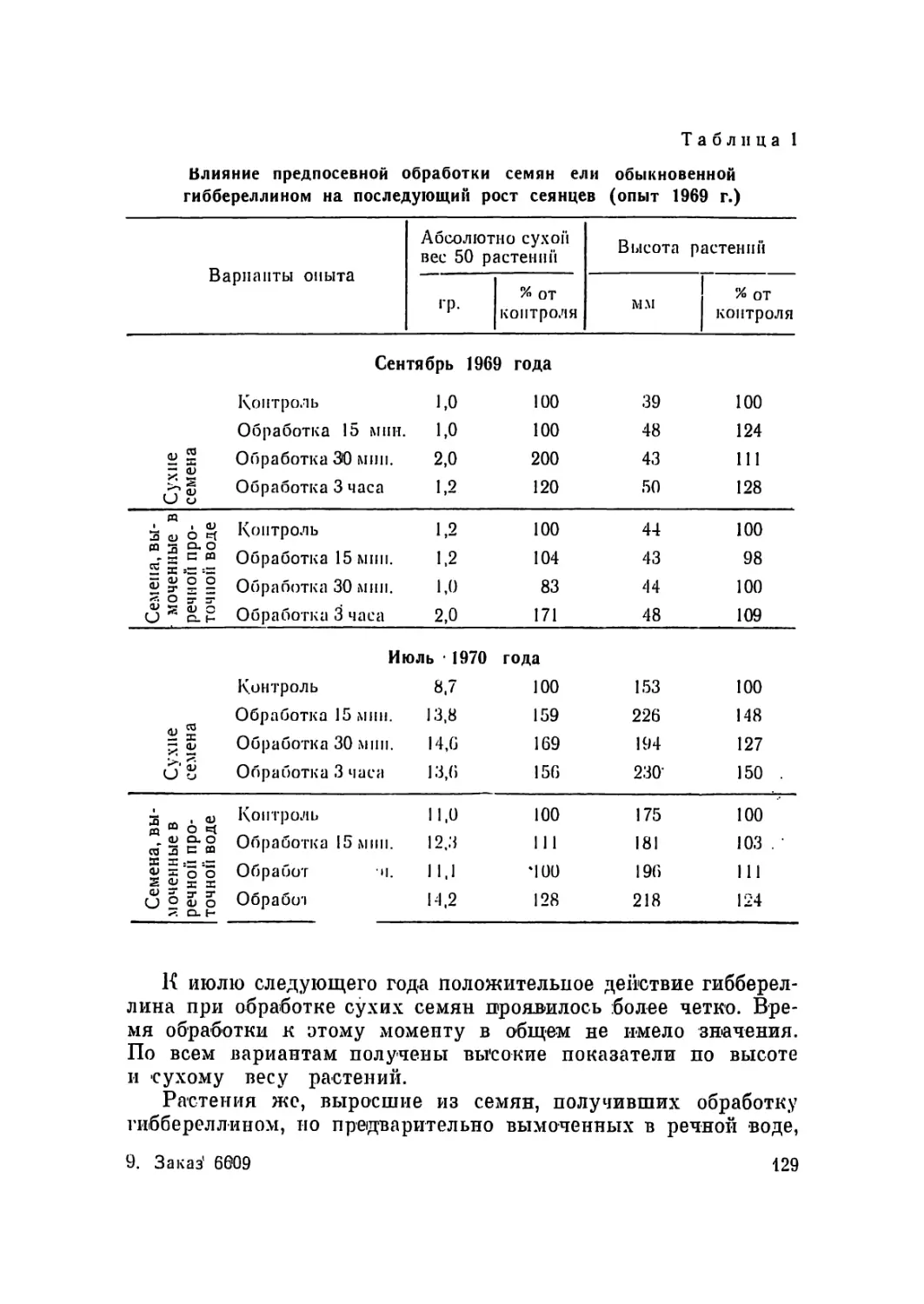
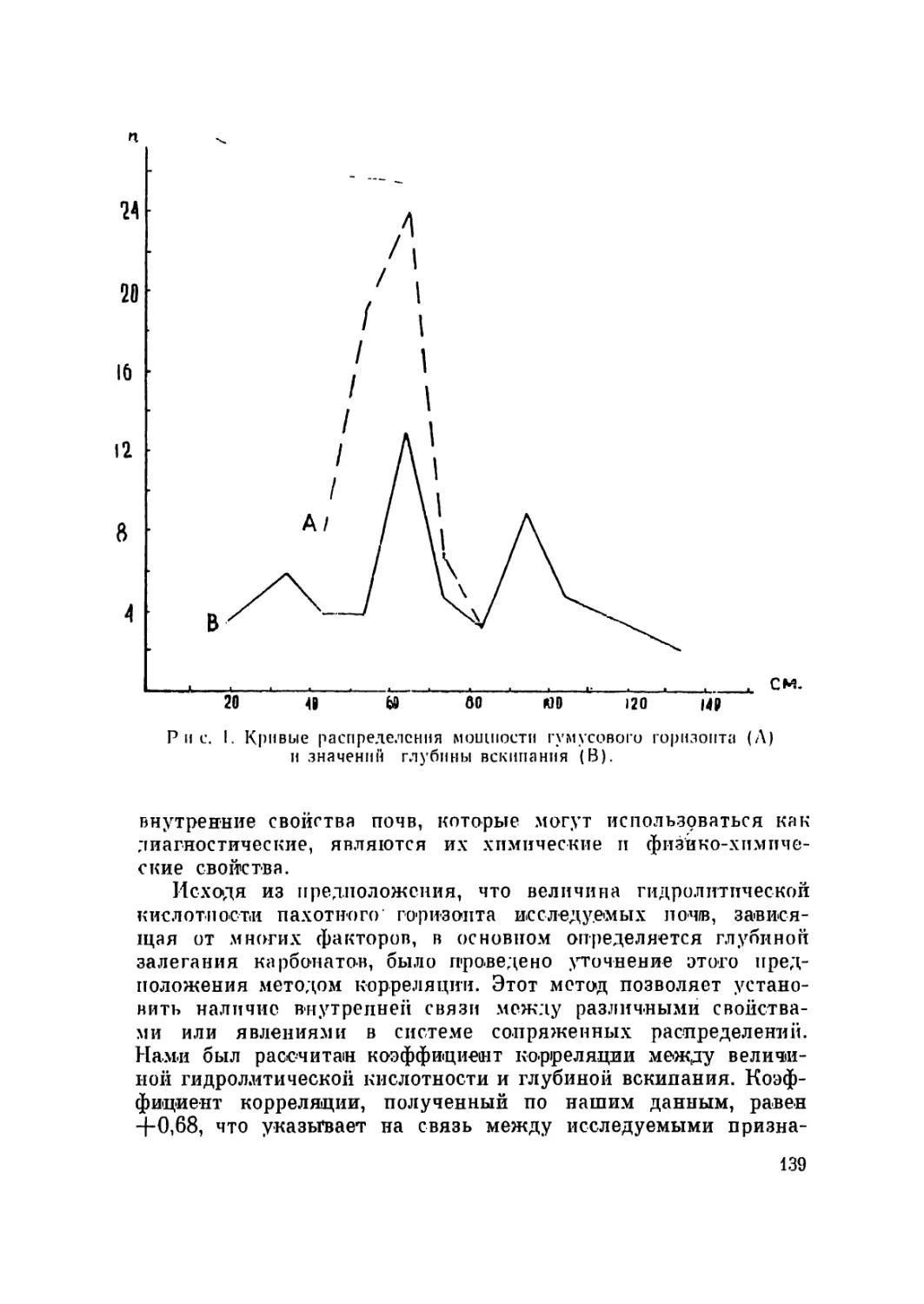
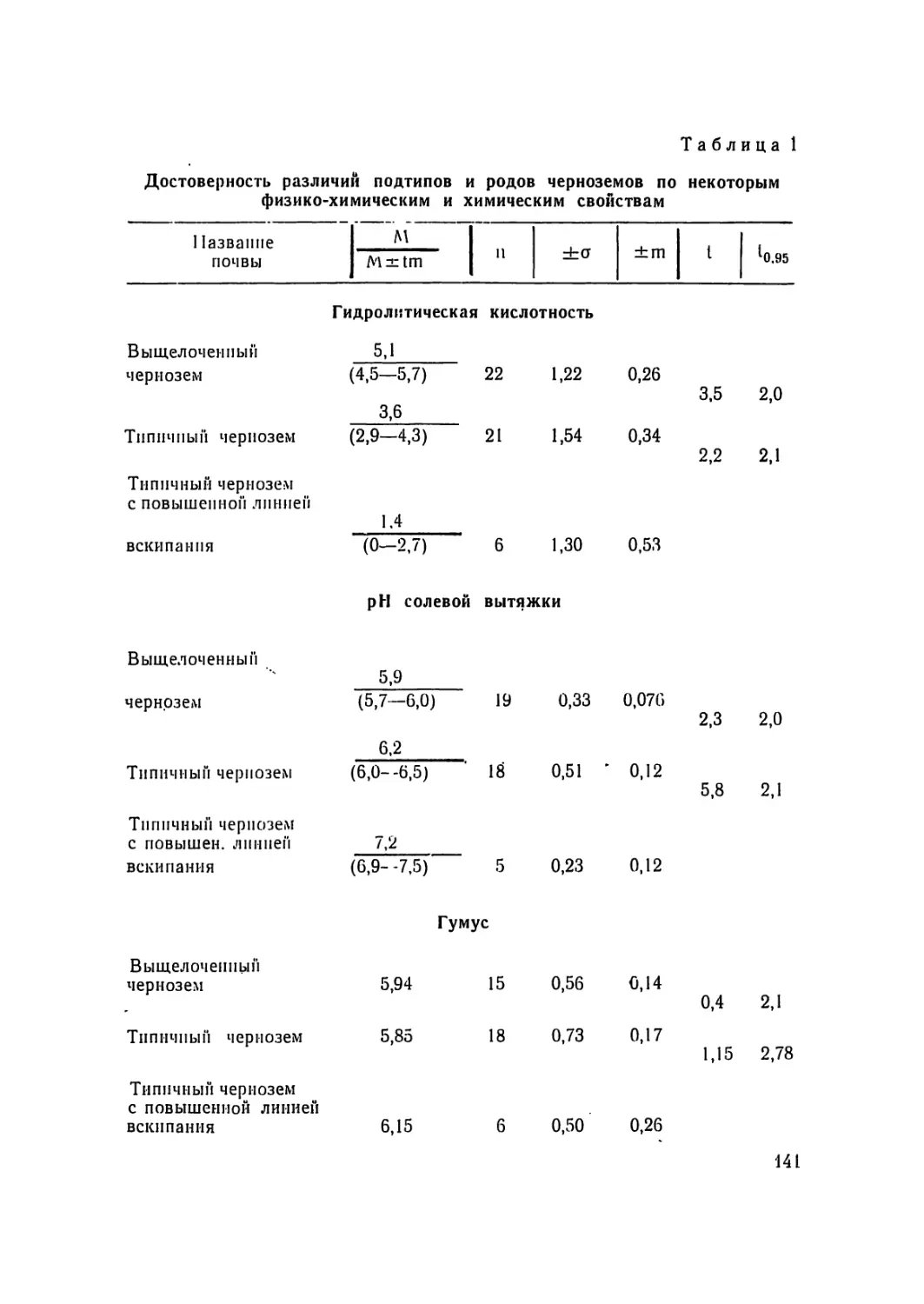
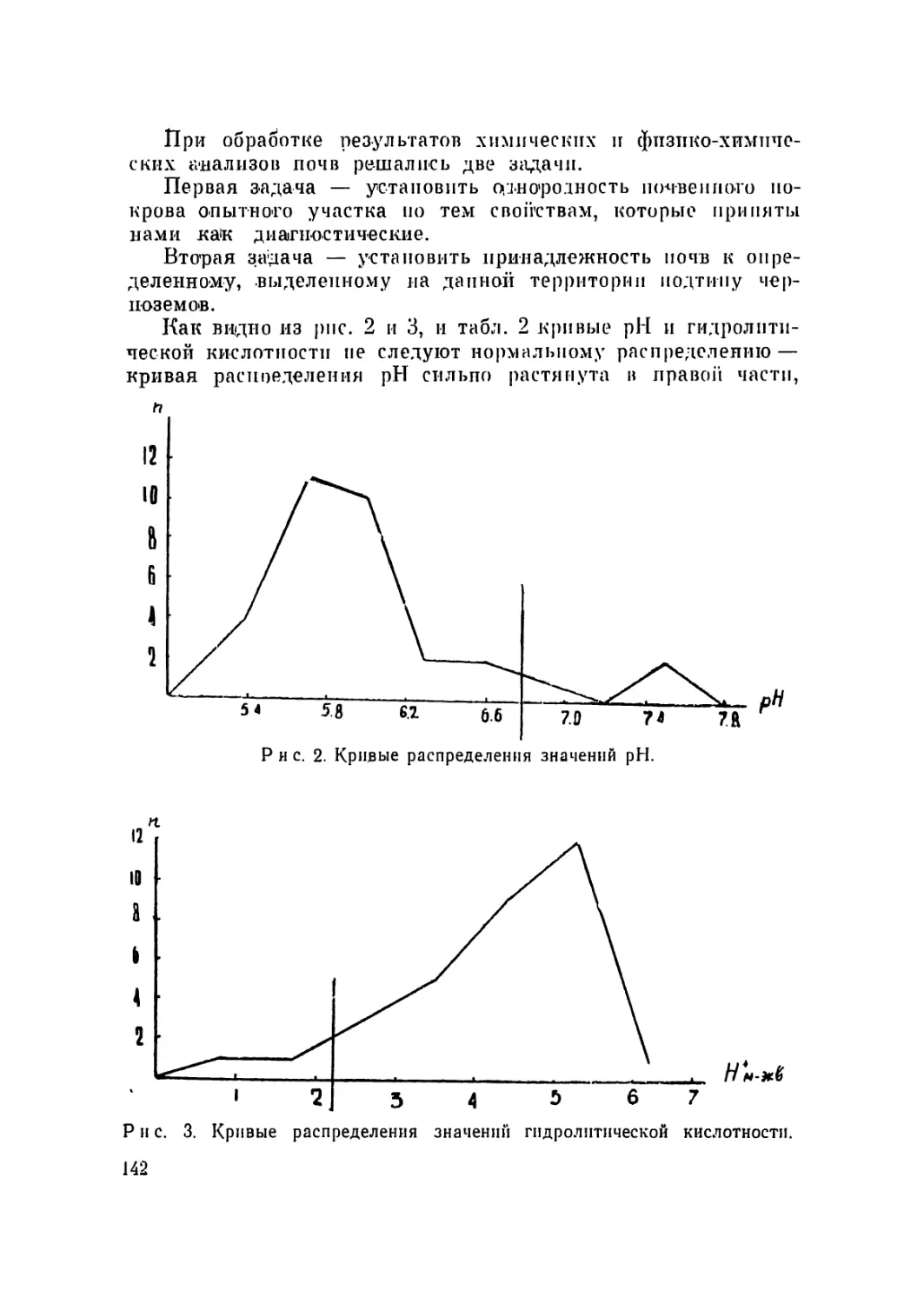
/






























































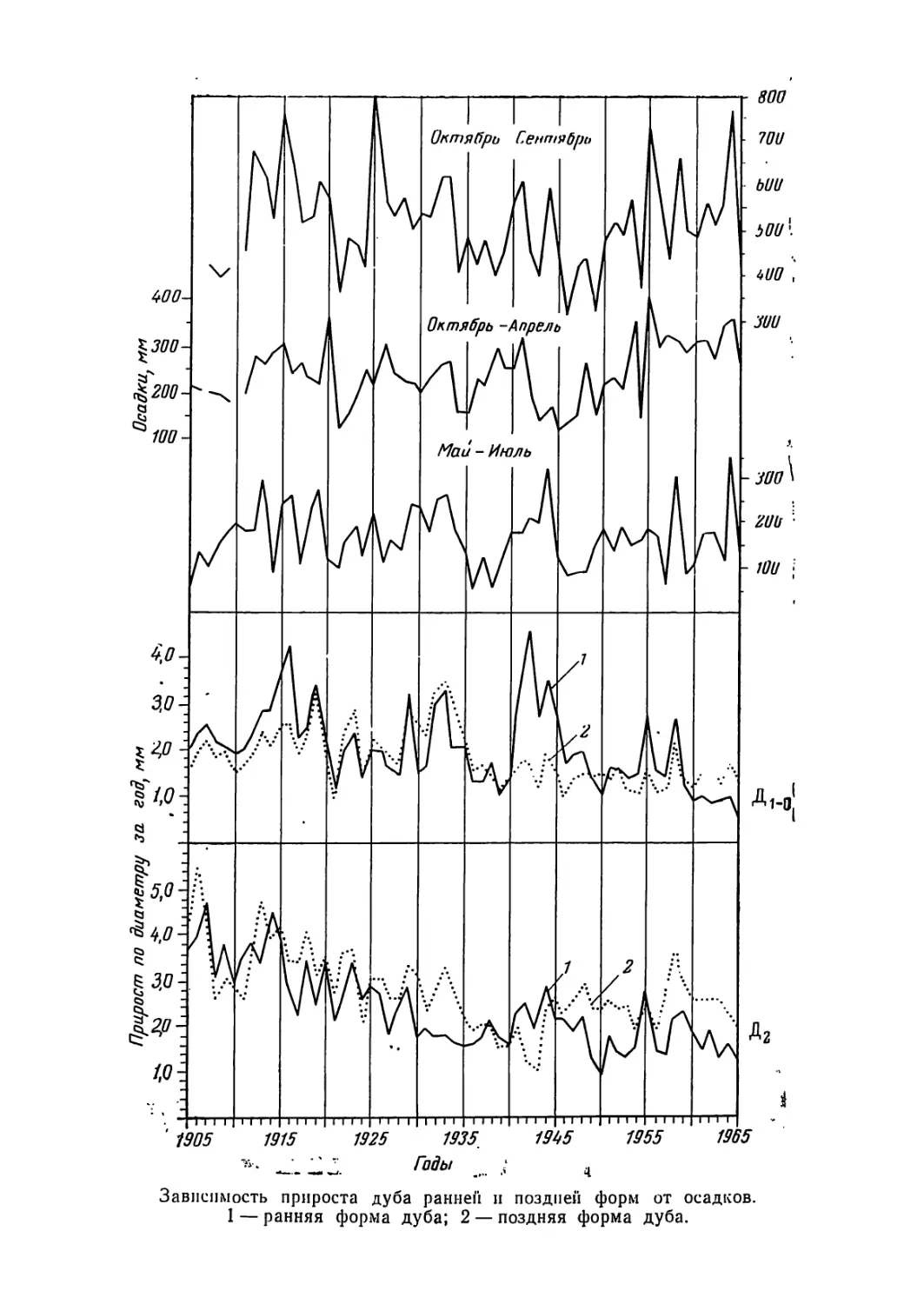



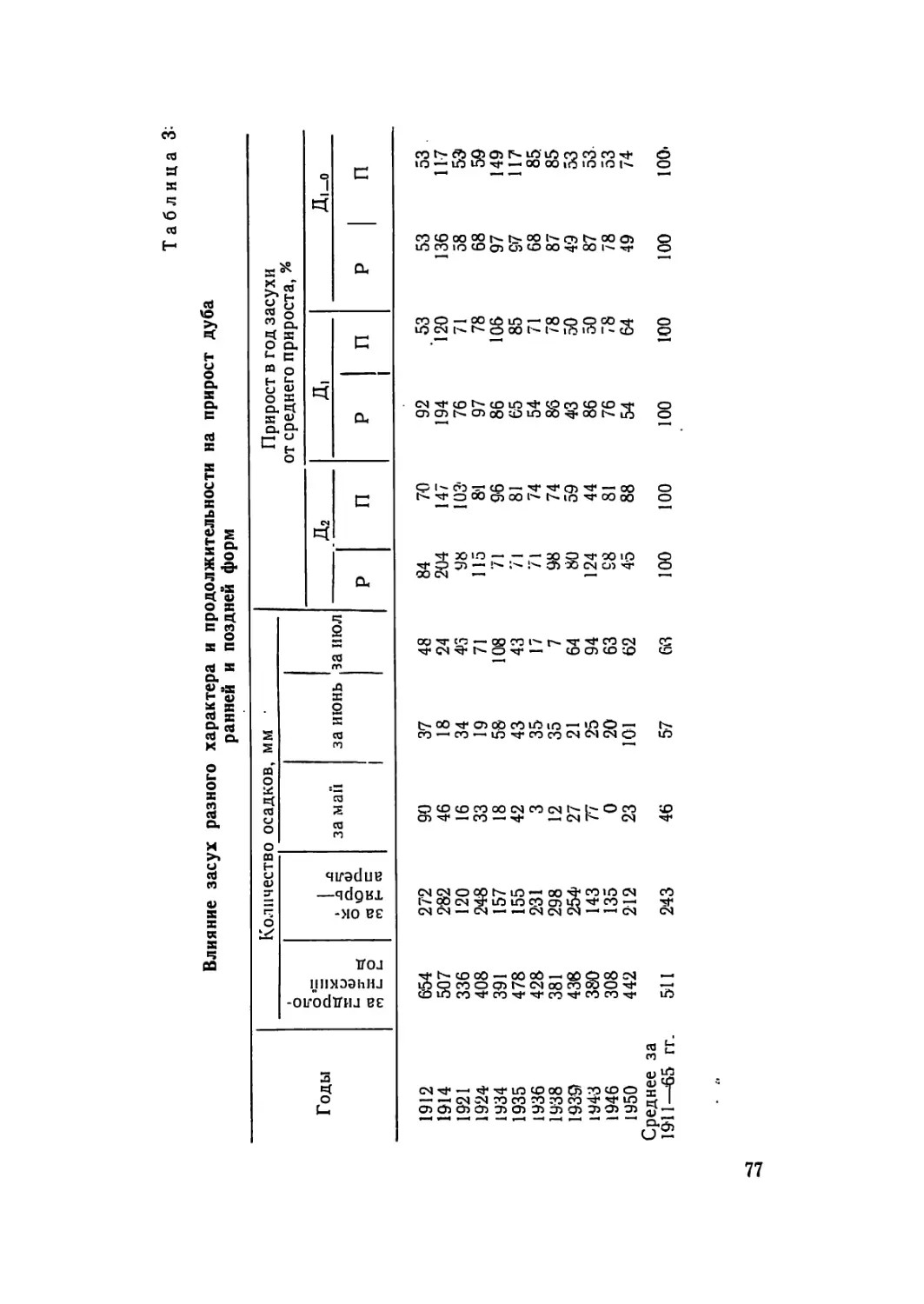



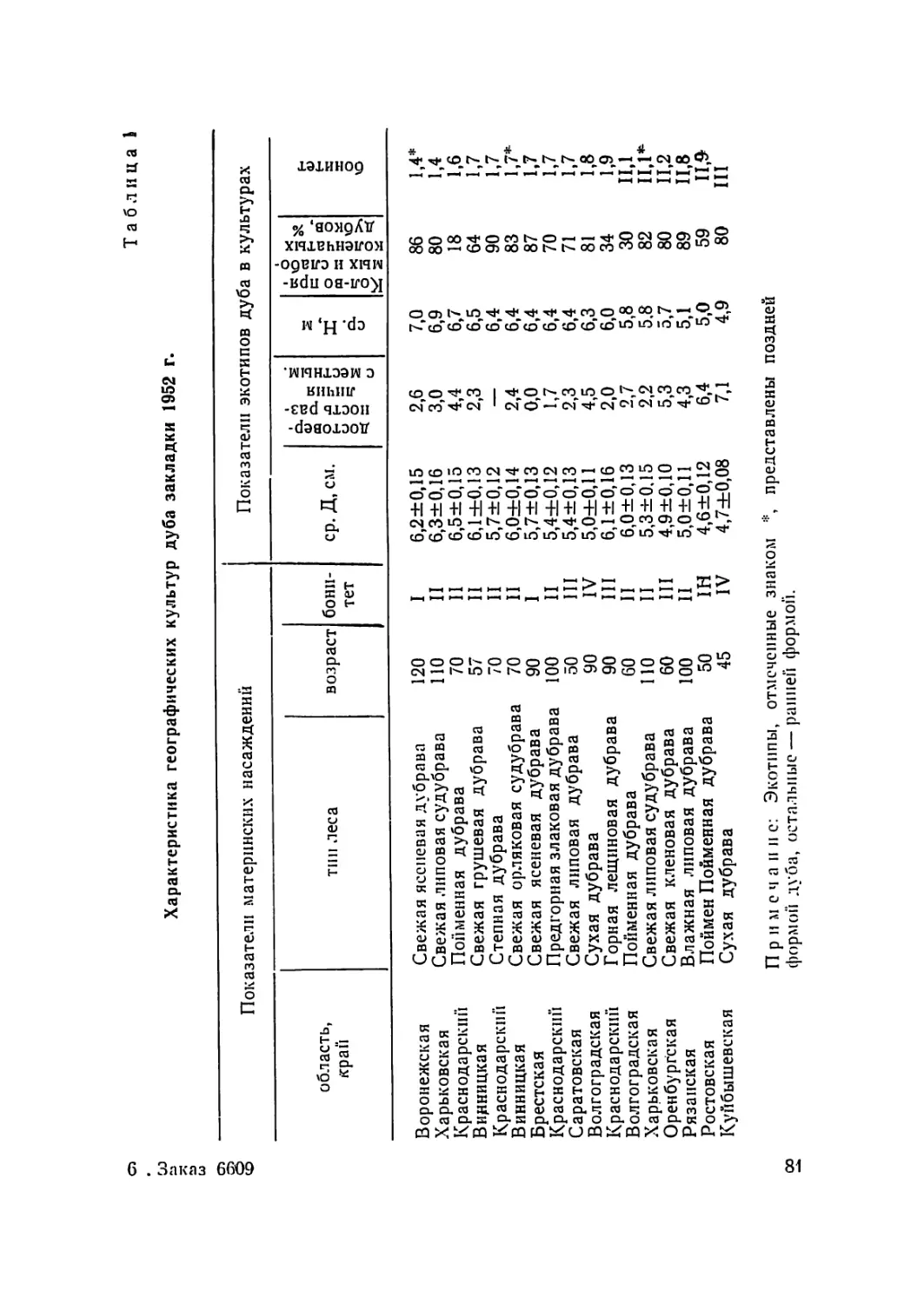









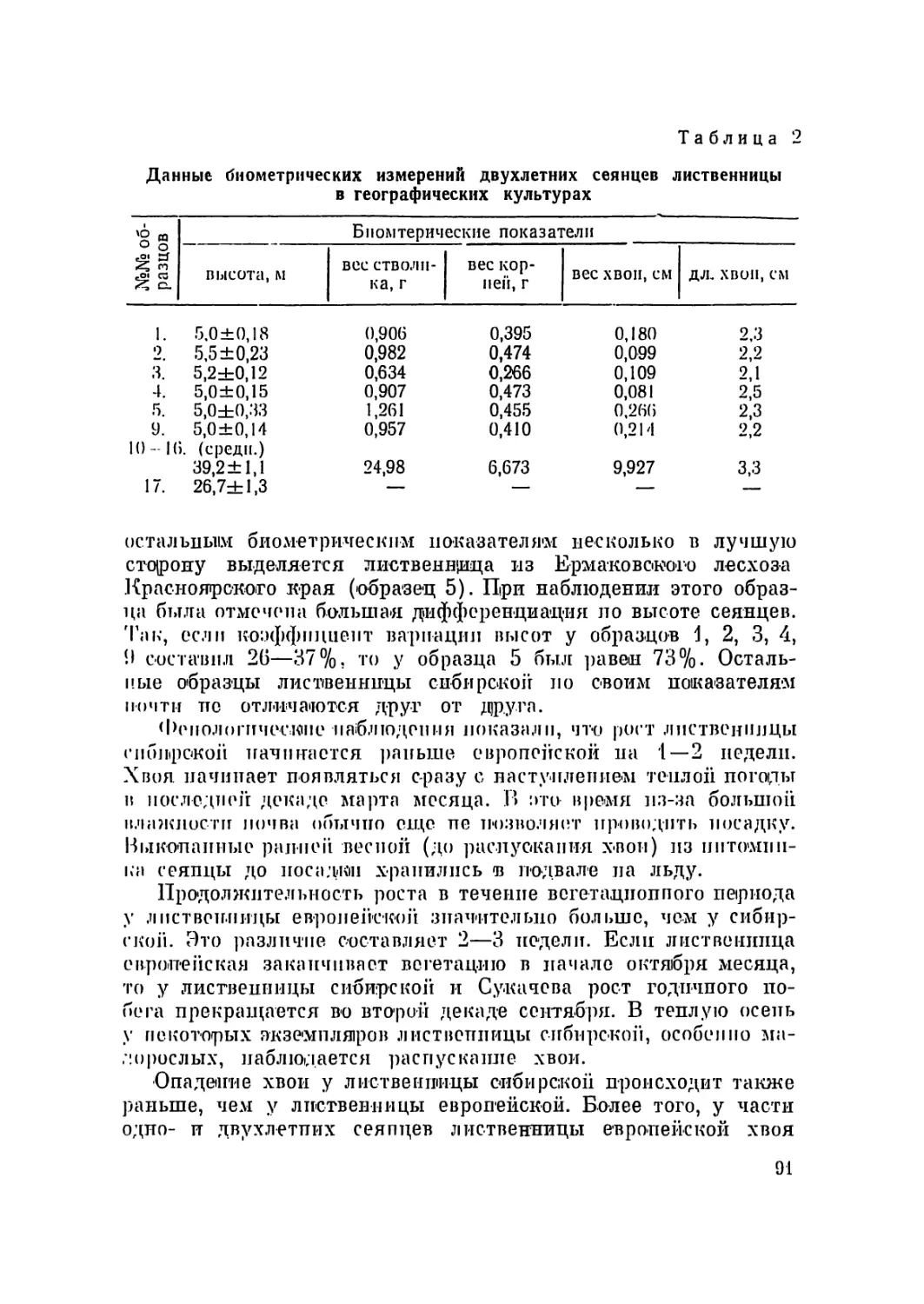














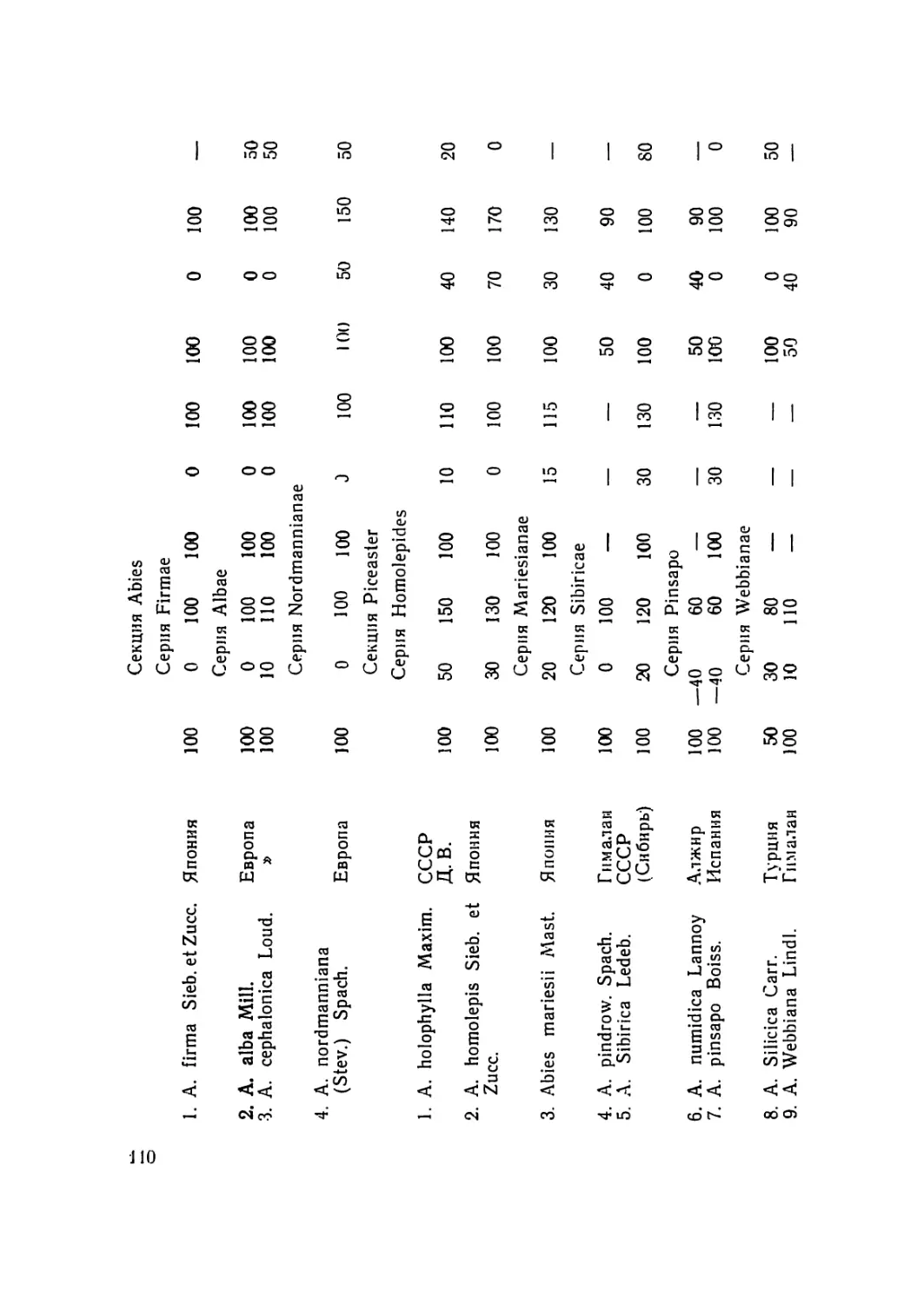



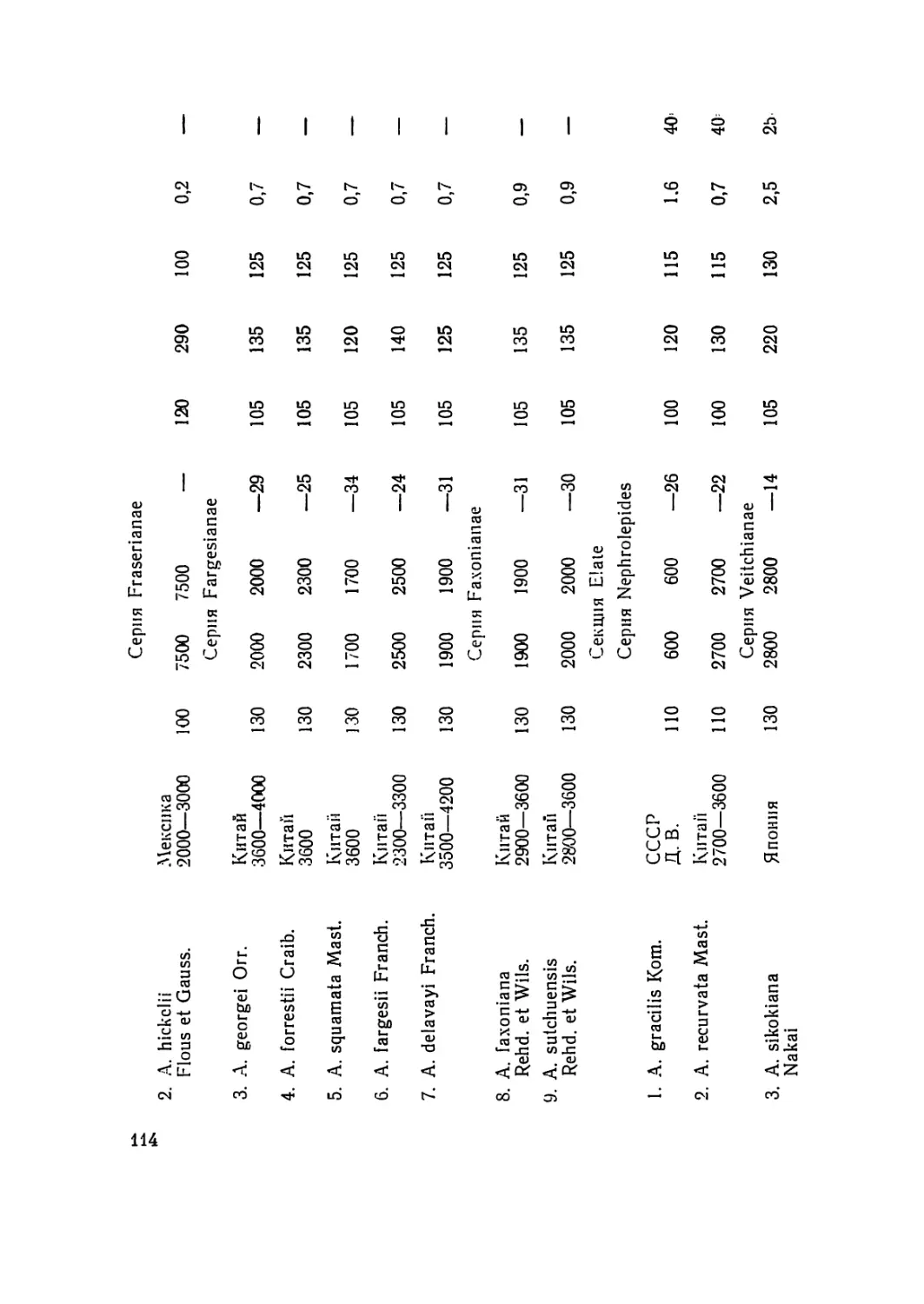
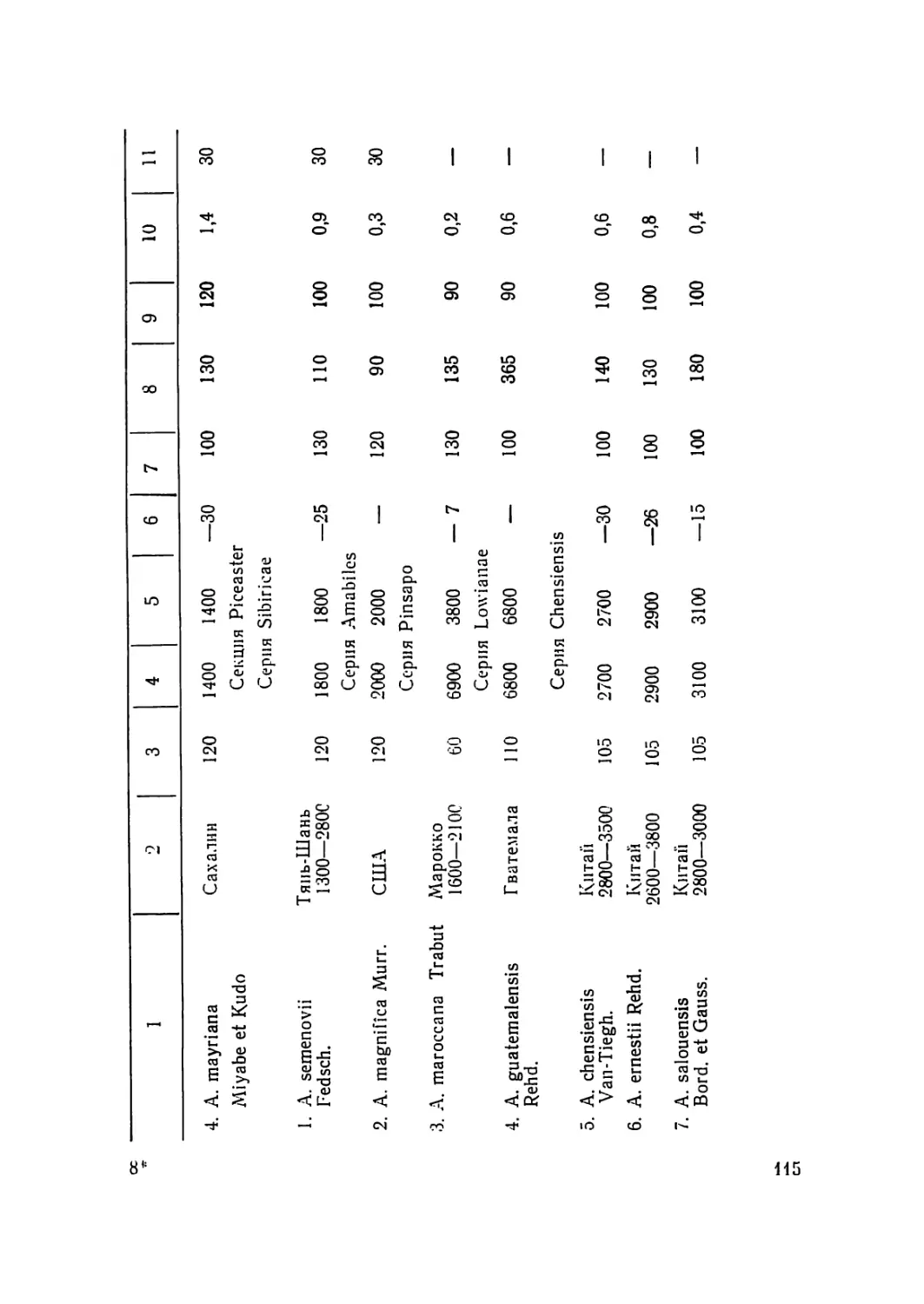








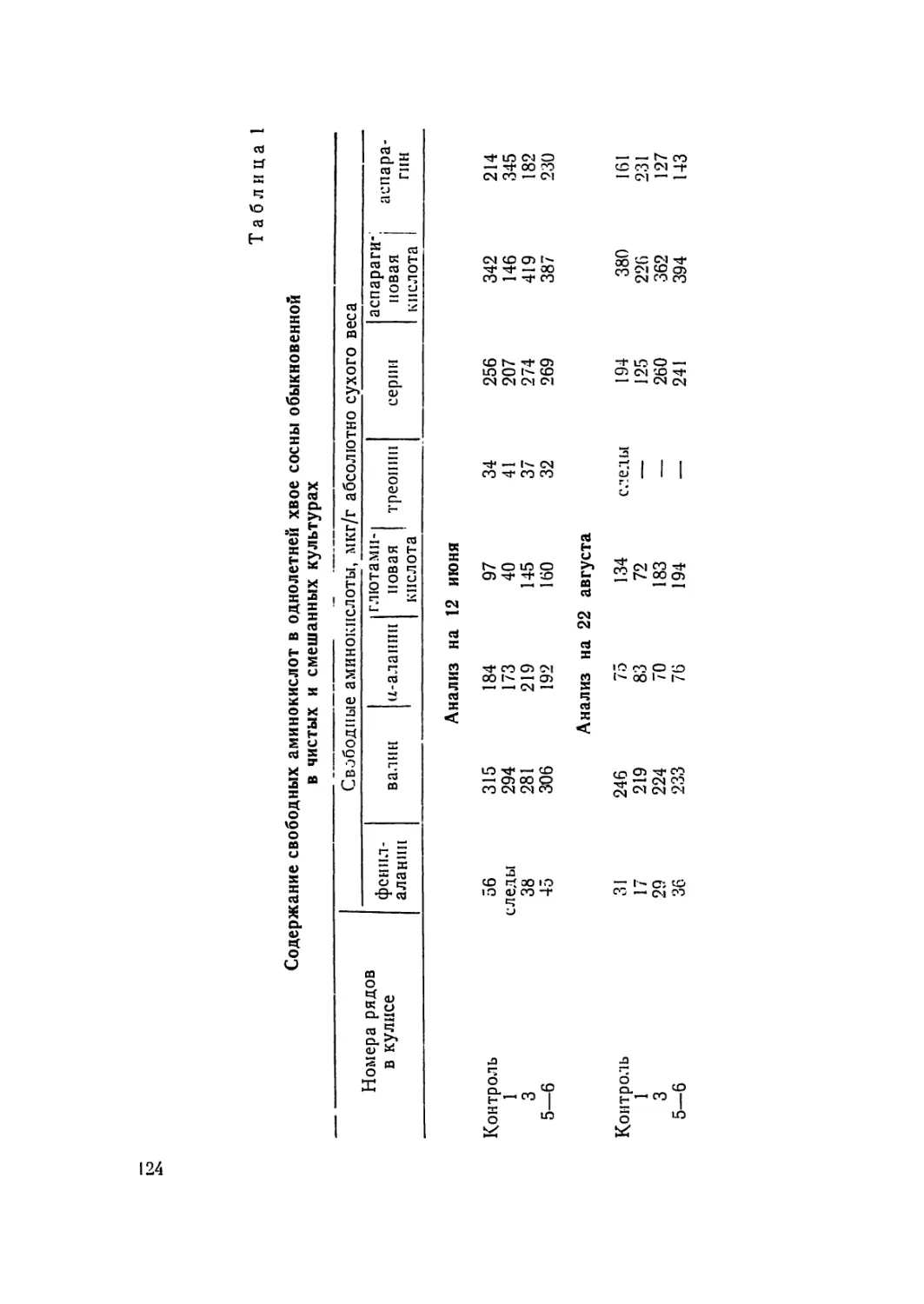




























































Текст
Государственный комитет лесного хозяйства
Совета Министров СССР
Всесоюзный научно-мсследовательоК)ИЙ институт
лесоводства и механизации лесного хозяйства
Генетика, селекция
и интродукция лесных пород
Тематический сборник научных трудов
Выпуск I
Воронеж, 1974 г.
УДК’ 634.030
В тематическом сборнике «Генетика, селекция и интродукция лесных
пород» представлены работы, выполненные научными сотрудниками
ЦИИИЛГиС и ведущими специалистами лесного хозяйства лесной зоны
страны.
Материал размещен в 5 разделах: селекция; семеноводство и лесные
культуры; экология, физиология и биохимия; анатомическое строение,
физико-механические и химические свойства древесины; защита ее
от вредителей и болезней.
Публикуемые научные статьи в сборнике имеют определенный за-
конченный характер и могут быть использованы производственниками
и научными работниками лесных и биологических институтов.
Библ, ссылок 228, илл. 25, табл. 50.
Печатается по решению Головного совета Всесоюзного научно-иссле-
довательского института лесоводства п механизации лесного хозяйства
(ВНИИЛМ).
Редколлегия:
Ефимов IO. II., Калуцкий К. К. (научи, ред.), Коспченко Н. Е.,
Пугач Е. А. (зам. науч, ред.), проф. Томчук Р. И., Харченко Н. А.,
Чусов И. В. (отв. секретарь).
К. К. КАЛУЦКИЙ
Основные направления научно-исследовательских работ
Центрального научно-исследовательского
института лесной генетики и селекции
Директивами XXIV съезда КПСС перед лесным хозяйством
страны поставлена задача повышения продуктивности и улуч-
шения качественного составе! лесов. Для успешного решения
этой задачи необходимо использовать прежде всего достиже-
нии лесной генетики и селекции. Мероприятия по реализации
этих достижений, с одной стороны, ио липин применения селек-
ции при выращивании леса (семеноводство, культуры, рубки
ухода и возобновления); с другой стороны, по линии улучше-
ния существующих и разработки повьгх методов лесной селек-
ции на основе современных достижений пауки о наследствен-
ности — генетики. Развитие этих исследований приобретает
все более важное значение для лесного хозяйства. В нашей
стране в атом направлении проводится значительная работа.
Многие научные, научно-производственные организации и вузы
страны участвуют в разработке отдельных проблем селекции
и генетики. Установлены основные принципы районирования
древесных растении, во многих частях СССР изучается формо-
вое разнообразие основных лесообразующих пород, получено
много перспективных гибридов, определены пути генетической
проверки селекционного материала.
Область лесного семеноводства характеризуется глубокими
исследованиями развития растений, а также методов создания
семенных плантаций для основных лесообразующих по-род.
Достижения в деле создания высокопродуктивных форм де-
3
ревьев и кустарников методом гибридизации, отбора плюсовых
деревьев и создания лесосемеииых плантации находят все более
широкое использование в .практике лесного хозяйства. Успешно
проходит внедрение результатов лесной селекции по отбору
плюсовых деревьев, селекционной оценке спелых насаждений
и созданию прививочных лесосемеииых плантаций в Латвий-
ской ССР (Институт лесохозяйственных проблем) и в Литов-
ской ССР (ЛитНИИЛХ). Комплексно решаются проблемы ге-
нетики, селекции и семеноводства в Карельской АССР (Инсти-
тут леса Карельского филиала АН СССР) и других. Организо-
вано сортоиспытание на Украине, в Белоруссии, Узбекистане
и в 22-х краях и областях РСФСР.
Теоретической основой селекции и семеноводства является
генетика. Положительным фактором является то, что все боль-
шее число научных учреждений СССР проводит исследования
по генетике и цитоэмбриологии древесных растений, по изу-
чению кариотипов основных лесообразующих пород.
Однако многие рекомендации еще не получили внедрения
в лесохозяйственную -практику. Научные исследования но лес-
ной генетике, селекции и семеноводству ведутся разрозненно,
малочисленными группами или же отдельными учеными. Во
многих учреждениях отсутствуют экспериментальные базы,
оснащенные необходимым оборудованием. Недостаточной явля-
ется подготовка лесных специалистов в этой области.
Для расширения и углубления работ по лесной генетике,
селекции, семеноводству п интродукции по постановлению Го-
сударственного комитета Совета Министров СССР по пауке и
технике от 24 августа 1970 года в системе Гослесхоза СССР
организовано Всесоюзное иаучио--нроизводственпое объедине-
ние лесной селекции древесных пород с Центральным научно-
исследовательским институтом леспой генетики и селекции
и с сетью лесных питомников по выращиванию сортового се-
лекционного материала для создания лесных культур и защит-
ных лесных насаждений. Установлены основные направления
научной деятельности института по лесной генетике, селекции,
семеноводству и интродукции древесных пород.
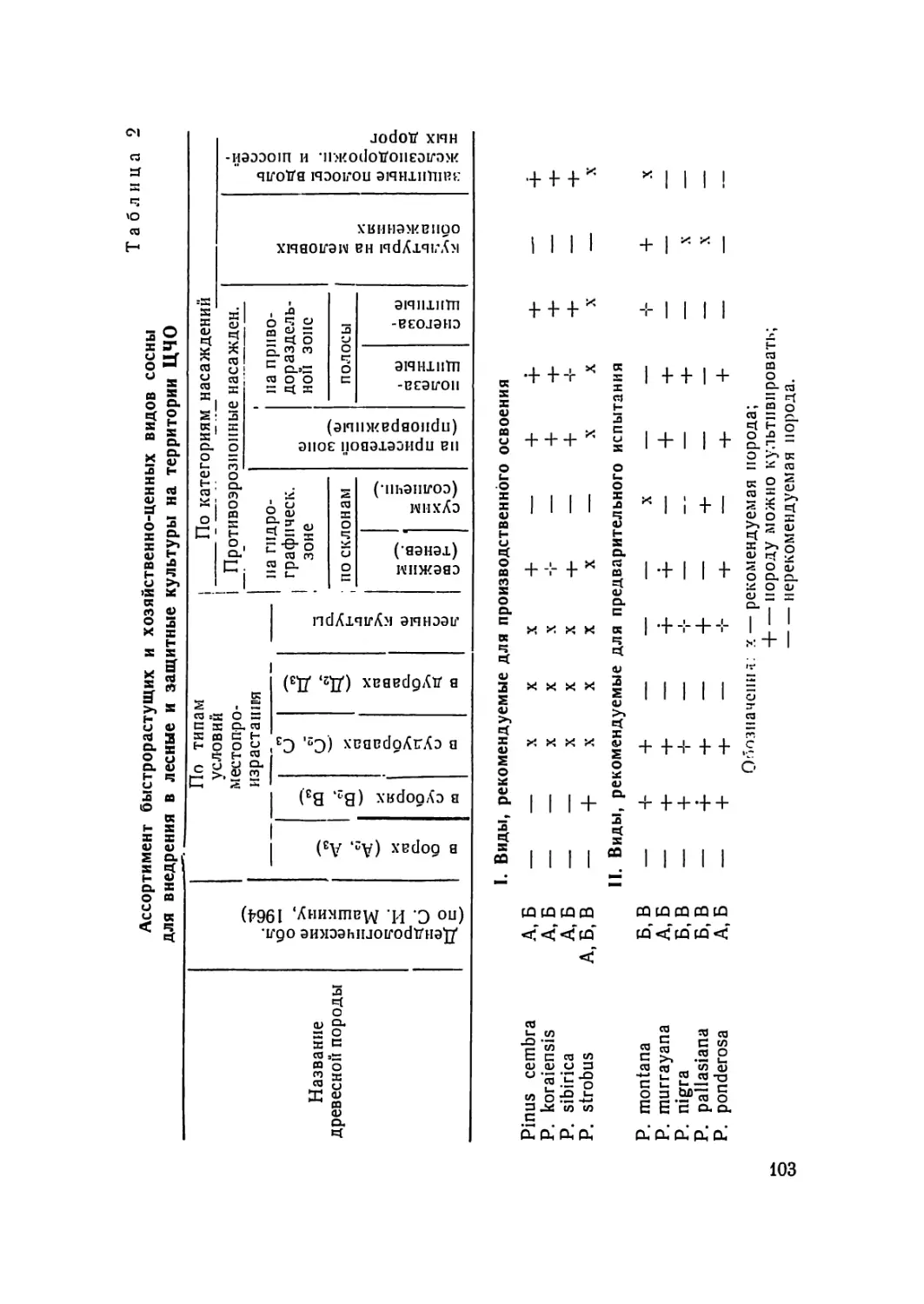
В области лесной селекции — теории и практики улуч-
шения лесных пород и выведения их форм и сортов, наиболее
полно отвечающим требованиям народного хозяйства, ведущи-
ми признаны следующие вопросы:
— изучение внутривидовой изменчивости лесных пород,
установление и изучение типичных форм, формового состава
природных популяций и его динамики в связи с возрастом
4
древостоя и изменением условии среды, выявление наиоолес
ценных в хозяйственном отношении форм с целью их исполь-
зования при семеноводстве, лесовыращиваиии и рубках ухода;
— разработка вопросов отбора, учета и сохранения гене-
тических фондов основных древесных пород, необходимых для
их селекции, семеноводстве! и вегетативного размножения. Сюда
относится разработка для разных пород и различных зон стра-
ны научно обоснованных технических указании по селекцион-
ной оценке и отбору плюсовых деревьев и насаждений (семен-
ных заказников), методики и правил их инвентаризации; со-
ставление региональных реестров-карточек лесного селекцион-
ного фонда; разработка вопросов о хозяйственном режиме, фор-
мах использования этого фонда и создание региональных «архи-
вов клопов», коллекций арборетумов, путем прививок и другими
способами вегетативного размножения лучших форм, для сохра-
нения этого невозместимого, в случае его утраты, фонда и более
удобного его использования при исследовании и размпоженип;
— разработка методов элитной апробации — ускоренной
оценки наследственных свойств плюсовых деревьев и насажде-
ний с целью повышения эффективности селекционного отбора
(аналитической селекции) на основе корреляции признаков,
ранней диагностики, контрольных прививок и культур в разных
экологических условиях, комплексного использования такса-
ционных, морфологических, анатомических, цитологических, фи-
зиологических, биохимических и биофизических показателей,
математического анализа и прогнозов;
— разработка новых эффективных методов синтетической
селекции на основе генетических исследований по эксперимен-
тальной полиплоидии и получению мутаций, а также широкого
применения гибридизации (разноэкологических скрещиваний);
выведение с их помощью быстрорастущих, технически ценных,
высокосмолопродуктпвиых, плодоппщевых и других форм лес-
ных пород, устойчивых к неблагоприятным условиям среды,
вредителям и болезням;
— разработка методов селекции новых ценных форм хвой-
ных и лиственных пород и организация их сортоиспытания в
культурах при вегетативном и семенном разведении, закладка
серии опытов в разных экологических условиях и различных
зонах страны в целях скорейшего выявления и внедрения в
производство лучших из этих форм, сортов и гибридов.
К задачам селекции и сортоиспытания непосредственно при-
мыкают проблемы интродукции и акклиматизации лесных
пород.
5
Выдающийся советский генетик и селекционер академик
И. И. Вавилов в своих работах обращал внимание ботаников и
лесоводов па практически безграничные возможности использо-
вания многих древесных экзотов. Оп указывал, что бедность
дендрологического состава Восточной Европы (в том числе тер-
ритории СССР) по сравнению с лесами Северной Америки, Ки-
тая, Дальнего ГЗостока обязывает лесоводов и дендрологов обра-
тить пристальное внимание па привлечение иноземных пород,
так как многие «из экзотов могут иметь серьезное лесоводст-
веяиое и хозяйственное значение» (И. И. Вавилов. Соч., т. V,
М.-Л, 1965 г.).
Однако, несмотря на то, что производственной практикой
как в нашей стране, так и за рубежом доказана огромная эко-
номическая эффективность интродукционных мероприятий, до
сих пор в общем объеме лесовосстановительных работ разведе-
тпте быстрорастущих и хозяйственно-ценных видов деревьев и
кустарников занимает пока еще недостаточное место.
Например, из 600 видов рода Quercus на обширных прост-
ранствах Европейской части СССР культивируются только
J0—15 видов, а всего в Советском Союзе интродуцировано
лишь немногим более 40 видов.
Из 120 видов рода Pinus в лесном хозяйстве и защитном
лесоразведении пашли более пли менее широкое применение
только 10—42 видов.
Исключительно медленно внедряются в производственные
н опытные культуры и многие другие весьма ценные перепек
тпвиыо виды хвойных и лиственных деревьев. Поэтому основ-
ным направлением работ ЦНИИЛГнС по интродукции па бли-
жайшие годы намечены:1
— Разработка методов интродукции-н проведение экспери-
ментальных исследований древесных пород для повышения
продуктивности лесов.
— Разработка ассортимента пород для дендрариев — своего
рода маточников для дальнейшего их размножения. Закладка
таких дендрариев также необходима для расширения базы
экспериментальных работ по интродукции и акклиматизации в
различных зонах страны.
В области лесного семеноводства актуальными п требую-
щими научной разработки являются следующие вопросы:
— Разработка лесосеменного районирования на основе
исследований имеющихся и закладки географических и эколо-
гических опытных культур; выявление ценных (сортовых) эко-
6
типов и возможности повышения продуктивности лесонасаж-
дении путем их широкого использования.
— Разработка наиболее эффективных методов зональной
технологии закладки и формирования постоянной лесоссмеипой
базы на селекционной основе: прививочных плантаций, план-
таций из отборного материала семенного происхождения, се-
менных участков, а также системы мероприятий по их рацио-
нальной эксплуатации.
— Разработка методов и схем массового производства гиб-
ридных семян первого поколения от скрещивания разных эко-
типов, межвидовой гибридизации, инцухт-гетерозисных скре-
щиваний в целях создания устойчивых и высокопродуктивных
лесонасаждений.
— Исследование биологии плодоношения лесных пород и
разработка па этой основе методов ускорения плодоношения,
повышения урожайности и качества семян, сокращения межсе-
мепмых периодов — путем применения стимуляторов, удобре-
ний, агротехнических, лесозащитных и других мероприятий.
