/








































































































































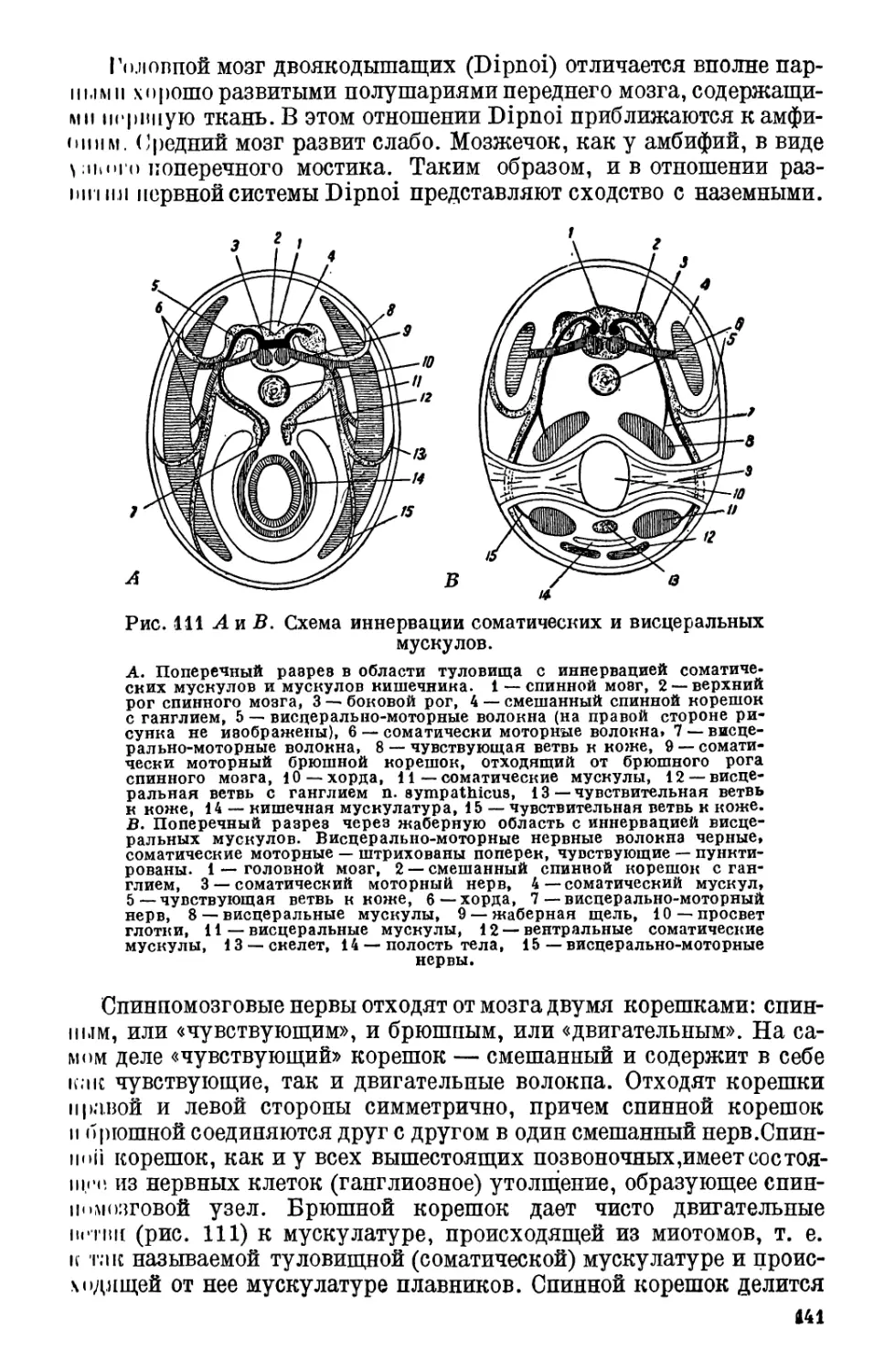




















































































































































































































































































































































































































































































































































































































































































































































































































































































































Текст
Кашкаров Д.Н., Станчинский В.В.
Курс зоологии
позвоночных животных
БИОМЕДГИЗ
1940
ПРЕДИСЛОВИЕ КО ВТОРОМУ ИЗДАНИЮ
Авторы полагают, что в университетском курсе изложение
должно итти с определенной точки зрения в связи с задачами со-
ветской школы. Кроме того, курс зоологии позвоночных — это
основной курс, который будут прорабатывать не только будущие
специалисты-зоологи, но и биологи других специальностей. С этим
багажом последние выйдут из университета, и многие из них пойдут
на педагогическую работу, где им придется преподавать и зоо-
логию.
Основная точка зрения авторов, проводимая в курсе, такова,
что изучение зоологии должно дать целостное представление о жи-
вотных и в первую очередь об их эволюции, зависимости от среды
и о практическом значении их для человека.
Некоторых товарищей несколько смущали размеры курса,
обилие преподносимого в нем материала. Но авторы полагают,
что учебник вовсе не должен служить конспектом для заучивания
курса к зачету. Студент должен работать с учебником, а по-
этому учебник должен быть интересен для студента, оп должен
быть таким, чтобы его можно было не заучивать, а читать и усваи-
вать; оп должеп быть рассчитан на подготовленного студента, от-
носящегося к изучению предмета активно и аккуратно слушаю-
щего лекции. То, что необязательно для студента, может быть вы-
несено петитом. Нет ничего скучнее, как выучивать морфологи-
ческие характеристики, когда данные морфологии преподносятся
без обобщающей, связывающей и осмысливающей их идеи. Поэтому
авторы излагают морфологические данные в эволюционном осве-
щении, связывая каждый факт с другими идеей развития. То же
самое относится и к систематике. Внесение в нее эволюционного
принципа, включение в систему животных ископаемых групп при-
дает ей гораздо больший интерес.
Опыт пяти лет преподавания с данным учебником показал,
что авторы не ошиблись. Учебник, несмотря на то, что был издан
i* з
плохо, со многими опечатками и редакторскими недосмотрами,
пользовался у студентов определенным успехом: они предпочитали
заниматься по этому учебнику, чем по другому; даже государствен-
ники не зоологи не пожелали воспользоваться разрешением сда-
вать экзамен по Холодковскому, которое пришлось дать им из-за
недостачи книг.
Опыт показал, что студенты хорошо справляются с книгой,
несмотря на ее объем, что книга возбуждает в них интерес к зооло-
гии. Поэтому авторы в настоящем издании не изменили принципа
построения курса, оставив прежний план. Зато, учтя сделанные
студентами и преподавателями замечания и свой опыт, авторы зна-
чительно подработали курс, освободив его от опечаток, погрешно-
стей стиля, уточнив формулировки, освежив некоторые места
новыми данными. Авторы надеются, что второе издание их курса
зоологии позвоночных, широко использовапного в первом издании
и преподавателями средней школы, будет так же тепло встречено
широкими кругами читателей, как и первое.
Д. Кашкаров
В. Станчинский
ПРЕДИСЛОВИЕ К ПЕРВОМУ ИЗДАНИЮ
Настоящий «Курс зоологии позвоночных» является перерабо-
танным и дополненным изданием «Курса биологии позвоночных»,
притом ие столько переработанным, сколько дополненным, именно
главами по морфологии и систематике. Биологическая или, точ-
нее, экологическая часть подверглась сравнительно небольшой
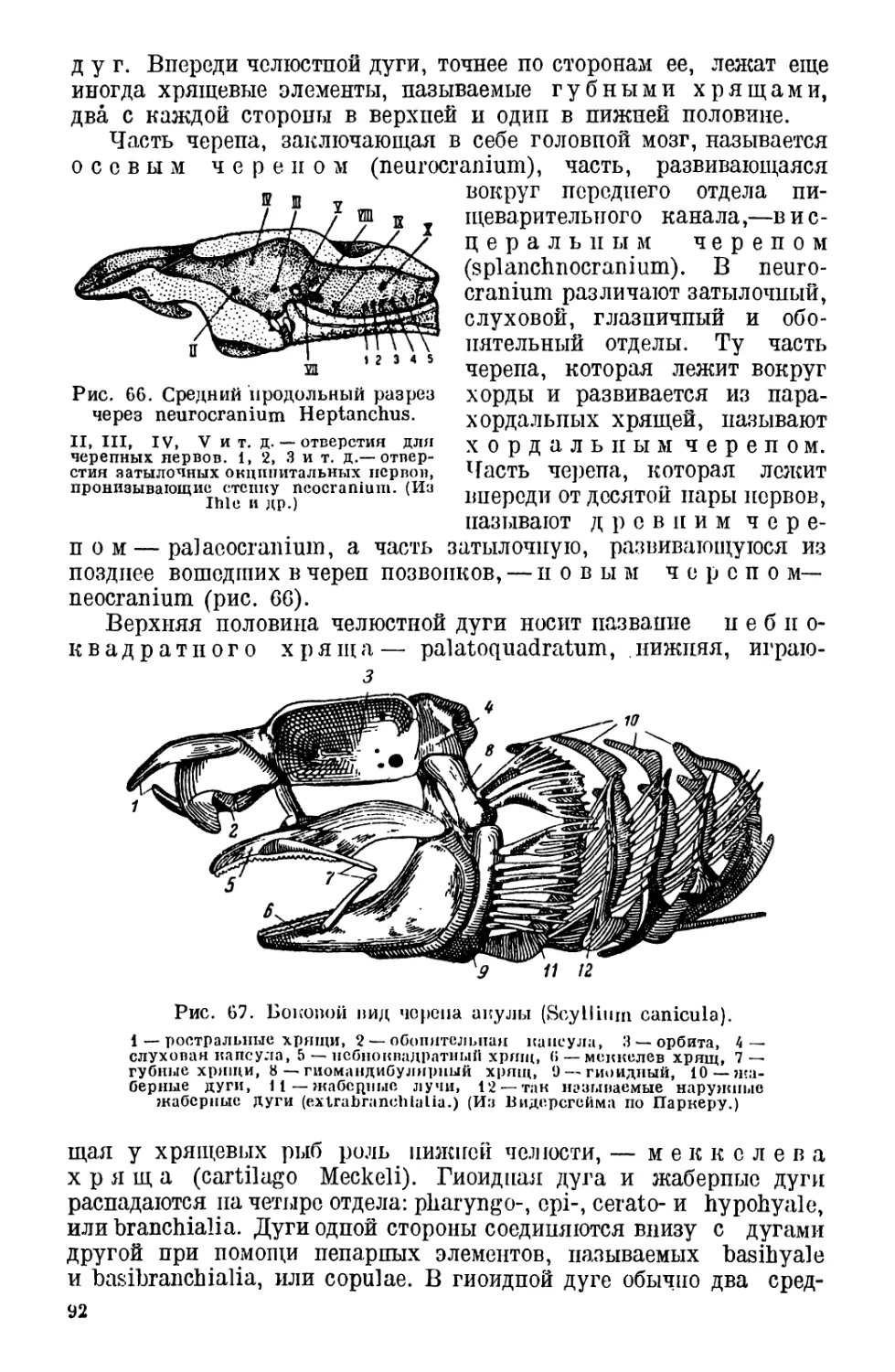
переработке.
Выпуская «Курс биологии позвоночных», авторы ставили себе
задачу заполнить существующий пробел в преподавании зоологии
в вузах, которая до сих пор строилась почти исключительно на
основах морфологии и систематики, предоставив в распоряжение
преподавателей и студентов систематический курс биологии, праг
вильнее, экологии позвоночных животных. Авторы исходили из
мысли, что по морфологии и систематике уже имеются соответ-
ствующие курсы и что, кроме того, морфология проходится в курсе
сравнительной анатомии, в котором опа может быть изложена го-
раздо лучше, систематика же проходится на практических занятиях.
Однако в такой оторваппости биологии от морфологии и систе-
матики оказался целый ряд неудобств, идущих в ущерб цельности
курса и затрудняющих усвоение студентом зоологии. К тому же
сравнитсльпая анатомия проходится далеко не во всех вузах и пе
всегда предшествует зоологии позвоночных. При прохождении
систематики па практикумах ускользает эволюционный принцип
и совершенно выпадают ископаемые формы.
Поэтому в настоящее издание решено было включить морфоло-
гию и систематику, проводя в той и другой, насколько возможно,
эволюционный принцип. А для этого необходимо было включить
в систематику (а также и в морфологический обзор) ископаемых
представителей классов. Это введение значительно усложнило систе-
матику в смысле объема, но внесло в нее несколько больше ясности.
Набор части систематических обзоров петитом облегчает студенту
изучение систематики, отделяя более важное от менее важного.
5
Говоря о филогенетической зависимости различных системати-
ческих групп, мы не боялись некоторой гипотетичности, совершенно
неизбежной в филогенетических построениях.
Объем курса, конечно, больше нормального. Однако авторы ис-
ходили из того, что уровень подготовки студенчества и уровень его
запросов непрерывно поднимаются. К тому же увеличились и тре-
бования к студенту. От пего требуются усвоение основ науки, из-
вестная культурность в избранной им области. И требования эти
все время возрастают. Курс, который давал бы лишь элементарные
сведения, сейчас не удовлетворял бы ни предъявляемым требова-
ниям, пи интересам преподавателей и студентов. Современный сту-
дент ищет более полного знания и хочет, кроме того, иметь инте-
ресную книгу, что очень трудно дать, применяя краткий схемати-
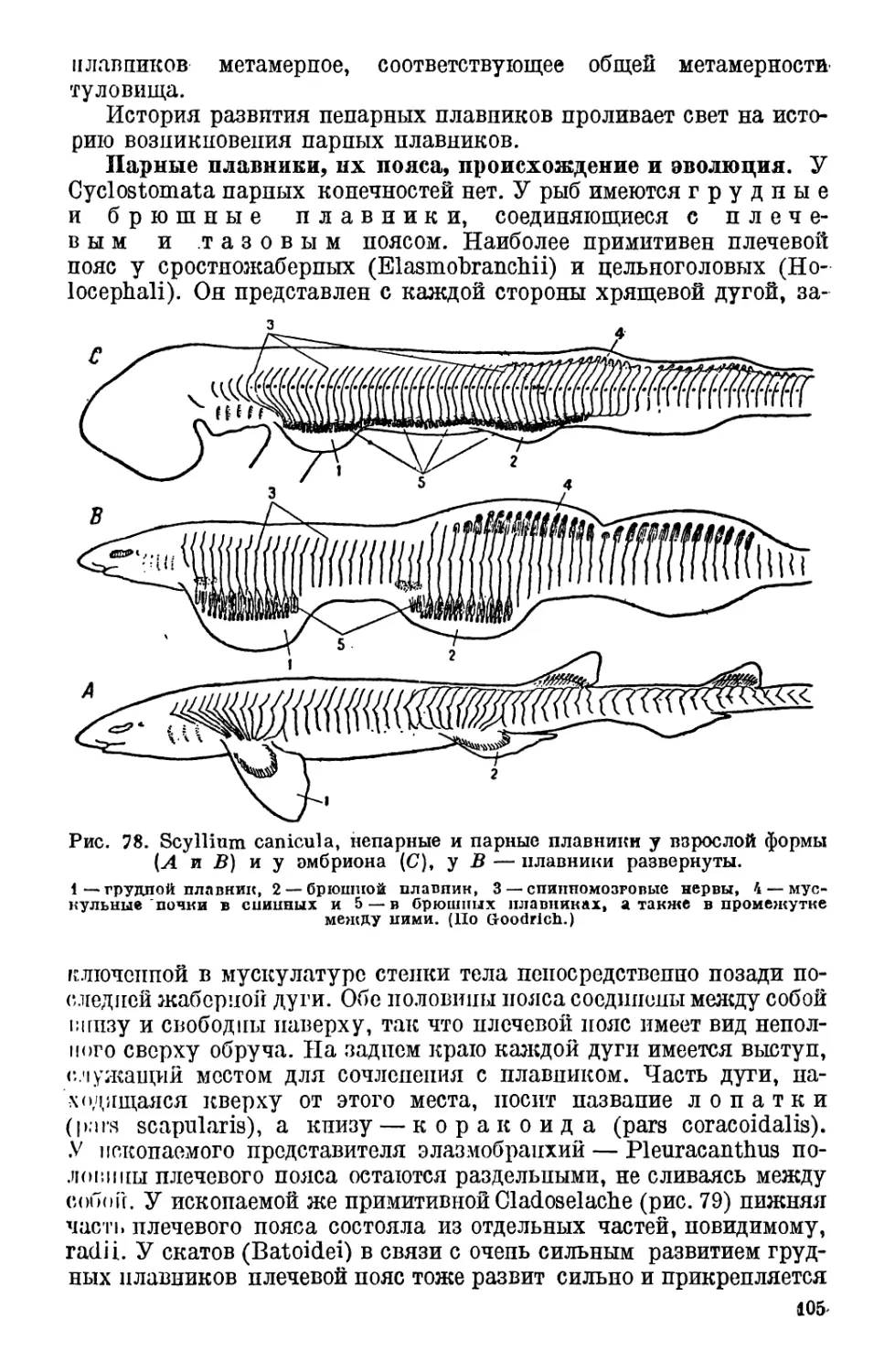
ческий курс.
Авторам желательно было бы богаче снабдить курс рисунками,
но сделать этого не удалось в настоящем издании.
Объектйвные условия, удаленность авторов друг от друга в те-
чение последнего года не позволяли согласовать написанные ими
части и совместно проредактировать курс. Этим объясняется неко-
торая невыдержанность размеров, а иногда и характер частей книги.
Д. Кашкаров
В. Статинский
ОГЛАВЛЕНИЕ
Предисловие ко второму изданию.................................. 3
Предисловие к первому изданию................................... 5
Часть первая
ОБЩАЯ ХАРАКТЕРИСТИКА ТИПА ХОРДОВЫХ. ПОЛУХОРДОВЫЕ.
ОБОЛОЧНИКИ. БЕСЧЕРЕПНЫЕ. КРУГЛОРОТЫЕ
Характеристика типа хордовых и обзор подтипов .... 13
Подтип 1. Полухордовые (Hemichordata).......................... 13
Класс 1. Кишечнодышащие (Enteropneusta) ................. 14
Класс 2. Перистожаберные (Pterobranehia)................. 18
Положение полухордовых в системе и филогенетические связи ... 21
Подтип 2. Личиночно-хордовые, или оболочники (Urochordata, или
Tunicata)................................................... 21
I. Строение личиночно-хордовых ........................... 22
II. Развитие личиночно-хордовых............................ 25
III. Систематический обзор личиночно-хордовых............... 27
Класс 1. Хвостатые, или личиночные (Copelata, или Lar-
vaeea).................................................... 28
Класс 2. Асцидии (Ascidiae) ............................. 28
Класс 3. Сальпы (Thaliacea) ............................. 30
IV. Значение личиночно-хордовых для эволюционного учения . 30
Подтип 3. Позвоночные (Vertebrate). Обзор подтипа. Общие напра-
вления в эволюции позвоночных................................ 31
Отдел А. Бесчерепные (Acrania)................................. 33
Класс 1. Головохордовые (Cephalochordata)................ 33
Строение ланцетника (33)—Развитие ланцетника (42).
Положение Acrania в системе и происхождение хордовых........... 47
Отдел В. Черепные (Craniata)................................... 50
Подотдел 1. Бесчелюстные (Agnathostomata)...................... 50
Класс 1. Круглоротые (Cyclostomata) .................... 51
Анатомическое строение (51). — Развитие миноги (60). — По-
ложение круглоротых в системе (62).
Классификация, распространение и экология круглоротых.......... 63
Класс 2. Панцырные (Ostracodermi) ....................... 66
Происхождение и ранняя эволюция черепных (Craniata)............ 69
Часть вторая
РЫБЫ
Подотдел 2. Челюстноротые (Gnathpstomata)...................... 72
Класс 3. Рыбы (Pisces)................................... 72
I. Характеристика и обзор класса............................ 72
7
II. Строение рыб............................................ 75
1. Форма тела и движение.................................. 75
2. Кожа рыб. Железы....................................... 78
3. Наружный скелет........................................ 79
4. Мышцы......................................•........... 82
5. Внутренний скелет...................................... 83
Позвоночник (83) — Череп. Сегментация головы (88). — Раз-
витие черепа. Хрящевой череп (90).—Типы окостенений
черепа (94).—Покровные окостенения в черепе рыб(95).—
Замещающие хрящ окостенения в черепе рыб (96). — Эво-
люция черепа в разных отрядах рыб (98).—Скелет конеч-
ностей. Непарные плавники (102). — Парные плавники,
их пояса, происхождение и эволюция (105). — Проис-
хождение парных конечностей (109).
6. Органы дыхания рыб.................................... 111
Жаберные щели, жабры и крышечный аппарат (111). — На-
ружные жабры (114). — Механизм дыхания у рыб и его
эволюция (115). —Добавочные органы и способы дыхания;
плавательный пузырь (118). — Дыхание рыб (123).
7. Органы кровообращения у рыб........................... 125
Сердце ( 125) —Артериальная система (127). — Венозная си-
стема (128). — Кровь рыб (131).
