАКТЕРИСТИКА
Тело лентовидное, сплюснутое в спинно-брюшном направлении. В
большинстве случаев оно состоит из отдельных члеников, или проглоттид,
совокупность которых составляет стробилу. Передний конец тела несет
особо модифицированный членик — головку, или сколекс (scolex), несу-
щий специальные органы фиксации к тканям хозяина — присоски и особые
крючья, часто расположенные на способном выдвигаться хоботке. Пище-
варительной системы нет. В члениках метамерно повторяется комплекс
органов, присущих каждому из них, а именно выделительные трубки, нерв-
ные тяжи, мощно развитые половые органы. Цестоды — гермафродиты.
Развитие сопровождается сменой хозяев и превращением. Если у сосаль-
щиков еще сохранились некоторые видимые связи со свободноживущими
177
предками (наружный паразитизм, наличие глазков, слабое развитие ку-
тикулы у моногенетических форм и другие признаки), то цестоды значи-
тельно глубже и полнее приспособились к паразитическому существова-
нию. Из перечисленных признаков особенно характерно полное исчезно-
вение кишечника. Изменен также и онтогенез. Так, у цестод нет явственно
обособленных зародышевых листков. В то время как личинки сосальщиков
все же претерпевают фазу свободного существования, личинки цестод
ведут паразитический образ жизни, и только зародыши короткое время
пребывают в наружной среде. Далее, в то время как часть сосальщиков
живет в качестве эктопаразитов, лентецам свойствен только эндопаразити-
ческий образ жизни. Все они (в половозрелом состоянии) обитают, как
правило, в кишечнике, реже в желчных каналах печени и в полости тела
различных животных; паразитируют ленточные черви и у человека. Всего
их описано свыше 1500 видов.
Представитель класса цестод
— бычий солитер (Taeniarhynchus saginatus)
Бычий цепень белого цвета. Он достигает огромных размеров, до 10—
12 м в длину.
Сколекс имеет до 2 мм в ширину и вооружен четырьмя мощными при-
сосками, снабженными мышцами. Задняя часть сколекса образует шейку
(рис. 159). Позади шейки тянется ряд члеников, общее число которых
достигает 2000. Форма члеников меняется по мере удаления от головки.
Ближайшие к сколексу членики короткие, причем их длина значительно
меньше ширины. Задние членики, напротив, удлинены, достигая в ширину
4—8 мм при 15—20 мм в длину.
Рис. 159. Головки цестод. / —
свиной цепень (Taenia soli-
um); // —бычий цепень (Tae-
niarhynchus saginatus). Видны
мощные присоски (acetabulae),
а у Т. solium — также венчик
крючьев на коротком хоботке
Рис. 160. Часть поперечного среза через членик
бычьего цепня (Taeniarhynchus saginatus):
/—кутикула, 2 — основание эпителиальных клеток,
3—кольцевая мускулатура, 4, 6—сечения волокон про-
дольной мускулатуры, 5 — эпителиальные клетки, 7—дор-
зовентралиная мускулатура. 8—мышечная клетка, 9 —
«известковые тельца»
Кожно-мускульный мешок. Головка (сколекс) и членики покрыты
многослойной кутикулой. Под кутикулой расположен эпителий (рис. 160).
Свободные концы эпителиальных клеток погружены во внутреннюю парен-
химу. Под кутикулой на уровне оснований эпителиальных клеток развита
довольно сильная кольцевая мускулатура. Под слоем кольцевой мускула-
туры расположен слой продольной мускулатуры. Кроме этой последней,
имеются волокна продольных мышц, которые равным образом проходят
178
между клетками эпителия. Описанный комплекс элементов образует кож-
но-мускульный мешок.
Внутренняя паренхима пронизана дорзо-вентральными мышцами и,
кроме того, продольными мышечными волокнами. Мышцы у бычьего цепня
очень сильные, чем объясняется значительная подвижность его в кишечни-
ке человека. Отдельные задние членики, оторвавшись от стробилы, могут
самостоятельно выползать из анального отверстия. Они могут ползать по
телу человека, по его белью и т. п. Системы органов, за исключением поло-
вой , развиты слабо.
Нервная система. Нервная система состоит из продольных нервных
стволов, самые крупные— боковые. В головке они соединены поперечными
Рис. 161. Гермафродитный членик бычьего цепня:
/ — двураздельный яичник. 2—яйцевод. 3— тельце Мелиса, 4—жел-
точник, 5 — ссмеприемник, 6—влагалище, 7 — клоака, 8 — начало
матки, идущее от оотипа, 9—матка (не имеет наружного отверстия),
10— мешок, в котором лежит циррус, 11— семяпровод, 12 — семен-
ники, 13—ствол выделительной системы, 14—нервный тяж
комиссурами, которые связаны с довольно сложным нервным головным
узлом. Органы чувств не развиты.
Пищеварительная система. Пищеварительная система отсутствует.
Питание происходит через поверхность тела. Паразит погружен в химус
(жидкое питательное содержимое) тонкой кишки человека. Этот питатель-
ный раствор диффундирует сквозь кутикулу в паренхиму. Проводниками
соков служат проксимальные (ближайшие, прилегающие к кутикуле)
плазматические части эпителиальных клеток кожи (рис. 160).
В паренхиматозных клетках происходит поглощение проникших сюда
веществ. Паренхима богата отлагающимся здесь животным крахмалом,
пли гликогеном, который играет большую роль в процессах анаэробного
дыхания.
Дыхательная и кровеносная системы отсутствуют.
Выделительная система. Состоит из многочисленных клеток с «рес-
ничным пламенем» и тонких каналов, которые впадают в боковые продоль-
ные выводные каналы. Эти каналы в каждом членике соединяются задним
поперечным каналом. Выделительная пора расположена на заднем членике,
в середине его свободного края. При отрыве этого членика выделительные
трубки открываются наружу своими отверстиями и новой общей выдели-
тельной поры не образуется. Из описания выделительной системы видно,
что она носит характер протонефридиев.
Половая система. Половая система достигает огромного развития
(рис. 161). Каждый членик имеет свою автономную половую гермафродит-
ную систему. В ближайших к головке члениках половой системы нет. На
179
расстоянии примерно 1 м от головки в члениках имеется уже явственно раз-
витая гермафродитная половая система. В задних члениках остается лишь
матка, наполненная яйцами. Остальные половые органы исчезают (кроме
выводных протоков половых органов). Членики, содержащие развитые
половые органы, называются гермафродитными, задние — зрелыми.
Мужские половые органы — семенники (рис. 161) —
имеют форму пузырьков, разбросанных в паренхиме членика. Число их
превышает тысячу. Семенные пузырьки имеют выводящие канальцы (vasa
efferentia), которые очень трудно видеть и можно обнаружить тогда, когда
они наполнены сперматозоидами.
Выводящие канальцы впадают в семяпровод (vas deferens). Последний
сильно извивается и выносит семя в совокупительный орган, или cirrus.
Он лежит в совокупительном мешке, откуда может выпячиваться. Мужское
половое отверстие открывается в половую клоаку.
Рис. 162. Часть стробилы (уменьшено) и зрелый членик
бычьего цепня. Ветвистая матка набита многочисленными
яйцами
В половую клоаку открывается и женское половое отверстие. Отверс-
тие клоаки открывается сбоку членика, то справа, то слева, причем пра-
вильного чередования нет.
Женские половые органы — яичник у бычьего цепня
парный, состоит из правой и левой долей. Обе доли связаны перемычкой,
от которой начинается яйцевод (oviductus). Проток яйцевода соединяется
с протоком оотипа, загибается кпереди (рис. 161) и впадает в расширенный
семеприемник (vesicula seminalis).
Сперматозоиды проникают в семеприемник по длинному участку
яйцевода, который образует влагалище. Последнее открывается наружу
женским половым отверстием в половую клоаку.
Яйца, выведенные из яичника и оплодотворенные в семеприемнике
семенем, в оотипе получают из желточников желточный материал и покры-
ваются скорлупой. Из оотипа яйца по короткому протоку попадают в матку.
Последняя расположена по медиальной линии членика.
По мере созревания члеников матка все более разрастается, причем
от ее срединного ствола в стороны — вправо и влево — образуются много-
численные (в среднем 15) боковые ветви. Все другие половые органы, кроме
яйцевода, семяпровода, совокупительного органа и клоаки, исчезают
(рис. 162).
180
Паренхима зрелого членика почти целиком занята огромной ветвистой
маткой, буквально набитой многочисленными зародышами в яйцевых обо-
лочках. Зрелая матка не имеет никакого сообщения с половыми путями
и наружной средой. Для того чтобы зародыши могли освободиться, отор-
вавшийся от стробилы зрелый членик должен попасть во внешнюю среду.
Здесь он разрушается, а яйца освобождаются и при соответствующих ус-
ловиях проделывают сложный путь развития.
Развитие. Развитие цепня сопровождается превращением и сменой
хозяев. Ко времени выделения наружу яйца развиваются в характерного
Рис. 163. Яйца цестод. 1, // — яйца бычьего цепня;
/// —зародыш широкого лентеца в .момент, выхода из
скорлупы, в которой остается эмбриональная оболочка;
/V — зародыш:
/ — шестикрючный зародыш, 2 — оболочка зародыша, 3—скор-
лупа яйца, 4 — нитевидные придатки (филаменты) скорлупы яйца,
5 — желточная клетка
шестикрючного зародыша, или онкосферу. Онкосфера имеет радиально
исчерченную оболочку, которая в свою очередь охвачена наружной эмбри-
ональной оболочкой, защищенной сверху скорлупкой яйца (рис. 163).
Последняя снабжена особыми нитевидными придатками — филаментами.
В благоприятных для паразита условиях, например, когда экскремен-
ты человека попадают в помещения для крупного рогатого скота, яйца
с онкосферой внутри легко попадают в желудок быка или коровы. Здесь
онкосферы освобождаются из яйцевых оболочек, выносятся в кишечник
животного и затем внедряются при помощи своих крючьев в слизистую
оболочку кишки, проникая этим путем в кровяное русло. Странствуя по
кровяному руслу быка или коровы, зародыши проникают обычно сначала
в печень, затем в сердце, легкие и другие органы. При этом онкосфера
развивается в своеобразную личинку, финку, называемую в данном случае
бычьим цистицерком (cysticercus bovis). Цистицерк представляет собой
пузырь величиной с горошину, причем внутри пузыря развивается го-
ловка, которая ввернута внутрь, в полость пузыря. Полость эта наполнена
жидкостью. Головка может выдвигаться из пузыря наружу. Этого можно
добиться механически, надавливая на пузырь, но головка выдвигается
наружу и самостоятельно под влиянием различных физических и химиче-
181
ских факторов, нормально под влиянием желудочного сока и соответствую-
щей температуры.
Дальнейшее развитие происходит в человеке. Если недостаточно про-
жаренная или проваренная говядина съедена человеком и попадает в желу-
док, произойдет следующее: финка выпячивает наружу головку и начи-
нает делать попытки фиксироваться на месте при помощи присосок. Обычно
ей это удается сделать в тонкой кишке, причем самый пузырь переваривает-
ся действием желудочного сока человека.
Присосавшаяся финка состоит первоначально из лишенной крючьев
головки и шейки. Затем начинается весьма своеобразный рост цепня,
выражающийся в образовании новых члеников.
Членики всегда растут от шейки, а не от члеников. Естественно поэто-
му, что самые молодые членики стробилы находятся около шейки, самые
старые — на заднем .конце тела. Следовательно, каждый членик, отпочко-
вавшись от шейки, затем отодвигается назад, так как от шейки отпочковы-
вается новый членик и т. д. Отодвигаясь все более назад, членик зреет,
в нем развиваются описанные половые органы, членик становится герма-
фродитным, наконец, зрелым, т. е. содержащим лишь матку, набитую си-
дящими в яйцевой оболочке онкосферами. Зрелый членик отрывается, на
его место становятся следующие членики и т. д.
Следовательно, стробила не есть нечто постоянное; напротив, она не-
прерывно образуется заново. Это — цепь постепенно сдвигающихся назад,
изменяющихся в положении (все дальше от головки), размерах, форме
и внутренней организации члеников, нечто вроде «живой конвейерной
системы».
Бычий цепень — космополит. Наносит довольно сильный вред чело-
веку, вызывает сильное малокровие и другие явления.
Циклы развития некоторых других представителей
класса цестод
У цестод, как и у сосальщиков, жизненные циклы осуществляются
далеко не однотипно. Разнообразие жизненных циклов цестод снова пока-
зывает приспособленность к существованию в различных условиях не только
половозрелых форм паразитов, но и их личинок. Рассмотрим жизненные
циклы некоторых цестод.
Цикл развития лентеца широкого Di phyllobothrium latum. Полово-
зрелая форма лентеца (рис. 164) обитает в тонких кишках человека, соба-
ки, кошки, песца.
Рис. 164. Широкий лентец (Diphyllobothrium latum). / — сколекс, видна
ботрия (/); II — гермафродитный членик; III —зрелые членики:
/ — ботрии, 2—набитая яйцами матка, 3— желточные фолликулы, 4 — яичник, 5 — ци-
руснын мешок, 6 — семенники, 7 — тельце Мелиса, 8 — наружное отверстие матки
182
Развивается широкий лентец с двумя промежуточными хозяевами.
Яйцо для дальнейшего развития должно попасть в воду, где вылупляется
личиночная форма — корацидий (рис. 165), покрытый ресничками; он
должен быть заглочен веслоногим рачком из рода Cyclops (см. рис.
301xi). В рачке корацидий превращается во вторую стадию— процеркоид
(рис. 165). Рачок заглатывается рыбой (промежуточным хозяином лентеца
могут быть многие пресноводные рыбы), где процеркоид превращается в
последнюю личиночную стадию—плероцеркоид (рис. 165), локализую-
щийся .в мышцах. Поедание недостаточно обработанной рыбы (непроварен-
ной, непрокопченной и пр.) ведет к заражению человека. Кроме человека,
окончательными хозяевами могут быть собака, кошка, лисица, песец.
Рис. 165. Развитие широкого лентеца. / — корацидий (/)
в оболочке (2), покрытой ресничками; II—процеркоид;
III — плероцеркоид (головкой вниз), видны ботрии (3)
Цикл развития эхинококка (Echinococcus granulosus). Эхинококк вы-
зывает заболевание, называемое эхинококкоз. Окончательным хозяином
эхинококка (рис. 166) являются собака и волк, в кишечнике которых
обитает половозрелая форма паразита. Промежуточными хозяевами слу-
жат человек и копытные. Человек заражается личинками эхинококка от
зараженных собак; копытные же, поедая траву; загрязненную испражне-
ниями зараженных собак или волков. Из кишечника личинки попадают в
кровь, разносятся по кровеносным сосудам и, оседая в легких, печени,
мозгу и других органах, вырастают в очень крупные, пузыревидные личи-
ночные формы — эхинококки. Внутри этих пузырей, которые могут до-
стигать размеров головы ребенка, лежат дочерние и внучатные пузыри
с головками будущих стробил. Рост материнского пузыря может привести
к смерти промежуточного хозяина, а его труп или внутренности могут стать
добычей волков и собак, которые, таким образом, и получают готовых
к дальнейшему развитию личинок. Материнские и дочерние пузыри раз-
рываются, головки прикрепляются к стенке кишечника и образуется ко-
роткая стробила.
ОБЩИЙ ОБЗОР КЛАССА
Внешнее строение. Большинство цестод характеризуется расчленен-
ностью тела на членики, хотя известны формы с нерасчлененным телом.
Размеры стробилы подвержены значительным колебаниям. Стробила взрос-
лой цестоды Mathevolepis petrotschenkovi достигает всего 0,6 мм, тогда
как другие цестоды имеют в длину несколько метров.
1.83
Число члеников (проглоттид) колеблется от 5—6 до 1000 и больше в
одной стробиле. Форма сколекса также очень изменчива (рис. 167) и в
значительной степени зависит от характера, величины и числа присосок;
Различают следующие типы органов фиксации.
1 Присоски (acetabulae) — обычно четыре (как у бычьего цепня).
Это сильные мышечные аппараты с внутренним глухим карманом (рис.
167). Они обычны у всех форм отряда Cyclophyllidea (цепни).
2. Ботридии — особые удлиненные защемляющие аппараты на ско-
лексе, часто вооруженные добавочными присасывательными аппаратами.
Они могут иметь очень сложное строение (рис. 167).
/ П
Рис. 166. Эхинококк (Echinococcus granulosus). /— половозрелая особь; //—строение
эхинококкового пузыря:
i — головка (с крючьями на хоботе и четырьмя присосками), 2— второй членик с гермафродитной
половой системой, — третий членике маткой, набитой многочисленными яйцами, 4 — кутикулярная
оболочка пузыря, 5 — герминативная (производящая) оболочка, 6—9— эндогенные (внутренние) до-
черние пузыри на разных стадиях развития, внутри образуются головки, 10 — сколекс, превращенный
в пузырь, 11 — 14 — экзогенные (наружные) пузыри на разных стадиях развития, 15 — вторичное раз-
множение пузырей
3. Присасывательные щели, или ботрии, свойственны довольно мно-
гим формам. Это примитивные присасывательные органы, более слабые,
чем присоски. Обычно их две (см. рис. 164, 167), но может быть одна.
4. У некоторых форм сколекс вооружен четырьмя втяжными щупаль-
цами, покрытыми загнутыми назад крючочками, как это мы видим у пред-
ставителей Tetrarhynchoidea (рис. 167).
5. Крючья у многих форм сопровождают описанные присасывательные
аппараты. Обычно они расположены на хоботке головки. Этот хоботок
подвижен. Когда червь присасывается, хоботок давит на слизистую обо-
лочку кишки хозяина, причем крючья впиваются в ткань и ранят ее. Хобо-
ток часто имеет явственную полость (рис. 167).
Обычно сколекс невелик, около 1 мм, но известны цестоды с крупным
сколексом; так у Anoplocephala gigantea он достигает 6 мм в ширину.
Позади сколекса у всех цестод лежит особый орган — шейка, которая
служит источником развития новых, от нее формирующихся члеников.
Проглоттиды (членики цестод) в высшей степени многообразны по ве-
личине и форме. По общей структуре членики бывают двух основных типов:
184
а) краспедотные членики характеризуются тем, что задний край каждого
предшествующего членика налегает в виде «паруса» на передний край
каждого последующего членика; б) в случаях акраспедотных члеников
они отделяются друг от друга простой перетяжкой, без «паруса». Эти раз-
личия имеют значение в систематике цестод. Далее обычно различают
Рис. 167. Головки цестод. / — Cyathocephalus truncatulus, животное по-
казано целиком (на головном конце — одна ботридия в виде воронки);
// — головка Polypobothrium vaccarii, вооруженная четырьмя ботри-
диями; /// —сколекс Taenia marshali с длинным хоботом и присоска-
ми (acetabulae), членики стробилы с острыми углами, выполняющими
роль зацепок; IV — сколекс Hymenolepis, четыре присоски (acetabulae)
и длинный хоботок, вооруженный крючьями; V — Hymenolepis cario-
са — сколекс, хоботок имеет явственную полость; VI — сколекс Р1а-
norothrium monticelli, на вершине сколекса присоска, шейка покрыта
обратно загнутыми щетинками; VII — Rybelinia lingualis в сколексе че-
тыре выдвижных хобота (/), связанных с мышцами (2)
анаполизические стробилы, задние членики которых не отделяются от стро-
билы к моменту их созревания, и аполизические стробилы, у которых
задние зрелые членики нормально отделяются от стробилы. К первому
типу относится стробила широкого лентеца, ко второму — бычьего цепня.
Каждый членик несет свой комплект метамерно повторяющихся органов
наподобие того, как мы это видели у бычьего цепня.
Кожно-мускульный мешок. Тело цестод покрыто кутикулой. У некото-
рых форм кутикула несет мелкие щетинки, обычно загнутые назад. Под
кутикулой лежит слой клеток погруженного эпителия, а под ним, и в сущ-
ности пронизывая его, залегает мускулатура — кольцевая и продольная
185
(рис. 160). Кроме этой субкутпкулярной мускулатуры, у цестод имеются
поперечные мышцы в самой паренхиме членика, причем они образуют слой,
лежащий несколько отступя от субкутикулярной мускулатуры, отделяясь
от него слоем паренхимы. Таким образом, эти мышцы как бы делят парен-
химу членика на два слоя — корковую паренхиму, лежащую между субку-
тикулярными мышцами кожно-мускульного мешка и указанными мыш-
цами, и сердцевинную, или центральную, паренхиму, отделенную от корко-
вой теми же мышцами (рис. 168). В этой сердцевинной паренхиме залегает
половая система цестод. Наконец, толщу паренхимы пронизывают дорзо-
вентральные мышцы, соединяющие спинную и брюшную стороны тела.
Обычно в паренхиме залегают многочисленные известковые тельца — про-
дукты обмена (рис. 160).
Нервная система. Нервная система состоит из нервного узла, залегаю-
щего в сколексе, и нервных стволов. Всего развито десять продольных ство-
лов, из них наиболее крупные — боковые.
Пищеварительная система. Пищеварительная система всегда отсут-
ствует. Как это уже было сказано выше, питательный субстрат, в котором
живет ленточный червь, вса-
сывается им всей поверхно-
стью тела, причем в тканях
цестод обнаружены специаль-
ные ферменты, которые обус-
ловливают переваривание
всосанной питательной массы
(химуса).
Выделительная система.
Выделительная система пос-
троена по типу протонефри-
дий с терминальными клет-
ками, несущими «ресничное
пламя». Экскреторные стволы
хорошо развиты прежде всего
в сколексе и затем, переходя
в проглоттиды, образуют бо-
Рис. 168. Схема соотношений между мышцами
и паренхимой у цестод (Amabilia lamelligera):
1 — субкутикулярные мышцы кожно-мускульного мешка,
2—корковая паренхима, 3— поперечные мышцы паренхи-
мы, 4 — срединная паренхима
ковые системы выделитель-
ных стволов, из них два дорзальных и два вентральных. Иногда эти
стволы усложнены, образуют разветвления, анастомозируют между собой,
дают ветви и т. п. В задней проглоттиде спинные и брюшные стволы сое-
диняются вместе, открываясь наружу задней порой, расположенной
обычно по середине заднего края последнего членика. Иногда в крупных
боковых экскреторных сосудах имеются клапаны, препятствующие движе-
нию жидких экскретов в обратном направлении.
Половая система. Половая система цестод в высшей степени многооб-
разна, хотя она и состоит, как правило, из органов, которые описаны выше
для бычьего цепня. Половое отверстие расположено либо по средней
брюшной линии (медиовентрально), либо на боку членика, либо (реже)
на спинной стороне его. Половое отверстие всегда ведет в половую клоаку,
принимающую мужское и женское половые отверстия. Яичник обычно
парный; чаще оба его компонента приближены друг к другу и лежат по
средней линии. У многих форм, однако, половые органы отодвинуты к
краям членика, образуя парные компоненты (рис. 169). Строение яичника
также многообразно. У тэний он имеет сетчатое строение (рис. 169) иногда
лучистое, чаще яичники компактны. Изменчиво и положение семенников;
они расположены то кпереди, то кзади от желточников и матки, то по бо-
кам от нее. Число их также изменчиво — от одного до нескольких сотен.
Соотношения между половыми органами у цестод показаны на рис. 169,
на котором очень ясно видно, что оотип принимает в себя протоки желточ-
ника, тельце Мелиса, яйцевод, проток семеприемника.
186
Размножение и развитие (онтогенез). Оплодотворение у цестод дости-
гается в результате совокупления, причем циррус выпячивается наружу и
вводится в отверстие вагины. Если в кишечнике имеется несколько особей,
Рис. 169. Половая система цестод. / — членик Catenotaenia dendritica;
II — Moniezia planissima; /// — Mosgovoyia pectinata; /V — Bertiella
elongata. Строение половой системы, а в частности и отношение оотипа
к другим половым органам, особенно явственны у Mosgovoyia:
1 — яичник. 2—яйцевод, 3—семеприемник, 4— влагалище, 5 — проток желточника,
6—желточник, 7 — тельце Мелиса, 8—матка, 9 — семенник, 10— оотип, 11— семяиз-
вергательный канал, 12— циррус
то может иметь место перекрестное оплодотворение. Однако, если в кишеч-
нике находится лишь одна особь, то наблюдается копуляция между члени-
ками одной и той же особи или даже самооплодотворение данного членика.
Оплодотворение яиц происходит в семеприемнике, а формирование заро-
187
дыша — в оотипе. Дробление яиц происходит либо в матке, либо во внеш-
ней среде. Характер дробления своеобразный. Он носит незакономерный
характер и приводит к развитию морулоподобной клеточной массы, кото-
рая вслед за тем дифференцируется на эктодермическую эмбриональную
и внутреннюю оболочку, у некоторых форм покрытую ресничками (рис.
163), у других — имеющую хитиноподобную консистенцию и радиально
исчерченную структуру. В результате дробления формируется зародыш,
Рис. 170. Яйца трематод и цестод и типы личинок цестод. / — яйца тре-
матод (/1— Fasciola hepatica; /2— Dicrocoelium lanceatum; /3— Clonorhis
sinensis; /4—Aragonimus ringeri; /5 —Schistosoma japonicum); харак-
терна крышечка, открывающаяся при выходе зародыша; II — яйца
цестод (//1—Taenia solium; //2—Taeniarhynchus saginatus; //3— Dip-
hyllobothrium latium; //4— Hymenolepis папа — карликовый цепень);
III—типы личинок цестод (///i—плероцеркоид; III2—цистицерк (финка);
II/з— цистицеркоид; ///4— ценурус)
вооруженный крючьями. Последних либо 6 — тогда это онкосфера,
либо 10 — тогда зародыш называется ликофорой. Матка зрелых члеников
набита соответственно онкосферами или ликофорами.
Если матка специального выводного отверстия не имеет, то описанные
зародыши выводятся наружу в члениках, которые отрываются и выделя-
ются во внешнюю среду вместе с фекалиями окончательного хозяина. Чле-
ники в наружной среде распадаются, а зародыши освобождаются. Если
матка имеет специальное отверстие, то зародыши выделяются наружу
через выводное отверстие матки в полость кишки хозяина, а оттуда вместе
с его фекалиями наружу. Если онкосфера или ликофора попадут в своего
промежуточного хозяина, то зародыш развивается в его органах в личинку.
Различают несколько типов личиночных форм (рис. 170).
1. Плероцеркоид. Имеет головку и плотное тело без пузырчатой зад-
ней части.
188
2. Цистицерк. Имеет головку, ввороченную внутрь пузыря. Головка
может выдвигаться наружу.
3. Цистицеркоид. Кроме пузыря, позади имеет хвостиковидный при-
даток.
4. Ценурус. Пузырь, внутри которого имеется несколько головок.
5. Эхинококк. Личиночная форма, которая состоит из материнского
пузыря с дочерними и внучатными пузырями внутри, причем в пузырях
образуются сколексы (рис. 166).
Личинки перечисленных типов могут перейти к дальнейшему разви-
тию и превращению в половозрелую форму лишь в том случае, если они
попадут в кишечник окончательного хозяина. В желудке пузырь распада-
ется, а головка закрепляется и начинает производить членики.
У ряда форм проникновению в окончательного хозяина предшествует
не один, а несколько промежуточных хозяев.
Сказанное выше можно обобщить следующим образом.
1. Чтобы развилась неполовозрелая стадия цестоды, т. е. финка,
промежуточным хозяином должна быть проглочена онкосфера (или лико-
фора).
2. Для развития половозрелой формы цестоды окончательным хозяи-
ном должна быть проглочена финка (например, цистицерк, ценурус и т. д.).
3. Хозяин, в котором живет неполовозрелая форма, всегда становится
промежуточным.
4. Хозяин, в котором живет половозрелый червь, всегда является
окончательным.
Экология и распространение. Цестоды, как мы видели, ведут исключи-
тельно паразитический образ жизни. Паразитическое существование дает
значительные преимущества. Прежде всего оно обеспечивает обильное
питание. Цестоды буквально погружены в питательный раствор. Во-вторых,
они защищены от каких-либо резких колебаний температуры. Если исклю-
чить воздействие приходящих факторов (например, глистогонных средств),
то в целом они обитают в относительно постоянной среде, подвергающейся
незначительным колебаниям. Однако они в своем индивидуальном суще-
ствовании всецело зависят от физиологического состояния хозяина.
Все цестоды находятся под постоянным воздействием перистальтических
движений стенок кишечника. Так как среда тела хозяина является постоян-
ной средой жизни эндопаразита, то естественно, что в этой среде паразит
не только питается, но и выделяет, например, в кишечник хозяина все
продукты собственного обмена веществ. При таких условиях цестоды долж-
ны обладать рядом приспособлений, которые позволяют им существовать
в своеобразной среде их обитания. Эти приспособления следующие.
1. Приспособления для прикрепления к телу
хозяина, как мы видели, присасывательные аппараты и крючья.
У длинных цестод и самые членики отчасти благодаря мощной мускулатуре
способны противостоять (в их взаимных связях) перистальтике кишечника.
У некоторых цестод членики имеют несколько заостренные задние края,
что также позволяет им цепляться за складки кишечного тракта.
2. Приспособления к бескислородной среде.
Все цестоды способны к анаэробному дыханию, происходящему за счет
расщепления гликогена. Поэтому всем цестодам свойственно энергичное
накопление гликогена в собственных тканях. Так, например, в тканях
собачьего цепня (Dipylidium caninum) содержится (в процентах к общему
количеству сухого вещества) 23,02% гликогена.
3. Приспособления к существованию в пита-
тельной среде. Существование в питательном субстрате делает
высоко полезной выработку приспособлений к максимальному использо-
ванию пищи. Особенно ярко выражена эта приспособленность у цестод.
Обитание в кишечнике, стенки которого энергично воспринимают пи-
тательный химус, делает питание цестоды через ротовое отверстие невыгод-
189
ным. Оно заменяется питанием всей поверхностью тела. Этот способ пита-
ния позволяет паразиту накапливать огромные количества питательных
веществ (углеводов и жиров). Эта способность накопления питательных
веществ имеет большое значение в жизни паразита, так как обеспечивает
существование основной его жизненной функции — продуцирования гро-
мадного количества зародышей.
4. Приспособления к сохранению вида. Огром-
ная плодовитость цестод является очень важным условием их существова-
ния. Выше было указано, что они выделяют продукты обмена в тело хо-
зяина. Это ведет к интоксикации хозяина. Кроме того, цестоды наносят
органам хозяина травматические повреждения и поражения. При таких
условиях переполнение хозяина паразитами могло бы вызвать быструю
гибель хозяина.
В связи с этим у цестод, как и у трематод, вырабатываются приспособ-
ления к размножению вне хозяина. Одним из них является смена хозяев —
переход из окончательного в промежуточного и обратно. Этот путь разви-
тия приводит к возможности проникновения в органы других окончатель-
ных хозяев и, следовательно, к распределению некоторого числа паразитов
в органах большого числа хозяев, а тем самым и к некоторому снижению
степени инвазии каждого из них в отдельности. Легко понять, что снижение
инвазии влечет за собой снижение смертности хозяев, а, следовательно,
и зависящих от них паразитов. Однако миграции из одного хозяина в дру-
гого связаны с многочисленными случайностями. Во многих случаях
эти миграции сопряжены с временным пребыванием во внешней среде.
Кроме того, и связи между промежуточными и окончательными хозяевами
далеко не всегда осуществляются. Отсюда понятно, что процесс смены хо-
зяев неизбежно влечет за собой большие потери. Гибнут зародыши и личин-
ки. Таким образом, огромная плодовитость паразитов (например, невоору-
женный ценень продуцирует ежедневно от 170 000 до 5 000 000 яиц на 1 г
фекалий) имеет значение фактора, компенсирующего сложные условия
смены хозяев и огромную гибель потомства. Без этой плодовитости смена
хозяев была бы невозможна (поголовная гибель немногих зародышей).
Следовательно, оставался бы лишь один способ размножения — в одном
и том же хозяине. Однако мы видели выше, что переобременение органов
хозяина в конечном счете невыгодно для самого паразита. Поэтому развитие
огромной плодовитости и смена хозяев оказалась биологически наиболее
выгодной формой паразитического существования.
Если распространение трематод ограничено распространением их про-
межуточных хозяев—моллюсков, то для цестод характерен значительно более
широкий круг промежуточных хозяев, а соответственно и более широкое
распространение. Так, среди промежуточных хозяев цестод известны пред-
ставители олигохет, пиявок, моллюсков, ракообразных, насекомых, много-
численных пресмыкающихся и млекопитающих. Это несомненно сыграло
большую роль в том, что фактически цестоды распространились по всему
земному шару, в том числе и в засушливых и пустынных областях, где
трематоды обычно представлены беднее или вовсе отсутствуют.
СИСТЕМАТИЧЕСКИЙ ОБЗОР КЛАССА
Класс ленточных червей делится на 2 подкласса: 1) нерасчлененные
(Cestodaria) и 2) расчлененные ленточные черви (Cestoda).
ПОДКЛАСС I. НЕРАСЧЛЕНЕННЫЕ ЛЕНТОЧНЫЕ ЧЕРВИ
(CESTODARIA)
Небольшая группа паразитических червей с нерасчлененным телом
с одним комплектом половых органов; зародыши с 10 крючьями (лико-
фора).
190
Сюда относится паразит полости тела осетровых рыб (Amphilina)
(рис. 171); паразит кишечника химеровых рыб — Gyrocotyle и некоторые
другие.
ПОДКЛАСС II. РАСЧЛЕНЕННЫЕ ЛЕНТОЧНЫЕ ЧЕРВИ
(CESTODA)
В этот подкласс входит основная масса ленточных червей, большин-
ство которых лентообразны, с расчлененным на отдельные членики телом.
Личинка с 6 крючьями (онкосфера). Сюда относятся несколько отрядов,
основные из которых следующие: 1) лентены (Pseudophyl-
lidea) и 2) цепни (Cyclophyllidea).
ОТРЯД ЛЕНТЕЦЫ (PSEUDOPHYLLIDEA) 1
Большей частью многочленистые цестоды. Сколекс
без крючьев и вооружен ботриями. Яйца с крышечкой,
как у трематод.
К нерасчлененным лентецам принадлежат несколько
видов гвоздичников (Caryophyllaeus, рис. 172). Они обла-
дают одним комплексом половых органов, сколекс отсут-
ствует. Паразиты карповых рыб. Промежуточными хо-
зяевами служат олигохеты (Tubifex, Limnodrilus).
У ремнецов тело также не
расчленено, но комплекты поло-
вых органов закономерно повто-
ряются. Паразиты водяных птиц
(гагар, чаек и др.).
Широкий лентец (Di phyllo-
bothrium latum) — наиболее извест-
ный представитель лентецов (см.
рис. 164, 165). Головка вооружена
двумя ботриями. Стробила имеет в
длину до 9 м и больше; члеников
3000 — 4000. Гермафродитные чле-
ники характеризуются тем, что
половые отверстия открываются
на брюшной стороне членика, а
не на боку. Матка имеет выводное
отверстие, яйца свободно выделя-
ются через это отверстие наружу,
отделения же отдельных члеников,
как у бычьего цепня, не происхо-
дит.
ОТРЯД ЦЕПНИ (СУСЬОРНУLLIDEA)
Сколекс имеет четыре присос-
ки. Половое отверстие всегда сбо-
ку проглоттиды. Имеется один жел-
точник, обычно лежащий позади
матки. Яйца без крышечки, онко-
сфера с шестью крючьями, без рес-
ничек.
Рис. 172. Гвоз-
дичник Caryo-
phyllaeus muta-
bi 1 is:
J—сколекс, 2—се-
менники, 3 —
желточники, 4 —
семяпровод, 5 —
сумка цирруса
Рис. 171. Нерасчленен-
ный ленточный червь
амфилина (Amphilina
ligu loides):
К этому отряду принадлежат
наиболее распространенные цесто-
/ — матка, 2—влагалище,
3 — семяпровод, 4 — семен-
ники, 5 — желточники
191
ды, в том числе описанный выше бычий цепень. Из числа других предста-
вителей назовем следующих.
Свиной цепень (Taenia solium) имеет сколекс, вооруженный венчиком
крючьев и четырьмя присосками (см. рис. 159). Стробила достигает 2 м
в длину. Половые отверстия сбоку членика, то справа, то слева, без пра-
вильного чередования. Яичник, как у бычьего цепня, двулопастной, но
с добавочной третьей долькой, расположенной под влагалищем и семяпро-
водом. К матке идет по девяти (основных) боковых ветвей с каждой стороны
маточного ствола (у бычьего цепня их 15, см. выше). Окончательным хозяи-
ном служит человек, промежуточными — свинья, собака, кошка и другие
млекопитающие, а также человек (см. ниже), в органах которого из онко-
сфер развиваются финки (рис. 173).
Эхинококк (Echinococcus granulo-
sus) — маленький цепень, стробила ко-
торого содержит 3—4 членика. Общая
длина стробилы 2,7—5,4 мм. Сколекс
несет 4 присоски и вооружен 38—34
крючьями. Обычно только задний чле-
Рис. 173. Финка (справа) и шестикрю- Рис. 174. Мозг овцы, поражен-
чный зародыш свиного солитера (Taenia ный ценурусом Multiceps mul-
solium) ticeps (/)
ник несет развитую продольно вытянутую матку с яйцами (см. рис. 166),
в которых развиваются онкосферы. Окончательным хозяином служат со-
бака и волк, в кишечнике которых можно обнаружить огромные количества
взрослых эхинококков. Промежуточным хозяином этой цестоды служат
копытные и человек.
Альвеококк (Alveococcus multilocularis)—близкая эхинококку цестода.
Стробила имеет 1,3—2,2 мм в длину, члеников 2—4. Задний членик несет
матку мешковидной формы. Сколекс с четырьмя присосками и 28—32
крючьями. Окончательные хозяева — лисица, песец, кошка, промежуточ-
ные — грызуны, человек.
Овечий мозговик (Multiceps multiceps) паразитирует в кишечнике
собаки, которая выполняет роль окончательного хозяина, тогда как про-
межуточным хозяином служат жвачные, в том числе овца. Ценурус часто
поражает мозг овцы (рис. 174).
Собачий цепень (Dipylidium caninum) — паразит кишечника собаки
(рис. 175) и кошки. Сколекс этого цепня несет хоботок, венчики крючьев
и четыре присоски. Длина стробилы 15—35 см. Матка сетчатая, в петлях
этой сетчатой матки лежат семенники. Половой аппарат двойной. Яичники
дольчатые. Личинка типа цистицеркоида, с хвостом.
Мониезия (Moniezia) — крупные цепни (рис. 175) с широкими члени-
ками. Сколекс округлый, без хоботка, с четырьмя присосками. Каждый
гермафродитный членик несет двойной комплект половых органов, почему
192
половые отверстия двусторонние. Матка в виде сложной сети тяжей и тру-
бок. Взрослые формы находятся в кишечнике жвачных, которые служат,
следовательно, окончательными хозяевами. Промежуточными хозяевами
служат клещи — орибатиды (Oribatoidea), в полости тела которых раз-
виваются цистицеркоиды. Сюда относится ряд видов, основные из них:
Moniezia expansa, стробила до 5 м и М. benedeni — цестода, достигающая
Рис. 175. Представители Cyclophyllidea. / — сколекс п шейка Dipylidium caninum,
видны хобот, присоски (acetabulae), длинная шейка и молодые членики; // — гер-
мафродитный членик того же вида; /// —сколекс Moniezia expansa; /V — стробила
A\oniezia benedeni:
1—левый и правый яичники, 2—желточники, 3—матка, имеющая вид ячеистой сетки, 4— влага-
лище, 5— циррусныи мешок, циррус торчит наружу. 6—семенные пузырьки, лежащие в петлях
(ячейках) матки
2,5—4 м в длину. Паразиты крупного рогатого скота, коз, овец, лосей и
других животных.
Из представителей других отрядов следует упомянуть своеобразных
четыреххоботных цестод (Tetrarhynchoidea), у которых, помимо приса-
сывательных органов, имеются четыре втяжных хоботка, усаженных загну-
тыми назад крючками (рис. 176). Эти хоботки могут втягиваться в особые
карманы при помощи мышц ретракторов (гл. retractores).
К этому отряду относится ряд родов, представленных многими видами,
паразитирующими во внутренних органах некоторых морских рыб.
7 Курс зоологии, т. I
193
ВРЕДОНОСНОСТЬ ЦЕСТОД
Цестоды из-за особенностей своего развития относятся к числу опас-
ных паразитов животных и человека. Рассмотрим здесь несколько приме-
ров. Так, у свиного цепня (Taenia solium) проглоттиды, оторвавшись от
Рис. 176. Представители отряда Tetrarhynchoidea. / — сколекс и начало
стробилы Nybellinia lingualis. видны присоски и хоботки; // — часть стробилы
того же вида; III — часть хоботка с многочисленными крючьями; IV — сколекс
Tentacularia bicolor; V — часть стробилы этого же вида
стробилы, выносятся наружу вместе с экскрементами человека и разру-
шаются. Попав в желудок промежуточного хозяина, шестикрючные онко-
сферы освобождаются от оболочек, проникают в кишечник, а через его
стенки — в кровяное русло промежуточного хозяина. Здесь онкосфера
развивается в финку типа цистицерка (Cysticercus cellulosae, рис. 173).
Финки застревают в различных органах, где и остаются. Дальнейшее раз-
витие их возможно, если промежуточный хозяин будет съеден человеком.
Таким промежуточным хозяином является свинья. Попав в желудок чело-
194
века, цпстицерк высовывает головку и делает ощупывающие движения.
Пузырь растворяется желудочным соком, а головка выносится в тонкую
кишку человека, где она укрепляется и начинает продуцировать чле-
ники.
Как сказано выше, человек может быть и промежуточным хозяином
свиного цепня. Это бывает в тех случаях, когда онкосферы попадают в
желудок человека либо через рот (при неопрятности, грызении ногтей
и т. д.), либо посредством рвоты, когда в результате обратной перисталь-
тики членики и онкосферы (яйца) попадают в желудок. Здесь онкосферы
освобождаются от оболочек, возвращаются в кишку и внедряются в кровя-
ное русло. В органах человека (в печени, сердце, легких, мозге, глазном
яблоке и т. д.) тогда развиваются финки.
Болезнь, вызываемая финками и известная под общим названием ци-
стицеркоза, смертельно опасна, так как в большинстве случаев пора-
жает мозг.
Другим опасным для человека цепнем нужно считать эхинококка.
И здесь опасность в том, что человек — промежуточный хозяин эхинококка.
Человек заражается от собаки онкосферами, сидящими в яйцевых обо-
лочках.
Онкосферы эхинококка развиваются в органах человека в эхинококко-
вый пузырь. Он имеет сложно устроенную стенку (рис. 166) состоящую
из толстой наружной кутикулы и герминативного слоя. В последнем идет
образование новых, дочерних, пузырей. Эти пузыри могут быть эндогенными,
растущими внутри материнского пузыря, и экзогенными, растущими на
наружной поверхности материнского пузыря.
Внутри дочерних пузырей могут образовываться внучатные пузыри.
В перечисленных пузырях образуются головки эхинококка. Окончательный
хозяин эхинококка (собака) заражается, поедая эхинококкозные органы
промежуточного хозяина — кошек, кроликов и т. д.
Эхинококкоз человека — очень опасная болезнь. Эхинококковые пу-
зыри локализуются прежде всего в печени, а затем и в других органах,
обусловливая огромные опухоли живота и тяжелую болезнь легких, мозга,
костей (трубчатых) и т. д.
Альвеококкоз (см. выше) — не менее опасная форма заболевания
человека. Альвеококкоз, в процессе которого человек становится проме-
жуточным хозяином этого цепня, характеризуется тем, что в печени чело-
века развивается сложный комплекс личиночных форм, имеющих вид густо
и плотно расположенных мелких пузырей.
Дефинитивным хозяином овечьего мозговика (Multiceps multiceps) слу-
жит собака. Рассеиваемые ею онкосферы для дальнейшего развития должны
попасть в кишечник жвачных. В их внутренних органах развивается
личинка этого цепня типа ценуруса. Ценурусы особенно опасны для
овец, у которых они часто локализуются в головном мозге, вызывая
так называемую вертячку овец. Вертячка овец кончается гибелью
животного.
Среди паразитов крупного рогатого скота и овец одним из наиболее
опасных нужно считать крупных мониезий (Moniezia, рис. 175). Зре-
лые членики этого цепня выпадают наружу. Если это произошло на паст-
бище, то по разрушении членика яйца мониезии попадают на траву
и на почву.
Здесь широко распространены маленькие клещи из группы орибатид.
Орибатиды очень мелки и многочисленны. Они заражаются зародышами
мониезии, и в результате в их теле развиваются личинки этой цестоды. Жвач-
ные, срывая траву, проглатываюторибатид, а вместе с ними личинок моние-
зий. В результате овцы и коровы становятся окончательными хозяевами со-
ответственных видов мониезий. Мониезиозы овец и крупного рогатого ско-
та — тяжелое заболевание, особенно для овец. В запущенных случаях
мониезиоз ведет животных к гибели.
7*
195
БОРЬБА С ЦЕСТОДАМИ
Болезни, вызываемые цестодами, известны под общим названием цесто-
дозов. Многие из них, как это было указано выше, опасны для жизни чело-
века и домашних животных. Ввиду этого с цестодами ведется систематиче-
ская борьба. Как и в отношении трематод, она основана на решении глав-
ной задачи — разорвать цепь развития цестоды. Разумеется, в каждом
отдельном случае это достигается различными путями в соответствии с
особенностями онтогенеза данной цестоды.
Так, в борьбе со свиным цепнем (Taenia solium) огромное значение при-
обретает воздержание от употребления в пищу свиных продуктов (окоро-
ков, грудинок, свиной колбасы) неизвестного происхождения, которые
могут содержать финки цепня. Ведь достаточно их проглотить, чтобы сде-
латься окончательным хозяином этого цепня. Опасность превращения
человека в промежуточного хозяина свиного цепня предотвращается содер-
жанием в чистоте жилого помещения, уничтожением тараканов, которые
могут на лапках заносить онкосферы на продукты питания, тщательным
мытьем овощей с огородов, своевременным изгнанием из кишечника (де-
гельминтизация) свиного солитера (если он есть), чтобы избежать опасности
попадания онкосфер из кишечника в желудок при рвоте, что делает чело-
века также и промежуточным хозяином цепня.
В основе борьбы с эхинококком лежит уничтожение бездомных собак —
окончательных хозяев этого паразита и лечение домашних, охотничьих
и служебных собак, больных кишечным эхинококкозом. Эти мероприя-
тия препятствуют превращению человека в промежуточного хозяина эхи-
нококка. Однако важно предотвратить и превращение собаки в оконча-
тельного хозяина этого паразита. Часто собака заболевает, поедая органы
домашних животных — промежуточных хозяев эхинококка, например
овец. Поэтому принимаются меры к недопущению собак на территорию
боен и т. п.
В борьбе с мониезиозами (см. выше) жвачных огромное значение имеет
пастбищный режим. Так, телят и овец в целях предотвращения контакта
с промежуточными хозяевами мониезий (клещи орибатиды), выпасают на
пастбищах, где до того не было выпаса жвачных.
Больные животные подвергаются специальному лечению, направлен-
ному на изгнание мониезий из их кишечника.
ФИЛОГЕНИЯ МОНОГЕНЕИ. ТРЕМАТОД И ЦЕСТОД
Трематоды безусловно происходят от турбеллярий и в частности пря-
мокишечных. Так, много сходных черт имеется между указанными группа-
ми в общем плане организации нервной системы. И там, и здесь имеется
головной нервный узел; продольные стволы трематод гомологичны соот-
ветственным нервным стволам прямокишечных турбеллярий. Выделитель-
ные органы построены также сходно. Имеются черты сходства в строении
кишечника. Некоторые паразитические турбеллярии необычайно похожи
на трематод; так, с достижением взрослого состояния они теряют реснич-
ный покров, а иногда, свободные личинки некоторых трематод, как мы
видели, несут реснички на поверхности эпителия. Различия связаны пре-
имущественно с чертами специализации трематод и развитием у них приспо-
соблений к паразитизму. Можно принять, что трематоды — это перешед-
шие к паразитизму прямокишечные турбеллярии.
Надо отметить, что в последнее время новое освещение получил вопрос
о происхождении моногеней. Их сближают с цестодами и считают, что они
произошли от рабдоцёл независимо от дигеней.
Значительно сложнее вопрос о филогении цестод. Они настолько глу-
боко изменены паразитизмом, что точек соприкосновения с соседними
группами у них меньше.
196
Однако у нерасчлененных цестодарий нервная система сходна по своей
организации с таковой дигенетических трематод. Этот факт, так же как и
черты сходства в организации половой системы трематод и цестод, был
причиной, по которой часть исследователей склонялась к мысли, что цесто-
ды произошли от трематод, выражая своей организацией более глубокую
специализацию в направлении паразитического существования. С другой
стороны, было показано, что сколекс цестод гомологичен переднему кон-
цу тела прямокишечных турбеллярий, тогда как выделительная система
первых весьма сходна с выделительной системой вторых. Поэтому не лишен
основания и другой взгляд, согласно которому цестоды происходят от пря-
мокишечных турбеллярий. Последняя теория более вероятна. Во многих
чертах своей организации цестоды резко отличаются от трематод. Так,
им свойственно асимметричное расположение половых отверстий (на боку
проглоттид). Нервная система их несет черты радиальной симметрии в рас-
положении основных стволов. Хотя эта черта частично имеет, по-видимому,
вторичное происхождение, но целиком к этому ее свести нельзя; ее можно
сопоставить с элементами радиальной симметрии у турбеллярий. Все пере-
численные факты позволяют считать, что цестоды отделились от древних
прямокишечных турбеллярий независимо от трематод и притом, вероятно,
раньше последних. Пройдя длинный исторический путь приспособления к
паразитизму, они подверглись значительно большей, чем трематоды
специализации, приобрели оригинальную организацию и утратили боль-
шинство признаков сходства со своими свободноживущими предками.
ТИП V. КРУГЛЫЕ, ИЛИ
ПЕРВИЧНОПОЛОСТНЫЕ, ЧЕРВИ
(NEMATHELMINTHES, ИЛИ
ASCHELMINTHES)
ХАРАКТЕРИСТИКА
К круглым червям относятся многообразные формы, весьма различные
в деталях организации, но обладающие следующими общими для них приз-
наками. Тело обычно удлиненное, более или менее округлое в поперечном
сечении. Ротовое отверстие обычно расположено на переднем конце тела.
На заднем конце тела на спинной или брюшной стороне расположено зад-
непроходное отверстие. Кожно-мускульный мешок состоит из кутикулы,
подлежащего эпителия и мышц. Для мышечной системы характерна ре-
дукция кольцевых мышц и наличие продольных мышц, либо лежащих
более или менее полными лентами, либо представленными только отдель-
ными мышечными тяжами. Внутренние органы лежат в первичной полости
тела, или протоцеле. Характернейший признак протоцеля — отсутствие
у него собственной выстилки. В протоцеле имеется полостная жидкость
с плавающими в ней клетками. Нервная система представлена различно
ориентированными продольными нервными стволами. Имеется полное или
неполное окологлоточное нервное кольцо. Кровеносной и дыхательной
систем нет. Дыхание либо полностью аэробное — через кожу, либо частич-
но анаэробное. Выделительная система представлена протонефридиями
или специальными выделительными каналами иного строения. Половая
система либо гермафродитная, либо раздельнополая. Ее строение у пред-
ставителей отдельных классов этого типа очень различно. Метамерии,
т. е. закономерной повторности органов, нет. Круглые черви очень широко
распространены, населяя моря, пресные воды, почву; многие из них перешли
к паразитическому образу жизни в органах животных, растений и человека.
КЛАССИФИКАЦИЯ
Тип круглые, или первичкополостные, черви (Nemathelminthes, или Ascheltninthes)
Класс 1. Брюхоресничные, или гастротрихи (Gastrotricha)
Класс 2. Собственно круглые черви, или нематоды (Nematoda)
Подкласс I. Аденофореи (Adenophorea)
Подкласс II. Сецерненты (Secernentea)
Класс 3. Волосатиковые (Gordiacea)
Класс 4. Коловратки (Rotatoria)
198
КЛАСС 1. БРЮХОРЕСНИЧНЫЕ, ИЛИ ГАСТРОТРИХИ
(GASTROTRICHA)
ХА РАКТЕРИСТИКА
Мелкие червеобразные животные. Тело покрыто кутикулой, но на
брюшной стороне у большинства форм имеется ресничный эпителий (от-
сюда и название класса). Полость тела первичная. Кишечник сквозной.
Кровеносная и дыхательная системы отсутствуют. Выделительная система
протонефридиального типа. Нервная система состоит из большого надгло-
точного ганглия и двух продольных стволов. Большинство гермафродиты.
Развитие без превращений. Большая часть обитает в пресных водах.
ОБЩИЙ ОБЗОР КЛАССА
Внешнее строение. Гастротрихи (рис. 177)—маленькие (0,05—1,5 мм)
животные, имеющие удлиненное, часто бутылкообразное, округлое в попе-
речном сечении тело. Кутикула обычно покрыта чешуйками и несет ще-
тинки. Брюшная сторона либо сплошь, либо частично покрыта ресничками.
Рис. 177. Гастротрихи. / — общий вид Chaetonotus maximus с брюш-
ной стороны; II — анатомия пресноводной гастротрихи; III — анато-
мия морской гастротрихи Macrodasys budenbrocki:
1 — пищевод, 2 —кишечник, 3— анальное отверстие, 4 — семенники (большое сход-
ство с Rhabdocoela), 5 — семяпровод, 6 — циррус, 7 — яичник, 8 — яйца, 9 — копу-
лятивная бурса, 10 — хвост, 11 — протонефридий
199
Под кутикулой лежит эпителиальная ткань (гиподерма). Мышцы
только продольные не образуют сплошного слоя и представлены разоб-
щенными узкими лентами. Поэтому у гастротрих нет настоящего кожно-
мускульного мешка. Головной конец несет ротовое отверстие; на заднем
конце тела либо на спине, либо на брюхе (что типичнее) расположен анус.
Задний конец тела у многих форм вильчато раздвоен. В этом случае вер-
шины вилки имеют поры, через которые выделяется клейкий экскрет
желез, лежащих в основании отростков вилки.
Внутреннее строение. Кишечная трубка состоит из ротовой полости,
снабженного радиальной мускулатурой пищевода, энтодермической сред-
ней и короткой задней кишок. Выделительная система представлена про-
тонефридиями (две трубки — одна справа и другая слева), заканчивающи-
мися типичными терминальными органами с «ресничным пламенем». Нерв-
ная система интересна тем, что так называемый мозг, лежащий в области
пищевода, не отделен от эктодермы. От мозга идут два нервных ствола.
Из органов чувств особенно интересны ресничные ямки — органы хими-
ческого чувства. Развитие изучено недостаточно. Известно около 175 видов.
Экология, представители, филогенез. Гастротрихи частью живут в
пресных водах, частью в море. Это типичные донные организмы. Они ловко
и быстро, часто скачками снуют в покрывающем дно бассейна иле или в дет-
рите в поисках добычи, которую они вовлекают в рот при помощи энергич-
ной работы сильных ресничек. В пресных водах весьма обычны виды рода
Chaetonotus, принадлежащие к отряду Chaetonotoidea. Среди морских
форм интересны представители второго отряда — Macrodasioidea, обладаю-
щие рядом своеобразных признаков.
Брюхоресничные представляют собой очень интересную группу в том
отношении, что сочетают признаки как турбеллярий (частичный ресничный
покров, протонефридии, строение половой системы), так и первичнополост-
ных (кутикулизация покровов, сквозной кишечник, первичная полость
тела). В связи с этим они имеют большое значение для понимания происхож-
дения нематод (см. ниже).
КЛАСС 2. СОБСТВЕННО КРУГЛЫЕ ЧЕРВИ,
ИЛИ НЕМАТОДЫ (NEMATODA)
ХАРАКТЕРИСТИКА
Нематоды, как показывает уже их название, образуют центральную
группу типа круглых червей. Тело удлиненное, нитевидное, веретено-
видное, обычно суженное на переднем и заднем концах. Размеры тела
варьируют от долей миллиметра до нескольких метров. Поперечное сече-
ние тела всегда правильно круглое. Ротовое отверстие терминальное, в
центре переднего конца тела. Анус лежит на брюхе. Участок тела позади ану-
са образует хвост. Все тело покрыто сплошной кутикулой, имеющей отверс-
тия только для ротовой полости, ануса, выделительного органа, половых
путей, немногих кожных желез. Мышцы — только продольные, в виде
более или менее сплошных лент, особенно заметно отделенных друг от
друга по боковым полям. Протоцель характеризуется высоким тургором
(полостной тургор), поддерживающим форму тела. Центральная нервная
система состоит из продольных стволов, среди них крупнейший брюшной.
Имеется полное окологлоточное нервное кольцо. Пищеварительная си-
стема включает в себя переднюю, среднюю и заднюю кишки. Просвет пи-
щевода трехгранный. Стенка средней кишки обычно состоит из одного
слоя эпителиальных клеток. В пищеводе имеются пищеварительные железы.
Выделительная система состоит из различно устроенных выделительных
клеток. Протонефридии отсутствуют. Половые органы трубчатые, парные;
200
в подавляющем числе случаев раздельнополые организмы. Самки имеют
обособленное половое отверстие. У самцов половые продукты и содержимое
кишечника выпадают в общую клоаку. Самцы имеют специальный аппарат
(спикулы) для расширения женского полового отверстия. Нигде и ни на
какой стадии развития у нематод не бывает мерцательного эпителия и под-
вижных ресничек. Дробление детерминированное, билатеральное, полное.
Развитие обычно с неполным превращением. Нематоды живут в морях,
в солоноватоводных бассейнах, в пресной воде, в минеральных источниках,
в горячих ключах, в почве, в различных гниющих и бродящих органиче-
ских остатках, а также паразитируют в растениях, животных и человеке.
Общая численность нематод огромна, по Коббу — около 500 000 видов.
Представитель класса нематод — лошадиная аскарида
(Parascaris equorum)
Лошадиная аскарида живет в тонкой кишке лошади, питаясь ее содер-
жимым. Нематода имеет белый или розоватый цвет и достигает довольно
крупных размеров. Самки имеют в длину до 37 см, самцы — до 25. Распо-
знаются они и по другому признаку. У самок тело выпрямленное, тогда
как у самцов хвостовой конец загнут на брюшную сторону (рис. 178).
П I
Внешнее строение. Форма тела веретеновидная, в
поперечном сечении круглая. Передний и задний концы
тела сужены. Тело червя обладает необычайной упру-
гостью. Впечатление такое, будто полость тела перепол-
нена жидкостью и его стенки испытывают значитель-
ное внутреннее давление. Так, оно и есть в действитель-
ности: с одной стороны, очень велико давление
полостной жидкости на стенки тела, ас другой — очень
велика прочность и эластичность покровов. Последним
свойством обладает кутикула, покрывающая все тело
нематоды. Ее эластичность определяется особенностями
химического строения и морфологической структурой.
Рис. 178. Лошадиная аскарида Parascaris equorum. / —
самец; 11 — самка (у самца, имеющего меньшие размеры,
хвост загнут на брюшную сторону); /// — кишка лошади,
прободенная аскаридами
Кутикула состоит из нескольких слоев, причем внутренний слой волокнис-
тый. Волокна имеют опорное значение.
Кожно-мускульный мешок. Под кутикулой лежит эпидермис, извест-
ный у нематод под названием гиподермы. Под гиподермой мышцы. Отдель-
ных клеток в гиподерме не видно, и она состоит из плазматической массы
201
с лежащими в ней ядрами. Следовательно, гиподерма представлена син-
цитием.
Мускулатура состоит из продольных мышц. Последние не образуют
сплошного слоя. Напротив, мышцы явственно разделены на четыре ленты,
из которых две принадлежат спинному сектору, а две другие — брюшному
(рис. 179). Спинные ленты отделяются от брюшных наиболее явственно,
так как по бокам тела гиподерма резко утолщена и образует два продоль-
ных боковых гиподермалъных валика. Эти боковые валики можно заметить
и снаружи по бокам тела, в виде продольных боковых линий. Обе спинные
Рис. 179. Строение аскариды. I — поперечный разрез через середину
тела самки; // —разрез яичника; III—эпителий кишечника:
1— рахис, 2— яичник, 3 — спинной нервный тяж, 4—кутикула, 5—гиподерма, 6 —
кишечник, 7 — начальный отдел яичника, 8— выделительный канал, 9 — яйцевод, 10—
сократимая (фибриллярная) часть мышечной клетки, 11— матка, 12 — пузыревидная
часть мышечной клетки (саркоплазма), 13— брюшной нервный тяж, 14—боковые вали-
ки гиподермы с боковыми (латеральными) нервными тяжами, 15 — оогоний, 16 — стенка
яичника, 17 — ворсинки на поверхности клеток, 18— зона поглощения питательных
соков, 19—секреторные зерна, 20— гликогеновые зерна (трофохопдрии), 21 — ядра
клеток эпителия, 22 — обкладка кишечного тракта
ленты и соответственно брюшные также явственно обособлены, хотя и
менее резко. Между спинными лентами лежит разделяющий их спинной
нервный ствол, тогда как обе брюшные ленты отделены друг от друга соот-
ветственным брюшным нервным стволом.
Нервная система. Окологлоточное нервное кольцо охватывает пищевод
вблизи головного конца и отсылает ряд нервных стволов. Наиболее мощный
среди них брюшной (вентральный). Вдоль спины тянется дорзальный нерв-
ный ствол, менее развитый. По бокам тела имеются латеральные нервные
202
стволы. Кроме этих стволов, у аскариды имеются еще субдорзальные и
субвентральные стволы (рис. 180). Перечисленные стволы соединяются ко-
миссурами и связаны с довольно многочисленными нервными узлами, или
ганглиями.
От центральной нервной системы по направлению к кутикуле идут
нервы, подходящие к органам чувств. Последние у аскариды представлены
многочисленными мелкими папиллами. У самок их относительно немного.
У самцов, напротив, в области анального отверстия, кпереди и кзади от
него, значительное количество мелких папилл, позволяющих им во время
совокупления ориентироваться в положении женского полового отверстия.
Обильно иннервирован также передний конец тела и в частности губы, окру-
жающие ротовую полость.
Полость тела. Полость тела представлена первичной полостью, или
протоцелем (псевдоцель). Эта полость не имеет собственных
стенок и огра-
связанные
с ним нервы аска-
риды:
/—окологлоточное нервное кольцо,
2 — брюшной нерв, 3—спинной
нерв, 4 — боковые нервы, 5— суб-
дорзальные нервы, 6 — субвентраль-
ные нервы
ничивается непосредственно мускулатурой.
Протоцель до отказа наполнен полостной
жидкостью, в которую погружены отдельные
различно ориентированные соединительно тка-
ные клетки. Жидкость эта у аскарид издает
запах, напоминающий валериановые капли.
Он обусловлен присутствием в полостной
жидкости масляных кислот — валериановой,
капроновой и др. Жидкость раздражает сли-
зистые оболочки (это всегда нужно помнить
при вскрытиях аскариды).
Пищеварительная система. Пищевари-
тельная система разделена на три отдела:
переднюю кишку, среднюю и заднюю. Рото-
вое отверстие, ведущее в переднюю кишку,
расположено строго терминально на переднем
конце тела и окружено тремя мощно развиты-
ми губами, которые вооружены губными па-
пиллами. Ротовое отверстие ведет в глотку,
имеющую трехгранный просвет и выстланную
кутикулой. Стенка глотки состоит из перифе-
рического эпидермального слоя и мощной
мышечной системы, состоящей из радиально
расположенных волокон. Она имеет значение насасывающего органа.
Глотка переходит в пищевод, покрытый поперечными складками. Пищевод
переходит в цилиндрическую среднюю кишку. Пищевод и средняя киш-
ка у аскарид не имееют мышечной обкладки. Стенка кишки состоит из
однослойного эпителия энтодермического происхождения, причем в клет-
ках содержится большое количество зерен гликогена (рис. 179), а внутрен-
няя поверхность клеток, обращенная к просвету кишки, несет палочко-
видный слой. Аскарида питается содержимым тонкой кишки лошади,
причем ей свойственно внеклеточное пищеварение (в отличие от плоских
червей, у которых пищеварение внутриклеточное). Ворсинчатый палочко-
видный слой на поверхности кишечных клеток имеет значение всасыватель-
ного аппарата, и через его посредство содержимое кишечника аскариды
(растворы питательных веществ) всасывается в стенку кишки. Аскариды
переваривают белки и углеводы. Последние отлагаются в кишечных клет-
ках в виде гликогена. Организм аскариды очень богат гликогеном (до
32% в сухом остатке).
Кровеносной и дыхательной систем нет. Раньше считали, что лошадиная
аскарида, по крайней мере во взрослом состоянии, не способна к дыханию
свободным кислородом. Старые опыты Вайнланда показали, что в кислороде
аскарида живет более короткие сроки, чем в среде, богатой углекислым
газом. Соответственно принималось, что дыхание аскариды идет по типу
203
Рис. 181. Пе-
редний конец
тела Parascaris
equorum, вид-
ны четыре
крупные фаго-
цитарные клет-
ки и боковые
выделительные
каналы
апаэрооного и сопровождается распадом гликогена, т. е. выражается в
процессах гликолиза, о которых мы уже знаем (см. «Общая характеристика
плоских червей»). Хотя это положение остается в силе и теперь, и хотя
отчасти этим объясняется тот факт, что тело аскариды весьма богато гли-
когеном (см. выше), тем не менее было выяснено, что дыхательные процессы
аскариды не ограничены анаэробными. Оказалось, что аскариды имеют
собственный гемоглобин со специфичным спектром поглощения, причем
этот гемоглобин в различных органах аскариды неравноценен.Так,в мыш-
цах имеется гемоглобин, отличный по своей структуре от гемоглобина
гиподермы. Гемоглобины аскариды обладают близким сродством с
кислородом и легко переходят в оксигемоглобин. Оказалось, далее,
что кщцечник животных — хозяев аскарид— вовсе не столь беден кислоро-
дом, как это принималось ранее. Во всяком случае он там
имеется в количествах, которые необходимы паразитичес-
кому червю для дыхания свободным кислородом. Стало
быть аскарида нуждается в кислороде и может сущест-
вовать при минимальном его количестве. Очевидно, те
количества кислорода, с которыми экспериментировал
Вайнланд, были чрезмерными для этих животных. Про-
цессы дыхания осуществляются у аскарид через кожные
покровы'. Именно этим и объясняется наличие гемоглобина
в гиподерме.
Выделительная система. Выделительная система аска-
риды своеобразна. Роль выделительных органов играют
продольные боковые каналы, включенные в боковые валики
гиподермы (рис. 181). В задней части каналы заканчивают-
ся слепо, тогда как впереди они соединяются в общий
выводной проток, который и открывается наружу выде-
лительной порой, расположенной на брюхе позади головной
области. Боковые каналы являются производными мощно
развитой клетки с крупным ядром. Эта клетка гомологич-
на шейной железе свободных нематод (см. ниже).
В связи с выделительной системой нужно упомянуть
о фагоцитарных клетках. Они лежат в полости тела как
раз на боковых валиках гиподермы (рис. 181) и находятся
в непосредственном соприкосновении с полостной жидко-
стью. Эти клетки вбирают в себя продукты распада, накап-
ливающиеся в полости тела. На фагоцитарную природу
этих клеток указывают следующие факты. Если впрыс-
нуть в полость тела аскариды тушь, сепию, кармин и другие витальные
краски или, например, кровь лягушки, то соответственные вещества
(например, эритроциты лягушки) захватываются фагоцитарными клетка-
ми. Клетки эти хорошо видны простым глазом и несут отростки, давшие
повод называть их звездчатыми органами. Природа связи этих клеток с
выделительными каналами неясна.
Половая система. Половая система представлена половыми трубками.
У самки (рис. 182) эта половая трубка парная, у самца непарная.
Женские половые органы состоят из правого и левого
яичников, правого и левого яйцеводов, правой и левой маток. Обе матки —
правая и левая — впадают в общее влагалище, которое открывается нару-
жу половым отверстием, лежащим на брюшной стороне.
Матки имеют крупный диаметр. Постепенно обе матки становятся
тоньше и переходят в еще более тонкие яйцеводы, а эти последние — в
нитевидные яичники.
Описанные части четко различаются гистологически (и цитологически).
В яичниках зреют и формируются яйца. Характерным гистологическим
признаком яичника является особый аксиальный (осевой) стержень,
проходящий внутри яичниковой трубки. Это — рахис (rachis). Рахис имеет
204
синцитиальную структуру; вокруг него радиально, как густые спицы
колеса вокруг втулки, расположены половые клетки (оогонии); часть труб-
ки, в которой рахис исчезает, должна считаться яйцеводом. В матке име-
ется явственный просвет канала, в полости которого лежат яйца и спер-
матозоиды. Последние проникают сюда после совокупления самки
с самцом.
Мужские половые органы представлены непарной
трубкой, вершинные части которой имеют нитевидный диаметр и соответст-
вуют семеннику. Далее трубка имеет более крупный диаметр, образуя
семяпровод. Последний переходит в семяизвергательный канал, который
открывается в заднюю кишку, выполняющую, следовательно, у самца
функцию клоаки. Сюда же у самца выводятся из особых мешков спикулы.
Рис. 182. Анатомия лошадиной аскариды. / — самка; // —
самец (кишка разрезана):
1 — ротовые сосочки, 2 — глотка, — пищевод, 4 — фагоцитарные
клетки, 5—средняя кишка, 6 — семенник, 7 — семяпровод и семя-
извергательный канал, 8—яичник, 9 — яйцевод, 10 — матка, 11—
влагалище, 12—боковые валики гиподермы, 13— задняя кишка,
14 — брюшной нервный ствол
Эти спикулы при помощи особой мускулатуры высовываются через аналь-
ное отверстие наружу. Во время совокупления самец вводит спикулы в
половое отверстие самки, раздвигая им просвет влагалища, и изливает
семя.
Оплодотворение яиц происходит в матке. Яйца аскариды —
классический объект, на котором изучаются процессы созревания и опло-
дотворения яйца. У аскариды, как у немертин, оба процесса совпадают
во времени. Дробление идет по билатеральному типу.
Развитие яиц мы рассмотрим на примере другого вида — человеческой
аскариды.
205
Другие важнейшие представители класса нематод
и их жизненные циклы
Человеческая аскарида (Ascaris lumbricoides). Этот червь имеет органи-
зацию, сходную с организацией лошадиной аскаридой, но он несколько мень-
ше по размерам: самки 20—40сл1, самцы 15—20 ел. Оплодотворенные яйца
выделяются через отверстие влагалища аскариды и оказываются таким
образом в просвете кишечника человека. Вместе с калом они попадают в
наружную среду, и только здесь начинается их дробление. Дробление
яиц происходит только в присутствии кислорода. Оптимальная температура
равна 33° С, при этом яйца аскариды развиваются в течение 9—13 дней;
при более низкой температуре развитие идет дольше. Яйца, содержащие
уже оформившуюся личинку, становятся инвазионными, т. е. способными
к дальнейшему развитию в теле хозяина (человека). Из этого видно, что
яйца, не развившиеся и, в частности, только что выделившиеся наружу,
не могут заразить хозяина.
Инвазионное яйцо, попавшее в рот человека, проникает в пищевод
и желудок, а затем в кишечник. Освободившиеся от оболочек личинки
проделывают необычайно интересные миграции. Личинки активно внед-
ряются в кровеносные сосуды кишечника. Попав в кровяное русло, личин-
ки проникают в кровеносную систему печени, отсюда в нижнюю полую
вену и далее в правое предсердие и желудочек сердца, а оттуда в легкие
и затем в трахею и рот. Изо рта они попадают в пищевод и желудочек чело-
века, а отсюда в тонкую кишку. За время этих странствований личинки
развиваются, становятся крупнее и, попав описанным выше образом вто-
рично в кишечник, остаются здесь и достигают половозрелого состояния.
Аскарида — довольно обычный паразит человека. Не имея сильных
органов прикрепления, аскарида тем не менее хорошо противостоит дви-
жениям кишечника, опираясь о его стенки своим изогнутым пружинистым
напряженным телом. Она в состоянии активно передвигаться в кишечнике.
Аскарида может пробуравить оболочку кишки и проникнуть в полость
тела человека, в легкие, оттуда в дыхательное горло, носоглотку, полость
носа, в евстахиеву трубу.
Так как человек заражается от человека же, то меры профилактики
состоят преимущественно в опрятности, в чистоте тела и жилища.
Трихина (Trichinella spiralis). Половозрелые трихины (рис. 183ц)
обитают в тонких кишках свиньи, крысы и человека. Самцы маленькие,
длина их вытянутого тела не превышает 1,6 мм, на заднем конце видны
две конические лопасти, окружающие анальное отверстие, и две пары со-
сочков. Самки крупнее, их тело достигает 3—4 мм, задний конец без лопас-
тей. В отличие от аскариды женская половая система трихины непарная.
Большой интерес представляет жизненный цикл трихины. Личинки
трихинеллы развиваются в известковых капсулах в мышцах свиньи или
других млекопитающих, в том числе крыс, мышей и т. п. Если свинья
съест мясо крысы и проглотит вместе с ним инкапсулированнььх личинок
трихинелл, то в желудке свиньи капсулы разрушаются, а личинки высво-
бождаются и проникают в тонкие кишки. Здесь они внедряются в ворсинки
эпителия слизистой оболочки. В толще ее личинки быстро развиваются и
достигают половой зрелости, превращаясь в самок и самцов. Самки опло-
дотворяются, самцы погибают, а самки здесь же, т. е. в ткани стенки тон-
кой кишки, родят личинок. Личинки попадают в лимфатические сосуды,
а оттуда в кровеносную систему. По кровяному руслу они достигают по-
перечнополосатых мышц преимущественно наиболее подвижных органов,
в том числе ног. Здесь личинки оседают, спирально скручиваются, инкап-
сулируются и превращаются в так называемых «мышечных трихинелл».
Цикл возобновляется, если свинья или ее труп будут съедены другими
животными или человеком, в организме которого трихинелла может про-
делать описанный путь онтогенеза. Фактически трихинелла — гельминт,
205
Рис. 183. Нематоды — паразиты животных. / — острица (Enterobius
vermicularis); II —трихина (Trichinella spiralis) в капсуле (в мышцах);
III — власоглав (Trichocephalus trichiunis); IV — кишечная угрица
(Strongyloides stercoralis); (/Vi— филяриевидная личинка; IV2— самка
из кишечника человека; IV3—рабдитовидная личинка; IV*—самец сво-
бодноживущего поколения; IV з—самка свободноживущего поколения)
V — яйца нематод (Vi—Enterobius vermicularis; V2— Trichocephalus
trichiurus; V3— V*—Ascaris lumbricoides; V5— Ancylostoma duodenale;
V6— Necator americanus; — Trichostrongylus instabilis; V8— Stron-
gyloides stercoralis):
1 — пищевод, 2—бульбус, 3—кишечник, 4 — анус, 5— матка, 6 — половое отверстие^
Z — семенник, 8 — спикула
характеризующийся если не сменой хозяев, то сменой локализации. Свинья
(или человек) — окончательный хозяин трихинеллы, когда последняя до-
стигает половозрелого состояния у нее в стенках тонкой кишки, но вместе
с тем свинья (или человек) — промежуточный хозяин трихинеллы, когда
она локализована у нее в мышцах. В данном случае приспособленность
к паразитизму достигла высшего своего выражения, когда паразит ни на
какой стадии развития не выходит из тела хозяина и когда смена локали-
зации достигается не путем смены особей хозяина, а путем смены локали-
зации в одном и том же хозяине.
ОБЩИЙ ОБЗОР КЛАССА
Внешнее строение. Размеры тела, как это мы видим и в других группах
животных, очень варьируют. Известны формы, длина которых достигает
всего немногих долей миллиметра, например морская нематода Trichoderma
minutum имеет 82 мк в длину. А вот огромная нематода Placentonema
gigantissima, паразитирующая в плаценте кашалота, достигает в длину
8—8,4 м. Различия в размерах тела во многом зависят от образа жизни
нематод. Свободноживущие формы (дальше они называются просто сво-
бодными) обычно невелики — от долей миллиметра и до 20—30 мм (не-
которые морские формы). Небольших размеров достигают и нематоды —
паразиты растений. Напротив, наиболее крупные размеры присущи немато-
дам — паразитам животных. Ширина тела (его диаметр) всегда относитель-
но невелика, поэтому форма тела нематод веретеновидная, а у очень тон-
ких форм даже нитевидная. У некоторых видов форма тела уклоняется от
типичной и противоречит общему названию этой группы (нематода —
слово, берущее свое начало от греческого пета — нить). У таких нематод
с уклоняющейся формой тела это последнее вздувается и становится
похожим то на лимон, то на тыкву или бутылку с длинным горлышком
и т. п. Для всех нематод характерен хвост, под которым понимается учас-
ток тела, лежащий позади анального отверстия. В огромном большинстве слу-
чаев ротовое отверстие расположено строго на переднем конце тела, анус —
всегда на брюшной стороне его. По средней продольной (медианной) брюш-
ной стороне тела обычно в пределах передней половины тела либо сзади
нее, ближе к анусу, лежит женское половое отверстие (вульва). В области
головного конца, вдоль тела, на брюшной и на боковых сторонах, а также
в области хвоста имеются отверстия кожных желез. В остальном кутикула
покрывает тело нематоды сплошным панцирем.
Кожно-мускульный мешок. Кожно-мускульный мешок состоит из
кутикулы, подлежащего эпителия, или гиподермы, и мышечного слоя,
состоящего из лент продольной мускулатуры. Эти элементы необходимо
рассмотреть подробнее.
Кутикула нематод — сложное образование. У паразитических
нематод, поселяющихся в кишечном тракте (например, у аскарид), она со-
стоит из нескольких слоев сложного химического строения. В жизни нема-
тод кутикула имеет огромное значение, которое будет выяснено ниже. Здесь
же укажем только на ее защитное значение, обусловленное тем, что во-
первых, кутикула обладает более или менее выраженными свойствами по-
лупроницаемости и, во-вторых, эластичностью, плотностью и вязкостью.
Кутикула нематод чаще кольчатая, причем каждое поперечное кольцо
обычно состоит из рядов точек, пластинок и т. д.
Г иподерма сплошным слоем подстилает кутикулу. Она относи-
тельно тонкая, но имеет важное значение в жизни нематод. Во-первых,
за ее счет образуется кутикула. Во-вторых, за счет гиподермы развиваются
различные железы кожного происхождения, играющие немаловажную
роль в физиологии нематод. В-третьих, у паразитических нематод, поселяю-
щихся в органах животных, гиподерма имеет, по-видимому, большое
значение в процессах кислородного дыхания, так как в гиподерме, как
203
уже указывалось, содержится гиподермальный гемоглобин. У свободных
нематод кислородное дыхание идет через гиподерму. По бокам тела нематод
гиподерма утолщена, и здесь образуются резко выраженные продольные
гиподермальные валики, которые глубоко вдаются в первичную полость
тела (протоцель). У части нематод в эти валики включены экскреторные
каналы.
Мышц ы нематод состоят только из продольных лент. Мышечные
клетки удлинены и состоят из двух продольно расположенных участков:
саркоплазмы и лежащей сбоку от нее зоны мышечных фибрилл. Мышцы
нематод отличаются значительной энергией сокращения, но движения этих
животных довольно ограничены. Так, нематоды способны изгибать свое
тело преимущественно в дорзо-вентральном направлении. Только головной
конец тела нематоды отличается более свободными движениями, изгибаясь
в любые стороны. Кроме мышечных лент, составляющих элемент кожно-мус-
кульного мешка, у нематод имеются различные специальные мышцы. Та-
ковы мышцы, управляющие зубами у хищников; мышцы-протракторы,
выдвигающие стилет у паразитов растений; мышцы, управляющие мужским
копулятивным аппаратом; мышцы женских половых выводных путей; мыш-
цы пищевода и т. д. Некоторые из этих мышц упоминаются ниже.
Полость тела. Полость тела первичная, или протоцель. Протоцель
не имеет какой-либо специальной характеристики, вернее, ее можно было
бы характеризовать «от противного»: это такая полость тела, которая не
имеет собственных стенок и не сообщается какими-либо отверстиями с внеш-
ней средой. Фактически это щели между стенкой тела, с одной стороны,
и внутренними органами — с другой. В протоцеле скапливаются различные
продукты обмена, подлежащие удалению.
Протоцель наполнен жидкостью. Эта жидкость обусловливает высокий
полостной тургор. Полостное давление передается на кутикулу, которая
сдерживает это давление, этим и обеспечивается типичная для данной не-
матоды форма тела.
Нервная система. Нервная система нематод состоит из шести нервных
тяжей: брюшного, спинного, двух субмедианных и двух боковых (см.
рис. 180). Все они лежат продольно, самый мощный брюшной. Эти тяжи
передними отрезками направлены вперед, к голове, задними — к хвостовой
области. Своего рода «демаркационной» линией между передними и задними
отрезками нервных тяжей служит полное нервное кольцо, охватывающее
пищевод. При этом, как правило, перед кольцом нервные стволы прилегают
к ткани пищевода, позади нервного кольца они отходят к гиподерме. Фак-
тически позади нервного кольца центральная нервная система целиком
прижата и даже связана с гиподермой, почему некоторые авторы говорят
о кожно-мускульно-нервном мешке у нематод. Близость центральной нерв-
ной системы к эпидермису (гиподерме) — признак примитивной организа-
ции. Нервное кольцо состоит из нервных волокон. Обычно оно лежит
где-нибудь в средней части пищевода, но может быть далеко передвинуто
вперед, близко от головного конца тела. Позади нервного кольца и в облас-
ти пищевода имеются скопления ганглиозных (нервных) клеток (рис. 184).
Органы чувств. Органы чувств наиболее явственно развиты у свобод-
ных нематод. У них наблюдаются осязательные органы, или тангорецеп-
торы, органы химического чувства, или хеморецепторьг, наконец, иногда
фоторецепторы, воспринимающие свет. _ Органов равновесия не найдено.
Осязательные органы сосредоточены преимущественно
в области головного конца, и прежде всего на голове, вблизи ротового
отверстия. В этой связи необходимо подробнее остановиться на структуре
головы. Головной конец тела нематод весьма многообразен по своей орга-
низации. В центре лежит ротовое отверстие. По его периферии имеются спе-
циальные образования. Прежде всего это губы, подвижные придатки,
которые прикрывают ротовое отверстие и могут плотно замыкаться. Губ
чаще шесть, иногда больше, изредка их три, даже две. На их поверхности
209
развиты губные сосочки, или губные папиллы (рис. 184). Обычно это корот-
кие образования, часто едва возвышающиеся над поверхностью кутикулы.
По оси папилл проходит нерв, и они имеют значение осязательных органов.
Такое же значение придается второму кругу головных органов — головных
щетинок, или головных папилл, которые сидят на головных буграх. Голов-
ные папиллы имеют примерно те же очертания, что и губные, тогда как
щетинки отличаются своей удлиненностью. Вдоль длинной оси щетинок
также проходит чувствующий нерв. Кроме того, осязательные щетинки
у части нематод разбросаны по поверхности кутикулы, сосредоточиваясь
преимущественно на переднем (пищеводном) отрезке тела и в области хвоста.
Рис. 184. Органы чувств нематод. I —Mononchus (голова); II — Ethmolai-
mus (голова); III — Ditylenchus (голова); IV — Monhystera (голова); 7 —
Dorylaimus (голова); VI — Ditylenchus (пищевод); VII — Rhabditis (хвостовой
конец тела самца); VIII — Ethmolaimus (хвост самца):
1 — губные сосочки, 2—головные щетинки, 3—круглые амфиды, 4—спиральные амфиды,
5—карманообразные амфиды, 6 — поровидные амфиды, 7 — нервное кольцо, 8— ганглиозные
клетки, 9 — бурсальные папиллы (сосочки), 10 — дополнительные органы
Вдоль оси этих щетинок также проходят чувствующие (осязательные) нер-
вы. Такова периферическая осязательная система нематод. К ней нужно
отнести и особые боковые осязательные органы, всегда расположенные
у части нематод по бокам пищевода и известные под названием боковых шей-
ных сосочков, или дейридов. Дейриды часто очень мощно развиты у многих
паразитических нематод. Осязательные органы в области хвоста особенно
сильно развиты у самцов, что связано с половыми функциями — ориента-
цией в положении женского полового отверстия.
Органы химического чувства расположены у нема-
тод по бокам головы или на боковых губах. Обычно это карман, погружен-
ный в кутикулу, причем внутри кармана, играющего роль «обонятельной
полости», торчит мощный пучок хеморецепторных нервных фибрилл. Хе-
морецепторы нематод получили название амфид, Амфиды бывают различ-
ной формы: карманообразные, спиральные, круглые, щелевидные, поро-
видные и т. п. (см. рис. 184). Амфиды особенно сильно развиты у самцов,
в чем нередко выражен половой диморфизм у свободных нематод. У парази-
210
Рис. 185. Фазмиды (/) у
Seinura winchesi
тических нематод амфиды хорошо развиты; обычно они лежат не по бокам
головы, а на губах, как это имеет место у паразитов растений.
Органы зрения. У некоторых нематод имеется явственно вы-
раженный пигментный бокал и лежащая спереди от него линза. В этих
случаях имеются основания рассматривать подоб-
ные образования как настоящие фоторецепторы.
Ф а з м и д ы, или боковые хвостовые желе-
зы, — очень маленькие одноклеточные органы,
имеющиеся у части нематод и расположенные по
бокам хвоста. Иногда они также приобретают зна-
чение осязательных аппаратов (рис. 185).
Пищеварительная система. Пищеварительная
система состоит всегда из передней, средней и зад-
ней кишок. Наиболее сложной дифференцировке
подвергается передняя кишка, начинающаяся ро-
товой полостью. В нее ведет ротовое отверстие.
Спереди обычно оно прикрыто губами. За губами
расположена губная полость. За ней следует соб-
ственно ротовая полость. Она может иметь глад-
кие стенки либо быть вооружена зубами, копьем
или стилетом (рис. 186).
За ротовой полостью следует пищевод. Иногда
он простой и состоит из мышечной ткани и пище-
водного просвета трехгранной формы. В других случаях он дифференци-
рован на отделы, имеющие специальные названия, и снабжен луковицами,
или бульбусами (рис. 187). Различают средний бульбус и задний, или карди-
альный, бульбус, который у многих форм снабжен специальным дробильным
Рис. 186. Строение головного конца и ротовой полости не-
матод. / — Caenorhabditis (без вооружения); II — Mesodip-
logaster (с зубом — /), III—Mononchus (с онхом —2); IV—
Ditylenchus (со стилетом —3)\ V — Dorylaimus (с копь-
ем —4)
аппаратом, перемешивающим сбившиеся в комки кусочки пищи. Бульбу-
сы обычно имеют сильную мускулатуру. У части нематод кардиальный
бульбус почти лишен мышц, и тогда сильно развиваются три пищеводные
железы, более слабо развитые у других форм. Одна из них — спинная,
две другие — субвентральные.
211
I л
Рис. 187. Бульбус нематод в движении (/, //):
1 — пищевод, 2—бульбус, 3— начало кишечника, 4—кардий
Рис. 188. Организация нематод:
1— головные щетинки, 2— стома, 3— амфида, 4 —
пищевод, 5—нервное кольцо, 6 — бульбус, 7 —
«желудочек», 8 — продолжение средней кишки,
9— задняя кишка, 10—анус, 11— хвост,/2—жен-
ское половое отверстие, 13—матка, 14— яйцо,
15 — яйцевод, 16 — яичник, 17 — спикулы, 18 — ру-
лек, 19 — семенник, 20— семяпровод, 21 —семя-
извергательный канал, 22—«дополнительные» органы
самца, 23—шейная (выделительная) железа, 24 —
ее проток, 25— выделительная пора, 26— хвостовые
железы
ьное отверстие обычно имеет форму
ганов нет. Процессы дыхания осуще-
и, вероятно, через кишечный тракт.
Рис. 189. Выделительная система нематод.
1 —лошадиная аскарида; // —Ascaris
ostroumovi; III —свободноживущая фор-
ма. Всюду явственно видно, что основу
выделительных каналов составляет выде-
лительная клетка
На границе между пищеводом и средней кишкой имеется отдел, кото-
рый получил название кардия (см. рис. 187). Кардий иногда вдается в
кишку и служит клапаном, который не пропускает жидкое содержимое
средней кишки обратно в пищевод. Таковы отделы передней кишки.
Средняя кишка состоит из однослойного эпителия энтодермического про-
исхождения. Она имеет цилиндрическую форму. В переднем отрезке она
обычно шире, часто образуя здесь «желудочек», в области половых орга-
нов она сужена и вновь расширяется в задней части тела. Не доходя до
анального отверстия, средняя кишка переходит в заднюю кишку, или
ректум, выстланный кутикулой. По бокам задней кишки у части нематод
имеются ректальные железы. Ана.
поперечной щели.
Дыхательных и кровеносных о
ствляются через кожные покровы
Свободные нематоды кислородолю-
бивы.
Выделительная система. Выде-
лительная система нематод свое-
образна. В связи с всеобщим
исчезновением ресничек у них
исчезли и протонефридии. Функции
выделения осуществляются особой
выделительной железой, получив-
шей название шейной железы. У
части свободных нематод она пред-
ставлена одной единственной клет-
кой, от которой вперед, к голове,
идет проток. Его выделительное
отверстие всегда расположено на
брюшной стороне тела, обычно в
области средних отделов пищевода
или против заднего бульбуса, на-
конец, позади него (рис. 188). В дру-
гих случаях выделительная систе-
ма представлена двумя железисты-
ми клетками, которые связаны с
экскреторными каналами, имеющи-
ми различное расположение.
Иногда эти каналы идут справа и
слева, располагаясь в боковых
полях (валиках) гиподермы. В других случаях имеется лишь один непарный
выделительный канал (рис. 189). Во всех случаях каналы неизменно ве-
дут к выделительному протоку, который на брюшной стороне тела, как это
было указано выше, заканчивается выделительной порой. Фактически вы-
делительные каналы являются производными выделительной клетки. У
ряда нематод, ведущих свободный образ жизни, кроме шейной железы,
имеются и другие кожные железы, которые выполняют функции выделе-
ния. Сюда относятся три терминальные хвостовые железы, обычные у мно-
гих пресноводных и морских форм и, как правило, отсутствующие у на-
земных форм и паразитов. Их протоки направляются к концу хвоста, где
они и открываются наружу. Экскрет этих желез (см. рис. 188) в воде зас-
тывает в вязкую нить. Фазмиды, о которых выше упоминалось, некото-
рыми учеными также рассматриваются как выделительные органы. С выде-
лительной системой связаны упоминавшиеся фагоцитарные клетки, которые
имеются и у многих свободных нематод.
Половая система. Половая система нематод в типичных случаях со-
стоит из парных половых трубок. Женская половая система чаще парная,
мужская очень часто ограничена одной половой трубкой. Женская половая
213
трубка состоит из яичника, занимающего вершину трубки, яйцевода, мат-
ки и выводных путей, по которым яйца выводятся наружу. К этим вы-
водным путям у некоторых нематод относится снабженный мышцами
яйцемет. Во всех случаях имеется вагина, которая заканчивается вуль-
вой — женским половым отверстием (см. рис. 188). По ходу половой трубки
у некоторых нематод наблюдается развитие особых желез. Иногда женская
половая трубка характеризуется явлением протерандрии. Под последней
понимается способность сначала производить в женской половой трубке
за счет специальных участков яичника сперму, а затем образовывать
и выделять яйца, которые этой спермой оплодотворяются. В других слу-
чаях оплодотворения яиц нет вовсе, а яйца развиваются партеногенети-
чески, т. е. без оплодотворения.
Мужская половая система (см. рис. 188) состоит
из семенника, семяпровода и семяизвергательного канала. В семеннике
образуются сперматозоиды, которые по созревании переходят в семяпровод
и затем в семяизвергательный канал. Он отличается более толстой мышеч-
ной стенкой. Конечный отдел мужской половой трубки (семяизвергательный
канал) связан с особым совокупительным аппаратом. Он состоит из одной
пары спикул и обычно непарного пулька. Спикулы более или менее изог-
нуты, состоят из очень плотной кутикулы и связаны с особой спи-
кулярной мускулатурой, которая выдвигает спикулы (мышцы-протрак-
торы) и втягивает их обратно (мышцы-ретракторы) в спикулярное влагали-
ще. Спикулы растягивают вульву и служат как органы, облегчающие про-
цесс осеменения самки. Рулек — это добавочный орган мужского полового
аппарата, который ведет спикулы в определенном направлении, не позво-
ляя им отклоняться в стороны. У многих нематод самцы имеют копулятив-
ную бурсу, представленную бурсальными крыльями, лежащими по бокам
хвоста. С помощью бурсальных крыльев самец прочно фиксируется на
теле самки в области расположения ее вульвы. Хвост самцов вооружен
чувствующими папиллами и щетинками, частично в виде «ребер»,
входящих в состав бурсы.
У других нематод чувствующие аппараты, ориентирующие самца в
положении вульвы самки, расположены по медиальной линии на брюшной
стороне тела преанально (перед анусом), иногда в виде длинного ряда ося-
зательных аппаратов, известных под названием дополнительных органов
(см. рис. 188).
Яйца нематод развиваются в сложной слоистой оболочке. Величина
яиц различна. У нематод — паразитов животных — яйца относительно к
их размерам тела мелкие и их много. У свободных нематод их в матке мало
(1—2), и они обычно по отношению к размерам тела очень крупные, иногда
едва уступают соответственному диаметру тела. У паразитов растений
яйца также относительно крупные, а плодовитость этих нематод несравнима
с плодовитостью нематод — паразитов животных. Сперматозоиды нематод
имеют различную форму, иногда шаровидную, причем характерным приз-
наком, отличающим их от сперматозоидов большинства других групп жи-
вотного царства, нужно считать отсутствие подвижного хвоста. Спермато-
зоиды нематод способны к амебоидным движениям.
Развитие яиц идет по билатеральному типу. В результате фор-
мируется плотный зародыш, имеющий сперва форму боба, затем неболь-
шого свернутого в яйцевой скорлупе червячка плоской формы; позднее
личинка приобретает цилиндрическую форму, обычно суженную на концах.
Чаще личинка не вполне похожа на взрослую форму. Во многих случаях
она имеет совершенно иной пищевод, форму хвоста, иную форму ротовой
полости, иные пропорции тела и т. п. В ходе формообразования личинка
линяет. У нематод описаны четыре линьки и соответственно пять возрастов.
Пятый возраст соответствует взрослому состоянию. В целом с полным ос-
нованием можно говорить о том, что у нематод развитие протекает с превра-
щением.
214
Рис. 190. Rhabdias buffonis. I — самка, общий вид;
// — передний конец тела самки
Развитие (онтогенез). Развитие нематод очень разнообразно. У сво-
бодных нематод онтогенетические циклы протекают по простой схеме.
Яйца дробятся обычно во внешней среде или в матке. Затем они выделяются
из половых путей наружу, и здесь из них вылупляются личинки, которые
в той же среде, где живут и взрослые формы, претерпевают линьку и дости-
гают взрослого состояния.
Однако с переходом к специфическим условиям существования у не-
матод осложняются и циклы развития. У видов, живущих в загнивающих
органических веществах и получивших наименование сапробиотических,
онтогенез протекает более сложно. Дробление идет очень быстро и приводит
к вылуплению личинок,
которые отличаются кути-
кулой, обладающей резко
выраженными свойствами
полупроницаемости. Фа-
ктически кутикула таких
личинок превращается в
цисту. Эти личинки могут
быть названы длительно-
стадиальными личинками.
Они легко переносят бла-
годаря свойствам своей
кутикулы неблагоприятные
условия и превращаются во
взрослые формы, когда
вновь попадут в условия
сапробиотической среды.
С переходом к парази-
тизму в органах животных
онтогенез еще более осло-
жняется. В случаях гете-
рогонии имеется два поко-
ления. Одно из них пара-
зитическое. Это поколение
развивает только герма-
фродитных самок. Такова,
например, Rhabdias buffo-
nis — паразит легких ля-
гушек (рис. 190). Самки
развивают протерандрически сперва сперматозоидов, а затем яйца. Опло-
дотворенные яйца попадают из легких в кишечник лягушки, а оттуда во
внешнюю среду. В почве развивается свободная генерация, состоящая из
самцов и самок. Самки этой генерации не столь плодовиты, продуцируют
немногие яйца, из которых выходят личинки. Последние вместе с пищей
и частицами почвы заглатываются лягушками, и в их легких вновь воспро-
изводится паразитическое поколение.
У кишечной угрицы (Strongyloides stercoral is), паразитирующей в
кишечнике человека, наблюдаются различные способы развития. Так,
часть личинок, выйдя из кишечника человека, развивается по типу гетеро-
гонии. В почве развиваются самцы и самки; от оплодотворенных самок рож-
даются личинки, которые через рот или через кожу попадают в организм
человека; в его кишечнике они вновь дают паразитическое поколение.
Другая часть личинок, оставаясь в почве, линяет, превращается в длинных
с тонким пищеводом так называемых филяриевидных личинок, которые
проникают в организм человека либо через рот, либо через кожу (перку-
танный путь проникновения) и развиваются в гермафродитных паразитных
самок (см. рис. 183).
215
Уже в этих случаях мы видели, что личинки требуют для своего разви-
тия соответственного пребывания во внешней среде. Многие паразитические
нематоды развиваются по этому типу онтогенеза,т. е. без участия промежу-
точных хозяев. При этом нужно различать две формы развития без проме-
жуточных хозяев. В одних случаях наружу выделяются яйца нематод.
Яйца испытывают разную судьбу. Например, яйца паразита человека —
острицы (см. рис. 183), выйдя наружу из кишечника хозяина (человека),
уже через 6 ч становятся способными заразить другого или того же челове-
ка, т. е. становятся инвазионными (способными к развитию в органах хо-
зяина). Объясняется это тем, что еще в кишке человека яйца успели пройти
эмбриональные стадии развития и попадают во внешнюю среду уже с ли-
чинкой.
В других случаях, например, яйца Ascaris lumbricoides в кишечнике
человека не развиваются и нуждаются для своего развития в кислороде
воздуха (см. выше). Поэтому их пребывание во внешней среде до достижения
инвазионного состояния становится более длительным. Затем они, достиг-
нув известной стадии развития, должны для завершения его до стадии
личинки проникнуть вновь в своего хозяина.
Наконец, многие нематоды характеризуются тем, что во внешнюю среду
попадают не яйца, а свободные личинки. Личинки для достижения инва-
зионного состояния должны некоторое время прожить в почве, где они
ведут сапробиотический образ жизни. В качестве примера рассмотрим
здесь развитие опасного человеческого паразита — анкилостомы (Апсу-
lostoma duodenale) (рис. 191). Паразит живет в тонкой кишке человека и
продуцирует яйца. Яйца выходят во внешнюю среду и проникают в почву.
Из яиц вылупляются личинки, которые имеют типичный рабдитоидный
пищевод, характеризующийся наличием двух бульбусов. Эти личинки
живут в почве в качестве сапробиотических форм, т е. питаются продукта-
ми органического распада. Они претерпевают линьку и затем подвергают-
ся метаморфозу, причем у них меняется строение пищевода. Личинка
приобретает новую форму, известную под названием филяриевидной ли-
чинки. Эта личинка снова линяет, оставаясь при этом в шкурке и превраща-
ясь в длительно-стадиальную форму, способную переживать неблагоприят-
ные условия примерно в течение года и больше. При наступлении благо-
приятных условий эта личинка, прошедшая, таким образом, две линьки
(следовательно, личинка 3-й стадии), высвобождается из своей шкурки и
становится способной заразить человека, т. е. инвазионной. Заражение
(инвазия) протекает перкутанно, т. е. через кожные покровы человека.
В дальнейшем личинка проникает в кровеносную систему и достигает
кишечника, предварительно проникнув в легкие. В других случаях личин-
ка может инвазировать человека, непосредственно попадая к нему в рото-
вую полость и оттуда в кишечный тракт Нематоды, которые развиваются
по типу «органы хозяина — почва — органы хозяина», называются гео-
гельминтами.
Другой большой биологической группой нематод нужно считать био-
гельминтов, которые нуждаются для завершения своего онтогенеза в про-
межуточных хозяевах. В качестве примера такого онтогенеза рассмотрим
здесь развитие легочной нематоды овец — протостронгилюса (Protostron-
gylus, рис. 191). Паразиты гнездятся в легких овец. Здесь они выделяют
яйца, которые тут же, в легких, претерпевают стадии развития и превра-
щаются в личинок. Вместе с кашлем личинки попадают в глотку овцы,
а отсюда в кишечник. Затем вместе с экскрементами (катышками фекалий)
личинки попадают во внешнюю среду, т е. на почву Если в воздухе до-
статочно влаги, то личинки активно выходят из катышков овцы и подни-
маются несколько вверх по стеблям травинок пастбища. Однако дальней-
шее их развитие возможно только в том случае, если они проникнут в мол-
люсков. Роль промежуточных хозяев синтетокаулюса могут играть многие
брюхоногие моллюски как раковинные, так и голые. Личинки активно
216
проникают в моллюска и локализуются в его ноге. Здесь они подвергаются
дальнейшему развитию, достигая инвазионной стадии. В дальнейшем эти
инвазионные личинки выходят из моллюсков, вновь попадают на пастби-
ще и вместе с травой заглатываются овцами, которых и заражают.
При более глубокой специализации биогельминт может вовсе не выхо-
дить из организма хозяина, проделывая все стадии онтогенеза в его орга-
нах, но меняя свою локализацию соответственно стадиям развития. По
7
Рис. 191. Нематоды — паразиты животных. / — передний конец тела
Synlhetocaulus hobmaieri; // —хвостовой конец тела самца того же
вида; видна бурса (/) и темные спикулы (2); III—хвостовой конец
самки того же вида; IV — передний конец тела Haemonchus contortus;
V — голова — Delafondia vulgaris; VI — голова кривоголовки (Апсу-
lostoma duodenale)
этому типу идет развитие трихинеллы Trichinella spiralis (см. рис. 183).
Экология и распространение. В отличие от трематод и цестод нематоды
имеют более широкую экологическую характеристику.
^Многочисленные виды нематод (описано не менее 25 000 видов) ведут
свободный образ жизни, обитая на дне океанов, морей, рек, лиманов и
других водоемов. Кроме того, представители многих видов нематод (вероят-
но, около 10 000 видов) живут в почве. Почвенная фауна нематод богата
экологическими группировками, так как нематоды заселяют решительно
все почвы, вплоть до песков пустынь, очень богатых ими. Многие виды
живут в чистой земле. Другие предпочитают почву, богатую органическими
гниющими частицами, остатками корневой системы растений и т. п. Эти
нематоды, находимые также на трупах животных, образуют группу сапро-
217
биотических нематод. Сапробиотические нематоды биологически связаны
с бактериями. Последние доводят углеводы и белки до растворимого состоя-
ния, и затем эти продукты распада (либо углеводы, либо белки) потребляют-
ся сапробиотическими нематодами.
Многие нематоды поселяются в ходах короедов, т. е. в коровой трухе.
Массы нематод заселяют мхи, болота, имеются виды, специфичные для
бродящих жидкостей, для фекальных масс, в том числе для навоза жвачных.
Уксусная угрица живет в уксусе. Почвенные нематоды приурочены к
Рис. 192. Spaerularia bombi. / — молодая самка, матка (/) слабо выпячена;
// —следующий этап развития, матка заметно выросла: III —взрослая
самка кажется придатком огромной матки
корневой системе растений, т. е. к ризосфере. Многие из них проникли в
органы растений. Известное число видов живет в растительных органах
в качестве паразитов. Эти нематоды вооружены стилетом и характеризуют-
ся специфичными онтогенезами (см. ниже). Наконец, очень большое число
видов нематод (тысячи видов) приспособились к паразитизму в органах
животных, поражая аннелид, ракообразных, насекомых, моллюсков, ам-
фибий, рептилий, птиц и млекопитающих, в том числе человека.
Нематоды распространены по всему земному шару, от Южного до
Северного полюса. Очень многие из них — космополиты. Известны немато-
ды горячих источников (до 40°С и выше). Зимой в самую стужу в слое листь-
ев лесных деревьев можно найти обильную фауну нематод, так как низкие
температуры зимы не препятствуют их активному существованию, если
218
вода не замерзла (что может иметь место, в частности, в глубоких слоях
лиственного покрова в лесу).
Паразитизм в органах животных вызвал у нематод ряд специфических
приспособлений. Для нематод — паразитов животных, с одной стороны, —
характерно увеличение размеров тела, резкое повышение плодовитости,
Рис. 193. Некоторые нематоды — паразиты растений. I—IV — галло-
вая нематода (/ — голова самца; II — взрослая самка; /// —личинка
в яйцевых оболочках в состоянии первой линьки; IV — инвазионная
личинка); V — стеблевая нематода картофеля (самка); VI — голова
той же нематоды; VII — гнилостная овсяная нематода (голова); VIII —
голова луговой нематоды:
1—губная полость, 2— стилет, <?—головка стилета, 4—мышцы: протракторы сти-
лета, выталкивающие его наружу, 5 — пищевод, 6— просвет пищевода, 7 —проток
спинной пищеводной железы, 8— пищеводные железы, 9 — средний бульбус, 10 —
средняя кишка, 11— нервное кольцо, 12—яичник, 13 — яйцевод, 14—предматоч-
ная железа, 15 — яйцо, 16— передняя матка (у стеблевой нематоды и обе матки у
галловой), 17 — женское половое отверстие, 18— задняя матка, 19 — анус
уменьшение размеров яиц, развитие способности к анаэробному дыханию
(гликолиз в частности), развитие специальных приспособлений к наруше-
нию целостности органов, в которых нематода паразитирует. Таковы, на-
пример, зубы у Ancylostoma. С другой стороны, у этих нематод подверга-
ются редукции амфиды, никогда не наблюдаются глазки, нет длинных ще-
тинок ни на голове, ни на теле, никогда не бывает хвостовых терминальных
219
Рис. 194. Луково-чесночная стеблевая не-
матода (Ditylenchus alii):
1— стилет, 2—пищевод, J—средний бульбус,
4 — железистая часть пищевода, 5 — кишечник,
6 — задняя кишка, 7. — нервное кольцо, 8 — выде-
лительное отверстие, 9 — яичник, 10 — матка, 11—
яйцо, 12 — железы, 13—передняя матка, 14 —
задняя матка, 15 — анальное отверстие, 15 — жен-
ское половое отверстие
желез, изменяется строение выделительной системы. Для нематод, парази-
тирующих в органах животных, характерно также явление смены хозяев
(для биогельминтов).
Нематоды — паразиты растений — характеризуются иными призна-
ками. Размеры их никогда не бывают крупными (не свыше 5 мм в длину).
Плодовитость относительно невелика. Виды, самки которых продуцируют
за индивидуальную жизнь несколько сотен или несколько выше 1000 яиц,
редки. У других число яиц колеб-
лется в среднем около 250 штук на
самку. Яйца обычно относительно
крупные. У всех фитопаразитичес-
ких нематод ротовая полость пре-
вращена в стилет Его острие ско-
шено как игла медицинского шпри-
ца, внутри стилета проходит тонкий
канал. Задний (проксимальный)
конец стилета несет большей
частью особые базальные головки.
Их всегда три. От головок вперед
к перегородкам головной капсулы
тянутся три пучка миофибрилл.
Это мышцы-протракторы, выдви-
гающие стилет наружу (рис. 193).
Фитопаразитическая нематода (фи-
тогельминт) «стреляет» своим сти-
летом, нанося неоднократные уколы
и пронзая стенки растительных
клеток. Одновременно с ударом
стилета в растительную ткань
выделяются особые пищеводные
ферменты, вытекающие из пище-
водных желез, в особенности из
спинной. Эти ферменты расщепляют
растительные белки, гидролизуют
углеводы и растворяют пектино-
вые оболочки растительных клеток,
разрушая растительную ткань.
Смены хозяев у фитопарази-
тических нематод не бывает. Мно-
гие из них неоднократно воспрои-
зводят поколения в органе расте-
ния, накапливаются в нем в
огромных количествах и ведут растения к гибели, как это наблюдается, на-
пример, при паразитировании луковой стеблевой нематоды, поражающей
луковицы различных луков и чеснока (рис. 194).
СИСТЕМАТИЧЕСКИЙ ОБЗОР КЛАССА
Класс нематод делится на 2 подкласса: 1) аденофореи (Adenophorea)
и 2) сецерненты (Secernentea).
ПОДКЛАСС I. АДЕНОФОРЕИ (ADENOPHOREA)
Этот подкласс, ранее называемый фазмидиа (Phasmidia), представлен
в основном свободноживущими формами, обитающими на дне морей, океа-
нов и пресных водоемов, а равно в почвенных водах. К этому подклассу
220
Рис. 195. Гигантский свайник (Dioctophy-
me renale) во вскрытой почке собаки. I
самец; // — самка
относятся 2 крупных отряда: 1) хромадориды (Chromadorida) и 2) эно
плиды (Enoplida). Основные признаки подкласса следующие:
1) мелкие или крупные формы — свободноживущие или паразиты
животных (меньшинство); частью паразиты растений;
2) с гиподермой связаны особые гиподермальные железы, у свободно-
живущих форм моря и пресных вод обычно развиты хвостовые железы,
выделяющие содержимое наружу через пору на конце хвоста;
3) фазмиды отсутствуют;
4) органы химического чувства хорошо развиты и обычно расположе-
ны по бокам головы — спиральные или карманообразные или редуцирован-
ные (у паразитов животных);
5) шейная железа в виде одной
клетки, боковых выделительных
каналов нет;
6) у самцов очень обычны до-
полнительные органы.
К паразитам животных отно-
сятся, например, Dioctophyme
renale (рис. 195) — паразит почек
собак и других млекопитающих,
власоглав Trichocephalus trichiu-
rus— паразит слепой кишки чело-
века (рис. 183ш), трихинелла —
Trichinella spiralis (см. рис. 183 ц)—
паразит млекопитающих в том чис-
ле свиней, многих грызунов (кры-
сы, мыши, морские свинки, кроли-
ки и др.), а равно человека,
хозяевами этой нематоды могут
также быть медведи, барсуки, кош-
ки, куницы, лисицы.
К паразитам растений среди
аденофорей относятся некоторые
представители отряда эноплид, вооруженные длинным ротовым копьем, с
помощью которого они высасывают соки из корешков различных растений
Эти формы опасны и как переносчики вирусных болезней растений.
ПОДКЛАСС II. СЕЦЕРНЕНТЫ (SECERNENTEA)
Этот подкласс, ранее называемый афазмидиа (Aphasmidia), характе-
ризуется следующими дифференциальными признаками-
1) большей частью мелкие формы, свободноживущие или паразити-
ческие;
2) гиподермальные железы редуцированы; хвостовых желез с порой
на конце хвоста нет (см. аденофорей);
3) фазмиды обычно присутствуют;
4) органы химического чувства (амфиды), как правило, не по бокам
головы, а на латеральных губах, обычно поровидные;
5) шейная железа с длинными боковыми каналами;
6) у самцов дополнительных органов нет.
В пределах этого подкласса огромного развития достиг паразитизм.
Свободноживущие сецерненты представлены семействами (например, Rhab-
dilidae), экологически связанными с гниющим (сапробиотическим) субстра-
том либо с почвой и корнями растений (например, семейство Cephalobidae).
Паразитические формы представлены многочисленными группами форм,
паразитирующих у различных животных. Большинство форм входит в
отряд Rhabditida, куда относятся и названные выше семейства сапробио-
221
тических нематод и многочисленные паразиты животных. Таковы предста-
вители подотряда стронгилят (Strongylata), среди которых назовем криво-
головку Ancylostoma duodenle, широко распространенную между 45° с. ш.
и 30° ю. ш.; некатора (Necator americanus и другие виды); опасный паразит
лошади — Strongylus vulgaris, легочные метастронгилиды овец и коз, в
том числе виды родов Dictyocaulus, Protosrongylus, Muellerius, Syntheto-
caulus, сингамиды (Syngamidae), в том числе Syngamus—паразиты дыхате-
льной системы птиц и млекопитающих; трихостронгилиды (Trichostrongy-
lidae), среди них Haemonchus contortus — опасный паразит желудка овец
и других жвачных, и многие другие формы.
В отряде аскаридид (Ascaridida) имеется ряд паразитов млекопитающих,
в их числе человеческая аскарида (Ascaris lumbricoides), лошадиная аска-
рида (Parascaris equorum), свиная аскарида (Ascaris suum), паразит кошек
Toxocara mystax и другие виды.
Из многих форм отряда спирурид (Spirurida) следует указать на нит-
чатки (Filariata). Все они имеют длинное нитевидное тело, ротовая полость
рудиментирована, рулька нет, женское половое отверстие близко к перед-
нему концу тела. Сюда относятся многие виды, в том числе нитчатка Банкроф-
та (Wucheria bankrofti) — возбудитель слоновой болезни; Loa 1оа, ми-
грирующая под роговицей глаз человека (Африка), ришта (Dracunculus
medinensis), паразитирующая в подкожной клетчатке человека, и дру-
гие виды.
К подклассу сецернентов относится также обширный отряд тиленхид
(Tylenchida), включающий большое число групп и видов, представленных
паразитами насекомых и растений. Таковы многочисленные паразитические
тиленхиды жуков короедов. Тиленхиды этих жуков имеют положительное
значение, так как способствуют снижению их численности. Тиленхиды —
паразиты растений, напротив очень опасные и вредоносные формы, о важ-
нейших особенностях организации которых выше было уже сказано. Сюда
относятся многочисленные группы фитопаразитических нематод, или фито-
гельминтов, поражающих большинство сельскохозяйственных и техниче-
ских культурных растений. Таковы стеблевые нематоды, паразитирующие
в клеверах, луках, чесноках, землянике, нарциссах и других растениях
(Ditylenchus dipsaci), картофельная стеблевая нематода (Ditylenchus des-
tructor), опасные галловые нематоды (род Meloidogyne), гетеродеры свеклы
(Heterodera schachtii), картофеля (Н. rostochiensis), злаков (H.avenae), при-
носящие огромные убытки различным культурам, луговые нематоды рода
Pratylenchus и др. (рис. 193).
ВРЕДОНОСНОСТЬ ПАРАЗИТИЧЕСКИХ НЕМАТОД
Паразитические нематоды животных в большинстве патогенны, т. е.
вызывают различные болезненные процессы в организме своих хозяев.
Объясняется это рядом причин. Нематоды отравляют организм хозяина сво-
ими выделениями, влияют на состав крови хозяина, механически раздра-
жают занятый ими орган и т. п.
Многие нематоды вредят здоровью человека. Таковы человеческая
аскарида (Ascaris lumbricoides), власоглав (Trichocephalus trichiurus), три-
хинелла (Trichinella spiralis), кривоголовка (Ancylostoma duodenale), ришта
(Dracunculus medinensis) и другие формы. Аскариды вызывают у человека
различные болезненные явления. Выше указывалось, что личинки аскари-
ды странствуют (мигрируют) по кровеносным сосудам. В этой «миграцион-
ной» стадии могут наблюдаться изменения в составе крови (эозинофилия),
повышение температуры; в период, когда личинки проникают в легкие и
бронхи, иногда наблюдаются явления воспаления легких и бронхита. В пе-
риод паразитирования аскарид в кишечнике наблюдаются потеря аппетита,
рвота, тошнота, явления анемии и т. п. Власоглав обычно локализуется в
слепой кишке человека, внедряясь в слизистую оболочку ее своим тонким
222
концом. Власоглав питается кровью, вызывая различные поражения слепой
кишки. Кроме того, власоглав вносит с собой бактериальные инфекции.
Он вызывает снижение работоспособности, аппетита и т. д.
Много опаснее трихинелла. В период, когда паразит локализуется в
кишечнике, и в особенности в его слизистой оболочке, могут наблюдаться
тяжелые кишечные расстройства, не менее тяжелы явления трихинеллеза
в период проникновения трихинелл в мышцы человека.
Кривоголовка также вызывает тяжелые симптомы. Когда личинки
этого паразита внедряются в кожу, развивается зуд и различные характер-
ные изменения поверхности кожи. Когда кривоголовка достигает кишечника,
возникают сложные желудочно-кишечные расстройства, иногда сопрово-
ждающиеся психическими явлениями.
Ришта вызывает в подкожной клетчатке человека воспалительные
процессы и нарывы. Паразит развивается со сменой хозяев. Из вскрывше-
гося нарыва высвобождаются личинки, которые, попав в воду, для даль-
нейшего развития должны быть проглочены циклопами (Cyclops). В теле
циклопа личинка линяет, а затем становится неподвижной. Если человек с
водой проглотит зараженного циклопа, то с ним в его кишечник попадает
и личинка ришты. В дальнейшем она странствует по кровеносной системе
человека, локализуясь, как было указано, в подкожной клетчатке, обычно
на ногах человека. Здесь же взрослая ришта и вызывает развитие нарывов.
Животные также страдают от паразитирования нематод. Большие убыт-
ки хозяйству приносят легочные метастронгилиды, а в особенности Dictyo-
caulus, вызывая у телят и овец тяжелое легочное заболевание (диктиока-
улез). Коневодству наносит ущерб Strongylus vulgaris; Haemonchus con-
tortus — паразит желудка овец, опасен для этих животных, вызывает
значительный падеж. Значительный ущерб свиноводству наносят метастрон-
гилиды (Metastrongylus) свиней, паразитирующие в легких этих животных
и т. д.
Большой вред сельскому хозяйству причиняют растительно-паразити-
ческие нематоды, выделяющие, как уже было сказано выше, особые ферменты
и разрушающие растительные органы. Многие фитопаразитические нема-
тоды наносят огромный ущерб хозяйствам. Так, картофельная гетеродера
резко снижает урожай картофеля. Галловые нематоды вредят культурам
закрытого грунта повсюду, а в южных районах — многочисленным куль-
турам открытого грунта. Огромные потери злакам наносит злаковая гете-
родера (см. выше). Стеблевые нематоды картофеля и лука могут быть насто-
ящим бичом этих культур. Так, потери урожая лука и чеснока могут до-
стигать 60% нормального урожая.
БОРЬБА С ПАРАЗИТИЧЕСКИМИ НЕМАТОДАМИ
Борьба с паразитическими нематодами всецело определяется их биоло-
гией и закономерностями циклов их индивидуального развития. Естествен-
но, что в отношении многих паразитических нематод, развитие которых
протекает со сменой хозяев, в основу методов борьбы кладется принцип,
с которым мы уже ознакомились при рассмотрении методов борь-
бы с фасциолой. Так, например, борьба с риштой включает в себя
борьбу с рачками-циклопами, поскольку они являются промежуточными
хозяевами этой нематоды (см. выше). При ликвидации дракункулеза очень
важную роль в общем комплексе мероприятий сыграли общие санитарно-
гигиенические мероприятия (устройство водопроводов и пр.), которые осво-
бодили население от необходимости пользоваться водоемами (хаузами) для
водоснабжения. С другой стороны, было предпринято поголовное лечение всех
больных дракункулезом. Этим был нанесен удар по взрослой риште, вместе
с тем был этим же ликвидирован важный этап в развитии паразита — вы-
хождение яичек ришты в водоем (во время купания больных в хаузах). Дра-
кункулез в нашей стране в настоящее время ликвидирован.
223
В отношении нематод геогельминтов, у которых нет промежуточных
хозяев и развитие идет по типу «хозяин — почва — хозяин», схема борьбы
другая. Так, в борьбе с диктиокаулюсом (Dictyocaulus) важнейшее значение
имеет пастбищный режим, предотвращающий возможность заражения жи-
вотных этим Опасным паразитом (см. выше) Наряду с этим в отношении не-
матод — паразитов животных —широкое применение получила специальная
химиотерапия, в ходе которой животные получают лекарственные препа-
раты, убивающие и изгоняющие нематод.
Что касается растений, то терапия растений приобретает некоторое
распространение лишь в самое последнее время, и основным мероприятием
в отношении многих фитонематод (нематод растений) остается уничтожение
больных растений, обеззараживание почвы высокой температурой (сухим
паром) и различными химическими препаратами. Еще большее значение на
широких пространствах колхозных полей, огородов и бахчей приобретает
севооборот, который основан на том, что круг хозяев большинства фитоне-
матод все же ограничен известными (правда, часто очень широкими) пре-
делами и что каждая из фитонематод имеет «излюбленный» круг растений —
хозяев. При севообороте выбираются, по возможности, культуры, стойкие
против данной нематоды, что позволяет победить врага, не давая ему воз-
можности размножаться в наиболее благоприятных для него условиях.
ФИЛОГЕНИЯ НЕМАТОД
Основной группой, о происхождении которой в первую очередь и пойдет
речь, являются морские нематоды. Море—колыбель нематод. Оригиналь-
ная организация нематод, отсутствие у них протонефридиев, подвижных
ресничек (где бы то ни было), своеобразие половой системы, мышц и т д.
делают их трудно сравнимыми с другими большими группами (типами) жи-
вотных. Их сравнивали и сближали с турбелляриями, сопоставляли с ан-
нелидами и даже с членистоногими.
Исследования над морскими гастротрихами (отряд Macrodasyoidea)
позволяют подтвердить и развить ранее высказывавшийся взгляд о близо-
сти нематод к гастротрихам. Черты сходства касаются очень многих призна-
ков. У гастротрих из отряда Macrodasyoidea (рис. 177ui) наблюдается хвост,
лежащий позади расположенного на брюхе ануса, терминальное расположе-
ние ротового отверстия, изумительно сходное строение ротовой полости,
пищевода и кишки. Пищевод гастротрих имеет радиальную мускулатуру и
трехгранный просвет, как у нематод. Кишка и там, и здесь состоит из одного
слоя энтодермических клеток. Необычайно сходны ресничные ямки некоторых
морских гастротрих с амфидами нематод. Велико сходство некоторых физи-
ологических особенностей, закономерностей роста и т д.
Вместе с тем гастротрихи более примитивны, чем нематоды, менее
специализированы, ближе к турбелляриям. Действительно, во многих чер-
тах организации (ресничный покров, обонятельные ресничные ямки, строе-
ние полового аппарата и другие признаки) гастротрихи, несомненно, близки
к турбелляриям.
Наиболее близко к истине представление, согласно которому гастро-
трихи отчленились от турбеллярий (прямокишечных) и в свою очередь от
основания их ствола отошли нематоды.
КЛАСС 3. ВОЛОСАТИКОВЫЕ (GORDIACEA, ИЛИ
NEMATOMORPHA)
ХАРАКТЕРИСТИКА
Свободноживущие (во взрослом состоянии) нитевидные черви (рис. 196).
Тело покрыто плотной кутикулой. Полость тела первичная, содержащая
нежную соединительную ткань. У взрослых форм пищеварительный
224
канал частично зарастает и не функционирует Кровеносная, дыхательная
и выделительная системы отсутствуют. Раздельнополы, развитие с превра-
щением. Обитают в большинстве в пресных водах. Личинки ведут паразити-
ческий образ жизни.
ОБЩИЙ ОБЗОР КЛАССА
Внешнее строение. Тело очень длинное, у самок достигает 160 см длины
при толщине 1—1,5 мм.
Кожно-мускульный мешок. Под кутикулой лежит эпителий, псд ним
продольная мускулатура.
Нервная система. Нервная система образована нервным кольцом, ох-
ватывающим пищевод, и брюшным нервным тяжем.
Пищеварительная система. Пищеварительная система представлена
длинным каналом; ротовое отверстие — на переднем конце, анальное —
Рис. 196. Волосатик (Gordius aquaticus)
Рис. 197 Строение и развитие
Nematomorpha. / и // головной
и задний концы тела самца воло-
сатика; /// и IV — личинка воло-
сатика в яйцевой оболочке и без
нее
на заднем. Замечательно, что у взрослых волосатиков пищеварительная
трубка частично зарастает и не функционирует.
Половая система. Половая система образована скоплениями клеток;
у самок они представляют яичники; у самцов — семенники. Половые про-
дукты выводятся наружу посредством двух каналов, которые впадают в
клоаку, куда открывается и задняя кишка.
Развитие. Развитие волосатиковых крайне интересно. В результате
эмбрионального развития формируется своеобразная личинка. Она имеет
вооруженный шипами выдвижной хобот (рис. 197), при помощи которого
личинка может передвигаться. Кутикула личинки кольчатая. У личинки
имеется кишка, которая заканчивается заднепроходным отверстием.
Личинки сперва свободно живут на дне водоема, а затем либо активно
внедряются через кожу в тело хозяина, либо (что справедливо для большин-
ства хозяев — насекомых) проглатываются им. Интересно, что личинки,
долго не попадающие в хозяина, по-видимому, инцистируются и в таком
виде могут быть проглочены не только водными, но и наземными насекомыми.
Попав в хозяина, личинка активно забирается в его полость тела, мышцы
8 Курс геологии, т. I
225
или жабры и подвергается превращению. Достигнув взрослого состояния,
волосатики — самцы и самки — выходят из тела хозяина и копулируют,
причем самки откладывают многочисленные яйца. Яйца повторяют описан-
ный выше цикл развития.
Интересно, что если хозяин, в которого попала личинка, очень мал, то
развитие личинки останавливается и может в дальнейшем идти лишь в том
случае, если этот хозяин будет съеден каким-либо хищным и притом более
крупным насекомым, например каким-нибудь плавунцом и т. п. Смена хо-
зяев и развитие волосатика, несомненно, не лишены широкого теоретиче-
ского интереса, поскольку они могут служить примером путей возникнове-
ния явления смены хозяев в процессе филогенеза.
Взрослые волосатики живут в воде; известно около 100 видов. Их длин-
ное тело выглядит как попавший в воду волос. Оно имеет коричневую окрас-
ку. Волосатики медленно движутся на дне водоема или обвиваются вокруг
подводных растений. На зиму волосатики уходят в грунт водоема. Наиболее
известный Gordius aquaticus, широко распространенный в пресных водах.
Самцы этого вида (и рода) имеют вильчато раздвоенный хвост (рис. 196).
Позади клоаки две полулунные складки. Хвост самки не разделен, на его
конце анус.
Волосатики, несомненно, близки нематодам, но представляют уклоняю-
щуюся группу.
КЛАСС 4. КОЛОВРАТКИ (ROTATORIA)
ХАРАКТЕРИСТИКА
Мелкие, в большинстве свободноплавающие (планктонные), в основном
пресноводные животные. На переднем конце имеется более или менее сложный
ресничный аппарат, состоящий из колец ресничек. Этот аппарат носит на-
звание коловратного, или коловращательного (отсюда и название «коло-
вратки»). Тело покрыто или мягким эпителием, или более или менее сложным
панцирем (рис. 198iv). Полость тела первичная. В передней части пищева-
рительного аппарата обычно имеется сложный «жевательный» аппарат
(рис. 198). Кровеносная и дыхательная системы отсутствуют. Выделитель-
ная система протонефридиального типа. Нервная система дифференцирована
и состоит из ряда узлов. Раздельнополы, причем у ряда форм резко выра-
жен половой диморфизм (рис. 198). В большинстве случаев имеются смена
двуполого и партеногенетического развития, а также сезонные изменения
(цикломорфоз).
ОБЩИИ ОБЗОР класса
Внешнее строение. Форма тела большей частью вытянутая, в попереч-
ном сечении либо круглая, либо сплющенная. Некоторые формы шарообраз-
ны (Trochosphaera). Размеры от 0,04 мм до 2,5 мм.
В типичных случаях легко различимы: головной конец, вооруженный
указанным коловратным аппаратом, туловище и хвостовой отдел, называе-
мый ногой. Нога обычно заканчивается двухвершинной вилкой (так назы-
ваемыми пальцами) (рис. 198). Концы вилки снабжены железами, выде-
ляющими клейкое содержимое. Это орган прикрепления (фиксации), при
помощи которого коловратка удерживается на месте.
Тело коловратки очень часто несет явственную кольчатость. Последняя
не имеет, однако, никакого отношения к членистости. Это скорее правильно
расположенные кольцевые складки наружных покровов, связанные со
способностью коловраток сокращать свое тело, причем складки входят друг
в друга как колена подзорной трубы. Коловратный аппарат устроен и функ-
ционирует по-разному.
226
Если коловратка свободно взвешена в воде, то действующий наподо-
бие винта коловратный аппарат приобретает главным образом значение
органа плавания (движения). Если коловратка прикрепляется к месту, то
этим усиливается действие коловратного аппарата как органа привлечения
Рис. 198. Коловратки. I—III —Brachionus urceolaris. (/ —сам-
ка; II — самец; /// — изолированная трубка — протонефридий
с «ресничным пламенем»); IV — планктонная коловратка (Noth-
olca longispina), длинные придатки обеспечивают «парение» в
воде; V — сидячая коловратка (Melicerta ringens), скопление
особей, сидящих друг на друге в своих защитных трубках:
1 — щупальца, под ними нервный ганглий и глазок. 2 — мастакс, 3 — «желу-
док», 4 — заднепроходное отверстие, 5 — слюнные железы, 6 — мочевой пу-
зырь, 7 — протонефридий, 8 — яичник, 9 — яйцо, 10 — семенник, 11 — сово-
купительный орган; задний конец самки с типичной вилкой
добычи; вихрь воды, созданный работой коловратного аппарата, устремля-
ется к головному концу тела коловратки, увлекая сюда и различные мелкие
организмы. Последние и захватываются коловраткой.
Кожные покровы состоят из однослойного эпителия, имеющего синци-
тиальный характер, и расположенных под ним тяжей мускулатуры.
Панцирь может быть весьма разнообразным по форме, структуре и ри-
сунку. Он часто снабжен выростами, особенно многочисленными у типичных
планктонных коловраток, у которых шипы увеличивают парящую поверх-
ность тела (рис. 198iv). У сидячих форм кожные покровы выделяют
8*
227
оболочку, принимающую форму трубки («домика»), в которой сидит живот-
ное. Трубка служит приспособлением для защиты. Коловратка может втя-
гиваться в трубку.
Мышцы не составляют сплошного слоя, а образуют систему кольцевых
продольных и поперечных мышечных тяжей; кроме указанных, имеются
мышцы, связывающие кожу с внутренними органами и, наконец, обслужи-
вающие эти последние.
Нервная система. Нервная система состоит из надглоточного узла,
представленного системой нервных клеток. От узла отходят нервы. Нер-
вы, направляющиеся к наружным покровам, связаны с органами чувств.
К последним принадлежат глазки и особенно осязательные органы.
Пищеварительная система. Пищеварительная система начинается рото-
вым отверстием, обычно вооруженным кругом (венчиком) ресничек. Это —
ротовой венчик ресничек коловратного аппарата (рис. 198). Ротовое
отверстие ведет в глотку. Глотка снабжена ресничками и переходит в особое
расширение, называемое у коловраток зобом, или мастаксом (mastax). Ма-
стакс имеет хитиновую выстилку и особое внутреннее хитиновое вооружение
(рис. 198,2), производящее движения, напоминающие жевание.
Ритмические движения хитиновых частей мастакса (различают обычно
два молоточка и одну наковальню) направляют пищу в пищевод, а отсюда в
объемистый желудок. Стенки последнего состоят из одного слоя клеток.
Содержимое желудка изливается в заднюю кишку, которая в типичных
случаях открывается наружу анальным отверстием. Последнее расположено
на спинной стороне, у основания хвостового отдела. Пищеварительный тракт
сопровождается так называемыми слюнными и желудочными железами.
Первые впадают в глотку, или зоб, вторые — в полость желудка.
Дыхание. Дыхание происходит через покровы. По-видимому, кислород
воды используется также через кишечник и выделительную систему Вода,
обильно заполняющая кишечник, проходит через полость тела и органы вы-
деления, отдавая свой кислород.