•— Разработка наиболее эффективных методов использова-
ния сортовых семян, в том числе способов долговременного их
хранения, предпосевной подготовки и других приемов; наибо-
лее рациональных методов семенного, вегетативного размноже-
ния и выращивания сортового посадочного материала в питом-
никах.
Обязательным для селекционеров и семеноводов надо счи-
тать участие в разработке и усовершенствовании специализи-
рованными институтами создания и усовершенствования машин
и приспособлений для сбора лесных семян с растущих деревьев
в насаждениях, на лесосеменных участках и плантациях, а так-
же технологии заготовки семян и переработки лесосеменного
сы;рья.
Для успешного решения изложенных выше проблем необхо-
дима разработка генетических основ лесной селекции — ее тео-
ретического фундамента. В качестве основных проблемных во-
просов лесной генетики, цитологии и эмбриологии, направлен-
ных на повышение эффективности селекционных работ и лес-
ного семеноводства в СССР, необходимо назвать следующие:
— Наркологическое п цитогенетическое изучение лесных по-
род в связи с проблемой повышения эффективности их селек-
ции: определение чисел хромосом у видов местных и интро-
дуцированных лесных пород; выявление спонтанных полиплои-
дов и лесохозяйственных особенностей их; исследование морфо-
логии хромосом и составление идиограмм основных кариотипов
7
древесных пород, знание которых необходимо для работ по
экспериментальному мутагенезу. • »i
— Разработка методов экспериментального получения rio.ni/;
плоидов в связи с использованием их в гибридизации па гете-
розис с целью «преодоления времени в лесоводстве»; получе-
ние нередуцированной (диплоидной) пыльцы путем колхпципп-
рования с последующим использованием ее в гибридизации с
диплоидным видом для синтеза авто- или аллотротриплоидов,
обладающих гетерозисом, создание коллекций полиплоидов лист-
венных и хвойных пород, их испытание, изучение по морфо-
логическим, эколого-физиологическим, анатомическим и лесохо-
зяйственным показателям с целью выявления ценных форм для
практической селекции и использования в лесном хозяйстве.
— Цитоэмбрнологическое изучение видов, разновидностей,
гибридов и форм главнейших лесных пород в связи с их селек-
цией, задачами повышения урожайности семян и сокращения
периодичности их плодоношения. Изучение закономерностей
сексуализации (половой дифференцировки) побегов и почек,
динамики заложения и развития генетических органов в зави-
симости от климатических факторов л погодных условий:
изучение жизнеспособности пыльцы и эффективности разных
форм опыления, выяснение причин отпада завязей и шишек,
пустосемянности и невсхожести семян; разработка практиче-
ских рекомендаций для повышения урожайности семян и со-
кращения периодичности плодоношения.
— Исследование внутривидового разнообразия н геиетиче-
ской структуры популяций основных лесных пород; изучение
генетики оплодотворения, наследования признаков у гибридных
сеянцев; разработка генетических методов ранней диагностики
наследственных свойств плюсовых деревьев; количественная
оценка относительной роли генотипа и среды в формировании
признаков и свойств; изучение генетической структуры попу-
ляций и разработка некоторых рекомендаций для аналитиче-
ской селекции и лесного элитного семеноводства.
В обосновании научных выводов и рекомендаций наряду
с отделами лесной генетики, селекции, семеноводства и интро-
дукции, существенное участие принимают также следующие
отделы и лаборатории: лесозащиты и иммунитета; экологии,
физиологии и биохимии растений; лесного почвоведения; фи-
зико-механических, химических и анатомических свойств дре-
весины; биокибернетики; экономики и научно-технической ин-
формации.
Целенаправленная координированная деятельность всех от-
8
дрлов и лаоораторип по разраоотке проолемных вопросов лес-
юй генетики и селекции обеспечит комплексную проработку
роблем на достаточно высоком научном уровне.
За истекшее время институтом начаты работы по всем ос-
новным научным направлениям. В отдельных случаях они
пока носят поисковый характер, поскольку серьезных исследо-
вании не проводилось ни в пашен стране, ни за рубежом. По
другим идет накопление фактического материала с целью по-
следующего обобщения и передачи производству для внедре-
ния в лесокультурную и лесоводствеипую практику.
Установлены деловые связи со многими научными учреж-
дениями страны, с которыми проводятся совместные научные
исследования (институты: химической физики АН СССР, фи-
зиологии растений АН СССР, физиологии растений АН Украин-
ской ССР им. Докучаева, Воронежский лесотехнический инсти-
тут и Воронежский госуниверситет).
Научно-исследовательская и производственная деятельность
объединения будет строиться в тесном контакте с широкой
сетью селекционных лабораторий, имеющих эксперимеитальио-
производствениые питомники, а также лесохозяйственными
предприятиями.
В настоящее время объединению переданы: Астраханская
лесная опытная станция, Шптювский опорный пункт и Семи-
лукский лесной опытпо-показательпый селекционный питом-
ник*. '
Гослесхозо-м СССР утвержден проект развития широкой
сети лабораторий и питомников в различных лесорастительных
зонах страны. В основу принципа размещения зональной сети
положено представительство основных лесорастительных зон
с учетом охвата исследованиями главных древесных пород
этих 'ЗОН.
В связи с этим встает необходимость разработки организа-
ционной и экономической основы и формы взаимоотношений
селекционных питомников с лесохозяйственными предприятия-
ми по производству п использованию улучшенного семенного
и посадочного материала, по изучению постоянной лесосемен-
пой базы па селекционной основе.
В дальнейшем появится необходимость в создании при объ-
единении контрольно-консультативной группы по лесному сс-
* Вновь организованы Белореченский научно-производственный се-
лекционный лесхоз с орехоплодовой селекционной лабораторией, Ново-
сибирская селекционная лаборатория и Брянская селекционная лабо-
ратория с лесным питомником.
9
мсповодству с возложением на нее обязанностей инспектиро-
вания деятельности селекционных питомников и лесхозов.
Широкое развертывание работ по лесной генетике и селек-
ции, связанное с образованием объединения и института, вы-
двигает в число первоочередных задач плановую подготовку
специалистов по этой отрасли в техникумах и вузах страны.
При институте планируется организация аспирантуры и посто-
янно действующих курсов повышения квалификации лесных се-
лекционеров-семеноводов.
Завершение комплекса намеченных организационно-техни-
ческих мероприятий по созданию паучнонпроиЛводствеииого
объединения создаст реальные предпосылки к переводу всего
лесосемеппого и лесокультурного дела в стране па генетико-
селекционную основу.
С. П. МАШКИН, А. К. БУТОРИНА,
М. М. БЕЛОЗЕРОВА, И. М. ПОЖИДАЕВА,
Л. С. МУРАЯ, Л. И. ХАТУНЦЕВА,
10. А. ДОРОНИН
Сравнительное изучение кариотипов сосны меловой
(Р. crctacea Kalen.) и сосны обыкновенной (Р. Silvestris L.)
в популяциях Центрального Черноземья
Введение. Для успешной селекционной работы необходи-
мо глубокое и всестороннее изучение исходного материала, фи-
логенетической связи видов и родов (И. И. Вавилов, 1960,
1965, 1965 а). В этом плане особого язлимания заслуживает изу-
чение материальных основ наследственности организмов — их
хромосомного аппарата.
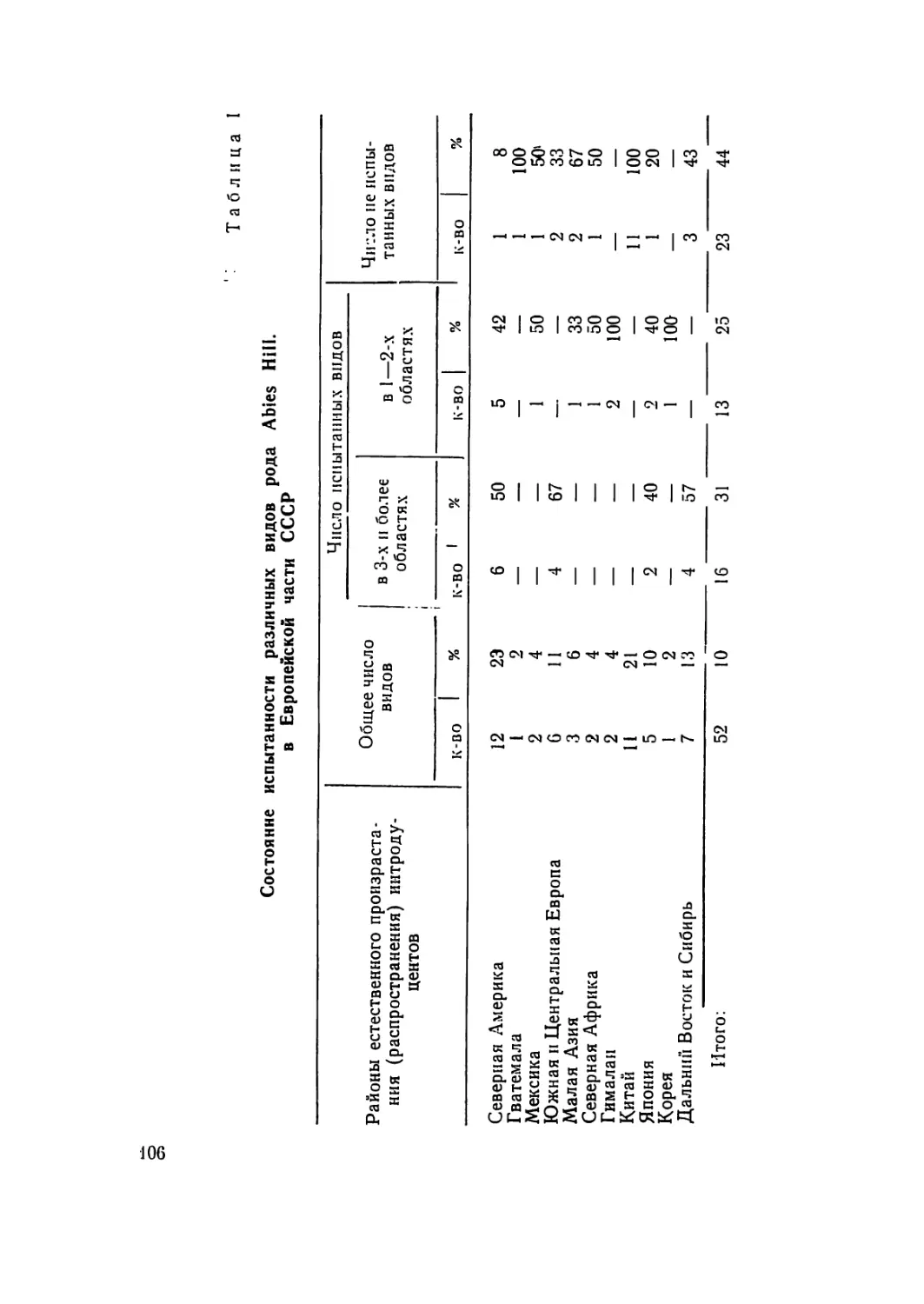
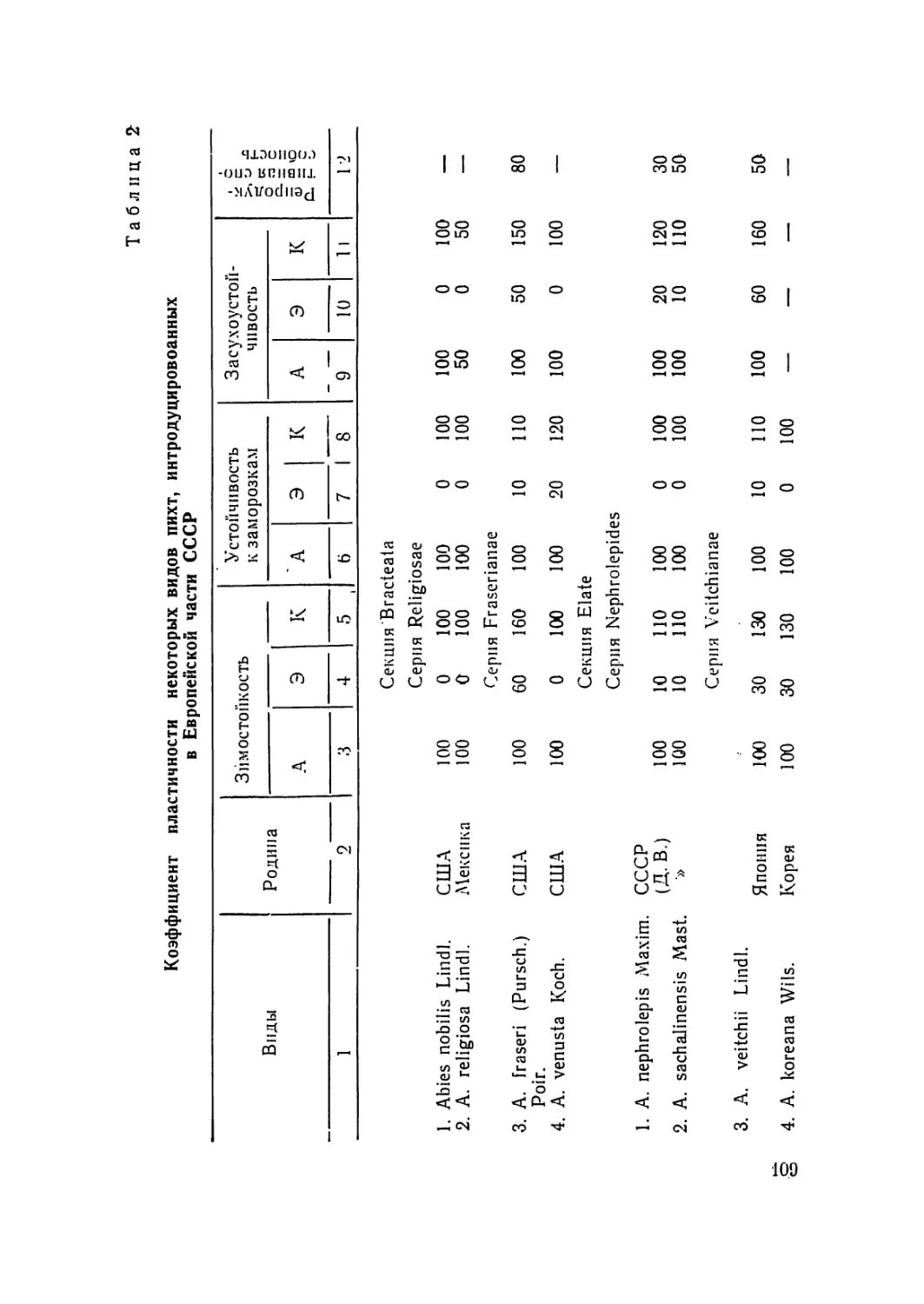
В наркологическом отношении род Pin us L. изучен далеко
недостаточно. В течение последних четырех десятилетий раз-
ными авторами установлены кариотипы 24 видов сосны. Всего
же род Pinus содержит около 100 видов, которые произрастают
в умеренных странах северного полушария, в более южных
шпротах — в горах и немногие — в тропическом поясе север-
ного полушария. Из них 75 впдов произрастают в СССР, боль-
шая часть которых падает на долю иитродуцентов (С. Я. Соко-
лов, 1949).
Абсолютное большинство видов сосны не изучено даже на
10
числа хромосом. Это относится и к тем видам сосыы, которые
дико произрастают в лесах СССР, что ограничивает их исполь-
зование в селекционных работах. 13 частности, Pinus crelacea К.
(сосна меловая) не изучена па числа хромосом ни в одной из
своих популяций; Pinus silvcsiris L. пе изучена всесторонне в
наркологическом отношении даже в основных популяциях свое-
го произрастания. Опа изучена с составлением иднограмм в
ряде популяций в Норвегии (Aass, 1957), в Швеции (Nata-
rajan, Ohba, Simak, 1961), в США (Saylor, 1964) и в СССР,
в Тувинской АССР (Л. Ф. Правдин, 1964).
Как отмечает Л. Ф. Правдин (1970), работы по изучению
кариотипов лесных пород на популяционном уровне только на-
мечаются. Начало наркологическим исследованиям рода Pinus
было положено К. Саксом и X. Саксом (К. Sax and И. Sax,
1933). Как К. Сакс и X. Сакс, так и другие исследователи:
Мехра и Хошоо (Mehra and Khoshoo, 1956), Аасс (Aass,
1967), Натараяи, Охба и Симак (Natarajan, Ohba, Simak,
1961), Сейлор (Saylor, 1961, 1964), ТОм (Yim, 1963),
Л. Ф. Правдин (1964), Кумар и др. (Kumar et al., 1966),
Шншииаш'вкли Р. М., 1968; Педерик (Pederic, 1967, 1970),
Хетти, Ападхауа и Кедхариат (Cheetly, Upadhaya, Kedharnath,
1970), Ападхауа п Кедхариат (Upadhaya, Kedharnath,
1970), LUaiMc ур Рехман и Пнрвлн (Shams-ur-Rehman and
Perveen, 1970), отмечали значительное однообразие кариоти-
пов всех видов рода Pinus, имеющих, ikuiK правило, 24 хромосо-
мы в соматических клетках (2п-^24). Полиплоидные формы
встречаются л пин» в единичных случаях (М. В. Комиссарова,
1947; Kanezawa, 1949; Zinnai, 1952; Morgen, 1958; Khoshoo,
1959). Полиплоидия у xiBioiiиых обычно сопровождается рез-
ким снижением жизненности, угнетенным состоянием дерева,
его отсталым ростом, причем, .как у естествен пых форм, так’
п у полученных зксперимеиталыю. Так*, в Швеции с -помощью
колхицина была получена тетрашюпдная форма сосны обыкно-
венной (2п=48). Возраст ее 25 лет, рост замедленный, дерево
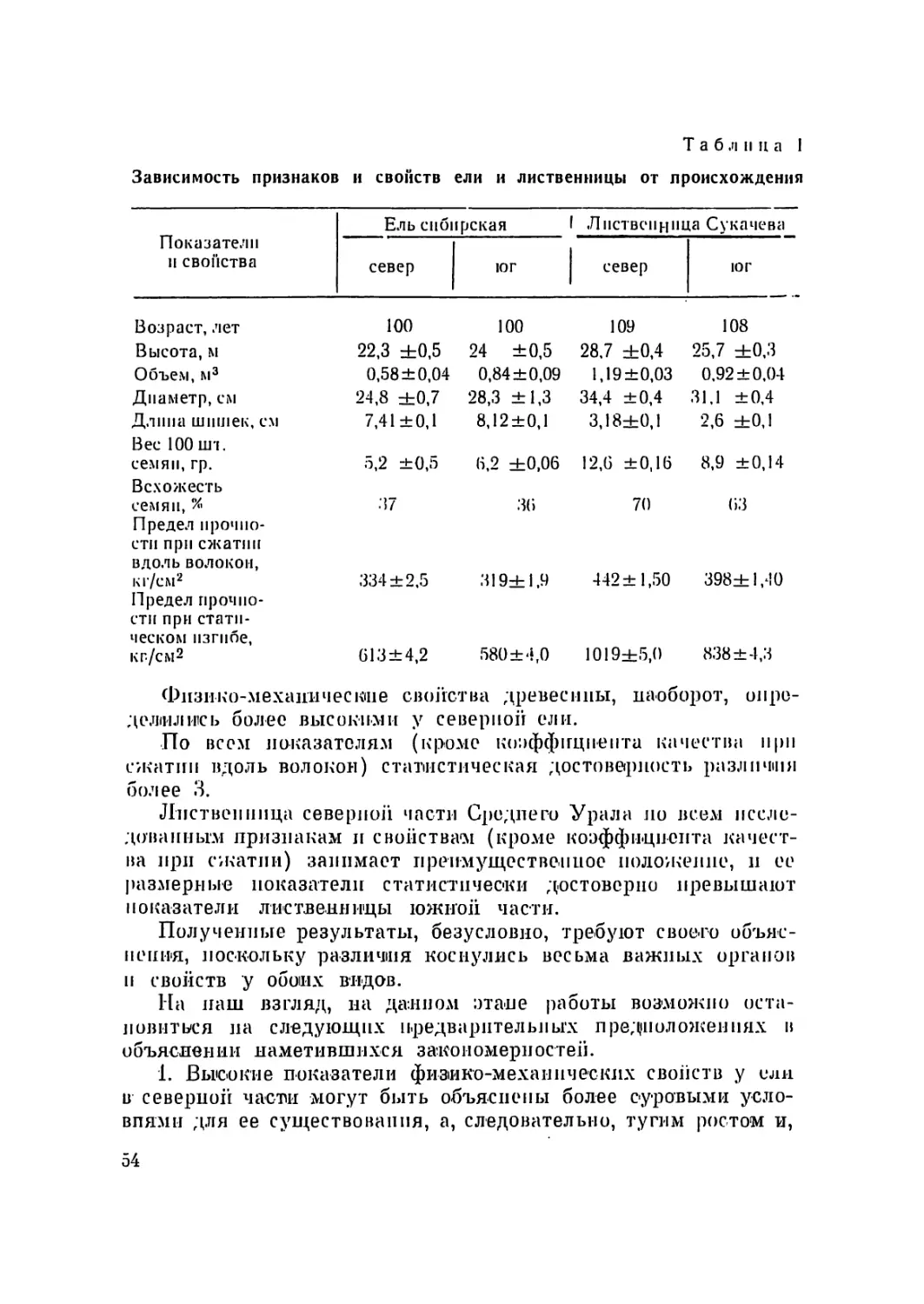
несколько искривлено (И. А. Картель, Е. Д. Мапцовпч, 1970).
Большинство авторов, изучавших кариотип различных видов
рода Pinus, указывали, что из 12 хромосом их основного набора
одна до некоторой стеисин гетеробрахпальиа, а другие имеют
приблизительно медианное положение центромеры. При атом
наблюдается также и значительное генетическое сходство хро-
мосом различных видов сосны, о чем свидетельствует, по дан-
ным Сакса (Sax, 1960) легкая скрещиваемость разных видов
ее между собой- и сравнительно нормальное поведение хромо-
11
сом б мепозе у межвидовых гиюридов первого поколения.
Но в ряде случаев между кариотипами отдельных видов сосны
наблюдаются незначительные различия по относительной длине
хромосом (которые, однако, можно приписать и к ошибкам из-
мерении), а также по положению центромеры в предпоследней
паре хромосом (Saylor, 1964; Upadhaya and Kedharnath, 1970).
Основное же различие между кариотипами разных видов
сосны проявляется по наличию и локализации вторичных пе-
ретяжек, хотя этот признак является достаточно сильно варь-
ирующим. Так, Аасс, Натараяи, Охба п Симак, Сейлор (Aass,
1957; Natarajan, Ohba, Simak, 1961; Saylor, 1964), изучав-
шие кариотип сосиьт обыкновенной, произрастающей в Норве-
гии, Швеции н США, установили у нее различное число вто-
ричных перетяжек: Аасс — 1 (в первой паре хромосом), На-
тараян, Охба и Симак — 5 (в 1, II, VI, VII и X нарах хромо-
сом), Сейлор — 3 (в I, II и X нарах хромосом). Л. Ф. Правдин
(1964) у сосны обыкновенной из Тувинской АССР обнаружил
также 3 вторичных перетяжки (в первой, второй и шестой па-
рах хромосом). Поэтому нельзя не согласиться с замечанием
Сейлора о том, что ценность вторичных перетяжек, как диагно-
стической черты кариотипов сосны, является сомнительной.
II тем не менее, па том уровне, па котором находятся в настоя-
щее время исследования кариотипов у хвойных, данный диагно-
стический признак продолжает оставаться главным.
Несмотря на то, что хромосомы сосны очень крупные (по-
рядка до 30 мк), исследование кариотипа сопряжено с рядом
трудностей, касающихся, главным образом, идентификации хро-
мосом, ввиду их значительной гомеоморфности, а также изуче-
ния их топкой структуры.
Возможно, что применение более топких методов исследова-
ния, а именно метода пахптепного анализа и метода построения
поликарпограмм, позволит выявить в кариотипах различных ви-
дов с оспы более определенные и постоянные различия, без чего
невозможно проводить глубокие цитогенетические исследова-
ния.
Особого внимания заслуживают работы, в которых прово-
дятся кариологические исследования гаплоидных наборов хро-
мосом (при изучении гаплоидных митозов в мужских и жен-
ских гаметофитах). Так, К. Сакс и X. Сакс (К. Sax and Н. Sax,
1933), Сантомоур (Santomour, 1960), Саркар (Sarcar, 1963),
Педерик (Pederic, 1967, 1970), Симан (Simak, 1969) изучали
хромосомы в тканях женского гаметофита, Аасс (Aass, 1957) —
в мужском гаметофите.
12
Точность таких исследовании повышается на 50% в резуль-
тате исключения возможных ошибок при подборе пар гомологов.
Для установления глубоких различий по морфологии хро-
мосом между различными видами сосны необходимо изучение
тонкой структуры хромосом, что достигается с помощью при-
менения метода пахитешгого анализа. На стадии пахииемы в
мейозе за счет значительной деопирализации хромосом можно
изучать такие морфологические признаки каждой пары конъ-
югирующих хромосом, которые недоступны для исследования
при обычном рассмотрении метафазных хромосом в митозе, на-
пример число и величину узелков, которые состоят из гетерохро-
матина и наследуются как мутантные гены, характерные осо-
бенности хромомер и степень гетеропикноза хромомер около
центромер, наличие хромосомных перестроек. В связи со слож-
ностью, метод пахитеииого анализа для изучения тонкой струк-
туры хромосом использовался пока на незначительном количе-
стве объектов (рис, кукуруза, картофель, малина, черешня).
Что касается лесных пород в этом плане следует отметить ра-
боту Педерпка (Pedcric, 1967), .изучавшего с помощью пахитеп-
ного анализа тонкую структуру хромосом у Pinus radiata
D. Don. Для получения наиболее полных сведений о строении
кариотипа у исследуемых нами объектов желательно использо-
вание всех вышеупомянутых методов. Настоящая работа явля-
ется первым этапом таких комплексных исследовании.
На этом этапе исследований были поставлены следующие
задачи:
I. Провести сравнительное карпологическое изучение сос-
ны меловой из разных популяций н сосны обыкновенной во-
ронежской популяции в целях установления возможных раз-
личий между ними на хромосомном уровне.
2. Разработать на основе данных наркологического анализа
предложения по использованию сосны меловой в генетико-се-
лекционных работах.
Объекты и методика исследований. Объектами исследова-
нии послужили: сосна меловая (Pinus cretacea Kalen.) из
урочищ Ржевка, Бекарюковка, Чураево Шебекинского лесхоза
Белгородской области и из урочища Мордва Острогожского
лесхоза Воронежской области, а также сосна обыкновенная
(Pinus silvestris L.) европейского подвида subsp. Silvest-
ris L.) (Л. Ф. Правдин, 1964) из Левобережного лесниче-
ства Воронежского ЛТП. Данные виды между собою фило-
генетически связаны, но имеют различия (Б. М. Козо-Полян-
скип, 1931; С. И. Машкин, 1959, 1971). Таким образом, иссле-
1.
дования .проводились на популяционном уровне. Хромосомы
изучались в копчиках корешков проростков семян сосны от
свободного опыления. Для анализа были взяты семена с де-
ревьев естественного произрастания. Шишки сосны меловой из
урочищ Бекарюковка, Ржевка, Чураево и Потудаиь собирались
в конце февраля — начале марта с нижней части южной сто-
роны кроны столетних деревьев. Шишки сосны обыкновенной
собирались аналогичным образом с деревьев 80-летиего воз-
раста.
Для проведения кариологических исследований готовились
давленно^постюяиньге препараты но методике Wittmann
(1962), модернизированной сотрудниками отдела цитологии
Никитинского государственного ботанического сада Е. Г. Шо-
феристовой.
Сухие иестратифнцнрованпыс семена сосны меловой и сосны
обыкновенной проращивались па влажной фильтровальной бу-
маге в чашках Петри при 23—25° в термостате. Процент про-
растания семян сосны обыкновенной был высоким (90—95%),
процент прорастания семян сосны меловой колебался от 80
(у семян сосны из урочища Чураево) до 50 (у семян сосны из
урочища Мордва), в последнем случае чаще наблюдалось за-
гнивание семян.
В целях максимального сокращения длины хромосом при
одновременном их утолщении и более равномерного распределе-
ния их па метафазных пластинках проростки семян, корешки
которых достигли длины от 1,5 до 2 см, подвергались пред-
обработке 1%-ным раствором колхицина в точение 5 часов при
комнатной температуре или 0,002 раствором 8-окснхнпо.ппиа
при комнатной температуре с предварительным выдержива-
нием в холодильнике (при 0°+'Г) в течение суток. Как пока-
зали паши исследования, предобра бот ка колхицином, как пра-
вило, дает лучшие результаты, чем обработка окспхпполппом,
поскольку при обработке колхицином хромосомы сокращаются
более равномерно.
После предобработки промытые в проточной воде кончики
корешков длиной 0,5 см фиксировались последовательно в
двух фиксаторах Карну а (фиксатор № 1 — уксусный алко-
голь — 3:1 — в течение 1—2-х суток; фиксатор № 2 — спирт:
хлороформ:ледяная уксусная кислота — 6:3:1 — 10—15 мин.).
После фиксации для лучшего прокрашивания хромосом ма-
териал помещался в 45%-ную уксусную кислоту со следами
железа на 10—15 мии., после чего переносился в гематоксилин,
где выдерживался в течение одних суток. После прокрашивания
14
лишняя краска удалялась 45%-ной уксусной кислотой, послед-
няя отсасывалась полосками фильтровальной бумаги и мате-
риал заключался в каплю смеси Гойера, накрывался покров-
ным стеклом и делался мазок. Просмотр препаратов осуществ-
лялся под микроскопом МБИ-6 при увеличении 20><7.
Затем избранные метафазиые пластинки фотографирова-
лись при увеличении 40X20 или 40X15, их негативное изобра-
жение вместе с масштабной линейкой, полученной с помощью
объектмикрометра, отбрасывалось на лист ватмаиовской бумаги,
и контуры хромосом по внутреннему краю обводились каран-
дашом.
Всего было исследовано 47 метафазиых пластинок (МП) по
сосне меловой и 8 МП по сосне обыкновенной. Вся документа-
ция по изучению МП велась отдельно по каждой пластинке
и хранилась в индивидуальных пакетах. Измерения хромосом
осуществлялись с помощью намоченной толстой нитки (№ 10),
которая накладывалась по оси хромосом. При этом измерялись
длина каждого плеча хромосомы и расстояние от вторичной пе-
ретяжки до центромеры. Результаты измерений наносились на
миллиметровую бумагу. Способ измерения хромосом, с по-
мощью инженерного циркуля-измерителя, наиболее часто при-
меняемый при наркологических последованиях, также исполь-
зовался нами. Так как при том it другом способе измерения
были получены очень сходные результаты, мы остановились на
первом способе, как более быстром и не менее точном, особенно
для измерения хромосом с резкими изгибами.
При проведении наркологического анализа определялись:
1) число хромосом;
2) абсолютная длина каждой хромосомы в микронах (как
сумма их плеч);
3) суммарная абсолютная длина гаплоидного набора;
4) относительная длина хромосом, как процентное отноше-
ние абсолютной длины каждой хромосомы к суммарной длине
гаплоидного набора;
5) центромерпый индекс, как отношение абсолютной длины
короткого плеча к абсолютной длине длинного;
6) относительная длина длинного плеча — определялась
по сумме плеч и их отношению;
7) относительная длина короткого плеча;
8) локализация вторичной перетяжки, как процентное отно-
шение расстояния от вторичной перетяжки до центромеры к
общей длине данного плеча.
Результаты работы. Впервые было установлено число хро-
15
мосом у сосны меловой (Pinus cretacea Kalenicz) в мет-афля-
ных пластинках меристемы кон чикав корешков, оно оказалось
равным 24 (2п = 24).
Ниже приводятся данные кариотипического анализа,"-, ос.»:.
меловой из разных популяций и сосны обыкновенной во§он•*.
ской популяции. н
Кариотипы популяций сосиы меловой:
а) кариотип сосиы меловой из урочища Бекарюковка. 1Кь-
бекииского лесхоза, Белгородской области.
Для определения кариотипа сосны меловой из урочища
Бекарюковка нами были использованы 12 метафазных пласти-
нок (МП).
В различных случаях при изучении хромосом сосиы данной
популяции использовалась предобработка 1% -пым раствором
колхицина или 0,002 раствором 8-окспхииолина на холоде.
В таблице 1 приводятся данные по абсолютной и относи-
тельной длинам хромосом, а также по центромерному индексу.
В отличие от абсолютной длины хромосом данные по относи-
тельной длине хромосом очень выровненные. Все величины
относительных длин каждом пары хромосом из разных пла-
стинок отличаются друг от друга и от своего среднего значе-
ния с вероятностью менее 5%, по таблице критерия Стыодента
(И. А. Плохинскпй, 1970).
Как видно из таблицы 1, по отдельным парам хромосом
(1, V, X) наблюдается некоторое варьирование показателей
центромерного индекса, что вызвано, по-видимому, ошибками
измерений. На возможность подобного рода ошибок указы-
вается в работе Bentzer, Bothmer, Engstrand, Guslafsson,
Snogerup (1971). Основными источниками таких ошибок слу-
жат: 1) измерения хромосом, производимые разными лицами
и в разных условиях; 2) разные уровни спирализации хро-
мосом; 3) способы .измерения хромосом па фотографиях или
рисунках. ।
Однако, в связи с тем, что указанные отклонения носили
единичный характер, мы ими на практике пренебрегли.
Вычисление относительной длины короткого плеча (а) и
длинного (Ь) производилось нами по формуле, исходя из зна-
чения их суммы (х=а+Ь) — относительной длины хромосо-
мы и отношения
д
(у = ^-)— центромерного индекса. Отсюда:
b = т-—, а = х—b
l+y
16
каз GG09
to
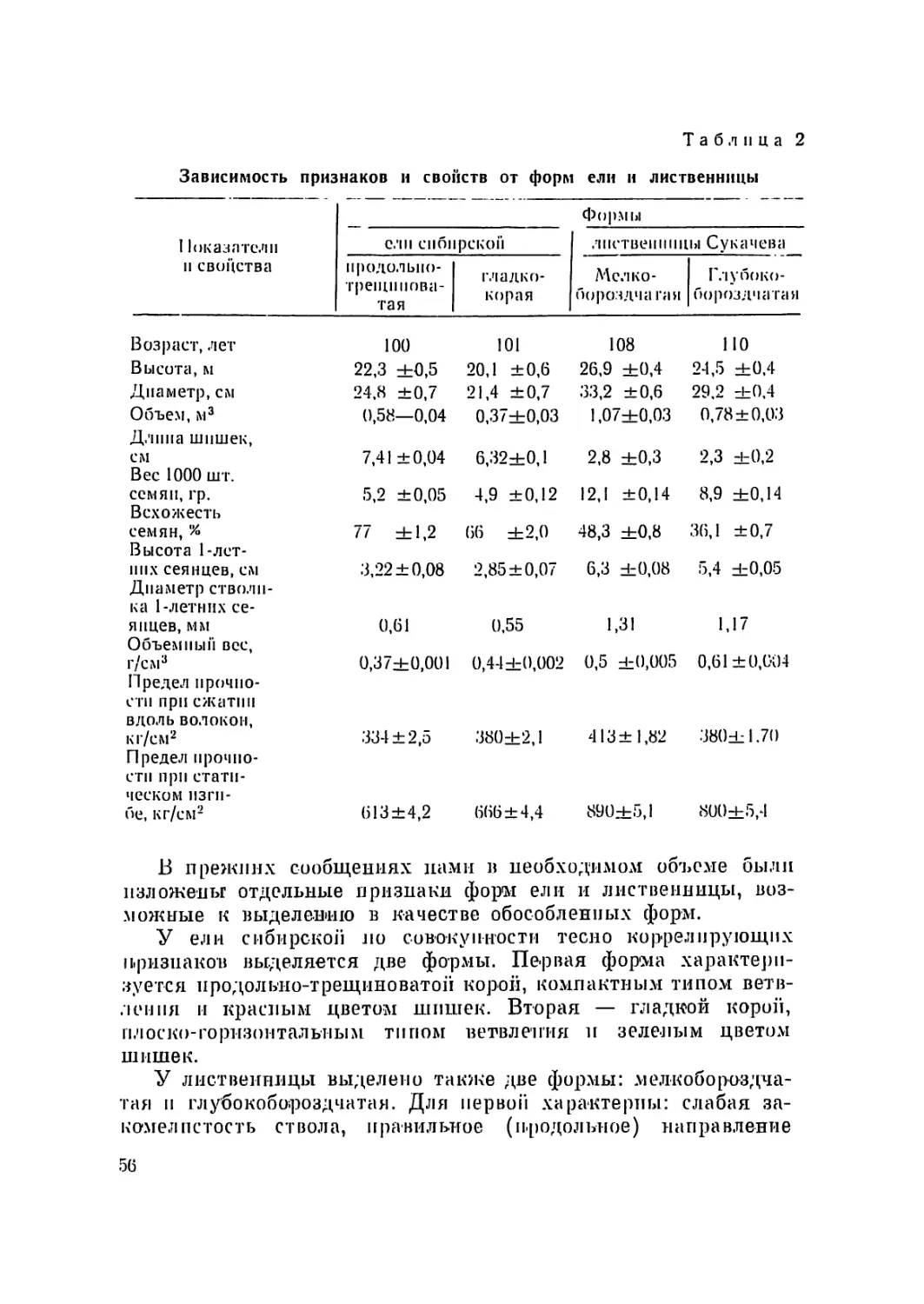
а 1
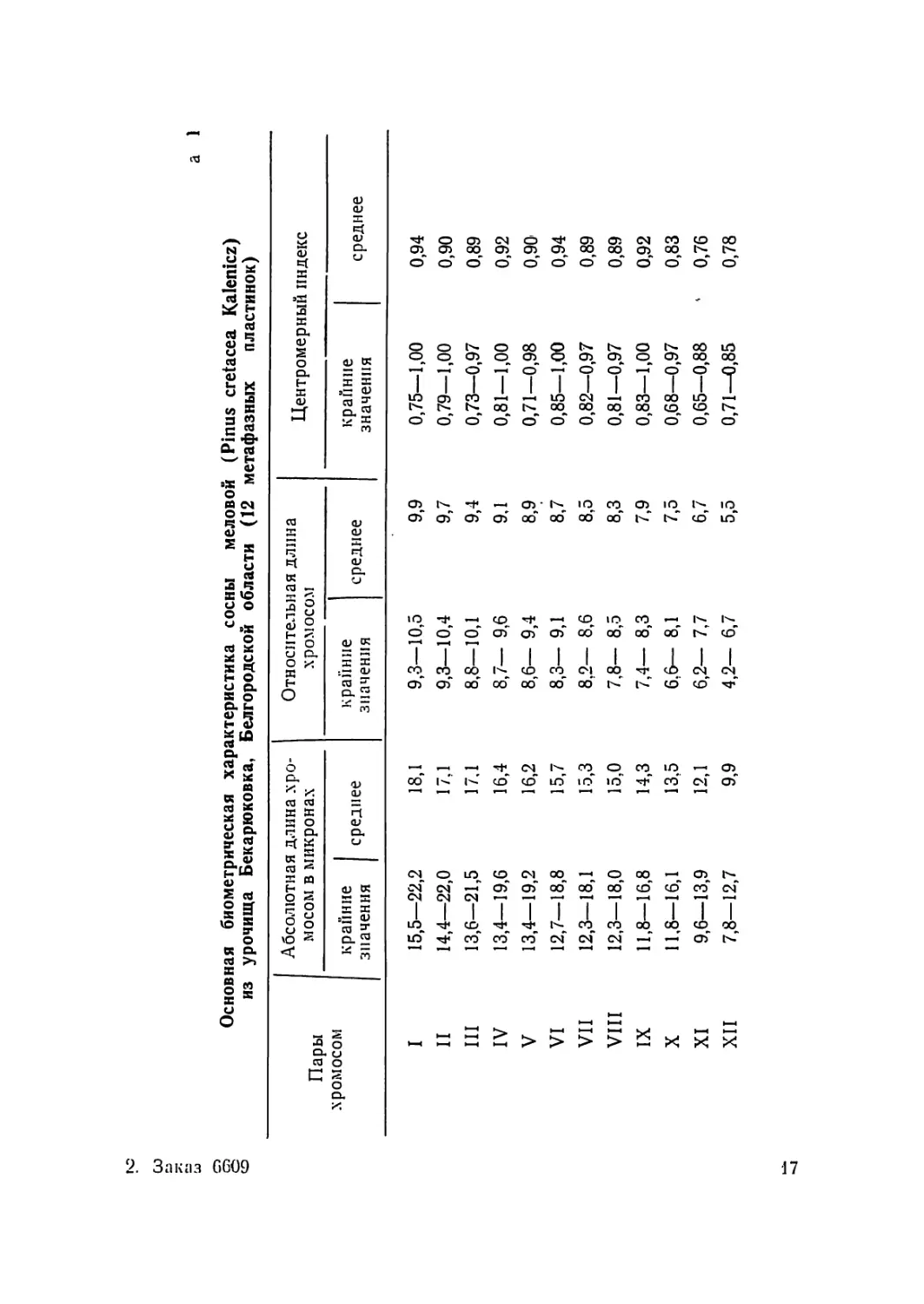
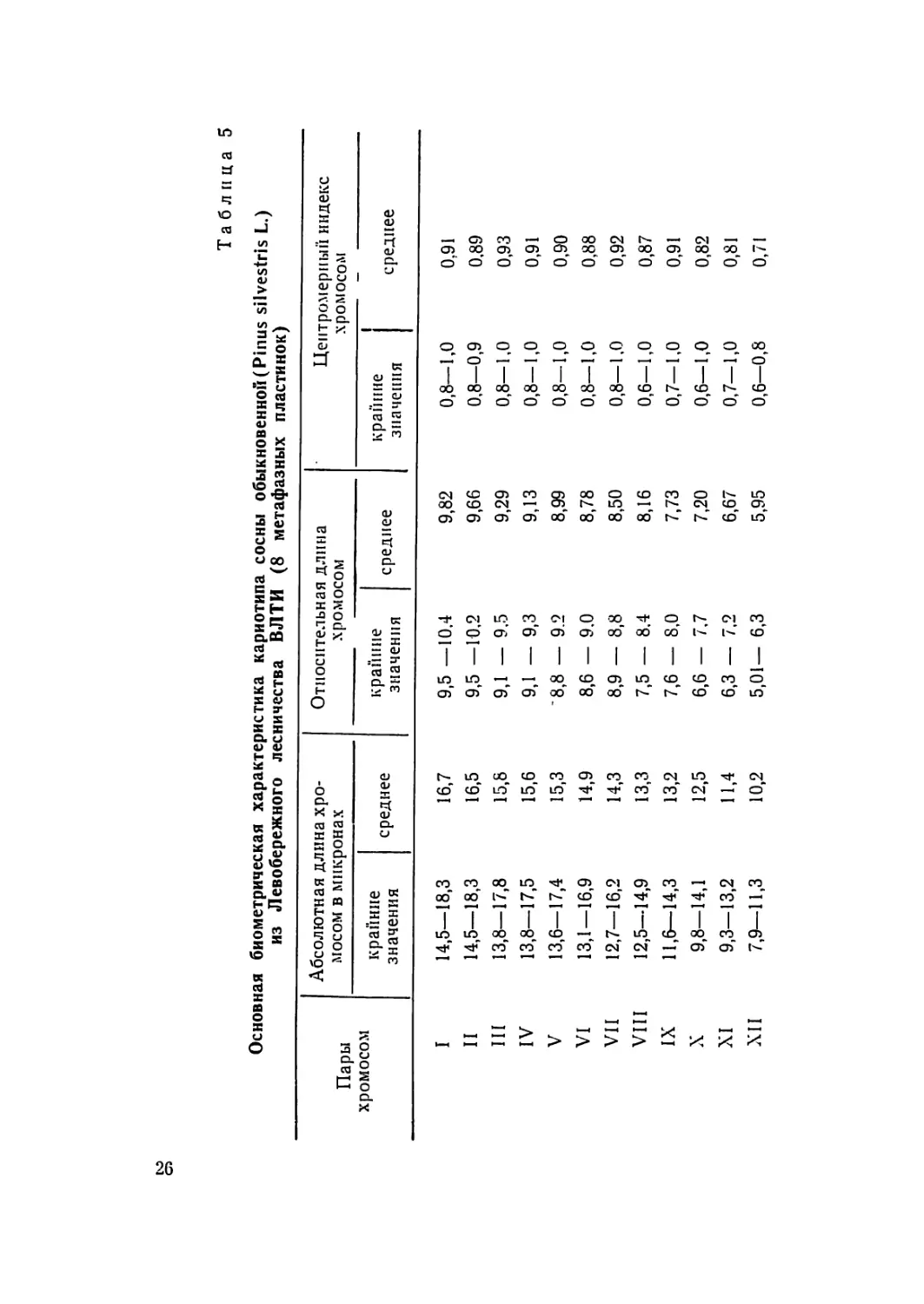
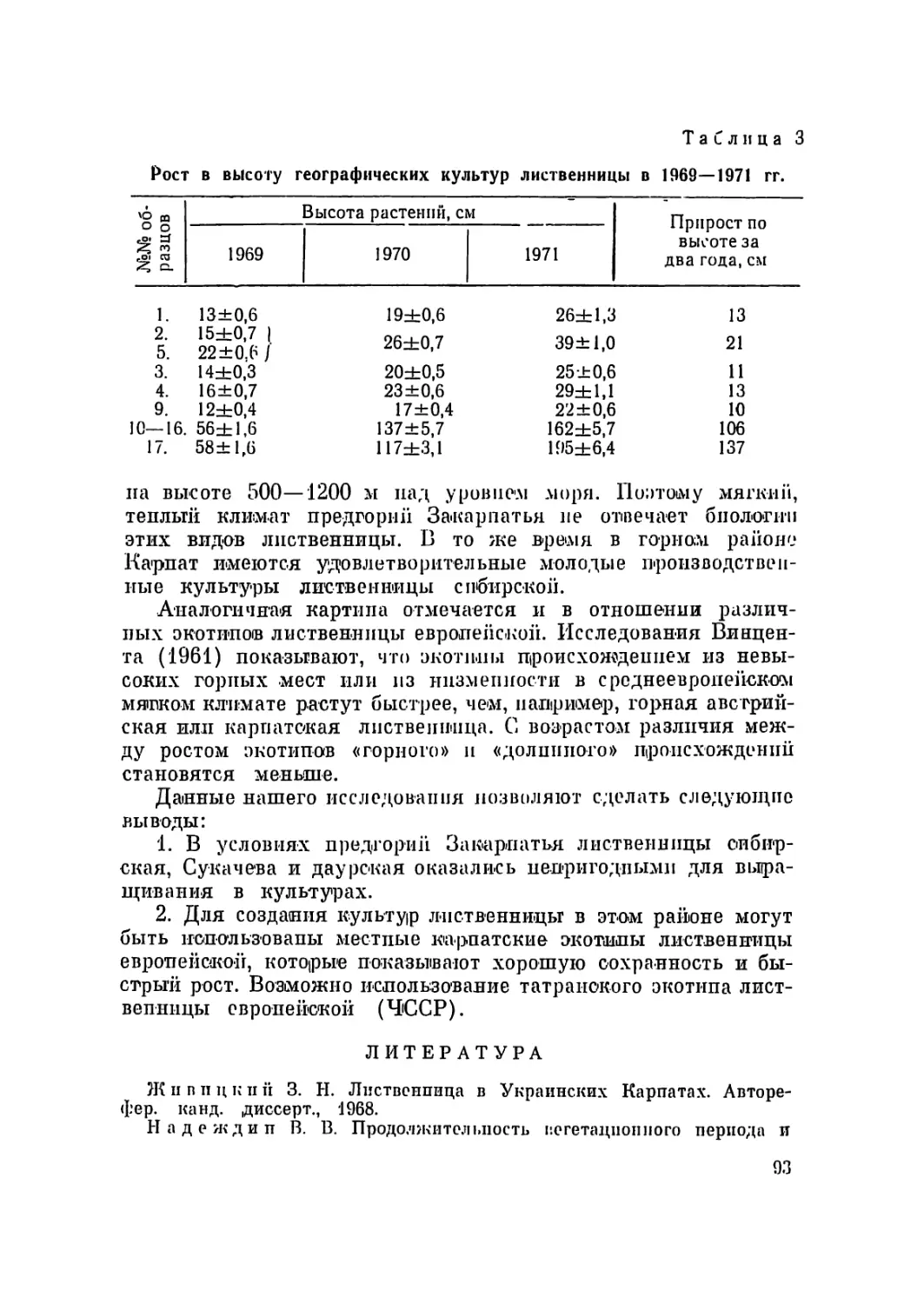
Основная биометрическая характеристика сосны меловой (Pinus cretacea Kalenicz)
из урочища Бекарюковка, Белгородской области (12 метафазных пластинок)
Пары .хромосом Абсолютная длина .хро- мосом в микронах Относительная длина хромосом Центромерный индекс
крайние значения среднее крайние значения среднее крайние значения среднее
I 15,5—22,2 18,1 9,3—10,5 9,9 0,75—1,00 0,94
II 14,4—22,0 17,1 9,3—10,4 9,7 0,79—1,00 0,90
III 13,6—21,5 17,1 8,8—10,1 9,4 0,73—0,97 0,89
IV 13,4—19,6 16,4 8,7— 9,6 9,1 0,81—1,00 0,92
V 13,4—19,2 16,2 8,6— 9,4 8,9 0,71—0,98 0,90'
VI 12,7—18,8 15,7 8,3— 9,1 8,7 0,85—1,00 0,94
VII 12,3—18,1 15,3 8,2— 8,6 8,5 0,82—0,97 0,89
VIII 12,3—18,0 15,0 7,8— 8,5 8,3 0,81—0,97 0,89
IX 11,8—16,8 14,3 7,4— 8,3 7,9 0,83—1,00 0,92
X 11,8—16,1 13,5 6,6— 8,1 7,5 0,68—0,97 0,83
XI 9,6—13,9 12,1 6,2— 7,7 6,7 0,65—0,88 ' 0,76
XII 7,8—12,7 9,9 4,2— 6,7 5,5 0,71—0,85 0,78
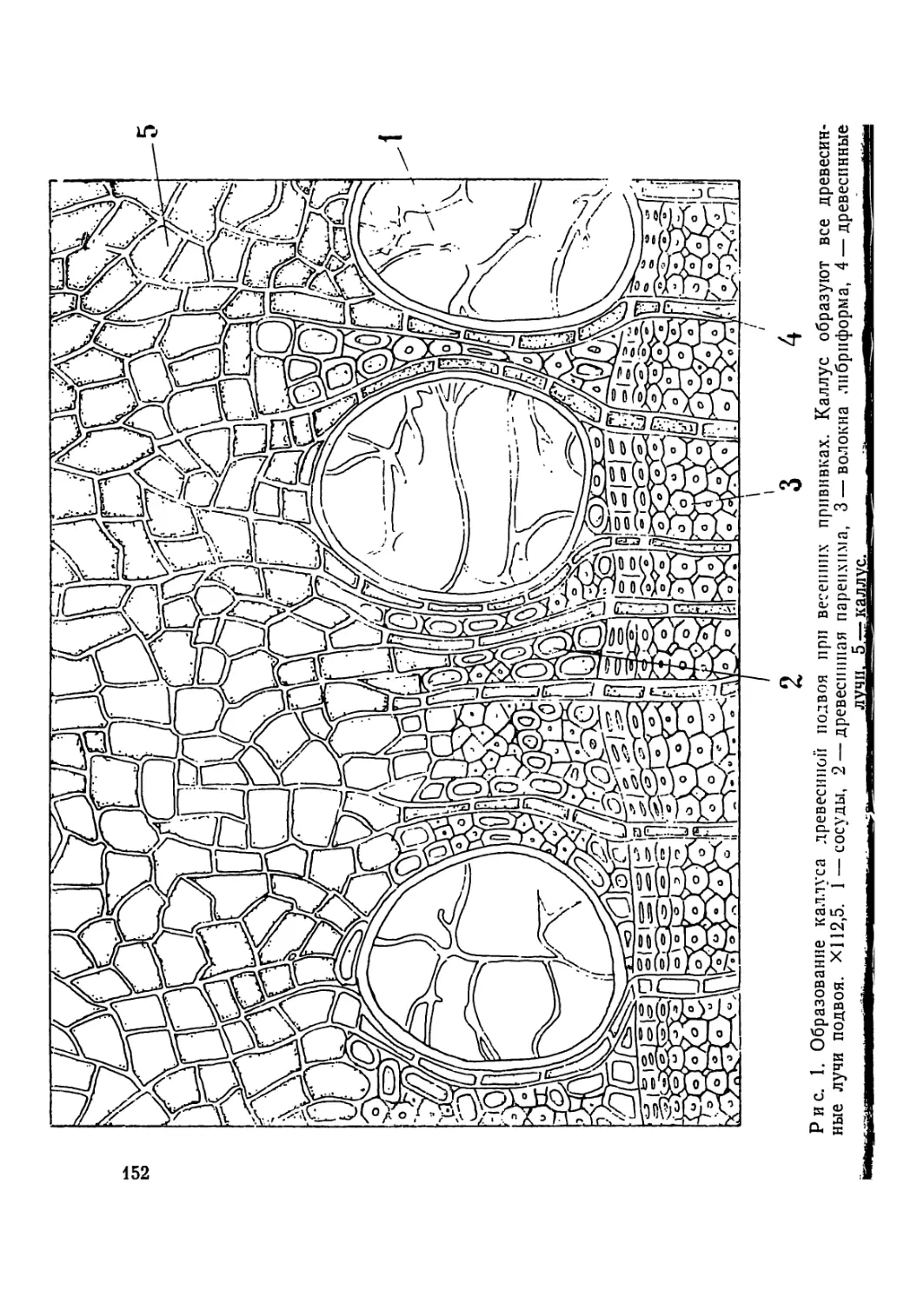
Рис. 1. Метафазпая пластинка сосны меловой из урочища Бекарюковка
(а, б) и се идиограмма (в). Предобработка произведена колхицином.
На рис. 1а приводится микрофотография метафазной пла-
стинки сосны меловой из урочища Бекарюковка. Съемка про-
изводилась при увеличении 40 <20. На рис. 16 — рисунок топ
же пластинки, выполненный с помощью рисовального аппара-
та РА-4 при увеличении 90X7. Как видно из рисунка и микро-
фотографии, хромосомы сосны имеют большое количество свет-
лых зон — гетерохроматиновых районов, которые в отдельных
случаях легко принять за вторичные перетяжки, с другой сто-
роны, нередко имеет место фенотипическая изменчивость хро-
мосом со вторичными перетяжками, благодаря чему последние
не всегда можно наблюдать. Всего в кариотипе сосны меловой
ни урочища Бекарюковка нами было установлено 5 вторичных
перетяжек (в I, II, III, IV и V парах хромосом). По средним
показателям относительной длины длинного и короткого плеча
хромосомы с учетом локализации вторичной перетяжки построе-
на идиограмма или схематическое изображение кариотипа сос-
ны меловой из уроч. Бекарюковка, представленная на рис. 1 в.
б) кариотип сосны меловой из урочища Чураево, ПТебе-
кииского лесхоза, Белгородской области.
Для определения кариотипа сосны меловой из урочища Чу-
раево было проанализировано и использовано 20 метафазных
пластинок.
В таблице 2 приводятся данные по биометрической харак-
18
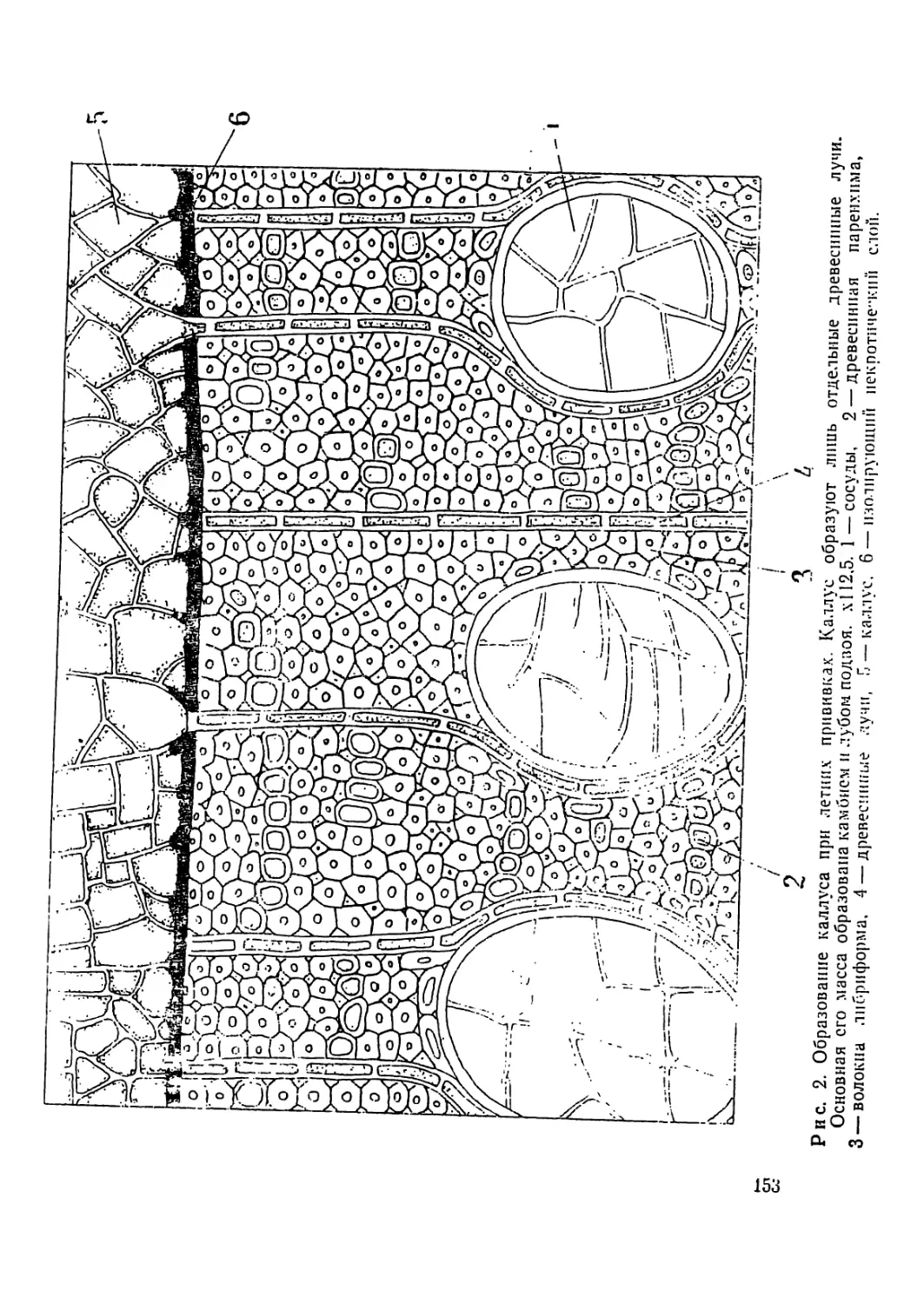
Р л с. 2. Метафазная пластинка сосны меловой из урочища Чураево
(а, б) н ее идпограмма (в). Предобработка произведена колхицином.
