/





























































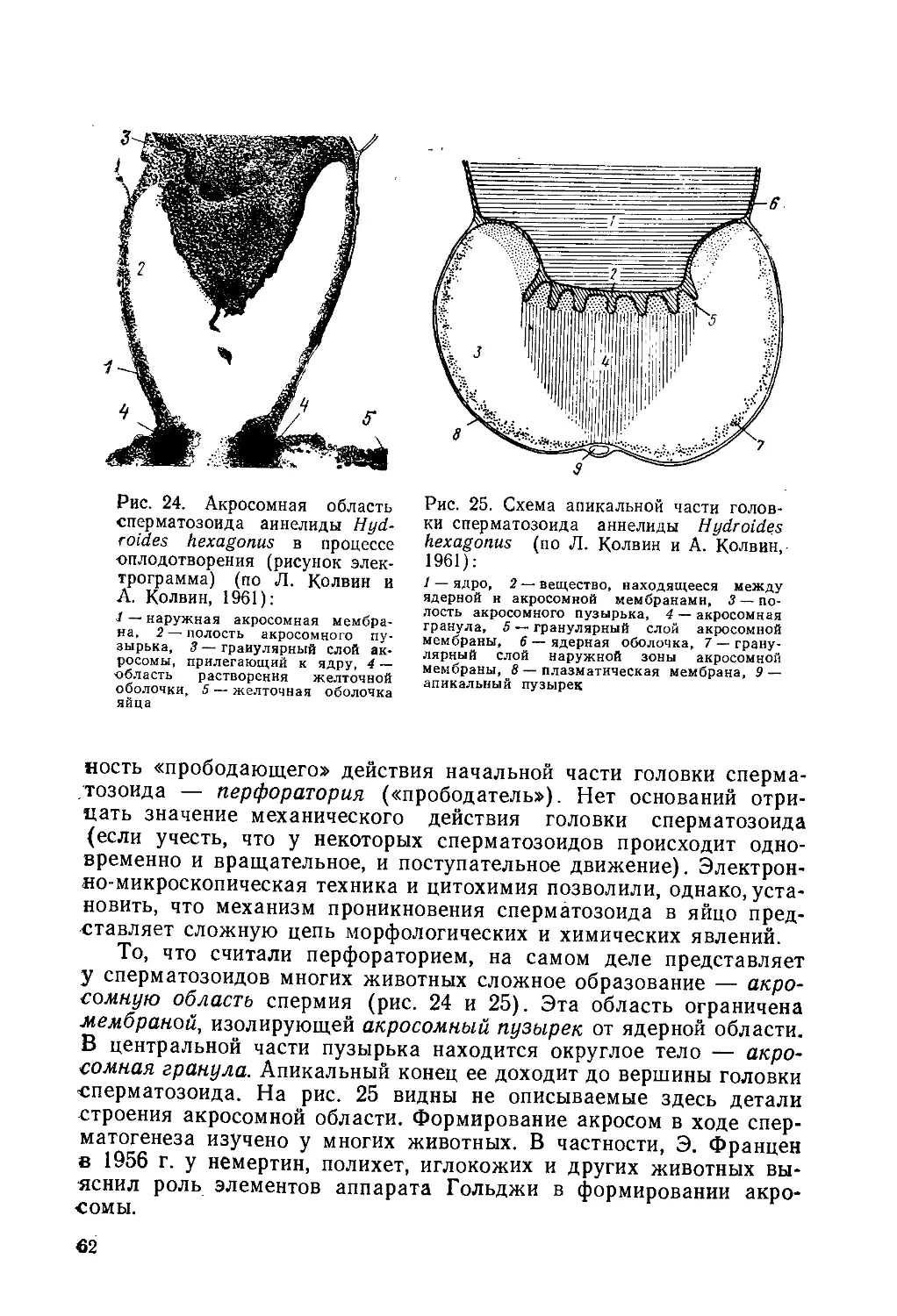






















































































































































































































































































































































































































Автор: Токин Б.П.
Теги: анатомия анатомия человека сравнительная анатомия общая биология медицина биология эмбриология
Год: 1987




Текст
Б.П.ТОКИН
ОБЩАЯ
ЭМБРИОЛОГИЯ
ИЗДАНИЕ ЧЕТВЕРТОЕ,
ПЕРЕРАБОТАННОЕ И ДОПОЛНЕННОЕ
ДОПУЩЕНО МИНИСТЕРСТВОМ
ВЫСШЕГО И СРЕДНЕГО
СПЕЦИАЛЬНОГО ОБРАЗОВАНИЯ СССР
В КАЧЕСТВЕ УЧЕБНИКА'
ДЛЯ СТУДЕНТОВ БИОЛОГИЧЕСКИХ
СПЕЦИАЛЬНОСТЕЙ УНИВЕРСИТЕТОВ
МОСКВА «ВЫСШАЯ ШКОЛА» 1987
ББК 28.03
Т 51
удк 611—013
Рецензенты:
кафедра эмбриологии Московского государственного универ-
ситета им. М. В. Ломоносова (зав. кафедрой проф. В. А. Го-
лнченков) и проф. Б. Г. Новиков (Киевский государственный
университет им. Т. Г. Шевченко)
Токин Б. П.
Т51 Общая эмбриология: Учеб, для биол. спец,
ун-тов.—4-е изд., перераб. и доп.—М.: Высш, шк.,
1987,—480 с.: ил.
В учебнике излагаются основные проблемы эмбриологии: вопро-
сы происхождения половых клеток, оплодотворения, эмбриогенеза,
регенерации и соматического эмбриогенеза,’ иммунитета зародышей
и другие вопросы; достижения описательной и экспериментальной
эмбриологии на основе использования новейших химических и физи-
ческих методик; различные теории индивидуального развития организ-
мов, пограничные между генетикой и эмбриологией вопросы.
Учебник предназначается для студентов университетов. Может
быть использован студентами педагогических, сельскохозяйственных,
медицинских и других высших учебных заведений.
2001000000—173 ББК 28.03
Т 001(01)—87 161—87 57.022
ф Издательство «Высшая школа», 1977
@ Издательство «Высшая школа», 1987, с изменениями
ПРЕДИСЛОВИЕ
Эмбриология — одна из важнейших ветвей биологии — пережива-
ет полосу нового расцвета, обусловленного прогрессом всего есте-
ствознания. Индивидуальное развитие организма, сущность и при-
чины формообразовательных процессов, характеризующих это
развитие, такие важнейшие вопросы эмбриологии, как проблемы
детерминации и дифференциации, — все это оказалось на передо-
вом плане исследований не только эмбриологов, но и генетиков,
цитологов, биохимиков и молекулярных биологов.
Дальнейший прогресс генетики немыслим без исследований це-
пи генотипически обусловленных процессов «развития признаков»,
а дальнейший прогресс цитологии — если клетка изучалась бы
абстрактно, вне развития организма.
Каждый год приносит новые факты, возникают новые идеи,
подчас объявляющие устаревшим то, что было сказано только
вчера. При такой ситуации было бы ошибкой не сообщать студен-
ту о новых фактах и новых гипотезах, но было бы не меньшей
ошибкой сообщать это новое в догматической форме, как бесспор-
ное слово в науке.
При подготовке книги к четвертому изданию в учебнике сох-
ранена дискуссионная манера обсуждения принципиальных теоре-
тических вопросов. Для привлечения внимания молодежи к реше-
нию народнохозяйственных задач внесен небольшой заключитель-
ный раздел, посвященный проблемам, в решении которых необхо-
димо участие специалистов-эмбриологов.
Срок, прошедший после выхода в свет третьего издания «Об-
щей эмбриологии», за которое Б. П. Токину была посмертно при-
суждена Государственная премия СССР, достаточно велик — 8
лет. За этот период появилось много новых фактических данных,
потребовавших внесения поправок в текстовую и иллюстративную
части книги. В настоящее время достаточно большое внимание
уделяется генетико-эмбриологическим проблемам в ущерб эмбрио-
лого-физиологическим, эмбриолого-экологическим и другим погра-
ничным с эмбриологией проблемам. Акцент в определении сущно-
сти индивидуального развития неоправданно сместился с разви-
вающегося организма на геном, поскольку онтогенез стал рассма-
триваться как процесс реализации наследственной информации,
закодированной в молекулах ДНК. Поэтому в учебник внесены
новые данные, касающиеся вопросов соотношения эмбриологии,
генетики и молекулярной биологии.
Поскольку преподавание основ биологии индивидуального раз-
вития введено во всех университетах СССР и курс лекций чита-
3
ется для всех студентов-биологов зоологических и ботанических
специальностей, в настоящее издание вйлючена глава, которая
посвящена основам эмбриологии растений. Эта глава написана
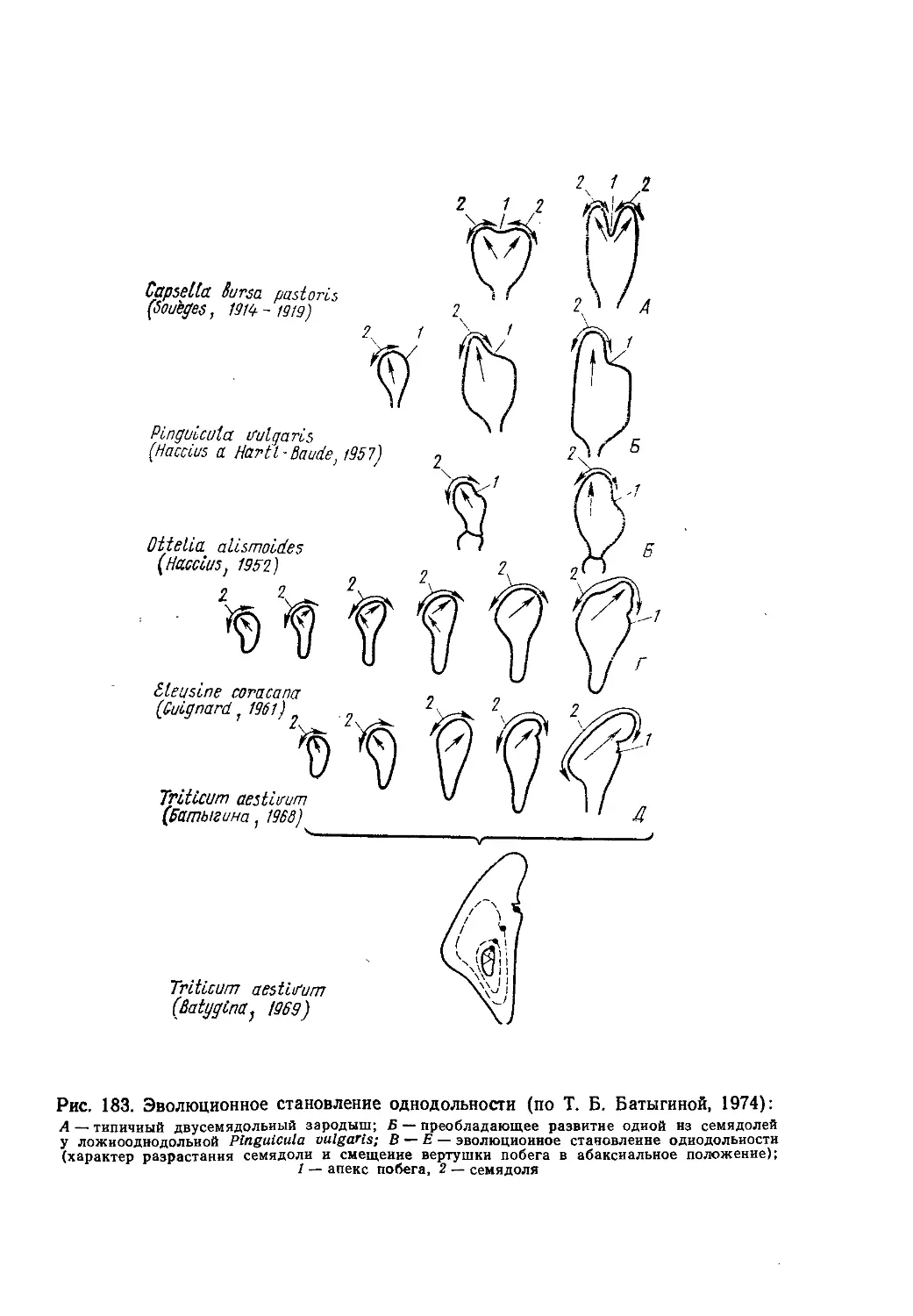
доктором биологических наук Т. Б. Батыгиной.
Доработка рукописи по замечаниям рецензентов осуществлена
Г. П. Коротковой. При доработке сохранены сложившаяся струк-
тура изложения материала, эволюционный аспект рассмотрения
основных эмбриологических проблем, включая и генетико-эмбрио-
логический. В связи с этим в книге сохранен очерк теории лар-
вальных сегментов П. П. Иванова, имеющий большое значение
для эволюционных построений. В учебнике по возможности сохра-
нен авторский стиль изложения, отличающийся образностью,
сохранена и неизбежная субъективная оценка некоторых идей,
о чем всегда предупреждал и сам автор.
Большую помощь в создании учебника оказали сотрудники ка-
федры эмбриологии Ленинградского государственного университе-
та: Н. С. Габаева, Д. Г. Полтева, Е. Б. Кричинская, С. М. Ефре-
мова, Е. Л. Авенирова — и сотрудники лаборатории эмбриологии
Ботанического института АН СССР. Всем им я приношу глубокую
благодарность.
Доктор биологических наук, профессор
Г. П. Короткова
Глава I
ПРЕДМЕТ, КРАТКАЯ ИСТОРИЯ И СОВРЕМЕННЫЕ НАПРАВЛЕНИЯ
В ЭМБРИОЛОГИИ
СОДЕРЖАНИЕ НАУКИ
Название эмбриология происходит от греческих слов эмбрион
— зародыш и логос — слово, учение. Оно не соответствует содер-
жанию современной науки. Эмбриология действительно описыва-
ет и выясняет причины всех процессов собственно зародышевого
развития — от оплодотворения яйца сперматозоидом до вылупле-
ния зародыша из яйцевых оболочек у яйцекладущих животных
или до выхода его из материнского организма у живородящих.
Однако она изучает и предзародышевый период—формирова-
ние половых клеток, и постэмбриональной период, потому что нет
таких животных, развитие которых совершенно заканчивается в
яйцевых оболочках или до рождения и которым оставалось бы
только расти. У млекопитающих животных некоторые системы
органов (например, половая, эндокринные железы и другие си-
стемы) приобретают дефинитивные, т. е. окончательные, свойст-
венные взрослому состоянию структуры и функции спустя тот или
иной промежуток времени после рождения. Зародыши многих
животных, освобождаясь от яйцевых оболочек, имеют строение,
мало сходное со строением взрослых организмов; у них развива-
ются провизорные (временные) органы, которые обеспечивают их
самостоятельное существование. Такие зародыши и личинки жи-
вут в сходной, а у ряда животных в совершенно отличной от има-
го внешней среде и имеют специальные приспособления к этой
среде. Впоследствии совершается метаморфоз, в ходе которого
личиночные органы (органы чувств и движения, пищеварения,
органы выделения и нервная система) преобразуются, прежде
чем достигнут своего дефинитивного состояния.
Эмбриологи растений изучают не только гаметогенез и разви-
тие семени, но и процесс прорастания семян, образование проро-
стков, а также постэмбриональные морфогенезы (см. гл. XVIII).
Предметом эмбриологии являются и восстановительные мор-
фогенезы, т. е. процессы развития, наступающие после случайных
травм или в результате автотомии (самокалечения). Сложные и
своеобразные процессы развития происходят у животных и расте-
ний при бесполом и вегетативном размножении, а также после
деструктивных процессов, связанных, например, с инцистировани-
ем, инволюцией (образованием редукционных тел). Эмбриологи
5
изучают также и патологические процессы, уродливые формооб-
разования.
Таким образом, предметом эмбриологии оказываются разно-
образные морфогенетические процессы, совершающиеся в ходе
индивидуального развития особи или колонии.
Издавна предлагались названия, более отвечающие содержа-
нию науки об индивидуальном развитии. В прошлом столетии
Э. Геккель стал употреблять термин «онтогения» — индивидуаль-
ное развитие и «филогения» — историческое развитие животных
и растений. В соответствии с ними науку об индивидуальном раз-
витии было бы правильнее называть онтогенетикой. С конца
прошлого столетия в связи с внедрением экспериментальных ме-
тодов в изучение морфогенетических процессов стали употреб-
лять такие названия, как «механика развития» (В.Ру), «динами-
ка развития» (М. Завадовский), «физиология развития» и др.
Однако они не вошли в быт научных лабораторий и практику
преподавания, поскольку оказались односторонними, не отражаю-
щими разнообразия задач и проблем эмбриологии.
В последние десятилетия науку о закономерностях индивиду-
ального развития стали называть «биологией развития». Предмет
науки об индивидуальном развитии должен включать различные
аспекты изучения морфогенетических процессов (морфологичес-
кие, физиологические, биохимические, биофизические, генетичес-
кие, экологические, филогенетические и др.). Содержание же по-
нятия «биология развития» в значительной степени определяется
молекулярно-генетическими аспектами изучения индивидуального
развития, что недостаточно для причинного анализа любого из
перечисленных ранее морфогенетических процессов. Широкое ис-
пользование термина «биология развития» — это дань уважения
успехам в области молекулярной генетики, биохимии и цитологии,
но вряд ли из-за этого надо отказываться от устоявшегося назва-
ния «эмбриология».
ИЗ ИСТОРИИ ЭМБРИОЛОГИИ — ОТ АРИСТОТЕЛЯ
ДО МЕЧНИКОВА И КОВАЛЕВСКОГО
Зачатки эмбриологических знаний о млекопитающих и птицах
были уже в Древнем Египте, Вавилонии, Ассирии, Индии и Китае.
С именами Гиппократа (IV в. до н. э.) и Аристотеля (384—
322 г. до н. э.) связывают не только определенные знания в об-
ласти развития организмов, но и формулирование важных эмбри-
ологических представлений. Аристотель анатомировал и изучал
зародышей многих животных, вскрывал куриные яйца на разных
стадиях развития, изучал развитие сердца куриного зародыша,
исследовал развитие акулы, каракатицы, обнаружил партеногене-
тическое развитие пчел, изучал, вероятно, абортивных человечес-
ких зародышей. Конечно, далеко не все наблюдения Аристотеля
6
были правильными, а некоторые не представляют и историческо-
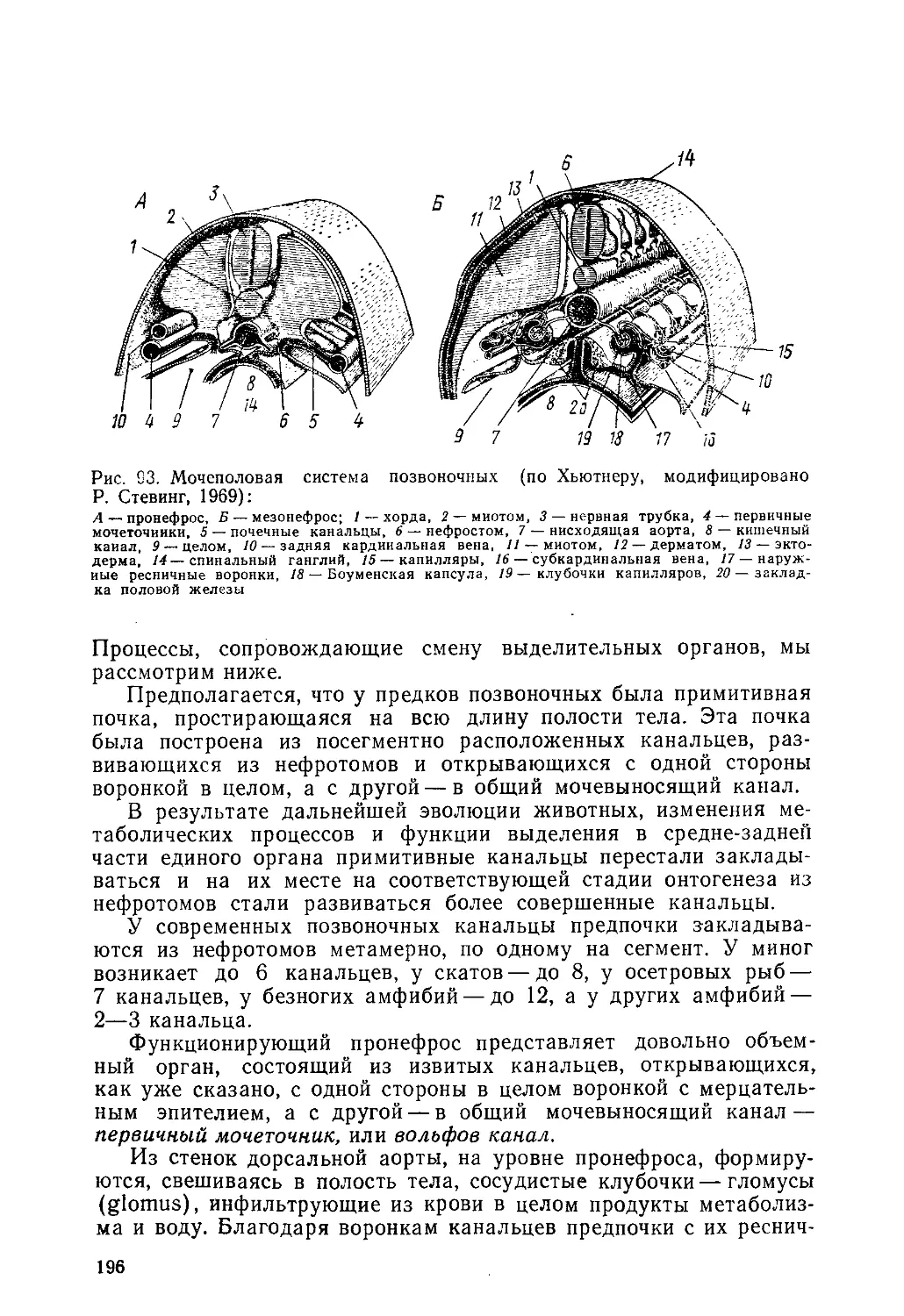
го интереса. Однако исследования, приписываемые Гиппократу, в
особенности теория индивидуального развития Аристотеля, оказа-
ли огромное влияние на развитие эмбриологии.
Согласно двусеменной теории Гиппократа плод образуется
через смешение мужского и женского «семени» (конечно, в то
время никакого представления о половых клетках не было). Вод-
ном из трактатов Гиппократа написано, что все части зародыша
образуются в одно и то же время, но те, которые по природе
своей толще, появляются прежде тонких. Эта идея — предвосхи-
щение господствовавшей в течение XVII—XVIII вв. теории пре-
формизма (от лат. ргае — впереди, перед и forma — форма). Со-
гласно этой теории каждый зародыш является уже вполне сфор-
мированным, имеющим все части организмом, которому остается
только расти. Выражаясь современным языком, в зародыше пре-
добразованы и преформированы все особенности будущего орга-
низма, происходит лишь рост без дифференциации. Наиболее
крайние и наивные преформисты представляли себе, что каждый
организм, в том числе и человеческий, заключает в себе громад-
ное количество поколений зародышей, вложенных друг в друга
в теле прародителей со времени сотворения мира (наподобие из-
вестной детской игрушки — вкладывающиеся один
в другой все более мелкого калибра шарики).
Между представителями двух направлений пре- ш к
формистов велись горячие споры. Анимальку- Ш Ж)
листы, или сперматики, считали, что зародыши—
это семязачатки. Овисты думали, что зародыши
в миниатюрном виде находятся не в мужском се-
мени, а в яйце, и семя, входящее в яйцо при опло- Jy
дотворении, составляет питательный материал за- F
родыша. Сторонниками преформизма были выдаю- II/
щиеся ученые XVII и XVIII вв. А. Левенгук, If
Я. Сваммердам, М. Мальпиги, А. Галлер, Ш. Бон- 7
не. Для состояния эмбриологии того времени очень
показателен рисунок — сатира на анималькулистов,
отстаивавших преформистские взгляды (рис. 1).
Аристотель сформулировал впервые теорию
эпигенеза (от греч. epi — после и genesis — проис-
хождение), более соответствующую современной
эмбриологии. Он, однако, внес в нее идеалистиче-
ское содержание. Аристотель думал, что зародыш
человека развивается из менструальной крови, яв-
ляющейся только материалом для развития, кото-
рому «семенная жидкость придает «форму»; ины-
t
Рис. 1. Фантастическое изображение строения сперматозоида
человека преформистами (из кн. Д. Нндхема, 1947)
7
ми словами, мужской динамический элемент придает форму жен-
скому пластическому элементу. В менструальной крови, по Ари-
стотелю, заложена лишь возможность развития, как в мраморной
глыбе — статуя, но должен быть фактор осуществления. Важно
не то, из чего развивается зародыш, а посредством чего. Мы дол-
жны представить нематериальный фактор осуществления—энтеле-
хию наподобие того, как идея статуи содержится в мысли скульпто-
ра. Энтелехия и есть истинная сущность. Если бы глаз был живым
существом, то истинной сущностью его был бы не материал, из
которого он устроен, а его зрение. Эта теория может быть опреде-
лена как идеалистический эпигенез. Несмотря на это эпигенетиче-
ские идеи Аристотеля сыграли выдающуюся роль в науке.
Сколь незначительным и медленным был прогресс эмбриоло-
гии в течение многих столетий, видно, из того, что только в 1600
и 1604 гг. появились описания и рисунки развития куриного и че-
ловеческого зародышей, принадлежащие Д. Фабрицию. Ему ка-
залось, что цыпленок развивается из завитков белка, халаз.
В 1652 г. В. Гарвей провозгласил: «Omne vivum ex ovo» — все
живое из яйца. Нельзя, однако, думать, что знаменитый созда-
тель теории кровообращения имел истинное представление о яйце.
В то же время Р. Грааф увидел в яичнике яйцевые мешочки, ко-
торые он принял за яйца. На самом деле это сложные образова-
ния, вошедшие в науку под названием граафовых пузырьков,
внутри которых только одна клетка является яйцом (рис. 2).
Я. Сваммердам также в середине XVII в. описал развитие яйца
лягушки. А. Левенгук и Гам увидели в «семени» животных мно-
жество маленьких подвижных телец, которые А. Левенгук
в 1690 г. назвал семенными животными (Spermatozoa).
Анатом М. Мальпиги в
1672 г. зарисовал и описал
куриный зародыш в разные
дни насиживания, но эти опи-
сания представляют лишь ис-
торический интерес. Эти и
другие эпизодические и боль-
шей частью ошибочные на-
блюдения не заложили, одна-
ко, фундамента эмбриологии
как науки. Занимались не раз-
витием зародыша, так как гос-
подствовали идеи префор-
мизма, а описанием отдель-
ных картин роста уже предоб-
разованных частей.
Натуралисты XVII в. эпи-
генетического направления
были лишь жалкими эпигона-
Рис. 2. Фолликул (яичник женщины):
1 — ооцит, 2 — фолликулярные клетки, 3 —
полость фолликула
8
ми Аристотеля с его глубокими, хотя и ошибочными, воззрениями.
Так, в 1620 г. Т. Фиенус, профессор в Лувене, не привнося ничего
нового, доказал, что «семя» свертывает менструальную кровь
в бесформенную лепешку, на что требуется три дня. После этого
разумная душа (энтелехия), вступившая в матку с семенем, вхо-
дит в бесформенную материю,.которую она там нашла, и начи-
нает придавать ей форму.
Важнейшей вехой в истории эмбриологии должен считаться
1759 г. В этот год была опубликована диссертация «Теория
развития» 26-летнего К- Вольфа, ставшего впоследствии академи-
ком Петербургской Академии наук. Вольф обосновал эпигенез,
проследил развитие зародыша курицы, опровергнув преформизм.
Дал описание развития кишечного канала с самого начала до пол-
ного формирования, открыл особые органы в человеческом и кури-
ном зародышах—вольфовы тела, или первичные почки. Вольф не
имел никаких представлений о клеточном строении организмов,
однако своими наблюдениями он доказал, что все части тела
животных и растений являются разнообразными группами ма-
леньких «пузырьков», «гранул», складывающихся в листки (то,
что впоследствии названо зародышевыми листками). Он впер-
ные высказал идею метаморфоза растений, рассматривая различ-
ные органы (чашечка, венчик, тычинки) как видоизменения листа.
Однако работы основателя научного эпигенеза обратили на себя
внимание лишь более чем полстолетия спустя.
Основателем современной эмбриологии является петербург-
ский академик Карл Максимович Бэр, подаривший миру свое
знаменитое сочинение «История развития животных», изданное
в 1828 г. Труд Бэра основан на фундаментальных наблюдениях
за развитием куриного зародыша и зародышей млекопитающих..
Он обосновал эпигенетическое представление в более глубокой
форме, чем это было сделано Вольфом. Творчество Бэра — яркое
доказательство несостоятельности преформистских взглядов в той
форме, какони выражались философами и натуралистами в течение
многих столетий — от Аристотеля до XVIII в.
К- М. Бэр создал учение о зародышевых листках. Он различал
два первичных листка — анимальный и вегетативный. Анималь-
ный листок, по Бэру, разделяется в конце развития на кожный
слой, дающий покровы, нервную систему и органы чувств, и на
мускульный, дающий мышцы и кости. Вегетативный разделяется
на сосудистый слой, который дает мезентерии и сосуды, и слизистый
слой, вместе с участками сосудистого образующий стенки кишеч-
ного канала. Эти слои дают первичные органы, преобразующиеся
в дефинитивные. Учение Бэра о «зародышевых листках» предше-
ствовало современным представлениям о зародышевых листках
или пластах, созданным А. О. Ковалевским и И. И. Мечнико-
вым.
Бэр сделал много важных открытий. Он описал хорду у ку-
9
риного и человеческого зародышей. Показал, что у зародышей
высших животных и человека в периоды их развития формируют-
ся органы, функционирующие у низших в течение всей жизни
(вольфовы тела, спинная струна). Он доказал, что человек разви-
вается по единому плану со всеми позвоночными животными и
впервые увидел яйцо млекопитающих и человека. В 1864 г. Пе-
тербургская Академия наук по случаю пятидесятилетия научной
деятельности Бэра заказала в его честь юбилейную медаль с
надписью: «Orsus ab ovo hominem homini ostendit» (Начав с яйца,
он показал человеку человека). Бэр сформулировал закон заро-
дышевого сходства, согласно которому у зародыша возникают
сначала общие признаки типа, затем появляются признаки класса,
рода, вида и, наконец, индивидуальные признаки (см. гл. XVII).
'Как ни велики заслуги гениального натуралиста Бэра, следу-
ет помнить, что он не был последовательным эволюционистом.
Удивительно, 'что, работая лишь на «надклеточном уровне» (ра-
боты Т. Шванна и М. Шлейдена, изложивших учение о клеточном
строении организмов, вышли в 1838— 1839 гг.), почти не прибе-
гая к микроскопу и не руководствуясь эволюционной идеей, Бэр
явился основателем современной эмбриологии. Творчество Бэра за-
вершает доклеточный и додарвиновский период в развитии эм-
бриологии. Современная эмбриология немыслима без учения о
клетке и без эволюционной идеи.
СОЗДАНИЕ ЭВОЛЮЦИОННОЙ ЭМБРИОЛОГИИ
Честь основания эволюционной эмбриологии, базирующейся на
принципах учения о клеточном строении организмов, принадлежит
А. О. Ковалевскому и И. И. Мечникову. Оба замечательных уче-
ных своими многочисленными сравнительными наблюдениями
над развитием большого числа животных создали фактическую
основу современной эмбриологии. До них хорошо изучено было
лишь развитие некоторых позвоночных и членистоногих. Почти
ничего не было известно о развитии кишечнополостных, мол-
люсков и иглокожих. В представлениях ученых беспозвоночные
и позвоночные животные представляли обособленные группы и
сильным было влияние идей Кювье и Бэра о не связанных между
собой типах животного мира. Ковалевский обратил особое внима-
ние на «переходные», «уклоняющиеся» формы. Среди его замеча-
тельных открытий особенно важным явилось установление род-
ства асцидий с позвоночными животными, в то время как их обыч-
но относили к моллюскам или моллюсковидным. Большое
значение для уничтожения «пропасти» между типами имело и его
исследование по развитию ланцетника. Ковалевский и Мечников,
отправляясь от дарвиновского учения, создавали именно сравни-
тельную эмбриологию, сопоставляя образование полости тела, пи-
10
щеварительного канала, нервной и сосудистой системы у разных
животных.
Значение творчества А. О. Ковалевского для развития науки
очень хорошо охарактеризовано словами его ученика, эмбриолога
К. Н. Давыдова: «А. О. Ковалевский по праву должен считаться
фактическим основателем современной эмбриологии. Он первый
констатировал у беспозвоночных зародышевые листки и доказал
существование единства плана в развитии всех животных. Совре-
менная теория зародышевых пластов — этот основной принцип
сравнительной эмбриологии — в значительной степени обязана сво-
им развитием именно Ковалевскому. Ковалевский, наконец, пер-
вый ясно понял и оценил значение эмбриологии для уяснения эво-
люции и своими трудами более чем кто-либо другой способство-
вал окончательному торжеству эволюционной идеи. Работы его
над развитием ланцетника и асцидий останутся вечным памятни-
ком мощности человеческой мысли ... »'.
Действительно, современными представлениями о зародышевых
листках мы обязаны А. О. Ковалевскому, обнаружившему экто-
дерму, энтодерму и мезодерму у всех типов животных. Термины
эти были предложены Э. Геккелем. Благодаря работам Мечнико-
ва, Ковалевского и других эмбриологов второй половины XIX в.
установлены единые принципы развития беспозвоночных и позво-
ночных животных. Глубокое изучение ранних стадий развития жи-
вотных стало возможно лишь после внедрения в эмбриологию уче-
ния о клеточном строении организмов, а также благодаря прогрес-
су микроскопической техники. Эмбриологические исследования
все теснее переплетались с исследованиями гистологического
строения зародышевых листков и развития тканей и органов. Вто-
рая половина XIX в. ознаменовалась необозримым количеством
гистоэмбриологических исследований зародышей разных животных,
проведенных зарубежными и русскими учеными. Многими рабо-
тами А. О Ковалевского, И. И. Мечникова и других отечествен-
ных ученых, ставшими классическими, пользуется без существен-
ных поправок современная эмбриология.
Эволюционная эмбриология сослужила немалую службу,
вместе со сравнительной анатомией и палеонтологией, для доказа-
тельства эволюции животных и выяснения исторического родства
одних групп животных с другими. Мысли Дарвина, работы
И. Меккеля и Бэра подготовили почву для формулирования
Ф. Мюллером и Э. Геккелем закономерности, известной под на-
званием биогенетического закона, согласно которому онтогенез
(развитие особи) есть краткое повторение филогенеза (развитие
вида). Идея взаимосвязанности онтогении и филогении сыграла
важную роль в развитии эмбриологии, общей зоологии и эволю-
1 Давыдов К. Н. Курс эмбриологии беспозвоночных. Петербург — Киев,
1914. С. 5.
11
ционной морфологии, хотя формулировки Мюллера и Геккеля
несовершенны. В нашем столетии вопросами соотношения онто- и
филогенеза особенно много занимался А. Н. Северцов — один из
крупнейших эволюционных морфологов. О его теории филэмбрио-
генезов говорится в гл. XVII. Там же дан анализ биогенетического
закона в свете современной биологии.
Проблемы эволюционной эмбриологии в нашей стране с успе-
хом развивали А. Н. Северцов, И. И. Шмальгаузен, П. П. Иванов,
П. Г. Светлов, А. А. Захваткин, Г. А. Шмидт, А. Г. Кнорре и дру-
гие эмбриологи. К эволюционной эмбриологии тесно примыкают
эколого-эмбриологические исследования, выясняющие адаптации
на различных стадиях индивидуального развития к той или иной
среде обитания (Г. А. Шмидт, С. Г. Крыжановский, Б. С. Матвеев,
А. Н. Трифонова и др.). В результате этих исследований были раз-
работаны различные принципы периодизации индивидуального
развития и даны их морфофизиологические характеристики.
Исследования, пограничные между эмбриологией и физиологи-
ей, обогатили науку об индивидуальном развитии фактами о ста-
новлении функций, обеспечивающих в совокупности интегратив-
ность и адаптивность организма (П. К. Анохин, И. А. Аршавский
и др.). Большое влияние на развитие эмбриологии в нашей стра-
не оказало творчество зоологов и сравнительных анатомов
И. И. Шмальгаузена, А. Н. Северцова, В. Н. Беклемишева,
В. А. Догеля, А. В. Иванова, а также гистологов А. А. Заварзи-
на, Н. Г. Хлопина и др.
ЭКСПЕРИМЕНТАЛЬНАЯ ЭМБРИОЛОГИЯ
Экспериментальные методы начали использоваться в эмбриоло-
гии и зоологии с середины XVIII в. Л. Спалланцани занимался
искусственным осеменением собаки. Р. А. Реомюр разрабатывал
вопросы инкубации куриных яиц и в 1749 г. выпустил книгу «Об
искусстве выводить цыплят». А. Трамбле обессмертил свое имя
опытами на гидрах; в 1744 г. вышли его знаменитые «Мемуары
о пресноводных полипах». Однако это были лишь эпизоды на фо-
не описательной эмбриологии, каковой она являлась до конца
прошлого столетия. Эмбриология долгое время отвечала на воп-
рос: как происходит развитие? По мере успехов эмбриологии и
всей науки о жизни исследователи все более настойчиво ставили
вопрос: почему развитие происходит так, а не иначе? В. Ру при-
надлежит честь основания экспериментального направления в эм-
бриологии, названного им механикой развития организмов. Одно-
временно с ним некоторые другие ученые стали использовать
различные экспериментальные методики в своих исследованиях.
Е. Пфлюгер, например, в 1883 г. изучал действие силы тяжести
на развитие яйца лягушки. Вскоре начали экспериментировать
Г. Дриш, Ж. Лёб и многие другие. В 1883 г. В. Ру провел исто-
12
рический опыт, имеющий целью выяснить, как происходит разви-
тие одного из бластомеров яйца лягушки в случае умерщвления
другого на двухбластомерной стадии развития (см. гл. XI). Увле-
ченный каузально-аналитическим методом (выяснением причин
развития), В. Ру основал в 1883 г. специальный журнал, ставший
идейно-научным и организационным центром для эмбриологов
экспериментального направления всех стран. В новом журнале,
с первых его номеров, начали сотрудничать Т. Морган — один из
основателей современной‘генетики, Э. Вильсон и Т. Бовери—вид-
нейшие цитологи, Ж. Лёб и Р. Лилли — создатели физико-хими-
ческого направления в биологии, Г. Шпеман — автор оригиналь-
ной теории индивидуального развития — теории организационных
центров, Г. Дриш, которому эмбриология обязана' выдающимися
экспериментами по изоляции бластомеров, ц др. (впоследствии
Дриш встал на ложную позицию в области философии, возглавив
неовиталистическое направление).
В разных главах настоящей книги дано изложение исследова-
ний и направлений основных школ экспериментальной эмбриоло-
гии XX в. Как и в любой науке, прогресс в эмбриологии определял-
ся созданием новых гипотез и теорий, открытием новых методик и
развитием смежных областей знаний. Начиная с 1940 г. большую
роль стали играть электронно-микроскопические исследования.
Для прослеживания судьбы тех или иных бластомеров, для изу-
чения передвижения клеточного материала в ходе развития важ-
ное значение имела разработанная В. Фогтом методика меток,
наносимых витальным красителем на отдельные части зародыша.
Эта методика дала возможность выяснить процессы гаструляцик
у амфибий и других животных.
Исключительное значение, притом все возрастающее в наше
время, имели разработка методик культивирования тканей и за-
чатков органов вне организма, соответствующие способы хирур-
гии, подбор питательных сред, способы их стерилизации. Многие
ученые — Ж. Лёб, А. Каррель, Н. Г. Хлопин, Э. Вольф, У. Фелл и
др. — внесли большой вклад в науку о культуре клеток, тканей
и органов вне организма. Однако честь открытия метода тканевых
культур принадлежит Р. Гаррисону.
Наибольшее влияние на экспериментальную эмбриологию в
XX в. оказала школа Г. Шпемана, предложившего свою теорию
индивидуального развития (см. теорию организационных центров
в гл. XI) и разработавшего прекрасные методики микрохирургии
на зародышах: снятие оболочек яиц животных, пересадка частей
одного зародыша другому, изготовление благоприятной жидкой
среды для развития и др. Школа Шпемана дала эмбриологии
много фактов о взаимозависимости «частей» развивающегося
зародыша (см. гл. XI). Влияние работ Шпемана было значитель-
ным до 40-х годов нашего столетия. Особенно большое значение
13
для эмбриологии имели работы учеников Шпемана — Г. Ман-
гольд и И. Гольтфретера.
Исключительная роль в развитии эволюционной эмбриологии
принадлежит отечественным эмбриологам Д. П. Филатову и
П. П. Иванову. Воспитанные на традициях эволюционной морфо-
логии, они явились создателями школ, влияние которых не осла-
бевает со временем. Д. П. Филатов сделал важные открытия, ка-
сающиеся развития органа слуха и глаза у позвоночных, разра-
ботал свои методики микрохирургии, а главное — положил нача-
ло сравнительной экспериментальной эмбриологии.
Ученик Филатова В. В. Попов своими исследованиями на
функционирующих глазах развивающихся позвоночных и на зачат-
ке внутреннего уха обосновал функциональное направление
в экспериментальной эмбриологии. Было установлено, что различ-
ные условия освещения влияют на функциональное состояние
сетчатки, а это отражается на ее способности к «индукции» рого-
вицы и на контроле за ростом и дифференциацией хрусталика.
Попов считает доказанным влияние на процесс онтогенеза «услов-
ного раздражения» (сочетание светового раздражения как «ин-
дуктора» нового хрусталика с вибрационными раздражениями).
Формативные связи и взаимные влияния между частями раз-
вивающегося организма — важнейший и неотъемлемый раздел
в современных эмбриологических исследованиях. Ученые продол-
жают исследования индукционных взаимодействий частей разви-
вающегося организма.
Особое направление исследований в эмбриологии берет свое
начало от работ Г. Дриша, который попытался осмыслить и
объяснить явления эмбриональных регуляций, развития частей
зародыша в новую особь, в новый полноценный организм того же
вида животного или растения. Объяснение этим процессам не мог-
ло быть дано на уровне близкодействующих зависимостей, напри-
мер индукционных взаимодействий или геномных регуляций. Тре-
бавались гипотезы, исходящие из представлений о наличии в целом
организме или его частях интеграционных механизмов, поддер-
живающих определенное состояние целостности. Эти очень важ-
ные для эмбриологии направления исследований привели к фор-
мулированию представлений о полях (А. Г. Гурвич, Н. К. Коль-
цов, Е. Гиено и др.), физиологических градиентах (Ч. Чайлд)
или градиент-полях. Позднее стали разрабатываться более част-
ные вопросы эмбриональных и постэмбриональных регуляций,
такие, как, например, «плюс-минус взаимодействия» М. М. Зава-
довского, многочисленные работы о гормональных регуляциях
гаметогенеза, эмбриогенеза, метаморфоза, регенерации и других
морфогенетических процессов.
Особое место в эмбриологии занимают исследования вегета-
тивного и бесполого размножения (в особенности у растений),
а также восстановительного морфогенеза. Начиная с Р. Реомюра
14
и А. Трамбле, выполнивших в первой половине XVIII в. первые
работы по регенерации, исследования восстановительных морфо-
генезов интенсивно развивались с привлечением широкого круга
объектов. Преобладание преформистских идей в объяснении реге-
нерационных способностей животных породило в это время до-
вольно распространенное направление исследований, а именно
проведение повторных ампутаций (например, восстановившихся
конечностей или отделов тела) для выяснения того, сколь много
запасных зачатков имеется в теле регенерирующего животного.
В XIX в. под влиянием творчества Ч. Дарвина восстановительные
процессы стали изучать с позиций их значения для прогрессив-
ной эволюции вида (Т. Морган, А. Вейсман, Г. Пржибрам).
Во второй половине XIX в. сформировались эксперименталь-
ная ботаника и физиология растений. Регенерационные процес-
сы, в особенности при черенковании, стали одними из главных
предметов исследования ботаников этого профиля (Г. Фёхтинг,
К. Гёбель, Ф. Ваккар, И. Сакс, Г. Клебс и др). Большой вклад
в развитие регенерационных исследований внесли советские био-
логи (П. П. Иванов, Э. Е. Уманский, Л. Н. Жинкин, М. А. Ворон-
цова, Л. Д. Лиознер и многие другие исследователи).
Экспериментальная эмбриология бурно развивается во многих
странах в связи с прогрессом всего естествознания и представле-
на многими оригинальными школами.
МЕСТО ЭМБРИОЛОГИИ СРЕДИ ДРУГИХ НАУК.
ОСНОВНЫЕ ЧЕРТЫ СОВРЕМЕННОЙ ЭМБРИОЛОГИИ
Современная эмбриология имеет характерные признаки. Большин-
ство лабораторий во всех странах занято экспериментальными
исследованиями, но наряду с ними продолжается описание гисто-
эмбриологическими'способами развития тех или иных животных,
не изученных или недостаточно изученных в эмбриологическом
отношении. Если эмбриология прошлого и в значительной мере
нашего века была чисто морфологической наукой, современная
эмбриология все более становится наукой морфофизиологиче-
ской, использующей новые методы работы, вызванные прогрес-
сом физики, химии и математики.
Отношение эмбриологов к своему объекту долгое время на-
поминало отношение матери к ребенку. Для матери сын или дочь
в любом возрасте — ребенок, у которого все в будущем. Так и
большинство эмбриологов считало единственной задачей своей
науки — изучение смены структур в ходе развития организма,
выяснение того, что в последующем возникает из тех или иных
клеток и участков зародыша. Эта задача остается в полной мере
и у современных эмбриологов. Однако выяснить причины онто-
генеза невозможно без изучения своеобразия физиологии и био-
15
химии зародышей на ранних стадиях их развития и без изучения
меняющегося в ходе развития отношения к среде. Поэтому вни-
мание современных эмбриологов все более привлекается к изу-
чению функций, свойственных собственно зародышам, физиологии
и биохимии зародышей на разных этапах развития изменению
функционирования генома и развитию интегративных систем
(нервной, эндокринной, иммунологической).
Современная эмбриология ставит своей задачей управление
развитием, организмов. Осуществление этой задачи возможно при
условии тесной связи эмбриологии с другими науками, в первую
очередь с гистологией, цитологией, биохимией (включая молеку-
лярную биологию) и генетикой. Цитологи и гистологи также за-
интересованы в связи с эмбриологией, так как они изучают не
абстрактные клетки и ткани, а клетки и ткани развивающихся
организмов или взрослых животных, но не как какие-то стабиль-
ные кирпичики, а в связи с состоянием органов и организма в це-
лом.
Эмбриология должна быть теснейшим образом связана с ге-
нетикой и цитогенетикой. Эту связь начал осуществлять еще
В. Ру. Т. Морган, изучая явления наследственности и изменчи-
вости, неоднократно пытался осуществить синтез генетики и
эмбриологии. Тесная связь эмбриологии с экологическими наука-
ми проявляется при изучении влияния внешней среды на разви-
тие организмов.
Задачи и объект исследований определяют и выбор соответ-
ствующих методик. Это могут быть прижизненные наблюдения,
микрокиносъемка развивающихся зародышей, процессов регене-
рации и других явлений, методики разрезов зародышей, их спе-
циальной окраски. Эмбриология, - как и другие биологические
науки, использует методы, диктуемые развитием физики, химии
и математики. Таковы, например, методики электронной микро-
скопии. Для изучения клеточного метаболизма как основы диф-
ференциации (см., гл. XI) используются радиоактивные изотопы,
методы ауторадиографии. Все более тесной становится связь
эмбриологии с физикой и химией. Во-первых, эти науки помогают
создавать новые методики исследования.. Во-вторых, многие био-
физические и биохимические процессы эмбрионального развития
вообще невозможно изучать без участия физика, химика и мате-
матика.
При выяснении причинных механизмов формообразователь-
ных процессов доминирующими исследованиями, особенно начиная
с 20-х годов нашего столетия, были биохимические. Вышедшая
- в 1931 г. известная сводка «Химическая эмбриология» Дж. Нид-
хема, не охватившая, конечно, все исследования, уже содержала
библиографию 8000 биохимических работ в области эмбриологии.
Начиная с 50-х годов в связи с успехами молекулярной биологии
и выяснением роли нуклеиновых кислот в синтезе белков и в яв-
16
лениях наследственности интерес к биохимии эмбрионального
развития возрос еще больше. В частности, появились многочис-
ленные данные об изменениях в синтезе белков и нуклеиновых
кислот в ходе развития организма, развития отдельнах органов,
в процессах дифференциации клеток и тканей. Но, к сожалению,
крайне мало биохимических исследований, в которых доказыва-
лось бы, что изменения в метаболизме являются действительно
причиной тех или иных морфологических явлений.
Еще труднее сообщить что-либо, «устоявшееся» в области фи-
зики эмбрионального развития. Между тем даже грубое физико-
химическое моделирование жизненных процессов в прошлом веке
(«искусственные клетки» и др.) помогало биологии, а в настоя-
щее время прогресса естествознания создание моделей, киберне-
тических устройств, имитирующих те или иные явления развития
организмов, приобретает еще большее значение.
Таким образом, закономерности развития организмов иссле-
дуются с использованием методов физики, химии и математики
на разных уровнях: молекулярном, клеточном и на уровне позна-
ния взаимодействия между клетками, тканями, частями зароды-
ша, т. е. на уровне развивающегося организма как целого.
На каком уровне проводится исследование, требуются ли биофи-
зические, биохимические, гистохимические, хирургические или
иные методики — все это зависит от изучаемого явления и от за-
дачи исследования. При изучении развития организмов наука
имеет дело как с процессами молекулярного и атомного характе-
ра, для познания которых требуется применение квантовой фи-
зики, так и с надмолекулярными, клеточными и надклеточными
процессами, которые не только не требуется, но и невозможно
изучать лишь на молекулярном уровне, — взаимные отношения
частей зародыша, специальные биологические структуры, влияние
биотических факторов среды на развитие зародыша (см. гл. XIV)
и обширный круг других биологических явлений.
Основой современной эмбриологии служит эволюционная идея.
Развитие зародыша может быть понято должным образом лишь
в том случае, если его рассматривать как результат исторического
развития организмов. Вот почему в настоящей книге многие про-
цессы эмбрионального развития рассматриваются в сравнитель-
ном, эволюционном аспекте.
Глава II
ПОЛОВЫЕ И СОМАТИЧЕСКИЕ КЛЕТКИ. ПРОИСХОЖДЕНИЕ
И СТРОЕНИЕ ПОЛОВЫХ КЛЕТОК
ПОЛОВОЕ И БЕСПОЛОЕ РАЗМНОЖЕНИЕ
Большинству многоклеточных животных свойственно половое
размножение, при котором женская половая клетка — яйцо— со-
единяется с мужской половой клеткой —сперматозоидом. При
этом образуется одна оплодотворенная яйцеклетка — зигота (от
греч. zygota— спаренная) в отличие от половых клеток — гамет
(games — брак). Зигота по существу уже является организмом
в самом начале его развития.
При бесполом размножении специализированные клетки не
формируются, новому организму дают начало соматические (те-
лесные) клетки. О различных формах бесполого размножения см.
гл. XVI.
Половое размножение объединяет обширный круг морфоло-
гических и физиологических явлений: различия между полами,
строение и развитие органов, в которых развиваются половые
клетки, физиологические особенности самцов и самок и т. д. Эти-
ми и связанными с ними вопросами занимаются цитология, ге-
нетика, эндокринология, биохимия, общая зоология и другие
науки. В этой главе рассматриваются вопросы происхождения
и развития половых клеток, их соотношение с соматическими
клетками. Из общебиологических проблем, интересующих эмбрио-
логов, остановимся только на связи полового процесса с размно-
жением.
Привычная мысль, без сомнения, связывает половой процесс
с размножением, так как действительно у многоклеточных живот-
ных эти процессы составляют единство, являясь причиной уве-
личения числа особей данного вида. У Protozoa связь между раз-
множением и половым процессом не столь очевидна, и проводить
четкую грань между половым и бесполым размножением у них
нелегко. В серию подчас очень многочисленных бесполых раз-
множений лишь иногда вклинивается половой процесс, который
непосредственно к размножению, т. е. к увеличению числа осо-
бей данного вида, не приводит. При половом процессе у простей-
ших может иметь место соединение двух клеток — копуляция, a
у других видов — конъюгация — временное взаимодействие двух
особей, приводящее к реорганизации ядра и других частей кле-
ток-организмов и заканчивающееся разъединением конъюгирую-
щих особей. При конъюгации происходит обмен микронуклеусами
18
и в некоторых случаях цитоплазмой. В случае эндомиксиса реор-
ганизация ядерного аппарата происходит у единичных особей.
Таким образом, эти виды полового процесса или процессы,
физиологически и с точки зрения генетики соответствующие ему,
не приводят непосредственно к размножению; некоторые из них
приводят даже к уменьшению числа особей (при копуляции).
Лишь при определенных способах репродуктивного деления
многоклеточных половой процесс оказался неразрывно соединен-
ным с размножением.
Жизненные циклы одноклеточных растений и животных вклю-
чают различные типы агамной репродукции, половой процесс, а
также стадии инцистирования и эксцистирования. Инцистирова-
ние может совпадать со стадией зиготы, но может происходить
и в другой период, чаще всего совпадающий с наступлением
неблагоприятных условий для жизнедеятельности организма. У по-
давляющего большинства высших растений происходит последо-
вательная смена полового и бесполого размножения, характе-
ризующаяся разным соотношением стадий гаметофита и споро-
фита (см. гл. XIX).
Э. Бауэр в 1935 г. обосновал предположение, что при любой
форме полового процесса за счет энергии, выделяемой в резуль-
тате гибели части живой материи, происходит перестройка остаю-
щихся живыми структур, имеющих отношение к наследственности,
перезарядка потенциалов, в результате чего изменяется жизне-
способность организма. В самом деле, все виды полового и беспо-
лого размножения сопровождаются теми или иными «трупными»
явлениями: гибель полярных телец при созревании яиц, гибель
части живой материи при конъюгации инфузорий, а также в
связи с их делением и т. д. Об этом мы будем еще говорить
в главах о бесполом размножении и о биогенетическом законе.
В этой связи заслуживает внимания закономерно наступаю-
щая дегенерация клеточных элементов в яичниках и семенниках,
а также явления гибели животных вскоре после размножения,
например горбуши. Эти процессы тесно связаны с явлениями
репродукции, так как если удалить у той же горбуши половые
железы, то она не погибает и живет не один год, а до девяти
лет.
Принципиально такие же явления встречаются в жизни расте-
ний. Пальма Corypha umbracylifera, расцветая, дает начало
огромному соцветию и умирает. Индивидуальную жизнь этого
растения можно продлить, задерживая цветение путем отрезания
ветвей, образующих бутоны.
ПОЛОВЫЕ И СОМАТИЧЕСКИЕ КЛЕТКИ
Длительный период в развитии биологии (конец прошлого века
и первая четверть нашего) характеризовался резким противопо-
ставлением половых и соматических клеток: «фетишизированием»
19
9
Рис. 3, Яйцо курицы (по А. Романову и А. Романовой, 1949).
Белковые оболочки: 1 — наружная жидкая, 2 — плотный (белковый) мешок, 3 — халазифер-
ная, 4 — внутренняя жидкая, 5 — белковая связка, 6 — халаза. Слои желтка: 7 — желтый
желток, 8 — белый желток, 9 — бластодиск, 10 — желточная мембрана, 11— паидерово яд-
ро, 12 — канал латебры, 13 — латебра, 14, 15 — подскорлуповые оболочки, 16 — воздушная
камера, 17 — скорлупа, 18 — восковая кутикула
половых клеток, наделением их свойствами, которые на самом
деле им не присущи. Каковы основные отличия половых клеток
ст соматических?
1. Сперматозоиды и яйцеклетки имеют не диплоидный набор
хромосом, как это свойственно соматическим клеткам, а гапло-
идный, т. е. в два раза уменьшенное число'хромосом. Так, сома-
тические клетки пчелы имеют 32 хромосомы, а сформированные
половые клетки—16. Соматические клетки человека имеют
46 хромосом, а сперматозоиды и яйца — 23.
2. У половых клеток резко измененное по сравнению с сома-
тическими ядерно-плазменное отношение. Это очевидно на при-
мере яиц птиц (рис. 3). Собственно яйцом, яйцеклеткой птицы,
считается только «желток». Объем желтка яиц птиц в миллионы
раз превышает объем клетки, исходной в его развитии. Объем
же ядра столь резко не изменяется. И если из массы цитоплазмы
яйца птицы исключить вещества, которые как бы включены в нее.
«про запас», для развития зародыша (желточные включения),
и говорить о «чистой цитоплазме» (что не совсем правильно),
все равно очевидно, что масса яйца сильно возрастает в объеме.
То же наблюдается у яиц любого размера у всех животных,
будь то большие яйца головоногих моллюсков, имеющих размеры
до 10—15 мм в диаметре 1,5—2-миллиметровые яйца гидры и
лягушки (рис. 4) или яйца млекопитающих, зародыши которых
уже на ранних стадиях развития получают все необходимое от
20
Рис. 4. Яйцо (икриика)
лягушки:
Z, 2 — студенистая оболоч-
ка, 3 — анимальный полюс,
4 — вегетативный полюс
Рис. 5. Яйцо женщины (процесс оплодот-
ворения) (по А. Мюнтцингу, 1963).
Яйцо (2) окружено фолликулярными клетками;
видны сперматозоиды (3); некоторые проникли
через «барьер» из фолликулярных клеток; слева
видно полярное тельце (Z)
материнского организма. Например, у яйца женщины (100—
300 мкм в диаметре) имеется также резко измененное соотноше-
ние ядра и цитоплазмы (рис. 5). У пиявки Nephelis ядерно-плаз-
менное отношение в яйцевых клетках равно 0,15/428, а в сомати-
ческих оно составляет 0,3/3 и менее. Значит, ядерно-плазменное
отношение яйцеклетки по сравнению с телесными клетками в этом
случае изменяется приблизительно в 300 раз. Впоследствии при
развитии зародыша ядерно-плазменное отношение делящихся кле-
ток восстанавливается до свойственного соматическим клеткам.
Это происходит у разных животных в неодинаковое время, но
большей частью к 5—6-му или 7-му делению яйца.
Не менее «уродливые» изменения в соотношении цитоплазмы
и ядра происходят при образовании мужских половых клеток.
Как правило, сперматозоиды имеют очень мелкие размеры: у кро-
кодилов, например, эти клетки около 20 мкм, у человека —50—
70 мкм (рис. 6). Об их ничтожных размерах можно судить, если
представить, что сперматозоиды, оказавшиеся исходными в разви-
тии всех современных людей (три миллиарда), по объему равны
горошине. Имеются, впрочем, исключения: сперматозоиды одной
из амфибий — Discoglossus pictus — около 2 мм длиной.
Объем ядер сперматозоидов несколько меньший, чем у сома-
тических клеток, а «чистой цитоплазмы» у них очень мало. Грубо
говоря, сперматозоид — это ядро клетки плюс различные струк-
туры, обеспечивающие его встречу с яйцом (см. об этом далее).
По рис. 6 можно легко создать представление о соотношении
21
Рис. 6. Общий вид (Д) и схема строения ти-
пичного сперматозоида (5); продольный срез
головки сперматозоида (В):
1 — головка, 2 — промежуточный отдел, 3 — хвост,
4 —- акросома, 5 — ядро, 6 — центросома, /—-мито-
хондриальная спираль, 8 — осевая нить, 9 — цент-
ральное кольцо, 10 — акросомная мембрана, И —
наружные структуры ядра, 12 — полость, образую-
щаяся при инвагинации акросомной мембраны, 13 —
базальный кольцевой материал, 14 — перегородка
(простенок), связывающая базальное кольцо, 15—16—
материал, прилегающий к акросомной мембране,
17— осевое сгущение, 18 — мембраноподобная «муф-
та», окружающая осевую нить, 19 — плазматическая
мембрана, 20 — неоформленный материал — пред-
шественник акросомной инти
ядра и цитоплазмы,4 ес-
ли учесть, что головка
сперматозоида почти
целиком состоит из
ядра.
3. Известно, что
нормальные проявле-
ния жизни клетки, и
прежде всего ее мета-
болизм, могут осуще-
ствляться лишь при
тесной взаимозависи-
мости ядра и цито-
плазмы. Если мужские
и женские половые
клетки оказываются
крайне измененными по
сравнению с жизнедея-
тельными телесными
клетками, то следует
думать, что обмен ве-
ществ у них необыч-
ный. Высказано пред-
положение, что яйце-
вая клетка в отношении
обмена веществ нахо-
дится в состоянии де-
прессии, которое мож-
но уподобить, как пред-
полагает Б. П. Токин
(1955), анабиозу. Дис-
симиляторные и асси-
миляторные процессы
минимальны. У спер-
матозоидов столь ма-
лое количество цито-
плазмы и питательных
веществ, что если бы
им и был свойствен
нормальный метабо-
лизм, то он не мог бы
продолжаться долго.
В половых железах или в половых протоках самца спермато-
зоиды находятся в неподвижном и анабиотическом состоянии.
Оказавшись вне мужской половой системы, они, как правило,
живут очень короткое время. Так, сперматозоиды форели погиба-
ют в воде уже через 30 с, а у колюшки — через полчаса.
22
В половых путях самок длительность жизни сперматозоидов
у разных животных неодинакова. В яйцеводах кур сперматозои-
ды петуха живут 30—40 дней, у кролика 8—12 ч, в матке и яйце-
водах женщины — 5—8 дней (а вне организма, в семенной жид-
кости— 2—3 ч). Имеются кажущиеся исключения. В семяприем-
нике самок пчел сперматозоиды сохраняют свою жизнеспособность
2—2,5 года, но они находятся в неактивном состоянии, с едва
«теплющимся» метаболизмом. У летучих мышей спаривание про-
исходит в начале осени. Оплодотворения, однако, не происходит,
так как животные вскоре впадают в спячку, а с этим связано
сильное понижение температуры тела и другие неблагоприятные
для развития зародыша явления. Сперматозоиды сохраняются в
течение всего зимнего периода в половых путях самки и весной,
с возобновлением нормальной жизни, оплодотворяют яйцо. Имеют
место и паузы в развитии зародышей. Так, спаривание у косули
Capreolus vulgaris происходит в августе, незамедлительно начи-
нается развитие зародыша, но через несколько дней (на стадии
бластоцисты — иначе бластодермического пузырька, см. гл. VI)
развитие останавливается до ноября, после чего возобновляется
и идет беспрерывно до конца. Подобное явление наблюдается и
у некоторых других животных (например, у соболя) и называет-
ся диапаузой. Причины этих явлений еще недостаточно выясне-
ны, однако несомненно, что большую роль при этом играют тем-
пературные условия.
Н. А. Трипольская (1967) создавала условия, при которых
температура тела белых крыс снижалась за 6—7 ч до 20 °C.
Охлаждению подвергались крысы на 4-й день беременности, ко-
гда зародыш находится на стадии бластодермического пузырька
и еще не имплантируется в стенку матки. Охлаждение в этот
период вызывает состояние, подобное диапаузе, наблюдаемой, как
уже сказано, у некоторых животных в естественных условиях.
Возникает таким образом значительная задержка в имплан-
тации.
4. Долгое время среди большей части биологов было распро-
странено мнение о том, что половые клетки в отличие от сома-
тических— это малодифференцированные «тотипотентные» клет-
ки и что по мере развития организмов новые и новые клеточные
поколения становятся все более и более дифференцированными.
Это утверждение неправильное (см. гл. XI). Яйца и спермин —
высокодифференцированные клетки, обладающие многими специ-
альными, выработавшимися в процессе эволюции приспособле-
ниями для выполнения специфических функций: встречи мужских
и женских половых клеток—осеменения, защиты зародыша от
инфекций и вредных абиотических факторов. Яйцевые клетки
каждого вида животных характеризуются особой структурой
(«проморфологией»), от которой зависят особенности развитая орга-
низма; яйцевые клетки содержат те или иные вещества, требую-
23
A
Рис. 7. Схема развития оболочки у
ооцита в яичнике тритона (по Вартен-
бергу, 1962):
Л — до начала образования оболочки; Б—В —
в период образования оболочки; Г — зрелое
яйцо перед овуляцией (отростки фолликуляр-
ных клеток н мнкроворсинкн ооцита втяну-
лись); / — отростки фолликулярных клеток с
десмосомными участками связи между ооци-
том и фолликулярными клетками. 2-мнкро-
ворсники ооцита, 3 — формирующаяся яйце-
вая оболочка
щиеся для метаболизма раз-
вивающегося эмбриона, и т. д.
(см. гл. XI).
5. Измененный по сравне-
нию с соматическими клетка-
ми обмен веществ, наличие спе-
циальных структур, мешаю-
щих нормальному протеканию
жизненного цикла клетки (ее
онтогении), — все это приво-
дит к тому, что сперматозои-
ды оказываются неспособны-
ми нормально, как это свой-
ственно большинству сомати-
ческих жизнедеятельных кле-
ток, развиваться и доходить
в своем развитии до итогового
этапа онтогении клетки — ми-
тоза. Делящихся сперматозои-
дов ни у одного вида живот-
ных никто не наблюдал. Яйце-
вые клетки также не могут
делиться без влияния особых
внешних факторов, а именно:
если не состоится оплодотво-
рение или если яйцо не будет
активировано к развитию пар-
теногенетическими агентами
(см. гл. III).
6. Иллюстрацией высокой
специализации и дифференциа-
ции половых клеток могут слу-
жить оболочки яйцевых кле-
ток. За редкими исключениями
(губки, некоторые кишечнопо-
лостные и моллюски) оболоч-
ки как морфологические обра-
зования имеются у яиц всех
животных.
В зависимости от происхо-
ждения различают первичные,
вторичные и третичные обо-
лочки.
Первичные оболочки яиц
возникают путем секреции
ооцитом веществ, формирую-
щих оболочку. Это, как пра-
24
вило, очень тонкая пленка, непосредственно контактирующая
с плазматической мембраной ооцита. Она может быть толстой и
прочной (у некоторых животных) . Чаще всего первичную оболочку
называют желточной.
Электронно-микроскопические исследования свидетельствуют
о большой общности в строении и процессах формирования жел-
точной оболочки у яиц разных позвоночных (рис. 7). С момента
возникновения и по мере ее развития желточная оболочка перфо-
рируется отростками фолликулярных клеток, соприкасающихся
с поверхностью яйца. Таким образом, несмотря на возникновение
между фолликулярными клетками и ооцитом желточной оболоч-
ки, контакт ооцита и фолликулярных клеток сохраняется. На по-
верхности ооцита в свою очередь образуется огромное количество
микроворсинок, вдающихся во внутреннюю часть желточной обо-
лочки и в 10—20 раз увеличивающих поверхность ооцита. Образо-
вание желточной оболочки обычно совпадает с тем моментом
оогенеза (период большого роста ооцита), когда происходит интен-
сивное поступление питательных веществ в ооцит (см. с. 44). Обра-
зование отростков фолликулярными клетками и микроворсинок
на поверхности ооцита служит своего рода адаптацией,
облегчающей транспорт в ооцит питательных веществ, несмотря
на образование в этот период желточной оболочки, пред-
ставляющей своеобразный барьер на пути веществ, поступаю-
щих в яйцеклетку. К концу периода большого роста незадолго
до овуляции (к этому времени интенсивность питания ооци-
та понижается) отростки фолликулярных клеток, как и микро-
ворсинки ооцита, втягиваются, пористость желточной оболочки
при этом либо исчезает, либо сохраняется в виде радиальной
исчерченности оболочки, столь характерной для оболочек яиц по-
звоночных.
Вторичные оболочки яиц, или хорион, возникают за счет фол-
ликулярных клеток яичника либо путем секреции ими вещества
оболочки, либо из самих фолликулярных клеток, сохраняющих-
ся на поверхности ооцита при овуляции. Примерами вторичных
оболочек могут служить хорион яиц насекомых, вторичные оболоч-
ки яиц рыб, птиц. У насекомых хорион хитиновый и нередко очень
толстый.
Третичные оболочки очень разнообразны по строению, хими-
ческому составу и значению. Образуются они при прохождении
яйца по яйцеводам за счет секреторной деятельности желез в
стенке яйцевода или желез, расположенных за пределами поло-
вого тракта. Примеры третичных оболочек яиц — это галлерта,
одевающая яйцевые клетки в кладках моллюсков, студенистые
оболочки яиц амфибий, белковая, подскорлуповая и скорлуповая
оболочки яиц птиц, коконы многих червей и моллюсков. К тре-
тичным оболочкам относится также и белковая жидкость внутри
кокона, в которой находится не одно, а несколько яиц. Присут-
25
ствие всех трех типов оболочек встречается не у всех яиц; яйц®
может иметь лишь одну или две из названных оболочек (первич-
ную и вторичную или вторичную и третичную).
Строение яйцевых оболочек животных обычно соответствует
тем условиям, в которых происходит развитие ооцита, а затем
и зародыша и в связи с этим разнообразно, в особенности когда
речь идет о вторичных и третичных, т. е. наружных, оболочках
яиц яйцекладущих животных (хорион насекомых, студенистые
оболочки яиц, развивающихся в воде, кожистые и скорлуповые
оболочки яиц рептилий и птиц и т. д.). Очень разнообразно, на-
пример, строение оболочек яйцевых клеток рыб: оно связано с
неодинаковыми экологическими условиями, в которых протекает
нерест и развитие зародыша у разных видов рыб. У плацентар-
ных млекопитающих, развитие зародышей которых происходит в
матке, в относительно сходных условиях, строение оболочки яйца
(zona pellucida) у представителей различных видов довольно
однообразно.
Функции, выполняемые оболочками яиц, многообразны.
У ооцитов и развивающихся зародышей через оболочки идет про-
цесс обмена, осуществляется связь с внешней средой. Значитель-
на роль оболочки яйца в связи с оплодотворением; они могут
предотвращать или ограничивать полиспермию (см. с. 58).
На ранних этапах эмбриогенеза оболочки яйца обеспечивают
сохранение зародышем присущей ему формы, что важно для
нормального хода формообразовательных процессов; у некоторых
животных вещества оболочек могут играть роль в питании за-
родыша в определенные периоды его развития, например, заро-
дыш курицы во второй половине эмбрионального развития за-
глатывает остатки белковой оболочки яйца.
Из всех многообразных функций, выполняемых оболочками
яйца, наиболее общая и значительная — защитная. Она особенно
важна у яйцекладущих животных, развитие которых протекает
вне организма матери. Яйцевые оболочки могут предохранять
зародыш от разнообразных повреждений физического порядка,
связанных с колебаниями в окружающей среде: температуры,
влажности, химического состава, от повреждений механического
характера, а также от неблагоприятного воздействия биотических
факторов среды, предотвращая возможные инфекции зародыша,
что особенно важно на ранних этапах развития. Так, у ряда живот-
ных, исследованных в этом отношении (амфибии, птицы, репти-
лии), оболочки яиц обладают антибиотическим действием в отно-
шении различных микроорганизмов, выполняя функцию защиты
эмбрионов от инфекций (см. гл. XV).
Таким образом, половые клетки — очень сложные, высоко-
дифференцированные клетки. Специальные особенности структу-
ры яиц способствуют реализации их возможности развиваться в
целый организм. Ядра яиц и сперматозоидов в генетическом
26
смысле обеспечивают развитие признаков, свойственных матери
и отцу.
Долгим заблуждением в науке было представление об этих
свойствах половых клеток, как о чем-то исключительном, таком,
что принципиально отличает их от соматических клеток. В гл. XVI
и XVII будут приведены многочисленные доказательства того,
что нормальные, жизнеспособные телесные клетки в генетическом
отношении не беднее половых. Наоборот, они богаче, так как
содержат диплоидное число хромосом. А с эмбриологической
точки зрения они при определенных экспериментальных или при-
родных условиях могут быть аналогами половых клеток и раз-
виваться в целый организм. Так происходит при бесполом раз-
множении, естественной и экспериментальной полиэмбрионии, со-
матическом эмбриогенезе. Эти явления будут описаны ниже. Пока
ограничимся ярким примером из мира растений. Клетки листьев
бегонии Begonia rex являются интегральной частью тканей, а
при особых условиях они могут развиваться в новый организм
с его воспроизводительными органами; при этом развившийся
организм окажется с теми видовыми и индивидуальными наслед-
ственными свойствами, какие характеризуют и исходное растение.
Таким образом, несмотря на многие отличия половых клеток
от соматических, между ними нет пропасти, о которой думали
многие ученые до 30-х годов нашего столетия, исходя из предвзя-
тых концепций наследственности и развития (А. Вейсман и др.),
ТЕОРИЯ ЗАРОДЫШЕВОГО ПУТИ: СТРАНИЦЫ ИЗ ИСТОРИИ
ЭМБРИОЛОГИИ, цитЬлогии И ГЕНЕТИКИ
Процессы формирования мужских половых клеток (сперматоге-
нез) и женских (оогенез) имеют много сходного в поведении
ядер клеток и очень большие различия в отношении преобразо-
вания других структур. Поэтому в дальнейшем будут затронуты
как общие вопросы гаметогенеза (происхождения и развития
половых клеток), так и некоторые специальные, касающиеся или
только яиц, или только сперматозоидов.
Чтобы выяснить важность вопросов происхождения половых
клеток для эмбриологии и генетики, остановимся сначала на ис-
тории вопроса.
А. Вейсман в 1885—1896 гг. создал теорию «непрерывности
зародышевой плазмы». Существуют якобы две плазмы: сомати-
ческая, участвующая в развитии тела организма, и зародышевая,
передающаяся непрерывно от половых клеток одного поколения
к половым клеткам каждого нового поколения. Носителями жиз-
ни являются, по Вейсману, «биофоры» — группы молекул органи-
ческих веществ. Они находятся в ядре клеток и через поры ядра
могут переместиться в цитоплазму. Ядру принадлежит активная
роль, а цитоплазма, ее поведение и дифференциация, структура и
27
отправления зависят от биофор, точнее — от групп биофор — де-
терминантов (определителей). В зародышевой плазме находится
полный набор детерминантов; их столько, сколько типов однород-
ных клеток у взрослого организма. Детерминанты, как и биофоры,
растут и размножаются (делятся), так что для любого числа од-
нородных клеток определенной ткани достаточно одного исход-
ного детерминанта. Все детерминанты, определяющие развитие
всех признаков организма, сгруппированы в ядре в иды. Во время
митоза иды слагаются в иданты, соответствующие структурам
хромосом. Таким образом, по Вейсману, наступающие в развитии
организма различия между клетками происходят путем сортиров-
ки наследственных единиц (детерминантов). В ходе эмбриональ-
ного развития в связи с делением клеток детерминанты распреде-
ляются по различным клеткам, происходит постепенная растрата
их, обеднение ими у новых и новых поколений специализирующих-
ся клеток. Только половые клетки имеют полный набор детерми-
нантов, а потому они и способны развиваться в целый организм.
Как увидим в гл. XII, некоторые идеи Вейсмана в той или
иной форме возрождаются и в новейшее время, а именно мысль о
неидентичности ядер клеток разного типа тканей. В целом же эта
чисто умозрительная теория с ее не основанным на эмпирических
фактах противопоставлении «сомы» и «половой плазмы» пред-
ставляет только исторический интерес. В 30-х годах один из осно-
вателей современной генетики Т. Морган, а до него эмбриологи
решительно отказались от этой концепции, так как было доказано,
что при митозе каждая из сестринских клеток получает полный
набор хромосом (а значит, и вейсмановских гипотетических опре-
делителей). Кроме того, была доказана возможность развития
целых организмов из изолированных бластомеров — стройная и
формально логичная теория Вейсмана оказывала длительное
время большое влияние на генетические и эмбриологические по-
строения; она импонировала и многим эмбриологам, например
В. Ру, создавшему представление о зародыше как о «мозаике
бластомеров». На рубеже нашего столетия теория Вейсмана, ка-
залось, получила и превосходное цитогенетическое подтверждение.
Перед Вейсманом вставали трудные вопросы. Соматические клет-
ки по мере развития организма все более обедняются потенция-
ми, растрачивают свои детерминанты. Каким же образом из обед-
ненных телесных клеток возникают в яичниках и семенниках
половые клетки с их полным набором детерминантов? В 1899 г.
появилось исследование Бовери, которое, казалось, разрешало
это тяжелое для теории Вейсмана противоречие. Бовери обнару-
жил, что уже первые два бластомера яйца лошадиной аскариды
Ascaris megalocephala отличаются друг от друга (рис. 8,А). При
втором дроблении в одном из бластомеров Si (АВ), исходном для
эктодермы, происходит диминуция (уменьшение) .хроматина бла-
годаря отбрасыванию дистальных концов хромосом в цитоплазму;
28
Рис. 8. Дробление яйца Ascaris megalocephala (по О. Цур-Штрассеиу):
Д— стадия двух бластомеров с веретенами второго дробления; Б, В, Г — стадии четырех
бластомеров и их последующего перемещения. Заштрихованы бластомеры, дающие пер-
вичную эктодерму, в инх видна диминуция хроматина (по Т. Боверн, 1899): Д, Е, Ж—ста-
дии 16 бластомеров (Д — вид с брюшной стороны, Е— справа. Ж — слева)
в другом бластомере Pi — родоначальнике будущих внутренних
органов и половых клеток — диминуции хроматина нет. И при
последующих дроблениях бластомеры Pi, Р2, Рз и Р4 оказывают-
ся без диминуции, и именно эти клетки являются исходными для
всех будущих сперматозоидов или яиц. Выходит, что уже на ста-
дии 16 бластомеров имеется половой зачаток — гонобласт. Это
было триумфальным цитогенетическим подтверждением теории
Вейсмана. Была обоснована теория особого зародышевого путы
(Keimbahn), разделявшаяся
ологов. Отголоски этой тео-
рии в той или иной форме
имеются и в новейшее
время.
Как справедливо обра-
тил внимание П. Г. Светлов,
грубый перевод с немецкого
Keimbahn — зародышевый
путь — лучше заменить на-
званием «линия половых кле-
ток». На рис. 9 дана схема
линии, половых клеток на
долгое время большинством эмбри-
примере развития аскариды. рис g схема «зародышевого пути» у аска-
ЧерНЫМИ кружками изобра- риды. Пояснения см. в тексте
29
жен ряд бластомеров, в которых на протяжении пяти первых дроб-
лений не происходит диминуции хроматина. После шестого дробле-
ния они образуют гоноциты. Другие бластомеры и их производ-
ные, изображенные на рисунке выше черных кружков, делятся
с диминуцией хроматина и являются поколениями соматических
клеток.
Исследования, касающиеся диминуции хроматина, продолжа-
ются и в настоящее время. Показано, что у некоторых насекомых
(Coleoptera и Diptera) в соматических клетках элиминируются
целые хромосомы (Е-хромосомы), и в ядрах, попадающих в об-
ласть «полярной плазмы» на вегетативном полюсе яйца, сохраня-
ется полный набор хромосом. Повреждение области полярной
плазмы (например, лучами Рентгена) ведет к развитию стериль-
ных особей.
Ж. Гердон и X. Вудленд (1968) не только подтверждают ди-
минуцию хроматина у лошадиной аскариды, но и доказывают, что
она происходит под цитоплазматическим контролем. К. Мориц
(1967) рентгенизовал цитоплазму (но не ядро) Pi клетки. Это при-
водит к диминуции в обеих дочерних клетках, т. е. и в презумп-
тивной половой клетке. Автор делает вывод: половая плазма
(плазма линии половых клеток) содержит фактор, защищающий
хромосомы от диминуции.
Представления об отсутствии принципиальных различий между
половыми и соматическими клетками прочно обоснованы фактами
эмбриологии и генетики. Однако дискуссии по этому поводу не
утихают на протяжении более чем ста лет со времени постановки
этой проблемы в работах В. Вальдейера (1870), М. Нуссбаума
(1880), А. Вейсмана (1885, 1896) и др. В современной биологии
имеются две точки зрения по вопросу о соотношении половых и
соматических клеток.
Одни исследователи считают, что формирование половых кле-
ток тождественно процессам дифференцировки клеток в любых
иных направлениях, а все особенности половых клеток связаны со
спецификой их дифференцировки, обусловленной репродуктивными
функциями: они отрицают непрерывность линии половых клеток
в ряду поколений, но не отрицают реального факта раннего обо-
собления их в онтогенезе многих животных, служащих единствен-
ным источником для образования половых клеток организма.
Другие исследователи придерживаются представления о непре-
рывности в ряду поколений линии половых клеток, считая их раз-
витие особым процессом, не имеющим подобия с другими явлени-
ями дифференциации. Эти исследователи утверждают о сущест-
вовании фундаментальных различий между соматическими и по-
ловыми клетками, состоящих в сохранении последними (как и их
предшественниками в онтогенезе) качества тотипотентности. Воз-
никновение половых клеток они связывают с теми клетками за-
родыша или организма, в которых в результате ооплазматической
за
сегрегации окажется сосредоточенной зародышевая плазма, де-
терминирующая линию половых клеток и их предшественников
в онтогенезе. (Зародышевая плазма по современным представле-
ниям в отличие от Вейсмановских определяется как совокупность
цитоплазматических факторов, детерминирующих линию непре-
рывных в ряду поколений тотипотентных клеток, в том числе и
половых.) Природа зародышевой плазмы пока не известна, хотя
ее маркеры — половые детерминанты, выявляются визуально
в ооцитах многих животных на светооптическом или ультраструк-
турном уровне. Однако даже у животных с наиболее четко выра-
женными половыми детерминантами (Diptera, Anura) структуры
эти не непрерывны в онтогенезе. Функция зародышевой плазмы
предположительно состоит в предохранении генома клеток
и их предшественников от соматизации, т. е. в обеспечении сохране-
ния первичными половыми клетками их исходных потенций. Этим
постулируется представление о непрерывности в ряду поколений
линии тотипотентных клеток, заменившее сегодня представление
А. Вейсмана о непрерывности «зародышевого пути», но по сущест-
ву возрождающее его в несколько измененной форме. Эта концеп-
ция доступна критике.
Прежде всего следует помнить, что под тотипотентностью кле-
ток понимают скрытую, но сохранившуюся ими способность дать
при развитии целый организм. Такими потенциями при половом
размножении Metazoa обладает зигота, а у животных с регуля-
тивным типом дробления — бластомеры в ходе нескольких первых
делений дробления. В эмбриогенезе по мере развития потенции
всех клеток, не исключая и половые, все более суживаются (хотя
утрата ими тотипотентности во многих случаях сопровождается
сохранением эквипотенциальности ядер). Ни первичные половые
клетки, ни половые клетки всех этапов дифференцировки (ни го-
нии, ни гаметоциты, ни, тем более, зрелые половые клетки — яйца
и сперматозоиды) не являются тотипотентными. С известной на-
тяжкой можно говорить о тотипотентности зрелых яиц у некоторых
животных в смысле их способности к партеногенезу; при этом
следует помнить, что партеногенез при половом размножении —
вторичное явление. Тотипотентность — это качество, воссоздавае-
мое каждый раз с появлением зиготы и утрачиваемое клетками
зародыша, в том числе и половыми, в ходе их специализации.
Половые клетки — это продукт организма, одноклеточного или
многоклеточного, а их детерминация и дифференцировка всегда
происходит под интегративным воздействием организма как це-
лого.
Из сказанного ясно, что для утверждения тезиса непрерыв-
ности тотипотентных клеток ни в онтогенезе, ни, тем более, в ряду
поколений не имеется достаточных оснований. Современные пред-
ставления о «зародышевой плазме», в известной степени возрож-
31
дающие гипотезу Нуссбаума—Вейсмана о непрерывности «заро-
дышевого пути» в трансформированной новыми данными форме,
спорны.
СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ О ПРОИСХОЖДЕНИЙ
ПОЛОВЫХ КЛЕТОК В ОНТОГЕНЕЗЕ
При описании процессов эмбрионального развития можно гово-
рить, конечно, о линии половых клеток, не вкладывая в это
выражение метафизического содержания, ие противопоставляя
«бессмертную половую плазму» «смертной соме». У некоторых
животных половой зачаток действительно образуется очень рано,
но столь же рано обособляются и зачатки других органов и си-
стем. На той же стадии развития аскариды, а именно на стадии
16 бластомеров (см. рис. 8,Д, Е, Ж), на которой обособляется по-
ловой зачаток, не менее отчетливо и обособление зачатка энто-
дермы (средней кишки). Бластомеры mst и рот при дальнейшем
развитии дают зачатки стомодеума, или эктодермальной глотки
(st, от); т и ц— первичные зачатки мезодермы; с дает эктодерму
хвостовой части. Более того, на стадии 4 бластомеров (см.
рис. 8,5, В, Г) судьба бластомеров А и В ясна: они дадут экто-
дерму зародыша, a EMSt явится родоначальником энтодермы, ме-
зодермы и глотки.
Таким образом, пет оснований придавать какое-то особое зна-
чение понятию «зародышевый путь» в отличие от «пути на экто-
дерму», от «пути на стомодеум» или от «пути на глаз».
Столь ранних случаев обособления полового зачатка, какое
наблюдается у аскариды, очень немного (Sagitta, циклоп, некото-
рые насекомые). У большинства животных (у всех позвоночных)
половой зачаток развивается сравнительно поздно. Формирование
половых клеток и половых желез — семенников у самцов и яични-
ков у самок — является не более таинственным, чем любой другой
органогенез.
Еще в XIX в. появились различные взгляды на вопрос о том,
из каких клеток происходят половые клетки, являются ли они но-
выми и новыми поколениями исходных первичных половых кле-
ток, которые обособляются на определенных этапах эмбриональ-
ного развития, или они могут развиваться из клеток каких-либо
тканей половых желез.
Как уже говорилось, вопросы происхождения половых клеток
у животных связаны с конкретными, неодинаковыми для разных
животных вопросами развития половых желез — семенников и
яичников. Зачаток половых желез у животных возникает по-раз-
ному. Во многих случаях он образуется вместе с перитонеальными
стенками целомических мешков. У животных с ранним обособле-
нием первичных половых клеток (оно происходит еще до возник-
новения мезодермы) перитонеальные клетки, покрывая слоем по-
32
ловые клетки, также принимают участие в развитии половой же-
лезы, являясь, в частности, источником образования фолликуляр-
ных клеток *.
Нет оснований думать, что вопрос о происхождении половых
клеток может быть решен однозначно для всех организмов. В сов-
ременной биологии имеются две точки зрения. Одни исследователи
считают, что формирование половых клеток тождественно процес-
сам дифференциации клеток в любых иных направлениях. Другие
стоят на точке зрения непрерывности линии половых клеток, счи-
тая это особым процессом, не имеющим подобия с другими явле-
ниями дифференциации. Считают даже, что «процесс зародышево-
го пути» — это не дифференциация, а сохранение первоначального
генома клетки без какой-либо существенной активации его частей.
Обратимся к фактам.
У губок нет непрерывности зародышевой плазмы: первичные
половые клетки могут развиваться у них из амебоцитов или хоа-
ноцитов. О. Тюзе (1962) утверждает, что половые клетки происхо-
дят у известковых и кремниевых губок или из хоаноцитов сфор-
мированной губки или из археоцитов (из амебоцитов с крупным
ядром). Процессы оогенеза и сперматогенеза происходят у губок
также, как и у других многоклеточных.
В отношении кишечнополостных также нет оснований говорить
о «линии половых клеток». У гидр и других кишечнополостных нет
постоянных семенников и яичников. Их половые клетки могут
возникать из интерстициальных клеток, а также из зимогенных
клеток гастродермиса (А. Бернет и др., 1966). Развившиеся (в эк-
спериментальных условиях) из фрагмента гастродермиса целые
гидры нормально почкуются; через 7 недель в популяциях подоб-
ных гидр появляются и особи с гонадами.
У полипа Clava squamata все половые клетки концентрируются
в гонофорах. Если отрезать зону гонофора «с гроздьями» половых
клеток под гипостомом, то при регенерации нового гипостома со
щупальцами восстанавливается и весь половой аппарат за счет
присутствующих в теле полипа интерстициальных клеток.
Л. М. Лосева (1970) доказала, что женские половые клетки
у актинии Bunodactis stella образуются заново в каждый период
половой активности непосредственно из эпителиальных клеток
гастродермиса (септ). В оогенезе отсутствует стадия оогониальных
делений. Зародыши и молодые животные не содержат клеток, ко-
торые можно было бы считать половыми. Созревание половых
клеток происходит асинхронно; в одном мезентерии можно встре-
тить ооциты, находящиеся на разных стадиях развития (рис. 10).
У некоторых червей возможна регенерация гонад из участков
тела, в которых отсутствуют половые элементы. Это наблюдается
1 Современные данные о происхождении половых клеток см. в кн.:
Айзенштадт Т. Б. Цитология оогенеза. М., 1984.
3»
Рис. 10. Часть мезентерия Bunodactis stella с
ооцитами (по Л. М. Лосевой, 1970)
у планарий, а среди
полихет — у серпу-
лид; наблюдается
это и у многих оли-
гохет. Принципи-
альный интерес пред-
ставляет то, что у
этих животных по-
ловые клетки, обо-
собляются иа ранних
стадиях эмбрио-
нального развития;
можно говорить о
«линии половых
клеток», но сомати-
ческие клетки спо-
собны восстанавли-
вать половые элементы, которые, безусловно, утрачиваются в ре-
зультате ампутации.
Д. Бьерн (1962) исследовал регенерацию гонад у немертины
L. ruber. У животного отрезали передние фрагменты на уровне
передней кишки (позади рта). На 7-й день после операции у реге-
нерирующих фрагментов L. ruber появлялись оогонии и спермато-
гонии. Источником их служат свободные клетки паренхимы.
У мшанок «обособление половых клеток» происходит на позд-
них стадиях развития. При бесполом размножении и регенерации
из соматических клеток у них возникают половые клетки.
Членистоногие дают нам многочисленные и яркие примеры
раннего обособления линии половых клеток.
В поверхностном слое цитоплазмы (периплазме), в области
заднего полюса имеется морфологически отличимый участок —
«оосомы». При делении ядер, перемещении их в периплазму и
формировании бластодермы особая судьба выпадает на долю
тех ядер, которые окажутся в оосоме. Окружающая их цитоплазма
разрастается, затем отделяется от остальной части яйца, образуя
полярные клетки. У Drosophila melanogaster образование поляр-
ных клеток происходит на этапе развития зародыша, когда он
состоит уже из 256 клеток и более. В цитоплазме заднего полюса
яйца имеются какие-то условия, способствующие формированию
гоноцитов (т. е. первичных половых клеток).
Пока не установлены надежные признаки первичных половых
клеток у рыб, их происхождение остается неясным. Большинство
авторов в настоящее время считают, что в течение всей жизни
рыбы в половой железе сохраняются потомки первичных половых
клеток, являющихся основным источником новых генераций как
оогоний, так и сперматогоний.
Невозможно сомневаться в наличии линии половых клеток
34
у всех позвоночных, однако споры по этому вопросу не прекра-
щаются, к тому же эмбриология не располагает достаточными
фактическими данными, например о рыбах.
Многие эмбриологи придерживаются «дуалистических» взгля-
дов на развитие половых желез у позвоночных. Они считают, что
соматические ткани яичника и семенника всегда мезодермального
происхождения, а первичные половые клетки являются дериватами
различных зародышевых листков и впоследствии мигрируют к за-
чатку половой железы (часто благодаря амебоидным движени-
ям). Об этом имелись данные еще в 30-х годах.
У амфибий первичные половые клетки, окруженные мезенхи-
мой, могут быть обнаружены в микроскопических участках над
формирующейся брыжейкой, откуда они перемещаются в мезодер-
мальный эпителий, участвующий в развитии половой железы.
У птиц и рептилий первичные половые клетки были найдены на
ранних стадиях развития, перед началом сегментации мезодермы
во внезародышевой желточной энтодерме. Клетки эпителиального
типа обрастают здесь нераздробившуюся массу желтка. Они круп-
нее, чем обычные мезодермальные, содержат большое, более или
менее вакуолизированное ядро, их протоплазма отличается от та-
ковой окружающих клеток своим отношением к красителям, фор-
ма их близка к сферической. Сам по себе факт присутствия пер-
вичных половых клеток во внезародышевой желточной энтодерме
еще не доказывает, что именно они или их потомки окажутся позд-
нее составной частью функционирующей половой железы.
Считается, однако, доказанным, что «первичные половые клет-
ки» мигрируют в мезенхиме и по кровеносным сосудам в область
целомической мезодермы, где происходит закладка гонад. В. Дан-
чакова в 1932 г. выжигала у куриного эмбриона зачатковый серп.
В результате операции в закладке гонады развивающегося эмбри-
она пролиферация зачаткового эпителия не обнаруживалась. Рас-
полагаются ли «первичные половые клетки» в пределах зачатко-
вого эпителия или во всей толще гонады? Гибнут ли вскоре все
эти клетки, сыграв свою роль при формировании половой железы,
или часть их становится родоначальником будущих половых кле-
ток? Эти вопросы в достаточной степени еще не выяснены. Наибо-
лее точными данными экспериментальная эмбриология располага-
ет в отношении птиц и амфибий, и нет сомнений в том, что без
мигрирующих в зачаток гонады «первичных половых клеток» яйца
или спермин не формируются.
Несмотря на обилие исследований о развитии яичников и се-
менников у млекопитающих, вопрос о происхождении половых
клеток у них далеко еще не ясен.
Согласно данным одних авторов с момента прекращения мито-
тического размножения первичных половых клеток у плода ново-
образования яйцевых клеток не происходит. Другие исследователи
утверждают, что оогенез продолжается в течение всего репродук-
35
тивного периода жизни самки. Подсчитано (Т. Бэкер, 1963), что
число половых клеток у двухмесячного плода человека около
600 000. К пятому месяцу развития — 6 800 000. Позднее происхо-
дит "массовая дегенерация клеток, так что ко времени рождения
остается около 1 млн. нормальных ооцитов. Из этого количества
примерно 300 000 сохраняется к семилетнему возрасту ребенка.
Многие ученые уверены в том, что первичные половые клетки
полностью или частично дегенерируют и происходит формирова-
ние половых клеток из клеток герминативного эпителия. Описано
новообразование яйцевых клеток из герминативного эпителия
в разные периоды жизни (после рождения) у мышей, крыс, мор-
ских свинок, кошки, собаки, обезьяны и человека. Некоторые ис-
следователи уверены в том, будто все клетки герминативного эпи-
телия являются потенциальными яйцеклетками.
Однако, по данным многих авторов, после закладки половых
желез ооциты не могут образовываться из клеток герминативного
эпителия, и абсолютно все ооциты происходят в результате раз-
множения первичных половых клеток. Оогенез у млекопитающих,
согласно мнению этих авторов, заканчивается на поздних стади-
ях развития зародыша или вскоре после рождения.
Исследования А. Г. Кнорре и А. Г. Семеновой-Тян-Шанской
(1972, 1973 и др.) на позвоночных, зародышах человека позволили
с полной уверенностью говорить о внегонадном возникновении
первичных половых клеток у высших позвоночных, о миграции их
в закладку гонады. Вследствие этого, по мнению авторов, в отно-
шении позвоночных надо отбросить понятие «зачатковый эпите-
лий» и отказаться от мысли, будто есть эпителий — источник об-
разования половых клеток. Имеет место раннее «обособление» ли-
нии половых клеток. Можно говорить о специальном половом
зачатке — гонобласте.
Действительно, существуют многочисленные данные о мигра-
ции первичных половых клеток, об активном движении гоноци-
тов через массу мезенхимных клеток (движение амебоидного ха-
рактера), а также достоверные данные о переносе половых клеток
с кровотоком.
Местом первичной локализации гоноцитов эмбриона человека
служит краниальная зона зародышевого щитка (А. Г. Семенова-
Тян-Шанская, 1973). Первичные половые клетки на разных ста-
диях развития человека оказываются в разных частях зароды-
ша — они попадают в мезодерму первичной полоски, оказываются
в составе кровяных островков, желточных сосудах, впоследствии
в выстилке задней кишки, эндотелиальной закладке сердца, и дру-
гих образованиях.
В гонадах позвоночных гонии располагаются скоплениями —
«гнездами». Позже вокруг каждой половой клетки за счет сома-
тических элементов гонады формируется фолликул (от лат.
folliculus — мешочек). Стенка фолликула состоит из фолликуляр-
36
ного эпителия, прилегающего к поверхности половой клетки и со*
единительно-тканной теки, в которой разветвляются кровеносные
капилляры, питающие заключенную в фолликуле половую клетку.
В оогенезе фолликул формируется по завершении гениальных де-
лений и, как правило, содержит одну половую клетку — ооцит I
порядка. В сперматогенезе формирование фолликула происходит
задолго до завершения гениальных делений, продолжающихся
внутри сформированного фолликула, благодаря чему семенной
фолликул содержит большое количество мужских половых клеток.
Кроме того, группы половых клеток внутри семенного фолликула
могут быть заключены в сперматоцисты; стенка каждой сперма-
тоцисты образована одной или несколькими фолликулярными
клетками. Гомологом семенных фолликулов низших позвоночных
у Amniota являются семенные канальцы. Находясь в контакте
с половыми клетками, фолликулярный эпителий выполняет роль
посредника в их взаимоотношениях с родительским организмомг
А
Рис. 11. Схема изменений трансформирующегося фолликулярного эпителия в
оогенезе пеляди Corregonus peled (по И. Д. Чииаревой, 1975):
А — период цитоплазматического роста ооцита; В — период трофоплазмэтического роста
ооцита; В —в конце вителлогенеза; / — базальная мембрана, 2 — фолликулярная клетка,
3 — субфолликулярное пространство, 4 — отростки фолликулярных клеток, 5 — микроворсин-
ки ооцита, 6 — zona radiata, 7 — поверхность ооцнта
37
регуляторных, трофических и иных. На определенных этапах га-
метогенеза фолликулярный эпителий несет гормональную фунК’
дню; в случае гибели половых клеток осуществляет их фагоцитоз;
продуцирует вторичные яйцевые оболочки.
Особенно велика роль фолликулярного эпителия в оогенезе
в связи со значительным ростом ооцита и формированием яйцевых
оболочек. В юных фолликулах фолликулярный эпителий состоит
из одного слоя сильно уплощенных, функционально малоактивных
клеток. На протяжении оогенеза фолликулярный эпителий позво-
ночных претерпевает значительные морфофизиологические пре-
вращения. Предложено различать четыре вида фолликулярных
эпителиев у позвоночных (Н. С. Габаева, 1970)—однослойный
статически плоский, в ходе оогенеза почти не изменяющийся (бес-
хвостые амфибии); трансформирующийся изоморфный, приобре-
тающий в ходе оогенеза строение кубического, столбчатого или
ложномногорядного (акулы, костистые рыбы, птицы); трансфор-
мирующийся анизоморфный, клетки которого в ходе оогенеза диф-
ференцируются в двух направлениях—на собственно эпителиаль-
ные и железистые (скаты, чешуйчатые рептилии), и многослойный
изоморфный — примером его служит фолликулярный эпителий
плацентарных млекопитающих. Предполагается, что наиболее
примитивным, исходным в эволюционных изменениях фолликуляр-
ного эпителия можно считать статически плоский, свойственный
низшим вторичноротым. При овуляции стенка фолликула разры-
вается и яйцо освобождается в полость гонады либо в полость
тела, откуда попадает в выводные пути половой системы. Иллюст-
рацией изменений транформирующегося фолликулярного эпителия
может служить рис. 11,Б, В.
СПЕРМАТОГЕНЕЗ И ООГЕНЕЗ
Начало онтогенеза связывают с моментом образования зиготы,
процессы же, предшествующие образованию зародыша — возник-
новение первично-половых клеток в организме, размножение и
дифференцировка половых клеток — гаметогенез, а также оплодо-
творение,— объединяют понятием проэмбриональное развитие,
или прогенез.
Гаметогенез — формирование яиц и сперматозоидов — изучает-
ся кроме эмбриологии в разных аспектах многими науками, в осо-
бенности цитологией и генетикой. Остановимся на некоторых воп-
росах гаметогенеза. Весь процесс развития половых клеток можно
разделить на ряд периодов (рис. 12), характеристика которых бу-
дет дана позднее.
Схема строения семенника человека и млекопитающих дана на
рис. 13,-4. В многочисленных извитых трубочках (семенных ка-
38
Сперматоцит
1-го порядка
Первое полярное
Сперматиды
Вторая метафаза
мейоза
Первая метафаза меойза
Конъюгация гомологичных
хромосом
да
Ооцит 2-го
порядка,
Рис. 12. Образование сперматозоидов (А) и яйцеклеток (5) у животных и
человека (по А. Мюнтциигу, 1963)
Вторичные
полярные
тельца
Сперматозоиды
Яйцеклетка
нальцах) происходит развитие сперматозоидов. Стенки канальцев
состоят из соединительно-тканной основы и слоя сертолиевых
клеток с включенными в него половыми клетками на различных
стадиях развития (рис. 13,5).
Исходные в формировании сперматозоидов клетки — сперма-
тогонии имеют округлую форму, относительно большое ядро и не-
39
Рис, 13. Схема строения семенника и эпидидимиса человека (4) и участок стен-
ки семенного канальца семенника человека (Б) (по И. Клермонту, 1955):
/ — головка эпидидимиса, 2 — tunica albuginea, 3 — перегородка, 4 — семенные канальцы,,
i — vas deferens, 6 — проток эпидидимиса, 7 — выносящие протоки, 8 — хвост эпидидимиса,
9— прямые канальцы, 10 — формирующийся сперматозоид, //—сперматиды, 12 — спермато*
циты 1-го порядка, 13— сперматогонии, 14— клетки Сертоли
большое количество цитоплазмы. Следует серия митотических
делений, в результате которых количество сперматогоний может
стать очень большим. В семенниках многих животных имеется
особая зона размножения.
Одна из характерных особенностей сперматогониальных деле-
ний — незавершение цитотомии. В результате потомки одного ис-
ходного гония образуют клон клеток, объединенных цитоплазма-
тическими мостиками. Число гениальных делений специфично для
вида, ио, как правило, большее, чем в оогенезе. Сперматогонии
разных генераций отличаются морфологически — по величине и
степени конденсации хроматина. В ходе гоииальных делений ме-
няются параметры клеточного цикла. Так, у мыши S-фаза мито-
тического цикла удлиняется от 7 до 14,5 ч, а фаза g2 укорачивает-
ся с 14 до 4,5 ч.
Первый период сперматогенеза называется периодом размно-
жения, когда сперматогонии митотически делятся.
Затем деление сперматогоний прекращается, они вступают во
40
второй период — фазу роста, во
время которой в ядрах клеток
осуществляется профаза мейоза.
Клетки, приступившие к росту,
называются сперматоцитами 1-го
порядка. Во многих семенниках
этому соответствует особая зона
роста.
Третий период развития муж-
ских половых клеток называется
периодом созревания, который
заключается в двух последова-
тельных делениях сперматоцитов
1-го порядка; в результате после
1-го деления образуются два
сперматоцита 2-го порядка, а за-
тем четыре сперматиды (см. рис.
12,А), отличающиеся от исходных
клеток меньшими размер-ами, от-
носительно большим количеством
цитоплазмы, иным отношением
к красителям, а главное — важ-
ными преобразованиями ядер,
связанными с осуществлением
мейоза. Таким образом, мейоз по
Рнс. 14. Схема спермиогеиеза (по
И. Клермонту и К- Леблоиду, моди-
фицировано А. Хэмом, 1961):
А — 3 — последовательные стадии преоб*
разования сперматиды в сперматозоид у
человека; / — зона Гольджи, 2— мито*
хондрин, 3 — ядро, 4 — центриоля, 5
жгутик, 6 — акросома, 7 — цитоплазматн*
ческая мембрана
15. Основную массу головки
времени совпадает с периодами
роста и созревания в ходе гаме-
тогенеза, о чем будет сказано
в дальнейшем.
Последний, четвертый, пери-
од— период формирования спер-
миев, или спёрмиогенез, представ-
ляет серию сложных преобразо-
ваний, изображенных на рис. 14,
спермия составляет ядро. Цитоплазма практически отсутствует.
У ряда животных в переднем конце головки имеется «акросола» —
важный аппарат для процесса оплодотворения, о чем будет ска-
зано в гл. III. Акросома происходит путем преобразования аппа-
рата Гольджи сперматид. Центросома сперматиды принимает
участие в формировании среднего участка сперматозоида — про-
межуточного или соединительного отдела. В его составе распола-
гаются клеточный центр и митохондрии. У млекопитающих между
головкой и средней частью спермия имеется перехват — шейка,
в котором локализован клеточный центр. Третий отдел спермия —
жгутик, или хвост, в нем различают основную, или главную, часть
и концевую часть, суженную в связи с тем, что не все фибриллы
осевого комплекса доходят до конца хвоста спермия.
41
Рис, 15. Сперматиды человека
(рисунок электроиограммы (по Д. Фовитту н
М. Бургосу, из ки. А. Хэма, 1961):
А — головной участок сперматиды иа стадии, соответствующей изображенной на рис. 14,Б;
Б — сперматида иа более поздней стадии, соответствующей изображенной на рис. 14,Ж;
/ — зона Гольджи, 2 — акросома, 3 — капсула головки, 4 — ядерная мембрана, 5 — ядро,
б — жгутик, 7 — митохондрия, 8 — клеточная мембрана сперматиды
Рис. 16. Нетипичные спермин. А,
Б, В. — Pinnotheres, Maja, Munida
(ракообразные, по Н. К. Кольцо-
ву, 1906); Г — Aglena (паук, по
К- Безенбергу, 1905); Д — Copris
(жук, по С. Баллоинцу, 1890)
Спермин разных животных име-
ют неодинаково устроенную хвосто-
вую часть (так называемые типич-
ные спермин, см. рис. 6); в редких
случаях сперматозоиды имеют не
хвост, а другого типа двигательный
аппарат — нетипичные спермин
(рис. 16), или двигательный аппа-
рат вообще отсутствует.
Представление о яичнике млеко-
питающих дает рис. 17. Это схема
разреза через яичник женщины.
Видны соединительно-тканная стро-
ма и разные стадии развития фол-
ликулов с ооцитами. На поверхно-
сти яичника герминативный эпите-
лий.
Как уже говорилось, Р. Грааф
принимал за яйцо образование,
впоследствии названное граафовым
пузырьком. На самом же деле это
многослойный фолликул с полостью
(что придает ему вид пузырька),
наполненной жидкостью и выстлан-
4?
Рис. 17. Схема строения яичника женщины. Последовательные стадии созрева-
ния фолликулов (по Б. Пэттену, 1946):
/ — зачатковый эпителий, 2 — первичный фолликул, 3 — многослойный фолликул, 4 — обра-
зование полости граафова пузырькй, 5 — зрелый граафов пузырек, 6—атрезня фолликула,
7 — выход яйцевой клетки, 8 — желтое тело, 9 — сосуды
ной фолликулярным эпителием. Участок утолщенного фоллику-
лярного эпителия, в котором расположено яйцо, называется яйце-
носным, бугорком. Во время овуляции целостность стенки фолли-
кула нарушается, яйцо выходит в брюшную полость и, как пра-
вило, попадает в воронку яйцевода.
Процесс оогенеза у разных животных, очень своеобразный
в деталях, вполне сопоставим с фазами сперматогенеза (см.
рис. 12,5). Во-первых, имеется период размножения мелких кле-
ток с относительно крупным ядром и небольшим количеством
цитоплазмы, называемых оогониями. У женщин и у всех самок
млекопитающих этот период заканчивается еще у эмбриона, до
рождения. У большинства других животных и низших позво-
ночных деления оогоний в яичнике продолжаются и у взрослых
форм. Так образуется первая генерация ооцитов, большинство из
которых пребывает в малоизмененном состоянии многие годы до
половой зрелости, когда начинаются процессы роста ооцитов.
Период роста в оогенезе длительнее, чем в сперматогенезе, и
подразделяется на несколько подпериодов или этапов. Вступле-
ние ооцита 1-го порядка в период роста совпадает с вступлением
его в профазу мейоза. Процесс роста в оогенезе разделяют на пе-
риоды малого роста и большого роста. В период малого роста
клетка вырастает незначительно, но в ее ядре в это время
осуществляются важные превращения первых этапов профазы
мейоза вплоть до диплотены. Период большого роста разделяют
на период цитоплазматического роста, характеризуемый интенсив-
ным ростом цитоплазмы и ядра, максимальным развитием хро-
43
Рис. 18. Функциональные отношения
между растущим ооцитом и вспомога-
тельными клетками (по Т. Б. Айзён-
штадт, 1984).
Источником рРНК служат ядро ооцита и яд-
ра трофоцитов (см. стрелки). Желточные
белки секретируются вне гонады и поступа-
ют в ооцит из кровяного русла через меж-
клеточные пространства фолнкулярного эпи-
телия. Небольшая часть желтка синтезирует-
ся эндогенно самим ооцитом.
Ц — рибосомы, 2 — вителлогенин, 3 — кровенос-
ный сосуд; 4 — фолликулярные клетки, 5 —
трофоциты, 6 — эндоплазматический ретику-
лум, 7 — аппарат Гольджи, 8 — ядрышко, 9 —
желток, 10 — кариосфера
мосом типа «ламповых ще-
ток» и период трофоплазмати-
ческого (или дейтоплазмати-
ческого) роста, в конце кото-
рого в ооплазме. осуществля-
ется накопление желтка; ин-
тенсивность синтеза РНК в
ядре в это время падает.
Одна из особенностей ооге-
неза животных состоит в спо-
собности ооцитов в период
большого роста накапливать
значительные количества ре-
зервных веществ—РНП и
желтка, необходимых для
развития зародыша. Процесс
накопления яйцеклеткой желт-
ка называют вителлогенезом.
Вителлогенез может осуще-
ствляться за счет синтеза
желтка внутри ооцита (эндо-
генный желток), либо синтези-
роваться вне яичника (экзо-
генный желток) (рис. 18).
Синтез эндогенного желтка
происходит в эндоплазматиче-
ском ретикулуме и аппарате
Гольджи; накопление желтка
у некоторых животных может
идти и в митохондриях, кото-
рые при этом перерождаются
в желточные гранулы. Эндо-
генный синтез желтка характерен для солитарного оогенеза.
У животных с алиментарным оогенезом эндогенное образова-
ние желтка дополняется экзогенным, при этом белки — предше-
ственники желтка — вителлогенины, синтезируются вне яичника,
например у позвоночных в печени. Вителлогенин поступает в кровь,
а в яичнике поглощается растущими ооцитами. Большую роль
в избирательном поглощении вителлогенина яйцевыми фолли-
кулами играет фолликулярный эпителий. Представление о сложной
системе регуляции экзогенного синтеза желтка, в которой прини-
мают участие гонадотропные и стероидные гормоны, можно по-
лучить из схемы на рис. 19. В ооцит вителлогенин поступает в со-
ставе пиноцитозных пузырьков; в дальнейшем он распадается на
более простые белки — липовителлин и фосвитин, которые входят
в состав экзогенного желтка. Пиноцитозные пузырьки с экзоген-
ным желтком при слиянии с пузырьками Гольджи, содержащими
44
Рис. 19. Схема гормональной регуляции синтеза вителлогенина и его поглощения
ооцятом (по Б. Фоле, Т. Ннколс, М. Редшоу, 1968)'
эндогенный желток, формируют гранулы смешанного желтка (см.
рис. 18). Т. Б. Айзенштадт (1984) предполагает, что в филоге-
незе по мере обогащения яиц желтком и увеличения их объема
уменьшалась роль эндогенного и увеличивалась роль экзогенного
синтеза желтка. Объем зрелого яйца, как уже отмечалось, может
возрастать в тысячи и десятки тысяч раз по сравнению с оого-
ниями. По окончании роста может произойти оплодотворение, и
то, что можно назвать (по аналогии с фазами сперматогенеза)
периодом созревания, начинается только после проникновения
в яйцо сперматозоида (например, у аскариды). У других живот-
ных вхождение сперматозоида происходит после первого деле-
ния созревания (например, у ланцетника), и, наконец, у многих
животных сперматозоид проникает в яйцо только после созрева-
ния яйца. Все указанные периоды оогенеза представляют сово-
купность сложных морфологических и биохимических преобразо-
ваний клетки, ее цитоплазмы, ядра, органоидов клетки, мембран.
Ооциты и их оболочки по сравнению с соматическими клетками
представляют по-особому дифференцированные образования.
Созревание яйца — это прежде всего особого порядка преобра-
зования в ядре. В связи с этим отметим своеобразие деления
ооцита 1-го порядка: одна сестринская клетка (ооцит 2-го поряд-
ка) окажется почти такой же величины, как и ооцит 1-го поряд-
ка, а другая сестринская клетка благодаря совершенно необыч-
ному поведению цитоплазмы в телофазе деления окажется очень
маленькой «почкой» в области будущего анимального полюса
яйца. Это первое редукционное, или первое полярное тельце.
45
Рис. 20. Половые и вспомога-
тельные клетки при разных ти-
пах оогенеза (схематизировано
по Н. С. Табаевой, 1984):
<4 — диффузный солитарный ооге-
нез — амебоидный ооцит губки
Leucandra gossei. Диффузный аго-
иадный алиментарный оогенез;
Б — примитивный яйцевой фолли-
кул губкн Hlppospongia communis;
В — ооцнты и питающие клетки
губкн Sycon raphanus-, Г—яйцевая
капсула трематоды Fasciola hepa-
tica, локализованный алиментарный
фолликулярный оогенез; Д — це-
почка клеток — ооцит и трофоциты
нз целома полнхеты Diopair а сир-
геа; локализованный (внутригонад-
ный) солитарный оогенез; Е—учас-
ток стенки яичника моллюска
Sphaerlum striatinum; Ж — попе-
речный срез яичника Parascaris
equorum, локализованный алимен-
тарный нутримеитарный оогеиез;
3 — поперечный срез яичника пияв-
ки Glosslphonia complanata; И —
участок стенки яичника многонож-
ки Geophilus с фолликулами; К —
фолликул нз яичника таракана
Perlplaneta; Л — участок яичника
млекопитающего, содержащий яй-
цевой фолликул; М — многослой-
ный аинзоморфиый фолликулярный
эпителий яичиика чешуйчатых реп-
тилий; Н— фолликул из яичника
шелкопряда, ооцит с группой тро-
фоцитов расположен внутри фол-
ликула (пример фолликулярио-иут-
римеитариого оогенеза): 1— ооцит,
2 — фолликулярные клетки, 3 —
фолликулярный эпителий, 4 — жел-
точные клетки, 5 — питающие клет-
ки, 6 — трофоциты, 7— яйцо, 3 —
герминативный эпителий яичника,
9— стенка яичиика, 10— строма
яичника, 11 — рахис
Ооцит 2-го порядка в ходе деления «отпочковывает» второе ре-
дукционное тельце, или второе полярное тельце. Только в этом
состоянии яйцо может считаться вполне зрелым.
При сравнении процессов сперматогенеза и оогенеза необхо-
димо обратить внимание, во-первых, на то, что размножение спер-
матогониев происходит дольше и интенсивнее, чем размножение
оогониев; во-вторых, рост ооцитов у многих животных идет не-
соизмеримо длительнее, чем рост сперматоцитов. Каждый сперма-
тоцит дает четыре сперматиды, формирующиеся в четыре зрелых
сперматозоида, в то время как каждый ооцит дает только одно
зрелое яйцо.
В заключение остановимся на некоторых вопросах эволюции
гаметогенеза, разработанных Н. С. Габаевой. '
Исходным типом в эволюции гаметогенеза можно считать диф-
фузный, солитарный (без участия вспомогательных клеток)
46
(рис. 20,Л), распространенный среди губок, кишечнополостных,
некоторых полихет, эхиурид. У некоторых многоклеточных диф-
фузный гаметогенез оеуществляется по алиментарному типу, т. е.
с участием вспомогательных клеток, хотя и без гонад (некоторые
губки, полихеты, рис. 20,5, В, Д). У ряда низших первичноротых
(некоторые кишечнополостные, приапулиды, некоторые черви и
моллюски), хотя уже имеются простые мешковидные гонады,
оогенез сохраняет черты солитарного типа, так как протекает
в гонаде без участия вспомогательных клеток (рис. 20,Е,Ж).
Следующий этап в эволюции гонад многоклеточных связан с воз-
никновением в них вспомогательных элементов гаметогенеза, об-
разующихся в большинстве случаев за счет соматических клеток
гонад (фолликулярные эпителии при фолликулярном типе али-
ментарного гаметогенеза, рис. 20,И, К, Л, М) либо за счет неко-
торой части половых клеток («трофоциты» и др. — при нутри-
ментарном типе алиментарного гаметогенеза, рис. 20,Г, 3,Н).
В эволюции алиментарного оогенеза происходило усовершен-
ствование трофических отношений женских половых клеток с ор-
ганизмом, обеспечивающих накопление яйцом резервных веществ.
В эволюции алиментарного сперматогенеза происходила все боль-
шая изоляция развивающихся гоноцитов от внутренней среды ор-
ганизма. Особенно наглядно изолирующая роль фолликулярного
эпителия в сперматогенезе может быть проиллюстрирована
на примере сперматогенного эпителия млекопитающих. Клетки
фолликулярного эпителия, выстилающие семенные канальцы, со-
единены межклеточными контактами такой структуры, что ока-
зываются непроницаемыми для большинства веществ, находя-
щихся в крови животного. Этот так называемый гематотестику-
лярный барьер имеется в семенниках млекопитающих, а также
некоторых рыб и насекомых.
В целом эволюция алиментарного гаметогенеза осуществля-
лась по пути все большей специализации и расширения функции
вспомогательных структур, достигших наибольшего развития у на-
секомых среди первичноротых и у позвоночных среди вторично-
ротых животных. О значении фолликулярного эпителия в гаме-
тогенезе можно судить по перечислению его основных функций:
это изолирующая, барьерная роль, функция транспорта, а иногда
и синтеза трофических веществ для гоноцитов; гормонально-регу-
ляторная, связанная с синтезом гормонов и гормоноподобных
веществ, регулирующих гаметогенез; способность к фагоцитозу
погибающих половых клеток; опорно-механическая и морфоге-
нетическая функции; в оогенезе функция выработки вещества вто-
ричной оболочки яйца и др.
Таким образом, характеризуя взаимоотношения половых кле-
ток с организмом в филогенезе, можно отметить две противоре-
чивые, но взаимодополняющие тенденции — все большую изоля-
цию половых клеток от внутренней среды организма и, с другой
47
стороны, все большую зависимость развития половых клеток от
вспомогательных элементов гаметогенеза
МЕЙОЗ. РЕДУКЦИОННОЕ ДЕЛЕНИЕ
В процессах оогенеза и сперматогенеза давно уже привлекало
внимание генетиков и эмбриологов, выработавшееся в ходе эво-
люции изменение митоза, происходящее в периодах роста и со-
зревания половых клеток. При нормальном митозе две сестрин-
ские клетки имеют точно такой же набор хромосом, какой был
в делящейся материнской клетке. Все нормальные соматические
клетки, в том числе и те, из которых развиваются половые клет-
ки, содержат парные, гомологичные хромосомы, одна из которых
привнесена при оплодотворении сперматозоидом,, а другая —
яйцом. В ходе же оогенеза и сперматогенеза происходят процес-
сы, в результате которых зрелые половые клетки оказываются
гаплоидными, т. е. с числом хромосом (haplos — одиночный),
в два раза меньшим по сравнению с соматическими клетками.
Редукция числа хромосом осуществляется в результате специали-
зированного клеточного деления — мейоза (meiosis — уменьше-
ние). При оплодотворении в результате слияния ядер гамет объ-
единяются гаплоидные наборы мужской и женской половых
клеток и тем самым восстанавливается диплоидность хромосом-
ного набора (diplos — двойной). Таким образом, в диплоидном
наборе хромосом любого организма каждая хромосома представ-
лена парой идентичных хромосом, ведущих свое происхождение
одна от матери, другая — от отца. Такие парные хромосомы на-
зываются гомологичными. Благодаря редукции, предшествующей
оплодотворению, гарантируется постоянство числа хромосом в ря-
ду поколений, несмотря на происходящий периодически процесс
кариогамии — слияния ядер двух гамет, который сопровождает
половой процесс.
У многоклеточных животных мейоз происходит в ходе гамето-
генеза (гаметическая редукция); он начинается по завершении
гениальных делений и по времени совпадает с периодами роста
и созревания половых клеток.
Мейоз — это процесс, включающий два последовательных кле-
точных деления, подготавливаемых общей для них профазой мей-
оза. Перед вступлением клетки в профазу мейоза в предмейотиче-
ской интерфазе (наступающей вслед за оогониальным делением)
и вплоть до созревания количество ДНК в хромосомах оказывает-
ся равным 4п, т. е. вдвое большим, чем в соматических диплоид-
ных клетках.
1 Габаева Н. С. О строении и функциях фолликулярного эпителия семен-
ников позвоночных // Современные проблемы сперматогенеза. М., 1982.
48
Профаза мейоза в отличие от таковой митоза сопровождается
более значительными преобразованиями ядерных структур, боль-
шей длительностью во времени, достигающей иногда многих лет
(например, у птиц и млекопитающих). Условно профазу мейоза
делят на несколько стадий (лептотенная, зиготенная, пахитенная,
диплотенная, диакинез).
На лептотенной стадии (от лат. leptos — тонкий) в ядрах поло-
вых клеток периода роста, т. е. ооцитов и сперматоцитов 1-го по-
рядка, выявляется диплоидное число тонких в связи со слабой
степенью спирализации хромосомных нитей. К концу лептонемы
по длине хромосом начинают выявляться небольшие утолщения —
хромомеры,' число, величина и расположение которых вдоль хро-
мосомы специфичны для каждой пары гомологов.
Зиготенная стадия (zeugos — пара) состоит в конъюгации, т. е.
в попарном соединении по длине гомологичных хромосом. Сбли-
жение гомологичных хромосом происходит за счет взаимодействия
комплементарных участков ДНК, которые выполняют здесь ме-
хано-химическую функцию. Пары конъюгирующих хромосом назы-
ваются бивалентами. Сдваивание хромосом в зиготенной стадии,
как будет видно далее, подготавливает редукционное деление.
Пахитенная стадия (pachis — толстый) характеризуется завер-
шением конъюгации гомологов; благодаря усиливающейся спира-
лизации хромосомы в этот период происходит тесное взаимное
закручивание хромосом в составе каждого бивалента. В конце этой
стадии в составе каждой хромосомы, образующей бивалент, дела-
ется различимой ее двухроматидная структура. С этого момента
бивалент может быть назван тетрадой. Однако в действительности,
и это доказано экпериментально, уже лептотенные хромосомы
имеют двухроматидную структуру. Итак, две гомологичные хро-
мосомы, конъюгируя, образуют бивалент, каждая из этих хромо-
сом расщепляется на две хроматиды, в результате чего бивалент
превращается уже в четырехнитчатую структуру — тетраду. Че-
тырехнитчатое строение пахитенных хромосом отражает подготов-
ленность хромосомного аппарата клетки к двум последующим
мейотическим делениям, в результате которых каждая из образую-
щих клеток получит по одному элементу из каждой тетрады.
На диплотенной стадии (diplos — двойной) начинается процесс
расхождения, расталкивания и раскручивания гомологичных хро-
мосом в составе каждого бивалента. При этом обнаруживается,
что несестринские хроматиды в биваленте могут быть соединены
между собой в одной или нескольких точках, образуя перекресты —
хиазмы. Хиазмы являются следствием происходящего в предшест-
вующий период профазы мейоза кроссинговера — обмена участка-
ми между несестринскими хроматидами, происходящего в резуль-
тате разрывов и воссоединений в соответствующих локусах
хроматид. Кроссинговер, приводящий к обмену генетическим ма-
териалом между хроматидами, служит одной из причин увеличе-
49
ния резерва наследственной изменчивости в потомстве при половом
размножении.
У ооцитов в ходе диплотенной стадии может наблюдаться та
или иная степень деспирализации хромосом, особенно в случаях
оогенеза солитарного типа («разрыхление» хромосом, образова-
ние «ламповых щеток» и т. п.); такое состояние ядра может быть
довольно длительным — охватывает весь период большого роста
ооцита. Это свидетельствует об активизации ядерного аппарата и
его участия в накоплении в яйцеклетке резерва нуклеиновых кис-
лот и запасных веществ, необходимых для обеспечения начальных
этапов эмбриогенеза. В случае алиментарного типа питания ооци-
та, напротив, можно наблюдать явления, свидетельствующие об
инактивации хромосомного аппарата ооцита — сильно спирали-
зованные хромосомы в этом случае образуют клубок той или иной
плотности — кариосферу. Как известно, при алиментарном типе
питания важную роль в снабжении ооцита запасными веществами
выполняют питающие клетки или фолликулярный эпителий
яичника.
Диакинез отличается все большей степенью спирализации хро-
мосом, все большим смещением хиазм к концевым участкам хро
мосом (терминализация хиазм). Диакинез завершает профазу
мейоза.
К началу метафазы первого мейотического деления исчезает
ядерная мембрана, формируется веретено деления, по экватору
которого располагаются биваленты, т. е. сдвоенные в результате
конъюгации гомологичные хромосомы. В анафазе от каждого би-
валента к полюсам веретена отходит по целой хромосоме, что при-
водит к числовой редукции хромосом в ядрах образующихся кле-
ток. Рассмотрим это на произвольном числовом примере:
если диплоидное число хромосом 8, то при конъюгации гомоло-
гичных хромосом образуются 4 бивалента, которые располагают-
ся по экватору веретена первого деления мейоза; в анафазе этого
деления одна хромосома из состава каждого бивалента отойдет
к одной клетке, другая — к другой. Каждая из клеток, таким
образом, получит по 4 хромосомы, что соответствует для взятого
примера гаплоидному числу хромосом. Таким образом, произойдет
числовая редукция хромосом. В связи со сказанным первое деле-
ние мейоза принято называть редукционным или уменьшительным
делением.
Расхождение хромосом к полюсам при первом мейотическом
делении происходит таким образом, что тетрады делятся в плос-
кости конъюгации (а не в плоскости продольного расщепления
хромосом), значит, к полюсам расходятся диады разного проис-
хождения: в одну клетку попадает диада отцовского происхожде-
ния, а в другую — диада материнского происхождения.
После непродолжительной интерфазы наступает профаза вто-
рого мейотического деления. Второе деление мейоза по форме на-
50
Рис. 21. Схема мейоза. А—В — конъюгация хромосом;
Г, Г'—Д, Д' — метафаза мейоза (по А. Мюнтцингу,
1963)
поминает митоз и не сопровождается изменением числового соста-
ва хромосом, в связи с чем его называют эквационным или
уравнительным делением. Однако оно принципиально отличаете»
от митоза тем, что отходящие к двум клеткам из состава каждой
хромосомы хроматиды могут быть не идентичны по генетическим
свойствам вследствие происшедшего кроссинговера.
В целом в ходе делений мейоза перекомбинация генетического
материала осуществляется дважды: при первом делении в связи с
расхождением к каждому из полюсов веретена части материнских
и части отцовских хромосом, при втором — вследствие расхожде-
ния разнокачественных по генетическому составу хроматид.
Все сложные преобразования хромосомного аппарата клетки,
сопровождающие мейоз, имеют два важнейших следствий:
1. Редукцию числа хромосом и соответственно количества
ДНК в ядре половых клеток.
2. Перекомбинацию генетического материала между форми-
рующимися клетками, увеличивающую резерв наследственной из-
менчивости будущего потомства.
На рис. 21 дана схема мейоза. Для большей простоты приво-
дится случай, когда в оогонии или сперматогонии имеются только
две пары гомологичных хромосом.
При сперматогенезе оба мейотических деления равномерные и
в результате получаются четыре одинакового размера клетки —
сперматиды, впоследствии преобразующиеся в сперматозоиды
(см. рис. 12). Они содержат гаплоидный набор хромосом. Мейоти-
ческие деления ооцита неравномерны, и в результате образуются
одно целое яйцо с гаплоидным набором хромосом и три малень-
51
ких редукционных, или полярных, дегенерирующих тельца
(см. рис. 12).
Различают несколько стадий (или фаз) жизненного цикла
клетки, или онтогении клетки. Вслед за делением наступает пост-
митотическая (пресинтетическая) фаза Gp Затем следуют фаза
синтеза S, во время которой происходит редупликация ДНК, пост-
синтетическая (премитот.ическая) стадия G2 и, наконец, послед-
ний этап онтогении клетки, обусловленный предыдущим развити-
ем,— митоз М.
Соотношение во времени между фазами онтогении клетки мо-
жет меняться при различных состояниях тканей и организма.
Вопросы о значении этих изменений в явлениях специализации
клеток, в нормальной жизни тканей и органов далеки от ясности,
однако, несомненно, они заслуживают большого внимания эмбрио-
логов.
Период синтеза ДНК, предшествующий первому мейотическо-
му делению при развитии женских половых клеток у мышей, за-
нимает 10—10,5 ч, т. е. более чем средняя продолжительность этой
стадии в соматических клетках эмбрионов и взрослых животных.
Профаза мейоза в ооцитах млекопитающих может продолжать-
ся в течение нескольких лет.
При сперматогенезе увеличивается продолжительность фазы
синтеза ДНК. У первых генераций сперматогоний мыши .S-фаза
имеет продолжительность 7—8 ч, в последней генерации — 14,5 ч.
БИОХИМИЧЕСКИЕ АСПЕКТЫ ГАМЕТОГЕНЕЗА
Процессы созревания ооцита, как и другие стадии оогенеза, осу-
ществляются под гормональным контролем. Так, например, гона-
дотропный гормон у позвоночных животных, выделяемый в кровь
самок, запускает процесс созревания ооцита, что было показано
не только в экспериментах in vivo, но и in vitro при непосредствен-
ном воздействии на изолированные фолликулы гонадотропными
гормонами. Фолликулярный эпителий, реагируя на гонадотропный
гормон, продуцирует стероидный гормон (прогестерон), который
непосредственно влияет на ооцит. Вся совокупность описанных ра-
нее событий, сопровождающих процесс созрерания яйца,—это си-
стемные, взаимосвязанные события, затрагивающие как организ-
менные интегративные системы (функционирование нейрогормо-
нальных систем), так и внутриклеточные процессы, совершающие-
ся на молекулярном уровне и связанные с функционированием
генома разных типов клеток и с транспортами биологически ак-
тивных веществ внутри клетки и в циркуляторных системах целого
организма.
В настоящее время выявлены факторы, участвующие в осуще-
ствлении различных моментов созревания. Так, например, в начале
созревания в цитоплазме ооцита появляется фактор, вызывающий
52
дезорганизацию оболочки ядра. Появление этого фактора по-ви-
димому связано с активностью прогестерона. Затем появляется
фактор, вызывающий конденсацию хромосом, фактор, способству-
ющий образованию пронуклеусов, и ряд других. Таким образом
внутри ооцита осуществляется скоординированная последователь-
ность структурно-биохимических процессов, в реализации которых
участвуют и ядро и цитоплазма. Различная последовательность
событий созревания и оплодотворения также обусловлена появле-
нием биологически активных веществ, блокирующих тот или иной
этап мейоза.
Выход яйцеклеток из яичников в яйцеводы и их откладка во
внешнюю среду или задержка в соответствующих отделах поло-
вых путей самки при внутреннем осеменении — это также гормо-
нально зависимые процессы. Благодаря раскрытию многих интим-
ных механизмов этих процессов в настоящее время удается успеш-
но управлять репродуктивной функцией у рыб и сельскохозяйст-
венных животных.
Сформированная яйцеклетка всегда поляризована. Полярность
проявляется не только в градиентном распределении различных
по размерам и структурно-биохимическим характеристикам жел-
точных включений, не только в положении ядра, но и в органи-
зации кортикального слоя и цитоскелета, а также в распределении
других цитоплазматических компонентов.
Форма яйцеклетки и ее общая структурная организация — это
такие же специфические видовые признаки, как и признаки взрос-
лых особей. От того, как организована яйцеклетка, а затем и зиго-
та, зависят последовательность и характер начальных этапов
развития. Окончательные структурно-биохимические перестройки
совершаются в яйцеклетке после оплодотворения. Они приводят
к сложным преобразованиям структур цитоплазмы, получившим
название сегрегации цитоплазмы. В результате этих процессов в
различных участках яйца оказываются локализованы особые фак-
торы, специфически влияющие на функции ядер, репродуцирую-
щихся в ходе дробления. К числу таких факторов относятся и
половые детерминанты, о которых уже говорилось.
Много интересных вопросов возникло в эмбриологии в послед-
ние десятилетия в связи с прогрессом физики и химии. Однако
«начало» онтогенеза — дозародышевый период, формирование
яйцевых клеток в химическом и биофизическом отношении изучено
недостаточно, несмотря на большое число исследований в 60—
80-х годах о белках и нуклеиновых кислотах. Пока невозможно
четко изложить биохимические и биофизические данные, касаю-
щиеся гаметогенеза. Ограничимся изложением лишь некоторых.
Нельзя сделать бесспорные выводы, касающиеся вопросов об-
мена веществ — ассимиляции и диссимиляции. Результаты иссле-
дований разных авторов противоречивы. На основании биохими-
ческих исследований гаметогенеза у лягушек, морских ежей и рыб
53
можно утверждать, что наиболее высокая интенсивность дыхания
свойственна ооцитам на ранних стадиях оогенеза.
Значительно больше биохимических работ по вопросам угле-
водного обмена на различных стадиях оогенеза. У большинства
животных в ходе оогенеза накапливаются резервные углеводы,
главным образом гликоген, интенсивное потребление которого и
других резервных соединений происходит в начале эмбрионально-
го развития. Вероятно, до этого, в конце оогенеза изменяются ре-
гуляторные механизмы энзиматических процессов. Эмбриология,
однако, не располагает необходимыми данными о ферментных си-
стемах ооцитов у разных животных. На ооцитах вьюна обна-
ружено, что имеет место увеличение всех ферментов углеводного
обмена в процессе роста ооцитов,
Известно, какое значение придается в современных генетиче-
ских теориях РНК и ДНК. Сообщим некоторые факты о них, ка-
сающиеся гаметогенеза.
В клетках тканей ДНК находится преимущественно в ядре —
в составе хромосом и ядрышка. Небольшое количество ДНК на-
ходится в составе митохондрий. У зрелых яйцевых клеток внеядер-
ная ДНК преобладает: находится в митохондриях (у яйцеклеток)
и в желточных пластинках. Количество цитоплазматической ДНК
в яйцах морских ежей приблизительно соответствует количеству
ее у 16 диплоидных ядер. У яиц амфибий резерв ДНК эквивален-
тен 5000—10 000 клеточных ядер.
Большая часть РНК — это рибосомальная (до 80—90 %). Ме-
сто ее синтеза — ядрышко. Транспортная РНК цитоплазмы состав-
ляет около 10 %. Во время оогенеза синтезируются все РНК; па-
раллельно осуществляется и синтез белков. Активный синтез
иРНК происходит на хромосомах типа «ламповых щеток». Ин-
формационная РНК (иРНК) запасается в цитоплазме яйца (ча-
стицах) в информосомах (комплексах иРНК и белка), поэтому
синтез белка может происходить некоторое время даже без ядра.
Скорость синтеза РНК в ооцитах намного выше, чем в бластоме-
рах дробящегося яйца. Зрелая яйцеклетка вьюна имеет значитель-
ный запас рРНК, а количество иРНК относительно небольшое.
У амфибий обнаружено, что рибосомальная РНК не синтезируется
до гаструляции. Это означает, что весь синтез белка в ходе обра-
зования зародыша, состоящего приблизительно из 30 000 клеток,
осуществляется только при участии рибосом, образовавшихся в хо-
де оогенеза. Морфологическим показателем синтеза рРНК «про
запас» может быть или увеличение размера ядрышка ( в случае,
если в ооците имеется одно гигантское ядрышко), или увеличение
количества ядрышек, их амплификация. На стадии диплотены в
кариоплазме ооцита у амфибий может оказаться несколько сот
отдельных «свободноплавающих» ядрышек. В них синтез рРНК
может происходить в 60 раз быстрее, чем в хромосомах.
Недостаточно исследований различных физических параметров
54
8 ходе оогенеза. Недавно стали появляться работы с использова-
нием микроэлектродной техники: введения внутрь клетки тончай-
шего микроэлектрода (заполненная электролитом пипетка диа-
метром менее 1 мкм, что практически не повреждает клетку). Та-
ким образом удается измерить разность электрических потенциа-
лов между внутренней средой клетки и внешней средой — слабым
раствором солей в воде. Эта методика позволяет прижизненно
анализировать работу клеточных механизмов, которые регулиру-
ют состав ионов и их концентрацию внутри клетки и обмен ионов
клетки с окружающей средой. Этот обмен у многоклеточных орга-
низмов опосредован высокопроницаемыми контактными мембрана-
ми, которые содержат щелевые контакты — скопления белков-кон-
нексонов. Последние образуют каналы диаметром около 2 нм,
соединяющие цитоплазму контактирующих клеток. Через щелевые
контакты проходят вещества с низкой молекулярной массой—ионы
и молекулы не более 900 Д для клеточных мембран млекопита-
ющих, 1500—2000 Д для насекомых. Изучение проницаемости для
более крупных молекул, чем ионы, переносящих электрический
ток и ответственных за явление электрической связи между клет-
ками, делается при помощи внутриклеточных микроинъекций ве-
щества с флуоресцентными метками (флуоресцеин, проционовый
желтый, родамин и др.), которые могут быть обнаружены после
фиксации и in vivo в ультрафиолетовом микроскопе.
Как видим, благодаря прогрессу физики и химии современные
эмбриологи изучают «начало» индивидуального развития — доза-
родышевый период — формирование яйцевых клеток и спермато-
зоидов с принципиально новых позиций.
Еще в 1935 г. Э. С. Бауэр обосновал необходимость термоди-
намических подходов в исследованиях закономерностей онтогене-
за, вплоть до явлений старения, продолжительности жизни инди-
видуума.
На основании экспериментальных данных изучения интенсив-
ности дыхания и содержания АТФ в онтогенезе вьюна А. И. Зо-
тин, Н. Д. Озернюк утверждают, что на ранних стадиях оогенеза
в период малого роста ооцитов происходит омоложение системы.
В конечном счете все процессы онтогенеза, включая и старение,
обусловлены генотипически. Однако только прогресс современной
физики и химии позволяет основательно ставить исследования по
выяснению морфофизиологических механизмов, определяющих вы-
сокие морфогенетические потенции зиготы или иных репродуктив-
ных тел, т. е. выяснить, за счет каких энергетических источников
становится возможной реализация тех или иных этапов морфоге-
нетического процесса.
Глава III
ОПЛОДОТВОРЕНИЕ И ПАРТЕНОГЕНЕЗ
ОСЕМЕНЕНИЕ
Совокупность явлений, благодаря которым происходит контакт
между женскими и мужскими половыми клетками, называется осе-
менением. Приспособления у организмов и у половых клеток, обес-
печивающих этот процесс, разнообразны у разных видов живот-
ных. В случаях наружного осеменения сперматозоиды и яйца вы-
деляются во внешнюю среду, например у большинства водных
животных. При внутреннем осемененци, благодаря копулятивным
органам и согласованному поведению самца и самки, сперматозоиды
вводятся в половые пути самки. Выбрасывание самцом семенной
жидкости со сперматозоидами — эйякуляция — представляет
сложный акт, обусловленный, особенно у млекопитающих и чело-
века, специальным строением и функциями мужского полового
органа, мускулатуры семявыносящего протока, строением влага-
лища, системой безусловных и условных рефлексов. Сколь разно-
образны явления, сопровождающие встречу мужских и женских
половых клеток, можно судить по ряду примеров.
Для некоторых пиявок характерно кожное осеменение, при ко-
тором сперматозоиды во время спаривания прикрепляются к гипо-
дерме тела на спинной стороне, затем они проникают между клет-
ками гиподермы в полость тела и в матку сквозь ее стенки. У сала-
мандр при очень сложном взаимном поведении самца и самки
откладывается сперматофор (мешочек с большим количеством спер-
матозоидов). Самка помещает сперматофор в свою клетку, где
впоследствии сперматозоиды освобождаются и оплодотворяют
яйцеклетки.
При гермафродитизме (когда у одного и того же индивидуума
имеются и яичники и семенники) в редких случаях происходит са-
моосеменение (например, у паразитических ленточных червей),
большей частью происходит осеменение одной особью другой (на-
пример, у дождевых червей). У некоторых гермафродитов разви-
тие и функционирование семенников и яичников происходит в раз-
ное время, так что один и тот же индивидуум при копуляции ока-
зывается то самцом, то самкой.
Копуляция, как правило, достаточно надежно (а у многих жи-
вотных совершенно гарантированно) обеспечивает оплодотворение.
Например, у всех заднежаберных, высших переднежаберных, голо-
воногих моллюсков в результате копуляции оплодотворение про-
56
«сходит в 100% случаев. У многих немертин самки и самцы откла-
дывают яйца и сперматозоиды в общую слизистую массу, что так-
же гарантирует оплодотворение.
Процессы осеменения, являясь объектом многочисленных зо-
ологических и физиологических исследований, во многих случаях,
однако, изучены недостаточно. При наружном осеменении многих
водных беспозвоночных (переднежаберных моллюсков, актиний,
иглокожих) яйца и спермин выбрасываются в воду лишь тогда,
когда самец и самка приближаются друг к другу, что зависит, не-
сомненно, от влияний друг на друга на расстоянии благодаря про-
дукции возбуждающих химических веществ, и благодаря особым
рецепторам, свойственным самцам и самкам. Более чем у 40 видов
актиний, моллюсков, немертин и других животных обнаружено,
что сначала самец отдает сперму и уже затем возбужденная
«спермовой средой» самка выталкивает свои яйца.
Интересные примеры «химических взаимоотношений» между
самцом и самкой можно наблюдать у актиний. Половозрелые сам-
ки актинии Sagartia troglodytes перемещаются по направлению
к отдающему сперматозоиды самцу. Это движение прекращается,
когда педальные диски приходят в тесный контакт. Тела животных
образуют почти замкнутый свод, под которым происходит слияние
выделенных в воду сперматозоидов и яиц. После этого самцы и
самки отъединяются, а масса оплодотворенных яиц растекается.
У другого вида актиний — Halcampa duodecinwirrata самец и
самка также возбуждают друг друга на расстоянии, по-видимому,
химическими (а может быть, и физическими) факторами, но пере-
мещения их не происходит. Они сильно вытягиваются в базиапи-
кальном направлении. Самец отдает сперматозоиды в воду, а сам-
ка при этом наклоняется им навстречу.
После процессов осеменения следуют процессы оплодотворения.