8. Органы пищеварения рыб............................... 131
9. Органы выделения и размножения (мочеполовая система)
рыб..................................................... <35
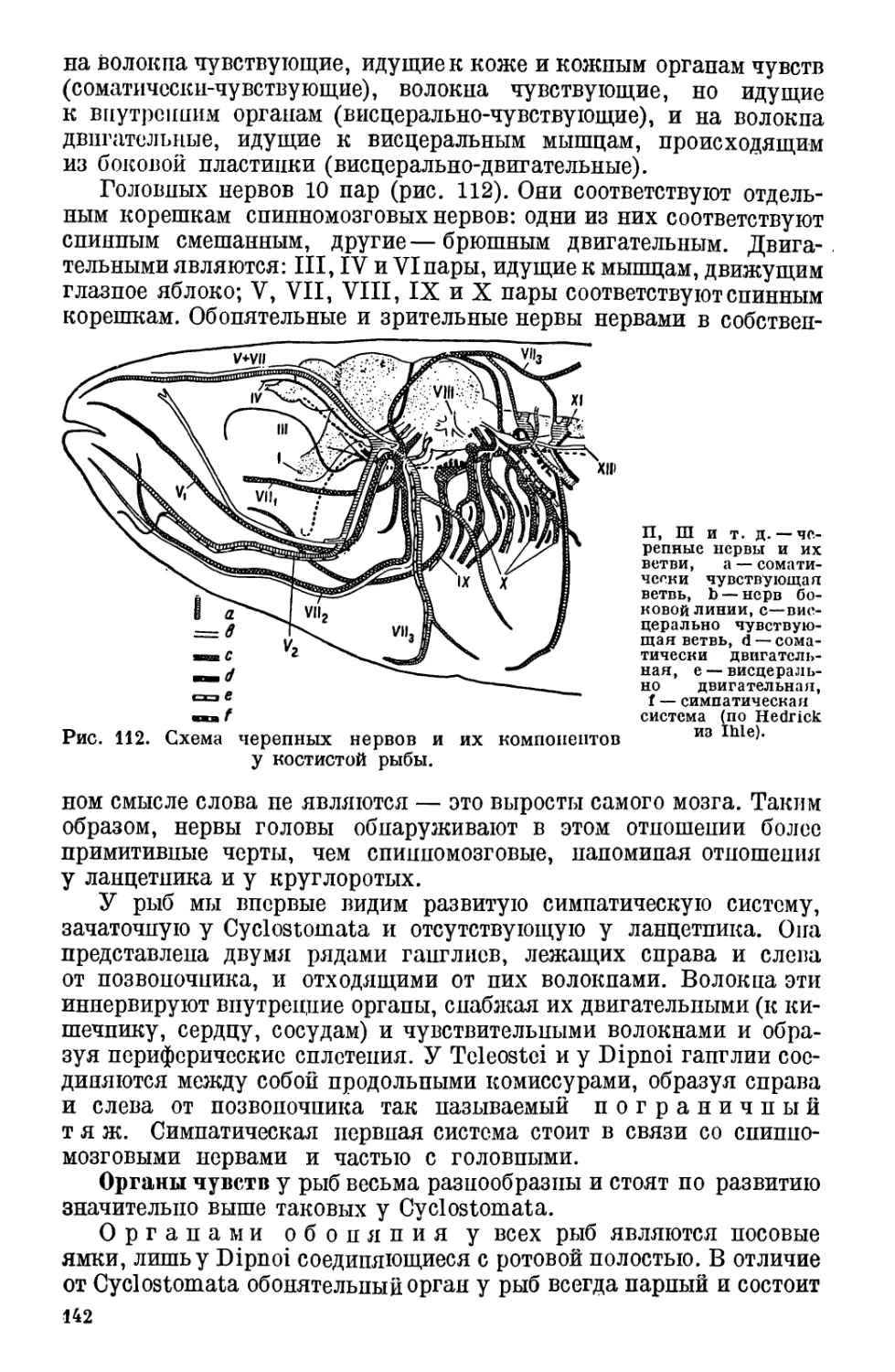
10. Нервная система и органы чувств у рыб................ 139
Нервная система (139).—Органы чувств (142).—Функция го-
ловного мозга у рыб (145).
III. Развитие рыб.......................................... 146
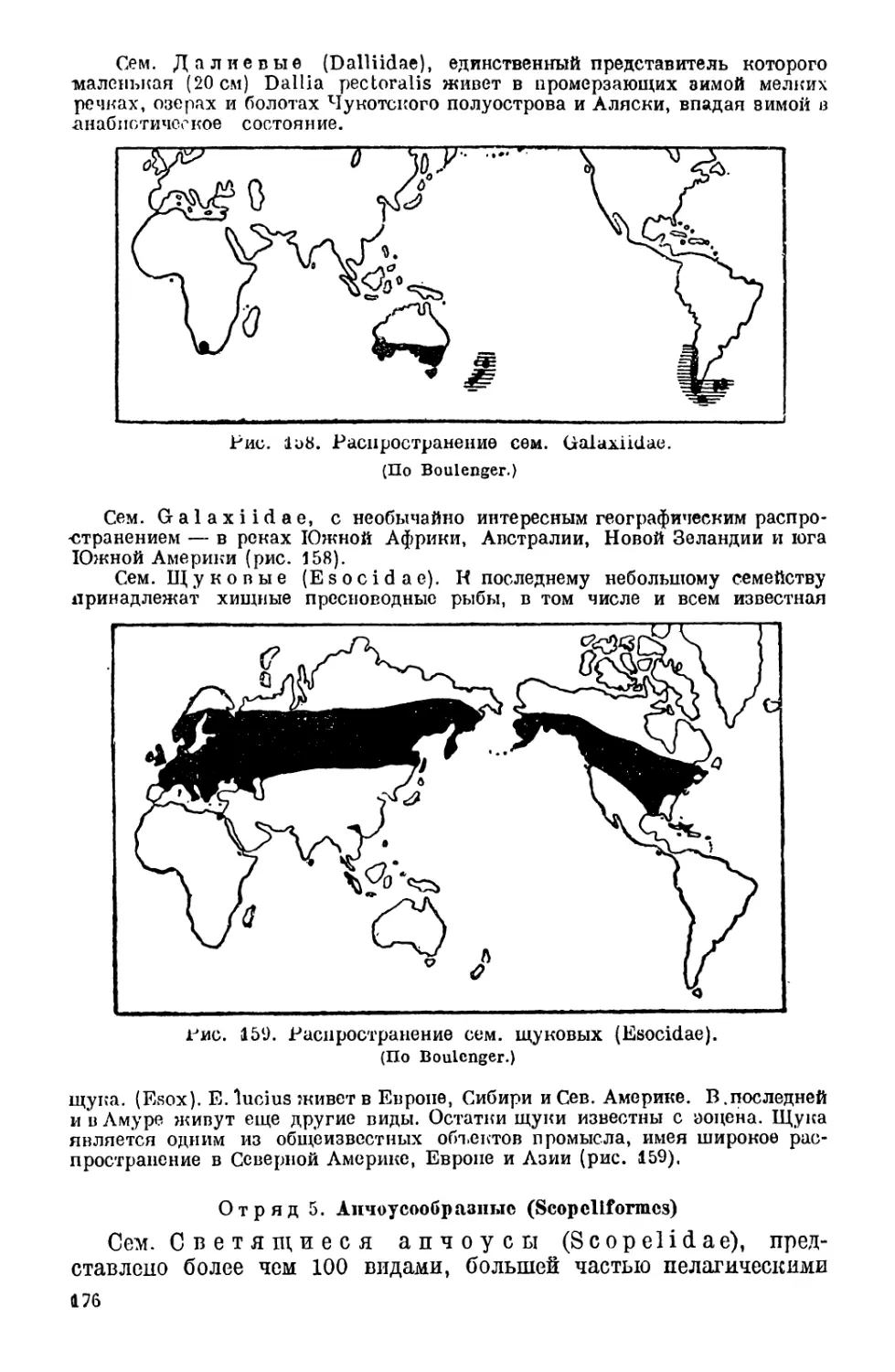
IV. Классификация рыб..................................... 148
V. Филогенетическое развитие рыб.......................... 193
VI. Экология рыб........................................... 200
1. Вода как среда для жизни рыб.......................... 201
Соленость и жесткость воды (201). — Содержание газов в во-
де (204). — Температура воды (205). — Глубина, давление
и освещение в воде (207).
2. Рыбы и водная среда. Приспособления к условиям обита-
ния, адаптация формы и движения. Типы приспособления. 208
Пелагические рыбы — нектон и планктон (209). — Придон-
ные рыбы, или рыбы бентоса (213).
3. Питание и приспособление к роду пищи у рыб............ 216
4. Окраска рыб........................................... 223
5. Защитные приспособления у рыб. Ядовитые железы, элект-
рические органы, панцыри и некоторые другие......... 226
6. Размножение рыб....................................... 231
7. Возраст и рост рыб. Продолжительность жизни........... 241
8. Изменчивость и наследственность. Образование рас .... 247
9. Миграции рыб.......................................... 252
10. Роль рыб в биоценозах и в круговороте веществ........ 271
11. Географическое распространение рыб................... 276
12. Поведение рыб (высшая нервная деятельность).......... 288
VII. Значение рыб для человека............................. 293
1. Рыбные промыслы СССР и главнейшие промысловые рыбы 294
2. Охрана рыб и интенсификация рыбного хозяйства .... 299
Ч асть третья
АМФИБИИ, ИЛИ ЗЕМНОВОДНЫЕ
Класс 4. Амфибии, или земноводные (Amphibia).......... 304
I. Характеристика и обзор класса....................... 304
II. Строение амфибий..................................... 306
1. Форма тела и движение............................. 306-
2. Общие покровы амфибий.............................. 311
3. Наружный скелет.................................... 313
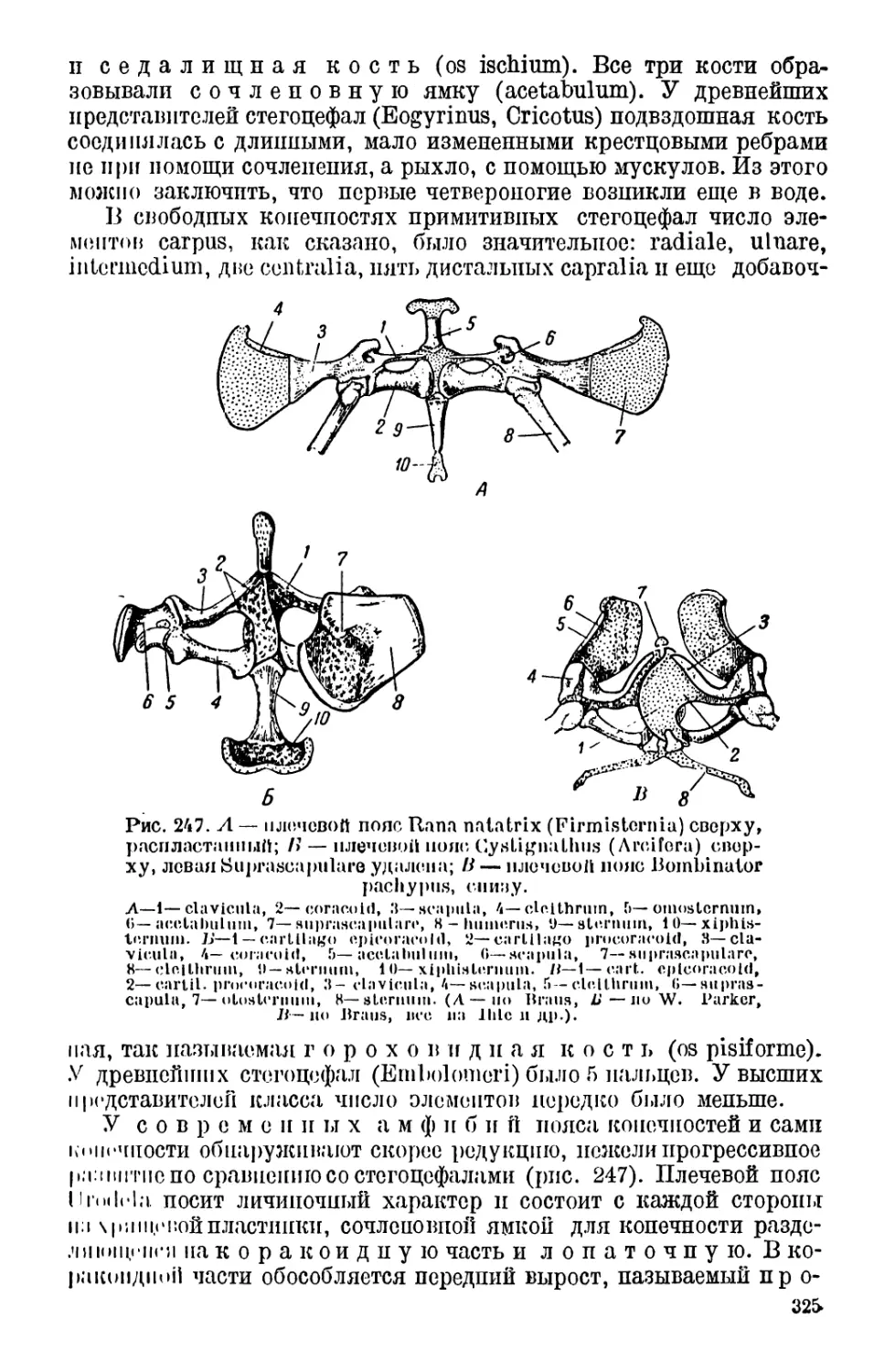
4. Внутренний скелет.................................. 314
Позвоночник (314). — Череп (318). — Конечности и их
пояса (323).
5. Мускулатура........................................ 327
6. Пищеварительная система амфибий.................... 329
7. Органы дыхания амфибий............................. 331
8. Органы кровообращения у амфибий.................... 334
9. Мочеполовая система амфибий........................ 339
40. Нервная система и органы чувств амфибий............ 341
Нервная система (341).—Органы чувств (344).
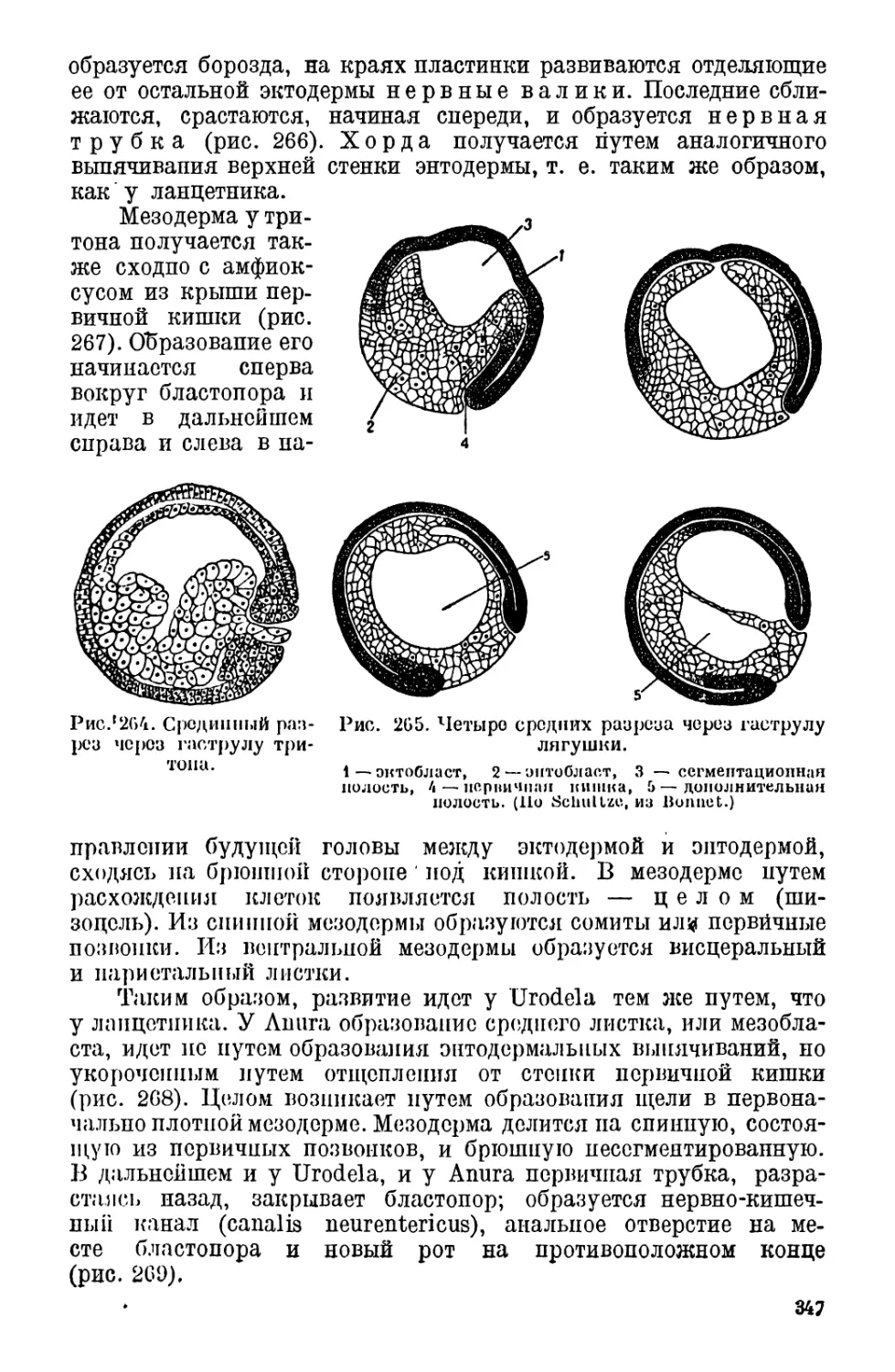
III. Дробление и развитие яйца у амфибий................. 345
IV. Классификация амфибий............................... 348
V. Происхождение амфибий............................... 361
VI. Экология амфибий.................................... 364
4. Приспособление амфибий к условиям среды обитания: ти-
пы — роющий, водный, наземный, древесный.............. 364
2. Питание амфибий. Приспособление к роду пищи........ 367
3. Окраска амфибий.................................... 369
4. Защитные приспособления. Регенерация............... 371
5. Размножение амфибий................................ 373
Половой диморфизм (373). — Выбор места для размноже-
ния (374). — Спаривание и кладка яиц (375). — Развитие и
метаморфоз (376). — Забота о потомстве (377). — Неоте-
ния (381).
6. Приспособление амфибий к климатическим условиям су-
ществования .......................................... 383
7. Роль амфибий в биоценозах и в круговороте веществ. Эко-
номическое значение амфибий....................... 385
8. Поведение амфибий.................................. 387
9. Географическое распространение амфибий............. 391
Часть четвертая
РЕПТИЛИИ, ИЛИ ПРЕСМЫКАЮЩИЕСЯ
Характеристика высших позвоночных (Amniota)...........• . . 395
Общий обзор (395).—Строение яйца, развитие зародыша и
зародышевые оболочки высших позвоночных (396).
Кла сс 5. Рептилии, или пресмыкающиеся (Reptilia) . . . 401
I. Общая характеристика и обзор современных и ископаемых
форм класса рептилий................................ 401
II. Строение рептилий.................................... 404
1. Форма тела рептилий и их движения.................. 404
2. Общие покровы рептилий............................. 409
3. Наружный, или кожный, скелет....................... 412
4. Внутренний скелет рептилий......................... 414
Позвоночник (414). — Череп (417). — Конечности и их поя-
са (425).
5. Мускулатура рептилий............................... 434
6. Органы пищеварения рептилий........................ 436
7. Органы дыхания рептилий............................ 438
8. Органы кровообращения рептилий..................... 438
• Сердце (438).—Артериальная система (439). — Венозная
система (440).
9. Мочеполовая система рептилий......................... 440
Выделительная система (440). — Половые органы (442).
10. Нервная система и органы чувств рептилий............. 443
Головной мозг (443). — Орган равновесия и слуха (444).
Дыхательная область носа (444). — Орган зрения (445).
III. Классификация рептилий (Reptilia).................... 445
IV. Филогенетическое развитие рептилий.................... 500
Изменения среды в эпоху появления рептилий. Отноше-
ние рептилий к амфибиям, адаптация к жизни на
суше (500). — Филогенез отдельных групп рептилий (502).
V. Экология рептилий..................................... 506
1. Приспособления к условиям обитания. Типы: наземный,
подземный, древесный, водный............................ 506
Приспособления к наземному и подземному образу жиз-
ни (506). — Приспособления к жизни на деревьях и к по-
лету (511). — Приспособления для жизни в воде (515).
2. Питание рептилий и приспособления к роду пищи .... 517
3. Окраска рептилий..................................... 521
4. Приспособления для защиты и нападения................ 523
5. Размножение рептилий . ............................. 525
6. Отношение рептилий к климатическим факторам.......... 529
7. Биоценотические отношения рептилий................... 531
Пища и роль рептилий в различных средах обитания . . . 532
8. Значение рептилий для человека...................... 533
Ядовитые змеи......................................, 534
9. Поведение рептилий (высшая нервная деятельность) .... 539
10. Географическое распространение рептилий.............. 542
Ч аслятая
ПТИЦЫ
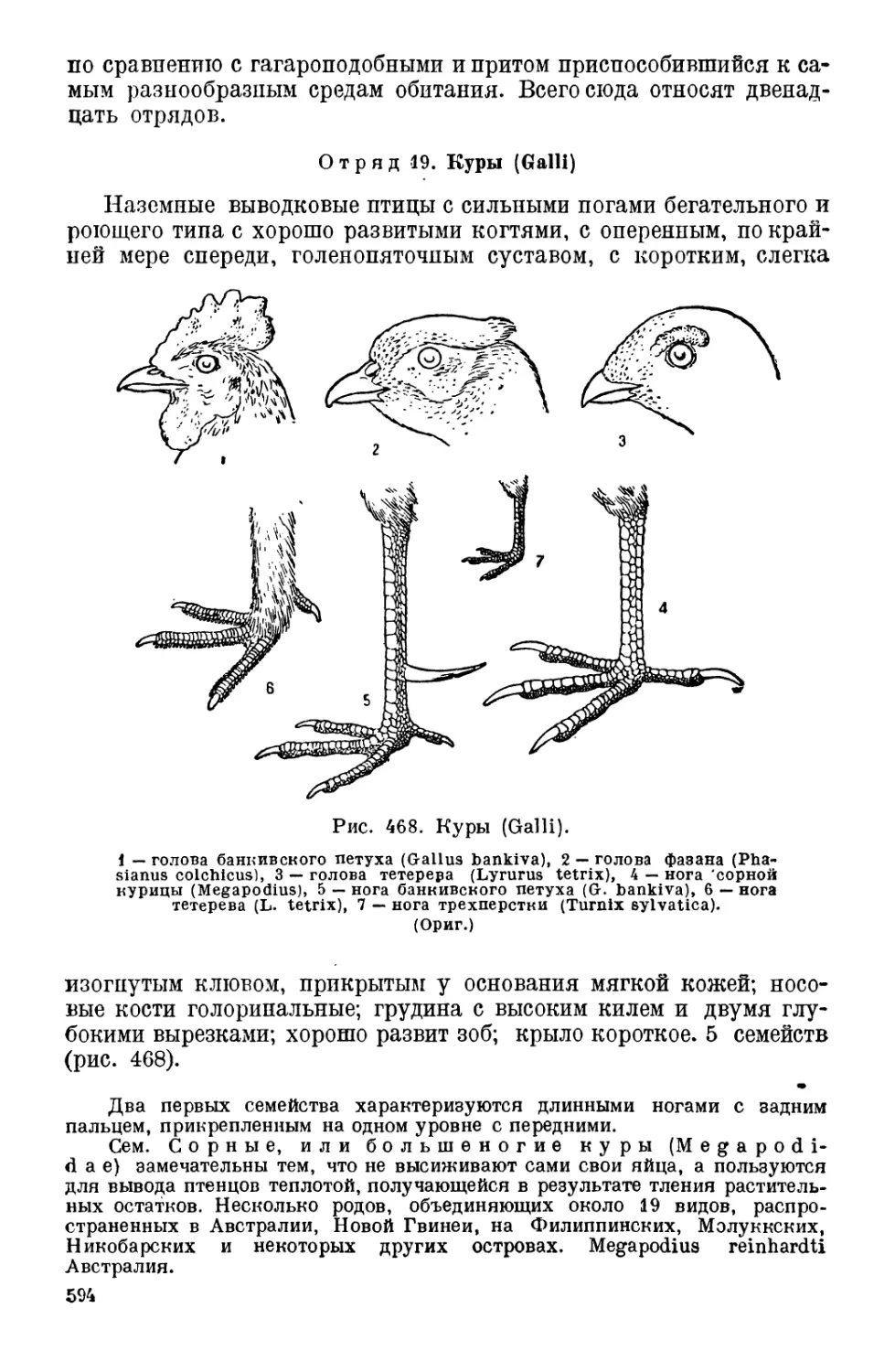
Класс 3. Птицы (Aves)............................... 550
I. Характеристика класса птиц.......................... 550
II. Строение птиц......................................... 551
1. Покровы.............................................. 551
2. Скелет птиц........................................ 556
Позвоночник (557).—-Грудина (558). — Череп (558).—Конеч-
ности (563).
3. Мышечная система..................................... 564
4. Органы пищеварения................................... 566
Пищеварительный канал птиц (566).—Печень (568).— Под-
желудочная железа (568).
5. Органы дыхания птиц.................................. 568
6. Кровеносная система...........*...................... 570
7. Мочеполовая система птиц............................. 571
8. Нервная система и органы чувств...................... 574
Головной мозг (574). — Кожные органы чувств (575). — Ор-
ган слуха и равновесия (575). — Глаза (576 ). — Орган
обоняния (576).
III. Происхождение птиц................................... 576
IV. Классификация птиц................................... 580
V. Эволюция птиц в свяаи с приспособлением к различным
средам обитания.......................................... 617
VI. Полет птиц......................................... 622
Скользящий, или планирующий, полет (624). Гребной, или
пропеллирующий, полет (625). — Парящий полет птиц,
или парение (627).
VII. Пища птиц........................................... 629
10
VIII. Размножение птиц.................................... 636
Вторичные половые признаки птиц (636). — Спаривание
птиц (640).—Гнезда птиц (641). — Яйца птиц (643). — Птен-
цы (647). — Птенцы-паразиты (650).
IX. Линька птиц........................................ 651
Возрастные и сезонные изменения оперения............ 652
X. Окраска птиц........................................ 654
XI. Приспособления птиц в целях защиты . . ............ 656
XII. Зависимость жизни птиц от условий среды.............. 659
1. Климатические факторы существования птиц........... 659
Влияние температуры (659). — Влияние света (662). — Влия-
ние влаги (664).—Влияние атмосферного давления (666).