терпстнке хромосом. Применялась предобработка колхицином.
В наборе хромосом сосиы меловой из урочища Чураево нами
не наблюдалось хромосом со вторичными перетяжками.
На рис. 2 а, б представлены микрофотография и рисунок МП
сосны из указанной популяции, па рис. 2 в дала ее идиопрамма.
в) кариотип сосиы меловой из урочища Ржевка, Шебекпп-
ского лесхоза, Белгородской области.
Для определения кариотипа сосны меловой из урочища
Ржевка было использовано 10 метафазных пластинок. При изу-
чении хромосом применялась предобработка колхицином и
8-оксихинолнном. Следует отметить, что предобработка 8-оксн-
хиполииом дает больший спектр изменчивости опирализации
хромосом по сравнению с колхицином.
Максимальная длина хромосомы при обработке 8-оксихи-
нолипом достигает примерно 36 микрон, при обработке колхи-
цином пемиого более 23 микрон, минимальная длина хромосо-
мы при обработке 8-оксихинолииом достигает 19 микрон, кол-
хицином — 15 микрон. В таблице 3 приводятся данные по
биометрической характеристике хромосом.
При изучении данного кариотипа сосны меловой нами йылп
2* 19
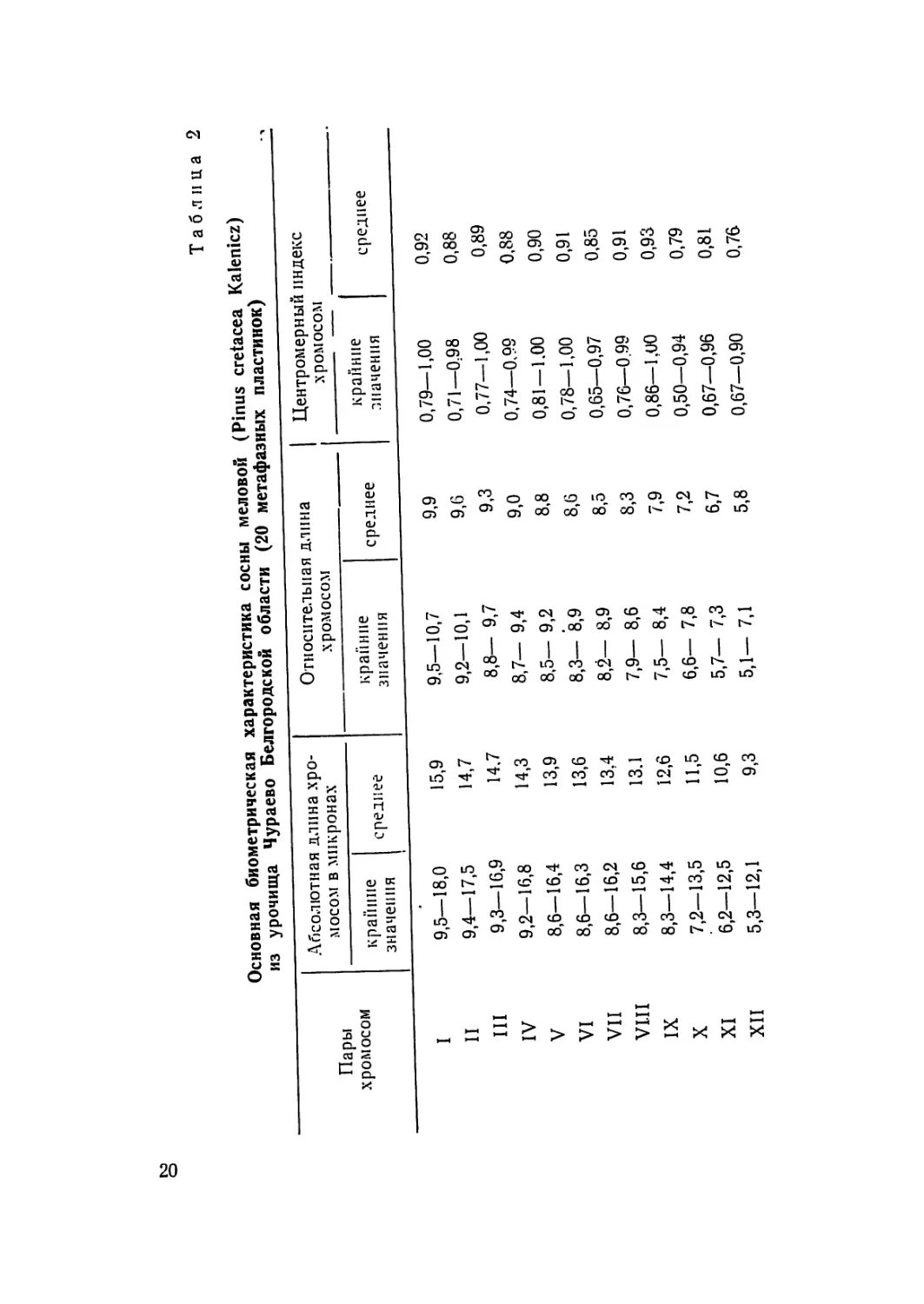
Таблица 2
Основная биометрическая характеристика сосны меловой (Pinus cretacea Kalenicz)
из урочища Чураево Белгородской области (20 метафазных пластинок)
Пары хромосом Абсолютная длина хро- мосом в микронах Относительная длина хромосом Центромерный индекс хромосом
крапине значения среднее крапине значения среднее крайние значения среднее
I 9,5—18,0 15,9 9,5—10,7 9,9 0,79—1,00 0,92
II 9,4—17,5 14,7 9,2—10,1 9,6 0,71—0:98 0,88
III 9,3—16,9 14.7 8,8— 9,7 9,3 0,77—1,00 0,89
IV 9,2—16,8 14,3 8,7— 9,4 9,0 0,74—0.99 0,88
V 8,6-16,4 13,9 8,5— 9,2 8,8 0,81 — 1.00 0,90
VI 8,6—16,3 13,6 8,3— 8,9 8,6 0,78—1,00 0,91
VII 8,6—16,2 13,4 8,2— 8,9 8,5 0,65—0,97 0,85
VI.II 8,3—15,6 13.1 7,9— 8,6 8,3 0,76—0.99 0,91
IX 8,3—14,4 12,6 7,5— 8,4 7,9 0,86—1.U0 0,93
X 7,2—13,5 И,5 6,6— 7,8 7,2 0,50—0,94 0,79
XI 6,2—12,5 10,6 5,7— 7,3 6,7 0,67—0,96 0,81
XII 5,3—12,1 9,3 5,1— 7,1 5,8 0,67—0,90 0,76
впервые обнаружены спутничные хромосомы (VII пара).
Имеется в виду спутник классической формы — округлое тель-
це на тонкой нити (Р. Ригер и Л. Михаэлис, 1967). В силу
значительной фенотипической изменчивости спутиичпых хро-
мосом, «классический» спутник обнаруживается не па всех
пластинках.
Р и с. 3. Метафазная пластинка сосны меловой из урочища Ржевка
(а, б, в, г) и ее идиограмма (д). Предобработка произведена 8-оксихиио-
лином (а, б) и колхицином (в, г).
г) кариотип сосны меловой из урочища Потудаиь, Острогож-
ского лесхоза, Воронежской области.
Для определения кариотипа сосны меловой из урочища По-
тудаиь было использовапо 5 метафазиых пластинок.
В таблице 4 приводятся данные по абсолютной и относи-
тельной длинам хромосом и центромерному индексу.
21
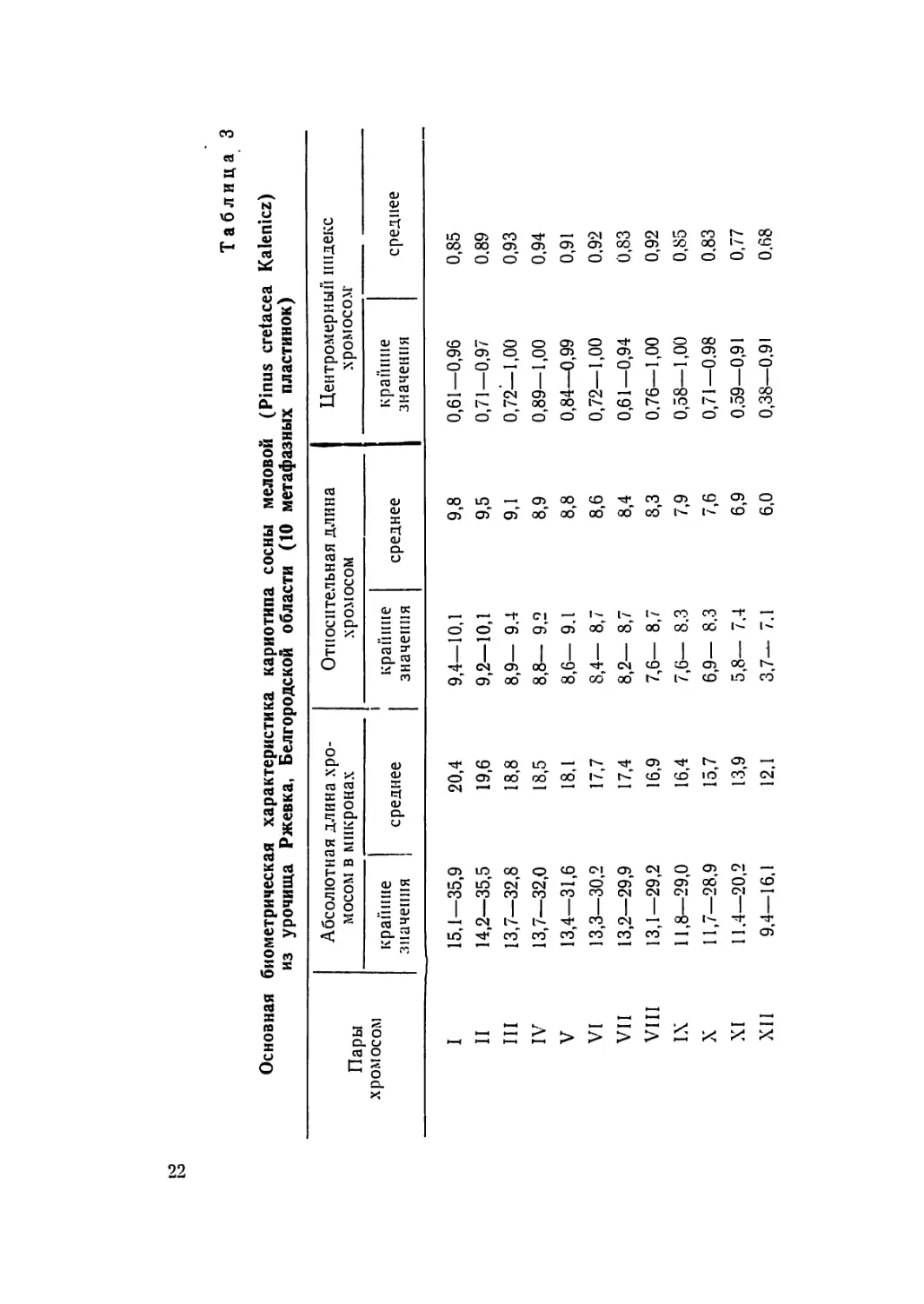
Таблица 3
Основная биометрическая характеристика кариотипа сосны меловой (Pinus cretacea Kalenicz)
из урочища Ржевка, Белгородской области (10 метафазных пластинок)
Пары хромосом Абсолютная длина хро- мосом в микронах Относительная длина хромосом Центромерный индекс хромосом-
крайние значения | I среднее 1 ! крайние значения среднее крапине значения среднее
I 15,1—35,9 20,4 9,4—10,1 9,8 0,61—0,96 0,85
П 14,2—35,5 19,6 9,2—10,1 9,5 0.71—0,97 0,89
III 13,7—32,8 18,8 8,9— 9.4 9,1 0,72—1,00 0,93
IV 13,7—32,0 18,5 8,8— 9,2 8,9 0,89—1,00 0,94
V 13,4—31,6 18,1 8,6— 9,1 8,8 0,84—0,99 0,91
VI 13,3—30,2 17,7 8,4— 8,7 8,6 0,72—1,00 0,92
VII 13,2—29,9 17,4 8,2— 8,7 8,4 0,61—0,94 0,83
VIII 13,1—29,2 16,9 7,6— 8,7 8,3 0.76—1,00 0,92
IX 11,8—29,0 16,4 7,6— 8.3 7,9 0,58—1,00 0,85
X 11,7—28,9 15,7 6,9— 8.3 7,6 0,71—0,98 0.83
XI 11.4—20,2 13,9 5,8— 7,4 6,9 0,59—0,91 0,77
XII 9,4—16,1 12,1 3,7— 7.1 6,0 0,38—0,91 0.68
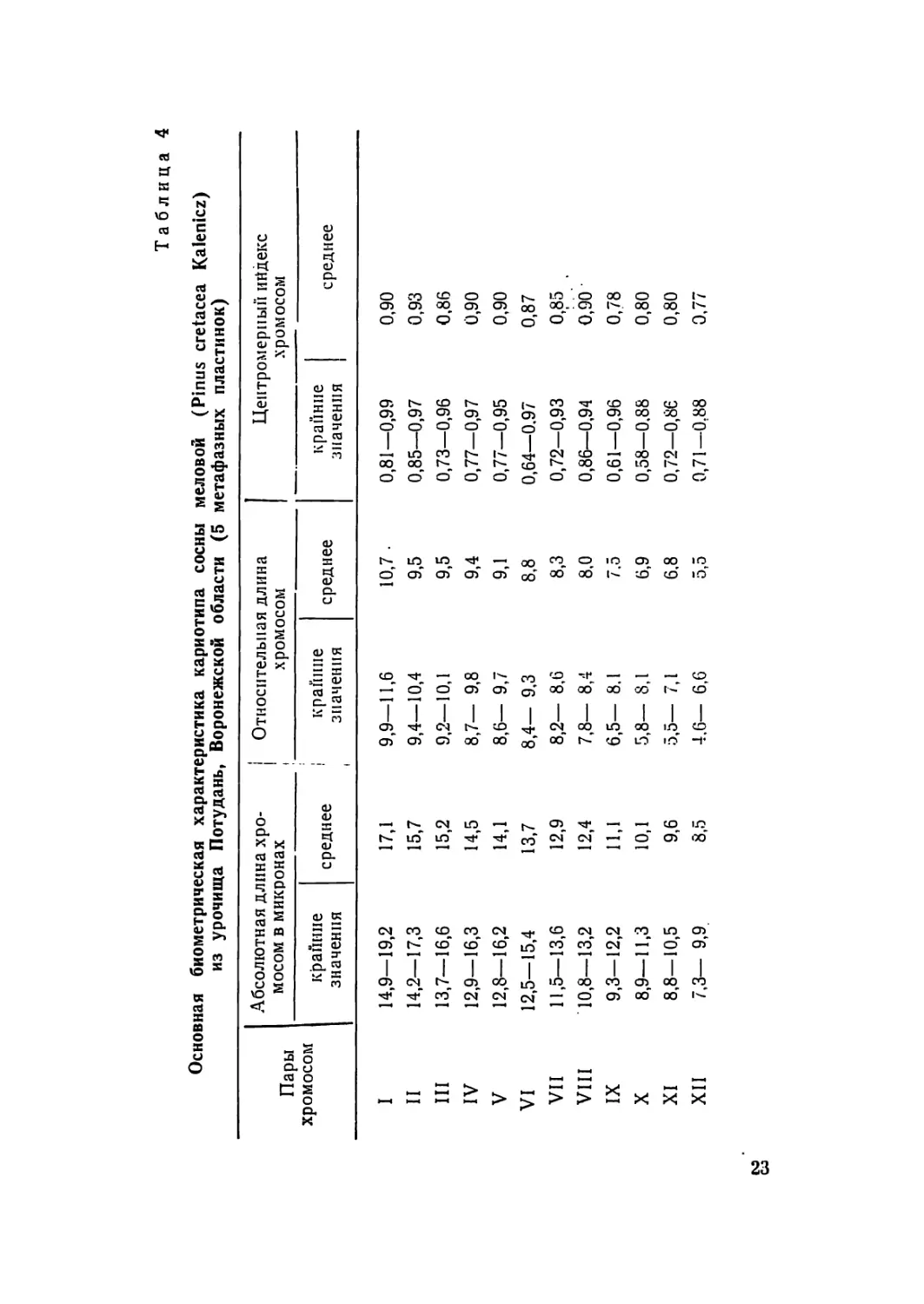
Таблица 4
Основная биометрическая характеристика кариотипа сосны меловой (Pinus cretacea Kalenicz)
из урочища Потудань, Воронежской области (5 метафазных пластинок)
Пары хромосом Абсолютная длина хро- мосом в микронах । Относительная длина , хромосом Центромерный ийдекс хромосом
крайние значения среднее I крапине значения среднее крайние значения среднее
I 14,9—19,2 17,1 9,9—11,6 10,7 • 0,81—0,99 0,90
II 14,2—17,3 15,7 9,4—10,4 9,5 0,85—0,97 0,93
III 13,7—16,6 15,2 9,2—10,1 9,5 0,73—0,96 0,86
IV 12,9—16,3 14,5 8,7— 9,8 9,4 0,77—0,97 0,90
V 12,8—16,2 14,1 8,6— 9,7 9,1 0,77—0,95 0,90
VI 12,5—15,4 13,7 8,4— 9,3 8,8 0,64—0,97 0,87
VII 11,5—13,6 12,9 8,2— 8,6 8,3 0,72—0,93 0,85
VIII 10,8—13,2 12,4 7,8— 8,4 8,0 0,86—0,94 0,90
IX 9,3—12,2 11,1 6,5— 8.1 7.5 0,61—0,96 0,78
X 8,9—11,3 Ю,1 5,8— 8,1 6,9 0,58—0,88 0,80
XI 8,8—10,5 9,6 5,5— 7,1 6,8 0,72—0,86 0,80
XII 7,3— 9,9. 8,5 4.6— 6,6 5,5 0,71—0,88 0,77
Так же, как и в кариотипе сосны меловой из урочища Ржев-
ка, в кариотипе сосны меловой и.з урочища Нотудань были
обнаружены1 спутничные хромосомы (IV пара) со спутником
классической формы. На рис. 4 (а, б) представлены микрофо-
тография и рпсупок МП сосны меловой из урочища Нотудань
с двумя спутничными хромосомами. На рис. 4 (в, г) представ-
лены микрофотография и рисунок метафазной пластинки, на
которой спутник по обнаруживается. Па рис. 4 (д )дана ндно-
грамма сосны меловой из урочища Нотудань.
д) кариотип сосны обыкповетгной европейского подвида из
Левобережного лесничества Воронежского ЛТИ.
Р и с. 4. Метафазные пластинки сосны меловой со спутником (а, б)
п без спутника (в, г) из урочища Потудаиь п их идиограмма (д). Пред-
обработка произведена колхицином.
24
Для определения кариотипа сосны обыкновенной европей-
ского подвида (было попользовано 8 метафазных пластинок
(всего же было проанализировано 22 МП).
В табл. 5 — данные по абсолютной и относительной длинам
хромосом и центромерному индексу. Так как для обработки под-
бирал псьметафавтые пластинки! со сходной степенью спиралнза-
цин, разница но абсолютной длине между одпими и темп же па-
рами хромосом в разных пластинках не превысила 3,8 микрон.
Па рис. 5 (а, б) представлены микрофотография и рису-
нок МП сосны обыкновенной, па рис. 5 (в) дана ее идиограмма.
Обсуждение полученных данных и общее заключение.
Как показали результаты математической обработки, вели-
чины абсолютной и относительной длины гаплоидных наборов
у обоих видов сосны совпадают друг с другом п со своим сред-
ним значением с вероятностью свыше 93% по таблице крите-
рия Стыодепта (И. А. Плохнпской, 1970); значительное сход-
ство наблюдается также и но относительной длине всех пар
хромосом и по их центромерному индексу. Однако, кариотипы
сосны меловой и сосны обыкновенной не тождественны. Раз-
ница между ними проявляется, прежде всего, по количеству
и локализации вторичных перетяжек. Так, у сосны обыкновен-
ной европейского подвида, полученной из Левобережного лесни-
чества Воронежского ЛТП, можно отметить 5 вторичных пере-
тяжек (в I, 11, 111, V и IX нарах хромосом). Однако эти дан-
ные можно рассматривать только как предварительные.
Рис. 5. Метафазная пластинка сосны обыкновенной (а, б) н сс нднограм-
ма (в). Предобработка произведена колхицином.
25
Таблица 5
Основная биометрическая характеристика кариотипа сосны обыкновенной(Pinus silvestris L.)
из Левобережного лесничества ВЛТИ (8 метафазных пластинок)
Пары хромосом Абсолютная длина хро- мосом в микронах Относительная длина хромосом Центромерный индекс хромосом
крайние значения среднее крайние значения среднее крайние значения среднее
I 14,5—18,3 16,7 9,5 —10,4 9,82 0,8—1,0 0,91
II 14,5—18,3 16,5 9,5 —10,2 9,66 0,8—0,9 0,89
III 13,8—17,8 15,8 9,1 — 9,5 9,29 0,8—1,0 0,93
IV 13,8—17,5 15,6 9,1 — 9,3 9,13 0,8—1,0 0,91
V 13,6—17,4 15,3 '8,8 — 9.2 8,99 0,8—1,0 0,90
VI 13,1—16,9 14,9 8,6 — 9,0 8,78 0,8—1,0 0,88
VII 12,7—16,2 14,3 8,9 — 8,8 8,50 0,8—1,0 0,92
VIII 12,5—14,9 13,3 7,5 — 8,4 8,16 0,6—1,0 0,87
IX 11,6—14,3 13,2 7,6 — 8,0 7,73 0,7—1,0 0,91
X 9,8—14,1 12,5 6,6 — 7,7 7,20 0,6—1,0 0,82
XI 9,3—13,2 Н,4 6,3 — 7,2 6,67 0,7—1,0 0,81
XII 7,9—11,3 10,2 5,01— 6,3 5,95 0,6—0,8 0,71
Шведские ученые, изучавшие кариотип сосны обыкновенной,
установили у нее разное число вторичных перетяжек.
Если сопоставить имеющиеся данные по изучению карио^
типа сосны обыкновенной с полученными нами, то можно
сказать, что признак вторичной перетяжки является кон-
стантным для I и II пар хромосом и варьирующим — для
остальных.
У соспы меловой из урочища Чураево, Шебекинского лес-
хоза Белгородской области не было установлено вторичных
перетяжек, у сосны меловой из урочища Бекарюковка, наобо-
рот, их было обнаружено 5 (в I, II, III, IV и V парах хромо-
сом), у сосиы меловой из урочища Мордва (Потудапь) Воро-
нежской области отмечено 2 вторичных перетяжки (в I и
IV нарах хромосом), у сосны меловой из урочища Ржевка,
Белгородской области — 5 (в I, III, IV, VII и IX парах
хромосом).
У сосны меловой из урочищ Ржевка и Мордва нами были
обнаружены спутничные хромосомы со спутником «классиче-
ской формы» (округлое тельце на тонкой нити), чего не было
обнаружено до сих пор ни одним автором ни для одного из
11 од ви до в с ос п ы обыкновенной.
Спутничные хромосомы наблюдались не на всех метафазных
пластинках сосны меловой из урочищ Ржевка и Мордва, а
лишь на отдельных, причем чаще у сосны меловой из Ржевки.
Такое явление, по-видимому, связано со значительной феноти-
пической изменчивостью спутничных хромосом, как это отмеча-
лось в работах Г. В. Дерягина и А. Б. Иорданского (1971).
На основе обобщения полученных данных можно сделать
следующие выводы и практические предложения:
1. В целях облегчения подсчета числа хромосом п изучения
их морфологии удобнее применять предобработку раствором
колхицина, так как он дает более равномерное сокращение
хромосом по сравнению с 8-оксихинолином.
2. Число хромосом у сосиы меловой равно 24 (2п^24),
так же, как и у сосны обыкновенной.
3. По абсолютной и относительной длине гаплоидных на-
боров, абсолютной и относительной длине хромосом и их плеч
между сосной меловой и сосной обыкновенной отсутствуют
достоверные различия. Различия между кариотипами сосны
меловой и сосны обыкновенной проявляются по количеству и
локализации вторичных перетяжек.
4. У сосны меловой из урочища Ржевка и Потудань обна-
27
ружены спутнпчпые хромосомы, у сосны обыкновенной спутни-
ки неизвестны.
5. В пределах популяций у сосны* меловой наблюдаются
значительные различия по количеству и локализации вторич-
ных перетяжек, наличию и отсутствию спутников. Это свиде-
тельствует о значительно большем ее полиморфизме по срав-
нению с сосной обыкновенной и что обусловлено, возможно,
большей древностью сосны меловой, как лиинеевского вида —
третичного реликта, чем сосны обыкновенной, как ее песчапо-
го деривата.
6. Сосна меловая, выделенная Калиниченко (1849) в осо-
бый вид (Р. cretacea Kai.) за свою морфологию (приплюснутая
крона, короткая хвоя, мельче шишки, чем у сосны обыкновен-
ной), географию (юг Европейской части СССР), экологию
(растет на меловом субстрате), имеет также фенологическую
изоляцию (по наблюдениям 10. А. Дороиииа, цветет иа несколь-
ко дней позже сосны обыкновенной), дает более качественную
древесину (Е. М. Кондратюк, 1957), а также, как показывают
настоящие исследования, имеет и кариологические особенно-
сти. т. е. свою структуру кариотипа и наследственную специфи-
ку. На наш взгляд, сосна меловая (Р. cretacea Kalenicz) вполне
удовлетворяет комплексу основных требований, предъявляе-
мых к критерию самостоятельного вида — морфологическому,
географическому, экологическому, историческому, популяцион-
ному, цитологическому и другим (К. М. Завадский, 1968), что
свидетельствует о биологической обособленности ее как вида.
7. Учитывая биологическую обособленность соспы меловой,
как вида, ее специфические экологические и лесохозяйствен-
ные особенности, необходимо дифференцированно использовать
ее в селекции и в практике облесения обширных пространств
меловых обнажений юга Европейской части СССР. С этой
целью следует выявить внутривидовое разнообразие, отобрать
цепные формы и плюсовые деревья.
Целесообразно также заложить опыты по географическим
и лесотппологическим культурам ее па Среднерусской п Волж-
ской возвышенности, подобно географическим культурам сосны
обыкновенной, созданным на песчаных почвах.
8. Необходимо расширить работы по созданию массовых про-
изводственных культур сосны меловой па меловых обнажениях
юга Европейской части СССР, где 'Насчитывается их свыше
1,5 млн. га, т. е. больше площади всех природных и искусствен-
но созданных лесов Центрального Черноземья. Все это имеет
огромное народно-хозяйственное значение, ггак как может
28
дать большой мелиоративный, лесохозяйственный и. экономиче-
ский эффект.
По ориентировочным подсчетам при облесении названной
площади меловых обнажений страна может ежегодно допол-
нительно получать прибавку урожая зерновых за счет мелио-
ративного влияния посадок сосны меловой но 2—3 центнера
зерна с гектара, что в пересчете по коэффициенту: 1 га поса-
док сосны — 15 га посевов зерновых, мелиоративный эффект
всей площади ее насаждений выразится в ежегодном дополни-
тельном сборе зерна в 30—45 млн. центнеров, не говоря уже
о водоохранном и санитарно-гигиеническом значении ее по-
садок, а также о возможностях прямого и побочного использо-
вания ее насаждений.
9. Все существующие остатки меловых боров на юге Евро-
пейской части СССР (Белгородская, Воронежская и Донец-
кая области) являются уникальными памятниками и эталона-
ми природы, а также резерватами генетических фондов ценных
редких растений. Необходимо усилить мероприятия, направ-
ленные на сохранение п восстановление сосновых насаждений
на мелах и шире использовать их как ценнейшие семенные
маточники.
ЛИТЕРАТУРА
Вавилов И. И. Селекция как паука. Избранные труды. Изд.
АН СССР. М.-Л., 1960-
Вавилов И. И. Вид как система. Избран, труды. Изд-во «Наука»,
М.-Л., 1965.
В а в п л о в Н. И. Закон гомологических рядов в наследственной
изменчивости. Избран, труды. Изд-во «Наука», М.,-Л., 1965.
Дерягин Г. А., Иорданский А. Б. 1971. Фенотипическая из-
менчивость спутничиых хромосом. Сообщение I. V. Allium сера
Z A, Tistulosum Z., A. altaicum Pull «Генетика», № 10, 13—22, 1971.
Завадский К. М. Вид и разнообразие. Изд-во «Наука», Л., 1968.
Коз о-П о л я и с к и й Б. М. В стране живых ископаемых. Очерк
из истории горных боров на степной равнине ЦЧО. Учпедгиз, М., 1931.
Кондратюк Е. М. До характеристики деревинидеяких вид в сос-
ни. Допов АН Укр. РСР, § 6, 1957.
Мацкевич Н. В., Лутков А. Н. Числа хромосом полиплоид-
ных видов лесных деревьев и кустарников. Со. Экспериментальная по-
липлоидия и селекция растений. Изд. СО АН СССР, Новосибирск, 1966.
Машкин С. И. Редкие реликтовые виды древесных растений
Центрального Черноземья и их значение для агролесомелиорации и
зеленого строительства. В сб.: «Охрана природы Центральпо-Черпо-
зомпоп полосы», № 2, Воронеж, стр. 170—182, 1959.
29
Машкин - С. Й. Дендрология Центрального Черноземья. Том I.
Нзд-во ВГУ, Воронеж, 1971.
Плохннский И. А. Биометрия. Изд. Московск. ун-та, М., 1970.
Паву л соне С. А., Иорданский А. В., Гипдилис В. М.
Сравнительный морфологический анализ хромосом Allium сера L. п
Allium tistulosum L. Генетика, т. VI, № 2, 40—55. 1970.
Павулсоне С. А., Иорданский А. В. Новый способ повы-
шения разрешающей способности кариограммного анализа. Генетика,
т. VII, № 4, 149,1971.
Правдин JT. Ф. — Сое па обыкновенная (Изменчивость, внутриви-
довая систематика н селекция). М., изд. «Наука», 1964.
Правдин Л. Ф. — Сосна обыкновенная (Изменчивость, внутриви-
довая генетика, селекция и семеноводство», изд-во «Карелия», Петроза-
водск, .1970.
Ригер Р., Михаэлис А. Генетический и цитологический сло-
варь. М., 1967.
Соколов С. Я. Сем. Pinaceae Lindb. — сосновые. Деревья и кустар-
ники СССР, т. 1, Изд-во АН СССР, М.-Л., 1949.
Соколов С. Я., Св язе в а 0. А. География древесных растений
СССР. Изд-во «Наука», М.-Л., 1965.
III и ш и н а ш в и л и Р. М. Карпотип сос.ны Сосновского. Цитология,
т. 10, № 2, 255, 1968.
Aass J. A cytological analysis of scots pine (Pinus Silvestris L.) from
Skyak Norway MaddeL Det. norske skogl — Porsoksvesen 14: 96—109. 1957.
Benz er B., Bolhmer В. V., Engstrand L„ Yastafsson M.,
Snogerup S. Some souices of error in the determination of arm rations
of chromosomes. Bot. Notis. Vol. 124, № 1: 65—67, 1971.
Khochoo T. N. Polyploidy in gymnospermes. Evolution v. 13 № 1, 1959.
Kanezawa R. Polyploids appeared jo 5ui|poos uia\j hi forest trees.
«Jap. For. Soc.», v. 31, 22—24, 1949.
L i n n a 1 J. Tetrapioid plants of Japancnslc red pine (Pinus densiflora
Sieb. Luce.) discovered in transplant beds. Jour. Jap. For., Sci., 34: 185—187,
1952.
Meh г a P. N. and Khoshoo T. N. Cvlologv of conifers. J. Genetic,
54(1): 165—180, 1956.
M er gen F. Natural polyploidy in Slash pine. For. Sci., 4: 283—293,
1958.
N a t a г a j a n A. T., Ohba K. and Sima k M. Karyotype analysis of
Pinus Silvestris. Hereditas, 47: 379—382, 1961.
Pederic L. A. Chromosome relationships between Pinus species.
Silvae Genetica, Bd. 19, H5/6: 171—180, 1970.
Santamour F. S. New chromosome counts in Pinus and Picea. Sii-
vae Genetica, 9: 87—88, 1956.
S arc ar P. Cromosome studies in Pinus species (abstr.). Can. J. Genet.
Cytol. 5: 107, 1963.
Sax K. and Sax H. J. Chromosome number and morphology in Coni-
fers. Jour. Arnold arboretum 14: 356—375, 1933.
Sa’x K. Meiosis in interspecific pine hybrids. For. Sci., 6(2): 135—138,
Saylor L. CC. A karyotypic analysis of selected species of Pinus.
Silvae Genetica, Bd. 10: H 3: 77—84, 1961*.
Saylor L. C. A karyotype analysis of Pinus group Lariciones. Silvae
Genetica, Bd. 13, H 6: 165—170. 1964.
30
Shams-ur-Rehman and Perveen. Karyotypic analysis of Pinus. Wallichiana
AB Jacks Syn. Pyriffithii. The Pakistan Journ. of Forestry. 20; 3: 283—292,
1970.
Wittmann W. Aceto-ircn--haematoxilin for stainig chromosomes in
squashes of plant material. Stain TechnoL 37, № 1, 1962.
Upadhana L. P. and Kedharnath S. Karyotype four species of
Pinus naturally occuring in Indian. Forestry, 96: 657—667, 1970.
Jim K. F. Karyotype analysis of Pinus rigida. Hereditas, 49, 274—276,
1963.
M. M. ВЕРЕСИН, А. П. ЦАРЕВ
К итогам сортоиспытания тополей в Воронежской области
Введение. Для успешного внедрения в производство
тополей чрезвычайно важное значение имеет правильный выбор
их ассортимента. В СССР выведено и интродуцировано много
ценных и перспективных гибридов, форм и сортов тополей, но
огромное разнообразие природных условий нашей страны тре-
бует дифференцированного подхода к выработке рекоменда-
ций для практики в пределах того или иного региона. Научно
обоснованный ассортимент может быть выявлен только в про-
цессе сортоиспытания. К сожалению, у нас еще не создана еди-
ная государственная сеть сортоиспытания, и эта работа про-
водится отдельными ведомствами и лицами по различным
программам п методикам (Ростовцев, 1963; Иванников, 1971;
Старова, 1971; Озолин, 1971 и др).
В Воронежской области (Центральная лесостепь) работы
по сортоиспытанию тополей были начаты в 1952 году под
методическим руководством М. М. Вереспна, которым был соз-
дан ряд испытательных участков (Вересии, 1967). Помимо них
в 1961 году в Новоусманском лесхозе (под Воронежем) был
заложен сортоиспытательный участок под методическим руко-
водством ВНИИЛМа. Всего в испытании в Воронежской обла-
сти находится 52 образца видов и гибридных форм тополей.
В настоящей статье сообщаются некоторые итоги этой работы.
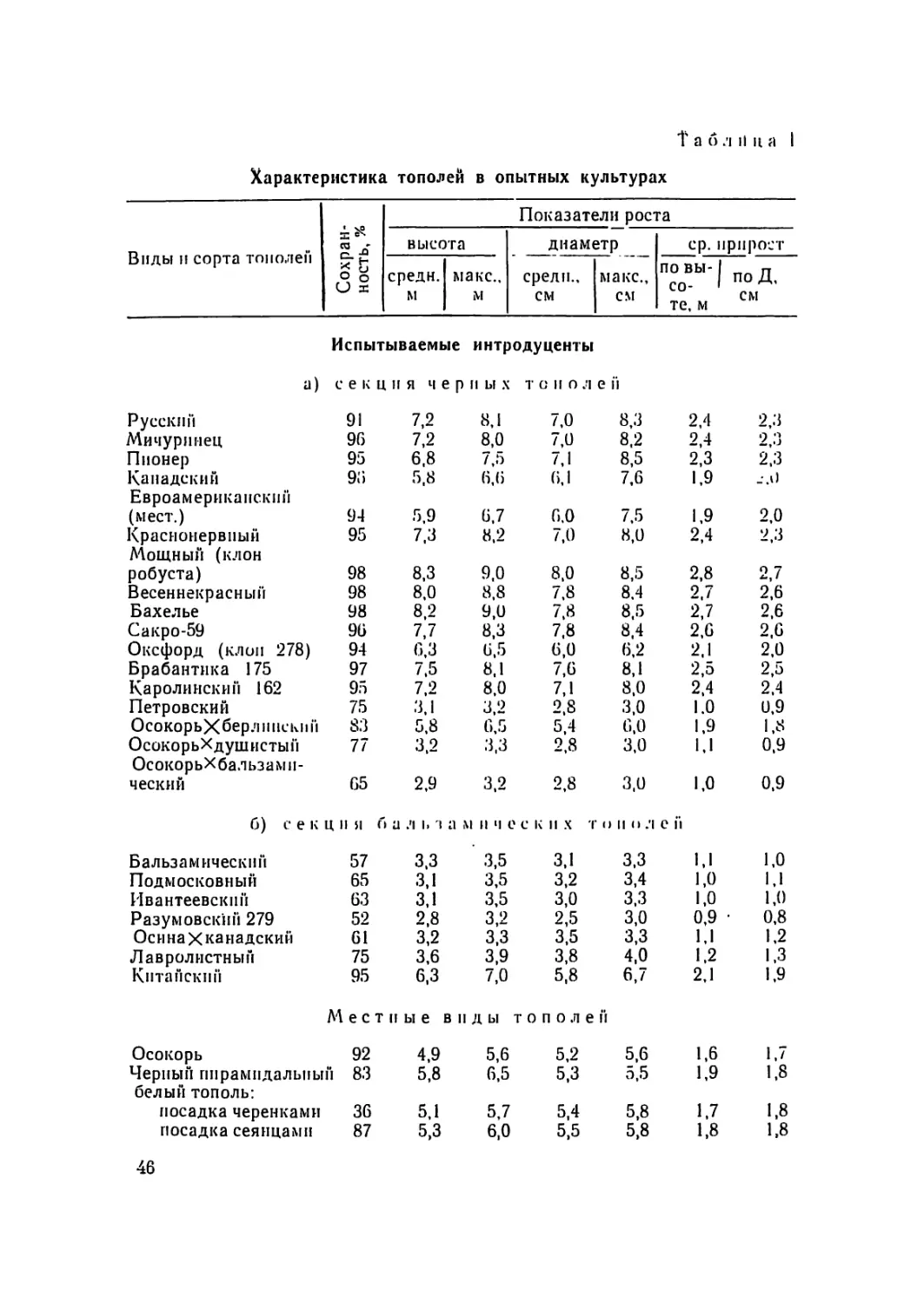
Объекты и объем исследований. Краткая ха-
рактеристика объектов исследования приведена в таблице 1.
Общая площадь сортоиспытательных участков составляет око-
ло 6,0 га. Они расположены в пойме на аллювиальных слои-
стых и зернистых почвах и в нагорных условиях иа водораз-
деле и приводораздельном склоне. Аллейные посадки находят-
ся в нагорных условиях. Общее количество учтенных деревьев
31
Таблица 1
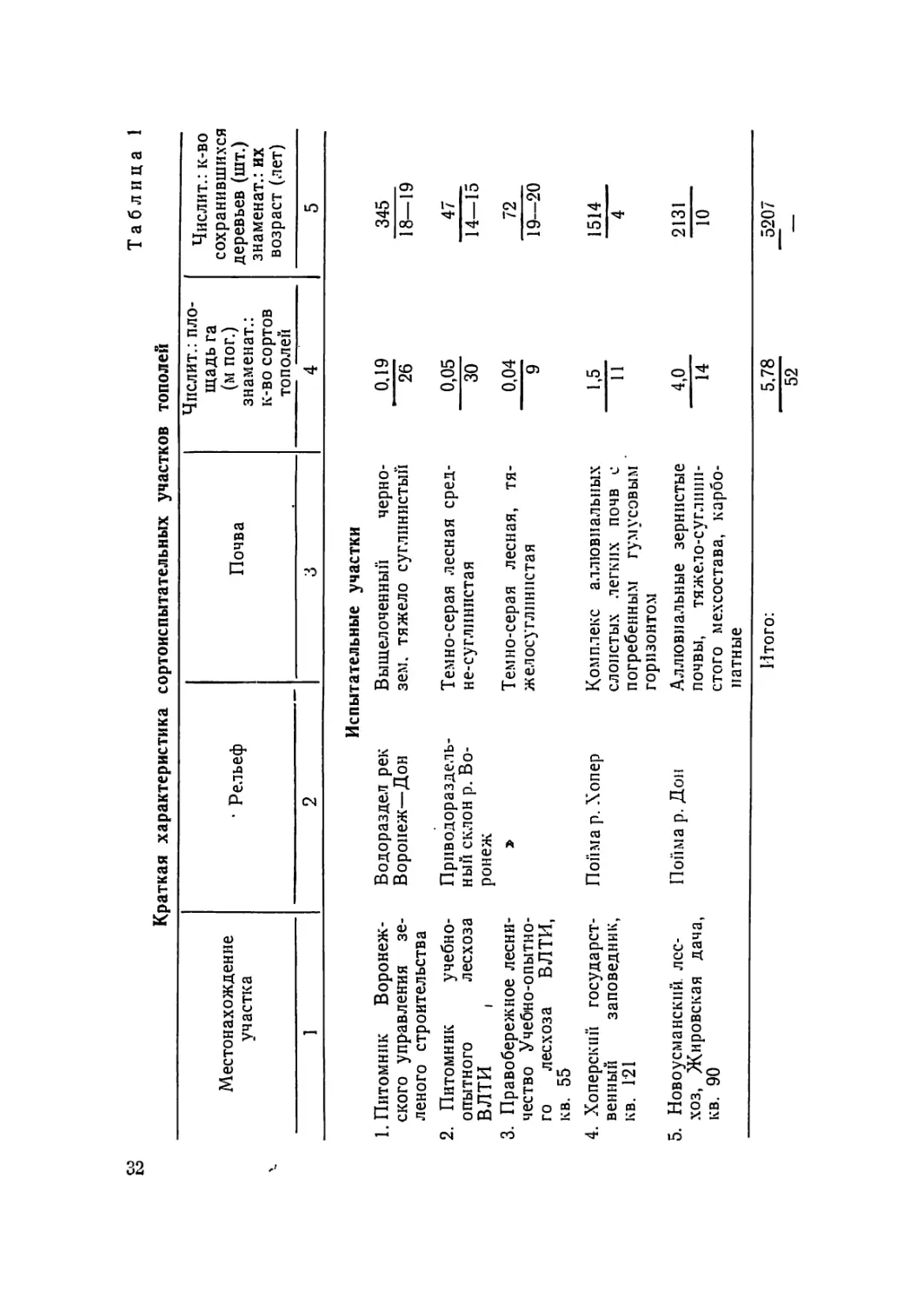
Краткая характеристика сортоиспытательных участков тополей
Местонахождение участка • Рельеф Почва Числит.: пло- щадь га (м пог.) знамен ат.: к-во сортов тополей Числит.: к-во сохранившихся деревьев (шт.) знаменат.: их возраст (лет) 5
1 2 3 4
Испытательные участки
1. Питомник Воронеж- Водораздел рек Выщелоченный черно- 0,19 345
ского управления зе- леного строительства Воронеж—Дон зем. тяжело суглинистый 26 18—19
2. Питомник учебно- Приводораздель- Темно-серая лесная сред- 0,05 47
опытного лесхоза ный склон р. Во- не-суглинистая 30 14—15
ВЛТИ । ронеж
3. Правобережное лесни- Темно-серая лесная, тя- 0,04 72
чество Учебно-опытно- го лесхоза ВЛТИ, кв. 55 жел осугллннстая 9 19—20
4. Хоперский государст- Пойма р. Хопер Комплекс аллювиальных 1,5 1514
венный заповедник, слоистых легких почв с 11 4
кв. Г21 погребенным гумусовым ’ горизонтом
5. Новоусманский. лес- Пойма р. Дон Аллювиальные зернистые 4,0 2131
хоз, Жировская дача, кв. 90 почвы, тяжело-суглини- стого мехсостава, карбо- 14 10
натные
Итого: 5,78 _5207
52
Заказ 6609
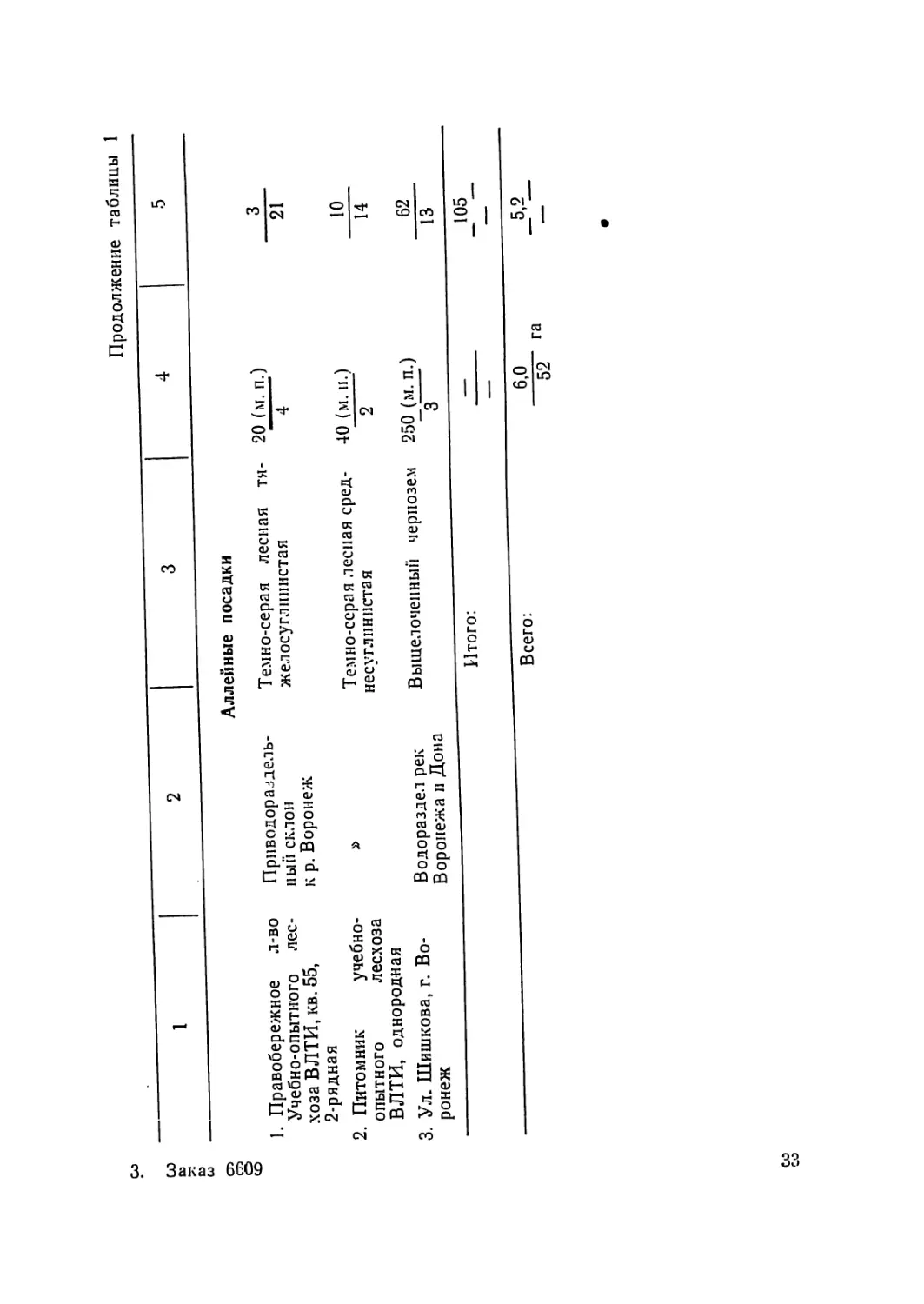
Продолжение таблицы 1
1 2 3 4 5
Аллейные посадки
1. Правобережное л-во Приводораздель- Темно-серая лесная тя- 20 (м. п.) 3
Учебно-опытного лес- хоза ВЛТИ, кв. 55, 2-рядная 11 ый склон к р. Воронеж желосуглнннстая 4 21
2. Питомник учебно- Темно-серая лесная сред- 40 (м. и.) 10
опытного лесхоза ВЛТИ, однородная несуглинистая 2 14
3. Ул. Шишкова, г. Во- Водораздел рек Выщелоченный чернозем 250 (м. и.) 62
ронеж Воронежа и Дона 3 13
Итого: — _105__
— —
Всего: 6,0 5,2
52 га “
5,2 тысячи. Возраст их колебался От 4 лет до 21 года. Пред-
ставлены и изучались следующие виды и гибриды тополей (не-
сколько тополей, из числа произрастающих в Жировской даче
отмечены в списке и в таблицах буквой «ж», поскольку не
исключена неточность в их наименовании при закладке опыта);
I. Виды: 1) белый (Р. alba L.), 2) осина (Р. tremula L.),
3) бальзамический — «ж» (Р. balsamifera L.), 4) волосисто-
плодный (Р. trichocarpa Torr et Gray).
II. Гибриды белых тополей (внутри- и межсекционные):
5) Волле 'камышинский (Р. alba L. х Р. Bolleana Lauche),
6) Советский (то же), 7) гибрид (Р. alba L. х Р. tremula L.).
111. Гибриды осин (межсекцпонные): 8) осина Веселобоко-
веньковская (Р. tremula L. < Р. Bolleana Lauche), 9) тополь
Яблокова (то же), 10) гибрид (Р. tremula L. X Р. alba L.),
11) гибрид — «ж» (Р. tremula L. X Р. Simonii Сагг.), 12) гиб-
рид - «ж» (Р. tremula L. а Р. canadensis Moench).
IV. Гибриды черных тополей (внутрисекционные): 13) Пио-
нер — «ж» (Р. pyramidalis Roz. А Р. nigra L.),14) Русский —
«ж» (то же), 15) Мичуринец (то же), 16) Пирамидально-осо-
коревый камышинский (то же), 17) гибрид № 180 — «ж»
(Р. nigra L :< Р. pyramidalis Roz.), 18) гибрид (Р. pyrami-
dal is Roz. X Р. deltoides Marsh), 19) Мариландика-543
(P. X euramericana (Dode) Guinier cv.’marilandica’), 20) Ma-
риландика-476 (то же), 21) Мариландика румынский (то же),
22) Робуста-162 (Р. X euramericana (Dode) Guinier cv. ’ro-
busta'), 23) Робуста-173 (то же).
V. Гибриды черных тополей (межсекционные): 24) гибрид
(Р. pyramidalis Roz. X Р. berolinensis Dipp.), 25) Краснонерв-
ный (Р. rubrinervis hort. Alb.), 26) Пирамидальный улучшен-
ный .№ 667 (Р. pyramidalis Roz. X Р. Simonii Carr.),27) Пира-
мидальный улучшенный № 673 (то же), 28) гибрид ЭС-38
(Р. deltoidts Marsh :< (Р. alba L. + Р. tremula L. + P. balsa-
mifera L.) — .по данным отдела генетики ЦНИИЛГиС
(10. А. Смирнов) тополь ЭС-38 является трип л ои дом; 29) гиб-
рид № 117 — «ж» (Р. nigra L. X Р. berolinensis Dipp.),
30) гибрид № 87 — «ж» (то же); 31) гибрид № 165 — «ж»
(Р. nigra L. X Р. berolinensis Dipp.), 32) Петровский — «ж»
(Р. Petrowskiana Schroed.).
VI. Гибриды бальзамических тополей (внутрисекцион-
ные): 33) Соболева (Р. Maximowiczii Henry X Р. suaveolens
Fisch).
VII. Гибриды бальзамических тополей (межсекционные):
34) Бальзамический улучшенный (Р. balsamifera L. X Р. Ье-
34
Т а б л п ц а 2
АвторЫ-сеЛекцИоНерЫ й МестоПроисХождение тополей
Авторы селекционеры №№ тополей, введенных в сортоиспытание Где получены гибриды Откуда прислан по- садочный материал для опытов
1 2 3 4
Альбепскпй А. В. 25 34 5, 16, 18, 26, 27 г. Москва г. Москва г. Камышин г. Москва г. Камышин г. Камышин
Альбенекий А. В. Делицнна А. В. 42, 43 г. Москва г. Камышин
Березин А. М. 11, 12, 17,29,30, 31,38, 40 г. Уфа г. Уфа
Бондаренко Н. 11. 24 г. Умапь г. Умань
Вересин М. М. 7, 10, 28, 35,36,41 г. Воронеж г. Воронеж
Соболев И. 11. 33 г. Минск г. Минск
Шумилина 3. К. 44, 45, 46, 47, 48, 49, 50,51,52 г. Москва г. Москва
Щепотьев Ф. Л. 8 п. Веселые Бокоп. Веселые Боко- веньки Кирово* веньки Кирово- градской обл. градской обл.
Яблоков А. С. 6, 9, 13, 14, 15, 37, 39 г. Пушкино г. Пушкино (Ивантеевка) (Ивантеевка) Московской обл. Московской обл.
— 1.2 — Местные, воронежские
— 4 — г. Рига
— 3 — г. Уфа
— 32 г. Москва г. Уфа
— 21 — Румыния, Констанца
— 19, 20. 22, 23 — Чехословакия
rolinensis Dipp.), 35) гибрид ЭС-53 (Р. balsamifera L. х Р. ру-
ramidalis Roz.), 36) гибрид ЭС-42 (то же), 37) Подмосков-
ный — «ж» (Р. suaveolens Fisch ' Р. tremula L.), 38) гиб-
рид № 188 — «ж» (Р. balsamifera L. X Р. canescens Smith),
39) Ивантеевский — «ж» (Р. suaveolens Fisch X Р. berolinen-
sis Dipp).
VI11. Сложные гибриды настоящих тополей: 40) гиб-
3
35
ряд № 161 — «ж» (Р. nigra L. X Р. berolinensis Dipp.) ><
P. balsamifera L.), 41) гибрид (тополь Самофала X P. pyrami-
dalis Roz.), 42) Московский улуч-шеииым № 33 (P. moskowien-
sis Schroecler X P. berolinensis Dip-p.), 43) Московский улуч-
шенный б/№ (то же), 44) гибрид № 1111 (Р. moskowiensis
Schroeder X Р. rubrinervis hort. Alb.), 45) гибрид № 1115
(то же), 46) гибрид № 1131 (то же), 47) гибрид № 1133 (то
же), 48) гибрид № 1416 (Р. moskowiensis Schroeder X Р. be-
rolinensis Dipp.) X Р. rubrinervis hort. Alb.), 49) гиб-
рид № 1420 (то же), 50) гибрид № 1429 (то же), 51) гиб-
рид № 1430 (то же), 52) гибрид № 1436 (то же).
В сортоиспытании представлены тополя, полученные из
11 пунктов. Авторы-селекционеры и местопроисхождение от-
дельных гибридов и видов тополей приведены в таблице 2..
Методика и с с ледова и и й. Изучались сохранность,
рост, устойчивость к биотическим и абиотическим факторам
среды н качество древесины испытываемых тополей. При раз-
работке методики учтены предложения по методике сортоиспы-
тания ВНI4PIJIM (Ростовцев, .1961) и УкрНИИЛХА (Старова,
1962). У каждого дерева, определялись диаметр, высота, раз-
мер и форма ствола, повреждение морозами, энтомовредите-
лямн, болезнями и животными, отмечался пол.
Форма кроны считалась узкой прими;!.ал иной, если отноше-
ние ее ширины к длине было менее ’А; среднепирампдаль-
ной — отношение колебалось от 7з до ’А и широкоппрамидаль-
iioii — было более 7з-
По форме ствола приняты следующие категории:
1) прямо!*! ствол — искривлений ствола нет или они очень
слабые от комля до вершины; 2) средио-искривленный ствол —
наблюдалась легкая кривизна ствола или до начала кроны он
был прямо]*!, а в кроне изгибался и разветвлялся; 3) кривой
ствол — значительная кривизна ствола наблюдалась до нача-
ла кроны.
Пораженность морозами определялась по наличию морозо-
бойиых трещин.
Степень поражения первичными энтомовредителями и фито-
болеапямн молодых побегов и листьев оценивалась по следую-
щей шкале: 0 — поражение отсутствует — пораженных листь-
ев или побегов пет; 1 — слабое — поражено до 10%; 2 — сред-
нее — 11—25%; 3 — сильное — 26—50%; 4 — очень силь-
ное — 51—75%; 5 — полное поражение — поражено свыше
75% листьев или побегов.
36
Степень поражения вторичными зптомовредителями и фи-
тоболезиямл стволов оценивалась по следующей шкале:
О — повреждений нет;
1 — повреждение слабое — растение имеет: 1 летное отвер-
стие или единичные вздутия, небольшие участки единичного
। го сел еа ги я щитовки;
2 — повреждение среднее — 2—5 летных отверстий или
вздутия на многих ветвях, на стволе и ветвях большие участ-
ки поселения щитовки пли повреждения болезнью (до поло-
вины окружности ствола);
3 — повреждение сильное — изломы в месте поражения
стеклянницей (6—10 летных отверстий) или вздутия на боль-
шинстве ветвей (па некоторых по несколько штук), в резуль-
тате поражения больших участков ствола и скелетных ветвей
щитовк-ой или болезнью наблюдается усыхание некоторых ске-
летных ветвей;
4 — повреждение очень сильное — растение отмерло пли
отмирает.
Анализ полевых материалов позволил выделить обобщенные
категории, характеризующие виды и гибриды тополей ио габи-
тусу кроны, форме ствола, производительности и устойчивости.
Габитус кроны тополей одного и того же вида или клопа
варьировал в зависимости от условий роста и возраста. С уче-
том амплитуды варьирования виды и формы, находящиеся в
испытании, были разделены по этому признаку на следующие
три категории (по преобладающему числу деревьев в насаж-
дениях более старшего возраста): узкокронные (пирамидаль-
ные), пол пирамидальные и ширококропные (тополя с раски-
дистой кроной).
Категории но форме ствола устанавливались соответственно
преобладанию растений с прямыми, слабопскривлеиными или
кривыми стволами.
По производительности все тополя были разбиты также на
три категории: 1) высокая производительность: средний при-
рост в 10-летием и старшем возрасте был выше 15 м3/га; рост
испытываемого сорта при 5%-пом уровне значимости досто-
верно лучше, чем у контрольного; и у единично сохранившихся
деревьев объем ствола выше или равен объему ствола одного
из лучших сортов, взятого за контроль в данных условиях
местопроизрастания. Таким эталоном был взят тополь красно-
нервный, деревья которого в 15-летнем возрасте имели средний
объем ствола 0,4 м3.