МОНО- И ПОЛИСПЕРМИЯ
Для оплодотворения яйца требуется один сперматозоид. Происходит
соединение его ядра с ядром яйцеклетки, и начинается развитие
организма. Это — моноспермия. Но еще в прошлом столетии были
известны случаи полиспермии.“По данным С. Рюккерта (1890), при
оплодотворении яиц голубя в цитоплазму проникают 15—25 спер-
матозоидов. Доказано, что при моноспермии проникновение в яйцо
«лишних» сперматозоидов вызывает уродства в развитии. Так, если
в яйцо лягушки проникает несколько сперматозоидов, то с его
ядром сливается, однако, ядро только одного сперматозоида. При-
внесенные с другими спермиями центросомы образуют также цент-
ры дробления, вследствие чего вместо нормального гтерВОНТ Дроб-
ления яйцо уродливо дробится сразу на несколько бластомеров.
Развивающийся зародыш оказывается нежизнеспособным.
57
Большинство эмбриологов (например, А. С. Гинзбург, 1963; на
основании работ на осетровых) убеждены в том, что для многих
животных полиспермия вредна, в норме после проникновения од-
ного сперматозоида возникает «барьер» против проникновения
других сперматозоидов, изменяются поверхностные слои цитоплаз-
мы и образуется оболочка оплодотворения (см. об этом далее).
А. С. Гинзбург (1967), подводя итоги многочисленным наблю-
дениям, утверждает, в частности, что оплодотворение у осетровых
рыб осуществляется по типу моноспермии, но в экспериментальных
условиях можно добиться полиспермного оплодотворения. В этом
случае оплодотворение носит явно патологический характер. Гинз-
бург наблюдала одновременное проникновение через разные ми-
кропилярные каналы нескольких — до четырех—сперматозоидов.
Ядра проникших спермиев соединяются с женским пронуклеосом.
При первом делении дробления в полиспермных яйцах возникают
многополюсные митозы, и анимальная область яйца обычно разде-
ляется сразу на 3, 4 и большее число бластомеров. При выращи-
вании 200 зародышей, развивавшихся из полиспермных яиц осетра
и севрюги, было установлено, что все они развиваются уродливо.
Около половины их погибает еще до достижения стадии вылуп-
ления.
Эти интересные факты, конечно, не противоречат тому, что у не-
которых животных безусловно установлена полиспермия как зако-
номерное явление: у некоторых членистоногих—насекомых и пауко-
образных, брюхоногих моллюсков, хордовых (акулообразных и хи-
меровых рыб), хвостатых амфибий, рептилий и птиц.
Хотя полиспермия свойственна сравнительно небольшому числу
животных, представляет большой интерес изучение ее физиологи-
ческого и эмбриологического значения. Имеется ли корреляция
между типом оплодотворения и строением оболочек яиц? По мне-
нию А. С. Гинзбург (1967), корреляции нет: полиспермия наблюда-
ется как у яиц, оболочки которых легко преодолеваются спермия-
ми, так и у яиц с оболочками, не проницаемыми для спермиев и
снабженными микропилярными каналами (насекомые). Моноспер-
мия имеет место у многих таких животных, яйцевые клетки кото-
рых проницаемы для сперматозоидов в любой точке поверхности
яйца, например у иглокожих, млекопитающих и др.
ВЗАИМНОЕ ВЛИЯНИЕ ЯИЦ И СПЕРМАТОЗОИДОВ
НА РАССТОЯНИИ И ПРИ КОНТАКТЕ
Давно уже многие эмбриологи отмечали влияние каких-то химиче-
ских веществ, выделяемых яйцом, на находящиеся от него на рас-
стоянии сперматозоиды. Ф. Р. Лилли (1912—1921) убедился в том,
что морская вода, в которой оказались зрелые яйцевые клетки
морского ежа («яичная вода»), может вызывать усиление подвиж-
ности сперматозоидов и их последующую агглютинацию. Вероят-
58
но, те же химические вещества влияют возбуждающе на самцов и
вызывают нерест. Такие свойства «яичной воды» описаны не толь-
ко для ежей, но и для многих других животных: морских звезд,
моллюсков, круглоротых (Latnpetra fluviatilis), рыб и амфибий.
Имеются данные о хемотаксическом, «притягивающем» влиянии
«яичной воды» на сперматозоиды.
Продуцируемые яйцами специфические вещества, без которых
невозможно оплодотворение, Ф. Р. Лилли (1919) назвал фертили-
зином. Эти вещества присутствуют на поверхности яйца и могут
также находиться в яйцевых оболочках. Считают, что фертили-
зин — это гликопротеин или мукополисахарид. Как протеин он
содержит аминокислоты, а как полисахарид имеет молекулы одно-
го или более моносахаридов — глюкозу, фруктозу или галактозу.
И аминокислоты, и моносахариды неодинаковы у разных видов
животных, так что различны и фертилизины. Молекулярная масса
одного из фертилизинов около 300 000.
Предполагается, что в поверхностном слое цитоплазмы сперма-
тозоидов содержатся особые вещества — антифертилизины. Это
кислые протеины с молекулярной массой около 10 000. А. Тайлер
(1958) рассматривает систему фертилизин — антифертилизин как
систему антиген — антитело и считает, что они подходят друг
к другу, как это принято образно говорить в иммунохимии, как
«ключ» к «замку».
Гипотетические вещества, продуцируемые яйцами и спермато-
зоидами или содержащиеся в поверхностных слоях цитоплазмы,
по инициативе М. Гартмана (1940) многие эмбриологи называют
гамонами (гормонами гамет), причем гиногамоны (гормоны яйце-
клеток) вырабатываются яйцами, андрогамоны (гормоны мужских
половых клеток) — сперматозоидами. Яйцо содержит гиногамон I,
активирующий движение сперматозоидов, и гиногамон II, или фер-
тилизин, агглютинирующий их. Сперматозоиды содержат также не
менее двух андрогамонов: андрогамон I, по-видимому, парализует
подвижность сперматозоидов, а андрогамон II растворяет оболоч-
ку яйца. Последний называют нередко лизином, и отождествляют
с энзимом гиалуронидазой, несомненно играющей важную роль
в оплодотворении: этот энзим разжижает студенистые вещества.
Влияние гиалуронидазы на мукополисахарид — гиалуроновую'
кислоту давно констатировали микробиологи при анализе прони-
цаемости микробов сквозь ткани многоклеточных животных. Пред-
полагают, что гиалуронидаза может играть важную роль в про-
никновении сперматозоидов между клетками фолликулярного эпи-
телия, которые у яиц некоторых млекопитающих оказываются
в большом количестве на поверхности зрелого яйца.
И. И. Соколовская (1947) в опытах на кроликах доказала, что
оплодотворения не происходит, если при осеменении самки вносит-
ся менее 1000 сперматозоидов. Но оно не может состояться и в том
случае, если в половых путях оказывается избыточное количество
59
Рис. 22. Сперматозоид Hemicentrotus pulcherrimus после обработки «яичной во-
дой» (по К. Дан, 1954):
2 — головка, 3 — промежуточная область, 4~ хвост
1 —- акросомная нить,
сперматозоидов (более 100 000 000). Это связывают с недостатком
гиалуронидазы в первом случае и с ее избытком во втором, т. е.
с недостаточным или избыточным количеством лизирующих суб-
станций.
Факт влияния «яичной воды» на спермий подтвержден и элек-
тронно-микроскопическими исследованиями. На рис. 22 видна
часть сперматозоида Hemicentrotus pulcherrimus, обработанного
«яичной водой». Произошли резкие изменения: набухание спер-
мин, выбрасывание акросомной нити.
В новейших исследованиях все большее внимание привлекает
характеристика поверхности взаимодействующих гамет. Так, у ас-
цидий показано избирательное взаимодействие спермиев с поверх-
ностью яйцеклетки, предшествующее акросомной реакции. «Узна-
вание» нужных спермиев происходит благодаря наличию специ-
альных гликопротеиновых молекул в оболочке яйца — рецепторов,
с которыми взаимодействуют поверхностные молекулы головки
спермия (А. Монрой, Ф. Розати, 1979).
При оплодотворении яиц, выметываемых гидромедузами, спер-
мий всегда входит в яйцо на анимальном полюсе, что связано со
спецификой оболочки в этом районе.
Свойства поверхности яйца, обусловливающие взаимодействие
со спермием, проявляются в процессе созревания и оплодотворения
(С. Р: Миллер, 1981). В начале второго мейотического деления
в плазматической мембране яйца возникают рецепторные зоны, от-
ветственные за узнавание поверхности спермия. После отделения
второго направительного тела эти зоны концентрируются в районе
анимального полюса. В этом процессе принимают участие микро-
трубочки.
Условно можно выделить следующие группы веществ, участву-
ющих в осеменении и оплодотворении: 1) собственно гамоны —
вещества, повышающие подвижность сперматозоидов; они видо-
специфичны; 2) гликопротеины — вещества, обеспечивающие взаи-
60
Рис. 23. Строение микропиле (по О. Ф. Сакун и Н. А. Будкой, 1964):
А — микропиле в зрелом ооците рыбы сырти; Б — микропиле у осетровых; 1 — соединитель-
но-ткаиная оболочка, 2— фолликулярная оболочка, 3 — бугорки, приклеивающиеся к суб-
страту, 4 — радиально исчерченная оболочка, 5 — замыкающая клетка, 6 — кортикальные
альвеолы, 7 — желток, 8— канал микропиле, 9 — студенистый слой, 10 — ампула микропиле,.
// — мелкозернистый желток, 12 — пигмент
модействие со специфическими поверхностными рецепторами и вы-
зывающие агглютинацию спермиев; 3) лизины — лизирующие ве-
щества, выделяемые при акросомной реакции.
ПРОНИКНОВЕНИЕ СПЕРМАТОЗОИДА В ЯЙЦО.
АКРОСОМНЫЙ АППАРАТ
Сформированные яйцевые клетки имеют разнообразные поверхно-
стно расположенные структуры, через которые сперматозоид про-
никает при оплодотворении. Это прежде всего оболочки, которые
есть, за редкими исключениями, у всех животных.
Как же происходит проникновение сперматозоидов через эти
оболочки? У некоторых яиц проникновение сперматозоида проис-
ходит только через особое «отверстие» микропиле — маленький
участок первичной или вторичной оболочки, где не произошло
обычное формирование структуры, свойственной всей поверхности
(рис. 23). Это тот участок поверхности ооцита, через который про-
никают питательные вещества. С прекращением питания на завер-
шающих стадиях роста ооцита одна из клеток фолликула остает-
ся в тесном соединении с поверхностью ооцита и отмирает после
полного сформирования яйца. Конечно, и в случае наличия микро-
пиле сперматозоиду требуется преодолевать «барьер», так как ми-
кропиле не представляет собой полого канала.
Методы электронной микроскопии позволили выявить у спер-
матозоидов морских звезд, некоторых моллюсков, аннелид, кругло-
ротых, осетровых рыб и других животных акросомную реакцию.
В доэлектронно-микроскопическую эпоху предполагали возмож-
61
Рис. 24. Акросомная область
сперматозоида аинелиды Hyd-
roides hexagonus в процессе
•оплодотворения (рисунок элек-
трограмма) (по Л. Колвин и
А. Колвин, 1961):
1 — наружная акросомная мембра-
на, 2— полость акросомного пу-
зырька, 3 — гранулярный слой ак-
росомы, прилегающий к ядру, 4 —
область растворения желточной
оболочки, 5 — желточная оболочка
яйца
Рис. 25. Схема апикальной части голов-
ки сперматозоида аннелиды Hydroides
hexagonus (по Л. Колвин и А. Колвин,
1961):
1 — ядро, 2 — вещество, находящееся между
ядерной н акросомной мембранами, 3 — по-
лость акросомного пузырька, 4— акросомная
гранула, 5 — гранулярный слой акросомной
мембраны, 6 — ядерная оболочка, 7 — грану-
лярный слой наружной зоны акросомной
мембраны, 8 — плазматическая мембрана, 9 —
апикальный пузырек
ность «прободающего» действия начальной части головки сперма-
тозоида — перфоратория («прободатель»). Нет оснований отри-
цать значение механического действия головки сперматозоида
(если учесть, что у некоторых сперматозоидов происходит одно-
временно и вращательное, и поступательное движение). Электрон-
но-микроскопическая техника и цитохимия позволили, однако, уста-
новить, что механизм проникновения сперматозоида в яйцо пред-
ставляет сложную цепь морфологических и химических явлений.
То, что считали перфораторием, на самом деле представляет
у сперматозоидов многих животных сложное образование — акро-
сомную область спермия (рис. 24 и 25). Эта область ограничена
мембраной, изолирующей акросомный пузырек от ядерной области.
В центральной части пузырька находится округлое тело — акро-
сомная гранула. Апикальный конец ее доходит до вершины головки
«сперматозоида. На рис. 25 видны не описываемые здесь детали
строения акросомной области. Формирование акросом в ходе спер-
матогенеза изучено у многих животных. В частности, Э. Францен
в 1956 г. у немертин, полихет, иглокожих и других животных вы-
яснил роль элементов аппарата Гольджи в формировании акро-
сомы.
62
При соприкосновении с яйцом или с «яичной водой» (в которой
находились яйца) сперматозоид быстро претерпевает изменения.
Оболочка акросом разрушается, и выбрасывается тонкая упругая
акросомная нить. У сперматозоидов морских ежей она всего около
1 мкм длиной, а у морских звезд — до 25 мкм, у осетра — 5—
8 мкм.
Электронная микроскопия позволила выявить ту структуру
акросомного аппарата у спермиев морских ежей и моллюсков, ко-
торая в первую очередь реагирует на близость яйца или его обо-
лочек, воспринимает воздействия, вызывающие акросомную реак-
цию. Это — тончайший слой плотного материала, лежащего непо-
средственно под плазматической мембраной в апикальной части
спермиев.
Акросомная нить, возникающая у сперматозоида еще до кон-
такта его с яйцом, проникает через студенистую оболочку яйца,
а затем и через желточную. В связи с этим образуется «конус»
оплодотворения, форма которого может меняться от довольно пра-
вильного конуса, расположенного вокруг акросомной нити, до псев-
доподиеподобных выростов. По мере проникновения головки спер-
матозоида акросомная нить постепенно исчезает. Далеко не у всех
животных сперматозоиды имеют такой акросомный аппарат
с акросомными нитями. Акросомных нитей может и не быть. Осо-
бенности организации спермиев связаны со строением яйцевых
клеток, в частности с тем, имеются ли микропилярные каналы
в оболочке яиц. Так, у костистых рыб в процессе сопряженной
эволюции строения сперматозоидов и яиц произошла, вероятно,
редукция, утрата акросомы. У яиц костистых рыб спермий в глуби-
не микропилярного канала вступает в непосредственный контакт
с ооплазмой; при этом акросома, являющаяся орудием для преодо-
ления барьера между гаметами, утрачивает свое значение
(А. С. Гинзбург, 1967).
Все высказанные ранее предположения о гамонах и о гиалуро-
нидазе сохраняют свое значение и при толковании электронно-ми-
кроскопических фотографий. На рис. 24 видно, что уже через 5 ми»
после осеменения яйца аннелиды Hydroides hexagonum участок
желточной оболочки оказывается лизированным, акросомная нить
исчезает и начальная часть головки сперматозоида уже проникает
в цитоплазму яйца.
При изучении оплодотворения у млекопитающих найдено,
что акросома кроме гиалуронидазы содержит протеолитические
ферменты типа трипсина, способные разрушать фолликулярные
клетки и блестящую оболочку. Это установлено в опытах in vitro.
А. Колвин и Л. Колвин в 1955 и 1956 гг. обнаружили, что яйца
реагируют только после того, как акросомные нити вступают в не-
посредственный контакт с мембраной яйца и с лежащей под ней
цитоплазмой. А. Тайлер (1959) представляет процесс оплодотво-
рения как «пиноцитоз», под которым подразумевается поглощение
63
клетками жидких веществ из внешней сре-
ды (рис. 26). Подобные представления воз-
никали еще в начале нашего столетия. Дейст-
вительно, сходство с фагоцитозом или пиноцито-
зом большое (возникновение «воспринимающе-
го» бугорка, проникновение акросомы и погру-
жение в цитоплазму яйца спермия), но схема
Тайлера не отражает данных электронной мик-
роскопии и цитохимии, согласно которым проис-
ходят сложные взаимоотношения между мем-
бранами яйца и спермия, локальный лизис жел-
точной оболочки и т. п.
Эксперименты показывают, что для актива-
ции яйца к развитию не обязательно проникно-
вение спермия в яйцо. Требуется лишь «пуско-
вой механизм», возникающий при начальных
(только что описанных) процессах взаимодей-
ствия спермия с яйцом. Для побуждения яйца
Рис. 26. Схема к развитию и для оплодотворения не требуется
оплодотворения
(по А. Тайлеру,
1959)
проникновения всего спермия. Нередко хвосто-
вая часть его остается вне яйца. Активация
яйца (но не истинное оплодотворение) может
произойти, если экспериментатор оттянет микро-
пипеткой сперматозоид только что начинающий проникать в яйцо.
Дробление может начаться. Если же ввести в яйцо сперматозоид с
помощью микропипетки и тем самым не допустить возниковения
процессов, совершающихся нормально в поверхностных частях
яйца при его контакте со сперматозоидом, активации яйца не Про-
исходит. Эти и другие многочисленные наблюдения говорят в поль-
зу важной роли процессов, совершающихся при оплодотворении
в кортикальных (поверхностных) структурах яйца.
ОБОЛОЧКА ОПЛОДОТВОРЕНИЯ. БИОХИМИЧЕСКИЕ ИЗМЕНЕНИЯ,
ПРОИСХОДЯЩИЕ В ЯЙЦЕ ПОСЛЕ ОПЛОДОТВОРЕНИЯ
На поверхности яйца в связи с проникновением сперматозоида
происходят две группы связанных между собой процессов: физико-
химические изменения поверхностного слоя цитоплазмы и образо-
вание оболочки оплодотворения. Особенно хорошо изучены эти
процессы Ж. Рунстрёмом (1950—1963) и Л. Ротшильдом (1956)
на яйцах морских ежей.
Поверхность зрелого неоплодотворенного яйца морского ежа
имеет структуру, изображенную схематично на рис. 27.
В месте проникновения сперматозоида незамедлительно начи-
наются физико-химические изменения, распространяющиеся бы-
строй волной по всей поверхности яйца. В течение двух секунд по-
сле соприкосновения со сперматозоидом происходят биохимические
64
Рис. 27. Схема строения не-
оплод отворенного (4) и
оплодотвореииого (5) яйца
морского ежа (по Ж- Рун-
стрем, 1952):
1 — оболочка оплодотворения,
2 — желточная оболочка (3—
5 мкм), 3 — слой гранул, 4~
слой пигментных гранул, 5 —
внутренний протоплазматиче-
ский слой, 6 — желток (эндо-
плазма), 7 — экстрагрануляр-
ная зона, 8 — наружная поверх-
ность цитоплазмы яйца, 9 — пе-
рнвнтеллиновое пространство
процессы, видимым выражением которых является исчезновение
кортикальных гранул. В результате экзоцитоза содержимое кор-
тикальных гранул (производных комплекса Гольджи) присоеди-
няется к материалу желточной оболочки, а одевающие их мембра-
ны оказываются в составе плазматической мембраны яйца. Так
возникает оболочка оплодотворения. Процесс формирования этой
оболочки протекает быстро и завершается уже в первую минуту.
Схема образования оболочки оплодотворения в яйце морского ежа
представлена на рис. 27. Часть кортикальных гранул, не участвую-
щих в образовании оболочки оплодотворения, превращается в тон-
кую гиалиновую мембрану. Образующаяся оболочка оплодотворе-
ния отделяется от поверхности цитоплазмы, вместе с чем возника-
ет заполненное жидкостью перивителлиновое пространство.
После образования оболочки оплодотворения другие спермато-
зоиды не могут проникать в яйцо. Так обстоит дело у яиц некото-
рых кишечнополостных, аннелид, иглокожих, костистых рыб, лягу-
шек.
Механизм предотвращения полиспермии изучался многими,
особенно на яйцах морских ежей. Согласно данным Л. Ротшильда
и М. М. Сванна (1951—1956) имеет место двухфазное блокирова-
ние полиспермии: в первые 1—2 с после установления контакта
между оплодотворяющим сперматозоидом и яйцом по поверхно-
сти яйца распространяются какие-то невидимые изменения, кото-
рые делают ее менее доступной для других сперматозоидов. Затем
следует вторая фаза полного блокирования: видимые изменения
кортикального слоя яйца, после завершения которых яйца стано-
вятся полностью непроницаемыми для сперматозоидов.
Ж. Рунстрём (1957) и другие предполагали, что основное пре-
пятствие для^проникновения дополнительных сперматозоидов у яиц
морских ежей обусловлено влиянием студенистой оболочки яйца
на спермин и образованием на поверхности яйца гиалинового слоя.
65
А. С. Гинзбург (1967) считал, что у яиц осетровых и лососевых
рыб, а также и у морского ежа блокирование полиспермии — это
одноактный процесс, основанный на секреции содержимого корти-
кальных телец, так как блокирование проникновения спермиев
в ооплазму совпадает по времени с процессом выделения содер-
жимого кортикальных гранул.
Как видим, существуют разные мнения по этому вопросу. Сле-
дует думать, что механизм блокирования у яиц разных животных
различен. Приведем еще один пример кортикальной реакции, изу-
ченной А. С. Гинзбург и Т. А. Детлаф (1960—1962) на яйцах осет-
ровых рыб.
Электронно-микроскопическое изучение оплодотворенных яиц
дало возможность определить, что уже через 5 с после осеменения
яиц севрюги содержимое кортикальных гранул начинает изливаться
под оболочку, но весь процесс секреции растягивается до 2 мин.
На поверхности яйца образуется тонкий слой выделенного содер-
жимого кортикальных гранул. Этот слой обусловливает разобще-
ние поверхности яйца от оболочки; перивителлиновое пространст-
во в это время еще неразличимо в световом микроскопе. Расши-
рение перивителлинового пространства происходит путем
выделения гидрофильных коллоидальных веществ из цитоплазмы
анимальной области яйца.
Как видим на примере яиц морских ежей и осетровых рыб, кор-
тикальным гранулам принадлежит большая роль. Это установлено
исследованиями на яйцах многих других животных. Различают
в связи с разной консистенцией содержимого кортикальные грану-
лы и кортикальные альвеолы. Электронно-микроскопические иссле-
дования яиц осетра и севрюги показали, что кортикальные грану-
лы имеют разную форму и размеры (от 0,3—0,5 до 5 мкм). Рас-
полагаются они в непосредственном контакте с клеточной оболочкой.
Кортикальные альвеолы, обнаруженные у 5—6 , видов костис-
тых рыб, представляют тельца более крупные — до 35 мкм в диа-
метре. Интересно, что строение кортикальных альвеол видоспеци-
фично. Основными компонентами кортикальных гранул и альвеол
являются полисахариды, по-видимому, связанные с белками. Мем-
браны кортикальных альвеол костистых рыб содержат лициды.
В связи с бурно протекающими при оплодотворении процесса-
ми в кортикальном слое происходят энергичные изменения в со-
стоянии всего яйца. У яиц амфибий на стороне, противоположной
месту вхождения сперматозоида, в области экватора происходит
видимое изменение в пигментации: пигмент отступает внутрь, и эта
часть поверхности яйца становится более светлой (серой, если пиг-
мент был черным, или желтоватой, если пигмент бурый), представ-
ляя серый серп или серый полумесяц.
Очевидно, что такие преобразования не могли бы наступить без
специфических биохимических процессов, без перемещения цито-
плазмы, вызванного проникновением сперматозоида. При оплодо-
66
творении изменяется состояние всего яйца. Процессы эти изучены
недостаточно даже у таких излюбленных эмбриологами объектов,
как амфибии. Так, имеют еще место споры о механизме образова-
ния «серого серпа».
И у яиц всех других животных при оплодотворении происходят
специфичные для каждого вида организмов процессы изменения
протоплазмы. Вскоре после проникновения сперматозоида в яйцо
морского ежа Paracentrotus livtdus пигментированный слой прото-
плазмы (а пигмент у ооцита распределен относительно однородно
во всем поверхностном слое цитоплазмы) приходит в состояние
возбуждения, происходит быстрое передвижение участков цито-
плазмы, так что ниже экватора образуется отчетливое пигменти-
рованное «кольцо». Процесс этот начинается в месте вхождения
сперматозоида в яйцо.
О больших физико-химических изменениях, происходящих во
всем яйце, свидетельствуют и передвижения в цитоплазме яйца как
собственного ядра, так и проникшего сперматозоида, точнее — его
головки и шейки. При вхождении сперматозоида впереди оказыва-
ется головка, а затем шейка, где локализована центросома. Близ-
лежащая цитоплазма яйца изменяется, возникают лучи уплотнен-
ной цитоплазмы, в связи с чем головка сперматозоида перемеща-
ется так, что впереди оказывается центросома. Вместе с движени-
ем головки сперматозоида происходит и движение ядра яйцеклет-
ки. Нельзя думать, что эти движения хаотичны. Наоборот, можно
говорить о закономерной «тропе оплодотворения». По мере дви-
жения ядер навстречу друг другу происходит изменение их орга-
низации. Наиболее характерным признаком при преобразовании
ядер в пронуклеусы считается начало репликации в них ДНК-
В конце концов оба пронуклеуса приходят в соприкосновение, и
наступает их соединение (рис. 28). Слияния женского и мужского
ядер (их хромосом) не происходит.
Рис. 28. Оплодотворение у
стерляди (по Г. М. Персову,
1955):
А — погружение в толщу желтка
мужского и женского пронуклеусов
(через 1 ч 43 мнн после осемене-
ния, температура воды 10 °C); Б —
сближение пронуклеусов (через 1 ч
30 мнн после осеменения, темпеоа-
тура воды 14,8 °C); В —метафаза
деления сегментационного ядра
(через 3 ч после осеменения, тем-
пература воды 14,8 °C)
67
Процесс оплодотворения можно рассматривать как выведение
яйца из состояния «анабиоза» (см. гл. II). В ходе первых 5—6
дроблений происходит восстановление нормального соотношения
между ядром и цитоплазмой и вместе с тем обмена веществ, свой-
ственного нормальным соматическим клеткам, не потерявшим спо-
собность к делению. Близкий к этому взгляд высказал Браше, ко-
торый предполагал, что неоплодотворенное яйцо находится в со-
стоянии «анестезии», «отравления». Оплодотворение возвращает
яйцеклетку к дыханию первичных ооцитов.
Можно отметить серию биохимических изменений, происходя-
щих в яйце в связи с оплодотворением.
1. Еще в 1908 г. О. Варбург обнаружил резкое возрастание по-
требления кислорода тотчас после вхождения сперматозоида. Это
имеет место, например, у яиц морского ежа, асцидий и некоторых
аннелид. Так, по данным Т. Накацавы и др. (1970), через 10—20 с
после оплодотворения интенсивность дыхания яиц морских ежей
резко повышается и сохраняется на высоком уровне 5 мин. Но
нельзя считать это общим явлением, так как, например, у амфибий
не доказано резкое возрастание потребления кислорода, а у мол-
люска Cumingia и аннелиды Chaetopterus, как показали Дж. Нид-
хем (1942) и Л. Ротшильд (1956), понижается потребление кис-
лорода после оплодотворения.
2. В первые 10 мин после оплодотворения отмечено усиление
углеводного обмена, заметное потребление гликогена.
3. В первые же минуты после оплодотворения увеличивается
содержание свободных аминокислот, что, вероятно, является пока-
зателем усиления диссимиляторных процессов.
4. Резко возрастает (в сто раз и более!) фосфатный обмен,
в десять раз и более возрастает калийный и кальциевый обмен.
5. Несомненно, изменяется активность протеолитических фер-
ментов.
6. Изменяется проницаемость яйцевых мембран, она резко воз-
растает, что доказано, например, в отношении фосфатов; изменя-
ются электрические свойства и др.
7. Зрелая яйцевая клетка имеет все необходимое для синтеза
важнейших соединений — белков и нуклеиновых кислот. В изо-
билии имеются типичные рибосомальные частицы, с которыми свя-
зан синтез белков. Доказано наличие в неоплодотворенной яйцевой
клетке иРНК и тРНК и всех необходимых для синтеза белка фер-
ментов.
Несмотря на то что яйцеклетка имеет необходимые аппараты
для синтеза белков, многочисленные исследования показывают,
что синтез белка в ней не происходит. Причины этого не ясны:
имеются ли в яйце какие-либо ингибиторы (тормозящие вещества)
или, несмотря на наличие всех компонентов белоксинтезирующего
аппарата, отсутствуют какие-то необходимые связи и сочетания
между этими компонентами.
68
При оплодотворении незамедлительно начинается интенсивный
синтез белков, матрицами для которого в течение первых стадий
развития служат в основном матричные (информационные) РНК,
синтезированные в процессе развития яйцевой клетки. Но тотчас
после оплодотворения начинается и синтез новых мРНК. Особенна
много доказательств этому дано в опытах Т. Гультина (1950) и
Д. Стаффорда (1964) на яйцах морских ежей.
Активацию белоксинтезирующего аппарата в яйцеклетках
в связи с оплодотворением можно доказать и в опытах in vitro.
Т. Гультин в 1961 г. обнаружил, что рибосомы, выделенные из не-
оплодотворенного яйца в бесклеточной среде, оказываются неак-
тивными, а рибосомы, выделенные из яйцеклетки через 10 мин по-
сле оплодотворения, уже синтезируют белок. Через час после опло-
дотворения (начало первого дробления) активность выделенных
рибосом достигает максимума.
ж. Браше (1963) и В. Гличин (1965) отмечали, что в зрелых
яйцевых клетках морских ежей до оплодотворения не происходит
синтеза РНК. Тотчас после оплодотворения начинается синтез не-
рибосомальных типа информационных РНК.
Следует отметить, что синтез белков активируется как при
оплодотворении, так и при партеногенезе.
Механизм активации яйца при оплодотворении не выяснен. Из-
вестно только, что у морских ежей сначала происходит усиление
интенсивности дыхания, разрушение кортикальных гранул, акти-
вация некоторых ферментов, а уже затем — лишь через 5—7 мин
после оплодотворения наблюдается возрастание синтеза белков.
Через 30 мин после оплодотворения яйца морского ежа уровень
белкового синтеза возрастает в несколько раз по сравнению с не-
оплод отворенным.
Интересный факт обнаружен при изучении рибосом. В резуль-
тате оплодотворения происходит быстрое образование полисом-
ных структур. В неоплодотворенном яйце морского ежа лишь не-
многие рибосомы объединены в полисомы, а через 40—120 мин
после оплодотворения 25—40% рибосом объединяются в полисом-
ные агрегаты. Благодаря этому постепенному изменению (форми-
рование полисом) и возрастает скорость синтеза белка.
Синтез ДНК, согласно данным Ж- Браше (1968), начинается
тотчас же после оплодотворения. Несколько позже начинается син-
тез РНК, а затем и синтез белков. Однако имеются данные о том,
что синтез иРНК начинается также тотчас после оплодотворения.
Из всех данных биохимии можно сделать заключение, что опло-
дотворение яйца — это начало работы всего аппарата, от которого
зависит синтез белка, начало функционирования нового ядра,
о чем свидетельствует синтез новых иРНК.
ЕСТЕСТВЕННЫЙ И ИСКУССТВЕННЫЙ ПАРТЕНОГЕНЕЗ
Активация яйца к развитию может происходить и без участия
сперматозоида. У некоторых животных развитие яиц без оплодо-
творения происходит строго закономерно и называется естествен-
ным партеногенезом (от греч. parthenos — девственница и genea—
развитие). Партеногенез распространен среди насекомых (тлей),
низших ракообразных, коловраток, встречается у моллюсков и дру-
гих животных. У коловраток в течение лета из отложенных яиц
развиваются только поколения самок, которые, в свою очередь,
несколько раз откладывают летние яйца, развивающиеся без опло-
дотворения. В последних партеногенетических поколениях появля-
ются самцы и самки, откладывающие зимние, оплодотворенные,
яйца. Своеобразное явление — так называемый факультативный
партеногенез — имеет место у пчел, ос и муравьев. После осемене-
ния в семяприемнике царицы оказывается много сперматозоидов.
Семяприемник устроен так, что самка как бы по произволу может
откладывать оплодотворенные или неоплодотворенные яйца. Опло-
дотворенные яйца развиваются в личинки рабочих пчел или цариц,
а неоплодотворенные — в личинки трутней.
Естественный партеногенез происходит и у некоторых живот-
ных (например, у сосальщиков), которые размножаются уже на
личиночных стадиях развития. Такой партеногенез называется пе-
догенезом.
Возникают два вопроса: 1) генетико-эмбриологический и
2) чисто эмбриологический. Первый вопрос — каков набор хромо-
сом у соматических клеток партеногенетически развивающихся
животных? У пчел и ос оказывается гаплоидный набор хромосом.
В соматических клетках других развивающихся партеногенетиче-
ским путем животных (тлей, коловраток, ракообразных) оказыва-
ется диплоидный набор хромосом. Последнее обеспечивается раз-
ными путями, один из которых заключается в том, что партено-
генетически развивающееся яйцо начинает развитие после того,
как первое редукционное тельце втягивается обратно и соединяет-
ся с ядром яйца. Так происходит у рачка Artemia salina. В других
случаях может задерживаться выделение 2-го редукционного тель-
ца (у тлей). Редукция числа хромосом может и не происходить
(у некоторых орехотворок).
Второй вопрос — каков механизм естественного партеногенеза?
Сравним ли он с процессами, происходящими при оплодотворении?
К сожалению, эмбриология еще не располагает достаточными дан-
ными, чтобы ответить на этот вопрос. Следует думать, что в случа-
ях естественного партеногенеза происходят такие же морфологи-
ческие и биохимические процессы, и прежде всего в кортикальном
слое яйца, какие имеют место при оплодотворении. Какие-то фак-
торы внешней среды (а в случаях, подобных факультативному пар-
теногенезу у пчел, какие-то эндогенные факторы) играют роль
70
сперматозоидов, обеспечивая «пусковой механизм» активации
яйца.
Для более полного понимания механизма активации яйца спер-
матозоидом или естественными партеногенетическими агентами
большое значение приобрели опыты по искусственному партеноге-
незу, начатые еще в прошлом столетии. А. А. Тихомиров в 1886 г.
впервые обнаружил, что неоплодотворенные яйца тутового шелко-
пряда начинают развиваться после обработки их концентрирован-
ной серной кислотой, при изменении температуры или после меха-
нического повреждения, вызываемого потиранием яиц щеткой. Уда-
валось выводить полноценных гусениц из активированных таким
образом яиц.
Б. Л. Астауров (1940—1960) разработал промышленный спо-
соб стимулирования развития неоплодотворенных яиц тутового
шелкопряда. Используя особые методы обработки яиц тутового
шелкопряда высокой температурой и на определенной стадии раз-
вития, ему удалось получить в большом количестве партеногенети-
ческие бабочки женского пола.
Многочисленные эксперименты по партеногенезу проведены
многими исследователями. О. и Р. Гертвиги сообщили о возмож-
ности партеногенетического развития яиц морских ежей после об-
работки их хлороформом или стрихнином.
Обширные наблюдения были проведены Ж. Лёбом (1913,1916).
В частности, он убедился в том, что если яйца морского ежа будут
находиться в течение нескольких минут в растворах масляной,
уксусной или валериановой кислот, а затем последовательно пере-
несены в подщелоченную морскую воду и нормальную воду, то
образуется оболочка оплодотворения. Такое же действие оказыва-
ют и другие жирные кислоты. Ж. Лёб считает, что во всех указан-
ных и иных случаях партеногенеза сначала происходит частичный
цитолиз кортикального слоя яйца. Последующая (после действия,
например, кислот) обработка яйца подщелоченной морской водой
корригирует повреждение поверхностного слоя яйца, в результате
чего усиливаются окислительные процессы.
Близки к взглядам Ж. Лёба мысли Ж- Рунстрёма (1949), кото-
рый утверждал, что «ключом» для активации яйца является
разрушение энзимо-ингибиторного комплекса, находящегося в кор-
тикальном слое яйца. Это разрушение вызывается факторами, про-
дуцируемыми сперматозоидом; они связывают ингибитор и осво-
бождают энзим. Ингибитором, по Рунстрёму, является мукополи-
сахарид, сходный или идентичный с веществом желточной оболоч-
ки. Гипотезу Ж. Лёба, как и ее дальнейшие видоизменения, нель-
зя еще считать вполне обоснованными.
И. Деляж в 1909 г. обрабатывал неоплодотворенные яйца тан-
нином и аммиаком (первое вещество вызывает коагуляцию, а амми-
ак разжижает). В результате наблюдался партеногенез. Деляж
рассматривал активацию яйца как чередование разжижения и
71
коагуляции. Большой интерес вызвали опыты Е. Батайона, про-
веденные в 1910 г. Он вызывал партеногенетическое развитие яиц
лягушек и жаб уколом иглы, однако процент положительных слу-
чаев в опытах оказывался значительно большим, когда он вводил
вместе с иглой капли крови или кровяных клеток взрослой лягуш-
ки (или соответственно жаб). Батайон думал, что в его опытах
удаляются какие-то токсические вещества, находившиеся в неопло-
дотворенном яйце. Процесс оплодотворения он рассматривал как
«реакцию очищения». Это объяснение опытов Батайона надо счи-
тать мало аргументированным.
Многими авторами испробованы разнообразные химические и
физические агенты, вызывающие партеногенетическое развитие, —
гипертоническая и гипотоническая морская вода; соли калия, каль-
ция, магния и др.; органические кислоты — молочная, олеиновая
и др.; бензин, ацетон; перемежающаяся температура; электриче-
ский ток; ультрафиолетовые лучи и т. д. Создается впечатление
полной неспецифичности партеногенетических агентов. Но это не-
правильно, так как, во-первых, далеко не всякое вещество и не
всякий физический агент может вызвать активацию яйца; во-вто-
рых, партеногенез яйцевых клеток разных видов животных вызы-
вается как общими для всех агентами, так и особыми, применимы-
ми в экспериментах только с данным видом яиц. В то же время
поражает разнообразие испытанных агентов. Все они, однако, вы-
зывают одно общее явление, а именно: они повреждают кортикаль-
ный слой яиц, при партеногенезе возникают процессы, подобные
тем, какие происходят и при оплодотворении. Не случайны, в част-
ности, данные Лёба о лизисе кортикального слоя, наступающем
при действии партеногенетических факторов.
Таким образом, данные по искусственному партеногенезу под-
тверждают гипотезы о важнейшей роли в оплодотворении корти-
кальных изменений в яйцевых клетках.
Проблема партеногенеза ждет новых исследователей, так как
представляет большой теоретический интерес и не безразлична
для практики.
К сожалению, к вопросам естественного и искусственного пар-
теногенеза у позвоночных не было привлечено необходимого вни-
мания. Лишь яйцо лягушки было предметом многочисленных ис-
следований.
Возможность естественного и искусственного партеногенеза
у рыб констатирована уже давно. Удавалось доводить мальков
карпа, развившихся из партеногенетических яиц, до возраста 8,5
месяца. Яйца одних видов рыб могут осеменяться сперматозоида-
ми других видов рыб. Истинного оплодотворения не происходит,
но имеет место активация яйца к совершенно нормальному разви-
тию; при этом развиваются только самки.
Многочисленные давние опыты скрещивания разных видов жи-
вотных между собой позволяют исследователям говорить о неспе-
72
цифичности агентов, побуждающих яйцо к развитию. Скрещивали
разных животных и активировали яйца сперматозоидами другого
вида животных: разные виды морских ежей, яйца морского ежа и
спермий морской лилии или спермий голотурий, кольчатых червей
и моллюсков; опыты по оплодотворению яиц тритона спермиями
саламандры; скрещивание зубра, бизона с другими видами рога-
того скота, лошади и зебры, оленей разных видов и т. д.
М. Олсен в 1954 г. доказал возможность естественного партено-
генеза у птиц. Выведена порода индеек, у которой приблизительно
в половине случаев наблюдается естественный партеногенез; при
этом из неоплодотворенных яиц индеек всегда развиваются самки.
Большой интерес представляют наблюдения И. С. Даревского
и В. Н. Куликовой (1957—1964) за скальной ящерицей Lacerta
saxicola. У трех подвидов этой ящерицы установлен естественный
партеногенез. Удалось, в частности, получить потомство от не-
скольких самок, выведенных в лаборатории и содержавшихся за-
тем на протяжении двух лет изолированно от самцов в террариуме.
Из партеногенетических яиц развиваются исключительно самки.
Т. Меслин в 1962 г. высказал предположение, что некоторым
североамериканским ящерицам — кнемидофорам свойствен полно-
ценный естественный партеногенез. Ж- Пинкусу в 1936 г. удалось
добиться партеногенетического развития яиц млекопитающих (кро-
ликов). Известны многие попытки вызвать партеногенетическое
развитие яиц млекопитающих. Ставились опыты на крысах
(Ж. Пинкус, 1930, 1939), кроликах (К. Аустин, 1956), хорьках
(Ц. Чанг, 1957) и других животных. Партеногенез вызывался
(в опытах in vivo и in vitro) под влиянием низких и высоких тем-
ператур, гипертонической и гипотонической сред, под влиянием
трипсина, суспензии сперматозоидов других видов и т. д. В неко'
торых случаях удавалось наблюдать, что активировалось до 90%
использованных в экспериментах яиц.
А. Витковская (1973) в большом проценте случаев вызывала
партеногенетическое развитие яиц мыши действием электрического
тока 20—50 В. Яйца мыши получали в результате спонтанной ову-
ляции у самок, спаренных с вазэктомированными самцами, или
после гормональной стимуляции. Витковская наблюдала развитие
вплоть до формирования бластоцист. Кариологический анализ за-
родышей (в предимплантационном развитии) показал, что в 60%
случаев клетки гаплоидны, в 16,7% диплоидны, в 1,7% тетрапло-
идны и в 21,6% представляют мозаику из гаплоидных и диплоид-
ных клеток.
А. Тарковский и другие исследователи вызвали партеногенети-
ческое развитие яиц у мышей электрическим воздействием (30—
50 В) на яичник и яйцеводы, при активированной гонадотропина-
ми или спонтанной овуляции после спаривания самок с вазэкто-
мированными стерильными самцами. Имплантирующиеся партено-
генетические зародыши погибали в течение 8—10 дней: из 87 им-
73
плантировавшихся зародышей к 10-му дню жизни живыми оста-
вались только два. Достоверных данных о возможности партено-
генетического развития у млекопитающих на более поздних ста-
диях, чем бластоциста, пока нет.
Почти все, что говорилось в предыдущем разделе о биохимиче-
ских изменениях, наступающих в яйце в связи с оплодотворением,
относится и к яйцу, активированному к развитию партеногенети-
ческими агентами. Сразу начинается интенсивный синтез белка.
По скорости включения меченых аминокислот яйцеклетки после
оплодотворения и активации партеногенетическими факторами не
отличаются друг от друга в течение первых часов развития.
Белковый синтез можно обнаружить не только у целых яиц,
активированных к развитию, но и в безъядерных фрагментах.
Э. В. Гарвей (1936) удалось партеногенетическими агентами акти-
вировать к развитию безъядерные участки яйца морского ежа.
П. И. Денни и А. Тайлер (1964) показали, что в таких безъядерных
фрагментах также начинается интенсивный синтез белка. Этот
•факт дополнительно свидетельствует о том, что в цитоплазме не-
оплодотворенных яиц имеются все необходимые компоненты для
белкового синтеза, включая и готовые мРНК.
На основании сказанного следует, что оплодотворение являет-
ся совокупностью процессов, начинающихся с контактирования
яйца и сперматозоида и до момента слияния пронуклеусов. В ре-
зультате этого оплодотворенная яйцевая клетка приобретает ди-
плоидный набор хромосом, обеспечивающий передачу наследствен-
ных свойств, химическими и морфологическими явлениями в кор-
тикальном слое яйца и во всей его протоплазме, что обусловливает
переход яйца из состояния анабиоза в активное состояние. Стано-
вится возможным онтогенез яйца как клетки, ее развитие, способ-
ность дойти в этом развитии до митотического деления.
Глава IV
ДРОБЛЕНИЕ ЯИЦ
ПРЕДВАРИТЕЛЬНЫЕ ПОНЯТИЯ О ДРОБЛЕНИИ
Оплодотворенное яйцо является клеткой и в то же время это уже
организм на ранней стадии развития. В результате клеточных де-
лений и других процессов одноклеточный организм превращается
в многоклеточный. Возникшее при оплодотворении путем соеди-
нения пронуклеусов ядро обычно через несколько минут начинает
делиться, вместе с этим происходит и деление протоплазмы. Кдеъ_
,ки_дробящегося яйца называют бластомерами^ процесс деления
называется дроблением или сегментацией. Вдавления протоплаз-
мы называют бороздами дробления. У одних животных (напри-
уйГёрттГягушек) “борозда дробления появляется в области анималь-
ного полюса и распространяется к вегетативному. Одновременно
борозда углубляется до центральной части яйца. Еще до оконча-
ния первого дробления на поверхности появляется вторая борозда
дробления. Таким образом, бластомеры во внутренних своих
частях некоторое время остаются неразъединенными. У других
\яиц борозда дробления почти одновременно опоясывает все яйцо.
Период дробления завершается стадией развития организма,
называемой бл астрлой._Тсм ,~гл7VI ГОд на из важнейших особенно-
стей периода дробления—отсутствие роста ра!звйва1ощетося~орга^
низма. несмотря на совершающиеся дкдения. Более того, к концу
дробления объем ^а^дыша несколько уменьшается. Деление кле-
ток взрослых организмов, как правило, интимно связано с ростом
клеток. В онтогении же бластомеров нет периода роста, бласто-
меры перед дроблением не увеличиваются в размерах.
Таблица 1. Изменение соотношений ядра и цитоплазмы в ходе дробления яиц
морского ежа
Стадии Объем Отношение объема плазмы к объему ядра
всей клетки, мкм* плазмы, мкм* ядра, мкм8
2 бластомера . 100 000 91653 8 347 10,9
4 43 ОСО 41 402 1598 25,9
8 бластомеров 22 343 21 387 956 22,3
16 , 9 361 8 783 578 15,2
32 бластомера 4814 4 314 5С0 8,6
64—132 1 633 1 438 215 5,8
Бластула 759 656 103 6,31
75
Ядра. Дррб^щихДлщшшеррв.ще уменьшаются в. размерах^ в то
время как объем протоплазмы в результате каждого очередного
дробления''уменьшается вдвое. Поэтому к шестому или седьмому
дроблению яиц (а у некоторых животных уже к четвертому или
пятому) восстанавливается нормальное соотношение объемов .ядра
и цитоплазмы в бластомерах, которое, как мы видели, оказывается
сильно нарушенным^ половых клеток (табл. 1).
С теми или иными отклонениями от ожидаемых теоретических
цифр такое изменение ядерно-плазменного отношения происходит
при дроблении всех яиц. Например, перед оплодотворением яйца
морского ежа отношение объема ядра к объему плазмы 1 : 550,
а к концу дробления, т. е. на стадии бластулы, это отношение 1 :6.
КЛАССИФИКАЦИЯ ЯИЦ ПО КОЛИЧЕСТВУ ЖЕЛТКА
И РАСПРЕДЕЛЕНИЮ ЕГО В ЦИТОПЛАЗМЕ
Разным группам животных свойствен свой способ дробления.
Вместе с эволюцией организмов происходит эволюция в структуре
и биохимических свойствах яиц, в их «проморфологии», опреде-
ляющей особенности дробления. Не созданы, однако, гипотезы об
эволюции дробления, которые удовлетворяли бы сравнительных
эмбриологов. Проблема крайне осложняется тем, что, с одной
стороны, в пределах отдельных типов существуют разные способы
дробления, с другой стороны, у очень далеких друг от друга жи-
вотных могут быть одни и те же способы дробления. В данном'
•разделе сначала дается чисто формальная морфологическая
классификация способов дробления яиц, а затем делается попыт-
ка причинного анализа.
Большинство вопросов о причинах, дррбдения. .остается еще .нет
ясным из-за недостатка биофизических и биохимических данных.
Далека от ясности центральная проблема — почему онтогенез
клеток и бластомеров завершается митозом. При такой ситуации
нельзя пренебрегать различными гипотезами, которые хотя бы
отчасти помогают объяснять процессы дробления. Важнейшей яв-
ляется гипотеза, высказанная_'О,. Гертвигом. Она связывает осо-
бенности'Дроблеция с количеством желтка _и разным распределе-
нием его в цитоплазме. Вот почему, прежде чем описывать мор-
фологию начальных стадий развития зародышей разных живот-
ных, мы должны остановиться на классификации яиц по количе-
ству и распределению желтка.
Алецитальные яйца. Это яйца почти без желткаиди „скачешь
незначительным количеством его. г 'Гаковы, например, яйцевые
клетки [млекопитающих, плоских червей_^рис. 29,А). Поскольку
яйцевые клетки млекопитающих все же содержат небольшое ко'
личество относительно равномерно расположенного желтка, то их
можно называть и изолецитальными яйцами.
76
Рис. 29. Типы яйцевых клеток по
количеству и расположению желт-
ка:
А — алецитальиое яйцо; Б — гомолеци-
тальное (изолецитальное); В—телоле-
цитальное; Г — центролецитальное
Гомолецитальные, или изоде-\
цитальные, яйца. В этих яйцах
желток относительно равномерно
распределен в цитоплазме, ^дро
у таких яиц расположено, в цен-
тре (рис. 29,Б). Такое строение
Имеют яйца многих моллюсков,
иглокожих и ланцетника. Гомо-
"лецитальные яйца могут иметь и
очень большое количество. желт-
ка, например яйца гидр или не-
мертины Lirieiis gesserensis.
Телолецитальные яйца (рис.
29,В). ЖеЛтОК режб неравномер -
но распределен'~~в' цитоплазме
б области аномального полюса
мало или почти совсем нет желт-
ка; большая часть его находится
в вегётативном полушарии, где
желточные зерна и•пластинки
лежат более плотно.
Примером телолецитальных яиц являются яйца амфибий, на-
пример лягушек. Ооцит и зрелое яйцо лягушки отчетливо поляр-
,ны: оци~~имёют~ длинную ось и, перпендикулярно к ней — более
короткую. В области вег.ет.ативного ‘лолюса"лицеи светлое Д серое
или желтоватое), ацимальная же половина пигментирована. У яиц
одних видов лягушек пигмент черный, а у других—-'бурый или
коричневый. Отмечен и градиент расположения РНК вдоль ани-
мально-вегетативноц оси__Положение ядра эксцентричное; оно
находится в анимальиой половинёГвыше экватора. В анимальиой
половине значительно .меньше желтаа~~^бКГ"К^ёгетативной7ПЕсли
мы повернем оплодотворенное яйцо так, чтобьг пигмента ров энный
полюс оказался внизу, а светлый наверху, то яйцо незамедлитель-
но возвратится к нормальному положению. Это обусловлено оби-
лием желтка в вегетативном полушарии, имеющем больший
удельный вес по сравнению с остальными частями яйца.
Такое же строение имеют яйца птиц (см. рис. 3), у которых,
однако, область желтка достигает огромных размеров по сравне-
нию с очень небольшим y4acTKQH^a^.2{S§^&fneBlJiif дйскд'м)»
менее загруженным желтком. Здесь расположено ядро. Не случай-
но собственно яйцо птицы в общежитии называется желтком,.
Зародышевый диск в первые дни развития зародыша находится
наверху. Это имеет, вероятно, большое физиологическое значение,
так как «верхняя часть» яйца под наседкой всегда имеет более
высокую температуру, чем нижняя (выше на 2—3 и более граду-
сов). Обычная создающаяся под наседкой температура оказывает-
ся оптимальной для собственно зародыша, но она является Крита-
77
ческой для различных ферментов, имеющихся в вегетативном по-
лушарии.
Телолецитальные яйца с огромным количеством желтка, зани-
мающим большую част^'яйца, свойственны и некоторым другим
животным, например головоногим моллюскам.
Центролецитальньге яйца имеют большое количество равно-
мерно расположенного желтка (поэтому могут быть условно на-
званы также гомолецитальными), однако у поверхности яйца име-
ется тонкий слой цитоплазмы, почти лишенной желтка (рис. 29,Г).
Расположенное ближе к центру яйца ядро также окружено слоем
относительно чистой цитоплазмы. Эти яйца характерны для чле-
нистоногих.
МОРФОЛОГИЯ ДРОБЛЕНИЯ ЯИЦ
Полное и частичное дробление. Различают дробление полное или,
голобластическое, и частичное — меробластическое.
а
Рис. 30. Развитие морского ежа:
Д—£ —стадии дробления; Ж, 3 — бластула;
X — гаструляция; Ji—O — личинки: 1 — первичная
мезенхима, 2 —вторичная мезенхима, 3 — бласто-
пор
и,
При полном дроблении
(рис. 30) вся яйцевая клет-
ка дробится на меньшие
клетки. Проникая до самых
глубоких частей яйца, бо-
розды дробления распро-
страняются в то же время
по всей поверхности яйца.
Это свойственно яйцам але-
цитального, изолецитальцо-
го типа й некоторым яйце-
вым клеткам телолёциталь-
ного типа, например яйцам
амфибий (см. рис. 33).
При частичном дробле-
нии борозды дробления не
проникают глубоко внутрь
яйца, большая часть яйца
оказывается вне сегмента-
ции. Частичное дробление,
в свою очередь, у разных
животных оказывается очень
своеобразным. Следует раз--
личать поверхностное иди-
скоидальное дробления. По-
верхностное дробление (рис.
31), характерное для насе-
комых и большинства дру-
гих членистоногих, отлича-
ется следующими особенно-
стями, связанными сострук-
78
А
Рис. 31. Поверхностное дробление яйца
жука Ну drophilus (по К. Хейдеру, 1890):
А, Б — раннее дробление; В — позднее дробле-
ние; Г — начало образования бластодермы (пе-
рибластула)
турой центролецитальных
яиц:"У"'ЭТйг-яиц‘ свободна
от желтка цитоплазма, ок-
ружающая расположенное
в центре ядро и ее перифе-
рический слой. В густой мас-
се желтка имеются тончай-
шие \ цитоплазматические
тяжи, благодаря которым
вся цитоплазма яйца (и пе-
риферийная ее часть, и уча-
сток вокруг ядра) оказыва-
ется единым целым. Ядро
делится на два, четыре
и т. д. После нескольких
делений (неодинаково у раз-
ных яиц) ядра с небольши-
ми количествами .ЦИтбплазмы передвигаются от центра к поверх-
ности яйца (рис. 31,А, В). Цитоплазма, сопровождающая-каждое
ядро, смешивается с поверхностной цитоплазмой яйца. Мигри-
рующие на поверхность яйца ядра располагаются на довольно,
близких расстояниях друг от друга, образуя синцитиальный
слой. Затем наступает процесс очень своеобразной сегментации
периферийной, цитоплазмы, происходит постепенное обособле-
ние вокруг ядер клеточных территорий^ (рис. 31,В, Б). Борозды
дробления появляются, но они не заходят глубоко в яйцо. В~рё-
зультате периферийная часть яйца превращается в сплошной эпш
телиепрдобньщ сдо-й. клеток—в бластодерму, внутренние участки
которой не ограничены от нераздробившейся центральной массы
яйца. На следующем этапе развития зародыша бластодерма мор-
фологически целиком обособляется от желтка и приобретает ис-
тинно^кле точную структуру. У большинства яиц, которым свойст-
венно такое дробление, ...о.1дельнь1еядр'а'С'небильшим участком
ц«топлазмь1...осташ.ся^В--.скюллениях желтка. Эти клетки — вител-
лрфаги («пожиратели желтка», желточные клетки), перерабаты-.
вающие желток, не участвуют непосредственно в построении за-
родыша, но выполняют важную физиологическую роль ^“эмбри-
ональном развитии.
При развитии центролецитальных яиц полость дробления не
возникает, и только на более поздних ста лй'ях~~ра~звитйя бласто-
Дерма. отделяется от лежащего.дод ней желтка.
У многих яиц, которым свойственно также частичное дробле-
ние, оно совершается не по всей__поверхности, а лишь в районе
анимального полюса, где расположено ядро. Такое ухробление
навываетсяТЭйскоцЗальмылг. Оно особенно распространено'"у по-
звоночных и характерно для акул, скатов, костистых рыбуггре-
смыкаюшихсши.птиц (рис. 32).
79
Рис. 32. Дискоидальное дробление куриного яйца (по Дж. Паттерсону, 1910):
А — вид с аннмального полюса области дробления; Б — разрезы дробящегося бластодиска
Равномерное и неравномерное дробление. Образующиеся в ре-
зультате дробления бластомеры могут казаться почти одинаковы-
ми по размерам. Такое дробление называется равномерным. Оно
свойственно, как.правило, гомолецитальным яйцам.
Более распространено неравномерное дробление. Типичным
примером является дробление
Рис. 33. Неравномерное дробление
яйца лягушка
яйца лягушки, начиная с треть-
его дробления (рис? 33).
Строго говоря, настоящего
равномерного дробления не су-
ществует. Уже первые лва Лла-
стомера при любом характере
дробления отличаются друг от
друга и по размерам и, особен-
но, по структуре й биохимиче-
ским свойствам (см. гл. XI).
В отношении некоторых живот-
ных, например ланцетника, гово-
80
пят, что дробление их яиц
почти равномерное. По внеш-
нему виду бластомеры кажут-
ся одинаковыми, на самом же
деле в области вегетативного
полюса бластомеры большего
размера и отличаются по фор-
ме. Яйцо одного и того же
вида животного на разных
этапах развития может дро-
биться равномерно или нерав-
номерно. Например, первые
два дробления яйца лягушки
(рис. 33), равномерные, а по-
Рис. 34. Радиальное дробление яйца
голотурии Synanta digitata (по Е. Зе-
ленка, 1936)
следующие — резко неравномерные.
Синхронное и несинхронное дробление. Последовательные дроб-
ления бластомеров могут наступать одновременно! например
у яиц морских ежей до 32-бластомерной стадии цли у амфибий на
ранних этапах дробления. Такое синхронное дробление отличается
от несинхронного дробления, которое наблюдается у низших мно-
гоклеточных животных, у некоторых червей и млекопитающих.
Классификация дробления, основанная на взаимном простран-
ственном расположении бластомеров. В этой классификации вы-
деляют несколько типов.
Радиальное дробление (рис. 34 и 35) в его наиболее типичном
виде характерйзуётся"'Длёдующими особенностями. Первое дроб-
ление меридиональное^ъ'ло проходит в меридиональной плоскости
яйца. Второе дробление также меридиональное, оно проходит че-
рез главную ось яйца, но под прямым углом к плоскости первого
дробления. Плоскость 3-го дроблениш лежит под прямьшжуглом
к 11лрскост^яя.щерш^.ддуд дрйбдений: и главной оси яйца. Это
дробление происходит в плоскости, параллельной" экватору, поче-
му и называется экваториальным или широтным. Четыре бласто-
мера анимального полушария будут лежать непосредственно под
четырьмя бластомерами вегетативного полушария, создавая кар-
тину радиальной симметрии.
Для аннелид, большинства моллюсков, немертин и некоторых
планарий (Polyclada) характерно спиральное дробление (рис. 36
и37|. При этом типе дробления бластомеры, входящие в состав
горизонтальных рядов (образовавшиеся в результате экваториаль-
^“Х делений), не располагаются один над другим: бластомеры
верхнего ряда как бы вклиниваются между бластомерами нижнего
ряда. При спиральном дроблении веретена дроблений после треть-
его дробления направлены косо (рис. 36). Борозды дроблений
проходят не меридионально или экваториально, а под углом в 45°
к~ меридиану и экватору; следующие борозды проходят под пря-
81
яйца ланцетника
Рис. 35. Радиальное дробление
(по А. О. Ковалевскому, 1867)
мым углом к преды-.
дущйм и, значит,
опять-таки под уг-
лом в 45° к эквато-
ру. Если мысленно
продолжить верете-
но дробления в обе
стороны по поверх-
ности яйца, то полу-
чится спиральная
линия (рис. 38). Та-
кая ситуация созда-
ется благодаря тому,
что в анафазе каждого деления
дробления два дочерних бластоме-
ра отклоняются от оси веретена в
противоположные стороны (В. Н.
Мещеряков).
После первых двух дроблений
образуются четыре бластомера, ко-
торые называются основным кварте-
том и обозначаются Л, В, С и D.
Бластомеры основного квартета мо-
гут быть примерно одинакового раз-
мера (рис. 37,£), тогда образую-
щиеся при последующих дробле-
в анимальиой области будут так-
(рис. 37,В). Такое дробление при-
Рис. 36. Схема первых стадий
спирального дробления:
А — переход к восьмиклеточиой ста-
дии; Б — переход к шестнадцатикле-
точной стадии
ниях более мелкие клетки
же одинаковой величины
нято называть гомоквадрантным. Если клетки основного квар-
тета не одинаковы по величине, то и их производные также раз-
личаются. Обычно самые крупные размеры имеет бластомер D и
соответственно микромеры, которые от него отделятся. Неравно-
мерное спиральное дробление называют гетероквад рантным
(рис. 39).
В зависимости от направления смещения мелких бластомеров
(микромеров) по отношению к крупным бластомерам (макроме-
рам) спиральное дробление подразделяется на два типа. При
дексиотропном дроблении смещение микромеров осуществляется
по часовой стрелке (если смотреть на дробящееся яйцо с анималь-
ного полюса). Такой тип дробления изображен на рис. 36 и 37.
При леотропном дроблении микромеры смещаются против часовой
стрелки. Первые четыре микромера называют первым квартетом
микромеров. Их обозначают малыми буквами латинского алфави-
та с коэффициентом 1 (la, lb, 1с, Id). Основной квартет после
выделения первого квартета микромеров называют первым квар-
тетом макромеров; обозначаются они соответственно 1А, IB, 1С
и 1D. При образовании квартетов микромеров происходит чередо-
82
Рис. 38. Схема положения веретен
дробления яйца при спиральном
дроблении:
А — дексиотропное; Б — леотропное дроб-
ление; 1 — анимальиый полюс, 2 — веге-
тативный полюс
Рис. 37. Схема спирального дробле-
ния. Вид с анимального полюса:
А — два бластомера; Б — четыре бласто-
мера; В — восемь бластомеров; Г —
16 бластомеров:
Д, В, С, D — макромеры; a, b, с, d — мик-
ромеры. Коэффициенты перед буквами
означают последовательность их образова-
ния
вание дексиотропных и леотропных дроблений. Для возникающих
после четвертого дробления 16 бластомеров приняты такие индек-
сы: бластомер первого квартета микромеров 1а дробится на бла-
стомеры 1а1 и 1а2, а бластомер основного квартета 1А дробит-
ся на бластомер 2а и 2А. Бластомеры 2а, 2Ь, 2с,и 2d составляют
второй квартет микромеров, а бла-
стомеры 2А, 2В, 2С и 2D образуют
квартет макромеров. Возникшие в
результате очередных дексиотроп-i
ных и леотропных дроблений 32 и
64 бластомера обозначаются по то-
му же принципу.
В ходе спирального дробле-
ния чаще всего выделяется четы-
ре квартета микромеров, которые
соответственно занимают все более
удаленное положение от ани-
мального полюса. Поскольку одно-
временно с делением макромеров
делятся и первые квартеты мик-
ромеров, то к концу дробления
на анимальном полюсе образуется
значительное количество клеток—
производных первого квартета
Рис. 39. Гетероквадрантное дроб-
ление яйца (по А. Пеннерсу, из
кн. П. П. Иванова, 1937):
Л — два бластомера; Б, В, Г — обра-
зование первого квартета микромеров
83
микромеров. Поскольку общее число клеток, образующихся при
спиральном дроблении, невелико, то удается проследить «судьбу»
каждого бластомера. При использовании буквенно-цифровой но-
менклатуры все бластомеры получают определенный индекс. Ока-
залось, что все бластомеры, обозначаемые малыми буквами и
имеющие коэффициент 1, т. е. все клетки, образовавшиеся из пер-
вого квартета микромеров, пойдут на построение предротового
отдела личинки. Из клеток второго квартета возникает венчик
ресниц — прототрох; третий квартет микромеров пойдет на обра-
зование мезодермальных и отчасти эктодермальных производных
личинки и, наконец, четвертый квартет микромеров и четвертый
квартет макромеров образуют кишечник. Особую роль при спи-
ральном дроблении играют бластомеры 2d и 4d. Первый называ-
ется соматобластом, и из него образуется эктодерма заднего отде-
ла личинки, а в дальнейшем покровы взрослого червя, 4d называ-
ется мезобластом. Этот бластомер делится на две клетки
телобласты — родоначальницы мезодермальных телобластических
полосок, из которых образуются целомы личинки, а в ряде слу-
чаев и целомическая мезодерма взрослого организма.
По мнению О. М. Ивановой-Казас, спиральное дробление раз-
вивалось постепенно. Элементы этого типа дробления встречаются
у низших многоклеточных. Так же постепенно происходило и его
«вырождение», т. е. замена другими формами дробления.
Дробление яиц нематод, коловраток, асцидий и некоторых дру-
гих животных имеет ту особенность, что уже на ранних этапах
дробления проявляется билатеральная симметрия в расположении
бластомеров (см. рис. 8). Такое дробление называется билате-
ральным или двустороннесимметричным. Вне зависимости от свое-
образия дробления в деталях (у разных животных) при билате-
ральном типе дробления каждый бластомер правой половины эм-
бриона соответствует подобному бластомеру левой половины.
Следует упомянуть об очень своеобразном двусимметричном
типе дробления, свойственном ктенофорам, у которых уже на
стадии четырех бластомеров определяются обе плоскости симмет-
рии будущего организма — продольная и поперечная.
Строго закономерный характер дробления яиц каждого вида
животных обусловлен сложившимся в ходе эволюции особым
морфофизиологическим механизмом. Кажущимся исключением
является анархическое дробление, которое иначе называют «бес-
порядочным» и «хаотическим», например дробление яиц некото-
рых метагенетических медуз. Анархическое дробление гидромеду-
зы Oceania .armata (рис. 40) описал впервые И. И. Мечников
в 1886 г. Как и у других гидромедуз, никакой «анархии» в начале
дробления нет: первая борозда дробления меридиональная, вре-
зающаяся; появляется она на анимальном полюсе; вторая борозда
тоже меридиональная, но появляется на внутренней стороне блас-
томеров и распространяется центробежно (рис. 40,В). Однако
84
Рис. 40. Развитие гидромедузы Oceania armata.
А—Г — дробление; Д—Е — бластула; Ж — личинка (по И. И. Мечни-
кову, 1886)
порядок взаимного расположения бластомеров после третьего
дробления может варьировать. Чем дальше идет дробление, тем
более проявляется «беспорядочный характер» (рис. 40,В, Г).
Такие своеобразные явления дали повод П. П. Иванову (1945)
утверждать, что яйца некоторых кишечнополостных не имеют ни-
какой детерминации и предварительной структуры. Это утверж-
дение трудно аргументировать, так как анархический тип дроб-
ления кажется хаотичным лишь при условии, если закономерным
считают только такое пространственное расположение бластоме-
ров, которое укладывается в те или иные принятые при описании
дробления геометрические схемы, в понятие о симметрии. Совер-
шенно очевидно, однако, что при кажущемся совершенно «беспо-
рядочным» дроблении яйца гидромедуз развиваются в животные
именно данного, а не другого вида. Не выяснены еще полностью
закономерности такого развития.
Кажущаяся хаотичность и простота анархического дробления
позволили Л. Н. Жинкину в 1951 г. высказать предположение
о том, что оно является примитивным, не установившимся и что
ют него произошли спиральный и радиальный типы дробления.
85
Черты спирального дробления у кишечнополостных отмечал ещ
И. И. Мечников в 1886 г., а впоследствии С. Берг (1941) и К. Ни
хольм (1949). По гипотезе Г. А. Шмидта (1951), радиально
дробление возникло из спирального.
У актинии Sagartia troglodytes дробление может варьировать
Оно может быть равномерным или неравномерным, бластомерь
могут располагаться как при радиальном, так и при спирально»
дроблении.
Вопросы эволюции дробления пока еще слабо разработаны
Вероятно, будут иметь значение исследования эволюции дробле
ния в пределах отдельных типов. При анализе анархическогс
дробления следует принять во внимание своеобразную способ-
ность к полиэмбрионии и соматическому эмбриогенезу (см
гл. XVI) зародышей многих кишечнополостных. Фрагменты заро-
дыша гидромедузы Oceania armata способны развиваться в нор-
мальные личинки уменьшенного размера. Подобные явления от-
мечены и у гидр. М. Н. Грузова, изучая развитие гидр, в 1956 г.
обнаружила, что зародыши, пройдя нормальное, закономерное
развитие вплоть до гаструляции, при формировании эмбриотеки
(особая оболочка вокруг зародыша) и все зимнее время оказы-
вается менее дифференцированным, анархизированным и напоми-
нает структуры геммул и сомитов (см. гл. XVI), хотя до форми-
рования эмбриотеки развитие происходит строго закономерно.
У гидр дробление почти полное и равномерное. Оно заканчивается
целобластулой (см. далее), и закономерно совершается гаструля-
ция. И вот при таком закономерном развитии зародыш теряет
свою нормальную интеграцию и в некоторых случаях вместо
одного нормального зародыша развиваются два. Предполагают,
что в результате такой полиэмбрионии начинается развитие по
типу соматического эмбриогенеза, еще недостаточно изученное
эмбриологами (см. гл. XVI).
Интересно в этой связи отметить, что у Oceania полиэмбриония
не обязательно имеет место, не все бластулы делятся. Форма не-
которых эмбрионов становится постепенно все более правильной,
зародыш удлиняется и, наконец, принимает вид языкообразной
планулы. Это означает, что нет анархии в дроблении, но имеется
стадия дезинтеграции зародыша, которая может привести к по-
лиэмбрионии.
Как видим, ничто не свидетельствует о хаосе анархического
дробдения, и этот термин нельзя считать удачным. Многие водные
низшие многоклеточные организмы обладают низким уровнем ин-
теграции; яйца их мало отличаются от соматических клеток (они
не имеют обычных яйцевых оболочек, у них относительно нормаль-
ное, как и соматических клеток, соотношение ядра и цитоплаз-
мы, нормальный метаболизм). У этих малоинтегрированных орга-
низмов и их зародышей выработался особый характер дробления
как адаптация к меняющимся внешним условиям, выработалась
86
возможность перехода к соматическому эмбриогенезу. Более под-
ходит к такому типу дробления название регуляционный тип
дробления.
СКОРОСТЬ ДРОБЛЕНИЯ. ПРИЧИНЫ ДРОБЛЕНИЯ ЯИЦ
И ДЕЛЕНИЯ СОМАТИЧЕСКИХ КЛЕТОК
Яйцам каждого вида животных свойствен свой особый ритм дроб-
ления. У золотой рыбки Carassius auratus дробление совершается
регулярно через каждые 20 мин. У яиц лягушки первые дробле-
ния совершаются приблизительно через каждый час. Яйца млеко-
питающих дробятся очень медленно: интервалы между дробле-
ниями достигают десяти часов и более.
Авторадиографические исследования позволили узнать инте-
ресные детали изменения времени протекания разных фаз жизнен-
ного цикла клеток (их онтогении). Так выяснено, что на ранних
стадиях дробления у бесхвостой амфибии Xenopus laevis фаза
удвоения ДНК (фаза S) занимает всего 10 мин. Во время гастру-
ляции фаза S составляет приблизительно 1 ч; в ранней нейруле
эта фаза замедляется, занимая до 5 ч. Короткая онтогения блас-
томера на ранних стадиях дробления обусловлена ускорением
времени процесса митоза, фазы (G2), следующей за синтезом
ДНК и отсутствием стадии Gi, следуемой за митозом.
У некоторых животных после совершившихся дроблений насту-
пает пауза, закономерно прерывающаяся в определенные сроки.
У летучих мышей половой акт (осеменение) происходит поздней
осенью, развитие же яйца начинается лишь весной. Пауза в раз-
витии характерна для многих низших ракообразных.
Скорость дробления яиц, естественно, изменяется под влияни-
ем температуры. Обычно закономерности изменения темпа дроб-
ления близки к правилу Вант-Гоффа, установленному для химиче-
ских реакций: при повышении (или понижении) температуры на
10 °C процесс ускоряется (или, соответственно, замедляется)
в 2—3 раза. Следует подчеркнуть, что изменение температуры
(если оно не выходит за определенные для каждого вида живот-
ного рамки нормальных температур) не приводит к качественным
изменениям в развитии, а лишь сказывается на темпе дробления.
Ранний период развития, как правило, характеризуется син-
хронным дроблением. Для митотического цикла дробящихся блас-
томеров характерно полное отсутствие фаз Gi и G2, а также не-
продолжительность стадий S и собственно митоза. Это связано
с тем, что синтетические процессы в бластомерах выражены слабо
из-за того, что почти все необходимое для этих процессов накоп-
лено в период оогенеза и сохраняется в бластомерах. Кроме того,
укорочение S-фазы обеспечивается синхронизацией начала репли-
кации в различных репликонах генома.
Наличие периода синхронного дробления послужило основани-
87
ем для использования продолжительности одного синхронного
дробления в качестве безразмерной единицы для характеристики
продолжительности различных периодов развития (Т. А. Детлаф).
В зависимости от количества желтка, его распределения,
а также неравномерности распределения других цитоплазматиче-
ских компонентов яйца период синхронного дробления раньше или
позже переходит в период асинхронных дроблений. Этот процесс
связан в первую очередь с появлением и удлинением фаз Gi и G2.
Наименее выражен период синхронных дроблений у яиц с мозаич-
ным дроблением, а также у некоторых примитивных многоклеточ-
ных, которые вообще могут иметь слабо упорядоченный в прост-
ранстве и во времени процесс дробления.
Поздний период дробления принято называть бластуляци-
ей, так как в этот период вся совокупность бластомеров приоб-
ретает характер эпителиоморфного пласта, ограничивающего
полость бластулы. Период бластуляции характеризуется удлине-
нием фазы S в митотическом цикле бластомеров. Повышение син-
тетических процессов в дробящемся зародыше связано с исполь-
зованием молекул мРНК, уже имеющихся в яйцевой клетке. Ге-
ном зародыша, возникший после оплодотворения, начинает функ-
ционировать не сразу, а лишь после прохождения определенного
количества делений дробления и нормализации ядерно-плаз-
менного отношения. Чаще всего это приурочено к стадии позд-
ней бластулы — началу гаструляции. По мере дробления вступают
в действие еще недостаточно изученные факторы взаимодействия
бластомеров, факторы интеграции зародыша, которые приводят
к тому, что темпы деления клеток в разных частях зародыша ста-
новятся неодинаковыми. «Угнетающее» действие одних частей
зародыша на другие сказывается и в появлении дегенерирующих
клеток уже на ранних стадиях онтогенеза животных.
Если бы темпы дробления сохранялись неизменными и дроб-
ление оставалось бы синхронным, то следовало бы, например, что
через 20 ч эмбрионального развития лягушки, т. е. приблизитель-
но на стадии начинающейся гаструляции, зародыш состоял бы из
миллиона клеток, чего, конечно не происходит. Через двое суток
на стадии поздней гаструлы имелось бы 25* 1013 клеток, к моменту
ранней хвостовой почки, т. е. к концу суток, количество клеток
превышало бы 1028, а размер зародыша оказался бы 5,2-1012 м3
(если радиус клетки считать постоянным и равным 5 мкм). Вступ-
ление в действие внутренних факторов развития, тормозящих ми-
тозы в одних частях зародыша и стимулирующих митозы в дру-
гих частях, играет важную роль в дифференциации клеток и
тканей в процессах развития. Об этом эмбриологи догадывались
давно, но убедительными экспериментальными доказательствами
этого фактора эмбриология стала располагать недавно.
К этому вопросу имеют отношения гипотезы, основанные на
признании межклеточных взаимодействий, осуществляющихся по-
88
средством специфически действующих веществ (В. С. Буллоу,
1965; П. Вейс, 1957; Г. Туманишвили, 1968). По Буллоу, деление
клетки — нормальное событие в ее жизни. Если бы не было регу-
ляторного контроля над делением, все клетки делились бы с оди-
наковой скоростью, обусловленной их генетическими особенно-
стями.
Таким образом, должны быть факторы, подавляющие деление
клеток. В 70-е и 80-е годы в межклеточной среде были открыты
/кейлоны, (хелоны)—вещества, тормозящие деление клеток. Кон-
центрация кейлонов зависит от числа клеток в данном объеме и
от количества кейлона, вырабатываемого одной клеткой, что обу-
словлено генетически и является особенностью данного клеточного
типа. Количество кейлона, вырабатываемого одной клеткой, веро-
ятно, постоянно и, значит, изменение числа клеток играет решаю-
щую роль в концентрации кейлона. Уменьшение числа клеток
в клеточной популяции приводит к повышению митотической ак-
тивности.
Г. Туманишвили (1968) считает, что в клетке существуют два
типа веществ: в связи с функцией ядер клеток вырабатывается
вещество, тормозящее митотическую активность, но есть в клетке
другое вещество, вырабатывающееся в цитоплазме; оно повышает
митотическую активность. По этой гипотезе концентрация ядер
зависит от митотической активности и находится в обратной за-
висимости от скорости роста цитоплазмы и продукции межкле-
точного вещества. При накоплении «избытка ядер» в ткани насту-
пает торможение клеточных делений. Косвенным подтверждением
наличия веществ является установленный Туманишвили факт, что
при введении в куриные зародыши экстрактов органов взрослой
курицы в соответствующем органе зародыша митотическая актив-
ность повышается.
Некоторыми исследователями придается значение поверхност-
ным мембранам как регуляторам деления животных клеток. По-
верхность клетки в течение жизненного цикла (онтогении) под-
вергается закономерным изменениям. Предполагается, что уста-
новление определенного контакта между клетками может изме-
нять прохождение этапов митотического цикла. Изменение внеш-
них воздействий может вызывать значительные изменения свойств
поверхности клетки, что влияет на прохождение процессов
митоза.
Нелегко применить какую-либо из этих гипотез к явлениям
неодинаковых темпов клеточных делений в разных частях зароды-
ша. Но эти гипотезы заслуживают внимания эмбриологов. Конеч-
но, должны быть факторы надклеточного порядка, возникающие
в результате взаимоотношений клеток развивающегося заро-
дыша.
Почему происходит сегментация яйца? Каков биохимический и
биофизический механизм дробления? Этот далекий от ясности
89
вопрос тесно связан с проблемой деления клеток, хотя дробление
имеет столь специфические черты, что маловероятно существова-
ние одной-единственной причины дробления яиц и деления клеток
тканей. Может быть, неосновательны и поиски единого механизма
дробления телолецитальных, центролецитальных и гомолециталь-
ных яиц. Конечно, должны существовать особые, по крайней мере
дополнительные факторы, которые определяли бы поведение де-
лящихся ядер в яйцах членистоногих и загадочное явление фор-
мирования клеточных перегородок вокруг ядер, оказавшихся на
периферии яйца. Однако в ходе дробления яиц всех животных мы
видим типичное кариокинетическое деление. Не раз отмечавшиеся
при изучении развития тех или других животных картины амитоза
или спорны, или относятся к тем поздним стадиям эмбрионально-
го развития, когда происходит специфическая дифференциация кле-
ток и тканей, при которой амитоз может иногда превалировать
над митозами или даже стать на время исключительной формой
деления ядер. В других случаях амитотическая перешнуровка
ядер, обнаруженная у ранних зародышей, происходит в клетках,
идущих по пути дегенерации. Амитозы наблюдаются на стадиях
гаструлы и нейрулы у амфибий, но, по-видимому, это связано, как
полагает А. Г. Кнорре (1959), с уже начинающимся отмиранием
отдельных клеток. Впрочем, в этом вопросе не все еще ясно. Не
понятны пока механизм и эмбриологическое значение возникнове-
ния у эмбрионов амфибий двуядерных и многоядерных бласто-
меров.
Помимо указанных ранее особенностей дробления, по сравне-
нию с митозом, а именно отсутствия роста бластомеров, прогрес-
сирующего уменьшения цитоплазмы можно отметить и ряд других
отличий. Если при нормальных митозах, как правило, телофазное
состояние ядер возникает параллельно известным процессам де-
ления протоплазмы, то при дроблении яиц, например амфибий,
деление ядра и цитоплазмы происходит отчасти асинхронно: ядро
переходит в стадию профазы еще до того, как заканчивается об-
разование борозды дробления.
Гипотеза А. Г. Гурвича (1923—1945) о митогенетических лу-
чах— об ультрафиолетовых лучах порядка 193—220 нм, продуци-
руемых клетками и играющих роль «фактора осуществления»
митоза «готовой» к делению клетки, к сожалению, оказалась оши-
бочной (Б. П. Токин, 1933, и др.). Если бы и было подтверждено
существование этих лучей, они оказались бы не «митозорождаю-
щими», а одним из стимуляторов клеточного деления. Этим не
умаляется значение творчества А. Г. Гурвича и, в частности, его
работ о митогенетических лучах; он приковал внимание биологов
и физико-химиков к проблеме излучений при биохимических ре-
акциях, что уже представляет огромный интерес. Теория «гормо-
нов клеточного деления» Г. Габерляндта (1918), фактически со-
вершенно доказанная, не является, однако, теорией клеточного
90
деления, а касается также явлений стимулирования делений и
к своеобразным процессам дробления она неприменима. При
анализе дробления может быть принята во внимание и интересная
гипотеза Р. Гертвига, согласно которой следует видеть причину
деления клеток в меняющемся ядерно-плазменном отношении
(Kernplasmarelation).
Объем ядра сначала отстает в своем росте от объема плазмы,
но незадолго до следующего деления резко возрастает. Это при-
водит к ядерно-плазменному напряжению. Когда оно достигает
наивысшей точки, ядро приобретает способность расти за счет
плазмы, возникают токи жидкостей, создаются и другие физико-
химические условия, вызывающие деление клетки. Этой старой
гипотезой, вероятно, не следует пренебрегать, хотя в биохимиче-
ском и физиологическом отношении гипотеза эта совершенно не
конкретна, а применить ее к дроблению яиц не представляется
возможным, так как при дроблении яиц отсутствуют явления рос-
та клеток, однако и в этом случае ядерно-плазменные отношения
изменяются.
Многочисленные гипотезы в последние десятилетия касаются
собственно деления ядер, а не дробления яиц в целом. Таковы
предположения о том, что пусковой механизм митоза — это явле-
ние удвоения ДНК, что хромосомы вырабатывают особое веще-
ство, вызывающее деление клетки, а также другие гипотезы.
Возможно (хотя и уродливое) дробление при полном отсутст-
вии ядра, что впервые наблюдала Е. Гарвей в 1936 г. Это сооб-
щение было встречено резким скептицизмом эмбриологов, но впо-
следствии оно полностью подтвердилось. Гарвей центрифугирова-
ла яйца морских ежей. Отрывающиеся при сильном
центрифугировании безъядерные фрагменты яйца активировались
к развитию партеногенетическими агентами. Несмотря на полное
отсутствие ядер происходило дробление (во всяком случае про-
цессы, напоминающие дробление). Дробление доходило до стадии
бластулы, и как будто возможны были случаи развития бластул
с ресничным аппаратом у клеток.
Представление о значении хромосом в делении клетки не под-
тверждается, например, таким опытом: если удалить ядро перед
делением инфузории стентор, деление ее все равно может совер-
шиться и образуются две новые клетки (В. Тартар, 1961).
Множество фактов говорит о том, что дробление яиц и размно-
жение клеток не может считаться только функцией самих единич-
ных клеток.
Заслуживают внимания следующие гипотезы о причинах появ-
ления борозд дробления.
1. Согласно «гипотезе роста звезды» «звезды» (хромосомы,
удаляющиеся к полюсам), возникающие при митозе, по мере уве-
личения обусловливают удлинение клетки, увлекают за собой кле-
точную поверхность, вызывая образование борозды дробления.
91
2. «Гипотеза удлинения веретена» основана также на явлениях
изменения митотического аппарата. Согласно ей лучи «звезды»
прикрепляются к поверхности клетки, веретено деления удлиня-
ется, а экваториальная поверхность втягивается внутрь.
3. «Гипотеза амебоидного движения» утверждает, что у каж-
дого полюса делящейся клетки или бластомера происходит обра-
зование псевдоподий, возникают амебоидные движения, направ-
ленные в противоположные стороны, что и вызывает разделение
клетки или бластомера. Эта гипотеза заслуживает серьезного
внимания, так как явления амебоидного движения бластомеров и
клеток, как увидим далее, играют, несомненно, большую роль
в различных морфогенетических процессах.
4. Суть «гипотезы сократимого кольца» состоит в кажущемся
убедительным и основанном на фактах предположении о наличии
особых уплотненных участков кортикального слоя яиц. Сокраще-
ние полоски желатинизированного кортикального слоя приводит
к разделению яйца или бластомера. Яйцо или бластомер можно
уподобить резиновому мячу, перетягиваемому шнурком. Одно из
возражений против этой гипотезы таково: приходится предпола-
гать невероятно сильное сокращение, которому нет аналогий в фи-
зиологии; «кольцо» должно сокращаться, например, у яиц морских
ежей до 0,05 своей первоначальной длины (первоначальная длина
«кольца» около 300 мкм, а при дроблении диаметр перетяжки
менее 5 мкм). Даже мышца способна сокращаться только до 0,1
своей длины, а при переходе золь^гель сокращение еще мень-
шее.
5. Представляет интерес эксперимент Д. М. Арнольда (1971)
на зародыше осьминога Loligo peulii. На стадии 1—4 борозд дроб-
ления подрезали стеклянной иглой основание борозд дробления
(область цитоплазмы, расположенной непосредственно под бороз-
дой дробления). Борозда исчезала или сокращалась. Автор пред-
полагает, что в основании борозды дробления имеется волокни-
стая полоска, сокращение которой ведет к втягиванию борозды
дробления.
6. «Гипотеза расширяющейся мембраны», высказанная
М. Сванном и Ж- Митчисоном (1952—1958), заинтересовала мно-
гих эмбриологов. Авторы придают решающее значение факту
увеличения, растяжения площади поверхности яйца. Если помес-
тить на поверхность дробящегося яйца морского ежа частицы
каолина или угольного порошка и измерять расстояние между
частицами в ходе дробления, то обнаружится растяжение корти-
кального слоя яйца. Оно возникает у полюсов яйца (у полюсов
будущей митотической фигуры) и распространяется по его по-
верхности в направлении к экватору митотической фигуры. Лежа-
щий у экватора участок сначала сокращается, а затем, когда
образуется борозда дробления, сильно растягивается. Кортикаль-
ный слой яйца благодаря особому расположению молекул белка
92
подобен эластической мембране, натяжение которой возрастает
при растяжении. При растяжении поверхностного слоя яйца у по-
люсов происходит соответствующее сокращение у экватора. В ре-
зультате яйцо сначала несколько уплощается, а затем приобрета-
ет форму гимнастической гири, начинается перетягивание яйца.
Сванн и Митчисон предполагают, что дочерними хромосомами
в анафазе выделяется Х-вещество, от которого зависят изменение
в расположении молекул и сокращение мембраны. Это вещество
достигает участков полюсов яйца скорее, чем участков поверхно-
сти, расположенных у экватора, так как первые находятся на бо-
лее близком расстоянии от источника Х-вещества (т. е. от хромо-
сом). Если гипотеза об этих веществах вызывает сомнение (хотя-
бы уже потому, что дробление возможно, как увидим далее, и без
ядра), то представления о сокращающихся мембранах можно
считать доказанными. Данные других исследователей прямо или'
косвенно подтверждают эти представления. Д. Марслянд и
Ж- Ландау в 1954 г. доказали, что в начале дробления яиц мор-
ского ежа Arbacia повышается вязкость вертикального слоя пре-
имущественно на месте будущей первой борозды, а на полюсах
вязкость уменьшается. Это приводит к удлинению полярной оси.
По мере стягивания экваториального пояса кортикального слоя
образуется борозда дробления. Сжатие экваториального пояса
кортикального геля, по мнению названных исследователей, — ре-
шающий фактор дробления.
Интересную гипотезу о механизме образования борозд дроб-
ления у яиц осетровых рыб и амфибий предложил А. И. Зотин
в 1962 г. Поверхностный слой яиц способен к возбуждению и со-
кращению в ответ на механическое раздражение. Раздражителем,
вызывающим сокращение поверхностного слоя яиц при образова-
нии борозды дробления, является внутренняя стенка (диастема),
которая возникает в ранней анафазе в средней части веретена и
к концу анафазы приближается к поверхности яйца. Диастема —
узкая зона слабо окрашивающейся, видоизмененной цитоплазмы,
расположенной на месте будущей «стенки», разделяющей бласто-
меры. Подобные структуры действительно найдены у яиц морских
ежей, амфибий, осетровых рыб. Зотин перемещал диастему внутри
цитоплазмы путем сжатия яйца, частично разрушая ее с помощью
тяжелой воды или механическим путем. Такими способами удается
вызвать образование дополнительных борозд дробления и появле-
ние избыточных бластомеров.
8. Интересные данные о механизме дробления яиц амфибий
были получены С. Сондерсом (1974). Оказалось, что в месте по-
явления врезающейся борозды поверхность яйца становится
складчатой, что является результатом локального сокращения
микрофиламентов поверхностного слоя цитоплазмы. В области
небольшой щели, возникающей на границе двух бластомеров
в анимальной части яйца, к поверхностным мембранам подходят
93-
микропузырьки, продуцируемые комплексом Гольджи. Мембраны,
ограничивающие эти пузырьки, сливаются с плазматической мем-
браной бластомеров, увеличивая тем самым ее поверхность. Со-
держимое пузырьков выливается в межклеточную щель. Это —
начало образования внутренней среды зародыша.
Таким образом, процесс дробления включает наряду с процес-
сами перестройки микрофиламентов и цитоскелета, ведущих к об-
разованию «сократимого кольца», также и новообразование
участков поверхностных мембран благодаря синтетическим и экс-
креторным цитоплазматическим процессам.
ПРАВИЛА О. ГЕРТВИГА И ИХ ПРИМЕНЕНИЕ К АНАЛИЗУ
РАЗВИТИЯ АМФИБИЙ, РЫБ, ПТИЦ И НАСЕКОМЫХ
Уже упоминалось о большом значении, которое придает эмбрио-
логия правилам О. Гертвига для понимания причин разных типов
дробления яиц.
Первое правило Гертвига: ядро занимает центр активной
протоплазмы (полностью или почти полностью свободной от
желтка). В алецитальных и изолецитальных яйцах ядро располо-
жено ближе к центру (рис. 41,Л), а у телолецитальных оно рас-
полагается эксцентрично, в анимальном полушарии.
Второе правило Гертвига: веретено деления ядра располага-
ется в направлении наибольшей протяженности чистой протоплаз-
мы (рис. 41,Б).
Ярким примером правильности этих утверждений служит
характер дробления телолецитальных яиц амфибий. Ядро у них,
как уже сказано, расположено в анимальном полушарии. Наи-
большая протяженность чистой протоплазмы параллельно эквато-
риальной плоскости (см. рис. 42,Л и Л'). В этом направлении и
окажется веретено первого деления ядра. Кариокинетический
процесс связан с передвижением раз-
личных составных частей клетки, ее
хромосом, частей цитоплазмы, корти-
кального слоя. Классическая описатель-
ная гистология и цитология, так много
давшие биологии с - их методиками
фиксации и окрашивания клеток, при-
вивают, однако, изучающему клетки, тка-
ни и развитие организма совершенно
не соответствующее действительности
представление о клетках, как о чем-то
неподвижном. Мы неоднократно будем
убеждаться в том, что такое представле-
ние о клетках необоснованно.
Различного рода закономерным дви-
жениям составных частей ядра и цито-
1 Б
Рис. 41. Образование пер-
вого веретена дробления яй-
ца круглого червя (по
О. Гертвигу, из. кн.
П. П. Иванова, 1937):
А — сближение пронуклеусов;
.Б — веретено дробления
^4
плазмы могут мешать зерна и пластинки желтка. Если желток
расположен очень компактно, то процесс дробления в определен-
ном участке яйца может подавляться.
Первая борозда дробления у яиц амфибий определяется ука-
занным направлением веретена деления ядра (параллельно эква-
ториальной плоскости, иначе — перпендикулярно к анимально-
вегетативной оси яйца). Поэтому дробление произойдет
в плоскости одного из меридианов яйца (рис. 42, А и А').
Положение веретен 2-го дробления и расположение новой бо-
розды дробления также подтверждают правила Гертвига: в каж-
дом из двух бластомеров веретена дробления располагаются, как
и при 1-м дроблении, параллельно экваториальной плоскости,
под прямым углом к бывшему направлению веретена 1-го дроб-
ления. Это происходит потому, что наибольшая протяженность
чистой протоплазмы остается параллельной экватору и плоскости
1-го дробления. В результате произойдет снова меридиональное
дробление. Борозды 1-го и 2-го дроблений появляются сначала
на анимальном полюсе или вблизи от. него и постепенно вреза-
ются в вегетативное, перегруженное желтком полушарие
(рис. 42, 5 и 5х). Итак, первые два дробления яиц амфибий —
полные и равномерные. Однако после двух дроблений наиболь-
шая протяженность чистой протоплазмы оказывается в направ-
лении анимально-вегетативной оси, и именно в этом направлении
расположены веретена третьего деления ядер во всех образовав-
шихся четырех бластомерах. Это направление перпендикулярно
к экваториальной плоскости и плоскостям первых дроблений,
в связи с чем очередное дробление окажется экваториальным
(рис. 42, Г и Г'). Дробление опять-таки происходит полное, но уже
Рне. 42. Положение веретен дробления сбоку и с анимального полюса:
ложеннё "бл астомеплв е!тлг л Б’ Б'~ вт°Р°е дробление, В, В'-третье дробление; Г, Г' - по-
диженне оластомеров после третьего дробления
9&
.неравномерное; третья борозда дробления разделяет каждый
бластомер на неодинаковые половины — верхняя (анимальная)
меньшего размера, а нижняя (вегетативная), содержащая жел-
ток, большего размера. Затем две меридиональные борозды при
четвертом дроблении сегментируют яйцо на 16 бластомеров,
а две экваториальные борозды пятого дробления дробят яйцо
на 32 бластомера. В дальнейшем меридиональные и экваториаль-
ные борозды чередуются, что отчасти также подтверждает пра-
вила Гертвига, но теперь уже вступают в действие не менее
мощные факторы: взаимное давление бластомеров, процессы
изменения желтка, различные явления взаимозависимости частей
развивающегося эмбриона. Дробление перестает быть синхронным,
и, как уже говорилось, в одних участках зародыша темп деления
Рис. 43. Дискоидальное дробление
яйца костистых рыб: форели (Л—Г,
по О. Коршу), мурены (Д, по
Г. Букке):
1 — парабласт, 2 — бластоцель, 3 — блас-
тодерма, 4 — желток
может усилиться, а в других—
замедлиться; появляются и
клетки, идущие по пути деге-
нерации. Эти и другие процес-
сы уже не являются функцией
единичных бластомеров, и для
их анализа требуется изучать
развивающийся зародыш как
интегрированное целое.
Не менее показательны
другие примеры, подтвержда-
ющие правила Гертвига: раз-
витие птиц (см. рис. 32), кос-
тистых рыб (рис. 43), а также
рептилий. У яиц костистых
рыб первое, второе, третье,
а нередко и четвертое дробле-
ния меридиональные. Каждый
из образовавшихся 16 бласто-
меров остается соединенным
с желтком, полного обособле-
ния клеточных территорий не
происходит. Зародышевый
диск имеет незначительный
размер по сравнению с мас-
сой желтка. При пятом дроб-
лении яиц костистых рыб
четыре центральных бластоме-
ра делятся экваториально,
вследствие чего один из каж-
дой образующейся пары бла-
стомеров отделяется от желт-
ка и приобретает вид клетки
с вполне обособленной кле-
'36
точной территорией, а другой из этой пары остается под ним
в протоплазме, соединенной с желтком (рис. 43, Б, В). В это
время каждый из 12 периферических бластомеров делится
меридионально, в направлении формирующегося бластодиска
(бластодерма), причем один из каждой пары сыновних бластоме-
ров оказывается «краевым» бластомером, связанным с желтком,
а другой — свободным, лежащим центрально. Однако в одном
участке бластодиска (будущая его задняя сторона) меридиональ-
ная борозда проходит параллельно поперечному диаметру
бластодиска. Связанные с желтком периферические бластомеры
образуют краевую зону бластодиска. В этой зоне самые крайние
бластомеры в течение всего периода дробления сохраняют связь
с желтком. По мере того как такие «синцитиальные» бластомеры
в краевой зоне, а также и подобного характера бластомеры под
всем бластодиском в результате очередных дроблений все более
беднеют цитоплазмой, образование новых клеток постепенно
прекращается, однако ядра размножаются. Совокупность этих
ядер называют перибластом или парабластом, или также мероци-
тами (рис. 43, Д,1). Они играют роль в переработке желтка.
Постепенно формируется бластула, состоящая приблизительно
из тысячи клеток, иногда с выраженной полостью, наполненной
жидкостью (бластоцель). Бластула снизу ограничена пара-
бластом.
Принципиально такую же картину в смысле применимости
правил Гертвига к анализу закономерностей дробления мы видим
и у птиц (см. рис. 32).
В бластуле птиц различают: наружный слой центральной
части бластодиска — это плотный слой мелких бластомеров; под
ними, в полости с жидкостью находится большое количество
бластомеров, также не соединенных с желтком; дно полости
и ее боковые стенки состоят из крупных периферических бласто-
меров. Постепенно, без деления клеток, происходит некоторое
изменение в строении бластулы, а именно: бластомеры полости
бластулы подходят к поверхностному слою бластодиска, клетки
приобретают вид эпителия, который надвигается и ложится
на периферическое поле бластодиска. Центральная часть зароды-
ша благодаря просвечивающей жидкости полости бластулы ока-
зывается более светлой; она называется area pellucida — светлое
поле, а периферическая часть бластодиска, лежащего на желт-
ке,— area ораса — темное поле.
Как видим, правила Гертвига вполне применимы к развитию
телолецитальных яиц. Некоторые эмбриологи (П. П. Иванов,
1945) считают, что и дробление центролецитальных яиц не про-
тиворечит этим правилам,хотя объяснение характера поверхност-
ного дробления центролецитальных яиц, исходя из принципов
Гертвига, не может быть сделано с такой же ясностью, как это
осуществлено в отношении телолецитальных яиц.
97
О влиянии распределения желтка на характер дробления
можно судить и на основании экспериментов. Так, центрифугиро-
ванием только что оплодотворенных яиц лягушки можно добить-
ся еще большего уплотнения желтка в вегетативном полушарии,
в результате чего яйцо начинает дробиться дискоидально.
Ниже показана зависимость характера дробления от того или
иного типа яиц.
Схема зависимости формы дробления от типа яиц
Итак, все факты в совокупности позволяют установить связь
между типами дробления яиц, их количеством и распределением
в них желтка, хотя биохимический и биофизический механизм
этой связи в сущности до сих пор неизвестен. Эмбриология
может говорить лишь о том, что желток как «инертный материал»
является механическим препятствием при дроблении. На самом
деле, как увидим далее, вопрос этот гораздо сложнее.
СЛУЧАИ, ПРОТИВОРЕЧАЩИЕ ПРАВИЛАМ О. ГЕРТВИГА
Изучая эмбриональное развитие паразитических перепончатокры-
лых, О. М. Иванова в 1954 г. обнаружила, что у наездников в свя-
зи с паразитическим образом жизни происходит уменьшение
размеров яиц и обеднение их желтком. Изменяется и тип дробле-
ния. Вместо поверхностного, сбойственного насекомым, дробление
становится настоящим тотальным (полным), например у Copido-
soma gelechiae. Имеются и случаи, не поддающиеся объяснению.
Яйца Prestwichia aquatica, Caraphractus reductus и Aphidius
fabarum совершенно лишены желтка. Несмотря на это, дробление
яиц первых двух' форм типичное поверхностное от начала до конца.
Своеобразно дробление яйца Aphidius fabarum. Это очень
маленькие яйца, всего 86X30 мкм. Несмотря на отсутствие
желтка, дробление сначала, как и обычно для насекомых, непол-
ное, деление ядер не сопровождается разделением цитоплазмы
(рис. 44^4), но через 5 ч после начала дробления происходит
обособление клеточных границ, яйцо сразу распадается на
98
несколько (до 10) бластомеров
(рйс. 44,5). О. М. Иванова
•(1954) полагает, что поверхност-
ный тип дробления насекомых,
возникший в связи с накоплени-
ем желтка и закрепленный на-
следственно, продолжает действо-
вать еще некоторое время и пос-
ле исчезновения желтка. Эти
суждения справедливы, если
предполагать, что эволюция
форм дроблений происходила не
только в связи с изменениями
в количестве и распределении в
яйцах желтка, но и в связи с из-
менением каких-то других неиз-
вестных механизмов.
Рнс. 44. Развитие зародыша Aphi-
dius fabarum на стадиях раннего
дробления (А, Б) и образования
бластодермы (В) (по О. М. Ивано-
вой, 1954)
Накопилось много парадоксальных фактов, требующих
создания новых гипотез. Так, например, установлено, что летние
яйца ракообразного Moina paradoxa беднее желтком, чем зимние,
но дробление их частичное, а богатые желтком зимние дробятся
по голобластическому типу.
Большой интерес представляют начатые А. Г. Гурвичем
в 1908 г. опыты по центрифугированию яиц. Можно подвергнуть
яйца морского ежа Arbacia punctulata вскоре после оплодотворе-
ния такому центрифугированию (в течение 2—5 мин), что ядро
разрушится, и ядерный материал можно обнаружить лишь в виде
глыбок, окрашивающихся по Фёльгену. Яйцо сохраняет сфериче-
скую форму, через несколько минут после центрифугирования
начинается дробление при полном отсутствии каких-либо следов,
веретена дробления. В небольшом проценте случаев такие яйце-
клетки развивались даже до хорошо плавающих личинок — плу-
теусов. Результаты опытов по центрифугированию трудно подда-
ются анализу. В. Циммерман и Д. Марслянд в 1960 г. доказали,
что центрифугирование и ультрацентрифугирование яиц нематод
(150 000 g в течение 3 дней и даже 400 000 g в течение получаса)
не препятствуют нормальному дроблению. Л. Я. Бляхер в 1947 г.
наблюдал дробление яйца тритона после 20-минутного центрифу-
гирования (1500—2000 об/мин), несмотря на то, что, казалось бы,
происходят катастрофические изменения в расположении желтка
и ограноидов яйца. Как будто существует какой-то не видимый
в световой микроскоп, не разрушаемый центрифугированием
каркас, от которого преимущественно зависит дробление. Эта
соблазнительная гипотеза, однако, очень уязвима: приходится
думать о таком каркасе, который не разрушается, несмотря на
резкие изменения в расположении частиц желтка и других
составных частей яйца.
99
Следует отметить, что А. Г. Гурвич и Л. Я. Бляхер объясняют
возможность дробления яиц после энергичного центрифугиро-
вания не тем, что не разрушается гипотетический каркас.
Их предположения основаны на представлении о природе прото-
плазмы, состоящей из неравновесных молекулярных констелля-
ций (см. гл. XIII).
Известны явления, которые необъяснимы с точки зрения пра-
вил Гертвига. Таковы, например, результаты опытов Гарвей
по партеногенетической активации лишенной ядра части цито-
плазмы яйца морского ежа, в результате которой все же проис-
ходит дробление.
Многие другие факты также требуют особого объяснения;
к ним, строго говоря, неприменимы правила Гертвига. Так, напри-
мер, равномерное дробление свойственно, как правило, яйцам
с равномерным распределением желтка, однако изолецитальные
яйца некоторых млекопитающих дробятся неравномерно. Вероят-
но, очень важную роль при всех типах дробления играют еще
недостаточно изученные типы контактов между бластомерами,
характер межбластомерных пространств, большая или меньшая
поверхность соприкосновения между бластомерами, явления вза-
имного скольжения клеток, образование на поверхности клеток
псевдоподий, выростов протоплазмы, возможность амебоидных
движений и иные явления. Неподвижность бластомеров — лишь
кажущееся явление. Вспомним, например, яйца нематоды. В раз-
витии этих яиц наблюдается перемещение бластомера Р2 и EMSt
и образование ромбовидной фигуры на стадии четырех бластоме-
ров (см. рис. 8). Активные перемещения бластомеров характерны
и для спирального типа дробления. Причинный механизм этих
явлений остается пока невыясненным.
ИНТЕГРАЦИЯ ЗАРОДЫШЕЙ. ФУНКЦИОНАЛЬНЫЙ, ДОФУНКЦИОНАЛЬНЫЙ,
ЦИТОТИПИЧЕСКИЙ И ОРГАНОТИПИЧЕСКИЙ ПЕРИОДЫ РАЗВИТИЯ ЯИЦ
На любой стадии развития зародыш представляет интегрирован-
ное целое, а не сумму бластомеров и клеток. Взаимодействие
бластомеров, обусловенное их стуруктурой и жизнедеятельностью,
а также жизнедеятельностью зародыша в целом, носит своеобраз-
ный характер у разных животных. У одних ранние зародыши
более интегрированы, чем у других; интеграция развивающихся
зародышей непрерывно меняется по мере развития. Зародыш на
любой стадии развития живет, функционирует. Представления
В. Ру о так называемых дофункциональном и функциональном
периодах развития представляют большой интерес, хотя в неко-
тором отношении ошибочны. Дофункциональным периодом Ру
называл все стадии развития до гаструляции, до закладки орга-
нов. Этим не отрицается значение основной идеи Ру и для совре-
менной эмбриологии. Он утверждал, что на определенных стадиях
100
Рис. 45. Дробление Bimastus (по
П. Г. Светлову, 1928):
А —стадия после третьего дробления;
Б — плакула с дорсальной стороны;
В —стадия 50 бластомеров с задней
стороны: 1 и 2 —бластомеры А и В,
дифференцированные в осморегулято*
рЫ> мезодермальные полоски, 4 —
мезотелобласт, 5 — энтодерма, 6— ле-
вые и правые эктотелобласты
развития органов их дефинитивная функция отсутствует.
Эт0 — очевидный факт. Развитие органов обусловлено историче-
ски, а не только их функционированием в онтогенезе, дефинитив-
'ная функция органа подготовляется предыдущими стадиями раз-
вития. Ру, к сожалению, не учитывал того, что и до возникнове-
ния дефинитивной функции на любой стадии развивающийся
орган играет какую-либо функциональную роль ка'к часть заро-
дыша, от взаимоотношения с которой так или иначе зависит
развитие других частей зародыша (см. гл. XI, в частности о роли
хордо-мезодермального материала в развитии нервной системы).
Более интересны взгляды П. П. Иванова на период до гаст-
руляции и период, начинающийся с гаструляции. Первый период
он называет цитотипическим, а последующий — органотшгическим.
«Яйцо... лишь комплекс однозначных и независимых друг
от друга единиц... Зародыш состоит... из самостоятельных одно-
клеточных единиц... И физиология их является суммой физиоло-
гических процессов этих самостоятельных клеток».1 Эти взгляды
не аргументированы, хотя многие мысли П. П. Иванова об особен-
ностях этапа, начинающегося гаструляцией, представляют боль-
шой интерес.
Приведем некоторые факты, убеждающие в неправильности
указанной характеристики зародышей как «суммы бластомеров».
В дроблении яйца дождевого червя Bimastus constrictus мы ви-
дим стадию, когда бластомеры А и В (рис. 45, А, 1, 2) (точнее—
три бластомера А, В' и В") оказываются «органами осморегуля-
ции и выделения», они не принимают участия в дальнейшем дроб-
лении яйца, в построении тела зародыша, но без них не могло
бы состояться развитие. Это связано с особой,экологией эмбрио-
нального развития животного: зародыш развивается в дождевой
влаге, постоянно поглощает воду вследствие деплазмолиза. Для
нормального развития требуется удаление избыточной воды, что
и делают бластомеры А и В. Эти бластомеры выступают в каче-
стве провизорных «огранов» зародыша. При появлении других
органов осморегуляции они погибают; весь зародыш развивается
из бластомеров С и D.
Существуют паразитические медузы Cunina и Cunoctantha
со слабо развитым зонтиком и длинным ротовым стебельком,
1 Иванов П. П. Общая и сравнительная эмбриология. М„ Л., 1937. С. 85.
101
Рис. 46. Зародыши паразитической ме-
дузы Cunina на стадиях дробления (4)
и бластулы (Б) (по И. И. Мечникову,
1886):
} — зародыш, 2 —• амебоидный фороцит
погруженным в пищеваритель-
ную полость другого вида ме-
дузы-хозяина. Уже первые два
бластомера у нее различны
(рис. 46,4).
Один бластомер продолжа-
ет развиваться и дает в конце
концов личинку — полипоид-
ную актинулу, а другой бла-
стомер, подобный амебе, не
дробится, он охватывает со
всех сторон дробящийся бла-
стомер и, образно говоря,ста-
новится «извозчиком»: благо-
даря амебоидпым движениям
он постепенно переносит развивающийся эмбрион в каналы гаст-
ральной системы медузы-матери. Этот бластомер называется фо-
роцитом. Сам зародыш, находясь в фороците, продолжает разви-
ваться; на стадии, предшествующей полной бластуле, он «освобо-
ждается» от фороцита и выходит на его поверхность. На этой
стадии формируется своеобразная «полубла’стула» в виде чаши,
соединенной с амебоидным фороцитом (рис. 46,5)..
В гл. XI будут приведены показатели состояния зародышей на
разных стадиях развития, взаимозависимости бластомеров и дру-
гих частей зародыша на более поздних стадиях. В то же время
будет показано, что бластомеры равнонаследственны, они потен-
циально одинаковы и с точки зрения генетики, цитогенетики и
эмбриологии содержат одинаковые ядра, одинаковые хромосомы
и гены. Будет также сообщено, что отдельные бластомеры, буду-
чи изолированными от интегрирующих влияний зародыша как
целого, могут изменить путь своего развития и дать начало
новому целому эмбриону. Это является, несомненно, одним из
показателей взаимозависимости в жизни бластомеров, влияния
их друг на друга при нормальной интеграции зародыша.
БИОХИМИЯ РАННИХ СТАДИИ ЭМБРИОНАЛЬНОГО РАЗВИТИЯ
Морфогенетические процессы, сопровождающие эмбриональное
развитие, несомненно, теснейшим образом связаны с биохими-
ческими и биофизическими процессами. К сожалению, мало мож-
но назвать определенных фактов, прямым образом устанавли-
вающих обусловленность морфологических преобразований в раз-
витии зародышей тем или иным химическим или физическим
процессом. Вот почему ряд интересных фактов о биохимии
эмбрионального развития приходится сообщать после описания
морфологии развития, что является (с точки зрения будущей
эмбриологии) нелогичным.
102
Некоторые факты фундаментального значения для физиологии
и биохимии развития были уже сообщены: 1) своеобразие инди-
видуального развития бластомеров в отличие от онтогении клеток
тканей, а именно: бластомеры не имеют периода роста, и с каж-
дым новым дроблением они прогрессивно уменьшаются в раз-
мерах; 2) к пятому-шестому дроблениям у бластомеров как кле-
ток восстанавливается нарушенное в ходе оогенеза и сперматоге-
неза соотношение ядра и плазмы, что обусловливает и изменения
в метаболизме.
Как ни своеобразен жизненный цикл бластомеров (онтогения
каждого бластомера), они являются клетками и подчиняются
закономерностям онтогении клеток, вскрытым биологией (см.
гл. XVII). Дробление — это процесс митоза, и при каждом
дроблении хромосомный материал должен удваиваться, так как
два брата-бластомера, возникающие в результате дробления,
будут иметь нормальный диплоидный набор хромосом. Как и
в жизненном цикле клеток тканей, в связи с дроблением количе-
ство ДНК в ядре должно удваиваться, приближаться к тетрап-
лоидному состоянию. Дупликация ДНК действительно подтверж-
дается. Р. Мак-Мастер в 1955 г. измерял количество ДНК в ядрах
бластомеров, готовых к очередному дроблению в ходе развития
эмбриона морского ежа Lytechinus variegatus, начиная с только
что оплодотворенной яйцеклетки до личиночных стадий; при этом
автор для измерения брал произвольные бластомеры — тем более
ценны его данные (табл. 2).
Было изучено общее количество ДНК и РНК у ручьевой фо-
рели на стадии синхронных делений (дроблений) и на стадиях
средней и поздней гаструлы. Количество РНК и ДНК в зародыше
Таблица 2. Среднее количество ДНК ядер на ранних этапах развития
морского ежа Lytechinus variegatus в условных единицах
Стадия ДНК Состояние ядра
Оплодотворенная яйцеклетка 1,11+0,6 Близкое к 4п
2 бластомера 1,02+0,4 То же
4 8 бластомеров 1,09+0,4 1,08+0,7 я
16
микромеры 0,82+0,6 2п<4п
мезомеры 1,04+0,4 Близкое к 4п
макромеры 28 бластомеров 1,02+0,4 То же
микромеры 1,10+0,5
мезомеры 0,95+0,6
макромеры Бластула Эктодерма гаструлы 0,01+0,3 0,68+0,4 0,60+0,3 2п<4п 2п+
Эктодерма раннего плутеуса 0,62+0,3 2п
103
за этот период увеличивается в 1,5—2 раза (Г. В. Донцова, Г. М.
Игнатьева, Н. Н. Ротт, И. И. Толсторуков, 1970).
В оогенезе накапливается обычно большой резерв негенетиче-
ской ДНК. Она содержится в основном в цитоплазме и расходуется
в период дробления. Содержание «запасной» ДНК в некоторых
яйцевых клетках составляет около 99 % всего количества ДНК.
Ядерная ДНК ооцита также превышает количество ее в диплоид-
ном наборе хромосом. В яйцах морских ежей и амфибий количе-
ство ДНК превышает содержание ДНК в диплоидных клетках
тех же видов от 200 до 25 000 раз. В курином яйце резерв ДНК
еще больший. В яйцах сверчка Grillus в 1600 раз больше ДНК,
чем в сперматозоиде.
Оплодотворение — это начало работы биохимического аппара-
та, от которого зависит синтез белков. С оплодотворением связано
и начало функционирования нового ядра — синтез новых иРНК.
СИНТЕЗ БЕЛКА И ВНУТРИКЛЕТОЧНЫЕ РЕГУЛЯЦИИ
В РАННЕМ РАЗВИТИИ
В раннем эмбриогенезе сразу же после оплодотворения наступает
активация синтетических процессов. В них вовлекаются различ-
ные РНК, ранее запасенные в ходе оогенеза. Образуются полисо-
мы. Синтезируются в основном белки такого же типа, как в ооци-
те, но в отличие от оогенеза эти белки не запасаются впрок, а сра-
зу же используются на строительство различных клеточных струк-
тур. Параллельно с этим расходуются и те белки, которые были
«отложены про запас» в ооците. Однако было бы неправильно
думать, что никаких новых белков в дробящемся яйце не возни-
кает. Небольшое количество белков регуляторного типа всегда
появляется в ранний период развития. Об их образовании и зна-
чении для развития судят на основании экспериментов по блоки-
рованию белкового синтеза. Если бы все необходимые для ранних
этапов развития белки были запасены в яйце, то добавление ве-
ществ, блокирующих синтез белков, не отразилось бы на ходе
развития. На самом деле, по данным ряда исследователей, разви-
тие в таких условиях тормозится.
Что касается транскрипционной активности в ранний период
развития, то этот процесс видоспецифичен и разные классы РНК
транскрибируются в определенной последовательности.
Исключительный интерес представляют вопросы энергетическо-
го обмена. Каков источник энергии, требующийся для всех видов
морфологических и физиологических процессов при дроблении
яиц? Общий, несомненно правильный ответ, что энергия образу-
ется за счет диссимиляторных процессов, недостаточен, так как
наблюдаются очень своеобразные случаи, еще не поддающиеся
точному анализу.
Уже говорилось о том, что процесс активизации яйца и его оп-
лодотворения выводит яйцо из состояния анабиоза, у некоторых
104
яиц наблюдается вспышка окислительных процессов, например, у
асцидий. Но у других яиц дыхание в результате оплодотворения
ослабляется (например, у моллюска Cumingia, полихеты Chaeiop-
terus и др.) или остается на том же незначительном уровне (нап-
ример, у полихеты Sabellaria). Не доказаны различия между ды-
ханием неоплодотворенного и оплодотворенного яиц лягушки. В
течение первых минут после оплодотворения происходит заметное
потребление гликогена.
Экспериментами на яйцах аскарид, трихостронгилид, оксиу-
рисов и других паразитических червей (М. М. Завадовский и др.,
1915, 1928) доказано, что если среду, в которой развивается яйцо,
лишить кислорода, дробление прекращается до тех пор, пока кис-
лород снова не будет поступать в яйцо. Лишить яйца аскарид
доступа кислорода в эксперименте легко, пользуясь особым строе-
нием оболочек яиц, не пропускающих многие химические вещест-
ва. Можно залить яйца аскариды, находящиеся в пробирке, на-
пример, ртутью, преграждающей доступ кислорода. Дробление
прекращается. Если через несколько месяцев ртуть слить и по-
местить яйца в воду, дробление продолжается. Также и яйцо мор-
ского ежа без кислорода во внешней среде не дробится. Яйцо же
лягушки при полном анаэробиозе достигает стадии ранней га-
струлы, яйцо форели также может дробиться.
Неоднократно изучалась ди-
намика потребления кислорода
дробящимися яйцами разных
животных. Получены строго за-
кономерные кривые дыхания в
ходе развития зародышей (рис.
47 и 48 и табл. 3).
Во время развития дыхание,
как видим из рисунков, усилива-
ется.
Минуты разбития
Рис. 48. Кривая дыхания
яиц лягушки (100 яиц) (по
Ж- Браше, 1961)
время, ч
Рис. 47. Потребление кислорода
нормально развивающимися яйца-
ми морского ежа нз двух разных
партий (по Ж. Браше, 1961):
I —• вылупление, 2 — конец гаструляции
105
Таблица 3. Потребление кислорода
в ходе дробления яйца лягушки
и в ходе гаструляции
(по Б. Керну, Ж. Барту и Л. Барт,
1954)
Стадия Поглощение Оз. мл/мг, сухого ве- щества за 1 ч
Дробление 0,054
Ранняя бластула 0,069
Поздняя С, 108
Средняя гаструла 0,141
Поздняя » 0,162
Еже Ж. Лёб проводил наблю-
дения за дыханием в ходе дро-
бления яиц. Если неоплодотво-
ренные яйца морского ежа в те-
чение часа поглощают 0,24 мг
кислорода, то в первый час пос-
ле оплодотворения они поглоща-
ют 0,94 мг О2, во второй час —
0,80, в третий час — 0,87, в чет-
вертый—0,91 и пятый—1,05 кг
О2.
В исследованиях динамики
окислительных процессов в ходе
дробления помимо строго законо-
мерных кривых, характерных
для яиц каждого вида животных, обнаружены факты, позволяю-
щие предполагать, что каждый бластомер, поскольку он развива-
ется между двумя очередными дроблениями, дышит с неодинако-
вой интенсивностью в разные моменты своей онтогении. Е. П. Лион
в 1902 г. заметил, что яйца морского ежа, находящиеся на одной
стадии развития (например, на стадии двух или 16 бластомеров),
в разные моменты этой стадии обладают неодинаковой чувстви-
тельностью к «дыхательному яду»—к KCN. Через 10—15 мин
после оплодотворения яйца наиболее чувствительны, но по мере
приближения к первому дроблению чувствительность к яду умень-
шается. Особенно интересно, что после первого дробления яйца
снова приходит состояние наибольшей чувствительности, которая
постепенно снижается по мере приближения второго дробления.
Ю. Г. Юровицкий и Л. С. Мильман (1975) изучили изменения
углеводного обмена в оогенезе в раннем эмбриогенезе у костистых
рыб. В период большого роста происходит запасание гликогена.
Такому характеру обмена ооцита соответствуют относительно вы-
сокая активность гликогенсинтетазы и очень низкая активность
фосфорилазы. Ооциты используют глюкозу, поступающую из кро-
ви самки. В раннем эмбриональном развитии изменяется направ-
ленность углеводного обмена зародыша, в десять раз возрастает
активность фосфорилазы, а активность гликогенсинтетазы вдвое
снижается.
Благодаря использованию микроэлектродной техники, о кото-
рой говорилось в гл. II, были получены интересные данные, ка-
сающиеся биоэлектрической характеристики внутренней среды,
возникающей в период дробления. Так, например, у зародышей
рыб мембранный потенциал микромеров оказался ^40 мВ со
знаком, обратным знаку наружной среды, т. е. электрохимический
градиент мембраны, покрывающей наружную поверхность заро-
дыша, отличается от градиента мембраны, выстилающей межкле-
точную полость. Это означает, что уже на стадии дробления кле-
106
точные поверхности, обращенные к внутренней среде, находятся
в иных условиях.
К стадии бластулы устанавливается барьер высокого сопро-
тивления^ связанный с появлением плотных контактов (или зон
замыкания) между бластомерами. Коэффициент электрической
связи (Кэс) или отношение напряжения на поверхностной мембра-
не одной клетки к соответствующему электрическому показателю
соседней клетки для разных стадий дробления оказался сущест-
венно различным. Так, например, у зародыша вьюна (5—6 пер-
овых'дроблений) и у зародыша аксолотля (первые восемь дробле-
ний) у всех бластомеров Кэс— 1. При переходе к бластуляции
значение этого коэффициента постепенно уменьшалось до 0,3—0,2.
Внутри зародыша наряду с бластомерами, устанавливавшими
плотные контакты с соседними клетками, встречались одиночные
клетки или их группы, характеризующиеся очень низким коэффи-
циентом связи, т. е. практически разобщенные друг от друга. Эти
данные позволили заключить, что в составе презумптивных заро-
дышевых листков еще на стадии бластулы возникают гетероген-
ные клеточные популяции, отличающиеся по свойствам образова-
ния контактов с другими клетками. Более того, оказалось, что и
механизмы образования мембранных потенциалов у разных групп
клеток были неодинаковыми, что свидетельствует о начальных
этапах мембранной дифференцировки клеток развивающегося за-
родыша.
Таким образом, в оплодотворенном яйце начинается цепь по-
следовательных событий, которые не зависят от генома яйца.
Сюда относятся перестройки мембран, активация ряда фер-
ментов, изменение соотношения ибиов К+ и Na+ и др. Эти измене-
ния коррелятивно , связаны с. уровнем цАМФ, содержанием ней-
ромедиаторов, изменением уровня дыхания и мембранных потен-
циалов.
Согласно представлениям К. А. Кафиани и А. А. Костомаро-
вой (1978) период раннего дробления характеризуется градуаль-
ными изменениями в системе малых молекул, следствием чего
оказываются сдвиги в системе макромолекул. Отмечается также,
что эти сдвиги совершаются очень быстро. Следовательно, неге-
иетические факторы контролируют временную последовательность
событий в раннем эмбриогенезе.
Как уже говорилось, в период раннего развития начинает
формироваться и внутренняя среда зародыша, которая оказыва-
ется причастной к процессам водного обмена. Мутации, нарушаю-
щие активный трансмембранный транспорт ионов, ведут к нару-
шению водного баланса, следствием чего оказываются тератоге-
незы, проявляющиеся в серьезных морфологических дефектах.
Глава V
БЛАСТУЛА. ГАСТРУЛА. ЗАРОДЫШЕВЫЕ ПЛАСТЫ
МОРУЛА И БЛАСТУЛА
Многими путями происходила эволюция животных. Очень разно-
образны и специфичны связи развивающихся организмов со сре-
дой. Несмотря на это и большие особенности в морфологии и фи-
зиологии дробления у разных видов животных, дробление у боль-
шинства организмов завершается сходным периодом развития —
формированием бластулы (от греч. Waste, blastos — росток, зача-
ток). Это один из многочисленных показателей общности проис-
хождения животного мира и один из примеров параллелизма
в эволюционном развитии структур. Но это не означает, что за-
родыши всех животных на стадии бластулы одинаково устроены;
наоборот, наряду с основными общими чертами имеются сущест-
венные, притом разнообразные особенности бластул разных жи-
вотных.
У некоторых гидроидных полипов (например, Clava) в ре-
зультате дробления образуется плотный, без полости шар взаимо-
связанных бластомеров, напоминающий тутовую ягоду или ягоду
ежевики (рис. 49,Д). В таких случаях говорят о моруле (от лат.
morum — тутовая ягода).
В зависимости от многих причин (наличие оболочек яиц, вяз-
кость и иные физические свойства цитоплазмы яиц и др.) дробя-
щееся яйцо в общем сохраняет свою первоначальную сфериче-
скую форму, а бластомеры могут оказывать большое давление
друг на друга, приобретать многогранную форму и не оставлять
между собой никаких щелей; при этом и образуется типичная
морула. В других случаях бластомеры, оставаясь шарообразными
клетками, более или менее сплющиваются в местах соприкосно-
вения и между ними в местах закругления остаются щели. Так,
эмбрион лягушки или тритона уже на стадии четырех бластоме-
ров имеет небольшую полость с жидкостью — продуктом жизне-
деятельности клеток. Полость — полость дробления (сегментаци-
онная полость) — по мере дробления постепенно увеличивается.
В честь Бэра, впервые описавшего эту полость, ее называют так-
же бэровской полостью. По мере дробления в центральной части
зародыша бластомеры удаляются друг от друга, на периферии же
они все теснее контактируют, так что слой бластомеров приоб«
ретает вид эпителиоидного пласта. Полость дробления, превра-
тившаяся постепенно в полость бластулы, называется бластоце-
лем (рис. 49, Б,Д), т. е. полостью зародыша, точнее — первичной
108
Рис. 49. Типы бластул:
Л — морула Clava; Б —бластула морского ежа; В — бластула лягушки; Г —бластула кос-
тистой рыбы- Д — бластодиск птицы; Е — плавающая амфибластула губки; Ж — стерро-
бластула Lucernaria; 3 — плакула Cuculanus elegans. Вид с поверхности (1) и сбоку (2)
полостью зародыша. Слои клеток, составляющие стенку бластулы,
называются бластодермой. Во многих случаях стенка бластулы
состоит только из одного слоя бластомеров, например, у зароды-
ша морского ежа (рис. 49,В). Хотя в вегетативной половине яйца
желтка несколько больше, происходит полное и почти равномер-
ное дробление; бластула оказывается такого строения, что бла-
стодерма в вегетативном полушарии состоит из бластомеров
большего размера.
В случае телолецитальных яиц, как мы уже отмечали, имеют
место различные типы дробления, приводящие к тому, что бла-
стодерма (по крайней мере на поздних этапах развития бластулы)
оказывается многослойной, с неравными размерами бластомеров
в анимальной и вегетативной половинах. Такова, например, бла-
стула у амфибий (рис. 49,В).
Однослойную бластулу с большим бластоцелем называют ти-
пичной бластулой или целобластулой. Понятие «типичная бласту-
ла не имеет особого сравнительно-эмбриологического смысла и
является условным. Бластула без выраженного бластоцеля
(рис. 49,Ж) называется стерробластулой (от греч. sterros — креп-
кий).
При поверхностном дроблении бластоцель заполнен желтком,
а не только продуктами жизнедеятельности бластомеров. Поэто-
му нет достаточных оснований говорить о «полости дробления» и
109
о «бластоцеле». Бластодерма у такой бластулы состоит из одного
слоя клеток. Бластулу столь своеобразного строения называют
перибластулой (см. рис. 31,Б).
Дискоидальное дробление приводит к формированию диско-
бластулы, бластоцель которой имеет вид сплющенной щели, под-
час очень узкой, находящейся под зародышевым диском
(рис. 49,Г, Д). Жидкость дискобластулы, видимо, не только про-
дукт жизнедеятельности бластомеров, она образуется и в резуль-
тате происходящих биохимических процессов в подстилающем
зародышевый диск желтке, который в этом месте разжижается.
Некоторые эмбриологи выделяют еще амфибластулу; это цело-
бластула с резко отличающимися бластомерами вегетативного и
анимального полушарий. Бластула лягушки, например, может
быть названа амфибластулой. Однако различия бластомеров ани-
мального и вегетативного полушарий бластулы могут касаться
не только морфологических признаков, но и своеобразных функ-
циональных особенностей бластомеров, отчетливо различимых
уже на стадии бластулы и определяющих своеобразие в дальней-
шем развитии зародыша. Это относится к амфибластуле губок
(рис. 49,Б). На 8-м дроблении яйца Sycon raphanus передняя по-
ловина образовавшейся удлиненной бластулы состоит из реснич-
ных клеток, а задняя — из крупных зернистых клеток. Выходя
из тела матери, бластула плавает при помощи ресничек, при-
крепляется на подходящем субстрате и развивается в губку.
Если воду освободить от ионов Са, амфибластула губок распа-
дается на две различные части. Изолированная половинка, со-
стоящая из ресничных клеток, быстро погибает, а состоящая из
зернистых бластомеров продолжает развиваться и дальше в пол-
ноценную губку. Если уже на первых стадиях дробления возни-
кают те или иные различия между бластомерами, то на стадии
бластулы любого вида животных клетки различных участков от-
личаются размерами, структурами и химическим составом. На-
чиная со стадии бластулы, следует говорить не о бластомерах,
а о клетках бластодермы.
Существует еще своеобразный тип бластулы—плакула, отли-
чающийся от других видов бластул тем, что это двухслойная
пластинка (рис. 49,3). Такая эмбриональная форма образуется,
когда бластомеры при дроблениях располагаются лишь в двух-
параллельных плоскостях, благодаря чему и образуются два пла-
ста сближенных бластомеров-клеток. Плакула характерна для
наземных OUgochaeta. Такое строение бластулы определяет даль-
нейшее своеобразное развитие, о чем будет сказано в связи с про-
цессами гаструляции.
Большого внимания эмбриологов заслуживает изучение физио-
логической и морфологической роли жидкости бластоцеля — внут-
ренней среды развивающегося зародыша (см. гл. XIV). По со-
ставу и консистенции она неодинакова у бластул разных живот-
но
ных (у морских ежей она студенистая, а у других видов - это
подвижная жидкость); всегда чем-либо отличается от воды, окру-
жающей эмбрион водных животных.
ГАСТРУЛА. ТИПЫ ГАСТРУЛЯЦИИ
В гл. II приводились примеры раннего обособления полового за-
чатка, как и зачатков других органов и систем. Отмечалось, в ча-
стности, что уже на стадии 16 бластомеров развития яйца лоша-
диной аскариды, когда обособляется половой зачаток, бластомеры
Е1 и Е11 (сдо. рис. 8) являются уже «обособившимся» зачатком
средней кишки. При следующем дроблении бластомер st на пра-
вой стороне вместе с от на левой представляют зачаток стомо-
деума, или эктодермальной глотки; т и ц, а также бластомер
D — зачатки мускулатуры; С — источник формирования наружных
покровов хвостовой части зародыша. Таким образом, на очень
ранних стадиях развития определяется судьба отдельных бласто-
меров и их групп — зачатков отдельных органов.
Эти и другие случаи, когда зачатки будущих органов обособ-
ляются на первых этапах развития в виде отдельных бластоме-
ров или маленьких их групп, в основном характерны для живот*
ных с мозаичным типом дробления. В большинстве случаев столь
раннего обособления зачатков не происходит. Бластула, состоящая
подчас из большого количества бластомеров (у асцидий—128,
у ланцентника — около 3000, у лягушки — много сотен и т. д.)
переходит в своем развитии в новую стадию — гаструлу (от греч.
gastre— выпуклость, чрево сосуда; gaster — желудок). Зародыш
на этой стадии состоит из ясно отличимых пластов клеток, так
называемых зародышевых пластов или листков — наружного, или
эктодермы (от греч. ectos — находящийся снаружи), и внутрен-
него, или энтодермы (entos — находящийся внутри). Возникает
и третий зародышевый листок — средний, или мезодерма (от
греч. mesos — находящийся посредине). Совокупность процес-
cofi, приводящих к гаструле, называется гаструляцией.
Несмотря на большое разнообразие процессов гаструляции,
можно дать вполне определенную классификацию гаструл и пу-
тей возникновения их из предыдущей стадии бластулы. К сожа-
лению, еще не создана классификация, которая исходила бы из
принципа эволюции, так как историческое происхождение разных
типовгаструляции изучено недостаточно, и эмбриология имеет
в этой области больше научных загадок, чем достоверных све-
дений.
Некоторые способы гаструляции встречаются во всех типах
животного мира.
Инвагинационная гаструла (рис. 50,Д) образуется путем ин-
вагинации (впячивания) части однослойной бластодермы (стенки
бластулы) внутрь бластоцеля. Грубое подобие такого типа гастру-
ляции результат впячивания стенки резинового мячика в ре-
111
Рис. 50. Способы гаструляции (по П. П. Иванову):
А — инвагинация у Phoronis; Б — однополюсная иммиграция у гидроида Clytia\ В, В' —
лямннация у Geronia\ Г, Г' — эпиболическая гастула у ВопеШа-. / — эктодерма, 2 — энто-
дерма, 3—бластоцель
зультате надавливания на нее. Впячивание можно довести вплот-
ную до внутренней стенки мячика (на противоположном от места
вдавления участке), или вдавление можно сделать не столь
значительным. Образуется двухслойный мешок, наружной стен-
кой которого является первичная эктодерма, а внутренней — пер-
вичная энтодерма. Впячивание образует архентерон (первичный
кишечник), а отверстие, при помощи которого он сообщается
с наружной средой, называется первичным ртом или бластопором
(рис. 50,Д).
Судьба бластопора у разных животных неодинакова. У мно-
гих животных первичный рот, соответственно развиваясь и диф-
ференцируясь, превращается в дефинитивный рот взрослого орга-
низма. Эти животные называются первичноротыми (Protostomia).
К первичноротым относятся черви, моллюски и членистоногие.
Не менее обширна группа вторичноротых (Deuterostomia),
у которых бластопор превращается или в анальное отверстие, или
(у хордовых) в нервно-кишечный канал, находящийся в заднем
конце эмбриона. Рот у таких животных возникает на переднем
конце, на брюшной стенке, в результате особых формообразова-
тельных процессов (впячивание эктодермы, прободение стенки
средней кишки). К Deuterostomia относятся: щетинкочелюстные,
плеченогие, иглокожие, кишечнодышащие и хордовые.
Указанными признаками отнюдь не исчерпывается необходи-
мость разделения животных на две группы: первично- и вторично-
112
ротые. В дальнейшем будут сообщены иные не менее важные при-
знаки обеих групп.
Очень многие вопросы являются спорными потому, что био-
логия еще не знает истинных предков современных многоклеточ-
ных (см. гл. XVII); нет единого мнения н о филогенетическом
значении разного типа гаструл. Многие исследователи под влия-
нием теории гастреи Геккеля (см. гл. XVII) считали и считают
исходной в эволюции процессов гаструляцин ннвагинационную
гаструлу, некоторые — плакулу. Нет убедительных аргументов
в пользу предположений об инвагинации как первичном типе га-
струляции. Следует согласиться с теми зоологами (Ю. В. Мам-
каев, 1968), которые думают, что на начальных этапах возник-
новения многоклеточных, т. е. на начальных этапах становления
их онтогенеза, имели место различные неустоявшиеся способы
гаструляцин н лишь позднее постепенно выработались и получили
распространение определенные способы гаструляцин.
Многие современные исследователи, присоединяясь к И. И. Меч-
никову, считают исходной формой гаструлы иммиграционную
гаструлу и выводят все способы образования энтодермы из муль-
типолярной иммиграции и деляминации.
Возвращаясь к характеристике инвагинационной гаструлы, от-
метим, что в ней имеются две полости: в большей или меньшей
степени сохраняется бластоцель (хотя в некоторых случаях эта
полость почти нацело вытесняется впячиванием) и возникает но-
вая полость — гастроцель, или гастральная полость, или полость
первичного кишечника (архентерона).
Иммиграционная гаструла характерна для многих кишечно-
полостных (рис. 50,Б). Она развивается вследствие активного
выселения части клеток стенки бластулы внутрь бластоцеля. Это
выселение клеток в случаях униполярной иммиграции происходит
лишь с одного полюса (вегетативного). Выселившиеся клетки об-
разуют позднее внутренний слой гаструлы (энтодерму). Таким
образом, как н при инвагинации, зародыш делается двухслойным.
Так происходит гаструляция, например, у гидроидного полипа
Glytia, гидромедузы Aequorea или у медузы Octorchis gegenbauri.
Униполярная иммиграция свойственна почти всем гидромедузам,
особенно тем, для которых характерна плавающая целобластула.
Клетки бластодермы могут проникать в бластоцель не в ка-
ком-либо одном участке, а по всей поверхности зародыша, при-
чем такая иммиграция может начинаться уже на стадии 16 бла-
стомеров. Эта так называемая мультиполярная иммиграция
свойственна, например, зародышу медузы Solmundella, что обна-
ружил впервые И. И. Мечников. Мультиполярная иммиграция,
однако, редкое явление.
У многих видов кишечнополостных, которым свойственна им-
миграционная гаструла, происходит столь массовое активное вы-
селение клеток- бластодермы, что бластоцель полностью исчезает,
113
так как заполняется плотной массой клеток. Как видим, у имми-
грационной гаструлы нет бластопора и, значит, нет характерного
для инвагинационной гаструлы сообщения гастроцеля с внешней
средой. Можно лишь, проводя далеко идущую аналогию (оправ-
данную представлением об общности животного мира), говорить
о месте бластодермы, соответствующем бластопору, что, однако,
доступно критике. В случае мультиполярной иммиграции еще
меньше оснований говорить о бластопоре или даже указывать
какой-либо соответствующий ему участок бластодермы.
Деляминационная гаструла, встречающаяся также у кишечно-
полостных, впервые описана И. И. Мечниковым (рис. 50,В,Д').
Морфогенетические процессы, сопровождающие этот редкий слу-
чай гаструляции, очень своеобразны и в типичном случае кажут-
ся простыми. Яйца некоторых сцифомедуз, например Geryonidae,
имеют различимые, концентрически расположенные участки ци-
топлазмы: один — плотная, зернистая эктоплазма и другой —
ячеистого строения эндоплазма, в который заходит наружный
эктоплазматический слой, содержащий полужидкий желток.
Структура яйца такова, что оно может быть отнесено к центро-
лецитальным яйцам.
У таких яиц происходит относительно равномерное синхрон-
ное дробление, так что образуется зародыш, состоящий из 2, 4,
8, 16 и, наконец, 32 бластомеров, кажущихся одинаковыми по
размерам и пространственному положению.
Дальнейшие дробления бластомеров происходят параллельно
поверхности зародыша, и в результате дроблений наружный слой
.бластомеров будет состоять из одной (бывшей) эктоплазмы,
а внутренний — из эндоплазмы и частично из эктоплазмы. Про-
исходит деляминация (расщепление) одного слоя клеток на два.
В дальнейшем дробятся только «внутренние» бластомеры, при-
том опять-таки параллельно поверхности зародыша; возникают,
таким образом, две легко различимые клетки: наружный, «экто-
плазматический» бластомер и внутренний, «эндоплазматический».
В результате такой своеобразной гаструляции зародыш оказы-
вается шаром, состоящим из 64 плоских клеток, образующих на-
ружный слой — эктодерму, и из 32 более высоких клеток внут-
реннего слоя — энтодермы.
У животных, для которых характерна типичная морула (на-
пример, у Clava squamata), процесс гаструляции происходит так,
что клетки, находящиеся на периферии, приобретают отличное от
других клеток строение. Пласт первых клеток образует эктодер-
му, а внутренний пласт — энтодерму. Такой тип гаструляции
можно рассматривать как один из вариантов деляминации.
Эпиболическая гаструла формируется у некоторых животных
с резко выраженным телолецитальным строением яиц, например
у ВопеШа (рис. 50,ГиГ')- Значительного размера медленно де-
лящиеся бластомеры (макромеры), с большим содержанием желт-
.114
ка не обнаруживают способности к каким-либо перемещениям.
Их обрастают (наползают на них) более быстро делящиеся мел-
кие, лежащие на поверхности клетки (микромеры). Выходит, что
у зародышей таких животных бластопор отсутствует, архентероа
не формируется. Только впоследствии, когда макромеры, делясь,,
становятся меньшего размера, образуется, по неясным биохими-
ческим и физическим причинам, полость, формируется зачаток
первичного кишечника.
Смешанный тип гаструляцин. Типы гаструляцин кажутся на-
столько различными, что их невозможно сравнивать. Несомненно,,
каждый тип гаструляцин требует причинного анализа, но все же1
в некотором отношении разные способы гаструляцин сравнимы..
Так, своеобразный процесс эпиболии можно рассматривать как
один из особенных вариантов инвагинации. Процессы делямина-
ции можно уподоблять процессам иммиграции, конечно, с боль-
шими оговорками. Важно отметить далее, что гаструляцин у мно-
гих животных происходит так, что имеют место и явления ин-
вагинации, и явления эпиболии, и другие морфогенетические
процессы. Такова, положим, гаструляцин у амфибий. Можно по-
этому безыскусственно выделить пятый тип гаструляцин — сме-
шанный.
ОБРАЗОВАНИЕ МЕЗОДЕРМЫ У
У всех животных, за исключением кишечнополостных, в связш
с гаструляцией (параллельно с ней или на следующем этапе,
обусловленном гаструляцией) возникает и третий зародышевый-
пласт — мезодерма. Это совокупность клеточных элементов, зале-
гающих между эктодермой и энтодермой, т. е. в бластоцеле. Таким,
образом, зародыш становится не двухслойным, а трехслойным.
У высших позвоночных трехслойное строение зародышей возни-
кает уже в процессе гаструляцин, тогда как у низших хордовых
и у всех других типов в результате собственно гаструляцин обра-
зуется двухслойный эмбрион.
Вопросы о путях образования мезодермы у разных животных,
издавна интересовали и сравнительных анатомов, и эмбриоло-
гов. В общем они могут считаться решенными, однако не в аспек-
те причин соответствующих морфогенетических процессов, а в пло-
скости формально-морфологического описания этих процессов,.
Если отвлечься от всех разнообразных деталей формирования1
мезодермы у разных животных, можно установить два принци-
пиально^ разных пути возникновения ее: телобластический, свой-
\fotosiom^ и энтероцельный, характерный для Deute-
s а. у первичноротых во время гаструляцин на границе-
ежду эктодермой и энтодермой, по бокам бластопора, уже име-
тся две большие клетки (или несколько таких клеток-тело-
115,
>1
Рис. 51. Грубая схема образо-
вания мезодермы у первичио-
ротых (А) и вторичноротых
(Б) (по В. М. Шимкевичу,
1925, модифицировано):
1 — эктодерма, 2 — мезенхима, 3 —
энтодерма, 4 — телобласт (Л) и це-
ломическая мезодерма (5)
бластов), отделяющие от себя (вследствие делений) мелкие клет-
ки (рис. 51,А, рис. 69). Таким образом формируется средний
пласт — мезодерма. Телобласты, давая новые и новые поколения
клеток мезодермы, остаются на заднем конце зародыша. По этой
причине такой способ образования мезодермы и называют тело-
бластическим (от греч. teios — конец).
При энтероцельном способе совокупность клеток формирую-
щейся мезодермы появляется в виде карманоподобных выступов
первичного кйшечника (выпячивание его стенок внутрь бласто-
целя, рис. 51,5,4). Эти выступы, внутрь которых входят участки
первичной кишечной полости, обособляются от кишечника и от-
деляются от него в виде мешочков. Полость мешочков превраща-
ется в целом, т. е. во вторичную полость тела, целомические меш-
ки могут подразделяться на сегменты.
Такая характеристика способов возникновения среднего заро-
дышевого листка не отражает всего многообразия вариаций и от-
клонений, строго закономерных для отдельных групп животных.