Выделительная система. Выделительная система представлена типичны-
ми протонефридиями. Они начинаются клетками с описанным выше «реснич-
ным пламенем». Обычно имеются два боковых выделительных канала. Эти ка-
налы изливают свое жидкое содержимое в выделительный (мочевой) пузырь.
Последний открывается в задний конец кишки. Прозрачность покровов коло-
враток позволяет видеть работу «ресничного пламени» выделительной си-
стемы с необычайной ясностью.
Половая система. Самцы обычно меньше самок и имеют более простую
организацию.
Мужская половая система состоит из семенника, семя-
провода и совокупительного органа.
Женская половая система состоит из яичника, желточ-
ника и яйцевода. Яйца одеты скорлупой. Созревание яиц (овогенез) проис-
ходит всегда в организме самок, тогда как эмбриональное развитие зародыша
происходит либо в наружной среде (яйцекладущие коловратки), либо в орга-
низме самки (живородящие самки). Дробление полное, видоизмененного
спирального типа.
Развитие. Развитие коловраток отличается многими интересными осо-
бенностями.
У части коловраток (у морских форм из семейства Seisonidae) размноже-
ние всегда совершается с участием самцов, столь же развитых, как и самки.
У большинства коловраток наблюдается чередование партеногенетического
и двуполого размножения. При этом партеногенетические самки производят
два рода яиц: амиктические и миктические яйца. Первые имеют двойной
(диплоидный) набор хромозом, вторые — одинарный (гаплоидный) набор.
Амиктические яйца развиваются только партеногенетически и дают только
самок. Миктические яйца, напротив, развиваются половым путем и дают
самцов. С появлением самцов миктические яйца оплодотворяются, полу-
228
чают, следовательно, диплоидный набор хромозом и превращаются в покоя-
щиеся миктические яйца. Эти яйца имеют более плотную скорлупу.
Отношения между этими двумя сортами яиц связаны с гетерогонией,
завершающейся в течение определенного цикла. Цикл проходит следую-
щим образом. Из покоящихся яиц развиваются самки. Эти самки дают ами-
ктические яйца, снова развивающиеся в самок. Так в течение известного пе-
риода возникает несколько поколений «амиктических» самок. Это — период
партеногенетического размножения. Затем наступает период, когда из ами-
ктических яиц развиваются «миктические» самки, дающие миктические яйца.
Из этих яиц партеногенетически возникают самцы, которые оплодотворяют
миктических самок. У последних развиваются покоящиеся амиктические
яйца. Из этих яиц всегда развиваются самки, которые снова продуцируют
амиктические яйца. Следовательно, цикл возобновляется. Итак, в течение
одного цикла происходит смена партеногенетического поколения поколени-
ем, полученным из оплодотворенных яиц, которое вновь сменяется партено-
генетическим поколением.
У различных видов коловраток и в зависимости от различий физико-
химических условий водоемов число таких циклов в течение года может
быть различным: у некоторых видов наблюдается в течение года один цикл
(моноцикличные формы), у других — два (двуцикличные формы) или более
(полицикличные формы).
Экология и представители. Коловратки распространены по всему зем-
ному шару. Описано их свыше 1500 видов. В значительной степени этот факт
объясняется тем, что ничтожные размеры коловраток облегчают перенос
этих животных, а особенно их покоящихся яиц на большие расстояния раз-
личными водными животными (например, водными птицами). Однако осо-
бенно легко осуществляется перенос на большие расстояния различных поч-
венных коловраток, которые могут высыхать (анабиоз) и в таком виде вет-
ром заноситься в новые места обитания. Многие коловратки являются в
связи с этим «космополитами». Исследования Н. В. Воронкова, одного из
лучших знатоков коловраток, показали, однако, что представления о «ко-
смополитизме» отдельных видов коловраток были сильно преувеличены.
Известны многие виды, приуроченные, например, только к холодным или,
напротив, к тропическим или субтропическим областям. Коловратки по
преимуществу пресноводные формы, но известны и морские. Очень многие
коловратки приспособились к почвенным условиям, населяя, например,
растительный перегной лесной почвы, мхи и т д.
Водные формы составляют важную составную часть планктона. Эти
планктические формы обладают обычно сильным коловращательным аппа-
ратом или длинными отростками кутикулы, обеспечивающими парение в
воде, тогда как нога у многих типичных планктических коловраток может
отсутствовать или, во всяком случае, быть уменьшенной. Типичной планк-
тонной формой нужно считать, например, коловратку Notholca longispina
с ее длинными выростами, обеспечивающими парение (рис. 198iV). У дру-
гих планктических коловраток имеются длинные придатки, при помощи
которых животные производят прыгательные движения в воде. Таковы
снабженные мощными придатками Polyarthra platyptera — представители
рода Triarthra, Pedalion и др. У многих коловраток, живущих в толще
воды, эти приспособления отсутствуют, но зато они обладают мощным коло-
вращательным аппаратом, который позволяет этим формам активно дер-
жаться в толще воды. Таковы представители родов (рис. 198i ), Synchae-
ta, Conochilus, Rattulus и множество других форм, входящих в отряд сво-
бодноплавающих коловраток (Ploima).
Очень многие коловратки ползают Ползание наиболее характерно
для донных, а равно почвенных форм, поселяющихся во мху или раститель-
ном перегное. Ползающие коловратки имеют длинное червеобразное тело
и ногу. Особенно типичны представители отряда Bdelloidea, например, не-
обычайно характерный, живущий на дне Rotifer neptunius и другие формы.
229
Почвенные формы коловраток обладают замечательной способностью высы-
хать, сохраняя жизнь и впадая таким образом в состояние анабиоза, от кото-
рого они освобождаются, попав во влажную среду.
Следующую экологическую группу образуют сидячие формы. Они при-
крепляются к субстрату своей ногой, причем животное сидит либо в тонкой
слизистой трубочке (например, Stenophanoceras fimbriatus, Floscularia
proboscidea и др.), либо в более плотных трубках, которые коловратка строит
из комочков различных материалов, привлекаемых действием коловра-
щательного аппарата (например, Melicerta, см. рис. 198). Такие сидячие
коловратки входят в состав отряда Rhisota.
Ряд форм (из семейства Notommatidae) приспособился к паразитизму.
Такова, например, интересная Diglena volvocicola, паразитирующая внутри
шаров Volvox aureus, и ряд других форм. Хозяевами паразитических коло-
враток являются некоторые олигохеты, мшанки, некоторые растения и т. п.
Коловратки питаются различными микроорганизмами и являются в
сущности микроскопическими хищниками. Размножаясь в больших массах,
коловратки способствуют биологическому очищению водных бассейнов,
поскольку уничтожают множество организмов и препятствуют переобре-
менению воды органическими продуктами. Наряду с этим, коловратки бла-
годаря своему массовому размножению составляют немаловажный корм
для других, более крупных, организмов, например мальков различных рыб.
ФИЛОГЕНИЯ КОЛОВРАТОК
О происхождении коловраток высказывались самые разнообразные
предположения. Наибольшим признанием пользовалась в свое время так
называемая трохофорная теория (Б. Гатчек), основывавшаяся на сходстве
коловратного аппарата коловраток с ресничными поясами личинки много-
щетинковых кольчатых червей — трохофоры (см. ниже, рис. 216j ). По этой
теории коловратки рассматривались как неотенические, т. е. достигшие по-
ловой зрелости, личинки кольчатых червей. Успеху этой теории способство-
вало нахождение своеобразной шарообразной коловратки (Trochosphaera),
внешне похожей на трохофору
В настоящее время трохофорная теория большинством исследователей
не разделяется. Во-первых, оказалось, что трохосфера — типичная коловрат-
ка, и ее кажущееся сходство с трохофорой — конвергентное сходство, свя-
занное с одинаковыми условиями существования (планктонными). Во-вто-
рых, доказано, что наиболее примитивными, т е. первичными, коловратками
являются ползающие (червеобразные), а не планктонные.
Ввиду этого гораздо естественнее сближать коловраток с первичнопо-
лостными червями, с которыми они имеют безусловно ряд общих признаков
(первичная полость тела, отсутствие настоящей метамерии и др.), и выводить
от турбеллярий как особую специализированную ветвь, эволюционировав-
шую в другом направлении, чем гастротрихи и нематоды.
Дополнение к типу Nemathelminthes
КЛАСС СКРЕБНИ (ACANTHOCEPHALA)
ХАРАКТЕРИСТИКА
Своеобразные паразитические черви, имеющие цилиндрическое тело с
втяжным хоботом, усаженным загнутыми назад крючьями (рис. 199). Тело
покрыто плотной кутикулой. Полость тела первичная. Пищеварительная,
кровеносная и дыхательная системы отсутствуют. Выделительная система
протонефридиального типа в некоторых случаях отсутствует. Нервная си-
230
стема развита слабо. Раздельнополы. Развитие с метаморфозом и сменой
хозяев. Паразиты различных позвоночных, промежуточные хозяева — ра-
кообразные и насекомые.
Рис. 199. Скребни. / —
анатомия скребня (Echino-
rhynchus); II — хобот
скребня (сильно увеличен-
ный); ///—личинка скребня
(Echinorhynchus proteus):
1— хоботок, вооруженный крючь-
ями, 2 — ретрактор хоботка, 3—
влагалище хоботка, 4—лемниски,
5 —мышца (ретрактор), идущая
от влагалища хоботка к стенке
тела, 6—лигамент (связка), 7 —
семенники, 8— семяпроводы,
9 — железы, 10 — семенной пу-
зырь, заканчивающийся островер-
шинным придатком (II), играю-
щим роль копулятивного аппарата
ОБЩИЙ ОБЗОР КЛАССА
Внешнее строение. Тело скребней в поперечном сечении круглое и обычно
несет наружную членистость, которая не имеет, однако, никакого отношения
к метамерии. Животные эти имеют различные размеры. Одни из них не
превышают в длину нескольких миллиметров, другие достигают 20 и
даже 40 см.
Кожно-мускульный мешок. Кожно-мускульный мешок состоит из ку-
тикулы, гиподермы и подлежащей мускулатуры. Гиподерма представлена
довольно своеобразной тканью, пронизанной волокнами и заключающей в
себе сложную систему каналов. Каналы эти из гиподермы проникают в осо-
бые выросты, так называемые лемниски (рис. 199), лежащие в полости
тела. Мускулатура тесно связана с гиподермой и состоит из наружного
кольцевого и внутреннего продольного слоев.
Нервная система. Нервная система представлена нервным узлом, рас-
положенным в основании хобота. От него отходят боковые нервные стволы,
направляющиеся к заднему концу тела.
Питание происходит через кожу. Скребни, живущие в кишечнике,
пользуются его питательным содержимым. Всасываемые кожей питательные
растворы, по-видимому, проникают вглубь по системе кожных каналов.
Выделительная система. Протонефридии изливают свое содержимое в
выводные половые протоки (самки или самца), около которых они и распо-
ложены в форме комочков извитых канальцев.
Половая система. Мужская половая система состоит
из двух семенников и семяпроводов, которые соединяются в один канал,
вливающийся в семенной пузырек. Последний заканчивается островершин-
ным придатком, играющим роль мужского совокупительного органа. Этот
орган лежит в соответствующей сумке (бурсе). Семя вводится самцом в
половые пути самки. Оплодотворение яиц происходит в полости тела самки.
Женская половая система довольно своеобразна. Яйца
развиваются в яичниках, лежащих внутри так называемого лигамента. Этот
лигамент представлен мышцей, идущей от основания хоботного влагалища
231
(см. ниже) к заднему концу тела. Выпавшие из лигамента группы яиц по-
падают в полость тела, где они плавают, претерпевая процесс созревания и
оплодотворения. Оплодотворенные и начавшие дробиться яйца обычно
имеют удлиненную форму. Выделение их наружу происходит при помощи
весьма своеобразных выводных путей. В полости тела самки примерно вдоль
длинной оси тела имеется особый аппарат, так называемый колокол. Коло-
кол производит глотательные движения, посредством которых яйца втяги-
ваются в его внутренний просвет В задней части колокола имеются два от-
верстия, ведущие в яйцеводы и оттуда в матку. По этим путям заглоченные
колоколом яйца попадают во влагалище и затем наружу, т е. в кишечник
или полость тела хозяина.
При этом описанный путь могут проделать лишь оплодотворенные
дробящиеся яйца, так как их меньший диаметр позволяет им проскользнуть
через узкие отверстия яйцеводов. Комки неоплодотворенных яиц, прогоняе-
мые глотательными движениями колокола, не имея возможности проникнуть
в яйцеводы, выталкиваются через особое более крупное отверстие в колоколе
обратно в полость тела. Таким образом, колокол играет роль своеобразного
сортирующего прибора.
Развитие. Обычно зародыш, попав из хозяина в наружную среду,
должен быть сперва проглочен своим промежуточным хозяином, а послед-
ний — окончательным. Здесь зародыш сперва развивается в похожую на
цистицерка личинку, которая затем приобретает черты организации взрослых
форм (см. рис. 199).
Образ жизни. Паразитируя в кишечнике различных животных (рыбы,
птицы, млекопитающие), скребни хорошо противостоят перистальтике ки-
шечника благодаря своему вооруженному крючьями хоботу
Этот хобот поэтому должен рассматриваться как приспособление для
прикрепления. Крючья хобота заострены и загнуты назад. Паразит внедря-
ется хоботом в слизистую оболочку, причем крючья обеспечивают прочную
фиксацию к тканям. Этому еще способствует подвижность хобота, который
при помощи своих мышц может оттягиваться назад, погружаясь в располо-
женное на переднем конце тела особое влагалище хобота (см. рис. 199)
Представители. Скребень-великан (Macracanthorhynchus hirundina-
ceus). Самец — до 15 см, самка — до 40 см длиной. Паразит кишечника сви-
ньи. Яйца, выпавшие из кишечника свиньи, для дальнейшего развития долж-
ны быть проглочены личинками некоторых пластинчатоусых жуков
(например, личинками бронзовки, майского жука и пр.). Свиньи заражаются,
поедая личинок. Впрочем, в полости тела личинок зародыши скребня-вели-
кана остаются и после метаморфоза личинок и превращения их во взрослых
жуков. Вследствие этого свиньи могут заразиться скребнем и при поедании
взрослых жуков.
Из других скребней назовем паразита рыб Echinorhynchus proteus,
Е. polymorphus, паразитирующего в кишечнике некоторых уток и т д.
Всего известно примерно 260 видов.
Филогения. Филогения скребней не выяснена. Однако наличие пер-
вичной полости тела служит основанием к тому, чтобы ставить эту группу
рядом с первичнополостными. Ряд исследователей выделяют скребней в
особый тип и выводят их непосредственно от прямокишечных турбеллярпй.
ТИП VI. НЕМЕРТИНЫ (NEMERTINI)
ХАРАКТЕРИСТИКА
Тело удлиненное, более или менее сплющенное в спинно-брюшном на-
правлении, гладкое, покрытое снаружи ресничным эпителием с большим
количеством желез. У некоторых форм довольно явственно обособлена голова
и имеется втяжной хобот, лежащий над кишечником. Под эпителием лежит
соединительнотканый слой и далее несколько слоев мускулатуры, обычно
очень сильно развитых. Полость тела отсутствует, и промежутки между орга-
нами заполнены паренхимой. Кишечник сквозной. Дыхательная система
отсутствует, дыхание происходит через кожу Кровеносная система хорошо
развита, замкнутая. Выделительная система протонефридиальная и тесно
связана с кровеносной. Нервная система состоит из головного узла, нервного
кольца, охватывающего влагалище хобота, и двух продольных боковых ство-
лов. Раздельнополы. Развитие прямое или с превращением. В громадном
большинстве — морские формы, несколько видов обитает в пресных водах,
некоторые — во влажных наземных условиях, ряд видов перешел к парази-
тизму. Всего описано около 1000 видов.
ОБЩИЙ ОБЗОР ТИПА
Внешнее строение. Многие немертины, особенно не имеющие иглы на
хоботе, обладают ярко окрашенным телом. Во
носит характер покровительственной. Не-
которые невооруженные (без стилета) формы
необычайно сходны с вооруженными (мими-
крия).
Кожно-мускульный мешок. Кожно-
мускульный мешок немертин развит очень
сильно. Он имеет следующее строение.
Снаружи расположен однослойный эпителий
(рис. 200), состоящий из цилиндрических
клеток с ресничками. В эпителиальном
слое много железистых клеток, выделяю-
щих на поверхность кожи слизь. Под эпи-
телием лежит так называемый основной,
или опорный, слой с разбросанными ядрами
клеток и тонковолокнистой структурой.
Глубже расположен слой кольцевой муску-
латуры. За ним обычно следует слой диаго-
нальных мышц и, наконец, слой продоль-
ной мускулатуры. У ряда форм эти отноше-
ния могут быть сложнее, а именно: имеются
два слоя продольных мышц, между кото-
многих случаях окраска эта
Рис. 200. Продольно-боковой
(парамедиальный) срез через
тело немертины (Drepanophorus
albolineatus) в области средней
кишки:
1 — эпителий кожи, 2 — опорный
слой, 3—слой кольцевых мышц, 4 —
диагональные мышцы, 5 — продоль-
ная мускулатура, 6 — боковой ствол
нервной системы, 7 — дорзо-вентраль-
ная мускулатура, 8 — сечение средней
кишки, 9—сечение ринхоцелома,
10— сечение кровеносного сосуда,
11 — яйцо
233
рыми включен один слой кольцевых мышц (трехслойный кожно-мускуль-
ный мешок).
Кроме перечисленных мышц, у немертин имеются п дорзовентральные
мышцы, наподобие таких же мышц у Platodes.
Нервная система. Нервная система немертин достигает довольно вы-
сокого развития. Центральная нервная система состоит из головных узлов
и продольных стволов. Головные узлы образуют так называемый головной
мозг немертин и достигают значительного развития. Узлы связаны комиссу-
рами (перемычками), причем дорзальная и вентральная комиссуры образуют
кольцо, через которое проходит влагалище хобота (рис. 201).
Рис. 201. Немертины. / — анатомия; // —деталь вы-
делительных стволов и их отношение к кровеносному
сосуду; /// — личинка немертины — пилидий:
1— кишечник, 2— карманы кишечника, 3— вентральный крове-
носный сосуд, 4 — спинной кровеносный сосуд, 5 — боковой сосуд,
6 — выделительные сосуды, 7 — яичники. 8 — хобот, 9— стилет,
/0 — церебральный орган, 11—глазки, 12—надглоточная нервная
комиссура, 13—подглоточная нервная комиссура, 14 — теменной
орган
Что касается продольных стволов центральной нервной системы, то
обычно они расположены вдоль боков тела, по одному стволу с каждой сто-
роны. Нервные стволы лежат либо в кожно-мускульном мешке, либо в па-
ренхиме.
От головного узла отходят многочисленные нервы к переднему краю,
к внутренним органам, к хоботу, к коже, к органам чувств. Наиболее сильно
развиты спинной нерв (часто двойной — верхний и нижний), брюшной нерв
и глоточный.
Органы чувств. Сюда относятся довольно многочисленные образования.
Глаза большей частью имеются и притом в большом числе. Организация глаз
очень примитивна. К ним подходит глазной нерв, а самый глаз имеет явст-
венные световоспринимающие клетки и соответствующую пигментную об-
кладку.
У некоторых форм имеются органы слуха и равновесия — статоцисты.
234
Весьма типичны так называемые церебральные органы, лежащие по бокам
головного конца и связанные с головным мозгом.
Полость тела. У взрослых особей полость тела отсутствует. Внутрен-
ние органы погружены в нежную паренхиматозную ткань, совершенно про-
зрачную у живых форм. Упоминавшаяся дорзо-вентральная мускулатура в
области средней кишки имеет явственно метамерное расположение. Фибрил-
лы этой мускулатуры связаны с кольцевыми мышцами (рис. 200).
Пищеварительная система. Пищеварительная система представлена
трубкой, которая начинается расположенным на головном конце тела рото-
вым отверстием. Последнее обращено к брюшной стороне. Ротовое отверстие
ведет в короткий пищевод. Пищевод переходит у некоторых форм в расши-
ренную часть, так называемый желудок. Пищевод и желудок образуют перед-
нюю кишку (рис. 201). За передней кишкой следует средняя, которая у многих
форм образует правильно повторяющиеся симметричные боковые карманы.
Задняя кишка короткая и заканчивается на заднем конце тела анальным
отверстием.
Хобот — характернейший орган немертин. Он представлен слепым,
имеющим форму трубки мешком, который лежит в хоботной полости, или
ринхоцеломе. Путем сжатия мускулатуры ринхоцелома хобот, втянутый в
полость ринхоцелома, может быть выдвинут наружу наподобие пальца пер-
чатки. Обратное втягивание его в ринхоцелом достигается сокращением
особой мышцы (ретрактор хобота).
У некоторых форм выводной канал ринхоцелома (rhynchodaeum)
открывается в начале передней кишки, и тогда хобот выталкивается наружу
через ротовое отверстие. Хобот у многих форм вооружен стилетом и ядо-
витой железой.
Немертины — хищники. При помощи хобота они хватают или убивают
(стилетом) своих жертв, преимущественно различных полихет. Однако
крупные немертины нападают даже на мелких рыб и некоторых асцидий.
Кровеносная система. Кровеносная система представлена обычно
одним спинным сосудом и двумя боковыми. От этих сосудов отходят много-
численные ответвления, образующие сложную сеть капилляров. Крове-
носная система замкнутая. Главные сосуды переходят друг в друга
на заднем и переднем концах тела. Кровь некоторых форм содержит гемо-
глобин. Кровь течет в спинном сосуде сзади наперед и в боковых — спе-
реди назад.
Дыхательная система отсутствует. Газообмен происходит через кожу.
Выделительная система. Состоит из двух основных стволов, иногда
коротких, иногда длинных и сильно извивающихся (рис. 201). Эти стволы
лежат по бокам тела. В их просвет вливаются многочисленные канальцы,
которые образуют густую сеть. На слепых концах канальцев имеются замы-
кающие их клетки с «ресничным пламенем». Эти слепые концы канальцев
обычно тесно связаны с кровеносными сосудами и погружены в их стенки.
При этом прямого сообщения между кровеносными и выделительными кана-
лами, разумеется, нет.
Половая система немертин обладает интересными чертами. Немертины
большей частью раздельнополые животные. Половые железы представлены
генитальными (половыми) мешочками, которые лежат у самцов и самок по
бокам кишечника (рис. 201). Иногда в расположении этих мешочков нет
особой правильности. Но у ряда форм с метамерными карманами кишки ге-
нитальные мешочки лежат между двумя соседними карманами кишечника и
носят характер метамерно повторяющихся органов. От этих генитальных
мешков отходят протоки, которые открываются наружу половыми порами.
Зрелые яйца обычно оплодотворяются после их выделения через поры в
наружную среду. Впрочем у некоторых форм наблюдается внутреннее опло-
дотворение. При этом самец, находящийся в близком соседстве с несколькими
самками, выбрасывает семя в воду. Сперматозоиды достигают половых от-
верстий самок и проникают в женские генитальные мешки.
235
Следует отметить, что оплодотворение яиц совпадает с процессом их
созревания.
Развитие. Дробление яиц полное, детерминированное, спирального
типа. Дальнейшее развитие происходит либо прямым путем, либо с превра-
щением. В последнем случае из яйца развивается личинка: либо так называе-
мый пилидий, либо дезоровская личинка.
Пилидий особенно характерен. Он имеет шлемовидную или зонтико-
видную форму с двумя лопастями. По краям зонтика расположены реснич-
ки, а на вершине — весьма характерный пучок ресничек (см. рис. 201) —
теменной орган. У пилидия имеются ротовое отверстие, пищевод и кишеч-
ник. Заднепроходного отверстия нет. Личинка ведет пелагический образ
жизни. Спустя некоторое время пилидий подвергается своеобразному пре-
вращению, в результате которого внутри его наружных покровов образуется
взрослая немертина, освобождающаяся наружу через разрыв покровов
пилидия.
Экология и распространение. Немертины, за исключением нескольких
пресноводных и наземных видов, живут в море, обитая преимущественно
на дне, хотя известны и пелагические формы. Последние встречаются на
больших глубинах. Немертины широко распространены во всех морях, от-
сутствуют они только в Каспийском и Аральском морях. В ряде тропиче-
ских стран известны некоторые почвенные формы, они — обитатели скры-
тых мест, их можно найти среди гниющих листьев, в трещинах коры и т. п.
Некоторые немертины перешли к паразитизму Таков один род, представ-
ленный видами, поселяющимися на жабрах крабов.
ФИЛОГЕНИИ НЕ МЕРТ ИН
Л4ногие исследователи присоединяют немертин к плоским червям,
включая и тех и других в особый (довольно плохо обоснованный) тип
несегментированных червей (Ашега). Это нельзя признать правильным.
Наличие у немертин анального отверстия и в особенности кровеносной си-
стемы заставляет признать, что организация немертин много выше, чем орга-
низация плоских червей. По-видимому, немертин надо рассматривать как
особую ветвь первичноротых, возникшую от турбеллярий. С последними
немертин сближают многие черты сходства — наличие ресничного покрова,
строение кожно-мускульного мешка, нервной системы, органов чувств и
т. д., а также некоторые моменты эмбрионального развития.
ТИП VII. КОЛЬЧАТЫЕ,
ИЛИ ВТОРИЧНОПОЛОСТНЫЕ,
ЧЕРВИ (ANNELIDES)
В организации кольчатых червей вместе с чертами, характерными для
круглых червей (передний конец тела, кожно-мускульный мешок, билате-
ральная симметрия, мезодерма), появляются и другие чрезвычайно важные
особенности. Некоторые из них, впервые возникнув у кольчатых червей,
переходят в другие типы, знаменуя собой новый подъем уровня организации
животных.
Эти новые черты организации следующие: а) вторичная полость тела—
целом, представляющая собой систему внутренних полостей тела, имеющих
собственную эпителиальную стенку; б) метамерия — расчлененность тела
на сегменты, одинаковые в своем внешнем строении и с повторением в них
жизненно важных структур — кровеносной, выделительной, нервной, пи-
щеварительной и половой систем; в) боковые выросты тела — параподии —
специальные органы движения.
ХАРАКТЕРИСТИКА
Тело у большинства форм состоит из отдельных колец — сегментов,
или метамер, закономерно повторяющихся от головного отдела до аналь-
ного членика, или пигидиума, на конце которого обычно вентрально поме-
щается заднепроходное отверстие. Этой метамерии соответствуют характер-
ные для многих кольчатых червей боковые подвижные выросты тела, так
называемые параподии и пучки щетинок.
Наружной метамерии соответствуют разделение внутренней полости
тела перегородками (диссепиментами) на отдельные участки (сомиты) и
метамерное расположение ряда внутренних органов. Правильно повторяют-
ся нервные узлы, кольцевые кровеносные сосуды, выделительные органы
(метанефридии) боковые карманы средней кишки и половые органы (гона-
ды) У некоторых форм сегментация как наружная, так и внутренняя, а
также придатки тела могут в связи с образом жизни редуцироваться. Кож-
но-мускульный мешок состоит из кутикулы, эпителия и кольцевой и про-
дольной мускулатуры, а также внутренней выстилки полости тела — цело-
телия.
Полость тела — вторичная (целом) у большинства хорошо развитая и
лишь у немногих форм редуцированная.
Центральная нервная система состоит из надглоточного нервного узла
(«головного мозга»), окологлоточного кольца и брюшной нервной цепочки с
посегментно расположенными нервными узлами.
Кишечник сквозной, сложный, делится на переднюю, среднюю и зад-
нюю кишки, иногда он осложнен боковыми выростами.
237
Кровеносная система замкнутая и состоит из сосудов, часть которых
обладает сокращающимися стенками, что обеспечивает правильную цирку-
ляцию крови. У некоторых групп кровеносная система редуцирована и за-
меняется системой лакун — остатков редуцированной полости тела. Кровь
у ряда форм содержит гемоглобин.
Дыхание осуществляется в большинстве случаев всей поверхностью
тела, у некоторых форм имеются специальные выросты — жабры.
Выделительная система представлена посегментно расположенными
метанефридиями, или сегментарными органами. Их воронка открывается
в полость тела (целом), а другой конец — наружу.
Размножаются половым и бесполым путем типа почкования. Раздельно-
полые или гермафродиты. У одних имеется довольно сложная половая си-
стема. У других специальных половых органов нет, половые клетки обра-
зуются из целотелия и выводятся наружу через нефридии — обычные или
специализированные. Дробление спиральное, развитие прямое или с пре-
вращением. В последнем- случае имеется характерная личинка — трохо-
фора.
Обитают в морях, пресных водах и почве. Некоторые формы — обита-
тели влажных мест и паразиты. Всего известно около 8000 видов.
КЛАССИФИКАЦИЯ
Тип Кольчатые, или вторичнополостные, черви (Annelides)
Класс 1. Многощетинковые кольчецы, или полихеты (Polychaeta)
Подкласс I. Свободно-подвижные полихеты (Errantia)
Подкласс II. Сидяче-прикрепленные полихеты (Sedentaria)
Подкласс III. Мизостомиды (Myzostomida)
Класс 2. Малощетинковые кольчецы, или олигохеты (Olygochaeta)
Класс 3. Пиявки (Hirudinea)
Отряд 1. Щетинконосные пиявки (Acanthobdellae)
Отряд 2. Хоботные пиявки (Rhynchobdellae)
Отряд 3. Челюстные пиявки (Gnathobdellae)
Отряд 4. Глоточные пиявки (Pharhyngobdellae)
Класс 4. Эхиуридовые (Echiuroidea)
КЛАСС 1. МНОГОЩЕТИНКОВЫЕ КОЛЬЧЕЦЫ,
ИЛИ ПОЛИХЕТЫ (POLYCHAETA)
ХАРАКТЕРИСТИКА
Типичные кольчатые черви с хорошо развитыми у большинства форм
боковыми выростами — параподиями, несущими пучки щетинок. У боль-
шинства форм также хорошо выражен головной отдел. В некоторых случаях
в связи с условиями существования (роющий или сидячий образ жизни,
паразитизм) эти признаки претерпевают регресс и животные внешне сильно
изменяются (см. рис. 210). Целомическая полость тела развита в громадном
большинстве случаев хорошо. Дыхание осуществляется через кожу или
имеются специальные выросты тела (жабры). Обычно раздельнополые;
половые продукты образуются из целомической выстилки и выводятся через
нефридии — обычные или видоизмененные. Развитие с превращением, у
большинства имеется личинка — трохофора. Почти все обитают в морях,
в пресных водах встречаются в единичных случаях (например, оз. Байкал).
Некоторые формы — паразиты. Полихеты играют существенную роль в
питании рыб, в частности промысловых. Известно около 5000 видов.
Представитель класса полихет—нереида (Nereis)
Род Nereis объединяет большое число видов, широко распространен-
ных во всех наших морях. Последующее описание относится главным обра-
зом к виду N. pelagica.
238
Внешнее строение. Нереида — буровато-зеленого цвета с металличе-
ским отливом, стройное тело, достигающее 120 мм в длину Задняя часть
тела несколько уже передней и средней. Спинная сторона тела выпуклая и
заметно темнее окрашена, чем брюшная. Тело состоит из 80—100 сегментов,
хорошо обособленных друг от друга, причем передние сегменты резко вы-
деляются от остальных и образуют голову нереиды (рис. 202). Голова со-
Рис. 202. Строение полихет. / — головной конец Nereis pelagica; // — анатомия
Nereis pelagica; /// — параподия Nereis pelagica; IV — клетки кишечника Aphrodite
aculeata; V — центральная нервная система Phyllodoce laminosa; VI — «мозг» Phyl-
lodoce laminosa; VII — разрез через глаз Vanadis ramosa (сем. Alciopidae):
1— простомиум (prostomium), 2—перистомиум (peristomium), 3 — щупальца (антенны), 4— пальпы,
5— усики, 6—глазки, 7 — обонятельные ямки, 8— параподии, 9— глотка, 10 — пищевод, 11 — желе-
зы пищевода, 12—кишечник, 13— спинной кровеносный сосуд, 14 — брюшной кровеносный сосуд,
15 — диссепимент, 16—метанефридии (начинаются воронками в полости тела; их каналы проходят
сквозь диссепименты и порами открываются наружу), 17 — брюшная нервная цепочка, 18—кольцевые
кровеносные сосуды, 19 — спинной циррус, 20 — брюшной циррус, 21 — спинной пучок щетинок, 22 —
брюшной пучок щетинок, 23 — клетки кишечника в различных стадиях экскреторной деятельности
(светлые и темные), 24 — жидкость, содержащаяся в просвете кишечника с каплями пищеварительного
экскрета, выделенного клетками, 25 — надглоточные ганглии, 26 — подглоточные ганглии, 27—ган-
глии брюшной нервной цепочки, 28—нервы, идущие от «мозга» к щупальцам, 29— глаза, 30— часть
комиссуры, соединяющей надглоточные ганглии с неизображенными здесь подглоточными, 31 — сет-
чатка, 32—чувствительные палочки зрительных клеток, 33 — так называемый греефовский орган
(сократимые волокна, укорочение которых меняет положение хрусталика и обеспечивает аккомода-
цию хрусталика к видению вдаль), 34— хрусталик, 35—роговица, 36 — железистая клетка, 37 —
стекловидное тело
239
стоит из двух хорошо различимых отделов: переднего — простомиума
(prostomium) и заднего — перистомиума (peristomium) Простомиум имеет
треугольную форму и несет важные органы чувств, а именно: а) осязатель-
ные органы — тонкие щупальца, или антенны, и более толстые и даже мас-
сивные пальпы; б) две пары глазков, лежащих в основании простомиума;
в) органы химического чувства, или обонятельные ямки, лежащие позади
задней пары глаз. Перистомиум несет длинные, обращенные вперед при-
датки — перистомиалъные усики, или циррусы (cirri). На брюшной стороне
перистомиума расположено ротовое отверстие.
Туловищные сегменты несут по бокам параподии — типичные для много-
щетинковых кольчецов придатки. Каждая параподия (см. рис. 202) состоит
из двух основных частей: спинной ветви, или нотоподия, и брюшной ветви,
или нейроподия. Обе эти ветви вторично делятся на лопасти. Дорзально от
спинной лопасти нотоподия расположен спинной циррус параподия, тогда
Рис. 203. Поперечный разрез через тело полихеты:
1—кожа, 2—кольцевые мышцы, 3 продольные мышцы, 4 — жабры,
5 — спинной циррус, 6 — нртоподий (спинная ветвь параподии), 7 — цир-
рус нейроподия, 8— целом, 9—кишечник, 10—спинной сосуд, 11—
брюшной сосуд, 12 — параневральные кровеносные сосуды, 13—нервный
ствол, 14 — пора метанефридия, лежащего в выделительной камере цело-
ма, 15—ацикула, 16—метанефридий, 17 — яйца, 18 — невроподий (брю-
шная ветвь параподии), 19 — целомический эпителий, 20—воронка мета-
нефридия
как нейроподий несет соответственный брюшной циррус. Пронизывая ткань
параподии, наружу торчат пучки характерных щетинок, среди которых две
отличаются особенно заметной толщиной и получили название ацикул (см.
рис. 211). Эти ацикулы имеют значение опорных элементов параподии. Во
время плавания параподии движутся и выполняют роль гребных органов.
Щетинки способны (благодаря особой мускулатуре) выдвигаться и втяги-
ваться. При выдвигании наружу они увеличивают гребущую поверхность
параподий.
Кожи о-мускульный мешок. Снаружи тело нереиды покрыто плотной,
но тонкой кутикулой. Под ней лежит сплошной слой эпителия, под которым
расположена мускулатура. Кутикулу вместе с лежащим над ней эпителием
называют кожей. Хорошо развитая мускулатура слагается из двух слоев.
Снаружи лежит кольцевой слой, под ним — продольный (рис. 203) Изнутри
стенка тела выстлана целомическим эпителием, или целотелием (см. ниже).
Продольная мускулатура образует четыре мощные мышечные ленты. Две
из них лежат на спинной стороне, разделенные небольшим промежутком, в
котором расположен спинной кровеносный сосуд. Две другие ленты, менее
развитые, расположены на брюшной стороне тела. Они также несколько
разобщены, так как здесь по средней (медианной) брюшной линии лежат
брюшная нервная цепочка и невральный кровеносный сосуд. Кроме этих
240
продольных мышечных лент, очень характерны диагональные, или транс-
версальные, мышечные пучки, которые одними концами прикрепляются к
стенке тела по бокам брюшной нервной цепочки, а другими — подходят к
параподиям и входят частично в состав мышц этих последних. Трансверсаль-
ные мышцы отделяют небольшие полости от общей полости тела, в которых
обычно расположены выделительные органы.
Полость тела. Внутренние органы Nereis лежат в полости тела. В от-
личие от первичной полости тела нематод у всех кольчецов и у Nereis в том
числе имеется вторичная полость тела, или целом. Важнейшим признаком
целома, отличающим его от первичной полости тела, нужно считать наличие
собственной целомической стенки, состоящей из основной бесструктурной
мембраны и лежащего на ней целомического эпителия, или целотелия. Эта
целомическая стенка плотно прилегает, с одной стороны, к стенкам тела,
образуя соматоплевру, а с другой — к стенкам кишечной трубки, образуя
Рис. 204. Развитие и строение целома аннелид (схемы поперечных разрезов).
/ — начальная стадия развития целома; // формирование целомических
мешков; /// сформировавшийся целом:
1 кишка. 2—мезодермические полоски — зачатки будущих целомических мешков (мезо-
бласты), 3—первичная полость тела, 4 — полость целомического мешка—целом (слева
так же), 5—собственная стенка целомического мешка, 6 — соматоплевра, 7 — спинной
кровеносный сосуд, 8— спланхноплсвра, 9 — брюшной кровеносный сосуд. Оба сосуда (7, 9)
лежат соответственно в спинной и брюшной мезентериальных перегородках
спланхноплевру. Сам целом состоит из отдельных парных камер, или цело-
мических мешков (рис. 204). Над кишечной трубкой и под ней стенки этих
мешков сближаются и образуют спинной мезентерий над кишечникОхМ и
брюшиной под ним. Спереди и сзади каждый целомический мешок отделен
от предыдущего и последующего поперечной перегородкой — диссепимен-
том (см. рис. 202). Целом имеет, следовательно, метамерное строение, так
как состоит из парных закономерно повторяющихся целомических мешков.
Положение диссепиментов строго соответствует наружной сегментации,
или метамерии. В целоме расположены метамерно расчлененные, или ме-
тамерно повторяющиеся внутренние органы, а сама полость целома запол-
нена целомической жидкостью, в которой находятся специфические клеточ-
ные элементы и жидкие и зернистые продукты обмена веществ.
Нервная система. Основу нервной системы составляют окологлоточ-
ное нервное кольцо и брюшная нервная цепочка. Окологлоточное нервное
кольцо слагается из следующих компонентов. На спинной стороне лежит
мощно развитый комплекс надглоточных ганглиев (рис. 205). Комплекс
этот парный. От него, охватывая глотку, идет нервное кольцо, состоящее из
двух — правого и левого — коннективов. Коннективы обращены на брюш-
ную сторону и здесь соединяются с первым парным нервным ганглием брюш-
ной нервной цепочки. Последняя состоит из метамерно расположенных
ганглиев, которые, повторяясь из метамера в метамер, тянутся вдоль всей
брюшной стороны тела, соединяясь между собой парными коннективами
(рис. 2056). От окологлоточного нервного кольца идут нервы к головным
241
органам чувств, в частности к пальпам и усикам, а от метамерных гангли-
ев — нервы, которые иннервируют тело и его периферию.
Пищеварительная система. Как уже было указано, ротовое отверстие
расположено на перистомиуме вентрально. Оно ведет в ротовую полость,
или буккальный отдел, отличающийся мускулистыми стенками и способ-
ный выворачиваться наружу. За буккальным отделом лежит глотка. Она
также мускулиста, причем на границе между ней и буккальным отделом
расположены острые и загнутые внутрь челюсти. Когда буккальный отдел
выворачивается наружу, то челюсти обнажаются (рис. 206, 207) и служат
органами захвата добычи. Весь этот аппарат мощно развит и достигает у
нереиды (Nereis pelagica) пятого метамера (сегмента). Глотка переходит в
относительно узкий пищевод, в который изливается содержимое слюнных
желез. Вся описанная часть кишечника составляет передний кишечник, или
стомодеум. За ним следует средняя кишка, или мезодеум. Строение ее до-
вольно однородное на протяжении всех метамер тела. В пределах каждого
целомического мешка кишка несколько расширяется, сжимаясь вновь дис-
сепиментами. В средней кишке происходит переваривание захваченной пищи.
Пищеварение всегда внеклеточное. За средней кишкой следует короткая
Рис. 205. Центральная нервная система:
1—нервы к пальпам. 2—нервы к усикам (ниррусам),
3—надглоточный ганглий, 4, 5 — окологлоточное нервное
кольцо, 6 — нервы брюшной нервной цепочки, 7.— начало
брюшной нервной цепочки
задняя кишка, или прок-
тодеум. Она соответствует
последнему отрезку тела —
пигидиуму, в области ко-
торого задняя кишка от-
крывается наружу аналь-
ным отверстием.
Кровеносная система.
Сосуды кровеносной систе-
мы несут красную кровь.
Спинной сосуд (см. рис.
203) обладает способностью
к пульсации, т. е. сокра-
тительными движениями
стенок и обычно гонит
кровь сзади наперед. Спе-
циальные сосуды, охваты-
вающие кишечную трубку
и расположенные метамер-
но в каждом сегменте
Рис. 206. Схема выпячи-
вания глотки и буккаль-
ного отдела. / — глотка
и буккальный отдел втя-
нуты; // — буккальный
отдел вывернут, глотка
выдвинута вперед:
1 — буккальный отдел, 2—
глотка
Рис. 207 Головной ко-
нец Nereis. Глотка (/) с
большими челюстями
выдвинута наружу
(кольцах тела), переводят
кровь в брюшной сосуд (см.
рис. 203), который не спо-
собен к самостоятельной
пульсации. Кровь движет-
ся в нем спереди назад.
Помимо этих направлений
кровеносных токов, важное
значение имеют сосуды,
которые несут кровь от
спинного сосуда к парапо-
диям. Это параподиальные
сосуды. В параподиях кро-
веносные сосуды приобре-
тают характер капилляров,
где происходит окисление
крови, близко соприкасаю-
щейся здесь с кислородом
наружной среды. Спинной
сосуд доходит до простоми-
242
ума, брюшной сосуд заканчивается на уровне глотки, т. е. он несколько
короче. Кожа также интенсивно снабжается капиллярными кровеносными
сосудами. Вместе с тем, кишечная трубка и все внутренние органы, а равно
диссепименты также обильно орошаются кровью.
Дыхательная система. Капилляры кровеносной системы параподий
и кожи имеют очень большое значение в дыхательных процессах Nereis,
тогда как специальных жаберных выростов у видов этого рода не имеется.
Выделительная система. Она состоит у Nereis из парных метанефридиев
(рис. 208). Их нет только в пяти передних метамерах и трех или четырех
задних. Метанефридии Nereis очень типичны. Каждый метанефридий со-
стоит из железистого метанефридиального тела, имеющего форму мешочка,
пронизанного извитым нефридиальным каналом. Канал этот начинается
еще вне тела метанефридия в полости соответ-
ствующего целомического мешка воронкой, или
нефростомой. Нефростома быстро сужается в
канал, который пронизывает стенку лежащего
перед ней диссепимента и входит в полость сле-
дующего целомического мешка (переднего по от-
ношению к предшествующему), где и лежит са-
мое тело метанефридия. Внутри метанефридиаль-
ного тела передняя часть нефридиального канала
Рис. 209. Nereis и его
эпитокная форма (Hete-
ronereis)
Рис. 208. Метанефридий Nereis:
1 — воронка, открытая в целом, 2— начало нефридиаль-
ного канала (в месте его прохождения сквозь диссепи-
мент), 3—часть канала, несущая мерцательные рес-
нички, 4 — выделительная пора
(ближайшая к нефростоме, рис. 208) несет тонкие реснички, которые согла-
сованно работают и гонят жидкость, находящуюся в трубке нефридиаль-
ного канала, вперед к наружному выходу, т. е. во внешнюю среду. Эта
задняя часть нефридиального канала не несет ресничек. Наружное отверстие
нефридиального канала называется нефропорой. Таким образом, жидкое
содержимое метанефридия, проникая в нефридиальный канал из целома, вы-
водится наружу через нефропору. Такова анатомия метанефридия. Что
касается его функции, то она состоит в удалении через нефридиальный
канал жидких продуктов обмена, которые накапливаются частью в виде
зерен в целомической полости.
Половая система. Половые железы, или гонады, образуются у Nereis
в целотелии (целомическом эпителии) в области брюшной стенки тела.
Nereis — раздельнополые полихеты. К моменту, когда в гонадах начинается
образование половых клеток (либо яиц, либо сперматозоидов), Nereis пре-
терпевает типичные изменения. Дело в том, что половые клетки формируются
только в задних метамерах. Эти последние в период половой деятельности
приобретают иную форму, чем передние членики (рис. 209). Явление это
называется эпитокией (с греческого «эпитокос» — рождение), а членики
(метамеры), несущие половые элементы, — эпитокными. Обычно животные
живут на дне моря, но эпитокные метамеры, отрываясь от тела нереиды,
плавают в верхних слоях морской воды. Эпитокные комплексы метамер
243
известны также под названием гетеронереидных, Nereis — живут в морях,
ведя придонный образ жизни.
Питание у нереид смешанное: они грызут при помощи челюстей подвод-
ную растительность, способны нападать на мелких беспозвоночных, или
они питаются, детритом, взрыхляя челюстями грунт В наших морях
виды рода Nereis очень обычны и широко распространены.
ОБЩИЙ ОБЗОР КЛАССА
Внешнее строение. Как и в пределах других групп животных, размеры
тела полихет подвержены значительным колебаниям. Наряду с мелкими фор-
мами, иногда едва превышающими 1 мм, известны крупные полихеты, до-
стигающие 3 м в длину. Большей частью тело полихет стройное и относи-
тельнотонкое. Однако известны формы с коротким и плотным, даже толстым
туловищем (рис. 210). У части форм тело на всем протяжении однородно.
У других оно, напротив, явственно дифференцировано на головной, туловищ-
ный и хвостовой отделы. Многие виды свободно плавают в воде. Другие
ведут сидяче-прикрепленный образ жизни, и у таких форм развиваются
трубки, выполняющие роль «домиков», в которых сидят их обладатели —
сидячие полихеты. Для этих животных очень обычны яркие окраски —
желтые, красные, фиолетовые, зеленые, черные и т. п., нередко с красивыми
металлическими оттенками.
Тело полихет, как и у Nereis, всегда расчленено на отдельные метамеры,
или сегменты, которые дифференцированы на три основных отдела: голов-
ной, туловищный и хвостовой, или пигидиальный. Головной отдел всегда
состоит из предротового метамера, или простомиума, и ротового метамера,
или перистомиума. Число туловищных метамер различно. Пигидиум всегда
состоит из одного членика. У многих форм, ведущих сидяче-прикрепленный,
или седентарный, образ жизни, тело явственно делится на голову, туло-
вище и хвост, причем хвостовые метамеры могут быть сильно изменены и
упрощены по сравнению 'с туловищными или головными (см. рис. 210).
Как уже указывалось, простомиум несет особые придатки усики
(или антенны) и пальпы. Эти органы, с которыми мы познакомились на при-
мере нереиды, у различных полихет подвергаются значительным измене-
ниям. Так, у живущих в песчаных норках Magelona papillicornis пальпы
превращены в длинные придатки, используемые для ловли добычи (рис. 211).
У Sabellaria alveolata (см. рис. 210) пальпы образуют нитевидные жабер-
ные выросты, сидящие на широких боковых лопастях. У Serpula (см. рис. 210)
пальпы представлены двумя лопастями, усаженными тонкими выростами,
покрытыми ресничками и образующими венчик ловчих органов. Один из
лучей этого венчика превращен при этом в крышечку (см. рис. 210), которая
закрывает вход в «домик» червя. У Amphitrite (см. рис. 210) ловчие щупаль-
ца образуют пышный венец, украшающий головной конец тела. Эти примеры
показывают различные формы превращений головных органов.
Параподии всегда состоят из спинной ветви, или нотоподия, (notopo-
dium) и брюшной ветви, или нейроподия (neuropodium). Спинная ветвь
несет спинной усик, или спинной циррус, тогда как нейроподий соответст-
венно вооружен брюшным циррусом. В основной части параподии развиты,
как это мы видели у Nereis, комплекты щетинок, которые принадлежат как
спинной, так и брюшной ветви параподий. У многих полихет имеются кроме
пучков тонких щетинок, толстые опорные щетинки, или ацикулы (рис. 212).
Параподии у различных полихет подвергаются многообразным изменениям.
На рис. 212 представлены некоторые примеры этих изменений. Вверху по-
казана типичная параподия такого же типа, как у Nereis, ниже — парапо-
дия Phyllodoce, у которой спинной циррус превращен в плавательный орган,
или плавник. Наконец, на нижнем рисунке показана параподия Eunice,
у которой спинной циррус превращен в жабру Жабра снабжена капилляр-
ными кровеносными сосудами и в ней происходит энергичное окисление
244
крови. Обычно каждый метамер тела полихеты имеет параподии по одной
паре на метамер. Однако в большом числе случаев параподии сохраняются
далеко не на всех метамерах. Довольно однородный ряд параподий имеет
Nereis. У многих других форм, в особенности седентарных (сидяче-при-
крепленных), параподии сохраняются на передних метамерах, но исчезают
на задних (см. рис. 210).
Различное строение параподий определяется их функциями. Вообще
параподии — органы движения. Двигательными, или локомоторными,
Рис. 210. Разные полихеты. I — Arenicola marina (роющая форма); II Nereis
pelagica (плавающая форма); /// Phyllodoce paretti (плавающая форма);
IV Amphitrite edwardsi (плавающая форма); V—Sabellaria alveolata; VI
Serpula (сидячие формы); VII—Aphrodite aculeata (ползающая форма):
1 — щупальца, 2 — жабры, 3 — крышечка, 4 — параподии
245
компонентами их являются нотоподий и нейроподий, тогда как спинной и
брюшной усики функционируют как органы осязания. При помощи пара-
подий полихета может ползать по дну моря, а выдвигая пучки щетинок,
усиливать гребущий эффект параподии и, таким образом, производить пла-
вательные движения. Превращение спинного цирруса в плавник (рис. 212)
усиливает плавательную функцию параподий у форм, для которых плавание
особенно характерно (например, Phyllodoce).
Естественно, что отсутствие или присутствие параподий, а равно форма
и размеры метамер влияют на общее строение тела полихеты. Когда*все ме-
тамеры приблизительно одинаковы, как
это мы видели у Nereis, принято гово-
рить о гомономной метамерии. У многих
полихет либо в качестве постоянного
признака их организации, либо в перио-
ды их полового размножения метамеры
тела оказываются неравнозначными,
неодинаковыми по своей морфологиче-
ской организации. В этих случаях сле-
дует говорить о гетерономной метаме-
рии. свидетельствующей о морфологи-
ческой неравнозначности метамер.
Рис. 211. Magelona papil-
1 icorni s:
1 — ловчая параподия, 2— обыч-
ные параподии
Рис. 212. Типы параподий.
/ — Nereis; // — Phyllod
осе; III —Eunice:
/ — нотоподий, 2 — невроподиГ!,
3 — спинной циррус, 4 — брюш-
ной циррус, 5 — жаберный вы-
рост, 6 — ацикулы
Кожно-мускульный мешок. Кожно-мускульный мешок состоит из
плотной, но тонкой, похожей на хитин кутикулы, подлежащего эпителия,
кольцевой и продольной мускулатуры и выстилающего ее изнутри целоте-
лия. принадлежащего целому. Кутикула полихет пронизана многочислен-
ными отверстиями — порами кожных желез, которыми очень богат подле-
жащий эпителий. Выделения этих желез имеют двоякое значение: частью
защитное (по-видимому, экскрет белковых желез), частью они идут на по-
строение «домиков», т. е. временных или постоянных трубок, в которых
живут многие полихеты. Обычно выделяемые экскреты, идущие на построе-
ние трубки, смешиваются с песчинками и застывают в прочную массу.
Кольцевая мускулатура, как правило, тонкая. Напротив, продольная
мускулатура образует четыре мощные продольные ленты (см. рис. 203).
В вертикальном направлении они разделены спинным и брюшным мезенте-
риями, по бокам тела — особыми косыми (трансверсальными) мышцами.
Спинные ленты, как правило, сильнее, чем брюшные. Мышечные волокна
принадлежат к типу гладкой мускулатуры. Косые, или трансверсальные,
246
мышцы тянутся по бокам тела от нервной цепочки к параподиям и отделяют
участки целома, в которых обычно располагаются выделительные органы
(метанефридии).
Целом и его функции. С элементами целома мы уже познакомились выше.
Еще раз только подчеркнем, что основным признаком его, который необхо-
димо твердо помнить, остается наличие собственной целомической стенки
(целотелия), за счет которой и образуются элементы целома — сомато- и
спланхноплевра, брюшной и спинной мезентерии, диссепименты. Напомним
еще раз, что сам целом состоит из независимых парных (правого и левого)
целомических мешков, которые метамерно повторяются из сегмента в сегмент
соответственно наружной метамерии. Камера целома, отделенная трансвер-
сальными мышцами, обычно называется метанефридиальной, а большая
камера, стенки которой охватывают кишку, — кишечной камерой. У мно-
гих полихет лежащие тотчас позади головы метамеры не разделены диссе-
пиментами, а образуют «грудные» или торакальные, метамеры. Эти послед-
ние отделены от расположенных позади «брюшных» сегментов крепким дпс-
сепиментом, иногда называемым «диафрагмой».
В тех случаях, когда диссепименты частично отсутствуют целомиче-
ская жидкость может свободно переливаться из одних метамер в другие,
поэтому тело в соответственных местах вздувается. У некоторых форм такое
вздувание тела в результате притока целомической жидкости имеет при-
способительное значение, например при рытье нор одной из полихет — пе-
скожила (Arenicola). Сказанное выше демонстрирует еще одну важную
функцию целома: полостная жидкость его обеспечивает полостной тургор,
придающий телу известную ригидность и сохраняющий его форму.
Целом — орган с очень важными и ответственными функциями. Цело-
мическая жидкость изотонична морской воде, в которой живут полихеты.
Целомическая жидкость содержит слабые концентрации растворенных аль-
буминоидов, сахара, жиров и других веществ. В целомической жидкости
взвешены специфические форменные элементы, в том числе лейкоциты,
лимфоциты и фагоциты. Кроме того, в полостной жидкости взвешены зер-
нистые лейкоциты, имеющие у ряда форм значение носителей резервных
питательных веществ, в частности альбуминоидов. Следует также указать,
что частично резервные жировые гранулы могут скопляться в целотел ин,
ограничивающем целомические полости. В других случаях целомическая
жидкость содержит особые клеточные включения, богатые жировыми ве-
ществами. Такие полостные клетки получили наименование элеоцитов.
Элеоцитов может быть очень много, и их совокупность приобретает зна-
чение полостной жировой ткани (аналогичной жировому телу насекомых),
имеющей большое значение при развитии половых продуктов. На-
конец, в целомической полости включены особые клетки, функционально
связанные с экскреторной (выделительной) функцией. В этой связи в клет-
ках целомической жидкости, в том числе в упомянутых элеоцитах и грану-
лоцитах, могут накапливаться экскреторные гранулы, которые имеются
также в особо измененных клетках целотелия, выстилающего полость сег-
ментов. Так возникает очень характерная хлорагогенная ткань, содержа-
щая окрашенные в желтый или бурый цвет продукты обмена. Все эти про-
дукты выносятся из организма полихеты через посредство экскреторной
системы. Наконец, укажем, что в целомической жидкости могут быть вклю-
чены эритроциты, имеющие большое значение в дыхательных процессах.
Наполнение целома внутриполостной жидкостью позволяет вторичной
полости тела выполнять важнейшую опорную функцию. Давление целоми-
ческой жидкости придает телу и параподиям упругость. Вместе с сегмента-
цией, усиливающей возможность изгибания тела и параподиями, опорная
функция вторичной полости тела позволяет многощетинковым существовать
в весьма различных условиях океана, вплоть до больших глубин.
Нервная система. Нервная система лежит либо в эпителии, что нужно
считать или более примитивным признаком, или чертой упрощения органи-
247
зации, либо (что чаще) — под эпителием. Как всегда, она состоит из около-
глоточного кольца и брюшной нервной цепочки. Окологлоточное кольцо
образовано за счет крупных спинных ганглиев и коннективов, которые охва-
тывают глотку и соединяются в области первого брюшного нервного узла.
У полихет спинные ганглии достигают мощного развития и часто называются
мозгом. На рис. 205 показан мозг Nereis, а на рис. 20 — мозг Phyl-
lodoce. От спинных ганглиев и близлежащих нервных элементов идут нервы
к хорошо развитым органам чувств — антеннам и пальпам.
Органы чувств. Полихеты обладают хорошо развитыми глазами, иног-
да достигающими значительной величины и сложности (см. рис. 202). У не-
которых форм глаза по своему строению напоминают глаза позвоночных.
Такие глаза характерны, например, для семейства Alciopidae. В других
случаях глаза представлены простыми пигментными бокалами, дно ко-
торых содержит зрительные клетки. У некоторых полихет имеются инвер-
тированные глаза (наподобие глаз турбеллярий). Глаза расположены либо
на головной лопасти (Nereis), либо на первом сегменте тела или у основания
щупалец (антенн) вблизи параподий (седентарные Serpulidae), или даже на
брюхе, как это имеет место в эпитокных члениках у палоло. Число глаз
варьирует от 2—4 до 200 и больше.
Щупальца на простомиуме и спинной циррус параподий, равно как раз-
бросанные в коже чувствующие клетки, имеют значение органов осязания.
К химическим органам принадлежат пальпы, а также особые нухальные
органы, представленные в простейшем случае ресничными ямками (рис. 213).
Кроме того, в коже имеются также чувствующие клетки, обладающие функ-
цией органов химического чувства. У седентарных (сидяче-прикрепленных)
форм имеются также статоцисты. Обычно они расположены на переднем
конце тела и имеют форму закрытых или открытых пузырьков. В последнем
случае в полости пузырька могут быть песчинки, раковинки диатомовых
водорослей ит. п., играющие роль отолита.
Пищеварительная система. Основные части передней кишки, т. е. бук-
кальный отдел, глотка и пищевод, описанные выше для Nereis, присущи
многим полихетам. Иногда этот отдел подвергается упрощениям. У боль-
шинства хищных полихет особенно мощного развития достигает выдвижная
глотка. У некоторых форм длина выдвижной глотки достигает половины
длины тела. Глотка вооружена «челюстями». Когда добыча схвачена, она
удерживается челюстями, а с помощью специальных мышц-ретракторов
глотка вводится обратно внутрь передней кишки вместе с добычей. Строение
пищевода различно, часто это простая трубка, в других случаях она услож-
нена. Весь передний отдел кишечника имеет эктодермическое происхо-
ждение.
Средняя кишка имеет форму трубки, иногда простой, иногда с карма-
нами, которые располагаются метамерно в соответствии с целомическими
мешками. У части полихет средняя кишка усложнена развитием глубоких
боковых выростов, которые увеличивают поверхность средней кишки. По-
следняя должна рассматриваться как орган, усваивающий захваченную
пищу. Здесь происходят ее химическая переработка и всасывание питатель-
ных растворов. В связи с функцией средней кишки для нее характерно на-
личие однослойного эпителия, функционирующего как усваивающий аппа-
рат и богатого железистыми клетками. Средняя кишка характеризуется
некоторыми усложнениями организации, которых мы здесь не будем ка-
саться. Вся средняя кишка имеет энтодермическое происхождение.
Задняя кишка очень короткая, заканчивается анальным отверстием.
Она имеет эктодермическое происхождение. Кишечник в целом всегда несет
на себе печать приспособления к тому роду пищи, который характерен
для соответственных групп полихет.
Кровеносная система. У всех типичных полихет она имеет примерно
то же строение, что и у Nereis, хотя она и варьирует в деталях. Спинной
кровеносный сосуд, обладающий способностью к пульсации, имеет кольце-
248
вую мускулатуру, которая и обеспечивает ток крови. Спинной сосуд дости-
гает головной лопасти, где превращается в капилляры, орошающие ее.
Кровь направляется в этом сосуде всегда вперед, а часть ее течет по сосудам
мезентерия к кишечному кровеносному сплетению. Это последнее связано с
брюшным кровеносным сосудом, по которому кровь течет спереди назад.
Вся эта система в действительности сложнее, так как на путях между спин-
ным сосудом, кишечным сплетением и брюшным сосудом
кровь омывает все элементы целома, в том числе мезен-
терии, диссепименты, а равно параподии. В параподиях
часть кровеносных сосудов также способна к самостоя-
тельной пульсации. Отметим, что в составе кишечного
сплетения имеются особые расширенные сосуды, извест-
ные под названием сердец пульсаторных аппаратов,
которые гонят кровь из кишечного сплетения к брюш-
ному сосуду.
Органы дыхания. Органами дыхания у полихет слу-
жат прежде всего жабры (рис. 212). Обычно они распо-
ложены на параподиях. Это — выросты тела, включающие
участок целома, соответствующий сомитам. Покрыты они
кутикулой, под которой лежит эпителий, обильно снаб-
женный капиллярными сосудами. Эти сосуды находятся
в близком контакте с внешней средой и, таким образом,
через их посредство обеспечивается газообмен, и, в част-
ности, кровь полихеты отдает углекислый газ и обога-
щается кислородом. Положение жаберных выростов
очень характерно. Часто они занимают лишь определен-
ные участки тела. Так, у пескожила (Arenicola) они
развиты лишь на средних метамерах тела. В других слу-
чаях жабры образуются на спинной ветви передних па-
раподий или за счет пальп. Подобное расположение жабер
Рис. 213. Ну-
хальные орга-
ны (/) у Polyg-
ordius
вполне понятно
у сидячих (седентарных) форм: жабры должны находиться вне трубки, в
которой сидит седентарная полихета, т. е. на переднем конце тела.
Выделительная система. У более примитивных полихет имеются прото-
нефридии. Однако эти последние
связаны с целомом, и в этом их
существенное отличие от прото-
нефридий плоских червей. Связь
с целомом осуществляется в про-
тонефридиях полихет специаль-
ными приспособлениями, кото-
рые получили название солено-
цитов. Каждый соленоцит (рис.
214) представлен клеткой, кото-
рая содержит ядро. От клетки
идет узкий то длинный, то более
короткий канал, в котором мер-
цает жгут Комплекс таких
соленоцитов расположен на вер-
шине протонефридия, обращен-
ной в целом данного целомичес-
кого мешка (рис. 214). Через
посредство соленоцитов целоми-
ческая жидкость фильтруется и
попадает в канал соленоцита.
Жидкость гонится жгутом даль-
ше и проникает из соленоцитов
в общий канал нефридия. Отсю-
да удаляемая жидкость прони-
Рис. 214. Протонефридии полихет. 1,11 —
соленоциты, реснички которых проникают в
нефридиальный канал; /// —протонефридий
Alciope:
/ — половая воронка, 2 — половая трубка, 3—нефри-
дий, идущий от розетки соленоцитов
249
кает через нефропору наружу. Усложнения такого протонефридия, снаб-
женного соленоцитами, указаны ниже. В более сложных и более типичных
случаях соленоциты исчезают и нефридиальный канал открывается в по-
лость целома нефростомой; выделительный канал становится сквозным.
Именно этот факт характеризует и отличает метанефридий от типичного
протонефридия. Строениеметанефридиев варьируету разных групп полихет,
но в принципе соответствует организации метанефридий, характерных для
Nereis (см. рис. 208).
Метанефридии выводят наружу продукты обмена, о которых говори-
лось выше при рассмотрении строения и функций целома.
Половая система. Половые желе-
зы, или гонады полихет, как уже
указывалось, расположены в целоте-
лии, оставаясь первоначально прикры-
тыми им. Как правило, их нет в пе-
редних метамерах. В типичных
случаях они следуют метамерно вдоль
тела, в виде парных образований
вплоть до пигидиума. В других слу-
чаях число гонад сокращается до
нескольких пар (2—5). У седентарных
форм гонады могут располагаться в
торакальном отделе (грудном), или
в брюшном (абдоминальном). Поло-
вые продукты всегда выпадают из
гонад в целомическую полость. Высво-
бождение половых элементов во внеш-
нюю среду достигается у разных
форм различными путями. Иногда
через разрыв стенки тела; иногда
путем отрыва всей задней части тела,
содержащей половые продукты; нако-
Рис. 215. Эпитокия и шизогамия у поли- нец, сплошь и рядом половые элемен-
хет. / — эпитокия палоло (Eunice viri- ты ВЫВОДЯТСЯ наружу через ПОСредст-
djs); // — шизогамия у Autoiytus com- в0 ПОЛОвых протоков. Эти последние
utus. В задней части тела образовался г
участок, развивающий голову не являются, однако, чем-то особым
в организации полихет; напротив, это
специально модифицированные участ-
ки мета- или протонефридиев. На рис. 214 показан протонефридий с
соленоцитами, причем рядом с комплексом протонефридий расположена
трубка полового протока. Он открывается в полость сомита половой ворон-
кой, переходящей в тело полового протока, по которому половые продукты
и проникают в нефридиальный канал, а затем во внешнюю среду. Иногда
половой проток и нефридий полностью обособляются друг от друга. Несо-
мненно, что филогенетически образование специальных половых протоков —
позднейшее явление.
Размножение. Размножение полихет протекает в двух формах — по-
ловой и бесполой.
Половое размножение. Полихеты в большинстве раздель-
нополы. Половой диморфизм выражен не всегда. У некоторых групп полихет
самцы отличаются от самок большим развитием органов чувств — антенн,
пальп. У ряда форм яйца освобождаются наружу, как говорилось выше,
через разрыв стенки тела и попадают в воду, где они и оплодотворяются.
То же происходит и у форм со специальными половыми протоками или в
случаях, когда яйца выбрасываются через метанефридий. Однако в некоторых
случаях наблюдается оплодотворение яиц и в материнском организме. У
части форм период размножения связан с явлением эпитоклш, или эпигамии.
В этих случаях, как это имеет место у Nereis, задние членики тела изменяют-
250
ся по структуре и образуют комплекс эпитокных члеников, содержащих
половые продукты. Обычно эти членики отличаются морфологически от
передних метамеров, несодержащих половых продуктов (рис. 215) и извест-
ных под названием атокных. В это же время внутренние органы эпитокных
участков подвергаются глубоким изменениям: кишечник дегенерирует,
стенки тела истончаются, сквозь них просвечивают половые продукты,
дающие окраску всему телу эпитокной формы. Половые особи всплывают к
поверхности моря, ведя пелагический образ жизни. Это способствует более
широкому распространению половых продуктов и полихет данного вида.
У многих форм эпитокные участки отрываются от атокных, причем именно
эпитокные участки носятся в пелагической зоне моря, тогда как атокные ос-
таются на дне, регенерируя утраченные метамеры. Когда половые продукты
попадают в воду, эпитокные участки погибают. Примером подобных явлений
может служить размножение при помощи эпитокии у Eunice viridis, извест-
ного под названием «палоло» (рис. 215). Эпитокные участки этой полихеты в
невероятных количествах появляются на поверхности моря у берегов ост-
ровов Фиджи и Самоа. У Nereis имеются аналогичные процессы эпитокии
(см. выше). По-видимому, биологическое значение эпитокии состоит в том,
что у форм, размножающихся этим способом, нет специальных половых
протоков, и образование эпитокных участков тела позволяет освобождать
половые продукты через разрыв стенки тела. При этом эпитокные участки
или особи гибнут, но атокные сохраняются и возобновляют размножение в
результате регенерации новых эпитокных участков.
У некоторых полихет наблюдается явление шизогонии, когда эпиток-
ный участок, прежде чем оторваться отатокного, образует в месте отрыва
голову, превращаясь, следовательно, в самостоятельную особь, несущую
половые продукты (рис. 215). Отложив в воду половые продукты, эти до-
черние зооиды, как их называют, гибнут, а атокная часть, или материн-
ский зооид, снова образует новый эпитокный участок.
Бесполое размножение. Легко понять, что в случаях
шизогонии мы имеем дело с новообразованием, которое вполне сравнимо с
Рис. 216. Столонизация и почкование у полихет. / — столонизация у
Myrianida fasciata. Рядом (//, ///) два отдельных зооида:; IV — почко-
вание у SylHs ramosa
251
Рис. 217. Трохофора Polygordius. / трохофо-
ра; II—IV — превращение трохофоры во взро-
слого червя:
1—ротовое отверстие, 2—кишка, 3—анус, 4 — прото-
нефридии с соленоцитами, 5 — теменная пластинка, 6 — про-
тотрох (пояс ресничек)
Рис. 218. Протонефридий трохофоры:
1— соленоциты, 2—их жгуты, 3— мембраны, 4 —
выделительная пора
почкованием. В данном слу-
чае почкование протекает в
продольном направлении.
Однако у полихет бывает и
почкование, протекающее в
поперечных направлениях.
Так, у Syllis ramosa (рис. 216)
в результате бокового почко-
вания образуется целая сеть
особей. Это почкование в дан-
ном случае не связывается с
половым размножением и с
эпитокией и приобретает са-
мостоятельное значение одно-
го из способов размножения.
У Myrianida (рис. 215)
процессы почкования идут
так быстро, что едва на заднем
конце тела успевает обосо-
биться новый зооид, как от
заднего метамера полихеты
начинает обособляться следу-
ющий зооид и т. д. В резуль-
тате образуется целая цепочка
зооидов — столон, или стро-
била. Процесс подобного поч-
кования известен под назва-
нием стробиляции, или сто-
лонизации. В конце концов
задние дочерние зооиды (бла-
стозооиды) отрываются, выде-
ляют половые продукты и
погибают
Полихетам свойственна
регенерация. Как мы видели
выше, регенерируют после
эпитокии новые метамеры: у
многих полихет регенерируют
антенны, щетинки параподий,
иногда головная лопасть со
всеми ее придатками, в осо-
бенности у седентарных
форм, легче подвергающихся
ранениям и калечению.
Развитие. Развитие либо
с метаморфозом, либо прямое.
Для форм, развивающихся с
метаморфозом, весьма харак-
терна личинка, называемая
трохофорой.
Мы здесь опишем весьма
типичную трохофору (рис.217)
Polygordius. Вполне развитая
трохофора этого червя имеет
грушевидную форму тела. По
экватору трохофоры проходят
два пояса ресничек: верхний,
или прототрох, и нижний,
252
или метатрох. Между ними лежит ротовое отверстие, которое ведет в
пищевод. Пищевод переходит в обширный желудок. Последний суживается
и переходит в короткую энтодермическую кишку. Кишка заканчивается
задней эктодермической кишкой, открывающейся наружу анальным отвер-
стием. Последнее расположено на нижнем полюсе тела трохофоры. Верхний
полюс трохофоры несет характерный орган— теменную пластинку, зачаток
будущих надглоточных узлов. Над теменной пластинкой возвышается темен-
ной султан ресничек. Трохофора имеет протонефридиальный выделительный
аппарат. У трохофоры Polygordius этот аппарат представлен одной парой
трубок.Слепые концы этих трубок заканчиваются в полости тела трохофоры
терминальными органами в виде своеобразных клеток, замыкающих канал.
Каждая такая клетка (терминальный орган) переходит в трубку (рис. 218).
Эти трубочки на свободных концах замкнуты, а внутри каждой такой трубки
колеблется длинная нить. Описанный терминальный орган — соленоцит.
Соленоциты открываются в общую полость нефридиального канала. У
трохофоры Polygordius таких каналов два. Они соединяются в один вывод-
ной проток, который открывается наружу порой.
Трохофора некоторое время плавает в верхних слоях воды, а затем
подвергается метаморфозу. Нижний полюс личинки вытягивается в черве-
образный членистый (сегментированный) придаток. Примерно в это время
трохофора опускается на дно. Верхний полюс личинки подвергается редук-
ции и превращается в головной конец червя. Внутренняя организация тро-
хофоры также изменяется. Часть личиночных органов атрофируется. Ис-
чезают протонефридии и личиночная мускулатура. Напротив, нервный узел
теменной пластинки и кишечный канал превращается в соответствующие
органы взрослого червя. За счет теменного нервного узла образуется над-
глоточный узел нервного окологлоточного кольца. В то же время в туловище
развивающейся личинки образуется брюшная нервная цепочка. Последняя
развивается за счет эктодермы и притом назависимо от надглоточного узла
из продольных зачатковых валиков эктодермы.
Выше упомянуто, что нижний конец трохофоры вытягивается и превра-
щается в туловищные сегменты. Параллельно с этим происходит и развитие
вторичной полости тела, или целома. На этом процессе ввиду его теорети-
ческой важности мы несколько задержимся.
У только что образовавшейся трохофоры можно видеть примерно по бо-
кам задней кишки две характерные клетки — мезобласты. Мезобласты
дают начало мезодермическим полоскам (см. рис. 204), которые ложатся у
трохофоры по бокам кишечника. Параллельно с ростом нижнего конца тро-
хофоры, превращающегося в тело червя, мезодермические полоски удли-
няются и при этом сразу разделяются в поперечном направлении на отдель-
ные группы клеток, которые расположены симметрично по бокам кишки.
Эти симметрично и метамерно расположенные участки мезодермы называют-
ся сомитами.
В дальнейшем внутри каждого из сомитов обособляется полость и со-
миты превращаются в уже нам известные целомические мешки, которые
являются не чем иным, как участками (камерами) вторичной полости тела,
или целома, тогда как границы соприкасающихся мешков соответствуют
диссепиментам. Каждая пара целомических мешков (см. рис. 204) разра-
стается, охватывает кишечную трубку и постепенно вытесняет первичную
полость тела. За счет соприкасающихся стенок правого и левого мешков
образуются спинная и брюшная брыжейки, тогда как сохранившиеся над
и под кишечником остатки первичной полости, редуцированные до степени
спинной и брюшной щелей, соответственно превращаются в спинной и брюш-
ной кровеносные сосуды.
Соответственно возникновению этих первых сомитов происходит и
наружная сегментация тела трохофоры. Следовательно, несколько первых,
так называемых личиночных, или ларвальных, сегментов, возникает сразу
благодаря одновременному разделению мезодермической полоски. Остальные
253
Рис. 219. Dinophilus
apatris:
1 — глаза, 2 — ротовое от-
верстие и глотка, 3—хо-
бот, 4 — пищеводные же-
лезы, 5 — кишка, 6 — зад-
няя кишка, 7 — яичник
постларвальные сегменты возникают в дальнейшем путем постепенного
отпочковывания от последнего ларвального сегмента.
Описанный процесс показывает роль мезобластов в образовании целома
и кровеносной системы. В то же время он подчеркивает зависимости, су-
ществующие между развитием целома и кровеносных
сосудов. Как правило, кровеносные сосуды образуются
у форм, имеющих целом (хотя бы и подвергшийся на
тех или иных стадиях развития вторичной редукции),
за счет остатков первичной полости тела. У некото-
рых примитивных кольчецов, а именно у представи-
телей рода Dinophilus (рис. 219) можно превосходно
наблюдать зависимость между развитием целома и
кровеносной системы. У этих форм имеется и целом, и
первичная полость тела, т. е. псевдоцель. Но целом
мал и представлен небольшими участками, в которых
находятся половые продукты. Напротив, псевдоцель
составляет наибольшую часть полости тела и играет
роль кровеносных лакун. Можно себе представить,
что в процессе исторического развития дальнейшая
редукция остатков первичной полости привела к раз-
витию обособленных кровеносных сосудов, как это и
показывает описанный выше процесс развития трохо-
форы.
Экология и распространение. Громадное большин-
ство полихет — морские формы. Только немногие ви-
ды приспособились к жизни в солоноватой и пресной
воде. В этом отношении особенно интересен род Мапа-
junkia (семейство Sabellidae), один вид которого М.
baikalensis обитает в Байкале.
В морях полихеты играют очень большую роль,
встречаясь повсюду и иногда в громадных количест-
вах. Типичная пх стация — дно, где они населяют
песок, ракушечник, илистый песок, галечник, зарос-
ли подводной растительности и т. п. Здесь можно
различить целый ряд экологических групп.
Таковы, например, роющие формы, представителем
которых может служить пескожил (Arenicola marina, рис. 210), делающий в
илистом песке U-образные норы. Пескожил подобно дождевому червю про-
пускает песок через кишечник. «Морская мышь», или афродита (Aphrodite
aculeata), может служить примером ползающих
форм (см. рис. 210). Многие виды, имеющие
длинное тело или веслоподобные ветви пара-
та подий, хорошо плавают или даже ведут пелаги-
/ \ ческий образ жизни и живут, следовательно, в
/ Хя \ толще воды, например Tomopteris (рис. 220),
/ \ Другие полностью или частично утратили спо-
/ \ собность к передвижениям и превратились в
/ \ СИДЯЧИХ животных. В этих случаях полихеты
/ \ сидят в «домиках» (рис. 210). Последние либо
л । сплошь построены за счет выделений кожи,
I либо состоят из разного рода материала,
\ 1 скрепленного выделениями цементных желез.
\ / На переднем конце у этих форм развивается
/ обычно султан щупалец, при помощи которого
они фильтруют воду.
Отношения полихет с другими организмами
Рис. 220. Tomopteris euchae- Довольно многообразны. По способу питания
ta — планктонная полихета у полихет можно наметить несколько групп.
254
Зоофаги питаются животной пищей. Среди зоофагов есть типичные
хищники (см. рис. 210), подвижные, с хорошо развитыми глазами, глотка
вооружена крепкими челюстями. Хищные полихеты питаются по преимущест-
ву полихетами же и другими мелкими организмами, хватая даже мелкую
рыбешку. Другие зоофаги, как, например, Aphrodite, ползая по дну, под-
бирают различных животных, живущих в илистом песке или детрите.
Многие полихеты — фитофаги, питаются подводными растениями.
Интересно, что, например, в пределах рода Nereis имеются как зоофаги
(в том числе хищники), так и фитофаги.
Большое число полихет питается органическими веществами заилен-
ного песка (пескожил) или детритом (органические вещества, остатки жи-
вотных и растений, лежащие на песке). Некоторые полихеты ведут себя как
комменсалы (нахлебники), поселяясь, например, внутри губок, на раковинах
моллюсков (в особенности полихеты из семейства Serpulidae), на панцирях
крабов и т. п.
Сидячие полихеты сплошь и рядом образуют многочисленные скопле-
ния особей, среди которых живут и другие организмы, например некоторые
моллюски, амфиподы, мшанки и т. п.
Известны, наконец, некоторые паразитические полихеты (Myzosto-
mida).
Полихеты имеют довольно большое значение в питании различных жи-
вотных. Полихетами питаются раки иглокожие, многочисленные, в том чи-
сле и важные промысловые рыбы (пикша, камбала и др.), а в пределах мелко-
водной прибрежной полосы полихет хватают различные птицы. Палоло в
период их размножения используют в пищу человеком.
СИСТЕМАТИЧЕСКИЙ ОБЗОР класса
Класс полихет разбивается на 3 подкласса: 1) свободноподвижные
(Errantia), 2) сидячеприкрепленные (Sedentaria), 3) мизостомиды (Myzo-
stomida).
ПОДКЛАСС I. СВОБОДНОПОДВИЖНЫЕ ПОЛИХЕТЫ
(ERRANTIA)
Представители этого подкласса в большинстве случаев ведут свободно
подвижный образ жизни, имеют большей частью длинное тело, состоящее
из гомономных члеников с хорошо развитыми параподиями. Простомиум
всегда явственный, нефридии метамерны.
Сюда принадлежит ряд семейств, основные из них (см. рис. 210): Aphro-
ditidae, например, Aphrodite aculeata; Phyllodocidae — Phyllodoce paretti,
Tomopteridae — Tomopteris euchaeta (рис. 220), Syllidae — Syblis ramosa;
Nereidae — Nereis pelagica и т. д. Многие Errantia ведут хищный образ
жизни, тогда как другие формы растительно- или детритоядные.
К этому же подклассу относятся и упрощенные формы полихет, при-
надлежащие к семействам Polygordidae, Protodrilidae и Dinophilidae1.
Типичным родом первого из этих семейств является Polygordius, второго —
Protodrilus. Формы эти интересны тем, что у них редуцируются параподии,
но имеются щетинки. В остальном у них типичная аннелидная организация.
Большой интерес представляют полихеты третьего семейства Dinop-
hilidae.
Представителем этого семейства может служить род Dinophilidae
рис. 219), у которого сохраняются мерцательные обручи, напоминающие
ресничные пояса трохофоры, имеются протонефридии, целом редуцирован
1 Ранее эти семейства объединялись в особый класс Archinnelides, т. е. примитив-
ные кольчецы.
255
до степени полости, в которой лежат гонады и имеются кровеносные лакуны—
гомолог протоцеля. Все динофилиды — мелкие микроскопические формы,
несомненно упрощенные в связи с более простыми условиями существования.
Все эти формы обитают в морях. Polygordius и Protodrilus известны в
Черном море, Dinophilus — в Белом.
ПОДКЛАСС II. СИДЯЧИЕ ПОЛИХЕТЫ
(SEDENTARIA)1
Сюда относятся частью свободные (обычно роющиеся в грунте), частью
сидячие формы. Простомиум развит слабо, параподии тоже более или менее
редуцированы. Нефридии у многих форм сосредоточены лишь в более круп-
ных передних члениках. У многих явственная гетерономность с хорошо
обособленными грудными сегментами. Питаются детритом, илом, мелкими
организмами. Семейства. Arenicolidae, например Arenicola marina, Tere-
bellidae, Amphitrite edwardsi, Sabellidae — Sabellaria alveolata; Serpuli-
dae — Serpula (рис. 210).
ПОДКЛАСС III. МИЗОСТОМИДЫ (MYZOSTOMIDA)
Тело плоское, овальное или дисковидное (рис. 221), у некоторых форм внешне
похоже на тело плоских червей. Внешняя метамерия не выражена. Параподии пред-
ставлены пятью парами брюшных выростов,
вооруженных щетинками, загнутыми
в виде крючков, или вообще отсутствуют. У не-
которых форм мышцы, двигающие параподии,
имеют поперечнополосатое строение. Цент-
ральная нервная система правильно расчле-
ненная, метамерная. Целом развит слабо,
внутренние органы лежат в паренхиме. Пе-
редняя кишка представлена мускулистой
глоткой, могущей выпячиваться наружу
Средняя кишка с боковыми мешковидными
выростами. Задняя кишка играет роль клоаки,
так как в нее впадают пара нефридий и яйце-
вод. Кровеносная и дыхательная системы
отсутствуют Гермафродиты. Половые клетки
формируются в стенке целома, скопления их
5
Рис. 221. Внешний вид мизостомиды:
1 — отверстие, через которое наружу выпячивается
глотка, 2 краевые циррусы, 3 — боковые органы,
4— параподии, 5 — анальное отверстие, 6 — мужское
половое отверстие
образуют яичник и семенники. Последние открываются наружу. Развитие с превра-
щением — образуется личинка, сходная с трохофорой.
Мизостомиды комменсалы и паразиты иглокожих, главным образом морских
лилий, а также морских звезд и офиур.
Организация мизостомид несет признаки упрощения под влиянием экто- и эндо-
паразитизма и их можно рассматривать, как полихет, измененных паразитическим
существованием.
КЛАСС 2. МАЛОЩЕТИНКОВЫЕ КОЛЬЧЕЦЫ,
ИЛИ ОЛИГОХЕТЫ (OLIGOCHAETA)
ХАРАКТЕРИСТИКА
Параподии отсутствуют, и щетинки сидят непосредственно в стенке
тела. Головной отдел выражен слабо. Щупалец и усиков нет. Целомическая
полость развита хорошо. Гермафродиты. Развитие без превращения. Пресно-
водные и почвенные формы. Известно примерно 2500 видов.
1 Название этого подкласса не совсем соответствует его содержанию в него
входят не только сидячие формы, обитающие в трубках, но и формы, обитающие в
грунте (роющие).
256
Представитель класса олигохет — дождевой, или
земляной, червь
(Lumbricus)
Под названием дождевые, или земляные, черви обычно понимают раз-
личные виды почвенных олигохет, относящихся к нескольким родам (Lumb-
ricus, Eisenia, Allolobophora и др.), которые все относятся к одному семей-
ству лумбрицид (Lumbricidae).
В нашем описании имеется в
виду преимущественно вид Lum-
bricus terrestris.
Внешнее строение. Особи
этого вида имеют в длину 15—
30 см. Тело типичной червеобраз-
ной формы и состоит из явст-
венных колец, число которых
достигает 140—180. Кольца пе-
реднего конца несколько уже,
чем остальные. Спинная сторона
тела округлая, брюшная нес-
колько уплощена. Примерно на
30-, 31- или 32-м члениках тела
имеется особое образование, так
называемый поясок (clitellum).
Это орган, выделяющий экскрет,
из которого строится кокон.
Червь имеет ржаво-бурую
окраску, более темную на спин-
ной стороне. Поясок окрашен
светлее. Кожа влажная и слизис-
тая В коже сидят щетинки. В
каждом кольце их 8: две па-
ры — субдорзальные, две па-
ры — субвентральные,
Кожи о-мускульный мешок.
Кожно-мускульный мешок име-
ет типичное строение. Под кути-
кулой находится эпителий (рис.
222), под ним двойной слой мы-
шечных волокон: кольцевые
мышцы, глубже — продольные.
Под продольной мускулатурой,
непосредственно примыкая к
ней, лежит тонкая пленка сома-
топлевры с явственными целоте-
лиальными клетками.
Нервная система. Централь-
ная нервная система вполне
типична. Она состоит из около-
глоточного нервного кольца и
брюшной нервной цепочки.
Нервное кольцо слагается из
Рис. 222. Строение стенки тела дождевого
червя (поперечный срез). / — кожно-мус-
кульный мешок; II — кутикула и эпителий:
1—кутикула, 2—эпителий, 3—кольцевые мыш-
цы, 4 — продольные мышцы, 5 — целомический
эпителий, 6 — белковая железа, 7 — слизистая же-
леза, 8 — базальная мембрана, 9 — эпителиальные
клетки
парного надглоточного узла,
кольцевых (глоточных) комиссур
и парного подглоточного нерв-
ного узла, который является
вместе с тем первым узлом брюш-
ной нервной цепочки (рис.
9 Курс зоологии» т. I
257
I 6
223щ). Сквозь окологлоточное кольцо
проходит глотка. Брюшная нервная це-
почка состоит из нервных узлов, по од-
ному узлу в каждом сегменте.
Органы чувств. Дождевые черви
очень резко реагируют на прикоснове-
ние, действие температуры, света и т д.
Кожа их богата нервными окончаниями,
чувствующими клетками.
Полость тела. Полость тела пред-
ставлена целомом (вторичной полостью
тела): она выстлана целотелием. Как и
у вышеописанных групп аннелид, по-
лость тела дождевых червей разделена
на камеры диссепиментами (рис. 223).
Косых мышц нет. Спинная мезентериаль-
ная перегородка отсутствует, и, следова-
тельно, полость тела не делится на пра-
вую и левую половины.
Следует, однако, отметить, что брю-
шная мезентериальная перегородка име-
ется. Она расположена между кишкой и
брюшным кровеносным сосудом (рис.
224).
Целом наполнен полостной жидкос-
тью, в которой взвешены различные
клеточные элементы, в том числе подле-
жащие удалению через метанефридии.
Полостная жидкость может выделяться
наружу через особые межсегментные
спинные поры.
Пищеварительная система. Пищева-
рительная система начинается ротовым
отверстием, которое лежит на переднем
конце тела. Оно несколько смещено на
брюшную сторону Первый сегмент тела
(prostomium) прикрывает ротовое отвер-
стие сверху Самое же отверстие откры-
вается во втором членике, называемом
и здесь перистомом.
Ротовое отверстие ведет в широкую
глотку, которая суживается и переходит
в довольно длинный пищевод. В пищевод
открываются своеобразные известковые
железы (морреновские железы), которые,
по-видимому, играют важную в физио-
логическом отношении роль. В них обра-
Рис. 223. Строение дождевого червя (Lumbri-
cus terrestris). / — головной конец (Л — сбо-
ку; Б — с брюшной стороны): // — анато-
мия; /// — передний отдел нервной системы:
/ — мужское, 2 — женское половое отверстие, 3 — по-
ясок (точками показано расположение щетинок), 4 —
глотка, 5 — пищевод, 5 —зоб, 7 —желудок, 8 — киш-
ка, 9 —- спинной кровеносный сосуд, 10 — «сердца»,
// — брюшной кровеносный сосуд, 12 — субневральный
кровеносный сосуд, 13— метанефридий, 14— воронка
метанефридия, 15—надглоточный нервный узел (го-
ловной мозг), 16 — ганглий брюшной нервной цепоч-
ки, 17 — окологлоточное кольцо, 18—боковые нервы,
19 — диссепименты, 20 — семеприемники
258
зуется известь, которая выводится (в 10-, 11 и 12-м сегментах) в просвет
пищевода. Дождевые черви, питаясь растительными остатками, заключен-
ными в проглатываемой ими почве, вводят в пищевод пищу, богатую гуму-
совыми кислотами. Выделения известковых желез нейтрализуют действие
этих кислот (впервые это мнение высказал Дарвин).
В 14-м сегменте пищевод расширяется в зоб. Из зоба захваченная пища
попадает в мышечный желудок (рис. 223ц), где пища, по-видимому, несколько
измельчается, чему способствуют захватываемые с ней песчинки.
Из «желудка» пища попада-
ет в кишку Строение кишки до-
вольно своеобразно. Диссепи-
менты образуют на ней явствен-
ные метамерные перетяжки.
Вдоль кишки тянется складка,
тифлозолис.
На поперечном разрезе ки-
шки видно (рис. 224), что
тифлозолис глубоко впячивается
вдоль всей кишки внутрь ее
полости. Этим достигается зна-
чительное увеличение всасыва-
тельной поверхности кишки, в
чем, конечно, и заключается
основное значение тифлозолиса.
Поверхность средней кишки
покрыта желтым налетом. Это
так называемые хлорагогенные
клетки (рис. 224). Кишка —
переваривающий и всасываю-
щий орган. В ней переваривают-
ся белки, углеводы и жиры. Пи-
Л 5 Ю 9
Рис. 224. Поперечный разрез через тело дож-
девого червя (Lumbricus terrestris):
1 — кутикула, 2 — эпителий, 3 — кольцевые мышцы,
4 — продольные мышцы, 5 — целомический эпителий,
6 — полость тела (целом). 7 — просвет кишечника, над
ним — канал тифлозолиса, кишечник покрыт хлораго-
генными клетками, 8— спинной кровеносный сосуд,
9 — брюшной кровеносный сосуд, 10— брюшная нерв-
ная цепочка, 11 субвентральные щетинки, 12— суб-
дорзальные щетинки, 13— мышцы, 14 — разрез метане-
фридня с выводным протоком, 15 — капилляры в
щеварение внеклеточное, проис- коже н кожно-мускульном мешке
ходит в просвете средней кишки,
куда выделяются пищеварительные соки. В эпителии средней кишки явст-
венно различимы экскреторные клетки, выделяющие в просвет кишки
пищеварительные ферменты, тогда как другие клетки выполняют всасы-
вательную функцию.
7 10 11
Рис. 225. Схема кровеносной системы олигохеты:
/ — спинной сосуд, 2 — капилляры головы, 3—невральные сосуды (для простоты показан один);
4 — париетальные сосуды, 5 — входящий сосуд мстансфридия, 6 — его выносящий сосуд, 7 — ла-
теральный сосуд, 8 — «сердце», 9 — брюшной сосуд, 10 — входящий кожный сосуд, 11 — выно-
сящий кожный сосуд. Схема упрощена — метанефридиальныс и кожные сосуды показаны лишь
частично, чтобы оставались ясными отношения латеральных сосудов и т. п. Стрелки показы-
вают направление кровяного тока, кишечная трубка показана пунктиром
Задней кишкой считается отдел, лишенный тифлозолиса. Сюда попа-
дают непереваренные частицы почвы, а равно остатки растительных тканей.
Дождевые черви пропускают сквозь кишечник землю, а следовательно,
9*
259
последней разветвляется
' 1
Рис. 226. Половая система дождевых
червей:
1 —брюшная нервная цепочка, 2'—семеприем-
ники, 3, 5, 7—семенные пузыри (мешки), 4,
6 — семенные капсулы (основание семенных пу-
зырей), 8 — семенники, 9 — воронка семенника,
от которой идет семяпровод (//), 10—воронка
семенника и соответствующая ей вторая пара
семенников (14) < 12—яичник, 13—яйцевод
и содержащиеся в ней растительные остатки. Но они способны затаскивать
в норки листья и молодые побеги различной травянистой растительности.
Кровеносная система. Кровеносная система состоит из замкнутых
кровеносных сосудов, а именно крупного спинного и брюшного сосудов и
лежащего под нервной системой неврального сосуда. По бокам неврального
сосуда (рис. 225) видны еще боковые околонервные сосуды.
Спинной сосуд связан с кольцевыми сосудами. На переднем конце (в
пределах 7—11 -го сегментов) эти последние образуют так называемые сердца.
Сердца имеют мускулистые стенки и пульсируют. Кровь течет в спинном
сосуде сзади наперед, в брюшном — в обратном направлении.
Кровь дождевого червя содержит растворенный в плазме гемоглобин
и имеет красный цвет.
Органы дыхания отсутствуют. Дождевой червь дышит через кожу,
личество кровеносных капилляров.
Кровь капилляров окисляется ки-
слородом, который диффундирует
сквозь влажные покровы тела.
Интересно, что упомянутые
выше морреновские (известковые)
железы играют, по-видимому, так-
же довольно важную роль в про-
цессах, связанных с дыханием.
Эти железы, по некоторым данным,
играют роль коллекторов углекис-
лой извести, накопляющейся в
результате жизнедеятельности
червя.
Органы выделения. Органы
выделения представлены метанеф-
ридиями, имеющими типичную
организацию. Нефростом лежит в
камере вторичной полости тела (це-
ломе), суживается, прободает дис-
сепимент, входит в состав тела
метанефридия (сегментарного орга-
на), уже в другой камере и закан-
чивается порой. Сегментарные
органы в каждой камере целома попарно повторяются: один — справа,
другой — слева, из сегмента в сегмент (рис. 223ц).
Выделительные процессы таковы. Жидкие отбросы жизнедеятельно-
сти, циркулирующие по кровяному руслу, в конце концов попадают в хло-
рагогенные клетки, которые покрывают поверхность средней кишки. Эти
собиратели экскретов, называемые также экскретофорами, затем отми-
рают, отпадают от поверхности средней кишки и погружаются в полостную
жидкость, откуда они и выводятся наружу через метанефридиальные
каналы.
Половая система. Половая система дождевых червей (рис. 226) обладает
рядом специфических особенностей. Их половой аппарат сосредоточен в не-
большом числе сегментов; у Lumbricus terrestris в пределах 9—15-го сег-
ментов тела. Дождевые черви — гермафродиты.
Мужской половой аппарат. Семенники представлены
маленькими и трудно обнаруживаемыми телами в 10-м и 11-м члениках тела.
Их четыре , по два в каждом из этих сегментов. Семенники прижаты к брюш-
ной стороне и связаны с эндотелиальной выстилкой стенки тела. От семен-
ников идут выводные протоки (vasa efferentia), которые впадают соответст-
венно в правый и левый семяпроводы (vas deferens). Последние открываются
в 15-м сегменте наружу, на брюшной стороне.
Семенники и воронки семявыносящих каналов охвачены семенными
260
Рис. 227.
Копуляция
у лумбрицид
капсулами, которые образуют большие мешкообразные выпячивания, из-
вестные под названием семенных пузырей, или семенных мешков (vesiculae
seminales). Эти мешки хранят скопляющееся, в них мужское семя.
Женский половой аппарат состоит из яичников и яйце-
водов. Яичников два; у Lumbricus terrestris они лежат в 13-м сегменте. Оба
яйцевода начинаются в этом же членике, пронзают диссепимент и откры-
ваются наружу половыми отверстиями в 14-м сегменте на брюшной стороне
тела. Яйцеводы начинаются воронкой. Они должны рассматриваться как
гомологи сегментарных (нефридиальных) органов.
К перечисленным органам принадлежит система семеприемников (ге-
ceptaculae seminales) в 9-м и 10-м сегментах. Семеприемники наполняются
во время совокупления сперматозоидами другого червя.
Оплодотворение и образование кокона. Два червя-партнера приклады-
ваются друг к другу своими брюшными сторонами и притом задними кон-
цами тела врозь (рис. 227). Оба червя выделяют капли семен-
ной жидкости. Эти капли попадают в область отверстий се-
меприемников партнеров. Отверстия открываются в 9-м и 10-м
члениках тела на брюшной стороне. Через эти отверстия капли
семенной жидкости всасываются в полость семеприемников
червей-партнеров. Другими словами, перекрестное совокуп-
ление заканчивается лишь обменом семенем, которое хранит-
ся в семеприемниках. Обменявшись семенем, черви-партнеры
расходятся.
Когда наступает процесс выделения яиц, наблюдается
следующее любопытное явление. Из эпителиальных желез
пояска (clitellum) происходит обильное выделение экскрета,
который образует вокруг пояска муфту. В дальнейшем червь
начинает производить волнообразные сократительные движе-
ния головного конца, постепенно сдвигая муфту к переднему
концу. В то же время из яйцеводов выделяются яйца, кото-
рые и откладываются в сдвигающуюся муфту, когда она про-
ходит мимо 16-го членика тела (половые женские отверстия),
шем , когда муфта с яйцами проходит мимо отверстий семеприемников
(в 9-м и 10-м сегментах), из них вытекает семенная жидкость и происходит
оплодотворение. Следовательно, только в этот момент имеет место оплодо-
творение яиц данного червя семенем его бывшего партнера.
Соскользнувшая с головного конца муфта еще влажна и вязка. Откры-
тые ее концы слипаются, закрываются, и муфта превращается в кокон,
внутри которого находится 1—3 яйца. Последний лежит в земле (где про-
исходят и описанные выше явления). В коконе яйца развиваются в молодых
червей. Личиночных стадий нет.
Организация дождевого червя обладает рядом особенностей, которые
надо рассматривать как приспособления к жизни в почве. Приспособления
эти следующие.
Обилие кожных желез и зависящая от них слизистая влажность кожи.
Эта особенность, во-первых, обеспечивает кожные дыхательные процессы,
во-вторых, облегчает скольжение в почве, в-третьих, защищает кожу от
возможных поранений об острые частицы почвы.
В условиях сухости почвы червь увлажняет поверхность тела полост-
ной жидкостью, выделяемой через спинные поры. Следовательно, спинные
поры обеспечивают защитную реакцию червя на недостаток влаги.
Сильная продольная мускулатура, в особенности в области простомиума
обеспечивает сильные роющие движения. Lumbricus terrestris, как и многие
другие его родичи, вбуравливается в землю при помощи мощно развитой
головной лопасти (prostomium) и затем отжимает землю в стороны сильным
головным концом.
Короткие, но крепкие щетинки (см. рис. 224) имеют значение при дви-
жении вперед и вместе с тем позволяют червю держаться в норке, что имеет
В дальней-
261
защитное значение (червя трудно вытащить из норки, так так щетинки
цепляются за почву).
Экология. Дождевой червь (Lumbricus terrestris) обитает в различных
почвах, но наиболее благоприятна для него почва огородных и садовых куль-
тур. В странах с влажным климатом дождевых червей больше. Однако в
почвах, пропитанных водой, а равно на болотах, в особенности торфяных,
Lumbricus terrestris не живет или во всяком случае эти почвы для него не
характерны. Норы его достигают 2—2,5 м глубины.
Почва, богатая растительными остатками, составляет главный источ-
ник питания червя. Пропуская через кишечник почву, червь выделяет ее
затем на поверхность земли в виде характерных кучек экскрементов. Кроме
того, по ночам черви наиболее активны, они затаскивают в свои норы опав-
шие листья, травинки, мелкие растения.
ОБЩИИ И СИСТЕМАТИЧЕСКИЙ ОБЗОР КЛАССА
Внешнее строение. Олигохеты представлены разнообразными формами,
отчасти довольно мелкими (в несколько миллиметров), отчасти крупными
формами. Так, один из бразильских представителей олигохет, Rhynodrilus
fafner, достигает 210 см.
Характерные для Chaetopoda щетинки развиты у олигохет не столь
типично, как у полихет. Они сидят в коже и часто явственно связаны с под-
ходящей к ним мускулатурой.
Членики тела — гомономны. Их число колеблется от 2—3 у мелких форм
и до 600 у крупных. Как у многощетинковых кольчецов, у олигохет
явственно различимы prostomium и peristomium. Задние сегменты у многих
форм уже и меньше, чем передние. К придаткам тела относятся щетинки, а
также сравнительно редко встречающиеся усики — головные, спинные,
брюшные или даже анальные. Щетинки довольно различны по форме, как
и у Poliychaeta, играют важную роль в систематике.
Интересно, что щетинки (в типичных случаях) образуют вентральные и
дорзальные пучки, что соответствует положению вентральной и дорзальной
ветвей параподий полихет.
Поясок, или clitellum, — образование весьма характерное для всех
олигохет
Кожно-мускульный мешок. Кожно-мускульный мешок состоит из
кутикулы, эпителия (гиподермы), мышц и тонкой перитонеальной выстилки
(соматоплевры).
Кутикула олигохет пронизана отверстиями, через которые открываются
протоки различных органов, выводящих жидкие экскременты. Таковы про-
токи слизистых одноклеточных желез кожи, а у почвенных форм спинные
поры, выводящие наружу полостную жидкость, наконец, отверстия половых
желез и органов, с ними связанных.
Гиподерма у низших олигохет состоит из одного, а у высших из одного-
двух слоев эпителиальных клеток.
Мышцы образуют два слоя: кольцевой и продольный. Кольцевая му-
скулатура состоит из волокон, погруженных в соединительно-тканевую
основу. Продольные мышцы развиты сильнее кольцевых. У многих форм
они состоят из мышечных волокон, которые ориентированы перпендику-
лярно к поверхности тела, напоминая мышечные элементы нематод. У дру-
гих — ив частности у дождевых червей — продольные мышцы образуют
волокна, разбитые соединительноткаными перегородками на отдельные
полоски (см. рис. 224). И в том и в другом случае продольные мышцы тя-
нутся продольными лентами вдоль тела. Обычно таких лент имеется 4: дор-
зальная, две латеральные, одна вентральная.
Нервная система. Центральная нервная система состоит из окологло-
точного кольца и брюшной нервной цепочки, от которой отходят нервные
тяжи, направляющиеся в различные части тела. Надглоточный узел около-
262
глоточного кольца содержит нервные (ганглиозные) клетки. У более при-
митивных форм надглоточный узел тесно связан с эктодермой, у остальных
вполне от нее обособлен. Окологлоточные комиссуры состоят из неврофи-
брилл. Первый нервный узел брюшной цепочки образует подглоточный
ганглий окологлоточного кольца. Кроме брюшной нервной цепочки, у оли-
гохет имеются два весьма характерных боковых нервных ствола, они лежат
по бокам тела в боковых лентах продольной мускулатуры и берут начало от
окологлоточного кольца.
Органы чувств. Органы чувств олигохет развиты слабее, чем у по-
лихет К ним принадлежат* ресничные ямки (органы химического чувства)
щупальца, представленные обычно непарным выростом головной лопасти
(prostomium), чувствующие щетинки, расположенные на головном конце
тела, статоцисты, встречающиеся в коже некоторых форм, наконец, глаза,
имеющие обычно форму пигментных бокалов. В коже олигохет разбросано
большое число чувствующих клеток. Все эти образования связаны с перифе-
рической нервной системой.
Полость тела. Целотелий выстилает всю полость тела (целом). Спин-
ная мезентериальная перегородка (брыжейка) отсутствует, брюшная наблю-
дается, но обычно недоразвита. Кишечная трубка и кровеносные сосуды
охвачены спланхноплеврой. Диссепименты хорошо развиты, однако у не-
которых форм частично отсутствуют. Целом наполнен полостной жидкостью.
У некоторых форм (почвенных) она, по-видимому, ядовита и при выпрыски-
вании играет защитную роль. В полостной жидкости содержатся клеточные
элементы, например, лимфоциты, а равно хлорагогенные клетки.
Пищеварительная система. Пищеварительная система начинается при-
крытым ротовой лопастью (prostomium) ротовым отверстием. Последнее
ведет в ротовую полость. У некоторых олигохет она вооружена.
Ротовая полость расширяется и переходит в мускулистую глотку. С
глоткой связаны лежащие над ней или позади нее железы Глотка переходит
в пищевод. Полость пищевода, по крайней мере в задней его части, выстлана
ресничным эпителием.
Задняя часть пищевода у примитивных олигохет расширяется в эзо-
фагеальный (от oesophagus — пищевод) желудок, достигающий иногда
огромных размеров. Кроме того, в качестве особых производных пищевода
нужно иметь в виду морреновские железы, или хилусные карманы. Это бо-
лее или менее глубокие железистые карманы, лежащие над пищеводом и
по бокам его.
Пищевод переходит в среднюю (энтодермическую) кишку Просвет
последней обычно выстлан мерцательным эпителием, а охватывающий кишку
перитонеальный эпителий (целотелий) превращен на протяжении средней
кишки в хлорагогенные клетки. У многих олигохет, как и у дождевых чер-
вей, передняя часть средней кишки вздута и образует интестинальный желу-
док (от intestinum — кишечник). Следовательно, энтодермический интести-
нальный желудок нужно отличать от эктодермического эзофагеального же-
лудка.
У многих олигохет кишка снабжена более или менее явственно выра-
женным и у разных форм различно устроенным тифлозолисом.
Стенка кишечника покрыта продольной и кольцевой мускулатурой, а
мышечный слой в свою очередь покрыт хлорагогенными клетками.
Кровеносная система. Кровеносная система у разных олигохет имеет
различное строение. У многих форм она состоит из следующих основных
частей. Над кишкой лежит длинный пульсирующий спинной кровеносный
сосуд. Этот сосуд имеет сложно устроенные протоки, соединяющие его с
кишечным синусом. Последний состоит из щелей между кишечным эпителием
и кишечной мускулатурой и наполнен кровью. Кровь из этого синуса на-
правляется в спинной кровеносный сосуд, течет вперед, в расположенные в
передней части его кольцевые сосуды и проникает в брюшной кровеносный
сосуд. Здесь кровь течет спереди назад и снова входит в кишечный синус.
263
Кровь, наполняющая синус, находится в тесном соприкосновении с эпи-
телием кишечника. Эпителиальный слой кишки содержит межклеточные или
внутриклеточные хилусовые канальцы, проводящие питательное содержи-
мое кишечника (хилус) в кровь.
Кровеносная система описанного типа более примитивна. У высших
олигохет (и у дождевых червей в том числе) кровообращение много сложнее.
Кровь течет сзади наперед по спинному сосуду (рис. 225); достигнув
головного конца, кровь течет по двум основным руслам: частью направляется
в капилляры головы, частью же в «сердца». Проследим первый путь. До-
стигнув капилляров головы, кровь вновь идет по двум путям: в латеральные
(боковые) сосуды и в невральные сосуды. Из латеральных и невральных
сосудов (орошающих большую часть тела) кровь направляется в большие
париетальные сосуды, которые впадают в спинной сосуд.
Проследим теперь второй путь, проходящий через «сердца». Из «сер-
дец» кровь проходит в брюшной сосуд, по которому течет отчасти вперед в
капилляры головы, а отчасти назад до заднего конца тела. В то же время
брюшной сосуд дает ряд ответвлений к внутренним органам, например ме-
танефридиям, и к стенкам тела. Омыв органы, обогатившись кислородом в ка-
пиллярах кожи, кровь попадает в выносящие сосуды, которые выводят ее
в уже упоминавшиеся париетальные сосуды (комиссуры). Последние впа-
дают в спинной сосуд. Следовательно, париетальные сосуды — это коллекто-
ры крови, идущей из латеральных, невральных, кожных и внутриполостных
кровеносных сосудов. Этот тип кровообращения свойствен и дождевым чер-
вям.
Дыхательная система. Дыхательной системы в виде специально обо-
собленных органов у олигохет, за малым исключением, нет. Однако, с дру-
гой стороны, кожа олигохет пронизана многочисленными капиллярами и
несомненно является многофункциональным органом, выполняющим роль
органа дыхания.
Выделительная система. Выделительная система представлена мета-
нефр идиями, имеющими в общем уже известное нам строение, несколько,
однако, усложненное. Наиболее интересны случаи, когда вместо более или
менее типичных метанефридиев развиваются меронефридии. В этом случае
в одном сегменте развивается (справа и слева) несколько метанефридиев.
Половая система. Половые органы олигохет представлены гонадами.
Сперматозоиды нитевидны, яйца примитивных олигохет богаты желтком. В
простейших случаях яйцо и сперма выпадают из гонад в полость тела и от-
сюда выводятся наружу через особые протоки. Однако , как это мы видели у
дождевого червя, половые органы могут достигать большой сложности.
Мужскими половыми протоками могут быть либо особо дифференцирован-
ные метанефридпи, либо обособленные от них половые протоки. Женские
протоки обычно являются видоизмененными метанефридиями. В связи с
этим, например, яйцевод у дождевого червя по своему положению вполне
соответствует метанефридию. Напротив, семяпровод дождевого червя явст-
венно обособлен от метанефридия и представлен каналом, который откры-
вается наружу порой не в соседнем членике (как это наблюдается в случае
настоящих метанефридиев), а, напротив, в 15-м членике, как это имеет место
у Lumbricus terrestris.
Развитие. Обычно имеет место копуляция (рис. 227). Дробление
яиц происходит по спиральному типу. Развитие эмбриона совершается в
коконе, тогда как свободноплавающая личинка, в частности трохофора,
отсутствует. Сформировавшиеся в коконе молодые червячки выходят из
кокона наружу, растут и достигают половозрел ости.
Бесполое размножение у олигохет довольно обычно и притом
встречается в различных формах.
В более простых случаях происходит архитомия, при которой тело
олигохеты распадается на несколько участков или даже отдельные сегменты,
из которых каждый может развиваться в нового червя. В случаях паратомии
264
на том месте, где должно произойти отделение заднего участка тела, разви-
вается регенеративная зона. В этой зоне образуются сегменты, часть которых
развивает задний участок материнской (передней) особи, тогда как другие
образуют передний (головной) участок дочерней (задней) особи (рис. 228).
Прежде чем образовавшаяся таким образом дочерняя особь отделится
от материнского тела, в регенеративной зоне начинается снова описанный
процесс. Так возникают целые цепочки этих особей (например, у Chaeto-
gaster).
Регенерация. Сказанное выше само по себе говорит о высокой регене-
ративной способности олигохет. Дождевой червь принадлежит к числу клас-
сических объектов по изучению регенерации. Несколько члеников, выре-
занных, например, из тела Lumbr-
icus rubellus, регенерируют в целого
червя. Наблюдения над другой
формой, а именно Allolobophora
longa показали, что уже из отрезка,
содержащего 9—13 члеников, вы-
растает целый червь. У представи-
телей рода Lumbriculus каждый
членик способен восстановить всего
червя, несмотря на то, что тело
последнего состоит из 100—150 чле-
ников.
Экология. Олигохеты в боль-
шинстве пресноводные формы. Мно-
гие из них не выносят даже не-
больших примесей соли (Lumbri-
culidae). Тем не менее среди них
имеются эвригалинные формы, спо-
собные переносить все переходы
от пресной к настоящей морской
(соленой) воде (Enchytraeus albi-
dus), тогда как другие являются
даже стеногалинными формами,
т е. способными жить только в
солоноводных бассейнах. Морские
олигохеты также известны. От прес-
новодных олигохет можно найти
Рис. 228. Размножение (паратомия) у Aelo
soma:
1 — граница между материнской и дочерней осо-
бями
переходы к амфибиальным формам,
живущим в почвах, обильно пропитанных водой, и, наконец, к формам,
приспособившимся к жизни в относительно сухих почвах (Lumbricidae).
Пределы приспособленности олигохет к температурным колебаниям
могут быть весьма значительными. Некоторые виды являются эвритермич-
ными, приспособленными к широким колебаниям температуры, тогда как
другие стенотермичны, т. е. приспособлены лишь к более узким колебаниям
температуры. Известны как тропические, так и полярные формы. Олигохеты
избегают прямого солнечного света. Многие из них безусловно оксифильны,
т е. нуждаются в обильных количествах кислорода, но, например, обычный
в наших водах Tubifex tubifex (рис. 229) очень не требователен и, по-види-
мому, способен даже к анаэробному дыханию.
Олигохеты питаются преимущественно растительной пищей, илом или
детритом. Многие формы питаются живыми растениями. Зоофаги редки.
К ним принадлежат, например, Chaetogaster (см. ниже) или байкальский
Agriodrilus vermivorus, поедающий червей (олигохет). Большинство олиго-
хет свободноживущие животные. Лишь немногие среди них ведут образ
жизни комменсал истов или паразитов. К комменсал истам следует отнести,
например, обычного пресноводного Chaetogaster limnaei. Он поселяется в
дыхательной полости некоторых брюхоногих моллюсков (например, Physa
265
fontinalis, Planorbis corneus и др.), используя этих животных лишь как свое-
образное местообитание, тогда как пищей этой интересной олигохеты яв-
ляются различные мелкие организмы, в частности церкарии трематод.
Олигохеты составляют очень важную составную часть фауны дна пресно-
водных водоемов. Формы, питающиеся за счет ила данного водоема (Tubi-
fex) (рис. 229), успевают в течение известного времени пропустить весь
ил водоема сквозь кишечник. В некоторых озерах они выносят на поверх-
Рис. 229. Tubifex
обитатель дна пресно-
водных водоемов
ность дна озера до 12,5 кг ила (сухой вес) на 1 м2
Там, где Tubifex встречается в больших массах, он
оказывает, следовательно, огромное влияние на дно.
Наряду с этим, олигохеты имеют очень боль-
шое значение как источник питания многих рыб.
Среди пресноводных олигохет нашей фауны
укажем на следующих. Очень обычны представите-
ли семейства Naididae, в частности представители
родов Nais (N blanci, N pseudoobtusa); Stylaria
(S. lacustris) (рис. 230), а также упоминавшийся
выше Chaetogaster limnaei. В пределах этого семей-
ства распространено размножение по типу парато-
мии. Интересно семейство Enchytreidae, частью
представленное пресноводными, частью почвенны-
Рис. 230. Stylaria lacustris—обитатель толщи
воды пресноводных водоемов
ми, реже морскими формами. Одним из самых обычных представителей
семейства Tubificidae является Tubifex (Т tubifex) и другие виды (рис.
229). Этот червь зарывается в ил головным концом тела, тогда как задний
конец его тела торчит наружу и производит ритмические колебания. Tubifex
хорошо уживается даже в плохо содержимых аквариумах.
Некоторые олигохеты обитают в морях (часть представителей семейства
Phreodrilidae). В почве, кроме упомянутых выше Enchytreidae, живут
представители семейства Lumbricidae, к которым относятся дождевые черви.
Значение дождевых червей в жизни почвы. Дождевые черви, роясь в
почве, оказывают значительное влияние на ее свойства, в известной мере на
растительность и на почвенные процессы.
266
Выделяя на поверхность свои экскременты, дождевые черви тем самым
перемешивают землю, вынося на поверхность глубжележащие слои. Это
перемешивание почвы способствует ее разрыхлению, что наряду с наличием
нор дождевых червей дает растениям возможность лучше и глубже укоре-
няться.
Огромная масса дождевых червей, живущая в почве, влияет на почвен-
ные процессы. Погребая в почве растительные остатки, дождевые черви
способствуют накоплению в ней органических веществ, образующих гумус.
Кроме роли собирателей гумуса, дождевые черви имеют известное зна-
чение в развитии тех процессов, которые в гумусе происходят Гумус под-
вергается разложению и окислению под влиянием деятельности почвенных
микробов, а минеральные вещества, возникающие этим путем, потребля-
ются корнями растений. Для развития упоминаемых здесь процессов нужны
два фактора — воздух и влажность. Дождевые черви, роясь в почве, ари-
зируют ее, а вместе с тем облегчают и доступ влаги в почву.
В конечном счете, дождевые черви ощутительно влияют на развитие
процессов нитрификации почвы (наряду с другими почвенными организ-
мами).
Роль дождевых червей в почве хорошо демонстрируется следующими
данными. Было показано, что один червь (из числа крупных видов, напри-
мер, Lumbricus terrestris) съедает в течение месяца свыше 0,61 г сухого орга-
нического вещества. Нетрудно рассчитать, что если на 1 м2 площади обитает
в среднем 30 червей, то за 6 месяцев они пропустят сквозь кишечник около
1000 кг земли на 1 га. А между тем число червей на 1 м2 площади может
превышать при благоприятных условиях 100 штук.
Некоторые данные подтверждают исключительно большое значение
дождевых червей в повышении плодородия почвы. Так, экспериментальные
исследования некоторых специалистов показали, что жизнедеятельность
дождевых червей улучшает структуру почвы, повышая ее прочность против
эрозии. Далее было показано, что в экскрементах дождевых червей содер-
жатся значительные количества важных для растений элементов. Так,
оказалось, что в почве, содержащей дождевых червей, было обнаружено
азота в пять раз, фосфора в семь раз, поташа в одиннадцать раз больше,
чем в почве, не содержащей дождевых червей. Экспериментальные наблю-
дения выяснили, что присутствие дождевых червей значительно повышает
урожай ряда культур, в том числе картофеля, ржи, гороха и др. Заслужива-
ют также внимания данные, согласно которым дождевые черви, пропуская
почву через кишечник, уничтожают некоторых вредоносных для растений
бактерий и снижают численность проволочных червей. Эти новые данные
были сообщены Пауэллом (1953). Неудивительно, что, по некоторым рас-
четам, в долине Нила, столь славящейся плодородием, дождевые черви
приносят своей жизнедеятельностью до 115 т удобрений на акр земли.
Были также получены следующие данные о влиянии дождевых червей
на урожай пшеницы (табл. 3).
Таблица 3
Влияние дождевых червей на урожай пшеницы
Количество червей в сосуде Количество червей в пересчете на 1 га Общий вес рас- тений в сосуде Вес собранных зерен Количество соб- ранных зерен
Без червей 0 110,6 27,6 873
4 880 000 116,5 31,1 882
8 1 790 000 175,5 50,7 1313
16 3 540 000 I 183,0 55,6 1422
Наряду с полезным значением дождевых червей, следует указать на
то, что они могут быть источником заражения домашних животных неко-
267
торыми паразитическими червями. Дождевые черви могут иметь значение
промежуточных хозяев метастронгилид (семейство нематод) свиней и неко-
торых паразитических нематод домашней птицы (куры, утки, индюки).
Тем не менее, полезное значение дождевых червей в жизни почвы и в сущест-
вовании растений неизмеримо выше вредного воздействия их в качестве
промежуточных хозяев паразитических червей.
Среди почвенных форм олигохет известны очень крупные и даже ги-
гантские формы семейства Glossoscolecidae, куда относятся огромный Rhi-
nodrillus, достигающий 210 см в длину, и не менее крупный австралийский
Megaloscolex australis, делающий в земле большие норы, напоминающие
миниатюрный вулкан с кратером.
ФИЛОГЕНИЯ ОЛИГОХЕТ
Олигохеты должны рассматриваться как формы, отделившиеся от по-
лихет и приспособившиеся к жизни в пресной воде и в почве.
КЛАСС 3. ПИЯВКИ (HIRUDINEA)
ХАРАКТЕРИСТИКА
Параподии и в большинстве случаев щетинки отсутствуют. Наруж-
ная кольчатость не соответствует внутренней сегментации. Полость тела
редуцирована и превращена в систему лакун. У многих форм эти лакуны
функционируют как кровеносные сосуды, а кровеносная система редуциро-
вана. Характерный признак — наличие присосок — одной или двух. Одна
присоска помещается обычно на переднем конце и связана с ротовым отвер-
стием, вторая — на заднем. Гермафродиты. Развитие без превращений.
Ведут обычно полупаразитический образ жизни. Обитают главным образом
в различных пресноводных водоемах, некоторые перешли к наземному образу
жизни, немногочисленные формы обитают в морях. Известно около 250
видов.
ОБЩИИ ОБЗОР КЛАССА
Внешнее строение. Форма тела пиявок довольно типична. Обычно они
более или менее сплющены в дорзо-вентральном направлении. Спинная
сторона тела выпуклая, брюшная—более плоская. Впрочем, известны
формы с цилиндрическим телом.
Размеры тела сильно колеблются от мелких форм с длиной в пределах
сантиметра и кончая относительно крупными формами, достигающими 20 см.
Тело пиявок покрыто тонкой кутикулой, которая имеет правильную
кольчатость. Однако эта кольчатость кутикулы не соответствует сегментам
тела. На каждый сегмент у разных пиявок приходится различное число
колец. Так, у Acanthobdellae на каждый типичный сегмент обычно прихо-
дится 4 кольца, у хоботных пиявок —2—14 колец, у челюстных — 5 колец
и т. д.
Границы сегментов можно определить по некоторым внешним призна-
кам, например по окраске, а равно по анатомическим признакам, например
по правильной повторяемости явственных узлов нервной цепочки, карма-
нов зоба, метанефридиев, половых органов.
Кожно-мускульный мешок. Кожно-мускульный мешок состоит из
эпителия и мускулатуры. Эпителий очень богат железами, причем обычно
у основания эпителиальных клеток скопляются пигментные клетки, обус-
ловливающие окраску тела пиявок. Мускулатура трехслойная: кольчатая,
диагональная и продольная. Кроме того, имеется дорзо-вентральная му-
скулатура. Мускулатура отличается очень большой силой и огромным раз-
витием.
268
Полость тела. Полость тела редуцировалась до степени лакун (сину-
сов) и щелей между органами. Редукция вторичной полости тела происхо-
дит благодаря сильному развитию паренхимы и мускулатуры.
Нервная система. Нервная система носит метамерный характер и сходна
с нервной системой прочих аннелид. Она состоит из окологлоточного
кольца и брюшной нервной цепочки. Последняя обычно состоит из гангли-
озных узлов и связывающих их
комиссур (коннективы). От узлов
отходят боковые нервы, которые
затем разветвляются в стороны
и входят в состав очень сложной
симпатической и периферической
нервной системы, на которой мы
останавливаться не будем.
Органы чувств. К органам
чувств относятся прежде всего
рассеянные в коже чувствующие
клетки. В коже пиявок имеются
также свободные нервные окон-
чания, а равно особые осязатель-
ные бокалы, которые состоят из
удлиненных эпителиальных кле-
ток. К комплексу этих клеток
подходит нерв. У всех пиявок
имеются глаза. У медицинской
пиявки глаза представлены пиг-
ментными бокалами. По оси
бокала проходит зрительный
нерв. Самый бокал наполнен
зрительными клетками. К каж-
дой зрительной клетке подходят
ответвления зрительного нерва.
Зрительные клетки рассеяны по
всей поверхности кожи. Число
глаз у различных пиявок раз-
лично.
Пищеварительная система.
Пищеварительная система до-
вольно своеобразна. Ротовое
отверстие находится на дне пе-
редней присоски и ведет в рото-
вую полость. Последняя у части
форм вооружена челюстями, у
других—лежащим в особой по-
лости хоботом, который может
всасываться наружу либо через
рот, либо через особое отверстие.
Ротовая полость переходит в
глотку (рис. 231). Последняя
Рис. 231. Анатомия медицинской пиявки
(Hirudo medicinalis). / — строение кишеч-
ника; // — при снятом кишечнике:
1— край ротовой присоски, 2 — верхняя челюсть,
3— глотка, 4 — ее мышцы, 5 — первая и последняя
пара слепых карманов зоба, 6— тонкая кишка;
7 — прямая кишка, 8—задняя присоска, 9 — мета-
нефридий, 10—11 — эпителий (кожа), 12—кольце-
вые мышцы, 13 — продольные мышцы, 14— парен-
хима, 15 — «мозг», 16 — брюшная нервная цепочка,
17 (внизу) — семяпровод, 18— проток метанефри-
дия, 19 — мочевой пузырь метанефридия, 20—се-
менной мешок, 21 — совокупительный орган, 22 —
яичник, 23 — первая пара семенников, 24 — боковой
кровеносный сосуд
охвачена пакетом одноклеточных желез. У медицинской пиявки протоки
этих желез открываются у краев ротового отверстия (так называемые
губные железы), частью по особым протокам непосредственно в челюсти.
При этом выделяется своеобразный секрет, так называемый гирудин, пре-
пятствующий свертыванию крови, высосанной пиявкой.
За глоткой следует объемистый зоб, снабженный боковыми карманами.
Спереди назад у медицинской пиявки эти слепые карманы становятся посте-
пенно длиннее. Задняя пара карманов удлинена и протянута до заднего
конца тела, располагаясь по бокам кишечной трубки.
269
Наблюдения над медицинской пиявкой показали, что кровь, попавшая
в зоб, изменяется очень медленно. Так, кровь лягушки оставалась без вся
ких гистологических изменений в течение первых четырех дней. Через 14
дней можно было различить довольно много измененных эритроцитов.
Лимфоциты оставались явственными. Заглоченные при сосании ранки мы-
шечные волокна оставались без изменений даже спустя месяц после сосания
крови.
Оказалось, что зоб пищеварительной функцией не обладает и служит
лишь резервуаром захваченной пищи, например насосанной крови (у меди-
цинской пиявки). Огромные размеры зоба, составляющего большую часть
кишечного тракта, имеют адаптивное значение, так как обеспечивают воз-
можность заглатывания больших запасов пищи. Мощное развитие зоба—
характерный признак пиявок.
Зоб переходит в короткий желудок, в котором происходят перева-
ривание и всасывание пищи. Внутренняя стенка желудка покрыта
спиральными складками, увеличивающими всасывательную поверхность.
Пищеварение происходит в просвете желудка. Последняя переходит в тон-
кую кишку, равным образом способную как к переваривающей, так и вса-
сывающей функциям и имеющую внутренние складки. Кишка заканчивается
расширенной прямой кишкой, которая открывается наружу заднепроход-
ным отверстием.
Кровеносная система. Кровеносная система у различных пиявок по-
строена различно. У Acanthobdellae она сходна с таковой у олигохет, замк-
нута и состоит из спинного и брюшного сосудов, лежащих в полости тела.
У хоботных пиявок тоже имеются спинной и брюшной сосуды, которые
лежат в спинном и брюшном синусах — остатках редуцированного целома.
Кроме того, у них имеются боковые синусы. Синусы не сообщаются с кро-
веносными сосудами.
У челюстных пиявок кровеносные сосуды исчезают, и их функции це-
ликом выполняют боковые кровеносные синусы (лакуны), имеющие мышечные
стенки и превращающиеся в боковые кровеносные сосуды. Более мелкие
лакуны, наполненные кровью, не имеют мышечных стенок. Над кишечником
и под кишечником у этих пиявок имеются не сосуды, а синусы. У медицин-
Рис. 232. Поперечный разрез через тело медицинской пиявки:
1— эпителий (кожа), 2 — кольцевая мускулатура, 3—мощная продольная мускулатура, 4 —
дорзо-вентральные мышцы, 5 — небольшие лакуны, несущие кровь, 6—разрез кишечника,
7 — слепой карман кишечника, 8— боковой кровеносный сосуд, 9—часть метанефридия, 101
11 — проток семяпровода, 12 — семенник, 13—семяпровод, 14 — пузырь метанефридия, 15 —
спинной кровеносный синус, 16— брюшной кровеносный синус, 17 — брюшная нервная цепоч-
ка, 18— проток метанефридия, 19— ботриондная ткань
270
ской пиявки те же отношения: по бокам тела происшедшие за счет синусов
пульсирующие кровеносные сосуды (рис. 232), а над и под кишечником —
синусы с значительно более тонкими стенками. Брюшной синус охватывает
при этом нервную цепочку как чехол.
Кровь у пиявок либо бесцветная, либо окрашена гемоглобином в крас-
ный цвет.
Дыхательная система. Дыхательная система отсутствует. Газообмен
происходит посредством кожи, богатой капиллярами кровеносных сосудов.
Выделительная система. Выделительная система представлена мета-
нефридиями. Последние обнаруживают метамерное расположение (рис. 231,
232). У медицинской пиявки их 17 пар. Тело метанефридия представлено
сложной системой извитых внутриклеточных канальцев. Они берут свое
начало от воронки.
Воронка каждого нефридия открывается в соответствующее боковое
ответвление брюшного синуса. Это подтверждает, что, синусы пиявок —
остатки целома, так как нефростом всегда открывается в целом. Противо-
положный конец нефридия открывается наружу посредством нефропоры,
лежащей справа и слева на брюшной стороне.
Интересно, что у пиявок воронка не соединяется с каналом метанефри-
дия. Жидкие продукты, захваченные воронкой, по-видимому, проникают
из воронки в канал метанефридия осмотическим путем. Это разобщение
воронки и метанефридиального канала — явление вторичное.
Половая система. Половая система гермафродитная (см. рис. 231).
Мужские половые органы состоят из метамерно распо-
ложенных семенников, коротких семявыносящих канальцев (vasa efferen-
tia), двух длинных продольных семяпроводов (vasa deferentia), впадающих
в правый и левый семенные пузыри (vesiculae seminales), которые выносят
семя в семяизвергательные каналы (ducti jeculatorii), вливающиеся в
канал совокупительного органа.
У многих пиявок совокупительного органа нет, и пучки сперматозоидов,
скрепленные железистыми выделениями выводных протоков, выделяются
наружу в сперматофоре. Это удлиненный мешочек, который приклеивается
к брюшку пиявки-партнера вблизи женского полового отверстия. Из
открывшегося сперматофора семя проникает в женское половое отверстие.
У других форм сперматофор целиком вводится во влагалище или прикрепля-
ется пиявкой к коже ее партнера.
Женские половые органы медицинской пиявки состоят
из одной пары яичников, коротких яйцеводов и влагалища, лежащего на
брюшной стороне позади мужского совокупительного органа. Яичники
имеют форму компактных шарообразных телец либо удлинены.
Оплодотворенные яйца откладываются в кокон. Последний, так же как
у олигохет, образуется за счет выделений пояска (clitellum), который ко
времени кладки яиц набухает вследствие сильного развития в нем кожных
желез. Число яиц, откладываемых в кокон, различно у разных пиявок, оно
колеблется от одного до двухсот.
Коконы откладываются на различные подводные предметы, в том
числе и на растения. Некоторые пиявки (Helobdella stagnalis, Protoclepsis
tesselata) носят коконы на брюхе. Коконы весьма различны по разме-
рам. Кокон медицинской пиявки достигает 2 см в длину и содержит
до 20 яиц.
Развитие. Пиявкам свойственно спиральное детерминированное дроб-
ление. У них нет свободноживущей личинки. Тем не менее зародыши, на-
ходящиеся в коконе, безусловно являются личинками, обладающими ли-
чиночными органами, в частности ресничным аппаратом, протонефридиями
ит. д. Личинка плавает в жидкости кокона при помощи ресничного аппарата.
Позднее происходит метаморфоз. Личиночные органы разрушаются, про-
исходит процесс перестройки тканей личинки, и она превращается в моло-
дую пиявку, освобождающуюся из кокона. Интересно, что у эмбрионов
271
пиявок закладывается настоящий сегментированный целом в виде метамер-
ных целомических мешков.