2) Средняя производительность: средний прирост в 10-лет-
37
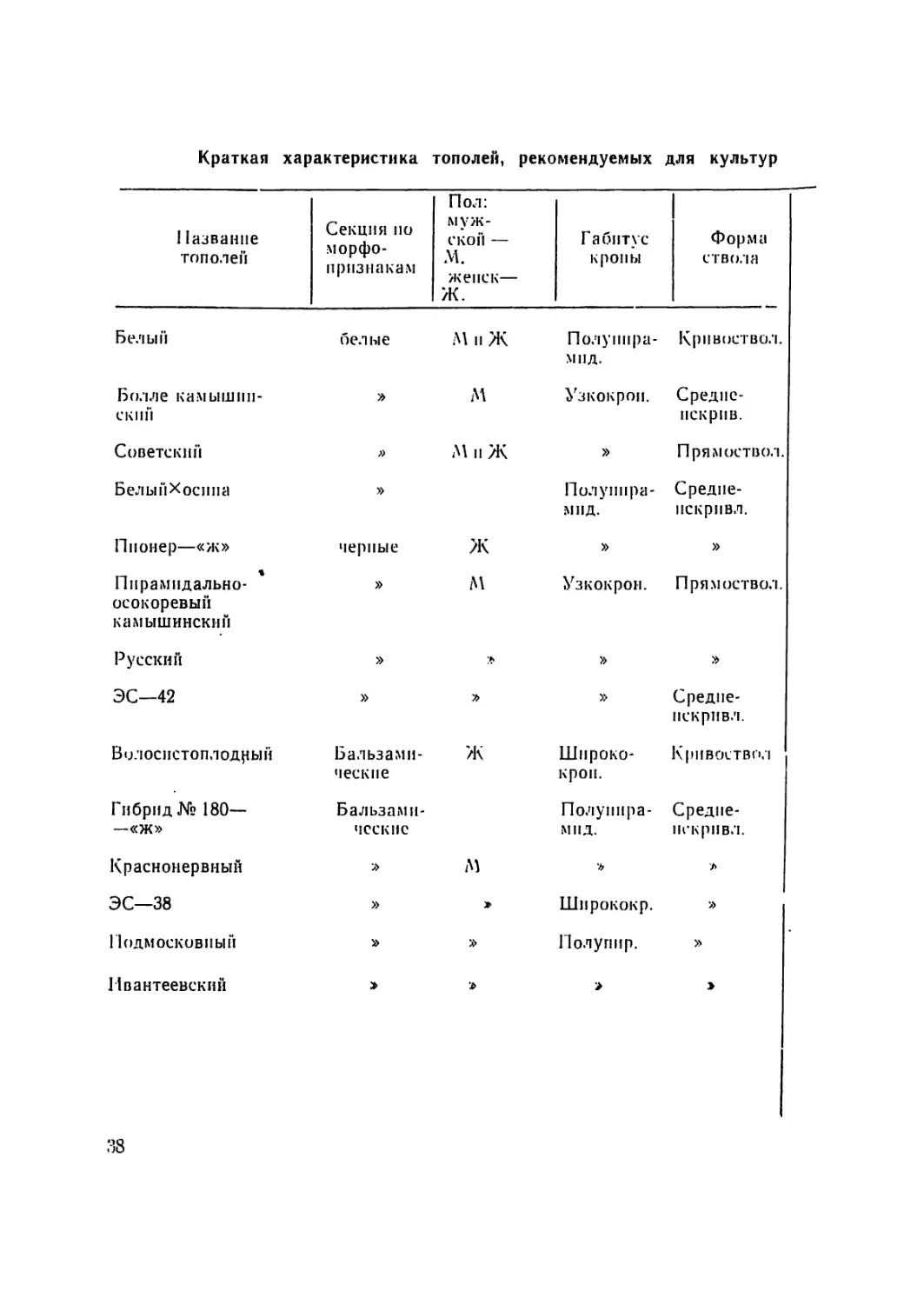
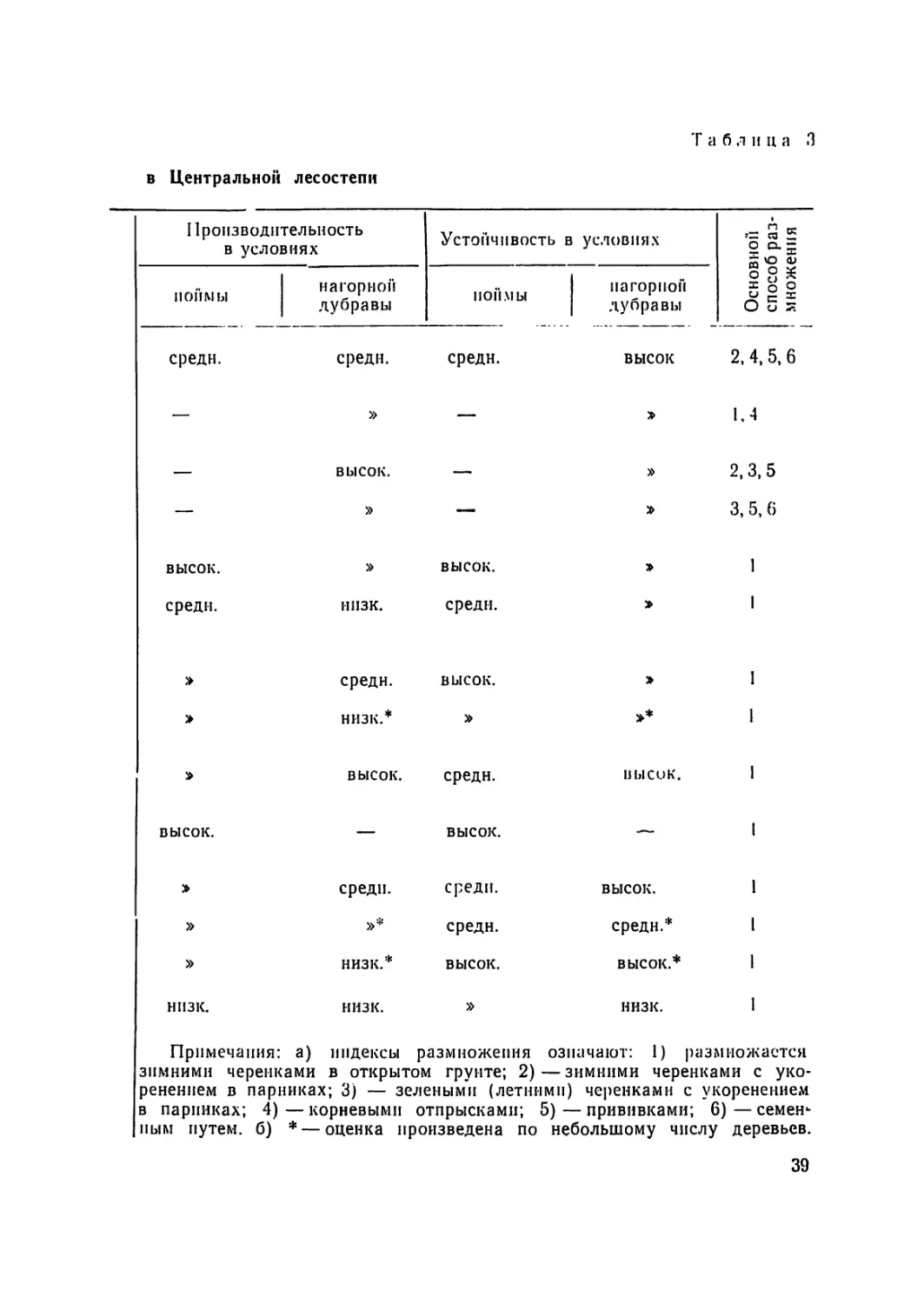
Краткая характеристика тополей, рекомендуемых для культур
11азванпе тополей Секция по морфо- ii ризнакам Пол: муж- ской — М. жепск— Ж. Габитус кроны Форма ствола
Белый белые М п Ж Полупнра- мпд. Кривоствол.
Болле камышин- ский » М Узкокрон. Средпе- нскрпв.
Советский » М п Ж » Прямоствол.
Белы и Хосп и а » Пол у пира- мид. Средпе- пскрпвл.
Пионер—«ж» черные Ж » »
Пирамидально- осокоревый камышинский » м Узкокрон. Прямоствол.
Русский » » »
ЭС—42 » » Средпе- пскривл.
Волоспстоплодцый Бальзами- ческие ж Широко- Кривоствол крои.
Гибрид № 180— Бальзами- Полупира- Средне-
—«ж» ческие мнд. нек рпвл.
Краснонервный м 'Ь л
ЭС—38 » > Шпрококр. »
Подмосковный » » Полупир. »
Ивантеевский х> >
38
Т а б л и ц а .3
в Центральной лесостепи
Производительность в условиях Устойчивость в условиях зной б раз- ения
поймы нагорной дубравы поймы | нагорной дубравы О 8 * = О ° о 5 §
средн. средн. средн. высок 2, 4, 5, 6
— » — 1,4
— высок. — » 2,3,5
— » — 3, 5, 6
высок. » высок. » 1
средн. ннзк. среди. 1
средн. высок. > 1
низк.* >* 1
высок. средн. высок. 1
высок. — высок. — 1
среди. среди. высок. 1
» »* средн. средн.* 1
» низк.* высок. высок.* 1
ннзк. низк. » низк. 1
Примечания: а) индексы размножения означают: 1) размножается
зимними черенками в открытом грунте; 2)—зимними черенками с уко-
ренением в парниках; 3) — зелеными (летними) черенками с укоренением
в парниках; 4) — корневыми отпрысками; 5) — прививками; 6) — семен-
ным путем, б) * — оценка произведена по небольшому числу деревьев.
39
нем и старшем возрасте колебался от 10 до 15 м3/га; рост испы-
тываемого сорта пе отличался достоверно от роста контрольного
сорта; у единично сохранившихся деревьев объем ствола меньше
па 1 — 50% по сравнению с объемом контроля.
3) Низкая производительность: средний прирост в принятом
возрастном диапазоне меньше .10 м3/га; но росту уступают досто-
верно контролю; и у единично сохранившихся деревьев объем
ствола на 50% (и более) ниже, чем .у тополя красиоверв-
но го.
По устойчивости к эитомо- и фитовредителям и факторам
абиотической среды были выделены следующие три категории:
1) высокая— до 25% слабо (балл «1») и средне (балл «2»)
пораженных деревьев или до 10% сильно и очень сильно
пораженных (балл «3», «4», «5»);
2) средняя — от 26 до 50% слабо и средае-норажепных
деревьев или от 11 до 25% сильно и очень сильно пораженных;
3) низкая — более 50% слабо и средне пораженных деревь-
ев или свыше 25% сильно и очень сильно пораженных.
Во всех случаях устойчивость оценивалась по отношению
деревьев к наиболее опасному повреждению для данных усло-
вий .произрастания.
Р е з у л ь т а т ы и с с л е д о в а н п й. Производительность
тополей резко варьировала в зависимости от сорта. Так, в пой-
менных условиях средний годичный прирост колебался от
7,7 м3/га (у гибрида осина х канадский) до 24,7 м3/га (у топо-
ля Пионер); в условиях нагорной дубравы (водораздельный
участок) от 1,4 м3/га (у т. Соболева) до 23,0 м3/га (у т. Крас-
лонервного).
Поражение первичными зпгомовредптелям'п в той или иной
степени наблюдалось па всех участках и почти у всех тополей.
Наиболее распространенными были повреждения от листоедов,
различных видов, трубковерта Bycliscus populi L., тополевого
слоника-блошки Rhynchaenus (Orchestes) populi F., различно-
го рода тлей. Однако степень поражения листвы не превы-
шала 10% ее количества у дерева.
Из вторичных вредителей ладо отметить большую тополе-
вую стеклянницу (Sessia apiformis CL), стеклянницу малую
(Sciapteron tabaniforme Rott.), яблоневую запятовидную щи-
товку (Lepidosaphes ulmi L.).
Наиболее устойчивы в пойменных условиях к вторичным
вредителям оказались тополь белый, Пионер, гибрид ЭС-38,
Робуста-173, волосистоплодный, Русский (поврежденных расте-
ний не более 10—14%).
40
В условиях пагорпой дубравы большинство тополей оказа-
лись непораженными вторичными вредителями. Наилучшимн
но устойчивости были Болле камышинский, Советский, Пира-
мидально-осокоревый камышинский, Красиолервиый.
Наиболее пораженными оказались тополя — гибриды № 33,
1416, 1111, Бальзамический улучшенным.
Из фнтоболезней в нагорных условиях наиболее распростра-
нены септориоз листьев (поражал тополя № 33, Бальзамиче-
ский улучшенный, № 673), цитоспорноз (тополь Ивантеев-
ский), камедетечение от разных причин (667, 673, 1133), ги-
ноксплоиовый рак (тополь Яблокова).
В пойменных и пагориых условиях встречаются поврежде-
ния стволов морозобойными трещинами. Более значительное
н'оврежде.ине ими (до 50—57% стволов) отмечено для тополей
Бальзамического улучшенного и Пирамидального улучшенного
.№ 673. Тополя Пирамидально-осокоревый камышинский, Крас-
нолервпый, Бальзамический улучшенный № 667, Пионер, Ро-
буста-173, белый — имели 10—30% пораженных стволов, у
остальных тополей поражение отсутствовало или не превышало
3—5% от общего количества стволов.
По показателям производительности и устойчивости топо-
лей из 52 находящихся в сортоиспытании видов п клопов выде-
лено И наиболее перспективных, рекомендуемых для разведе-
ния в условиях ЦЧР. Краткая характеристика этих тополей
нрнводится в таблице 3.
В основном ото растения, показавшие высокую пли среднюю
производительность при высокой устойчивости. Некоторые то-
поля ле отличаются высокими показателями роста (Подмосков-
ный, Ивантеевский п др.), но включены в перспективный ассор-
тимент для озеленения, т. к. представлены только мужскими,
(не дающими «пуха») особями п отличаются декоратив-
ностью.
С учетом установленных по данным исследований особен-
ностей выделенных лучших видов и сортов тополей, можно
дать следующие рекомендации по их внедрению, в зависимо-
сти от условий местопроизрастания (пойменные пли нагорные)
и категории насаждений (лесные культуры, защитные посадки,
озеленение).
Для лесных культур и защитных лесонасаждении в пой-
менных условиях могут быть рекомендованы тополь белый,
гибрид № 180 «ж», Пионер, тополь ЭС-38, Краснонервный и
волостгстоплодпый; в нагорных условиях для этих же целей —
тополь белый, волосистоплодиый, гибрид тополь белый X оси-
41
на, Пионер, Краспопервный. В защитные насаждения в нагор-
ных условиях могут быть рекомендованы также тополи Болле
к а м ы и j -1111 с к11 й и П т । р а м 11 д а л ы । ()- о с (> к ор е в ы н к а м ыш и и с к и й.
В озеленительные посадки в пойме н в условиях нагорных
дубрав могут быть рекомендованы следующие тополи, представ-
ленные только мужскими клопами: Болле камышинский, Пи-
рамидально-осокоревый камышинский, Русский, Краевоиерв-
ный, гибрид ЭС-42, гибрид ЭС-38, Подмосковный и Ивантеев-
ский; для озеленения могут быть также рекомендованы муж-
ские клоны тополя белого, Советского и гибрида тополь бе-
лы и X осина.
В качестве временной быстрорастущей породы в культурах
дуба, но нсныта.нням в нагорных условиях, могут быть рекомен-
дованы узкокронные, не заглушающие дуб тополя: Болле камы-
ши помпй. Пирамндалыю-осокоревый камышинский, Русский,
гибрид ЭС-42.
Некоторые тополя были представлены в опытах недоста-
точным количеством деревьев или еще молодыми экземпляра-
ми. Для этих тополей не дается рекомендаций по внедрению;
однако предварительные данные говорят в пользу дальнейшего
испытания некоторых из них. К таким перспективным тополям
надо отнести тополи Мариландпка-476, 543 и Марпландмка ру-
мынский, ЭС-53 и ЭС-38 (для лесных культур и защитных ле-
сонасаждении). Тополь Соболева, № 1115, пирамидальный
улучшенный № 667 н № 673, гибриды ЭС-38 и ЭС-53 (для озе-
ленения).
ЛИТЕРАТУРА
Верес пн М. М. Итоги опытных работ ВЛТИ по интродукции ц
селекции тополей в Воронежской области ла 1951 — 1966 it. (Научпый
отчет за 19G6 г.). Машинопись, Воронеж, 1967.
И в а и и и к о в С. И. Выведение и использование тополей в СССР
и за рубежом (Обзор). Москва, ЦБНТИ Гослесхоз,, 1971.
О з о л и и Г. И. Степень изученности и практического использования
тополей в СССР и зарубежных странах. Труды ВНИИ агролесомелиора-
ции, вып. I (61), 1970 (1971). а
Ростовцев С. А. Методика сортоиспытания тополей. Пушкино,
1961.1
Ростовцев С. А. Районирование культуры сортовых тополей
в Европейской части РСФСР. Пушкино, 1963.
Старова П. В. Биологические основы селекции тополей. Авторе-
ферат дисс. на сопск. уч. степени доктора биологических паук. М.,
J971.
Старова П. В. Методика сортоиспытания тополей. Харьков, 1962.
42
А. И. СИДОРОВ
Сортоиспытание тополей в пойменных условиях Кубани
Пойменные леса в Краснодарском крае занимают площадь
свыше 50 тыс. га. В основном эти насаждения малопродуктив-
ны (запас около 100 м3 на 1 га, выход деловой древесины
в пределах 20—25%, средний годичный прирост 3,4—4,2 м3).
В то же время условия поймы Кубани (осадки 550 мм, средне-
годовая температура +'10сС, число дней вегетационного перио-
да до 250, уровень грунтовых вод 1,5—2,5 м) позволяют вы-
ращивать высокопроизводительные тополевые насаждения.
Вопросами подбора ассортимента сортовых тополей и их
выращивания в нашей стране занимались многие исследова-
тели: П. Л. Богданов (1936, 1958, 1965), А. М. Березин (1939),
А. С. Яблоков (1940, 1962, 1967), А. В. Альбенский (1946,
1956, 1959), М. М. Вересин (1953, 1959), Г. П. Озолии (1956),
Л. Ф. Правдин (1960), С. П. Иванников (1961, 1966), Ф. Л. Ще-
потьев (1962), Г. И. Редько (1963, 1966), Н. В. Старова (1964),
Д. Д. Лавриненко (1966) и др. Вместе с тем по указанным
вопросам исследований на Кубани проведено было крайне не-
достаточно.
С целью уточнения ассортимента тополей, наиболее при-
годных для выращивания в этой зоне, с 1961 года в ряде лес-
хозов, размещенных в пойме Средней Кубани (Армавирский,
Кропоткинский и др.), ВНИИЛМ и Министерство лесного хо-
зяйства РСФСР организовали ведомственное сортоиспытание
гибридных тополей, непосредственно осуществлявшееся Северо-
Кавказской ЛОС (В. Н. Какушкпн) и автором настоящей
статьи.
Опытные объекты были заложены на площади 95 га. Иссле-
дование . проводилось по единой методике ВНИИЛМ
(С. П. Иванников и С. А. Ростовцев, 1961). В сортоиспытание
включено 24 перспективных вида и сорта тополей отечествен-
ной и зарубежной селекции, рекомендованных ВНИИЛМ. Для
сравнения дополнительно включены местные тополи: осокорь,
черный пирамидальный н белый тополь.
В течение 1967—1970 гг. нами была проведена инвентари-
зация сортоиспытательных культур, включавшая в себя фено-
логические наблюдения и снятие биометрических характеристик
по росту и продуктивности, а также устойчивости против энто-
мовредителей и фитоболезией. Ниже приводятся основные ито-
ги этих исследований,
43
По фенологии отмечается более раппее начало разверзаппя
почек (начало роста) у тополей бальзамической секции
(22—24 марта). Окончание роста и заложение верхушечных
почек у этих тополе]”! происходит в первой декаде сентября.
Продолжительность периода роста побегов составляет
170—172 дня. В чти же сроки начитают и заканчивают рост
тополя: Петровский, осокорь X душистый, осокорь X бальзами-
ческий из секции черных тополей.
Тополи селекции А. С. Яблокова (русский, мичуринец,
пионе])) начинают рост с 9 апреля и закладывают верхушеч-
ную почку в конце сентября, имея продолжительность роста
170 дней. Виды и сорта из секции черных тополей (мощный,
весен нек-рас и ый, Бахелье, с-акро-59, Прибалтика-175, Каролип-
ский-162, Канадский) начинают вегетацию с 11 —12 апреля и
закладывают верхушечную почку в конце первой декады ок-
тября, продолжительность роста 183 дня. Примерно те же
сроки начала и окончания роста имеют местные виды тополей
(осокорь, тополь белый и черный пирамидальный, признанный
международной комиссией по тополю в качестве стандарта для
фенологических наблюдений).
Последовательность наступления других фаз развития —
созревания семян, пожелтения и опадения листьев и др. зави-
сит в большей мере от погодных условий, особенно от количе-
ства и частоты осадков, а потому не всегда является постоян-
ной. Раньше желтеют (начало ссптября) н полностью опадают
к 25 сентября листья у бальзамических тополей. Иногда опаде-
ние листьев у этих тополей начинается в августе и уже закан-
чивается в начале сентября. Долго стоят в облиственном со-
стоянии (до 25—27 октября) тополи: русский, мичуринец,
пионер, осокорь X берлинский, китайский, краспопервный.
Очень долго удерживают листья черные тополи (мощный, ве-
сеп/нскраспый, бахелье, брабаптина-175, каролпнский-162, сак-
ро-59, канадский) и местные виды тополей (осокорь, тополь
белый и черный пирамидальный). Опадение листьев у этих
тополей заканчивается 9—12 поября. У тополя мощного листья
до конца вегетации сохраняют зеленую окраску. Это имеет
значение при оценке декоративных качеств разных видов то-
полей.
Исследования показывают, что продолжительность вегета-
ции тополей в условиях Кубани не одинакова. У бальзамиче-
ских тополей период вегетации равен 181—187 дням, у черных
тополей — 197—209 дпям, у местпых видов тополей 210—
214 дням. Все эти особенности могут существенно облегчить
44
или уточнить определение видов и сортов тополей, установле-
ние сроков сбора и высева семян, а также оптимальных перио-
дов проведения различных уходов за тополями.
Различные виды п сорта тополей в разной степени пора-
жаются вредителями и болезнями. Так, исследования показы-
вают, что тополь Краснонервный в наибольшей степени под-
вержен различным заболеваниям. Его молодые стволики и вет-
ви сильно повреждаются бактериальным язвенно-опухолевид-
ным раком, возбудителем которого на тополях является бакте-
рия Micrococcus populi Del. (А. И. Воронцов и В. П. Гречкин,
1962).
В культурах посадки 1967 г. выявлена зараженность в сла-
бой степени (до 10%) стволиков тополя краснонервиого доти-
щицей тополевой. Это заболевание вызывается сумчатым гри-
бом, коиидиальпая стадия которого имеет название — Dot-
hichiza populea Sacc. Листья тополя краснонервиого поража-
ются в сильной степени ржавчинным грибом и преждевремен-
но опадают (начало сентября).
Сорта из секций черных тополей (петровский, осокорь X
берлинский, осокорь X душистый, осокорь X бальзамический)
имеют повреждения в пределах 10—15% бактериальным язвен-
но-опухолевидным раком и в большей степени (15—20%)
повреждаются вредителями — стеклянницей темпокрьглой
(Paranthreno tabanifarinis Ratt.) и малым тополевым усачем
(Saperda populea L.). /
Тополи бальзамической секции (подмосковный, ивантеев-
ский, разумовскпй-279, осина X канадский, бальзамический)
также повреждаются темпокрьглой стеклянницей п малым то-
полевым усачом. Листья этих тополей в сильной степени по-
вреждаются тополевым листоедом тг ржавчинным грибом. Из
бальзамических тополей более устойчивыми к вредителям ока-
зались тополи китайский и лавролистпый. Они имеют еди-
и г । ч и ы е пив р еж де и и я.
Высокую устойчивость против болезней и вредителей пока-
зали виды и сорта из секции черных тополей: брабантина-175,
каролинский-162, мощный, весениекрасный, бахелье, сакро-59,
канадский, евроамерикаиский и др. Эти тополи не имеют по-
вреждений. Также не имеют повреждений местные виды тополей
(осокорь, тополь белый, черный пирамидальный).
В результате 5-лет.исго сортоиспытания выявились тополи,
непригодные для разведения в условиях Кубани. К ним отно-
сятся виды и сорта из бальзамической секции тополей, за
исключением китанекого.
43
t а б л и ц a I
Характеристика тополей в опытных культурах
Виды п сорта тополей Сохран- ность, % Показатели роста
высота диаметр ср. прирост
средн, м макс., м средн., см макс., см под- те.. “
Испытываемые интродуценты
а) с е к ц и я ч е р и ы х т о и о л е й
Русский 91 7,2 8,1 7,0 8,3 2,4 2,3
Мичуринец 96 7,2 8.0 7,0 8.2 2.4 2,3
Пионер 95 6.8 7,5 7,1 8,5 2,3 2,3
Канадский 95 5,8 6,6 6,1 7,6 1,9 _•.<)
Евроамерикаискип
(мест.) 94 6.9 6,7 6.0 7,5 1,9 2.0
Краснонервпый 95 7.3 8,2 7.0 8.0 2.4 2,3
Мощный (клон
робуста) 98 8.3 9.0 8.0 8,5 2.8 2,7
Весеннекрасный 98 8,0 8.8 7,8 8.4 2.7 2.6
Бахелье 98 8.2 9.0 7.8 8.5 2,7 2.6
С акр о-59 96 7,7 8.3 7.8 8.4 2.6 2.6
Оксфорд (клоп 278) 94 6,3 6.5 6.0 6.2 2.1 2,0
Брабантика 175 97 7,5 8.1 7,6 8,1 2,5 2.5
Каролинский 162 95 7.2 8.0 7,1 8.0 2,4 2.4
Петровский 75 3.1 3.2 2.8 3.0 1.0 0.9
ОсокорьХ берлински и 83 5.8 6,5 5,4 6.0 1,9 1.8
ОсокорьХдушистып 77 3,2 з.з 2,8 3,0 1,1 0.9
ОсокорьХбальзами-
ческий 65 2,9 3.2 2,8 з.о 1,0 0.9
б) с е к ц и я б а Л 1. 1 а м и ч о с к и X Т О II о л с й
Бальзамический 57 3,3 3,5 3.1 з.з 1.1 1,0
Подмосковный 65 3,1 3,5 3,2 3,4 1,0 1,1
Ивантеевский 63 3,1 3,5 3,0 3.3 1.0 1,0
Разумовский 279 52 2.8 3,2 2,5 3,0 0,9 • 0,8
Осина X канадский 61 3,2 з.з 3.5 3,3 1,1 1.2
Лавролистнып 75 3,6 3.9 3,8 4,0 1.2 1.3
Китайский 95 6,3 7,0 5,8 6,7 2,1 1,9
Местные виды тополе й
Осокорь 92 4.9 5,6 5,2 5.6 1.6 1.7
Черный пирамидальный 83 5,8 6.5 5.3 5,5 1,9 1.8
белый тополь:
посадка черенками 36 5,1 5,7 5,4 5,8 1,7 1,8
посадка сеянцами 87 5,3 6,0 5,5 5,8 1,8 1,8
46
Рис. 1. Культуры тополя свроамсрпкапского в коридорах. Возраст
15 лет, средняя высота 21,5 м. диаметр 21 см. запас 387,6 м-ч/га
(Армавирское л-во. кв. 3).
47
Р и с. 2. Опытные культуры тополя мощного (клон 236) в пойме Кубани
(Армавирское лесничество, кв. 25). Возраст 4 года, средняя высота
11,2 м. диаметр 12 см, запас 104 м3/га.
В таблице 1 приводятся осповиые показатели, характери-
зующие рост тополей по высоте и даметру в 3-летнем возрасте
в опытных культурах Армавирского лесничества, кв. 25 (пой-
ма р. У рун, на аллювиальных мощных легко суглинистых поч-
вах с уровнем грунтовых вод 1 ,5—2 м). Посадка культур
произведена в 1966 г. черенками по лесному типу с размеще-
нием 3X2 м. Уходы проводились ежегодно по 4—5 культива-
ции в междурядьях и ручная прополка в рядках.
Из таблицы 1 видно, что некоторые сорта из секции черных
тополей (петровский, осокорь X душистый, осокорь X бальза-
мический, также как и тополш из бальзамической секции,
растут неудовлетворительно, к вредителям и болезням неустой-
чивы.
Тополи краснонервный оксфорд-278 и осокорь X берлин-
ский па сортоиспытательных объектах показали хороший рост,
но отмечена их слабая устойчивость против болезней и энто-
мовредителей.
Вполне удовлетворительный рост ио высоте и диаметру, но
меньший в сравнении с лучшими гибридными сортами черных
тополей, показали местные тополи (осокорь, черный пирами-
дальным и тополь белый). В среднем за год они имеют прирост
по высоте 1,6—1,9 м, ио диаметру 1,7—1,8 см.
Лучшие результаты по сохранности, росту и общему раз-
витию имеют растения из секции черных тополей: мощный,
бахелье, вернирубепс, сакро-59, брабантп.иа-175, каролин-
ский-162, русский, мичуринец, пионер, канадский, евроамерп-
капский (местный) и др. Эти тополи за 3 года достигли высоты
5,8—8,3 м с диаметром па высоте груди — 6—8 см, ие повреж-
даются морозами, выращиваются без полива, прямоствольны
с хорошо развитыми здоровыми кронами. У отдельных видов
(сортов) тополей выход целлюлозы 46,1—48,8% (С. П. Иван-
ников и С. А. Ростовцев, 1961).
Для разведения и выращивания в лесорастительных усло-
виях Кубани наиболее перспективные виды и сорта тополей
следующие. В лесные культуры рекомендуются тополи: мощ-
ный, весенпекраапый, бахелье, сакро-59, пионер, каролин-
ский-162, брабаптпна-175, канадский, евроамериканский (ме-
стный), осокорь и тополь белый. Для озеленения рекоменду-
ются тополи: русский, мичуринец (селекции А. С. Яблокова),
бахелье, мощный, китайский, черный пирамидальный, евро-
америкаискнй (местный).
Исследования показывают, что тополевые насаждения из
лучших гибридных сортов в оптимальных условиях выращива-
4. Заказ 6609
49
пня в пойме Кубани дают исключительно высокую продуктив-
ность. Так, например, в тине условий произрастания Дз на бо-
гатых луговых почвах (Армавирское лесничество, кв. 3) куль-
туры тополя ев'роамерикапского в коридорах в возрасте 15 лет
имели среднюю высоту 21,5 м, диаметр 21 см, запас древесины
па 1 га до 400 м3 (по таблицам Л. В. Вицина, 1967), средний
годичный прирост 25,8 м3 (рис. 1).
В культурах лесного типа (Кавказское лесничество, кв. 4)
па плодородных лугово-аллювиальных почвах с уровнем грун-
товых вод 1,5—2 м насаждения тополя канадского в во-зрасте
16 лет имели среднюю высоту 24 м, диаметр 28 см, запас
древесины на 1 га 470 м3, средний годичный прирост 29,4 м3.
Опытные культуры из тополя мощного в аналогичных усло-
виях (Армавирское лесничество, кв. 25) в 4-летнем возрасте
имеют среднюю высоту 11,2 м, диаметр 12 см, запас древеси-
ны на 1 га 104 м3 (рис. 2). Насаждения гибридных тополей в
оптимальных условиях поймы Кубани уже к 20—25 годам (ре-
комендуемый возраст рубки) могут иметь запас до 700 и более
кубометров древесины на 1 га.
ЛИТЕРАТУРА
Альбе некий А. В. Культура тополей. Гослестехиздат, 1946. Новые
гибриды тополей, ж-л Л. х., № 9, 1956. Селекция древесных пород и
семеноводство. Гослесбумиздат, 1959.
Бог д а н о в П. Л. Тополя и их культура. Изд. Лесная промышлен-
ность. М., 1936, 1965. Новые гибриды тополей. Ж. Л. х.. № 3, 1958:
Березин А. М. Описание гибридных тополей. Тр. ВНИИЛХ, 1939.
Б и ци п Л. В. Таблицы для таксации тополевых культур. Изд.
Лесн. пром., 1967.
В ере си и М. М. Семеноводство и размножение тополей. Ж. Л. х.,
№ 4, 1953. Создание семенных тополевых насаждений (стр. 258—260)
в кн. Альбепский А. В. «Селекция лесных пород п семеноводство»,
1959.1
Воронцов А. И., Гречки и В. П. Вредители и болезни тополей!
и меры борьбы с ними. Гослесбумиздат, М., 1962.
В в а н и и к о и С. П. и Ростовцев С. А. Методика сортоиспытания
тополей, ВНИИЛМ, М., 1961. Некоторые итоги испытаний сортовых то-
полей, ж. Л. х., № 8, 1961.
Иванников С. П. Разведеппе и выращивание тополей и осины
в лесостепи. Изд. ЦНИИТЭИ леспром., М., 1966.
К а к у ш к и н В. Н. Выращивание сортовых тополей на Северном
Кавказе. Сб. «Выращивание лиственных пород», ЦНИИТЭИ леспром.,
М., 1967ч
Л а в р и н е п к о Д. Д., Редько Г. И., Прилуцкий А. В. и др.
Создание тополевых насаждений. Кн. изд. Лесн. пром.. М., 1966.
О з о л и н Г. П. Выращивание тополей в Узбекистане. Ташкент,
Госиздат УзССР, 1956.
50
Прав д и н Л. Ф. п Ф нлп моиов а В. Д. Культура тополя в
Европейской части СССР. Сб. «Проблемы повышения продуктивности
лесов», т. III, гл. 6, изд. АН СССР, I960.
Старова Н. В. Селекция и сортоиспытание тополей на Украине.
Изд. «Урожаи», Киев, 1964j
Ф и л и м о и о в а В. Д. Культура тополей за границей. Гослесбумиз-
дат, М., 1962.
Щепотьев В. Л. и Павленко Ф. А. Быстрорастущие древес-
ные породы. М., 1962w
Яблоков А. С. Селекция тополей, дуба, ясеня, осины и ореха.
Гр. ВНИИЛХ, вып. VII, 1940.
Селекция древесных пород, Сельхозиздат, 1962.
Некоторые итоги работ по селекции лесных пород в РСФСР. Сб. «Ле-
сохозяйственная и лесомелиоративная наука в СССР», М., 1967.
Е. А. ПУГАЧ, М. С. НЕКРАСОВ
Идеи Г. Ф. Морозова о развитии селекции в лесоводстве
на примере некоторых древесных пород среднего Урала
Г. Ф. Морозов одним из первых лесоводов выдвинул идею
развития селекции в лесоводстве. В статье «О лесоводствснных
устоях» (1962) он писал: «Как в сельском хозяйстве возникла
целая отрасль селекций или искусственного отбора, так и у нас
в лесоводстве должна возникнуть особая глава о сознательном
отборе, представляющем собою регулирование естественного
отбора в условиях п в порядке хозяйственной дисциплины».
Рассматривая принципы вмешательства человека в лес,
Г. Ф. Морозов неоднократно обращал внимание на то, чтобы
они, увеличивая производительность природного леса, как мож-
но меньше ослабляли его природную устойчивость.
Отдавая должное искусственному отбору, как методу по-
вышения производительности древостоев, он раскрыл значение
для леса открытого Дарииным закона наследственности и нз-
мепчивости. Г. Ф. Морозов, касаясь влияния среды на измен-
чивость свойств, писал: «...что особенности внешней среды мо-
гут в известных пределах видоизменить наследственные био-
логические свойства растений, отражаясь на быстроте и пре-
дельности роста, величине растений, их формах, на плодоноше-
нии, на степени теневыносливости и т. д.».
Идеи Г. Ф. Морозова об улучшении лесных насаждений
и повышении их производительности селекционными метода-
ми получили широкое развитие в трудах крупных ученых как
в нашей стране (В. Н. Сукачев, 1922, 1928, 1938; Н. П. Кобра-
4*
51
нов, 1925; М. К. Турский, 1929; IT. С. Нестеров, 1935; С. С. Пят-
иицкнй, 1951, 1954; II. А. Коновалов. 1949, 1960; Л. Ф. Прав-
дин, 1960, 1964. и’др.), так и за рубежом (в Англии — Франц,
1956; в Германии — Мюнх, 1928; Ромедер и Шенбах, 1962; в
Чехословакии — Гейтманек, 1956; в Швеции — Килланд ер,
1955; Л и ндк вист, 1956).
Особое внимание Г. Ф. Морозов уделял связи признаков и
свойств у древесных пород с географическом средой. Ои писал:
«11 светолюбив, и быстрота роста, и форма ствола и корней,
и требовательность корней к составу почвы, и отношение к
насекомым, и качеству древесины, и семенная производитель-
ность, и долговечность,* и устойчивость породы — все это и
другое находится в он редел ениой и закономерной связи с геогра-
фической средой» (Учение о лесе, стр. 379, 1949). Это указа-
ние Г. Ф. Морозова явилось толчком к постановке целого ряда
региональных научных исследований изменчив ост и морфо-био-
логических признаков и свойств у древесных пород Союза в
узких рамках отдельных географических зон и районов. По ели
обыкновенной можно отметить работы: Гаврися (1938);
Г. Л. Тышкевич (1958); В. А. Панина (1959); Д. С. Голод
(1960); Пчелина (1961); Л. 14. Милютина (1963); П. И. Вой-
чал ь (1955, 1959) п но лиственнице сибирской н Сукачева —
В. И. Сукачева (1924), 11. В. Дылнса (1947, 1961), Б. П. Колес-
никова (1946), II. А. Лагова (1959), А. В. Альбенского (1959)
и других. Работы указанных авторов убедительно подтверждают
высказанные Г. Ф. Морозовым предположения о наличии в при-
роде индивидуальной и географической изменчивости ели и
лиственницы в пределах отдельных частей ареала. Это в пер-
вую. очередь относится к ели обыкновенной и лиственнице
’сибирской. Менее изучена изменчивость ели сибирской и лист-
венницы Сукачева па Урале. Особенно отрывочны сведения но
этим породам в пределах Среднего Урала, на восточном и за-
падном склонах, северной п южной части его.
Данное сообщение и преследует цель восполнить этот про-
бел — изложить основные результаты наших исследований
по обеим породам в пределах указанных районов Среднего
Урала.
Территория Среднего Урала представляет собой обширную
естествепно-гео1гра'фи1чес1кую область, с резко обособленными
районами как в широтном, так и долготном направлениях.
Различия в природном комплексе западного склона цент-
ральной части и восточного склона наложили свой отпечаток
52
и на древесную растительность. Европейский склон н водораз-
дельная полоса являются зоной господства еловых лесов. На
азиатском же склоне господствуют сосняки. Кроме того, внутри
каждой части наблюдается неоднородность климатических и
почвенно-грунтовых условий в зональном аспекте (Колесников,
1960; Горчаков-скин, 1965).
Указанный комплекс лесорастительиых условий отдельных
частей Среднего Урала не мог не оказать существенного влия-
ния на биолого-морфологическую природу ели и лиственницы
и, как будет показано ниже, сыграл решающую роль в качест-
венном обособлении южпых и северных популяций этих пород.
С целью выявления этих различий нами были подобраны
и соответствующие для исследования объекты. Для ели — на
западном склоне в Чусовском районе и на восточном склоне —
в Артннском районе.
Для лиственницы Сукачева — в южной части — Полевской
рапой, в северной — Карпинский район. Указанные пункты
по своим л-есорастительпым комплексам достаточно резко очер-
чены и являются наиболее типичными для своих естественных
районов.
Характеристика объектов и методика полевых и лаборатор-
ных исследований нами излагались ранее (М. С. Некрасов, 1965,
1966; Е. А. Пугач, 1962, 1964, 1964-а), поэтому ограничимся
приведением пунктов сбора материала и ссылкой па прежде
опубл и ко ванные материалы.
Обсуждение результатов исследования
Географическая изменчивость
Рассмотрим полученные результаты исследования географи-
ческой изменчивости основных морфо-биологических признаков
и свойств у ели сибирской и лиственницы Сукачева. Для удоб-
ства разбора материала изложение результатов приводится с де-
лением на северную и южную части, под которыми понимаются
соответственно пункты, расположенные в конечных частях огра-
ни ченного исследованием ареала.
Сводная таблица 1 иллюстрирует качественную неоднород-
ность по указанным признакам ели и лиственницы и четко
обособляет северные и южные популяции этих пород. Еловые
древостои южной части превосходят древостои северной по так-
сационным показателям и биометрическим характеристикам ге-
неративных органов. Во всех случаях достоверность статисти-
ческого различия установлена высокой (t>3).
53
Таблица 1
Зависимость признаков и свойств ели и лиственницы от происхождения
Показатели п свойства Ель сибирская 1 Лиственница Сукачева
север юг север юг
Возраст, лет 100 100 109 108
Высота, м 22,3 ±0,5 24 ±0,5 28,7 ±0,4 25,7 ±0,3
Объем, м3 0,58 ±0,04 0,84 ±0,09 1,19±0,03 0,92 ±0,04
Диаметр, см 24,8 ±0,7 28,3 ±1,3 34,4 ±0,4 31,1 ±0,4
Длина шишек, см Вес 100 ни. 7,41 ±0,1 8,12±0,1 3,18±0,1 2,6 ±0,1
семян, гр. Всхожесть 5,2 ±0,5 6,2 ±0,06 12,6 ±0,16 8,9 ±0,14
семян, % Предел прочно- сти при сжатии вдоль волокон, 37 36 70 63
кг/см2 Предел прочно- сти при стати- ческом изгибе, 334 ±2,5 319± 1.9 442 ±1,50 398± 1,40
кг/см2 613±4,2 580 ±4,0 1019±5,0 838 ±4,3
Физико-механическше свойства древесины, наоборот, опре-
делил гись более высокими у северной ели.
По всем показателям (кроме коэффициента качества при
сжатии вдоль волокон) статистическая достоверность различит
более 3.
Лиственница северной части Среднего Урала но всем иссле-
дованным признакам и свойствам (кроме коэффициента качест-
ва при сжатии) занимает преимущественное положение, и ее
размерные показатели статистически достоверно превышают
показатели лиственницы южной части.
Полученные результаты, безусловно, требуют своего объяс-
нения, поскольку различит коснулись весьмгх важных органов
п свойств у обоих видов.
На наш взгляд, на данном этапе работы возможно оста-
новиться на следующих предварительных предположениях в
объяснении наметившихся закономерностей.
1. Высокие показатели фи-эико-механических свойств у или
в северной части могут быть объяснены более суровыми усло-
виями для ее существования, а, следовательно, тугим ростом и,
54
естественно, платной древесиной. Замедленный рост в высоту и
по диаметру определил более плотное формирование годичных
слоев (у северной ели их в 1 см 5,6; у южной — 4,4), что и
объясняет ее высокие показатели по объемному весу, пределу
прочности при сжатии и статическом изгибе.
Лучший рост в высоту и по диаметру ельников южной ча-
сти отражает более оптимальные условия для ели лесорастн-
тельного комплекса этого района в сравнении с северным (бо-
лее продолжительный вегетационный период, лучше почвенные
условия и др.). Этим же можно объяснить размеры генератив-
ных органов и посевные качества семян.
2. Лиственница северной части Среднего Урала на многих
позициях занимает довольно устойчивое положение, не усту-
пая своим основным соперникам — ели и сосне.
Резко выраженный континентальный климат этой части и
почвенные условия более благоприятны для лиственницы, не-
жели в южной.
Это и определило преимущественное положение по изучен-
ным свойствам лиственницы северной части в сравнении с
южиой.
Не лишено смысла и предположение о более динамичной
природе лиственницы северной части, связанное с историческим
расселением этой породы на Урале. Лиственница южной части
ведет свою историю с более ранних времен, нежели северная.
Оледенение не затронуло в южной части лиственницы и ее
возраст значительно больший, чем у лиственницы северной
части, формирование древостоев которой протекало путем рас-
селения лиственницы из севера Европейской части и Сибири
(К. Н. Игошина, 1963; И. В. Дылпс, 1947). Такой контакт от-
дельных рас н популяций лиственницы не мог не вызвать
к жизни более динамичных и устойчивых форм, каковыми и
отмечаются памп северные популяции лиственницы.
Высказанные предположения безусловно не претендуют на
исчерпывающую полноту и необходимы дальнейшие исследо-
вания в подтверждение их или замене более доказательными.
Наши исследования по географической изменчивости обеих
пород, приведенные выше, и являются вескими аргументами
в пользу дальнейших работ в этом направлении.
Индивидуальная изменчивость
Сложный комплекс почвенно-грунтовых, климатических и
фитоцеиотических условий, совместно с историческим разви-
тием ели и лиственницы определили довольно обширный на-
бор морфологических форм в рассматриваемом районе.
55
Таблица 2
Зависимость признаков и свойств от форм ели и лиственницы
11оказателп и свинства Формы
елн сибирской ли ственнпцы Сукачев а
продольно- тая 1 Мелко- Глубоко- бороздча гая бороздчатая
Возраст, лет 100 101 108 ПО Высота, м 22,3 ±0,5 20,1 ±0,6 26,9 ±0,4 24,5 ±0,4 /Диаметр, см 24,8 ±0,7 21,4 ±0,7 33,2 ±0,6 29,2 ±0,4 Объем, м3 0,58—0,04 0,37±0,03 1,07±0,03 0,78±0,03 Длина шишек, см 7,41 ±0,04 6,32±0,1 2,8 ±0,3 2,3 ±0,2 Вес 1000 шт. семян, гр. 5,2 ±0,05 4,9 ±0,12 12,1 ±0,14 8,9 ±0,14 Всхожесть семян, % 77 ±1,2 66 ±2,0 48,3 ±0,8 36,1 ±0,7 Высота 1-лет- них сеянцев, см 3,22±0,08 2,85±0,07 6,3 ±0,08 5,4 ±0,05 Диаметр стволи- ка 1-летних се- янцев, мм 0,61 0,55 1,31 1,17 Объемный вес, г/см3 0,37±0,001 0,4-1±0,002 0,5 ±0,005 0,61 ±0,004 Предел прочно- сти при сжатии вдоль волокон, кг/см2 334±2,5 380±2,1 413±1,82 380±1.70 Предел прочно- сти при стати- ческом изги- бе, кг/см2 613±4,2 666±4,4 890±5,1 800±5,4
В прежних сообщениях нами в необходимом объеме были
изложены отдельные признаки форм ели и лиственницы, воз-
можные к выделению в качестве обособленных форм.
У ели сибирской ио совокупности тесно коррелирующих
n-ризиаков выделяется две формы. Первая форма характери-
зуется продольно-трещиноватой корой, компактным типом ветв-
ления и красным цветом шишек. Вторая — гладкой корой,
плоско-горизонтальным типом ветвления и зеленым цветом
шишек.
У лиственницы выделено также две формы: мелкобороздча-
тая и глубокобороздчатая. Для первой характерны: слабая за-
немел и с тость ствола, правильное (продольное) направление
56
борозд, темно-серый цвет коры и узкая (5—7 м) пирамидаль-
ная по форме крона. Для второй — сильная закомелистость,
неправильное (угловатое или ступенчатое) расположение бо-
розд, пластинчатое строение светло-коричневой коры и широкая
(6—J2 м) яйцевидная по форме крона.
Проведенное комплексное изучение выделенных форм ио
основным морфолого-биологическим признакам и свойствам
подтвердило их не только морфологическую, но и глубокую био-
логическую разпокачествепность.
В таблице 2 приведены основные результаты этого изуче-
ния в разрезе форм, которые позволяют сделать следующие
выводы:
Формы ели. 1. Наиболее высокие таксационные показатели
(высота, диаметр, объем ствола) при почти одинаковом воз-
расте имеет ель сибирская с продольпотрещиноватой корон,
превосходя ель с гладкой корон по высоте на 9,9%, по диамет-
ру — на 13,7% и объему на 36,1 % • Причем, это различие ока-
залось существенным (t > 3).
2. Более длинные шишки по сравнению со второй имеет
первая форма. Различие достоверно (t>3). Лучшие физиче-
ские показатели (’объем 1000 шт. семян) и посевные качества
семя и имеет также первая форма.
3.. Размеры однолетних сеянцев оказались меньшими у вто-
рой формы'. Разница с первой формой составляет по высоте
11,3%, длине стержневого корпя — 40,3%, диаметру стволи-
ка - 98%. I
4. Древесина ели с гладкой корой тяжелее па 15,9% и проч-
нее на 8—12,1% древесины ели, имеющей продол ьнотрещпно-
ва тую кору.
Ф о р м ы л ш с т в е и и п ц ы
1. Мелкобороздчатая форма опережает глубокобороздчатую
но всем таксационным показателям.
2. Более высокий рост в высоту при относительно меньшей
закомелистости объясняется высокими физико-механическими
свойствами древесины (объемный вес и пределы прочности).
3. Существенное различие прослежено и па размерах гене-
ративных органов и свойствах семян.
4. Наиболее доказательно различие рассматриваемых форм
при анализе роста и развития однолетних сеянцев.
По высоте стволика, длине корней и диаметру преимуще-
ство сохраняется за мелкобороздчатой формой, что дает
57
основание предполагать наследственный характер указанной
м о р ф о л о г । р । е с ко i i и зм е и ч и в о с ти.
В ы в о д ы. Изложенные выше материалы убедительно под-
тверждают необходимость проведения селекционных исследо-
ваний в каждом естественно-историческом районе ареала той
или Hiiioii породы.
Глубокая генетическая изментовость ели и лиственницы на
небольшой по площади территории Урала — наглядный при-
мер этому.
Выделенные формы ели и лиственницы различны не только
по морфологическим признакам, по и по основным биологиче-
ским свойствам и хозяйственная ценность их неодинакова.
Оставление лучших форм при рубках ухода, в качестве се-
менников при главном пользовании, использовании их в качест-
ве маточников для создания прививочных плантаций позволит
знач<ител1.по поднять продуктивность и жн-зпенность уральских
еловых н лиственных лесов.
Результаты наших исследований еще раз подтвердили гени-
альные высказывания корифея отечественного лесоводства
Г. Ф. Морозова о тесной связи древесной породы и среды, слож-
ной внутри видовой изменчивости, имеющей важное значение
в хозяйственной практике.
Л II Т Е Р А Т У Р А
Г о р ч а к о в с к п й II. Л. История развития растительности Урала.
Свердловск, 19zi7.
Д ы л и с Н. В. Сибирская лиственница. М., I9zi7.
Il coin и и а К. Н. Лиственница на Урале. Материалы ио истории
флоры и растительности (.’ССР. 1963.
К о и о в а л о в II. А. Пути применения селекции в лесосеменпом
деле. Сб. статей но обмену опытом на предприятиях лесной промыш-
ленности н лесного хозяйства Свердловской области. I960.
Колесников Т>. П. Естественно-историческое районирование ле-
сов (па примере Урала). Сб. вопросов лесоведения и лесоводства. Доклад
па V мировом лесном конгрессе. 1960.
Морозов Г. Ф. Учение о лесе. М.—Л., 1949.
Морозов Г. Ф. О лссоводствеппых устоях. М., 1962.
Некрасов М. С. К изучению формового разнообразия ели па
Среднем Урале. ИВУЗ (лесной журнал), № 6, 1965.
Некрасов М. С. Селекциопно-лесоводствеппые особенности и тех-
нические свойства древесины ели сибирской па Среднем Урале. Научно-
техническая конференция по итогам паучпо-псслсдовательских работ за
1964—65 гг. Сб. аннотации и докладов. Свердловск, 1966.
П р а в д и н Л. Ф. Быстрорастущие и хозяйствеппо-цспные древес-
ные породы п их значение для повышения продуктивности лесов.
М.-Л., 1960.
58
Пугач Е. А. Изменчивость таксационных показателей у деревьев
лиственницы Сукачева с различными морфологическими признаками.
Сб. тр. по леей. пром, и леей, хоз-ву. ВНИИЛП, 1964.
Пугач Е. А. Изменчивость морфологических признаков у листвен-
ницы на Среднем Урале. Сб. «Лиственница», т. II, 1964.
С у к а ч е в В. И. Очередные задачи русской дендрологии. Тр. Все-
рос. леев. конф. Вып. I, 1922.
А. В. КОЗЬМИН
О березе с декоративной текстурой древесины
Береза с декоративной текстурой .древесины издавна при-
влекала внимание лесоводов. Линдквист различал в Шве-
ции по текстуре следующие типы древесины березы: ледя-
ную (die Eisbirke), пламенную (die Flammbirke), коричневую
узорчатую березу (die Braunmaserbirke). Пламенную березу
за рубежом находили на севере и в средней части Европы
(Линдквист, 1951, 1954; Шрок и Шольц, 1953; Руден, 1954;
Гардинер, 1962 и др.).
Ледяную березу Линдквист считает экстремальным типом
пламенной березы. На поверхности ствола ледяной березы,
по его описанию, имеются частые бессистемно расположенные
бугры.
В лесах Архангельской, Кировской и Воронежской областей
единично встречаются стволы березы с буграми! н свилеватым
расположением волокон. У дерева, найденного нами в Архан-
гельской области (рис. .1), бугорки па поверхности ствола
были. размером от еле заметных до величины 5X4X1 см и
располагались друг от друга па расстоянии от I до 6 см. При
продольном распиле ствола поверхность среза имела красивый
блеск полосами. Ствол слегка извилистый неровный. Кора
серовато-белая, толщиной 0,6 см, у комля отслаивающаяся.
Произрастала береза в типе леса ельник — черничник свежий.
Состав насаждения до рубки был 9Е1Б, полнота — 0,6.
Коричневая узорчатая береза (в СССР носит название «ка-
рельская», «узорчатая») за рубежом имеется на юге Швеции,
Финляндии, единично ее встречали в Польше, Чехословакии,
ГДР (Боллапд, Шольц, 1960; Шольц, 1963; Пехман, 1961 и др.).
В СССР се находили кроме Карельской АССР п Белорус-
сии, где имеются ее наибольшие запасы, в Латвии, Ленинград-
ской и Костромской областях (Соколов, 1950; Яблоков, 1962;
Сакс и Банде*]), 1969; Багаев, 1965 и др.) (рис. 2).
59
Рис. 1. Ствол бугорчатой березы. Кнземскпп лесхоз Архангельской
области. Фото автора.
Работами Н. О. Соколова, А. С. Яблокова и других исследо-
вателей доказана возможность выращивания карельской (узор-
чато]’!) березы из семян от свободного опыления, собранных с
деревьев, имеющих морфологические признаки «узорчатой»
березы. В настоящее время культуры карельской березы с хо-
рошо выраженны-миг морфологическими признаками «узорча-
тости» па стволах в виде «перехватов», бугров имеются в Ка-
рельской АССР, Ленинградской, Московской, Воронежской,
Кировской, Омской, Ташкентской областях, Латвийской ССР,
Украинской ССР и других мостах (Соколов, 1950; Яблоков,
1962; Любавская, 1966; Вереей н, Литвак, .1968; Судохольский,
1971; Яскина, .1971 и др.).
Изучение десятилетних культур узорчатой березы, выра-
щенных Кировским лесхозом (Кировская область) из семян
от свободного опыления, собранных в Карелии с деревьев,
имеющих морфологические признаки «узорчатости» показало,
что из 50 сохранившихся экземпляров березы, которые росли
двумя рядами в условиях достаточного освещения, у 15 стволов
имелись хорошо выраженные морфологические признаки узор-
чатой березы (рис. 3). У 6 стволов эти признаки были слабо
60
Р и с. 2. Карельская береза в заказнике Петрозаводского лесхоза Карель-
ской АССР. Фото автора.
Р л с. 3. Десятилетняя карельская береза из культур, зало-
женных Кировским лесхозом Кировской области. Фото автора.
выражены, у остальных деревьев их не было. 13 экземпляров
березы с ггрнзпаким «узорчатости» имеют кустовндную форму,
один экземпляр высокоствольную, остальные деревья были
короткоствольной формы. Деревца кустовидной формы мпого-
ветвистые, разветвление их стволов начинается почти от самой
земли. На стволиках хорошо видны «перехваты», «вздутия»,
бугры.
Средним диаметр десятилетних карельских берез — 3 см,
средняя высота — 3 м. У обычных берез, произрастающих в
G2
Рис. 4. Гнездо из 4 порослевых стволов каповой березы, произошедших
от одного дерева. На всех стволах имеются капы. Вятско-Поляпскип
лесхоз Кировской обл. Фото автора.
этих же культурах, средний диаметр — 5 см, средняя высота —
6 м. Следовательно, карельская (узорчатая) береза уступает
в росте обычной березе, произрастающей в тех же условиях.
Высоко ценятся изделия из наростов другой разновидности
березы' — каповой (рис. 4).
В СССР ка»по1В'ая береза шмеется в Новосибирской области,
Башкирской АССР, в небольшом количестве на юге Кировской
области, единично встречается в Московской!, Горьковской,
Архангельской, Воронежской и других областях.
Самый большой капокорень у березы имел средний попереч-
ник 193 см и высоту над землей 50 см. Самый большой
ствольный кап имел размеры 55X32X15 см. Стволовые капы
большей частью встречаются по несколько штук на дереве,
на отдельных стволах их число достигает нескольких десятков.
В Вятско-Поляпском лесхозе Кировской области в кв. 81
Бурецкого л-ва (пойма реки Вятки) 'произрастает насаждение
березы, в котором как примесь имеется береза со стволовыми
капами и капокорнями. Самый большой стволовый кап достигал
размеров 53X45X10 см, небольших капов (в среднем 7X6X3 см)
у отдельных деревьев было до 60 шт. на одном стволе.
Наиболее часто капокорни крупной величины можно встре-
тить у отдельно растущих деревьев березы пушистой в местах,
оптимальных для ее произрастания. Рост капов у каповой бе-
резы начинается с образования колоний спящих почек. Число
спящих почек в колонии постепенно увеличивается и обра-
зуется нарост, который особенно быстро растет в тот период,
когда дерево хорошо разовьет крону п корневую систему.
Иногда путают наросты типа кап с сувелем, тоже наростом
большей частью одиночным па стволе или у корневой шейки
дерева (редко) и имеющего другую природу образования
(рис. 5). Сувель значительно менее ценен, чем кап, но может
употребляться на мелкие поделки. Основным отличием капа
от оувеля является то, что на поверхности капа имеется боль-
шое количество близко расположенных друг к другу спящих
почек. Природа образования капа связана с этими почками.
Если сделать разрез через кап, то можно увидеть, что шейки
почек берут начало из середины дерева и постепенно разветв-
ляются к поверхности.
На сувеле нет почек, он часто имеет более неправильную
форму чем стволовый кап. Сувель достигает больших размеров
па дереве, чем стволовый кап. Редко встречаются стволовые
капы, отходящие от поверхности ствола до величины радиуса
64
Рис. 5. Нарост па березе типа сувель.
Зплапрскпй лесхоз Башкирской АССР.
Фото автора.
его возле ка.па, сувель же нередко отходит от ствола па вели-
чину больше радиуса, до величины диаметра дерева в этом
месте (иногда и больше).
Как показали разрезы через сувель, его древесина в два-
три и даже более раз прирастает быстрее, чем обычная возле
него. Образование сувеля начинается не с первых лет жизни
дерева, а в более поздние годы, по рост может продолжаться
в течение нескольких десятилетий.
Отмеченные образования у деревьев березы имеют разную
природу, что следует иметь в виду при разведении березы с
декоративной текстурой. древесины.
ЛИТЕРАТУРА
Козьмин А. В. Каповая береза. Изд-во ЦБНТИ лесхоз., М., 1970.
Литвак П. В. Карельская береза в Украинском полесье. «Украин-
ский ботанический журнал», 25, № J, 1968.
Л ю б а в с к а я А. Я. Селекция и разведение карельской березы.