Сходен с телобластическим, но лишь внешне, способ образования
мезодермы не путем деления телобластов, а путем возникновения
на краях бластопора непарного плотного зачатка (группы кле-
ток), впоследствии разделяющегося на две симметричные полоски
клеток. При энтероцельном способе зачаток мезодермы может
быть парным или непарным; в одних случаях формируются два
симметричных целомических мешка, а в других — сначала обра-
зуется один общий целомический мешок, впоследствии разделя-
ющийся на две симметричные половины.
Уже говорилось о своеобразных процессах развития нематод
и других животных, в отношении которых было бы искусственно
применять понятие «зародышевые пласты»,— у них, минуя фор-
мирование клеточных зародышевых пластов, зачатки будущих
органов обособляются в виде отдельных бластомеров.
Ввиду важности для эмбриологии в целом и для понимания
процессов развития органов в следующей главе будет дан срав-
нительно-эмбриологический очерк процессов гаструляции у раз-
личных животных, внесены соответствующие коррективы в слиш-
ком упрощенные классические представления о зародышевых ли-
стках, в частности об энтероцельном способе образования
мезодермы.
116
ПРОИЗВОДНЫЕ ЗАРОДЫШЕВЫХ ПЛАСТОВ. ЦЕЛОМ
Гаструляции — это этап в развитии зародыша. Зародышевые пла-
сты не являются чем-то обособленным друг от друга, их возник-
новение и дальнейшие изменения происходят благодаря взаимной
зависимости частей эмбриона. Зародышевые пласты как совокуп-
ности клеток отличаются друг от друга не только своим положе-
нием в общей системе зародыша, но уже и некоторыми цитологи-
ческими особенностями. В то же время эксперименты убеждают,
что их судьбу можно еще изменить, заставляя идти на «строи-
тельство» необычных для них клеточных систем и органов (см.
гл. XI и XVII).
При нормальном развитии зародышей зародышевые пласты,
взаимодействуя между собой под воздействием интегрирующих
влияний зародыша как целого, продолжают дифференцироваться
в определенном направлении, и каждый из них принимает участие
в возникновении зачатков определенных органов и систем орга-
нов. Можно говорить о новом этапе в развитии зародышей — об
органогенезе.
Во всем животном мире определенные органы берут начало от
одного и того же зародышевого пласта. Исключения из этого за-
кона, о которых будет сказано в дальнейшем, обусловлены изме-
нениями в онтогенезе, в связи с теми или иными своеобразными
путями эволюции животных. Их рассматривают как гомологич-
ные образования в зародыше. О зародышевых пластах см.
в гл. VII—IX.
Производные эктодермы. Большая часть клеток, составляющая
наружный пДаст, размножаясь и соответственно дифференцируясь,
так и остается на поверхности, принимая участие в развитии по-
кровов тела. Из них формируются: наружный эпителий, кожные
железы, поверхностный слой зубов, роговых чешуйит. д. Почти
всегда -каждый орган развивается из клеточных элементов двух,
а то и всех трех зародышевых пластов. Например, кожа млеко-
питающих развивается из экто- и мезодермы.
Как правило, обширная часть первичной эктодермы (до трети
и более всей поверхности зародышей амфибий) вследствие осо-
бых морфогенетических процессов «погружается» внутрь, под на-
ружный эпителий, и дает начало всей нервной системе. У многих
животных эктодерма на переднем и заднем концах тела впячи-
вается по направлению к переднему и заднему концам развива-
ющегося из энтодермы кишечника (средняя кишка). Впячения эти
прорываются в полость средней кишки и образуют stomodeum
(передняя кишка) и proctodeum (задняя кишка).
Производные энтодермы. Внутренний зародышевый пласт, диф-
ференцируясь сопряженно с другими частями зародыша, разви-
вается в эпителий средней кишки и ее пищеварительные железы.
Развитие эпителия дыхательной системы (жаберного отдела и
117
легких) у разных позвоночных прослежено не одинаково полно
и не столь еще ясно в деталях. Бесспорно, что этот эпителий раз-
вивается из переднего отдела кишечника. Однако нельзя катего-
рически сказать, что он весь эктодермального происхождения, так
как в его происхождении несомненно участвует клеточный мате-
риал прехордальной пластинки (см. с. 126 и др.).
Производные мезодермы. Все остальные, не перечисленные до
этого органы развиваются из мезодермы: все мышечные ткани,
где бы они ни находились (стенка тела, кишечника и других об-
разований), все виды соединительной, хрящевой и костной тканей,
каналы выделительных органов, перитонеум полости тела, кро-
веносная система, часть тканей яичников и семенников. При раз-
витии соответствующих органов происходит и специфическая диф-
ференцировка клеточных элементов мезодермы. У большинства
животных средний пласт появляется не только в виде совокуп-
ности клеток, образующих компактный эпителиеподобный слой,
т. е. собственно мезодерму, но и в виде рыхлого комплекса раз-
розненных, амебоподобных клеток. Эта часть мезодермы назы-
вается мезенхимой. Мезодерма и мезенхима отличаются друг от
друга по своему происхождению, между ними нет прямой связи,
они не гомологичны. Мезенхима большей частью эктодермально-
го происхождения, начало же мезодерме дает энтодерма. У поз-
воночных, однако, мезенхима в меньшей своей части эктодермаль-
ного происхождения, основная же масса мезенхимы имеет общее
с остальной мезодермой происхождение. У многих животных со
спиральным дроблением мезенхима появляется еще в ходе дроб-
ления. У иглокожих источником мезенхимы являются микромеры
и энтодерма. Клетки дна формирующегося первичного кишечни-
ка мигрируют внутрь бластоцеля.
Несмотря на отличное от мезодермы происхождение, мезенхи-
ма может рассматриваться как часть среднего листка. Она играет
большую роль в формировании личинки и дефинитивных органов.
Для уяснения разбираемых в дальнейшем вопросов требуется
иметь представление о важном образовании — целоме, вторичной
полости тела. У всех животных, которым свойствен целом, начало
полым целомическим мешкам дает мезодерма. Было уже сказано,
что при энтероцельном происхождении мезодермы целомические
мешки образуются путем изменения, дифференцирования карма-
ноподобных выпячиваний первичного кишечника. При телобла-
стическом и ему подобном способах, когда образуются мезодер-
мальные тяжи, внутри них появляется щель, превращающаяся
в конце концов в целом. Целомические мешки формируются сим-
метрично по бокам кишечника. Стенка каждого целомического
мешка, обращенная в сторону кишечника, называется спланхно-
плеврой. Стенка же, обращенная в сторону эктодермы зародыша,
называется соматоплеврой.
Таким образом, в ходе развития зародыша формируются раз-
118
Рис. 52. Схема органогенеза заро-
дышей высших позвоночных (по
К. Уоддингтону^ 1957):
I — нервная трубка, 2 — сомит, 3 —
хорда, 4 ~ кишка, 5 — боковая мезо-
дерма, в —целом, 7 —эпидермис, в —
глотка, 9 — жаберные щели, 10 —
глазной пузырь, 11 — мозг
личные полости, имеющие важное морфогенетическое или функ-
циональное значение. Сначала появляется полость Бэра, превра-
щающаяся в первичную полость тела — бластоцель, затем в связи
с процессами гаструляцин возникает гастроцель (или гастральная
полость), наконец, у многих животных — целом. При образовании
гастроцеля и целома бластоцель все более уменьшается, так что
от бывшей первичной полости тела остаются лишь щели в проме-
жутках между стенками кишки и целома. Эти щели у некоторых
животных превращаются в полости кровеносной системы. Гастро-
цель со временем превращается в полость средней кишки.
При энтероцельном способе обособления мезодермы и целома
за счет гастроцеля, кроме того, возникает н вторичная полость
тела.
Процессы гаструляцин непосредственно приводят к периоду
органогенеза. У одних животных закладываются органы и си-
стемы органов, приобретающие постепенно дефинитивное зна-
чение, а у других животных формируются сначала органы, свой-
ственные личинке, затем совершается метаморфоз (см. гл. X) и
происходят процессы формирования дефинитивных органов взрос-
лого организма.
В связи с отсутствием единого плана в строении эмбрионов
всех беспозвоночных невозможно дать схему абстрактного эмб-
риона беспозвоночного животного. Схема же позднего эмбриона
позвоночного животного изображена на рис. 52.
ТЕОРИЯ ЗАРОДЫШЕВЫХ ПЛАСТОВ
На существование у зародышей курицы двух пластов обратил
внимание еще в «доклеточный» период развития эмбриологии
К. Вольф (11759). Позднее X. Пандер в 1817 г. также описал за-
родышевые пласты у куриного эмбриона. К. Бэр создал основу
для теории зародышевых листков, так как распространил понятие
о зародышевых листках на всех позвоночных (1828). Но, конечно,
основателем современной теории зародышевых листков должен
считаться А. О. Ковалевский, который в 1865—1871 гг. вместе со
своими современниками И. И. Мечниковым (1886) и Э. Геккелем
119
(1866,1871) обнаружили зародышевые листки у беспозвоночных жи-
вотных и внесли коррективы в их описание, наполнив понятие
новым содержанием, обусловленным эволюционной идеей, дарви-
низмом.
Геккель создал гипотезу о происхождении многоклеточных, со-
гласно которой предком их является гастрея — двухслойное жи-
вотное, по строению подобное зародышам современных живот-
ных на стадии гаструлы (см. гл. XVII). Эктодерма и энтодерма,
по мнению Геккеля, рекапитулируют (см. гл. XVII) «первичные
органы» этого гипотетического предка современных многоклеточ-
ных.
Эмбриологи прошлого столетия рассматривали зародышевые
листки как основные первичные органы, наделяя их определен-
ными функциями. Эктодерму называли Hautsinnesblatt — кожно-
чувствующим листком, а энтодерму Darmdrusenblatt — слизистым
или кишечно-железистым слоем, листком первичного кишечника.
Зародышевые листки всех животных стали рассматриваться
как наследие первичной двухслойной формы предка Metazoa, как
гомологичные образования. В этом суть теории зародышевых ли-
стков.
Теория зародышевых пластов — самое крупное морфологиче-
ское обобщение за всю историю эмбриологии. Она сыграла выда-
ющуюся роль в сравнительной, эволюционной эмбриологии.
И в настоящее время, несмотря на сомнения в ее правильности
и на явную ошибочность ряда утверждений основателей ее, по-
нятия об эктодерме и мезодерме, применяемые к развитию почти
всех многоклеточных животных, никем из эмбриологов не оспа-
риваются, хотя многие ученые вкладывают в них разное со-
держание. Все, однако, согласны с тем, что эти понятия характе-
ризуют определенные этапы развития зародышей, отражают по-
следовательность образования зачатков органов, историческую
непрерывность функционального характера листков, их органо-
образовательную специфичность. Все эволюционисты-эмбриологи
не отрицают, что зародышевые листки, возникающие в ясной фор-
ме у подавляющего большинства животных, являются одним из
многочисленных свидетельств единства животного мира.
Cnidaria и Ctenophora — многоклеточные организмы, тело ко-
торых строится из двух зародышевых пластов — эктодермы и эн-
тодермы. Между ними находится мезоглея. Это эластичный не-
клеточный материал сложного состава, выделяемый эпителиаль-
ными клетками. Основной его компонент — коллагеновые фиб-
риллы.
Труды А. О. Ковалевского обогатили эмбриологию многими
фактами и идеями. Естественно, в теорию зародышевых листков
XIX в. эмбриология XX в. не могла не внести существенных по-
правок. Уже во время создания этой теории И. И. Мечников от-
вергал гипотезу Геккеля о гастрее, считая, что рассматривать
120
инвагинационную гаструлу как первичную эмбриональную форму
нет оснований. Более примитивной, по мнению Мечникова, надо
считать мультиполярную иммиграцию, и предок современных Me-
tazoa не мог быть геккелевской гастреей (см. гл. XVII). Образо-
вание гаструлы путем инвагинации не характерно для низших
многоклеточных. Мечников установил большое распространение
внутриклеточного пищеварения у низших многоклеточных: губок,
кишечнополостных, низших животных. На основании сопоставле-
ния большого числа эмбриологических, сравнительно-анатомиче-
ских и физиологических данных Мечников создал, в противовес
геккелевской теории о гастрее, теорию фагоцителлы (или парен-
химеллы), согласно которой предком Metazoa было первобытное
животное, не имеющее пищеварительной полости. Пищевые ча-
стицы у них захватывались поверхностными клетками благодаря
фагоцитарной способности и оказывались достоянием всех клеток,
так как эти подвижные клетки внедрялись внутрь. Вот почему и
примитивные формы гаструляции ныне живущих организмов сле-
дует искать в развитии тех низших многоклеточных, у которых
непосредственно в результате гаструляции не возникает первичная
пищеварительная полость. Такова гаструляция, идущая по пути
иммиграции клеток. Процессы же инвагинации — более позднее
явление в эволюции гаструляции.
Естественно, нельзя оспаривать факт рекапитуляции в разви-
тии современных многоклеточных возникновения двухслойности
у предков (см. гл. XVII). В то же время надо отказаться от мно-
гих наивных, упрощенных представлений как об индивидуальном
развитии животных, так и об эволюции явлений эмбрионального
развития. Перечислим некоторые факты, заставляющие критиче-
ски относиться к утверждениям теории зародышевых листков в ее
классической форме.
1. Классическая описательная эмбриология сводила образова-
ние зародышевых листков к неравномерному росту частей зароды-
ша, образованию складок, выпячиваний, врастаний и т. п. Эмб-
риологи могли видеть лишь внешние проявления гаструляции,
последовательный ряд состояний. Методами описательной эмбрио-
логии, в особенности в «доклеточный» ее период, даже талантли-
вый исследователь не мог бы изучить сложную клеточную и
биохимическую динамику гаструляцйонных движений.
Следует отказаться от представлений о зародышевых пластах
как о стойких определенных образованиях, о первичных «орга-
нах» зародыша. Гаструляция — это процесс. Зародышевые пла-
сты— это органическая часть интегрального целого зародыша;
они не имеют самостоятельных функций, а в связи с взаимодей-
ствием частей в их развитии «функция» состоит в способности,
благодаря интегрирующим влияниям зародыша как целого,
дать в ходе развития ряд производных (см. гл. VII, VIII
и IX).
121
2. Образование мезодермы у зародышей многих позвоночных
животных происходит, по существу, не по энтероцельному спосо-
бу. Так, в частности, в ходе развития амфибий нельзя говорить об
образовании чего-либо подобного «целомическим мешкам», отда-
ленно напоминающего энтероцельный способ образования мезо-
дермы у ланцетника. Уже в начале гаструляции формирующаяся
мезодерма вворачивается в полость гаструлы независимо от энто-
дермы. Вместо движения мезодермы от средчедорсальной линии
вниз и вбок, как думали ранее, мезодерма, появляющаяся в об-
ласти боковых и нижней губ (отнюдь не в виде «карманоподоб-
ных выступов» энтодермы), стягивается к спинной стороне тела.
В дальнейшем, в главах о гаструляции у позвоночных, мы убе-
димся в том, что при сохранении представлений о двух типах об-
разования мезодермы (телобластический и энтероцельный) тре-
буется вносить очень существенные поправки в старые представ-
ления о гаструляции разных животных и в сущности требуется
новая классификация процессов гаструляции. Нет оснований ис-
кать у всех животных гаструляцию как образование обязательно
двухслойного зародыша, рассматривать образование мезодермы
как какой-то особый процесс, оторванный от процессов самой га-
струляции. У кишечнополостных, у которых нет столь развитого
среднего пласта, как у других животных, гаструляция заверша-
ется формированием только экто- и энтодермы. В ходе эволюции
животных и, значит, эволюции эмбрионального развития появ-
ление и развитие третьего зародышевого пласта было связано
с изменениями гаструляции. У амфибий и других животных обра-
зование мезодермы — это одно из явлений самой гаструляции, а не
обособленный, вторичный процесс. Нельзя противопоставлять про-
цесс гаструляции как образование экто- и энтодермы образованию
мезодермы.
3. Не следует на основании исключений и отклонений от схе-
мы органогенеза, продиктованной теорией зародышевых листков,
отказываться от самой теории, но не следует и пренебрегать эти-
ми исключениями, чтобы правильно представлять процесс разви-
тия зародышей и не отклонять вполне аргументированную мысль
о необходимости реформы теории.
Сравнительный эмбриолог классического направления, создан-
ного А. О. Ковалевским, П. П. Иванов указывал (1945), что при
ранней закладке органов характерные признаки зародышевых
листков исчезают, о них говорить нельзя. У турбеллярий, сосаль-
щиков и других животных образование различных органов проис-
ходит из однородной клеточной массы, без каких-либо признаков
обособления зародышевых пластов. У коловраток и скребней
(Acanthocephala) о зародышевых пластах вообще говорить нель-
зя, так как эмбриональный клеточный материал непосредственно
«распадается» на отдельные зачатки органов. Также уже отме-
чалось, что и о зародышевых листках аскарид можно говорить
122
лишь условно; у зародышей аскарид очень рано обособляются
эктодерма, половой зачаток и т. д. Интересна деталь: сразу по-
является по нескольку отдельных зачатков для нервной системы
и глотки, на что обращал внимание П. П. Иванов в связи с ана-
лизом теории зародышевых листков.
По теории зародышевых листков хорда развивается из энто-
дермы, нервная система—-из эктодермы и т. д. Между тем обна-
ружены не единичные случаи, резко противоречащие этому пра-
вилу. Так, у асцидйй определенные группы бластомеров дают и
хорду, и нервную систему; другие бластомеры дают кишечник,
третьи — наружный покров, четвертые — мышечные и мезенхимные
элементы.
О. М. Иванова и А. Г. Кнорре (1963) считают, что можно рас-
сматривать все эти и иные подобные случаи у разных живЬтных
как вторичное изменение ранних. стадий развития, что зароды-
шевые листки, как и вообще весь онтогенез, способны до неузна-
ваемости изменяться в ходе эволюции под влиянием определен-
ных, вполне объяснимых причин. Тем самым все подобного рода
«исключения» лишь подтверждают общее правило.
4. В сказанное ранее об органогенетической специфичности
зародышевых пластов надо внести существенные дополнения и
коррективы. В гл. XI о детерминации указаны результаты много-
численных экспериментов, согласно которым клеточный материал
эктодермы, энтодермы и мезодермы можно заставить изменить
развитие, участвовать в формировании совершенно не свойствен-
ных данному листку органов. Это говорит о том, что судьба кле-
точного материала каждого зародышевого пласта, строго говоря,
еще не предопределена. Развитие и дифференцировка каждого
пласта, их органогенетическая специфичность обусловлены взаи-
имовлияниями частей целостного зародыша и возможны лишь при
его нормальной интеграции.
5. И в нормальном развитии некоторых животных закономер-
но «нарушается» установленная теорией зародышевых листков
специфичность. Так, мышечные волокна волосяных сумок кожи,
ириса глаза, некоторые хрящевые элементы развиваются не из
мезодермы, а из эктодермы. Клеточный материал ганглионарной
пластинки, т. , е. производное эктодермы, образует в норме че-
люстную дугу, подъязычную и все жаберные дуги, т. е. образо-
вания, которые должны возникать из мезодермы. Задняя часть
нервной пластинки, возникающей из эктодермы, образует у за-
родышей амфибий в норме не нервную ткань, а хвостовую мус-
кулатуру. J
6. Развитие организма в результате бесполого размножения и
экспериментально вызванного соматического эмбриогенеза (см.
гл. XVI) пока совершенно невозможно не только объяснять, но
даже описывать в понятиях «зародышевые листки».
Некоторые экспериментальные эмбриологи (например, О. Ман-
12»
гольд, 1925), не довольные фанатизмом приверженцев теории за-
родышевых листков, готовы считать зародышевые листки лишь
чисто топографическим понятием, удобным для описания явлений
развития. Эта точка зрения также излишне фанатична и не мо-
жет считаться обоснованной. Несмотря на важность всех указан-
ных противоречий и сомнений, нельзя отрицать реальность су-
ществования зародышевых листов как гомологичных образований.
Эволюция явлений эмбрионального развития, как и вообще эво-
люция любого биологического явления, не происходила прямо-
линейно, и, естественно, имеется много факторов, противоречащих
теории. Критически оценивающий теорию зародышевых листков
П. П. Иванов определяет зародышевый пласт как сумму всех за-
чатков частей будущих органов, находящихся на начальной, од-
ной и той же стадии морфологической дифференцировки.
Прогресс эмбриологии столь значителен, что нельзя сомне-
ваться в возможности радикального пересмотра теории зародыше-
вых листков с сохранением ее основы — представления о листках
как гомологичных образованиях.
Как уже было сказано, эмбриология располагает доказатель-
ствами гомологии одноименных зародышевых листков у всех
многоклеточных животных. Теория зародышевых листков имеет
большое общебиологическое значение как одно из доказательств
единства всех многоклеточных. Однако не следует считать под-
рывом эволюционной эмбриологии и возвращение к теории типов
Кювье и Бэра мысли о радикально ином понимании процессов
гаструляции и формирования зародышевых листков. Вопрос о за-
родышевых листках неразрывно связан с гипотезами о происхож-
дении многоклеточных животных. Нельзя игнорировать большое
число фактов, не укладывающихся в теорию зародышевых листков.
Можно говорить о гомологии зародышевых листков, если стоять
на позиции монофилетического представления об эволюции живот-
ного мира организмов и рисовать обычные схемы родословного
дерева. Методологически вполне оправданы и подкрепляются ог-
ромным числом эмбриологических фактов мысли Н. К- Кольцо-
ва (1936) о том, что основные типы отделились друг от друга так
давно, что связать их между собой и отвести от одного корня
не представляется возможным.
В случае правильности идей о полифилетическом происхожде-
нии организмов данные эмбриологии о зародышевых листках мож-
но рассматривать как свидетельство явлений параллелизма в
онтогенезе животных с разными «установками развития»
(П. П. Иванов), «типами развития» (Бэр). При этом так мучаю-
• щие эмбриологов многочисленные отклонения у разных животных
от обычных представлений о зародышевых листках, приобрели бы
значение как доказательства разных типов развития. Особый ин-
терес представляют воззрения Г. П. Коротковой (1979), которая
полагает, что яйцевые клетки, бластулы, гаструлы, личинки, за-
124
родышевые листки полностью гомологичны только у тех видов,
которые относятся к одному и тому же типу организмов. Сравне-
ние онтогенеза животных или растений, относящихся к далеко
отстоящим типам, позволяет выявлять не столько гомологичные,
сколько аналогичные структуры и процессы. Эти выводы вытекают
из новой теории о фазной эволюции онтогенеза, в основе которой
лежит утверждение об обязательности чередования полового и
бесполого размножения.
Вопрос о зародышевых листках рассматривается и в главе XVII
в связи с проблемами о соотношении онтогенеза и филогенеза.
КАРТЫ ПРЕЗУМПТИВНЫХ ОРГАНОВ
В изучении процессов гаструляции большую роль сыграла пред-
ложенная В. Фогтом в 1925 г. методика маркировки частей за-
родыша. Кусочки агара необходимой величины помещаются в
растворы витальных красителей (метиленовый синий, нейтральный
красный, коричневый Бисмарка и Др.). Пропитавшиеся красите-
лями кусочки агара прикладывают к поверхности эмбриона в
пунктах, интересующих экспериментатора. Благодаря диффузии
красителя определенная группа клеток оказывается окрашенной
(рис. 53, А). Если на стадии бластулы сделать метки в разных ее
участках, можно проследить меняющееся в ходе гаструляции по-
ложение этих меток, описать гаструляционные перемещения частей
зародыша (рис. 53, Б, В, Г). Благодаря методу Фогта превосходно
изучен процесс гаструляции у амфибий. Конечно, методику Фогта
невозможно применить к очень маленьким яйцам, а также к тем,
оболочки которых непроницаемы для витальных красителей. В
этих случаях прибегают к другим способам изучения: маркировке
Рис. 53. Методика В. Фогта:
— сп<е<пи1<1Ма₽КИ₽ОВКИ витальиымн красителями, эксперимент Фогта; Б — ранняя гаструла;
/ ц III /уГааспУост (участки V'—VH инвагинировалнсь); Г — поздняя гаструла (участки
полпжаийрУпраспространились иа большую поверхность по сравнению с их первоначальным
ляннТя пластинка ^₽г„аструляции): "Р°"итанный краской агар, 2-зародыш, .7 - стек-
сальная губа бластопоп?И?аЮЩаЯ эаР°Дыш в определенном положении, 4 - воск, 5 — дор-
пробка У° аст°пора, 6 — аиимальный полюс, 7 — вегетативный полюс, « — желточная
125
тушью, частичками мела, инъекцией некоторых веществ внутрь
•отдельных бластомеров й т. п.
На стадии поздней бластулы амфибий и других животных
можно точно указать участки, из которых разовьются те или иные
органы. Принято говорить о картах презумптивных органов (от
лат. praesumptio — предположение, основанное на вероятности).
Англичане говорят о fate map — о карте судьбы. Некоторые эм-
бриологи предпочитают говорить о картах презумптивных зачат-
ков, или презумптивных областей.
На стадии поздней бластулы амфибий можно безошибочно
указать границы обширной области анимального полушария заро-
дыша, клеточный материал которой разовьется в эпидермис кожи
(рис. 54, /). В этой области можно указать участки эктодермаль-
ного материала для органа обоняния, слуха и эктодермальной
части рта, а также участки презумптивной линзы глаза, слизи-
стых желез.
Известны и границы не менее рбширной области анимального
полушария, являющейся источником формирования нервной систе-
мы (рис. 54, 4). В свою очередь в пределах презумптивной нерв-
ной системы уже на стадии бластулы можно указать участки,
которые пойдут на развитие соответствующих частей глаза. В
промежуточной, или маргинальной, зоне бластулы (маргинальной,
или краевой, зоной называются области, в которых формируются
края бластопора) на дорсальной стороне имеется обширная зона
презумптивной хорды (рис. 54, 6). Рядом с ней находится область
прехордальной пластинки (от лат. ргае — перед, прехордальный,
находящийся перед хордой).
В ходе гаструляцин этот материал первым инвагинирует, пе-
редвигается к анимальному полюсу и затем становится частью
крыши архентерона, располагаясь впереди переднего конца хорды.
Далее по направлению к вегетативному полюсу, но все еще в
составе промежуточной зоны находится клеточный материал,
дифференцирующийся впоследствии в часть рта, жабр и глотки
энтодермального происхождения. Бластопор возникнет в этой об-
ласти. Части промежуточной зоны, лежащие по обеим сторонам
от презумптивной хорды, служат материалом для мышечных сег-
Рис. 54. Карта презумптивных
органов (бластула амфибий).
Вид сбоку (4) и со спинной
стороны (5) (по Ж- Пастелсу,
1942):
/ — эпидермис, 2 — боковая мезо-
дерма, 3 — место замыкания буду-
щего бластопора, 4 — нервная пла-
стинка, 5 — хвостовые сомиты, 6 —
хорда, 7 — туловищные сомиты,
8 — прехордальная пластинка, 9 —
головная кишка, 10 — туловищная
энтодерма
126
ментов тела (рис. 54, 7). Боковые и брюшные части промежуточ-
ной зоны участвуют как мезодермальный материал в формировании
полости тела, почек и других образований. Этот участок может
быть назван латеральной (или боковой) презумптивной мезодер-
мой или, еще точнее, вентральной мезодермой. Наконец, в районе
вегетативного полюса лежит материал презумптивной средней и
задней кишок (рис. 54, 10).
Пространственное расположение презумптивных органов та-
ково, что части будущей головы — глаза, органы слуха и обоня-
ния— находятся вблизи анимального полюса бластулы, соответ-
ствующего переднему концу зародыша. В центре среднеспинной
части'зародыша, там, где промежуточная зона наиболее протяжен-
ная, находится материал хорды. Анимальный полюс яйца амфибии
почти соответствует переднему концу позднего зародыша и взрос-
лого животного. Область вегетативного полюса будет задним кон-
цом зародыша. Самый широкий участок промежуточной зоны —
А
Рис. 55. Карта презумптивных зачатков:
А— расположение презумптивных зачатков на серии поперечных срезов через бластодиск
радужной форели (по В. Боллард, 1973); Б — презумптнзные зачатки в эпнбласте курицы
(по Г. Розенквнст, 1966): 1 — презумптивиая мезодерма, 2 — презумптнвиая хорда, 3—пре-
зумптивная энтодерма, 4 — перидерма, 5 — презумптнвиая нервная система, 6 — внезароды-
шевая эктодерма, 7 — зародышевая эктодерма, 8 — нервная пластинка, 9 —хорда, 10 —
зародышевая энтодерма, // — сомиты, /2 — спланхнотом, стрелкой обозначена передне-зад-
няя ось симметрии
127
ам, где расположен клеточный материал хорды, соответствует
пине зародыша, а противоположный — брюху.
Следует, однако, обратить внимание на очень важное явление:
। ходе гаструляцин презумптивные органы, расположенные на по-
1ерхности зародыша, окажутся после закрытия бластопора лежа-
цими внутри взрослого организма (хорда, нервная система, кишка
I др.). Презумптивный эпидермис кожи, занимающий хотя и об-
пирную, но вполне определенную часть только анимального полу-
пария бластулы, разовьется впоследствии в эпидермис кожи, по-
рывающий всю поверхность взрослого животного. Происходит это
з результате сложных морфогенетических движений пластов в ходе
'аструляции.
Не менее точные исследования позволили создать карту пре-
зумптивных органов у зародышей других животных. Так, напри-
мер, у костистых рыб презумптивные зачатки располагаются в
бластодерме не в одной плоскости, как у амфибий, а в разных
плоскостях (рис. 55, А). Презумптивные материалы эктодермы,
хордомезодермы и кишечной энтодермы лежат друг над другом,
образуя многослойную структуру. Гаструляционный процесс не
связан у костистых рыб с подворачиванием края бластодиска и
осуществляется по способу расслоения — деляминации.
У птиц карта презумптивных органов имеет следующие осо-
бенности (рис. 55, Б). Передняя часть эпибласта area pellucida—
это область презумптивного эпидермиса кожи, а также эмбрио-
нальных оболочек. В area pellucida находится в виде серпа пре-
зумптивная нервная пластинка. Позади нее лежит презумптивна?
хорда. По средней линии, позади хордальной области, расположен
клеточный материал презумптивных кишечной энтодермы и мезо-
дермы.
Карты презумптивных областей ранних зародышей можно со-
ставить в отношении развития любого вида животных. Полнота
и точность их зависят, конечно, от степени изученности развития
того или иного животного.
Глава VI
СРАВНИТЕЛЬНЫЙ ОБЗОР ПРОЦЕССОВ ГАСТРУЛЯЦИИ
ГАСТРУЛЯЦИЯ У АСЦИДИЙ, ЛАНЦЕТНИКА И МИНОГИ
Процессы дробления у различных представителей хордовых, как
мы видели, очень своеобразны и зависят от проморфологии яиц,
в особенности от количества и распределения желтка. Процессы
гаструляции также варьируют в пределах Chordata. Однако воз-
никающие в результате гаструляции главные или осевые зачатки
группируются в одинаковый у всех хордовых осевой комплекс
{хорда, нервная трубка, осевая мезодерма). Дальнейшие процес-
сы органогенеза и связанная с ним дифференциация тканей и кле-
ток приводят снова к большому разнообразию организации разных
животных.
У асцидий гаструляция начинается со стадии 128 бластомеров.
Особенности проморфологии яйца таковы, что карту презумптив-
ных зачатков можно нарисовать уже в начальных этапах разви-
тия. Цитоплазма оплодотворенного яйца — гетерогенная система
{рис. 56, А, Б). Анимальная половина состоит из прозрачной ци-
топлазмы, вегетативная — из зернистой. Ясно различаются и раз-
ные участки области экватора: протоплазма будущей передней
половины зародыша расположена в виде подковы, она темно-се-
рого цвета, а задняя половина желтая, Темно-серая протоплазма —
материал хорды и нервной пластинки, желтая — мускулатуры и
Рис. 56. Развитие асци-
дии Cynthia (по Э. Кон-
клину, 1905).
Стадии: А — двух бластоме-
ров; Б —• восьми бластоме-
ров; В —• 64 бластомеров;
г — раиияя гаструла — вид
с вегетативного полюса;
Д — поздняя гаструла, вид
с дорсальной стороны; 1 —
темно-серая цитоплазма, 2 —
цитоплазма, богатая желт-
ком, 3 —• «желтая» цито-
плазма, 4 — презумптнвиая
нервная пластинка, 5 — пре-
зумптнвиая хорда, 6 — энто-
дерма, 7 — мезодерма, 8 —
мускульные клетки, 9 — эк-
тодерма
129
мезенхимы. На стадии 64 бластомеров (рис. 56, В) темно-серый
серп делится на два ряда бластомеров; передний ряд развивается
в нервную систему, а задний — в хорду. В области «желтых бла-
стомеров» происходит также обособление двух рядов; задний из
них (или наружный ряд) — источник развития мускулатуры, а пе:
редкий (или внутренний) ряд дает мезенхиму. Участок протоплаз-
мы в вегетативном полушарии, расположенный между материалом
хорды и материалом мезенхимы,— будущая энтодерма (кишеч-
ник). Светлая протоплазма анимального полушария — это буду-
щая эктодерма.
Итак, задолго до гаструляцин судьба разных частей зародыша
(при нормальных условиях развития) предопределена.
На стадии 128 бластомеров начинается гаструляцин, которая
осуществляется по способу инвагинации в сочетании с эпиболией.
Нарастание передней (дорсальной) губы бластопора, а также сбли-
жение боковых губ бластопора приводят к образованию спинной
поверхности зародыша. Эктодермальная группа бластомеров, а за
ней хордальные и мезодермальные бластомеры погружаются
внутрь в результате инвагинации.
Гаструляцин у ланцетника была предметом многочисленных
исследований и изучена во всех подробностях.
В бластуле ланцетника ясно различим энтодермальный мате-
риал, находящийся в районе вегетативного полюса; в одном уча-
стке краевой (маргинальной) зоны находится материал мезодермы,
в другой области — материал хорды. Анимальное полушарие за-
нято клетками презумптивного эпидермиса кожи и презумптивной
нервной системы (рис. 54, А, Б).
Одна из особенностей гаструляцин ланцетника по сравнению
с асцидиями заключается в том, что она начинается тогда, когда
зародыш (после 10-го дробления) состоит из большого числа (бо-
лее 1000) клеток, так что материал каждого осевого зачатка пред-
ставлен большим их количеством.
Гаструляцин у ланцетника типично инвагинационная, начинает-
ся она выпячиванием презумптивной энтодермы. Вслед за энтодер-
мой инвагинирует в бластоцель материал хорды, а через боковые
в вентральную губы бластопора погружается мезодерма. В районе
дорсальной губы бластопора перемещается материал будущей
хорды. Гаструла имеет вид двухслойной чаши, бластоцель почти
нацело вытесняется инвагирующим клеточным материалом. Затем
изменяется конфигурация зародыша в целом, он удлиняется
(рис. 57).
Как только энтодермальный пласт приходит в контакт с вну-
тренней стороной эктодермального пласта, начинаются процессы,
приводящие к формированию зачатков осевых органов.
У миноги можно отметить свои особенности в процессах гаст-
руляции и формировании осевых зачатков по сравнению с лан-
цетником.
130
1
Рис. 57. Ранние стадии развития ланцетника (по Э. Коиклииу, 1932):
А—Е— поздняя бластула и гаструляции (медиальные срезы); 1—материал эктодермы; 2 —
<5ластоцель, 3— материал энтодермы, 4 — материал мезодермы, 5 — хорда, 6—архентерон,
7 — бластопор, 8— дорсальная губа бластопора
О. В. Чекановская (1933—1936) установила, что стенка бласту-
лы становится многослойной. В ее вегетативной части находятся
крупные, плотно прилежащие друг к другу клетки. Они образу-
ют эктодермальный зачаток. Анимальная часть бластулы, состоя-
щая из мелких клеток, оказывается «раздутой» в результате на-
копления жидкости в полости, смещенной в эту часть зародыша.
На границе мелкоклеточной анимальиой и крупноклеточной веге-
тативной частей бластулы располагается кольцевидная зона сред-
них по размеру клеток — краевая зона. Это район будущего бла-
стопора. В передней части краевой зоны появляется в виде косой
щели бластопор — место инвагинации. Клеточный материал дор-
сальной губы бластопора нарастает на эктодермальную подушку
(рис. 58, 5). Ближайшие клетки промежуточной зоны перемещаются
через боковые края бластопора в бластоцель. Стенки постепенно
удлиняющегося гастроцеля состоят из клеток различного проис-
хождения. Верхняя стенка образована слоем клеток, инвагиниро-
вавшим через переднюю губу бластопора. Этот клеточный- мате-
риал развивается в хорду. Дно полости гаструлы состоит из
крупных клеток энтодермы, возникшей из клеток эктодермальной
подушки (рис. 58, Г). Из этого клеточного материала развивается
кишечник.
Клетки промежуточной зоны — это материал мезодермы
£рис. 58, В), который оказывается в составе архентерона между
материалом хорды и энтодермы.
В отличие от ланцетника анальное отверстие у миноги обра-
зуется не на заднем конце тела, а гораздо ближе к середине.
Карты презумптивных органов бластул ланцетника и миноги
почти идентичны. Презумптивная хорда и нервная система миноги
131
Рис. 68. Гаструляция у миноги:
Л — внешний вид бластулы; Б — сагиттальный разрез гаструлы (из кн. П. П. Иванова,
1945); В - схема топографии материалов иа одной из стадий гаструляцин (по П. Гатта,
О. В. Чекаиовской, 1945); Г — фронтальный разрез поздней гаструлы (но П. Гатта, 1915);
9 — анимальный полюс, 2 — вегетативный полюс, 3 — бластоцель, 4 — бластопор, 5 — дор-
сальная губа бластопора, 6 — полость архентерона, 7 — хорда, 3—нервная пластинка, Р —
эктодерма, 10 — энтодерма, 11 — мезодерма
расположены так же, как у зародышей ланцетника. Вокруг мно-
гослойной энтодермальной пластинки находится мезодермальный
материал, за исключением переднего участка, где находится хор-
да. У ланцетника отсутствует прехордальная пластинка, которая
при гаструляцин у миноги оказывается в составе головной кишки,
ГАСТРУЛЯЦИЯ У АМФИБИЙ
Вследствие исключительного значения амфибий как объекта экс-
периментальной эмбриологии здесь дается подробный очерк га-
струляции у этих животных, а также начальных этапов форми-
рования нервной и других систем органов.
Карта презумптивных зачатков бластулы амфибий сходна с
расположением презумптивного материала зародышей асцидий,
ланцетника и миноги. Как у ланцетника и миноги, области бласто-
132
дермы у амфибий, являю-
щиеся источником формиро-
вания мускулов, хорды,
кишки и других органов, в
результате гаструляции ока-
зываются внутри зародыша.
Инвагинация у амфибий не
может происходить так, как
у ланцетника, потому что
вегетативное полушарие
яйца очень перегружено
желтком.
Первый заметный при-
знак начинающейся гастру--'
ляции у лягушек и трито-
нов — это появление бла-
Рис. 59. Изменение формы бластопора и
закрытие его в ходе гаструляции заро-
дыша лягушки (по Б. И. Балийскому,
1961):
стопора, т. е. вдавления или щели в середине серого серпа
(рис. 59). Первоначально небольшая щель все более удлиняется
по окружности—по границе краевой зоны и вегетативной области
зародыша. В конце концов окружность замыкается. Ограниченный
кольцевидным бластопором клеточный материал вегетативной
области имеет вид подушечки и называется желточной пробкой
(лучше было бы назвать ее эктодермальной пробкой).
Формирование бластопора происходит благодаря последова-
тельному вовлечению в процессы инвагинации эктодермального и
мезодермального клеточного материала. В полость бластулы с по-
верхности перемещаются клетки, лежащие на границе пигменти-
рованного и светлого полушарий, и около щели оказываются с
одной стороны темные (пигментированные) клетки, с другой —
светлые клетки. Поэтому желточная пробка выглядит светлой, как
бы включенной в темную раму. Гастральное впячение образуется
в районе дорсальной губы бластопора благодаря встречному дви-
жению в сторону бластопора хордо-мезодермального материала
со стороны анимального полюса и эктодермального — со стороны
вегетативного. Оно постепенно углубляется, и к моменту образо-
вания кольцевидного бластопора почти достигает своим дном
анимального полюса. У боковых и заднего краев желточной проб-
ки полного гастрального впячения не наблюдается, происходит
лишь подворачивание края, обрастание анимальными клетками
вегетативных. Инвагинация и связанные с ней у амфибий процес-
сы эпиболии (нарастание одних клеточных масс на другие) при-
водят к постепенному закрытию бластопора. Эктодермальная
пробка оказывается внутри зародыша. Окончательного закрытия
бластопора, впрочем, не происходит, все еще остается очень узкий
канал, идущий с поверхности внутрь зародыша.
Инвагинация у амфибий резко отличается от инвагинации у
ланцетника. В начале формирования бластопора имеет место дей-
133
ствительно инвагинация, но вскоре, а затем в течение всего про-
цесса гаструляции происходит очень своеобразный процесс пере-
мещения с поверхности в бластоцель клеточного материала
будущих прехордальной пластинки, хорды и мезодермы. Новые и
новые участки бластодермы заворачиваются внутрь, оказываются
в бластоцеле. Суть явления состоит в том, что губы бластопора,
особенно верхняя (иначе передняя или дорсальная),— это непре-
рывно меняющееся в смысле клеточного состава образование. Губы
бластопора можно уподобить участку реки; новые и новые порции
воды (клеток) протекают в этом месте, причем самый энергичный
поток клеток — это дорсальная губа бластопора. Речка клеток
здесь как бы суживается, течение сильнее, а «спокойные воды»
текут, как увидим, с обширной поверхности зародыша.
Каковы пути и характер гаструляционных перемещений кле-
точного материала? На этот вопрос дан точный ответ еще в 1929 г.
Фогтом, использовавшим свой метод маркировки частей зародыша
витальными красителями. Рис. 59 и 60 дают представление о пе-
ремещениях клеточного материала.
Следует отметить, что помимо замыкания края бластопора по
поверхности зародыша в кольцо происходит процесс эпиболии,
выражающийся в перемещении губ бластопора к вегетативному
полюсу, в связи с чем диаметр желточной пробки постепенно со-
кращается.
По мере гаструляционных передвижений клеточного материала
формируется гастроцель — полость, окруженная со всех сторон
инвагинировавшим клеточным материалом (рис. 60). Гастроцель
увеличивается, вовлекаемый внутрь клеточный материал все более
вытесняет бывшую полость бластулы. В конце гаструляции дном
гастроцеля является энтодерма, а крыша — это материал хорды
Материал мезодермы примыкает к хордальному зачатку с боков
(см. рис. 59, Е).
Достойно особого внимания поведение презумптивного клеточ-
ного материала нервной системы и эпидермиса кожи. В конце
концов будущий эпидермис и материал нервной системы покрыва-
ют всю поверхность зародыша (рис. 60, 1, 2). Зависящий от ин-
тегрирующих влияний всего зародыша процесс распространения
эпителиального покрова на всю поверхность зародыша склады-
вается из активного перемещения клеточных пластов и их истон-
чения.
Презумптивный эпидермис кожи перемещается и истончается
во всех направлениях. Совокупность клеток презумптивной нерв-
ной системы — нервная пластинка перемещается почти исключитель-
но в меридиональных направлениях — к дорсальной губе бласто-
пора. Кроме того, он сокращается в поперечном направлении. В
результате презумптивная область нервной системы оказывается
вытянутой в анимально-вегетативном направлении (рис. 61, <?).
Как уже сказано, материал презумптивной хорды составляет в кон-
134
Рис. 60. Последовательные этапы гаструляции у амфибий (по Г. Шпеману, 1936):
А, Б, В, Г, Д — медиальные сагиттальные срезы; Е— стадия, соответствующая изображен-
ной иа Д в поперечном разрезе; 1 — эктодерма, 2 — нервная пластинка, 3 — хордомезодер-
ма, 4 — энтодерма, 5 — мезенхима
це гаструляции часть крыши первичной кишки и в виде более уз-
кого удлинившегося пласта подстилает находящуюся над ним
более широким пластом клеток будущую нервную систему
(рис. 61, 3, 4). На переднем конце зародыша, впереди материала
хорды, составляя как бы часть его, лежит материал ввернувшейся
сюда прехордальной пластинки, которая в поздней бластуле лежит
на ее поверхности ближе всего около будущего бластопора.
Рис. 61. Схема гаструляции у амфибий
(по В. Фогту, 1929):
А — ранняя гаструла; Б — средняя гаструла;
В — поздняя гаструла (стадия желточной
пробки); Г —конец гаструляции; 1 — пре-
зумптивный эпидермис, 2 — вентральная мезо-
дерма, 3— презумптнвиая нервная система,
4 — презумптивная хорда, 5 — прехордальная
пластинка, 6 — презумптивная энтодерма, 7 —
бластоцель, 8 — бластопор, 9 — вентральная
губа бластопора, 10 — желточная пробка,
11 — дорсальная губа бластопора, 12 — археи-
терои (крестом обозначено положение вегета-
тивного полюса)
135
Рис. 62. Сагиттальный разрез гаструлы трито-
на (по В. Фогту, 1929):
/—дорсальная граница энтодерма, 2 — хордомезо-
дермальная мантия, 3 — граница мезодермальной
«мантии>. Видно обособление от энтодермы хордо-
мезодермальной «маитии>
Большая часть мезодермального материала вворачивается
внутрь бластоцеля через боковые и брюшную губы бластопора.
Клеточный пласт мезодермы при перемещении в переднем направ-
лении оказывается между экто- и энтодермой (рис. 62). На этом
этапе гаструляцин следует говорить о хордомезодерме или, точ-
нее, о едином эпителиальном пласте хордомезодермальной ман-
тии, состоящей из инвагинировавшей через дорсальную губу пре-
зумптивной хорды и с боков — мезодермы. Мезодермальный мате-
риал не достигает, однако, самого переднего конца, т. е. того места,
где формируется впоследствии рот. Он сильно утолщен вблизи
хорды.
Остается сказать о поведении эктодермального материала, ко-
торый в бластуле амфибий находится частично в вегетативной
области и частично в краевой зоне (см. рис. 60, 4 и 61, 6).
Энтодермальный материал, первоначально составляющий ямку
гастрального впячения, позднее примет участие в образовании
передней части гастроцеля и разовьется в переднюю кишку.
Энтодерма вегетативной области, оказавшаяся внутри зароды-
ша после закрытия бластопора, составит впоследствии толстый
слой клеток дна и боковых стенок архентерона, и из нее разовьется
средняя кишка зародыша.
ГАСТРУЛЯЦИЯ У КОСТИСТЫХ РЫБ
Дискоидальное дробление, как уже говорилось, приводит к фор-
мированию дискобластулы. Между бластодиском и желтком у не-
которых рыб имеется небольшая полость — бластоцель. Иногда
бластоцель совершенно не выражен. Бластодиск соответствует
крыше и краевой зоне бластулы амфибий и ланцетника. Дно бла-
стулы— нераздробившийся желток, покрытый парабластом.
Процесс гаструляцин у костистых рыб начинается тогда, когда
многослойный бластодиск покрывает только небольшую часть
желтка яйца, а заканчивается вместе с полным обрастанием всего
«желточного шара». Это означает, что гаструляция включает и
136
разрастание бластодиска. Карта презумптивных органов дискобла-
стулы изображена на рис. 55, А.
До недавнего времени считалось, что гаструляция у костистых
рыб осуществляется подворачиванием краевого бластодиска, гомо-
логичного дорсальной и боковым губам бластопора позвоночных
с голобластическим типом развития; в этом участке под бластодиск
перемещаются презумптивные материалы хордомезодермы и ки-
шечной энтодермы. По будущей краниокаудальной оси зародыша
в результате подворачивания формируется осевой комплекс зачат-
ков, дальнейшая дифференцировка которого имеет ряд общих черт
с таковым у других позвоночных.
В последние годы гаструляция костистых рыб получила иную
трактовку благодаря работам американского исследователя В. Бол-
ларда (1973—1982) на лососе и чукучане (карпообразные). Поль-
зуясь методом прижизненной маркировки бластодиска с помощью
частичек мела, вводимых на разную глубину бластодиска в раз-
личных его участках, и прослеживая судьбу меченых клеток, Бол-
лард показал, что при образовании зародышевых листков у ло-
сося и чукучана подворачивания бластодиска не происходит.
Презумптивные материалы эктодермы, хордомезодермы и кишеч-
ной энтодермы располагаются в зародышевой части бластодиска
слоями, налегающими один на другой. Гаструляция осуществляется
в результате двух процессов: деляминации, т. е. обособления этих
двух слоев друг от друга, и конвергенции, т. е. «схождения» или
перемещения материала этих пластов к медиальной оси зародыша,
что приводит к формированию осевого комплекса зачатков. Про-
цесс гаструляции в этом случае отличен от такового у других поз-
воночных, а карта презумптивных зачатков оказывается трёхмер-
ной (см. рис. 55, А)<
Периферические участки бластодиска становятся краями обра-
стания и постепенно вместе с перибластом распространяются по
желтку: завершение этого процесса приводит к окончательному
образованию желточного мешка. Желточная энтодерма у костистых
рыб образована перибластом и, следовательно, имеет иное по
сравнению с кишечной происхождение. По мере обрастания желт-
ка по периферии бластодиска идет выселение кровяной мезенхимы,
распространяющейся по перибласту и впоследствии формирующей
сосуды желточного мешка.
Клеточный материал всех трех зародышевых пластов по перед-
нему и боковым краям бластодиска начинает нарастать на жел-
ток. Таким образом формируется желточный мешок. Энтодерма,
из которой развивается эпителий желточного мешка, возникает из
глубоких слоев бластодиска (парабласт). У многих рыб кишечная
и желточная энтодермы имеют разное происхождение.
Желточный мешок — это орган зародыша, в функциональном
отношении нераздельная его часть, так как без желточного мешка
жизнь зародыша и дальнейший онтогенез невозможны. Значит, тра-
137
диционное разделение зародыша на собственно зародыш и «вне-
зародышевые» части в сущности неправильно и лишь отчасти
оправдывается тем, что зародышевый мешок-—временный прови-
зорный орган. Однако можно указать многие образования в ходе
онтогенеза разных животных, составляющие интегральные части
зародыша, исчезающие впоследствии, но которые в эмбриологии
не считаются внезародышевыми частями.
Желточный мешок как часть зародыша выполняет многообраз-
ные функции.
1. Это орган с трофической функцией, так как дифференцирую-
щийся энтодермальный пласт продуцирует ферменты, помогаю-
щие расщеплять вещества желтка, а в дифференцирующемся ме-
зодермальном пласте образуются кровеносные сосуды, находящие-
ся в связи с сосудистой системой собственно зародыша.
В результате благодаря изменению ферментами составных частей
желтка, всасыванию эпителием желточного мешка и поступлению
их в сосуды желток становится пищей собственно зародыша и од-
новременно «внезародышевых» частей. У костистых рыб эктодер-
мальный эпителий участвует в формировании кожи. Из этого
также видно, сколь условно разделение зародыша рыб на заро-
дыш и «внезародышевые» его части.
2. Желточный мешок — орган дыхания. Газообмен зародыша
с внешней средой происходит через стенки сосудов желточного
мешка и наружный (эктодермальный) эпителий.
ГАСТРУЛЯЦИЯ У РЕПТИЛИЙ и птиц
Яйцевые клетки у птиц и рептилий гигантские, с большим количе-
ством желтка, резко выраженного телолецитального типа. Собст-
венно яйцом является желток (см. рис. 3), покрытый желточной
оболочкой. Белковая оболочка («белок»), которой одевается яйцо
птиц по мере продвижения его по яйцеводу, а также скорлуповач
и подскорлуповые оболочки — третичные, они возникают благода-
ря секреторной деятельности желез яйцевода. У яиц черепах так-
же имеется белковая оболочка, а у ящериц и змей она отсутствует
или представлена чрезвычайно тонким слоем.
Дробление и формирование бластулы у птиц происходит в яй-
цеводе. Дробится лишь небольшой участок яйца, расположенный
в районе анимального полюса. Первые бластомеры распластаны
на поверхности желтка. После второго-третьего делений дробле-
ния бластомеры делятся асинхронно. В центре формирующегося
бластодиска появляются борозды, проходящие в плоскости, па-
раллельной поверхности яйца, в результате чего возникают изоли-
рованные от желточной массы клетки. Краевые бластомеры про-
должают сохранять тесную связь с желтком. По мере увеличения
количества клеток в составе бластодиска его поверхностный слой
138
все более приобретает эпителиоморфную организацию, а под ним
появляемся подзародышевая полость. Тело зародыша будет раз-
виваться ^3 части бластодиска — зародышевого щитка (рис. 63).
Вся средняя часть бластодиска вместе с зародышевым щитком
расположена над зародышевой полостью и не соприкасается
с желточной массой яйца. Эта зона, как уже отмечалось, называ-
ется area pellucida (прозрачная зона). Она окружена по краям
зоной клеток, прилегающих к желтку,— area ораса (темная зона).
Процессы гаструляции у птиц были предметом многочислен-
ных исследований и до сих пор трактуются эмбриологами неоди-
наково. К моменту откладки яйца в средней части area pellulcida
клетки сгущаются, располагаются плотнее и эта часть бластодис*
ка становится толще. Ее называют зародышевым щитком. В зад-
ней части зародышево-
го щитка на стороне,
обращенной к подзаро-
дышевой полости, на-
чинается обособление
путем деламинации
внутреннего слоя кле-
ток— первичного гипо-
бласта (материала
внезародышевой энто-
дермы).
Первичный гипо-
бласт оказывается- ле-
жащим на дне подза-
родышевой полости,
образуя непрерывный
слой клеток, контакти-
рующих с желтком.
Позднее на заднем кон-
це зародышевого щит-
ка образуется второе
поколение клеток гипо-
бласта — вторичный ги-
побласт, который по-
степенно вытесняет
первичный гипобласт
к периферии бласто-
диска. Некоторые ав-
торы относят образо-
вание гипобласта к яв-
лениям, предшествую-
щим собственно гаст-
руляции. Было пока-
зано, что гипобласт
Рис. 63. Гаструляция у птиц:
А — вертикальный разрез бластодиска куриного яйца
в начале гаструляции; Б — ранияя первичная полос-
ка; В — дефинитивная первичная полоска (по
А. Г. Кнорре, 1959); Г, Д — стадии головного отрост-
ка (по В. Гамбургеру и X. Гамильтону, 1951); 1 —
средняя часть наружного листка (эктобласта), заро-
дышевый щиток, 2— внутренний листок (эндобласт),
5 — внезародышевая эктодерма периферических
участков area pellucida, 4 — желточная оболочка, 5—
желток, 6— area ораса, 7 — area pellucida, 8 — пер-
вичная полоска, 9 — геизеновскнй узелок, 10 — голов-
ной отросток
139
имеет отношение к детерминации полярности зародыша.
Позднее в области зародышевого щитка образуется первичная
полоска, располагающаяся вдоль средней линии зоны. Она посте-
пенно удлиняется и занимает 2/3 или 3/4 этой зоны. На переднем
конце первичной полоски образуется утолщение — ёензеновский
узелок, в центре которого появляется ямка (углубление). По всей
длине первичной полоски, начиная от гензеновского узелка к краю
бластодиска, формируется первичная бороздка.
Еще до образования гензеновского узелка клеточный матери-
ал первичной полоски становится источником формирования ки-
шечной энтодермы зародыша, которая мигрирует внутрь под эпи-
бласт и распространяется латерально, постепенно оттесняя пер-
вичный и вторичный гипобласт на периферию. Со дна первичной
бороздки мигрируют затем внутрь клетки мезодермы. Впереди
гензеновского узелка в составе эпибласта располагаются клетки,
которые погружаются в районе гензеновского узелка внутрь и за-
тем оказываются лежащими под эпибластом по средней линии
первичной полоски. Гензеновский узелок рассматривают как го-
молог дорсальной губы бластопора амфибий, а материал, распо-
ложенный до погружения впереди его,— презумптивная хорда.
Правые и левые края первичной бороздки — гомологи боковых
губ бластопора амфибий.
ОСОБЕННОСТИ РАЗВИТИЯ МЛЕКОПИТАЮЩИХ
До тех пор, пока не будут выяснены более детально вопросы про-
исхождения млекопитающих и их эволюция, любое предположе-
ние. об историческом происхождении особенностей их развития
неизбежно будет носить спорный характер.
Предками млекопитающих являлись, вероятно, яйцекладущие
рептилии. Наиболее примитивны из современных млекопитающих
также яйцекладущие животные. В подклассе первозверей (Proto-
theria) яйцо утконоса Ornithorhynchus развивается вне материн-
ского организма; у ехидны Echidna яйцо развивается в кожной
складке на брюшной стороне тела, но питание от матери зародыш
не получает. Яйца этих животных содержат большое количество
желтка, дробление частичное, свойственное телолецитальным яй-
цам. Эмбриональное развитие во всех существенных чертах сход-
но с развитием рептилий.
В подклассе сумчатых (Metatheria) зародыши также почти не
получают питание от матери. У некоторых сумчатых развивается
примитивная плацента, функционирующая непродолжительное
время. Зародыши у сумчатых, например, рождаются недоразви-
тыми, очень маленькими (1—2 см) и донашиваются матерью в
сумке, где устанавливается связь пищевода зародыша с соском
матери: ее молоко впрыскивается в пищевод. Яйца у этих живот-
ных содержат небольшое количество желтка.
140
У в'Цсших млекопитающих, а значит, и у человека яйцо прак-
тически\не содержит желточных включений и у него нет в доста-
точном количестве ни органических, ни неорганических веществ
даже для^дачальных стадий развития. Все необходимое для свое-
го развитий зародыш получает с самого начала от материнского
организма. Яйцевые клетки плацентарных микроскопического
размера (1010—200 мкм в диаметре), алецитального или изолеци-
тального типа. Как происходила эволюция яиц и каков характер
развития млекопитающих по мере прогресса установления связи
зародышей с материнским организмом, эмбриология не знает,
хотя и высказаны важные предположения. Очень интересна и
вполне аргументирована смелая гипотеза зоолога В. А. Фаусека,
согласно которой эволюцию установления все более тесных связей
зародыша с материнским организмом надо рассматривать в свете
явлений паразитизма: у зародыша, как у паразита, развиваются
приспособления для использования материнского организма в ка-
честве места обитания и как источника пищи.
Для. анализа причин характера дробления яиц млекопитаю-
щих недостаточно тех гипотез, которые изложены в гл. IV о дроб-
лении. Казалось бы, раз яйца млекопитающих практически лише-
ны желтка, их дробление должно быть таким же, как у изолеци-
тальных яиц беспозвоночных и низших хордовых. На самом деле
у многих высших млекопитающих дробление кажется неправиль-
ным, оио несинхронное, зародыш проходит, например, стадию
трех, пяти бластомеров и т. д. Если учесть сложность организации
млекопитающих, а также большие видовые особенности в дробле-
нии, закономерно встречающиеся у разных видов млекопитающих,
то надо признать, что говорить о дроблении яиц млекопитающих
как о «неправильном» имеется еще меньше оснований, чем об
«анархическом» дроблении некоторых кишечнополостных.
У большинства млекопитающих в результате дробления возни-
кает морула, но у некоторых видов (например, у слоноземлеройки
или хоботкового прыгунчика) уже на четырехбластомерной ста-
дии появляется бластоцель. Эта стадия развития соответствует
бластуле других позвоночных животных.
Внутренняя масса бластомеров морулы отличается от поверх-
ностнолежащих своей дальнейшей судьбой. Внутренняя группа
бластомеров — эмбриобласт — это источник развития зародыша,
его оболочек и желточного мешка, а поверхностно лежащие бла-
стомеры являются трофобластом («питающим зачатком»). Эти
клетки участвуют в установлении связи зародыша с тканями мат-
ки. Многие эмбриологи предпочитают вместо названия «эмбрио-
бласт» употреблять название «внутренняя клеточная масса», а
вместо «трофобласт» — «трофэктодерма».
Морула млекопитающих превращается в бластоцисту — бла-
стодермический пузырек (рис. 64), размеры которого у разных
животных неодинаковы (у кролика около 7 мм в диаметре). По-
141
Рис. 64. Разрез ранней (Д) и позд-
ней (5) бластоциты и дифферен-
цирующего эмбриобласта (В) (по
К. Уоддингтону, 1957):
1— трофобласт, 2 — эмбриобласт, 3 —
энтодерма, 4 — эктобласт, 5 — раубе-
ров слой трофобласта
лость его наполнена жидкостью,
вероятно, продуктом жизнедея-
тельности клеток зародыша.
К одному из участков однослой-
ной стенки бластоцисты приле-
гает скопление клеток эмбрио-
бласта — зародышевый узелок
(рис. 64,2).
Участок трофобласта, с кото-
рым соприкасается эмбриобласт,
называют рауберовым слоем
(рис. 64,5). В ходе дальнейшего
развития зародышевый узелок
превращается в зародышевый
щиток. Начинаются процессы
гаструляции. В зародышевом
щитке обособляется энтодерма
(рис. 64,3), в результате чего он становится двухслойным, состоя-
щим из эпи- и гопобласта (рис. 64, 3,4). В зародышевом щитке,
как и у птиц, будут происходить процессы формирования первичной
полоски и гензеновского узелка, т. е. тех же структур, с которыми
связано образование системы осевых органов зародыша.
На стадии гаструляции зародыш имплантируется в стенку мат-
ки. Это — сложный в морфологическом и физиологическом отноше-
ниях процесс, подробности которого здесь не описываются, а отчасти
сообщаются в гл. VI. Зародыш внедряется в слизистую оболочку
матки целиком или только отростками трофобласта. Выросты
трофобласта называются первичными ворсинками. Впоследствии
развиваются вторичные ворсинки с соединительной тканью и со-
судами. Так формируется хорион — ворсинчатая оболочка, гомоло-
гом которой является серозная оболочка зародышей рептилий и
птиц.
У млекопитающих, как у птиц и рептилий, формируются амни-
он, аллантоис, а также и желточный мешок, хотя яйцеклетки мле-
копитающих лишены желтка. Функции этих образований, однако,
изменились.
Не касаясь большого круга вопросов об особенностях разви-
тия млекопитающих и человека, остановимся только на одном
вопросе — о способах связи эмбриона с материнским организмом.
Эта связь обеспечивается формированием специального органа —
плаценты (детского места). По строению плаценты разделяются
на несколько типов.
Плацента эпителиохориального типа, или полуплацента, ха-
рактерна для некоторых видов сумчатых, свиньи, лошади и дру-
гих млекопитающих (рис. 65,4). Ворсинки хориона, контактируя
со слизистым эпителием матки и входя в его углубления, не раз-
рушают материнские ткани. Маточные железы секретируют ма-
142
точное «Молоко (эмбриотроф), которое всасывается ворсинками
хориона. \Гаким образом материнский организм обеспечивает за-
родыш питательными веществами и кислородом. При родах вор-
синки хориона отъединяются от эпителия матки без разруше-
ния ее. \
Десмохорцальная плацента, или соединительно-тканная хори-
альная (у жвачных, рис. 65,5), развивается так, что ворсинки
внедряются в Атенку матки, эпителий ее слизистой оболочки раз-
рушается; тканй зародыша оказываются в контакте с соедини-
тельной тканью &атки; однако более интимная связь между мате-
ринским организмом и зародышем не устанавливается и при этом
типе плацентации. Область контакта зародыша с тканями мате-
ри осуществляется в районах котиледон, небольших площадок,
разбросанных по поверхности обширного плодного пузыря, пре-
Г
Рис. 65. Схема плацент (ориг.):
А — эпителиохориальная; Б — соединительно-тканная хориальная; В — эндотелнохориаль-
ная; Г — гемохориальная; / — эпителий хорнона, 2 — эпителий стенки матки, 3 — соедини-
тельная ткань ворсннкн хориона, 4 — соединительная ткань стенки маткн, 5 — кровеносные
сосуды ворсинок хорнона, 6 — кровеносные сосуды стенкн матки 7 — материнская кровь
143
восходящего в сотни раз объем самого зародыша. После родов в
стенке матки остаются участки, лишенные эпителия, последую-
щая регенерация происходит очень быстро. .7
Вазохориальная плацента, или эндотелиохориальная' (у хищ-
ных), обеспечивает еще более тесную связь зародыша с матерью
(рис. 65,В). При плацентации происходит не только разрушение
слизистого эпителия матки, но и деструктивные процессы в соеди-
нительной ткани: трофобласт ворсинок хориона оказывается в не-
посредственном контакте с эндотелием сосудов. Этот способ пла-
центации обеспечивает более совершенное снабжение зародыша
питательными веществами и кислородом.
Гемохориальная плацента развивается у некоторых насекомо-
ядных, летучих мышей, грызунов, обезьян и у человека (рис.
65,Г). Благодаря плацентам этого типа устанавливается еще бо-
лее интимная связь зародыша с материнским организмом. Сово-
купность очень сложных процессов (лизис эпителия матки, разру-
шение соединительной ткани и сосудов) приводят к тому, что
трофобласт непосредственно соприкасается с кровью матери, вор-
синки хориона «купаются» в материнской крови.
Строение гемохориального типа плацент варьирует у разных
животных. У обезьян и человека ворсйнки хориона сложно вет-
вятся, и тем самым создается огромная поверхность соприкосно-
вения ворсинок с кровью. Поэтому такая плацента называется
ворсинковой гемохориальной плацентой. Увеличение поверхности
контакта трофобласта с кровью в других случаях (у грызунов,
например) достигается путем сильного разветвления выпячива-
ний трофобласта, образующих сложный лабиринт каналов, по ко-
торым течет материнская кровь; поэтому такого типа плацента
называется лабиринтной гемохориальной плацентой.
Естественно, что при такой связи зародыша с матерью роды
сопровождаются сильным кровотечением, отпадает эпителий сли-
зистой оболочки матки. Стенка матки обладает хорошей регене-
рационной способностью, и происходит быстрое восстановление ее.
БИОХИМИЧЕСКИЕ ПРОЦЕССЫ ПРИ ГАСТРУЛЯЦИН
Биохимические процессы при гаструляцин изучены, к сожалению,
недостаточно и притом на ограниченном круге объектов. Сравни-
тельного очерка биохимических явлений при гаструляцин дать
невозможно. К тому же биохимические данные приобретают цен-
ность для эмбриологии только при условии, если удается устано-
вить их причинную связь с явлениями формообразования. Таких
данных, однако, мало, к тому же они спорны (см. гл. XI).
Эмбрион в ходе гаструляцин, как и до нее, не растет. Для
происходящих при гаструляцин процессов клеточного деления, пе-
ремещения пластов клеток н других явлений требуется затрата
энергии. Она возникает при диссимиляции. Происходят процессы
144
распадД запасных веществ эмбриона или же (например, у млеко-
питающих) необходимая энергия получается в конечном счете от
материнского организма. Период гаструляции — период бурных
морфогенетических процессов, которые, конечно, сопровождаются
и энергичным метаболизмом; при этом он приобретает существен-
ные особенности по сравнению с предыдущими стадиями развития
зародыша.
Потребление кислорода заметно возрастает на стадии гастру-
лы. Так, по данным Б. Коэна, на стадии дробления за час яйцо
лягушки поглощает 0,054 мл3 Ог на миллиграмм сухой массы,
на стадиях средней и поздней гаструлы — соответственно 0,141 в
0,162 мл3.
Происходят и качественные изменения диссимиляторных про-
цессов, однако эти изменения недостаточно изучены, а имеющиеся
разрозненные данные пока трудно интерпретировать. У зародышей
амфибий и морских ежей на стадии дробления, вероятно, не про-
исходит распад белков. Очень заметно уменьшается в ходе дроб-
ления количество углеводов, в особенности гликогена. Эксперимен-
тально доказано (о чем говорилось ранее), что яйца амфибий,
помещенные в атмосферу азота (в анаэробные условия) развива-
ются нормально; при этом происходит гликолиз, что очевидно из
появления молочной кислоты. Кроме того, в аэробных условиях
происходит также нормальное развитие; при этом на стадии гаст-
рулы потребление кислорода увеличивается. Вероятно, у эмбри-
онов некоторых животных возможны компенсаторные изменения
в метаболизме, при развитии эмбрионов в меняющихся условиях.
Как уже говорилось, по мнению А. С. Спирина и др. (1964),
исследовавших биохимические процессы по ходу развития вьюна»
на всех стадиях развития происходит синтез белка и рибонуклеи-
новых кислот. Однако до стадии поздней бластулы не происходит
программирования рибосом новосинтезированной мРНК. Полири-
босомные белок-синтезирующие структуры цитоплазмы до стадии
поздней бластулы содержат относительно стабильную матричную
РНК, синтезированную не ядрами зародыша, а материнским яд-
ром в процессе оогенеза. По мнению авторов, перед гаструляцией
наступает период, когда информосомы (комплексы мРНК с бел-
ком), синтезированные ядрами клеток зародыша, репрограмми-
руют рибосомы. При этом возможен частичный или полный рас-
пад старой материнской мРНК.
Транскрипция генетической информации, необходимой для
морфогенетических процессов на стадии гаструлы, у морской звез-
ды Asterias forbesi происходит между 6 и 11 ч после оплодотво-
рения, т. е. в период от стадии бластулы до стадии выселения ме-
зенхимных клеток. Гаструляция начинается через 18 ч с неболь-
шими отклонениями в разных случаях.
Большое количество фактов говорит о том, что вместе с гаст-
руляцией начинается и более энергичное образование белков.
145-
В 50-е годы было распространено убеждение, что у зародышей до
гаструляцин вообще не происходит синтез белков, однако последу-
ющие многочисленные исследования этого не подтвердили. Важ-
нейший источник для синтеза белков — желток. Новообразующи-
еся белки отличаются качественно, что доказано в опытах на яй-
цах амфибий и морских ежей. Много интересных данных получено
в работах эмбриологов стокгольмских школ. Т. Густафсон и
М. Хьелт в 1951 г. изучали относительное количество 18 различ-
ных аминокислот в ходе развития морских ежей. Оказалось, что
во время дробления до поздней бластулы соотношение кислот
почти стабильно, а с начала гаструляцин до формирования личин-
ки количество одних кислот уменьшается, других — возрастает.
Т. Густафсон и И. Хассельберг в 1951 г. нашли, что одна груп-
па ферментов остается количественно неизменной во все время
развития до плутеуса включительно, а количество, точнее актив-
ность, другой группы ферментов, не изменяясь во время дробле-
ния, возрастает с начала гаструляцин (ферменты апираза, сукци-
натдегидрогеназа, глутаминаза и др.).
Во время гаструляцин осуществляется синтез новых белков.
Какова их морфогенетическая функция и ферментативная роль,
еще не ясно. Нет полной ясности и в некоторых чисто биохими-
ческих вопросах. Возникают ли новые белки или же происходит
активация ферментов, сохранившихся со времени оогенеза? Однако
в некоторых случаях удалось установить, что новые специфические
белки, характерные для развития определенных тканей, синтези-
руются тогда, когда следует дифференциация тканей. Например,
ферменты, необходимые для синтеза меланина (в клетках нервно-
го гребня у амфибий), появляются в ходе нейруляции, когда оче-
редные процессы развития полностью зависят от генной актив-
ности, от белков, возникающих под влиянием генома.
Иммунохимическими методами П. Перлман и Т. Густафсон
в 1948 г. доказали изменение антигенной характеристики прото-
плазмы клеток во время гаструляцин. Вновь образующиеся анти-
гены (белки) вызывают появление новых антител.
Привлекает особое внимание эмбриологов факт наступающих
цитохимических отличий клеток, сопровождающих переход от
бластулы к гаструле, на что указывал П. П. Иванов (1945). Не
меньшего внимания заслуживают факты химических различий
клеток в разных участках гаструлы, повышение интенсивности
метаболизма в районе спинной губы бластопора и другие
явления. О них говорится в разных главах книги, в частности
в гл. XI.
На основании молекулярно-биологических исследований можно
сделать следующие главные выводы о проявлении генной актив-
ности в эмбриональном развитии амфибий. Неэнергичный синтез
иРНК происходит уже на стадиях дробления и медленно увели-
чивается до стадии средней бластулы. В жизни бластулы насту-
146
пает момент, когда во всех клетках зародыша число генов, нахо-
дящихся в состоянии активности, сильно возрастает, синтез
иРНК увеличивается раз в 20. На стадии формирования поздней
бластулы возрастает и синтез тРНК. Количество иРНК, оказыва-
ющейся на стадии средней гаструлы, остается приблизительно та-
ким же и на протяжении нейруляции.
Синтез рибосомной РНК начинается во время гаструляции. Как
уже говорилось, процессы развития до гаструляции, зависящие от
рибосомной РНК, происходят только благодаря рибосомам, воз-
никшим в оогенезе. Новая рибосомная РНК синтезируется в зна-
чительном количестве накануне вылупления зародыша. Как уже
неоднократно говорилось, процессы развития до гаструляции не
зависят от синтеза новой РНК, совершающегося в ядрах бласто-
меров.
У зародышей иглокожих иные сроки начала синтеза разных
РНК. У некоторых видов синтез рибосомной РНК начинается еще
до гаструляции, на стадии мезенхимной бластулы, а синтез
тРНК — во время гаструляции. Выходит, что у амфибий —обрат-
ные соотношения: рибосомная РНК синтезируется во время гаст-
руляции, а транспортная —уже на стадии бластулы (между
средней и поздней бластулой). Молекулярные генетики отмечают
и сходство зародышей амфибий и иглокожих: у тех и других син-
тез иРНК начинается еще во время дробления, а синтез рибосом-
ной и транспортной РНК задерживается.
У рыб иРНК синтезируется только на стадии ранней бластулы.
В яйцах млекопитающих синтез РНК, как и в яйцах амфибий,
начинается во время дробления, даже на стадии двух бластоме-
ров (у яиц мыши).
В табл. 4 на основании исследований многих авторов представ-
лены данные о содержании РНК в яйцах животных и о начале
синтеза РНК геномом зародыша.
В заключение следует еще раз остановиться на вопросах мо-
лекулярной биологии и генетики развития организма. На основа-
нии данных многих авторов можно подвести такой итог.
Генная активность проявляется уже тотчас после оплодотво-
рения, но имеет место лишь незначительная транскрипция «ин-
формации». Обусловленная генной активностью продукция не
вступает тотчас же в соответствующие биохимические и морфоло-
гические связи и используется впоследствии, на поздних стадиях
развития.
Поведение бластомеров определяется «программой» синтезиро-
ванных в оогенезе материнских матриц и процессами, происходя-
щими при оплодотворении (формирование полисом и др.). В пе-
риод бластулы в ядрах бластомеров синтезируются новые матри-
цы, с которыми связано дальнейшее развитие. Синтез белков
в период бластуляции у многих животных определяется материн-
скими матрицами, но в это же время происходит ц образование
147
"Таблица 4. Первоначальное содержание РНК в яйцах различных животных
и начало синтеза РНК геномом зародыша (по Э. Дэвидсону, 1969)
Животные Содержание РНК в яйце, мкг1 Стадия, иа которой начинается синтез РНК
рибосомные РНК | иРНК | тРНК
Вто ричн о ротые
Иглокожие (боль- шинство родов) 0,С02 Гаструла; у некоторых видов мезенхимная бластула Раннее дроб- ление Гаструла
Асцидии (Asci- dia) — — Раннее дроб- ление —
Костистые рыбы (Misgurnus) 2,2 После гаст- руляцни Бластула —
Амфибии ;(Xenopus) 4,0 Гаструла Дробление Средне- поздняя бластула
Птицы (Gallus) -Ч.20002 Дробление ——
Млекопитающие .-(Mas) 0,0016-0,0020 4— 8-клеточ- ная стадия Раннее дроб- ление или перед первым дробле- нием
.Первично ротые
Нематоды -(Ascaris) 0.0С0С60 До слияния пронуклеусов Перед первым дроблением —
Моллюски (Пу anas sa) 0, С04 До постгаст- руляцнонного периода, вероят- но, уже на ста- дии 16 клеток До постгаст- руляцнонного периода, веро- ятно, уже на стадии дробле- ния
Насекомые -.(Oncopeltus) 0,31 Гаструла Перед нача- лом гаструляции
1 Более трети общего количества РНК, экстрагированной из яйца, имеет молекулярную
'Массу, характерную для молекул рибосомной РНК.
а Содержание РНК во всем яйце (в белке н желтке). До сих пор невозможно установить,
какая часть этой РНК используется зародышем.
новых иРНК, без которых не смогли бы состояться морфогенети-
ческие процессы на стадии гаструляции, они определяют особен-
ности белкового синтеза. В свете многих исследований молекуляр-
ные биологи не сомневаются, что именно так происходит в ходе
развития амфибий и иглокожих. У зародышей млекопитающих
участие новых генных продуктов происходит раньше.
Сколь долго играют роль образованные в оогенезе материн-
ские рибосомные и иРНК, неизвестно; предполагают, что они фун-
кционируют длительное время и после гаструляции.
ПРИЧИНЫ ГАСТРУЛЯЦИИ
Тщетно искать одни и те же причины морфогенетических движе-
ний 1 при гаструляциях разного типа, например при образовании
1 Термин предложен В. Фогтом в 1925 г.
Д48
деляминационной и инвагинационной гаструл. Однако не исклю-
чено наличие и общих черт у кажущихся совершенно различных
явлений.
Имеются исследования процессов гаструляции, осуществленные
многими авторами. К сожалению, за единичными исключениями,
все исследования проведены в основном на двух объектах — яйцах
морских ежей и, в меньшей мере, на зародышах амфибий. Это
означает, что экспериментальная эмбриология далеко не исчер-
пала разнообразие явлений гаструляции при разных ее типах и
еще не в состоянии формулировать общие закономерности. По
этой причине можно дать лишь иллюстрации подчас довольно про-
тиворечивых фактов и гипотез, касающихся гаструляции.
1. Причиной гаструляции некоторые эмбриологи считают не-
равномерный рост в разных областях зародыша, точнее—неоди-
наковые темпы деления клеток в разных частях зародыша. В слу-
чае типичной эпиболической гаструлы, вероятно, следует прини-
мать во внимание эту гипотезу, так как одной из причин обраста-
ния анимальными бластомерами вегетативных несомненно явля-
ется факт сильно замедленных темпов деления вегетативных блас-
томеров. Также и при анализе механизма смешанных типов гаст-
руляции, например у амфибий, одним из факторов гаструляции
является, вероятно, неравномерный «рост». Однако эта давняя
в эмбриологии гипотеза совершенно бессильна объяснить многие
явления. Например, может и не быть делений клеток, а процессы
гаструляции происходят. Из-за этого не следует вообще отказы-
ваться от мысли о значении митотического градиента во время
гаструляции, иллюстрацией чему служат наблюдения И. Агреля
митотического градиента у зародыша морского ежа (Psammechi-
nus miliaris) 8—22-часового возраста (подсчитывались митозы
в эктодерме, первичной мезенхиме, энтодерме и во вторичной ме-
зенхиме) .
У 10-часовых зародышей обнаружено уменьшение числа мито-
зов по направлению от анимального полюса бластулы к ве-
гетативному. Через новые 3 ч в экваториальной области зародыша
митозов оказывается меньше, чем у полюсов. Это распределение со-
храняется до конца гаструляции. Автор считает, что инвагинация
(энтодермальное впячивание) связана с локальным усилением
митотической активности в области бластопора. Образование же
первичной и вторичной мезенхимы зависит не от размножения
клеток, а от миграции их.
2. Л. Румблер в 1902 г. предложил объяснить гаструляцию
чисто механическим путем: клетки впячивающейся части изменя-
ют форму, становятся «клиновидными», обращенные в полость
бластулы концы клетки становятся более широкими, а концы, об-
ращенные к внешней среде, суживаются. Вследствие этого и в ре-
зультате давлений клеток друг на друга происходит втягивание
данного участка пласта клегок внутрь. Изменение формы клеток
149
обусловлено разницей в поверхностном натяжении с той и другой;
стороны клеток.
Близкие к этому представлению мысли высказал в 1941 г.
А. Р. Мур в связи с анализом формы клеток при нейруляции
(рис. 66).
3. Предполагают, что неодинаковая интенсивность поглощения
воды различными клетками поздней бластулы может быть причи-
ной морфогенетических движений. Наблюдения показывают, од-
нако, что эти различия невелики.
4. Предполагают, что во внутренней цитоплазме клеток форми-
руются параллельно расположенные волоконца, в связи с чем:
клетки удлиняются, а это имеет значение при впячивании энто-
дермы при гаструляцин. Действительно, при гаструляцин, напри-
мер амфибий, инвагинирующие клетки оказываются длинными,
бутылкообразными (рис. 67). Однако неизвестно еще, почему клет-
ки удлиняются и почему они принимают бутылковидную форму.
Это явление, кажущееся причиной, нуждается в цитологическом и
физико-химическом объяснении.
5. Интересную гипотезу предложил И. Гольтфретер в 1943—
1944 г. для объяснения причин «втягивания» клеток в районе
дорсальной губы бластопора при гаструляцин у амфибий. Соглас-
но его наблюдениям у яиц амфибий, помимо студенистой и жел-
точной оболочек, имеется еще тончайшая пленка, видимая в све-
товой микроскоп, если на пленке адсорбируется краситель. Пленка
эта — coat (от англ, coat,— одежда) покрывает клетки анимально-
го полушария (рис. 67,А, Б). Таким образом, дистальные концы
клеток как бы прикреплены к coat, а большая, проксимальная
Рис. 66. Механизм инвагинаций (по
А. Муру, 1941):
А— схематический разрез гаструлы; Б— фор-
ма клеток при нейруляции; 1 — первичная ак-
тивная зона гастральной пластинки, 2 — пла-
стическая зона, 3 — новое активное кольцо.
В связи с деформацией при, образовании бо-
роздки индивидуальные клетки (одна из них
показана изолированной) имеют форйу клина,
две стороны которого наклонены друг к другу
Рис. 67. Представления Гольтфре-
тера о coat (по И. Гольтфрете-
ру, 1943):
А, Б — группы клеток из анимального
полушария, одетые coat; В — разрез
ранней гаструлы (видны бутылкооб-
разные клетки); 1 — анимальный по-
люс, 2 — бластоцель, 3 — вентральная
губа бластопора, 4 — желточная проб-
ка, 5 — дорсальная губа бластопора >
150
часть клеток свободна от нее. В результате большой комплекс
клеток как бы связан. Эластичная, растяжимая пленка — основ-
ная структура, соединяющая поверхностно лежащие клетки в еди-
ное целое и поддерживающая совокупность клеток, как «спаян-
ную» массу.
При гаструляции отмечаются энергичные амебообразные дви-
жения клеток в области дорсальной губы бластопора, но изолиро-
ванными эти движения быть не могут, так как дистальные концы
всех клеток связаны с их общей «одеждой». Сила натяжения и
сократимость пленки противодействуют амёбоидному движению
и зависящему от него впячиванию клеток. Если клетка не отде-
ляется от coat, то она, вытягиваясь, превращается в бутылкооб-
разную (рис. 67,В) и даже нитевидную. Пленка заставляет клет-
ки вести коллективный образ жизни. Как слагаемое их совмест-
ных действий, при регулируемом влиянии coat происходит неиз-
бежное впячивание групп клеток внутрь.
Логично предположение Гольтфретера и о том, что некоторые
участки coat в ходе гаструляции окажутся внутри зародыша, coat
будет препятствовать слипанию клеток и тем самым играть фор-
мообразовательную роль при развитии просвета кишечника, нерв-
ной трубки и почечных канальцев.
Агенты, уменьшающие силу натяжения поверхностной пленки
или совершенно удаляющие ее (лишение Са, растворы с pH выше
9,6 или ниже 4,4 и др.), обусловливают экзогаструляцию. Так, по
данным Гольтфретера, лимоннокислый натрий растворяет coat
у амфибий, и происходит экзогаструляция.
Эти мысли очень подкупают своей ясностью. Однако Б. Балин-
ский в 1961 г., изучая электронно-микроскопические снимки заро-
дышей амфибий, не обнаружил ни coat, ни вообще каких-либо
мембран, помимо известных оболочек яйца амфибий. Грустно от-
казываться от оригинальной гипотезы Гольтфретера. Может быть,
в этом нет надобности, так как и Балинский утверждает, что ди-
стальные концы клеток очень прочно соединены между собой бла-
годаря каким-то «цементирующим» субстанциям, являющимся
экскрецией самих клеток. Координирующим механизмом морфо-
генетических движений могут быть, вероятно, и другие клеточные
структуры, например нитевидные отростки клеток. Таким обра-
зом, их поведение в районе дорсальной губы бластопора можно
объяснить исходя из предположений Гольтфретера, но заменив
слишком прямолинейные представления о coat как пленке и о ме-
ханизме движения более физиологической теорией.
6. Для анализа процессов гаструляции, как и других морфо-
генетических движений клеток и клеточных масс, следует все
больше привлекать многочисленные цитологические данные о раз-
ных типах связей между клетками, обнаруживаемых химически-
ми и электронно-микроскопическими исследованиями. Одним из
инициаторов изучения взаимодействия между клетками является
151
английский исследователь М. Эберкромби, а также Дж. Картис,
Характер сцепления между клетками, разная степень слипа-
ния, структура межклеточных пространств и другие факторы игра-
ют, по-видимому, огромную роль во всех явлениях формообразо-
вания, в том числе и в процессах гаструляцин. Связи между
клетками могут быть разнообразными: выросты мембран клеток,
выпускание псевдоподий, ундулирующие мембраны (пластинча-
тые выросты, способные волнообразно колебаться) и т. п.
Между клетками даже таких плотных тканей, как эпителии,
никогда не бывает полного контакта, всегда существует между
их мембранами пространство (60 нм и более). Большие простран-
ства между клетками (до долей мк) наблюдаются в мезенхим-
ных тканях. Пространства между клетками — не пустота, они за-
полнены веществами — продуктами жизнедеятельности клеток.
Старые представления о клетках как стабильных кирпичиках
и тканях как стабильных сооружениях из них следует целиком
оставить. Клетки могут скользить своими поверхностями,
они никогда не бывают в покое, если они живы, и стабильность
сооружений — тканей и органов — объясняется не тем, что клетки
находятся в мертвом, неподвижном состоянии, а иными надкле-
точными закономерностями. В этом отношении возможна анало-
гия архитектоники живых существ с физическим состоянием твер-
дых тел, «твердость» которых отнюдь не объясняется неподвиж-
ностью молекул. Последние находятся всегда в движении. Но
архитектоника пластов клеток может сильно меняться при нор-
мальных процессах формообразования, и клетки, освобожденные
из общих связей, могут проявлять скрытые до того потенции.
7. Одной из биологических особенностей клеток является их
способность к амебоидным движениям и фагоцитозу. Эти две
черты — отражение древнейших свойств клеточных существ.
Б. П. Токин в 1955 г. в связи с проблемой иммунитета зародышей
предполагал, что следует придавать значение важному обстоя-
тельству: потенциально всем жизнедеятельным клеткам любых
тканей, принадлежащих организмам любой степени сложности,
свойственны фагоцитарная способность и амебоидная подвиж-
ность (см. гл. XV). Эта способность находится под «арестом» ин-
теграции тканей благодаря взаимным влияниям клеток и, веро-
ятно, еще неизвестным факторам биологической и физико-хими-
ческой природы. Взаимное влияние клеток имеется уже на са-
мых ранних стадиях развития зародыша в эпителиального типа
пластах, как и в любом скоплении клеток. В случае нарушения
состояния интеграции и освобождения их от обычных корреляций
клетки начинают проявлять способность к амебоидному движению
и к фагоцитарным реакциям.
Впервые Гольтфретером показано, что если зародыши амфибийг
на стадии бластулы, гаструлы или нейрулы дезинтегрировать (до-
биться химическими способами диссоциации зародыша на. отдель-
на
ные клетки и группы их), то проявляется фагоцитарная актив-
ность одиночных клеток.
Таким образом, способность клеток зародыша к амебоидной
подвижности является прочно установленным экспериментами
фактом. Это помогает объяснять поведение одиночных клеток и их
групп, наблюдаемое в ходе гаструляции, происходящей по типу
униполярной и мультиполярной иммиграции. Одним из моментов
морфогенетических движений при гаструляции у морского ежа яв-
ляется активное перемещение клеток внутрь бластоцеля. Несом-
ненно, и при смешанных типах гаструляции, например у амфибий,
способность к амебоидным движениям клеток играет важную роль.
Несколько искусственно сближать явления фагоцитоза с про-
цессами обволакивания анимальными бластомерами вегетатив-
ных, наблюдающихся при гаструляции, которая происходит по типу
эпиболии, хотя не исключена возможность того, что эти процессы
имеют общую морфофизиологическую основу в жизни клеток.
8. В эмбриологической литературе не раз дискутировался воп-
рос — являются ли морфогенетические движения, наблюдаемые
в ходе гаструляции, функцией зародыша в целом или к моменту
перехода от бластулы к гаструле происходит уже такая дифферен-
цировка частей зародыша, что эти части способны к автомати-
ческим, не зависящим от целого преобразованиям. Однако столь
резкое противопоставление целого его частям и соответственно
противопоставление предполагаемых двух групп факторов может
быть несколько метафизично и заслуживает внимания лишь в по-
рядке рабочей гипотезы, дающей возможность экспериментиро-
вать для получения ориентировочных данных.
Эксперименты в таком направлении оказались очень полезны-
ми, так как результаты их хотя и не объясняют процессы гастру-
ляции, но ставят ряд новых неожиданных проблем, что уже зна-
менует прогресс знаний. Если поместить зародыши амфибий
в подщелоченную воду (pH 9,6 или 9,8), то они распадаются на не
связанные между собой клетки. Масса диссоциированных таким
образом клеток приготовлялась из эктодермы, энтодермы и мезо-
дермы со стадии гаструлы, из эпидермиса и нервной пластинки
со стадии нейрулы и т. п. Массы клеток смешивались в различ-
ных комбинациях и помещались в растворы с нормальным для них
pH (8,0). Клетки начинали соединяться в агрегаты соответственно
их свойствам (рис. 68). Эктодермальные клетки и клетки нервной
пластинки со стадии нейрулы оказывались на поверхности комоч-
ков, а мезодермальные клетки и клетки энтодермы, находивши-
еся в общей массе с клетками эпидермиса, имели явную тенден-
цию расположиться внутри комочков. При комбинации диссоции-
рованных клеток всех трех зародышевых пластов эктодермальные
клетки концентрировались на поверхности и мезодермальные
клетки оказывались между эктодермальными и энтодермальными
клетками.
153
Рис. 68. Поведение агрегатов клеток диссоциированного зародыша амфибий (по
Т. Тауну и И. Гольтфретеру, 1955).
Клетки: 1 — эктодермы, 2 — энтодермы, 3 — мезодермы
Эти и подобные им факты следует взять на учет при анализе
процессов гаструляции. П. П. Иванов и другие исследователи
обращали внимание на то, что на стадии поздней бластулы ам-
фибий клетки в разных частях зародыша неодинаковы по своим
физиологическим потребностям: эктодермальные клетки «требу-
ют».одних условий, а клетки, омываемые внутренней средой—•
жидкостью бластоцеля, проявляют свою физиологическую актив-
ность при иной среде.
Каковы бы ни были физико-химические особенности - клеток,
побуждающие их к определенным типам взаимосвязи (будь то
особенности клеточных мембран, своеобразие слипания клеток
или иммунохимические свойства типа антиген — антитело), эти
особенности несомненно играют роль в морфогенетических движе-
ниях клеток и клеточных масс. В дальнейшем эти вопросы будут
дополнительно рассмотрены в гл. XV.
Поведение клеточных элементов и частей эмбриона не авто-
номно: 1) намечающаяся клеточная дифференцировка в разных
частях зародыша есть результат развития зародыша в целом; 2)
в дальнейшем развитии поведение зародышевых пластов и разных
участков зародыша, как увидим в гл. XI, определяется не только
приобретенными морфогенетическими потенциями частей, но и
взаимным влиянием частей. К тому же не следует переоценивать
результаты указанных опытов, так как в лучшем случае их ре-
зультаты лишь отдаленно напоминают действительные процессы
нормального развития — формирование соответствующих пластов
клеток, полостей и т. д. В гл. XI о детерминации будут сообщены
многие факты якобы независимого развития частей зародыша
в условиях эксплантации и дан анализ этих фактов.
ГИПОТЕЗА Т. ГУСТАФСОНА
Работы Т. Густафсона и его коллег могут служить примером пло-
дотворных поисков связи между биохимией и морфологией раз-
вития. Густафсон справедливо утверждает, что в эмбриологии
154
почти нет гипотез и фактических данных о причинных зависимос-
тях между биохимическими изменениями (например, в синтезе
нуклеиновых кислот и белков), с одной стороны, и морфологичес-
кими изменениями — с другой. Необходимо искать «мосты». В фи-
зиологии стандартной практикой является объяснение процессов
в терминах физических наук, например объяснение биения сердца
и циркуляции крови на основе гидродинамики. В эмбриологии
также требуется изучение электрических, гидростатических и иных
внутриклеточных и межклеточных сил. Это поможет найти недо-
стающие звенья связи между химическими и морфологическими
'Процессами.
Густафсон пытался объяснить морфологию развития морского
•ежа, в частности процессов гаструляцин, с использованием в ос-
новном двух групп закономерно наблюдаемых явлений: по-первых,
степени слипания, степени контактов между клетками и, во-вто-
рых, амёбоидной подвижности клеток.
Используя метод цейтраферной микрокиносъемки, он изучил
такие явления, которые не были известны эмбриологии. В зре-
лом, еще не дробящемся яйце имеются три зоны с определенным
значением (рис. 69,А): клетки анимального полушария — это бу-
дущая эктодерма (3-я зона): клетки вегетативного полушария —
это клетки первичной мезенхимы (1-я зона) и, наконец, клетки,
находящиеся между названными зонами, — это энтомезодерма
(2-я зона). За первые 10 ч (приблизительно после десятого дроб-
ления) образуется сферическая однослойная бластула, состоящая
примерно из 1000 клеток.
Через несколько часов около 40—50 клеток из 1-й зоны про-
двигаются в полость бластулы (рис. 69,Д). Это первичные мезен-
химные клетки. Оказавшись в бластоцеле, они в конце концов
займут определенное положение в форме кольца с двумя ветвями,
распространяющимися по внутренней поверхности стенки блас-
тулы, по направлению к анимальному полюсу (рис. 69,Ж).
Затем следует стадия гаструляцин, формирования архентерона.
Это происходит в результате инвагинации клеток 2-й зоны, напо-
добие пальцевидного вдавления в стенку мячика. Образующееся
отверстие (бластопор) впоследствии превратится в анус. Вершина
впячивающегося вдавливания приходит на вентральной стороне в
контакт с эктодермой (рис. 69,Л). В результате происходящей пер-
форации этих двух пришедших в контакт слоев клеток образуется
рот. Архентерон не остается одинаковой на всем протяжении труб-
кой, в разных участках он сужается, подразделяется на целом,
пищевод, желудок и проктодеум. Происходит изменение формы
зародыша, в результате чего он приобретает характерную форму
личинки (рис. 69, Л, М). Скелет, начавший образовываться пер-
вичными мезенхимными клетками, простирается - до эктодермы
в пяти пунктах — в четырех на вентральной стороне и в одном на
дорсальной. Таким образом, вместе с другими гистогенетическими
155
Рис. 69. Последовательные стадии развития морского ежа
(Л— М) (по Т. Густафсону, 1963)
процессами формируются руки, а личинка-плутеус приобретает
дефинитивный вид (рис, 70). Это происходит уже через двое суток
после оплодотворения.
Рис. 70. Схема плутеуса (по Т. Густаф-
сону, 1963);
1 — рот, 2 «—скелет, 3 —желудок, 4 — аналь-
ное отверстие, 5 — целомкческая мезодерма,
6 — ресничный шнур, 7 — руки
Итак, можно отметить
четыре основные категории
морфогенетических процес-
сов: 1) формирование поло-
го шара — бластулы; 2) из-
менение кривизны стенок;
3) направленное движение
клеток, как это видно по
поведению первичных ме-
зенхимных клеток, и 4) пе-
ремещения пластов клеток
на значительную дистан-
цию, что имеет место при
инвагинации.
Почему происходят все
эти процессы? Т. Густафсон
предлагает учесть амебоид-
ную активность клеток,
формирование ими псевдо-
подий и изменения в кон-
тактных отношениях между
клетками, закономерно на-
ступающие при их слипа-
нии.
156
Рис. 71. Модель изменения объема бластулы (по Т. Густафсо-
ну, 1963)
Почему в результате дробления формируется4 не компактная*
масса клеток, а полый шар (бластула)? По Густафсону, это под-
дается объяснению в терминах механики и геометрии. Примем во-
внимание радиальный план дробления яйца ежа, а также то, что-
клетки очень сильно прилипают к гиалиновой мембране, имеющей-
ся на наружной поверхности бластулы (на схеме рис. 71 она вы-
делена линией). Гиалиновый слой (мембрана) достаточно хорошо-
изучен. Он может быть отделенют клеток. Этот слой необходим для
развития; гаструляция у морских ежей может блокироваться, если,
нарушить целостность гиалинового слоя.
Как видно из схемы, внешний и внутренний объемы бластулы по
мере делений клеток по геометрическим причинам должны возра-
стать.
Дальнейшие изменения формы, все более локальные, не зави-
сят от делений клеток. Например, ко времени инвагинации проис-
ходят различные утолщения стенки и ее кривизны. Сферическая^,
форма преобразуется так, что становится треугольной. Если кон-
такт между клетками будет возрастать, а контакт их с мембра-
ной останется константным, то слой клеток будет неизбежно ис-
кривляться. Если контакт с мембраной уменьшается, то образу-
ется слой столбчатых клеток.
Однако вскоре в развитии зародыша вступают в силу новые-
факторы не меньшего значения. Это — «псевдоподиальная актив-
ность» клеток, возможность активных перемещений клеток. На
стадии поздней бластулы происходит миграция первичных мезен-
химных клеток в бластоцель (см. рис. 69,Д). В области вегетатив-
ного полюса связь между клетками и с гиалиновой мембраной-
ослабляется, клетки приобретают амебоидную подвижность и про-
тискиваются в бластоцель. Это сопровождается энергичными пуль-
сациями клеток на их внутренней поверхности (естественно, что
на внешней стороне, где они связаны с мембраной, пульсаторные
движения исключены). Псевдоподиальная активность первичных
мезенхимных клеток усиливается, энергично образуются длинные
псевдоподии, которые как бы «выстреливаются» клетками в блас-
тоцель (рис. 72). Концы псевдоподий контактируют со стенкой
157
Рис. 72. Контактирование псевдоподий клеток
первичной мезенхимы с внутренней поверхно-
стью стенки бластулы морского ежа (по
Т. Густафсону, 1963)
бластулы. Сокращение
псевдоподий вызывает
«подтягивание», переме-
щение клеток. Если же
не произошло контакта
концов псевдоподий со
стенками, то они втяги-
ваются в клетку. Клетки
рассеиваются в нижней
половине бластулы. Т. Гу-
стафсон делает предполо-
жения об особенностях со-
стояния разных участков
стенки бластулы, о свое-
образии контактов меж-
ду клетками и в связи
с этим о неодинаковых
возможностях слипания концов псевдоподий с разными клетка-
ми. Для понимания образования первичной мезенхимы оказыва-
ются ненужными старые представления о хемотаксисе клеток.
Густафсон дает объяснение и процессам инвагинации и обра-
зования вторичной мезенхимы. При первой фазе инвагинации от-
мечается энергичное «кипение», и «вспузыривание» клеток, подоб-
ное тому, что обнаружено и для клеток первичной мезенхимы. Это
опять-таки связано с ослаблением контактов между клетками, но
связь с гиалиновой мембраной при этом остается. По чисто меха-
ническим и геометрическим причинам должна происходить инваги-
нация. Если концы пласта фиксированы, клетки имеют тенденцию
к округлению и контакт их друг с другом уменьшается, то слой
должен изменяться так, как это и происходит в начале инва-
гинации. Продвижению пласта способствует «выстреливание»
псевдоподий клетками вершины инвагинирующей области. Концы
псевдоподий слипаются с клетками эктодермы, и это содействует
«подтягиванию» первичной кишки. Клетки на вершине конуса,
наконец, теряют связь между собой и с гиалиновой мембраной и
внедряются в бластоцель, образуя вторичную мезенхиму. Есть
указания на то,- что вещества, связанные с функционированием
нервной системы, играют важную роль в морфогенетических про-
цессах уже на ранних стадиях развития организмов. Утверждают,
что синтез медиаторов нервной системы происходит уже на ран-
них стадиях развития зародышей, задолго до возникновения нерв-
ной -системы. Медиаторы нервной системы —это низкомолекуляр-
ные физиологически активные вещества — ацетилхолин, серотонин,
адреналин и др., — благодаря которым осуществляется передача
нервных влияний от клетки к клетке. Предполагают, что некото-
рые из этих веществ, вырабатываясь уже на ранних стадиях раз-
вития, играют роль своеобразных внутриклеточных гормонов,
158
регулирующих процессы деления клеток; затем они приобретают
функции гормонов, играющих роль в движении клеток, и, наконец,
становятся медиаторами нервной системы. Удалось обнаружить,
синтез медиаторов на очень ранних стадиях развития. Так, яйце-
клетки вьюна синтезируют серотонин уже на 12 мин после опло-
дотворения. Синтез ацетилхолина, серотонина и адреналина обна-
ружен в яйцеклетках морских ежей, он резко усиливается сразу
после оплодотворения, за 1,5—4 ч до первого клеточного деления,
и значит, за 10—15 сут до начала формирования нервной системы.
(Г. А. Бузников, 1968). Предполагают, что «до нервный» синтез
медиаторов — универсальное явление. Эти вещества играют важ-
ную роль в регуляции процессов деления клеток.
К изложенным представлениям о значении физических явле-
ний и амебоидного движения в морфогенетических процессах
Т. Густафсон в 1966—1969 гг. внес существенно новое, именно-
в связи с представлениями о медиаторах. Изучая влияние на про-
цессы развития морского ежа многих фармакологических веществ,
блокирующих нервногуморальную трансмиссию (1968), он обна-
ружил функционирование примитивного нейросекреторного меха-
низма, в котором большая роль принадлежит серотонину. Благо-
даря псевдоподиальной активности на стадии гаструляции из
вершины архентерона, а позже из эктодермальных ресничных
шнуров мигрируют клетки, которые можно назвать «первичными
нейронами»; в них локализованы медиаторы нервной системы.
Эти клетки играют роль в нервной трансмиссии и стимулируют
морфогенетическое движение, активируя группы эктодермальных
клеток. Густафсон предположил, что серотонин служит веществом,
обеспечивающим гаструляционные передвижения клеточных плас-
тов. Серотонин нейтрализует влияние различных веществ, способ-
ных остановить гаструляцию.
Исследования влияния нейрофармакологических веществ не-
сомненно интересны.
Г лава VII
РАЗВИТИЕ ПРОИЗВОДНЫХ ЭКТОДЕРМЫ
КЛАССИФИКАЦИЯ ОРГАНОВ КАК ПРОИЗВОДНЫХ
ЗАРОДЫШЕВЫХ ПЛАСТОВ
Органогенез — предмет исследования эмбриологии, гистологии,
анатомии, физиологии и других наук, так как он является сово-
купностью процессов изменения формы, тканевого строения, воз-
никновения и изменения биохимических и физиологических яв-
лений и т. д. Эмбриология должна изучать процессы, сопровож-
дающие развитие того или иного органа в их совокупности и
при разных способах репродукции. Однако это пока идеал, к ко-
торому стремятся. На самом деле развитие органов изучается в
разных аспектах и подчас очень односторонне, в отрыве морфо-
логии от физиологии. Невозможно сообщить в книге необозримое
количество данных о развитии всех органов животных. В даль-
нейшем будут даны только отдельные примеры. Выбор их не
произволен: он диктуется или особой важностью этих органов
для жизни организмов, или возможностью на развитии данного
органа познакомиться с общими закономерностями формообра-
зования, вскрытыми эмбриологией, или, наконец, большей изу-
ченностью развития некоторых органов излюбленных эмбриоло-
гами объектов. К сожалению, состояние эмбриологии не позво-
ляет дать какой бы то ни было сравнительный очерк органогенеза
-беспозвоночных и позвоночных животных. Потребности меди-
цины и ветеринарии обусловили большее внимание к органоге-
незу позвоночных и особенно млекопитающих. Вот почему в
книге дается описание органогенеза именно позвоночных животных.
При изучении органогенеза надо учесть, что классификация
органов по их генезису (производные эктодермы, энтодермы и
мезодермы) вполне оправдана эмбриологическими данными и
теорией зародышевых пластов в том смысле, что возникновение
зачатка (закладки) органа связано с локальными изменениями
определенных участков того или иного зародышевого пласта.
Однако в большинстве случаев органы позвоночных — производ-
ные двух или всех трех зародышевых пластов (например, яичник,
кишечная трубка, орган слуха, нос и т. д.), при этом подчас
^невозможно выделить какой-либо главенствующий клеточный ма-
териал, от которого зависел бы механизм развития органа. По-
этому классификация органов на эктодермальные, эктодермаль-
ные и мезодермальные столь же доступна сомнениям, критике,
160
ограничениям, как и теория зародышевых пластов, на которой
эта классификация основана.
Нельзя придавать этой классификации тот смысл, будто кле-
точный материал первоначальной закладки органа приобретает
какие-то особые гистотипические и «органные» свойства. Это
неверно, во-первых, потому, что развитие органа, по крайней мере
вначале, не есть автономный процесс, оно происходит при тесней-
шем взаимодействии составляющих зачаток клеток и взаимодей-
ствии различных зачатков. Во-вторых, при бесполом' размноже-
нии и экспериментальном соматическом эмбриогенезе развитие
органов той же структуры и с теми же функциями может про-
исходить существенно по-иному, в результате иных гистогенети-
ческих, анатомических и физиологических процессов.
НЕЙРУЛЯЦИЯ. РАЗВИТИЕ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Следующая за гаструляцией стадия в развитии позвоночных —
нейруляция. Зародыш на этой стадии называют нейрулой. Мате-
риал презумптивной нервной системы у позвоночных животных —
это обширный пласт клеток, покрывающий хорду и сомиты. По
краям нервной пластинки образуются два нервных, или медул-
лярных, валйка— утолщения пластов клеток презумптивной
нервной системы (рис. 73). Эти утолщения возникают сначала на
переднем конце зародыша, а затем постепенно появляются в
средней или задней частях его. Впоследствии валики становятся
все более высокими, сближаются сначала в срединной части, а
затем и в задней; наконец, смыкаются своими гребнями, образуя
нервную, или медуллярную, трубку. Эта трубка, развивающаяся
в спинной мозг, приблизительно одинакового диаметра на всем
протяжении. Передний отдел нервной трубки образуется позд-
нее, также путем смыкания валиков, которые, однако, здесь бо-
лее высокие, поэтому головной отдел медуллярной трубки ока-
зывается более расширенным. Из него развиваются передний и
средний мозговые пузыри.
В передней части нервной
трубки полость ее шире, чем
в задней.
Позднее головной отдел
медуллярной трубки превра-
щается в головной мозг. По-
лость задней части нервной
трубки превращается в канал
спинного мозга.
Рис. 73. Нейрула амфибий:
А—вид с дорсальной поверхности; Б — попе-
речный разрез; 1—нервный валик, 2 — нерв-
ная пластинка, 3— хорда, 4 — эктодерма, 5 —
мезодерма, 6 — энтодерма
Медуллярная трубка пол-
ностью отъединяется от покры-
вающего ее будущего эпидер-
миса. Края эпидермиса соеди-
161
няются над нервной трубкой. Важно отметить, что часть клеток
не включается ни в нервную трубку, ни в эпидермис. Они распо-
ложены в виде полоски по всей длине нервной трубки. Образуется
нервный гребень, или ганглиозная пластинка. После того как нерв-
ная трубка отделяется от эпидермиса, эта полоска располагается
между нервной трубкой и покрывающим ее эпителием, но впо-
следствии она разделяется на правую и левую половины, которые
залегают дорсолатерально по отношению к нервной трубке. Пе-
редний участок презумптивного головного мозга довольно долго
не закрывается. Отверстие на переднем конце нервной трубки
называется нейропором (или невропором). Задний отдел нервной
пластинки при замыкании в трубку прикрывает своим сводом
бластопор. Он находится теперь на заднем конце дна медулляр-
ной трубки и становится нервно-кишечным каналом. (У некото-
рых позвоночных, в частности у хвостатых амфибий, нервно-ки-
шечный канал не возникает.) В области шеи и туловища нервная
трубка дифференцируется в спинной мозг, а передняя часть
нервной трубки превращается в головной мозг. Передний уча-
сток нервной пластинки у всех позвоночных и в особенности у
человека значительно шире, и с самого начала формирования
нервной трубки диаметр ее неодинаков в разных участках. Зна-
чительно более расширен передний, головной участок, и стенки
трубки здесь толще, чем в задней части. Дорсальный и вен-
тральный участки стенки медуллярной трубки более тонкие, чем
разрастающиеся боковые участки. Канал нервной трубки пре-
вращается в центральный спинно-мозговой канал развившегося
мозга.
В разных участках переднего отдела нервной трубки благо-
даря неравномерному росту очень рано становятся заметными
в определенных участках утолщения, образуются карманоподоб-
ные выпячивания, особенно значительные в участках будущих
мозговых полушарий. Одновременно по той же причине снаружи
рано образуются в определенных местах неглубокие сжатия
нервной трубки и явственно обозначаются три мозговых пузыря:
prosencephalon — передний, mesencephalon — средний и rombence-
phalon — задний (рис. 74). Передний мозговой пузырь в даль-
нейшем подразделяется в свою очередь на telencephalon — зача-
ток большого мозга, иначе концевого, и diencephalon — зачаток
промежуточного мозга. Mesencephalon, развиваясь и дифферен-
цируясь, превращается в средний мозг. Rombencephalon при даль-
нейшем развитии подразделяется на metencerphalon — зачаток
cerebellum (мозжечка и моста) и myelencephalon — зачаток про-
долговатого мозга (medulla oblongata, рис. 75).
Продолговатый мозг переходит в спинной. Верхняя стенка
myelencephalon остается при дальнейшем развитии мозга тонкой,
и в ней не образуются нервные клетки. Большая часть верхней
стенки впоследствии врастает вместе с сосудистой оболочкой в
162
Рис. 75. Развитие головного мозга человека (ста-
дия пяти мозговых пузырей) (по Б. Пэттеиу,
1944):
/ — обонятельная доля, 2 —перекрест зрительных пуч-
ков, 3 — полушарие большого мозга, 4 — контур проме-
жуточного мозга, 5 — средний мозг, 6 — мозжечок, 7 —
продолговатый мозг, 8 — спинной мозг, 9 — гортань.
Римскими цифрами обозначены черепно-мозговые нервы
Рис. 74. Развитие голов-
ного мозга у двухднев-
ного куриного эмбриона
(по Б. Пэттену, 1944):
1 — передний мозг, 2—сред-
ний мозг, 3— глазной пу-
зырь, 4—-задний мозг
полость четвертого желудочка мозга (см. далее), образуя сосу-
дистое сплетение четвертого желудочка (plexus chorioideus чет-
вертого желудочка). Продолговатый мозг образуется в резуль-
тате разрастания и развития боковых и нижних стенок myelen-
cephalon. Так как при развитии продолговатого мозга у всех
позвоночных на его поверхности образуются желобки (бороздки),
то он разделяется на
большое число сегмен-
тов — нейромеров'. Осо-
бенно отчетлива сегмен-
тация у зародышей рыб.
Она, однако, не связана
с метамерным развитием
черепных нервов, а носит
временный характер, йот
нее не сохраняется следа
в дефинитном мозге.
Еще до возникновения
отчетливого подразделе-
ния переднего мозгового
пузыря на зачаток боль-
шого и промежуточного
мозга на его боковых
стенках выпячиваются
глазные пузыри (см. рис.
74.5).
Дорсальная часть
нервной трубки вытягива-
Рис. 76. Изменения топографии зачатков в
заднем конце зародыша амфибий и превраще-
ние заднего конца нервной трубки в хвостовые
сомиты (по И. Бойтелю, 1931):
А — ранняя хвостовая почка; Б, В — поздняя хвосто-
вая почка; / — нервная трубка, 2— хорда, 3 — энто-
дерма, 4 — просвет кишки, 5—постанальная кншка,
6 — сомиты
163
ется сильнее, чем вентральная вследствие чего задний конец труб-
ки выносится за бластопор. Затем происходит изгибание трубки на
некотором расстоянии от бластопора. Вершина образующегося
изгиба становится зачатком хвоста (рис. 76). Участок, лежащий
между вершиной зачатка хвоста и бластопором, дифференцирует-
ся как мышцы хвостовой области.
У разных животных, в зависимости от степени сложности нерв-
ной системы, имеются значительные анатомические и гистологиче-
ские особенности, а также особенности в топографии различных
участков центральной нервной системы позвоночных.
У низших позвоночных, например у акуловых рыб, форма
дефинитивного мозга мало отличается от мозга их зародышей.
У высших позвоночных, в особенности у человека, развивающийся
мозг ранних зародышей по своей морфологии мало отличается
от мозга взрослых рыб или амфибий, но дефинитивный мозг,
особенно по клеточной и тканевой структуре, резко отличается
от мозга низших представителей позвоночных. Прогрессивная
эволюция организмов не обязательно сопровождалась усложне-
нием в устройстве всех органов. Так, в некотором отношении
глаза у птиц или некоторых насекомых более совершенны, чем
глаза млекопитающих. Что касается эволюции головного мозга,
то у человека он достиг поразительного совершенства, и, грубо
говоря, бывший и дальнейший большой прогресс в его струк-
туре и функциях не коррелирован со сравнительно медленными
и менее радикальными преобразованиями других органов, напри-
мер кишечного тракта.
РАЗВИТИЕ СПИННОГО МОЗГА И ПЕРИФЕРИЧЕСКОЙ
НЕРВНОЙ СИСТЕМЫ
Спинной мозг развивается так, что клетки его стенок, размно-
жаясь, дифференцируются в двух направлениях—одни в ней-
роглию (поддерживающие клетки), а другие — в нервные. Под-
держивающие клетки, располагающиеся вокруг нервного канала
и в толще боковых стенок, образуют отростки, которые как бы
поддерживают, скрепляют нервные клетки. Клеточные тела
нейробластов образуют серое вещество спинного мозга, а нерв-
ные волокна и их миелиновые оболочки — бе.лое&<вещество.
Часть отростков выходит из спинного мозга. Это — брюшные
корешки спинно-мозговых нервов. Вследствие разрастания брюш-
ных и спинных частей серого вещества возникают передние и
задние рога спинного мозга. В связи с развитием конечностей
вместе с отхождением нервов конечностей в спинном мозге раз-
виваются шейное и поясничное утолщения. В книге даются лишь
общие сведения о развитии периферической нервной системы.
Двигательные нервные волокна нервной трубки выходят сег-
ментно расположенными пучками и дают начало двигательным
164
корешкам спинно-мозговых нервов. Позднее после образования
ганглиозной пластинки дифференцируются в сегментные спинно-
мозговые узлы с двумя отростками: одни отростки растут по
направлению к спинному мозгу и прорастают в него — это бу-
дущие чувствительные корешки спинно мозговых нервов, другие
отростки растут в брюшном направлении и соединяются с во-
локнами двигательных корешков в спинно-мозговые нервы. Эти
смешанные нервы отделяют от себя задние ветви, которые иннер-
вируют спинную мускулатуру двигательными волокнами и кожу
спины — чувствительными волокнами. Затем спинно-мозговые
нервы продолжаются в виде передних ветвей, иннервирующих
брюшную мускулатуру (двигательные нервы) и кожу (чувстви-
тельные нервы).
От передних ветвей вырастают висцеральные ветви (с дви-
гательными и чувствительными волокнами). Впоследствии в кон-
цевых частях этих ветвей из нервных клеток закладываются по
обеим сторонам нисходящей аорты два стврла симпатической
нервной системы.
Развивающиеся головные нервы при описании можно объеди-
нить в три группы: 1) чувствующие нервы, из которых упомянем
обонятельный, слуховой и глазную ветвь тройничного нерва;
2) двигательные нервы — это нервы глазодвигательных мышц и
подъязычный; 3) смешанные нервы, формирующиеся из двига-
тельных волокон головного мозга и чувствительных нервных во-
локон, так называемых головных узлов, возникающих за счет
ганглиозных валиков, образующихся впереди заднего мозгового
пузыря. К этой группе нервов относится тройничный и лицевой,
языкоглоточный и блуждающий.
РАЗВИТИЕ ГЛАЗА
Развитию органа зрения позвоночных посвящено необозримое ко-
личество работ. Глаза амфибий — излюбленный объект в экспе-
риментальной эмбриологии. В СССР многочисленные исследова
ния проведены Д. П. Филатовым, В. В. Поповым, Т. В. Лопа-
шовым, В. А. Голиченковым, Д. В. Поповым, - Г. Г. Квинихидзе
и др. Если миновать детали развития этого сложного органа и
почти не касаться гистогенетических процессов, изучаемых обыч-
но в гистологии, развитие глаза представляется в следующем
виде.
Как уже было сказано, первоначальные закладки глаз воз-
никают в виде глазных пузырей — выпячиваний стенок промежу-
точного мозга (рис. 77,5). Размер глазных пузырей относительно
остальной части prosencephalon у разных позвоночных сильно
варьирует. Так, по данным Б. И. Балинского (1958), у различ-
ных видов лягушек глазные пузыри составляют от 10 до 50 %
всего переднего мозга (включая и промежуточный). У костистых
165
Рис. 77. Схема развития глаза (по
Г. Г. Щеголеву, 1923):
/ — эктодерма, 2 — утолщение эктодермы,
дающее хрусталик, 3 — глазной пузырь,
4 — впячивание утолщенной эктодермы,
5— зачаток пигментной оболочки, 6 —за-
чаток сетчатой оболочки, 7 — передняя
стенка хрусталика, 8 — задняя стейка
хрусталика, 9 — зачаток стекловидного те-
ла, 10— глазной стебелек, //—верхнее
веко, 12 — конъюнктивный мешок, 13 —
роговица, 14 — эпителий хрусталика, 15 —
передняя камера глаза, 16— передний
листок сосудистой оболочки хрусталика,
17 — радужная оболочка, 18 — ресничное
тело, 19 — стекловидное тело, 20 — реснич-
ная круговая пленка, 21 — тело хрустали-
ка, 22 — задний листок сосудистой обо-
лочки хрусталика, 23— радужная часть
сетчатки, 24 —ресничная часть сетчатки,
25 — сетчатка, 26 — пигментная оболочка,
27 — сосудистая оболочка, 28 — белочная
оболочка
рыб, рептилий и птиц возникают зачатки глаз значительно боль-
шего размера по сравнению с глазными пузырями амфибий.
У зародышей млекопитающих они небольшие. По мере развития
глазные пузыри все более обособляются от мозга и остаются
соединенными с ним тонкими глазными стебельками (рис. 77,70).
Увеличиваясь, глазной пузырь приходит в непосредственный
контакт с эктодермой и одновременно в нем начинают происхо-
дить сложные морфологические изменения. Наружная поверх-
ность глазного пузыря уплощается, а затем инвагирует внутрь,
так что глазной пузырь превращается в конце концов в двух-
слойную чашу — глазной бокал. Инвагинированная стенка бокала
значительно толще, чем наружная. Внутренний слой клеток глаз-
ного бокала — это закладка сетчатки (retina), наружный слой —
закладка пигментной оболочки (stratum pigmenti или tapetum
nigrum). Край глазной чаши впоследствии превращается в край
зрачка. Полость глазной чаши — это будущая задняя камера
глаза, заполняющаяся стекловидным телом, которое развивается
из внутреннего листка глазного бокала, т. е. оно эктодермального
происхождения. Однако в формировании стекловидного тела при-
нимают участие и клеточные элементы мезодермального проис-
хождения, врастающие через сосудистую щель.
Сначала очень широкое отверстие глаза постепенно сильно
сужается, но так, что это сужение происходит неодинаково по
всей окружности. На вентральном крае глазного бокала отвер-
стие остается более открытым. Здесь образуется желобок, дости-
гающий глазного стебелька (fissura chrioidea). По желобку в
заднюю камеру глаза входят кровеносные сосуды и мезенхим-
ные клетки, которые затем и окажутся в стекловидном теле.
Впоследствии края сосудистой щели зарастают. У человеческого
зародыша это происходит уже на седьмой неделе.
Задняя (большая) часть внутреннего слоя бокала дифферен-
те
цируется в зрительную часть сетчатки (pars optica retinae), а пе-
редняя (меньшая) часть — в радужную и ресничную части. Рес-
ничная часть оболочки вместе с прилегающей в этом месте пиг-
ментной оболочкой образует концентрические складки, в которые
врастает клеточный материал мезодермального происхождения.
Так возникает ресничное тело (corpus ciliaris), в котором раз-
виваются ресничные мышцы. К ресничному телу с помощью во-
локонец— ресничной круговой пленки—прикрепляется хруста-
лик. Из радужной части сетчатки при участии сигментной и сосу-
дистой оболочек возникает радужная оболочка (iris). Клеточные
элементы сетчатки дифференцируются и в нервные волокна, из
которых развивается зрительный нерв', он заполняет просвет глаз-
ного стебелька. Из клеточного материала мезодермального про-
исхождения, врастающего в сосудистую щель глазного стебелька,
развиваются кровеносные сосуды, питающие сетчатку.
Глазной бокал не составляет еще глаза. Для нормальной функ-
ции зрения должны развиваться добавочные важные структуры.
Это прежде всего хрусталик (линза), благодаря которому вхо-
дящие в глаз лучи света преломляются. Линза развивается из
эктодермального эпителия, который в месте контакта с эктодер-
мой наружного участка глазного пузыря утолщается, а клеточ-
ные элементы претерпевают специфическую дифференцировку, они
становятся призматическими и превращаются впоследствии в хру-
сталиковые волокна. Клеточная масса будущего хрусталика от-
шнуровывается, превращаясь постепенно по своей структуре и
физическим свойствам в хрусталик.
У разных групп позвоночных развитие хрусталика и отде-
ление его от эпидермиса варьирует. У птиц и млекопитающих
возникают утолщения всего эпидермального слоя (в месте кон-
такта с глазным пузырем). Из складывающихся утолщений обра-
зуются на короткое время открытые кнаружи карманы; эти утол-
щения впоследствии и превращаются в пузырек — формирующую-
ся линзу.
У амфибий и костистых рыб только внутренний листок эпи-
дермиса принимает участие в утолщении; это утолщение отде-
ляется от эпидермиса в виде очень плотной массы клеток, ко-
торые, перегруппировываясь, образуют хрусталик. При любом
типе формирования хрусталиков в них происходят сложные ци-
тологические преобразования. Клетки внутренней стороны пу-
зырька будущей линзы удлиняются. и постепенно преобразуются
в длинные волокна. Ядра клеток дегенерируют, цитоплазма де-
лается плотной и прозрачной. Волокна располагаются не беспо-
рядочно, а в строго закономерном направлении; при этом и
формируется сферическое, или эллипсоидное, преломляющее свет
тело линзы. Но часть линзового эпителия таким образом не из-
меняется и покрывает измененную часть. Возникает точка роста
линзы — место соединения неизменного эпителия с массой во-
167
локон линзы. Это, фигурально выражаясь, мастерская линзооб-
разования. В этом месте эпителиальные клетки непрерывно пре-
вращаются в волокна, линза растет.
На примере развития глаза экспериментальные эмбриологи
еще в начале нашего столетия убедились, что в ходе органогене-
за происходит взаимозависимость развивающихся клеток, тка-
ней, частей органов. Эта взаимозависимость, однако, такого рода,
что тем или иным клеточным и тканевым компонентам принад-
лежит «ведущая» роль. Они, образно говоря, индуцируют раз-
витие соседнего клеточного материала (см. гл. XI). Этот факт,
имеющий большое принципиальное значение, был установлен
Г. Шпеманом и В. Льюисом еще в 1901 г. В. 1904 г. было обна-
ружено, что между развитием глазной чаши и развитием линзы
существует тесная связь: глазной пузырь индуцирует развитие
из прилежащего к нему эктодермального эпителия хрусталика.
От глазной чаши, вершина которой контактирует с эпидермисом,
как бы исходит какое-то «организующее» влияние — стимул пре-
образования клеток эпидермиса в хрусталик. Огромным количе-
ством экспериментов большого числа исследователей, в частно-
сти советских ученых, было доказано, что любые эпидермальные
клетки при определенных условиях побуждаются к формирова-
нию линзы или линзоподобных образований. Доказательства пра-
вильности этого положения получены разными эксперименталь-
ными способами, i
Если удалить 'зачаток глаза (глазной пузырь или начинаю-
щий формироваться глазной бокал) и предотвратить контакт глаз-
ного пузыря с эпидермисом, то, как правило, линза не возникает.
Эксперименты такого рода сами по себе еще не убеждают пол-
ностью в «индукционной» способности глазного пузыря. Во-пер-
вых, эти эксперименты грубые, так как резко нарушают нор-
мальное состояние клеточных масс в зародыше. Во-вторых, дело
осложняется тем, что не всегда достигается положительный ре-
зультат в таких опытах. К тому же у некоторых амфибий (Rana
esculenta, Xenopus laevis и др.) обнаружена некоторая степень
«независимого» развития хрусталика, когда предварительно уда-
лялся презумптивный материал глаза на стадии нейрулы. Раз-
витие в этих случаях никогда не бывает нормальным, но нередко
формируются довольно типичные линзы с соответствующими ци-
топлазматическими преобразованиями клеток в волокнистые
структуры. Однако эти вселяющие сомнение факты требуют осо-
бого анализа и не опровергают основных данных о влиянии
глазного пузыря на эпидермис, из которого возникает хрусталик.
Демонстративные результаты дают опыты, в которых уда-
лялся презумптивный эпидермис линзы и на его место переса-
живался эпидермис, взятый с какой-либо другой части зародыша,
например с головы или брюха. Эпидермис побуждался в этих
опытах к развитию линзы.
168
Наконец, убедителен и третий тип экспериментов. Если вы-
резать глазной пузырь и трансплантировать его под эпидермис
в другой части тела зародыша, то иногда эпидермис развивает-
ся в линзу.
Считается, что для индукции хрусталика необходим непосред-
ственный контакт эпидермиса с глазным пузырем: Мэк Кийен
в 1951 г. показал, что помещение между глазным пузырем и эпи-
дермисом тонкого листочка целлофана (в опытах на курином
зародыше) предотвращает индукционное действие глазного пу-
зыря. Однако тот же исследователь в опытах 1958 г. (также на
курином зародыше) доказал, что частичное экранирование эпи-
дермиса от глазного пузыря тонкой полоской агара не предот-
вращает полноценного развития хрусталика. Если индуцирующий
агент химической природы, то он должен был в этих опытах или
проходить через агар, или обойти вокруг него.
Химическая природа индуцирующего действия глазного пузы-
ря пока неясна. Есть предположения, отчасти подтверждаемые
экспериментами, что когда глазной пузырь приходит в контакт
с эпидермисом, он содержит большое количество, РНК, а пре-
зумптивный хрусталиковый эпидермис имеет мало нуклеиновых
кислот. После контакта глазного пузыря с эпидермисом появ-
ляется большое количество РНК и в эпидермисе. Может быть,
молекулы нуклеиновых кислот проходят от клеток глазного пу-
зыря в клетки презумптивной линзы. Однако если бы это и ока-
залось так (что не удается пока доказать), нелегко дать хими-
ческую интерпретацию индукционной роли глазного пузыря в
образовании линзы.
Некоторые исследователи (например, Б. И. Балинский, 1961)
предполагают, что индукционное воздействие на презумптивный
линзовый эпителий исходит не только из глазного пузыря, но и
из головной мезодермы, и делают попытку объяснить причины
обнаруженных многими авторами отклонений в результатах опы-
тов по индукции хрусталика глазным бокалом.
Согласно данным Н. Джакобсона (1966), индуцирующее влия-
ние на эктодерму, подготовляющее детерминацию линзовой за-
кладки, осуществляется не только глазным зачатком, но и энто-
дермой, и презумптивной сердечной мезодермой.
Столь подробное изложение опытов, касающихся развития
хрусталика, важно потому, что, как увидим в гл. XI, упоминав-
шиеся интересные данные классических работ Г. Шпемана и
В. Льюиса послужили толчком для создания в 30-х годах
Г. Шпеманом теории индивидуального развития, главенствовав-
шей в эмбриологических исследованиях более двух десятилетий.
Другие добавочные структуры глаза — сосудистая оболочка,
склера, или, иначе, белочная оболочка, и роговица — обеспечи-
вают его функцию.
Наружные оболочки глаза (сосудистая и склера) образуются
169
из мезенхимы, окружающей развивающийся глазной бокал. Внут-
ренний слой клеток мезенхимы развивается в сеть кровеносных
сосудов, окружающих пигментный эпителий. Это и есть закладка
сосудистой оболочки. Наружная масса мезенхимных клеток —
источник формирования склеры — преобразуется в фиброзную
капсулу, играющую защитную роль и служащую местом прикреп-
ления мышц глаза.
У птиц и рептилий капсула становится хрящевой и даже кост-
ной, у других позвоночных она остается фиброзной.
Сосудистая и белочная оболочки вначале прилегают друг к
другу по всей поверхности глазного яблока. Впоследствии между
ними, в передней части, возникает пространство — передняя ка-
мера глаза. Склера становится в этом месте прозрачной и уча-
ствует в образовании роговицы - (cornea). Соответствующий же
участок сосудистой, оболочки идет на построение наружного со-
единительно-тканного слоя радужной оболочки и покрывающей
хрусталик зародыша сосудистой оболочки.
Как уже сказано, роговица развивается из эпидермального
эпителия и мезенхимы. Эпителий и соединительная ткань буду-
щей роговицы в ходе развития глаза становятся прозрачными.
Покрывающий глаз эпителий пигментирован.
Многочисленными экспериментами Г. Шпемана (1901) и
О. Мангольда (1931) доказано, что превращение участка кожи
в роговицу зависит- от причинных» влияний, идущих от хруста-
лика и всего глазного бокала. Если удалить глазное яблоко до
образования роговицы, то она не развивается. Зависимость рого-
вицы от всех остальных частей глаза сохраняется все время.
Если удалить глаз у личинки амфибий или у взрослого живот-
ного, то роговица вскоре теряет прозрачность и постепенно при-
обретает структуру кожи.
У различных позвоночных неодинаковое строение различных
добавочных структур глаза. У двухмесячного зародыша человека
по верхнему и нижнему краям роговицы возникают две складки
кожи. Это будущие верхнее и нижнее веки. На третьем месяце
эти складки, разрастаясь, временно срастаются. Полость между
веками и роговицей глаза называется конъюнктивальным меш-
ком.
РАЗВИТИЕ ЭПИДЕРМИСА И ОРГАНОВ, В ОБРАЗОВАНИИ КОТОРЫХ
ОН УЧАСТВУЕТ
Большая часть эктодермы, остающейся на поверхности зароды-
ша после нейруляции, развивается в эпидермис кожи. Но мы
уже знаем, что и различные другие структуры являются дери-
ватами эктодермы. Она участвует в формировании глаза, так
как из нее развиваются хрусталик и роговица. Большое число
других органов и частей органов развиваются в связи с диф-
170
ференциацией эпидермиса: перья, чешуи, потовые и сальные же-
лезы, млечные железы у млекопитающих.
Эпидермис раннего эмбриона птиц и млекопитающих снача-
ла состоит из одного слоя клеток. У куриного зародыша он, диф-
ференцируясь взаимозависимо с подлежащей соединительной
тканью, на 7-й день развития становится двухслойным. Лишь
на поздних стадиях развития птиц и млекопитающих возникает
внутренний листок, состоящий из ряда слоев размножающихся
клеток, — так называемый мальпигиев слой эпидермиса. У ку-
риного зародыша на 17-й день эпидермис становится многослой-
ным. В теснейшей взаимозависимости дифференцируются эпите-
лий и соединительно-тканная часть кожи — дерма. Вместе они
и составляют кожу как орган. Соединительно-тканная часть кожи
развивается из мезенхимы, являющейся производной мезодер-
мального пласта зародыша (дерматом), и нервного гребня. Нерв-
ный гребень в начале своего появления после смыкания нервной
пластинки в медуллярную трубку представляет массу клеток,
лежащих на месте бывшего смыкания краев медуллярной пла-
стинки. Этот клеточный материал, мигрируя от места происхож-
дения в латеральном и вентральном направлениях, у разных
позвоночных участвует в образовании многих структур (жабр,
частей лицевого скелета и т. д.). Висцеральный скелет, напри-
мер, полностью развивается из клеточного материала нервного
гребня.
У амфибий эпителий ранних эмбрионов двухслойный. Наруж-
ный листок называется перидермой, а внутренний — сенсорным,
потому что он принимает участие в образовании некоторых орга-
нов чувств.
Начальные этапы развития многих структур — производных
эпидермиса — сходны в том отношении, что сначала образуются
локальные утолщения эпидермального эпителия. Эти пластинко-
образные утолщения называются плакодами. В соответствующих
местах возникают парные плакоды, производными которых явля-
ются обонятельные мешки, черепные нервы, слуховые пузырьки
и др. Из плакод, находящихся в соседстве со слуховыми, раз-
виваются органы чувств боковой линии. У рыб боковая линия
тянется вдоль всего тела, однако вся она образована плакодами,
возникающими в области развития органа слуха.
Зачатком обонятельной выстилки органа обоняния служит пе-
редняя, впоследствии обособляющаяся часть нервной пластинки.
В результате особых морфогенетических перемещений клеток,
и прежде всего вследствие впячивания центрального участка, пла-
коды превращаются в обонятельные мешки, сообщающиеся с на-
ружной средой посредством отверстий — ноздрей. Клеточные эле-
менты стенок обонятельных мешков превращаются в обонятель-
ный эпителий с его специфической цитологической дифференци-
ровкой. У чувствительных клеток эпителия вырастают нервные
171
отростки, которые образуют обонятельный нерв. Нерв врастает
в стенку telencephalon. Внешне морфологическое строение обо-
нятельного органа зародыша у большинства рыб мало отличает-
ся от дефинитивного состояния этого органа. Но у наземных по-
звоночных, а также у некоторых рыб развиваются внутренние
«ноздри» — хоаны, а у млекопитающих, кроме того, с развитием
органа обоняния связано образование носослезного канала.
РАЗВИТИЕ ОРГАНА СЛУХА
Основные черты развития органа слуха таковы. Появляющиеся
в эктодерме на уровне заднего мозга- слуховые плакоды
(рис. 78,/) инвагинируют и превращаются в слуховые пузыри,
отъединяющиеся от эпидермиса (рис. 78,2). В ходе эмбриональ-
ного развития они трансформируются во внутреннее ухо.
У амфибий слуховые плакоды возникают как утолщения
внутреннего слоя эпидермиса, наружный же слой клеток совер-
шенно не участвует в их образовании. У всех амниот соответ-
ствующие участки всего эпидермиса (обоих слоев) участвуют
в образовании плакод. Слуховые «мешочки», или слуховые пу-
зыри, образуются вследствие инвагинации плакод. Сначала пу-
зырек имеет грушевидную форму; верхняя и нижняя стенки его
начинают образовывать полые выросты (рис. 79). Верхний вы-
Рис. 78. Четыре стадии развития внутреннего уха — от слуховой плакоды до
сформированного слухового пузырька человеческого зародыша (по К. Стритеру,
1942, 1945):
1 — слуховая плакода, 2 — слуховой пузырь, 3 — центральная нервная система, 4 — спин*
ная аорта
172
Рис. 79. Схема развития внут-
реннего уха зародыша челове-
ка (по К. Стритеру, 1906):
/ — зачаток эндолимфатического
протока, 2 — зачаток улиткового
протока, 3 — зачаток улитки, 4—6—
три полукружных канала,- 7 — эндо-
лимфатический мешок, 8, 9 — за-
чатки эллиптического и круглого
мешочков
рост — это зачаток эндолимфатического протока. Нижний вы-
ступ— зачаток улиткового протока, который, сильно удлиняясь,
как бы спирально закручивается. Улитковый проток трансфор-
мируется в улитку (cochlea). Почти одновременно с началом
образования эндолимфатического протока на верхней стороне
пузыря начинают формироваться три полукружных канала. Воз-
никающая борозда подразделяет пузырек на овальный и круг-
лый мешочки.
В результате указанных процессов слуховой пузырек превра-
щается в перепончатый лабиринт, весь эпителий которого экто-
дермального происхождения. Снаружи лабиринт окружен массой
мезенхимных клеток мезодермального происхождения.
Судьба окружающей лабиринт мезенхимы сложна. Часть ме-
зенхимной массы сильно изменяющихся клеток превращается в
жидкость, и вокруг перепончатого лабиринта возникает напол-
ненное этой жидкостью перилимфатическое пространство. Впо-
следствии слои «нерастворившихся» мезенхимных клеток, окру-
жающих лабиринт, превращаются в хрящевую слуховую капсу-
лу, которая окружает и защищает внутреннее ухо. Еще позднее
происходит окостенение, и образуется костный лабиринт, по всей
конфигурации почти соответствующий перепончатому лабиринту.
Уже говорилось о классических опытах В. Льюиса и Г. Шпеманэ
о взаимоотношении в развитии глазного пузыря и хрусталика.
Не меньший интерес представляют опыты Льюиса (1907) о влия-
нии слухового пузырька на развитие слуховой капсулы. Если
слуховой пузырек эмбриона вырезать и трансплантировать на
стадии хвостовой почки другому зародышу амфибий того же
вида, то окружающие трансплантат клетки могут развиться в
дополнительную хрящевую капсулу: слуховой пузырек как бы
индуцирует образование хрящевой капсулы. Тщательный анализ
этого явления дан классическими опытами московского эмбрио-
лога Д. П. Филатова и его школой (1916, 1924, 1927). Д. П. Фи-
латов (1916) удалял слуховой пузырек у зародышей жабы.
Хрящевая капсула слухового пузырька на оперированной сто-
роне не развивается. При пересадке слухового пузырька в раз-
ные области головы и в другие области тела вокруг них форми-
руется хрящевая капсула из любой мезенхимной ткани.
173
Важнейшая часть органа слуха — барабанная полость, или
полость среднего уха, возникает из полости жаберного кармана.
Полость среднего уха отъединяется от полости глотки, но связь,
однако, остается благодаря протоку — закладка евстахиевой
трубы. Барабанная полость разрастается и развивается далее;
формирующиеся вначале вне ее слуховые косточки оказываются
внутри барабанной полости, отделяясь от нее слизистой оболоч-
кой среднего уха.
Детали процессов развития внутреннего и среднего уха у раз-
ных животных своеобразны, но общий принцип развития остает-
ся таким же.
Наружное ухо у млекопитающих и человека развивается из
жаберного кармана. Средний участок последней образует ямку
ушной раковины, превращающуюся постепенно в полость ушной
раковины. Возникает наружный слуховой проход. Мезодермаль-
ного происхождения клеточный материал, находящийся на гра-
нице между барабанной полостью и полостью наружного уха,
дифференцируется в барабанную перепонку.
Путем разрастания бугорков и складок на глоточных дуж-
ках (на первой и второй) постепенно развивается ушная раковина.
В заключение следует упомянуть, что у разных животных
имеются особые органы, в развитии которых также принимает
участие наружный зародышевый пласт — эктодерма. Различные
органы развиваются так, что их первоначальной закладкой яв-
ляются не утолщения типа плакод, а выступы. Таковы, напри-
мер, первоначальная закладка плавников и наружных жабр (у
рыб, у личинок амфибий), закладка балансер у личинок сала-
мандр, закладка парных конечностей и других органов. Началь-
ные этапы развития конечностей у жабр сходны. Происходит
выпячивание эпидермиса, которое наполняется массой мезенхим-
ных клеток. Однако развитие конечностей — сложный процесс,
в котором принимают участие производные других зародышевых
пластов; при этом эктодерме во многих отношениях доминирую-
щая роль не принадлежит.
В этом разделе далеко не исчерпаны вопросы о развитии
производных эктодермы. Почти не рассмотрены гистогенетические
и цитологические вопросы, составляющие специальный предмет
других наук.
ГАНГЛИОЗНАЯ ПЛАСТИНКА И ДРУГИЕ ПРОИЗВОДНЫЕ ЭКТОДЕРМЫ
На основании всех данных этой главы можно сделать заключе-
ние о том, что в развитии нервной системы и всех органов чувств
единственным или доминирующим источником клеточного мате-
риала является эктодерма. Нервная система — целиком произ-
водное эктодермы. Нервная система и органы чувств развива-
ются из разных зачатков. Более того, лишь условно можно го-
174
ворить о зачатке нервной систёмы, так как нервная система и
органы чувств обязаны в своем\развитии, во-первых, нервной
трубке, развивающейся из нервнощ пластинки, во-вторых, пла-
кодам и, в-третьих, ганглиозной пластинке. У них общий генезис,
все они производные эктодермы, но соотношение каждого из этих
зачатков с другими частями развивающегося зародыша, их мор-
фологические особенности и разная судьба в развитии не позво-
ляют без существенных оговорок объединять общим названием
нервного зачатка. К тому же ганглиозная пластинка принимает
участие не только в формировании элементов нервной системы,
но и других образований (скелетных, например).
В этой связи ганглиозная, или ганглионарная, пластинка
(Neural crest, по определению английских авторов) заслуживает
особого внимания. Собственно, это не резко обособленное мор-
фологическое образование, по крайней мере в начале ее появ-
ления, т. е. вскоре после нейруляции. Когда у зародышей амфи-
бий края нервных валиков сливаются у средней линии, масса
эктодермальных клеток (не только «выбитых» из общей связи
пласта клеток нервных валиков, но и клеток прилежащей экто-
дермы) располагается дорсально от нервной трубки. Эта масса
однородных клеток не отграничена от покрывающей нервную
трубку кожной эктодермы. Клетки ганглиозной пластинки яв-
ляются предшественниками меланофор и эктомезенхимы. Этот
термин указывает на эктодермальное происхождение мезенхим-
ных клеток в отличие от мезенхимы, развивающейся из мезодер-
мы. Факт образования мезенхимы из эктодермы считается неко-
торыми эмбриологами прямым противоречием доктрины специ-
фичности зародышевых листков (С. Герстадиус, 1967).
Клетка ганглиозной пластинки.
Симпатический нейуобласт
ХрящеЬая
Клетка элдстической_дболочки
Клетка симпатического
ганглия
Рис. 80. «Судьба» клеток ганглиозной пластинки
Биполярный 'нейробласт
Униполярная клетка
дорсального корешка
ганглия
Хромаффинная
клетка
Хроматофор
Одонтобласт
Остеоцит
Мезенхимная ~
клетка голодной
области
175
Neural crest существует у всех позвоночных, но исследован
хорошо только у амфибий, Ц связи с особенностями формиро-
вания нервной трубки у равных позвоночных, конечно,, и меха-
низм образования ганглиозной пластинки и морфология ее при-
обретают свои особенности. Клеточный материал ганглиозной
пластинки участвует в разных образованиях. В эмбриологии
дискутируется вопрос и о том, каковы собственные «внутренние»
потенции клеточных масс в разных участках ганглиозной пла-
стинки.
Вскоре после образования ганглиозной пластинки клетки ее
начинают мигрировать между нервной трубкой и эпидермисом
сначала в виде пласта, затем «прядью» клеток или как единич-
ные клетки в вентральном и латеральном направлениях. Образ-
но говоря, ганглиозная пластинка — не стабильное образование,
а очень «беспокойное»: от нее направляются потоки клеток к
некоторым презумптивным органам. Более заметны потоки кле-
ток в области головы и шеи, чем в туловищной области.
Клетки ганглиозной пластинки перемещаются главным обра-
зом между мезодермой и эпидермисом, вдоль внутренней стенки
последнего, но они мигрируют и между нервной трубкой и ме-
зодермой и достигают средней линии тела.