2. Сезонные явления в жизни птиц в зависимости от клима-
тических факторов .................................... 666
Отношение к месту обитания (667). —Жизнь птиц в тропи-
ческих странах в сухое время года (668).—Жизнь птиц
зимой (668). — Перелет птиц} (671). — Условия весенней,
летней и осенней жизни птиц (679).
3. Экотопические факторы.............................. 681
Воздух как среда обитания птиц (682). Вода как среда оби-
тания птиц (683).—Болота как среда обитания птиц (684).—
Открытые пространства как среда обитания птиц (684) —
Древесная растительность как среда обитания птиц (687).
4. Биоценотические факторы............................ 688
Взаимоотношения между птицами и растениями (688).—
Взаимоотношения между птицами и другими живот-
ными (689). — Взаимоотношения между особями одного
и того же вида (693).
XIII. Экономическое значение птиц ........................ 694
1. Домашние птицы . •................................. 694
2. Промысловые птицы.................................. 702
3. Полезные в хозяйстве птицы......................... 706
4. Вредные в хозяйстве птицы.......................... 711
Часть шестая
МЛЕКОПИТАЮЩИЕ
Класс 7. Млекопитающие (Mammalia)...................... 712
I. Характеристика класса млекопитающих................ 5.12
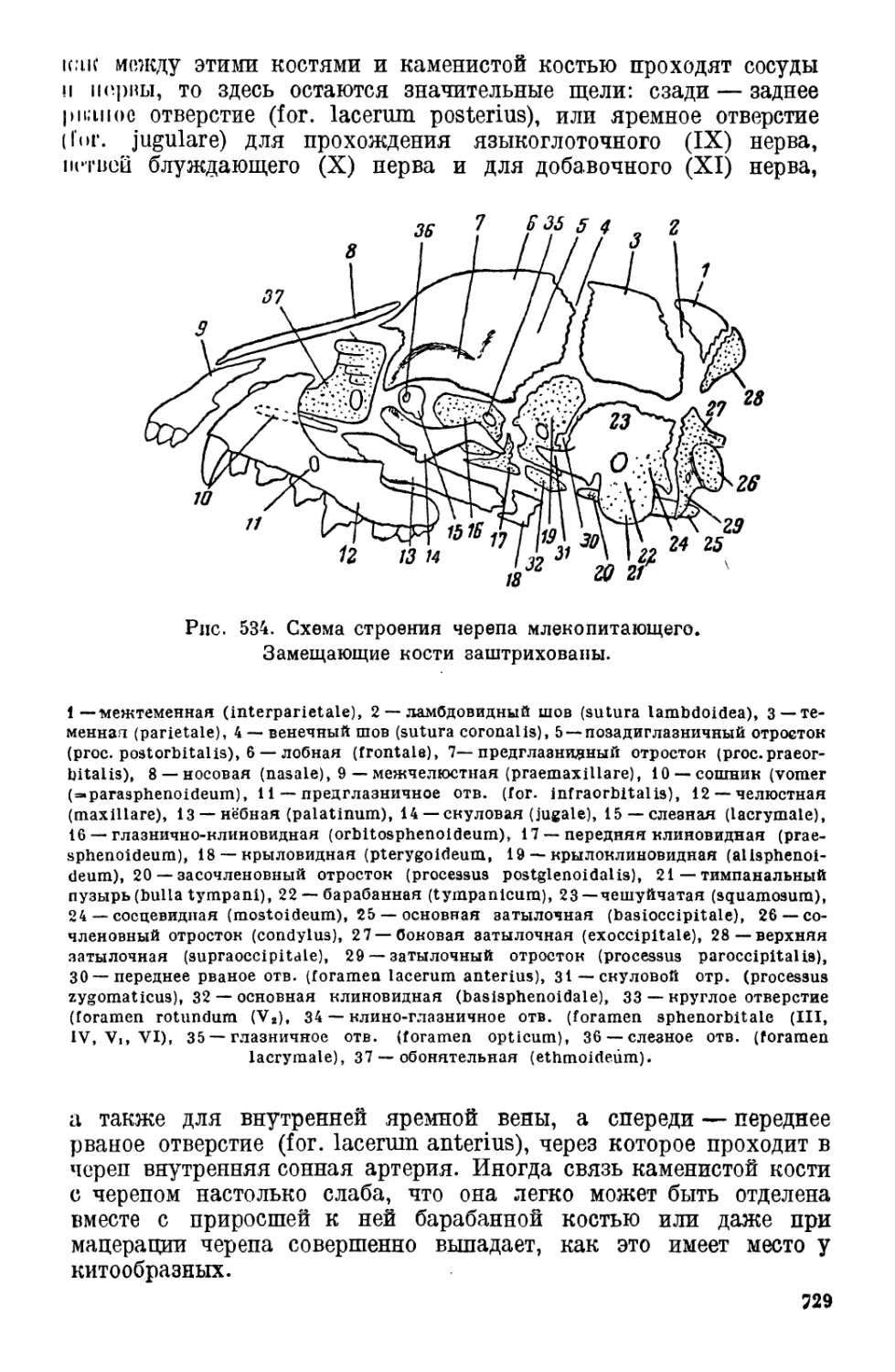
II. Строение млекопитающих............................. 713
1. Покровы............................................ 713
Кожа (71*3); — Волось/(715).— Когти, могтии копыта (717).—
Кожные железы (719).
2. Скелет............................................. 724
Позвоночник (725).—Грудина (726). — Череп (728). — Пояса
конечностей (736). — Скелет конечностей (737).
3. Мускулатура........................................ 747
4. Органы пищеварения................................ 750
Рот (751). — Ротоцыежелезы (752 ). — Зубы',(752). — Кишеч-
ный канал (766). — Печень (766).
5. Органы дыхания..................................... 766
6. Кровеносная система................................ 768
7. Органы выделения................................... 769
8. Органы воспроизведения............................ 770
9. Железы внутренней секреции......................... 775
11
ЙО. Нервная система..................................... 777
li. Органы чувств....................................... 780
Органы осязания (780).—Органы равновесия и слуха (781). —
Глаза (782).—Органы обоня лия (783).—Органы вкуса (783).
111. Происхождение млекопитающих.......................... 784
IV. Классификация млекопитающих........................... 786
V. Эволюция млекопитающих в связи с приспособлением к
среде .................................................. 857
VI. Зависимость млекопитающих от условий среды. Экология
млекопитающих............................................ 858
1. Приспособление к условиям обитания среды как субстрата.
Приспособление к биотопическим условиям............... 858
Млекопитающие открытых мест обитания (859). —Млеко-
питающие роющие и подземные (861). — Лесные млеко-
питающие (865). — Летающие млекопитающие (869).—
Водные млекопитающие (872).
2. Приспособления к климатическим особенностям среды . . 876
Приспособления к температурным условиям (876). — Зимняя
и летняя спячка (878). — Кочевки млекопитающих (880). —
Зависимость отсвета (881). — Зависимость от влаги (883).
3. Биоценотические факторы существования млекопитающих . 884
Пища млекопитающих. Приспособление млекопитающих
к разного рода пище (885).—Зависимость млекопитающих
от растений (889).
Взаимоотношения млекопитающих с другими животными
биоценоза.......................................... 892
Взаимоотношения между самцами и самками и отношение
к детенышам........................................ 895
VII. Высшая нервная деятельность млекопитающих...........' 898
VIII. Экономическое значение млекопитающих.................. 900
1. Домашние млекопитающие, их история и экология. . . . 901
Собака (902),—Домашняя кошка (905).—Крупный рога-;
тый скот (905),—Мелкий рогатый скот (911).—Домашние
свиньи (918), — Домашние лошади (919).—Домашний
осел (923).—Северный олень (925).—Кролики (925).
2. Промысловые и охотничьи млекопитающие............. 926
3. Вредные млекопитающие............................. 942
4. Роль млекопитающих в распространении заболеваний . . 944
5. Влияние человека на изменение фауны млекопитающих . 945
Заповедники СССР (949).
IX. Географическое распространение птиц и млекопитающих . 951
I. Нотогейское царство.................................. 953
Новозеландская область (954). — Австралийская область
(956).
2. Неогейское царство................................... 959
Неотропическая область (959).
3. Арктогейское царство................................. 964
Мадагаскарская область (965).—Эфиопская область (966).—
Восточная, или Индийская, область (968). —Голарктиче-
ская область (971).
Рекомендуемая литература...................................... 978
Указатели..................................................... 979
ЧАСТЬ ПЕРВАЯ
ОБЩАЯ ХАРАКТЕРИСТИКА ТИПА ХОРДОВЫХ.
ПОЛУХОРДОВЫЕ. ОБОЛОЧНИКИ.
БЕСЧЕРЕПНЫЕ. КРУГЛОРОТЫЕ
ХАРАКТЕРИСТИКА ТИПА ХОРДОВЫХ
И ОБЗОР ПОДТИПОВ
К типу хордовых животных (Chordata) относятся животные,
характеризующиеся следующими признаками: 1) у всех хордовых
па ранних стадиях развития или в течение всей жизни имеется
осевой скелет в виде спинной струны, или хорды
(chorda dorsalis), представленной шнуром из клеток,
обычно сильно вакуолизированных, одетых эластической оболоч-
кой; 2) в переднем отделе пищеварительного канала имеется боль-
шее или меньшее количество жаберных щелей, посред-
ством которых полость глотки сообщается с наружной средой:
подобно хорде жаберные щели имеются или в течение всей жизпг
или только в зародышевом состоянии; 3) нервная система в виде
трубки находится на спиппой стороне тела пад хордой; 4) сердце
если есть, находится па брюшпой стороне животного под пищева
рительным каналом.
Тип хордовых состоит из трех подтипов. Сюда принадлежат вес
животные, называемые п о з в о п о ч п ы м и (V е г t е b г a t а)
ланцетник, круглоротые, рыбы, земноводные, пресмыкающиеся
птицы и млекопитающие. Кроме того, к типу хордовых припадле
жат так называемые полухордовые (Adelochor
data, Н е m i с h о г d a t а): червеобразные морские формы
у которых признаки, характерные для хордовых, не явствешп
и могут быть замечены лишь при внимательном изучении, иль
чипоч но-хордовые (Urochordata), или обе
л о ч п и к и (Т u n i с a t а). Последние во взрослом состояпи
совершенно пе похожи па хордовых животных, но в личипочно
состоянии обнаруживают явственные признаки припадлежпост
к Chordata. Hemichordata и Urochordata живут, как сказаш
в морях.
ПОДТИП 1. ПОЛУХОРДОВЫЕ (НЕМЮПОЕВЛТА
К этому подтипу принадлежат морские животные более пт
мепее червеобразной формы, долгое время считавшиеся за беси
звопочпых, но обнаруживающие как в строении, так и в развит!
ряд черт, сближающих их с позвоночными и личиночно-хордовым:
Представлен подтип небольшим количеством форм: Balanoglossus,
Dolichoglossus, Ptychodera, Rhabdopleura, Cephalodiscus и еще
некоторыми другими. Подтип подразделяется на два класса: ки-
шечнодышащих (Enteropneusta), к которым относятся первые три
рода, и перистожаберных (Pterobranchia), с родами Rhabdopleura
Рис. 1. Glosso-
balanus. Целое
животное.
1 — жаберная об-
ласть. 2 — ошей-
ник, 3 — половые
выросты, 4 — вы-
пячивания сле-
пых печеночных
выростов, 5—хо-
боток, 6 —задне-
проходное отвер-
стие.
(По Шпенгелю,
ив Паркера.)
и Cephalodiscus. Иногда всех этих животных на-
зывают кшпечпо-дышащими, так как передняя
часть кишечника служит у них для дыхания, и
подразделяют их на червевидных (Enteropneusta
vermiformia) и перистожаберпых (Enteropneusta
pterobranchia).
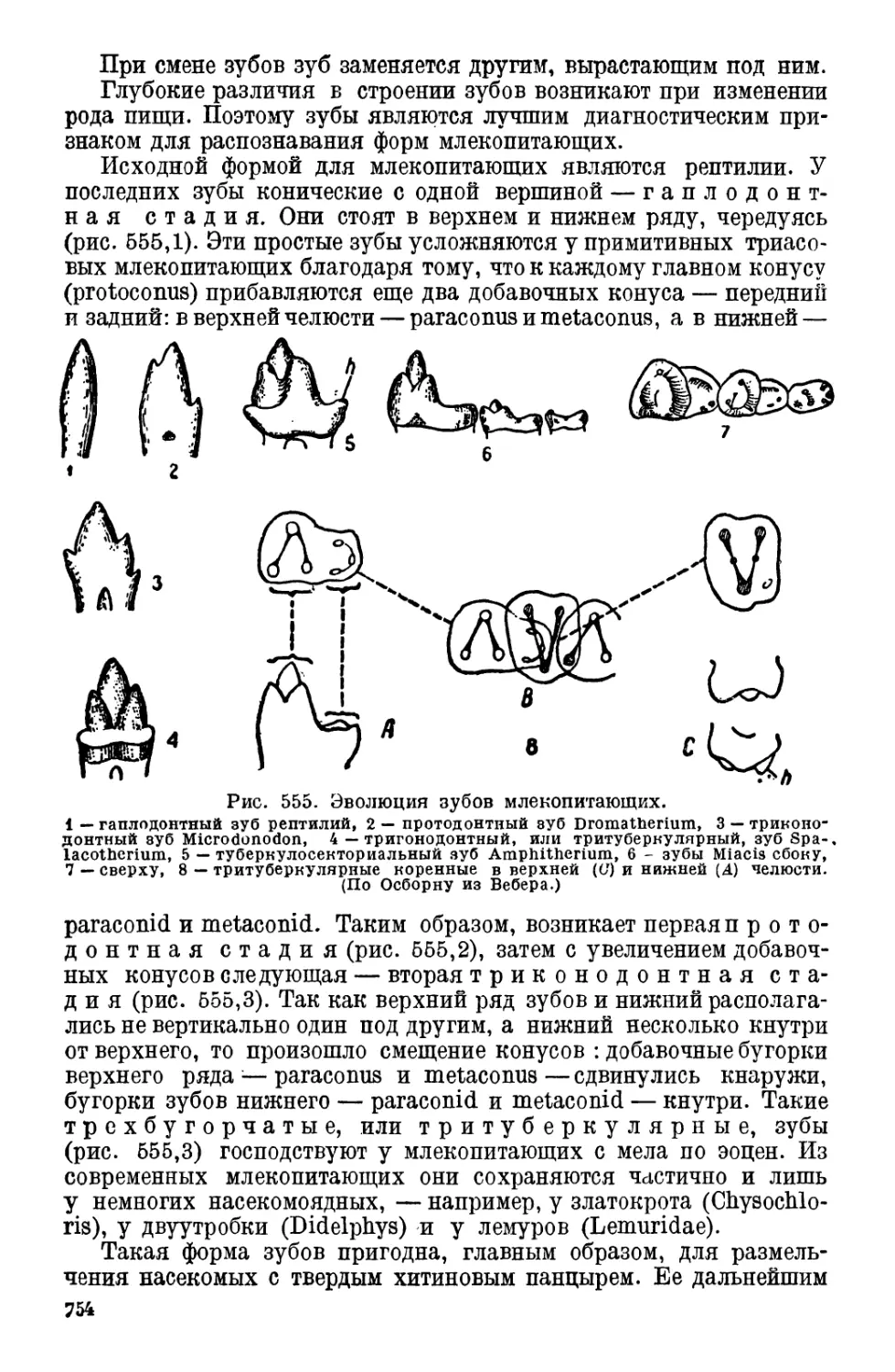
Класс 1. Кишечнодышащие (Enteropneusta)
Представителем этого класса является род Ва-
lanoglossus, сперва считавшийся единственным в
классе, но впоследствии разбитый на ряд родов,
отличающихся несколько по внешнему виду, но
весьма сходных по строению: Balanoglossus, Glos-
sobalanus, Dolichoglossus, Ptychodera и др.
О внешней форме этих животных дает пред-
ставление рис. 1, изображающий Glossobalanus
nrinutus.
Glossobalanus (рис. 1) имеет червеобразное тело,
длиной от 2.5 до 1.5 см, разделенное на три отдела:
вытянутый в конус хоботок (proboscis), по-
зади которого находится рот. За ртом следует
вздутый цилиндрический ошей-
ник, а далее идет длинное заостряющееся кзади
туловище, на заднем конце которого находится
заднепроходное отверстие (anus).
Встречается это животное в морях, где прячется
под камнями или роется в песке и иле.
Своеобразный по внешности Glossobalanus еще
более своеобразен по своему внутреннему устрой-
ству. Прежде всего покровы его, как у многих
беспозвоночных, представлены на всем протяже-
нии ресничными клетками, а также очепь богаты
одноклеточными железами, выделяющими слизь.
Слизь эта вместе с прилипающими к ней зернами
песка образует иногда вокруг тела трубку. Му-
скульный слой, лежащий под покровами, пред-
ставлен гладкими мускульными волокнами. Дви-
гается Glossobalanus очень медленно, при содей-
ствии ресничек, как некоторые беспозвоночные,
и хоботка, при постоянно открытом рте. В песок
животное зарывается при помощи хоботка и ошей-
ника, способных вздуваться и становиться плот-
ными благодаря наполнению водой через особые —
14
хоботковую и ошейниковую — поры. Здесь имеется известное-
сходство с амбулакральными ножками иглокожих.
Туловище делится на жаберную область, пеленочную и брюшную.
На переднем крае ошейника, на брюшной стороне, лежит р о-
товое отверстие, ведущее в ротовую полость. Верхняя
стенка последней образует выпячивание, направленное вперед,
к основанию хоботка. Это выпячивание, состоящее из крупных ва-
куолизированных клеток, занимает положение, сходное в известной
мере с положением спинной струны хордовых животных, с которой,
его многие и сравнивают. Под выпячи-
ванием лежит скелет хоботка, охваты-
вающий рот.
В жаберной области находится ряд
жаберных отверстий, веду-
щих в кишечный канал.
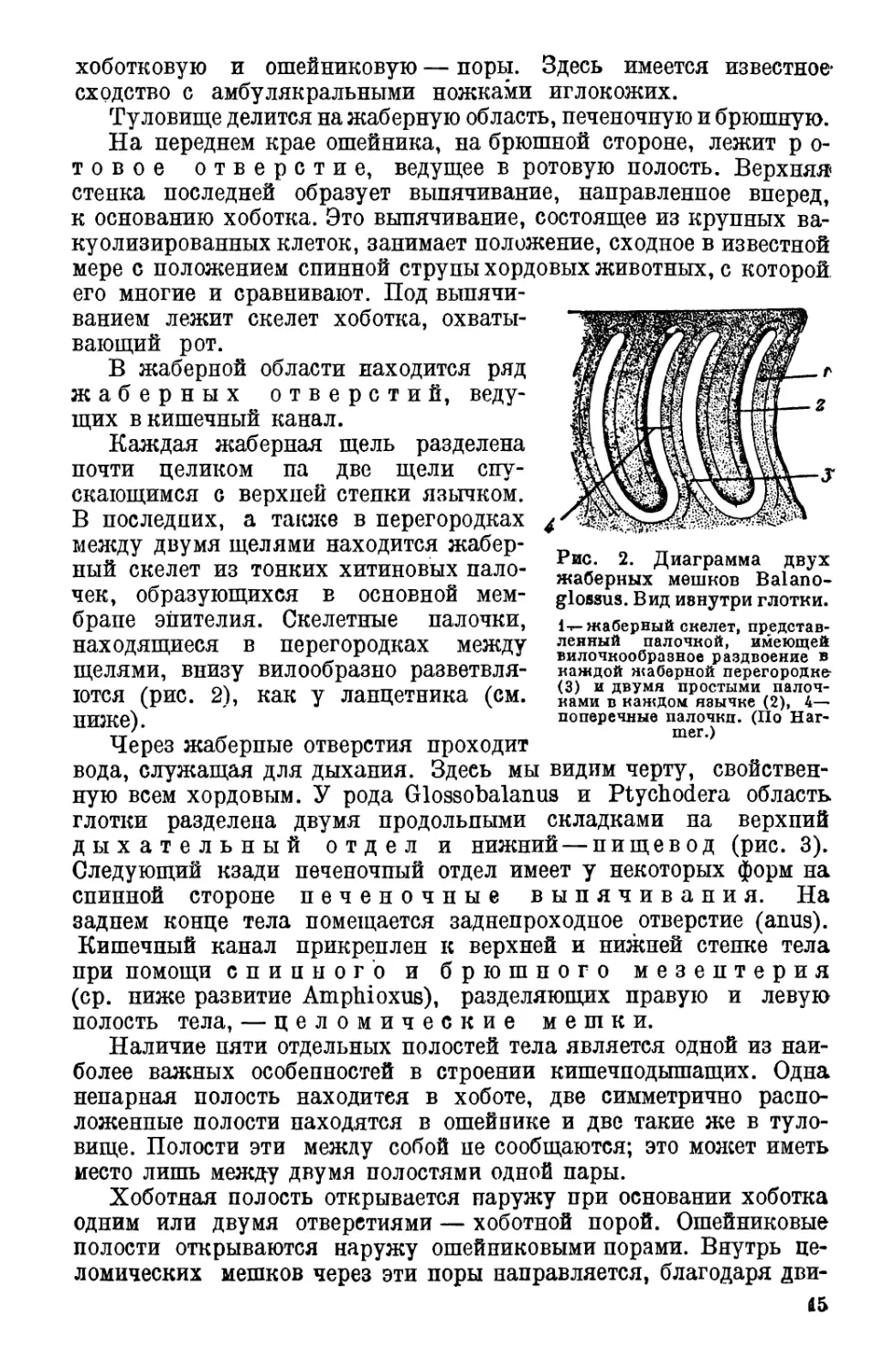
Каждая жаберная щель разделена
почти целиком па две щели спу-
скающимся с верхней стенки язычком.
В последних, а также в перегородках
между двумя щелями находится жабер-
ный скелет из тонких хитиновых пало-
чек, образующихся в основной мем-
бране эпителия. Скелетные палочки,
находящиеся в перегородках между
щелями, внизу вилообразно разветвля-
ются (рис. 2), как у ланцетника (см.
ниже).
Через жаберные отверстия проходит
Рис. 2. Диаграмма двух
жаберных мешков Balano-
glossus. Вид изнутри глотки.
1т-жаберный скелет, представ-
ленный палочкой, имеющей
вилочкообразное раздвоение в
каждой жаберной перегородке-
(3) и двумя простыми палоч-
ками в каждом язычке (2), 4—
поперечные палочки. (По Har-
mer.)
вода, служащая для дыхания. Здесь мы видим черту, свойствен-
ную всем хордовым. У рода Glossobalanus и Ptychodera область
глотки разделена двумя продольными складками на верхний
дыхательный отдел и нижний — пищевод (рис. 3).
Следующий кзади печеночный отдел имеет у некоторых форм на
спинной стороне печеночные выпячивания. На
заднем конце тела помещается заднепроходное отверстие (anus).
Кишечный канал прикреплен к верхней и нижней стенке тела
при помощи спинного и брюшпого мезентерия
(ср. ниже развитие Amphioxus), разделяющих правую и левую
полость тела, — целомические мешки.
Наличие пяти отдельных полостей тела является одной из наи-
более важных особенностей в строении кишечподышащих. Одна
непарная полость находится в хоботе, две симметрично распо-
ложенные полости находятся в ошейнике и две такие же в туло-
вище. Полости эти между собой не сообщаются; это может иметь
место лишь между двумя полостями одной пары.
Хоботная полость открывается наружу при основании хоботка
одним или двумя отверстиями — хоботной порой. Ошейниковые
полости открываются наружу ошейниковыми порами. Внутрь це-
ломических мешков через эти поры направляется, благодаря дви-
45
жениго ресничек, ток воды. Хоботок и ошейпик вздуваются, на-
полняясь водой, и образуют годный для бурения аппарат, при по-
мощи которого Balanoglossus зарывается в песок.
Кровеносная система образована из спинного и брюшного
продольных стволов, соединяющихся между собой дугообраз-
ными сосудами, находящимися по бокам ошейника. Спинной сосуд
способен сокращаться, как и брюшной сосуд и пара боковых сосу-
дов в области печеночных выростов. Кровь в спинном сосуде дви-
жется сзади наперед. Лежит спинной сосуд над спинной струной.
Переднюю хоботковую часть спинного сосуда называют «сердцем».
Однако движение крови по телу осуществляется при помощи осо-
бого органа — сердечного пузыря. Это — полый му-
скулистый мешок, замкнутый и не наполняющийся кровью. Он
Рис. 3. Сагиттальный разрез Glossobalanus. Схематично.
1 — хоботок, 2 — ошейник, 3 — туловище, 4 — полость хоботка,
5—-ротовое отверстие, 6 —центральная нервная система, 7 —
спинная струна, 8 — жаберный отдел кишечника, 9 — пищевод-
ный отдел кишечника, 10—жаберные щели, 11 —полость ошей-
ника, 12 —спинной кровеносный сосуд, 13—брюшной крове-
носный сосуд. (Из Шиплея и Мак-Брайда.)