«Лесная промышленность», М., 1966. •
Сакс К. А. и Б а н д е р В. Л. Карельская береза в Латвийской
ССР. В сб. «Вопросы лесной селекции и семеноводства в Латвийской
ССР». «Зипатне», Рига, I960.
5. Заказ 6609
65
Соколов Н. О. Карельская береза. Гос. изд. Карело-Фиископ ССР.
Петрозаводск, 1950.
Суходольский Д. А. Опыт разведения и акклиматизации березы
карельской в Сибири. Лесное хозяйство, № II, 1971.
Яблоков А. С. Селекция древесных пород. Изд-во сельскохозяй-
ственной литературы, журналов и плакатов. М., 1962.
Я с кип а А. В. Выращивание карельской березы в Ташкентском
оазисе. Лесохозяйственная информация. Реф., выпуск ЦБНТИ лесхоз,
№ 3, 1971.
Boland und Scholz. Rassenfragcn der wichtigsten Holzarlen in der
Volksrepublik Polen. Forst und Jagd, Sonderheft «Forstliche Samenplanla-
gen». II Berlin, Juni, 1960.
Gardiner A. A Note on the Variation of Flamy Figure in silver
Birch. Silvae Genetica, II, Heft 5, 6, 1962.
Lindquist B. Forstgenctik in der Schwcdischen Waldbaupraxis. Ber-
lin, 1951.
Pec maun. Die Struktur des Holzes a!s Qualitalsmerkmal. Allgemeine
Forstzeitschrift .№ 40—41, 1961.
Ruden G. Ober die Maserbirke und einige andere abnorme Holzbil-
dungen bei Birke. Meddelelser Norsche Skogsfozoksvesen, 12, 1954.
Scholz E. Das VerbreitLingsgebiel der Braunmaserbirke. Archiv fur
Forstwesen, 12, Heft 12, 1963.
Schrock 0. und Scholz E. Einiges uber Furnierbirken insbesondere
Flammbirken und deren Erkcnnon im Bestandc. Der Wa Id (Zeitschr. lur
Forst — Holzwirtschaft), № 6, 1953.
В. А. ГОРОХОВ, Б. В. СКОРИКОВ
О состоянии лесного семеноводства
в лесхозах Воронежской области
и перспективах его развития
Директивами XXIV съезда КПСС по девятому пятилетнему
плану предусмотрено более полное и рациональпое использо-
вание ресурсов и земель государственного лесного фонда, по-
вышение продуктивности и качественного состава лесов.
В текущей пятилетке лесоводам Воронежской области пред-
стоит выполнить большой объем работ по лесовосстановлению
и защитному лесоразведению. Всего будет посажено и посеяно
леса па площади 63000 га, в том числе по основным лесообра-
зующим породам — сосне и дубу — соответственно 4000 и
26000 га. Для выполнения этих работ потребуется значитель-
ное количество высококачественных семян, ценных по наслед-
ственным свойствам. Повышение продуктивности и качества
создаваемых лесонасаждении невозможно без организации лес-
66
ноге свхУгеноводства на nayni-ioii основе, па достижениях лесной
генетики ;и селекции.
Основной задачей, которая стоит сейчас перед лесоводами
пашен области., является создание постоянной лесосеменной
базы, полностью обеспечивающей предстоящие работы по вос-
становлению лесов в государственном лесном фонде,, а также
посадку защитных лесных насаждений на землях колхозов и
совхозов семенами с высокими наследственными качествами.
Создание постоянной лесосеменной базы па селекционной
основе начато в предприятиях лесного хозяйства с 1960 года.
Эти работы концентрируются в наиболее ценных лесных
массивах области, где сохранились •вы,сококачестве1нные и про-
дуктивные насаждения сосны и дуба. Это Ус-манский и Хре-
ИОВ1С.КОЙ боры, Шипове кая и Теллермановская дубравы. Здесь
.па значительных площадях проведена селекционная инвента-
ризация древостоев и выделены плюсовые насаждения и де-
ревья, являющиеся базой для организации сортового семеновод-
ства. Всего в настоящее время в лесхозах области отобрано
<60 плюсовых деревьев сосны и 227 плюсовых деревьев дуба.
(Основные работы по сосне ведутся в Сомовском лесхозе,
.насаждения которого являются южной оконечностью массива
«Усмаиский бор». Общая цлощадь лесов лесхоза — 9043 га.
Преобладающей породой является сосна, занимающая 71% всей
.лесопокрытой площади. .Средний бонитет сосняков — 1, полно-
та — 0,79,
(Семенная баз.а создавалась как путем отбора и формирова-
ния постоянных лесосеменных участков в наиболее производи-
тельных и высококачественных естественных насаждениях и
культурах, так и путем закладки искусственных семенных
участков и плантаций. В 1960 году в лесхозе на площади
10 га в одном из лучших сосновых насаждений отведен семен-
ной заказник с целью получения лучших по наследственности
семян для закладки постоянных лесосеменных участков. Воз-
раст насаждения — 60—80 лет, средняя высота — 29 м, сред-
ний диаметр — 34 см, бонитет — 1, полнота — 0,7. В 1961 го-
ду в заказнике вырублены все минусовые деревья.
Большая работа проведена в лесхозе по закладке семенных
плантаций сосны. Основным методом при этом было создание
плантаций материалом семенного происхождения. За десяти-
летие с 1961 по 1971 г. посадкой отборных сеянцев, выращен-
ных из семян с лучших деревьев, заложено более 95 га лесо-
семенных плантаций. При этом плантации создавались различ-
ными способами: квадратно-одиночной редкой посадкой с раз-
5*
G7
мощением 2X2 (по И. К. Вехову); площадками редкого разме-
щения 5X5 м (-по М. М. Вересину), при этом в площадку
размером 1X1 м высаживалось -по 5 сеянцев; аллейной посад-
кой с размещением в ряду через 0,5 м и между рядами — через
5 м (по П. И. Дементьеву).
В последующем на плантациях проводились и проводятся
мероприятия по постепенному переживанию насаждений. Ко-
нечной целью будет формирование семенных насаждений «са-
дового» типа, с редким размещением (5X5 м, 8X8 м), на кото-
рых будут проводиться мероприятия по ускорению и усилению
плодоношения (уход за почвой, внесение удобрений, обрезка
крон и т. п.).
На площади 6,3 га в лесхозе заложены плантации методом
прививки черепков с лучших деревьев. Прививки производи-
лись в обычных лесных культурах с шириной междурядий
1,5 м. Прививался каждый третий ряд, в ряду прививки про-
изводились через 4 м. Таким образом, па 1 га было привито
в среднем 550 деревьев. Смешение клопов на плантациях по-
рядное, т. е. каждый ряд прививался черепками с одного дере-
ва. Представительство клопов на плантациях — от 10 до 20 и
выше. Некоторые плантации уже вступили в пору плодоноше-
ния, и с них ужо собирали семена с улучшенными наследст-
венными свойствами.
Приведенные данные позволяют говорить, что в Ооновском
лесхозе по существу организовано семенное хозяйство по сосне.
Большое участие в проведении этой работы приняли работники
Воронежской зональной лесосемеппой станции.
-Всего в лесхозах области на 1 ноября 1971 г. числилось
514 га постоянных лесосемеииых участков и плантаций, из
них заложено методом изрежпван-ия лучших лесных культур —
369 га, посадкой отборных сеянцев — 106 га, прививкой че-
ренков — 39 та.
Работа по созданию постоянной лесосеменной базы дуба —
ценнейшей породы нашей области — ведется в основном в
Воронцовском леспромхозе. Леса его представляют часть извест-
ного лесного массива «Шипов лес». Здесь организовано спе-
циализированное лесо-семенное хозяйство по проекту, состав-
ленному в 1963—64 гг. институтом «Союзгипролесхоз». На пло-
щади свыше 9 тыс. га проведена селекционная инвентаризация
насаждений, выделены нормальные лучшие (2654 га), нормаль-
ные средние (6489 га) и минусовые (330 га) насаждения.
Отобрано 212 плюсовых деревьев дуба. В насаждениях раз-
68
личного класса возраста заложены постоянные лесосеменныо
участки на площади 2021 га.
В настоящее время леспромхозом выполняются намеченные
проектом работы. Все семенные участки остолблены; продол-
жаются работы по изреживанию древостоев.
В 1968 году заложена семенная плантация дуба на пло-
щади 3 га посевом желудей, собранных с плюсовых деревьев.
Всего в лесхозах области заложен о постоянных лесосемен-
ных участков дуба на площади 2579 га.
Приведенные данные показывают, что в предприятиях Во-
ронежского управления лесного хозяйства за последнее десяти-
летие проведена значительная работа по созданию постоянной
лесосеменной базы основных лесообраэующих пород, способной
обеспечить потребности в семенах для лесокультурного фонда.
Однако задача состоит не только в том, чтобы получить
достаточное количество семян. Необходимо, чтобы эти семена
были высоких наследственных качеств. Поэтому в настоящее
время разрабатываются мероприятия по постепенному переводу
лесного семеноводства на сортовую, селекционную основу.
В ближайшие годы в основных лесных массивах закончится
селекционная инвентаризация насаждений. Отобранные плю-
совый деревья и насаждения явятся базой для закладки в ши-
роких масштабах семенных плантаций сосны и дуба.
Учитывая большую перспективность плантаций, созданных
прививкой, намечается продолжать закладку таких плантаций
по сосне в предприятиях управления в 1971 — 1975 гг. на пло-
щади 75 га. Кроме того, проектируется создание плантаций
посадкой сеянцев, выращенных из семян с улучшенной на-
следственной основой. В Новоу сманском, Сомове ком, Давыдов-
ском лесхозах и Хреновском лесхозе-техникуме таких планта-
ций за пятилетие будет заложено общей площадью 95 га.
Закладка плантаций дуба сеянцами с последующей привив-
кой и лесосемеиньгх участков в предприятиях управления про-
ектируется на пятилетие на площади 195 га.
IO. II. ЕФИМОВ
Влияние осадков иа прирост дуба черешчатого раиией
и поздней форм в центральной лесостепи
Основное свойство ранней (var. ргаесох Czern.) и поздней
(var. tardiflora Czern.) форм дуба черешчатого — неодновре-
менное начало 'вегетации — предопределяет неодинаковые
возможности использования ими почвенной влаги. Это
наряду с другими факторами лежит в основе различий в тер-
риториальном распределении н продуктивности дуба данных
форм. Однако сведения о приурочен ноет,и дуба ранней и позд-
ней форм к почвам различной влажности и об энергии их роста
в зависимости от экологических условий противоречивы (Кобра-
по-в, 1925; Харнтонов1пч, 1930; Плотников, 1937; Пятницкий,
1941; Еиькова, .1950; Карандина, 1966 и др.). Последнее при-
вело к разным выводам относительно степени златолюбия или
засухоустойчивости данных форм, что нередко затрудняет их
11 ра в ил ьн ое 11 он ол ьзова пне.
Настоящая статья посвящена одной из сторон затронутого
вопроса: выяснению зависимости прироста дуба ранней и позд-
ней форм от количества выпадающих осадкою. Исследования
проведены в лесостепи Воронежской области, в насаждениях
Морозовского лесничества Бутуряиновского лесхоза. (Шипов
лес).
В условиях центральной лесостепи, характеризующейся не-
устойчивым увла?кпе1пие1м, осадки являются одним из ‘важней-
ших факторов, влияющих па рост древесной растительности.
Известно, что дуб очень чутко реагирует изменениями прироста
на колебания осадков (Костин, 1960). В соответствия с целями
нашей работы мы считали возможным и целесообразным рас-
сматривать влияние па прирост дуба осадкою вне связи с други-
ми факторами — температурой воздуха, эптомо1И'Нвазия-
ми. и т. п.
При выборе объектов исследования основное внимание об-
ращалось па сходство экологических условий для сравнивае-
мых насаждений дуба обеих фо*рм, так как нередко именно
игпорирсмвание этого ирл-пципа являлось основной причиной
противоречивой оценки их биологии. Пробные площади с пре-
обладанием дуба ранней и поздней форм отводились в сравни-
мых условиях произрастания (насаждения одного возраста, со-
става и полноты). Краткая характеристика проб приводится
в таблице 1.
70
Таблица 1
Рост и производительность насаждении дуба ранней и поздней форм
в различных условиях произрастания
Тип усло- вии произ- растания Возраст Фенологи- ческая фор- ма дуба Средине размеры дуб Бонитет 1 Полнота Запас на 1 га, м3
диаметр, с.м высота, м
85 Ранняя 34,0 ±0.75 26,5 I 0.72 298
Дс 90 Поздняя 35.8±0.74 28.8 I 0.80 371
Ранняя 27.2±0.42 19.8 III 0.84 252
Д| 85 Поздняя 26.2 ±0.60 21.6 III 0.76 224
Д.-С 1 85 Ранняя 25.1 ±0,91 15,8 IV 0.92 211
Поздняя 23.9±0.67 17.3 IV 0.88 196
На пробных площадях, заложенных в свежей (Дг), сухой
(Д|) и солонцовой (Д1-0) дубравах, осенью 1965 г. срублено по
3 средних модельных дерева. Для анализа хода роста были
выпилены кружки с высоты 1,3 м. С помощью измерительной
лупы с десятикратным увеличением измерена величина годич-
ных приростов с точностью до 0,1 мм. Замеры производили
но четырем радиусам, ориентированным по странам света;
величину радиального прироста за каждый год вычисляли как
среднюю из 12 определений.
Сведения о количестве осадков по месяцам за 1906—1965 гг.
взяты на ближайших к пробным площадям метеостанциях —
Красный Кордон н Бутурлпнов-ка.
При оценке влияния осадков на прирост дуба обеих форм
необходимо исходить из особенностей динамики их вегетацион-
ного развития. Лнстораспусканпе у дуба ранней формы на-
ступает в Шиповом лесу в среднем 4—6 мая, позднего —
22—23 мая (Плетминцева, 1966). Окончание вегетации у них
происходит примерно одновременно. Поэтому продолжитель-
ность периода -вегетации у дуба поздней формы заметно короче.
Активная деятельность камбия у дуба начинается, по одним
данным, за несколько дпей до л и сто распускания (Харитонович,
1939; Вихров и Епькова, 1953; Самцов, 1966), по другим —
спустя некоторое времш после раскрытия листовых почек
(Раскатов, 1946,1948). Наибольшие величины, по данным этих
авторов, прирост дуба имеет в мае — июле. В последующем
деятельность 'камбия ослабевает и отложения годичного слоя
в августе и сентябре незначительны, особенно на сухих поч-
71
вах. В соответствии с этим осадки гидрологического года
( с октября предыдущего года но сентябрь последующего) раз-
биты нами на несколько периодов с целью выяснения значения
каждого из пих для прироста дуба.
Между годичным приростом и количеством осадков нахо-
дили графическую зависимость (рис. .1) н коэффициенты кор-
реляции. Корреляционная связь устанавливалась для 60-летпе-
го (1906—1965 гг.) и 20-летнего (1920—1939 гг.) периодов роста
’ дуба. При анализе за основу взят последний период (табл. 2),
так как в это время относительно стабилизировался прирост
дуба (возраст деревьев был равен 40—60 годам) и возрастные
изменения прироста в меньшей стеиепи отражались на коэффи-
циентах корреляции. Для 60-летпего периода абсолютные зна-
чения коэффициентов корреляции ниже, по по характеру связи
он аналогичен апдлизируемому периоду, в связи с чем мы по
приводим этих венных.
Таблица 2
Зависимость прироста дуба ранней (Р) и поздней (П) форм
ио диаметру от количества осадков
Период Коэффициенты корреляции между годичным приростом и осадками
свежая дубрава (Д2) сухая дубрава (ДО солонцовая дубрава (Д|_о)
р 1 п 1 Р | П Р | П
Октябрь—
сентябрь +0.24 +0.38 + 0.41 + 0.35 +0.36 +0.45*
Октябрь—июль + 0.29 + 0.51* +0.58** +0.77** +0.71** +0.60**
Октябрь—
апрель +0.33 + 0.07 + 0.30 +0.23 —0.02 +0.03
Май—июль +0.07 +0.57** + 0.47* + 0.74** +0.66 +0.79**
Май—июнь + 0.10 + 0.60** +0.53* +0.70** +0.79** +0.78**
Июнь—июль +0.10 +0.51* + 0.40 +0.63** + 0.49* +0.71**
Май —0.19 +0.27 + 0.15 +0.43 +.054* + 0.47*
Июнь +0.18 +0.52* + 0.48* +0.73** + 0.63** +0.69**
Июль +0.03 + 0.28 +0.19 +0.32 +0.21 +0.45*
Август —0.11 —0.10 —0.08 —0.08 —0.13 +0.01
Примечание: * Коэффициенты корреляции, достоверные при уровне зна-
чимости Р=0.05.
** Коэффициенты, достоверные при Р=0.01.
72
Зависимость прироста дуба ранней и поздней форм от осадков.
1 — ранняя форма дуба; 2 — поздняя форма дуба.
Рост дуба определяется уровнем влатообеспеченностп в ве-
гетационный период, что связано с поступлением воды в виде
осадков и их расходом (па транспирацию, испарение с поверх-
ности п внутриничвепны'й отток). Последнее в значительной
степени обусловливается водным режимом почв.
Темпо-серые (Д2), светло-серые (Д|) п солонцеватые
(Д1-) почвы существенно различаются по характеру водного
режима.
В painieBOceiiiiiin период, после снеготаяния, запасы влаги в
почве в дубравах всех типов достаточны. Последующая дина-
мика влажности зависит от физико-химических свойств данных
почв. Темпо-серые почвы благодаря хорошо выраженной струк-
।урпостп п высокой водоудерживающей способности характе-
ризуются постепенным расходом влаги. В засушливые годы он
усиливается, во влажные нередко запасы воды в почве (в зоне
расицространеИ'ПЯ корней) превосходят весенний уровень. Со-
лонцеваты? почвы отличаются быстрой потерей весенней влаги.
Недостаток водил в период вегетации, восполпяеамый лишь ча-
стично в ыш а дающим п осадками, усугубляется физиологической
сухостью, недоступностью воды растениям вследствие высокой
кочще пт рации обменного натрия (Зопн, 1951).
Светло-серые пич»зы по характеру водного режима зани-
мают как бы* промежуточное положение между двумя назван-
ными выше типамн почв.
По данным В. Е. Вихрова п Е. II. Еньковон (1953), отло-
жения годичного слоя дуба ранней формы в свежей дубраве
ла май составляют до 60% от общего прироста за год. Форми-
рование его, как видно из наших данных, происходит в основном
за счет запасов весенней влаги (коэффициент корреляции
с осадками октября — апреля составляет +0,33). В результа-
те колебание сумм выпадающих в теплый период осадков поч-
ти по отражается па его приросте (рис. I и табл. 2). По-ввдимо-
му, на характер вегетации ранней формы в этих условиях
большее влияние оказывают другие факторы, в частности,
температура воздуха. Некоторым подтверждением этому слу-
жит отрицательная корреляция между приростом и осадками
мая (—0,19). Выпадение осадков в мае сопровождается, как
правило, понижением температуры, что снижает энергию роста
дуба.
Максимум прироста дуба поздней формы, по данным тех же
авторов, падает на июнь — июль. Вегетация его начинается
при меньших запасах весенней влаги. Поэтому осадки летних
месяцев оказывают на его рост существенное влияние. Наибо-
74
лее тесную связь годичный прирост дуба поздней формы имеет
в свежей дубраве с суммарными осадками мая — нюня (+0,60).
На светло-серых почвах (Д1), быстро теряющих весеннюю
влагу, дуб ранней формы уже более зависим от летних осадков.
Наивысшую же сопряженность его прирост имеет здесь с осад-
ками октября — июля, определяющими баланс влаги в почве
в период вегетации (+0,58). Последнее характерно в еще боль-
шей степени для дуба поздней формы (+0,77). Зависимость
роста дуба Поздней формы от осадков летнего периода прояв-
ляется па этих почвах более четко, чем ранней формы.
На солонцеватых почвах (Д1-0), с их своеобразным вод-
ным режимом, прирост дуба обеих форм почти в равной степе-
ни тесно связан с осадками весенне-летнего периода. Следует
лишь отметить, что осадки июля для дуба поздней формы более
важны, чем для ранней. Впрочем, это проявляется во всех
условиях произрастания и обусловлено динамикой вегетацион-
ного прироста данных форм.
Связь годичного прироста с осадками августа практически
отсутствует у дуба обеих форм во всех условиях произраста-
ния.
Таким образом, в центральной лесостепи для роста дуба в
толщину определяющую роль играют весенне-летние осадки
(кроме ранней формы на темно-серых почвах).
Дуб ранней н поздней форм, как показал анализ, по-разно-
му реагирует па осадки, выпадающие в отдельные периоды
.гидрологического года, что связачго с различиями в динамике
их сезонного развития. Раньше начиная вегетацию, дуб раиной
формы успевает сформировать определенную часть годичного
прироста за счет запасов влаги, накопленных после снеготая-
ния. Поэтому он менее зависим от количества оса'дков, выпа-
дающих в период вегетации. Последнее особенно характерно
для свежен дубравы. С увеличением же сухости почв возрастает
связь прироста дуба ранней формы с летними осадками.
Дуб поздней формы во всех условиях произрастания постав-
лен, как правило, в худшие условия водоснабжения, так как
начинает вегетацию при меньших запасах почвенной влаги.
Отсюда и более высокая степень связи прироста дуба этой фор-
мы с осадками вегетационного периода.
Более резкие колебания прироста дуба поздней формы,
коррелирующие с колебаниями суммы осадков периода макси-
мального роста (рис. 1), создают впечатление большего злато-
любия дуба этой формы по сравнению с ранней. Известно,
что более заметное снижение прироста позднего дуба в отдель-
75
ные засушливые годы нередко является основанием считать
его менее засухоустойчивым (Харитонович, 1930; Топчиев,
Травлсев, Апостолов, 1961 и др.). В связи с этим интересно
проследить, как реагируют фенологические формы дуба на за-
суху изменениями прироста.
Влияние засухи на рост дуба мы выразили в степени изме-
нения величины годичного прироста по сравнению со средним
приростом (табл. 3). В таблице анализируются годы, охаракте-
ризованные С. II. Кост-иным. (1963) как засушливые для цеит-
]) ал ь по й л есо с т ей н.
Полученные данные показывают, что при оценке влияния
на прирост дуба ранней и поздней форм засухи необходимо
учитывать ее характер и продолжительность. Так, при доста-
точной влагообеопегченности весной (количество осадков за
октябрь — апрель и за май близкое к средним многолетним
данным) от летней засухи дуб поздней формы теряет прирост
в большей степени (1912, 1914, 1924, 1938, 1943 гг.). Если за-
суха захватывает лишь весенний период (май), а в последую-
щем выпадает достаточное количество осадков, во всех типах
дубрав более заметно падение прироста у дуба ранней формы
(1934, 1935, 1936, 1950 гг.). В некоторые годы при засухах,
продолжающихся длительный период (май — июнь), различия
между фенологическими формами прослеживаются менее четко
(1921, 1939, 1946 гг.).
Анализ показывает, что наиболее неблагоприятно отражает-
ся на росте дуба засуха, захватывающая начальный период,
его вегетации. Разный ритм сезонного развития дуба ранней
и поздней форм обусловливает и разную реакцию их на засуху
в зависимости от характера и продолжительности последней.
Для условий центральной лесостепи характерны засухи,
захватывающие в основном поздневесенний (конец мая) и
летний период. Поэтому поздний дуб, в силу своей биологиче-
ской особенности — позднего распускания листьев — чаще
попадает под их действие. Однако в благоприятные по увлаж-
нению периоды он формирует повышенные приросты, компен-
сируя потери его при засухе, что свидетельствует о способ-
ности дуба этой формы полнее использовать выпадающие в
период вегетации осадки.
Наши исследования проведены над деревьями, растущими
в насаждениях с господством дуба одной из форм при отсутст-
вии конкуренции за влагу. В случае совместного произраста-
ния дуба обеих форм, особенно в условиях неблагоприятного
водного режима почв, можно предположить более жесткие усло-
76
Таблица 3;
Влияние засух разного характера и продолжительности на прирост дуба
ранней и поздней форм
Годы Количество осадков, мм • Прирост в год засухи от среднего прироста, %
за гидроло- гический год за ок- тябрь— апрель за май 1 за июнь 1 .за июл
д2 Д| Д1—0
р П р 1 1 П р 1 П
1912 654 272 90 37 48 84 70 92 53 53 53
1914 507 282 46 18 24 204 147 194 120 136 117
1921 336 120 16 34 43 98 103- 76 71 58 53
1924 408 248 33 19 71 115 8’1 97 78 68 59
1934 391 157 18 58 108 71 96 86 106 97 149
1935 478 155 42 43 43 71 81 65 85 97 117
1936 428 2'31 3 35 17 71 74 54 71 68 85
1938 381 298 12 35 7 98 74 86 78 87 85
1939 438 254 27 21 64 80 59 43 50 49 53
1943 380 143 77 25 94 124 44 86 50 87 53-
1946 308 135 0 20 63 98 81 76 78 78 53
1950 442 212 23 101 62 45 88 54 64 49 74
Среднее за 1911—65 гг. 511 243 46 57 68 100 100 100 100 100 100.
вия водоснабжения для -дуба поздней формы ввиду лучшего
использования дубом ранней формы весенней влаги. Это долж-
но обеспечить дубу ранней формы перевес в конкуренции
ina сухих почвах. Именно поэтому в естественных древостоях
Шипова леса, как показали .наши исследования, в сухих п
«солонцовых типах дубрав редко встречаются участки с совме-
стным произрастанием дуба обеих форм. Решающее влияние
на их распределение оказывает здесь отбирающее действие
весенних заморозков. Дуб ранне]! формы господствует в этих
типах на возвышенных местоположениях, поздней — в морозо-
бойны'х понижениях, по склонам сухих балок. При этом дуб
поздней формы не уступает здесь в росте по диаметру раннему,
а по высоте даже превосходит его (табл. 1). Это говорит о
возможности успешного роста дуба поздней формы па сухих
почвах. Несомненно, здесь мы имеем дело с особыми экотипа-
ми дуба ранней и поздней форм, сформировавшимися в про-
цессе эволюции в этих неблагоприятных условиях произраста-
ния.
Результаты нашей работы позволяют сделать вывод о том,
что различия в характере прироста дуба ранней и поздней
форм в зависимости от количества осадков, как и в террито-
риальном распределении их на почвах разной влажности, не
являются показателем засухоустойчивости или златолюбия той
или другой формы. Эти различия связаны лишь со сроками
листораспускания феноформ дуба, обусловливающими разную
возможность использования ими осадков в период вегетации.
Л II Т Е Р А Т У Р А
Вихров В. Е., Е и ь к о n а Е. И. Динамика вегетационного при-
роста древесины рано- и позднораспускающихся форм дуба в связи
с условиями произрастания. Тр. ин-та леса АН СССР, т. 9, 1953.
Е п ь к о в а Е. И. Территориальное размещение рано- и позднорас-
пускающихся форм черешчатого дуба. Докл. АН СССР, т. 74, № 1, 1950.
3 о н н С. В. Водный режим почв дубовых лесов. Тр. ин-та леса
АН СССР, т. 7, 1951.
Карандина С. Н. Рост и развитие дуба в падинах Западного
Казахстана. «Наука», М., 1966.
К об рано в Н. П. Селекция дуба. «Новая деревня», М., 1925.
Кост и и С. И. Колебания климата в Центральной лесостепи Рус-
ской равнины. Научи, зап. ВЛТИ, т. 21, 1960.
Костин С. И. Повторяемость засушливых и влажных периодов в
Центральной части лесостепи Русской равнины. Сб. «Вопросы повы-
шения продуктивности лесного хозяйства». Изд. ВГУ, Воронеж, 1963.
Плетминцева Т. И. Селекция позднораспускающейся разновид-
ности дуба черешчатого для массового выращивания. Изд. ЦНИИТЭИ-
леспром, М., 1966-
78
П л о т п и ков Н. С. Сравнительная ценность каштаполистного, ар-
мянского и рас обыкновенного дуба в степных условиях Азово-Черно-
морского края. Лесомелиорация и лесное хозяйство. Вып. I, 1937.
Пят-пицкий С. С. — Экологические типы обыкновенного дуба и
их использование в лесокультурной практике. Леспое хозяйство, № 3,
1941.
Р а с к а т о в П. В. Ход прироста древесины дуба в течение вегета-
ционного периода в различных экологических условиях. Научи, зап.
Вороне?кск. лесохоз. ин-та, т. 9, 1946.
Р а с к а т о в П. Б. К вопросу о формировании годичных колец дре-
веси1иы дуба. Науч. зап. Воропежск. лесохоз. ии-та, т. 10, 1948.
С а м ц о в А. С. Периодичность роста и формирования структуры
древесины рано- и поздпораспускающихся форм дуба черешчатого в
БССР. Автореф. каид. дис., Минск, 1966.
Топчиев А. Г., Травле ев А. П., Апостолов Л. Г. Биоло-
гические особенности ранней и поздней форм дуба черешчатого. Лес-
пое хозяйство, № 7, 1961.
Харитонович Ф. Н. Сезонный прирост ранораспускающегося
(Quercus pedunculata v. ргаесох) и нозднораспускающегося (Quercus ре-
dii culata v. tardiflora) дуба. Tp. по лесному опытному делу Украины,
вып. 15, Харьков, 1930.
А. М. ШУТЯЕВ
Географические культуры дуба черешчатого
в степных условиях Краснодарского края
Большой объем работ по созданию полезащитных насажде-
ний в стенных условиях Северного Кавказа вызывает необ-
ходимость использования желудей из различных пунктов Евро-
пейской части СССР. Однако завоз семян из областей с резко
различными лесорастптел иными условиями по всегда приводит
к положнтельпо-му лесокультурному эффекту (Ерусалимский,
1969; Колданов п Траве нь, 1970). Одной из причин гибели
дуба является нарушение принципов районирования перевозок
желудей.
В 1952—1953 гг. в Армавирском лесхозе были заложены
географические культуры дуба с целью выявления областей,
откуда можно завозить желуди для степной части Краснодар-
ского края. Закладку опыта осуществляла Северо-Кавказская
ЛОС совместно с Армавирским лесхозом в порядке создания
сети географических культур дуба ВНИИЛМ. Район опыта
относится к первой агролесомелиоративной зоне Краснодарско-
го края (Барышман, 1968). Годовое количество осадков —
550 мм, среднегодовая температура воздуха 10,2°, почва —
79
лредкавказский слабокарбонатный чернозем на бурых глинах.
Поверхность участка ровная.
Лесная полоса состоит из 28 рядов площадок дуба. Расстоя-
ние между площадками в ряду — 3 м, между рядами — 5 м.
На метровых площадках желуди высевали под лопату в 5 лу-
нок. С целью получения сравнительно одинаковой густоты по-
севов количество высеваемых желудей в одну лунку зависело
от их доброкачественности (при 100% доброкачественности в
лупку высевали 7 желудей). Культуры заложены но схеме:
сопутствующая порода — кустарник — дуб — кустарник —
сопутствующая порода. В качестве сопутствующей породы вве-
дена яблоня лесная, в качестве кустарнп-ка — акация желтая.
Схема не выдержана, и в некоторых рядах вместо яблони
растет ясень зеленый, вместо акации желтой — бирючина, клен
татарский. Общая площадь участка (5,78 га) разбита на квар-
талы* размером 0,25 та.
В 1967—1969 гг. нами установлена фенологическая форма
экотипов дуба, обмер культур проведен в июле 1969 г. На мо-
мент обмера высота яблони достигала 5 м, клена татарского —
4 м, акации желтой — 3 м, поросли вырубленного в 1967 г.
ясеня — 3 м. Лесоводственного ухода в площадках дуба не
было. От каждого варианта намечали два ряда дуба, средних
по сохранности дубков и с примерно одинаковым окружением
сопутствующими породами, на которых обмеряли от 150 до
200 деревьев. Среднюю высоту определяли по графикам высот,
построенным для каждого варианта. Для этого шестиметровой
рейкой измеряли высоту у 25—30 деревьев разного диаметра.
Сведения о материнских насаждениях заимствованы у Е. II. Бу-
дянского (1953) и Р. А. Поповой (1952).
В литературе данные географические культуры (секция
1952 г. по обмерам 1958 г.) описаны Е. Н. Будянским (1963).
Полного отпада дуба в условиях опыта нет. Однако геноти-
пическая изменчивость желудей разного географического и эко-
логического происхождения обусловила различия в таксацион-
ных показателях экотипов дуба (табл. 1). В настоящее время
среднее количество деревьев на одной площадке по вариантам
колеблется в пределах 3,4—4,6 шт. Отпад происходит за счет
отстающих дубков центральных лунок.
Количество сухостойных* деревьев достигает 13%. Диаметр
сухостоя колеблется в пределах 2,4—4,6 см.
В культурах 1952 г. средняя высота худшего экотипа дуба
(ранняя форма из сухой дубравы Куйбышевской области) на
30% меньше высоты лучшего экотипа (поздняя форма из
80
о
Таблица 1
. Заказ 6609
Характеристика географических культур дуба закладки 1952 г.
Показатели материнских насаждений Показатели экотипов дуба в культурах
область, край тип леса возраст бони- тет ср. Д, см. достовер- ность раз- личия с местным. ср. Н, м Кол-во пря- мых и слабо- коленчатых дубков, % бонитет
Воронежская Свежая ясеневая дубрава 120 I 6,2±0,15 2,6 7,0 86 1,4* Харьковская Свежая липовая судубрава ПО II 6,3±0,16 3,0 6,9 80 1,4 Краснодарский Пойменная дубрава 70 11 6,5±0,15 4,4 6,7 18 1,6 Видницкая Свежая грушевая дубрава 57 II 6,1 ±0,13 2,3 6,5 64 1,7 Краснодарский Степная дубрава 70 II 5,7±0,12 — 6,4 90 1,7 Винницкая Свежая орляковая судубрава 70 II 6,0±0,14 2,4 6,4 83 1,7s* Брестская Свежая ясеневая дубрава 90 I 5,7±0,13 0,0 6,4 87 1,7 Краснодарский Предгорная злаковая дубрава 100 II 5,4±0,12 1,7 6,4 70 1,7 Саратовская Свежая липовая дубрава 50 III 5,4±0,13 2,3 6,4 71 1,7 Волгоградская Сухая дубрава 90 IV 5,0±0,11 4,5 6,3 81 1,8 Краснодарский Горная лещиновая дубрава 90 III 6,1 ±0,16 2,0 6,0 34 1,9 Волгоградская Пойменная дубрава 60 II 6,0±0,13 2,7 5,8 30 11,1 Харьковская Свежая липовая судубрава НО II 5,3±0,15 2,2 5,8 82 11,1* Оренбургская Свежая кленовая дубрава 60 III 4,9±0,10 5,3 5,7 80 11,2 Рязанская Влажная липовая дубрава 100 II 5,0±0,11 4,3 5,1 89 11,8 Ростовская Поймен Пойменная дубрава 50 IH 4,6±0,12 6,4 5,0 59 II,9^ Куйбышевская Сухая дубрава 45 IV 4,7±0,08 7,1 4,9 80 III
Примечая п с: Экотипы, отмеченные знаком *, представлены поздней
формой луба, остальные — ранней формой.
с!вежеп ясеневой дубравы Шипова леса). Средняя высота
J 8-летних экотипов дуба колеблется в пределах 4,9—7 м, сред-
ний диаметр — 4,6—6,5 см, класс бонитета 1,4-1 II, число
прямых и слабоколенчатых деревьев — 18—90%.
Самая плохая форма стволов у дуба пойменного пропс хож-
дения, хотя их размеры л могут быть больше, чем у нагорных
экотипов. Так, краснодарские пойменные дубки превосходят
местные нагорные но размерам, но по качеству стволов значи-
тельно уступают им. В основном, они коленчатые, колено-виль-
чатые и наклонные. Искривление ствола отмечается через каж-
дые 40—70 см, величина прогиба достигает 50 см. Большая
крпвоствольность дуба, выращенного из местных пойменных
желудей, отмечена памп н в производственных культурах Арма-
вирского лесхоза, в лесных полосах Каневского района (Ба-
рышмап, 1968) и совхоза «Кубань» (Волкова, 1962) Краснодар-
ского края.
Лучше местного степного экотипа дуба растут дубки позд-
ней формы из воронежской свежей ясеневой дубравы (Шипов
лес), ранней формы из харьковской свежей липовой дубравы,
винницкой грушевой дубравы. Одинаковые высоты с местными
имеют дубки поздней формы из свежей грабовой дубравы Вин-
ницкой области, ранней формы из предгорной дубравы Красно-
дарского края, раппей формы из свежей ясеие-вой дубравы
Брестской области, ранней формы из свежей липовой дубравы
Саратовской области. Существенные различия по диаметру в
сравнении с местным экотипом имеются у ранней формы из
Оренбургской, Куйбышевской, Рязанской, Волгоградской об-
ластей.
В культурах 1953 г. экотипов дуба, растущих лучше мест-
ного, нет. Близок к местному экотип из брянской свежей дуб-
равы, остальные существенно отстают по диаметру, их высоты
составляют 83—71% от высоты местного.
В опыте имеется пять экотипов дуба из трех областей
БССР. Разница но высоте между лучшим экотипом ранней
формы из свежей ясеневой дубравы Брестской области и худ-
шим экотипом поздней формы из пойменной дубравы Моги-
левской области в 17-летнем возрасте составляет 42,8%, по диа-
метру 107%. Таким образом, на белорусские дубравы нельзя
смотреть, как па единое целое. И в производственном масштабе
результат от использования белорусских желудей получается
различным.
В степных лесничествах Краснодарского края имеются хоро-
шие насаждения, выращенные из белорусских желудей (Ба-
82
(У
раиораспускающейся формы из желудей
Р н с. 1. Крпвоствольиые дубки р ‘
местной пойменной дубравы.
82.
Р п с. 3. Культуры дуба нтдиораснускающейся формы из желудей све-
жей ясеневой дубравы (Шипов лес) Воронежской области. Экотип
по диаметру и высоте занимает первое место.
Рис. 4. Культуры луба ранораспускающенся формы из желуден липовой
дубравы Рязанской области. Экотип по диаметру и высоте занимает
одно из последних мест.
рытпман, 1969). В то же время в соседней Ростовской области
на предкавкаэских черноземах отмечено плохое состояние бе-
лорусских дубков (Колданов, Травень, 1970). К сожалению,
как в первом, так и во втором случаях не говорится о типо-
логическом и формовом происхождении желудей.
О росте ранней и поздней форм дуба в степных условиях
Краснодарского края и смежных с ним областей сведений
очень мало. О худшем росте дуба поздней формы на предкав,-
казских черноземах сообщает В. А. Мельчанов (1963). В 1'0-лет-
ием возрасте дуб ранней формы превосходит дуб ноздрей
формы по высоте на 22,6%, по диаметру на 42,5%. И. В. Суха-
нова (1958) в тех же условиях лучший рост и качество стволов
наблюдала у дубков поздней формы.
И в данных случаях не сообщается о происхождении же-
лудей.
В армавирском опыте мало климатипов, представленных
одновременно ранней и поздней формами дуба. Ранняя форма
из харьковской липовой судубравы имеет такие же показатели
роста, как и поздняя форма из воронежской свежей дубравы.
Но дубки харьковской поздней формы из того же типа леса,
что рапняя форма, значительно отстают от последних. У позд-
ней формы больше сухостойных деревьев. Дубки ранней и
поздней форм могилевского экотипа из свежей лещиновой
судубравы по росту и качеству стволов не отличаются, а у
пойменного экотипа той же области раипяя форма превосходит
позднюю по высоте на 9,5%, по диаметру на 15,4%. Винницкая
область представлена ранней и поздней формами дуба, но опп
относятся к разным почвенным экотипам: поздняя форма — из
свежен судубравы, ранняя — из свежей грушевой дубравы.
Различий по показателям роста почти пет.
Лучшим среди экотипов дуба Краснодарского края являет-
ся степной. У предгорного экотипа незначительно уменьшается
диаметр (показатель различия 1,7). У горного экотипа по
сравнению со степным на 6,6% меньше высота и хуже качест-
во стволов. Пойменный равнинный экотип по высоте и диамет-
ру занимает первое место, по качеству стволов — последнее
(82% кривых и вильчатых стволов).
Опытные культуры в Краснодарском крае, так же как и по-
добные опыты в Центральной лесостепи Европейской части
СССР свидетельствуют о том, что дуб черешчатый в пределах
своего ареала -имеет ряд климатипов и эдафотипов, исторически
сформировавшихся в результате естественного отбора под влия-
нием разнообразных физико-географических условий. Эти эко-
87
типы отличаются потребностью в свете, влаге, тепле, ритмом
сезонного развития, быстротой роста и производительностью
древостоев. При перемещении желудей из одних условий в
другие их наследственные особениостй проявляются в различ-
ной степени в зависимости от характера новых условий роста.
Существующее наставление по лесосеменному делу (1963)
не учитывает агролесомелиоративные зоны Северного Кавказа.
В частности, для равнинной части Краснодарского края выде-
ляются три зоны (Барьгшман, 1968): I — входят районы с
количеством годовых осадков 450—500 мм, II — 550—600 мм,
III — 630—710 мм. Неоднородность агроклиматических усло-
вий края предопределяет в свою очередь дифференцированный
подход к переброске желудей из других областей.
На основании изучения географических культур дуба под
Армавиром, анализа лесорастительных условий инорайонных
дубрав и лесомелиоративных зон для полезащитного лесоразве-
дения в первой зоне Краснодарского края можно использовать
желуди из наиболее продуктивных свежих нагорных дубрав
Волгоградской, Саратовской, Воронежской, Винницкой и дру-
гих областей, расположенных ниже южной границы лесостепи
или в ее южной части. Для второй и третьей агролесомелиора-
тивных зон, менее подверженных засухам, желуди можно за-
возить из нагорных дубрав центральной лесостепи, северных
районов левобережья УССР, белорусских ясеневых дубрав на
суглинках.
В степных районах возможно использование желудей из
предгорных дубрав дуба черешчатого до 400—500 м над уров-
нем моря. Наставление (1963) рекомендует заниженные вы-
сотные пределы для предгорного экотипа. Из желудей горного
экотипа дуба вырастают культуры пониженной продуктивности
н качества. Из желудей поименного происхождения любого
географического пункта, в том числе и из местных, вырастает
дровяной лес. •
Изложенные придержки по районированию семян дуба по-
зволят повысить сохранность, продуктивность и качество стен-
ных насаждений на Северном Кавказе.
ЛИТЕР АТУРА
Барыш мая Ф. С. Основы защитного лесоразведения па Кубани.
Краснодар, кн. изд-во, 1968.
Будянский Е. Н. Создание географических культур климатиче-
ских и почвенных экотипов дуба в степпых районах лесоразведения.
Паучп. отчет, Майкоп, 1953.
88
Будянский Е. Н. Географические культуры дуба летнего в
Краснодарском крае. Лесн. хоз-во, № 3, 1963.
В о л к о в а А. В. Леспыс полосы совхоза «Кубань». Лссп. хоз-во,
№ 1, 1961.
Е р у с а л п м с к и ii В. И. О причинах усыхания дубрав в степной
зоне. Лссп. хоз-во, № JI, I960.
Кол да нов В. Я., Травспь Ф. П. Чему учит опыт создания
дубрав промышленного значения. Лесн. хоз-во, № 7, 1970.
М ел ьчап о в В. А. Исследование роста дуба летнего в лесных куль-
турах па предкавказскйх черноземах. Лесоэксплуатация и лесн. хоз-во,
№ 22, 1963.
Наставление но лесосемепному делу. М., 1963.
Попова Р. А. Создание географических культур климатических
и почвенных экотипов дуба в степных районах лесоразведения. Научи,
отчет, Майкоп, 1952.
Суханова И. В. Некоторые биологические и лесоводственпыс осо-
бенности ранней и поздней форм дуба черешчатого в Каменской обла-
сти. Автореферат канд. диссерт., 1958.
Н. И. МАМОНОВ
Географические культуры лиственницы
в 'условиях предгорий Закарпатья
Лиственница на Украине является ценной интродуцирован-
ной породой. Лишь в высокогорьях Карпат имеется несколько
автохтонных участков лиственницы польской. В Закарпатье
имеется ряд старовозрастпых культур лиственницы европей-
ской, некоторые из них очень продуктивны. Так, культуры
лиственницы европейской, созданные в 1814 году в Раховс-ком
лоспичестгоо, имеют запас 1200 м3 на 1 га. В последнее время
стали создаваться культуры лиственницы сибирской, но опыт
их производства пока небольшой. В связи с этим изучение куль-
тур, созданных в условиях предгорий Закарпатья из привоз-
ных семян различных экотипов некоторых видов лиственницы
имеет научное и практическое значение.
Семена лиственницы сибирской, Сукачева и даурской бьгл.и
получены в 1965—1966 гг. из восточных районов СССР, семепа
экотипов лиственницы европейской собраны в различных лес-
ничествах Закарпатья (табл. 1). Все образцы семян были
стандартными и относились к первому и второму классам ка-
чества.
Посев семян был произведен 31 марта 1966 г. в питомнике
Закарпатской ЛОС, расположенном в предгорном районе на
высоте над уровнем моря 150 м. Климат района мягкий, влаж-
89
Таблица 1
Пункты сбора семян Larix, использованных для создания
географических культур
№№ об- разцов Вид Пункты сбора
1. Larix sibirica Ldb 2. » 3. » 4. » 5. » 6. Larix dahurica Turcz 7. » 8. » 9. Larix Sukaczcwii Djil 10—16. Larix decidua Mill 17 » Качугскпй лесхоз Иркутской области Усть-Капскнй лесхоз Алтайского края Сонскнй лесхоз Красноярского края Чунскпй лесхоз Иркутской области Ермаковскпи лесхоз Красноярского края Горный лесхоз Якутской АССР Усть-Алданскнн лесхоз Якутской АССР Канский лесхоз Якутской АССР Н.-Лялннскпй лесхоз Свердловской обл. Различные экотипы из Закарпатья УССР Татралскнй экотип
ный, среднегодовая температура 9,3°С. Среднегодовое количе-
ство осадков 850 маг, в теплый период выпадает 500 мм. Поч-
ва — суглинистый оподзолепный бурозем. Перед посевом се-
мена замачивали в воде в течение 20 часов.
После посева прядай были политы водой и замульчированы
слоем древесных опилок толщиной 1—3 см.
Начало появления всходов было отмечено 26 апреля. При
атом различные образцы вели себя по-разному. Так, всходы у
образцов 3, 4, 5 лиственницы сибирской и большинства образ-
цов лиственпицы европейской были недружными. Наоборот,
образцы 1, 2, 9 взошли дружно, а образцы л-пствеиницы даур-
ской (6, 7, 8) появились в единичном количестве. В последую-
щее время всходы всех образцов лиственницы даурской погиб-
ли, сеянцы лиственницы сибирской и европейской имели впол-
не удовлетворительный вид.
Осенью 1967 года у двухлетних сеянцев были проведены
измерения некоторых показателей (табл. 2).
Данные таблицы 2 показывают, что между двухлетними
сеянцами лиственницы европейской и сибирской имеется значи-
тельное различие. Так, по высоте лиственница европейская
превышает сибирскую в 4—8 раз, но весу стволика аз 20—27 раз,
но весу корней в 14—25 раз, по весу хвои в 37—122 раза и
по длине хвои в 1,3—1,6 раза.
Среди образцов лиственницы сибирской по своему росту и
90
Таблица 2
Данные биометрических измерений двухлетних сеянцев лиственницы
в географических культурах
№№ об- разцов Б11омтерические показатели
высота, м вес стволи- ка, г вес кор- ней, г вес хвои, см дл. хвои, см
1. 5.0 ±0,18 0,906 0,395 0,180 2,3
2# 5,5 ±0,23 0,982 0,474 0,099 2,2
3.’ 5,2±0,12 0,634 0,266 0,109 2,1
4. 5,0±0,15 0,907 0,473 0,081 2,5
5. 5,0±0,33 1,261 0,455 0,266 2,3
9. 5,0±0,14 0,957 0,410 0,214 2,2
10- 1( ). (средн.) 39,2± 1,1 24,98 6,673 9,927 3,3
17. 26,74=1,3 — — — —
остальным биометрическим показателям несколько в лучшую
сторону выделяется лиственница из Ермаковокого лесхоза
.Красноярского края (образец 5). При наблюдении этого образ-
ца была отмечена большая дифференциация по высоте сеянцев.
Так, если коэффициент вариации высот у образцов 1, 2, 3, 4,
9 составил 26—37%, то у образца 5 был равен 73%. Осталь-
ные образцы лиственницы сибирской по своим показателям
почти тте отличаются друг от друга.
Фенологические наблюдения показали, что рост лиственницы
сибирской начинается раньше европейской на 1—2 педели.
Хвоя начинает появляться сразу с наступлением теплой погоды
в последней декаде марта месяца. В это время из-за большой
влажности почва обычно еще пе позволяет проводить посадку.
Выкопанные ранней весной (до распускания хвои) из питомни-
ка сеянцы до посадки! хранились в подвале па льду.
Продолжительность роста в течение вегетационного периода
у лиственницы европейской значительно больше, чем у сибир-
ской. Это различие составляет 2—3 недели. Если лиственница
европейская закапчивает вегетацию в начале октября месяца,
то у лиственницы сибирской и Сукачева рост годичного по-
бега прекращается во второй декаде сентября. В теплую осень
у некоторых экземпляров лиственницы сибирской, особенно ма-
лорослых, наблюдается распускание хвои.
Опадеигие хвои у лиственницы сибирской происходит также
раньше, чем у лиственницы европейской. Более того, у части
одно- и двухлетних сеянцев лиственницы европейской хвоя
91
сохраняется всю зиму и имеет зеленый цвет. У лиственницы
аибщэакой после окончания вегетации хвоя желтеет, затем
быстро опадает. Большая продолжительность вегетационного
периода ,у лиственницы европейской по сравнению с сибирской
отмечается и для других географических районов (Надеждин,
1964).
Посадка лиственницы па лесокультурную площадь произ-
ведена 3 апреля 1968 года. Участок площадью 1,12 га был
разбит на секции. Числю секции было таким, чтобы каждый
образец имел трахкратную повторность
Расстояние между рядами было принято 2 м, в рядах 1 м.
Посадка была произведена вручную под лопату. С питомника
•для посадки брались наиболее рослые сеянцы. Маленькие,
слабые сеянцы были распикированы в школе и использовались
в последующие годы для дополнения.
В культурах за 3 года произошел большой отпад, однако
у лиственницы европейской сохранность оказалась значитель-
но выше (71—77%) по сравнению с лиственницей сибирской
(33—60%). Отпад произошел, в основном, в результате по-
вреждения растений личинками майского хруща и выжимания:
более развитая корневая система лиственницы европейской ока-
залась более устойчивой к этим факторам по сравнению с лист-
венницей сибирской.
Рост в высоту различных образцов лиственницы является
одним из лучших показателей состояния географических куль-
тур, соответствия биологических особенностей этих образцов
-данным условиям лесорастительной среды. Измерения высот
лиственницы в культурах проводили в течение 1969—1971 гг.
(табл. 3).
Из таблицы 3 видно, что различие по росту в высоту .между
образцами лиственницы европейской и сибирской ие только
сохранилось, ио стало более отчетливым. Это различие за 3-лет-
ний период достигает 5—10-жратной величины. Особенно хоро-
шо растет татранскпй экотип лиственницы европейской
(ЧССР). 13 мягком климате предгорий Закарпатья листвеппица
сибирская и Сукачева растут плохо. Следует отметить, что это
не противоречит научным данным по биологии Larix. Листвен-
ница является древесной породой гор п континентального
климата и это служит основным фактором ее природного про-
израстания (Тимофеев, 1961).
Особенно относится это к даурской и сибирской лиственни-
цам. В частности, присланные нам семена лиственницы сибир-
ской и даурской были собраны в насаждениях, расположенных
92
Таблица 3
Рост в высоту географических культур лиственницы в 1969—1971 гг.
№№ об- разцов Высота растений, см Прирост по высоте за два года, см
1969 1970 1971
1. 13±0,6 19±0,6 26±1,3 ! 13
2. 5. 15±0,7 | 22±0.6 f 26±0,7 39 ± 1,0 • 21
3. 14±0,3 20±0,5 25 ±0,6 i 11
4. 16±0,7 23±0,6 29±1,1 13
9. 12±0,4 17±0,4 22 ±0,6 i 10
10—16. 56±1,6 137±5,7 162±5,7 106
17. 58 ± 1,6 117±3,1 195±6,4 137
па высоте 500—1200 м над уровнем моря. Поэтому мягкий,
теплый климат предгорий Закарпатья не отвечает биологии
этих видов лиственницы. В то же время в горном районе
Карпат имеются удовлетворительные молодые производствен-
ные культуры лиственницы сп‘бирс-кой.
Аналогичная картина отмечается и в отношении различ-
ных экотипов лиственницы европейской. Исследования Винцен-
та (1961) показывают, что экотипы происхождением из невы-
соких горных мест или из низменности в среднеевропейском
мягком климате растут быстрее, чем, например, горная австрий-
ская или карпатская листвеипшца. С возрастом различия меж-
ду ростом экотипов «горного» и «долинного» происхождений
становятся меньше.
Данные нашего исследования позволяют сделать следующие
выводы:
1. В условиях предгорий Закарпатья лиственницы сибир-
ская, Сукачева и даурская оказались непригодными для выра-
щивания в культурах.
2. Для создания культур лиственницы в этом районе могут
быть использованы местные карпатские экотипы лиственницы
европейской, которые показьнвают хорошую сохранность и бы-
стрый рост. Возможно использование татраиского экотипа лист-
венницы европейской (ЧССР).
ЛИТЕРАТУРА
Ж и в п ц к п й 3. Н. Лиственница в Украинских Карпатах. Авторе-
фер. канд. диссерт., 1968.
Надеждин В. В. Продолжительность вегетационного периода и
93
сроки роста разных видов и экотипов лиственницы в Московской об-
ластщ
Тимофеев В. П. Роль лиственницы в поднятии продуктивности
лесов. Изд. АН СССР, М., 1961.
4. Vincent G—.VUtteleiiropalscbe Larc iea und ihre /\uslesc Praha, 1961
P. Г. МОИСЕЕВ
Особенности прививок хвойных пород в Карпатах
Разработке методов прививок хвойных пород для условии
Карпат положили начало 10. Ю. Боберский (1966) и В. Б. Лог-
гинов (1967). С 1966 г. эти работы были продолжены нами
с включением в исследования дополнительно целого ряда хвой-
ных как местных, так и экзотов.
В настоящей статье анализируется материал пятилетних ис-
следовании по методам прививок хвойных пород. За этот период
было проведено свыше 20 тыс. прививок различными метода-
ми 20 видов хвойных растений. Исследованиями преследовалась
цель уточнения лучших способов, сроков прививок и их изоля-
ции, подбора подвоев, первичного ухода за прививками и т. д.
На каждый вариант опыта ежегодно приходилось до 100 при-
вивок.
Для уточнения лучшего способа прививок нами были опро-
бованы па пяти хвойных породах прививки в приклад сердце-
виной на камбий по Проказипу (1960), в приклад камбием
па камбий по Гтргидову и Долголпкову (1964), в расщеп вер-
хушечной почки, улучшенная копулировка и в боковой зарез
(табл. 1). Как показали опыты, приживаемость в первую оче-
редь, зависит от качества проведенных работ. Выявить пре-
имущества того пли иного способа оказалось трудно.
Наиболее простыми в исполнении и надежными оказались
способы прививок в приклад камбием на камбий и сердцевиной
па камбий. В дальнейшем при опытных работах использова-
лись только эти способы.-
Причем, выбор одного из этих способов зависел от диаметра
привоя. Черенки диаметром более 0 4—0,5 см лучше прививать
в приклад сердцевиной на камбий, меньшего диаметра — кам-
бием на камбий.
Улучшенная копулировка требует более высокой квалифи-
кации прививальщиков. Лучше этим способом проводить при-
вивки пород с плотной древесиной (псевдотсуга, лиственница).
9-1
Таблица 1
Приживаемость хвойных пород в зависимости от способа прививки, %
Порода
Камбием
на камбии
Способ прививки
о
ш
Сосна кедровая
европейская 9.3
Пихта белая 90
Пссвдотсуга 93
Лиственница
европейская 90
Ель обыкновенная 93
85 72 — 31
80 80 — 50
92 — 90 —
90 — 90 —
90 87 — —
В целом все из перечислепных способов можно использо-
вать при прививках за исключением способа в боковой зарез.
Однако благодаря простоте выполнения, надежности лучше
всего прививать хвойные породы впрпклад сердцевиной па кам-
бий и камбием па камбий.
При изоляци прпвпвок применялись полиэтиленовая пленка,
нитки, изоляционная лента, топкая резина, полиэтиленовая
пленка с последующей изоляцией полиэтиленовыми мешочками
(табл. 2).
Большой разницы в приживаемости при изоляции прививок
различным материалом не наблюдается.
Более низкая приживаемость получена при обвязке нитка-
ми. Это связано в первую очередь с тем, что у части прививок
запоздали со снятием ниток, что привело к перетяжкам и ги-
бели прививок.
Для медленно растущих по диаметру пород вполне пригодна
1} качестве обвязки тонкая резина. Для изоляции прививок сос-
ны кедровой этот материал мало пригоден. Вследствие быстро-
го роста ио диаметру прививаемых компопептов разрушение
резины происходит раньше, чем наступает полное срастание
привоя с подвоем. Так, при весенних прививках сосиы кедро-
вой и изоляции резиной полностью срослось всего 34%. У
остальных прививок срастание произошло участками и привой
имел угнетенный вид и незначительный прирост в первые два
года.
Наилучшие результаты дала обвязка полпэтплеповой плеп-
95
Таблица 2
Зависимость приживаемости хвойных пород от применяемого
изоляционного материала, %
Изоляционный материал
Привой нитки полиэтиле- новая пленка тонкая резина изоляцион- ная лента полиэтиле- новые ме- шочки
Сосна кедровая евр. 94 89 92 97 98
Пихта белая 78 82 80 85 —
Ель обыкновенная 85 90 90 87 80
Лиственница
европейская 75 93 90 90 —
Псевдотсуга 90 92 90 — —
кон и полиэтиленовой -пленкой с последующей изоляцией ме-
шючками.
При изоляции! полиэтиленовыми мешочками приживаемость
прививок не возрастает, но улучшается качество срастания и
рост привоя в первый год. На следующий год разницы в росте
практически не обнаружено. Так в первый год при применении
полиэтиленовых мешочков прирост привоя составлял 5,0±0,2 см,
без мешочков 3,5±0,1 см, па следующий год соответственно
21,0±1,00 и 19,8±0,84 см.
В других районах страны отмечают значительное возраста-
ние приживаемых при прививках сосны с изоляцией полиэти-
леновыми мешочками. В Карпатах благодаря влажному не-
жаркому климату необходимость в применении полиэтилено-
вых мешочков отпадает. Также незначительно влияет обма-
зывание места обвязок пластилином пли парафином.
Применение парафина давало некоторый положительный
результат при прививках лиственницы и сосны кедровой при
изоляции нитками. Парафинирование прививок пихты дало
отрицательный результат. Приживаемость парафинированных
прививок упала до 50%. Успешность срастания прививок во
многом зависит от техники их исполнения и в первую очередь
от плотности соприкосновения прививаемых компонентов и
своевременного удаления обвязки.