Как уже говорилось, клетки нервного гребня принимают уча-
стие в формировании многих структур. Клетки ганглиозной пла-
стинки принимают участие в формировании спинальных ганглиев,
ганглиев симпатической нервной системы, большей части висце-
рального скелета, мозгового вещества надпочечников, пигмент-
ных клеток (рис. 80). За исключением пигмента ретины, вероятно,
все хроматофоры у рыб, амфибий, птиц и млекопитающих — про-
изводные нервного гребня. Пигментные клетки находятся в эпи-
дермисе, волосах, перьях и других образованиях.
Глава VIII
РАЗВИТИЕ ПРОИЗВОДНЫХ ЭНТОДЕРМЫ
РАЗВИТИЕ ПИЩЕВАРИТЕЛЬНОГО КАНАЛА У НИЗШИХ ПОЗВОНОЧНЫХ
Об эктодермальных органах можно говорить лишь с большими
оговорками, так как в развитии органов пищеварения и дыхания
принимают участие производные не только энтодермы, но всех
трех зародышевых листков.
В зависимости от типа гаструляции у позвоночных развитие
органов пищеварения происходит разными путями. Поэтому сле-
дует рассмотреть сначала развитие пищеварительного канала
у низших позвоночных с характерным для них полным дробле-
нием, а затем у животных с частичным дроблением яиц.
Дном первичного кишечника у ланцетника служит клеточный
материал энтодермы, крышей его—материал презумптивных
хорды и мезодермы (рис. 81). Дефинитивный пищеварительный
канал у ланцетника начинает формироваться после того, как хор-
да и мезодерма отъединяются в ходе развития эмбриона от энто-
дермы и возникает полость, выстланная только клетками экто-
дермального происхождения.
У миног и миксин, у ганоидных и двоякодышащих рыб, амфи-
бий происходит то же: крыша архентерона, образованная пре-
зумптивной хордой и отчасти мезодермой, из-за морфогенетиче-
ских перемещений клеточного материала обособляется от энто-
дермы; края энтодермального пласта клеток соединяются,
и в результате образуется полость, выстланная эктодермальными
клетками (рис. 82).
Возникающая кишка, однако, неодинакова в разных участках.
Передняя часть — передняя кишка, более расширена и выстлана
тонким пластом клеток. Задний участок — средняя кишка не-
Рис. 81. Формирование осевых органов у ланцетника. Поперечные срезы заро-
дыша (по Э. Конклину, 1932):
/ — нервная пластинка, 2 — эпидермис, 3 — первичная кишка, 4 — хорда, 5 —энтодерма,
6 — мезодермальные сомиты, 7 — нервная трубка, 8 — кншка, 9— целом
177
Рис. 82. Развитие эктодермальных ор-
ганов Triturus taeniatus от нейрулы до
плавающей личинки (по Б. И. Балинско-
му, 1947):
А, Б, Д, И — сагиттальные срезы; В, Г —
фронтальные срезы; 1— нервная пластинка,
2 — архентерон, 3 — бластопор, 4 ~ полость
передней кишки, 5 — полость средней кишки.
6 — жаберные карманы, 7 — нервная трубка,
8 — хорда, 9 — печеночный вырост, 7(9— по-
лость средней кишкн, // — глотка, 12 — фор-
мирующийся головной мозг. 13 — желудок,
14 — печень, /5 — двенадцатиперстная кишка,
16— поджелудочная железа, /7 — спинной
мозг
сколько уже, ее дорсальная
(обращенная к спине) стен-
ка состоит также из тонко-
го пласта эктодермальных
клеток, но вентральная
стенка очень толстая, она
образована массой больших
клеток с обильным количе-
ством желтка.
Помимо обычных морфо-
логических исследований,
развитие кишечной трубки
у амфибий прослежено
Б. И. Балинским (1947) и
другими исследователями
способами маркирования
разных участков эктодер-
мального эпителия виталь-
ными красителями. При
этом, естественно, требова-
лось прибегать к операциям:
делали разрезы через ме-
дуллярную пластинку у ран-
них зародышей, а у позд-
них— через стенку тела; к
внутренней поверхности эк-
тодермального пласта на
короткое время приклады-
вали кусочки агара, пропи-
танного красителем. После
операции происходит быст-
рое слипание краев раны и
не обнаруживаются резкие
отклонения от нормального
развития.
Благодаря образованию
складок вентральной стенки
передней кишки, что проис-
ходит на стадии нейрулы,
она подразделяется на бо-
лее широкую, переднюю,
часть, лежащую под формирующимся головным мозгом, и на зад-
нюю, суженную часть. Передняя часть развивается в полость рта
и в полость глотки, а задняя часть — в печеночный дивертикулум
(полый слепой вырост), который у лягушек является зачатком
печени и поджелудочной железы, а у саламандр, по данным Ба-
линского, участвует в формировании желудка и двенадцатиперст-
178
ной кишки. Этот исследователь тщательно проследил развитие
пищеварительного канала у лягушек. Выяснено, что энтодерма
передней кишки принимает участие наряду с эктодермой в разви-
тии энтодермального эпителия рта, а также глотки, пищевода,
желудка, передней половины двенадцатиперстной кишки (duode-
num) , легких, печени и части поджелудочной железы.
Энтодермальные клетки средней кишки— источник формирова-
ния эпителия задней половины duodenum, части поджелудочной
железы, а также всего остального кишечника и клоаки.
Представляющие интерес процессы развития и гистологической
дифференциации пищеварительных органов у низших позвоноч-
ных здесь не описываются. Однако с различными иллюстрациями
морфогенетических процессов, сопровождающих развитие пище-
варительных органов (причины формирования трубок, изгибов,
удлинений, утолщений и т. п.), можно познакомиться на с. 140,
где говорится о морфогенетических движениях клеток и клеточ-
ных пластов.
Представление о развитии «эктодермальных органов» у низ-
ших позвоночных и об их топографии можно создать по схеме
Б. И. Балинского, которому эмбриологи обязаны знанием многих
подробностей органогенеза у амфибий (рис. 82).
РАЗВИТИЕ ОРГАНОВ ПИЩЕВАРЕНИЯ У ПТИЦ,
МЛЕКОПИТАЮЩИХ И ЧЕЛОВЕКА
При меробластическом дроблении яиц птиц процессы развития
пищеварительной системы и связанных с ней генетически органов
оказываются очень своеобразными и более сложными, чем у низ-
ших позвоночных.
Источником для развития энтодермального эпителия пищева-
рительного канала служит участок эпибласта в составе первич-
ной полоски. Этот материал в ходе гаструляции мигрирует под
эпибласт и оказывается в составе гипобласта, содержащего, кро-
ме того, клетки желточной энтодермы. Впоследствии полоска
презумптивной кишечной энтодермы удлиняется и образуется
складка — «кишечный желобок». Срединный участок полоски ле-
жит под презумптивной хордой и презумптивными сомитами.
Лежащие рядом, справа и слева, участки сгибаются книзу, схо-
дятся и соединяются по средней линии (рис. 83). Внутренняя по-
верхность складки идет на формирование энтодермы кишки;
наружная поверхность складок является продолжением энтодер-
мального слоя, выстилающего желточный мешок.
Замыкание трубки происходит в направлении от переднего,
а затем и заднего концов к срединному участку. Передний и зад-
ний слепые концы формирующейся кишечной трубки прилегают
непосредственно к соответствующим участкам кожной эктодермы.
Пищеварительная трубка замыкается сначала на переднем
179
Рис. 83. Стадия развития передней
кишки куриного зародыша (по
Р. Беллеру, 1953):
/ — утолщенная стенка передней кншки,
2 —передний конец головной кишкн, 3 —
крыша передней кншкн, 4 — утолщенная
мезодерма (спланхноплевра), 5 — утолщен-
ная энтодерма, 6 — кишечная энтодерма.
На переднем конце зародыша эктодерма,
нервные складки и париетальная мезодер-
ма удалены
конце зародыша. Она соответствует передней кишке амфибий,
а замыкающийся впоследствии задний участок кишечного желоб-
ка соответствует средней кишке. На границе между передней и
средней кишками остается участок кишечного желобка, в котором
он не замыкается полностью и где образующийся кишечник про-
должает быть в связи с желточным мешком. Сначала очень ши-
рокое отверстие постепенно уменьшается, превращаясь в узкое
отверстие стебелька, соединяющего полость »кишки с полостью
желточного мешка.
Начальный, очень широкий (по сравнению с просветом сред-
ней кишки) участок передней кишки участвует в развитии пище-
вода, большей части duodenum, печени, поджелудочной железы.
Участки, соответствующие полости рта и глотки, уплощены и
растянуты в поперечном направлении, причем., участок глотки
в виде глоточных мешков вытягивается вперед. Участки же, соот-
ветствующие пищеводу, желудку и duodenum, округляются; в по-
перечном направлении они не уплощены. Округляется также и
средняя кишка. Диаметр ее с самого начала развития уже про-
света передней кишки.
Как и развитие любого органа, формирование желудочно-ки-
шечного тракта позвоночных не происходит обособленно от раз-
вития других частей зародыша. Это накладывает отпечаток на
рост и форму разных участков пищеварительного канала. Пище-
вод у высших позвоночных сильно удлиняется по сравнению
с таковым у амфибий. В связи с неравномерным ростом и влия-
нием соседних частей тела происходят изгибы и перемещения
разных участков развивающегося пищеварительного канала. Так,
передний конец желудочной области (будущая основная часть
желудка) перемещается на левую сторону, а пилорический отдел
желудка перемещается вниз и к середине. Передний участок
двенадцатиперстной кишки принимает поперечное положение.
Длина кишечника и спиральное закручивание его у высших позво-
ночных и даже у разных видов млекопитающих сильно варьиру-
ет. Передняя и средняя кишки, как уже сказано, оканчиваются
слепыми выростами.
180
Анальное, или клоакальное, отверстие у млекопитающих и
птиц образуется так. У высших позвоночных нет бластопора как
отверстия, посредством которого архентерон сообщался бы
с внешней средой. Клоакальное отверстие у них—новообразова-
ние; оно появляется вследствие «прободения» стенки тела в месте
контакта с ней заднего конца средней кишки. Место прободения
определяется уже на стадии первичной полости, у ее заднего
конца. На стадии поздней гаструлы на заднем конце обособления
зародышевых пластов не происходит, между массой эктодермаль-
ных и эктодермальных клеток мезодерма не проникает, так что
в этом участке образуется двухслойная пластинка. Это и есть
клоакальная мембрана. В ходе развития зародыша, когда проис-
ходит обособление средней кишки от желточного мешка, кло-
акальная мембрана оказывается в непосредственном соседстве
со стенкой киш^и. Она расположена сначала дорсально, но поло-
жение ее изменяется в связи с развитием впереди мембраны хво-
стовой почки, и в конце концов клоакальная мембрана оказыва-
ется у основания хвоста обращенной вниз.
Контактирующаяся с клоакальной мембраной часть кишки
расширяется, формируется • клоака. В месте соприкосновения
клоакальной мембраны с эктодермой в последней образуется не-
большая ямка: формируется" proctodeum — образование эктодер-
мального происхождения. После перфорации клоакальной мем-
браны проктодеум становится частью пищеварительного тракта.
РАЗВИТИЕ РТА
Начальный отдел пищеварительного тракта эктодермального
происхождения. В месте контакта передней кишки с кожной экто-
дермой на последней образуется ротовая ямка, или впадина
(stomodeum). Часть ротового'впячения (на ее дорсальной сторо-
не) развивается в зачаток передней доли гипофиза. Этот зача-
ток— карман Ратке, проникает через соединительную ткань и
соединяется с задней долей гипофиза, формирующейся в связи
с процессами развития промежуточного мозга.
Формирование рта и ротовой полости, варьируя у разных по-
звоночных, представляет очень сложный процесс, в который во-
влекается клеточный материал всех трех зародышевых пластов.
Схематично можно представить себе развитие рта и частей ли-
цевого отдела млекопитающих (рис. 84) таким образом. Отвер-
стие ротовой ямки у зародыша млекопитающих сначала представ-
ляет дыру, по окружности которой образовалось пять выступов:
верхний непарный — лобный, два парных верхних выступа —
верхнечелюстные отростки и два нижних — нижнечелюстные. Ниж
нечелюстные выступы, срастаясь, образуют нижний край ротового
отверстия.
В дальнейшем одновременно с формированием лобного отро-
стка обонятельных плакод на нем возникает средний лобный и
181
Рис. 84. Последовательные стадии развития
лицевых частей зародыша свиньи (Д—В) (из
кн. Б. И. Балинского, 1961):
1— глазной пузырь,' 2 — слезно-носовая щель, 3 —
отросток верхней челюсти, 4 — мандибулярная дуга,
5 — гиоидная дуга, 6 — обонятельная ямка, 7 — назо-
латеральный отросток, 8 — назо-меднальный отрос-
ток, 9 — медиально-фронтальный отросток, 10 — глаз,
11 — язык, 12 — ннжняя челюсть, 13 — наружное но-
совое отверстие
два боковых носовых от-
верстия (рис. 84, 7, 8).
По мере роста верхнече-
люстные отростки сраста-
ются с боковыми носовы-
ми и со средним лобным
отростками, образуя
верхний край ротового
отверстия.
На среднем лобном
отростке возникает попе-
речный носовой валик —
зачаток кончика носа.
Отверстия обонятельных
ямок преобразуются в на-
ружные носовые отвер-
стия. Боковые носовые
отростки превращаются в
крылья носа. Общая кон-
фигурация лица — много-
образный процесс разра-
станий и перемещений
разных зачатков, про-
цесс, который оканчива-
ется лишь в постэмбрио-
нальном периоде.
Первичная ротовая
полость постепенно под-
разделяется на носовую и
собственно ротовую.
Верхняя стенка ротовой
полости — первичное мяг-
кое нёбо. Задние концы
упоминавшихся ранее обонятельных ямок, достигая мягкого нёба,
открываются в первичную ротовую полость благодаря образую-
щимся отверстиям — первичным хоанам, также уже упоминав-
шимся ранее.
Глоточная перепонка, являющаяся задней границей первичной
ротовой полости, перфорируется, вследствие чего эта полость при-
соединяется к средней кишке.
На внутренних сторонах обоих верхнечелюстных отростков
образуются нёбные отростки, срастающиеся и разделяющие пер-
вичную ротовую полость на собственно ротовую и носовую, в ко-
торую открываются первичные хоаны. Параллельно с этими
процессами на внутренней поверхности среднего лобового от-
ростка образуется носовая перегородка, разделяющая носовую
полость на правую и левую. Края нёбных отростков срастаются
182
между собой и с нижним краем носовой перегородки, вследствие
чего нёбо приобретает окончательное строение. В дальнейшем про-
исходит формирование носо&ых раковин из складок, возникающих
на поверхности носовых полостей.
Зубы развиваются из клеточного материала эктодермы (или
энтодермы у других животных), являющегося источником эмали,
и из мезенхимы ганглионарной пластинки, идущей на развитие
«сосочка» зуба'.
Язык (lingua) возникает в виде небольшого выступа нижней
поверхности глоточной кишки между челюстной и подъязычной
дужками (рис. 84, 11).
РАЗВИТИЕ ЖАБЕРНОЙ ОБЛАСТИ
За первичной ротовой полостью находится глоточный, или жа-
берный, отдел кишечной трубки. Часть ее принимает участие
в образовании окончательной ротовой полости, эпителий которой,
таким образом, составлен из участков эктодермального и эндо-
дермального происхождения. В развитии глоточной области важ-
ную роль играют энтодермальные производные, связанные с про-
изводными других зародышевых пластов. О ведущей роли энто-
дермы в некоторых из процессов формообразования можно
говорить лишь условно.
У амфибий жаберный отдел кишки со свойственными ему
структурами развивается из передней, расширенной, кишки.
В конце нейруляции возникают попарно, одна вслед за другой,
начиная е участка, лежащего позади нижнечелюстной дуги, вы-
пячивания боковых стенок глоточной области. Это жаберные
карманы. Разрастаясь, они проникают через массу мезодермаль-
ных клеток до презумптивного эпидермиса, на поверхности кото-
рого возникают борозды (желобки), соответствующие приблизив-
шимся эндодермальным мешкам. Происходит соединение наруж-
ных слоев клеток эндодермальных выпячиваний и внутреннего
клеточного пласта желобка. В результате перфорации образовав-
шейся пластинки появляются жаберные щели. Благодаря этим
отверстиям полость глоточной кишки сообщается с внешней
средой. Закладка жаберных щелей наблюдается у всех позвоноч-
ных. У водных жабродышащих животных формируются органы
дыхания — жабры, на стенках жаберных щелей развиваются жа-
берные нити — внутренние жабры. Однако даже у некоторых
водных позвоночных (у костистых рыб и амфибий) жаберные
мешки первой пары остаются замкнутыми, и соответствующие
жаберные щели и жаберные структуры не развиваются.
У рептилий, птиц, млекопитающих и человека жабры никогда
не функционируют как органы дыхания, но жаберные мешки и
1 Развитие зубов рассматривается в учебных руководствах по гистологии.
183
Рис. 85. Полусхема развития «эктодермальных» органов зародыша человека (по
К. Стритеру, 1942):
/ — головной мозг, 2 — сердце, 3 — легкие, 4 — желточный мешок, 5 — аллантоис, 6 — щи-
товидная железа, 7 — венозный сниус, 8 — печень, 9 — дорсальная часть поджелудочной
железы, 10 — желчный пузырь, И — глоточные мешки, 12 — рот, 13 — пищевод, 14 — желу-
док, 15—ямка для венозного синуса, 16 — эпителий печени, 17 — мезонефрический проток
жаберные щели развиваются. У зародыша человека с каждой
стороны возникают четыре жаберные щели, чаще же всего закла-
дывается еще и слабо развивающаяся пятая щель. Однако даль-
нейшего развития жаберного аппарата, образования жаберных
нитей не происходит. Это характерно для всех высших позвоноч-
ных, причем у некоторых даже не возникают и жаберные отвер-
стия. На это явление давно обратили внимание зоологи, и оно
считается одной из ярких иллюстраций рекапитуляции (см. гл.
XVII). Однако этот случай нельзя рассматривать просто как
«воспоминание» эмбрионов высших животных о происхождении
от низших, дышащих жабрами форм. Хотя почти вся система
структур начинающего развиваться жаберного аппарата редуци-
руется, развитие некоторых органов обусловлено этим кажущим-
ся бесполезным формированием жаберного аппарата. У зароды-
шей человека и у всех высших позвоночных первая пара жабер-
ных мешков участвует в возникновении наружных слуховых
отверстий, полости среднего уха и евстахиевой трубы. Вторая
пара глоточных щелей у зародыша человека принимает участие
в образовании нёбных миндалин. За счет эктодермальных клеток,
отделяющихся от стенок третьего, четвертого и пятого жаберных
мешков, развиваются железы с внутренней секрецией: зобная
(тимус), околощитовидные (паратиреоидные) железы или эпите-
лиальные тельца.
С глоточной кишкой связано и развитие щитовидной железы,
а именно зачаток ее возникает в виде кармана на вентральной
стенке глотки, отделяется от нее и перемещается в вентральном
направлении к трахее (у наземных животных; рис. 85,6).
184
РАЗВИТИЕ ЛЕГКИХ, ПЕЧЕНИ И ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ
Органы дыхания — гортань (larynx), дыхательная трубка (bron-
chus) и легкие (pulmo) — возникают общим зачатком — кармано-
подобным выпячиванием на вентральной стороне пищеваритель-
ного канала, позади жаберной области (рис. 85,3). Конец выпя-
чивания, расширяясь и дифференцируясь, разветвляется; две
ветви, вырастающие в сторону и назад, развиваются в бронхи и
легкие. Срединная (между этими ветвями) часть развивается
в трахею. На верхнем конце выпячивание, преобразовавшись
в трубку, остается связанным с кишкой; при этом передний рас-
ширенный участок ее дифференцируется в гортань. У низших
позвоночных зачатками легких являются мешкообразные расши-
рения на концах бронхов.
У птиц и млекопитающих в результате обильного ветвления
бронхов, развития на их концах многочисленных легочных пузырь-
ков и благодаря специфической тканевой дифференциации созда-
ется орган дыхания очень сложной организации. Хотя у чело-
века легкие начинают функционировать лишь после рождения,
развитие легких с легочными пузырьками происходит на ранних
стадиях развития. На восьмой неделе закладывается и хрящевой
скелет дыхательной трубки.
Печень (hepar) у всех позвоночных возникает из эктодермаль-
ного эпителия брюшной стороны duodenum (рис. 85, 8\. Зачаток
желчного пузыря возникает на заднем конце первичного задат-
ка печени (рис. 85,10). Параллельно развивается и поджелудоч-
ная железа (pancreas), один из двух зачатков которой возникает
из брюшной стенки duodenum. Образующееся здесь выпячивание
развивается в проток поджелудочной железы. Второй дорсальный
зачаток поджелудочной железы у амниот образуется на спинной
стороне duodenum, впереди зачатка печени (рис. 85,9). Структура
поджелудочной железы и система протоков варьируют у разных
позвоночных.
Глава IX
РАЗВИТИЕ ПРОИЗВОДНЫХ МЕЗОДЕРМЫ
ПЕРВЫЕ ЭТАПЫ ДИФФЕРЕНЦИАЦИИ МЕЗОДЕРМЫ
В результате процессов гаструляции у хордовых мезодерма при-
обретает вид плотных толстых клеточных слоев, расположенных
справа и слева от хорды (см. рис. 86,6). Вскоре совершаются
морфогенетические процессы, приводящие к сегментации мезодер-
мы и к образованию внутри клеточных масс мезодермы полостей
(см. рис. 81,9).
Мезодермальные клеточные массы, прилежащие к хорде и
нервной трубке, утолщаются и дифференцируются постепенно
спереди назад на метамерно расположенные сегменты — сомиты
(см. рис. 52,2). Обособление сомитов происходит сначала в голов-
ном конце тела, затем эта дифференциация распространяется и
на другие участки мезодермы в каудальном направлении. У пяти-
недельного человеческого зародыша уже становится 43—44 пары
Рис. 86. Схема органогенеза и гистогенеза у зародышей высшего позвоночного
(по А. Г. Кнорре, 1959):
1— кожная эктодерма (эпидермис), 2 — ганглиозная пластинка (чувствительные нейроны,
периферическая нейроглия, хроматофоры), 3 — нервная трубка (нейроны, нейроглия), 4 —
хорда, 5 — дерматом (соединительно-тканная основа кожи), 6 — мнотом (скелетно-мышеч-
ная ткань), 7 — склеротом (хрящевая н костная ткань), 8 — нефротом (почечный эпителий),
9 — париетальный листок спланхнотома (мезотелий), 10 — висцеральный листок спланхното-
ма (мезотелий, ткань сердечной мышцы), 11 — кишечная эндотерма (кишечный эпителий),
12 — мезенхима (соединительная ткань, кровь, гладкомышечная ткань), 13 — внезародыше-
вая эктодерма (эпителий амниона), 14 — эндотелий аорты, 15— желФочная энтодерма (эпи-
телий желточного мешка), 16 — целом
186
сомитов. У млекопитающих сегментация начального головного
участка мезодермального пласта происходит с опозданием: вто-
рая пара сомитов, а затем первая возникают по времени после
третьей и последующих пар. П. Г. Светлов и другие усматривают
в этом иллюстрацию правильности теории метамерии П. П. Ива-
нова и называют первые два сегмента ларвальными, гомологич-
ными сегментами личинок предков хордовых (см. гл. XVII).
У ряда животных (в частности, у ланцетника) масса мезодер-
мальных клеток, составляющих сомит, имеет в середине неболь-
шую полость, так что он выглядит пузырьком с толстыми стенка-
ми. Вместе с начинающейся гистологической дифференциацией
изменяется и форма сомитов: они вытягиваются в дорсовентраль-
ном и уплощаются в срединно-боковом направлении.
Продолжением мезодермы сомитов являются более рыхлые
клеточные массы, не подразделяющиеся на сомиты и не утолщаю-
щиеся. Эти более тонкие краевые участки называются спланхно-
томами, боковыми пластинками или пластинками латеральной
мезодермы (см. рис. 52,5). Внутри них возникают полости — за-
чатки будущей полости тела. Таким образом, клеточный мате-
риал боковой мезодермы подразделяется на два листка: наруж-
ный — париетальный, соматоплевру, и внутренний—висцеральный,
спланхноплевру (рис. 86, 9, 10). Названия «соматоплевра»—стен-
ка тела и «спланхноплевра» — стенка внутренностей образованы
от греч. soma — тело, splanchnon — внутренности, pleura—бок,
боковая стенка.
Целомический эпителий происходит из клеточного материала
обоих листков. Клеточный материал разных участков сомитов
служит источником развития органов и систем органов. Хотя
между этими участками в начальный период дифференцировки
сомитов нет резких границ, топография их строго закономерна.
Наружный участок сомита, прилегающий к кожной эктодерме,
является источником развития соединительно-тканной части
кожи (дермы) и называется дерматомом (рис. 86. 5). Сред-
ний слой клеток (рис.
перечно-полосатых
сомита — склеротом (от греч. scleros — твердый, скелетный и
tomos — отрезок, отдел), имеющий вид рыхлой мезенхимы, разо-
вьется в хрящевую и костную ткани осевого скелета (рис. 86,7).
Участки спланхнотомов — «стебельки сомитов», находящиеся ря-
дом с ними, развиваются в нефротомы — источник формирования
выделительной системы. Нефротомы — это полоски клеток, связы-
вающие сомиты с латеральным листком мезодермы (рис. 86,8).
Гладкая мышечная ткань кишечника развивается из висцерально-
го листка боковой мезодермы, мускульные волокна стенок крове-
носных сосудов — из висцерального и париетального листков,
мышцы сердца — из висцерального листка. Клетки миотомов диф-
ференцируются в мышечные волокна. У низших животных миотомы
86, 6) пойдет на построение по-
мышц тела, а внутренний участок
187
по сравнению с другими участками сомитов очень большие,
а у высших позвоночных очень большими участками являются
склеротомы.
Дифференцируясь, мышечные сегменты разрастаются книзу
в пространствб между кожей и париетальным листком спланх-
нотома, пока в конце концов не встретятся на брюшной стороне.
В таком виде морфологическая дифференцировка оказывается
в общем сходной у рыб и водных личинок амфибий. У различных
групп позвоночных дальнейшее развитие мышечной системы име-
ет свои особенности в зависимости от характера строения и ме-
' стоположения околомоторных органов, развития парных конечно-
стей, плавников, хвоста. Мускульные -ткани, формирующиеся из
сомитов головной области, дифференцируются в шесть пар глаз-
ных мышц.
РАЗВИТИЕ ОСЕВОГО СКЕЛЕТА
Наиболее важные черты сложных морфологических преобразова-
ний, происходящих входе развития осевого скелета у позвоночных,
таковы. Опорной системой тела в онтогенезе разных позвоночных
служит хорда. На следующих этапах онтогенеза развиваются
частично (в связи с хордой и независимо от нее) хрящевые струк-
туры. Хрящевой скелет оказывается дефинитивным у круглоротых
(Cyclostomata) и у хрящевых рыб (Elasmobranchii). У большин-
ства позвоночных на более поздних"стадиях онтогенеза хрящевой
скелет дополняется и в большей своей части замещается костным
скелетом.
Источником для развития осевого скелета, как уже сказано,
служит склеротбм. Эта часть сомитов состоит из массы мезен-
химных клеток. Размножаясь, они заполняют пространство, окру-
жающее хорду,-покрывают ее и оказываются источником развития
хрящевых тел и дуг позвонков (рис. 87). Мезенхима склеротома
распространяется таким образом, что покрывает развивающийся
спинной мозг на всем его протяжении, хвостовые артерии и вены.
С каждой стороны хорды развиваются треугольные массы, углы
Рис. 87. Схема разви-
тия осевого скелета (по
Б. И. Балийскому, 1961):
Д — спинная н брюшная
пары дуг позвонка; Б —
дейральные дуги н ребра
туловищного отдела; В —
нейральная н гемальная
дуги хвостового отдела; 1—
нервная трубка, 2 — хорда,
3 — нейральная дуга, 4 —
гемальная дуга
188
которых постепенно вытягиваются в спинном, брюшном и средин-
ном направлениях. Так возникают зачатки невральных, попереч-
ных и хордальных отростков позвонков. Хордальные отростки од-
ной стороны срастаются с хордальными отростками другой сторо-
ны, таким образом начинают формироваться позвонки. В образо-
вании позвонка участвует задняя часть одного склеротома и
передняя часть следующего за ним склеротома; так, образую-
щиеся позвонки не соответствуют «первичным позвонкам» (соми-
там) и их миотомам: каждый миотом соприкасается с задней
половиной одного и передней половиной следущего за ним позвон-
ка’ т. е. они располагаются на стыках двух миотомов. Вскоре
начинается охрящевение позвонков. Заключенная в образующийся
позвоночный столб хорда впоследствии почти целиком дегенери-
рует, точнее редуцируется до межпозвоночных прослоек. Хряще-
вой, а позднее костный позвоночный столб замещает функцио-
нально хорду.
Хрящевые ребра развиваются независимо от позвоночника
в межмышечных связках, в их спинных отделах. Они растут
в брюшном направлении, концы выросших хрящевых ребер сли-
ваются друг с другом, образуя зачаток грудины.
Сложные процессы происходят при развитии скелета головы,
т. е. развитии черепа и висцерального скелета. Череп образуется
по бокам и впереди переднего конца хор-
ды. Сначала развивается и дифференци-
руется соединительная ткань, окружаю-
щая головной мозг и развивающиеся
органы слуха, зрения и обоняния. Этот
так называемый перепончатый череп
постепенно охрящевевает, а впоследст-
вии происходит окостенение.
У ранних эмбрионов позвоночных хря-
щевой череп состоит из нескольких неза-
висимых зачатков: трабекул («черепных
перекладин»), парахордальных хрящей
(от греч. para — подле, chorde — струна;
парахордальный — находящийся рядом,
по бокам хорды) и хрящевых капсул во-
круг органов зрения, слуха и обоняния
(рис. 88).
Передняя часть трабекул развивается
из клеточного материала, мигрирующего
из ганглиозной пластинки, значит, она
эктодермального происхождения. Задняя
часть их развивается из мезенхимы скле-
ротомов. Хрящевые капсулы органов
чувств развиваются из мезенхимы разно-
f
Рис. 88. Зачатки хрящевого
черепа позвоночных (по
Б. И. Балансному, 1961):
/ —* трабекулы, 2 — обонятель-
ная капсула, 3 — глазная кап-
сула, 4 — парахордальные хря-
щи, 5 —слуховая капсула, б —
первый позвонок, 7 — хорда;
черные области — хрящ на пер-
воначальной стадии; места сли-
яния разных компонентов обо-
значены точками на правой
стороне рисунка
189
го происхождения: слуховая капсула — из мезенхимы склерото-
мов, хрящ носовой капсулы, как показал в 1939 г. И. И. Шмаль-
гаузен, развивается из эмбрионального зачатка носа. Неясно про-
исхождение соединительно-тканной капсулы, окружающей глазное
яблоко и превращающейся впоследствии в хрящевую.
РАЗВИТИЕ СЕРДЦА
Вся кровеносная и лимфатическая системы — производные мезо-
дермы. В этом разделе сообщаются лишь основные факты, касаю-
щиеся развития важнейшего органа сосудистой системы—сердца,
и почти опускаются вопросы развития сосудов.
Развитие сердца можно рассмотреть на примере амфибий.
В ходе гаструляции происходит распространение мезодермального
пласта клеток между экто- и энтодермой. На стадии нейрулы
разрастание спинного и спинно-бокового участков мезодермальной
мантии доходит до головной области зародыша. Разрастание это,
однако, неравномерное: на брюшной стороне зародыша, в ораль-
ной и глоточной областях, остается в виде треугольника участок,
в который мезодерма не проникает (рис. 89).
В период, следующий за нейруляцией, происходит постепенное
схождение (в середине свободной от мезодермы области) краев
разрастающейся мезодермальной мантии (спланхнотомов). Слева
и справа, к середине свободного от мезодермы участка, распро-
страняются мезенхимного типа рыхло расположенные клетки,
которые являются парным зачатком сердца. Точнее, этот клеточ-
ный материал — зачаток эндокардия, т. е. эндотелия полости
сердца (рис. 90, 4). Клетки мезенхимы в виде продольной поло-
ски накапливаются, и в результате формативных перемещений
клеточного материала формируется тонкостенная трубка, развет-
вляющаяся на обоих концах: две ветви на переднем конце — это
будущие брюшные аорты, а две ветви на заднем конце — это
две желточные вены. Как и эндокардиальная трубка, ветви ее —
будущие сосуды — состоят также только из одного слоя клеток
эндотелия, возникшего вследствие дифференциации мезенхимы.
Одновременно с этими процессами края мезодермального пласта
клеток доходят друг до друга и срастаются под зачатком сердца
(т. е. под эндокардиальной трубкой), который затем покрывается
висцеральным листком мезодермы.
При слиянии мезодермальных листков под и над зачатком
сердца образуются перегородки — спинной и брошной мезокардии
(брыжейки сердца, рис. 90,6). Брюшная перегородка вскоре исче-
зает, и благодаря этому целомические полости (правой и левой сто-
рон) оказываются неразделенными. Впоследствии рассасывается
и спинной мезокардий. Расширяющийся участок целомической по-
лости в районе сердца образует околосердечную полость (рис.
90,9), которая являясь сначала частью общей полости тела, впо-
190
следствии развивается в полость, обусловленную от
целома. Эпикардий и миокардий (сердечная мыш-
ца) возникают из клеток висцерального листка ме-
зодермы, дифференцирующихся в наружный эпи-
телий и мускульную ткань сердца (рис. 90,8).
Перикардий является производным париетально-
го листка спланхнотома.
Вначале развития сердце — это полая, почти
прямая, без всяких подразделений трубка. Путем
не описываемых здесь изгибаний и поворотов, сжи-
маний в одних местах и расширений в других
происходит изменение его формы, и оно подразде-
ляется на четыре основные части: венозный синус
(sinus venosus), предсердие (atrium), желудочек
(ventriculus) и артериальный конус (conus arte-
riosus).
Задолго до полного сформирования сердца, еще
образуются периферические сосуды, начинается его
Рис. 89. Поло-
женне зачатка
сердца заро-
дыша тритона
на стадии ней-
рулы (по К.
Гертлеру)
прежде чем
ритмическая
пульсация. Оно находится, образно говоря, в состоянии готовно-
сти играть свою функциональную роль.
Развитие сердца амфибий явилось объектом многих экспери-
ментальных исследований; данные этих исследований могут слу-
жить иллюстрацией влияния одних частей зародыша на другие
(см. гл. XI).
В связи с не изученными до конца взаимовлияниями клеток
Рис. 90. Развитие сердца зародышей амфибий (из О. Гертвига, 1906):
А- Д —тритон; Б — саламандра; В — лягушка; / — энтодерма, 2 — париетальный листок
латеральной мезодермы, 3 — висцеральный листок латеральной -мезодермы, 4 — зачаток
эндокардия, 5 — эндокардиальная трубка, 6 — брюшной мезокардий, 7 — перикардий, 8 —
мнокарднй, 9— околосердечная полость, 10— полость сердца
191
мезодермы и прилегающей энтодермы задолго до сформирования
сердца со структурой, свойственной его дефинитивному состоянию,
уже имеет место специфическая клеточная и тканевая дифферен-
циация презумптивной мезодермы сердца. Такое заключение
можно сделать на основании вырезания презумптивной сердеч-
ной мезодермы и культивирования ее вне зародыша. По К. Герт-
леру (1928), такая складывающаяся тканевая система способна
развиваться в свойственном ей направлении: дифференцируется
характерная для сердца мышечная ткань, начинающая ритмично
сокращаться.
При развитии сердца несомненно происходит взаимодействие
всех трех зародышевых листков. О. Мангольду в 1936 г. и
Б. И. Балинскому в 1939 г. удалось осуществить операции полно-
го удаления энтодермы у зародышей амфибий. Техника операций
была такова, что мезодерма, производной которой является
сердце, оставалась неповрежденной. Однако у таких безэнтодерм-
ных зародышей сердце совершенно не развивалось.
П. Штер в 1924 г. обнаружил, что, достигнув определенной
стадии развития, зачаток сердца может некоторое время в усло-
виях эксплантации продолжать развиваться, значит, независимо
от окружающих его частей зародыша развиваются «пульсирую-
щие» мышечные ткани, формируется сердечная трубка. Подобные
явления наблюдаются и в экспериментах на зачатках других
органов (см. гл. XI).
Согласно опытам Г. Экмана (1925), если вырезать не весь
парный зачаток сердца, а только правую или левую половины
его и культивировать их вне зародыша, каждая половина (как
оставшаяся в зародыше, так и удаленная) может дифференциро-
ваться в ритмично сокращающиеся ткани. Нельзя считать удиви-
тельным явлением, что развитие сердца в условиях культивиро-
вания вне организма половины его зачатка вскоре останавлива-
ется, так как в норме для развития сердца требуется его
функционирование, чтобы была циркуляция крови.
Можно добиться развития двух сердец у одного эмбриона,
если сделать надрез перед тем, как соединятся презумптивные
зачатки сердца с правой и левой сторон. Можно поместить между
двумя презумптивными зачатками сердца чужеродную ткань,
в результате не произойдет срастания зачатков правой и левой
сторон. В этих условиях у одного зародыша могут развиваться
два пульсирующих сердца. Считают, что на стадии до слияния
двух зачатков сердца они не «строго детерминированы». Интер-
претация подобных явлений дается в гл. XI.
РАЗВИТИЕ ВЫДЕЛИТЕЛЬНОЙ СИСТЕМЫ
Строение различных типов выделительных органов у животных
сопоставлено на рис. 91. Эта схема дает представление об эво-
люции выделительной системы, а по рис. 92 можно составить
192
органов (А — Д — по
Рис.
А. Г.
Л — протонефридии (низшие черви); Б — метанефрнднн, или целомодукты (кольчатые чер-
ви); Г 4 г, _ .... х г. .... /
шие позвоночные, зародыши высших позвоночных); ,
ночные, изображена развивающаяся вторичная почка зародыша); Е,
типа экскреторных «единиц» с внутренними н наружными клубочками;
ветвления .протонефридиев, 2— то же, при большем увеличении (видна крупная клетка —
соленоцит, замыкающая слепой конец веточки протонефридия), 3 — мерцательное пламя,
4 — выводные отверстия протонефридиев (одна пара на все тело червя), 5— каиалец мета-
нефридия, 6—'его мерцательная воронка, открытая в целом, 7—выводные отверстия мета-
нефрнднев (по паре на сегмент), 8—'Стенка целома (вторичная полость тела), 9— кожный
эпителий, 10— границы между сегментами тела, 11— каналец предпочки, 12 — его мерца-
тельная воронка, открытая в целом, 13 — проток предпочки, 14 — гломус (клубок кровенос-
ных капилляров, прилегающий снаружи к стенке целома). 15— аорта, 16 — ее сегментар-
ные веточки (приносящие артерии), /7 — выносящие артерии, 18 — мальпигиево тельце, со-
держащие мальпигиев капиллярный клубочек, 19 — срез протока первичной почки, 20 —
мочеточник. 2/ —лоханка, 22 — метанефрогенная ткань, 23 — развивающиеся канальцы вто-
ричной почки, 24 — почечная артерия, 25 — нефроцель, 26 — нефростом
91. Строение различных
Кнорре, 1959; Е — Ж — по Р. Фразеру, 1950):
типов
выделительных
В— предпочка (зародыши н личинки низших позвоночных); Г — первичная почка (низ-
‘ Д — вторичная почка (высшие позво-
Ж — два основных
1 — концевые раз-
Рис. 92. Отношения пронефроса, мезонефроса и метанефроса в ходе развития
выделительной системы млекопитающих: А — предпочка; Б, В—первичная почка;
Г — вторичная почка (по Б. П. Пэттену, 1959):
1— канальцы пронефроса, 2 — проток пронефроса, 3— канальцы мезонефроса, 4 — клоака,
5 — канальцы мезонефроса с нефростомами, 6 — канальцы мезонефроса без нефростомов,
7 — проток мезонефроса, 8 — аллантоис, 9 — проток метанефроса, 10 — мюллеровы протоки,
11— семенник, 12 — аллантоис, 13 — канальцы метанефроса, 14 — яичник, /5 — дегенерирую-
щие канальцы и проток мезонефроса. Пронефрос у эмбрионов человека появляется в кон-
це третьей недели на стадии 9—10 сомитов и регрессирует уже в конце четвертой недели:
'мезонефрос достигает наибольших размеров в конце второго месяца
представление о развитии выделительной системы в онтогенезе
млекопитающих и человека. Сложная перестройка выделительной
системы в онтогенезе этих органов с основанием сопоставляется
с ее филогенетической историей и является одним из демонстра-
тивных примеров рекапитуляций (гл. XVII).
У губок и кишечнополостных нет выделительных органов;
клетки у этих организмов могут освобождаться от ненужных про-
дуктов обмена благодаря своим клеточным механизмам.
Наиболее примитивные органы выделения — протонефридии
(рис. 91,Л), характерны для низших червей, у которых нет кро-
веносной системы, забирающей продукты обмена из тканей и
выносящей их к почкам. Это сильно разветвленные, проникающие
во все участки тела трубочки, открывающиеся на поверхности
кожи порами. Концы разветвления не имеют отверстий, а состоят
из одной или нескольких клеток с ресничным аппаратом. Благо-
даря колебательным движениям длинных жгутиков ток жидкости
направляется к выходному отверстию.
У кольчатых червей, которым свойствен целом и кровеносная
система, имеются метанефридии — трубочки, воронкообразный
конец которых открывается в полость тела, а другой конец —
на поверхность кожи (выделительные поры) (рис. 91, Б). Реснич-
ный аппарат воронки обеспечивает направление тока жидкости
из целомической полости во внешнюю среду. Через метанефридии
(в тех сегментах тела, где в стенке целома развиваются половые
клетки) выводятся наружу и половые клетки.
У позвоночных развиваются почки — совокупность мочевыно-
сящих, секретирующих канальцев. В отличие от метанефридий
кольчатых червей канальцы почки позвоночных связаны с крове-
носными сосудами в единое морфофизиологическое целое.
Отражением эволюции выделительной системы у позвоночных
являются различной степени сложности процессы, приводящие
к смене органов выделения в ходе онтогенетического развития.
У низших позвоночных (рыбы и амфибии) в зародышевом
периоде развивается относительно примитивно построенная голов-
ная почка или предпочка (pronephros), сменяющаяся потом ту-
ловищной или первичной почкой (mesonephros). Только у мик-
син из круглоротых и у некоторых низших рыб предпочки фун-
кционируют во взрослом состоянии. У зародышей высших
позвоночных (рептилий, птиц и млекопитающих) закладывается
зачаточный, вскоре редуцирующийся пронефрос (см. рис. 91, В\
92, Л). Каудальнее пронефроса развивается и функционирует
некоторое время первичная почка (mesonephros) (см. рис. 91, Г;
92, Б, В; 93, 4), которую можно назвать в связи с ее локализа-
цией туловищной почкой; затем у ее задних границ образуется
и функционирует окончательная наиболее совершенного строения
вторичная почка (metanephros). Это тазовая почка (см. рис.92, Г).
195
/4
Рис. 03. Мочеполовая система позвоночных (по Хьютнеру, модифицировано
Р. Стевинг, 1969):
Л — пронефрос, Б — мезонефрос; / — хорда, 2 —миотом, 3 — нервная трубка, 4 — первичные
мочеточники, 5 — почечные канальцы, 6 — нефростом, 7 — нисходящая аорта, 8 — кишечный
канал, 9 — целом, 10 — задняя кардинальная вена, 11 — миотом, 12 — дерматом, 13 — экто-
дерма, 14— спинальный ганглий, 15 — капилляры, 16 — субкардинальная вена, 17 — наруж-
ные ресничные воронки, 18— Боуменская капсула, 19—клубочки капилляров, 20—заклад-
ка половой железы
Процессы, сопровождающие смену выделительных органов, мы
рассмотрим ниже.
Предполагается, что у предков позвоночных была примитивная
почка, простирающаяся на всю длину полости тела. Эта почка
была построена из посегментно расположенных канальцев, раз-
вивающихся из нефротомов и открывающихся с одной стороны
воронкой в целом, а с другой — в общий мочевыносящий канал.
В результате дальнейшей эволюции животных, изменения ме-
таболических процессов и функции выделения в средне-задней
части единого органа примитивные канальцы перестали заклады-
ваться и на их месте на соответствующей стадии онтогенеза из
нефротомов стали развиваться более совершенные канальцы.
У современных позвоночных канальцы предпочки закладыва-
ются из нефротомов метамерно, по одному на сегмент. У миног
возникает до 6 канальцев, у скатов — до 8, у осетровых рыб —
7 канальцев, у безногих амфибий — до 12, а у других амфибий —
2—3 канальца.
Функционирующий пронефрос представляет довольно объем-
ный орган, состоящий из извитых канальцев, открывающихся,
как уже сказано, с одной стороны в целом воронкой с мерцатель-
ным эпителием, а с другой — в общий мочевыносящий канал —
первичный мочеточник, или вольфов канал.
Из стенок дорсальной аорты, на уровне пронефроса, формиру-
ются, свешиваясь в полость тела, сосудистые клубочки — гломусы
(glomus), инфильтрующие из крови в целом продукты метаболиз-
ма и воду. Благодаря воронкам канальцев предпочки с их реснич-
196
ним аппаратом моча — продукты метаболизма и вода — вбирается
и выводится из тела во внешнюю среду первичным мочеточником.
И канальцы, и протоки у всех позвоночных—производные
мезодермы, они развиваются из нефротомов, о которых упомина-
лось в связи со строением сомитов. Почечные канальцы развива-
ются так. Полоска клеток (нефротом)—связующее звено между
сомитом и боковой пластинкой мезодермы, расслаивается, как и
вся боковая мезодерма, на париетальный и висцеральный листки;
таким образом возникает полость нефротома — нефроцель (см.
рис. 91, 25). Связь этой щели с полостью сомита (миоцелем)
вскоре исчезает, но связь с полостью между двумя листками ла-
теральной мезодермы (целомом или спланхноцелем) в случае
примитивного типа выделительных органов остается. Спиннобоко-
вая стенка нефротома вытягивается в полую трубочку (каналец),
сообщающуюся с полостью тела воронкой — нефростомом (см.
рис. 91,26).
Первичный мочеточник у всех позвоночных развивается из
собственного зачатка, закладывающегося у задних границ про-
нефроса в виде плотного клеточного скопления на париетальном
листке. Клетки — источники для развития мочеточника активно
перемещаются в каудальном направлении и клоаке, вдоль нижних
границ сомитов. По мере перемещения клеточный тяж приобре-
тает строение канала — мочеточника.
При образовании мезонефроса устанавливается непосредствен-
ная связь почечных канальцев с кровеносной системой. Происхо-
дит впячивание (в виде чаши) «слепых» расширенных концов
канальцев, образуются вместилища — боуменовы капсулы, в кото-
рых возникают клубочки капилляров (glomerulus) благодаря
врастанию приносящих артерий. Так образуется мальпигиево
тельце, имеющее вид капсулы с клубочком (см. рис. 91, Г; рис. 92).
Это строение и характерно для' типа почки, называемой первич-
ной почкой, вольфовым телом, иначе mesonephros. Мочевые ка-
нальцы открываются в пару выводных (вольфовых) каналов,
являющихся продолжением пронефрических протоков.
Главное отличие первичных почек от предпочек заключается,
как уже сказано, в том, что подлежащие выделению продукты
метаболизма поступают в экскреторные органы не из целома,
а непосредственно из крови. Первичная почка развивается из
большого числа туло’вищных нефротомов, канальцы ее не образу-
ют воронок и не имеют связи с целомом.
Такое строение выделительной системы, конечно, прогрессивно.
Отражением эволюции выделительной системы является, в част-
ности, то, что у примитивных низших позвоночных, например
у акул, наряду с капсулами и сосудистыми клубочками сохраня-
ются еще и воронки, связанные с целомом.
Мезонефрос, как и пронефрос, является производным нефро-
томов, но канальцы образуются не путем трансформации, нефро-
197
томов, а более сложным путем. У селяхий же и некоторых амфи-
бий мочевые канальцы формируются непосредственно из нефро-
томов. У большинства же амфибий и всех высших позвоночных
мезодермальные клеточные массы нефротомов превращаются
в нефрогенный шнур мезенхимных клеток, тянущийся вдоль дор-
сального края латеральной мезодермальной пластинки. Уплотняю-
щиеся группы мезенхимных клеток превращаются в эпителиаль-
ные пузырьки, преобразующиеся в почечные канальцы, впадающие
в вольфов проток. В области каждого сегмента развивается
несколько канальцев. У каждого канальца образуется бокаловид-
ное впячивание — упоминавшаяся ранее боуменова капсула.
В этой полости формируется клубочек тончайших сосудов, кото-
рые при выходе из капсулы собираются в отводящий кровеносный
сосуд (vas defferens). Клубочек сосудов с боуменовской кап-
сулой называют почечным или мальпигиевым тельцем (см.
рис. 91, 18).
На рис. 93 дана схема мочеполовой системы у позвоночных
животных.
Метанефрос — дефинитивная почка (у птиц и млекопитающих)
развивается из несегментированной массы нефротомов задних
сегментов туловища и из эпителиального зачатка вторичного мо-
четочника. Почечные канальцы образуются из тяжей нефротом-
ного клеточного материала, в котором возникают просветы. Про-
исходит развитие канальцев, они соприкасаются с внедряющи-
мися разветвлениями почечных артерий, возникают мальпигиевы
тельца. Еще до этих процессов начинается образование мочеточ-
ников. У человека в начале четвертой недели концевые отделы
вольфовых каналов утолщаются и на дорсальной стороне утол-
щений образуются небольшие выпячивания, которые врастают
в нефрогенные массы клеток и превращаются в каналы — зачатки
мочеточников, а возникающие на их концах расширения дают
начало формированию почечных лоханок. В нефрогенной массе
клеток происходит тканевая дифференциация, развиваются два
слоя. Внутренний слой, облегчающий вросшие зачатки почечных
лоханок, дифференцируется затем в извитые канальцы и маль-
пигиевы тельца. Клетки наружного слоя дифференцируются в
соединительную ткань — строму и волокнистую оболочку дефи-
нитивной почки (см. рис. 91, Д).
Возникшие из стенок вольфовых каналов правый и левый мо-
четочники после разделения клоаки на прямую кишку и моче-
половой синус открываются в последний благодаря отверстиям
вольфовых каналов. Когда происходит развитие и обособление
от мочеполового синуса мочевого пузыря и мочеиспускательного
канала, мочеточники обособляются от вольфовых каналов, на
спинной стороне мочевого пузыря образуется самостоятельное
отверстие мочеточников.
Развитие органов размножения очень тесно связано с разви-
198
тием выделительной системы. Это единый комплекс развивающих-
ся структур.
Сложные процессы развития мочеполовой системы у млекопи-
тающих— появление пронефроса, мезонефроса и метанефроса —
сопровождаются тем, что некоторые органы появляются, а затем
полностью исчезают, так и не начав специфически функциониро-
вать (рудиментарный пронефрос). При редукции предпочки, раз-
витии первичной и затем вторичной почек и половых органов
происходят разнообразные морфогенетические движения, дегене-
рация тех или иных клеточных масс, фагоцитарные явления и
другие процессы. Эмбриология не располагает еще вполне удов-
летворительным анализом этих явлений, в частности в аспекте
иммунологическом (см. гл. XV). Неясны также вопросы о зна-
чении . для развития зародыша формирования рудиментарных
органов. То, что предпочка у млекопитающих не выполняет своей
специфической выделительной функции, еще не означает, что ее
развитие и резорбция не играют никакой функциональной, фор-
мообразовательной роли. Предпочка у высших позвоночных — не
только «воспоминание» о предках. Впоследствии (гл. XI) мы
узнаем, например, что не выполняющая специфической функции
хорда, развивающаяся у зародышей амфибий, имеет большое
«функциональное значение», так как развитие нервной системы
не может происходить без взаимодействия с этим зачатком.
РАЗВИТИЕ ПОЛОВОЙ СИСТЕМЫ
Строение и развитие половой системы, в частности органов со-
вокупления, рассматривается в руководствах по анатомии, ги-
стологии, общей зоологии и по другим наукам. Процессы раз-
вития половых органов с точки зрения механики развития не
представляют особого интереса, а служат лишь добавочными
иллюстрациями морфогенетических процессов, рассматриваемых
на примерах других органов. Вследствие этого многие вопросы
развития половой системы здесь опускаются.
Мужские и женские гонады позвоночных развиваются в зад-
ней половине тела из клеточного материала латеральной пла-
стинки мезодермы (рис. 93). Зачаток гонад появляется в виде
продольной полоски перитонеального эпителия. На поверхностях
первичных почек, вблизи кишечной брыжейки, образуются неболь-
шие утолщения — половые валики. Они представляют собой ком-
пактную массу клеток, состоящую из нескольких слоев и вы-
дающуюся в полость тела. Впоследствии связь между развиваю-
щейся гонадой и перитонеальным листком латеральной мезодер-
мы сужается, и гонады оказываются как бы подвешенными к
перитонеальной стенке.
Половые валики состоят из двух родов клеток. Большинство
клеток очень сходно с клетками перитонеального эпителия. Они,
199
однако, уже на начальных стадиях развития половых валиков
принимают характер столбчатого эпителия — это зачатковый эпи-
телий (рис. 93,20). Клетки другого типа — с пузыревидными
ядрами значительно большего размера, чем обычные мезодер-
мальные клетки. Цитоплазма их отличается от первого типа
клеток своим отношением к красителям (по данным исследова-
ний на птицах). У зародышей амфибий они характеризуются
большим содержанием желтка, чем окружающие клетки. Эти
клетки — первичные половые клетки в составе зачаткового эпи-
телия.
Как уже говорилось в гл. II, эти клетки имеют особое про-
исхождение, они мигрируют в зачатки половых желез из жел-
точного мешка и внедряются в эпителий целома, входящий в
состав развивающейся железы. Размножаясь, они становятся
мельче и неотличимы от других клеток. Клеточная масса же-
лезы как бы «пересыпана» этими мигрировавшими первичными
половыми клетками, которые и являются источником для разви-
тия спермиев в семенниках и яиц в яичниках. Клетки же гонад
мезодермального происхождения дифференцируются в поверхност-
ный эпителий гонады, клетки стромы гонады и фолликулярный
эпителий.
РАЗВИТИЕ КОНЕЧНОСТЕЙ
Конечности амфибий, птиц и млекопитающих являлись предме-
том многочисленных исследований. Классическая эмбриология
занималась вопросом — как происходит их развитие? Экспери-
ментальная эмбриология выясняла взаимозависимость клеток и
Тканей в развитии конечностей. Конечности хвостатых амфибий —
излюбленный объект специалистов по регенерации. Вот почему
здесь дан краткий очерк развития конечностей амфибий.
Начало развития конечности связано с париетальным лист-
ком латеральной пластинки мезодермы (см. рис. 85 и 94), в ко-
тором под верхним краем происходит утолщение. Это, однако,
не есть разрастание участка боковой мезодермы, а скорее ми-
грация клеток и скопление их в массе мезенхимы, располагаю-
щейся между боковой мезодермой и эпидермисом. Целость (не-
прерывность) структуры париетального листка мезодермы не на-
рушается.
У зародышей амфибий такое скопление мезенхимы происхо-
дит почти одновременно в районе будущих передних и задних
конечностей. У рептилий, птиц и млекопитающих скопления ме-
зенхимы происходят не только в районах развития конечностей,
но и по всей длине в виде гребней (вольфовы гребни). Однако
в ходе развития зародыша в большей своей части эти сгущения
мезенхимы рассасываются, а дифференцировка мезенхимы про-
исходит только на «территориях конечностей».
В этих областях эпидермис, покрывающий мезенхиму, не-
200
Рис. 94. Схема происхождения
мезодермального зачатка ко-
нечности зародыша амфибий
(по Б. И. Балийскому, 1961):
/ — миотом, 2 — почечный канал,
3, 8 — зачаток конечности, 4 — па-
риетальный листок латеральной
мезодермы, 5 — висцеральный лис-
ток латеральной мезодермы, 6
энтодерма, 7 — спинной мозг
сколько утолщается и выпячивается.
Образующаяся таким образом почка
конечности состоит из клеток мезо-
дермального происхождения и клеток
эпидермиса. Интересно, что по степени
дифференциации и по своим морфоге-
нетическим потенциям два компонента
почки конечности (мезенхима и эпи-
дермис) не равнозначны. Ведущая
роль на первых этапах развития при-
надлежит мезенхиме. Экспериментами
Б. И. Балинского еще в 1931 г. было
доказано, что если вскоре после обра-
зования нервной трубки у зародыша
амфибий вырезать кусочек из боковой
мезодермы и трансплантировать его
под эпидермис бока или головы, ино-
гда развивается на не свойственном
ей месте добавочная конечность. Уда-
лив лоскут кожи на «территории» ко-
нечности, можно трансплантировать
вместо него кусок эпидермиса, экстир-
пированный из любой области тела.
Пересаженный материал входит в состав формирующейся почки
конечности.
Поведение эпидермиса почки конечности отличается особен-
ностями у разных позвоночных. У рептилий, птиц и млекопи-
тающих ранняя почка конечности уплощается, а пи краям упло-
щения эпидермис утолщается в виде гребня у одних позвоночных
(птиц) и в виде плотной складки у других (рептилий). Отмече-
ны патофизиологические отличия клеток эпидермальных утолще-
ний по сравнению с клетками рядом лежащих частей эпидермиса:
они содержат больше РНК, гликогена и щелочной фосфатазы.
Это свидетельствует о высокой метаболической активности.
На стадиях, предшествующих формированию видимой почки
конечности амфибий, указанными экспериментами Б. И. Балин-
ского и других авторов доказана «ведущая роль» мезенхимы,
ее индуцирующее влияние на прилегающий эпидермис. Мезенхи-
ма выполняет роль как бы стимула специфической дифференци-
ровки эпидермиса. В свою очередь эктодермальный гребень почки
конечности влияет на дифференцировку клеточных элементов
мезодермального происхождения, на все развитие почки конеч-
ности. В этом убеждают опыты на зародышах птиц, проведенные
Ж. Сондерсом в 1948 г. Если удалить эктодерму эпидермального
гребня почки трехдневного куриного эмбриона, то дистальная
часть крыла не формируется, хотя подлежащая мезодерма в
этих опытах не повреждается.
201
Однако считается, что ведущую роль в дифференциации, по
крайней мере на первых этапах развития конечности, у птичьих
зародышей играет мезодерма. В этом убеждают опыты Е. Цвил-
линга, проведенные на курином зародыше в 1955 г. Химическим
способом (действие «раствора версена») он удалял эпидермис
почки конечности, трансплантировал на его место лоскуты эпи-
дермиса из разных мест другого зародыша (например, из области
зачатка крыла) и пересаживал почку конечности на бок третьего
зародыша. Развитие продолжалось, причем развивалось или
крыло, или нога, в зависимости от того, какого происхождения
была взята мезодерма — из почки крыла или из почки ноги. Эти
же опыты дали возможность заключить, что происхождение эпи-
дермиса не играет ведущей роли, эта роль принадлежит ме-
зодерме.
Результаты этих и подобных им опытов интерпретируются
разными исследователями неодинаково в зависимости от теории
индивидуального развития (см. гл. XI).
Интерес представляют опыты Б. И. Балинского (1925), а
позднее и других авторов, в которых удавалось получать
развитие добавочных конечностей у хвостовых амфибий. Меж-
ду зачатками передней и задней конечностей под кожу вво-
дили инородные ткани — слуховой пузырек (в опытах Балинско-
го), зачаток гипофиза или обонятельного мешка. В результате
возникали добавочные конечности; при этом Балинский в 1933 г.
убедился в том, что если один и тот же стимулятор всаживать
ближе к нормальной передней или к задней конечности, проис-
ходит соответственно развитие передней или .задней конечности.
Анализ дается на основе представлений о морфогенетических
полях.
Внешне морфологическая картина роста и развития почки
конечности такова. По мере удлинения почки дистальная часть
ее оказывается более плоской и более широкой. На этой стадии
«лопатки», или «пластинки», длина всего зачатка превышает его
ширину. Края лопатки становятся неровными: определяются не-
большие выросты — зачатки пальцев, которые продолжают расти,
а в промежутках между ними заметного роста не наблюдается.
Интересно, что у всех высших позвоночных с пятипалыми ко-
нечностями зачатки всех пяти пальцев возникают одновременно,
а у амфибий сначала появляются первые два пальца, а затем
последовательно появляются зачатки 3-го, 4-го и 5-го пальцев.
Эмбриологии еще неизвестны какие-то надклеточные и надтка-
невые факторы морфогенеза. По мере роста конечности проис-
ходят строго закономерные, не описываемые здесь процессы сги-
бания и вращения, которые обусловливают формирование трех
отделов конечности и заложения суставов. Эти морфологические
изменения представляют в сущности единый с тканевой диффе-
ренцировкой частей развивающейся конечности процесс.
202
«Намек» на развитие костного скелета можно усмотреть й
том, что первоначально относительно одинаковой плотности мас-
са мезенхимы на определенном этапе развития почки конечности
оказывается в виде уплотнений (куч) клеток в одних местах и
в очень разрыхленном состоянии в других местах. Участки уплот-
нений мезенхимы дифференцируются в хрящ. Нельзя представ-
лять процесс формирования скелетных частей так, что одно-
временно на всем протяжении, положим, презумптивной бедрен-
ной кости происходит охрящевение. Этот процесс начинается в
нескольких местах. Очаги охрящевения впоследствии объединя-
ются в единый скелетный комплекс. Проксимальные части кост-
ного скелета закладываются, в общем, ранее, чем дистальные.
У хвостовых амфибий сначала можно заметить оформление sty-
lopodium (humerus или femur), затем закладываются части zeu-
gopodium (radius и ulna; tibia и fibula) и позднее всего autopO-
dium. Зачатки пояса конечностей появляются ранее, чем зачатки
костей autopodium. В пальцах проксимальные фаланги заклады-
ваются ранее, чем дистальные.
Кровеносные сосуды возникают на ранних стадиях развития
конечности. Заслуживает особого внимания факт большой инди-
видуальной изменчивости в количестве и общем рисунке сосуди-
стой системы как развивающейся конечности, так в сущности и
во взрослом состоянии любого органа.
Огромное количество экспериментов проведено на развиваю-
щихся конечностях, особенно у амфибий и птиц, для познания
закономерностей механики их развития. Заслуживают особого
внимания эксперименты, в которых производилось расщепление
зачатка конечности на два участка и предотвращение их слипа-
ния и срастания путем, например, вставки между ними пластин-
ки инородных тканей. В этих опытах каждая половина может
развиваться в целую конечность. Это говорит о том, что факторы
развития и дифференциации конечности отнюдь не находятся
только в начальной системе почки конечности, что развитие
почки зависит от окружения. Результаты опытов по расщепле-
нию конечности не представляют сенсации в экспериментальной
эмбриологии. Принципиально те же результаты получены во
многих экспериментах по расщеплению зачатков и других орга-
нов (например, зачатка глаза).
В природных условиях можно наблюдать явления множествен-
ного развития конечностей, особенно у амфибий. Может быть,
в основе этих явлений лежат и механические травмы, и опреде-
ляемые патологическими внутренними факторами развития про-
цессы, аналогичные экспериментальному расщеплению зачатков
или физиологической изоляции частей. Возможно, происходят
процессы повреждения внутренними факторами в «территориях
конечностей», аналогичные тем, которые имеют место, например,
при введении под кожу разных тканей, о чем можно судить по
203
упоминавшимся опытам Б. И. Балийского и других авторов.
Источником для развития пояса конечностей является та же мас-
са мезенхимы, от которой начинается и развитие почки конеч-
ности, а именно — периферическая часть этой мезенхимной
массы.
Итак, началом развития конечности служит обособление ме-
зенхимы, мигрирующей из париетального листка боковой ме-
зодермы. Это не означает, что все структуры конечности мезодер-
мального происхождения развиваются из этого источника.
Мышечные ткани конечности в своей массе развиваются из ме-
зодермы нижних краев миотомов. У зародышей рыб видны «поч-
ки» презумптивных мускулов, образующихся на нижних краях
миотомов, входящих впоследствии в состав развивающейся ко-
нечности на начальных этапах развития, В почке конечности
мускульные почки миотомов соединяются в общую массу клеток,
которая дифференцируется в мускульные ткани.
У амфибий, рептилий, птиц и млекопитающих образования
таких мускульных почек миотомов не происходит, однако клеточ-
ные элементы миотомов могут мигрировать в почки конечностей.
Развитие нервов конечностей тесно связано с развитием всей
нервной системы.
ПОВЕДЕНИЕ КЛЕТОК И КЛЕТОЧНЫХ КОМПЛЕКСОВ
В РАЗЛИЧНЫХ ФОРМООБРАЗОВАТЕЛЬНЫХ ПРОЦЕССАХ
В эмбриологии должны содружествовать разные типы исследо-
ваний: молекулярный, клеточный, надклеточный и др. Какой
аспект и какие методики должны доминировать — зависит от
конкретных проблем и явлений. Однако почти во всех явлениях
эмбрионального развития прежде всего бросаются в глаза раз-
личные преобразования клеток и клеточных масс, в связи с чем
возникает много цито- и гистоэмбриологических проблем, а также
вопросов о динамике и архитектонике клеточных масс.
Ни одна часть эмбриона не бывает стабильной и не занимает
определенного положения, как это кажется по препаратам фик-
сированных зародышей и их частей. От изменения формы и по-
ложения первых бластомеров дробления до органогенеза вклю-
чительно происходят строго закономерные изменения клеток, их
индивидуальные и совокупные перемещения и другие подобные
явления.
Кажется, что в явлениях эмбрионального развития (разных
стадий его и у разных животных) существует столь большое
разнообразие, что они не поддаются какой-либо классификации.
Конечно, механизм процессов каждого типа гаструляцин — само-
стоятельная проблема и на первый взгляд ничего общего не
амеет, например, с проблемами, возникающими при анализе фор-
мирования нервной трубки или кишки. Однако между многими
204
явлениями, кажущимися совершенно различными, можно устано-
вить и общие черты. Живая природа многообразна, но не хао-
тична.
Многие исследователи делали попытки классифицировать мор-
фогенетические процессы, происходящие в эмбриональном раз-
витии. Рассмотрим одну из таких классификаций, предложенную
Б, И. Балинским (1961). Согласно ей различают следующие
типы морфогенетических движений клеток и клеточных масс.
1. Местные утолщения эпителиальных пластов (например, в
случае образования нервной пластинки у амфибий). Механизм
утолщения может быть различным: удлинение клеток эпители-
ального пласта в направлении, перпендикулярном к поверхности
пласта, увеличение числа клеточных слоев и т. д. Форма клеток
может меняться: из уплощенных они могут превращаться в ку-
бовидный, столбчатый эпителий.
2, Обособление эпителиальных пластов. Параллельно или пер-
пендикулярно к поверхности эпителиального пласта может появ-
ляться щель. В образовавшейся щели могут скапливаться жидкие
продукты жизнедеятельности клеток. Примером расщепления
(обособления) пластов путем возникновения и развития щелей-
полостей может служить развитие висцерального и париетального
листков боковой мезодермы. Мезодермальные сомиты возникают
благодаря появлению щелей, перпендикулярных к поверхности
пласта.
3. Возникновение складок пластов эпителиальных клеток. В ре-
зультате этого формируются желоб.ки, карманоподобные склад-
ки. Края складок (желобков) могут смыкаться, образуются
трубки.
Механизм образования желобков может быть различным. Не-
редко изгибание связано с локальными утолщениями эпители-
ального пласта или даже является их следствием. Так происхо-
дит образование желобка нервной пластинки. Впоследствии
образующаяся трубка обособляется от рядом лежащих клеточ-
ных масс. В случае обособления карманоподобных впячиваний
образуются полые пузырьки. Самые различные зачатки возни-
кают этим способом: слуховые пузырьки, многие железы. При-
мером этого типа морфогенетических движений является и воз-
никновение первичной пищеварительной полости у многих жи-
вотных. В свою очередь могут иметь место самые различные
механизмы образования складок, впячиваний (см. гл. VI).
При локальном утолщении эпителиального пласта кнаружи
образуются выросты, наросты. Механизм образования выростов
может быть различным. Б. И. Балинский (1961) предполагает,
что выросты никогда не возникают вследствие локального уси-
ления размножения клеток или увеличения размеров самих кле-
ток, они образуются в результате каких-то механизмов концен-
трации и перегруппировок клеток. Таковы ворсинки хориона у
205
млекопитающих, жаберные нити у амфибий и рыб. Кончики вы-
ростов могут быть плотными, но если возникает полость, в нее
могут проникать соединительная ткань и кровеносные сосуды.
4. Соединение (срастание, слипание) обособленных клеточных
масс. Пример: «сплавление» краев нервной пластинки, а значит,
превращение ее в трубку.
5. Распад участков эпителиальных пластов. В результате
образуется рыхлая мезенхимного типа ткань — материал для воз-
никновения зачатков тех или иных органов. Локальный распад
эпителиальных пластов — это нарушение (и подчас резкое) це-
лостности пласта клеток. Такое разрушение происходит, напри-
мер, при образовании «нервного гребня» — ганглиозной пластин-
ки: часть эпителия (край складки нервной пластинки) распадает-
ся на отдельные клетки, заполняющие пространство между
нервной трубкой и покрывающим ее эпидермисом. В создании
ганглиозной пластинки могут участвовать и клетки дорсальной
стенки самой нервной трубки.
Распространенное явление — отделение клеток и групп их от
общего эпителиального пласта без повреждения его. Пример: от-
деление клеток от висцерального листка латеральной мезодер-
мы. Образующаяся мезенхима оказывается вокруг пищевари-
тельной трубки, из нее развивается гладкая мышечная и соеди-
нительная ткани кишечника.
6. Возникновение сгущений мезенхимных клеток (уплотняю-
щихся масс мезенхимы), дифференцирующихся в хрящ, костную
или мышечную ткань.
7. Концентрация мезенхимы вокруг различных эпителиальных
образований — пузырьков, «карманов», трубок. Мезенхима мо-
жет служить источником образования хрящевых и костных кап-
сул. Примеры: скелетные капсулы при развитии органов слуха
позвоночных, капсула вокруг носового мешка, череп (мозговая
коробка) и др. Из скоплений мезенхимы могут развиваться со-
единительно-тканные капсулы (окружающие почку, печень, селе-
зенку и др.).
8. Вторичная эпителизация мезенхимы, возникшей из эпите-
лиальных пластов. Примеры: превращение клеток, мигрирующих
из первичной полоски птиц в слои эпителиевидных мезодермаль-
ных пластов, дающих сомиты и другие образования; возникнове-
ние эпителиальных пластов из масс мезенхимных клеток при
формировании эндотелия кровеносных сосудов.
В подавляющем большинстве случаев причинный анализ мор-
фогенетических движений еще ждет своих исследователей. Однако
важная задача экспериментальной эмбриологии — выяснение при-
чин тех или иных изменений в поведении индивидуальных кле-
ток, скоплений клеток, а также клеточных пластов в каждом
конкретном формообразовательном процессе — будь то процесс
гаструляции или образование зачатков органов и т. п. Направле-
206
ние исследований в этой области определяется теориями инди-
видуального развития, которым посвящены последующие гла-
вы XI—XVI.
Анализ явлений гаструляцин относится и ко многим другим
морфогенетическим движениям, только что перечисленным по
схеме Давенпорта—Балинского. По-видимому, следует придать зна-
чение важному факту: всем жизнедеятельным клеткам любых
тканей любой степени сложности организмов свойственна фаго-
цитарная способность и амебоидная подвижность (см. гл. VI и
XVI). Эта способность проявляется при дезинтеграции тканей,
эпителиальных пластов и плотных скоплений мезенхимы. Особен-
ности перемещения клеток определяются своеобразием физико-
химических свойств поверхностей клеток, межклеточных субстан-
ций, характером амёбоидного движения и т. п.
Несомненно важное формообразовательное значение имеют
перемещения не одиночных клеток, а целых пластов, частей за-
родышей. В культурах тканей на периферии растущих кусочков
можно наблюдать разнообразные движения клеточных масс, ве-
роятно, обусловленные отчасти неравномерным размножением
клеток (выростов, «языков» и т. п.). Перемещения пластов кле-
ток и их особенности обусловлены субстратом, на котором воз-
можна эта подвижность. Эпителии могут распространяться толь-
ко на плотных субстратах, мезенхимные же клетки могут дви-
гаться и по волокнам. Направление движения клеток опреде-
ляется ориентацией фибрилл.
Движениями клеток эпизодически интересовались многие ис-
следователи. Еще классик экспериментальной эмбриологии В. Ру
изучал поведение изолированных бластомеров яиц лягушки и
сделал предположение о значении «цитотаксисов» в эмбриональ-
ном развитии. Для объяснения движения сперматозоидов неко-
торые эмбриологи использовали понятие хемотаксиса.
Большое число исследований было посвящено явлениям пере-
мещения клеток и клеточных пластов в процессах гаструляцин.
Ранее были изложены представления Густафсона о значении
амёбоидных движений клеток в гаструляцин морского ежа. В но-
вейшее время уделяется большое внимание явлениям слипания
и отталкивания клеток.
Совершенно очевидно, что имеются различные механизмы мор-
фогенетических движений. Методом цейтраферной киносъемки
изучали на зародыше тритона поведение клеток нейрального эпи-
телия от момента появления нервной пластинки до контакта меж-
ду нервными валиками (М. Барнсайд, А. Джакобсон, 1968). При
длине зародыша 2,5 мм клетки нейрального эпителия смещаются
на 1 мм со скоростью от 4 до 95 мкм/ч. Движущиеся клетки
сохраняют свой контакт. Согласно этому исследованию одной из
главных причин «перемещения» клеток (а может быть, един-
ственной) является деформация пласта, которая обусловлена раз-
207
личиями в изменении формы составляющих его клеток (изме-
няется высота клеток).
Как уже говорилось (см. с. 159), в 1960—1968 гг. появились
работы о влиянии на движение клеток физиологически активных
веществ — медиаторов, благодаря которым осуществляется пере-
дача нервных влияний от клетки к клетке. На разных стадиях
развития обнаружены вещества (серотонин и др.), играющие
роль своеобразных клеточных гормонов. Т. Густафсон и М. То-
неби (1970) изучали влияние на морфогенетические процессы
у эмбрионов морского ежа 223 нейрофармэкологических веществ
и сделали вывод о стимулирующем действии серотонина и об
угнетающем действии его ингибиторов на образование первичной
мезенхимы. Они считают, что это связано с влиянием серотонина
на подвижность клеток. Ацетилхолин влияет преимущественно на
тот последний период гаструляции, когда устанавливаются
псевдоподиальные контакты между клетками архентерона и кры-
шей гаструлы.
Помимо изучения движения клеток, клеточных пластов и бо-
лее сложных комплексов клеток, требуются исследования и внут-
риклеточных движений, а также движений на молекулярном
уровне, вероятно, играющих важную роль в процессах деления
клеток, в их взаимодействии и дифференциации. Начаты иссле-
дования о закономерностях внутриклеточных движений, выясне-
но, что в процессах деления растительных клеток происходит за-
кономерная смена одних типов движения цитоплазмы другими.
В интерфазе движение цитоплазмы активное, типично циркуля-
ционное. В профазе совершается переход от циркуляционного к
броуновскому. В анафазе происходят обратные изменения в ха-
рактере движения. В конце телофазы, при полном восстановле-
нии ядерно-плазменных соотношений, движение цитоплазмы ста-
новится снова типично циркуляционным. Удалось выявить зако-
номерности в движении хромосом, волнообразно-извилистые дви-
жения хромосомных плеч (В. Ф. Боженко, 1973).
Молекулярная биология открыла своеобразное явление «само-
сборки» — образования биологических структур из однородных
молекул путем их сближения. Вероятно, такого рода движения,
основанные на физико-хичических закономерностях и сходные
с теми, какие наблюдаются и в неживых системах, также играют
роль в формообразовательных процессах. Представляют интерес
исследования регуляции в морфогенетических движениях темпов
клеточного деления в разных участках зародыша.
ПРОВИЗОРНЫЕ ОБРАЗОВАНИЯ ЗАРОДЫШЕЙ РЕПТИЛИЙ И ПТИЦ
О провизорных органах уже упоминалось. Указывалось, что тра-
диционное разделение зародыша на собственно зародыш и внеза-
родышевые части неправильно, так как в функциональном отно-
208
шении внезародышевые образования — неотделимая часть заро-
дыша на определенных стадиях его развития. Эти образования,
сопровождающие развитие многих животных, представляют боль-
шой интерес для эмбриологии, эволюционной морфологии и фи-
зиологии.
Желточный мешок. У зародышей рептилий и птиц, так же
как у низших позвоночных животных с меробластическим типом
яйцевых клеток, формируется желточный мешок. Однако, как об
этом уже говорилось, обрастание бластодиском желтка у репти-
лий и птиц не совпадает топографически с процессом формиро-
вания осевых органов, который осуществляется в средних участ-
ках бластодиска. В обрастании желтка принимают участие все
зародышевые пласты, однако мезодермальный пласт несколько
отстает от энтодермального и эктодермального, так как и по
времени он возникает позднее. Боковая мезодерма разделяется
на париетальный (наружный) и висцеральный (внутренний)
листки, которые продолжаются за пределами тела зародыша и
таким образом оказываются в составе периферических участков
бластодиска, участвующих в обрастании желтка. Собственно
стенка желточного мешка будет образована желточной энтодер-
мой и висцеральным листком мезодермы. Эктодерма с парие-
тальной мезодермой окажутся в составе серозной оболочки или
хориона, который будет покрывать зародыш вместе с желточным
мешком (см. далее). Пространство между хорионом и стенкой
желточного мешка называется экзоцеломом. В экзоцеломе раз-
растается аллантоис (рис. 95).
В отличие от рыб желточный мешок у рептилий и птиц дол-
гое время остается незамкнутым. По мере расходования желтка
он сморщивается и становится складчатым. Впоследствии перед
вылуплением зародыша из яйцевых оболочек желточный мешок
с небольшим количеством желтка перемещается в брюшную по-
лость и в течение месяца после вылупления полностью исчезает.
В течение второго и третьего дней развития куриного заро-
дыша во внуренней части area ораса развивается сеть крове-
носных сосудов. Эта область называется сосудистым полем area
vasculosa, а внешняя часть area ораса превращается в желточ-
ное поле area vitellina (рис. 96). Появление кровеносных сосудов
неразрывно связано с возникновением эмбрионального кроветво-
рения. В мезодерме будущего сосудистого поля появляются кро-
вяные островки, представляющие скопления клеток. Периферийная
часть этих плотно лежащих групп клеток дифференцируется в тон-
кий эпителиальный пласт, который окажется эндотелием сосудов,
а внутренние клетки, наоборот, обособляются друг от друга и
дифференцируются в кровяные клетки. Таким образом, клетки
крови оказываются внутри сосудов. Между развивающимися в раз-
ных кровяных островках сосудами устанавливается связь, разви-
вается сложная, густая, кажущаяся очень неправильной сеть сосу-
209
Рис. 95. Схема вполне сформирован-
ных амниона и аллантоиса Sauropsi-
da (из кн. П. П. Иванова):
/ — экзоцелом, 2 — зародыш, 3 — кишеч-
ник, 4 — полость амниона, 5 — эктодерма
амниона, 6 — мезодерма амниона, 7 — ме-
зодерма серозной оболочки, 8 — эктодер-
ма серозной оболочки, 9 — выстилка ал-
лантоиса, 10 — мезодерма аллантоиса,
И — желточный мешок, (жирной линией
обозначена энтодерма)
Рис. 96. Куриный зародыш, окружен-
ный сосудистым полем (из кн. Б. Й. Ба-
линского, 1961):
I — слуховой пузырек, 2 — терминальный си-
нус, 3 — «шов» вдоль линии соединения ам-
ниотических складок, 4 — глаз, 5 — сердце,
6 — край амниона, 7 — желточная артерия,
8—'Задняя амниотическая складка
дов (рис. 96). Эта сеть распро-
страняется и в area pellucida,
устанавливается связь с соб-
ственно зародышем. Из ска-
занного ясно, что одна из
функций желточного мешка
зародышей птиц — эмбрио-
нальное кроветворение. В са-
мом зародыше лишь впослед-
ствии образуются кроветвор-
ные органы — печень, селезен-
ка, костный мозг.
Связь сосудистой системы
желточного мешка с сосудис-
той системой собственно заро-
дыша устанавливается благо-
даря развитию правой и левой
желточно-брыжеечных вен, со-
единяющихся в непарный ве-
нозный проток, который, в свою
очередь, соединяется с веноз-
ным синусом развивающегося
сердца. Связь с сосудистой си-
стемой собственно зародыша
устанавливается и посредством
правой и левой желточных ар-
терий, являющихся разветвле-
ниями спинной аорты. Сердце
зародыша начинает функцио-
нировать (сокращаться) в кон-
це второго дня; с этого времени
возникает кровоток, и кровь
поступает из желточного мешка
в собственно зародыш. Как уже
говорилось, на внутренней по-
верхности стенки желточного
мешка образуются глубоко
проникающие в желток склад-
ки, в которые заходят и кро-
веносные сосуды. Клетки вну-
треннего листка желточного
мешка (клетки желточного
эпителия) выделяют фермен-
ты, помогающие изменять
желток; эти клетки всасывают
продукты расщепления желтка
и он, таким образом, становит-
210
ся достоянием развивающегося зародыша. Весь желток не исполь-
зуется в эмбриональный период; на поздних этапах развития втя-
гивающийся желточный мешок (в значительной мере уже редуци-
рующийся) еще содержит небольшое количество желтка, которое
идет на питание зародыша в течение первых четырех дней после
вылупления.
Таким образом, желточный мешок — важнейшая интегральная
часть зародыша на определенных этапах его развития. Это про-
визорный орган с мультифункциональным значением: он обеспе-
чивает переваривание и всасывание желтка, одновременно слу-
жит органом эмбрионального кроветворения и органом дыхания
эмбриона.
Морфологическое обособление собственно зародыша от «вне-
зародышевых частей» (без потери функциональной связи всех
частей развивающегося организма как целого) начинает проис-
ходить еще в начале развития желточного мешка. Зародыш на-
чинает возвышаться над поверхностью бластодиска, обособляется
головной конец, затем хвостовой и все остальные части тела.
Тело собственно зародыша птицы или рептилии морфологи-
чески обособляется от «внезародышевых образований» благодаря
образующейся туловищной подзародышевой складке, которая по-
том становится кольцевидной и отделяет все тело эмбриона. Раз-
личные участки складки появляются неодновременно: сначала
появляется часть складки непосредственно впереди головы — го-
ловная складка, так что первыми возвышаются над поверхностью
желточного мешка голова и передняя часть туловища; вскоре
образуются и боковые и задняя части складки, благодаря чему
возвышается хвостовой конец эмбриона и задняя часть его туло-
вища. Туловищная складка под эмбрионом все более сокращает-
ся, так что возникает очень узкий стебелек — упомянутый уже
пупочный шнур (канатик), посредством которого собственно за-
родыш продолжает быть связанным со своими «внезародышевы-
ми» частями. Канатик состоит из всех трех зародышевых слоев:
из наружного листка — эктодермы, из двух пластов среднего
листка — мезодермы и из внутреннего листка — энтодермы. Экто-
дермального происхождения канал пупочного канатика связывает
полость кишки зародыша с полостью желточного мешка. Здесь же
находятся, как мы видели, кровеносные сосуды и часть аллан-
тоиса.
Зародышевые оболочки. У зародышей рептилий и птиц кроме
желточного мешка образуются еще три провизорных органа, ко-
торые принято называть зародышевыми оболочками, — амнион,
сероза и аллантоис. Эти органы можно рассматривать как вы-
работавшиеся в процессе эволюции адаптации эмбрионов, кото-
рые вместе с желточным мешком «эмансипируют» зародыши реп-
тилий и птиц от превратностей разнообразных влияний внешней
среды (см. гл. XV). Эти эмбриональные адаптации возникли и
211
совершенствовались путем естественного отбора — в конечном
счете, значит, в результате влияния изменяющихся условий абио-
тической и биотической среды (см. гл. XIX).
Амнион и сероза возникают в теснейшей взаимосвязи. Их
закладка появляется в area pellucida на некотором расстоянии
от головного конца зародыша как поперечная складка, которая,
нарастая, изгибается над передним концом головы зародыша и
покрывает его, как капюшон (рис. 96,6). В дальнейшем боковые
участки амниотических складок нарастают по обеим сторонам
собственно зародыша спереди назад, все более сближаясь, и в
конце концов соединяются друг с другом и срастаются. Зародыш
все более и более оказывается заключенным в образующуюся
амниотическую полость (см. рис. 95). Эта полость сначала вы-
глядит узкой щелью между зародышем и внутренней поверхностью
амниотической складки. Однако уже вскоре образовавшаяся по-
лость наполняется и растягивается амниотической жидкостью —
продуктом жизнедеятельности клеток. Эмбрион оказывается со
всех сторон окруженным амниотической жидкостью. С «внезаро-
дышевыми частями» эмбрион теперь связан только пупочным ка-
натиком-.
Каково тканевое строение стенки амниотической полости?
Амниотические складки состоят из эктодермы и париетального
листка мезодермы. Мезодермальный клеточный материал затем
проникает в переднюю часть складки и после этого стенка ам-
ниона становится на всем протяжении одинакового гистологиче-
ского строения.
Сопряженно со стенкой амниотической полости развивается
еще одно важное провизорное образование — хорион, серозная
оболочка, или сероза (см. рис. 95). Как уже говорилось, амнион
возникает в виде складки, значит контактируются два пласта,
при этом каждый состоит из эктодермального листка и листка
париетальной мезодермы. Таким образом, следует представить
себе четырехслойное образование. Поэтому возникают сопряжен-
но две оболочки: амниотическая, состоящая из эктодермального
листка, «смотрящего» на зародыш, и мезодермального, «смотря-
щего» наружу, и серозная, которая состоит также из эктодермы,
обращенной кнаружи, и из мезодермы, обращенной внутрь. На-
ружная оболочка разрастается по всей поверхности под скорлу-
пой, над экзоцеломом. Это и есть сероза.
Амнион и сероза являются, конечно, «оболочками», так как
действительно покрывают и отъединяют собственно эмбрион от
внешней среды. Однако это — органы, части зародыша с важны-
ми функциями. Амниотическая жидкость создает водную среду
для эмбрионов животных, в ходе эволюции ставших сухопутными.
Она предохраняет развивающийся зародыш от высыхания, со-
трясений, прилипания к оболочкам яйца. Интересно отметить,
что роль амниотической жидкости у млекопитающих отметил
212
еще Леонардо да Винчи. Вероятно, амнион играет и еще какую-
либо неизвестную роль, может быть, в формообразовании. Ам-
ниотическая жидкость не является обычной водой. Ее химиче-
ский состав гораздо сложнее, и он меняется в ходе развития
зародыша. Может быть, амниотическая жидкость не безразлична
для дифференциации поверхностных слоев клеток зародыша.
Поскольку помимо яйцевых оболочек развились в эволюции
животных такие эмбриональные приспособления, сопряженно с
ними должны были появляться и другие особенности в морфо-
логии и физиологии зародышей, хотя бы уже потому, что стенка
амниона является дополнительным препятствием, помимо белко-
вой и других оболочек яйца, к поступлению кислорода из внешней
среды.
Вместе с аллантоисом, о котором речь пойдет далее, сероз*
ная оболочка принимает участие в дыхании и резорбции остатков
белковой оболочки. Загустевшие остатки белка скапливаются в
районе острого конца яйца, куда подрастают и края серозной
оболочки. Некоторые исследователи считают, что под действием
выделяемых хорионом ферментов может происходить расщепле-
ние белков; образующиеся низкомолекулярные соединения могут
поступать через серозную оболочку в кровеносные сосуды ал-
лантоиса и затем в зародыш. С 12—14-го дней развития остав-
шаяся часть белка начинает перемещаться по серозо-амниоти-
ческому каналу в полость амниона (М. Н. Рагозина, 1961). Этот
белок заглатывается зародышем, попадает в кишечник и под-
вергается перевариванию. Интересно, что кишечник развиваю-
щейся птицы начинает функционировать еще до вылупления из
оболочек. Может быть, кроме того, оказавшийся в кишечнике за-
родыша «белок» благодаря ясно выраженным антибиотическим
свойствам продолжает играть иммунологическую роль (см.
гл. XV) и влиять на процессы дифференциации стенки кишки.
Развивается и еще один провизорный орган — аллантоис, вы-
полняющий сначала функцию зародышевого мочевого пузыря (см.
рис. 95). Продукты азотистого обмена не выводятся зародышем
во внешнюю среду, а скапливаются в аллантоисе. Его происхож-
дение отлично от происхождения амниона и хориона — он по-
является как вентральный вырост задней кишки, врастающий
в кишку из области клоакальной перегородки проктодеума. В-со-
ставе аллантоиса находится эпителий кишки и висцеральный
листок мезодермы с сосудами. У куриного зародыша этот выступ
появляется уже на третий день развития. Он быстро растет и
вскоре проникает во внезародышевый целом, в пространство
между желточным мешком, амнионом и хорионом. Аллантоис
продолжает быть связанным с собственно зародышем посред-
ством узкого стебелька. В середине эмбрионального развития птиц
и рептилий аллантоис разрастается под хорионом по всей по-
верхности зародыша с желточным мешком.
213
Прилегающий к хориону висцеральный листок мезодермы
аллантоиса срастается с его париетальным листком. Образуется
хорио-аллантоис (см. рис. 95). Теперь аллантоис благодаря выхо-
ду сосудов на поверхность может выполнять и функцию снабже-
ния зародыша кислородом.
В конце эмбрионального развития птиц и рептилий провизор-
ные органы зародыша постепенно прекращают свои функции, ре-
дуцируются, зародыш начинает дышать воздухом, имеющимся
внутри яйца (в воздушной камере), пробивает скорлупу, осво-
бождается от яйцевых оболочек и оказывается во внешней среде.
Все сообщенное о «внезародышевых» образованиях относится
не только к птицам, но и к рептилиям. Однако в развитии неко-
торых рептилий есть ряд особенностей, из которых укажем сле-
дующие. У ящериц и змей нет белковой оболочки, яйцеклетка
(«желток») контактируется непосредственно с подскорлуповой
оболочкой. В процессе обособления зародыша от стенки желточ-
ного мешка он не поднимается над поверхностью бластодиска
(ему и некуда подниматься, так как он находится вблизи скор-
лупы), а, наоборот, погружается в желток. Хорион и амнион
вначале состоят только из эктодермальных пластов клеток, позд-
нее между ними врастает мезодерма. Функциональное значение
всех «внезародышевых» органов у рептилий и птиц одинаково.
Глава X
МЕТАМОРФОЗ
ОБЩИЕ СВЕДЕНИЯ О ПОСТЭМБРИОНАЛЬНОМ РАЗВИТИИ
И МЕТАМОРФОЗЕ
После выхода молодого животного из яйцевых оболочек или из те-
ла матери начинается постэмбриональный период развития. Раз-
витие может быть прямым или сопровождаться превращением
(метаморфозом). При прямом развитии новорожденное животное
уже обладает всеми основными чертами организации взрослого
и отличается главным образом меньшими размерами и не вполне
развитыми половыми железами, так что постэмбриональное раз-
витие сводится в основном к росту и достижению половозрелости.
Прямое развитие наблюдается, например, у пресноводной гидры,
многих нематод, головоногих моллюсков и у большинства позво-
ночных.
При развитии с метаморфозом из яйца выходит личинка, обыч-
но в то же время имеющая специальные личиночные органы, от-
сутствующие во взрослом состоянии. П. П. Иванов характеризу-
ет личинку как организм, вынужденный на ранних стадиях разви-
тия приспособиться к условиям свободного существования путем
ранней дифференцировки необходимых для этого специально личи-
ночных органов. Поэтому в процессе последующего развития на-
ряду с прогрессивным развитием одних органов происходит ре-
дукция других. Если эти перестройки происходят постепенно
(например, .у кольчатых червей или ракообразных), метаморфоз
называют эволютивным, если же они совершаются за сравнитель-
но короткий срок — катастрофическим. Иногда дегенеративные
процессы особенно велики и преобладают над прогрессивными
(у асцидий, паразитического ракообразного саккулины), такой
метаморфоз называется некробиотическим. Он сопровождается
массовой дегенерацией личиночных структур и паузой в двига-
тельной активности животного.
Как правило, метаморфоз связан с переменой образа жизни
(переход от планктонного образа жизни к сидячему у губок, ки-
шечнополостных, мшанок, асцидий или от свободного существо-
вания к паразитизму у саккулины) или со сменой среды обитания
(переход из воды на сушу у амфибий). Эта смена экологических
отношений в некоторых случаях несомненно отражает события,
имевшие место в филогенезе (например, у паразитических раков,
асцидий, амфибий), но в то же время имеет большое биологиче-
ское значение, так как планктонные личинки сидячих животных
215
и свободноживущие у паразитических играют роль расселительной
стадии. Поэтому в организации личинок (как и на всякой другой
стадии развития) могут быть черты как палингенетического, так
и ценогенетического характера (см. гл. XVII). Так, например,
характерная для личинок иглокожих билатеральная симметрия и
наличие у них трех пар целомических мешков не имеют приспо-
собительного значения и могут расцениваться только как свиде-
тельство происхождения этого типа от билатеральносимметричных
трехсегментных животных. А развитие длинного ресничного шну-
ра, образующего многочисленные петли и изгибы у бипиннарий
морских звезд, а также аурикулярий голотурий и длинных рук,
поддерживаемых скелетными иглами, у личинок морских ежей и
офиур — несомненно являются вторично возникшими (в связи
с планктонным образом жизни) структурами.
Развитие с метаморфозом широко распространено в животном
мире. Поскольку такие примитивные животные, как губки и гидро-
идные полипы, развиваются с метаморфозом, существует мнение,
что метаморфоз является изначальной особенностью онтогенеза
Metazoa (И. И. Ежиков, 1939; А. А. Захваткин, 1949). В процессе
эволюции многие животные перешли к прямому развитию. Этот
процесс «выпрямления» развития можно шаг за шагом проследить
в таких группах животных, как гидроиды, брюхоногие моллюски,
ракообразные и др. Он осуществляется таким образом: в яйцах
начинает накапливаться больше желтка, что дает возможность
животному больший отрезок своего онтогенеза пройти под покро-
вом яйцевых оболочек. В результате личинка «эмбрионизируется»,
т. е. из активной свободноживущей стадии превращается в одну
из стадий эмбрионального развития. Все специально личиночные
структуры, обеспечивавшие раньше питание личинки, ее движение
и другие процессы, теперь утрачивают свое значение и постепен-
но устраняются из развития. Весь ход развития упрощается, так
как из него выпадают все процессы перестройки, связанные с ме-
таморфозом. Из яйца выходит маленькое животное, похожее на
взрослое. Но известны и такие случаи, когда метаморфоз возник
вторично. Так, общепризнано, что полное превращение насекомых
имеет вторичное происхождение. Согласно известной теории Бер-
лезе — Ежикова, оно возникло в связи с уменьшением количества
желтка в яйцах. Впрочем, по этому частному вопросу высказыва-
лись и другие точки зрения.
МЕТАМОРФОЗ ГИДРОИДНЫХ полипов
Наиболее простые формы имеет метаморфоз у гидроидных поли-
пов. Яйца гидроидов лишены оболочек, поэтому началом постэм-
брионального развития условно считается образование целоблас-
тулы, клетки которой несут жгутики (рис. 97,А). Такая бластуло-
образная личинка плавает, но не питается. Ее строение постепенно
усложняется. Путем миграции части клеток внутрь бластоцель
216
Рис. 97. Метаморфоз Hydrozoa (по И. И. Мечникову, 1886):
А — бластула Tiara leycostyla; Б, В, Г — образование паренхимулы у
Clytia flavidula\ Б, В — однополюсная иммиграция; Г — паренхимула;
Д— прикрепление планулы Clytia к субстрату; Е, Ж — образование гид-
ранта Clytia
Рис. 98. Превращение плаиулы S/o-
motheca в сидячую полипоидную ста-
дию (по С. Риттеихаузу, 1910):
/ — планула, прикрепившаяся плашмя к суб-
страту, 2 — появление выроста на ее верхней
стороне, 3—8 — последующие стадии оформле-
ния полипа — основателя колонии
заполняется аморфной массой клеток энтодермы. Эта личинка
называется паренхимулой (рис. 97,5, В, Г). Затем клетки энтодер-
мы расходятся (или частично дегенерируют), так что возникает
центральная полость (гастроцель), и эпителизируются; эта стадия
называется планулой. На стадии планулы уже различаются нерв-
ные, интерстициальные, иногда железистые и стрекательные клет-
ки. Затем планула прикрепляется передним концом, утрачивает
жгутики и распластывается по субстрату в форме лепешечки, от
которой кверху вырастает стебелек (рис. 97,Д, Е). На конце сте-
белька образуется вздутие, которое преобразуется в гидрант: на
его вершине прорывается ротовое отверстие, вокруг которого
формируется венчик щупалец (рис. 97,Ж)- Молодой полип начи-
нает питаться, растет и дает начало колонии.-
При метаморфозе гидроидных полипов дегенеративные явления
незначительны (исчезновение жгутиков), и постепенное усложне-
ние организации происходит непрерывно. Однако в физиологиче-
ском смысле кульминационным пунктом метаморфоза является
момент прикрепления планулы. В это время животное переходит
от свободноплавающего образа жизни к сидячему, начинается
активное питание и происходит смена физиологической полярно-
сти: паренхимула и планула плавают одним концом вперед,
а после прикрепления доминирующим полюсом становится про-
тивоположный (вегетативный) полюс, на котором образуется рот.
Иногда личинки прикрепляются боком (Microcoma, Stomotheca)
и сперва образуют разветвленный зачаток гидроризы, от которой
позднее начинают расти кверху гидранты (рис. 98). В этом слу-
чае ось окончательной полярности полипа располагается под пря-
мым углом к первоначальной продольной оси личинки. А. А. За-
хваткин (1949) предполагает, что метаморфоз гидроидов сопро-
вождается временной деполяризацией организма.
Как уже отмечалось, у многих гидроидов наблюдается тенден-
ция к сокращению метаморфоза, проявляющаяся в том, что ста-
дии бластулы, паренхимулы, а иногда и планулы превращаются
в пассивные эмбриональные стадии.
МЕТАМОРФОЗ ИГЛОКОЖИХ
У многих иглокожих (морских звезд и морских ежей) постэмбри-
ональное развитие тоже начинается со стадии жгутиконосной
бластулы. Только после выхода зародыша из яйцевых оболочек
218
Рис. 99. Метаморфоз Asterias vulgaris (по Е. Мак Брайду, 1914):
А — образование целома; Б — диплеврула; В, Г — бипиннария около трехнедельного воз-
раста. Вид с оральной (В) и аборальной (Г) сторон; 1 — мезеихнма, 2 — обособляющийся
целом, 3 — ресничный шнур, 4 — стомодеум, 5—пищевод, 6 — желудок, 7 — посторальиый
ресничный шнур, 8 — преоральный ресничный шнур, 9 — рот, 10 — целом, 11 — задняя киш-
ка, 12 — анальное отверстие
происходит гаструляция, причем образовавшийся путем впячива-
ния первичный кишечник подразделяется поперечной перетяжкой
на более глубокую часть, зачаток целомической мезодермы, и
эктодермальный зачаток кишки (рис. 99,А). Одновременно путем
выселения отдельных клеток из вегетативной части бластулы или
из стенок первичного кишечника происходит обособление мезен-
химы. Бластопор превращается в анальное отверстие, а на одной
из сторон личинки (будущей брюшной), ближе к переднему кон-
цу, прорывается рот. С этого момента личинка начинает самосто-
ятельно питаться. Затем образуется околоротовая впадина, по
краю которой проходит кайма особенно длинных жгутиков — рес-
ничный шнур. Такая личинка называется диплеврулой (рис. 99,5).
Дальнейший рост личинки сопровождается образованием ло-
219
пастей и выростов, на которые переходит и ресничный шнур. Для
каждого класса иглокожих характерна своя форма личинки; би-
пиннария — для морских звезд (рис. 99,В, Е), аурикулярия— для
голотурий, плутеусы — для морских ежей и офиур и своеобразная
боченковидная личинка — для морских лилий. Однако, несмотря
на разнообразие и сложную форму, все личинки характеризуются
двусторонней симметрией, которая распространяется н на внутрен-
нюю организацию. Непарный целомический мешочек успевает
к этому времени разделиться на три пары целомов, лежащих по
бокам от кишечника. Впрочем, целомы правой стороны могут
сильно запаздывать в развитии, поскольку в формировании взрос-
лого иглокожего главную роль играют органы левой стороны и,
в частности, левый средний целом — гидроцель.
Переход от билатеральной трехсегментной организации личин-
ки к радиальной симметрии взрослого животного происходит так.
Гидроцель окружает пищевод и замыкается в кольцо — зачаток
околоротового кольца амбулакральной системы. На его поверх-
ности появляется пять выпячиваний — зачатки радиальных кана-
лов, по ходу которых начинается формирование амбулакральных
ножек. Радиальную симметрию принимают нервная и кровеносная
системы. У морских ежей метаморфоз усложнен тем, что все фор-
мообразовательные процессы сосредоточиваются на дне особой
амниотической полости, образующейся путем впячивания кожных
покровов личинки в области гидроцеля.
Из изложенного видно, что при метаморфозе иглокожих ради-
альная симметрия проявляется впервые в строении гидроцеля.
Гидроцель играет важную роль во взаимодействии зачатков, на-
поминающую роль «организатора» (см. гл. XI). В связи с этим
представляет интерес тот факт, что у морских ежей иногда (как
аномалия) встречаются плутеусы с одинаково хорошо развитыми
целомами слева и справа; тогда образуются два гидроцеля, две
амниотические полости и метаморфоз приводит к образованию
уродливых ежей с удвоенными органами оральной стороны.
МЕТАМОРФОЗ АСЦИДИЙ
Личинки асцидий по внешнему виду напоминают головастиков:
они состоят из вздутого переднего конца и узкого хвоста, благо-
даря колебаниям которого личинки плавают (рис. 100,А). В теле
личинки резко различаются две группы органов или зачатков:
чисто личиночные органы и органы дефинитивные. К числу первых
относятся: хорда, нервная система, состоящая из нервной трубки
и переднего ганглиозного утолщения, содержащего глазок и ста-
толит, и две расположенные по бокам от хорды мышечные ленты.
К личиночным органам относится также находящийся на перед-
зем конце прикрепительный аппарат, состоящий из чувствитель-
зых и железистых органов.
!20
10
Рис. 100. Метаморфоз асцидии Clone intestinalis (из ки. Е. Коршельта, 1936):
1— прикрепительным орган, 2 — эндостиль, 3 — статолит, 4 — глаз, 5 —глотка, 6 — хорда,
7 — головной мозг, 8 — нервная трубка, 9 — хвост, 10— оральный сифон, 11 — ганглий, 12 —
атриальный сифон, 13 — жаберный отдел, 14 — пищевод, 15 — желудок, 16 — кишка
Дефинитивными частями личинки являются зачатки дыхатель-
ного аппарата, кишечника, кожных покровов, сердца и других ор-
ганов. Личиночные органы лежат преимущественно в хвосте,
а дефинитивные целиком сосредоточены в туловищном отделе. Так
как личиночные органы начинают функционировать сразу после
рождения личинки, то к этому времени их развитие уже заверше-
но. Дефинитивные органы, наоборот, начинают функционировать
только после метаморфоза; поэтому у личинки они находятся
в более или менее недоразвитом состоянии. Личинки асцидий не
способны воспринимать пищу извне и имеют в основном рассели-
тельное значение.
Период плавания личинки обычно не превышает 2—3 дней.
Часто уже через несколько часов личинка прикрепляется к суб-
страту передним концом, хвост ее втягивается в туловище, хорда,
мышцы и нервная система распадаются на отдельные клетки и
флагоцитируются. От нервной системы личинки остается лишь не-
большая группа клеток, дающая начало дефинитивному нервному
ганглию. Комплекс внутренних органов будущей асцидии изменя-
ет свое положение в пространстве: на стадии личинки ротовой и
клоакальный сифоны расположены на спинной стороне, т. е. об-
ращены кверху, а в первый момент после прикрепления они ока-
221
зываются обращенными в сторону (рис. 100,5). За1ем в течение
2—3 дней весь комплекс внутренних органов поворачивается так,
что сифоны снова обращены вверх. Дефинитивные органы завер-
шают свое развитие и начинают функционировать иногда в первые
же часы метаморфоза, а иногда только через несколько недель.
Это зависит от того, насколько хорошо они были развиты у ли-
чинки.
В метаморфозе асцидий обращает внимание резкое разграни-
чение и относительная независимость личиночных и дефинитивных
органов. Развитие дефинитивных органов происходит равномерно,
и наступление метаморфоза не вызывает резкого его ускорения.
Личиночные же органы подвергаются уничтожению сразу после
прикрепления личинки. Поэтому метаморфоз асцидий можно на-
звать катастрофическим и некробиотическим. При этом надо от-
метить, что вместе с хвостом личинки исчезают и наиболее харак-
терные черты организации хордовых. Таким образом, основное
содержание метаморфоза асцидий состоит в переходе к сидячему
образу жизни и связанным с ним упрощением организации.
МЕТАМОРФОЗ НАСЕКОМЫХ
У насекомых рост и изменение формы тела при постэмбриональ-
ном развитии сопряжены с периодическими линьками. Типы пост-
эмбрионального развития высших насекомых (Pterygota) разно-
образны, но все же можно выделить два основных, которые при-
нято называть неполным и полным превращением.
У насекомых с неполным превращением (Hemimetabola) из
яйца выходит молодое животное, очень похожее на взрослое на-
секомое (имаго), но отличающееся размерами, относительно
крупной головой и сильно недоразвитыми крыльями (рис. 101,А).
Однако с каждой линькой размеры и пропорции тела все больше
приближаются к окончательным. Последняя линька отличается от
предыдущих более значительными изменениями — только после
нее крылья и гонады завершают свое развитие и начинают функ-
ционировать. В этот период развитие происходит постепенно, зна-
чительных дегенеративных явлений не наблюдается, и весь про-
цесс можно назвать метаморфозом лишь с некоторой натяжкой.
Но иногда у Hemimetabola незрелые стадии живут в иной среде
обитания, чем имаго, или отличаются образом жизни. Тогда могут
появиться специальные личиночные органы. Например, живущие
в воде личинки стрекоз имеют органы водного дыхания, а нижняя
губа превращена у них в своеобразный хватательный орган —
маску. Смена среды обитания и редукция личиночных органов
тоже обычно бывают приурочены к последней линьке. В этом
случае термин «метаморфоз» применяется с большим основанием.
Более глубокие изменения происходят при метаморфозе у на-
секомых с полным превращением (Holometabola). Здесь из яйца
222
л
Рис. 101. Метаморфоз насекомых (из Б. Н. Шванвича, 1949).
Постэмбриональное развитие таракана (яйцо, шесть возрастов личинки и взрослые насеко-
мые) (Д) и сосновой совки (пять возрастов гусеницы, куколка и взрослое насекомое) (Б)
выходит личинка, совершенно не похожая на имаго. Гусеницы
бабочек, например, отличаются гомономной сегментацией тулови-
ща (рис. 101,5), ротовым аппаратом примитивного грызущего типа
(в то время как у взрослых бабочек ротовые части образуют
высоко специализированный сосущий хоботок), отсутствием слож-
ных глаз, не вполне развитыми грудными конечностями и наличи-
ем ложных ножек на брюшных сегментах. Еще больше отличают-
ся от имаго личинки мух — они имеют червеобразную форму,
голова у них сильно недоразвита и втянута внутрь, а конечности
вообще отсутствуют. Наружных зачатков крыльев у личинок
Holometabola никогда не бывает.
Личинки Holometabola проделывают ряд линек, значительно
увеличиваются в размерах, но по форме меняются мало. Затем
происходит линька, приводящая к наступлению стадии куколки.
Последняя очень похожа на имаго формой тела и строением его
придатков. У куколки различаются зачатки крыльев, но они еще
не расправлены и плотно прижаты к телу (см. рис. 101,5). Ха-
рактерная черта куколки — ее пассивность: она не питается и поч-
ти неподвижна, но внутри нее в это время происходят интенсивные
формообразовательные процессы разрушения личиночных органов
и тканей и замены их имагинальными. По окончании этих пере-
строек происходит последняя линька, и вылетает имаго.
Внутренние процессы при метаморфозе насекомых впервые
были описаны в ставших классическими исследованиях А. О. Ко-
223
Рис. 102. Развитие има-
гинальных дисков у мух
Muscidae (из кн.
Б. Н. Шванвича, 1949):
А — личинка; Б—Г — кукол-
ка; / — атрнум, 2—4 — за-
чатки грудных ног, 5 —
глотка, 6 — зачаткн аитеин,
7 — глаз, 8 — пищевод, 9 —
головной мозг. 10 — зачаток
хоботка, I, 11, 111 — груд-
ные сегменты
валевского (1887). Он показал, что происходящие при линьке на
куколку и кажущиеся внезапными изменения общей формы тела
и появление зачатков крыльев, ног, яйцеклада и других органов
на самом деле подготавливаются еще у личинки. Наружные при-
датки тела, отсутствующие у взрослой мухи, закладываются у ли-
чинки в форме дисковидных утолщений гиподермы, состоящих из
мелких клеток. Эти утолщения, названные имагинальными диска-
ми, растут благодаря размножению составляющих их клеток, но
при этом погружаются внутрь и не видны снаружи (рис. 102).
Только во время линьки на куколку имагинальные диски «выво-
рачиваются»— выходят на поверхность и принимают форму тех
органов, зачатками которых являются. Остальная гиподерма ли-
чинки может превратиться в кожные покровы имаго, подвергшись
большим или меньшим гистологическим преобразованиям, но у мух
личиночная гиподерма полностью дегенерирует и замещается но-
вой, образующейся за счет специальных имагинальных дисков.
Подобные же процессы дегенерации личиночных тканей и
замещения их имагинальными происходят и во внутренних орга-
нах. Источником образования передней и задней кишки взрослой
мухи служат скопления имагинальных клеток, имеющие форму
колец. В средней кишке личинки мелкие имагинальные клетки
рассеяны диффузно между крупными личиночными клетками. Раз-
множаясь, они образуют небольшие пла-
стинки, которые разрастаются поверх ли-
чиночной кишки и, сливаясь, образуют
новый эпителий кишки, а личиночные
клетки разъединяются и оказываются
лежащими внутри имагннальной кишки,
где и перевариваются.
Аналогичные процессы происходят
в мальпигиевых сосудах, жировом теле,
мускулатуре и т. д. Таким образом, во
время метаморфоза весь организм насе-
комого в большей или меньшей степени
обновляется. Обычно при этом меняются
образ жизни (появляется способность
к полету), характер питания, поведение,
инстинкты. Разрушение личиночных ор-
ганов (гистолиз) часто совершается при
участии фагоцитов (см. гл. XV).
Большой интерес представляет физио-
логический механизм метаморфоза насе-
комых. Раньше полагали, что причиной
разрушения личиночных органов являет-
ся старение, снашивание их клеток. Дру-
гие авторы были склонны сильно пре-
увеличивать роль фагоцитов, которые
224
нападают якобы на здоровые жизнеспособные ткани личинки и
уничтожают их. В настоящее время имеется огромное количество
экспериментальных данных, которые показывают, что все пост-
эмбриональное развитие насекомых находится под контролем гор-
мональных факторов.
Наиболее важными инкреторными структурами насекомых яв-
ляются нейросекреторные элементы мозга, торакальные железы и
прилежащие тела (corpora allata), расположенные в голове. Функ-
циональные отношения между этими органами схематически мож-
но представить следующим образом. Нейросекреторные клетки
выделяют вещества, стимулирующие деятельность торакальных
желез. Последние выделяют гормон линьки. Если удалить тора-
кальные железы, насекомое утрачивает способность линять и, сле-
довательно, дальнейший рост и развитие становятся невозможны-
ми. После реимплантации торакальных желез линьки возобновля-
ются. Торакальные железы достигают максимального развития
накануне метаморфоза, а у взрослого насекомого дегенерируют,
в результате чего имаго больше не линяет.
Секрет прилежащих тел получил название ювенильного гормо-
на. Пока в крови содержится достаточно ювенильного гормона,
линьки не сопровождаются значительным изменением формы и
имеют личиночный характер. Накануне метаморфоза активность
прилежащих тел падает, содержание ювенильного гормона в крови
резко снижается и становится возможным превращение. Это до-
казывается опытами с клопом Rhodnius: удаление прилежащих
тел у молодой личинки вызывает преждевременный метаморфоз и
образование карликового взрослого клопа. Наоборот, трансплан-
тация прилежащих тел от молодой личинки к личинке последнего
возраста, которая должна превратиться в имаго, делает эту по-
следнюю линьку личиночной и приводит к появлению лишней
личиночной стадии.
МЕТАМОРФОЗ У НИЗШИХ позвоночных
Личинки круглоротых, ганоидных, двоякодышащих и костистых
рыб, а также амфибий в большей или меньшей степени отличаются
от взрослых форм, и у них в ходе роста и развития происходят очень
существенные преобразования. Так, у ганоидных и двоякодыша-
щих рыб образуется своеобразная личинка, характеризующаяся
присутствием наружных жабр. У миноги освободившаяся от яйцевых
оболочек маленькая личинка — пескоройка отличается от взросло-
го организма. Стомодеум у нее еще не сообщается с кишечной
трубкой, и личинка до прорыва дна стомодеума в кишку питается
за счет остатков желтка. К этому времени устанавливается нор-
мальное жаберное дыхание: жаберные щели становятся сквозны-
ми отверстиями глотки (наружу), образуются жаберные складки
на стенках мешковидного вздутия эктодермального участка жа-
берной щели (канала). Глаза у пескоройки маленькие, недораз-
225
витые, они скрыты под ко-
жей. В течение личиночного
состояния, продолжающего-
ся 3—4 года, пескоройка
сильно вырастает и в ее ор-
ганизации происходят и
другие преобразования.
Пронефрос сменяется мезо-
нефросом. Первичная почка
выполняет главную роль в
функции выделения как
личинок, так и взрослой
миноги.
При метаморфозе зача-
ток щитовидной железы
становится функционирую-
щим органом с внутренней
секрецией. Эта железа обо-
собляется от кишечника, ут-
рачивается сообщение с
глоткой, продуцируемый ею
гормон начинает попадать
в кровь. Преобразуется пе-
редний отдел тела песко-
ройки: увеличиваются гл-аза,
дифференцируется хряще-
вая ткань частей черепа,
разрастается ротовая часть
головы, внутри рта появля-
ются новые кожные сосоч-
ки, превращающиеся в рого-
вые зубы миноги. Одновре-
менно разрастается гипо-
физ. Большие изменения
происходят в переднем от-
деле кишечника, он прини-
мает дефинитивное, очень
характерное для миноги
Рис. 103. Метаморфоз бесхвостых (Rana pipiens) (/) н хвостатых (Pleurodeles
Waltlii) (II) амфибий:
/ — А — головастик перед метаморфозом; Б — начальная фаза метаморфоза головастика
(рост задних конечностей); В— прорезывание передних конечностей, уменьшение хвостово-
го плавника; Г, Д -— заключительные стадии метаморфоза, постепенное появление органи-
зации лягушки (по Е. Битчи, 1956); II — А — личиночная стадия в завершенной форме;
Б — Г — постепенное исчезновение наружных жабр, изменение хвостового плавника (по
Л. Галльену и М. Дюроше, 1957)
226
строение: образуются два лежащих друг над другом отдела —
трубчатый пищевод и жаберный отдел, причем оба они открыва-
ются в ротовую полость. При метаморфозе сплошные спинной и
брюшной плавники дифференцируются в спинной, хвостовой и
брюшной. Желчный пузырь и его проток исчезают.
Очень тщательно изучен метаморфоз у амфибий — как проте-
кание морфологических и физиологических преобразований, так и
причины их. Процессы метаморфоза у разных групп амфибий име-
ют особенности, что отражает своебразия их онтогенеза, возник-
шие в эволюции. У личинок бесхвостых амфибий морфофизиологи-
ческие перестройки при метаморфозе более интенсивные; они со-
ответствуют глубоким изменениям в связи с переходом от водной
жизни животного к наземной.
-При метаморфозе головастика в лягушку все системы органов
подвергаются той или иной перестройке. Происходящие внешние
изменения личинки общеизвестны (рис. 103). Длинные, развет-
вленные наружные жабры закрываются развивающимися жабер-
ными крышками, они атрофируются, и головастик начинает ды-
шать внутренними жабрами. Ко второй половине личиночной жиз-
ни уже функционируют легкие. После образования жаберной
крышки у основания хвоста головастика появляются зачатки зад-
них конечностей, а затем позади задней пары жаберных дуг воз-
никают зачатки передних конечностей. До метаморфоза они оста-
ются закрытыми жаберной крышкой.
При метаморфозе происходит развитие легких, а вместе с ни-
ми— легочных артерий и вен. Затем атрофируются внутренние
жабры, исчезает жаберная крышка. Открываются и быстро раз-
виваются передние конечности, ускоряется рост и задних конеч-
ностей. Вследствие изменения в притоке крови к разным частям
тела, особенно вследствие усиленного роста задних конечностей
(в связи с чем кровь в задней части тела направляется главным
образом в бедренную аорту и по бедренным венам возвращается
в нижнюю полую вену), происходит отмирание клеток хвостового
плавника, хвостовых мышц и хорды, атрофия личиночного хвоста.
При этом разыгрываются фагоцитарные и воспалительные про-
цессы (см. гл. XV).
Большие преобразования совершаются в кишечнике, он укора-
чивается, происходят гистогенетические преобразования. Реконст-
руируется голова, формируется среднее ухо, развиваются слезные
железы век. Изменяется строение челюстей и черепа, отпадают
клюв и роговые зубы, сильно вырастают глаза.
Скелет подвергается также большим, но более медленным пре.-
образованиям — происходит окостенение хрящевых частей и обра-
зование накладных костей. Большие изменения наблюдаются
в мочеполовом аппарате. Происходят структурные изменения
кожи.
Среди хвостатых амфибий особый интерес представляет аксо-
227
лотль, являющийся личиночной стадией амбистомы (мексиканской
хвостатой амфибии). Аксолотль, сохраняя личиночную организа-
цию, приобретает способность к размножению. Это явление полу-
чило название неотении.
У подвергающихся метаморфозу хвостатых амфибий перестрой-
ки тканей и органов менее резкие. Однако регрессивные явления
имеют место. Так происходят атрофия наружных жабр, резорб-
ция части хвостового плавника, изменения в головной области,
особенно в висцеральном скелете, в структуре кожи.
Вопросы о механизмах регуляции процессов метаморфоза здесь
не рассматриваются. Они крайне сложны и своеобразны у разных
животных.
Путем многочисленных и разнообразных опытов доказано, что.
метаморфоз амфибии связан с деятельностью щитовидной железы.
Этот процесс возможен только при условии функциональной зре-
лости всей системы: гипоталамус — гипофиз — щитовидная железа.
В какой-то момент преметаморфоза (когда уровни тиреоидного
и тиреотропного гормонов очень низкие) гипоталамус становится
чувствительным к тиреоидному гормону и стимулирует освобож-
дение тиреотропного гормона, который повышает уровень тиреоид-
ного гормона, обеспечивая метаморфоз (М. С. Мицкевич, 1978).
Таким образом, метаморфоз всегда связан с изменениями
в образе жизни животного и соответственно имеет приспособи-
тельное значение. В то же время в его осуществлении участвуют
очень сложные внутренние механизмы.
Глава XI
ДЕТЕРМИНАЦИЯ ЗАЧАТКОВ ОРГАНОВ
И ДИФФЕРЕНЦИАЦИЯ КЛЕТОК И ТКАНЕЙ
ПРОБЛЕМА ДЕТЕРМИНАЦИИ И ДИФФЕРЕНЦИАЦИИ
Одни из важнейших понятий в эмбриологии — понятия дифферен-
циации и детерминации, отражающие основные явления преемст-
венности, последовательности процессов развития организма. В он-
тогенезе непрерывно происходят процессы дифференциации, т. е.
появляются новые и новые различия между бластомерами, разны-
ми участками зародыша, клетками и тканями, возникают разные
органы. По сравнению с исходной в развитии яйцевой клеткой ор-
ганизм кажется необычайно сложным. Дифференциация — это
такое структурное, биохимическое или иное изменение развиваю-
щегося организма, при котором относительно однородное превра-
щается во все более различное, касается ли это клеток (цитоло-
гическая дифференциация), тканей (гистологическая дифференци-
ация) или органов и организма в целом, идет ли речь
о морфологических или физиологических изменениях. Если стоять
на позициях более широких определений сущности дифференциа-
ции, то нет оснований для сужения понятия дифференциации
только до клеточного уровня. Дифференциация — это и возникно-
вение новых ферментных систем, специфических белков, новых
свойств отдельных локусов мембран, изменения различных физи-
ческих параметров, появление новых структурных элементов и но-
вых внутренних сред, разнообразных неклеточных структур. В по-
нятие «дифференциация» должны быть включены субмикроскопи-
ческие изменения в клетках, связанные с их поведением как ин-
тегральных частей тканей.
Вопрос о последовательности становления в онтогенезе разли-
чий, относящихся к субмолекулярному или надмолекулярному
уровням, очень сложный и дискутируется разными школами со-
временной эмбриологии. Успехи биохимии и морфологии индиви-
дуального развития дают основание утверждать, что такие явле-
ния, как внешняя форма и симметрия какой-либо биологической
дискретности (мембраны, митохондрии, клетки и т. п.), неотдели-
мы от особенностей и типа связей составляющих их компонентов
(молекул, молекулярных ассоциаций и т. п.); изменчивость тех и
других тесно коррелирована. Можно говорить о различных про-
явлениях единого процесса дифференциации, затрагивающего мно-
гие структурные уровни зародыша — развивающейся целостной
системы.
229
Основная задача описательной и экспериментальной эмбрио-
логии и состоит в изучении преемственности процессов дифферен-
циации. При выяснении причинного механизма тех или иных диф-
ференцировок употребляется термин «детерминация». Многие
эмбриологи удовлетворялись представлением о детерминации как
о совокупности факторов, определяющих формообразовательные
процессы. Понятие детерминации возникло не в связи с вейсма-
новским понятием «детерминанты» (см. гл. II и XII), хотя не-
которые авторы (В. Ру) детерминирующими факторами считали
«наследственную субстанцию».
Взгляды некоторых современных авторов близки к этой мысли,
поскольку под детерминацией понимают предопределенность и
необратимость тех или иных процессов дифференциации. Используя
вейсмановские идеи, такое представление о детерминации можно
характеризовать утверждением: раз клетка имеет определенные
детерминанты и она лишилась других, значит, судьба ее пред-
определена.
Многие ученые понимали под детерминацией ограничение или
утрату тех или иных потенций, благодаря чему могут осущест-
вляться остающиеся потенции, приобретается возможность само-
дифференцировки. Если приобретается способность какой-либо ча-
сти к самодифференцировке (даже и при изменении условий разви-
тия), то говорят, что эта часть детерминирована.
В связи с учениями о влиянии одних частей зародыша на дру-
гие и вместе с обнаружением явлений лабильной детерминации
(о чем м,ы будем говорить впоследствии) в понятие детерминации
разные исследователи стали вносить разное содержание в зависи-
мости от того, как они интерпретируют факты о влиянии «частей»
зародыша друг на друга.
Г. Шпеман (1936) считал, что какая-либо часть зародыша ста-
новится детерминированной с того момента, когда она может раз-
виваться путем самодифференцировки.
В связи с обнаружением факта возможности передифференциро-
вок и с наличием этапов в развитии зародыша и органов «нестой-
кого определения судьбы» многие исследователи предпочитали вы-
брасывать понятие «стойкости», «определенности судьбы» из содер-
жания термина «детерминация». Так, Д. П. Филатов (1934) под
детерминацией понимал такое воздействие одной части развиваю-
щегося организма на другие части, благодаря которому эти послед-
ние при наличии определенных условий проходят часть пути своего
развития. Близкое этому определение дал в 1959 г. Б. П. Токин, го-
воривший о детерминации как об установлении в ходе онтогенеза
организма, развивающегося в данных конкретных условиях, таких
взаимосвязей между частями зародыша, при которых клеточные
комплексы проходят определенный путь развития.
Согласно определению Б. И. Балинского, данному в 1940 г., де-
терминация — это устойчивость начавшихся процессов дифферен-
та
циации, их тенденция развиваться в раз намеченном направлении,
несмотря на изменение условий.
Как увидим далее, это определение относится далеко не ко всем
случаям детерминации и дифференциации, и следует предпочитать
определения более общего характера. Мы убедимся в существова-
нии многочисленных явлений, при которых развитие органов и це-
лых организмов возможно путем утраты специфических структур
тканей и клеток, и в том, что новым процессам дифференциации
не предшествует то, что разные авторы называют детерминацией.
Это тем более заставляет придерживаться более общего характе-
ра определения терминов «детерминация» и «дифференциация».
Г. П. Короткова (1979) подчеркнула, что появление различий —
нечто всегда конкретное: это либо различия, видимые в микроскоп,
либо выявляемые биохимическими или иными методиками, но они
касаются не всех компонентов клетки, а определенных ее частей
(ядра, ядрышка, митохондрий и т. п.). В любом организме или
клетке существует большое число гомологичных единиц, подлежа-
щих сравнению.
Надо рассматривать дифференциацию как процесс появления
пространственных—временных различий в совокупности гомологич-
ных «дискретных» единиц, входящих в состав одной и той же био-
логической системы. С точки зрения Г. П. Коротковой, понятие
«дифференциация» должно отражать многообразие преобразова-
ний состояния дифференцированности разных типов гомологичных
дискретных единиц. Нельзя понимать осуществление этого процес-
са как однонаправленную линейную изменчивость состояния гете-
рогенности различных гомологичных компонентов клетки или орга-
низма только в сторону ее повышения или понижения. В онтогене-
тическом цикле имеет место непрерывный процесс сложных и мно-
жественных изменений и передифференцировки разных типов дис-
кретных единиц (органоидов, клеток, тканей, органов, метамеров
и др.). Мы не можем говорить об уровне или степени дифферен-
циации клетки или организма вообще, а лишь применительно
к строго определенным ее элементам, связям или показателям.
Такое представление о дифференциации предостерегает от из-
лишне прямолинейных, небиологичных высказываний.
В XX в. выдающуюся роль в разработке проблемы детермина-
ции и дифференциации сыграла школа немецкого ученого Г. Шпе-
мана. Им обнаружено явление «организационных центров» у заро-
дышей животных, осуществлен экспериментальный анализ разви-
тия нервной системы и органов чувств, разработаны новые микро-
хирургические методики, в особенности способы трансплантации
частей одного зародыша другому, что связано со специальной тех-
никой снятия оболочек яиц и приготовлением специальных сред.
Говорят об аутотрансплантации (аутопластике) в случае пересад-
ки частей на другие м,еста того же самого зародыша. Но на 'заро-
дышевых стадиях развития относительно легко удаются и гомо-
231
трансплантации (т. е. пересадки от одного зародыша другому за-
родышу того же вида животного), и гетеротрансплантации (пере-
садки от одного вида другому в пределах одного рода), и даже
пересадки частей эмбрионов более отдаленных видов — ксенопла-
стика. Животное, от которого берется трансплантат, называется
донором; животное, которому пересаживается трансплантат, на-
зывают хозяином или реципиентом. Разработка методик транс-
плантации дала возможность обнаружить многие важные явления.
Огромный фактический материал шпемановской школы и ряд
идей сохранили свое значение до настоящего времени, хотя, есте-
ственно, прогресс эмбриологии, генетики и всей биологии заставил
отказаться от его теории индивидуального развития, в чем мы убе-
димся в дальнейшем.
Большую роль в развитии представлений о дифференциации и
детерминации сыграли работы многих исследователей — Д. П. Фи-
латова, Т. А. Детлаф, П. П. Иванова, П. Г. Светлова, Г. А. Шмидта,
К. Уоддингтона, Дж. Нидгемд, П. Вейса, Л. Барта, Ж- Рунстрёма,
Т. Густафсона, Ж- Браше, Э. Вольфа, С. Герстадиуса, С. Тойвонена
и многих других эмбриологов, генетиков, биохимиков.
ОПЫТЫ ПО ИЗОЛЯЦИИ БЛАСТОМЕРОВ. ПРИМЕНЕНИЕ
ПОНЯТИЙ «ДЕТЕРМИНАЦИЯ» И «РЕГУЛЯЦИЯ» К ЯЙЦЕВЫМ КЛЕТКАМ
Еще в 1869 г. Э. Геккель, наблюдая за развитием яиц сифоно-
фор, а в 1883 г. В. Ру, изучая яйца амфибий, обнаружили возмож-
ность развития целых организмов из изолированных бластомеров.
Впоследствии обнаружили это явление и многие другие ученые
(Г. Дриш, 1895; Г. Шпеман, 1900; А. Герличка, 1905; Е. Конклин,
1906; Г. А. Шмидт, 1933, и др.) в опытах на большом числе беспо-
звоночных и позвоночных животных.
Изоляция бластомеров достигается разными механическими и
химическими способами: перетягиванием с помощью волоса или
освобождением морской воды от кальция и т. п.
Рис. 104. Перешнуровка яйца тритона после оплодотворения
(по Г. Шпеману, 1919):
А — схема перешнуровки; Б, В — развивающиеся зародыши
232
Рис. 105. Опыт изоляции бластомеров у кролика (по Ф. Зейделю, 1959):
Д— стадия двух бластомеров; Б — тотчас после умерщвления одного из бластомеров; В,
/' — Дробление оставшегося в живых бластомера (убитый бластомер справа); Д — развив-
шиеся из изолированных бластомеров кролики (20-дневного возраста) с их матерью (в
центре)
Г. Шпеман перетягивал оплодотворенное яйцо тритона волося-
ной петлей и оно приобретало вид гимнастической гантели
(рис. 104,Л) - Можно добиться, чтобы ядро оказалось в одной поло-
вине «гантели». В ядерной половине начинается дробление, а безъ-
ядерная не дробится. При ослаблении волосяной плети перетяну-
тый участок может быть расширен и одно из ядер (на стадии 2, 4, 8,
16 и 32 бластомеров) может оказаться в безъядерной половине.
233
Если затем волосяную петлю снова затянуть, каждая половина
может развиться в нормальный зародыш (рис. 104, Б, В).
Ф. Зейделю в 1956 и 1959 гг. удалось осуществить изоляцию
бластомеров зародышей кролика. Оплодотворенное яйцо вымыва-
лось из половых путей самки. На стадии двух бластомеров одного
из них убивали иглой (рис. 105,5). После этого яйцо вводилось
в матку, удавалась имплантация зародыша, происходило полноцен-
ное его развитие (рис. 105,5, Г). Аналогичные операции удавалось
производить и на стадии четырех бластомеров.
Не менее интересные результаты получили А. К. Тарковский,
Б. Минц и др. Тарковский осуществил изоляцию одного из четырех
бластомеров яйца мыши. Происходило развитие вплоть до стадии
возникновения сомитов и даже более поздних стадий. Осущест-
влены также опыты изоляции бластомеров яйца крысы со стадии
двух, четырех и восьми бластомеров.
Развитие однояйцевых близнецов у человека — принципиально
то же явление. Если близнецы имеют общие оболочки, то есть осно-
вание предполагать, что они произошли в результате изоляции
бластомеров или других частей одного зародыша. Такие близнецы
оказываются одного пола и обычно очень сходны, вплоть до оди-
накового расположения линий на коже ладонной поверхности руки,
считающегося индивидуальным свойством. Рождение преимущест-
венно разнояйцевых двоен у человека — не очень редкое явление:
одно на 80 родов. Разнояйцевые тройки рождаются в одном случае
из 6400 родов, четверни — один случай на 512 000 родов. Известны
единичные случаи рождения и пяти, и шести детей. У различных
позвоночных — птиц, рептилий и рыб — описывались случаи двух
или нескольких относительно нормальных зародышей, развивших-
ся, несомненно, из одного яйца.
Опыты по изоляции бластомеров, а также другие данные цито-
логии, генетики и эмбриологии давно позволили сделать вывод
большого принципиального значения: несмотря на начинающуюся
с самого начала дробления дифференциацию (см. гл. IV), первые
дробления равнонаследственны. Бластомеры, находясь в системе
целого зародыша, подвергаясь взаимным формативным влияниям
и так или иначе дифференцируясь, не теряют, однако, своих видо-
вых свойств. При определенных условиях бластомеры и соматиче-
ские клетки на поздних стадиях развития организма и даже клетки
и комплексы клеток взрослых и старческих организмов могут на-
чать развиваться в целый организм, стать аналогом половых кле-
ток. Возможны далеко идущие регулятивные явления. Каждый
бластомер содержит все хромосомы, одинаковые гены. Иные пред-
ставления (идеи В. Ру о «мозаичном» развитии, гипотеза Вейсмана
о детерминантах и «зародышевом пути» и т. п.) оказались несо-
стоятельными в решении проблем дифференциации (см. гл.
II, IV).
Не на всех зародышах удаются успешные опыты по изоляции
234
бластомеров. Это дало повод эмбриологам в начале XX в. выделить
два типа яиц: «мозаичные» яйца или, иначе, яйцевые клетки с де-
терминированным характером дробления и регулятивные яйцевые
клетки. К числу первых причисляли прежде всего яйцевые клетки
большинства моллюсков и червей, характеризующихся спираль-
ным и билатеральным типом дробления (см. гл. IV). Мы видели
на примере развития аскарид, коловраток и других животных
очень раннее различие между бластомерами и очень раннее пред-
определение их судьбы (см. гл. II).
При детерминированном дроблении изолированные бластомеры
способны к «самодифференцировке». Однако и у регулятивных яиц
имеет место раннее предопределение судьбы бластомеров и можно
говорить о карте презумптивных органов бластулы (см. гл. V).
Опыты по изоляции бластомеров на яйцах с детерминированным
характером дробления или совсем не удаются, или не приводят
к развитию целых организмов. При разделении бластомеров на
двухклеточной стадии червя Lattice один из них дает сегментиро-
ванную личинку без переднего конца, а другой, наоборот, не обра-
зует сегментированного туловища. Если на восьмибластомер-
ной стадии яйца гребневика (имеющего восемь ребер с мерцатель-
ными пластинами) изолировать бластомер, то из него разовьется
личинка с одним валом гребных пластинок.
Несмотря на кажущуюся убедительность этих и других приме-
ров, традиционное разделение яиц на детерминированные и неде-
терминированные, или регулятивные, следует считать условным.
Детерминированные яйца на той или иной стадии развития прояв-
ляют способность к регуляции. Яйцо ланцетника причислялось
к яйцам с детерминированным характером дробления, но оно спо-
собно к регуляции с первых стадий дробления. Уродливые личинки
гребневиков, полученные из изолированных бластомеров, способны
к регуляции (могут развиться в нормальных особей). Можно гово-
рить о регулятивных возможностях трохофор червей и моллюсков,
так как наступает стадия метаморфоза, стадия очень существен-
ных морфологических и физиологических преобразований, которые
не могли бы совершаться без способности к регуляции.
Сколь правильно сопоставлять явления детерминации и регу-
ляции? Ответ на этот вопрос различен, он зависит от того или иного
понимания самой проблемы детерминации. Многие считают, что
в ходе развития зародышей по мере дифференциации клеток проис-
ходит прогрессирующее сужение их потенций, так что, наконец,
формируются ткани, состоящие из клеток «окончательно диффе-
ренцированных», и не может быть превращения тканевых элемен-
тов, происшедших из определенной эмбриональной закладки, в эле-
менты, берущие начало из другой эмбриональной закладки. По-
добные взгляды излишне персонифицируют единичные клетки и не
согласуются с фактами.
В дальнейшем (особенно при анализе явлений соматического
235
эмбриогенеза и регенерации, см. гл. XVI) мы встретимся с много-
численными фактами, находящимися в противоречии с точками
зрения, сводящими проблему детерминации и дифференциации
к единичным клеткам. Яйца асцидий или кольчатых червей и не-
мертии причислялись к строго детерминированному типу. Есте-
ственно предположить, что все новые и новые поколения клеток,
дифференцируясь и специализируясь, становятся, наконец, необ-
ратимо детерминированными. Но, как увидим в дальнейшем, взро-
слым асцидиям свойствен соматический эмбриогенез (см. гл. XVI),
при котором целый организм может развиться из телесных клеток.
Не надуманным парадоксом, а действительным и до сих пор за-
гадочным противоречием самой природы является то, что дробле-
ния яиц равнонаследственны, что митоз — идеальное выравнивание
двух сестринских клеток в отношении их ядер, что никакого обед-
нения одних бластомеров или клеток по сравнению с другими не
происходит, и в то же время уже первые бластомеры абсолютно
у всех яиц чем-либо отличаются друг от друга, дифференцируются.
Во взрослом организме нельзя встретить двух тождественных кле-
ток, но это нельзя рассматривать как следствие митоза (см. гл. XII
и XVI).
Понятие детерминации и дифференциации клеток необходимо
связывать не только и даже не столько с единичными клеткамл, но
с взаимодействием клеток, с устанавливающейся в развитии заро-
дыша новой и новой его интеграцией, с функцией новых возни-
кающих в ходе развития систем.
Различия в уровне интеграции, в характере взаимодействия
клеток (в том числе и биохимического и физического) на одних
и тех же стадиях развития (например, на стадии бластулы) у раз-
ных видов животных могут быть очень существенными. Не исклю-
чены случаи, когда на более молодых стадиях развития зародыш
более интегрирован, чем на более поздних стадиях. Так, личинки
асцидий, вероятно, более интегрированы, чем взрослые формы.
У зародышей многих моллюсков и червей, по-видимому, большая
взаимозависимость бластомеров, чем у клеток более поздних за-
родышей.
Изучение взаимозависимости клеток и частей зародышей, осо-
бенно в биохимическом аспекте, составляет важнейшую главу
эмбриологии. Взаимодействия клеток многообразны: электриче-
ский, механический контакт, на почве излучений, изменений про-
ницаемости мембран и т. п. Многообразны химические взаимоот-
ношения— изменения в среде концентрации ионов, обмен молеку-
лами, выделение в среду продуктов жизнедеятельности клеток
и др.
Начиная с 70-х годов стали умножаться исследования о взаи-
моотношении клеток в ходе онтогенеза животных, особенно бурно
стала развиваться наука о мембранах клеток, их проницаемости,
межклеточных субстанциях.
236
Рис. 106. Опыт В. Ру:
А — «полуэмбрнон», развившийся из одного бластомера после умерщвления другого (на
двухбластомерной стадии); Б — зародыш на стадии ранней иейрулы; / — мертвая полови-
на, 2 — живая половина, 3 — бластоцель, 4 — археитерон, 5 — нервный валик, 6 — хорда
Обратимся снова к фактам. Уже из сказанного ясно, что слу-
чаи неудач опытов по изоляции бластомеров, невозможность по-
лучить из изолированных бластомеров целые зародыши сами по
себе не оправдывают еще необходимости разделения яиц на де-
термированные и регуляционные. Исследователь встает перед по-
ка непреодолимыми техническими трудностями в опытах по изо-
ляции бластомеров у многих животных. Невозможно, например,
освободиться от оболочек яиц некоторых животных без разруше-
ния зародышей, находящихся в норме под давлением внутрияйце-
вой жидкости.
Не преодолены технические трудности в опытах по изоляции
бластомеров, например, у яиц нематод. Один из первых двух бла-
стомеров яйца аскариды убивали ультрафиолетовым лучом (дру-
гой бластомер затемняли и, значит, сохраняли живым). Развития
живого бластомера не наблюдалось. Такой результат, однако, мо-
жет быть объясним техникой опыта, что следует из анализа инте-
ресной дискуссии конца прошлого столетия, разгоревшейся меж-
ду В. Ру и другими исследователями.
В. Ру в 1887 г. укалывал горячей иглой один из первых блас-
томеров зародыша лягушки. Оставшийся живым бластомер раз-
вивался, но разрушение другого ведет за собой недоразвитие всей
половины тела зародыша (рис. 106). Ру сделал вывод в пользу
своего представления о зародыше как мозаике бластомеров, судь-
ба которых предопределена.
О, Гертвиг в 1893 г. и другие исследователи, возражая Ру, до-
казывали, что из каждого бластомера развивается целый нор-
мальный зародыш. Эксперименты обеих спорящих сторон были
безупречны. Особенности же в результатах объяснялись, с.одной
стороны, разной техникой опытов, а с другой — излишне односто-
ронней оценкой результатов, продиктованной предвзятой точцой
зрения. В опытах Ру в сущности не было изоляции бластомеров:
237
оставался убитый «привесок», и это отразилось на формообразо-
вательных процессах. Происходила задержка регуляции вследст-
вие механического препятствия регулятивным перемещениям со
стороны прижатой к зародышу массы мертвой протоплазмы уби-
того бластомера. Регуляция, однако, наступает: происходит, по
выражению Ру, «постгенерация» и развивается целый организм.
Аналогичная ситуация может быть отмечена и в отношении
яиц аскариды. Если оплодотворенное яйцо аскариды подвергнуть
центрифугированию, то дробление изменится таким образом, что
первые два бластомера окажутся равноценными и в обоих бласто-
мерах не произойдет диминуции хроматина, т. е. начнется как бы
развитие двойников. Это свидетельствует о том, что до начала
дробления яйца аскариды обладают высокой регулятивной спо-
собностью?-Вероятно, при несовершенных способах изоляции
бластомеров у моллюсков и червей могут быть еще большие по-
мехи в развитии целого организма из отдельных бластомеров.
Из всех изложенных данных вытекает вывод: разграничение
яиц с детерминированным развитием и яиц регулятивных относи-
тельно.
ВЗАИМОЗАВИСИМОСТЬ ЧАСТЕЙ ЗАРОДЫШЕЙ
НА РАННИХ СТАДИЯХ ИХ РАЗВИТИЯ
Как мы видели, яйцевые клетки имеют строгую видовую специ-
фичность (cm:^mt»4V, XI). Какое бы яйцо ни брать, оно с самого
начала не является гомогенным, а имеет в морфологическом, био-
химическом и физиологическом смысле сложную структуру. Это
в одинаковой мере относится как к яйцам с детерминированным
характером дробления, так и к регуляционным яйцам. Развитие
любого--яйца, обусловленное его «проморфологией» и внешними
условияМ; строго детерминировано с самого начала развития.
Приведем дополнительные доказательства. У яиц со спираль-
ным типом дробления, как мы видели, судьба бластомеров пред-
определена на самых ранних этапах развития.
Принципиально то же мы вправе утверждать и в отношении
регуляционных яиц любых животных. Можно, например, с полной
достоверностью сделать мысленную проекцию различных частей
плутеуса на соответствующие части яйца морского ежа. На ста-
дии 16 бла-етомеров мы вполне точно предусматриваем, что из
верхних восьми бластомеров (мезомеров) сформируется эктодер-
ма гаструлы, из средних четырех макромеров — стенка первичной
кишки, а микромеры дадут клетки первичной мезенхимы. В этом
смысле каждый участок любого яйца и каждый из бластомеров
двуклеточной стадии уже строго детерминированы: их судьба
при определенных условиях развития и при нормальной интегра-
ции зародыша строго предопределена.
Как только в результате дробления образуются два первых
238
бластомера, каждый из них становится
неразрывной частью новой биологической
системы, и его поведение определяется этой
системой. Каждая стадия развития орга-
низма есть новое состояние целостности,
интеграции. Зародыш и на стадии двух бла-
стомеров— интегрированное целое, и пове-
дение каждого бластомера обусловлено
этой интеграцией.