Экология и распространение. Подавляющее большинство пиявок —
пресноводные организмы. Морские известны только из семейства Ichtyob-
dellidae. Рыбья пиявка, по-видимому, способна проникать и жить в солоно-
ватой воде лиманов. Пресноводные пиявки эвритермны, в большинстве спо-
собны к земноводному образу жизни, выходя иногда на сушу (например,
для кладки яиц). Однако долгое пребывание на солнце для этих животных
обычно губительно. Тем не менее известны наземные тропические пиявки,
живущие во влажных местах. Все пиявки — хищники, переходящие,
однако, в сторону паразитизма. Некоторые пиявки превращаются
даже в эндопаразитов, забираясь, например, в глотку и дыхательное горло
теплокровных животных. Такова, например, пиявка Limnatis nilotica,
Рис. 233. Общий вид пиявок. / — медицинская пиявка
(1а — вскрытый с брюшной стороны передний конец тела
медицинской пиявки, видны три челюсти; 16 — челюсть
медицинской пиявки, сильно увеличено); // конская пи-
явка (Haemopis sanguisuda); III —Glossiphonia compianata;
IV — Herpobdella octoculata
которая забирается в гортань лошади (Африка). Близкая к ней, встречаю-
щаяся в Туркмении и Узбекистане, Limnatis turkestanica забирается в
носоглотку человека и сосет кровь, вызывая кровотечения. Иногда она
проникает даже в трахею. Попавшую туда пиявку удаляют хирургическим
путем во избежание удушения. Заражение пиявкой происходит при питье
сырой воды из водоемов, где они живут.
Медицинское значение пиявок. Применение пиявок в медицине для
кровопусканий известно с глубокой древности. Для этой цели используются
различные виды челюстных пиявок, но наибольшее значение имеет так
называемая медицинская пиявка Hirudo medicinalis (рис. 233). Эта крупная
пиявка широко распространена в пресных водах и в естественных условиях
питается кровью различных позвоночных животных. Такой характер пита-
ния выработал у этой пиявки ряд приспособительных черт как морфологи-
ческих, так и физиологических, уже отмеченных при описании пиявок.
Сюда прежде всего относятся боковые карманы зоба (см. рис. 231), дающие
возможность медицинской пиявке сразу всосать большое количество крови
и затем долгое время оставаться без пищи. Опыты показали, что медицин-
ская пиявка может не питаться несколько месяцев. Насосавшаяся пиявка
272
в несколько раз превосходит по объему «голодную». Приспособлением физи-
ологического характера является наличие в слюнных железах пиявки
особого вещества — гирудина, благодаря которому кровь, находящаяся в
зобе пиявок, очень долгое время не свертывается, остается свежей и может
постепенно усваиваться пиявкой.
Особенно много пиявок потреблялось медициной в прошлом веке. Так,
например, считается, что в сороковых годах во Франции использовали в
год около 25000000 пиявок! В то время медицина употребляла их почти от
всех болезней! В дальнейшем использование пиявок сильно упало. В настоя-
щее время они опять очень прочно вошли в арсенал медицины, но уже
используются только при определенных заболеваниях. Употребляются
пиявки главным образом при повышенном кровяном давлении (гипертонии),
когда некоторое уменьшение крови в организме способствует снижению
давления. При «укусе» пиявка пропиливает очень характерную треуголь-
ную ранку, что объясняется наличием у неё трех челюстей (рис. 233).
Вводя в ранку гирудин, пиявка лишает кровь способности свертываться,
поэтому ранка, оставляемая пиявкой, долгое время кровоточит.
Усиленный спрос на пиявок со стороны медицинских учреждений,
естественно, не может быть покрыт только выловом пиявок из естественных
водоемов. В связи с этим в настоящее время имеются специальные питом-
ники, где пиявок разводят.
СИСТЕМАТИЧЕСКИЙ ОБЗОР КЛАССА
ОТРЯД ЩЕТИНКОНОСНЫЕ ПИЯВКИ (ACANTHOBDELLAE)
Передней присоски нет, тело состоит из 30 сегментов, из которых 5
сегментов (от 2 до 6) несут по 2 пары щетинок с каждой стороны. Целом
имеется и разделен на метамерные отрезки.
Всего лишь одно семейство Acanthobdellidae с одним представителем—
Acanthobdella peledina (рис. 234). Река Енисей, Онежское озеро и другие
водоемы Кольского полуострова. Паразит лососевых.
Рис. 234. Акантобделла (Acanthobdella peledina).
/ — общий вид; //— головной конец
ОТРЯД ХОБОТНЫЕ ПИЯВКИ (ЯНУNCHOBDELLAE)
Вооружены хоботом. Зоб и кишка с боковыми метамерными карманами.
Оплодотворение при помощи сперматофоров, которые либо прикрепляются
к любому месту тела, либо вводятся в женское половое отверстие.
Представители:
Piscicola geometra — рыбья пиявка, широко распространена в пресных
водах, паразит рыб (щука, окунь, налим и др.). Пиявка иногда покидает
хозяина и сидит на растениях. Коконы— на растениях или различных под-
водных предметах.
Hemiclepsis marginata реках и озерах. Питается кровью уток, пога-
нок и других водных птиц. Коконы прикрепляются к брюшной стороне тела.
Hemiclepsis marginata нападает на рыб, тритонов, головастиков, лягу-
шек. Обычно достигает 25—30 мм в длину. Эта пиявка, как впрочем и другие
273
пиявки этого отряда, проявляет заботу о потомстве. Кучки яиц, прикреп-
ленные к подводным предметам, пиявка прикрывает собственным телом и,
совершая ритмичные волнообразные движения, обеспечивает приток свежей
воды.
Glossiphonia complanata/— обитает в стоячих и текучих водах.
Питается кровью моллюсков и червей. Обычна на растениях или под кам-
нями (рис. 233).
ОТРЯД ЧЕЛЮСТНЫЕ ПИЯВКИ (GNATHOBDELLAE)
Типичные формы имеют челюсти. У европейских форм их три. Челюсти
действуют как пилы. Семя вводится посредством сперматофора во влагалище.
У медицинской пиявки — посредством совокупительного органа.
Центральное семейство — Hirudinidae. Эти пиявки хорошо плавают
при помощи волнообразных движений. Кладут яйца в землю. Личинки вы-
ходят наружу вполне сформированными. Примеры: Haemopis sanguisuga —
конская пиявка (рис. 233). Весьма обычная форма. В прудах, канавах,
стоячих водах, реже в текучих водах. Пиявка нападает на беспозвоночных
животных, глотая червей и личинок несекомых. Сосать кровь она не может
Hirudo medicinalis — медицинская пиявка (рис. 233). Обитает в
торфяных болотах, заводях, речных плавнях, тростниковых озерах и т д.
Пиявка эта на севере не живет, но очень обычна в южных реках (например,
в Днепре), а также на Кавказе, по Сыр-Дарье, известна на Алтае.
ОТРЯД ГЛОТОЧНЫЕ ПИЯВКИ (PHARYNGOBDELLAE)
Челюсти отсутствуют. Глотка — огромная, трехгранная в поперечном
сечении. Сюда относятся виды рода Herpobdella (очень обычны Н. nigrico-
lis и Н. octooculata) (рис. 233) и Trochaeta (Т. subviridis).
ФИЛОГЕНИЯ пиявок
Пиявки произошли от олигохет. Связующим звеном является отряд
Acanthobdellae. У единственного представителя этого отряда A. peled i па
еще сохранился целом, а щетинки на головном конце гомологичны щетинкам
олигохет, кроме того, у Acanthobdellae имеется замкнутая кровеносная
система. Некоторые авторы рассматривают даже эту группу как изменен-
ных паразитизмом олигохет. Acanthobdellae отличаются сборной органи-
зацией, что показывает родственные связи олигохет и пиявок. В противо-
положность этому, олигохеты из сем. Branchiobdellidae во многом напоми-
нают пиявок.
КЛАСС 4. ЭХИУРИДОВЫЕ (ECHIUROIDEA)
Небольшая группа кольчатых червей, сильно измененная в связи с роющим
образом жизни. Тело мешкообразное. Метамерия во взрослом состоянии* полностью
отсутствует Параподий нет, сохраняется только несколько пучков щетинок. Обычно
имеется более или менее развитой невтяжной хобот (головная лопасть). Полость тела
развита хорошо. Кишечник сквозной. Кровеносная система сильно редуцирована.
Выделительные органы у большинства представлены особыми связанными с кишеч-
ником анальными мешками с многочисленными мерцательными вороночками, откры-
вающимися в полость тела. 1—4 пары нефридиев обычно функционируют как вывод-
ные половые протоки. Раздельнополые, иногда с резко выраженным половым ди-
морфизмом. Личинка типа трохофоры.
274
Исключительно морские формы, ве-
дущие роющий образ жизни. Особый ин-
терес в этом классе представляет Bonnelia
viridis (Средиземное море и Атлантический
океан), у которой резко выражен половой
диморфизм. Самки — крупные черви с
длинным хоботом, достигающие в длину
(с хоботом) до 100 см (рис. 235). Самцы —
очень мелкие, длиной в 1—3 мм, покрыты
ресничками, паразитируют в половых не-
фридия х самок. Следует отметить, что
впервые по-настоящему расшифровал этот
своеобразный половой диморфизм А. О. Ко-
валевский (1870); до его исследования
самцы боннелии считались просто парази-
тическими инфузориями.
Рис. 235. Bonnelia viridis. / самка (силь-
но уменьшено); // — самец (сильно уве-
личено)
ФИЛОГЕНИЯ КОЛЬЧАТЫХ ЧЕРВЕЙ
В отношении филогении кольчатых червей (в основном — полихет)
было выдвинуто несколько теорий. Одна из них турбеллярная теория
происхождения аннелид (А. Ланг и Э. Мейер). По этой теории, прежде всего
интересен тот факт, что у некоторых турбеллярий намечается правильная
повторяемость некоторых внутренних органов боковых ветвей кишечни-
ка, гонад, нервов и т д. (ложная метамерия). Это явление в известной мере
бросает свет на происхождение настоящей метамерии кольчецов. Можно
думать, что последняя является дальнейшим развитием такой повторяемости
комплексов органов. Эта повторяемость сперва касалась лишь немногих
систем органов, но в дальнейшем распространилась и на остальные, а
затем в связи с усилением движения охватила и кожно-мускульный мешок.
Кроме того, основания для турбеллярной теории следующие: 1)у некоторых
турбеллярий поперечные нервные тяжи отходят от продольных стволов
через более или менее равные промежутки, вследствие чего возникает
известное сходство с брюшной нервной цепочкой аннелид; 2) метанефридии
аннелид легко выводятся из протонефридиев турбеллярий, в частности,
как уже указывалось, многим аннелидам свойственны видоизмененные
протонефридии, снабженные соленоцитами, а у трохофоры тоже имеются
соленоциты, которые затем заменяются метанефридиями; 3) известное
сходство между мюллеровской личинкой турбеллярий и трохофорой анне-
лид, а равно тот факт, что турбелляриям свойственно спиральное дроб-
ление яиц, столь характерное и для аннелид.
К турбеллярной теории довольно близко примыкают представления о
происхождении аннелид от немертин (Н. А. Ливанов), обладающих некото-
рой метамерностью внутренних органов и кровеносной системой.
Менее приемлема трохофорная теория происхождения аннелид (Б. Гат-
чек). Согласно этой теории, трохофора аннелид прообраз гипотетического
предка трохозоона, который возник от форм, близких гребневикам. Следо-
вательно, тот факт, что аннелиды проходят в своем развитии стадию трохо-
фор,с точки зрения этой теории, говорит в пользу предположения о про-
исхождении аннелид от трохозоона. В соответствии с этими взглядами прини-
мается, что формы в роде Dinophilis (см. рис. 218) являются ближайшими
275
потомками трохозоона и имеют значение промежуточных форм, от которых
следует считать происхождение остальных аннелид. Такое положение весьма
спорно, более правильно предположить, что динофилиды — вторично упро-
щенные кольчецы, а не исходная форма этой группы.
Наконец, крупнейший советский специалист по сравнительной анато-
мии и зоологии В. Н. Беклемишев склоняется к мысли о происхождении
вторичнополостных червей непосредственно от форм, более близких к ки-
шечнополостным.
В настоящее время еще трудно вынести окончательное суждение в
отношении справедливости той или иной теории. Среди рассмотренных
нами классов аннелид наименее вторично изменены полихеты, благодаря
чему они и должны считаться исходной группой в этом типе.
Заканчивая обзор паренхиматозных червей, первичнополостных чер-
вей, немертин и кольчатых червей целесообразно рассмотреть сравнитель-
ную таблицу 4, в которой сведены важнейшие черты организации названных
типов:
Таблица 4
Сравнительная таблица
Признаки или системы органов Плоские черви Круглые черви Немертины Кольчатые черви
Полость тела Her Первичная Нет Вторичная
Состав кожно- Эпителий, Гиподерма и про- Эпителий, Эпителий, коль-
мускульного мешка кольцевые, косые и про- дольные мышцы дольные мышцы кольцевые, косые и про- дольные мышцы цевые и про- дольные мышцы
Центральная Несколько про- Несколько про- Продольные Брюшная нерв-
нервная сис- тема дольных стволов, крупнейшие боковые дольных ство- лов, крупней- ший брюшной боковые стволы ная цепочка
Окологлоточное Имеет ганг- Не имеет гангли- Имеет ганг- Имеет ганглии,
нервное коль- лии, охваты- ев, охватывает лии, охва- охватывает
цо вает глотку пищевод тывает кар- ман хобота глотку
Пищеваритель- Передняя и Передняя, сред- Передняя, Передняя, сред-
ная система средняя кишки няя и задняя кишки средняя и задняя киш- ки няя и задняя кишки
Анальное от- верстие Нет Есть Есть Есть
Кровеносная си- стема Нет Нет Есть (замкну- тая) Есть (замкнутая)
Дыхательная система Нет Нет Нет У части форм имеются жабры
Выделительная Протонефри- Протонефридии Протонефри- Протонефридии и
система дии или особые вы- делительные клетки или ка- налы эктодер- мического про- исхождения дии метанефридии
Половая система Большинство— гермафроди- ты Большинство раз- дельнополы , имеется поло- вой диморфизм Большинство раздельно- полы Типичные формы раздельнополы, но имеются и гермафродиты
ТИП VIII. МШАНКИ (BRYOZOA, ИЛИ
POLYZOA)
ХАРАКТЕРИСТИКА
Сидячие в громадном большинстве, колониальные животные (табл.У)
иногда с массивным известковым скелетом. Отдельные особи помещаются в
ячейках, из которых могут выпячиваться их передние концы с ротовым от-
верстием. Рот окружен венцом щупалец. Нервная система в связи с сидячим
образом жизни сильно редуцирована и состоит только из одного узла и от-
ходящих от него нервов. Гермафродиты. Развитие с метаморфозом — име-
ется свободноплавающая личинка. В настоящее время известно около 3500
видов. Обитают мшанки главным образом в морях, в пресных водах пред-
ставлены небольшим количеством видов.
Формы, имеющие скелет, хорошо сохраняются в ископаемом состоянии,
где их известно, начиная с ордовика, около 5000 видов.
ОБЩИЙ ОБЗОР ТИПА
Внешнее строение. Колонии мшанок очень разнообразны по форме
(табл. V). Одни образуют приподнимающиеся с субстрата тонкие деревца
или более массивные образования, внешне похожие на гидроидов или корал-
лов. Другие покрывают субстрат тонкой сеткой или сплошной коркой.
Размеры колоний очень различны — от нескольких миллиметров до несколь-
ких десятков сантиметров. Они могут быть студенистые, хитинообразные
или известковые. Колонии состоят обычно из очень большого числа особей,
так, например, листообразная колония Flustra (табл. V) весом в 13 кг со-
стоит из 1 330 000 отдельных особей. У некоторых колоний все особи оди-
наковы — мономорфные колонии, у других наблюдается довольно резко
выраженный полиморфизм особей — полиморфные колонии.
Каждая особь помещается в особой ячейке (рис. 236). Размеры ячеек
очень небольшие и обычно не превышают 1 мм. Стенки ячейки различного
строения и от их состава зависит и характер всей колонии, они могут быть
толстыми студенистыми, тонкими хитинообразными или могут быть пропи-
таны более или менее сильно известью. Ячейка имеет отверстие, через кото-
рое может выпячиваться передняя часть особи, или лофофор. Лофофор несет
ротовое отверстие, окруженное венцом щупалец, расположенных кольцом
(большинство мшанок) или подковообразно (большинство пресноводных
форм). Лофофор выпячивается обычно гидростатически, а втягивается осо-
быми мышцами-ретракторами. Отверстие ячейки может закрываться раз-
личными способами. У некоторых форм оно стягивается специальными
мышцами, у других закрывается особой крышечкой (operculum).
277
3
Таблица V. Пресноводные и морские мшанки СССР:
1 — Eucrata loricata — северные и дальневосточные моря, 2 — Flustra foliacea—
Белое море и дальневосточные моря, <7 — Alcyonidium gclatinosum— северные и
дальневосточные моря, 4— Ногпега lichenoides — северные и дальневосточные
моря, 5 — Retepora cellulosa — северные и дальневосточные моря, 6 — Defrancia
lucernaria—северные моря, 7 — Membranoipora — все моря, 8— Plumatelia re-
pens— пресные воды, 9 — Cristatella mucedo — пресные воды
сперва направляется назад
14
Рис. 236. Схема строения от-
дельной особи голоротой мшан-
ки в разрезе:
1 — венец шупалец, 2 стенки ячей-
ки, 3— вторичная полость тела, 4 —
целомический эпителий, 5 — мышцы
ретракторы, 6 — ротовое отверстие,
7 — пищевод, 8—желудок, 9 — зад-
няя кишка, 10 — анальное отверстие,
11 — нервный ганглий, 12 яичник,
13 — семенники, 14 — канатик
Полость тела. Полость тела вторичная (рис. 236), имеет мезодерми-
ческую выстилку и заполнена полостной жидкостью с клеточными элемен-
тами. В полости тела находятся внутренние органы.
Пищеварительная система. Пищеварительная система начинается ро-
товым отверстием. У большинства пресноводных мшанок (покрыторотые
мшанки) над ротовым отверстием имеется особый вырост — эпистом, при-
крывающий рот, у всех других мшанок эпистома нет (голоротые мшанки)
Ротовое отверстие ведет в глотку, которая переходит в пищевод и в объеми-
стую среднюю кишку (желудок). Последняя
(рис. 236), а затем резко загибается впе-
ред и переходит в заднюю кишку, которая
открывается заднепроходным отверстием
на переднем конце — вне венца щупалец.
От конца средней кишки к стенке ячейки
идет особый мезентериальный тяж — кана-
тик (funiculus), который прикрепляет пи-
щеварительный канал к стенке.
Кровеносная и дыхательная системы.
Кровеносная и дыхательная системы у
мшанок отсутствуют Функцию органов
дыхания выполняют щупальца, функцию
крови — полостная жидкость.
Выделительная система. Выделитель-
ная система у большинства тоже отсутст-
вует; у некоторых мшанок она, однако, име-
ется и представлена двумя короткими кана-
лами. Один конец такого канала откры-
вается в полость тела мерцательной ворон-
кой, другой наружу, причем обычно
оба канала в своих конечных частях
сливаются вместе и открываются единым
отверстием в передней части. Наличие
мерцательной воронки, открывающейся в
полость тела, делает право считать выдели-
тельные органы мшанок метанефридиями.
Нервная система. Нервная система
состоит из одного нервного узла, который
лежит между ротовым отверстием и анусом,
и отходящих от этого узла нервов.
Половая система. Половая система
большинства мшанок гермафродитная.
Скопление половых клеток, возникающих,
как у аннелид, из целотелия, образует го-
нады, которые находятся в полости тела.
Они сосредоточены обычно на канатике.
Сперматозоиды выводятся наружу через
выделительные трубки либо через особые
отверстия в стенке тела. Яйца, напротив,
остаются в полости тела и оплодотворяются сперматозоидами другой особи.
Развитие. Первые стадии развития оплодотворенных яиц протекают в
полости тела материнской особи или в особых измененных особях (см. ниже).
Личинки мшанок довольно разнообразны как по внешней форме, так и по
внутренней организации. Проплавав некоторое время в воде, личинка опу-
скается на дно, прикрепляется, претерпевает превращение и становится
первичной особью, которая затем путем почкования образует колонию.
Превращение сопровождается сложными явлениями. У многих мшанок
после прикрепления ко внутренней организации личинки происходят глу-
бокие изменения. Все ее внутренние органы подвергаются полному разру-
279
шению; сохраняются лишь эктодерма и мезодермические элементы. Кишеч-
ник взрослой особи образуется из эктодермы, а за счет мезодермы образуются
гонады, мускулатура и выстилка полости тела (рис. 237).
Кроме полового размножения, у мшанок очень большое значение имеет
бесполое, а именно почкование. В результате последнего и разрастаются
большие колонии. Почки образуются в определенных местах тела, что и
определяет форму колонии. Интересно отметить, что в образовании почек
участвуют только эктодерма и мезодерма.
Особая форма бесполого размножения, которое имеет место у пресно-
водных мшанок, — это внутреннее почкование. При этом в канатике обра-
зуются особые обычно дисковидные тела — статобласты, которые затем
выпадают в полость тела. Статобласты (рис. 238) покрыты двустворчатой
Рис. 237 Личинка мшанок Cyph- Рис. 238. Статобласт Cristatella muce-
onautes: do. Створки раскрылись, и из статоб-
/ — теменная пластинка# 2 — ротовое ласта выходит молодая мшанка
отверстие, 3—грушевидный орган, 4 —
прототрох, 5 — анальное отверстие, 6
присоска# 7. — кишечник
плотной скорлупой и в большинстве случаев окружены особым ячеистым,
так называемым плавательным кольцом, а иногда имеют и хитиновые крю-
чочки. Когда осенью колония мшанок погибает и распадается, статобласты
выпадают наружу и благодаря плотной оболочке перезимовывают Таким
образом статобласты представляют собой приспособление к переживанию
неблагоприятных условий (см. геммулы губок). В умеренных широтах
это происходит в зимний период, в тропиках — в засушливый период, когда
многие водоемы пересыхают.
Весной (или в начале периода дождей) скорлупа статобласта лопается
и половинки ее раскрываются, находящийся внутри зародыш выходит (см.
рис. 238) и развивается в молодую мшанку, которая затем путем обычного
почкования образует колонию. Плавательное кольцо и хитиновые крючочки
статобластов служат для пассивного разноса статобластов и тем способст-
вуют расселению их. Так, например, статобласты индо-китайской мшанки
Lophopodella, имеющие на концах хитиновые крючочки, были неожиданно
найдены в водоемах дельты Волги (Абрикосов и Косова, 1963), куда они
были, по-видимому, занесены перелетными птицами, к которым пассивно
прицепились при помощи упомянутых крючочков.
Полиморфизм. У полиморфных колоний, кроме основной массы обыч-
ных особей, несущих питающую функцию и образующих половые продукты,
имеется и ряд видоизмененных особей. Эти особи бывают обычно трех родов.
Авикулярии (рис. 2390, имеющие вид птичьего клюва, представляют собой
особи, у которых все внутренние органы редуцируются, но и сильно разви-
ваются крышечка ячейки (которая превращается в своеобразную «челюсть»),
280
и мышцы-ретракторы, приводящие ее в движение. Назначение авикулярий—
защита и очищение колоний от мелких животных и посторонних предметов.
Вибракулярии (рис. 239ц) тоже служат для защиты колонии, они имеют
длинный жгут, находящийся благодаря действию сильно развитых мышц
все время в колебательном движении. Оеции (рис. 240) представляют собой
выводковые камеры, куда после оплодотворения переходят яйца и где про-
исходит развитие личинок.
Кроме этих обычных форм полиморфизма, у некоторых мшанок имеются
еще измененные особи, служащие для прикрепления колоний к субстрату.
Рис. 240. Полиморфизм мшанок-
1 — обычная особь, 2 — оеция с различ-
ными стадиями развития личинки (<3)
П
Рис. 239. Полиморфизм мша-
нок. / —авикулярия; // ви-
бракулярия:
1 — неподвижная ветвь «клюва»,
2— подвижная ветвь «клюва», 3 —
мышцы, двигающие подвижную
часть (челюсть), 4— подвижной бич,
5—мышцы, приводящие его в дви-
жение
Экология и распространение. Большинство мшанок обитает в морях.
В пресных водоемах известно около 100 видов, относящихся главным обра-
зом к группе покрыторотых (см. выше). В морях мшанки обитают, начиная
с полосы отлива и до больших глубин, встречаются они как в арктических,
так и в тропических морях; в последних, правда, они более многочисленны.
В наших морях мшанки представлены довольно богато, особенно в северных
и дальневосточных.
В пресных водоемах мшанки обитают как в стоячих, так и в текучих
водах, причем главным образом по берегам, на водных растениях. В нашей
стране пресноводные мшанки хоть и не многочисленны, но распространены
широко во всех водоемах; наиболее обычны виды рода Plumatella (табл. V).
Кроме того, особенно надо отметить Cristatella mucedo (табл. V), колонии
которой в отличие от всех других мшанок подвижны и могут медленно пол-
зать по субстрату на широкой подошве, образующей основание колонии.
Колонии мшанок очень сильно изменяются по форме в зависимости от
условий существования—движение воды, глубины, характера субстрата
281
и пр. У одного и того же вида могут быть колонии различной формы — от
стелющихся по субстрату до образующих плотные массы на различных
подводных предметах.
Питаются мшанки путем фильтрации воды при помощи щупалец. От-
фильтрованные частицы детрита и мелкие животные и растительные орга-
низмы попадают в рот и далее в кишечный тракт
Образование колонии путем почкования идет у мшанок при благоприят-
ных условиях очень интенсивно. Разрастаясь на различного рода гидро-
технических и водопроводных сооружениях, мшанки могут приносить
иногда довольно существенный вред, закупоривая и засоряя их.
ПОЛОЖЕНИЕ В СИСТЕМЕ
Положение мшанок в системе животного мира долгое время остава-
лось неясным, что обусловлено значительными особенностями их строения
и сложностью развития. Старые авторы (Линней и др.), основываясь на
чисто внешнем сходстве, относили их к кишечно-полостным, причем имею-
щих мягкий скелет — к гидроидным, а обладающих известковым скеле-
том — к коралловым.
Более детальное изучение мшанок показало, что они организованы зна-
чительно сложнее полипов (сравните рис. 82 и 236). Тогда их стали объеди-
нять в один тип с плеченогими под различными названиями: червеобраз-
ные, моллюскообразные, щупальцевые. В настоящее время зоологи рассмат-
ривают мшанок как самостоятельный тип (Bryozoa или Polyzoa). Мшанки,
как и плеченогие (см. ниже), представляют особую древнюю ветвь живот-
ного мира, занимающую промежуточное положение между первичнороты-
ми и вторичноротыми (см. табл. XVI).
ТИП IX. ПЛЕЧЕНОГИЕ (BRACHIOPODA)
ХАРАКТЕРИСТИКА
Сидячие одиночные животные, имеющие двустворчатую раковину,
(рис. 241), створки которой в отличие от двустворчатых моллюсков покры-
вают тело животного со спинной и брюшной стороны. От тела, около рта
отходит пара особых придатков, так называемые руки, которые поддержи-
ваются скелетными выростами спинной створки, так называемым ручным
аппаратом (рис. 242). Кишечный канал у большинства форм замкнутый, не
имеющий анального отверстия. Выделительная система представлена выде-
лительными каналами с воронками (целомодуктами), открывающимися во
вторичную полость тела. Нервная система состоит из надглоточного и
подглоточного ганглиев с отходящими от них нервами. Развитие с мета-
морфозом, причем закладка мезодермы происходит энтероцельным путем
(рис. 243), что сближает плеченогих со вторичноротыми.
Плеченогие исключительно морские животные. В настоящее время
известно только около 200 видов, тогда как в ископаемом состоянии, начи-
ная с докембрия, около 7000.
ОБЩИЙ ОБЗОР ТИПА
Внешнее строение. Все плеченогие имеют раковину. У боль-
шинства животных створки раковины неодинаковые, брюшная створка
обычно более вогнутая и образует
макушку, в которой имеется глубо-
кая вырезка, так называемый дель-
тирий. Через эту вырезку выходит
особый вырост — стебелек, при
помощи которого большинство
плеченогих прирастает к субстрату
Стебелек занимает не всю вырезку,
а только ее вершину, основание же
ее закрыто особой пластинкой (или
двумя пластинками), так называ-
емыми дельтидиальными пластин-
ками (рис. 242). У некоторых пле-
ченогих стебелек не прикрепляется
к субстрату, а превращен в муску-
листую ножку, при помощи кото-
рой животное делает норки в пес-
чаном дне (Lingula).
Створки раковины или просто
соединяются мышцами — беззамко-
Рис. 241. Плеченогие на субстрате. Ви-
дны стебельки, при помощи которых
они прикрепляются
283
еые плеченогие, или на створках имеются особые выросты — зубы, образую-
щие замок, при помощи которого створки и соединяются, — замковые плече-
ногие. Кроме того, имеется довольно сложная система мышц как замы-
кающих створки, так и открывающих их.
Спинная створка имеет различной сложности выросты, поддерживаю-
щие руки, так называемый ручной аппарат. Он может состоять просто из
двух крючковидных выростов, но у большинства плеченогих образует слож-
ный петлеобразный каркас. Особой сложности ручной аппарат достигал у
некоторых ископаемых плеченогих, например, у Spirifer.
Рис. 242. Схема строения плеченогого. / продольный разрез; // —фрон-
тальный разрез:
/ — сосочки мантии,- входящие в канальцы раковины, 2— спирально завитой участок руки,
3—мышцы замыкателя раковины, 4 — мышцы, раскрывающие раковину, 5 — стебелек, 6 —
сердце, 7 — рот, 8 — пищевод, 9 — желудок, 10— замкнутая кишка, 11 и 12 — прямые участки
рук, 13—цирры рук, 14 — щетинки по краю мантии. 15 — перепонка, соединяющая руки, 16—
печень, 17— органы выделения (целомодукты), 18— дельтидиальные пластинки
По своему строению раковина плеченогих состоит из наружного рого-
вого слоя и двух известковых — пластинчатого и призматического. Харак-
терной особенностью раковины плеченогих являются тонкие слепые каналь-
цы, пронизывающие призматический слой.
Створки раковины выстланы изнутри мантией, которая представ-
ляет собой выросты тела. В отличие от мантии двустворчатых моллюсков
мантия плеченогих имеет полость, которая представляет собой выросты
полости тела животного. Мантия образует многочисленные сосочки, которые
входят в канальцы раковины. Мантия ограничивает довольно обширную
мантийную полость.
Тело животного занимает только сравнительно небольшую их часть ман-
тийной полости. Большая часть ее занята руками — парными, полыми
выростами тела, отходящими по бокам ротового отверстия и поддерживае-
мыми скелетными выростами спинной створки (см. выше). Руки усажены
многочисленными усиками, покрытыми ресничками. Движение этих ресни-
чек вызывает ток воды в раковину, с которым плеченогое получает пищу и
кислород для дыхания.
Полость тела. Полость тела — вторичная, довольно хорошо выражен-
ная, выстланная целомическим эпителием.
284
Нервная система. Нервная система представлена небольшим надгло-
точным ганглием, более крупным подглоточным ганглием, соединенным с
первым окологлоточными коннективами, и отходящими от обоих ганглиев
нервными тяжами.
Органы чувств отсутствуют.
Пищеварительная система. Пищеварительная система начинается ле-
жащим между руками щелеобразным ртом, который ведет в пищевод,
желудок и заднюю кишку. В желудок у плеченогих открываются протоки
особых желез, которые именуются печенью. Заднепроходное отверстие
имеется не у всех форм.
Рис. 243. Развитие плеченогих (по А. О. Ковалевскому). /
образование целомических пузырьков в виде боковых вырос-
тов первичного кишечника. II целомические пузырьки
отделились от первичного кишечника; III —личинка
Дыхательная и кровеносная системы. Кровеносная система не замкнута
и состоит из лежащего на спинной стороне сердца и сосудов, которые откры-
ваются в полость тела. Функцию органов дыхания выполняют руки. Они
постоянно омываются водой, что обеспечивает доступ кислорода к кровенос-
ным сосудам.
Выделительная система. Выделительная система представлена целомо-
дуктами в числе одной или двух пар.
Половая система и развитие. Раздельнополые животные. Половые про-
дукты развиваются в целотелии и образуют гонады, находящиеся в полости
тела. Оплодотворение внутреннее. Развитие сопровождается превращением.
Развивающаяся личинка плавает в воде, оседает на дно, прикрепляется,
претерпевает сложный метаморфоз и превращается в молодую брахиоподу
(рис. 243.)
Экология и распространение. Плеченогие — исключительно морские
животные, требующие для своего существования высокой солености, в
связи с чем они не встречаются в таких слабо соленых морях, как Балтий-
ское, Черное и Каспийское. К температуре они менее требовательны и
встречаются как в теплых, так и в холодных морях, хотя в первых пред-
ставлены богаче. Современные плеченогие — обитатели сравнительно боль-
ших глубин и не встречаются в прибрежных частях моря. В наших морях
плеченогие представлены немногими видами, наиболее обычна в северных
морях Rhynchonella psittacea.
Плеченогие — фильтраторы, они получают пищу, состоящую из ча-
стиц детрита и микроорганизмов, из тока воды, вызываемого движением
ресничек рук.
285
В прошлые геологические эпохи плеченогие были значительно разно-
образнее как по форме тела, так, видимо, и по биологии. Являясь массовы-
ми формами палеозойских морей, они имеют очень большое значение при
геологических исследованиях как одни из важнейших руководящих форм.
ПОЛОЖЕНИЕ В СИСТЕМЕ
Положение плеченогих в системе животного мира до настоящего вре-
мени недостаточно выяснено. Долгое время их объединяли вместе с мшан-
ками в один тип (см. выше). Многими зоологами и палеонтологами плече-
ногие рассматриваются в качестве самостоятельного типа. В строении и
развитии их имеются как признаки, сближающие плеченогих с первичноро-
тыми (сходство целомодуктов с целомодуктами кольчатых червей, ганглиоз-
ная нервная система, способ образования рта взрослого животного), так и
черты, напоминающие вторичноротых (недетерминированное дробление,
энтероцельный способ возникновения целома и мезодермы, некоторое
сходство с вторичноротыми в расчленении целома). Наряду с этим в орга-
низации плеченогих имеются черты своеобразия. На этом основании пола-
гают, что плеченогие, как и мшанки, занимают промежуточное положение
между первичноротыми и вторичноротыми (см. табл. XVI). На особенности
строения плеченогих, безусловно, большое влияние оказал прикрепленный
образ жизни, в связи с которым у них конвергентно выработались некоторые
признаки, сводные с другими сидячими животными.
ТИП X. МОЛЛЮСКИ (MOLLUSCA)
ХАРАКТЕРИСТИКА
Моллюски представляют собой резко отграниченный тип первично-
ротых животных. Их тело в большинстве случаев состоит из головы, внут-
ренностного мешка и ноги и покрыто твердой известковой раковиной. Ра-
ковина формируется при участии особой складки кожи-мантии. Мантия
свисает со спинной стороны внутренностного мешка и между ней и стенками
внутренностного мешка имеется то более, то менее обширная полость —
мантийная полость (см. рис. 266). В мантийной полости лежат органы ды-
хания, концевые отделы выделительных, половых органов и кишечника,
совокупность этих структур называют мантийным комплексом органов.
Органы дыхания — жабры у большинства моллюсков построены одно-
типно: по обеим сторонам оси жабры сидят многочисленные треугольные
лепесточки, а сама ось пронизана кровеносными сосудами и нервом. У одних
моллюсков ось жабры свободно свисает в мантийную полость, у других она
приросла к стенке мантийной полости, вследствие чего лепестки могут быть
лишь с одной стороны оси. И в том и в другом случае жабры такого типа
называются ктенидиями.
На голове расположены ротовое отверстие, органы чувств, а у неко-
торых специальные органы для схватывания добычи (см. рис. 278).
Нога моллюсков — это сильно утолщенная за счет мускулатуры брюшная
сторона внутренностного мешка (см. рис. 249, 270). Разрастаясь в разных
направлениях эта мускулизированная сторона тела образует весьма различ-
ные типы ног* широкие — ползательные, клиновидные — вбуравливаю-
щиеся в грунт, похожие на плавники —для плавания, округлые приса-
сывающиеся и др.
В глотке большинства моллюсков лежит своеобразное образование —
радула (терка) Это хитинойдная лента, на которой сидят многочисленные
хитинойдные же клювовидные зубцы. Как правило, радула служит для
соскабливания растительной пищи и лишь в редких случаях для активного
захвата ее, при хищничестве. В среднюю кишку впадает обширная печень —
орган переваривания пищи и место накопления питательных веществ.
У моллюсков есть вторичная полость тела (целом). В большинстве
случаев, однако, вторичная полость тела представлена лишь околосердеч-
ной сумкой — перикардом и полостью половых желез. Кровеносная система
незамкнутая, с обширными лакунами или синусами, занимающими много
места в голове, внутренностном мешке и ноге. Есть сердце, состоящее из
двух предсердий и желудочка. В некоторых случаях одно из предсердий
редуцируется, в других имеется четыре предсердия. Органы выделения
моллюсков — почки. Одним отверстием почки соединены с перикардием,
другим открываются наружу Такое положение почек по отношению к вто-
287
ричной полости тела (перикарду) позволяет считать их целомодуктами,
однако в сравнении с кольчатыми червями сильно усложненными.
Центральная нервная система состоит или из нервных тяжей, или из
нервных узлов, соединенных между собой нервными комиссурами или коннек-
тивами. У некоторых моллюсков нервные узлы концентрируются в рас-
положенный в голове мозг У большинства видов органы чувств развиты
хорошо.
Среди моллюсков есть как раздельнополые, так и гермафродитные виды.
Дробление оплодотворенного яйца в большинстве случаев происходит
по спиральному типу. Развитие либо со свободноплавающей личинкой, либо
без нее, т. е. прямое. На ранних стадиях личинка весьма похожа на тро-
хофору (см. рис. 246, 267).
Некоторых моллюсков добывают и разводят с целью использова-
ния в пищу (устрицы, гребешки, каракатицы и др.) или для получения жем-
чуга, раковина других идет для получения перламутра.
Многие моллюски являются вредными животными. Наземные слизни
повреждают сельскохозяйственные культуры, морской корабельный червь
разрушает находящиеся в море деревянные сооружения. Велика роль
пресноводных и наземных моллюсков как промежуточных хозяев парази-
тических червей домашних и диких животных и человека.
В настоящее время известно более 100 000 видов моллюсков. Таким
образом, моллюски —второй по величине тип в мире животных, уступаю-
щий по количеству видов лишь членистоногим.
Моллюски обитают в морях, пресных водах и на суше, небольшое число
их — паразиты иглокожих.
КЛАССИФИКАЦИЯ
Тип Моллюски (Mollusd)
Класс 1. Панцирные (Loricata)
Класс 2. Бороздчатобрюхие (Solenogastres)
Класс 3. Моноплакофоры (Monoplacophora)
Класс 4. Брюхоногие (Gastropoda)
Подкласс I. Переднежаберные (Prosobranchia)
Отряд 1 Двупредсердные (Diotocardia)
Отряд 2. Однопредсердные (Monotocardia)
Подкласс II. Заднежаберные (Opisthobranchia)
Отряд 1. Покрытожаберные (Tectibranchia)
Отряд 2. Голожаберные (Nudibranchia)
Отряд 3. Крылоногие (Pteropoda)
Подкласс III. Легочные (Pulmonata)
Отряд 1. Сидячеглазые (Basommatophora)
Отряд 2. Стебельчатоглазые (Stylommatophora)
Класс 5. Лопатоногие (Scaphopoda)
Класс 6. Двустворчатые, или Пластинчатожаберные (Bivalvia, или
Lamellibranchiata)
Отряд 1. Первичножаберные (Protobranchia)
Отряд 2. Нитежаберные (Filibranchia)
Отряд 3. Настоящие пластинчатожаберные (Eulamellibranchia)
Отряд 4. Перегородчатожаберные (Septibranchia)
Класс 7. Головоногие (Cephalopoda)
Подкласс I. Наружнораковинные (Ectocochlia)
Отряд 1. Кораблики (Nautiloidea)
Отряд 2. Аммониты (Ammonoidea; только ископаемые формы)
Подкласс II. Внутреннераковинные (Endocochlia)
Отряд 1. Десятиногие (Decapoda)
Отряд 2. Восьминогие (Octopoda)
288
КЛАСС 1. ПАНЦИРНЫЕ (LORICATA)
ХАРАКТЕРИСТИКА
К этому классу относятся моллюски, раковина которых состоит из
восьми отдельных пластинок, налегающих друг на друга и покрывающих
тело со спинной стороны. В длинной мантийной борозде, идущей по бокам,
расположены многочисленные жабры (рис. 244). Обитают в морях. Известно
около двухсот видов.
ОБЩИЙ ОБЗОР КЛАССА
Внешнее строение. Тело состоит из головы, внутренностного мешка
и удлиненной ноги. Срединная часть спинной стороны внутренностного меш-
ка занята пластинками раковины (см. рис. 244). Свободные от раковины
1 П
Рис. 244. Хитон (Tonicella marmorea), обычная форма наших северных
морей. / —со спинной стороны; 11 —с брюшной стороны:
/—а пластинки раковины, 9 — мантия, 10 — голова, 11 — нога, 12 — жабры, 13 — рото-
вое отверстие, 14 — анальное отверстие> 15 — отверстие почек, 16— половое отверстие
края спинной стороны представляют собой участки мантии, пронизанные
мышцами. Этот крышеобразно нависающий над ногой край мантии называет-
ся перинотумом.
С внутренней стороны между перинотумом и ногой справа и слева
имеется узкое пространство, называемое мантийной бороздой (см. рис. 244).
Раковина панцирных образуется из восьми пластинок, каждая
из которых состоит из двух слоев: наружного — органического и внутрен-
него — известкового. Пластинки налегают одна на другую как черепицы
и не сращены между собой, благодаря чему моллюск может сворачиваться
в кольцо.
Кроме пластинок, в коже имеются иногда многочисленные известковые
чешуйки, иголки и зернышки разнообразной формы.
У некоторых панцирных, например у крупного, достигающего 18 см
длины дальневосточного криптохитона (Cryptochiton stelleri), пластинки
могут обрастать окружающей кожей и не видны.
Мышечная система. Мускулатура развита сильно особенно на спинной
стороне, где отдельные мышцы поддерживают пластинки раковины, а дру-
гие, прикрепляясь к ним, идут в ногу.
10 Курс ЗООЛОГИИ, 7. I
289
Нервная система. Центральная нервная система (рис. 245) состоит из
головного (церебрального) нервного тяжа, расположенного впереди глотки,
и отходящих от него боковых (плевральных) и ножных (педальных) нервных
тяжей. Плевральные тяжи идут вдоль боков тела близко к поверхности,
переходя сзади друг в друга. Педальные стволы лежат в толще мускула-
туры ноги и друг в друга не переходят. Соединения церебральных тяжей с
педальными и плевральными образуют нервное кольцо, охватывающее глот-
ку. Интересно отметить, что у панцирных есть все же ганглии. Эти малень-
кие ганглии лежат на глотке и посылают нервы к мышцам глотки и к ра-
дуле. Таким образом, у моллюсков концентрация нервных клеток впервые
возникает при необходимости иннервации наиболее динамичных и подвиж-
ных органов — глотки и радулы, которые в связи со соскребыванием пищи
с субстрата находятся в постоянном движении.
Органы чувств. Органы чувств представлены особыми образованиями,
так называемыми эстетами, — чувствительными выростами эпителия,
прободающими раковину. У некоторых представителей часть эстетов прев-
ращается в органы зрения.
Пищеварительная система. Ротовое отверстие ведет в ротовую полость,
которая переходит в обширную полость глотки. Со дна глотки поднимается
валик — «язык» — несущий радулу. В следующий за глоткой пищевод впа-
дают сахарные железы, перерабатывающие содержащийся в пище крахмал
в сахар.
Средняя часть кишечного канала начинается желудком, который затем
переходит в довольно длинную кишку. Имеется объемистая печень. Аналь-
ное отверстие помещается на заднем конце тела (см. рис. 244).
Дыхательная система. Многочисленные жабры (ктенидии) располагают-
ся по бокам тела — в мантийной борозде. Видимо, лишь задняя пара жа-
Рис. 245. Внутреннее строение хитона. I — нервная система; // — выде-
лительная, кровеносная и половая системы:
1 — окологлоточное нервное кольцо, 2—радулярные ганглии, 3— ножные нервные
стволы, 4 — туловищные нервные тяжи, 5 — жабры (ктенидии), 6 — анальное отверстие,
7—рот, 8 — жаберная вена, 9 — почки, 10 — половая железа, 11— протоки половой же-
лезы, 12— наружное отверстие почки, 13— перикардиальная полость, 14 — сердце, 15—
реноперикардиальное отверстие
П
290
ужу по бокам тела (см. рис. 245ц).
Рис. 246. Развитие хитона. / — свободно-
плавающая личинка; II — ее метаморфоз;
/// — молодой хитон:
1 — ресничное кольцо, 2— зачаток ноги, 3 — зачаток
скелета
бер гомологична жабрам других моллюсков, в то время как остальные —
это вторичные, адаптивные жабры, т. е. развивающиеся из кожи (при не-
обходимости усиления газообмена). Количество этих адаптивных жабр у
разных видов панцирных моллюсков разное.
Кровеносная система. Сердце лежит в перикардии на спинной стороне
тела, в его задней половине (рис. 245ц). Оно состоит из одного желудочка
и двух предсердий, симметрично расположенных по его бокам, соединенных
с желудочком атриовентрикулярными отверстиями. От желудочка отхо-
дит аорта, в предсердия впадает по одному сосуду, приносящему окислен-
ную кровь из’ жабр. Периферическая система сосудов развита слабо и во
многих частях тела заменяется лакунами.
Выделительная система. Пара почек, представляющих собой сложно
разветвленные древовидные образования, открывается одним концом в пе-
рикардий, а другим концом ь
Половая система. Все пан-
цирные, за исключением одного
вида, раздельнополы. Гонады,
часто сливающиеся в непарный
орган по средней линии тела,
лежат над кишечником (см. рис.
245ц). Открываются они по бо-
кам тела вблизи наружных от-
верстий почек. У некоторых
форм наблюдается половой ди-
морфизм, выражающийся в раз-
личной окраске ноги, жабр и
других мягких частей тела.
Развитие. Оплодотворение
наружное, в результате дробле-
ния образуется свободноплаваю-
щая личинка трохофорного типа
с сильно развитым предротовым
ресничным кольцом и теменной
пластинкой с султаном длинных
ресничек (рис. 246). У личинки
довольно рано появляются за-
чаток ноги, пластинки раковины и чешуйки кожного скелета. Она все чаще
и чаще опускается на дно, а затем вовсе перестает плавать и превращается
во взрослое животное. Такое постепенное превращение личинки во взрос-
лый организм называется эволютивным метаморфозом.
Экология и распространение. Панцирные моллюски — обитатели глав-
ным образом полносоленых и теплых морей, поэтому у нас они
встречаются главным образом в дальневосточных морях, где их известно
в настоящее время 37 видов, в то время как в северных — всего 7. В Чер-
ном море панцирные представлены двумя видами, а в Каспийском, Азовском
и Балтийском морях они вообще отсутствуют.
Панцирные моллюски обитают в большинстве своем в прибрежной зоне.
Широкой ногой они могут очень крепко присасываться к береговым камням.
Благодаря плоской форме тела панцирные иногда совершенно сливаются
с субстратом, чему способствует и окраска многих форм, сходная с цветом
субстрата. Питается большинство панцирных растительной пищей, соскаб-
ливая при помощи радулы водоросли с камней. Некоторые из них — хищ-
ники, питающиеся преимущественно фораминиферами или губками.
Филогения. Панцирные — примитивная группа моллюсков. Это видно
из строения нервной системы, центральные отделы которой лишены ган-
глиев. Многие исследователи считают их даже самыми примитивными
моллюсками, ближе других, стоящими к кольчатым червям — общим пред-
кам всех моллюсков.
10*
291
КЛАСС 2. БОРОЗДЧАТОБРЮХИЕ (SOLENOGASTRES)
ХАРАКТЕРИСТИКА
Рис. 247. Nematome-
nia flavens на гидрои-
де. видна брюшная
бороздка
Моллюски с червеобразным телом (рис. 247). Тело почти полностью об-
растает мантией. На брюшной стороне имеется бороздка (отсюда и название
класса), в которой расположен покрытый ресничным эпителием валик (ру-
димент ноги). Раковина отсутствует, но в наружных покровах тела есть ма-
ленькие известковые спикулы. Нервная система со-
стоит из четырех продольных стволов, соединенных
между собой спереди и сзади. Развитие у большин-
ства с метаморфозом. Исключительно морские формы.
ОБЩИЙ ОБЗОР КЛАССА
Внешнее строение. Бороздчатобрюхие — неболь-
шая группа животных, охватывающая всего около
120 видов. Размеры тела обычно небольшие, достигаю-
щие 15 см. Тело покрыто кутикулой, в которой
располагаются иногда многочисленные известковые
тельца, образующие в некоторых случаях сплошной
чешуйчатый покров.
Мышечная система. Под кутикулой расположен
мускульный мешок, состоящий из нескольких различ-
ных слоев. Движение моллюска происходит при по-
мощи этих мышц путем изгибания тела, как у червей.
Когда кутикула очень плотная и изгибание тела невоз-
можно, моллюск передвигается скользящим движени-
ем при помощи ресничек брюшного валика.
Нервная система и органы чувств. Нервная си-
стема состоит из нескольких ганглиев, лежащих в пе-
редней части тела, и четырех нервных стволов, иду-
щих вдоль тела и соединенных между собой. Органы
чувств развиты слабо.
Пищеварительная система. Пищеварительный
ротовым отверстием на переднем конце тела, прохо-
дит через все тело и открывается на заднем конце в клоаку, куда откры-
ваются и протоки перикардия. В ротовой полости у некоторых форм
имеется, как и у других моллюсков, радула.
Дыхательная и кровеносная системы. Специальных органов дыхания
нет, дыхательные функции выполняют кожные складки в стенках клоаки.
Кровеносная система состоит из двухкамерного сердца, лежащего в пери-
кардии, и системы синусов.
Половая система. Бороздчатобрюхие в большинстве гермафродиты.
Половые продукты из гонад выпадают в перикардий и затем выводятся на-
ружу через протоки перикардия.
Экология и распространение. Бороздчатобрюхие — обитатели исклю-
чительно полносоленых морей, где они встречаются на различных глубинах
и различных грунтах. Часть из них — активные хищники, обитающие среди
кораллов и гидроидов и объедающие их полипы; часть — обитатели илистых
грунтов, пассивно питающиеся илом. В наших морях бороздчатобрюхие
представлены на Севере и на Дальнем Востоке.
Филогения. Бороздчатобрюхих надо считать очень специализирован-
ной группой, имеющей общие черты с Loricata, но рано отошедшей от обще-
го с ними ствола развития и приобретшей в связи со сходными условиями
существования ряда черт, сходных с червями (форма тела, кожно-мускуль-
ный мешок).
канал открывается
292
КЛАСС 3. МОНОПЛАКОФОРЫ (MONOPLACOPHORА)
ХАРАКТЕРИСТИКА
До 1957 г. были известны только ископаемые представители этого клас-
са из кембрийских, силурийских и девонских отложений. Первый ныне
живущий представитель Neopilina galatheae найден набольших глубинах
'около 3600 м) в Тихом океане у берегов /Мексики (датская океанологическая
экспедиция).
Билатеральносимметричные моллюски с колпачковидной раковиной,
вершина которой наклонена вперед, в сторону головы. Имеется обширная
вторичная полость тела; метанефридии, жабры, предсердия и многие мышцы
расположены метамерно.
ОБЩИЙ ОБЗОР КЛАССА
Тело состоит из переднего отдела — нерезко обособленной головы.
довольно высокого внутренностного мешка и дисковидной ноги. В окружаю-
дей ногу мантийной борозде лежат пять пар оригинально устроенных жабр
и остальные органы мантийного комплекса (рис. 248).
Рис. 248. Neopilina galatheae. / — вид сверху; // — вид с брюшной сто-
роны:
1 — передний отдел, 2 — нога, 3 — мантия, 4 — рот, 5 — анальное отверстие, 6 — жабры
(по Ле мхе)
Раковина состоит из трех слоев: наружного органического (периост-
ракум), срединного — призматического, состоящего из кристаллов угле-
кислого кальция, и подстилающего раковину перламутрового слоя, тоже
состоящего из кристаллов углекислого кальция. На внутренней стороне
раковины хорошо заметны места прикрепления шести и более пар мускулов
ноги и головы. Прикрепление мускулов — ретракторов ноги и головы
к раковине убедительно показывает, что раковина выполняет опорную
функцию.
Важно отметить, что вершина колпачковидной раковины загнута
вперед и немного нависает над головой. Это резко отграничивает монопла-
кофор от тех современных брюхоногих моллюсков, которые имеют такую
же колпачковидную раковину.
Полость тела. Вторичная полость тела хорошо развита. Она состоит из
обширного парного туловищного отдела, парного околосердечного (пери-
кардиального) отдела и полового отдела в виде двух пар половых желез.
293
Нервная система. Центральная часть нервной системы состоит из
окологлоточных нервных тяжей, боковых туловищных тяжей и ножных
тяжей. Боковые и ножные нервные тяжи огибают тело и ногу, переходя
друг в друга. Таким образом, туловищная часть центральной нервной
системы представлена двумя длинными дугами. На окологлоточных тяжах
развиты небольшие ганглии.
Пищеварительная система. Пищеварительная система начинается глот-
кой с радулой, переходит в петлеобразно извитой кишечник, открывающий-
ся сзади в мантийной борозде анальным отверстием. В средние отделы ки-
шечника впадает печень.
Кровеносная система. Кровеносная система состоит из сердца и слабо
развитых сосудов; большее развитие получают артериальные и венозные
синусы. Сердце с двумя парами желудочков, которые расположены по бокам
задней кишки; в каждый желудочек впадает свое предсердие. Таким образом,
имеются четыре предсердия, лежащих по бокам тела, в его задней
части.
Выделительная система. Органы выделения представлены шестью па-
рами целомодуктов, одни из которых соединены с туловищным, а другие с
околосердечным отделом вторичной полости тела. В целомодукты централь-
ной части тела впадают протоки половых желез. Наружные — выводные —
отверстия метанефридиев открываются в мантийную борозду.
Образ жизни и распространения. Немногочисленные виды ныне живущих
моноплакофор обитают, видимо, только на больших глубинах (более 2000,и)
и только в одном районе Тихого океана — у берегов Центральной и Южной
Америки. В кишечнике некоторых экземпляров были обнаружены радио-
лярии.
Филогения. Толкования организации моноплакофор противоречивы.
Одни исследователи считают, что характерная для этой группы метамерия
метанефридиев и других структур, так же как и сильное развитие вторичной
полости тела свидетельствуют о глубокой примитивности моноплакофор.
Эти исследователи склонны видеть в них не только предков всех моллюсков,
но и предков членистоногих. Таким образом, по этой концепции монопла-
кофоры связаны в своем происхождении непосредственно с кольчатыми
червями. Другие ученые полагают, что большинство черт метамерии моно-
плакофор вторичны. Истинная, наследованная от кольчатых червей, мета-
мерность проявляется лишь в наличии двух-трех целомических полостей.
В этом отношении они не отличаются принципиально от других моллюсков,
Не отрицая примитивности моноплакофор, эти исследователи считают их
глубоко специализированной группой, которая нс может считаться исход-
ной для всех других моллюсков.
КЛАСС 4. БРЮХОНОГИЕ (GASTROPODA)
ХАРАКТЕРИСТИКА
К этому классу относятся моллюски, у которых мантийная полость со
всем комплексом ее органов (анальное отверстие, отверстие почек, органы
дыхания и др.) лежит впереди, над головой, или сбоку. Кишечник в глуби-
не висцерального мешка образует петлю. У многих нервные тяжи, соот-
ветствующие плевральным (т. е. боковым) нервным тяжам панцирных мол-
люсков и моноплакофор, образуют перекрест, так называемую хиастоневрию
(см. рис. 253). Для большинства брюхоногих характерна асимметрия
(см. рис. 253), которая выражается в том, что имеется только один желудо-
чек сердца, одна почка, одна жабра, одна половая железа; эти органы на
другой стороне тела редуцированы. Раковина либо конусовидная с верши-
ной, склоненной назад, либо завита в спираль.
294
У части наземных и пресноводных брюхоногих, образующих подкласс
легочных, жабра заменена «легким» — участком мантии, обильно снабжен-
ным ветвящимися кровеносными сосудами.
Известно около 100 000 морских, наземных и пресноводных видов.
Весьма большое количество ископаемых форм.
Представитель класса брюхоногих — виноградная
улитка
(Helix pomatia)
Внешнее строение. Виноградная улитка (рис. 249) относится к под-
классу легочных, характерная черта которых отсутствие жабр и наличие
легкого. Тело улитки достигает 6—8 см в длину, на спинной стороне поме-
щается большая спирально завитая раковина
диаметром до 5 см. В нормальном состоянии
хорошо видна голова с двумя парами щупа-
лец: короткими — осязательными и длинны-
ми — несущими глаза. На переднем конце
помещается ротовое отверстие. Вблизи рото-
вого отверстия под основанием правого глаз-
ного щупальца помещается половое отверстие.
У края устья раковины также на правой
стороне тела находится дыхательное отвер-
Рис. 250. Центральная нервная
система («головной мозг») вино-
градной улитки:
1 — головные ганглии, 2 — ножные
ганглии, 3—плевральные ганглии,
4 — париетальный ганглий, 5 — ту-
ловищный ганглий, 6 —паллиальный
ганглий
Рис. 249. Виноградная улитка (Helix poma-
tia) в естественную величину:
1—раковина, 2—голова, 3—верхние и нижние щу-
пальца, 4 — нога, 5 — дыхательное отверстие, 6—по-
ловое отверстие, 7 — анальное отверстие, 8— глаза
стие, ведущее в мантийную полость, а рядом с ним — анальное и отверстие
мочеточника.
При неблагоприятных условиях улитка втягивает голову и ногу в
раковину, а устье раковины закрывает слизью.
Нервная система (рис. 250). Нервная система состоит из около-
глоточных ганглиев и периферических нервов. В состав окологлоточных нерв-
ных углов входят лежащие над глоткой головные (церебральные) ганглии и
лежащие под глоткой тесно сближенные парные ножные (педальные), пар-
ные плевральные и непарные паллиальный, париетальный и туловищный
(абдоминальный) ганглии. Церебральные ганглии соединены с педальными
и плевральными ганглиями хорошо заметными коннективами, остальные
коннективы и комиссуры короткие.
Органы чувств. Имеются органы осязания, обоняния, равновесия (стато-
цистами) и зрения. Глаза (рис. 251), расположенные на концах длинных
щупалец, представляют собой отшнуровавшееся впячивание покровов —
глазной пузырь. Полость глаза занята сферической линзой — хрусталиком.
Передняя стенка глазного пузыря прозрачна, боковая и задняя стенки пиг-
ментированы. Ко дну глаза подходит зрительный нерв.
295
1
2.
Рис. 251. Разрез
через глаз виноградной улит-
ки:
1— наружная роговица, 2 — внутренняя роговица, 3—
хрусталик, 4 — сетчатка, 5—зрительные клетки, 6 —
пигментные клетки
Пищеварительная система. Ротовое отверстие ведет в довольно обшир-
ную ротовую полость. За ротовой полостью следует глотка, в которой поме-
щаются челюсть и радула с многочисленными зубчиками. Челюсть и терка
служат для отгрызания и перетирания частей растений, которыми улитка,
как и большинство брюхоногих,
питается. В глотку открывается
также пара слюнных желез.
После глотки идет короткий
пищевод, расширяющийся в
объемистый зоб, который затем
переходит в небольшой желу-
док. Желудок окружен долями
большой печени, занимающей
верхние обороты спирали рако-
вины и повторяющей их по
форме.
От желудка отходит кишка,
в глубине завитка она делает
петлю и идет в обратном направ-
лении, переходя в заднюю киш-
ку Анальное отверстие лежит
рядом с дыхательным впереди,
на правой стороне. Печень улит-
ки функционирует не только как
пищеварительная железа, в ее
полости происходит и всасыва-
ние пищи.
Дыхательная система. Лег-
кое представляет собой участок мантийной полости, богато снабженный
кровеносными сосудами. Стенки этих сосудов очень тонкие
происходит обмен газов из воздуха, входящего в мантийную
дыхательное отверстие.
и через них
полость через
Рис. 252. Внутреннее строение виноградной улитки (схема):
/ _ рот, 2 — глотка, 3 — слюнные железы, 4 — кишечник, 5 — печень, 6 — анальное отверстие,
7— сердце, 8— почка, 9— наружное отверстие почки, 10—легкое, 11— дыхательное отвер-
стие, 12 — окологлоточные ганглии, 13—гермафродитная железа, 14 — семя-яйцепровод, 15 —
яйцевод, 16—семяпровод, 17 — жгут (флаггелум), 18— мешок «любовных стрел», 19 — белко-
вая железа, 20 — проток и резервуар семеприемника (пальчатые железы не изображены), 21 —
половое отверстие, 22 — перикардиальная полость, 23—реноперикардинальное отверстие.
Стрелками показано направление движения крови
296
Кровеносная система. Кровеносная система, как у всех моллюсков,
незамкнутая. Сердце лежит у стенки мантийной полости в околосердечной
сумке, или перикардии, и состоит из одного предсердия и желудочка. В
предсердие поступает окисленная кровь из сосудов легкого, которая затем
переходит через отверстие, снабженное клапанами, в желудочек. От по-
следнего берет начало аорта, которая ветвится и распадается в конце концов
на капилляры. Из капилляров кровь поступает в систему мелких полостей,
лишенных собственных стенок. Из этих полостей кровь затем вновь соби-
рается в сосуды, но уже венозной системы, которая доставляет кровь к лег-
кому Кровь виноградной улитки имеет слабосиний пигмент — гемоцианин,
содержащий медь и связывающий кислород в небольших количествах. Ос-
новная масса кислорода находится в крови в растворенном виде.
Выделительная система. Непарная почка помещается около перикар-
дия, куда открывается небольшой покрытой ресничками воронкой. По-
лость почки сильно сужена многочисленными складками, в которых про-
ходят кровеносные сосуды. Из этих сосудов почечный эпителий и извлекает
подлежащие удалению вещества. Продукты выделения откладываются в
почке в виде комочков мочевой кислоты, которые периодически, каждые
14—20 дней, выводятся наружу Выводное отверстие почки открывается
около анального отверстия.
Половая система. Виноградная улитка, как все легочные брюхоно-
гие, гермафродит. Половая система очень сложная, главным образом из-за
наличия целого ряда дополнительных желез. Основная часть половой
системы — гермафродитная железа, в которой образуются и яйцеклетки и
сперматозоиды. От этой железы отходит гермафродитный проток, быстро,
однако, делящийся на мужской и женский протоки, которые на большом
протяжении внешне слиты. В начальную часть женского протока открывает-
ся белковая железа, выделяющая слизистое вещество, которое одевает яйцо,
в конечную часть — проток семеприемника и пальчатые железы, образую-
щие скорлупу яйца.
Мужской проток впадает в совокупительный орган, который открывает-
ся в небольшую полость перед наружным половым отверстием. В эту же
полость открывается и конечная часть женского
С совокупительным органом связан тонкий труб-
чатый придаток — бич, секрет которого облекает
большие массы сперматозоидов, образуя сперма-
тофоры. Кроме того, в конечную часть половой
системы открывается мешок «любовных стрел»,
в котором образуются известковые тельца — «любов-
ные стрелы». При копуляции они втыкаются в
кожу партнера и служат, по-видимому, для раз-
дражения. Каждая из участвующих в копуляции
особей вводит сперматофор во влагалище другой
особи.
Экология и распространение. Виноградная
улитка распространена в южных и западных рай-
онах европейской части СССР и встречается в
садах, виноградниках и на пастбищах. В цен-
тральных районах она обитает только местами,
как завезенная форма.
Питаются улитки растительностью, скобля ее
протока — влагалища.
Рис. 253. Откладывание
яиц виноградной улит-
кой
радулой.
Размножаются они, откладывая яйца, имеющие вид мелких горошинок.
Яйца откладываются в землю, в ямку, которую улитка предварительно
выкапывает ногой (рис. 253). Развитие прямое — из яйца вылупляется ма-
ленькая, но полностью сформированная улитка, лишь половая система
которой еще не развита.
297
При массовом размножении виноградная улитка может наносить боль-
шой вред садовым культурам, объедая почки и листья.
На зиму улитки впадают в спячку, закапываясь для этого в почву.
При надлежащем уходе виноградные улитки хорошо живут в терра-
риумах.
Положение в системе. Виноградная улитка относится к подклассу
легочных и отряду стебельчатоглазых.
ОБЩИЙ ОБЗОР КЛАССА
Раковина у громадного большинства брюхоногих спирально завита
вправо, но у представителей некоторых родов и видов, а также у отдельных
индивидуумов спираль может быть закручена влево1. Примером влево за-
крученного брюхоногого могут служить пресноводные роды физа (Physa)
и аплекса (Apalexa).
Высота раковины различна, она может быть и очень высокой, например
черноморская церициум (Cerithium), и очень уплощенной, например прес-
новодная катушка (Planorbis). У некоторых форм раковина не спирально
завитая, а имеет вид конического колпачка. Примером такого рода моллюс-
ка могут служить обычные на морских побережьях морские блюдечки
(Patella, Acmaea), а также пресноводная чашечка (Ancylus) (табл. VI).
Раковина состоит из трех слоев — слаборазвитого органического (пе-
риостракум) и двух известковых, из которых особенно сильно развивается
средний — фарфоровидный. Внутренний — перламутровый — слой у брю-
хоногих обычно развит слабо, за несколькими исключениями, например, у
морского ушка (Haliotis) он развит сильно, и этого моллюска добывают
именно из-за его раковины (см. ниже). У большинства переднежаберных
имеется еще одно дополнительное скелетное образование — крышечка, рас-
положенная на верхней поверхности задней части ноги. При втягивании
моллюска в раковину крышечка плотно закрывает ее отверстие. Примером
брюхоногого, имеющего крышечку, может служить обычная наша пресно-
водная живородка (Vivipara) и ряд морских форм.
Многие брюхоногие имеют редуцированную раковину или совершенно
лишены ее. Такое явление распространено главным образом среди задне-
жаберных, например у плавающего морского ангелочка из крылоногих
Clione (см. рис. 260), а также у различных представителей голожаберных
Dendronotus и др. (см. рис. 261). Помимо заднежаберных, раковина
может редуцироваться или отсутствовать и у некоторых переднежаберных,
например у плавающих киленогих (см. рис. 262), а также у некоторых ле-
гочных, например у слизней (см. рис. 263).
Нога. Нога у большинства брюхоногих имеет такое же строение, как
и у виноградной улитки. Резко отличаются в этом отношении плавающие
формы. У них нога или превращается в два боковых придатка — плавника
(крылоногие, см. рис. 260), или часть ноги, сплющенная с боков, образует
выдающийся книзу непарный плавник (киленогие).
Нервная система. Нервная система построена различно у разных
групп брюхоногих, причем можно проследить постепенное усложнение этой
системы от низших форм к высшим. Усложнение это, во-первых, выражается
в том, что имеется постепенный переход от системы, состоящей из нервных
тяжей, равномерно усеянных нервными клетками, к системе нервных узлов,
содержащих нервные клетки и соединенных волокнистыми тяжами без нерв-
ных клеток. Таких далеко отстоящих друг от друга нервных узлов у брюхо-
ногих четыре пары и один непарный узел: пара головных, пара ножных,
пара плевральных, пара туловищных и непарный — внутренностный узел.
Во-вторых, усложнение нервной системы состоит в сближении нервных уз-
лов. Это достигает максимума у наземных легочных брюхоногих и у неко-
1 Чтобы определить, в какую сторону завита раковина, надо расположить ее
вершиной вверх, а устьем к себе. Если при таком положении устье лежит вправо от
оси раковины, то раковина завита вправо, если — влево, то раковина завита влево.
298
у части заднежаберных (см.
9
8
Рис. 254. Пресноводный брюхоногий моллюск—
лужанка Vivipara (переднежаберные, однопред-
сердные). ~
торых заднежаберных. В этом случае нервные узлы не только сконцентри-
рованы так, что не видны соединяющие их нервные тяжи (комиссуры и кон-
нективы), но и так, что некоторые ганглии внешне слились друг с другом
(см. рис. 251).
Перекрещивание основных нервных тяжей, т. е. хиастоневрия (рис.
254), выражена у всех переднежаберных,
далее) и весьма немногих
легочных. У остальных зад-
нежаберных нет и следов
перекреста нервов, а у осталь-
ных легочных есть лишь ос-
таточные явления этого утра-
ченного перекреста. Ликвида-
ция хиастоневрии есть след-
ствие все большего и боль-
шего укорочения коннективов
и следствие концентрации
нервных узлов.
Органы чувств. Органы
чувств развиты у брюхоно-
гих, как это видно на при-
мере виноградной улитки,
довольно хорошо. У водных
форм имеются также органы
химического чувства — осфра-
дии, отсутствующие обычно
у наземных форм.
П ищеварительная систе-
ма. У ряда брюхоногих имеет-
ся втягивающийся подвижной
хоботок, на конце которого
помещается ротовое отверстие.
При помощи такого хоботка
Раковина удалена и мантийная по-
лость вскрыта:
2 — сердце, 3 — кишечник, 4 — выводной про-
5—расширение яйцевода, в котором разни-
7 — нервные
9 — анальНое
/ — жабра,
ток почки, . _
вается молодь, 6 — окологлоточные ганглии,
стволы, (видна хиастоневрия), 8—печень,
отверстие
моллюск может захватывать не только
растительную пищу, но и мелких животных, а также высасывать других
моллюсков (см. ниже). Наличие печени и радулы характерно для всех
брюхоногих. Многие из них обладают слюнными железами.
Дыхательная система. Лишь у очень небольшого числа брюхоногих
имеются две жабры (ктенидия) на переднем конце — часть переднежабер-
ных. У большинства имеется одна перистая жабра (см. рис. 254). В ряде
случаев, например у голожаберных, жабры редуцируются, а вместо них
по краям мантии или по всему телу образуются адаптивные жабры (уме. 261).
У наземных и большинства пресноводных представителей брюхоногих,
как у виноградной улитки, жабры редуцируются и заменены легким.
Кровеносная система. У большинства брюхоногих, как и у виноградной
улитки, сердце состоит из одного желудочка и одного предсердия. Исклю-
чение составляют двупредсердные примитивные брюхоногие, у которых или
два нормальных предсердия соответственно двум жабрам (рис. 255), или
одно нормальное и одно рудиментарное.
Выделительная система. Выделительная система обычно представлена,
как у виноградной улитки, одной почкой, лежащей в непосредственной
близости к перикардию. У двупредсердных брюхоногих — две почки.
Половая система. Среди брюхоногих имеются как гермафродиты,
так и раздельнополые. К первым относятся легочные и заднежаберные, ко
вторым — переднежаберные. У низших форм (двупредсердных) нет спе-
циальных половых протоков и гонады открываются в почку. Половая
система раздельнополых форм сравнительно проста, гермафродитных же,
как это имеет место у виноградной улитки, очень сложная.
Развитие. Дробление оплодотворенного яйца у брюхоногих спираль-
299
ное. У одних из яйца выходит трохофорообразная свободноплавающая
личинка, как и у полихет называемая трохофорой. В отличие от трохофоры
полихет у трохофоры брюхоногих не происходит сегментации зачатков ме-
зодермы и трохофора не вытягивается в длину. Вместо этого у трохофоры
брюхоногих образуется сильный спинной горбообразный вырост, в который
1
Рис. 255. Брюхоногий моллюск Puncturella (пе-
реднежаберные, двупредсердные, парножабер-
ные). Раковина удалена и мантийная полость
вскрыта:
1—голова, 2—головные щупальца, 3— нога, 4—внут-
ренностный мешок, 5—мантия, разрезанная и отведенная
в стороны, 6 — жабры, 7—анальное отверстие
заходит кишка и в кото-
ром формируется печень.
Этот горб — будущий вну-
тренностный мешок (рис.
256). Снаружи на вершине
горба образуется первич-
ная колпачковидная рако-
вина, впоследствии заме-
няющаяся окончательной
раковиной. Вокруг перед-
ней — головной — части
личинки развивается мощ-
ный пояс ресничек. На этой
стадии личинка брюхоно-
гих называется велигер
(Veliger). Брюшная сторо-
на велигера, противолежа-
щая спинному выросту,
превращается в ползатель-
ную поверхность ноги, на
которую и оседает личинка.
С этого времени перед на-
ми уже почти закончивший
свое развитие брюхоногий
моллюск. Он продолжает
расти, при этом интенсивно
растущая раковина растет
по спирали.
Важно отметить, что у личинок некоторых примитивных брюхоногих
перед оседанием на дно колпачковидная раковина — уже сильно развитый
внутренностный мешок и мантийная полость, лежащая сзади, поворачивают-
ся на 180°. В результате этого поворота мантийная полость с комплексом
ее органов (жабры, анальное отверстие и др.) перемещается в переднее по-
ложение и оказывается лежащей над головой моллюска. Этот присущий лишь
личинкам брюхоногих поворот раковины и внутренностного мешка назы-
вается личиночным закручиванием, или торсионным процессом (см. рис. 256).
Рис. 256. Личинки морского брюхоного моллюска. 1 — стадия трохофоры;
11 — стадия велигера (до личиночного закручивания); III — стадия ве-
лигера после личиночного закручивания:
1 — вершина эмбриольной раковины, 2—зачаток головы, 3—зачаток ноги£ 4 —анальное
отверстие, 5—внутренностный мешок
300
Рис. 257. Схема происхожде-
ния переднего положения ман-
тийного комплекса и хиасто
неврии. / исходная, гипоте-
тическая форма; // — форма с
законченным скручиванием:
/ — жабры, 2 — нервные тяжн
У большинства других брюхоногих из яйца выходит уже почти пол-
ностью сформированный брюхоногий моллюск. Он отличается от взрослого
лишь меньшими размерами тела, раковиной с меньшим количеством оборо-
тов и неразвитой половой системой. Таким образом, в этом случае развитие
идет без свободноплавающих личиночных стадий — трохофоры и велпгера,
а торсионный процесс происходит на очень ранних этапах развития заро-
дыша, в яйце.
ПРОИСХОЖДЕНИЕ ЛИЧИНОЧНОГО СКРУЧИВАНИЯ
И АСИММЕТРИИ
Как было отмечено, перемещение мантийной полости в переднее поло-
жение и асимметрия — характерные особенности брюхоногих моллюсков.
Вопрос о происхождении этих явлений — один из сложных вопросов мор-
фологии беспозвоночных, для его объяснения было выдвинуто немало ги-
потез. Многие из них совершенно устарели, из других составилось совре-
менное представление о происхождении этих
особенностей организации брюхоногих моллю-
сков.
По мнению всех исследователей, предпо-
лагаемый предок брюхоногих был двусторонне
симметричным. Сердце с двумя боковыми
предсердиями парные жабры и почки были
расположены сзади (рис. 257). Здесь же нахо-
дилось и анальное отверстие. Такой моллюск
имел раковину в виде низкого колпачка. Да-
лее, в ходе эволюционного развития брюхо-
ногих объем внутренностного мешка стал
увеличиваться, что повлекло за собой увели-
чение высоты и конусовидности раковины.
Вряд ли такая форма раковины была удобна
при передвижении животного. Предпочтитель-
ней оказалась форма внутренностного мешка
с вершиной, склоненной вперед, как у иско-
паемых и современных Monoplacophora. Однако и такое расположение
вершины внутренностного мешка над головой не вполне удобно ни для
свободноплавающей личинки, ни для осевшего на дно взрослого моллюска.
И в том, и в другом случае раковина, нависая, давит своим краем на голо-
ву, затрудняя и ползание и быстрое втягивание под раковину головы с
органами чувств. Несомненно выгоднее обратное расположение раковины
с висцеральным мешком, при котором над головой будет больше свободного
пространства и, следовательно, меньше помех при движении.
Поворот внутренностного мешка и связанной с ним раковины на 180°
вначале происходил от случая к случаю. Однако дальнейшее увеличение
объема висцерального мешка и, следовательно, высоты завитка привело,
видимо, к необходимости «узаконить» этот поворот и сделать его обязатель-
ным явлением в онтогенезе брюхоногих моллюсков. Как уже упоминалось,
такой поворот можно наблюдать при развитии примитивных современных
брюхоногих. Этот поворот на 180°, перемещающий мантийную полость с
комплексом ее органов в переднее положение, называется личиночным за-
кручиванием, или торсионным процессом. Он же вызывает петлеобразное
изгибание кишечника и своеобразный перекрест нервных тяжей (хиасто-
неврию).
Дальнейшее прогрессивное увеличение объема внутренностного мешка
привело к необходимости выработать наиболее выгодную форму раковины.
Вначале это были плоскоспиральные раковины, т. е. раковины с завитком,
скрученным в спираль, лежащую в одной плоскости (рис. 258). Такими были
раковины ископаемых беллерофонтид. Они заменились еще более совершен-
301
ными — турбоспиральными раковинами с конусовидным спиральным завит-
ком. Такая форма висцерального мешка и раковины лучше всех других
обеспечивает возможность дальнейшего увеличения объема внутренност-
ного мешка при минимальном увеличении внешних размеров тяжелой ра-
ковины. Именно такие турбоспиральные раковины и получили наибольшее
развитие в классе брюхоногих.
Однако торчащий в сторону завиток создавал не меньше неудобств для
движения моллюска. Неудобства от неравновесного положения внутрен-
ностного мешка должны были возрастать при дальнейшем увеличении высо-
ты турбоспирали. Видимо, для ликвидации этого неравновесного располо-
жения брюхоногие стали носить свою раковину в положении, изображенном
Рис. 258. Образование асимметрии раковины и мантийного комплекса (схема).
I — V — ряд последовательных гипотетических стадий:
1 — жабры, 2— желудочек, 3— предсердия, 7— рудимент предсердия, 5—задняя кишка
на рис. 258ш, т. е. при сильно косом положении оси турбоспирали. Такое
постоянное положение раковины привело к значительному сокращению объе-
ма правой части мантийной полости и, как следствие этого, к редукции пра-
вой жабры, правого предсердия и правой почки, т. е. взаимосвязанных ор-
ганов. В этом и проявляется столь характерная для большинства брюхо-
ногих асимметрия.
Следует отметить, что степень редукции названных органов у современ-
ных моллюсков находится в прямой зависимости от степени развития турбо-
спирального завитка. Так, современные примитивные брюхоногие с колпач-
ковидной раковиной совершенно симметричны, у других — с низким завит-
ком и малым количеством оборотов раковины — далеко не полная редукция
этих органов правой стороны.
СИСТЕМАТИЧЕСКИЙ ОБЗОР КЛАССА
ПОДКЛАСС I. ПЕРЕДНЕЖАБЕРНЫЕ (PROSOBRANCHIA)
ХАРАКТЕРИСТИКА
Брюхоногие моллюски, у которых мантийный комплекс лежит в перед-
ней части тела и ктенидии помещаются впереди сердца (см. рис. 254, 255).
Раковина, за редким исключением, хорошо развита, обычно спиральная,
реже колпачковидная. В большинстве случаев имеется крышечка. Морские
формы, лишь небольшое число приспособилось к обитанию в пресных водах
или вышло на сушу.
Подкласс делится на 2 отряда: 1) двупредсердные и 2) однопредсердные.
ОТРЯД ДВУПРЕДСЕРДНЫЕ (DIOTOCARD1A)
К этому отряду относятся наиболее примитивные брюхоногие. У одних
форм мантийный комплекс симметричный, т. е. состоит из парных ктени-
диев, почек и предсердий (парножаберные, рис. 258). У других — комплекс
302
несимметричный, остается один ктенидий и нормально развито одно пред-
сердие, тогда как второе рудиментарно (непарножаберные). В некоторых
случаях ктенидии вообще отсутствуют и заменены кольцом вторичных
жабр по краю мантии (кругложаберные). Раковина или спирально завитая,
или колпачковидная, в некоторых случаях уплощенная с рядом отверстий
(морское ухо). В этот отряд входит небольшое число видов, обитающих
главным образом в морях. В пресных водах и на суше встречаются очень
немногие. Сюда относятся из морских форм Puncturella с симметричной ра-
ковиной в виде колпачка, с вершиной, загнутой назад, и отверстием на
вершине; морское ухо (Haliotis); морские блюдечки (Patella, Acmaea)
(табл. VI) и ряд форм со спирально закрученной раковиной (Trochus,
Margarites и др.). Из пресноводных сюда относится Theodoxus — небольшой
моллюск с низкой раковиной, обитает в водоемах юга нашей страны.
ОТРЯД ОДНОПРЕДСЕРДНЫЕ (MONOTOCARDIA)
К этому отряду относится основная масса переднежаберных. Имеется
одна жабра и одно предсердие. Раковина обычно спирально завитая, лишь
у немногих (киленогие) она редуцируется (рис. 262). К этому отряду
принадлежит большое число морских форм, богато представленных во всех
морях: Littorina, Natica, Buccinum (табл. VI) и др.
В пресных водах представителей этого отряда меньше, и они сильно
уступают в числе легочным (см. ниже). Наиболее обычны: живородка, или
лужанка (Vivipara, Bithynia) (табл. VI), и др. Много видов из этого отряда
обитает в оз. Байкал, где имеется несколько эндемичных родов (Baikalia,
Benedict!а и др.).
ПОДКЛАСС П. ЗАДНЕЖАБЕРНЫЕ (OPISTHOB RANCHI А)
ХАРАКТЕРИСТИКА
Брюхоногие, у которых мантийная полость со всеми ее органами лежит
на боку справа, а единственное предсердие и единственная жабра — позади
желудочка (рис. 259). Это положение мантийной
полости и жабры почти соответствует исходному,
т. е. их положению до торсионного процесса. Та-
кое расположение этих органов возникает благо-
даря так называемому регулятивному раскручи-
ванию — перемещению мантийной полости из
положения, достигнутого торсионным процессом
в почти обратное. В наибольшей степени это
достигается у заднежаберных, способных к бы-
стрым поступательным движениям, или у задне-
жаберных, утративших раковину и тоже движу-
щихся очень активно. Естественно, что при
активном поступательном движении удобнее
жабра, направленная назад, тем более если она
не прикрыта раковиной. Неудобства, которые
возникали у предков переднежаберных с их зад-
ним положением мантийной полости, у современ-
ных заднежаберных не проявляются вследствие
Рис. 259. Морской брюхо-
ногий моллюск Philine (зад-
нежаберные), раковина и
часть мантии удалены, пи-
щеварительная система не
показана:
/ — голова, 2 — нога, 3 — жабра,
4— сердце, 5 — почка, 6 — гер-
мафродитная железа
редукции раковины и развития асимметрии,
предшествовавшей регулятивному раскручива-
нию.
Исключительно морские формы — некоторые
ползают по дну, другие свободно плавают в тол-
ще воды.
Подкласс делится на 3 отряда: 1) покрытожа-
берные, 2) голожаберные и 3) крылоногие.
303
ОТРЯД ПОКРЫТОЖАБЕРНЫЕ (TECTIBRANCHIA)
Ползающие формы. Раковина развита слабо, в некоторых случаях она
более или менее полно закрыта мантией. Сюда относятся* Cylichna, Philine
(см. рис. 259) (северные моря), морской заяц Aplysia (теплые моря).
ОТРЯД ГОЛОЖАБЕРНЫЕ (NUDIВRANCHIA)
Ползающие формы. Раковина редуцирована.
Настоящие жабры отсутствуют и заменены наруж-
ными выростами кожи различной сложности. Сюда
относятся: Dendronotus (рис. 261), Cadlina, Согу-
phella и др.
Рис. 260. Крылоногий моллюск — морской ангелок (Cli-
one limacina). Обычная форма наших северных морей
ОТРЯД КРЫЛОНОГИЕ (PTEROPODA)
Планктонные формы. Ра-
ковина в большинстве слу-
чаев редуцирована, если
имеется, то очень тонкая,
полупрозрачная. Нога пре-
вращена в два боковых вы-
роста. Сюда относятся: мор-
ской ангелок Clione limacina
(северные моря) и др. (рис.
260).
Рис. 261. Голожаберный мол-
люск — дендронотус (Dendronotus
arborescens, Японское море)
Рис. 262. Киленогий мол-
люск — каринария (Carin-
aria mediterranea, Средизе-
мное море):
I — рудиментарная раковина, 2 —
нога
304
ПОДКЛАСС III. ЛЕГОЧНЫЕ (PULMONATA)
ХАРАКТЕРИСТИКА
Жабры редуцированы и заменены сложно разветвленной в мантии
сетью кровеносных сосудов. Раковина у большинства представителей раз-
вита хорошо, спирально завитая, различная по высоте, иногда совершенно
плоская. В некоторых случаях раковина редуцирована. Наземные и вторич-
новодные— пресноводные формы. Подкласс делится на 2 отряда: 1) сидя-
чеглазые и 2) стебельчатоглазые.
ОТРЯД СИДЯЧЕГЛАЗЫЕ (BASOMMATOPHORA)
Глаза расположены при основании щупалец. Раковина обычно хорошо
развитая, спирально завитая, редко колпачковидная. Главным образом прес-
новодные формы. Сюда относится основная масса наших пресноводных брю-
хоногих моллюсков — Limnaea, Planorbis, Physa, Ancylus (с колпачковид-
ной раковиной) и др. (табл. VI).
ОТРЯД СТЕБЕЛЬЧАТОГЛАЗЫЕ (STYLOMMATOPHORA)
Глаза расположены на вершине второй пары щупалец. Раковина у
большинства форм хорошо развита, спирально завитая, в некоторых слу-
Рис. 263. Слизень (Arion):
/ — редуцированная раковина, обросшая мантией^ 2— дыха-
тельное отверстие
чаях (слизни) редуцирована. Наземные формы. Сюда относится большое
число наземных брюхоногих моллюсков, так называемых улиток — Helix,
Bradibaena, Vallonia, Clausilia и др. и слизней — Arion, (рис. 263), Limax,
Agriolimax и др.
ОБРАЗ ЖИЗНИ И РАСПРОСТРАНЕНИЕ БРЮХОНОГИХ
Брюхоногие в большинстве свободноживущие, подвижные животные.
Паразитических форм среди них очень немного.
Паразитизм столь сильно изменяет строение тела (рис. 264), что
узнать в этом паразите моллюска можно лишь в результате изучения
эмбрионального развития, которое до определенных стадий идет у них
характерным для моллюсков путем. Паразитируют исключительно на игло-
кожих — морских звездах.
Большинство брюхоногих ведут донный ползающий образ жизни и от-
носятся, таким образом, к бентосу. Некоторые из них—плавающие формы,
относящиеся к макропланктону; сюда относятся все киленогие (см. рис.
262), все крылоногие и некоторые другие. У этих форм наблюдается обычно
редукция или исчезновение раковин.
305
Таблица VI. Брюхоногие пресных вод (7—12) и морей (13—19) СССР:
1— малый прудовик (Limnaea truncatula), 2— ушковый прудовик (L. auricula-
ria), 3— обыкновенный прудовик (L. stagnalis), 4— Ancylus fluviatilis, 5—Phy-
sa fontinalis, 6— Valvata piscinalis, 7 — Amphipeplea glutinosa, 8 — живородка
обыкновенная (Vivipara vivipara) (крышечка отдельно), 9— Bythinia leachi^
JO — Aplexa hypnorum, 11 — Baicalia stidae — Байкал, 12 — катушка обыкновен-
ная (Planorbis corneus) (a — сверху, 6 — сбоку), 13—морское блюдечко (Acmaea
testudinalis)— северные моря, 14—Natica clausa — северные моря, 15 — Rha-
рапа bezoar — Японское и Черное моря, 16 — Cerithium ponticus — Черное море,
17— литторина (Littorina rubis) — северные моря, 18— Nassa reticulata — Черное
море, 19 — Buccinum undatum — северные моря
306
Рис. 264. Паразитический брю-
хоногий моллюск (Parenteroxenos
dogieli) из полости тела голоту-
рии, естественная величина до
130 см\
1 — передний конец, 2—задний конец
тела
Движение брюхоногих происходит при помощи ноги или ее придатков.
У ползающих форм нога хорошо развита и движение совершается благодаря
пробегающим по подошве волнам сокращения мускулатуры.
Питаются брюхоногие в большинстве растительной пищей, которую они
соскабливают при помощи радулы. Некоторые из них—хищники, захваты-
вающие при помощи хобота мелких животных. Другая небольшая группа
хищников захватывает добычу сильно измененными зубами радулы, выво-
рачивая для этого свою глотку. Третьи имеют сложные слюнные железы,
выделением которых они растворяют створки раковин двустворчатых и
высасывают их мягкие части. Примером такого хищника может служить
довольно обычная в наших северных морях натика (Natica) (табл. VI).
Дышат брюхоногие или при помощи
жабр, или при помощи легкого. К легоч-
ным брюхоногим относятся не только на-
земные формы, но и многие пресноводные.
Последние могут рассматриваться как
вторичноводные формы, происшедшие от
наземных. Дышат они, забирая в мантий-
ную полость воздух, для чего регулярно
поднимаются на поверхность воды. Назем-
ные формы могут существовать только в
таких местах, где имеется хотя бы неболь-
шая влажность. При наступлении жаркого
времени дня они выделяют большое коли-
чество слизи, которая как бы заклеивает
отверстие раковины. Прикрепившись в
таком состоянии к растительности, моллюс-
ки пребывают так до ночи, когда они вновь
делаются активными.
Размножаются брюхоногие, как и все
моллюски, только половым путем. Оплодо-
творение внутреннее, причем у гермафро-
дитных форм оно взаимное. Яйца отклады-
ваются обычно группами в виде различных
по форме кладок. У водных брюхоногих
они слизистые в виде колбасок или кучек.
У морских форм образуется личинка парус-
ник, или велигер (см. рис. 256), представ-
ляющая собой измененную трохофорную
личинку с несколькими предротовыми лопастями. Велигер парит в толще
воды. У него довольно рано образуются характерные для брюхоногих ра-
ковина и нога и, как уже было сказано, он опускается на дно и превра-
щается во взрослого моллюска.
Наземные формы откладывают одиночные яйца, покрытые плотной обо-
лочкой. Развитие у наземных прямое, без личиночных стадий и протекает
до самого конца внутри яйцевых оболочек. У некоторых форм оплодотворен-
ные яйца развиваются в особой камере, представляющей собой расширение
яйцевода. В таких случаях из материнского организма выходят уже вполне
сформировавшиеся маленькие моллюски. Примером такого живородящего
моллюска может быть наша обычная пресноводная живородка (Vivipara)
(см. рис. 254).
Распространены брюхоногие очень широко. В морях они обитают, на-
чиная с прибрежных камней до больших глубин. На береговых камнях се-
верных морей постоянно можно встретить большое количество мелких перед-
нежаберных литторин (Littorina), остающихся при отливе вне воды, крепко
присосавшихся к камням. Здесь встречаются и морские блюдечки (Астаеа).
Зона водорослей очень богата брюхоногими, здесь встречаются разнообраз-
ные переднежаберные и заднежаберные. Глубже встречаются крупные, до-
307
стирающие 10 см в высоту, трубачи (Buccinum и Neptunea), хищные моллюс-
ки, имеющие хорошо развитый хобот.
Распространены брюхоногие по всем морям, причем особенно обильны
и разнообразны они в тропиках, где встречаются очень крупные формы, до-
стигающие 30—40 см высоты. Во всех наших морях брюхоногие достаточно
разнообразны. Всего в морях СССР известно более 500 видов.
Пресные водоемы также обильно населены брюхоногими. Они встре-
чаются в больших озерах и в маленьких весенних лужах, в бурных горных
речках и на заболоченных лугах. Всего в пресных водах СССР известно
160 видов брюхоногих, причем особенно богато ими оз. Байкал, где имеется
особая эндемичная фауна, насчитывающая 72 вида, из которых отдельные
виды очень оригинальны. Некоторые пресноводные брюхоногие встречаются
иногда в громадном количестве, так, например, малый прудовик Galba
truncatula (см. ниже) достигает в благоприятных условиях нескольких
миллионов экземпляров на 1 га.
Наземные брюхоногие встречаются по всему земному шару, за исклю-
чением Крайнего Севера; особенно богато представлены они в тропиках,
где имеются очень крупные формы, достигающие 15 см в высоту Обитают
они как на равнине, в полях и в лесу, так и в горах до высоты 5000м. Осо-
бенно многочисленны они во влажных местах с известковой почвой. В пре-
делах СССР наземные брюхоногие разнообразны, особенно в южных районах
(Крым, Кавказ, Средняя Азия). Всего у нас их известно около 600 видов.
Так же как и пресноводные формы, наземные брюхоногие иногда встречаются
массами, например распространенная у нас на юге зебрина (Zebrina).
ПРАКТИЧЕСКОЕ ЗНАЧЕНИЕ БРЮХОНОГИХ
Некоторых морских и наземных брюхоногих добывают и даже разводят
(например, виноградную улитку) как объекты, пригодные в пищу. Это рас-
пространено в странах Дальнего Востока, США и в Западной Европе, в
меньшей степени у нас в Прибалтике.
Несомненно значительнее вред, причиняемый брюхоногими. Из мор-
ских брюхоногих следует отметить рапану (Rapana) — крупного моллюска,
проникшего в Черное море из Средиземного. Этот моллюск сильно выедает
устриц — разводимых двустворчатых моллюсков. Из наземных брюхоно-
гих следует назвать безраковинных слизней, которые в некоторые годы
сильно вредят всходам озимых (полевой слизень Agriolimax agrestis) и
огородным культурам, поедая их стебли, листья и плоды.
Следы повреждений, оставляемые моллюсками, очень характерны: на
листьях — неправильно округлые дыры, а на плодах и клубнях — широкие,
неглубокие ямы. Действуют они при этом радулой, следы от которой и бы-
вают видны на повреждениях. Полевой слизень размножается обычно все
лето, откладывая до 500 яиц кучками по 20—30 яиц в землю или под мох.
Из отложенных яиц через 2—3 недели выводятся молодые слизни, которые
уже через 1,5 месяца делаются половозрелыми. Таким образом, темп раз-
множения полевых слизней велик, но вместе с тем он очень зависит от внеш-
них условий — температуры и влажности. Слизни отмирают уже при повы-
шении температуры до 30°С и при падении влажности почвы до 10—15%
Поэтому массовое развитие слизней обычно бывает в прохладное и сы-
рое лето.
Лучшим средством борьбы со слизнями является применение отравляю-
щих приманок (метальдегид).
Другой способ борьбы со слизнями, вредящими в садах и огородах,
сгруживание в кучи опавших листьев с последующим сжиганием их. При
сгруживании листьев важно как можно лучше оголить почву, тогда слизни
непременно соберутся в массе.
Ряд наземных и пресноводных брюхоногих — промежуточные хозяева
паразитических червей человека и домашних животных. Большое значение
308
в этом отношении имеет малый прудовик (Galba truncatula) — промежуточ-
ный хозяин печеночной двуустки (Fasciola hepatica) (см. выше). Этот мол-
люск очень широко распространен и населяет главным образом мелкие
водоемы, а также сырые болотистые луга иногда в громадном количестве.
Первым промежуточным хозяином кошачьей двуустки (Opistorchis
felineus) служит пресноводный моллюск Bithynia leachi, а на Дальнем Во-
стоке пресноводные Melania — промежуточные хозяева для шистозом —
кровепаразитов человека.
Велико значение брюхоногих как пищи некоторых промысловых
рыб и морских млекопитающих — китов. Киты питаются одним из массо-
вых планктических видов Clione limacina (морской ангелок) (рис. 260).
ФИЛОГЕНИЯ БРЮХОНОГИХ
Несомненно, что брюхоногие с характерным для всех них передним
или боковым положением мантийной полости происходят от форм, у которых
она располагалась сзади, как у панцирных и моноплакофор. На это указы-
вает прежде всего эмбриональное развитие примитивных брюхоногих (На-
liotis — морское ухо, Patella — морское блюдечко и некоторых других).
У свободноплавающих личинок этих моллюсков мантийная полость с аналь-
ным отверстием и жабрами до торсионного процесса лежит сзади, а личиноч-
ная раковина своей вершиной нависает над головой. Организация совре-
менных моноплакофор во многом соответствует этим «доторсионным» ли-
чинкам примитивных брюхоногих и вследствие этого, видимо, соответствует
и их предку. Но в то же время организация моноплакофор многим и от-
личается от него. Следует, видимо, предположить, что брюхоногие имели
много общего с моноплакофорами и панцирными, однако эти три группы
рано «разошлись» и каждая по-своему специализировалась. Основное
в специализации брюхоногих — это выработка торсионного процесса, раз-
витие турбоспиральной раковины и асимметрии.
Среди брюхоногих наиболее изменены и специализированы по сравни
нию с примитивными переднежаберными легочные и заднежаберные. У
первых это связано с выходом на сушу (редукция жабры и развитие лег-
кого), у вторых в основном связано с развитием большой подвижности и
редукцией раковины.
КЛАСС 5. ЛОПАТОНОГИЕ (SCAPHOPODA)
ХАРАКТЕРИСТИКА
Двустороннесимметричные моллюски с трубкообразной, открытой на
обоих концах, несколько суженной к одному концу, раковиной (рис. 265).
Мантия срастается в цельную трубку. Имеются нога
и небольшой головной вырост без глаз, несущий обыч-
но многочисленные щупальца. В ротовой полости
имеются роговые челюсть и терка. Жабр нет, их заме-
няют выросты задней кишки. Кровеносная система
сильно редуцирована — сердце без предсердий, сосу-
дов нет. Почки не сообщаются с перикардием. Нерв-
ная система с ясно обособленными нервными узлами.
Раздельнополые, половая железа непарная и откры-
вается в правую почку. Развитие с трохофорообразной
щчинкой.
Экология и распространение. Лопатоногие — ис-
ключительно морская группа, охватывающая всего
Рис. 265. Денталиум (Dentalium), схематический продольный
разрез-
/ — раковина, 2—нога,-5— голова, 4—головные щупальца; 6—мантия,
6 — мантийная полость^ 7 — кишечник, 8 — гонада
309
около 150 видов. Обитают в грунте, куда вкапываются при помощи ноги,
высовывающейся из широкого конца раковины. Суженный конец обычно
торчит из грунта. Лопатоногие — хищники, питающиеся преимущественно
фораминиферами и молодыми двустворчатыми моллюсками. Распростране-
ны лопатоногие во всех морях. В наших северных морях наиболее обычен
морской зуб (Dentalium entale), достигающий 2—3 см в длину.
Филогения. Происхождение лопатоногих к настоящему времени еще
не выяснено.
КЛАСС 6. ДВУСТВОРЧАТЫЕ,
ИЛИ ПЛАСТИНЧАТОЖАБЕРНЫЕ
(BIVALVIA, ИЛИ LAMELLIBRANCHIA)
ХАРАКТЕРИСТИКА
Билатсральносимметричные моллюски, раковина которых состоит из
двух створок, охватывающих тело с боков. Обширная мантийная полость с
комплексом мантийных органов лежит по бокам тела (рис. 266). Обособлен-
ные. 266. Мантийная полость и строение жабр у различных двустворча-
тых моллюсков, схематизированный поперечный разрез тела. / —первич-
ножаберные; // —нитежаберные; III —настоящие пластинчатожаберные;
IV — перегородчатожаберные
ной головы нет, органы чувств развиты довольно слабо. Нет также челюстей
и радулы. Последнее стоит в связи
с тем, что двустворчатые питаются путем
отфильтровывания органической взвеси
из воды, засасываемой в рот через ман-
тийную полость. Билатеральная симмет-
рия отчетливо выражена и во внутрен-
ней организации — двупредсердное серд-
це, две почки и т. д. Свободноплаваю-
щие личинки трохофорного типа (рис.
267), у многих свободноплавающей ли-
чинки нет.
Обитают в морях и пресноводных во-
доемах. Известно около 15000 видов.
Рис. 267. Личинка морского двустворчатого
моллюска:
1 — венчики ресничек, 2 — зачаток раковины, 3 — рот,
4 — желудок, 5 — анальное отверстие, 6 — теменная
пластинка
'ЭЮ
Представитель класса двустворчатых —
беззубка (Anodonta)
Внешнее строение. Живая беззубка вне воды плотно смыкает створки
раковины парой замыкательных мускулов; причем все мягкие части втя-
гиваются в раковину. В воде раковина приоткрыта и из нее высовываются
на переднем конце клинообразная нога, а на заднем два небольших выроста
мантии — нижний вводящий и верхний выводящий сифоны (рис. 268).
Раковина состоит из двух боковых, вполне симметричных створок с
несколько эксцентрично расположенной вершиной, сдвинутой к переднему
концу. На спинной стороне раковины имеется эластическая связка — ли-
гамент, которая связывает обе створки и автоматически открывает их,
когда ослабевает действие замыкательных мускулов. Лигамент образуется
за счет наружного органического слоя раковины. Раковина беззубки со-
стоит из трех слоев — наружного, органического — конхиолинового1, так
Рис. 268. Беззубка Anodonta с обычным
положением ноги и сифонов, уменьшено:
1—нога, 2—вводящий сифон, 3—выводящий си-
фон, 4 — вершина раковины с обнаженным фарфо-
ровидным слоем, 5 — лигамент
Рис. 269. Правая створка беззубки с
внутренней стороны:
/ — мантийная линия, 2 — отпечатки замыка-
тельных мышц, 3 — опечаток протрактора но-
ги, 4— отпечатки ретракторов ноги
называемого периостракума, и двух известковых — среднего — фарфоро-
вого—и внутреннего—перламутрового. Периостракум на вершине раковины
обычно стирается и на ней поэтому обычно обнажен толстый фарфоровый
слой. На поверхности раковин проходят концентрические линии, отражаю-
щие рост моллюска и являющиеся, таким образом, линиями роста.
С внутренней стороны раковина выстлана перламутровым слоем с ха-
рактерным переливающимся блеском. На внутренней стороне обеих створок
имеются отпечатки некоторых мягких частей: двух хорошо видных замы-
кательных мускулов и несколько более слабо выраженных отпечатков края
мантии, двух мускулов, втягивающих ногу (ретракторов), и одного муску-
ла, выдвигающего ногу (протрактора) (рис. 269).
Мантия. Мантия подстилает обе створки своей формой соответствует
форме раковины. Ее наружный эпителий принимает участие в формирова-
нии всех трех слоев раковины, что происходит при восстановлении повреж-
денных участков. При нормальном росте животного раковина строится при
участии края мантии по всей его длине.
На заднем крае складки мантии не полностью срастаются и образуют
два сифона, причем нижний — вводящий — имеет бахромчатый край, со-
стоящий из ряда отростков, или папилл.
Мантийная полость. Под мантией расположена обширная мантий-
ная полость, в которой помещаются жабры (рис. 270). В мантийную
полость открываются: на переднем конце ротовое отверстие, а на заднем,
около выводящего сифона, анальное отверстие, отверстия почек и половых
желез.
1 Конхиолин —белковое вещество, содержащее до 50% углерода, 16,5% азота,
0,8% серы и др.
311
Вся поверхность жабр покрыта ресничным эпителием, постоянное дви-
жение которого вызывает непрерывный ток воды через вводящий сифон
внутрь мантийной полости. Вместе с водой приносятся пищевые частицы и
Рис. 270. Внутреннее строение беззубки (схема):
] — мантия, 2 — вога, 3—вводящий сифон, 4—выводящий сифон, .5—замыкатель-
ные мышцы, 6 — ротовые щупальца, 7 — ротовое отверстие, 8 — желудок, 9 — пе-
чень, кишечник, И — анальное отверстие, 12—сердце,- 13—перикардий, 14 —
почка, 15—кеберов орган, 16—головной ганглий, 17 — ножной ганглий, 18— вис-
церальный ганглий, 19— гонада, 20— мантийная полость
необходимый для дыхания кислород. Вода проделывает по мантийной по
лости сложный путь и выходит через выводящий сифон, унося углекислоту,
продукты обмена и экскременты.
Нога. Нога представляет собой мускулистый клинообразный вырост
брюшной стенки внутренностного мешка, направленный острой вершиной
Рис. 271. Нервная систе-
ма беззубки (схема):
1 — головные ганглии, 2 — ту-
ловищные ганглии, 3—нож-
ные ганглии
вперед. Нога имеет систему мышц, управляющих ее
движением, в частности мышцы выдвигающие и
мышцы, втягивающие ее. В верхнюю часть ноги,
прилежащую к собственному телу, входят некото-
рые внутренние органы. Нога иннервируется осо-
бой парой ножных ганглиев (см. ниже).
Полость тела. Полость тела сильно редуци-
рована и представлена только околосердечной
сумкой (перикардием) и полостями половых желез
(рис. 270).
Нервная система. Имеются три пары нервных
узлов — головные, ножные и туловищные (рис. 270,
271). Ганглии каждой пары соединены между собой
короткими комиссурами; кроме того, головные узлы
соединены с туловищными и ножными.
Органы чувств. Органы чувств развиты слабо
и представлены главным образом отдельными чув-
ствительными клетками, рассеянными в эпителии
ротовых лопастей, папиллах вводящего сифона и
мантии. Имеется также пара небольших статоцис-
тов, лежащих недалеко от ножных ганглиев, и ос-
фрадии.
Пищеварительная система. Ротовое отверстие
расположено в передней части животного. С обеих
его сторон лежит по паре ротовых лопастей. Рот
открывается непосредственно в короткий пищевод,
переходящий в довольно объемистый желудок.
312
между собой соединительноткаными
Рис. 272. Глохидий беззубки. / — отдельный
глохидий; II глохидий на плавнике рыбы.
/ — створки раковины, 2 зубцы ее, 3 — замыкатель-
ный мускул, 4 — чувствительные щетинки
Вокруг желудка расположена пищеварительная железа, или печень. На
брюшной стороне желудка помещается своеобразное образование, так
называемый кристаллический стебелек — стекловидно-прозрачная студени-
стая палочка, он сортирует поступающие в желудок частицы и выделяет
вещества, склеивающие их.
От желудка начинается длинная кишка, образующая несколько петель
и открывающаяся анальным отверстием около выводящего сифона.
Дыхательная система. Органы дыхания, или жабры, расположены по
бокам ноги и представлены с каждой стороны ее двумя полу жаб рами —
внутренней и наружной. Каждая полужабра слагается из ряда изогнутых
жаберных нитей, тесно соединен
перекладинами и образующих
две решетчатые жаберные плас-
тинки — нисходящую и восхо-
дящую, переходящие внизу одна
в другую и соединенные между
собой перекладинами.
Таким образом, каждая по-
лужабра представляет собой ре-
шетчатое двуслойное образо-
вание.
Кровеносная система. Кро-
веносная система незамкнутая.
На спинной стороне помещается
сердце, состоящее из желудоч-
ка, лежащего по средней линии
и прободенного кишкой, и двух
предсердий, лежащих по бокам
его. От желудочка вперед и назад
отходят два основных сосуда —
аорты, распадающиеся далее на
артерии и капилляры. Капилля-
ры изливают кровь в лакуны,
расположенные среди паренхи-
мы. Кровь, насыщенная углекис-
лотой, собирается в вены, про-
ходит через жабры и органы
выделения; затем поступает в
предсердия, а из них в желу-
дочек.
Выделительная система. Органами выделения является пара почек,
лежащих по одной с каждой стороны перикардия и носящих название боя-
нусовых органов. Почки представляют собой сильно видоизмененные цело-
модукты (см. выше) и открываются с одной стороны в перикардий, с другой—
в мантийную полость. Кроме того, органом выделения служит кеберов ор-
ган, представляющий собой железистое утолщение передней стенки пери-
кардия.
Половая система. Беззубки обычно раздельнополы, но внешне по полу
почти неотличимы. Парная половая железа, или гонада, лежит в верхней
части ноги между петлями кишки (рис. 270), открывается она в мантийную
полость недалеко от отверстия почек. Гонады развиты не всегда одинаково
и обычно сильно увеличиваются в размере во время половозрелости беззуб-
ки, которая наступает в условиях средней полосы СССР обычно
осенью.
Развитие. Оплодотворение происходит в мантийной полости, куда
откладываются яйца и приносятся с водой сперматозоиды. Дальнейшее
развитие оплодотворенных яиц происходит в жабрах, причем как выводко-
вые сумки используются обычно обе наружные полужабры. В жабрах раз-
313
виваются личиночные формы — глохидии, имеющие вид маленького моллюс-
ка уже с двустворчатой раковиной (рис. 272).
Количество глохидий в одной беззубке может достигать несколько сот
тысяч. Глохидии остаются в жабрах матери довольно долгое время, обычно
до весны. Для дальнейшего развития глохидии должны попасть на плавники
рыб.
Таблица VII. Двустворчатые пресных вод СССР:
1 — перловица (Unio pictorutn), 2— дрейссена (Dreissena polymorpha), 3 —
беззубка (Anodonta cygnea), 4 — Cristaria plicata — водоемы Дальнего Восто-
ка, 7 — шаровка (Sphaerium corneum), 6—горошинка (Pisidiuni amnicum),
7 —речная жемчужница (Margaritana margaritifera)— водоемы севера евро-
пейской части СССР
Это происходит, когда рыба проплывает около беззубки с готовыми
глохидиями — они выталкиваются через сифоны наружу и с помощью осо-
бых клейких нитей (биссуса) и шипов личиночной раковины прикрепляются
к жабрам или плавникам. Через некоторое время глохидии погружаются в
ткань плавника. Они не имеют развитого кишечника, питание их происхо-
дит за счет тканей хозяина при помощи клеток личиночной мантии. Парази-
314
тический период продолжается от 1 до 2 месяцев, после чего выросший уже
маленький моллюск покидает хозяина, опускается на дно и переходит к
нормальному образу жизни.
Экология и распространение. Различные виды рода Anodonta широко
распространены и встречаются в водоемах различного типа как текучих,
так и стоячих. В СССР известно 12 видов беззубок. Наиболее обычные —
крупная, достигающая 20 см, Anodonta cygnea (табл. VII), обитающая в стоя-
чих водоемах, и более мелкая A. piscinalis, обитающая в реках.
Беззубки, как большинство двустворчатых, — фильтраторы и питаются
главным образом отфильтрованным из воды детритом. Размножаются без-
зубки обычно осенью, когда происходит откладка яиц в мантийную полость
и оплодотворение. Выход глохидий наружу бывает весной, а в летние ме-
сяцы происходит созревание половых продуктов.
Рост беззубок происходит обычно более интенсивно в первые годы
жизни, а затем замедляется. Беззубки очень изменчивы в связи с условиями
существования (характер водоема, химизм воды, условия питания и пр.).
Систематическое положение. Род Anodonta относится к отряду Eula-
mellibranchia, т. е. настоящих пластинчатожаберных.
ОБЩИЙ ОБЗОР КЛАССА
Раковина. Створки раковины охватывают тело моллюска с боков, но
у форм, лежащих на грунте, например у морского гребешка (Pecten)
(табл. VIII), или прирастающих одной створкой, например у устриц (рис.
273), так же охватывая тело с боков, они занимают нижнее и верхнее поло-
жение и обозначаются как брюшная и спинная. У таких форм имеется только
один замыкательный мускул, занимающий срединное положение.
Створки раковины скрепляются у большинства двустворчатых
не только лигаментом и за-
мыкательными мышцами, как у
беззубки, но еще и замком.
Замок состоит из развивающих-
ся на верхнем крае одной створ-
ки выступов, или зубов, и соот-
ветствующих углублений на
другой, в которые зубы входят.
Замки устроены разнообразно.
Наиболее обычны два типа их —
таксодонтный, состоящий из
многочисленных мелких одина-
ковых зубов, и гетеродонтный,
состоящий из немногочисленных
различных по форме зубов
(табл. VIII).
Редукция раковины наблю-
дается у двустворчатых очень
редко. Примером может служить
корабельный червь (Teredo nava-
lis) — очень своеобразный мор-
ской моллюск, сверлящий дре-
весину и приносящий этим боль-
шой вред (см. ниже). Тело этого
моллюска сильно вытянуто, при-
чем раковина покрывает только
небольшой участок переднего
конца (рис. 274). Служит она не
для защиты, а помогает моллюс-
ку сверлить дерево.
Рис. 273. Группа устриц на
свае
Рис. 274. Корабельный червь
(Teredo navalis):
1— редуцированная раковина, 2—
рудиментарная нога, 3 — сифоны, 4 —
мантийная полость, 5 — жабры, 6 —
кишечник, 7 — сердце, 8 — гонады
315
Таблица VIII. Двустворчатые морей СССР:
/ — мидия (Mytilus edulis) — северные моря, 2— Portlandia arctica — северные моря,- 3 — фазе-
олина (Modiola phaseolina) — Черное море, 4 — белая ракушка (Spisula sachalinensis) — Япон-
ское море (а — снаружи, б— с внутренней стороны; 1 — замок гетеродонтный), 5 — сердцевид-
ка (Cardium edule)—Черное море, Балтийское, Каспийское и Аральское моря, Мурманское
побережье, о — Leda pernula — северные моря (а — снаружи, б—с внутренней стороны; 1 —
замок гаксодоытный), 7— гребешок (Pecten islandica) — северные моря
солеи также регулируется организмом
Рис. 275. Nucula. Левая створка раковины и
мантия удалены. Вид с левой стороны:
I — выросты мантии, 2— внутренностный мешок, 3 —
ротовое отверстие, 4 — нога с сосочковидными вырос-
тами, 5 — подошва ноги, 6 — ротовые лопасти с при-
датками, 7 — ктенидий (левый)
Формирование раковины происходит следующим образом. Кровь при-
носит к клеткам края мантии (или к участку мантии, лежащему под повреж-
денным участком раковины) сложный комплекс белково-липоидных веществ.
Эти вещества «выпотевают» на поверхности мантии и накапливаются в узком
пространстве между мантией и раковиной, т. е. там, где происходит образо-
вание жемчужин. Из этих веществ особые клетки мантии строят так назы-
ваемые матрицы — органические структуры, на которых и происходит
кристаллизация сначала фосфорнокислого кальция, замещаемого потом
углекислым кальцием. Углекислый кальций откладывается в его наименее
растворимой форме — в форме арагонита. Важно отметить, что матрицы,
находясь под контролем организма, могут регулировать процесс кристалли-
зации. Кровью же приносятся к месту формирования раковины и соли
кальция; количество поступаюшд
моллюска. Таким образом, в
строительстве раковины участву-
ет весь организм, а не только
мантия, как считали ранее.
Нога. Нога у различных
форм может быть развита по-
разному. У большинства дву-
створчатых она имеет такое же
строение, как у беззубки, у не-
которых, например у сердце-
видки (Cardium), она более узкая
и вытянутая. У ряда форм она
теряет свои локомоторные функ-
ции и подвергается редукции.
Так, например, у мидии (Mytilus)
нога имеет вид тонкого языка
(табл. VIII), у морского гребеш-
ка — небольшого выроста, а у
устрицы вообще отсутствует. У
примитивной Nucula нога имеет
ползательную плоскую подошву
(рис. 275).
Нервная система и органы
чувств. Нервная система и орга-
ны чувств имеют общий для всех
двустворчатых план строения, типично выраженный у анодонты. У некоторых
форм, например у гребешков, на краю мантии имеются светочувствительные
органы — глазки, имеющие вид замкнутых пузырьков.
Пищеварительная система. Построена у большинства двустворчатых
так же, как и у беззубки.
Дыхательная система. Жабры построены у различных форм по-разно-
му, что и служит основой систематизации двустворчатых (см. рис. 266).
У примитивных форм жабры представлены двумя рядами коротких плоских
жаберных лепестков. У более сложных жаберные лепестки удлиняются и
превращаются в петлеообразные жаберные нити, не связанные спайками,
свободно свисающие в мантийную полость. У настоящих пластинчатожабер-
ных, например у беззубки, жаберные нити соединяются спайками и обра-
зуют сложные жабры.
Кровеносная и выделительная системы. Кровеносная и выделитель-
ная системы построены у большинства форм так же, как и у беззубки.
Половая система. Большинство двустворчатых раздельнополы, но
имеется и ^^гермафродитных видов. Внешне половой деморфизм у раздель-
нополых форм выражен очень слабо. Половые железы, или гонады, обычно
парные, симметрично расположенные по бокам тела. Определенной формы
они не имеют и состоят из массы сильно разветвленных трубочек, лежащих
317
среди паренхимы. Полость гонад представляет собой целомическую по-
лость.
Развитие. У большинства двустворчатых развитие протекает иначе
нежели у беззубки. Из яйца выходит свободноплавающая трохофора, прев-
ращающаяся во вторую личиночную стадию — велигер. Велигер двуствор-
чатых отличается от велигера брюхоногих тем, что на его спине двулопаст-
ная мантия формирует двустворчатую раковину. Личиночный поворот
(торсионный процесс) у двустворчатых отсутствует.
СИСТЕМАТИЧЕСКИЙ ОБЗОР КЛАССА
ОТРЯД ПЕРВИЧНОЖАБЕРНЫЕ (PROTOBRANCHIA)
Немногочисленная группа примитивных двустворчатых. Примитив-
ность выражена в целом ряде признаков. Раковина с таксодонтным зам-
ком, нога с ясно выраженной подошвой, т. е. ползательная (см. рис. 275),
рядом с церебральными ганглиями лежит пара плевральных ганглиев (у
других двустворчатых плевральные ганглии внешне не обособлены), строе-
ние жабер (см. рис. 266) одинаковое с таковым у моллюсков из других клас-
сов — это ктенидии (см. стр. 317), полость гонад соединена с почками и они,
таким образом, служат протоками половых желез, т. е. в данном случае
почки — это типичные целомодукты, такие же, как у кольчатых червей.
К этому отряду относятся обычная для северных морей Nucula, Nu-
culana, Joldia (J. arctica, обитающая в арктических морях) (табл. VIII).
ОТРЯД НИТЕЖАБЕРНЫЕ (FILIBRANCHIA)
Большая группа двустворчатых, у которых лепестки жабр вытянуты в
длинные сложенные пополам нити (см. рис. 266), у некоторых между нис-
ходящим и восходящим коленами нитевидных листков есть перемычки. Ши-
роко известны представители: морской гребешок (Pecten), который может
плавать с помощью створок раковины, мидия (Mytilus) (табл. VIII), съедоб-
ный моллюск, нити биссуса которого находили применение у древних — из
них приготовляли шелковистые ткани (и ныне это делают, но в меньшем ко-
личестве, используя для этого биссус крупной пинны). Сюда же принад-
лежит Area с таксодонтным замком раковины (у большинства других ните-
жаберных замок либо более совершенный, либо он отсутствует и створки
скрепляются лишь лигаментом, как, например, у Ostrea — устриц).
ОТРЯД НАСТОЯЩИЕ ПЛАСТИНЧАТОЖАБЕРНЫЕ
(EULAMELLIBRANCHIA)
Наиболее многочисленная группа двустворчатых. Нисходящие и вос-
ходящие нити слиты в цельные пластинки. Эти нисходящие и восходящие
пластинки соединены друг с другом многочисленными перекладинами,
благодаря чему возникает решетчатая структура соединенных пластинок—
полужабр (см. рис. 266). У большинства замок сложный — гетеродонтный,
т. е. с неодинаковыми зубами правой и левой створок.
В пресных водах широко распространены беззубка—Anodonta, Unio —
перловица, шаровки и горошинки (Sphaeridae) (табл. VII). Многочисленны
морские сердцевидки — Cardium (табл. VIII), у которых червеобразная
пружинящая нога; с ее помощью они могут совершать резкие прыжки.
Тридактны, вес раковины которых достигает многих десятков килограммов,
и множество других весьма разнообразных двустворчатых.
318
ОТРЯД ПЕРЕГОРОДЧАТОЖАБЕРНЫЕ (SEPTIBRANCHIA)
Небольшая группа глубоководных двустворчатых, у которых основная
часть жабр редуцирована. Для дыхания служит часть мантийной полости,
отграниченная от остальной ее части остатками жабры в виде перегородки
с отверстием (рис. 266). Благодаря тому, что перегородка прикреплена к
мантии и к ноге она может то подниматься, то опускаться. Этими движения-
ми обеспечиваются захват и засасывание в дыхательные участки мантийной
полости довольно крупных придонных планктических животных, которые
служат пищей. Эти оригинальные двустворчатые — Cuspidaria и другие —
лежат на дне, погрузившись в ил спинной стороной раковины.
ОБРАЗ ЖИЗНИ И РАСПРОСТРАНЕНИЕ ДВУСТВОРЧАТЫХ
Двустворчатые — малоподвижные животные, живущие на дне пресно-
вэдных и морских водоемов. Большинство двустворчатых зарывается на раз-
личную глубину в грунт, выставляя свой задний конец с сифонами, и лишь
немногие лежат на грунте на боку, например морской гребешок. Некоторые
виды совершенно неподвижны и прикреплены к подводным предметам ниж-
ней створкой, как, например, устрицы (см. рис. 273).
Другие — мидии (табл. VIII), пинны—удерживаются на камнях или
даже передвигаются по отвесным стенкам подводных скал с помощью
биссуса. Биссус — это множество тонких, очень прочных, эластичных нитей,
выходящих из расположенного на ноге сосочка. Они представляют собой
клейкие секреторные выделения специальной биссусной железы.
Некоторые формы вбуравливаются в твердый субстрат (скалы, камни),
например фолас (Pholas), в днища кораблей и другие деревянные сооруже-
ния, находящиеся в море (корабельный червь).
Передвигаются зарывающиеся двустворчатые при помощи ноги, дви-
жение их очень медленно. Своеобразно передвигаются лежащие на грунте
морские гребешки: они вспрыгивают со дна благодаря толчку от сильного
выбрасывания воды при резком закрывании створок раковины.
Почти все двустворчатые — фильтраты. Пропуская через свою ман-
тийную полость большое количество воды, они отфильтровывают содер-
жащуюся в ней взвесь как живую (микропланктон), так и мертвую (детрит)
и выпускают воду наружу чистой. Фильтрация воды двустворчатыми прохо-
дит порой весьма интенсивно; так, например,мидия, имеющая длину 5—6 см,
пропускает через свою мантийную полость за час около 3,5 л воды. Отфиль-
трованные частицы ослизняются и либо поступают в ротовое отверстие и далее
в кишечник, либо частично выделяются прямо наружу в виде так называемых
псевдоэкскрементов. Поскольку частицы эти ослизнены, они не рассеиваются
в воде, а откладываются в виде рыхлых отложений вокруг поселений мол-
люсков. Таким образом, двустворчатые моллюски, с одной стороны, освет-
ляют окружающую воду, а с другой, — образуют донные отложения, иног-
да достаточно мощные. Это явление в целом носит название биофильтрации,
а организмы, вызывающие это явление, главным образом двустворчатые
моллюски, биофилътраторами.
У форм, глубоко зарывающихся в грунт, имеются длинные сифоны,
которые они выставляют на поверхность грунта; через эти сифоны и идет
ток воды в мантийную полость и из нее.
Постоянная фильтрация воды через мантийную полость служит и
для дыхания двустворчатых — вода омывает жабры, где и происходит
газообмен.
Размножаются двустворчатые, как и все моллюски, только половым
путем.
Оплодотворение происходит или в мантийной полости, или в наружной
среде. Дробление спиральное. У всех морских двустворчатых гаструла пре-
319
вращается в типичную трохофорообразную личинку (рис. 267) и развитие,
следовательно, идет с метаморфозом,
У трохофорообразных личинок на спинной стороне образуется зачаток
раковины, сперва непарный, а потом перегибающийся и образующий две
створки. После этого личинка опускается на дно и постепенно превращается
во взрослую форму.
У многих пресноводных видов личинки нет совсем и развитие идет без
метаморфоза, У других пресноводных оно идет так, как это было описано
для беззубки. В этих случаях паразитизм глохидиев обеспечивает расселение
вида. Интересно отметить, что рыбы могут заносить глохидиев в верховья
рек, т. е. против течения, что было бы не под силу свободноплавающим ли-
чинкам двустворчатых.
Среди пресноводных двустворчатых лишь у дрейссены (табл. VII)
(Dreissena) есть свободно плавающая трохофорообразная личинка. Как
полагают, она сохранилась еще у этого вида потому, что дрейссена сравни-
тельно недавно вселилась в пресные воды.
У двустворчатых можно хорошо проследить рост по тем линиям нара-
станий, которые имеются на раковине. По этим линиям подобно тому, как
это делается на древесине, можно установить возраст, а также характер
роста моллюска.
Двустворчатые широко распространены как в морях, так и в пресных
водах. В морях они обитают, начиная с отливной зоны до больших глубин,
населяя разнообразные грунты как мягкие, так и твердые. Некоторые из
них образуют массовые скопления. Известны, например, массовые скопле-
ния устриц и мидий. На банках моллюски обитают иногда сотнями тысяч
экземпляров. Наиболее обильны двустворчатые в тропических морях, где
имеются громадные, достигающие 1,5 м в длину моллюски. Примером таких
гигантских двустворчатых может служить обитающая главным образом в
Индийском океане гигантская тридакна (Tridacna). Размеры этой ракушки
настолько велики, что прибрежное население использует их створки для
хозяйственных целей (хранилища для воды и пр.). Вес одной такой створки
достигает 125 кг.
Во всех наших морях двустворчатые многочисленны и разнообразны.
В прибрежной зоне наших северных морей массовыми формами являются:
мидия (см. выше) и макома (Масота); последняя достигает нескольких
сотен экземпляров на 1 м2 дна; особенно характерна она для Балтий-
ского моря.
В пресных водах двустворчатые моллюски распространены также ши-
роко. Часть из них приурочена к текучим водам,например перловица (Unio),
другие, наоборот, более характерны для стоячих, например беззубка.
В пресных водоемах нашей страны известно около 50 видов двустворчатых,
причем наиболее богата фауна пресных водоемов Дальнего Востока, где
имеется ряд очень интересных и оригинальных форм, например очень круп-
ная кристария (Cristaria plicata) (табл. VII), имеющая большой киль на верх-
ней части раковины. Интересна по своему распространению дрейссена.
Основной район обитания этого моллюска — Каспийское море. Отсюда
он проник в пресные воды и по речным системам распространился до Бал-
тийского моря. Такому распространению способствовало то, что дрейссена
прикрепляется при помощи биссуса к днищам судов. Местами дрейссена
развивается в громадных количествах. Проникая в трубы водопроводных
сооружений, она иногда совершенно закупоривает их
ПРАКТИЧЕСКОЕ ЗНАЧЕНИЕ ДВУСТВОРЧАТЫХ
Целый ряд двустворчатых моллюсков добывают и даже разводят как
пищевые объекты или для использования раковин, причем в некоторых ме-
стах этот промысел развит очень интенсивно. Главным объектом как про-
мысла, так и разведения является устрица (Osterea). Этот моллюск исполь-
320
зуется человеком очень давно. Уже в стоянках первобытного человека в так
называемых кухонных остатках часто попадаются створки устриц. В настоя-
щее время устриц не столько добывают, сколько разводят. Разведение устриц
2Г
Рис. 276. Промысловые двустворчатые. / раковина речной перловицы
после выреза кружков для пуговиц; // створка настоящей жемчу-
жницы (Pteria) с образующимися на ней жемчужинками
производится в особых питомниках, куда помещают молодь и где она выра-
стает Развито устричное хозяйство главным образом в Голландии, Франции
и США, устрицы используются как в свежем, так и в консервированном виде.
Общий улов устриц составляет около 1,6 млн. центнеров в год. У нас про-
мысел устриц развит слабо, так как в наших морях устрицы встречаются
редко. На севере их нет совсем, в Черном море они хотя и имеются, но в
небольших количествах. В последние годы некоторое развитие получил
промысел устриц на Дальнем Востоке.
Вторым промысловым объектом на морских побережьях является ми-
дия (Mytilus), широко распространенная и образующая местами громадные
поселения, главным образом в прибрежной полосе.
11 Курс зоологии, т. I
321
Объектами промысла являются также морские гребешки (Pecten), бе-
лая ракушка (Spisula), прибрежная ракушка (Муа arenaria), сердцевидка
(Cardium edule) и некоторые другие (табл. VIII).
У нас промысел двустворок развит на Дальнем Востоке, где фауна их
очень обильна и где они представлены крупными формами, например Myti-
lus dunkeri, Pecten jessoensis, Spisula sachalinensis, а также в небольшом
размере на Черном море.
В некоторых количествах промышляются и пресноводные двустворки,
раковины которых используются как сырье для изготовления различных
перламутровых изделий, главным образсм пуговиц (рис. 276). Помимо про-
мысла пресноводных ракушек как перламутрового сырья, их добывают и
Рис. 277. Разрушенные деревянные свайные строения,
подточенные корабельным червем (Калифорния)
используют в качестве корма для домашней птицы. Дело в том, что в нор-
мальный рацион кур всегда должно входить некоторое количество извести,
необходимой для правильного образования скорлупы яйца. Лучшей по
усвояемости служит известь створок пресноводных двустворок. Створки
перемалывают, превращая в кормовую муку и в определенном количестве
прибавляют в корм птиц. Мягкие части обычно используются на корм
свиньям.
Некоторых пресноводных двустворчатых, а именно пресноводную жем-
чужницу (Margaritana margaritifera) (табл. VII), распространенную у нас
на севере (Карелия, Кольский полуостров, Архангельская область), добы-
вают из-за жемчуга, который образуется у нее довольно часто. Основной
объект добычи жемчуга — настоящая жемчужница (Pteria margaritifera),
распространена в тропических морях Индийского и Тихого океанов (см.
рис. 276). Встречается эта ракушка на сравнительно небольших глубинах
и добывается специальными ловцами — «искателями жемчуга».
Жемчужины образуются в узкой щели между мантией и раковиной в
результате откладывания арагонита — перламутра с особым расположе-
нием кристаллических пластинок — вокруг попавших туда инородных ча-
стиц, чаще всего песчинок.
Наряду с этими полезными двустворчатыми, имеются и вредныеформы,
а именно корабельные черви, или шашни (роды Teredo (рис. 274), Bankia
и др.), наносящие очень большие повреждения деревянным сооружениям в
море и на морских побережьях (рис. 277). Моллюски эти широко распростра-
нены во всех теплых морях. У нас древоточцы встречаются в Черном море
и в морях Дальнего Востока, где они особенно опасны. Борьба с ними ведет-
ся путем пропитывания древесины перед погружением в воду особым со-
ставом, который мешает личинкам моллюска осесть на нее.
Большой вред приносят двустворчатые, входящие в группу пресновод-
ных и морских организмов — обрастателей. Так, митилястр, дрейссена и
322
другие двустворчатые поселяются в массовом количестве в водоводных тру-
бах различных гидротехнических сооружений, со временем нацело забивая
их собой. Обрастатели поселяются и на днищах кораблей, сильно снижая
их мореходные качества. Борьба с двустворчатыми-обрастателями очень
затруднительна.
ФИЛОГЕНИЯ ДВУСТВОРЧАТЫХ
Прямые нервные тяжи и заднее положение анального отверстия сви-
детельствует о том, что у двустворчатых нет торсионного процесса, свойствен-
ного брюхоногим. Принимая во внимание, что нога примитивных двуствор-
чатых имеет плоскую ползательную подошву, можно предположить, что
предок двустворчатых обладал организацией, близкой к организации пан-
цирных и моноплакофор. Таким образом, двустворчатые, видимо, такая же
специализированная ветвь моллюсков, как и брюхоногие, происходящая от
одного с ними предка. Своеобразное строение двустворчатых — их ракови-
на, клиновидная нога, редуцированная голова (след которой—церебральные
и плевральные ганглии) — возникает в связи с сугубо бентическим образом
жизни, в состоянии полупогруженности или полной погруженности в грунт
Именно при таком образе жизни необходима двустворчатая раковина, слу-
жащая надежной защитой при давлении с боков.