Одним из основных вопросов при прививках являются сроки
проведения работ. Выделяют два срока прививок: весенние и
лстппе (Проказил Е. Н., 1960; Коновалов И. А. и Пугач Е. А.,
9G
1968 и др.). Весенние прививки рекомендуют проводить с сере-
дины апреля по июнь, летние — в конце июля, начале августа,
когда сформированы почки, одревеснели побеги, но жизнедея-
тельность камбия не ослаблена.
Рекомендации проводить прививки с апреля по июнь слиш-
ком неконкретны и пользоваться трудно ими, особенно в Кар-
патах, где в связи с вертикальной зональностью наступление
весны* происходит неодновременно.
Для установления оптимальных сроков, npiiBinBKiii проводи-
лись в трех пунктах на различных высотах: 400 м, 700 м и
1000 м над уровнем моря, в течение всего вегетационного пе-
риода при этом регистрировались фенофазы подвоя и привоя и
погодные условия.
По всем породам для различных высот над уровнем моря
получены аналогичные результаты, только па больших высотах
сроки начала работ были сдвинуты па более позднее время.
Оптимальных календарных сроков даже для одного лесо-
растителытого пояса установить не удалось вследствие разно-
временного наступ л сипя весны по годам.
Ранее полученные В. Б. Логгпновым (1967) данные о рез-
ком снижении процента приживаемости прививок в более
поздние весенние ершей нами не подтвердились. При правиль-
ном хранении привойного материала >п благоприятных погодных
условиях высокая приживаемость (до 90%) наблюдалась даже
при прививках в двадцатых числах июня. Вопрос о продлении
сроков прививок зимними черепками определяется в первую
очередь условиями их хранения. Лучшие результаты были
получены,- когда черенки хранились в полиэтиленовых мешках
в ледниках пли в холодильной установке.
Если оптимальные календарные сроки пе выявлены, то за-
висимость приживаемости прививок от фенофаз подвоя п при-
воя прослеживается хорошо (табл. 3).
Наилучшую приживаемость прививки имели в период ак-
тивной деятельности клеток камбия прививаемых компонентов.
При слабой активности жизнедеятельности клеток камбия пе-
риод срастания затягивается. Так, в начале апреля, когда
деятельность клеток только начиналась, полное срастание ком-
понентов наступило в среднем через 40 дней, а при прививках
в начале мая на 25—30 день. Удлинение периода срастания
приводило к значительному отпаду и плохому росту привоя в
первые два года. Низкий процент приживаемости осенних при-
вивок, когда жизнедеятельность камбия уже заканчивается,
объясняется тем, что, по-видимому, значительная часть их пе
7. Заказ’ 6609
97
Таблица 3
Зависимость приживаемости прививаемых растений от их фенофаз, %
Фснофаза Привой
сосна кед- ровая ев- ропсйск. пихта белая ель обык- новенная
Начало деятельности камбия подвоя, привой в состоянии покоя 80 88 70
Подвой тронулся в рост, стадия — вытягивания верхушечной почки, нача ло деятельности камбия привоя 95 90 90
Активный рост подвоя в высоту, рас пускание почек у привоя. 96 97 92
Однолетние пооеги подвоя и привоя нс одревеснели, верхушечная почка за- ложена 90 89 93
Однолетние побеги одревеснели у под- воя н привоя, активность камбия не снижена 92 90 90
Окончание деятельности камбия, кора от древесины отстает плохо 50 45 45
успевает срастись и гибнет в зимнее время. Оставшиеся при-
вивки в первые два года -шмели прирост 2—5 с.м в год, в то
время, как у весенних и летних прививок на второй год он
достигал 12—15 см.
Следует отметить, что -в Карпатах особенно в горнолесном
поясе, прививки неодревеснев-шш-ш черенками дают положи-
тельный результат. Так, в 1971 г. из 200 прививок ели, прове-
денных 1 июля, погибло к 15 сентября всего 6 штук.
Значительную роль в приживаемости прививок играют по-
годные условия. При наступлении засушливых, жарких дней
в период приживания значительная часть их гибнет.
Например, у ели приживаемость в засушливые периоды па-
дает до 60%. Особенно сильно на приживаемости сказываются
засушливые периоды при летних прививках неодревесневшими
черенками. В этом случае она падала до 50%.
С целью выявления •влияния возраста дерева на приживае-
мость и рост привоя были сделаны прививки черенками, взя-
тьями с деревьев возрастом 50—60 лет и 200 лет. Полученные
данные показали, что возраст дерева, с которого заготавливались
черенки, существенного влияния на приживаемость не оказы-
вает (90—98% и 90—95%). Различие же наблюдалось в даль-
нейшем росте привоя. Если на второй год прирост в высоту
98
прививок ели в первом случае составлял 45,0± 1,00 см, то во
втором — 22,0±0,38 см. Разница в приросте продолжала наблю-
даться и на третий год после прививок. Аналогичные данные
о влиянии возраста дерева на рост и приживаемость прививок
сосны получены1 Шульгой В. В. и Мосиным В. И. (1970) для
условий Казахстана. По нашим данным место заготовки че-
ренков в кроне не играет роли в приживаемости, но рост
привоев несколько меньший в высоту выполненных черенками,
заготовленными с нижней части кроны. Особенно значительная
разница наблюдалась у прививок ели обыкновенной. Если че-
ренки, заготовленные из верхней части кроны дерева, имели
прирост на второй год 30—35 см, то из нижних ветвей лишь
17—25 см. Кроме того, черенки из нижних ветвей давали
значительный процент горизонтально растущих побегов (до
40%).
У лиственницы была отмечена также тенденция давать
горизонтально растущие побеги, по при подвязывании к кольям
побег на следующий год рос вертикально. Ike привои кедро-
вой сосны независимо от места заготовки в кроне черенков
росли вертикально.
При закладке прививочных плантаций не всегда возможно
найти необходимое количество подвоев. В связи с этим были
проведены опыты по межвидовым и межродовым прививкам.
Для кедровых сосен в качестве подвоев кроме сосны обык-
новенной испытывались сосны веймутова, черная, баикса, ель
обыкновенная, лиственница европейская, пихта. На всех соснах
приживаемость сосны кедровой составляла 80^-95%. При при-
вивка! сосны кедровой на сосне банкса уже на 3—4.год прп-
вой перерастает по диаметру подвой. При межродовых привив-
ках приживаемость кедровых сосен не превышала 10%. Пихту
белую прививали на пихте белой, ели обыкновенной, листвен?
лице европейской. Приживаемость свыше 70% была получена
на подвоях пихты и ели. Однако на следующий год на еловом
подвое большинство прививок погибло. Причину гибели приви-
вок не удалось установить. С подобным явлением сталкивалась
Яковлева Л. В. (1967). При межвидовых прививках на пихте
белой хорошо приживались пихты кавказская, бальзамическая,
одноцветная, испанская, сибирская и др.
Псевдотсуга тмссолистная прививалась на ель, пихту, лист-
венницу. Наиболее хорошим подвоем, исключая псевдотсугу,
являлась лиственница, на что указывал также и Ковали-
шин В. П. (1966). На лиственнице приживалось свыше 90%
черепков псевдотсуги, а рост привоев мало чем отличался
7* 99
ат роста прпвгпвок псевдотсуги на псевдотсугу. Размеры при-
ростов па второй год были соответственно 25,6 и 29 см.
Высокая приживаемость (90%) псевдотсуги была и на
подвое ели. Здесь щрп.рост в высоту был значительно ниже —
15,7 см. На пихте приживаемость была около 90%, но при-
вивки имели угнетенный вид и на следующий год гибли.
У оставшихся прив1пвок даже на 4 год прирост не превышал
10—12 см. На ель обыкновенную было привито более 15 видов
и разновидностей елей. Во всех случаях приживаемость была
пе ниже 75%. Рост в высоту большинства прививок был зна-
чительно выше, чем, когда этот вид рос на собственных корнях.
Например, на 4-й год после прививки, привой ели колючей
(форма голубая) давал прирост до 65 см.
В связи с вышеизложенным при прививках хвойных пород
можно расчитывать на успех, в первую очередь, при межвидо-
вых прививках. Межродовые прививки во многих случаях дают
отрицательный результат и для рекомендации их производству
требуются дополнительные исследования.
ЛИТЕРАТУРА
Воверськпй 10. 10. Вирощувапня щеплен1х саджанщв модрины.
Ужгород, 1966. ।
Г и р г и д о в Д. Я., Д о л г о л и к о в В. И. О прививках ели обыкно-
венной. Сб. научно-исследовательских работ по лесному хозяйству.
ЛенНИПЛХ, М., 1964.
Коновалов Н. А., П у г а ч Е. А. Основы лесной селекции и сор-
тового семеноводства. М., 1968.
Ковалишин В. П. Межвидовые и межродовые прививки хвойных
пород. Лесоэксплуатация и лесное хоз-во, № 4, ЦНИИОТИ, Леспром,
1966.
Л о г г и н о в В. Б. Испытание некоторых способов прививки ели в
Карпатах. Лесоводство и агролесомелиорация. Вып. 9, Киев, 1967.
Прок а зин Е. П. Новый метод прививки хвойных для создания
семенных участков. Лесное хоз-во, № 5, 1960.
Шульга В. В., Мосин В. И. Некоторые вопросы элитного семе-
новодства сосны обыкновенной (Pinus silvestris L.) в Казахстане. Сб.
Лесная генетика, селекция и семеноводство. Петрозаводск, 1970.
Яковлева Л. В. Опыт межвидовой и межродовой прививки хвой-
ных в открытом грунте. Лесное хозяйство, № 6, 1967.
А. В. ЛУКИН
Основные итоги интродукции видов рода Pinus L.
в Центрально-Черноземных областях
История интродукции видов рода Pinus L. в Центрально-
черноземных областях (ЦЧО) имеет более чем полуторавеко-
вую давность. Нашими исследованиями, а также по литератур-
ным данным (О. Г. Каппер, 1933, 1954; И. К. Вехов, 1937,
1949, 1953; С. И. Машкин, 1952, 1964, 1971; В. Н. Вехов, 1958;
А. В. Лукин, 1961, 1966; И. Г. Акимочкин, 1961; Н. К. Вехов,
В. Н. Вехов, 1962; Е. С. Павловский, 1962; М. К. Кузьмин.,
1969 и др.) установлено, что в различного рода культурах на
территории ЦЧО в настоящее время произрастают 22 вида,
5 разновидностей и форм и 1 гибрид сосны, в том числе 2 вида
местных и 20 видов интродуцированных.
В систематическом отношении все имеющиеся здесь сосны
относятся к двум подродам, различающимся между собой по
количесту пучков в хвое: сосны однопучковые (подрод Haplo-
xylon Koehne) — 9 видов и сосны двухпучковые (подрод
Diploxylon Koehne) — 13 видов.
Из числа интродуцированных сосен в лесорастительных
условиях ЦЧО 19 видов растут в форме деревьев и только 1 вид
имеет кустарниковую 'биоморфу.
В пределах описываемой территории 4 вида сосны встре-
чаются в разного рода культурах часто, 8 видов — редко и
8 видов — единично.
Наибольшее количество видов сосны интродуцировано в
ЦЧО из Северной Америки (9 видов, или 45%), 6 видов (30%)
являются уроженцами Европы и 5 видов (25%) происходят пз
Азии.
Наблюдения пад зимостойкостью и морозоустойчивостью
интродуцированных сосен показывают, что в ЦЧО сильно под-
мерзает только Pinus monticola Dougl., недостаточно зимо-
стойкими оказались Pinus funebris Кот., Pinus excelsa Wall.,
Pinus resinosa Ait. и Pinus rigida Mill. Все остальные сосны
(15 видов, или 75%) являются здесь вполне зимостойкими
и морозоустойчивыми.
Наиболее засухоустойчивыми из испытанных сосен явля-
ются Pinus montana Mill., Pinus nigra Arn., Pinus scopulorum
Lemm., а наименее требовательными к плодородию почвы ока-
зались Pinus flexilis James., Pinus funebris Korn., Pinus ha-
mata D. Sosn., Pinus montana Mill., Pinus murrayana Balf.,
101
Tinuspumila (Pall.) Regel, Pinus resinosa Ait, и Pinus scopulo-
rum Lemm.
Из общего числа интродуцированных сосен 2 вида только
вегетируют и лишь изредка цветут, 9 видов дают всхожие семе-
на и 9 видов образуют устойчивый, жизнеспособный самосев и
подрост, что является показателем высокой степени пх аккли-
матизации (В. П. Малеев, 1933).
В то же время продолжительный интродукционный опыт
показывает, что подавляющее большинство иноземных и ипо-
раноиных видов сосны в условиях ЦЧО значительно уступает
но быстроте роста и продуктивности образуемых ими насажде-
ний местному «эталону» — сосне обыкновенной (Pinus silvest-
ris L.). Только сосна веймутова (Pinus strobus L.) в куль-
турах на достаточно плодородных и удовлетворительно увлаж-
ненных почвах способна образовывать здесь насаждения, кото-
рые растут в высоту и по диаметру быстрее культур обыкно-
венной сосны (таблица 1).
Табл п ца 1
Сравнительные показатели роста культур интродуцированных сосен в ЦЧО
Соотношение в % %
Высот Диаметров
Виды сосны
Pinus silvcstris L. 100 100
Р. banksiana Lamb. 70— 93 77— 85
Р. cembra L. 36— 49 48— 52
Р. flcxilis James. 70 102
Р. funebris Korn. 33 75
Р. hamata D. Sosn. 79— 86 84—101
Р. koraiensis Sieb. ct Zucc. 59— 63 54— 57
Р. montana Mill. 40— 53 35— 42
Р. monticola Dougl. 69 95
Р. murrayana Balf. 76— 99 81— 90
Р. nigra Arn. 62—100 58—103
Р. pallasiana Lamb. 48- 54 93— 97
Р. рейсе Gris. 51— 98 50- 79
Р. ponderosa Dougl. 65— 77 73— 99
Р. pumila (Pall.) Regel 19— 26 —
Р. resinosa Ait. 27 54
Р. rigida Mill. 29 32
Р. scopulorum Lem. 51— 60 63— 89
Р. sibirica (Rupr.) Mayr. 48— 82 30—112
Р. strobus L. 85—112 82—120
102
Таблица 2
Ассортимент быстрорастущих и хозяйственно-ценных видов сосны
для внедрения в лесные и защитные культуры на территории ЦЧО
Название
древесной породы
По типам
условий
местопро-
израстания
1____По_категориям_насаждени2£
I Противоэрозионные насажден.
па приво-
дораздель-
ной зоне
па гпдро-
графпческ.
зоне
полосы
защитные полосы вдоль
железподорожи. и шоссей-
ных дорог
I. Виды, рекомендуемые для производственного освоения
Pinus cembra А, Б — — X X X 4 — 4 4 _|_ — 4
Р. koraiensis А, Б — — X X X — 4 — н-
Р. sibirica А, Б — — X X X н- — 4 4 — 4
Р. strobus А, Б, В — + X X X X — X X X — X
II. Виды, рекомендуемые для предварительного испытания
р. montana Б, В — 4 4 — — — X — — 4 4 X
р. murrayana А, Б — н- 4 — -f- 4 — + н- — — —
р. nigra Б, В — 4 — — — — •4- — X —
р. pallasiana Б, В — 4 — + — -f- — — — X —
р. ponderosa А, Б — 4 4 — н- — 4 4 — — —
Обозначение: х— рекомендуемая порода;
Н---породу можно культивировать;
---- нерекомендуемая порода.
Выводы и предложения
1. В лесораст.птельпых условиях ЦЧО некоторые иноземные
и пиорайонные виды сосны благодаря быстрому росту и выдаю-
щимся техническим качествам древесины и других продуктов,
ценным водо — охранио — защитным свойствам и высокой
устойчивости к неблагоприятным факторам внешней среды мо-
гут иметь большое значение в деле существенного и быстрого
повышения продуктивности лесов.
2. На основании тщательного и всестороннего изучения по-
ведения интродуцированных видов сосиы в новых условиях
местопроизрастания для лесора-стительных условий ЦЧО авто-
ром разработай и рекомендуется ассортимент быстрорастущих
и хозяйственно-ценных сосен-экзотов для различного рода лес-
ных и защитных культур (таблица 2).
3. В целях расширения работ по интродукции и акклимати-
зации хвойных древесных растений в ЦЧО, опираясь при этом
на многочисленные методы предварительного выбора интроду-
цеитов, нами разработан рекомендательный перечень видов
сосны, включающий 13 названий, а именно: Pinus albicaulis
Engelm., Р. aristata Engem., P. bungeana Iffrei, P. bungeana
Zucc., P. contorta Dougl., P. gerarcliana Wall., P. heldreichii
Christ., P. kochiana Klolzch., P. lambertiana Dougl., P. palust-
ris Mill.. P. parviflora Sieb. et Zucc., P. pungens Michx., P. vir-
giniana Mill.
ЛИТЕРАТУРА
А к цм оч кип II. Г. Итоги интродукции древесных и кустарниковых
пород на Лесостепной опытпо-сслекциоппой станции за 35 лот. Ефре-
мов, 1961.
Вехов В. И. Особенности роста некоторых видов сосен в условиях
лесостепи. Научные доклады высшей школы (биологические пауки),
№ 2, 195В.
Вехов И. К. Быстрота роста экзотов в условиях лесостепи. Курск,
19374
Вехов И. К. Быстрота роста экзотов в условиях степи. Гослес-
бумиздат, М., 1949.
Вехов Н. К. Деревья и кустарники Лесостепной селекционной
опытной станции. Изд.во МКХ РСФСР, М., 1953.
Вехов Н. К., Вехов В. Н. Хвойные породы Лесостепной стан-
ции (Итоги интродукции). Изд-во МКХ РСФСР, М., 1962.
Каппер О. Г. Экзоты ЦЧО. ВНИАЛМИ, 1933.
Капп ер О. Г. Хвойные породы. Гослесбумиздат, М.-Л:, 1954:
Кузьмин М. К. Деревья и курстарники Лесостепной опытно-селек-
циоипой станции. Воронеж, 1969.
•104
Лебедев Г. И. Акклиматизация древесных и кустарниковых пород
(из опыта Лесостепной степи). Изд-во МКХ РСФСР, М., 1953.
Л у к и п А. В. Морозоустойчивость хвойных пород в условиях цент-
ральной лесостепи. ИВУЗ, «Лесной журнал», № 4, 1961.
Лукин А. В. Ассортимент быстрорастущих и хозяйственно ценных
дровеспо-кустарпиковых пород для лесных и защитных культур (реко-
мендации). Липецк, 1966.
Малеев В. П. Теоретические основы акклиматизации растении.
Приложение к «Трудам по прикладной ботанике, генетике и селек-
ции», Л., 1933.
Машкин С. 1Г. Дикорастущие и разводимые деревья и кустраники
Воронежской области, в Вкп. «Деревья и кустарники Воронежской об-
ласти», Воронеж, 1952.
Маш кип С. И. Депдрофлора Центрально-Черноземной полосы
('ССР и перспективы ее овогащепня. Научные записки Воронежского
отделения Всесоюзного ботанического общества, Воронеж, 1964.
М а ш к и н С. И. Дендрология Центрального Черноземья. Изд-во ВГУ,
Воронеж, 1971.1
Павловский Е. С. Таксационное описание лесных насаждений
Каменной степи. Изд-во «Коммупа», Воронеж, 1962.
Н. А. БОЛОТОВ
Перспективы дальнейшей интродукции видов пихты
(Abies Hill) '
в Европейской части СССР
Интродукция ппхтьг и Европейскую часть СССР насчи-
тывает полуторавековую давность. За ото время- накоплен до-
статочный опыт, позволяющий подвести итоги и наметить
перспективы дальнейшего интродукционного испытания видов
рода Abies Hill, произрастающих в различных районах земного
шара.
К настоящему времени в Европейскую часть страны из
52 известных видов пихт интродуцировано 29. Опыт их аккли-
матизации освещен в справочных материалах по деревьям и
кустарникам и в специальной научной литературе (II. Д. Не-
стерович, 1950; А. Л. Лыпа, 1952; О. В. Соколова, 1952;
В. В. Ухапо®, 1952; Н. М. Андронов, 1953; М. А. Железнова-
Каминская, 1953; Ф. Б. Орлов, 1953; А. М. Маурнпь, 1957;
Б. М. Алимбек, 1958; И. А. Кауров, 1961; II. К. Бехов, В. ТТ. Ве-
хов, 1962; А. С. Лаитратова, 1969; К. А. Андреев, 1971;
С. И. Машкин, 1971 и др.).
Состояние испытаппости интродуцентов рода Abies Hill в
Европейской части СССР- отражено в таблице 1.
105
Таблица I
Состояние испытанности различных видов рода Abies Hill,
в Европейской части СССР
Районы естественного произраста- ния (распространения) интроду- центов Общее число видов Ч11ело испытанных видов Число не испы- танных видов
в 3-х и более областях в 1—2-х областях
к-во 1 % к-во 1 % к-во 1 % к-во | %
Северная Америка 12 23 6 50 5 42 1 8
Гватемала 1 2 — — — — 1 100
Мексика 2 4 — — 1 50 1 50'
Южная и Центральная Европа 6 11 4 67 — — 2 33
Малая Азия 3 6 — — 1 33 2 67
Северная Африка 2 4 — — 1 50 1 50
Гималаи 2 4 — 2 100 — —
Китай 11 21 — — — — 11 100
Япония 5 10 2 40 2 40 1 20
Корея 1 2 — — 1 100 —
Дальний Восток и Сибирь 7 13 4 57 — — 3 43
Итого: 52 10 16 31 13 25 23 44
Приведенные данные показывают, что наиболее испытан-
ными в Европейской части СССР являются североамериканские
и европейские виды; совершенно не испытывались пихты, про-
израстающие в Китае.
Для сравнения степени адаптации видов пихт, интроду-
цированных в различных областях Европейской части СССР,
известные нам многочисленные исследования, выполненные
разными авторами, в разное время и различными методами,
были приведены к небольшому числу показателей, достаточно
полно отражающих этот процесс. Такими показателями являют-
ся: зимостойкость, устойчивость против заморозков, засухо-
устойчивость и репродуктивная способность.
Степень устойчивости интродуцентов по тому или иному
признаку выражалась в процентах: у вполне устойчивых в
данных природных условиях она оценивалась 100%, у стра-
дающих в различной степени, но сохраняющих жизнеспособ-
ность — 50%, У гибнущих — равной нулю.
Репродуктивная способность, как дополнительный признак
адаптации у видов, имеющих жизнеспособный самосев и под-
рост, оценивалась 100%, У видов, дающих всхожие семена —
50%, и не продуцирующих всхожих семян — равной нулю.
Сопоставление показателей адаптации с агроклиматически-
ми показателями местообитания интродуцентов показало, что
большая корреляция наблюдается между суммой положитель-
ных температур свыше 10°С, обеспеченной осадками, и сте-
пепыо зимостойкости; продолжительностью безморозного перио-
да и повреждаемостью интродуцентов заморозками;- гидротер-
мическим коэффициентом (Г. Т. К.) и степенью засухоустойчи-
вости.
Эти три основных агроклиматических показателя пункта
интродукции сравнивались с темп же показателями естествен-
ного ареала иптродуцента, на основании чего вычислялось мак-
симальное (экстремальное) отклонение между ними (в процен-
тах) при наивысшей степени адаптации иптродуцента (в боль-
шинстве случаев 100%).
Величина экстремального отклонения (э) выражается фор-
мулой:
Э = 100о/о<
1 !сст
где Пест- и Писк. — соответствующий агроклиматический по-
казатель естественного и искусственного
ареалов.
107
Коэффициент пластичности определяется >по формуле
К=А+Э, где А — процент адаптации интр оду цента в данных
климатических условиях, Э — величина экстремального откло-
нения.
Следовательно, коэффициент пластичности вида отражает
не только степень адаптации иптродуцента в новых условиях
среды, по и особенности филогенеза вида, экстремально про-
являющиеся в новых условиях произрастания.
На основе приведенной методики расчетов для всех интро-
дуцированных в Европейской части СССР видов пихт нами
определен коэффициент пластичности (табл. 2).
Данные таблицы 2 показывают, что общей закономерности
в варьировании пластичности внутри рода ле обнаруживается.
Однако для наиболее близкородственных серий и видов, дли-
тельное время развивающихся в сходных климатических усло-
виях, такая закономерность заметно проявляется. Особеппо
близки коэффициенты пластичности в секции Abies, которая
считается одной из древнейших (А. Е. Маценко, 1964). Две
ее серии Nordmannianae и Albae представляют собой тупико-
вую ветвь эволюции. Изучение их палеоареалов показывает,
что виды этой секции, пережившие оледенение в южных ре-
фугиумах и эндемически ограниченные в постоянных ареалах,
имеют малую пластичность. Исключение составляет только
Abies nordmanniana (Stev.) Spach. Однако дело здесь не в
высокой пластичности этой пихты в засушливых условиях, а
в изменении самих условий естественного ареала в голоцене
в сторону увеличения влажности. Другими словами, высокий
коэффициент пластичности (К) в данном случае подчеркивает
только консерватизм наследственности данной секции. Отсюда
можно сделать вывод, что пластичность новых иптродуцентов
секции Abies будет также невысокой.
Секция Bracleala объединяет наиболее древние пихты, про-
израстающие в районах земного шара, в наименьшей степени
подвергавшихся геологическим и климатическим катаклизмам.
Пихты серий Religiosae и Fraserianae, видимо, также будут
иметь наиболее низкую пластичность. И все же потенциальные
возможности этой секции в отношении интродукции достаточ-
но велики, примером чего может служить A. fraseri (Pursch.)
Poir., испытавшая в Аллеганских горах сильное воздействие
ледников. Подобные условия, по мнению китайских геологов
Ли-Сы-Чуана и Яп-Цынь-Шапя (Чень Ши Сюнь, 1961), могли
существовать в плейстоцене или несколько ранее в высокогор-
108
Таблица 2
Коэффициент пластичности некоторых видов пихт, интродуцировоанных
в Европейской части СССР
Виды Родина 9 Зимостойкость Устойчивость к заморозкам Засухоустой- чивость Репродук- тивная спо- собность
А Э к А э | к А Э к
1 3 4 5 , б 7 1 8 10 11 12
Секция Bracteata Серия Religiosae
1. Abies nobilis Lindl. США 100 0 100 100 0 100 100 0 100 —
2. A. religiosa Lindl. Мексика 100 0 100 100 0 100 50 0 50 —
Серия Fraserianae
3. A. fraseri (Pursch.) США 100 60 160 100 10 110 100 50 150 80
Poir.
4. A. venusta Koch. США 100 0 100 100 20 120 100 0 100 —
Секция Elate
Серия Nephrolepides
1. A. nephrolepis Maxim. СССР
(Д. В.) 100 10 ПО 100 0 100 100 20 120 30
2. A. sachaJinensis Mast » 100 10 110 100 0 100 100 10 110 50
Серия Vcitchianae
3. A. veitchii Lindl.
Япония 100 30 130 100 10 110 100 60 160 50
4. A. koreana Wils. Корея 100 30 130 100 0 100 — — — —
Секция Abies Серия Firmae
1. А. firma Sieb. etZucc. Япония 100 0 100 100 Серия Albae 0 100 100 0 100 —
2. А. alba Mill. Европа 100 0 100 100 0 100 100 0 100 50
3. А. cephalonica Loud. » 100 10 110 100 0 100 100 0 100 50
Серия Nordmannianae
4. A. nordmanniana
(Stev.) Spach. Европа 100 0 100 100 Э 100 100 50 150 50
1. A. holophylla Maxim. СССР Д.В. 100 Секция Piceaster Серия Homolepides 50 150 100 10 по 100 40 140 20
2. A. homolepis Sieb. et Zucc. Япония 100 30 130 100 0 100 юо 70 170 0
3. Abies mariesii Mast. Япония 100 Серия Mariesianae 20 120 100 15 115 100 30 130
4. A. pindrow. Spach. Гималаи 100 Серия Sibiricae 0 100 — 50 40 90
5. A. Sibirica Ledeb. СССР (Сибирь) 100 20 120 100 30 130 100 0 100 80
6. A. numidica Lannoy Алжир 100 Серия Pinsapo —40 60 — 50 40 90
7. A. pinsapo Boiss. Испания 100 —40 60 100 30 130 100 0 100 0
8. A. Silicica Carr. Турция 50 Серия Webbianae 30 80 — 100 0 100 50
9. A. Webbiana LindL Гималаи 100 10 110 — — — 50 40 90 —
1 2 3 4 5 6 7 8 9 10 11 12
Серия Lowianae
10. A. concolor Lindl. et США
Gord. 100 0 100 100 0 100 100 0 100 80
11. A. grandis Lindl. США 100 30 130 100 20 120 50 -10 40 30
12. A. lasiocarpa Nutt. США 100 0 100 100 0 100 100 0 100 50
13. A. lowiana Murr. США — — — 100 0 100 — — — —
Серия Arizonicae
14. A. arizonica Merr. США 100 50 150 100 0 100 100 —60 40 30
15. A. balsamea Mill. США 100 30 Cepi 130 100 !я Amabilcs 0 100 0 100 100 80
16. A. amabilis Ford. США 100 20 120 100 20 120 100 0 100 30
пой зоне пихтовых лесов Центрального Китая. В связи с этим,
видимо, будет логичным принять значение коэффициента пла-
стичности китайских пихт средним для секции Bractoata.
Для неиспытанных пихт секции Elate, произрастающих дл и-
тельное время в сходных климатических условиях муссонного
климата Японии и Дальнего Востока, п имеющих достаточное
количество близкородственных видов, уже прошедших -первич-
ное испытание, видимо можно принять коэффициент пластич-
ности -средний для каждой серии, а для A. mayriana Miyabe el
Kudo средний между сериями Nephrolepides и Veitchianae.
Секция Piceaster содержит самое большое разнообразие мо-
лодых видов, имеющих и широкие, и узкие ареалы, распола-
гающиеся от субтропиков до северной тайги. Поэтому предпо-
лагаемые коэффициенты пластичности пптродуцептов этой се-
рии необходимо брать по аналогии с наиболее близкими вида-
ми, произрастающими в сходных климатических условиях.
Агроклиматические показатели вероятных ареалов интро-
дуцентов рассчитывались памп по следующей формуле:
- Пест (100-К)
1 bin 1 -^ест 1 । QQ
где Пин и Пет — соответственно агроклиматический показа-
тель зоны культивирования пнтродуцента п
его естественного ареала;
К — предполагаемый коэффициент пластичности
пнтродуцента, который коррелируется с дан-
ным агроклиматическим показателем.
Анализ данных таблицы 3 показывает, что из 24 возмож-
ных для интродукции новых видов пихт лишь менее половины
могут быть использованы в лесостепной и лесной зонах Евро-
пейской части СССР. Наиболее перспективными здесь будут
виды китайских пихт из серий Fargesianae и Faxonianae (за
исключением явно теплолюбивых A. fargesii Franch. и A. for-
restii Craib.).
В северо-западной части лесной зоны, в условиях повышен-
ной влажности, могут быть успешно испытаны виды пихт,
имеющие естественные ареалы на Советском Дальнем Востоке,
такие как камчатская (A. gracilis Кот.) и сахалинская
(A. mayriana Miyabe et Kudo).
Перспективными в районах засушливой лесостепи -могут
оказаться высокогорные экотипы Северо-американской A. mag-
nifica Murr., а для юго-западной части лесной и лесостепной
зон — A. semenovii Fedtsch.
lt2
Таблица 3
Предполагаемые коэффициенты пластичности и агро-климатические
показатели ареалов вероятных интродуцентов
Название вида М если I а х о ж де икс осте - ствспиого ареала и вы- сота над уровнем моря в м Коэффициент пластич- ности -К» зимостой- кости Агро-] показа гаем of о Л климатиче» тел н пред г i зоны кул рования £ § ° 3 О о 2 2 А = средний из абсо- ~ ° £ лютных мпнпму- g Q3 ?5 м о в 7 Коэффициент «К» устой- чивости к заморозкам Продолжительность безморозного периода предполагаемой зоны культивирования (дней) Коэффициент «К» засу- хоустойчивости Минимальный гидротер- мический коэф. (ГТК) предполагаемой зоны культивирования Репродукционная спо- собность предполагае- мого пптродуцента
1 о 3 4 5 ! 6 । 7 8 9 19 11
Секция Abies
Серия Albac
1. A. borisii—regis Mattf. Балканы
1000—1700 105 3100 3100 —18 100 205 100 0,6 50
2. A. nebrodensis Mattf. Сицилия 2000
105 2900 1900 —18 100 206 100 0,6 50
3. A. bornmillleriana Mattf. Малая Азия
1100—210С 100 3400 2100 —15 100 190 100 0,3 50
4. A. equi-trojani Малая
Aschers. et Sint. Азия
1300—1700 100 5700 1800 —13 100 260 100 0,2 50
Секция Bracteata
Серия Religiosae
1. A. shastensis Lemmon США 100 4300 1000 — 100 — — — —
Серия Fraserianae
4^ 2. A. hickclii Мексика
Flous et Gauss. 2000—3000 100 7500 7500 — 120 290 100 0,2 —
Серия Fargesianae
3. A. georgei Orr. Китай
3600—4000 130 2000 2000 —29 105 135 125 0,7 —
4. A. forrestii Craib. Китай
3600 130 2300 2300 —25 105 135 125 0,7 —
5. A. squamata Mast. Китай
3600 130 1700 1700 —34 105 120 125 0,7 —
6. A. fargesii Franch. Китай
2300—3300 130 2500 2500 —24 105 140 125 0,7 —
7. A. delavayi Franch. Китай
3500—4200 130 1900 1900 —31 105 125 125 0,7 —
Серия Faxonianae
8. A. faxoniana Китай
Rehd. et Wils. 2900—3600 130 1900 1900 —31 105 135 125 0,9 —
9. A. sutchuensis Китай
Rehd. et Wils. 2800—3600 130 2000 2000 —30 105 135 125 0,9 —
Секция Elate
Серия Nephrolepides
1. A. gracilis Kom. СССР
д.в. по 600 600 —26 100 120 115 1.6 4О>
2. A. recurvata Mast. Китай
2700-3600 по 2700 2700 —22 100 130 115 0,7 40:
Серия Veitchianae
3. A. sikokiana Япония 130 2800 2800 —14 105 220 130 2,5 2b
Nakai
1 2 3 4 5 6 7 8 9 10 11
4. A. mayriana Сахалин 120 1400 1400 —30 100 130 120 1,4 30 Miyabe et Kudo Секция Piceaster Серия Sibiricae 1. A. semenovii Тянь-Шань Fedsch. 1300—280C 120 1800 1800 —25 130 110 100 0,9 30
Серия Amabilcs
2. A. magnifica Murr. США 120 2000 2000 — 120 90 100 0,3 30
Серия Pinsapo
3. A. maroccana Trabut Марокко 1600—2100 60 6900 3800 — 7 130 135 90 0,2 —
Серия Lowianae
4. A. guatemalensis Rehd. Гватемала ПО 6800 6800 — 100 365 90 0,6 —
Серия Chensiensis
5. A. chensiensis Van-Tiegh. Китай 2800—3500 105 2700 2700 —30 100 140 100 0,6 —
6. A. ernestii Rehd. Китай 2600—3800 105 2900 2900 —26 100 130 100 0,8 —
7. A. salouensis Bord. et Gauss. Китай 2800—3000 105 3100 3100 —15 100 180 100 0,4 —
Наименее перспективными для интродукции в Европейскую
часть СССР, видимо, будут консервативные виды секции Abies
л Л. niaroccana Trabut., а также теплолюбивые китайские пихты
серии Chensienses п Nephrolepides п субтропические виды
пихт из Мексики и Гватемалы.
Перспективность высокогорных пихт Центрального и Юж-
ного Китая для интродукции в Европейской части СССР
косвенно подтверждается данными китайских лесоводов
(Ма-Цзи, 1955 и др.), сообщающих об успешном произраста-
нии самой южной из пихт Китая A. georgei Огг. в лесных
культурах Северного и Северо-Восточного Китая.
Таким образом, еще 8—9 новых цепных в лесохозяйствеи-
1Ю1М отношении видов пихт успешно могут выдержать интро-
дукционное испытание п произрастать .в Европейской части
СССР.
«II II Т Е Р А Т У Р /\
А л и м б о к В. М. Основные результаты опыта интродукции древес-
ных и кустарниковых пород в Поволжье. Со.: «Быстрорастущие и хоз.
ценные породы». М., 1958.
А н Дрее в К. А. Морозоустойчивость древесных экзотов в условиях
Петрозаводска. Уч. зап. Петр, университета, т. XVII, вып. 6, 1969.
Андропов П. М. О зимостойкости деревьев и кустарников в
Ленинграде. Сб.: «Интродукция растений и зеленое строительство»,
БИН, АН СССР, вып. 3, 1953.
Бехов Н. К., Бехов В. Н. Хвойные породы Лесостепной станции
(итоги интродукции), Изд-во МКХ РСФСР, М., 1962.
Деревья и кустарники. Тр. Никитского ботанического сада. Ялта,
1939. i
Деревья и кустарники. Т. 1, Голосеменные, All СССР, М., 1949.
Железнов а-К а м и н с к а я М. А. Результаты интродукции хвой-
ных экзотов в Ленинграде и его окрестностях. Тр. ИНН, вып. 3,
серия 6, «Интродукция и зеленое строительство», 1953.
It а у р о в И. А. Итоги интродукции древесных и кустарниковых
пород в районе Ленинграда. Бюлл. Главн. бот. сада АН СССР, вып. 41,
1961.
Качалов А. А. Деревья и кустарники. Пзд.во «Лесная промыш-
ленность», М.т 1970.
Л а и т ратова А. С. Интродукция древесных растений Восточно-
Азиатского происхождения в условиях Карелии». Уч. зап. Петрозавод-
ского университета, т. XVII, вып. 6, 1969.
Л ы п а А. Л. Дендрологические богатства УССР и их использование.
Со.: «Озеленение населенных мест», 1952, Киев.
Соколова О. В. Зимостойкость древесных и кустарниковых пород
па питомниках Ботан. сада БИНа. Сб.: «Интродукция и зеленое строи-
тельство», вып. 2, 1952, серия 6.
У х а н о в В. В. Результаты перезимовки хвойных деревьев и кустар-
ников в зиму 1939—1940 гг. в районе Ленинграда. Тр. БИНа, вып. 2,
«Интродукция растений и зеленое строительство», 1952.
116
Чень Ши-Сюнь. Климат Китая. Изд-во ипостраи. лит., .М., 1961.
М а цепко А. Е. Обзор рода Abies. Ботан. матер, гербария БПНа
АН СССР, 1963.
М ацен к о Л. Е. Пихты восточного полушария. Сб.: «Флора и
систематика высших растении», серия 1, вып. 13, 1964.
Машкин С. И. Дендрология Центрального Черноземья. Т. 1,
издательство Воронежского университета, Воронеж, 1971.
Маурипь А. М. Хвойные экзоты Латвийской ССР. Рига. 1957.
Ма-Цзи. Леса Китая. Сб.: «Леса и почвы Китая». Изд-во иност-
ранной лит., М., 1955.
Мировой агро-климатический справочник. Л.—М., 1937.
Нестерович Н. Д. Акклиматизация древесных растении в зеленом
строительстве и леспом хоз-ве БССР. Минск, 1950.
Орлов Ф. Б. Интродукция древесной растительности па Севере.
Ж. «Лрспое хозяйство», 12, 1953.
А. Г. КАСЮКОВ
К вопросу о газоустойчпвостп п фптонцидпостп
древесных пород в условиях юго-востока РСФСР
Положительная роль зеленых насаждений в системе го-
родской застройки (промышленной п жилой) заключается в
оздоровлении атмосферы, улучшении микроклимата, защите от
солнечной радиации, ветра, пыли, шума, дыма, копоти. Оздо-
ровление атмосферы происходит за счет фотосинтеза, т. е.
процесса превращения световой энергии, поглощенной хло-
рофиллом, в химическую энергию органических соединений,
синтезируемых в зеленых клетках из углекислого газа и волы
с выделением большого -количества кислорода.
Наряду с выделением кислорода поглощается углекислый
газ. Следовательно, озеленение играет совершенно незамени-
мую роль в природных процессах, жизненно важных для че-
ловека.
Зеленые растения выделяют не только кислород. В 1928 г.
советские ученые Б. П. Токин, А. Филатова и А. Тебякина
открыли повое свойство растений — способность вытелять фи-
тонциды. Эти летучие вещества создают невосприимчивость
или природный иммунитет растений к различным заразным
болезням.
Учеными было замечено, что выделяемые растениями фи-
тонциды снижали «количество микробов в воздухе: фитонциды
туи — на 67%, хризантемы мелкоцветной на 66%, райграса
117
пастбищного на 58%, хризантемы крупноцветной — па 55%,
герани и бегонии — на 43%, аспарагуса па 38%.
Если учесть, что в 1 см3 молока бывает 30 000 бактерии,
а в I г почвы до 10—15 млн. микробов, то становится попят-
ным значение фитонцидов растении в оздоровлении среды.
Из декоративных растений, рекомендуемых для озеленения
юго-востока, наиболее фитонцидными являются: пион дикий
(корень), кедр атласский, клен татарский и сахарный, дуб
пушистый, каштан калифорнийский, апельсиновое дерево,
черемуха обыкновенная, можжевельник крымский и казац-
кий, чубушник, граб европейский, смородина черная, айва
обыкновенная, сосна кедровая, береза бородавчатая, тополь
серебристый (Токин, 1967).
Зеленые листья растений выделяют в воздух массу лету-
чих веществ: водород, метан, этан и этилен, бутап и бутилен,
пропан, ряд альдегидов, спиртов, эфирных масел и других
газообразных соединений. Созревающие плоды также выделяют
определенные летучие соединения. Как правило, летучих ве-
ществ -в воздухе немного — тысячные доли граммов на 1 кг
воздуха. Однако летучше вещества растений способствуют
ионизации воздуха, они биологически активны. При попадай и и
в организм они действуют как стимуляторы и актнвизаторы
жизненных процессов.
Этот факт особенно важен для здоровья человека, потому
что тяжелые ноны (заряженная пыль, копоть, дым, разные
испарения) вредны, а легкие ионы, особенно попы отрицатель-
ного знака молекул кислорода оказывают благотворное и це-
лебное действие на живые организмы. Легкие отрицательны'?
попы рекомендуют вводить в состав коплициопированпого воз-
духа. Применение такой •ионизации снижает у человека утом-
ляемость и резко сокращает заболеваемость.
Число легких отрицательных попов увеличивает сосна обык-
новенная, дуб красный, туя западная, лиственница сибирская,
дуб черешчатый, ель обыкновенная, пихта одноцветная, липа
мелколистная. Неблагоприятное влияние оказывает дуб болот-
ный, каштан конский, орех грецкий, черный и серый, клеи
остролистный (Лахно, 1960).
Известно, что насаждения существенно влияют па темпе-
ратуру и влажность. Они -задерживают атмосферные осадки,
уменьшают испарение почвенной воды. Испарение влаги с
поверхности почвы среди деревьев п кустарников в полтора-
два раза меньше, челг с поверхности открытых мест. Причи-
на — более низкая температура, ослабленная сила ветров,
118
наличие подстилки. В свою очередь, отмеченные условия более
бла гоприятно влияют на произрастание насаждении.
Таким образом, перечисленные свойства зеленых растении:
выделение в атмосферу кислорода, поглощение углекислого га-
за, увеличение количества легких отрицательных ионов в воз-
духе, смягчение перепадов температуры и влажности, а также
бактерицидные свойства растений в воздухе и почве — позво-
ляют сделать вывод, что в руках человека деревья, кустарни-
ки и газоны могут быть незаменимыми инструментами но улуч-
шению микроклимата жилой и производственной среды. В про-
изводственной среде озеленение может иметь такие же функ-
ции и назначение, как и на городской территории. Одна ко
специфика промышленного предприятия отражается на качест-
венной стороне озеленения, в частности, на выборе пород де-
ревьев, кустарников и цветов. Большой интерес представляет
сумма качеств и свойств самих растений. В связи с тем, что в
данном случае рассматривается создание максимально устой-
чивых посадок в условиях промышленных предприятий с вред-
ными выделениями и в их защитных зонах, остановимся па
отношении растений к некоторым химическим соединениям.
Наибольший вред зеленым насаждениям наносят сернистый
газ, хлорнстып водород, хлор, фтористо-водородная и кремне-
фтористоводородная кислота, сероводород, синильная кислота,
азотная и азотистая кислоты (Бойченко, 1952). Убытки США
в 1950—51 гг. от загрязнения воздуха составили 1500 млн. дол-
ларов, в Англии в 1947 г. — 100 млн. фунтов стерлингов,
во Франции — 240 млн. франков (Габинова, Попов, 1962).
В настоящее время борьба с промышленными загрязнения-
ми все еще пе достигла требуемого уровня. Поэтому наиболее
устойчивыми средствами остаются градостроительные меро-
приятия, в частности, средства озеленения. Однако далеко не
все растения выносят действие газов. Газоустойчивость расте-
ний изменяется -в зависимости от многих факторов: характера
производства и степени загрязненности, климатических и поч-
венных условий, агротехники, возраста растений, характеров
насаждений и их расположения по отношению к источнику
загрязнения (Бойченко, 1952; Колесников, 1960; Спр. «Озеле-
нение городов», 1960; Галактионов и др., 1967).
На основе целого ряда литературных источников в условиях
загрязнения промышленными газами возможно применение сле-
дующих деревьев, кустарников и вьющихся растений: де-
ревья — абрикос обыкновенный, акация белая, вяз мелколист-
ный, гледичия трехколючковая, ива белая, ива вавилонская,
119
тополя — канадский, пирамидальный, китайский, черный,
белый, софора японская; кустарники — айва низкая, айва
японская, акация желтая, бирючина обыкновенная, бу-
зина красная, калина гордовина, лох узколистый, скум-
пия, бирючина обыкновенная, тамарикс одесский, виноград ди-
кий, плющ, можжевельник виргинский (Колесников, 1962).
Благодаря морфологическому строению листа вяз мелколист-
ный задерж.ивает в 6 раз больше пыли, чем тополь (Кудинов,
1968). Сирень венгерская и бузина красная также эффективно
задерживают пыль и сами при этом не повреждаются.
При обследовании ряда промышленных предприятий юго-
востока автором отмечена хорошая выносливость акации ново-
мексиканской, дерезы ромбовидной, абрикоса обыкновенного,
ивы вавилонской, акации белой (шаровидная форма), которых
нет в перечне газоустойчивых растений.
Приведенные свойства ряда деревьев, кустарников и цве-
точных растений могут стать исходным моментом для озеле-
нения огромных пространств санитарно-защитных зоп крупных
промышленных -предприятий юго-востока РСФСР.
Л II Т Е Р А Т УРА
Бойченко Е. 11. Защитные насаждения городов степных районов.
М.,. 1952.
Галактионов И. М., Бу А. В., Осин В. А. Декоративная
дендрология, М.(> 1967.
Г а би но в а Ж. Л., Попов Г>. В. Основные направления борьбы
за чистоту атмосферного воздуха. «Городское хозяйство Москвы», №
1962j
-Колесников А. И. Декоративная дендрология. М., P.Mill. Сира ноч-
ник «Озеленение городов», т. Ill, второй полутом. М., 196(1.
Кудинов О. В. О планировочной организации санитарно-защитных
зон. Киев, 1968.
Л ахи о Е. Санитарно-гигиеническая po.ni» зеленых насаждений.
«Озеленение городов», Киев, 1966.
То кин Е. Целебные яды растений. Ленинград, 1967.
Ю. М. СПАХОВ|
Особенности роста и содержания свободных аминокислот
у древесных растений в смешанных культурах
Интенсивность роста и направленность физиолого-биохими-
ческих процессов у растении в смешанном и -простом сообщест-
вах различны (Spurr, 1964; Гродзинский, 1970). Этому в зна-
чительной степени способствуют особенности взаимоотношений,
складывающиеся между растительными организмами при сов-
местном произрастании (Чернобривепко, 1956; Гродзинский,
1965; Колесниченко, 1968). В то же время известно, что интен-
сивность роста и развития .растений, активность синтетических
процессов в них тесно связана с динамикой азотистых соедине-
ний, в частности, с содержанием свободных аминокислот
(Плешков, Фоуден, 1959; Хавкин, 1964; Сытин к, 1966; Кара-
петян. 1970 и др.). Образуясь в процессе фотосинтеза или в
результате синтетической деятельности корней, свободные ами-
нокислоты в дальнейшем являются основными исходными
соединениями, обеспечивающими синтез белков. Качественный
и количественный состав свободных аминокислот в раститель-
ном организме может служить критерием обеспеченности ра-
стений элементам it питания и показателем их жизнеспособно-
сти (Плешков и др., 1959; Церлияг, Зинкевич, 1967). В связи
с этим изучение особенностей роста и содержания свободных
аминокислот у совместно произрастающих древесных растений
разных видов может быть использовано по только для выясне-
ния характера и механизма их взаимоотношений, по для раз-
работки системы физиолого-биохимических показателей, тесно
коррелирующих с энергией роста растений. В дальнейшем эти
показатели могут быть использованы в качестве физиолого-
биохимических признаков ранней диагностики интенсивности
роста древесных растений.
В настоящем сообщении приведены результаты исследова-
ния особенностей роста и некоторых сторон аминокислотного
обмена у совместно произрастающих сосны обыкновенной (Pi-
ous sibveslis L.) и березы бородавчатой (Betula verrucosa
FJirh.). Эти породы широко распространены в естественных
лесах и лесных культурах центральной лесостепи, а взаимоот-
ношения и особенности физиолого-биохимических процессов их
изучены недостаточно.
Объектами исследования служили незатронутые рубками
ухода 16-летние сосново-березовые культуры, расположенные
121
в зеленой зоне г. Воронежа. Местоположение участка ровное,
почвы средпегумусспроваппые (с признаками оподзоливания)
песчаные, второй ладлуговой террасы г. Дои«а. Смешение
пород в культурах произведено кулисами: 10 рядов сосны че-
редовались с 5 рядами березы, ширила междурядий 1,5 м.
При таком способе смешения только 1-й и 10-й ряды сосны
находятся в непосредственном контакте с рядами березы.
Исследования проведены на 6 пробных площадях: одна (конт-
роль) в массиве чистых культур сосны и пять в смешанных
культурах — три сосновые кулисы и две разделяющие их бе-
резовые кулисы. Таксационные обмеры и биохимические ана-
лизы на пробных площадях проведены отдельно для каждого
ряда деревьев. Содержание свободных аминокислот в хвое
(листьях) однолетних побегов древесных растений определяли
методом распределительной хроматографии на бумаге (Бояр-
кип. 1956; Трофимова, Солопепко, 1967).
Результаты исследований показывают, что существует зна-
чительная разница в росте 1 и 10 рядов деревьев соспы, при-
мыкающих к .рядам березы, п средних (5, 6) рядов. Деревья
сосны крайних (стыковых) рядов имеют средний диаметр па
21—30%, среднюю высоту на 17—20% и средний объем на
•18—52% меньше, чем деревья центральных рядов. Указанные
различия статистически достоверны. Деревья сосновой кулисы
в пограничных с березой рядах также значительно уступают
ио показателям роста (на 10—31%) деревья аг сосны в чистых
культурах. Снижение средних высот и диаметров деревьев сос-
ны в сравнении с контролем распространяется и па вторые от
березы ряды кулисы, начиная же с третьего ряда, наблюдается
их увеличение. В центральных рядах кулисы средний объем
деревьев сосны наибольший, па 32—12% выше, чем в чистых
посадках.
Снижение показателей роста па стыках кулис, по сравне-
нию с центральными рядами, наблюдается пе только у соспы,
по и у березы. Береза в рядах на стыках с соспоп имеет
диаметр на 6—7%, высоту на 13—16% и средний объем де-
рева па 19—22% меньше, чем в центральном (третьем) ряду
кулисы. Различия, в средних высотах деревьев статистически
достоверны.
Переход взаимного угнетения сосны и березы при близ-
ком произрастании в стимулирование роста по мере удаления
друг от друга хорошо укладывается в известную закономер-
ность действия физиологически активных веществ — так на-
зываемой «доза-эффект». Очевидно, в исследуемых нами куль-
122
турах большое значение во взаимоотношениях сосны и бере-
зы ‘имеет их биохимическое взаимовлияние, осуществляемое
посредством фитонцидов, концентрация которых уменьшается
по мере удаления одного растения от другого. Это предполо-
жение подтверждается результатами наших лабораторных и
вегетационных опытов (Колесниченко, Спахов, Опалова, 1971;
Спахов, .1971; Спахова, Колесниченко, Спахов, 1971).
В таблицах 1 и 2 приведены данные о содержании и ди-
намике в течение вегетации свободных аминокислот в листьях
и хвое древесных растении в сосново-березовых культурах.
В течение вегетационного периода в однолетней хвое сосны
как в чистых, так и в смешанных с березой культурах обна-
ружен большой набор свободных аминокислот (табл. 1). В наи-
большем количестве найдены валин, серин, астра ратиновая
кислота и аспарагин. В незначительном количество содержит-
ся фенилаланин, треопип (в начале вегетации). Наблюдается
изменение количественного содержания в хвое свободных ами-
нокислот в зависимости от физиологического состояния де-
ревьев. Суммарное количество свободных аминокислот в хвое
ослабленных деревьев сосиы (на стыке с березой) па 22% мень-
ше, чем у хорошо растущих деревьев в средних рядах сосно-
вой кулисы. У сосиы, растущей на стыке с березой, по срав-
нению со средними рядами в 3—4 раза надает содержание
в хвое глютаминовой кислоты, в 2 раза— серина (22/VIIT)
и в 1,5—3 раза — аспарагиновой кислоты. Изменение балан-
са свободных аминокислот в растениях не может не отразить-
ся на их росте (Haylor, 1959). Увеличение содержания сво-
бодных аминокислот усиливает движение протоплазмы и по-
вышает темп обмена веществ (Генкель, Окпппа, 1964). В свя-
зи с этим, повышенное содержание свободных аминокислот в
хвое деревьев средних рядов сосновой кулисы свидетельствует
об их высокой жизнеспособности п интенсивности роста ((Бо-
ровикова, 1968).
В то же время, у деревьев сосны, растущих па стыке с бере-
зой, наблюдается повышен ное количество в хвое аспарагина.
Можно предположить, что направленность азотного обмена
в сторону накопления аопарагппа связана с отмеченным ра-
нее недостатком углеводов в хвое деревьев этих рядов (Спахов,
1971). При наличии достаточного количества углеводов аммиак,
образующийся в растениях при дезаминировании аминокислот,
вступает в реакцию прямого аминирования с кетокислотамп,
которые образуются из углеводов. В случае же недостатка
123
Таблица 1
Содержание свободных аминокислот в однолетней хвое сосны обыкновенной
в чистых и смешанных культурах
Номера рядов в кулисе Свободные аминокислоты, мкг/г абсолютно сухого веса
фенил- аланин валин а-аланпн гл ютами-! новая | кислота треонин серин аспараги-; новая кислота 1 аспара- гин
Контроль 56 315 Анализ на 184 12 июня 97 34 256 342 214
1 следы 294 173 40 41 207 146 345
3 38 281 219 145 37 274 419 182
5—6 45 306 192 160 32 269 387 230
Контроль 31 246 Анализ на 22 августа 75 134 следы 194 380 161
1 17 219 83 72 — 125 226 231
3 29 224 70 183 — 260 362 127
5-6 36 233 76 194 — 241 394 143
углеводов аммиак обезвреживается путем образования амидов,
в данном случае аспарагина.
По качественному составу и количественному содержанию
свободных аминокислот береза несколько уступает сосне
(табл. 2). Всего в листьях березы обнаружено 7 свободных
аминокислот. Одна из них, двигающаяся на хроматограмме за
глютаминовой кислотой, не идентифицирована. В наибольшем
количестве в листьях березы содержится серин и аспараги-
новая кислота, в наименьшем — лейцин и аспарагин. В целом
содержание свободных аминокислот в листьях несколько сни-
жается к концу вегетации. Вначале вегетационного периода
наблюдается значительное снижение (в 2 раза) количества
глютаминовой кислоты в листе ях слаборастущих деревьев бе-
резы (т. е. на стыке с сосной) по сравнению с деревьями сред-
него ряда кулисы. 11а количественное содержание других сво-
бодных аминокислот в листьях березы присутствие сосны
практически не оказало влияния.
Результаты наших исследований свидетельствуют о том,
что в процессе взаимовлияния сосны обыкновенной и березы
бородавчатой в смешанных культурах наблюдаются сущест-
венны? н. мененпя содержания свободных аминокислот (особен-
но глютаминовой гмелоты) в хвое и листьях деревьев этих по-
род. Указанные изменения тесно связаны с ослаблением росто-
Таблпца 2
Содержание свободных аминокислот в листьях березы бородавчатой
в смешанных сосново-березовых культурах
Номера рядов в кулисе Свободные аминокислоты, мкг/г абсолютно сухого веса
лей- цин .валпп глюта- миновая кислота ненден- тнфпцн- ровапная кислота серин аспараги- новая кислота аспа- рагин 1
1 следы Анализ на 278 125 12 июня + 298 305 78
3 следы 256 287 + 283 289 65
5 следы 249 143 + 287 271 71
1 85 Анализ на 22 августа 104 172 + 167 375 следы
3 92 129 203 + 135 342 следы
5 76 122 187 + 154 350 следы
125
hbix процессов у сосны п березы на стыках и усилением их в
средних рядах кулис. Это дает возможность использовать со-
держание свободных аминокислот в листьях (хвое) как одни из
диагностических признаков интенсивности роста древесных
растений.
Л 11Т Е Р А Т У Р А
Боровикова А. М. Содержание свободных аминокислот в хвое
деревьев соспы разных классов роста. Лесоведение, 6, 1968.
Бояркин А. И. Разноцветное проявление аминокислот на бумаж-
ных хроматограммах. Физиол. растений, т. 3, вып. 4, 1956.
Генкель П. А., О к пи на Е. 3. Состояние покоя и морозоустой-
чивость плодовых растений. «Наука», М., 1964.
Г р о д з и н с к и й А. М. Аллелопатия в жизни растений и их сооб-
ществ. «Наукова думка».-, Киев, 1965.
Г р о д з и п с к н й А. М. Изучение физиологических и биохимиче-
ских процессов в растительных сообществах, Сб.: Физиолого-биохими-
ческие основы взаимодействия растении в фи то ценозах, выв. I, «Нау-
кова думка», Киев, 197CL
Карапетян К. А. Динамика содержания углеводов и аминокис-
лот в различных органах и тканях древесных в течение годичного
цикла развития. Ки.: Онтогенез высших растений. АП Арм. ССР, Ере-
ван, 1970.
К о л е с и и ч е н к о М. В. Биохимические взаимовлияния древесных
растений. «Лесная промышлниость», М., J 968.
К о л е с н п ч е п к о М. В., С и ахов 10. КГ., С п а х о в а А. С. Осо-
бенности роста п фосфорного обмена у соспы и березы при совместном
произрастании. Физиол. растений, т. 18, вып. 5, 1971.
Плешков Б. П., Ф о уде н Л. Содержание свободных аминокислот
и аминокислотный состав белков листьев ячменя в зависимости
от условий минерального питания и возраста. Пзв. Тимирязевск. с.-х.
акад., т. 5, вып. 30, 1959j.