лежит в хоботке, между расширенным передним концом спинного
сосуда и зачатком хорды. Расширение пузыря выдавливает кровь
из расширения сосуда (лакуна) назад в окологлоточные сосуды.
При сокращении сердечного мешка кровь из спинного сосуда посту-
пает в лакуну. Таким образом, кровеносная система построена не-
сколько иначе, нежели у остальных хордовых.
Полы раздельны. Яичпики и семенники в виде двух рядов
мешковидных органов расположены вдоль жаберной области тела
и дальше кзади' открываясь наружу рядами отверстий.
Нервная система, однако, построена по-иному, чем у других хор-
довых. В глубине эпителиального слоя лежит слой нервных воло-
кон. Этот слой образует на спинной и брюшной сторонах тела ган-
глиозный продольный ствол, тянущийся вдоль всего туловища.
Спинной тяж можно проследить до хоботка. В области ошейника он
отделен от эпидермиса, чего пет в туловище , и содержит гигантские
нервные клетки (рис. 3), Замечательно, что у некоторых видов
Balanoglossus в этой части ствола имеется канал — невро-
цель, у других — лишь отдельные полости. Эту часть нервной
системы можно рассматривать как центральную. На границе ошей-
ника и туловища спинной и брюшной стволы соединяются между
собой кольцевидным утолщецйем. С нервными стволами не связано
16 .''
никаких органов чувств. Чувствующие клетки разбросаны в эпи-
дермисе хоботка и переднего края ошейника.
В качестве выделительного органа действует находящееся
па задней стенке полости хоботка, между ним и лагуной, сосу-
дистое сплетение — glomerulus. Выделенные
в полость хоботка продукты при сокращении степок последнего
выгоняются с водой наружу через хоботковую пору.
Развитие полухордовых очень интересно и помогает нам вы-
яснить их положение в системе. Опо больше изучено у червевидных
кишечподышащих, нежели у перистожаберных.
Оплодотворение у Balanoglossus происходит вие тела матери.
Яйца мелки, мелкие зерна желтка равномерно распределены в про-
топлазме. Дробление яйца—полное и равномерное, идет по радиаль-
ному типу и очень сходно с дроблением яйца у иглокожих (Echi-
nodermata). В результате получается бластула с небольшим бласто-
целем и с клетками, вытянутыми в вышину, напоминающими эпи-
телий. На одной стороне бластулы, вследствие изменения формы
клеток, получается впячивание, т. е. образуется гаструла путем
инвагинации. Властопор замыкается, и на его месте образуется
^впоследствии заднепроходное отверстие. Часть первичной кишки,
шли архентерона, начинает отшнуровываться у слепого его конца,
и из отшнуровавшейся части образуется хоботпый целом, открываю-
щийся затем на спинной стороне личинки хоботной порой наружу.
Вся личинка покрыта в это время мерцательными волосками, кото-
рые на верхней стороне личинки, на так называемом теменпом поле,
достигают значительной длины. Кишечник двумя перехватами раз-
деляется па пищевод, желудок и кишку. Пищевод загибается
к брюшной стороне личинки, где и открывается наружу, образуя
ротовое отверстие. Анальное отверстие, как сказано, открывается
на месте бластопора. То же происходит и у иглокожих. Дальше,
опять-таки как у иглокожих, на брюшпой стороне тела, возле рта
образуется широкое околоротовое вдавлепие. Последнее, окаймлен-
ное по краю мерцательным шнуром, разрастается таким образом,
как это наблюдается у бипиппарии — личинки иглокожих.
В таком виде личинка освобождается от яйцевой оболочки и на-
зывается торпарией. По своему строению опа совершенно
сходна с только что вышедшей из яйца личинкой иглокожих, на-
зываемой диплеурулой. У торпарии образуется сердце
и две пары целомов. Последние развиваются путем выпячивания
первичпой кишечной полости: впереди пара средних, или воротни-
ковых, целомов, позади — пара задних, или туловищных Благо-
даря этому личинка становится двусторонне симметричной. Сердце
образуется из задней стенки хоботного целома, от которой отде-
ляется сперва плотное скопление клеток, превращающееся затем
в пузырек — перикардий или околосердечную сумку —и в сердце,
способное пульсировать. Здесь имеется сходство с позвоночными.
Из первичпой же кишки, а имеппо на боровых стенках пищевода,
путем выпячивания и прорыва последййх наружу, образуются
1—4 пары жаберных щелей. ЗатеМчп4&ледпие языковидным вы-*
ростом дорзальной стенки делятся на две щели. В образовании жа-
берных щелей обнаруживается огромное сходство с развитием
вышестоящих подтипов хордовых — личиночно-хордовых (Uro-,
chordata) и позвоночных (Vertebrata). В дальнейшем наступает
превращение. Два передние мерцательные шнура утрачиваются,
личинка перестает плавать и опускается на дно, где ползает при
помощи волосков заднего шнура. Двумя перехватами определяется
воротниковый отдел. Передний отдел тела вытягивается в хобот,
задний сильно растет, образуя туловищный отдел. В задней стенке
головного целома развивается выделительный орган — головная
почка. На спинной стороне переднего отдела пищевода образуется
выпячивание со стенками из упругих вакуолизированных клеток.
Это выпячивание соответствует по месту своего развития и по гисто-
логическому строению спинной струне или хорде позвоночных.
Нервная система в виде трубки образуется в воротниковом отделе
личинки из впячивающейся вниз эктодермальной пластинки по-
добно тому, как это имеет место у всех хордовых животных.
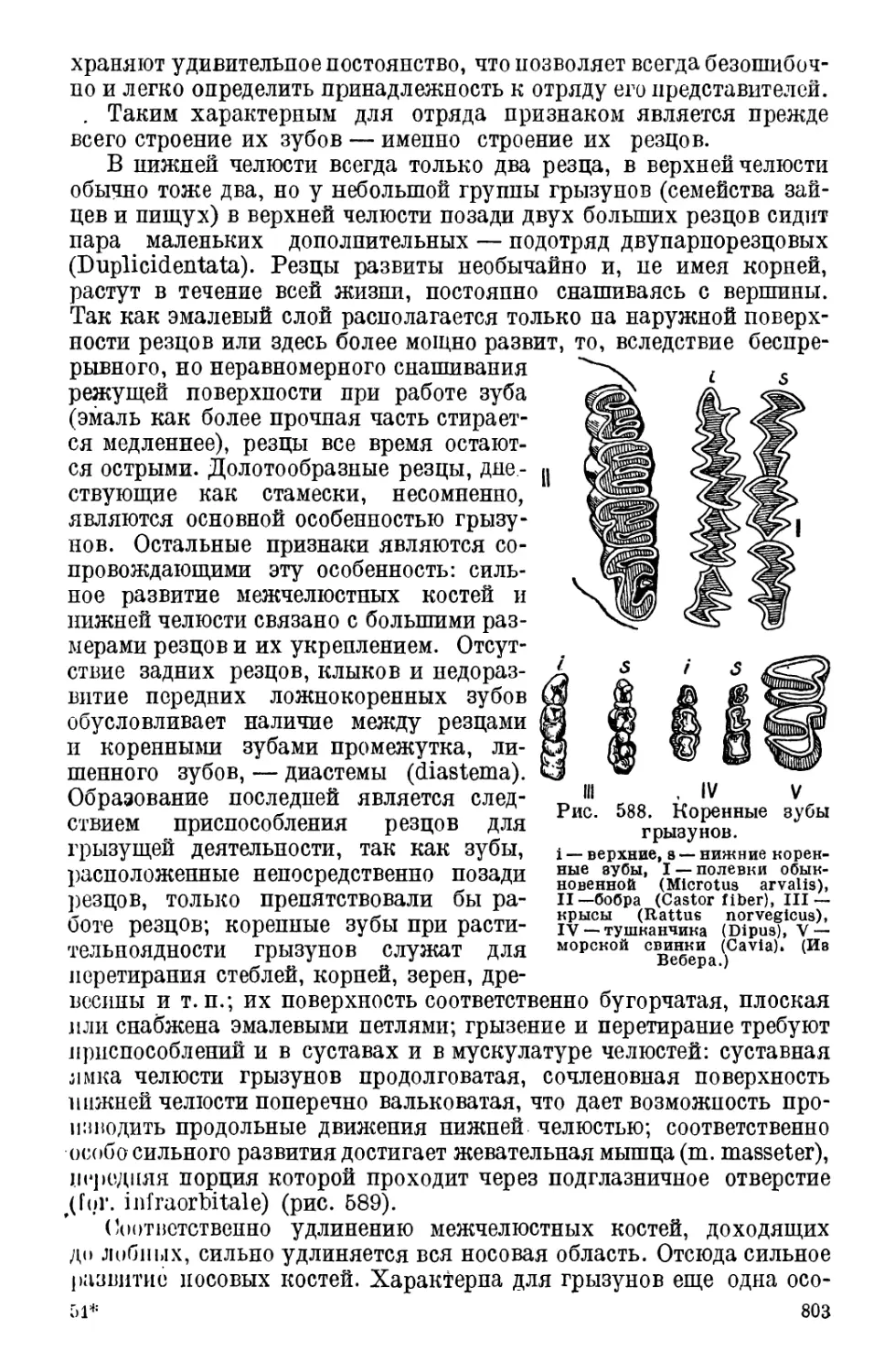
Класс 2. Перистожаберные (Pterobranchia)
К Hemichordata относятся еще роды Cephalodiscus и Rhab-
dopleura, выделяемые в отдельный класс. В основном их организа-
ция сходна с организацией кишечнодышащих, но есть и существен-
ные отличия помимо формы.
Рис. 4. Cephalodiscus
Л. Маленькая часть общего ствола
в натуральную величину. В. Отдель-
ная особь колонии, увеличенная в
65 раз. (По McIntosh, из Harmer.)
dodecalophas.
В. Сагитальный разрез:
I —анальное отверстие, 2, 2В, —
первая, вторая .4 третья полости тела,
3— кишка, 4 --ротовое отверстие,
5 — спинная струна, в — централь-
ная нервная система, 7 — пищевод,
8 — яичник, 9 —пигментированный
яйцевод, 10 — глотка, 11 — хоботко-
вая пора, 12 —хобо^эк, 13—желудок,
14 — стебель (по На *пег).
18
। rph.ilodiscus (рис. 4) представляет собой мелкое животное,
их трех миллиметров длиной, обладающее, однако, всеми важ-
ih'IIiiiiimii органами, которые имеются у кишечнодышащих Тело
• hi ризделено па хобот, ошейпик и туловище. Хобот уплощен
н прикрывает расположенный позади пего рот. Передняя полость
и1 и открывается наружу двумя хоботковыми порами; в ошейнике
н< • к и,ть тела парная и открывается наружу двумя порами; третья
in• 'вють находится в туловище. От ошей-
.... отходят па спиппой стороне 4—6
пгрообразных «рук» с огромным числом
in у ни л(‘Ц. По нижней стороне рук, име-
iniiipil вогнутую форму, пища продви-
। эгтся ко рту при помощи ресничек. От
гиннной стороны переднего отдела пище-
варительного капала отходит выпячива-
ние гомолог спинной струны. Пара
жаберных отверстий открывается на глот-
ки наружу, будучи прикрыта выростом
воротника. За глоткой идут; пищевод,
объемилтыП желудок и кишка. Анальное
отпоротое открывается вблизи от рта
благодаря S-образному изгибу кишеч-
ника.. Центральная нервная система пред-
ставлена значительной массой нервной
ткани в спинном эпидермисе воротника,
не погружающейся в глубину в виде
трубки, как у кишечнодышащих. Яич-
ники расположены между глоткой и кон-
цом кишки и открываются наружу между
анусом и нервной системой.
Живет Cephalodiscus (как и Rhabdop-
leura) па большой глубине в море коло-
ниями, в которых осбби, не связанные
между собой органически, сидят в общей
студенистой оболочке. Rhabdopleura (рис. 5)
еще меньше Cephalodiscus. Вероятно,
вследствие малой величины этой формы у
Рис. 5. Часть коло-
нии Rhabdopleura
normanni.
1 — две руки со щупаль-
цами, 2 —анальное от-
верстие, 3 — желудок,
4 — кобот. (По Sars 413
Harmer.)
нее не были найдены
некоторые органы, папример, хоботковая пора и жаберные щели.
Анатомически этот род обнаруживает теспое сходство с Cephalo-
discus. Это животное живет колониями, в которых отдельные
особи, или зооиды, находясь в системе ветвистых перепончатых
трубок, соединены между собой.
Каждая особь в колонии Rhabdopleura состоит из длинного сте-
белька, мускулистого и способного сокращаться; при сокращении
стебелька животное увлекается в прозрачную наружную выделяе-
мую хоботком трубку. Ошейник песет лишь пару рук с двойным
рядом щупалец.
Как Cephalodiscus, так и Rhabdopleura способны размножаться
почкованием.
s*
19
оиЛатеральные или трехслойные
Кишечно-
дышащие
Oeuterostomia
Позвоночные
бесчерепные черепные
Оболочники
Первичные по-
звоночные
Иглокожие
Prn*03t0fn.’Q
Немертины
5ра,иопо- Ншант
Нематоды
Ресничные
черви '
I
f
J'
Насекомые
Пачки Мечехвост
Ракоскорпи-
оны
Многощетин-
ковые
Щупальцевые
X Мапощетин-
' ковые ,
Олигомерные
vePeo Пиявки
Моллюски \
Многоножки
Перипатус
Раки
Первично-.
трахейные
Полимерные
черви
шикиЬ~ Пен™чы I Первичные иерви
Гребневики I
Коралловые полипы ’
Простейшие
Первичные биченосцы
Кишечнопо-
лостные
Сцифомедузы
.Гидроиды
Губки
Филогенетические взаимоотношения полухордовых с другими типами
и группами.
Развитие перистожаберпых при половом размножении известно
меньше, нежели развитие кишечподышащих, по и здесь дробление
полное и равномерное, за бластулой образуется гаструла, причем
разными способами. Властопор и здесь становится заднепроходным
отверстием. Целомы, числом пять, образуются из первичпой кишки
20
Г
_1
J
выпячиванием ее стенок, как и у Balanoglossus. Так же как и у по-
следнего, у Rhabdopleura образуются нервная система на спинной
стороне шейки и орган гомологичный спинной струне.
ПОЛОЖЕНИЕ ПОЛУХОРДОВЫХ В СИСТЕМЕ И ФИЛОГЕНЕТИЧЕ-
СКИЕ СВЯЗИ
Таким образом, несмотря на внешнее отличие формы, ряд при-
знаков сближает полухордовых с позвоночными и заставляет
отнести и тех и других к типу хордовых:
1) наличие жаберных щелей, ведущих из глотки наружу и раз-
дёленпых язычком надвое, как у Amphioxus (см. ниже); 2) положе-
ние главной части нервной системы с внутренней полостью на
спинной стороне ошейника; 3) наличие на спинной стороне глотки
выроста, гистологически напоминающего хорду; 4) гистологическое
разделение глотки на верхнюю, дыхательную, часть с жаберными
щелями и нижнюю, выстланную слизистыми и мерцательными
клетками и служащую для проведения пищи, как у личипочпо-
хордовых и ланцетника (см. пилсе); 5) наличие и способ образо-
вания полостей тела; (J) наличие сердца и околосердечной полости;
7) наличие печеночных выростов в средней части кишечника; 8) об-
разование в задней стенке головного целома выделительного ор-
гана — головной почки, или glomerulus, подобного тому, что
имеется у низших позвоночных; 9) метамерпое расположение жа-
берных щелей, печеночных выростов тела и половых желез.
Все эти признаки заставляют считать, что между полухордовы-
ми и позвоночными существует известное родство.
С другой стороны, эмбриональное развитие полухордовых об-
наруживает близость их с иглокожими. Дробление яйца, образо-
вание и характер бластулы и гаструлы, образование целома из
переднего конца первичной кишки идут одинаково у тех и других.
Сходство молодой торнарии с личинкой иглокожих совершенно по-
ра: ительно. Эти факты следует объяснять таким образом, что игло-
кожие и полухордовые имеют общее происхождение, что полухор-
доьые являются близкими потомками тех двусторонне-симметрич-
ных форм, от которых произошли иглокожие. Так как подтип
полухордовых очень древен, то мы видим в пределах его различные
виды специализации, вызванные особенностями образа жизни.
ПОДТИП 2. ЛИЧИНОЧНО-ХОРДОВЫЕ,
ИЛИ ОБОЛОЧНИКИ (VROCHORDATA
ИЛИ TUNIC АТ А)
Сюда относятся сидячие или свободноплавающие морские формы,
имеющие мешкообразное или бочепкообразное тело, иногда с хво-
стовым придатком. Тело одето большей частью толстой кутику-
лярной мантией. Хорда не всегда есть у взрослых животных, обычно
имеется лишь в личиночном состоянии. Широкая глотка служит для
дыхания. Сердце имеется.
21
1. СТРОЕНИЕ ЛИЧИНОЧНО-ХОРДОВЫХ
За представителя подтипа мы можем взять асцидию (Ascidia)
(рис. 6). Тело ее обычно прочно прикреплено к твердому субстрату
(дно, камни, раковина моллюска) и имеет вид мешка с двумя отвер-
стиями- вводной и выводной сифон, ротовое и клоакаль-
ное отверстия. Прибавив к воде аквариума туши, мы
видим, что вода входит в первое и выходит из второго. Время от
времени тело более или менее резко сокращается, благодаря чему
происходит выбрасывание воды. Это происходит всегда, если жи-
вотное вынуть из воды.
Вскрывая животное, мы видим следующее (рис. 7). Туника,
наружный слой стенок тела, состоит из плотного прозрачного ве-
Рис. 6. Ascidia men-
tula с правой стороны.
1 — ротовое отверстие,
2—клоакальное отвер-
стие. (По Herdman.)
Рис 7. Разрез через тунику и мантию Ascidia,
показывающий отношение эктодермы к стенке
тела и кутикуле.
1 — пузырчатые клетки туники, 2 — кровеносные ка-
налы, 3 — клетки соединительной ткани мантии, 4 —-
эктодерма, 5 — блуждающие клетки, 6—мускульные
волокна, разрезанные вдоль и поперек. (По Herdman.)
щества, в котором имеется большой процент целлюлозы.
Это единственный случай нахождения целлюлозы в теле животных.
Туника является продуктом выделения клеток эпидермиса. Внутри
пее находятся так называемые пузырчатые клетки, затем различ-
ные веретенообразные' и звездчатые клетки, пигментные и др.
Первые скопляются главным образом во впешнем слое туники.
Все эти клетки являются выселившимися в тупику мезодермальны-
ми, так называемыми блуждающими клетками, а также клетками
эктодермы и энтодермы. Кроме того, в тупике паходятся ветвящиеся
каналы с концевыми расширениями. Каждый канал разделен про-
дольной перегородкой. Эти каналы образуются врастанием в ту-
нику эпидермы с сосудами. По ним движется кровь: по одной сто-
роне канала в одну сторону, по другой — в другую
Под туникой паходятся мягкие стенки тела, или мантия
(рис. 8). Тело не срастается с туникбй. Срастание имеет место только
22
в круг щупальцами.
з
Рис. 8. Послойно вскрытая
асцидия
1 — туника, 2 — мантия, 3 — рото-
вое отверстие, 4 — стенка глотки
или жаберной полости, 5 — около-
жаберная, клоачная или атриаль-
ная полость, 6 — эндостиль, 7 —
отверстие пищевода, 8 — шел удои,
9 — кишка, 10—анальное отвер-
стие, 11—клоачное отверстие,
12 — половая железа, 13 — отвер-
стие полового протока, 14—нерв-
ный ганглий, 15 — выделительная
железа. (По Herdman.)
вокруг ротового и вокруг клоакального отверстий. Остальное тело
свободно висит внутри туники. Мантия состоит из эпидермиса,
соединительной ткани и мышечных волокон. Последние па краях
м;штни, где она срастается с туникой, образуют сильные мышечные
замыкатели — так называемые сфинктеры. В других местах маптии
волокна перекрещиваются в беспорядке. Ротовое отверстие ведет
в обширную глотку или жаберную полость, вход
в которую окружен расположенными
глотки пронизаны множеством ма-
леньких отверстий, называемых сти-
гмами (stigmata); они рас-
положены правильными рядами и
ведут из полости глотки в так на-
зываемую клоачную, или а т-
риальную, полость. Эта
полость со всех сторон окружает
глотку, кроме одной, где стенка глот-
ки срастается с мантией. Здесь вдоль
внутренней поверхности глотки име-
ются две продольные складки, раз-
деленные бороздой. Борозда высти-
лается клетками двоякого рода. Одни
из них — ресничатые, другие — же-
лезистые. Все это образование носит
название э н д о с т и л я. На проти-
воположной эпдостилю спинной сто-
роне идет срединный продольный вы-
рост — так называемая спинная
борозд а, — клетки которого снаб-
жены респичками. Спереди, около
начального конца глотки эпдостиль
переходит в респич п о е око-
логлоточное кольцо, ок-
ружающее этот конец. Вперед от
этого кольца лежит другое окологло-
точное кольцо. Назначение эпдостиля
следующее: различные микроскопи-
ческие организмы, приносимые в глот-
ку с водой, склеиваются той слизью,
которую выделяют клетки эндостиля, в комочки, и последние про-
гоняются ресничками эпдостиля кпереди к окологлоточ-
ной борозде и отсюда переходят па спипную борозду,
реснички которой гопят пищевые комочки к отверстию пищевода.
Пищевод переходит в короткий желудок. Последний переходит
в кишку, образующую двойную петлю и открывающуюся задне-
проходным отверстием в атриальную полость Вдоль внутренней
поверхности кишки имеется складка, подобная той, которую мы
наблюдаем у дождевых червей, и называемая тифлозол ь—
typhlnsolis.
23
Кровеносная система у асцидий развита лучше, нежели у Ва-
lanoglossus (рис. 9). На брюшной стороне глотки находится
сердце. Окружающее его пространство называется около-
сердечной, перикардиальной полостью.
Сердце это представляет утолщение расположенного под эн-
достилем брюшного сосуда. При посредстве Сети сосудов
в глоточной области брюшной сосуд соединяется со спинным кро-
веносным сосудом. Сокращения сердца идут от одного конца сердца
к другому, затем наступает пауза, а последующее сокращение идет
в обратном направлении. Таким образом кровь течет то от сердца
Рис. 9. Схема продольного
разреза через асцидию пос-
ле удаления оболочки и
мантии.
1 — сердце, 2 — брюшной со-
суд, 3 — спинной кровеносный
сосуд, 4 — нервный ганглий,
5 — поперечные сосуды, 6 —
половая железа. (По Herdman.)
к спинному сосуду, то от последнего к
сердцу. Сосуды, как и у Adelochordata,
не имеют собственных стенок. Их имеет
только сердце.
Нервная система представлена у
асцидий единственным нервным узлом,
лежащим между ротовым и клоакаль-
ным отверстиями. От этого узла отходят
нервы к различным органам.
Орган выделения представлен не-
парным скоплением прозрачных пу-
зырьков, помещающемся в изгибе петли
кишечника. Пузырьки нередко содер-
жат мочевую кислоту.
Полы не раздельны, асцидии гермаф-
родиты. Яичник и яйцевод образуют
вместе ветвистый пучок трубочек, ле-
жащий на левой стороне тела в петле
кишечника. Яйцевод и семяпровод тесно
прилегают друг к другу и открываются
в атриальную полость рядом с задне-
проходным отверстием, образуя таким
образом клоаку. Вода, выходящая из
жаберных щелей и через атриальную
полость, выносит с собой наружу и по-
ловые продукты.
Таким образом, в- строении взрослой асцидии мы пе находим
почти никаких признаков строения хордовых животных: пи хорды,
пи нервной системы в виде трубки, пи настоящих жаберных щелей.
Пассивный, сидячий образ жизни и кутикулярная туника также
не свойственны ни одному хордовому животному. Но принадлеж-
ность асцидий к последним становится совершеппо очевидной, если
мы исследуем развитие асцидии. Изучение этого развития Кова-
левским в 1866 г. явилось важным момептом в истории эволюцион-
ного учения и в истории понимания генетических взаимоотношений
хордовых животных.
24
II. РАЗВИТИЕ ЛИЧИНОЧНО-ХОРДОВЫХ
Яйца асцидий мелки, бесцветны и почти не имеют желтка.
Оплодотворение происходит или впе тела или у других видов в ат-
риальной полости, откуда выходит уже личинка. Перекрестность
оплодотворения обыкновенно обеспечивается неодновременным
созреванием яиц и спермы у одного и того же животного. В неко-
торых случаях происходит самооплодотворение. Яйцо окружено
Рис. 10. Развитие асцидии.
А — зрелое яйцо. 1 — калиммоциты, 2 — фолликулярные клетки, 3 — прото-
плазма, 4 — ядро. В —спермин. С—разрез, показывающий бластоцель.