Пассивный способ добывания пищи — фильтрация, при котором жи-
вотное не перемещается совсем или мало подвижно, привел к редукции го-
ловы как места сосредоточения органа чувств.
Большое разнообразие двустворчатых объясняется тем, что ими освоены
весьма разнообразные условия в пределах дна моря и в пресноводных водо-
емах. Некоторые из них смогли все же «выйти из грунта» и стать несколько
более подвижными.
КЛАСС 7. ГОЛОВОНОГИЕ (CEPHALOPODA)
ХАРАКТЕРИСТИКА
Билатеральносимметричные моллюски, у которых рот окружен много-
численными ловчими щупальцами. Эти щупальца представляют собой сильно
измененную ногу, которая потеряла у головоногих свою обычную функцию.
У одной крайне малочисленной группы донных головоногих есть весьма
своеобразная наружная раковина, у большинства других раковины нет или
она рудиментарна и скрыта под кожей спины.
Движение совершается реактивным способом благодаря толчку от вы-
брасываемой из мантийной полости воды.
Наружнораковинные головоногие обладают двумя парами жабр и
двумя парами предсердий, внутреннераковинные — одной парой пред-
сердий. Развитие прямое, без личинки.
Обитают только в морях. Известно около 200 ныне живущих видов,
ископаемых известно около 8000.
Представитель класса головоногих —
тихоокеанский кальмар (Ommastrephes sloanei
pacificus)
Внешнее строение (рис. 278) Тело кальмара удлиненное, цилиндри-
ческое с хорошо выраженной головой и заостренным задним концом. По
бокам тела в задней части имеются треугольные плавники. Общая длина те-
ла достигает 40—50 см.
Голова несет на переднем конце ротовое отверстие, окруженное 10 щу-
пальцами, или руками, причем 8 из них короткие, а 2 длинные, так называе-
11*
32а
мые ловчие. Восемь коротких щупалец несут по всей длине 2 ряда присосок.
Каждая присоска имеет вид чашечки, прикреплена к щупальцу коротким
стебельком и имеет роговое колечко с зубчиками. Ловчие руки несут при-
соски только на расширенных концах.
На брюшной стороне головы имеется воронка, открывающаяся широким
концом в мантийную полость, а более узким наружу. По бокам головы на-
Рис. 278. Кальмар тихоокеанский (Огп-
mastrephes sloanei pacificus) с брюшной
стороны:
/ — голова, 2 — туловище, 3 — плавник,
4 — край мантии, 5 — вход в мантийную по-
лость, 6 — воронка, 7 — ловчие щупальца,
8 короткие щупальца, 9 — присоски, 10— рот,
11 — глаза
ходится пара глаз сложного строе-
ния. Под каждым глазом помещает-
ся обонятельная ямка.
Мантия и мантийная полость
(рис. 279). Пронизанная большим
количеством мышечных элементов
Рис. 279. Самец тихоокеанско-
го кальмара с вскрытой ман-
тийной полостью, мантия раз-
резана по средней линии и раз-
вернута в стороны:
1 — голова, 2 — воронка, 3 — мантия,
4 — валики запонок. 5 — желобки за-
понок, 6 — жабры, 7 — мешок внут-
ренних органов, 8— чернильный ме-
шок, 9— прямая кишка, 10—аналь-
ное отверстие, 11 — отверстие почек,
12— мужское половое отверстие,/<3—
задняя аорта, 14 — звездчатый ман-
тийный нервный узел
кожная складка — мантия — как воротничком охватывает голову, но так,
что между ними остается узкая щель, ведущая в мантийную полость. Кро-
ме того, что мантия сращена с кожей спины, она еще «пристегнута» в двух
точках на брюшной стороне к внутренностному мешку, чуть ниже головы.
Это скрепление осуществляется особыми структурами туловища и мантии,
так называемыми запонками. На туловище — это довольно сложно устро-
енные углубления близ воронки, в то время как на внутренней стороне
мантии — это хрящеватые выпуклости. Сокращением мышц хрящеватые
выпуклости втискиваются в ямки, в результате чего верхний край мантии
плотно прижимается к голове и вход в мантийную полость закрывается.
Расслабление мышц ведет к тому, что щель между мантией и головой зияет
и мантийная полость наполняется водой.
324
нозную кровь из задней части тела. Сокращением венозных сердец кровь
проталкивается в жабры, где и окисляется. Задний участок головной вены
и полные вены покрыты особыми выпячиваниями, так называемыми веноз-
ными придатками, которые вдаются в полость почек и способствуют извле-
чению из крови продуктов обмена.
Выделительная система. Органы выделения представлены парными
почками, которые в виде мешков неправильной формы лежат в полости
тела. Их рыхлая масса тесно окружает венозные придатки. Открываются
почки с одной стороны в полость тела, с другой — наружу, в мантийную
полость.
Половая система. Кальмар, как все головоногие, раздельнопол. Внешне
самцы отличаются от самок только одним измененным коротким щупальцем
Рис. 281. Нервная система осьми-
нога:
1—головной ганглий, 2— глазные ганг-
лии, 3 — звездчатые ганглии, 4 — желудоч-
ный ганглий, 5 — нервы рук
Рис. 282. Кровеносная система головоногого
моллюска (схема):
1 — желудочек, 2—аорта, 3— большая головная вена,
4 — жаберные сердца, 5 — жабры, 6— предсердия, 7 —
слепые выпячивания венозных сосудов, вдающиеся в
почки (стрелки указывают направление тока крови)
(см. рис. 278), которое служит для перенесения сперматофора из мантийной
полости самца в мантийную полость самки. Семенник непарный, лежит в
задней части тела, от него отходит длинный извитой семяпровод, образую-
щий небольшое расширение, в котором образуются сперматофоры. Откры-
вается семяпровод в мантийную полость.
Яичник тоже непарный, зрелые яйца выпадают в полость тела, откуда
выводятся уВ мантийную полость через парные яйцеводы. В связи с женской
половой системой находится и пара скорлуповых желез, самостоятельно
открывающихся в мантийную полость.
Экология и распространение. Тихоокеанский кальмар — очень под-
вижное животное, которое может быстро передвигаться и хорошо ориенти-
роваться в пространстве.
Движение кальмара происходит путем выбрасывания из мантийной
полости через воронку сильной струи воды, т. е. реактивным способом. Быст-
рое, стремительное движение нападения кальмар совершает так, что конец
тела с плавником обращен вперед. Этим достигается большая маневренность.
В иных случаях воронка может отгибаться в любом направлении, что поз-
воляет кальмару двигаться даже боком.
326
Плывущий кальмар складывает щупальца так, что они не мешают дви-
жению, и тело легко рассекает воду. Плавники служат рулями глубины.
Кальмары плавают очень хорошо и могут совершать длительные миграции,
проплывая тысячи километров.
Кальмар, как и большинство головоногих, способен быстро менять
окраску в зависимости от фона окружающей среды и состояния животного.
Изменение окраски связано с наличием в соединительнотканом слое кожи
пигментных клеток — хроматофор. Плазма этих клеток наполнена тем-
ными зернами пигмента, к клеткам прикрепляются многочисленные мускуль-
ные волокна, иннервируемые особыми нервными окончаниями. При сок-
ращении мышц клетки сильно растягиваются и превращаются в темные
пятнышки, а вся кожа темнеет. При раслаблении мышц клетки сжимаются,
и вся кожа светлеет В спокойном состоянии окраска кальмара обычно свет-
лая, особенно на брюшной стороне. При раздражении, которое передается
через центральную нервную систему нервным окончаниям, связанным с
мускульными волокнами пигментных клеток, клетки растягиваются, и
кальмар темнеет. Эта способность к перемене окраски имеет для кальмара
очень большое значение, давая ему возможность оставаться незаметным
среди окружающих предметов (покровительственная окраска).
Кальмар — хищник, с этим связана его большая подвижность, хоро-
ший ловчий аппарат и крепкие челюсти. Тихоокеанский кальмар обычно
стаями нападает на косяки рыб и может уничтожать их в большом количе-
стве. Активному нападению и одновременно и защите очень помогают выде-
ления чернильного мешка. Будучи чем-либо раздражен, кальмар выпускает
содержимое мешка в воду и под этой «дымовой завесой» может назаметно
подплывать к жертве и также незаметно скрываться от врага. Кальмар схва-
тывает рыбу ловчими щупальцами, челюстями прогрызает ей затылок и вые-
дает обычно только мозг и мясо передней части спины. В этом отношении
тихоокеанский кальмар — вредное животное, так как уничтожает и портит
довольно большое количество рыб. Вместе с тем этот кальмар является про-
мысловым объектом и его добывают в наших дальневосточных морях (см.
ниже).
Тихоокеанский кальмар — теплолюбивая форма — широко распро-
странен в Японском море, по побережьям Японии и в Желтом море.
Положение в системе. Тихоокеанский кальмар относится к подклас-
су внутреннераковинных и отряду десятиногих.
ОБЩИЙ ОБЗОР КЛАССА
Внешнее строение. Головоногие — самые крупные из всех беспозво-
ночных и одни из самых крупных животных вообще. Известен представи-
тель головоногих — кальмар архитеутис (Architeutis princeps), достигаю-
щий в длину вместе с вытянутыми щупальцами 18 м. Некоторые ископаемые
головоногие — аммониты (см. ниже)—достигали в поперечнике 2 м.
По своему внешнему виду оба подкласса — наружнораковинные и
внутреннераковинные — резко отличаются друг от друга. Первые, пред-
ставленные в настоящее время несколькими видами рода Nautilus — кораб-
лики (рис. 283) — имеют наружную спирально завитую раковину, разде-
ленную на камеры. Само животное занимает последнюю камеру, а все ос-
тальные наполнены газом и представляют собой своеобразный гидростати-
ческий аппарат. От спинной стороны животного отходит отросток — сифон,
который пробадает все перегородки и доходит до самого конца спирали
раковины.
Внутреннераковинные, основная группа современных головоногих, не
имеют наружного скелета. Тело состоит из головы с венцом, щупалец, или
рук, окружающих ротовое отверстие, и туловища, различного по форме.
Щупалец может быть 10 (см. рис. 276) или 8 (рис. 284), по этому признаку
и происходит деление подкласса на 2 отряда. В первом случае два щупальца
327
более длинные. Щупальца несут присоски, а иногда и роговые крючки для
захвата добычи. Кроме щупалец, на голове имеются два глаза, по одному с
каждого бока. Глаза обычно ярко окрашены, чаще всего они изумрудно-
зеленые.
Туловище может быть или стройное, удлиненное, имеющее торпедооб-
массивное, несколько сплюснутое в
спинно-брюшном направлении (ка-
ракатицы), или, наконец,округлое,
мешковатое (осьминоги). У кальма-
ров обычно хорошо развит хвосто-
вой плавник, у каракатиц — два уз-
ких боковых плавника по бокам тела
Форма тела головоногих тесно
связана с их образом жизни.
На брюшной стороне тела у
всех головоногих имеется воронка.
Как уже сообщалось, руки и
воронка — это измененная нога.
Доказательством этому служит то,
что и руки и воронка иннервиру-
ются от нервного узла, соответст-
разную форму (кальмары), или более
Рис. 283. Кораблик (Nautilus), ракови- вующего ножным — педальным —
на в разрезе: узлам Других МОЛЛЮСКОВ.
1 — жабры Окраска головоногих очень
разнообразна. Некоторые формы,
живущие в толще воды, почти совершенно бесцветны и прозрачны, у боль-
шинства же имеется пигмент. Головоногие могут быстро и резко менять
окраску. Особенно эта способность развита у осьминогов: будучи в спокой-
ном состоянии, они обычно молочно-белые, при раздражении делаются
темно-красными или коричнево-лиловыми.
Рис. 284. Осьминог (Octopus siIbertianus), восточное побе-
режье Камчатки. Естественная величина до 60 см
Внутренний скелет. У большинства ныне живущих форм скелет со-
стоит из-более или менее хорошо развитой спинной пластинки. В.некоторых
случаях (у каракатиц) она массивная, хорошо обызвествленная (так назы-
ваемая кость каракатицы) (рис. 285i ), в других (у кальмаров) — тонкая
роговая. Эта пластинка представляет собой сильно измененный рудимент
328
скелета живших в прошлые геологические периоды внутреннераковинных
головоногих — белемнитов.
Кроме спинной пластинки, у большинства внутреннераковинных
имеются головные хрящи (рис. 285п), защищающие подобно черепу позвоноч-
ных головной мозг Состоят эти хрящи действительно из хрящевой ткани.
Это единственный случай наличия хрящевой ткани у беспозвоночных.
Полость тела. Полость тела вторичная, находится у головоногих в
различной степени редукции. Сравнительно хорошо развит целом у кораб-
ликов. У десятиногих он редуцирован сильнее, а
у осьминогих еще больше, даже сердце лежит у
них вне целома, что является единственным ис-
ключением среди всех моллюсков.
Нервная система. Нервная система внут-
реннераковинных наиболее сложная среди бес-
позвоночных. Состоит из центральной части, или
головного мозга, и периферической части.
Головной мозг (см. рис. 282) представляет
собой мощное скопление нервных ганглиев, рас-
положенных вокруг пищевода. Верхнюю часть
мозга составляют два головных ганглия, от кото-
рых отходят слуховые, обонятельные и зритель-
ные нервы. Последние образуют при основании
два больших глазных, или оптических, ганглия.
По бокам головного мозга расположены два
плевральных ганглия, посылающие нервы к ман-
тии. Нижняя часть мозга образована в передней
части двумя педальными ганглиями, а в задней—
двумя туловищными. Таким образом, головной
мозг внутреннераковинных головоногих состоит
из четырех пар слитых между собой ганглиев.
Периферическая нервная система состоит из сис-
темы нервов и немногих ганглиев, а именно пары
звездчатых ганглиев, расположенных на внутрен-
ней поверхности мантии, и желудочного ганг-
лия, расположенного в стенке желудка.
У корабликов, т е. наружнораковинных,
нервная система состоит из окологлоточных нерв-
ных тяжей без нервных узлов.
Органы чувств. Лучше всего развиты глаза,
представлющие собой очень сложные образова-
ния. У корабликов они имеют форму бокала с
Рис. 285. Внутренний ске-
лет каракатицы. / спин-
ная пластинка; // — голов-
очень суженным наружным отверстием, выстлан- ной хрящ
ного сетчаткой; роговицы, хрусталика и стекло-
видного тела нет.
У внутреннераковинных глаза устроены значительно сложнее (рис. 286),
имеются хрусталик, радужина и роговица и по остроте зрения, способности
к аккомодации они могут быть сравнимы с глазами позвоночных.
У некоторых форм глаза достигают громадных размеров, так, например,
у гигантских кальмаров (Architeutis) они достигают 30 см в диаметре.
Имеются органы обоняния (осфрадии)и органы равновесия (статоцисты).
Пищеварительная система. Пищеварительная система начинается ро-
товым отверстием, расположенным на голове внутри венца щупалец. В ро-
товой полости всегда имеется пара мощных роговых челюстей, имеющих
вид птичьего клюва, и радула, представляющая собой тонкую хитиновую
пластинку с острыми зубами. В ротовую полость впадает одна или две пары
слюнных желез. Далее идет пищевод, переходящий в желудок. В желудок
открываются протоки объемистой, иногда сложно устроенной печени. От
329
желудка отходит кишка, образующая более или менее сложную петлю,
открывающаяся в мантийную полость.
Таким образом, у головоногих, как и у брюхоногих, анал ное отверстие
лежит не сзади, а впереди и пищеварительный тракт образует резкое колено.
Рис. 286. Аргонавт (Argonauta argo). I —самка, несколько уменьшенная; П —
самец, сильно увеличенный:
/ — ложная раковина — выводковая сумка, 2 — два щупальца, ее образующие, 3—гектокотильl
развернутом виде
Однако у головоногих это следствие простого подгибания длинного внутрен-
ностного мешка, но не следствие торсионного процесса, как у брюхоногих.
Недалеко от анального отверстия в кишечник у многих головоногих
впадает чернильная железа, в которой вырабатывается черный секрет.
Дыхательная система. Дыхательная система представлена двумя или
четырьмя жабрами, расположенными в мантийной полости. Циркуляция
воды в ней обеспечивается ритмическими сокращениями самой мантии при
движении.
Кровеносная система (см. рис. 281). Очень сложная, почти замкну-
тая, так как ряд капилляров артерий непосредственно переходит в капил-
ляры вен. Из жабр окисленная кровь направляется в предсердия, оттуда в
желудок и далее разносится по всему телу, изливаясь местами в кровеносные
синусы.
Выделительная система. Выделительная система представлена изме-
ненными метанефридиями, у большинства соединяющимися с полостью тела
или ее рудиментом и открывающимися в
мантийную полость. У корабликов их две
пары, у остальных — одна. Характерна
довольно тесная связь между почками и
кровеносной системой (см. выше).
Половая система. Все головоногие
раздельнополы, причем иногда половой
диформизм выражен очень резко. Примером
может служить встречающийся в Среди-
земном море аргонавт (Argonauta argo) (рис.
287), у которого самка довольно крупная
и имеет особую ложную раковину (см. ниже),
а самец — маленький обычного вида осьми-
ног. Очень часто одна рука самца голово-
ногих превращена в особый придаток —
гектокотиль, при помощи которого проис-
ходит оплодотворение. Наполненный спер-
матофорами гектокотиль может иногда
Рис. 287. Глаз каракатицы (про-
дольный разрез):
1 — хрусталик, 2—роговица, 3 — ра-
дужная оболочка, 4— сетчатка, 5— глаз-
ной ганглий, 6 — глазной нерв
330
желудка отходит кишка, образующая более или менее сложную петлю,
открывающаяся в мантийную полость.
Таким образом, у головоногих, как и у брюхоногих, анал ное отверстие
лежит не сзади, а впереди и пищеварительный тракт образует резкое колено.
Рис. 286. Аргонавт (Argonauta argo). I —самка, несколько уменьшенная- П
самец, сильно увеличенный:
/ — ложная раковина — выводковая сумка, 2— два щупальца, ее образующие, 3—гектокотиль е
развернутом виде
Однако у головоногих это следствие простого подгибания длинного внутрен-
ностного мешка, но не следствие торсионного процесса, как у брюхоногих.
Недалеко от анального отверстия в кишечник у многих головоногих
впадает чернильная железа, в которой вырабатывается черный секрет.
Дыхательная система. Дыхательная система представлена двумя или
четырьмя жабрами, расположенными в мантийной полости. Циркуляция
воды в ней обеспечивается ритмическими сокращениями самой мантии при
движении.
Кровеносная система (см. рис. 281). Очень сложная, почти замкну-
тая, так как ряд капилляров артерий непосредственно переходит в капил-
ляры вен. Из жабр окисленная кровь направляется в предсердия, оттуда в
желудок и далее разносится по всему телу, изливаясь местами в кровеносные
синусы.
Выделительная система. Выделительная система представлена изме-
ненными метанефридиями, у большинства соединяющимися с полостью тела
или ее рудиментом и открывающимися в
мантийную полость. У корабликов их две
пары, у остальных — одна. Характерна
довольно тесная связь между почками и
кровеносной системой (см. выше).
Половая система. Все головоногие
раздельнополы, причем иногда половой
диформизм выражен очень резко. Примером
может служить встречающийся в Среди-
земном море аргонавт (Argonauta argo) (рис.
287), у которого самка довольно крупная
и имеет особую ложную раковину (см. ниже),
а самец — маленький обычного вида осьми-
ног. Очень часто одна рука самца голово-
ногих превращена в особый придаток —
гектокотиль, при помощи которого проис-
ходит оплодотворение. Наполненный спер-
матофорами гектокотиль может иногда
Рис. 287. Глаз каракатицы (про-
дольный разрез):
1 — хрусталик, 2 — роговица, 3 — ра-
дужная оболочка, 4— сетчатка, 5— глаз-
ной ганглий, 6—глазной нерв
ззо
отрываться, самостоятельно плавать, змееобразно извиваясь, и заползать в
мантийную полость самки. Такие действия гектокотиля дали основание
старым исследователям первоначально рассматривать его как самостоятель-
ный паразитический организм.
Половая железа обычно непарная, занимает самый задний конец тела.
Имеется особый орган, где происходит формирование сперматофоров. Спер-
матофоры различны по форме, они могут иметь вид длинных бутылочек или
просто трубочек, в которых помещается сперма.
Развитие. Развитие головоногих сильно отличается от развития других
моллюсков. Чрезвычайно большое количество желтка в яйце приводит к
тому, что дроблению подлежит лишь поверхностная его часть. Следов спи-
рального дробления у большинства головоногих нет. На поверхности яйца
формируется зародыш, который постепенно растет, потребляя в качестве
пищи желток своего яйца. Из яйца выходит маленький головоногий моллюск,
быстро затем растущий.
СИСТЕМАТИЧЕСКИЙ ОБЗОР КЛАССА
ПОДКЛАСС I. НАРУЖНОРАКОВИННЫЕ (ECTOCOCHLIA)
Этот класс характеризуется наличием наружной раковины, разделен-
ной перегородками на камеры, а также наличием четырех жабр. В связи с
последним признаком этот подкласс часто называется четырехжаберным
(Tetrabranchiata) в отличие от другого подкласса — двужаберных (Dibran-
chia).
Подкласс делится на 2 отряда: 1) кораблики и 2) аммониты.
ОТРЯД КОРАБЛИКИ (NAUTILOIDEA)
Из современных форм к этому отряду относится очень небольшое число
видов, объединяемых в один род Nautilus (см. рис. 283), имеющих очень уз-
кий ареал распространения. Они встречаются только в тропических морях
между юго-восточной Азией и
Австралией.
В прошлые геологические
эпохи кораблики обильно насе-
ляли моря, начиная с кембрия,
причем наиболее древние из них
имели прямую почти цилиндри-
ческую раковину (Orthoceoros).
ОТРЯД АММОНИТЫ
(AMMONOIDEA)
Этот отряд содержит исклю-
чительно ископаемые формы,
очень богато представленные
особенно в мезозое, где они были
одной из самых многочисленных
групп морской фауны (рис. 288).
Рис. 288. Ископаемые головоногие.
/ белемнит (реконструкция); // —
чертов палец; III — аммонит (рекон-
струкция); IV — аммонит (Phyllo-
сега):
1 — фрагмакон, 2 — спинной вырост, 3 — ро-
струм. 4— шовная линия
33’
Характеризуется этот отряд тем, что перегородки раковины соприка-
саются с ее стенками не простыми линиями, как у корабликов, а очень слож-
ными — извилистыми, получившими название шовных линий. Всего извест-
но около 5000 видов аммонитов, крайне разнообразных как по строению,
так, видимо, и по биологии. В связи с этим аммониты являются одними из
важнейших руководящих ископаемых.
ПОДКЛАСС II. ВНУТРЕННЕРАКОВИННЫЕ (ENDOCOCHLIA)
Представители этого подкласса не имеют наружной раковины, которая
изменена у них во внутренний скелет, имеющий в основном опорные функ-
ции. Жабр — две.
Подкласс делится на 2 отряда. 1) десятиногие и 2) восьминогие.
ОТРЯД ДЕСЯТИНОГИЕ (DECAPODA)
Число рук 10, причем 2 из них более длинные. К этому отряду относят-
ся наиболее известные современные головоногие (кальмары и каракатицы),
а также ископаемые — белемниты.
Белемниты были внешне, по-видимому, похожи на кальмаров, но имели
более сложный внутренний скелет. Скелет их (см. рис. 288) состоял из кону-
са, разделенного на камеры (фрагмакон). Спинная сторона этого конуса вы-
давалась вперед в виде плоского выроста, а книзу он имел придаток с мас-
сивным известковым футляром (рострум) Рострумы белемнитов хорошо
сохраняются в ископаемом состоянии и принадлежат к одним из самых рас-
пространенных окаменелостей (так называемые «чертовы пальцы»).
Из современных форм необходимо отметить, кроме тихоокеанского каль-
мара, обыкновенную каракатицу (Sepia officinalis), широко распространен-
ную в теплых морях; россию (Rossia), наиболее обычную в наших северных
морях; гигантского кальмара (Architeutis); обыкновенного кальмара (Lo-
ligo vulgaris) и глубинного светящегося кальмара (Thaumatolampus diade-
ma) и др.
ОТРЯД ВОСЬМИНОГИЕ (OCTOPODA)
Число рук восемь, тело массивное, почти округлое (см. рис. 284). В от-
личие от десятиногих — донные формы, ползающие по грунту.
Сюда относится ряд осьминогов, встречающихся главным образом в
теплых морях. Следует особо отметить северного осьминога (Bathypolypus
arcticus), встречающегося в наших северных морях; илистого осьминога
(Cirrotauma murrayi) с перепонками между руками и аргонавта (Argonauta
argo) (см. рис. 286).
ОБРАЗ ЖИЗНИ И РАСПРОСТРАНЕНИЕ ГОЛОВОНОГИХ
Все головоногие — подвижные животные, хорошо передвигающиеся
или по дну (осьминоги) или в толще воды (кальмары). Менее подвижны при-
донные кораблики.
Движение головоногих происходит при помощи рук (донные формы —
осьминоги) или реактивным способом, т е. путем выбрасывания воды из
мантийной полости через воронку (плавающие формы — кальмары). Осьми-
ноги передвигаются сравнительно медленно. Обычно они неподвижно сидят
в подводных гротах, среди скал, в куче камней и подкарауливают добычу.
Такому образу жизни помогает способность осьминогов быстро менять свою
окраску, приспосабливаясь к окружающей среде. Активному нападению, а
одновременно и защите очень помогают выделения чернильного мешка.
332
Все головоногие—хищники. Питаются они, захватывая жертву ру-
ками и «расклевывая» ее мощными челюстями, в виде птичьего клюва.
Оплодотворение у головоногих внутреннее. Самец переносит спермато-
форы в мантийную полость самки при помощи гектокотиля. Яйца сравни-
тельно крупные, богатые желтком и покрытые плотной оболочкой. Отклады-
ваются они обычно группами. У некоторых форм, например у аргонавта, две
сильно измененные руки самки выделяют особую ложную раковину (см.
рис. 286), в которую откладываются яйца и которая, таким образом, являет-
ся особого рода выводковой сумкой. Развитие головоногих прямое и целиком
проходит внутри яйца. В связи с большим количеством желтка у головоно-
гих дробление проходит не по спиральному типу, характерному для всех
других групп моллюсков, а по дискоидальному Вылупившаяся молодь у
большинства форм отличается от взрослых только величиной и слабой пиг-
ментацией.
Головоногие—теплолюбивые и соленолюбивые животные. Это обстоя-
тельство определяет их распространение. В громадном большинстве они
обитатели теплых и полносоленых морей. В связи с этим в наших краях они
представлены очень небольшим числом представителей. В южных морях
они полностью отсутствуют в северных представлены несколькими ви-
дами, среди которых в первую очередь необходимо указать небольших пе-
лагических россий.
Более многочисленны головоногие в дальневосточных морях, где их
известно 27 видов, среди которых имеется несколько очень крупных, дости-
гающих 3 м в длину осьминогов и ряд кальмаров.
ПРОМЫСЛОВОЕ ЗНАЧЕНИЕ ГОЛОВОНОГИХ
Целый ряд головоногих (кальмары и осьминоги) является объектом про-
мысла. Их добывают главным образом в странах Дальнего Востока, а также
в США и в Средиземном море. Заготовка их производится примерно так же,
как и двустворчатых моллюсков; их потрошат и затем высушивают обычно
на солнце. Всего добывается в год около 1 700 000 ц головоногих. У нас, в
Японском море, за последние годы развился промысел кальмара (Ommast-
rephes sloanei pacificus) и осьминогов.
В Китае развит промысел каракатиц для добывания содержимого чер-
нильного мешка, из которого изготовляется так называемая китайская тушь.
ФИЛОГЕНИЯ ГОЛОВОНОГИХ
Ископаемый материал показывает, что у обеих групп головоногих —
наружнораковинных и внутреннераковинных — был общий предок с прямой
цилиндрической наружной раковиной, разделенной внутри на камеры. Мож-
но считать, что этот общий предок всех современных головоногих обладал
двумя парами жабр и двумя парами предсердий. На то указывает организа-
ция современных наружнораковинных — вымирающих примитивных го-
ловоногих.
Эти следы метамерности многие исследователи расценивают как свиде-
тельство того, что предки головоногих происходят от древнейших и прими-
тивнейших моллюсков, обладавших чертами внутренней и наружной мета-
мерии. Видимо, группа моллюсков с видоизмененной ногой — будущие
головоногие — раньше других отклонилась от общего для всех моллюсков
предка и стала специализироваться на основе хищничества. Необходимую
при таком образе жизни большую подвижность эта группа моллюсков ста-
ралась совместить с наличием наружной раковины, превратив ее в напол-
ненный газом гидростатический орган. Однако возрастающая подвижность и
общая активность головоногих привела не к усилению раковины как гид-
ростатического аппарата, а к еередукции; те жегруппы, которые сохраняли
наружную поддерживающую раковину, почти вымерли.
333
филогения моллюсков
Наличие в развитии большинства моллюсков черт, сходных с кольчаты-
ми червями (спиральное дробление, трохофорообразные личинки), а также
некоторые общие черты в строении взрослых форм (целомическая полость
тела, характер органов выделения) дают право считать, что моллюски и
кольчатые черви — родственные типы вторичнополостных животных, при-
чем моллюски представляют собой ветвь, рано отклонившуюся от древней-
ших и примитивнейших кольчатых червей.
Последующая эволюция моллюсков и кольчецов шла по-разному
Кольчатые черви развивались, вырабатывая активный образ жизни,
связанный с большой подвижностью. В связи с этим у них выработалась
внешняя и внутренняя сегментация, появились органы движения — па-
раподии, сформировался головной отдел, снабженный комплексом органов
чувств.
Моллюски, наоборот, развивались в основном как малоподвижные,
донные, формы. Строение ноги, особенно у примитивных групп разных
классов, показывает, что нога — не столько орган активного движения,
сколько орган прочного прикрепления к субстрату. Развитию активного
движения моллюсков всегда препятствовала раковина. Прогрессивное раз-
витие раковины весьма характерно для моллюсков; ранняя ее закладка в
период эмбрионального развития свидетельствует о том, что раковина
возникла, видимо, на самых ранних этапах их эволюции. Часто наблюдае-
мая у моллюсков редукция раковины — всегда вторичное явление, сменяю-
щее период ее сильного развития.
Что касается соотношения отдельных классов внутри типа, то более
примитивными следует считать панцирных, бороздчатобрюхих и монопла-
кофор с их нервной системой в виде простых тяжей без ганглиев и обшир-
ными отделами вторичной полости тела (у Neopilina). В то же время все
эти три класса представляют собой по-разному специализированные группы
(вторичная метамерия наружных и внутренних органов, структура рако-
вины и т. п.) и поэтому ни один из них не может считаться исходным для
всех остальных.
Двустворчатые и лопатоногие — наименее подвижные моллюски, в
основном развивались, вырабатывая приспособления к фильтрации и к
закапыванию на дне водоема. Двустворчатая раковина обеспечивает воз-
можность и того и другого.
Брюхоногие — более подвижная группа, в основном развивалась, вы-
рабатывая приспособления к питанию путем соскабливания растений с суб-
страта; хищники среди них — вторичные и немногочисленные формы. Асим-
метрия и торсионный процесс делают их весьма своеобразной группой, но
вместе с этим и несомненно близкой по основным чертам к предкам моллюс-
ков (шапочковидная раковина и билатериальная симметрия примитивных,
нога). Важно отметить, что те брюхоногие, которые стали обитать на суше,
по-видимому, были одними из первых наземных животных вообще.
Головоногие самые подвижные из всех моллюсков. Наряду с чертами
большого прогресса организации (строение нервной системы, органы чувств,
почти замкнутая кровеносная система) головоногие сохранили и некоторые
черты метамерных предков — две пары жабр и предсердий у наружнора-
ковинных и более развитую вторичную полость тела, чем у брюхоногих и
двустворчатых.
Древние (ископаемые) представители этого класса — аммониты и белем-
ниты — были в определенные периоды истории Земли основными группами
морской фауны. Дальнейшее изменение их заключалось в постепенном осво-
бождении от скелета, который из наружного защитного образования прев-
ращается во внутреннее, опорное, что и способствовало усилению подвиж-
ности головоногих.
ТИП XI. ЧЛЕНИСТОНОГИЕ (ARTHROPODA)
ХАРАКТЕРИСТИКА
Членистоногие — двустороннесимметричные сегментированные жи-
вотные с метамерно расположенными членистыми конечностями. Тело их
в большинстве случаев состоит из трех отделов — головы, груди и брюш-
ка, состав которых у разных групп неодинаков. Тело покрыто сплошной
хитинизированной кутикулой', кутикула распадается на более твердые щит-
ки (склериты) и на перепончатые участки, соединяющие их.
Органы дыхания членистоногих разнообразные в соответствии со сре-
дой, в которой обитают эти животные. Это или органы водного дыхания —
жабры, или органы атмосферного дыхания—легкие—или трахеи', в случае
малой величины членистоногого и нежности его покрова специальные орга-
ны дыхания могут отсутствовать и дыхание осуществляется всей поверх-
ностью тела.
Центральная нервная система, первоначально парная, лестничного
типа. Передний отдел ее («головной мозг») расположен над кишечником,
остальная часть образует брюшную нервную цепочку. Полость тела смешан-
ная (миксоцель), происшедшая от слияния вторичной полости тела (цело-
ма) с первичной. Мускулатура частично поперечнополосатая.
Кровеносная система не замкнута, кровь приводится в движение сок-
ращениями сердца, расположенного над кишечником и обладающего пар-
ными щелями — остиями.
Органы выделения представлены связанными с кишечником мальпи-
гиевыми сосудами и сходными с ними придатками средней кишки. Нефри-
диальная система (сегментарные органы) сохранилась весьма неполно, она
представлена некоторыми грудными и головными железами и выводящими
половыми протоками.
Членистоногие в громадном большинстве раздельнополы.
Тип членистоногих — самый многочисленный и многообразный тип
животных. Это связано с необычайным разнообразием условий существова-
ния членистоногих — нет такой подходящей для существования животных
среды, где они не встречались бы.
Атмосфера, поверхность и глубины самых разнообразных водоемов,
поверхность и глубокие слои почвы, снежные поля и вечные льды — все
эти биотопы населены членистоногими. Среди членистоногих имеются и
паразиты животных и растений, живущие как на поверхности тела хозяина,
так и в различных органах и тканях.
Паразитизм может привести к тому, что животное утрачивает признаки
членистоногого (см. рис. 311) и лишь его онтогенетическое развитие указы-
вает на принадлежность к членистоногим.
Общее число членистоногих около 900 000 видов.
По своему строению и онтогенезу тип членистоногих является дальней-
шим развитием типа кольчатых червей (см. «Общий обзор типа членистоно-
гих»).
335
КЛАССИФИКАЦИЯ
Тип членистоногие (Arthropoda)
Подтип I. Жабернодышащие (Branchiata)
Класс 1. Ракообразные (Crustacea)
Подтип II. Трилобиты (Trilobita)
Подтип III. Хелицероносные (Chelicerata)
Класс 1 Меростомовые (Merostomata, или Palaeostraca)
Класс 2. Паукообразные (Arachnida)
Подтип IV Первичнотрахейные (Protracheata, или Onychophora)
Подтип V Трахейнодышащие (Tracheata)
Класс 1 Многоножки (Myriopoda)
Класс 2. Несекомые (Hexapoda, или Insecta)
Подтип I. Жабернодышащие (Branchiata)
ОБЩАЯ ХАРАКТЕРИСТИКА
Подтип жабернодышащих представлен одним классом ракообразных
(Crustacea), вследствие этого характеристика подтипа соответствует харак-
теристике класса.
Этот подтип включает преимущественно водных членистоногих, дыша-
щих жабрами. Жабры представляют собой тонкостенные выросты конечно-
стей грудного или реже брюшного отделов тела. Немногочисленные жаберно-
дышащие, приспособившиеся к сухопутному существованию, например мок-
рицы, сохраняют дыхание при помощи жабр. У мелких форм жабры могут
исчезать и газообмен совершается через покровы тела.
Голова жабернодышащих состоит из головной лопасти (акрона), на
которой помещаются глаза, и пяти сегментов, несущих каждый по паре
придатков. Все придатки, кроме первой пары, представляют собой в той
или иной степени видоизмененные конечности. Придатки двух передних сег-
ментов выполняют функцию органов чувств, реже также двигательную, ос-
тальные служат для размельчения и отфильтровывания пищи и помещаются
по бокам и позади рта. Сегменты головы в большей или меньшей степени
сливаются между собой.
Помимо определенного числа головных сегментов для жабернодыша-
щих характерно строение органов выделения. Они представляют собой ви-
доизмененные метанефридии, но в отличие от метанефридиев кольчатых чер-
вей имеются обычно в количестве только одной пары (редко двух), откры-
вающейся при основании второй или последней пары головных придатков.
КЛАСС РАКООБРАЗНЫЕ (CRUSTACEA)
КЛАССИФИКАЦИЯ1
ч Класс ракообразные (Crustacea)
Подкласс I. Листоногие (Branchiopoda)
Отряд 1. Жаброноги (Anostraca)
Отряд 2. Собственно листоногие (Phyllopoda/
Подотряд щитни (Notostraca)
Подотряд ветвистоусые (Cladocera)
Подкласс II. Челюстеногие (Maxillopoda)
Отряд 1 Веслоногие (Copepoda)
Отряд 2. Усоногие (Cirripedia)
Подкласс III. Ракушковые (Ostracoda)
Отряд 1 Ракушковые (Ostracoda)
Подкласс IV. Высшие ракообразные (Malacostraca)
Отряд 1. Тонкопанцирные (Leptostraca)
Отряд 2. Мизиды (Mysidacea)
Отряд 3. Бокоплавы (Amphipoda)
Отряд 4. Равноногие (Isopoda)
Отряд 5. Десятиногие (Decapoda)
1 Приводятся только наиболее важные подклассы и отряды.
336
Таблица IX. Речной рак Astacus astacus. / — общий вид со спинной стороны
вскрытого рака; // — вскрытый желудок рака:
/ — клешни, 2—ветви антеннулы, 3— лобный шип (rostrum), 4—усик (эндоподит) антенны, 5—слож-
ный глаз, 6 — желудок, 7 — шейная бороздка, 8— железы средней кишки, 9 — передние артерии,
10— яичник, 11— сердце, 12 — ходильная ножка, 13—жабра, 14 — бранхиостегит, 15—верхняя ар-
терия брюшка, 16— нервная цепочка, 17 — задняя кишка, 18— тельсон, 19— чешуйка (экзоподит)
антенны, 20 — уроподы
До недавнего времени подклассы листочогих, челюстеногих и ракуш-
ковых раков объединяли в один подкласс низших раков. В настоящее вре-
мя признано, что такое объединение невозможно, так как эти группы раков
различны по своему происхождению.
Представитель класса ракообразных — речной рак
(Astacus leptodactylus и Astacus astacus)
(табл. IX)
Внешнее строение. Передняя часть тела — головной и грудной отделы—
покрыта головогрудным щитом — карапаксом, и ее сегментация заметна
только с брюшной стороны. Задняя часть тела — брюшко — разделяется на
шесть подвижных друг по отношению к другу сегментов и заканчивается
тельсоном, по краям которого расположена пара пластинчатых задних
брюшных конечностей — уроподов, Уроподы вместе с тельсоном образуют
хвостовой веер.
Поперек карапакса проходит шейная, или цервикальная, борозда,
отделяющая головную его часть от грудной. Передний его край вытянут впе-
ред в заостренный на конце клювик, или рострум. при основании которого
прикрепляются сложные фасеточные глаза. Боковые края карапакса опу-
скаются вниз и прикрывают боковые края тела и расположенные на осно-
ваниях и частично на боках тела жабры. Между стенкой тела и боковыми
краями карапакса с обеих сторон остаются широко сообщающиеся с наруж-
ной средой жаберные полости.
Голова, как у всех ракообразных, несет пять пар придатков — антенну-
лы. антенны, мандибулы, максиллулы и максиллы. Антеннулы прикрепляют-
ся под рострумом между глаз и имеют трехчлениковый стебель и два много-
Рис. 289. Конечности речного рака. / —антенна (конец жгута —эндоподита—сре-
зан): // — мандибула; /// — максиллула; IV — максилла; V — ногочелюсть первой
пары; VI — ногочелюсть второй пары; VII — предпоследняя ходильная ножка;
VIII —ножка третьего брюшного сегмента; IX —ножка первого брюшного сег-
мента самца; X — то же, второго брюшного сегмента; XI — уропод
338
Рис. 290. Центральная нер-
вная система, кишечник и
эндоскелет речного рака:
/ — надглоточный нервный узел,
2 — подглоточный нервный узел,
J — брюшная нервная цепочка,
4 — глаз, 5 — нерв антенны,
6 — симпатическая нервная си-
стема, 7 — желудок, 8 — сред-
няя кишка (разрезана), 9 — зад-
няя кишка, 10 — эпдоскелет гру-
ди, 11 — бранхиостегит, 12—уро-
поды (ножки шестого абдоми-
нального сегмента)
члениковых жгута. Антенны расположены с наружной стороны от глаз.
Они, так же как все последующие конечности, подходят под схему двувет-
вистой конечности (рис. 289) и состоят из двучленикового протоподита,
многочленикового эндоподита и чешуевидного экзоподита. Антеннулы и ан-
тенны богаты чувствительными щетинками и служат органами чувств.
Мандибулы, максиллулы и максиллы находятся по бокам рта и служат для
размельчения и отфильтровывания пищи. Соответственно этому у них силь-
но развиты эндиты, а экзоподиты редуцированы. Эндоподиты сохранились.
Максиллы снабжены с наружной стороны сильно развитой лопастью, об-
разовавшейся в результате слияния экзо- и эпиподита, с помощью которой
рак вызывает движение воды в жаберной полости, нужной для дыхания.
Грудь состоит из восьми сегментов, но три передние сегмента у десяти-
ногих ракообразных срастаются с головой. Их конечности превращены в
ногочелюсти, функционирующие как дополнительные ротовые придатки.
Все три пары ногочелюстей имеют наруж-
ную и внутреннюю ветви—экзо-и эндоподиты
и снабжены измененными в жабры придат-
ками — эпиподитами. Конечности осталь-
ных пяти сегментов выполняют двигательную
и дыхательную функции. Они лишены экзопо-
дитов, но несут эпиподиты — жабры. Три пер-
вые пары этих ходильных ног заканчиваются
клешнями, при помощи которых раки могут
хватать пищу и передавать ее ротовым придат-
кам.
Конечности передних пяти брюшных сегмен-
тов называются плеоподами. У самца первая и
вторая пары плеоподов преобразованы в копуля-
тивный аппарат, приспособленный для переноса
сперматофоров из половых отверстий самца к
половым отверстиям самки. У самки первая па-
ра плеоподов отсутствует. Остальные плеоподы
представляют собой типичные двуветвистые ко-
нечности. Самка прикрепляет к ним выметанную
икру. Последняя пара брюшных конечностей —
уже упоминавшиеся уроподы.
Нервная система и органы чувств (рис. 290).
Надглоточный ганглий лежит в голове близ
основания рострума. От него отходят пять пар
нервов — к глазам и их мышцам, к антеннулам,
роструму и антеннам. Окологлоточные коннекти-
вы огибают пищевод, направляясь вниз и назад,
и впадают в подглоточный ганглий, представляю-
щий собой результат слияния ганглиев сегмен-
тов мандибул, максиллул, максилл и трех пар
ногочелюстей. Далее вдоль брюшной стороны
тела тянется брюшной нервный тяж, состоящий
из двух сближенных продольных нервных ство-
лов, с парными ганглиями в каждом сегменте. Соответственно числу сег-
ментов на грудной отдел приходится пять (не считая подглоточного), а
на брюшной шесть пар ганглиев. Между предпоследней и последней па-
рами грудных ганглиев нервные тяжи раздвинуты, так как между ними
проходит нисходящая артерия. От окологлоточных коннективов отходят
нервы симпатической нервной системы, обслуживающей желудок, сердце и
другие внутренние органы.
Стебельчатые глаза имеют типичное для членистоногих строение. Ор-
ганы равновесия — статоцисты — представляют собой полости, находя-
щиеся в основном членике стебля антеннул. Эти полости сообщаются с
12*
339
наружной средой треугольным отверстием, прикрытым щетинками. В по-
лости статоциста находятся песчинки, играющие роль статолитов. На дне
полости располагаются чувствительные щетинки, на которые давят статоли-
ты. При изменении положения тела по отношению к горизонтальной плос-
кости статолиты начинают оказывать давление на боковые чувствительные
щетинки и рак быстро реагирует на это, приводя свое тело в нормальное
положение. Органами обоняния, осязания и химического чувства служат
специальные щетинки антеннул, антенн и частично ротовых придатков.
Рис. 291. Продольный разрез через тело речного рака:
1 — пищевод, 2—кардиальный отдел желудка, 3— пилорический отдел желудка, 4—пищевари-
тельная железа, 5 — семенник, 6 — сердце, 7 — верхняя брюшная артерия, 8 — нисходящая арте-
рия, 9— задняя кишка, 10—окологлоточная коннектива, 11— эндоскелет груди, 12 — копулятив-
ные органы самца, 13 — антеннула, 14 — жгут антенны, 15 — поднервная продольная артерия
Пищеварительная система (рис. 291, 292). Короткий пищевод, начинаясь
от ротового отверстия, поднимается почти вертикально и расширяется в
объемистый желудок. Желудок разделен на две камеры — переднюю кар-
диальную и заднюю пилорическую. К стенкам кардиальной камеры прикреп-
лены три направленных внутрь мощных хитиновых зубца — один сверху и
два с боков. Они образуют так называемую желудочную мельницу, разма-
Рис. 292. Поперечный разрез
через головогрудь речного ра-
ка в области сердца:
1 — жаберная покрышка, 2 — основа-
ние ножки, 3 — жабра, 4 — сердце,
5 — пищеварительная железа,
6 — кишка, 7 — нисходящая артерия,
8 — нервная цепочка, 9 — поднервная
продольная артерия
лывающую пищу, уже частично размельчен-
ную мандибулами. Снаружи к желудку под-
ходит сложная система мышц, приводящих в
движение желудочную мельницу. В боковых
стенках кардиальной камеры имеются углуб-
ления, в которых помещаются богатые изве-
стью дисковидные жерновки, или гастролиты.
После линьки они перевариваются, а содержа-
щаяся в них известь идет на затвердение
нового панциря. В меньшей по объему пило-
рической камере находится сложная система
складок с щетинками по краям, образующая
весьма совершенный фильтр. Он пропускает
в среднюю кишку и пищеварительную железу
только полужидкую и достаточно размельчен-
ную пищу. Средняя кишка очень короткая, в
13 раз короче задней. На ее границе с пило-
рической камерой желудка двумя отверстиями
открывается большая двулопастная пищева-
рительная железа, иногда неправильно на-
зываемая печенью. Она состоит из массы
тонких трубочек, эпителиальные стенки кото-
рых выделяют пищеварительные ферменты и
усваивают переваренную пищу. Пищевари-
тельные ферменты попадают из пищеваритель-
ной железы в желудок, где происходит не
340
только размельчение и фильтрация, но и переваривание пищи. Задняя
кишка имеет вид прямой трубки и открывается анальным отверстием на
брюшной стороне тельсона.
Кровеносная система. Мешковидное приблизительно пятиугольное
сердце помещается близ спинной стороны тела внутри околосердечной сум-
ки, или перикардия. Стенка сердца прободена тремя парами остий, через
которые кровь поступает в его полость из полости перикардия. От сердца
отходят шесть артерий', одна, непарная, — вперед
к голове, две — также вперед к антеннам, одна,
непарная, — назад, над задней кишкой, две — на-
зад и в стороны к обеим лопастям пищеваритель-
ной железы и, наконец, одна, нисходящая артерия—
почти вертикально вниз, к брюшной стороне тела,
где она впадает в брюшную артерию. Из артерий
кровь изливается в просветы между органами —
лакуны — и двигается по ним. Она попадает в жаб-
ры, где окисляется, а затем по жаберно-сердечным
каналам переходит в полость перикардия, а оттуда
в сердце. Поскольку значительную часть своего
пути кровь проходит по лакунам, а не внутри кро-
веносных сосудов, кровеносную систему рака надо
считать незамкнутой.
Выделительная система (рис. 293). Основны-
Рис. 293. Антеннальная
железа речного рака:
/ — железистая часть, 2 — на-
ружное отверстие железы в
основании антенны, 3— ветви
антенны
ми органами выделения служит пара видоизменен-
ных метанефридиев, так называемые антеннальные, или зеленые, железы.
Это относительно крупные округлые железы, расположенные в головном
отделе и открывающиеся короткими протоками в основных члениках антенн.
Каждая железа состоит из маленького целомического мешочка, извитого,
расширенного в начале канала, и обширного мочевого пузыря.
Рис. 294. Мужская половая систе-
ма речного рака:
/ — семенник, 2 — семяпровод, 3— на-
ружное половое отверстие
Рис. 295. Женская половая си-
стема речного рака.
1 — яичник, 2 — яйцевод, 3—наруж-
ное половое отверстие
Половая система (рис. 294 и 295). Семенник и яичник имеют сходное
строение. Парные по своему происхождению, они, однако, срастаются в
своей задней части, так что передняя часть каждой железы двулопастная,
а задняя — непарная. Семенник относительно гораздо меньше яичника.
От границы между парной и непарной его частями отходит пара длинных
341
/ /7
Рис. 296. Головогрудь. / —
Astacus leptodactylus; II—
Astacus astacus
распространения за счет
Биология обоих виде
сильно извитых семепроводов, открывающихся половыми отверстиями в ос-
новных члениках задней пары грудных ног. Также парные яйцеводы, нао-
борот, короткие, прямые и открываются в основных члениках третьей пары
ходильных ног (шестой пары грудных ног).
При спаривании самец двумя парами передних плеоподов перекосит
сперму из своих половых отверстий в район расположения половых отвер-
стий самки, после чего оба партнера расходятся. Масса перенесенных спер-
матозоидов склеена вязким застывающим в воде веществом и представляет
собой сперматофор. Только через несколько дней самка начинает отклады-
вать икру, причем выделяется секрет, растворяющий связующее вещество
сперматофора. Освобожденные сперматозоиды оплодотворяют яйца. Оп-
лодотворенные яйца прикрепляются к плеоподам. Там и происходит их
развитие.
У речного рака развитие прямое. Из яиц выходят маленькие рачки,
отличающиеся от взрослых только отсутствием уроподов и некоторыми дру-
гими второстепенными признаками. Рачки оста-
ются на плеоподах матери, где несколько раз
линяют, после чего приступают к самостоятель-
ному существованию.
Биология. Оба вида обычных в СССР речных
раков (рис. 296) —длиннопалый (Astacus lepto-
dactylus) и широкопалый (A. astacus) — живут
на дне пресноводных водоемов. Особый подвид
длиннопалого рака обитает в Каспии.
Длиннопалый рак широко распространен в
бассейнах притоков наших южных морей, а так-
же в Западной Сибири, широкопалый рак встре-
чается в бассейнах рек, впадающих в Балтий-
ское море; он обычен в прибалтийских республи-
ках и в некоторых районах Белоруссии и
Украины. Длиннопалый рак вытесняет широко-
палого, постепенно расширяя область своего
области распространения последнего.
)в сходная. Они живут только в водоемах с бла-
гоприятным кислородным режимом и в случае наступления заморов быстро
погибают. Представители этих видов населяют прибрежные части водоемов.
Днем, а также зимой они укрываются в норах, которые вырывают в откосах
берегов, под камнями и т. д. Ночью они выходят из нор в поисках пищи.
Раки всеядны, но основной их пищей служит водная растительность.
Если представляется возможность, они охотно поедают живых животных и
их трупы.
Размножение происходит осенью. В зависимости от размера самка от-
кладывает от 60 до 500—600 яиц. В первый год жизни рак линяет 8 раз,
во второй — 5 раз, а затем самки обычно линяют 1, а самцы 2 раза в год.
Раки достигают половозрелости на третыо-четвертую осень жизни, а живут
до 20 лет.
ОБЩИЙ ОБЗОР РАКООБРАЗНЫХ
Ракообразные по числу видов и особей занимают одно из первых мест в
фауне морских и пресных водоемов. Они обитают как на дне, так и в толще
воды, начиная от прибрежных мелководий и кончая наибольшими океани-
ческими глубинами (около 11 км). Они приспосабливаются к жизни в под-
земных водах и в самоосадочных соленых озерах. Подавляющее большин-
ство ракообразных ведет свободноподвижное существование, но среди них
есть и паразиты, а также виды, перешедшие к сидячеприкрепленному обра-
зу жизни. Некоторые ракообразные выработали приспособления к обита-
нию на суше. Всего известно более 20 000 видов ракообразных.
342
Размеры этих животных колеблются в широких пределах — от мил-
лиметра и менее и до 60—80 см длины (некоторые лангусты и омары).
Внешнее строение. Расчленение тела. Тело ракообразных обычно (за
исключением некоторых паразитических и сидячеприкрепленных видов)
ясно сегментировано и разделяется на три отдела — голову, грудь и брюшко.
Голова состоит из акрона и пяти следующих за ним сегментов. Акрон
всегда срастается с антеннулярным сегментом и сегментом антенн, образуя
протоцефалон. Следующие за ними три задних головных сегмента — сег-
менты мандибул, максиллул и максилл — также срастаются между собой в
так называемый гнатоцефалон. У некоторых ракообразных (жаброноги
(рис. 297), мизиды, десятиногие и др.) прото- и гнатоцефалон сохраняют
Рис. 297 Жаброног Branchipus спинкой вниз:
1 — глаз науплиуса, 2 — антеннула, 3— антенна, 4—ман-
дибула, 5—максиллярная железа, 6—сердце, 7 — ки-
шечник, 8— penis, 9— сложный глаз
свою самостоятельность, у других (веслоногие, бокоплавы и т. д.) они сли-
ваются без всяких следов шва в сложную голову, или синцефалон.
Если число головных сегментов у всех ракообразных постоянно, то
число грудных и брюшных сегментов чрезвычайно изменчиво. Так, напри-
мер, у щитней их нередко больше 50, а у ракушковых ракообразных не
больше двух. Только высшим ракообразным свойственно постоянное число
грудных (8) и брюшных (6 или изредка 7) сегментов, а челюстеногим —
грудных (6) сегментов. Брюшной отдел заканчивается хвостовой пластин-
кой, или тельсоном, гомологичным пигидию полихет и часто снабженным
парой придатков — фуркой, или вилочкой.
У многих ракообразных один, три или даже четыре передних грудных
сегмента срастаются с головой, образуя вместе с нею головогрудь. Граница
между головой и грудью может быть не всегда четкой благодаря присутствию
у многих ракообразных так называемого головогрудного щита, или кара-
пакса.
Карапакс представляет собой двойную хитинизированную складку пок-
ровов заднего головного сегмента (максиллярного), покрывающую сверху
грудной отдел и часто спускающуюся вниз по его бокам. Карапакс может
иметь форму плоского щита у щитней (рис. 298), двустворчатой раковины
(у ветвистоусых, ракушковых и др.) или приблизительно цилиндрическую
(у креветок, речных раков, мизид и др.). У многих ракообразных (жабро-
ногие, веслоногие, бокоплавы, равноногие и др.) он отсутствует.
Каждый сегмент тела в принципе несет пару конечностей. Однако так
обстоит дело только у большинства высших ракообразных. У остальных
брюшные сегменты, как правило, лишены конечностей.
Конечности ракообразных, за исключением антеннул, гомоло-
гичны параподиям полихет — вернее гомологичны брюшной части парапо-
дий — невроподиям. Антеннулы соответствуют пальпам полихет и в отли-
чие от остальных конечностей, связанных нервами с брюшным нервным
тяжем, иннервируются от надглоточного ганглия. Исходной формой всех
конечностей ракообразных (кроме антеннул) считается одноеетвистый
стержень с рядом выростов на внутренней (эндиты) и наружной (экзиты)
343
стороне. У большинства ракообразных часть этих эндитов и эк-
зитов редуцировалась, а другие, наоборот, гипертрофировались, в резуль-
тате чего сформировалась типичная двуветвистая конечность, состоящая
из основной части—двух или трехчленикового протоподита (членики его
Рис. 298. Щитень Triops (Apus) cancriformis
называются прэкоксоподит, который присутствует довольно редко, коксо-
подит и базиподит) и двух расположенных на его конце ветвей — наружной,
или экзоподита, и внутренней, или эндоподита. Кроме того, часто имеется
экзит коксоподита, или эпиподит, превращенный у многих ракообразных
в жабру (см. рис. 299).
Основными функциями конечностей ракообразных следует считать
двигательную, дыхательную и функцию улавливания пищи и передачи ее к
Рис. 299. Ножка Le-
pidarus:
/ — эпиподит (жабра),
2 — экзоподит, 3 — эндо-
подит и эндиты
ных сегментов их
ротовому отверстию. У наиболее примитивных рако-
образных, например у жаброногов, щитней и других,
каждая из грудных конечностей выполняет все три
функции и имеет соответственно служащие для пла-
вания экзо-иэндоподит, дыхательные отростки преэпи-
подит и эпиподит и эндиты, подгоняющие пищу ко
рту. Однако у большинства ракообразных наблюдает-
ся специализация их конечностей в каком-либо
определенном направлении, приводящая к утрате
тех или иных частей (экзоподита, эндитов и т. д.).
Ротовые придатки всех ракообразных узко специали-
зированы как органы размельчения и фильтрации
пищи, а в случае слияния с головой передних груд-
конечности приобретают промежуточное строение
между ротовыми придатками и ходильными ногами и получают название но-
гочелюстей.
Скелет. Тело и конечности ракообразных покрыты хитиновым панци-
рем, твердость которого у многих форм увеличивается за счет присутствия
в покровах извести.У крупных ракообразных имеется и внутренний скелет,
состоящий из отдельных хитинизированных пластинок и тяжей.
Наружный скелет препятствует беспрерывному росту ракообразных.
Поэтому их рост связан с периодически повторяющимися линьками. После
сбрасывания старого панциря ракообразные быстро растут до тех пор,
пока не затвердеет новый панцирь, после чего рост прекращается до следую-
щей линьки.
Окраска ракообразных зависит от пигментных зерен, находящихся в
толще покровов. Из нескольких обычных присутствующих пигментов наи-
344
более стоек красный. Поэтому при варке раков только он сохраняется, а
остальные разрушаются. Кроме того, у многих ракообразных под хитино-
вым покровом располагаются звездчатые пигментные клетки хроматофоры.
Некоторые виды креветок, крабов и равноногих способны менять окраску
благодаря концентрации пигментных зерен в центре одних хроматофоров и
распределению пигмента иного цвета по всем отросткам других хроматофо-
ров. Передвижения пигмента в хроматофорах зависят от гормонов, выделяе-
мых в кровь железами внутренней секреции, расположенными близ глаз
или в глазном стебельке. Гормональная деятельность контролирует и часто-
ту линек.
Мышечная система. Как у всех членистоногих, мускулатура рако-
образных представлена отдельными, метамерно расположенными мышцами.
Нервная система. Центральная нервная система, так же как у других
членистоногих и кольчатых червей, состоит из надглоточного ганглия,
окологлоточных коннективов и двойного брюшного нервного тяжа с парой
сближенных между собой ганглиев в каждом сегменте. В случае слияния
или даже укорочения сегментов сливаются и сближаются и их ганглии.
Так, например, ганглии сегментов гнатоцефалона образуют единый подгло-
точный ганглий, к которому присоединены и ганглии сегментов ногочелю-
стей. У крабов и морских желудей с их укороченным телом все ганглии
брюшной нервной цепочки срастаются в единый мощный нервный центр.
Органы чувств. В качестве органов чувств у ракообразных функци-
онируют глаза, статоцисты, а также расположенные главным образом на
антеннулах и антеннах многочисленные и разнообразные видоизмененные
щетинки, служащие органами обоняния и осязания. Обычно на протоцефа-
лоне находится пара сложных фасеточных глаз. У некоторых отрядов (жаб-
роноги, мизиды, десятиногие) они стебельчатые, у других (щитни, бокопла-
вы, равноногие) — сидячие. В отдельных случаях эти глаза объединяются в
один непарный глаз, как, например, у ветвистоусых. Наряду со сложными
фасеточными глазами многие ракообразные обладают простым непарным
лобным или, как его называют, науплиальным глазом. Последнее название
объясняется тем, что этот глаз присущ первой личиночной стадии ракообраз-
ных — науплиусу, но нередко он переходит и ко взрослым формам, а у
веслоногих является единственным органом зрения. Статоцисты встречаются
не у всех ракообразных. Они расположены либо при основании антеннул
(у десятиногих), либо в основании эндоподита задних брюшных ног (у
мизид).
Пищеварительная система. Кишечник начинается ротовым отверстием,
расположенным на брюшной стороне головы, обычно на некотором расстоя-
нии от переднего ее конца. Передний отдел кишечника состоит из прямого
пищевода, расширяющегося в желудок, подразделенный у высших ракооб-
разных на кардиальную и пилорическую камеры. Далее следует относитель-
но очень короткая средняя кишка, в которую открываются протоки пище-
варительной железы. Эта железа особенно сильно развита у высших рако-
образных, а у остальных она представлена небольшим выростом или даже
совсем отсутствует. Она выделяет пищеварительные ферменты, но, кроме
того, в ней происходит частично всасывание пищи. Наконец, она у некото-
рых ракообразных участвует в процессе выделения и осморегуляции. Сред-
няя кишка переходит в заднюю, которая в виде прямой трубки доходит до
тельсона и на его брюшной поверхности открывается наружу анальным
отверстием.
Кровеносная система. У разных ракообразных кровеносная система
построена по-разному. У большинства есть сердце, иногда имеющее вид
длинного спинного сосуда, снабженного многочисленными парными остия-
ми (у жаброногов и ротоногих), а иногда оно короткое, мешковидное, с не-
большим количеством остий. У многих мелких ракушковых и веслоногих
сердце отсутствует. У высших ракообразных от сердца отходит 7 или 8 арте-
рий, из которых кровь выливается в лакуны, у остальных кровеносных сосу-
345
дов нет. Сердце лежит внутри околосердечной сумки. В крови растворен
гемоцианин, играющий такую же роль в связывании кислорода, как гемогло-
бин. Железо заменено в нем медью. У некоторых ракообразных, например
у дафнии, в крови растворен гемоглобин. Кровь иногда синяя, иногда крас-
новатая, иногда бесцветная и богата различного рода лейкоцитами. Окис-
ленная в жабрах или у поверхности тела, кровь поступает в околосердечную
сумку, потом в сердце и оттуда в артерии или в полость тела.
Дыхательная система. Ракообразные в большинстве случаев живут в
воде и дышат кислородом, растворенным в воде. У некоторых ракообраз-
ных специальных органов дыхания нет — они дышат всей поверхностью
тела, покрытого тонким нежным покровом. Этот тип дыхания наблюдается
преимущественно у особенно мелких ракообразных (например, у веслоно-
гих — Copepoda). У большинства ракообразных дыхательный процесс лока-
лизован: его может обслуживать внутренняя поверхность головогрудного
щита (некоторые низшие раки) или брюшные ножки (равноногие). Однако
обычно роль органов дыхания выполняют специальные нежные выросты —
жабры, сидящие то на основании грудных ножек (эпиподиты), то на брюш-
ных ножках (ротоногие). В случае развития жабр на груди головогрудной
щит может служить защитным покровом, а для поддержания тока воды в
образуемой полости служит вырост конечностей, способный к самостоятель-
ному движению (скафогнат). В тех случаях, когда ракообразные живут на
суше (равноногие), у них могут быть слабо развитые органы атмосферного
дыхания — трахеи — в брюшных ножках.
Выделительная система. Выделение осуществляется при помощи фаго-
цитов крови, эпителия пищеварительной железы и видоизмененных метане-
фридиев—антеннальных и максиллярных желез. Антеннальные железы
свойственны большинству отрядов высших ракообразных, за исключением
равноногих, ротоногих и некоторых других, у остальных имеются максил-
лярные железы. У некоторых примитивных мизид функционируют одновре-
менно как антеннальные, так и максиллярные железы.
Половая система. В подавляющем большинстве случаев ракообразные
раздельнополы, причем обычно самцы даже внешне отличаются от самок (по
величине, форме тела, присутствию специальных приспособлений для удер-
живания самок и т д.). Только для отряда усоногих характерен гермафроди-
тизм. Кроме того, отдельные случаи гермафродитизма известны среди не-
которых высших ракообразных, в частности среди некоторых креветок,
бокоплавов и клешненосных осликов. Первым свойственна протерандрия:
при наступлении половозрелости рачок становится самцом, а затем превра-
щается в самку У некоторых клешненосных осликов наблюдается обратное
явление, или протогиния, вообще очень редко встречающаяся в животном
мире. Довольно широко распространена среди ракообразных способность к
девственному размножению, т. е. к партеногенезу. Партеногенез свойствен
щитням, ветвистоусым, некоторым жаброногам и ракушковым.
Половые железы — яичники и семенники — первично парные образо-
вания и располагаются между кровеносной и пищеварительной системами
органов. Обе железы иногда частично сливаются между собой, образуя не-
парный отдел. У высших ракообразных яйцеводы открываются парным от-
верстием на брюшной стороне или при основании конечностей шестого груд-
ного сегмента, семяпроводы —восьмого грудного сегмента. У других рако-
образных положение половых отверстий варьирует
Спаривание заканчивается прикреплением сперматофора близ половых
отверстий самки, после чего оба партнера расходятся. После этого самка
приступает к икрометанию, причем вместе с яйцами выделяется секрет, рас-
творяющий оболочку сперматофора. Таким образом осуществляется наруж-
ное оплодотворение. Лишь у немногих ракообразных сперма вводится в по-
ловые пути самки, что обеспечивает внутреннее оплодотворение.
Развитие. Количество желтка в яйцах различных ракообразных раз-
лично, и это определяет разный характер дробления яиц. Для богатых желт-
346
ком яиц характерно поверхностное дробление, для бедных желтком яиц —
полное дробление. У некоторых ракообразных с полным дроблением яйца
описаны признаки спирального дробления, столь типичного для кольчатых
червей. В яйце развивается общая для всех ракообразных личинка — науп-
лиус (рис. 310, 311, 317).
Тело науплиуса внешне не сегментировано и состоит из акрона и трех
сегментов — антеннул, антенн и мандибул. Эти три пары конечностей
уже сформированы, но антенны располагаются не впереди, как у взрослых
ракообразных, а позади рта. По своему строению науплиус соответствует
метатрохофоре полихет.
У части ракообразных (веслоногие, усоногие, ракушковые, некоторые
креветки) науплиус покидает яйцевую оболочку и приступает к самостоя-
тельному существованию. У других эта личиночная стадия проходит в яйце
и свободное существование начинается в какой-нибудь более поздней личи-
ночной стадии. Наконец, есть много ракообразных с прямым развитием, у ко-
торых из яйца выходит уже вполне сформировавшийся, но еще маленький
молодой рачок.
Метаморфоз науплиуса очень напоминает метаморфоз личинок полихет.
Перед задним концом тела у него обосабливается зона роста, последователь-
но образующая перед собой сегмент за сегментом с зачатками соответ-
ствующих им конечностей. В процессе метаморфоза задняя часть тела
вместе с зоной роста отодвигается назад и в конце концов превращается
в тельсон.
СИСТЕМАТИЧЕСКИЙ ОБЗОР КЛАССА
ПОДКЛАСС I. ЛИСТОНОГИЕ (BRANCHIOPODA)
Голова не срастается с грудными сегментами и несет фасеточные глаза и
науплиальный глаз. Грудные конечности листовидные, служащие для дви-
жения, дыхания и подачи пищи ко рту. Ротовые придатки слабо развиты, ман-
дибулы лишены эндоподита. Число грудных и брюшных сегментов непо-
стоянно.
Исключительно свободноподвижные ракообразные, обитающие, за не-
многим исключением, во внутренних водоемах.
ОТРЯД ЖАБРОНОГИ (ANOSTRACA)
Характеризуется отсутствием карапакса. Тело вытянуто, длина его обыч-
но около 1 см и более. Антеннулы тонкие, антенны более мощные, особенно у
самцов. Фасеточные глаза сидят на стебельках. Грудь состоит из 11 (реже
17 или 19) одинаковых сегментов, несущих одинаково устроенные конечно-
сти. Брюшко длинное тонкое, лишено конечностей (см. рис. 297). Развитие
с метаморфозом. Около 150 видов.
Жаброноги — обычные обитатели временных весенних водоемов. Неко-
торые виды приспособились к жизни в необычных условиях. В этом отно-
шении особенно интересна Artemia salina, населяющая соленые озера
вплоть до самоосадсчных. Из всех многоклеточных животных Artemia
выдерживает наиболее высокую соленость (почти до 300о/оо)- Будучи
единственным многоклеточным животным в таких водоемах, Artemia
развивается в них в огромном количестве. В последнее время ее искус-
ственно разводят на рыбоводных заводах в качестве корма для рыб.
Интересно, что соленость влияет на строение рачка: особи, живущие
при очень высокой солености, отличаются укороченными ветвями фурки и
слиянием двух задних брюшных сегментов. В последнее время доказа-
на полиплоидность этого вида.
347
ОТРЯД СОБСТВЕННО ЛИСТОНОГИЕ (PHYLLOPODA)
Карапакс имеется, но у разных подотрядов устроен по-разному.
Подотряд щитни (Notostraca)
Карапакс в виде единого спинного щита. Фасеточные глаза сидячие,
сближенные, перед ними есть науплиальный глаз. Число сегментов тела
достигает 40, из них на передних 12 по одной паре ног, на остальных по
нескольку (до 6 пар). Задние сегменты лишены конечностей. Антеннулы и
антенны редуцированы. Тельсон с парой длинных нитевидных расчлененных
фуркальных ветвей (см. рис. 298). Крупные ракообразные, длиной до
6 у см. Развитие начинается с метанауплиуса.* Всего 9 видов.
Эти своеобразные крупные рачки подобно многим жаброногим обитают
главным образом во временных весенних водоемах и только на севере живут
в постоянно существующих озерах и прудах. Большую часть своей жизни
они проводят в стадии покоящегося яйца, легко переносящегося ветром, на
лапках водоплавающих птиц и другими случайными путями из водоема в
водоем. Замечательна необычайная консервативность щитней. Наиболее
обычный и широко распространенный вид Triops cancriformis в ископаемом
состоянии известен из триасовых отложений. Это единственный пример та-
кой долговечности современного вида.
Подотряд ветвистоусые (Cladocera)
Мелкие (до 2—3 мм длины) ракообразные, в массе населяющие всевоз-
можные пресноводные водоемы. Карапакс имеет вид прозрачной двустворча-
той раковины, у некоторых видов сохраняющейся только в виде редуцента
на спинной стороне тела. Фасеточные глаза объединяются в один непарный
глаз. Грудь состоит из 4—6 неясно разделенных сегментов, несущих листо-
видные конечности, брюшко укороче-
но, загнуто на брюшную сторону и
заканчивается двумя когтевидными
придатками. Антеннулы очень корот-
кие, антенны, наоборот, сильно раз-
виты и служат органами плавания
(рис. 300, 301). Развитие без метамор-
фоза (за исключением одного вида).
Ветвистоусые составляют обычно
основную часть планктона пресных
водоемов. Среди них есть и немало
видов, живущих только на дне или
среди подводных растений.
Большая часть видов питается,
отфильтровывая водоросли, бактерии
и частицы органического вещества,
взвешенные в воде. Это совершается
путем движения ног, на которых есть
выросты со щетинками. Переплетаясь,
эти щетинки образуют фильтрующую
сеть, на которой оседают пищевые
частицы. Другие выросты и щетинки
Рис. 300. Daphnia:
/ — антеннула, 2 — антенна, 3 — непарный глаз на-
уплиуса, 4 — сложный глаз, 5 — раковинная (мак-
силлярная) железа, 6 — сердце, 7 — вторая ножка,
8—коготки, 9 — выводковая камера, 10 — яичник,
11 — кишечник, 12 — печеночный отросток
348
Рис. 301. Ветвистоусые (/—IX) и веслоногие (X—XII) пресных вод.
/ — Polyphemus pediculus; II — Holopedium gibberum; /// — Sida cris-
tallina; IV — Diaphanosoma brachyurum; V — Daphnia longispina; VI —
Daphnia cucullata; VII —Scapholeberis mucronata; VIII —Bosmina
longirostris; IX —Chydorus; X —Cyclops; XI — Diaptomus coeruleus;
(самка, справа антеннула самца); XII — Canthocamptus staphylinus
проталкивают этот осадок к ротовому отверстию. Только несколько видов с
редуцированной раковиной ведет хищный образ жизни (рис. 302). Грудные
ножки этих видов специально приспособлены для захвата различных мел-
ких животных.
Пока условия существования благоприятны, ветвистоусые размножают-
ся партеногенетически и в водоемах обитают исключительно самками. Яйца
Рис. 302. Leptodora:
1 — антеннула, 2 — сложный глаз, 3—антенна, 4— пер-
вая грудная ножка, 5—выводковая сумка (редуциро-
ванная раковина)
вынашиваются в выводковой
камере, помещающейся на спин-
ной стороне тела под раковиной.
При ухудшении условий суще-
ствования, например осенью,
появляются самцы, более мел-
кие, чем самки, и последние
начинают откладывать гаплоид-
ные, нуждающиеся в оплодотво-
рении, яйца. Таких яиц немного
(обычно одно или два), и после
оплодотворения из них образу-
ются стойкие покоящиеся яйца.
У многих видов покоящиеся
яйца окружаются в выводковой
камере хитиновой оболочкой и
возникает так называемое сед-
лышко, или эфиппиум (рис. 303).
Покоящиеся яйца ветвистоусых
свободно переносят высыхание и
низкую температуру и служат
основным средством расселения.
Многим планктонным ветви-
стоусым свойственно явление
цикломорфоза, т е. закономер-
ных циклических изменений формы раковины. У озерной Daphnia cucul-
lata летом постепенно вырастает остроконечный шлем (рис. 304), у Bosmina
coregoni—«горб» и увеличивается длина антеннул.
Рис. 303. Седлышко дафний. / —
Ceriodaphnia quadrangula; // —
Daphnia pulex
ТШУШДХтПЕУЯУ! ш ш
Рис. 304. Сезонная изменчивость у Daphnia на
протяжении года (внизу обозначения месяцев)
Ветвистоусые служат важнейшим объектом питания пресноводных рыб,
в особенности их молоди. На рыбоводных заводах разводят некоторые виды
ветвистоусых, в первую очередь Daphnia и Moina, для кормления получае-
мых из икры мальков.
ПОДКЛАСС II. ЧЕЛЮСТЕНОГИЕ (MAX1LLOPODA)
Голова лишена сложных фасеточных глаз и у свободноживущих видов
срастается с одним или двумя передними грудными сегментами. Грудь
состоит из шести сегментов и несет расчлененные двуветвистые конечности,
350
лишенные жаберных придатков и эндитов. Жабр нет. Ротовые придатки
свободноживущих видов сильно развиты и вызывают специальный направ-
ленный ко рту ток воды. Развитие начинается со стадии науплиуса.
Помимо свободноживущих видов, в подкласс входит значительное ко-
личество сидячеприкрепленных и паразитических видов с соответственно из-
мененной организацией.
ОТРЯД ВЕСЛОНОГИЕ (COPEPODA)
Мелкие (большей частью 1—2 мм, реже до 1 см, паразиты более крупные)
пресноводные и морские ракообразные, лишенные карапакса, с одним или
двумя передними грудными сегментами, срастающимися с головой. Орга-
нами плавания служат длинные антеннулы и двуветвистые грудные ножки.
Имеется только науплиальный глаз. Брюшко короткое, сегментированное, с
фуркальными ветвями на конце. Известно около 2000 видов.
Отряд разделяется на шесть подотрядов, из которых три объединяют
паразитические виды. Три подотряда, представители которых ведут свобод-
ный образ жизни, различны по своей биологии.
К подотряду Calanoida (рис. 305) принадлежат планктонные виды, оби-
тающие преимущественно в море. Их антеннулы очень длинные, брюшко
Рис. 305. Calanus. / —рачок сбоку; // —ножка; /// —мандибула:
/— антеннула, 2 — антенна, 3 — мандибула, 4— максилла, 5 — ногочелюсть, 6 — брюшко^ 7 — экзопо-
дит, 8 — эндоподит, 9 — протоподит, 10 — плавательные ноги
укорочено, самка носит на себе один яйцевой мешок. Calanoida — господ-
ствующая группа в морском планктоне. На их долю приходится обычно не
менее 90% массы всего морского планктона. Ими питаются многие рыбы, в
том числе такие важные промысловые объекты, как сельдь, иваси, килька и
многие другие. В пресных водах подотряд представлен широко распростра-
ненным родом Diaptomus. Как морские, так и пресноводные Calanoida
питаются преимущественно водорослями, которые они отфильтровывают
при помощи щетинок ротовых придатков и размельчают мандибулами.
В подотряд циклопов (Cyclopoida) входят главным образом донные и
придонные виды (есть и паразитические), отличающиеся гораздо более ко-
роткими антеннулами, сравнительно длинным брюшком, резко обособлен-
ным от грудного отдела, и присутствием у самки двух яйцевых мешков.
Циклопы особенно многочисленны в пресных водах. В отличие от предыду-
щего подотряда это хищники.
Третий подотряд — Harpacticoida — представлен как в море, так и в
пресных водах очень мелкими донными червеобразными рачками. Их антен-
нулы короткие, грудные сегменты подвижны по отношению друг к другу,
брюшко по ширине почти не отличается от грудного отдела, самка вынаши-
вает один яйцевой мешок. Harpacticoida нигде не достигают такого массо-
вого развития, как представители двух первых подотрядов, но зато встре-
чаются и в таких своеобразных условиях, как условия подземных вод, мо-
ховых болот и больших океанических глубин.
351
Многочисленные паразитические веслоногие (например, Lernaea) па-
разитируют на различных рыбах и водных беспозвоночных. Организация
многих из них упрощена, некоторые полностью утеряли сегментацию и ко-
Рис. 306. Метаморфоз у Lernaea. / — метанауплиус;
// —копеподидная стадия; /// —сидячая куколочная
стадия; IV — спаривание самца с самкой; V — послед-
няя стадия паразитирующей самки:
1 — антеннула, 2—антенна, 3—грудные ножки, 4 — орган при-
крепления паразита к хозяину, 5— шнур с яйцами
нечности и превратились
в бесформенные колбасо-
видные тела. Однако из
яйца даже таких реду-
цированных существ
выходит типичный сво-
бодный науплиус, по
строению которого мож-
но судить о системати-
ческом положении взро-
слой формы (рис. 306).
Рис. 307. Карповая вошь
(Argulus foliaceus), снизу:
/ — шип, 2— антеннула, 3—ан-
тенна, 4 — присоска (максил-
лула), 5—хоботок с двумя па-
рами челюстей, 6—максилла,
7 — кишечник, 8 — плавательные
ножки, 9 — брюшко с семенни-
ками
К отряду Copepoda тесно примыкает отряд карповых вшей (Вгап-
chiura) — эктопаразитов рыб (рис. 307).
ОТРЯД УСОНОГИЕ (CIRRIPEDIA)
Все усоногие — морские обитатели, ведущие во взрослом состоянии
сидячеприкрепленный или паразитический образ жизни и в связи с этим
обладающие весьма своеобразной и упрощенной организацией. Известно
около 800 видов этих ракообразных. Непаразитические усоногие — морские
желуди и морские уточки — прикрепляются к всевозможным подводным
предметам — к камням и скалам, сваям, днищам судов, к другим животным.
Тело их неясно сегментировано и окружено складкой кожи — мантией,
соответствующей карапаксу других ракообразных. Мантия выделяет разное
количество пропитанных известью пластинок. Эти пластинки соединяются
между собой так, что образуют домик или раковину, целиком покрывающую
тело животного. Две или четыре пластинки соединены между собой подвижно
и снабжены приводящими их в движение мышцами. Благодаря этому рако-
вина может приоткрываться и вода поступает в мантийную полость жи-
вотного.
Головной отдел тела, которым животное прикрепляется к субстрату,
очень сильно видоизменен. Маленькие антеннулы соединены с субстратом,
352
антенны отсутствуют. Рот помещается на конце ротового конуса и окружен
ротовыми придатками упрощенного строения. Грудь несет шесть пар дву-
ветвистых ножек с многочлениковыми ветвями. Эти ножки совершают одно-
временно характерные взмахи, причем вода загоняется в мантийную полость.
Рис. 308. Морская уточка (Lepas) со вскрытой раковиной, сбоку. / —
взрослая стадия; // — циприсовидная личинка:
1—антеннула и цементная железа, 2—сводящая щитки (scuta) мышца, 3—кишечник,
4 — яичник, 5 — семенник, 6 — penis, 7 — carina, 8 — terga, 9 — scuta (щитки раковины),
10— брюшко, 11 — глаз (парный)
На конце грудного отдела находится длинный жгутовидный мужской копу-
лятивный придаток, при помощи которого сперма переносится в мантий-
ную полость другой особи. Брюшко сильно редуцировано.
В отличие от подавляющего большинства ракообразных усоногие — гер-
мафродиты, однако у них происходит перекрестное оплодотворение. У не-
которых видов, помимо гермафродитных особей, имеются дополнительные
самцы, открытые Ч. Дарвином, много занимавшимся усоногими. Они сильно
редуцированы и живут в мантийной полости гермафродитных особей.
Из яйца выходит свободно плавающий науплиус (рис. 310), после ряда
линек превращающийся в характерную для усоногих циприсовидную личин-
ку. Она снабжена двустворчатой раковиной и плавает при помощи шести пар
грудных ножек, похожих на плавательные ножки
веслоногих. Своими антеннулами, снабженными це-
ментными железами, она прикрепляется к субстрату,
после чего подвергается сложному метаморфозу,
заканчивающемуся формированием маленького усо-
ногого рачка.
Морские уточки (рис. 308) отличаются от мор-
ских желудей присутствием стебля, соответствую-
щего удлиненной части головы. Как те, так и
другие широко распространены во всех морях и
Рис. 309. Наружный вид морского желудя Balanus (сбоку)
1 и 2 — крышечка
353
океанах от полосы прибоя до глубин примерно 7 км. Некоторые виды посе-
ляются на скалах выше уровня моря, довольствуясь только брызгами, ко-
торые до них долетают.
Усоногие, в особенности морские желуди (рис. 309), имеют большое от-
рицательное практическое значение. Они составляют основную массу об-
растаний судов. Судно с обросшим днищем теряет свою скорость, благодаря
чему возрастают затраты на топливо. О массе морских желудей на днищах
судов можно судить по тому, что с одной китобойной матки, проплававшей
7 месяцев в Тихом океане, сняли 40 т крупных морских желудей. Приходит-
ся периодически очищать от обрастаний морские суда, изобретать необра-
стающие краски и т д.
Пелагические личинки усоногих (рис. 310) усиленно потребляются
планктоноядными рыбами.
Рис. 310. Личиночные фазы Balanus. / — науплиус; // — метанауплиус:
антеннула, 2 — антенна, 3—мандибула, 4 — максиллула, 5 — брюшко
Паразитические усоногие ракообразные поражают главным образом де-
сятиногих ракообразных. Представители одной из групп этих паразитов
обтают в толще раковины некоторых моллюсков, а также в скелете ко-
раллов.
Среди паразитов ракообразных интересен паразит крабов (рис. 311).
Во взрослом состоянии мешковидный паразит помещается под брюхом кра-
ба, а многочисленные разветвленные выросты его тела оплетают все внутрен-
ние органы хозяина, в частности гонады. Если самец краба заражается сак-
кулиной в молодом возрасте, он постепенно теряет признаки самца и приоб-
ретает признаки самки. Описаны случаи полного перерождения семенников в
яичники, способные продуцировать нормальные яйца.
Личинки саккулины существенно не отличаются от личинок непарази-
тических усоногих. Циприсовидная личинка прикрепляется к щетинкам
брюшных ножек краба, отбрасывает раковину, грудной отдел и науплиаль-
ный глаз, и оставшаяся часть головного отдела проникает в тело краба. Там
паразит начинает расти вдоль кишечника, развивает корневидные выросты и,
достигнув определенных размеров, при очередной линьке хозяина выпячи-
вается наружу. Таким образом, саккулина сначала оказывается эктопарази-
том, потом переходит к эндопаразитизму и, наконец, возвращается к эктопа-
разитизму.
3 54
Рис. 311. Sacculina. / — взрослый паразит, продольный разрез; // —науплиус;
/// — циприс; IV, V, VI, VII —дальнейшие фазы развития после прикрепления к
щетинке хозяина:
/ — нервный узел, 2—массы яиц ь толще мантии, 3 — яичник, 4 — семенник, о — придаточная поло-
вая железа, 6 — корневидные отростки
ПОДКЛАСС III. РАКУШКОВЫЕ (OSTRACODA)
В этом подклассе имеется единственный отряд с тем же названием.
Ракушковые — мелкие (редко более
Рис. 312. Cypridina:
1 — антеннула. 2 — антенна, 3 — .мандибула#
4 — максиллула, 5 — максилла, 6 — первая ножка,
7 _ вторая ножка, 8 — глаз науплиуса, 9— слож-
ный глаз, 10—сердце, 11— раковина
мм) морские и пресноводные пре-
имущественно донные ракооб-
разные с укороченным телом,
целиком заключенным в обычно
богатую известью двустворчатую
раковину, которая соответствует
карапаксу (рис. 312). Голова
снабжена пятью парами придат-
ков, грудь — всего двумя пара-
ми. На границе между грудью и
брюшком поперек тела проходит
мощный мускул — замыкатель
створок раковины. Укороченное
брюшко у некоторых заканчива-
ется фуркой. Развитие начина-
ется со стадии науплиуса, уже
имеющего раковину. Известно
1500 видов.
Ракушковые передвигаются
по дну или водорослям, переби-
рая антенной и задними грудны-
ми ножками. В составе морской
фауны, помимо донных, имеются
и планктонные виды.
Благодаря твердой раковине, ракушковые хорошо сохраняются в иско-
паемом состоянии. Они широко используются в геологии как руководящие
ископаемые.
ПОДКЛАСС IV. ВЫСШИЕ РАКООБРАЗНЫЕ
(MALACOSTRACA)
Грудной отдел состоит из восьми, брюшной из шести или изредка из
семи сегментов. Брюшные сегменты несут конечности. Желудок подразде-
лен на две камеры, пищеварительная железа сильно развита. Помимо серд-
ца, имеются кровеносные сосуды. Карапакс у некоторых отрядов есть, у
других отсутствует. Развитие с метаморфозом или прямое. В подкласс вхо-
дит около 15 000 видов, распределяющихся между 5 надотрядами и 13 от-
рядами. Здесь рассматриваются только важнейшие отряды.
ОТРЯД ТОНКОПАНЦИРНЫЕ (LEPTOSTRACA)
Карапакс имеет вид двустворчатой раковины, срастающейся только
с передней частью головы. Ростру м подвижно сочленен с карапаксом. Восемь
пар одинаково построенных грудных двуветвистых конечностей с пластин-
чатыми эпи- и экзоподитами. Брюшко состоит из семи сегментов и заканчи-
вается тельсоном, несущим фуркальные ветви. Имеются как антеннальные,
так и максиллярные железы.
Тонкопанцирные могут считаться наиболее примитивными из высших
ракообразных. Об этом свидетельствуют гомономная сегментация (у боль-
шинства высших ракообразных 6-й и 7-й брюшной сегменты сливаются
между собой) и присутствие фурки. Это очень древняя группа. В верхнепа-
леозойских отложениях найдены формы, очень сходные с современными.
356
В современной фауне отряд представлен всего четырьмя родами с еди-
ничными видами. Они обитают исключительно в море у дна (за одним исклю-
чением), в области мелководья. Наиболее широко распространена Nebalia
bipes (рис. 313, 314).
Рис. 313. Nebalia:
1 — рострум, 2 — антенну ла, 3 — антенна, 4 — сводящая створки мышца; 5 — сердце;
$ — грудные ножки, Z — первая ножка брюшка, 8 — шестая ножка брюшка, 9 — щупик
максиллулы
Рис. 314. Грудная ножка Nebalia:
/ — протоподит, 2 — эпиподит, 3 — экзопо-
дит, 4 — эндоподит
ОТРЯД МИЗИДЫ (MYSIDACEA)
По форме тела напоминают кре-
веток. Имеют стебельчатые глаза.
Карапакс только прикрывает сверху
задние грудные сегменты, но не сра-
стается с ними. Брюшко состоит из
шести цилиндрических подвижно со-
члененных сегментов и заканчивается
пластинчатым тельсоном. У большин-
ства видов грудные конечности дву-
ветвисты, но лишены жабр, брюшные
конечности слабо развиты, за исклю-
чением последней пары — уроподов.
В основании эндоподита уроподов
расположен крупный статоцист. Сам-
ки вынашивают яйца в выводковой
сумке, образованной выростами осно-
ваний грудных ног. Развитие прямое.
Известно около 500 видов. Большин-
ство имеет длину около 1 см, но некоторые глубоководные виды достигают
20 см.
Мизиды живут преимущественно в морях, где ведут придонный образ
жизни, встречаясь от приливо-отливной полосы до глубин 7 км. Известно
также много планктонных морских видов. Некоторые мизиды, обитающие в
Каспийском море и в опресненных частях Черного и Азовского морей, про-
никают в реки и подымаются по ним вверх по течению. В холодных северных
озерах и в устьях северных рек встречается реликтовая мизида (Mysis re-
lic ta). Есть пресноводные мизиды и на Дальнем Востоке.
Многие мизиды служат излюбленной пищей рыб. Для увеличения кормо-
вой базы рыб некоторые виды мизид успешно акклиматизированы в ряде
водохранилищ южной части СССР, а также в озере Балхаш.
357
ОТРЯД БОКОПЛАВЫ (AMPHIPODA)
около 1—z см, немногие до
Рис. 315. Gammarus:
/ — антеннула, 2 — антенна, 3 — ногочелюсть (груд-
ная ножка), 4—жабра пятой грудной ножки, 5 —
восьмая грудная ножка, 6 четвертая брюшная
ножка
Тело обычно сжато с боков. Глаза сидячие. Карапакс отсутствует.
Первый грудной сегмент слит с головой, остальные семь подвижно сочленены
друг с другом. Первая пара грудных конечностей превращена в ногочелю-
сти, следующие две пары хватательные: последний членик каждой — ко-
готь — может пригибаться к расширенному предпоследнему, образуя вместе
с ним так называемую ложную клешню. Остальные грудные конечности хо-
дильные. Три передние пары брюшных коаеч юстей плавательные (плеопо-
ды), а три задние пары — прыгательные (уроподы). Благодаря такой диф-
ференциации конечностей отряд и получил свое латинское название, которое
в переводе означает разноногие. Жабры расположены на грудных конеч-
ностях. Размножение, как у мизид. Описано около 3000 видов. Длина тела
см длиной.
Подавляющее большинство бо-
коплавов живет в море, приспосо-
бившись к существованию в самых
различных условиях и на всех
глубинах. Помимо множества дон-
ных видов, есть и планктонные.
Небольшая группа бокоплавов па-
разитирует на китах (китовые вши).
Есть бокоплавы, перешедшие к
полусухопутному обитанию. Они
массами встречаются в прибрежной
части южных и восточных морей.
Сравнительно менее богата пресно-
водная фауна бокоплавов. Только
озеро Байкал выделяется необы-
чайным изобилием и разнообрази-
ем этих ракообразных. Оттуда
известно 240 видов, причем все они свойственны только Байкалу и рассе-
лились лишь по вытекающей из него Ангаре и дальше по Енисею. Много
бокоплавов морского происхождения населяет реки, впадающие в Каспий-
ское, Черное и Азовское моря. В остальных пресноводных водоемах ши-
роко распространены виды рода Gammarus (рис. 315), обитающие в озе-
рах, реках и различных горных потоках и ручьях. В очень мелких
водоемах бокоплавы передвигаются на боку, от чего и происходит их наз-
вание, но в более крупных водоемах они ползают и плавают спинной сто-
роной вверх.
Бокоплавами питаются очень многие рыбы, в том числе и промысловые.
Сами рачки всеядны. Они способствуют очищению водоемов, поедая труппы
погибших животных, но иногда приносят и некоторый вред, так как портят
рыболовные сети и пойманных в них рыб.
ОТРЯД РАВНОНОГИЕ (ISOPODA)
Подобно бокоплавам равноногие лишены карапакса, имеют такое же
расчленение грудного отдела и такие же сидячие глаза. Однако тело их
обыкновенно уплощено в спинно-брюшном направлении, грудные конечно-
сти устроены более однообразно, а часть брюшных превращена в жабры.
Часто некоторые или все брюшные сегменты сливаются с тельсоном. Органа-
ми выделения служат максиллярные, а не антеннальные железы. Развитие
прямое, как у мизид и бокоплавов.
К этому отряду относится около 1500 видов, обитающих в море, в прес-
ных водах и на суше. Размеры равноногих колеблются от одного или несколь-
ких миллиметров до 27 см. В морях разноногие населяют преимущественно
дно или водоросли и встречаются от приливо-отливной зоны до наибольших
358
океанических глубин. Среди них есть и зарывающиеся и ползающие по суб-
страту формы, есть хищники, растительноядные виды, потребители органиче-
ского вещества грунта и паразиты рыб и беспозвоночных. По внешнему виду
морские равноногие весьма разнообразны. В пресных водах очень обычны
водяные ослики, живущие во всевозможных стоячих водоемах. Особый ин-
терес представляют многочисленные сухопутные равноногие, известные под
названием мокриц. Они обитают не только во всевозможных сырых местах —
в домах, сараях, оранжереях, а также в почве, но и в пустынях. Замеча-
тельно, что эти сухопутные ракообразные сохранили унаследованное ими
от водных предков жаберное дыхание. Они используют кислород, растворен-
ный в тонкой пленке воды, покрывающей их жаберные листочки. Однако у
многих видов в порядке приспособления к сухопутному существованию воз-
никли ложные трахеи — тонкие трубочки, вдающиеся в толщу жаберного
листочка. При помощи этих ложных трахей мокрицы могут дышать атмосфер-
ным воздухом. От настоящих трахей они отличаются тем, что их стенки ли-
шены спиральных хитиновых утолщений.
Водные равноногие в меньшей степени используются в пищу рыбами,
чем бокоплавы. Среди мокриц есть вредители культурных растений, но,
известны и пустынные мокрицы, повышающие там плодородие почв, подобно
тому, как это делают дождевые черви.
ОТРЯД ДЕСЯТИНОГИЕ (DECAPODA)
Наиболее крупные из ракообразных. Три передних грудных сегмента
срастаются с головой, карапакс срастается со всеми грудными сегментами.
Глаза стебельчатые. Самки прикрепляют выметанную икру к брюшным нож-
кам. Развитие обычно с метаморфозом, только у многих пресноводных и глу-
боководных видов прямое.
Я ш
Рис. 316. Ракообразные. / — Камчатский краб (Paralithodes camtschatica),
уменьшено в 15 раз // — прибрежный травяной краб (Carcinus maenas);
/// —рак-отшельник (Pagurus setosus)
359
К десятиногим относятся такие на первый взгляд несходные между собой
ракообразные, как креветки, речные раки и омары, раки-отшельники, кра-
бы(рис. 316). Они различаются главным образом строением брюшного отде-
ла. Всего известно около 8500 видов десятиногих.
У креветок, речных раков и некоторых других брюшной отдел хорошо
развит, превосходит по длине голову и грудь и снабжен полным набором
конечностей. У раков-отшельников брюшко спирально закручено, лишено
хитинового покрова и обычно помещается в пустой раковине брюхоногих
моллюсков, которая служит надежной защитой для них. Представители
близких к ним видов, например камчатский краб (неправильно называемый
Рис. 317. Penaeus. I — науплиус; // — протозоэа
крабом), подворачивают свое брюшко под головогрудной отдел, но оно сох-
раняет следы асимметрии, что свидетельствует о происхождении таких кра-
бовидных форм от настоящих раков-отшельников. Брюшные конечности ра-
ков-отшельников и их родичей развиты обычно только с левой стороны.
Наконец, у настоящих крабов брюшко всегда подогнуто под головогрудь,
укорочено и сужено, карапакс расширен и уплощен в спиннобрюшном на-
правлении и имеет форму овала, треугольника, четырехугольника или мно-
гоугольника. Брюшные ножки сильно редуцированы, особенно у самцов.
Метаморфоз у десятино?их ракообразных протекает различно. Лишь у
немногих креветок из яйца выходит типичный науплиус (рис. 317). У боль-
шинства личинка покидает яйцо на более поздней стадии развития — на ста-
дии зоэа, имеющей карапакс, сегментированный брюшной отдел, зачатки
задних грудных ног и заднюю пару брюшных ног (рис. 18); зоэа плавает пре-
имущественно при помощи хорошо развитых ногочелюстей. Зоэа крабов обла-
дают вздутым карапаксом со спинным, боковыми и ростральным шипами,
карапакс зоэа раков-отшельников с глубокой задней выемкой и т. д. Эта
стадия затем переходит в мизисную стадию (отсутствующую у некоторых
групп), за которой следует последняя личиночная стадия —декаподитная,
по форме карапакса сходная со взрослыми. Декаподитные стадии разных
групп десятиногих настолько различны, что имеют разные названия (у кра-
бов — мегалопа, у раков-отшельников и близких форм — глаукотоэ). Как
360
Рис. 318. Penaeus. / — стадия зоэа;
// — мизисная стадая:
1 — вторая грудная ножка, 2— шестая
брюшная ножка
уже указывалось, у большинства пресноводных (речные раки, крабы) и
глубоководных видов развитие прямое, без свободных личиночных стадий.
По способу передвижения десятиногих ракообразных можно разбить на
две группы — ползающих и плавающих. К последним принадлежат кревет-
ки с их удлиненным брюшком и сильно развитыми плавательными конеч-
ностями — плеоподами, к первым — все остальные, плеоподы которых не
служат для плавания.
Десятиногие ракообразные обитают главным образом в море. Здесь они
встречаются как на дне, так и в толще воды до глубины 5—6 км. Особенно бо-
гата этими ракообразными область
морских мелководий тропиков, в
частности коралловые рифы, изоби-
лующие самыми разнообразными кра-
бами. В тропиках живут и интересные
сухопутные десятиногие, так называ-
емые манящие крабы, обладающие
резко асимметричными клешнями и
роющие выше уровня моря очень
сложные ходы, и близкий к ракам-
отшельникам пальмовый вор, питаю-
щийся кокосовыми орехами. Его
жабры редуцированы, а вместо них в
качестве органов дыхания атмосфер-
ным воздухом функционируют распо-
ложенные под краем карапакса тонко-
стенные складки наружных покровов.
Когда личинкам этих ракообразных
приходит время выходить из яйца,
родители направляются к морю и
выпускают личинок в родную стихию.
Число видов пресноводных деся-
тиногих ограничено. Это различные
виды речных раков, встречающиеся
преимущественно в умеренных широ-
тах обоих полушарий и на Мадагаска-
ре, пресноводные крабы и креветки, приуроченные, наоборот, главным обра-
зом к тропической зоне. В СССР пресноводные крабы обычны на Кавказе,
в Крыму и в Средней Азии, а креветки — в реках и озерах Дальнего Восто-
ка и в пещерах Закавказья.
Благодаря крупным размерам многие десятиногие ракообразные (омары,
лангусты, креветки, крабы, речные раки) используются человеком в пищу и
служат важным объектом промысла. В СССР важное промысловое значение
имеет камчатский краб Paralithodes camtschatica, добываемый в Охотском
море с крупных судов — краболовов, на которых производится его консер-
вирование. Гораздо меньшее промысловое значение имеют креветки и реч-
ные раки.
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ РАКООБРАЗНЫХ
Многих ракообразных, в первую очередь десятиногих, добывают
во многих странах в большом количестве и используют в пищу. Как
уже указывалось, в СССР развит промысел камчатского краба, некото-
рых креветок и речных раков. В других странах интенсивно промыш-
ляют омаров, лангустов, различных крабов и креветок. В странах юго-
восточной Азии креветок разводят в специальных прудах, соединенных
каналами с морем. Широко практикуется изготовление консервов из
крабов и креветок.
Очень велико значение ракообразных в качестве пищи промысло-
вых рыб и китов. Молодь почти всех рыб как в море, так и в пресных
361
водах откармливается преимущественно планктонными ракообразны-
ми — веслоногими и ветвистоусыми, а многие рыбы, такие, как сельдь,
шпрот, килька и другие планктоноядные рыбы, питаются ими всю
жизнь. Высшие ракообразные, например, бокоплавы и мизиды, состав-
ляют существенную часть пищи многих донных рыб, использующих
донную фауну. На рыбоводных заводах для питания выведенных из
икры мальков рыб и подросшей молоди разводят кормовых ракообраз-
ных, в первую очередь некоторых ветвистоусых (Daphnia, Moina), а
также жабронога Artemia salina. Мизид и бокоплавов перевезли в >
многие вновь созданные водохранилища южной части СССР, а также
в озеро Балхаш, с тем, чтобы улучшить условия откорма живущих там рыб.
Важную роль играют ракообразные в процессах очистки водоемов.
Они поедают трупы погибших животных, а также их экскременты, пре-
пятствуя, таким образом, загрязнению воды продуктами разложения
органического вещества. Мокрицы перерабатывают растительный мате-
риал, обогащая почву органическими веществами, улучшают ее струк-
туру и, благодаря этому способствуют повышению ее плодородия
Среди ракообразных есть и вредные. Паразитические веслонсгге
и карповые вши могут вызвать гибель рыб, особенно молодых. Прес-
новодные веслоногие служат промежуточными хозяевами опасных пара-
зитов человека — широкого лентеца и ришты. Морские желуди, масса-
ми нарастая на днищах судов, снижают их скорость и увеличивают
расходы на их эксплуатацию. Рачки древоточцы — равноногий рачск
Limnoria и бокоплав Chelura — разрушают деревянные сооружения в море.
ФИЛОГЕНИЯ РАКООБРАЗНЫХ
В ископаемом состоянии ракообразные известны, начиная с ниж-
него кембрия, причем уже тогда существовали некоторые современные
отряды, например остракоды. Очевидно, формирование всего класса ра-
кообразных и даже его основных подклассов происходило в далекое до-
кембрийское время, от фауны которого не сохранилось никаких остатков.
Сравнение ракообразных с другими современными и вымершими
группами позволяет предполагать происхождение ракообразных от об-
щего предка с трилобитами и хелицеровыми. Эти древнейшие неизвест-
ные нам членистоногие произошли, в свою очередь, от каких-то коль-
чатых червей.
Ныне живущие подклассы ракообразных непосредственно не род-
ственны между собой. Все они представляют боковые ветви общего
ствола, отделившиеся от него в глубокой древности.
Подтип И. Трилобиты (Trilobita)1
ХАРАКТЕРИСТИХА
Трилобиты — вымершие, палеозойские членистоногие, исключительно
морские формы, обладавшие примитивными признаками: однородность
(гомономность) строения конечностей и сегментов тела, примитивный мета-
морфоз и др. Этими признаками они напоминают предков членистоногих —
аннелид.
Длина тела 2—10 см, реже до 75 см. У примитивных форм тело состо-
ит из двух отделов: слитной головы и явственно расчлененного многосег-
ментного туловища (рис. 319). Тело заканчивается небольшой анальной
1 Наряду с трилобитами в палеозойских морских отложениях встречаются не-
которые другие похожие на них членистоногие. Эти малоизученные формы относятся
к особым классам; вместе с классом собственно трилобитов их иногда объединяют
в подтип трилобитообразных (Trilobitomorpha).
362
лопастью, называемой у членистоногих тельсоном. У большинства трило-
битов тельсон и задние сегменты в разном числе сливаются в хвостовой от-
дел — пигидий (рис. 320). Тело делится продольными спинными бороздами
на выпуклую срединную часть (ось, или рахис) и уплощенные боковые час-
ти (так называемые плевры). С тройным продольным и поперечным делением
тела связано название трилобитов, что значит трехдольчатые. Покров спин-
ной стороны был очень плотный, а брюшной — тонкий перепончатый.
Голова и пигидий покрыты щитами, сегменты тела, расположенные
между ними, соединялись подвижно, что позволяло трилобиту свертываться
на брюшную сторону. В состав головы входят головная лопасть, которая
4
Рис. 319. Трилобит Triarthrus becki
с брюшной стороны
1 — антеннулы, 2 ротовое отверстие,
3 — конечности головы, 4— анальная ло-
пасть
Рис. 320. Трилобит Granularia proto-
lenorum со спинной стороны:
1 — 4 — спинки сегментов головы, 5 — части
головного щита, образованные акроном,
6 — глаза, 7 рахис, 8 — плевры, 9 — пи-
гидий
соответствует простомиуму аннелид и называется у членистоногих акрон,
и четыре послеротовых сегмента. Акрон образует переднюю и боковые
части полулунного головного щита, ему же принадлежит на дорзальной
стороне головы пара крупных фасеточных глаз и иногда имевшиеся простые
глаза, а на вентральной стороне — пара нитевидных членистых усиков —
антеннул, прикрепленных впереди рта. Позади рта расположены четыре
пары двуветвистых конечностей сегментов головы. Эти сегменты образуют
выпуклую срединную часть головного щита, на которой обычно различи-
мы их поперечные границы. Конечности головы полностью подобны ту-
ловищным. Все они построены по двуветвистому типу и состоят из осно-
вания и двух членистых ветвей (рис. 320) Считается, что конечности
такого типа произошли от параподий полихет; эти конечности удлинились
и расчленились. Однако у трилобитов и ракообразных (см. выше) они устро-
ены различно. У трилобитов от основного членика отходит неветвистая
расчлененная ножка, а вторая ветвь конечности представлена жаброй —
членистым придатком основного членика с жаберными листочками Нож-
ки имеют жевательные отростки — эндиты.
Трилобиты развивались с метаморфозом, во многом напоминающим
развитие аннелид. Из яйца выходила очень мелкая (до 1 мм) личинка про-
зы
Рис. 321. Ранние стадии послезародышевого
развития трилобитов Olenellidae. / — личин-
ка-протаспис; //—/// —образование новых
сегментов на переднем крае анальной лопас-
ти (анаморфоз):
1 — головная лопасть, 2—личиночные сегменты, 3 —
глаза, 4—анальная лопасть, 5 — новые, постларваль-
ные, сегменты
таспис. Самые ранние известные стадии имеют цельное тело. Далее оно
сразу делится на головную лопасть с глазами и антеннулами, на три или
четыре личиночных (ларвальных) сегмента и анальную лопасть. В этом
состоянии протаспис соответствует расчленившейся трохофоре аннелид.
Дальнейшее развитие сопрово-
ждалось появлением туловищ-
ных сегментов, длительным рос-
том и линьками. Новые пост-
ларвальные сегменты, как у
аннелид, появлялись в процессе
анаморфоза, т. е. один за другим
обособлялись кпереди пред-
анальной зоной роста (рис. 321).
Известно несколько тысяч
видов трилобитов. Они были ши-
роко распространены в морях в
течение кембрия, силура и дево-
на и вымерли к концу палеозой-
ской эры. Большинство обитало
в мелководье, ползая или плавая
у дна; некоторые виды зарыва-
лись в грунт; были, по-видимому, и обитатели больших глубин. Вслед-
ствие многочисленности трилобиты широко используются как руководящие
ископаемые для установления возраста содержащих их отложений.
Подтип III. Хелицеровые (Chelicerata)
ХАРАКТЕРИСТИКА
Наиболее существенные признаки хелицеровых — редукция антеннул
и характер расчленения тела, связанный со специализацией конечностей
(рис. 322, Л). К голове присоединяются два туловищных сегмента, образуя
передний отдел тела — головогрудь (просому), несущую шесть пар конеч-
ностей, которые выполняют жевательную и двигательную функции. Ос-
тальные сегменты туловища образуют брюшко (опистосому), конечности
которого выполняют половую и дыхательную функции и в значительной
части атрофируются. У всех хелицеровых головогрудь и брюшко соеди-
няются посредством седьмого (так называемого предполового) сегмента,
который в той или иной степени редуцирован.
Передняя часть головогруди образована акроном, на котором имеются
глаза и срединный вырост—эпистом —с верхней губой, нависающей над ро-
товым отверстием (рис. 322, б)- Близ рта расположены передние две пары
конечностей, участвующие в захвате и пережевывании пищи: хелицеры,
обычно прикрепленные перед ртом и снабженные клешней1, и педипальпы
(ногощупальца), основные членики которых (тазики, или коксы) имеют
жевательные отростки — эндиты (рис. 322). Остальные четыре пары ко-
нечностей головогруди—ноги — служат для передвижения, иногда сохраняя
при этом жевательные эндиты на тазиках. Головогрудь покрыта сверху
обычно цельным щитом, но иногда сохраняется разделение на голову и два
туловищных сегмента, несущих две задние пары ног. Между основаниями
ног у части форм имеется грудина — стериум; у наземных Arachnoidea
грудная пластинка (стернит) педипальп ограничивает сзади ротовое от-
верстие.
1 Chelicerata значит клешнеусые: антеннулы исчезли и на их место передвину-
лась первая пара послеротовых конечностей — клешневидные хелицеры.
364
В составе брюшка объединяются две группы сегментов: семь передних
(считая предполовой) несут видоизмененные конечности и образуют перед-
ний отдел брюшка —мезосому—и шесть задних, которые всегда лишены ко-
нечностей и образуют задний отдел — метасому, которая оканчивается
анальным придатком (тельсоном) с анальным отверстием. Второй брюшной
сегмент (восьмой по общему счету) половой, на нем всегда расположено
Б
Рис. 322. Схема строения Chelicerata. А—вид сбоку; Б —вид спе-
реди: / —голова; // —головогрудь (просома); /// —брюшко
(опистосома), IV — переднебрюшье (мезосома); V — заднебрюшье
(метасома):
1 — 19 — сегменты тела, 20 — акрон с глазами, 21 — эпистом. 22 — анальная ло-
пасть (тельсон), 23—хелицеры, 24 — педипальпы, 25 — ноги, 26 —- конечности
полового сегмента, 27— дыхательные конечности брюшка, 28 — акрон с глазами,
29 — эпистом с верхней губой, 30— хелицеры (левая удалена), 31 — педипальпы,
32 — тазик педипальп с жевательным отростком (эндитом), 33— ротовое отвер-
стие
половое отверстие, а его конечности у части форм образуют половые крыш-
ки. Конечности остальных пяти сегментов у водных форм представлены
жаберными ножками, а у наземных превращены в легкие, которые по числу
и положению различны и могут исчезать, заменяясь дыхательными труб-
ками — трахеями.
Хелицеровые в отличие от трилобитов в большинстве развиваются
без метаморфоза. Яйца богаты желтком и вылупляется полносегментная
маленькая особь, в общем похожая на взрослую. К Chelicerata относятся
первично водные формы класса Merostomata и большое число наземных,
объединенных в класс Arachnoidea.
КЛАССИФИКАЦИЯ
Подтип хелицеровые (Chelicerata)
Класс I. Меростомовые (Merostomata, или Palaeostraca)
Отряд 1. Мечехвосты (Xiphosura)
Отряд 2. Ракоскорпионы (Eurypteroidea)
Класс 2. Паукообразные (Arachnoidea)
Отряд 1. Скорпионы (Scorpiones)
Отряд 2. Жгутоногие (Pedipalpi)
Отряд 3. Ложноскорпионы (Pseudoscorpiones)
Отряд 4. Сольпуги (Solifugae)
Отряд 5. Сенокосцы (Opiliones)
Отряд 6. Пауки (Агапеае)
Отряд 7. Клещи (Acarina)
365
КЛАСС 1. МЕРОСТОМОВЫЕ (MEROSTOMATA, ИЛИ
PALAEOSTRACA)
ХАРАКТЕРИСТИКА
Меростомовые — морские хелицеровые, дышащие жабрами. Голово-
грудь цельная, покрыта сплошным щитом с простыми медиальными гла-
зами и более сложными боковыми. Хелицеры небольшие, с клешней на
конце, расположены перед ртом. Остальные пять пар конечностей голово-
груди расположены по сторонам щелевидного ротового отверстия, снабжены
жевательными отростками и либо несут на концах клешню, либо заострены.
Они служат для передвижения и захвата пищи. Иногда задние ноги пла-
вательные. Брюшко расчленено или слитное; анальная лопать превращена
в хвостовой шип. Конечности мезосомы пластинчатые; восьмой сегмент не-
сет половые крышки, следующие пять сегментов — жаберные ножки —
направленные назад пластинки с жаберными листочками на задней приле-
гающей к телу стороне. Сегменты метасомы, лишенные конечностей, либо
образуют суженный задний отдел, либо сливаются с метасомой и редуци-
руются.
К меростомовым относятся вымершие палеозойские ракоскорпионы
(Eurypteroidea) и мечехвосты (Xiphosura), известные с силура и частью
ныне живущие. Это крупные формы, среди вымерших были гиганты более
2 м длины.
СИСТЕМАТИЧЕСКИЙ ОБЗОР КЛАССА
ОТРЯД МЕЧЕХВОСТЫ (XIPHOSURA)
лых морях у побережий
Мечехвосты представляют большой эволюционный интерес как един-
ственная сохранившаяся группа первично водных хелицеровых, во мно-
гом очень примитивная; насчитывается всего 5 видов встречающихся в теп-
". Тело 50—90 см в длину, состоит из массивной
щитовидной головогруди и слитного
брюшка, несущего на конце длинную
острую иглу, причлененную подвиж-
но (рис. 323). Головогрудь и брюшко
также сочленены подвижно посредст-
вом первого брюшного сегмента. На
головогрудном щите имеются простые
срединные и более сложные боковые
глаза. Последние состоят из многих
глазков, напоминая фасеточные глаза
трилобитов, но покрыты общей рого-
вицей. Конечности головогруди груп-
пируются вокруг рта на вентральной
стороне. Хелицеры более короткие;
педипальпы и ноги сходного строения,
длинные; все, кроме последней пары
ног, имеют на конце клешни; на тази-
ках педипальп и трех пар ног развиты
эндиты, на тазиках четвертой пары ног
7
6
Рис. 323. Мечехвост Limulus polyphemus
с брюшной стороны:
1 — головогрудной щит, 2 — брюшной щит, 3 — хво-
стовая игла, 4 — хелицеры, 5 — педипальпы, 6 но-
ги, 7 — рудименты конечностей 7-го сегменте! (хи-
ларии), 8 — половые крышки, 9 — жаберные крыш-
ки, 10 — жаберные ножки
366
имеется придаток (так называемый флабеллум), отвечающий жаберной ветви
ножки трилобитов. Задняя пара ног приспособлена для ползания и выка-
пывания ямки в песке при откладке яиц. Брюшко имеет шесть пар конеч-
ностей; на втором сегменте — половые крышки, наследующих пятижабер-
ные ножки, которые служат для дыхания и плавания. Они состоят из
двух ветвей — внутренней узкой, собственно ножки, и плоской наружной,
несущей жаберные листочки и отвечающей жаберной ветви трилобитов.
Передняя пара жаберных ножек образует крышки, прикрывающие осталь-
ные четыре пары. Рудименты конечностей имеются и на первом брюшном
сегменте (так называемые хиларии). ?
Пищеварительная система мечехвостов представлена пищеводом, жевате-
льным желудком”, средней и задней кишкой; в среднюю кишку впадают две
Рис. 324. Развитие мечехвоста Limulus mollucanus. / —
зародыш на стадии образования личиночных (протаспи-
совых) сегментов; // —вылупившаяся личинка с брюшной
стороны
пары протоков ее придатка очень крупной печени, состоящей из многих
железистых долек. Органами выделения служат коксальные железы,
расположенные в головогруди; четыре пары желез соединяются общим
потоком, который переходит в длинный извитой проток, открывающийся
наружу. Эти железы соответствуют целомодуктам полихет. Сердце длин-
ное трубчатое с восемью парами щелевидных отверстий — остий, передней
и несколькими парами боковых артерий, которые сливаются в боковые
стволы. Кровь через артерии изливается в полость между органами, соби-
рается в систему лакун-синусов, поступает к жабрам, возвращается в око-
лосердечный синус — перикардий и оттуда через остии в сердце. Нервная
система представлена надглоточным ганглием (мозгом), от которого отходят
глазные и антеннулярные нервы (последние сохранились среди хелицеро-
вых только у мечехвостов, хотя антеннулы исчезли и у них), и сомкнутыми
ганглиями нервной цепочки, из которых шесть посылают нервы к голово-
грудным сегментам, а пять иннервируют брюшко. Половые железы парные
с многочисленными анастомозирующими разветвлениями; сохраняется пар-
ность и полового отверстия, расположенного в основании половых ножек.
Оплодотворение наружное. Самка откладывает яйца в ямку, вырытую
в песке в полосе отлива, а самец поливает их спермой. Яйца богаты желт-
ком и вылупляется полносегментная молодая особь, которая отличается
короткой хвостовой иглой (рис. 324,//). Дальнейшее развитие сопровожда-
ется длительным ростом и линьками. В эмбриональном развитии мечехвостов
обнаружена четырехсегментная стадия, соответствующая личинке трилоби-
тов—протаспису (рис. 324,/ ). Последующие туловищные сегменты у зародыша
мечехвостов формируются один за другим преданальной зоной роста, т е.
так же, как они появились у трилобитов, но в послезародышевом развитии.
На примере мечехвостов видно, что процесс анаморфоза (появления иосле-
личиночных сегментов) переместился у хелицеровых в эмбриональный пе-
риод, что связано с большим запасом желтка в яйце.
367
Мечехвосты в основном хищники, питаются моллюсками и другими
беспозвоночными. Современные формы (род Limulus и др.) распространены
у берегов Центральной Америки, юго-восточной Азии, Малайских, Филип-
пинских, Японских островов. Вымершие представители отряда известны в
силуре. Мечехвосты по побережью юго-восточной Азии служат объектом
промысла.
ОТРЯД РАКОСКОРПИОНЫ (EURYPTEROIDEA)
Целиком вымерший отряд морских хелицеровых. Крупные, частью ги-
гантские (более 2 м) членистоногие. Тело состоит из головогруди, покрытой
цельным щитом с глазами, и крупного 12-сегментного брюшка с постаналь-
ной иглой на конце (рис. 325). Хелицеры короткие с клешней. Педипальпы
14
13
12
10
11
П
Рис.
cheri
8
9
325.
с брюшной стороны; II — Palaeophonus
nuncius со спинной стороны:
1 — 6 — конечности просомы, 7 — 11 — пластинчатые жа-
берные ножки мезосомы, 12 — половые придатки 8-го сег-
мента, 13 — эндиты задних ног, 14 — грудина (стернум)
Ракоскорпионы. / — Eurypterus fis-
и ноги ходильные с жевательными эндитами на тазиках; задние ноги у части
форм веслообразные, плавательные. На втором сегменте мезосомы имеются
половые крышки, на остальных (обычно кроме третьего) — жаберные нож-
ки, устроенные, как у мечехвостов, но сросшиеся попарно в виде жаберных
крышек; шесть сегментов метасомы лишены конечностей.
Ракоскорпионы населяли мелководье морей, некоторые, по-видимому,
проникали и в пресные воды. Это были хищные донные животные, частью
способные плавать у дна. Они находимы в морских и континентальных от-
ложениях от кембрия до перми и многочисленны в силуре (роды Eurypterus,
Slimonia, Mixopterus и др.). С эвриптеридами через силурийских водяных
скорпионов (Palaeophonus, рис. 325//) связаны по происхождению современ-
ные скорпионы.
368
Таблица X. Паук-крестовик Araneus, самка (общий вид снизу и сбоку):
7 —хелицера, 2 челюсть ногощупальца, 3—ногощупальце, 4 — нижняя губа, 5—тазик второй пары
ножек, 6—грудина, 7 — щиток, прикрывающий легкое, 8— вскрытое листовидное легкое, 9— стиг-
ма легкого, 10—половой вырост самки, 11— печень, 12—средняя кишка, 13— сердце, 14 — яич-
ник, 15 — мальпигиева трубочка, 16 — паутинные железы, 17— трахеи, 18— передняя паутинная
бородавка, 19—медиальная паутинная бородавка, 20—задняя паутинная бородавка, 21 — анальное
отверстие, 22— срез через стенку тела, 2<3—дорзо-вентральная Mbimuaj 24 — коготки
13 Курс зоологии, т. I
369
КЛАСС 2. ПАУКООБРАЗНЫЕ (ARACHNOIDEA)1
ХАРАКТЕРИСТИКА
Паукообразные — наземные хелицеровые, имеющие органы воздуш-
ного дыхания — листовидно-складчатые легкие, спрятанные в углублениях
на сегментах брюшка, и трубчатые трахеи, причем у ряда форм легкие
полностью заменяются трахеями. Головогрудь обычно цельная, несет шесть
пар конечностей и покрыта сплошным щитом, но может быть разделена на
голову с хелицерами, педипальпами и двумя парами ног и два туловищных
сегмента, несущих остальные две пары ног Хелицеры короткие 2—3-чле-
никовые с клешней, когтевидным концевым члеником или стилетовидные
(у некоторых паразитических клещей). Педипальпы с эндитами на тазик; х
щупальцевидные или с клешней на конце. Стернит педипальп ограничивает
сзади ротовое отверстие. Иногда эндиты сохраняются на тазиках передних
пар ног. Число сегментов брюшка варьирует в связи с постепенной атро-
фией сегментов метасомы; в ряде случаев сегменты брюшка слиты. К пау-
кообразным относится ряд ядовитых форм; наибольшее значение имеют кле-
щи, как паразиты, переносчики возбудителей болезней человека и живот-
ных, а также вредители растений и запасов продуктов (зерна, муки и проч.).
Представитель класса паукообразных —
паук-крестовик (Araneus diadematus)
Паук-крестовик (табл. X, рис. 326) принадлежит к отряду пауков (Ага-
пеае), подотряду высших двулегочных пауков (Araneomorpha, или Dip-
neumones).
Внешнее строение. Тело взрослой самки около 20 мм длины, самца —
заметно мельче. Головогрудь и брюшко приблизительно яйцевидной формы,
соединяются друг с другом подвижно коротким тонким стебельком, пред-
ставляющим собой первый брюшной сегмент. Головогрудь покрыта цельным
спинным щитом, в передней части которого в два поперечных ряда распо-
ложены восемь простых глаз. Снизу между основаниями ног расположен
выемчатый по краям щиток — грудина. К грудине примыкает небольшой
стернит педипальп, ограничивающий сзади ротовое отверстие. Спинной-
щит и грудина не сегментированы и о составе головогруди из 6 слившихся
сегментов внешне можно судить по наличию 6 пар конечностей. Из них
13 9
Рис. 326. Схема строения двулегочного паука:
1—хелицера, 2 — ядовитая железа, 3—глаза, 4 мозг, 5 — ротовое отверстие,
6—сосательный желудок, 7— средняя кишка, 8— выросты средней кишки,-
9—протоки печени, 10—ректальный пузырь, 11— анальное отверстие, 12—
мальпигиевы сосуды, 13 сердце, 14 — легкие, 15 — стигма, 16 — легочный си-
нус. 17— яичник, 18— яйцевод, 19 — паутинные железы. 20—паутинные боро-
давки
1 Арахна—по-гречески паук; в древней Греции — имя девушки, которая по
преданию, достигла такого высокого ткацкого искусства, что вызвала на состязание
Афину; побежденная, но не признавшая своего поражения Арахна разгневанной бо-
гиней была превращена в паука.
370
передние — хелицеры двучленистые—направлены вниз. Крупный вздутый
основной членик имеет на медиальной стороне бороздку, в которую может
вкладываться когтевидный концевой членик. Близ вершины последнего
находится отверстие протока ядовитой железы.
За хелицерами расположены педипальпы (ногощупальца). Их основ-
ные членики (коксы) имеют уплощенные челюстные отростки (эндиты), ко-
торые снизу и с боков прикрывают ротовое отверстие и изнутри покрыты
жесткими волосками. Дистальные пять члеников педипальп образуют щу-
пальца. Ротовое отверстие щелевидное поперечное, спереди ограничено верх-
ней губой, имеющей вид мясистого бугорка.
Паук убивает добычу хелицерами, вонзая в нее когтевидные членики
хелицер и впрыскивая секрет ядовитых желез. Содержимое добычи высасы-
вается, причем щетка волосков на челюстных отростках педипальп препят-
ствует попаданию твердых частиц в ротовое отверстие. У самцов концевой
членик щупальца педипальп снабжен копулятивным аппаратом. Его стро-
ение у пауков различно. В простейшем случае это полый грушевидный при-
даток, суживающийся в иглу с отверстием на конце. У паука-крестовика он
устроен сложнее и внутри также заключает резервуар, проток которого
открывается небольшой порой на конце короткой иглы. Самец наполняет
совокупительные резервуары спермой, вытекающей из семенного протока
на брюшке. При спаривании он вводит иглу копулятивного аппарата в от-
верстие семеприемника самки и впрыскивает сперму в его полость.
Ноги паука состоят из 7 члеников: тазика (соха), вертлуга (trochanter),
бедра (femus), двучленистой голени (tibia) и двучленистой лапки (tarsus).
Лапка заканчивается гребенчатыми коготками, используемыми при по-
стройке паутинной сети. Крупное брюшко крестовика не сегментировано
и покрыто кожистой эластичной кутикулой. Следы сегментации усматри-
ваются только в расположении элементов характерного крестообразного
рисунка брюшка и в наличии видоизмененных конечностей — легких и па-
утинных бородавок. Судя по строению эмбриона, брюшко высших пауков
образовано пятью слитыми сегментами (не считая 1-й—стебелек); пара лег-
ких принадлежит 2-му сегменту, а две пары паутинных бородавок—4-му и
5-му сегментам; третья (медиальная) пара бородавок обособляется от второй.
Наружный покров паука состоит, как обычно у членистоногих, из со-
держащей хитин кутикулы и подстилающего ее эпителиального слоя кле-
ток — гиподермы. Под наружным покровом имеется слой мускулатуры.
Богато развита и сложно дифференцирована более глубокая мускулатура
головогруди и конечностей.
Нервная система. Центральная нервная система сосредоточена в голо-
вогруди; здесь расположена уплощенная нервная масса, образованная
слившимися ганглиями нервной цепочки. Сегментация этой подглоточной
нервной массы обнаруживается в расположении внутренних скоплений ганг-
лиозных клеток, а внешне намечена утолщенными основаниями пяти пар
нервов, из которых передняя пара иннервирует педипальпы, а четыре дру-
гие— ноги. В заднем направлении отходит нервный ствол, делящийся на два
нервных тяжа, которые проходят через сочленовный стебелек и иннервиру-
ют органы брюшка. Надглоточный узел (мозг) соединен с головогрудной
массой короткими и толстыми коннективами, между которыми проходит
горизонтальный участок узкой глотки. От надглоточного узла вперед об-
щим основанием отходят глазные нервы, разветвляющиеся по числу глаз.
Под глазными нервами отходит пара нервов хелицер.
Органы чувств. Глаза крестовика, каки других паукообразных, устрое-
ены по типу простого глаза, имеется хрусталик и стекловидное тело, а под
ним ретина, состоящая из зрительных и пигментных клеток; пара передних
медиальных глаз в деталях гистологического строения отличается от ос-
тальных. Органами осязания и восприятия колебаний служат многочис-
ленные волоски или щетинки, у основания которых имеются нервные клет-
ки, посылающие отростки в центральную нервную систему. Имеется также
13*
371
несколько типов более сложно устроенных кожных органов чувств на паль-
пах, ногах, паутинных бородавках, близ наружного полового аппарата и
в других местах. Часть из них выполняет обонятельную и вкусовую функ-
цию (хеморецепторы), часть, по-видимому, регистрирует изменения влаж-
ности воздуха и т. п. Паук чутко реагирует на внешние воздействия, меха-
нические и звуковые колебания, изменения освещенности, влажности воз-
духа и др. Богато развитые органы чувств обеспечивают сложнейшие ин-
стинкты, связанные с постройкой ловчей сети, ловлей добычи, поведением
при спаривании и др.
Пищеварительная система. Пищеварительная система состоит из перед-
ней, средней и задней кишки (см. рис. 326). Передняя кишка представлена
глоткой, которая, суживаясь, проникает сквозь центральную нервную мас-
су и переходит в сосательный желудок. Глотка выстлана кутикулой, стенки
сосательного желудка образованы четырьмя кутикулярными пластинками,
к которым прикрепляются мощные радиальные мышцы. Этот орган служит
для высасывания жертвы. В глотку близ ротового отверстия открываются
небольшие железы, расположенные в основаниях педипальп и верхней
губы и обычно называемые слюнными. Средняя кишка в головогруди имеет
пять пар железистых слепых выростов, из которых четыре пары загнуты
на брюшную сторону и проникают в полости тазиков ног. Секрет око-
лоротовых желез и слепых отростков обладает протеолитическими свой-
ствами (интенсивно растворяет белки); он вводится в жертву, превращая
ее содержимое в полужидкую кашицу, которая всасывается, т. е. у паука
наблюдается частичное внекишечное пищеварение.
В брюшке средняя кишка дугообразно изогнута, и в нее открываются
несколько пар ветвящихся слепых железистых придатков, которые плотной
массой заполняют верхнюю половину полости брюшка. Это так называе-
мая печень, которая выделяет пищеварительные ферменты и служит для
всасывания питательных веществ. В печени пища частично переваривается
и внутриклеточно. Средняя кишка переходит в небольшую заднюю, обра-
зующую расширение — ректальный пузырь. На границе средней и задней
кишок открываются трубчатые выделительные органы — мальпигиевы со-
суды. Остатки пищи и секрет мальпигиевых сосудов скапливаются в рек-
тальном пузыре и выводятся через прямую кишку, которая открывается на
анальном бугорке. Средняя кишка и мальпигиевы сосуды образуются в
эмбриональном развитии из эктодермального зачатка, передняя и задняя
кишки — из эктодермы.
Кровеносная система. Кровеносная система не замкнутая, представ-
лена кровеносными сосудами, имеющими собственные стенки, и системой
лакун, или синусов, — полостей между органами. Пульсирующий спинной
сосуд — сердце — помещается в брюшке и имеет форму трубки, в стенках
которой имеются кольцевые мышцы и три пары щелевидных отверстий —
остий, снабженных клапанами. Вперед от сердца отходит передняя аорта,
ветвящаяся на артерии, по бокам — несколько пар артерий, назад — зад-
няя аорта, из артерий кровь изливается в систему лакун. Омывая все ор-
ганы, кровь попадает затем в окололегочный синус, обогащается кислоро-
дом, возвращается в перикардиальный синус и через остии в сердце.
Дыхательная система. Органы дыхания у крестовика — пара листо-
видно-складчатых легких и трубчатые трахеи. Легкие расположены в ос-
новании брюшка по сторонам от полового отверстия (табл. X), где имеются
две поперечные щели — стигмы легких. Стигма ведет в полость легкого,
на стенке которой имеется ряд плоских карманов, расходящихся веерооб-
разно (рис. 327). Карманы связаны перемычками и не спадаются, так что
воздух свободно проникает между ними. Кровь циркулирует в полостях
карманов, обмен газами происходит через их тонкие кутикулярные стенки.
Трахейная система состоит из двух неветвящихся трубок, которые на-
правлены вперед от общего кармана, открывающегося малозаметной попе-
речной щелью перед паутинными бородавками (табл. X).