Плешков Б. П., Шмырева Т. В., II в а н к о 1П. Изменение
содержания свободных амипокпелот в листьях и корнях кукурузы в
зависимости от условий питания растений. Физиол. растений, т. 6,
вып. 6, 195,9.
С и а х о в 10. М. О роли аллелопатии в изменении физиологического
состояния молодых древесных растений при совместном произраста-
нии. Автореф. капд. диссерт., Воронеж, 1971.
С п а х о в а А. С., КолеснпчепкоМ. В., С п а х о в 10. М. Влия-
ние фитонцидов на физиолого-биохимические процессы в древесных
растениях. Сб.: Физиологически активные соединения биогенного про-
исхождения. Изд. МГУ, 1971.
Сытиик К. М. (редактор). Физиолого-биохимические основы роста
растений. «Наукова думка», Киев, 1966.
Трофимова О. С., С о л о н е н к о Л. П. Количественное определе-
ние содержания аминокислот с помощью бумажной хроматографии
(методика). СО АН СССР. Новосибирск, 1967.
Хавкин Э. Е. Изменение азотного обмена в процессе роста расте-
ний. Агрохимия, 6, 1964.
126
Ц е р л и и г В. В., Зинкевич А. С. Содержание свободных амино-
кислот в тканях как показатель обеспеченности растопил азотом. Агро-
химия, 9, 1967.;
Черной р и и с и к о С. И. Биологическая роль растительных выде-
лений и межвидовые взаимоотношения в смешанных посевах. «Совет-
ская паука», М., 1956.
А. Е. САМСОНОВА
Гиббереллин и рост сеянцев ели обыкновенной
Многие исследователи еще недавно считали, что гиббереллин
синтезируется только микроорганизмами. Затем было установ-
лено наличие этого стимулятора роста в вегетативных частях
растений, а несколько лет назад и в семенах. Приведенные в
литературе материалы (Усовскин, 1958; Овчаров,1969), ука-
зывают на высокую отзывчивость многих семян на дополни-
тельное обогащение их гиббереллином. Хотя механизм дей-
ствия этого стимулятора роста вскрыт еще не полностью,
однако имеющиеся в настоящее время данные говорят о
том, что гиббереллин участвует в важнейших процессах
обмена веществ, способствующих нарушению периода покоя
семян п стимулированию их прорастания.
В целях испытания действия гиббереллина на ускорение
роста сеянцев в высоту на питомнике Макспмец-Гладыщанско-
го лесничества Надворняпскоги лесокомбината были поставле-
ны опыты с предпосевным намачиванием семян елн обыкно
венной .в водных растворах гиббереллина.
Первый опыт заложен 19 мая 1969 г. Обрабатывали сухие
семепа и семена, предварительно вымоченные в течение трех
суток в проточной речной воде, как обычно делается в про-
изводственных условиях. Испытывали действие раствора гиб-
береллина 0,01%-ной концентрации. Время намачивания се-
мян — 15—30 минут и 3 часа. Контролем в этом опыте слу-
жили сухие семепа и семепа, вымоченные в воде.
Второй опыт поставлен 15 мая 1970 г. Сухие семепа обра-
батывали гиббереллином концентрации 0,01%, 0,1%, 0,5%,
1,0% в течение 3-х, 12-ти и 24-х часов. Контролем являлись се-
мепа, вымоченные соответствующее время в дистиллированной
воде. Большие концентрации гиббереллина испытывали в целях
установления оптимальной дозы, необходимой для лучшего
прорастания семян и лучшего роста растений.
При приготовлении растворов точно взвешенное кол-пче-
127
ство порошка гпобереллнпа растворяли в 10 мл этилового спир-
та и разбавляли водой до нужного объема. Так как в водных
растворах активность гиббереллина надает, то растворы гото-
вили непосредственно перед опытом. Семена вымачивали в
стеклянных стаканах на свету при комнатной температуре.
Посев семян сделан па глубину 1 — 1,5 см. Повторность в
опытах трехкратная.
Наблюдения за опытом 1969 г. показали, что раньше всего
взошли семена, предварительно вымоченные в речной воде и
обработанные гиббереллином в течение трех часов. Вслед за
ними взошли сухие семена, получившие такую же обработку.
Всходы то другим вариантам опыта начали появляться только
через 4—6 дней. Через две педели разница между первыми
двумя вариантами и другими внешне была большая, ио затем
она постепенно начала сглаживаться. Учет растеши! проведен
в сентябре 1969 г. и в июле 1970 г. Сеянцы к атому времени
по всем вариантам опыта значительно выравнялись.
Из таблицы 1 видно, что растения, семена которых перед
посевом выдерживали в гиббереллине, за небольшим исключе-
нием, имели лучшие показатели по общей длине, сухому весу
по сравнению с копт рольными. Вымачивание семян в речной
проточно!! воде без воздействия гиббереллина улучшало всхо-
жесть их, а затем увеличивались сухой вес и высота надземной
части растений.
Намачивание семян в водных растворах гиббереллина
0,01% в течение трех часов повышало энергию их про-
растания и начальную скорость роста растений. Хотя резкое
ускорение в первые дни ростовых процессов постепенно сни-
жалось, растения в сентябре по всем показателям были лучше
контрольных.
Вымачивание семян в растворах гиббереллина в течение
15 и 30 минут пе повышало энергию С^ .^оастанпя, но расте-
ния'к сентябрю шмели хорошие показ;.,,-.- .. f»;,o высоте и сухому
весу. -----
Большой положительный эффект получен уже в первый год
исследования при обработке гиббереллином сухих семян. Оче-
видно, это явление объясняется тем, что само вымачивание в
проточной воде значительно уменьшает содержание ингибито-
ров в кожуре семян (Овчаров, 1969) и эффект от действия гиб-
береллина при расчете на вымоченный контроль снижается.
Следует однако отметить, что обработка предварительно вымо-
ченных в речной воде семян в течение трех часов гибберелли-
ном дала нанлучшие результаты по энергии прорастания.
128
Таблица 1
Влияние предпосевной обработки семян ели обыкновенной
гиббереллином на последующий рост сеянцев (опыт 1969 г.)
Варианты опыта Абсолютно сухой вес 50 растений Высота растений
гр. % от контроля мм % от контроля
Сентябрь 1969 года
Контроль 1,0 100 39 100
Обработка 15 мни. 1,0 100 48 124
(У со си Обработка ЗЮ мни. 2,0 200 43 111
U о Обработка 3 часа 1,2 120 50 128
m 3 QJ 6 си е( Контроль 1,2 100 44 100
СП 3 сх о
Е m Обработка 15 мни. 1,2 104 43 98
7?. о 'о о Обработка 30 мин. 1,0 83 44 100
<У О * су с. н Обработка 3 часа 2,0 171 48 109
Июль * 1970 года
Контроль 8.7 100 153 100
со Обработка 15 мни. 13.8 159 226 148
<У X <у Обработка 30 мни. 14,6 169 194 127
и aj Обработка 3 часа 13,6 156 230- 150 .
(У Контроль н.о 100 175 100
о
► со 3 Е О CQ Обработка 15 мин. 12,3 111 181 юз. •
Я X и X о О Обработ и. Н.1 '100 196 111
S о X X
су 52 о 2 0J У О Обрабо! 14.2 128 218 124
К июлю следующего года положительное действие гибберел-
лина при обработке сухих семян проявилось более четко. Вре-
мя обработки к этому моменту в общем не имело значения.
По всем вариантам получены высокие показатели по высоте
и сухому весу растений.
Растения же, выросшие из семян, получивших обработку
гиббереллином, но предварительно вымоченных в речной воде,
9. Заказ' 6609 129
отличались от контрольных в меньшей степени, чем первые.
Причем, лучшими в этом опыте снова оказались растения в ва-
рианте с 3-часовой обработкой гиббереллином.
В опыте 1970 года первые всходы появились 3 июня в ва-
риантах с 24-часовой обработкой; 8 июня начали появляться
редкие всходы в вариантах с 12-часовой обработкой. К 12 июня
в основном по этим двум вариантам появились групповые всхо-
ды, а в вариантах с 3-часовой обработкой — единичные.
Учет растений 17 июля показал (табл. 2), что при 24-часо-
вой обработке семян положительное действие гиббереллина на
последующий рост сеянцев в высоту снижалось с повышением
его концентрации. Сухой вес растений, наоборот, возрастал со-
ответственно повышению концентрации гиббереллина. Однако
положительное действие однопроцентного раствора гибберелли-
на по отношению к сухому весу сеянцев несколько снижалось.
При 12-часовой обработке отмечено увеличение высоты и
сырого веса сеянцев с повышением концентрации гибберелли-
на до 0,5%. Положительное действие однопроцентного раство-
ра было несколько слабее.
При 3-часовой обработке с повышением концентрации гиб-
береллина наблюдалось некоторое увеличение веса и довольно
значительное — высоты сеянцев (исключение составляет ва-
риант, с 0,5%-ной концентрацией).
Как показывают данные таблицы 2, обработка семян раст-
вором гиббереллина различной концентрации (0,01 % —1 %)
в течение трех часов дает хорошие положительные результаты.
Увеличение врвхмени обработки до 12 и 24-х часов хотя и по-
вышает начальную всхожесть семян и энергию их прорастания,
но последующее состояние сеянцев остается в тех же пределах
пли даже несколько хуже.
Полученные данные указывают на довольно высокую отзыв-
чивость семян ели обыкновенной на дополнительное обогаще-
ние их гиббереллином.
Намачивание семян в растворах гиббереллина концентрации
от 0,01 % до 1,0% повышает энергию их прорастания и началь-
ную скорость роста растений. Интенсивность роста сеянцев рез-
ко увеличивается на второй год.
Наиболее отзывчивы на обработку гиббереллином сухие се-
мена. Вымачивание семян в проточной воде уже повышает
энергию прорастания их, улучшает всхожесть и способствует
лучшему росту, большему накоплению сухой массы сеянцев.
Повышение концентрации раствора гиббереллина требует
130
Таблица 2
Влияние предпосевной обработки семян ели обыкновенной гиббереллином
на последующий рост сеянцев (опыт 1970 г.)
Варианты опыта С\хой вес 50 растений Высота растений
время об- работки концентрация гиббереллина гр. % от контроля мм % от контроля
3 часа Контроль 0,47 100 80 100
» 0,01 0,62 131 104 129
» 0,1 0,67 142 108 134
» 0,5 0,65 138 102 128
» 1,0 0,60 128 НО 137
12 часов Контроль 0,50 100 79 100
» 0,01 0,60 120 100 126
» 0,5 0,70 140 ПО 139
» КО 0,60 120 103 130
24 часа Контроль 0,46 100 79 100
» 0,01 0,55 120 104 132
» 0,1 0,57 124 98 123
» 0,5 0,62 135 94 119
» 1.0 0,55 120 79 100
уменьшения времени обработки, хотя начальная скорость про-
растания семян и увеличивается в обратном порядке.
Довольно хороший устойчивым положительный эффект в
условиях наших опытов дает намачивание семян раствором гиб-
береллина 0,01%-ной концентрации. Время обработки — от
15 минут до 3 часов.
ЛИТЕРАТУРА
Овчаров К. Е. Физиологические основы всхожести семян. «Наука»,
М., 1969.
У с о в с к и и Б. И. Гиббереллин в овощеводстве. Сад и огород, №11,
1958.;
И. Я. КАЗАНЦЕВ
Лесорастительные условия в Волго-Ахтубинской пойме
и дельте Волги после зарегулирования стока реки
В Астраханской области промышленность, сельское хозяй-
ство и население ежегодно потребляет 1,3—1,5 млн. м3 древе-
сины, а заготовляется па месте пемпотим больше 10% этого
9*
131
количества. Поэтому повышение продуктивности лесов области
имеет важное хозяйственное значение.
По данным Астраханского управления лесного хозяйства на
1/1-71 г. покрытая лесом площадь гослесфонда составляет
80,2 тыс. га, находится опа в основном в Волго-Ахтубинской
пойме и дельте Волги. Кроме того, 16,9 тыс. га покрытой лесом
площади в пойме н дельте имеют колхозы, совхозы и запо-
ведник (Лесной фонд СССР, 1968).
Преобладающая часть лесопокрьттой площади гослесфонда
занята ветлой (57,1%) и осокорем (13,0%). Эти древесные
породы в зоне умеренного климата являются самыми быстро-
растущими, ио продуктивность осокорников и ветляпиков на
Нижней Волге очень низкая и составляет в спелом и пере-
стойном возрасте всего лишь 116 м3/га.
Дело в том, что в пойменных лесах исторически сложилась
экстенсивная форма ведения хозяйства, влекущая за собой низ-
кополиотпость, перестойность и низкотоварность насаждений.
Следовательно, повышение продуктивности пойменных лесов
возможно путем снижения возраста рубки п своевременного
восстановления лесосек наиболее быстрорастущими древесны-
ми породами в данных экологических условиях.
Зарубежный и отечественный опыт показывает, что в пой-
менных условиях наиболее высокопродуктивными являются гиб-
ридные сорта тополей, обладающие гетерозисом (Филимонова,
1962, Иванников, 1971).
Сортоиспытание тополей в Волго-Ахтубинской пойме ведет-г
ся с 1959 г. (Ростовцев, 1965). Продолжительный срок сорто-
испытания позволил установить, что наиболее высокопродук-
тивными и устойчивыми к факторам внешней с ре .ы являются
евроамерпканские сорта тополей (№ 175 и №176 «Брабантский»,
№ 59 «Сакро», № 236 «Мощный», «Весенпекраспый», «Ба-
хелье», № 162 «Каролинский» (Ивашников, 1971; Казанцев,
1971). Так, в 9-летних опытных культурах лесного типа при
размещении 3X3 м сортовые тополя образуют насаждения с за-
пасом 200—350 м3/га и даже больше, а делянка местного осо-
коря в этих условиях имеет запас 125 м3/га. Перспективные
сорта тополей лесхозы области в настоящее время широко вво-
дят в лесные культуры.
Поскольку основным экологическим фактором, определяю-
щим ареал тополей в пойме, является продолжительность за-
топления участков паводковыми водами, то возникает необходи-
мость выявления количества площадей, которые возможно за-
нять под культуры тополей.
132
В связи с зарегулированием стока р. Волги продолжи-
тельность паводков и максимальная высота подъема паводко-
вых вод уменьшились, по специфика пойменных условий со-
хранилась.
По Астраханскому водомерному песту средняя высота мак-
симальных уровней полой воды с 1935 г. по 1955 г. была
285±3,3 см, а после зарегулирования стока реки с 1956 г. но
1970 г. она стала 259=^3,4 см. Количество паводков, соответст-
вующих по уровню средним и выше, составляло в 1935—55 гг.
53,4%, при современных условиях это соотношение составляет
45%, а но сравнению с прежними уровнями подъема полой
воды составляет 36,7%, подобные изменения водного режима
произошли на всей территории Волго-Ахтубинской поймы и
дельты Волги. Количество незатопляемых и редко затопляемых
площадей в связи с этим увеличилось.
Наряду с высотой подъема полой воды также уменьшилась
продолжительность паводков. Раньше она на границе ветля-
ников с кустарниковыми ивами доходила до 100 дней, а в на-
стоящее время больше 80 дней ®етляпп1К|И низких уровней
не затапливаются. В зимнее время в связи с повышенными
расходами В'одьг ГЭС они часто затапливаются и повреж чаются
льдом, что существенно ограничивает лесопригодность таких
участков.
Наши наблюдения (Казанцев, 1970, 1971) показывают, что
тополя из секции черных хорошо выдерживают затопление
в условиях Волго-Ахтубинской поймы и дельты Волги до
60—70 дней. Поэтому .в места, занятые ветлой, но’^затапливае-
мые зимними паводками, появилась возможность продвижения
тополей.
Почвы поймы являются продуктом деятельности текучих
вод, что обусловливает им большую пестроту в механическом
составе и слоистость (профиля.
Однако, сходный лесорастительны'й эффект на первый
взгляд, казалось бы, различных почвенных разностей позволяет
классифицировать пойменные почвы по типам лесораститель-
ных условий. Используя материалы лесоустройства, покрытую
лесом площадь мы распределили по типам лесорастительиых
условий (табл. 1). При этом за основу приняты классифика-
ции И. В. Трещевского (1961, 1966) и В. Г. Шаталова (1965),
разработанные ими для лесов поймы р. Дона и Волги. Наша
классификация отличается тем, что имеет большую дробность
зон по продолжительности затопления. Зоны выделены в соот-
133
Таблица 1
Распределение покрытой лесом площади (в га) Астраханского
Управления лесного хозяйства (леса в Волго-Ахтубинской пойме
и дельте Волги) по типам лесорастительных условий
Режим и длительность затопления
Группы почвогрунтов со
сходным лесораститель-
ным эффектом
в основном проточное, местами застойное
от нескольких до 15 дней
ПС ПП ППЗ ПД
Спльиозасолепные почвы
всех почвенных подтипов
и всех зон затопления
Малосформированные
супесчано-песчаные неза-
соленные п слабосолеи-
ные
Маломощные и резко-
слоистые незасоленные
и слабозасолеиные
Слоистые и слоисто-
.зсрнистые незаселенные,
слабо и средпезасолеп-
пые
Зернисто-слоистые и зер-
нистые незаселенные,
слабо и срсднезасолеи-
ныс
О 890 — — — — — —
1 3447 2889 723 — — — —
2 — 341 7622 13Q661 — — —
3 — 318 1710 20937 8598 16000 9523
4 — 282 2110 1644 _ — _
Примечание: ПН — пойма пезатонляемая; ПР — пойма резкого
затопления (О—20 дней); ПК — пойма кратковременного затопления
(0—35 дней); ПС — пойма средиеиродолжнгелыюго затопления (15—45
дней); ПП — пойма продолжительного затопления (25—70 дней); ППЗ--
пойма продолжительного летнего и зимнего затопления (28—80 дней);
ПД— пойма длительного затопления (30—100 дней).
вествии с экологическими нишами основных древесно-кустар-
никовых пород поймы.
Выделенные в таблице тины лесорастительпых условий со-
ставляют площадь 38 665 га н являются экологически пригод-
ными для выращивания евроамерпканскнх тополей без ороше-
ния. Практически возможно освоить 30—35 тыс. га, т. к. часть
площади из-за раздробленности будет теряться. Поскольку ука-
занная площадь является .покрытой лесом, то культуры перспек-
тивных видов л сортов тополей на пей возможно создавать
только путем постепенной реконструкции существующих вет-
ляннмков и осокорников.
134
Претворение в жизнь этого мероприятия позволит при
15—20-летнем обороте рубки довести ежегодную заготовку
тополевой древесины до 500—600 тыс. м3.
ЛИТЕРАТУРА
Иванников С. II. Выведение и использование тополей в СССР
и за рубежом. Изд. ЦБНТИ Лесхоза, М., 1971.
Казанцев И. Я. Селекция и интродукция тополей па Нижней
Волге. Материалы паучпой конференции по вопросам лесного хоз-ва
(секция генетики и селекции), Пушкино, ВННИЛМ, 1970.
Казанцев И. Я. Сортовые тополя в условиях Волго-Ахтубинской
поймы, Лесное хоз-во, № 12, 1971.
Лесной фонд СССР. Изд. Лесная промышленность, М., 1-969.
Ростовцев С. А. Тополя в культурах Волго-Ахтубинской поймы.
ЦНИПТЭИ Леспром, М., 1965. . *
Т р е щ е в с к и й И. В. К вопросу о лесорастительных условиях
поймы Нижнего Дона. Науч. зап. ВЛТИ, т. XXIV, 1961.
Трещевский И. В. Лесоразведение в поймах засушливых районов
Волго-Донского бассейна. Лесное хоз-во, № 7, 1966.
Филимонова В. Д. Культура тополей за границей, Гослесбум-
издат, М., 1962.
Шаталов В. Г. Защитное лесоразведение в пойме Среднего Допа.
Автореф. кандидатской диссертации. Воронеж, 1965.
И. П. ЧИГИРИНЦЕВ, А. Б. БЕЛЯЕВ,
В. Н. СТЕБАКОВА, П. Б. БУДАНЦЕВ
Опыт уточнения диагностических признаков
подтипов черноземов методом математической обработки
В настоящее время методы математической статистики как
наиболее объективные, точные и универсальные находят все
более широкое применение в почвенных исследованиях (Важе-
нин, 1963; Сорокина, 1965, 1971; Крупенников и др., 1968;
Счастиая, 1969; Фридланд и др., 1969; Орешкина, 1971 и др.).
Использование математической статистики особенно необходимо
в тех случаях, когда приходится дифференцировать близкие
в морфологическом отношении почвы.
Объектом нашего исследования была территория Семилук-
ского лесного селекционного опытно-показательного питомни-
ка, расположенного в 25 км к северо-западу от города Воро-
нежа и входящего в состав Белгородско-Касторен1ского почвен-
но-климатического района типичных и выщелоченных чернозе-
мов (Адерихин, Тихона, 1963).
135
Территория питомника является базой для научно-исследо-
вательской работы отделов и лабораторий ЦНИИЛГпС, поэтому
в наших исследованиях было особенно важно установить сте-
пень варьирования морфологических, химических и физико-
химических свойств почв и найти среди них наиболее достовер-
ные признаки для выделения почвенных разностей.
Почвообразующими породами па обследуемой территории'
являются бурые покровные карбонатные тяжелые суглинки.
Территория характеризуется в основном равнинным рельефом,
местами переходящим в слабопологие склоны крутизной
0,5—1°.
По ныне действующе!! классификации («Указания по диаг-
ностике п классификации почв», 1967) выделение подтипов
черноземов в данной зоне производится по степени выщелочен-
ности (глубина вскипания от НС1), мощности гумусового гори-
зонта и содержанию гумуса.
Почвенный покров питомника представлен типичными и
выщелоченными черноземами, о морфологии которых можно
судить по описанию следующих разрезов.
Разрез 71
Почва — типичный чернозем среди емощпый тяжел ос у гли-
нистый. Разрез заложен на ровном месте широкого слабоволни-
стого водораздела в северо-восточно]’! части питомника.
Угодье — пашня.
Гор. Апах 0—30 см —
темно-серый, сухо!!, структура пылевато-крупноком-
коватая, уплотнен, продольные трещины, масса мел-
ких корешков, тяжел осу гл ин истый, переход посте-
пенный по цвету (заметный по структуре).
Гор. Ап/п 30—45 ом —
темно-серый, сухой, структура крупнокомковато-зер-
п иста я, уплотнен, продольные трещины более мелкие,
чем в верхнем горизонте, корней меньше, тяжело-
суглинистый, переход постепенный.
Гор. В| 45—70 см —
темпо-серый со слабым буровато-коричневым оттен-
ком, сухой, структура комковато-зернистая, уплот-
нен, мелких корешков еще меньше, чем в гор. Ап/п,
переход заметный.
Гор. Вг 70—96 см —
грязпо-бурып, более пли менее однородно окрашен,
136
сухой, уплотненность слабее, чем в вышеперечислен-
ных горизонтах, структура комковатая с единичны-
ми зернами, встречаются светло-бурые кротовины,
карбонаты в виде легкой припудрепиости и псевдо-
мицелия, единичные корешки, переход заметный.
Гор. В/С 96—140 см -
грязно-бурый, неравномерно окрашен, с бурыми зате-
ками гумуса, слабо уплотнен, карбонаты по всему
нрофилю, тяжелосуглинистып.
Гор. С 140-180 см —
бурый карбонатный лессовидный тяжелый суглинок.
Вокипа пне от соляной кисл оты с 70 см.
Разрез .Ni 8
Почва — выщелоченный чернозем средпемощпый тяжело-
суглинистый. Разрез на слабо пологом склоне юго-восточной
экспозиции в центральной части питомника. Угодье — пашня.
Гор. Апах 0—30 см —
темпо-серый, почти черный, свежий, тяжел осу глини-
стый, структура пылевато-комковатая, слабо уплот-
нен, переход незаметный по цвету, ио заметный по
структуре.
Гор. Ап/п 30—54 см —
темно-серый, почти черный, свежий, структура ком-
ковато-зернистая; глинистый, уплотнен, переход по-
степенный.
Гор. В| 54—77 см —
темно-серый с буровато-коричневым оттенком, све-
жий, структура зернисто-комковатая, глинистый, уп-
лотнен, обратные кротовины, ходы червей, переход
заметный.
Гор. Ва 77—125 см —
грязио-бурый с красноватым оттенком, неравномерно
окрашен, с затеками гумуса, структура комковатая,
прямые и обратные кротовины, глинистый.
Гор. С 125—150 см —
светло-бурый, свежий, уплотнен, карбонаты в виде
псев1домицелия.
Вскипание от соляной кислоты со 125 см.
Как следует из морфологического описания, изучаемые поч-
вы близки между собой: они имеют однотипный характер ге-
нетического профиля, одинаковую мощность гумусового горн-
137
зонта, в них полностью отсутствуют признаки иллювиалыюстп
в горизонте В (В/С), и различаются они лишь положением кар-
бонатного горизонта. Следовательно, в пашем случае, основ-
ным морфологическим признаком подтипов черноземов являет-
ся глубина вскипания от соляпой кислоты.
Для проверки этих выводов методами математической ста-
тистики морфологические характеристики почвенных профилей
были представлены в виде вариационных рядов. По данным
вариационных рядов были рассчитаны следующие параметры:
Мо — среднее арифметическое; m — ошибка среднего ариф-
метического; с — среднее квадратическое отклонение; %2 —
критерий достоверности различия эмпирического п теоретиче-
ского распределений.
Построенные вариационные кривые анализировались с целью
установления их соответствие нормальному распределению.
Как показали результаты математической обработки мощности
гумусового горизонта (А+-В), полученная величина %2 состави-
ла 3,78 и оказалась меньше стандартной величины (Ро,95= 11,1).
Это значит, что различие между нормальным п эксперимен-
тально полученным распределением незначимо, оно несу-
щественно и полученное распределение можно считать нор-
мальным. Это видно также из рисунка 1, где кривая А рас-
пределения мощностей гумусового горизонта близка к нор-
мальной, что свидетельствует об однородности рассматривае-
мой группы почв по этому признаку. Следовательно, по мощ-
ности гумусового горизонта, как одного из диагностических
признаков, 'исследованные почвы не могут быть разделены на
подтипы.
Анализ кривой распределения глубин вскипания (рис. 1,
кривая В) указывает на ненормальное распределение этой
величины. Несоответствие эмпирической кривой распределения
теоретической (трехвершинпая кривая) свидетельствует о том,
что в рассматриваемую группу почв включены профили, отно-
сящиеся к разным классификационным группам.
На полученной нами кривой распределения глубин вскипа-
ния отчетливо выделяются три вершины, соответствующие
вскипанию на глубине 30—40 см, 60—70 см п 90—110 см. Это
дает нам возможность выделить три группы почв с амплиту-
дой колебания глубины вскипания до 50 см (вы'соковскипаю-
щпе), 50—85 (типичные), 85—130 см (выщелоченные). Эти
]раницы установлены по объективным показателям — мини-
мальным частотам на кривой распределения.
Достаточно точными показателями, характеризующими
138
Рис. I. Кривые распределения мощности гумусового горизонта (А)
и значении глубины вскипания (В).
внутренние свойства почв, которые могут использоваться как
диагностические, являются их химические п физико-химиче-
ские свойства.
Исходя из предположения, что величина гидролитической
кислотности пахотного горизонта исследуемых поч/в, завися-
щая от многих факторов, в основном определяется глубиной
залегания карбонатов, было ггро-ведено уточнение этого пред-
положения методом корреляции. Этот метод позволяет устано-
вить наличие внутренней связи между различными свойства-
ми или явлениями в системе сопряженных распределений.
Нами был рассчитан коэффициент корреляции между величи-
ной гидролитической кислотности и глубиной вскипания. Коэф-
фициент корреляции, полученный по нашим данным, равен
-f-0,68, что указывает на связь между исследуемыми призна-
139
ками средней степени сопряженности. Чтобы оцепить досто-
верность коэффициента корреляции, сравниваем его с ожидае-
мым, равным 0,37 (при уровне значимости Р = 0,01 и числе
степеней свободы п = 48). Рассчитанный пами коэффициент
превосходит это значение. Следовательно, вопреки утвержде-
нию некоторых авторов (Денисова, 1968), получена для одно-
родной территории коррелятивная достаточно высокая зави-
симость величины гидролитической кислотности с глуби или
вскипания, и эта величина может служить в нашем случае объ-
ективным диагностическим признаком для выделенных подти-
пов черноземов. Это подтверждается и достоверными разли-
чиями величии гидролитической кислотности выделенных под-
типов (табл. I). Достоверные различия подтипов обнаружены
так же по величине pH (табл. 1). Особенно резко по физико-
химическим свойствам выделяется группа высоковскипающпх
черноземов.
Одним из самых употребительных диагностических пока-
зателей внутренних свойств почв является содержание гумуса.
Учитывая это, мы попытались выяснить, имеются ли досто-
верные различия между подтипами черноземов по содержанию
гумуса в пахотном слое для наших почв. Как видно из табли-
цы I, разницы между подтипами по содержанию гумуса пе
обнаружено: все черноземы относятся к среднегумусиым
(t=0’,4; 1,15; при t(0,95) =2,12; 2,78).
Не обнаружено разницы по подтипам и в запасах гумуса
в слое 0—40 см (Мо для типичного чернозема равно
261 т/га, для выщелоченного — 261 т/га и для. типичного вы-
соковскппающсго — 273 т/га).
Установленные закономерности, показывающие взаимо-
связь морфологических и физико-химических свойств почв,
позволили выявить для изученной территории характеристи-
ки подтипов по некоторым химическим и физико-химическим
свойствам. При наличии таких характеристик представляется
возможным уточнять границы почвенных подтипов при круп-
номасштабном картировании опытных участков без закладки
тт описания дополнительных почвой пых разрезов и бурения
глубоких скважин, применяя лишь агрохимическую характе-
ристику почв.
Нами выло л немо агрохимическое обследование опытного
участка отдела семеноводства. Смешанные образцы почв от-
бирались с каждой опытной делянки из пахотного, горизонта
с глубины 0—30 см. Закладка разрезов и глубокое бурение
были нежелательны из-за возможной порчи делянок.
140
Таблица 1
Достоверность различии подтипов и родов черноземов по некоторым
физико-химическим и химическим свойствам
11азваппе 1 М ±m I
почвы 1 Mnztm п t l0.95
Гидролитическая кислотность
Выщелоченный 5,1
чернозем (4,5—5,7) 3,6 22 1,22 0,26 3.5 2,0
Типичный чернозем (2,9—4,3) 21 1,54 0,34 2,2 2,1
Типичный чернозем с повышенной линией 1,4
вскипания (0—2,7) 6 1,30 0,53
pH солевой вытяжки
Выщелоченный 5,9
чернозем (5,7—6,0) 19 0,33 0,076 2,3 2,0
6,2
Типичный чернозем (6,0--6,5) 18 0,51 ‘ ’ 0,12 5,8 2,1
Типичный чернозем с повышен, линией 7,2
вскипания (6,9- -7,5) 5 0,23 0,12
Гумус
Выщелоченный чернозем 5,94 15 0,56 0,14 0,4 2,1
Типичный чернозем 5,85 18 0,73 0,17 1,15 2,78
Типичный чернозем с повышенной линией вскипания 6,15 6 0,50 0,26
141
При обработке результатов химических и физико-химиче-
ских анализов почв решались две задачи.
Первая задача — установить о^тнородность почвенного по-
крова опытного участка по тем свойствам, которые приняты
нами ка‘к диагностические.
Вторая задача — установить принадлежность почв к опре-
деленному, выделенному на данной территории подтипу чер-
ноземов.
Как видно из рис. 2 и 3, и табл. 2 кривые pH и гидролити-
ческой кислотности пе следуют нормальному распределению —
кривая распределения pH сильно растянута в правой части,
Р и с. 3. Кривые распределения значений гидролитической кислотности.
142
а кривая распределения гидролитической кислотности имеет
левую асимметрию.
Используя критерий «доброкачественности» выборок
М±2а(Ро.95), мы установили, что делянки с pH 7,0 и выше
и с гидролитической кислотностью меньше 1,7 мг-экв. на
100 г почвы выходят за вероятный предел границ -вариацион-
ных рядов. Эти три делянки были выбракованы как нети-
пичные.
Xi2 — критерий достоверности и различия эмпирического и
теоретического распре деления для всей совокупности,
Х22 — для совокупности после выбраковки сомнительных ва-
риантов.
Оставшиеся после выбраковки сомнительных вариантов де-
лянки принадлежат к одной совокупности и подчиняются за-
кону нормального распределения.
Принадлежность почв опытного участка к определенному
подтипу черноземов устанавливали по величине М — довери-
тельной границе среднего арифметического для гидролитиче-
ской кислотности и pH, сравнивая ее с этими же показателя-
ми, установленными для каждого подтипа черноземов всей
территории питомника (табл. 2).
Из сопоставления этих данных вытекает, что почвы опыт-
ного участка относятся к подтипу выщелоченных черноземов.
Таблица 2
Результаты математической обработки величин гидролитической
кислотности и pH опытного участка
Свойства почв М п ±а ±111 у. % Pj% х.= 1 Х22
Гидролит. кислотность 4,5 (4,1- 29 0,95 -4,9) 0,18 21,0 4,1 14,75 (14,1)* 6,9 (И 1)
pH солевой вытяжки 5,9 29 о,з 0,056 5,1 1,0 44,4 9,6
(5,8—6,0) (15,5)* (П 1)*
П р и м е ч а н и е: * в скобках приведены критические значения X2
при Р = 0,95.
143
Выводы
1. Проведенная математическая обработка некоторых морфо-
логических характеристик, химических и физико-химических
свойств поив показала:
а) при выделении подтипов черноземов глубина вскипания
является надежным показателем, определяющим некоторые
важные химические и физико-химические свойства почв;
б) величина plI солевой вытяжки и пиролитическая кис-
лотность пахотного горизонта находятся в коррелятивной с-вязп
с глубиной вскипания и являются диагностическими показате-
лями подтипов черноземов.
2. Выделение величины pH и гидролитической кислотности
в качестве диагностических признаков подтипов черноземов
дают возможность уточнить почвенную карту опытного участ-
ка, сделать выбраковку нетипичных делянок и установить при-
надлежность почв к определенному подтипу.
Однако следует оговориться, что эти признаки не могут
быть универсальными для данного подтипа в различных геогра-
фических зонах, па разных почшообравующих породах, раз-
личных элементах рельефа и т. п.
Л 11 Т ЕРА Т У Р А
А д е р е х и н П. Г., Т и х о в а Е. П. Агрохимическая характеристика
почв ЦЧП. В кн.: «Агрохимическая характеристика почв СССР», т. 2,
«Наука», М., 1963. /
Важенин И. Г. Применение метода вариационной статистики а
почвенно-агрохимических исследованиях. Почвоведение, № 2, 1963.
Денисова Н. В. К вопросу о разделении типичных и выщелочен-
ных черноземов «Химия, генезис и картография почв», М., 1968.
Крупенников И. А., Лунева Р. И., Рябинина Л. Н. Иссле-
дование корреляционных зависимостей между свойствами почв и их
производительностью. «Химия, генезис и картография почв». М., 1968.
Орешкина Н. С. О некоторых закономерностях варьирования
влажности в модельной дерново-подзолистой почве. Почвоведение, № 9,
197L
Сорокина Н. П. Статистический метод оценки смытости на при-
мере типичных черноземов Курской опытной станции. Почвоведение,
№ 2, 1965.
Сорокина Н. П. Применение статистических методов при уточне-
нии диагностики черноземов. В сб.: «Крупномасштабная картография
почв», «Наука», М., 1971.
С ч а с т н а я Л. С. О диагностических признаках серых лесных почв
Среднерусской лесостепи. Почвоведение, № 3, 1969.
144
Указания но диагностике и классификации почв, вып. 3. Почвы
стопных областей СССР. Изд-во «Колос», М., 1967.
Фрндладд В. М., Белобров В. П., Дайнеко Е. К. Опыт
стастпческого анализа морфологических свойств черноземов целинной
степи. Почвоведение, № 4, J969.
Ю. Е. БУЛЫГИН
Представления о древесных растениях
как о саморегулирующихся автоматически управляемых
живых системах природы
За сравнительно небольшой исторический отрезок времени
кибернетика как наука о законах управления сложными дина-
мическими системами развилась в широкое и содержательное
научное направление современности. Ее «магическая» сила
заключена во всеобщности идей связи и управления, имеем ли
мы дело с живыми организмами пли с автоматами в технике.
Процессы управления этими динамическими системами подчи-
нены некоторым общим закономерностям. И в том и другом
случае задача управления заключается в поддержании регули-
руемых параметров системы па строго заданном, программиро-
ванном уровне, независимо от действия возмущающих факто-
ров среды или, как говорят инженеры, помех.
Работа живого автомата (биоэкоса), как и всякого другого
регулирующего устройства, в математической форме может
быть описана системой нелинейных дифференциальных уравне-
ний вида:
ср(х, х', х", ... у, у', у", ...) - 0 (1)
где х', х", ... у', у", ... — производные по времени от входного
и выходного сигналов.
Разложив в ряд Тэйлора, можем записать
есть оператор диффренцирования.
10. Заказ1 6609
145
Перейдя к операторной форме, кат: это принято в теории
регулирования, будем иметь?
(Т (2р2 г Т.,р+1 )у = (к3р2-;- к2р 4- к()х (3)
где Ti, Т2 — физически имеют смысл постоянных линий, не
строго
/ d? \ / d<p \
Т 2 = V "сТу^/ц . т = \ dyZ ) о .
1 I d(p ) ’ 2 ( d? ’
\ dy А) \ dy /о
К, Кг, Кз — имеют смысл коэффициента передачи
Тогда уравнение (3) можно записать в следующем виде:
_ Xi + k2p + k3p
- T/V + bpfl *Х
Выражение (4) называют передаточной функцией системы
и его обозначают буквой W(P) или К(Р).
Но на какое-то звено системы может воздействовать не одно,
а несколько возмущений, последнее характерно для всех жи-
вых систем природы, включая и древесные растения.
Тогда исходное уравнение (3) можно представить в виде:
Q(p)y = SRi(p)x! (5)
=1
или, переходя к передаточным функциям, можем записать:
11 К|(Р) П / ч /сч
y=,iwx-=,E=r(p)x' ()
Здесь Xi — входные воздействия на звено (i = l, 2, ...
п);
146
Q(P) и R’iCP) — полиномы относительно Р;
(n) — — передаточная функция звена для i-ro
' Q (р) входного воздействия.
В этом смысле древесные растения, хотя и отнесены к раз-
ряду низших регулянтов в природе [3, 5], тем не менее они
также являют собой образчик совершенства регулирования и
саморегулирования.
Древесные растения не только поддерживают гомеостазис
(оптимальное состояние системы), но еще и обладают широ-
ким диапазоном приспособляемости (адаптирования) к различ-
ным условиям их произрастания. Процесс адаптирования в ма-
тематическом выражении можно представить в виде получения
изображения выходящего сигнала по входящему с помощью
прямых и обратных преобразований по Лапласу и Фурье,
имеющих следующий вид:
По Фурье: ’
Г —iwt
F |f (t) |=h(t)e • dt = S(io)) (7)
oo
— ] P icot
F|S(jw)j= -97- S(]w) c • d<»=f(l)
— 20
По Лапласу:
p -P‘
S(p)= f(t)e -dt
0 с+гл pt (8)
f(t)= "^rjslpje -dp
c—i00
Управление, таким образом, обеспечивает целенаправленное
приспособление системы к действию возмущающих факторов.
Конечно, в этой аналогии имеется одно то важное обстоя-
тельство, которое мы должны учитывать при моделировании
растений и целых биогеоценозов, а именно: растения, как пра-
вило, дают нелинейные осциляторы, то есть эти системы обыч-
но нелинейны и описываются системой нелинейных дифферен-
циальных уравнений [1, 2]. Однако, нелинейность системы' не
10*
147
означает, что она не может быть описана математическим пу-
тем, п что поэтому невозможно осуществить ее моделирование.
В этом случае для определения параметров системы нередко
используют прием линеаризации, то есть приведение нелиней-
ных систем к линейным (линейной модели), которые с успе-
хом могут быть описаны системой линейных дифференциаль-
ных уравнений.
Существующий математический аппарат позволяет описать
некоторые нелинейности иа аналоговых вычислительных ма-
шинах.
Более того, такая особенность исследования динамических
систем позволяет изучать не только свойства, но и, например,
приступить уже сейчас к массовому составлению карт типов
леса и моделированию будущих лесов нашей страны [4, 6J,
а также широко использовать оптимальное программирование
для оптимизации генетического фонда хозяйственно-ценных
хвойных и лиственных древесных пород.
Автоматизм живых систем природы осуществляется и воз-
можен только при наличии так называемой обратной связи в
системе, то есть направленного действия с проверкой резуль-
тата контролем от управляемого объекта к управляющему
устройству.
Без обратной связи пет управления. С нарушением сигнала
подачи обратной связи система не может функциониро-
вать [7].
Передаточную функцию для сигнала связи можно предста-
вить как
_ w(pl , .
"•с’~ l+w(p) 3 1 ’
и для ошибки регулирования
We (р)—— = -/ (10)
V ' Х3 l+w(p)
Здесь:
х0. с- — величина сигнала обратной связи;
Хз — задающее воздействие.
Итоговый эффект определяется суммой воздействия
Хз-J-f
где f — возмущение.
х= [ WS3X(P) + [ Wfx(p) |Lk. (П)
148
Здс‘сь х — регулируемый параметр, пли
Wx:tx(p)X3+Wfx(p)f
1-1-w(p)
Но древесные растения представляют как правило много-
контурные системы. Для таких систем величина регулируемого
параметра выразится формулой:
y — wfix(PXi
l+w(p)
Кибернетический взгляд па природу древесных растений
обещает много нового и полезного в лесоводстве.
На повестке дня таком важный вопрос, как моделирование
оптимальных лесо-семенных хозяйств и разработка автоматизи-
рованной системы управлениями этими хозяйствами.
Не менее важен и такой вопрос, как разработка систем жиз-
необеспечения в процессах лесовыращпвання высокосортного
посадочного материала в лесных питомниках и их автомати-
зация.
Такая постановка вопроса во многом будет способствовать
улучшению качественного состава лесов и повышению их про-
дуктивности.
ЛИТЕРАТ УРА
Гудвин Б. Временная организация клетки. Изд. «Мир», 1966. Пе-
ревод с английского А. М. Жаботинского под редакцией С. Э. Шполя.
Гродинз Ф. Теория регулирования и биологические системы. М..
«Мир», 1966. Перевод с английского В. Л. Наппельбаума и Тепенбаума.
Нестеров В. Г. Кибернетика живой природы. М., 1962.
Нестеров В. Г. Программные леса и модели будущих лесов.
М., -1963.
Нестеров В. Г. Кибернетика и сельское хозяйство. Изд. «Зна-
ние», М., 1966.
Нестеров В. Г. Опыт применения оптимального программирова-
ния в лесном хозяйстве. М., 1970.
Ш м а л ь г а у з -е и И. И. Кибернетические вопросы биологии. Изд.
«Наука», Сибирское отделение. Новосибирск, 1968.
149
II. Е. КОСИЧЕНКО, В. К. ШИРНИН
Анатомия мест срастания прививок дуба,
выполненных различными способами
В связи с большим объемом работ по созданию леоосемен-
ных плантации важно установить наиболее простой способ
прививок, дающий падежные результаты срастания привоя и
подвоя. Большую помощь в этом отношении может оказать
анатомический анализ мест срастания прививочных компонен-
тов, поскольку он позволяет изучить возникновение и характер
развития тканей в местах срастания н сделать выводы о на-
дежности И ПРИГОДНОСТИ ТОГО ИЛИ ИНОГО СПОСОба H'pHiBM'BIKH.
Микроскопическим изучением прививок хвойных и листвен-
ных пород занимались Ф. С. Груздов (1954), А. И. Северова
(1951, 1958), В. М. Пнр-ага (1962), Л. А. Лебеденко (1964),
В. Н. Не.нюхин (1966) и др. Сведения о раневых процессах
имеются в трудах Н. П. Кренке (1928), Л. А. Иванова (1939),
В. Ф. Раздорского (1949), А. А. Яцеико-Х(мелеbciko-го (1954,
1961), В. Г. Александрова (1966), К. Эзау (1969). По сраста-
нию прививок дуба известны только работы М. М. Котова
(1967) и В. И. Белоуса (1970). М. М. Котов производил при-
вивки зелеными черенками за кору боковых побегов 11 -летних
подвоев дуба. В. И. Белоус исследовал прививки в «мешок» на
высоком штамбе с полиэтиленовыми пакетами. Эти работы, од-
нако, не дают сравнительной анатомической оценки наиболее
распространенных способов прививок дуба, в связи с чем и ка-
честве их на практике судят обычно но проценту приживае-
мости п характеру роста привоя.
Мами была поставлена задача на основе анатгмнческого ана-
лиза мест срастания сравнить процессы образования и разви-
тия тканей при наиболее распространенных способах прививок
и выделить лучш1не из них.
Объектом исследования являлись опытные прививки дуба
черешчатого, выполненные к. с.-х. н. Ефимовым 10. П. Изуча-
лись следующие способы прививок: в «мешок» камбием привоя
к древесине подвоя, а также камбием привоя к камбию подвоя
и «за кору» камбием привоя к камбию подвоя. Изоляция мест
срастания производилась полиэтиленовыми пакетами в два сро-
ка: весной (18—21 мая) и летом (15—19 июля) 1971 года.
Привоем при весенних прививках служили зпмпие черенки дуба
поздней формы, а подвоем — 7-летпие дубки рапней формы.
150
Лотппе прививки производились зелеными черенками поздней
формы дуба, заготовленными в день прививки.
В конце сентября было взято но 4—5 прививочных участков
кащдого варианта, которые фиксировались в спирте (96°).
Окрашивание микросрезов производилось по общепринятой ме-
тодике (Наумов и Козлов, 1954; Прозина, 1960). Анатомиче-
ский анализ 96 постоянных мшкроиреиаратов прививок дуба,
проведенный в конце вегетацнош-юго периода, позволил сопоста-
вить срастание тканей на различных сезонных этапах.
Исследованием установлено, что сравниваемые способы при-
вивок имеют как общие стороны, так и существенные различия
процесса срастания прививочных компонентов. Известно, что
в первые дни контакт между компонентами прививки осущест-
вляется путем диффузного поступления воды в привой (Неию-
хнп, 1966). Прививочное ранение вызывает активизацию всех
живых клеток древесины и луба. Неповрежденные клетки вбли-
зи поверхностей срезов сильно разрастаются (гипертрофия) и
энергично делятся (гиперплазия), образуя промежуточную па-
ренхимную ткань — каллус (Эзау, 1969), в образовании которо-
го принимают участие живые клепки как подвоя, так и привоя.
У древесных пород роль подвоя в образовании каллуса является
преобладающей (Mergen, 1955; Лебеденко, 1964; Ленюхин,
1966). Наши наблюдения также показали, что основная масса
каллуса возникает в результате интенсивных делений и роста
клеток камбия п луба подвоя, плотно прилегающих к привою.
Каллус первоначально возникает в нижней части «мешка» и
в углах рапы. Затем он постепенно заполняет крупные проме-
жутки между лубом и древесиной подвоя. Древесина подвоя
пе остается безучастной к образованию каллуса. Клетки еще не
полностью дифференцированных древесинных лучей, прилегаю-
щие к оголенной поверхности, также активно делятся и растут,
пополняя промежуточную ткань. Степень участия древесины
подвоя в образовании каллуса неодинакова и зависит от време-
ни прививки. Весенняя древесина подвоя образует каллус па
всем протяжении раны. В его образовании принимают участие
как широкие, так и узкие древеспнпые лучи (рпс. 1). При лет-
ящих прививках поздняя древесина подвоя образует меньше кал-
луса. Здесь он возникает местами в результате делений клеток,
главным образом, широких древесинных лучей (рис. 2).
Вначале каллус состоит из однородных паренхиматических
клеток, которые обеспечивают контакт между привоем и под-
воем. Затем, с началом срастания постоянных проводящих тка-
ней прививочных компонентов, клетки каллуса подвергаются
151
Рис. 1. Образование каллуса древесиной подвоя при весенних прививках. Каллус образуют все древесин-
ные лучи подвоя. XI 12,5. 1—сосуды, 2 — древесинная паренхима, 3 — волокна либрпформа, 4 — древесинные
----------—-----------------лучи. _5 —каллус. _ _ . ---------------------------;------
Рис. 2. Образование каллуса при летних прививках. Каллус образуют лишь отдельные древесинные лучи.
Основная его масса образована камбием и лубом подвоя, х! 12,5. 1—сосуды, 2 — древесинная паренхима,
3 — волокна лнбриформа, 4 — древесинные лучи, 5 — каллус, 6 — изолирующий иекпотиче-кий слон.
изменениям. Первоначально топкие целлюлозные оболочки их
сильно утолщаются и одревесневают, при этом часть клеток
каллуса начи-пает отмирать. Одревеснение и отмирание каллуса
начинается на 15—18 день прививки от коры подвоя, где пер-
вый контакт проводящих тканей устанавливается раньше (Бе-
лоус, 1970).
Ряд исследователей отмечают дифференциацию клеток кал-
луса на водоироводятцпе элементы у хвойных пород и у тра-
вянистых растений (Grafts, 1934; Лебеденко, 1964; Александ-
ров, 1966; Ненюхпн, 1966). В противоположность этому
В. И. Белоус (1970) указывает, что промежуточная паренхи-
матическая ткань между привоем и древесиной подвоя не пре-
вращается в древесину.
Наблюдения, проведенные нами, показали, что каллус, бу-
дучи разнородным по происхождению, состоит из участков, ха-
рактеризующихся различной степенью дифференциации клеток.
Участки каллуса, возникшие от древесинных лучей подвоя,
состоят пз однородных паренхиматических клеток, которые за-
тем не дифференцируются в водопроводящие элементы. Ана-
логичное строение имеет также каллус, образованный лубяной
паренхимой подвоя тг привоя. По иному развиваются участки
каллуса, возникшие из окончаний дуг пли остатков камбиаль-
ных клеток подвоя и привоя. В этом случае образуется слабо-
диффсрепцированяая паренхиматическая ткань, в которой
встречаются проводящие и механические элементы, короткие
но длине и крупные в поперечнике. Таким образом, дифферен-
циацию каллуса надо рассматривать в связи с происхождением
отдельных ого участков.
При любом способе прививок па оголенной части подвоя
образуется защитная древесина, которая препятствует проник-
новению возбудителей заболеваний. При этом клетки лучей и
тяжей древесинной паренхимы через поры оболочек проникают
в полости сосудов, сильно разрастаются и закупоривают их.
Клетки разросшейся паренхимы внутри сосудов называют тил-
лями, которые со временем о.чревосневают и отмирают. Вслед-
ствие образования тплл в сосудах древесина теряет водопрово-
чящуго функцию. Одновременно отмирают также клетки дре-
весинной паренхимы и лучей. Этот процесс при прививках
аналогичен образованию защитной древесины при зарастании
ран. Как указывает А. А. Яценко-Хмелевский (1954), интен-
сивность процесса рапе вой реакции увеличивается с повыше-
нием температуры п снижается прп низких температурах. Из
154
числа факторов, обусловливающих этот процесс, основным так-
же является кислород (Prael, 1888).
Как показали паши наблюдения, у летних прививок дре-
весина отмирает быстрее и меньше образует каллуса. Поэтому
важным техническим условием прививок, особенно летних,
является хорошая изоляция поверхности среза подвоя от окру-
жающей среды (покрытие раны пластилином, применение
полиэтиленовых пакетов).
Из разрушенных оболочек клеток прираневой зоны, а также
пз клеток, отмерших вследствие высушивания и раздавливания,
могут возникнуть участки изолирующего некротического слоя
(изолирующей прослойки). Как указывает К. Эзау (1969),
процесс образования изолирующего слоя аналогичен рубцева-
нию ран. По данным Н. П. Крейкс (1928). Л. А. Лебеденко
(1964), В. Н. Нешохииа (1966) некротический слой образуется
не всегда, одпако авторы не останавливаются на причинах это-
го явления.
Наблюдения, проведенные нами, показали, что участки не-
кротического слоя могут появляться и в холе спастания привоя
с подвоем (повторные некротические слои). Они образуются
в результате надвигания одной ткани на другую л смятия пер-
вых слоев соприкасающихся клеток.
Важным моментом является тот факт, что если при привив-
ке достигнуто полное совмещение камбиальных вон привоя п
подвоя, то повторные пекротпческие слои не образуются, так
как в этом случае происходит совместный прирост но-вы'х тка-
ней. Чаще всего повторные некротические слан встречаются
при прививке в «мешок» камбием привоя к древесине подвоя.
Изолирующий слой в ходе роста прививки разрывается и
рассасывается. Разрыв изолирующего слоя осуществляется, оче-
видно, вследствие роста при.вив1ки, а также в результате раство-
рения его продукта лги жизнедеятельности клеток соседних тка-
ней. А. И. Северова (1951, 1958) указывает, что если слой изо-
лирующей прослойки 'остается ненарушенным, то он может
явиться причиной гибели прививки. Толщина изолирующего
некротического слоя зависит от числа разрушенных клеток
прираневой зоны. Отсюда следует, что прививку необходимо
производить остроотточепным инструментом.
На внутренней части луба подвоя восстанавливается непре-
рывный слой камбия. В его регенерации принимают участие па-
ренхимные клетки, расположенные между разобщенными
участками камбия. Камбий подвоя в месте прививки начинает
откладывать вначале слабо, а затем все более дифференциро-
155
ванные элементы древесины и луба. По своему строению древе-
сина, образуемая камбием в месте прививки, напоминает ране-
вую древесину открытых ран. Доля участия трахеид, сосудов,
лмбриформа в ней уменьшается, а живых •паренхимпых клеток
увеличивается. Особенно много здесь образуется шириних мпо-
горядных лучей, количество 'которых в раневой древесине в
40—50 раз больше, чем в допрививочной. Они составляют боль-
шую часть объема древесины. Рапевый луб также отличается
высокой степенью паренхиматпзацпп: в нем 'почти отсутствуют
группы волокон и имеется большое число широких многоряд-
пых лучей.
Послепрививочный слой древесины привоя по содержанию
т-капей и размерам их элементов отличается от ранее отложен-
ного слоя. В нем, как и в раневой древесине подвоя, очень мно-
го лучей. Кроме того, про-светы сосудов здесь сильно увеличи-
ваются. Размер их в тапгентальном направлении составляет в
среднем 50—52 мк, в то время как в ранее отложенном слое —
23—24 мк. Значительное и быстрое ра-звитие .водопроводящей
системы позволяет привою более полно использовать усиленный
приток воды и минеральных веществ, создаваемый хорошо раз-
витой 'Корневой системой подвоя. Видимо этим и объясняется
то обстоятельство, что при благоприятных условиях однолетние
прививки достигают высоты 1,5—2,0 м.
Кнаружи от камбия привоя дифференцируются обычные
элементы луба: ситовидные трубки, волокла, лубяная паренхи-
ма и радиальные лучи. Однако паренхимные клетки здесь дли-
тельное время сохраняют способность к делениям и росту, при-
нимая участие в дальнейшем срастании привоя и подвоя.
Для раневой послепрпвпвочной древесины подвоя характер-
но аномальное строение. Здесь нарушается вертикальная ориен-
тация элементов. В частности, трахеальные элементы могут
располагаться под различными углами к осп ствола, иногда да-
же горизонтально.
Рассмотрим особен пости срастания привоя с подвоем п рн
различных способах прививки.
1. Прививка в «мешок»
а) Камбием привоя к древесине по'.твоя
В. М. Спдорчспко (1949) предполагал, что срастание приви-
вочных компонентов при данной орнентацип привоя будет нап-
лучшим. Он писал, что наиболее активная часть камбия оста-
ется, очевидно, па отделяемой коре тт обеспечивает сращение с
древесиной привоя, с другой стороны, камбиальный слой древе-
сины подвоя легко срастается с заболонью привоя. Проти.вопо-
156
ложного мнения придерживается В. И. Белоус (1970). На осно-
вании проволенпьгх исследований он пришел «к выводу, что нет
необходимости поворачивать nepenoiK обратной -стороной, так
как древесина подвоя в «мешке» постепенно отмирает и ие
участвует в срастании прививочных компонентов.
Наши исследования показали, что срастание черепка с забо-
лонью подвоя зависит от времени прививки. У весенних приви-
вок к моменту образования защитной древесины клетки радиа-
льных лучей интенсивно делятся п их производные органичес-
ки сливаются с общей тканью каллуса, образуемого камбием
привоя. Дифференцирующиеся вслед за этим элементы луба
привоя тга значительном протяжении имеют непосредственный
контакт с еще не отмершей заболонной древесиной подвоя. При
летних прививках участие древесины подвоя в образовании кал-
луса ограничено и здесь обычно наблюдается слабое фрагмен-
тарное срастаппе прививочных компонентов. Таким образом,
этот способ обеспечивает быстрое срастание привоя с подвоем
только при весенних прививках и не дает желаемого эффекта
при летних. РЬз особенностей этого способа, которые положитель-
но выделяют его среди других, необходимо также отметить
быстрое соединение древесины привоя с послепрививочпой
древесиной подвоя, а также активпую роль камбия и луба при-
воя в заполнении пустот «мешка» паренхимной тканыо. Окон-
чания камбиальной дуги привоя крыловидно разрастаются по
щели «мешка», а их производные, дифференцируясь в проводя-
щие ткани, обеспечивают контакт и срастание с прово.чящпмп
тканями подвоя (рис. 3.). Луб привоя сильно вытягивается в
тангеита л ином направлении и сливается с каллусом. У летних
прививок, у которых со стороны защитной древесины каллус
почти отсутствует, па периферии луба возникает слой пробки.
Однако прививка в «мешок» камбием привоя к древесине
подвоя имеет ряд недостатков: поскольку выпуклая (камбиаль-
ная) часть черенка прилегает к выпуклой заболонной части
подвоя, то в результате образуется крупная щель «мешка», ко-
торая должна заполняться новообразованными тканями; кроме
того, при этом способе прививки невозможно совместить камби-
альные поверхности привоя и подвоя, так как они имеют проти-
воположную ориентацию, что несколько усложняет п замедля-
ет срастание постоянных проводящих тканей прививочных
компонентов.
При благоприятных условиях температуры, влажности воз-
духа и водоснабжения (при весенних прививках) эти недостат-
ки сглаживаются интенсивным каллусообразовапием, быстрым
157
Рис. 3. Схематическое изображение срастания летней
прививки в «мешок» камбием привоя к древесине под-
воя. 1 — перидерма, 2 — вторичная кора, 3 — вторичный
луб, 4—раневып луб, 5 — раневая древесина, 6 —
нослспрививочная древесина привоя, 7 — луб привоя,
8 — каллус, 9 — защитная д| свесила подвоя.
установленном контакта меж,ту компонентами нрптгвкп н нача-
лом ранней жп-зиедеятелыпости привоя.