В этом отношении большой принципи-
альный интерес представляют опыты сдав-
ливания яиц. Если яйцо лягушки Rana tem-
poraria сдавить двумя пластинками в ани-
мально-вегетативном направлении, то по-
сле двух меридиональных дроблений, про-
исходящих как и при нормальных услови-
ях, следует не горизонтальное, а опять
Рис. 107. Яйцо лягушки
на стадии восьми блас-
томеров. Результат сдав-
ливания яйца в ани-
мально-вегетативном на-
правлении (по Г. П.
Святогору, 1975)
меридиональное, приводящее к расположе-
нию восьми бластомеров в один слой в виде пластинки (рис. 107).
То же наблюдается и на стадии 16 и 32 бластомеров. После удале-
ния давящей силы из таких яиц могут развиться нормальные за-
родыши.
Опыты сдавливания яиц между стеклянными пластинками,
опыты центрифугирования, изменение температурных условий и
другие могут привести к тому, что у форм с гетероквадратным
дроблением (Chaetopterus, Nereis, Sabellaria) первое деление
окажется равномерным. В таких экспериментальных условиях
каждый из двух бластомеров, будучи изолированным, может раз-
виваться в почти нормальную личинку. Таким образом, у яйца со
строгой детерминированностью дробления возможна регуляция,
экспериментальное вызывание развития двойников.
ЛАБИЛЬНАЯ И СТАБИЛЬНАЯ ДЕТЕРМИНАЦИЯ
В гл. -V говорилось, что благодаря разработанной Фогтом методи-
ке витального окрашивания на ряде животных, особенно на амфи-
биях, хорошо прослежена судьба частей зародыша (см. рис. 53),
и говорилось о картах презумптивных органов бластулы амфибий
и других животных.
Рисуются участки клеточных комплексов на стадии бластулы
тритона, являющиеся источником энтодермы, кожной эктодермы,
нервной пластинки, хорды, сомитов, боковой мезодермы, хвоста,
жабр, передней конечности. Это означает одновременно, что мы
предусматриваем, какие группы клеток являются исходными для
формирования нервной системы, эпителия кишечника, эпителия
кожи, мускулатуры и других органов будущего взрослого орга-
239
Рис. 108. Схема трансплантации частей зародыша:
Презумптивные зачатки: 1 — нервной системы, 2 — сомитов,
3 — энтодермы, 4 — бластоцель
низма. Такие карты презумптивных органов не менее точно могут
быть нарисованы на стадии бластулы птиц и других животных.
Означает ли это, что клетки на стадии бластулы необратимо де-
терминированы и приобретают такие структуры и функции, кото-
рые не позволяют изменять предустановленный путь их развития?
Г. Шпеман взял у поздней бластулы в начале гаструляции
пигментированного зародыша тритона кусочек клеточного матери-
ала будущей медуллярной пластинки, а у непигментированного
зародыша тритона — кусочек будущей кожной эктодермы и пере-
садил их взаимно зародышам (рис. 108). Пересаженные клеточ-
ные комплексы развивались не в том направлении, в каком они
были «детерминированы», не по месту происхождения, а согласно
новому окружению и вошли гармонически в состав зародыша хо-
зяина: кусочек будущей кожной эктодермы принял участие в раз-
витии нервной системы, и наоборот. Г. Мангольд пересаживала
кусочки эктодермы в различные места зародыша, и в зависимости
от места приживления пересаженный материал принимал участие
в образовании органов, на развитие которых он, конечно, не был
«детерминирован», например на развитие сомитов, почечных ка-
нальцев, хорды, стенок кишечника и т. п. Дальнейшими опытами
выяснено, что не только будущая эктодерма в опытах с транс-
плантацией может пойти на построение любого органа, в зависи-
мости от места пересадки, но так же ведут себя и будущие мезо-
дерма и энтодерма. Г. Шпеман установил для стадии бластулы
понятие лабильной детерминации.
Классические опыты шпемановской школы свидетельствуют о
том, что поведение клеток не только их собственная функция.
Источником детерминации служат не единичные клетки, их ядра
или цитоплазма. Ни одно явление дифференциации не может быть
исследовано исходя из единичных клеток. Поведение каждой клет-
ки обусловлено специфическими связями между клетками, связя-
ми, возникающими в ходе развития организма в определенных ус-
ловиях существования, при непрерывно меняющихся состояниях
целостности. Поведение каждой клетки (ее размеры, форма, плос-
240
кости и темпы делений и т. п.) зависит не только от свойств еди-
ничных клеток, но от всего организма, представлен ли он стадией
двух бластомеров, бластулой или разнообразными тканями и
органами поздних стадий развития. Многие факты говорят, прямо
или косвенно, в пользу этих представлений.
Мы переходим теперь к фактам, на первый взгляд противоре-
чащим сказанному. Перед нами зародыш на стадии поздней гаст-
рулы или нейрулы. Появилась медуллярная пластинка. При пере-
садке участка ее в любое место другого зародыша образуется
часть нервной системы. Пересаженный кусочек кожной эктодермы
образует эпидермис кожи или соответствующие органы, генетиче-
ски связанные с эктодермой (слуховой пузырек, хрусталик и др.),
значит, он ведет себя по месту происхождения, а не нового поло-
жения. _
Шпемановская школа утверждала, что теперь наступает окон-
чательная, стабильная детерминация в отличие от лабильной де-
терминации на стадии бластулы или ранней гаструлы. Означает
ли это, однако, что теперь клетки, наконец, так дифференцирова-
ны и детерминированы, что произошло сужение их потенций, что
окончательно предопределена судьба единичных клеток?
И на стадиях поздней гаструлы, а также нейрулы мы не впра-
ве говорить об окончательной детерминации. Явление передетер-
минации казавшихся необратимо детерминированными участков
зародыша доказано на стадиях поздней нейрулы и хвостовой поч-
ки. Особенно показательны опыты по развитию глаза и органа
слуха у амфибий (работы Шпемана, Льюиса, Филатова и др.).
Об этом говорилось в гл. VII. Шпеман в 1901 г. разрушал при по-
мощи горячей иглы зачатки глазных бокалов у зародыша лягуш-
ки на стадии нейрулы еще до момента образования из эпидер-
мального слоя клеток хрусталика. В этом случае хрусталик сов-
сем не развивался. Значит, для развития хрусталика нужен кон-
такт эпидермального слоя с глазным бокалом. Если участок кле-
ток, из которого мы ожидаем развития хрусталика, заменить
участком эктодермы с другого места, то последний развивается
в хрусталик.
Поведение клеток на любых стадиях развития зародыша пред
определено, они действительно детерминированы, если продолжи
ют быть в определенной биологической системе клеток, составля
ют часть ее, находятся под замком корреляций и если организм
развивается в определенных условиях.
Под детерминацией следует понимать, как об этом говорилоа
в начале главы, установление в ходе онтогенеза организма, раз
вивающегося в данных конкретных условиях, таких взаимосвязе!
между клетками, при которых клеточные комплексы проходят оп
ределенный путь развития. Сами клетки при этом изменяются
приобретают и утрачивают те или иные структуры и функции, эт<
24
не подлежит сомнению. Более того, развитие организма приводит
к таким стадиям, когда начинаются процессы дегенерации отдель-
ных клеток и тканей, прекращается способность некоторых клеток
к делению и тому подобные явления, зависящие опять-таки от за-
кономерностей целостности организма. Клетки, специфически диф-
ференцируясь, все более отличаются друг от друга. Но в то же
время было бы чудом, если бы клетка какого-либо эпителия на-
цело теряла свои основные видовые черты при сохранении способ-
ности к делению.
«ОРГАНИЗАЦИОННЫЕ ЦЕНТРЫ» РАЗВИВАЮЩИХСЯ ЗАРОДЫШЕЙ.
ПОНЯТИЕ «ИНДУКЦИИ»
Г. Шпеман срезал у зародыша тритона на стадии ранней гастру-
лы всю верхнюю половину (анимальное полушарие), повернул ее
на 180° и снова срастил с нижней половиной. Нервная пластинка
развивалась на той стороне, на которой находился материал быв-
шего серого серпа, а теперь формирующейся губы бластюпора,
иначе говоря, нервная пластинка развивалась не из того мате-
риала, который участвует в нормальном развитии, а из другого.
Г. Шпеман решил, что от верхней губы бластопора по направ-
лению к анимальному полюсу распространяется какое-то влияние,
которое заставляет клетки эктодермального слоя развиваться
в сторону нервной пластинки, индуцирует ее образование. Эту
область он назвал организационным центром, а материал, из ко-
торого исходит индуцирующее влияние, — организатором, иначе—
индуктором.
Отметим некоторые основные факты, полученные в дальней-
ших исследованиях. Г. Шпеман и Г. Мангольд производили ре-
ципрокную пересадку дорсальной губы бластопора от одного вида
тритона (пигментированного) к другому (непигментированному).
Это позволило определить, за счет каких клеток (донора или ре-
ципиента) формируются нервная система, хорда и мезодерма.
Было установлено, что губа бластопора (в случае удачного экс-
перимента) превращалась в хордомезодермальные зачатки (до-
норского происхождения), которые индуцировали развитие из
эктодермы нервной пластинки (реципиентного происхождения).
Поведение самого трансплантата было не всегда одинаково.
В большинстве случаев, инвагинируя под ткани хозяина, транс-
плантат развивается в общем в том же направлении, как и при
нормальном морфогенезе, — он образует большую часть хорды и
сомитов. Отсюда был сделан вывод, что клеточный материал са-
мого организатора детерминирован и развивается путем само-
дифференцировки, а клеточный материал, окружающий транс-
плантат, преобразуется в направлении, которое ему нормально
не свойственно.
242
Для анализа явлений индукции- вторичной медуллярной пла-
стинки в опытах трансплантации дорсальной губы бластопора
в другой зародыш и для решения вопроса о том, можно ли пред-
полагать, что дорсальная губа бластопора является забежав-
шим вперед (в смысле дифференцировки) участком зародыша,
первостепенное значение имеют многие исследования И. Гольт-
фретера. В 1938 г. он культивировал в стандартных средах ма-
ленькие кусочки, вырезанные из гаструлы аксолотлей и тритонов.
Оказалось, кусочки из эктодермальной области (как проспектив-
ной эпидермальной, так и проспективной медуллярной) в условиях
эксплантации не формируются в типичные ткани. Когда выреза'
ется большая пластинка, она может принять форму эпителия, но
быстро дезинтегрируется на отдельные клетки.
Эксплантаты из различных участков эндотермальной области
(вегетативная половина зародыша) в условиях таких опытов ока-
зываются локально более детерминированными: формируются
различные эпителии, напоминающие эпителий кишечного тракта,
печени, поджелудочной железы. Эти структуры, выражаясь язы-
ком шпемановской школы, развиваются в отсутствие организа-
тора.
Особый интерес представляют в условиях тех же опытов ре-
зультаты эксплантации клеточного материала из различных уча-
стков краевой зоны. Эксплантаты способны развиваться в раз-
личные ткани, но судьба их не соответствует точно их проспек-
тивным (в ходе нормального развития) потенциям. Маленькие
кусочки из будущей хорды не только дают развитие хорды, но
и сомитов, возникают нередко невральные и эпидермальные
структуры. Вполне убедительный вывод из этих фактов сделали
в 1955 г. И. Гольтфретер и В. Гамбургер: все участки краевой
зоны продуцируют в условиях эксплантации более широкое раз-
нообразие тканей, чем они дали бы, находясь в системе зародыша
(рис. 108, Д,В). Однако из данных Гольтфретера напрашиваются
еще более значительные выводы.
Кусочки из верхней губы бластопора и из боковых губ фор-
мируют в условиях эксплантации сложную структуру, начинает-
ся как бы развитие целого организма (рис. 108,В). Формируется
аксиальная система с «голово-хвостовой» полярностью и била-
теральной симметрией. В этом сложном образовании имеются хор-
да и спинной мозг, которые окружены миобластами. Впереди мы
видим мозгоподобное расширение нервной трубки, рядом — эпи-
дермис, мезенхимные и «гигантские клетки», которые как бы
представляют мозговую мезоэктодерму.
Таким образом, будущий хордомезодермальный материал
в районе губы бластопора на стадии ранней гаструлы способен
давать в условиях изоляции от целого разнообразные производ-
ные. Этот клеточный материал не детерминирован по какому-
либо одному пути развития. Опыты не позволяют делать вывод
24,
Рис. 109. Опыт И. Гольтфретера:
А — распределение презумптивных зачатков на стадии ранней гаструлы амфибий; Б — диф-
ференциация, выявленная в опытах эксплантации маленьких кусочков из разных областей
зародыша со стадии гаструлы. Результаты экспериментов проецируются иа ту же стадию
(ранняя гаструла); В — эксплаитированные кусочки из области будущих сомитов ранней
гаструлы амфибий; / — боковая пластинка, 2 — эктодерма, 3 — нервная пластинка, 4 —
хорда, 5 — сомиты, 6 — энтодерма
о том, что клеточный материал организатора, т. е. дорсальной
губы бластопора, является забежавшим вперед в смысле диф-
ференцировки участком, а скорее наоборот, так как судьба кле-
ток будущей энтодермы на стадии ранней гаструлы более «опре-
делена». Клеточный же материал краевой зоны (в том числе и
дорсальной губы бластопора) очень лабилен в формообразова-
тельном отношении и в условиях изоляции от целого зародыша
имеет тенденцию начать развиваться в целый организм, а не
только давать структуры, свойственные хордомезодерме.
Исследования лаборатории Г. Шпемана о роли спинной губы
бластопора в развитии амфибий вызвали, особенно в 1925—
1940 гг., огромное количество работ. Многие исследователи пока-
зали наличие организационных центров у различных эмбрионов:
у бесхвостых амфибий, рыб и птиц. Понятие «организационный
центр» стало вскоре применяться и в отношении взрослых орга-
низмов в связи с процессами регенерации. Стали говорить об
организаторе у гидры (гипостом), нашли организаторы у плана-
рий, насекомых, иглокожих. Одновременно стала формироваться
теория индивидуального развития Шпемана. Эмбриональное раз-
витие стало представляться как включение в действие все новых
244
и новых «индукторов», «организаторов». Этому способствовали
превосходные экспериментальные исследования органогенеза, ре-
зультаты которых, к сожалению, стали трактоваться крайне од-
носторонне. Губа бластопора стала называться организатором
1-го порядка, глазной бокал (индуцирующий хрусталик) и слу-
ховой пузырек (индуцирующий слуховую капсулу) — организато-
рами 2-го порядка и т. д.
Ценные результаты многочисленных исследований школы Шпе-
мана и его последователей из других лабораторий, давших эмб-
риологии блестящее доказательство взаимозависимости частей
зародыша, его интеграции'на любой стадии развития, стали трак- '
товаться все более односторонне, как действие организаторов
на якобы индифферентный клеточный материал. Это была свое-
образная полоса в развитии эмбриологии, когда, казалось, най-
дено было основное в объяснении процессов формообразования
и критические замечания отдельных ученых (Д. П. Филатов,
1935; Б. П. Токин, 1934) против односторонних увлечений рас-
сматривались как что-то мешающее развитию науки. Создавшая-
ся в то время «теория организационных центров», несомненно,
содержала излишне односторонние и даже фанатичные взгляды,
которые потерпели поражение перед лицом новых, не менее увле-
кательных фактов, обнаруженных впоследствии самой же шпе-
мановской школой.
ФЕНОМЕН «МЕРТВЫХ ОРГАНИЗАТОРОВ»
Перед исследователями встал вопрос: сколь специфично дейст-
вие «организаторов», «индукторов»? При пересадке организатора
от бесхвостой амфибии (лягушка-жерлянка) зародышу хвостатой
амфибии (тритон) была обнаружена индукция медуллярной пла-
стинки. В случае пересадки от зародыша птицы в зародыш три-
тона «организатор» также оказывает индуцирующее действие.
Это происходит и в случае пересадки зародышу кролика органи-
затора тритона.
Было обнаружено, что организатор способен индуцировать и
после полного разрушения его структуры, даже после полного
разрушения клеток. Перемешивали раздавленные (между двумя
стеклянными пластинками) кусочки верхней губы бластопора,
делали комочки из них и всаживали в полость бластулы друго-
го зародыша. Индукция имела место. Индукционные свойства ор-
ганизатора не утрачивались также после кипячения, обработки
спиртами и т. п.
В настоящее время ясно, что все эти попытки найти специ-
фическое формообразовательное вещество не достигли цели. Хи-
мические вещества, принадлежащие к различным группам соеди-
нений, дают одинаковый эффект. Ф. Ж. Фишеру и Е. Веймейеру
удалось обнаружить положительный результат с тимонуклеино-
245
вой и адениловой кислотами. Положительные результаты полу-
чены с жирными кислотами из растительных масел, с эфирными
вытяжками из печени и сала свиньи, с синтетическими органиче-
скими веществами — масляной кислотой и оксилалкоголем. По-
ложительные результаты дала нуклеиновая кислота зобной и под-
желудочной желез. Л. Барт в 1934—1939 гг. обнаружил индук-
ционное влияние ядовитого для животных тканей вещества —
цефалина и продуктов его расщепления. Е. А. Финкельштейн
получил положительный результат с помощью кусочков агара,
пропитанных продуктами гидролиза тканей (1937), или дипепти-
дом карнозином (1938). К- Уоддингтон получил индукцию с по-
мощью нафталина и сквалена.
В понятие «организатор» стали вкладывать представление
о химических формообразовательных веществах. Развивающийся
организм представился мозаикой микролабораторий, вырабаты-
вающих формообразовательные вещества, а развитие организма —
включением в действие разных органообразующих веществ. Стали
толковать о веществах, индуцирующих глаза, уши, жабры и пр.
Возвратимся к теории организаторов. В обычной схеме об
индукционных влияниях организаторов клеточный материал, ко-
торый реагирует, индуцируется, подразумевается как нечто ин-
дифферентное. Однако это не так. Было показано, что закладка
осевой мезодермы представляет не только индуктор нервной труб-
ки, но и сама нуждается для своей дифференциации в форматив-
ном воздействии со стороны закладки нервной системы. При этом
имеет место не односторонняя индукция, а взаимодействие частей
развивающегося зародыша. Мезодерма, будучи изолированной от
нервной трубки и хорды, не превращается в мышечные клетки
или в скелетные элементы, но развивается по двум путям: пре-
вращается в рыхлую мезенхимную ткань или в избыточные ка-
нальцы предпочки. Один и тот же индуктор может индуцировать
различные образования. Так, слуховой пузырек при пересадке на
бок зародыша хвостатых амфибий индуцирует добавочную конеч-
ность, тот же слуховой пузырек в ином месте и на другом этапе
онтогенеза индуцирует слуховую капсулу. Он же может быть
в роли индуктора добавочного волокнистого ядра хрусталика
в. случае соприкосновения с зачатком хрусталика. При пересадке
в туловищную и головную части гаструлы верхней губы бласто-
пора со стадии поздней гаструлы (т. е. когда головной участок
хордомезодермы уже подвернут внутрь, продвинулся в направле-
нии будущего переднего конца зародыша и губой бластопора яв-
ляется участок, который попадает под эктодерму туловища на
следующей стадии) в головной части хозяина трансплантат ту-
ловищного отдела хордомезодермы вызывает в эктодерме обра-
зование дефектных головных частей, а такой же трансплантат,
пересаженный в туловищную часть, вызывает в ее эктодерме
только туловищные закладки.
246
Оказалось, что искать в единичных клетках «регулирующее»
вещество, которое могло бы дать ключ к пониманию дифферен-
цировки, бесполезно. Причину дифференцировки следует искать
в реагирующей ткани, в которой дифференцировка, собственно,
и происходит. Только в определенные периоды эта ткань способ-
на отвечать на воздействия организатора, т. е. обладает тем, что
называют компетенция. Кстати сказать, термин «компетенция»,
введенный К. Уоддингтоном и Дж. Нидхемом, конечно, не дает
объяснения явлениям, но очень полезен, так как предостерегает
от упрощенной антропоморфной точки зрения об организаторах,
•двдб^я Действующих на индифферентный материал.
Следует отметить попытки Изучения взаимных влияний частей
развивающихся зародышей, сочетая изложенные ранее экспери-
ментально-морфологические методики с методиками радиобиоло-
гии, биофизики и молекулярной биологии. В. В. Попов (1967)
экспериментировал на функционирующих глазах развивающихся
позвоночных и на зачатке внутреннего уха этих животных. Уста-
новлена связь между функциональной активностью развивающе-
гося органа и его «индуцирующей способностью». Попов (1966),
в частности, приводит следующий пример. Условия освещения по-
разному влияют на функциональное состояние сетчатки, возбуж-
дая или, наоборот, угнетая ее функции. А функциональное со-
стояние сетчатки, в свою очередь, отражается на ее способности
к индукции роговицы и на ее влиянии на дифференциацию хру-
сталика.
АНАЛОГИЧНО ЛИ ДЕЙСТВИЕ «ОРГАНИЗАТОРА» В ХОДЕ НОРМАЛЬНОГО
РАЗВИТИЯ ЗАРОДЫША И В СЛУЧАЕ ПЕРЕСАДКИ ЕГО В ДРУГОЙ ЭМБРИОН!
Этот вопрос должен быть дополнен другим (который будет ра-
зобран ниже): аналогично ли действие живого и мертвого орга-
низаторов в случае трансплантации их в эмбрион? Ответы на эти
вопросы имеют решающее значение для интерпретации всех дан-
ных шпемановской школы.
Такая постановка вопросов была сделана Б. П. Токиным еще
в самом начале увлечений открытием мертвых организаторов и
тогда же им был дан отрицательный ответ на них. Филатов
в 1935 г. также дал отрицательный ответ на второй вопрос.
Требуется вспомнить, как протекает процесс гаструляции и
как возникает закладка нервной системы у амфибий (см. гл. VII
и рис. 73). Нами уже обращалось внимание на тот факт, что пер-
вичный организатор, т. е. дорсальная губа бластопора, не явля-
ется какой-либо хотя бы относительно стабильной структурой.
В нормальном развитии организатор — это подвижное, меняющее-
ся клеточное образование. Некоторые эмбриологи очень удачно
называют губу бластопора «речкой клеток». Через губу бласто-
247
пора проходит клеточный материал обширной поверхности, на-
ходящийся между нею и анимальным полюсом.
Если пересаживать дорсальную губу бластопора со стадии
ранней гаструлы, то она индуцирует комплекс осевых зачатков
переднего отдела зародыша, а если пересаживать эту же губу со
стадии более поздней гаструлы, то тогда она индуцирует комп-
лекс осевых зачатков туловищно-хвостового отдела. Это озна-
чает, что в зависимости от того, какой клеточный материал ока-
зывается в момент трансплантации в области дорсальной губы,
его индукционные способности окажутся неоднотипными. На ос-
новании этих и аналогичных экспериментов были созданы пред-
ставления о существовании двух типов индуцирующих воздейст-
вий первичного организатора: передне-мозгового и спинно-хвосто-
вого. Как увидим далее, это дало основание для поиска так
называемых «нейрализующих» и «мезодермализующих» веществ
(Л. Саксен, С. Тойвонен, 1963).
Вернемся к анализу действия живых и мертвых «организато-
ров». При действии мертвых и живых «организаторов» наблюда-
ются явления, которые невозможно отождествлять с теми, что
происходят при взаимодействии частей зародыша в ходе нор-
мального развития.
Эффект индукции живым организатором, особенно чужерод-
ным, выражается не в развитии какой-либо ткани, органа или
части органа, а в развитии (или в начальных попытках развития)
прилежащего к трансплантату клеточного материала или самого
пересаженного материала в сторону целого организма. Все раз-
нообразные опыты с живыми и особенно с мертвыми организато-
рами свидетельствуют о возможности экспериментальной поли-
эмбрионии на стадиях бластулы и ранней гаструлы амфибий, рыб,
птиц и млекопитающих.
Г. Шпеман и Г. Мангольд в одном из опытов на месте пере-
садки кусочка верхней губы бластопора (взятого от зародыша
гребенчатого тритона) на боку зародыша хозяина (обыкновен-
ный тритон) наблюдали развитие целого добавочною зародыша
(рис. 1!10). Последний имел хорду, сомиты, длинную мозговую
трубку, почечные канальцы, развились два слуховых пузырька,
глазные бокалы, сзади — зачаток хвоста, из энтодермы образо-
валась кишечная полость.
Не менее интересны случаи экспериментальной полиэмбрио-
нии, зарегистрированные в подобных опытах по трансплантации
на эмбрионах рыб, птиц и млекопитающих (рис. 111).
Еще больший интерес представляют результаты транспланта-
ции организаторов у птиц. В 1933 г. в опытах К. Уоддингтона и
Г. А. Шмидта донорами были куриные эмбрионы, а хозяевами —
эмбрионы утки; в других опытах, наоборот, участки эмбриона
утки трансплантировались эмбриону курицы. Пересаживались раз-
личные участки первичной полоски (область гензеновского узелка
248
Рис. 110. Экспериментальная полиэмбриония у
Triturus taeniatus (поперечный разрез через средин-
ную область зародыша) (по Г. Шпеману и Г. Ман-
гольд, 1924).
Первичный эмбрион иа левой стороне рисунка, вторичный
(индуцированный) — на правой; 1 — первичная нервная
трубка, 2 — вторичный пронефрический проток, 3 — вторич»
ная кишка, 4 — вторичный сомнт, 5 — индуцированная
нервная трубка, 6 — хорда
Рис. 111. Эксперимен-
тальная полиэмбрио-
ния у Fundulus (по
В. Лютеру, 1935):
1 — первичный эмбрион,
2 — индуцированный эм-
брион
и др.). Описываемые авторами результаты и приводимые ими
фотографии внешнего вида зародышей и гистологических препа-
ратов не оставляют никаких сомнений в том, что здесь имеет
место полиэмбриония. Приведем иллюстрацию из этих опытов.
На рис. 112,А представлены эмбрионы хозяина и индуцирован-
ный через 26,5 ч после трансплантации передней половины пер-
вичной полоски (включая и гензеновский узелок) куриного эмб-
риона (15-часовой инкубации) эмбриону утки (26-часовой инку-
бации). Мы видим, что прекрасно развившийся индуцированный
эмбрион (его головной отдел) находится на той же стадии раз-
вития, какой достиг и эмбрион-хозяин.
На рис. 112,5 показаны результаты другого опыта, в кото-
ром хозяином был также утиный зародыш 28-часовой инкубации,
а донором — куриный зародыш 15-часовой инкубации. Фотосни-
мок сделан через 26,5 ч после трансплантации. Налево от эмб-
риона хозяина виден вторичный эмбрион. Изучение срезов за-
родышей показало, что индуцированный эмбрион имеет вполне
сформированную голову, хорду, сомиты, сердце, формирующийся
орган слуха и другие образования.
Сходны с этими явлениями случаи экспериментальной поли-
эмбрионии, вызываемые механическими травмами зародышей
птиц (рис. 112,В).
249
Рис. 112. Экспериментальная полиэмбриония у птиц:
Л, Б — результаты двух опытов К. Уоддингтона и Г. А. Шмидта (/ — эмбрион индуцирован-
ный, 2 — эмбрион основной); В — пять утиных зародышей, развившихся в результате меха-
нического разделения бластодермы на кусочки (из кн. А. Далька, 1957)
Не требуется доказывать полную искусственность попыток
привлечь результаты подобных опытов для объяснения индукции
нервной пластинки хордомезодермой, имеющей место в ходе нор-
мального развития животного. В одном случае наблюдается вза-
имное влияние частей развивающегося зародыша, а в другом —
принципиально иное явление — развитие (или попытка развития)
целого организма.
При экспериментальном изучении формирования нервной труб-
ки у амфибий (опыты эксплантации и др.) превосходно доказано
влияние на этот процесс других участков зародыша. Ф. Е. Леман
удалял проспективную хорду у зародыша тритона, а Ж- Пастелс
подавлял ее дифференциацию воздействием LiCl или посредством
центрифугирования. В этих опытах сомиты формировались непра-
вильно («сплавлялись»). При таких условиях стенка нервной
трубки вместо того, чтобы сформироваться в тонкую пластинку,
образует толстую массу (рис. 113,Д). Когда же нервные ткани
дифференцировались только в контакте с хордой (без нормаль-
ных сомитов), то участок нервной трубки, контактирующийся
с хордой, формируется в тонкую пластинку (рис. 113,Б). В слу-
чае наличия хорды и сомитов, находящихся лишь на одной стороне
от нервной трубки, происходит формирование резко асимметрич-
ной нервной трубки (рис. 113,В). Когда спинной мозг развивает-
ся, будучи окруженным мезенхимой (т. е. без контакта с хордой
и сомитами), стенки нервной трубки оказываются одинаковой
толщины во всех участках (см. рис. 112,Г). И. Г. Гольтфретер и
В. Гамбургер (1955) предполагали, что в нормальном развитии
кажущееся (на основании указанных опытов) антагонистическое
действие хорды и сомитов дополняет друг друга, в результате
250
Рис. 113. Формативное влияние сомитов, хорды и мезодермы на форму
нервной трубки у амфибий. Пояснения см. в тексте (по И. Гольтфрете-
ру и В. Гамбургеру, 1955)
чего формируется нормальная нервная трубка (рис. II13,Д). Если
такое объяснение и не кажется очень убедительным, нельзя не
усмотреть в приведенных материалах прекрасное доказательство
взаимодействия частей развивающегося эмбриона. Что же обще-
го имеют с этими фактами данные об индукции вторичного эмб-
риона в случае пересадки губы бластопора в чужой эмбрион?
Понятия «ассимиляторной индукции» (установленное Г. Шпе-
маном и Б. Гейницем в 1927 г.) и «компенсаторной индукции»
(установленное О. Мангольдом в 1932 г.) указывают на тот ин-
тересный факт, что при трансплантации губы бластопора различ-
ным образом может вовлекаться в формирование вторичного за-
родыша клеточный материал хозяина, или, наоборот, вовлекает-
ся клеточный материал трансплантата в формирующийся из кле-
точного материала хозяина вторичный эмбрион.
Таким образом, необходимо отличать две принципиально раз-
ные группы явлений: взаимозависимость частей развивающегося
эмбриона и экспериментальную полиэмбрионию. Особенно неар-
гументированно отождествлять явления индукции медуллярной
пластинки в нормальном развитии с явлениями развития вто-
ричного зародыша в опытах с мертвыми организаторами.
251
ИНДУКЦИЯ ВТОРИЧНЫХ ЭМБРИОНОВ — ЧАСТНЫЙ СЛУЧАЙ
ЭКСПЕРИМЕНТАЛЬНОЙ ПОЛИЭМБРИОНИИ
В результате воздействия пересаженных мертвых организаторов
развиваются два зародыша: один — вследствие обычных процес-
сов развития, другой — из клеточного материала того же зароды-
ша под влиянием организатора. Конечно, не всегда наблюдается
столь отчетливая картина, но даже и в случае образования не
всех осевых органов вторичного эмбриона, а лишь вторичной ме-
дуллярной пластинки имеется одно и то же явление — развитие
в сторону целого организма.
В самом деле, что означает индукция вторичной медуллярной
пластинки? Она образуется на определенной стадии развития за-
родыша, на стадии, когда и сам нормальный первичный эмбрион
представлен лишь медуллярной пластинкой как наиболее диф-
ференцированной частью целого.. На этом этапе онтогенеза жи-
вотного нет еще ни органов слуха, ни глаз, ни конечностей, ни
сердца и т. д.; характеристикой продвинувшейся дифференциров-
ки эмбриона на данной стадии развития служит именно медулляр-
ная пластинка. Вторичный эмбрион столь же полноценен, он так-
же представлен самой дифференцированной на этой стадии ча-
стью — медуллярной пластинкой. Выходит, что если бы дальней-
шего развития индуцированного эмбриона и не происходило (что
вполне естественно, если принять во внимание условия экспери-
ментов), то мы обязаны были бы говорить о развитии эмбриона,
о развитии целого. То, что часто развивается дефектный зародыш,
не должно смущать биологов, ибо иного результата трудно ожи-
дать в условиях опытов.
Мы делаем общий вывод: клеточный материал хозяина под
влиянием индуктора вместо дифференциации в определенные тка-
ни и органы и образования частей зародыша развивается в целый
организм, в добавочный эмбрион. Явления биологически различ-
ные нельзя оценивать как идентичные. С одной стороны, устанав-
ливается взаимозависимость частей развивающегося эмбриона:
слуховой пузырь — слуховая капсула, глазной бокал — хрусталик,
дорсальная губа бластопора — медуллярная пластинка и т. д.
С другой стороны (в опытах с трансплантацией индуктора в чу-
жой эмбрион), мы имеем явление нарушения нормальных взаи-
мосвязей частей развивающегося организма, дезорганизацию, луч-
ше сказать, дезийтег^ацийт нормальной клеточной системы, раз-
витие некоторых клеточных комплексов не в сторону отдельных
частей, а по линии целого организма.
Факты и анализ и& принуждают прийти к выводу: если антро-
поморфное понятие «организатор» имеет какой-либо смысл в при-
менении к развитию организма, поскольку делается ударение на
связи клеток и их комплексов, то понятие «организатор» в опы-
тах трансплантации в чужой эмбрион (в живом или мертвом со-
стоянии этот материал пересаживается, взят ли он от раздавлеи-
252
ных дафний или от трупа человека) лишено вообще всякого био-
логического смысла.
Роль подсаженного организатора заключается в резком нару-
шении обычных взаимных индукций. Значит, организатор не ор-
ганизует и не индуцирует, а выступает в роли дезорганизатора,
лучше сказать дезинтегратора, структурных и функциональных
соотношений клеток и клеточных систем нормального эмбриона.
Происходит изоляция клеточных комплексов из-под формативно-
го влияния целого.
Следует особо подчеркнуть, что в абсолютном большинстве
опыты по индукции вторичных эмбрионов не удается осуществлять,
на более поздних стадиях. Это объясняется тем, что на стадиях
поздней гаструлы и нейрулы начинает уже появляться способ-
ность к регенерации, создаются новые иммунологические возмож-
ности и зародыш становится еще более интегрированным, чем на:
предшествующих стадиях. Вследствие этого обусловливается воз-
можность репарации после повреждения чужеродными-
трансплантатами, перестройка начавшего развиваться в нервную-
пластинку клеточного материала под влиянием интегрального це-
лого.
Таким образом, явления, анализом которых мы столь долго
занимались, вероятно, сходны по своему механизму с явлениями:
полиэмбрионии у других животных, возникающей благодаря ка-
ким-то спонтанно происходящим нарушениям нормальной инте-
грации зародыша.
Индукции вторичных эмбрионов у амфибий, рыб, птиц и мле-
копитающих, как мы убедились, служат примерами эксперимен-
тальной полиэмбрионии. Нельзя, конечно, это утверждение рас-
пространять абсолютно на все случаи индукции вторичной ме-
дуллярной пластинки, описанные в литературе.
Теория индивидуального развития Шпемана представляет уже
только исторический интерес, но огромный фактический материал:
его школы далеко не весь ис-
пользован эмбриологией, он
требует дальнейшего анализа.
Из работ последнего времени
следует обратить внимание на
интереснейшие факты, обна-
руженные Хасаном Насером
Эль-Дином и Н. С. Табае-
вой (1980). Авторами отмече-
ны случаи формирования
нервной трубки в отсутствии
хорды у эмбриона вьюна
(возникших в результате экс-
периментальной полиэмбрио-
нии). Развивались, в частно-
Рис. 114. Срез через нервные трубки,,
развившиеся в отсутствии хорды у за-
родышей-близнецов вьюна
253:
сти, и эмбрионы с несколькими нервными трубками при отсут-
ствии хорды (рис. 114). Эти факты как будто несопоставимы даже
с кажущимися бесспорными утверждениями о значении хордоме-
зодермы в «индукции» нервной системы в ходе онтогенеза.
ХИМИЧЕСКИЙ АСПЕКТ ИЗУЧЕНИЯ ДЕТЕРМИНАЦИИ
И ДИФФЕРЕНЦИАЦИИ КЛЕТОК И ТКАНЕЙ
Обратимся вновь к тому, что совершается в ходе нормального
развития, — к явлениям дифференциации и к анализу процессов,
названных Шпеманом индукцией, и разберем, что, собственно,
является взаимозависимостью частей в развитии.
В 50-х и 60-х годах XX в. в связи с возрастающим взаимо-
влиянием биологии, физики и химии и с использованием новых
методик (ауторадиография, электронная микроскопия и др.) сно-
ва повысился интерес к химии индукторов, хотя содержание это-
го понятия резко изменилось. Во-первых, считается неоснова-
тельным искать какое-либо одно формообразовательное вещество,
вызывающее индукцию. Во-вторых, все меньшее число исследо-
вателей уподобляет явления индукции, наблюдаемые в ходе
нормального развития зародышей, феномену мертвых организато-
ров. В-третьих, вместо гипотезы Шпемана об индукционных влия-
ниях организатора на «индифферентный» клеточный материал
утвердилась мысль о взаимозависимости частей в развитии за-
родышей.
В течение длительного периода, последовавшего после увле-
чения и разочарования химическими исследованиями (40-е и 50-е
годы), некоторые эмбриологи продолжали изучать природу ин-
дукторов. К ним относятся прежде всего С. Тойвонен и Т. Ямада,
которые вместе с другими учеными создали качественную гипо-
тезу индукторов. Тойвонен испытал действие сотен тканей разных
животных на их способность к индукции осевых зачатков у ам-
фибий. В 1938 г. он обнаружил, что некоторые индукторы обла-
дают качественно различным действием: ткань печени морских
свинок индуцирует почти исключительно передний мозг и его
производные (сначала передний мозг, затем глаза и балансеры).
Костный мозг морской свинки индуцируют туловищные и хвосто-
вые структуры (миотомы, хорду, почечные канальцы и т. д.).
Ф. Леман в 1950 г. предложил гипотезу, принятую Тойвоне-
ном, Ямадой п другими исследователями. Согласно ей, первичная
индукция (сущность действия организатора — дорсальной губы
бластопора) может быть вызвана всего двумя агентами, образу-
ющими два взаимоперекрывающихся градиента. Одно вещество
индуцирует исключительно переднеголовные (архенцефалические)
структуры, а другое—туловищно-хвостовые (дейтеренцефаличес-
кие) структуры. Если много архенцефалического агента и мало дей-
теренцефалического, то индуцируется передний мозг; если первого
агента относительно мало, а второго больше, то возникает
254
заднеголовная область; если мало
переднеголовного агента и много туло-
вищно-хвостового, то индуцируются
туловищно-хвостовые отделы. Все это
и имеет место, согласно гипотезе, в
нормальном развитии амфибий; надо
представить себе наличие определен-
ных индуцирующих веществ в соответ-
ствующих количественных комбина-
циях в разных участках зародыша.
С. Тойвонен и Л. Саксен в 1955 г.
проверили эту гипотезу эксперимен-
тально, поставив опыты на зародышах
амфибий с одновременной и раздель-
ной индукцией — печенью и костным
мозгом. Ткани печени и костного моз-
га или то и другое вместе пересажи-
вали в гаструлу тритона или завер-
тывали в. эктодерму гаструлы тритона
(метод «сэндвичей»). Как видно из
схемы на рис. 115, ткань печени инду-
цирует преимущественно образование
переднего мозга и его производных и
изредка также заднего мозга со слу-
ховыми пузырьками; индукция мезо-
дермы не наблюдается. Костный мозг
индуцирует главным образом мезодер-
мальные структуры; головные струк-
туры не образуются. В опытах с одно-
временным действием печени
Рис. 115, Схема действия полей
индукторов (по С. Тойвонену
и Л. Саксену, 1955):
А — два перекрывающихся поля
действия индукторов, вызванных
одновременно пересаженными в
гаструлу тканями костного мозга
(7) и печени (2); между ними по-
ле, на котором эти два действия
перекрываются (комбинированное
действие); Б — соответствующий
опыт, проведенный иа «эктодер-
мальном сэндвиче» с двусторон-
ним полем действия
и костного мозга возникли
структуры всех уровней тела
нормальной личинки. В ряде
случаев на брюшной стороне
хозяина развивались полные
вторичные зародыши, подоб-
ные тем, которые возникают
нередко и при трансплантаци-
ях верхней губы бластопора
или при действии мертвых ор-
ганизаторов.
Тойвонен предполагает, что
каждый из двух индукторов
образует свое активное поле,
при одновременном их дейст-
вии возникнет комбинирован-
ное поле (рис. 115).
Рис. 116. Схема взаимодействия двух,
градиентов (по С. Тойвонену, 1955):
N — нейрогенное влияние, М — мезодермали-
зующее влияние, А — переднеголовные (ар-
хенцефалические) индукции, D — заднеголов-
ные индукции, С — спинно-хвосговые индук-
ции; / — передний мозг, 2 —глаза, 3 — плако-
ды, *4 — балансеры
255-
На основании подобных опытов Тойвонен представляет себе
существование такого индукционного механизма у зародышей.
Должен существовать нейрогенный агент (N-агент), образующий
дорсолатеральный градиент, и мезодермальный (или М-агент)—
краниокуадальный и одновременно дорсолатеральный (рис. 116).
В передней части спинной области зародыша, т. е. в районе дей-
ствия нейрогенного агента, возникают головной мозг и его произ-
водные. Дальше назад по спине, где сильный нейрогенный агент
комбинируется со слабым мезодермальным, возникают спинной
мозг и слуховые пузырьки. В задней части спинной области более
сильно действие М-агента, поэтому центральная нервная система
развивается в спинной мозг, формируются сомиты, хорда и весь
комплекс мезодермальных структур.
Предположения и данные разных исследователей о химиче-
ской природе N- и М-агентов очень противоречивы. Предполага-
ют, что М- и N-агенты — это белковые соединения. X. Тидеман
(1968) извлекал из гомогенатов девятидневных куриных зароды-
шей индуцирующий мезодермальный фактор белковой природы
(относительная молекулярная масса 25000—30 000). Он инакти-
вируется тиогликолевой кислотой или меркаптоэтанолом, значит,
содержит сульфидный мостик цистина. При имплантации этого
вещества в гаструлу амфибии происходило во многих случаях
формирование мезодермальных структур. Трудно, однако, гово-
рить об этом веществе как о специальном «мезодермальном фак-
торе», так как в опытах кроме формирования мышщ- хорды и
почечных трубочек образовывались и энтодермальные ткани.
Считают, что индукция архенцефалических структур может быть
получена рибонуклеопротеинами. Однако чистые протеины из ко-
стного мозга и куриного эмбриона действуют как каудально-мезо-
дермальные индукторы.
Анимализации можно добиться уменьшением хирургическим
путем вегетативного материала или обработкой перед оплодотво-
рением некоторыми химическими веществами, такими, как KCN,
или обработкой в период дробления иодобензойной кислотой или
2,5-тиометилцитозином.
Вегетализация, т. е. более интенсивное развитие энтодермаль-
ных зачатков, может быть вызвана обработкой дробящихся яиц
литием или хлорамфениколом. К 70-м годам химия «индукторов»
оказалась столь же неясной, как и в период односторонних хи-
мических увлечений эмбриологов в 30-е годы.
Подробно изложенная интересная гипотеза Тойвонена не дает,
к сожалению, ничего принципиально нового по сравнению со ста-
рыми односторонне химическими схемами сущности индукторов и
организаторов, только вместо одного вещества думают о двух
или нескольких. Должны быть приняты во внимание следующие
очевидные недостатки гипотезы Тойвонена, на которые отчасти
указывает и сам автор. Во-первых, эта гипотеза говорит только
:256
Рис. 117. Вариации проявления тенденций «анимализации» и «вегетализации»
развивающихся яиц морских ежей (по Т. Густафсону, 1965):
А—Г — различные случаи «анимализированных» личинок; Д — нормальная гаструла; Е—
Ж — различные случаи «вегеталнзированных» личинок. В случае «Ж» эктодерма редуциро-
вана до маленькой «почкн». Анимализация (редукция вегетативного материала) производи-
лась или обработкой химическими веществами перед оплодотворением (например, SCN),
или обработкой в ходе дробления яйца иодобензойиой кислотой или 2,5-тиометилцитозином.
Вегеталнзацня производилась обработкой солями лития или хлорамфениколом по ходу
дробления яиц
об «индукторах» и совершенно не касается главного вопроса —
о «реагирующих» системах. Во-вторых, экспериментальное обос-
нование ее дано на основании действия каких-то веществ тканей
млекопитающих, а делается попытка объяснений явлений нор-
мального развития зародышей амфибий. Требуется доказать, что
выделенные Тойвоненом, Ямада и другими «индуцирующие веще-
ства» действительно присутствуют в нормальной гаструле. Если
они присутствуют, то каково их местоположение? Поэтому к ги-
потезе Тойвонена и сходным с ней относятся все те критические
замечания, которые были уже сделаны о шпемановской школе.
С данными Тойвонена перекликаются результаты опытов по экзо-
гаструляции (рис. 117).
О ВОЗМОЖНОСТИ РАЗЛИЧНОЙ ДИФФЕРЕНЦИАЦИИ КЛЕТОК
ВСЛЕДСТВИЕ РАЗНЫХ ХИМИЧЕСКИХ ВЛИЯНИЙ
И. Гольтфретер склоняется к мысли о том, что ключ для объяс-
нения индукции скорее следует искать в реагирующих клетках,
чем в индукторах. Конечно, любой формообразовательный про-
цесс имеет ту или иную биохимическую основу, но совершенно
необязательно представлять эту основу в виде специфических ин-
дуцирующих веществ. На примере партеногенеза мы знаем, что
агенты различной природы могут вызвать одно и то же явление.
Интересные данные о химических влияниях на дифференциацию
клеток презумптивного эпидермиса гаструлы лягушек (Rana pipi-
ens) в условиях культуры in vitro получены Л. Бартом и Л. Барт
в 1960, 1962 и 1963 гг. Доказано, в частности, что различные
концентрации LiCl обусловливают дифференцировку клеток пре-
257
зумптивного эпидермиса в разные типы клеток: нервные клетки,
пигментные, мускульные. В зависимости от концентрации, про-
должительности действия, температурных и иных условий и другие
вещества (NaHCOs, MgSO4, СаСЬ) могут вызывать разный эф-
фект —• разного типа дифференциацию клеток презумптивного
эпидермиса. Эти и другие интересные данные предостерегают от
излишне упрощенных представлений о химической природе ин-
дукторов и пока, к сожалению, не дают возможности судить об
истинной причине гистологической дифференциации в ходе раз-
вития организма, в частности о возможном химическом влиянии
хордомезодермы на обширную область эктодермы зародышей
амфибий, развивающуюся в нервную систему.
Б. П. Токин (1955) предполагает, что в ходе развития заро-
дыша возникают вещества, тормозящие клеточные деления и по-
вреждающие клетки эктодермы. В формировании нервной системы
могут иметь значение какие-либо альтерирующие воздействия
внешней среды, разность внешней и внутренних сред (см.
гл. XIV).
Л. Барт в 1941 г. убедился в том, что эктодерма Ambystoma
punctatum может развиваться в нервного типа структуры в усло-
виях эксплантации, в отсутствие каких бы то ни было организа-
торов. Особо интересен факт пропорциональной зависимости
«нейрализации» клеточного материала от степени дезагрегации
кусочков зародышей, вызываемой искусственной средой. Гольт-
фретер подтвердил и значительно расширил эти данные, связав
вопрос о «нейрализации» с изменением под влиянием различных
агентов поверхностей клеток, со способностью к «слипанию» и,
наоборот, к дезагрегации.
При снижении pH до 4,5 происходит уплотнение клеточных
мембран эксплантатов из зародышей A. punctatum, и в связи
с этим клетки дифференцируются по типу эпидермальных обра-
зований. Наоборот, если подвергать кусочки зародышей кратко-
временному действию неорганических кислот или щелочей, алко-
голя, дистиллированной воды или растворов без ионов кальция,
наступает набухание клеток, их дезагрегация, возникает амебоид-
ная подвижность клеток и, в случае более длительного действия,
цитолиз.
При перенесении эксплантата (после кратковременного дей-
ствия повреждающих веществ) в более нормальную среду проис-
ходит «реинтеграция клеток» и их «нейрализация». Чем больше
клетки приводятся к границе смерти, утверждает Гольтфретер,
тем больше приобретают тенденцию развиваться по типу нерв-
ной ткани. Следует подчеркнуть, что в опытах воздействия по-
вреждающими агентами наблюдается иногда не просто «нейра-
лизация», но и уродливое эмбриональное развитие, появление раз-
нообразных структур. Изложенные факты и предположения Гольт-
фретера отчасти сходны с мыслями о значении дезинтегрирую-
258
щих факторов при индукции вторичных зародышей (в опытах
с живыми и мертвыми организаторами).
Проникают ли индуцирующие субстанции в реагирующие клет-
ки или эффект индукции обусловливается контактом индуциру-
ющих и реагирующих тканей?
Этот естественно напрашивающийся вопрос дискутируется
начиная с 1932 г. Шпемановская школа не сомневалась
в необходимости тесного контакта индуктора и реагируклцего ма-
териала. Однако многие исследователи, среди них О. Мангольд
(11932) и Ж. Браше (1961), говорили о возможности диффузии
индуцирующих агентов. П. Вейс (1949) считал, что сущность ин-
дукционного действия —это реакции клеточных поверхностей.
Молекулы индуктора являются своего рода штампами, по шаб-
лону которых возникает новый набор молекул на поверхности
реагирующей клетки. Этот новый набор молекул и определяет
начало дифференциации клеток. Доказать правильность этой ги-
потезы Вейса химическим путем не удалось.
Электронно-микроскопические исследования не принесли че-
го-либо нового в споры о механизме индукционного действия.
Предполагают возможность пиноцитоза, ссылаясь на наблюдае-
мые благодаря электронной микроскопии маленькие пузырьки,
включающиеся в мембраны клеток презумптивной эктодермы.
С очень большими сомнениями их идентифицируют с веществами
индуктора. Сторонники перехода, проникновения индуцирующих
веществ прибегали к разного типа экспериментам. Пытались не
допускать контакта клеточных поверхностей, создавая различные
преграды для гипотетических химических веществ (слой желточ-
ной оболочки, целлофановая или агаровая пластинки, фильтры
из эфира целлюлозы); при этом использовались в опытах мембра-
ны с известными диаметрами пор (порядка от 230 до 3,4 мкм).
Результаты всех опытов не позволяют сделать выводов, хотя в ря-
де случаев вызывалась индукция. Толкование этих положитель-
ных случаев осложняется неясностью возможного влияния самих
использованных мембран (химически или механически дезинте-
грирующих реагирующие ткани).
В пользу проникновения химических индуцирующих веществ
косвенно говорят положительные результаты опытов с бесклеточ-
ными экстрактами из индукторов. Использовались, например,
фракции экстракта из куриного эмбриона, содержащего «мезо-
дермальный индуктор»: возникали нервные клетки, меланофоры
или мышечные волокна и клетки хорды.
Ж. Сирлин (1956), С. Брама и К. Уоддингтон (1956, 1959) и
другие авторы использовали метод радиоактивных меток. Для
этого вводили в ткани зародыша-донора меченные по S или С
соединения (метионин, глицин и др.). Затем индуцирующие части
трансплантировали в зародыш-реципиент, вызывая индукцию либо
нервной пластинки, либо других зачатков. Исследовалось, содер-
259
жатся ли радиоактивные атомы в индуцированных из тканей хо-
зяина образованиях? В результате была обнаружена высокая ра-
диоактивность в индуцированной нервной пластинке, и нередко
менее радиоактивными оказывались мезодермальные и эктодер-
мальные ткани хозяина. Эти эксперименты, однако, не дают до-
казательства того, что имеет место проникновение «индукторов»
именно в реагирующие клетки, так как большая концентрация
радиоактивных веществ в индуцированной нервной пластинке мо-
жет еще проще объясняться более высоким обменом веществ в ней.
Эти опыты также не дают доказательств в пользу предположений
о концентрации радиоактивных веществ на поверхности клеток
индуцированных структур. Основной недостаток метода — полная
неясность вопроса, проходят ли меченые субстанции в их исход-
ной форме (аминокислоты или нуклеиновые кислоты), или же они
окажутся включенными в другие макромолекулярные соединения,
которые и будут иметь какое-либо значение в дифференциации
клеток? Значит, если в опытах такого рода и убеждаешься, что
те или иные меченые вещества проникнут через клеточные мем-
браны, нельзя, однако, решить, имеет ли это отношение к индук-
ции.
Многие исследователи использовали иммунохимические методы.
Д. Раундс и К. Фликинжер (1958) культивировали химерные экс-
плантаты, состоявшие из мезодермы лягушки и эктодермы три-
тона. Через 4—5 дней культивирования ткани отделяли и изуча-
ли антигенные свойства эктодермы тритона: содержатся ли в ней
антигены лягушки? Результаты опытов позволили сделать за-
ключение о переходе белков лягушки в ткани тритона. Конечно,
вывод этот не очень обоснован, так как возможны «загрязнения»
энтодермы клетками (или их обрывками) мезодермы.
С. Тойвонен на основании своих опытов и данных других ав-
торов предположил, что антигенный материал не только может
прилипать к поверхности клеток, но и проникать в протоплазму,
несмотря на то, что речь идет о макромолекулярных частицах,
подобных белкам. Таким образом, по-прежнему химия индукции
неясна.
ДЕГЕНЕРАЦИЯ И СМЕРТЬ КЛЕТОК В ХОДЕ ПРОЦЕССОВ
ФОРМООБРАЗОВАНИЯ
Заслуживают большого внимания различного рода деструктивно-
го характера процессы, закомерно наступающие в онтогенезе жи-
вотных. Как уже отмечалось, на ранних стадиях развития заро-
дышей вступают в действие факторы, тормозящие митозы в одних
частях зародыша и стимулирующие их в других частях. В резуль-
тате взаимодействия клеток и тканевых систем, при непрерывно
меняющихся состояниях интеграции зародыша закономерно на-
ступают явления дегенерации и смерти клеток в тех или иных
260
частях эмбриона. Особенно демонстративны деструктивные процес-
сы, происходящие при метаморфозе животных—насекомых, асци-
дий, амфибий и др. И процессы регенерации, как правило, сопро-
вождаются гибелью клеточных элементов.
При развитии мочеполовой системы у млекопитающих проис-
ходит редукция предпочки и другие процессы, при которых имеет
место дегенерация и смерть значительных клеточных масс.
Деструктивным процессам в эмбриогенезе в современной эмб-
риологии уделяется большое внимание. Так, Ж. Фросберг и
Б. Кёллен (1968) считают, что процессы клеточной гибели в он-
тогенезе имеют не меньшее значение, чем клеточное размножение
и дифференциация клеток.
Удовлетворительной теории этих явлений, однако, не создано,
и каждый исследователь рассматривает деструктивные процессы
в рамках своих, подчас односторонних научных интересов. Так,
Ж. Сондерс и Ж. Фаллон (1966) предлагают рассматривать ги-
бель клеток в эмбриогенезе как явление, запрограммированное
генетически. Это утверждение ничего не объясняет, ибо в конеч-
ном счете все определяется генотипически.
Приведем примеры наблюдающихся деструктивных процессов.
Дегенерация и смерть клеток в яркой форме наблюдаются при
развитии нервной системы и органов чувств у позвоночных. Ги-
бель клеток неизбежно происходит при резорбции провизорных
органов, как уже указывалось, например, при редукции пронефро-
са, при резорбции Мюллерова протока эмбрионов — самцов. Дест-
руктивные процессы имеют место у позвоночных животных при
изменениях формы зачатков органов, разделении слоев клеточных
масс, при слиянии закладок органов, образовании полостей ит. п.
При развитии плеча и предплечья куриного зародыша, особенно
локтевого сустава, возникают строго локализованные участки не-
кроза клеток. Выяснено, когда и какая группа клеток дегенери-
рует. Если клетки, неизбежно подвергающиеся гибели в опреде-
ленное время и в определенном месте, пересадить в.иные части за-
родыша, они не погибают и участвуют в формообразовательном
процессе. Придается значение тому факту, что умирающие клетки
выделяют какие-то вещества, стимулирующие митоз в соседних
клетках.
Некоторые авторы создали конкретные предположения о том,
какую роль играет гибель клеток в формообразовательных про-
цессах. Так, А. Глюксман (1951) считает, что гибель части клеток
создает условия для морфогенетических перемещений клеток (на-
пример, при инвагинации, разделении слоев клеток и т. п.).
Б. Менкесом и другими исследователями (1964) обнаружено, что
дегенерация в спинальных ганглиях, в сомитах, склеротоме, мио-
томах, зачатках глаза и в других местах зародышей разных мле-
копитающих мышей, крыс, кроликов, человека — происходит на
определенных стадиях онтогенеза.
261
Гибель клеток в определенных местах необходима, чтобы про-
изошло соединение или разделение частей органов или образова-
ние просвета в какой-либо плотной структуре. Так, гибель клеток
происходит при «разделении» пальцев на руках и ногах, при от-
делении губы от десен, при формировании век и образовании
центральных каналов тех или иных протоков.
Вряд ли следует искать какую-то одну закономерность, кото-
рая давала бы объяснение всем разнообразным случаям деструк-
тивных процессов, втречающихся на стадиях дробления, форми-
рования зачатков органов, при метаморфозе и т. д. Может быть,
помимо специальной формообразовательной роли деструктивных
процессов, в каждом частном случае имеются все же и какие-то
общие морфофизиологические черты.
Так, если верна теория Э. С. Бауэра, то следует думать, что
при гибели части живой материи (клеток, межклеточных веществ,
тканей) освобождается энергия, которая служит для поддержа-
ния неравновесного состояния остающейся живой материи.
ТЕОРИЯ М. М. ЗАВАДОВСКОГО О ПРОТИВОРЕЧИВОМ
ВЗАИМОДЕЙСТВИИ МЕЖДУ ОРГАНАМИ В ТЕЛЕ
РАЗВИВАЮЩЕГОСЯ ЖИВОТНОГО
Среди многих теорий, касающихся взаимного влияния различных
систем развивающегося организма, особый интерес представляет
теория М. М. Завадовского, экспериментально обоснованная еще.
в 30—40-х годах. Интерес эмбриологов к этой теории не умень-
шается, чему способствует, вероятно, родство представлений За-
вадовского современным понятиям кибернетики об обратной свя-
зи. На теории Завадовского следует остановиться еще и потому,
что она, касаясь главным образом явлений постнатального раз-
вития, убеждает эмбриологов в необходимости изучать взаимо-
действия «частей», а не односторонних гипотез об «индукции».
По Завадовскому, разввдие любого признака на любой стадии
онтогенеза осуществляется именно на основе взаимодействия «ча- -
стей». Конечно, развивающийся организм — это целостная систе-
ма, но, чтобы понять отдельные явления, мы должны «вырывать»
их из всеобщей связи. Развитие особи осуществляется в единстве
превращений формы и функции, друг друга обусловливающие.
Автор использовал при создании своей теории преимущественно
данные о формообразовательной роли желез с внутренней секре-
цией и особенно результаты своих классических исследований
развития половых признаков у птиц и млекопитающих. Зависимые
половые признаки, например гребень, сережки, бородка, голосовой
аппарат, половой инстинкт и другие признаки петуха, развивают-
ся под воздействием половых гормонов семенника, так же как
зависимые половые признаки курицы и женских особей других
видов животных развиваются под влиянием яичника (рис. 118).
262
Рис. 118. Форма перьев петуха и курицы куропатчатой итальянской породы (по
М. М. Завадовскому, 1922):
1 — гребень, 2 — бородка, 3 — сережки, 4 — щека, 5 — грива, 6 — бахрома, 7 — большие сер-
повидные, 8 — малые, 9 — гузковые, 10— плечевые, // — поддерживающие, /2 — покрываю-
щие, 13 — маховые первого порядка, 14 — маховые второго порядка, /5 —грудь, /6 —живот
Половые признаки касаются формы и пигмента пера, шпор,
размеров и окраски гребня, бородки и сережек, посадки корпуса,
пения, спаривания, насиживания яиц, половых проводящих путей,
половых желез.
Рост бороды и усов, огрубение голоса и пробуждение инстин-
кта у юноши и формирование вторично-половых признаков совер-
шается в период полового созревания. Кастрация мужчцн приво-
дит к выпадению у кастрата ряда вторично-половых признаков —-
не развиваются борода и усы, голос сохраняет детские интонации.
У стареющих женщин вместе с угасанием функции яичника не-
редко начинается рост волос наподобие бороды и усов.
Завадовский удалял у неполовозрелых петухов оба семенника.
В результате — резкие отклонения в развитии гребня, бородки и
сережек, они малого размера и сохраняют ювенильную блед-
ность. Если кастрировать половозрелого петуха с хорошо разви-
тым головным убором, то гребень, бородка и сережки
постепенно сморщиваются, уменьшаются в размерах. Это
заметно уже на третий-четвертый день после кастрации. Кастри-
рованный петух теряет половой инстинкт. Если пересадить на шею
кастрированного петуха (в период, когда вторично-половые приз-
наки еще не отчетливо выражены) семенник, взятый от другой
263
Рис. 119. Опыт «превращения полов» (по М. М. Завадовскому, 1922):
Л —нормальный петух; В —нормальная курица; В — кастрированный петух; Г —кастриро-
ванная курица; Д — кастрированный петух после имплантации ему яичника; Е — кастриро-
ванная курица после имплантации ей семенников
особи той же породы, то через 4 месяца обнаруживается усилен-
ный рост и порозовение гребня, до того очень бледного. Через
8 месяцев гребень значительно превосходит в размере гребни кон-
трольных. нормальных петухов. Гистологическое исследование
приживленного семенника показало, что крупные кровеносные со-
суды хорошо снабжают ткани семенника.
Итак, формирование головного убора, петушиного голоса и
инстинкта совершается при участии гормона семенника.
При кастрации курица после первой же линьки надевает пету-
264
шиный наряд как по форме, так и по окраске, на ногах выраста-
ют шпоры (рис. 119). Если пересадить семенники кастрированным
курочкам, то в случае хорошего приживления гормоны семенника
могут вызвать образование зависимых самцовых признаков из
тканей самки: развивается петушиный гребень, головной убор,
курица начинает петь по-петушиному. Замечательно, что Зави-
довский на основании многих своих опытов в 40-х годах сделал
вывод о том, что ткани самца тождественны тканям самки, что
под влиянием полового гормона самца они дифференцируются
в ткани самца, а под влиянием женского полового гормона —
в ткани самки. У птиц самка гетерозиготна, у нее оба гормона.
Не касаясь исследований последних десятилетий химии половых
гормонов и их формообразовательной роли, следует отметить, что
выводы Завадовского о гетеросексуальности кур вполне соответст-
вуют представлениям генетики о гетерогаметности самки (они не-
сут ХУ хромосомы, а самцы — XX хромосомы).
Подобные описанным эксперименты проведены Завадовским
на фазанах и других птицах, а также на млекопитающих.
Представление о том, что тот или иной вторично-половой при-
знак развивается из тканей покровов головы при участии полово-
го гормона семенника, например петушиный гребень развивается
из тканей покровов головы, Завадовский выразил формулой:
X + Y—*А, где X — ткани покровов головы; У — половой гормон
семенника; А — символ петушиного гребня. Это «алгебраическое»
выражение того, что между X и У существует взаимодействие,
которое необходимо для развития признака.
Было показано, что, как правило, там, где одна система оказы-
вает влияние на другую, то и эта другая оказывает влияние иа пер-
вую, образуя взаимодействующую систему: если У оказывает вли-
яние на X, то и X оказывает в свою очередь влияние на У. Если
отрезать гребень у петуха, то можно убедиться в том, что гребень,
развившийся под влиянием семенника, в свою очередь оказывает
влияние на жизнь семенника: у петухов со срезанными гребнями
семенники развиваются значительно сильнее, чем у нормальных,
контрольных петухов, происходит увеличение массы семенников
в 1,5—2 раза. Гребень оказывает какое-то формативное влияние
на развитие семенника, а семенник стимулирует гребень. Взаимо-
влияние этих органов имеет взаимно противоречивый характер.
Завадовский выражает это взаимодействие как «плюс — минус
взаимодействие». Семенникч^гребень. В опытах на следующих
системах автор убедился в правильности своих представлений.
1. Семенник — гребень. 2. Яичник — матка. 3. Семенник — се-
менные пузырьки. 4. Гипофиз (гонадотропный) —яичник. 5. Гипо-
физ (гонадотропный) семенник. 6. Гипофиз (тиреотропный) —
щитовидная железа. 7. Гипофиз (лактогенный)—яичник.
Приведем один пример. Яичник млекопитающих стимулирует
развитие матки. Если удалить яичник, то матка атрофируется.
265
Опыт кастрации коров и свиней, а также наблюдения в гинеколо-
гических клиниках подтверждают это.
В результате опытов по влиянию гонадотропных (гонада —
половая железа, тропность — направленность действия) гормонов,
а именно: гонадотропного гормона гипофиза, гонадотропного
агента, находящегося в крови и сыворотке жеребой кобылы, в мо-
че беременных женщин и других препаратов, — происходит гипер-
трофия матки и всей системы вторично-половых признаков. Если
удалить у молодых крыс матку, то оказывается, что их яичники
развиваются сильнее, чем у нормальных крыс. Выходит, что мат-
ка как-то тормозит функцию яичника.
Завадовский сделал вывод о том, что вследствие противоречи-
вого взаимодействия между частями развивающегося организма он
«представляет собой саморегулирующуюся систему, с высокой
степенью устойчивости, так как регуляция присуща не только
организму как целому, но и каждому звену в отдельности».
Как видим, теория М. М. Завадовского обосновывается мно-
гими фактами. Однако, конечно, нельзя считать формулу
Х-|-У—>-А выражением любого формообразовательного процесса.
М. М. Завадовский, как обычно происходит в творчестве каждо-
го ученого, вносящего реальный вклад в науку, распространил
свою теорию на больший круг явлений, чем она способна объяс-
нить.
Основным недостатком этой теории следует считать представ-
ление о реактивном характере всех процессов формообразования
(см. гл. XIV). Однако основанная преимущественно на явлениях
формообразования в послезародышевом периоде развития высших
позвоночных, она может претендовать на неменьшую научность,
чем теория развития Г. Шпемана, которая также выхватывает из
целого организма те или иные единичные системы. Завадовский
вправе применять свою теорию «плюс-минус взаимодействия» и
к результатам опытов Шпемана и Льюиса по пересадке глазного
бокала тритона в различные части тела, в результате чего «инду-
цируется» образование роговицы, а также к результатам опытов
Д. П. Филатова—пересадка зародышевого слухового пузырька,
что вызывает индукцию формирования хрящевой капсулы из сое-
динительной ткани.
Согласно современным данным, как об этом говорит Б. Г. Но-
виков (1975, 1981), приводимые М. М. Завадовским примеры для
подтверждения правильности теории не совсем удачны. Прежде
всего для взаимодействия вторично-половых признаков с гонада-
ми нет каналов для отрицательной обратной связи. Принцип об-
ратных связей в регуляции функции гонад замыкается на уровне
гипоталамуса. Точно так же существуют три уровня регуляции
функции щитовидной железы: авторегуляция (базальная актив-
ность), регуляция на уровне гипофиза и на уровне гипоталамуса.
Гормональная жё бисексуальность на самом деле присуща обоим
266
полам, поэтому она не связана с различиями в генетической ор-
ганизации самца и самки. Установленный Завадовским принцип
эквипотенциальности тканей самца и самки не носит универсаль-
ного характера. Даже в пределах относительно узких системати-
ческих групп можно построить ряд постепенного перехода от за-
висимого к независимому, от гормона, типу развития одного и того
же признака. Следовательно, если у одних форм генная актив-
ность, определяющая развитие полового признака, контролирует-
ся половыми стероидами, то у других их индуцирующая роль от-
сутствует. Б. Г. Новиков высказал предположение о том, что воз-
никновение половых признаков и их превращение в видовые обу-
словлено появлением особых систем рецепторных клеток, которые
в ходе эволюции испытывают соответствующие преобразования
(т. е. изменяется их компетенция).
Можно внести и другие поправки и замечания, касающиеся
аргументации теории М. М. Завадовского, однако она, несомнен-
но, внесла большой вклад в понимание взаимодействия между
частями развивающегося организма.
ЖИЗНЕННЫЕ ЦИКЛЫ КЛЕТОК В ХОДЕ РАЗВИТИЯ ОРГАНИЗМА
Для оценки важности биохимического аспекта исследований эм-
брионального развития следует вспомнить сообщенные ранее
данные молекулярной биологии. В дальнейшем (гл. XII) будут
рассмотрены вопросы о значении молекулярно-биологических и
генетических данных, касающихся проблемы дифференциации кле-
ток и тканей. В 60—80-х годах под влиянием прогресса биохимии,
генетики и цитологии появились новые аспекты исследования
дифференциации клеток и тканей. Интересные исследования, свя-
занные с представлениями об онтогении клетки, ее жизненном
цикле. Большинство авторов предпочитают термин «митотический
цикл», но, конечно, этот термин более узкого содержания, чем
термины «жизненный цикл» или «онтогения клетки».
В индивидуальной жизни клетки отмечают постмитотическую
(Gi) фазу, фазу редупликации (-синтетическую, S), постсинтети-
ческую (G2) и стадию митоза (М). Утверждают, что в случае
дифференциации клетки образования ДНК не происходит, а имеет
место синтез различных специфических белковых соединений.
В отличие от биохимических процессов, совершающихся при обыч-
ном «митотическом цикле», когда «интерфаза» именуется автосин-
тетической, в случае дифференциации клетки «интерфазу» назы-
вают гетеросинтетической. Считается, что клетки в этом случае
выходят из цикла размножения. Обосновывается целесообраз-
ность выделения «особого периода неустойчивого состояния кле-
ток, когда они как бы «принимают решение»: или вступить на
путь дифференциации, или начинать новый митотический цикл.
267
Этот период принято обозначать Ci»1. Описанная схема клеточной
дифференциации — только частный случай специализации клеток,
требующий дальнейших исследований.
Несмотря на существование противоречивых данных об анта-
гонизме между синтезом ДНК и синтезом специфических белков,
исследования особенности клеточных циклов в ходе эмбриональ-
ного развития представляют большой интерес для эмбриологии.
Доказано, что может быть и совпадение (одновременное протека-
ние) процессов синтеза ДНК и специфических белков. Мыслима
дифференциация клеток без потери хотя бы и временной способ-
ности к размножению. В ходе нормального развития организма,
благодаря интеграционным механизмам тканей и зародыша как
целого, закономерно возникает локализация митотической актив-
ности клеток, подавление митозов в определенных участках, деге-
нерация клеточных элементов и т. д. Таким образом, исследова-
ние «митотических циклов» в эмбриональном развитии выходит
за рамки изучения соотношения синтеза ДНК и синтеза специфи-
ческих белков.
Обширные исследования ряда цитологов (О. И. Епифанова,
А. А. Заварзин, А. К. Дондуа и др.), естественно, интересуют эм-
бриологов. Приведем примеры некоторых исследований. Л. Ф. Ан-
дреева (1970) методом авторадиографии с применением Н3-тими-
дина изучала синтез ДНК и длительность «клеточных циклов»
(онтогении клеток) в эмбриогенезе морского ежа Strongylocentro-
tus droebachiensis на стадиях поздней бластулы, гаструлы и
плутеуса. В ходе развития эмбрионов продолжительность «клеточ-
ного цикла» непрерывно увеличивается. На стадии поздней бла-
стулы включение Н3-тимидина в ДНК наблюдается в течение
всей интерфазы. На стадии средней-поздней гаструлы происходит
разделение интерфазы на три периода: Gi, S и G2. Доля периода
интенсивного синтеза ДНК непрерывно уменьшается. От стадии
поздней гаструлы до плутеуса продолжительность клеточного
цикла все время увеличивается, а доля периода синтеза ДНК в
клеточном цикле сокращается.
В период дробления основной пик синтеза ДНК приходится на
телофазу и раннюю интерфазу. На стадии средней бластулы наи-
более активный синтез ДНК происходит в середине интерфазы.
Во время гаструляцин — во второй половине интерфазы. Таким
образом, утверждает Л. Ф. Андреева, в ходе эмбрионального раз-
вития на стадии ранней интерфазы синтез ДНК становится все
меньшим и возрастает количество ДНК, редуплицирующейся на
поздних стадиях интерфазы.
Какой биологический смысл в этой закономерности, сказать
пока трудно. Ряд исследователей считают, что имеется корреляция
1 Заварзин А. А. Синтез ДНК и кинетика клеточных популяций в онтоге-
незе млекопитающих. Л., 1967. С. 6.
268
между временем редупликации генов (ДНК) и его функциональ-
ной активностью. Происходит становление характерного для сома-
тических клеток клеточного цикла с разделением интерфазы на
три периода (Gi, S и G2).
Некоторые данные имеются о митотических циклах в ходе
сперматогенеза и оогенеза. Общая продолжительность «митоти-
ческого цикла» на разных этапах сперматогенеза 27—30 ч. У спер-
матогоний первых генераций имеется продолжительная фаза G2
около 11—14 ч, и в соматических клетках она 2—3 ч. В последу-
ющих генерациях сперматогоний время G2 сокращается до 4,5 ч.
Отмечается увеличение продолжительности фазы синтеза ДНК: У
первичных генераций сперматогоний время S 7—8 ч, а время син-
теза ДНК сперматогоний последней генерации 14,5 ч. Такие же
факты обнаружены в отношении оогоний.
На определенных этапах гаметогенеза синтез ДНК прекраща-
ется. В связи с процессом оплодотворения в женском и мужском
пронуклеусах (еще до их соединения) синтез ДНК возобновляет-
ся. Редупликация ДНК у пронуклеусов морских ежей наступает
через 15—20 мин после оплодотворения.
О том, происходит ли синтез ДНК на стадиях дробления, гово-
рилось ранее. Установлено, что синтез ДНК имеет место у эмбри-
онов всех животных с самых первых этапов развития.
Дондуа на основании данных разных исследователей делает
следующие обобщения об особенностях митотических циклов в пе-
риод дробления: 1) отсутствие премитотической фазы Gi; 2) не-
большое удельное значение фазы синтеза ДНК в цикле (10—
20%); 3) высокое удельное значение фазы G2 и митоза и значи-
тельная скорость протекания синтеза ДНК- Автор и его коллеги
провели обширные исследования митотических циклов в период
формирования зачатков различных органов на зародышах птиц
и млекопитающих. Наиболее важный вывод заключается в том,
что в период, когда при формировании тканей зачатки органов
вступают в дифференциацию, при которой синтезируются специ-
фические для данной ткани белки, митотические циклы приобре-
тают особые черты: снижается пролиферативная активность, клет-
ки переходят из авто- в гетеросинтетическую фазу, происходит
выключение синтеза ДНК. Однако приводятся случаи сочетания
и синтеза ДНК и синтеза специфических белков.
Изложенные материалы, несмотря на дискуссионность их ин-
терпретации разными исследователями, как уже сказано, пред-
ставляют интерес для эмбриологии.
И. И. ШМАЛЬГАУЗЕН ОБ ОРГАНИЗМЕ КАК ЦЕЛОМ
В ИНДИВИДУАЛЬНОМ РАЗВИТИИ
В заключение этой главы остановимся на некоторых идеях
И, И. Шмальгаузена, внесшего большой вклад в эволюционную
морфологию, сравнительную анатомию и эмбриологию.
269
Вопросы об интегрирующих факторах онтогенетического и фи-
логенетического развития до исследований Шмальгаузена совер-
шенно не разрабатывались. Были лишь эпизодические высказыва-
ния многих авторов. Проблемой интеграции занимались физиоло-
ги, а эмбриологи особое внимание уделяли проблеме дифферен-
циации. Однако форма и функция неразрывно связаны друг с дру-
гом и морфологические явления имеют и свою физиологическую
сторону. Резко критикуя идеалистические построения Г. Дриша и
других о целом как о чем-то предшествующем частям и опреде-
ляющем до конца все свойства клеток, тканей и органов, Шмаль-
гаузен дает строго научную, материалистическую интерпретацию
понятия «целое» (см. гл. XIII). Он ставил и разрешал вопросы об
историческом становлении взаимозависимостей, о возникновении
и эволюции корреляционных систем.
Ни на одной стадии развития организм не есть мозаика час-,
тей, органов или признаков. Организм на данной стадии развива-
ется всегда как специфическое целое. Сложная система связей,
т. е. корреляций, обусловливает развитие организма как целого.
Прогрессивное усложнение организации (как в филогении, так
и в онтогенезе) означает дифференциацию целого и специализа-
цию частей. Отдельные «части» приобретают свои самостоятель-
ные функции. Однако любая специализированная часть связана с
остальным организмом, и—подчеркивает Шмальгаузен — чем
более специализирована часть, тем более она становится в зави-
симость от других частей организма, обеспечивающих выполнение .
основных жизненных функций всего организма (питания, дыха-
ния, выделения и др.). Следовательно, «дифференциация и спе-
циализация неизбежно связаны с соподчинением частей, с инте-
грацией».
«... Любой орган имеет свои специфические функции. Однако,
с одной стороны, эти функции подчинены всему организму в це-
лом и, с другой стороны, они осуществляются через посредство
функциональной деятельности составляющих данный орган тканей
и клеток. Каждая высшая система контролирует и координирует
работу низшей. В этом выражается и взаимосвязь циклов регуля-
ции, которые не изолированы друг от друга, а соединены всегда
входными и выходными каналами связи с другими регуляторны-
ми циклами. Вместе с тем в функциях высшей системы осущест-
вляется не суммирование деятельности низших систем, а их инте-
грация. В каждой высшей системе проявляется своя качественная
специфика, которая создается только организацией этой высшей
системы. Поэтому мы и при рассмотрении формообразовательных
систем должны учитывать уровень их интеграции»1.
Центральной мыслью Шмальгаузена о закономерностях онто-
1 Шмальгаузен И. И. Регуляция формообразования в индивидуальном раз-
витии. М., 1964. С. 122.
270
генеза является следующая: «... движущие силы индивидуально-
го развития создаются по мере дифференцировки зародыша
в результате взаимодействия продуктов этой дифференцировки.
Взаимодействие разных частей ведет к новым дифференцировкам
и дальнейшим взаимодействиям. Устойчивость организации поко-
ится не на прочности каких-либо структур, а на сложности систе-
мы взаимозависимостей (корреляций) и на регуляторном их ха-
рактере. В результате этих взаимозависимостей организм разви-
вается на всех стадиях как одно согласованное целое,.. Вместо
прежнего, недостаточно определенного термина корреляция я
употребляю теперь гораздо более выразительный русский термин
«взаимодействие»1.
Из этой важной идеи вытекает много выводов, в частности
отношение Шмальгаузена к школе Шпемана, данные которой из-
ложены в настоящей главе. «Мы можем лишь условно говорить
об индукторе и реакторе. В процессе взаимодействия они постоян-
но меняются местами. Всегда реагирующая часть оказывает и об-
ратное влияние на индуктор». «... В большинстве формоообразо-
вательных систем обоюдный характер зависимостей совершенно
ясен, и выделение индуктора как более активной части системы
имеет в значительной мере условный характер»2.
Эмбриологам важно подчеркнуть, что Шмальгаузен говорил
о корреляции не в статическом смысле сосуществования призна-
ков, а о закономерностях в соотносительном их изменении.
Шмальгаузену было чуждо какое-либо одностороннее — хими-
ческое или иное — представление о корреляциях. Конечно, можно
строить классификацию корреляций (взаимодействий) на основа-
нии различных критериев; например, по характеру связи — хими-
ческие, физические и иные. Можно классифицировать взаимосвя-
зи по характеру осуществляемых изменений (стимуляция, депрес-
сия и т. п.) или по специфическим биологическим факторам онто-
генеза, при посредстве которых реализуется данная взаимозависи-
мость. Оговаривая, что нельзя провести твердые грани между раз-
ными типами корреляций, Шмальгаузен в 1938 г. предложил клас-
сификацию, не потерявшую значения и в настоящее время. Он
говорил о геномных, морфогенетических и арготических взаймо-
зависимостях.
Геномные зависимости могут осуществляться, по-видимому,
непосредственно генами, через какие-то биохимические процессы,
совершающиеся в клетках, в которых реализуется изменение; это
зависимости не двух или более частей друг от друга, а от одного
или нескольких связанных генов.
Так как новые и новые движущие силы индивидуального раз-
вития создаются по мере дифференцировки зародыша, а значит,
1«^vfta'iaRA/3eH И' И' Регуляция формообразования в индивидуальном раз-
ВИТИИ» IУ04.
2 Там же. С. 59.
271
не обязательно проводить в каждом случае прямую линию от того
или иного гена ко всем возникающим в онтогенезе взаимодейст-
виям (хотя все события в онтогенезе в конечном счете обусловле-
ны генотипически), Шмальгаузен выделяет морфогенетические
корреляции. Это взаимозависимости между двумя (или более)
морфогенетическими процессами.
Огромное число таких корреляций, как мы видели, установле-
но школой Шпемана: хордомезодерма и развитие нервной систе-
мы из прилегающей эктодермы; преобразование эктодермы в на-
ружную часть роговицы под влиянием хрусталика глаза; зачаток
глаза определяет развитие хрусталика и т. д.
Шмальгаузен выделяет эргонтические корреляции, иначе —
функциональные. Это соотношения между частями (например, ор-
ганами), обусловленные функциональными зависимостями между
ними. «При изменении одного элемента корреляционной пары его
функция изменяется, и это изменение сказывается и в изменени-
ях строения связанного с ним другого элемента. Я подчеркиваю,
что здесь говорится не просто о зависимости в функциях, а о по-
бочном формообразовательном эффекте этой зависимости»1.
Эргонтические корреляции — это зависимости, устанавливаю-
щиеся через дифинитивные функции, характерные для различных
органов уже сформированного организма. Примеры таких корре-
ляций: зависимость между развитием нервных центров, нервов и
развитием периферических органов; зависимости между развити-
ем мышцы, нервов и кровеносных сосудов, снабжающих мышцу;
зависимость между развитием мышцы и формированием скелет-
ных частей, к которым эти мышцы прикрепляются, и т. п.
Различные типы корреляций характерны для этапов индивиду-
ального развития. «Так как морфогенетические процессы сами
приводят к новым соотношениям, новым связям, то они вместе
с тем вносят такие изменения в процессе развития, которые вы-
зывают новые морфогенетические процессы. Таким образом раз-
вертывается целая цепь взаимно обусловленных процессов формо-
образования» 1 2.
Как видим, анализ явлений дифференциации и детерминации,
который был дан в настоящей главе, не противоречит высказы-
ваниям выдающегося естествоиспытателя.
В настоящей главе мы познакомились с большим количеством
фактов, гипотез. Этим далеко не исчерпываются ни факты, ни ги-
потезы, относящиеся к проблеме детерминации. Отчасти этой про-
блемы мы касались, давая анализ ранних стадий развития заро-
дышей (гл. IV, V и др.). В дальнейшем мы в сущности будем про-
должать изучать центральную проблему эмбриологии почему раз-
вивается организм и почему происходит дифференциация.
1 Шмальгаузен И. И. Организм как целое в индивидуальном и историческом
развитии. М., Л., 1938. С. 20—21.
2 Там же. С. 23.
272
Глава XII
ЭМБРИОЛОГИЯ И ГЕНЕТИКА
ПОГРАНИЧНЫЕ МЕЖДУ ЭМБРИОЛОГИЕЙ И ГЕНЕТИКОЙ ВОПРОСЫ
Генетика и эмбриология должны быть родными сестрами. В своем
прогрессе и в своих заблуждениях они влияли друг на друга.
Связь между- этими науками в первой четверти века персонифи-
цировалась в лице Т. Моргана — экспериментального зоолога и
эмбриолога и одного из основателей современной генетики.
В 20—30-е годы генетика удовлетворялась такими представле-
ниями, согласно которым от генов, как от определенных мате-
риальных точек хромосом, проводились прямые линии к призна-
кам организма, минуя процессы его развития. Все основные гене-
тические гипотезы того времени были созданы фактически вне
связи с эмбриологией. У генетиков того времени не было потреб-
ности в изучении эмбрионального развития даже основного объ-
екта исследований — плодовой мухи дрозофилы. Оперируя поня-
тиями «ген» и «признак взрослого организма», генетика в сущ-
ности игнорировала онтогенез и все эмбриологические данные.
Устанавливались математически точные корреляции между тем,
что обнаруживалось в хромосомном аппарате половых клеток и у
взрослых организмов. О соотношении половых и соматических
клеток господствовали представления, столь ярко выраженные
у одного из наиболее физиологично мыслящих генетиков того вре-
мени Р. Гольдшмидта. Он сравнивал (1923) положение половых
клеток в организме с положением паразита, с положением цари-
цы в государстве термитов: сама она не принимает активного уча-
стия в жизни государства, ее содержат рабочие для того, чтобы
она могла дать новые поколения и выполнить этим свое единст-
венное назначение. Эти уцрощенные неправильные идеи не могли
способствовать связи генетики с эмбриологией, и эти две науки
во многих проблемах оказывались не «родными сестрами»,
а «троюродными тетками»: генетика сама по себе и эмбриология
сама по себе. Однако вместе с бурным прогрессом генетика все
более сближалась с эмбриологией, физиологией и биохимией. Это-
му способствовали следующие открытия и обобщения генетики и
эмбриологии.
«Любой признак организма определяется многими генами, в
конечном счете всем генотипом. С другой стороны, каждый ген
обладает множественным, т. е. плейотропным, эффектом»1.
1 'Лобашев М. Е. Генетика. Л., 1963. С. 411—412.
273
«... Развитие признака как детали многоклеточного организ-
ма определяется всей системой генотипа в процессе многоступен-
чатых и взаимосвязанных актов»1. «Гены отнюдь не являются за-
чатками признаков, которые растут и распределяются при разви-
тии особи ... Признаки организмов не передаются через мате-
риальную наследственную систему в готовом виде. Они в каждом
новом поколении развиваются заново. По наследству передаются
только клеточные структуры, которые представляют собой мор-
фофизиологическую систему наследственных элементов»1 2.
Было бы проще для эмбриолога и генетика думать, что все
признаки взрослого организма имеют непосредственное матери-
альное представительство в ядре исходной оплодотворенной яй-
цевой клетки, что является абсурдом с химической и физической
точек зрения. Наследственность, выраженная в «проморфологии»
яиц (см. гл. IV и XI), обусловливает совершенно специфический
для данного вида организма путь развития, и нет оснований про-
ецировать со взрослого состояния организма все его признаки на
зиготу, отыскивая соответствующие атомы, молекулы или участ-
ки нуклеиновых кислот и белков, «ответственные» за каждый
признак, так же как не требуется эволюционисту пытаться нахо-
дить «материальных представителей» всех признаков человека в
«первобытной амебе» — родоначальнице животного мира.
Исключительную роль в развитии генетики сыграл прогресс
биохимии, ее раздела, именуемого молекулярной биологией, от-
крытие роли нуклеиновых кислот в синтезе белков и в явлениях
наследственности. Эти открытия переводят генетику все более
на рельсы эмбриологии, физиологии и биохимии. Наследственность
все более вырисовывается как «процессы воспроизведения опре-
деленных форм обмена веществ между ядром и цитоплазмой клет-
ки и внешней средой в ряде поколений»3.
Генетику сблизил с эмбриологией также отказ от первоначаль-
ных вейсманистского характера иллюзорных представлений, буд-
то исходные эмбриональные клетки являются недифференцирован-
ными, тотипотентными, а в ходе развития происходит потеря тех
или иных первоначальных свойств. В гл. II й XI мы убедились
в ошибочности этих взглядов.
Развитие любого организма — это не авторепродукция моле-
кул, клеток, тканей, а непрерывные закономерные изменения, воз-
никновение нового и нового в химическом, физическом, структур-
ном и физиологическом отношениях. В ходе развития организма
меняются белковые субстанции, реактивность протоплазмы, уро-
вень и характер интеграции зародыша, антигенная характеристи-
ка новых поколений клеток и другие свойства организма. Так,
1 Лобашев М. Е. Генетика. Л., 1967. С. 553.
2 Дубинин И. П. Проблемы радиационной генетики. М. 1961. С. 111.
3 Там же.
274
представления о том, что свойства миллионов разным образом
дифференцированных клеток взрослого организма непосредствен-
но представлены в исходных в развитии организмов половых или
соматических клетках — эти представления, сыгравшие прогрес-
сивную роль в генетике начала нашего века, являются теперь
слишком упрощенными и не соответствуют данным эмбриологии.
Наконец, важнейшее значение для развития генетики и эмбри-
ологии имел отказ от вейсманистского противопоставления сома-
тических и половых клеток (см. гл. II)'. Оказалось, что в резуль-
тате митоза из одной клетки возникают две, ядра которых идеаль-
но воспроизводят свойства ядра исходной клетки. Имеет место
авторепродукция, обусловленная спецификой ДНК в хромосомах.
Каждая из сестринских клеток несет всю полноту наследственной
информации, характерующей данный организм.
Клетки разных тканей могут радикально отличаться друг от
друга по многим особенностям обмена веществ, морфологии и
функциям, но если они способны совершать свою онтогению и до-
ходить до очередного митоза, они содержат одно и то же коли-
чество ДНК и вся эта ДНК принадлежит к одному типу. Каждая
клетка содержит полный геном организма, полный комплект «ге-
нетического материала». Исключением являются случаи полиплои-
дии и политении клеток, которые можно рассматривать как выхо-
дящие за норму.
В гл. XI отмечалось уже, что это бесспорное для эмбриологов
и генетиков положение выдвинуло ряд еще далеко не разрешен-
ных обеими науками проблем. Требуется расшифровать «пара-
докс природы»: все более дифференцирующиеся в ходе эмбрио-
нального развития клетки сохраняют, однако, благодаря митозам
все свои видовые свойства, несут всю полноту наследственной ин-
формации, характеризующей данный организм. Клетка выглядит
и как интегральная часть целого, и как потенциальный организм.
Вопросы связи генетики и эмбриологии подробно обсуждают-
ся в руководствах по генетике. Здесь указываются лишь основные
принципиальные проблемы связи этих наук.
ГЕНЫ И ОРГАНОГЕНЕЗ
В конечном счете все явления развития организма и возможность
отклонений от этого развития имеют основу в генотипе; гены «кон-
тролируют» развитие. Развитие наследственных признаков, реали-
зация их генотипа в фенотипе, зависит от взаимодействия различ-
ных наследственных факторов и от условий развития. Данный
признак может проявиться неполно или совсем не проявиться в
онтогенезе, если не будет соответствующих условий.
Известно более .1000 наследственных болезней человека.
С точки зрения эмбриологии и биохимии эти болезни можно рас-
275
сматривать как аномалии развития, вызываемые наследственно
закрепленным нарушением различных биохимических процессов
на разных стадиях индивидуального развития организма, наруше-
нием нормальной интеграции зародыша на той или иной стадии
развития. Естественно, чем ранее в эмбриональном развитии на-
рушаются те или иные стороны метаболизма, тем больше наблю-
дается дефектов в развитии.
Состояние генетики и эмбриологии не таково, чтобы можно
было вполне удовлетворенно объяснить биохимический и иной
вероятный механизм действия генов на процессы развития орга-
низма, на органогенез. В то же время генетическая обусловлен-
ность различных отклонений от нормального развития не вызы-
вает сомнений, и в некоторых случаях вскрыто, что те или иные
аномалии связаны с определеными хромосомами и зависят или
от мутаций, или от условий развития, так что нет оснований со-
мневаться в возможности генетического и цитогенетического
объяснения любой наследственно обусловленной болезни. Благо-
даря работам. С. Райта и К. Вагнера (1934) стала известна му-
тантная линия у морских свинок, для которой характерны законо-
мерные, поддающиеся генетическому анализу аномалии в развитии
головы: происходит в большей или меньшей степени недоразвитие
передней части головы, приводящее к циклопии. Голова даже
вообще может не развиться, хотя все в области туловища разви-
вается более или менее нормально. Подобные мутации отмечены
и у мышей. У мышей же зарегистрирована мутация, влекущая за
собой укорочение или даже полное отсутствие хвоста. Обнаружены
такие мутации, как отсутствие ног у телят, деформация клюва
у птиц и многие другие. Молекулярно-генетический анализ этим
явлениям не дан — не выяснено, какие моменты последовательных
стадий развития подобных патологических процессов следует ста-
вить в причинную зависимость от генной активности.
Зарегистрированы мутации, выражающиеся в увеличении чис-
ла пальцев на передних или задних конечностях. Такие мутации
обнаружены Т. Картером в 1954 г. у ряда животных, но генети-
чески изучены у немногих (например, мутация у мышей, называе-
мая luxate и обозначиваемая 1х).
Генетически изучена аномалия в развитии дрозофилы, выра-
жающаяся в том, что вместо развития из соответствующего имаги-
нального диска антенны развивается маленькая конечность с дву-
мя коготками. Из имагинальных дисков, предназначенных для раз-
вития глаза или крыла, формируется конечность. Доказывается,
что эти ненормальности в развитии обусловлены изменением ге-
нетической конституции — присутствием мутантных генов вместо
нормальной аллели этого гена.
Одним из убедительных примеров плодотворного генетического
анализа процессов развития является анализ механизма определе-
ния пола, связанного с особенностями состава хромосомного аппа-
276
рата. Так, у прямокрылых насекомых и у самок млекопитающих
две половые Х-хромосомы (XX), а у самца — только одна Х-хро-
мосома и одна У-хромосома (ХУ), отсутствующая у особей жен-
ского пола. Таким образом, в этом случае самцы являются гете-
розиготными в отношении определения пола. Имеются два типа
•сперматозоидов: оплодотворение одним приводит к развитию самок,
оплодотворение другим — к развитию самцов.
У птиц, бабочек и некоторых рыб гетерогаметные (в отноше-
нии определения пола), наоборот, самки. В отношении определения
пола у ряда животных оказалось превосходное совпадение гене-
тических, цитологических и эмбриологических данных. Однако
генетический механизм определения пола для многих животных
остается еще неясным. У рыб не наблюдается видимых различий
между половыми хромосомами и обычными аутосомами. У земно-
водных и пресмыкающихся также не удалось выявить половые
хромосомы. У некоторых животных определение пола зависит
только от среды. В этих случаях оплодотворенные яйцеклетки по-
тенциально бисексуальны: в зависимости от тех или других усло-
вий развивается самец или самка. Примером этого может слу-
жить Boneilia viridis.
О «ГЕНЕТИЧЕСКОМ КОНТРОЛЕ» НАД ДИФФЕРЕНЦИАЦИЕЙ КЛЕТОК И ТКАНЕЙ
Большое количество фактов в эмбриологии и генетике, касаю-
щихся роли ядра цитоплазмы не поддается еще объяснению.
Все явления эмбрионального развития генотипически обусловле-
ны, но обычно не анализируется участие генов в нормальном
эмбриональном развитии, включая гаструляцию и развитие орга-
нов. Это можно видеть и по настоящей книге, в которой дается
анализ процессов дробления,- гаструляции и других без упомина-
ния о генетической обусловленности всех явлений и используются
различные факты, касающиеся, например, количества и располо-
жения желтка и т. п. Это происходит, с одной стороны, потому,
что эмбриология пока почти не располагает генетическими и ци-
тогенетическими данными, касающимися механизма развития.
С другой стороны, экспериментальная эмбриология располагает
большим количеством фактов, которые как будто исключают
«контроль» хромосом и генов в развитии.
Поскольку возможно нормальное развитие при партеногенезе,
наличие отцовских генов необязательно для всех процессов раз-
вития, за исключением тех признаков, которые могут быть
обусловлены генами сперматозоида. Но и хромосомы матери так-
же не необходимы для основных процессов развития. Об этом
говорит возможность получения андрогенетических Зародышей, т. е.
зародышей, развивающихся из яйца с ядром исключительно отцов-
ского происхождения. Так, Б. А. Астауров и В. П. Острякова-Вар-
шавер в 1957 г. убивали ядро яйца тутового шелкопряда одного
277
вида; яйцо оплодотворялось затем сперматозоидами другого вида.
Так как у этих видов возможна полиспермия, то удалось добить-
ся слияния ядер двух сперматозоидов, в результате чего осу-
ществляется диплоидный гетероспермный андрогенез. Происходит
нормальное развитие.
Некоторые исследователи (например, Ж. Браше, Ю. А. Филип-
пенко и др., 1929) считали, что существует наследственность,
контролирующая ранние стадии развития (обусловленная и ядром
и цитоплазмой), и наследственность менделевского типа, обуслов-
ленная ядром. Некоторые эмбриологи высказывают мысль, что
гены не участвуют в образовании основных органов зародыша,
а контролируют последние этапы дифференциации. Так, с этой
точки зрения цвет глаз находится под контролем генов, а разви-
тие самого глаза происходит без участия генов. В качестве агру-
ментов для обоснования этой точки зрения могут быть использо-
ваны многие факты. В гл. XI говорилось о роли области серого
серпа в развитии амфибий. Если оставить неповрежденным ядро
яйцеклетки, но локально разрушить серый серп, резко нарушается
дифференцировка хорды и нервной системы. Можно эксперимен-
тально, по желанию, устанавливать плоскость симметрии у яиц
амфибий, вызывать появление серого серпа в желаемом месте.
Возможно (хотя и уродливое) дробление при полном отсутст-
вии ядра, что впервые наблюдала Е. Гарвей в 1936 г. Это сообще-
ние было встречено резким скептицизмом эмбриологов,
но впоследствии, однако, оно полностью подтвердилось.
Э. Вильсон (1896) сдавливал оплодотворенные яйца Nereis
между двумя стеклянными пластинками, при этом первые, три
деления происходили в меридиональном направлении. Зародыш
оказывался в виде пластинки из восьми клеток. Затем давление
ослаблялось, дробление продолжалось, зародыш теперь состоял
из восьми микромеров и восьми макромеров. В дальнейшем раз-
вивалась. свободноплавающая личинка, у которой зачаток кишки
состоял не из четырех, а из восьми макромеров. Четыре ядра
макромеров первого квартета (из них развивается апикальный
орган и прототрок) в указанных экспериментальных условиях
становятся ядрами -энтобластов. Вильсон еще в конце прошлого
века заключил, что у полихет дифференциация клеток зависит
не от ядра, а от особенностей цитоплазмы.
Дробление в отсутствие ядра у амфибий отмечалось многими
авторами. Е. Штауфер в 1945 г. разрушал мужские и женские
пронуклеусы в яйцах аксолотлей. В таких опытах отмечен случай
развития почти нормальной безъядерной бластулы.
С другой стороны, имеется ряд достоверных фактов о несом-
ненном участии генов на ранних этапах развития зародышей.
Д. Ф. Паульсон в 1940, 1945 гг. обнаружил, что в результате пол-
ной утраты Х-хромосомы происходит нарушение ранних стадий
развития дрозофиллы (изменяется продвижение ядер к периферии,
278
уродливо развивается бластодерма). Потеря отдельных участков
хромосом может повлечь уродливое развитие нервной системы,
кишечника, производных мезодермы.
Уже говорилось об ошибочности представлений многих моле-
кулярных биологов и генетиков о половых клетках, как, якобы,
недифференцированных. Эмбриологи не имеют оснований поддер-
живать эту устаревшую точку зрения. Причина сохранения ста-
рого взгляда на структуру и функции яйца и спермия заключа-
ются в излишне прямолинейном, хотя и увлекательном представ-
лении многих исследователей, согласно которому все морфогене-
тические процессы эмбрионального развития обусловлены систе-
мой геномных переключений. Излишне оптимистически думают,
что для понимания того, как происходит дифференциация, нужно
узнать только, когда и какие гены активируются. Однако все
факты, касающиеся ранних этапов эмбрионального развития,
заставляют утверждать, что система генетических переключений
не может играть решающую роль в ранних стадиях эмбриональ-
ного развития.
Выражаясь генетическим и молекулярно-биологическим язы-
ком, вся информация, необходимая для ранних стадий развития,
содержится в цитоплазме яйца еще до оплодотворения. Когда
в развитии появляются новые генетические «указания», опреде-
ляющие дифференцировку и развитие, при теперешних возможно-
стях эмбрио-генетического анализа мыслимо узнать лишь путем
определения времени появления новых «инструкций» от аппарата
наследственности сперматозоида. Молекулярная биология распо-
лагает достаточно точными методами определения ферментов.
Доказано, что отцовские ферменты появляются на сравнительно
поздних стадиях развития: у амфибий — после того как у заро-
дыша начинается сердцебиение и мышечные сокращения. Фермен-
ты, участвующие в изменении оболочки яйца морского ежа, имеют
материнское происхождение и т. д. Общий вывод бесспорен:
на ранних этапах развития дифференциация осуществляется
не путем включения транскрипции, а за счет генетико-эмбриоло-
гической программы, имеющейся уже в цитоплазме яйца.
В настоящее время в области молекулярной биологии интен-
сивно разрабатываются вопросы, касающиеся установления вре-
мени начала синтезирования и функционирования тех или иных
видов РНК и белков (ферментов). Было показано, что в ооплазме
содержится все необходимое для ранних этапов развития, т. е.
материнский геном обеспечивает запас необходимых строитель-
ных материалов и энергии для осуществления дробления. Выявле-
ны и изучены разнообразные мутации, влияющие на процессы
раннего развития.
Можно сделать вывод: на ранних стадиях развития все слож-
ные морфогенетические процессы (дробление, формирование
бластулы, а в некоторых случаях и формирование ранней гастру-
279
лы) происходят на фоне слабой активности генома зародыша.
Установлено, что все изменения в функционировании генома
бластомеров и клеток более поздних зародышей зависят от взаимо-
действия ядер с цитоплазмой. Распределение качественно различ-
ных участков цитоплазмы, как уже отмечалось ранее, зависит
от проморфологии зиготы и от сегрегации цитоплазмы. В этом
смысле можно говорить, что программа ранних периодов разви-
тия определяется проморфологией зиготы и связанной с ней
сегрегацией цитоплазмы. К сожалению, мало исследований цито-
генетического и генетико-эмбриологического характера по вопро-
сам роли разных структур сперматозоида в начальных этапах
эмбрионального развития. Естественно, все внимание в нашем
столетии сосредоточивалось на изучении роли ядра. Однако име-
ется много косвенных данных, позволяющих решать вопросы —
принадлежит ли ядру и другим структурам сперматозоида роль,
так или иначе дополняющая роль яйцеклетки в развитии.
Л. Д. Удалова (1975) установила, что у ранних зародышей крыс,
отцы которых подвергались действию рентгеновского облучения,
наблюдаются разного рода хромосомные аберрации. Есть основа-
ния полагать, что отцовская наследственность у млекопитающих
важна для нормального развития трофобласта и последующего
контакта со стенкой матки (развития плаценты). Это обстоятель-
ство многие авторы связывают с неудачами получения поздних
стадий партеногенетически развивающихся зародышей.
Обнаружено, что раковины некоторых моллюсков (Limnaea)
обычно закручены в правую (декстральную) спираль, но изредка
попадаются особи с раковиной, завернутой влево (синистральный
завиток). Доказано действие генов на очень ранних стадиях
развития. Направление завитка раковин коррелировано с направ>
лением наклона веретена митоза при третьем дроблении. Это
направление веретена предопределяется до оплодотворения вли-
янием организма матери на еще незрелое яйцо в яичнике. Отцов-
ский ген направления завитка, привносимый сперматозоидом,
не изменяет характера завитка раковины, обусловленного мате-
ринской наследственностью. Фенотипическое проявление генети-
чески обусловленного направления завитка раковины зависит
не от генотипа самой особи, а от генотипа ее матери. Этому де-
монстративному примеру материнской наследственности дан
вполне удовлетворительный генетический анализ и выяснено,
почему в природе наблюдаются определенные числовые отноше-
ния— редкие случаи особей с синистральными завитками по срав-
нению с декстральными. Ген декстрального направления домини-
рует над геном синистрального.
Для обрисовки трудностей, встающих перед генетикой и
эмбриологией, следует возвратиться к тому диалектическому
противоречию, о котором говорилось в гл. XI: бластомеры вслед-
ствие механизма митоза равнонаследственны, и в то же время
280
с самого начала развития происходит дифференциация. Нет ни
одного факта в эмбриологии, подтверждающего теорию Вейсмана,
согласно которой хромосомы в разных бластомерах содержат
качественно неодинаковые «детерминанты» (гены), которые
ответственны за дифференциацию. Речь может идти лишь о раз-
личном функционировании генома.
Заслуживают внимания идеи А. А. Заварзина, который отме-
чал, что в связи с бурным развитием генетики и установлением
непосредственной связи генов с хромосомами изучение ядерных
структур свелось преимущественно к изучению этих структур
в половых клетках. Поэтому кариология в значительной степени
сводится к цитогенетике. По традиции ядро изучается вне клеток
и тканей, й ткани (их клетки) изучаются вне кариологии и гене-
тики. Каждая соматическая клетка обладает видовой специфич-
ностью: клетки кошки, лягушки, карася, жужелицы и т. д. С дру-
гой стороны, учитывая физиологическую и генетическую роль ядер
и развитие разных тканей с большими особенностями их клеток
(в отношении структур и функций), А. А. Заварзин не видит
другого, более логичного выхода из противоречий, чем необходи-
мость признания различий в кариотипе разных тканей. Он реши-
тельно отвергает теорию детерминант Вейсмана: «... факты снова
ставят проблему неравнонаследственного деления не в той, конеч-
но наивной форме, как это пытался делать Вейсман, а на новом
этапе развития наших знаний. Мне кажется, ничто иное, кроме
этой идеи, не выдерживает даже самой поверхностной критики.
Действительно, при тканевых дифференцировках происходит
какое-то ограничение возможностей при сохранении видовых
признаков»1.
Некоторые исследователи, не зная статьи А. А. Заварзина,
высказали родственные мысли и сделали попытки эксперимен-
тальных доказательств их правильности. Г. Штерн, В. Олфрей,
А. Мирский и Г. Ситрен в 1952 г. установили, что ядра разных
тканей отличаются друг от друга по содержанию в них фермен-
тов.
Ц. Паван в 1955 г. и другие исследователи изучали тонкую
структуру политенных хромосом личинок насекомых (типа гигант-
ских хромосом клеток слюнных желез). В гигантских или поли-
тенных хромосомах в результате эндомитозов происходит много-
кратное (до 1000 раз) удвоение хроматид хромосом, без
их расхождения, в связи с чем деспирализованная хромосома
утолщается, так как наращиваются все новые и новые хромати-
ды. Длина гигантских хромосом превышает' обычные метафазные
хромосомы в 100—250 раз. Последовательность дисков одинакова,
однако наблюдаются закономерные особенности в «набухании»
• Заварзин А. А. Кариология и гистология // Жури, общей биологии,
т. IX. 1948. № 4. С. 285.
281
Рис. 120. Левый конец хромосомы XII ооцита Triturus crista-
tus. Имеется много нормальных пар петель ламповых щеток и
одна пара гигантских петель (1) (по И. С. Галлу, X. Г. Кал-
лану, 1962)
разных участков хромосом, в образовании и развитии «пуфов»—
«вздутых» участков. В различных тканях или на разных стадиях
развития одной и той же ткани признаки активности проявляют
разные локусы хромосом; при этом морфологические изменения
хромосом строго закономерно совпадают с определенными перио-
дами в развитии тканей. Это установлено при изучении развития
слюнной железы. Когда период активности заканчивается, вздув-
шийся участок почти полностью возвращается в свое исходное
состояние.
Интересны наблюдения, проведенные X. Калланом в 1960 г. над
гигантскими хромосомами типа «ламповых щеток», которые обна-
руживаются в ядрах ооцитов рыб, амфибий, птиц и других позво-
ночных и у некоторых беспозвоночных (рис. 120).
От хромомер хромосом в сторону от оси отходит пара или
несколько петель разной величины — от гигантских до очень
небольших. Вероятно, эти петли содержат белок и участки непре-
рывной ДНК. Предполагают, что петли эти — продукт деятель-
ности генов и что в разных локусах хромосом проявляется разная
активность. Действительно, в соответствующих локусах гомоло-
гичных хромосом чаще всего петли имеют одинаковую форму,
но есть и гомологичные локусы с петлями различной формы,
что объясняют гетерозиготностью, непосредственно проявляющей-
ся в самих хромосомах. Такое объяснение подтверждается
результатами опытов по скрещиванию.
В последнее время усиливается интерес к вопросам «функцио-
. нальной организации» хромосом, и, в частности к активным
участкам хромосом — к пуфам, о которых говорилось ранее и
с которыми многие исследователи связывают дифференциацию
282
клеток и тканей. Пуфы рассматриваются' как участки локальной
деспирализации хромосом, обеспечивающей в сочетании с биохи-
мическими механизмами переход генов, локализированных в них,
в активное состояние. Следует поэтому подробнее остановиться
на соответствующих цитогенетических и эмбриологических вопро-
сах. Генетика считает, что роль хромосом состоит в «кодирова-
нии» и «реализации» «наследственной информации». Этой роли
соответствует структура хромосом, а именно строго закономерное
расположение внутри хромосомы «наследственного материала».
Организация хромосомы должна обеспечивать закономерную
редупликацию ДНК. Эти и другие вопросы освещаются в посо-
биях по генетике и цитогенетике.
И. И. Кикнадзе провела обширные исследования на гигантских
хромосомах слюнных желез хирономид (Chironomus dorsalis
Meig), а также и на других клетках животных и растений. Эти
данные приводятся в дальнейшем. Она утверждает, что в хромо-
сомах слюнных желез функционируют около 220 пуфов, это
составляет около 26% от общего числа дисков хромосомы. Пуфы
связаны в своем развитии только с дисками политенных хро-
мосом, но не с междисковыми пространствами. Основой для обра-
зующегося пуфа является, как правило, единичный диск. Диски
могут содержать разное количество ДНК. Могут образовываться
и сложные пуфы, в формировании которых принимает участие
несколько дисков (рис. 121,Л, Б). Диски хромосом идентичны
хромомерам и объединяют комплексы генов, связанных в своем
действии. Кикнадзе утверждает, что пуфы являются единствен-
ными центрами синтеза РНК в клетке. Пуфы — это специфиче-
ские дискретные единицы, играющие роль в синтезе РНК и
редупликации ДНК. В работе некоторых пуфов отмечается опре-
деленная периодичность. Крупные пуфы функционируют непре-
рывно во всех клетках слюнной железы, они способны к синтезу
РНК. Таких пуфов около 80% от общего числа клеток железы,
остальные функционируют лишь в части клеток.
Не следует думать, что функционирование основных пуфов
в ядрах Ch. dorsalis обязательно связано с локальной редуплика-
цией ДНК в их области, так как оказалось, по данным Кикнадзе,
что образование многих крупных пуфов происходит в ядрах,
потерявших способность к редупликации ДНК (в клетках пред-
куколки и куколки).
Имеет ли место белковый метаболизм в пуфах? Вопрос этот
не выяснен, но имеются данные, что количество гистонов в пуфе
не меняется при его развитии.
Исследования и гипотезы И. И. Кикнадзе могут служить иллю-
страцией состояния спорных вопросов, интересующих эмбриоло-
гию, о функционировании хромосом в онтогенезе животного. Счи-
тается, что единая хромосомная нить состоит из- хромомер,
объединяющих группы генов. Хромомеры, изменяясь структурно
283
Рис. 121. Формирование пуфов в хромосомах клеток слюнных желез Chironomus
dorsalis (по И. И. Кикнадзе, 1967):
А — онтогенез пуфа, 2— 7А3. Пуф. развивается нз одного диска; Б — общая схема возмож-
ных путей формирования пуфов; / — за счет нескольких дисков (сложный пуф); 2— за
счет тонких и 3 — более крупных дисков с разным количеством ДНК; 4 — «движение» при
образовании сложного пуфа
и биохимически, могут быть в активном или в неактивном состоя-
нии. Предполагается, что дифференциация клеток связана с акти-
вацией разных частей генома в разных типах клеток.
На ранних этапах дробления яйца хромосомы отличаются от
хромосом клеток бластулы и последующих стадий развития. Ядра
первых бластомеров очень крупные, хромосомы большой длины,
спирализация их слабая. Налицо все признаки высокой актив-
ности хромосом, но синтеза рибосомной РНК в это время не про-
исходит. На стадии бластулы в клетках, по данным Кикнадзе,
происходит уменьшение ДНК («диминуция»), и хромосомы уко-
рачиваются, так что на стадии 33—64 бластомеров они в 2,5—3
раза короче хромосом первого деления дробления и в 3—4 раза
тоньше их. Меняется и структура ядер в периоде между деления-
ми. Эти явления нестабильного состояния ДНК—«материальных
носителей наследственности» — трудно объяснить.
284
Формирование пуфов Кикнадзе обнаружила на разных стадиях
метаморфоза личинок Ch. dorsalis. Общее количество пуфов
увеличивается в течение четвертого личиночного возраста, но спе-
цифических пуфов, характеризующих каждую стадию метамор-
фоза, немного: четыре — у предкуколки и три — у куколки. С чем
связано образование этих пуфов, каково их значение? Эмбриоло-
ги могут относиться к высказываниям цитогенетиков лишь как
к предположениям, так как не доказана причинная зависимость
между формированием пуфов и какими-либо определенными мор-
фофизиологическими явлениями. Кикнадзе считает эти пуфы
«эффекторами, регулирующими изменение общей картины функ-
ционирования хромосом при смене стадий метаморфоза». Дейст-
вительно, какая-то связь между формированием пуфов и стадиями
метаморфоза есть, но ничего неизвестно о том, что означают эти
корреляции. Происходит в ходе метаморфоза изменение соотно-
сительной активности разных пуфов, например, многие слаба
функционирующие у личинок пуфы, у предкуколки и куколки
функционируют энергично, часть пуфов, наоборот, прекращает
свою деятельность. Кикнадзе утверждает, что изменения картины
пуфов в ходе метаморфоза коррелированы с определенными
физиологическими процессами в клетках слюнной железы —
с усилением синтеза мукополисахаридов, с появлением в цито-
плазме белковых гранул.
Интересен, но не поддается объяснению тот факт, что основ-
ное количество пуфов, характеризующих геном клеток слюнных
желез, является устойчивым в разных популяциях и у мутантных
организмов. Доказано влияние на активность хромосом факторов
внешней среды, например температуры. Таковы интересные дан-
ные исследований Кикнадзе, которые эмбриологи, конечно, берут
на учет в своих гипотезах. Однако эмбриология пока не распола-
гает доказательствами того, что причиной тех или иных формо-
образовательных процессов являются именно возникновение it
функционирование пуфов. Это относится и к «ламповым щеткам».
Многие генетики и эмбриологи вынуждены быть пока осто-
рожными в наделении пуфов и ламповых щеток морфогенетиче-
скими функциями. Может быть, формирование пуфов — одно из
проявлений реагирования клеток на биохимические изменения
в клеточных системах, а не причинный момент формообразова-
тельных процессов. Имеются указания (И. Клевер, 1965) на то,
что образование пуфов в клетках слюнных желез можно вызвать
различными воздействиями, например изменением соотношения
ионов Na+ и К+ в ядре.
ЯДРО И ЦИТОПЛАЗМА В РАЗВИТИИ
Эмбриологи и генетики занимались исследованиями взаимодей-
ствия между ядром и цитоплазмой клеток. ДНК не может проя-
вить свои биохимические возможности, если она окажется
28S
изолированной от других химических веществ и структур хромо-
сомы. Изолированная ДНК, как и изолированнная белковая
молекула,—мертвые образования. Кроме ДНК в состав хромосом
входят белки, РНК и Другие вещества. Выяснено, что соотноше-
ние гистоновых и негистоновых белков неодинаково в клетках
разных тканей и варьирует на разных стадиях дифференциации
клеток одного и того же типа. В хроматине клеток бластулы мор-
ского ежа содержится по отношению к ДНК (принятого за еди-
ницу) 1,04 гистонов; 0,48 негистоновых белков; 0,039 РНК и дру-
гих веществ. Соответственно цифры для плутеуса морского ежа:
Ю,86; 1,04; 0,078.
Многообещающие опыты были проведены Р. Бриггсом и
Т. Кингом в 1955 г. Неоплодотворенное яйцо лягушки активиру-
ется к партеногенетическому развитию (уколом), затем микрохи-
рургическим путем из клетки удаляется ядро. В микропипетку
всасывают какую-либо клетку зародыша с интересующей экспе-
риментатора стадии развития — со стадии бластулы или гаст-
рулы. При всасывании клетка разрушается, но ядро с окружаю-
щей цитоплазмой остается. Посредством той же микропипетки
ядро с остатками цитоплазмы инъецируется в энуклеированную
яйцеклетку. Оказалось, что при пересадке в энуклеированное
яйцо ядер клеток, взятых из анимального полушария бластулы,
приблизительно в 30% случаев яйца дробятся нормально. В боль-
шинстве случаев (в пределах этих 30%) происходит нормальная
гаструляция. Данные ряда последующих исследований этих и
других авторов показали, что в некоторых сериях опытов до 80—
100% яиц-реципиентов достигали стадии бластулы. В общем тот
же результат был обнаружен в опытах, в которых йнъецировались
ядра, взятые из клеток анимальной половины и ранней гаструлы.
Был сделан вывод: ядра клеток на стадии бластулы и ранней
гаструлы еще не дифференцированы, «тотипотентны», каждое
ядро может полноценно заменить ядро зиготы.
Аналогичные опыты на других видах амфибий, в частности на
жабах, также дают право говорить о «тотипотентности» ядер
клеток стадии бластулы и ранней гаструлы. Этого и ожидали
авторы, разделяя традиционное (хотя и необоснованное:
см. гл. XI) убеждение, будто и сами клетки еще «не дифферен-
цированы».
Если брать для инъекций энуклеированное яйцо ядра клеток
с более поздних стадий гаструляции, то оказывается, что ядра,
взятые из клеток будущей хорды и мезодермы, не обеспечивают
полного развития: возникают уродливые нейрулы, а чаще разви-
тие останавливается на стадии бластулы или нейрулы. В других
опытах Кинг и Бриггс выяснили прогрессирующее ограничение
потенций ядер, взятых из эктодермальных клеток, расположенных
в области презумптивной средней кишки (из клеток дна первич-
ной кишки). Результаты этих опытов были пестрыми; ядра неко-
286
торых клеток при пересадке их в энуклеированные яйца еще
обеспечивали нормальное развитие, но часто развитие останав-
ливалось на стадии бластулы или гаструлы. Только одна треть
ядер энтодермальных клеток средней нейрулы' при транспланта-
ции их в энуклеированное ядро обеспечивает развитие зародышей
до стадии бластулы и только в шести случаях из ста такие
бластулы продолжают нормальное развитие. То же относится и
к ядрам клеток, закладок хорды и нервной пластинки на стадиях
поздней гаструлы и нейрулы. Авторами был сделан вывод, что
ядра одних клеток энтодермы еще остаются тотипотентными,
но ядра большинства других клеток подвергаются на этой стадии
дифференциации.
Из всех опытов был сделан общий вывод, что в ядрах бласто-
меров в ходе дробления и на последующих стадиях развития
зародыша происходят изменения: они теряют свою эквипотен-
циальность.
Независимо от Р. Бриггса и Т. Кинга, Л. А. Никитина и
О. Г. Строева (1960—1969) провели опыты по гомотранспланта-
ции ядер эктодермы и ее производных в энуклеированные яйце-
клетки четырех видов амфибий — Bufo viridis, B.bufo dsiaticus^
Rana arvalis и R. temporaria. Результаты опытов принципиально
те же, что и в опытах Кинга и Бриггса. Однако в опытах Ники-
тиной для пересадки брали ядра из клеток глазного пузыря и
глазного бокала (со стадии хвостовой почки). Оказалось, что и
ядра дифференцирующихся клеток закладки глаза сохраняют
потенции, присущие ядру зиготы. С увеличением возраста зароды-
шей — доноров ядер — наблюдается уменьшение процента нор-
мально развивающихся зародышей — реципиентов. Были сделаны
выводы, что происходит возрастное ограничение потенций ядер
у все большего числа дифференцирующихся клеток.
Изложенные данные, подтвержающие указанную гипотезу
Заварзина, к сожалению, не могут считаться убедительными и,
конечно, не решают проблем о роли ядра и цитоплазмы клеток
в развитии зародышей и о вероятном различии ядер дифференци-
рующихся клеток. В этих опытах, к сожалению, нельзя было-
инъецировать чистые ядра, неизбежно вводилась и цитоплазма
клеток. Конечно, относительное количество инъецированной цито-
плазмы (по сравнению с цитоплазмой яйца) незначительно,
особенно когда клетки-доноры были очень мелкими (со стадии
поздней гаструлы). Но пренебрегать возможной биохимической
ролью этих небольших количеств цитоплазмы нельзя, зная, сколь
значительный эффект могут обусловливать микродозы тех или
иных веществ в биохимических процессах (ферменты и др.).
В опытах Кинга и Бриггса наносились ощутимые травмы зароды-
шам (и при удалении ядра яйца, и при иньекции чужих ядер),
поэтому трудно решить, за счет каких причин могли быть случаи
аномалий в развитии.
287
Изящные опыты Кинга и Бриггса, давшие указанный резуль-
тат, не удалось с тем же успехом провести на зародышах хвоста-
тых амфибий. В точно таких же опытах на яйцах тритона заро-
дыши не развивались нормально со стадии бластулы даже
в случаях, когда инъецировались ядра клетки со стадии ранней
бластулы. Выходит, что у зародышей тритона какие-то иные
закономерности дифференциации ядер, а это кажется сомнитель-
ным. Данные других исследователей заставляют более насторо-
женно относиться к выводам, сделанным Кингом и Бриггсом.
Дж. Гердон в 1962 г. доказал способность ядер клеток кишеч-
ного эпителия питающихся головастиков Xenopus laevis обеспе-
чивать эмбриональное развитие: в 10 случаях из 726 оперирован-
ных зародышей (в энуклеированные яйца инъецировались ядра
клеток кишечного эпителия) развились головастики.
Результаты опытов дают право автору делать вывод о том, что
ядро может обусловить дифференциацию клеток и в то же время
сохранять генетическую информацию, необходимую для нормаль-
ного процесса развития. Т. Кинг и Мэк-Кинкель (1962) пересажи-
вали в яйцеклетку лягушки Rana pipiens ядра клеток опухоли
(аденокардициномы), возникающие при образовании опухоли из
дифференцированных клеток почки. В этих опытах были получе-
ны плавающие головастики с дифференцированными хордой, ко-
жей, мозгом, мышцами и т. д.
Трудно сделать определенные выводы из опытов Р. Бриггса
и др. (1964), проведенных на яйцах аксолотля. Яйца освобождали
•от оболочек, хромосомы их разрушали действием ультрафиолето-
вых лучей. Активация яиц к развитию вызывалась действием
электрического тока. Ядра для пересадок брали от других заро-
дышей со стадии нейрулы, из клеток хорды, боковой мезодермы,
медуллярной пластинки и энтодермы. В большинстве случаев
развитие не шло дальше стадии бластулы и ранней гаструлы,
но при пересадке ядер из хорды развитие иногда продолжалось
до стадии нейрулы (6% случаев), хвостовой почки (4%) и даже
до вылупления (0,6%)- При инъекции ядер, взятых из клеток
•нервной пластинки, в 3% случаев нормальное развитие продол-
жалось до стадии нейрулы.
Исследователями дается противоречивая оценка всех опытов
по пересадке ядер. Одни считают, что, по-видимому, происходят
какие-то необратимые изменения клеточных ядер, другие сомне-
ваются в реальных возрастных ограничениях ядерных потенций
на таких поздних стадиях, как хвостовая почка.
Интересны опыты ряда авторов, пытавшихся разрешить
проблему роли ядра и цитоплазмы в развитии путем убиения
ядер яйца или бластомеров действием ионизирующей радиации.
Они исходили из гипотезы, будто при действии ионизирующего
излучения можно убить ядро клетки, а цитоплазму оставить нор-
мальной.
288
Б. В. Конюхов (1973) так представляет контроль клеточной
дифференциации. Дифференцировка клеток связана прежде всего
с синтезом специфических белков, появление которых определя-
ется дерепрессией локусов ДНК и синтезом соответствующих
иРНК. Многие думают, что процессы дифференциации и морфо-
генеза осуществляются так, что те или другие гены включаются
и выключаются в одной и той же клеточной системе. Конюхов
считает, что дифференциация связана только с дерепрессией ге-
нома «тотипотентных недифференцированных» клеток раннего
зародыша. Сначала дерепрессируются гены, контролирующие раз-
множение и процессы общего метаболизма, затем активируются
первые «тканеспецифические гены», детерминирующие специали-
зацию бластомеров в стволовые клетки, а в период гисто- и орга-
ногенеза дерепрессируются гены, которые контролируют синтез
белков, специфичных для тех или иных клеточных систем. Эта
гипотеза возникла в связи с иммуноэмбриологическими исследо-
ваниями Конюхова (см. гл. XV) антигенов, возникающих в ходе
развития организма (видоспецифические, стадиеспецифические,
органоспецифические). Она связана с популярным в 60—80-е годы
представлением о стволовых клетках. Определенный клон клеток
образуется в результате многократных делений первичной инициа-
торной, стволовой клетки, у которой дерепрессируются и затем
сохраняются в активном состоянии соответствующие тканеспеци-
фические локусы ДНК. Стволовые клетки, думают Конюхов и
другие авторы, возникают на стадии гаструлы. Таковы миогенные,
хондрегенные, фиброгенные и иные.
ЭМБРИОЛОГИЯ, ГЕНЕТИКА И МОЛЕКУЛЯРНАЯ БИОЛОГИЯ
Как уже сказано, между эмбриологией и генетикой существует
тесная связь, которую пытались наполнять новым содержанием
несколько поколений генетиков и эмбриологов, начиная с Т. Мор-
гана и В. Ру. В 60—80-е годы огромное число генетиков, молеку-
лярных биологов и эмбриологов, вдохновленные превосходными
успехами в познании синтеза белка, увлеченно работали над
вскрытием преимущественно биохимических закономерностей,
эмбрионального развития, теснейшим образом коррелированных
с морфогенетическими процессами. Как и при возникновении вся-
кой новой, бурно развивающейся науки, в молекулярной биологии
и генетике индивидуального развития имеется много противоре-
чивых исследований, спорных представлений. Интерпретация мно-
гих фактов еще неясна. Совершенно неизвестно еще эмбриологи-
ческое, морфогенетическое значение многих биохимических дан-
ных. Эмбриологи, однако, не вправе игнорировать даже и
не «устоявшиеся» еще представления.
Впоследствии, после изложения всех основных проблем
эмбриологии, будут рассмотрены спорные и бесспорные вопросы
289
А. С. Спирин (1965) говорил: «Многим казалось (а иным ка-
жется и по сей день), что открытие информационной РНК
(мРНК) и связанных с ней генных регуляторных механизмов
обещает легкий путь к решению проблем клеточной дифференци-
ровки. Уже родилось немало красивых схем, где простой игрой
активности генов, простым включением и выключением синтеза
различных мРНК в процессе развития и дифференцировки якобы
можно объяснить и сами эти процессы. Схему регуляции Жакоба
и Моно (Jacob, Monod, 1961), столь плодотворную в объяснении
ряда процессов у недифференцирующихся одноклеточных, сдела-
ли почти аксиомой и в объяснении процессов клеточной диффе-
ренцировки. Переход от одной стадии развития к другой стал
рассматриваться как довольно несложный акт «индукции» на ген-
ном уровне: некий «индуктор», подготовленный предыдущей ста-
дией, активирует новую порцию ранее «репрессированных» генов,
последние начинают продуцировать новые мРНК, которые опре-
деляют синтез новой серии белков, а уже последние-то и опреде-
ляют всю специфику новой стадии.
Хотелось бы, однако, предупредить о том, что никаких экспе-
риментальных подтверждений применимости схемы Жакоба и
Моно к процессам клеточной дифференцировки пока нет. Надо
помнить о той возможности, что схема регуляции Жакоба и Моно
может быть ограничена вполне конкретным, определенным кругом
явлений, общих как для недифференцирующихся, так и для диф-
ференцирующихся клеток, но не имеющих отношения к самой
дифференцировке. С другой стороны, регуляция на генном уровне
по Жакобу и Моно может быть лишь вырванной частью, случай-
ным фрагментом сложной общеклеточной регуляторной системы,
и в этом случае также было бы крайне наивно пытаться
объяснить такие общеклеточные процессы, как развитие и диф-
ференцировка, на основании знания одного маленького известного
нам фрагмента (хотя, разумеется, знания этого может быть
вполне достаточно для объяснения некоторых частных явлений),
например, индукции или репрессии синтеза данного фермента»1.
Из открытий Менделя, апробированных победным развитием
генетики в течение восьми десятилетий нашего века, вытекает
бесспорный принцип о дискретности наследственной детерминации
признаков. Однако бесспорно и то, что эмбриология не имеет
основания говорить о независимом возникновении и развитии при-
знаков в онтогенезе. Нормальный онтогенез — это совокупность
процессов изменения состояния целостности. Что означает проти-
воречивость двух бесспорных утверждений?
Уже говорилось, что блестящие успехи генетики оказались
без всякой связи с эмбриологией. «С этой стороны, существова-
1 Спирин Д. С., Белицина Н. В. Информационные рибонуклеиновые кислоты
в раннем эмбриогенезе / / Успехи современной эмбриологии. Т. 59, вып. 2.
1965. С. 187.
292
ли алгебраические символы генов А, В, С, за которыми еще
не было вскрыто никакой физико-химической или биологической
реальности, а с другой,— были сформированные признаки»1.
Между «геном» и «признаком» оказалась «пропасть». «Я твердо
уверен, что именно решение проблемы наследственного осущест-
вления информации в процессе индивидуального развития, про-
блем генетики развития, стало сейчас направлением главного
удара не только генетики, но и всей современной общей биоло-
гии»1 2.
Несмотря на усилия многих талантливых ученых по заполне-
нию «пропасти» между геном и признаком, успехи пока черезмер-
но скромны. Прогресс генетики, биохимии, эмбриологии обеспе-
чивает в настоящее время возможность заполнения «пропасти»
между геном и признаком.
Основной метод генетики — гибридологический анализ, скре-
щивание самцов и самок с альтернативными признаками, регист-
рация признаков у потомков. Объектом же эмбриологии являются
процессы развития, становление в онтогенезе типовых, семейст-
венных, родовых, видовых признаков; это закономерности про-
цессов дробления, гаструляции, органогенеза. Почему, к примеру,
развиваются два глаза, четыре конечности, та или иная структура
у семяпочки растений и т. п.? Как видим, объекты исследований
у эмбриологов (процессы развития) и генетиков (сопоставление
«начала» развития с итоговыми этапами) были существенно раз-
личными.
Понятия «ген», «генотип», «фенотип» предложил в 1909 г.
В. Иоганнсен. Он выступил одним из первых против представле-
ния об организме как о сумме признаков, каждому из которых
соответствует самостоятельный наследственный зачаток. Фенотип
особи он определил как совокупность доступных непосредствен-
ному наблюдению или анализу признаков. Описательно он
представлял его крайне расчлененным, морфологически — как
элементы при постройке, физически — как отдельные свойства
или функции, химически — в виде отдельных составных
частей и пр. Живой организм он понимал как целое,
как единую систему не только во взрослом состоянии, но и в те-
чение всего развития.
Естественное соотношение генетики и эмбриологии не может
складываться на основе гегемонии взглядов и гипотез, вытекаю-
щих из анализа фактов, полученных только генетическими или
только эмбриологическими методами исследования.
Развитие альтернативных признаков, конечно, не какая-то
категория явлений, обособленная от развития «основных»
1 Астауров Б. Л. Генетика и
генез. Т. 3. 1972. № 6. С. 549.
2 Там же. С. 547.
проблемы индивидуального развития // Онто-
293
соотношения генетики и эмбриологии и сделаны предположения
о дальнейшем совместном прогрессе обеих наук.
Огромное количество фактов, накопившихся в области моле-
кулярной биологии, генетики, биохимии, не оставляет сомнений
в том, что генетический аппарат включает хромосомы (содержа-
щие линейно расположенные гены в ДНК и регуляторные белки)
и полирибосомные комплексы (включающие иРНК, рибосомы,
гРНК и факторы трансляции). Генетическая информация о син-
зете специфических белков закодирована в виде определенной
последовательности азотистых оснований в цепи ДНК. Гены
кодируют ферменты, а с последними связаны все жизненно важ-
ные функции организма.
Генетические и молекулярно-биологические представления об
эмбриональном развитии, о детерминации излагаются в руковод-
ствах по генетике. Многие исследователи считают, что существу-
ет «антагонизм» между синтезом ДНК и синтезом специфических
белков, ответственных за дифференциацию. Дифференцированные
тканевые клетки, завершившие процесс специализации и синтези'
рующие специфические белки, как будто не способны к синтезу
ДНК (А. А. Заварзин, 1964, и многие другие исследователи).
Однако не менее убедительны и противоположные взгляды. Так,
при исследовании изменения содержания ДНК и РНК в скелет-
ной мускулатуре куриного эмбриона в ходе ее развития установ-
лено, что могут протекать одновременно и синтез ДНК и синтез
специфических белков. Об этом говорят и другие данные.
Как же трактуются генетикой и эмбриологией процессы детер-
минации и дифференциации?
Все последовательные процессы онтогенеза находятся под
генетическим контролем. В ходе онтогенеза происходит синтез
тех или иных белков в результате последовательного активирова-
ния тех или иных участков генетического аппарата и блокирова-
ния других, иначе говоря, происходит индукция или репрессия
образования специфических РНК. Существует мнение, что на ран-
них стадиях развития гены, имеющие отношение к дифференциа-
ции различных клеток, еще не активны, не образуют иРНК,
не синтезируются белки, имеет место репрессия. Соответствующие
гены становятся активными в определенное время в соответствую-
щих клеточных популяциях в связи с теми или иными морфо-
генезами.
Остановимся несколько подробнее на цитогенетических гипо-
тезах и фактах, касающихся явлений эмбрионального, развития.
Каждый тип специализированных (т. е. по-особому дифферен-
цированных) клеток многоклеточного организма содержит харак-
терные ферменты. Молекулярная биология и генетика без каких-
либо сомнений обосновывает тезис: тканевые особенности основы-
ваются на различиях белкового аппарата клеток. В связи с этим
290
логично утверждается, что процессы дифференциации связаны
с различиями в синтезе многих индивидуальных белков.
Раз все полноценные клетки имеют совершенно идентичные
геномы, а дифференциация клеток в ходе развития организма
имеет место и при этом синтезируется только часть тех фермен-
тов, для которых имеется информация в его геномной ДНК,
значит, в ядре должен быть какой-то аппарат, от которого зави-
сит, в каких клетках, на какой стадии развития организма опре-
деленный ген активен, т. е. обеспечивает синтез специфической
иРНК, и в каких клетках и когда — неактивен, репрессирован.
В свете этой гипотезы последовательные процессы детермина-
ции и дифференциации могут мыслиться таким образом, что
в ядре имеется «программа», определяющая строго закономер-
ную последовательность репрессии и депрессии отдельных струк-
турных единиц генома, в результате чего строятся определенные
белки, характерные для данных клеток, на данных стадиях онто-
генеза и не свойственные другим клеткам на других стадиях
онтогенеза.
Не касаясь деталей организации и функционирования генов,
следует сказать, что информативная емкость генома в ходе эво-
люции повышалась. Она наименьшая у вирусов и наибольшая
у эукариот. Кроме того, процессы транскрипции и трансляции
у эукариот усложнились за счет их разделения во времени и
пространстве. Появились особые этапы, связанные с посттранс-
крипционной модификацией, обеспечивающейся процессингом.
Ряд генетиков обсуждают вопросы о «эпигенетических» фак-
торах. Существует много «внешних» воздействий в отношении
хромосом и клеток. Эти воздействия возникают в процессе разви-
тия организма и влияют на «генетическую» управляющую систему.
Думают, что даже внутри ядра и каждой отдельной хромосомы
по отношению к индивидуальным генам и группам функциональ-
но связанных генов (оперонов) другие гены, их продукции в дан-
ной хромосоме и в других хромосомах могут выступать как внеш-
ний, не только репрессивный, но и индуцирующий фактор и
внехромосомные условия могут влиять на дифференцировку всего
генома через эту систему.
НЕРЕШЕННЫЕ ГЕНЕТИКО-ЭМБРИОЛОГИЧЕСКИЕ ВОПРОСЫ
Каждый год в современной биологии появляется большое коли-
чество новых факторов, касающихся наследственности и измен-
чивости. Фундаментальные же понятия классической менделев-
ской генетики остаются такими, какими они были формулирова-
ны при ином состоянии знаний в области генетики и эмбриоло-
гии. Нельзя не видеть своеобразного кризиса роста генетики,
частую смену различных гипотез, что знаменует, конечно, прогресс
генетики и эмбриологии.
291
признаков (видовых, типовых). Менделевский анализ, положим,
зубчатости или гладкого контура листа какого-либо вида расте-
ния должен привести к представлениям о соответствующих генах
и о дискретности. Однако нельзя сомневаться в том, что «глад-
кость» и «зубчатость» появляются не обособленно от развития
листа в целом. Генетик скажет то же, что и эмбриолог. «Когда
мы. изучаем наследование того или иного признака (например,
красно-белая окраска венчика у растений, черно-белая окраска
шерсти у мышей, комолость-рогатость у скота и т. д.) и уста-
навливается моногенное расщепление по одной аллельной паре,
то называем соответствующие гены по определяемым признакам
лишь условно, на самом же деле такое определение гена отно-
сится лишь к одной из замеченных нами сторон его действия:
учитываемый нами признак является лишь его частным проявле-
нием. Для того, чтобы иметь представление о полном проявлении
гена, необходимо изучить его действие на всех этапах развития
организма, что не представляется пока возможным». «... Разви-
тие наследственно определяемых признаков всегда осуществля-
ется в целостной системе процессов формирования всего организ-
ма, так как оно обусловливается системой генов — всем генотипом.
Отсюда другой стороной взаимодействия геиов будет плейотроп-
ное действие одного и того же гена на многие признаки и
свойства»1.
Отметим еще одну важную генетико-эмбриологическую проб-
лему. Теория гена и все гипотезы области генетики развития
созданы на основе анализа явлений, наблюдаемых при половом
эмбриогенезе. Гаметы и зигота — исходные клетки в генетическом
анализе; цитогенетический анализ основан на процессах митоза
и мейоза. В меньшей степени изученными оказались различные
формы бесполого размножения, соматический эмбриогенез,
полиэмбриония, при которых исходными в онтогенезе являются
единичные соматические клетки или совокупности их, а не зигота.
Начальные морфогенетические процессы при бластогенезе не мо-
гут быть уподоблены дроблению яиц, процессам гаструляции и др.
Открытие кода синтеза белковой молекулы стало началом но-
вой эпохи не только в биофизике и биохимии, но и в генетике и
эмбриологии, ибо наука сказала подлинно великое слово о важ-
нейших веществах, без которых нет ничего живого. Однако воз-
никло, как это всегда и бывает при истинном прогрессе, много
новых вопросов и трудностей.
В большинстве генетических и эмбриологических лабораторий
стихийно возникли две «аксиомы». 1. Суть индивидуального раз-
вития можно свести к явлениям дифференциации клеток. Морфо-
генез — это процесс изменения от недифференцированного со-
стояния к дифференцированному. 2. Явления дифференциации
1 Лобашев М. Е. Генетика. Л., 1967. С. 176—177.
294
можно объяснить генотипически обусловленными закономерностя-
ми последовательных синтезов специфических белков.
За двадцать лет «торжества» этих идей произошло много на-
учныу событий, заставляющих пересмотреть эти «аксиомы».
К. Уоддингтон — генетик и эмбриолог, десятилетиями мечтавший
о синтезе двух наук, с энтузиазмом встретивший формирование
молекулярной биологии, говорил: «Как объяснить появление тц-
ких структур, как митохондрии, хлоропласты, рабдомеры и т. п.?
Ясно, что к этому вопросу чисто генетические методы, например,
рекомбинационный анализ, почти не имеют отношения... Необ-
ходимо задать один вопрос, который звучит весьма непривычно:
когда молекула не является молекулой?. . Силы, обеспечивающие
образование из жиров, углеводов таких элементов, как, например,
ядерная оболочка, вовсе не похожи на силы, с которыми имеет
дело классическая химия макромолекул. Существует целый ряд
проблем, таких, как проблема генотропных веществ, компетенция
клеточной ультраструктуры, морфогенеза в целом, которые, по-
видимому, нельзя решить с помощью генетических и биохимиче-
ских методов... Я хотел бы дополнить модную в настоящее время
молекулярную генетику основами эмбриологии»1.
Эмбриолог П. Г. Светлов писал: «Биохимический метод в на-
стоящее время по преимуществу ограничивается вопросами, свя-
занными с редупликациями ДНК и специальными синтезами бел-
ков, т. е. лишь сектором одной из многочисленных форм диффе-
ренциации .. ,»2
И: И. Шмальгаузен, способствовавший прогрессу эмбриологии
и генетики, говорил: «Молекула ДНК—это, конечно, настоящий
кибернетический код. Однако это не значит, что он сам по себе
определит" все развитие (онтогенез. — Б, Т.) ... Молекулярная
генетика и биохимия, конечно, не разрешат проблемы развития.
Для этого нужен подход на более высоком уровне интеграции».
Представления о триаде «ДНК-^РНК—^-специфический белок»
не могут быть единственным путем познания процессов дифферен-
циации. Обходится факт, что сами биохимические компоненты со-
здаются и преобразуются под эгидой системы, именуемой клеткой,
под эгидой биологической организации, а не сами по себе, явля-
ются всегда зависимыми структурами и процессами, а не само-
регулирующимися системами. Процессы развития принципиально
отличаются от «самосборки», развитие предполагает изменение
самих структурных элементов.
1 Уоддингтон К. Морфогенез и генетика. М., 1966. С. 98.
Светлов П. Г. Тезисы VII Всес. съезда анатомов, гистологов и эмбриоло-
гов. Тбилиси, 1966. С. 26.
Глава XIII
ГИПОТЕЗЫ ОБ ЭМБРИОНАЛЬНЫХ ПОЛЯХ.
ТЕОРИЯ ФИЗИОЛОГИЧЕСКИХ ГРАДИЕНТОВ
РАЗВИВАЮЩИЙСЯ ОРГАНИЗМ — МЕНЯЮЩЕЕСЯ СОСТОЯНИЕ
ЦЕЛОСТНОСТИ
Изучая состояние разных проблем, мы убедились, что в зави-
симости от задачи эмбриологи избирают тот или другой способ
работы и тот или иной «уровень» исследования: молекулярный,
клеточный, надклеточный, — и все эти пути исследований не всту-
пают в противоречие друг с другом. Хотя ни одно биологическое
явление нельзя объяснить свойствами индивидуальных атомных
процессов или свойствами отдельных молекул, но ясно, что неко-
торые проявления жизни, особенности надмолекулярных структур
и энергетики живой материи получат объяснение именно в связи
с прогрессом квантовой физики. Этому не противоречит то, что
многие процессы развития организма заведомо следует изучать на
уровне надклеточном, оперируя понятиями «интеграция», «орга-
низм как целое», «взаимодействие частей» и т. п. В этом мы убе-
дились при анализе процессов дифференциации. Целостность раз-
вивающихся организмов проявляется прежде всего во взаимоза-
висимости частей в развитии зародыша, что было подвергнуто
анализу в гл. XI.
Развивающийся организм — это закономерная непрерывная ди-
намика структурированных процессов, новое и новое состояние
целостности. Клетки зародыша находятся под «замком корреля-
ций», под «арестом» интеграционных механизмов, обусловливаю-
щих сужение их проспективного значения. С этим связаны их био-
химические свойства, появление или потеря тех или иных струк-
тур, изменение размеров и формы клеток, темпы деления и т. п.
Это относится абсолютно ко всем организмам, в частности и к тем,
дробление которых неточно называется «анархическим». Развива-
ющийся организм — не сумма развивающихся признаков и, конеч-
но, не сумма белков, а динамика преобразования взаимозависи-
мостей дискретных единиц, в свою очередь являющихся целост-
ными системами.