372
Выделительная система. Органы выделения двоякого типа: мальпи-
гиевы сосуды и коксальные железы. Кроме того, выделительную функцию вы-
полняют специальные клетки (нефроциты и гуаноциты), лежащие в полости
тела. Мальпигиевы сосуды представлены четырьмя ветвящимися слепо зам-
кнутыми на концах трубками, которые впадают в ректальный пузырь по
его сторонам на границе средней и задней кишок. Мальпигиевы сосуды вы-
стланы плоским эпителием, в клетках которого образуются зерна гуанина—
основного продукта выделения. Коксальные железы, представляющие у
паукообразных остатки системы целомодуктов, находятся у крестовика
при основании первой пары ног У взрослого паука они не функционируют.
Ядовитые железы. Ядовитые железы расположены в передней час-
ти головогруди при основании хелицер Это пара довольно крупных
цилиндрических желез, заходящих в полости основных члеников хелицер.
Наружную обкладку железы образует спирально завитая лентообразная
мышца, при сокращении которой яд изливается через тонкий проток,
открывающийся на конце когтевидного членика хелицер.
Прядильный аппарат. Прядильный аппарат представлен тремя парами
паутинных бородавок и паутинными железами. В покое паутинные боро-
давки вместе с анальным бугорком
На вершинах бородавок имеются
многочисленные паутинные трубоч-
ки, через которые выделяется сек-
рет — паутина, застывающая при
соприкосновении с воздухом. Па-
утинные железы заполняют ниж-
нюю часть полости брюшка самки.
Их строение и величина неодина-
ковы (рис. 326); различают трубко-
видные, ампуловидные, древовидные
и грушевидные железы. Последние
особенно многочисленны и соедине-
ны в пучки по числу бородавок
(табл. X). Роль различных желез и
бородавок различна, трубковидные :
вого кокона, ампуловидные — для постройки сети, грушевидные — для оп-
летания добычи; древовидные выделяют липкий секрет, покрывающий сеть.
Половая система. Непарное половое отверстие у самки и самца распо-
ложено в основании брюшка между легочными стигмами. Самец лишен на-
ружных половых придатков близ полового отверстия и имеет копулятив-
ные органы на пальпах. У самки перед половым отверстием расположен не-
парный половой вырост (эпигина), во вздутом основании которого имеется
пара семеприемников (табл. X). Яичники лежат в брюшке, парные, в от-
личие от большинства других пауков, у крестовика не срастаются концами.
Это две широкие трубки, покрытые многочисленными яйцевыми фоллику-
лами и имеющие гроздевидную форму. Вперед от яичников отходит пара
яйцеводов, сливающихся в непарный отдел. Расширенную часть последнего
называют маткой', его суженная концевая часть — наружная матка — от-
крывается щелевидным яйцевыводным отверстием. Каждый семеприемник
соединяется протоком с наружной маткой и открывается отверстием на
эпигине. Через эти отверстия при спаривании вводится игла копулятивного
аппарата, сперма попадает в семеприемник и поступает далее в матку.
У самца семенники расположены в передней трети брюшка, парные,
продолжаются вперед парными семепроводами, которые сливаются в не-
парный проток, открывающийся наружу отверстием между стигмами.
Распространение и биология. Паук-крестовик широко распространен
в СССР Излюбленные места его обитания — опушки леса, поляны, порос-
шие редким кустарником. Самка строит колесовидную вертикальную сеть,
которая состоит из неправильной рамы, из лучей и двух спиральных нитей.
373
образуют общую сомкнутую группу.
Рис. 327. Схема легкого паукообразных:
1 — стигма, 2— щели между карманами,
3 — карманы
^лезы выделяют паутину для яйце-
Одна из нитей вспомогательная, к концу постройки сети разрушается и за-
меняется новой, покрытой липким секретом. Самка сидит либо в центре сети,
либо в убежище, к которому от центра сети протянута сигнальная нить.
При попадании добычи и подаче «сигнала» паук направляется в центр сети
и постукивая по нитям пальпами и ногами, определяет положение добычи.
Последняя густо оплетается паутиной и высасывается. Изношенная сеть
заменяется новой. Самцы не строят сеть и обычно держатся под листьями
близ сети самки. После спаривания самец стремится скрываться, так как
оплодотворенные самки склонны поедать самцов. Отложенные яйца опле-
таются коконом. Вылупившиеся паучки сначала не питаются, существуя
за счет остатков желтка в кишечнике; держатся многочисленным скоплени-
ем среди нитей паутины. Далее паучки расходятся и начинают строить
маленькие сети, размеры которых по мере роста пауков возрастают
Рис. 328. Ixodes ricinus, самка со спинной
стороны; / голодная; // насосавшаяся
крови (увеличение одинаковое):
/ — спинной щиток, 2 — хоботок
Второй представитель класса паукообразных —
аксодовый клещ (Ixodes ricinus)1
Иксодовый клещ (Ixodes ricinus) относится к группе паразитиформных
клещей (Parasitiformes), семейству иксодовых клещей (Ixodidae). Это вы-
соко специализованный кровососущий паразит наземных позвоночных,
служащий среди паукообразных одним из примеров крайнего видоизменения
организации в связи с паразитизмом.
Внешнее строение. Тело клеща цельное, овальное, уплощено в спинно-
брюшном направлении. Разделение на сегменты и отделы исчезло. Хелице-
ры и педипальпы соединены в хобо-
ток, расположенный спереди и слу-
жащий для прикрепления к живот-
ному-хозяину и сосания его крови.
Четыре пары ног прикрепляются
уплощенными тазиками на вент-
ральной стороне в передней поло-
вине тела. Между ними примерно
по середине тела расположено
половое отверстие, а ближе к зад-
нему краю — анальное. По бокам
тела имеются пластинки пери-
тремы, на которых расположены
дыхательные отверстия — стигмы.
У клеща резко выражен половой
диморфизм. Самка крупнее самца,
насасывает много крови и сильно
увеличивается. Длина тела голод-
ной самки около 4 мм, напитавшей-
ся — 10—12 мм. Длина тела самца
около 2,5 мм. Диморфизм выражен
также и в развитии щитков на теле.
У самки тело покрыто эластичной кутикулой и имеется только небольшой
спинной щиток (рис. 328). У самца щиток покрывает всю спину и имеется
несколько щитков на брюшной стороне (рис. 329).
Хоботок (рис. 330) имеет трубчатое склеритное основание, образованное
сросшимися тазиками педипальп. От основания отходит срединный вырост—
гипостом, усаженный направленными назад шипиками. По бокам основа-
ния прикрепляются четырехчлениковые пальпы (видоизмененные щупаль-
ца педипальп). Внутри основания хоботка над гипостомом помещаются в
1 Клеща Ixodes ricinus иногда называют собачьим клещом. Однако это название
более укрепилось за другим видом иксодовых клещей — Rhipicephalus sanguineus,
который распространен на юге СССР, часто паразитирует на собаках и передает чело-
веку возбудителей марсельской лихорадки.
374
кожистых футлярах хелицеры. Они имеют на конце режущие зубчатые пла-
стинки и могут выдвигаться вперед. Попав на тело животного-хозяина, клещ
прорезает кожу хелицерами и вводит в ранку гипостом, с помощью которого
прочно прикрепляется и длительно (обычно несколько дней) сосет кровь.
У самцов хоботок служит также для копуляции.
Нервная система. Нервная система концентрирована. Центральная
нервная масса расположена на брюшной стороне несколько впереди от по-
Рпс. 329. Ixodes ricinus, са-
мец с брюшной стороны:
/ — гипостом, 2—пальпы, 3 —
основание хоботка, 4 — тазик пе-
редней ноги (концевые части ног
не показаны), 5 — половое отвер-
стие, 6 — перитрема со стигмой,
7 — щитки брюшной поверхности,
8 — анальное отверстие
лового отверстия и пронизана пищеводом.
Она образована слиянием надглоточного, под-
глоточного и брюшных ганглиев. От нервной
массы отходит 11 пар нервов, иннервирующих
хоботок, ноги и внутренние органы.
Органы чувств. Органы чувств главным
Рис. 330. Ixodes ricinus, хоботок самки, сле-
ва со спинной, справа с брюшной сторо-
ны:
/ — гипостом, 2 хелицеры, 3 — пальпы, 4 — футляры
хелицер, 5 — основание хоботка, 6 — поровые поля.
7. — 10 членики пальп
образом обонятельные и осязательные. У Ixodes ricinus глаз нет, но у
некоторых других иксодовых клещей имеется пара простых глаз по бокам
спинного щитка. Органом обоняния, играющим важную роль при подсте-
регании хозяина, служит так называемый орган Галлера, расположенный
на лапках передних ног Это ямка, на дне которой имеются тонкостенные
выросты (сенсиллы), иннервируемые отростками группы нервных клеток.
Подстерегая хозяина, клещи сидят на растительности в характерной позе,
выставив передние ноги с обонятельными органами (рис. 331).
Пищеварительная система. Ротовое отверстие расположено в основании
гипостома под хелицерами, ведет в глотку, которая выстлана плотной кути-
кулой и имеет мускулатуру для насасывания крови. В глотку открываются
протоки крупных слюнных желез (рис. 332), секрет которых обладает ана-
стезирующим действием и вводится в ранку при укусе. От глотки тянется
узкий пищевод, который пронизывает нервный узел и открывается в цент-
ральную часть средней кишки. Последняя имеет шесть пар длинных слепых
выростов, служащих резервуарами для всосанной крови и особенно раз-
витых у самок. Самка за один прием высасывает до 400 мг крови, что в
300—400 раз превосходит ее вес в голодном состоянии. Средняя кишка,
резко суживаясь, переходит в ректальный пузырь, куда открываются и
мальпигиевы сосуды. Ректальный пузырь через короткую заднюю кишку
открывается анальным отверстием.
Дыхательная система. Органы дыхания—трубчатые трахеи, в которые
воздух поступает через две стигмы, расположенные по бокам тела на щитках
375
перитремах. Через стигму воздух поступает в округлую полость, от которой
отходит главный ствол, делящийся на два пучка трахей. Стенки трахей
укреплены хитиновой спиральной выстилкой, трахеи сильно ветвятся и
оплетают все органы.
Кровеносная система. Кровеносная система развита слабо. На спинной
стороне под покровом расположено овальное сердце с двумя остиями. От
Рис. 331. Клещи Ixodes, слева — самка, подстерегающая
хозяина, справа — самка, откладывающая яйца
Рис. 332. Ixodes ricinus, вскрытая упитанная самка:
1—хоботок, 2—глотка. 3 — слюнная железа, 4 — орган Женэ, 5 средняя кишка,
6—слепые выросты средней кишки, 7—ректальный пузырь, 8 — мальпигиевы сосуды^
9 — яичник, 10 — яйцевод, 11 — нервный узел, 12 — трахеи
376
него вперед отходит аорта, расширяющаяся вокруг нервного узла в нерв-
ный синус, от которого через сосуды кровь изливается в полость тела. По-
ступает кровь в сердце из полости тела через остии.
Выделительная система. Органы выделения — пара очень длинных
мальпигиевых сосудов, открывающихся в ректальный пузырь.
Половая система. У самки яичник непарный и вместе с отходящими от
его концов яйцеводами образуется замкнутое кольцо; яичник сильно уве-
личивается по мере переваривания крови и созревания яиц. Яйцеводы от-
крываются в непарную матку, которая переходит в объемистое влагалище
и через так называемое преддверие открывается наружу половым отверсти-
ем. Во влагалище впадает пара придаточных желез и непарный семеприем-
ник. У самца семенники парные, лежат вдоль тела. От них отходят семяпро-
воды, сливающиеся в семяизвергательный канал, открывающийся наружу.
В канал открываются четыре пары придаточных желез и семенной пузырь.
Секрет придаточных желез служит для образования сперматофора (мешочка
со сперматозоидами), который самец при спаривании вводит в половое от-
верстие самки.
Биология, развитие. Ixodes ricinus — пастбищный кровосос, подсте-
регающий животных-хозяев на растительности в открытой природе. Насо-
савшаяся крови самка отпадает, переваривает кровь и откладывает яйца
обычно на поверхность почвы среди растительности. Яйца при откладке
обволакиваются клейким секретом, выделяемым железистым органом (так
называемый орган Женэ), который расположен близ глотки и может выпя-
чиваться наружу (рис. 332). Самка кладет яйца раз в жизни и очень плодо-
вита, в кладке насчитывается несколько тысяч яиц. Из яиц вылупляются
мелкие личинки, у которых нет четвертой пары ног, стигм и полового от-
верстия. Личинка, линяя, превращается в нимфу, которая имеет четыре
пары ног и перитремы со стигмами, но лишена полового отверстия. Нимфа,
линяя, превращается во взрослого клеща. Клещ перед каждой линькой дол-
жен попасть на животное-хозяина и всосать определенную порцию крови.
Личинки и нимфы сосут кровь мелких животных (грызунов, птиц, ящериц),
взрослые клещи паразитируют на более крупных диких животных, на скоте,
собаках, охотно присасываются и к человеку. Большая часть клещей в про-
цессе метаморфоза гибнет, так и не встретив животного-прокормителя Гибель
компенсируется высокой плодовитостью клеща и способностью длительно
голодать на всех фазах развития.
Распространение. Медицинское значение. Ixodes ricinus широко рас-
пространен в европейской части СССР, за исключением северных и юго-
восточных районов, в Западной Европе и Северной Африке. Он обитает на
пастбищах с кустарником, в лиственных и хвойно-лиственных лесах. Служит
переносчиком возбудителя пироплазмоза крупного рогатого скота и пере-
дает человеку вирус западной формы клещевого энцефалита. Близкий к
Ixodes ricinus вид Ixodes persulcatus, распространенный по всей таежной
зоне, имеет большое эпидемиологическое значение как природный храни-
тель и переносчик человеку вируса весенне-летнего энцефалита.
ОБЩИЙ ОБЗОР КЛАССА
Расчленение тела, конечности. Для Arachnoidea характерна возрастаю-
цая концентрация сегментов вплоть до их полного слияния, редукция зад-
них сегментов брюшка — метасомы, высокая и довольно однотипная специ-
ализация конечностей головогруди и постепенная атрофия видоизмененных
конечностей брюшка — легких в связи с переходом к трахейному дыханию.
Голова обособлена у сольпуг и немногих других. У большинства пауко-
образных головогрудь цельная. Число сегментов брюшка сокращается за
счет редукции метасомы: скорпионы имеют 13 сегментов брюшка (из них
1-й внешне не выражен), жгутоногие и ложноскорпионы — 12, членисто-
брюхие пауки — 11, сольпуги и сенокосцы — 10, большинство клещей —
377
7, высшие пауки — 6, некоторые специализованные клещи — 4—2. В по-
следнем случае редуцируются и концевые сегменты мезосомы. Атрофиру-
ются и брюшные конечности. У скорпионов они представлены на мезосоме
половыми крышками, гребневидными органами чувств и четырьмя парами
легких, у жгутоногих и четырехлегочных пауков — двумя парами легких,
у двулегочных пауков — одной парой легких, у остальных паукообразных
легкие исчезают и заменяются трахеями. Для большинства пауков и клещей
характерно исчезновение границ между сегментами брюшка.
Нервная система. Нервная система также концентрируется. У скор-
пионов (рис. 333) имеется парный надглоточный ганглий, связанный кон-
нективами с головогрудной ганглиозной массой, дающей нервы к педипаль-
пам и 4 парам ног, и 7 ганглиев брюшной нервной цепочки. У сольпуг, кро-
ме общей нервной массы, сохраняется один узел, у пауков вся нервная це-
почка сливается в головогрудную массу; у большинства клещей исчезает
разграничение головного мозга и головогрудной массы.
Рис. 333. Внутреннее строение скорпиона, схема:
1—мозг, 2—подглоточная нервная масса, 3—медиальный глаз, 4—боковые глаза, 5 —
6—сердце, 7 передняя аорта, 8 — задняя аорта, 9 — боковые артерии, /0 — вентральная
артерия, 11— легкие, 12 — средняя кишка, 13 — протоки печени, 14 — анальное отверстие,-
15 — ядовитая железа (печень и половая система не показаны)
Органы чувств. Органы чувств представлены осязательными волос-
ками и более специализованными кожными органами: вкусовые и обоня-
тельные сенсилы (хеморецепторы), органы, воспринимающие колебания
(сейсморецепторы), влажность воздуха (гигрорецепторы) и др. Глаза име-
ются у большинства паукообразных в числе 8, 6, реже 2 пар на головогруди;
они устроены по типу простых глаз и несколько различаются в деталях
гистологического строения. Имеются и слепые формы.
Пищеварительная система. Передняя кишка обычно образует расши-
рение— снабженную мышцами глотку, которая служит для насасывания
полужидкой пищи. В переднюю кишку открывается пара желез, называе-
мых слюнными. Средняя кишка в большинстве случаев образует слепые
выросты, увеличивающие ее вместимость и всасывающую поверхность.
Брюшной отдел кишечника обычно имеет парный железистый вырост, назы-
ваемый печенью, которая выделяет пищеварительные ферменты и служит
для всасывания питательных веществ. В печени происходит также внутри-
клеточное пищеварение. Задняя кишка часто образует выпячивание — рек-
тальный пузырь. Большинство Arachnoidea — хищники, причем обычно
содержимое жертвы перед высасыванием растворяется протеолитическими
ферментами слюнных желез и кишечника, т е. имеет место частичное вне-
кишечное пищеварение. Среди клещей, помимо хищников, имеются расти-
378
тельноядные формы и разнообразные паразиты позвоночных и насекомых,
питающиеся кровью, роговыми частями покровов и др.
Дыхательная система. Органы дыхания представлены легкими (см.
рис. 326), которые в пределах класса постепенно редуцируются и заменя-
ются трахеями. Легочный мешок вдается внутрь от щелевидной стигмы. От
его внутренней стенки отходят многочисленные листовидно-складчатые кар-
маны. В карманах циркулирует кровь, а между ними проникает воздух.
Трахеи представляют собой трубки, неветвистые или ветвящиеся; их стен-
ки изнутри выстланы продолжением наружной кутикулы, которая на про-
тяжении трахеи имеет спиральное утолщение, препятствующее спадению
стенок. Легкие—видоизмененные жаберные конечности, унаследованные
от водных хелицеровых; трахеи — новообразование, органы, более при-
способленные к воздушному дыханию. Легкие сохранились у более прими-
тивных форм: у скорпионов их четыре пары (на 4—7-м сегментах брюшка),
у жгутоногих и четырехлегочных пауков — две пары (на 2-м и 3-м сегмен-
тах), у двулегочных пауков — одна пара (на 2-м сегменте). Последние од-
новременно с легкими имеют и пару трахей. Остальные Arachnoidea имеют
только трахеи, которые могут открываться разным числом стигм на различ-
ных участках тела, обычно на брюшке, но иногда близ оснований ног или
хелицер (у клещей).
Кровеносная система. Кровеносная система по степени развития свя-
зана с характером дыхания, степенью расчленения и величиной тела. Серд-
це наиболее развито у крупных расчлененных форме локализованным легоч-
ным дыханием. Так, у скорпионов сердце длинное трубчатое с 7 парами
остий, передней и задней аортами и метамерными артериями; у жгутоногих
остий 5 пар, у пауков — 3—4 пары, у сенокосцев — 2 пары; у клещей серд-
це имеет вид мешочка с парой остий или отсутствует Через аорту и боковые
артерии кровь из сердца изливается в систему лакун — пространств между
органами. У форм, имеющих легкие, она проходит через легочные синусы
и, обогатившись кислородом, возвращается в околосердечный синус и через
остии в сердце. При трахейном дыхании кровь собирается к сердцу из всех
полостей между органами. Кровь паукообразных бесцветна и содержит
клетки нескольких типов.
Выделительная система. Выделительная система представлена маль-
пигиевыми сосудами, являющимися у Arachnoidea новообразованием, и кок-
сальными железами, которые соответствуют целомодуктам. Мальпигиевы
сосуды — пара ветвящихся, слепо замкнутых на концах трубок, открыва-
ются на границе средней и задней кишок. Они эктодермального происхож-
дения, т е. принадлежат средней кишке. В эпителии и просвете мальпигие-
вых сосудов накапливаются зерна гуанина — главного продукта выделения
паукообразных. Коксальные железы образованы мешковидной частью ме-
зодермального происхождения, извитым протоком (лабиринтом), резервуа-
ром и наружным выводным протоком. Они имеются в числе одной-двух
пар, открываются у оснований ног и у взрослых форм функционируют
редко.
Половая система (рис. 334). Паукообразные раздельнополы. Половые
железы расположены в брюшке и в исходном состоянии парны. В ряде слу-
чаев наблюдается слияние правой и левой гонад. Так, у самца скорпионов
семенники парные и каждый состоит из двух трубок, связанных перемыч-
ками (рис. 334/); у самок скорпионов яичник один и состоит из трех тру-
бок (рис. 334//), из которых средняя, очевидно, есть результат слияния двух
медиальных трубок, подобных таковым самца. У многих пауков, сенокосцев
и клещей парные гонады срастаются концами в кольцо (рис. 334///). Парные
яйцеводы и семепроводы открываются непарным половым отверстием всегда
на втором сегменте брюшка. Строение выводной части половой системы и
копулятивные приспособления самцов весьма разнообразны. У самок обыч-
но имеется расширение яйцеводов — матка и семеприемники. У самцов ко-
пулятивные органы либо связаны с половым отверстием, либо таковыми
379
служат педипальпы (пауки) или хелицеры (некоторые клещи). В ряде слу-
чаев оплодотворение сперматофорное — с помощью пакетов со спермиями.
Развитие. Большинство паукообразных откладывает яйца, но имеют-
ся и живородящие формы (скорпионы, некоторые клещи и др.).Яйца богаты
Рис. 334. Внутренние половые органы паукообразных. / — самца
скорпиона; // —самки скорпиона; /// —самки паука:
/ — семенник, 2—семяпровод, 3—копулятивный орган, 5 — яичник, 5—яйцевод,
6 — семеприемник
желтком, благодаря чему дробление частичное, поверхностное, все сегмен-
ты тела и конечности формируются в эмбриональном развитии, и из яйца
вылупляется маленькая полносегментная особь, похожая на взрослую. После-
зародышевое развитие прямое, сопровождается преиму-
щественно ростом. Только у клещей в связи с малыми
размерами яиц вылупляется шестиногая личинка и имеет
место метаморфоз. Изучение зародышей примитивных
паукообразных позволяет полнее понять строение взрос-
лых. Так, у зародыша скорпионов (рис. 335) на всех сег-
ментах мезосомы закладываются брюшные конечности, из
которых первая пара затем исчезает, вторая — превра-
щается в половые крышки, третья — в гребневидные ор-
ганы, а остальные четыре пары — в легкие.
Рис. 335. Зародыш скорпиона:
1 — хелицеры, 2— педипальпы, 3—ходильные ноги, 4—конечности мезосо-
мы
СИСТЕМАТИЧЕСКИЙ ОБЗОР КЛАССА
В классе Arachnoidea насчитывается до 12 современных отрядов и не-
сколько ископаемых. В настоящем курсе рассмотрено 7 отрядов.
ОТРЯД СКОРПИОНЫ (SCORPIONES)
Скорпионы (рис. 336) средних размеров или крупные более 15 мм па-
укообразные, живут в областях с теплым климатом. Расчленение тела хо-
рошо выражено. Щитки тела и конечности сильно склеротизованы. Голо-
вогрудь покрыта цельным спинным щитком. Хелицеры небольшие клешне-
видные, педипальпы с крупной клешней, передние две пары ног имеют
380
жевательные отростки. Сегментов брюшка 13, но первый внешне не различим,
они группируются в более широкое переднебрюшъе и узкое гибкое задне-
брюшье1. На переднебрюшье полно представлены производные конечностей:
на втором сегменте — половые крышки, на третьем — гребневидные органы
чувств, на следующих четырех — легочные мешки. Заднебрюшье несет на
конце вздутый постанальный членик, в котором имеется парная ядовитая
железа, открывающаяся на конце когтевидной иглы. Это орган нападения
и защиты. Скорпионы питаются насекомыми, мокрицами, пауками и т п.
Рис. 336. Скорпион Buthus eupeus со спинной и брюшной
стороны. // — VII—сегменты мезосомы (/ не выражен):
1 — 6 — сегменты метасомы, 7 — хвостовой придаток (тельсон), 9 — голо-
вогрудной щит, 10 — хелицеры, 11— педипальпы, 12—эндиты ног*
13 — грудина, 14 половые крышки, 15 — гребневидные органы*
16 — стигмы
Они схватывают добычу клешнями педипальп и, загнув хвост на спину,
жалят ее ядовитой иглой.
Днем скорпионы прячутся под камнями, в трещинах почвы, норах.
На охоту они выходят ночью. Живородящи, молодь держится некоторое
время на теле самки. В южных районах СССР широко распространены
виды родов Buthus и Euscorpius.
Ужаление представителями наших видов для человека не смертельно,
но вызывает сильную боль, опухоль, температурную реакцию. Дня через
2—3 эти явления проходят. В тропиках известны случаи ужаления мест-
ными крупными видами скорпионов со смертельным исходом.
Современные скорпионы по строению близки к ископаемым силурий-
ским морским скорпионам Palaeophonus (см. рис. 325//), которые в свою
очередь сближаются с эвриптеридами. Отличия сухопутных скорпионов
заключаются главным образом в том, что жаберные ножки превратились
у них в легкие.
ОТРЯД ЖГУТОНОГИЕ (PEDIPALPI)
Довольно крупные формы, внешне несколько напоминают скорпионов,
но брюшко не разделено на передне- и заднебрюшье, не имеет ядовитой
иглы и обычно заканчивается членистой нитью (рис. 337). Педипальпы хва-
1 Последний конический членик расширенной части брюшка есть первый сегмент
.метасомы.
381
Рис. 337. Жгутоногие Thelyphonus
caudatus
Рис. 339. Сольпуга Galeode- aranoides
тательные, передние ноги обычно
длинные жгутовидные, играют
роль усиков. Имеют две пары
легких. Хищники. Встречаются
в тропических странах.
ОТРЯД ЛОЖНОСКОРПИОНЫ
(PSEUDOSCO RPIONES)
Мелкие (1—6 мм) паукооб-
разные, внешне похожие на скор-
пионов, но с широким 11-сег-
ментным брюшком без хвостовой
иглы. Дышат трахеями. На кон-
це подвижного членика клешни
хелицер открываются протоки
паутинных желез, секрет кото-
рых служит для постройки гнез-
да. Хищники, питаются мелкими
членистоногими, которых ловят
клешневидными педипальпами.
Живут в лесной подстилке, дре-
весной трухе, под корой пней.
«Книжный» скорпион Chelifer
cancroides (рис. 338) встречается
в домах.
382
ОТРЯД СОЛЬПУГИ, ИЛИ ФАЛАНГИ (SOLIFUGAE)
Сравнительно крупные, до 5—1см, паукообразные. Голова и сегменты,
несущие две задние пары ног, обособлены. Хелицеры крупные с мощной
зубчатой клешней, педипальпы похожи на ноги и участвуют в передвиже-
нии. Брюшко из 10 члеников. Дышат трахеями. Ночные хищники. Встреча-
ются в жарких и сухих местностях, в СССР — в южных районах, особенно
в Средней Азии; обычны виды родов Galeodes (рис. 339), Karschia nRhago-
des. Укус не ядовит, но остатки пищи на хелицерах могут загрязнить рану
и вызвать воспаление.
ОТРЯД СЕНОКОСЦЫ (OPILIONES)
Небольшие (2—7 мм) паукообразные с укороченным телом и обычно
очень длинными ногами (рис. 340), педипальпы и клешневидные хелицеры
Рис. 340. Сенокосец Liobunum rotundum, самка, откладывающая
яйца в почву
также заметно удлинены. Брюшко расчленено и широким основанием сое-
диняется с головогрудью; его концевые тергиты сдвинуты на нижнюю сто-
рону Самки некоторых форм имеют длинную выпячивающуюся яйцеклад-
ную трубку. Дышат трахеями. Питаются личинками насекомых, слизня-
ками и другими мелкими беспозвоночными. Распространены очень широко,
живут в опавшей листве, среди травянистой растительности, на стволах
деревьев и т п. Обычны виды рода Phalangium.
ОТРЯД ПАУКИ (ARANEAE)
Обильный видами отряд, насчитывающий более 20 000 видов. Пауков
делят на три подотряда. Самые примитивные пауки — небольшой тропиче-
ский подотряд членистобрюхих (Liphistiomorpha) — отличаются расчле-
ненным 11-сегментным брюшком (рис. 341/). Представители двух других
подотрядов — пауков-птицеядов (Mygalomorpha) и еысших двулегочных
пауков (Araneomorpha) имеют слитное брюшко. По числу легких пауков
делят на четырехлегочных, к которым относятся первые два подотряда, и
двулегочных — третий подотряд. У последних кроме легких, имеется пара
трахей, а у одного семейства высших пауков (Caponiidae) легкие исчезают
и заменены двумя парами трахей. Строение пауков рассмотрено выше на
примере двулегочного паука-крестовика (стр. 370).
Пауки распространены очень широко, заселяют самые различные места
обитания и представляют ряд интересных биологических особенностей. Все
они хищники, питаются главным образом насекомыми и другими членисто-
383
Рис. 341. Пауки (Агапеае). Тропические пауки. / —членистобрюхий паук (Liphis-
tius malayanus); // — паук-птицеяд (Grammostola actaeon), длина тела до 7,5 си;
/// —Micrathena cyanospina; IV — Gasteracantha curvispina, самка и самец (при-
мер крайней степени полового диморфизма); V — Myrmecium gounlli, похожий на
муравья. Пауки нашей фауны: VI — самка каракурта (Lathrodectes tredecimgut-
tatus); VI! —бокоход (Coriarachne depressa); VIII —скакун (Salticus cingulatus);
IX — самка Lycosa с яйцевым коконом
ногимп, которых убивают хелицерами и высасывают; большинство имеет
ядовитые железы, секрет которых вводится хелицерами в жертву, но есть и
неядовитые формы. Некоторые крупные тропические пауки-птицеяды (Avi-
cularia и др.; рис. 341//), помимо членистоногих, охотятся на мелких птиц,
грызунов, рептилий. По образу жизни пауков делят на бродячих, которые
ловят добычу на бегу или прыгая на нее, и тенетных, которые строят ловчие
сети. Ловчие устройства пауков очень разнообразны.
Часть пауков, например тарантулы (Lycosa), обычные у нас в степной
полосе, подстерегают добычу в отвесной норке, которую роют в почве и вы-
стилают паутиной; попавших в норку насекомых высасывают, а остатки вы-
брасывают наружу. У некоторых пауков норка или гнездо продолжается в
Рис. 342. Atypus piceus в своей норке Рис. 343. Laseola tristis при ловле
муравья
трубчатую паутинную галерею (рис. 342). Многие пауки устраивают на зем-
ле или среди растительности плоский ловчий полог, у края которого рас-
положено отверстие, ведущее в гнездо. Такие тенета характерны и для до-
мового паука Tegenaria domestica. Многие лесные виды укрепляют рыхлые
горизонтальные пологи на многочисленных нитях (рис. 344). К наиболее
сложным относятся различные варианты колесовидной сети, в частности
крестовиков (рис. 345). Иногда такая сеть оттягивается центральной нитью
и имеет вид широкой воронки. У паука Hyptiotes сеть представлена сектором
из 4 лучей и поперечных липких нитей; от места соединения лучей отходит
нить, на которой сидит паук, подобрав нить петлей и натягивая сеть
(рис. 346). При попадании добычи паук повторно отпускает и натягивает пет-
лю, благодаря чему добыча прилипает к смежным нитям; затем паук бежит
к ней, оплетает паутиной и высасываег.
Взрослые самцы обычно ловчих сетей не строят, но пользуются паути-
ной для наполнения спермой копулятивных органов педипальп. Капля спер-
мы выпускается на специально сотканную сеточку (см. рис. 346), откуда
вбирается копулятивными органами. При спаривании проявляются весьма
сложные инстинкты. В ряде случаев самки агрессивны по отношению к сам-
цам, которые после спаривания спасаются бегством, так как могут быть съе-
дены самкой. Самцы некоторых пауков, например видов Pisaura, отвлекают
внимание самки при спаривании, принося ей оплетенное паутиной насеко-
мое. Пока самка его высасывает самец наполняет ее семеприемники спермой,
385
вводя копулятивные органы поочередно в правое и левое отверстия эпиги-
ны. У пауков обычно выражен половой диморфизм, иногда очень резкий
(см. рис. 341 /у).
Самки всех пауков пользуются паутиной для постройки яйцевого ко-
кона. Яйца откладывают на плоскую или мешковидно сотканную паутину,
которую затем оплетают в виде
плотного кокона. У большинства
пауков самка либо носит кокон при
себе, прикрепив его под брюшком к
паутинным бородавкам или держа
хелицерами, либо охраняет кокон в
гнезде. Вылупившиеся паучки не-
которое время держатся в гнезде
или на теле самки (у бродячих па-
уков сем. Lycosidae, тарантула и
др.), затем расходятся и начинают
Рис. 345. Колесовидная сеть кресто-
вика (показана половина)
Рис. 344. Ловчий полог Linyphia
triangularis
жить самостоятельно. Молодые паучки разносятся на паутинках ветром,
особенно в ясные осенние дни. Рост пауков сопровождается несколькими,
чаще четырьмя, линьками. Многие пауки живут около года. В этом случае
обычно зимуют яйца в коконе. При двухлетнем жизненном цикле зимуют
либо подросшие молодые пауки, либо половозрелые. Крупные тропические
пауки-птицеяды, достигающие 6—8 см, живут несколько лет.
Пауки многочисленны в природе и
составляют характерный компонент фауны
различных зон и биотопов, но еще мало
изучены. Среди примерно 1500 видов пау-
ков фауны СССР около 900 видов обитает в
лесной зоне. Здесь обильны мелкие пауки
сем. Erigonidae, Linyphiidae и других, кото-
рые строят сети в кроне деревьев и кустар-
ников в виде пологов и многочисленных
перекрещенных нитей; обычны и крестовики
сем. Argiopidae. На цветах и стволах деревь-
Рис. 346. Сеть паука Hyptiotes
cavatus; под ней паук, натягива-
ющий сеть; внизу сперматическая
сеточка самца паука
ев живут не строящие сетей уплощенные пау-
ки бокоходы (Thomisidae; рис341уп), кото-
рые подстерегают добычу в неровностях ко-
ры, и прыгуны (Salticidae, рис. 341 viii), пры-
гающие на свою жертву. Последние высоко
специализованные хищники, обладают под-
вижными глазами. Многие виды живут в
травянистой растительности и лесной под-
386
стилке, например многочисленные бродячие хищники сем. Lycosidae. Близ
водоемов обычны крупные зелено-коричневые пауки Doloniedes, которые
охотятся на водяных насекомых, бегая по воде и ныряя. В водоемах встре-
чается водяной паук Argironeta aquatica, который строит под водой воздуш-
ный колокол, принося воздух с поверхности на несмачивающемся опушенном
брюшке. У пауков нередко явление покровительственной окраски и подра-
жательной формы — мимикрии. Так, пауки, охотящиеся на цветках (Misu-
mena и др.), окрашены под цвет венчика в белый, желтый, зеленый и другие
тона, некоторые пауки удивительно похожи на муравьев (рис. 341 у) Осо-
бенно причудлива форма некоторых тропических пауков (рис. 341 ш).
Среди пауков есть ядовитые формы, опасные для человека и животных.
Укус тарантулов и крупных крестовиков нашей фауны не опасен и может
вызвать только небольшое местное воспаление. Очень опасен укус каракурта
(Lathrodectus tredecimguttatus), встречающегося в степной полосе СССР Это
паук около 1 см длиной, черный, с красными точками на брюшке (рис. 341 Vi).
Самка устраивает гнезда в траве и углублениях почвы; питается насекомыми
и другими пауками. Укусы, наносимые животным, часто смертельны даже
для лошади, коровы, верблюда. Человек при укусе чувствует сильную
боль, появляется пот, рвота, судороги. Эти явления продолжаются 3—4 дня,
после чего наступает медленное выздоровление. Бывает и смертельный исход.
ОТРЯД КЛЕЩИ (ACARINA)
Клещи — мелкие паукообразные (0,1—0,5 мм, реже до 15 мм), очень раз-
личные по образу жизни и строению. Существует две основные систематиче-
ские группы клещей: настоящие клещи (Acariformes) и паразитиформные кле-
щи (Parasitiformes). Эти группы представляют самостоятельные отряды, да-
лекие по происхождению. Здесь они рассматриваются как один отряд Acari-
па в целях краткости изложения. В отличие от большинства других пауко-
образных клещи имеют огромное медицинское и хозяйственное значение.
Их интенсивно изучают. Наука о клещах — акарология — по своим зада-
чам параллельна науке о насекомых — энтомологии.
Сегменты тела сливаются в отделы двумя способами: либо имеется
головогрудь, несущая шесть пар конечностей, либо голова с хелицерами,
педипальпами и двумя парами ног и туловище, в составе которого объе-
диняются брюшко и сегменты, несущие две задние пары ног (рис. 347i ).
У примитивных клещей тело сегментировано, и в составе брюшка различи-
мы 7 сегментов рис. (347к щ); у некоторых высоко специализованных
клещей, напротив, исчезает и разделение на отделы. Ротовые органы
(эпистом, хелицеры и педипальпы) часто объединяются в хоботок или
«головку», строение которой бывает различно. Хелицеры, клешневидные
у хищных и многоядных форм, у клещей, питающихся кровью живот-
ных или соком растений, превращены в колющие стилеты или более слож-
ные режущие образования. У паразитов ротовые органы иногда имеют при-
крепительные приспособления (гипостом и др.). Примитивные клещи не
имеют щитков, кутикула очень тонкая и дыхание кожное. У многих дру-
гих имеются щитки, которые иногда сливаются в сплошной панцирь. Парал-
лельно в разных группах клещей формируется трахейная система, причем
стигмы могут открываться по бокам брюшка, у оснований ног или хелицер.
Благодаря малым размерам клещей их осязательные волоски (щетин-
ки) относительно велики, обычно постоянны по числу и часто выполняют спе-
циальные функции. Они бывают уплощены и образуют дополнительный
покров или торчат в виде крепких игл, охраняя клеща от хищников, или
очень длинны и опущены, обеспечивая парусность при переносе ветром и
т. п. Особенно разнообразно строение щетинок у паразитов, которые помога-
ют им удерживаться на теле хозяина.
Строение наружного полового аппарата примитивных клещей сложно
и очень сходно у обоих полов, что связано с оплодотворением без копуля-
387
Рис. 347. Клещи (Acarina). / — примитивный сегментированный клещ сем.
Terpnacaridae; II — панцирный клещ Galumna mucronata; III —мучной клещ
Tyroglyphus farinae; IV —расселительная нимфа — гипопус Т. farinae; V —
перьевой клещ Analgopsis; VI — чесоточный зудень Acarus siro; VII —
паутинный клещ Tetranychus telarius; VIII—галлообразующий клещ (справа—
образуемые галлы); IX—личинка краснотелки Trombicula autumnalis; X —
пресноводный клещ Arrhenurus neumani; XI — куриный клещ Dermanyssus gal-
linae; XII — аргасовый клещ Alectorobius papillipes
ции: самец оставляет на субстрате сперматофоры, а самки собирают их на-
ружными гениталиями. У других клещей формируются приспособления для
спаривания, причем копулятивным органом самцов могут служить хели-
церы, специальный придаток брюшка и др. Яйцевыводное отверстие самок
также занимает различное положение и иногда разобщено со специальной
копулятивной порой.
Внутреннее строение примитивных клещей в общем довольно ти-
пично для паукообразных, за исключением недоразвития дыхатель-
ной и кровеносной систем в связи с малыми размерами и кожным
дыханием. У более специализованных форм наблюдается ряд аномалий.
Например, обособление задней кишки и превращение ее в экскреторный ор-
ган у некоторых форм, сосущих сок растений, усиленное развитие половой
системы у самок некоторых паразитов и т. п.
Клещи в большинстве откладывают яйца, но есть и живородящие,
вплоть до таких, которые рождают взрослых особей. Развиваются с ме-
таморфозом. Из яйца вылупляется шестиногая личинка, лишенная чет-
вертой пары ног, стигм и полового отверстия. Личинка, линяя, превращается
в нимфу с четырьмя парами ног и обычно со стигмами, но без полового от-
верстия. Нимфа обычно имеет 2—3 возраста, т. е. линяя 2—3 раза, превра-
щается во взрослую форму У настоящих клещей при этом наблюдается час-
тичный анаморфоз: последовательное образование трех сегментов в пред-
анальной части брюшка. Циклы развития клещей разнообразны. Широко
распространено, особенно у паразитов, явление неотении', исчезновение
взрослой фазы и размножение в фазе нимфы или личинки. В некоторых груп-
пах, напротив, взрослая фаза (имаго) специализируется — взрослый клещ
покрыт панцирем, не боится пересыхания и очень долговечен. Иногда ним-
фа становится расселительной (так называемый гипопус)', она имеет плотный
покров, не питается и расселяется, прикрепившись к насекомым или поз-
воночным. В ряде групп метаморфоз усложнен: личинка паразитирует на
насекомых или позвоночных, избыточно питается и превращается в покоя-
щуюся нимфу, подобную куколке насекомых, в которой путем сложного
гистолитического метаморфоза протекает дальнейшее развитие.
ОСНОВНЫЕ ГРУППЫ КЛЕЩЕЙ, ИМЕЮЩИЕ
МЕДИЦИНСКОЕ И ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
К настоящим клещам (Acariformes) относятся панцирные, амбарные,
перьевые, волосяные и чесоточные, паутинные и галлообразующие клещи,
краснотелки, водяные клещи и многие другие.
Панцирные клещи (Oribatei) — обширная группа обитателей
почвы. Тело взрослых клещей защищено плотным панцирем (рис. 347ц).
Питаются органическими остатками, грибами, микрофлорой. Развитие в
почве, личинки и нимфы влаголюбивы; взрослые клещи менее чувствитель-
ны к пересыханию, долговечны и могут расселяться. Oribatei служат про-
межуточными хозяевами ленточных глист (Anoplocephalata), поражающих
диких и домашних млекопитающих, особенно овец — так называемый мо-
ниезиоз, глистное заболевание, вызываемое Moniezia expansa и др. Клещи,
поедая помет зараженных животных на пастбищах, заглатывают яйца
глист, которые развиваются в организме клещей до стадии цистицеркоида.
Заражение животных происходит при поедании клещей с травой на
пастбищах. В борьбе с мониезиозом применяется ряд мер, в первую оче-
редь рациональный выпас с учетом зараженных пастбищ.
Амбарные, или зерновые, клещи (Tyroglyphoidea)
мелкие белесоватые с тонким глянцевитым покровом. Живут в скоплениях
растительных остатков, в почве, очень влаголюбивы. Питаются гниющей
растительной пищей и микрофлорой. В развитии имеется расселительная
нимфа — гипопус (рис. 347iv), которая не питается, очень устойчива к
внешним факторам и расселяется на насекомых и других животных. Попадая
389
в олагоприятные условия, гипопусы линяют и дают начало колонии кле-
щей, которые при высокой влажности и наличии пищи очень быстро раз-
множаются. Некоторые виды приспособились к жизни в запасах зерна и
продуктах его переработки: муке, крупах и др., которые сильно повреж-
дают
Клещи проникают в склады с зерном с полей, где встречаются в почве,
соломе, гнездах грызунов. Через повреждения оболочки они вгрызаются
в зерно, поедают зародыш, а при повышенной влажности и эндосперм. Зер-
но теряет всхожесть и товарные качества, засоряется шкурками и экскре-
ментами клещей; пораженная мука слипается в комки и загнивает Защита
запасов от клещей основана на создании в элеваторах и складах условий,
исключающих их размножение — понижении влажности зерна, его провет-
ривании, охлаждении и др., а также мерах, препятствующих попаданию
клещей с полей. В ряде случаев применяют ядохимикаты — акарициды.
Наиболее вредные виды: мучной клещ Tyroglyphus farinae (см. рис. 3471 и)
и удлиненные клещи Tyrophagus noxius и Т perniciosus. Помимо зерна,
представители некоторых видов вредят в сыроварении, виноделии, повреж-
дают силос и корнеплоды в хранилищах.
Чесоточные'клещи (Sarcoptoidea) относятся к обширной груп-
пе паразитических клещей, куда входят также перьевые клещи (Analgesoi-
dea), паразитирующие в оперении птиц (см. рис. 347v ), волосяные клещи
(Listrophoroidea), паразитирующие в шерсти млекопитающих, и др. Чесо-
точный зудень Acarus siro (см. рис. 347vi) вызывает у человека кожное
заболевание — чесотку Самка зудня 0,3 лш, самец вдвое мельче. Самка
питается кожей, прогрызая в ее роговом слое извитые ходы до 15 мм длиной,
которые различимы через поверхность кожи в виде сероватых линий. Яйца
откладываются в ходах, причем над ними самка обычно прогрызает венти-
ляционные отверстия. Личинки и нимфы первого возраста живут в ходах,
где питаются остатками кожи, изгрызенной самкой. Сами они ходов не
прогрызают Нимфы второго возраста и самцы выходят на поверхность ко-
жи, где спариваются. Оплодотворенная нимфа прогрызает новый ход, где
превращается в самку Заражение происходит при соприкосновении с
больным чесоткой в редких случаях через платье больного. Человека мо-
гут заражать и другие чесоточные клещи, которые паразитируют на до-
машних животных: собаках, овцах, свиньях, лошадях и других—и сильно
вредят в животноводстве. Лечат чесотку химическими препаратами, кото-
рые применяют в виде мазей, растворов; животных купают в ваннах или
обрабатывают газообразными препаратами в специальных камерах.
Тетраниховые, или паутинные, клещи (Tetrany-
choidea) живут на листьях и других частях растений, сосут сок. Многие
виды выделяют паутину, покрывая ею нижнюю сторону листьев, где скоп-
ляются клещи. Обычно окрашены в зеленые, желтые, красноватые и другие
тона. Покровы мягкие, ротовой аппарат колюще-сосущий. Яйца приклеи-
вают к нижней поверхности листьев. Развитие укорочено (личинка, 1—2
нимфальных возраста) Наиболее известен обыкновенный паутинный клещ
Tetranychus telarius (синоним Т uricae) (см. рис. 347Vn), который живет
на очень многих видах растений и особенно сильно повреждает хлопчатник,
бахчевые, тепличные и комнатные растения. Летом клещи зеленоватые, к
осени самки краснеют и уходят на зимовку, прячась в почве, растительных
остатках. С весны до осени на растениях развивается несколько поколений
клещей. В оранжереях клещи размножаются и зимой Листья пораженных
растений покрываются пятнами, сливающимися в сплошное побеление,
и засыхают При массовом размножении клещей растения гибнут Другой
широко распространенный вид — красный плодовый клещ Panonychus
ulmi—сильно вредит плодовым; у этого вида зимуют яйца в трещинах коры.
Наряду с прямым вредом некоторые виды тетраниховых клещей переносят
вирусы растений. В борьбе с клещами применяют различные акарициды.
ДДТ мало эффективен, так как уничтожает хищников — других клещей
390
и насекомых, поедающих тетранихид, и может даже стимулировать размно-
жение последних. Хорошие результаты дают яды системного действия,
т. е. проникающие в растение и делающие его сок ядовитым для клещей.
Галлообразующие, или четырехногие, к л е-
щ и (Eriophyoidea, или Tetrapodili) вызывают на листьях и других частях
растений патологические разрастания в виде подушек из волосков или
мешковидных галлов, в которых они живут (см. рис. 347уш) Клещи очень
малы, имеют удлиненное тело, колюще-сосущие ротовые органы и только
две пары ног. Они по происхождению связаны с тетраниховыми клещами,
но представляют крайне неотенические формы, выходящие из яйца и
размножающиеся в состоянии, более раннем, чем личинка (активные полуэм-
брионы). Среди клещей они служат примером наиболее высоко специали-
зованных паразитов растений.
Клещи-краснотелки (Trombea) — обильная видами груп-
па клещей, для которых особенно характерен сложный метаморфоз. Из
отложенных в почву яиц выходят мелкие паразитические личинки, которые
сосут лимфу позвоночных или гемолимфу насекомых. Отпав от животного-
хозяина, они линяют и превращаются в покоящихся нимф, подобных ку-
колке насекомых. Из покоящейся нимфы выходит нимфа следующего воз-
раста, которая хищничает. Линяя, она вновь превращается в покоящуюся
нимфу, из которой выходит взрослый клещ, по образу жизни похожий на
вторую активную нимфу Паразитические шестиногие личинки (рис. 347ix)
имеют колюще-сосущий ротовой аппарат, мягкий покров и сильно разду-
ваются при сосании. Активные нимфы и взрослые клещи крупнее, густо
опушены волосками, имеют мощные хелицеры с когтевидными придатками;
часто они окрашены в красный цвет, откуда и пошло название этих клещей.
Среди краснотелок есть формы, личинки которых нападают на человека и
передают ему от диких животных опасное риккетсиозное заболевание —
японскую речную лихорадку, распространенную в Восточной Азии, на Япон-
ских и Индо-Малайских островах.
Водяные клещи (Hydracarina) — несколько обильных видами
групп клещей, обитающих в пресных и морских водах. Взрослые пресно-
водные клещи 1—2 мм длиной, иногда до 8 мм, часто имеют яркую окрас-
ку красные, зеленые и др. Тело покрыто кожистой кутикулой, ноги обычно
плавательные с длинными щетинками (см. рис. 347х ). Яйца откладывают
на подводные предметы. Развитие сходно с таковым краснотелок: личинки
паразитируют на насекомых, имеются покоящиеся нимфы; хозяевами
личинок клещей служат водные насекомые и их личинки, взрослые кле-
щи-хищники. Некоторые виды в течение всей жизни паразитируют в мантий-
ной полости и жабрах пластинчатожаберных моллюсков. Гидракарины ши-
роко распространены, составляют характерный элемент фауны водоемов
и один из компонентов пищи рыб.
К паразитоформным клещам (Parasitiformes) относятся
гамазоидные, иксодовые и некоторые другие. Это формы, имеющие наи-
большее медицинское значение.
Гамазоидные клещи (Gamasoidea) частью свободноживущпе,
частью паразитические формы, сосущие кровь рептилий, птиц и млеко-
питающих. Некоторые виды нападают на человека. Размеры — 0,1—1,5 мм\
на теле имеются щитки, по бокам пара стигм. Ротовые органы жующе-
сосущие (хелицеры с клешней) или колюще-сосущие (хелицеры в виде сти-
летов). Откладывают яйца, некоторые живородящи. Свободноживущпе
клещи-хищники, питаются нематодами, мелкими насекомыми, другими
клещами, а также микрофлорой; живут в почве, гниющих остатках,
гнездах грызунов, муравейниках, пчелиных ульях и т. п. Клещи, живущие
в навозе и на трупах, расселяются на жуках-навозниках и могильщиках,
на мухах. Среди гамазоидей можно проследить все стадии перехода к
паразитизму на позвоночных. Имеются формы, использующие последних
как средство расселения; другие, будучи хищниками, при случае сосут кровь,
391
вытекающую из поранений кожи; наконец, многие окончательно стали па-
разитами и питаются только кровью, прокалывая кожу.
Различны и взаимоотношения с животными-хозяевами: одни живут
и размножаются в норах и гнездах, периодически нападая на хозяев только,
чтобы насосаться крови; другие подстерегают хозяев в открытой природе:
наконец, есть постоянные паразиты, размножающиеся на теле хозяина или
даже живущие в полостях — дыхательных путях, слуховых проходах.
Гамазоидные клещи причастны к распространению возбудителей ряда ин-
фекционных болезней человека и животных — патогенных вирусов, риккет-
сий, бактерий, спирохет и др. Птичьи клещи рода Dermanyssus (см.
рис. 347xi) — паразиты многих видов диких, домашних и синантропных
птиц, в массе размножаются в гнездах. Из гнезд голубей, скворечников и
птичников они проникают в жилые помещения и нападают на людей, осо-
бенно ночью. Они прокалывают кожу и сосут кровь, вызывая покраснение
кожи в местах уколов, сыпь. Считают, что птичьи клещи причастны к рас-
пространению некоторых вирусных заболеваний. Крысиный клещ Ornit-
honyssus bacoti и мышиный клещ Allodermanyssus sanguineus сосут кровь
крыс и мышей. Они размножаются в жилых помещениях, нападают на че-
ловека и могут передавать ему от грызунов риккетсий крысиного тифа и
риккетсиозной оспы.
И кс од о вы е клещи (Ixodoidea) включают два целиком парази-
тичхжих семейства — аргасовых клещей (Argasidae) и собственно иксодо-
вых (Ixodidae) клещей. Сюда относятся важнейшие переносчики возбуди-
телей целого ряда опасных болезней человека и животных.
Apr азов ы е клещи (Argasidae) в СССР распространены главным
образом в Средней Азии и Закавказье. Они приурочены к засушливым
пустынным и предгорным районам, где живут в естественных убежищах —
пещерах, трещинах скал, норах и гнездах зверей и птиц, а также поселяют-
ся в жилых и хозяйственных постройках. Клещи сосут кровь млекопитающих
и птиц, могут нападать и на человека. Обитание в убежищах, подолгу
пустующих, привело к выработке ряда приспособлений. Аргазиды способ-
ны очень долго голодать (личинки около года, нимфы и взрослые до несколь-
ких лет), благодаря чему цикл развития может затягиваться до 25 лет. При
появлении животного или человека клещи наползают на него и сосут кровь
быстро и в большом количестве.
Тело голодных клещей дорзо-вентрально уплощено, кутикула соб-
рана в складки. При наполнении кишечника кровью складки расправляют-
ся и клещ принимает почти шаровидную форму. Хоботок аргазид, как и у
иксодид, снабжен гипостомом, но они не присасываются к хозяину надол-
го и, напитавшись, остаются в убежище, где происходит откладка яиц и
последующее развитие. Поселяясь в жилище, они нападают на людей обыч-
но ночью, подобно постельным клопам. Аргазиды служат переносчиками
спирохет клещевого возвратного тифа. Спирохеты сохраняются в организме
клещей пожизненно, причем самка заражает ими и свое многочисленное
потомство. Спирохеты передаются клещами при укусе от животного живот-
ному и человеку. Последнего заражают клещи рода Alectorobius (синоним —
Ornithodoros), особенно A. papillipes (см. рис. 347хп). Стойкие очаги кле-
щевого тифа были в старых глинобитных поселениях Средней Азии, которые
теперь по большей части ликвидированы. Представители рода Argas вредят
в птицеводстве, распространяя куриный спирохетоз.
Собственно иксодовые клещи (Ixodidae) распространены в раз-
личных ландшафтных зонах. В СССР известно более 50 видов, относящихся
к 6 родам. Строение и развитие иксодид рассмотрено на примере Ixodes ricinus
(стр. 374). Почти все они пастбищные паразиты, подстерегающие животных-
хозяев в открытой природе, чаще на растительности. Взрослые клещи обыч-
но сосут кровь крупных животных, личинки и нимфы питаются на грызу-
нах, птицах, рептилиях. Многие виды во взрослой фазе присасываются к
человеку При подстерегании хозяина на пастбище встреча с ним маловеро-
392
ятна,что компенсируется у иксодид рядом приспособлений. Клещ, попав на
животное, крепко присасывается и отпадает, только полностью напитавшись.
Самка кладет яйца один раз и очень плодовита (7—10 тыс. яиц и более).
Клещи на всех фазах способны длительно голодать. Большинство иксодид
в течение цикла развития трижды меняет хозяина. У некоторых видов ли-
чинки, напитавшись, не покидают хозяина, а превращаются в нимф, которые
вновь присасываются и затем отпадают. Наконец, есть виды, у которых все
развитие от личинки до самки проходит на одном обычно крупном животном,
и отпадает сытая самка для откладки яиц.
Иксодовые клещи служат переносчиками возбудителей многих инфек-
ционных болезней человека и животных. Они не только передают возбудите-
лей болезней при сосании крови от животного животному и человеку, но
длительно сохраняют инфекцию в своем организме и заражают свое много-
численное потомство, поддерживая таким образом существование природных
очагов болезней.
Клещи Ixodes persulcatus передают вирус клещевого энцефалита, рас-
пространенного в таежной зоне; Hyalemma marginatum заражает человека
вирусом крымской геморрагической лихорадки, Rhipicephalus sanguineus
передает риккетсий марсельской лихорадки, распространенной в Средизем-
номорье, a Dermocentor nuttali — риккетсий клещевого сыпного тифа Азии;
клещи рода Dermacentor сохраняют возбудителя туляремии — болезни
грызунов, опасной для человека; виды родов Ixodes, Hyalemma и другие
передают гемоспоридиальные заболевания животных. Иксодовые клещи
причастны также к распространению чумы, бруцеллеза и других болезней.
Советскими учеными академиками Е. Н. Павловским и В. Н. Бекле-
мишевым и их многочисленными учениками проведена большая работа по
выяснению значения клещей как переносчиков болезней и разработке мер
борьбы с ними. Результаты этих исследований вошли в общую систему борь-
бы с паразитами, проводимую в нашей стране.
ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ХЕЛИЦЕРОВЫХ
Среди членистоногих Chelicerata представляют единую систематичес-
кую группу, связанную по происхождению с трилобитами, которые в свою
очередь по ряду признаков сходны с аннелидами. С примитивными трило-
битами хелицеровых сближает сегментальный состав головы, способ форми-
рования сегментов тела, строение жаберных конечностей. В состав головы
тех и других, помимо головной лопасти — акрона—входят 4сегмента, пред-
ставленные у личинки трилобитов протасписа; среди хелицеровых соответ-
ствующая стадия имеется в эмбриональном развитии мечехвостов. Обособ-
ленная четырехсегментная голова сохраняется у части хелицеровых и во
взрослом состоянии (сольпуги, настоящие клещи и др.). У мечехвостов и
трилобитов сходно формируются и туловищные сегменты — один за другим
преданальной зоной роста с тем только различием, что этот процесс, назы-
ваемый анаморфозом, у трилобитов происходил в постэмбриональном раз-
витии, а у мечехвостов наблюдается в эмбриогенезе. Среди хелицеровых
известны и такие, у которых часть сегментов брюшка представлена постэм-
бриональными сегментами (вымершие ракоскорпионы, среди современных —
настоящие клещи). Составные части двуветвистых конечностей трилобитов
и жаберных ножек водных хелицеровых также считают гомологичными
(соответственными).
В отличие от трилобитов Chelicerata утратили антеннулы, их конеч-
ности дифференцировались, сегменты тела сократились в числе и объеди-
нились в отделы соответственно функциям конечностей. Первая пара
конечностей головы превратилась в хелицеры, вторая — в педипальпы,
две другие — в ноги; конечности двух туловищных сегментов объединились
с головой в головогрудь — просому Остальные сегменты образовали
брюшко, причем конечности его переднего отдела — мезосомы — у ряда
393
форм сохранили дыхательную функцию, а конечности заднего отдела —
метасомы — исчезли. Функцию соединения головогруди и брюшка взял
на себя 7-й (предполовой) сегмент, за которым всегда следует сегмент с
половым отверстием.
Chelicerata возникли в морской среде. Древнейшие водные хелицеровые
населяли палеозойские моря одновременно с трилобитами, от которых отде-
лились, вероятно, еще в докембрийское время. Эти крупные хищные членис-
тоногие жили в мелководье теплых морей и, судя по наличию их в континен-
тальных отложениях, обитали также в солоноватых и пресных водах. Рако-
скорпионы известны с кембрия до перми, мечехвосты — с силура, мало
изменившись, дожили до нашего времени. Непосредственными потомками
силурийских эвриптерид (водяных скорпионов Palaeophonus и др.) среди
наземных хелицеровых (Arachnoidea) считают скорпионов.
Arachnoidea сформировались в процессе перехода к сухопутному об-
разу жизни. Большинство отрядов известно из карбоновых отложений,
но возникли они, по-видимому, значительно раньше; клещи, подобные сов-
ременным низшим Acariformes, найдены в девоне. Главные приспособления
при выходе на сушу направлены на защиту от высыхания. Жаберное ды-
хание сменилось легочным, а далее трахейным. Жаберные ножки мезосомы
погрузились в полости и преобразовались в легкие, которые полнее пред-
ставлены у примитивных отрядов: 4 пары у скорпионов, 2 пары у жгуто-
ногих и четырехлегочных пауков, пара у высших двулегочных пауков. У
других паукообразных легкие заменены трахеями, которые у членистоно-
гих представляют новообразование и более приспособлены к воздушному
дыханию. В разных отрядах Arachnoidea трахейная система возникла более
или менее независимо, на что указывает различное расположение стигм: на
разных сегментах брюшка, а также у оснований ног или хелицер (у клещей).
Трахеи не гомологичны легким, но иногда образуются на их месте, напри-
мер у пауков.
С переходом к сухопутной жизни связана и дальнейшая концентра-
ция отделов тела и систем органов. Характерна постепенная редукция
метасомы и образование слитного брюшка, особенно у пауков и клещей.
У части клещей сливаются и отделы тела. Параллельно идет концен-
трация нервной системы и упрощение кровеносной. Последнее обусловлено
переходом к нелокализованному трахейному дыханию.
Существенно изменяется биология размножения и развитие. Наружное
оплодотворение, свойственное водным хелицеровым (мечехвосты), сменяется
у наземных сперматоформным оплодотворением и копуляцией. Защитой
спермы от пересыхания служат сперматофоры, У примитивных клещей,
живущих во влажной почве, сохранилось оплодотворение без спаривания
(самки подбирают сперматофоры, оставляемые самцами). У большинства
других паукообразных самец тем или иным способом переносит спермато-
фор в половое отверстие самки. Наконец, в некоторых случаях сперматофоров
нет и сперма изливается в семеприемники самки из копулятивных органов,
например резервуаров педипальп у пауков. Яйца хелицеровых, в том числе
водных, богаты желтком, и обычно вылупляется маленькая полносегмент-
ная особь, похожая на взрослую. У водных хелицеровых дальнейшее раз-
витие сопровождается длительным ростом и многими линьками. У наземных
форм рост и число линек сокращаются, чем обусловлены относительно ма-
лые размеры взрослой особи, быстрое достижение ею половой зрелости и спо-
собности размножаться. Эта выгодная во всех отношениях форма жизни
достигает крайней степени у самых маленьких хелицеровых — клещей.
Среди других наземных членистоногих она особенно характерна для насе-
комых (см. ниже).
На первых этапах перехода к сухопутному существованию возник тот
образ жизни, который свойствен большинству отрядов паукообразных и
во многом еще примитивен. Это животные, тесно связанные с почвой и с
влажными местами обитания, бродячие хищники, охотящиеся ночью, а
394
днем прячущиеся в укрытиях, трещинах почвы, под камнями, в норах и т. п.
или постоянно живущие в толще почвы и влажных растительных остатках.
Таковы в подавляющем большинстве скорпионы, жгутоногие, сольпуги,
сенокосцы, примитивные пауки, ложноскорпионы и клещи. Связь с почвой
порывают лишь немногие группы, главным образом высшие пауки и часть
клещей. Решающее значение в эволюции пауков имела способность выделять
паутину, которая первоначально использовалась для устройства яйцевых
коконов и выстилания убежищ, а далее стала применяться и для построй-
ки разнообразных ловчих сетей. Некоторые семейства пауков специализи-
ровались как бродячие дневные хищники (бокоходы, скакуны). Перейдя
к открытому существованию и заняв в природе всевозможные места обита-
ния, высшие пауки достигли расцвета, возникли различные экологические
группы, разнообразие формы и окраски, сложнейшие инстинкты и т. д.
Наибольшего разнообразия достигли клещи. Благодаря малым разме-
рам они заселили на суше буквально все мыслимые места обитания, в том
числе такие, которые недоступны более крупным членистоногим; они про-
никли также в пресные и морские воды. Существенное значение имел
метаморфоз клещей, который видоизменялся в разных направлениях соответ-
ственно условиям и образу жизни, особенно при паразитизме. Непаразити-
ческие формы в изобилии населяют почву, скопления всевозможных орга-
нических остатков, норы и гнезда позвоночных и насекомых. Многие группы
клещей перешли к паразитизму, тесно связавшись в своей эволюции с поз-
воночными, членистоногими и растениями. Паразитизм клещей разнооб-
разен: формы, питающиеся кровью или роговыми частями кожи позвоноч-
ных, гемолимфой насекомых, живущие в норах и гнездах, в шерсти и коже
млекопитающих, оперении птиц, под чешуями рептилий, в дыхательных
путях позвоночных-хозяев, на теле и в трахеях насекомых, в жабрах мол-
люсков; паразиты растений, сосущие сок, образующие галлы и т. д.
СРАВНЕНИЕ ЖАБЕРНОДЫШАЩИХ, ТРИЛОБИТОВ
И ХЕЛИЦЕРОНОСНЫХ
Можно предполагать, что три рассмотренных подтипа членистоногих
связаны друг с другом общими предками, причем связующим звеном между
жабернодышащими и хелицероносными в известной мере служат трило-
биты. По гомономности сегментации туловища и по наличию жабр на
всех конечностях они сходны с низшими раками (листоногимп), а по типу
строения конечностей и расположению их на головном отделе — с ракооб-
разными вообще. Однако, форма отделов тела и онтогенез связывают трило-
битов с низшими хелицероносными (мечехвостами) Это дает основание ряду
исследователей признавать их родство с меростомовыми (Palaeostraca) и
включать их в подтип хелицероносных.
Подтип IV. Первичнотрахейные
(Protracheata)
ХАРАКТЕРИСТИКА
Первичнотрахейные, или онихофоры (вооруженные коготками),—
червеобразные формы с неясной внешней сегментацией: на теле отсутствуют
границы сегментов; сегментация наружно намечается лишь парами ножек.
Голова неясно обособлена от туловища. Она несет три пары придатков —
впереди рта пару кольчатых усиков (рис. 348), в околоротовом углублении,
округленном кольцевой складкой, пару челюстей, по сторонам кольцевой
складки пару приротовых сосочков; челюсти и сосочки представляют ре-
зультат изменения двух пар ножек: челюсти представляют видоизмененные
395
парные коготки, сосочки представляют ножки, лишившиеся коготков.
Пара простых глаз расположена позади усиков. Туловище на подотделы не
делится. Покров содержит хитин, но
склеризованы лишь челюсти и пары
коготков на конце ножек. Ножки,
расположенные по двум боковым кра-
ям тела на границе выпуклой спинки
и плоской грудки, коротки и мягки,
со слабо выраженной членистостью
суженной дистальной части (рис. 349).
При каждой паре ножек по паре от-
верстий сегментальных органов (ме-
танефридий). Трахеи представлены
неветвящнмися тончайшими трубоч-
ками; на поверхности тела они откры-
ваются многочисленными отверстия-
ми различного положения. Половое
отверстие на брюшной стороне у зад-
него конца тела. Наружные половые
придатки у обоих полов отсутствуют.
склеризованных щитков на нем нет:
Рис. 349. Peripatopsis capensis
(справа полость тела вскрыта):
1 — усик, 2 — ножка, 3 — кишечнику
4 — слюнная железа, 5 — слизеотде-
лительная железа, 6 — сегменталь-
ный орган, 7 — нервный ствол#
8 — надглоточный узел, 9— семенник
Рис. 348. Peripatopsis
edrnardsii (голова снизу)
I—усик, 2—челюсть, 3—при-
ротовый сосочек
ОБЩИЙ ОБЗОР ПОДТИПА
Внешнее строение. Тело первичнотрахейных гусеницеобразное, ци-
линдрическое (рис. 349) с уплощением на брюшной стороне. От границы
нижней спинно-боковой поверхности отходят от 14 до 43 пар ножек, посте-
пенно суживающихся к свободному концу; основная часть ножки покрыта
мелкими кольцевыми складками, дистальная часть делится на 5—6 корот-
ких члеников, которыми животное опирается на субстрат так же, как на-
секомые на членики лапки; ножка оканчивается парой коготков. Только
ножки и связанные с их основанием отверстия сегментальных органов ука-
зывают на сегментацию тела: перетяжки как границы сегментов
отсутствуют. На переднем конце тела, т. е. на голове, расположена пара
умеренно длинных кольчатых усиков, за ними на тыльной стороне — пара
простых глаз. На нижней поверхности головы находится обнесенное бо-
родавками ротовое углубление с парой челюстей, а по сторонам от него —
по сосочку, расположенному на линии ножек туловища (рис. 348).
396
Эти сосочки представляют рудименты ножек, на конце сосочка находится
отверстие мощно развитой слизеотделительной железы (рис. 350), секрет
которой служит для обволакивания добычи хищника, он с силой выбра-
сывается через пору сосочка на насекомое (например на термита),
служащее пищей первичнотрахейному.
Нервная система. Центральная нервная система туловища (рис. 350)
состоит из пары широко расставленных нервных стволов, тянущихся на
всем протяжении туловища в парном боковом отделе полости тела на уров-
не основания ножек и сливающихся над задней кишкой. Утолщения
стволов по паре на сегмент намечены очень слабо, так как нервные клетки
распределены по стволу весьма равномерно, не образуя значительных скоп-
лений. Многочисленные комиссуры (до 10 на сегмент) соединяют оба ствола
на брюшной стороне друг с другом на всем их протяжении от глотки до
анального отверстия; приблизительно то же число поперечных нервов
Рис. 350. Peripatoides novae — zelandiae, поперечный срез
через туловище:
1— кутикула, 2—кольцевая мускулатура, 3— косая мускулатура,
4 — продольная мускулатура, 5 — слюнная железа, 6 — трансверзальная
мускулатура, 7 — сегментальный орган (нефридий), 8 — нервный ствол,
9—мускулатура ножки, 10 — коготки, 11 — лапка, 12 — средняя кишка,
13— выделительная пора, 14—слизеотделительная железа, 15 — диаф-
рагма, 16 — сердце, 17 — средний отдел полости тела, 18 — боковой отдел
полости тела
отходит в стороны, частично соединяясь с соответствующими нервами дру-
гой стороны под спинной поверхностью. Впереди оба продольных нервных
ствола соединяются с надглоточным узлом, своей нерасчлененностью напо-
минающим надглоточный узел кольчецов (характерное для ракообразных
и трахейнодышащих деление мозга на 3 отдела у первичнотрахейных от-
сутствует). Надглоточный узел иннервирует глаза, усики и мышцы челюс-
тей. Первый простой глаз первичнотрахейных строением напоминает глаза
кольчецов (Alciopidae)
Пищеварительная система. Кишечник первичнотрахейных очень
прост: это прямая трубка, почти на всем протяжении туловища представ-
ленная средней кишкой, передняя и задняя кишки образуют лишь очень
короткие отделы. Выпячиваний, вырабатывающих секрет, кишечная труб-
ка не образует; пара же слюнных желез открывается в околоротовое углуб-
ление позади челюстей. Мальпигиевых трубочек у первичнотрахейных нет.
Кровеносная система. Кровеносная система слагается из спинного
сосуда и системы полостей тела. Спинной сосуд сердце — не имеет ответ-
влений: кровь непосредственно поступает из полости тела в сердце через
парные боковые отверстия — остии, которых имеется по паре на каждый
сегмент туловища, и выносится из спинного сосуда в полость тела через
397
переднее отверстие. Как отсутствием артерий и вен, так и наличием спинной
диафрагмы — горизонтальной мышечной перепонки, расположенной под
сердцем и с ним связанной,— сердце первичнотрахейных напоминает спин-
ной сосуд насекомых.
Полость тела и выделительная система. Полость тела первичнотра-
хейных, как и типичных членистоногих,— миксоцель. Парная мышечная
перепонка дорзо-вентрального направления (см. рис. 350) разделяет по-
лость тела на 3 отдела — парные боковые и непарный средний; парный
боковой заключает в себе парный ствол нервной системы, сегментальные
органы и слюнные железы; в непарном среднем отделе помещаются кишеч-
ник, слизеотделительные железы, сердце и половая система. Сегментальные
органы расположены во всех сегментах тела и открываются наружу на
нижней поверхности ножек, как правило, при их основании; однако в
крайних передних и задних сегментах тела они превращены или в рудимен-
ты, или в органы особого назначения, как-то: слюнные железы, половые
протоки, анальные железы.
Половая система. Строение мужской и женской половой системы на-
поминает ее строение у членистоногих, в некоторых отношениях достигая
Рис. 351 Aysheaia pedunculata (реставрация)
еще более высокой специализации. Половые системы, мужская и женская,
парны; непарно лишь половое отверстие и ближайший отдел половых про-
токов. Яйца развиваются в парном отделе половых протоков, причем, как
правило, имеет место живорождение, что обязательно в случае мелких яиц;
развитие таких яиц происходит при участии плаценты.
Экология и географическое распространение. Первичнотрахейные
(Protracheata) — наземные животные, как правило, водятся в сырых мес-
тах под корой, под камнями и листьями; лишь немногие виды встречаются
в сухих местностях. Распространены они в тропическом поясе, где встре-
чаются во всех частях света. Ранее все известные виды объединялись в род
Peripatus, ныне различают целый ряд родов, среди которых наиболее из-
вестен род Peripatopsis с видом Peripatopsis capensis (Южная Африка).
Первичнотрахейные — исключительно ночные животные; к свету они
относятся отрицательно, и глаза им служат лишь для светоощущения; при
передвижениях они руководятся осязанием, причем главным органом ося-
зания им служат усики. Это исключительно хищники, питающиеся главным
образом насекомыми.
Организация и распространение первичнотрахейных позволяют пред-
полагать, что они — формы древние. Действительно, в последние десяти-
летия в древних отложениях кембрийской эпохи найдены ископаемые,
близко напоминающие современных первичнотрахейных. Более точные
сведения дало изучение ископаемого из среднекембрийских отложений Се-
верной Америки — Aysheaia (рис. 351). Это была морская форма, не имею-
щая, очевидно, трахей, как современные Onychophora, это ископаемое
обладало неясной сегментацией; покровы также несли кольца мелких бу-
горков, а ножки, напоминавшие ножки первичнотрахейных, были не от-
членены; в отличие от первичнотрахейных конец ножек не был расчленен,
а вместо пары коготков имел 6 щетинок; не было ни челюстей, ни сосочков
со слизеотделительной железой; усики имели гребневидную форму Таким
образом, строение этой ископаемой формы было проще строения первично-
трахейного.
398
ФИЛОГЕНИЯ ПЕРВИЧНОТРАХЕИНЫХ
Первичнотрахейные соединяют в себе признаки как членистоногих,
так и кольчатых червей; в зависимости от того, каким признакам сходства
приписывали преобладающее значение, первичнотрахейных относили то
к кольчецам, то выделяли их в промежуточную группу между кольчецами
и многоножками. С того времени, когда у первичнотрахейных обнаружили
трахейное дыхание, их сближают с Tracheata.
Противоречивость признаков первичнотрахейных выражается во
многом: так, наружный покров у них мягок и не несет склеритов, как и
у кольчецов, но он хитинизирован, как у членистоногих. Ножки первично-
трахейных не обособлены от сегмента, и основание их не расчленено, но
конец ножки состоит из мелких члеников и несет пару коготков, как у на-
секомых. Сегментальные органы, как у червей, имеются почти на всех сег-
ментах туловища, но полость сегментального органа не сообщается с полостью
тела, которая у первичнотрахейных, как и у всех членистоногих, смешанная
(миксоцель), а несет на внутреннем конце целомический мешочек (рис. 350),
представляющий собой обособившуюся часть вторичной полости. Как у
типичных членистоногих, половые протоки и слюннле железы первично-
трахейных преобразуются из сегментальных органов.
Надглоточный узел, как у кольчецов, наружно не расчленен, но внут-
ренним строением более напоминает мозг членистоногого.
Мощное развитие и анатомо-гистологическое строение туловищной
мускулатуры, а именно: ее слоистость, состав слоев и отсутствие поперечной
полосатости мышечных волокон — напоминают мускулатуру кольчатых
червей.
Подтип V. Трахейнодышащие (Tracheata)
ХАРАКТЕРИСТИКА
К этому подтипу относятся наземные членистоногие, дышащие возду-
хом при помощи трахейной системы. Для них характерна одна пара усиков,
расположенных впереди рта. Голова всегда ясно отделена от туловища,
которое состоит из многих сегментов. Наружные половые придатки у обоих
полов — видоизмененные ножки. Дыхальца (стигма) распределены сегмен-
тально.
КЛАССИФИКАЦИЯ
Подтип трахейнодышащие (Tracheata)
Класс 1. Многоножки (Myriopoda)
Подкласс I. Губоногие (Chilopoda)
Подкласс II Двупарноногие (Diplopoda)
Подкласс III. Сколопендреллы (Syniphyla)
Подкласс IV Пауроподовые (Pauropoda)
Класс 2. Насекомые (Insecta)
Подкласс I. Первичнобескрылые (Apterygota)
Подкласс II. Крылатые (Pterygota)
КЛАСС 1. МНОГОНОЖКИ (MYRIOPODA)
ХАРАКТЕРИСТИКА
Многоножки — наземные трахейнодышащие членистоногие с тулови-
щем, состоящим из большого количества сегментов или из несколько от-
личающихся друг от друга сегментов (т е. из гомономных или гетерономных
сегментов). В последнем случае большие метамеры чередуются с малыми.
У многоножек нет ясного деления на грудь и брюшко. Все сегменты туло-
399
вища несут многочисленные, в общем одинаковые конечности. Усики (ап
tennae) разнообразны по форме; как правило, они одноветвисты и нитевидны
или булавовидны. Число челюстей две или три пары. Органами выделения
служат отростки кишечника — мальпигиевы сосуды и нижнегубые железы—
нефридиального происхождения. Непарные или парные половые отверстия
у одних групп (например, у двупарноногих) расположены на одном из
передних сегментов тела (3-м или 4-м), а у других (например, у губоногих)
на предпоследнем сегменте тела.
Трахейная система в виде парных пучков в каждом сегменте туловища.
В голове этих пучков нет, и она снабжается трахеями, идущими от туловищ-
ных сегментов. Нервная система представлена головными ганглиями,
слившимися в одну общую массу—мозг—и брюшной нервной цепочкой,
соединенной с мозгом. Брюшная цепочка лестницевидная, ее парные узлы
по числу и положению соответствуют сегментам тела.
Все многоножки откладывают яйца. Постэмбриональное развитие мо-
жет быть прямым (эпиморфоз), обычно же развитие с личиночной фазой
(метаморфоз).
ПОДКЛАСС I. ГУБОНОГИЕ (CHILOPODA)
ХА РАКТЕРИСТИКА
Губоногие — хищные многоножки; охотятся за различными членис-
тоногими и червями обычно ночью, а днем прячутся в укрытиях и вообще
избегают сильного освещения. Губоногие очень подвижны, в особенности
мухоловки (Scutigera). Усики длинные, нитевидные из сходных члеников.
Челюстной аппарат состоит из пары нерасчлененных верхних челюстей и
двух пар сложных нижних челюстей (табл. XI). Нижние челюсти второй
пары похожи на ходильную ножку; их дистальная часть функционирует
как щупальце. Фасетки сложных глаз расположены обычно не плотно друг
к другу. Первый сегмент туловища несет характерные для губоногих
сильные острые клещи — измененные конечности (табл. XI), оканчиваю-
щиеся сильно хитинизированным острием с отверстием ядовитой железы.
Эти клещи служат для захвата и умерщвления добычи, а также для защиты.
Туловище состоит из большого количества сегментов: от 19 до многих
десятков. Сегментация в одних случаях гомономная (сегменты одинаковые);
в других случаях сегментация туловища гетерономная (сегменты неоди-
наковые) с чередованием больших и укороченных, не имеющих стигм сег-
ментов. На всех сегментах туловища хорошо развиты боковые склериты —
плевры, на которых обычно расположены стигмы. Трахейная система
сильно развита. Половое отверстие расположено у обоих полов на предпос-
леднем, половом, сегменте. Сегмент, лежащий перед половым, несет умень-
шенные ножки, превращенные в половые придатки.
Представитель подкласса губоногих — костянка
(Lithobius forficatus)
Костянка (табл. XI) часто встречается под камнями, под корой пней
и лежащих стволов, вообще там, где живут мелкие членистоногие и их
личинки, которыми питается этот хищник.
Внешнее строение. Костянка достигает 2—2,5 см длины. Тело, как
и у остальных членистоногих, покрыто щитками из толстого хитина, сое-
диненными тонким перепончатым хитином. Тело состоит из ясно обособ-
ленной головы и 19 сегментов туловища.
Голова. Плотная головная капсула впереди имеет наличник и
трехлопастную верхнюю губу (табл. XI), под которой расположено ротовое
отверстие, окруженное челюстным аппаратом. Верхние челюсти — манди-
булы — состоят только из одного членика, и их свободный край несет ряд
400
7
Таблица XI. Костянка (Lithobius forficatus), самка. / —общий вид с брю-
шной стороны вскрытого животного; II — голова снизу; III — вскрытая
ногочелюсть:
/— усик, 2— глаз, 3— верхняя челюсть, 4 — нижняя челюсть первой пары, 5 — нижняя
челюсть второй пары (ее щупальце), 6 — ногочелюсть, 7 — грудная пластинка, 8 — стиг-
ма, 9—нервная цепочка, 10—мальпигиева трубочка, 11 — яичник, 12—анальное отвер-
стие, 13—половые ножки, 14— ядовитая железа
зубцов. Лишь эта верхняя челюсть служит для измельчения пищи. Нижняя
челюсть первой пары состоит из трех члеников. Основания второй пары
нижних челюстей слились в непарный отдел, остальные три членика вытя-
нуты в короткий щупик. Обе пары нижних челюстей служат для ощупывания
и придерживания пищи.
На передней поверхности головной капсулы расположена пара длин-
ных многочленистых нитевидных усиков, или антенн, а по бокам лежат
сложные глаза, фасетки которых неплотно прилегают друг к другу.
Туловище. Сегменты туловища развиты неравномерно. Первый и
три последних сегмента резко отличаются от остальных. Первый сегмент
несет шестичленистые ногочелюсти, оканчивающиеся острым сильно
хитинизированным шипом, несущим отверстие ядовитой железы. Основные
членики этих ногочелюстей срослись в непарную пластинку.
Три задних сегмента отличаются от остальных туловищных сегментов
еще более. У самки первый из этих трех сегментов несет пару трехчленистых
половых ножек. Половое отверстие лежит на предпоследнем сегменте. Ос-
тальные сегменты туловища также неодинаковы: большие сегменты че-
редуются с меньшими. Только шесть больших сегментов (4-й, 6-й, 9-й,
11-й, 13-й и 15-й) сохранили дыхальца, расположенные же между ними
малые сегменты дыхалец не имеют.
Ходильная, или бегательная, ножка состоит из восьми члеников.
Самая задняя пара бегательных ножек потеряла свою локомоторную функ-
цию — она принадлежит укороченному сегменту, отличается особенной
длиной и служит органом осязания.
Нервная система. Центральная нервная система состоит из надглоточ-
ного и подглоточного ганглиев, сходных с соответствующими нервными
узлами насекомых. В туловище все сегменты, за исключением анального
сегмента (telson), имеют по парному нервному узлу, которые парными же
продольными коннективами связаны с узлами соседних сегментов. Таким
образом, брюшная цепочка состоит из семнадцати узлов, включая подгло-
точный узел, т. е. комплексный узел трех головных сегментов, несущих
челюсти. От надглоточного нервного узла иннервируются глаза и усики, от
узлов брюшной цепочки — ножки и мускулатура сегментов. По спинной
стороне над сердцем расположен нервный тяж с нервными клетками.— это
симпатическая нервная система, иннервирующая сердце.
Пищеварительная система. Кишечный канал костянки (рис. 352)
состоит из передней, средней и задней кишок. Передняя и задняя кишки
выстланы хитином и очень коротки; широкая средняя кишка тянется почти
на всем протяжении туловища. На границе средней и задней кишок в ки-
шечник впадает пара длинных неветвящихся мальпигиевых трубок это
выделительная система костянки. Рядом с ротовым отверстием открываются
протоки парных желез, принимаемых за слюнные.
Кровеносная система. Кровеносная система костянки, как и у прочих
трахейнодышащих, в том числе и у насекомых, незамкнутая. Однако эта
система у костянки подвергалась меньшей редукции, чем у насекомых.
Кроме спинного сосуда, расположенного над кишечником (сердце), имеется
подкишечный продольный сосуд, впереди связанный со спинным сосудом,
а сзади открывающийся в полость тела. Сердце тянется на протяжении всего
туловища, образуя 15 сегментально расположенных сердечных камер, от-
сутствующих лишь в последних задних сегментах. Каждая камера имеет
два отверстия — остии и две крыловидных мышцы, а также клапаны, обес-
печивающие движение крови от заднего тела к переднему. Около головы
от сердца отходит аорта, которая распадается на три сосуда—средний
идет в голову, а два боковых впадают в подкишечный продольный сосуд.
Кровь в сердце течет от заднего конца тела к переднему, по ветвям аорты
поступает в голову и в подкишечный сосуд, по которому движется обратно
к заднему концу тела. Поднимаясь в околосердечную полость, кровь через
остии вновь поступает в сердце.
402
Дыхательная система. Наружных отверстий трахейной системы шесть
пар, расположенных на плейритах шести больших сегментов тела. Сразу
за стигмой расположена камера, покрытая волосками и щетинками, пре-
пятствующими проникновению посторонних предметов. От камеры отходит
пучок ветвящихся трахей, выстланных хитином и оплетающих все органы.
Половая система. Костянки раздельнополы. Имеется один семенник
у самцов; у самок — один яичник и система придаточных желез (рис. 352).
У самца половое отверстие находится на вершине маленького коническо-
го выроста (penis), расположенного на последнем сегменте тела. У самки
функцию яйцеклада исполняет пара укороченных ножек 17-го сегмента.
Рис. 352. Lithobius. / — самец, вскрытый со спины; П — женская половая система:
1 — усик (обрезан), 2 — передний край головы, а—надглоючный узел, 4 — ядоносная ногочелюсть,
5 — пищевод, 6—слюнная железа, 7 — брюшная нервная цепочка, 8— основание ножки, 9—сред-
няя кишка, 10—мальпигиевы трубки, 11 — большая придаточная половая железа, 12— семенник,
13— малая придаточная половая железа, 14 — семяпроток, 15—спинной щиток, 16—наружные
половые придатки, 17 — яичник, 18—яйцеклетка, 19 и 20 — придаточные железы, 21—мальпигиева
трубочка, 22—граница яичника и яйцевода, 23—яйцевод, 24—вздутие мальпигиевой трубочки,
25 — расширение яйцевода, 26— семеприемник, 27 — ветви яйцевода, охватывающие прямую киш-
ку, 28 — протоки придаточных половых желез, 29—резервуар придаточной железы, 30— кроющий
щиток, 31 — прямая кишка, 32—части последнего сегмента тела, 33—задняя ножка, 34 — щиток,
отделяющий половой сегмент от анального, 35 — анальное отверстие, 36—наружные половые при-
датки, 37—спинка анального сегмента
Развитие. Из яиц костянки вылупляются личинки, сходные по строе-
нию со взрослым животным, но маленького размера и с меньшим количе-
ством сегментов и ножек. Первый период постэмбрионального развития кос-
тянки можно обозначить как анаморфоз, т е. от одного возраста личинки к
другому, разделенных линьками; в процессе пяти возрастов происходит
постепенное увеличение сегментов и ножек. Второй период постэмбриональ-
ного развития костянки может быть обозначен, как эпиморфоз, во время
которого происходят изменения строения в деталях: изменяется число чле-
14* 403
ников наружных органов, появляются щетинки, шпоры, развиваются
половые признаки.
Систематический обзор подкласса. По особенностям в постэмбриональ-
ном развитии губоногие делятся на 2 отряда: 1) эпиморфные губоногие
(Epimorpha) и 2) анаморфные губоногие (Anamorpha). К первым относятся
сколопендры (Scolopendromospha) с 25—27 сегментами туловища и гзофи-
лообразные (Geophilomorpha), имеющие длинное туловище со множеством
однообразно построенных члеников. Эпиморфные губоногие покидают яйцо,
имея полное число сегментов и ножек. Анаморфные губоногие, к которым
относятся наши костянки и мухоловки южной части СССР, покидают яйцо
с сокращенным числом ножек, с семью их парами. В первый период постэм-
брионального развития анаморфные губоногие последовательно с линьками
увеличивают число сегментов и ножек, т. е. проходят анаморфоз; в следую-
щий же период органы изменяются лишь в форме и величине, как это имеет
место в течение всего периода развития эпиморфных губоногих.
Филогения. Четыре отряда губоногих, а именно: 1) Geophilomorpha
(представитель — род Geophilus), 2) Scolopendromorpha (представитель
род Scolopendra), 3) Lithobiomorpha (представитель — род Lithobius) и
4) Scutigeromorpha (представитель — род Scutigera) дают весьма поучитель-
ный пример последовательных этапов развития целого ряда органов. Такая
последовательность эволюции органов позволяет уверенно говорить о
филогении губоногих. Филогенетический ряд представлен перечисленными
отрядами в том именно порядке, в каком они приведены выше, а именно
геофилообразные и сколопендры представляют низшие отряды в подклассе
губоногих; мухоловки (Scutigeromorpha) представляют высший отряд, осу-
ществляя высшую ступень в развитии органов.
В пределах филогенетического ряда губоногих наблюдается совершен-
ствование ходильных ножек и постепенный переход от змеевидного пол-
зания к быстрому бегу. Соответственно этому постепенно исчезает локо-
моторная функция туловища.
У геофилообразных и сколопендр незначительные размеры двух ос-
новных отделов ножки, тазика и вертлуга и их состав из мелких склеритов
указывают на раннюю стадию в развитии двух данных отделов за счет скле-
ритов плейры; у костянок эти два отдела ножек, особенно тазик, мощнее,
состав же их становится проще, и в соответствии с этим бег становится
быстрее, змеевидное изгибание туловища, т. е. участие туловища и его
мускулатуры в передвижении, слабеет; наконец, у мухоловок ножка
приобретает исключительную длину, тазик достигает огромных размеров,
бег становится стремительным, а локомоторная роль туловища сходит на
нет — утрачивается змеевидное изгибание тела.
Параллельно развитию локомоторной роли конечностей туловище
укорачивается, число его сегментов сокращается. Относительно всего
короче туловище у костянок и мухоловок, вместе с тем и число сегментов
туловища у них минимальное, лишь 19. Дальнейшее параллельное изме-
нение туловища заключается в гетерономизации его сегментов: у геофило-
образных сегментация туловища гомономная; у сколопендр сегментация то
близкая к гомономной, то отчетливо гетерономная; гетерономизация сег-
ментов прогрессирует от сколопендр к костянковым и далее к мухоловкам;
у губоногих гетерономизация захватывает в первую очередь тыльную
часть сегмента и выражена в том, что спинной щит каждого второго сегмента,
укорачиваясь, уступает место разрастающемуся спинному щиту впереди
лежащего сегмента; в связи с этим у мухоловок спинной щит каждого вто-
рого сегмента превращается в едва заметный рудимент, а щит предшествующе-
го прикрывает оба сегмента — простые сегменты превращаются в двойные.
Параллельно исчезновению спинных щитков сокращается и число стигм.
У геофилообразных стигмы имеются почти на всех сегментах, у костянок
их число сокращается до шести пар, у мухоловок в связи с исчезновением
тергитов шесть непарных стигм располагаются по средней части тела.
404
ПОДКЛАСС II. ДВУПАРНОНОГИЕ (DIPLOPODA)
ХАРАКТЕРИСТИКА
Двупарноногие (рис. 353) — многоножки, питающиеся за редким
исключением растительной пищей, главным образом продуктами гниения.
Формы малоподвижные. Усики развиты у двупарноногих слабо: они коротки,
состоят из небольшого числа члеников. Имеется пара сложных глаз, фасетки
Рис. 353. Кивсяк Tachypodojulus
которых обычно расположены рыхло. В связи с приспособлением к однооб-
разной растительной пище значительно изменился челюстной аппарат:
совершенно выпала первая пара нижних челюстей. Верхние челюсти (един-
ственная активная, измельчающая пищу пара челюстей) отличаются исклю-
чительно сложностью своего строения: это массивный орган, состоящий из
отделов. Нижняя губа (гнатохиларий), соответствующая второй паре ниж-
них челюстей (рис. 354ш), представляет широкую и длинную пластинку
сложного строения, образующую всю нижнюю стенку головы.
Рис. 354. Кивсяк. I —четвертый сегмент туловища спереди; II сегмент
снизу /// — передний конец тела (голова и грудь) снизу;
/ —диафрагма, к которой прикрепляется продольная мускулатура, 2— трахейный ме-
шок, 3 и 4 — стерпиты двойного сегмента, 5 — нижняя губа (гнатохиларий)
Туловище не подразделено отчетливо на отделы. По строению нижней
стернальной части сегментов можно различить передний отдел из четырех
сегментов, который называют грудью (thorax), и задний, состоящий из
неопределенного числа сегментов, называемый брюшком (abdomen). Каждый
из четырех сегментов груди соответствует одному метамеру: первый, второй
и четвертый сегменты несут по паре ножек (см. рис. 354), третий — их
лишен; сегменты же брюшка имеют по две пары ножек. Каждый сегмент
брюшка имеет два расположенных друг за другом «стернита» (нижние щиты),
с которыми связаны две пары ножек и две пары стигм с трахейными мешка-
ми (рис. 354ц ); от последних отходят пучки очень длинных равномерно
тонких трахейных трубочек. Из сравнения сегмента двупарноногого и гу-
боногого, что подчеркивается нахождением ножек и стигм у двупарноногих
на «стернитах», видно, что эти последние представляют сросшиеся по сред-
ней линии плейральные (боковые) участки двух соседних сегментов.
405
ПОДКЛАСС III. СКОЛОПЕНДРЕЛЛЫ (SYMPHYLA)
ХАРАКТЕРИСТИКА
Сколопендреллы (рис. 355) — подвижные мелкие беловатые, лишенные
органов зрения многоножки. Они встречаются как на поверхности почвы,
под камнями и листвой, так и в ее поверхностных слоях. Ведут подвижный
хищный образ жизни. По своим морфологическим особенностям сколопен-
дреллы являются высшим подклассом многоножек; сколопендреллы служат
звеном между нижестоящими многоножками — губоногими и насекомыми.
насекомыми сколопендрелл связывает строение челюстного аппарата
Рис. 355. Склопен-
дрелла (Scutigerel-
1а) с брюшной сто-
роны.
/ — выпячиваемый ме-
шочек, 2—грифелек
(рис. 356); наличие на голове за усиками
специального органа чувств, связанного с
процеребральной частью мозга (орган Лейди-
га многоножек, постантеннальный орган пер-
вичнобескрылых насекомых); распространение
на сегментах туловища рудиментов ножек
выпячиваемых мешочков и грифельков (styli);
форма тергитов туловища, близко напомина-
ющая тергиты личинок крылатых насекомых,
т. е. тергиты с зачатками крыльев. С низши-
ми губоногими сколопендрелл связывает н^
доразвитие базального отдела ножки (тазика),
состав спинок из двух друг за другом распо-
ложенных отделов. С двупарноногими, в
Рис. 356. Челюстной
аппарат Scolopendrella:
1 — верхняя челюсть, 2 — ни-
жняя челюсть, 3—нижняя
губа
частности, сколопендрелл сближает строение нижней губы и расположе-
ние полового отверстия на передних сегментах туловища.
Те признаки, которые появляются и развиваются у губоногих, полу-
чают дальнейшее развитие у сколопендрелл. К таким признакам относится
сокращение числа сегментов туловища, о чем свидетельствует уменьшение
у сколопендрелл числа пар развитых ножек до 12 (а в результате редукции
передней пары ножек, которая может превращаться в щетинки,— до 11
пар) и появление за каждой парой развитых ножек пары рудиментов в
виде двух выпячиваемых мешочков.
Такая пара мешочков представляет собой последний остаток сегмен-
та, подавлению которого впереди лежащим сегментом положено начало
еще у низших губоногих.
406
ПОДКЛАСС IV. ПАУРОПОДОВЫЕ (PAUROPODA)
ХАРАКТЕРИСТИКА
Весьма мелкие многоножки подобно двупарноногим, обладающие
лишь двумя парами челюстей, а именно верхними челюстями и нижней гу-
бой, напоминающей гнатохиларий. Усики в виде трехконечного жгута.
Туловище, включая анальный сегмент, 12-сегментное; 1-й сегмент с руди-
ментами ножек; каждый из следующих 9 сегментов несет пару развитых
ножек, последние два их лишены. Число тергитов меньше, чем сегментов,
а именно: 6 или 10. Половое отверстие — между 2-й парой ножек. Развитие
с анаморфозом: известны личинки, имеющие 3, 5, 6, и 8 пар ножек. Пауро-
подовые встречаются в пнях и, по-видимому, питаются жидкой пищей.
КЛАСС 2. НАСЕКОМЫЕ (INSECTA)
ХАРАКТЕРИСТИКА
Из всех классов животного и растительного мира класс насекомых
самый большой по количеству входящих в него видов. Число видов насе-
комых далеко превосходит число видов остальных классов животного мира,
вместе взятых, но точно определить количество этих видов весьма трудно.
По одним данным, оно приближается к миллиону, а по другим — значи-
тельно больше.
Насекомые подобно другим трахейнодышащим обитатели суши, и
только небольшое количество их перешло с суши в водную среду, а именно
в пресные водоемы.
Плодовитость многих насекомых определяется сотнями, а иногда и
тысячами яиц, откладываемых одной самкой. К тому же многие насекомые
Рис. 357 Жук-плавунец (Macrodytes marginalis). / вид со спинной стороны (спин-
ки брюшка частично вырезаны для показания сердца и крыловидных мышц); //
вид с брюшной стороны:
1 — усик, 2—переднеспинка, J—элитра, 4 — щиток, 5 — заднсспинка, 6— стигма, 7—-сердце,
8—крыловидная мышца, 9 — челюстное щупальце, 10 — нижнегубное щупальце. 11 — горловой щиток,
/2 — тазик, 13— вертлуг, 14 — бедро, 15 — голень, 16 — лапка, 17 — тазик задней ножки
407
дают по нескольку поколений в год. Если эти качества касаются насекомых,
кормовой базой которых оказываются растения, возделываемые человеком,
или предметы, нужные человеку, то становится понятным, сколь опасны
они для людей и их хозяйства. Тоже самое относится к насекомым, перено-
сящим различные заболевания человека или скота.
Тело взрослого насекомого (рис. 357) обычно отчетливо делится на три
отдела, голову, грудь и брюшко. Голова ясной перетяжкой отделена от ту-
ловища. Головная капсула несет пару сложных глаз, три простых глазка
и пару одноветвистых антенн. Ротовое отверстие окружено тремя парами
ротовых придатков — одночленистыми верхними челюстями, или мандибу-
лами, пары сложно устроенных нижних челюстей, или максилл, и нижней
губой — второй парой нижних челюстей, сросшихся в единое образова-
ние, или лабиум.
Грудной отдел состоит из трех сегментов, каждый с одной парой одно-
ветвистых членистых ножек. У высших крылатых насекомых (Pterygota)
на 2-м и 3-м сегментах обыкновенно расположено по паре крыльев.
Брюшко обычно состоит из 10 сегментов, однако первичным для насе-
комых надо считать 12-сегментное брюшко. У некоторых насекомых коли-
чество сегментов в результате редукции может снижаться до шести. Половое
отверстие самки расположено на 8-м сегменте брюшка, а у самцов на 9-м.
Исключение составляют только некоторые бескрылые насекомые — дву-
хвостки ибессяжковые, отличающиеся от всех остальных насекомых и ря-
дом других особенностей.
Яйцеклад образован конечностями 8-го и 9-го сегментов, а параметры
половые придатки самцов — конечностями 9-го.
КЛА ССИФИКАЦИЯ
КЛАСС НАСЕКОМЫЕ (Insecta)
Подкласс I. Первичнобескрылые (Apterygota)
Отряд 1. Ногохвостки (Collembola)
Отряд 2. Бессяжковые (Protura)
Отряд 3. Двухвостки (Diplura)
Отряд 4. Щетинкохвостые (Thysanura)
Подкласс II. Крылатые (Pterygota)
Отряд 1. Поденки (Ephemeroptera)
Отряд 2. Стрекозы (Odonata)
Отряд 3. Таракановые (Blattodea)
Отряд 4. Богомолы (Mantodea)
Отряд 5. Прямокрылые (Orthoptera)
Отряд 6. Термиты (Isoptera)
Отряд 7 Привидениевые (Phasmodea)
Отряд 8. Веснянки (Plecoptera)
Отряд 9. Уховертки (Dermatoptera)
Отряд 10. Членистохоботные (Rhinchota)
Отряд 11. Сеноеды (Copeognatha)
Отряд 12. Пухоеды (Mallophaga)
Отряд 13. Вши (Anoplura)
Отряд 14. Пузыреногие (Thysanoptera)
Отряд 15. Жуки (Coleoptera)
Отряд 16. Сетчатокрылые (Neuroptera)
Отряд 17 Перепончатокрылые (Hymenopfera)
Отряд 18. Скорпионовые мухи (Mecoptera)
Отряд 19. Двукрылые (Diptera)
Отряд 20. Блохи (Aphaniptera)
Отряд 21. Ручейники (Trichoptera)
Отряд 22. Чешуекрылые (Lepidoptera)
Представитель класса насекомых — черный таракан
(Blatta orientalis)
Черный таракан — представитель отряда таракановых (Blattodea).
Отряд таракановых относится к древнейшим отрядам подкласса крылатых
насекомых (Pterygota); типичные представители данного отряда встречаются
408
в отложениях каменноугольного периода. Они существовали, таким обра-
зом, уже в ту эпоху, которая дала нам древнейших ископаемых крылатых
насекомых.
Родиной черного таракана (Blatta orientals) считается тропическая
Азия. Оттуда с товарами по торговым путям, как полагают, он проник в
средиземноморские страны, а к концу XVI в. в Западную Европу (Англию,
Голландию). В настоящее время он распространен повсеместно в Европе,
будучи тесно связан с жильем человека.
Внешнее строение. По расчленениютела на отделы черный
таракан—типичное насекомое. Голова у него шейным перехватом ясно
обособлена от туловища. Грудной отдел туловища слагается из трех сег-
ментов, именуемых передне-, средне-, и заднегрудью (pro-, meso- и metat-
horax), которые несут по паре ножек. Средне- и заднегрудь, кроме того,
несут у взрослого самца по паре хорошо развитых крыльев (рис. 358), у
Ю
Рис. 358. Самец и самка черного таракана (Blatta orient al is), сверху / —
самец; II — самка:
1 — усик, 2 — челюстное щупальце, 3 — переднегрудь, 4—надкрылье, 5 — бедро, 6 —
голень, 7 — лапка, 8 —десятый сегмент брюшка, 9 — церки, 10 — голова, 11 — средне-
грудь, /2 — заднегрудь
взрослой же самки на их месте лишь рудименты крыльев. Задний отдел ту-
ловища, десятичленистое брюшко, несет лишь одну пару развитых конеч-
ностей, церки (cerci), расположенных на заднем конце тела по сторонам
анального отверстия.
Голова таракана прячется под передней частью переднегруди:
из-под переднеспинки выступает лишь верхняя часть ее. На нижнем конце
головы, вокруг глоточного отверстия, сосредоточены ротовые части. На
боковых поверхностях расположены крупные сложные глаза. На передней
поверхности (рис. 359) находится пара длинных щетинковидных усиков
(antennae), состоящих из большого числа мелких члеников. У самца усики
несколько длиннее тела, у самки короче. Усики служат органом осязания
и обоняния. Ротовые части представлены верхней губой, подглоточником и
тремя парами челюстей. Полукруглая верхняя губа ограничивает ротовое
отверстие спереди. Непосредственно позади глоточного отверстия свисает
языкообразный вырост — подглоточник, за которым расположена нижняя
губа (рис. 360), состоящая из непарного основания и из парной дистальной
части с трехчленистыми щупальцами (рис. 361). В складке между подгло-
точником и нижней губой расположено отверстие парной слюнной железы
(рис. 367). С помощью двух пар лопастей на конце нижней губы принимается
жидкая пища. Нижняя губа представляет собой вторую пару нижних че-
люстей. По сторонам глоточное отверстие и расположенный за ним подгло-
409
1
Рис. 360. Вид головы черного тара-
кана сзади:
Рис. 359. Вид головы черного тара-
кана спереди:
1— темя, 2—сложный глаз. — усик,
7 — лоб, 5—наличник, 6—верхняя губа,
7 - верхняя челюсть, 8 — челюстное щу-
пальце. -9—ннжнегхбнос щупальце,
10— простой // — щека
/ — темя, 2 — затылочное отверстие, — ви-
сок, 4 — позадищечная область, 5—подве-
сок, 6—стволик, 7— челюстное щупальце,
8 и 9 — внутренняя и наружная лопасть ниж-
ней челюсти, 10 — ппжнегубное щупальце,
11 и 12— язычок и придаточный язычок, 13—
подподбородок, 14— подбородок
Рис. 361. Челюстной аппарат черного таракана.
/ — верхняя челюсть; II верхняя губа, Ill
нижняя челюсть; IV нижняя губа:
/ — подвесок, 2 — стволик, 3 — челюстное щупальце, / —внут-
ренняя лопасть, 5—наружная лопасть, 6—подподбородок,
7 подбородок, 8—предподбородок (лигула). 9 — нижнегуб-
ное щупальце, 10—язычок, // — придаточный язычок
410
точник ограничены парой верхних челюстей, следующих непосредственно за
верхней губой и первой парой нижних челюстей. Верхние челюсти пред-
ставляют пару зазубренных на медиальном крае нерасчлененных пласти-
нок. Нижние челюсти обладают сложным расчленением и несут пятичленис-
тое щупальце. Последнее, как и нижнегубное щупальце, служит для ося-
зания. Вместе с верхними челюстями нижние челюсти измельчают твердую
пищу; для этого на конце нижней челюсти имеется коготкообразная внут-
ренняя лопасть, прикрываемая мягкой наружной лопастью (см. рис. 361).
Рис. 362. Поперечный разрез грудного сегмента
насекомого, схема. / — в момент подъема крыла;
II в момент опускания крыла.
1 — спинка, 2—бочок, ^—грудка, 4 — крыло. 5 — дорзо-всн-
тральная мышца, 6 — продольная мышца
Г р у д ь. Мягкое туловище черного таракана дорзо-вентрально сплюс-
нуто. То и другое помогает таракану прятаться в узкие щели. Грудь состоит
из трех хитиновых колец, или сегментов. Каждый сегмент (рис. 362) состоит
Рис. 363. Передняя
ножка таракана
склериты передне-
груди и шейки:
1 — тазик, 2 — верт-
луг, 3—бедро, 4 — го-
лень, 5 — лапка
Рис. 364. Крылья левой стороны черного
таракана:
1 — надкрылье, 2 — крыло, 3 — срсднеснинка,
4 — заднсслннка
из четырех участков: спинного (спинка), двух боковых (бочки) и нижнего
(грудка). На всех трех сегментах на границе бочка и грудки расположены
ножки. У самца на 2-м и 3-м сегментах груди на границе бочка и спинки
имеется по крылу. Таким образом, три сегмента груди (передне-, средне- и
411
заднегрудь) несут по паре ножек, а среднегрудь и заднегрудь, кроме того,
и по паре крыльев. Переднегрудь прикрыта большой спинкой, под которой
прячется и голова. Все ножки у таракана бегательные. Они слагаются из
пяти отделов (рис. 363). От основания ножки к свободному концу располо-
жены следующие отделы: тазик, или ляжка (соха), вертлуг (trochanter),
бедро (femur), голень (tibia) и лапка (tarsus). Основной отдел —тазик —
очень большой, заключает внутри себя сильную мускулатуру, приводящую
в движение остальную часть ножки, служит и для прикрытия мягкой ниж-
1
Рис. 365. Цент-
ральная нервная
система черного та-
ракана:
1— надглоточный узел,
2—подглоточный узел,
3 — 5 — грудные узлы,
6—11 — брюшные узлы
ней поверхности груди. Самые длинные отделы ножки
бедро и голень — друг с другом образуют коленное
сочленение. Лапка состоит из пяти члеников, придающих
гибкость данному отделу, которым ножка ступает на
субстрат. На конце лапка несет еще коготковый членик
с парой коготков, с помощью которых ножка цепляется
за неровности субстрата. Между коготками находится
присоска, позволяющая таракану бегать по поверхности
в любом положении.
Крылья (рис. 364) представляют складки наружного
Рис. 366. Головные узлы
центральной нервной систе-
мы черного таракана сбоку:
/ — прото церебральная часть мо-
зга, 2—дейтоцеребральная часть
мозга с усиковыми нервами,
3 — тритоцеребральная часть мо-
зга, 4 — возвратный нерв (симпа-
тическая система), 5—окологло-
точная коннектива, 6—подгло-
точный узел с челюстными нерва-
ми, 7 — эндоскелет (tentorium),
8 — глотка
покрова, сплюснутые в тон-
кую пластинку Утолщения,
так называемые жилки, или
нервы, проходящие вдоль и
поперек крыла, придают кры-
лу прочность. Передняя пара
крыльев, принадлежащая
среднегруди, более груба, ко-
жиста. Это так называемые
надкрылья. Пара крыльев зад-
негруди перепончата. Задние
крылья частично могут скла-
дываться в складки. В покое
крылья складываются на
спинную поверхность брюш-
ка, причем задние крылья
прикрываются надкрыльями.
Самка несет лишь пару руди-
ментов крыльев на средне-
груди.
Брюшко состоит из
десяти сегментов. От грудных
сегментов они отличаются
отсутствием конечностей:
лишь по бокам последнего
сегмента имеется пара коротких мелкочленистых конечностей — это церки,
на заднем конце тела играющие ту же роль, что усики на переднем. Наибо-
лее развиты сегменты брюшка, начиная со 2-го и кончая 7-м. По строению
задних сегментов брюшка взрослые самки ясно отличаются от самцов
(см. «Половая система»).
Под задним краем десятой спинки по средней линии расположено аналь-
ное отверстие.
Нервная система. Центральная нервная система черного таракана
состоит из 11 узлов, из которых два узла, надглоточный и подглоточный
расположены в голове, три— в груди и шесть— в брюшке (рис. 365). Все узлы
соединены коннективами, ясно парными в голове и груди и соприкасающи-
мися в брюшке. Развиты узлы нервной цепи неодинаково, самый крупный —
надглоточный узел, или мозг (рис. 366), снабжающий нервами органы
зрения (парные сложный и простой глаза), усики и их органы
обоняния и осязания и верхнюю губу. Также крупные подглоточный
412
и три грудных узла (рис. 365) иннервируют ротовые части, ножки, крылья и
мускулатуру головы и груди. Из шести узлов брюшка значительно больше
остальных крайний задний, снабжающий половую систему и церки и проис-
шедший от слияния ряда нервных узлов.
Органы чувств. Отожные глаза таракана типичного строения, состоя-
щие приблизительно из 1800 фасеток. Нервными окончаниями богаты лопас-
ти нижне-челюстного аппарата и их щупальцы (органы осязания и вкуса);
Рис. 367. Вскрытый черный таракан (<5). / —вид сверху; // —вид
сбоку:
1 — глотка, 2 — пищевод, 3—зоб, 4—мускулистый желудок, 5— слепые кишки,
6—средняя кишка, 7—задняя кишка, 8—мальпигиевы трубочки, 9—спинной сосуд,
10 — надглоточный узел, 11— брюшная нервная цепочка, 12 — семенник 13—непарная
придаточная железа, 14—семенной пузырек с парной придаточной железой, 15—боко-
вые трахейные стволы, 16 — слюнная железа, 17—резервуар слюнной железы, 18 — об-
щий проток, 19—стигма
нервными окончаниями богаты и усики. Роль усиков как органов осязания
и обоняния явствует из обычного их направления при движении вперед и
в стороны, а также из непрерывных колебательных движений жгута.
Пищеварительная система. Кишечник черного таракана, как обычно
у членистоногих, делится на три основных отдела: переднюю, среднюю
и заднюю кишки. От глоточного отверстия передняя кишка поднимается
вверх и, дугообразно изгибаясь, направляется назад, сохраняя это на-
правление до области брюшка. Задний конец передней кишки занимает
переднюю часть полости брюшка, большая же, задняя, часть брюшка вме-
щает в себя среднюю и заднюю кишки, которые образуют в ней по петле
(рис. 367). Передняя кишка состоит из пяти отделов, из которых лишь че-
тыре передних различимы внешне, пятый же в виде кольцевой складки
вдается в начальную часть средней кишки наподобие горлышка воронки,
вставленной в бутыль. Эта заслонка препятствует пище, поступившей в
413
полость средней кишки, вернуться обратно в переднюю кишку. Передний
отдел передней кишки — глотка (pharynx), начинающаяся впереди глоточ-
ным отверстием, втягивает и всасывает грубо измельченную челюстным
аппаратом и смоченную слюной пищу в свою полость. Она обладает сильной
мускулатурой, способной энергично расширять и сужать эту полость.
Глотка проходит через окологлоточное кольцо центральной нервной сис-
темы (см. выше) и у затылочного отверстия переходит в следующий отдел
передней кишки, в пищевод (Olsophagus), постепенно расширяющийся
в зоб (ingluvies). Задняя, мешкообразная, часть зоба помещается уже в
брюшке. В ней принятая пища задерживается и частично подвергается пере-
вариванию и всасыванию. Из зоба пища попадает в среднюю кишку не сразу
Небольшие порции пищи, поступив из зоба в следующий отдел жеватель-
ного, или мускулистого, желудка (proventriculus, рис. 368), подвергаются
Рис. 368. Продольный разрез через мускулистый желудок таракана
/ — зоб. 2 — мускулистый желудок, «V — средняя кишка, 4 — слепая кишка
там дальнейшему измельчению и фильтрации. Такое дополнительное из-
мельчение пищи способствует ее перевариванию. Жевательный желудок,
имеющий форму бокала или чаши, обращенный отверстием вперед, служит
не только измельчающим пищу аппаратом, но и фильтром, пропускающим
пищу, лишь достаточно подготовленную к поступлению в среднюю кишку
Для измельчения в передней, более широкой, части жевательного желудка
имеется кольцо из шести сильно хитинизированных продольных складок-
пластин (рис. 368).
Средняя кишка, или желудок (ventriculus), черного таракана — самый
короткий отдел пищеварительного канала, собранный к тому же в петлю
(рис. 367). Эпителий средней кишки сплошь железистый. Наряду с ролью
железы, выделяющей пищеварительный секрет, средняя кишка функцио-
нирует и как отдел, всасывающий продукты пищеварения. Кольцо слепых
кишок, впадающих в передний конец средней кишки, служит для увели-
чения секреторной и всасывающей поверхности средней кишки. Эпителий
средней кишки лишен хитиновой выстилки.
Граница средней и задней кишок наружно намечается местом впадения
трубкообразных мальпигиевых желез (см. рис. 367) — органов выделения
продуктов распада в теле таракана. Мальпигиевых трубочек у черного
таракана насчитывается около ста, они расположены в шесть пучков и
представляют собой цилиндрические выпячивания кишечника. Кзади от
места их впадения начинается задняя кишка, анальным отверстием откры-
вающаяся на заднем конце брюшка.
Кровеносная система. Кровь у таракана, как и насекомых вообще,
движется по участкам полости тела между внутренними органами. Лишь на
очень ограниченном протяжении кровь течет по сосуду, обладающему соб-
ственными стенками: это расположенный под наружными покровами в
полости тела так называемый спинной сосуд, состоящий из сердца и аорты.
Трубкообразное сердце тянется по средней линии на протяжении брюшка.
В груди его продолжением служит аорта. Сзади сердце у таракана окан-
414
чивается слепо. На своем переднем конце аорта открывается в полость тела
между надглоточным узлом и глоткой. Стенку спинного сосуда образует
мышечная ткань. В отличие от аорты трубки равномерной ширины, сердце
имеет расширения, расположенные друг за другом,— камеры. У таракана
наибольшее число таких сердечных камер, встречаемое у крылатых насе-
комых: их насчитывается тринадцать, из которых две расположены в груди,
остальные же одиннадцать камер — в брюшке. Таким образом, камеры рас-
пределены приблизительно посегментно (см. рис. 367). У зародыша тарака-
на они расположены строго сегментально. Каждая сердечная камера обла-
дает парой боковых щелевидных отверстий — остий, через которые кровь
из окружающей полости тела поступает в полость камеры. Оба края щелевид-
ного остия, вдаваясь в полость камеры, образуют клапан, мешающий обрат-
ному оттоку крови из камеры в полость тела. Последовательное сокращение
сердечных камер гонит кровь из камеры в камеру от заднего конца к перед-
нему Расширение полости камер происходит при участии крыловидных мышц.
В связи с развитием дыхательной (трахейной) системы у таракана,
как у огромного большинства насекомых, кровь сохранила функцию пере-
носа лишь питательных веществ и продуктов распада. Этим именно и объяс-
няется редукция системы кровеносных сосудов.
Дыхательная система. Дыхание черного таракана обеспечивается си-
стемой древовидно ветвящихся в теле трубок — системой трахей, опле-
тающих все внутренние органы. По своему происхождению трахейная
система является системой впячиваний наружного покрова, чем объясняется
и ее строение из тонкого слоя хитина, образующего внутреннюю выстилку
полости грудки, и тонкого же слоя гиподермы, окружающего хитин. От
основных трахей отходят ветви, последовательно ветвящиеся на все более
мелкие ветви. У таракана и огромного большинства насекомых трахеи сли-
ваются в общую систему, основная часть которой состоит из продольных
и поперечных стволов (см. рис. 367). С окружающей средой полость трахей-
ной системы таракана сообщается, как у большинства насекомых, десятью
парами стигм. Из десяти пар стигм две пары находятся на грудном отделе
по бокам, остальные же восемь пар — на сегментах брюшка.
Выделительная система. Органами выделения служат мальпигие-
вы сосуды, или трубки, расположенные в полости тела (см. рис. 367). Они
омываются кровью и, принимая в себя из крови путем осмоса продукты рас-
пада, выделяют их через заднюю кишку наружу.
Жировое тело. На внутренней поверхности стенки брюшка и
между внутренними органами (например, в петлях кишечника) можно
наблюдать скопление белого вещества. Это жировое тело — ткань, содер-
жащая большое количество запасных питательных веществ — жира, гли-
когена, белка. Жировое тело имеется у всех насекомых, и заключающиеся
в нем запасные питательные вещества используются организмом насекомого
при голодании и при созревании яиц. Жировое тело несет и выделительную
функцию, в нем накопляются продукты распада тканей тела.
Половая система. Самцы и самки Blatta orientalis во взрослом состоя-
нии легко отличимы друг от друга. Половые отличия — половой димор-
физм — в основном сводятся, во-первых, к различному развитию крыльев
(а также связанных с крыльями спинок) и, во-вторых, к различной степени
редукции восьмого и девятого сегментов брюшка. Оба сегмента у самки
редуцированы в большей степени, чем у самца, и скрыты под частями седь-
мого сегмента брюшка.
У самца нижний щиток девятого сегмента брюшка увеличен, несет
на заднем краю пару одночленистых придатков, грифельков (styli). Он слу-
жит защитным приспособлением для мужских наружных половых придат-
ков — субгенитальной пластинкой. Наружный половой аппарат самца
слагается из ряда хитиновых выростов и пластин.
Внутренняя половая система самца (рис. 369) состоит из пары семенни-
ков (testiculi), расположенных под пятой спинкой брюшка, парного семя-
415
протока (vas deferens), двух семенных пузырьков (vesiculae seminales),
непарного семяизвергательного канала (dictus ejaculatorius), который от-
крывается наружу при основании penis, и двух придаточных желез, одна
из которых парная. Каждый семенник состоит из 30—40 пузыревидных до-
лек, собранных в общую гроздь.
Рис. 369. Половые органы самца черного таракана. 7 —сбоку; /7 —снизу:
/, 2 и 3 — придаточные железы, 4—семенник, 5 — семяпроток, 6 — семяизвергательный канал,
7 — наружный половой (копулятивный) аппарат
Половая пазуха самки расположена под седьмым-десятым спинными
щитками брюшка. В половой пазухе после спаривания образуется яйце-
вой кокон (ootheca, рис. 372), в котором протекает зародышевое развитие
Рис. 370. Наружные половые органы
самки черного таракана:
1—рудименты яйцеклада, 2—наружное по-
ловое отверстие, 3 — церк, 4 — субгениталь-
ный щиток
таракана. Три пары коротких выростов
сообща образуют усеченный конус — ру-
диментарный яйцеклад (рис. 370). Меж-
ду основаниями рудиментов яйцеклада
находится отверстие придаточной желе-
зы, дающей секрет для построения
кокона. Отверстие непарного яйцевода,
или влагалища (vagina), расположено в
слепом конце половой пазухи под вы-
ростами яйцеклада.
Внутренние половые органы самки
таракана (см. рис. 371) состоят из пары
яичников, пары яйцеводов, влагалища
(непарного яйцевода), семеприемника и
придаточных половых желез, дающих
секрет для изготовления оболочки коко-
на. Каждый яичник (ovarium) состоит
из восьми яйцевых трубочек, которые
являются местом постепенного роста и
формирования яйцеклеток. Пара яйце-
водов (oviductus) у взрослой самки
впадает в короткий непарный яйцевод.
Внутри кокона черного таракана
поперек его продольной оси в два ряда
расположены яйца. Содержимое кокона
представляет результат одной кладки:
при кладке каждая яйцевая трубочка
416
дает одно яйцо и так как яйцевых трубочек в каждом из двух яичников
насчитывается восемь, то в коконе черного таракана имеется всегда шест-
надцать яиц.
Экология и развитие таракана. Экология и развитие таракана будут
рассмотрены нами на примере другого вида — прусака (Blattella germani-
Рис. 371 Внутренний половой аппарат самки черного
таракана:
/—яйцевая трубочка яичника, 2 — зрелое яйцо, 3— яйцевод,
4 — семеприемник, 5 — придаточная железа
са, рис. 372). Этот вид семейства таракановых (Blatticlae) — космополит.
Он встречается в СССР значительно чаще, чем Blatta orientalis. Прусак
встречается в СССР исключительно как домашнее насекомое. Предпола-
гают, что его родина — юго-вос-
ток Азии. По Европе он распро-
странился, по-видимому, значи-
тельно раньше черного таракана
в связи с легкостью его заноса и
скоростью размножения. Его
отличительными от черного та-
ракана признаками служат:
меньшая длина (от 1,1 до 1,5 см)
имаго, буро-желтая окраска,
наличие развитых крыльев каку
самца, так и у самки. Нормаль-
ными условиями для прусака
являются температура 20°С и
некоторая влажность среды. При
температуре 4°С прусаки еще
способны к движению. Окочене-
ние от холода начинается при
+2°С, жизненный минимум
при — 5°С. Прусак проводит
день в щелях и других укрытых
Рис. 372. Blattella germanica (прусак). / —
самец сверху; //—самка с яйцевым коко-
ном снизу /// — нижняя губа
417
местах. С наступлением же сумерек покидает их, отправляясь на поиски
корма.
При беге прусак одновременно перемещает две крайние ножки одной
стороны и среднюю ножку другой, ступая при этом на все пять члеников
лапки. Положение поверхности для него безразлично: прусаки бегают
столь же легко по вертикальным поверхностям, как и по горизонтальным,
и по обращенным вниз, например по потолку. Это осуществляется благода-
ря наличию между двумя коготками на конце лапки присоски. Прыжки,
которые прусак может делать в горизонтальной плоскости, вверх и вниз,
не превышают 4—10 см. При прыжке с большой высоты прусак пользуется
и крыльями, кожистые надкрылья отводятся при этом вперед и в стороны,
сложенные же в складки перепончатые задние крылья расправляются и
производят быстрые колебательные движения. Крылья — не активный ор-
ган передвижения, они служат лишь для направления движения тела и
притормаживания.
Прусак обычно питается плодами и хлебом. Принятие пищи происхо-
дит в начале и в конце ночи, периодически, продолжается примерно около
минуты и прерывается на четверть часа и более. Имеет место и канниба-
лизм, причем поеданию подвергаются более слабые особи, особи в линьке,
коконы.
Отделенные при помощи верхних челюстей и внутренних лопастей ниж-
них челюстей частицы твердой пищи с помощью гипофаринкса препровож-
даются в переднюю кишку. Слюнной аппарат состоит из парной дольчатой
железы, дольки которой с помощью системы парного и непарного протоков
изливают свой секрет и содержимое парного резервуара на внутреннюю по-
верхность нижней губы (см. рис. 367). Секрет слюнной железы не только
облегчает глотание, но и служит пищеварительным секретом, переводя
крахмал в глюкозу
Через короткий промежуток времени после спаривания у самки разду-
вается средняя часть брюшка — местонахождение яичника. Через не-
сколько дней начинается кладка и построение кокона, на что требуется не
больше суток.
Одновременно с оставлением кокона молодой прусак переживает
линьку Последующих линек насчитывается шесть; они отделяют друг от
друга семь возрастов, из которых шесть молодых и один имагинальный. При
линьке происходит образование щели по средней спинной линии на протя-
жении груди и головы. Через эту щель постепенно выступает покрытое бес-
цветным хитином белое тело. Развитие от момента выхода из кокона и до има-
гинальной фазы длится от 4 до 6 месяцев или даже более. Длительность воз-
раста колеблется индивидуально; она различна и для разных возрастов
одной особи.
Молодые возрасты прусаков отличаются от имагинальной фазы главным
образом размерами, отсутствием крыльев и половых придатков или их раз-
витием. Кроме того, имеются отличия количественного порядка и в разви-
тии других органов, как-то: членистости усиков, церк. От первого возраста
и до имагинального все изменения протекают постепенно, от возраста к
возрасту Такое постепенное развитие при общем сходстве первого молодого
возраста с имагинальным и при отсутствии специальных личиночных (про-
визорных) органов характерно для эпиморфоза.
За 2—3 дня до линьки молодь прекращает питаться. Подыскав себе
подходящее для линьки место, прусак впадает в оцепенение на 4—6 ч,
по истечении которых образуется трещина вдоль средины спинной линии
в области груди и головы; через трещину молодь освобождается Весь
процесс линьки тянется часа четыре. Освободившееся от экзувия, т е. ста-
рого покрова, насекомое совершенно бело и сильно вздуто; вздутость
через несколько часов пропадает, хитин темнеет По прошествии 10 ч
слинявший прусак совершенно не отличается по своей окраске от нели-
нявших.
418
Второй представитель класса насекомых —
медоносная пчела (Apis mellifera)
Медоносная пчела — представитель семейства пчелиных (Apidae) в
отряде перепончатокрылых (Hymenoptera).
Медоносная пчела среди пчел, разводимых человеком, имеет наиболее
широкое применение (Европа, Северная Америка, часть Азии и Африки).
По особенностям внешнего строения и по особенностям инстинкта разли-
чают ряд разновидностей (пород) данного вида.
Перепончатокрылые (Hymenoptera), к которым относится медоносная
пчела, отличаются высокой специализацией в строении всех отделов тела:
специализированы голова, грудь, брюшко, усики, челюстной аппарат,
грудные ножки и придатки брюшка. По степени развития инстинкта род Apis
и ближайшие к нему роды, образующие группу настоящих пчел (подсем.
Apinae), относятся к наиболее высокоразвитым перепончатокрылым; они
образуют одну из наиболее сложных форм сообщества. В состав сообщества
пчел или пчелиной семьи входят три касты (фазы): матка, самцы (трутни)
п рабочие пчелы (самки с недоразвитыми половыми органами).
Внешнее строение, (рис. 373). Голова пчелы резко обособлена от туло-
вища шейной перетяжкой, допускающей свободное вращение головы вокруг
продольной оси. Рассматриваемая спереди голова рабочей пчелы (рис. 374)
представляется яйцевидной, с ротовым полюсом, направленным вниз и
несущим длинный хоботок. Верхнебоковые части головы несут большие
сложные глаза; простые же глаза в числе трех расположены наверху, на
темени у средней линии. Оба усика, сидящих на лбу непосредственно над
лицевым щитком на уровне нижней границы сложных глаз, коленчаты. Ос-
новной отдел усика представлен лишь одним весьма длинным члеником;
остальная часть усика, состоящая из 11 коротких члеников, образует коле-
но с основным отделом. По сравнению с головой рабочей пчелы голова
матки меньше и хоботок значительно укорочен. У трутня голова крупнее
головы матки и рабочей пчелы; причина этому огромные сложные глаза,
которые сходятся на темени, оттесняя
простые глаза вперед. Хоботок трут-
ня укорочен. Пчела питается некта-
ром и пыльцой, и поэтому измельче-
ние твердой пищи излишне. Верхние
челюсти получают новое назначение:
у рабочей пчелы это органы, придаю-
щие форму воску при изготовлении
сот у трутня и матки они служат для
выхода из запечатанной ячейки.
Нижнечелюстной аппарат пчелы,
как и перепончатокрылых вообще,
представляет анатомически и функци- /2
онально единое целое. Это аппарат
для принятия жидкой пищи, для че-
го дистальные части нижней челюсти
н нижней губы (рис. 375) складыва-
ются вместе, образуя трубочку
Г р у д ь. Две основные особен-
ности служат отличием грудного отде-
Рис, 373. Apis mellifera:
/—усик, 2—сложный глаз, 3—глазок,
4 — верхняя челюсть, 5 — хоботок, 6 — пер-
вый членик лапки, 7 — переднеспинка,
8 — среднеспинка, 9 — переднее крыло,
10 — заднее крыло, 11 — тергит первого
брюшного сегмента, /2 — седьмой сегмент
брюшка
ла пчелы, как и высших перепончато-
крылых вообще: 1) грудной отдел
вместе с первым сегментом брюшка
обособляется от остальной части брюш-
ка резко выраженной перетяжкой,
благодаря чему, а также обособлению
419
Рис. 374. Apis mellifera. / — матка; // —ее голова, /// —
рабочая пчела; /V ее голова; V — трутень; VI — его голова
Рис. 375. Голова пчелы. / — голова в продольном разрезе (хи-
тиновые части); II хоботок в расправленном виде:
1 _ глотка, 2 — подвесок, 3— уздечка. 4 — подподбородок, 5 — подбородок,
6 — лопасть (наружная) нижнем челюсти, 7 — нижнегубное щупальце,
£— язычок (медиальная лопасть нижней губы), 9— верхняя челюсть, 10 — на-
ружная лопасть нижней губы, 11 — нижнегубное щупальце
от головы, грудной отдел получает почти шаровидную форму; 2) среднегрудь
значительно больше передне- и заднегруди, что связано с преобладанием
передних крыльев над задними и расположением летной мускулатуры в
среднегруди. Как задние, так и передние крылья перепончаты. Задние кры-
лья значительно меньше передних; на переднем краю задние крылья несут
ряд крючочков, которыми задние крылья цепляются за утолщение на
заднем краю передних крыльев и благодаря этому буксируются послед-
ними. Пчелы относятся к лучшим летунам среди насекомых.
Ножки пчелы не только локомоторные органы: они служат и для сбора
пыльцы с поверхности всего тела. Для выполнения этой роли на голени и
первом членике лапки ножек имеются аппараты из длинных или коротких
щетинок. Первый членик лапок отчасти в связи с его новой ролью расширен
Рис. 376. Apis melli-
fera. / —задняя нож-
ка снаружи; II — то
же с внутренней сто-
роны
1 — корзиночка, 2 — обно-
жка (комочек пыльцы),
•? — первый членик лапки
со щеточкой
Рис. 377. Жалоносный аппарат
пчелы в расправленном виде:
1 — основание стилета( нижняя створ-
ка), 2 — влагалище жала, 3 — уголь-
ник, 4 — квадратный щигок, 5 — про-
токи ядовитых желез
и удлинен. Передние ножки собирают пыльцу с головы и передней части
туловища, средние — со средней части туловища, а задние — с брюшка.