D — ранняя стадия гаструлы. В—более поздняя стадия гаструлы. В —
более поздняя стадия зародыша, показывающая зачатки хорды и нервной
трубки. G—поперечный разрез тела зародыша, показывающий метабласт
и образование нервного канала. Н —более поздний зародыш, показываю-
щий тело, хвост, хорду, нервный канал и кишечный канал. J — молодая
личинка, готовая к вылуплению. поперечный разрез хвоста личинки.
1—эктодерма, 2 —энтодерма, 3 — бластоцель, 4—бластопор, 5 — спинная
струна, 6 — первичная кишечная полость. 7 *— canalis neurentericus, 8 —
нервный канал, 9 —глаз, 10 — слуховой пузырек, 11—мозговой пузырь.
(По Ковалевскому» н другим, из Herdman.)
крупными фолликулярными клетками. Часть их мигрирует в яйцо,
размножается и образует в поверхностном слое последнего так на-
зываемые «калиммоциты», образующие testae cellae. Сегментация
яйца полная (рис. 10). В результате дробления получается бла-
стула с небольшой сегментационной полостью. Уже с восьми-
25
клеточной стадии клетки, находящиеся на будущей брюшной сто-
роне, становятся меньше и являются зачатком будущей эктодермы.
Более крупные и зернистые клетки будущей спинной стороны обра-
зуют энтодерму. Клетки последней принимают цилиндрическую
форму. Более быстрый рост на переднем конце зародыша ведет
к искривлению последнего, в результате чего происходит как бы
впячивание внутрь клеток будущей энтодермы, полость бластулы
уничтожается, и получается гаструла. Последняя принимает про-
долговатую форму, и отверстие бластопора определяет задний конец
тела. Спинная поверхность зародыша становится более плоской.
Клетки, ограничивающие бластопор, становятся кубическими и об-
разуют иа спинной стороне так называемую медуллярную
пластинку. В последней появляется борозда, справа и слева
ограниченная медуллярными складками. Складки
растут кзади, встречаясь между собой позади бластопора. Кроме
того, они растут вверх и встречаются между собой, замыкая канал—
невроцель. В задний конец невроцеля открывается бластопор,
т. е. первичпокишечная полость соединяется с полостью нервной
системы при посредстве нервно-кишечного канала (canalis пенгеп-
tericus). Впереди невроцель некоторое время остается открытым.
Отверстие здесь называется невропором.
Под задним концом медуллярной пластинки клетки энтодермы,
начиная от бластопора, образуют пластинку, растущую вперед.
Эта пластинка затем отчленяется от энтодермы. Из средней ее
части образуется так называемая хорда, или спинная
струпа, из боковых и задней частей обособляется м е з о д е р-
м а. Из остающейся полости первичной кишки получается жаберпый
мешок, а при дальнейшем росте и весь пищеварительный капал.
Задний конец тела личинки быстро удлиняется, образуя хвост.
Он состоит из эктодермы, нервной трубки, хорды под пей и из тяжа
эктодермальных клеток под хордой, представляющего кишечный
канал. Позднее в хвост врастает и мезодерма, образующая по сто-
ронам хвоста продольные тяжи поперечнополосатых мышечпых
клеток. Клетки эктодермы начинают секретировать кутикулярную
оболочку. Последняя образует на спинной стороне тонкие про-
дольные складки: спинной и брюшной плавники. В передней части
тела эктодерма образует присасывательные сосочки. На переднем
конце нервной трубки образуется расширение с тонкими степками.
От этого расширения отходит пузырчатый вырост — так называе-
мый чувствующий пузырек. В последнем образуются
зачатки пигментированного глаза и отоцисты (слухового органа).
В переднем отделе кишечного канала (в глотке) образуется рес-
ничатый выворот, вступающий в тесную связь с мозговым пузырем.
В этом вывороте видят гомолог гипофизы высших позвоночных.
Рот образуется па верхней стороне переднего конца тела. Пара
эктодермальных заворотов, атриальные и перибронхиальные кар-
маны сливаются по средпеспинной линии, образуя атриальную
полость Первичные жаберные щели (protostigmata) образуются
растущими друг другу навстречу выпячиваниями глотки и впячи-
26
на пнями стенок атриальной полости. У рода Ascidia их шесть
пар, подразделяющихся потом на stigmata.
Спустя два-три дпя после оплодотворения уже вылупляется
личинка. В течение короткого времени она ведет свободный образ
лсизни. В это именно время у нее сильпо развиваются нервная си-
стема, органы чувств, причем глаз имеет сетчатку, пигментный слой,
хрусталик и роговицу, развиваются мускулы по бокам хорды.
Сто — наивысший пункт развития личинки асцидии. В это время
признаки Chordata выражены у нее наиболее ярко: форма тела,
хорда, нервная система в виде трубки, жаберные щели. Сердце
и околосердечная сумка в мезобласте между нижней стороной
глотки и желудком, вероятно, не имеют генетического родства
с таковыми высших хордовых.
После нескольких часов, самое большее — одного дпя по вы-
луплении, личинка прикрепляется передними железистыми сосоч-
ками к какому-либо предмету и подвергается регрессивному мета-
морфозу, приводящему ее ко взрослому состоянию. Сами сосочки,
которыми личинка прикрепилась, начинают атрофироваться, и
функцию их выполняет теперь быстро развивающаяся тупика.
Хвост быстро разрушается и рассасывается, хорда и первпая трубка
исчезают, так же как чувствующий пузырек. Лишь задняя утол-
щенная стенка его сохраняется в виде нервного узла, под которым
сохраняется и зачаток гипофизы. Уничтожение хорды, нервной
трубки и мускулов происходит при помощи особых клеток — фа-
г о ц и т о в. Кишечпый капал, наоборот, быстро разрастается.
Жаберный мешок увеличивается в размерах, в стенки его прони-
кают кровеносные каналы, число жаберпых отверстии значительно
увеличивается благодаря распадению щелей на поперечные ряды
stigmata. Желудок и кишка удлиняются, и последняя изгибается.
В мезодерме между желудком и кишкой образуются гонады и их
продукты. Так как передний конец тела растет очепь быстро, то
рот, поворачиваясь, перемещается наверх от точки прикрепления.
Атриальное отверстие образуется позади ротового путем впячи-
вания покровов и слития дпа получающейся ямки с наружными
частями двух жаберных щелей.
Развитие асцидии в высшей степени замечательно. Здесь мы
видим, как личинка, сперва чрезвычайно сходная с эмбрионом лан-
цетника или амфибии, являясь типичным хордовым животным,
с развитием подвергается дегенерации и становится животным,
совершеппо на животных хордовых пе похожим Развитие асци-
дии показывает настоящее положение Urochordata в системе.
Если бы мы пе зпали этого развития, мы поместили бы асцидий
безусловно, среди беспозвоночных. Факты онтогенеза асцидий,
рассматриваемые с точки зрения биогенетического закона, опре-
деленно указывают паи па родство их с Chordata.
III. СИСТЕМАТИЧЕСКИЙ ОБЗОР ЛИЧИНОЧНО-ХОРДОВЫХ
Характеристика подтипа дана была выше. Подтип распадается
па три класса, представляющие различные степени специализации.
27
Класс 1. Хвостатые, или личиночные (Copel ata, или Larvacea)
Рис. 11. Продольный оптический разрез через Oi-
kopleura. Часть хвоста отрезана (По Herdman.)
1—туника, 2 —эктодерма, 3 — рот, 4 — жаберный ме-
шок, 5 — наджаберный валин, 6 — пищевод, 7—желу-
док, 8 — кишка, 9 — анальное отверстие, 10 — отоциста,
Hi 11 а и 113 — нервный ствол, 12 и 12j — нервные ган-
глии, 13 —сердце, 14—орган чувств, 15 —ресничная
воронка, 16 —ротовая железа, 17 — эндостиль, 18 —
подглоточный валик, 19 — стигмы, 20 — атриальное
отверстие, 21 — мускулы хвоста, 22 — хорда, 23 — се-
менник, 24 — яичник, 25 — отверстие гонад наружу.
К классу Larvacea относится только одно семейство:
аппендикулярии (Appendiculariidae) с не-
сколькими родами, из которых важнейшими являются А р р е п-
dicularia и Oikopleura, всего 40 видов (рис. 11).
Это мелкие формы, остающиеся в течение всей своей жизни на ли-
чиночной стадии развития, ведущие свободный пелагический образ
жизни и сохраняющие хвостовой придаток с хордой или спинной
струной Жаберных
отверстий только два,
причем открываются
они прямо наружу,
так как атриальной
полости у Larvacea
нет. От главного нерв-
ного узла по спинной
стороне хвоста до са-
мого его конца идет
нервный ствол. Тупи-
ка развита слабо и
представлена так на-
зываемым «домиком»,
временной оболочкой,
лишенной кровенос-
ных сосудов и час-
то сбрасываемой. На
месте ее быстро об-
разуется другая.
Развитие происходит
без превращепия.Поч-
кования не наблю-
дается.
Этот класс обнаруживает, таким образом, черты организации,
(хвост, хорда, отсутствие атриальной полости, нервный ствол и т. д.),
которые можно па основании того, что мы знаем о развитии асци-
дии, считать древними, примитивными. Представители этого класса
подверглись дегенерации лишь в очень незначительной степени или
вовсе ей не подверглись, а потому соответствуют личиночной ста-
дии других, более специализированных групп. Хвостатые оболоч-
ники живут в морях большей части света, от Арктики до Антарк-
тики, придерживаясь обычно поверхности, причем движутся при
помощи хвоста.
Класс 2. Асцидии (Ascldiae)
Формы большей частью сидячие, колониальные или простые.
Во взрослом состоянии никогда не имеют пи хвоста ни следов хорды.
К асцидиям относится главная масса Urochordata. Одиночные формы
всегда являются сидячими; свободно плавающие встречаются исклю-
28
чительпо среди колониальных. Всегда хорошо развита туника,
образование вполне постоянное, увеличивающееся в течение всей
жизни, притом имеющее более сложную структуру вследствие
иммиграции в нее клеток тела и врастания кровеносных сосудов.
Объемистая глотка пронизана массой жаберных отверстий — стигм,
открывающихся не наружу, а в особую атриальную полость. Раз-
витие у большинства происходит с превращением, как описано
выше. Многие асцидии способны с помощью почкования образо-
вывать колонии, плавающие или сидячие.
Делятся на три подкласса: 1. Простые асцидии
(Ascidiae simplices) с родами Clavellina, Ci опа, Ascidia, Cynthia,
Molgula и другими. 2. Сложные асцидии (Ascidiae сош-
positae) (рис. 12), сидячие формы, размножаю-
щиеся почкованием, благодаря чему образуются
колонии, в которых отдельные особи зооиды
внедрены в общую студенистую массу без от-
дельных туник («синасцидии»). От простых асци-
дий их отделяют лишь последний признак и
способность почковаться. В некоторых случаях
атриальные отверстия нескольких особей сое-
диняются, образуя общее клоакальное отвер-
стие. Сложные асцидии происходят от простых.
Наиболее известный род — Botryllus. Почкова-
ние у этого рода происходит путем образования
у родительской особи маленьких выпячиваний
«си-
Рис. 12. Две
стемы» из колонии
Botryllus viola-
ceus..
1 — общая клоака
системы, 2 — жабер-
ные отверстия асци-
диозооидов. Увели-
чено. (По Milne-Ed-
wards из Herdman.)
атриальпой полости. Обычно же почкование
происходит путем образования полого пальце-
видного выроста глотки, называемого столо-
ном, делящегося па отрезки, каждый из ко-
торых дает почку. 3. Светящиеся ас-
цидии (Ascidiae luciae, или Ascidiae salpae-
formes, сальпоподобные). Сюда относится одно
только семейство Pyrosomatidae с родом Py-
rosoma и 6 видами. Формы колониальные, свободно плавающие,
рот и клоака па противоположных концах тела. Отдельные зооиды
заключены в общую тупику таким образом, что ротовые отверстия
направлены кпаружи, а атриальные внутрь к центральной полости
колонии, являющейся общем клоакальной полостью и открываю-
щейся наружу одним общим отверстием. Вся колония походит па
цилиндр с отверстием па одном конце. Колонии пиросом ведут
пелагический образ жизни, имеют размеры от 5—8 см до 4 м.
Они обладают способностью сильно и красиво фосфоресцировать -
Развитие прямое, без хвостатой личиночной стадии. В яичнике
достигает зрелости лишь одно яйцо, развивающееся затем внутри
родительского организма. Очень рано у асцидиеподобного зооида
позади эпдостиля образуется столон, распадающийся иа четыре
части. Эти тетр а зооиды дают начало целой колонии.
29
Класс 3. Сальпы (Thalicea)
Сюда принадлежат свободно плавающие формы небольшой
величины, прозрачные, как стекло, боченковидпой формы со ртом
и клоакой, лежащими на противоположных концах. Во взрослом
состоянии никогда не бывает хвоста и хорды. Жаберных отверстий
два пли два ряда. Туника всегда есть, более или менее развитая.
Мышцы лежат в виде обручей на бочке. Сокращением этих мышц
года выбрасывается из тела, благодаря чему животное движется.
В развитии наблюдается чередование поколений; половое поколе-
ние сменяет бесполое. Половым путем возникают одиночные саль-
з
5
Рис. 13. Боченок (Doliolum). Бес-
полая стадия сбоку. Видны почки
на ранних стадиях развития.
1 — ротовое отверстие, 2 — атриаль-
ное отверстие, 3 — нервный увел,
4 — спинной столон, 5 — брюшпой
столон.
пы, бесполым — цепи из сальп.
Развитие особи сальпы, происшед-
шей половым путем, папример, так
г называемого бочепочка (Doliolum),
во всех основных чертах сходно
с развитием простых асцидий. Но
затем от тела личинки вырастают
два отростка — спинной и
брюшной столоны (рис. 13).
На брюшном образуются почки в
виде маленьких клеточных скоп-
лений. Эти почки, оторвавшись от
столона, ползут кверху, к спинно-
. му столону, и на нем укрепляются.
Спипной столон растет в длину,
укрепившиеся па нем почки развиваются в зооиды, цепь их с ро-
стом столона и образованием новых все удлиняется. Родительская
же особь подвергается ряду своеобразных изменений: она утра-
чивает жабры, эндостиль, кишечный капал; мышечные лепты ее
утолщаются, нервная система, необходимая для мышечной дея-
тельности, также прогрессивно развивается, и весь родительский
организм превращается в оргап движения всей колонии, зооиды
которой являются особями половыми. У других сальп развитие
и чередование поколений происходят несколько иначе. Сальпы
распространены в морях чрезвычайно широко, ведут пелагический
или сидячий образ жизни. Среди сидячих многие встречаются на
очень больших глубинах. Наиболее известны роды: Doliolum
и Salpa.
IV. ЗНАЧЕНИЕ ЛИЧИНОЧНО-ХОРДОВЫХ для эволюционного
УЧЕНИЯ
На примере оболочников (а также полухордовых) мы показали,
каким путем можно разбираться в вопросах происхождения жи-
вотных, в их родственных взаимоотношениях Иллюстрация, ко-
нечно, далеко пе полная, но пример этот достаточно хорошо пока-
зывает значение изучения индивидуального развития для понима-
ния места животного в системе, которая должна отражать процесс
исторического или филогенетического развития. Развитие асци-
зо
дни показывает нам, что последние должны быть отнесены к хор-
довым животным, но что они значительно уклонились от своей ро-
доначальпой формы, подвергшись регрессивному метаморфозу,
утратив подвижность, став сидячими формами. С другой стороны,
сравнение развивающейся личинки асцидий с организацией взрос-
лых хвостатых тупикат, Larvacea, приводит к заключению, что
последние являются классом, наименее удалившимся от общих
оболочникам предков. Сложные асцидии развились, конечно,
из простых. В этом убеждает пас сравнение организации тех и дру-
гих. От асцидий типа Clavellina могли произойти и Ascidiae luciae
и Thaliacea — сальпы. Древние унаследованные признаки обна-
руживаются в эмбриональных и личиночных стадиях, у взрослых
же они исчезают и появляются новые признаки. Эти последние
являются адаптациями, т. е. приспособлениями к усло-
виям существования: к жизни, например, у берегов, в открытом
море, к свободному плаванию и к сидячему образу жизни.
Мепее ясна принадлежность к хордовым полухордовых. Но
и там, па основе сравнительного изучения строе-
ния, а также и д д и в и д у а л ь п о г о р а з в и т и я, мы
можем все же сделать известные заключения о весьма вероятном
родстве этого подтипа с хордовыми животными. Наши заключения
были бы гораздо более убедительными, если бы мы имели ископаемых
представителей самых древнейших хордовых, т. е. свидетельства
палеонтологии. К сожалению, этого нет: палеонтология ничего не
говорит нам о происхождении типа хордовых, так как древнейшие
его представители не имели еще скелета, который мог бы до нас
сохраниться. Мы вынуждены довольствоваться заключениями
о происхождении, основанными па косвенных доказательствах
сравнительной анатомии и эмбриологии.
ПОДТИП 3. ПОЗВОНОЧНЫЕ (VERTEBBATA)
ОБЗОР ПОДТИПА
ОБЩИЕ НАПРАВЛЕНИЯ В ЭВОЛЮЦИИ ПОЗВОНОЧНЫХ
У позвоночных особенности хордовых выражены наиболее
явственно. Те признаки, которые у полухордовых только намечены
и толкования которых могут вызывать сомнения (например, за-
чаток хорды в виде выпячивания верхней степки переднего отдела
пищеварительного канала), а у личиночно-хордовых проявляются
лишь в личиночном состоянии, — у позвоночных развиты совер-
шенно отчетливо от низших представителей до высших. Хорда,
или спинная струна, имеется или в течение всей жизни или в заро-
дышевом состоянии, а потом вытесняется хрящевым или костным
позвоночником, развивающимся частью из оболочек хорды, частью
из ‘ окружающей хорду перихордальной соедини-
тельной ткани. Нервная система лежит над хордой
в виде трубки с каналом или невроцелем внутри, занимая все про-
31
тяжепие тела, от переднего конца до заднего. Жаберные щели
в числе нескольких пар пронизывающие глотку, всегда имеются
у зародыша, а у многих форм сохраняются и в течение всей жизни.
Наконец, сердце, имеющееся у подавляющего большинства позво-
ночных, лежит на брюшной стороне тела. У одной группы (Acrania,
Amphioxus) сердца еще нет, ио ему соответствует так называемая
брюшная аорта, лежащая также па брюшной стороне, под глоткой.
Всем позвоночным свойствен еще один орган, развивающийся
как выпячивание брюшной стенки кишечника, образующее мас-
сивный железистый орган — печень.
Позвоночные составляют высшую ступень эволюционного раз-
вития в сравнении с ранее описанными подтипами полухордовых
и личиночно-хордовых. Они приобрели ряд признаков, которые под-
нимают на большую высоту общий физиологический уровень их
организации: скелет дает более прочную опору мышцам — органам
движения; нервная система более развита, а, следовательно, реакции
поведения становятся более четкими и совершенными; сердце де-
лает более совершенным кровообращение, а с ним вместе и общий
обмен веществ; увеличение энергии движения влечет за собой уси-
ление сгорания, т. е. увеличивает запас энергии животного. Та-
кого рода изменения, поднимающие организм па большую функцио-
нальную высоту, носят название ароморфозов (от грече-
ского слова alp© — поднимаю и gopcpojai; изменения формы).
Позвоночные появляются па земле позднее других животных.
Кроме того, уже у низших из позвоночных, кроме Acrania, развился
твердый скелет, наружный и внутренний. Благодаря этим двум
обстоятельствам древние вымершие позвоночные гораздо полнее
сохранились до нас р ископаемом состоянии, чем представители
других типов. Это дает возможность в вопросах происхождения раз-
личных классов позвоночных опираться на данные палеонтологии,
и филогенез (историческое развитие группы) позвоночных,
восстанавливаемый на основе параллельного изучения сравнительно
анатомическим, эмбриологическим й палеонтологическим мето-
дами, известен нам гораздо лучше и с бблыпими деталями, нежели
филогенез других типов. Некоторые страницы эволюции позвоноч-
ных известны с большими деталями. Эволюция позвоночных шла,
как и всякая эволюция, в тесной взаимосвязи со средой, которая
в свою очередь менялась. Естественный отбор, отбор из бесчислен-
ного множества случайных изменений вызывал к жизни все более
и более приспособленные, адаптивно измененные формы. Начав
свою эволюцию в воде, невидимому, в пресной, позвоночные при-
способились к различным условиям водной среды, а затем, выйдя
па сушу, дали формы, приспособленные к самым разнообразным
условиям существования на земле. Большие изменения в устрой-
стве поверхности последней сопровождались большими скачками,
«революциями» в развитии позвоночных (выход на сушу, появление
новых классов и т. д.). Перевороты эти, совершавшиеся, как мы
дальше увидим, под контролем среды, подготовлялись изнутри.
Каждая новая стадия эволюции покоилась на изменениях, воз-
32
нпкших раньше, в предыдущей стадии. Каждая эпоха имела свое
направление в развитии позвоночных, соответствующее в целом из-
менениям физической среды [эпоха панцырпых (Ostracodermi),
эпоха рыб, эпоха стегоцефал, рептилий, млекопитающих]. Разви-
тие в направлении слишком специальпом (так называемые идио-
адаптации) давало группе на некоторое время преимущества
в борьбе за жизнь, и группа процветала, по при изменении среды
эта же специализация являлась источником гибели (например,
динозавров). Каждый новый класс в эволюции позвоночных
приобретал признаки, дающие ему большую относительную неза-
висимость от среды, пока пе появился человек, освобождаю-
щийся от власти последней.
Подтип позвоночных распадается на два отдела. Первый отдел
представлен так называемыми бесчерепными (Acrania),
куда относится знаменитый ланцетник (Amphioxus). Бесчереп-
ные являются наиболее примитивными из позвоночных, в известной
мере связывающими последних с личиночно-хордовыми беспозво-
ночными. Второй отдел — черепные (Craniata) — подразде-
ляется па дна подотдела: па животных без челюстей — Agnatho-
stomata (у них одна ноздря, почему их называют также Monorhi-
ла), куда из современных животных относятся круглоротые
(Cyclostomata), лишенные челюстей миноги и миксины, и на живот-
ных с челюстями и с парными ноздрями — челюстноротые
(Gnathostomata). К последним принадлежат классы: рыбы
(Pisces), амфибии, или земноводные (Amphibia),
рептилии, или пресмыкающиеся (Reptilia), п т и-
ц ы (Aves) и млекопитающие (Mammalia), к которым
относится и человек.
ОТДЕЛ А. БЕСЧЕРЕПНЫЕ (ACRANIA)
Класс 1. Головохордовые (Cephalochordata)
Строение ланцетника. К отделу бесчерепных и классу голово-
хордовых принадлежат два семейства: Branchiostomatidae с двумя
родами — Branchiostoma и Asymmetron и небольшим числом видов,
н Amphioxididae. Одним из подродов рода Branchiostoma является
наилучше известный ланцетник (Amphioxus). Он был сперва
описан как моллюск и назван Limax lanceolatus, по затем, значи-
тельно поздпее, он был признан за позвоночное животное и назван
родовым именем Branchiostoma. Видовое название за ним сохра-
нилось: Branchiostoma lanceolatum. Название Amphioxus было
дано поздпее и теперь относится к одному из подродов. Ланцетник
принадлежит к наиболее популярным животным. Хотя оп пе имеет
никакого практического значения, он привлекал и привлекает к себе
внимание исследователей. Им занимались крупнейшие ученые, над
ним работает масса молодых зоологов.
Причина такого внимания заключается в том значении, кото-
рое он имеет для понимания эволюции позвоночных, их организа-
ции и онтогенеза этого подтипа.
3 Кашкаров а Ставчиаский 33
Организация ланцетника проста в одном и сложна в другом
отношении, так лее просты и его функции: пищеварение, крово-
обращение, и т. д., проста и история развития.
Амфиоксус является маленьким (4—10 см)* прозрачным живот-
ным, имеющим вид заостренного с обоих концов ланцета. Живет
он у берегов, зарываясь обычно с большой быстротой в песок,
откуда выставляет передний конец тела, со щупальцами. Находясь
на поверхности песка, он лежит на одной стороне тела. Плавает по
ночам (рис. 14).
Рис. 14. Branchiostoma lanceolatum.
А — с брюшной стороны, В —о боковой стороны, 1 — атриальное отверстие,
2 —усики, 3 —спинная струна, 4 — миомеры, 5 — ротовая воронка,6— аналь-
ное отверстие, 7 — метаплевральная складна, 8 — спинной плавник, 9 — брюш-
ной плавник, 10 —хвостовой плавник.
Вдоль всего тела у лапцетника по спинной стороне идет продоль-
ная складка — спинной плавник. На заднем конце
тела оп загибается вокруг последнего и, образуя хвостовой
плавник, переходит на брюшную сторону, как брюшной
плавник (точнее: подхвостовой). Последний занимает заднюю
треть тела. На передней границе ее находится отверстие, называе-
мое атриальным. Не доходя до него, брюшной плавник переходит
в парные складки, правую и левую, идущие па границе между бо-
ковыми и брюшной сторонами тела. Это — так называемые боко-
вые, или метаплевральпые, складки (metapleurae).