б) Камбием привоя к намбню подвоя
Этот способ прививки при соответствующих размерах черен-
ка и подвоя обеспсчп.вает плотное прилегание прививочных
компонентов друг к другу, что уменьшает размеры щели «меш-
ка». Наиболее существенной 'отличительной его чертой являет-
ся возможность совмещения камбия привоя с регенерируемым
па лубяной части камбием подвоя. Это возможно при условии,
если «яа-чистку» коры черепка производить с особой тщатель-
ностью, пе оставляя толстого слоя луба. Конечно, данное усло-
вие трудно выдержать по всей длине прививаемой части череп-
ка, поэтому в отдельных местах камбиальные золы прививоч-
ных компонентов не совпадают и между ними откладываются
древесина подвоя и луб привоя. С последующим ростом привив-
ки разобщенные участки камбия посредством паренхимы сли-
ваются между собой in к концу вегетационного периода форми-
руется общее, почти правильное кольцо камбия. Уже в первый
158
гоп жизни прививки общий для привоя и подвоя камбий обра-
зует очень широкий слои древесины с большим числом широ-
ких радиальных лучей и крупными сосудами (рис. 4).
При совместном отложении проводящих тканей прививоч-
ных компонентов повторные некротические слои не образуются
и пе препятствуют сообщению между привоем и подвоем. Отме-
ченная особенность этого способа прививки и составляет его
главное преимущество перед другими. Наиболее заметно оно мо-
жет проявиться в неблагоприятных условиях при летних при-
вивках, когда быстрое достижение прямого контакта между •про-
водящими тканями прпвивки является необходимым условием
их приживаемости и высоко]"! устойчивости к механическим
повреждениям. Прививка камбием привоя к камбию подвоя мо-
жет уступать предыдущему лишь при весенних прививках
вследствие более замедленного каллусообразовапия и отсутствия
непосредственного контакта между лубом черепка и заболон-
ной частью подвоя.
Рис. 4. Схематическое изображение срастания летней
прививки в «мешок» камбием привоя к камбию под-
воя. 1 — перидерма, 2 — вторичная кора, 3 — вторич-
ный луб, 4 — раневый луб, 5—раневая древесина,
6—носленривнвочная древесина привоя, 7 — луб при-
воя, 8 — каллус, 9 — защитная древесина нодвоя,
10 — общин послепрививочный годичный слои древе-
сины.
159
2. Прививка «за кору» камбием привоя к камбию подвоя.
В тех случаях, когда «мешок» на подвое выполнить невоз-
можно, приводят прививку «за кору». При тгом способе привив-
ки в .коре подвоя делается два открытых раневых разреза. В
местах надрезов коры интенсивно образуется каллус, *в «котором
затем дифференцируется .камбий. В лубе подвоя ;в местах пора-
нения закладывается феллоге.н, который формирует мощный
слон раневой .пробки, заполняющей рану. Клетки пробки круп-
ные, равномерно разросшиеся, в радиальном .поперечнике они
значительно больше плоских клеток обычной пробки (рис. 5).
Срастание прививочных компонентов в этом случае идет также,
как и .при прививке в «мешок» камбием привоя к камбию
подвоя.
В сравнении с прививкой в «мешке» камбием прнг.оя к
камбию подвоя этот способ имеет недостатки, обусловленные
отделением полосы коры у .поз воя: заболонная часть древесины
подвоя в .месте поранения отмирает быстрее, усиливается воз-
Р и с. 5. Схематическое изображение срастания летней
прививки «за кору» камбием привоя к камбию подвоя.
1—перидерма, 2— вторичная кора, 3 — вторичный
луб, 4—раневый луб, 5 — раневая древесина, 6 —
носленрививочиая древесина привоя, 7 — луб привоя,
8 — каллус, 9 — защитная древес ина подвоя, 10 —
общий послепрпвпвочный годичный слой древесины, И —
раневая перидерма.
IG0
ложность 1троникнове1нпя инфекции и ослабляется прочность
прививки.
Неблагоприятные условия, имеющие место при летних при-
вивках, будут усиливать отрицательные стороны этого способа.
Однако эти недостатки могут быть значительно ослаблены и да-
же полт постною исключены, если привику производить весной и
принять при этом ряд дополнительных мер для прочного соеди-
нения привоя с подвоем и для тщательной изоляции места
прививки.
На 'основании проведенных ц-ьссл вдеваний можно сделать
следующие выводы и предложения:
1. Срастание прививочных комтюментов начинается с воз-
никновения каллуса. Наиболее энергичное образование каллу-
са наблюдается при прививке в «мешок» камбием привоя к
древесине подвоя. Степень дифференциации каллуса зависит от
его происхождения. Впоследствии клетки каллуса утолщаются,
одревесневают и отмирают, способствуя скреплению ггривоя с
jподвоем. Однако прочность соединения каллуса с защитной
древесиной подвоя невелика, поэтому подвязку прививок к
опорам пеибхолпмо сохранять до полного зарастания раневых
мест.
2. По своему строению поелепрививочные (раневые) прово-
дящие ткани подвоя отлнчаюются от обычных. Раневые древе-
сина и луб имеют .много широких лучей, обеспечивающих связь
с привоем. Механические ткани здесь слабо развиты, а в лубе
отсутствуют почти полностью. Вертикальная ориентировка этих
тканей часто нарушается.
3. В результате встречного .прироста тканей прививочных
компонентов и смятия первых соприкасающихся слоев клеток
возникают послерапевые ’некротические слои, которые могут
ухудшать (чюбщаем-ость между привоем и подвоем. При привив-
ках в «Минск» камбием привоя к древесине подвоя 1по1вторных
11 ек р о т 1i чес к и х с л о-св образу етс я бол ьше.
4. Весенние прививки в «мешок» камбием привоя к древе-
сине п.олв1оя отличаются быстрым заполнением «мешка» каллу-
сом и уста ио влепи ем контакта проводящих тканей прививочных
компонентов. Летние 'прививки, вследствие быстрого отмирания
заболонной древесины подвоя, подобным преимуществом перед
другими способами нс -обладают.
5. Прививка в «мешок» камбием привоя к камбию подвоя
характеризуется более полным совмещением камбиальных зон
прививочных компонентов, что ведет к отложению общих слоев
11. Заказ 6609
161
проводящих тканей. Этот метод более аффективен при летних
прививках, иежел.и iupii весенних.
6. Процесс срастания проводящих тканей при прививках
«за кору» и в «мешок» в основном одппа.ков. Однако, наруше-
ние целостности коры подвоя влечет за собой ускорение ране-
ной реакции у подвоя и ослабление механической прочности
прививки. В связи с этим пр1гв1И1В1К.п «1за кору» лучше проводить
весной с обязательной изоляцией раневы-х участков, добиваясь
прочного скрепления привоя с подвоем.
Л И Т Е Р А Т У Р А
Александров В. Г. Анатомия растений. «Высшая школа», М.,
1966.
Белоус В. И. Срастание прививок дуба. Сб. «Лесоводство и агро-
лесомелиорация», вып. 23, «Урожаи», Киев, 1970.
Груздов Ф. С. Прививки растений. Сельхозгиз, М., 1954.
Иванов Л. А. Анатомия растений. Гос-лестехиздат. Л., 1939.
Кренке Н. П. Хирургия растений. «Новая деревня», Л., 1928.
Котов М. М. Срастание прививочных компонентов дуба (каллюсно-
иочковые побеги). Изв. вузов «Лесной журнал», № 4, Архангельск,
1967. I
Лебедев к о Л. А. Анатомия прививок ели обыкновенной. Сб. науч-
но-исследовательских работ по лесному хозяйству, ЛенННИЛХ, вып. VIII,
Л., 1964.
Наумов II. А. и К о з л о в В. Е. Основы ботанической микротехни-
ки. «Советская наука», М., 1954.
Пеню хин В. II. Анатомия прививок некоторых видов сосен. Изд.
вузов, «Лесной журнал», № 3, Архангельск, 1966.
П п р а г а В. М. Некоторые особенности прививок сосны в открытом
грунте. «Лесное хозяйство», № 3, 1962.
П ризина М. II. Ботаническая мпкротехпика. «Высшая школа», М.,
1960.
Раз дор ск ий В. Ф. Анатомия растений. «Советская наука», М.,
1949. (
Северова А. И. Вегетативное размножение хвойных. Изд-во
АН СССР, М., 1951.
Северова А. II. Вегетативное размножение хвойпых древесных
пород. Гослесбумиздат. М.-Л., 1958.
Сидор чен ко Б. М. Прививка дуба. «Лесное хозяйство», № 7,
1949;-
Э з а у К. Анатомия растений. «Мир», М., 1969.
Я ц е н к о-Х м е л е в с к и й А. А. Основы и методы анатомического
исследования древесины. Изд-во АН СССР. М.-Л., 1954.
Я ц е н к о-Х м е л с в с к и й А. А. Краткий курс анатомии растений.
«Высшая школа», М„ 1961.
Crafts A. S. Phloem anatomy in two species of Nicotiana, with notes
on the interspecific graft union. Bot. Gaz., 95, 1934.
Mergen F. Anatomical study of slash pine graft unions. Fla. Acad. Sci.
Quart Jour., 17, 1955.
Ргаё! E. Vergleichende Untersuchungen uber Schutz — und Kernholz der
Laubbaume. Jahrb. Wiss. Bot., 19, 1888.
162
ПОЛОЖЕНЦЕВ П. А., АРТЮХОВСКИЙ А. К.,
ХАРЧЕНКО Н. А.
Таксономическая принадлежность мермитпд фауны СССР
В систематическом отношении мермитиды (Me rm i th i cl ae,
Nematoda) сложная и мало изученная группа гельминтов. Это
обстоятельство, в сочетании с открывающимися перспективами
использования мермнтид в биологической борьбе с вредными
бес’П'031ва-но1Члы'ми, привлекает к их изучению вое большее чис-
ло исследователей как в СССР, так и за рубежом (США, Кана-
да, Франция и др.). Особенно интенсивные исследования этой
группы гельминтов проводятся в нашей стране, лдс за послед-
ние годы значительно возросло число публикаций во фауне,
систематике и биологии мермитпд. По сборам, проведенным
в СССР, описано 60 новых для науки видов, обосновано 2 под-
семейства и 10 новых родов Mermithidae. Фауна мермитпд на-
шей страны обогатилась большим количеством новых находок
из различных географических районов н экологических усло-
вий. Правильность определения родовой принадлежности обна-
руженных видов чрезвычайно затруднена вследствие неупоря-
доченности отдельных родовых критериев и описания ряда ви-
дов но единственным экземплярам постпаразнтическпх и даже
паразитических личинок. Отмеченное вызвало необходимость
уточнения с п стема'Т1гчес1;|()1Г() пол иже Мия мермитпд, зарегн-
стрировапных на территории страны. В настоящей статье при-
водится систематический список мермнтпд фауны СССР с ука-
занием их хозяев н мест распространения.
Класс NEMATODA.
Отряд ENOPLIDA Chitw, 1937.
Подрод DORYLAIMATA, Chitw, 1933.
Надсемейство MERM1THO1DEA Wiilker, 1934.
Семейство MERMITHIDAE Braun, 1883.
Подсемейство MERM1TH1NAE (Filipjev, 1934) Artjukhov-
sky, 1971.
*]. Род MERMIS Dujardin, 1842.
1. M. nigrcscens Dujardin, 1842.
Хозяин: Orthoptera (различные виды) по Быстрицкому,
1933.
Место обнаружения: Куйбышевская область (Положенцев,
1941); Воронежская область (Положенцев, Артюховскпй, 1958);
Среднее Поволжье (Положенцев, Артюхове кий, Харченко,
1Р 163
1964)); Северный Кавказ и Крым (Положениев, Харченко,
1965).
2. М. kirgisica Kirjanova, Karawaeva, Romanenko, 1959.
Хозяин: неизвестен.
Место обнаружения: Южная Киргизия (Кирьянова, Кара-
ваева, Романенко, 1959).
Подсемейство PARAMERMITHINAE Artjukhovsky, 1971.
I. Род AGAMERMIS Cobb, Steiner a. Christie, 1923.
1. A. decaudata Cobb, Steiner, Christie. 1923.
Хозяин: Lepidoptera (боярышница, пяденица плодовая и др.)
но Артюховскому, 1955.
Место обнаружения: Воронежская область (Артюхове кип,
1955).
2. A. angusticephala Steiner, 1924.
Хозяин: неизвестен.
Место обнаружения: Брянская область (Ипатьева, 1966).
II. Род ALLOMERMIS Steiner, 1925.
1. A. trichotopson, Steiner, 1925.
Хозяин: неизвестен.
Место обнаружения: Воронежская облает!» (Положенцев,
Артюховскпй, 1958).
Ш. Род AMPH1B1OMERM1S Artjukhovsky, 1969.
1. A. kirjanovae Pologenzev, Artjukhovsky, 1958. (спн. Ие-
xamermis kirjanovae Polog, Art., 19.58).
Хозяин: неизвестен.
Место обнаружения: Воронежская область (Положенцев,
Артюховскпй, 1958); Брянская область (Ипатьева, 1966).
2. A. ghilarovi Polog., Art., 1958. (спи. Amphimermis
ghilarovi Polog., Ari., 1958).
Хозяин: Lepidoptera (различные виды совок) ио Иегробову,
1962.
Место обнаружения: Луганская область (сборы М. С. Гиля-
рова), Воронежская область (Негробов, 1962, Харченко, 1966);
Северный Кавказ и Крым (Положенцев, Харчепко, 1965); Кур-
ская область (Артюховский, Харченко, 1965).
3. A. silvatica Artjukhovsky, Khartshenko, 1971.
Хозяин: неизвестен.
Место обнаружения: Воронежская область (Артюховский,
Харченко, 1971).
4. A. rivalis Art., Khartsh., 1971.
Хозяин: неизвестен.
Место обнаружения: Воронежская область (Артюховский,
Харченко, 1971).
1С4
5. A. hydrophilis Art., Khartsh., 1971.
Хозяин: неизвестен.
Место обнаружения: Воронежская область (Артюховскпп,
Харченко, 1971).
IV. Род AMPHIDOMERMIS Fili.pjev, 1933.
1. A. tenuis Hagmcier, 1912.
Хозяин: Coleoptera (майский хрущ и др.) по Артюховско-
му, 1955.
Место обнаружения: бассейн р. Печоры (Охотипа, 1953);
Воронежская область (сборы Артюховского).
2. A. pologenzevi Ipatjeva, 1963.
Хозяин: Coleoptera (.майский хрущ) по Ипатьевой, 1963.
Место обнаружения: Брянская область (Ипатьева, 1963).
V. Род AMPHIMERMIS Kaburaki, Jamainura (спн. Comple-
xoinermis Filipjev, 1933).
1. A. elegans Hagmeier, 1912 (син. Mermis elegans Hagm.,
1912; Complexomermis elegans Hagm.. 1912).
Хозяин: Lepidoptera, Coleoptera, Orthoptera (различные ви-
ды) по Артюховс.кому, 1955.
Место обнаружения: Воронежская область (Артюховскпп,
1955; Харченко, 1966). Ростовская область (сборы М. С. Ги-
лярова), Латвийская ССР (сборы В. К. Эглптис), Южная Кир-
гизия (Кирьянова, Караваева, Романенко, 1959). Среднее По-
волжье (Положенцев, Артюховский, Харченко. 1964), Курская
область (Артюховский. Харченко, 1964), Курская область
(Артюховский, Харченко, 1965), Крым и Северный Кавказ
(Положенцев, Харченко, 1965), Украинские Карпаты (Хар-
ченко, 1970), Алтайский край (сборы А. И. Черепанова).
2. A. litoralis Art., Khartsh., 1971.
Хозяин: неизвестен.
Место обнаружения: Воронежская область (Артюховский,
Харченко, 1971).
VI. Род BATHYMERMIS Daday, 1911. (
1. В. Helvetica Daday, 1911.
Хозяин: неизвестен.
Место обнаружения: Витебская область (Кипенварлиц,
1954).
VII. CAPITOMERMIS Rubzov, 1968.
1. С. crassiderma Rubzov, 1968.
Хозяин: Diptera (.различные виды мошек) по Рубцову, 1968.
Место обнаружения: Ленинградская область (Рубцов, 1968).
VIII. Род EURYMERMIS МйПег, 1931.
1G5
1. E. tuberculata var., var., tuberculata, latebrosa Art.,
Khartsh., 1971.
Хозяин: Diplera (личинки слепней) по Артюховскому, Хар-
ченко, .1971.
Место обнаружения: Воронежская область (Артюховский,
Харченко, 1971).
2. Е. krasnitskyi Art., Khartsh, 1971.
Хозяин: Diplera (личинки слепней) по Артюховскому, Хар-
ченко, 1971.
Место обнаружения: Воронежская область (Артюховский,
Харченко, 1971.
IX. Род EUMERM1S Daday, 1911.
1. Е. behningi Steiner, 1929.
Хозяин: незвестен.
Место обнаружения: озера поймы р. Волги (сборы Бенинга).
X. Род GASTROMERMIS (Micoletzky, 1923), Filipjev, 1934.
1. G. aqualilis Hagm., 1912.
Хозя1пп: Diplera (различные виды мошек) по Рубцову, 1963.
Место обнаружения: Ленинградская область (Рубцов, 1963).
2. G. gastrostoma. Steiner, 19'18. (спи. Paramcrmis gastros-
toma, St., 1918).
Хозяин: Diplera (различные вады мошек) ио Рубцову, 1963.
Место обнаружения: Ленинградская область (Рубцов, 1963).
3. G. acroamphidis Steiner, 1929 (син. Limnomermis
acroamphidis Steiner, 1929).
Хозяин: неизвестен.
Место обнаружения: бассейн р. Камы (сборы Бенинга);
Куршский залив Балтийского моря (Артюховскпй, Вахидов,
1971).
4. G. euvaginata var. simplex Steiner, 1929 (син. Limno-
mermis euvaginata var. simplex St., 1929).
Хозяин: неизвестен.
Место обнаружения: бассейн р. Камы (сборы Бенинга).
5. G. viridis Welch, 1963.
Хозяин: Diptera (различные виды мошек) по Рубцову, 1963.
Место обнаружения: Ленинградская область (Рубцов, 1963).
6. G. boophthorae var., var. boophthorae, glaucescens, coeru-
lescens, cinerea, vittata, distoma, minifrons Welch, Rubzov, 1965.
Хозяин: Diptera (мошки) по Вел чу и Рубцову, 1965.
Место обнаружения: Ленинградская область (Волч, Руб-
цов,. 1965).
7. G. rosalbus Rubzov, 1966.
Хозяин: Diptera (мошки) по Рубцову, 1966.
1G6
Место обнаружения: Ленинградская область (Рубцов, 1966).
8. G. virescens van, var. virescens, acutipens, Rubzov, 1966.
Хозяин: Diptera (мошки) по Рубцову, 1966.
Место обнаружения: Ленинградская область (Рубцов, 1966).
9. G. minuta Rubzov, 1966.
Хозяин: Diptera (мошки) но Рубцову.
Место обнаружения: Ленинградская область (Рубцов, 1966).
10. G. odagmiae Rubzov, 1966.
Хозяин: Diptera (мошки) по Рубцову, 1966.
Место обнаружения: Ленинградская область (Рубцов, 1966).
11. G. crassifrons Rubzov, 1966.
Хозяин: Diptera (мошки) по Рубцову, 1966.
Место обнаружения: Лениградская область (Рубцов, 1966).
12. G. clinogaster Rubzov, 1966.
Хозяин: Diptera (мошки) по Рубцову, 1966.
Место обнаружения: Ленинградская область (Рубцов, 1966).
13. G. crassicaudata Rubzov, 1966.
Хозяин: Diptera (мошки) но Рубцову, 1966.
Место обнаружения: Ленинградская область (Рубцов, 1966).
14. G. longispicula Rubzov, 1966.
Хозяин: Diptera (мошкп) по Рубцову, 1966.
Место обнаружения: Ленинградская область (Рубцов, 1966).
XI. Род HEXAMERMIS Steiner, 1924.
1. Н. albicans Siebold, 1848 (спн. Mermis albicans Sieb,
1848).
Хозяин: Lepidoptera, Coleoptera, Orthoplera (многие виды)
no Assmus, 1858; Васильеву, 1904; Положенцеву и Артюхов-
скому, 1953, 1958; Артюховскому, 1955; Харченко, 1966.
Место обнаружения: Московская область (Assmus, 1858);
Киевская область (Васильев, 1904); Куйбышевская область
(Положенцев, 1952); Воронежская область (Положенцев, Артю-
ховский, 1953, 1958; Артюховский, 1955; Харченко, 1966);
Ростовская область (сборы М. С. Гилярова); Южная Кирги-
зия (Кирьянова, Караваева и Романенко, 1959); Новосибирская
область и Горно-Алтайская автономная область (сборы
А. И. Черепанова); Среднее Поволжье (Положенцев, Артю-
ховский, Харченко, 1964); Северный Кавказ и Крым (Поло-
женцев, Харченко, 1965); Украинские Карпаты (Харченко,
1970); Ленинградская и Закарпатская области (Рубцов, 1971).
2. Н. brevis Hagmeier, 1912 (син. Mermis brevis Hagm.,
1912).
Хозяин: Coleoptera, Diptera (различные -виды) по Артю-
167
ховскому, 1955; Артюховскому, Харченко, 1966; Харченко.
1966; .Рубцову, 1971. ।
Место обнаружения: Воронежская область (Артюховский,
1955; Положенцев, Артюховскии, 1958; Харченко, 1966); Ро-
стовская и Луганская области (сборы М. С. Гилярова); Горно-
Алтайская автономная область (сборы А. И. Черепанова);
Южная Киргизия (Кирьянова, Караваева, Романенко, 1959);
Среднее Поволжье (Положенцев, Артюховскии, Харченко,
1964); Северный Кавказ и Крым (Положенцев, Харченко,
1965); Латвийская ССР (сборы В. К. Эглитис); Украинские
Карпаты (Харченко, 1970); Курская область (Артюховскщу,
Харченко, 1965); Ленинградская и Закарпатская области (Руб-
цов, 1971).
3. Н. pussardi Baylis, 1933 (сип. Mermis pussardi Baylis,
1933).
Хозяин: неизвестен.
Место обнаружения: Воронежская область (Положенцев,
Артюховскии, 1958); Луганская и Ростовская области (сборы
М. С. Гилярова); Среднее Поволжье (Положенцев, Артюхов-
скин, Харченко, 1964); Северный Кавказ и Крым (Положенцев,
Харченко, 1965); Украинские Карпаты (Харченко, 1970).
4. Н. slepposis Art., Khartsh., 1965.
Хозяин: неизвестен.
Место обнаружения: Курская область (Артюховский, Хар-
ченко, 1965); Украинские Карпаты (Харченко, 1970).
5. Н. pratensis Polog., Art., Khartsh., 1965.
Хозяин: неизвестен.
Место обнаружения: Воронежская область (Положенцев,
Артюховский, Харченко, 1965).
6. Н. artjukhovskii Khartsh., 1966.
Хозяин: неизвестен.
Место обнаружения: Воронежская область (Харченкс,
1966).
7. Н. latioesophaga Art., Khartsh., 1971.
Хозяин: неизвестен.
Место обнаружения: Воронежская область (Артюхов^;;:;•
Харченко, 1971).
8. Н. cornuta Gleiss, 1955.
Хозяин: Coleoptera (колорадский жук) по Ковалю, 1971. ,х
Место обнаружения: Черновицкая область (Коваль, 197
9. Н. sujdae Rubzov, 1971.
Хозяин: Coleoptera по Рубцову, 1971.
168
Место обнаружения: Ленинградская область (сборы Г. 13. Ве-
зем чу к).
XII. Род HYDROMERM1S Corti, 1902.
I. Ы. contorta (Linstow, 1889). Hagmei-er, 1912. (спи. Мег-
mis contorta Linstow, 1889; Paramermis contorta (Linstow)
Kohn, 1905; Hydromermis rivicola Corti, 1902).
Хозяин: Diptera (мошки) по Рубцову, 1963.
Место обнаружения: Ленинградская область (Плотников,
190'1; Рубцов, 1963).
2. Н. fluviatilis Hagmeier, 1912 (син. Paramermis flu-
. ialilis Hagm., 1912; Limnomermis fluviatilis Hagm., 1912).
Хозяин: неизвестен.
Место обнаружения: Бассейн реки Печоры (Охотииа, 1953).
3. Н. boslrycoides Steiner, 1918 (сии. Paramermis bostrycoi-
des Steiner, 1918).
Хозяин: Diplera (мошки) по Рубцову, 1963.
Место обнаружения: Ленинградская область (Рубцов, 1963).
И. leptoposlhia Steiner, 1929.
Хозяин: неизвестен.
Место обнаружения: Бассейн рек Вятка и Юва (сборы Бе-
нинга); бассейн реки Печоры (Охотина, 1953).
5. II. ас и lea ta Rubzov, 1967 (син. Limnomermis aculeata
Rubzov, 1967).
Хозяин: Diptera (мошки) по Рубцову, 1967.
Место обнаружения: Ленинградская область (Рубцов, 1967).
XIII. Род 1SOMERM1S Coman, 1953.
1. I. solienamphidis. Steiner, 1929 (син. Eumermis. sole-
nampliidis Sbeiner, 1-929). ' .
Хозяин: неизвестен.
Место обнаружения: бассейн р. Камы (сборы Бешшга). :
2. 1. wisconsiensis Welch.
Хозяин: Diptera (мошки) но Рубцову, 1963.
Место обнаружения: Ленинградская область (Рубцов, 1963).
3. I. rossica Rubzov, 1968.
'эзяип: Diptera (мошки) по Рубцову, 1968.
,1'есто обнаружения: Ленинградская область (Рубцов, 1968).
XIV. Род LANCEIMERM1S Artjukhovsky, 1969.
1. L. zschokkei Schmassman, 1914 (син. Paramermis
4‘hokkei schmass., 1914; Limnomermis zschokkei Schmass.
. J14).
Хозяин: Diptera (мошки) по Рубцову, 1963.
Место обнаружения: Ленинградская область (Рубцов, 1963).
169
2. L. trachelata Steiner, 1929 (ciih. Limnomermis trachelata,
St., 1929).
Хозяин: неизвестен.
Место обнаружения: бассейн реки Камы (сборы Бенинга).
3. L. prolata Coman, 1961 (син. Paramermis prolata Com.,
1961).
Хозяин: Diptera (мошки) ио Рубцову, 1963.
Место обнаружения: Ленинградская область (Рубцов, 1963).
4. L. lanceicapita Rubzov, 1967 (сии. Limnomermis lanceica-
pila Rubzov, 1967).
Хозяин: Diptera (мошки) по Рубцову, 1967.
Место обнаружения: Ленинградская область (Рубцов, 1967).
5. L. ivashtschenkovae Rubzov, 1970.
Хозяин: Diptera (мошки) по Рубцову и Иващенко, 1970.
Место обнаружения: Ивановская область (Рубцов и Ива-
щенко, 1970).
XV. Род LIMNOMERMIS Daday, 1911.
1. L. distoicha Steiner, 1929.
Хозяин: неизвестен.
Место обнаружения: бассейн р. Камы (сборы Бенинга).
2. L. potamophila Steiner, 1929.
Хозяин: неизвестен.
Место обнаружения: бассейн р. Камы (сборы Бенинга).
3. L. cryophila Rubzov, 1967.
Хозяин: Diptera (мошки) но Рубцову, 1967.
Место обнаружения: Ленинградская область (Рубцов, 1967).
4. L. macronuclei Rubzov, 1967.
Хозяин: Diptera (мошки) но Рубцову, 1967.
Место обнаружения: Ленинградская область (Рубцов, 1967).
XVI. Род MESOMERMIS Daday, 1911.
1. М. охусегса Steiner, 1929.
Хозяин: неизвестен.
Место обнаружения: бассейн р. Камы (сборы Бенинга).
2. М. flumenalisj Welch, 1963.
Хозяин: Diptera (-мошки) по Рубцову, 1963.
Место обнаружения: Ленинградская область (Рубцов, 1963).
3. М. melusinae var. var., melusinae, biseriata, baicalen-
sis. canescens, Rubzov, 1966.
Хозяин: Diptera (мошки) по Рубцову, 1966.
Место обнаружения: Ленинградская область, Коми АССР,
оз. Байкал (Рубцов, 1966).
4. М. vernalis Rubzov, 1966.
Хозяин: Diptera (мошки) по Рубцову, 1966.
170
Место обнаружения: Ленинградская область (Рубцов, 1966).
5. М. albicans Rubzov, 1966.
Хозяин: Diptera (мошки) по Рубцову, 1966.
Место обнаружения: Ленинградская область (Рубцов, 1966).
6. М. ornata Rubzov, 1966.
Хозяин: Diptera (мошки) по Рубцову, 1966.
Место обнаружения: Ленинградская область (Рубцов, 1966).
7. М. acuticaudata Art., Khartsh., 1971.
Хозяин: неизвестен.
Место обнаружения: Воронежская область (Артюховский,
Харченко, 1971).
XVII. Род NEOMERMIS Linstow, 1904.
1. N. macrolaimus Linstow, 1904.
Хозяин: неизвестен.
Место обнаружения: Саратовская область (сборы Скорико-
ва).
XVIII. Род OESOPHAGOMERM1S Artjukhovsky, 1969.
1. О. lerricola Hagmeier, 1912 (син. Merniis terricola Hagm.,
19.12).
Хозяин: неизвестен.
Место обнаружения: Воронежская, Курская области (Хар-
ченко, 1966).
2. О. paramonovi Polog., Art., 1958 (спп. Filipjevimermis
paramonovi Polog., Art., 1958).
Хозяин: неизвестен.
Место обнаружения: Воронежская область (Положенцев,
Артюховский, 1958); Курская область (Артюховский, Харчен-
ко, 1965); Украинские Карпаты (Харченко, 1970).
3. ,0. ferghancnsis Kirjanova, Karavaeva, Romanenko, 1959
(сии. Hexamermis ferghancnsis Kirjan., Karav., Roman., 1959).
Хозяин: Lepidoptera но Кирьяновой, Караваевой, Романен-
ко^ 1959.
Место обнаружения: Южная Киргизия (Кирьянова, Кара-
ваева, Романенко, 1959).
4. О. brevivaginala Art., Khartsh., 1971.
Хозяин: неизвестен.
Место обнаружения: Воронежская область (Артюховский,
Харченко, 1971).
XIX. Род OCTOMERMIS.
1. О. paludicola Linstow, 1883 (син. Mermis paludicola
Linst., 1883, Bathymermis paludicola Linst., 1883).
Хозяин: неизвестен.
171
Место обнаружения: Туркестан (Федченко, 1874, 1886).
XX. Род PARAMERMIS Linstow, 1904.
1. Р. lepnevae Filipjev, 1933.
Хозяин: неизвестен.
Место обнаружения: Алтай, Телецкое озеро (сбиры Лепне-
вой, 1932, 1949).
2. Р. ascaroides Art., Khartsh., 1971.
Хозяин: неизвестсчь
Место обнаружения: Воронежская область (Артюховский,
Харченко, 1971).
XXI. Род POLOGENZEV1MERM1S Kirjanova, Karavaeva,
Romanenko, 1959.
1. Р. kulagini Pologenzev, 1941 (син. Psammomermis ku-
lagini, Polog., 1941).
Хозяин: Coleoptera (майский жук) no Положенцеву, 1941.
Место обнаружения: Куйбышевская область (Положенцев,
1941); Среднее Поволжье (Положенцев, Артюховский, Харчен-
ко, 1964); Курская область (Артюховский, Харченко, 1965);
Украинские Карпаты (Харченко, 1970).
XX1J. Род PSAMMOMERMIS Pologenzev, 1941.
1. Р. korsakovi Polog., 1941.
Хозяин: Coleoptera (майский жук) по Положенцеву, 1941.
Место обнаружения: Куйбышевская область (Положенцев,
1941; Ипатьева, 1966); Среднее Поволжье (Положенцев, Артю-
ховский, Харченко, 1964).
2. Р. busuluk Poliog, 1941.
Хозяин: неизвестен.
Место обнаружения: Куйбышевская область (Положенцев,
1941), Воронежская область (Положенцев, Артюховский, 1958),
Среднее Поволжье (Положенцев,- Артюховский, Харченко,
1964).
3. Р. alechini Art., Khartsh., 1965.
Хозяин: неизвестен.
Место обнаружения: Курская область (Артюховский, Хар-
ченко, 1965); Украинские Карпаты (Харченко, 1970).
XXIII. Род PSEUDOMERMIS de Mann, 1903.
1. Р. zykoffi de Mann, 1903.
Хозяин: неизвестен.
Место обнаружения: бассейн р. Волги (сборы Зыкова, 1903,
1904).
172
XXIV. Рол SKRJABINOMERMTS Pologcnzov, 1952.
1. S. sukatshevi Polog., 1952.
Хозяин: неизвестен.
Место обнаружения: Куйбышевская область (Положенцев,
1952), Воронежская область (Положенцев, Артюховский, 1958);
Среднее Поволжье (Положенцев, Артюховский, Харченко,
1964).
2. S. tolski Polog., 1952.
Хозяин: неизвестен.
Место обнаружения: Куйбышевская область (Положенцев,
1952), Воронежская область (Положенцев, Артюховский, 1958).
Среднее Поволжье (Положенцев, Артюховский, Харченко,
1964).
XXV. Род SPICULIMERM1S Artjukhovsky, 1963.
1. S. bursata Steiner, 1929 (сии. Mesomcrmis bursala
Steiner, 1929).
Хозяин: неизвестен.
Место обнаружения: бассейн р. Камы (сборы Бенинга).
XXVI. Род STRELKOVIMERMIS Rubzov, 1970. (син.
Kurshymermis Sakhidov, Poinar, 1970).
1. S. singularis Strelkow, 1964 (син. Filipjevimermis
singularis Strelkow, 1964).
Хозяин: Diptera (мотыль) по Стрелкову, 1964.
Место обнаружения: Рыбинское водохранилище (Стрелков,
1964).
2. S. viridis Sakhidov, Poinar, 1970 (син. Kurshymermis
viridis Sakhidov, Poinar, 1970).
Хозяин: Diptera (хирономиды) по Захидову и Пойиару,
1970.
Место обнаружения: Калининградская область, Куршский
залив (Захидов, Пойнар, 1970).
Виды с неясным систематическим положением
1. Allomermis lasiksi Rubzov, 1970 — описан тго одной па-
разитической личинке, найденной на берегу р. Облы Ленин-
градской области.
2. Hexamermis coccinelae var. var. coccinelae, byssoides,
adaliae, exochomi Rubzov, 1971 — описапы по одной или не-
173
скольким паразитическим личинкам, добытым из жуков Закар-
патской, Воронежской и Ленни градской областей.
3. Limnomermis tenuicauda Rubzov, 1967 — описай по
постпаразитическим личинкам из мошек Ленинградской об-
ласти.
4. Mermis pentatomiae Rubzov, 1970 — описан по пост-
паразитической личинке из клопа Новосибирской области.
5. Mesomermis bistrata Rubzov, 1966 — описан по пост-
паразитическим личинкам из мошек Ленинградской области.
6. Agamomermis heleis Rubzov, 1967 — описан по постпа-
разптическим личинкам из мокрецов, собранным в Казахстане
Глуховой.
Отсутствие важных диагностических признаков у перечис-
ленных видов и? позволяет определить их родовую принадлеж-
ность.
Несостоятельные виды
1. Mermis acuminata Linstow, 1883;
2. Mermis drassi Linstow, 1883;
3. Mermis rotundata Linstow, 1883 — описаны по коллек-
ции Федченко (1874) из Туркестана. .В диагнозе не указыва-
ются многие существенные видовые и родовые -признаки.
Рисунки отсутствуют.
4. Mermis explicans Fedschenko, 1874;
5. Mermis longissima Fedschenko, 1874 — обнаружены Фед-
ченко (1874, 1886) в Туркестане, как паразиты Orlhoptera
(Pachytylis migratorius). Описания червей очень неполные,
а рисунки изображают строение гиподермальиых хорд на по-
перечных срезах тела.
6. Mermis piscinalis Linstow, 1904 — отмечен па терри-
тории СССР Штейнером (Steiner, 1929) по сборам Бенинга
из бассейна р. Волги. В диагнозе вида указаны лишь некото-
рые размеры тела (длила, ширина).
Таким образом, в составе фауны мермптид СССР к настоя-
щему времени (1971) насчитывается 84 вида, относящиеся к
26 родам. Для 45% паразитических червей хозяева остались по
выявленными. Наиболее распространенными мермнтмдами явля-
ются Amphimermis elegans, Hexamermis albicans, Hexamermis
brevis, Hexamermis pussardi u Mermis nigrescens. Несостоя-
тельными признаны 6 видов и 5 видов медмитид отнесены
к группе с неясным систематическим положением.
174
РЕФЕРАТЫ
УДК 634.0.30.
«Основные направления научно-исследовательских работ Централь-
ного научно-исследовательского института лесной генетики и селекции».
К. К. Калуцкий. «Генетика, селекция и интродукция лесных пород»,
бып. 1, Воронеж, 1974, стр. 10.
В статье изложены основные направления научной деятельности
Центрального паучпо-псслсдовательского института лесной генетики
и селекции, организованного в системе Государственного комитета лес-
ного хозяйства Совета Министром СССР Всесоюзного научно-производ-
ственного объединения «Союзлесселекцття»^
Библ, ссылок и табл. нет.
УДК 634.0.1 G5.4:674.032.16 (470.32).
Сравнительное изучение кариотипов сосны меловой (Pinus cretacea
Kalen) и сосны обыкновенной (Pinus silvestris L.) в популяциях Цент-
рального Черноземья». С. II. Магакип, Л. К. Буторипа, М. М. Белозе-
рова, И. М. Пожидаева, Л. С. Мурая, Л. Н. Хатунцова, Ю. А. Доронин.
«Генетика, селекция и интродукция лесных пород», вып. 1, Воронеж-,
1974, стр. 10—31.
Проведено сравнительное кариологическое изучение четырех попу-
ляций сосны меловой из Белгородской и Воронежской областей и воро-
нежской популяции сосны обыкновенной^
Впервые установлено число хромосом у соспы меловой (2п = 24)
п выявлены некоторые различия в кариотипах сосны меловой и обыкно-
венной, в частности, по количеству и локализации вторичных перетя-
жек, а также наличию и отсутствию хромосом с «классическим» спут-
ником (последний обнаружен в двух популяциях сосны меловой). Вы-
явлен также полимормизм на хромосомном уровне между разными
популяциями сосиы меловой. Составлены идиограммы.
Сделан вывод о биологической самостоятельности вида соспы ме-
ловой и о необходимости дифференцированного использования ее в се-
лекции и лесомелиорации. Освещено современное состояние меловых
боров и указано на необходимость их строгой охраны и восстановления
как памятников природы и ценных маточников-семенников.
Библ, ссылок 44. илл. 5, табл. 5.
175
УДК 634.0.232.11:674.031.623.23(470.324).
К итогам сортоиспытания тополей в Воронежской области. М. М. Пе-
респи, А. П. Царев. «Генетика, селекция и интродукцпз лесных пород»,
вып. 1, Воронеж, 1974, стр. 30—4.2.
Изучено 52 вида и клона тополей, полученных из 11 пунктов и вве-
денных в испытательные насаждения Воронежской области. На основа-
нии исследования роста, устойчивости и качества 5,2 тыс. деревьев
4—21-летнего возраста в условиях поймы и нагорных дубрав выделено
14 перспективных тополей и даны рекомендации но их внедрению в
насаждения различного целевого назначения. Подробно излагается ме-
тоди ка и с сл едов алия.
Библ. ссылок 7, табл. 3.
УДК 634.0.232.13:674.031.32 (470.62).
«Сортоиспытание тополей в пойменных условиях Кубани». А. И. Си-
доров. «Генетика, селекция и интродукция лесных пород», вып. I, Во-
ронеж, 1974, стр. 42—5 L
Излагаются результаты сортоиспытания гибридных тополей в пойме
Кубани. Опытные объекты заложены в I960 году на 95 га, с использо-
ванием 24 образцов гибридных тополей отечественной и зарубежной
селекции п 3-х образцов местных видов тополей.
Приводятся показатели роста, фенологическое развитие п устойчи-
вость к вредителям п болезням гибридных тополей. Указывается па
непригодность для разведения в условиях Кубани бальзамических и
некоторых гибридных сортов из секции черных тополей. Рекомендуются
для лесных культур наиболее перспективные виды и сорта гибридных,
а также местных тополей. Приводятся примеры высокой продуктивности
тополевых насаждений, созданных в пойме Кубани из перспективных
гибридных сортов.
Библ, ссылок 20, нлл. 2, табл. I.
УДК 634.0.165,6(470.54).
«Идеи Г. Ф. Морозова о развитии селекции в лесоводстве на примере
некоторых древесных пород Среднего Урала». Е. А. Пугач, М. С. Некра-
сов. «Генетика, селекция и интродукция лесных пород», вып. I, Воро-
неж, 1974, стр. 51—59.
В статье рассматриваются материалы но индивидуальной и географи-
ческой изменчивости лиственницы Сукачева п ели Сибирской в При-
уралье и на Среднем Урале. Возросла сложная генетическая природа
популяций лиственницы и ели в указанных районах, что хорошо под-
тверждает основные тезисы, высказанные но лесной генетике и селек-
ции корифеем отечественного лесоводства Г. Ф. Морозовым.
Библ, ссылок 13, табл. 2.
УДК 634.0.811.6:674.031.632.134.
О березе с декоративной текстурой древесины. А. В. Козьмин. «Ге-
нетика, селекция и интродукция лесных пород», вып. I, Воронеж, 1974,
стр. 59—66j
В статье говорится о карельской! (узорчатой), каповой и пламенной
разновидностях березы, о их морфологических признаках, о встречае-
мости в естественных насаждениях.
Приведены результаты исследований 10-летних культур узорчатой
березы в Кировской области, на основании которых сделан вывод о
наследуемости у потомства признака «узорчатости» в этих условиях, а
также данные об условиях местопроизрастания каповой березы.
Библ, ссылок 17, нлл. 5.
176
УДК 634.0.232.3(470.324).
«О состоянии лесного семеноводства в лесхозах Воронежской обла-
сти и перспективах его развития». В. А. Горохов, Б. В. Скориков. «Гене-
тика, селекция и интродукция лесных пород», вып. 1, Воронеж, 1974,
стр. 66—69.
Приводятся данные о работах по созданию постоянной лссосемепноп
базы в лесхозах Воронежской области. За десятилетие (1961 — 1971 гг.)
отобрано 287 плюсовых деревьев сосны и дуба, заложено свыше 3 тыс.
га постоянных лесоссмепных участков и плантаций. Разработаны ме-
роприятия по постепенному переводу семеноводства на селекционную
основу. В 1971—75 гг. планируется заложить сортовым материалом
365 га плантаций.
УДК 6.‘14.0.111:674.031.632.264.2 (470.32).
«Влияние осадков на прирост дуба черешчатого ранней и поздней
форм в Центральной лесостепи». 10. П. Ефимов. «Генетика, селекция и
интродукция лес-пых пород», вып. 1, Воронеж, 1974, стр. 69—78.
Диализируется связь годичного прироста дуба раппеп и поздней
форм с количеством осадков за различные периоды года. Показаны
особенности влияния засух разного характера и продолжительности
па прирост форм дуба. Отмечено, что различия в характере связи
прироста раннего и позднего дуба с осадками обусловлены разной
динамикой из сезонного развития.
Библ, ссылок 15, илл. I, табл. 3.
УДК 6.34.0.232:674.ОУ» 1.632.264.2(470.62).
«Географические культуры дуба черешчатого в степных условиях
Краснодарского края». А. М. Шутяев. «Генетика, селекция и интродук-
ция лесных пород», вып. 1, Воронеж, 1974, стр. 78—88.
Рассмотрены количественные и качественные показатели роста дуба
черешчатого разных климатипов, почвенных экотипов и фенологических
форм в 17-летних географических культурах, заложенных в Арма-
вирском лесхозе. Даны нридержки но пере.мещепию семян дуба из
Европейской части СССР и северо-кавказских дубрав в степную часть
Краснодарского края с учетом его агролесомелиоративных зон.
Библ, ссылок 9, илл. 4, табл. 1.
УДК 634.0.232:674.032.14(477.87).
«Географические культуры лиственницы в условиях предгории За-
карпатья». П. II. Мамонов. «Генетика, селекция и интродукция леспых
пород», вып. I, Воронеж, 1974, стр. 89—94.
Излагаются результаты исследований географических культур, соз-
данных в 1968 г. в предгорьях Закарпатья с использованием 18 образ-
цов лиственницы европейской, сибирской, Сукачева и даурской.
Приводятся показатели роста и фепология различных видов лист-
венницы, отпад в культурах за 3-летний период и причины отпада.
Указывается па непригодность для лесных культур в условиях Закар-
патья лиственницы сибирской, Сукачева и даурской из восточных рай-
онов СССР. Рекомендуется местный экотип лиственницы европейской,
а также использование привозной лиственницы европейской из Татр.
Библ, ссылок 4, та-бл. 3.
• УДК 634.0.232.328.5:674.031.032:
«Особенности прививок хвойных пород». Р. Г. Моисеев. «Генетика,
селекция и интродукция лесных пород», вып. 1, Воронеж, 1974,
стр. 94—100.
Обобщен материал пятилетиях исследований по прививкам хвой-
12. Заказ 6609 177
ных пород в Карпатах. Анализируются применяемые способы, сроки
прививок, методы их изоляции. Показана зависимость приживаемости
от фснофаз прививаемых компонентов и погодных условий в момент
прививки. Рассмотрены вопросы влияния возраюта маточного дерева,
места заготовки черенков в кроне на приживаемость и дальнейший рост
привоев.
Библ, ссылок 8, табл. 3.
УДК 634.0.181.28:674.032.475.4 (470:32).
«Основные итоги интродукции видов рода Pinus L. в Центрально-
черноземных областях». Лукин А. В. «Генетика, селекция п интродук-
ция лесных пород», вып. 1, Воронеж, 1974, стр. 100—105.
Исследовались эколого-биологические особенности, рост и продук-
тивность иптродуцироваппых видов сосеп (20 видов) в культурах па тер-
ритории Центра л ьпо-Черпоземных областей.
Приведены сравнительные показатели (в процентах) роста в высоту
и ио диаметру сосен-экзотов и местной сосны обыкновенной в одинако-
вом возрасте и в сходных условиях местопроизрастания.
Разработап и рекомендуется ассортимент сосен-экзотов для широ-
кого производственного освоения (4 вида) и для предварительного
испытания (5 видов) в различного рода лесных и защитных куль-
турах. Дан рекомендательный перечень видов сосеп для интродукции,
включающий 13 названий.
Библ, ссылок 17, табл. 2.
УДК 634.0.181.28:674.032.475.242(47).
«Перспективы дальнейшей интродукции видов пихты (Abies Hill)
в Европейской части СССР». Болотов Н. А. «Генетика, селекция и интро-
дукция лесных пород», вып. I, Воронеж, 1974, стр. 105—117.
На основе изучения опыта интродукции видов р. (Abies Hill) в Евро-
пейской части СССР и анализа юстественпо-климатпческих условий со-
временных ареалов и палеоареалов рассчитан коэффициент пластичности
интродуцированных видов пихт. С использованием гомологических за-
кономерностей в развитии систематических близких видов и общих
для них коэффициентов пластичности выявлена перспективность для
первичного испытания в Европейской части СССР группы наиболее
ценных в лесохозяйственном отношении видов пихт Центрального и
Западного Китая.
Библ, ссылок 22, табл. 3.
УДК 632.151:581.524.1.
К вопросу о газоустойчпвостп п фитонцидностп древесных пород
в условиях юго-востока РСФСР. Ка шоков А. Г. «Генетика, селекция и
интродукция лесных пород», Воронеж, 1974, стр. 117—120.
В статье частично обобщены имеющиеся в настоящее время све-
дения о га.зоустойчивости и фитонцидностп некоторых древесных и тра-
вянистых растений, произрастающих на юго-востоке РСФСР. Литератур-
ные данные по газоустойчпвостп древесных растений дополнены мате-
риалами, полученными автором при обследовании промышленных пред-
приятий. Приведен перечень наиболее фитонцидных п газоустойчивых
древесно-кустарниковых пород, пригодных для озеленения промышлен-
ных предприятий исследуемого региона.
Библ, ссылок 7.
УДК 581 (143.133.5+524.1).
Особенности роста и содержания свободных аминокислот у древес-
178
ных растений в смешанных культурах. Спахов Ю. М. «Генетика, се-
лекция и интродукция лесных пород», Воронеж, 1974, стр. 120—127.
В 16-летних сосновых и сосново-березовых культурах кулисного
способа смешения (10 рядов сосны обыкновенной чередуются с 5 ря-
дами березы бородавчатой) отдельно по рядам проведены таксационные
обмеры деревьев и биохимические анализы на содержание свободных
аминокислот в листьях (хвое) древесных растений. При совместном про-
израстании у сосны и березы наблюдаются существенные изменения в со-
держании свободных аминокислот (особенно глютаминовой кислоты) в
хвое и листьях. Указанные изменения тесно связаны с ослаблением
ростовых процессов у сосны и березы на стыках и усилением их в
средних рядах кулис. Это даст возможность использовать содержание
свободных аминокислот в листьях (хвое) как один из диагностических
признаков интенсивности роста данных древесных пород.
Библ, ссылок 19, табл. 2.
УДК 581.143.
Гиббереллин и рост сеянцев ели обыкновенной. Самсонова А. Е.
«Генетика, селекция и интродукция лесных пород», Воронеж, 1974,
стр. 127—131.
В целях испытания действия гиббереллина на ускорение роста
сеянцев в высоту па питомнике Максимец-Гладышанского лесничества
Надворняпского лесокомбината были поставлены опыты с предпосевным
намачиванием семян ели обыкновенной в водных растворах гибберел-
лина.
Установлена довольно высокая отзывчивость семян ели на дополни-
тельное обогащение их гиббереллином. Намачивание семян в 0,01%-пом
растворе гиббереллина повышает эпергшо пх прорастания п начальную
скорость роста растении. Интенсивность роста растений резко увели-
чивается на второй год. Наибольший эффект получен при обработке
сухих семян (длила опытных растений по отношению к контролю
на 50—80% выше). Повышение концентрации раствора требует умень-
шения времени обработки, хотя начальная скорость прорастания семян
и увеличивается в обратном порядке. В условиях постановки опытов
хорошей концентрацией является 0,01 %-пая концентрация гибберел-
лина, а время обработки семян — от 15 минут до 3-х часов.
Библ, ссылок 3, табл. 2.
УДК 634.0.116.28. i
Лесорастительиые условия в Волго-Ахтубинской пойме и дельте
Волги после зарегулирования стока реки. Казанцев И. Я. «Генетика,
селекция и интродукция лесных пород». Выпуск 1, Воронеж, 1974,
стр. 131—135.
Приводится распределение площадей Волго-Ахтубипской поймы но
градациям затопления, засоления и плодородия.
Наиболее пригодные для разведения тополей площади находятся
в условиях редкого (0—20 дней), кратковременного (0—35 дней),
средпепродолжптелыюго (15—45 дней) и, частично, продолжительного
(25—75 дней) затопления. На незаселенных и слабозасоленных (до
среднего засоления) слоистых и зернистых почвах. Общая площадь
этих земель равна 30—35 тыс. га.
УДК 634.0.114.4(470.324).
«Опыт уточнения диагностических признаков подтипов черноземов
методом математической обработки». Чпгнриицев И. П., Беляев А. Б.,
12* 179
Стебакова В. И., Буданцев П. Б. «Генетика, селекция и интродукция
леспых пород», вып. 1, Воронеж, 1974, стр. 135—145.
В статье приводятся результаты математической обработки морфоло-
гических и физико-химических свойств выщелоченных, типичных
(«нормальных» и пысоковскнпающих) черноземов, залегающих в одина-
ковых условиях рельефа па территории Семилукского лесного селек-
ционного опытно-показательного питомника. Основными диагностиче-
скими признаками подтипов являются: глубина вскипания, гидролити-
ческая кислотность и величина pH солевой вытяжки. Не получено до-
стоверных различии между подтипами по мощности гумусового гори-
зонта, содержанию и запасам гумуса в слое 0—40 см.
Библ, ссылок 10, табл. 3.
УДК 634.0.232.32+634.0.181.28-1-634.0:165.
Представления о древесных растениях как о саморегулирующихся
автоматически управляемых живых системах природы». Булыгин Ю. Е.
«Генетика, селекция и интродукция леспых пород», вып. I, Воронеж,
1974, стр. 145—148.
Изложен взгляд автора па многолетние древесные растения как
па живые автоматы на основе бноэкоса в свете современной теории
регулирования и оптимального управления. Живые автоматы рассмат-
риваются как сложиые мпэгоконтурные системы, для которых харак-
терно наличие глубокой обратней связи и направленное приспособле-
ние (адаптирование системы) к действию возмущающих факторов сре-
ды. Приведен ряд формул, с помощью которых может быть онисап
этот сложный процесс механизма саморегуляции и адаптирования.
Такой подход к изучению жизни древесных растении позволяет
уже сейчас приступить к моделированию генетического фонда лесов
будущего и разработке автоматизированных систем управления в про-
цессе лесовыра щи вания, в первую очередь, выращивания сортового се-
менного и посадочного материала в лесных питомниках и плантациях,
а также ряд других важных вопросов лесной генетики, (’(‘лекции и
интродукции древесных и кустарниковых пород.
Бибч. ссылок 7j
УДК 581.824:634.0.232.328.5.
Анатомия мест срастания прививок дуба, выполненных различны-
ми способами. И. Е. Косичепко, В. К. ТПнрпнн. «Генетика, селекция и
интродукция лесных пород». Воронеж, 1974. стр. 149—162.
На основании анатомических исследовании мест срастания прививок
дуба в «мешок» камбием привоя к древесине подвоя (к/д) и камбием
привоя к камбию подвоя (к/к), а также «за кору» к/к установлено, что
успешность срастания прививочных компонентов при условии надежной
их изоляции зависит от способа прививки, ориентации черенка и от
сроков прививки. Всегда предпочтительны весенние прививки в «меток»,
которые обеспечивают длительную жизнеспособность паренхимных тка-
ней прививочных компонентов и их высокую меристематическую актив-
ность. Весной наиболее быстрое и падежное срастание привоя с под-
поем имеет место при прививках в «мешок» к/д вследствие длительной
жизнеспособности и активной роли в срастании клеток древесинных лу-
чей подвоя и камбия привоя. Летом участие древесины подвоя
в срастании ограничено, в этом случае способ прививок в «мешок» к/к
обеспечивает надежное срастание привоя с подвоем, так как происходит
слияние их камбиальных зоп и образование общего слоя древесины.
Библ, ссылок 21, рис. 5.
180
УДК 634.0.13.
«Таксономическая принадлежность мермитид фауны СССР». Поло-
женцев П. А., Артюховскпп А. К., Харченко Н. А. «Генетика, селекция и
интродукция», вып. 1, Воронеж, 1974, стр. 162—174. Фауна мермитид (Мег-
mithidac, Nematodes) СССР к настоящему времени (197'1) насчитывает 84 ви-
да, относящихся к 27 родам. Для 45% паразитических червей хозяева оста-
лись не выявленными. Наиболее распространенными мермитидами явля-
ются Amphimermis elegans, Hexamermis ablikans, Hexamermis brevis, He-
xamermis pussardi, Mermis nigrescens.
Несостоятельными признаны 6 видов (Mermis acuminata Linstov, 1833,
Mermis drassi Linstow, 1883, Mermis rotundata Linstow, 1883, Mermis expli-
cans Fedscelmko, 1874, Mermis longissima Fedschenko, 1874) и 5 видов мер-
митид отнесены к группе с неясным систематическим положением (АПо-
mermis lasiusi Rubcov, 1970, Hexamermis coccinelae var. coccindae, byssoi-
dcs, adaliae, elochomi Rubcov, 1971, Zimnomermis tenuicauda Rubcov, 1967,
Mermis pentatomis Rubcov, 1970, Mesomermis bistrata Rubcov, 1966, Ago-
momcrmis hclcis Rubcov, 1967).
СОДЕРЖАНИЕ
К. К. К а луцкий. Основные направления научно-исследова-
тельских работ Центрального научно-исследовательского
института лесной генетики и селекции ....
Генетика, цитология и эмбриология
С. 11. Машкпп, /V К. Буторина, М. М. Белозерова,
И. М. П о ж и д а е в а, Л. С. М ура я, Л. Н. Хат у и ц е-
в а, Ю. А. Доронин а. Сравнительное изучение карио-
типов сосны меловой и сосны обыкновенной в популяциях
Центрального Черноземья ........................................10
Селекция
М. М. В е р е с и н, А. П. Царев. К итогам сортоиспытания то-
полей в Воронежской области ...... 31
А. И. Сидоров. Сортоиспытание тополей в пойменных условиях
Кубани ...................................43
Е. А. Пугач, М. С. Н е к р а с о в. Идеи Г. Ф. Морозова о разви-
тии селекции в лесоводстве на примере некоторых древес-
ных пород Среднего Урала..................................51
А. В. Козьмин. О березе с декоративной текстурой древесины 59
Семеноводство и лесные культуры
В. А. Горохов, Б. В. Скориков. О состоянии лесного семе-
новодства в лесхозах Воронежской области и перспективах
его развития .............................. Об
Ю. П. Ефимов. Влияние осадков на прирост дуба черешчатого
ранней и поздней форм в Центральной лесостепи ... 70
А. М. Ш у т я е в. Географические культуры дуба черешчатого в
степных условиях Краснодарского края ..... 79
Н. И. М а м о н о в. Географические культуры лиственницы в
условиях предгорий Закарпатья..............................89
Р. Г. Моисее в. Особеппости прививок хвойных пород в Кар-
патах .............................. 94
А. В. Лук и и. Основные итоги интродукции рода Pinus L. в
Цептральио-Черпоземпых областях ........................101
Н. А. Болотов. Перспективы дальнейшей интродукции видов
пихты в Европейской части СССР..........................105
Экология, физиология и биохимия
А. Г. К а с ю к о в. К вопросу газоустойчпвости и фитонцидио-
стн древесных пород в условиях юго-востока РСФСР . 117
|l(). М. С п а х о bJ Особенности роста и содержания свободных
аминокислот и древесных растений в смешанных культурах 121
А. Е. Самсонов а. Гиббереллин и рост сеянцев ели обыкно-
венной .................................127
«1. Я. Казанцев. Лесорастптельпые условия в Волго-Ахтубпн-
ской пойме и дельте Волги после зарегулирования стока
реки .....................; . . . 131
II. П. Ч и г и р и и ц е в, А. Б. Беляев, В. Н. Стеб а ко в а,
П. Б. Буда п ц с в. Опыт уточнения диагностических при-
знаков подтипов Черпозсчмов методом математической обра-
ботки ..................... < . . 135
Ю. Е. Б у л ы г и п. Представления о древесных растениях как о
саморегулирующихся автоматически управляемых живых
системах природы ............................145
Н. Е. К о с и ч е и к о, В. К. III и р и и п. Анатомия мест сраста-
’ нпя прививок дуба, выполненных различными способами 150
П. А. П о л о ж е н ц е в, А. К. Артюхове к и й, Н. А. Харчо п-
ко. Таксономическая принадлежность мермнтпд фауны
СССР ............................163
Рефераты ......................... 175
ГЕНЕТИКА, СЕЛЕКЦИЯ И ИНТРОДУКЦИЯ
ЛЕСНЫХ ПОРОД
Выпуск I
(Тематический сборник научных трудов)
Л65747 Формат 60_x84I/i6, объем 11,5 п. л. Заказ 6609-73, т. 1000.
Воронеж, Областная типография Управления издательств, полиграфии
н книжной торговли, ул. 20-летня Октября, 73-а.