Процессы формообразования нельзя объяснить исходя из еди-
ничных клеток. О. Гертвиг (1898) был прав, утверждая, что меж-
ду понятием агрегата клеток и понятием биологического соедине-
ния клеток такая же большая разница, как между понятием смеси
двух объемов водорода и одного объема кислорода, с одной сто-
роны, и химического соединения — молекул воды, с другой.
296
Поведение клеток и клеточных комплексов в той или иной сте-
пени зависит от состояния зародыша в целом: клетки имеют боль-
шие потенции, чем те которые реализуются в развитии. Зароды-
ши— гетерогенные системы, разные части которых отличаются
веществами, интенсивностью метаболизма, темпами деления кле-
ток, степенью дифференциации и т. д. Применительно к поздним
Стадиям развития и к взрослым организмам интеграция означает
объединение клеток в более сложные и работоспособные едини-
цы — органы и аппараты. Для характеристики интегрированности
важно учитывать: полифункциональность органов; способность или
неспособность изолированных от всего организма частей к продол-
жению жизни: отсутствие, наличие и особенности сложно устроен-
ных органов и аппаратов, от которых зависит жизнь всех других
органов и тканей—характер строения и функционирования
единого для всего организма нервного аппарата; наличие общих
для всего организма пищеварительной, инкреторной, выделитель-
ной и других взаимодействующих систем.
Многие явления вынуждают думать о надклеточных факторах.
1. Перережем поперечно планарию. Хвостовая половина будет
восстанавливать недостающий головной конец. Значит, ткани, на-
ходящиеся вблизи раневой поверхности (положим, протяжен-
ностью 2—3 мм), примут участие в построении головной полови-
ны червя. Представим теперь, что мы разрезали червя на 2—3 мм
ближе к хвосту, чем это было в первом опыте. Те же самые ткани
(в области этой 2—3-миллиметровой полоски), которые ранее
принимали участие в построении головного конца, теперь оказа-
лись в головной половине и в зависимости от каких-то механизмов
интеграции принимают участие в построении уже не головного
отдела червя, а хвостового. Одних молекулярных и клеточных па-
раметров для объяснения этих явлений недостаточно.
2. Изменим путем операции положение плечевого нерва три-
тона, отведем его в направлении
к задней конечности, т. е. в об-
ласть действия поля конечности:
может вырасти добавочная ко-
нечность (рис. 122,А). Мы мо-
жем, однако, отвести нерв в новое
положение так, что он окажется
в поле спинного плавника или
гребня. Соответственно развива-
ется, образование, подобное
плавнику или гребню (рис.
122,5).
Поступим подобным образом
с седалищным нервом. Отведем
его от нормального положения
в направлении к передней ко-
8 М
т! Г
Рис. 122. Действие отведенных нер
вов, свободно кончающихся в поля,
органов (по Е. Гиено, 1927):
А — отведение в поле конечности привс
дит к образованию конечности; Б — отве
дение в поле спинного гребня образуе
спниной гребень; В — отведение в пол
хвоста приводит к образованию добавоч
ного хвоста
29
нёчности. Может образоваться добавочная конечность, но если
тот же нерв будет приживлен в области действия поля хвоста, воз-
никнет добавочное хвостоподобное образование (рис. 122, В).
Выходит, что на один и тот же раздражитель, на один и тот
же нерв реакция тканей и их систем будет различной; это зави-
сит, по Вейсу, от специфического влияния полей.
3. Пересадим переднюю конечность тритона, клетки тканей
которого имеют гаплоидный набор хромосом, на тело «диплоид-
ного тритона». После приживления ампутируем ее на каком-либо
уровне. Регенерат будет весь гаплоидным.
Р. Гертвиг в 1927 г. имплантировал гаплоидную почку перед-
ней конечности личинки хвостатой амфйбии в тело диплоидной
личинки, в область действия поля конечности. Пересаженная поч-
ка может не приживаться и дегенерировать, но иногда, распа-
даясь, она стимулирует ткани хозяина, и возникает добавочная
конечность; при этом клетки ее имеют не гаплоидные, а диплоид-
ные ядра.
Отсюда ясно, что изучение закономерностей онтогенетического
развития требует разносторонних исследований на разных уров-
нях — молекулярном и электронном, клеточном, тканевом и орга-
низменном. Поэтому достойны уважения, но научно неверны одно-
сторонние увлечения каким-либо одним уровнем, например моле-
кулярным, каким-либо один аспектом исследования, например
биохимическим.
Кажущиеся резко отличными взгляды разных авторов, в сущ-
ности, не противоречат друг другу. Так, В. А. Энгельгардт (1971)
под влиянием успехов биохимии утверждает: «Молекулярная био-
логия изучает явления жизни, оперируя неживыми (лишенными
жизни) объектами ... Редукционизм означает принцип исследова-
ния, основанный на убеждении, что путь познания сложного лежит
через расчленение этого сложного на все более и более простые
составные части и изучение их природы и свойства» *.
Другой биохимик, также увлеченный молекулярной биологией,
А. Сент-Дьердьи (1972) пишет, «молекулярная биология вовсе не
последнее слово в науке... Нам необходимо получить максимум
сведений относительно молекул, квантов и электронов, чтобы
приблизиться к пониманию жизненных явлений. Мы не должны,
однако, забывать, что молекулярный уровень представляет лишь
один из многих уровней организации, в то время как слово
«жизнь» подразумевает совокупность всех функций и всех реак-
ций» 1 2.
Правильно и утверждение Г. Селье: «Жизнь клетки в ее нерас-
члененном веществе. Чем дальше вы расчленяете эти живые ком-
1 Энгельгардт В. А. Часть и целое в биологических системах // Природа.
1971. № 1. С. 26.
2 Селье Г. На уровне целого организма. М., 1972. С. 5.
298
плексы, тем дальше вы уходите от биологии, в конце концов вам
остаются только величественные, вечные и всеобъемлющие физи-
ческие законы неживой природы»1.
До 20-х годов нашего столетия проблема целостности разви-
вающегося организма была монополией витализма. Но сторонники
этой ненаучной философии не затрудняли себя причинным анали-
зом явлений целостности организации живых существ, создавая
понятия, дававшие лишь иллюзию объяснения. Однако некоторые
термины, предложенные виталистами (например, «проспективная
потенция», «перспективное значение» и др.), широко использова-
ны эмбриологами и уже не имеют идеалистического содержания.
Один из самых воинственных виталистов Г. Дриш создал свою те-
орию индивидуального развития. Он придал идеалистический
смысл высказыванию Гертвига о том, что судьба бластомеров
есть функция положения их в целом. Это положение, если его не
абсолютизировать, представляет интерес.
Г. Дриш считал, что все клетки зародыша и их ядра эквипотен-
циальны, одинаковы по своей «проспективной потенции» и обла-
дают одинаковой способностью развиваться в любую часть орга-
низма. Проспективная потенция гораздо шире, чем ее проспектив-
ное значение, т. е. то, что на самом деле реализуется. Проспек-
тивные потенции эквипотенциальных частей при развитии
зародыша всегда ограничиваются, так как зародыш является гар-
монической эквипотенциальной системой, которой подчинены все
части.
Экспериментальные «доказательства» витализма он пытался
находить, в частности, в явлениях регенерации и в опытах по раз-
витию новых целых организмов из изолированных (изъятых из
систем целого зародыша) бластомеров полагая, что оставшиеся
части зародыша перестраиваются и создается новая гармониче-
ская эквипотенциальная система.
Дришем была создана виталистическая философия биологии,
которую можно назвать идеалистическим эпигенезом (Б. П. То-
кин, 1933). Он заимствовал у Аристотеля понятие «энтелехия»
(см. гл. I). Это, якобы, истинная сущность явления в отличие от
«динамис», т. е. от возможности его осуществления. «Энтелехия» —
особый витальный внепространственный, вневременной, нематери-
альной природы фактор. Его нельзя уподобить и энергии, так как
он не может иметь количественную характеристику. Это фактор,
стоящий над клетками, тканями и материей. Энтелехия — «инже-
нер развития». Представление о развивающемся организме Дриш
выразил формулой B(x)—,f(S, I, Е), где проспективное значение
какой-либо части х организма (то, что реально формируется из
этой части в ходе развития организма) обозначается В; это про-
Селье Г. На уровне целого организма. М., 1972. С. 10.
299
спективное значение части х есть функция / величины этой части
3, ее положения в целом I и энтелехии Е. Энтелехия, не будучи
многообразием в пространстве и времени, создает таковое, подоб-
но тому как акт художественного творчества, сам по себе не про-
странственный, выливается в создание пространственного харак-
тера. В связи с результатами опытов по изоляции бластомеров
Дриш утверждал, что идея целого находится в его частях; разви-
тие новых целых организмов нельзя объяснить свойствами мате-
рии. Попробуйте, говорил он, разрезать на две части автомобиль.
Мы не получим из него два автомобиля. А зародыш является та-
кой машиной, у которой каждая часть может стать полноценной
машиной такого же рода. Основная идея Дриша, касающаяся раз-
вития организма и неизбежно приводящая к идеализму, такова.
В ходе индивидуального развития происходит увеличение прост-
ранственного многообразия (речь идет о всех «элементах» заро-
дыша). Однако в окружающем зародыш пространстве сколько-
нибудь значительного разнообразия нет. Значит (?), многообразие
возникает как бы из ничего. Таким образом, нельзя обойтись без
представления об особом виталистическом факторе, обеспечива-
ющем это многообразие, без понятия «энтелехия».
Другим примером идеалистических взглядов на развитие орга-
низмов может служить теория В. Брандта, созданная в 1931 г.
Согласно его взглядам главные явления в онтогенезе — осущест-
вление «типа», «формы как таковой». Форма противопоставляется
конкретным процессам дифференциации, обусловленным наслед-
ственностью. Осуществление «типа развития» происходит через
процесс детерминации, который является самостоятельным, неза-
висимым от наступающей позднее дифференциации. Дифференци-
рующаяся материя пассивна, в нее возможно «вкладывать» при
помощи процесса детерминации различные формы. Форма как та-
ковая — «субъект», могущий быть отождествленным с кантовской
«вещью в себе». Детерминация — акт, при котором нематериаль-
ное начало («тип», «воля к природе») воздействует на материю.
Эти взгляды, как и более сложная концепция Дриша, имеют в
основе два утверждения: пассивность развивающегося материала
и наличие внешних нематериальных «факторов», определяющих
развитие.
Существовало много причин для возникновения виталистиче-
ских концепций в биологии; одна из них — примитивность и бес-
помощность механистического материализма в объяснении явле-
ний целостности живых существ. Дриш в молодости был сторон-
ником «механистического» объяснения жизненных явлений, но
впоследствии стал лидером неовитализма.
Эксперименты Дриша внесли существенный вклад в проблему
детерминации и регенерации. Ряд предложенных им терминов и
понятий, очищенных от идеалистического истолкования соответ-
ствующих этим терминам явлений, стал достоянием эмбриологии.
300
Таковы понятия «проспективная потенция» и «проспективное зна-
чение».
Некоторые эмбриологи (например, Ю. Шаксель), считая себя
материалистами, объявили проблему «организм как целое» лож-
ной, чуждой научной эмбриологии и вместо разработки этой проб-
лемы отстранились от нее, поверхностно критикуя витализм. Меж-
ду тем давно известно множество явлений, свидетельствующих об
интеграции зародышей. Давно уже стала очевидной несостоятель-
ность представлений о зародыше как о мозаике частей. Неудов-
летворенность теориями индивидуального развития, игнорирующи-
ми бесспорные факты интеграции зародышей и не считающихся
с проблемой организма как целого, явилась одной из причин для
создания гипотез об эмбриональных полях.
ГИПОТЕЗЫ ОБ ЭМБРИОНАЛЬНЫХ ПОЛЯХ
Понятие «поле» заимствовано биологами у физиков, использу-
ющих его при характеристике зависимости изменения .частиц ма-
терии от свойств пространства, в котором находятся эти частицы.
В биологию это понятие было введено Бовери в 1901 и 1910 гг.
Гипотеза Л. Г. Гурвича разрабатывалась с 1913 г. и была за-
вершена в 1944 г. Первоначальная концепция поля (1922—1930)
была чисто идеалистической. Процессы развития зародыша мыс-
лились как материализация «динамически преформированной
морфы».
Изучая гистологию формирующегося хрусталика и роговицы,
а также растущего корешка лука, Гурвич решил, что распределе-
ние митоза беспорядочно, хаотично, не детерминировано строго,
подчинено закону случая. Он решил создать концепцию, согласно
которой поведение «однородных элементов» всех микроморфоге-
нетических процессов регулировалось бы «единым фактором».
Морфогенез определяется этим единым фактором — «преформи-
рованной морфой». Морфогенез, по Гурвичу, — это перемещение
клеток, упорядочивание их пространственного расположения и
движение клеток в каждый данный момент определяется законом,
связанным с еще не существующей в этот момент структурой, с
окончательной конфигурацией зачатка. Первоначальная концепция
поля мыслилась им как нематериальный фактор, стоящий над
клетками, над материей.
В 1944 г. А. Г. Гурвич написал о новой «теории биологиче-
ского поля», в которую включил понятие «поле клетки». Область
действия этого поля выходит за пределы клетки и клетки оказы-
вают своими «полями» влияние друг на друга. Этим определя-
ются движения находящихся поблизости клеток. Происходит синтез
клеточных полей в единое «актуальное поле». Поля складываются
подобно правилам векторного сложения, но свойства «общего поля»
нельзя объяснить только свойствами «полей» клеток. По мере уда-
301
ления клеток их взаимодействия быстро ослабляются, так как ин-
тенсивность действия поля обратно пропорциональна квадрату рас-
стояния. В .ходе эмбрионального развития все время происходит
эволюция «поля целого», целое в данное мгновение развития опре-
деляет ход дальнейшего развития. ,
По мнению Гурвнча, клеточное поле анизотропно, оно непре-
рывно и преемственно, имеет радиальное строение, при делении
клетки делится и ее поле. «Поле», «целое», как и в первона-
чальных представлениях Гурвича, оказывается и в новой интер-
претации биологического поля чем-то стоящим над клетками, над
зародышем.
Некоторые эмбриологи полагают, что всю глубину идей Гурви-
ча и их плодотворность можно по достоинству оценить только те-
перь, что он опередил свое время, ие был правильно понят со-
временниками (Л. В. Белоусов, Д. С. Чернавский, 1983). Эти ис-
следователи защищают мысли о хаотичности митотической актив-
ности клеток, беспорядочном их расположении, например в бла-
стулах, о том, что «порядок» создается полем. Представить воз-
никновение различий в комплексе якобы однородных элементов
можно, если допустить, что они «соподчинены» некоторому «по-
лю». Морфогенез представляется ими как перемещения «кинема-
тически независимых элементов» зародыша. С их точки зрения
процессы онтогенеза могут быть разделены на две независимые
категории — процессы морфогенеза (обусловленные полем переме-
щения клеток или клеточных комплексов) и процессы внутренней
дифференцировки клеток (изменения клеток по любым показате-
лям, за исключением их локализации). Утверждается, будто клет-
ки должны отталкиваться друг от друга, вводится понятие «от-
талкивающее поле» и т. д. и т. п.
А. Г. Гурвич — одна из ярких звезд в нашей отечественной
науке. Вопреки своей виталистической идеологии он высказал ряд
интереснейших идей, с которыми должны согласиться биологи и
в эпоху молекулярной биологии, бурного развития естествознания:
он привлек внимание биологов к вопросам о лучистой энергии,
сопровождающей биологические процессы, предложил интересную
теорию «протоплазмы», независимо от Э. С. Бауэра создал близ-
кую к Бауэровскому всеобщему принципу об устойчивом нерав-
новесии биологических систем гипотезу с неравновесных молеку-
лярных констелляциях и сделал много ценного для теоретической
биологии.
Гурвич привлек также внимание к закономерностям изменения
формы зачатка и зародыша, что очень важно для разработки то-
пологии дефинитивных живых систем и топологии эмбриогенеза
(Д’Арси Томпсон, 1916—1948; Г. П. Короткова, 1968, 1979; В. М.
Маресин, 1982; Е. В. Преснов, В. В. Исаева, 1985, и др.). Эти ас-
пекты морфогенеза связаны с описанием преобразования целост-
ных клеточных систем, целостных зачатков, и они не могут быть
302
поняты исходя нз поведения отдельных клеток или исходя из био-
химических процессов, совершающихся в этих клетках.
В понятие «поля» исследователи вкладывали разное содержа-
ние. Дж. С. Гексли и Г. Р. де Бер (1934) думали о поле как об
, области, в пределах которой определенные факторы действуют
одинаково. Внутри поля, по их представлениям, существует состоя-
ние равновесия. Поле — это единая система, а не мозаика, где
одни части можно было бы удалить или заменить так, чтобы при
этом система не изменилась. Внутри поля-системы может быть
разная концентрация химических веществ, могут быть градиенты
метаболизма в смысле физиологических градиентов Чайлда.
По К. Уоддингтону, «поле» можно представить как систему,
организация которой обусловливается тем, что положение, заня-
тое неустойчивыми ее частями в одном участке системы, порож-
дает определенные отношения к положению, занятому подобными
же частями в других участках. Их уравновешенное состояние и
обусловливает эффект поля. Уодингтон считает, что это пред-
ставляет близкую аналогию с равновесием, наступающим в части-
це стали, когда сама она ориентируется в направлении магнит-
ных сил.
Г. Шпеман также применял понятие поля. Его поле организато-
ра (Organizationsfeld) по существу равнозначно «доминантной об-
ласти» Чайлда. Как мы увидим, Чайлд внес физиологическое со-
держание в морфологические представления Шпемана об органи-
зационных центрах. По Чайлду, организационные поля — это
участки зародыша, которые доминируют, определяют развитие
других участков, отличаясь более высокой физиологической ак-
тивностью.
Гипотеза П. Вейса. По П. Вейсу (1925, 1927, 1930), поле—
это система сил, имеющих определенное направление, т. е. систе-
ма векторов. Первоначальные представления о поле Вейс создал
на основании явлений регенерации, анализируя роль остатка ам-
путированного органа. Регенерация, например, конечности трито-
на — это не сумма тканевых регенераций, а целостный процесс,
зависящий от системы влияний, которая может быть определена
как морфогенетическое поле. Поле присуще остатку органа как
целому, оно имеет динамический характер. После ампутации орга-
на поле действует на регенерационную бластему, которая явля-
ется нуллипотентной. Формообразовательные воздействия на нее
не локализованы в тех или иных частях остатка органа. Более
того, между тканями остатка органа и тканями регенерата нет
никакой генетической связи. В числе доказательств этого утверж-
дения были опыты, согласно которым удаление из остатка органа
кости или мускулатуры не препятствует нормальному развитию
регенерата. Сколь правильны, с точки зрения современных дан-
ных, факты, на которые опирается Вейс, — об этом будет гово-
риться в гл. XVI о регенерации.
303
Вейс применил понятие поля и к эмбриональному развитию.
Яйцо характеризуется единым полем. В результате расчленения
поля происходит дифференциация частей и автономизация их.
Части зародыша индифферентны и организуются под влиянием
целого. Поле, по Вейсу, материально, но оно не химической при-
роды, так как химические факторы могут активировать, а не соз-
давать поле.
Итак, Вейс, не удовлетворяясь одними клеточными параметра-
ми, создал представление о поле как факторе развития, факторе
детерминации, предполагая, что клеточный материал, на который
действует поле (регенерационная бластема, части зародыша), яв-
ляется сам по себе индифферентным. В развивающемся организме
образуются новые и новые поля, так что весь организм оказыва-
ется разбитым на ряд сфер действия, соответствующих полям раз-
личных органов. Эти представления очень близки к предложениям
Е. Гиено о. существовании регенерационных территорий — хвоста,
правой задней конечности и т. п.
Процесс регенерации представлялся П. Вейсу таким образом:
из морфогенетического регенерационного поля—«поля органа»
возникают подчиненные поля: кожи, скелета, мускулатуры. Эти
поля осуществляют дальнейшую детерминацию, и создаются но-
вые: плечевой кости, локтевого сустава и т. д. Поля находятся в
иерархической зависимости друг от друга, развитие регенерата
следует представить себе как субординацию полей.
Наиболее уязвимо в теории Вейса — представление о пассив-
ности, нуллипотентности клеточных систем, на которые действует
поле. Эта теория является в сущности крайним выражением взгля-
дов на явления формообразования как на реактивные процессы.
Гипотеза Н. К. Кольцова. Ряд ученых пытался создать свое
представление о поле. Среди них несомненно интересна и в на-
стоящее время гипотеза Н. К. Кольцова. Представления Н. К.
Кольцова (1934) о целостности организма и его теории поля чуж-
ды витализму. Это попытка рассмотрения данных эксперименталь-
ной эмбриологии и генетики в физико-химическом аспекте.
Ооцит и яйцо — организованные системы с определенно выра-
женной полярностью, с определенным расположением клеточных
структур. Уже в ооцитах находятся разнообразные вещества и
структуры, дающие своеобразную реакцию на кислые и основные
красители в зависимости от их pH. Это означает, что различные
части клетки могут иметь те или иные положительные или отри-
цательные заряды. В целой клетке поверхность ее, как правило,
заряжена отрицательно, а поверхность ядра и хромосом —поло-
жительно. При созревании ооцита создается соответственно его
строению электрическое силовое поле, «закрепляющее» это стро-
ение. Под влиянием силового поля в клетке должны возникать
определенные катафорезные токи перемещения веществ, объясня-
ющиеся разностью потенциалов. При активации яйца, вызванной
304
сперматозоидом и партеногенетически, происходит изменение ды-
хания, pH, иногда резкое, проницаемости мембран и передвиже-
ния веществ. Ядра спермия и яйцевой клетки (у голобластических
яиц) после сложных передвижений сходятся в центре или по глав-
ной оси ближе к анимальному полюсу. По Кольцову, эти явления
обусловлены, очевидно, напряжениями перезаряжающихся сило-
вых полей, разностью потенциалов. Таким образом, начинающий
развиваться зародыш — это силовое поле. В ходе развития разные
пункты силового поля характеризуются разностью потенциалов.
Речь идет не только об электрических потенциалах, но и о хими-
ческих, температурных, гравитационных, диффузионных, капилляр-
ных, механических и др.
Даже такой сравнительно простой фактор, как уменьшение
или увеличение проницаемости оболочек яиц.в каком-либо участ-
ке, неизбежно вызывает изменения токов жидких веществ яйце-
клетки. Благодаря тому, что между бластомерами существуют
определенные связи, можно представить себе, что изменение то-
ков жидких веществ может влиять и на пространственное распо-
ложение бластомеров. Различного характера потенциалы, их из-
менения не только сопровождают развитие зародышей и являются
отражением состояния его интеграции, но и играют важную роль
в развитии, определяя поведение отдельных бластомеров и все-
го зародыша. В ходе развития силовое поле зародыша изменяет-
ся: оно усложняется, дифференцируется, но остается единым.
Вероятно, не без влияния идей Чайлда о доминантных обла-
стях зародыша и представлений Шпемана об организаторах, во
всяком случае не вступая в противоречие с ними, Кольцов говорит
о главных центрах с высокой разницей потенциалов, о центрах
второй, третьей степени и т. д. Он говорит о градиентах с напря-
жением, убывающим от одного потенциала к другому. По мнению
Кольцова, однако, теория градиентов Чайлда — аналитическая
теория — теория расчленяющая (ибо чайлдовские градиенты могут
явиться лишь слагаемыми силового поля), тогда как силовое поле
Кольцова отражает развитие яйца как единого целого.
От каждого центра распространяются градиенты, определя-
емые всем силовым полем. Область дорсальной губы бластопора
у амфибий — это, вероятно, самый важный комплекс центров си-
лового поля гаструлы, и этот комплекс определяет дальнейший
этап развития.
Н. К- Кольцов с предельной ясностью говорит о физическом
характере силового поля, причем его теория поля выгодно от-
личается от изложенных ранее «реактивных» теорий тем, что
поле выводится именно из состояния клеток, а не оказывается
фактором, стоящим над зародышем, фактором, приходящим к не-
му извне. При состоянии биофизики 30-х годов Кольцов не мог
создать более конкретных физических представлений о поле за-
родыша. Поведение каждого участка зародыша зависит от его
305
предварительной структуры, от влияния общего силового поля и
влияния близлежащих областей этого поля. Представления Н. К-
Кольцова в этом отношении выгодно отличаются от шпеманов-
ских, что особенно наглядно, когда он прибегает к понятию «ин-
дукция». «Конечно, в силовом поле развивающегося яйца и заро-
дыша нет участков, которые мы могли бы назвать исключительно
организаторами или исключительно индуцируемыми: в каждом
участке выражены обе функции, только в разной пропорции. Это
положение непосредственно вытекает из самого понятия о едином,
но расчлененном силовом поле». При развитии зародыша все бо-
лее усложняется его структура и структура разных его частей —
происходит дифференциация. «Развитие яйца как единого целого,
несмотря на распадение его на отдельные бластомеры, зароды-
шевые листки и зачатки органов, обеспечивается тем, что сило-
вое поле его остается единым, постепенно усложняясь, диффе-
ренцируясь с течением развития. Изменяются потенциалы в полю-
сах этого единого силового поля, причем число полюсов по ме-
ре дифференцировки возрастает. Потенциалы в полюсах силового
поля могут быть электрические, гравитационные, механические,
капиллярно-активные, химические» *.
Н. К- Кольцов вводит понятие «силовое поле внешней среды»
(гравитационное, световое и химическое), приписывая ему важ-
ное значение, так как оно влияет на силовое поле внутри заро-
дыша, например определяет направление роста у сидячих живот-
ных.. Как видим, Кольцов при создании своей теории поля после-
довательно проводит принцип физико-химического рассмотрения
явлений эмбрионального развития.
В новейшей эмбриологии Н. К. Кольцов нашел бы много фак-
тов, не противоречащих его теории поля.
По данным эндокринологии зародышевого развития, разные
участки зародыша, и не находящиеся в контакте, могут химически
влиять друг на друга, и уже в эмбриональной развитии вступают
в действие регуляторные биохимические механизмы. Начиная с
40-х годов получено много данныхfо том, что функциональная ак-
тивность желез устанавливается еще в зародышевый период и что
ряд зародышевых гормонов играет формообразовательную роль
на строго определенных Этапах развития эмбриона.
В гл. X говорилось о значении гормона щитовидной железы
в метаморфозе амфибий. Исходное состояние генитальных струк-
тур у всех зародышей (у млекопитающих) одинаково. На этой
ранней стадии половой системы имеются как женские (мюллеро-
вы), так и мужские (вольфовы) протоки. Если удалить у таких
зародышей закладки гонад, происходит развитие, характеризую-
щее женский пол: исчезают вольфовы протоки, мюллеровы прото-
ки сохраняются. Этот факт и другие показывают, что развитие
1 Кольцов Н. К. Организация клетки М., Л., 1936. С. 564.
306
женской половой системы у млекопитающих не нуждается в дей-
ствии половых гормонов и в наличии яичников. Опыты показыва-
ют, что половая же система мужского типа может развиваться
только при наличии полового гормона семенника, иначе не проис-
ходит дифференцировка вольфовых протоков и редукция мюлле-
ровых протоков. Роль гормона семенника в развитии генитально-
го тракта проявляется в самом начале нормальной половой диф-
ференцировки гениталиев. У кролика, например, это происходит
на 19—20-й дни развития. М. С. Мицкевич (1947, 1949) доказал,
что. если не допускать нормальной функции щитовидной железы
у зародышей птиц, происходит задержка окостенения скелета, не-
доразвитие эмбрионального оперения и другие патологические
явления. Зародышевое развитие у млекопитающих становится не--
нормальным в случаях отсутствия или недостатка тиреоидного
гормона.
Имеются данные и о значении других желез. Н. К. Кольцов
предполагал, что и на очень ранних этапах эмбрионального раз-
вития гормоноподобные вещества, нормально возникающие в ходе
развития, играют регуляторную роль. Близкие к взглядам Коль-
цова мысли высказывали и другие исследователи. Б. Вейсберг
(1958) предложил единую, физическую трактовку разных морфо
генетических процессов, создав представление о «колебательных
полях». Он изучал колебания электрических потенциалов у мик-
сомицетов, сходство некоторых органических форм, например ко
лоний шампиньонов, с расположением мелких частиц в акустиче-
ском поле. Вейсберг предположил, что колебательные поля при-
водят к тому, что клеточные комплексы должны разделяться на
территории, внутри которых колебания синхронизируются по фа-
зам, а между территориями создается разность фаз. Происходя-
щее вследствие этого пространственное разъединение может при-
водить к морфогенетическим движениям: впячивание клеток при
гаструляции, расположение полукружных каналов внутреннего
уха, формирование гребных пластинок у ктенофор и т. п.
ОСНОВЫ ТЕОРИИ ФИЗИОЛОГИЧЕСКИХ ГРАДИЕНТОВ Ч. ЧАЙЛДА
Как мы неоднократно убеждались (гл. IV, V, XI и др.), развива-
ющийся организм на любой стадии представляет собой не сумму
независимых частей, а является целостной системой, причем со-
стояние интеграции, присущее зародышу с самого начала, меня-
ется в ходе развития, так как возникают новые структуры с новы-
ми функциями. Изучение динамики целостности развивающегося
организма невозможно только морфологическими методами:
нужны физиологические, биохимические и биофизические иссле-
дования.
Большой интерес для эмбриологии представляет теория физи-
ологических градиентов американского физиолога, эмбриолога и
307
общего биолога Ч. Чайлда. Ее называют также теорией «аксиаль-
ных» градиентов. Согласно этой теории, обоснованной многочис-
ленными экспериментами ее создателя и его последователей,
интенсивность жизненных процессов неодинакова в разных частях
тела: она закономерно понижается по какой-либо оси тела или
его органов. Основным показателем интенсивности жизненных
процессов, по Чайлду, является уровень метаболизма, изучаемый
по интенсивности окислительно-восстановительных процессов. По
мнению автора, количественные различия в уровне метаболизма,
или градиенты1, имеют значение простейших систем, определяю-
щих интеграцию тех организмов, у которых в эволюции не выра-
ботались еще или находятся в примитивном состоянии интеграци-
онные механизмы более высокого порядка, как-то: нервная систе-
ма, железы внутренней секреции и др. Свою теорию Чайлд при-
менял преимущественно для анализа интеграции у растительных
организмов, протозоа, кишечнополостных, низших червей и др.
Однако физиологическими градиентами, как увидим далее, харак-
теризуются и эмбрионы высокоорганизованных животных, а так-
же формирующиеся органы этих животных.
Каким образом можно убедиться в различии физиологической
активности разных участков тела, в наличии физиологических
градиентов? Основной способ, сыгравший решающую роль в раз-
витии теории градиентов,— это метод определения дифференци-
альной чувствительности разных частей тела к различным по-
вреждающим агентам: кислотам, щелочам, веществам, подавляю-
щим дыхание, лучистой энергии.
Поместим туфельку Paramecium caudatum в растворы НС1,
метиленового синего высокой концентрации, KCN, подвергнем ее
действию ультрафиолетовых лучей или будем содержать в усло-
виях недостатка кислорода (рис. 123). Хотя организм будет омы-
ваться ядами или подвергаться действию лучистой энергии со
всех сторон, умирание его будет происходить в строго определен-
ном направлении: мы зарегистрируем передне-задний градиент
чувствительности, т. е. отмирание начинается с переднего конца,
что выражается нередко, и очень заметно, в отторжении умираю-
щих частей от остающихся еще живыми и способными к обычной
раздражимости, сохранению работы ресничного аппарата и т. д.
Принципиально тот же результат мы получим в опытах действия
повреждающих веществ на гидру. Прежде других частей тела бу-
дет, как правило, распадаться гипостом со щупальцами, и умира-
ние распространится в передне-заднем (апико-базальном) направ-
лении. В какой-то момент к этому присоединяется распад в про-
тивоположном направлении, со стороны подошвы и стебелька.
Планарии имеют такой же градиент повреждения: животное
1 Градиент — различие, ступенчатость, постепенность.
308
Рис. 123. Дифференциальная чувствительность Paramecium (по Ч. Чайлду и Де-
виии, 1926):
А—Г — действие ультрафиолетовых лучей; Д, 3 — метиленовый синий высокой концентра-
ции; И, К—KCI; Л, М — недостаток кислорода
начинает умирать «с головы». Эти удивительные явления Чайлд
объясняет наличием градиентов физиологической активности: чем
интенсивнее метаболизм в каком-либо участке тела, тем энергич-
нее реагирует он на альтерирующие воздействия. Добавим к это-
му, что если концентрация ядов настолько незначительна, что
животное не умирает, то, по Чайлду, участки тела с максималь-
ной интенсивностью обмена веществ переносят альтерирующее
влияние ядов лучше; они, по выражению Чайлда, акклиматизи-
руются, быстрее выздоравливают после прекращения их действия.
Очень важно утверждение Чайлда о неспецифичности действия
повреждающих факторов любой химической и физической приро-
ды, о принципиально одинаковых «градиентах повреждения» под
влиянием любых ядовитых веществ, высокой температуры, уль-
трафиолетовых лучей, витальных красителей в высоких концент-
рациях и т. д. Эти факты, конечно, говорят о наличии особых
состояний целостности испытуемых животных, о том, что происхо-
дит не локальный ответ на действие вредных агентов, обусловлен-
зый, например, реакцией яда с теми или иными веществами по-
верхностных частей клеток или одноклеточных организмов.
309
К числу важнейших положений теории градиентов Чайлда
принадлежит также утверждение: между различными частями те-
ла у примитивных животных существуют различия лишь количе-
ственного характера. В дальнейшем мы увидим, что Чайлд и его
последователи также думают и о зародышах высокоорганизован-
ных животных. Физиологические градиенты можно выявить у за-
родышей до тех стадий, пока в результате прогрессирующей диф-
ференциации клеток и тканей не возникнут качественные разли-
чия между разными частями тела и когда явление физиологиче-
ских градиентов можно наблюдать не на целом эмбрионе, а на
закладках тех или иных органов.
ЯВЛЕНИЯ ЭМБРИОНАЛЬНОГО РАЗВИТИЯ В СВЕТЕ
ТЕОРИИ Ч. ЧАЙЛДА. ДОМИНАНТНЫЕ ОБЛАСТИ
Ч. Чайлд и его последователи (А. В. Беллами и др.) применили
теорию градиентов к анализу явлений эмбрионального развития.
Процессы формообразования на разных стадиях развития и иные
формообразовательные процессы, например сопровождающие ре-
генерацию, ставятся в зависимость от различных этапов изменя-
ющейся системы физиологических градиентов. Исходным поло-
жением служит утверждение, что прежде чем появляются качест-
венные отличия между различными частями тела, последние от-
личаются лишь количественно, а именно уровнем физиологиче-
ской активности. Возникновение различного рода структур, про-
цессы морфологической дифференциации происходят на основе
количественных отличий в уровнях физиологического градиента.
На разных его уровнях возникают и разные дифференцировки.
Участки с наивысшей активностью доминируют над другими, ока-
зывают влияние на дифференциацию соседних участков.
По Чайлду, возникновение полярности и симметрии в их пер-
воначальной простейшей форме обусловлено внешними фактора-
ми. Под влиянием внешних факторов возникают чисто количест-
венные различия в уровне обмена веществ по какой-либо оси, и
эти количественные различия переходят в различия качественно-
го порядка. Яркий пример этого Чайлд видит в поведении яйце-
клеток бурой водоросли Fucus. Чем объясняется их полярность?
До начала развития яйцеклетка имеет форму шара. Первый при-
знак развития — небольшая деформация, которая выражается
первоначально в небольшом выпячивании определенного участка,
не отличающегося по своей структуре от других частей яйца
(рис. 124,А). Удлинение яйца в этом участке приводит к образо-
ванию выроста (рис. 124,5). Яйцо делится в плоскости, перпенди-
кулярной к оси, по которой происходит удлинение (рис. 124,5).
Клетка с выростом становится первичным ризоидом, органом при-
крепления, а другая клетка дает начало таллому. Градиенты ин-
310
A
Рис. 124. Градиенты индофеноловой реакции на различных стадиях развития
водоросли Fucus (по Ч. Чайлду, 1941):
А — ранняя и Б — поздняя стадии развития с градиентом, уровень которого понижается,
начиная от кончика ризоида; В — стадия двух клеток. Кроме^указанного обнаруживается
второй, более слабый градиент, уровень которого понижается в направлении от апикальной
области к ризонду; Г, Д — более поздние многоклеточные стадии развития. Направление
градиентов указаны стрелками
дофеноловой реакции обозначены на рисунке стрелками; видно,
что при разделении на две клетки появляется второй градиент
чувствительности (рис. 124,Г, Д).
Чайлд установил, что различные внешние условия ответствен-
ны за возникновение градиентов яйца Fucus: при неодинаковом
освещении частей яйцеклетки ризоид развивается на менее осве-
щенной стороне; при использовании в экспериментах электриче-
ского тока ризоид образуется на стороне, обращенной к аноду;
развитие ризоида при разной концентрации водородных ионов
среды, омывающей разные участки яйца, обеспечивается на той
стороне яйцеклетки, где кислотность выше.
При изучении оогенеза'полихет, немертин, морских ежей Чайлд
убедился, что возникновение полярности ооцитов объясняется
действием неодинаковой среды на разные участки ооцита: центр
участков, подвергающихся действию овариальной жидкости, ста-
новится анимальным полюсом; в этом районе обмен веществ наи-
более интенсивен; противоположный полюс (которым яйцо при-
крепляется к стенке яичника) становится вегетативным полюсом.
Анимальный полюс у ооцита лягушки развивается в том участ-
ке, где входят в фолликул артериальные сосуды. На рис. 125 изо-
бражены ооциты полихеты Sternaspis и лягушки. У полихеты
ооцит прикреплен при помощи узкой ножки, содержащей отвер-
стие для сосудов, а ядро ооцита расположено в противоположном
конце, который выдается в овариальную полость. Полярность
ооцита отражает результат различий в транспорте питательных
311
веществ к яйцу. Они поступают из стенки яичника или благодаря
мигрирующим «кормящим» клеткам, покрывающим свободную
поверхность растущего ооцита. На растущий ооцит могут оказать
влияние локальные различия в содержании кислорода и углекис-
лоты. Одно из видимых показателей полярности — образование
полярных телец у анимального полюса. У яиц морских ежей обна-
ружен градиент в расположении митохондрий и региональные
различия в расположении рибонуклеиновых кислот и в синтезе
белков.
В ооцитах гидромедузы Phialidium направление анимально-ве-
гетативной оси определяется градиентом окислительно-восстано-
вительных процессов в яйце, направление которого в свою очередь
зависит от градиента аэрации (концентрация О2), закономерно
устанавливающегося между прикрепленным к бластостилю полю-
сом ооцита (низкая аэрация — вегетативный полюс) и полюсом,
обращенным к внешней среде (высокая аэрация — анимальный
полюс).
Как уже упоминалось ранее, при действии различных ядов вы-
является отчетливый анимально-вегетативный градиент чувстви-
тельности у еще не оплодотворенного яйца лягушки (рис. 126).
Отмирание яйца начинается в области анимального полюса (по
Чайлду, это область наивысшей физиологической активности) и
постепенно распространяется по направлению к вегетативному
полюсу (рис. 126,Л). После оплодотворения обнаруживается тот
же анимально-вегетативный градиент, но особенно быстрое уми-
рание происходит вдоль спинной стороны яйца, в направлении
к области серого серпа (рис. 126,Б). У бластулы наряду с преж-
ним градиентом появляется новый градиент — дорсовентральный,
от середины серого серпа, т. е. от района будущей дорсальной
губы бластопора. Здесь образуется второй центр высокой физио-
логической активности. Во время гаструляции анимальный
Рис. 125. Возниковение первичной полярности у ооцитов (по А. Беллами, 1919):
А — ранний и Б — сформированный ооциты у полихеты Sternaspis (по Ч. Чайлду, 1915);
В — сформированный ооцит Rana pipiens; f — артериальные сосуды и 2 —венозные сосуды
312
Рис. 126. Дифференциальная чувствительность
яиц Rana pipiens на ранних стадиях развития
(по А. Беллами, 1919):
Четыре стадии умирания: А — недробящегося яйца;
Б — яйца на ранних стадиях дробления; В — на ста-
дии поздней бластулы; Г — на стадии гаструлы
центр высокой физио-
логической активности
ослабляется и, наобо-
рот, усиливается вто-
рой центр, находящий-
ся в районе бластопо-
ра (рис, 126,В, Г).
В начале нейруля-
ции полюс высо-
кой физиологической
активности остается в
области замкнувшихся
губ бластопора и пере-
ходит на образующий-
ся здесь зачаток хво-
ста. Система градиен-
тов все более усложня-
ется. По длиннику фор-
мирующейся медулляр-
ной пластинки, а имен-
но в ее передней ча-
сти, возникает новый
более высокий центр
физиологической ак-
тивности. Это проявле-
ние интеграции в пределах зачатка формирующейся относительно
автономизированной нервной системы (см. гл. XI).
Механизмы влияния доминантных областей на дифференци-
ровку рядом находящихся участков могут быть, очевидно, неоди-
наковыми на разных стадиях развития.
Сущность теории Чайлда состоит, таким образом, в том, что
какие-то «внешние» факторы (например, разность сред: овариаль-
ная жидкость и ткани яичника) вызывают количественные разли-
чия в яйце и эмбрионе, на основе которых возникают в дальней-
шем качественные (структурные и функциональные) различия.
Особенности дифференциации связаны с уровнями физиологиче-
ской активности. Участки наивысшей активности — доминантные
области.
Физиологические представления Чайлда не вступают в проти-
воречие с экспериментально-морфологическими исследованиями
школы Шпемана. Первоначальная анимально-вегетативная по-
лярность представляет очевидный и важный фактор развития яиц
амфибий.
Биохимические исследования, несмотря на некоторую противо-
речивость их результатов у разных авторов, в общем подтвержда-
ют теорию градиентов Чайлда. Так, гаструла (амфибий) разреза-
313
лась на несколько частей и измерялось дыхание каждой части.
Максимальная интенсивность дыхания оказалась в районе дор-
сальной губы бластопора (т. е. организатора Шпемана или доми-
нантной области Чайлда). Эти данные, полученные Л. Сце в
1953 г., подтверждают более ранние исследования Ж- Браше. Еще
большего внимания заслуживают утверждения Р. Фликинжера и
Р. Блаунта (1958), использовавших полярографический метод,
позволяющий более точно измерить дыхание частей яйца. Резуль-
таты этих опытов также свидетельствуют о существовании в яйце
градиента окислительно-восстановительных процессов.
Методом восстановления витальных красителей в анаэробных
условиях, которыми были окрашены живые зародыши (Чайлд
и др.), установлено наличие анимально-вегетативного и (в ходе
развития зародыша) дорсовентрального градиента дыхания..
П. П. Иванов в 1949 г. отметил усиленное потребление углево-
дов в начале гаструляцин амфибий. Углеводный обмен в дорсаль-
ной губе бластопора наступает ранее и протекает более интенсив-
но, чем в брюшных частях зародыша. Сравнивалось включение
14СОг в нуклеиновые кислоты и белки в различных участках за-
родыша. Фликингер в 1954 г. установил наличие градиентов:
включение 14СО2 в нуклеопротеиды происходит по двум градиен-
там — анимально-вегетативному и дорсовентральному. Синтез
белка и РНК во время дробления не удается, обнаружить, но он
четко регистрируется с наступлением гаструляцин. Синтез рибо-
нуклеопротеидов за счет желтка преобладает в дорсальной поло-
вине развивающегося яйца, обнаруживая дорсовентральный гра-
диент. Анимально-вегетативный градиент РНК можно обнару-
жить уже в только что оплодотворенном яйце. Во время гаструля-
ции к этому градиенту в результате синтеза новых РНК прибав-
ляется дорсовентральный градиент.
Таблица 5. Содержание РНК и ДНК в разных областях гаструлы амфибий
Область РНК ДНК
Triturus alpestris Ambyatoma mexicanum Triturus alpestris Ambystoma mexicanum
Участки эктодермы без подлежащей мезодермы Участки эктодермы с при- лежащей мезодермой Дорсальная губа бласт- опора Крыша первичной кишки 3,25+0,04 4,46+0,27 2,08+0,1 1,78+0,07 2,51+0,12 4,48+0.22 2,37+0,09 2,5+0,22 1,47+0,25 1,18+0,07 2,45+0,25 5,45+0,92
Примечания: 1. Количества выражены в мкг/мг. 2. Средние цифры из пяти опреде-
ленна.
314
Рис. 127. Дифференциальная редукция кра-
сителя (янус зеленый) на ранних стадиях
развития куриного зародыша (по О. Руло-
ну, 1935):
Л—В — увеличение редукции на стадии первич-
ной полоски; Г, Д — стадия головного отростка;
Е — Ж — стадия первого межсомитного желобка.
Повреждение головной области и замедление ре-
дукции обозначены крупными черными точками.
Редукция обозначена точками
Неожиданные и труд-
но объяснимые результа-
ты дали цитохимические
исследования М. Пфаут-
ча (1960) о распределе-
нии нуклеиновых кислот
в разных частях зароды-
шей тритонов и аксолот-
лей на стадиях гаструлы
и нейрулы. Он обнару-
жил в эктодерме более
высокое содержание
РНК, чем в дорсальной
губе бластопора. Методи-
ки, которыми пользовал-
ся автор, были более
совершенны, чем гисто-
химические способы об-
наружения ДНК другими
авторами, получившими
противоположные данные
(табл. 5). Таким обра-
зом, приходится отказать-
ся от мысли, что в дор-
сальной губе бластопора
происходит более интен-
сивный синтез белка, чем
в эктодерме. Можно, ко-
нечно, предполагать, что дорсальная губа как-то передает эктодер-
ме способность к белковому синтезу. Есть предположение, что эта
роль выполняется посредством РНК. Действительно, Пфаутч обна-
ружил более высокую концентрацию РНК в уйасТках эктодермы,
подстилкой которых был мезодермальный материал, по сравнению
с иными участками эктодермы.
Многочисленные иные биохимические данные также подтверж-
дают гетерогенность развивающегося зародыша именно в том
смысле,, какой придавал этому явлению Чайлд.
Дезорганизация градиента химическими и физическими спосо-
бами сопровождается ненормальностями развития.
На зародышах других животных обнаружены принципиально
те же явления. У куриных эмбрионов в гензеновском узелке пер-
вичной полоски (по теории Шпемана, тождественной с дорсаль-
ной губой бластопора) наблюдается максимальная биохимическая
активность (рис. 127).
315
К ЧЕМУ ПРИВОДЯТ НАРУШЕНИЯ СИСТЕМЫ ГРАДИЕНТОВ ЗАРОДЫША!
Чайлд и его последователи пытались экспериментально доказать,
что высокий полюс градиента координирует формообразователь-
ные процессы и что нарушение системы градиентов приводит к
нарушению нормальных процессов формообразования. Изучалось
влияние на эмбриональное развитие гидромедузы (Phialidiutn
gregarium KCN, LiCl), фенилуретана и других ядов. При нор-
мальном развитии имеет место униполярная иммиграция. При
действии ядов происходит нечто вроде мультиполярной иммигра-
ции; при этом число иммигрирующих клеток возрастает, и вместо
формирования однослойной энтодермы образуется плотная масса
клеток. У формирующихся планул теряется полярность, личинка
не вытягивается в длину. Если личинка прикреплялась, то выра-
стал не один стебель, а одновременно несколько.
Были предприняты и другие попытки, изменяя систему гради-
ентов, доказать, что физиологические градиенты являются осно-
вой развития. В этом отношении представляют интерес опыты
Ф. Г. Джилькриста 1929, 1933 гг. по усилению или, наоборот,
противодействию исходному градиенту яйца амфибий. Считая, что
изменение температуры ускоряет или замедляет процессы разви-
тия, т. е. вызывает различия не качественного, а лишь количест-
венного порядка, Джилькрист воспользовался температурой для
вмешательства в систему градиентов. Еще до опытов Джилькри-
ста было установлено, что «прибавление» на ранних стадиях раз-
вития к существующему
анимально - вегетативному
градиенту температурного
градиента приводит к раз-
личиям в величине бласто-
меров, к аномалиям при
гаструляции, но в конечном
счете—к появлению эм-
брионов и молодых личинок
с увеличенной или, наобо-
рот, уменьшенной головой
в зависимости от того, ка-
кую область — анимальную
или вегетативную—будет
экспериментатор сильнее
обогревать.
Джилькрист помещал
бластулы тритона между
двумя алюминиевыми пла-
стинками, из которых одна
была нагрета, а другая под-
вергалась охлаждению. Та-
Рис. 128. Опыты Ф. Джилькриста (1929)
по влиянию температурного градиента на
хвостатых амфибий (на стадии бластулы
создавалась разница в 5° в нагреве двух
ее сторон):
А, Б — нагревалась левая сторона (А — вид со
спины, Б — внд спереди; левая нервная складка
большего размера и более дифференцирована);
В—Д — внд со спины; Е — вид сбоку (видны за-
кладки вторичной нервной системы на стороне,
подвергшейся более сильному обогреву)
316
кой неравномерный обогрев продолжался в течение двух суток,
а затем зародыши развивались в нормальных условиях. Если тем-
пературный градиент создавался так, что к нагретой пластинке
была обращена правая (или левая) половина тела зародыша,
а другая половина тела соприкасалась с охлажденной пластинкой,
то вызывались резкие нарушения нормального развития медулляр-
ной пластинки (рис. 128). Медуллярная пластинка оказывается
резко асимметричной, а именно: на подогретой стороне она ненор-
мально сильно развита, на охлажденной же — недостаточно. В не-
которых случаях на обогретом боку медуллярная пластинка рас-
пространяется за свои обычные пределы, в других случаях обра-
зуются добавочные нервные пластинки, которые могут возникнуть
и независимо от основных. Гистологическое строение добавочных
нервных пластинок или добавочных участков, подобно нормаль-
ному.
Могут быть и менее наглядные факты, но всегда на подогре-
той стороне нервные валики, а позже и соответствующие полови-
ны головного и спинного мозга оказываются более утолщенными.
Эти результаты можно объяснять так: температурный гради-
ент потому вызвал развитие добавочных структур, что обусловил
изменение нормального градиента физиологической активности.
Именно созданная разность температур целостного зародыша вы-
звала это явление, так как повышение или понижение температу-
ры всего зародыша повлекло бы ускорение или замедление разви-
тия, а не качественные различия в дифференциации. Приводи-
лось возражение против такой интерпретации опытов Джилькри-
ста. Некоторые исследователи думают, что вызванные в этих
опытах нарушения нейруляции зависят от нарушений в процессах
гаструляцин, в результате чего необычно распределяется клеточ-
ный материал крыши первичной кишки. Другие исследователи
считают, что это возражение не имеет значения, так как в силе
остается факт, что созданный температурный градиент вызвал из-
менения характера дифференциации.
Принципиально тот же результат получен В. Фогтом. В его
опыте специальная камера разделяется на две части серебряной
пластинкой с круглыми отверстиями, куда помещают зародышей
амфибий, препятствуя тем самым смешиванию воды в камерах.
Через одну половину камеры протекает холодная вода (2—
5°С), а через другую — теплая (19—22°С). В результате искусст-
венно созданного температурного градиента развитие одной поло-
вины зародыша задерживается. Через 3—4 дня, когда появляются
нервные валики, такие ненормальные зародыши помещают в
обычные условия. Развитие продолжается так, что, положим, на
одной стороне образовалась уже нервная пластинка с валиками,
а на другой — процесс нейруляции еще не начался. Условия опы-
та Фогта таковы, что в отличие от экспериментов Джилькриста
граница между нагретой и охлажденной половинами зародыша
317
была слишком резкой; поэтому разница в развитии половин очень
велика. Несмотря на это, прибегнув к методике маркировки ви-
тальными красителями зародыша, еще не подвергнутого темпера-
турному воздействию в указанной «камере Фогта», удалось уста-
новить, что на обогретой половине гаструляционные передвижения
материала оказались нормальными, а на охлажденной стороне
инвагинировало меньше материала. Сторонники приложения тео-
рии физиологических градиентов к анализу процессов дифферен-
циации ,считают, чтр и в опытах Фогта имело значение изменение
активности физиологических процессов, так как равномерный
обогрев не вызвал бы качественных изменений в явлениях диф-
ференциации.
Для оценки теории Чайлда следует привлечь и другие много-
численные факты. Так, опыты Джилькриста видоизменялись та-
ким образом, что создавалась разность температур не в дорсовент-
ральном, а в анимально-вегетативном направлении. Если созда-
вать более высокую температуру в области анимального полюса
и пониженную температуру в области вегетативного полюса, т. е.
если усиливать естественный анимально-вегетативный градиент
яйца, то развиваются зародыши с ненормально большими голо-
вами. Если же ставить эксперимент так, что разность температур
препятствует нормальному градиенту физиологической активно-
сти, т. е. если охлаждать область анимального полюса и, наобо-
Рис. 129. Подавление развития переднего отдела головы Fundulus heteroclitus:
А — нормальный зародыш; Б — циклопин; В — циклопин с уменьшением размера глаза;
Г—аиофтальмия; Д—Л —вид спереди нормальной головы и головы на различных стадиях
подавления ее развития (Л; Б; Д—Л — по И. Стоккарду; В, Г—по Дж. Гексли и Г. де Бе-
ру, 1934)
318
рот, обогревать область вегетативного полюса, то зародыши раз-
виваются с ненормально маленькими головами. Вероятно, к этим
данным, говорящим в пользу теории Чайлда, следует добавить
о результатах экспериментов, в которых развивающиеся зароды-
ши подвергались такого рода химическим воздействиям, когда по-
ражались или стимулировались области высокой физиологической
активности. Такие эксперименты проведены на зародышах рыб и
амфибий до гаструляцин или во время нее. Подбиралась такая
концентрация раствора хлористого магния или некоторых других
веществ, при которой ни одна часть зародыша не умирает; ни
о каком разрушении органов не может быть и речи, так как в мо-
мент опытов на стадиях бластулы или гаструлы никаких органов
еще нет. Речь может идти, таким образом, о поражении физиоло-
гической активности. В результате таких опытов передние части
головы недоразвиваются, глаза личинки сидят ненормально близ-
ко друг к другу или совсем сливаются, может возникнуть даже
одноглазие — циклопия (рис. 129,В—И). Могут слиться зачатки
органа обоняния, недоразвивается рот животного (рис. 129,Г, К,
Л). Подобные уродства наблюдаются в природе при разных на-
рушениях нормальной среды для развития. Сторонники теории
Чайлда имеют основание в орбиту своего анализа взять, уродства
и человеческих младенцев и эмбрионов домашних животных, хотя
конкретный химический механизм этих явлений и не выяснен:
случаи недоразвития головы, полное отсутствие головы, циклопия
и т. п. ’ ,
ЗНАЧЕНИЕ ТЕОРИИ Ч. ЧАЙЛДА ДЛЯ ЭМБРИОЛОГИИ
В предыдущих параграфах рассмотрены попытки применения тео-
рии Ч. Чайлда к анализу явлений эмбрионального развития. Ко-
личественные различия в уровне физиологической активности воз-
никают в ооците в связи с действием внешних факторов. Эти
различия превращаются затем в качественные различия. Каждый
зачаток образуется на определенном уровне градиента; дело, од-
нако, не просто в интенсивности жизненных процессов в этом
участке, а в градиентной системе, в относительном местоположе-
нии данного участка. Теория градиентов, на первый взгляд, явля-
ется физиологическим подтверждением теории индукторов Шпе-
мана, во всяком случае кажется, что эти теории не противоречат
друг другу. Этот вывод особенно напрашивается при сопоставле-
нии взглядов Шпемана на дорсальную губу бластопора как на
организатор и взглядов Чайлда на этот участок зародыша как на
доминантную область. Но из теории физиологических градиентов
не вытекает необходимость понятия «индуктор». Повышение фи-
зиологической активности в том или ином участке зародыша мо-
жет происходить без участия индуктора. Вспомним в этой связи
теорию трофического эпигенеза Далька (см. гл. XIV). Система
319
градиентов все время меняется в ходе развития зародыша, созда-
ются новые и новые градиентные системы (в пределах дорсальной
губы бластопора, в пределах формирующейся нервной системы,
почки конечности и т. д.).
Совершенно очевидно, что опыт применения теории Чайлда
к процессам индивидуального развития явился прогрессом и свое-
образной реакцией на одностороннее морфологическое направле-
ние в эмбриологии. Длительная история удачных и неудачных по-
пыток использования теории Чайлда в эмбриологии дает основа-
ние считать, что меняющаяся в ходе развития зародышей система
физиологических градиентов,— один из показателей изменения
состояния целостности зародышей. Это не означает, что теория
градиентов не нуждается в развитии и что она одна сама по себе
является теорией индивидуального развития. И до сих пор неяс-
ных вопросов и противоречий в этой теории слишком много, чтобы
она могла считаться бесспорной.
Одно из основных утверждений Чайлда о неспецифичности
градиентов повреждения при действии агентами различной при-
роды нуждается в существенных поправках. Картины морфологи-
ческих изменений, сопровождающие смерть, разнообразны и зави-
сят от природы повреждающего' агента. Особенно резкие откло-
нения от обычно описываемых схем «градиента распада» наблю-
даются при действии биологически активных веществ (фитонци-
ды, антибиотики животного происхождения).
В резкое противоречие с теорией Чайлда вступают результа-
ты некоторых опытов, согласно которым отчетливые физиологи-
ческие градиенты у многих зародышей удается выявить лишь со
стадий, когда уже имеется определенная степень интеграции раз-
вивающегося организма и когда разные участки зародыша разли-
чаются, конечно, не только количественно. Если в основе физио-
логических градиентов лежат только различия количественного
порядка и если на базе градиентов возникают качественные раз-
личия, то анимально-вегетативный градиент лучше всего должен
был бы выявляться на самых ранних стадиях. Однако в опытах
Н. И. Вержбицкой (1958) с эмбрионами морского ежа Strongilo-
cecentrotus droebachiensis не удалось наблюдать ясного градиен-
та до стадии восьми бластомеров, т. е. когда достигается опреде-
ленная степень интеграции и когда в разных частях зародыша
имеются не только количественные, но и качественные различия.
Основное представление о градиентах как о системе частей,
имеющих лишь количественную характеристику (уровень мета-
болической активности), представление, что нет никаких отличий
в структуре,— крайне упрощено и нуждается в изменении в соот-
ветствии с данными морфологии, биохимии и биофизики. Впро-
чем, и Чайлд, конечно, не думал столь примитивно.
Много и других сомнений и возражений возникает при оценке
значения теории Чайлда для эмбриологии, но при всех своих не-
320
достатках и недоработанности она остается одной из интересных
теорий хотя бы уже потому, что она не односторонне морфологи-
ческая, а базируется на большом количестве физиологических
экспериментов и является теорией широкого общебиологического
значения.
Теория доминантных областей Чайлда и теория организацион-
ных центров Шпемана — одна преимущественно физиологического
содержания, а другая морфологическая — как бы дополняют
друг друга. Чайлд, однако, считал бы такое сопоставление теорий
неправильным. Представление об организаторах он считал оши-
бочным в том отношении, что даже первичный организатор нельзя
назвать первичным. Действие его, по Чайлду,— лишь этап в про-
цессе усложнения организации эмбриона. Он не только не вызы-
вает появления первых признаков организации, но, напротив, сам
оказывается производным существовавшей ранее организации.
Чайлд, вопреки взглядам Шпемана, присоединяется к тем, кто
считает, что при индукции мертвыми или чужеродными тканями
характер индукционного эффекта если не целиком, то во всяком
случае в большей степени зависит от состояния реагирующей си-
стемы. В условиях нормального развития даже детерминация про-
дольной и спинно-брюшной оси зародыша не зависит от органи-
затора.
Как видим, теория Чайлда представляет большой интерес для
эмбриологии. К сожалению, эта теория пока оторвана от совре-
менной генетики, хотя несомненно, что системы физиологических
градиентов возникают на генетической основе.
Нельзя сомневаться в том, что и генетики заинтересуются тео-
рией Чайлда, а методиками современной биохимии удастся углу-
бить ее и освободить от возможных ошибок, обусловленных несо-
вершенством прежних способов исследования.
МЫСЛИМОЕ СОЗДАНИЕ НЕСКОЛЬКИХ «ПОЛЕЙ»
Любая конструкция поля, основанная на предположении об од-
ном якобы «решающем» факторе формообразовательных процес-
сов, в лучшем случае односторонне отразит индивидуальное раз-
витие. При меняющихся состояниях целостности, изменениях во
взаимоотношениях частей вступают новые и новые механизмы ин-
теграции. Претензии на описание и тем более анализ любого фор-
мообразовательного процесса исходя из какой-либо одной конст-
рукции поля или градиента иллюзорны.
Возможно и необходимо создать ряд теорий полей, отражаю-
щих различные стороны явлений интеграции: это могут быть поля
электрических потенциалов, поля лучистой энергии, продуцируе-
мой клетками, поля адгезии, поля денатурации белков, поля
свойств мембран, поля межклеточных субстанций и т. д. Эти поля
могут способствовать углублению понимания развивающегося
321
организма как целого. Здесь следует напомнить высказывания
эмбриологов разных направлений о клеточном поле.
Уже упоминались гипотезы Н. К. Кольцова, Дж. Гексли и
Г. де Бера, П. Вейса и др. Теория физиологических градиентов
Ч. Чайлда может быть названа теорией поля. Пространство, за-
нятое организмом, представляет собой поле, скаляром которого
является физиологическая активность, а градиентами — оси тела
(П. Г. Светлов). Поле и градиент — сопряженные понятия, оба
связаны с представлением о целостности организма, его интегра-
ции (И. А. Аршавский).
Теория поля Чайлда может быть названа (в соответствии со
взглядами Аршавского) теорией динамики целостности развива-
ющегося организма. Количественные различия в уровне метабо-
лизма, или градиенты, мыслятся как простейшие системы, опре-
деляющие интеграцию таких организмов, у которых в эволюции
еще не выработались или находятся в примитивном состоянии ин-
теграционные механизмы другого порядка.
П. Г. Светлов — единственный эмбриолог, обосновывающий
необходимость введения в эмбриологию таких физиологических
понятий, как раздражение, возбуждение, торможение. Он гово-
рит о химическом взаимодействии частей развивающегося орга-
низма, о возможном выделении клетками веществ, которые могут
морфогенетически воздействовать на другие клетки или зачатки
или на организм в целом. Таковы гормоны — вещества, вызываю-
щие эмбриональную индукцию, химические воздействия нервной
системы. По Светлову, влияние веществ одной части зародыша на
другую можно рассматривать как влияние раздражителей, кото-
рые могут и не быть компонентами метаболизма. Много доказа-
тельств его правоты можно найти в фактах эмбриологии. Таковы,
например, факты, обнаруженные Г. А. Бузниковым и Т. Густафсо-
ном, касающиеся медиаторов и их роли на ранних этапах эмбрио-
нального развития. Вводя в круг рассмотрения явлений развития
возбуждения и торможения, нет надобности ограничиваться це-
лостным изучением объекта на клеточном уровне: передача раз-
дражений от клетки к клетке у безнервных многоклеточных орга-
низмов (в частности, у растительных) хорошо известна. Ясно, что
возбуждаться и тормозить могут и зачатки органов, и области
тела, и весь зародыш в целом. Автор предлагает радикально ре-
формировать эмбриологию, настаивая на том, чтобы она не чуж-
далась физиологии. Поле Светлова — это такой надклеточный
фактор, который возникает и определяется на основе жизни кле-
ток, их взаимодействия. С его точки зрения в основе всякой тео-
рии развития должны лежать три главных принципа: целостность
организма на любой стадии его развития; представление о разви-
тии как о реактивном процессе и принцип фаз или периодичность
любого морфогенетического процесса.
322
В заключение остановимся на взглядах К. Уоддингтона, кото-
рый сдержанно и скорее скептически относится к использованию
понятия поля в эмбриологических исследованиях. Он считает, что
«...биологи не смогли придумать ничего лучшего, как постулиро-
вать наличие морфогенетического поля, которое определяет облик
образуемой структуры. Слово «поле», разумеется, представляет
собой расплывчатое понятие. В его обычном смысле оно означает,
что внутри участка развивающейся структуры действует какая-то
сила (или простой ряд сил), правильно распределяющаяся в
пространстве. В случае биологических явлений трудность заклю-
чается в том, чтобы определить что это за сила»1. «Мы знаем
очень мало о материальной природе причинных агентов тех про-
цессов, для объяснения которых мы употребляем термин «поле»1 2.
Появление в эмбриологии гипотез о полях, градиентах или
градиент-полях следует рассматривать как неизбежный резуль-
тат того, что морфогенетические процессы не могут быть поняты
только на путях аналитических исследований. Рассмотрение раз-
вивающихся зародышей как целостных систем требует особых ме-
тодов и особых моделей для их описания и объяснения. И в со-
временной эмбриологии создаются понятия о ближайших и ди-
станционных контролирующих механизмах (Т. Ямада), о позици-
онной информации (Л. Вольперт) и связанных с ней двумерных
моделях органных полей (П. Френч и Б. Брайент). Последние
с успехом используются для анализа регуляционных процессов,
совершающихся в имагинальных дисках насекомых и для объяс-
нения особенностей восстановительных процессов после разных
типов повреждения конечностей членистоногих и амфибий.
Концепция позиционной информации Вольперта основывается
на предположении о существовании в развивающемся организме
градиента некоего морфогена — гипотетического вещества, влияю-
щего на развитие той или иной структуры. Морфоген распростра-
няется между определенными точками отсчета. Процесс «усвое-
ния» клетками информации о своем положении назван Вольпер-
том спецификацией. В зависимости от характера предшествующей
дифференцировки клетки «интерпретируют» позиционную инфор-
мацию в конкретные структуры («паттерны») того или иного ор-
гана.
Природа и само существование «морфогенов» остаются пока
еще очень неясными, так же как и природа индуцирующих аген-
тов, о которых говорилось в связи с анализом теории организато-
ров Шпемана.
В последние годы делаются попытки создания новых концеп-
ций векторных полей, влияющих на морфогенезы. В основу этих
1 Уоддингтон К. Морфогенез и генетика. М., 1974. С. 184—185.
2 Там же. С. 202.
323
концепций кладутся представления математиков об устойчивых и
неустойчивых структурированных процессах. Для анализа гради-
ентно и волнообразно распространяющихся морфогенетических
процессов используют, в частности, модели математика А. Тью-
ринга, которые объясняют, на основе каких взаимодействий абст-
рактных веществ в физико-химическом процессе могут возникать
диссипативные процессы. Предложенные им ' уравнения могут
быть использованы для объяснения возможных механизмов обра-
зования, например мезодермальных сомитов, позвонков, периоди-
чески повторяющихся рисунков или структур на поверхности тела
и т. п. В качестве взаимодействующих веществ в развивающихся
системах называют гипотетические активаторы и ингибиторы, мо-
гущие влиять на развитие какого-либо конкретного признака. Эти
идеи близки к гипотезе С. Роуза (1952) об ингибиторных разме-
жеваниях, согласно которой раз возникшая структура выделяет
фактор, тормозящий развитие аналогичной структуры по-соседст-
ву с ней.
Делаются попытки приложения к онтогенезу синергетики
(К. Хакен,-1980)—научного направления, развивавшегося на ос-
нове математической теории динамических систем. Создаются
различные модели, объясняющие скачкообразные эмбриологиче-
ские процессы, а также предлагаются гипотезы «параметрическо-
го» управления онтогенезом (Л. В. Белоусов, Д. С. Чернявский,
Г. И. Соляник, 1985). В этой теории в основу разделения регули-
рующих факторов на устойчивые и динамические" кладется вре-
менная иерархия морфогенетических процессов.
Таким образом, в связи с прогрессом математики в области
топологии, поведения и управления различными динамическими
системами и т. п. создаются возможности для более адекватного
описания ряда эмбриологических событий в их нерасчлененном
состоянии.
Анализ разных теорий поля не убедил нас в необходимости
признать какую-либо из них теорией индивидуального развития,
могущей удовлетворить эмбриолога. Мы могли лишь убедиться в
том, что понятие поля не является случайным. Вне зависимости
от методологии исследователей оно отражает тот очевидный факт,
что любые представления о зародыше как мозаике частей, как
сумме бластомеров и т. д. несостоятельны, что организм на любой
стадии развития так или иначе интегрирован, представляет це-
лостную систему.
Глава XIV
РАЗВИТИЕ ОРГАНИЗМА И СРЕДА
Классическая общая эмбриология в лице К. Ф. Вольфа, X. Пан-
дера, К- М. Бэра и сравнительная эмбриология Ковалевского—
Мечникова созданы вне каких-либо идей, наблюдений и экспери-
ментов по вопросам о роли внешней среды в развитии организмов.
Экспериментальные эмбриологические школы конца XIX в. и
нашего столетия или совсем не уделяли внимания внешней среде
(например, школа Шпемана, Герстадиуса и др.), или проводили
лишь эпизодические исследования (В. Ру, П. П. Иванов и др.).
В начале XX в. многие исследователи интересовались влиянием
силы тяготения на эмбриональное развитие, значением осмотичес-
кого давления, газового состава среды, химического состава мор-
ской и пресной воды.
Представляют исторический интерес взгляды одного из осново-
положников «механики развития»—В. Ру. Он различал находя-
щиеся в яйце «детерминирующие факторы» и «факторы реализа-
ции», существующие в среде. По его представлениям, можно гово-
рить о четырех периодах в развитии животного организма. Первые
стадии эмбрионального развития определяются исключительно на-
следственными факторами; во втором (промежуточном) периоде
развитие обусловлено частично наследственными факторами, час-
тично раздражителями внешней среды. В этом периоде органы про-
должают развиваться под влиянием внутренних факторов, но
в связи с началом функционирования те или иные дифференци-
ровки возникают в организме под влиянием трофических функци-
ональных раздражителей. Характер последующего развития (тре-
тий период) определяется только внешними раздражениями —
гипертрофией одних частей (вследствие деятельности) и атрофией
других (от бездействия). Он придавал исключительное значение
для осуществления этого периода трофическим функциональным
раздражителям. Наконец, он предлагал отличать и четвертый пе-
риод— увядание или старость.
В гл. XIII излагалась теория Чайлда, который объяснял явле-
ния возникновения первичной полярности ооцитов, явления сим-
метрии яйцевых клеток и ранних зародышей действием внешних
факторов. Эти взгляды не потеряли значения для современной
эмбриологии.
Большой интерес к проблемам влияния внешней среды на раз-
витие животных проявили советские эмбриологи, особенно начи-
ная с 40-х годов. Этот интерес стимулировался общим для всей
биологии направлением активного вмешательства в развитие ор-
325
ганизмов, стремлением к управлению онтогенезом. Предложено
много интересных теорий периодизации онтогенеза животных
(В. В. Васнецов, Б. С. Матвеев, Г. А. Шмидт, И. А. Аршавский
и др.). Сформулированы новые поставленные развитием науки
и потребностями практики задачи и возникли новые главы эм-
бриологии, в которых предметом изучения являются факторы сре-
ды эмбрионального развития. Такова, например, проблема имму-
нитета эмбрионов (см. ниже). Большое значение приобрели воп-
росы «эмбриологической» радиобиологии.
ВНЕШНЯЯ СРЕДА И НЕОБХОДИМЫЕ УСЛОВИЯ РАЗВИТИЯ
Эволюционное возникновение того или иного вида сопровождалось
приспособлением организмов к определенным условиям обитания,
появлением мутаций, закрепленных естественным отбором. Раз-
ные виды характеризуются различным диапазоном наследственно
закрепленной изменчивости, размахом физиологической адаптации
к изменениям среды.
В течение по крайней мере 8 миллионов лет рыба лещ сущест-
венно не изменялась. С другой стороны, известно, что лещ при-
надлежит к видам, легко акклиматизируемым в разных условиях.
Свойства широкой адаптации к меняющимся условиям у этого
вида животных наследственно закреплены.
В одном и том же водоеме в одинаковой среде развиваются
разные животные. В инкубаторе при одинаковых условиях из яйца
курицы развивается цыпленок, а из утиного яйца — утенок. Одна-
ко далеко не всегда кажущаяся одинаковой среда действительно
такова. Организм любого вида животных — исторический «сгус-
ток» эволюции. Яйцеклетки и ранние зародыши всех животных
различны. У каждого вида животных создались в ходе эволюции
свои отношения к внешней среде, свой диапазон приспособлений
к меняющимся условиям и т. д. То, что для эмбрионов одного ви-
да животных составляет необходимейшее условие развития, яв-
ляется нейтральным или вредным для других видов. В ходе
эволюции изменялись отношения организмов к среде, изменялись
закономерности наследственности, изменчивости, естественного
отбора и закономерности самой эволюции. Отражением этого
сложного и извилистого исторического процесса является большое
разнообразие живой природы, особенности изменчивости и на-
следственности у разных групп современных животных и растений.
Попытки навязать природе абсолютно одинаковую схему измен-
чивости и наследственности являются, может быть, очень важной
ошибкой биологии.
К- А. Тимирязев говорил о «требованиях», которые организм
предъявляет к среде в ходе своего развития. Из одних и тех же
веществ среды строится протоплазма разных организмов. Отно-
326
шение животных к одним и тем же окружающим веществам и ви-
дам энергии неодинаково. У многих организмов на разных этапах
онтогенеза закономерны переходы из одной среды в другую. У ря-
да насекомых личинки и куколки развиваются в воде или почве,
а взрослые обитают в воздухе; у большинства амфибий происхо-
дит также смена водной и воздушной сред.
Своеобразная смена сред имеет место и в развитии рыб, птиц
и млекопитающих. У последних, например, смена сред развития
эмбриона связана с имплантацией, рождением. Выясняется, что
каждому этапу свойствен особый приспособительный характер
строения, физиологии, поведения. В ходе развития животных
имеет место подчас сложная биологическая смена тех или иных
функций и на этой основе меняется отношение к внешней среде.
Первичным органом дыхания у зародышей костистых рыб явля-
ется сосудистая система на желточном мешке. Затем наступает
редукция этой системы (в связи с использованием желтка); у со-
мовых (Callichtys), у донной личинки функцию дыхания берет
на себя грудной плавник с мощной сетью кровеносных сосудов,
лишенный в этот период скелетных образований. После вылупле-
ния, когда зародыш переходит к свободному образу жизни, дыха-
тельная функция грудного плавника замещается постепенно жа-
берным дыханием.
Первичной формой дыхания у личинок амфибий является жел-
точная сосудистая сеть, замещающаяся впоследствии наружными
жабрами личинок амфибий. Затем функцию дыхания принимают
на себя жабры, сходные с таковыми у рыбы, наконец — легкие.
Меняющиеся отношения к внешней среде в ходе развития ор-
ганизмов есть отражение адаптаций, имевших место в эволюции
вида. Видовые адаптации развиваются в процессе эволюции не
как отдельные частные приспособления, а как элементы системы
взаимозависимых адаптаций. Функциональная и экологическая
специфичность этапов онтогенеза связана с возникновением и сме-
ной провизорных корреляций (временных, обеспечивающих жиз-
ненные проявления на данном этапе онтогенеза). Эти провизорные
корреляции Н. Л. Гербильский в 1963 г. рассматривал как про-
цессы реализации филогенетических адаптаций в онтогенезе. При-
меры провизорных корреляций: степень развития эмбриона и фун-
кция желез вылупления, выделяющих вещества, изменяющие
оболочки и дающие эмбриону возможность освобождения от них;
железы сформировываются в определенный этап развития; сущест-
вуют корреляции между эмбриональным кровообращением, прови-
зорными органами дыхания и желточным питанием.
В ходе эмбрионального развития животного не прерывается
связь организма с внешней средой, постоянно происходит обмен
веществ и преобразование энергии. Развивающийся организм ни-
когда не бывает застывшим морфологическим сооружением, в нем
непрерывно осуществляются процессы деструктивные и структуро-
327
образовательные, идущие на разных уровнях организации (атом-
ном, молекулярном, клеточном и т. п.).
Какие условия среды прежде всего привлекают внимание эм-
бриолога? Это обыденные, массовые, естественно встречающиеся
в природе: 1) температура; 2) вода — степень влажности воздуха
для яиц сухопутных животных, химический состав воды, соотно-
шение и концентрация солей для яиц водных животных; 3) содер-
жание кислорода, углекислого и других газов в среде, окружаю-
щей эмбрион; 4) атмосферное давление; 5) лучистая энергия и др.
В отношении всех необходимых условий можно говорить о ми-
нимуме, максимуме и оптимуме, значение которых для разных
эмбрионов может быть очень различным. Это относится, напри-
мер, к температуре.
Температура оказывает влияние прежде всего на обмен ве-
ществ. В пределах минимума и максимума она не вызывает ка-
чественных изменений в развитии, а способна ускорить или за-
медлить развитие. Так, при изменении температуры на 10 °C
скорость развития изменяется вдвое (ускоряются или замедляют-
ся химические процессы по правилу Вант-Гоффа). При 20 °C яйца
лягушки развиваются в зародыши с хорошо развитым хвостом, а
другие яйца из той же яйцеклетки при 15 °C к этому времени
достигнут только стадии нейрулы. Каждому виду животных свой-
ственна своя оптимальная температура окружающей среды. Ло-
сось мечет икру в ноябре, мальки вылупляются в мае. Темпера-
турный минимум для них —4 °C, а максимум +9, -(-10 °C. Более
высокая температура препятствует нормальному развитию. В ес-
тественных условиях развитие лососей, начинающееся при +8,
4-2 °C, происходит затем при —2 °C.
Для яиц разных видов птиц температурный оптимум различен.
Интересны особенности отношения развивающегося куриного за-
родыша к температуре. Температурный оптимум — около 38 °C.
Некоторые ученые-птицеводы считают, что температура в те-
чение всего периода эмбрионального развития не должна быть
одинаковой, что необходимы периодические охлаждения яиц кур
на разных этапах эмбриогенеза. То же доказано в отношении за-
родышей уток и гусей. Птицеводы таким образом имитируют тем-
пературные условия, создающиеся при естественном насиживании
курицей. Существует взгляд, согласно которому и в другом отно-
шении целесообразно «имитировать» природу: должен быть неоди-
наковым обогрев разных частей яйца, как это происходит при
естественном насиживании. Под наседкой верхняя половина яйца
имеет более высокую температуру — градуса на 2—3 (рис. 130).
Создание искусственного температурного градиента внутри
яйца Других животных (например, амфибий) может повлечь, од-
нако, появление уродств, как это мы видели в связи с изложением
теории физиологических градиентов Чайлда (см. гл. XIII).
На примере температурных условий эмбрионального развития
328
мы видим, сколь своеобразны истори-
чески сложившиеся отношения к внеш-
ней среде у разных видов животных.
То же можно сказать и о других обы-
денных, всегда встречающихся в при-
роде факторах внешней среды. Напри-
мер, лучи разных длин волн оказыва-
вают влияние на развитие; как прави-
ло, развитие ускоряется при действии
фиолетовой и синей частей спектра
солнечных лучей и замедляется при
действии лучей красной части спектра.
Но это не есть общая закономер-
ность. Достаточно вспомнить развитие
яиц, нормально развивающихся в тем-
ноте.
Еще более разнообразны и порой
своеобразны связи развивающихся за-
родышей с «химической средой»: для
одних (например, морских форм) оп-
тимальны одни комбинации веществ и
их концентрации, а для других эти
вещества гибельны (например, для
пресноводных животных). Для одних
животных наличие кислорода во внеш-
ней среде—абсолютно обязательное
условие развития, а у других процес-
сы диссимиляции идут по-иному. Де-
монстративным примером значения
кислорода являются яйца аскарид,
Рис. 130. Температурный
градиент развивающихся
яиц птиц (по Ф. Г. Джиль-
кристу, 1929)
оксиуриусов, трихостронгилид и некоторых других паразитических
червей. Если лишить развивающийся эмбрион этих животных ки-
слорода, развитие останавливается. Это можно проверить экспе-
риментальным путем, если поместить, например, яйца лошадиной
аскариды (на любой стадии развития) под слой ртути или содер-
жать в герметических сосудах в бескислородной среде.
Принципиальным вопросом в эмбриологии является сравнение
роли в эмбриональном развитии обыденных, естественно встречаю-
щихся условий — на суше, в водоемах, воздухе, почве и таких
факторов, как ионизирующая радиация, соли тяжелых металлов
и т. п. Мощный с точки зрения физики й химии фактор может
оказаться значительно менее эффективным при действии на раз-
вивающийся организм, чем обыденные факторы, о которых мож-
но думать, что в ходе эволюции организм на них уже отреаги-
ровал.
Существует вполне обоснованный взгляд: инструментом управ-
ления развитием организмов должны скорее быть не агенты, ко-
329
торые не встречаются в среде жизни данного организма, а скорее
варьирование какого-либо важного условия жизнедеятельности,
нормально включенного в основные жизненные процессы разви-
вающегося зародыша (Б. Л. Астауров). Такова, например, темпе-
ратура окружающей среды.
ЭВОЛЮЦИЯ ЯИЦ. ОБ ОТНОСИТЕЛЬНОЙ ЭМАНСИПАЦИИ
ОТ ВНЕШНЕЙ СРЕДЫ ЭМБРИОНОВ РАЗНЫХ ЖИВОТНЫХ
Любой организм на любой стадии развития в норме обычно так
или иначе связан с внешней средой. Организм может некоторое
время обходиться без притока пищи извне и заниматься «самоед-
ством», но и в этом случае связь с внешней средой поддержива-
ется благодаря процессам диссимиляции.
Все современные организмы в конечном счете являются про-
дуктом внешней среды. Эволюция организмов — совокупность про-
цессов взаимосвязи организмов между собой и с неорганической
средой. Изменчивость организмов и естественный отбор — явления,
по самой свой сути связанные с отношением организмов к среде.
Однако верно и другое: почти вся современная внешняя среда,
как указывал еще в 1926 г. В. И. Вернадский, оказалась продук-
том жизни. Газовый состав атмосферы (весь кислород воздуха)
биогенного происхождения. Океаны, вся гидросфера пронизана
жизнью; концентрация солей, pH и т. д. обусловлены в значитель-
ной мере организмами. История литосферы (30—80 км в глубь
земли) связана также с жизнью.
Не менее важно учесть и другое соображение. Принято гово-
рить о «взаимодействии» организмов, их эмбрионов и среды на
почве метаболизма. При ассимиляции, однако, живые организмы
уподобляют внешнюю среду самим себе, из одних и тех же эле-
ментов внешней среды строится разная протоплазма у разных
организмов.
В ходе эволюции происходила, образно говоря, эмансипация
организмов от внешней среды. В самых разнообразных направ-
лениях изменялась реактивность протоплазмы, вплоть до воз-
никновения механизма антигенной реактивности; вместе с тем
в историческом процессе естественного отбора приспособлений,
в частности отбора приспособлений у ранних эмбрионов, создава-
лись и механизмы, отгораживающие развивающиеся организмы от
внешней среды. Таковы, например, оболочки яиц животных. Ярким
примером «эмансипации» от среды служит эмбриональное разви-
тие многих паразитических червей. Возьмем яйца лошадиной,
свиной или человеческой аскариды. Они исключительно резистент-
ны ко многим повреждающим агентам. Яйца лошадиной аскариды,
как это выяснил М. М. Завадовский еще в 1914 г., способны жить
3 дня в формалине, 14—20 дней в глицерине, 14—80 дней в ксило-
ле, 9 дней в бензоле, сутки в дымящейся НС1, 5 дней в крепкой
330
HNO3 и т. д. Лишь концентриро-
ванный раствор H2SO4 разруша-
ет яйцо мгновенно. В растворах
HgCl2, C11SO4, НС1 и КОН яйца
аскариды развиваются даже не-
сколько быстрее, так как эти ве-
щества убивают микроорганиз-
мы, могущие «перехватывать»
кислород, совершенно необходи-
мый для яиц. Какие химические
вещества могут свободно прони-
кать в зародыши или выходить во
внешнюю среду? Н2О, СО2 и О2.
Для развития яйца аскариды
вплоть до личинки не требуется
проникновения из внешнего мира
ничего, кроме воды и кислорода.
Такая эмансипация эмбриональ-
ного развития от среды очень вы-
годна для паразитических форм.
Оплодотворенное яйцо, лошади-
ной, свиной или человеческой ас-
кариды окажется в кишечном
тракте животного-хозяина в сосед-
стве со многими ядовитыми для
зародыша веществами. В кишеч-
нике без кислорода развития не
происходит. По выходе из кишеч-
ника яйцо обычно попадает в сре-
ду со многими повреждающими
веществами, поэтому в ходе эво-
люции выработались своеобраз-
ные биохимические и иные при-
способления, предохраняющие
зародыши. Основное приспособ-
ление— структура оболочек.
По современным данным, по-
сле оплодотворения в яйце аска-
риды возникает многослойная
оболочка (3—5 мнм толщиной).
Сначала образуется поверхно-
стный, вителлиновый слой, затем
Рис. 131. Схема строения яйцевой
оболочки аскариды Ascaris lum-
bricoides (по А. Бёд, 1971):
1 — трехслойная мембрана, 2 — вител-
лнновый слой, 3 — хитиновый слой,
4 — липидный слой. 5 — клеточная мем-
брана, 6, 7 — шероховатый ЭПР и
плотные гранулы кортикального слоя
цитоплазмы ооцита
хитиновый и еще позднее самый
внутренний, липидный слой. Все они имеют фибриллярную струк-
туру. На рис. 131 дано изображение всех слоев яйцевой оболочки
после их окончательного формирования у Ascaris lumbricoid.es.
Структура и степень выраженности всех слоев оболочки водоспе-
цифичны. Оболочка яйца аскариды предохраняет зародыш от
331
Таблица 6. Эволюция яиц (по Дж. Нидхему, 1931, модифицировано)
Животные Органические вещества: белки, угле- воды, жиры, ЛИПОИДЫ, стеролы и т. п. Соли Вода Кислород
Водные
Кишечнополостные, ктеиофоры + — — —
Иглокожие, иапример Echinus + — —
Моллюски, например Sepia + . — •
Ракообразные, иапример Emerita + Г““ — —
Костистые рыбы, иапример Salmo + + — —
Селяхии, например Scyllium + + —
Наземные
Амфибии, иапример Rana + + — —
Рептилии, например Tropidonous + + — —
Thalassochelys „
Амфибии, иапример Hyla + + + —
Рептилии, иапример Tropidonous + + + —
Птицы, иапример Gallus + + + +
Водные птицы, например Podiceps + + + ±
Млекопитающие, иапример Ното 0 0 0 0
Водные млекопитающие, например 0 0 0 0
Balaenoptera
Примечание. Знак плюс означает, что яйца снабжены данным веще-
ством от родителей в достаточном количестве для эмбрионального развития;
знак минус — не снабжены этими веществами в достаточном количестве; знак
плюс-минус — вещество вырабатывается внутри яйца и поступает извне; нуль —
яйца получают необходимые вещества иным путем (например, от материнского
организма). Жирная линия отделяет группы веществ, имеющихся в яйце, от
веществ, получаемых извне.
механических воздействий. Она пропускает воду, кислород и угле-
кислый газ, но непроницаема для солей и многих неорганиче-
ских и органических соединений.
Какими веществами, необходимыми для развития, эмбрион
обеспечивается, родительским организмом и какие вещества им бе-
рутся из внешнего мира? Это видно из табл. 6. Яйца морских бес-
позвоночных получают от родительских организмов в достаточ-
ном для развития эмбриона количестве органические вещества —
белки, жиры, углеводы и т. д. Вода, кислород и соли берутся из
внешней среды. Зависимость эмбрионов от солевого состава среды
является, вероятно, одной из причин того, что в пресной воде зна-
чительно меньше беспозвоночных, чем в морской: нет иглокожих,
головоногих моллюсков, асцидий, очень мало кишечнополостных,
червей и моллюсков. В этом, вероятно, главная причина Трудности
332
акклиматизации многих форм. Известно, что взрослые моллюски
некоторых морских видов превосходно выживают в пресной воде,
но яйца их не развиваются в ней и гибнут.
Яйцевые клетки рыб оказываются достаточно снабженными
органическими и минеральными веществами, однако воду и кис-
лород они воспринимают извне. Таковы и яйца амфибий. У всех
высших беспозвоночных и почти у всех Anamnia зародыш обес-
печен не только органическими веществами, но и солями.
Новый этап «эмансипации» яиц от внешней среды наступил в
эволюции вместе с появлением первых наземных позвоночных —
рептилий, яйца которых содержат достаточное для развития за-
родыша количество органических субстанций, солей и воды.
К такому же типу яиц относятся и яйца некоторых амфибий,
например Rana opisthodon. Метаморфоз у этого вида амфибий
происходит внутри яйца, и появляющаяся молодь не отличается
от взрослых. Яйца другой древесной лягушки Megalophrys longi-
pes подобны яйцам рептилий. Однако и в этих случаях эмбрио-
нальное развитие указанных амфибий и рептилий зависит от на-
личия воды во внешней среде.
Яйца птиц имеют достаточное количество органических ве-
ществ, солей и воды. Впервые мы видим у птиц некоторую «эман-
сипацию» зародышей на ранних стадиях развития и от кислорода
внешней среды. Любопытно, что яйца водяных птиц, развивающи-
еся в воде или близко от нее, содержат, однако, достаточно воды
для развития.
Таким образом, в ходе эволюции действительно имеет место
установление таких отношений к среде, которые можно определить
несовершенным термином «эмансипация». Птица снесла яйцо.
Сформированы оболочки. Начавшееся в ходе продвижения яйца
по яйцеводу эмбриональное развитие приостанавливается, и в этом
состоянии яйцо может находиться в течение длительного времени.
Что необходимо для дальнейшего развития? Определенная (в до-
статочно широком диапазоне изменений) температура, наличие
кислорода во внешней среде и определенная влажность. Те же
факторы внешней среды необходимы и для яиц тысяч других ви-
дов птиц и животных, например для яйца лошадиной аскариды.
Кислород, воспринимаемый из внешнего мира, не играет специ-
фической формообразовательной роли. К яйцу птицы на прс^тя-
жении всего периода развития совершенно неприменимо понятие
«взаимодействие» (с внешней средой). Столь совершенная «эман-
сипация» от внешней среды явилась результатом эволюции отно-
шений со средой.
Яйцевые клетки млекопитающих не имеют в достаточном для
эмбрионального развития количестве ни органических веществ,
ни солей, ни воды, ни кислорода: яйцо находится в полной зависи-
мости от материнского организма в отношении жидкостей, газов
и органических веществ. Их поступление в развивающийся заро-
333
дыш происходит, начиная с ранних стадий развития. До имплан-
тации бластоциты в стенку матки внешней средой дробящегося
яйца следует считать среду половых путей, продукты жизнедея-
тельности тканей яйцеводов и матки.
ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ И ВНУТРЕННЯЯ СРЕДА
Представление о внутренней среде развивающегося эмбриона ка-
жется нелогичным, так как любая возникшая в ходе развития за-
родыша структура или жидкость является частью зародыша, и
можно говорить лишь о взаимодействии частей целостной системы,
однако можно рассматривать одну часть зародыша как внешнюю
среду для другой части.
До последнего времени в эмбриологии почти не обращали вни-
мания на различные возникающие и меняющиеся в ходе эмбри-
онального развития жидкости — продукты жизнедеятельности кле-
ток. Уже на стадии восьми бластомеров (у амфибий) между ними
возникает полость дробления, заполненная жидкостью. Имеет ли
она какое-либо значение для дальнейшего развития? Какое зна-
чение имеют жидкости полости бластулы, гаструлы? Каково зна-
чение перивителлиновой жидкости? Каков состав и каково значе-
ние жидкости бластодермического пузырька у млекопитающих?
Каково значение жидкости яйцеводов и матки для развития яиц
Млекопитающих до имплантации зародыша в стенку матки?
За редкими исключениями подобные образования й зародыше-
вом развитии животных не интересовали исследователей. Недо-
статочно даже выяснено значение амниотической жидкости мле-
копитающих и человека, несмотря на заинтересованность ею не
только науки, но и практики медицины и ветеринарии.
Вследствие неразработанности и спорности большинства отно-
сящихся к этой проблеме вопросов следует ограничиться приве-
дением лишь отдельных фактов, которые невозможно объяснять
без понятия «внутренняя среда».
Одни эмбриологи отрицают постановку вопроса о внутренней
среде, другие создают теории эмбрионального развития, факти-
чески основанные на признании ее. Такова теория трофического
эпигенеза бельгийского эмбриолога А. М. Далька (1953).
Анизотропия ооплазмы, полярность, билатеральная симметрия
яйца возникают, по Дальку, в связи с неравномерным распреде-
лением кровеносных сосудов вокруг формирующегося ооцита. Эти
мысли близки теории Чайлда (см. гл. ХШ). Направление диффе-
ренцирования и характер роста каждого зачатка в значительной
мере определяются степенью васкуляризации окружающей его
ткани. Пока ветвление кровеносных сосудов зародыша не достиг-
ло полного развития, разные части зародыша снабжаются кровью
в различной степени, и в связи с этим разные зачатки находятся
в неодинаковых трофических условиях. А. Дальк отрицает пред-
334
ставления об органообразующих участках, якобы существующих
в яйце; клетки раннего зародыша изопотентны, но они могут раз-
виваться в разных направлениях в зависимости от васкуляризации
рядом лежащих частей тела. Биохимические особенности разных
участков тела, например разные количества нуклеопротеидов, об-
разование и активация ферментных систем, накопление энергети-
ческих ресурсов—гликогена, мукополисахаридов и т. д. — все это
связано с условиями кровообращения.
Большое внимание внутренней среде уделял П. П. Иванов. По
его представлениям, для нормального прохождения процессов
гаструляции у амфибий необходимо различие в составе и свойст-
вах жидкости бластоцеля, с одной стороны, и воды как внешней
среды, с другой. На стадии многослойной бластулы у амфибий,
перед гаструляцией, клетки в разных частях зародыша уже имеют
различную «среду»: часть клеток анимальной половины имеет не-
посредственный контакт с внешней средой (они окажутся источ-
ником кожной эктодермы и нервной системы), часть клеток нахо-
дится под влиянием внутренней среды, омывается жидкостью бла-
стоцеля (они дадут энтодерму и мезодерму).
В ходе гаструляции возникают, в свою очередь, различные
условия для разных клеток; одни, уходя в бластоцель, все-таки
образуют поверхность, соприкасающуюся с «внешней средой» че-
рез бластопор, т. е. со «средой» гастроцеля. Эти клетки дают
энтодерму. Часть же клеток окажется вне непосредственного кон-
такта с внешней средой. Эти клетки явятся источником мезодермы
и половых клеток.
Таким образом, почти с самого начала развития существуют
две среды: наружная и внутренняя (жидкость бластоцеля). При
достижении стадии гаструлы в энтодерме и эктодерме возникают
новые свойства, создаются биохимические и иные отличия между
клетками; клетки эктодермы и энтодермы начинают по-разному
реагировать на внешние воздействия. Выяснено, например, что
энтодерма гаструлы лягушки обладает большей кислотностью,
чем эктодерма.
Для процессов гаструляции необходима разность сред—внеш-
ней й внутренней. Для правильной дифференциации наружного
эпителия и правильного его разрастания, как пласта клеток, тре-
буется, чтобы эпителий с двух своих плоских сторон омывался
неодинаковыми по Свойствам жидкостями: с одной стороны, он
должен соприкасаться с жидкостью внешней среды, а противопо-
ложная сторона должна омываться жидкостью внутренней среды,
т. е. жидкостью бластоцеля, лимфой, кровью и т. п. Такой вывод
сделан на основании опытов с культивированием изолированных
зародышевых тканей амфибий. Предполагается, что свойства и
«потребности» двух противоположных концов каждой клетки эпи-
телия неодинаковы, что в биохимическом отношении она биполяр-
на. Видимым выражением биполярности в нормальном эпителии
335
зародыша лягушки служит концентрация пигмента на стороне
клеток, обращенных во внешнюю среду.
П. П. Иванов (1939) помещал в кровяную плазму яйца Rana
temporaria и Triturus taeniatus (на стадиях дробления, бластулы,
гаструлы, нейрулы и на более поздних стадиях). Оболочки яйца
перед этим удалялись. Тем самым нарушалась обычная разность
внутренней и внешней среды. Плазма крови, вероятно, более
близка по своим свойствам к нормально создающейся внутренней
среде зародыша. Результаты опытов таковы. Если в опыт берутся
зародыши до гаструляцин, она совсем не происходит ни у лягуш-
ки, ни у тритона. Если же помещать в кровяную плазму заро-
дыши со стадии гаструлы, нормальное развитие продолжается.
Формируется нормальный комплекс органов. Значит, раз начав-
шись, гаструляция не изменяет своего хода развития под влия-
нием кровяной плазмы. Таким образом, разность сред имеет осо-
бое значение для начала процессов гаструляцин.
Бластомеры бластулы лягушки после пересадки их в кровяную
плазму проявляли большую активность, амёбоидно изменяли
форму, уползали внутрь бластулы, частью же беспорядочно пе-
ремешивались на поверхности. Эти факты интересны в связи
с изучением возможности передвижения клеток и клеточных плас-
тов в ходе гаструляцин, о чем говорилось ранее (гл. V и VI).
При помещении в кровяную плазму яиц лягушки на стадии
дробления или бластулы бластоцель обычно исчезает, клетки об-
разуют плотное скопление.
Кровяная плазма резко изменяет поведение эктодермальных
клеток; она изменяет условия, необходимые для нормального рас-
положения клеток; эктодерма приобретает правильное эпителиоид-
ное расположение клеток, когда одна сторона соприкасается
с внешней средой, а другая с внутренней средой. Нарушение по-
лярности и дифференцировки происходит поэтому как в случае
удаления мезенхимы (опыты Гольтфретера), так и в случае со-
хранения контакта внутренней стороны эктодермы с мезенхимой,
но когда дистальные участки клеток находятся не в условиях
обычной внешней среды, а имеют контакт с кровяной плазмой
или другими жидкостями, имитирующими внутреннюю среду. Не-
обходимая для процессов формообразования разность сред унич-
тожается. При этом клетки утрачивают полярность, пигмент бес-
порядочно рассеивается по всей протоплазме. Отдельные клетки
погружаются в более глубокие слои, начинаются процессы, при-
водящие к образованию маленьких цист. Если цисты образуются,
то каждая клетка восстанавливает свою биполярную ориентиров-
ку, так как снова появляется разность сред: один конец клетки
теперь соприкасается с внутренней средой зародыша, а другой —
с иной по составу жидкостью полости цисты, являющейся продук-
том жизнедеятельности клеток.
336
При длительном (в течение нескольких недель) наблюдении-
над состоянием зародышей, культивируемых в жидкостях внут-
ренней среды, оказалось, что, хотя ткани оставались живыми и
росли, взаимное их расположение носило хаотический характер,,
никаких признаков организации целого не наблюдается. Такая
анархия объясняется именно тем, что уничтожение нормальной
разности сред обусловило нарушение нормального состояния це-
лостности.
ЭКЗОГАСТРУЛЯЦИЯ
Из опытов получения экзогаструл видно, что изменение внешней
и внутренней среды может сильно изменять процессы развития.
Еще Дриш в 1893 г. обратил внимание на то, что если яйца
морского ежа подвергать действию повышенной температуры^
(30 °C), то первичный кишечник вместо инвагинации выпячива-
ется наружу, — образуется экзогаструла. На ненормальном ки-
шечнике появляются, однако, перетяжки, соответствующие под-
разделению кишечника нормальной личинки.
Ц. Гербсту в 1892 г. удалось вызвать экзогаструляцию у яиц
морского ежа, изменяя концентрацию литиевых солей в морской
воде. Если поместить дробящееся яйцо морского ежа или бластулу
в морскую воду, где часть солей заменена изотоническим коли-
чеством солей лития, то гаструляционные передвижения клеточ-
ного материала нарушаются, инвагинации не происходит, но клет-
ки дифференцируются по типу эктодермальных, что заметно по
величине коротких мерцательных ресничек; реснички эктодер-
мальных клеток значительно длиннее. Чем дольше зародыш на-
ходится под влиянием солей лития, тем больше становится энто-
дермическая часть. После длительного пребывания в растворах
солей лития почти вся стенка гаструлы оказывается по своему
строению энтодермой и только небольшая группа клеток в обла-
сти анимального полюса оказывается эктодермой (рис. 132). Чи-
сло мезенхимных клеток нормальное. П. П. Иванов считает это
естественным,, так как мезенхимные клетки и в случае экзогаст-
руляции оказываются в нормальной среде, т. е. в жидкости бла-
стоцел я.
Изменяя внешнюю среду, можно, наоборот, получить зароды-
ши с подавленной энтодермой и даже чисто эктодермические за-
родыши (Дриш), — например повышением содержания солей
кальция и выключением из морской воды аниона SO4.
Удовлетворительного объяснения явлениям экзогаструляции
до сих пор не дано. П. П. Иванов считает, что сам процесс гаст-
руляции, переход клеток будущей энтодермы в бластоцель, вызы«
вается тем, что они нуждаются в той среде, которую могут найти
только в жидкости бластоцеля; для эктодермальных же клеток
337
А Б
В
Рис. 132. Экзогаструляция у морского ежа Sphaerechinus granularis. А; Б; В —
результаты, зависящие от концентрации солей лития (по Ц. Гербсту, из ки.
Дж. Гексли и Г. де Бера, 1936):
/ — эктодерма, 2— промежуточная часть лнчннкн, соединяющая вывороченный кишечник
<с эктодермальной частью, 3 — клетки мезенхимы, 4 — участок, соответствующий первичной
«кишке
Рис. 133. Экзогаструляция лягушки. А; Б—внешний вид; В — разрез (по
И. Гольгфретеру, 1933):
1 — эктодерма, 2 — задняя кишка, 3 — средняя кишка, 4 — пронефрос, 5 — мускульный сег-
мент, 6 — хорда, 7 — передняя кишка; стрелками показано передвижение клеточного мате-
риала
наиболее благоприятным условием является соприкосновение их
с внешней средой.
Экзогаструляцию вызывали и у других животных: Конклин в’
1906 г. — на яйцах моллюска Crepidula, Гертвиг и Вильсон — на
яйцах лягушки.
Укажем интересные случаи экзогаструляции у лягушки, вы-
званные Гольтфретером помещением бластул в 0,35%.-ный раствор
NaCl (рис. 133). Хордомезодермальный материал сначала лежит
на поверхности, а затем погружается внутрь и со всех сторон
окружается кишечным эпителием. У «экзоэмбриона» в той или
иной степени развиваются: пронефрос и мезонефрос, гонады, глад-
кая мускулатура, хорда, пульсирующее сердце (без клеток крови),
целомическая полость, соединительная ткань, кишка. Все эти об-
разования не имеют никаких нервных связей, так как нервная
система не развита. В эктодерме не обнаруживается дифферен-
цировки: отсутствуют нервная трубка, роговица и хрусталик, глаз,
слуховые пузырьки, зубы и т. д.; она состоит из массы недиффе-
ренцированных относительно однородных клеток.
ЭМБРИОН И БИОТИЧЕСКИЕ ФАКТОРЫ СРЕДЫ
Развитие любого организма происходит в биотической среде;
большинство организмов развивается в нестерильных условиях
(в почве, водоемах, на суше) и в контакте с другими животными
или растениями, которые могут оказать непосредственное дейст-
вие и играть заметную роль в изменении солевого и газового сос-
тава среды развивающегося эмбриона, в изменении температуры
и т. д.
Многие органические вещества, составляющие естественную
среду для эмбрионов и являющиеся продуктами жизнедеятельно-
сти находящихся в этой среде животных и растений, несомненно
играют не меньшую роль в жизни зародышей, чем неорганические
соединения.
Многие моллюски, черви, амфибии, все рыбы и ряд других
животных откладывают яйца в водную среду, в которой может на-
ходиться большое количество бактерий, грибов и простейших.
Многие насекомые развиваются в сильно инфицированной среде,
даже в разлагающихся трупах животных или растений (мухи- из
родов Phormia, Calliphora и др.).
Остановимся здесь только на взаимоотношении эмбрионов жи-
вотных с высшими растениями. Эта область исследований была
вызвана открытием фитонцидов (Б. П. Токин, 1928—1930)—про-
дуцируемых растениями веществ (в атмосферу, почву, воду), об-
ладающих способностью убивать или тормозить размножение и
развитие микро- и макроорганизмов. Фитонциды — главный фак-
тор иммунитета растений. Как правило, они представляют собой
комплекс летучих органических веществ, обладающих способно-
339-
стью проникать через оболочки многих яиц. Так, летучие фитонци-
,ды лука Allium сера или манника Glyceria aquatica убивают на
.расстоянии зародышей моллюсков.
Развитие зародышей моллюсков Physa fontinalis L. тормозится
.или они убиваются под влиянием летучих фитонцидов ряда при-
брежноводных, плавающих и подводных (погруженных) расте-
ний.
Возьмем отложенные яйца Rana temporaria. Половину яиц по-
местим в чашку Петри с аквариумной водой, а другую —в такую
же воду, но внесем в нее нетравмированную синезеленую водо-
росль осциллярию. На пятый день контрольные зародыши (в ак-
вариумной воде) уже освобождаются от яйцевых оболочек,
а в опытной чашке (с водорослью) зародыши оказываются еще
на стадии гаструлы и нейрулы. На девятый день они погибают.
Фитонциды некоторых растений стимулируют развитие моллюс-
ков. Опыты с аиром (Acorus calamus) убедительны: когда конт-
рольные зародыши продвигаются в своем развитии до стадии
начала формирования ноги, зародыши, развивающиеся в присут-
ствии аира, доходят до стадии сформированных моллюсков, каки-
ми они оказываются незадолго перед освобождением из яйцевых
оболочек.
Некоторые исследователи (Ф. А. Гуревич, 1949) говорят об
избирательной способности моллюсков откладывать яйца на опре-
деленных частях определенных видов растений. Яйцекладки Р1а-
norbis corneus на ряде растений встречаются очень редко, (напри-
мер, на маннике), хотя эти растения не представляют «механиче-
ских препятствий» для откладывания яйца. Несколько чаще клад-
ки встречаются на Caltha palustris, Equisetum palustre, Carex
acuta. На других растениях, таких как Acorus calamus, Nuphar
luteum, Nymphea alba, Polygonum amphibium, яйцекладки встре-
чаются наиболее часто. Возможно, что в этих явлениях играют
роль фитонциды водных растений. Предположения о значении
фитонцидов возникают и при изучении развития многих насекомых
и клещей, судьба которых тесно связана с жизнью тех или иных
растений.
Вспомним явление галлообразования. Летучие фитонциды
листьев дуба являются мощным повреждающим фактором для
протоплазмы клеток многих организмов, а в случае орехотворки
мы имеем идеально пригнанную друг к другу жизнь эмбриона и
тканей листьев дуба, причем речь, конечно, идет об их взаимодей-
ствии, в частности об активном изменении биохимических особен-
ностей тканей листа, окружающих животное на личиночных стади-
ях его развития.
Эмбрионы клещей и мясных мух чувствительны к фитонцидам
многих высших растений, между тем они проявляют исключитель-
ную стойкость в отношении антибиотиков низших растений, про-
дуцируемых бактериями и грибами (пенициллин, грамицидин,
340
стрептомицин и др.). Это, несомненно, приходится ставить в связь
с тем, что в ходе эволюции эмбрионы указанных животных адап-
тировались к токсическим веществам низших растений, с которы-
ми эмбрионы находятся неизбежно всегда в контакте.
В 60—70-е годы появился ряд научных сообщений, приковыва-
ющих особое внимание эмбриологов. Эмбриональное развитие
некоторых животных, по-видимому, невозможно без какого-то
своеобразного симбиотического участия микроорганизмов. Так,
К. Д. Спиндлер и В. А. Мюллер (1972) выяснили, что для мета-
морфоза планул Hidractinia echinata требуется участие бактерий:
без стимулов, исходящих от них, метаморфоз не происходит.
Экспериментально доказано влияние микроорганизмов на про-
цессы оседания, прикрепления и метаморфоза личинок ряда бес-
позвоночных животных: кишечнополостных, кольчатых червей,
щупальцевых, иглокожих. Обнаружено значение симбиотических
микроорганизмов в процессах бесполого размножения сцифистом.
Так, для стробиляции Mastigias papia необходимо присутствие
симбиотических зооксантелл. У лишенных симбионтов полипов
медузы Cassiopea andromeda стробиляция прекращается. Если
полип снова инфицировать зооксантеллами, то через несколько
недель стробиляция возобновляется. Почкование же не нуждается
в присутствии симбионтов. (У этой медузы нормальное бесполое
размножение протекает двумя способами — стробиляцией и почко-
ванием сцифистом.)
Таким образом, назрела необходимость разработки новой про-
блемы: эмбриональное развитие животных и микроорганизмы.
ЗАМЕЩЕНИЕ В ХОДЕ ЭВОЛЮЦИИ ВНЕШНИХ ФАКТОРОВ
ОРГАНООБРАЗОВАНИЯ ВНУТРЕННИМИ
Характер закладки нервной системы и ее развития у разных по-
звоночных существенно различен. У амфибий, а из рыб у акул
нервная система закладывается как пластинка на спинной сторо-
не зародыша, края которой сходятся и образуют трубку. У кос-
тистых же рыб нервная система закладывается как утолщение
эктодермы. Сначала это сплошной тяж, потом вследствие расхож-
дения клеток образуется канал нервной системы.
О наличии существования разных механизмов органообразова-
ния у амфибий говорят исследования А. А. Машковцева, который
помещал личинки Rana temporaria, R.esculenta, Pelobates fuscus и
Siredon pisciformis в аквариумы до их вылупления из оболочек и,
закрывая поверхность воды сетками, не давал возможности личин-
кам подниматься на поверхность воды и вдыхать воздух. Личинки
указанных амфибий начинают дышать легкими через несколько
дней после вылупления из оболочек. Оказалось, что у аксолотлей,
проживших много месяцев в аквариумах под сетками, легкое было
не дифференцировано и оставалось похожим на гладкостенный
341
Рис. 134. Схема развития легкого Siredon (Л); Pelobates (5); Bufo (В) и Lacer-
ta (Г) (по А. А. Машковцеву, 1936).
Заштрихованы структуры легкого, возникающие под влиянием функции дыхания. Не за*
штрихованы структуры, возникающие под влиянием эндогенных факторов внутренней средь»
организма. I, II, III — этапы дифференциации легкого
плавательный пузырь рыб. У контрольных же особей легкое в это
время имело сложную ячеистую структуру. Значит, у аксолотля
ячеистая структура и сложная сеть кровеносных сосудов возника-
ют лишь в связи с дыхательной функцией — вдоха и выдоха
(рис. 134,Л).
Опыты с головастиками лягушки и чесночницы показали: если
легкое не функционирует (головастики находятся под сетками),
оно в своей дифференцировке достигает первой стадии поперечной
фрагментации (рис. 134,5), а полная фрагментация происходит
под влиянием комплекса явлений, связанных с вдохами и выдоха-
ми. Совершенно такие же опыты проведены с личинками жаб, т. е.
амфибий, наиболее приспособившихся к сухопутному образу
жизни.
342
У головастиков жаб (рис. 134,В) легкое закладывается поздно,
оно не имеет связи с ротовой полостью и наполнено серозной жид-
костью. Полная фрагментация легкого происходит еще до первого
вдоха; легкие имеют в это время вид виноградной грозди. Первый
вдох воздуха происходит через 5—20 мин после выползания мо-
лодых жаб из воды. Они интенсивно дышат на суше легкими, и
под влиянием усиленного дыхания количество альвеол легкого
в несколько дней увеличивается в два раза, затем легкое растет
медленно. А. А. Машковцев делает вывод: у жаб вся типичная
структура легких возникает под влиянием внутренних факторов
до начала функции органа (под влиянием гормона щитовидной
железы).
У чесночницы, а в большей мере у лягушки и жабы, при раз-
витии легкого повышается роль внутренних факторов в дифферен-
циации этого органа, происходит замещение внешних факторов
внутренними.
Что касается ящериц и черепах, у них все типичные структуры
легкого возникают так же, как и у жаб, до первого вдоха, но
«процесс автоматизации эмбрионального развития легкого репти-
лий» зашел значительно дальше, чем у жаб (рис. 134,Г).
У клоачных и сумчатых большая часть структур легкого воз-
никает под морфогенным влиянием вдохов и выдохов, а у плацен-
тарных млекопитающих — под влиянием эндогенных факторов.
Гипотеза А. А. Машковцева, несмотря на спорность, представ-
ляет интерес, однако требует анализа на основе данных современ-
ной генетики, особенно в вопросе о противопоставлении эргонти-
ческих корреляций геномным (см. с. 271).
КРИТИЧЕСКИЕ ПЕРИОДЫ В РАЗВИТИИ ОРГАНИЗМОВ
Термин «критические периоды» еще в 1897 г. ввел ботаник
П. И. Броун. Он говорил о периодах наибольшей чувствительности
сеянцев злаков, всходов картофеля и иных растений. Он считал,
что в эмбриональном и постэмбриональном развитии происходит
смена периодов, во время которых организм стоек к различным
внешним воздействиям на периоды, когда он чувствителен в рав-
ной мере к действию разнообразных агентов.
Ц. Р. Стоккард в 1921 г. положил начало представлениям
о критических периодах развития животных организмов. Он пред-
ставил себе онтогенез как ряд последовательных этапов, разли-
чающихся скоростями развития. Критические периоды характери-
зуются наибольшей скоростью развития. На ранних стадиях эм-
бриогенеза эти периоды являются критическими для всего орга-
низма в целом, на более поздних стадиях имеются критические
периоды в развитии отдельных органов. Внешние факторы, к ко-
торым особенно велика чувствительность в эти периоды, могут
ускорять, замедлять или совсем приостанавливать развитие.
343
Некоторые исследователи предполагают, что периоды разви-
тия, характеризующиеся интенсивно идущими процессами диффе-
ренциации, являются высоко чувствительными к разнообразным
внешним воздействиям. Опыты на эмбрионах позвоночных живот-
ных (лягушек, рыб, млекопитающих) действительно показывают,
что на разных стадиях развития эмбрионы характеризуются раз-
личной стойкостью к тем или иным повреждающим воздействиям.
Летальная доза рентгеновского излучения для эмбрионов кроли-
ка первых десяти дней беременности равна (2,6—3,9) • 10~2 Кл/кг
(100—150 Р), для эмбрионов 10—14 дней — 5,16-Ю-2 Кл/кг
(200 Р), для плодов 17—20 дней — 0,14 Кл/кг (550 Р) и для пло-
дов 23—27 дней — 0,23 Кл/кг (900 Р). В некоторые периоды раз-
вития зародыши стойки или чувствительны к ряду внешних фак-
торов, причем реакция их на воздействие бывает однотипной (так
же как реакция мускула или нерва глаза оказывается однотипной
при действии многих раздражителей). Но таких критических пе-
риодов, во время которых зародыш одинаково чувствителен к дей-
ствию всех внешних факторов среды, не существует.
В. М. Коровина в 1953 г. показала, что к концу нейруляции
появляются различия в реакции на кислоты и щелочи: стойкость
к кислотам продолжает повышаться, к щелочам же зародыши де-
лаются чувствительными.
Многие другие факты не позволяют говорить о чередовании
стойких и нестойких этапов в индивидуальном развитии живот-
ных. Вместе с тем нельзя отрицать своеобразия реагирования за-
родышей на разных этапах эмбриональной жизни. Если не
абсолютизировать взгляд на критические периоды и не утверж-
дать, что господствует однотипность ответа на агенты самой раз-
личной химической, физической и биологической природы, то
нельзя игнорировать утверждение многих авторов о наличии кри-
тических периодов, так как они основаны на фактах из развития
животных. Таковым является, например, начало гаструляцин
у амфибий и рыб, у млекопитающих — стадия бластоцисты в пе-
риод ее имплантации в стенку матки.
Представляют интерес взгляды на эмбриональное развитие
Р. Гольдшмидта (1927, 1932). Он рассматривал онтогенез как
«координированную систему скоростей развития». Критические
периоды — это «узловые точки» пересечения кривых скоростей
различных реакций. В эти периоды гармонически сочетаются, «пе-
ресекаются» независимые до того процессы развития, что и
обусловливает нормальное развитие. Эти периоды схождения ра-
нее независимых процессов развития совпадают со временем де-
терминации. Повышенная в эти периоды чувствительность обуслов-
ливается большей скоростью и разнообразием «перекрещивающих-
ся» процессов развития, ибо координация этих процессов лабиль-
ная и может быть легко нарушена при действии внешних агентов.
Эти мысли близки к некоторым современным высказываниям.
344
Остановимся на оригинальной гипотезе критических периодов
П. Г. Светлова. Он различает три группы воздействий внешней
среды: 1) повреждающие воздействия, приводящие к смерти или
вызывающие патологические явления; 2) модифицирующие воз-
действия внешней среды, вызывающие отклонения от нормы, — эти
отклонения не носят патологического характера («морфозы», му-
тации); 3) закономерное действие среды, обеспечивающее «норму
развития». Эти воздействия (наличие кислорода, питание, темпе-
ратура и т. д.) не бросаются в глаза, но они-то -и представляют
наибольший интерес для эмбриологии. По Светлову, в клетках
имеются высокочувствительные рецепторы, «настроенные на вос-
приятие строго определенных импульсов». Минимальные дозы
каких-либо воздействий могут играть большую роль в формооб-
разовании.
Повреждаемость и смертность зародышей неодинаковы на про-
тяжении развития. Существуют критические периоды, в которые
эмбрион более восприимчив к действию внешних агентов. Подъем
•чувствительности клеток в критические периоды развития связан
с пониженной репаративной способностью в эти периоды. Детер-
минация, являясь «ступенчатым процессом», связана с этими за-
кономерно наступающими в онтогенезе периодами. Критические
периоды — это и есть периоды детерминации, в течение которых
происходят высокая повреждаемость и пониженная регуляторная
деятельность. Под детерминацией Светлов понимает ступенчатый
процесс, являющийся актуальной причиной следующих за ним
цепных процессов дифференциации. Развитие организма и его
элементов и состоит, по его мнению, из небольшого числа крупных
периодов, каждый из которых начинается актом детерминации,
включающим развертывание событий дифференциации до опреде-
ленного состояния. По достижении этого состояния может после-
довать новый акт детерминации. Внешние воздействия являются
импульсами, приводящими те или иные части зародыша в состоя-
ние лабильной детерминации. Затем следуют процессы роста и
дифференциации, а потом опять наступает критический период —
период детерминации. Согласно этим взглядам онтогенез в целом
реактивный процесс, и его основные закономерности должны быть
выражены в понятиях физиологии раздражения. Зародыш в опре-
деленные периоды как бы «прислушивается» к тому, что делается
вокруг него. Поэтому он и восприимчив в этот период к действию
гормонов, индукторов, давлению, растяжениям, электрическому
току, к тому, что оказывается во внешней среде, а внешней средой
для любой части зародыша являются и соседние клетки, и наличие
кислорода, и т. д.
Преимущество теории П. Г. Светлова состоит в том, что она
ставит проблему детерминации на физиологическую основу и не
довольствуется односторонне химическими или генетическими ме-
тодами исследования.
345
Критические периоды не рассматриваются как периоды, в ко-
торых зародыш был бы более чувствительным вообще к факторам
среды. Автор гипотезы считает, что в один критический период
зародыш более чувствителен, например, к температурным воздей-
ствиям, а в другой — к химическим факторам и т. д. Эта интерес-
ная теория требует дальнейшей разработки.
Взгляды о реактивности морфогенетических процессов кажут-
ся убедительными, если принимать без сомнения одно из основных
положений генетики и генотипической однородности всех клеток
развивающегося организма. В связи с этим тезисом логично сде-
лать вывод о том, что развитие — не автономическое развертыва-
ние деятельности геномов клеток, а последовательные, реактив-
ного характера процессы, определяемые внешними факторами.
Однако необязательно мыслить взаимоотношения клеточных эле-
ментов и их производных как реактивный процесс, при котором
причиной должен быть «внешний» для клетки фактор. Развиваю-
щийся организм — меняющееся состояние целостности: требуется
искать новые и новые механизмы интеграции, взаимодействия эле-
ментов организма, а не одностороннего действия «индукторов» на
клетки. Впрочем, следует ожидать дальнейшего развития теории
П. Г. Светлова и создания более конкретных представлений
о «реактивных процессах». Может оказаться, что споры о крити-
ческих периодах — споры не о существе явлений, а скорее терми-
нологического характера.
Можно предположить, что в аспектах теории критических пе-
риодов отдельные моменты развития оказываются связанными
с перестройкой интеграционных механизмов, наступающей в опре-
деленные периоды в зародыше в целом и в отдельных его частях.
Все рассмотренные гипотезы о критических периодах имеют
серьезную опору в фактах, так как действительно в ходе онтоге-
неза наступают периоды, отличающиеся большим своеобразием.
Таков период гаструляции, с которым связан переход к новому
состоянию зародыша — формированию зародышевых листков, за-
кладыванию органов, к начальным стадиям формирования тканей.
Существуют и иные точки зрения, согласно которым представ-
ление о критических периодах является искусственным. Так,
А. П. Дыбан на основании многолетних экспериментов утвержда-
ет, что в эмбриогенезе нет общих критических периодов к агентам
с разным механизмом действия (рис. 135). Эмбриофизиолог
И. А. Аршавский считает, что каждый период в развитии организ-
ма характеризуется своей особой формой адаптации к определен-
ным специфическим условиям среды и своим уровнем резистент-
ности к разнообразным альтерирующим воздействиям.
Единодушия в оценке различных периодов (или стадий, этапов
и т. п.) как более или менее резистентных не существует. Многие
считают, что наиболее чувствительны ранние стадии и это они от-
личаются наибольшей однотипностью реагирования, а по мере
346
Эмбриональная смертность
SO
SO
SO
JO
\\
20
10
•-----Тмридин
Оранил
Актиномицин Д
Дмшкттерин
СалицилоВокислый
натрий
Гексен ал
SO
SO
70
•i\
Дробление
Органогенез
зародыш 8 матке
Зародыш
яйиеВоде
Дни
беременности
Ооразооание зарооыиае-
ЛиСТПКС'5
Рис. 135. Чувствительность эмбрионов крыс к повреждающим воздействиям на
разных стадиях развития (по А. П. Дыбану, 1968).
Периоды максимальной чувствительности эмбрионов к разным агентам не совпадают, т. е.
в эмбриогенезе крыс нет общих критических периодов к агентам с разным механизмом дей-
ствия
0-----Нефрогтритческая
сыВоротка
—— Б-неркатпопурин
развития специфичность ответов на действие внешней среды уси-
ливается. Однако эволюционисту трудно представить, чтобы эм-
брионы, являющиеся аккумуляторами эволюционного процесса,
были особенно нежны и податливы в отношении влияния факто-
ров, вызывающих наследственную изменчивость или морфозы.
Эмбрионы животных на ранних стадиях мало податливы к из-
менениям, но если происходит даже незначительное изменение
каких-либо структур и функций на начальных этапах развития
зародышей (например, небольшие изменения характера борозд
дробления или изменение состава жидкости бластоцеля), то они
могут повлиять многообразно и, так сказать, в геометрической
прогрессии на формирование тысяч признаков взрослых организ-
347
мов. В этом отношении организм можно уподобить очень сложно-
му и большому вееру: достаточно немного раздвинуть его у осно-
вания, чтобы вызвать большие видимые изменения во всем веере.
Самый «чувствительный» период — проэмбриональный — про-
цессы ово- и сперматогенеза. Вероятно, этот период заслуживает
наибольшего внимания эмбриологов, ставящих перед собой задачу
управления развитием.
Проблема чувствительных периодов в эмбриогенезе имеет боль-
шое практическое значение. Так, в частности, сведения о наиболее
чувствительных периодах развития необходимы врачам при уста-
новлении дозировки лекарственных препаратов или химиотерапев-
тических воздействий, назначаемых пациентам, особенно беремен-
ным женщинам. Эти знания необходимы также при разработке
мероприятий по борьбе с вредителями полей, с паразитами в дру-
гих аналогичных ситуациях.
Из всех данных- о влиянии внешней среды на развитие организ-
мов можно сделать такой вывод. Эмбриолог, желающий не пас-
сивно, не созерцательно описывать развитие животных, а ставить
перед собой смелые задачи управления развитием организмов,
перевода эмбриологии на уровень, образно говоря, биотехники
эмбрионального развития, должен вместе с генетиками наряду
с применением методов селекции все более использовать факторы
внешней среды. Современная генетика, которую иногда несправед-
ливо упрекают в автогенетических взглядах^ считает, что свойства
организмов обусловлены не только их генетической конструкцией,
но и внешними влияниями, которым подвергается развивающийся
организм. Фенотип есть результат взаимодействия между геноти-
пом и средой. Следует расширить понятие «среда», и, в частности,
вместе с физиками и химиками выяснять, нет ли таких факторов
среды, которые были скрыты от взора эмбриолога из-за несовер-
шенства наших знаний о космических излучениях, о гравитации,
о взаимосвязанности явлений биологических, химических и физи-
ческих, но которые играют существенную роль в эмбриональном
развитии животных.
В связи с развитием космических полетов особое значение
приобрел фактор гравитации. Влияние силы тяжести на развитие
зародышей оказалось неодинаковым для разных типов эмбриоге-
незов. Уже отмечалось ранее, что насильственное поворачивание,
например, яйца амфибий анимальным полюсом вниз вызывает из-
менение в локализации серого серпа. Центрифугирование ранних
зародышей амфибий может привести к нарушению дробления,
гаструляцин, а в некоторых случаях к образованию двойниковых
уродств. Если имитировать состояние невесомости путем клиноста-
тирования (равномерное и медленное вращение вокруг горизон-
тальной оси), то наблюдается широкий спектр нарушений, наблю-
даемых, например, у амфибий, начиная со стадии ооплазматиче-
ской сегрегации, а затем на стадии дробления и гаструляцин
348
(Я- Г. Дорфман, 1974; Я- Г. Дорфман, В. Г. Черданцев, 1977),.
Аналогичные эффекты были получены на птицах (Clavert, 1962)
и рыбах (А. А. Костомарова, 1965; Г. М. Игнатьева, 1974). Многие
виды беспозвоночных оказались мало чувствительными к измене-
нию силы тяжести в период эмбрионального развития.
Таким образом, факторы внешней среды могут оказывать в за-
висимости от дозы различное влияние: 1. Ускорять или замедлять-
развитие (слабое температурное воздействие). 2. Вызывать ано-
малии развития (гормональные воздействия). 3. Оказывать по-
вреждающие воздействия, в результате чего развитие останавли-
вается на той или иной стадии онтогенеза (влияние ингибиторов
синтеза нуклеиновых кислот или белков). 4. Вызывать морфозы-
и мутации, затрагивающие структуру гамет или генетический ап-
парат (ионизирующие воздействия).
Среди биотических факторов среды, влияющих на процессы-
эмбрионального развития, особое место занимают гормоны. Из
медицинской и ветеринарной практики можно привести много при-
меров, свидетельствующих о том, что атипическое развитие плодов-
млекопитающих животных и человека может быть связано с эндо-
кринными нарушениями, вызванными пороками материнского
организма. Так, например, при избытке в крови матери кортикоидов
происходят определенные изменения в гипоталамо-гипофизарной
системе зародыша, что вызывает редукцию у него коры надпочеч-
ника. У больных диабетом самок рождаются детеныши, страдаю-
щие ожирением. Нарушения у матери тиреоидных функций вызы-
вают у ребенка аномалии в развитии скелета.
Гормон, проникающий в кровяное русло плода через плаценту,,
может оказывать непосредственное действие на органы-мишени,
но может осуществлять и воздействие, опосредованное через гипо-
таламо-гипофизарную систему эмбриона.
По наблюдениям Л. Г. Лейбсона и его сотрудников, гормо-
нальная регуляция метаболизма развивающегося зародыша оказы-
вается возможной на такой стадии, когда регулируемый процесс
оказывается неотъемлемой частью (звеном) целостной функцио-
нальной системы.
По современным представлениям в дефинитивном состоянии
у позвоночных животных могут быть выделены пять взаимосвя-
занных звеньев (рис. 136): I — ткань или орган-мишень; II —эндо-
кринная железа; III — аденогипофиз; IV—система гипоталамиче-
ских рилизинг-факторов и V — система регуляции этих факторов..
Каждый из уровней гормональной регуляции не развивается
в онтогенезе изолированно, что создает сложность в анализе пря-
мых и опосредствованных влияний факторов внешней среды, за-
трагивающих эти системы (М. С. Мицкевич, 1978).
Внутренние нейрогуморальные регуляторные системы, контро-
лирующие гаметогенез и эмбриогенез, существуют не только
у позвоночных, но также и у беспозвоночных животных.
34ft
Рис. 136. Схема механизмов дефинитивной гормональной функции млекопитаю-
щих животных, включая пять основных уровней регуляции (I—V) (по
М. С. Мицкевич, 1978):
1 — внегипоталамические регуляторные центры, 2 — преоптическая область, 3 — область ар-
куатного и вентромедиального ядер, 4— оптическая хиазма, 5 — яичник, 6 — семенник, 7 —
надпочечник, 8 — щитовидная железа, 9 — матка, 10— семенные пузырьки, // — простата,
12 — печень, 13 — кость, 14 — мозг; АКТГ — адренокортикотропный гормон, ГРГ — гонадо-
тропный рилизинг-гормон, ГТГ — гонадотропный гормон, КРГ — кортикотропный гормон,
.Л/Г — лютеинизирующий гормон, ЛТГ — лактотрофный гормон, СТГ — соматотропин, ТРГ —
тиротропин-рилизииг-гормон, ТТГ — тиреотропный гормон, ФСГ — фолликулостимулирую-
щий гормон
35(1
Глава XV
ЭМБРИОЛОГИЯ И ИММУНОЛОГИЯ. ЗАЩИТНЫЕ РЕАКЦИИ
ЗАРОДЫШЕЙ, антигенная реактивность
Необходимость тесной связи зоологии с иммунологией впервые
обосновал И. И. Мечников в 1883 г. «Если явления нападения со-
ставляют предметы зоологического исследования, то и столь близ-
кие им явления инфекции и сопротивления ей входят в ту же об-
ласть исследования» *.
И. И. Мечников и А. О. Ковалевский (1887) обратили внима-
ние на то, что такие иммунологические механизмы, как фагоци-
тоз, играют важную роль в формообразовательных процессах, осо-
бенно в процессах метаморфоза амфибий и насекомых. Эти и дру-
гие классические исследования Мечникова впоследствии послужил»
отправным пунктом Б. П. Токину (1955) для формулирования
проблемы иммунитета эмбрионов как главы сравнительной эмбри-
ологии и общей зоологии.
Организм неистово защищает свою целостность многими путя-
ми.' Среди них большое значение принадлежит защитным реакци-
ям: фагоцитозу и воспалению, выработке «антибиотических» суб-
станций и др. Однако первенствующую роль играет антигенная
реактивность.
Остановимся прежде всего на иммунной реакции — антигенной
реактивности. Это эволюционно выработавшийся ответ организма
на вторжение (парентерально) бактерий, низших грибов, вирусов,
паразитических простейших и других чужеродных веществ в ос-
новном чужих белков. Все вещества, могущие вызвать иммунную
реакцию, называются антигенами. Существуют и аутоиммунные
реакции, вызываемые веществами, принадлежавшими данному ор-
ганизму, если на протяжении значительной части его жизни они
были изолированы от реагирующей системы, а впоследствии ока-
зались освобожденными в результате патологического процесса.
Суть антигенной реакции — образование антител, т. е. циркулиру-
ющих в крови белков, строение которых таково, что при встрече
их с антигенами могут происходить реакции, уничтожающие анти-
ген или его действие: агглютинация (склеивание, если антигенами
служат клетки), разрыв клеточных оболочек, детоксикация (если-
антиген — токсин, положим, вырабатываемый бактерий), лизис
(разрушение антигенных клеток). Может иметь место и фагоци-
1 Мечников И. И. Избранные биологические произведения. М., 1950.
С. 484—485.
35 Ь
тоз— захватывание, иногда переваривание чужеродного агента
клетками фагоцитами, макрофагами.
Общепризнано, что иммунная реакция может осуществляться
.путем активирования иммунокомпетентных клеток-лимфоцитов, их
деления, миграции и дифференцирования сначала на Т- и В-лим-
фоциты, а затем специализации в плазматические клетки, килле-
ры и др.
Главным действующим фактором гуморального иммунитета
являются антитела. Клеточный и гуморальный иммунитет помимо
защиты от чужеродных включений вызывает отторжение тканей
при гомо- и гетеротрансплантациях, прекращает жизнедеятель-
ность паразитов и др.
Волнующим’ вопросом в иммунологии был и еще в значитель-
ной мере остается — как же организм отличает «чужое»? Орга-
низм представляется чувствительным инструментом, в котором
под влиянием антигенных раздражителей, даже никогда не нахо-
дившихся в контакте с организмом, вырабатываются все новые и
новые «клавиши». Ничтожные количества чужого белка вызыва-
ют антигенную иммунную реакцию. «Гениальными» клетками
оказываются клетки лимфатической системы. Число лимфоцитов
в организме человека огромно: находясь в циркулирующей лимфе,
они попадают в венозную систему числом около десяти миллиар-
дов ежедневно.
Благодаря циркуляции в кровотоке (из лимфатических узлов —
в лимфатические сосуды, затем в венозную систему) совершается
их круговорот. Лимфоциты на начальной стадии активации отли-
чаются друг от друга: В-лимфоциты, вырабатывающие антитела,
и Т-лимфоциты, от которых зависит клеточный иммунитет, а так-
же регуляция процессов антителообразования. Буква Т взята от
слова тимус, где скапливаются Т-лимфоциты, а В — от слова
bursa fabricii — фабрициева сумка, имеющаяся только у птиц.
Среди первых, кто обратил внимание на необходимость исполь-
зования в эмбриологии понятий антиген — антитело, был
И. Л. Кричевский (1913—1923), пытавшийся использовать реакции
иммунитета в изучении биогенетического закона и установивший
отличия антигенов зародыша от антигенов взрослого животного,
а также изменяемость антигенов в ходе эмбрионального развития.
Начиная с 40-х годов нашего столетия взаимная связь эмбрио-
логии и иммунологии все более возрастает. Ряд эмбриологов и
иммунологов обнаружили важные закономерности и создали ин-
тересные гипотезы.
П. Вейс в 1950 г. уподоблял отношения между поверхностями
(молекулами) взаимодействующих клеток эмбриона отношениям
между молекулами антигена и соответствующего антитела.
А. Тайлер в 1935 г. предполагал, что из клеток зародыша выходят
аутоантитела, влияющие на дифференциацию тканей и органов.
Исследования П. Б. Медавара и Ф. М. Бернета (1945), а также
352
М. Гашека (1955) посвящены явлениям совместимости и несовме-
стимости тканей при пересадках, но они имеют большое значение
для анализа явлений эмбрионального развития в аспекте иммуно-
логии.
СОВМЕСТИМОСТЬ И НЕСОВМЕСТИМОСТЬ ТКАНЕЙ ПРИ ПЕРЕСАДКАХ
Микробиолог Ф. М. Бернет в 1959 г. высказал очень важную для
эмбриологии гипотезу, а зоолог П. Б. Медавар экспериментально
подтвердил ее правильность. Суть ее сводится к тому, что способ-
ность организма отличать «свой» белок от «чужого» приобретает-
ся в процессе эмбрионального развития. Чужой белок, введенный
зародышу, должен создать у него «привычку», и организм, раз-
вившийся из такого зародыша, должен обладать толерантностью
к данному типу белка и не реагировать на него выработкой анти-
тел.
Эти идеи возникли в связи с явлениями несовместимости тка-
ней при трансплантации. Лоскут кожи, перемещенный из одной
области тела в другую у одного и того же человека (млекопитаю-
щего или птицы), обычно приживляется. Если же трансплантиро-
вать лоскут кожи от одного индивидуума другому, то он при-
живляется лишь временно, не более чем на неделю. Вскоре после
трансплантации мобилизуются иммунологические силы организма
для отторжения инородного лоскута кожи: начинается воспаление,
нормального кровоснабжения не создается, трансплантат омертве-
вает и в конце концов отторгается. Организм в силу своих имму-
нологических свойств «рассматривает» белки трансплантата как
антигены и вырабатывает против них антитела. Люди разделены
по типу крови на четыре группы. При переливании крови от одно-
го индивидуума другому, принадлежащему к той же группе по
крови, не происходит выработки антител. Но всегда опасно пере-
ливание крови от индивидуума с одной группой индивидууму
с другой группой. Эти известные явления также имеют в основе
антигенную реактивность. Организм принимает за «свое» только
то, что в отношении белковых субстанций генетически идентично
заменяемой части.
Идеи П. Б. Медавара и Ф. М. Бернета были продолжены
М. Гашеком. Толерантность к тому или иному антигену можно
создать, если ввести его в ткани эмбриона. Возьмем две чистые
высокоинбредные линии мышей А и Б. Внутри каждой группы
возможны пересадки кожи, заканчивающиеся приживлением, но
пересадки между А и Б не дают положительного результата
вследствие антигенной реактивности. Можно, согласно идее Бер-
нета, ввести в эмбрионы беременных самок А порцию живых кле-
ток, например из селезенки или почек, взятых от эмбрионов того
же возраста линии Б. Эмбриональное развитие продолжается, и
роды оказываются нормальными. Пересадим таким 6—8-недель-
353
ним мышатам А кожу от мышат Б. Трансплантат приживляется,
кожа оказывается химерной: если мыши Б имели черную шерсть,
а мыши А белую, то белая мышь будет иметь белую кожу с при-
жившимся черным лоскутом. Толерантность'к чужой коже может
сохраниться месяца четыре, а то и в течение всей жизни.
М. Гашек в 1953 г. существенно развил теорию толерантности.
Он установил, в частности, для эмбрионов птиц период, в течение
которого можно вызвать адаптацию к чужеродному антигену пу-
тем инъекции в этот адаптивный период соответствующих антиге-
нов. Это тот период, когда зародыш или новорожденный организм
еще не способен к выработке антител, когда иммунологическая
система эмбриона или очень молодого организма еще не разви-
лась, когда встреча с чужеродным антигеном может вести к из-
менению его дальнейшей иммунологической реактивности.
Согласно клонально-селекционной теории иммунокомпетентные
клетки становятся неспособными осуществлять синтез определен-
ных антител. Возникшая толерантность может быть полной и не-
полной. В последнем случае антигенная реактивность, т. е. специ-
фический ответ на влияние антигена, может полностью восстано-
виться.
Автор провел серию экспериментов по иммунологическому
сближению для выработки толерантности к тем или иным анти-
генам. Это особенно успешно достигалось в опытах парабиоза
птичьих эмбрионов: на тех или иных стадиях развития у двух яиц
(индейки и цесарки или утки и курицы и в других сочетаниях)
удаляются участки скорлупы и сближаются хорио-аллантоисные
мембраны (рис. 137,А). При помощи зародышевого диска третьего
эмбриона (взятого у 20—40-часового эмбриона) два зародыша
сращиваются. Для этого зародышевый диск помещают на хорио-
аллантоис одного из яиц (рис. 137,В); яйца прикладываются друг
к другу просверленными в скорлупе отверстиями, место соедине-
ния заливается парафином. Вскоре в ткани зародышевого диска
врастают кровеносные сосуды обоих эмбрионов, устанавливается
связь между кровеносной системой обоих зародышей, обмен
Рис. 137. Иммунологическое сближение эмбрионов птиц. Пояснения см. в тексте
(по М. Гашеку, 1957)
354
кровью (рис. 137,5). После такого иммунологического сближения
создавалась иммунологическая толерантность: пересадка кожи от
одного вида птицы другому виду (осуществленные после рожде-
ния) давала положительный результат. На почве этих и многих
других экспериментов возникли, вероятно, излишне оптимистиче-
ские надежды хирургов. Почему бы, думают некоторые из них, не
вводить каждому ребенку в первый день жизни по нескольку мил-
лионов отцовских лейкоцитов и выработать тем самым толерант-
ность к антигенам отца. Впоследствии в случае ожогов сына можно
пересадить ему кожу отца. К сожалению, подобный путь подавле-
ния антигенной реактивности и создания условий для пересадок
тканей и органов нереален.
Огромная практика хирургов по пересадке кожи, суставов и
костей, роговицы глаза, почек и сердца, все данные эксперимен-
тов по трансплантации тканей и органов птиц и млекопитающих
убеждают в том, что помехой является не.только антигенная реак-
тивность. При пересадке, например, кожи имеет большое значение
то обстоятельство, что пока кровеносные сосуды прорастают в пе-
ресаженную кожу, в ее тканях могут происходить изменения,
ведущие к дегенерации. Иные трудности встречает хирург, поло-
жим, при пересадке почек. Большой комплекс биологических яв-
лений сопутствует приживлению или отторжению трансплантата:
сколь велика способность к репаративной и физиологической ре-
генерации и к срастанию данных тканей и частей органов (напри-
мер, в случае сращивания сосудов), столь энергичны фагоцитар-
ная активность и воспалительные реакции и т. п.
Антигенная реактивность отсутствует у губок и кишечнополост-
ных. Однако межвидовые, а иногда и межклональные пересадки
у этих животных в конце концов приводят к отторжению транс-
плантатов. Не получается срастания и в случае выращивания
личинок близкородственных видов в близком соседстве друг от
друга. На границах соприкасающихся губок, например, развивает-
ся зона контакта, в которой образуется толстая прослойка колла-
гена, препятствующая объединению их в одну общую колонию.
Таким образом, реакция распознавания «своего» и «чужого» име-
ет место уже у самых просто организованных организмов. Если
смешать соматически клетки двух видов губок или гидр, то они
легко рассортировываются, образуя однородные в видовом отно-
шении конгломераты. В ряде опытов были получены химерные
организмы (соматические гибриды), однако спустя несколько
месяцев в них начинали преобладать клетки какого-либо одного
вида за счет постепенного отторжения или фагоцитоза гибнущих
клеток другого вида.
Удаются пересадки различных частей эмбрионов (на стадиях
бластулы, гаструлы, нейрулы и хвостовой почки) от одного вида
амфибий другому виду. У амфибий приживаются без признаков
дегенерации кожа, глаза, конечности, сердце и иные органы, взя-
355
тые даже от других видов. Пересаженный глаз тритона прижи
ляется и регенерирует зрительный нерв. В 1945 г. Н. Б. Синиц!
впервые разработал методику пересадки сердца у лягушек. Of
жили с пересаженным сердцем по многу лет. Если произвест
сращение передней части эмбриона одного вида тритона с задне
частью тритона другого вида (на той стадии, когда уже сформ:
рованы голова, туловище и хвост), то создается химерное живо
ное. Можно получить химеру между альпийским и мексикански
аксолотлями.
ИММУНОБИОЛОГИЧЕСКИЕ ВЗАИМООТНОШЕНИЯ ПЛОДА И ОРГАНИЗМА
К обсуждаемым проблемам тесно примыкают иммуноэмбриолог:
ческие вопросы, волнующие теоретическую и практическую мед:
цину,— иммунобиологические взаимоотношения плода и органи
ма женщины-матери. Медицина и эмбриология не могут полность
объяснить ряд явлений. Почему в периоде внутриутробного ра
вития и особенно на ранних стадиях развития зародыша человек
происходит гибель очень большого числа зародышей? С какиь
факторами связаны ненаследственные заболевания и ранняя пос
натальная гибель детей?
Среди различных факторов, обусловливающих указанные явл
ния, большое внимание исследователей приковано к антигеннс
реактивности. Каждый человек и каждое млекопитающее живо
ное уникальны: в составе' клеток и тканей организма имеете
огромное чцело различных антигенов. Относительное исключен!
составляют близнецы. Также установлено, что антигенный состг
клеток и тканей эмбрионов характеризуется огромным разнообр
зием. Мать и эмбрион отличаются своими изоантигенами. Все т
что сказано о значении антигенной реактивноти в явлениях н
совместимости тканей при трансплантации, казалось бы, долж!