По бокам тела видны прогнуты^ вперед линии, обязанные суще-
ствованию соединительнотканных перегородок, разделяющих про-
дольные тяжи мускулов, лежащих по бокам тела. Это — так на-
зываемые миосепты. Миосепты правой и левой стороп лежат
не друг против друга, а чередуются, т. е. имеется некоторая асим-
метрия.
На переднем конце тела имеется отверстие, окруженное щу-
пальцами (cirri). Это отверстие ведет в полость, на дне которой
лежит ротовое отверстие, окруженное перепонкой — парусом
(velum), играющим роль сфинктера и несущим несколько щупалец.
Заднепроходное отверстие расположено несимметрично, с левой
34
стороны брюшного плавника. Впереди от пего лежит другое, более
крупное атриальное отверстие (atrioporus), через
которое сообщается с внешним миром особая полость — атри-
альная полость (atrium).
Общие покровы ланцетника представляют интерес
и том отношении, что они связывают позвоночных с беспозвоноч-
ными. Эпидермис однослойный, как у последних и у Hemichordata
и Urochordata. Среди цилиндрических клеток эпидермиса лежат
одноклеточные слизистые железы и чувствующие клетки. Снаружи
эпидермис одет тонкой кутикулой (черта беспозвоночных,
исчезающая у позвоночных) и покоится па нежпой основной мем-
бране. Под эпидермисом находится кожа (cutis, derma) — слабо
развитый слой студенистой соединительной ткани.
Мышечный слой состоит из двух продольных мышеч-
ных лент, разделенных соединительнотканными перегородками —
мносептами, или мио коммам и, приблизительно па 60 ми о-
м е р о в. Миосепты, или миокоммы, имеют форму горизонтально
лежащей буквы V, вершиной направленной вперед. Между каж-
дыми двумя мносептами находятся прикрепленные к ним поперечно-
полосатые мышечпые пластинки. Сокращение по-
следних по очереди то па одной стороне тела, то на другой влечет
за собой изгибы тела, имеющие место при плавании и зарыва-
нии в песок. Слой поперечнополосатых мускулов идет еще по-
перек брюшной стороны передних двух третей тела: попе-
речный мускул (musculus transversus). В отличие от
червей и других беспозвоночных, у которых кожно-мускульный
мешок имеет равномерную толщину по всему телу, у ланцет-
ника, как и у высших позвоночных, верхняя спипная часть
мускулатуры значительно толще брюшной.
Скелет ланцетцика представлен хорошо развитой хордой,
пли спинной струной (chorda dorsalis), идущей от са-
мого переднего конца головы до конца хвоста. Хорда лежит над
кишечником и под нервной системой между миомерами правой и ле-
вой сторон, ближе к спинной стороне животного, чем к брюшпой.
Состоит хорда из волокнистых дисков. Диски эти, как увидим
дальше, представляют остатки клеток, из которых первоначально
состоит хорда. По верхнему и нижнему краю хорды располагаются
остатки этих клеток. Снаружи хорда одета топкой бесструктурной
оболочкой, так называемой наружной эластической
(elastica externa). Эта оболочка является продуктом выделения
клеток хорды. Вокруг хорды располагается слой соединитель-
ной ткани — перихордальная соединительная
ткан ь; опа связана с соединительной тканью,одевающей нервную
систему, и с соединительной тканью, образующей миосепты и пе-
реходящей в подкожную соединительную ткань. Благодаря такому
устройству получается прочный остов, каркас, поддерживающий
тело и передающий сокращение мышц на все тело. Кроме того,
в cirri находится опорная ткань, напоминающая клеточный хрящ.
Также и спинной и брюшпой плавники поддерживаются уплот-
з* 35
ненным участком соединительной ткани. Из скелетных элементов
у ланцетника имеется еще жаберный скелет, построен-
ный из волокнистого бесклеточного вещества, являющегося выде-
лением прилежащих эпителиальных клеток, т. е. образованием
кутикулярным.
Пищеварительный капал. Ротовое отверстие,
находящееся на дне ротовой воронки и окруженное венчиком из
10—16 щупалец, ведет в глотку. Последняя очень объемиста и про-
стирается назад до половины тела. Служит глотка не столько для
питания, сколько для функций дыхания. Стенки глотки пронизаны
более чем 100 парами жаберных щелей, косо расположенных сверху
и спереди вниз и назад. Благодаря такому косому расположению
(рис. 15) на поперечном разрезе через область глотки мы видим
Рис. 15. Схема Покрытого Branchiostoma lanceolatum.
1 — анальное отверстие, 2 — 2'— атриальная полость, 3 —атриопор, 4—головной
мозг, 5 — жаберные щели, 6 — первичные, 7 —вторичные межжаберные перегородки,
8 — хвостовой плавник, 9 — центральный канал нервной системы, 10—усики, 11 —
целом, полость тела, 12 —спинной плавник, 13 —лучи спинного плавника, 14 —по-
ловые железы, 15—кишка, 16 —скелет ротовой воронки и щупалец, 17 —печень,
18 —рот, 19 —миомеры, 20 — хорда, 21—нефридий, 22 — обонятельная ямка, 23 —
ротовая воронка, 24 — глотка, 25 — спинной мозг, 26 — анальный плавник, 27 —
парус. (По Паркеру.)
целый ряд жаберных щелей. Последние отделяются друг от друга
узкими промежутками, укрепленными особым скелетом из тонких
палочек кутикулярного характера. Они двоякого рода: одни —
первичные, другие — вторичные, развивающиеся
у личинки позднее, как выросты, разделяющие первичную жаберную
щель на две вторичных. У молодой личинки жаберные щели рас-
положены метамерпо.
Жаберные* щели ланцетника открываются пе
наружу, а в особую атриальную полость (рис. 16).
Однако — это явление вторичное. Первоначально жаберные щели
открываются наружу, а затем уже развивается атриальная полость,
охватывающая жаберную область. Таким образом, жаберные
щели ланцетника по положению и существу дела подобны жабер-
ным щелям рыб.
В перегородках между жаберными щелями проходят крове-
носные сосуды. Вдоль средней линии глотки по брюшной ее сто-
роне проходит желобок, называемый подглоточным, или
эндостилем. Желобок этот выстлан мерцательным эпителием
с четырьмя рядами железистых клеток. Посередине находится ряд
клеток с очень длинными ресничками. На переднем конце эндостиль
36
разделяется на две ветви, охватывающие передний конец глотки
с боков и переходящие на спинной стороне в наджаберный желобок,
идущий до заднего конца пищевода, где он переходит в кишку.
Весь кишечник ланцетника, ротовая воронка и жаберные щели
выстланы мерцательным эпителием. Благодаря деятельности рес-
ничек, сгибающихся в одном направлении, вода входит в рот и идет
в глотку, неся с собой микроскопические пищевые частицы. Слизь,
выделяемая клетками эндостиля, действием ресничек последнего
гонится вперед, склеиваемые ею частицы пищи (микроорганизмы,
например) оседают на дно подглоточной борозды и продвигаются
Рис. 16. Branchiostoma lanceolatum.
А —поперечный разрез глоточной области, 1 — спинной мозг, 2—спин-
но-мозговые нервы, 3 — хорда, 4 — полость глотки, 5 — полость печени,
6 — лндостиль, 7 — межжаберные перегородки, 8—атриальная полость,
9 — гонады, 10 — мломеры, И —целом, 12 —нефридий, 13 — миосепты.
В — поперечный разрез кишечной области: 1 — оноложаберная полость,
2 —целом, 3 —спинная аорта, 4 — кишка, 5 — миотом, 6 — хорда,
7—спинной мозг, 8 — подкишечная вена. (По Герковичу и Рольф из
Паркера и Гасвеля.)
к окологлоточным ее разветвлениям, а оттуда к спинпой борозде.
По последней пища идет назад к кишечнику. Вода же действием
ресничатого эпителия прогоняется через жаберные щели. Такт!
образом, и дыхание, и питание ланцетника происходят при содей-
ствии ресничатого энителия; иначе говоря, эти акты у него пас-
сивны. Эндостиль сравнивают со щитовидной железой высших
позвоночных. Кишка у ланцетника прямая, и дифференцировка
отделов в пей едва намечена. Передний более широкий отдел лишь
с натяжкой может быть назван желудком. От брюшной стороны его
отходит вперед слепой вырост, сравниваемый с печенью высших
позвоночных, которая развивается, как выпячивание брюшной
стенки кишки. В атом выросте, как и в печени, имеется так
называемое воротное кровообращение (см. ниже).
37
Далее кзади кишка суживается и оканчивается назади а н а л ь-
отверстием.
п ы м,
К р
или заднепроходным,
овеносная система
ланцетника еще очень при-
митивна, сердца нет, сосудов не-
много, и они построены просто.
В этом отношении ланцетник сбли-
жается с червями. Но план строе-
ния кровеносной системы — это
тот план, который мы видим и у
высших позвоночных (рис. 17). Под
пищеварительным каналом, на-
правляясь вперед к глотке, лежит
сократимый брюшной сосуд —
брюшная аорта (aorta ven-
tralis). Под глоткой она отдает
справа и слева приносящие
жаберные сосуды, иду-
щие к первичным жаберным пере-
городкам; от этих сосудов отходят
поперечные ветви к сосудам, лежа-
щим во вторичных жаберных пе-
регородках. Эти сосуды не сооб-
щаются с брюшной аортой. Сосу-
ды всех жаберных перегородок от-
крываются на спинной стороне че-
рез посредство выносящих
жаберных артерий с
каждой стороны в одну из пар-
ных спинных аорт, идущих
над глоткой. Эти парные аорты
соединяются пазади в одну непар-
ную спинную аорту (aorta
v dorsalis). Последняя идет назад не-
посредственно под хордой до зад-
него конца тела, отдавая по пути
ветви к кишечнику, мышцам, степ-
кам тела и т. д. Впереди правая
верхняя аорта продолжается даль-
ше, чем левая, до конца тела, где
расширяется.
В сосудах жаберных перегоро-
док кровь окисляется кислородом,
заключающимся в воде, проходя-
щей через жаберные отверстия,
так что в аорте течет уже окислен-
ная кровь. Пройдя по отходящим
от нее сосудам к органам тела, кровь отдает тканям свой кислород,
а взамен обогащается углекислотой и различными продуктами об-
мена веществ, собираясь в венозные сосуды. Кровь,
38
прошедшая стенки кишечника, собирается в так называемую лод-
ки ш е ч j( у ю вену (vena subintestinalis), несущую кровь
вперед к печеночному выросту. Здесь она распадается на сеть
к а и и л л я р о в, образующих воротную систему пе-
чепп. Из печени кровь собирается в печеночную вену
(vena liepatica), которая переходит в упомянутую выше брюшную
порту, предварительно приняв в себя справа и слева парные вены —
к и» ньеровы протоки (ductus Cuvieri). Последние сла-
гаются из двух вен с каждой стороны: из вены, несущей кровь из
передней половины тела — передней кардинальной
в е п ы — и из вены, несущей кровь из задней половины — зад-
ней кардинальной вены.
Кровь у ланцетника бесцветная. Прогоняется она по сосудам
сокращением стенок брюшной аорты и сокращением расширенных
оснований жаберных сосудов, т. е. так называемых
жаберных сердец (bulbilli).
Таким образом, у ланцетника намечены уже ос-
новные черты кровообращения типичных позвоноч-
ных: 1) у пего имеется главный артериальный ствол —
аорта, по которому кровь течет спереди назад,
2) хотя сердца нет, по брюшная аорта, по которой
редуцированная кровь течет к жаберным щелям,
функционально выполняет роль сердца и замещает
его топографически, 3) имеется воротная система
печени.
Нервная система ланцетника еще очень
примитивна. На спинной стороне, непосредственно
лежит нервная трубка, окруженная соединительной
зади опа доходит почти до самого конца хорды, впереди не до-
ходит до переднего конца хорды. Передний конец нервной труб-
ки значительно расширен. В этом расширении можно видеть пер-
вую ступень развития головного мозга. В связи с этой расширен-
ной частью нервной системы стоят два органа чувств:
глазное пятно и обонятельная ямка. Вдоль
центрального канала всего мозга паходятся скопления пигмента,
связанные с особыми клетками. Это — так называемые глазки
Гессе (рис. 18).
* От переднего расширенного конца центральной первпой системы
отходят две пары «головных нервов», снабжающих предротовые
части тела. От остальной части мозга в каждом сегменте отходит
пара спинных нервов; вептральпо — чисто двигательный
(моторный) корешок к боковой мускулатуре туловища и дор-
залыю — чувствующий корешок, содержащий чув-
ствующие ветви — к коже и к внутренним органам, и двигатель-
ные ветви, по не к туловищной мускулатуре, а к мышцам внутрен-
ностей кишечника, сосудов и к поперечной мышце брюха (рис. 19).
Между спинными и брюшными корешками у лаицетпика пет ника-
кой связи, они свободны. Это — примитивная черта, свойственная
зародышевому состоянию высших хордовых. Кроме того, нервы
39
Рис. 18. Гла-
зок Hesse из
спинного моз-
га Amphiosus.
(По Camillo
Schneider.)
над хордой,
тканью. На-
Рис. 19. Вгап-
chiost ош а
lanceolatum.
Передняя
часть цент-
ральной нерв-
ной систе-
мы сверху.
Видны спин-
ные и брюш-
ные нервы.
правой и левой сторон расположены пе симметрично, а чередуются
друг с другом, как и миомеры.
Система органов чувств так же проста и при-
митивна. На поверхности тела, на краю паруса и на его щупальцах
и па щупальцах (cirri) переднего конца тела имеются связанные
с нервной системой чувствующие, видимо осязательные
клетки. На спинной стороне передней части тела слева имеется
выстланная мерцательным эпителием ямка, связанная волокнами
с передним выступом центральной нервной системы. Это — ямка
Келликера, принимаемая за обонятельный орган.
У личинки она сообщается с полостью центральной
нервной системы невропором. На верхней сто-
роне ротовой воронки имеется еще мерцательная
ямка — ямка Гатчека, принимаемая за о р-
ган вкуса. Орган зрения представлен
пигментным пятпом на передней стенке мозгового
пузыря. Орган слуха отсутствует вовсе.
Полость тела, или целом (см. рис. 16).
Полость тела у взрослого ланцетника развита слабо,
так как ее вытесняет развивающаяся здесь особая
полость — атриальная. Представлена полость тела в
жаберной области парой спинных целомических ка-
налов, находящихся с каждой стороны над глоткой по
бокам верхне-глоточной борозды. От этой пары по-
лостей вниз идут узкие каналы, спускающиеся по
первичным жаберным перегородкам; внизу они сое-
диняются в непарный капал — эндостил-
лярный целом, идущий под глоткой. Позади
глотки спинные каналы и брюшной соединяются в
узкую полость, охватывающую желудок и печень, а
в области кишки, позади атриопора полость тела
расширяется, охватывая кишку; лишь с правой сто-
роны она несколько стеснена продолжением сюда
выроста атриальной полости. Вся полость тела вы-
стлана плоским эпителием, каждая клетка которого
снабжена одним мерцательным волоском.
Органы выделения очень своеобразны и сближают
ланцетника с беспозвоночными. Весьма вероятно, что различные
части тела лапцетпика способны к выделению азотистых продук-
тов обмена. Но главная роль принадлежит нефридия м.
Нефридии являются сегментально расположенными в жаберпой
области короткими изогнутыми трубочками, лежащими по бокам
глотки между атриальной полостью и парной полостью тела
(рис. 20). Каждая трубочка открывается несколькими отверстиями —
нефростомами — в полость тела и одним — в атриаль-
ную полость. Так как атриальную полость мы можем считать за
часть наружной среды, обособившуюся вторично (см. ниже), то
следовательно, нефридии представляют сегментально располо-
женные трубочки, открывающиеся нефростомами в полость тела,
40
еще
Рис. 20. Branehiostoma lan-
ceolatum. Нефридий левой сто-
роны с частью стенки глотки.
Значительно увеличено.
(По Herdman.)
а выводным отверстием наружу. Такое точно расположение имеют
нефридии, или сегментальные органы, коль-
чатых червей. Сходство с червями увеличивается
тем, что нефростомы у ланцетника
усажены особыми булавовидными
клетками—с о л еноци т а м и, или
пламенными клетками.
Это — единственный случай нахож-
дения соленоцитов у позвоночных.
Называются они пламенными клет-
ками потому, что внутри эти клетки
имеют капал, а в нем — длинный
мерцательный волосок, отходящий
от тела клетки, представленного бу-
лавовидным вздутием. Волосок нахо-
дится в движении наподобие пламени.
Клетки эти выделительные. На уров-
не нефридиев жаберные сосуды обра-
зуют сосудистую сеть (glo-
merulus), напоминающую то, что мы
впоследствии увидим у высших поз-
воночных.
Половые железы, или
гонады (рис. 16), располагаются
по бокам тела сегментально, выда-
ваясь в атриальную полость по
сторонам глотки и кишки. Построены они одинаково у самцов
и самок. У рода Asymmetron гонады есть только на правой сто-
роне тела. У ланцетника их около 26 пар. Когда половые продукты
в гонадах созревают, эпителий гонад разрывается, яйца и спермин
Рис. 21. Развитие ланцетника.
А — бластула, В, С, D — образование гаструлы, Ьр — бластопор, д — первичнокишечная
полость, В, F —зародыш, пр — невропор, сп—canalis neurentericus, mes— мезодерма,
coel — целомические мешки, п — полость нервной системы, ch — хорда. (По Щмаль-
гаузену.)
41
мелки (и.1 мм) и содержат очень
попадают в атриальную полость, откуда выходят с водой через
атриопор.
Развитие ланцетника (рис. 21, стр. 41) исключительно инте-
ресно. Оно показывает нам в наиболее типичном, упрощенном
виде развитие позвоночного животного.
Оплодотворение происходит в воде, обычно вечером, эмбрио-
нальное развитие происходит ночью, а утром вылупляется личипка.
го желтка. Дробление
поэтому полное и поч-
ти равномерное и за-
канчивается образо-
ванием бластулы.
Клетки нижнего по-
люса лишь немного
крупнее клеток верх-
него. Из бластулы пу-
тем выпячивания, и п-
вагинации об-
разуется г а с т р у-
л а. Полость бласту-
лы, или бласто-
цель, исчезает, а
получается новая по-
лость — первич-
ная кишка (аг-
chenteron), открываю-
щаяся наружу пер-
вичным ртом,
или бластопо-
ром. Впутренпий
слой, или энтодер-
м а, гаструлы обра-
зован более крупными
клетками бластулы,
наружный — экто-
дерма — более мел-
кими верхней стороны. У клеток эктодермы развиваются рес-
нички, благодаря которым зародыш уже в оболочке может дви-
гаться внутри нее, а по выходе отсюда — плавать.
Гаструла вытягивается, отверстие бластопора суживается, и на-
чинается развитие основных систем тела. Нервная система обра-
зуется из эктодермы верхней стороны гаструлы. Здесь по средней
линии образуется так называемая медуллярная бороз-
д а. Края ее растут навстречу один другому и замыкаются над по-
грузившимся глубже слоем цилиндрических клеток. Последний,
заворачиваясь и замыкаясь в трубку, становится нервной
трубкой (рис. 22). Края медуллярной борозды, начав смыкаться
по середине спинной стороны, продолжают смыкаться дальше па-
зади, замыкаются позади бластопора, обойдя его. Таким образом,
4
Рис. 22. Четыре стадии развития хорды, нерв-
ной системы и мезодермы у Amphioxus.
1 — эктодерма, 2 — хорда, 3 — полость первичной киш-
ки, 4 — энтодерма, 5 —целом, 6 — целомический кар-
ман, 71 — кожно-волокнистый листок, 7а—кишечно-
волокнистый листок, 8 — медуллярная пластинка, 9 —
нервный канал, 10—первичный позвонок (сомит).
(По Herdmann.)
42
Рис. 23. Поперечный
разрез через личинку
ланцетника.
л —спинной мозг, ch —
хорда, ту — миотом,
до — зачатой гонады,
вс — склеротом, coel —
целом, int — кишечник.
кишечный капал оказывается теперь в соединении с полостью
нервной системы посредством нервнокишечного к а на-
л a (canalis neurentericus). Впереди края медуллярной борозды
смыкаются позже, и здесь долгое время остается открытым отвер-
стие — и европор.
Одновременно происходит образование хорды из энто-
дерм ы. На верхней стенке первичной кишки образуется выпя-
чивание в виде продольного желобка. Края последнего сходятся, и
от кишечника таким образом отделяется клеточный тяж — х о р-
д а, или спинная струна (chorda dorsalis). По сторонам
последней одновременно с ней образуются парные, сегментально
расположенные выпячивания — закладка
мезодермы. Эти парные выпячивания
обособляются от кишечной полости и обра-
зуют метамерно расположенные целоми-
ческие мешки с полостью внутри
(энтероцель). Это — вторичная
полость тела. Целомические мешки
разрастаются между эктодермой, с одной сто-
роны, нервной системой, хордой и кишечни-
ком — с другой. Затем они делятся на спин-
ной отдел, лежащий по бокам от хорды и
нервной системы, и па брюшной, лежащий
по сторонам кишечника. Верхний отдел на-
зывается сомитом, или первичным
позвонком, нижний — боковой
пластинкой. Полости сомитов (м и о-
ц е л ь) остаются раздельными, полости бо-
ковых пластинок вскоре сливаются между со-
бой, образуя полость тела (сплапхпоцель).
В сомите различают следующие части (рис. 23): м и о т о м,
из которого разовьется туловищная мускулатура, часть, прилегаю-
щая к хорде; с к леротом — зачаток перихордальной соеди-
нительной ткани и ткани плавниковых лучей, складка, врастающая
от пижпе-внутренпсго угла сомита между хордой и кишечником,
и кожный листок — наружная стенка миоцеля. Из этого
листка развивается впоследствии соединительная ткань, находя-
щаяся под эпидермисом. В боковой пластинке отличают наружный
кожно-волокнистый листок — сомато-плевру, и внутрен-
ний — кишечпо-волокпистый — сплапхпоплевру. Из
него развивается гладкая мускулатура кишечного канала. Правая
и левая боковые пластинки сходятся между собой, образуя единую
полость тела. В виде слепых пальцевидных выпячиваний стенок
полости тела развиваются нсфридиальные канальцы. Позже они
открываются в образующуюся околожаберную полость. Гонады
так же развиваются, как выпячивания полости миотома, именно
нижпей его части, где было соединение с боковой пластинкой.
На левой стороне переднего конца впячиванием снаружи и вы-
пячиванием изнутри образуется рот. На брюшной же стороне,
43
передвигаясь потом на правую сторону, образуется первая жабер-
ная щель, устанавливающая прямое сообщение обширной глоточ-
ной полости с наружной средой. Сзади нервно-кишечный канал
закрывается, а заднепроходное отверстие (anus)
образуется подобно рту впячиванием снаружи.
Личинка вытягивается в длину, на левой стороне одна за дру-
гой образуются 14 лежащих друг за другом жаберных щелей, ко-
торые затем перемещаются на правую сторону. Здесь же, несколько
дорзальнее образуется второй ряд жаберных щелей. Затем нижний
ряд их перемещается обратно на левую сторону, число уменьшается
до 8, рот передвигается на брюшную сторону, и ланцетник стано-
Рис. 24. Поперечные разрезы трех личинок Ampbioxus.
1 —полость в спинном плавнике, 2 — кишка, 3 —спинной мозг, 4 —
хорда, 5 — соединительная ткань, 6 —миотом, 7 —аорта, 8 —целом,
9 — подкишечная вена, 10 — метаплевральная складка, 11 —податри-
альный валик, 12 — атриальная полость, 13 — лимфатическое простран-
ство в метаплевральной складке. (По Lankaster и Willey из Herdman.)
вится симметричным животным. В дальнейшем число жаберных
щелей увеличивается путем образования новых щелей позади ста-
рых и разделения уже образовавшихся путем вырастания сверху
каждой щели «язычка», разделяющего щель пополам, как у Hemi-
chordata,
Атриальная полость получается путем образования двух скла-
док, ограничивающих эктодермальный желобок.
Внутренние края этих складок срастаются между собой, огра-
ничивая небольшую околожаберную, или атриальную, полость.
Эта полость затем увеличивается в размерах и оттесняет полость
тела кверху, охватывая глотку (рис. 24). Из развития околожабер-
ной полости видно, что она выстлана эктодермой и является частью
наружной среды. Этой полости нет ни у одного высшего позвоноч-
ного. Она является, видимо, приспособлением для защиты жаберных
щелей у животного, зарывающегося в песок. То обстоятельство, что
на известной стадии развития ланцетника рот и жаберные щели на-
ходятся на одной стороне, вызывает предположение, что предки
44
л;пщ(‘Т1гика были асимметрично построенными животными, кото-
рис подобно современным камбаловым рыбам лежали на дне на
<»/i.iioii стороне тела; тогда нахождение жаберных щелей на одной
< 11>1 ><>пе тела было приспособлением к такому образу жизни. Когда
ihr животные стали зарываться в песок, развились путем отбора
казанные складки и атриальная полость.