В конечном итоге пыльца щеточкой, расположенной на внутренней поверх-
ности первого членика задних лапок, собирается со всей поверхности тела,
поступает в углубление на внешней поверхности голени задних ножек в
корзиночку (рис. 376). Хорошо развитый собирательный аппарат имеется
только у рабочей пчелы; у матки он недоразвит.
Для сбора пыльцы тело пчелы покрыто густым покровом щетинок, ча-
стью ветвистых: пыльца, попавшая на покров щетинок при соприкосновении
с пыльниками, задерживается на теле пчелы.
Брюшко. Особенности в строении брюшка пчелы тесно связаны с
ролью жала на конце брюшка пчелы. Истинный первый сегмент брюшка
соединен тесно с грудным отделом, замыкая его сзади (см. рис. 373). Брюшко,
обособленное от груди перетяжкой, образовано шестью сегментами, которые
соответствуют второму — седьмому сегментам брюшка первоначального
состава. Первый из данных шести сегментов образует передний тупой полюс
яйцевидного брюшка. Он суживается, образуя стебелек для сочленения с
задним концом грудного отдела. В пазухе между стернитом и тергитом
последнего наружного сегмента располагается жалоносный аппарат
(рис. 377). Этот последний представлен редуцированными сегментами
брюшка (8—10), само же жало с непосредственно прилегающими к нему
421
что стилеты скользят по
Рис. 378. Нервная система
Apis mellifera:
/—надглоточный узел, 2—узел
переднегруди, 3— узел средне- и
заднегруди первого и второго
сегментов брюшка, 4— узел тре-
тьего сегмента брюшка
частями не что иное, как яйцеклад, изменивший свою первоначальную функ-
цию на роль органа защиты и нападения. Наиболее существенной частью
всего аппарата, т. е. жала, являются три выроста, верхний из которых пред-
ставляет длинную суженную кзади пластинку, желобчато изогнутую,
передняя часть этого желобка луковицеобразно вздута. В этот вздутый
отдел выделяют секрет две ядовитые железы (см. рис. 380), из которых боль-
шая выделяет кислый секрет (муравьиную кислоту), а меньшая — щелоч-
ной. Пластинка, обращенная желобком вниз, служит влагалищем и веду-
щим аппаратом для двух стилетов жала, с которыми желобок соединен так,
нему, причем их зазубренный конец, вонзаясь в
тело, выступает за конец желобка.
То искусство, с которым пчела действует
жалом, обусловлено необычайной подвижностью
брюшка, его способностью изгибаться в любом
направлении, чему способствует, во-первых,
резко выраженная перетяжка в основании брюш-
ка, причем второй сегмент брюшка образует
стебелек и, во-вторых, способ соединения друг
с другом сегментов брюшка, дающий им возмож-
ность свободно двигаться относительно друг
друга.
Каждый впереди лежащий сегмент накры-
вает переднюю половину последующего, соеди-
няясь с ним хорошо развитой перепонкой. Эта
перепонка и допускает изгибание брюшка в
любом направлении. Накрытая часть нижней
поверхности сегмента брюшка несет пару круг-
лых участков с тонким хитином (зеркальце). На
зеркальце железистая гиподерма выделяет пла-
стинку воска, которая извлекается пчелой из
кармана между двумя следующими друг за дру-
гом стернитами с помощью щеточки задних но-
жек. Выделяется воск стернитами четырех пос-
ледних наружных сегментов брюшка. К выделе-
нию воска способна только рабочая пчела.
Нервная система. У зародыша пчелы наблю-
дается максимальное число узлов центральной
нервной системы, встречаемое у крылатых
насекомых вообще, а именно: 2 головных, 2 груд-
ных и 11 брюшных узлов. В течение личиночного периода развития число
брюшных узлов сокращается, к концу личиночного развития число голов-
ных и грудных узлов остается таким же, число же брюшных сокращается
до 8. В течение куколочной фазы происходит дальнейшее сокращение
числа узлов, притом как брюшных, так и грудных (см. рис. 378). Органы
чувств см. стр. 428.
Пищеварительная система. Разделение кишечного канала пчелы на от-
делы в общем соответствует разделению, описанному для таракана. В связи
с кормлением рабочей пчелой детки особое значение получают передняя
кишка, связанные с ней железы и те изменения, которые претерпевает в ней
пища у рабочей пчелы. Пищеварительную роль играет секрет нижнегубной
слюнной железы, упомянутой выше. Секрет слюнной железы содержит фер-
мент инвертазу, вызывающий расщепление тростникового сахара на ви-
ноградный и плодовый сахар, легко усвояемые организмом. Под влиянием
слюны нектар, содержащий наряду с виноградным и плодовым сахаром и
тростниковый, превращается в мед, что происходит в зобу, который по-
мещается в передней части брюшка. Тростникового сахара в меду относи-
тельно меньше. Из зоба мед может отрыгиваться обратно, поступая в ячей-
ки сот. Из других ферментов слюны пчелы особенно существенна диастаза^
422
переводящая крахмал в сахар. Крахмал имеется в содержимом пыльцы —
твердой пище пчелы. Пыльца дает рабочей пчеле и белковую пищу. Мед,
которым рабочие пчелы кормят матку, трутней и личинок, не содержит
белковых веществ. Белковое вещество будущие матки получают в течение
всего периода их развития, личинки же рабочих пчел и трутней — лишь в
течение первых трех дней развития в виде молочка или кашки, секрета гло-
точных желез, причем личинка матки получает более густую массу. Как
результат такого различия в составе пищи первоначально одинаково оп-
лодотворенные яйца в одном случае дают рабочую пчелу, в другом — матку.
Рис. 379. Кровеносная, трахейная и нервная системы
пчелы, сбоку:
1—трахейные мешки, 2—центральная нервная система, 3 — аор-
та, 4 — сердце, 5— спинная диафрагма, 6— брюшная диафрагма
Дыхательная система. Как хорошие летуны пчелы обладают богатой
системой трахейных мешков (рис. 379). Подвижность пчел, связанная с
большой затратой энергии, вызывает усиленное дыхание. Энергия дыха-
ния внешне сказывается в быстрых дыхательных движениях брюшка, при-
чем брюшко при вдыхании удлиняется и расширяется.
Половая система. В пчелиной семье различают матку и рабочих пчел.
Это как бы две касты самок. Нормально к размножению способны лишь
матки, которых, как правило, в пче-
линой семье лишь одна. Рабочие пче-
лы обычно к размножению неспособ-
ны, внутренние половые органы их
сильно редуцированы: у них парные
части половой системы — яичники и
яйцеводы — представлены парой поло-
сок, лишенных развитых яйцевых
трубочек и соединяющихся сзади в
короткий непарный отдел, соответст-
вующий непарному яйцеводу. Лишь
при известных условиях у рабочих
пчел наблюдается развитие небольшо-
го числа (обычно 4—5) яйцевых тру-
бочек. Такие рабочие пчелы (трутов-
ки) способны откладывать яйца, но
эти яйца развиваются лишь в трутней,
так как остаются неоплодотворенны-
Рис. 380. Половая система пчелиной матки:
/— яичник, 2— парный яйцевод, 3— семеприсмпнк,
4 — придаточная железа семеприемпика, 5—влага-
лище, 6 — резервуар большой ядовитой железы.
7 — малая ядовитая железа, 8— жало
423
ми: трутовки не спариваются с трутнями и аппарата для оплодотворения
яиц — семеприемника — не имеют.
Каждый яичник (рис. 380) матки содержит в среднем 150 яйцевых тру-
бочек. Основание каждой трубочки имеет по одной зрелой яйцеклетке, так
что матки в любой момент готовы к кладке 300 яиц. Парный яйцевод сое-
диняет яичники с влагалищем, с начальной более узкой частью которого
связан узким каналом шарообразный семеприемник. Канал снабжен аппа-
ратом, с помощью которого матка может оплодотворять яйца, выпуская
из семеприемника несколько сперматозоидов; оплодотворенные яйца дают
самок, неоплодотворенные развиваются в трутней. Матка живет 3—4 года.
Особенной плодовитостью она отличается на первом году и в начале второ-
го года жизни: в этот период в разгар сезона она может класть до 1500—
2000 яиц ежедневно.
Развитие. Яйцо, отложенное маткой в ячейку сот, развивается в тече-
ние трех суток, после чего из яйца вылупляется червевидная ясно сегменти-
рованная безногая личинка с небольшой головой (рис. 381); голова лишена
Рис. 381. Apis mellifera. / —личинка; II — куколка трутня
глаз и усиков и вооружена функционирующими верхними челюстями и
рудиментами нижнечелюстного аппарата, который выпускает перед заку-
кливанием при плетении кокона нить.
Упрощенному внешнему строению личинки соответствует и упрощен-
ная внутренняя организация: так, кишечник у личинки — прямая трубка;
полость средней кишки замкнута сзади, и непереваренные остатки пищи
остаются в ней до наступления периода окукления.
По окончании личиночного развития ячейка с личинкой запечатывает-
ся рабочей пчелой, а личинка, устроив себе в ячейке кокон, превращается
в куколку (рис. 381).
Таким образом, пчела в своем развитии проходит фазу личинки и
куколки, т. е. ее развитие полное, или голометаморфоз.
Биология. Жизнедеятельность пчелиной семьи зависит от времен года.
Весной начинается усиленное размножение, увеличение состава семьи,
с началом лета связано размножение семей — роение, за которым следует
усиленный сбор меда и перги; с падением осенью сбора меда состав семьи
сокращается: прекращается размножение, вымирает значительная часть
рабочих пчел, удаляются из улья все трутни; к зиме сохраняется лишь
молодой состав рабочих пчел и матка, прерывающая кладку, которая, од-
нако, может возобновиться в конце зимы в условиях жизни в омшаннике.
Весеннее размножение является подготовкой к роению и усиленному
сбору меда. Сначала самка увеличивает рабочий состав семьи, откладывая
оплодотворенные яйца в более мелкие пчелиные ячейки, размер которых
в среднем равен 5,37 мм. Позже откладываются неоплодотворенные яйца
в трутневые соты, ячейки которых в среднем равны 6,91 мм. Подготовкой
к наступающему роению служит постройка на краю сота мисочкообразных
ячеек большого диаметра, которые после откладки оплодотворенных яиц
будут надстроены и превращены в маточные ячеи. В отличие от шести-
гранных пчелиных и трутневых ячеек, обладающих трехгранным дном, эти
маточные ячейки конусовидны с округленной направленной вниз вер-
шиной.
424
Роение есть уход из родного улья старой матки с летным (более старым,
вылетающим за взятком) населением улья. В старом улье остаются лишь
нелетные рабочие пчелы и молодь, не покинувшая ячеек. Таким образом,
состав населения старого улья оказывается омоложенным. Первая матка,
вышедшая из маточной ячеи, должа заместить старую, ушедшую с роем.
Однако в улье есть маточные ячейки. Часто между вышедшей маткой и
матками в ячеях происходит перекликание, называемое на языке пчеловода
пением маток. Наличие невышедших из ячей маток служит поводом к даль-
нейшему роению. Таких повторных роев может быть несколько. Иногда до
ухода роя вышедшие матки присоединяются к рою, между матками проис-
ходит бой, в результате которого живой остается одна матка.
Лишь первый вылетевший рой, рой оплодотворенной матки, обеспечен
для дальнейшего размножения. Задача этого роя заключается в отыскании
нового жилья, постройке сот, засеве маткой ячеек этих сот и сборе меда
и перги. Однако не исключено в этом случае вторичное оплодотворение
матки.
Постройка сот ведется рабочими пчелами сверху вниз в два слоя. Как
правило, ячейки получают форму шестигранной призмы и трехгранное пи-
рамидальное дно. Нижняя часть сот идет на засев маткой, медовые ячейки
занимают верхнее положение. Засев маткой ячеек ведется от центра к пери-
ферии сота.
Для обеспечения потомством роя с молодой маткой, а равно семьи,
оставшейся в старом улье, необходимо оплодотворение молодой матки. Для
этого матка делает вылет, во время которого в воздухе и происходит половой
акт, в котором принимает участие лишь один трутень.
В течение своей жизни, которая в разгар сезона длится Р/2—2 месяца,
рабочая пчела переходит от одной работы к другой. Первая по времени
работа состоит в очистке ячеек, в которые матке предстоит класть яички,
и в обогревании детки. В дальнейшем молодыми рабочими пчелами произ-
водится кормление детки, сначала взрослой, а с активизацией глоточных
желез, дающих молочко, более молодой. Во второй период ими производит-
ся приемка меда от пчел-сборщиц. Они выносят сор из улья, несут стороже-
вую службу и пр. Одновременно эти «нелетные» пчелы совершают первые
пробные полеты. Срок наступления первого вылета за взятком не строго
определенный; он может быть через 11—34 дня по выходе имаго.
Широко поставленные над рабочей пчелой опыты и наблюдения по-
казали, что пчела способна различать цвета и запахи и воспринимать об-
разы предметов. С рабочей пчелой были проведены опыты по ее дрессировке
на различные цвета, запахи и формы. Особенно интересны ее первые проб-
ные полеты, во время которых она последовательно знакомится с окружаю-
щей улей обстановкой, очевидно, запечатлевая формы предметов. Эти по-
леты удлиняются по мере изучения окрестностей.
ОБЩИЙ ОБЗОР КЛАССА
Конечности головы. Усики в ряду головных придатков насекомых за-
нимают крайнее переднее положение и обычно значительно выступают.
Форма усиков весьма разнообразна; наиболее обыкновенны нитевидные и
суживающиеся к свободному концу щетинковидные усики. Будучи сущест-
венным органом осязания и обоняния, усики редко отсутствуют
Первичный челюстной аппарат насекомых — грызущий, описанный
выше у таракана (см. рис. 409 и 410): это универсальный аппарат, прини-
мающий как твердую, так и жидкую пищу; последняя слизывается с по-
мощью нижней губы и подглоточника. Огромное большинство отрядов на-
секомых обладает грызущим челюстным аппаратом (таракановые, богомолы,
привиденьевые, прямокрылые, стрекозы, жуки и др.).
У тех насекомых, которые питаются преимущественно жидкой пищей,
относительно большее развитие и особую дифференцировку получает ниж-
425
нечелюстной аппарат, т. е. нижние челюсти (maxillae) и нижняя губа (labi-
um). Это имеет место у перепончатокрылых (медоносная пчела — стр. 420,
рис. 375). Еще более измененным может быть челюстной аппарат в тех слу-
чаях, когда он приспособлен для питания исключительно жидкой пищей
(клопы, двукрылые, чешуекрылые и некоторые другие отряды). У таких
сосущих насекомых челюстной аппарат в целом или отдельные его части
Рис. 382. Голова двукрылого, схема. / —спереди; // сзади:
/—верхняя губа. 2 — верхняя челюсть, ^ — нижнечелюстное щупальце,
4 — нижнечелюстная щетинка, 5 — нижняя губа, 6 — подушечка (labella)
нижней губы, 7 стволик нижней челюсти, 8 — усик, 9 — простые глаза
(верхние и нижние челюсти, верхняя губа, гипофаринкс) могут быть пред-
назначены для укола. Примером простого сосущего аппарата, иногда на-
носящего и укол, может служить таковой у чешуекрылых, у которых соса-
тельный хоботок образуется нижними челюстями, составляющими правую
и левую половины сосательной трубочки (табл. XIIv)- Большое разнооб-
разие в строении челюстного аппарата дают двукрылые: сосательным ап-
1 паратом двукрылых служит трубочка,
Рис. 383. Голова клопа, вскрытая
сбоку
I — глотка, 2 — верхнечелюстная щетинка,
3 — ее рычажок, 4 — нижнечелюстная щетин-
ка, 5— ее рычажок, 6— слюнной проток,
7—слюнной резервуар (насосик), 8 — верх-
няя губа. 9 — нижняя губа, 10 — усик
образуемая верхней губой (labrum) и
подглоточником (hypopharynx); эта тру-
бочка дополняется нижней губой (рис.
382), которая подводит к концу сосатель-
ной трубочки жидкость, служащую пи-
щей; вместе с тем нижняя губа выпол-
няет и роль футляра для трубочки. У
колюще-сосущего аппарата двукрылых
колющим аппаратом помимо сосательной
трубочки могут служить стилетообраз-
ные верхние челюсти и такой же формы
части нижних челюстей, (рис. 3822 и 4).
Другой случай колюще-сосущего че-
люстного аппарата дают хоботные (кло-
пы, цикады, тли и др ). Ив данном
случае нижняя губа, обладающая харак-
терной для хоботных членистостью (рис.
3839), образует футляр для колюще-
сосущего аппарата; однако сосательную
трубочку, а вместе с тем и проток для
направления слюны в ранку жертвы
образуют не верхняя губа и подглоточ-
426
3 5
6 и 2 1
Рис. 384. Поперечный раз-
рез через хоботок клона-,
гладыша.
1 — верхнечелюстная щетинка
2— нижнечелюстная щетинка.
3— пищеприемнын канал.
4—слюнной проток, 5—верхняя
губа, 6—нижняя губа
ник, а пара длинных стилетов нижних челюстей, расположенных по оси
хоботка (рис. 384), которые вместе с парой верхнечелюстных стилетов, рас-
положенных по обе стороны нижнечелюстных стилетов, образуют колю-
щий аппарат.
Как правило, в тех случаях, когда челюстной аппарат взрослого на-
секомого (имаго) специализирован для принятия жидкой пищи, у личинки
челюстной аппарат грызущего типа (перепончатокрылые, двукрылые, че-
шуекрылые) (табл. XII). У насекомых встречается и полная или почти
полная редукция челюстного аппарата; она
встречается у взрослого насекомого, не прини-
мающего пищи (поденки, некоторые чешуе-
крылые), а также у насекомого на личиночной
фазе, когда личинка — паразит; в этом случае
редукция может распространяться и на имаго.
Конечности груди. Грудные ножки насеко-
мого обладают характерным для них расчлене-
нием (см. рис. 363).
Первоначальной формой грудной ножки
насекомых следует считать бегательную ножку.
Такая ножка может специализироваться; она
может приспособляться к плаванию. Обычно в
плавательную (рис. 357) превращается только
задняя пара ножек. Ножка может быть прыга-
тельной. Ею обычно бывает также только третья
пара; у прыгательной ножки удлинены бедро и
голень (см. рис. 398). Ножка может быть хватательной (хищной). У такой
ножки бедро и голень также увеличены и способны прикладываться вплот-
ную друг к другу Между бедром и голенью ущемляется добыча. Хвата-
тельной бывает передняя ножка (рис. 397). Передняя ножка может быть и
роющей, у роющей ножки отделы укорочены, расширены и утолщены.
Конечный аппарат ножки — коготковый членик (praetarsus) — пред-
назначен для удержания тела на поверхности и остановки движения, для
чего имеется парный или непарный коготок.
Крылья. Крылья представляют собой складки наружного покрова на
границе спинок и бочков средне- и заднегруди. В тех случаях, когда крылья
могут складываться на брюшко, весьма часто из двух пар крыльев передняя
пара бывает сильнее хитинизированной, кожистой в отличие от задней
пары, которая сохраняет перепончатость. Кожистые крылья, служа защитой
задним перепончатым, получают наименование надкрыльев. Чтобы умес-
титься под более узким надкрыльем, задняя часть перепончатого крыла
ложится в продольные складки. Крылья обладают системой продольных
и поперечных утолщений, которые называются жилками (рис. 385). В жил-
ках проходят каналы, по которым располагаются нервы и трахеи. Эти кана-
лы сообщаются с полостью тела. Жилкование крыла может быть сведено
к определенной схеме, изображенной на рис. 385. Жилки ветвятся, причем
Рис. 385. Схема жилкования крыла:
1—костальная жилка, 2—субкостальная жил-
ка, 3— радиальная жилка, «/—медиальная
жилка, 5 — кубительная жилка, 6 — анальные
жилки, 7 — ячейки
Рис. 386. Конец брюшка и яйцеклад
кузнечика:
1— нижняя створка, 2 — верхнебоковая створка,
3 — медиальная створка. 4— восьмой сегмент
брюшка
427
5
Рис. 387 Мозг кузне-
чика
/ — зрительные лопасти,
2— фронтальная лапасть с
грибовидным телом, 3— про-
тоцерсбралъная доля, 4— дей
тоцеребральная доля с усико-
вым нервом, 5 — нерв парного
простого глаза, 6 — фрон-
тальный узел с отходящим
от него назад непарным сим-
патическим нервом (nervus
recurrens), 7 — окологлоточ-
ная коннектива
ветвление жилок дихотомическое. У большинства крылатых насекомых
имеются как передние, так и задние крылья. Распространены случаи ре-
дукции как передних, так и задних крыльев или полного отсутствия одной
пары при недоразвитии или отсутствии другой пары. Так, у двукрылых
(Diptera) задние крылья превращаются в рудименты. Полное отсутствие
крыльев может быть не только вторичным, но и первичным (Apterygota).
Как показали исследования, полет насекомых связан со сложным
движением крыла, которое сводится не только к опусканию и поднятию
крыла, но и к ряду иных движений. Сложность движения вызывается
способом сочленения крыла с частями сегмента и деформациями, которые
претерпевают части сегмента вследствие сокращения летательных мышц.
Сокращение летательных мышц влияет на форму сегмента, а деформация
последнего влияет на положение крыла.
Конечности брюшка. На брюшке насекомых встречаются выросты —
гомологи грудным ножкам и конечностям головы; таковы различного вида
рудименты на брюшке первичнобескрылых насеко-
мых, ложные ножки гусениц (личинок чешуекры-
лых и некоторых перепончатокрылых); сюда
относится пара придатков на заднем конце брюшка
(церки), наружный женский, а частично и мужской
половые аппараты. Наружный женский половой
аппарат — яйцеклад — представляет конечности
восьмого и девятого брюшных сегментов (рис.
386). Назначение яйцеклада — пробуравливать ход
внутрь растения, животного или в почву для от-
кладывания яиц. Во многих случаях, когда кладка
производится на поверхности какого-либо предмета
или в воду, яйцеклад утрачивается. В других слу-
чаях с исчезновением яйцеклада роль его могут
взять на себя непосредственно задние сегменты
брюшка самки, сужаясь и превращаясь в трубку;
таков яйцеклад многих мух. С другой стороны, у
высших перепончатокрылых яйцеклад превраща-
ется в жало-, сохраняя схему строения яйцеклада,
жало может служить исключительно орудием
защиты (рис. 377).
Нервная система. В строении центральной
нервной системы (см. рис. 365 и 378) у насекомых
встречаются те же видоизменения, что и у ра-
со случаями сильного ее расчленения (надглоточ-
ный, подглоточный, три грудных и восемь брюшных узлов) и ясно парного
строения, что имеет место у примитивных насекомых, встречаются
случаи крайней концентрации нервной системы; вся брюшная цепоч-
ка может быть сведена к сплошной ганглиозной массе, что особенно часто
имеет место у личинок и личинковидных взрослых особей в случае отсут-
ствия конечностей и слабого расчленения тела. В надглоточном узле (рис.
387) привлекает внимание развитие внутренней структуры протоцеребраль-
ной части мозга, в частности грибовидных тел. Отмечено, что строение гри-
бовидных тел, которые занимают место в верхней части мозга, образуя
здесь одну или две пары бугров (рис. 388) по сторонам средней линии,
стоит в тесной связи с развитием инстинкта насекомого.
Органы чувств. Органы чувств у насекомых дифференцированы и хоро-
шо развиты. Преобладают по своему значению органы осязания и обоняния.
Органы осязания наружно представлены щетинкой. Органы обоняния так-
же обладают формой типичной щетинки, которая, видоизменяясь, может
превращаться в отчлененные тонкостенные выступы и неотчлененные паль-
цевидные выступы и тонкостенные плоские участки покрова. Важнейшим
местонахождением окончаний обонятельных нервов служат усики.
428
кообразных. Наряду
1акова, например, роль усиков как органов оооняния у мух и чешуе-
крылых, которые на огромном расстоянии различают даже слабые запахи.
Лучше изучено обоняние пчел; оказалось, что их способность воспринимать
запахи близка к нашей: те запахи, которые воспринимаем мы, воспринимают
и пчелы, те запахи, которые смешиваем мы, смешиваются и пчелами; орга-
ны обоняния сосредоточены также преимущественно на усиках. Вкусы
Рис. 388. Мозг рабочей пчелы. Справа — внешнее строе-
ние; слева — фронтальный разрез, внутреннее строение:
1 — грибовидное (стебельчатое) тело, 2—центральное тело, 3—зри-
тельная лопасть, 4 — обонятельная дейтоцеребральная доля с двумя
усиковыми нервами, 5—подглоточный узел с нервами трех челюстей
сладкий, горький, кислый и соленый насекомыми также различаются;
органы вкуса находятся на щупальцах ротовых частей, на лапках; острота
вкусового ощущения у различных органов одного и того же насекомого
может быть различной; она бывает гораздо выше, чем у человека. Сложные
глаза насекомого воспринимают движение предметов, могут в некоторых
случаях воспринимать и форму предметов; высшие перепончатокрылые
(пчелы) могут воспринимать и цвета, в том числе и такие, которые челове-
ком не воспринимаются («ультрафиолетовый»); однако цветное зрение не
столь разнообразно, как у человека: так, пчела в левой части спектра ощу-
щает желтый цвет, прочие же цвета—как оттенки желтого; правую сине-фио-
летовую часть спектра пчелы ощущают также как один цвет. Острота зре-
ния пчел гораздо ниже остроты зрения человека.
В некоторых отрядах, как-то в отряде прямокрылых (Orthoptera), к
которым относятся кузнечики, сверчки и саранчовые, распространены так
называемые тимпанальные органы^ строение тимпанальных органов, а
равно и то, что виды, обладающие ими, имеют самцов со звуковыми органами,
заставляют предполагать в тимпанальных органах органы слуховые. Тим-
панальные органы у кузнечи-
ков и сверчков распо-
ложены на голени под ко-
ленным суставом, а у саранчо-
вых и цикад на боках первого
брюшного сегмента наружно
представлены углублением,
иногда окруженным складкой
покрова и с тонкой натяну-
той перепонкой на дне; на
внутренней поверхности пе-
репонки или в ближайшем с
ней соседстве расположено
нервное окончание своеобраз-
ного строения.
Пищеварительная систе-
ма. Кишечник насекомых,
Рис. 389. Схема строения насекомого:
1—граница передней и средней кишок, 2—граница сред-
ней и задней кишок, 3— верхняя челюсть, 4 — подглоточ-
ник, 5 — нижняя губа, 6 — глотка, 7 — пищевод, 8 — сред-
няя кишка, 9—мальпигиева трубочка, 10 — тонкая кишка,
11 — прямая кишка, 12—аорта, 13—сердце, 14—слюн-
ная железа, 15 — надглоточный узел, 16—подглоточный
узел, 17 — брюшная цепочка, 18— симпатическая нервная
система, 19 — яичник, 20 — непарный яйцевод, 21 — груд-
ной отдел брюшной нервной цепочки
429
как и других членистоногих, состоит из трех отделов — передней, сред-
ней и задней кишок (рис. 389).
Центральным наиболее существенным отделом кишечника насекомых,
как и членистоногих вообще, служит средняя кишка, вырабатывающая
пищеварительный секрет; секрет содержит ферменты, необходимые для
переваривания всех составных частей пищи данного насекомого; процесс
переваривания и всасывания пищи происходит здесь же в средней
кишке. Некоторые представители насекомых могут переваривать такие ве-
щества, как кератин, воск, клетчатку, вообще трудно поддающиеся изме-
нению. В отдельных случаях в переваривании пищи принимают участие
микроорганизмы, симбионты насекомого; это наблюдается у некоторых тара-
канов, термитов, личинок пластинчатоусых жуков и др. Образуемые сре-
дней кишкой слепые придатки увеличивают поверхность кишки, выполняя
ту же функцию: они выделяют пищеварительные соки и всасывают пищу
У бескрылых (низших) насекомых средняя кишка представляет широкий,
прямой и лишенный придатков отдел, как у многоножек; передняя и задняя
кишки низших насекомых простотой своего строения напоминают соответ-
ствующие отделы многоножек; в дальнейшем появляется расчленение пе-
редней и задней кишок на ряд подотделов (рис. 367). Развитие кишечника
и его расчленение на отделы зависят от рода пищи и степени ее использо-
вания Наиболее длинной кишка бывает тогда, когда пища малопитательна
(растительноядность, копрофагия); так, кишечник короедов в 2т/4 раза пре-
восходит длину их тела. Однако, у личинок некоторых отрядов насекомых,
принимающих жидкую питательную пищу, кишка не только выпрямлена,
но и не соединена с задней кишкой, оканчиваясь слепо.
Кровеносная система. Кровеносная система насекомых, как всех чле-
нистоногих, открытая, но ограничивается спинным сосудом: утрачена сис-
тема как венозных, так и артериальных сосудов, она заменена полостью
тела, движение крови в которой поддерживается сократимыми перегород-
ками (диафрагмами) и другими приспособлениями. Спинной сосуд (см.
рис. 379, 389) представлен сердцем — отделом, несущим первоначально
расположенные посегментно пары остий — отверстий, через которые кровь
поступает в полость сердца, и аортой, лишенной остий. Сердце составляет,
как правило, заднюю, расположенную в брюшке часть спинного сосуда,
аорта же его переднюю, грудную часть. Однако у низших насекомых
(например, таракана) сердце может распространяться и на грудь (рис. 367),
тогда как у высших насекомых сердце может занимать лишь часть брюшка.
У особенно мелких насекомых сердце может и отсутствовать.
Дыхательная система. Трахейная система насекомых не только вводит
в тело насекомого воздух, но и распределяет его по всем органам тела: сво-
ими разветвлениями она пронизывает все его части. Все тело представляет
как бы легкое: дыхательная функция крови если и не исключена вполне,
все же чрезвычайно снижена. Воздух распространяется по трахейной систе-
ме путем диффузии. Параллельно со снабжением органов и тканей кислоро-
дом по трахейной системе происходит и выделение воды; чем сильнее про-
цесс дыхания тем сильнее и испарение; для предотвращения чрезмерного
испарения каждое дыхальце обладает замыкательным аппаратом, который
дает доступ воздуху в трахейную систему лишь постольку, поскольку ор-
ганизму требуется кислород: при росте активности насекомого и повышении
температуры повышается обмен веществ, повышается потребность в кисло-
роде и соответственно дольше трахейная система остается открытой. При
особенно значительной активности открытой трахейной системы оказывается
мало: появляются приспособления, проветривающие трахейную систему;
продольная и дорзо-вентральная мускулатура тела, сокращаясь и расслаб-
ляясь, ведет к сокращению и увеличению объема тела, а вместе с тем и
трахейной системы, что в свою очередь ведет к выдыханию и вдыханию
воздуха; количество выдыхаемого воздуха увеличивается в тех случаях,
когда в трахейной системе имеются участки с тонкими стенками; всего чаще
430
Рис. 390. Личинка
вислокрылки Sialis
lut ari а
такие участки принимают форму объемистых мешков, наполняющихся при
вдыхании и спадающихся при выдыхании; такие трахейные пузыри (мешки)
имеются у всех хороших летунов: чем энергичнее полет насекомого, тем
сильнее развиты пузыри трахейной системы (см. рис. 379).
У водных насекомых, дышащих атмосферным воздухом, вырабатыва-
ются дополнительные приспособления в виде углублений и выростов на
поверхности тела и покрова из волосков, которые служат резервуарами
запаса воздуха и путями, проводящими воздух. Так, у водного клопа
гладыша (Notonecta) вдоль нижней поверхности брюшка имеется слабо выра-
женное парное вдавливание, отороченное рядами щетинок, что является
приспособлением для удержания запаса воздуха.
Однако у насекомых распространены и случаи
дыхания исключительно кислородом, растворенным в
воде. При этом, если некоторые стигмы и остаются
открытыми, они служат исключительно для вывода
использованных при дыхании газов; обычно же все
каналы трахейной системы, примыкающие к стигмам,
зарастают; воспринимают же воздух из окружающей
воды соответственно измененные участки поверхности
тела. Дыхание кислородом, растворенным в воде,
весьма распространено у личинок. В этом случае
обычно появляются специальные тонкостенные выро-
сты, преимущественно на брюшке, которые получают
общее название жабр. Особым распространением
пользуются трахейные жабры, т. е. тонкостенные
выросты тела с разветвлениями трахейной системы
внутри них (рис. 390) В таких случаях трахейная
система, очевидно, лишь распределяет воздух, не
воспринимая его извне (многие жуки, поденки, стреко-
зы, веснянки, сетчатокрылые, двукрылые, ручейники).
Выделительная система. Органы выделения на-
секомых представлены мальпигиевыми сосудами —
обычно очень длинными трубочками, впадающими в
кишечник на границе средней и задней кишок (рис.
389). Число их весьма различно. Мальпигиева тру-
бочка выделяет как твердый, так и жидкий экскрет.
Дополнительным органом выделения у низших насекомых (Apterygota),
как и у многоножек, служат нижнегубные железы. Продукты распада тка-
ней частично не выносятся наружу и накопляются в жировом теле.
Половая система. Как правило, половая система самца и самки со-
стоит из парной половой железы (семенника, яичника), парного протока
(выводящего протока, яйцевода) и непарного протока (семяизвергательного
канала, влагалища), оканчивающегося наружным отверстием.
Как у самца, так и у самки имеются придаточные половые железы.
Придаточные половые железы самца (рис. 369) вырабатывают среду для
спермы и материал для построения сперматофор. Такой сперматофор
приклеивается к половому отверстию самки (прямокрылые). У самки
придаточные железы образуют секрет для смазки яйцеклада или построе-
ния кокона.
Развитие. Эмбриональное развитие насекомых в ти-
пичном случае начинается с поверхностного дробления яйца, что связано
с богатым содержанием у последнего питательного желтка. Образовавшаяся
на поверхности яйца бастодерма на некотором участке поверхности толще;
это утолщение дает зародышевую полоску и определяет брюшную сторону
зародыша. Образующаяся вдоль края зародышевой полоски кольцевая
складка, обрастая и срастаясь над зародышевой полоской, дает два слоя за-
родышевых оболочек — внутреннюю (амнион) и наружную (серозу). От
пласта клеток зародышевой полоски на стороне, обращенной к желтку,
431
Рис. 391. Яйцо и
зародышевая полос-
ка жука-плавунца:
1—усик, 2—верхняя
челюсть, 3—нижняя
челюсть, 4 — нижняя
губа, 5 — вторая груд-
ная ножка, 6 — первая
брюшная ножка, 7—
нервная цепочка, 8
анальное отверстие
обособляется первичная энтодерма — общий зачаток энтодермы и мезо-
дермы. В дальнейшем зародышевая полоска сегментируется (см. рис. 391).
Постэмбриональное развитие насекомых весьма
разнообразно в связи с разнообразием условий существования молоди насе-
комого и степени ее приспособления к ним. Можно различать два основных
типа постэмбрионального развития: неполное превращение—гемимета-
болию и полное превращение — голомепгаболию. Голометаболия характери-
зуется глубокими изменениями молоди, в результате которых молодь по
своему строению резко отлична от взрослого насекомого (имаго). Поэтому
переход от состояния молоди, т. е. от личиночной фазы к фазе имагиналь-
ной, требует коренной перестройки органов; это достигается при участии
промежуточной фазы между фазами личиночной и има-
гинальной, в течение которой молодь сохраняет покой;
эта промежуточная фаза — куколочная. В течение ку-
колочного состояния внутренние органы насекомого
либо подвергаются большей или меньшей перестройке,
либо подвергаются полному разрушению (гистолизу) и
строятся заново; в последнем случае образование органа
заново происходит за счет так называемых имагиналъ-
ных дисков, состоящих из клеток эмбрионального харак-
тера (гистобластов). Таким образом, куколочный пери-
од — период деятельного гистолиза и гистогенеза (ново-
образования тканей). Внешне куколка представляет
имагинальную фазу с характерными для нее органами,
которые, однако, остались недоразвитыми.
Голометаболично развитие в высших отрядах на-
секомых у жуков, чешуекрылых, ручейников, блох,
перепончатокрылых и двукрылых.
В случае неполного превращения — гемиметабо-
лии — столь глубокого различия в строении молодой и
взрослой фазы нет: молодь то более, то несколько менее
напоминает по строению имагинальную фазу Наконец,
в тех случаях, когда переход от молодой фазы к взрос-
лой сводится лишь к росту и сбрасыванию (в связи с
ростом) старого покрова (экзувия), говорят об аметабо-
лии, т е. отсутствии превращения; примером аметаболии
служит развитие Collembola среди Apterygota. Однако
обычно с последовательными возрастами, которые отде-
лены друг от друга линьками, т е. сбрасыванием покро-
ва предшествующего возраста, происходят и изменения
в форме органов: так, например, относительно увели-
чивается длина усиков и церок, увеличивается число
и вновь появляться некоторые органы, как, например, на-
ружные половые придатки. Таково развитие у некоторых Apterygota, как-
то: Thysanura и Diplura и у большинства гемиметаболичных Pterygota.
У некоторых Pterygota, развивающихся с гемиметаболией (стрекоз, рис. 401,
поденок, рис. 402; веснянок и части членистохоботных), развитие осложня-
ется в связи с тем, что молодь живет в иных условиях, чем имаго; так, молодь
стрекоз, поденок и веснянок развивается в водоемах и в связи с этим при-
обретает органы водного дыхания — жабры, которые представляют пример
личиночных (провизорных) органов. Условия существования отражаются
и на развитии других органов личинки, которые приспосабливаются к
обстановке, например превращение в копательный орган передних ножек
молоди цикады в связи с ее подземной жизнью.
Со сбрасыванием куколочной оболочки и переходом к имагинальной
фазе, способной к размножению, рост и линька насекомого прекращаются;
лишь среди Apterygota встречаются виды (Collembola, Thysanura), которые
с переходом к размножению продолжают линять и расти.
члеников:
432
СИСТЕМАТИЧЕСКИЙ ОБЗОР КЛАССА
Класс насекомых делится на два подкласса: 1) подкласс первичнобеск-
рылых насекомых (Apterygota), которые крыльев не имеют и предки кото-
рых крыльями, по-видимому, не обладали; 2) подкласс крылатых насекомых
(Pterygota), которые либо обладают крыльями, либо произошли от крылатых.
Тот и другой подклассы в свою очередь делятся на отряды, которых в
общем насчитывается свыше 30; из них лишь четыре относятся к подклассу
первичнобескрылых.
ПОДКЛАСС I. ПЕРВИЧНОБЕСКРЫЛЫЕ НАСЕКОМЫЕ
(APTERYGOTA)
ХАРАКТЕРИСТИКА
Подкласс объединяет четыре отряда насекомых, а именно:
1. щетинкохвостых (Thysanura, рис. 392), 2) двухвосток (Diplura),
3) бессяжковых (Protura) и 4) ногохвосток (Collembola, рис. 393), обладающих
исконной бескрылостью. Помимо признака бескрылости, данные отряды
Рис. 392. Machilis. I — вид
сбоку; // —три сегмента
брюшка снизу
обладают рядом других общих для них приз-
наков; сюда относится наличие рудиментов
ножек на брюшке, в некоторых случаях (рис.
392) имеющихся почти на всех сегментах
брюшка, эти рудименты несут выпячиваемые
мешочки, которые представляют исчезнувшую
в своем первоначальном виде дистальную
часть ножки. Рудименты брюшных ножек
могут сохранять локомоторную роль ножек
или вторично приобретать ее, превращаясь в
прыгательные и останавливающие прыжок
органы; это имеет место у ногохвосток, у ко-
торых рудименты пары ножек четвертого
брюшного сегмента превращаются в прыга-
тельную вилочку (рис. 393), а рудименты пер-
вого брюшного сегмента в орган, останавли-
вающий прыжок, — в двулопастный выпячи-
ваемый мешочек. Широко распространенным
признаком всех первичнобескрылых служит
Рис. 393. Achrutes:
1 — прыгательная вилочка
1— усик, 2 — челюстное щупаль-
це, 3—рудимент абдоминальной
ножки, 4 — хвостовая щетинка
и наличие грифельков (styli), которые также могут превращаться в ры-
чажки для прыжка. Такое распространение у первичнобескрылых руди-
ментов ножек на брюшке, выпячиваемых мешочков и грифельков тесно
связывает первичнобескрылых с многоножками и притом именно с выс-
шими многоножками —сколопендреллами. Родство многоножек и первично-
15 Курс зоологии, т. I
433
бескрылых становится тем более вероятным, что и у первичнобескрылых
насекомых намечается слияние двух метамер в общий сегмент. У махилид
наличие двух пар рудиментов ножек в виду двух пар выпячиваемых мешоч-
ков на одном брюшном сегменте указывает на то, что сегмент брюшка
состоит из двух метамер; у сколопендрелл на это указывает пара разви-
тых ножек и пара рудиментов ножек на одном и том же сегменте брюшка.
На первичность бескрылого состояния Apterygota и на их близость к много-
ножкам указывает и тесное сходство представителей двух групп в эко-
логическом отношении: как многоножки, так и первичнобескрылые живут
в почве, под корой пней, в щелях, в подстилке и проч.
Об исключительной древности первичнобескрылых свидетельствует то,
что даже столь измененные (вторично) представители этого подкласса,
как ногохвостки, существовали уже в девонский период, для которого кры-
латые насекомые неизвестны. На древность первичнобескрылых указывает
и то, что Collembola и Thysanura среди них питаются грибками, водорос-
лями и лишайниками.
ПОДКЛАСС II. КРЫЛАТЫЕ НАСЕКОМЫЕ (PTERYGOTA)
ХАРАКТЕРИСТИКА
К крылатым насекомым относят тех, которые обладают одной или двумя
парами крыльев или утратили крылья вторично. Во взрослом состоянии
локомоторных органов на брюшке они не имеют. В современной фауне в
этом подклассе насчитывают 28 отрядов. Ниже описываются отряды
наиболее интересные в биологическом и экономическом отношениях.
ОТРЯД ПОДЕНКИ (EPNEMEROPTERA)
Поденки (рис. 394) получили свое наименование благодаря чрезвычай-
ной кратковременности жизни взрослых особей, в то время как личинки
могут жить несколько лет В тесной связи с короткой жизнью имагиналь-
Рис. 394. Поденка. I личинка С1оёоп dipterum; II — линька субимаго на
имаго
ной фазы во взрослом состоянии поденки не питаются: челюстной аппарат
их атрофирован и кишечник, полость которого прервана на границе сред-
ней и задней кишок, наполнен воздухом, что имеет значение при полете.
Две пары перепончатых крыльев, которые поденка в покое держит над
грудью приложенными друг к другу, отличаются, как и у стрекоз, богатым
жилкованием.
434
Подобно стрекозам, поденки развиваются с неполным метаморфозом,
личинка развивается в воде. Происходящее в воде личиночное развитие
длится несколько лет и насчитывает много возрастов (у Cloeon — 23).
Личинка несет трахейные жабры на передних сегментах брюшка и в допол-
нение к ним хвостовые нити, которые в числе 2—3 имеются и у имаго. По-
следняя нимфальная фаза, линяя, дает первую имагинальную фазу, так
называемый субимаго, за которой следует вторая имагинальная фаза. Среди
ныне живущих крылатых насекомых лишь поденки обладают двумя имаги-
нальными возрастами. У вымерших насекомых из отряда Palaeodicty ор-
tera (каменноугольный период) их было несколько. Представитель —
Ооёоп dipterum.
ОТРЯД СТРЕКОЗЫ (ODONATA)
На имагинальной фазе стрекозы (рис. 395) — подвижные хищники,
особенно крупные неравнокрылые стрекозы (Anisoptera),— обыкновенная
стрекоза (Libellula) и коромысло (Aeschna), ловящие добычу на лету. Мел-
кие равнокрылые стрекозы (Zygoptera) хватают пищу с прибрежных расте-
Рис. 395. Aeschna. I — имаго; II — нимфа; /// — го- Рис. 396. Личинка Ag-
лова нимфы с вытянутой маской rion (Coenagrion)
ний. Обе пары крыльев стрекоз перепончаты и отличаются тем, что не
имеют анальной части, ложащейся в складки. Это связано с тем, что крылья
стрекоз не складываются на брюшке, а в покое держатся либо распростер-
тыми в стороны (неравнокрылые стрекозы), либо, прикладываясь друг
к другу, направляются вершинами назад и вверх (равнокрылые стрекозы).
Стрекозы отличаются тем, что, обладая типичной личинкой, т. е. мо-
лодой фазой с провизорными органами, не имеют куколочной фазы; таким
образом, развитие происходит с неполным метаморфозом. Важнейшие
провизорные органы личинок данного отряда функционально связаны с
окружающей средой, резко отличной от среды имагинальной фазы; личинки
ведут водный образ жизни и в связи с этим имеют трахейные жабры, у
взрослой особи отсутствующие.
Личинки (нимфы) (рис. 395 и 396) стрекоз малоподвижны, несмотря на
свои длинные и тонкие ножки, которыми они держатся за подводные пред-
15* 435
меты. Малоподвижность личинок стрекоз возмещается подвижностью их
нижней губы, образующей хватательный аппарат этого хищника (рис. 395).
Это так называемая маска в покое прикрывает остальной челюстной аппа-
рат, в действии же вытягивается, хватая добычу
ОТРЯД ТАРАКАНОВЫЕ (BLATTODEA)
ОТРЯД БОГОМОЛЫ (MANTODEA)
Тараканы и богомолы (рис. 358 и 397) относятся к древнейшим насе-
комым; первые существовали уже в каменноугольный период палеозойской
эры. Это два близких друг другу отряда, их родственная близость выра-
жается в сходном строении грызущего челюстного аппарата, увеличенной
переднеспинке, пятичленистости лапки, всеми члениками опирающейся
на субстрат в строении двух пар крыльев, в расчленении брюшка. Эта
близость двух отрядов выражается в том, что представители обоих отрядов
откладывают яйца в изготовляемый из секрета придаточных половых же-
лез кокон, который формируется в особой половой пазухе самки с помо-
щью специально приспособленных укороченных створок яйцеклада, бо-
гомолы (рис. 397) приклеивают коконы к растениям.
Рис. 397 Кладка Mantis religiosa
Постэмбриональное развитие в обоих отрядах неполное.
От таракановых богомолы отличаются тем, что тело их цилиндричес-
кое и удлиненное, передние ножки хватательные, тазики двух задних пар
ножек умеренной величины. Богомолы в отличие от всеядных тараканов —
типичные хищники; первые ведут дневной образ жизни, вторые — ночной,
в связи с чем у богомолов органы зрения (сложные и простые глаза) раз-
виты значительно лучше, чем у тараканов.
Большая часть таракановых и богомолов — насекомые тропического
пояса. Черный таракан (Blatta orientalis) как типичный представитель на-
секомых описан выше. Представитель богомолов южной полосы СССР Man-
tis religiosa изображен на рис. 397.
ОТРЯД ПРЯМОКРЫЛЫЕ (ORTHOPTERA, ИЛИ SALTATORIA)
Крылатые насекомые с прыгательной третьей парой ножек, парой тим-
панальных органов на передних ножках или первом сегменте брюшка, ор-
ганами стрекотания и грызущим челюстным аппаратом. Развиваются
с неполным превращением.
Сюда относятся три подотряда: кузнечики (Tettigoniodea), сверчки
(Gryllodea) и саранчовые (Acridodea). Саранчовые — преимущественно оби-
436
татели открытых пространств, лугов и степей. Кузнечики встречаются
чаще на пространствах, поросших кустарником или покрытых редкими
деревьями. Сверчки представлены в СССР небольшим числом видов и мас-
сами встречаются только в южной полосе Союза.
Все прямокрылые обладают прыгательной третьей парой ножек
(рис. 398). Самцы всех прямокрылых стрекочут, причем одни из них (куз-
нечики и сверчки) издают звук трением надкрыльев друг о друга, другие
(саранчовые) — трением заднего бедра о надкрылье. Со способностью изда-
вать звук связано наличие органа слуха (тимпанального органа), который
ш
Рис. 398. Азиатская перелетная саран-
ча. / взрослая фаза (А —самка, Б —
самец); // личиночная фаза; ///
кубышка
у кузнечиков и сверчков находится
на передней голени, у саранчовых же
сбоку, на переднем сегменте брюшка.
У кузнечиковых и сверчковых усики
длинные щетинковидные; у саранчо-
вых они коротки и нитевидны. Все
прямокрылые обладают грызущим
челюстным аппаратом. Саранчовые
растительноядны, хотя при случае не
отказываются от животной пищи.
Среди кузнечиковых преобладают
плотоядные, хотя многие питаются
преимущественно растительной пи-
щей.
Экология. Кузнечики и сверчки
имеют значительно развитый в длину
яйцеклад. Откладывают яйца то в
одиночку, то небольшими кучками.
Кузнечики откладывают их в почву
или в части растений. Саранчовые кла-
дут яйца в почву Для кладки самка
выкапывает с помощью яйцеклада и
вытягиваемого в длину брюшка глу-
бокую ямку, в которую одновременно
с яйцами впускает секрет придаточ-
ной железы в виде пенистой массы,
затвердевающей в кубышку (см. рис.
398). Благоприятные метеорологичес-
кие условия могут способствовать мас-
совому размножению саранчовых, что влечет за собой уничтожение ими
окружающей растительности, в том числе и сельскохозяйственных куль-
тур. Особенно вредны для сельского хозяйства саранчовые, ведущие стад-
ный образ жизни. К стадным саранчовым в СССР относятся перелетная,
или азиатская, саранча (Locusta migratoria), степная саранча, или марок-
кская кобылка (Dociostaurus maroccanus), и прус, или итальянская саран-
ча (Calliptamus italicus). Мароккская кобылка встречается в южной поло-
се СССР, перелетная саранча и прус, начиная с лесостепной полосы к югу.
Для перелетной саранчи основными гнездилищами являются плавни боль-
ших южных рек СССР, где саранча находит излюбленную свою пищу—
тростник. Перелеты взрослой саранчи служат причиной распространения
саранчи далеко за пределы ее гнездилищ. Однако в местах залетного рас-
пространения саранча является временным гостем, исчезающим через ряд
лет Инстинкт стадности у саранчовых проявляется вскоре после рожде-
ния — с момента, когда они начинают передвигаться. Особенно длитель-
ные передвижения у перелетной саранчи начинаются с третьего личиноч-
ного возраста. Передвигаясь в определенном направлении, кулига пешей
саранчи, т е. саранчи, не обладающей развитыми, функционирующими
крыльями, образует поток, состоящий сплошь из особей молодых возрастов.
Миграции вызываются потребностью в корме.
437
Скорость передвижения у личинок третьего и четвертого возрастов
перелетной саранчи достигает 7 м в минуту, у личинок пятого возраста —
12 м, становясь особенно значительной между 10 и 15 ч. К 18 ч передвиже-
ние обычно прекращается. Личиночных возрастов у перелетной саранчи,
мароккской кобылки и самцов пруса насчитывается пять. У самок пруса —
шесть. Они различимы по размерам личинки, развитию крыловых зачатков
и числу члеников усика (см. рис. 398ц).
Массовое размножение и последующее снижение численности обуслов-
ливаются не только метеорологическими факторами, но и параллельно про-
исходящим размножением хищников и паразитов, связанных с саранчой.
Особенно деятельно уничтожается взрослая саранча и ее личинки розовым
скворцом. Не избегают уничтожения и кубышки, которых выкапывают и
поедают птицы (грачи, стрепеты и др.), мелкие грызуны, как, например,
тушканчики. Существенному уничтожению подвергаются кубышки и со
стороны жуков-нарывников (Meloidea), личинки которых, а равно и мух-
жужжал (Bombyliidae), питаются яйцами саранчовых.
ОТРЯД ТЕРМИТЫ (ISOPTERA)
Термиты обладают двумя парами почти одинаковых перепончатых
крыльев, прикорневая часть которых швом отделяется от остальной части
крыла. Образуют сообщества, в которые входят нормальные самцы и сам-
ки, добавочные половые особи, рабочие и солдаты. Термиты (см. рис. 418)
подобно муравьям, пчелам и осам ведут общественную жизнь. Число особей
сообщества колеблется от нескольких сот до миллионов. Термиты питаются
всевозможной пищей, переваривая и клетчатку. Способность переваривать
клетчатку связана с присутствием в их задней кишке паразитических жгу-
тиковых. Термиты распространены главным образом в тропическом поясе.
В СССР термиты встречаются в Закавказье и в южной части Средней Азии.
В тропических странах они известны разрушением построек и унич-
тожением запасов.
ОТРЯД ЧЛЕНИСТОХОБОТНЫЕ (RHYNCHOTA)
Для данного отряда характерен колюще-сосущий челюстной аппарат
с верхне- и нижнечелюстными щетинками, расположенными в одной плос-
кости, и с нижней губой в виде членистого хоботка, служащего для щетинок
футляром (см. стр. 382 и рис. 383). Этот аппарат устроен и функционирует
одинаково как у имаго, так и у личинки.
К данному отряду относятся клопы, или полужесткокрылые (Hete-
roptera), с неоднородно устроенным надкрыльем, кожистым в основании
и перепончатым в вершинной части; сюда же относятся цикадообразные,
или равнокрылые, хоботные (Homoptera) с однородными передними крыль-
ями. К равнокрылым хоботным относятся певчие цикады (Cicadidae), пен-
ницы (Cercopidae), цикадки (lassidae и другие семейства), листоблошки
(Psyllidae), тли (Aphidae) и червецы (Coccidae).
Развитие у членистохоботных обычно эпиморфное, реже с метаморфозом
(самцы червецов).
Среди клопов преобладают растительноядные формы. Среди плотояд-
ных клопов нередки хищники. К ним относятся клопы наших пресноводных
водоемов; как-то: гладыш (Notonecta), плавт (Naucoris), водяной скорпион
(Nepa) и другие. Многие растительноядные клопы серьезные вредители
сельскохозяйственных культур, как-то: черепашки, (Eurygaster), огородные
клопы (род Eurydema).
Общеизвестен постельный клоп (Cimex lectularius); к тому же роду
Cimex относится и птичий клоп (С. hirundinis). Любопытно то, что постель-
ный клоп — космополит. Он питается кровью только теплокровных, ак-
438
тивен в узких пределах температуры и весьма чувствителен к сырости.
Его широкое распространение объясняется завозом с мебелью и другими
вещами; всюду в человеческом жилье он находит себе пищу, тепло и необ-
ходимый ему сухой воздух.
Певчие цикады (Cicadidae) — крупные представители цикадообраз-
ных, отличающиеся парными певчим и тимпанальным аппаратами самца,
расположенными в передней части брюшка. Самки откладывают яйца под
кору древесных растений. Личинка свою часто многолетнюю жизнь (суще-
ствуют цикады, живущие до 17 лет) проводит под землей, питаясь соком
подземных частей растений. В СССР цикады встречаются преимущественно
на юге.
Мелкие цикадообразные пенницы (Cercopidae) и цикадки (lassidae)
распространены широко, часто встречаясь на лугах ив кустарниках. Пенни-
цы известны своими личинка-
ми, которые окружают свое
тело пеной, взбиваемой из
выделений кишечника и сек-
рета кожных желез.
Весьма серьезными вре-
дителями культурной и дикой
растительности являются ли-
стоблошки, тли и червецы.
Листоблошки (Psyllidae)
часто встречаются на листьях
деревьев. Они сильно вредят
яблоне и груше. На молодых
побегах ольхи часто встреча-
ются покрытые восковым
пушком нимфы листоблошки.
Как у прямокрылых, задние
ножки листоблошек прыга-
тельные.
Бичом наших огородных
и садовых культур являются
тли (Aphidae). Таковы, на-
пример, гороховая тля, бах-
чевая тля, кровяная тля,
Рис. 399. Злаковая тля Toxoptera graminis. I —
крылатая девственница; // — ее личинка; ///
ее нимфа; IV — бескрылая девственница
филлоксера.
Во внешнем строении тли (рис. 399) характерно образование конце-
вого сужения последнего членика усика. Усики состоят из трех-шести
члеников. Сложные глаза имеют всегда особую трехфасеточную группу,
часто сидящую на особом бугорке. Брюшко на спинной стороне шестого
сегмента часто несет пару трубочек, или бугорков, называемых соковыми.
У тлей, как и у всех перечисленных членистохоботных, развитие с эпимор-
фозом.
Сосанием органов своего хозяина-растения тля часто вызывает отмира-
ние или уродство органов и образование на них наростов — галл.
Тли интересны двумя биологическими особенностями: периодической
миграцией и связанной с миграцией сменой поколений.
Подобно тлям червецы и щитовки (Coccidae) длительно сосут различные
органы растений, являясь вредителями, часто вызывающими гибель рас-
тения. Хотя от щитовок страдают и деревья средней и северной полос СССР,
но особенно вредят они плодовым деревьям южной полосы Союза, где фауна
щитовок гораздо богаче. Густо покрывая стволы деревьев, щитовки исто-
щают своего хозяина, нарушают движение соков и влекут отмирание всего
растения или его части. В отличие от тлей червецы полиморфизмом поко-
лений и миграцией не обладают. У них резко выражен половой диморфизм.
Самка червеца бескрыла, личинковидна и обладает развитием с эпиморфо-
439
зом, тогда как самец претерпевает типичный метаморфоз с фазой куколоч-
ного покоя и заканчивает свое развитие крылатой фазой.
Как взрослая самка, так и молодые фазы покрыты спинным щитом,
состоящим из воскоподобных и смолоподобных веществ секрета кожных
желез.
ОТРЯД СЕНОЕДЫ (COPEOGNATHA)
ОТРЯД ПУХОЕДЫ (MALLOPHAGA)
ОТРЯД ВШИ (ANOPLURA)
Сеноеды, пухоеды и вши — преимущественно мелкие насекомые.
Первые — свободноживущие и питаются главным образом низшими расте-
ниями, встречаясь на коре и под корой деревьев, на листьях, во мху, в
гнездах, часто в книгах и в коллекциях растений и насекомых. Среди них
имеются как крылатые (рис. 400), так и бескрылые формы. Представители
Рис. 400. Сеноеды. I — Atropos pulsatoria; // Peripsocus
сеноедов, часто встречаемые в гербариях и книгах. Atropos pulsatoria
(рис. 400i ) и Troctes divinatorius.
Пухоеды внешне напоминают вшей. Это также мелкие наружные па-
разиты теплокровных, встречаемые, однако, не только на млекопитающих,
но и на птицах. Питаются пухоеды, как правило, различными продуктами
эпидермиса, как-то: роговыми чешуйками, пухом и др. Сообразно с пищей
челюстной аппарат пухоедов грызущий. Представитель — собачья вошь
(Trichodectes latus, рис. 401).
Вши — наружные паразиты исключительно млекопитающих, кровь
которых они сосут. Обладая подобно рассмотренным выше отрядам непол-
ным превращением, они отличаются от них своим колюще-сосущим челюст-
ным аппаратом, расположенным внутри головы. В момент укола этот ап-
парат, состоящий из ряда щетинок, выступает через поровидное ротовое
отверстие. Грудь подобно голове мала; она не расчле-
нена на сегменты. Поражают величиной концы ножек
с единственным коготком, с помощью которого вши
прикрепляются к волосам. Различные вши приурочены
к определенным видам млекопитающих. На человеке
паразитируют три вида.
Головная вошь (Pediculus capitis) живет обычно
на волосах головы, платяная вошь (Pediculus vestime-
nti) — в складках белья и платья и лобковая вошь
Рис. 401 Trichodec- (Phthirus pubis) — преимущественно в волосах лобка,
tes latus иногда подмышек и даже на ресницах (в последнем
440
случае они могут вызывать воспаление век). Вши откладывают яйца и
прикрепляют их или к волосам, или к одежде. Яйца обладают очень плот-
ной оболочкой и имеют на тупом свободном конце крышечки с отверсти-
ями (рис. 402).
Головная и платяная вши явля-
ются переносчиками возвратного и
сыпного тифов. Сыпной тиф передает-
ся при сосании крови, возвратный —
путем втирания в тело полостной
жидкости вши при расчесывании мес-
та укуса и раздавливании вши на
теле человека.
Рис. 402. Платяная вошь. Pediculus vesti-
nent. / яйцо; // —взрослая особь
П
ОТРЯД ЖЕСТКОКРЫЛ ЫЕ, ИЛИ ЖУКИ (COLEOPTERA)
Самый богатый видами отряд: некоторые семейства насчитывают де-
сятки тысяч видов (слоники — около 50 тыс. видов). Жуки обычно имеют
твердые наружные покровы и твердые, лишенные жилок надкрылья —
элитры. Мягки лишь спинки отдела брюшка, прикрываемые элитрами.
Перепончатые задние крылья складываются под элитрами. Челюстной ап-
парат у жуков типично грызущий. Основание нижней губы сросшееся с
черепом. Переднеспинка большая. Развитие с голометаморфозом (типичным
метаморфозом).
Экология. Условия существования и пища жуков крайне разнообразны; парази-
тизм, однако (бобровая блоха Platypsyllus castoris),— исключение.
Типичными хищниками являются как взрослые, так и личинки жужелиц (Са-
rabidae, рис. 403) и плавунцов (Dytiscidae, рис. 357). Жужелицы ночные жуки,
днем скрывающиеся под камнями, под корой и пр. В общем считаются полезными,
так как уничтожают много вредителей. Примером особенно полезного жука может
служить красотел (Calosoma sycophanta), водящийся на деревьях (рис. 403).
К типичным хищникам относятся божьи коровки (Coccinellidae). К менее ти-
пичным хищникам — коротконадкрылые жуки (Stahylinidae), которые частично
питаются и гниющими веществами. К ним по образу жизни примыкают мертвоеды
и могильщики (Silphidae), питающиеся падалью, иногда хищничающие.
Хищничество личинок и растительноядность взрослой фазы наблюдаются у
водолюбов (Hydrophilidae), внешне близко напоминающих плавунцов, и у нарывни-
ков (Meloidae). «Личинки некоторых нарывников поедают яйца саранчовых. Уничто-
жая кубышки саранчовых, поедая находящиеся в кубышках яйца, нарывники при-
носят пользу; однако те же виды во взрослом состоянии являются вредителями сель-
скохозяйственных культур.
Чисто растительноядные ряд семейств жуков. Таковы чернотелки (Tenebrio-
nidae), внешне напоминающие жужелиц, отличающиеся чрезвычайной твердостью
покрова. Их личинки, встречаемые в почве — опасные вредители подземных частей
полевых культур. По сходству с личинками щелкунов — проволочниками они
получили наименование ложнопроволочников.
Представители щелкунов (Elateridae, рис. 403) известны своей способностью,
лежа на спине, подпрыгивать. Как взрослые щелкуны, так и их личинки, живущие
в поле, проволочные черви вредят культурной и дикой растительности, протачивая
корневую систему растения. Борьба с этими вредителями весьма сложна.
Усачи, или дровосеки (Cerambycidae, рис. 403), относительно крупные жуки
с дугообразно загнутыми, направленными в сторону и назад усиками. Их белые мя-
систые личинки обладают увеличенной переднегрудью, в которую прячется головка,
вооруженная мощными верхними челюстями, с помощью которых они протачивают
ходы. Эти ходы выгрызаются в коре и в дереве, причем буровая мука, использованная
как пища, заполняет ходы. Одни усачи протачивают ходы в мертвых стволах, другие
живут в живых деревьях. Генерация длится два года или больше.
441
У листогрызов (Chrysomelidae), личинки которых питаются листвой деревя-
нистых или травянистых растений, тело укорочено, часто полу шаровидно. Обычное
явление — металлический блеск покрова.
У взрослых особей слоников, или долгоносиков (Curculionidae рис. 404 и 431),
лицевая часть головы более или менее вытянуты в так называемую головотрубку.
По сторонам головотрубки бороздки для вкладывания основания усика. Форма
головотрубки соответствует той роли, которую она играет при кладке. В растении
выгрызается с помощью головотрубки и верхних челюстей углубление, в которое
откладывается яйцо. В тех случаях, когда яйцо откладывается на поверхности расте-
ния, головотрубка укорочена. Личинка слоника белая, безногая, с поперечными
складками на туловище.
Рис. 403. Жуки. I — красотел (Calosoma sycophanta); II щелкун
Elater, имаго (//, а) и личинка щелкуна Elater (II, б); /// — усач Мо-
nochamus (III, а) и его личинка (III, б); IV — майский хрущ Melolo-
ntha melolontha
Родственны слоникам короеды (Ipidae), личинки которых напоминают личинок
слоников, взрослые же особи отличаются от слоников отсутствием ясной головотруб-
ки. Самка короеда прогрызает под корой ход, от которого в стороны направляются
ходы, прогрызаемые деткой, постепенно расширяющиеся и оканчивающиеся куколоч-
ной колыбелью (рис. 405). Живут короеды под корой или в древесине преимущественно
хвойных деревьев, но встречаются и в лиственных, особенно часто в вязе. Часть коро-
едов — древесинники, питаются древесиной, пробуравливая глубоко проникающие
в нее ходы, где происходит кладка яиц и развитие молоди. В связи с малопитатель-
ностью древесины особое значение получает для вскармливания молоди древесинника
грибок, растущий по стенкам ходов. Любопытно, что различным видам древесинни-
ков соответствуют различные виды грибков. Благодаря определенному уходу со сто-
роны жука грибок образует плотный налет, состоящий из конидий (спорообразующих
побегов) на стенке хода; клетки конидий отличаются особенно богатым содержанием
питательных веществ, используемых молодью.
Пластинчатоусые (Scarabaeidae) имеют усики с пластинчатой, веерообразно
расправляющейся булавой, плотным телосложением и роющими передними ножками
442
(рис. 403 и 424). Их толстые личинки, туловище которых покрыто поперечными склад-
ками и обладает лишь слабыми ножками, живут обычно в почве. В одних случаях
они питаются гниющими веществами (навозниками), в других — корнями растений
(хрущи). Взрослые растительноядные, питаются листьями и цветками. Навоз исполь-
зуется в качестве пищи как взрослыми навозниками, так и их личинками. Общеиз-
вестный навозный жук Geotrupes выкапывает в земле отвесный ход, от которого
Рис. 404. Долгоносик
Hylobius abietis
Рис. 405. Схема гнез-
да короеда:
/ — маточный ход.
личиночный ход
на различном уровне отходят боковые ответвления. Все ходы самцом и самкой сов-
местно набиваются навозом, причем в самой широкой части отвесного хода заклады-
вается яйцо. Другой навозник — священный скарабей (Scarbaeus sacer) катает из
навоза шарики, которые он затем откатывает в заготовленные им норки. Запасы навоза
частью поедаются взрослым жуком, частью служат пищей для молоди, развивающей-
ся из яйца, заложенного в толщу скатанного шарика.
ОТРЯД СЕТЧАТОКРЫЛЫЕ (NEUROPTERA)
Сетчатокрылые из насекомых с полным превращением имеют ряд при-
митивных признаков. Перепончатые крылья их характеризуются густой
сетью жилок, ротовые органы измененного грызущего типа. Тело удлинен-
ное, голова небольшая. Сетчатокрылые плотоядны. К этому отряду отно-
сятся муравьиные львы (Myrmeleontidae) по общему виду похожие на стре-
коз, но отличающиеся от них булавовидными усиками. Личинки муравьиных
львов питаются муравьями и другими мелкими насекомыми. Личинки вы-
рывают воронкообразную ямку в песке, на дне которой они закапываются,
выставляя наружу челюсти, и подстерегают добычу. Они живут преи-
мущественно в жарких странах, но встречаются в южной и средней Европе
и СССР. Кроме муравьиных львов, к сетчатокрылым относятся златоглазки
(Chrysopidae) — нежные насекомые с длинным и тонким телом, личинки
которых питаются тлями.
ОТРЯД ПЕРЕПОНЧАТОКРЫЛЫЕ (HYMENOPTERA)
Перепончатокрылые наряду с двукрылыми относятся к насекомым,
эволюция которых пошла наиболее далеко. Эволюция в пределах отряда
перепончатокрылых выступает вполне отчетливо не только в изменении всех
отделов тела в определенном направлении, но и в изменении их поведения,
что выразилось в образовании сообществ, в способах добывания пищи, в
обеспечении ею потомства.
Общими признаками всех перепончатокрылых служат сосущий или
грызуще-лижущий челюстной аппарат, а также наличие двух пар крыльев,
причем обе пары перепончаты. Передние крылья всегда больше задних.
Все перепончатокрылые переживают в своем развитии голометаморфоз.
443
К семействам отряда перепончатокрылых относятся наездники (Ich-
neumonidae, Braconidae и Chalcididae), орехотворки (Cynipidae), настоящие
осы (Vespidae), пчелы (Apidae) и муравьи (Formicidae).
К наездникам (Ichneumonidae и Braconidae) относятся очень подвижные строй-
ные перепончатокрылые, с более или менее выраженной перетяжкой в основании
брюшка. Личинка наездника червевидна и, как правило, лишена ножек, усиков и
глаз. Ведет паразитический образ жизни и бывает то эндо-, то эктопаразитом: самка
откладывает яйца на поверхности тела или в тело хозяина. Для этого самка обладает
соответствующей длины яйцекладом, с помощью которого она может иногда откла-
дывать яйца глубоко в древесину дерева, в котором живет хозяин-гусеница. Опреде-
ленные наездники придерживаются определенных хозяев. Так, Ichneumonidae при-
держиваются гусениц чешуекрылых, Aphidiidae тлей. Цикл паразита может быть
строго приурочен к циклу развития хозяина. Весеннее поколение самок Apanteles
glomeratus откладывает яйца в первое поколение гусениц капустницы; самки, разви-
вающиеся из этих яиц, откладывают яйца во второе поколение гусениц капустницы.
Используя своего хозяина, паразит начинает с пожирания менее существенных орга-
нов. Этим достигается то, что хозяин остается живым до момента окукления паразита.
Паразитический же образ жизни ведут личинки хальцидид (Chalcididae), к ко-
торым относятся мелкие наездники. Хозяевами им служат чешуекрылые, жуки, пере-
пончатокрылые и двукрылые на различных фазах развития, причем яйцом паразита
часто заражается яйцо хозяина. Хальцидиды, как яйцееды, развивающиеся в некото-
рых случаях в 2—3 недели и дающие семь поколений в сезон, могут иметь практиче-
ское значение в борьбе с вредителями.
Среди хальцидид имеются и вторичные паразиты, т. е. паразиты паразитов
(иногда называемые сверхпаразитами), уничтожающие полезных первичных парази-
тов.
Орехотворки (Cynipidae) мелкие перепончатокрылые с сидячим или стебель-
чатым брюшком, личинки которых паразитируют в растениях и насекомых. У расте-
ний они часто вызывают образование галлов. Самые типичные орехотворки образуют
галлы на листьях дуба (чернильные орешки).
Настоящие, или складчатокрылые, осы (Vespidae) в покое складывают
передние крылья вдоль. Они бывают одиночными или общественными.
В последнем случае, как и все прочие общественные перепончатокрылые,
они имеют касту рабочих ос, которыми могут быть лишь самки. Для по-
стройки гнезда роется полость в почве или в растении, которая у одиночных
ос штукатурится минеральными веществами. Общественные же осы строят
ячейки-гнезда из бумажной массы. Бумажные массы изготовляются ими
из древесинных волокон с помощью секрета желез. Ячейки располагаются
горизонтальными слоями — сотами, отверстиями вниз (см. рис. 416). Как
правило, ячейки служат только для вывода детки, пища же в ячейки скла-
дывается в исключительных случаях. Накопление пищи для детки произ-
водится лишь у одиночных ос. Общественные осы снабжают детку непрерыв-
но свежей пищей, состоящей преимущественно из разжеванной мускула-
туры насекомых.
Пчелиные (Apidae) отличаются по способу питания от прочих перепон-
чатокрылых: они питаются исключительно нектаром и пыльцой. Этим
определяется тесная связь пчелиных с высшими растениями. Они состав-
ляют приблизительно половину всех опылителей растений.
В интересах сбора пищи значительным изменениям по сравнению с
другими перепончатокрылыми подверглись челюстной аппарат и покровы:
первый значительно удлинился, что дало возможность более широко ис-
пользовать медоносные растения. Изменившиеся же покровы дают возмож-
ность собрать больше пыльцы. В составе покрова появляются перистые,
собирательные волоски, захватывающие при посещении цветка пыльцу
Покровы могут смазываться у высших пчел медом, что также способствует
приставанию к ним пыльцы. Это позволяет собрать пыльцу в дальнейшем
в комочек. Наиболее развит собирательный аппарат шмелей и медоносных
пчел (род Apis).
Способ постройки гнезда у пчел весьма разнообразен. В основном,
однако, можно различить два типа гнезд: гнезда, выкапываемые в земле
или выгрызаемые в древесине, и гнезда, возводимые из земли, из частиц
444
растений или из секрета, выделяемого покровами, т. е. из воска. Второй тип
гнезд, очевидно, совершеннее первого.
По образу жизни пчелы распадаются на две основные категории на
одиночных и общественных пчел. Одиночные пчелы представлены лишь сам-
цами и самками. Общественные имеют, кроме того, и рабочих пчел, которые
представляют недоразвитых самок. Как третью категорию можно принять
пчел-кукушек\ это нахлебники, не имеющие своих рабочих пчел и не имею-
щие собирательного аппарата, это формы деградировавшие, откладывающие
яйца в чужие гнезда часто близко родственных форм.
Из примеров, иллюстрирующих различные ступени эволюции пчел,
приведем два первый иллюстрирует самую низкую ступень, второй са-
мую высокую, наблюдаемую у пчел.
Одиночная пчела Prosop is имеет почти голое тело. Перистые, т е. со-
бирательные, волоски весьма редки, настоящего собирательного аппарата
нет Как нектар, так и пыльца заглатываются и отрыгиваются затем в ячей-
ке. Гнезда строятся по первому типу и представлены лишь рядом располо-
женных друг за другом ячеек в стебле растений. Таким образом, экологи-
чески Prosop is весьма напоминает роющую осу.
Примером общественных пчел могут служить шмели и медоносная
пчела.
Муравьи (Formicidae) от прочих перепончатокрылых отличаются тем,
что второй и третий сегменты брюшка сужены в тонкий стебелек (стр. 455).
Будучи плотоядными муравьи уничтожают огромное количество вре-
дителей и должны считаться полезными. Кроме того, они приносят пользу,
уничтожая мертвые организмы.
Постройки муравьев разнообразны: один (Tetramorium, Formicina)
строят подземные гнезда, другие (крупные Camponotus) устраивают ходы
в древесине и вызывают этим гибель живых стволов; третьи устраивают му-
равьиные кучи.
ОТРЯД ДВУКРЫЛЫЕ (DIPTERA)
Этот отряд характеризуется тем, что имаго обладает лишь одной парой
развитых крыльев и имеет челюстной аппарат сосущий или колюще-сосу-
щий. Развитие с голометаморфозом. Уже данная краткая характеристика
указывает на специализацию как органов взрослой особи, так и молодой
фазы.
Двукрылые делятся на длинноусых (Nematocera) с типичными усиками
нитевидной формы (комары, мош-
ки, бабочницы и др.) и на корот-
коусых (Brachycera) с усиками,
превращенными в трехчленистый
короткий орган (домашние мухи,
слепни, оводы и др.)
К двукрылым относятся насе-
комые с наиболее развитым поле-
том: это искусные летуны, обла-
дающие самым быстрым полетйм.
Как показывают опыты, развитие
полета зависит не только от формы
и строения крыла, развития муску-
латуры среднегруди, которой при-
надлежат функционирующие кры-
лья, но и от высокой специализа-
ции рудиментов задней пары
крыльев, которые превратились в
орган, направляющий полет, в
жужжальца, на поверхности и
внутри которых расположен ряд
Рис. 406. Мясная муха (Calliphora eryth-
rocephala). / имаго; II кокон; III
личинка
445
органов чувств. В связи с развитием мускулатуры среднегруди последняя
подавляет развитие передне- и заднегруди и достигает огромных размеров
(рис. 406).
Большая часть двукрылых яйцекладна, однако не редки случаи живо-
родности, причем личинки родятся на разных стадиях развития. Так, ли-
чинки серой мясной мухи (Sarcophaga) родятся на ранней стадии, кровосос-
ки (Hippoboscidae) — на поздней стадии, причем личинка тут же окукли-
вается.
Личинка двукрылых червевидна и обыкновенно лишена конечностей
(рис. 407). Ножки могут быть на крайних сегментах туловища, типа ложных
Рис. 407. Личинка перистоусового комара Chaoborus:
1 — усик, 2 — верхняя челюсть, 3 — трахейные пузыри, 4 — жабры
ножек (рис. 408). Яйца откладываются в ту среду, которая служит источ-
ником питания* в воду, в навоз, на поверхность органов животных или рас-
тений, служащих пищей, и т. д. Следствие этого — неразвитость не только
локомоторных органов, но и органов чувств и даже челюстного аппарата.
Более примитивны личинки длинноу-
сых (Nematocera). Личинка комара рода
Culex (рис. 409) имеет ясно выраженную
головку с боковыми глазами, короткими
усиками и грызущим челюстным аппаратом
полного состава, хотя и измененного строе-
Рис. 409. Culex. 1 —личинка;
II — куколка
Рис. 408. Личинка комара-звонца Tendipes:
1 — голова, 2—ложные ножки, 3— жабры
ния. Грудные сегменты срослись без сохранения границ в общий шаровид-
ный отдел, лишенный ножек. Роль локомоторного органа взяло на себя
девятисегментное брюшко, гибкое, подвижное и покрытое длинными щетин-
ками. На восьмом сегменте брюшка сидит вырост — стигмофор, несущий
пару стигм, с помощью которых личинка дышит, подвешиваясь к по-
верхности воды.
Хорошо развит головной отдел также у личинки Chaoborus (рис. 407),
относящейся к тому же семейству комариных (Culicidae), и личинки звонца
Tendipes (Chironomus, рис. 408). Обе личинки ведут также водный образ
жизни.
Другой тип личинки, тип личинки безголовой, весьма широко распро-
странен у короткоусых (Brachycera) и осуществлен у синей мясной мухи
(Calliphora, рис. 406). За небольшой лишенной органов зрения и усиков го-
ловкой, вооруженной лишь парой крючковидных верхних челюстей, сле-
дует одиннадцатисегментное туловище, лишенное ножек. Условия, в ко-
торых развиваются личинки двукрылых, самые разнообразные. Весьма
широко распространено питание продуктами распада.
446
Много личинок двукрылых, питающихся живыми растительными
тканями, в том числе и минеров. Эти личинки вредят многим культурным
растениям; такова шведская муха, повреждающая зерна злаков и их побеги,
яровая и ростковая мухи, минирующие стебли, капустная муха, прота-
чивающая корни капусты и др.
Имеются и хищные личинки двукрылых. Таковы все личинки слепней
(Fabanidae), питающиеся личинками насекомых и червями, личинки жуж-
жал, питающиеся в некоторых случаях яйцами саранчовых, пиявкообраз-
ная личинка журчалки (Syrphus), которая водится на листьях и хвое и
пожирает тлей.
Как куколка насекомых вообще, так и куколка двукрылых обнаружи-
вает во всей своей организации сходство с имаго: сосущий челюстной аппарат,
длинные усики, ножки и крылья являются лишь недоразвитыми по срав-
нению с соответствующими органами имаго. В результате приспособления
к среде куколка, однако, может также обладать провизорными органами.
Так, куколка обыкновенного комара (Culex), обладающая подвижностью,
имеет (рис. 409) на спинной поверхности груди пару рожковидных стигмо-
форов со стигмами для дыхания атмосферным воздухом.
У так называемых круглошовных короткоусых (Brachycera cyclorhapha),
к которым относятся самые обыденные представители мух, появляется
новое для фазы куколки защитное приспособление, прикрывающее ку-
колку снаружи. Это именно ложный кокон (пупарий, рис. 406), который
в отличие от настоящего кокона — продукта секрета желез личинки насе-
комых — является последней личиночной шкуркой, не сброшенной при
линьке, затвердевшей и прикрывающей куколку до момента выхода имаго.
При освобождении имаго из куколочной оболочки (которая здесь остается тон-
кой и нежной) и из кокона передние кольца ложного кокона отскакивают в виде кры-
шечки по кольцевой линии. У длинноусых (Nematocera) ложный кокон встречается
как исключение (у некоторых галлиц Cecidomyidae).
Из комариных (Culicidae) три европейских рода, а именно Anopheles,
Culex и Аёбе8 распространяют возбудителей болезней. Anopheles
заражает человека всеми формами малярии. Culex распространяет возбу-
дителей птичьей малярии. Один вид Аёбез argenteus, или Stegomyia, зара-
жает человека желтой лихорадкой. У всех трех родов личинки и куколки
живут в воде.
Рис. 410. Голова самца (/) и самки (II) Culex:
1 — глаз, 2 — усик, 3 — щупальце, 4 — нижняя губа
Во взрослой фазе — представители всех трех родов имеют длинные усики (рис.
410 и 41 1), ясно отличные у самца и самки; у самки они несут мутовки коротких ще-
тинок, у самца же щетинки мутовок длинны и становятся короче к концу усика. Сам-
ки в основном питаются кровью, самцы-нектаром, В связи с питанием кровью у самок
имеется длинный хоботок со щетинковидными, колющими верхними и нижними че-
люстями, верхней губой и подглоточником. У самцов челюстные щетинки недоразвиты.
Грудь в связи с развитием летательной мускулатуры горбообразно вздута; крылья
447
узкие и длинные, с чешуйками; ножки тонкие и длинные. Брюшко узкое и длинное,
из девяти сегментов. Личинки всех трех родов червевидны (рис. 409 и 412). Их голова
обособлена от туловища перехватом и несет на верхней губе пару приводимых в дви-
жение специальной мускулатурой кистей, направляющих ток воды с населением по-
верхностных слоев водоема к ротовому отверстию. Туловище лишено ножек. Передви-
жение личинки происходит с помощью гибкого брюшка и длинных щетинок на груди
и брюшке; на восьмом сегменте брюшка единственная пара стигм, с помощью
которых личинка дышит атмосферным воздухом. Наряду с органами дыхания
атмосферным воздухом имеются в окружности анального отверстия и жабры.
Куколки — не принимающие пищи, но подвижные, в виде запятой, «головка» которой
соответствует голове и груди насекомого (рис. 409). На спинной стороне грудной части
пара сигмофоров, несущих по стигме.
Рис. 411. Голова самки (/) и самца (//) Anopheles:
1 — глаз, 2 — усик, 3 — щупальце, 4 — нижняя губа
Рис. 412. Личинка комара Anopheles
Отличительные признаки яйца, личинки, куколки и крылатой фазы Anopheles —
малярийного комара, с одной стороны, Culex и Аёбез—сдругой, см. на рис. 409—413.
Биология малярийного комара (Anopheles). Покинув на поверхности водоема
куколочную оболочку, Anopheles ищет себе убежище. При наступлении вечерних
сумерек самцы первыми вылетают из убежища для роения. В рой влетает самка и уле-
тает с одним из самцов для спаривания. Повторяется ли в жизни самца или самки
спаривание, неизвестно. Как правило, самка сначала спаривается, затем напивается
крови После того как самка напилась крови, она отыскивает себе на несколько дней
убежище, где переваривает кровь. В это же время в яичниках развиваются яйца.
Для созревания яиц требуется при температуре в 20°С дней 14. По созревании яиц
448
16
ш
Таблица XII Капустница (Pieris brassicae). / — общий вид вскрытой гусеницы;
// —конец головы гусеницы; III линька гусеницы на куколку; IV —линька
куколки на имаго; V голова взрослой бабочки:
/ — усик, 2—верхняя губа, 3—верхняя челюсть, 4 — нижняя челюсть, 5—нижнегубное щупаль-
це, 6—прядильная трубочка, 7 — передняя ножка, 8—стигма, 9 — крыло, 10 — шелкоотделитель-
ная железа, // — трахеи, /2 — мальпигиевы трубочки, 13—ложная ножка, 14 — средняя кишка,
15 — задняя кишка, 16 — половая железа
самка летит к водоему, где и откладывает яйца, после чего значительная часть самок
погибает, часть же питается повторно и продолжает класть яйца.
При нежности покровов комара влажность атмосферы имеет большое влияние
на продолжительность жизни малярийного комара: оптимальная влажность 80—90%
В связи с этим комар ищет себе наиболее подходящую обстановку. Различной
влажностью следует объяснить, что комар днем ищет убежища в постройках, в траве,
избегает особенно мест с высокой температурой, ищет защиты от ветра и т. д. и пьет
кровь преимущественно вечером, в сумерки и ночью (днем относительная влажность
атмосферы значительно снижается). Там же, где влажнось значительна и днем, Ano-
pheles будет кусать и в это время суток> Самка Anopheles питается кровью как чело-
века, так и других млекопитающих. Она может питаться кровью представителей и
других классов позвоночных. Наконец, наблюдались случаи сосания крови и у пред-
ставителей беспозвоночных. Выбор объекта в общем —дело случая: чем крупнее
животное, тем чаще оно служит объектом питания. При выборе, однако, играет роль
и тепло, излучаемое животным,
(/) и
Рис. 413. Кладка и яйца Culex
Anopheles (//)
опаснее. Различные виды Anopheles
наконец, запах, им издаваемый. Как показывают
наблюдения над комарами в неволе, Anopheles
может питаться и соками растений (напри-
мер, плодов). По мере усвоения пищи разви-
ваются яички. Когда яйца в яичках разви-
лись, комар ищет водоем для кладки. Число
яиц в кладке колеблется то 60 до 350, причем,
как правило, каждая предшествующая кладка
больше последующей. Кладка изменяется и в
связи с сезоном: особенно велика плодовитость
весной (перезимовавшие самки), к середине
лета она уменьшается, после чего опять воз-
растает. Дальность полета комара может
быть значительной, опыты с окрашенными ко-
марами показали, что они могут улетать на
расстояние до 5 км. Однако их находили и за
десятки километров, что приписывается случа-
ям пассивного заноса (ветер, завоз с транспор-
том). С повышением температуры интенсив-
ность жизненных проявлений комара повыша-
ется, он больше питается, чаще откладывает
яйца. Вместе с тем уменьшается и срок его
жизни Средняя продолжительность жизни
имаго комаров летом около двух недель (вре-
мя, недостаточное для развития малярийного
плазмодия). Однако часть комаров выживает
дольше: максимальная продолжительность
жизни около одного месяца. В связи с уве-
личением продолжительности жизни осенью
комар как переносчик малярии становится
зимуют на различных фазах: то в виде яйца,
то личинки, то взрослого. Господствующий в средней полосе СССР вид комара Anop-
heles maculipennis зимует в фазе взрослого насекомого. С первыми же холодами пища
прекращает идти на созревание яиц, а идет на накопление жирового тела. С накоп-
лением жирового тела комары осенью начинают переходитть к зимовке, отыскивая
себе убежище. Обычный для средней полосы СССР Anopheles maculipennis перези-
мовывает за счет запасенного осенью жирового тела и поэтому может зимовать в
холодном помещении.
Вылупляющаяся из яйца Anopheles личинка проходит четыре возраста, отделен-
ных друг от друга линьками. Личинка питается частью организмами поверхностной
пленки водоема, частью организмами (живыми и мертвыми) на дне водоема и на по-
верхности растений водоема. Личинка то поднимаете» к поверхности воды, то погру-
жается на глубину водоема. Находясь на поверхности воды, личинка с помощью
двух подвижных кистей из волосков на верхней губе (рис. 412) приводит во враща-
тельное движение участок поверхностной пленки вместе с организмами в ней. При
этом колебательное движение кистей гонит ток воды назад к ротовому отверстию и
челюстному аппарату, где происходит отфильтровывание пищевых частиц, всасывае-
мых далее в глотку. Пища на дне водоема соскребывается кистями и челюстями.
Активным является поднятие личинки со дна. Так как поднятие требует усилий,
то личинки населяют участки водоема не глубже 1 м, если водоем не зарос; однако
при зарастании водоема роль второгодна могут играть водные растения. Весьма чутки
личинки к движению поверхности воды. Движение, вызванное ветром, дождем, тече-
нием воды, губит личинок; таким образом, местом заселения являются преимуществен-
но водоемы со стоячей водой. Значительное влияние на плотность населения личинок
в водоеме имеет кормность водоема. На выжимаемость личинок и на скорость их раз-
вития влияет температура воды. Оптимальная температура для личинки около
25° С. С повышением температуры срок развития личинки сокращается; минималь-
ный срок всего развития около 15 дней. С повышением температуры от 35°С развитие
450
задерживается. В связи с ускорением развития и удлинением периода активности
комара (вылет с зимовки при средней температуре в 20° С, кладка при средней темпе-
ратуре в 7—8°С) растет и возможное число поколений. В средней полосе СССР число
поколений два-три.
ОТРЯД БЛОХИ (APHANIPTERA)
Блохи — наружные паразиты теплокровных — млекопитающих и
птиц, кровь которых они сосут. Их челюстной аппарат колюще-сосущий;
по строению и отправлению он напоминает аппарат членистохоботных:
как и у последних, колющим и сосущим аппаратом служит пара нижне-
челюстных щетинок, к которым в роли колющего аппарата присоединяется
верхняя губа; и у блох нижняя губа со своими щупиками служит футляром,
в который вкладывается колюще-сосущий аппарат.
Все три пары ножек довольно
длинны (рис. 414); особенно разви-
ты тазики и лапки. При прыжке,
очевидно, преобладающую роль
играют задние ножки, особенно
длинные, крыльев у блох нет.
Личинка блохи — удлиненный
безногий червячок (рис. 414), пок-
рытый длинными щетинками и не-
сущий пару коротких церк, с по-
мощью которых личинка переме-
щается. Личинка питается гнию-
щими органическими остатками
животного происхождения, для
чего обладает слабо развитым
грызущим аппаратом. Перед окук-
ливанием личинка окружает себя
коконом. Срок развития человечес-
кой блохи (Pulex irritans) при бла-
гоприятных условиях от 2 до 4
недель, однако развитие может
значительно затянуться (период
развития куколки до 220 дней).
Блохи питаются главным обра-
зом кровью млекопитающих, реже кровью птиц. Виды блох часто при-
урочены к определенному виду хозяина, однако при случае блохи могут
переходить и на другие виды.
Питание блох периодическое и происходит в течение короткого времени
или более длительно; последнее переходит в постоянный эктопаразитизм.
Первый способ питания связан с жизнью блохи в гнезде хозяина, послед-
ний— с жизнью хозяина блохи на открытых пространствах, как это имеет
место у копытных. От своего специфического хозяина блохи при случае
легко переходят на других животных, которые тесно соприкасаются с хо-
зяином. Такой переход от одного хозяина к другому способствует широко-
му распространению тех заболеваний, переносчиками которых служат
блохи. Блохи — основные переносчики чумы. Как переносчик чумы извест-
на блоха Xenopsylla cheopis — паразит грызунов: чумные бактерии, всо-
санные с кровью в кишечник блохи, не только не гибнут в ее кишечнике,
но и размножаются в нем. Бактерии заносятся в ранку хозяина при пита-
нии блохи.
ZF
Рис. 414. Ctenocephalus canis (превраще-
ние). I —личинка; // — куколка^ III —
имаго; IV — кокон
ОТРЯД РУЧЕЙНИКИ, ИЛИ ПУХОКРЫЛЫЕ (TRICHOPTERA)
Ручейники внешне напоминают бабочек и им, несомненно, родствен-
ны, ясно отличаясь от последних лижуще-сосущим челюстным аппа-
ратом.
16*
451
Личинки, питающиеся растительной пищей, червевидны, с мягким
брюшком, которое обычно прикрывается чехликом (рис. 415), построенным
из растительных или минеральных частиц. Для дыхания брюшко несет
нитевидные трахейные жабры. Личинки ручейников живут в воде.
Рис. 415. Личинка Phryganea в чехлике
ОТРЯД ЧЕШУЕКРЫЛЫЕ, ИЛИ БАБОЧКИ (LEPIDOPTERA)
Два признака, обычно присутствующие одновременно, характерны
для данного отряда, это чешуйчатый покров крыльев, обусловливающий
те роскошные цвета и переливы, которыми отличаются крылья бабочки,
и сосущий челюстной аппарат, в образовании которого участвуют лишь
лопасти нижних челюстей. В случае отсутствия одного из двух признаков
имеется налицо другой, например, у бабочки тутового шелкопряда (Вот-
byx mori) отсутствует хоботок, у стеклянниц (Aegeriiclae) крылья почти
лишены чешуй и стекловидны. Длина хоботка у бабочек весьма различна;
она иногда превышает длину тела (некоторые бражники).
Червевидную личинку чешуекрылых называют гусеницей (табл. XII,
рис. 425 и 431). Челюстной аппарат гусеницы грызущий.
Нижняя губа гусеницы несет отверстие, из которого выделяется шел-
ковина, у чешуекрылых употребляемая на построение кокона, в котором
происходит закукливание; иногда закоконирования не происходит, шелко-
вина же идет на прикрепление куколки к постороннему предмету пояском
(табл. XII).
Гусеница — фаза роста чешуекрылых; она питается исключительно
растительной пищей и отличается большой прожорливостью, при массовом
размножении она может быть очень серьезным вредителем в сельском и
лесном хозяйстве.
Из чешуекрылых человеком используется и искусственно им разво-
дится для получения шелка тутовый шелкопряд (Bombyx mori). Тутовый
шелкопряд на имагинальной фазе — средней величины белая бабочка,
имеющая в размахе крыльев 4—6 см, Передние крылья обладают серпо-
видной вершиной. Хоботок у бабочки отсутствует В отличие от гребенчатых
усиков самки усики самца перистые. Гусеница серого или белого цвета с
рогом на спинке восьмого сегмента брюшка.
Тутовый шелкопряд в диком состоянии неизвестен. В связи с одомаш-
нением бабочка утратила способность полета. За отсутствием хоботка ба-
бочка неспособна питаться.
В Китае тутовый шелкопряд культивировался с незапамятных времен
(По преданиям, в Китае шелководство возникло за 2700 лет до н. э.).
К выкормке шелковичных червей (гусениц тутового шелкопряда) приступают
весной с появлением листвы на шелковице. Грена — яички тутового шелкопряда
оживляется при температуре в 22—25°С. Вылупляющиеся гусеницы на кусочках
резаного тутового листа переносятся на кормовую этажерку в условии хорошей вен-
тиляции. Через промежутки в несколько часов гусеницам сменяется корм, который
задается сначала в виде резаного, позже цельного листа шелковицы. В течение 30—
40 дней гусеница проходит пять возрастов, прерываемых линьками, после чего окру-
жает себя плотным коконом белого, желтого или зеленоватого цвета и окукливается.
Для закоконирования по краям этажерки ставятся коконники— прутья деревьев.
Выход бабочки наступает на 15—18-й день после начала завивки кокона. Так как
при выходе бабочки цельность нити кокона нарушается, то бабочка своевременно
452
замаривается горячим паром. В настоящее время предпринимаются шаги к разведе-
нию и других шелкопрядов, как, например, китайского дубового шелкопряда (Ап-
theraea pernyi).
НЕКОТОРЫЕ ВОПРОСЫ БИОЛОГИИ НАСЕКОМЫХ
ОДИНОЧНЫЕ И ОБЩЕСТВЕННЫЕ НАСЕКОМЫЕ
Огромное большинство насекомых живет одиночно и даже самцы и
самки держатся обособленно друг от друга; лишь на время размножения
они объединяются в пары, после чего снова расходятся. У таких насекомых
забота самки-матери о потомстве ограничивается кладкой. Имеется, однако,
около 6000 видов насекомых, ведущих общественную жизнь; сообщество
насекомых по происхождению своему — семья; по своему составу это слож-
ная семья, члены которой выполняют различную роль; для сообщества
характерно деление на стазы, т. е. на группы особей, выполняющих опре-
деленную роль, причем имеется стаза насекомых-рабочих, нормально не
способных размножаться, а приспособленных для выполнения иного спе-
циального назначения; обычно в стазу рабочих входят только недоразвитые
самки, у термитов и недоразвитые самцы.
От настоящих сообществ насекомых следует отличать подготовитель-
ную ступень, когда нет еще характерного для сообщества насекомых разде-
ления на стазы, но родители или по меньшей мере самка-мать живет с
потомством. Во всех остальных случаях, т. е. когда нет ни деления на
стазы, ни родителей данного поколения, имеет место стадность. Для иллю-
страции всех трех явлений — стадности, подготовительной ступени к
сообществу и настоящего сообщества — приведем следующие примеры.
Стадность широко распространена у саранчовых. Низшую ступень
стадности можно наблюдать у бескрылых саранчовых и у саранчовых, не
совершающих дальних вылетов с места отрождения. Это так называемые
кобылки (например, сибирская кобылка). Кобылки лишь в исключительных
случаях размножаются в больших количествах. Пример второй ступени
стадности дает прус\ в годы умеренного размножения он ведет себя подобно
кобылкам, в годы же массового размножения прус на фазе личинки предпри-
нимает странствования, на имагинальной фазе — перелеты. Отсюда можно
предположить, что основной причиной массовых передвижений служит
чрезмерная плотность населения, что влечет за собой недостаток пищи.
К третьей, а именно высшей ступени стадности относятся такие саранчовые,
как азиатская саранча и мароккская кобылка, которые совершают регуляр-
но странствования и перелеты. Эти саранчовые могут размножаться в ог-
ромных количествах.
Случаи, которые относятся к подготовительной ступени к сообществу,
не все одинаково сложны.
Простой случай подготовительной ступени можно проследить на обык-
новенной уховертке Forficula auricularia из отряда кожистокрылых
(Dermatoptera). Самка заблаговременно выкапывает в земле пещерку,
куда откладывает яйца, мать охраняет яйца до выхода из них молоди, после
чего молодь остается при матери в течение еще нескольких дней, а затем
разбредается,.
Более сложный случай наблюдается у жуков-навозников. Так, самец
и самка Copris lunaris держатся парами; они выкапывают в земле углубле-
ние, которое наполняют округлыми комочками навоза с яйцом в каждом
из них; родители остаются в выкопанной в земле ямке, пока вышедшая из
яиц молодь не съест своего комочка, после этого самец, самка и молодь,
развившаяся до имагинальной фазы, совместно оставляют свое убежище
и разбредаются. В двух приведенных случаях проявляется забота матери
или даже обоих родителей о своем потомстве, в последнем случае вплоть до
времени превращения потомства в имаго.
453
Рис. 416. Гнездо осы Vespa
germanica (продольный раз-
рез)
погибают, остаются в живых
Сообщество с делением на стазы встречается лишь у перепончатокры-
лых (осы, пчелы и муравьи) и термитов.
К одиночным осам относится вид рода Eumenes. Самка-мать ведет
типичную одиночную жизнь; однако она обеспечивает свое потомство гнез-
дом и пищей.
У представителей одиночных ос рода Belonogaster (Африка) самка
строит гнездо обширнее. Личинок кормит сначала одна самка-мать; с появ-
лением первых самок-дочерей к уходу за деткой приступают эти последние;
с возрастом они начинают охотиться за гусеницами и добывать материал
для построения гнезда; наконец, самые взрослые особи самок-дочерей на-
чинают класть яйца. Самцы живут паразитически, не выполняя никакой
работы и не покидая гнезда. С перенаселением гнезда небольшие рои самок
оставляют гнездо и устраивают новое.
Общественные осы умеренного пояса (рр.
Vespa, Polistes) обладают определенным, из
года в год повторяющимся циклом развития
сообщества. Оплодотворенная осенью самка
перезимовывает под мхом в одиночестве. В
следующем году она основывает гнездо пос-
тройкой нескольких ячей (рис. 416), куда
откладываются яйца; выкармливаемые самкой-
матерью личинки дают ос-рабочих. С момента
появления рабочих ос работа по дальнейшему
расширению гнезда и по добыванию пищи
отходит к рабочим осам; а самка-мать посвя-
щает себя размножению. К осени рабочие
осы начинают строить ячеи большего размера,
в которых выводятся нормальные самки и
самцы. Самки оплодотворяются самцами,
вскоре после чего сообщество распадается:
самка-мать, рабочие осы, самцы и личинки
лишь молодые оплодотворенные самки, кото-
рые перезимовывают в защищенном месте с тем, чтобы весной приступить к
основанию новой семьи.
Среди пчел также встречаются одиночные пчелы и общественные пчелы.
Сообщество медоносных пчел (Apis mellifera) представляет наивысшую
ступень у пчел вообще. Высокое развитие его у медоносных пчел выражает-
ся в том, что все стазы и физически и функционально высоко специализи-
рованы; так, рабочие пчелы, как правило, не откладывают яиц и обладают
весьма специализированным аппаратом для сбора пыльцы и способностью
давать воск. Пчелиная матка имеет длинное брюшко, укороченный хоботок
и жало, не способна выделять воск и у нее отсутствует собирательный
аппарат. Таким образом, она только откладывает яйца, в разгар сезона до
3—3,5 тыс. в сутки; при такой плодовитости одной самки достаточно для
обеспечения улья потомством, в связи с чем в семье пчел, как правило, име-
ется всего одна матка. /Латка при своей специализированной организации
не способна к самостоятельной жизни; в связи с этим нет момента, когда
матка остается одинокой, без окружения рабочими пчелами. Одна и та же
матка может обслуживать улей в течение 3—4 лет, причем общая плодо-
витость матки достигает 1 000 000 яиц. Расселение медоносной пчелы, раз-
множение семей происходит исключительно путем роения.
Все муравьи ведут общественную жизнь. Как правило, в сообществе
муравьев имеется нормальная кладущая самка и рабочие муравьи одного
и того же вида; кроме того, могут быть молодые крылатые самки и самцы.
Наиболее простой случай имеется у представителей подсемейства
Ponerinae. Они образуют небольшие сообщества, ведут хищнический образ
жизни, живут в простых земляных гнездах; рабочие муравьи Ponerinae не
полиморфны и мало отличаются от нормальных самок. Вообще же муравьи
451
(pp. Camponotus, Formica) дают примеры наиболее сложных обществ с
полиморфным составом муравьев-рабочих, а нередко и плодущих самок.
Муравьи-рабочие, так же как и рабочие особи у других перепончатокрылых,
представляют собой не способных к размножению самок; от нормальных
самок, сбросивших свои крылья, они отличаются более простым строением
груди и более слабо развитыми глазами, а также менее развитым брюшком.
Полиморфизм рабочих муравьев сводится в одних случаях к изменению
в размерах и к непропорциональности отдельных частей тела, особенно часто
бывает увеличена голова; такие особи предназначены для защиты колонии
или для нападения на чужие гнезда
(рис. 417); кроме того, встречаются
и промежуточные по своим приз-
накам самки, связывающие типич-
ного рабочего муравья с нормаль-
ной самкой; такие промежуточные
формы получили название эргато-
идных, псевдогин и пр. В образо-
вании таких промежуточных форм
существенная роль приписывается
пище, которой их кормили при
развитии. Иногда отклоняющаяся
форма приобретается рабочими
муравьями временно; такова «гине-
коидность» рабочих, которые за
отсутствием нормальной самки бе-
рут на себя роль половых особей.
В отличие от рабочих особей нор-
мальные самки и самцы первона-
чально имеют крылья, которые ими
отламываются после спаривания.
Самка, оборвавшая себе после
спаривания крылья, отыскивает
убежище, в котором она себя за-
муровывает. Молодь, вылупившая-
ся из отложенных самкой мелких
яиц, вскармливается ее секретом;
и поэтому называются «солдатами».
Рис. 417. Муравей Pheidole instabilis.
/ —самец; II —солдат; III, IV —про-
межуточные фазы рабочих; V — рабочий
этот секрет появляется за счет жи-
рового тела самки и распадающейся у нее летательной мускулатуры. Лишь
после появления первых взрослых рабочих муравьев, что может произойти
и через год после спаривания, самка начинает питаться той пищей, которую
ей приносят рабочие муравьи. С течением времени сообщество разраста-
ется. Ежегодно оно дает самцов и самок, которые основывают новые ко-
лонии. Колония продолжает существовать до гибели самки, ее основав-
шей, что может произойти лет через 10—15.
Однако основание самкой новой колонии может происходить и иначе:
она может попадать в сообщество другого вида, лишившегося своей самки,
где и принимается рабочими муравьями данного сообщества. Так, напри-
мер, самка рыжего лесного муравья (Formica rufa) может попадать к темно-
коричневому муравью (Formica fusca). Образуется временное сообщество
смешанного состава, которое становится однородным по вымирании рабо-
чих Formica fusca, адаптировавших самку Formica rufa. Такое внедрение
самки может быть и насильственным: так, кровавокрасный муравей (Formica
sanguinea) может внедриться в общину Formica fusca, убив самку этого
вида. Несмотря на состав сообщества исключительно из рабочих Formica
fusca, оно долго может оставаться смешанным благодаря набегам Formica
sanguinea на соседние сообщества Formica fusca с захватом у них куколок
для пополнения состава рабочих муравьев. Примером обязательного «ра-
бовладельчества» может служить муравей-амазонка (Polyergus rufescens).
455
Способность строить гнездо, вскармливать молодь и самостоятельно питать-
ся утрачена муравьем-амазонкой. Его челюстной аппарат служит ему ис-
ключительно орудием в его набегах и похищении «рабов».
Совместное существование с муравьями ведут и насекомые других
отрядов. Сюда относятся в числе других и тли, которые в виде экскрементов
или секрета поставляют муравьям сахаристую жидкость. Известны случаи
разведения грибов муравьями (Atta). В основном же муравьи плотоядны
и лишь немногие растительноядны.
Очень сложно устроены колонии термитов (Isoptera). В колониях всег-
да имеются две группы особей: половые особи и рабочие термиты. Назна-
чение первых — служить размножению, вторых — выполнять всевозмож-
П УШ
Рис. 418. Termes lucifugus. / кладущая самка, // —
крылатая фаза (половая особь); /// добавочная по-
ловая особь; IV рабочий V солдат
ные работы. Функция размножения лежит на особях с хорошо развитыми
крыльями, которые после лёта теряют крылья и попарно основывают новую
колонию. Половые железы у рабочих термитов недоразвиты. В колонии
имеются также и «солдаты», роль которых обычно сводится к защите сооб-
щества. Вскоре после роения крылатые самцы (рис. 418) и самки сбрасы-
вают крылья, отделяя их по поперечному шву и сохраняя от крыла лишь
треугольный обрубок, так называемую крыловую чешуйку. По форме
чешуйки этих самцов и самок нетрудно отличить от добавочных половых
особей, обладающих зачаточными крыльями в виде округлых чешуек.
С развитием внутренних половых органов брюшко матки увеличивается
в несколько, иногда в 9 раз, и может превратиться в белый мешок с темными
поперечными полосками (см. рис. 418) —тергитами и стернитами брюшка.
При отсутствии в колонии половых особей рабочие-термиты заменяют их
добавочными половыми особями. Последние, очевидно,— недоразвитые
особи, нимфы. У рабочих-термитов, покров туловища которых отличается
мягкостью и белизной, глаза и крылья абсолютно отсутствуют и соответ-
ственно с отсутствием крыльев недоразвиты средне- и заднегрудь. Недораз-
виты и половые железы. Голова у «солдат» (см. рис. 418) соответственно
исполинскому развитию их челюстей сравнительно велика. В отличие от
общественных перепончатокрылых у термитов рабочими особями и «сол-
датами», а также добавочными половыми особями могут быть как самки,
456
Таблица XIII Предохранительная окраска и форма насекомых. 1—2, 3—4
случаи предохранительной окраски и формы, 5—6 — 7—8 — случаи мимикрии:
/ — Kailima с расправленными крыльями сверху, 2 — она же, сидящая на ветке среди листьев,
3— Libythea ccltis с расправленными крыльями, 4 —они же со сложенными крыльями, 5 — Bombus
semenowi (перепончатокрылые), 6 — Portschinskia przewalski (двукрылые), 7 — Lycorea halia (гелико-
нида), 8 Disniorphia astynonico
так и самцы. Термиты-рабочие строят термитник, чистят его и всех чле-
нов сообщества до «солдат» включительно, добывают пищу, кормят и уха-
живают за молодью.
Образование новых колоний происходит главным образом путем рое-
ния, реже путем ухода части термитов-рабочих с молодью. Огромное
большинство особей роя, состоящего из самцов и самок, гибнет, становясь
жертвой хищников. Половозрелость и половой акт наступают несколько
спустя после роения. Первое время совместной жизни половые особи все
необходимые работы выполняет самостоятельно. Только после того, как
первое потомство выросло, они постепенно начинают освобождаться от ра-
бот и сосредоточиваются на размножении. В последующем новые особи
производятся на свет исключительно в маточной ячейке, где происходит
непрекращающаяся кладка яиц. Маточная ячейка занимает центральное
положение в термитнике и отличается особой прочностью.
ПОЛИМОРФИЗМ НАСЕКОМЫХ
Рис. 419. Chermes viridis, цикл поколений
и развитие. / —основательницы; // —
крылатой галловой тли; /// —ложной ос-
новательницы; IV — полоноски; V—обое-
полого поколения
О полиморфизме, связанном с общественной жизнью насекомых, сооб-
щено было выше. Другие случаи полиморфизма связаны с сезоном и ус-
ловиями питания. Наиболее яркий пример такого полиморфизма дают
тли. Полиморфизм тлей тесно связан со сменой у них поколений и периоди-
д ческой миграцией. Дело в том, что
часть определенного периода вре-
мени тля проводит на одном хозя-
рц ——. \ ине, часть—на другом. Миграция
7 6 часто связана с сезонными переме-
нами: осень, зиму и весну данный
вид тли проводит на каком-нибудь
древесном растении, а лето — на
другом (оно может быть и травя-
нистым). Первый хозяин считается
основным, второй — промежуточ-
ным. Однако миграция может быть
и не сезонной: так, хермес зеленый
один год проводит на одном хвой-
ном дереве, другой — на другом
(рис. 419). Смена поколений и мор-
фологические особенности каждого
поколения связаны с миграцией, с
переселением и с пребыванием на
том или другом хозяине; поколе-
ния отличаются друг от друга на-
личием и отсутствием крыльев,
той или иной степенью развития
глаз, усиков, соковых трубочек,
челюстного аппарата, а также плодовитостью; одни живородящие, другие
кладут яйца; одни поколения имеют самцов, другие партеногенетичны.
Обычно в полном цикле поколений тлей яйценосность наблюдается
только однажды: яйца кладут самки обоеполого поколения. Партеногенети-
ческая самка, развивающаяся из оплодотворенного яйца, получила назва-
ние самки-основательницы. Это поколение самок-основательниц считают
первым в цикле, последним же считается поколение обоеполое. Как поко-
ление самок-основательниц, так и обоеполое поколение развиваются на ос-
новном растении.
Переход с основного хозяина на промежуточного и обратно осущест-
вляется путем перелета. Поэтому в цикле имеются два поколения, особенно
подвижных, вооруженных крыльями и с хорошо развитыми ножками и
органами чувств (усиками, глазами). Промежуточные партеногенетические
458
поколения, неразлучные с питающим их растением, обладают слабо разви-
тыми локомоторными органами и органами чувств, но обычно более пло-
довиты.
Таким образом, цикл, как правило, слагается из следующих поколе-
ний: 1) бескрылая самка-основательница, 2) крылатая партеногенетическая
самка (эмигрант), 3) бескрылая партеногенетическая самка (поселенец),
4) крылатая партеногенетическая самка (реэмигрант), 5) самец и нормаль-
ная самка. В ряду указанных поколений четыре первых размножаются пар-
теногенетически и обычно живородящи. Число поколений может быть и
больше пяти.
У бабочек различные поколения одного и того же вида могут обладать
различной общей окраской и рисунком. Такое различие явно стоит в связи
с условиями сезона и поэтому получило название сезонного диморфизма.
Примером сезонного диморфизма может служить бабочка араьиниа (Arasch-
nia levana), перезимовавшие куколки которой дают рыжее с черными пят-
нами весеннее поколение, тогда как поколение, появляющееся в конце
лета, черное с белыми перевязями. Опыты показывают, что при воздействии
низкой температурой на куколок летнего поколения арашниа можно полу-
чить весеннюю форму бабочки (A. levana), а при воздействии высокой тем-
пературы на куколку весеннего поколения можно получить летнюю форму
(A. prorsa).
ЗАЩИТНЫЕ ПРИСПОСОБЛЕНИЯ НАСЕКОМЫХ
Встречаясь в природе в огромном числе видов и особей, насекомые
служат пищей хищникам. Ясно, что в тех случаях, когда насекомые мало-
подвижны, как, например, листотелы, или палочники, либо велики, ярко
окрашены (бабочки) и т. п., они хорошо заметны для хищника и поэтому
рискуют подвергнуться полному уничтожению. Понятно, что именно у
таких насекомых в результате отбора должны выработаться защитные при-
способления, которые должны им помочь в борьбе за существование; таким
защитным приспособлением служит покровительственная окраска, совпа-
дающая с окраской окружающей их природы, «подражание» форме расте-
ний и их частей, «подражание» окраске и форме других насекомых (миме-
тизм), хорошо защищенных особенностями в строении тела. Среди насе-
комых встречаются яркие примеры развития предохранительной окраски
и «подражания» другим организмам. Эти признаки могли, очевидно, выра-
ботаться лишь в условиях, указанных Дарвином, при наличии широкой
способности данного насекомого к изменчивости и в условиях остро про-
текающего отбора. Весьма частое явление в природе — это зеленая окраска
насекомого на фоне зелени или серовато-коричневая окраска на соответ-
ствующем фоне. Такова окраска прямокрылых, богомолов, палочников,
клопов и других насекомых. Не в меньшей степени затрудняет распознава-
ние насекомого пестрая окраска с мелкой штриховатостью и пятнистостью.
Пестрая окраска из серых менее ярких тонов характерна для нижней по-
верхности крыльев булавоусых (дневных) бабочек, которая у сидящей ба-
бочки обращена наружу (см. рис. 1 и 2, 3 и 4 на табл. XIII), тогда как
верхняя поверхность крыльев, прилегающая к крылу с другой стороны, об-
ладает яркой окраской. У сидящей же ночницы (Noctuidae) крылья рас-
полагаются двускатно, с наклоном в обе стороны, и соответственно этому
верхняя наружная поверхность крыльев окрашена в серые или бурые тона.
Весьма любопытный пример предохранительной окраски крыльев сидящего
насекомого дает палочник Tropidoderus children! (рис. 420). Когда он в
покое, надкрылья и крылья налегают у него частично друг на друга,
причем неприкрытые кожистые участки передних и задних крыльев ок-
рашены в темно-зеленый цвет (на рисунке изображены мелкой сеткой),
прикрытые же кожистые участки крыльев — в желтый (на рисунке изобра-
жены точками).
459
Другим средством защиты является покровительственная форма, т е.
подражение форме частей растений или других предметов. Примеры такого
подражания дают в изобилии палочники и листотелы (рис. 421, 422 и 423),
а также гусеницы пядениц. Классический пример покровительственной
Рис. 420. Tropidoderus children!
формы дают бабочки Kalli-
ma Libythea, «подражаю-
щие» листу (табл. XIII).
Мимикрия представля-
ет частный случай предо-
хранительной окраски,
когда один вид животного
подражает другому виду
животного. В стаях бабоч-
ки Lycorea из сем. Heli-
conidae, летающих в до-
лине р. Амазонки, встреча-
ются отдельные особи Dis-
morphia astynome — ба-
бочки из сем. Pieridae,
рисунком поразительно
похожие на Lycorea (табл.
XIII), хотя по анатомичес-
ким признакам оба вида
сильно различаются. Бро-
сающаяся в глаза окраска,
стадность и медленный полет геликонид должны бы привлекать внимание
хищников, а между тем птицы и другие хищники не преследуют гелико-
нид, потому что они несъедобны. Таким образом, съедобная пиерида Dis-
morphia, становясь сходной с несъедобными геликонидами, скрывается от
преследования. То, что Dismorphia встречается в единичных экземплярах
Рис. 421 Diapheromera femo-
rata
в стае геликонид, делает эффект реальным;
при большом числе съедобных Dismorphia
стая не была бы обеспечена от нападения
хищника. При несъедобности геликонид их
медленный полет, стадность и яркая окра-
ска получают смысл как знаки, предосте-
регающие, предупреждающие хищника. Та-
ким образом, при мимикрии объект подра-
жания должен обладать и действительно
обладает особыми средствами самозащиты.
Рис. 422. Phyllium scy-
the, самка (вид сверху)
Рис. 423. Листооб-
разное надкрылье
кузнечика Pteroc-
hroza
460
В приведенном случае мимикрии самка Dismorphia обладает большим
сходством с Lycorea, чем самец Dismorphia, что особенно существенно
для сохранения вида. Примеры подражания другим насекомым дают
мухи-журчалки (Syrphidae), формой и окраской напоминающие воору-
женных жалом пчел и шмелей. Пример прямого использования сход-
ства в форме и окраске дают пчелы-кукушки (рода Psithyris), близко на-
поминающие тех пчелиных, в гнездах которых они развиваются и нах-
лебничают.
Особенно интересны те случаи мимикрии бабочек, когда вид в раз-
личных местах своего распространения подражает различным образцам,
относящимся не только к различным видам, но и к различным родам дру-
гого семейства бабочек.
Примером может служить самка Papilio тегоре; этот вид встречается от гор
Эфиопии до южной оконечности Африки. Самец Papilio тегоре — большая желтова-
тая бабочка с небольшими черными пятнами; самки сильно отличаются от самцов
и окраской и формой крыльев; однако в Эфиопии, наряду с такими отличающимися
от самцов и мимикрирующими самками Р тегоре, встречаются иногда и такие, у
которых сохранилось полное сходство с самцом. На юге Африки, в Натале и Капской
провинции самка Р тегоре образует две формы самок, подражающие двум иммунным
формам данаиды Amauris echeria, причем в Натале Р. тегоре подражает натальской
форме A. echeria, в Капской Земле — капской форме последней; мало того, между
обеими разновидностями Amauris имеются переходные формы, которым подражает
и Papilio тегоре. В той же Капской провинции Papilio тегоре обладает еще двумя
разновидностями, которые подражают двум другим данаидам — Amauris niavius и
Danais chrysippus. И в данном случае, как в случае Papilio тегоре мимикрия осно-
вана на том, что данаиды, служащие Papilio тегоре образцом, несъедобны.
Не только образец может быть несъедобным, несъедобным может быть и подра-
жатель. Такие случаи, которые, казалось бы, лишают мимикрию ее смысла, объяс-
няются просто: несъедобность вида еще не гарантирует его от попыток поедания хищ-
ником; в случае двух несъедобных и исходных видов тот и другой вид потерпят от
хищников меньший урон.
МАССОВОЕ РАЗМНОЖЕНИЕ НАСЕКОМЫХ
Наблюдения показали, что плотность населения какого-либо вида
насекомых на данном участке не остается из года в год одной и той же —
она меняется. Эта неустойчивость особенно ярко проявляется при периоди-
ческом массовом размножении. Обратить на это внимание заставило массо-
вое размножение вредных насекомых. Периодичность массового размноже-
ния была выяснена для перелетной саранчи; оказалось, что за годами на-
растания плотности населения саранчи наступали годы максимума, после
чего плотность населения падала (табл. 5).
При сопоставлении с ме- теорологическими данными Таблица 5
оказалось, что годы прогрес- сивного увеличения ПЛОТНО- Годы прогрессив- сти населения отличались у повышенной средней темпера- Годы максимума Годы начала угасания
турой и пониженным количе- 1890—1891 ством осадков; этим доказы- 1897—1898 валась связь массового раз- 1920—1921 множения с благоприятны- ми для саранчи метеороло- гическими условиями. Сущность явления в 1892-1893 1899 1922—1923 конечном счете 1894 1900 1924 > сводилась к
тому, что в годы прогрессивного увеличения плотности населения саран-
чи повышенная температура способствовала более раннему отрождению
и окрылению саранчи; поэтому увеличивался и период яйцекладки.
Метеорологические условия, температура и влажность влияют на пло-
довитость насекомых: достаточно высокая температура и влажность могут
вызвать значительное повышение плодовитости насекомых; с другой сторо-
ны, засуха может повлечь полное бесплодие самок и соответственно сниже-
ние численности насекомого.
461
Однако метеорологические условия не являются единственной при-
чиной, влияющей на размножение насекомых; на него должны влиять и
обилие или недостаток пищи, наличие или отсутствие природных врагов —
хищников и паразитов данного насекомого. Наглядный пример влияния
обилия пищи на массовое размножение дает лес. В местах порубок леса
скопляется большое количество неиспользованного лесного материала и
отдельных пострадавших и оставшихся стоять деревьев; в ближайшие
же годы здесь наблюдается массовое размножение короедов, которое может
перерасти в поражение короедами и-здорового леса (см. выше). Наконец,
для хищников и паразитов могут наступить неблагоприятные условия,
вследствие чего их влияние на численность насекомого снижается. На
наездников, этих основных паразитов насекомых, погода оказывает особое
влияние: пасмурная погода в дни их лета препятствует откладке яиц, что
может повлечь уменьшение численности данных паразитов. Все указанные
условия могут способствовать массовому размножению насекомых-вреди-
телей. Однако со временем условия могут измениться: условия питания
могут сложиться невыгодно для насекомого, достигшего массового разм-
ножения; параллельно усиленному размножению насекомого будет
расти и размножение хищников и паразитов, усиленно снижающих его
численность.
Слишком плотное заселение насекомыми ограниченного пространства
при массовом размножении ведет и к развитию болезней, причиной кото-
рых служат различные микроорганизмы, грибы и бактерии. Примером
болезни, вызываемой бактериями, может служить фляшерия, поражающая
гусениц бабочек; фляшерия, часто поражающая разводимых человеком
шелкопрядов (Bombyx, Antheraea), вызывается последовательным действием
двух бактерий (Streptococcus bombycis и Bacillus bombycis).
За максимумом следует более или менее быстрое снижение численности
и период депрессии впредь до нового наступления исключительно благо-
приятных условий.
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ НАСЕКОМЫХ
Роль насекомых в природе очень велика. Как уже было сказано, боль-
шая плодовитость видов, питающихся культурными растениями, запасами
или предметами, нужными человеку, делает эти виды опаснейшими вреди-
телями, нередко приносящими серьезнейший ущерб хозяйству.
От вредителей болезней растений и от сорняков человечество теряет
от 30 до 50% урожая продовольственных культур. Мировые годовые потери
зерна от вредителей и болезней в среднем равны 83 млн. тонн. Этим количе-
ством зерна можно было бы прокормить около 400 млн. человек в течение
года. Очень большой процент этих потерь должен быть отнесен за счет на-
секомых-вредителей. Не меньший ущерб наносится и техническим культу-
рам, а также лесоводстну. Особое значение многие насекомые имеют как
паразиты человека и сельскохозяйственных животных и главное как пере-
носчики различных заболеваний — чумы, желтой лихорадки, малярии,
сонной болезни, тифов у человека и наганы, сурры, суауру и ряда других
заболеваний у домашних животных.
Однако, помимо вреда, насекомые приносят и большую пользу. Яв-
ляясь опылителями высших растений, они совершенно необходимы для
существования этих растений. Целый ряд видов рыб, птиц и других живот-
ных питается исключительно насекомыми. Ряд видов насекомых одомашнен
человеком ради вырабатываемых этими насекомыми продуктов (мед, воск,
шелк и др.). В Японии доходы от шелкообрабатывающей промышленности
оказываются одной из главных приходных статей государственного бюд-
жета.
Многие насекомые являются ассенизаторами, поедающими гниющие
вещества, гнилостные микроорганизмы и т. д.
462
Наконец, значительное количество насекомых — хищников и па-
разитов — уничтожает различных вредителей и нередко используется че-
ловеком для борьбы с вредителями полевых культур, в особенности с вре-
дителями цитрусовых плантаций и т. д.
Насекомые — опылители растений
Огромное значение имеют насекомые как опылители цветов; велика
роль чешуекрылых и особенно перепончатокрылых — обладателей доста-
точно длинного хоботка. Чешуекрылые и перепончатокрылые опыляют
именно те цветки, которые обладают особенно скрыто расположенными
нектарниками (гвоздичные, губоцветные, мотыльковые, первоцветные,
вьюнковые и многие другие). Средством привлечения насекомого является
не только накопленный в соответствующих органах (шпорцах, трубочках,
венчиках) нектар, но и окраска, форма и, наконец, запах цветка. Опре-
деленным цветкам соответствует определенный опылитель — насекомое.
Так, гвоздика (Dianthus carthusianorum), имеющая красный цвет, опыляет-
ся дневными бабочками; белая ночная фиалка (Platanthera bi folia) — ноч-
ными бабочками. Полевой вьюнок (Convolvus sepium) опыляется бражником.
Цветы, опыляемые пчелиными, окрашены преимущественно в синий и фи-
олетовый цвета. В зависимости от строения цветка при всасывании нектара
в цветок погружается то лишь хоботок, то вся головка, соответственно
этому при перекрестном опылении пыльники приходят в соприкосновение
и пыльца переносится то той, то другой частью тела. Об интенсивности
роли чешуекрылых и перепончатокрылых можно судить по наблюдениям
над бражником Macroglossum, который в одну минуту поспевает посетить
27—29 цветков фиалки.
Пчеловодство и шелководство
Пчела (строение, размножение, экология, сбор меда и выделение воска)
разобрана выше. Здесь отметим только, что пчеловодство среди сельско-
хозяйственных отраслей занимает гораздо большее место, чем обычно при-
нято думать. Еще до 1941 г. в США средняя стоимость годового урожая
меда по оптовым ценам превышала 30 млн. долларов, а средняя стоимость
годового сбора воска превышала 2 млн. долларов. Однако главное значение
пчел — это опыление плодовых растений и трав, которое пчела выполняет
надежнее других насекомых. По развитию пчеловодства первое место в
мире занимает Советский Союз. Особенно развито пчеловодство в колхозах
и совхозах Дальнего Востока, Северного Кавказа, Сибири и Поволжья.
Из 30 млн. семей, насчитываемых на всем земном шаре, около 9 млн. нахо-
дится в СССР В последние годы развилась новая отрасль пчеловодства —
получение «молочка» — белкового, очень витаминизированного вещества,
употребляемого плечами для питания матки и совсем молодых личинок.
Молочко используется в фармакологии и парфюмерной промышленности.
Неменьшее хозяйственное значение, чем пчеловодство, имеет и шел-
ководство. Кривая развития шелководства во всех странах, за исключением
Испании, неуклонно шла вверх до конца тридцатых годов нашего века.
Однако, с 1938 по 1952 г. мировое производство шелка сократилось пример-
но в 2 раза. Так в 1938 г. на земном шаре было получено 439 000 т коконов,
а в 1952 г. только 217 000 т. Основными производителями шелковичных
коконов являются Китай и Япония, на долю которых приходилось более
70% мирового сбора коконов.
СССР занимает в мире третье место, а в Европе первое. За годы Совет-
ской власти сбор коконов увеличился почти в 3 раза. Из Советских респуб-
лик по производству шелковичных коконов первое место занимает Узбек-
ская ССР, на долю которой приходится более 50% всего сбора. Главным
образом разводится тутовый шелкопряд, но за последнее время стали
463
разводить и другие виды: китайского дубового шелкопряда, из коконов
которого вырабатывается чесуча, клещевинного, айлантового и некоторых
других шелкопрядов. '
Насекомые — вредители сельского
и лесного хозяйства
Технические, зерновые, овощевые культуры, ягодники, плодовые на-
саждения, леса поражаются многочисленными вредителями из класса
насекомых. Личинки многих жуков — хрущей (рис. 424), долгоносиков,
щелкунов (так называемые проволочники), чернотелок (ложнопроволочники),
гусеницы совок (рис. 425) — вре-
дят корневой системе; гусеницы
чешуекрылых, в основном моли,
личинки двукрылых минируют ли-
стья, проделывая в листовой пла-
стинке ходы. Взрослые пыльцееды,
некоторые нарывники, пластинча-
тоусые, уховертки и другие сгры-
зают тычинки и пестики. Гусеницы
некоторых молей и других бабочек
поедают мякоть плодов и семена.
Некоторые короеды (рис. 426) губят
молодые деревья, выедая сердцеви-
ну побегов. Другие короеды пита-
ются корой или древесиной. Размер
потерь от вредных насекомых ис-
ключительно велик.
Вредители полевых культур.
Вредители полевых культур отно-
сятся к большому количеству отря-
дов насекомых — к прямокрылым,
хоботным, жукам, бабочкам, двук-
рылым и т. д.
Серьезными вредителями ока-
Рис. 424.Майский хрущ Melolontha hippo- зываются саранчовые, как стадные
castani. / —самка, // —самец; /// —ли- так и одиночные формы. В СССР
чинка; IV — куколка примерно 12 видов саранчовых
должны рассматриваться как се-
рьезные и опасные вредители. В свое время страшнейшим вредителем была
азиатская, или перелетная, саранча (Locustra migratoria), приносящая
неисчислимые беды, уничтожая растительность на огромных площадях
нашего юга и в соседних государствах. Этот вредитель упоминался в древ-
нейших мифах и легендах и рассматривался как бич и несчастье наро-
дов. Однако рационально поставленная борьба, основанная на хорошем
знании экологии, поведения и распространения перелетной саранчи,
значительно снизила ее численность, а также и вред, причиняемый ею. По-
стоянным местом обитания (резервацией) этой саранчи в СССР в основ-
ном оказываются плавни по дельтам рек юга и юго-востока, по берегам Чер-
ного, Азовского, Каспийского и Аральского морей и озера Балхаш. В
этих местах проводились основные истребительные работы.
Гораздо большее значение в настоящее время стали иметь другие
виды саранчовых, такие, как итальянский прус, сибирская кобылка, шисто-
церка и некоторые другие.
Сибирская и темнокрылая кобылки (Gomphccerus sibiricus и Stauro-
derus sibiricus) распространены преимущественно в степной и лесостепной
частях Сибири, но встречаются и за пределами СССР. В начале века (1903—
1905 гг.) сибирская кобылка уничтожила в Западной Сибири около 870 000 га
464
посевов. На юге этот вид встречается только в горах Кавказа, Средней Азии
и Западной Европы, где серьезно вредит высокогорным посевам и паст-
бищам.
Итальянский прус (Calliptamus italicus) распространен в Крыму,
Среднем и Нижнем Поволжье, в Предкавказье, в Казахстане, Киргизии,
Рис. 425. Капустная совка (Barathra brassicae). I — бабочка; // —кладка яиц;
/// — яйцо сверху и сбоку; IV — гусеница, вид сверху; V — гусеница, вид
сбоку; VI — куколка; VII — повреждение листа гусеницей
Рис. 426. Короед-древесник Xyloborus. / —самка и самец; II —абро-
зия (культура грибка); /// — ходы в древесине
во всей Средней Азии и на юго-востоке Западной Сибири. Вредит большому
количеству полевых и технических культур. Съедает листья, а часто и стебли.
Большой ущерб хлебным злакам наносят клопы-черепашки (рода Еигу-
gaster, Aclia и др.). Среди них особое значение имеет вредная черепашка
17 Курс зоологии, т, I 465
(Eurigaster integriceps). Повреждая в основном пшеницу, ячмень, овес
и рожь, вредная черепашка наносит этим культурам весьма серьезный
вред. Основными районами обитания вредителя являются южные респуб-
лики Средней Азии, Кавказ, Крым, а также прилегающие к ним с юга
страны. Отсюда
Рис. 427. Яб-
лонная тля
Aphis poini на
ветви яблони
черепашка распространилась по степной и лесостепной
зонам Европейской части СССР и встречается иногда
даже в районах Куйбышева и Воронежа. Для черепаш-
ки, как и для многих других вредных насекомых, очень
характерны резкие колебания численности, причем пе-
риоды массового размножения, длящиеся иногда по мно-
гу лет, сменяются периодами депрессии, когда числен-
ность вредителя, а одновременно и причиняемый им вред
резко падают.