относиться и ко взаимоотношению материнского организма и з,
родыша. Однако в большинстве случаев взаимоотношения эти i
перерастают в патологические процессы.
Таким образом, после имплантации бластоцисты и установл
ния плацентарной связи плода и матери беременность продолж
ется, несмотря на то, что зародыш в антигенном отношении чуж
роден материнскому организму. Почему «гомотрансплантат
«паразит»-зародыш не отторгается? На эти очень важные вопр
сы при современном состоянии знаний ответить нельзя, гипотез х
существует много.
В первые месяцы развития в крови зародыша человека мог;
формироваться групповые и типовые антигенные факторы, ере;
них резус-фактор. Эритроциты большинства людей агглютинир
ются сывороткой крови кроликов, иммунизированных кровь
обезьян макака-резус, но эритроциты некоторых людей не аггл!
тинируются. Резус-фактором называется антиген, ответственнь
356
за выработку антител (при иммунизации кролика) и вызывающий
агглютинацию эритроцитов у человека и обезьян. Обладающих
этим антигеном называют резус-положительными (Rh+), а не
обладающих — резус-отрицательными (Rh—). Резус-фактор опре-
деляется доминантным геном Rh, а отсутствие его — рецессивным
геном rh. У 85% людей содержится этот фактор; их называют ре-
зус-положительными. У 15% людей этого фактора нет; их назы-
вают резус-отрицательными.
В чем суть болезни? Предположим, что эмбрион гетерозиготен
по резус-фактору, он обладает способностью вырабатывать анти-
ген, который через плаценту попадает в кровь матери. В крови
резус-отрицательной матери против антигенов резус-положитель-
ного эмбриона (унаследовавшего Rh+ от отца) вырабатываются
антитела. Антитела, попадая в кровь эмбриона, вызывают у него
частичную агглютинацию эритроцитов, что обусловливает анемию
у ребенка.
Далеко не в ста процентах случаев происходит катастрофа
вследствие несовместимости. Как правило, в случае Rh-несовмес-
тимости первый ребенок рождается нормальным, так как антитела
у матери или не образуются, или титр их низкий. С каждой по-
следующей беременностью может возрастать сенсибилизация ма-
тери, тем самым опасность выработки антител возрастает, а зна-
чит, и опасность для развития плода. Но сенсибилизация может
не состояться. Выяснено к тому же, что не во всех случаях
Rh-фактор отца передается ребенку. Если отец гомозиготен по
Rh-фактору, то в этом случае все дети будут резус-положительны-
ми и вероятность гибели зародыша окажется наибольшей.
Существует мнение, что мать и плод постоянно находятся в со-
стоянии взаимно иммунологической несовместимости. Зародыш,
согласно этому взгляду, можно рассматривать как гомотранс-
плантат. Это созвучно очень смелой, крайне спорной идее, вы-
сказанной Б. А. Фаусеком. Он предполагал, что в ходе эволюции
внутриутробного развития между организмом матери и зародышем
создавались условия, характерные для явления паразитизма, и
зародыш может быть уподоблен паразиту.
Как бы то ни было, действительно антигенная характеристика
тканей зародыша и материнских тканей может оказаться суще-
ственно различной. Иммунологические реакции, возникающие
в организме женщины (выработка антител, воспалительные про-
цессы), могут оказаться факторами, приводящими к гибели яйце-
вые клетки и зародыш на стадии бластоцисты, приводить к само-
произвольному аборту, мертворождению или к возникновению ге-
молитической болезни и к другим заболеваниям.
Отмечено, что иммунная реакция на антигены, содержащиеся
в тканях зародыша и плаценты, может обусловить развитие у бе-
ременной женщины заболевания токсикозом. Эти явления можно
объяснить, если принять во внимание установленные факты о про-
357
ницаемости плаценты как для клеточных элементов крови плод,
так и для антител, содержащихся в материнской крови. Однаи
указанные патологические проявления иммунологической нес<
вместимости организма матери и плода — редкие исключения,
они неизмеримо более редки, чем явления несовместимости ткане
и органов при пересадках. Это требует своего объяснения, кг
ковое и дается исследователями в виде различных гипотез. Tai
думают, что во время беременности, в связи с изменением де5
тельности желез внутренней секреции, снижается иммунология!
ская реактивность женского организма. Предполагают, что у мг
теринского организма вырабатывается в связи с беременность!
толерантное состояние в отношении антигенов тканей зародыш
и он становится «ареактивным». Многие исследователи решающе
значение придают барьерной функции плаценты и плодных об(
лочек. Считают, что плацента — прочная перегородка между мг
терью и плодом. Это, однако, противоречит ряду фактов. Неке
торые иммунологи думают, что ткани плаценты могут избирателе
но пропускать совместимые и задерживать несовместимы
изоантитела материнской крови. Высказываются предположени
и о том, что защита матери от антигенов плода и защита плод
от антигенов матери обеспечивается благодаря антигенной нее
тральности тканей плаценты.
Более детальное ознакомление с этими важными медико-эм(
риологическими вопросами не входит в задачу общей эмбриолс
гии. Л. С. Волкова (1970) на основе исследований пришла ксл<
дующему заключению. Несмотря на то, что между плодом
материнским организмом, по-видимому, во всех случаях беремег
ности происходит двусторонняя иммунологическая реакция и зг
родыш может быть уподоблен гомотрансплантату, он, как правг
ло, выживает во «враждебном» материнском организме. Сред
многих причин выживаемости, вероятно, большое значение имее
то, что клетки трофобласта, непосредственно контактирующи
с материнскими тканями, вероятно, не имеют чужеродных дл
материнских тканей антигенов. Между матерью и плодом пре
исходит постоянный обмен клеточных элементов в незначител!
ных количествах, что вызывает возникновение толерантности к не
совместимым антигенам.
Изучая взаимодействие иммунологических факторов органи;
ма матери, плода, плаценты, плодных оболочек и околоплодно
жидкости, автор пришла к убеждению, что нормальное эмбрш
нальное развитие обеспечивается своеобразной регуляцией кол!
чества несовместимых элементов (изоантигенов, изоантител) кр(
ви матери и плода. Возможность такой регуляции обусловлен
различной антигенной дифференцировкой пограничных межд
матерью и плодом тканей и жидкостей и благодаря наличи,
в околоплодных водах большого количества групповых антигено!
способных нейтрализовать антитела материнской крови.
358
При беременности, осложненной резус-сенсибилизацией или
возникновением позднего токсикоза, групповая дифференцировка
тканей плаценты, плодных оболочек и околоплодной жидкости
нарушается: в них устанавливается «смешение» антигенных фак-
торов, принадлежащих матери и плоду, или «исчезают» антигены,
свойственные этим тканям и жидкостям. Эти изменения создают
условия для более свободного проникновения к плоду материн-
ских антител и для развития у него гемолитического заболева-
ния.
ГИПОТЕЗЫ И ЭКСПЕРИМЕНТЫ ПО ВОПРОСАМ МОРФОГЕНЕТИЧЕСКОЙ РОЛИ
АНТИГЕННОЙ РЕАКТИВНОСТИ
Как уже говорилось, многие исследователи пытались перекинуть
мост между иммунологией и эмбриологией. А. Тайлер в 1955 г.
и другие исследователи утверждали, что во взаимоотношении
сперматозоида и яйца при оплодотворении играет роль наличие
на их поверхности веществ, реагирующих друг с другом по типу
реакции антиген — антитело (фертилизин и антифертилизин). По
П. Вейсу, в процессах дифференциации тканей и органов наблю-
даются отношения, подобные системе антиген — антитело. Пове-
дение клеток регулируется такими отношениями между клеточ-
ными поверхностями, какие можно наблюдать между антигенами
и антителами: поверхность одной клетки играет роль антигенов,
а поверхность другой—антител. Вейс предполагает, например,
что антигены клеток глазной чаши развивающихся амфибий при-
тягивают из протоплазмы эпидермальных клеток вещества, комп-
лементарные для антигенов клеток глазной чаши. Это и лежит
в основе формирования хрусталика (см. гл. XI и XV). Родствен-
ные взгляды в 1939 г. высказывал И. Гольтфретер, который на-
блюдал, что диссоциированные клетки бластулы и ранней гаст-
рулы амфибий, помещенные в физиологический раствор Рингера,
беспорядочно «агглютинируются», ио затем конгломерат клеток
распадается на эктодерму и энтодерму. Выходит, что клетки каж-
дого зародышевого пласта имеют сродство и «находят» друг
друга.
Сходные взгляды высказывались в связи с опытами по про-
тиранию губок сквозь сито (см. гл. XVI). В настоящее время
показано, что если губки разных видов диссоциировать на клет-
ки, то клетки, принадлежащие разным видам, расходятся в раз-
ные стороны и образуются скопления.
Доказано, что по определенным физико-химическим законам и,
вероятно, благодаря тем или иным биологическим свойствам (сли-
пание и т. п.) клетки одних и тех же тканей могут «предпочти-
тельней» агрегироваться. Вероятно, подобные взгляды несколько
односторонни, ио ведь при защите всякого нового оригинального
представления ученые неизбежно односторонни и даже фанатич-
359
ны, и это нельзя ставить им в упрек. Что касается известны:
опытов по протиранию губок сквозь сито, а также подобных опы
тов М. Айзупет (1935) с гидрами, то, конечно, предрассудков
является утверждение о «сборе», «агрегации» организмов, «о ре
конструировании» прежних организмов из диссоциированного кле
точного материала (см. гл. XVI). Большая часть клеток поел*
протирания может погибнуть, но развитие (соматический эмбрио
генез) происходит. Из протертой губки может развиваться мног<
губок. Еще менее применимы предположения о «сборе» диссо
циированных клеток в старый организм к опытам по протирании
гидр. Если физико-химические явления «слипания» клеток даю-
повод думать о восстановлении «прежней» эктодермы и «преж
ней» энтодермы, то, конечно, нет никаких оснований говорит
о восстановлении прежней мезоглеи и прежнего организма. В<
всех случаях происходят своеобразные процессы развития (см
гл. XV и XVII).
Большое число исследований вызвала гипотеза, согласно ко
торой выходящие из клеток аутоантитела служат причиной соот
ветствующей дифференцировки тканей и органов.
Бесспорным фактом является закономерное появление новы:
и новых белков в ходе эмбрионального развития. Происходи'
непрерывное изменение антигенных свойств. Создаются ли меж
ду белками предыдущих и последующих этапов развития отноше
ния по типу антиген — • антитело и, если создаются, имеют ли этт
своеобразные иммунологические реакции морфогенетическое зна
чение? Этим вопросам посвящены многие исследования.
ИММУНИТЕТ ЭМБРИОНОВ
Б. П. Токиным в 1959 г. и ранее была обоснована проблема им
мунитета эмбрионов как глава сравнительной эмбриологии, вклю
чающая исследования в трех аспектах:
1. Каковы защитные силы эмбрионов различных животных?
2. Когда в ходе онтогенеза появляются те или иные иммуно
логические свойства? Какова динамика изменений фагоцитарно!
активности клеток различных частей зародышей? Когда появля
ется возможность воспаления? Происходит ли (и на каких ста
днях развития) продукция антибиотических субстанций? Когд.
возникает антигенная реактивность в онтогенезе птиц и млекопи
тающих?
3. Третий аспект исследований, касающийся связи иммуноло
гических реакций и морфогенетических процессов, обосновываете,
следующей гипотезой. В индивидуальном развитии иммунологи
ческие свойства появляются и развиваются не в качестве «шпаг:
для дуэлей» будущего взрослого организма с патогенными микро
организмами, но как необходимейшие факторы развития. Как в хо
де эволюции организмов различные иммунологические свойств
360
возникали и развивались на базе более общих функций (напри-
мер, фагоцитарные реакции возникали на базе внутриклеточного
пищеварения), так и большинство иммунологических свойств,
возникающих и меняющихся в ходе эмбрионального развития,
являются теми или иными сторонами закономерностей эмбрио-
нального развития и необходимы для процессов формообразова-
ния, в органогенезе, в процессах метаморфоза, регенерации, бес-
полого размножения, соматического эмбриогенеза и в других яв-
лениях формообразования. Все иммунологические свойства следует
рассматривать как факторы, могущие поддерживать определен-
ные состояния интеграции организма на разных стадиях разви-
тия.
ИЛЛЮСТРАЦИИ ЗНАЧЕНИЯ ИММУНОЛОГИЧЕСКИХ РЕАКЦИЙ
В ЯВЛЕНИЯХ ФОРМООБРАЗОВАНИЯ
Значение фагоцитарных реакций. В организмах трудно, а может
быть, и нельзя найти структуры и функции, имеющйе лишь ка-
кое-либо одно-единственное значение. Так обстоит дело и с фа-
гоцитозом. Еще И. И. Мечников в 11883 г. высказывал мысль
о формообразовательном значении фагоцитарных реакций у взрос-
лых организмов и в ходе развития животных. Он обратил внима-
ние на участие фагоцитов в разнообразных процессах: в инволю-
ции матки и яичника у млекопитающих, дегенерации нервных
центров при старческой атрофии, поседения волос. «Во время эмб-
рионального развития различных и в том числе очень развитых
организмов фагоцитарная функция имеет далеко простирающееся
влияние. Использование питательного желтка для развития за-
родыша возникло благодаря фагоцитозу. В продолжение долгого
периода развития, например при метаморфозе различных живот-
ных, фагоциты вызывают глубочайшие изменения как во внешнем
виде, так и во внутренней организации, пожирая и уничтожая
некоторые органы целиком и без остатка»'. И. И. Мечников, изу-
чая метаморфоз голотурий и морских звезд (1883), а также ме-
таморфоз амфибий (1884), установил, что многие личиночные
органы подвергаются уничтожению, что в этих процессах играют
роль клетки с фагоцитарными свойствами. Таким путем осуще-
ствляется резорбция хвоста головастика лягушки при метамор-
фозе. Впоследствии это было подтверждено многими авторами.
Б. В. Алешин (1936) считает, что резорбция личиночного хвоста
протекает по типу воспалительного процесса: имеет место альте-
рация тканей, сосудистые реакции с энергичной иммиграцией лей-
коцитов, фагоцитоз отмирающих клеток.
1 Мечников И. И. Учение о фагоцитах н его экпериментальные осно-
мы // Собр. соч. Т. 7. М., 1952. С. 496.
361
Рис. 138. Скопление фагоцитов возле сосудов туники (А) и фагоциты с захва-
ченными частицами кармина возле сосудов (Б) (по О. М. Ивановой, 1966)
А. О. Ковалевский в 1885 и 1887 гг., исследуя метаморфоз
насекомых, убедился в важной роли фагоцитарных реакций: во
время метаморфоза мух выселяющиеся в большом количестве лей-
коциты разрушают и переваривают личиночные ткани и органы
(за исключением нервной системы, полового зачатка и сердца).
А. О. Ковалевский в 1890 г., а в 1940 г. О. М. Иванова-Казас от-
метили роль фагоцитов в метаморфозе асцидий: мезенхимные
клетки-фагоциты захватывают и переваривают мышечные и нерв-
ные клетки хвоста личинки, клетки хорды.
О. М. Иванова-Казас (1966) изучала роль фагоцитарных реак-
ций в жизни асцидии Distaplia unigermis и убедилась, что унич-
тожение всякого рода дегенерирующих частей тела (хвоста и
нервной системы личинки при метаморфозе, торакса и сосудов
туники отмирающих зооидов) происходит при активном участии
фагоцитов. Если ввести инородное тело — частички кармина, уже
через сутки все они оказываются заглоченными клетками, а более
крупные комки частиц кармина инкапсулированными (рис. 138).
Фагоцитарные реакции происходят и в процессах регенера-
ции, на определенных этапах которых неизбежно происходит ги-
бель клеточных элементов, пожирание их фагоцитами. Хотя эта
функция фагоцитов выглядит внешне только как «санитарная*
(освобождение организма от ненужного), но если без этой функ-
ции невозможен процесс регенерации, значит, фагоцитоз играет
формообразовательную роль. Формообразовательная роль фаго-
цитарных и воспалительных реакций доказана и при изучении
различных процессов нормального развития организмов, напри-
мер при формировании органов слуха у позвоночных (Д. П. Фи-
латов, 1935), при формировании мочеполовой системы и т. д.
Е. Б. Кричинская (4965) изучала процессы развития выдели-
тельной системы куриного зародыша. Процесс деструкции мезо-
нефроса начинается приблизительно на 13-й день инкубации. На-
блюдаются явления, которые автор уподобляет «хроническому
362
воспалению». Лизис клеточных элементов при деструкции мезо-
нефроса, фагоцитарные реакции, осуществляемые клетками эндо-
телия кровеносных сосудов, и вставочной соединительной ткани,—
эти и другие явления совершаются значительно медленнее, чем
при воспалительных процессах. В случае, например, резорбции
хвоста головастика, но и к процессам деструкции мезонефроса
применимо утверждение Б. В. Алешина: «...Воспаление, принимая
участие в формировании дефинитивной структуры организма, ока-
зывается морфогенетическим фактором».
Явления фагоцитоза при некоторых формообразовательных
процессах у губок и гидр. Очень интересным примером значения
фагоцитарных реакций для процессов формообразования являют-
ся своеобразные явления, наблюдаемые при формировании яйце-
Рис. 139. Процесс оогенеза у гидры:
А — находящаяся в состоянии дезинтеграции эктодерма (в районе временного яичника) с
энергично размножающимися i-клетками; Б — два сливающихся ооцита; В — сформирован-
ная яйцевая клетка (Б — по И. Б. Токниу, 1955; В — по Н. Клейненберг, 1872)
363
вых клеток у гидр. И. Б. Токин в 1954 г. убедился, что описанные
Н. Клейненбергом и А. Коротаевым еще в прошлом столетии яв-
ления фагоцитоза при формировании яйца гидры не теряют своей
ценности и в настоящее время (рис. 139).
В яичнике благодаря размножению i-клеток возникает много
потенциальных яйцеклеток. В начале оогенеза невозможно найти
какие-либо различия между многими клетками-оогониями. Лишь
один оогоний впоследствии превращается в яйцо, остальные идут
на питание. Будущая яйцевая клетка постепенно фагоцитирует
другие клетки-оогонии и переваривает их. В результате электрон-
но-микроскопического анализа этого процесса Т. Б. Айзенштадт
(1975) установила, что в i-клетках, вступивших на путь оогенеза,
появляются специфические для оогоний маркеры (скопления в ци-
топлазме тонковолокнистого материала). В период роста ооцитов
и образования желточных включений происходит слияние оого-
ний и ооцитов с будущей яйцеклеткой. Цитоплазма и ядро «ас-
симилированных» клеток превращаются в яйцеклетке в желточного
типа включения. Фагоцитарные реакции происходят при форми-
ровании яиц губок.
Процесс образования геммул — внутренних почек у губок со-
провождается полной дезинтеграцией губок, умиранием большей
части клеток, повышением амебоидной подвижности остающихся
живыми клеток, фагоцитарными реакциями.
Таким образом, фагоцитарные реакции, являющиеся потен-
циально иммунологическими, играют формообразовательную роль:
без них не было бы оогенеза у гидры и губок, не происходило
бы образование геммул и т. д.
Почему бластоциста имплантируется в стенку матки? Боль-
шинство исследователей рассматривают процесс имплантации как
взаимодействие бластоцисты и тканей материнского организма.
С момента овуляции в матке происходят изменения, обуслов-
ленные деятельностью гормонов яичника и гипофиза. Развиваю-
щееся яйцо, пройдя в яйцеводах период дробления и достигнув
стадии бластодермического пузырька, попадает в полость матки.
В это время, помимо изменений матки, вызванных гормональной
деятельностью, в тканях матки происходят и локальные измене-
ния, возникающие в связи с взаимодействием с бластоцистой.
Клетки соединцтельно-тканной стромы в районе контакта или
приближения зародыша к стенке матки быстро преобразуются
в децидуальные клетки, располагающиеся параллельными про-
свету матки рядами. Они характеризуются большими размерами
и многоядерностью. В децидуальной ткани наблюдается большое
число митозов. Впоследствии в децидуальную ткань врастают
кровеносные капилляры, происходит расширение сосудов, повы-
шение проницаемости их стенок, усиливается приток лейко-
цитов.
364
Бластоцисты, как уже сказано, оказываются в матке и неко-
торое время пребывают как бы в «свободном» положении, взве-
шенными в жидкости просвета матки, без контакта с эпителием
матки. У разных млекопитающих период «свободного» располо-
.жения бластоцисты различен — от нескольких часов (у хомяков)
и 2—3 дней (у человека) до нескольких месяцев (у соболя, гор-
ностая, медведя, броненосца и др.). В последнем случае говорят
о паузе в развитии бластоцисты. В определенный момент бласто-
циста приходит в контакт со слизистой оболочкой стенки матки
и начинает в нее погружаться. Происходит имплантация бла-
стоцисты и затем начинается процесс формирования плаценты.
Единого мнения о механизме имплантации в эмбриологии до сих
пор еще нет.
Вероятно, наряду с другими факторами иммунологические ре-
' акции играют первенствующую роль в процессах имплантации
и формирования плаценты.
Б. П. Токин и А. Г. Филатова в 1953 г. убедились в том, что
развивающиеся яйцевые клетки млекопитающих до их имплан-
тации в стенку матки обладают антибиотическими свойствами:
они убивают находящиеся в контакте с ними многие бактерии.
Бактерицидными свойствами обладает и жидкость полости бла-
стоцисты (рис. 140). Эти свойства, имеющие потенциально им-
мунологическое значение, играют, по-видимому, существенную
роль в процессах им-
плантации. Антибиоти-
ческие вещества, про-
дуцируемые бластоци-
стой, воздействуют, ве-
роятно, на сенсибили-
зированную гормона-
ми матку, участвуют
в лизисе эпителия, яв-
ляются одним из фак-
торов, вызывающих де-
цидуальную реакцию
со стороны тканей
стенки матки.
Е. В. Зыбина в
1957 г. изучала на-
чальные стадии им-
плантации зародышей
крысы? На 4—5-е сутки
после оплодотворения
бластодермический пу-
зырек свободно лежит
в просвете рога матки.
Стенка рога матки вы-
Рис. 140. Подавление размножения Вас.
megatherium яйцевыми клетками кролика
на ранних стадиях развития. А—кон-
троль, обильный рост бактерий; Б—Г —
опытные висячие капли (в физиологиче-
ском растворе с бактериями находились
вымытые из половых путей яйцеклетки) (по
Б. П. Токину и А. Г. Филатовой, 1953)
365
Рис. 141. Развитие зародыша крысы и его
имплантация в стенку матки (по Е. В. Зы-
биной, 1958):
А — зародыш крысы на 5,5 сут после осеменения.
Зародыш находится в обособившейся импланта-
ционной камере, не касаясь ее стенки. Видно ло-
кальное уплощение эпителия; Б — зародыш кры-
сы на стадии 6,5 дней после осеменения. Заро-
дыш прикреплен к стенке имплантационной каме-
ры. Эпителиальный пласт, находившийся в кон-
такте с зародышем, <исчез». Клетки соедини-
тельной ткани, окружающие имплантационную
полость, превратились в децидуальные
стлана однослойным эпите-
лием. Зародыш в это время
представляет пузырек с не-
большой полостью внутри.
На 5-е сутки зародыш еще
не соприкасается с эпители-
ем стенки матки, но уже
можно отметить изменения
эпителия и 3—4 слоев со-
единительно-тканных кле-
ток, принадлежащих к эпи-
телию (рис. 141,Д). Как
будто зародыш, выделяя ка-
кие-то вещества, активно
действует на соответствую-
щий участок рога матки.
На 6-е сутки бластоци-
ста приходит в контакт с
эпителием матки (рис.
141,5). Количество клеток
трофобласта к этому време-
ни сильно возрастает в ре-
зультате митотических деле-
ний немногих исходных кле-
ток. В месте контакта бла-
стоцисты со слизистой
оболочкой образуется отек; в отечную строму и погружается заро-
дыш. В процессе имплантации клетки трофобласта полностью ли-
зируют прилегающий к ним эпителиальный слой. Клетки трофо-
бласта становятся гигантскими, образуют синцитий, соединяясь
друг с другом своими отростками. К этому времени разыгрывают-
ся фаготицарные реакции. Клетками трофобласта фагоцитируются
ядра и продукты распада лизирующихся эпителиальных клеток и
лейкоцитов. Предполагают, что трофобласт при имплантации осу-
ществляет питание зародыша в значительной мере за счет разру-
шаемых им клеток эпителия и подлежащих децидуальных клеток.
Немного позднее, когда зародыш оказывается окруженным со всех
сторон материнской кровью, клетки трофобласта активно фагоци-
тируют в большом количестве также эритроциты и лейкоциты.
Эти и другие данные исследований Е. В. Зыбиной не оставля-
ют сомнений в важной роли фагоцитарных реакций в процессах
имплантации. Очень вероятна также серьезная роль антибиоти-
ческих субстанций. В процессах имплантации в стенке матки ра-
зыгрывается комплекс явлений (отек, изменения сосудов, выхож-
дение лейкоцитов, отмеченные уже дегенеративные процессы
в эпителии и ярко выраженная фагоцитарная активность), кото-
рый является в сущности процессом воспаления. И. Г. Михайлова
366
(1964), исследуя фагоцитарные и воспалительные явления в про-
цессах формирования плаценты у мышей, крыс и кроликов, убе-
дилась в том, что в ходе беременности первичные гигантские
клетки трофобласта заменяются вторичными гигантскими клетка-
ми трофобласта, обладающими меньшей фагоцитарной активно-
стью. Автор пришла к убеждению, что в ходе развития эмбриона
в сформированной плаценте фагоцитарные и воспалительные яв-
ления, сыграв свою формообразовательную роль в процессах им-
плантации и образовании плаценты, должны подавляться. В про-
тивном случае те же иммунологические реакции окажутся вред-
ными, они приводили бы к элиминации эмбриональных тканей.
Б. П. Токин в 1955 г. предположил, что не только при про-
цессах имплантации, но и при различных других нцпатологических
процессах (например, при развитии различных органов и систем)
воспалительные явления оказываются необходимыми и законо-
мерными. Это относится, например, к развитию мочеполовой си-
стемы позвоночных, к процессам возникновения мезонефроса и
метанефроса.
Особый интерес представляет изучение роли иммунологиче-
ских реакций в нормальном функционировании семенников и
яичников. Так, например, Н. А. Пахомова (1975) изучила состоя-
ние яичников в разные периоды до и после нереста у краба Hyas
araneus. Через 7 суток после нереста часть ооцитов на II стадии
роста подвергается разрушению, резорбируются ооциты III ста-
дии, не овулировавшие во время нереста. Фолликулярные клетки,
проникающие в цитоплазму ооцитов, и незернистые амебоциты
фагоцитируют некротизированные структуры (рис. 142,А, Б, В, Г).
На 14—21-е сутки после нереста состояние яичника таково, что
к нему вполне приложимо понятие «воспалительный процесс».
В яичнике одновременно идут процессы лизиса разрушающихся
структур, процессы фагоцитоза и (по-видимому, сопряженно
с этими явлениями) процессы размножения оогоний и развития
ооцитов. Число митозов в участках пролиферации увеличивается.
Фагоцитозу подвергаются зрелые ооциты, оставшиеся в яич-
нике после нереста, небольшое число ооцитов III стадии роста
и опустевшие фолликулы. В этот период строение яичника кажет-
ся хаотичным, это как бы «депо» амебоцитов; все промежутки
между растущими половыми клетками в строме яичника
заполнены элементами гемолимфы. Одновременно интенсивно
протекают формообразовательные процессы в ооцитах старой и
новой генерации. Воспалительные и восстановительные процессы
в яичнике заканчиваются через 35 суток после нереста.
Аналогичные процессы деструкции половых клеток и их фаго-
цитоза вспомогательными клетками или фагоцитами имеют место
при периодическом функционировании гонад не только в ооге-
незе, но и в сперматогенезе других животных.
Д. П. Филатов (1916, 1927), изучая развитие органа слуха
367
Рис. 142. Яичник краба Hyas araneus (по Н. А. Пахомовой, 1975):
Л — ооцит на начальной стадии вителлогенеза. В яичнике через сутки после нереста; Б —
«атретический фолликул» в янчнике через сутки после нереста; В — резорбция ооцита
111 стадии роста; Г — яичник через 14 сут после нереста. Строма янчника заполнена вие-
сосудистыми элементами гемолимфы. Преобладают иезернистые амебоциты
у амфибий, обратил внимание на то, что в мезенхиме вокруг слу-
хового пузырька разыгрываются процессы, очень сходные с вос-
палительными. Без них не может происходить формирование
слуховой капсулы. Если в мезенхиму вводить инородные тела,
например целлоидиновый или парафиновый кусочек, т. е. если
вызывать экспериментальное воспаление, то вокруг инородных
тел происходит образование тех же зачатков, которые наблюда-
ются при типическом развитии слухового пузырька. Таким обра-
зом, природа была очень «экономна» в своем эволюционном раз-
витии: одни и те же реакции (воспалительные) оказались лежа-
щими в основе разнообразных процессов развития.
ДИНАМИКА ИММУНОЛОГИЧЕСКИХ РЕАКЦИЙ В ХОДЕ
ЭМБРИОНАЛЬНОГО РАЗВИТИЯ
Исследования роли иммунологических реакций в формообразова-
тельных процессах. находятся пока лишь на стадии накопления
фактов; невозможно поэтому предусмотреть, какие аспекты даль-
нейших исследований представят в будущем наибольший интерес.
368
Поскольку становится очевидной роль иммунологических реакций
в разных процессах эмбрионального развития, приобретают осо-
бое значение вопросы динамики иммунологических свойств в он-
тогенезе. В ходе эмбрионального развития животного происходит
сложная смена разных иммунологических свойств, о чем, в част-
ности, свидетельствуют экспериментальные данные, полученные
при изучении развития птиц: защитное значение белковой обо-
лочки яиц, возникновение возможности фагоцитарных и воспали-
тельных реакций в тканях зародыша, защитное значение амниона
н аллантоиса, возникновение антигенной реактивности.
Много данных по этим вопросам в экспериментах на амфи-
биях, птицах и млекопитающих получено А. К. Дондуа (1956,
11958, 1959), Е. Б. Кричинской (1955, 1959), И. Г. Михайловой
(1958, 1962). В кровяное русло и в различные ткани эмбрионов на
разных этапах их развития вводились инородные тела (тушь, кар-
мин, кизельгур, целлоидиновые иголочки). Изучались фагоцитар-
ные реакции и возможность вызывать воспаления.
Фагоцитоз желтка клетками желточной энтодермы куриного
зародыша наблюдается уже в первые сутки инкубации, у 15—
18-часовых эмбрионов. А. К. Дондуа установил, что воспалитель-
ная реакция в ответ на введение инородного тела возникает ра-
но, по крайней мере уже на 2-й день инкубации яиц птиц. Фаго-
цитарные реакции у 2—4-суточных эмбрионов осуществляются
мезенхимными клетками, первичными лимфоцитами и эндотели-
альными клетками сосудов различных органов. В зависимости от
величины инородных тел наблюдается активность или одиночных
фагоцитов, или «групповой фагоцитоз», образование мезенхима-
тозной капсулы, изолирующей инородное тело от тканей зароды-
ша. Характер воспалительных реакций, однако, изменяется в ходе
развития в связи с дифференциацией клеток и тканей. Вместе
с клеточной и тканевой дифференциацией, формированием сосу-
дов, началом функционирования нервной системы воспалительные
процессы (возникшие на базе фагоцитоза) усложняются и при-
обретают, наконец, свой «взрослый характер». Интересны детали:
у 6—8-суточного эмбриона формирующаяся сначала вокруг ино-
родного тела мезенхиматозная капсула затем дегенерирует и
образуется фибробластическая капсула. 11 — 14-суточный эмбрион
реагирует на повреждение прежде всего сосудистой реакцией
(выселяется большое количество специальных лейкоцитов и лим-
фоцитов), образуется фибробластическая капсула.
На рис. 143,А видна реакция мезенхимных клеток двухсуточ-
ного зародыша в ответ на введение инородных тел. Она выра-
жается в фагоцитарных реакциях. Это прообраз будущих про-
цессов воспаления. На рис. 143,В представлена картина воспале-
ния при введении целлоидиновой иголочки в ткани 10-суточного
куриного эмбриона, когда дифференциация клеток зашла уже да-
леко. В конце концов и в этот период основным компонентом
369
Рис. 143. Фагоцитарные и воспалительные реакции у зародышей птиц. А— фа-
гоциты из мезенхимы двухсуточного куриного эмбриона; Б — фагоцитоз крупных
частиц путем обволакивания; В — асептическое воспаление в соединительной
ткани десятисуточного куриного эмбриона (12 ч после операции) (по А. К. Дон-
дуа, 1955):
1 — кровеносный сосуд, 2 — эритроциты, 3 — специальный лейкоцит, 4 — фибробласт, 5 —
некротическая зона, 6—фагоцитирующий лимфоцит, 7 — место растворившейся целлоиди-
новой иголочки, 8 — гистиоцит, 9 — зона свободных элементов, 10 — фнбропластическая
капсула
воспаления является фагоцитоз, но вместе с тем в соединитель-
ной ткани и крови происходят сложные процессы, в результате
которых вокруг инородного тела образуется фибробластическая
капсула, изолирующая инородное тело от организма.
Е. Б. Кричинская в 1954 г. инъецировала в одну из вен жел-
точного мешка эмбриона курицы бактерии или взвесь туши. В ре-
зультате гистологических исследований ей удалось обнаружить
энергичную фагоцитарную активность клеток эндотелия сосудов
развивающейся печени, селезенки, сердца, желточного мешка, ал-
лантоиса. На 4—7-е сутки развития цыпленка эндотелиальные
клетки всех названных органов обладают приблизительно оди-
наковой фагоцитарной активностью. На поздних стадиях разви-
тия эмбриона наибольшая фагоцитарная активность наблюдается
в эндотелии сосудов печени и селезенки.
Все данные о развитии фагоцитарных и воспалительных реак-
ций в онтогенезе позволили А. К. Дондуа в 1957 г. заключить,
что «защитный фагоцитоз» как явление иммунитета возникает
в онтогенезе на базе «физиологического фагоцитоза».
В заключение следует отметить, что, по-видимому, всем клет-
кам любых организмов (без всякого исключения) потенциально
свойственны амебоидная подвижность и фагоцитарная способ-
370
Рис. 144. Клетки, изолированные со стадии гаструлы
амфибий. Видны псевдоподии и фагоцитированные кап-
ли жира (по И. Гольтфретеру, 1948)
ность. Она проявляется ясно в условиях дезинтеграции тканей,
на что обращалось внимание в других главах этой книги
(гл. VI). Отмечались уже, в частности, опыты И. Гольтфретера
(гл. VI), который изолировал клетки со стадий бластулы, гастру-
лы и нейрулы амфибий. Автор убедился в том, что клетки обла-
дают амебоидной подвижностью и выраженными фагоцитарными
свойствами (рис. 144).
ЗАЩИТНОЕ ЗНАЧЕНИЕ ЯЙЦЕВЫХ ОБОЛОЧЕК
Яйцевые оболочки — пограничные между зародышем и внешней
средой структуры, вероятно, играют гораздо большую роль, чем
предполагали ранее. Выясняется их важное значение для обес-
печения стерильных условий для развивающегося организма. Ил-
люстрацией этой важной роли оболочек в явлениях формообразо-
вания могут служить данные исследователей об оболочках яиц
птиц и пресмыкающихся. Известно, что куриное яйцо не загни-
вает и не плесневеет при хранении в течение длительного време-
ни, между тем скорлупа и подскорлуповая оболочка проницаемы
для бактерий и грибов. Если вылить «белок» свежеснесенного
куриного яйца в нестерильную посуду, он, как правило, не под-
вергается ни гниению, ни плесневению. На это явление обратил
внимание еще в 1909 г. П. Н. Лащенков, доказавший бактерицид-
ные свойства белка. В 1922 г. А. Флемминг предложил назвать
вещества куриного яйца, обусловливающие антибиотические
свойства, лизоцимом. Обнаружен широкий спектр антибиотиче-
371
Рис. 145. Aspergillus niger на третий день после внесения спор в белок куриного
яйца (А); воду (5) и питательную среду (В) (по Г. П. Коротковой, 1956)
ского действия натуральной белковой оболочки: она обладает не
только бактерицидными свойствами, а также фунгицидным и фун-
гистатическим действием (рис. 145). «Белок» .сохраняет свои анти-
биотические свойства (и, вероятно, свою роль в пассивном имму-
нитете) и после 11 дней инкубации, когда он смешивается
с амниотической жидкостью и заглатывается зародышем.
Н. А. Мовчан в 1964 г. обнаружила, что белок яиц черепахи
(Testudo horsfieldi) обладает также выраженными бактерицид-
ными и фунгицидными свойствами. К сожалению, значительные
материалы о роли микроорганизмов в эмбриональном развитии
животных (особенно насекомых) не систематизированы.
Несомненно, вопросы о физиологическом, защитном значении
оболочек яиц животных, об их значении для процессов формооб-
разования составляют важную главу эмбриологии. Особенно
большой интерес вызывают оболочки яиц насекомых, имеющих
подчас очень сложную структуру, состоящие из нескольких слоев,
в промежутках между которыми — сеть воздухоносного слоя. Га-
зообмен между яйцом и средой осуществляется через аэропиле.
У яиц некоторых паразитических насекомых развиваются до-
полнительные оболочки. Их функция — не только обеспечить про-
никновение в зародыш питательных веществ «хозяина», но и за-
щитная: эти оболочки дополнительно предохраняют зародыш от
иммунологических реакций пораженного животного или растения.
Благодаря использованию электронной микроскопии удалось вы-
явить детально структуру дыхательных пор. 3 яйцах насекомых
есть тончайшие канальцы — гидропиле, по ним проходит вода
(в определенные периоды развития), а микропиле — система ка-
налов, по которым к яйцу проходят сперматозоиды. Вероятно,
все эти поры мультифункциональны.
Глава XVI
БЕСПОЛОЕ РАЗМНОЖЕНИЕ, СОМАТИЧЕСКИЙ ЭМБРИОГЕНЕЗ
И РЕГЕНЕРАЦИЯ
РАСПРОСТРАНЕННОСТЬ БЕСПОЛОГО РАЗМНОЖЕНИЯ
В ЖИВОТНОМ МИРЕ
Несмотря на огромное разнообразие, формы размножения могут
быть объединены в две группы: половое и бесполое. Бесполое, или
.вегетативное (от лат. vegeto — расту), размножение — размно-
жение без участия яйцевых клеток и сперматозоидов: новый орга-
низм развивается из соматических клеток. Такой вид размножения
эмбриологи называют бластогенезом. Термин «бластогенез» ис-
пользуют и для обозначения наиболее ранних стадий развития
млекопитающих и человека. Применяется и термин «бластозооид»,
что означает развивающийся в результате бесполого размножения
организм.
Ботаники выделяют из явлений вегетативного размножения раз-
витие организмов из спор, называя этот вид размножения беспо-
лым. Бесполое размножение столь распространено в растительном
мире, что его можно считать одним из признаков растительных
организмов.
Бесполое размножение широко распространено в мире Proto-
zoa, а также у низших многоклеточных — губок, кишечнополост-
ных, у гребневиков и червей; оно свойственно мшанкам (Bryozoa)
и форонидам (Phoronis), некоторым иглокожим, характерно для
оболочников (Tunicata) (рис. 146).
По своему биологическому механизму к бесполому размноже-
нию близки явления нормальной полиэмбрионии, встречающиеся
у кишечнополостных, червей, членистоногих, а также и у позвоноч-
ных.
ПОЛИЭМБРИОНИЯ
Полиэмбрионию следует считать одной из форм бесполого размно-
жения. Это случай бесполого размножения, на стадиях эмбриональ-
ного развития, т. е. так же, как и при других видах бесполого раз-
множения, развитие новых организмов происходит из соматических
клеток, не достигших еще специализированного состояния. Это яв-
ление было обнаружено сначала ботаниками. Честь открытия по-
лиэмбрионии у животных принадлежит И. И. Мечникову, который
наблюдал в 1886 г. расщепление бластул у медузы Oceania armata
и развитие из каждого агрегата клеток целого организма. Важно
373
Рис. 146. Распространенность бесполого размножения в животном мире (п<
О. М. Ивановой, 1970):
1 — классы, в которых бесполое размножение отсутствует, 2— встречается редко, 3— сво.С
ствеиио целым семействам и отрядам, 4 — свойственно всем представителям
подчеркнуть, что у Oceania не все бластулы делятся, это не обяза
тельная полиэмбриония. У некоторых кишечнополостных могу
«почковаться» планулы (Chryosaora, Alcyonaria); у Lumbriculu
trapezoides может иметь место полиэмбриония на стадии бластуль
Закономерно наступающие в ходе развития случаи полиэмбри
374
«нии характеризуют
некоторых насекомых,
особенно наездников
(Encyrtus, Polygnotus,
JJthomastis и др.).
Рассмотрим в каче-
стве примера зародыш
паразитического наезд-
ника Ageniaspis fus-
.ciollis, откладывающе-
го яйца в кладки насе-
комого Hyponomeuta.
Происходит развитие
злррулоподобных групп
клеток (по 12—15 кле-
ток в каждой группе,
а иногда и до 60—80).
Морулы могут, в свою
очередь, разделиться
на две группы, каждая
из которых может раз-
виться в самостоятель-
ный организм (рис.
147).
Хорошо известна
полиэмбриония у мша-
нок. Морулообразный
зародыш (рис. 148,А)
принимает неправиль-
ную лопастную форму
(рис. 148,5) и затем
почкуется, давая нача-
ло многим вторичным
зародышам (рис.
148,5). Вероятно, еще
не все случаи полиэм-
брионии описаны. По-
тенциальные возмож-
ности полиэмбрионии
у многих животных
могут быть доказаны,
например, результата-
ми опытов по изоляции
бластомеров, по экспе-
риментальной полиэм-
брионии у амфибий и
птиц, о чем говорилось
в гл. XI.
Рис. 147. Полиэмбриония у наездника Agenias-
pis fusciollis:
А — общий вид; Би В — начало процесса полиэм-
брионии и последующий этап (Л — по Е. Мартину,
1914; Б и В —схема с препарата, по К. Кастельской,
1974); 1 — соедииительио-ткаииая капсула, 2 —внут-
ренний слой трофамниона, 3 — наружный слой тро-
фамниока, 4 — зародыши, 5 — парануклеус
Рис. 148. Полиэмбриония у мшанок (из
кн. Е. Коршельта и К. Хайдера, 1910):
А — морулообразиый первичный зародыш Cri-
sia occiaentalis, окруженный фолликулярны-
ми клетками; Б — распадение первичного за-
родыша иа вторичные у С. eburnea\ В — вто-
ричные зародыши н личинки на различных
стадиях развития у С. eburnea
Рис. 149. Полиэмбриония у Tatu-
sia novemcicta. Видны четыре
нормальных зародыша (по
X. Ньюмену и Д. Ж- Паттерсону,
1910)
Даже у млекопитающих встречается или закономерно наступа-
ющая полиэмбриония (например, у броненосцев), или вызывае-
мая экспериментально. К случаям полиэмбрионии можно отнести
и развитие однояйцевых однополых близнецов у человека, несом-
ненно развивающихся из частей раннего эмбриона.
У броненосцев «делится» бластоциста, возникают не одна (как
обычно), а несколько зародышевых полосок, из которых развива-
ются новые зародыши (у Tatusia hybrida — 8, у Т. novemcincta.
их 4). Новые эмбрионы имеют общую плаценту при самостоятель-
ном для каждого зародыша амнионе (рис. 149).
МОРФОФИЗИОЛОГИЧЕСКАЯ ОСНОВА ПРОЦЕССОВ БЕСПОЛОГО
РАЗМНОЖЕНИЯ И ОСОБЕННОСТИ РАЗВИТИЯ ПРИ БЕСПОЛОМ РАЗМНОЖЕНИИ
Разнообразие форм бесполого размножения у животных разной
сложности и своеобразия организации очень велико; соответствен-
но своеобразны и морфогенетические процессы, сопровождающие
бесполое размножение, а потому нелегко установить общие биоло-
гические закономерности. У ряда животных бесполое размножение
возникло как вторичное явление.
376
Наступление бесполого размножения связано либо с наруше-
нием нормальной интеграции организма, либо тех или иных его си-
стем.
О. М. Иванова-Казас (1976) доказывает, что имеется принци-
пиальное сходство закономерностей индивидуального развития при
бесполом и половом размножении. Анализируя явления бесполого
размножения у Cnidaria, Bryozoa, Tunicata, автор считает, что
к бластогенезу применимы такие понятия, как «рекапитуляция»,
«филэмбриогенез», «палингенез», «ценогенез» и др. С этим трудно
согласиться. Решающим является факт, что исходная клетка при
эмбриогенезе — зигота, а при любой форме бластогенеза и сома-
тического эмбриогенеза — это соматические клетки, входившие до
начала размножения в состав определенных органов и тканей ма-
теринского организма-, т. е. клетки, определенным образом специ-
ализированные. Начальные морфогенетические процессы при бес-
полом размножении не могут быть уподоблены дроблению яиц,
формированию бластул, процессам гаструляции и др. Понятие
«зародышевые листки» лишь с большими оговорками можно при-
менять к бластогенезу и далеко не во всех случаях. Бластогенез не
может рекапитулировать прошлую историю вида хотя бы по той
причине, что его закономерности уникальны для каждого конкрет-
ного случая. В бластогенезах не существует каких-либо общих
повторяющихся стадий развития. Эти морфогенезы более разнооб-
разны, чем половые эмбриогенезы.
Некоторые исследователи (Р. Бриен, 1956) утверждают, что
бластогенез — это совершенно новый, более поздний в филогене-
тическом смысле тип развития, возникший без связи с эмбриогене-
зом и имеющий другие закономерности. Однако это противоречит
тому, что большинство просто организованных животных размно-
жается бесполым путем.
При бесполом размножении дезинтеграция может затрагивать
весь организм или происходить локально. Паратомия v Oligochaeta
состоит в том, что в результате каких-то внутренних процессов
(без повреждения внешними факторами) и определенной части те-
ла появляется зона деления. В этой зоне происходит частичный не-
кроз клеток, изменяются клетки наружного эпителия, эпителия ки-
шечника, клетки мезодермального происхождения, происходит де-
Дифференциация эктодермальных и мускульных клеток. Прежние
структуры тканей в зоне деления радикально изменяются.
П. П. Иванов, в 1903 и 1907 гг. тщательно изучавший регенерацию
и бесполое размножение у Lumbriculus и других олигохет, сделал
заключение: при делении одной особи на две организм должен
пройти момент временного нарушения нормальных физиологиче-
ских связей между частями тела, а этот момент сопровождается
отмиранием одних клеток, дедифференцировкой других. Таким об-
разом происходит локальная или относительно локальная дезин-
теграция тканевых систем.
377
Вероятно, состояние местной дезинтеграции может наступать и?
при тех или иных формах почкования, например при почковании
гидры. Б. П. Токин (1959) предполагает, что в зоне почкования
нарушаются нормальные морфофизиологические взаимоотношения
между клетками в связи с процессами пищеварения. Особенно оче-
видная дезинтеграция или дезорганизация тела животного, сопро-
вождающая бесполое размножение, на примере губок. При гемму-
лообразовании большая часть тела губок распадается и без того-
мало интегрированные организмы нацело теряют свою нормальную
организацию.
О. М. Иванова-Казас (1970, 1976) говорит о морфофизиологи-
ческих причинах наступления бесполого размножения и лишь в
отношении некоторых случаев находит возможным применить тер-
мин «дезинтеграция». Действительно, далеко не всегда имеют ме-
сто видимые процессы, предшествующие бесполому размножению,,
которые можно было бы обозначить словом «дезинтеграция» тка-
ней. Одиако при любом типе бесполого размножения требуется вы-
яснить, как и почему наступает развитие новой особи из клеток,,
принадлежащих до момента почкования или деления материнско-
му организму.
Почкование асцидии Clavelina связано с явлениями местной
(летом) или общей (осенью) дезинтеграции; новый организм стро-
ится заново из соматических клеток; никакой преемственности
между полярностью материнского организма и таковой почки нет.
Многие асцидии обладают способностью к «регенерации», точ-
нее — к соматическому эмбриогенезу. Экспериментаторы ие раз;
убеждались, что практически любой участок тела асцидий может
дать целый организм и что развитие новых особей может происхо-
дить из участков тела, лишенных производных того или иного за-
родышевого листка или даже состоящих из клеток только одного-
зародышевого листка.
В заключение следует отметить, что, вероятно, всегда развитие
новой особи в результате бесполого размножения происходит не из
одной соматической клетки, а из групп клеток, которые можно
представить себе как подобие зародышей на стадии бластулы.
Впрочем, вопрос этот еще далек От ясности. Так, некоторые иссле-
дователи предполагают, например, что клетки почек медуз явля-
ются продуктом деления одной омоложенной клетки (Б. Брэм,
1908, 1911).
КЛАССИФИКАЦИЯ РЕГЕНЕРАЦИОННЫХ ЯВЛЕНИЙ, ПОНЯТИЕ
О СОМАТИЧЕСКОМ ЭМБРИОГЕНЕЗЕ
Следует отличать две группы явлений; собственно регенерацию и
соматический эмбриогенез. Регенерация — восстановление орга-
низмами утраченных частей: клеток, тканей, частей органов или
целых органов и систем. Таким образом, регенерацией можно на-
378
звать, например, восстановление части удаленной конечности три-
тона, удаленного участка тела гидры или инфузории, заживление
кожной раны у млекопитающих и т. п. Если бы часть отрезанного
листа растения могла восстановиться, это было бы регенерацией.
Соматический эмбриогенез — экспериментально вызванное раз-
витие новых организмов из отдельных соматических клеток или из
комплексов их. Эти явления имеют совершенно иные, чем при ре-
генерации, генезис и характер формообразования. При соматиче-
ском эмбриогенезе теряется прежняя индивидуальность, поляр-
ность, симметрия. Необходимость разграничения процессов реге-
нерации сознавалась многими учеными, в связи с чем предполага-
лись различные названия: «морфаллаксис», «реституция», «муль-
типолярные формы» и т. п.
Процессы регенерации в свою очередь нуждаются в классифи-
кации. Различают физиологическую и репаративную регенерацию.
Физиологическая регенерация — это процессы обновления
функционирующих тканей и органов, как-то: замена слущиваю-
щихся поверхностных слоев кожи новыми, регенерация эпителия
матки в связи с менструацией, смена перьев или шерстного покро-
ва и т. п.
Репаративной регенерацией называют процессы восстановления
частей тела, происходящие после различного рода повреждений.
Внутренние, а также некоторые наружные органы регенерируют по
типу регенерационной и компенсаторной гипертрофии. Регенераци-
онная гипертрофия — это процесс морфофункциональной пере-
стройки поврежденного или патологически измененного непарного
органа. В результате этого процесса увеличиваются объем и мас-
са органа без нарастания тканей на раневой поверхности. Исход-
ная форма органа при этом не восстанавливается. Компенсатор-
ной гипертрофией называют морфофизиологические перестройки
интактного парного органа (например, почки, семенника) после
удаления или повреждения другого.
Процессы соматического эмбриогенеза близки к некоторым
формам бесполого размножения. Экспериментально легче вызвать
соматический эмбриогенез у менее интегрированных организмов,
у тех, которым свойственно бесполое размножение. Соматическому
эмбриогенезу обязательно предшествует нарушение организменной
интеграции или появление участка тела, в котором часть клеток
оказывается изолированной от интеграционных влияний целого1.
Это обязательная основная биологическая предпосылка возможно-
сти перевода соматических клеток на путь развития целых орга-
низмов.
Несмотря на далеко зашедшую дифференциацию, способные
к размножению соматические клетки сохраняют свои видовые
1 Уточненная классификация восстановленных морфогенезов дана в статье
Коротковой Г. П. и Токина Б. П. О закономерностях эволюции востановитель-
ных морфогенезов // Биологические науки. 1979. № 11. С. 5—17.
379
свойства, в частности пол
неценный хромосомный ап
парат. В связи с делениям!
клетки частично дедиффе-
ренцируются, становятся
более лабильными в формо-
образовательном отноше-
нии. Если они (при дезинте-
грации тканей) не входят в
нормальное формообразова-
тельное русло тканевых за-
кономерностей, то могут
проявить свойственную им
потенцию развития целого
организма, дать начало про-
цессам . соматического эм-
бриогенеза (см. гл. II и XI).
Однако это может произой-
ти только при условии фи-
зиологической изоляции.
Процессы регенерации»
т. е. процессы развития не-
достающих частей организ-
Рис. 150. Образование побегов иа листе
Begonia rex, не отделенном от материн-
ского растения (по X. Гирфановой и
Б. П. Т окину, 1942)-.
А — лист, превращенный в решето; Б — образо-
вание побегов
ма, процессы типического строительства специализированных тка-
ней или органов как интегральных частей организма, напротив, не
могут нормально осуществляться, если организм или остаток орга-
на окажутся дезинтегрированными, чем-то хаотическим.
Регенерация и соматический эмбриогенез у растений. Растения
ио сравнению с животными — менее интегрированные организмы.
Им широко свойственно бесполое размножение и легче экспери-
ментально вызвать соматический эмбриогенез.
X. Н. Гирфанова и Б. П. Токин (1942) провели следующие экс-
перименты. На одном или двух (немолодых) листьях Begonia, rex
металлической трубочкой делаются близкие друг другу многочис-
ленные вырезы диаметром 3—5 мм, тем самым лист превращается
в решето (рис. 150,Л). При благоприятных температурных усло-
виях и условиях влажности по краям вырезов могут появляться
новые побеги (рис. 150,Б).
Морфогенетические процессы, сопровождающие соматический
эмбриогенез у растений, изучены крайне недостаточно. В большин-
стве случаев новый организм начинает развиваться не из одной,
а из группы телесных клеток. Однако возможно развйтие целых
организмов и из одной клетки. Примером могут служить резуль-
таты опытов Ф. Ц. Стюорда (1957), Р. Г. Бутенко (1964) и других
ботаников. В их работах даются иллюстрации различных этапов
культивирования вне организма клеток моркови, табака и
других растений. На рис. 151 мы видим, что при культивирова-
380
нии на специально приго-
товляемой агаровой сре-
де из единичных свобод-
ных клеток тканей мор-
кови возникает зароды-
шеподобная структура
(Д), которая может раз-
виваться в целое расте-
ние (Б). Схематически
.процесс развития из еди-
ничных клеток целого рас-
тения изображен на рис.
151,В.
Благодаря сходству
развития соматического
зародыша в условиях
культуры изолированных
клеток и эмбриона того,
же вида растения, обра-
зующегося при половой
репродукции, было пред-
ложено называть эту раз-
новидность соматического
эмбриогенеза — эмбрио-
идогенез (по Т. Б. Баты-
гиной и др., 1978).
Развившийся зародыш
можно изолировать и пе-
ренести в среду, благо-
приятную для выращива-
ния. Формируется проро-
сток, который можно пе-
ресадить в почву, он раз-
вивается далее в полно-
ценное растение, которое
дает цветы и образует
семена.
Восстановление типи-
чески организованных ча-
стей на раневой поверх-
ности, как правило, у ра-
стений не происходит.
Они либо сводятся к за-
Рис. 151. Соматический эмбриогенез у
растений (по Р. Г. Бутенко, 1964):
А -1 образование зародышеобразной структуры из
отдельной клетки моркови иа агаровой среде;
Б— развившееся из иее целое растение; В — схе-
ма культивирования тканей моркови и процесса
соматического эмбриогенеза; / — исходный кор-
неплод моркови, 2 — получение каллуса иа ку-
сочке ткани корнеплода, 3 — пассированная
культура; 4 — культура клеточных суспензий, 5,
6 — этапы соматического эмбриогенеза, 7— за-
родышеобразиые структуры, полученные из от-
дельных клеток, 8 -- развитие целого растения
щитным процессам («заживление» ран, формирование пробковой
ткани и т. п.), либо происходят компенсаторные замещения отсут-
ствующих частей за счет активизации спящих почек или меристе-
матических очагов. На раневых поверхностях растений часто обра-
381
зуется слабо упорядоченная каллусная ткань, за счет которой
могут формироваться новые побеги или корни.
Регенерация и соматический эмбриогенез у губок. X. Вильсон
в 1907 г. обнаружил сенсационное для того времени явление —
развитие губок из комочков соматических клеток, полученных пос-
ле продавливания тела губок через плотную мелкоячеистую ткань.
Позднее этот процесс наблюдали у других видов губок К. Мюл-
лер (1911) и Дж. Гексли (1923). Это дало повод говорить об ис-
ключительной регенерационной способности просто организован-
ных многоклеточных животных. Сам процесс развития губок после
диссоциации называли «рекомбинацией», «реституцией», «конъюнк-
цией» и др. Много неясного оставалось в отношении участия различ-
ных типов клеток как в образовании ранних конгломератов, так и
в процессах развития дефинитивных структур. Согласно классифи-
кации, предложенной Б. П. Токиным, процесс развития новых гу-
бок после их диссоциации был назван наряду с другими аналогич-
ными морфогенезами соматическим эмбриогенезом. В результате
обширных исследований Г. П. Коротковой оказалось, что восста-
новительные морфогенезы, протекающие у губок по типу сомати-
ческого эмбриогенеза, очень разнообразны. Менее интегрированные
многооскулумные губки при одном и том же способе опера-
тивного вмешательства отвечают на повреждение морфогенетиче-
ской реакцией, идущей по типу развития новых особей или коло-
ний и у них в меньшей мере наблюдаются собственно регенераци-
онные явления (восстановление утраченных частей при сохранении
исходной индивидуальности). У наиболее интегрированных (одно-
оскулумных) губок регенерационные процессы, наоборот, превали-
руют над соматическим эмбриогенезом. Г. П. Короткова полагает,
что клеточные механизмы осуществления того или иного вида ре-
генерации и соматического эмбриогенеза определяются не столько
уровнем организменной (или колониальной) интеграции, сколько
анатомическими и гистологическими особенностями организации
губок, водоносная система которых может быть устроена по типу
аскон, сикон или лейкон.
Если разрезать в продольном направлении представителей двух
видов Sycon (S. lingua, S. raphanus), из которых первый — потен-
циальная колония, а второй — одиночная губка, то их реакция на
повреждение будет существенно различной. «Половинки» и «чет-
вертушки» потенциальных колоний развиваются так, что старый
оскулум утрачивает свою функцию, частично подвергается деструк-
ции и зарастает регенерационной тканью одновременно со сраста-
нием краев раны. Новый оскулум разовьется как новообразование
в средней части оперированной губки. У этого же вида губки легко
можно получить развитие новых особей из небольших фрагментов
тела (рис. 152,А). Все эти разновидности восстановительного мор-
фогенеза относятся также к соматическому эмбриогенезу, так как
происходит развитие новой индивидуальности, о чем свидетельст-
382
Рис. 152. Восстановительные процессы у губок (по Г. П. Коротковой, 1963)’:
А — развитие Sycon lingua из небольших (около 2 мм) кусочков стенки тела спустя три
недели после операции; Б—регеиерация у S. raphanus, половинки губки на 15-й день после
операции; произошло срастание краев раны на значительном протяжении; В — формирова-
ние новых оскулумов у S. lingua
вует закладка новой морфологической оси и нового оскулума. Чер-
ты старой индивидуальности в ходе развития утрачиваются.
При таком же типе операции на S. raphanus (однооскулумная
губка) происходит изгибание боковых стенок тела, между краями
раны образуется регенерационная ткань, закрывающая раневое от-
верстие. Через 3—4 недели тело губки снова принимает цилиндри-
ческую форму, полярность не изменяется, новые оскулумы не воз-
никают. Можно говорить в этом случае о регенерации (рис. 152,Б).
Если вырезать кусочки 5. lingua из срединного участка тела, то<
совершаются преобразования, которые приводят к формированию
двух оскулумов (рис. 152,В).
У колониальной кремнероговой губки Halichondria panicea лег-
ко экспериментальным путем вызвать извращение полярности всей
колонии и добиться формирования новых оскулумов на бывшей
базальной стороне. Соматический эмбриогенез можно вызвать,
точечными ожогами, укалыванием иглой, т. е. в результате местной
дезинтеграции клеточных систем. Губкам свойственна и регенера-
ция, особенно более интегрированным формам. У Halichondria
panicea (слабо интегрированная многооскулумная губка) регене-
рационные процессы выражены слабо. При удалении части оску-
38а
лярного возвышения полноценное восстановление не завершается
даже через 2 месяца, а в 10—15% случаев губка вообще не реге-
нерирует. Хорошо выраженная регенерация наблюдается у более
интегрированных губок, например у Sycon, raphanus. Наконец, от-
метим очень интересный факт: если тому или иному виду губок
свойственны и регенерация и соматический эмбриогенез, экспери-
ментатор может по своему желанию увеличить или уменьшить
«удельный вес» той или другой реакции. Если желательно полу-
чить больший процент явлений соматического эмбриогенеза, то
нужно усилить дезинтеграцию оперируемого животного. Этого
можно добиться у низших многоклеточных животных очень простым
способом — наносить добавочные травмы, изолировать от целого
организма небольшие фрагменты или диссоциировать ткани до со-
стояния .клеточных взвесей. Если же требуется получить больший
процент случаев регенерации, то необходимо осуществить опера-
цию таким образом, чтобы не повредить функционирование
водоносной системы, не нарушить целостность, т. е. наносить незна-
чительные повреждения. Конечно, у некоторых губок исходная по-
лярность и функция водоносной системы сохраняются и при пере-
резке животного в продольном или поперечном направлении (одно-
оскулумные губки).
Возвращаясь к анализу развития губок из конгломератов кле-
ток после их диссоциации, необходимо отметить, что этот процесс
•ошибочно рассматривают некоторые исследователи как самосбор-
ку. В действительности всегда происходит гибель части клеток и
трансформация либо амебоцитов, либо хоаноцитов в другие клеточ-
ные типы. Это было убедительно показано в экспериментах с кон-
гломератами, составленными из клеток одного типа, например, из
архоцитов у пресноводных губок (Н. С. Никитин, 1974). Эти клет-
ки способны трансформироваться в пинакоциты, склеробласты,
хоаноциты и другие специализированные клеточные типы. В экспе-
риментах с маркировкой хоаноцитов и других типов клеток у мор-
ской губки Halisarca М. А. Волкова (1981) получила доказа-
тельства трансформации хоаноцитов в пинакоциты и амебо-
циты.
Явления клеточных трансформаций наблюдались и при разви-
тии известковых губок из небольших фрагментов тела. На рис. 153
показана трансформация хоаноцитов в пинакоциты, которая совер-
шается в составе эпителиоидного пласта при реконструкции водо-
носной системы у Sycon lingua в ходе соматического эмбриоге-
неза.
Получены многие доказательства того, что вещества, способст-
вующие регенерации, тормозят процессы соматического эмбриоге-
неза и наоборот. Так, 0-меркаптоэтаниол тормозит восстановление
удаленного оскулума у пресноводных губок, но одновременно не
мешает реализации соматического эмбриогенеза (А. Н. Суходоль-
ская, 1974).
384
Рис. 153. Различные стадии преобра-
зования хоаноцитов в уплощенные
клетки (по Г. П. Коротковой с соавт.,
„1965)
Регенерация и соматический
эмбриогенез у кишечнополостных
и ктенофор. В опытах с кишечно-
полостными можно констатиро-
вать принципиально те же зако-
номерности, которые обнаружены
у губок (Б. П. Токин, Д. Г. Пол-
тева и др.).
Кишечнополостные в целом от-
личаются хорошей регенерацион-
ной способностью. По мере ус-
ложнения организации кишечно-
полостных в ряду Hydrozoa-^-
-^Scyphozoa-^Anthozoa регенера-
ционные явления, как правило,
не затухают. У высших предста-
вителей типа они выражены в
превосходной степени. Соматиче-
ский эмбриогенез, наоборот, лег-
ко удается вызвать у низших ки-
шечнополостных. Что же касает-
ся ктенофор, то соматический
эмбриогенез, по-видимому, возможен только у аберрантных
форм из отряда Platyctenidea, которым свойственно бесполое
размножение, о чем можно судить по опытам И. Окада (1932),
X. Танака (1931), Г. Фримена (1967).
Соматический эмбриогенез не свойствен многочисленным пе-
лагическим гребневикам, размножающимся исключительно поло-
вым способом, в частности Вегоё и Bolinopsis, характеризующимся
высокой регенерационной способностью.
На рис. 154 даны схемы операций, проведенных на гребневиках
Вегое cucumis (А) и Bolinopsis infundibulum (Б). При всех типах
операций происходила полноценная регенерация.
И. В. Пылило провела наблюдения за регенерацией взрослых
и личинок ктенофор. Она впервые изучила и гистогенетические
процессы, сопровождающие регенерацию ктенофор. На
рис. 155,А, Б видна регенерация полностью и частично удаленного
гребного ряда Bolinopsis infundibulum.
Пресноводным гидрам свойственна регенерация (заживление
ран, регенерация гипостома и подошвы и т. п.). Кроме того, гидры
могут развиваться по типу соматического эмбриогенеза. Если про-
тирать Hydra oligactis сквозь шелковую ткань с отверстиями око-
ло 0,02 мм2, то, как и в опытах с губками, может образоваться кон-
гломерат соматических клеток, из которого разовьется целая
гидра. Фрагменты тела гидры могут регенерировать гипостом со
щупальцами и стебелек, но могут развиваться в целую гидру и по
типу соматического эмбриогенеза. Это зависит от того, подвергает-
385
Рис. 154. Схема операций на Вегоё cucumis (А) и Bolinopsis infundibulum (Б)
(по Б. П. Токину, 1962):
линиями (1—6) указаны типы разрезов в разных опытах. Объяснения в тексте.
Рис. 155. Регенерация гребного ряда Boli-
nopsis infundibulum (по И. В. Пылило,
1973):
А — регенерация части удаленного гребного ряда;
Б — регенерация полностью удаленного гребного
ряда
ся ли отрезанная часть те-
ла радикальной реоргани-
зации (без чего не может
произойти соматический
эмбриогенез). Стебелек
Н. oligactis считался
частью тела, не способной
развиться в целую гидру.
Б. П. Токину в 1934 г.
удалось, дезинтегрируя
отрезанные стебельки
(центрифугирование, мно-
гократное укалывание иг-
лой и т. п.), наблюдать
и из стебельков развитие
целых гидр (рис. 156,Л).
М. Я. Теплякова в 1937 г.
и независимо от нее
Г. С. Стрелин в том же
386
Рис. 156. Соматический эмбриогенез у гидры:
Л — развитие гидры из сращенных стебельков (по Б. П. Токниу, 1934); Б, В — развитие
почки на месте точечного ожога (по М. Я. Тепляковой, 1937); Г—Е — схема операции (по
Б. П. Токнну, 1958); Ж, 3 — уродливое развитие многих гидр из куска гастрального отдела
году вызвали соматический эмбриогенез путем точечных ожо-
гов в таких участках тела гидры (ближе к гипостому), где никогда
в норме не происходит почкование (рис. 156,В), и в результате
иных оперативных вмешательств (рис. 156,Г, Д, Е, Ж, 3).
Д. Г. Полтева (1965) вырезала полоску по окружности подо-
швы актинии Metridium senile fimbriatum (шириной 0,3—0,5 см),
в состав которой входили ткани подошвы и стенки колонки с участ-
ками оснований перегородок. Через неделю после операции края
разреза соединялись и срастались по всей длине полоски. Свобод-
ные концы закручивались внутрь и частично срастались с прилежа-
щими участками полоски. В этих местах формировались две акти-
нии со щупальцами (рис. 157,Д). На «стебельке», соединяющем
две новообразованные актинии, в дальнейшем наблюдалось иногда
Рис. 157. Соматический эмбриогенез у
актиний:
А—Г — различные случаи множественного
формирования оральных участков у Нагепас-
tis attenuata; объяснение в тексте (по
Ч. Чайлду, 1909, 1910); Д — две актнннн, раз-
вившиеся иа концах полоски, вырезанной из
педального диска Metridium. senile fimbriatum
(по Д. Г. Полтевой, 1965)
387
формирование дополнительных маленьких актиний. Порядок за-
кладки септ, щупалец, глотки и другие особенности этого сомати-
ческого эмбриогенеза были сходны с половым эмбриогенезом.
Регенерационные процессы у полипов, медуз и актиний иссле-
дуются во многих лабораториях. Установлено, что эти животные
обладают способностью восстанавливать значительные отделы те-
ла по типу регенерации. Кроме того, наблюдаются регенерацион-
ные способности и у колониальных форм. Так, например, колонии
гидроидных способны восстановить стелющуюся часть колонии, что
свидетельствует о наличии у них высокой колониальной интегра-
ции, доказанной также в специальных экспериментах Н. Н. Мар-
фениным.
Регенерация и соматический эмбриогенез у червей, асцидий и
других животных. Много исследований выполнено на планариях.
У некоторых видов '/зе часть всей длины червя и даже '/soo способ-
на развиваться в целый организм. Существует еще много неясных
вопросов, касающихся оценки результатов отдельных эксперимен-
тов и механизмов восстановительных морфогенезов. Не случайно,
что в экспериментах на планариях исследователи часто констати-
ровали гетероморфозы, развитие планарий-монстров и другие
«уродства» (рис. 158), которые следует рассматривать как непол-
ноценно протекающие явления соматического эмбриогенеза (Кри-
чинская, 1978).
Интересные факты обнаружила В. В. Исаева (1968) в исследо-
ваниях восстановительных процессов у планарии Dugesia tigrina
Girard (бесполая раса). При действии на планарий «версена»
(влияющего на межклеточные связи и способного диссоциировать
ткани на клетки), а также канцерогенных веществ провоцируется
Рис. 158. Гетероморфозы у плаиарий
(из ки. Е. Коршельта, 1927):
А—Е — различные случаи множественного
формирования головных и отчасти туловищ-
ных частей тела планарий в результате раз-
ных операций
Рис. 159. Гетероморфоз у пла-
иарии Dugesia tigrina, вызван-
ный p-меркаптоэтанолом (по
Б. П. Токину и Т. Н. Борисо-
вой, 1975)
388
соматический эмбриогенез. Б. П. Токин и Т. Н. Борисова (1975),
используя ингибитор регенерации p-меркаптоэтанол, а также кол-
хицин, вызывали соматический эмбриогенез у планарии Dugesia
tigrina (рис. 159).
Представляет интерес выяснение причины значительных коле-
баний регенерационных способностей, выявленных у представите-
лей разных родов планарий (Т. Лендер, 1965). Существуют плана-
рии, практически не способные восстанавливать утраченные части
тела, тогда как другие регенерируют передние и задние отделы
тела после их разрезания на разных уровнях тела по переднезад-
ней оси. Аналогичные наблюдения имеются и в отношении немер-
тин (В. Ку, 1930), кольчатых червей (О. Хес, 1963) и других бес-
позвоночных животных.
Известный, бывший когда-то сенсационным, пример соматиче-
ского эмбриогенеза дают асцидии. Это особенно интересно потому,
что асцндии занимают в эволюционном ряду высокое место. По
сравнению с другими хордовыми они, однако, примитивны в своей
организации, особенно взрослые животные. При метаморфозе ли-
чинок происходит упрощение нервной системы, исчезают глаза и
статоцист, атрофируется хвост вместе с хордой и т. п. Им свойст-
венно бесполое размножение и на этой основе — ярко выраженный
соматический эмбриогенез. Г. Дриш, один из первых открывший и
описавший это явление — явление «реституции», показал, что вы-
резанные в любом месте кусочки жаберной корзины или столона
Clavellina могут развиться в целую асцидию. Вырезанные кусочки
сначала превращаются в комок кажущихся однородными клеток,
происходит радикальная морфофизиологическая перестройка ис-
ходных тканей.
Не умножая примеры сомати-
ческого эмбриогенеза у разных
животных, отметим, что может
возникать соматический эмбрио-
генез и близкие к нему явления и
у более сложных организмов при
определенном их физиологическом
состоянии и на определенных эта-
пах онтогенеза. К этим явлени-
ям близко примыкают процессы
развития целых организмов из
изолированных бластомеров, экс-
периментальная полиэмбриония
у амфибий и птиц при действии
«мертвых организаторов» (см.
гл. XI) и др. Тератомы, или эм-
бриомы, у млекопитающих (опу-
холи, состоящие нередко из про-
изводных всех зародышевых лист-
Рис. 160. Инфузория-монстр, полу-
ченная в результате ианесеиия много-
кратных надрезов (по Калкинсу,
1911)
389
ков), может быть, следует рассматривать также в аспекте гипотез
о соматическом эмбриогенезе.
В заключение следует вспомнить Protozoa, организменность
многих из которых очень высока. Им широко свойственны процес-
сы регенерации, но в жизни простейших имеют место явления, ко-
торые, вероятно, следует уподобить соматическому эмбриогенезу.
У Stentor */б4 часть тела, у Dileptus и Dldinium — 1/75, а у Noctilu-
са miliaris — еще меньшего размера части могут развиваться в це-
лый организм. Ряду видов Protozoa свойственно наружное или
внутреннее почкование, в том числе множественное почкование
(например, до 24 почек у солнечников) и другие способы бесполого
размножения. На этой основе возможны явления соматического
эмбриогенеза при освобождении какого-либо участка от форма-
тивных влияний всего организма (рис. 160).
Регенерация и соматический эмбриогенез у позвоночных живот-
ных. Регенерации позвоночных животных уделено много внимания,
особенно в связи с обнаружением прекрасных регенерационных
возможностей печени, костной мышечной ткани, о чем будет гово-
риться ниже. Гораздо меньше внимания уделялось явлениям, близ-
ким к соматическому эмбриогенезу. Как уже говорилось, полиэм-
бриония свойственна и позвоночным животным. Тема об экспери-
ментальном вызывании полиэмбрионии не нова, однако она приоб-
ретает особый аспект в связи с теорией соматического эмбриогене-
за. Иллюстрацией этих исследований могут служить работы
Г. П. Святогора и Л. Н. Святогор. Исследования Г. П. Святогора
(1972—1974) по экспериментальной полиэмбрионии у птиц под-
твердили данные Э. Вольфа, Г. Лутца (1947) и других о возмож-
ности вызывания начальных стадий развития нескольких зароды-
шей из одного бластодиска птиц. На яйцах ряда птиц — перепела
Coturnix coturnix, курицы Gollas gallus, индейки Meleagris gallo-
pavo, гуся Anser anser —- в результате разрезания бластодиска
in ovo вызвано развитием нескольких эмбрионов (рис. 161).
В экспериментах Л. Н. Святогор и Г. П. Святогора по влиянию
ненормального положения дробящегося яйца лягушки Rana tem-
poraria в гравитационном поле выявлена возможность около 25%
случаев развития близнецов и около 40% зародышей с различны-
ми выростами на теле. В результате центрифугирования зароды-
шей лягушки R. temporaria на стадиях поздней бластулы — ран-
ней гаструлы при 600g в течение 10 мин наблюдается развитие
близнецов и зародышей с выростами на теле. Авторы вызывали
экспериментальную полиэмбрионию у травяной лягушки, перево-
рачивая дробящееся яйцо, т. е. создавая ненормальное положение
его в гравитационном поле. Процент развития близнецов был зна-
чителен— у 25% всех подопытных яиц. Те же исследователи вы-
зывали полиэмбрионию у травяной лягушки центрифугированием
яиц — при 600g в течение 10 мин. Нередко возникают выросты,
кажущиеся лишь локальным уродством. Однако гистологическое
390
Рис. 161. Экспериментальная полиэмбриония у птиц (по Г. П. Святогору, 1974):
А — близнецы индейки в опытах разрезания бластодиска на четыре части; Б — близнецы
курицы; В — трн зародыша утки в опытах разрезания бластодиска иа две части; Г —
близнецы перепела в опытах разрезания бластодиска иа две части
исследование выявило дифференциацию зачатков — производных
всех трех зародышевых листков.
Основное отличие регенерации от соматического эмбриогенеза.
Затухает ли регенерационная способность в ходе эволюции орга-
низмов? Неоднократно подчеркивалось, что соматическому эмбрио-
генезу обязательно предшествует дезинтеграция нормально корре-
лированных клеточных систем, тканей, органов и организма в це-
лом. Наоборот, процессы развития недостающих частей организ-
ма, процессы типического строительства специализированных тка-
ней или органов как интегральных частей организма не могут нор-
мально осуществляться, если организм или остаток органа окажут-
ся дезинтегрированными, анархическими, хаотичными. Что являет-
ся самым характерным для явления соматического эмбриогенеза?
Соматический эмбриогенез — это такая разновидность морфогене-
тической реакции на повреждения, в ходе которой происходит фор-
мирование морфологической оси нового организма (или несколь-
ких морфологических осей), а морфогенетические процессы выхо-
дят из-под контроля старой индивидуальности. Соматические клет-
ки способны в этих условиях стать исходными в развитии нового
индивидуума. К соматическому эмбриогенезу относятся: развитие
391
новых особей из комплексов соматических клеток после диссоциа-
ции тканей; развитие новых организмов из фрагментов тканей, ор-
ганов или организма; развитие организмов из участков личиноч-
ных или эмбриональных тканей, изолированных от развивающего-
ся организма и др.
Восстановительные процессы, вызванные повреждением, в той
или иной форме и степени свойственны абсолютно всем организ-
мам. У всех организмов в той или иной форме наблюдаются явле-
ния физиологической регенерации, совершающиеся на тканевом,
клеточном и субмикроскопическом уровнях. Обмен веществ с дис-
симиляцией и ассимиляцией — это не только химический, но и
структурный процесс. При диссимиляции окисляются не какие-либо
чужеродные включения в клетку, а составные ее части. Происхо-
дят непрерывные разрушения структур и регенерация их в связи
с обменом веществ. Это один из видов физиологической регенера-
ции, которая в ходе эволюции организмов совершенствовалась,
видоизменялась, приобретая своеобразные черты у разных живот-
ных в связи с особенностями их организации. Самообновление кле-
ток и тканей имеет место, вероятно, у всех животных. Кажущимся
исключением являются коловратки и нематоды, которым как буд-
то свойственно постоянное число клеток. Неизвестно, что дадут
специальные исследования регенерации этих животных, но бесспор-
но, что и им свойственная регенерация на субмикроскопическом
уровне, а также восстановление внутренних органов в условиях
неповрежденной кутикулы (Г. П. Короткова, Л. А. Агафонова,
1976).
В отношении регенерации можно установить принципиально
иную закономерность, чем для соматического эмбриогенеза. Мень-
шую регенерационную способность следует ожидать у слабо интег-
рированных организмов. Вместе с усложнением организации жи-
вотных в ходе эволюции не происходило затухание способности
к регенерации. Если говорить об основной тенденции, регенера-
ционная способность, будучи полезным свойством, подвергаясь дей-
ствию естественного отбора, усиливалась; приобретала своеобраз-
ный характер у разных животных. Это не означает, что совершен-
ствование физиологической и репаративной регенерации происхо-
дило прямолинейно. Эволюция регенерации происходила коррели-
рованно с другими явлениями, от которых зависят благополучие
и прогресс вида: иммунитет, особенности размножения и т. д.
Конечно, каждую точку зрения можно довести до абсурда. Ре-
генерационные способности млекопитающих и человека далеки от
совершенства не только с узко-утилитарной, медицинской точки
зрения, но и в сравнительно зоологическом плане. Любое млеко-
питающее не может идти ни в какое сравнение с хвостатыми ам-
фибиями, обладающими превосходной способностью восстанавли-
вать конечности. А когда констатируется большая возможность ре-
генерации тех или иных тканевых систем у млекопитающих, не сле-
392
дует забывать, что, например, при регенерации кожи с образова-
нием рубцовой ткани восстановление утраченного явно неполно-
ценное.
Регенерационные явления у человека изучались преимуществен-
но медиками, а не биологами. В 40—60-е годы проведены биоло-
гические исследования явлений регенерации у млекопитающих.
Особенно значительны работы школы М. А. Воронцовой и Л. Д. Ли-
ознера. Считалось, что у позвоночных животных, за исключением
хвостатых амфибий, регенерация выражена в незначительной сте-
пени. В свете новейших данных и изложенной теории соматическо-
го эмбриогенеза и регенерации привычные взгляды следует при-
знать ошибочными. Недоразумением является, в частности, утверж-
дение, будто птицам и млекопитающим свойственна регенерация
только тканей, а не органов. Л. Д. Лиознер в 1962 г. отмечал, что
ткани — части органов, и, если происходит восстановление повреж-
денных тканей как частей органов, нельзя говорить о гистогенети-
ческих процессах, происходящих при регенерации органа, как
о чем-то изолированном от них.
У млекопитающих, как правило, имеется хорошо выраженная
регенерация кожи, эпителия стенки кишечного тракта, матки, ро-
говицы глаза, печени, части яичников, почек, селезенки, легких,
эндокринных желез и других органов. Абсолютно всем тканям
млекопитающих и человека, включая и ткани нервной системы,
свойственна регенерация. Во всяком случае эта регенерация мо-
жет быть констатирована на клеточном уровне, в связи со струк-
турными процессами, сопровождающими обмен веществ. Некото-
рые восстановительные процессы у млекопитающих, по-видимому,
выражены в более яркой форме, чем у низших позвоночных.
Это относится к роговице глаза, тканям кишечного тракта
и др.
Большой интерес представляют исследования А. Н. Студит-
ского (.1954) по вылущиванию надкостницы бедренной и боль-
шеберцовой костей кроликов и собак. Происходит их полная ре-
генерация. За последние десятилетия были установлены факты
о механизмах регуляции регенерационного процесса со стороны
нервной и эндокринной систем, показано значение функциони-
рования иммунокомпетентных клеток в регенерационном процес-
се, выявлены разнообразные факторы регуляции роста тканей
и регуляции митотического цикла (см. А. Г. Бабаева, 1972;
В. Я. Бродский, И. В. Урываева, 1981; О. И. Епифанова, 1965).
Значение остатка органа при регенерации. Восстановление
клеткой, тканями, органом, организмом частей может совершать-
ся только при условии сохранения остатка данных систем, при-
чем сохранения не в хаотическом состоянии, а с совершенно типи-
ческими «замками» клеточных и тканевых корреляций.
Конечности тритона, аксолотля или головастика бесхвостых
амфибий, а также хвост этих организмов, ампутированные на
393
Рис. 162. Регенерация конечности сибирского четырехпалого тритона Hynobius
Keyserlinii (по А. И. Здруйковской, 1944)
А — схема операции; Б — тритон с регенерирующими конечностями: 1 — 4 — линии ампута-
ции, 5 — конечность, ампутированная после локальной рентгенизации с оставлением диска
необлучениых тканей, 6 — контрольная иерентгенизованиая конечность, ампутированная на
том же уровне. Заштрихованный участок правой конечности реитгеинзоваи
любом уровне, восстанавливают недостающее. При удалении поя-
са конечности или, в случае опытов с хвостом, при удалении его
на уровне клоаки регенерации не происходит.
Ряд экспериментов свидетельствует о том, что требуется со-
хранение остатка органа с его нормальной, а не анархизирован-
ной 'структурой. При облучении части бедра задней конечности
хвостатых амфибий, например 8000 Р (эксперимент А. И. Здруй-
ковской, 1944), а затем ампутации, произведенной по середине
облученного участка, регенерации не происходит, (рис. 162,3—4).
В случае удаления тех или иных тканей в остатке органа также
регенерация ухудшается или происходит атипично.
Развивается ли при регенерации каждая ткань от соответ-
ствующей предыдущей? Этот вопрос не решается однозначно для
всех животных, так как гистогенетические процессы у разных
организмов неодинаковы. Можно говорить, что для восстановле-
ния утраченных частей нередко характерен эпиморфоз, т. е. как
бы надставка недостающего. Однако в чистом виде эпиморфоз
никогда не встречается. Он сочетается чаще всего с морфал-
лаксисом — способом регенерации, предполагающим переплавку,
перегруппировку всех клеток и тканей, участвующих в восста-
новлении. Оба термина были предложены в начале нашего сто-
летия Т. Морганом. Так, например, при перерезании бедра ко-
нечности тритона процесс регенерации происходит не с восста-
новления недостающей части бедра, а затем голеии и стопы.
Происходит удивительное явление: сначала дифференцируются
дистальные части. В этом смысле регенерация конечности тоже
не «чистая» надставка.
Морфаллаксические перестройки бывают сильно выражены
при регенерации конечности у членистоногих. Этот механизм
регенерации присущ многим немертинам, планариям и кольча-
тым червям.
394
Несмотря на различие гистогенетических процессов, происхо-
дящих при регенерации у разных животных, можно все же отме-
тить важные общие черты, касающиеся поведения клеток. В пер-
вое время после травмы в районе регенерации неизбежны про-
цессы дезинтеграции, затрагивающие больший или меньший
участок тканей, наблюдается гибель клеточных элементов, воз-
можно инфицирование раны, имеет место фагоцитарная актив-
ность клеток, выполняющих «санитарную функцию» освобожде-
ния от, дегенерирующих клеток и чужеродных частиц, которые
могут оказаться в районе раны.
Как правило, после периода деструктивных процессов и вре-
менной утраты черт специализации клетками, прилегающими к
раневой поверхности, наступают явления редифференцировки,
т. е. возобновления у временно дедифференцированных клеток
черт исходной специализации. Наряду с этим в регенерации мо-
гут принимать участие и особые клетки, которые активируются
после травмы и мигрируют в область повреждения. Таковы
клетки-сателлиты в мышечной ткани у позвоночных животных;
необласты у червей, i-клетки у кишечнополостных и др.
ПРОИСХОЖДЕНИЕ КЛЕТОК ПРИ РЕГЕНЕРАЦИИ
Широкое распространение в различных областях биологии по-
лучили гипотезы о существовании среди дифференцированных
клеток взрослых организмов особых «тотипотентных», «эмбрио-
нального типа», «резервных», «вечно юных» и с другими назва-
ниями клеток, играющих роль в регенерационных процессах, при
развитии опухолей и т. д. Еще в 1887 г. М. Нуссбаум создал
представление об /-клетках гидры как тотипотентных, резервных
для регенерации. В 1892 г. X. Рандольф создала родственные
представления о резервных клетках у олигохет, назвав их нео-
бластами. Особенно энергично развивал гипотезу о резервных
клетках Ю. Шаксель (1914, 1915). Хотя действительно необласты
олигохет, i-клетки гидры или археоциты губок могут играть
очень важную роль в нормальных процессах регенерации и сома-
тического эмбриогенеза, гипотезы о «резервных клетках» как
единственных источниках клеток при регенерации оказались оши-
бочными, о чем говорилось еще в 1934 г. Б. П. Токиным.
Огромное число наблюдений проведено за необластами пла-
нарий. Эти крупные клетки с базофильной цитоплазмой, богатой
РНК, рассеяны по всему телу планарии, они оказываются в боль-
шом числе в районе раны и по всеобщему убеждению играют
важную роль при регенерации, образуют «регенерационную бла-
.стему». Споры о роли необластов происходят и в настоящее вре-
мя (Е. Б. Кричинская, 1980).
В настоящее время установлено, что кроме участия необла-
стов в образовании регенерационной почки они играют важную
395
роль в формировании гонад и железистых клеток кишечника,
т. е. обеспечивают физиологическое обновление. У планарий, раз-
множающихся бесполым путем, происходит накопление необла-
стов в зоне деления. В теле планарий необласты — единственные
клетки, способные к митотическому делению. Совокупность всех
этих фактов заставляет думать о существенной роли необластов
в восстановительных морфогенезах. Однако мнения исследовате-
лей относительно роли необластов расходятся. Так, например,
Р. Шандебуа (1960—1965) считает, что главную роль в регене-
рации играют фиксированные клетки паренхимы, а необласты
лишь накапливают и транспортируют в район регенерата РНК.
Вопрос об источниках необластов также далек от ясности.
Их характерная особенность — присутствие в цитоплазме хрома-
тоидных тел, которые по своей природе сходны с цитоплазма-
тичными маркерами первичнополовых клеток. В связи с этим
высказывается мнение о происхождении необластов за счет го-
ноцитов. В ряде работ доказывается возможность трансформации
секреторных клеток кишечника в необласты. Таким образом, во-
прос происхождения и роли этих клеток в морфогенезах остается
еще не решенным.
Дискуссия о роли интерстициальных клеток в регенерации
гидроидных имеет также длительную историю. Было получено
немало фактов как в пользу их важной роли в регенерации, так
и в пользу того, что регенерация может происходить и без их
участия. Наиболее убедительные данные об ограниченной роли
i-клеток в естественных и искусственно вызванных морфогенезах
были получены в 70—80-е годы нашего столетия1. При воздей-
ствии некоторыми веществами (колхицином, азотоипритом, окси-
мочевиной) гидры лишаются i-клеток и становятся состоящими
только из двух эпителиоидных платов эпидермиса и гастродер-
миса. Таких гидр называют «эпителиальными». В отсутствие
i-клеток у эпителиальных гидр не было нервных клеток, стрекаль-
ных клеток, гоноцитов. Они не способны были самостоятельно
питаться. Но если их искусственно кормить, то они начинали
почковаться и после удаления гипостома со щупальцами могли
их восстановить. Так было бесспорно установлено, что эпите-
лиоидные пласты тела гидры обеспечивают восстановление струк-
тур, в состав которых они входят. Однако эпителиальные гидры
не могли становиться половозрелыми, у них не появлялись нерв-
ные и стрекательные клетки. Значит, и у нормальных гидр в вос-
становлении утраченных структур принимают участие разные
типы клеток. Правда, в особых экспериментальных условиях
можно вызвать реализацию более широких формообразователь-
ных потенций у клеток, входящих в состав гастродермиса. Так,
1 Полтева Д. П. Дифференциация клеток и формирование плана строения
у гидры и гидроидов / / Онтогенез. 1983. Т. 14. № 2. С. 117—128.
396
например, при развитии гидры из фрагмента гастродермиса, ли-
шенного i-клеток, секреторные зимогенные клетки освобождаются
от секреторных гранул, уменьшаются в размерах и трансформи-
руются в i-клетки нового полипа.
Та или иная степень дедифференциации клеток в районе
травм, в районе нарушения нормальных корреляций между клет-
ками и тканями — важнейший процесс при регенерации всех жи-
вотных.
У амфибий эпителиальные клетки радужины могут превра-
щаться в клетки хрусталика. Клетки жаберного эпителия или
эпидермиса кожи могут превращаться в эпителий роговицы.
Экспериментально доказана возможность метапластических
превращений одной ткани глаза в другую (у позвоночных). Так,
возможна стимуляция метаплазии пигментного эпителия глаза в
сетчатку у головастиков, сеголеток и взрослых лягушек; напри-
мер, пигментный эпителий головастиков и сеголеток без мезен-
химных оболочек, помещенный в безлинзовый глаз головастика,
закономерно превращается в сетчатку (А. А. Сологуб, 1970).
При развитии немертин из предротовых фрагментов тела, не
содержащих производных энтодермы, возникает кишечник, зано-
во формируются половые железы. При регенерации олигохеты
Criodrilus из сегментов, не содержащих половых клеток, наряду
с другими системами заново формируются гонады. Стебелек асци-
дии Clavellina, которым она прикрепляется к столону, кишечника
не содержит. Между тем развивающаяся из этого кусочка асци-
дия имеет нормального устройства кишечник.
При травмировании тканей создаются условия для выхода
клеток из-под «замка» нормальных корреляций, а значит, и воз-
никает возможность появления относительно дедифференцирован-
ных и более лабильных в формообразовательном отношении кле-
ток. Конечно, не следует доводить эту мысль до абсурда, предпо-
лагая, что любая клетка любой ткани может превратиться в лю-
бую другую клетку любой другой ткани. Что при регенерации
происходит дедифференциация клеток и в то же время не имеет
места такого рода метаплазия, можно убедиться на примере реге-
нерации конечностей у амфибий.
Цитологическая картина регенерации начинает становиться
более ясной благодаря электронно-микроскопическим исследова-
ниям. Прежде всего подтверждены и уточнены процессы дедиф-
ференциации клеточных элементов, при этом значительные, что
дало повод Е. Хэй (1962) в шутливой форме сказать: «Ранняя ре-
генерационная бластема подобна быстро растущей тканевой куль-
туре, в которой мышечные и другие клеточные элементы, так ска-
зать, отказались от своей структуры и функциональной сложно-
сти, чтобы отдаться полностью удовольствию пролиферации».
В клетках бластемы видны большие ядра и отчетливые ядрышки,
397
происходит активный синтез белков и ДНК, рибонуклеопротеидов,
характерна базофилия цитоплазмы. При дедифференциации кле-
ток изменяется строение эргастоплазматической сети, комплекса
Гольджи. По мере редифференциации бластемных клеток на суб-
микроскопическом уровне происходят изменения в органеллах
клеток, в результате чего клетки приобретают типическую струк-
туру клеток данной ткани.
Прослежена дедифференцировка и редифференцировка хряще-
вых и мускульных клеток. И те и другие превращаются в клегки
бластемы; последние вместе с клетками, происходящими из дру-
гих соединительных тканей культи, формируют регенерационную
почку. Из клеток регенерационной почки формируются новые мус-
кульные и хрящевые клетки, что благодаря электронной микро-
скопии обнаруживается уже на 10-й день после ампутации; нали-
Рнс. 163. Изменения в тонкой структуре, которые
происходят в мезодермальных клетках бластемы;
их дифференциация в миобласты и мускульные
клетки, а также в первичные хрнщевые клетки и
хрящ. После ампутации конечности происходит
дедифференцнацня клеток (по Е. Хей, 1962):
/—скелетная мускулатура, 2 — хрящ, 3 — мезенхимные
клетки, 4 — миобласты, 5 — первичные хрящевые клетки
чие мускульных кле-
ток на 13-й день обна-
руживается и световой
микроскопией.
На рис. 163 дается
схема дедифференциа-
ции и редифференциа-
ции клеточных элемен-
тов скелетной мускула-
туры и хряща в ходе
регенерации конечно-
сти амфибий.
Созвучны с утверж-
дениями Е. Хэй резуль-
таты исследования
Т. Стин (1970) по ре-
генерации конечности
саламандр. Маркиро-
вались клетки регене-
ционной почки и транс-
плантировались на не-
обычное место или
культивировались in
vitro. Соединительно-
тканные клетки, в ча-
стности клетки хряща,
могут в определенных
условиях участвовать
как в образовании хря-
ща, так и мышечной
ткани или соединитель-
ной т. е. возможна
метаплазия тканей.
398
Следует, однако, обратить внимание на многие работы, осо-
бенно 80-х годов, в которых на основании электронно-микроско-
пических и иных наблюдений создаются новые гипотезы и утверж-
дения, идущие вразрез со всем сказанным о «резервных» клетках.
Некоторые исследователи не считают, что понятие «резервная
клетка» имеет лишь историческое значение. Созданному новому
понятию «стволовые клетки» придают нередко содержание, по-
добное тому, какое вкладывалось в понятие «резервные клетки».
Название «стволовые» присвоено клеткам эпителия кожи, нахо-
дящимся в нижнем, базальном слое кожи: на мембране, отделяю-
щей эпителий от соединительной ткани; в криптах стенки кишки,
а также исходным клеткам при кроветворении (в костном мозге,
селезенке). Стволовыми стали называть /-клетки гидры, необла-
сты олигохет, называвшиеся «резервными». Все перечисленные
клетки объединяют в группу «покоящихся клеток». Их отличает
то, что они вышли из репродуктивного цикла, но в то же время
сохранили жизнеспособность и потенции возобновить цикл деле-
ния и дифференцировки. «Покой» таких клеток не означает их ме-
таболической инертности (О. Е. Епифанова и соавт., 1983).
ЭЛЕКТРОННО-МИКРОСКОПИЧЕСКИЕ И ЦИТОХИМИЧЕСКИЕ
ИССЛЕДОВАНИЯ РЕГЕНЕРАЦИОННЫХ ПРОЦЕССОВ.
ВНУТРИКЛЕТОЧНАЯ РЕГЕНЕРАЦИЯ
В связи с использованием методов авторадиографии и электрон-
ной микроскопии оказалось возможным зарегистрировать восста-
новительные процессы, происходящие в ядре и цитоплазме клеток.
Д. С. Саркисов (1978) выделяет три формы внутриклеточной ре-
генерации (молекулярную, внутриорганоидную и органоидную) и
отмечает сходство проявления восстановительных процессов после
различных типов повреждений. Что касается репарации макро-
молекул, то в настоящее время интенсивно исследуются законо-
мерности репарации ДНК. В зависимости от характера повреж-
дений восстановительный процесс может осуществляться по-раз-
ному (Н. В. Томилин, 1977). Выявлены ферменты, участвующие
в репарации. Их набор уже достаточно велик и становится оче-
видным, что все последовательные этапы восстановительного про-
цесса от расчистки поврежденных или измененных участков ДНК
до их заполнения нормальными нуклеотидами выполняются при
участии ферментов. Эти же ферменты играют важную роль и в хо-
де нормальной жизнедеятельности клетки. Многие из ферментов,
участвующих в репарации ДНК, с успехом используют в области
генной инженерии.
Обширные электронно-микроскопические исследования измене-
ний клеточных структур в ходе восстановления клеток после иони-
зирующих повреждений проведены И. Б. Токиным (1974). Наибо-
лее интересные для цитоэмбриологии данные этих исследований
399
Рис. 164. Схема ультраструктурной организации клеток кишечного эпителия (по
И. Б. Токину, 1974):
Л — контроль; Б — при лучевом поврежденнн (дедифференцнрованное состояние); В — реге-
нерация (репарация)
таковы. Процессы организации клеток различных тканей и орга-
нов при нормальных и патологических состояниях своеобраз-
ны (рис. 164), однако найдены и достаточно общие закономерно-
сти дифференциации: происходит «.уменьшение относительных
объемов ядер и ядрышек и усложнение субмикроскопической ор-
ганизации, сопровождаемые количественными изменениями соот-
ношения «свободные рибосомы» — элементы гранулярной эндо-
плазматической сети и уровня удельной насыщенности цитоплаз-
мы мембранами органоидов»'.
Общей тенденцией при дифференциации является увеличение
числа митохондрий, повышение ферментативной активности. Про-
исходит усложнение комплекса Гольджи, состоящего, как прави-
ло, из пластинчатых компонентов и микропузырьков: появляются
крупные вакуоли и межцистернальные структуры. Увеличивается
численность и протяженность мембранных элементов эндоплазма-
тической сети. В процессах дифференциации большую роль игра-
ют межклеточные взаимодействия. В ходе дифференциации раз-
1 Токин И. Б. Проблемы радиационной цитологии. Л., 1974. С. 225.
400
виваются специализированные структуры — клеточные поверхно-
сти (W-образные складки, десмосомы и др.), устанавливается бо-
лее тесная связь между клетками. Происходит увеличение общей
площади поверхности мембран в клетке, уменьшается число сво-
бодных рибосом.
При дедифференциации клеток прооисходит ультраструктур-
ная реорганизация клеток. Важные факты о дедифференциации
обнаружены электронно-микроскопическими исследованиями.
В упоминавшемся исследовании Хэй (1962) обнаружено, что при
дедифференцировке мышечных и хрящевых клеток (в ходе про-
цесса регенерации конечности аксолотля) повышается содержа-
ние свободных рибосом и уменьшается количество элементов эн-
доплазматической сети.
И. Б. Токин, изучая восстановительные процессы в эпителии
кишечного тракта, клеток печени, других органов млекопитающих,
нашел в противоположность описанным ультраструктурным изме-
нениям при дифференциации, что при посттравматической регене-
рации происходит «.увеличение относительных объемов ядер и яд-
рышек, увеличение содержания гранулярного компонента ядрышка
и ядрышкосвязанного хроматина, возрастание числа свободных
рибосом и полисом и уменьшение площади мембранной поверхно-
сти клетки»1. Редукция специализированных структур клеточной
поверхности свидетельствует о снижении специфических функций
клетки (например, всасывающей в кишечном .эпителии), ослабле-
нии межклеточных связей, изменении уровня интеграции тканей и
межклеточных , взаимоотношений, значение которых неоднократно
подчеркивалось в этой книге.
Светооптические и электронно-микроскопические исследования
дают возможность говорить об изменении состояния клеток, про-
исходящем на начальных этапах регенерации, причем эта дедиф-
ференциация обычно наступает очень быстро, протекает кратко-
временно. В полном соответствии с морфологическими изменения-
ми происходят изменения и биохимические. Показано, что
свободные рибосомы осуществляют интрацеллюлярный синтез ве-
ществ (необходимых для вступления в митотический цикл и про-
хождения периода S и обеспечивающих энергетику митоза). При-
крепленные рибосомы осуществляют экстрацеллюлярный синтез,
поэтому увеличение числа свободных рибосом при дедифференци-
ации связано с усилением биосинтеза клетки, направленного на
удовлетворение собственных потребностей. При изучении природы
различий в синтезе разных типов белков в полисомах оказалось,
что интенсивность включения меченых соединений в мРНК из сво-
бодных полисом выше, чем в мРНК из связанных с мембранами
полисом. Увеличение объемов ядрышек и содержание гранулярно-
го компонента отражает усиление биосинтетических процессов,
1 Токин И. Б. Проблемы радиационной цитологии. Л., 1974. С. 257.
401
происходит активация синтеза РНК. И. Б. Токин показал, что
имеет место «конкуренция» между интрацеллюлярными и экстра-
целлюлярными биосинтезами в ходе дифференциации и дедиффе-
ренциации клеток: в ходе дедифференцировки клеток, как-уже го-
ворилось, происходит смена экстрацеллюлярного синтеза белков
интрацеллюлярным.
По-видимому, указанные закономерности носят универсаль-
ный характер. Так и при изучении дедифференциации клеток
растений отмечены черты, общие с изменениями животных клеток.
Дедифференциация растительных клеток связана с увеличениями
белкового синтеза, числа митохондрий и структур аппарата
Гольджи, ядрышка. Проявляются и специфические особенности
растительных клеток, например уменьшение толщины клеточной
оболочки. Р. Г. Бутенко (1970), изучая пролиферацию культиви-
руемых растительных клеток и процессы соматического эмбриоге-
неза у растений, отмечает ряд закономерностей, связанных с пе-
редифференцировкой специализированных тканей. Синтез белков,
репрессированный в период культивирования и дедифференциров-
ки клеток, снова дерепрессируется в ходе дифференциации тканей
при дальнейших процессах соматического эмбриогенеза.
На основании всех имеющихся в мировой литературе данных
следует заключить, что та или иная степень дедифференциации
высокоспециализированных клеток, а также активизация «покоя-
щихся» стволовых клеток очевидный факт. Длительное время су-
ществовавший в эмбриологии и цитологии критерий дифференциа-
ции клеток — утрата способности к митотическому делению —
следует считать ошибочным или имеющим значение лишь для ог-
раниченного круга формообразовательных процессов.
РЕГЕНЕРАЦИЯ И ОНТОГЕНЕЗ
Нельзя ожидать высокой регенерационной способности у организ-
мов в старческом состоянии, когда затухают функции различных
интеграционных механизмов, регулирующих жизнь целого орга-
низма и функции органов, без которых немыслим обмен веществ
в любой части тела (функция нервной системы, работа желез
внутренней секреции, деятельность дыхательной, кровеносной и
пищеварительной систем). В старческом организме развиваются
различного рода дистрофии, нарушается нормальная интеграция
тканей, прогрессирует ослабление процессов самообновления тка-
ней, вместе со всем этим ослабляется и способность к репаратив-
ной регенерации.
М. А. Воронцова в 1944 г. заметила, что если конечность годо-
валого тритона и аксолотля регенерирует полноценно уже через
1,5—2,5 месяца, то в старческом возрасте (после 6—10 лет) на ме-
сте ампутации происходит лишь заживление раны.
402
Заживление ран и другие регенерационные явления у млекопи-
тающих и человека к старости ослабляются, что является пред-
метом больших забот теоретика-биолога и практика-хирурга.
Закономерность падения регенерационной способности по ме-
ре старения относится, по-видимому, ко всем животным, размно-
жающимся только половым путем. Ее можно отметить и для про-
стейших. Деление простейших — итоговый этап их онтогенеза.
В результате достижения границы роста и ассимиляции наступает
старение, дезинтеграция и смерть индивидуума. Не случайно, что
на одно и то же оперативное воздействие (отрезание, например,
Vs части тела) инфузория (Onychodromus grandis и др.) отвечает
неодинаково в разные этапы онтогенеза: молодая инфузория реге-
нерирует, а у старой (незадолго перед делением) особи происхо-
дит реконструкция ядерного аппарата, имитирующая процессы
деления и вместе с тем происходит удлинение индивидуальной
жизни. Это наблюдали М. Гартман, Э. С. Бауэр и А. М. Гранов-
ская в 1935 г., В. И. Карелина и Б. П. Токин в 1949 г.
. Нельзя, однако, создавать утрированные схемы, касающиеся
однотипного изменения восстановительных морфогенезов в онто-
генезе. Закономерности изменения регенерации и соматического
эмбриогенеза зависят от структуры жизненного цикла, от наличия
или отсутствия в этом цикле бесполого размножения. Поэтому
«старческая» планария, готовящаяся к делению, будет регенери-
ровать лучше, чем молодая планария этого же вида или этой же
расы, не имеющая половой репродукции. Однако старческая пла-
нария, размножающаяся только половым путем, регенерирует
хуже, чем молодая особь того же вида.
СООТНОШЕНИЕ ПРОЦЕССОВ РЕГЕНЕРАЦИИ И БЛАСТОМАТОЗНОГО РОСТА
Предметом эмбриологии являются и патологические морфогенезы,
вызванные изменениями нормального состояния клеток, тканей и
органов, приводящие к бластоматозному росту, т. е. к возникнове-
нию новообразований (бластом), называемых злокачественными
^опухолями.
Очень сложная, волнующая человечество проблема злокачест-
венных опухолей в этой книге не рассматривается. Сообщаются
лишь некоторые факты о взаимоотношении процессов регенерации
и опухолевого роста.
Несмотря на многочисленные попытки цитологов, биохимиков,
генетиков и патологоанатомов, не удается обнаружить специфиче-
ские особенности биохимии и морфологии опухолевых клеток.
” В связи с прогрессом молекулярной биологии возникают под-
час оригинальные гипотезы «малигнизации», о злокачественном
перерождении клеток. По единичным клеткам нельзя опреде-
лить— принадлежат ли они нормальной или опухолевой ткани.
403
Решающим в понимании причин возникновения и развития опухо-
лей должно быть не столько исследование «злых» свойств еди-
ничных клеток, сколько изучение нарушения нормальной инте-
грации тканей, в результате чего размножающиеся клетки выхо-
дят из-под «замка» нормальных корреляций и не подчиняются
нормальным формативным влияниям. В случае, например, рака
кожи млекопитающих происходит дезинтеграция пластов эпите-
лия, нарушаются нормальные взаимоотношения между ними и
соединительной тканью.
Большую роль играют мембраны клеток, их адгезивные свой-
ства. Если эти свойства уменьшаются, изменяется взаимосвязь
между клетками, облегчается и отделение клеток друг от друга и
их миграция в другие органы.
Конечно, нарушения взаимоотношений между клетками и тка-
нями не являются процессами, обособленными от цитологических
изменений. Поэтому оправданы и крайне необходимы молекуляр-
но-биологические и электронно-микроскопические исследования
изменений, происходящих в клетках при канцерогенизации тка-
ней. Однако нет оснований надеяться найти какие-то особые,
«злые», «раковые» ультраструктурные особенности единичных,
«перерождающихся» клеток.
Нельзя увлекаться поэтому односторонне биохимическими (мо-
лекулярно-биологическими) исследованиями, создавая новые и
новые представления о перерождении нормальных клеток в опухо-
левые.
В организме происходят непрерывные формообразовательные
процессы в коже, молочной железе женщин, тканях стенки матки
и т. д. Во всех тканях в той или иной степени имеют место про-
цессы нормальной физиологической регенерации, обусловленной
определенной интеграцией тканей, органов и всего организма. При
нарушении регуляторных механизмов, и в особенности функции
нервной, эндокринной и иммунной систем, нормальные процессы
формообразования и самообновления тканей, а значит, и «строи-
тельство» тканей при физиологической и репаративной регенера-
ции происходить не могут.
Стойко дезинтегрировать ткани, нарушать типические процес-
сы формообразования и тем самым создавать потенциальную воз-
можность бластоматозного роста могут агенты разнообразной
природы: вирусы, бактерии, лучи Рентгена, ароматические. угле-
водороды, гормональные факторы и т. д.
Бластоматозный рост может возникнуть только при резком из-
менении нормальных процессов формообразования. Так называе-
мое предраковое состояние, анархизированное, атипическое раз-
витие тканей не обязательно переходит в бластоматозный рост.
Если ткани, находящиеся в физиологической и структурной связи
с пораженным участком, нормальны, то анархизированный кле-
404
точный материал снова входит в обычное формообразовательное
русло.
Все, способствующее нормальному формообразованию, долж-
но противодействовать опухолевому росту, и наоборот. Если во
всем организме или локально понижается регенерационная спо-
собность и нарушаются нормальные процессы формообразования,
то тем самым создаются условия для атипического, автономного,
опухолевого роста. В этом смысле, образно говоря, процессы реге-
нерации и процессы бластоматозного роста являются антагони-
стическими. Многие факты подтверждают это предположение.
1. Канцерогенные вещества (т. е. вещества, которые могут вы-
звать развитие опухолей) любой химической природы тормозят
регенерацию. А. И. Здруйковская в 1944 г. смазывала кожу крыс,
мышей и кроликов каменноугольным дегтем и раствором 9,10-ди-
метил-1,2-бензантрацена. Торможение регенерации происходило
как в опытах предварительного смазывания (один раз в день в те-
чение 17 дней) и последующего ранения, так и при подаче канце-
рогенного вещества на раневую поверхность. В других опытах она
прививала крысам рак или саркому Йенсена, а мышам — рак Эр-
лиха. А. Г. Филатова в 1947 г. прививала на боках мышей адено-
карциному. Оба автора производили затем ранения (на коже) в
районе опухоли (когда последняя была размером с фасоль). Об-
наруживалось торможение регенерации кожи по сравнению с кон-
трольными животными. В других опытах отмечено падение реге-
нерационной способности кожи мышей в районе спонтанного рака
грудной железы.
2. В медицине давно отмечены факты пониженной регенераци-
онной способности больных злокачественными опухолями: у них
наблюдается более частое расхождение швов при операциях, ос-
ложненное, неполноценное заживление кожных ран и т. п. Гово-
рилось уже, что у старческих организмов слабее их регенерацион-
ные возможности. В то же время известна более частая поражае-
мость опухолями стареющих организмов.
3. Известно, что рентгеновы лучи и другие виды ионизирую-
щей радиации могут явиться канцерогенными агентами. Логично
сопоставить это с тем, что рентгеновы лучи и радиоактивные ве-
щества тормозят процессы регенерации.
Однако общеизвестно превосходное лечебное противораковое
действие рентгеновых лучей. Это противоречие заключено в самом
биологическом явлении. Рентгеновы лучи, несомненно, во многих
случаях излечивают. Но как хирург, вырезая опухоль, еще не
уничтожает тем самым этиологических факторов, так и рентгено-
лог, «вырезая» опухоль лучом, не создает в организме или в соот-
ветствующих органах нормальных формообразовательных явле-
ний и нормальных структурных связей. Наоборот, поскольку всег-
да абсолютно неизбежно рентгенизуется не только опухоль, но
405
частично и здоровые ткани, рентгенолог, вылечивая рак, может
помочь рецидиву.
Таким образом, анализируя и в ином аспекте факты, обнару-
живаемые при рентгенизации, мы приходим к тому же заключе-
нию: поскольку рентгеновы лучи могут затормозить или нацело
подавить регенерационную способность, то при определенных ус-
ловиях они способствуют бластоматозному росту.
Следовало бы остановиться на вопросах — у всех ли животных
могут возникать спонтанно опухоли? У всех ли животных можно
экспериментально вызвать бластоматозный рост? К сожалению,
для ответа на эти вопросы нет ни достаточных фактических дан-
ных, ни обоснованных гипотез, так как эволюция опухолевых яв-
лений оказалась пока вне интереса биологов и медиков и имеются
только единичные наблюдения над беспозвоночными и низшими
позвоночными. Лишь некоторые факты не вызывают сомнений,
например превосходная регенерационная способность хвостатых
амфибий и крайне редкие случаи естественного образования опу-
холей у этих животных. Трудно вызвать у них и эксперименталь-
ные опухоли. Е. Г. Агапова и Б. П. Токин в 1962 г. получили ати-
пическое разрастание тканей конечностей аксолотля в результате
сочетания рентгенизации с последующим введением канцероген-
ных веществ.
Конечно, изложенная гипотеза не отвечает на многие встаю-
щие перед онкологией вопросы. Межклеточные взаимодействия,
интеграционные механизмы тканей и органов, которым эта гипо-
теза придает решающее значение, могут нередко нарушаться и
при нормальном состоянии организма. Организмы — не абсолютно
стабильные сооружения, в которых бы оказывались неизменными
проницаемость мембран клеток, концентрация ионов в межклеточ-
ных субстанциях, выделение клетками продуктов их жизнедея-
тельности и т. и. Напротив, происходят значительные изменения
в указанном и иных направлениях — на почве физиологической и
репаративной регенерации, на почве воспалительных процессов, в
ходе нормального функционирования органов. Молочная железа,
например, или стенка матки в разные этапы своей жизнедеятель-
ности находятся, так сказать, в состоянии нормальной «дезинте-
грации», например стенка матки во время менструаций.
Многие из этих изменений быстропроходящие. Если вступают
в силу регуляторные возможности тканей и органов (в частности
и в особенности способность к физиологической и репаративной
регенерации), указанные дезинтеграции не могут создать прочный
«предраковый фон». Указанные «дезинтеграции» закономерно
наступают в жизни тканей и органов. Сам процесс репаративной
регенерации, например любого органа хвостатых амфибий, вклю-
чает совершенно необходимый для регенерации этап «дезинтегра-
ции» тканей, без которого невозможно формирование регенераци-
406
онной бластемы. Это кажется видимым противоречием изложен-
ной гипотезе о соотношении регенерационных и опухолевых явле-
ний, и действительно необходимо углубление ее, дополнение но-
выми предположениями и экспериментами. Большое число фактов
говорит, однако, в пользу гипотезы.
ПРЕДПОЛОЖЕНИЯ ОБ ЭВОЛЮЦИИ РЕГЕНЕРАЦИОННЫХ ЯВЛЕНИЙ
И СОМАТИЧЕСКОГО ЭМБРИОГЕНЕЗА
Восстановительные морфогенезы следует рассматривать как одно
из древнейших свойств живых систем вообще, ибо в ходе обмена
веществ протоплазмы с его диссимиляцией и ассимиляцией всег-
да происходит разрушение и восстановление структур. Возникшая
на основе обмена веществ у первобытных живых систем физиоло-
гическая регенерация в дальнейшем совершенствовалась и разно-
образно видоизменялась на уровне клеток, тканей и органов.
Эволюция репаративной регенерации и явлений соматического
эмбриогенеза не могла происходить изолированно от эволюции
физиологической регенерации, от эволюции других защитных
свойств, изолированно от различных форм размножения, явлений
редукции и т. д.
Основные закономерности эволюции восстановительных мор-
фогенезов с учетом разграничения регенерации и соматического
эмбриогенеза могут быть сформулированы следующим образом.
Эволюция регенерации и соматического эмбриогенеза происходи-
ла дивергентно. Все факторы внешней среды и внутренние мута-
ционные изменения, которые ведут к общему ослаблению и нару-
шению интеграционных механизмов, независимо от стадии инди-
видуального развития особи, вызывают повышение потенциальных
возможностей возникновения в жизненном цикле либо полиэмб-
рионии, либо особой формы бесполого размножения (близкого по
своей природе к соматическому эмбриогенезу) и способствуют
появлению на той же стадии морфогенетической реакции, осуще-
ствляющейся по типу соматического эмбриогенеза.
Все факторы внешней среды и внутренние мутационные изме-
нения, которые не нарушают общей системы организменных ин-
теграционных механизмов, а способствуют возникновению лишь
локальных повреждений, либо бесполого размножения близкого
по своей природе к регенерационным процессам (например, архи-
томия, паратомия у червей), и одновременно оказываются причи-
ной появления на тех же стадиях онтогенеза явлений удвоения
крупных отделов тела и хорошо выраженных регенерационных
свойств. Обособляющиеся при бесполом (вегетативном) размно-
жении крупные участки тела сохраняют исходную полярность и
бластогенез осуществляется так же, как и регенерация удаленных
частей по типу эпиморфоза или морфоллаксиса с сохранением
407
основных черт организации развивающегося фрагмента. Состоя-
ние организменной интеграции не остается в онтогенезе постоян-
ным и всегда на любых этапах исторического развития сохраняет-
ся ситуация, когда особь в разные периоды жизненного цикла
имеет более и менее интегрированные связи макроотделов тела.
Следовательно, многообразие ответных реакций организма на по-
вреждение в ходе филогенеза всегда сохраняется. Если не сравни-
вать восстановительные морфогенезы только у взрослых особей,
а сопоставлять динамику этих процессов в жизненном цикле, то
тогда исключается всякая возможность для построения схем их
линейной изменчивости (в сторону повышения или понижения).
В пользу сохранения разнообразных ответных реакций на по-
вреждение высказываются в последние годы многие биологи, за-
нятые проблемами регенерации у животных и растений (М. А. Во-
ронцова, Л. Д. Лиознер, Р. Госс, А. Г. Юсуфов, Д. С. Саркисов,
Л. К. Романова и др.).
Глава XVII
ОНТОГЕНЕЗ И ЭВОЛЮЦИЯ. БИОГЕНЕТИЧЕСКИЙ ЗАКОН
ИДЕИ Ч. ДАРВИНА О СВЯЗИ ОНТОГЕНЕЗА С ЭВОЛЮЦИЕЙ
Еще в додарвиновский период анатомы и эмбриологи, особенно
И. Меккель в 1911 г., обратили внимание на то, что в развитии ор-
ганов у высших позвоночных, прежде чем они примут окончатель-
ное строение, наблюдаются стадии, на которых они очень сходны
с окончательно сформированными органами взрослых низших
представителей той же группы животных. Так, в эмбриональном
развитии птиц и млекопитающих образуются жаберные щели и
соответствующие артериальные дуги, чем зародыши этих живот-
ных напоминают взрослых рыб.
К. Бэр в 1828 г. установил, что зародыши разных позвоночных
на ранних стадиях развития очень сходны между собой, и сформу-
лировал свой знаменитый «закон зародышевого сходства» (см.
с. 423). В бэровском законе Дарвин видел наиболее удивительный
факт всей естественной истории. Дарвин придавал большое зна-
чение этим фактам в связи с разработкой своего эволюционного
учения. Он соглашался с тем, что древние животные так или ина-
че сходны с зародышами новых животных того же самого класса.
Взрослые формы отличаются от своих зародышей вследствие тех
изменений, которые наступают не в самом раннем возрасте,
а позднее и которые наследуются от родителей. «Это процесс,—
говорит Дарвин,— оставляя зародыш почти без изменений, посто-
янно накопляет в последовательных поколениях все большие и
большие изменения взрослой формы. Таким образом, зародыш
представляет собой как бы сохраняемый природный портрет преж-
него и менее измененного состояния вида... Зародыш... является
как бы свидетелем прошлых состояний, через которые прошел
вид... Эмбриология значительно выигрывает в интересе, если мы
будем видеть в зародыше более или менее потускневший портрет
общего прародителя всех членов одной большой группы в его
взрослом или личиночном состоянии»1. Нельзя думать, рассуждал
Дарвин, что образование у зародышей позвоночных жаберных
щелей и артериальных дуг в этой области связано со сходными
условиями и у млекопитающего, питающегося в утфобе матери,
1 Дарвин Ч. Сочинения. М., Л., 1959. Т. III. С. 458.
409
и в яйце птицы, насиживаемом в гнезде, и у икры лягушки в воде.
Дарвин был уверен, что в этих случаях повторяются признаки, ко-
торые были свойственны отдаленным предкам.
ОСНОВНОЙ БИОГЕНЕТИЧЕСКИЙ ЗАКОН
Ф. Мюллер и Э. Геккель (1866—1874) под влиянием эволюцион-
ной теории Ч. Дарвина сформулировали положение, сыгравшее
большую роль в эмбриологии и всей зоологии. Геккель назвал его
основным биогенетическим законом. Так как идеи закона близки
к мыслям Дарвина об отражении в онтогенезе истории развития
животных, то правильнее называть биогенетический закон зако-
ном Дарвина — Мюллера — Геккеля. Особенно много в обоснова-
ние закона сделано Ф. Мюллером. Он показал, что личинки раз-
личных ракообразных, резко отличающиеся друг от друга во
взрослом состоянии, очень похожи друг на друга. Особенно на-
глядно эта закономерность обнаруживается у паразитических
форм, например у Sacculina, паразитирующей на крабах. Взрос-
лое животное совершенно не похоже на ракообразных, но личинка
его в ходе развития как бы повторяет строение, характерное для
разных низших раков. Мюллер сделал заключение, что все низ-
шие раки произошли от одной общей формы, напоминающей по
своим признакам личинку современных низших ракообразных.
У Геккеля и Мюллера созревала мысль о том, что последователь-
ные изменения формы особи на протяжении индивидуального раз-
вития обусловлены филогенезом, т. е. развитием того рода, к ко-
торому принадлежит данный вид животного. Онтогенез является
кратким и быстрым повторением филогенеза, обусловленным на-
следственностью и приспособлением. Эта закономерность и была
названа Геккелем основным биогенетическим законом, более пол-
ная формулировка которого такова, что развитие зародыша (ин-
дивидуальный, или онтогенетический, ряд развития, онтогенез) —
это сжатое, сокращенное повторение развития всей группы форм,
возникших из одного общего корня (т. е. повторение развития
филетического, или палеонтологического; филогенез); повторение
тем полнее, чем больше сохраняется благодаря постоянному на-
следованию первоначальное развитие предков (палингенез). По-
вторение же тем менее полно, чем больше благодаря различным
приспособлениям было введено позднейших нарушений развития
(ценогенез). Согласно этому закону зародыши вышестоящих жи-
вотных похожи на взрослых нижестоящих предков. Яйца всех жи-
вотных «идентичны». Эволюция осуществляется благодаря при-
бавлению новых стадий в конце развития. Если бы ничто не ме-
шало, то в индивидуальном развитии точно повторялась бы эво-
люционная история вида, а значит, за часы, недели или месяцы
развития зародыша того или иного животного мы могли бы про-
смотреть, образно говоря, «кинофильм» о его историческом прош-
410
лом. При просмотре «фильма» об индивидуальном развитии мле-
копитающих мы увидели бы стадии, соответствующие взрослым
формам беспозвоночных животных, а затем рыбам, амфибиям и
рептилиям.
Однако проявлению в неизменном виде первобытных, древ-
них, палингенетических признаков (от греч. palaios — древний;
палингенезы — признаки, перешедшие в онтогенез животного из
его филогенеза) мешают ценогенезы — разнообразные признаки,
возникшие в онтогенезе как приспособление к условиям жизни за-
родышей личинок и взрослых животных. Примерами ценогенезов
могут служить амнион, хорион, аллантоис Amniota, громадные
прядильные железы личинок насекомых.
Из биогенетического закона следует, что древние (палингене-
тические) признаки раньше других должны появляться у зароды-
шей. Однако такая последовательность очень часто нарушается.
Сторонники биогенетического закона объясняют подобные нару-
шения следующими причинами. В ходе эволюции организмов про-
исходит сокращение времени развития и упрощение самого разви-
тия, изменение способов развития органа и т. д. Изменения во
времени закладки и развития тех или иных органов называются
гетерохронией. Некоторые ученые (Менерт) думают даже о зако-
не, согласно которому закладка прогрессирующих органов пере-
носится на более ранние стадии. При этом ссылаются на развитие
животных, имеющих детерминированный тип дробления, при ко-
тором закладка органов совершается очень рано. Примером гете-
рохроний являются более ранняя закладка и развитие головного
мозга, органов чувств и сердца у высших позвоночных по сравне-
нию с низшими и, наоборот, запаздывание в формировании поло-
вой железы и кишечника. Геккель утверждал, что могут изменять-
ся и пространственные отношения закладок органов, перенесение
зачатка из одной части зародыша в другую. Легкие наземных жи-
вотных и плавательный пузырь рыб являются видоизменением
задней пары жаберных мешков, но жаберные мешки лежат на бо-
ковой стороне кишечника, а легкие — на брюшной; плавательный
пузырь находится большей частью на спинной стороне кишечника.
Смещаются и зачатки этих органов у зародышей: легкие заклады-
ваются на брюшной стороне, а плавательный пузырь у большин-
ства рыб — на спинной стороне.
Биогенетический закон был использован Геккелем для «восста-
новления» картины филогенеза, для построения родословного дре-
ва органического мира. Геккель сдёлал много предположений, ка-
сающихся первых этапов эволюционного развития живых существ,
о которых нам ничего не может сказать палеонтология. Так, он
создал свою знаменитую теорию гастреи о предполагаемом предке
всех ныне живущих многоклеточных — Gastraea.
Форме и структуре этого гипотетического предка соответствует
411
Таблица 7. Параллелизм онтогении и филогении
Филогения Онтогения Какими современными формами иллюстрируется
Cytaea Древнейшее простейшее животное — первобытная амеба Могаеа Cytula Оплодотворенная яйцеклетка Morula Амебы
Сообщество однородных Шаровидная кучка Синамёбнй — сообще-
клеток — Protozoa одинаковых «сегмента- ционных шаров» ство амеб или других однородных клеток (Pro- tozoa)
Blastaea Blastula Volvox, Pandorina и
Пузыревидное первичное животное с полостью, со- держащей студень нлн жид- кость. Стенка состоит нз одного слоя ресничных кле- ток (псевдоподии преврати- лись в жгутики) Пузыревидный заро- дыш; состоит из одного слоя клеток с полостью внутри другие колониальные жгутиконосцы
Gastraea Многоклеточное живот- ное с кишечником и ртом, стенка двухслойная. Перво- начальная общая форма всех современных много- клеточных животных Gasfrula Двухслойный зародыш с первичным кишечни- ком и ротовым отвер- стием Гидра
стадия гаструлы современных организмов. Представление об ото-
бражении в онтогении животных их филогении дает табл. 7, со-
ставленная на основании работ Геккеля.
Как видно из данных таблицы, каждой стадии индивидуаль-
ного развития соответствует определенная предковая форма, ко-
торая сходна с некоторыми представителями современных живот-
ных. Например, стадия морулы соответствует предковой форме
морее, а ее современный аналог синамебий.
РЕКАПИТУЛЯЦИИ
Э. Геккелем было создано представление о гипотетических пер-
вобытных животных — бластее и морее (соответствующих бла-
стуле и моруле). Бластею он сопоставил о современными расти-
тельными колониальными организмами (Volvox). Стадию гастреи
он усматривал в типе кишечнополостных, состоящих из эктодер-
мы и энтодермы. Согласно биогенетическому закону названные
животные в ходе своего онтогенеза рекапитулируют строение
взрослых предков (от лат. recapitulatio — краткое повторение ска-
412
занного). Морфологи, особенно XIX в., описали много рекапитуля-
ций, которые далеко не всегда оказывались бесспорными.
1. Все многоклеточные организмы развиваются (при половом
размножении) из одной яйцеклетки, оплодотворенной спермием.
2. В эмбриональном развитии большинства животных образу-
ются зародышевые листки (см. гл. V). Эмбриологи прошлого
столетия рассматривали их как первичные органы, наделяли их
способностью к гисто-физиологической деятельности, что соответ-
ствовало теории гастреи Геккеля. Эктодерму называли кожно-
чувствующим слоем (Hautsinnesblatt), энтодерму — слизисто-же-
лезистым слоем (Darmdriisenblatt). Эти гомологичные органы
у разных животных получены как наследство от предполагаемых
Геккелем и другими учеными форм, у которых они существовали
в виде постоянных органов взрослых животных. Следует, однако,
вспомнить те споры, которые существуют вокруг теории зароды-
шевых листков (см. гл. V).
3. У всех позвоночных, включая и высших их представителей,
закладывается хорда, а у их предков, если судить по ланцетнику,
хорда оставалась на всю жизнь.
4. В ходе эмбрионального развития птиц и млекопитающих,
включая и человека, появляются жаберные щели и соответствую-
щий им скелетный и кровеносный аппарат—кровеносные сосуды,
пересекающие жаберные перегородки. Строение сердца человече-
ского зародыша в этот период напоминает строение такового
у рыб: оно с одним предсердием и одним желудочком. Выдели-
тельная система представлена пронефросом, также характерным
для рыб. Имеется и хвост, содержащий сегменты мезодермы,
хорду, нервную трубку с зачатками ганглиев и участок кишечни-
ка, впоследствии атрофирующегося (позади заднего прохода).
Образуются и позвонки, которые потом сливаются; к четырем
месяцам утробной жизни хвост уже не заметен, но при редких
патологических случаях остается на всю жизнь. На более поздних
стадиях развития человеческого зародыша жаберные щели и
пронефрос исчезают, образуется новая почка — мезонефрос; пред-
сердие теперь разделено на два — правое и левое. Можно видеть
на этой стадии черты строения рептилий. Наконец, формируется
четырехкамерное сердце, мезонефрос также исчезает, и развива-
ется новая почка, свойственная млекопитающим.
5. Демонстративным примером подтверждения биогенетическо-
го закона считается развитие Birgus latro — пальмового вора,
обитающего на островах Тихого и Индийского океанов (семейство
Coenobita). Его иначе называют крабом-разбойником или кокосо-
вым крабом, по внешнему сходству во взрослом состоянии с кра-
бами, хотя это интересное животное близко к ракам-отшельникам.
Нижняя мягкая сторона брюха защищена хитиновыми бляшками.
Пальмовый вор — сухопутное животное, до 30 см в длину и мас-
сой до 3 кг — живет в сухих местах, в лесу, питается кокосовыми
413
орехами. Дышит легкими и погибает, если его погрузить на 4—
5 ч в воду. Спаривание происходит вдали от воды. Яйца прикреп-
ляются к волоскам брюшных ножек самки. Ко времени выхода
личинок самки приближаются к морю, входят в него, личинки
оказываются в воде. Из яиц выходит планктонная личинка зоэа,
сходная с личинками раков-отшельников. Следует ряд пелагиче-
ских личиночных стадий и наконец развивается «рак-отшельник»,
прячущий свое мягкое брюшко в раковину брюхоногого моллю-
ска. Сначала стадия «рака-отшельника» протекает в море у бере-
га, затем животное переходит в стадию Coenobita. После одной
из линек пальмовый вор оставляет раковину, постепенно утрачи-
вает особенности рака-отшельника и превращается во взрослого
Birgus latro. Головогрудь разрастается до 10—12 см.
Во взрослом состоянии это животное дышит легкими, но в ходе
онтогенеза имеет место ряд сложных преобразований дыхательной
системы. У личинок функционирует 14 пар жабр, как у раков-
отшельников. По мере роста жаберные полости постепенно увели-
чиваются, их части на головогрудном панцире разрастаются
в большие выпуклые вздутия. На внутренней поверхности поло-
сти развиваются гроздевидные складки, пронизанные кровенос-
ными сосудами, тем самым поверхность полостей значительно
увеличивается; жаберные полости превращаются в легочные меш-
ки. Жабры не исчезают, хотя в опытах с удалением их доказано,
что никакой роли в дыхании они не играют. Сторонники биогене-
тического закона рассматривают индивидуальное развитие паль-
мового вора как яркую иллюстрацию повторения филогенеза
в онтогенезе.
6. Еще один из примеров рекапитуляции — онтогенез зубной
системы некоторых животных. Зародыши черепах и некоторых
птиц, для которых исходными в историческом развитии явились
зубастые рептилии, имеют зубы. Некоторые млекопитающие, на-
пример беззубые киты, во взрослом состоянии не имеют зубов.
Они задерживают во рту пищу с помощью сидящих на нёбе ро-
говых пластинок («китового уса»). Но у беззубых китов в эмбри-
ональный период появляются зубы (до 53 с каждой стороны).
Зубы эти не прорезаются, они разрушаются и рассасываются
тканями.
7. Индивидуальное развитие амфибий считается ярким приме-
ром рекапитуляций. Признаки взрослых хвостатых амфибий сдела-
лись личиночными признаками у бесхвостых. Головастики лягушек
пмеют хвост, напоминающий хвост рыб и хвостатых амфибий.
Имеются наружные жабры — гребенчатые придатки с каждой
стороны на голове; челюсти одеты роговым клювом, а впереди
него внутренняя поверхность губ усажена роговыми зубами;
позади рта на брюшной стороне находится подковообразная при-
соска. В таком виде по своему строению головастик сходен с по-
стоянножаберными амфибиями (Perennibranchiata), в течение
414
Рис. 165. Развитие кожи человека:
А — поперечный разрез через однослойный эпидермис на ранней стадии развития; Б—
поперечный разрез на несколько более поздней стадии развития, когда эпидермис состоит
из двух слоев эпителия: нижиего — цилиндрического и верхнего — плоского (по Ж. Вер-
шлюсу, 1927); В — поперечный разрез через кожу взрослого человека, состоящую из много-
слойного специализироваииого эпидермиса, в котором виден ряд дифференцированных слоев
(по Бему и Давыдову); / — клетки цилиндрического эпителия, 2 — кориум, 3— роговой
слой; 4 — блестящий слой, 5 —зернистый слой, 6 — мальпигиев слой
всей жизни имеющими жаберные щели, наружные жабры и хвост.
Роговые зубы напоминают таковые круглоротых (миног), присо-
ски придают сходство с личинками двоякодышащих рыб. В ходе
дальнейшего развития у головастика лягушек зарастают жабер-
ные щели, дыхание становится целиком легочным, но сохраняется
хвост, как у саламандр и тритонов. Затем исчезает хвост, и живот-
ное приобретает признаки Anura.
8. Эволюция скелета трехпалой конечности — также хороший
пример рекапитуляции. Шестинедельные зародыши современных
лошадей имеют три Пальца, боковые пальцы состоят из трех фа-
ланг, обнаруживаемых даже на пятом месяце беременности.
Позднее происходит срастание фаланг и превращение их в гри-
фелевидные косточки.
9. У эмбрионов современных рыб, которые произошли от хор-
довых, имевших 9 или более жаберных щелей и сосудов, обыкно-
венно появляются передние две жаберные щели и передние два
жаберных сосуда, отсутствующие у взрослых рыб в таком -виде,
в каком они были у предков.
10. При развитии тканей, как и при развитии органов, могут
появиться признаки, свойственные клеткам и тканям предков
этого животного, что дает основание говорить о гистогенетических
рекапитуляциях. Это можно наблюдать в развитии кожи млеко-
питающих и человека. На ранних стадиях онтогенеза эпидермиса
зародыша человека он состоит из однослойного цилиндрического
эпителия, как бы напоминая строение эпидермиса взрослого лан-
цетника; затем эпителий превращается в многослойный, но не оро-
говевающий (как у взрослых костистых рыб), затем в многослой-
415
ный слабо ороговевающий (как у Salamandra maculosa) и, нако-
нец, с такой дифференцировкой и характером ороговевания, как
yAmnita (рис. 165).
ТЕОРИЯ ФИЛЭМБРИОГЕНЕЗА
А. Н. СЕВЕРЦОВА. ЭВОЛЮЦИЯ ОНТОГЕНЕЗА
Более столетия прошло после того, как был сформулирован
биогенетический закон. Ввиду исключительной важности он не
раз подвергался критике и пересмотру. Выдающийся вклад в уче-
ние о соотношении онтогении и филогении сделал А. Н. Северцов.
Он изучал изменения пути развития отдельных органов потомков
по сравнению с тем, как эти органы развивались у взрослых,
Рис. 166. Онтогенез гидроида (по А. Н. Северцову, 1935):
А— зрелое яйцо; Б — оплодотворенное яйцо, прикрепленное ножкой к телу материнского
организма; В — начало дробления яйцеклетки; Г — однослойная бластула; из бластодермы
в полость бластулы выселяются отдельные клетки, нз которых образуется энтодерма заро-
дыша (образование энтодермы будущей гаструлы путем миграции клеток); Д— внутри за-
родыша образовался слой энтодермы (7), а на поверхности эктодерма (2) и наружная эм-
бриональная оболочка (5); Е — зародыш выползает из оболочки и переходит к свободному
образу жизни; 2К— зародыш ползает по дну бассейна; 3 — развитие щупалец (4); И —
молодая гидра со щупальцами и ртом (5); X —взрослая гидра
416
а также пути изменения в развитии организмов вообще. Была
создана теория эмбриональных новообразований, имеющих фило-
генетическое значение. Признаки крупных систематических групп
возникают путем образования и суммирования отклонений в ходе
эмбрионального развития. Такие эмбриональные изменения, отра-
жающиеся на дальнейшем строении взрослого животного, назва-
ны филэмбриогенезами. Стойко передающиеся потомству измене-
ния в индивидуальном развитии организмов или в развитии тех
или иных органов, новые характерные признаки могут возникнуть
в начале эмбрионального развития, в начале формирования соот-
ветствующих органов. Этот тип филэмбриогенеза назван архаллак-
сисом. В других случаях изменения могут затронуть какие-либо
срединные стадии в развитии органов (или всего организма).
Такой тип изменений называется девиацией. Наконец, изменения
могли происходить на конечных стадиях развития организмов
или на конечных стадиях развития органов. Эти изменения назва-
ны анаболией (т. е. надставкой к конечным этапам развития).
Таким образом, согласно Северцову, различные признаки взрос-
лого животного сформировываются в различное время на протя-
жении его онтогенеза и онтогенеза органов и переходят затем
в неизменном виде во взрослое состояние в качестве дефинитив-
ных признаков; надставки происходят над дефинитивным состоя-
нием признаков, которое, складываясь в онтогенезе, является и
«эмбриональным» и «взрослым» одновременно. Северцов создал
гипотезу происхождения и эволюции онтогенеза многоклеточных
животных, причем большое значение в ней имеют представления
об анаболиях и основанных на них рекапитуляциях. Являясь сто-
ронником гипотез происхождения многоклеточных от колоний од-
ноклеточных, Северцов предполагает, что у предков современных
многоклеточных, именно у таких, которые подобны ныне живущим
растениям Pandorina тогит или Volvox, онтогенез был коротким
и заключался в надставках стадий при развитии колоний: из
одноклеточной стадии произошла двухклеточная (первая надстав-
ка), из двухклеточной — четырехклеточная стадия (вторая над-
ставка) и т. д. У вольвокса путем ряда таких надставок организм
развивается до шарообразной колонии, могущей состоять более
чем из 20000 клеток, затем происходит стадия (надставка)
гистологической дифференцировки (некоторого разнообразия ти-
па клеток). У таких животных, как низшие кишечнополостные,
онтогенез удлинился (рис. 166). Гидра, по Северцову, имеет в он-
тогенезе стадии, соответствующие стадиям онтогенеза колониаль-
ных форм, — период дробления (рис. 166, А, Б, В). К этим ста-
диям прибавляются стадии образования двухслойного зароды-
ша— гаструлы, затем формирования полипа с эктодермой,
гастральной полостью, щупальцами (рис. 166, Г — И). К таким
стадиям — «надставкам» прибавляется только стадия роста и
гистологической дифференцировки (образование стрекательных и
417
* 2 J 4 5
Рис. 167. Гипотетический ход эволюции онтогенеза билатерально симметричных
Metazoa (по А. Н. Северцову, 1935):
/ — размножение свободно живущих-одноклеточных животных. Дочерние клетки после каж-
дого деления расходятся и начинают^ жить самостоятельно; 2 — онтогенез колонии однокле-
точных (по Volvox)', происходит дифференциация ^клеток на половые (черные) и соматиче-
ские (светлые), 3 — онтогенез многоклеточного организма типа гидры; к стадии оитогеиезз
колониальных форм прибавилась стадия бластулы и гаструлы (в действительности прибави-
лось больше стадий), 4 — онтогенез первичного двусторонне симметричного животного, у ко-
торого кроме энтодермы и эктодермы путем архаллаксиса развилась еще и мезодерма, 5 —
онтогенез высшего двусторонне симметричного многоклеточного животного, у которого эм-
бриональное развитие еще удлинилось путем новых надставок
иных клеток). Таким образом, онтогенез гидроида удлинился не-
сколькими новыми стадиями, добавленными к дефинитивному
состоянию колониальных форм одноклеточных.
Весь гипотетический ход эволюции онтогенеза билатерально
симметричных многоклеточных Северцов представляет себе так,
как это показано на рис. 167.
биогенетический закон в свете современной биологии
Нет сомнений в выдающихся заслугах Мюллера и Геккеля и
в большой исторической роли биогенетического закона. Идея о
взаимной обусловленности онтогении и филогении остается бес-
спорной, однако это не означает бесспорность биогенетического за-
кона. Некоторые эмбриологи считают, что ни у одного вида живот-
ных онтогенез не подчиняется этому закону; большинство призна-
418
ков в онтогенезе любого вида животного несет черты ценогенетиче-
ских изменений, и никакой рекапитуляции признаков быть не мо-
жет. Даже в случае анаболий эмбриологу и генетику нельзя пред-
ставить себе, чтобы наследственно изменялись лишь конечные ста-
дии онтогенеза или развития какого-либо органа и чтобы при этом
способе филэмбриогенеза были бы совершенно не затронуты
исходные половые клетки и ранние стадии развития. Ученые, ко-
торые не разделяют крайних взглядов, ведущих к полному отри-
цанию биогенетического закона, думают, однако, о его устарело-
сти и необходимости создания нового обобщения о соотношении
онтогении и филогении. Высказывались многочисленные критиче-
ские замечания.
1. Следует избегать взгляда, будто зародыши рекапитулируют
в своем развитии взрослых предков. Зародыши человека, напри-
мер, никогда не повторяют амфибий или рыб. Более соответству-
ет истине утверждение о том, что развитие органов вышестоящих
животных так или иначе повторяет эволюцию этих органов у ни-
жестоящих животных. И это правильно лишь при той оговорке,
что при окончательной функциональной дифференцировке орган
приобретает своеобразное, специфическое только для данного
вида животного строение. Нельзя проводить сравнение между
вполне развившимся органом низшего- представителя и эмбри-
ональным органом высшего, можно сравнивать только эмбри-
ональные формы, стадии развития высших и низших представи-
телей данного типа животных.
2. В ходе эволюции организмов эволюционировали и половые
клетки. Яйцевые клетки всех животных различны. Из этого вы-
текает, что истинных анаболий, при которых можно ожидать
рекапитуляций, не существует. Из представлений Геккеля вытека-
ет, что будто яйцевые клетки всех животных должны быть иден-
тичными. Поэтому неосновательно искать, как это делал Геккель,
для яйцевой клетки и для каждой ранней стадии развития заро-
дышей соответствующих гипотетических- предков: цитею, морею,
бластею, гастрею. Вот почему некоторые выдающиеся эмбриологи,
например П. П. Иванов, не придавали филогенетического значе-
ния исходным в развитии клеткам и ранним стадиям развития.
Эволюционная систематика и эмбриологические данные говорят
в пользу этих сомнений. Так, у кишечнополостных есть и бласту-
лы, и морулы; выходит, что одни произошли от Blastaea, а другие
от Могаеа. Кишечнополостным вообще свойственно многообразие
типов развития.
3. Привлекать для доказательства правильности биогенетиче-
ского закона теорию зародышевых листков как гомологичных
образований приходится с большой осторожностью в связи
со спорностью ряда ее положений (см. гл. V).
4. Биогенетический закон не является основным законом уже
потому, что как это доказал А. Н. Северцов, он может отражать
419
только закономерности изменений по типу анаболий, однако воз-
никает много трудностей для полного согласия с теорией филэм-
бриогенеза; в частности, трудно проводить границу между при-
знаками, проявляющимися в разные стадии онтогенеза, и трудно
объяснить возможность анаболий без изменений в яйцеклетках и
в ранних стадиях развития.