По поведению ланцетник стоит па очень низкой ступени разви-
тия. Деятельность его очень примитивна и не выше уровня червей
и тому подобных животных. Чувствительность к свету мала. Обус-
ловливается она не наличием глазного пятна, которое можно уда-
лить, не лишая ланцетника способности ощущать свет, а клетками
Гессе; быть может, и кожа чувствительна к свету. Обычно ланцет-
ник плывет от света в темноту. Это стоит в связи с тем, что он —
животное роющее. Поднятие температуры стимулирует ланцетника
к движению, температура в 40° вызывает смерть. Убивает лапцет-
пика и продолжительное действие температуры в 4°. Повидимому,
у ланцетника существуют разные рецепторы для восприятия тепла
и холода. Чувствительность к механическим стимулам велика,
в особенности чувствительны ротовая воронка и cirri. Зарывание
в песок ланцетника происходит пе столько вследствие отрицатель-
ного действия на него света (отрицательный фототропизм), сколько
благодаря стереотропизму, стремлению приводить свое тело в кон-
такт с твердыми телами. Есть у ланцетника чувствительность и к хи-
мическим стимулам. Вообще это животное мало деятельное, пассив-
ное. У пего имеются в потенции реакции и ощущения, свойствен-
ные позвоночным, но уровень его нервно-психической жизни стоит
очень низко, гораздо ниже, нежели у многих беспозвоночных.
Все это вместе взятое заставляет пас считать, что Acrania (Ce-
phalochordata) и Urochordata находятся между собой, быть может,
и в отдаленном, но всё же несомненном родстве, что пи те ни дру-
гие, конечно, не могут считаться предками друг друга, что Cepha-
lochordata и Urochordata происходят от одного общего корпя и ра-
зошлись в разные стороны, что от этого же корня произошли и
вышестоящие позвоночные (Craniata). Палеонтология ничего пе
говорит нам об этом корне, по па'осн&ве тщательного изучения исто-
рии развития и анатомии Acrania и низших Craniata (миноги, низ-
шие рыбы) мы можем его себе представить (рис. 25). Этот предок
был по ряду признаков близок к современным Acrania, но-хорда
у него пе доходила до переднего конца тела, у пего пе было около-
жаберной полости, число жаберных щелей было гораздо меньше
(около 17), нефридии открывались наружу, животные были по-
строены симметрично, свободно плавали, придерживаясь дна.
Время, когда жили эти примитивные Acrania, следует от-
нести, вероятно, к кэмбрийской эпохе. Существование таких жи-
вотных пе есть только гипотеза. Оно настолько обосновывается
сравпительноапатомическими и эмбриологическими данными, что
мы можем.быть уверенными в их реальности, хотя в ископаемом
состоянии они до нас не сохранились. От них произошли современ-
ные Acrania и Craniata. Первые приобрели свои особенности, при-
45
способившись к роющему образу жизни, к жизни в песке. В этой
среде они оказались в известной мере в стороне от борьбы за суще-
ствование и дожили до нашего времени в неизмененном почти виде.
Другой ряд потомков первичных Acrania привел к Craniata, кото-
рые затем эволюционировали двумя путями: один привел к Cyc-
lo s t о m at а и Ostracodermi, другой — к рыбам и че-
рез них к высшим черепным.
Наблюдающаяся в течение индивидуального развития Amphio-
xus несимметрия и положение жаберных щелей па одной стороне
объясняются предположением, что на известной стадии филоге-
нетического развития предки Amphioxus вели образ жизни, по-
добный образу жизни камбал, т. е. лежали на дне на одном боку,
тетического представителя примитивных Acrania, т. е.
предков современных Acrania (Branchiostomatidae) и че-
репных позвоночных (Cyclostomata, Ostracodermi и рыбы).
1 — анальное отверстие, 2 —брюшная аорта, 3 — спинная аорта,
4—жаберные щели, 5 — спинная струна, 6 — ротовое кольцо с
cirri, 7 — эндостиль, 8 — печеночный вырост, 9 — кишка, 10 —
ротовое отверстие, 11—-спинной мозг, 12 — мускульные сегмен-
ты, 13 — невроцель. (По А. Н. Северцову.)
и тогда жабры перешли на одну, верхнюю, сторону. Впоследствии
эти формы стали зарываться в песок, и в качестве защитного при-
способления у них развилась атриальная полость.
Асгашаявляются главным образом жителями тропических морей,
встречаясь между 40° северной и 40° южной широты. Больше всего
представителей бесчерепных в Индийском и Тихом океанах. Но ряд
видов является широко распространенным в мелких водах. Так,
Amphioxus найден в Атлантическом океане, в Средиземном и Чер-
ном морях, у западных берегов Северной и у восточных берегов
Южной Америки, у Южной Азии, Австралии, Новой Зеландии.
Такое широкое распространение вполне согласуется с примитив-
ностью организации, т. е. с древностью группы, указывая, что у нее
было много времени и возможностей к широкому распространению.
Acrania представлены двумя семействами — Branchiostomati-
dae с двумя родами: Branchiostoma и Asymmetron. У первого рода,
куда относится в качестве подрода и Amphioxus, гонады двусторон-
ние и метаплевральпые складки симметричны, у второго — гонады
есть лишь на правой стороне тела и метаплевральные складки не-
симметричны. Найденные в океане глубоководные Acrania — сем.
Amphioxididae с родом Amphioxides не имеют атриальной полости,
печеночного выроста, рот у них на левой стороне, жаберные
отверстия непарные и на брюшной стороне.
46 .
ПОЛОЖЕНИЕ ACRANIA В СИСТЕМЕ И ПРОИСХОЖДЕНИЕ
ХОРДОВЫХ
A< r.inin представляют большой интерес с эволюционной точки
tipriiiDi, так как позволяют сделать некоторые обоснованные пред-
нил<1жгпия о происхождении позвоночных.
Прежде всего ланцетник — настоящее хордовое животное, так
как оц имеет хорду, нервную систему в виде трубки на спинной
« тороне тела, ряд жаберных щелей, соответствующую сердцу брюш-
ную аорту, сегментированное тело. С другой стороны, Amphioxus
и ряде признаков еще сближается с беспозвоночными. Такими
признаками являются: однослойный эпителий, железы в эпидерми-
се одноклеточные, кутикула па его поверхности, долго сохраняю-
щийся ресничный покров, при помощи которого эмбрион движется,
кутикулярный скелет в жаберной области, слабое развитие соеди-
нительной ткани, отсутствие хряща или кости, сегментальные
выделительные органы, соленоциты, или пламенные клетки, от-
сутствие сердца и слабое развитие сети сосудов, жаберные сердца
(bulbilli). Эти признаки заставляют причислить ланцетника к по-
звоночным, но позвоночным примитивным. К примитивным приз-
накам принадлежат: слабое развитие нервной системы в области
мозга, раздельность спинных и брюшных корешков, эмбриональ-
ный характер расположения нервных клеток в мозгу, присутствие
хорошо развитой хорды, большое количество жаберных щелей, сег-
ментация мышц на всем протяжении тела, сегментально располо-
женные гонады, отсутствие плавников, глаз, уха, селезенки, под-
желудочной железы, выводпых половых протоков, пассивное пита-
ние при помощи ресничного эпителия. Все эти существенные приз-
наки организации явственно и неоспоримо говорят о прими-
тивности Acrania.
За эту примитивность говорит и значительное сходство органи-
зации и развития ланцетника с личиночно-хордовыми. И тут,
и там мы видим глотку, пронизанную многочисленными жаберными
отверстиями; тут и там в глотке имеются эндостиль, окологлоточ-
ное кольцо и надглоточная борозда. Общим признаком является
наличие атриальной полости. Но, с другой стороны, между Acrania
и Tunicata имеется целый ряд важных отличий, свидетельствую-
щих о том, что Acrania—не потомки оболочников. У последних
имеются туника, сердце, выводные протоки гонад, отсутствующие'
у ланцетника; по зато у пих пет пефридиев, нет сегментации,
у взрослых форм пет даже следов полости тела. Образ жизни со-
вершенно различен. Если мы сравним развитие ту ни кат с развитием
ланцетника, то увидим, что ранние стадии сходны у этих организ-
мов, но лишь потом развиваются значительные отличия.
Происхождение хордовых. Где же искать общий
корень Acrania и Urochordata, т. е. где искать корень, от которого
произошли позвоночные? Здесь перед нами встают все трудности
филогенетических построений. В особенности трудно ответить па
поставленный вопрос потому, что палеонтология здесь молчит.
До некоторой степени ее данные заменяет недавно найденный Аш-
47
phioxides. On найден на больших глубинах; а в глубинах часто
сохраняются архаические формы. На него можно смотреть как на
живое ископаемое, обладающее такими примитивными для Acrania
чертами, как отсутствие атриальной полости и печеночного выро-
ста и асимметрия. Но находка эта все же не дает ключа к понима-
нию возникновения хордовых.
Прежде казалось, что между позвоночными животными и бес-
позвоночными существует непроходимая пропасть. Однако знаком-
ство с развитием асцидий и с развитием ланцетника определенно
показало связь между хордовыми и беспозвоночными в целом ряде
признаков и заставило искать связь хордовых с той или иной груп-
пой беспозвоночных. Само собой попятно, что чем более далекие*
родственные связи мы стараемся выяснить, тем более встает перед
нами трудностей, тем более гипотетичными становятся
наши предположения. Палеонтологические данные в таких слу-
чаях отсутствуют, эмбриональное развитие более уклоняется от
филогенетического пути, пройденпого группой, и организацию
более уклоняющихся форм сравнивать труднее. Поэтому неуди-
вительно, что существует ряд гипотез о происхождении типа хор-
довых.
Прежде чем изложить кратко существующие гипотезы, следует
напомнить, что мы имеем две группы животных: первичноротых
(Protostomia) и вторичноротых (Deuterostomia). К первым от-
носятся черви, членистоногие и моллюски. Роту них развивается
па месте бластопора, мезодерма развивается из телобластов, кро-
веносная система не замкнута; эпидермис однослойный с кутикулой,
нервная система отделена от эпидермиса, личинка—трохофора,
выделительные органы протонефридии. У вторичноротых, к ко-
торым относятся хордовые, иглокожие, щетинкочелюстпые, па
месте бластопора развивается anus, а рот образуется заново; мезо-
дерма и полость тела развиваются из первичной кишки, крове-
носная система замкнутая, эпидермис часто многослойный, нерв-
ная система связана с эпидермой. Личинка может походить на тро-
хофору, но без протонефридиев. Эти две группы животных хорошо
обособлены и связываются с трудом.
Наиболее распространенной гипотезой долго была «анпелидпая»
гипотеза. Основой этой гипотезы является наличие явственно
выраженной сегментации как у аппелид, так и у позвопочпых жи-
вотных, у которых опа выражена в мускулатуре (миомеры), в ске-
лете (позвонки), в сосудистой системе (межсегмептальпые артерии
и т. п.), в первоначальной выделительной системе, в нервной си-
стеме. Весьма вероятно, что хордовые произошли от животных,
тело которых состояло из сегментов или метамер, подобно тому, как
у современных аннелид. Сегментальные выделительные органы по-
следних изумительно сходны с пефридиями амфиоксуса и с почеч-
ными трубочками у высших позвопочпых. Но трубочки впадают у
последних в продольный капал, открывающийся далее па заднем
конце тела. Одпако и у некоторых аппелид сегментальные органы
па каждой стороне тела открываются в один общий продольный про-
48
ток, идущий через ряд сегментов. При развитии выделительной
<1К'тсм1.1 некоторых позвоночных образование выводящего протока
<ч'1м<'нтальных органов происходит в связи с эктодермой. Однако
и настоящее время аннелидную теорию нельзя принять. Препят-
ствием к принятию аннелидной гипотезы является прежде всего
io обстоятельство, что расположение нервной системы и сердца
у аннслид как раз обратно тому, что имеется у позвоночных. По-
следние являются как бы перевернутыми аннелидами. Рот аннелид,
который при перевертывании оказался бы наверху мозга, должен
<>ыл исчезнуть, а у позвоночных образоваться новый. Чтобы выйти
из этого затруднения, некоторые допускали, что нейропор являлся
первоначально ртом, и кишка тянулась через весь мозг. По другому
автору, рот был на месте нейропора, ротовая полость — на месте
первичного желудочка мозга, а сообщение с кишкой происходило
через так называемую «воронку» (см. ниже). Кроме того, рот апне-
лпд образуется из бластопора, а у хордат — заново, т. е. первые
относятся к Protostomia, вторые — к Deuterostomia. Из указан-
ных затруднений стремится выйти «немертинная теория». Послед-
няя основана на том, что ряд признаков немертин, с точки зрения
анатомической, функциональной и эмбриологической, является
общим с таковыми миног и ланцетника или даже рыб. От немертин
произошли по этой гипотезе и так называемые Hemichordata (кишеч-
подышащие и др.), оболочники и ланцетник. Эта теория принимает,
что у предка позвоночных два нервных ствола срастаются, как
у немертин, па спипе, а не па брюшной стороне. Таким образом,
пет надобности допускать перевертывание и образование вторичного
рта. Выделительная система у немертин с соленоцитами, полость
тела, отсутствующая у взрослых, имеется при развитии и является
энтероцелей, т. е. образуется из первичной кишки.
Есть еще ряд гипотез о происхождении Chordata. Выводили
позвоночных от мечехвостов, от паукообразных, от ктенофор.
Наиболее вероятной в настоящее время является гипотеза, связы-
вающая хордовых с иглокожими, от одного корпя с которыми опи
произошли. Основывается эта гипотеза прежде всего па развитии
Balanoglossus, которое протекает удивительно сходным образом
с тем, как идет развитие у иглокожих. Этот общий с иглокожими
предок должен был походить па ту стадию развития иглокожих,
которая носит название дпплеурулы (Dipleurula). Этот предок был
двусторонне симметричным животным, с тремя парами целомиче-
ских мешков, ртом и анальным отверстием на брюшной стороне
тела. Иглокожие уже с самого начала перешли к сидячему образу
жизни, в связи с чем у пих развилась радиальная симметрия и
кожный папцырь (защита). К свободному образу жизни они (кроме
лилий) перешли вторично. Предки же хордовых пошли по пути
развития свободно живущих форм В связи с возрастающей актив-
ностью питания стала развиваться способность к активному дви-
жению, что достигалось развитием мускулатуры, ее сегментацией
па миомеры, развитием в связи с этим скелета для прикрепления
мышц, нервной системы, функционально связанной с мышечной
•1 - Кашкаров в Станчввскпй 4?
системой, развитием органов чувств, системы кровообращения,
выделительной системы, связанных с усилением обмена веществ.
Порвичиочереппые Современные Acrania
Protocraniata
Низшие бесчерепные
Acrania primitiva
Таким образом, па основе сравнительно анатомического изу-
чения низших хордовых и низших позвоночных, а также на основе
изучения их развития мы можем связать тип беспозвоночных с по-
звоночными и представить себе первые ступени их эволюции, хотя
палеонтология в данном вопросе нам ничего пе говорит. Один из
аргументов противников эволюционного учения, а именно, что мы
не можем связать между собой типы, которые-де были созданы не-
зависимо друг от друга, отпадает.
ОТДЕЛ В. ЧЕРЕПНЫЕ (CRANIATA)
К черепным (Craniata) принадлежат относившиеся прежде к ры-
бам круглоротые (Cyclostomata, миноги и миксины) и иско-
паемые примитивные позвоночные, объединяемые под названием
папцырпых (Ostracodermi), до самого последнего времени отно-
сившиеся также к рыбам; далее — рыбы (Pisces), амфибии
(Amphibia), рептилии (Reptilia), птиц bi(Aves) и млеко-
питающие (Mammalia). У всех Craniata в той или иной мере
развиты череп и головной мозг, в большей или меньшей степени
сегментирован осевой скелет, имеется сердце, кровь краспая,
с окрашенными кровяными тельцами. Отдел распадается па два
подотдела: бесчелюстных — Agnathostomata (или одпо-
ноздревых — Monorhina) и челюотпоротых — Gnathosto-
mata (или парпоноздревых — Amphirhina).
ПОДОТДЕЛ 1. БЕСЧЕЛЮСТНЫЕ (AGNATHOSTOMATA)
Сюда относятся хордовые животные, у которых рот не имеет
челюстей, жаберные мешки в средней части вздуты и снабжены
•50
... Ht
ндссь жаберными лепестками эптодермальпого происхождения;
им..гея одно посовое отверстие, нет парпых конечностей, слухо-
iioii ;шпарат с двумя или одним полукруглым каналом. Кроме ныне
живущих миног и миксин, составляющих класс круглоротых (Cyclo-
Bi.oinata) сюда относятся вымершие панцырпокожие, или просто
панцирные (Ostracodermi).
Класс 1. Круглоротые (Cyclostomata)
Крутлоротые, называемые еще мешкожаберными
(Marsipobranchii), характеризуются, кроме непарного носового
мешка, отсутствием парных конечностей, наличием жаберных меш-
ков, жаберным скелетом в виде коробки, отсутствием челюстей,
всю жизнь существующей хордой и примитивным хрящевым скеле-
том, отсутствием воротной системы почек и перекреста зрительных
нервов.
Анатомическое строение. По форме тела круглоротыс прибли-
жаются к рыбам. Одпако по организации они стоят гораздо ниже
и обладают вдобавок совершенно свеобразпыми чертами, говоря-
щими о том, что круглоротые проделали свой особый эволюционный
путь, очень рано ответвившись от общего ствола Craniata.
Тело круглоротых цилиндрическое, утреобразпое. Парпых
плавников пет. Непарные плавники, слабо развитые у миноги,
поддерживаются лучами из мягкого хряща. Кожа всегда голая.
Эпидермис многослойный и очень богат одпоклеточпыми железами,
причем, кроме обычных мелких слизистых бокаловидных клеток
имеются еще большие клетки, в плазме которых содержится зер-
нистый секрет. У мипог имеются еще особые крупные колбовидпые
клетки, в плазме которых находится длинная, спирально свернутая
пить, время от времепи продвигающиеся па поверхность. Зер-
нистые клетки мппог тоже, в конце концов, выталкиваются наружу.
У миксин все клетки эпидермиса становятся железистыми. На по-
верхности эпидермиса еще имеется cuticula. Такое обилие желези-
стых клеток, с одной сторопы, защищает тело выделяемой слизью,
с другой — служит для выделительной функции, для удаления из
организма продуктов обмепа.
Мускулатура круглоротых еще весьма примитивна.
Туловищная, или соматическая, мускулатура, происходящая из
миотомов, представлена правым и левым мускульными тяжами,
состоящими из миомеров, разделенных миосептами. Разделения
при помощи горизонтальной миосепты па спинную и брюшную
мускулатуру еще пет. Миосепты у миног лежат косо, подобно тому
как у ланцетпика. В жаберной области миомеры прерываются при-
сутствием жаберных щелей, и здесь получается разделение туло-
вищной мускулатуры па наджаберную, эпибрапхи-
а л ь н у ю и поджаберную, г и п о б р а я х и а л ь-
п у то. Туловищная мускулатура простирается па голову до глаза.
Из трех самых передних миотомов эмбриона образуются мускулы
глаза. Выросты миомеров пропинают в непарные плавники, но осо-
бых мускулов здесь еще не образуется.
Висцеральная мускулатура, происходящая из боковой пластин-
ки, лежит под туловищной, соматической мускулатурой Вокруг
жаберной области в целом (у миксин) или вокруг каждого жабер-
ного мешка лежит мускул, iазываемый общим сжимате-
лем (musculus constricor communis). Особый мускульный аппа-
рат висцерального т роисхождепия связан со скелетом языка.
Таким образом, мускулатура круглоротых еще очень примитивна
и не дифференцирована.
Внутренний скелет представлен прежде всего
х о р д о й с хорошо развитыми пузырчатыми клетками. По пери-
ферии хорды клетки менее изменены и носят название э п и т е-
Рис. 26. Pe^romyzon marinus. Боковой
вид части спинной струны и соседних
органов; левая половина удалена в
передней части путем медиального про-
дольного среза.
лия хорды. Эпителий
этот выделяет сперва тонкую
бесструктурную оболочку,
как у ланцетпика — на-
ружную эластичес-
кую оболочку (mem-
brana elastica externa), а за-
тем более мощную волок-
нистую. Хорда окруже-
на, как у ланцетника, пери-
хордальной соединительной
тканью. Однако в этой ткани
уже развиваются хрящи, со-
ответствующие зачатку поз-
вонков высших Craniata
1 — краниальная дуга, 2 — каудальная дуга,
3 — брюшной корешок, 4 — спинной коре-
шок, 5 — сегментальная артерия, 6—сег-
ментальная вена, 7 — хорда, 8 — спинной
мозг, 9 —аорта, 10 —левая задняя карди-
нальная вена. (Из Goodrich.)
(рис. 26). Хрящи эти лежат
по две пары в каждом сег-
менте, образуя в туловище
верхние дуги, краниаль-
ную (от слова cranium —
череп) и каудальную (cauda — хвост), а в хвостовой об-
ласти у миног и нижние дуги. У миксин хрящевых дуг нет вовсе.
Но и дуги миног не дают еще надлежащей защиты лежащей над
хордой нервной системе. Защита эта достигается тем, что часть
плотной перихордиалыюй ткани образует общий чехол вокруг
нервной системы, хорды и сосудов, лежащих под последней.
Внутрь от этой плотной ткани лежит рыхлая соединительная
ткань, а в пей уже — над хордой — лежит нервная система, под
хордой — артерия и вена. Парность дуг в сегменте носит назва-
ние первичной диплоспондилии. У круглоротых
мы видим ее в примитивнейшем виде.
Череп круглоротых также весьма примитивен. Та часть
черейа, в которой бывает заключен головной мозг и которая на-
зывается осевым черепом — neurocraniuni, представлена у
круглоротых хрящевой пластинкой, лежащей под мозгом и по
бокам переднего конца хорды (рис. 27). Пластинка эта имеет от-
52
Рис. 27. Petromyzon marinus. Схематическое изо-
бражение скелета передней области с черепными
нервами и брюшными корешками спинных нервов.
1—кольцевой хрящ, 2 — стилевидный хрящ, 3 — пе-
редний верхний хрящ, 4 — задний верхний хрящ,
5 — носовая капсула, 6 — слуховая капсула, 7 — 1—4-е
преджабсрные висцеральные дуги, 8 — наджаберные
комиссуры, 9 — поджаберные комиссуры, 10—жабер-
ные шсли, 11—передний боковой хрящ, 12—брюш-
ные комиссуры, 13 — подхордальные комиссуры, VII,
IX, X—головные нервы. (Но А. И. Северцову.)
иг । »cTiie (fenestra basicranialis) для назогипофизарного
к л и а л а (см. ниже) У миног от основания черепа поднимаются
\ p)iшгвые боковые стенки, крыша же черепа остается перепонча-
т<|Ц, лишь в одном месте от стенки к стенке перекинут хрящевой
мистик. У миксин перепончатыми остаются и бока черепа. С зад-
ним концом основания черепа соединяются хрящевые слухо-
вые капсул ы. Впереди с ним при помощи соединительной ткани
рыхло связана обонятельная капсула. Она непар-
ная, но пронизана парными отверстиями для обонятельных нер-
вов. В этом имеется указание на то, что у предков круглоротых
обонятельные органы были парные. Заднюю границу черепа опре-
деляет отходящий здесь от головного мозга блуждающий нерв
(nervus vagus). Так называемой затылочной части черепа, образо-
ванной врастанием в
череп передних туло-
вищных позвонков,
здесь еще нет.
Такой neurocrani-
um, как у круглоро-
тых, мы видим на
ранних стадиях раз-
вития черепа высших
Craniata. Это очень
примитивный череп.
Впереди от черепного
дна у миног лежит
тонкая непарная хря-
щевая пластинка —
задний верх-
ний хрящ, — он
налегает па задний
край переднего
верхнего хряща. Впереди лежит еще губной, или
кольцевидный, хрящ, а справа и слева этой области
находятся палочковидные передний (11) и задний (74)
боковые хрящи.
От основной пластинки вниз отходит хрящевая дуга, называемая
подглазничной (73). От нее же отходит стилевидный
отросток (72), соединенный с рожковидными хрящом (9J.
Позади стилевидного отростка лежит первая преджаберпая дуга
(7J. Язык поддерживается длинным непарным подъязычным
хрящом и может двигаться вперед и назад. Челюстей у кругло-
ротых нет . Вместо них сосательный аппарат из хрящей, называемых
губными. Как показали новейшие исследования, задний боковой
хрящ, подглазничная дуга и стилевидный хрящ являются перед-
ними частями висцерального скелета или жаберного (см. ниже).
(\'ш жаберный скелет представлен у миног коробкой из семи
изогнутых дуг, соединенных между собой над жаберными отвер-
стиями, под жаберными отверстиями, под хордой и па брюшной
53
Рис. 28.