Из жуков широко распространенными вредителями
полевых культур оказываются многочисленные виды
щелкунов и чернотелок. Они вредят главным образом
в стадии личинок — проволочников (личинки щелкунов)
и ложнопроволочников (личинки чернотелок). Эти личинки
живут в почве и вбуравливаются в подземные части раз-
личных полевых овощных культур. Вредоносность про-
волочников очень велика и из-за их присутствия (до
600 000 личинок на 1 га) — некоторые участки земли
оказываются совершенно неподходящими для полевых
и овощных культур. На юге СССР встречается хлебный
жук — кузька (Aniseplia austriaca), выедающий зерна
злаков. Этот жук нередко значительно повреждает
злаки, размножаясь в очень больших количествах. Так,
в Харьковской области, в 1924 г. было собрано и уни-
чтожено более 276 т этого вредителя.
Наиболее страшным вредителем полевых культур
оказывается колорадский картофельный жук (Leptinotar-
sus decemlineata) (рис. 428). В 1823 г. американский
энтомолог Томас Сэй в скалистых горах и у подножья в
прериях нашел жука на диком черном паслене. Лет за
десять до «золотой горячки» в штат Колорадо начали
проникать колонисты. Со своим хозяйством они привезли
культурный картофель. Посевы картофеля привлекли
этого жука и через несколько лет он настолько размножился, что стал
самым серьезным вредителем картофеля. Картофельный жук стал быстро
распространяться из штата в штат и вскоре заселил все Соединенные Штаты
и Канаду Жук размножался в таких количествах, что не только уничто-
жал посевы картофеля, но в некоторых местах останавливал поезда, так
как колеса буксовали в массе раздавленных жуков.
Рис. 428. Колорадский кар-
тофельный жук (Leptino-
tarsa decemlineata)
Около 1920 г. жука случайно, несмотря на
строжайшие карантинные меры, завезли во
Францию. С каждым годом жук распространял-
ся все на новых и новых территориях и к 1938 г
он уже расселился по всей Франции, перебрался
в Бельгию, Германию, Швейцарию, Испанию.
Каждая самка колорадского жука может отло-
жить до 400—600 яиц, а некоторые экземпляры
больше 2000, через 5—10 дней после откладки
из яиц вылупляются маленькие мясокрасные
личинки, набрасывающиеся на листья картофе-
ля. Уже через 16—25 дней они достигают своего
предельного размера, спускаются на землю, за-
рываются в почву и превращаются в куколку.
Через 6— 15 дней куколка превращается в мо-
466
Рис. 429. Амбарный долгоносик Calandra
granaria. I —имаго; Il —личинка; /// —
куколка; IV — повреждения
лодого жука. В зависимости от климата страны картофельный жук дает
от одного до трех поколений.
Серьезными вредителями зерна являются личинки жуков амбарных
долгоносиков (рис. 429).
Из бабочек полевым культурам наносят значительный ущерб многие
совки и огневки, особенно те, которым свойственно массовое размножение.
Этот ущерб наносится гусеницами. Из совок наибольшее значение имеет
озимая совка (Euxoa segetum), размножающаяся на озимых посевах. Озимая
совка распространена очень широко: по всей Европе, значительной части
Азии, Северной и Южной Африке, а также Австралии. Из огневок особенно
вредит луговой мотылек (Loxostege sticticalis). Луговой мотылек повреждает
не только злаки, но и технические культуры (свекловица, хлопчатник и др.).
Размеры вреда, причиняемого
луговым мотыльком, связаны с
числом поколений в сезон. В
средней полосе СССР луговой
мотылек дает лишь одно поко-
ление в год, в южных же райо-
нах число поколений достигает
четырех. При благоприятных
для лугового мотылька услови-
ях он может появиться в огром-
ных количествах. При значи-
тельном количестве гусениц у
повреждённых растений от лис-
тьев остаются лишь черешки и
крупные жилки, а иногда рас-
тения просто съедаются цели-
ком. В 1929 г. массовое появ-
ление лугового мотылька на-
несло убыток около 200 млн.
рублей, причем свеклы было
уничтожено этим вредителем
1. 278 000 tn.
Из двукрылых, вредящих
полевым культурам, наиболее
важны широко распространен-
ные гессенская и шведская мухи.
Гессенская муха (Mayetiola de-
structor) (рис. 430) — небольшое
насекомое, около 3 мм дли-
ны, похожее на комара. Вредит
зерновым хлебам (пшенице, ржи,
ячменю). Ее червеобразная личинка сосет междоузлия злаков, забрав-
шись в листовое влагалище. Этот вредитель нередко вызывает полное
бесплодие злака. Гессенская муха отличается чрезвычайно резкими колеба-
ниями численности. Так, на Украине, в зоне наибольшей ее вредности,
гессенская муха с 1913 по 1922 гг. встречалась редко, но с 1923 по 1927 гг.
наступил период массового ее размножения, принесший большие потери
урожая.
Шведская муха (Oscinosoma frit) величиной в 1,5—2 мм, черного цвета.
В пределах СССР встречается от самых южных до самых северных границ
произрастания злаков. Ее личинка проникает в стебель злака к зачатку
колоса и высасывает его. Число поколений шведской мухи может достигать
пяти в год. Урожай в некоторых случаях снижается этим вредителем на-
половину.
Большой вред лесам наносят прожорливые гусеницы шелкопрядов, на-
пример кольчатого (рис. 431).
17*
467
Насекомые — паразиты животных
Среди насекомых встречаются как экто-, так и эндопаразиты; отдельные
случаи паразитизма имеются в отрядах, представители которых, как прави-
ло, растительноядны или хищники. Имеются, однако, отряды, состоящие
сплошь из паразитов, таковы пухоеды (Mallophaga) — паразиты птиц и
млекопитающих, питающиеся частицами эпидермиса кожи. К отрядам,
состоящим сплошь из эктопаразитов, относятся также вши (Anoplura) —
Рис. 430. Гессенская муха Mayetiola destructor. / —самец; II —
самка; /// —яички на листе пшеницы; IV —личинка; V —
поврежденное растение с ложным коконом (/)
паразиты млекопитающих, питающиеся исключительно кровью. К крово-
сосущим эктог^разитам относится также отряд блох (Aphaniptera) и часть
клопов. Кровососущими эктопаразитами является и большое число дву-
крылых, то более, то менее тесно связанных со своим хозяином (комары,
слепни, жигалки, кровососки, никтерибии и др.). Наружные паразиты то
длительно связаны со своим хозяином (пухоеды, вши), то посещают хозяи-
на лишь на время кровососания (большинство клопов, блох, двукрылых).
К категории собственно эндопаразитов относятся различные мухи-тахины,
кожные, полостные и желудочные оводы. Бычий овод (Hypoderma bovis)
468
откладывает яйца на ноги крупного рогатого скота, вылупившаяся из яйца
личинка тут же вбуравливается в кожу, мигрирует по подкожной клетчатке
и доходит до пищевода, затем до кожи спины; здесь она достигает своего
полного развития. В коже появляются нагноения и образуется свищ, через
который взрослая личинка выпадает на землю, где и окукливается. Полост-
ные оводы, как, например, овечий овод (Oestrus ovis), на фазе личинки пара-
зитируют в носовых полостях млекопитающих. Яйца желудочных оводов
Рис. 431. Кольчатый шелкопряд Malacosoma neustria. / —
яйца; //—гусеница; III—куколка; IV и V—имаго; VI —
повреждения
откладываются самкой на волосы хозяина и вылупившиеся из них личинки
в итоге попадают в желудок. У желудочного овода (Gastrophilus intestinalis)
этот процесс, по-видимому, происходит следующим образом: из отложенного
самкой овода яйца вылупляется личинка, которая вбуравливается в кожу
лошади и, протачивая ход, вызывает зуд. Лошадь расчесывает зубами кожу
и при этом проглатывает личинку, которая основывается в желудке, при-
крепляясь к его стенке. По окончании личиночного развития личинки по
кишечнику выносятся наружу, после чего окукливаются. В коже млекопи-
тающих паразитируют личинки и других мух сем. Tachinidae (Larvivori
dae).
К насекомым эндопаразитам-хищникам относятся некоторые пред-
ставители мух-тахин, а равно и мух некоторых других семейств. Яйца мух-
тахин откладываются на растение, посещаемое хозяином паразита, и вылу-
пившиеся личинки нападают на хозяина, которым обычно является насе-
комое; в других случаях мать откладывает яйцо непосредственно на поверх-
469
ность тела хозяина, после чего вылупившаяся из яйца личинка вбуравли-
вается в тело хозяина. Большое число таких мух-тахин паразитирует в
гусеницах бабочек, в ложных гусеницах пилильщиков, личинках жуков,
в саранчовых. Важнейшими паразитами-хищниками являются представи-
тели перепончатокрылых — наездников. На личиночной фазе наездники
паразитируют в насекомых. Особенно велика роль наездников собственно
(Ichneumonidae) и блестящих наездников (Chalcididae). Наездники собст-
венно — относительно крупные формы, которые откладывают яйца на по-
верхность или внутрь личинок различных насекомых, особенно часто в
Рис. 432. Наездник-паниск и гусеница бабочки
гусеницах бабочек (рис. 432). Блестящие наездники — очень мелкие пере-
пончатокрылые, которые в большинстве случаев свои яйца откладывают
в яйца различных насекомых, преимущественно же бабочек; паразит разви-
вается в яйце хозяина, откуда и общее название паразита «яйцееды». Осо-
бое значение имеют яйцееды потому, что поражают насекомых вредителей
на стадии яйца, т. е. до нанесения ими вреда растениям; яйцееды могут
дать в течение сезона целый ряд поколений, размножаясь, таким образом,
в огромном количестве. Они могут сильно снизить численность вредителя
при массовом его размножении.
Насекомые — переносчики болезней
Вред, наносимый насекомыми растениям, не ограничивается лишь
поеданием различных органов растения: насекомое может прививать рас-
тению-хозяину различные болезни. Так, некоторые членистохоботные
{тли, цикадки) и трипсы прививают растениям вирусные заболевания.
Такова мозаичная болезнь, прйвиваемая картофелю тлей Macrosiphum.
Очень часто насекомые переносят на растения грибки и бактерии, вызываю-
щие гниение растения. Так, личинка мухи Phorbia brassicae переносит
бактерий, вызывающих гниение корней капусты. Личинка другой мухи
того же рода (Phorbia fuscipes) переносит бактерии на клубни картофеля,
вызывающие загнивание клубня. Сладкий продукт, образующийся при
гниении клубня, служит при этом пищей личинке. Сходное явление наблю-
дается для короеда Dendroctonus micans, который нападает на вполне
здоровые ели и прогрызает в слое луба широкие ходы. При этом он вводит
под кору грибок синей гнили, развивающейся на древесине. Данным гриб-
ком короед и питается. Питание грибком, развивающимся на стенке
ходов, распространено у короедов-древесинников. Жук ухаживает за
мицелием грибка, вследствие чего на мицелии развиваются спорообразую-
щие конидии (амброзия). Споры грибка, используемые на заражение дре-
весины, всегда хранятся у жука в мускулистом желудке. Усвоению древе-
сины личинками дровосеков, также прогрызающими себе ходы под корой,
470
помогают дрожжи, которые у личинки имеются в слепых выростах начала
средней кишки.
Целый ряд насекомых является переносчиком серьезных заболеваний
человека и животных. Приведем только важнейшие. Вши (платяная, воз-
можно и головная) переносят сыпной и возвратный тифы; блохи переносят
чумную инфекцию. Комнатная муха механически передает ряд кишечных
заболеваний. Тяжелые инфекции переносят мухи из рода Glossina, распро-
страненного в Африке; так, Glossina morsitans — муха цеце — передает
скоту смертельную трипанозомную инфекцию (болезнь нагана); другой
африканский вид того же рода — Glossina palpalis передает Trypanosoma
gambiense, вызывающую сонную болезнь. В слепнях предполагают перенос-
чиков сибирской язвы. Виды рода Phlebotomus — москиты — передают
кожный лейшманиоз (пендинку) и внутренностный лейшманиоз (калаазар).
На юге СССР (Средняя Азия, Кавказ, Крым) распространен москит Phle-
botomus papatasii — переносчик вирусного заболевания летнего гриппа
(москитки), по времени своего распространения совпадающего с периодом
лёта москита (с начала июня до середины октября). Комары из рода Anop-
heles, как уже говорилось выше, переносят малярию.
Борьба с вредными насекомыми
Для борьбы с насекомыми — вредителями сельского и лесного хозяй-
ства, амбарными вредителями, паразитами и переносчиками болезней людей
и скота, разработан целый ряд мер, применяемых порознь или в различных
сочетаниях. Эти меры могут быть сведены в следующие методы: агротехни-
ческий, механический, физический, химический и биологический. К этим
мерам надо отнести и карантин, т. е. систему мер, препятствующих проник-
новению вредителей в страну из зарубежных стран и из одной области стра-
ны в другую область.
Карантин. Карантинные мероприятия подразделяются на меро-
приятия внешнего карантина, направленные на охрану территории СССР
от завоза и проникновения сельскохозяйственных и лесных вредителей и
растений, отсутствующих в СССР или встречающихся, но имеющих огра-
ниченное распространение; и на мероприятия внутреннего карантина, в
задачи которого входит не допустить вредителей из зараженного района
в район, где вредители отсутствуют.
Внешний карантин касается многих вредителей различных культур,
например хлопка. В СССР карантинных вредителей хлопчатника нет.
Тем более опасно завезти в районы нашего хлопководства таких вредителей
хлопчатника, как моль «розовый червь» (Pectinophora gossipiella), хлопковый
долгоносик (Anthonomus grandis) и ряд других, причиняющих в зарубежных
странах (Объединенная Арабская республика, Мексика, Индия и др.)
чрезвычайный вред хлопковым плантациям.
Большое значение имеет садовый карантин, направленный на охрану
плодово-ягодных, лесодекоративных и субтропических культур. Среди
объектов этого карантина следует упомянуть кровяную тлю (Eriosoma
lanigerum) — одного из серьезнейших вредителей яблонь, австралийского
желобчатого червеца (Icerya purchasi) — вредителя цитрусовых культур,
калифорнийскую щитовку (Aspidiotus perniciosus) — вредителя плодовых,
семечковых и косточковых культур. К этому же карантину следует отнести
и противофиллоксерный карантин, направленный против распространения
филлоксеры (Dactylosphaera vastatrix). К карантинным объектам относится
и описанный выше колорадский картофельный жук.
Агротехнический метод борьбы заключается в улучшении условий про-
израстания культурных растений, одновременно служит фактором по-
давления и уничтожения вредителей. Так глубокая пахота и лущение уни-
чтожают почвенных вредителей — личинок хрущей, щелкунов, чернотелок,
долгоносиков, совок и т. д. Очистка посевов от сорняков, очистка полей от
471
стерни и других остатков после уборки урожая сжиганием этих остатков.
Сорняки — места резервации вредителей, а стерня служит убежищем и
иногда местом зимовок этих вредителей. Дальнейшими агротехническими
мерами подавления вредителя могут служить севооборот и срок посева:
чередование культур при севообороте оказывается губительным для моно-
фагов и олигофагов, особенно для тех из них, которые развиваются дольше
одного вегетационного периода и не способны к значительным перемещени-
ям. Изменение сроков посева (ранние посевы) может также нарушить пита-
ние вредителя, развитие которого приурочено к определенным фазам раз-
вития растения-хозяина. Особенное значение в борьбе с вредителями имеет
селекция имунных форм растений.
Механический метод охватывает ряд разнообразных ис-
требительных приемов борьбы с вредителями. Создаются преграды переме-
щению вредителей по посевам; сюда же надо отнести бредни для вылавли-
вания насекомых (например, бабочек лугового мотылька), ручные, конные
или тракторные уловители (например, черепашкоуловитель при борьбе с
клопом — вредной черепашкой), клейкие ловчие кольца, накладываемые
на деревья в садах, плантациях и т д.
Физический метод заключается в применении высоких и
низких температур, смертельных для вредителя, в применении перегретого
пара или в просушивании пораженного вредителем материала; применяется
против амбарных вредителей. Против вредителей применяется и электричес-
кий ток. Механический и физический методы используются для непосред-
ственного уничтожения вредителя. Для той же цели служат методы хими-
ческий и биологический.
Химический метод уничтожает вредителя, вызывая его от-
равление. Метод получил широкое применение, так как связан с использо-
ванием высокой техники (самолеты, мощная аппаратура в виде опылителей
и опрыскивателей, камеры для окуривания и проч.). Яды применяются
в пылевидном, жидком и газообразном состояниях. По своему действию
на органы вредителя яды делятся на кишечные, контактные (действующие
на покровы) и дыхательные (действующие через трахейную систему). При-
меняемые яды — инсектисиды — разнообразны по химическому составу;
широко применяются органические синтетические яды, такие как ДДТ
(дихлордифенилтрихлорэтан) и ГХЦГ (гексахлорциклогексан).
Биологический метод борьбы заключается в использо-
вании хищников и паразитов данного вредителя для его уничтожения или
подавления К применению биологического метода борьбы с насекомыми
склоняет их поражаемость в природных условиях паразитами и хищни-
ками; так, за счет личинки шведской мухи питаются и развиваются более
30 видов паразитов, за ©чет гусеницы лугового мотылька — более 30 видов
паразитов, за счет гусеницы капустной белянки — 37 видов паразитов.
Имеется ряд наблюдений, когда при массовой яйцекладке насекомого-вре-
дителя (капустная совка, кукурузный мотылек, непарный шелкопряд и
др.) его размножение сводилось на нет при заражении яиц вредителя яй-
цеедом. Роль паразитов-хищников заключается не только в непосредственном
истреблении вредителей, но и в прививке им болезнетворных микроорганиз-
мов, вызывающих среди них эпизоотии; кроме того, паразиты истощают
организм вредителей и снижают их половую продукцию. Гусеницы Leu-
cania, пораженные наездником Apanteles, едят вдвое меньше, чем здоровые;
если перелетная саранча, зараженная тахиной Blaesoxipha, остается жить,
то плодовитость ее резко снижается.
Среди насекомых-хищников, участвующих в уничтожении вредных насе-
комых, особое значение имеют некоторые жуки, как-то: красотел (жужелица),
божьи коровки и нарывники. Последние — лишь в личиночной фазе. Кра-
сотел (Calosoma sycophanta), живущий на кронах деревьев, деятельно уни-
чтожает гусениц (рис. 433) Божьи коровки хищничают подобно красотелу
как на фазе имаго, так и личинки. Представление о роли божьих коровок
472
как истребителей дает следующий пример: семиточечная божья коровка
за сутки поедает до 270 тлей.
Жуки-нарывники (Zonabris, Epicauta) откладывают яйца в местах,
пораженных кубышками саранчовых. Развивающиеся из яиц личинки внед-
ряются в кубышки и уничтожают яйца саранчовых. Хищником, питаю-
щимся специально тлями, является личинка сетчатокрылого Chrysopa
Рис. 433. Хищные насекомые. / — муха-журчалка и ее личинка; // златоглазка
(Chrysopa); ее личинка и яйца; /// и IV — красотел (Calosoma) и его личинка; V
божьи коровки (Coccinellidae) и их личинка; VI — жужелица (Carabus); VII —
коротконадкрылый жук
(златоглазка). Стебельчатые яйца златоглазки откладываются самкой в
местах нахождения тлей. За счет тех же тлей хищничают пиявкообразные
личинки мух-журчалок (Syrphidae) (см. рис. 433).
Для использования насекомых — паразитов и хищников — в' борьбе
с вредителями их ввозят из других стран и акклиматизируют, а в тех слу-
473
чаях, когда они не размножаются в достаточной мере в условиях природы,
стремятся создать такие условия или организуют их массовое разведение
в искусственных, лабораторных, условиях.
Из числа таких полезных ввезенных божьих коровок особенного
внимания заслужила Rodolia (Novius) cardinalis, применяемая для истреб-
ления червеца Icerya. Щитовка Icerya была завезена с растениями в Сухуми
и стала сильно вредить мандариновым деревьям; с помощью Rodolia удалось
подавить размножение Icerya.
Таким же образом австралийский хищный жук Cryptolaemus (с. Сосе-
inellidae) успешно подавляет другого вредителя цитрусовых — червеца
Pseudococcus gahani, завезенного впервые в СССР в 1931 г.; виды рода
Chilocorus поедают калифорнийскую щитовку (Aspidiotus perniciosus),
распространившуюся в СССР на Черноморском побережье Кавказа.
Опыты по борьбе с вредителями с использованием эндопаразитов-
хищников проводятся за последние годы в СССР весьма деятельно и дали
хорошие практические результаты. Так, в целях борьбы с кровяной тлей,
завезенной к нам еще около половины XIX в. и распространившейся по
территории Украины, Крыма, Кавказа и Средней Азии, был в 1930 г. вве-
зен наездник Aphilinus mali, который на родине регулирует размножение
кровяной тли, не допуская ее массового размножения. Проведенные опыты
дали заражение тли в местах выпуска Aphelinus на 80—100%. Из яйцеедов
самое широкое применение за последние годы получила трихограмма
(Trichogramma evanescens) — многоядный наездник из сем. Chalcididae.
Трихограмма любопытна тем, что уничтожает яйца вредителя, не только
заражая свои яйца, но и производя укол яйца яйцекладом; этим очень уве-
личивается эффективность полезной деятельности трихограммы самки,
которая может отложить всего до 40 яиц. Однако ограниченное число яиц
в кладке возмещается быстрым развитием яиц трихограммы (8—11 дней)
и соответственно большим числом поколений паразита в сезон. Трихограм-
ма имеет большое значение в борьбе с яблонной плодожоркой, озимой сов-
кой, луговым мотыльком, кукурузным мотыльком, хлопковой совкой.
Значение наездников-яйцеедов, к которым относится трихограмма,
заключается в их эффективности как средства борьбы, так как они размно-
жаются быстро и уничтожают вредителя на фазе яйца, когда он еще не на-
нес культуре вреда.
С той же целью уничтожения вредных насекомых используют насеко-
моядных птиц, создавая им благоприятные условия (например, искусствен-
ные гнездовья в лесах.)
Наконец, для борьбы с вредителями используют также грибы и бак-
терии, вызывающие у насекомых заболевания.
ОБЩИЙ ОБЗОР ТИПА ЧЛЕНИСТОНОГИХ
Предками членистоногих считают кольчецов; с ними членистоногих
связывает то, что те и другие покрыты кутикулой, сегментированы, обла-
дают специальными органами передвижения (локомоторными) и сходным
расположением систем внутренних органов; имеется сходство и в строении
отдельных органов. От кольчецов членистоногие отличаются более сложной
организацией, стоящей на более высоком уровне.
Наружный покров членистоногих, как и кольчецов, состоит из кутику-
лы и подстилающего ее клеточного слоя гиподермы. Кутикула членисто-
ногих содержит вещество, называемое хитином1; она гораздо сильнее
развита, чем у кольчецов, и распадается на систему слоев (рис. 434). Кути-
кула членистоногих служит не только покровом, но и наружным скелетом,
к которому прикрепляется мускулатура. Хитинизированная кутикула (хи-
1 Хитин — полисахаридное соединение, разные исследователи дают для него раз-
личные формулы.
474
7 4
Рис. 434. Разрез через хитин и гиподерму
гусеницы Cnetocampa, проведенный отвес-
но к поверхности тела:
1 — поверхностный слой хитина, 2 — глубокий слой
хитина, 3—гиподерма, 4 — пигмент в хитине
тин) может давать выросты и складки, направленные внутрь тела, которые
служат также местом прикрепления мышц и опорой для различных внут-
ренних органов; эти выросты образуют так называемый эндоскелет (см.
рис. 290). Поверхность кутикулы несет часто неотчлененные выросты и
отчлененные придатки — щетинки — и их видоизменения — чешуйки.
Внешнее строение. В отличие от кольчецов, у которых различают лишь
два отдела тела — слабо обособленные голову и туловище,— у членисто-
ногих группы сегментов тела образуют, как правило, три отдела — голову,
грудь и брюшко; однако дифференцировка тела на отделы может быть
различной; брюшко может быть не обособлено от груди (многоножки); с
другой стороны, отделы тела могут вторично сливаться друг с другом.
Конечности тела у членистоногих, как и у кольчецов, расположены
посегментно. Конечность членистоногого—первоначально локомоторный ор-
ган; однако с дифференцировкой
тела передние конечности, располо-
женные на голове, становятся орга-
нами, направляющими движение,
и носителями органов чувств (уси-
ки) или же органами, хватающими
и перерабатывающими пищу (челю-
сти); при тесном срастании головы
с грудью передние ножки грудно-
го отдела превращаются в челюсти
(ногочелюсти); в основном же ко-
нечности груди или конечности
всего туловища остаются локомо-
торными. Часто некоторые брюш-
ные ножки превращаются в поло-
вые ножки (копулятивные органы,
яйцеклад) и другие придатки.
Конечности кольчецов — па-
раподии — нерасчленены; они не
образуют с туловищем сустава и
не разделены на части — членики, связанные друг с другом сочленениями;
поэтому размах движений параподии ограниченный. Конечность же чле-
нистоногих подвижно сочленена с сегментом, которому она принадлежит,
и разделена на членики, которые соединены друг с другом сочленениями,
допускающими то более, то менее сложные движения. Форма конечности
и степень ее расчленения характерны для отдельных групп членистоно-
гих; для Branchiata характерна двуветвистая конечность.
Мускулатура членистоногих в отличие от кольчецов не образует под
наружным покровом сплошного кожно-мускульного мешка, а разбита на
отдельные мышечные тяжи. К тому же мускулатура членистоногих, в от-
личие от гладкой мускулатуры кольчецов поперечнополосатая, вследствие
чего она может быстро сокращаться.
Полость тела. Полостью тела у кольчецов служит вторичная полость
тела, т е. целом, У членистоногих вторичная полость тела сливается с пер-
вичной, образуя окончательную полость тела (миксоцель). Однако отдельные
участки целома могут сохраняться обособленными {органы выделения,
половые железы).
Нервная система. Центральная нервная система (рис. 435) членисто-
ногих в основных чертах имеет то же строение, что и кольчатых червей.
В первичном виде она парная и слагается из двух стволов, расположенных
параллельно друг другу вдоль брюшной стенки тела внутрь от гиподермы
(наружного покрова). В каждом сегменте тела ствол образует утолщение —
нервный узел, связанный с узлом другой стороны простой или двойной по-
перечной перемычкой. Часть продольного ствола, соединяющая два сле-
дующих друг за другом узла, получила название коннективы, поперечные
475
же перемычки — комиссур. По общему строению центральная нервная
система напоминает, таким образом, лестницу. Передняя пара коннектив
охватывает с обеих сторон переднюю часть пищевода так, что первая пара
или две первые пары узлов расположены над кишечником, остальная же
часть центральной нервной системы лежит под кишечником; поэтому перед-
ний конец в составе одного или двух пар узлов, слившихся в одно целое,
получил название надглоточного узла, или мозга, подкишечная же часть
называется брюшной цепочкой.
Центральная нервная система такой типичной лестничной формы имеет
у членистоногих весьма ограниченное распространение. Обычно же пара
продольных нервных стволов сближается друг с другом по средней линии
брюшной
стенки, причем каждая
Рис. 435. Типы строения нервной системы рако-
образных. / —Phyllopoda; // —Leptostraca;
/// — Stomatopoda; IV — Decapoda brachyura;
V Copepoda isokerandria
пара узлов сливается в общий непарный
узел (см. рис. 290). Парность
при этом внешне сохраняется
лишь коннективами; однако и
пара коннективов может
слиться во внешне непарную
коннективу. Дальнейшее из-
менение центральной нервной
системы заключается в том,
что узел приближается к впе-
реди лежащему узлу, причем
соединяющие их коннективы
укорачиваются и могут совсем
исчезнуть. В последнем слу-
чае два или более узлов сли-
ваются в общую нервную
массу; такое слияние узлов
происходит вместе с укороче-
нием или слиянием друг с
другом сегментов тела. Узлы
могут и перемещаться вдоль
коннектив; такое перемещение
узлов вперед наблюдается в
течение онтогенеза членисто-
ногих, именно благодаря
перемещению вперед передних
узлов брюшной цепочки мозг
членистоногих может слагать-
ся из двух пар узлов, слившихся в одну общую массу. Частью благода-
ря этому слиянию узлов, частью благодаря дальнейшей дифференцировке
центров надглоточного узла в нем различают три (см. рис. 387 и 388) от-
дела. Срастание расположенных друг за другом узлов с образованием
общего узла (подглоточного) наблюдается и в передней части брюшной це-
почки.
Органы чувств. В наиболее простом виде орган чувств (рецептор) чле-
нистоногих, как и орган чувств кольчатых червей, представлен восприни-
мающей раздражение специализированной гиподермальной клеткой (чув-
ствующей клеткой). Эта клетка связана с одной стороны непосредственно
с каким-либо кутикулярным образованием, а с другой — с центральной
нервной системой при участии своего отростка. В органах осязания чувствую-
щая нервная клетка связана часто со щетинкой; в случае органов обоняния
и вкуса щетинку обыкновенного строения замещают тонкостенные образо-
вания разнообразной формы — видоизмененные щетинки. Особенно слож-
ного строения достигают высшие органы чувств — органы слуха и зрения.
У членистоногих различают две категории органов зрения: сложные,
или фасеточные, глаза и глаза простые, иначе глазки. Только сложным, или
фасеточным, глазам принято приписывать значение органов собственно
476
Рис. 436. Продольный
разрез через два омма-
тидия сложного глаза
речного рака. Налево —
омматидий освещен, на-
право — в темноте:
I — роговица, 2 — гиподер-
мальные клетки роговицы,
3— клетки хрусталика, 4а и
и 46—пигментные клетки,
5а и 56— клетки сетчатки,
6 — рабдом, 7 — основная пе-
репонка
зрения, органов восприятия формы и движения предметов. В основном
строение глаз той или другой категории сходно. Составные части как про-
стого, так и сложного глаза располагаются послойно, причем наружные
слои, представленные частью слоями кутикулы, частью клеточными слоями,
играют роль преломляющей (оптической) среды, а глубже расположен
слой воспринимающих раздражение (зрительных) клеток, т. е. слой сетчат-
ки, или ретины.
От простого глаза сложный отличается тем, что разбита на участки не
только воспринимающая раздражение часть глаза,
но и часть преломляющая; так, в сложном глазу
хитиновый покров разбит на участки — фасетки.
Сложный глаз по всей своей толщине от по-
верхности до дна распадается на столбики, или
клинья, составляющие элементы глаза (рис. 436).
Каждый такой элемент сложного глаза называется
омматидием. В составе каждого омматидия имеется
группа из 7—9 клеток ретины (сетчатки); часть
каждой клетки ретины, ближайшая к оси всей
группы, обладает особым строением; на эту часть
непосредственно воздействует свет; эти части всех
клеток ретины омматидия образуют рабдом.
Пищеварительная система. Кишечный канал
членистоногих (см. рис. 389), как и кольче-
цов, имеет форму трубки с передним, глоточным,
или ротовым, отверстием и задним, анальным.
Глоточное отверстие расположено на брюшной
стороне у переднего конца тела, а заднепроход-
ное — на заднем конце тела или с брюшной сторо-
ны у заднего конца тела. Как и у кольчецов, име-
ются три отдела кишечника, из которых передняя и
задняя кишки развиваются как впячивания экто-
дермы и в связи с этим обладают кутикулярной
(хитиновой) выстилкой. Средняя кишка развивает-
ся из энтодермы. Средняя и задняя кишки членис-
тоногих в отличие от кишки кольчецов, могут
образовывать выпячивания, функционирующие как
железы (пример — мальпигиевы трубочки).
Кровеносная система. Кровь членистоногого,
состоящая из лимфы и гемоцитов (клеток крови),
движется частью по системе трубок, обладающих
собственными стенками, частью по синусам, или
лакунам, представляющим участки полости тела,
сообщающиеся друг с другом.
Таким образом, в отличие от замкнутой крове-
носной системы кольчецов кровеносная система чле-
нистоногих всегда не замкнутая. Наиболее разви-
та система кровеносных сосудов в тех случаях, когда дыхание локализова-
но, когда органами дыхания служат жабры или легкие. Когда же органы
дыхания распространяются по всему телу (трахейная система) или специаль-
ных органов дыхания вообще нет, тогда система кровеносных сосудов ре-
дуцируется, ограничиваясь одним спинным сосудом, но и он может исчезать.
Наиболее развитая кровеносная система (см. рис. 367) состоит из спин-
ного сосуда, образующего сердце, и отходящих от него вперед, назад и по-
сегментно в стороны артериальных сосудов. Кровь из сердца поступает в
полость тела.
Из всех частей полости тела кровь поступает в околосердечный синус
полости тела и далее через отверстия (остии) сердца в полость последнего.
Каждой паре остий соответствует отдел сердца — камера, сокращение стен-
477
Рис. 437. Антеннальная железа Mysis:
1 — целомический мешочек, 2— лабиринт, 3 —
выносящий проток (наружное отверстие)
ки которой дает движению крови определенное направление: из данной
камеры кровь поступает в одну из соседних камер (обычно лежащую впе-
реди). Ток крови направляется системой клапанов.
Дыхательная система. Членистоногие в зависимости от того, живут ли
они в воде или на суше, имеют различные органы дыхания. Для дыхания
в воде наряду с общей поверхностью тела служат специальные органы вод-
ного дыхания — жабры.
Для дыхания атмосферным воздухом членистоногим служат впяченные
внутрь тела участки покрова; сюда относятся трахеи и легкие; трахеи пред-
ставлены трубками, обычно ветвящимися.
Личинки насекомых, живущие в воде, имеют жабры, обычно комбини-
рующиеся с трахеями; в этом случае трахейная система не сообщается с
наружной средой, ее стигмы замкну-
ты и часть трахейной системы прони-
кает в жабру.
Выделительная система. Органа-
ми выделения у кольчецов служат
нефридии. К органам выделения чле-
нистоногих относятся: 1) трубкообраз-
ные слепые выпячивания средней или
задней кишки, так называемые маль-
пигиевы сосуды; 2) железы, связан-
ные с основанием конечностей головы
или груди,— антеннальная (см. рис.
293, 437), нижнегубная и коксальная.
Происхождение этой второй катего-
рии желез связывают с нефридиями
кольчецов.
Половая система. Половые желе-
зы кольчецов тесно связаны с целоми-
ческими мешками и заложены в их
стенке; они, таким образом, могут быть
многочисленными и расположены по-
сегментно. От половой системы коль-
чецов половая система членистоногих
отличается тем, что половых желез
и их протоков имеется не больше одной
пары. Внутренние половые органы —
семенники, яичники и их протоки —
занимают у самцов и самок в тулови-
ще сходное положение. С половыми
протоками связаны придаточные орга-
ны: семенной пузырек (vecicula semi-
nalis) простатическая железа самца, семеприемник (receptaculum seminis)
самки.
У самца членистоногих очень часто имеется совокупительный аппарат,
иногда очень сложный, включающий систему выростов около наружного
полового отверстия, и вырост (penis), обычно несущий половое отверстие.
У самки в связи с кладкой яиц в твердую среду бывает иногда сложно по-
строенный яйцеклад. Считают, что как совокупительный аппарат самца,
так и яйцеклад самки образовались из локомоторных ножек, так что есть
основание назвать наружный половой аппарат многих членистоногих по-
ловыми ножками — гоноподами.
Внутренний половой аппарат первоначально парный; в случае непар-
ного семенника или яичника железа образуется через слияние пары желез.
Развитие. Эмбриональное развитие членистоногих весьма разнообразно:
дробление яйца может быть как полным, притом равномерным и нерав-
номерным, так и частичным. Обычно яйцо членистоногого богато желтком,
478
в связи с чем дробление частичное, поверхностное; получившаяся от дроб-
ления бластодерма окружает бластоцель, заполненный желтком. Гастр ул я-
ция лишь редко представлена инвагинацией, обычно на определенном месте
поверхности яйца происходит иммиграция клеток.
В постэмбриональном развитии членистоногих можно различать пря-
мое развитие (эпиморфоз), не имеющее личиночных фаз, и метаморфоз —
развитие с личиночными фазами. Длительность эмбрионального и постэм-
брионального развития крайне варьирует.
ФИЛОГЕНИЯ ЧЛЕНИСТОНОГИХ
В очерке общей организации членистоногих было указано, что на ос-
новании сходства строения членистоногих со строением кольчецов пос-
ледних считают предками членистоногих; в этом вопросе разногласий не
существует. В свою очередь, общее сходство в строении ракообразных, три-
лобитов, хелицероносных и трахейнодышащих указывает на то, что это
сходство основано не только на происхождении всех трех подтипов от коль-
чатых червей, но и на непосредственном родстве этих подтипов, иначе го-
воря, по мнению большинства авторов, членистоногие в своей совокупности
представляют естественную систематическую группу.
Однако в вопросе, монофилетичен или полифилетичен тип членисто-
ногих, авторы расходятся.
Поскольку кольчатые черви — морские организмы, для приверженцев
монофилетического происхождения членистоногих естественно было искать
общего предка всех членистоногих среди морских форм, каковым являются
трилобиты, большая часть ракообразных и меростомовые. Авторы, придер-
живавшиеся этого взгляда, искали предков для прочих членистоногих
среди ракообразных. Так, Фриц Мюллер, ссылаясь на сходство в строении,
пытался обосновать происхождение насекомых от десятиногих раков и имен-
но от их зоёаобразной личиночной фазы, однако сходство это очень поверх-
ностное.
Последующие авторы стремились опереться на более подробный и точ-
ный анализ черт сходства, в том числе и на сходство в членистости ножки.
Сравнительное изучение строения членистой ножки, этого основного приз-
нака всех членистоногих, однако, вряд ли может дать положительный ответ.
При общем сходстве членистой ножки у различных типов в подробностях
ее строения имеется существенное расхождение. Это расхождение получает
тем большее значение, что сравнительное изучение ножки трахейнодышащих
показывает, что у последних эволюция членистости ножки шла путем, от-
личающимся от других членистоногих. Притом расхождение в формирова-
нии ножки ракообразных и трахейнодышащих заключается не только в
том, как развивалась членистость ножки, но и в ее первоначальном по-
ложении на сегменте, а также и в общем строении сегмента туловища. Осо-
бенно следует отметить то, что у ракообразных отсутствует самостоятельная
плевральная область сегмента, с которой теснейшим образом связана нож-
ка и ее формирование у трахейнодышащих. Эти и другие черты расхождения
в строении ракообразных и трахейнодышащих приводят к заключению, что
непосредственного родства между ракообразными и трахейнодыиющими
не существует.
Столь же неудачными следует считать попытки связать происхождение
трахейнодышащих непосредственно с трилобитами или с паукообразными.
Результаты анатомического исследования первичнотрахейных заста-
вили Геккеля еще в конце минувшего века отказаться от своей первоначаль-
ной (1866 г.) монофилетической родословной членистоногих и дать дифиле-
тическую схему (1896 г.), в которой филогения всех дышащих трахеями
членистоногих толковалась независимой от филогении водных членисто-
ногих, т. е. ракообразных, трилобитов и меростомовых. Таким образом, в
своей второй родословной Геккель высказался за дифилию членистоногих.
479
Еще дальше в этом направлении пошел наш эмбриолог П. П. Иванов, он
высказался в пользу трифилии членистоногих, т е. за самостоятельное,
независимое друг от друга происхождение от кольчецов трех основных
групп членистоногих — ракообразных, хелицероносных и трахейнодыша-
щих.
Как указано выше, все же можно предполагать, что жабернодышащие
и хелицероносные произошли от кольчецов одним общим корнем. С другой
стороны, не может быть сомнения, что трахейнодышащие, т е. многоножки
и насекомые, состоят друг с другом в непосредственном родстве и что насе-
комые произошли от многоножек (стр. 399—407).
В свою очередь, указания, что сформирование основных отделов чле-
нистой ножки произошло у многоножек, что базальные отделы ножки впер-
Рис. 438. Спинки Tracheata. /—/2-й и 14- тергиты тулови-
ща Lithobius (Chilopoda); // —тергиты 2-го и 3-го сегмен-
тов туловища Symphylella (Symphyla); /// тергиты 2-го и
3-го грудного и 1-го брюшного сегментов нимфы Baetis
(Ephemeroptera) с зачатками крыльев и жабры
Прямокрылые
и им
родственные
Веснянки
Поденки
Стрекозы
Цпенисто -
хоботные
и пузыреногие
Таракановые
и им
родствен -
ные
Жуки [
Сетчатокрылые
и им
родственные
Первичные
крылатые
Перепончато-
крылые
Первичнобескрылые
Двупарноногие
Сколопендрелловые
Первичные
сколопендрелловые
Губоногие
Пербичнотрахейные
Полихеты
Рис. 439. Схема филогенетического развития Tra-
cheata
480
вне сформировались у низших губоногих, свидетельствуют о том, что если
не Onychophora собственно, то близкие к ним формы послужили предками
многоножек. Таким образом, можно предполагать дифилию членистоногих.
Какова же наиболее вероятная родословная трахейнодышащих по
имеющимся ныне данным?
Как указывалось уже выше, можно предполагать, чтотрахейнодыша-
щие собственно, т. е. многоножки и насекомые, произошли от онихофорооб-
разных предков, близких к современным Onychophora. Средитрахейноды-
шащих (Tracheata) наиболее примитивной группой являются, несомненно,
низшие губоногие, в пределах которых, как описано выше (стр. 433), про-
исходил процесс эволюции ножки и сегментации туловища, высшую ступень
которой можно видеть у высших губоногих (мухоловок), двупароногих и
сколопендрелловых. У последних и у Thysanura среди низших насекомых
мы видим последнюю фазу слияния двух соседних сегментов туловища
в один. Те же сколопендрелловые связывают многоножек не только с пер-
вичнобескрылыми насекомыми, но и с крылатыми насекомыми. У низших
губоногих прототипом крыла служит парный выступ на заднем краю спин-
ного щита (рис. 438i ), который служит опорой для дорзо-вентральной мус-
кулатуры, в том числе и для мышц ножки. У сколопендрелловых (рис. 438ц)
этот выступ приобретает форму зачатка крыла в том виде, как мы его на-
ходим у нимф (личинок) насекомых, например поденок (рис. 438щ). Пре-
вращение опорного для мышц приспособления в подвижной орган — крыло,
которое имело место в эволюции насекомых, является заключительным
актом этого процесса.
Схема филогенетического развития трахейнодышащих представлена
на рис. 439.
ТИП XII. ИГЛОКОЖИЕ (ECHINODERMATA)
ХАРАКТЕРИСТИКА
Исключительно морские животные, во взрослом состоянии построенные
в большинстве случаев по лучевой симметрии (табл. XIV). В личиночных
стадиях хорошо выражена двусторонняя симметрия (рис. 440), и поэтому
лучевая симметрия взрослых должна рассматриваться как вторичная,
выработавшаяся в связи с переходом к сидячему или малоподвижному об-
разу жизни. У большинства форм хорошо выражен внутренний — мезо-
дермический — скелет, состоящий из отдельных известковых элементов
Рис. 440. Личинки иглокожих (схема). / —
голотурий (аурикулярия); II — морских
звезд (бипиннария); III — морских ежей
(эхиноплутеус)
Рис. 441. Разрез через стенку тела голотурии:
1—эпидермис, 2—соединительнотканый слой кожи, 3 —
отдельные скелетные пластинки, 4— мускулатура, 5—эпи-
телий, выстилающий целом
(рис. 441), в некоторых случаях
сливающихся в сплошной пан-
цирь. Полость тела вторичная —
целомическая — хорошо разви-
тая. Имеется особая система —
воднососудистая, или амбуля-
кральная, выполняющая глав-
ным образом двигательную функ-
цию.
Следует особо отметить, что
иглокожие вместе с хордовыми
относятся к группе вторично-
ротых животных: у личинки
первичный рот—бластопор — за-
мыкается, а новый рот, кото-
рый и будет ртом взрослого
животного, прорывается на
другом месте, часто на проти-
воположном полюсе личинки.
Мезодерма и вторичная
полость тела иглокожих за-
кладываются энтероцельным
путем в виде двух выпячи-
ваний стенок первичной
кишки. Далее эти выпячива-
ния отшнуровываются от
кишки и распадаются на три
4 пары пузырей, преобразую-
щиеся в участки вторичной
полости тела. Такой способ
формирования мезодермы и
вторичной полости тела ха-
рактерен также для хордовых.
482
Интересной особенностью иглокожих является то, что у большинства
из них имеются три нервные системы — эктоневральная, гипоневральная
и энтоневральная.
Все три системы связаны с прилежащим эпителием. Первая система
образуется из кожного эпителия, вторая из эпителия псевдогемальной си-
стемы (см. ниже), третья из эпителиальной выстилки целома. Последнее
обстоятельство особенно интересно, так как представляет собой исключи-
тельный случай формирования нервной системы за счет мезодермы.
В настоящее время известно всего около 6000 видов иглокожих. Игло-
кожие — очень древняя группа; благодаря хорошо выраженному скелету
они очень богато представлены в ископаемом состоянии. В прошлые геоло-
гические периоды были более разнообразны.
Практическое значение иглокожих небольшое. Некоторые из них яв-
ляются объектами промысла как пищевые объекты, многие служат пищей
ряда промысловых рыб, например камбалы.
КЛАССИФИКАЦИЯ
Тип иглокожие (Echinodermata)
Класс морские лилии (Crinoidea)
Класс морские звезды (Asteroidea)
Класс змеехвостки (Ophiuroidea)
Класс морские ежи (Echinoidea)
Класс морские огурцы, или голотурии (Holothurioidea)
КЛАСС 1. МОРСКИЕ ЗВЕЗДЫ (ASTEROIDEA)
ХАРАКТЕРИСТИКА
Основной класс иглокожих, у представителей которого все признаки
типа выражены очень хорошо. У взрослых форм как снаружи, так и в рас-
положении внутренних органов хорошо выражена лучевая симметрия
(рис. 442), у личинок бипиннарий — двусторонняя (рис. 440). Скелет из
известковых пластинок, расположенных главным образом на нижней по-
верхности животного. Амбулякральная система развита хорошо и служит
преимущественно для движения.
Представитель класса морских звезд
— морская звезда acme риас (Asterias rube ns)
Внешнее строение. Тело животного (табл. XIV, рис. 442) состоит из
центральной части, или диска, и отходящих от него пяти лучей. Легко от-
личается нижняя, или оральная, сторона и верхняя, или аборальная. В
центре оральной стороны расположено ротовое отверстие, а в бороздках,
идущих по лучам,— конечные части амбулякральной системы, так назы-
ваемые амбулякральные ножки (см. ниже), каждая с присоской на конце.
На аборальной стороне расположены: начальная часть амбулякральной
системы — мадрепоровая пластинка (см. ниже), анальное отверстие и от-
верстия половых желез.
Asterias rubens достигает в поперечнике 20 см, цвет ее лиловато-корич-
невый, иногда красноватый. Поверхность тела покрыта кожей (рис. 443)
с многочисленными короткими и тупыми иглами, между которыми располо-
жены особые образования — педицеллярии, имеющие вид щипчиков, слу-
жащих для очищения поверхности морской звезды от посторонних частиц
483
Рис. 442. Вскрытая морская звезда Asterias rubens (вид с
аборальной стороны), естественная величина до 20 см в диа-
метре (обычная форма наших северных морей)*
/— желудок, 2— печеночные выросты, 3—ректальные железы, 4— ка-
менистый канал с ыадрепоровой пластинкой, 5—ампулы амбулякральных
ножек, 6 гонады
/
Рис. 443. Участок кожи Asterias
rubens при большом увеличении:
/ — основания игл (сами иглы удалены),
2— педицеллярни, 3 — кожные жабры
Рис. 444. Скелет луча морской
звезды:
J амбулякральные пластинки, 2 —
адамбулакральные пластинки, 3 — ниж-
ние краевые пластинки, 4 — верхние
краевые пластинки, 5 — аборальная
стенка луча, 6 — амбулякральные ножки
и кожные жабры. Последние представляют собой полые конические, тонко-
стенные выросты, в которые заходят выросты полости тела.
Скелет. Скелет (рис. 444) состоит из известковых пластинок, располо-
женных несколькими рядами по оральной стороне и отчасти по бокам лучей.
По средней линии луча лежит двойной ряд амбулякральных пластинок,
образующий углубленную бороздку. Между этими пластинками выходят
наружу амбулякральные ножки. К амбулякральным пластинкам примы-
кает с каждой стороны по ряду адамбулякральных пластинок, а к послед-
ним — по два ряда краевых, образующих боковую стенку луча. Один набор
этих пластинок образует как бы отдельное звено, подвижно сочленяющееся
с соседними — передним и задним.
Мышечная система. Мускулатура развита довольно хорошо, что стоит
в связи со сравнительно слабым развитием скелета. Лучи звезды могут
Рис. 445. Разрез через диск и луч морской звез-
ды, схема (ср. с рис. 442):
1 — рот, 2 — желудок, 3 — анальное отверстие,/ — пече-
ночный вырост, 5 — мадрепоровая пластинка, 6 — осевой
комплекс, 7—каменистый канал, 8—кольцевой канал
амбулякральной системы, 9 — радиальный канал этой
системы, 10— амбулякральные ножки, // — их ампулы,
/2 — осевой орган, 13 — кольцевой канал кровеносной
системы, 14 — радиальный канал этой системы, 15 — ораль-
ное кольцо эктоневральной системы, 16 — радиальный
ствол этой системы, 17 — гонада
сгибаться как вверх, так и вниз. Сгибание осуществляется тремя мышеч-
ными лентами, идущими вдоль каждого луча и соединяющимися в центре
аборальной стороны. Кроме того, имеются мышцы, связывающие амбуля-
кральные пластинки одной пары, амбулякральные пластинки одного звена
с соответствующими пластинками впереди и сзади, а также амбулякральные
пластинки с адамбулякральными. Такая система мышц делает луч морской
звезды достаточно подвижным.
Полость тела, или целом. Под кожей, скелетными элементами и мус-
кулатурой расположена обширная полость тела, входящая в лучи
(рис. 445) Целом закладывается энтероцельно сперва в виде парного цело-
мического пузырька по обеим сторонам первичного кишечника. При даль-
нейшем развитии каждый из этих пузырьков распадается на три. Из обоих
задних участков образуется целом, из левого среднего — гидроцеля —
амбулякральная система (см. ниже). Правый средний участок редуцируется,
а оба передних образуют осевые синусы, составляющие часть псевдогемаль-
ной системы (см. ниже). Полость тела содержит полостную жидкость с мно-
гочисленными амебоидными клетками и выстлана ресничным эпителием.
Амбулякральная, или воднососудистая, система. Амбулякральная си-
стема (рис. 445) построена строго по лучевой симметрии и состоит из цен-
тральной части и радиальных ответвлений, расположенных в лучах. В цен-
тре расположен кольцевой канал, окружающий пищевод. От этого канала
отходят радиальные каналы в каждый луч, а также каменистый канал,
который идет к аборальной стороне и соединяет всю амбулякральную си-
485
стему с внешней средой посредством мадрепоровой пластинки (см. выше),
представляющей собой пластинку, пронизанную мельчайшими отверстиями.
На кольцевом канале, в промежутках между местами отхождения радиаль-
ных каналов, или в интеррадиусах располагаются попарно особые желе-
зистые придатки — тидемановы тельца, в которых образуются амебовид-
ные клетки целома. От радиальных каналов соответственно звеньям
скелетных пластинок ответвляются попарно многочисленные веточки, от ко-
торых внутрь полости луча отходят сократимые пузырьки, или ампулы,
7
Рис. 446. Схема строения осевого комплекса морской звезды (соотноше-
ния целома, амбулякральной, псевдогемальной и кровеносной систем):
/—мадрепоровая пластинка, 2—каменистый канал, 3—кольцевой канал амбуляк-
ральной системы, 4 — осевые каналы псевдогемальной системы, 5—кольцевые каналы
этой системы, 6 — половой канал этой системы, 7 — кольцевой канал кровеносной
системы,- 8— осевой орган, 9— аборальный кровеносный сосуд
а наружу — тоже сократимые амбулякральные ножки. Ножки эти могут
вытягиваться и имеют на конце присасывающуюся подошву.
Основной функцией амбулякральной системы является двигательная,
осуществляемая амбулякральными ножками. Через мадрепоровую плас-
тинку в амбулякральную систему поступает вода, которая и наполняет ее.
Ножки вытягиваясь, присасываются своими присосками к какому-либо
предмету, а затем, сокращаясь, подтягивают звезду в этом направлении.
При сокращении ножек избыток жидкости переходит в ампулы и далее в
радиальный, кольцевой и каменистый каналы и через мадрепоровую плас-
тинку выходит наружу.
Кроме двигательной функции, амбулякральной системе принадлежит
выделительная и в некоторой степени и дыхательная.
486
Рис. 447. Конец луча Asterias
rubens:
/ — амбулякральные ножки, 2 — кон-
цевое щупальце, 3 — глазок
По своему происхождению амбулякральная система представляет со-
бой обособившийся и разросшийся участок целома (см. выше).
Псевдогемальная система. Псевдогемальная система так же, как амбу-
лякральная, является частью целома. Она состоит из двойного сосудистого
кольца, окружающего пищевод и лежащего кнаружи от кольцевого канала
амбулякральной системы. От этого двойного кольца в лучи отходят ради-
альные каналы. Кроме того, в состав псевдогемальной системы входят осе-
вые синусы — полости, окружающие осевой орган кровеносной системы,
и аборальные синусы (рис. 446).
Нервная система, (рис. 445) построена тоже по лучевой симметрии.
Имеются нервное кольцо вокруг пищевода и отходящие от него в лучи ра-
диальные нервные стволы. От этих стволов отходят нервы к амбулякраль-
ным ножкам. Кроме этой поверхностной, или
эктоневральной, нервной системы, под ней
имеется еще глубинная, или гипоневральная,
вполне повторяющая по своему расположению
первую. На аборальной стороне имеется и
третья — аборальная, или энтоневральная,
нервная система, состоящая из пяти нервов,
проходящих по аборальной стороне лучей и
сходящихся в центре.
Органы чувств. Органы чувств развиты
довольно слабо. Органами осязания служат
амбулякральные ножки и короткие щупальца
на концах лучей (рис. 447). В связи с послед-
ними находятся просто устроенные светочув-
ствительные органы, или глазки, представляю-
щие простые глазные ямки.
Пищеварительная система. Пищевари-
тельная система (рис. 445) начинается
широким круглым ртом, который сильно
растягивается и вбирает в себя крупную до-
бычу. Короткий пищевод идет в объемистый
тонкостенный желудок, занимающий всю цен-
тральную часть диска. Желудок может выво-
рачиваться через ротовое отверстие наружу и обволакивать жертву. Вверх
от желудка к аборальному полюсу идет короткая прямая кишка, окру-
женная группой ректальных желез, от желудка отходят во все лучи пар-
ные отростки — «печеночные» выросты, имеющие вид трубочек, окружен-
ных в лучах придатками (рис. 442). Последние являются железистыми
образованиями, выделяющими пищеварительные соки в центральную часть
желудка, где и происходит переваривание захваченной пищи.
Кровеносная система. Кровеносная система также построена строго
по лучевой симметрии и расположена внутри перегородок псевдогемальной
системы. Внутри перегородки между кольцевыми каналами псевдогемаль-
ной системы проходит кольцевой канал кровеносной системы (рис. 446),
дающий радиальные каналы в лучи. Кольцевой канал соединяется с соот-
ветствующими каналами на аборальной стороне тела — осевым органом,
идущим внутри осевого комплекса и представляющим собой соединительно-
тканый тяж, пронизанный кровеносными сосудами. Аборальная часть
осевого органа имеет способность сокращаться. Осевой орган кровеносной
системы, окружающие его синусы и располагающийся по соседству с ними
каменистый канал амбулякральной системы входит в состав осевого комп-
лекса органов. От аборального сплетения кровеносной системы отходят
кровеносные сосуды к гонадам, а от аборального конца осевого органа
ответвляются сосуды, направляющиеся к кишечнику.
Наполняющая кровеносную систему жидкость не содержит никаких
пигментов и не несет дыхательных функций. В стенках осевого органа,
487
так же как в тидемановых тельцах, образуются амебоидные клетки целома.
Дыхательная система. Дыхание морской звезды осуществляется глав-
ным образом через кожные жабры (см. выше) (рис. 443), в которые входят
выросты целома. Кроме того, некоторую роль в газообмене играет и амбуля-
кральная система.
Выделительная система отсутствует. Образовавшиеся в теле звезды
продукты обмена выводятся при помощи амебоидных клеток целома. Эти
клетки нагружаются продуктами обмена и затем выходят из тела наружу
обычно через тонкие стенки кожных жабр или через амбулякральную
систему. Часть продуктов обмена не выводится из тела, а откладывается
в коже или других тканях тела в виде зерен и других образований.
Половая система. Половая система устроена очень просто и состоит из
пяти парных гонад, расположенных на аборальной стороне тела в интерради-
усах (рис. 442 и 445). При разрастании гонады заходят в лучи, причем в
один луч заходят отростки двух гонад с одной и с другой стороны. Звезды
Рис. 448. Регенерация морской звезды из одного отрезанного луча
раздельнополы, но определить пол можно только при вскрытии звезды по
цвету или микроскопическому строению гонад.
Развитие. Зрелые яйца откладываются самкой прямо вводу Туда
же в воду выпускает сперму и самец. Оплодотворение и развитие происходят
в воде. В яйце формируется и личинка звезды — микроскопическая бшшн-
нария (рис. 440), имеющая строго двубокую симметрию, причем парные
отростки, образующиеся по бокам тела, являются приспособлениями к
планктическому образу жизни. После некоторого времени плавания бипин-
нария претерпевает метаморфоз, опускается на дно и превращается в ма
ленькую звездочку. Следует упомянуть, что маленькие прозрачные личин
ки морских звезд послужили объектом, на котором И. М. Мечников открыл
явление фагоцитоза.
Экология и географическое распространение. Обитает Asterias на раз
личных глубинах. Ее можно найти, и у самого уреза воды на береговых кам-
нях и на глубине до 400 м. Передвигается звезда сравнительно быстро, дей-
ствуя амбулякральными ножками. При движении она обычно сперва нес-
колько приподнимает конец луча, как бы ощупывая им окружающую сре-
ду, затем вытягивает ножки, присасывается ими к субстрату и, сокращая
их, подтягивается вперед. Скорость передвижения звезды по горизонталь-
ной поверхности достигает 15 см в час.
Asterias — хищник. Питается главным образом моллюсками, захваты-
вая их целиком. Происходит это следующим образом: звезда выпячивает
свой тонкостенный желудок, охватывает им жертву и затем втягивает ее
внутрь, причем переваривание начинается еще до того, как желудок втянул-
ся внутрь тела. Раковина моллюска выбрасывается затем через ротовое
отверстие
488
У Asterias, так же как и у других морских звезд, сильно развита спо-
собность к регенерации. Один оторванный луч может восстановить недо-
стающие части, причем получается несколько странный по форме орга-
низм (рис. 448)
Распространена Asterias очень широко и встречается по всему побе-
режью Атлантического океана, начиная от Сенегала в Африке до Северной
Норвегии. В наших морях она встречается по всему Мурманскому побе-
режью и в Белом море, являясь наиболее обычной и часто встречающейся
морской звездой этих районов.
Систематическое положение. Asterias rubens относится к ряду педи-
целляриевых морских звезд, т. е. звезд, у которых всегда имеются педицел-
лярии.
ОБЩИЙ ОБЗОР КЛАССА
(табл. XIV). Это крупные звезды,
Рис. 449. Морская звезда, пожирающая
двустворчатого моллюска
Кроме описанной Asterias rubens, типичной пятилучевой морской звез-
ды, имеются многолучевые звезды, число лучей у которых достигает иногда
двух десятков. Примером таких звезд могут служить встречающиеся у нас
на севере представители рода Solaster
имеющие до 16 лучей и поражающие
своей яркой окраской — малиново-
красной или красно-оранжевой.
У других звезд лучи, наоборот,
выражены слабо, и они имеют вид
пятиугольников только с несколько
вытянутыми углами. Примером таких
звезд может служить встречающаяся
в Баренцевом море звезда церамстер
Ceramster granularis (табл. XIV).
Многие морские звезды питаются
моллюсками, причем не только брю-
хоногими, но и двустворчатыми. Пос-
ледних они охватывают лучами (рис.
449) и постепенно открывают, растя-
гивая створки амбулякральными
ножками. Раскрыв раковину, звезда
запускает внутрь выпяченный желу-
док и выедает мягкие части моллюска,
морские звезды — опасные вредители
выше) и первые приходится время от времени очищать от наползших на них
звезд. Иногда морские звезды нападают даже на небольших рыб. В отли-
чие от этих звезд другие звезды — грунтоеды.
Некоторые морские звезды живородящи: молодь развивается у них в
особой полости на спинной стороне.
Распространены морские звезды очень широко и встречаются во всех
морях, особенно в теплых и тропических. Благодаря своей оригинальной
форме и обычно яркой окраске они сразу бросаются в глаза на морском
побережье и придают ему особый колорит. Морские звезды, как и большин-
ство иглокожих, очень чувствительны к солености. Моря со слабой соле-
ностью, как наши южные моря и Балтийское море, непригодны для их
существования, и они там не встречаются. В наших северных и дальневос-
точных морях они разнообразны и многочисленны: на севере известно
33 вида их, на востоке более 60.
Кроме названных выше сравнительно небольших представителей
нашей фауны, необходимо упомянуть еще очень крупную дальневосточ-
ную звезду Evasterias echinosoma, достигающую в поперечнике лу-
чей 80 см.
В связи с таким способом питания
устричных и мидиевых банок (см.
489
КЛАСС 2. ЗМЕЕХВОСТКИ, ИЛИ ОФИУРЫ (OPHIUROIDEA)
ХАРАКТЕРИСТИКА
Иглокожие, с хорошо выраженной пятилучевой, симметрией. Тело со-
стоит из диска и пяти резко отграниченных от него очень подвижных рук
(табл. XIV). Анальное отверстие отсутствует. Амбулякральные ножки ру-
Рис. 450. Личинка офиуры-офиоплутеусз
/ — ресничный шнур, 2 — рот, 3 — анальное отверстие
диментарны и служат только для дыхания. Личинка двустороннесиммет-
ричная, с сильно развитыми боковыми отростками — офиоплутеус
(рис. 450).
ОБЩИЙ ОБЗОР КЛАССА
Рис. 451. Схематический поперечный
разрез через руку офиуры:
1 — 4—наружные скелетные пластинки,
5—позвонок, 6 — радиальный канал амбу-
лякральной системы, 7 — то же псевдоге-
мальной системы, 8 — радиальный нервный
ствол, 9 — амбулякральные ножки, 10 — це-
лом
Внешнее строение. Тело офиур состоит из центрального, в большинст-
ве случаев плоского диска и отходящих от него пяти лучей, или рук. У не-
которых офиур руки многократно дихотомически ветвятся (ветвистые офи-
уры), в результате чего получается очень сложная форма тела (табл. XIV).
На оральной стороне диска расположе-
ны ротовое отверстие и маленькая мад-
репоровая пластинка только с одним
отверстием. Все тело покрыто скелетными
пластинками, особенно хорошо выра-
женными на аборальной стороне диска.
Скелет. Скелет развит в руках и
состоит из ряда отдельных известковых
члеников, или позвонков, занимающих
почти всю полость руки (рис. 451). Они
соответствуют амбулякральным пластин-
кам морских звезд, ушедшим в глубь
тела и слившимся в единое образование.
Позвонки свободно соединены между
собой при помощи мышц, и поэтому ру-
ки очень подвижны и являются органа-
ми передвижения.
Полость тела. Полость тела развита
в диске, а в лучи продолжается только
в виде небольшого канала (рис. 451).
Амбулякральная система. Амбуляк-
ральная система построена по лучевой симметрии. Имеются околоротовое
кольцо и радиальные каналы, от которых отходят рудиментарные амбу-
лякральные ножки, не имеющие присосок и служащие для дыхания и от-
части осязания. У некоторых офиур на кольцевом канале имеются особые
резервуары, так называемые полиевы пузыри (рис. 452).
Нервная система и органы чувств. Нервная система образует околоро-
товые кольца и радиальные стволы в руках.
Органами осязания служат амбулякральные ножки. Других органов
чувств нет.
490
Пищеварительная система. Пищеварительная система состоит только
из большого мешковидного желудка, занимающего почти весь диск. Аналь-
ного отверстия нет.
Псевдогемальная и кровеносная системы. Также имеются околорото-
вые кольца и радиальные каналы. Кроме того, имеется и аборальное коль-
цо. В связи с тем, что мадрепоровая пластинка расположена на оральной
стороне, осевой комплекс идет не к аборальной, а, наоборот, к оральной
поверхности.
Половая система. Офиуры раздельнополы. Гонады образуются в ин-
террадиусах в особых эктодермических впячиваниях, или бурсах (см.
рис. 451). У некоторых форм оплодотворение происходит в этих бурсах
и там же протекает часть развития.
Рис. 452. Вскрытый диск Ophiura sarsi, сверху:
1 — рот, 2 — ротовые щупальца, 3 — желудок (на правой
половине он снят), 4 — кольцевой канал амбулякральноЙ
системы, 5 — полнев пузырь, 6 — гонады
Экология и географическое распространение. Офиуры — хищники, пи-
таются они различными мелкими животными, которых активно захватывают
своим большим ртом. Передвигаются они при помощи рук, которыми могут
очень хорошо действовать. В этом отношении особенно выделяются ветвис-
тые офиуры, которые своими сложными руками могут охватывать различ-
ные подводные предметы.
Офиуры распространены по всем морям на различных глубинах, начи-
ная с прибрежных горизонтов и кончая большими глубинами. В наших
северных морях встречается 49 видов офиур. У некоторых местах (Баренце-
во море) они встречаются в массовых количествах, например мелкая Ophi-
ura robusta достигает почти 3000 экз. на 1 м2 дна, буквально покрывая все
дно моря. Один вид офиур встречается и в Черном море.
Многие офиуры отличаются яркой и пестрой окраской и приспособле-
ны к цвету окружающей среды, например красномраморная офиофолис
(Ophiopholis aculeata) (табл. XIV) — одна из самых обычных офиур наших
северных морей, хорошо гармонирует с цветом береговых гранитных скал,
на которых часто встречается. Наиболее эффектны из наших офиур пред-
ставители ветвистых, например «голова Горгоны» (Gorgonocephalus eucne-
mis, табл. XIV) — крупная оранжевая офиура, достигающая 10 см, в
поперечнике диска с многократно ветвящимися руками. Обитает она на
средних глубинах во всех наших северных морях.
491
Таблица XIV Морские звезды и змеехвостки морей СССР:
1—Solaster papposus— северные и дальневосточные моря, 2 — Asterias rubens—
— Мурманское побережье. Белое море, 3 — Ceramster granularis — Мурманское
побережье, 4 — Ophiopholis aculeata — северные и дальневосточные моря, 5 —
голова Горгоны (Gorgon ocephalus) —северные и дальневосточные моря
КЛАСС 3. МОРСКИЕ ЛИЛИИ (CRINOIDEA)
ХАРАКТЕРИСТИКА
Прикрепленные иглокожие (табл. XV), ведущие постоянно сидячий
образ жизни или могут иногда отрываться от субстрата, на котором сидят,
и свободно плавать, двигая своими лучами, как плавниками.
ОБЩИИ ОБЗОР КЛАССА
Внешнее строение. Тело морской лилии состоит из чашечки (рис. 453) и
отходящих от нее вверх длинных разветвленных пяти лучей, или рук. От
нижнего конца чашечки отходят либо более или менее длинный стебелек
(стебельчатые морские линии), либо многочисленные усики или цирри
492
Рис. 453. Чашечка мор-
ской лилии сверху:
/ — рот, 2 — анальное отвер-
стие на бугорке, 3— амбу-
лякральные бороздки
(табл. XV), служащие для прикрепления лилии к субстрату. Лучи лилий
при самом основании ветвятся (см. рис. 453), и таким образом от чашечки
отходят как бы десять лучей. Подобно рукам офиур они складываются из
ряда члеников, состоящих из скелетных позвонков, подвижно соединенных
между собой, и могут поэтому достаточно хорошо изгибаться. К каждому
членику обычно попеременно то с правой, то с ле-
вой стороны причленяются боковые веточки — пин-
нулы.
В центре верхней поверхности чашечки нахо-
дится ротовое отверстие, от которого отходят пять
выстланных ресничками амбулякральных бороздок,
или желобков, переходящих далее на лучи и иду-
щих от их внутренней поверхности. В этих борозд-
ках располагаются амбулякральные ножки, или
щупальца. На верхней поверхности чашечки име-
ется также особый бугорок, на котором помещается
анальное отверстие (рис. 453).
Амбулякральная система. Амбул якрал ьна я
система состоит из околоротового кольцевого кана-
ла и отходящих от него пяти радиальных каналов,
идущих под амбулякральными бороздками и соответственно ветвящихся
по лучам. От этих каналов отходят веточки в пиннулы и амбулякральные
ножки, или щупальца, служащие у лилии для захвата пищи, осязания и
дыхания (рис. 454).
Несколько по-иному, чем у других иглокожих, сообщается у лилий
амбулякральная система с внешней средой. От кольцевого канала отходят
пять или более каменистых каналов, которые своими свободными концами
открываются в полость тела. Полость же тела сообщается с внешней средой
через многочисленные водяные поры, которые пронизывают верхнюю по-
верхность чашечки.
Рис. 454. Схематический поперечный
разрез через руку морской лилии:
/ — тело позвонка, 2 — амбулякральное щу-
пальце, 3— радиальный канал амбулякраль-
ной системы, 4 — каналы полости тела,
входящие в руки, 5 — радиальный ствол эк-
тоневральной системы, 6 — то же гипонев-
ральной системы, 7 — то же энтоневраль-
ной системы
Рис. 455. Развитие морской лилии.
/ —личинка (сбоку); // —стадия
цистоидеи:
/ —ресничные обручи, 2 — ротовое углуб-
ление, 3 — прикрепительная ямка, 4— об-
разующиеся скелетные пластинки
493
Нервная система. Нервная система представлена тремя видами систем.
Эктоневрольная образует околоротовое кольцо, от которого отходят радиаль-
ные стволы, проходящие по дну амбулякральных бороздок, причем залегает
оно очень близко к поверхности. Гипоневральная проходит несколько глуб-
же. В отличие от других групп иглокожих очень сильно развита эндонев-
ральная система, образующая нервное кольцо в аборальной части чашечки,
от которого отходят радиальные стволы в лучи, проходящие внутри поз-
вонков (рис. 454).
Пищеварительная система. В отличие от большинства иглокожих у
морских лилий ротовое отверстие направлено вверх. От него внутрь чашечки
идет пищевод, переходящий далее в желудок. От последнего отходит кишка,
которая заворачивает кверху и открывается анальным отверстием (см.
выше). Питаются морские лилии мелкими планктическими организмами, ко-
торых захватывают амбулякральными щупальцами. Попав в амбулякраль-
ные бороздки пищевые частицы направляются ко рту током воды, который
образуется движением ресничек, выстилающих бороздки.
Половая система. Морские лилии раздельнополы. Гонады образуются
в пиннулах, ближайших к основанию лучей. Половые продукты выходят
наружу, прорывая стенки тела.
Развитие. Оплодотворение наружное: в результате развития образуется
двусторонне симметричная личинка (рис. 455i ), свободно плавающая в
планктоне. Личинка не имеет ротового отверстия, а лишь ротовое углуб-
ление, не сообщающееся с кишечником. В связи с этим личинка не питает-
ся, а живет и развивается за счет запасов желтка в своем теле. Плавает она
короткое время и затем прикрепляется к субстрату особой прикрепительной
ямкой. Через некоторое время у нее развивается стебелек, и она превращает-
ся в стадию цистоидеи (рис. 455ц), получившей свое название от сходства
с вымершим палеозойским классом шаровиков (Cystoidea), у которых лучи
были развиты слабо или даже отсутствовали. При дальнейшем развитии
образуются лучи, и у бесстебельчатых лилий отпадает стебелек.
Экология и географическое распространение. Современные морские
лилии представлены небольшим количеством видов, причем обитают они
на сравнительно больших глубинах, а стебельчатые морские лилии принад-
лежат даже к характерным представителям глубоководной фауны. В наших
морях морские лилии очень немногочисленны: на севере их известно всего
3 вида, на Дальнем Востоке — 5, в наших южных морях они полностью
отсутствуют. В северных и дальневосточных морях наиболее часто встре-
чается гелиометра (Heliometra glacialis), достигающая 20 см высоты и оби-
тающая на глубинах от 10 до 1300 м. Кроме этой обычной лилии в глубоких
частях наших морей встречается стебельчатая лилия батикринус (Bathyc-
rinus) (табл. XV), достигающая 30 см высоты и обитающая обычно на глуби-
нах от 1000 до 2800 м.
Морские лилии хорошо представлены в ископаемом состоянии, причем
в прошлые геологические периоды, особенно в палеозое, были значительно
многочисленнее и разнообразнее, чем сейчас. Известны они с кембрия. Таким
образом, морские лилии наиболее древняя группа среди современных иг-
локожих.
КЛАСС 4. МОРСКИЕ ЕЖИ (ECHINOIDEA)
ХАРАКТЕРИСТИКА
К этому классу относятся иглокожие, имеющие сплошной панцирь,
образованный тесно соединенными между собой известковыми пластинками
(рис. 456). Снаружи тело обычно покрыто подвижными иглами, иногда
очень крупными (отсюда и название морские ежи). Большинство имеет
строго выраженную лучевую симметрию, но у некоторых она нарушается
(и животное приобретает двустороннюю симметрию, см. рис. 458).
494
Амбулякральная система служит у ряда форм только для дыхания.
У большинства имеется особый жевательный аппарат (аристотелев фонарь).
Личинка эхиноплутеус (рис. 440) двустороннесимметричная, свободно
плавающая в планктоне.
Рис. 456. Морской еж Strongylocentrotus droebachiensis, естествен-
ная величина до 9 см в диаметре (обычная форма наших северных
морей). Часть игл удалена, чтобы показать скелет с бугорками (/),
к которым прикрепляются иглы
ОБЩИЙ ОБЗОР КЛАССА
Внешнее строение. Большинство морских ежей имеет правильную ок-
руглую форму тела (рис. 456) при довольно большой высоте. Наряду с эти-
ми типичными, или правильными, морскими ежами имеются неправильные.
К таковым относятся плоские морские ежи, имеющие округлое, но очень
сплющенное в спинно-брюшном направлении тело (рис. 457), и сердцевидные
морские ежи, имеющие двустороннесимметричную форму тела (рис. 458).
Панцирь состоит обычно из 20 рядов известковых пластинок, причем
одни из них сплошные, а другие прободены мелкими отверстиями для вы-
хода амбулякральных ножек. Панцирь отсутствует обычно только на двух
участках тела: на оральной стороне вокруг ротового отверстия и на абораль-
ной — вокруг анального1. Особые скелетные пластинки располагаются у
правильных ежей вокруг последнего участка (рис. 459). Здесь имеются пять
крупных пластинок, из которых одна мадрепоровая. На этих пластинках,
в том числе и мадрепоровой, имеются довольно крупные отверстия, а име-
но половые, через которые выводятся половые продукты. Эти пластинки
соответствуют рядам непрободенных скелетных пластинок,- т. е. интерра-
диусам. Кнаружи от этих половых пластинок расположены тоже пять более
мелких пластинок, соответствующих радиусам, т. е. рядам прободенных
скелетных пластинок. Через отверстия, имеющиеся на этих пластинках,
1 Имеется небольшая группа морских ежей, так называемых мягких, у которых
скелетные пластинки не пропитываются известью, и поэтому весь скелет мягкий.
495
выходят наружу конечные веточки радиальных каналов амбулякральной
системы. Веточки эти содержат пигментные клетки и в связи с этим рассмат-
риваются обычно как светочувствительные органы, или «глазки» (ср. с «глаз-
ками» морских звезд). Снаружи панцирь покрыт тонким слоем кожи. Плас-
тинки панциря имеют обычно особые выпуклости, к которым подвижно
Рис. 457. Плоский морской еж. I — с аборальной стороны;
// — с оральной стороны; /// — в разрезе:
/ — петалоиды, 2—ротовое отверстие, 3—анальное отверстие
I и
Рис. 458. Сердцевидный морской еж. 1 — с аборальной сторо-
ны; II — с оральной стороны:
1,— петалоиды, 2 — рот, 3 — анальное отверстие
причленяются торчащие во все стороны иглы. Последние сильно развиты
у правильных ежей, достигая у некоторых тропических форм нескольких
сантиметров длины. У неправильных ежей они обычно короткие, лежащие.
Кроме игл, на поверхности морских ежей, так же как и у морских
звезд, имеются многочисленные мелкие разнообразные педицеллярии. Они
подвижны, имеют вид щипчиков и служат главным образом — для очище-
ния поверхности морских ежей от посторонних тел. Некоторые педицелля-
рии снабжены ядовитыми железками и, таким образом, служат защитными
органами. Размеры ежей очень различны, среди них имеются и сравнитель-
но мелкие формы и очень крупные, достигающие в поперечнике 20 см и более.
496
Примером крупного морского ежа может служить встречающийся у нас на
Западном Мурмане съедобный морской еж (Echinus esculentus), достигаю-
щий в поперечнике 17 см.
Амбулякральная система. Построена в общем также, как и у морских
звезд. От мадрепоровой пластинки вниз, к оральной стороне, направляется
заключенный в осевой комплекс каменис-
тый канал, впадающий в околоротовое
кольцо амбулякральной системы, располо-
женный над аристотелевым фонарем. От
кольца отходят радиальные каналы, иду-
щие вверх по внутренней поверхности
панциря по радиусам. От этих каналов
отходят амбулякральные ножки, выходя-
щие наружу через отверстия прободенных
пластинок и имеющие каждая по ампуле.
В интеррадиусах на кольцевом канале име-
ются особые придатки губчатого строения,
соответствующие тидемановым телам мор-
ских звезд. Основная функция амбуляк-
ральной системы правильных морских ежей
двигательная.
Рис. 459. Аборальная часть пан-
циря морского ежа:
1 — радиусы, 2—интеррадиусы, 3—
главные пластинки, 4 — половые плас-
тинки, 5—мадрепоровая пластинка,
6 — анальное отверстие
Нервная система и органы чувств.
Нервная система также сходна с системой
морских звезд. Имеется околоротовое коль-
цо с отходящими от него пятью радиаль-
ными стволами, причем параллельно рас-
положены две системы — эктоневральная
и гипоневральная. Органы чувств развиты слабо. Органами осязания слу-
жат амбулякральные ножки в особенности те, которые расположены на
оральной стороне близ ротового отверстия. Светочувствительными органами
являются амбулякральные ножки на концах лучей.
Пищеварительная система. Ротовое отверстие расположено у правиль-
ных и плоских ежей строго в центре оральной стороны тела. У сердцевидных
ежей оно смещено, что рез-
ко нарушает лучевую сим-
метрию этих ежей (рис.
459). Вокруг ротового от-
верстия у правильных ежей
имеется мягкая кожа, на
которой часто располагают-
ся многочисленные своеоб-
разные педицеллярии, по-
могающие ежу захватывать
пищу. У правильных и
плоских ежей из ротового
отверстия обычно выстав-
лены пять небольших зуб-
чиков. Эти зубчики — ко-
нечная часть сложного и
крайне своеобразного же-
вательного аппарата —
аристотелева фонаря1.
Аппарат этот имеет вид
конуса, построен по строго
пятилучевой симметрии и
Рис. 460. Схема строения морского ежа
/ — рот, 2 — анальное отверстие, 3— зуб аристотелев.! фона-
ря, 4—каменистый канал, 5 — кишечник, 6 — радиальный
канал амбулякральной системы, 7 — радиальный ствол нерв-
ной системы, 8—ампула, 9—радиальный канал сосудисто)!
системы, 10—амбулякральная ножка, 11—осевой орган 12 —
педицеллярия, 13—скелетная пластинка, 14—игла, 15 — го-
нада
1 Название происходит оттого, что это образование было описано еще Аристоте-
лем, который сравнивал его по форме с греческим светильником.
’/о 19 Курс зоологии, т. I
497
состоит из пяти основных трехгранных зубов, каждый из которых заканчи-
вается упомянутым зубчиком. Кроме этих пяти основных зубов, аристо-
телев фонарь имеет еще ряд скелетных образований и сложную муску-
латуру. При помощи последней он как бы подвешивается на пяти П-образ-
ных внутренних выростах панциря и может несколько выпячиваться
наружу, втягиваться внутрь, а также у него могут раздвигаться и сдви-
гаться зубы. При помощи аристотелева фонаря морские ежи соскребывают
различные водоросли, а иногда и мелких животных, например корненожек,
и перетирают их. От верхней части аристотелева фонаря внутрь тела
отходит кишечный канал, в связи с растительностью морских ежей очень
длинный (ср. с морской звездой), во много раз превышающей по длине вы-
соту ежа. Ввиду этого он образует сложные петли в полости тела ежей и
затем только подходит к анальному отверстию (рис. 460).
На протяжении части кишечного канала параллельно с ним идет
особая тонкая трубка — сифон, обеими своими концами впадающий в киш-
ку. В эту трубку входит заглатываемая вместе с пищей вода, и, по-види-
мому, она выполняет дыхательные функции.
Органы дыхания. У многих морских ежей имеются специальные органы
дыхания — кожные жабры, располагающиеся обычно в числе пяти пар
вокруг ротового отверстия на окружающей его мягкой площадке. Кроме
того, дыхательные функции выполняют амбулякральная система и упомя-
нутый выше сифон кишечника.
Псевдогемальная и кровеносная системы устроены в общем так же,
как и у морских звезд , т. е. имеются соответствующие кольцевые сосуды на
оральной и аборальной сторонах, связанные вместе осевым органом. От
кольцевых сосудов отходят радиальные сосуды. Кроме того, у морских ежей
сильно развиты кровеносные сосуды кишечника. Особенно развиты из них
два продольных, проходящих по обеим сторонам кишечника и соединяю-
щихся с оральным и аборальным кольцами.
Половая система. Гонады расположены на аборальной стороне в ин-
террадиусах и открываются наружу вокруг анального отверстия (см. выше).
Ежи обычно раздельнополы, но по внешнему виду определить пол невоз-
можно.
Развитие. Оплодотворение наружное; в результате дробления и даль-
нейшего развития образуется свободноплавающая личинка эхиноплутеус
(рис. 440).
Экология и географическое распространение. Морские ежи обитают
главным образом в теплых морях, где очень разнообразны и многочислен-
ны. В наших морях на севере они представлены всего пятью видами, причем
четыре из них встречаются редко. Часто встречается обыкновенный широко
распространенный морской еж сгпронгилоценпгропгус (Strongylocentrotus
droebachiensis), достигающий 8—9 см в диаметре. Обитает он, начиная с
прибрежных горизонтов и до больших глубин. Местами, например на Мур-
манском побережье, этот еж встречается в массовых количествах. В запад-
ных частях Мурманского побережья обитает упомянутый выше более круп-
ный, достигающий 17 см в диаметре съедобный морской еж, получивший свое
название от того, что по побережью Европы, где он обычен, его промыш-
ляют из-за икры, которую употребляют в пищу. В морях Дальнего Востока
фауна ежей значительно богаче, в наших южных морях они отсутствуют.
КЛАСС 5. МОРСКИЕ ОГУРЦЫ, ИЛИ ГОЛОТУРИИ
(HOLOTHUROIDEA)
ХАРАКТЕРИСТИКА
Своеобразный класс иглокожих, заключающий в себе червеобразные
организмы, внешне совершенно не похожих на других иглокожих и скорее
напоминающих толстых червей (табл. XV). На переднем конце обычно име-
498
ется венец щупалец, окружающих ротовое отверстие. Скелет в виде микро-
скопических известковых телец (спикул) различной формы (рис. 461). Лу-
чевая симметрия выражена нетипично. Амбулякральная система развита
слабо, в некоторых случаях она даже
рудиментарна. Личинка аурикулярия
(см. рис. 440) несколько напоминает
личинку морских звезд.
Рис. 461. Спикулы различных голотурий
ОБЩИЙ ОБЗОР КЛАССА
Внешнее строение. Червеобразное или бочонковидное тело голотурий
обычно мягкое. Наружная поверхность на ощупь часто шероховатая от
лежащих в подвижном слое спикул. В большинстве случаев кожа толстая,
темноокрашенная, но у ряда форм тонкая, полупрозрачная и через нее про-
свечивают внутренние органы. Венец щупалец в большинстве случаев
имеется, и у голотурий с древовидными щупальцами развит очень мощно
(табл. XV).
Амбулякральные ножки у некоторых голотурий имеются п распо-
ложены в виде пяти продольных рядов вдоль всего тела; у других они пол-
ностью отсутствуют (безногие голотурии, табл. XV).
Размеры голотурий очень различны: мелкие — до 2—3 см в длину,
некоторые крупные тропические формы — до I м. Цвет тела также разно-
образен, преобладают коричневато-лиловые тона.
Скелет. Скелет расположен под кожей и состоит из многочисленных
спикул. Форма спикул разнообразна, иногда очень причудлива (рис. 461).
Мышечная система. Мускулатура в связи со слабым развитием скелета
развита очень хорошо и состоит из нескольких слоев. Наиболее сильно раз-
вит кольцевой слой, сжимающий тело голотурии, и продольный, располо-
женный обычно пятью лентами, укорачивающий тело. Кроме мышц, распо-
ложенных в стенках тела, имеются еще мышцы, втягивающие венец щупалец
в передний отдел тела. Задним концом эти мышцы прикрепляются к продоль-
ным мускульным лентам в заднем конце тела, а передним — к особому из-
вестковому кольцу, окружающему глотку (рис. 462).. Кольцо это состоит
обычно из десяти пластинок — пяти радиальных и пяти интеррадиальных.
Полость тела. Очень обширная, наполненная целомической жидкостью.
Нервная система и органы чувств. Нервная система представлена до-
вольно типично для иглокожих. Имеются эктоневральная и гипоневральная
системы, состоящие каждая из околоротового кольца и пяти радиальных
стволов (рис. 463). Органы чувств развиты слабо и представлены только ор-
ганами осязания, которыми являются щупальца.
Амбулякральная система. Амбулякральная система состоит из около-
глоточного кольца, от которого отходят пять радиальных каналов, идущих
сперва к переднему концу, затем поворачивающих назад и проходящих по
радиусам под лентами продольных мышц (рис. 463). От передних отделов
этих каналов отходят вперед каналы в щупальца, которые таким образом
связаны с амбулякральной системой и представляют собой как бы разрос-
шиеся амбулякральные ножки. От радиальных каналов отходят амбуля-
кральные ножки с их ампулами. У безногих голотурий радиальных каналов
нет, и каналы в щупальца отходят непосредственно от околоротового кольца.
От окологлоточного кольца отходит также каменистый канал, обычно нс
связанный с внешней средой и открывающийся в полость тела, а также
один или несколько полиевых пузырей, свободно свешивающихся в полость
тела (см. рис. 462).
Псевдогемальная и кровеносная системы. Развиты у голотурий довольно
сильно. Помимо околоротового кольца и радиальных каналов, идущих по
радиусам, имеется сложная сеть кровеносных сосудов около кишечника и
вокруг левого «легкого».
Пищеварительная система. Ротовое отверстие ведет в длинную кишку,
изгибающуюся внутри тела и оканчивающуюся на заднем конце анальным
Таблица XV. Голотурии и морские лилии морей СССР:
1 — морской огурец (Cucumaria frondosa) — северные моря, 2 — трепанг (Stichopus
japonicum) — Японское море, 3— безногая голотурия (Caudina)—Японское море,
4—морская курочка (Psolus phantapus) северные моря, 5 — обыкновенная морская
лилия (Helioinetra gracialis) — северные и дальневосточные моря, 6 — стебельча-
тая морская лилия (Bathycrinus) — глубины дальневосточных морей и северных
частей Карского моря
отверстием. Последний отдел кишечника носит название клоаки, так как в
него открываются водные легкие (см. ниже), а у некоторых голотурий еще
особые образования — кювьеровы органы (рис. 462), представляющие
собой многочисленные слепые железистые трубочки. Функция этих образо-
ваний, по-видимому, защитная. При раздражении голотурии они выбрасы-
500
ваются через анальное отверстие и превращаются в липкие нити, обвола-
кивающие прикоснувшийся к голотурии предмет
Дыхательная система. Открывающиеся в клоаку водные, или ки-
шечные, легкие представляют собой пару длинных разветвленных тонко-
стенных образований, доходящих почти до переднего конца тела
(рис. 462). Путем сокращений клоаки в них входит вода и через их стенки
происходит газообмен. Левое «легкое» богато опутано сетью кровеносных
сосудов, и поэтому кислород поступает не в полостную жидкость, а непо-
средственно в кровь. Через стенки «лег-
ких» выходят наружу и амебоциты с
продуктами выделения.
Половая система. В отличие от всех
других иглокожих половая система у
голотурий представлена только одной
гонадой, находящейся в полости тела и
открывающейся наружу на переднем
конце тела (рис. 462). Большинство
голотурий раздельнополы, но имеются
ю
и
те-
з-
Рис. 462. Внутреннее строение голоту-
рии (безногой).
/ — венец щупалец, 2 известковое кольцо во-
круг глотки, 3—кишечник, 4 — клоака, 5 —
железистые трубки, впадающие в клоаку, так
называемые кювьсрсвы органы, 6—анальное
отверстие 7 — кишечные легкие, 8 амбуляк-
ральный кольцевой сосуд, 9 — каменистые ка-
налы, 10 — полнев пузырь, 11 — гонада, 12 ен-
ты продольных мышц
Рис. 463. Поперечный разрез через
ло голотурии, схема:
1 — стенка тела, 2 кольцевые мышцы,
ленты продольных мышц, 4—амбулякральные
ножки, 5 — их ампулы, 6 — радиальные каналы
амбулякральной и кровеносной систем и ра-
диальные стволы нервной системы, 7 — разре-
зы кишечника, 8— водные «легкие», 9 — кро-
веносные сосуды, 7 — гонада
гермафродиты. В последнем случае гонада попеременно продуцирует то
мужские, то женские половые продукты.
Развитие. Оплодотворение наружное. В результате дробления обра-
зуется свободноплавающая личинка аурикулярия (см. рис. 440), сходная с
личинками морских звезд.
Среди голотурий очень распространено явление самокалечения, или
автотомии. При раздражении, например вынимании из воды, голотурия
выбрасывает обычно через анальное отверстие большую часть своих внут-
ренностей: кишечник, правое «легкое» и пр. Выкинутые части могут затем
довольно быстро регенерировать.
Экология и географическое распространение. Голотурии — донные
животные, ползающие по дну при помощи амбулякральных ножек, щупа-
лец или просто благодаря мышечному сокращению тела. Многие из них за-
капываются в грунт, выставляя наружу только передний конец с щупаль-
цами. Цитаются голотурии обычно при помощи щупалец, захватывая ими
добычу и затем заглатывая ее. У некоторых щупальца служат своеобразным
18 Ку>с зоологии, т< I 501
Рис. 464. Глубоководная
голотурия Elpidia. Ку-
рило-Камчатская впади-
на, глубина около 10 000
м (по бокам тела сильно
развитые амбулякраль-
ные ножки, которые по-
могают держаться в мяг-
ком иле)
ловчим аппаратом — голотурия сильно расправляет их и благодаря слизи,
которая их покрывает, к ним пристает добыча; голотурия по очереди запус-
кает их в рот. Некоторые голотурии питаются органическими остатками
и микроорганизмами, находящимися в грунте, заглатывая его в больших
количествах.
Голотурии встречаются во всех морях на различных глубинах и грун-
тах. Они наиболее глубоко из всех иглокожих опускаются в глубины океа-
нов, и некоторые из них встречены в больших количествах в северной части
Тихого океана на глубинах до 9600 м (Elpidia, рис.
464). В наших морях они представлены достаточно
богато, на севере их известно 44 вида. Наиболее
часто встречается у нас на севере крупная, дости-
гающая полуметра длины бурая кукумария (Cucu-
maria frondosa), живущая в прибрежной зоне до
200 м на различных грунтах, особенно среди при-
брежных водорослей. Из безногих голотурий у нас
встречается длинная, червеобразная, белая, полу-
прозрачная хиродота (Chirodota), живущая в или-
стых грунтах. По форме интересна морская курочка
(Psolus, табл. XV), имеющая плоскую подошву для
ползания и вытянутые вверх концы: передний в виде
головки и задний в виде хвоста. Встречаются
голотурии и в Черном море (ср. с офиурами), где
представлены тремя, правда, небольшими и редко
встречающимися формами.
Некоторые голотурии обитают в планктоне,
например Pelagothuria natatrix.
Практическое значение. Многих голотурий в
странах Дальнего Востока употребляют в пи-
щу и в связи с этим их промышляют. Особенно в
этом отношении следует отметить трепанга (Sticho-
pus japonica, табл. XV), являющегося объектом
промысла и у нас на Дальнем Востоке, в Японском
море. Добывают трепангов обычно водолазы; их
очищают от внутренностей, варят, солят, затем
высушивают.
ИСКОПАЕМЫЕ ИГЛОКОЖИЕ
В прошлые геологические периоды иглокожие
были представлены очень богато, причем своеоб-
разными полностью вымершими классами: морские
бутоны (Blastoidea), шаровики (Cystoidea), текоидеи
(Thecoidea), карпоидеи (Carpoidea) и некоторые дру-
гие, известные из кембрия, силура, девона и карбо-
на. Наиболее древние формы имели более или менее
выраженную двустороннюю симметрию (рис. 465),
которая затем в связи с прикрепленным образом
жизни превратилась в лучевую.
Рис. 465. Реконструкция силурского иглокожевого Rhipido-
cystis. Хорошо видна почти правильная двухсторонняя сим-
метрия
502
филогения иглокожих
Иглокожие обладают настолько своеобразным строением, что истол-
кование их организации крайне затруднительно. Данные палеонтологии,
сравнительной анатомии, эмбриологии проливают некоторый свет на их
эволюцию и помогают понять особенности их строения.
В индивидуальном развитии иглокожих обращает на себя внимание
наличие двустороннесимметричной личинки. Эту билатеральность личин-
ки рассматривают как указание на то, что предки иглокожих были сво-
бодноподвижными двустороннесимметричными животными; радиальное
строение выработалось впоследствии в результате перехода предков игло-
кожих к сидячему образу жизни.
Подтверждением данного предположения служат сохранение сидячего
образа жизни у наиболее примитивных современных иглокожих морских
лилий — и факт временного перехода личинок морских звезд к сидячему
образу жизни, что совпадает с развитием в организации личинки пятилу-
чевой симметрии.
Постепенная выработка радиальной симметрии в типе иглокожих
хорошо прослеживается на ископаемых формах из класса Cystoidea (цистои-
деи). В кембрийских и силурийских отложениях обнаружен Aristocystis.
Тело его имело форму неправильного мешка. Суженным концом аристоцис-
тис прикреплялся ко дну. Ротовое отверстие находилось на противополож-
ном конце, здесь же на некотором расстоянии от рта открывалось анальное
отверстие.
Между ротовым и анальным отверстиями находились непарное по-
ловое и мадрепоровое отверстия. Скелет состоял из пластинок неправиль-
ной формы, расположенных без определенного порядка. Элементы радиаль-
ной симметрии в строении аристоцистиса отсутствовали. Отпечатки кишеч-
ника показывают, что форма его была петлеобразной подобно кишечнику
современных морских лилий.
Из нижнего силура известен другой представитель из класса цистпдей —
сферонис (Sphaeronis). Во многом он был сходен с аристоцистисом; суще-
ственное отличие от последнего заключалось в появлении на панцире мер-
цательных бороздок, сходившихся к ротовому отверстию. Функция их —
привлечение мелких пищевых частиц. Точно такой же способ питания свой-
ствен и современным морским лилиям. Новой ступенью в эволюции цистидей
надо считать удлинение приводящих борозд, увеличение их до пяти борозд
(вследствие разделения двух борозд из прежних трех на две ветви) и из-
вестное упорядочение в расположении известковых пластинок. Названные
признаки обнаружены у нижнесилурийского Proteroblastus (протероблас-
тус). Пластинки на его скелете разграничены на две группы: а) часть пла-
стинок приобретает правильную форму и располагается в пять двойных
рядов); б) другая часть пластинок, располагающаяся между рядами пра-
вильных пластинок, все еще не имеет правильной формы. На дальнейших
ступенях развития и остальные пластинки скелета стали располагаться
по радиальной симметрии и приобрели правильную форму.
Следует заметить, что первоначально радиальная симметрия возникла
у цистидей в виде приводящих борозд. Впоследствии радиальная сим-
метрия распространилась на нервную систему, возникшую амбулякральную
систему, половой аппарат и другие органы.
Возникновение радиальной симметрии у цистидей показывает, каким
образом развивалась радиальная симметрия у иглокожих.
Примитивные цистидеи — прообраз того предка, от которого произошли
современные морские лилии и остальные иглокожие.
Иглокожие разделяются на два подтипа: 1) стебельчатые Pelmatozoa
в составе одного ныне живущего класса — морские лилии (Crinoidea) и
2) Eleutherozoa, к которому относятся морские звезды (Asteroidea), офиуры
(Ophiuroidea), морские ежи (Echinoidea), голотурии (Holothuroidea).
18*
503
Стебельчатые отличаются от остальных классов положением в про-
странстве (ротовое отверстие направлено кверху); многие из них ведут си-
дячий образ жизни. Такое положение рта соответствует способу питания:
мелкие пищевые частицы механическим движением мерцательного эпителия
рук вовлекаются в ротовое отверстие. Все Eleutherozoa имеют другое поло-
жение в пространстве: ротовое отверстие направлено книзу (за исключением
голотурий). Такое положение связано с активным способом добывания пи-
щи; у голотурии оральный и аборальный полюсы располагаются по гори-
зонтальной оси.
На иглокожих можно видеть правило выработки радиальной симмет-
рии при переходе животных к сидячему образу жизни. Свободноподвиж-
ный образ жизни у всех Eleutherozoa у части Pelmatozoa явился результа-
том вторичного перехода от сидячего образа жизни к подвижному Свобод-
ноподвижный образ жизни не оказал влияния на изменения в радиальной
симметрии морских звезд и офиур. Объяснение этого факта заключается,
по-видимому, в медленном передвижении названных животных, при котором
не могла выработаться билатеральная симметрия, хотя у некоторых игло-
кожих наряду с сохранением элементов радиальной симметрии вторично
развиваются признаки двусторонней симметрии (морские ежи, голотурии).
ТИП XIII. ПОГОНОФОРЫ (POGONOPHORA)’
ХАРАКТЕРИСТИКА
Нитевидные, очень длинные животные (рис. 466), живущие в роговых
трубочках различной плотности. На переднем конце имеется различно
устроенный щупальцевый аппарат. Полость тела вторичная, образующаяся
энтероцельным путем (рис. 467). Пищеварительная и дыхательная системы
отсутствуют. Нервная система лежит на спинной стороне. Раздельнополы.
Развитие с превращением. Исключительно морские формы, обитающие глав-
ным образом на больших глубинах.
Рис. 466. Lamellisabella zachsi (большие глубины Охотского моря):
1 — передний отдел тела со щупальцевым аппаратом, 2—средний отдел тела,- 3 — задний отдел
тела, прикрепительные подушечки
Погонофоры были впервые открыты французским ученым Коллери
в 1914, однако до последнего времени они оставались неизученными. В
настоящее время в Тихом океане советскими экспедициями на судне
«Витязь» собрано около 90 видов, из них 44 описаны А. В. Ивановым в
специальной монографии. Они составили класс погонофор, который в
1944 г выделен в особый тип животных, промежуточный между беспозвоноч-
ными и хордовыми. Класс погонофор расчленяется на два отряда Atheca-
nephria с двумя семействами, в составе 27 видов, и отряд Thecanephria с
тремя семействами, в составе 17 видов.
ОБЩИЙ ОБЗОР ТИПА
Внешнее и внутреннее строение. Длинатела достигает у некоторых форм
нескольких десятков сантиметров при толщине всего в несколько десятых
миллиметра. На переднем конце расположен сложный щупальцевый аппа-
рат различного строения. Число щупалец колеблется от 1 до 220. Они или
полностью свободны, или спаяны вместе в трубку или спираль. Тело состо-
1 От греческого «погонос»— борода, т. е. несущие бороду
505
ит из трех отделов: переднего, несущего щупальцевый аппарат и головную
лопасть, среднего—небольшого — и заднего, образующего основную часть
тела. На последнем отделе имеются особые прикрепительные подушечки с
хитиноидными пластиночками, при помощи которых животное, по-види-
мому, цепляется за внутреннюю поверхность трубочки и передвигается в
достаточно мощная продольная мускулатура. Вторичная
полость тела в соответствии с расчленением его состоит
тоже из трех отделов — переднего непарного и среднего и
заднего — парных. Ни ротового и анального отверстий,
ни кишечника не обнаружено. Питание происходит при
помощи щупалец, которые выполняют и дыхательную
функцию. Кровеносная система замкнутая, на брюшной
стороне, в среднем отделе тела, имеется сердце. Выдели-
тельная система представлена парой целомодуктов передне-
го, непарного отдела целома. Нервная система примитив-
ная, представлена скоплением нервных клеток в головной
лопасти и спинным нервным стволом, идущим вдоль тела.
Погонофоры раздельнополы, у самцов образуются довольно
сложные сперматофоры. Развитие с личинкой, которая,
по-видимому, не выходит из материнской трубки.
Рис. 467. Схема развития Siboglinum — образование второй по-
лости тела (зачернена). Ясно видно, что полость образуется энте-
роцельным путем (ср. с рис. 10)
и распространение. Как показали исследования последних
лет, погонофоры — широко распространенная группа. Большинство их —
глубоководные животные, встречающиеся на глубинах до 9600 м, вместе с
тем среди них имеются и формы, обитающие на глубинах в несколько десят-
ков метров. Местами погонофоры встречаются в массовых количествах.
В настоящее время известно уже несколько десятков видов, объединяемых
в ряд родов (Siboglinum, Lamellisabella, Polybrachia и др.), изученных
главных образом советскими исследователями (А. В. Иванов).
Положение в системе и филогения. Положение погонофор в системе и
их филогения еще не совсем ясны. Целый ряд признаков (щупальцевый
аппарат, прикрепительные подушечки, упрощенное внутреннее строение),
безусловно, связан с их сидячим образом жизни. Наряду с этим у пого-
нофор имеется ряд черт (положение нервной системы, способ образования
целома, рис. 467), сближающих их с низшими хордовыми—полухордовыми
НЕКОТОРЫЕ ГРУППЫ БЕЗПОЗВОНОЧНЫХ
НЕЯСНОГО СИСТЕМАТИЧЕСКОГО ПОЛОЖЕНИЯ
Среди большого числа разнообразных групп беспозвоночных имеется ряд клас-
сов, систематическое положение которых до сих пор не совсем ясно.
Рассмотрим один такой класс.
КЛАСС ЩЕТИНКОЧЕЛЮСТНЫЕ (CHAETOGNATHA)
ХАРАКТЕРИСТИКА
Очень подвижные, с удлиненным телом животные, имеющие хорошо выражен-
ную голову (рис. 468). На голове ротового отверстия расположены две группы щетинок,
служащих для захвата пищи (отсюда название группы). Кровеносная, дыхательная
506
и выделительная системы отсутствуют. Нервная система разви-
та довольно хорошо и представлена головным и брюшным нерв-
ными узлами. Гермафродиты, развитие прямое. Исключитель-
но морские формы.
ОБЩИЙ ОБЗОР КЛАССА
Внешнее и внутреннее строение и развитие. Щетинкочелю-
стные — очень небольшая группа, охватывающая около 30 ви-
дов. Все они очень подвижные, сравнительно небольшие, дости-
гающие 9 см, обитатели толщи морской воды. Тело их удлинен-
ное с резко выраженной головой и плавниками —обычно с двумя
боковыми и хвостовыми. Покрыто тело однослойным эпителием,
который в области перехода тела в голову образует многослой-
ное утолщение. Под эпителием находится хорошо развитая
поперечно-полосатая мускулатура, особенно сложная в голове
в связи с головными щетинками. У щетинкочелюстных имеется
хорошо выраженная вторичная полость тела, разделенная
продольной перегородкой на правую и левую половины и двумя
поперечными — на три отдела.
Кишечник начинается на переднем конце. Вокруг ротового
отверстия имеется сложный челюстной аппарат. При помощи
этого аппарата щетинкочелюстные могут очень активно захва-
тывать пищу. Они резко выраженные хищники.
Нервная система развита хорошо и состоит из головного
узла, лежащего над кишечником, и брюшного, лежащего под
кишечником в середине тела. Узлы соединены между собой
комиссурами. Из органов чувств хорошо развиты глаза. Щетин-
кочелюстные — гермафродиты. Развитие без метаморфоза,
причем оно идет по типу вторичноротых животных, т. е.
мезодерма образуется типичным энтероцельным путем (рис. 10)
и рот взрослого животного образуется не из первичного рта за-
родыша.
Экология и распространение. Все щетинкочелюстные —
очень подвижные обитатели толщи морской воды. Питаются
планктонными беспозвоночными, а иногда нападают и на маль-
ков рыб. Обитают щетинкочелюстные во всех морях, иногда
скопляясь в массовых количествах. Представлены они широко и
в наших морях, за исключением Каспия.
Рис. 468. Схема строения Sagitta (с брюшной стороны)-
/—мозг, 2—окологлоточные нервы, 3—брюшной ганглий, 4— перифери-
ческие нервы, 5 — целом, 6 — желудок, 7 — яичники, 8 — яйцевод, 9 —аналь-
ное отверстие, 10—семенники, 11— семяпровод, 12—хвостовой плавник,
13 — головные железы, 14 — щетинки, 15 — мужское и 16 — женское половые
отверстия
Наиболее известный представитель щетинкочелюстных — морская стрелка.
(Sagitta), встречающаяся во всех наших морях. В периоды массового развития са-
гитты служат пищей промысловым планктоноядным рыбам.
Положение в системе и филогения. Положение щетинкочелюстных в системе
беспозвоночных пока неясно, так же как неясен их филогенез. По своему эмбриональ-
ному развитию они должны быть отнесены, безусловно, к вторичноротым, но среди
последних нет ни одной группы, с которой их можно было бы сблизить. С другой
стороны, по форме тела, характеру движений (изгибание тела) и полости тела щетин-
кочелюстные сходны с некоторыми червями.
Видимо, эту группу надо рассматривать как особую самостоятельную ветвь
вторичноротых.
ЗНАЧЕНИЕ БЕСПОЗВОНОЧНЫХ
В НАРОДНОМ ХОЗЯЙСТВЕ
Миллиарды беспозвоночных животных, населяющие буквально все
уголки земного шара, естественно, оказывают влияние на весь живой мир,
как животный, так и растительный, а в некоторых случаях и на неживую
природу. Постоянно сталкивается с беспозвоночными и человек.
Одни беспозвоночные являются полезными животными, которых человек
507
использует, а иногда и разводит (медоносная пчела, шелковичный червь»
устрица и некоторые другие).
Другие беспозвоночные (а таких большинство) приносят вред, и с ними
постоянно приходится вести упорную борьбу
Беспозвоночных с точки зрения их полезности можно рассматривать
в следующих направлениях.
1 Беспозвоночные или продукты, ими вырабатываемые, используемые
в пищу.
Сюда относится, в первую очередь, медоносная пчела, разводимая
человеком с древнейших времен и ставшая, по сути дела, настоящим домаш-
ним животным. Разведение пчел дает не только такой первоклассный пи-
щевой продукт, как мед, но и технически ценный продукт — воск.
Из других групп сюда относится ряд моллюсков, главным образом дву-
створчатых, ряд ракообразных, некоторые иглокожие и пр., причем не-
которых моллюсков и ракообразных не только промышляют, но и разводят
(например, устрицы и омары).
2. Беспозвоночные или продукты, ими вырабатываемые, используемые
как техническое сырье.
Сюда относятся в первую очередь некоторые бабочки, гусеницы которых
вырабатывают для закукливания шелковую нить (тутовый и дубовый шелко-
пряды). Бабочки эти с древних времен разводятся человеком. Кроме того,
сюда относится ряд моллюсков, раковины которых используются как
сырье для производства перламутровых изделий или добычи жемчуга, губ-
ки (туалетная губка), красный коралл, некоторые насекомые, например
кошениль, из которых вырабатывается краска — кармин. Сюда же мож-
но отнести и медицинскую пиявку, в настоящее время не только добываемую
в природе, но и искусственно разводимую для медицинских целей.
3. Беспозвоночные — опылители различных сельскохозяйственных, ого-
родных и садовых культур.
Сюда относится большое число различных насекомых, причем роль их
в этом отношении огромна. Надо отметить, что большую роль в опылении
некоторых сельскохозяйственных растений, например красного клевера,
играет медоносная пчела.
4. Беспозвоночные — кормовые объекты для промысловых рыб, птиц
и зверей.
Сюда относятся (как кормовые объекты для рыб) коловратки, различные
ракообразные, моллюски, свободноживущие черви, некоторые иглокожие,
личинки насекомых, а для наземных форм главным образом различные на-
секомые, их личинки и отчасти земляные черви.
5. Беспозвоночные — истребители и паразиты вредных беспозвоночных.
Сюда относятся главным образом некоторые насекомые, в первую
очередь наездники, откладывающие яйца в гусениц вредных бабочек и
таким образом снижающие численность вредных видов. Некоторую роль
в уничтожении вредных насекомых играют и хищные многоножки, а так-
же паразитические круглые черви.
6. Беспозвоночные, играющие роль в жизни почвы.
Сюда относятся главным образом земляные, или дождевые, черви, раз-
рыхляющие почву и вносящие в нее органические остатки. На громадную,
хотя и малозаметную роль этих червей указал еще в свое время Ч Дарвин,
посвятивший наблюдению над образом жизни червей многие годы и написав-
ший специальный труд «Образование растительного слоя земли деятель-
ностью дождевых червей». В жизни почвы довольно существенную роль
играют также ряд личинок насекомых, некоторые взрослые насекомые и
почвенные одноклеточные (амеба, жгутиковые и инфузории).
Вред, приносимый человеку и его хозяйству беспозвоночными, идет по
следующим направлениям:
1 Беспозвоночные — непосредственные паразиты, человека, вызывающие
у него ряд заболеваний.
508
Сюда относятся некоторые одноклеточные, в первую очередь возбудитель
малярии, которой на земном шаре ежегодно болеет несколько миллионов
человек. Далее следует ряд паразитических плоских и круглых червей, а
также ряд наружных паразитов из клещей и насекомых (вши, блохи, клопы).
2. Беспозвоночные — паразиты домашних и промысловых животных.
Сюда относится ряд одноклеточных, очень многие плоские и круглые
черви, отчасти пиявки, некоторые ракообразные (для промысловых рыб),
клещи и многие насекомые. Все эти паразиты наносят существенный вред
животноводству и требуют постоянной и планомерной борьбы с ними.
3. Беспозвоночные — паразиты сельскохозяйственных культур.
Сюда относится ряд круглых червей, например свекловичная нематода,
пшеничная нематода и некоторые другие, а также клещи и некоторые на-
секомые.
4. Беспозвоночные — переносчики и промежуточные хозяева возбудите-
лей различных болезней человека, домашних животных и сельскохозяйствен-
ных растений.
В этом отношении роль беспозвоночных очень велика. Не будучи в
данном случае сами паразитами, они являются переносчиками возбудите-
лей ряда болезней, причем самого разнообразного характера. Среди них
мы имеем и просто механических переносчиков различных инфекций и про-
межуточных хозяев, в которых происходит та или иная часть жизненного
цикла паразита.
Сюда относятся брюхоногие моллюски, некоторые ракообразные, мно-
гие клещи и большое количество насекомых (комары, мухи и т. д.). Борьба
с этими беспозвоночными исключительно важна, так как всегда лучше
предотвратить болезнь, чем лечить ее.
5. Беспозвоночные — вредители сельского и лесного хозяйства.
Сюда относится главным образом громадное количество различных на-
секомых, клещи и некоторые брюхоногие моллюски, например слизни.
6. Беспозвоночные — вредители различного рода запасов.
Сюда относятся различные насекомые и клещи (долгоносик, мучной
хрущак, мучные клещи), которые вредят запасам зерна и муки. Широко
известна обыкновенная моль, гусеницы которой уничтожают шерсть
и изделия из нее.
7. Беспозвоночные, засоряющие и повреждающие различные сооружения
и обрастающие суда.
Сюда относится ряд сидячих беспозвоночных (губки, гидроиды, мшанки,
прикрепленные моллюски и др.), которые, поселяясь в водопроводных
и других гидротехнических сооружениях, могут засорять, а в некоторых
случаях даже совсем закупоривать их.
Непосредственными разрушителями деревянных сооружений в море,
а также деревянных кораблей является сильно измененный двустворчатый
моллюск, так называемый корабельный «червь», и некоторые ракообразные.
Деревянным сооружениям на суше, а также деревянной мебели вредят
некоторые насекомые — жуки и термиты.
8. Ядовитые беспозвоночные.
Сюда относится ряд паукообразных (скорпионы, пауки), многоножки
(сколопендры), некоторые насекомые и кишечнополостные (медузы). Эти
животные могут своим ядом даже убивать человека и домашних животных
(так действуют главным образом крупные формы тропических стран) или
вызывать более или менее тяжелые поражения.
Изложенные выше данные более наглядно представлены на прилагае-
мой таблице (табл. 6), из которой ясно видно многообразие практического
значения беспозвоночных и разнообразие важных в практическом отноше-
нии групп.
Таким образом, мы видим, что значение беспозвоночных для человека
очень велико и, главное, разнообразно. Отрицательное значение беспозвоноч-
ных усугубляется тем, что вредные беспозвоночные очень многочисленны
509
Полезные
Группа беспозвоночных разводимые промышляемые как продукт пи- тания промышляемые как техническое сырье кормовые объек- ты промыс. овых животных опылители сель- скохозяйственных растений
Тип. Простейшие . — — — — —
Тип. Губки — — + — —
Тип. Кишечнополостные — + + — —
Тип. Плоские черви . . Тип. Круглые черви: — — — — —
Класс. Собственно круглые черви — — — — —
Класс. Коловратки — — — + —
Класс. Скребни Тип. Кольчатые черви: — —* — — —
Класс. Многощетинковые — + — + —
Класс. Малощетинковые — — — + —
Класс. Пиявки + — + — —
Тип. Мшанки Тип. Моллюски: — — — — —
Класс. Брюхоногие — + + + —
Класс. Двустворчатые + + + + —
Класс. Головоногие Тип. Членистоногие: — + + + —
Класс. Ракообразные + + + + —
Класс. Меростомовые (мечехвосты) . — + + —— —
Класс. Паукообразные ...... — —— — — —
Класс. Многоножки — —— — — —
Класс. Насекомые + + + +
Тип. Иглокожие + +
Таблица 6
Вредные
истребители и паразиты вреди- телей полезные в жиз- ни почвы паразиты че-~о- 1 века 1 1 паразиты про- мысловых и до- машних живот- ных паразиты сель- скохозяйственных растений переносчики бо- лезней и проме- жуточные хозяе- ва паразитов | вредители сель- скохозяйствен- ных культур вредители лес- ного хозяйства вредители запа- сов повреждающие сооружения и об- растающие суда ядовитые
— + 4- + — — — — — — —
— — — — — — — — — + —
— — — — — — — — — 4- +
— — + + — — — — — — —
+ — + + + — — — — — —
— — — + — — — — — — —
+ — — — — — — — —
— — + +. — — — — — — —
—- — — — — — — — — + —
— — — — — 4- 4- — — — —
— — — — — — — — — + —
— 4- — + — + — — — + —
i — — + 4- 4- + + + “Г — +
+ — —— — •— *— —— +
4- 4- + + + 4- + 4- +
как в качественном отношении (число видов), так и в количественном (число
особей того или иного вида). Для иллюстрации укажем, что число насеко-
мых, вредящих сельскому и лесному хозяйству, исчисляется сотнями
видов, и некоторые из них в момент массового развития появляются бук-
вально в миллионах экземпляров (например, саранча). Число паразити-
ческих червей тоже велико. У человека, например, паразитируют 22 вида
сосальщиков, 26 видов ленточных червей и 39 видов круглых. Количествен-
но паразитические черви могут быть тоже очень многочисленны. Извест-
ны случаи нахождения у человека в печени сразу 25 000 экземпляров дву-
устки (Opistorchis felineus).
Использование полезных беспозвоночных и борьба с вредными являют-
ся проблемами большой важности. Невозможно повысить по-настоящему
урожайность сельскохозяйственных культур без освобождения их от вре-
дителей из мира насекомых, а организовать эффективную борьбу с насеко-
мыми-вредителями нельзя без знания биологии.
Животноводство очень много теряет от гельминтов, которые пора-
жают всех домашних животных, внедряясь во все органы их тела, и умень-
шают выход мяса, удои молока и т. д. Борьбу с гельминтами можно поста-
вить рационально, только зная их биологию, в частности их циклы раз-
вития.
Для дальнейшего увеличения вылова рыбы в наших морях, озерах и
реках, т. е. увеличения рыбной продуктивности водоемов, надо знать, ка
кими беспозвоночными питаются эти рыбы и каких беспозвоночных надо все-
лить в водоемы, чтобы промысловые рыбы могли нормально питаться и
нагуливать вес.
Кожевенная промышленность терпит ущерб от тех изъянов на коже
крупного рогатого скота, которые наносят личинки кожных оводов, про-
буравливая кожу скота в наиболее ценных в техническом отношении час-
тях. Борьба с оводами — биологическая проблема, решить которую можно
только зная хорошо биологию оводов.
Сказанное ясно показывает, какие громадные задачи стоят перед био-
логами и, в частности, зоологами.
Надо добиться, чтобы наши поля и леса были освобождены от вреди-
телей, чтобы ни один грамм зерна или муки на наших складах не уничтожал-
ся жуком-долгоносиком или мучным клещом, чтобы человек и все домашние
животные были полностью избавлены от солитеров, аскарид, остриц и
прочих червей-паразитов. С другой стороны, необходимо, чтобы наши во-
доемы изобиловали рыбой. Необходимо более широко использовать такие
первоклассные пищевые продукты наших морей и пресных вод, как рако-
образные (креветки и крабы) и моллюски (мидии, устрицы, гребешки и др.).
Необходимо широко продвигать на север наших полезных беспозвоночных —
медоносную пчелу и шелкопрядов.
Остановимся более подробно только на некоторых вопросах, связанных
с хозяйственным значением беспозвоночных.
БЕСПОЗВОНОЧНЫЕ КАК КОРМОВЫЕ ОБЪЕКТЫ
ПРОМЫСЛОВЫХ РЫБ
В этом направлении значение беспозвоночных очень велико, причем
некоторые рыбы питаются бентическими беспозвоночными (двустворчаты-
ми моллюсками, некоторыми ракообразными, различными червями и игло-
кожими), другие планктическими, главным образом веслоногими рачками.
Примером бентосоядных рыб могут служить треска, камбала, осетровые
и др., планктоноядных — сельди. У трески беспозвоночные составляют
около 30% от всей поглощаемой пищи, у пикши около 50% и у камбалы и
ерша даже более 50%.
Пища сельдей, например всем известной каспийской сельди-пузанка,
511
состоит почти исключительно из беспозвоночных — веслоногих рачков
(около 70%) и мизид (около 20%).
Что касается пресноводных рыб, то в их питании большую роль играют
личинки насекомых, особенно двукрылых (хирономид).
Таким образом, непосредственно не используя тех или иных беспозво-
ночных, человек косвенно использует их через добываемую им рыбу
В связи с задачей значительно увеличить вылов рыбы в наших водоемах
вопрос о кормовых запасах приобрел особенно большое значение, так как
без достаточного количества кормовых беспозвоночных, естественно, не
может нормально развиваться и рыбное население, а следовательно, и
повышаться в нужной степени рыбная продуктивность водоемов.
Особенно остро встал вопрос на Каспии, где ввиду ряда причин,
связанных с геологической историей этого водоема, фауна донных беспоз-
воночных сравнительно бедна. Необходимо было усилить кормовую базу
этого моря, в первую очередь для таких ценных рыб, как осетровые. После
ряда экспериментальных исследований, проведенных нашими учеными
(проф. Л А. Зенкевич и его ученики), был намечен объект для акклимати-
зации в Каспии, а именно живущий в большом количестве в Азовском море
многощетинковый червь нереис (Nereis). В 1939 г был осуществлен массо-
вый перенос этого червя из Азовского моря в Каспийское. Нереис в Каспии
хорошо прижился, размножился и вошел в пищевой рацион осетровых рыб.
Это мероприятие наглядно показало, как человек может активно переделы-
вать природу
ПРОМЫСЛОВЫЕ БЕСПОЗВОНОЧНЫЕ
Как уже указывалось, беспозвоночные промышляются как пищевые
объекты и как источники технического сырья, причем промышляются в
основном водные беспозвоночные. Промысел наземных беспозвоночных ог-
раничивается очень небольшим числом форм из насекомых, например коше-
ниль.
По статистике мирового промысла водных животных, всего беспозво-
ночных промышляется ежегодно около 12 000 000 ц, что составляет пример-
но 7% общего улова водных животных. Из этого количества примерно
7 200 000 ц, или 60%, падает на различных моллюсков, примерно 3 600 000 ц,
или 30%,— на различных ракообразных; остальная часть приходится на
иглокожих, губок и кишечнополостных. Из моллюсков промышляются
главным образом устрицы, затем мидии, морской гребешок, сердцевидка
и ряд головоногих. Из ракообразных — омары, лангусты (крупные, дости-
гающие 0,75 м морские раки, по общему виду похожие, особенно омар, на
обыкновенного речного рака), различные крабы и креветки. Устрицы не
столько промышляются на естественных банках, сколько главным образом
разводятся, для чего существуют специальные «устричные заводы», где мо-
лодь выдерживается в особых садках, пока не достигнет промысловых раз-
меров. Всего устриц добывается в мире ежегодно примерно 1 600 000 ц.
Из ракообразных в некоторых странах разводятся омары.
В дореволюционной России беспозвоночных почти совершенно не до-
бывали, если не считать небольшого кустарного промысла мидий, креветок
и устриц в Черном море. После Октябрьской революции начал развиваться
промысел беспозвоночных преимущественно в Японском море (различные
беспозвоночные, главным образом гигантский, достигающий в размахе
ног 1 м, камчатский Kpa6(Paralithodes camtschatica), головоногие моллюс-
ки — кальмары и осьминоги, ряд двустворчатых — морской гребешок и
«белая ракушка», трепанги из иглокожих и некоторые другие).
Промысел камчатского краба представляет в настоящее время особую
ветвь нашей дальневосточной промышленности. Основным районом крабо-
вого промысла является западное побережье Камчатки. Добывают крабов
особые суда — краболовы, представляющие собой плавучие заводы, на
512
которых из выловленных крабов тут же в море изготавливаются консер-
вы. Такой краболов уходит из Владивостока на несколько месяцев и
возвращается груженный готовой продукцией. Ловят крабов крупно-
ячеистыми ставными сетями, в которые крабы, передвигаясь по дну,
запутываются.
На других наших морях промысел беспозвоночных развит еще очень
слабо, хотя сырьевые запасы промысловых форм, например мидий и креве-
ток, в них громадны. Взять у моря эти ценные в питательном отношении
ресурсы, безусловно, можно.
Кроме морских ракообразных, у нас достаточно широко промышляются
и пресноводные раки, причем особо ценным считается так называемый широ-
копалый рак (Astacus astacus), обитающий в водоемах Белоруссии, Смолен-
ской и Ленинградской областей и частично в водоемах Украины.
Что касается беспозвоночных, промышляемых как техническое сырье,
то здесь наибольшее значение имеют двустворчатые моллюски, раковины
которых используются при изготовлении различных перламутровых изде-
лий. В этом отношении у нас в настоящее время довольно широко исполь-
зуются пресноводные ракушки. До Октябрьской революции перламутр
ввозился к нам из заграницы.
НАСЕКОМЫЕ — ВРЕДИТЕЛИ СЕЛЬСКОГО
И ЛЕСНОГО ХОЗЯЙСТВА
Ряд насекомых, как уже было сказано,— опасные враги как сельского,
так и лесного хозяйства. В годы массового развития они могут опустошить
громадные площади посевов и лесов.
В царской России борьба с вредными насекомыми велась кустарно и
потому не могла дать нужного результата. Только в условиях планового
социалистического хозяйства это было признано государственно важным.
В дело разработки мер борьбы с насекомыми-вредителями включился ряд
наших крупных ученых — академик Кулагин, проф. Богданов-Катьков,
проф. Римский-Корсаков и многие другие. Борьба с вредными насекомыми
дала ощутимые результаты. В СССР азиатская саранча, например, не яв-
ляется таким бичом, каким она была в царской России. Так обстоит и с
другими вредителями (подробно об отдельных вредителях насекомых см.
в главе о насекомых).
Большим достижением в деле борьбы с вредными насекомыми является
применение так называемого биологического метода борьбы, т е. активного
использования различных животных, главным образом насекомых, в борьбе
с вредителями. В этом отношении на первом месте стоят наездники (из пе-
репончатокрылых), которые откладывают яйца в личинки различных на-
секомых, особенно бабочек, и даже в их яйца. Развивающиеся из этих яиц
личинки наездников уничтожают соответствующие личинки или яйца вреди-
телей. Особенно ценен в этом отношении наездник-яйцеед трихограмма
(Trichogramma evanescens). У нас в СССР биологический метод борьбы с
вредителями применяется очень широко; развито искусственное разведение
трихограммы для ее массового выпуска там, где наблюдается большое коли-
чество вредителей. Трихограмма имеет большое значение в борьбе с ябло-
невой плодожоркой, озимой совкой, луговым мотыльком, кукурузным мо-
тыльком, хлопковой совкой и другими чешуекрылыми.
КЛЕЩИ
Клещи — одна из наиболее вредных групп среди беспозвоночных.
Вред, приносимый клещами, очень разнообразен. Здесь и непосредственные
паразиты человека и домашних животных, так называемые зудни, вызываю-
щие изнурительную и неприятную болезнь — чесотку, и паразиты расте-
ний, и вредители запасов, так называемые «мучные клещи», и, наконец, пе-
реносчики различных опаснейших болезней человека, домашних животных
513
и сельскохозяйственных растений. Эта сторона «деятельности» клещей была
особенно полно выявлена ценными научными работами академика
Е. Н. Павловского и В. Н. Беклемишева и их учениками. Установлено, что
ряд заболеваний человека передается ему клещами. Так, например, тяжелая
болезнь, чаще встречающаяся в таежных районах, так называемый таеж-
ный, или весенне-летний, энцефалит, вызывается фильтрующимся вирусом,
а передается рядом иксодовых клещей, представителями рода Ixodes и др.
Клещи из рода Alectorobius являются переносчиками так называемого
клещевого возвратного тифа. Клещи также переносят туляремию. Возбу-
дители всех этих болезней в природе широко распространены среди млеко-
питающих и отчасти птиц, которые являются, таким образом, резервуарами
вируса. Высасывая кровь диких животных, а затем нападая на человека,
клещи заражают его соответствующей болезнью. Исследования Е. Н. Пав-
ловского природных очагов этих болезней и путей заражения ими человека
дали возможность совершенно по-иному поставить вопрос о борьбе с этими
болезнями, в частности с таежным энцефалитом. Борьба с клещами стала в
данном случае в центре внимания. Чтобы защищать человека от клещей
были разработаны и применены особые отпугивающие вещества, защитные
комбинезоны и пр. Вместе с тем микробиологами были выработаны предо-
хранительные прививки против энцефалита. Одна из опасностей, угрожаю-
щих человеку при завоевании тайги, была ликвидирована общими усилия-
ми биологов и микробиологов.
Эти работы показали, что может добиться наука в социалистическом
государстве, когда она поставлена на службу народному хозяйству и когда
научные- исследования проходят строго целеустремленно, комплексно, по
определенному плану.
ПАРАЗИТИЧЕСКИЕ ЧЕРВИ
Паразитические черви являются иногда очень опасными паразитами
человека и домашних животных. Если они обычно и не приводят к непосред-
ственной гибели, то сильно истощают организм. В некоторых случаях ки-
шечные паразиты своими прикрепительными крючочками (например, во-
оруженный солитер) или «зубами» (например, анкилостома) нарушают це-
лость кишечного эпителия, вызывая тем самым кишечные кровотечения и
«открывая ворота» различным инфекциям, а также способствуя возникнове-
нию кишечных язв.
В царской России зараженность различными гельминтами была очень
большой. В некоторых районах она охгатывала 100% населения. Особенно
страдали от гельминтов народности Севера, у которых заражение глиста-
ми было одной из причин их вымирания. С первых же лет Советской власти
у нас была развернута большая работа по борьбе с гельминтами. В этом
отношении надо особо отметить деятельность академика К. И. Скрябина
и его учеников, проведших большие работы по гельминтологическому об-
следованию населения нашей страны. Было установлено сильное заражение
некоторыми гельминтами: кошачьей двуусткой — для населения низовьев
Оби, например, до 85,2% заражения, широким лентецом—в некоторых
районах Сибири до 80% заражения и особенно аскаридой и острицей — по-
чти повсеместно большой процент заражения, причем последняя встречает-
ся особенно часто у детей.
Эти данные указали на необходимость организации серьезной и плано-
мерной работы с паразитическими червями. Борьба стала возможной бла-
годаря поднятию материального и культурного уровня населения и прове-
денным в нашей стране работам по улучшению бытовых условий жизни —
строительству водопроводов, канализации и пр. В связи со всеми этими ме-
роприятиями зараженность гельминтами значительно сократилась.
514
Хорошей иллюстрацией проведенной у нас борьбы с гельминтами яв-
ляется полная ликвидация заражения риштой (Dracunculusmedinensis) —
подкожным паразитом человека. До Октябрьской революции этот пара-
зит в большом количестве поражал население старой Бухары, что было
связано с исключительно низкими санитарно-гигиеническими условиями,
существовавшими тогда там. Дело в том, что промежуточным хозяином
ришты является веслоногий рачок — циклоп. Отсутствие водопровода за-
ставляло население пользоваться уличными бассейнами, так называемыми
хаузами. Больные риштой, совершая в хаузе омовение (ришта локали-
зуется обычно под кожей рук или ног), заражали бассейны личинками
ришты. Последние внедрялись в обитающих в хаузах циклопов. При питье
воды из этих же хаузов человек заглатывал вместе с водой зараженных цик-
лопов и тем самым заражался риштой. Все это усугублялось почти полным
отсутствием медицинской помощи. Проведенные мероприятия по строитель-
ству водопровода, по охране хаузов от загрязнений, а также организация
медицинской помощи населению полностью ликвидировали у нас дракун-
кулез.
Если биология некоторых гельминтов полностью изучена (эхинокок-
киоз и тениидоз), то в биологии многих из них, особенного ряда гельминтов
домашних животных, еще далеко не все ясно. Задача зоологов — разрешить
эти проблемы и дать в руки медицинских и ветеринарных работников твер-
дые основы для ликвидации этих гельминтозов.
БРЮХОНОГИЕ МОЛЛЮСКИ — ПРОМЕЖУТОЧНЫЕ ХОЗЯЕВА
ПАРАЗИТИЧЕСКИХ ЧЕРВЕЙ ЧЕЛОВЕКА
И ДОМАШНИХ ЖИВОТНЫХ
Ряд как пресноводных, так и наземных брюхоногих моллюсков явля-
ются промежуточными хозяевами сосальщиков и нематод человека и до-
машних животных. В них проходит значительная часть жизненного цикла
этих паразитов (см. соответствующие главы). Борьба с моллюсками — важ-
ное звено в борьбе с гельминтозами.