5. Биогенетический закон мало применяется в ботанике.
6. Многие мысли основателей биогенетического закона, особен-
но Э. Геккеля, в свете данных современной эмбриологии оказы-
ваются слишком упрощенными и неосновательными. Геккель
рассматривал эмбриональное развитие не как самостоятельную
сторону жизни, а как придаток филогенеза, как «филогенетиче-
ский справочник» (Н. К. Кольцов). Мы видели (см. гл. IV — XI),
что на всех стадиях развития эмбрион характеризуется своими
особенностями в отношении структур, функций, интеграции.
По Геккелю, на эмбриональных стадиях организм представляет
«склад», где в неприкосновенности хранятся утратившие значение
старинные, палингенетические структуры, якобы бесполезные для
организма. Геккель определял онтогенез как «краткое извлечение
или перечень филогении». Ученые, принимающие без возражений
биогенетический закон, должны предполагать, что естественному
отбору подвергаются только признаки взрослых организмов,
будто процесс эволюции происходит в порядке приспособления
(к изменяющимся условиям среды) взрослых форм, будто начи-
нают функционировать только сформировавшиеся органы, а части
ранних зародышей, закладки органов лишены функций, что не
соответствует действительности. Помимо приведенных ранее при-
меров развития функций в онтогенезе, остановимся еще на неко-
торых явлениях.
Хорда функционирует в течение всей жизни только у наиболее
древних групп позвоночных, редуцируясь у остальных во взрослом
состоянии организма. Но нельзя делать заключения о бесполез-
ности хорды у этих животных. Анализ явлений взаимозависимости
частей в развитии организмов (гл. XI) показывает важную фун-
кциональную роль хорды как «индуктора» нервной системы.
Может быть, у предков современных хордовых первая функция
хорды была также не скелетной.
Те же рассуждения применимы к жаберным щелям позвоноч-
ных (А. Седжвик, Н. К. Кольцов). Неизвестно, каковой была их
первоначальная функция. Может быть, при своем возникновении
в эволюции эти образования еще не были по своей функции жа-
берными щелями, а играли особую роль в процессах морфогенеза.
Жаберные дуги, сосуды, нервные плакоды, головные ганглии, же-
лезы внутренней секреции развиваются в связи с жаберными
щелями. Если жабры и утрачивают функцию органов дыхания,
то они играют важную функциональную роль как структуры,
420
во взаимодействии с которыми происходят нормальные процессы
морфогенеза.
7. Если следовать биогенетическому закону без должного кри-
тического анализа его на основе данных палеонтологии и всей
зоологии, то можно иногда впасть в иллюзии, на что обращал
внимание П. П. Иванов.
Сравним некоторые признаки зародышей человека и антропо-
идов. В отличие от последних на теле человека отсутствует
сплошной густой волосяной покров. У взрослого человека тело
покрыто редкими волосами, густые волосы имеются на голове и
около рта (у мужчин). В последний месяц утробной жизни эмбри-
он человека покрыт сплошным и равномерным покровом из тон-
ких и коротких волосков (lanugo, первичный пух), сбрасываемых
до рождения. Волосы на теле взрослого и lanugo — два разных
поколения волос.
Каков волосяной покров у зародышей гориллы и шимпанзе?
Первичный пух (lanugo) есть у эмбрионов обезьян; их кожа ка-
жется голой, так как волосы короткие и редкие, за исключением
волос на голове и верхней губе. Они появляются на 5-м месяце
эмбрионального развития и расположены подобно тому, как рас-
полагаются они в усах и бороде человека, на подбородке (рис. 168),
особенно у монгольской расы. Волосы у эмбриона шимпанзе кур-
чавые, сидят пучками. У новорожденной гориллы волосы имеются
только на голове, а - голое тело покрывается волосами только
на 2-м месяце после рождения.
Какой вывод можно сделать из этих фактов с точки зрения
биогенетического закона? Гомологом волосяного покрова шимпан-
зе и гориллы у человека являются волосы на голове, усы, борода
и редкие волоски на теле, но не lanugo. Характерное для взрос-
лого человека расположение волос на голове и около рта у горил-
лы и шимпанзе — временная зародышевая фаза волосяного покро-
ва. Следует сделать вывод: не человек (существованием у его
зародыша первичного пуха) повторяет в ходе эмбрионального
развития стадию сплошного волосяного покрова антропоидов,
а, наоборот, горилла и шимпанзе в зародышевой жизни рекапи-
тулируют состояния, которые у человека остаются на всю жизнь:
распределение волос у эмбрионов гориллы и шимпанзе сохраня-
ется у человека на всю жизнь. Выходит, что не человек произошел
от обезьяноподобных предков, а они произошли от человека,
скажем ради шутки, от мужчин (а не от женщин) с характерным
для них расположением волос на голове. Этот вывод, конечно,
совершенно неправилен, он не совпадает со всеми известными
данными о происхождении человека.
Как видим, описание явлений с позиции биогенетического
закона без учета всех других данных может привести к недоразу-
мениям.
Против упрощенных взглядов на соотношение онтогении и фи-
421
г д
Рис. 168. Головы зародышей человека (Д), гориллы (5, В) и шим-
панзе (Г, Д) (по Л. Больку, 1926)
логении выступал еще Бэр в 20-х годах прошлого столетия, когда
мысли о рекапитуляциях уже высказывались. «...Вообразим се-
бе,— писал Бэр, — что птицы изучили историю своего развития и
изучают теперь строение взрослых млекопитающих и человека.
Разве не могли бы их учебники физиологии утверждать следую-
щее: эти четвероногие и двуногие животные имеют в зародыше-
вом состоянии много сходства, потому что их черепные кости
разделены, у них не имеется клюва, как и у нас на пятый или
шестой день насиживания; их конечности довольно сходны между
собой и почти так же длинны, как наши. У них на теле нет ни еди-
ного настоящего пера, но только тонкий пушок, так что наши
птенцы уже в гнезде ушли дальше, чем они; их кости менее хруп-
ки и не содержат, как наши в юности, никакого воздуха; вообще
у них отсутствуют воздушные мешки, и легкие у них не разраста-
ются, как наши в раннем возрасте; зоб у них совершенно отсут-
ствует; железистый и мускульный желудки у них слились более
или менее в один мешок; способностью летать у них обладают
только летучие мыши, которые, по-видимому, являются наиболее
совершенными, остальные же не летают. И вот эти млекопитаю-
щие животные, которые в течение столь долгого времени после
рождения не могут самостоятельно отыскивать себе пищу и никог-
да не поднимаются над землей, претендуют на более высокую
организацию, чем мы?»1
1 Бэр К. М. История развития животных. Наблюдения и размышления.
Л., 1950. С. 292—293.
422
8. В ходе эволюции организмов, принадлежащих к очень отда-
ленным друг от друга формам, могут развиваться одинаковые или
сходные структуры и функции. У отдаленных друг от друга орга-
низмов могут развиваться сходные признаки внутреннего и внеш-
него строения, причем, такой- параллелизм в развитии обусловли-
вается не родством данных организмов, не общностью происхож-
дения, а приспособлением к одинаковым экологическим условиям.
Явление это называется конвергенцией. Примеры конвергенции
многочисленны. Так, например, форма тела у некоторых рыб
(акул), рептилий "(ихтиозавров), млекопитающих (дельфинов)
очень сходна. Живая природа в> ходе эволюции оказалась «эко-
номной». Наряду с изумительным разнообразием мира живого
через все ряды животных (и растений) проходят одни и те же
структуры и функции: единая структурная единица — «клетка»,
параллельное развитие.тканей животных (А. А. Заварзин). Пора-
зительно монотонна структура ресничек Protozoa, ресничек клеток
многоклеточных и хвоста спермиев. Митоз обнаруживается почти
у всех клеток. Хлорофилл свойствен - всем зеленым растениям.
Параллелизм (Гомологичных рядов в наследственной изменчи-
вости) исследовал и Н. И. Вавилов. Явления параллельного раз-
вития различных структур и функций следует учитывать при
анализе тех или иных рекапитуляций.
ЗАКОН К. БЭРА
Как уже говорилось, идеи, лежащие в основе биогенетического
закона, и идеи о связи онтогении и филогении возникали давно,
еще до Ч. Дарвина. Очень важные недооцененные современной
наукой мысли высказывал еще К. Бэр (1828). Он сформулировал
закон зародышевого сходства, или закон Бэра:
1. То, что является общим для большой группы животных,
образуется в зародыше раньше, чем более частное.
2. Из более общего в соотношении форм образуется менее об-
щее и так далее, пока, наконец, не образуется самое специальное.
3. Каждый зародыш определенной животной формы вместо
того, чтобы проходить стадии других определенных животных
форм, все более отдаляется от них.
4. По существу зародыш высшей животной формы никогда не
бывает похож на другую животную форму, но лишь на ее заро-
дыш.
Эмбрион позвоночных есть с самого начала позвоночное жи-
вотное. При развитии любого позвоночного, утверждал Бэр,
сначала закладываются признаки класса, к которому этот заро-
дыш принадлежит; только после того как появились в ходе раз-
вития, например, признаки птицы, начинают развиваться признаки
водяной или наземной птицы; еще позже появляются в ходе
эмбрионального развития признаки, например, кур (сем. Gallinae),
423
затем признаки рода Gallus, впоследствии формируются признаки
Gallus domestica и, наконец, позже всего появляются индивидуаль-
ные признаки данного цыпленка.
Эти давние утверждения Бэра не противоречат данным совре-
менной эмбриологии. Действительно, например, у всех хордовых
животных сначала формируются общие, всем свойственные при-
знаки— развивается хорда, формируется нервная трубка, му-
скульные сегменты.
В начале этой главы уже говорилось, какое значение придавал
Дарвин обобщению Бэра, называя явления зародышевого сход-
ства самыми удивительными фактами всей естественной истории.
Нелегко, однако, и в наше время определить отношение к зако-
нам Бэра и решить, в чем они правильны и что надо в них
изменить. Бэр думал, что каждому типу животного мира свой-
ственны свои закономерности развития, особый характер разви-
тия. Конечно, в ходе эволюции отдельных ветвей животного мира
на основе естественного отбора совершенствовались и выявлялись
определенные способы развития, включая морфологию и физи-
ологию половых клеток, тип дробления и т. д. А. Н. Северцов,
оценивая закон Бэра, считал, что признаки предков, которые
образуются на ранних стадиях филогенеза, в течение последующих
геологических эпох не изменяются. Эти древние константные
признаки—признаки крупных систематических групп появляются
в эмбриональной жизни прежде других.
Разбирая разные пути изменений, имеющих эволюционное
значение (архаллаксис, девиация, анаболия), А. Н. Северцов при-
ходит к выводу, что при эволюции, идущей по пути архаллаксиса,
не имеют места ни «надставки» конечных стадий, ни удлинения
морфогенеза. При архаллаксисе не может быть никаких рекапи-
туляций анцестральных признаков у потомков. Таким образом,
признаки, развивающиеся этим способом, не подчиняются закону
Бэра. Выходит, что древние (общие) признаки не образуются в он-
тогенезе раньше новых (менее общих признаков).
Некоторые эмбриологи считают «Закон Бэра» идеалистиче-
ским, едва заслуживающим упоминания; при этом объясняется
неправильным утверждение о том, что развитие происходит от
общего к частному, что имеется направленность развития. Такая
оценка мыслей спорна. Развитие организма, в свете данных эм-
бриологии и генетики, происходит, как думал и Бэр, от общего
к частному, а не от частного к общему. Организм как целое де-
терминирован в зиготе. Зигота — начальное состояние организма—
это целостная система, а не какие-либо частности в дискретном
состоянии. Определенная, исторически возникшая проморфология
яйца данного вида животных обусловливает строго определенную
цепь морфогенетических процессов. Он говорил о разных типах
развития разных групп животных, и эта мысль, конечно, не содер-
жит ничего идеалистического.
/
424
Бэр различал «степень (ступень) развития» и «тип развития».
Понятие «ступень развития» близко к современным понятиям
«высота организации» или «сложность организации». «Более вы-
сокой будет та организация, где различные отделы всей системы
органов несходны друг с другом и каждая часть имеет более
выраженную индивидуальность, чем там, где вся организация
в целом более однородна». «Чем однообразнее вся масса тела,
тем ниже ступень образования», «...чем более развита животная
жизнь в различных своих направлениях, тем более гетерогенны
те элементы, в которых выступают проявления этой жизни».
Что такое «тип организации» по Бэру? «Это — характер рас-
положения органических элементов и органов». «Один и тот же
тип может охватывать разные ступени развития и, наоборот,
та же самая ступень развития может быть достигнута в различ-
ных типах». Сочетание ступени развития с типом и дает большие
группы животных, названные классами.
Нет оснований эти высказывания называть идеалистическими.
Он обосновывал мысль о том, что каждому типу животного мира
свойственны свои закономерности развития. Даже идеалистиче-
ские выражения вроде «тип каждого животного с самого начала
фиксирован в зародыше и управляет всем развитием», «эмбрион
позвоночного есть с самого начала позвоночное животное» и т.п.,
по существу содержат мысли, являющиеся родственными пред-
ставлениям современных эмбриологов. П. П. Иванов также гово-
рил в сущности о типах развития, употребляя выражение «уста-
новка развития». Так, у большинства моллюсков имеется уста-
новка на личинку трохофору, или veliger, но у головоногих
моллюсков развитие идет с установкой на взрослое животное.
Эти выражения при желании можно упрекнуть в телеологичности,
что сознавал и П. П. Иванов, но они не более телеологичны, чем
множество других современных понятий, например «программи-
рование р.азвития» и т. п.
Приходится сожалеть, что эмбриологи недостаточно уделили
внимания закону Бэра и классификации «установок развития».
Попыткой систематизации «типов развития» было исследование
зоолога И. И. Ежикова (1939), на основании которого он делает
вывод о том, что онтогенез изменяется с течением эволюции
не только в связи с эволюцией взрослых форм и личинок, но со-
вершенствуется и сам по себе как процесс, ведущий к формиро-
ванию дефинитивного организма.
Закон Бэра должен привлечь внимание современных эмбри-
ологов. Г. П. Короткова и Б. П. Токин (1978) отметили ошибки,
допущенные исследователями в оценке закона Бэра. Как уже
было отмечено А. С. Северцов утверждает, будто закон Бэра —
следствие «анаболической эволюции» онтогенеза, сходство заро-
дышей есть результат того, что признаки общие—это признаки
древних анцестральных форм, т. е. организмов, относящихся
425
к другому типу организмов. Однако утверждение Бэра о зароды-
шевом сходстве ограничено пределами одного и того же типа жи-
вотных. Будучи сторонником теории типов Кювье, Бэр не мог
создать гипотез, касающихся перехода от одних типов к другим.
Не учитывается и то, что закон Бэра, строго говоря, не относится
к начальным стадиям развития—к яйцу, бластуле, гаструле.
Если учесть все работы Бэра и состояние микроскопической тех-
ники его времени, становится очевидным, что он говорил о сход-
стве зародышей (позвоночных) на стадиях органогенеза. Зароды-
шевое сходство обусловлено не сходством зародышей со взрослы-
ми особями предков, а, вероятно, параллелизмом организации
зародышей.
ПЕРВИЧНАЯ И ВТОРИЧНАЯ МЕТАМЕРИЯ ТЕЛА
Эмбриологи и эволюционные морфологи все большее значение
придают крупному морфологическому обобщению, сделанному
П. П. Ивановым в первой четверти нашего века и развиваемому
советскими и иштстранными исследователями (В. Н. Беклемишев,
1952; П. Г. Светлов, 1926—1957, и др.; Е. Коршельт, 1936, и др.).
Эта теория называется теорией первичной и вторичной метамерии
тела, теорией первичной гетерономии тела, или теорией ларваль-
ных сегментов. Хотя эта теория специально не сопоставлялась
с биогенетическим законом, она, однако, основана на фактах,
имеющих прямое отношение к доказательствам единства живот-
ного мира и отражения в индивидуальном развитии филогении.
Основы теории, выходящей за рамки эмбриологии, кратко изло-
жены ниже.
Метамерное строение тела животных и особенности метамерии
у разных животных зоологи издавна рассматривали как приспо-
собление к определенному образу жизни, к определенным формам
движения (возникновение придатков для ползания при змеевид-
ных движениях), как приспособление к увеличению плодовитости
(повторное расположение гонад), как полезное совершенствова-
ние кровообращения (повторное расположение анастомозов меж-
ду главными продольными сосудами) и т. п. Одни зоологи дума-
ли, что тело первоначальна несегментированных животных бла-
годаря появлению тех или иных органов разделялось на метамеры,
так как это было полезно. Согласно таким предположениям,
логично рассматривать метамерию мезодермы, разделение ее
в ходе эмбрионального развития на сомиты как следствие возник-
ших в эволюции приспособлений; метамерия мезодермы зависит
от метамерии других органов. Другие зоологи рассматривали ме-
тамерию как способность животного пролиферировать на своем
заднем конце однородные участки, метамеры, каждый из которых
содержит определенный комплекс органов или участков общего
органа. Согласно этому взгляду в образовании метамеров глав-
426
ную роль играют мезодермальные сомиты; с появлением сомита
начинается формирование нового метамера тела.
Функциональные особенности сегментов (функциональная ге-
терономность) разнообразны у разных животных: особенности
функционирования головы, частей туловища, особенности голов-
ных придатков. У некоторых животных резко выражены отличия
в строении передних и последующих сегментов. У кольчатых чер-
вей и членистоногих особенно бросаются в глаза эти отличия.
У Arthropoda передние — головные и челюстные — сегменты рез-
ко отличны от туловищных. Они специализированы на схватыва-
нии пищи, на механической обработке пищи. Это антенны, ман-
дибулы, максиллы и другие придатки (до семи пар головных при-
датков) .
Суть теории П. П. Иванова заключается в установлении зако-
номерности, по-видимому, относящейся ко всему животному миру:
известная «функциональная» гетерономность сегментов и их при-
датков является вторичной; наряду с ней имеется особая первич-
ная гетерономность состава тела, особый способ возникновения
полимерного строения тела в онтогенезе. Своеобразие первичной
гетерономности — результат происхождения каждой группы жи-
вотных.
Следует различать ларвальные и постларвальные сегменты.
Ларвальные, передние, головные сегменты отличаются от осталь-
ных (а также и от головных, которые возникают путем процессов
вторичной цефализации их) своей закладкой, способом метаме-
ризации и дефинитивным строением. Эти ларвальные сегменты
закладываются в эмбриональном развитии животного первыми.
Остальные сегменты — постларвальные — возникают из заднего
участка ларвального тела путем нарастания. Такое типично вы-
раженное явление наблюдается у большинства аннелид, многих
членистоногих. Сначала возникает первичный отдел тела, кото-
рый представляет минимум организации для получения способ-
ности к образованию тела. Он сегментирован, и от него начина-
ется образование остальных сегментов тела. У полихет, ракооб-
разных и трилобитов остальные сегменты тела образуются уже
после вылупления эмбриона из яйца, на стадии личинки.
У аннелид и членистоногих исходной и в филогенетическом и
в онтогенетическом отношении формой метамерных животных яв-
ляется водная личинка аннелид метатрохофора, или нектохета
(рис. 169), возникающая путем сегментации тела и мезодермы не-
сегментированной трохофоры. Возникновение всего ряда метамер
у аннелид идет спереди назад: первым возникают ларвальные сег-
менты, последними — самые задние постларвальные (так происхо-
дит у большинства метамерных животных, но по различным при-
чинам в связи с преобразованием в ходе эволюции это явление
у разных групп животных изменяется). Образование постларваль-
ных сегментов, однако, начинается только после возникновения
427
Рис. 169. Схема расположения ларвальной и постларвальной
мезодермы у олигохет (по П. П. Иванову, 1945):
1 — рот, 2 — ларвальные сомиты, 3 — постларвальные сомиты, 4 — ки-
шечник
и дифференциации на метамеры ларвального кон-
ца. Мезодерма в этом начальном, ларвальном теле
сначала не сегментирована, «ведущей» в сегмента-
ции является эктодерма. Мезодерма в трохофоре
не сегментирована. Это пара сплошных мезодер-
мальных полосок. Нижнее полушарие трохофоры,
вырастая в длину, дает определенное число сегмен-
тов, каждый из которых несет пару параподий
(эктодермального происхождения). Соответственно
этой «эктодермальной сегментации» затем и мезодермальные по-
лоски разделяются на соответствующее число сегментов (3—7).
После полного сформирования сегментированного личиночного те-
ла (после возникновения ларвальных сегментов) в области особой
зоны роста, около анального отверстия начинается развитие но-
вой серии сегментов — постларвальных. Сегментация в постлар-
вальной области происходит так, что мезодерма в этой области
обособляется в виде парных групп клеток и каждая группа дает
сомит, или серия постларвальных сегментов отделяется от конце-
вой первичной мезодермальной клетки в виде отдельных крупных
клеток, из которых каждая дает сомит.
Таким образом, у полихет происходит ясное разделение об-
разования ларвальных и постларвальных сегментов во времени.
У олигохет личинка отсутствует, и личиночное тело не является
исходным организованным отделом (как трохофора у полихет),
от которого бы развивались впоследствии постларвальные сегмен-
ты. У олигохет в яйце еще до вылупления зародыша возникает
более сложная организация, состоящая из зачатков и ларвальных
и постларвальных сегментов. Ларвальная мезодерма и в этом
случае обособляется раньше постларвальной, но дифференцировка
ее потом сильно задерживается, и она начинает сегментировать-
ся уже после того, как возникает серия передних постларвальных
сегментов. В связи со способом развития (смещение ларвальных
мезодермальных полосок в промежуток между кишечником и со-
митами, загибание полосок передними концами назад) у олиго-
хет взаимное положение ларвальных и постларвальных сомитов
отличается от того, что мы видим у полихет: в каждом головном
сегменте есть и ларвальная и постларвальная мезодерма
(рис. 170).
У ракообразных ясна первичная гетерономность принципиаль-
но такого же характера, как у полихет; многие ракообразные име-
ют сегментированную личинку — науплиус. Эти сегменты
П. П. Иванов называет науплиальными. Их всегда три. На заднем
428
Рис. 170. Метаморфоз полихеты Hydroides
(по П. П. Ииаиоиу, 1937):
Л — сегментация ларвального тела; Б — образо-
вание лараподий ларвальных сегментов; В — об-
разование постларвальных сегментов
конце нНуплиуса из зо-
ны нарастания возника-
ют постларральные мета-
науплиальные сегменты.
Теорию первичной и
вторичной метамерии
П. П. Иванов распростра-
нил и на хордовых. Эту
теорию применительно к
позвоночным развивал
П. Г. Светлов (1926,
1957).
У вторичноротых ис-
ходной онтогенетической
и, вероятно, филогенети-
ческой формой их мета-
мерии является, по
П. П. Иванову и П. Г.
Светлову, трехсегментная
морская личинка типа
диплеврулы кишечножаберных-и иглокожих. Передние сегменты го-
ловы ланцетника и позвоночных рассматриваются как гомологич-
ные сегментам диплеврулы. Они составляют ларвальное тело вто-
ричноротых. Остальная же часть головы, туловище и хвост состав-
ляют постларвальное тело. Первичная гетерономия сегментации
тела характерна для хордовых не в меньшей степени, чем для ан-
нелид и членистоногих (П. Г. Светлов, 1957). Первичная гетеро-
номия тела хордовых обосновывается авторами гомологией перед-
них сегментов тела хордовых с целомами кишечножаберных и их
личинок, двойственностью главной составной части головы — моз-
га. ‘К ларвальному телу П. П. Иванов относит весь передний и
большую часть среднего мозга.
Таким образом, формируется теория метамерии, охватываю-
щая весь животный мир. Конечно, многое еще в этой теории, осо-
бенно касающееся позвоночных, далеко от полной ясности. Для
эмбриологов, в связи с вопросами о соотношении онтогении и фи-
логении, спорным является вопрос о том, возникла ли первичная
гетерономия тела всех метамерных беспозвоночных и позвоночных
как последовательные общие ступени эволюции или это явление
возникло у первично- и вторичноротых независимо. П. Г. Свет-
лов считает, что это яркий пример морфогенетического паралле-
лизма. Если это так, то процессы образования ларвальных сег-
ментов и явление первичной гетерономии тела можно рассматри-
вать как рекапитуляции лишь отдельно, в пределах первичноро-
тых, с одной стороны, и вторичноротых — с другой.
Вероятно, теория первичной и вторичной метамерии тела Ива-
нова— Светлова каким-то образом будет сязана с большим мор-
429
фологическим обобщением В. А. Догеля (1954), известным как
теория олигомеризации гомологичных органов. Установление свя-
зи этих двух теорий диктуется уже тем, что необходимо внести
новые моменты в эволюционные представления о самой метаме-
рии. Как уже говорилось, некоторые зоологи думают о разделе-
лении первоначально несегментированного тела на метамеры, в
частности в связи с необходимостью увеличения плодовитости,
так как у некоторых групп животных в связи с этим возникает
повторное расположение гонад. Может быть, однако, метамерия
(или прообраз ее) в этом смысле является исторически первичным
явлением.
Как ни спорны многие, относящиеся к теории П. П. Иванова
вопросы, нельзя не видеть, что она отражает реально существую-
щие в эмбриогенезе и филогенезе явления.
ОНТОГЕНЕЗ PROTOZOA. СПОРНЫЕ ВОПРОСЫ ПРОИСХОЖДЕНИЯ
И ЭВОЛЮЦИИ ОНТОГЕНЕЗА МНОГОКЛЕТОЧНЫХ
Созданию новых представлений о соотношении онтогении и фило-
гении препятствует неясность вопроса происхождения многокле-
точных. От принятия той или иной точки зрения по этой проблеме
зависит отношение к биогенетическому закону, трактовка теории
зародышевых листков, признание или непризнание филогенети-
ческого значения ранних стадий эмбрионального развития, гипо-
тезы о возникновении и эволюции онтогенеза.
Стройная гипотеза возникновения и эволюции онтогенеза, соз-
данная А. Н. Северцовым (см. с. 417), приобретает значение, если
принять за истину его исходное, крайне спорное предположение, что
у Protozoa нет онтогенеза, что от «одноклеточных» путем анабо-
лий (надставок стадий в виде увеличения числа клеток) произош-
ли первобытные колониальные формы, подобием которых являют-
ся современные организмы типа Pandorina, Volvox и др., и что
истинные Metazoa развились из этих колониальных форм. Но
нельзя согласиться с тем, что материнская клетка дает после де-
ления начало не зародышу, а сразу двум «взрослым» дочерним
клеткам. Понятие онтогенез Protozoa, введенное Б. П. Токиным
в 1934 г., обосновано фактами эмбриологии и общей зоологии.
Рассмотрим морфологические и физиологические показатели
онтогенеза простейших, их жизненный цикл, «возрастные измене-
ния». Если не осложнять вопрос анализом различных своеобраз-
ных циклов жизни у разных Protozoa и анализом явлений поло-
вого размножения, то под онтогенезом простейших следует пони-
мать совокупность процессов развития, начиная с формирования
нового индивидуума в результате бесполого размножения мате-
ринской клетки до очередного деления. Конечным этапом онтоге-
неза, подготовленным развитием организма, является очередное
деление. У простейших бесполое размножение составляет в отно-
430
щении численности вида ведущий способ размножения. Лишь из-
редка в серию бесполых размножений вклинивается половой про-
цесс; при этом такие явления, как конъюгация инфузорий, строго
говоря, не связаны с размножением, так как в результате их чис-
ло индивидуумов не увеличивается. Лишь в ходе эволюции жи-
вотных половой процесс оказался соединенным с размножением,
а конъюгации и родственные процессы играют какую-то физио-
логическую роль. В результате митотического деления обе воз-
никающие сестринские клетки являются «омоложенными». . При
делении происходит повышение потенциала новых особей за счег
частичного умирания живой материи; освобождающаяся при этом
энергия передается оставшейся живой материи. Размножение од-
ноклеточных происходит, таким образом, за счет структурной
энергии тех частей клетки, которые при этом неизбежно умирают
(Э. С. Бауэр). Утверждение А. Вейсмана о бессмертии Protozoa
(справедливое лишь в том смысле, что никому не удалось дока-
зать естественного закономерного прекращения образования но-
вых особей) ошибочно, если учесть, что нельзя проблему смерти
индивидуума (кролика, березы, амебы и т. д.) заменять пробле-
мой длительности размножения при каких-либо условиях: гибель
березовой рощи, гибель всех кроликов в виварии, гибель
культуры одноклеточных и т. д. Смерть одноклеточного организ-
ма и исчезновение прежней индивидуальности связаны с размно-
жением простейшего; при этом всегда происходит умирание части
живой материи, «трупные явления», заметные у некоторых одно-
клеточных при исследовании и обычными морфологическими спо-
собами (например, у грегарин).
В результате деления (у разных Protozoa неодинаково) про-
исходит потеря многих структур и их развитие в онтогенезе се-
стринских особей: многие из органоидов материнской особи резор-
бируются во время или непосредственно после деления и заменя-
ются образовавшимися заново. У разных видов это может от-
носиться к аксостилю, жгутикам, основанию ундулирующей мем-
браны и т. п. Любой вид инфузорий, делящихся поперечно, явля-
ется примером этого. У заднего конца стентора в связи с делени-
ем образуется заново перистом, преобразуется ресничный аппа-
рат, структура пелликулы, формируются заново вакуоли.
У брюхоресничных инфузорий передний конец резко отличает-
ся от заднего. На нем имеются «ротовое отверстие» и околорото-
вая спираль мембранелл. Реснички на переднем и заднем концах
разные. В связи с делением «ротовое отверстие» у задней особи
образуется заново, происходят сложные преобразования в обоих
особях — в цитоплазме, органеллах, ресничном аппарате.
Инфузория Coleps hirtus одета поясками кутикулярных пла-
стинок. Резко отличается передней конец от заднего. При делении
между рядами пластинок возникает светлая полоса, лишенная
431
Рис. 171. Coleps hirtus во время деления (из кн.
Е. Коршельта и К- Хайдера, (1910):
А — видна образовавшаяся светлая кольцевая борозда —
протоплазма, лишенная кутикулярных пластинок; Б — рас-
ширение пояса голой протоплазмы, намечается борозда де-
ления; В — появление ресничек на увеличившемся в объеме
протоплазматическом теле; Г—появление рядов пластинок
на голой протоплазме новых двух организмов, удлинение
ресничек; Д и Е —передний и задний отделившиеся инди-
видуумы; пластинки более развиты, у передней особи сфор-
мирован задний конец, у задней особи сформирован перед-
ний конец
кутикулярных образований (рис. 171). Площадь «голой» прото-
плазмы быстро увеличивается, деление продолжается 20 мин.
В течение последующих 2 ч совершаются глубокие преобразова-
ния, связанные с восстановлением структур, потерянных в резуль-
тате деления.
В.различные этапы онтогении простейшие реагируют неодина-
ково на одни и те же дозы химических и физических агентов.
Вскоре после деления молодые особи находятся в состоянии по-
вышенного обмена, а по мере старения, при приближении к смер-
ти, т. е. к новому делению, интенсивность обмена падает. В ходе
онтогении простейших происходят закономерные процессы роста,
изменения интенсивности дыхания, чувствительности к K.CN, из-
менения в синтезе белков, в отношении к ядам, к витальным кра-
сителям и т. д. Старые, достигшие границы роста инфузории ды-
шат интенсивнее своих нормальных сестер, молодые — слабее, хо-
тя в норме старческие организмы
сивной диссимиляцией (табл. 8).
Таким образом, простей-
шие, как и все животные, име-
ют онтогенез, и взгляды
А. Н. Северцова о происхож-
дении онтогенеза путем ана-
болий следует признать недо-
статочно аргументированны-
ми. Может быть, разные ветви
многоклеточных произошли
разными путями, и нельзя дать
характеризуются менее интен
Таблица 8. Поглощение кислорода
инфузориями разного возраста
в норме и после операции
(по А. М. Грановской)
Возраст Нормальные, мк8 Оперирован- ные, мк8
Старые 70 728 116 101
Молодые 104 017 63 630
единой схемы возникновения и эволюции онтогенеза.
На основе открытий в области биохимии, молекулярной гене-
тики и в других науках за последние 10—15 лет исследования по
онтогенезу простейших (как и исследования онтогении клеток
многоклеточных организмов) приобрели широкий размах. Вошли
в быт лабораторий новые термины, например «митотический
цикл», жизненный цикл». Каждый из этих терминов имеет свои
преимущества. Первый из них отражает направленность исследо-
ваний по вопросам редупликации ДНК, матричного синтеза, син-
теза белков. Этот подход в исследованиях онтогенеза простейших
и онтогении клетки тканей должен входить важнейшей состав-
ной частью (но лишь частью) цитологических, морфологических
и иных исследований. Термин «онтогенез» простейших, включаю-
щий всю совокупность процессов индивидуального развития Pro-
tozoa,„ и термин «онтогения клетки» (применительно к клеткам
тканей) более широкие, чем термины, привлекающие внимание
к какой-либо одной стороне жизни клеток. Термин «онтогенез
простейших» вошел прочно в быт научных лабораторий.
Ряд исследователей выдвинули вопросы, существенно обога-
433
щающие этот термин. Так, В. Д. Калинникова и А. С. Северцов
(1983) предлагают включать в этот термин подчас очень слож-
ные циклы простейших в ряду поколений, например, имеющих
место у жгутиковых простейших Trypanozoniatidae.
Все гипотезы о происхождении многоклеточных основаны на
учении о клеточном строении организмов, на признании простей-
ших «клетками», и большинство гипотез исходит из положения о
происхождении первобытных Metazoa от колониальных форм про-
стейших. Весь путь эволюции, согласно этим гипотезам, пред-
ставляется так.
Неорганический мир ->• живая материя ' в доклеточиой форме ->•
-» клегки -> простейшие организмы -» колонии одноклеточных -»
-> многоклеточные животные.
Эту схему эволюции принимали Э. Геккель, И. И. Мечников
и многие другие. Отличия во взглядах касались лишь конкретных
представлений о предках современных Metazoa. Геккель, как уже
указывалось, говорил о «гастрее». Этот организм развился из
более древнего — «бластеи» и представлял «мешок» с отверстием,
состоящий из двух слоев клеток. Появился он в эволюции, подоб-
но тому как происходит впячивание в одном месте бластулы у жи-
вотных, для которых характерна гаструляция по типу инвагина-
ции (см. гл. V). Полость «мешка» гастреи стала кишкой, а отвер-
стие— первичным ртом. Наружный, эпителиального типа слой
клеток — это эктодерма, а внутренний, также эпителиального ха-
рактера, стал кишечным листком, первичной энтодермой. Подоб-
ными организмами, по Геккелю, являются современные кишечно-
полостные.
И. И. Мечников в 1886 г., отрицая теорию гастреи, создал свое
представление о предках многоклеточных — теорию фагоцителлы
(см. стр. 121).
Эти предложения с физиологической и эмбриологической то-
чек зрения более обоснованы, хотя, конечно, трудно представить
себе механизм погружения клеток с поверхности и возвращения
их на поверхность, причины сбрасывания жгутов и т. д. По Гек-
келю, внутренняя полость гастреи была пищеварительной. Несом-
ненно, однако, что у всех одноклеточных, из колоний которых воз-
никла гастрея, пищеварение было внутриклеточным. Согласно
теории Мечникова пищеварительная полость (кишка) фагоцител-
лы произошла не путем инвагинации (как думал Геккель о гаст-
рее), а вторичным путем, из массы центральных клеток. Тем са-
мым совершился переход к внеклеточному полостному пищеваре-
нию. Бластопор, который, по Геккелю, явился «первичным ртом»,
согласно взглядам Мечникова, — также вторичное образование.
С эмбриологической точки зрения теория Мечникова более обос-
нована. Геккель ссылался на кишечнополостных, строение кото-
434
рых сходно с гастреей. Согласно биогенетическому закону
в развитию. кишечнополостных, которые во взрослом состоя-
нии являются как бы гастреей, должна быть стадия
инвагинационцой гаструлы, но именно для кишечнополостных та-
кой способ образования энтодермы совершенно нехарактерен,
а имеют место гаструляционные процессы, совершающиеся путем
мультиполярной иммиграции, деляминации и эпиболии (см. гл.У).
Оригинальный, но спорный взгляд на происхождение много-
клеточных был высказан А. А. Захватанным в 1919 г. У некото-
рых жгутиконосцев происходят палинтомические (повторные) де-
ления. Это не простые деления, в результате которых образуются
две особи, а цепь следующих друг за другом делений; при этом
в промежутках между делениями дочерние особи не вырастают
до размеров материнской клетки. Промежуточные продукты па-
линтомии — спороциты, а заключительные—зооспоры. Последних
может быть очень много (сотни и тысячи). Палинтомические де-
ления могут быть разных типов. У всех Metazoa многоклеточность
чередуется с одноклеточностью (яйцо). Среди простейших (типа
современных колониальных Volvocoidea) наблюдается «многокле-
точность»: это колонии, возникающие путем палинтомических де-
лений. А. А. Захваткин считал возможным сравнивать палинто-
мические деления простейших с дроблением яиц животных. В част-
ности, было установлено, что по своим геометрическим формам
процесс образования колоний Volvocoidea сходен с гомоквадрат-
ным спиральным дроблением многоклеточных. Истинная много-
клеточность с дифференциацией в структуре и функциях клеток
возникла на основе того, что в результате палинтомических деле-
ний простейших оказалось возможным формирование колоний
свободно плавающих жгутиковых «бластул», бластулоподобных
организмов типа Volvox. '
Таким образом, А. А. Захваткин установил филогенетическую
связь некоторых стадий развития, именно бластулы низших Meta-
zoa со стадиями жизненного цикла Protozoa; он представил себе
эмбриональное развитие всех первичных Metazoa как процесс ме-
тагамного палинтомического размножения, как процесс зооспоро-
образования. У современных Mastigophora нередко возникают
синзооспоры, т. е. «постоянные или временные объединения мно-
гих или нескольких зооспор, возникших в результате нерасхож-
дения и сохраняющих свои прямые расселительные функции
(функции расселения вида)». Более прочное объединение особей
таких синзооспор привело к большей интеграции их, к гистологи-
ческой и функциональной дифференцировке составляющих клеток.
В ходе эволюции могли создаться две ветви: вольвоксообразные
колонии простейших и первобытные низшие истинные многокле-
точные.
Согласно всем изложенным гипотезам колониальные формы
являются звеном в происхождении Metazoa от первобытных Рго-
435
tozoa. Существуют и новые предположения, считающие простей-
ших особой ветвью животного мира и отрицающие происхожде-
ние многоклеточных от колоний одноклеточных. Подвергается
сомнению право называть Protozoa одноклеточными организмами.
Это организмы, лишь сходные по своей структуре с клетками
многоклеточных. Считается, что первые организмы были гетеро-
трофами, и нет оснований сравнивать предков Metazoa с колони-
альными формами ныне живущих растительных организмов (типа
Volvox). Странно, что среди современных животных организмов
невозможно найти такие колониальные формы Protozoa, которые
служили бы иллюстрацией правильности гипотез о происхожде-
нии многоклеточных из колониальных форм одноклеточных. С дру-
гой стороны, ботаники в своих гипотезах о происхождении мно-
гоклеточных не имеют оснований ссылаться на организмы типа
Volvox.
Некоторым зоологам кажется неаргументированным предполо-
жение, что возникшие в эволюции одноклеточные организмы с их
интеграцией, метаболизмом и особым отношением к среде как бы
пошли вспять в своем развйтии, и их интеграция, организмен-
ность, самостоятельность стали исчезать по мере развития- мно-
гоклеточности.
Рис. 172. Инфузория Calloscolex (Л) и коловратка Hydatina (Б) (из кн.
А. А. Заварзина, 1932):
/ — околоротовые реснички, 2 — различные сократиглые системы тела, 3 — мионема глотки,
4 — туловищный венчик ресничек, 5 — сократительная вакуоль, 6 — макронуклеус, 7 — зад-
няя кишка, 8 — ротаторный орган, 9 — мышцы коловратки, 10 — желудок, 11— нервная си-
стема, 12 — половая железа, 13— анальное отверстие, 14 — кишка
436
Если бйать современных Protozoa, то даже самые низшие из
них имеютючень сложное строение и ярко выраженую организ-
менность. Бесспорно, что некоторые инфузории по уровню инте-
грации могут превосходить многих многоклеточных. Примером
может служить- инфузория Cctloscolex, по сложности не уступаю-
щая коловратке Hydatina. Внешне морфологически эти организ-
мы сходны (рис. 172). Некоторые эмбриологи и до сих пор разде-
ляют старую гипотезу зоологов, обоснованную еще И. Деляжем
в 1896 г. Согласно ей предки Metazoa приобрели многоядерную
структуру (наподобие современных опалин), и различия в разных
частях их тела достигли высокой степени. У них и без клеточной
структуры могли быть выраженными органы — proctodeum, глот-
ка и т. п. Достаточно предположить возможность обособления
клеточных территорий вокруг ядер, чтобы представить себе по-
явление тканей и органов.
ПОИСКИ НОВОГО В ТЕОРИИ ЭВОЛЮЦИИ ОНТОГЕНЕЗА
С 30-х и 40-х годов, когда у эволюционных морфологов доминиро-
зали замечательные идеи А. Н. Северцова, в биологии произошло
иного событий, возникли и новые гипотезы, касающиеся проблем
эволюции, в частности проблем происхождения и эволюции онто-
генеза.
Все понятия, связанные с проблематии соотношения индивиду-
эльного и исторического развития, до недавнего прошлого созда-
эались на основе изучения половой репродукции многоклеточных,
гх гамет, бластул и гаструл, зародышевых листков, органогене-
за и т. п. Эти понятия не могли быть применены к развитию про-
стейших и тем более прокариотических организмов. В гипотезах
эб эволюции онтогенеза сопоставлялись почти исключительно
только явления, происходящие при половой репродукции.
Фактически обособленными от излучения закономерностей ин-
дивидуального развития оказались бластогенезы, тератогенезы,
юлиэмбриония, соматический эмбриогенез, физиологическая и
эепаративная регенерация. Оторванными друг от друга оказались
штегральные «части» онтогенеза, отдельно изучались эволюция
зроцессов дробления, эволюция восстановительных морфогене-
юв и т. д. 'Конечно, эмбриология обогащалась новыми фактами
1 идеями и в такого рода исследованиях. Однако давно уже на-
|рела необходимость в синтезе данных об эволюции различных
юрфогенетических процессов, встречающихся в жизненном цик-
ie одной и той же особи.
Г. П. Короткова (1979) обосновывает возможность и необхо-
[имость чередования в историческом развитии разных способов
зепродукции. Необходимость изменения характера репродуктив-
юго деления возникает тогда, когда изменившиеся абиотические
температура, соленость, загрязнения и др.) или биотические (ви-
437
русные или бактериальные инвазии, паразитизм и др.) факторы
среды блокируют сложившийся способ репродукции. В арсенале
вида и особи всегда имеется морфогенетический резерв, который
может быть использован для восполнения необходимой числен-
ности вида и для выживания особи. Переходы от одного типа ре-
продукции к другой естественно возможны не для каждого пред-
ставителя вида или рода, но тем не менее среди близкородст-
венных видов и родов такие возможности всегда имеют место за
счет природного полиморфизма популяций и видов. Частным слу-
чаем такого полиморфизма оказывается внутривидовая и внутри-
родовая вариабельность регенерационных свойств. Для того что-
бы осуществлялось чередование половой и агамной репродукции,
необходима преемственность тенденций к образованию гамет или
их аналогов, а также преемственность к возобновлению агамного
размножения. Возможность образования гамет не может исчез-
нуть у агамно репродуцирующихся организмов, потому что и у
растений и у животных геномы всех клеток сходны, а существую-
щие в организме растений меристематические клетки, у живот-
ных стволовые клетки, обладающие достаточно широкими формо-
образовательными потенциями, всегда обеспечивают возможность
переключения таких клеток на путь гаметогенеза.
Тенденция к возобновлению агамной репродукции сохраняет-
ся у растений и животных благодаря резервам восстановительных
морфогенезов или эмбриональных регуляций.
Появляющееся в ходе эволюции на какой-то исторический пе-
риод времени бесполое размножение или полиэмбриония накла-
дывают определенный отпечаток на возобновляющийся после это-
го периода половой эмбриогенез, на способы обособления гоноци-
тов, организацию гонад, различные особенности эмбриогенеза.
Г. П. Короткова доказывает наличие коррелятивной изменчи-
вости половых и соматических морфогенезов. Она впервые привле-
кает серьезное внимание эмбриологов, генетиков и эволюционных
морфологов к этому явлению в связи с созданием теории фазной
эволюции онтогенеза.
Создавая представление о фазном характере эволюции онто-
генеза, при котором неизбежно допущение чередования агамной
и половой репродукций и учитывая факты коррелятивной измен-
чивости анатомической организации, способа агамной репродук-
ции (или восстановительного морфогенеза) и характера половой
репродукции, Короткова предположила, что фазы эволюции он-
тогенеза не случайны, а закономерно связаны с переходами на но-
вый уровень структурной организации. Согласно этой гипотезе
этапы макроэволюции онтогенеза, в течение которых осущест-
влялось возникновение более сложных структурных единиц в со-
ставе организмов (например, переход от прокариот к эукариотам,
от одноклеточных к многоклеточным, от одиночных к колониаль-
ным, от однослойных к многослойным и т. п.), происходили бла-
438
годаря включению в жизненный цикл агамной репродукции. Имен-
но соматический морфогенез в виде незавершенной агамной ре-
продукции, незавершенной регенерации и т. п. был той реальной
основой, на которой был возможен переход к новому, более слож-
ному структурному уровню организации, а также осуществление
биологической полимеризации. Логическим следствием предпола-
гаемого характера эволюции онтогенеза был отказ от линейного
типа преобразования индивидуального развития и от надставоч-
ного способа усложнения полового эмбриогенеза.
Возникает вопрос — как регулируется и как осуществляется
переход от одного способа репродукции к другому? «Виновника-
ми» являются и внутренние (гормональные, зараженность пара-
зитами и др.), и внешние факторы (механические раздражения,
загрязненность воды и др.). Ответить на этот вопрос в настоящее
время можно лишь в предварительной форме.
Г. П. Короткова предполагает, что кратковременные и непо-
стоянные переходы, скажем, от половой репродукции к бесполой
с возобновлением в следующем цикле прежнего типа эмбриогене-
за, конечно, не должны затрагивать генома и они не влияют на ха-
рактер гамето- и эмбриогенеза. Это репродуктивные модификации.
Однако как гормональные, так и иммунологические сдвиги могут
повлечь за собой и мутации, те могут вызывать длительные, на-
следственно закрепленные изменения в структуре жизненного
цикла, например появление устойчивого метагенеза, что обяза-
тельно повлечет за собой изменения в гаметогенезе и половом эм-
бриогенезе по сравнению с таковыми гаметически репродукциро-
вавших предков.
В соответствии с развиваемой концепцией фазного преобразо-
вания индивидуального развития онтогенез рекапитулирует не
только черты полового эмбриогенеза, но и черты бластогенеза
предковых форм. Естественно, что при таких закономерностях
эволюции онтогенеза не может осуществляться прямая рекапи-
туляция черт развития отдаленных предков, вернее она ограниче-
на лишь пределами того исторического периода развития, в тече-
ние которого совершалось прохождение одной и той же фазы
филогенеза. Явления дивергенции и параллелизма как общие за-
кономерности эволюционного процесса находят, по мнению Ко-
ротковой, свое отражение и в эволюции онтогенеза. Параллелизм
в эволюции морфогенетических процессов обусловлен сходством
закономерностей перестройки целостного репродуктивного мор-
фогенеза, а дивергенция — расхождением морфогенетических про-
цессов из-за несовместимости условий для их реализации. Несовме-
стимость условий для осуществления половой и агамной репродук-
ции— частое явление в жизненном цикле растений и животных.
Поэтому не удивительно, что половой эмбриогенез и бластогенез—
это чаще всего чередующиеся события в жизненном цикле особи или
в ходе исторического развития вида.
439
Глава XVIII
НЕКОТОРЫЕ ВОПРОСЫ ЭМБРИОЛОГИИ РАСТЕНИИ
Эмбриология растений — наука о закономерностях зарождения и
первых этапах развития растений. Однако эмбриология рассмат-
ривает не только развитие зародыша (эмбриогенез), но и фор-
мирование мужских (тычинки) и женских (завязи и семяпочки)
структур, а также процессы, происходящие в цветке. Первые
этапы онтогенеза, т. е. эмбриогенез, трудно понять без постэмб-
рионального развития (формирование проростка). Вот почему
необходимо использовать структурно-функциональный подход
к изучению структур, развивающихся в семени. Развитие всех
структур, формирующихся в цветке, обычно взаимообусловлено
и тесно связано с экологией. Задача эмбриологии заключается
в выяснении закономерностей процессов воспроизведения (поло-
вого и бесполого) и в разработке способов управления этими про-
цессами. ,
Эмбриология растений — наука относительно молодая, как на-
учная ботаническая дисциплина сформировалась лишь в конце
XIX — начале XX в. В современной эмбриологии растений до-
вольно отчетливо выявляются два основных направления: 1 —
выявление закономерностей эволюционного процесса и решение
вопросов филогении и систематики (основным методом этих ис-
следований все еще остается сравнительно-описательный); 2 —
выявление закономерностей индивидуального развития, касающих-
ся воспроизведения растений и имеющих широкое применение
в практике (методы исследования — сравнительно-описательный и
экспериментальный). В подавляющем большинстве эмбриологиче-
ских исследований в качестве объекта избраны покрытосеменные,
или цветковые, растения. Покрытосеменные—самый большой отдел
растительного мира. Все важнейшие культурные растения относятся
к цветковым растениям. Они отличаются от остальных групп ра-
стений наличием в цветке камер, образованных тканями завязи,
в которых развиваются семена. Оказавшись в этом своеобразном
влажном вместилище, семена могут формироваться быстро, с ми-
нимальной затратой строительных материалов, что оказалось ис-
ключительно важным биологическим преимуществом, позволив-
шим цветковым существенно повысить семенную продуктивность.
Улавливание пыльцы у покрытосеменных осуществляется специа-
лизированными участками плодолистика — рыльцами, что также
является их характерной особенностью. Кардинальное отличие
покрытосеменных от других отделов растений — двойное оплодо-
440
творение. Эндосперм покрытосеменных в отличие от голосеменных
образуется в результате полового процесса.
ЦВЕТОК
Цветок — совокупность стерильных и фертильных структур, раз-
вивающихся на цветоложе. Стерильными структурами цветка яв-
ляются чашелистики и лепестки. К фертильным относятся ты-
чинки и плодолистики, или карпеллы. Границы между фертиль-
ными и стерильными структурами условны и относительны.
В гинецее обычно различают нижнюю фертильную часть —за-
вязь, и верхнюю стерильную — столбик. Верхняя часть столбика
преобразуется в рыльце. В завязи различают гнездо (или гнезда)
и стенку. В гнезде завязи формируются семяпочки (семязачатки),
место их возникновения называется плацентой. Существуют раз-
личные типы плацентации (А. А. Тахтаджян, 1945).
В тычинках и плодолистиках формируются мужские и жен-
ские половые клетки — гаметы. В завязи происходят процессы
опыления, оплодотворения и развития зародыша.
СПОРОГЕНЕЗ, ГАМЕТОФИТОГЕНЕЗ И ГАМЕТОГЕНЕЗ
В индивидуальном развитии покрытосеменного растения, размно-
жающегося половым путем, различают два этапа — спорофит и
гаметофит. Мейоз обеспечивает чередование этих этапов и при-
водит к образованию гаплоидных половых клеток (гамет). Орга-
низм, развивающийся от зиготы до образования спор, называет-
ся спорофитом. Гаплоидный этап развития растения называется
гаметофитом (от микро- и мегаспор до зиготы). В эволюции от
мхов к покрытосеменным происходят постепенная редукция гаме-
тофита и развитие спорофита, достигающие у покрытосеменных
максимального выражения. Спорофит у высших чаще всего пред-
ставлен деревьями, кустарниками и травами. Гаметофиты реду-
цированы и образуются в результате небольшого числа митоти-
ческих делений. Развитие женского гаметофита (зародышевого
мешка) у большинства сведено к трем делениям, а мужского
(пыльцевого зерна) —к двум (рис. 173).
Строение семяпочки. Семяпочка состоит из нуцеллуса (мега-
спорангия), окружающих его покровов (интегументов), пыльце-
входа (микропиле), халазы и семяножки (фуникулуса), с по-
мощью которой она связана с плацентой (рис. 174). В бугорке
семяпочки дифференцируется нуцеллус. В зависимости от степе-
ни развития нуцеллуса выделяют различные типы семяпочек.
В основании нуцеллуса появляются покровы. Семяпочки бывают
однопокровные и двупокровные, реже встречаются три или че-
тыре интегумента (ариллусы, ариллоиды). Встречаются семяпоч-
ки без интегументов (некоторые паразитные растения). Однопо-
441
A
Материнские клетки
микроспор (Zn)
• Диады
Трехклеточные пыльцевые зерна
Тетрадь!
Б
Мегаспоры (л)
I
/о\ Двуядерный заро-
дышевый мешок
/ р | Чртырехьядгоный
1 v---LJ 'зародышевый мешок
Семиклеточный весь-
Г°о I - миядерный зароды-
* шевый мешок
Рис. 173. Образование пыльцевых зерен (А) и зародышевых мешков (Б) у по-
крытосеменных растений (В. П. Банникова, О. А. Хведынич, 1982, с допол-
нениями):
/ — зародышевый мешок, 2 — вегетативная клетка, 3 — спермин, 4 — генеративная клетка
кровность считается прогрессивным признаком, она характерна
для большинства спайнолепестных. В верхней части семяпочки
интегументы (один или несколько) образуют микропилярный ка-
нал— микропиле. У разных видов растений микропиле по строе-
нию разное (экзостом, эндостом), а в случае недоразвития инте-
гументов оно практически не образуется. Часто микропиле запол-
нено сосочками или железистыми волосками, выделяющими слизь,
что способствует прохождению пыльцевой трубки. В процессе раз-
вития семяпочки могут образовываться и другие структуры: эпи-
стаза, оперкулюм, нуцеллярный колпачок, гипостаза, подиум, по-
стамент и обтуратор. Из плаценты в семяпочку через фуникулюс
проходит проводящий пучок в халазальную часть семяпочки, где
442
часто разветвляется. Ино-
гда он входит в нуцеллус
и интегументы. Существу-
ют виды растений, в се-
мяпочке которых отсут-
ствует проводящая систе-
ма, например у орхидных.
Мегаспорогенез. В апи-
кальной части нуцеллуса
в субэпидермальном или
более глубоких слоях
дифференцируется одна
или несколько археспори-
альных клеток (рис. 175).
Образование археспория
сопровождается увеличе-
нием размеров клеток,
числа органелл и мем-
бранных систем. Харак-
терной особенностью ар-
хеспориальных клеток
является сосредоточение
пластид, митохондрий и
эндоплазматического ре-
тикулума в апикальной
части, что говорит об их
полярности, которая наи-
более явственно проявля-
ется в мегаспороците.
Существуют два типа
Рис. 174. Строение анатропной семяпочки на
примере лотоса Nelumbo nucifera (по Т. Б. Ба-
тыгиной, Колесовой, В. Е. Васильевой, 1983):
1 — наружный интегумент, 2 — внутренний интегу-
мент, 3— зона обтуратора, 4 — нуцеллус, 5 — нуцел-
ляриый колпачок, 6 — гипостаза, 7 — зародышевый
мешок, 8 — яйцевой аппарат, 9 — сииергнды, 10 —
полярные ядра, // — центральная клетка, 12— анти-
поды, 13 — пыльцевая трубка, входящая в микропиле
археспория: одноклеточный и многоклеточный. Археспориальная
клетка может непосредственно дифференцироваться в мегаспоро-
цит (материнскую клетку мегаспор) или может претерпевать одно
или несколько делений, образуя париетальную (кроющую) клетку
или париетальную ткань. Редукция археспория идет по линии
уменьшения числа археспориальных клеток или уменьшения числа
делений в них; в последнем случае археспориальные клетки непо-
средственно становятся мегаспороцитами. Процесс мейоза сопро-
вождается интенсивным накоплением нуклеопротеидов, крахмала и
других включений. В результате первого мейотического деления,
как правило неравного, образуется диада, следующее деление при-
водит к образованию тетрады мегаспор (рис. 175). По характеру
протекания этих делений и расположению клеточных перегородок
определяют линейные, Т-образные, квадратные, тетраэдрические и
тетрады с косым расположением перегородок. У большинства по-
крытосеменных три мегаспоры разрушаются, а из одной развивает-
ся женский гаметофит (зародышевый мешок типа Polygonum).
443
19
Рис. 175. Мегаспорогенез и развитие женского гаметофита (зародышевого меш-
ка) пшеницы (по Т. Б. Батыгиной, 1974):
1—3 — верхняя часть нуцеллуса с археспориальной клеткой, 4—12 — мейоз н образование
тетрады мегаспор, 13 — дегенерация микропилярных мегаспор, 14 — 15 — двухъ- и четырехъ-
ядерные зародышевые мешки 16, 17 — сформированный зародышевый мешок, 18, 19 — зре-
лый зародышевый мешок
Классификация типов зародышевых мешков, предложенная
И. Д. Романовым (1945, 1981), на наш взгляд, в настоящее время
наиболее полная из всех имеющихся. В ее основу были взяты
следующие критерии: 1 — число мегаспор, образующих зароды-
шевый мешок; 2 — число митозов, происходящих после мейотиче-
ских делений; 3 — поведение ядер, определяющее особенности
конструкции (организации) зародышевого мешка. Согласно пер-
вому критерию (т. е. пр происхождению) выделяют три типа за-
444
родышевых мешков: моноспорические, биспорические и тетраспо-
рические. Мегаспорогенез у большинства покрытосеменных харак-
теризуется заложением перегородок при обоих делениях в мейозе
и образованием обособленных спор. Эта особенность, как полагает
И. Д. Романов (1981), указывает на его примитивность у семен-
, . Тип ЗюрооышеВого мешка MKM деление деление деление деление деление 30«AbtU ЗоИды- шеБый мешок
ИЯШ5П1 1 8HJIS8W1t|oUa0H0^ Polygonum-тип 0 © I О Q ©
Oenothera-тип 0 © 0 ©
Бисгофические вд[зодышс6ы<! мешки ДIlium тип 0 © 0 © 0 ©
Dodostemum- nodmun 0 © 0 0 0 ©
Balsam ita (Anthem is)-тип 0 0 0 Q 0 ©
Drusa-mun 0 © 0 Q
CEentonla подтип 0 0 Q © ©
3 X 3 fritiWaria-mun 0 0 Q © ©
Of S су Plumbagella-mun 0 0 0 0 ©
3 О 3 Adoxa-mun 0 0 0 ©
а Tulipa -tetraphytta -тип 0 0 0 ©
ал 3 Ж Eriostemones-mun 0 0 0 0
ОТ 2Т 3 о Penaea-mun 0 0 Q © 0^
о Е" & Plumbago-тип 0 0 0
Peperomla-mun 0 0 0 ©
AcalyphaEanceolaia -подтип 0 0 G ©. Ф
Рис. 176. Классификация типов зародышевых мешков (по
И. Д. Романову, 1971, с дополнениями)
445
ных растений. Предпосылками развития зародышевого мешка,
образуемого несколькими мегаспорами, является подавление за-
ложения перегородок при мейозе. И. Д. Романов выделил 13 ти-
пов зародышевых мешков. Наиболее распространенный тип
Polygonum-тип (рис. 176).
Гаметофитогеиез. Рассмотрим развитие и строение зародыше-
вого мешка Polygonum-типа. Моноспорический семиклеточный,
восьмиядерный зародышевый мешок образуется обычно из хала-
зальной мегаспоры (материнской клетки зародышевого мешка).
Первое деление материнской клетки зародышевого мешка служит
началом формирования женского гаметофита. В результате деле-
ния образуется двуядерный ценоцит — двухъядерный зародышевый
мешок. Два последующих деления приводит к образованию вось-
миядерного зародышевого мешка, на двух полюсах которого рас-
полагаются по четыре ядра. Далее одно из ядер каждой группы
двигается в центр зародышевого мешка, после чего начинается
процесс заложения клеточных перегородок. Образуется сформи-
рованный зародышевый мешок, состоящий из яйцевого аппарата,
центральной клетки с двумя ядрами и антиподального аппарата.
В результате дальнейшей дифференциации и специализации фор-
мируется зрелый зародышевый мешок, варьирующий у разных
видов по размеру, форме, положению в семяпочке, а также по
характеру строения его элементов (см. рис. 172, 175). Яйцевой
аппарат в зрелом зародышевом мешке обычно состоит из яйце-
клетки (см. рис. 173, 175), лежащей между двумя синергидами и
всегда имеет более крупное ядро, чем синергиды. Обычно ее ядро
располагается в апикальной части клетки, в базальной — круп-
ная вакуоль. В синергидах расположение обратное — ядро в ба-
зальной части и вакуоли — в апикальной. В базальной части си-
нергид дифференцируется нитчатый аппарат, нередко образуются
крючкообразные или зубовидные выросты (см. рис. 175). Яйце-
клетка имеет оболочку, пронизанную плазмодесмами, лишь в ба-
зальной части, а в апикальной части она представлена только
плазмалеммой. В яйцеклетке и синергидах имеются различные
клеточные органеллы. В яйцеклетке некоторые из органелл слабо
развиты, рибосомы обычно не сгруппированы в полисомы, в мито-
хондриях слабо выражены кристы и т. д. Такая морфология кле-
точных органелл позволила исследователям считать яйцеклетку
малоактивной. Синергиды боЛее богаты клеточными органеллами
и считаются более активными клетками. Нитчатый аппарат пред-
ставляет выросты оболочки клетки синергиды и состоит из геми-
целлюлозы и пектиновых веществ. Синергиды служат главным об-
разом для проведения пыльцевых трубок. Центральная клетка —
самая большая сильновакуолизированная клетка зародышевого
мешка (см. рис. 173, 175). Она как бы охватывает весь яйцевой
и антиподальный аппараты. Обычно в ее центре расположены два
гаплоидных полярных ядра (при развитии зародышевого мешка
446
по Polygonum-типу). По состоянию органелл цитоплазмы она явля-
ется метаболически активной. Характерная черта центральной клет-
ки — большое количество диктиосом. Центральная клетка после
оплодотворения дает эндосперм. Количество и форма антипод у
разных видов неодинаковы. У большинства покрытосеменных они
эфемерны, хотя известны случаи, когда в процессе развития за-
родышевого мешка они увеличиваются в числе и размерах (на-
пример, у бамбука, мятлика и др.). Ядра антипод часто бывают
высокой степени плоидности. Присутствие и число антипод зави-
сит в определенной степени от типа зародышевого мешка; имеют-
ся зародышевые мешки, в которых антиподы не образуются
(Oenothera-тип, Acalypha-тип и др.). Большинство исследователей
в настоящее время склоняются к тому, что зародышевый мешок
имеет собственную оболочку на протяжении всего развития вплоть
до образования в нем зрелого зародыша и эндосперма. Это под-
тверждается данными по выделению целых зародышевых мешков
из семяпочек путем мацерации. Существуют различные точки зре-
ния относительно происхождения зародышевого мешка (В. А. Под-
дубная-Арнольди, 1976; А. Г. Смирнов, 1982, и др.).
Развитие и строение тычинки. Формирование тычинки, пыль-
ника— это единый процесс развития сложной интегрированной
системы. На первых этапах развития тычиночный бугорок со-
стоит из меристематических клеток. Одновременно с обособлени-
ем пучка происходит образование обычно четырех лопастей
(гнезд). Каждая лопасть преобразуется в микроспорангий. Два
латеральных микроспорангия составляют теку, теки соединены
между собой связником (рис. 177). У некоторых видов (например,
злаки) в субэпидермальном слое меристемы каждой лопасти диф-
ференцируются археспориальные клетки, отличающиеся от осталь-
ных величиной, формой и структурой. Далее деление археспория
приводит к образованию париетального слоя и спорогенной ткани.
Париетальный слой в результате деления образует стенку пыль-
ника, представленную у разных видов различным числом слоев
клеток. Спорогенная ткань в результате сложных преобразований
(мейоз и др.) дает пыльцу. В развитии пыльника различают три
периода: премейотический, мейотический и постмейотический.
В премейотический период пыльник характеризуется митотиче-
ской активностью, в результате чего формируются стенка пыль-
ника и пыльцевые гнезда (сформированный пыльник). В мейоти-
ческий период идет дальнейшая дифференциация слоев стенки
пыльника и мейоз в микроспороцитах (материнских клетках мик-
роспор), в результате чего образуются тетрады гаплоидных мик-
роспор. В постмейотический период созревает пыльца и, как пра-
вило, идет дегенерация отдельных слоев стенки пыльника (зре-
лый пыльник) (см. рис. 174, 177).
Формирование стенки микроспорангия. Развитие спорогенной
ткани тесно связано с формированием отдельных слоев стенки и
447
Рис. 177. Развитие пыльника у молочайных (Euphorbiaceae) (поТ. Б. Батыгиной,
Т. Е. Колесовой, 1983):
А — сформированная стенка пыльника н спорогенная ткань; Б—Г — фрагмент стенки пыль-
ника на стадии: Б — синапсиса в микроспороцитах, В — тетрад микроспор; Г — одноклеточ-
ной пыльцы (в тапетуме появляются орбикулы); Д— стенка зрелого пыльника с кутннизи-
рованным экзотецием, эндотецием с фиброзными утолщениями, зрелые, двуклеточиые пыль-
цевые зерна с крахмалом; Е — поперечный срез зрелого пыльника: теки, состоящие из двух
гнезд каждая, соединены связником; 1— археспорий, 2 — микроспороциты, 3 — тетрады
микроспор, 4 —- одноклеточные пыльцевые зерна, 5— двуклеточные пыльцевые зерна, 6 —
экзотеций, 7 — эндотеций, 8 — средние слои, 9 — тапетум, /0 —орбикулы, // — связник,
12 — тека, 13 — гнездо пыльника
поэтому их изучение необходимо проводить сопряженно. Экзотеций,
(эпидермис) — самый наружный слой пыльника, в основном он
несет защитную функцию. Эндотеций характеризуется наличием
фиброзных утолщений (поясков) для вскрывания пыльника. Под
эндотецием формируются один или несколько средних слоев. Та-
петум — самый внутренний слой, непосредственно примыкающий
к спорогенной ткани. Клетки тапетума изначально одноядерные,
но в процессе развития пыльника они становятся двухъ- и много-
ядерными; иногда происходит слияние ядер. Реорганизация и раз-
рушение тапетума происходят на разных этапах развития пыль-
ника. В клетках тапетума по сравнению с другими слоями обна-
ружено много органелл. Они богаты нуклеопротеидами, жирами,
различными ферментами, витаминами и включениями. Все это
говорит о высокой метаболической активности этой ткани. При
нарушении нормальной дифференциации тапетума образуется сте-
рильная пыльца. Существуют различные типы тапетума. О. П.Ка-
мелина (1980) предложила классификацию тапетума, в которой
448
за основу принята и несколько дополнена классификация К. Кар-
ниеля (1963). Основными типами тапетума являются клеточный
секреторный тапетум и периплазмодиальный тапетум.
Классификация типов развития стенки пыльника. В 1963 г.
Т. Б. Батыгина, Э. С. Терёхин, Г. К. Алимова, М. С. Яковлев
предложили морфогенетическую классификацию типов развития
стенки пыльника и выделили четыре основных типа формирова-
ния ее у цветковых растений. Критериями при выделении типов
явились центробежное (Е. Варминг, 1873) и центростремительное
(К- Карниель, 1961) развитие стенки спорангия и происхождение
тапетума. Имеется и другая классификация (Дэвис, 1966), с на-
шей точки зрения не совсем удачная (Т. Б. Батыгина, И. И. Шам-
ров, 1981).
Микроспорогенез. В результате развития спорогенной ткани
в пыльнике (в премейотический период) образуются материнские
клетки микроспор (микроспороциты). Микроспороциты по своему
строению (плотности цитоплазмы, наличию в ней пластид, мито-
хондрий, рибосом, диктиосом и т. д., размерами ядер) отличают-
ся от клеток спорогенной ткани. Микроспороциты связаны между
собой плазмодесмами. Во время мейоза происходит дальнейшее
разрастание пыльника и сильное изменение микроспороцитов.
Профаза мейоза, как правило, является наиболее продолжитель-
ной стадией. В результате мейоза микроспороциты дают начало
тетрадам микроспор с гаплоидным набором хромосом (см.
рис. 172, 1173, 177). В зависимости от характера заложения пере-
городок в микроспороцитах различают три типа образования
тетрад: сукцессивный (последовательный), симультанный (одно-
временный) и промежуточный. Тетрады микроспор вскоре после
их образования начинают обособляться, а в дальнейшем они рас-
падаются на микроспоры. Характерная особенность микроспоро-
генеза у большинства видов растений — наличие каллозных обо-
лочек (полисахаридной природы) у археспориальных клеток, мик-
роспороцитов, клеток диад и тетрад. Во время мейоза происходят
значительные изменения и в стенке пыльника, выражающиеся
в сильной дифференциации и дегенерации отдельных ее слоев.
Развитие пыльцевого зерна. Развитие пыльцевого зерна (муж-
ского гаметофита) начинается с деления микроспоры (либо одно,
либо два деления). В результате первого деления, чаще диффе-
ренцирующего, образуются генеративная, меньших размеров, и
вегетативная, больших размеров, клетки. Особенностью первого
деления является асимметричность веретена (см. рис. 173). Ге-
неративная клетка в отличие от вегетативной характеризуется
малым количеством цитоплазмы, в которой, однако, имеются раз-
личные органеллы и включения. Одной из интересных особенно-
стей генеративной и вегетативной клеток является движение их
в пыльцевом зерне, особенно постепенное «внедрение» генератив-
ной клетки в цитоплазму вегетативной.
449
Гаметогенез (спермиогенез). В результате деления генератив-
ной клетки образуются два гаплоидных спермия. Это деление мо-
жет происходить либо в пыльцевом зерне, либо в пыльцевой
трубке, что определяет, будут лн зрелые пыльцевые зерна двух-
или трехклеточными (см. рис. 173). Спермин различаются по фор-
ме и величине, они бывают бедно- и богатоплазменные, в созрев-
ших пыльцевых зернах органеллы находятся в неактивном со-
стоянии, в состоянии телофазы или интерфазы. Зрелые пыльцевые
зерна имеют оболочку (спородерму) и аппертуру (поры, борозды,
щели), которая служит, для прорастания пыльцевой трубки.
В мужском гаметофите покрытосеменных (пыльце) в отличие от
голосеменных наблюдается последняя степень редукции, которая
приводит иногда лишь к одному делению. Вопрос о гомологиза-
ции отдельных структур пыльцевого зерна остается дискуссион-
ным.
ОПЫЛЕНИЕ И ОПЛОДОТВОРЕНИЕ
Подавляющему большинству покрытосеменных свойственно поло-
вое размножение, определяющим моментом которого является
оплодотворение. Оплодотворение — это сложный процесс, включа-
ющий две основные фазы: прогамную (от попадания пыльцы на
рыльце до вхождения пыльцевой трубки в зародышевый мешок)
н постгамную — двойное оплодотворение.
Различают два основных типа опыления: перекрестное (алло-
гамия, ксеногамия), когда рыльце одного растения опыляется
пйльцой другого; самоопыление (автогамия) — рыльце опыляется
пыльцой своего цветка. Перекрестное опыление имеет большое
биологическое значение, обеспечивая гетерозиютность популя-
ции— основу естественного отбора. Приспособления, препятству-
ющие самоопылению: двудомность, дихогамия, гетеростилия, са-
монесовместимость. Способы перекрестного опыления: анемофи-
лия, гидрофилия, энтомофилия, орнитофилия, анималофилия.
Прорастание пыльцы на рыльце, рост пыльцевой трубки
в столбике и завязи. Начало прорастания происходит в резуль-
тате взаимной активации пыльцы и рыльца. Путем выпячивания
через аппертуру и дальнейшего разрастания интины образуется
пыльцевая трубка, в которую переходит генеративная клетка или
спермин и ядро вегетативной клетки. Пыльцевые трубки растут
по поверхности стенок канала столбика, в самом канале или меж-
ду клетками проводниковой ткани. Далее, перейдя в завязь, они
растут в сторону зародышевых мешков, что, возможно, обуслов-
лено хемотропизмом (см. рис. 173, рис. 178). Цитоплазма пыль-
цевой трубки находится в ротационном движении, образуя два
тока: восходящий (к кончику пыльцевой трубки) и нисходящий.
Пыльцевая трубка ритмично растет своим кончиком. Во время по-
коя в кончике пыльцевой трубки идет синтез веществ, из кото-
450
рых во время роста стро-
ится ее оболочка (М. С.
Навашин, 1973). Базаль-
ные участки пыльцевой
трубки периодически от-
секаются каллозными
пробками и позднее деге-
нерируют. Основная мас-
са активной цитоплазмы
пыльцевой трубки вместе
со спермиями или генера-
тивной клеткой и вегета-
тивным ядром (если оно
выходит в пыльцевую
трубку) сосредоточены
вблизи ее кончика.
Движение генератив-
ной клетки, спермиев и
вегетативного ядра в
пыльцевой трубке. Спер-
мин преодолевают путь
от рыльца до женских
половых клеток, находясь
Рис. 178. Пыльцевые трубки в пестике пше-
ницы (по Т. Б. Батыгиной, 1962)
внутри пыльцевой трубки. Дискус-
сионным является вопрос: активно или пассивно они пере-
мещаются. Следует учесть, что органов движения — жгутиков,
ресничек — спермин не имеют. Наиболее убедительными являют-
ся представления С. Н. Коробовой (1977), полагающей, что кон-
чик пыльцевой трубки, отграниченный каллозной пробкой, следует
рассматривать как своеобразную клетку, в которой движение ге-
неративной клетки или спермиев, совершающееся аналогично дви-
жению ее собственного — вегетативного — ядра, происходит на
основе тех же внутриклеточных закономерностей, которые обус-
ловливают перемещение ядра в любой клетке при ее росте и раз-
витии. Любые формы движения биологических структур связаны
с наличием микрофибрилл, микрофиламентов при условии, если в их
состав входят белки актомиозинового типа. Подобного рода фибрил-
лярный материал обнаружен с помощью электронного микроскопа
в цитоплазме пыльцевой трубки около ядер вегетативной и генера-
тивной клеток, а также спермиев. Пыльцевая Трубка подходит
к зародышевому мешку через микропиле (порогамия) или внед-
ряется через халазу (апорогамия, халазогамия).
Вхождение пыльцевой трубки в зародышевый мешок. Пыльце-
вая трубка, как правило, входит в одну из синергид, изливая
в нее свое содержимое и разрушая ее. Содержимое пыльцевой
трубки под большим напором быстро проходит через синергиду
и попадает вместе со спермиями в «щель» — промежуток между
яйцеклеткой и центральной клеткой зародышевого мешка.
451
ДВОЙНОЕ ОПЛОДОТВОРЕНИЕ
Долгое время оставалась неясной судьба одного из двух спер-
миев, образующихся при делении генеративной клетки. Этот во-
прос был разрешен С. Г. Навашиным (1898), который, исследуя
лилейные, обнаружил, что кроме оплодотворения яйцеклетки в
зародышевом мешке происходит еще одно оплодотворение —
слияние второго спермия с ядром центральной клетки (рис. 179).
Это явление получило название двойного оплодотворения. Клас-
сическая работа С. Г. Навашина открыла новую эру в изучении
эмбриологии растений. - В настоящее время двойное оплодотво-
рение описано более чем у 300 видов растений и, по общему при-
знанию, является характерным признаком покрытосеменных.
Участие мужской цитоплазмы в оплодотворении вызывает
большой интерес в связи с проблемой неядерной наследственно-
сти. Некоторые авторы полагают, что спермин теряют свою ци-
топлазму, проходя через разрушающуюся синергиду, по мнению
других, они. остаются клетками вплоть до контакта с яйцеклеткой
и центральной клеткой.
Нет единого мнения о причинах и механизмах расхождения из
«щели» спермиев в разные женские клетки (основа двойного опло-
дотворения) .
Рис. 179. Двойное оплодотворение
пшеницы (по Т. Б. Батыгиной, 1961)
Различают два основных ти-
па оплодотворения: премитоти-
ческий и постмитотический
(рис. 180). При премитотиче-
ческом типе объединение муж-
ского и женского половых ядер
происходит сразу же после
установления их контакта, до
первого митоза зиготы (злаки,
сложноцветные). При постми-
тотическом типе объединение
происходит после начала ми-
тоза зиготы (у большинства
лилейных). Мужские и жен-
ские половые ядра к моменту
слияния сильно отличаются
друг от друга. Женское ядро
имеет структуру, типичную для
покоящейся клетки: крупное
ядрышко, сильно деконденси-
рованный хроматин. Ядро
спермия мельче, ядрышко
обычно отсутствует, хроматин
сильно конденсирован. При
премитотическом типе после
исчезновения контактирующих
452
Типы оплодотворения
Рис. 180. Типы оплодотворения (по Е. Н. Герасимовой-Навашииой, 1960)
участков оболочки ядра они объединяются в одно ядро зиготы (см.
рис. 173, рис. 181). Хромосомы спермия постепенно деконден-
сируются, выделяется и начинает увеличиваться мужское ядрыш-
ко, которое затем, сливаясь с женским, образует ядрышко ядра
зиготы. Аналогично, но значительно быстрее Ироисходит слияние
453
I
Рис. 181. Последовательные фазы изменения формы и структуры спермиев пше-
ницы от момента их вхождения в зародышевый мешок до полного объединения
с ядром яйцеклетки (I, А—С) и ядрами центральной клетки (II, А—Л) (по
Т. Б. Батыгиной, 1974)
половых ядер в центральной клетке. У пшеницы оно занимает
1,5 ч, а в яйцеклетке — 5—6 ч. При постмитотическом типе про-
исходят такие же превращения мужского хроматина и ядрышка,
но происходят они в ядре спермия, сохраняющего свою индиви-
дуальность вплоть до профазы деления зиготы. Стимулом к де-
лению оплодотворенных клеток является, как полагают некото-
рые авторы, внесение в женское ядро ДНК спермия, другие
отводят эту роль цитоплазме пыльцевой трубки, излившейся в
зародышевый мешок.
С. Г. Навашин указывал, что результатом двойного оплодо-
творения является своеобразная полиэмбриония, полагая, что
эндосперм также представляет собой зародыш, правда, идущий
на питание зародыша, развивающегося из зиготы. Существует и
иное мнение, отказывающее эндосперму в равноправии с зиго-
тическим зародышем (А. Л. Тахтаджян, 1980). Считают, что в
отличие от голосеменных развитие женского гаметофита здесь
разделено на две фазы. До оплодотворения с максимумом эко-
номии материала развивается предельно упрощенный гамето-
фит-— зародышевый мешок. Вторая фаза развития начинается
после слияния второго спермия с ядром центральной клетки,
которое является стимулом к бурному развитию эндосперма,
обеспечивающего быстрое создание необходимой для зародыша
пищи. Возникновение гибридного зародыша значительно повы-
сило жизнеспособность растений, их приспособляемость к меняю-
щимся условиям существования и обеспечило покрытосеменным
победное распространение по планете. В этом биологический
смысл двойного оплодотворения.
ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ
Большой вклад в изучение зародыша внесли Ганштейн (1870),
С. М. Розанов (1840—1870), Ф. М. Каменский (1851 —1912),
А. С. Фаминцын (1835—1918) и др.
Зигота — оплодотворенная яйцеклетка — образуется в резуль-
тате слияния гаплоидных гамет (рис. 182). Она представляет
одноклеточный зародыш — новый организм (спорофит) на самой
ранней стадии развития. Зигота — сложная интегрированная си-
стема с высокой метаболической активностью, что подтверждает-
ся электронно-микроскопическими исследованиями, выявившими
в ней наличие аппарата Гольджи, митохондрий, рибосом, пластид,
эндоплазматического ретикулума и различных включений (жи-
455
Рис. 182. Развитие и строение двудольного за-
родыша пастушьей сумки Cruciferae:
А—Г — образование четырехклеточного зародыша,
Д—— квадрантов, 3, И — октантов, К—-М — много-
клеточного зародыша и начало дифференциации се-
мядолей, // — семяпочка с молодым зародышем,
подвесочным гаусторнем н эндоспермом, О — зрелый
подковообразный зародыш с дегенерирующим подве-
сочным гаусторнем (А—Н — по Р. Суэжу, 1914—
1919; О — по И. Шаффнеру, 1906)
ры, белки и др.). Пос-
ле оплодотворения зи-
гота вступает в покой.
Продолжительность пе-
риода покоя у разных
растений неодинакова
(от минут до месяцев)
и в определенной сте-
пени зависит от внеш-
них условий. Во время
периода покоя в зиго-
те происходят глубо-
кие морфогенетические
изменения (X. Линс-
кенс 1969; В. Йенсен,
1972; Р. Капил, С. Бат-
нагар, 1981, и др.).
Форма и величина зи-
гот варьирует. Оболоч-
ка зиготы целлюлозная.
Т. Б. Батыгина
(1968, 1974) полагает,
что в развитии заро-
дыша можно выделить
две фазы: бластомери-
зацию и органогенез.
В 1-й фазе (прозм-
брио), особенно на первых ее этапах (в течение 3—4 клеточных
генераций), можно легко проследить происхождение и положение
каждого бластомера. Вторая фаза начинается с вычленения
эмбриодермы, дифференциации и морфологического обособления
отдельных органов.
Бластомеризация зародыша. В зиготе осуществляется первое
деление, как правило, неравное, дифференцирующее (рис. 182),
обычно сопровождающееся образованием клеточной стенки. Вслед-
ствие неравномерной вакуолизации зиготы фигура первого деле-
ния обычно оказывается сдвинутой к ее апикальному концу так,
что дочерние клетки получаются неравными. Апикальная клетка
оказывается меньшей, чем базальная, которая сильнее вакуоли-
зирована. Как отмечают многие исследователи, апикальная клет-
ка и ее дериваты становятся зоной активного протеинового
синтеза, сосредоточением органелл, различных включений, яв-
ляясь полюсом роста и морфогенеза. В зависимости от типа раз-
вития зародыша первая перегородка, как и последующие, распо-
лагается по отношению к оси проэмбрио по-разному. У большин-
ства изученных зародышей цветковых растений первое деление
бывает поперечным. Второе и третье происходят в апикальной
456
и базальной клетках, что ведет к образованию четырех клеток
квадранта. Последующие деления дают восемь клеток октанта.
Дальнейшие деления приводят к образованию многоклеточного
зародыша.
Органогенез. В ходе дальнейшего развития формируется
глобулярный зародыш, в котором начинается гистогенез — обра-
зование эмбриодермы. Дальнейший рост и развитие зародыша
связаны с сильным изменением его формы. В этих изменениях,
как и во всех сложных преобразованиях в течение органогенеза,
определяющую роль играет последовательное изменение ритма и
темпа активности в определенных областях зародыша (М. С. Яков-
лев, 1946; И. Рот, 1955; Т. Б. Батыгина, 1968, и др.). Эти разли-
чия обусловливают своеобразное положение и строение отдель-
ных органов у зародыша у различных видов растений. Некото-
рые исследователи делят зародыши на расчлененные и нерасчле-
ненные, между ними существуют переходные типы. Расчлененные
зародыши характеризуются дифференциацией зародыша, форми-
рованием структур и органов: семядоли, апекса побега (иногда
с дальнейшей дифференциацией примордиев листьев — почечки),
гипокотиля — под семядольного колена и апекса корня. Нерасчле-
ненные зародыши до полного созревания семян остаются недиф-
ференцированной клеточной массой. Иногда нерасчлененные ре-
дуцированные зародыши к моменту созревания семени, как по-
казали детальные исследования Э. С. Терёхина и 3. И. Никити-
чевой по паразитным растениям (1962—1983), состоят всего лишь
из нескольких клеток. На ранних стадиях развития зародышу
большинства покрытосеменных присуща радиальная симметрия,
которая на поздних стадиях развития сменяется на билатераль-
ную симметрию. Некоторые виды покрытосеменных имеют заро-
дыши с дорсовентральным строением с самых ранних стадий
развития (например, злаки). В зависимости от числа семядолей
в зародыше различают дву- и односемядольные зародыши. Число
семядолей у двудольных растений может варьировать (от одной
до нескольких). Среди однодольных выделяются своим развитием
и строением зародыши злаков. Для них характерно наличие сле-
дующих структур: щитка, колеоптиля, эпибласта, колеоризы, ли-
гулы— образований, отсутствующих у зародышей других покры-
тосеменных. Существуют различные гипотезы относительно трак-
товки разных структур зародыша злаков.
По своей форме зародыши могут быть прямыми (табак), со-
гнутыми (гвоздика), скрученными в спираль (повилика) и др.
По отношению к эндосперму или перисперму зародыш семени
занимает центральное положение (молочай), периферическое (ку-
коль) или боковое (злаки).
В зародышах некоторых растений был обнаружен хлорофилл.
М. С. Яковлев и Г. Я- Жукова (1971 —1973) разделили все заро-
457
дыши покрытосеменных по этому признаку на две большие груп-
пы: хлороэмбриофиты и лейкоэмбриофиты.
Электронно-микроскопическое исследование выявило, что при
формировании проэмбрио в его клетках увеличивается число
диктиосом, митохондрий и других органелл, тогда как в клетках
зрелых зародышей происходит исчезновение внутренних струк-
тур некоторых органелл. Так, в. хлоропластах исчезают граны,
в лейкопластах — строма и ламеллы и т. п. Биохимическое изу-
чение говорит о том, что в ходе эмбриогенеза идут значительные
изменения синтеза нуклеиновых и аминокислот. При созревании
семян содержание в зародыше физиологически активных веществ
и ферментов снижается.
Классификации эмбриогенеза. Существуют различные класси-
фикации типов эмбриогенеза (К. Шнарф, 1927—1929; Р. Суеж,
1936—1939; Д. Джогансен, 1950). Исходя из представлений о един-
стве путей прохождения начальных стадий развития у двудоль-
ных и однодольных было выделено шесть типов эмбриогенеза
двудольных. Эти типы основываются на различии в направлении
деления зиготы и первых бластомеров и их участии в построении
зародыша. М. С. Яковлев и М. Д. Иоффе (1957, 1961) показали,
что у различных представителей пеона ранние этапы развития
его зародыша не сопровождаются цитокинезом — образуется це-
ноцит, и только на определенном этапе его развития начинают
закладываться клеточные перегородки. Затем на многоклеточном
предзародышевом образовании возникают многочисленные заро-
дыши, но впоследствии развивается только один из них. Это дало
основание в 1958 г. дополнить классификацию Джогансена но-
вым седьмым типом эмбриогенеза (Paeoniaceae). Своеобразное
течение эмбриогенеза злаков не укладывалось ни в один из имею-
щихся типов развития зародыша. Детальное исследование Т. Б. Ба-
тыгиной (1968—1974) по эмбриогенезу разных видов пшеницы
выявило характерное наклонное положение перегородок по отно-
шению к оси зародыша и определенное участие бластомеров в
построении зародыша. Такой своеобразный способ развития и
строение раннего проэмбрио (его дорсовентральность) наряду с
особенностями органогенеза и строение зрелого зародыша позво-
лило выделить развитие зародыша злаков в особый восьмой тип
эмбриогенеза (Graminad-Poaceae).
Проблема эволюционного становления однодольности. Понима-
ние нормального эмбриогенеза невозможно без учета филогене-
тического положения исследуемой группы растений и особенно-
стей эволюционного становления зародыша однодольных. Подав-
ляющее большинство исследователей приходит к заключению, что
однодольные растения произошли от двудольных и что однодоль-
ный зародыш является эволюционно преобразованным зародышем
двудольного растения (рис. 183, А—Е). Существует несколько
принципиально различных гипотез эволюционного становления
458
Capselta Bursa, pastons
(Soueges, 1919-t9l9)
2„ 1
2
V. /
2
Plnguicuta vulgaris
(Hacclus a Hartl-Baude, >957)
Bteustne coracana
(fiulgnard; 1961)
OtteUa. alismobdes
(Haccius, 1952)
Trlticum aesttvum
(Батыгина, 1968)
2 1 2
Trlticum aesthfum
(Batygina, 1969)
Рис. 183. Эволюционное становление однодольности (по Т. Б. Батыгиной, 1974):
Д — типичный двусемядольный зародыш; Б — преобладающее развитие одной нз семядолей
у ложиооднодольиой Ptnguicula vulgaris; В — Е — эволюционное становление однодольности
(характер разрастания семядоли и смещение вертушки побега в абаксиальное положение);
1 — апекс побега, 2 — семядоля
семядоли у зародыша однодольных растений. Ряд авторов счи-
тает, что односемядольность возникла в результате редукции од-
ной семядоли двудольного зародыша (Abort Hypothese); другие
поддерживают теорию синкотилии, согласно которой происходит
слияние двух семядолей в единую семядолю зародыша. Для вы-
яснения конкретных эволюционных путей происхождения одно-
дольного зародыша существенное значение имеют данные о при-
роде его органов. Большое внимание уделяется происхождению
и эволюционному преобразованию семядолей и тесно связанному
с ним апексу побега. Особого внимания заслуживают представ-
ления, высказанные Ж. Гиньяром и Ж- Местром (1969, 1971),
о том, что в обеих группах цветковых а-пикальная часть проэм-
брио может быть уподоблена апикальной меристеме стебля, функ-
ционирование которой дает либо одну, либо две семядоли, по-
добно тому, как на апексе стебля возникают примордии листьев;
семядоли можно рассматривать как первую пару листьев. Де-
тальные исследования по эмбриогенезу выявили у одной группы
растений относительно медленное формирование точки роста, что
позволяет наблюдать «смещение» ее в онтогенезе зародыша,
у другой (злаки) это происходит так быстро, что создается лож-
ное впечатление о боковом формировании точки роста изначаль-
но (рис. 183). Терминальность семядоли и латеральность точки
роста — явление кажущееся, обусловленное изменениями хода
развития зародыша (в частности, неравномерностью темпов де-
ления клеток) и является у однодольных (в частности, злаков) —
вторичным как в онтогенетическом, так и в эволюционном планах
(М. С. Яковлев, 1946; Т. Б. Батыгина, 1968—1974, и др.). Та-
ким образом, по происхождению апекса побега и морфологиче-
скому значению семядоли, по-видимому, не существует глубоких
различий между однодольными и двудольными. Следует также
отметить, что нельзя провести резкую границу между проэмбрио
и зрелым зародышем и проростком, и если, таким образом, строе-
ние проростка связано с его образом жизни, то строение заро-
дыша является не более чем ранней стадией строения проростка.
Действительно, морфологическую природу органов нельзя понять
в отрыве от их функции. Различные модификации эмбриогенеза
в значительной степени должны быть, по-видимому, связаны с
эволюционным изменением способов прорастания.
РАЗВИТИЕ И СТРОЕНИЕ ЭНДОСПЕРМА
Эндосперм у покрытосеменных образуется в результате поло-
вого процесса. В большинстве случаев он триплоидный (при Po-
lygonum-типе зародышевого мешка). Он имеет большое значение
для развития зародыша как основной источник питания и акку-
мулятор различных физиологически активных веществ, необходи-
мых для нормального протекания процесса эмбриогенеза (рис. 184).
460
Рис. 184. Схема строения семяпочки лотоса на разных стадиях развития (по
Т. Б. Батыгиной и др., 1983):
/ — зародыш, 2 — эндосперм
Есть виды растений, у которых эндосперм практически не обра-
зуется (орхидные). В последнее время в культуре in vitro из ткани
эндосперма были получены регенеранты, что говорит о тотипо-
тентности его клеток и подтверждает половую природу, посколь-
ку растения-регенеранты имели клетки, содержащие триплоидное
число хромосом. Характерной особенностью эндосперма является
полиплоидизация его ядер, которая осуществляется различными
путями (М. Д. Иоффе, 1968). Это, несомненно, говорит о физио-
логической активности эндосперма. Различают три основных типа
эндосперма: нуклеарный (ядерный), целлюлярный (клеточный) и
гелобиальный (базальный). Они различаются в основном харак-
тером протекания начальных этапов. Темп развития эндосперма
неодинаков у разных видов: у одних с момента оплодотворения
проходит несколько дней (Asteraceae), у других — месяцы (Che-
nopodiaceae). Различна и судьба эндосперма: у некоторых видов
(злаки) он представлен тканью, существующей вплоть до про-
растания зерновки, у других (тыква, горох и др.) он потребляется
в процессе развития зародыша, и часто в зрелом семени он от-
сутствует или представлен тонкой пленкой. В связи с этим раз-
личают два типа семян: с эндоспермом и без эндосперма. В связи
с развитием эндосперма в семени следует сказать о формирова-
нии перисперма — структуры, функционирующей как эндосперм и
развивающейся чаще в семенах, где слабо развит эндосперм, и
по происхождению являющийся нуцеллусом, в клетках которого
откладываются запасные питательные вещества. Развитие эндо-
сперма, перисперма и зародыша происходит сопряженно и яв-
ляется единым сложным процессом, который обычно кончается
«поглощением» эндосперма и перисперма в процессе эмбриогенеза
461
или развития проростка. Все эти структуры развиваются в тес-
ной связи с тканями семяпочки, завязи, цветка и в целом всего
организма.
Семя и плод. По мере развития зародыша семяпочка значи-
тельно увеличивается в размерах и превращается в семя, а за-
вязь— в плод. Типы развития и строения семян и плодов разно-
образны.
МЕТАМОРФОЗ У ЦВЕТКОВЫХ РАСТЕНИЙ
В индивидуальном развитии некоторых цветковых, как установил
для паразитных растений Э. С. Терёхин (1976), имеет место ме-
таморфоз (рис. 185). Обычно этот процесс как у паразитных,
так и у других растений (орхидные) сопровождается некрозом
исчерпывающих свою функцию соматических зон и образованием
новых меристематических очагов развития. Метаморфоз может
проявляться на разных этапах онтогенеза и имеет разные формы.
Например, у орхидных, начиная с недифференцированного на
органы зародыша, весь онтогенез представлен своеобразными
структурами (протокормами, туберидиями и т. д.), которые по-
явились, как «вставки»,
обеспечивающие возмож-
ность перехода индиви-
дуума на вегетативный
способ размножения. Ме-
таморфоз у орхидных и
появление таких структур
обусловлено в основном
экологическими условия-
ми (Т. Б. Батыгина,
В. Е. Васильева, 1980).
АПОМИКСИС
И ПОЛИЭМБРИОНИЯ
У многих цветковых рас-
тений наряду с половым
и бесполым размножени-
ем существует апомик-
сис. Одни исследователи
приравнивают апомиксис
к половому размноже-
нию, другие—к бесполо-
му. Обнаружены различ-
ные типы апомиксиса:
регулярный (постоянный
и нерегулярный. Его так-
же делят на стимулятив-
Рис. 185. Метаморфоз в онтогенезе пара-
зитных растений. Внутренний круг — раз-
витие автотрофного растения; наружный—
развитие с метаморфозом (по Э. С. Тере-
хину, 1976)
462
ный (индуцированный), т. е. связанный с опылением, и автономный
(независимый от опыления). У растений различают частичный
апомиксис, когда у одного и того же таксона наряду с половым
размножением встречается и апомиктичное (различные виды мят-
лика), и полный, когда вся система размножения исключительно
апомиктична (ряд видов одуванчика). В настоящее время наи-
более распространенной является классификация и терминология,
предложенная В. А. Поддубной-Арнольди (1976). I. Партеноге-
нез—образование зародыша из неоплодотворенной яйцеклетки с
редуцированным или нередуцированным числом хромосом. Сюда
относятся гиногенез, андрогенез и партеногенез. II. Апогаметия —
возникновение зародыша не из яйцеклетки, а из других клеток
женского гаметофита, с редуцированным или нередуцированным
числом хромосом. III. Апоспория — развитие гаметофита не из
споры, а из диплоидной клетки спорофита. IV. Нуцеллярная эм-
бриония—образование зародышей из клеток нуцеллуса семяпоч-
ки; различают индуцированную и автономную, неиндуцированную
эмбрионию. V. Интегументальная эмбриония — образование заро-
дыша из клеток покровов семяпочки; различают индуцированную
и автономную интегументальную эмбрионию.
Несмотря на большое число имеющихся классификаций, к на-
стоящему времени нет ни одной приемлемой классификации, ко-
торая в соответствии с биологической сущностью апомиксиса
разграничила бы его формы. Понятие апомиксис трактуется неод-
нозначно. Мы полагаем, что нуцеллярная и интегументальная по-
лиэмбриония может рассматриваться как один из способов ве-
гетативного (бесполого) размножения, поскольку неполовые
зародыши (эмбриоиды) 1 образуются не из гамет, как это про-
исходит в случае партеногенеза, а из соматических клеток спо-
рофита. В этом аспекте образование многих эмбриоидов на«пред-
зародыше» у пеона также можно рассматривать как один из
способов вегетативного размножения, поскольку они образуются
из соматических клеток. Все эти явления в настоящее время
нужно рассматривать в свете новых данных по получению целых
растений in vitro из соматических клеток. Интересно отметить,
что нуцеллярная и интегументальная эмбриония по своему про-
исхождению очень сходна с явлением образования эмбриоидов
из клеток эпидермиса стеблей, корней, гипокотиля и других
структур (рис. 186), а также в некотором плане, с вивипарией
(живорождением): возникновением луковичек в пазухах листьев
вместо генеративных структур в соцветии или цветке. Несмотря
на общность происхождения этих явлений (из соматических кле-
ток спорофита), пути морфогенеза воспроизведения растений раз-
1 Эмбриоид — зародышеподобная биполярная с первых этапов развития
структура, образующаяся асексуально из соматической клетки.
463
Рис. 186. Развитие эмбриоидов из клеток
эпидермиса стеблей Ranunculus sceleratus
- (К. Конар, К. Натарайя, 1965):
А, Б, В — общий вид проростка с эмбриоидами;
Г — % — последовательные стадии развития эм-
бриоида; Л — М — сформированный эмбриоид;
Н — поперечный срез стебля с эмбриоидами
личны (Т. Б. Батыгина
и др., 1978): эмбриоидо-
генез— в случае интегу-
ментальной, нуцеллярной
эмбрионии, а также обра-
зования эмбриоидов, на-
пример у Ranunculus,
органогенез (гемморизоге-
нез)—в случае вивипа-
рии у Роа и Polygonum.
Интересны работы по изу-
чению ультраструктуры
инициальных клеток ну-
целлярных и интегумен-
тальных эмбриоидов (рис.
187) (Т. Н. Наумова,
1983). В последнее время
получило широкое рас-
пространение экспери-
ментальное исследование
апомиксиса. Существует
много гипотез, пытаю-
щихся объяснить апомик-
сис и его эволюционное
значение. Наиболее убе-
дительной, на наш взгляд,
является точка зрения
Д. Ф. Петрова (1964), по-
лагающего, что широкое
наследственное многооб-
разие различных форм
апомиксиса и способность их к сравнительно быстрой эволюции
(тип адаптации) обусловлены сочетанием полового и апомиктич-
ного способов размножения. Апомиктичный способ размножения
является одним из лучших способов достижения гомозиготности,
что обеспечивает высокую жизнеспособность потомства и практи-
чески исключает расщепление за счет образования клонов.
Полиэмбриония. Полиэмбриония — образование зародышей
(эмбриоидов) из различных клеток зародышевого мешка, а также
соматических клеток спорофита на разных стадиях его развития.
Впервые она была обнаружена А. Левенгуком в 1719 г. в семени
апельсина. Наиболее интересная и малоизученная форма поли-
эмбрионии — монозиготическая, или расщепленная, полиэмбрио-
ния. К ней относится образование одного или более зародышей
из зиготы, из различных клеток самого зародыша, которые могут
возникать на разных стадиях его развития (например, орхидные).
Дополнительные зародыши могут быть поли-, ди- и гаплоидными.
464
Полиэмбриония может быть постоянной, ре-
гулярной или случайной, нерегулярной. Обра-
зование многих зародышей на ранней стадии
развития зародышевого мешка еще не гово-
рит о том, что все они дадут нормальные
зародыши в зрелом семени. Большей частью
развивается один-два зародыша. М. С. Яковлев
(1957) предложил классификацию полиэм-
брионии, в основе которой был заложен гене-
тический принцип, учитывая при этом предше-
ствующую историю развития генеративных
структур у высших растений.
К сожалению, в кратном очерке нет воз-
можности сопоставить явления апомиксиса и
полиэмбрионии у растений с явлениями сома-
тического эмбриогенеза и полиэмбрионии у
животных (см. гл. XVI).
ПРОБЛЕМЫ СОВРЕМЕННОЙ ЭКСПЕРИМЕНТАЛЬНОЙ
ЭМБРИОЛОГИИ
Рис. 187. Нуцелляр-
ная полиэмбриония у
Sarcococca humilis.
Микропиллярная часть
развивающегося семе-
ни с многочнслеины-
ми нуцеллярными за-
родышами, эндосперм
клеточный (по Т. Н.
Наумовой, 1980)
Описательный метод не позволяет вскрыть
механизмы отдельных процессов, из которых
складывается сложная картина эмбриональ-
ного развития. В связи с этим все большее
значение приобретает экспериментальное направление в эмбриоло-
гии растений. Известно, что у многих цветковых в естественных
условиях могут образовываться кроме половых и неполовые заро-
дыши— эмбриоиды. Вопрос о том, как происходит развитие
эмбриоидов и сходно ли оно с развитием половых зародышей,
особенно стал интересовать биологов в связи с работами
Ф. Стюарда (1958) и Р. Г. Бутенко (1954—1984) и др., в которых
показана возможность экспериментальной индукции образования
эмбриоида из соматической клетки и получения из него целого
растения. Это и последующие открытия (получение гаплоидного
растения из пыльника, завязи, парасексуальная гибридизация)
имеют общебиологическое значение для изучения вопросов диффе-
ренциации клеток, тканей и органов, а также морфогенеза.
Независимо от того, получает исследователь растение из сома-
тической клетки или гаплоид из пыльцы, или отдаленный гибрид
в результате слияния протопластов, он сталкивается с проблемой
морфогенеза, прежде всего с такими вопросами: каковы морфо-
генетические потенции эмбриональных структур; каковы пути
морфогенеза in vivo и in vitro.
Открытие описанных явлений вновь привлекло внимание к
некоторым вопросам классической эмбриологии. До сих пор не
465
установлено, например, чем характеризуются инициальные клетки
эмбриоидов, какие факторы индуцируют их развитие, чем отли-
чаются половые зародыши и эмбриоиды, образованные in vivo
и in vitro. До сих пор еще не удалось вырастить нормальное
растение в условиях культуры из зиготы. Поэтому культивирова-
ние зиготы и зародыша на ранних стадиях эмбриогенеза остается
одной из сложнейших и интереснейших проблем современной эм-
бриологии.
Среди модельных систем для изучения закономерностей мор-
фогенеза особого внимания заслуживает пыльник, поскольку в
нем находят отражение основные морфогенетические процессы
растений в целом (например, чередование двух основных фаз в
развитии — спорофита и гаметофита, сопровождающееся мейо-
зом). Такие модели, как культура изолированных половых заро-
дышей (эмбриокультура) и эмбриоидокультура, представляют
интерес для изучения закономерностей дифференциации зародыша
и явления апомиксиса. Пристального внимания для практических
целей заслуживает разработка теоретических основ культивиро-
вания in vitro различных генеративных структур (завязи, семя-
почки, нуцеллуса, интегументов, зародышевого мешка, эндо-
сперма) .
Большой фактический материал, накопленный к настоящему
времени в сравнительной эмбриологии, и экспериментальные дан-
ные позволяют сделать заключение о том, что в эмбриогенезе
растений существуют критические периоды (Т. Б. Батыгина,
В. Е. Васильева, 1976—1986), например стадия автономности
(рис. 188). Изучение всех этапов эмбриогенеза и особенно ста-
дии автономности является теоретической предпосылкой для
прикладных исследований (сокращение периода покоя семян за
счет получения проростков из незрелых зародышей, преодоление
нескрещиваемости, установление оптимального срока сбора уро-
жая и др.). Каллус, полученный из недифференцированного на
органы зародыша, а также из отдельных его органов и других
генеративных структур, может быть использован как один из
способов получения новых форм растений и как метод клональ-
ного микроразмножения.
Дальнейшее всестороннее изучение закономерностей диффе-
ренциации зародышей у различных растений и моделирование
эмбриогенеза даст возможность управлять эмбриогенезом, что
является одной из главнейших задач не только эксперименталь-
ной эмбриологии, но и биологии развития в целом.
* *
*
Филогения и взаимоотношения между отдельными группами
растений — одна из сложнейших проблем ботаники. Эмбриология
дает материал для развития и уточнения наших представлений
о взаимоотношениях ряда порядков, семейств, родов и даже ви-
466
Антиподы
Зндослерм
Нуцеллус
Внутренний интегумент
Наружный интегумент
0] 1 234-567 8 3 )0 11
Оплодотворение
30
Дни после опыления
Рис. 188. Корреляции роста и развития зародыша и структур семяпочки лотоса
(по Т. Б. Батыгиной, В. Е. Васильевой, 1981):
А— динамика роста зародыша (1), семяпочки (2), завязи (3) и содержания растворимых
углеводов в зародыше in vivo (4); Б— динамика развития структур зародышевого мешка
н семяпочки. Стрелкой указана стадия начала отложения крахмала в зародыше. Период
формирования зародыша лотоса длится 30 дней с момента опыления: 1-й день — бластоме-
ризация, 2-й день — заложение э.мбриодермы, 3 — 4-й дни — инициация семядолей и точки
роста, 5-й день — заложение примордия 1-го листа почечки зародыша, 8-й день — заложе-
ние примордия 2-го листа почечки, 10-й день — появление пластинки 1-го листа почечки,
И-й день — заложение примордия 3-го листа почечки, 30-й день — сформированная почеч-
ка зародыша с двумя зелеными листьями и прнмордием 3-го листа. Зона X — период ста-
новления автономности зародыша (7—11-й дни)
30*
дов, положение которых в системе вызывало сомнение (П. Ма-
гешвари, 1950; В. А. Поддубная-Арнольди, 1982, и др.). Эмбрио-
логические признаки являются относительно консервативными по
сравнению с морфологическими, комплекс которых характери-
зует определенным образом различные систематические таксоны
покрытосеменных.
При обсуждении основных закономерностей эволюции при-
менительно к растениям необходимо учитывать специфику их
онтогенеза. В онтогенезе растений в сравнении с животными
имеется ряд фундаментальных отличий, среди которых основные —
неподвижный образ жизни, постоянство и сезонная цикличность
процессов роста и развития и др. Из всего этого следует и боль-
шая эволюционная пластичность растений. В их эволюции одни
способы эволюционных преобразований проявляются в очень сла-
бой форме (например, явления рекапитуляции), другие — распро-
странены широко (например, редукции и неотении) (А. Л. Тах-
таджян, 1943—1984, Э. С. Терёхин, 1977, А. С. Смирнов, 1982).
ПЕРСПЕКТИВЫ РАЗВИТИЯ ТЕОРЕТИЧЕСКОЙ
И ПРИКЛАДНОЙ ЭМБРИОЛОГИИ
Эмбриология стала за сравнительно короткий период развития
очень разветвленной областью биологии.
Одно из бурно развивающихся направлений, сформировавшее-
ся на стыке генетики, молекулярной биологии и эмбриологии, —
это биология развития. Исследователи, работающие по пробле-
мам биологии развития, в первую очередь видят свою задачу в
том, чтобы выяснить молекулярно-генетические закономерности
морфогенетических процессов. Эти задачи тесно смыкаются с
проблемами генной инженерии. При «имплантации» в геном по-
ловой или соматической клетки того или иного гена создаются
возможности преобразования признаков развивающегося зароды-
ша на основе исторически сложившегося морфогенеза, который
присущ данному виду животного или растения.
Особой областью теоретической и прикладной эмбриологии
становится клеточная инженерия, когда объединяются бластоме-
ры разных видов зародышей или создаются иным путем сома-
тические химеры. В 60-е годы такие химеры были получены у
мышей, а позднее и у других млекопитающих. Перспективный
метод анализа морфогенетических процессов — изучение разви-
тия мутантных животных и растений, установление корреляций
тех или иных пороков развития с нарушениями в структуре хро-
мосом и изменении в функционировании генома.
Другим не менее важным направлением эмбриологических
исследований должна стать в будущем популяционная эмбриоло-
гия, в задачу которой входит изучение вариабельности морфоге-
неза и фенотипа в популяции (изучение вариаций жизненного
цикла). Это направление смыкается с экологической эмбриоло-
468
гией, значение которой трудно переоценить. Установление влияния
различных абиотических и биотических факторов среды на раз-
личные морфогенетические процессы, непрерывно осуществляю-
щиеся в жизненном цикле животных и растений — важнейшее
звено в экологических исследованиях и анализе чувствительных
периодов, о чем говорилось ранее.
За последние годы возобновляется интерес к проблемам эво-
люции онтогенеза. Эволюционная эмбриология как раздел эво-
люционной морфологии в целом и эволюционной систематики —
важнейшее звено в филогенетике. Достаточно сказать, что во-
просы соотношения онтогенеза и филогенеза, так успешно раз-
рабатывавшиеся в прошлом и в начале нашего столетия, ока-
зались забытыми и в настоящее время нуждаются в дальнейшем
изучении с учетом новых данных, накопленных в морфологии
развития, экспериментальной эмбриологии и биологии развития.
Не менее важное место в системе эмбриологических знаний
занимает изучение индукционных взаимодействий, особенно в
плане сравнительной, эволюционной эмбриологии. Интересные
идеи и факты рождаются при анализе проблем, пограничных
между эмбриологией и другими науками: физиологией, иммуно-
логией, термодинамикой, физикой, математикой. Это не только
молекулярно-биологические аспекты решения проблем, погранич-
ных между этими разделами естествознания, но и иные аспекты
и, в частности, те, которые адекватны для тканевого, органного
и организменного уровней, т. е. решаемые на основе представ-
лений о системогенезе, о полях и градиентах, с учетом анализа
динамических автоколебательных процессов, теории регуляции и
многих других.
Прикладные аспекты современной эмбриологии также широки
и разветвленны, как широки и разветвленны связи этой науки с
другими дисциплинами, с различными областями практики. Об
отдельных проблемах прикладной эмбриологии речь уже шла в
различных главах учебника. Перечислим главнейшие проблемы,
решаемые при участии эмбриологов.
1. Медицинские проблемы, связанные с предотвращением и
лечением патологий гаметогенеза и эмбриогенеза. Регуляция ре-
генерационных процессов и лечение злокачественных образова-
ний. Разработка методов преодоления бесплодия.
2. Сельскохозяйственные проблемы повышения продуктивно-
сти культур в сельском хозяйстве и пород в животноводстве, кло-
нирование перспективных сортов, растений и пород животных,
улучшение ценных растений на основе соматического эмбриогенеза
и путем соматической гибридизации.
3. Экологические проблемы — установление возможностей пе-
реключения тех или иных видов животных и растений на рельсы
соматического воспроизводства (в условиях нарушения половой
репродукции). Разработка эффективных доз веществ, блокирую-
469
щих те или иные этапы индивидуального развития вредных орга-
низмов и паразитов. Установление предельно допустимых доз
воздействия ряда повреждающих факторов, встречающихся в есте-
ственной среде, производстве и быту. Создание банка генетиче-
ского материала, половых клеток, зародышей и взрослых особей.
Проблемы космической эмбриологии.
4. Биотехнологические проблемы — разработка условий и осу-
ществление регулируемого воспроизводства полезных микроорга-
низмов, беспозвоночных и позвоночных животных, а также во-
дорослей и высших растений (марикультуры, рыборазведение,
звероводство и др.).
5. Бионические — разработка аналоговых моделей, имитирую-
щих некоторые разновидности морфогенезов (регенерационные
модели, модели бластогенеза и др.).
ЛИТЕРАТУРА
Айзенштадт Т. Б. Цитология оогенеза. М., 1984.
Анохин П. К. Биология и нейрофизиология условного рефлекса. М., 1968.
Аршавский И. А. Физиологические механизмы и закономерности индивиду-
ального развития. М., 1982.
Бабаева А. Г. Иммунологические механизмы восстановительных процессов.
М„ 1972.
Банникова В. П, Хведынич О. А. Основы эмбриологии растений. Киев, 1982.
Батыгина Т. Б. Эмбриология пшеницы. Л., 1974.
Белоусов Л. В. Введение в общую эмбриологию. М., 1980.
Бесполое размножение, соматический эмбриогенез и регенерация / Под. ред.
Б. П. Токина. Л., 1972.
Биология развития растений. М., 1975.
Бляхер Л. Я. История эмбриологии в России (с середины XVIII до середя-
ны XIX в.). М„ 1955.
Бляхер Л. Я. Очерк истории морфологии животных. М., 1962.
Боннер Дж. Молекулярная биология развития. М., 1967.
Браше Ж- Биохимическая эмбриология. М., 1961.
Бэр К. М. История развития животных. Наблюдения и размышления. Т. 1.
М., 1950; т. 2. М„ 1953.
Внешняя среда и развивающийся организм. М., 1977.
Волкова Л. С. Иммунобиологические взаимоотношения организмов матери
и плода. М., 1970.
Вольф К. Ф. Теория зарождения. М., 1950.
Воронцова М. А. Регенерация органов у животных. М., 1949.
Воронцова М. А., Лиознер Л. Д. Физиологическая регенерация. М., 1955.
Газарян К.. Г., Белоусов Л. В. Биология индивидуального развития живот-
ных. М., 1983.
Гексли Дж. С. де Бер Г. Р. Основы экспериментальной эмбриологии. М.,
Л., 1936.
Герасимова-Навашина Е. Н. К цитолого-эмбриологическому пониманию
процесса опыления. // Тр. ин-та. Ботан. ин-т АН СССР, сер. VII,
вып. 3, 1952.
Гердон Дж. Регуляция функции генов в развитии животных. М., 1977.
Гормональные факторы индивидуального развития / Под. ред. Б. Л. Астау-
рова. М., 1974.
Детлеф Т. А., Гинзбург А. С., Шмальгаузен О. И. Развитие осетровых рыб.
М., 1981.
Догель В. А. А. О. Ковалевский. М., Л. 1945.
Догель В. А. Олигомеризация гомологичных органов. Л., 1954.
Дондуа А. К.. Андреева Л. Ф., Лукина Н. А. Клеточное размножение н
процессы дифференциации. Л., 1983.
Дыбан А. П. Очерки патологической эмбриологии человека. Л., 1959.
Дэвидсон. Э. Действие генов в раннем развитии. М„ 1972.
Епифанова О. И., Терских В. В., Полуновский В. А. Покоящиеся клетки.
М., 1983.
Ефимов Е. А. Посттравматическая регенерация кожи. М., 1975.
Завадовский М. М. Противоречивое взаимодействие между органами в те-
ле развивающегося животного. М., 1941.
Захваткин А. А. Сравнительная эмбриология низших беспозвоночных М
1949.
Зотин А. И. Термодинамический подход к проблемам развития роста и ста-
рения. М., 1974.
471
Иванов А. В. Происхождение многоклеточных животных. Л., 1968.
Иванов П. П. Общая н сравнительная эмбриология. М., Л., 1937.
Иванов П. И. Руководство по общей и сравнительной эмбриологии. Л., 1945.
Иванова-Казас О. М. Очерки по сравнительной эмбриологии перепончатокры-
лых. Л., 1961.
Иванова-Казас О. М. Бесполое размножение животных. Л., 1977.
Иванова-Казас О. М. Сравнительная эмбриология беспозвоночных живот-
ных. Новосибирск, 1975. Т. I; 1977. Т. II; 1978. Т. Ill, IV; 1979. Т. V;
1981. Т. VI.
Калинникова В. Д., Северцов А. С. Жизненные циклы паразитических про-
стейших как форма онтогенеза // Журнал общей биологии, т. XIV, 1983. № 2.
Карлсон Б. Основы эмбриологии по Пэттену. М., 1983.
Кафиани К. А., Костомарова А. А. Информационные макромолекулы в ран-
нем развитии животных. М., 1978.
Клеточное размножение и процессы дифференциации. М„ 1983.
Кнорре А. Г. Краткий очерк эмбриологии человека. Л., 1967.
Константинов А. В. Биология индивидуального развития. Минск. 1978.
Конышев В. А. Стимуляторы и ингибиторы роста органов и тканей живот-
ных. М„ 1974.
Короткова Г. П. Принципы целостности. Л., 1968.
Короткова Г. Л. Происхождение и эволюция онтогенеза. Л., 1979.
Короткова Г. П., Токин Б. П. Бэр и соверменная эмбриология // Бэр и
развитие естествознания. Талин, 1978.
Короткова Г. П., Токин Б. П. Эмбриология н генетика. Дискуссионные воп-
росы. Л., 1977.
Корочкин Л. И. Взаимодействие генов в развитии. М., 1977.
Ковалевский А. О. Избранные работы. М., 1950.
Лагучев С. С. Гормоны и митотический цикл клетки. М., 1975.
Лобашев М. Е. Генетика. Л., 1963.
Магешвари П. Эмбриология покрытосеменных. М., 1954.
Методы биологии развития. М., 1974.
Мечников И. И. Избранные биологические произведения. М., 1950.
Мина М. В., Клевезаль Г. А. Рост животных. М., 1976.
Мицкевич М. С. Гормональные регуляции в онтогенезе животных. М., 1978.
Меттсон П. Регенерация — настоящее и будущее. М„ 1982.
Мюллер Ф„ Геккель Э. Основной биогенетический закон. М., Л., 1940.
Нейфах А. А., Тимофеева М. Я- Молекулярная биология процессов разви-
тия. М., 1977.
Нидхем Дж. История эмбриологии. М., 1947.
Объекты биологии развития. М., 1976.
Поддубная-Арнольди В. А. Цитоэмбриология покрытосеменных растений.
М„ 1976.
Полежаев Л. В. Регенерация путем индукции. М., 1977.
Преснов В. В., Исаева В. В. Перестройки топологии при морфогенезе. М„
1985.
Пэттен Б. М. Эмбриология человека. М., 1959.
Романова Л. К- Регуляция восстановительных процессов. М., 1984.
Рузен-Ранге Э. Сперматогенез у животных. М., 1980.
Саксен Л., Тойвонен С. Первичная эмбриональная индукция. М., 1963.
Саркисов Д. С., Втюрин Б. В. Электронная микроскопия декструктивных и
регенераторных внутриклеточных процессов. М., 1967.
Светлов П. Г. Физиология (механика) развития. Т. I, И. Л., 1978.
Северцов А. Н. Морфологические закономерности эволюции. М., Л., 1934.
Сидорова В. Ф. Возраст и восстановительная способность органов у млеко-
питающих. М.. 1976.
Современные проблемы оогенеза. М., 1977.
Современные проблемы сперматогенеза. 1982.
Сравнительная эмбриология цветковых растений. М.. 1981, 1983, 1985.
472
Токин Б. П. Иммунитет зародышей. Л., 1935.
Токин Б. П. Регенерация и соматический эмбриогенез. Л., 1959.
Токин И. Б. Проблемы радиационной цитологии. Л., 1974.
Туманишвили Г. Д. Некоторые вопросы регуляции роста животных тканей.
Тбилиси, 1965.
Уоддингтон К. Морфогенез и генетика. М., 1964.
Филатов Д. П. Сравнительно-морфологическое направление в механике раз-
вития, его объект, цели и пути. М., 1935.
Шмальгаузен И. И. Организм как целое в индивидуальном и историческом
развитии. М., 1938.
Шмальгаузен И. И. Регуляция формообразования в индивидуальном разви-
тии. М„ 1964.
Шмидт Г. А. Эмбриология животных. Ч. I. Общая эмбриология; ч. И.
Частная эмбриология. М., 1951, 1953.
Яковлев М. С. Эмбриогенез 'И его значение для филогении растений. Кома-
ровские чтения. Вып. 13. М., 1960.
Embryology of Angiosperms. Ed. В. M. Johri. Springer — Verlag. Berlin, Hei-
delberg, New Jork, Tokye, 1984.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Автогамия 450
Автотомия 5
Агглютинация 58, 61, 359
Акросома 41, 62
Акросомная гранула 62
— нить 60, 63
— область 62
— реакция 60, 61
Акросомный аппарат 61, 63
— пузырек 62
Аллантоис 142, 210, 213, 214
Аллогамия 450
Амитоз 90
Амнион 210, 212
Амниотическая жидкость 212, 334,372
Амплификация генов 54
Амфибластула 110
Анаболия 417, 424
Андрогамоны 59
Андрогенез 278, 463
Андрогенетические зародыши 277
Анимальио-вегетативный градиент 312,
314, 316
Анимальный полюс 60, 77, 82, 83, 95,
126, 127, 133, 138, 155, 242, 311, 312
Антиген 289, 351, 352
Антигенная реактивность 351, 359,
369
Антитело 351
Антифертилизин 59
Апекс 457, 460
Апорогамия 451
Апомиксис 462, 463
Апоспория 463
Area op аса 95, 139
— pellucida 95, 128, 139, 210
— vasculosa 209
— vitellina 209
Ариллоиды 441
Ариллусы 441
Архаллаксис 417, 424
Археспорий 443
Архентерон 113, 115, 159, 177, 181,
208
Ассимиляция 53, 403
Аурикулярия 220
Аутотрансплантация 231
Аэропиле 372
Биогенетический закон 11—12, 19,
409—411
Биология развития 468
«Бнофоры» 27, 28
Биппинария 220
474
Бластема 303, 304, 397, 398
Бластея 412, 434
Бластогенез 294, 373, 377, 470
Бластодерма 79, 97, 99, 109, ПО, 114
133 ’
Бластодиск 97, 128, 136—139 209
390
Бластоматозный рост 403—406
Бластомер 13, 29, 54, 57, 76 80—103
108-111, 114, 116, 123, 126, 130
138, 141, 146, 207, 232—235, 287
299, 320, 334, 468
Бластопор 112, 114, 115, 126, 128 130
133, 135, 137, 140, 146, 151, 164’
181, 242, 251, 254, 313—315
Бластоцель (полость Бэра) 97 109
110, 113, 115, 118, 119, 130, 131, 134’
136, 154, 155, 157, 158, 216, 335
336
Бластоциста 73, 74, 141, 142 357 364
376
Бластула 75, 76, 86, 91, 97, 107, ПО,
124, 126, 128, 131, 132, 135, 138,
145—147, 149, 150, 152, 155—157,
235, 239, 268, 279, 284, 286, 287,
312, 334, 336, 390, 435
Бластуляция 88, 107
Боуменова капсула 197, 198
Вант-Гоффа правило 87
Вегетативный полюс 113, 126, 127,
130, 133, 218, 311, 312
Вителлогенез 44
Вителлогенин 44, 45
Вителлофаги 79
Вольфов канал 196, 198
Восстановительные морфогенезы 5, 14
Гаметогенез 5, 14, 27, 36, 37, 40, 46,
47, 52—54, 269, 441, 469
— алиментарный 46, 47
— солитарный 46
Гаметофит 19, 441, 443, 444, 466
Гаметофитогепез 441
Гамета 18, 294, 437, 441, 455
Гамоны 60, 63
Ганглионарная пластинка 123, 174,
175, 176, 183, 189, 206
Гаплоидный набор хромосом 20, 449
Гастрея 113, 120, 121, 434
Гастродерма 33
Гастроцель 113, 114, 119, 134, 136
Гаструла 90, 103, 108, 111, 113, 120,
122, 124, 145, 146, 148, 152, 241—
243, 253, 257, 268, 279, 286, 287,
289, 313, 334, 371, 417
— делямннацнонная 113, 114
— иммиграционная 113, 114
— ннвагннацнонная 111, 113
— эпнболнческая 114
Гаструляция 13, 54, 86, 101, 106,
110—117, 121, 122, 124, 125, 129,
130, 132—139, 142, 144—154, 159,
177, 179, 204, 206—208, 219, 240,
286, 293, 294, 307, 312, 319, 336,
344, 346, 377, 434
Геммула 86, 364
Гензеновскнй узелок 140, 249 .
Генная инженерия 468
Геном 3, 16, 31, 52, 87, 107, 146, 148,
275, 280, 281, 289, 346, 468
Гермафродитизм 56
Гертвнга правила 94—98, 100
Гетеростилия 450
Гетеротрансплантацня 232, 352
Гетерохрония 411
Гиалиновая мембрана 65
Гидропнле 372
Гидрофилия 450
Гидроцель 220
Гинецеи 441
Гнногамоны 59
Гнногенез 463
Гипертрофия компенсаторная 379
— регенерационная 379
Гнпобласт первичный н вторичный
139, 140, 142, 179
Гиподерма 56
Гипокотиль 457
Гипостаз 441
Гипотезы об эмбриональных полях
301
Гистогенез 186
Гнстолнз 224
Глазной бокал 166, 169, 170, 241, 245,
252
Глазные пузыри 163, 165, 167—169
— стебельки 166, 167
Головная складка 211
Гомологичные образования 118, 124
Гомотрансплантацня 352, 358
Гонады 33, 35, 37, 200, 222, 266, 397
Гонобласт 29, 36
Гоноцнты 34, 36, 47, 396
Гормон 47, 158, 225, 226, 265, 306
343, 349, 364
— гонадотропный 43, 52, 266
— тиреоидный 228, 307
— тиреотропный 228
— стероидный 44, 52
— ювенильный 225
Граафов пузырек 8, 42
Двудомность 450
Девиация 417, 4Й4
Дегенерация 36, 90, 199, 242, 260
Деднфференцнацня 395, 401
Делямннация 114, 128, 137
Дерма 171
Дерматом 171, 187
Детерминанты 28
Детерминация 3, 140, 154, 230—232,
241, 254, 272, 292, 300, 345
— лабильная 239—241
— стабильная 239, 241
Дефинитивные структуры н функции
5
Диада 443, 449
Диапауза 23
Диастема 93
Днктносома 447
Днплеврула 219, 429
Диплоидный набор хромосом 20, 48,
74
Днскобластула 110, 136
Днсснмнляцня 53, 144, 330
Дифференциация 3, 7, 16, 30, 33, 186,
229, 231, 254, 257, 261, 267, 270,
272, 279, 284, 288—291, 294, 295,
306, 309, 310, 317, 335, 359, 400
Дихогамия 450
Доминантные области 313
Дробление яиц 75, 78, 87, 177
— анархическое 84, 86
— билатеральное 84
— гетероквадрантное 82, 83, 239
— гомоквадрантное 82
— двуснмметрнчное 84
— декснотропное 82, 83
— днскондальное 78—80, 96, 98, 136
— леотропное 82, 83
— неравномерное 80, 81, 86, 98
— несинхронное 81
— поверхностное 78, 79, 98
— полное 78, 98
— равномерное 80, 81, 86, 98
— радиальное 81, 82, 85, 86
— синхронное 81
— спиральное 81—83, 85, 86, 100
— частичное 78, 98
Жаберные мешки 183
— шелн 413, 415
Железа щитовидная 184, 306
— поджелудочная 185
Железы кожные 117
— околощитовидные 184
— половые 33—35
— торакальные 225
Желток 44, 45, 76, 79, 88, 95—98,
109, 110, 114, 129, 136—138, 140,
142, 146, 200, 210, 211, 216
475
Желточный мешок 137, 138, 141, 179,
180, 209, 211, 213, 214
Желточная оболочка 25, 138
— пробка 133, 134
Завязь 440, 441, 450, 465, 466
Закон зародышевого сходства 10, 409,
423
Зародышевые листки 9, 11, 35, 107,
111, 119—125, 137, 175, 192, 413
— оболочки 211
Зародышевый мешок 138, 441, 442,
444, 446, 447, 451, 464
---- биспорический 445
— — моноспорический 445, 446
---- тетраспорический 445
— путь 27, 29, 31, 32
— узелок 142
— щиток 139, 140, 142
Зигота 18, 19, 287, 294, 455, 466
Имагинальный диск 276
Имаго 223
Иммиграция мультиполярная 113—
115, 153, 316
— униполярная 113, 153
Иммунитет эмбрионов 360
Имплантация 142, 365
Инвагинация 113, 115, 121, 131, 133
134, 155, 157, 158, 337, 434
Инволюция 5, 361
Индукция 242
Интеграция 86, 100, 236, 270, 295
Инцистирование 19
Каллозные оболочки 449
Каллус 466
Кариогамия 48
Кариосфера 50
Карман Ратке 181
Карты презумптивных органов 125,
----зачатков 126, 127, 137
Кейлоны 89
Клетки апикальные 456
— археспориальные 446, 447, 449
— базальные 456
— зимогенные 33
— иммунокомпетентные 393
— интерстициальные (i-клетки) 33
395, 396, 399
— меристематические 438
— мезенхимные 36, 174, 206, 243
— париетальные 443
— перитонеальные 32
— половые 19, 24, 27, 30—38 46 48
51, 56, 76, 279, 281, 419
---- первичные 36
— резервные 395, 399
— сертолиевые 38
— соматические 18, 19, 23, 26—30
34, 36, 45, 47, 48, 68, 70, 86, 87, 464
— «стволовые» 289, 399
— типотентные 31
— фолликулярные 24, 25, 33, 37, 62
— эпителиальные 33, 120, 205
— яйцевые 24, 26, 58, 69, 72, 104,
138, 235
Клеточное поле 301
Колеоптиль 457
Колеориза 457
Конвергенция 137, 423
Конъюгация 18, 19, 49
Копуляция 18, 19
Критические периоды в развитии
организма 343
Ксеногамия 450
Ксенопластнка 232
Куколка 224, 285, 327
Ламеллы 458
Лейкоэмбриофиты 458
Личинка 84, 119, 124, 215, 220, 319,
327
Макромеры 82—84, 103, 114, 278
Мацерация 447
Мегаспора 443
Мегаспорогенез 443, 444
Мегаспороцит 443
Медуллярная пластинка 240, 241, 243,
245, 252, 313
— трубка 162
Мезентерий >3
Мезенхима 35, 118, 155, 158, 171, 183,
188—190, 200, 201, 206, 336, 368
Мезоглея 120, 360
Мезодерма 11, 32, 35, 84, 111, 115—
123, 127, 130, 131, 134, 136, 153—
160, 175, 186, 190, 192, 199, 200,
201, 214, 219, 242, 246, 251, 260,
335, 413, 428
Мезокардий 190
Мезонефрос 194, 197, 199, 339, 363,
367
Мейоз 41, 43, 46, 48—52, 443
Мероциты 97
Метаболизм 23, 24, 86, 103, 145, 146,
196, 283, 297, 322, 349
Метамерия первичная и вторичная
Метаморфоз 5, 9, 14, 119 215—228,
261, 285, 341, 361, 429, 462
Метанефридии 195
Метанефрос 194, 198, 199
Метаплазия 397
Мнокардий 191
476
Микромеры 82—84, 103, 106, 118, 278
Микропиле 61, 372, 441, 442, 451
Микроспорангий 447
Микроспороциты 449
Миобласт 243
Миотом 187, 189
Митоз 24, 28, 48, 51, 52, 76, 87—
91, 103, 236, 280, 364
Мозговые пузыри 162
Моноспермия 57, 58
Морея 412
Морула 108, 114, 141
Морфоген 5, 323
Морфогенез 15, 202, 302, 323, 456, 468,
470
Морфогенетические движения клеток
205
Мюллеров проток 306, 307
Нейробласт 164
Нейроглия 164
Нейромер 163
Нейропор 162
Нейрула 90, 152, 161, 168, 178, 190,
241 253 328
Нейруляция 146, 147, 161, 175, 190,
313, 344
Необласты 396
Неотения 228
Нервная трубка 161
Нервные валики 161
Нефротом 187, 196—198
Нефроцель 197
Нитчатый аппарат 446
Нуцеллус 441, 443, 466
Нуцеллярный колпачок 442
Оболочка оплодотворения 64, 65
Оболочки яиц 63, 138
----вторичные 24, 25
----первичные 24, 25
----третичные 24, 25
Овуляция 25, 73
Онтогенез 12, 14, 31, 32, 53, 54, 76,
117, 123, 125, 138, 195, 196, 229,
254, 260, 271, 272, 295, 300, 326,
349, 402, 409, 414—418, 425, 430,
462, 468, 469
Онтогенетика 6
Онтогения клетки 6, 11, 24, 52, 87,
89, 103, 106, 267, 433
Оогенез 32, 33, 35, 37, 40, 42, 45—48,
54, 55, 104, 106, 145—147, 188, 363
Оогонии 34, 43, 46, 51, 364
Оосомы 34
Ооцит 24—26, 33, 34, 36, 43—46, 49,
50, 52, 54, 61, 106, 334, 367
Оперкулюм 442
Оплодотворение 23, 26, 43, 56—60,
62—70, 74, 89, 99, 104, 105, 232,233,
279, 450
— двойное 440, 450, 452
Опыление 450, 463
— перекрестное 450
— самоопыление 450
Органогенез 117, 119, 122, 160, 179,
186, 204, 245, 276, 293, 426, 437,
451, 456
Оргаиотипический период 101
Орнитофилия 450
Осеменение 23, 66, 87
— внутреннее 56
— наружное 56
Оскулум 382
Палингенез 377, 410
Парабласт 97, 136, 137
Паренхима 34
Паренхимула 121, 218
Партеногенез 6, 24, 56, 69—73, 257,
463
— естественный 70—72
— искусственный 70—72
— факультативный 70
Педогенез 70
Первичная бороздка 140
— полоска 140
Перибласт 97, 137
Перибластула 110
Перивителлиновая жидкость 334
Перивителлиновое пространство 65, 66
Перидерма 171
Перикардий 191
Период развития пыльника мейотиче-
ский 447
— постмейотический 447
— премейотический 447
--- яиц 100
------- дофункциональиый 100
-------органотипический 100, 101
---функциональный 100
— цитотипический 100, 101
Перисперм 457, 461
Перфораторий 62
Пестик 451
Пиноцитоз 63, 64, 259
Плакоды 171, 172, 181
Плакула 110, 113
Планула 86, 218
Пластиды 443
Плацента 140, 142, 143, 357, 365
— гемохориальная 144
--- ворсинковая 144
--- лабиринтная 144
— десмохориальная 143
— эндотелиохориальная 144
477
— эпителиохориальная 142
Плод 462
Плодолистики 441
Плутеус 99, 146, 156, 220, 286
Подиум 442
Полиспермия 26, 57, 58, 65, 66
Полиэмбриония 27, 86, 248—250, 252,
253, 373—376, 391, 464
Полярное тельце 45, 46
Порогамия 451
Постамент 442
Почечка 457
Почка конечности 201
Презумптивные органы 126, 128, 131
Прехордальная пластинка 118, 126
Провизорные органы 5, 138, 208
Прогенез 37
Проктодеум 155, 213
Проморфология 23
Пронефрос 194, 195, 197, 199, 226, 339
Протонефридии 195
Прототрох 84, 269
Профаза мейоза 39, 48, 49, 50
----диакинез 49, 50
----диплотенная 49, 50
---- зиготенная 49
---- лептотенная 49
---- пахитенная 49
Псевдоподиальная активность клеток
157
Пуфы 282—285
Пыльник 447, 448, 465
— зрелый 447
Пыльцевая трубка 446, 450, 451
Пыльцевое зерно 441, 442, 449
Пыльца 450
Размножение бесполое 14, 19, 27,
361, 373, 374, 385
— половое 19, 373
Рауберов слой 142
Регенерация 15, 34, 144, 200, 253, 261,
303, 361, 378—408
— репаративная 379
— физиологическая 379
Регуляция 232, 235, 350
Редукционное деление 48
Рекапитуляция 121, 184, 412, 415, 468
Репродукция 19
Реципиент 232, 242, 287
Роговица 169, 170, 339, 397
Рыльце 440, 441, 450
Самонесовместимость 450
Связник 447
Сегментация мезодермы 186
Сегрегация цитоплазмы 52
Семенник 32, 38, 40, 47, 118, 264, 265,
307
478
Семя 441
Семяножка 441
Семяпочка 441, 442, 447, 461, 466
Семядоля 457
Серозная оболочка 212, 213
Серый серп 66, 133, 312
Сетчатка 166, 167, 247
Синергиды 446
Склера 169, 170
Склеробласты 384
Склеротом 187—190
Слуховой пузырек 172, 173, 202, 245,
252, 256, 339
Соматобласт 84
Соматоплевра 118, 187
Сомиты 83, 163, 186, 187, 239, 240
248—250, 324
Сперматиды 41, 42, 51
Сперматогенез 33, 36, 37, 43, 45, 47,
48, 103, 269, 348
Сперматогоний 34, 39, 40, 51, 269
Сперматозоид 18—24, 39, 42, 45, 48,
56—64, 280
Сперматофор 56
Сперматоцит 41, 46, 49
Спермин 42, 57, 60, 61, 63, 65, 450,
451
Спермиогенез 41
Спланхноплевра 118
Спланхнотом 187, 188
Спорофит 19, 441, 455, 466
Спорогенез 441
Стекловидное тело 166
Стерробластула 109
Столбик 441, 450
Стомодеум 32, 111, 226
Стробилляция 341
Строма 200, 458
Тапетум 448, 449
— клеточный секреторный 449
— периплазмодиальныц 449
Тека 447
Телобластнческий способ образования
мезодермы 115, 116, 122
Телобласты 84, 116
Теория физиологических градиентов
Чайлда 307
Тератогенез 107
Тотипотентность 30, 31, 286
Толерантность 353
Трансплантация 231, 240, 247, 249, 353
Трофобласт 141, 142, 144
Трохофора 427
Туловищная подзародышевая складка
211
Тычинка 441
Фагоцителла 121, 434
Фагоцитоз 38, 47, 64, 152, 351, 361,
367, 369
Фаза роста 39
Фенотип 293
Фертилизин 59
Физиологический градиент 14, 307—
310, 320
Филогенез 12, 45, 47, 125, 410, 411,
414, 469
Филогения 6, 11, 270, 412, 439, 466
Филэмбриогенез 12, 377, 416, 417
Фитонциды 339, 340
Фолликул 36—38, 43
Фороцит 102
Фуникулус 441, 442
Халаза 441, 451
Хназмы 49, 50
Хлороэмбриофиты 458
Хоаноциты 33, 384, 385
Хоаны 172, 182
Хорда 128—134, 177, 239, 242, 250,
254, 259, 413
Хордомезодерма 128, 133, 136, 137,
244, 246, 250
Хорио-аллантонс 214
Хорнон 26, 142, 144, 209, 212, 213
Хрусталик 167, 168, 241, 339
Цветок 441
Целобластула 86, 109, 113
Целом 91, 116, 118, 119, 130, 155,196,
197, 213, 220, 229
Ценогенез 377, 410
Ценоцит 446
Центросома 67
Цитокинез 458
Цитотипический период 101
Чашелистики 441
Щиток 457
Эйякуляция 56
Экзогаструла 337
Экзогаструляция 151, 337, 339
Экзостом 442
Экзотепий 448
Экзоцель 209
Экзоцитоз 65
Эксплантация 243, 250
Эксцнстирование 19
Эктодерма 11, 32, 103, 112
117, 120, 123, 128, 137
160, 174, 239, 258, 341
Эктоплазма 114
Эмбриобласт 141, 142
Эмбриогенез 14, 31, 86, 87,
114, U5,
153, 155,
106, 107.
268, 302, 328, 377, 388, 438, 458,
465, 469
— соматический 27, 86, 123, 161, 232,
360, 361, 373, 378, 469
Эмбриодерма 457
Эмбриоид 463—466
Эмбриония 463
— интегументальная 463
— нуцеллярная 463
Эмбриология 3, 5, 6, 10, 16, 27, 120,
269, 351, 469
— популяционная 468
— эволюционная 10, 12, 120, 469
— экологическая 468
— экспериментальная 12, 14, 203, 206,
465
Эмбриотека 86
Эндокардий 190
Эндомиксис 19
Эндоплазма 114
Эндоплазматический ретикулум 443
Эндосперм 447, 457, 460, 466
— гелобиальный 461
— нуклеарный 461
— целлюлярный 461
Эндостом 442
Эндотеций 448
Энтелехия 8, 9, 299
Энтероцельный способ образования
мезодермы 112, 118
Энтодерма 11, 32, 35, 112—115, 117,
118, 120, 122, 128, 130, 131, 137,
139, 153, 160, 177, 179, 239, 335
Энтомезодерма 155
Энтомофилия 450
Энуклеация 286
Эпибласт 457
Эпиболия 115, 134, 153
Эпигенез 9, 299, 319
Эпидермис 128, 130, 134, 162, 168—
174, 176, 200, 201, 206, 258, 448
Эпикардий 191
Эпиморфоз 394
Эпистаз 442
Эпителий 35—38, 42—44 117, 138,
168, 169, 205, 397
Яичник 32, 42, 43, 53, 56, 118, 262
Яйца алецитальные 76, 90, 94, 98, 141
— гомолецитальные 77, 78, 80, 90
— телолецитальные 77, 78, 90, 94, 98,
114, 140
— центролецитальные 77—79, 97, 98.
114
Яйцевые оболочки 26, 86, 214 216,
371
Яйцевой аппарат 446
Яйцеклетка 20, 36, 50, 53, 54, 74, 99,
103, 159, 287, 310, 328, 420
ОГЛАВЛЕНИЕ
Предисловие........................................................... 3
Глава /. Предмет, краткая история и современные направления в эмбрио-
логии .............................................................. 5
Глава II. Половые и соматические клетки. Происхождение и строение по-
ловых клеток.................................................... . 18
Глава III. Оплодотворение и партеногенез..............................56
Глава IV. Дробление яиц...............................................75
Глава V. Бластула. Гаструла. Зародышевые пласты.......................Ю8
Глава VI. Сравнительный обзор процессов гаструляцин..................129
Глава VII. Развитие производных эктодермы............................160
Глава VIII. Развитие производных энтодермы...........................177
Глава IX. Развитие производных мезодермы.............................186
Глава X. Метаморфоз..................................................215
Глава XI. Детерминация зачатков органов и дифференциация клеток и
ткаией.............................................................229
Глава XII. Эмбриология и генетика....................................273
Глава XIII. Гипотезы об эмбриональных полях. Теория физиологических
градиентов.........................................................296
Глава XIV. Развитие организма и среда................................325
Глава XV. Эмбриология и иммунология. Защитные реакции зародышей.
Антигенная реактивность............................................351
Глава XVI. Бесполое размножение, соматический эмбриогенез и регенерация 373
Глава XVII. Онтогенез и эволюция. Биогенетический закон..............409
Глава XVIII. Некоторые вопросы эмбриологии растений..................440
Литература...........................................................471
Предметный указатель.................................................474
Учебное издание
БОРИС ПЕТРОВИЧ ТОКИН
ОБЩАЯ ЭМБРИОЛОГИЯ
Заведующий редакцией А. Г. Гаврилов. Редактор Т. А. Рыкова. Младшие редакторы
Е. И. Попова, И. М. Павлова. Художник Ю. Д. Федичкин. Художественный редактор
В. И. Мешалкии. Технический редактор Т.А. Новикова. Корректор С. К. Завьялова
ИБ № 6444
Изд. № Е-467. Сдано в набор 05.09.86. Подп. в печать 02.02.87. Т —08314.
Формат 60Х88'/16. Бум. тнп. № 2. Гарнитура литературная. Печать офсетная.
Объем 29,40 усл. печ. л. 29,40 усл. кр.-отт. 33,35 уч.-изд. л.
Тираж 16000 экз. Зак. № 5298. Цена 1 р. 50 к.
Издательство «Высшая школа», 101430, Москва, ГСП-4, Неглннная ул., д. 29/14.
Ордена Октябрьской Революции и ордена Трудового Красного Знамени МПО «Первая
Образцовая типография имени А. А. Жданова» Союзполиграфпрома при Государственном
комитете СССР по делам издательств, полиграфии и книжной торговли, 113054, Москва,
Валовая, 28.