Без знания биологии промежуточного хозяина невозможна рацио-
нальная борьба с паразитом. Чтобы бороться с врагом, надо хорошо
знать все его повадки и нащупать наиболее уязвимые места.
Пока из всех моллюсков — промежуточных хозяев гельминтов —
наиболее полно изучена биология промежуточного хозяина печеночной
двуустки — малого прудовика (Galba truncatula). Наблюдения и опыты
показали, что этот моллюск может существовать в самых различных ус-
ловиях на экологической грани между сушей и водой. Удалось найти одно
«уязвимое место» этого моллюска, а именно его отношение к щелочности
воды; опыты показали, что если довести активную реакцию воды (pH) в
водоеме, где обитают моллюски, до величины больше 10, они гибнут. Эти
опыты дали возможность биологам предложить способ борьбы с моллюска-
ми путем известкования мест их обитания.
В Казахстане, Киргизии и других районах, где развито овцеводство,
очень важной проблемой является изучение биологии наземных моллюсков—
промежуточных хозяев трематод и круглых червей. Знание биологии мол-
люсков даст возможность предложить более рациональные способы выпасов
овец с тем, чтобы допускать возможно меньший контакт их с моллюсками.
Для гельминтозов человека также важно изучение биологии некоторых
пресноводных моллюсков — промежуточных хозяев гельминтов. Это от-
носится к гельминтозам: клонорхоз, метагонимоз, парагонимоз и шестизо-
моз; последнее — тяжелое заболевание, вызываемое так называемой кровя-
ной двуусткой (Schistosoma japonicum).
515
БЕСПОЗВОНОЧНЫЕ, ВРЕДЯЩИЕ ГИДРОТЕХНИЧЕСКИМ
СООРУЖЕНИЯМ И ВОДНОМУ ТРАНСПОРТУ
Большой вред различным гидросооружениям и кораблям, кроме древо-
точцев, приносят различные сидячие беспозвоночные, входящие в состав
так называемых обрастаний. Сюда относятся губки, гидроидные и корал-
ловые полипы, сидячие кольчатые черви, мшанки, прикрепляющиеся дву-
створчатые моллюски, усоногие рачки и пр. Они обрастают самые различные
сооружения и предметы, находящиеся в воде: части пристаней, гидротех-
нические сооружения и корабли. Некоторые организмы проникают и в
водопроводные сооружения и при массовом развитии могут почти закупори-
вать их. Они могут развиваться в полной темноте при давлении в нес-
колько атмосфер. Вред обрастания состоит не только в том, что задерживает-
ся ток воды, но при этом портится и сама вода, приобретая плохой вкус и
запах.
Особенно сильно развиваются обрастания в морях, главным образом
в теплых, где рост организмов и образование колоний идут очень быстро.
Для всякого рода кораблей обрастания представляют большие затрудне-
ния — они увеличивают трение корабля о воду, из-за чего снижается его
скорость.
Борьба с обрастаниями ведется путем покрытия подводных сооруже-
ний и частей кораблей особыми красками, содержащими ядовитые примеси
мели и ртути. Почти все организмы, входящие в обрастание, имеют свобод-
ноплавающую личинку Ядовитые частицы, выделяющиеся из краски в
воду, отравляют личинок и таким образом предотвращают их оседание и
образование нового обрастания. Важной задачей биолога при изучении
обрастаний является изучение именно личинок — их биологии и сроков
развития с целью выявления наиболее эффективно действующих на них
веществ для включения их в покровные краски.
Работа над изучением беспозвоночных животных в нашей стране ве-
дется очень активно. Мы исходим из общего принципа: усиливать и разви-
вать все полезное и бороться со всем вредным, мешающим нашему хозяй-
ству и угрожающим здоровью человека.
ЭВОЛЮЦИОННОЕ РАЗВИТИЕ БЕСПОЗВОНОЧНЫХ
Эволюционное развитие представляет собой процесс исторического
преобразования организации животных и растений, идущий от простого к
сложному, от менее дифференцированного к более дифференцированному
В зависимости от условий существования в одних случаях происходит
процесс усложнения организации, возникают новые органы или усложняется
строение старых; это вызывает коррелятивное изменение в других органах:
одни из них соответственно также усложняются, другие, наоборот, соот-
ветственно редуцируются.
В других случаях, когда животные попадают в особые специфические
условия существования, переходят, например, к сидячему образу жизни
или паразитизму, наступает процесс односторонней специализации, ко-
торый сопровождается развитием одних органов и сильной редукцией
других.
Таким образом, эволюционный процесс состоит не только в постоянном
прогрессивном развитии организации животных, а представляет собой
сложный процесс, включающий и прогрессивные и регрессивные из-
менения.
Современный животный мир представляет собой систему конечных
разветвлений общего родословного древа, отчленившихся от главного ство-
516
ла на разных этапах исторического развития. Низшие ветви представляют
собой древнейших животных, сохранивших свойственную им примитив-
ную организацию, однако их современные представители не являются груп-
пами, остановившимися в своем эволюционном развитии. Они прошли соб-
ственный путь развития, и у них наряду с признаками, сохранившимися
от общего родоначального предка, развились иные, приспособительные к
новым условиям существования качества, которых не было у предков.
К этому нужно еще прибавить, что новые эволюционные изменения
возникают не только у взрослых форм, но и на самых различных этапах
индивидуальной жизни организма, при этом чем более ранней стадией яв-
ляется стадия возникновения нового эволюционного изменения, тем силь-
нее нарушается общий путь дальнейшего развития организма. Отсюда
следует, что при построении родословного древа животного мира нужно учи-
тывать не только организацию современных и ископаемых взрослых живот-
ных и условия окружающей их среды, но и данные по истории их индиви-
дуального развития.
Большинство зоологов держится монофилитической точки зрения и
утверждает, что разнообразные группы животного мира произошли от
общей родоначальной группы, лежащей в основе родословного древа
(табл. XVI). Своими основаниями корни животного и растительного царства
тесно переплетаются друг с другом, причем первичными организмами надо
считать растительные формы, питающиеся неорганическими веществами.
Животные формы, питающиеся обычно либо растениями, либо другими жи-
вотными, развились позднее.
Много примитивных черт имеет класс корненожек (Rhizopoda, 2)1.
Класс этот включает в себя, с одной стороны, голых амеб, имеющих весьма
примитивное строение, с другой,— более сложных одноклеточных, как
фораминиферы, солнечники и радиолярии, имеющих очень сложный ске-
лет, развивающийся в тесной связи с условиями их существования: сравни-
тельно тяжелые раковины у донных, ползающих форм и ажурный скелет с
различного рода придатками у свободнопарящих в воде форм.
Промежуточное положение в системе одноклеточных простейших (Pro-
tozoa) занимает класс жгутиковых (Flagellata), которые сохраняют некото-
рые черты растительных форм, так как имеют способность автотрофного
питания неорганическими веществами. Предки первичных жгутиковых
(Proflagellata) долгое время считались родоначальной группой, от которой
произошли все остальные классы современных одноклеточных животных.
Среди них встречаются и колониальные многоклеточные формы (Volvo-
cidae и др.). В настоящее время после новейших данных по вопросу о проис-
хождении жизни первичными животными существами считают предков
корненожек (Rhizopoda).
Класс споровиков (Sporozoa, 3), объединяющий одноклеточных, приспо-
собившихся к паразитическому образу жизни и характеризующихся очень
сложными циклами развития, имеет, по-видимому, двойственное проис-
хождение. Одна группа (телоспоридии) связана со жгутиковыми, другие
(неоспоридии) — ближе к корненожкам.
Высшая группа среди одноклеточных, безусловно, подтип ресничных
(Ciliophora). Оба класса, входящие в него (Giliata, 4 и Suctoria), характери-
зуются чрезвычайно сложным строением одноклеточного тела. Органеллы
движения — реснички — все же сближают их со жгутиковыми, от которых
их и выводят в виде особой филогенетической ветви.
Важным новым этапом в эволюции мира животных явилось возник-
новение многоклеточных животных (Metazoa), когда между клетками, со-
ставляющими их тело, произошло разделение труда и они стали выполнять
разные жизненные функции целого организма.
1 Эти цифры соответствуют цифрам на схеме эволюционного развития беспозво-
ночных (табл. XVI)
517
IF6
Среди одноклеточных встречаются колониальные формы, которые
могут считаться первым этапом на пути происхождения многоклеточных
животных. Среди жгутиковых, например, имеются колонии (Volvox и др.),
которые могут дать нам некоторое представление о первичных многоклеточ-
ных животных. Они характеризуются тем, что после многократных деле-
ний дочерние клетки не расходятся, а остаются вместе и образуют новый
единый организм. У него происходит дифференциация клеток в связи с
функцией, ими выполняемой (вегетативные клетки и клетки размножения).
Относительно происхождения первичных многоклеточных существует
несколько гипотез. Э. Геккель на основании существования в эмбриональном
развитии многих многоклеточных животных стадии однослойной бластулы
и двуслойной гаструлы считал эти стадии эмбрионального развития повто-
рением (рекапитуляцией) стадий эволюционного развития. Первым этапом
этого развития Геккель считал однослойный организм и назвал его блас-
теей, вторым — двуслойный организм из эктодермы и энтодермы с полостью
первичного кишечника, развившейся путем впячивания, и назвал его
гастреей. Эта гастрея, по мнению Геккеля, была родоначальной формой
для всех многоклеточных животных.
И. И. Мечников оспаривал эту гипотезу: он считал гаструлу, образую-
щуюся путем впячивания, вторичным образованием. Первичным процессом
превращения однослойного организма в двуслойный, по его мнению, яв-
ляется иммиграция части клеток внутрь однослойного зародыша (см.
рис. 9). Гипотетическое первичное двуслойное животное он называл парен-
химеллой, или фагоцителлой.
Первым этапом в эволюционном развитии многоклеточных являются
двуслойные животные, т. е. губки и кишечнополостные, тело которых со-
стоит только из двух слоев — эктодермы и энтодермы.
Тип губок (Spongia, 5) с их своеобразным строением и развитием
настолько отличается от всех других многоклеточных, что обычно выделяет-
ся в особую группу Parazoa или Enantiozoa, тогда как все остальные мно-
гоклеточные объединяются под названием настоящие многоклеточные —
Eumetazoa, или Enterozoa. Губки — очень древняя группа, отделившаяся
от основного ствола многоклеточных очень давно и представляющая осо-
бую боковую ветвь их эволюционного развития.
Таблица XVI. Схема эволюционного развития беспозвоночных:
1—одноклеточные; II— двуслойные; III— бесполостные и первичнополостные:
А — первичноротые; Б — вторичноротые
/. Одноклеточные (Protozoa); / — жгутиковые (Flagellata), 2 — корненожки (Rhizopoda), 3— спо-
ровики (Sporozoa), 4 — ресничные (Ciliata)
II. 5 — губки (Spongia). Кишечнополостные (Coelenterata): 6— гидроидные (Hydrozoa),- 7— сци-
фоидные (Scyphozoa), 8 — коралловые (Actinozoa), 9 — гребневики (Ctenophora)
ill. Плоские, или бесполостные, черви (Plathelminthes): 10— ресничные черви (Turbellaria), II —
сосалыцики (Trematodes), 12 — ленточные черви (Cestodes).
Круглые или первичнополостные, черви (Nemathelminthes): 13— брюхоресничные (Gastrotrichia),
14—коловратки (Rotatoria), 15—собственно круглые черви (Nematodes); 16 — волосатиковыс
(Gordiacea), 17 — колючеголовые, или скребни (Acanthocephali); 18— немертины (Nemertini). IV А .
Кольчатые черви (Annelides): 19 — многощетинковые (Polychaeta), 20— малощетинковые (Oli-
gochaeta): 21 — пиявки (Hirudinea), 22 — эхиуридовые (Echiuroidea).
Членистоногие (Arthropoda); 23—трилобиты (Trilobita), 24 — мечехвосты (Xiphozura), 25 — пауко-
образные (Arachnoidea), 26— ракообразные (Crustacea), 27— первичнотрахейные (Protracheata),
28 — многоножки (Myriopoda), 29 — насекомые (Insecta).
Моллюски (Mollusca): 30 — бороздчатобрюхие (Solenogastres), 31 — панцирные (Loricata), 32 —
двустворчатые (Bivalvia), 33 — лопатоногие (Scaphopoda), 34 — брюхоногие (Gastropoda), 35—голо-
воногие (Cephalopoda); 36—Мшанки (Bryozoa); 37—Плеченогие (Brachiopoda).
IV Б. 38 — Щетинкочелюстные (Chaetognatha). Иглокожие (Echinodermata): 39 — морские лилии
(Crinoidea), 40—морские ежи (Echinoidea), 41 — змеехвостки (Ophiuroidea), 42 — морские звезды
(Asteroidea), 43 — морские огурцы (Holothuroidea); 44 — Погонофоры (Pogonophora); 45 — Кишеч-
нодышащие — полухордовые (Enteropneusta — Hemichordata); 46 — Хордовые (Chordata); 47.—Мо-
ноплакофоры (Monoplacophora).
Примечание: пунктирные линии указывают, что филогения группы еще не совсем ясна;
черными кружками обозначены исходные гипотетические группы; цифрами в кружках отмечены
целиком паразитические группы.
519
Основное направление развития настоящих многоклеточных шло по
пути обособления двуслойных многоклеточных животных и дало прежде
всего кишечнополостных (Coelenterata, 6—9) — типичных дву-
слойных многоклеточных. Среди них наиболее простой группой надо счи-
тать гидроидных (Hydrozoa, 6), у которых уже имеются две характерные
для кишечнополостных жизненные формы — прикрепленный полип и сво-
бодноплавающая медуза. Дальнейшее эволюционное развитие кишечно-
полостных шло по линии усложнения двуслойного типа строения (образо-
вание эктодермической глоточной трубки, усложнение кишечной полости без
общего повышения организации). Развитие таких высших кишечнополост-
ных шло по двум направлениям: у представителей класса коралловых (Ас-
tinozoa, 8) по линии развития полипа как типичного сидячего организма, у
представителей класса сцифоидных (Scyphozoa, 7) по линии развития ме-
дузы как типичного свободноплавающего организма. Несколько особо
шло, по-видимому, развитие класса гребневиков (Ctenophora, 9) — свобод-
ноплавающих организмов. Активный образ жизни вызвал у них появление
зачатков двубокой симметрии и третьего зародышевого листка, или мезо-
дермы, как основы для формирования мускулатуры. В связи с наличием
этих признаков гребневики занимают несколько особое положение средн
кишечнополостных и рядом исследователей выделяются даже в особый тип.
Переход к ползанию, естественно, вызвал дифференцировку переднего
(головного) конца с нервным центром, более резко выраженную двубоко-
вую симметрию и развитие настоящей мезодермы, обеспечивающей разви-
тие мускулатуры и других внутренних органов. Совокупность всех этих
признаков есть те качественные изменения, которые имели место при пе-
реходе от низшего (двуслойного) этапа в эволюционном развитии многокле-
точных к высшему (трехслойному)
Первичные трехслойные дали начало нескольким эволюционным вет-
вям живого мира, из которых две—первичноротые (Protostomia, 10—35,
36, 37) и вторичноротые (Deuterostomia, 38—46) — достигли наибольшего
развития.
Первичноротые характеризуются рядом признаков: кутикулой, или
наружным эктодермическим скелетом, телобластическим способом образо-
вания мезодермы, преобразованием личиночного рта в рот взрослого живот-
ного, ганглиозной нервной системой и некоторыми другими.
К первичноротым относится ряд типов, а именно плоские черви (Plat-
helminthes, 10—12), немертины (Nemertini, 18), круглые черви (Nemathel-
minthes, 13—17), кольчатые черви (Annelides, 19—22), членистоногие (Art-
hropodo, 23—29) и моллюски (Mollusca, 30—35, 47).
Наиболее примитивным типом первичноротых, безусловно, надо счи-
тать плоских червей и среди них класс свободноживущих
ресничных червей (Turbellaria, 10). Представители этого класса имеют еще
ряд признаков, связывающих их с кишечнополостными. Сюда относятся:
отсутствие анального отверстия, элементы лучевой симметрии в строении
ряда органов, ресничный покров. Часть плоских червей перешла к пара-
зитизму, сперва, по-видимому, в качестве наружных паразитов, затем —
внутренних (классы сосальщиков (Trematoda, 11) и ленточных червей (Ces-
toidea, 12). Переход к такому специфическому образу жизни вызвал ряд
регрессивных изменений, например полное исчезновение кишечника у лен-
точных червей. Вместе с тем выработался ряд адаптивных признаков, тесно
связанных с жизнью на хозяине или внутри него, например органы при-
крепления, плотная кутикула.
Организмы, близкие к современным турбелляриям (их можно назвать
первичными трехслойными), были, по всей вероятности, исходными форма-
ми для ряда более высокоорганизованных групп первичноротых.
Прогрессивные изменения их выразились в появлении качественно
новых признаков, поднявших организацию этих животных на новую сту-
пень. Среди этих признаков одними из важнейших были: образование аналь-
520
ного отверстия, большая концентрация нервной системы и усиление мус-
кулатуры.
Безусловно, прогрессивным, но вместе с тем еще очень тесно связанным
с плоскими червями является тип немертин (Nemertini, 18), представ-
ляющий особую ветвь развития первичноротых. У них имеется сквозной
кишечник, хорошо развитый головной нервный узел, замкнутая кровенос-
ная система и очень мощная мускулатура, достигающая у некоторых из
них до 60% всей массы тела (Зенкевич, 1944). Вместе с тем у немертин име-
ются еще признаки плоских червей — ресничный покров, отсутствие на-
стоящей полости тела и протонефридиальная выделительная система.
Наличие этих признаков дает основание ряду авторов рассматривать немер-
тин даже только как подтип плоских червей.
Другой прогрессивной ветвью, идущей от первичных плоских червей,
является тип круглых червей (Nemathelminthes, 13—17). Связь
входящих в этот тип групп с ресничными червями долгое время была очень
неясна, и круглые черви стояли как бы особняком. В настоящее время эта
связь устанавливается довольно ясно через группу брюхоресничных (Gastrot -
richa, 13), имеющих признаки как ресничных (частичный ресничный покров
протонефридии), так и круглых червей (сквозной кишечник, кутикуляри-
зация покровов). У собственно круглых червей (Nematodes, 15), по-види-
мому, в связи со специфическими условиями существования — жизнь в
грунте водоемов, в почве или переход к паразитизму — произошло даль-
нейшее развитие признаков, имеющихся уже у гастротрих, а именно полное
выпадение ресничного покрова, образование мощной кутикулы и усиление
мускулатуры. Близка к собственно круглым червям небольшая группа воло-
сатиковых (Gordiacea, 16). В другом направлении эволюционировали к о-
ловратки (Rotatoria, 14). Если круглые черви перешли к жизни в
грунте и почве или к паразитизму, то коловратки, наоборот,— к жизни в
толще воды, в связи с чем ресничный покров превратился у них в сравни-
тельно мощный коловращательный аппарат — орган активного движения
в воде.
Несколько особняком среди круглых червей стоят колючеголовые, или
скребни (Acanthocephala, 17). С одной стороны, они имеют ряд признаков,
сходных с круглыми червями (полость тела, строение мускулатуры), с дру-
гой,— ряд особых, специфических признаков (отсутствие кишечника). Ряд
авторов в связи с этим считает более правильным выделить их в особый тип
и рассматривать как самостоятельную ветвь, происходящую непосредствен-
но от первичных плоских червей и сильно измененную в связи с пара-
зитизмом.
Наиболее прогрессивной ветвью, происходящей от исходных форм пер-
вичноротых, безусловно, является ветвь, давшая кольчатых чер-
вей (Annelides, 19—22) и далее членистоногих (Arthropoda, 23—29). Ка-
чественные отличия этой высшей группы червей от нижестоящих низших
типов (плоские черви, немертины, круглые черви) заключаются в том, что
у них появляются особые органы движения (боковые выросты тела), настоя-
щая (вторичная) полость тела и разделение тела на отдельные сегменты. Все
эти признаки, безусловно прогрессивные, и появление и развитие их сильно
повысило организацию высших червей сравнительно с низшими.
Среди кольчатых червей центральное место занимают многощетинко-
вые (Polychaeta, 19) с их громадным морфологическим разнообразием, тес-
но связанным с различными условиями существования. От этой централь-
ной группы в связи с особым образом жизни отошло несколько групп, часть
которых сохранила основные признаки типа, часть утеряла их. Сюда от-
носятся малощетинковые (Oligochaeta, 20), перешедшие к жизни главным
образом в почве, и пиявки (Hirudinea, 21), перешедшие к полупаразитичес-
кому образу жизни. Обе эти группы сохранили основные признаки кольче-
цов, например сегментацию. Значительно дальше отошли от многощетинко-
вых так называемые эхиуридовые (Echiuroidea, 22), которые в связи с ро-
20 Курс зоологии, т. I
521
ющим образом жизни утеряли ряд признаков, свойственных кольчатым
червям.
Дальнейшее усложнение придатков тела и распадение их на отдельные
элементы (членики) вновь качественно повысило организацию животных
и дало такую прогрессивную и исключительно богатую в современной эпохе
группу, как членистоногие (Arthropoda, 23—29).
Кроме членистых конечностей, у них хорошо отграниченная голова с
головным мозгом и целой системой различных рецепторов, дающих возмож-
ность животному хорошо воспринимать окружающую среду и все изменения,
в ней происходящие. Снаружи тело членистоногих покрыто хитиновым ске-
летом, который, естественно, лучше защищает животное, чем мягкая кути-
кула кольчатых червей.
Развитие различных групп членистоногих шло разными путями, но
предками их была, по-видимому, одна примитивная группа кольчатых
червей. Можно предполагать, что эволюционное развитие членистоногих
шло по трем основным направлениям: во-первых, ракообразные (Crustacea,
26); во-вторым, трилобиты (Trilobita, 23) — хелицеровые (Chelicerata),
мечехвосты (Xiphosura, 24) и паукообразные (Arachnoidea, 25); в-третьих,
первичнотрахейные (Protracheata, 27) — многоножки (Myriopoda, 28) и
насекомые (Insecta, 29).
Развитие этих трех ветвей шло по линии превращения гомономной сег-
ментации аннелид в гетерономную. Каждая ветвь начинается формами с
длинным гомономно расчлененным телом (листоногие, трилобиты, первично-
трахейные), затем происходит уменьшение числа сегментов, возрастает их
дифференцировка и появляется разделение тела на отделы. В процессе эво-
люционного развития членистоногих, кроме морфологических изменений,
произошли коренные изменения и в их биологии, а именно переход к на-
земному образу жизни. Этот переход произошел в ветви трилобиты — хели-
церовые, по-видимому, в карбоне, когда появились первые наземные скор-
пионы, очень близкие к морским формам из ракоскорпионов.
В ветви первичнотрахейные — насекомые этот переход произошел
возможно еще раньше, так как формы, подобные первичнотрахейным, из-
вестны уже из среднего кембрия.
Первичнотрахейные представляют очень большой интерес; по сути
они еще не настоящие членистоногие, но все же имеют ряд характерных для
них признаков (конечности с коготками, хитиновый покров, характер по-
лости тела), тесно сплетенных с признаками кольчатых (кожно-мускульный
мешок, нефридий), а в некоторых случаях (строение нервной системы) даже
плоских червей.
Высшими представителями членистоногих, безусловно, являются на-
секомые с их очень хорошо развитой нервной системой и органами чувств,
резко выраженным разделением тела на три отдела (голова, грудь, брюш-
ко), небольшим числом ходильных конечностей и появлением особых ор-
ганов движения — крыльев. Внутри класса насекомых произошла диффе-
ренцировка отдельных групп (отрядов) в связи с различными условиями
существования, характера питания. Надо отметить, кроме того, что часть
насекомых вторично перешла к водному образу жизни, сохранив при этом
воздушное дыхание, а часть стала паразитами, вторично утеряв в связи с
этим крылья.
Особой ветвью первичноротых являются моллюски (Mollusca.
30—35, 47). По некоторым признакам они близки к кольчатым червям
(характер полости тела, строение органов выделения, наличие кровеносной
системы), но вместе с тем резко отличаются от них по внешнему виду (от-
сутствие сегментации и парных придатков тела). Такое резкое отличие свя-
зано, по-видимому, с тем, что моллюски эволюционировали не в сторону
повышения активности, а в сторону малоподвижного, а в некоторых случаях
совершенно неподвижного образа жизни. В связи с этим у них развился та-
кой характерный для них признак, как сравнительно тяжелый наружный
522
скелет — раковина. Таким образом, можно предполагать, что моллюски
имеют общий корень с кольчатыми червями, но очень рано (т. е. геологи-
чески очень давно) от них отделились и эволюционировали как особая ветвь
первичноротых.
Внутри моллюсков наиболее примитивным классом надо считать моно-
плакофор (Monoplacophora, 47), панцирных, или боконервных (Loricata,
31), у которых сохранились некоторые примитивные признаки. Близки,
но имеют ряд специфических признаков бороздчатобрюхие (Solenogastres,
30). Очень типичными моллюсками как по строению, так и по образу жизни
надо считать двустворчатых (Bivalvia, 32). Дальнейшая эволюция моллюсков
пошла по линии увеличения активности и усиления в связи с этим голов-
ного отдела, большей концентрации нервной системы и усиления отдела
тела, связанного с передвижением,— ноги. Сюда относятся три класса —
брюхоногие (Lastropoda, 34), лопатоногие (Scaphopoda, 33) и особенно
высшая группа моллюсков — головоногие (Cephalopoda, 35). В связи с
большей активностью этих моллюсков раковина стала для них органом не
полезным, а, наоборот, мешающим прогрессивному развитию, и она стала
претерпевать регрессивное развитие. Это имеет место у ряда групп брюхо-
ногих (киленогие, крылоногие, слизни), а особенно у головоногих, боль-
шинство современных форм которых в отличие от ископаемых не имеют на-
ружного скелета. Скелет у них превратился во внутреннее образование,
несущее главным образом опорную функцию. К этому надо еще прибавить
то, что у брюхоногих произошел важный биологический сдвиг — переход
части их к наземному образу жизни.
Другая ветвь трехслойных — вторичноротые — характеризуется сле-
дующими признаками: наличием внутреннего мезодермического скелета,
образованием мезодермы энтероцельным путем, замыканием бластопора и
вторичным образованием рта взрослого животного, нервной системой не
ганглиозного типа, а в виде тяжей или трубки.
Ко вторичноротым относятся щетинкочелюстные (Chaetognathar
38), иглокожие (Echinodermata, 39—43), погонофоры (Pogonophora, 44),
кишечнодышащие (Enteropneusta, 45) и хордовые (Chordata, 46).
В отличие от первичноротых среди вторичноротых неизвестны предста-
вители первой эволюционной ступени, т. е. соответствующей низшим червям,
а все представители относятся к более высокоорганизованным формам,
имеющим сквозной кишечник и целомическую полость тела.
Своеобразной группой вторичноротых, которая занимает среди них
несколько особое положение, являются щетинкочелюстные
(Chaetognatha, 38). Эти свободноподвижные животные, имеющие ряд основ-
ных признаков вторичноротых, например энтероцельное образование ме-
зодермы, по некоторым признакам сходны с первичноротыми. По-видимому,
их надо рассматривать как особую ветвь вторичноротых, очень рано отде-
лившуюся от основного пути их развития.
Большая группа вторичноротых — иглокожие (Echinodermata,
39—43) — крайне своеобразна в связи с тем, что большинство представителей
имеют лучевую симметрию, что как будто бы роднит их с кишечнополостны-
ми. Как показывают исследования ископаемых иглокожих, а также их раз-
витие (личиночные формы), лучистая симметрия иглокожих — явление
вторичное, связанное с переходом к прикрепленному или малоподвижному
образу жизни. Таким образом, иглокожие в своем эволюционном развитии
несколько уклонялись от основного прогрессивного направления вторич-
норотых. При дальнейшей эволюции у некоторых иглокожих (голотурии,
сердцевидные ежи) вновь выработалась двубоковая симметрия.
Основное направление эволюции вторичноротых пошло по линии фор-
мирования подвижных форм. Сперва это были, видимо, формы, роющиеся
в грунте, подобные полухордовым — кишечнодышащим (Enteropneusta,
45) с их зачатком хорды и жаберными щелями, т. е. с признаками, характер-
ными для хордовых; затем появились более подвижные формы, подобные
20*
523
ланцетнику (46), уже относящиеся к настоящим хордовым (бесчерепным)
(о филогенезе хордовых см. в конце второго тома настоящего курса).
Кроме двух основных ветвей развития трехслойных (первично- и вто-
ричноротые), от первичных трехслойных, по-видимому, происходит еще
несколько небольших ветвей. К таким ветвям относятся мшанки (Вгуо-
zoa, 36) иплеченогие (Brachiopoda, 37). Обе эти совершенно различ-
ные группы очень древние, известные с кембрия. Современные их предста-
вители по своим признакам стоят на уровне высших червей (наличие целома
и метанефридиев). Характерной чертой обеих групп, многое объясняющей
в их строении, является то, что они прикрепленные животные. В связи с
этим у них развился наружный скелет и фильтрационный аппарат. По ряду
признаков (энтероцельное образование мезодермы) плеченогие ближе к
вторичноротым, мшанки — к первичноротым. В общем обе эти группы надо
рассматривать как самостоятельные, занимающие промежуточное положение
между первично- и вторичноротыми, развившиеся под сильным влиянием
специфического (прикрепленного) образа жизни и приобретшие в связи с
этим ряд сходных, конвергентных, черт: плеченогие — с двустворчатыми
моллюсками, мшанки — с кишечнополостными (полипами).
ЛИТЕРАТУРА
Общие руководства по всем разделам
Руководство по зоологии (под ред. Л. А. Зенкевича) т. I (одноклеточные, губки,
кишечнополостные, низшие черви); т. II (высшие черви, моллюски) 1940; т. III
(первичнотрахейные, многоножки, насекомые, иглокожие, щетинкочелюстные и др.)»
изд. Ин-та зоологии МГУ, 1951.
Догель В. А. Сравнительная анатомия беспозвоночных, т. I, 1938;
т. II, 1940.
Догель В. А. Общая протозоология, изд. АН СССР, 1962.
Беклемишев В. Н. Основы сравнительной анатомии беспозвоночных,
изд. 3-е, 1964.
Ливанов Н. А. Пути эволюции животного мира, 1955.
Иванов П. П. Общая и сравнительная эмбриология, 1937
Шмидт ГА. Эмбриология животных, т. I — Общая эмбриология, 1951;
т. II Беспозвоночные, 1953.
Коштоянц X. С. Основы сравнительной физиологии, ч. I, изд. 2-е, 1950;
ч. II, 1957
Бобринский Н. А., Зенкевич Л. А. и Бирштейн Я. А.
География животных, 1945.
Наумов Н. П. Экология животных, 1963.
Давиташвили Л. Ш. Краткий курс палеонтологии, 1958.
Kaestner A. Lehrbuch der Speziellen Zoologie. Band I, II, 1965.
Traite de Zoologie, publie par P. Grasse (1948—1963). Paris.
По паразитологии
Догель В. А. Общая паразитология, изд. 3-е, 1963.
Скрябин К. И. и Шульц Р. С. Основы общей гельминтологии, 1940.
Павловский Е. Н. Руководство по паразитологии человека, т. I, 1946;
т. II, 1948.
Павловский Е. Н. Природная очаговость трансмиссивных болезней
в связи с эпидемиологией зооантропонозов, изд. Наука, 1964.
Скрябин К. И. и др. Ветеринарная паразитология, 1934.
По гидробиологии
Зернов С. А. Общая гидробиология, изд. 2-е, 1949.
Зенкевич Л. А. Фауна и флора морей СССР, изд. 2-е, 1955.
Иванов А. В. Промысловые водные беспозвоночные, 1955.
По энтомологии
3-е,
Ш в а н в и ч Б. Н. Введение в энтомологию, изд-во ЛГУ, 1959.
Шеголев В. А. Сельскохозяйственная энтомология, Сельхозгиз, 1960.
Римский Корсаков М. Н. и др. Лесная энтомология, изд.
1949.
Розов С. А., Губин А. Ф. и др. Пчеловодство, изд. 3-е, 1955.
Яхонтов В. В. Экология насекомых, изд -во «Высшая школа», 1964.
525
Руководство к практическим занятиям
Иванов А. В., Полянский Ю. И., Стрелков А. А. Боль-
шой практикум по зоологии беспозвоночных, т. I, изд. 2-е, 1958; т. II, 1946.
Аверинцев С. В. Малый практикум по зоологии беспозвоночных, 1947.
Павловский Е. Н. Практикум по зоологии, изд. 2-е, 1938.
Роскин Г. И. и Левинсон Л. Б. Микроскопическая техника,
изд. 3-е, 1957.
Райков Б. Е. и Р и м с к и й - К о р с а к о в М. Н. Зоологические
экскурсии, изд. 6-е, 1956.
Руководство по определению беспозвоночных
Определитель насекомых Европейской части СССР, под ред. Тарбинского С. П.
и Плавил ыцикова Н. Н., 1948.
Плавильщиков Н. Н. Определитель насекомых, изд. 2-е, 1950 (более
краткий, охватывающий только наиболее обычных насекомых).
Жизнь пресных вод СССР, под. ред. Жадина В. И., т. I, 1940; т. II, 1949
(содержит определительные таблицы всех групп пресноводной фауны, начиная от одно-
клеточных и кончая млекопитающими).
Липин А. Н. Пресные воды и их жизнь, изд. 3-е, 1950 (содержит опреде-
лительные таблицы пресноводных беспозвоночных, за исключением одноклеточных).
X е с с и н Е. М. Краткий определитель пресноводной фауны, 1951.
Определитель фауны и флоры северных морей СССР, под. ред. Гаевской
Н. С., изд. 2-е, 1948 (содержит определительные таблицы всех групп морских беспоз-
воночных, начиная от одноклеточных и кончая личиночнохордовыми).
Атлас беспозвоночных дальневосточных морей СССР, изд. Зоол. ин-та АН СССР
1955.
«Фауна СССР» и «Определители по фауне СССР», издаваемые Зоол. ин-том
АН СССР (отдельные выпуски по беспозвоночным).
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
483,
497,
483,
485,
499.
486,
486,
489,
Аппарат челюстной грызу ще-лижущий 443
-----колюще-сосущий 426, 438, 450
— — лижуще-сосущий 450
— — сосущий 426, 443, 447
— парабазальный 58
Аргазиды 392
Аргонавт 330
Ареал 28
Аристоцистис 503
Археоциты 100, 101
Архитомия 264
Арцелла 43
Асимметрия 301, 309
Аскарида лошадиная 201—205, 222
— свиная 222
— человеческая 206, 222
Аскариды 222, 515
Аспидогастроидеи 144, 175
Ассимиляция 13
Аурикулярии 499, 501
Аутогамия 83
Афазмидиа 221
Ацикулы 240, 244
43, 96, 97
41
Авикулярии 280, 281
Автогенез 33
Автотомия 501
Автотрофные формы 82
Адамбулакральные пластинки 485
Аденофореи 198, 220, 221
Адолескария 166, 167, 168, 173
Акантобделла 273
Акарология 9
Акрон 336, 347, 363
Аксоподии 46
Аксостиль 58, 59
Актиния конская, или обыкновенная
133—135
Актиномиксидии 36, 73, 75
Альвеококк 192
Альвеококкоз 195
Алярия 169
Амбулякральная система
487, 488, 493, 495,
Амбулякральные ножки
490, 497, 499
— пластинки 485
Амебоциты 100
Амеба дизентерийная 42,
— предцистная форма
— просветная форма 42
— тканевая форма 42
Амебы 36, 37, 98, 509, 517
— раковинные 36, 37, 43
Амитоз 17, 78
Аммониты 288, 331, 332, 334
Амнион 431
Амфибластула 102
Амфидиски 104
Амфиды 210, 211, 219
Амфилина 191
Амфиподы 362
Анаморфоз 364, 367, 403
Анизогамия 49, 53
Анкилостома 216, 514
Аннелиды 275, 362
Антеннулы 338, 343, 347
Антенны 338, 347
Антомедузы 120, 121
Аппарат гидростатический 327
— Гольджи 17
— коловратный 226, 229
— коловращательный 520
— тимпанальный 439
— челюстной грызущий 427, 436,
441
440,
Бабезииды 72, 73
Бабочка арашниа 458
Бабочки 451, 458, 464, 508
Бабочкины 445
Бадяга 103, 104
Базальные зерна 48, 51, 54, 78
Балантидий 88
Батикринус 494
Беззубка 311, 312, 313
Беззубки 317, 318, 320
Белемниты 329, 331, 332, 334
Беллерофонтиды 301
Бесполостные 519
Бессяжковые 408
Бесщупальцевые 111
Бинарная номенклатура 10, 24
Биогельминты 216, 217, 220
Биотоп, 26
Биофильтраторы 319
Бипиннария 483, 488
Бластея 519
Бластозоиды 252
Бластомеры 21
Бластостиль 118
Бластоцель 23
Бластула 23
Блефаропласт 54, 55, 57
Блохи 408, 468, 471, 450, 509
Богомолы 408, 425, 436, 458
Бокоплавы 337, 358
Бокоход 384, 395
Боннелия 275
Бороздчатобрюхие 288, 292
Ботридии 184
Ботрии 184, 191
Бродяжка 92
Брюхоресничные 36, 198, 199
Бульбус 211, 212
Буцефалидидеи 175
Вакуоль пищеварительная 35, 38, 79,
80, 148
— пульсирующая 50, 52, 97
— сократительная 35, 38, 80, 87, 88, 92
Велигер 300, 307, 318
«Венерин пояс» 124
Веслоногие 337, 351
Веснянки 408, 431
Ветвистоусые 336
Вибрикулярия 281
Виды стенобатные 27
— стеногалинные 27
— стенотермные 27
— эвригалинные 27
— эвритермные 27
Вилочка прыгательная 433
Вислокрылки 431
Власоглав 207, 221, 222, 223
Внутроппораковииныо 288
Водолюбы 441
Возбудитель амебиаза человека 41
— малярии человека 65, 66
Волосатик 225, 226
Волосатиковые 198, 224, 225
Вольвенты 114
Вольвокс 52
Восьминогие 288, 332
Вторичноротые 282, 283, 507, 523, 524
Вошь головная 440
— карповая 352
— лобковая 440
— платяная 440, 441
— собачья 440
Вши 408, 440, 468, 471, 509
Гаметогония 61, 70
Гаметоциты 61, 64, 72
Гаметы 20, 37, 61
Гаплоспоридии 36, 61, 76
Гастрея 519
Гастро-васкулярная система 117, 126
Гастрозооид 123, 125
Гастролиты 340
Гастротрихи 199, 200, 224
— морские 224
Гаструляция 23
Гвоздичник 191
Гектокотиль 330
Геликониды 459, 460
Гелиометра 494
Гельминтология 8
Гемиметаболия 432
Геммулы 104, 105
Гемоспоридии 36, 61, 67, 98, 72
Генетика 8
Геогельминты 216, 224
Герпетология 8
Гетерогония 215, 229
528
Гетеродера картофельная 222
— свекловичная 222
Гетерохлоридиновые 36
Гидра пресноводная 113
Гидракарины 391
Гидрант 117, 118, 120
Гидробиология 8
Гидроидные 111, 112, 509
Гидроиды 111, 116, 119, 120, 509
Гидрокораллы 121, 122
Гидромедузы 112
Гидрообразные 111, 112, 509
Гидрополипы 112
Гидротэка 118, 120
Гидры 111
Гипопус 389, 390
Гипостом 374, 375, 387
Гидродактиль 158, 159
Гистология 7, 10
Гладыш 438
Глаз науплиальный 345, 351
Глаза стебельчатые 339
Глазки 228, 240, 317, 476, 487
Глохидии 314, 315, 320
Глютинанты 114
Глаукотоэ 360
Гнатоцефалон 343, 345
«Голова горгоны» 491
Головоногие 288, 333, 523
Голожаберные 288
Голометаболия 432
Голомстаморфоз 424, 445
Голотурии 482, 483, 498, 499
Голотурия безногая 501
Гонангии 119
Гонозоиды 125
Гоноподы 478
Гонотэка 119
Гонофор 121
Горошинки 314
Гребневики 111, 275, 522
— ползающие 140
Грегарины 36, 61, 67, 68, 96, 97, 98
Грена 75, 451
Губка сверлящая 108
— туалетная 99, 108, 508
Губки 99, 106, 121, 509, 521
— известковые 101, 106, 109
— классификация 101
— кремнероговые 101, 105, 108, 109
— пресноводные 105
— стеклянные 106, 107, 109
— трехосные 101, 106
— четырехосные 101, 108, 109
Губоногие 399, 400, 404
Гусеницы 451, 508
Дафния 350
Двужаберные 331
Двукрылые 408 , 426, 427, 428 , 431, 445, 446
Двупарноногие 399, 405
Двупредсердные 288
Двустворчатые 288, 319, 320, 323
Двуустка гигантская 175
— китайская 176
— кошачья, или сибирская, 169, 175,
514
— кровяная 169, 174, 175, 176, 516
— ланцетовидная 175
— легочная 176
— печеночная 175
Двухвостки 408
Дейриды 210
Дейтомерит 68
Деление редукционное 81
Дельтирии 283
Дендриты 18
Дендронотус 304
Денталиум 309
Десятиногие 288, 332, 337, 359
Дидиниум 85
Диморфизм половой 26, 83, 84, 317, 439
— сезонный 26, 458
Диски имагинальные 432
Диссепименты 241, 247, 258
Диффлюгия 43
Длинноусые 445
Долгоносик 443, 464, 471, 472, 509
— хлопковый 471
Дрейссена 314, 320, 322
Дробление 21, 22, 347
Дровосеки 470
Жуки коротконадкрылые 441
— пластинчатоусые 442
Журчалка 447
Заднежаберные 288
Замок, таксодонтный тип 315
— гетеродонтный тип 315, 318
Зебрина 308
Златоглазки 443, 473
Змеехвостки 483, 490, 492
Зоогеография 8, 13
Зооиды дочерние 251
— материнские 251
Зоомастигины 36, 54
Зоохлореллы 83
Зоофаги 255
Зоэа 360
Зудень чесоточный 388
Зудни 514
Жабернодышащие 336, 395
Жаберные сердца 325
Жаброног 343
Жаброногие 336, 344
Жабры кожные 485, 488, 489
— трахейные 431 435, 451
Жало 428
Жгутиковые 36, 509, 517
— классификация 48, 49
Жгутоногие 365, 379, 381, 382
Железы антеннальные 356, 478
— белковые 297
— биссусные 319
— гермафродитные 297
— гиподермальные 221
— зеленые 341
— кожные 261
— коксальные 367, 372, 379, 478
— максиллярные 356
— мальпигиевые 414
— морреновые 258, 260
— нижнегубные 431
— паутинные 373
— прядильные 151
— пищеварительные 340
— сахарные 290
— чернильные 330
— шейные 213, 221
— ядовитые 370, 373, 381, 385
Жемчужница настоящая 332
— пресноводная 322
Жесткокрылые 441
Жиардия 59
Живородка обыкновенная 306
— пресноводная 307
Жигалки 468
Жужелица 441, 472, 473
Жук австралийский 474
— долгоносик 467
дровосек 438
— колорадский 466, 467
— коротконадкрылый 473
— красотел 441
— кузька 466
— навозник 391, 453
— нарывник 438, 473
— плавунец 407, 432
Жуки 408, 425, 431, 441, 464, 466, 470,
472, 509
Иглокожие 482, 503, 523
классификация 483
Изогаметы 45, 49, 53
Инфузории безротые 97
— брюхоресничные 84, 90
— кругоресничные 84, 90
— малоресничные 84, 88
— равноресничные 84, 85, 86
— разноресничные 84, 87
— ресничные 36, 79, 81
— сосущие 36, 91, 92, 93
— хищники 85, 86
Ископаемые руководящие 30
Ихтиология 8
Ихтиофтириус 85
Кальмар гигантский 329, 332
— обыкновенный 332
— светящийся 332
— тихоокеанский 328, 332, 333, 515
Кампанелла 91
Капсула полярная 73
— стрекающая или крапивная 73, 111,
112, 115, 134
— хрящевая 325
Капустница 444, 452
Каракатица 328, 329, 330, 332, 333
Каракурт 384, 387
Карапакс 337, 343, 347, 348, 356, 359,
360
Кардий 213
Каринария 304
Кариогамия 82
Кариосомы 81
Кариокинез 17
Карпоидеи 502
Катушка обыкновенная 306
Кивсяк 405
Киленогие 523
Кинетопласт 54, 55, 57
Кишечнодышащие 523
Кишечнополостные ПО, 111, 521
— классификация 111
Классификация животных 24
Классы 11
Клетки воротничковые 99
— жгутиковоротничковые 101, 105, 108
529
Клетки стрекательные 11, 112, 114
— с «ресничным пламенем» 179, 227
— фагоцитарные 204
— хлорагогенные 259
Клеточная теория 10, 15
Клещ амбарный 389
— аргасовый 338
— водяной 391
— волосяной 390
— краснотелка 388, 391
— крысиный 392
— куриный 388
— мучной 388, 509
— мышиный 392
— паутинный 388, 390
— перьевой 388, 390
— плодовый 390
— пресноводный 388
— птичий 392
— чесоточный 390
Клещи 72, 73, 365, 376, 377, 378, 387, 388,
390, 392, 393, 394, 395, 505, 514
— глллообрлзующие или четырехногие
391
— гпмазоидные 391
— иксодовыс 392, 393, 514
— панцирные 388, 389
- - тётраипхАвые или паутинные 390
Клоп гладыш 427, 431
— огородный 438
— постельный 438
— птичий 465
— черепашка 465
Клрпы 426, 438, 458, 4G8, 509
Книдобласты ПО
Кмидоспоридпи 36, 61, 73, 75
Книдоциль 111
Кобылка марокская 437, 438, 453
— сибирская 453, 464
— темнокрылая 464
Кожно-мускульный мешок 145, 151, 162,
170, 178, 185, 201, 208, 231, 233,
240, 246, 262
Кокцидии 36, 61, 71, 72, 95, 96, 97, 98
Коловратки 105, 198, 227, 229, 508, 52СУ
— планктонные 227
— ползающие 229
— сидячие 227
Колонии мономорфные 227
— полиморфные 227
Кольчецы 255, 334, 397, 474, 475
— малощетинковые 238, 256
— многощетинковые 238
Комар звонец 446
— малярийный 448
— перистоусый 446
Комменсализм 27
Комменсалисты 96, 265
Конъюгация 78, 81, 83, 84, 89, 90, 92, 98
Кораблик португальский 124, 125
Кораблики 228, 327, 328, 331, 332
Коралл морское перо 131, 133, 137
— кожистый 132
— красный, или благородный, 131, 132,
133, 508
— органный 132
Кораллы восьмилучевые 111, 131, 132, 137
— мадрепоровые 135
— четырехлучевые 136
— шестилучевые
Коралловые 111, 129
Корацидии 183
Корнежгутиковые 36, 60
Корненожки 36, 37, 73, 98, 517, 519
— раковинные 98
Корнероты 128
Коровки божьи 441, 472, 473, 474
Короед древесник 465, 470
Короеды 430, 442, 464, 470
Короткоусые 445
Костянка 400, 401
Кошениль 508
Краб камчатский 359, 360, 513
— манящий 361
— травяной 359
Крабы 345, 354, 361, 511, 512
Красотел 442, 472, 473
Креветки 345, 360, 361, 362, 511, 512
Крестовик 386
Крестовичек 123
Кривоголовка 217, 222, 223
Криптомопады 36
Крпстария 320
Кровенлрачиты 72
Кровососки 468
Кругорссничные 36, 90
Крылатые 399, 408
Крылоногие 288, 523
Ктенидии 287, 299, 303
Кубомедузы 128
Кубышки 438
Кузнечики 427, 428, 429, 436, 437 459
Кукумария 502
Лангусты 3G1, 512
Ларпальпые сегменты 253, 364
Лауреров канал 165
Легочные 288
Лейшмапии 55, 57, 58
Леитец широкий 182, 183, 185, 191, 515
Лентецы 144, 191
Лептомедузы 120, 122
Лептоплана 157
Лигамент 311
Ликофора 189
Лилия морская 492, 493, 494
— стебельчатая 492, 494
Листки зародышевые 23
Листоблошки 438, 439
Листогрызы 442
Листоногие 336
Листотелы 458, 459
Литторина 307
Личинка мюллеровская 154
— неотеническая 230
— рабдитовидная 207, 216
— филяревидная 207, 215, 216
Ложнопроволочники 441, 466
Ложноскорпионы 365, 377, 381, 382
Лопатоногие 288, 523
Лужанка 297
Лумбрициды 257
Лучевики 36, 37, 47, 94, 95
Макома 320
Макрогаметоциты 61, 64, 70
Макрогаметы 49, 52, 64, 69, 84
Макроконъюганты 89, 90, 91
Макромеры 21, 139
Макронуклеус 78, 81, 82, 88, 92
Макросклеры 104
Максиллулы 338, 343
530
Максиллы 338. 343
Малакология 8
Малоресничные 36
Малощетинивые 521
Малый прудовик 308, 309, 515
Мальпигиевы сосуды 379
Малярийный плазмодий 62—67
Маммалогия 8
Мандибулы 338, 343, 347
Мантийная полость 287, 303, 310, 311. 313,
319, 324, 326
Мантия 284, 311, 317, 324
Мантийный комплекс органов 287, 302
Маритогония 173
Мастакс 228
Медузоиды 121
Медуза аурелия 126—128
— обелия 116—119
— сидячая 129
Медузы 111, 112, 116, 119, 121, 122, 123,
128, 509
А\езобласты 253
Мезосома 365, 366, 368, 393
Мембранеллы 77, 78, 87
— адоральные 87, 88, 90
Мембрана кожная 103
— ундулирующая 54, 79, 96
Мерозоиты 61, 64, 66, 70
Меростомовые 336, 365, 366, 479
Метамеризация 20
Метамерия 237, 256, 275
— гетерономная 246
— гомономная 246
Метамеры 20, 237
Метаморфоз 161, 214, 231, 277, 283, 285,
291, 320, 347, 356, 359, 389, 391, 479
Метанауплиус 348, 352, 354
Метанефридии 238, 243, 250, 279, 330, 336,
341, 523
Метасома 365, 366, 377, 394
Метастронгилиды 222, 223
Метатрохофоры 347
Метацеркарий 173
Мечехвосты 365—367, 393, 395, 521
Мидия 316—321, 511—512
Мизиды 337, 346, 357, 362, 512
Мизостомиды 255, 256
Микрогаметоциты 61, 64, 65, 70
Микрогаметы 49, 52, 53, 64, 65, 69, 84
Микроконъюганты 89—91
Микромеры 21, 139
Микронуклеус 78, 81, 82, 88, 92
Микросклеры 104
Микроспоридии 36, 73, 75, 76
Миксоспоридии 73, 74, 95, 96
Миксотрофные формы 52
Мимикрия 459, 460, 461
Мирацидии 166, 167, 173
Митилястр 322
Многоветвистокишечные 144
Многожгутиковые 36, 58
— паразитические 59, 60
Многоножки 336, 399, 430, 431, 433, 475,
521
Многощитинковые 521
Мозговик овечий 192
Мокрицы 359, 381
Моллюски 11, 287, 288, 318, 319, 320, 488,
508, 509, 522
— брюхоногие 216, 288, 293, 294, 302—
305, 334, 515, 523
— внутреннераковинпые 327, 332, 333
Моллюски головоногие 323, 326, 332, 334
— двустворчатые 175, 310, 318, 334
— десятиногие 327
— классификация 288
— лопатоногие 309, 310, 334
— наружнораковинные 331, 333
— нижнежаберные 318
— панцирные 289, 291, 294
Моллюсков черт 334
Моль обыкновенная 464, 509
— «розовый червь» 471
Мопиезия 192, 195
Моногенеи 196
Моноплакофоры 288, 293, 294. 309, 522
Морская звезда астериас 483—488
— — церамстер 489
Морская курочка 500
Морская мышь 254
Морская стрелка 507
Морские блюдечки 303, 306, 307
— бутоны 502
— гребешки 315, 316, 318, 319, 322,
511, 512, 513
— ежи 483, 484, 494—498, 503, 523
— желуди 345, 352—354
— звезды 483—486, 488, 489, 492, 503
— лилии 483, 492—494, 500, 503
— огурцы 483, 498, 500
— уточки 352, 353
Морское перо 131, 137
Морской ангелок 304
Морской заяц 304
Морфология 7, 13
Москиты 57, 471
Мотылек луговой 467, 472, 474, 514
— кукурузный 472, 474, 514
Мошки 445
Муравей амазонка 455, 456
— кровавокрасный 455
— лесной рыжий 455
— темнокоричневый 455
Муравьи 444, 445, 454, 455
Муравьиные львы 443
Муха гессенская 467, 468
— капустная 447
— комнатная 471
— мясная 445, 446
— шведская 467, 472
— це-це 471
Мухи 429, 509
— журчалки 460, 473
— кровососки 446
— скорпионовые 408
— тахины 468, 469
Мухоловки 400, 404, 481
Мшанки 105, 121, 277—281, 282, 509, 523,
524
— голоротые 279
Наездники 444, 470
Наркомедузы 123
Наружнораковинные 288
Нарывники 441, 464, 472
Насекомые классификация 408
— хищные 472—474
Науплиус 345, 347, 352, 353, 355, 356,
360
Некатор 222
Нематоды галловые 219, 220, 222
— луговые 222
— лука 220, 223
531
Нематоды морские 224
— овсяные 219
— сапробиотические 218
— стебля картофеля 219, 222
Немертины 233—235, 275, 276, 520
Неоспоридии 519
Нервная система гипоневральная 483, 487
— — эктоневральная 483, 487
— — энтоневральная 483, 487
Нереиды 238, 239, 512
Нестрекающие 111, 138
Нефропора 243
Нефростом 243
Нижнежаберные 318
Никтерибин 468
Нимфа 389, 390, 391
Нитчатка Банкрофта 222
Нитежаберные 288
Ногохвостки 408
Ножки бе га тельные 427
— грудные 427
— плавательные 427
— прыгатольиыо 427, 437
— роющие 427
— хнятнтольныр 427
Ноземы 75
Ночницы 458
ОПопятрльняя ямка 127, 240
Овод бычий 468
— желудочный 468, 469
— кожный 468
овечий 469
— полостной 468
Однопредсердные 288
Олигохеты 238, 256, 262—266, 274
Омары 360—362, 512
Омматидий 477
Онихофоры 395
Онкосферы 181, 188, 189, 191, 192, 194,
195
Онтогенез 21, 23, 178, 187, 217, 218
Ооциста 61, 65, 70, 71
Опалина 83—84
Опалиниды 83
Оплодотворение 21, 64
— внутреннее 21, 333
— наружное 21, 346, 367, 394
Органеллы движения 35
— питания 35
Организмы автотрофные 14, 48
— гетеротрофные 14, 48, 49
— миксотрофные 49, 50
Орган звездчатый 204
Органы аборальные 138—140
— Бэра 170, 176
— боянусовые 213
— Галлера 375
— Женэ 377
— Кювьера 500
— осевые 487
— тимпанальные 429, 436
Орехотворки 444
Орибатиды 193, 195
Орнитология 8, 13
Ослик водяной 359
— клешненосный 346
Острица 207, 216, 515
Осьминог 326, 328, 332, 333, 513
— илистый 332
— северный 332
Осы 454
— настоящие 444
Отбор естественный 32, 33
— искусственный 32
Офиоплутеус 490
Офиофолис красномраморная 491
Офиуры 490, 491, 502, 503
Офриосколециды 88, 89
Палеозоология 8
Палеонтология 13, 30
Палоло 255
Пальпы 240, 244
Палочки стрекательные 79
Палочники 458, 459
Пальмовый вор 361
Пандорины 53
Панспоробласты 74, 75
Панцирные 36
Паразитология 8
Парамеции 40, 78—83
Параподии 238, 240, 245, 24G, 25G, 268,
274, 334, 475
Паратомия 264
Паренхимелла 519
Партеногенез 21. 107. 229. 340
Партспогонпя 173
Парусник 123, 125
Паук бокоход 384, 386
— подиной 387
— крестовик 309, 373, 383
— лесной 384
скакун 384
Пауки 365, 381, 383, 384, 395, 509
бокоходы 386
дву легочные 379
— жгутоногие 379, 381, 382
— члепистобрюхие 384
четырехлегочные 379, 383
Паукообразные 336, 365, 369, 377, 380,
521
Пауроподовые 399, 407
Пебрины 75
Педипальпы 364, 366, 374, 380, 381, 387
Педицеллярии 483, 489
Педогамея 46
Пелликулы 48, 49, 77, 79, 81, 92
Пенетранты 114
Пенницы 438, 439
Первичнобескрылые 393, 408, 434
Первичножаберные 288, 318
Первичномонадные 36, 54
Первичноротые 282, 519, 522, 524
Первичнотрахейные 336, 395, 396, 398,
521, 522
Перегородчатожаберные 288
Переднежаберные 288
Перепончатокрылые 427, 428, 429, 443
Перисарки 118, 120
Перистом 78, 79, 88
Перловица 314, 318, 320, 321
Пескожил 249, 254
Пигидиум 237, 250, 363
Пиериды 459
Пилильщики 470
Пинны 318, 319
Пиноцитоз 38, 39
Пиреноид 52
Пироплазма 72
Питание автотрофное 51
— гетеротрофное 55, 198
532
Питание голозойное 49
голофитное 48, 49
— миксотрофное 51
— сапрофитное 49, 54
Пиявка кожная 272
— медицинская 272, 274
— рыбья 273
Пиявки глоточные 238, 273
— хоботные 238, 273
— челюстные 238, 274
— щетинконосные 238, 273
Плавт 438
Плавунцы 441
Плазматические классификация 36
Плазмодии 65
Плазмодий малярийный 96, 97
Планария молочная 145—150
Планктон 122
Планула 117, 119, 127
Пластинки гребневые 138
— дельтидиальные 283
— опорные 113
Пластинчатожаберные 288, 318
Пластинчатоусые 464
Плеоподы 338, 342, 361
Плероцеркоид 183, 188
Плеченогие 283—286, 523, 524
— беззамковые 283, 284
— замковые 284
— ископаемые 284
Плодожорка яблоневая 474, 514
Пневматофор 123, 125
Погонофоры 505, 506, 523
Поденки 408, 427, 431, 434, 435
Покрытожаберные 288
Покрыторотые 281
Полиморфизм 24, 118, 120, 125, 455, 457
Полипы 111, 112, 117, 118, 121, 132
— колониальные 129
Полихеты 238, 245, 250—252
— кольчатые многощетинковые 238
— паразитические 255
— свободноподвижные 238, 255
— сидячеприкрепленные 238, 255, 256
Полиэмбриония 159
Половой диморфизм 274
Полость гастральная ПО, 117, 123, 129,
131
мантийная 300, 301
тела вторичная 237, 241, 284, 287, 293
— — первичная 254
Почкование 251
— множественное 93
Привидениевые 408
Примит 69
Проволочники 431, 464, 466
Проглоттиды 184
Проктодеум 242
Простейшие классификация 36
Простомиум 240, 244
Протаспис 363, 364
Протерандрия 214
Протеробластус 503
Протомерит 68
Протонефридии 149, 153, 249, 250
Протоподиты 344
Прототрох 252
Протоцель 209
Процеркоид 183
Протоцефалон 343
Протоцилиаты 36, 78, 83
Прудовик малый 306
Прудовик обыкновенный 306
— ушковый 306
Прус 437, 438
— итальянский 464, 465
Прыгуны 384, 395
Прэкоксоподиты 344
Прямокрылые 408, 425, 429, 437, 464
Псевдоподии 36, 37, 40, 43, 44, 46, 48,
73, 98, 116
Псевдоцель 254
Пузыреногие 408
Пупарий 447
Пухокрылые 450
Пухоеды 408, 440, 468
Пчела медоносная 418, 424, 426, 444, 445,
454, 508, 511
Пчелы 419—423, 429, 444, 454, 460, 463,
508
— общественные 445
— одиночные 445
— кукушки 445, 461
Пяденицы 459
Рабдиты 151
Рабдом 477
Равноногие 337, 346, 358
Равноресничные 36
Радиолярии 517
Радула 287, 290
Развитие прямое 252, 342, 356, 357 361
партеногенетическое 226
— с превращением 161, 214, 277 283,
285, 320, 356, 359, 389
— эпиморфное
Размножение бесполое 20, 21 45, 49,
104, 116, 250, 251
— половое 20, 104, 116, 250, 280
— почкованием 20, 118, 238, 280
— партеногенетическое 228
Разноресничные 36
Рак речной 337—342, 360, 361
— длиннопалый 342
— — широкопалый 342
— отшельник 134, 135, 359, 360
Ракообразные 336, 342, 361—363, 397,
476, 479, 521
— классификация 336, 337
Ракоскорпионы 365, 366, 368
Ракушка белая 316, 322
Ракушковые 337, 356
Рапана 308
Рахис 204
Редии 166, 167, 168, 173
Реликты 28
Ремнецы 191
Ринхоцелом 235
Ришта 222, 223, 515
Россия 322
Рострум 332, 338, 356
Ротоногие 345
Ручейники 408, 431
Рулек 214, 222
Саккулина 354
Саранча азиатская, или перелетная, 437,
438, 453, 464
— прус, или итальянская, 437
— степная, или мароккская кобылка,
437, 438
533
Саранчовые 436
Саркоспоридии 36, 61, 76, 96
Сателлиты 69
Свайник гигантский 221
Сверчки 429, 436, 437
Сегментация гомономная 356
Сеноеды 408, 440
Сенокосцы 365, 377, 383
Септы 133
Сердцевидка 322, 512
Сетчатокрылые 408, 431, 443
Сецерненты 220—222
Сидячеглазые 288
Сизигия 69
Сикон 101 —103
Симбиоз 27, 96, 134
— внутриклеточный 83
Симбионты 96
•Симметрия билатеральная 19, 132. 142,
143, 482, 502, 504
— лучевая 482, 494, 502
— радиальная 19, 131, 133, 142, 503.
504
Сингамиды 22'2
Синкариои 82, 83
Синтетика у л юс 216
Синцсфалон 343
Система амбулякральная 463. 485. 488.
493, 495, 497, 499
— выделительная 19
— гэстро-наскулярная 117, 126
— дыхательная 19
— животною мира 24
— кровообращения 19
— мышечная 19
— нервная 19
— нервная диффузная НО
— органов размножения 19
— пищеварительная 19
— покровная 19
— скелетная 19
— эндокринная 19
Систематика 8, 10, 24
Сифоноглифы 131, 133
Сифонофоры 111, 112, 123, 125
Скарабей священный 443
Скафогнат 346
Склеробласты 100, 101
Сколекс 177, 178, 184, 192, 197
Сколопендреллы 399, 406, 433
Сколопендры 404, 509
Скорпион водяной 394, 438
— книжный 381
Скорпионы 365, 369, 377—381, 509
— сухопутные 381
Скребень великан 232
Скребни 230—231, 520
Слепни 447, 471
Слизень полевой 308
Слизни 523
Слоники 442
Совка капустная 465, 472
— озимая 467, 474, 514
— хлопковая 474, 514
Совки 464, 472
Соленоциты 249, 250, 253
Солитер бычий 178, 179
— вооруженный 514
Солнечники 36, 37, 46, 517
Сольпуги 365, 377, 382, 383
Сосальщики дигенетические 161
— моногенетические 144, 158
Сосательные палочки 91, 92, 93
Сперматофор 346, 394, 431
Спикулы 100, 101, 102, 106, 108, 109, 131,
214
Спируриды 222
Спланхноплевра 241
Споробласты 61, 65
Спорогония 61, 74, 75, 76
Споровики 36, 97, 98, 517
— слизистые 36, 61, 73
Спорозоиты 61, 62, 64, 65, 70, 73
Споросак 121
Спороцисты 71, 167, 168
Споры 61, 71, 74—76, 97
Статобласт 280
Статолит 139
Статоцисты 117, 122, 127, 152, 234, 312,
339, 345, 357
Стебелек несократимый 91
— сократимый 91
Стебел М1ятогл1пыо 288
Стебельчатые 183
Стентор 87
Стереолипы 114
Стигма 50
Стигмофор 446
Стилет 218, 220
Стнлонмхпя 90
Столон 252
Стомодеум 242
Стрекающие 111
Стрекоза обыкновенная 435
— коромысло 435
Стрекозы 408. 425, 431, 435, 436
Стробила 177, 183, 192, 252
— ананолизическая 185
— аполизическая 185
Стробиляция 127, 252
Стронгилоцентротус 498
Стронгиляты 222
Сувойка 91
Суктории 92
Сумка выводковая 333
Сцифистома 127
Сциооидные 111, 125, 522
Сцифомедузы 125—128
Сферонис 503
Тангорецепторы 209
Таракан черный 408—413, 416, 417, 436
— прусак 417, 418
Таракановые 408
Тарантул 385—387
Тахина 472
Тейлирииды 72, 73
Текоидеи 502
Телоспоридии 36, 61, 66, 519
Тельсон 337, 347, 348, 363, 365
Тельце выделительное 81
— Мелиса 165, 172, 186
— тидельмановы 486
Териология 8
Термиты 408, 438, 454, 456, 509
Тиленхиды 222
Типы 24
Тифлозолис 259
Тли 426, 238, 439, 444, 456, 457, 470
Тля бахчевая 439
— гороховая 439
— кровяная 439, 471, 474
Тонкопанцирные 337, 356
534
Торсионный процесс 309, 330
Трахейнодышащие 336, 397, 399
— классификация 399
Трахелиус 85
Трахелиды 111, 119, 122
Трахимедузы 123
Трематоды 144, 161, 196, 197
— брюхоротые 144, 175
— дигенетические 144
Трепанг 500, 502, 513
Трехветвистокишечные 144
Тридакна 320
Трилобиты 336, 362, 363, 364, 393, 395,
479, 521
Трипанозомы 54—56, 95—97
Трипсы 470
Трихина 206, 407
Трихинелла 221—223
— мышечная 206
Трихограмма 474, 513, 514
Трихостронгилиды 222
Трихоцисты 79
Трофозоиты 61, 72
Трохозоон 275
Трохофора 230, 238, 252, 253, 274, 275.
288, 300, 318, 320
Трубач 308
Турбеллярии 145, 196, 200, 224, 232, 520
— бескншечные 144, 155, 156
— многокишечные 157
— прямокишечные 144, 157
— трехветвистокишечные 157
Угрица кишечная 207, 215
— уксусная 218
Улитка виноградная 295—297
Улитки 305
Умбеллула 133
Уроподы 337, 342, 357
Усачи 441
Усоногие 337, 352
Устрицы 315, 317—321, 508, 511, 512
Уховертки 408, 453, 464
Фагоцителла 519
Фагоцитоз 38
Фазеолина 316
Фазмидиевые 220
Фазмиды 211, 213, 221
Факторы абиотические 26
— биотические 26
Фасциола печеночная 162—164
Физофора 123
Филаменты 181
Филлоксера 439, 471
Филогенез 23
Финка 189, 194, 195
Фитомастигины 36, 51
Фитомонады 36, 49
— автотрофные 52
— гетеротрофные 52
— миксотрофные 52
Фитонематоды 224
Фитофаги 225
Флабеллум 367
Фляшерия 462
Фолас 319
Фораминеферы 36, 37, 94, 98, 517
— макросферические 44
— микросферические 44
Фораминеферы многокамерные 44
— однокамерные 44
Формы моноцикличные 22
— полицикличные 229
— стеногамные 265
— эвригамные 265
Фрагмакон 332
Хальцидиды 444
Хелицеровые 364, 365, 394, 521
— классификация 365
Хелицероносные 336, 395
Хелицеры 364, 366, 368, 380, 381, 383,
385, 387, 391
Хермес зеленый 457
Хилярии 367
Хиродота 502
Хирономиды 512
Хитон 290, 291
Хламидофрис 43
Хлоромонады 36
Хоаноциты 55, 99, 100, 101, 106, 108
Хозяин дефинитивный 195
— окончательный 167, 177, 189
— основной 457
— промежуточный 167, 177, 195, 457
Хондриозомы 17
Хризомонады 36, 95
Хромадориды 221
Хроматофоры 48, 50—52. 327, 345
Хрущ майский 442, 464
Хрущак мучной 509
Хрущи 467, 471
Целом 237, 241, 247, 249, 254, 287, 475
485, 487, 523
Целомические мешки 241, 272
Целомодукты 285, 294, 318, 367
Целотелий 237, 241
Центропиксис 43
Ценурус 188, 189, 192, 195
Цепень бычий 178, 179, 180—182, 192
— карликовый 188
— свиной 178, 192, 196
— собачий 188
Цервикальная борозда 337
Церкария 166—168, 173
Церки 409, 428
Цестоды 177, 178, 191, 192, 196, 197
— нерасчлененные 144
— расчлененные 144
— четыреххоботные 193
Цикадки 438, 470
Цикады 426, 438, 439
— певчие 439
Цикадкообразные 439
Циклоз пищеварительный 79, 80
Цикломорфоз 350
Циклоп 93, 223, 351, 515 %
Циклопостииды 88, 89
Циприс 355
Цирры 90, 492
Циррус 240, 246
Цистицерк 181, 188, 189, 194, 195
Цистицеркоид 188, 189, 192
Цистогония 173
Цистозоид 123, 125
Цистоидеи 494, 503
Цисты 42, 43, 59, 69, 76, 77, 84, 85, 88
93, 94, 97
535
Цитостом 78, 79
Цитофаринкс 79
Челюстеногие 336
Червеобразные 282
Червец австралийский желобчатый 471
Червецы 438, 439, 474
Черви вторичнополостные 237
— кольчатые 142, 237, 238, 275, 276,
318, 334, 399, 520
— — классификация 238
круглые 198, 200, 208, 276, 520
классификация 198
ленточные 144, 177 178, 183,
187 519
— малощетинковые 75
паразитические 514
первичнополостные 142, 276
плоские 142—144, 276, 519
— —классификация 144
— ресничные, или турбеллярии 145
Чсриь дождевой, или земляной 257, 266
Чородооаннс поколений 45, 169
Черепашка продпая 4G5, 4GG, 472
Чернильный мешок 325, 327, 332, 333
Чернотелки 441, 464, 466, 471
Чертой палец 331, 332
Четырехжаберные 331
Чешуекрылые 408, 420, 427, 429, 251
Членистоногие 224, 335, 350, 393, 395,
ЗУ 9, 474, 520, 521, 522
— классификация 336
Членистохоботные 408, 438
Шаровики 494, 502
Шаровка 314, 318
Шелкопряд алантовый 464
— китайский дубовый 464
— клещевинный 464
— кольчатый 467, 469
— непарный 472
— тутовый 451, 463, 511
Шизогония 61, 64, 66, 70, 72, 75, 76, 251
Шизонт 64, 66, 70
Шмели 444, 445, 460
Щелкуны 441, 442, 464, 466, 471
Щетинкохвостые 408
Шетинкочелюстные 507, 523
Щитень 336, 344
Щитовка калифорнийская 471, 474
Щупальцевые 282
Эвглена 40, 49, 50, 51
Эвгленовые 36, 52
Эвдорина 53
Экзиты 344
Экзопаразиты 344
Экология 7
Эксконъюгапты 82
Эктопаразиты 95, 159
Элеоциты 247
Эмбриология 7, 10
Эмбриогония 173
Эндемизм 28
Эндиты 344, 370
Эндопаразиты 95
Эндоподиты 338, 344, 345, 357
Эпоплиды 221
Энтомология 8
Эпимернт 68
Эпиморфоз 479
Эпнподиты 338, 344
Эпистом 279, 364, 387
Эниморфоз 400
Эпитокия 243, 250
Эстеты 290
Эфиры 127
Эхинококк 189, 192, 195, 196
Эхинококковый пузырь 184, 195
Эхиноплютеус 495
Эхиурдовые 238, 474
Яйца миктические 228
— амиктические 228
— нематод 207
— трематод 188
Яйцевой кокон 416
Acanthobdella peledina 273
Acanthobdellae 268, 270, 273, 274
Acanthobdellidae 273
Acanthocephala 230, 519,* 521
Acanthometra elastica 47
Acariformes 387, 389, 394
Acarina 365, 387 388*
Acarus siro 388*, 390
Acelis arctica 157
Achrutes 433*
Acmeaea 29j^ 303, 307
Acmadaea testudinalis 306*
Acnidaria 111, 138
Acoela 144, 152, 153, 154, 155
Acridodea 436
Actinia equina 133, 134*
Actiniaria 135
Actinomonas mirabilis 60
Actinomyxidia 36, 73, 75
Actinophrys 46
— sol 46, 95
Actiaosphaerium 46
536
— eichhornii 46, 95
Actinozoa 111, 129, 519*
Adenophorea 198, 220
Aedes 447
Aegeriidae 452
Aeginopsis 123, 130*
Aelosoma 265
Aeschna 435
Aglantha 123, 130*
Agriolimax 305
— agrestis 308
Agriodrilus vermivorus 265
Agrion 435
Alaria alata 169
Alectorobius 392, 514
— papillipes 388*, 392
Alciope 249*
Alciopidae 239*, 248
Alcyonidium gelatinosum 278*
Alcyonium 132, 137*
Allocreadium isoporum 171
Allodermanyssus sanguineus 392
Allolobophora 257
longa 265
Allopora stejnegeri 121*
Alveococcus multilocularis 192
Amabilia lamelligera 186*
Amauris niavius 461
— echeria 461
Amera 236
Ammodiscus incertus 44
Ammonoidea 288
Amoeba limax 40
— polypodia 40
— proteus 37, 38, 39, 40, 41, 95
— radiosa 40
— terricola 41
— verrucosa 40
Amoebina 36, 37, 40
Amphilina 191
— liguloides 191*
Amphipoda 358
Amphitrite 244
— edwardsi 245*, 256
Analgesoidea 390
Anamorpha 404
Ancylostoma 219
— duodenale 207*, 216, 222
Ancylus 298, 305
— fluviatilis 306*
Anisoplia austriaca 466
Anisoptera 435
Annelides 25, 237, 238, 519*, 520, 521
Anodonta 176, 311, 315
cygnea 314*, 315
— piscinalis 315
Anopheles 64, 67, 447, 448*, 471
— maculipennis 66, 450
Anoplocephala gigantea 184
Anoplocephalata 389
Anoplura 408, 440, 468
Antheraea 462
— pernyi 453
Anthonomus grundis 471
Anthozoa 111, 112
Apelexa 298
Aphaniptera 408, 451, 468
Aphasmidia 221
Aphidae 438, 439
Aphidiidae 444
Aphilinus mali 474
Aphis pomi 466
Aphrodite aculeata 239*, 245*, 254, 255
Apidae 419, 444
Apinae 419
Apis 444
mellifera 419, 420*, 421*, 422*, 424*,
454
Aplexa hypnorum 306*
Aplysia 304
Apolemia 123
Apterigota 399, 408, 428, 432, 433, 434
Arachnoidea 365, 370, 377, 378, 379, 380,
394, 519*, 521
Agaronimus ringeri 188*
Araneae 365, 370, 383, 384*
Araneomorpha 383
Araneus 369*
— diadematus 370
Araschnia levana 459
— prorsa 459
Area 318
Arcella 43, 44
Arcella dentata 43
— vulgaris 43
Arenicola marina 245*, 247, 249, 254, 256
Arenicolidae 256
Argas 392
Argasidae 392
Argriopidae 386
Argrioneta aquatica 387
Arion 305
Aristocystis 503
Arrhenurus neumani 388*
Artemia salina 362
Arthropoda 25, 519*, 520, 521
Ascaridida 222
Ascaris lumbricoides 206, 207*, 216, 222
— ostroumovi 213*
— suum 222
Aschelminthes 195
Ascon 105, 106*
Aspidiotus perniciosus 471, 474
Aspidogaster conchicola 176
— limacoides 175
Aspidogastrodidea 144, 161, 175
Astacotrema cirrigerum 173
Astacus astacus 513
Astasia 95
Asterias rubens 483, 484*, 487*, 488, 489,
492*
Asteroidea 483, 503
Astomata 97
Astree 137*
Atentaculata 111
Athecanephria 505
Athecata 119, 120*
Atropos pulsatoria 440*
Atta 456
Atypus piceus 385
Aurelia aurata 126, 127*, 128*, 130*
Autolytus cornutus 250*
Aysheaia pedunculata 398*
Babesia bigemina, 72, 73
Babesesta canis 72*
Babesiidae 72
Bacillus bombycis 462
Baicalia 303
— stidae 306*
Balantidium coli 87*, 88, 96
suis 88
Balanus 353*, 354*
Barathra brassicae 465*
Basommatophora 288, 305
Bathycrinus 500*
Bdelloidea 229
Belonogaster 454
Benedictia 303
Beroe cucumis 124*, 140
Bertiella elongata 187*
Bistadiidae 37, 40
Bithynia 176, 303, 309
— leachi 169
Bivalvia 288, 310, 519*, 522
Blatta orientalis 408, 409*, 415, 417, 436
Blattella germanica 417
Blattidae 417
Blattodea 408, 436
Bombus semenovi 457*
Bombyliidae 438
Bombyx 462
— mori 75, 452
537
Bonnelia viridis 275*
Boophilus 72
Bradibaena 305
Brachionus urceoalris 227*
Brachiopoda 25, 283, 519*, 524
Brachycera 445
— cyclorhapha 447
Braconidae 444
Brachiata 475
Bryozoa 25, 277, 519*, 524
Buccinium undatum 306*
Buccinum 303, 308
Bucephalidae 175
Bucephalididea 144, 161, 175
Bucephalus polymorphis 174*, 175
Busaria truncatella 87
Buthus 381
— eupeus 381
Bythynia leachi 306*
Cadlina 304
Caenorhabdilis 211*
Calcartn 101 100
Calliphora crytlirocrpliala 44Г/ 446
Calliptamus italicus 465
Campanella umbellaria 91
Camponotus 455
Calonympha grassii 59, 60
Calonymphidae 60
Calosoma syncophanta 441, 442*, 472, 473*
Canidae 24
Canis lupus 24
Caponiidae 383
Carabidae 441
Carabus 473*
Cardium 317, 318
edule 316*
Carchesium polypinuni 91
Carcinus maenas 359*
Carinaria mediterranea 304*
Carnivora 24
Carpoidea 502
Caryophyllaeus 191
— mutabilis 191*
Catenotaenia dendritica 187*
Cecidomyidae 447
Centropyxis 43, 44
Cephalobidae 221
Cephalopoda 288, 519* 522
Cerambycidae 441
Ceramster granularis 489, 492*
Ceratomyxa 74*
Cercopidae 438, 439
Cerithium 298
— ponticus 306*
Cestoda 144, 190, 191
Cestodaria 144, 190
Cestoidea 144, 177, 520
Cestodes 519*
Cestoplana rubrocineata 156*
Cestus veneris 124*, 138
Chaetogaster limnaei 265, 266
Chaetognatha 25, 506, 519*, 523
Chaetonotoidea 200
Chalcididae 444, 470, 474
Chaoborus 446*
Chelicerata 364, 365*, 393, 394, 521
Chelifer cancroides 382
Chelura 362
Chermes viridis 458
Chilocorus 474
Chilodonella cyprini 86
Chilomastix mesnili 59
Chimera monstrosa 176
Chilomopas 38
Chilopoda 399, 400
Chirodota 502
Chironomus 150, 446
Chlamydomonas 95
— braunii 53
— steinii 53
Chlamydophris 43
Chlandra granaria 467*
Chloromonadina 36, 49
Chordata 24, 25, 519* 523
Chromadorida 221
Chrysomelidae 442
Chrysomonadina 36, 49, 95
Chrysopa 473*
Chrysopidae 443
Cicadidae 438, 439
Ciliata 36. 77, 78, 79, 517, 519*
Ciliophora 36, 77. 517
Clathrina 100*
Clausilia 305
Clione 108, 298
— Hmacina 304*, 309
Clonorchis sinensis 176, 188*
Cnetocampa 475
Cnidaria 111
Cnidosporidia 36, 61, 73, 98
Coccidae 438, 439
Coccidia, 36, 61, 67, 70
Coccidiomorpha 72
Coccinellidae 441, 473, 474
Codonisigia alloides 55
Codonisigia botrytis 55
Coelenterata 25, 110, 111, 519*=
Coeloplana 140
gonoctena 140* —
— kowalevski 140
— metschnikovi 140
Coenagrion 435
Coleoptera 408, 441
Coleps 85
Collembola 408, 432, 433, 434
Colpes hirtus 86*
Colpidium colpoda 86*, 95
Colpoda 93
— parviformis 92*
Conochilus 229
Convoluta convoluta 156*
— rosscoffensis 155
Convolvus sepium 463
Copeognatha 408, 440
Copepoda isokerandria 476*
Copris lunaris 453
Corallium rubrum 131, 132*
Cordylophora caspia 121*, 122
Coriarachne depressa 384
Cornacuspongia 101, 108, 109
Cornuspira violacea 35
Corycella armata 68*
Coryphella 304
Costia 55
— necatrix 55, 95
Craspedacusta 122
— sowerbii 122*
Craspedomonadinae 55, 109
Crenobia alpina 157
Crinoidea 483, 492, 519*
Cristaria plicata 314*, 320
538
Cristatella mucedo 278*, 280*, 281
Crustacea 519*, 521
Cryptochiton stelleri 289*
Cryptolaemus 474
Cryptomonadina 36, 49
Cryptomonas 95
Ctenocephalus canis 451*
Ctenophora 111, 138, 519*
Ctenoplana 140
Cucumaria frondosa 500*, 502
Culex 67, 446*, 447, 450*
— hirundinis 438
— lectularius 438
Culicidae 72, 446, 447
Curculionidae 442
Cuspidaria 319
Cyanea arctica 126, 128, 130*
Cyathocephalus trunculatus 185*
Cyclidium 92*
— glaucoma 85*
Cyclophyllidea 144, 184, 191, 193’
Cyclopostiidae 88
Cyclopostium bipalmatum 89
Cyclops 183
Cyclichna 304
Cynipidae 444
Cyphonautes 280*
Cypridina 356
Cysticercus cellulosae 194
Cystoidea 494, 502, 503
Dactylogyrus 160
Dactylosphaera vastatrix 471
Danais chrysippus 461
Daphnia 362
Decapoda 288, 359
— brachyura 476*
Defrancia lucernaria 278*
Demalitum 309
Delafondia vulgaris 217*
Dendrocoelum lacteum 145, 146*, 147*,
150, 153, 155, 156*, 157
Dendroctonus micans 470
— arborescens 304
Dentalium entale 310
Dermanyssus 392
— gallinae 388*
Dermatoptera 408, 453
Dermocentor nuttali 393
Deuterostomia 520
Diapheromera femorata 460*
Dicrocoelium 176
Dicrocoelium lanceatum 16b, 170, 188*
— voluptarium 171*
Dictyocaulus 222, 223, 224
Didinium 85, 86*, 95
— nasutum 86*
Dientamoeba fragilis 41
Difflugia 43, 44
— corona 43
— pyriformis 43
Digenea 172
Diglena volvocicola 230
Dimophyes 124*, 125
Dinoflagellata 36, 49
Dinophilidae 255
Dinophilus 254, 255, 256, 275
— apartis 254*
Dioctophyme renale 221
Diotocardia 288. 302
Diphyllobothrium latum 182*, 188*, 191
Diplopoda 399
Diplozoon paradoxum 160
Diplura 408, 432, 433
Diptera 408, 428, 445
Dipylidium caninum 192, 193*
Discomedusae 128
Discocotyle salmonsis 172
Dismorphia astynome 457*, 460, 461
Ditylenchus 210*, 211*
Ditylenchus alii 220*
— destructor 222
dipsaci 222
Dolomedes 387
Dorylaimus 210*, 211*
Dracunculus medinensis 222. 515
Dreissena polymorpha 314~
Drepanophorus albolineatus 233
Dytiscidae 441
Echinococcus granulosus 184, 192
Echinodermata 25, 482, 483, 519* 523
Echinoidea 483, 494, 503, 519*
Echinorhynchus polymorphus 232
— proteus 231*, 232
Echinus esculentus 497
Echiuroidea 238, 274, 519*
Ectocochlia 288
Eicomonas 93
Eimeria 70
— avium 71
— clupearum 71
— magna 70*. 71
— oxyspora 71*
— perforans 71*
— sardinae 71
sniydersi 71*
Eisenia 257
Elater 442*
Elateridae 441
Eleutherozoa 503, 504
Elpidia 502*
Enantiozoa 100, 519
Enchytraeus albidus 265
Enchytreidae 266
Endocochlia 288
Enoplida 221
Entamoeba coli 41
histolytica, var. magna 42, 94
— — var. minuta 41., 42
Enterobius vermiculatus 207*
Enteropneusta 519*, 523
Enterozoa 519
Ephelota gemmipara 92*
Ephemeroptera 408, 434
Ephydatia 103, 104
Epicauta 473
Epimorpha 404
Erigonidae 386
Eriophyoidea 391
Eriosoma lanigerum 471
Errantia 255, 238
Ethmolaimus 210*
Euciliata 36, 78, 84
Eucrata loricata 278
Eudorina 53, 95
Euglena acus 51
— oxyuris 51
— spirogyra 49
— viridis 49, 50, 51, 95
Eu^lenoidina 36, 49, 52
Euglypha 44
539
Euglipha alveolata 43
Eulamellibranchia 288, 315, 318
— Eumenes 454
Eunaetazoa 519
Eunice 246*
— viridis 250*, 251
Euplanaria gonocephala 156*, 157
— lugubris 157
Euplectella 107
— oweni 107
Euplotes 90
— charon 90*
Eurydema 438
Eurygaster 438
— intergriceps 466
Eurypteroidea 365, 366, 368
Eurypterus fischcri 368* 368
Euscorpius 381
Euspongia offi'cinalis 99*. 108
Evasterias echinosoma 489
Fasciola gigantica 175
— hepatica 162, 163* 164*, 165*, 166*,
176, 188*, 309
Filibruiicliia 288, 818
Plagollata 36, 48, 517, bio45
Floscularin proboscidoa 230
Elustra foliacea Z7fi'‘
Forarninifcra 36, 37, 44, 95
Fprficula auricularia 453
Formien 1G8, 455
— fusca 455
— rufa 455
— sanguinea 455
Formicidae 444, 445
Formicina 445
Franceata 399
Galba truncatula 309, 515
Galeodes aranoides 382
Galumna mucronata 388*
Gamasoidea 391
Gammarus 358*
Gasteracantha curvispina 384*
Gastrophilus intestinalis 469
Gastropoda 288, 294, 519*, 522
Gastrotricha 198, 199
Gastrotrichia 519
Geophilomorpha 404
Geophilus 404
Geotrupes 443
Girardia intestinalis 58, 59
Globigerina 45
Globotruncana area 44
Glossina morsitans 56, 471
Glossina palpalis 55, 471
Glossiphonia complanata 272*, 274
Glossoscolecidae 268
Gnathobdellae 274, 238
Gomphocerus sibiricus 464
Gonionemus vertens 123
Gonium 52, 95
Gordiacea 198, 224, 519*, 520
Gordius aquaticus 225*, 226
Gorgona 137*
Gorgonacephalus eucnemis 491, 492*
Gastrostyla 93
Grammostola actaeon 384*
Granularia protolenorum 363*
Gregarina 36, 61, 67
— blattarum 68*
— cuneata 69*
— polymorpha 69*
— steini 69*
Gryllodea 436
Gyrocotyle 191
Gyrodactilus elegans 158, 159*
Haemonchus contortus 217*, 222, 223
Haemopsis sanguisuga 274
Haemosporidia 36, 61, 67, 72
Haliclystus 129
Haliotis 298, 303, 309
Haplosporidia 36, 61, 76
Haplosporidium 76
Ileficella 168
Hcliconidac 460
Heliometra gracialis 494, 500*
Helix 305
— permit in 295
lleliozoa .30, .37. 45
Ilclobdclla atagiialis 271
I Icmichordata 519
HcmickpsH murgiiiata 273
llennegya 74w
Horpobdella 274
— n igricol is 274
— ocloculata 272*, 274
11cIcrochloridina 36
Hctcrodora avenac 222
— rostochiensis 222
— schachtii 222
Heteronereis 243*
Heteritricha 84, 87
Heterotrichia 36
Hexacorallia 111, 131, 133
Hippobocidae 446
Histrio pelinella 90*
Hirudinea 238, 268, 519*, 521
Hirudinidae 274
Hirudo medi-cinalis 269*, 272, 274
Hoferellus 74
Holothuroidea 483, 498, 503, 519*
Holophrya discolor 85*
Holotricha 78, 84, 85
Holotrichia 36
Homoptera 438
Hoplytophrya 97
Hornera lichenoides 278*
Hyalemma marginatum 393
Hyalomma 73
Hydra oligactis 113, 115*
Hydracarina 391
Hydrida 111, 116, 119
Hydroidea 111, 112
Hydrophilidae 441
Hydrozoa 111, 112, 140, 519*
Hylobius abietis 443*
Hymenolepis carioca 185*
— папа 188*
Hymenoptera 408, 443
Hypermastigidae 98, 60
Hypoderma bovis 468
Hyptiotes 385, 386*
Hypotricha 36, 84, 90
Hypotrichia 36
Icerya purchasi 471, 474
Ichneumonidae 444, 470
540
Ichthyophtirius multifilis 77*, 85
Ichtyobdellidae 272
Inperforata 44
Insecta 399, 407, 408, 519*, 521
Ipidae 442
Isopoda 358
Isoptera 408, 438, 456
Isospora belli 71*, 72
— rivolta 71*
Ixodes 514
— persulcatus 393
— ricinus 73, 374, 375*, 376*, 377, 392
Ixodidae 374, 392
Ixodoidea 392
Jassidae 438, 439
Jodamoeba biitschlii 41
Joldia arctica 318
Jungia aurantiaca 155*
Kallima 457*
Kerona 90
Liphistius malayanus 384*
Listrophoroidea 390
Lithobius 403*, 404
— forficatus 400, 401*
Lithobiomorpha 404
Lithoptelera egans 47
Littorina 303, 307
— rubis 306*
Loa loa 222
Locusta migratoria 464
Lonabris 473
Lophopodella 280
Loricata 288, 289, 292, 519*, 522
Loxostege sticticalis 467
Lubomirskiidae 105
Lucernaria 129
Lucernariida 129
Lumbricidae 257, 265
Lumbriculidae 265
Lumbriculus 265
Lumbricus 257, 265
— terrestris 257, 258*, 259*, 260, 261,
262, 264, 267
Lycorea 460, 461
— halia 457
Lycosidae 386, 387
Lynx 24
Lyperosia 56
Lacrimaria sp., 85*, 95
Lagena stricta 44
Lamblia intestinalis 59
Lamellibranchia 310
Lamellibranchiata 288, 310
Lamellisabella rachsi 505
Larvivoridae 469
Laseola tristis 385
Lathrodectes tredecimgluttatus 384, 387
Leda pernula 316*
Leischmania 55, 57
— donovani 57
— tropica 57
Lentospora 74
— cerebralis 74
Lepas 353*
Lepidoptera 408, 452
Leptinotarsa decemlineata 466*
Leptolida 111, 119
Leptoplana borealis 157
— tremellaris 157
Leptostraca 356, 476*
Lesquereusia modesta 43
Leucandra 105, 106
Leucon 105
Libellula 435
Libythea celtis 457*
Limnaea 305
— auriculata 306*
— stagnalis 174, 305*
truncatula 306*
Limnatis nilotica 272
— turkestanica 272
Limnodrilus 191
Limnoria 362
Limulus 368
— mollucanus 368
— polyphemus 366*
Lingula 283
Linyphia triangularis 386*
Linyphiidae 386
Liobunum rotundum 383*
Lionotus fasciola 85*
Liphistiomorpha 383
Madreporaria 27
Magellona papillicornis 244, 246*
Machilis 433*
Macracanthorhynchus hirudinaceus 232
Macraspis elegans 176
Macoma 320
Macrodasioidea 200, 224
Macrodasys budenbrocki 199
Macrodytes marginalis 407*
Macroglossum 463
Macrosiphum 470
Malacosoma neustria 469*
Malacostraca 356
Mallophaga 408, 440, 468
Mammalia 24
Manajunkia baikalensis 254
Mantis religiosa 436*
Mantodea 408, 436
Margaritana margaritifera 314*
Margarites 303
Margaropus annulatus calcaratus 73
Mastigamoeba aspera 60
Mathevolepis petrotschenkovi 183
Mayetiola destructor 467, 468*
Megaloscolex australis 268
Mecoptera 408
Melania 176, 309
Melicerta 230
— ringens 227*
Meloidea 438, 441
Meloidogyne 222
Melolontha hippocastani 464*
— melolontha 442*
Membraniopora 278*
Merostomata 365, 366
Mertensia 124*
Mesodinum rubrum 97
Mesodiplogaster 211*
Mesostoma ehrenbergi 151, 153*, 154*, 157*,
158
Metastrongylus 223
Metazoa 109, 517
541
Micrathena cyanospina 384*
Microhydra ryderi 122*
Microsporidia 36, 73, 75
Microstomum lineare 155, 157, 158
Misumenga 387
Mixopterus 368
Modiola phaseolina 316*
Moina 362
Mollusca 25, 287, 288, 519*, 520, 522
Monas vulgaris 55
Monhystera 210*
Moniezia 192, 195
— benedeni 193
— expansa 193*
— planissinia 187*
Monochamus 442*
Monogenea 144, 158
Mononchus 210*, 211*
Monoplacophora 288, 293, 301, 519*, 522
Monothalamia 44
Monotocardia 288
Mosgovoyia pcclinata 187*
Mullcrius 222
Multiceps multiceps 192, 195
Muslolidae 24
Mygalomorph.’i 383
Myrianida 252
— fasciata 251*
Myriopoda 399, 519*. 521
Myrmclcotiilidae 449
Myrmocium gounlii 384
Mysidacea 357
My sis 478*
— relicta 357
Mytilus 317
— edulis 316*, 318
Myxobolus 74
— bramae 74*
— fuhmani 74
— pfeiferi 74*
Myxosoma 74*
Myxosporidia 36, 61, 73, 98
Myzostomida 238, 255, 256
Naididae 266
Nais blanci 266
— pseudoobtusa 266
Nassa reticulata 306*
Natica 303
— clausa 306*, 307
Naucoris 438
Nautiloidea 288
Nebalia 357*
Necator americanus 207*, 222
Nemathelminthes 25, 142, 196, 230, 519*,
520
Nematobothrium filarina 169, 170
Nematocera 445, 446, 447
Nematoda 198, 200
Nematodes 519*, 520
Nematomenia flavens 292*
Nematomorpha 224
Nemertini 25, 142, 233, 519*, 520
Neopilina galatheae 293*
Nepa 438
Neptunea 308
Nereidae 255
Nereis 244, 245, 246*, 248, 250, 251, 512
— pelagica 238, 239*, 242, 243*, 245*,
255
Neuroptera 408, 443
Noctuidae 459
Nodomorphinia compressiuscula 44
Nosema apis 75
Nosema bombycis 75
Notholca longispina 227*, 229
Notommatidae 230
Notonecta 431, 438
Novalis 88
— cordiformis 88
Novius cardinalis 474
Nucula 317, 318
Nuculana 318
Nudibranchia 288, 304
Nutallia equi 73
Nybellina lingualis 194*
Nycthoterus faba 88
— ovalis 87*
Obelia 116, 117*, 118*. 119, 120
Octocorallia 111, 131
Octopoda 288
Odonata 408, 435
Oestrus ovis 469
Olonellidno 364
Olygochaeta 75, 238, 256. 519* 521
Oligotrichia 84, 88
Opalina raiiaruni 63, 64 ‘
Opalinidac 83
Opalinina 83
Ophiopholis aculeata 491, 492*
Ophiura sarsi 491*
Ophiuroidea 483, 490, 503, 519*
Ophryoscolecidae 88
Ophrioscolex caudatus 89*
— purkinjei 89*
Opiliones 365, 383
Opistobranchia 288, 303
Opistorchis felineus 169, 175, 309, 511
Opistotrichium janus 89
Orbitolites 38
Oribatei 389
Oribatoidea 193
Ornithodoros 392
Ornithonyssus bacoti 392
Orthoptera 408, 436
Oscillaria 38
Oscinosoma frit 467
Ostracoda 356
Ostrea 320
Oxytricha fallax 90*
Pagurus setosus 359*
Palaeophonus 381, 394
— nunicius 368*, 368
Palaeostraca 365, 366
Pandorina 53
Panonychus ulmi 390
Papilio merope 461
Paracyneta 92
Paragonimus westermani 175, 176
Paralithodes camtschatica 359*, 361, 512
Paramaecium 83
— bursaria 83
— caudatum 78, 79, 81, 82, 83, 95
— putrinum 95
Paramphistomum 169, 170
Parasitiformes 374, 387, 391
Parascaris equorum 201, 204*
542
Parazoa 109
Pardus 24
Parenteroxenos dogiel 307*
Patella 298, 303, 309
Pauropoda 399, 407
Pecten 315
— islandica 316*, 318
Pectinophora gossipiella 471
Pedalion 229
Pediculus capitis 440
— vestimenti 440, 441*
Pedipalpi 365, 381
Pelagothuria natatrix 502
Pelmatozoa 503, 504
Pelomyxa binucleata 40
Penaeus 360*, 361*
Peneroplis planatus 44
Pennatula 137*
Pennatularia 131
Perforata 44
Peridinium cincutum 93
Peripatoides novaezelandiae 397*
Peripatopsis capensis 396*, 398
Peripatopsis edwardsii 396*
Peripatus 398
Peripsocus 440*
Peritrichia 84, 90, 36
Phalangium 383
Pharyngobdellae 238. 274
Phasmidia 220
Phasmodea 408
Pheidole instabilis 455*
Pheronema 107
Philine 303, 304
Phlebotomus 57
— papatasii 471
Pholas 319
Phorbia brassicae 470
— fuscipes 470
Phoronoidea 25
Phryganea 452*
Phthirius pubis 440
Phyllium scythe 460*
Phyllodocidae 255
Phyllodoce laminosa 239*
— paretti 245*, 255
Phyllopoda 476*
Physophora 123, 125
Physophora hydrostatica 124*
Phytomastigina 36, 51
Phytomonadina 36, 49, 51, 52
Physa 298, 305
— fontinalis 266, 306*
Physalia 123, 124*
Pieridae 460
Pieris brassicae 449*
Pisaura 385
Piscicola geometra 273
Pisidium amnicum 314
Placentonema gigantissima 208
Planaria graffi 156*
Planootrichium monticelli 185*
Planorbis 298, 305
— corneus 266, 306*
Platanthera bifolia 463
Plathelminthes 25, 142, 144, 519*, 520
Plasmodium 62, 72
— falciparum 63, 66
— malariae 63, *66
— ovale 66
Plasmodium vivax 62, 63, 64, 65, 66
Plasmodroma 36
Platoda 158
Platodes 234
Platypsyllus castoris 441
Plecoptera 408
Pleodorina 52
Pleurobrachia 124*
— rhododactyla 140
Ploima 229
Plumatella 281*
— repens 278*
Pogonophora 25, 505, 519*, 523
Polistes 454
Polyartha paltyptera 229
Polybrachia 506
Polycelis cornuta 156*
— nigra 156*, 157
Polvchaeta 238, 262, 519*, 521
Polycladida 144, 151, 152*, 153. 154. 156*.
157
Polycystidea 68
Polyergus rufescens 455
Polygordidae 255
Polygordius 249, 252*, 253, 255, 256
Polymastigina 36, 49, 58
Polypobotrium vaccarii 185*
Polypodium hydriforme 119
Polystomella crispa 45
Polystomum integerrinum 159, 160
Polythalamia 44
Polytoma 95
Polyzoa 277
Poneurinae 454
Porifera (Spongia) 99
Portlandia arctica 316*
Portschinskia przewalski 457*
Poterion neptuni 108
Pratylenchus 222
Priatoma megista 56
Proflagellata 517
Proporus 155
Proteroblastus 503
Prosopsis 445
Prosobranchia 288, 302
Prosostomadidea 144, 161
Prosthiostomum siphunculus 156*
Protobranchia 288, 318
Protociliata 36, 78, 83
Protoclepsis tesselata 271
Protodrilidae 255
Protodrilus 255, 256
Protomonadina 36, 49
Protostrongylus 222
Protozoa 25, 35, 36, 98, 519*
Protura 408, 433
Prototracheata 395, 398, 519* 521
Pseudococcus gahani 474
Pseudophyllidea 144, 191
Pseudoscorpiones 365, 382
Pseudotrychonympha 48, 59
Psithyris 461
Psolus phantapus 500*, 502
Psyllidae 438, 439
Pterochroza 460*
Pteropoda 288, 304
Pterygota 399, 408, 432, 433, 434
Pulex irritans 451
Pulmonata 288, 305
Puncturella 299*, 303
540;
Pyroplasma ovis 73
Pyroplasmoidea 72
Radiolaria 36, 37, 47, 95
Rapana 308
Rattulus 229
Retepora cellulosa 278*
Rhabdammina abyssoruin 44
Rhabditidae 221
Rhabditis 210*
— buffonis 215*
Rhabdocoela 144, 151, 152*, 153, 154, 157
Rhapana bezoar 306*
Rhinosporidium seeberi 76
Rhipicephalus 73
— sanguineus 374, 393
Rhipidocysiis 502*
Rhisota 230
Rhizoniastigina 26, 37, 48, 49, 60
Rhizopoda 36, 517, 519
Rhizostoma piilmo 128. ISO*
Rhopilema esculanta 124
Rhyuchobdellae 238. 273
Rhynchonella pslttacca 285
Rhynchota 438, 408
Rhynchodrilus fafner 262
Rodolia oardinalis 474
Rotatoria 198, 226, 519” 520
Rotifer iicptuniuq 229
Rybelliniu inguilis 185?
Sabellaria alveolata 244, 245*, 256
Sabellidae 254, 256
Sacculina 28, 355*
Sagitta 507*
Sanatoria 436
Salticidae 386
Salticus cingulatus 384*
Sarcoptoidea 390
Sarcosporidia 36, 61, 76
Sarcocystis lindemanii 76
— mescheriana 76
— muris 76
Sarcocystis tenella 76
Sarcophaga 446
Sarsia 120, 122, 130*
Scaphopoda 288, 309, 519*, 522
Scarabaeida 442
Scarabaeus sacer 443
Schisostoma 169, 175
— haematobium 174*
— japonicum 176, 188*, 515
Schizotrypanum cruzi 96
Sciaenia aquila 169
Scolopendra 404
Scolopendrella 406*
Scolopendromorpha 404
Scorpiones 365, 380
Scutigera 400, 404
Scutigerella 406*
Scutigeromorpha 404
Scyphozoa 111, 112, 125, 519*
Secernentea 198, 220, 221
Sedentaria 238, 255, 256
Seinura winchesi 211*
Seisonidae 228
Septibranchia 288, 319
Serpula 244, 245*, 256
Serpulidae 248, 256
Siboglinum 506*
Silphidae 441
Siphonophora 111, 112, 123
Slaurophora 130*
Slimonia 368
Solaster 489
— papposus 492*
Solengastres 288, 292, 519*, 522
Solifugae 365, 383
Spaerularia bombi 218*
Spathiclium spatula 86*
Sphaeridae 318 <
Sphaerium corneum 314*
Sphaerospora 74*
Sphaerophrya stentoris 93
— magna 92*
Spirifer 284
Spiroloculina depressa 44
Spirostomum 35, 87, 95
— ambiguum 87
Spirurida 222
Spisula sachaltnonsis 316*
Spongta 25, 99, 101, 519*
Spongilla 103, 104
Spongillidae 195
Sporozoa 36, 60, 98, 517, 519*
Stahylinidac 441
Siaurudcrus sibiri<?u§ 464
Staurophora 120
Stenophanoceras fimbriatus 230
Stentor 35, 67. 95
— polymorphic 87A
Stephanopogon 98
Steptococcus bombycis 462
Stichopus japonicum 500*, 502
Stomoxys 56
Strongylocentrotus droebachiensis 495*
Strongyloides stercoralis 207*, 215
Strongylus vulgaris 222, 223
Styeocephalus longicolis 68
Stylaria lacustris 266
Styli 433
Stylochoplana maculata 156*
Stylochus tauricus 157
Stylommatophora 288, 305
Stylonichia 90
— mytusil 90*
Suctoria 36, 77, 78, 91, 93, 98, 517
Sycon 101, 102, 103, 105
— raphanus 101, 102*, 103
Syngamidae 222
Syllidae 255
Syllis ramosa 251*, 252, 255
Symphyla 399, 406
Synchaeta 229
Syncoryne 120
Syngamus 222
Synthetocaulus 216, 222
— hobmaieri 217*
Syrphidae 461, 473
Syrphus 447
Tabanidae 56, 447
Tachinidae 469
Tachypodojulus 405*
Taenia marshali 185*
— solium 178*, 188*, 192, 194
Taeniarhynchus saginatus 178, 188*
Tectibranchia 288
Tegenaria domestica 385
Telosporidia 36, 61, 67, 98
Telyphonus caiidatus 382*
544
Tenebrio molitor 69*
Tenebrionidae 441
Tendipes 446
Tentacularia bicolor 194*
Tentaculata 111
Terebellidae 256
Teredo navalis 315
Termes lucifugus 456*
Terpnacaridae 388*
Testacea 37, 43
Tetracorallia 136
Tetramorium 445
Tetranychoidea 390
Tetranychus telarius 388*, 390
— uricae 390
Tetrapodili 391
Tetrarhynchoidea 193, 194*
Tetraxonida 101, 108, 109
Tettigoniodea 436
Textullaria sagittula 44
Thecaphora 119
Thecoidea 502
Theileriidae 72
Theodoxus 303
Thomisidae 386
Thusanoptera 408
Thusanozoon brocchii 156*, 157
Thysanura 408, 432, 433, 434, 481
Tintinnidae 88
Tocophrya cyclopum 92*, 93
Tomopteridae 255
Tomopteris euchaeta 254*, 255
Tonicella marmorea 289
Toxocara mystax 222
Toxoptera graminis 439*
Tracheata 399, 480, 481
Trachelius 85
— ovum 85*
Trachylida 111, 119, 122
Trematoda 144, 161, 520
Trematodes 519*
Triarthra 229
Triarthrus becki 363*
Triaxonida 101, 106, 109
Trichinella spiralis 206, 207*, 217, 221,
222
Trichocephalus trichiurus 207* 221, 222
Trichodectes latus 440*
Trichoderma minutum 208
Trichogramma evanescens 474, 513
Trichomonas angusta 58
— hominis 58, 59
Trichoptera 408, 451
Trichostrongylidae 222
Trichostrongylus instabilis 207*
Tricladida 144, 151, 152*, 153, 155, 156*,
157
Tridacna 320
Trilobita 362, 519*, 521
Trilobitomorpha 362
Trochaeta subviridis 274
Trochosa 385
Trochosphaera 230
Trochus 303
Troctes divinatorius 440
Trombea 391
Trombicula autumnalis 388*
Tropidoderus 459
Tropidoderus children! 460*
Trypanosoma 54, 55
— brucei 56, 57
— cruzi 54
— evansi 56
— equiperdum 56
— gambiense 55, 56, 471
Trypanosomidae 54
Tubifex 191, 266*
— tubifex 265, 266
Tubificidae 266
Tubipora musica 132
Tubularia 120
— larynx 120*
Turbellaria 144, 145, 165, 519*, 520
Tylenchida 222
Tyroglyphoidea 389
Tyroglyphus farinae 388*, 390
Tyrophagus noxius 390
— perniciosus 390
Umbellula encrinus 133, 137*
Unio 320
— pictorum 314*
Uroleptus pisces 90*
Ursidae 24
Vallonia 305
Vanadis ramosa 239*
Velella 123, 124*
Veliger 300
Vermes 142
Vertebrata 24
Vespa 454
— germanica 454*
Vespidae 444
Vivipara 176, 297, 298, 303, 307
— vivipara 306*
Volvocidae 52, 517
Volvox 52, 95, 518
— aureus 53, 230
Vorticella 91
— microstoma 95
— nebulifera 91*
Wucheria bankrofti 222
Xyloborus 465*
Xyphosura 365, 366, 519, 521
Zachvatkinia sternae 388*
Zebrina 308
Zoochlorella 113
Zoomastigina 36, 49, 54
Zygoptera 435
ОГЛАВЛЕНИЕ
Предисловие к седьмому изданию . .......................................5
Введение ....... . .... 7
Зоологии как система наук . .... ... .... 7
История зоологии ........... -.,«. 9
Организм как целое . .......................... 13
Строение организма животных и функции его органов................. 15
Органы и системы Оргацуи животных . . ... 10
План строения и симметрия .......................... 10
Размножение и развитие .......................... 20
Система мира животных . ........ 24
Йаанмоотцршуннс животных с окружающей средой . . ... । 26
Географическое распространение животных ... . . 28
Геологическая история животного мира .... . . 30
Учение об эволюции ...................... 30
Тип I. Простейшие (Protozoa)
Характеристика ...............................35
Подтип I. Плазматические (Plasmodroma) .........36
Класс 1. Корненожки (Rhizopoda) ......... 36
Характеристика .... 36
Представитель класса корненожек — амеба (Amoeba proteus) ... 37
Систематический обзор класса ... .... 40
Отряд амебы (Amoebaina) ..........................40
Отряд раковинные амебы (Testacea) .....................................43
Отряд фораминиферы (Foraminifera) . ...................................44
Отряд солнечники (Heliozoa) .............................. . 46
Отряд лучевики (Radiolaria) 47
Класс 2. Жгутиковые (Flagellata) ............................................48
Характеристика 48
Представитель класса жгутиковых — эвглена (Euglena viriolis) • . 49
Систематический обзор класса . ..............51
Подкласс I. Фитомастигины (Phytomastigina)...................................51
Отряд эвгленовые (Euglenoidina) 52
Отряд фитомонады (Phytomonalina) . 52
Подкласс И. Зоомастигины (Zoomastigina) ... ...........54
Отряд первичномонадные (Protomonadina) ........................ . 54
Отряд многожгутиковые (Polymastigina) 58
Отряд корнежгутиковые (Rhizomastegina) ................................60
Класс 3. Споровики (Sporozoa) ............... ... 60
Характеристика ......................... . 60
Представитель класса споровиков — малярийный плазмодий (Plasmodium
vivax) . ...........................62
Систематический обзор класса 67
Подкласс I. Телоспоридии (Telosporidia) .....................................67
Отряд грегарины (Gregarina) 67
Отряд кокцидии (Cocci di а) ....................... 70
Отряд гемоспоридии (Haemosporidia) ... ..... 72
Подкласс II. Книдоспоридии (Cnidosporidia) ..................................73
Отряд слизистые споровики (Myxosporidia) ......... 73
546
Отряд актиномиксидии (Actinomyxidia) . . . . 75
Отряд микроспоридии (Microsporidia} . . . . 75
Подкласс III. Саркоспоридии (Sarcosporidia) . . 76
Подкласс IV. Гаплоспоридии (Haplosporidia) . . 76
Подтип II. Ресничные, или инфузории (Ciliata) 77
Характеристика ........................................................ 77
Класс 1. Ресничные инфузории (Ciliata) .... 78
Характеристика ........................................................ 78
Представитель класса ресничных инфузорий — парамеция (Paramaecium
caudatum) ........................................................... 78
Систематический обзор класса .................................... 83
Подкласс I. Протоцилиаты (Protoci 1 i at а) .................................83
Подкласс II. Собственно инфузории (Euciliata) ... ...........84
Отряд равноресннчные (Holotricha) . . . . ... 84
Отряд разноресничные (Heterotricha) . .... 87
Отряд малоресничные (Oligotricha) . . . . . . 88
Отряд брюхоресничные (Hypotricha) . . . . . . . 90
Отряд круглоресничные (Peritricha) ... . . . . . . 90
Класс 2. Сосущие инфузории (Suctoria) ... . . 91
Характеристика ....................... .................. . . . 91
Географическое распространение и экология простейших . . . . 93
Филогения простейших .............................................. . . 9&
Тип II. Губки (Spongia, или РогИега)
Характеристика ................................................... ... 99
Представитель типа губок — сикон (Sycon) ......................... ... 101
Второй представитель типа губок — бадяга ......................... ... 103
Общий обзор типа ................................................ ... 105
Систематический обзор типа ... ............ ... 106
Отряд известковые губки (Calcarea) ... 106
Отряд трехосные губки (Triaxonida) ... 106
Отряд четырехосные губки (Tetraxonida).......................... ... 108
Отряд кремнероговые губки (Cornacuspongia) ... ... 108
Филогения губок .................. ... ... ... 109
Тип III. Кишечнополостные (Coelenterata)
Характеристика .... ..................................1Ш
Систематический обзор типа ....................................111
Подтип I. Стрекающие (Cnidaria) . .111
Характеристика ............... ... .........................Ш
Класс I. Гидроидные (Hydrozoa) ... .........................112
Характеристика ....................................................... 112
Подкласс I. Гидрообразные (Hydroidea) ..............................112
Представитель сидячих гидрообразных — пресноводная гидра (Hydra
oligactis) ......................................................... 113
Представитель свободноплавающих гидрообразных — медуза обелия (ОЬе-
lia) .................................................................116
Систематический обзор подкласса ... .........................119
Отряд гидры (Hydrida) ..................................119
Отряд гидроиды (Leptolida) ....................................119
Отряд трахилиды (Trachylida) . . .........................122
Подкласс II. Сифоноформы (Siphonophora) ....................................123
Характеристика . ...........*....................................123
Общий обзор подкласса ..................................................123
Класс 2. Сцифоидные (Scyphozoa) ........................................ ... 125
Характеристика 125
Представитель класса сцифоидных — медуза аурелия (Aurelia aurita) 126
Общий обзор класса .....................................................128
Класс 3. Коралловые (Actinozoa) ............................................129
Характеристика .........................................................129
Подкласс I. Восьмилучевые кораллы (Octocorallia) ................131
Характеристика . . ........................................131
Общий обзор подкласса ....................................132
Подкласс II. Шестилучевые кораллы (Hexacorallia) ................133
Характеристика .........................................................133
Представитель подкласса шестилучевых кораллов — обыкновенная, или
конская, актиния (Actinia equina) ................................133
Общий обзор подкласса ..................................135
547
Подтип II. Нестрекающие (Acnidaria) •
. . 138
Характеристика .......................... . . ... 138
Общий обзор класса ............................. .... .... 138
Филогения кишечнополостных .... ....................140
Паренхиматозные черви, первичнополосатые черви и немертины
Тип IV. Плоские, или паренхиматозные, черви (Plathelminthes)
Характеристика . .... ... ...............144
Класс I. Ресничные черви, или турбеллярии (Turgbellaria) .................145
Характеристика . ...............145
Представитель класса ресничных червей молочная планария (Dendro-
coelum lacteum) .......................... . .... . 145
Общий обзор класса ............................................... .151
Систематический обзор класса . . . .... . 155
Отряд бескишечные турбеллярии (Acoeda) ............................ 155
Отряд многоветвнстокншечпые турбеллярии (Polycladida) . . 157
О1ряд трехветвистокишечные турбеллярии (Tricladida) . 157
Отряд прямокишечные турбеллярии (Rhabdocoela) . . . 157
Филогения турбеллярии ...... .... . . 156
К л а с с 2. Моиогсистичсскис сосальщики (Monogenca) . . . 158
I1родстапнтель класса мопогспс-и — гиродактмль (Gyrodactylua clcgana) 158
Общий обзор класса ............................................. , . 159
Клас с 3. Трематоды, или дигрпотиисскио еооллыцпки (Trematoda) . . . 1GI
Характеристика ..................................................... 151
Подкласс I. Дигснстичсскис сосальщики (Prosostoniadidea) .... 161
Представитель подкласса днгенстических сосальщиков — печеночная фас-
циола (Paeciola hopatica) ............... 162
ЦИКЛЫ рЯЛИПТИИ ПРКОТОрЫХ ДруГПХ ДПГОИОТПЧеСКТТХ Сосальщиков . 168
Общий обзор класса . . .................................. 169
Подкласс П. Пуцефалндиден (Biiccpiialidldea) .................... . . 175
Подкласс 111. Аспидогастродидсн (Aspidogadrodidea) .... 175
ПрсДопоспосТь сосальщиков. ........... 176
Борьба с сосальщиками . .................................. 176
Класс 4. Ленточные черви, или цестоды (Ce$toidea) 177
Характеристика . ......................... 177
Представитель класса цестод—бычий солитер (Taeniarhynchus saginatus) 178
Циклы развития некоторых других представителей класса цестод . . .182
Общин обзор класса ... . . . ......................183
Систематический обзор класса . . . . .... . 190
Подкласс I. Нерасчлененные ленточные черви (Cestodaria)....................190
Подкласс II. Расчлененные ленточные черви (Cestoda) .......................191
Отряд лентецы (Pseudophyllidea) . . . .................191
Отряд цепни (Cyclophyllidea) ................................. 194
Вредоносность цестод.................................................194
Борьба с цестодами . ......................................196
Филогения моногеней, трематод и цестод..............................196
Тип V. Круглые, или первичнополостные, черви
(Nemathelminthes, или Aschelmintes)
Характеристика .....................................................198
Класс 1. Брюхоресничные, или гастротрихи (Gastrotricha) .... 199
Характеристика .....................................................199
Общий обзор класса . . . .............................. 199
Класс 2. Собственно круглые черви, или нематоды (Nematoda)............200
Характеристика . . ...... . ........... 200
Представитель класса нематод — лошадиная аскарида (Parassaris equorum) 201
Другие важнейшие представители класса немертин и их жизненные циклы 206
Общий обзор класса ................................. . ............208
Систематический обзор класса ................................... .... 220
Подкласс I. Аденофореи (Adenophorea) ‘.................................. 220
Подкласс II. Сецерненты (Secernentea) . . .......... . . 221
Вредоносность паразитических нематод . . ..........- . . 222
Борьба с паразитическими нематодами ............................... 223
Филогения нематод .... ... ...............224
Класс 3. Волосатиковые (Gordiacea, или Nematomarpha) ....................224
Характеристика ... ......... ....................... 224
Общий обзор класса . . ................... .... 225
Класс 4. Коловратки (Rotatoria) .................................. 226
Характеристика ................................. 226
Общий обзор класса ... 226
548
Филогения коловраток . 230
Дополнение к типу Nemathelminthes 230
Класс скребни (Acanthocephala) . 230
Характеристика .... . 230
Общий обзор класса . . 231
Тип VI. Немертины (Nemertini)
Характеристика
Общий обзор типа
Филогения немертин
. 233
. 233
. 236
Тип VII. Кольчатые, или вторичнополостные, черви
(Annelides)
Характеристика . . . . . . . .... 237
Класс 1. Многощетинновые кольчецы, или полихеты (Polychaeta) . . 238
Характеристика .... ...... 238
Представитель класса полихет — нереида (Nereis) ....................238
Общий обзор класса . . .....................244
Систематический обзор класса . . ........................255
Подкласс I. Свободноподвижные полихеты (Errantia) ...... 255
Подкласс II. Сидячие полихеты (Sedentaria)................. ... 256
Подкласс III. Мизостомиды (Myzostomida) . ... 256
Класс 2. Малощетинковые кольчецы, или олигохеты (Oligochaeta) 9 . . 256
Характеристика ... . . . 256
Представитель класса олигохет—дождевой, или земляной, червь (Lum-
bricus) ... . . . . . . . . 257
Общий и систематический обзор класса .... .... . 262
Филогения олигохет . . ........... .................. 268
Класс 3. Пиявки (Hirudinea) ............................................. 268
Характеристика . ......................................... 268
Общий обзор класса . .... . ......................268
Систематический обзор класса .... ...................... 273
Отряд щетинконосные пиявки (Acanthobdellae) ........................ 273
Отряд хоботные пиявки (Rhynchobdellae) ..............................273
Отряд челюстные пиявки (Gnathobdellae) ..............................274
Отряд глоточные пиявки (Pharyngobdellae) .... .... 274
Филогения пиявок . .................................274
Класс 4. Эхиуридовые (Echiuroidea) ........................................274
Филогения кольчатых червей .............................. 275
Тип VIII. Мшанки (Bryozoa, или Polyzoa)
Характеристика
Общий обзор типа
Положение в системе
277
277
282
Тип IX. Плеченогие (Brachiopoda)
Характеристика ........................ 283
Общий обзор типа ...................................283
Положение в системе ...................... .... 286
Тип X. Моллюски (Mollusca)
Характеристика ... .........................287
Класс 1. Панцирные (Loricata) ... .........................289
Характеристика . . 289
Общий обзор класса ... .... .............. 289
Класс 2. Бороздчатобрюхие (Solenogastres) ................................292
Характеристика 292
Общий обзор класса ...................................................292
Класс 3. Моноплакофоры (Monoplacophora) . . ..............293
Характеристика . .........................................293
Общий обзор класса . . ................................. 293
Класс 4. Брюхоногие (Gastropoda) .................................... . . 294
Характеристика . . ................... ................294
Представитель класса брюхоногих — Лшоградная улитка (Helix pomatia) 295
549
Общий обзор класса......................................
Происхождение личиночного скручивания и асимметрии
Систематический обзор класса .........................
одкласс I. Переднежаберные (Prosobranchia) . .
Характеристика . . ........................
Отряд двупредсердные (Diotocardia) ...........
Отряд однопредсердные (Monotocardia) . ...
одкласс II. Заднежаберные (Opisthobranchia) . .
Характеристика . . . . . .
Отряд покрытожаберные (Tectibranchia)
Отряд голожаберные (Nudibranchia)
Отряд крылоногие (Pteropoda)
одкласс III. Легочные (Pulmonata)
Характеристика
Отряд сидячеглазые (Basommatophora)
Отряд стебельчатоглазыс (Styloinniatophora)
Образ жизни и распространение брюхоногих
Практическое значение брюхоногих
Филогения брюхоногих . ..................
Класс 5. Лопатоногие (Scaphopoda) .................. ,
Характеристика . .
К л а с с б. Двустворчатые, или пластинчатожаберные (Eivalvia.
libiandna) . .
Характеристика . . . . . ...
Представитель класса двуствурчатых —беззубка (Anodonta)
Общий обзор класса . . . , , .
Систематический обзор класса ...
Отряд
Отряд
Отряд
Отряд
Образ
Практическое значе14ис дпустворчатых
Филогения двустворчатых
Класс 7 Головоногие (Cephalopoda)
Характеристика
Представитель класса головоногих — тихоокеанский кальмар (Ommastre-
phes sloanei pacificus) ........... . ...........
Общий обзор класса ......................................
Систематический обзор класса ... ......................
Подкласс I. Наружнораковинные (Ectocochlia) ..........................
Отряд кораблики (Nautiloidea) ..................
Отряд аммониты (Ammonoidea) ..................
с II. Внутреннераковинные (Endocochlia) .... . .
десятиногие (Decapoda) ........................
п
п
п
плп Lamd-
298
301
302
302
302
302
302
302
302
304
304
304
305
305
305
305
305
30ft
309
309
309
первичножаберные (Protobranchia) . . . . .
пнтгжабгриыс (rilibrandiin) ......................
настоящие пластничатоткабсрншс (Eul a nielli bi andiia)
перегородчатожаберпые (Sept ibranchia) . , .
жииш п распространение двустворчатых . . ,
310
. 310
. 311
. 315
. 218
. 318
. 318
. 316
. 319
. 319
. 320
. 323
. 323
323
П о д к л а с
Отряд
Отряд восьминогие (Octopoda)
Образ жизни и распространение головоногих
Промысловое значение головоногих
Филогения головоногих ......................
323
327
331
331
331
331
332
332
332
332
333
333
Филогения моллюсков .......................................... 334
Тип XI. Членистоногие (Arthropoda)
Характеристика
. 335
Подтип I. Жабернодышащие (Branchiata) .... 336
Общая характеристика . . ...............................336
Класс ракообразные (Crustacea) ......................................336
Представитель класса ракообразных — речной рак (Astacus leptodactylus
и Astacus astacus) .... .............................338
Общий обзор ракообразных .................................................342
Систематический обзор класса ........................................... 347
Подкласс I. Листоногие (Branchiopoda) ................................... . 347
Отряд жаброноги (Anostraca) ....................................... . 347
Отряд собственно листоногие (Phyllopoda) .......................... . 348
Подотряд щитни (Notostraca) . . .......................348
Подотряд ветвистоусые (Cladocera) . . .......................348
Подкласс II. Челюстеногие (Maxillopoda) ......................................350
Отряд веслоногие (Copepoda) 351
Отряд усоногие (Cirripedia) .......................................... 352
Подкласс III. Ракушковые (Ostracoda) .... ..............356
550
Подкласс IV. Высшие ракообразные (Malacostraca) • 356
Отряд тонкопанцирные (Lepostraca) . . . 356
Отряд мизиды (Mysidacea) ............................................ 357
Отряд бокоплавы (Amphipoda) .... . 358
Отряд равноногие (Isopoda) ..... . 358
Отряд десятиногие (Decapoda) .... . 359
Хозяйственное значение ракообразных . . 361
Филогения ракообразных ... . . . 362
Подтип II. Трилобиты (Trilobita) . 362
Характеристика . 362
Подтип 111. Хелицеровые (Chelicerata) . 364
Характеристика .............. .............364
Класс I. Меростомовые (Merostomata, или Palaeostraca) .... 366
Характеристика . . . . . ................366
Систематический обзор класса 366
Отряд мечехвосты (Xiphosura) .........................................366
Отряд ракоскорпионы (Eurypteroidea) ..................................368
Класс 2. Паукообразные (Arachnoidea) ................................ . 370
Характеристика . . ......................370
Представитель класса паукообразных — паук-крестовик (Araneus dia-
dematus) 370
Второй представитель класса паукообразных — иксодовый клещ (Ixodes
ricinus) 374
Общий обзор класса 377
Систематический обзор класса ...........................................380
Отряд скорпионы (Scorpiones) . . ......................380
Отряд жгутоногие (Pedipalpi) 381
Отряд ложноскорпионы (Pseudoscorpiones) ............................382
Отряд сольпуги, или фаланги (Solifugae) ............................383
Отряд сенокосцы (Opiliones) . ............................383
Отряд пауки (Агапеае) ............................383
Отряд клещи (Acarina) .... . ......................387
Основные группы клещей, имеющие медицинское и хозяйственное значение 389
Происхождение и эволюция хелицеровых .... 393
Сравнение жабернодышащих, трилобитов и хелицероносных .... 395
Подтип IV. Первичнотрахейные (Protracheata) 395
Характеристика . ............................395
Общий обзор подтипа . . ...................................396
Филогения первичнотрахейных ............................................399
Подтип V. Трахейнодышащие (Tracheata) . . 399
Характеристика 399
Класс 1 /Многоножки (Myriopoda) ...................................... 399
Характеристика 399
Подкласс I. Губоногие (Chilopoda) ... ..............400
Характеристика . . .... ..............400
Представитель подкласса губоногих — костянка (Lithobius forficatus) . . 400
Подкласс II. Двупарноногие (Diplopoda) ........... ... 405
Характеристика ....................................................... 405
Подкласс III. Сколопендреллы (Symphyla) .............................406
Характеристика ................. . . 406
Подкласс IV Пауроподовые (Pauropoda) ...............................407
Характеристика ... ..............................407
Класс 2. Насекомые (Insecta) . ........... .... 407
Характеристика . . . ..............................407
Представитель класса насекомых — черный таракан (Blatta orientalis) . 408
Второй представитель класса насекомых — медоносная пчела (Apis mel-
lifera) . ......................... .............419
Общий обзор класса . .........................................425
Систематический обзор класса ......................................... 433
Подкласс I. Первичнобескрылые насекомые (Apterygota) ....................433
Характеристика . . ............................433
Подкласс II. Крылатые насекомые (Pterygota) .........................434
Характеристика .........................................................434
551
Отряд поденки (Ephemeroptera)
Отряд стрекозы (Odonata)
Отряд таракановые (Blattodea)
Отряд богомолы (Mantodea) ....
Отряд прямокрылые (Orthoptera, или Saltatoria)
Отряд термиты (Isoptera)
Отряд членистохоботные (Rhynchota)
Отряд сеноеды (Copeognatha)
Отряд пухоеды (Mallophaga)
Отряд вши (Anoplura)
Отряд жесткокрылые, или жуки (Coleoptera)
Отряд сетчатокрылые (Neuroptera)
Отряд перепончатокрылые (Hymenoptera)
Отряд двукрылые (Diptera)
Отряд блохи (Aphaniptera)
Отряд ручейники, или пухокрылые (Trichoptora)
Отряд чешуекрылые, или бабочки (Lepidoptora)
Некоторые вопросы биологии насекомых
Одиночные и общественные насекомые .................
Полиморфизм насекомых . . ...........
Защитные приспособления насекомых ....
Массовое размножение насекомых .......................
Хозяйственное значение насекомых
Насекомые — опылители растений
I Ьюлоподстор и шелководство ....
Насекомые — вредители сельского и лесного хозяйства
Насекомые — паразиты животных . .
Насекомые — переносчики болезнен ...................
Борьба с вредными насекомыми
Общий обаир тина членистоногих ... ...........
Филогения члоппстоцргих . . . ..............
Тип XII. Иглокожие (EcliInodcrmata)
Характеристика . ......................
Класс 1 Морские звезды (Asteroidea) ............................
Характеристика . . .
Представитель класса морских звезд — морская звезда астериас
rias rubens) . ..................
Общий обзор класса . ...........
Класс 2. Змеехвостки, или офигуры (Ophiuroidea)
Характеристика .....
Общий обзор класса . .
Класс 3. Морские лилии (Crinoidea)
Характеристика ...
Общий обзор класса . . ...
Класс 4. Морские ежи (Echinoidea) . ...
Характеристика . ....
Общий обзор класса . . ... ....
Класс 5. Морские огурцы, или голотурии (Holothuroidea) . .
Характеристика ... ........... . .
Общий обзор класса . . ... . . . .
Ископаемые иглокожие .... ... . .
Филогения иглокожих ..................... ..................
. 434
435
. 436
. 436
. 436
. 438
. 438
440
440
. 440
441
. 443
. 443
. 445
451
451
. 452
453
. 453
458
459
461
. 462
463
. 463
464
. 4G8
470
. . 471
. . 474
479
482
. . 483
483
(Aste-
483
. . 489
. 490
. 490
490
. . 492
. . 492
492
. 494
. . 494
495
. . 498
493
499
. . 502
. . 503
Тип XIII Погонофоры (Pogonophora)
Характеристика . .................505
Общий обзор типа . ... .............. . . 505
Некоторые группы беспозвоночных неясного систематического положения 506
Класс щетинкочелюстные (Chaetognatha) .... .... . 506
Характеристика ............... ................... ... . 506
Общий обзор класса . . ... . ... . 507
Значение беспозвоночных в народном хозяйстве . . . . 508
Беспозвоночные как кормовые объекты промысловых рыб . 512
Промысловые беспозвоночные ... . 512
Насекомые вредители сельского и лесного хозяйства . . 513
Клещи ....................................... . .... . 513
Паразитические черви . .... . ............. . 514
Брюхоногие моллюски — промежуточные хозяева паразитических червей
человека и домашних животных . . ... . 515
Беспозвоночные, вредящие гидротехническим сооружениям и водному
транспорту ................................... .... . . 516
Эволюционное развитие беспозвоночных ..................................516
Литература .... . ................................. 525
Предметный указатель........................................ . . • 527
552