А — разрез развивающегося зуба: черный —
функционирующий роговой эуб, под ним—смен-
ный эуб, начинающий ороговевать. В. Ротовая
воронка с роговыми зубами. [Из Goodrich, по
Warren (А) и Heckel и. Кпег (В)].
стороне. Задняя дуга соединена с хрящом, поддерживающим около-
сердечную сумку, или перикардий. У миксин жаберный скелет
сильно редуцирован.
Плавники поддерживаются тонкими лучами из эластического
хряща — так называемыми птеригиофорами.
В органах пи-
щеварения мы ви-
дим у миног также черты
примитивные и своеобраз-
ные. Ротовое отверстие ок-
ружено у мипог губами, а
у миксин усажено щупаль-
цами и ведет в воронкооб-
разную полость, воору-
женную зубами. В глубине
воропки лежит также во-
оруженный зубами, вы-
двигающийся язык. Зубы
роговые, а не костные
(рис. 28, А и В). Это — со-
сочки соединительной тка-
пи, одетые ороговевшим эпидермисом. Расположение зубов слу-
признаком, отличающим различные формы
жит систематическим
А
В
Рис. 29. Диаграмма головы
(Л) Bdellostoma и (В) Му-
xine.
1 — усики, 2 — каналы, веду-
щие пз жаберных мешков на-
ружу, 3 — общее наружное жа-
берное отверстие, 4 — жабер-
ные мешки, 4Х — вскрытый жа-
берный мешок, 5 — глотка. (Из
Goodrich, по Dean.)
миног. Рот ведет в глотку (pha-
rynx). Последняя у Petromyzon разде-
лена па верхний отдел, служащий для
проведения пищи — пищевод (oeso-
phagus), и пижний — жаберный
отдел. Пищевод переходит в кишку,
идущую прямо к анальному отверстию
и снабженную внутри спиральной
складкой слизистой оболочки — зачат-
ком спирального клапана.
Желудка нет. Перед входом в дыха-
тельный отдел глотки лежит занавесо-
подобная складка — парус (velum),
напоминая то, что мы видели у ланцет-
ника. Печень, развивающаяся как
выпячивание нижней стороны кишеч-
ника, представляет массивный орган
и пе имеет желчного пузыря. Поджелу-
дочная железа едва развита, селезенки
пет.
С каждой стороны глотки у кругло-
ротых находятся жаберные м е ш-
к и (рис. 29), числом от 6 (Myxine) до
14 (некоторые виды Bdellostoma). У
миног их 7. У Bdellostoma и Myxine эти
мешки сообщаются прямо с полостью
54
in'I и 71 пего отдела кишечной трубки; у Petromyzon жаберные
мгипсп открываются в верхний дыхательный отдел глотки, под
которым лежит пищевод. Но это образование вторичное, так
как у личинки миноги — Aminocoetes — разделения переднего
отдела пищеварительного канала па пищевод и дыхательный
отдел пет. От жаберных мешков кнаружи идут жаберные ходы,
которые у мипог и у Bdellostoma открываются наружу, каждый
самостоятельно, а у миксин (Myxine) они соединяются в один
общий канал (ductus branchialis communis), открывающийся на
брюшной стороне позади жаберного аппарата.
Внутренняя оболочка жаберных мешков имеет э н т о д ъ р-
мальное происхождение и образует многочислен-
ные тонкие листочки, богато снабженные сосудами. Вода входит
Рис. 30. Диаграмма сосудистой системы Bdellostoma.
1 — atrium, 2 —sinus venosus, 3, 4 — ventriculus, 5 — брюшная аорта, 6 — спинная
парная (правая) аорта, 7—непарная спинная аорта, 8—хвостовая вена, 9 — правая
задняя кардиальная вена, 9> — правая передняя кардиальная вена, 10 — воротная
вена печени, 11 — печень, 12 — подкишечпая вена. (Из Goodrich.)
в жаберные мешки через наружные жаберные отверстия, а у мик-
сины и через носовой ход; при сокращении общего сжимателя
глотки (muSculus constrictor communis) выходит обратно, а у мик-
син еще через особый непарный канал левой стороны наружу.
Органы кровообращения (рис. 30) построены
у круглоротых по тому же типу, как у ланцетника, по несколько
более развиты, хотя в общем еще примитивны. Под задним концом
жаберной коробки находится сердце. Последнее лежит в осо-
бой полости — перикардиальной. Сердце состоит из
предсердия (atrium), принимающего кровь из веноз-
ного синуса (sinus venosus), и из желудочка
(ventriculus). Между желудочком и предсердием имеются клапаны,
не позволяющие крови течь обратно. Приносящие сосуды
отходят от брюшной аорты и несут кровь к жабрам. Отсюда отходят
отводящие сосуды, соединяющиеся в продольную спин-
ную аорту. Туловищные артерии, отходящие от последней,
снабжают сегменты тела. Соответствующие вены соединяются в
кардиальные вены, передние и задние. В зад-
ние кардиальные вены переходит хвостовая вена. Подкишеч-
пая вена образует в печени воротную систему
печени. Воротной системы почек нет: почки получают кровь
113 аорты, а отдают через задние кардиальные вены. Передние
55
2
Рис. 31. Схема мальпигиева тельца
1 — воронка (нефростом), 2 — боума-
нова капсула, 3 — нефридиальная тру-
бочка, 4 — клубочек (glomerulus), 5 —
вена, 6 — артерия (По Кингсли.)
кардиальные, парные у личинок миноги, соединяются с задними
в протоки Кювье, но затем кювьеров проток у миног на
левой стороне исчезает, а у миксин исчезает правый.
Выделительные органы Апатомия низших по-
звоночных и изучение развития высших дают нам ясное представ-
ление об основном плане строения и функции выделительных и по-
ловых органов Craniata. Вся выделительная система может быть
сведена к системе парных трубочек, расположенных справа и слева
от позвоночника, от головы до хвоста, и открывающихся с одной
стороны в полость тела, с другой — в продольно расположенный
выводной проток. Отверстие трубочки, открывающееся в полость
тела, представляет выстланную
ресничным эпителием воронку.
Воронка эта ведет в короткую тру-
бочку, расширенную в виде тонко-
стенной камеры — боумено-
вой капсулы, в которую
с одной стороны вдается клубо-
чек сосудов (glomerulus).
Капсула и клубочек вместе обра-
зуют так называемое «мальпи-
гиево тельце» (corpus Mal-
pighii) (рис. 31).
Дальше за боуменовой капсу-
лой идет извитая железистая часть
выделительной трубочки, открывающаяся в выводной канал.
Функция воронки заключается в том, чтобы продвигать целоми-
ческую жидкость в трубочку; функция клубочка — фильтровать
жидкость из крови в полость боуменовой капсулы, а в железистой
части канальца выделяются азотистые продукты. Таков основной
план строения выделительных органов Craniata. Однако в различ-
ных классах и отрядах он подвергается различным видоизменениям.
Воронки (нефростомы) могут быть или не быть, glomerulus может
иметь различное положение (рис. 32), проток может подвергаться
различным видоизменениям, выделительные трубочки и протоки
могут вступать в различную связь с половыми железами.
Изучая развитие выделительной системы у низших позвоноч-
ных, мы видим, что первоначально закладываются трубочки с во-
ронками в самой передней части полости тела — головная
почка, pronephros, — но затем передние трубочки, развивающиеся
в самые ранние периоды жизни, атрофируются и уступают свое ме-
сто трубочкам, развивающимся позади них — первичной
почке (mesonephros), или вольфову телу. Glomerulus
располагается против воронки, в первичной почке он вдавливается
в трубочку, образуя мальпигиево тело. Первичная
почка сохраняется и функционирует у многих низших позвоноч-
ных. У высших первичная почка заменяется вторичной —
metanephros, развивающейся из заднего отдела общей закладки
выделительной системы (рис. 33).
56
Первично почечный проток у акул расщепляется вдоль на
вольфов канал, который остается в связи с канальцами
первичной
леров
почки и служит ее выводным
канал, который у самки
У других позвоночных
мюллеров канал обра-
зуется заново, а не пу-
тем расщепления.
На основании целого
ряда сравнительно ана-
томических и сравни-
тельно эмбриологичес-
ких данных установле-
но, что древнейшие Cra-
niata имели вдоль всего
туловища одну непре-
рывную серию посег-
ментно расположенных
выделительных трубо-
чек, открывавшихся в
полость тела. Рост их
назад и слияние внеш-
них концов привели к
образованию продоль-
ного выделительного ар-
протоком, и на м ю л-
стаповится яйцеводом.
/
Рис. 32а. Стереограмма головных почек.
1 — спинной мозг, 2 — ганглий спинного чувствую-
щего корешка, 3—брюшной двигательный корешок,
4 —хорда, 5—аорта, 6 — задняя кардинальная ве-
на, 7 —полость нефротома (10), 8 —клубочек, 9 —
воронка, 10 — нефротом, 11 — пронефрическая тру-
бочка, 12 — мезонефрическая трубочка, 13 —про-
нефрический проток, 14 — перитонсум, 15 — бры-
жейка (По Кингсли.)
Рис. 326. Стереограмма первичных почек.
1 — миотом, 2 — хорда, 3 — аорта, 4 — гломерулус,
5 — задняя кардинальная вена, 6 — половой валик,
7 — мезонефрические трубочки, 8 — вольфов про-
ток, 9 — перитонеум, 10 — нефростом (воронка), 11—
мальпигиево тельце. (По Кингсли.)
хинефрического капала,
идущего назад к клоа-
ке. В дальнейшем эта
древнейшая поч-
ка — archinephros — с
его архинефрическим
протоком дифференци-
ровалась в pro-, meso-
и metanephros.
Выделитель-
ные органы круг-
лор отых представлены
в виде двух длинных
тел, прикрепленных к
верхней стенке тела.
Выделительные органы
Bdellostoma имеют поч-
ти сегментированное,
строение, и в каждом
сегменте в почечный про-
ток открывается один
почечный каналец. У Myxinoidea и во взрослом состоянии
сохраняется головная почка — pronephros в виде значительной
величины органа, трубочки которого открываются в полость тела,
57
Рис. 33. Схема развития мочеполо-
вых органов Craniata
А — развитие головных почек (pronephros)
и сегментального протока. В —атрофия
пронефроса, развитие первичных (meso-
nephros) почек. С—дифференцировка про-
и мезонефрических протонов. D — разви-
тие вторичных почек (metanephros) у сам-
ца, Е— у самки. 1—pronephros, 2 — неф-
ридиальная трубочка, 3—половая железа,
в В —семенник, в Е — яичник, 4—не-
фростом, 5 — клоака, 6 — mesonephros,
7 — мальпигиево тельце, 8 — мезонефри-
ческий проток, 9 — про- или архинефри-
ческий проток, 10 — metanephros, 11 —
метанефрический проток. (По Паркеру.)
прсдстаплсппую впереди околосердечной полостью. Далее назад
лежит мезонефрическая часть почки. У миног передний отдел
ломки зачаточен, а развит задний отдел. От головной почки (pro-
nephros) наблюдаются лишь следы в виде нескольких канальцев,
сохраняющих свои воронки и открывающихся в околосердечную
полость.
Выводные протоки почек сливаются назади в мочеполо-
вой синус, а последний открывается наружу позади задне-
проходного отверстия мочеполо-
вым сосочком; в синус открыва-
ются особые отверстия — по-
ловые отверстия или
брюшные поры (pori genitales).
Через них выводятся полостная
жидкость и половые продукты.
Полы у миног раздельны,
миксины—гермафродиты. У сам-
ки миноги имеется большой не-
парный яичник (ovarium), у
самца — семенник (testiculus)
Половые продукты попадают в
полость тела, оттуда через по-
ловые отверстия в мочеполовой
синус и наружу. У миксин 'по-
ловая железа функционирует то
как семенник, то как яичник.
Нервная система.
У круглоротых имеется голов-
ной мозг. Но организован ой
еще примитивно, Он состоит уже
из тех отделов, из которых со-
стоит головной мозг вышестоя-
щих Craniata: переднего, сред-
него и заднего мозга с подразде-
лением переднего на полушария
и на промежуточный мозг, и
заднего — па мозжечок и про-
долговатый мозг (рис. 34). Все
эти отделы имеют внутри боль-
шие полости, свод их состоит не
из нервной, а из чисто эпители-
альной ткани, и расположены
все отделы по прямой линии, пе
покрывая друг друга. Это черты примитивные, повторяющиеся при
развитии мозга высших форм. Обонятельные доли (lobi olfactorii)
мозга очень велики и едва обособлены от очень небольших полуша-
рий (prosencephalon). Основание последних образовано так назы-
ваемым полосатым телом (corpus striatum). Промежу-
точный мозг (diencephalon) песет на своей верхней поверхности
68
чувствующих, ганглиозных
Рис. 34. Petromyzon marinus.
Головной мозг (А) сверху и (В)
снизу. 1 — обонятельные доли,
2 — полушария, 3 — промежуточ-
ный мозг, 4 — правый и 4» — левый
ganglion habenulae, 5 — пинеаль-
ный орган, 6 — воронка, 7 — сред-
ний мозг, зрительные доли, 8 —
мозжечок, 9—продолговатый мозг.
(По Альборну из Паркера.)
I пи \ in i iih'ii и и нервного вещества. Эти ганглии связаны с темен-
п 11 м г и и и о м. Последний состоит пз теменного глаза собствен-
н" । и и и г а л в п ы й орган) и лежащего под ним п а р а-
п и и га 1 в и ого (париетального) органа. Теменной орган
> си inn с г.1 игл нем посредством церва (nervuspinealis). Сам он лежит
ни । кожистой крышей черепа, прозрачной вэтом месте. Его дисталь-
но и чисть построена из эпителия, который в некоторых случаях
\ го I и 1.1 стел, образуя подобие хрусталика. Проксимальная часть
। иi;ia - сетчатка — состоит из
и опорных клеток. В полости глаза
<1ть из тонких волокон образует
с т г к ловидное те л о.
Па нижней стороне промежуточ-
ного мозга имеются так называемая
и о р о н к a (infundibulum) и плос-
I..UI гипофиза. Средний
м о з г (mesencephalon, lobi optici)
развит сравнительно слабо, м о з-
ж е ч о к (cerebellum) совершенно за-
чаточен; продолговат.ый мозг
переходит в спинной. У миксин мозг
несколько уклоняется от мозга миног,
и теменной глаз менее развит. Спин-
ной мозг лентовидный. Как и у лан-
цетника, спинные смешанные кореш-
ки чередуются с брюшными двига-
тельными и не сливаются с ними для
образования общего нерва. Головных
нервов десять пар: 1) обонятельный
нерв (п. olfactorius), 2) зрительный
(п, opticus), 3) глазодвигательный (п.
oculomotorius), 4) блоковый (n. troch-
learis), 5) тройничный (n. trigeminus),
с) отводящий (n. abducens), 7) лице-
вой (n. facialis), 8) слуховой (n. acu-
s(icus), 9) языкоглоточный (n. glossopharyngeus) и 10) блуждаю-
щий (n. vagus). У селахий, ганоидов и Dipnoi найден еще нерв,
< отходящий от переднего конца мозга к обонятельному эпителию —
концевой нерв (n. terminalis). .
Органы чувств круглоротых также обнаруживают
примитивные черты. Орган слуха у миксин представлен
перепончатым лабиринтом с одним единственным полу-
кружным каналом, у миног —два полукружных канала. Глаза
в существенном построены, как у других позвоночных, но у миног
"ни имеют ряд примитивных черт, а у миксин они дегенерируют от
неупотребления. Орган обоняния непарный, хотя нервы
и обонятельные доли — парные. Особенностью круглоротых яв-
лнется наличие обонятельного мешка, от которого
отходит большой питуитарный вырост, идущий назад
59
ние микромеры на верхнем
па нижнем полюсе, содержащие же
Рис. 35. Развитие миноги.
А и В — две стадии сегментации, С — ран-
няя зародышевая стадия сзади, В — разрез
через бластулу, Е — разрез через гаструлу,
F, G — две стадии развития хорды и нервной
системы. 1 — микромеры, 2 — макромеры,
3 — бластопор, 4 — бластоцель, 5 — образо-
вание нервной системы из эктодермы, 6 —
полость в энтодерме, 7 — хорда, 8 — спинные
узлы. (По Шиплею и Купферу из Паркера.)
между основанием мозга и дном черепа. В коже миноги имеются
свободные нервные окончания и специальные органы в виде поче-
чек. Кроме того, имеются особые органы — органы боковой линии,
представленные неглубокими ямками, на дне которых помещаются
чувствующие клетки. Эти органы иннервируются темп же нервами
что и органы боковой линии рыб, но построены мпого примитивнее.
Развитие миноги (рис. 35). Желток в яйце распределен неравно-
мерно, т. е. яйца телолецитальные. Сегментация пол-
ная, но неравномерная. В бластуле различаются более мелкие верх-
и крупные м а к р о м е р ы
ток. Растру ла развивается пу-
тем образования впячивания
между макромерами и микро-
Омерами, т. е. несимметрично.
I Полость впячивания носит на-
1 звание первичной к и ш-
к и (archenteron), а отверстие
ее — бластопора. Со вре-
менем бластопор становится
заднепроходным отверстием.
Рот образуется путем впячи-
вания передней стенки кишки
позднее.
Способы первоначальной
закладки главных зачатков у
миноги обнаруживают столь
большое сходство со способа-
ми их закладки у лапцетника
и у асцидий, что могут быть
сведены к ним. Наблюдающи-
еся отличия зависят от того,
что в яйце миног имеется боль-
шое количество желтка. Отсю-
да происходят различия в га-
струляции. Более поздние ста-
дии развития отличаются от
таковых у ланцетника, так как у миног дифференцируется головной
мозг, из целомической мезодермы образуются элементы хрящевого
скелета, головная и первичная почка и т. д. Следует отметить, что в
области четырех передних пар жаберных щелей на дне глотки обра-
зуется впячивапие в виде борозды, вполне соответствующее как по
способу закладки, так и по месту последней—эндостилю ланцетника
и асцидий и являющееся зачатком щитовидной железы (glandula
thyreoidea). Другое впячивапие образуется из энтодермы на перед-
ней стороне головы впереди от ротового впячивания; последнее
остается на месте, а первое, благодаря разрастанию верхней губы,
передвигается наверх и своим слепым концом направляется к концу
хорды, прилегая к нижней поверхности промежуточного мозга.
Впоследствии это впячивание становится тонкостенным мешком,
60
inn ii;rii.iii.’ieMbiM питуитарным. Его наружное отверстие
• • ।нилгтгя с отверстием обонятельной ямки (рис. 36).
Ih.i Iшляющаяся из яйца молодая минога по многим признакам
...... т(’я от взрослой формы и называется личинкой. Взрослой
фпрмои она становится, лишь подвергаясь метаморфозу через
। I года. В состоянии личинки она настолько уклоняется от взрос-
ши формы, что не только обыватели, но и зоологи считали ее за
особое животное — пескоройку (Ammocoetes).
Весьма вероятно, что Ammocoetes соответствует низшей стадии
люлюции позвоночных, так как в некоторых отношениях она
Рис. 36. Сагиттальный разрез через голову четырех ста-
дий развития Petromyzon.
1—эктодерма, 2—крыша мозга, 3 —полость мозга, 4—кишка,
5 —отверстие, ведущее в рот, 6 —гипофиза, 7 — носовая ямка,
8 — хорда. 9—верхняя губа, 10 — отверстие носовой ямки и гипо-
физы, будущая срединная ноздря, 11—воронка, 12 —гипофи-
зальный мешок, 13 —нижняя дуга. (По Goodrich.)
занимает промежуточное место между ланцетником и высшими по-
звопочпыми. Непарный плавник у пескоройки является непрерыв-
ным. Рот ее окружен своеобразно построенными губами; зачаточ-
ные и неспособные функционировать глаза глубоко скрыты под
кожей, теменной глаз хорошо развит. Семь жаберных щелей откры-
ваются непосредственно в глотку, пе разделенную на верхний и ниж-
ний каналы. Имеется желчный пузырь, отсутствующий у миноги.
И череп и жаберная коробка зачаточны. По образу жизни и по
способу питания Ammocoetes очень похожа на Amphioxus и даже
на асцидий. У пее имеется в глотке эндостиль. Питается, однако,
пескоройка не пассивно, а мелкими червями и ракообразными. В
коже пескоройки существует замечательное приспособление, защи-
61
щающсе се от бактерий, грибов и паразитов, живущих в песке, в
котором роется пескоройка: в коже ее содержится пищеваритель-
ный пептический фермент, переваривающий белки.
При наступлении метаморфоза глаза пескоройки увеличи-
ваются, эпдостиль обособляется от кишечника в замкнутый мешок,
распадающийся впоследствии па замкнутые пузырьки. Этот мешок
становится щитовидной железой, т. е. железой внутренней секре-
ции, выделяющей свой гормон в кровь. Весьма вероятно, что сам
метаморфоз начинается под влиянием обособления и начала функ-
ционирования щитовидной железы. Далее развивается хрящевой
череп, роговые зубы, глотка разделяется на дыхательный, или жа-
берпый, отдел и на пищевод, сплошной непарный плавник разде-
ляется па спинной, хвостовой и брюшной.
Развитие миксин известно гораздо меньше, нежели развитие
миног. Яйца у них крупны (2—3 см), имеют эллиптическую форму
и прицепки па полюсах, которыми они сцепляются друг с другом.
Яйца очень богаты желтком. Активная протоплазма сосредоточена
на одпом полюсе яйца в виде небольшого диска. Дробление цйца
дискоидальное, поверхностное.
Положение круглоротых в системе хорошо выяснено в недав-
нее время. Круглоротые пе могут быть отнесены к рыбам, как их
относили раньше (а некоторые относят и теперь): отсутствие челю-
стей и наличие сосательного аппарата из «губных хрящей», иепар-
Мцноги.
Petmryzontes
Предни круглоротых
Piotocycfostomaia
внутрижаберные
EntoSianchLata
BianchLostomatldae
Мамины,
MyxinoideL
Панцыряве
Ostiacocbeiml
Н рыбам
Наружножаберные
EctoEiartchiata
Пербичночерепные
Piotocianiato.
Низшие бесчерепные
Aczania piLtndJjfOL
пый обонятельный орган, отсутствие парных плавников, своеоб-
разная жаберная коробка, многочисленные жаберные мешки с жа-
брами энтодермальпого происхождения, отсутствие настоящих
зубов, примитивный еще череп (neurocranium), примитивный,
в виде хорды и отдельных дуг, осевой скелет, примитивный голов-
ной мозг, раздельно существующие спинные и брюшные корешки
сппппомозговых нервов, примптивпое архаическое строение почек,
примитивная недифференцированная мускулатура; наличие темен-
ного глаза — все эти признаки говорят, с одной стороны, за то,
что круглоротые являются весьма древними существами с прими-
тивными чертами организации, которые вероятно, были свойствен-
ны общим предкам черепных позвоночных, с другой стороны —
ряд признаков следует признать за признаки особой специализа-
&
Ц11Н, которые развились у них и некоторых ископаемых форм, па-
iimiiiiomi.ix панцирными (Ostracodermi) и объединяемых вместе
И круглоротыми в подотдел бесчелюстных (Agnathosto-
IlWitu). 11оследине противопоставляются всем остальным черепным
11011|1о||о<|цым как челюстпоротым (Gnathostomata).
КЛАССИФИКАЦИЯ, РАСПРОСТРАНЕНИЕ И ЭКОЛОГИЯ
КРУГЛОРОТЫХ
Круглоротые, таким образом, могут быть охарактеризованы
следующими признаками. Орган обоняния непарный, хотя спаб-
>1(о11П1.||| парными обонятельными нервами. С ним связан питуи-
тарный вырост, про-
инаыш1 ющнй основа-
нии черепа сверху.
Чолюстой пет. Рот ле-
жит па дно присасы-
ПНТОЛ1.ПОЙ воронки.
Кожи голпн, богатап
ЖПЛ1НН1МИ. Парных
IIЛИ«НИКОИ Нот. Ого-
ИОЙ околот представ-
лен ШЮТОПННоП хор-
дой и норпхордаль-
1ЮЙ соодпнительпой
ТИЛП1.Ю, в которой мо-
гут развиваться зача-
Л — Petromyzon fluviatilis, речная минога. В — Bdel-
lostoma dombeyi. С — Myxine glutinoaa. 1—жаберное
отверстие, 2 — жаберная щель» 3 —хвостовой плавник,
4—клоачное отверстие, 5t и 5з — спинные плавники»
6 —глаз', 7 — рот, 8 — отверстие ноздри. (По Goodrich.)
точные верхние дуги.
Череп хрящевой в основании и с боков, но с перепончатой крышей.
Висцеральный скелет представлен жаберной коробкой. Кишечный
канал прямой. Органы дыхания представлены жаберными мешками.
Головной мозг примитивен. Перекреста зрительных нервов (хиазмы)
нот. Воротной системы почек нет. Нет и артериального копуса.
< >ргак слуха с одним или двумя полукружными каналами. Почки —
мезонефрос, еще имеется рудимент пронефроса. Половые железы
непарные, не имеющие выводных протоков. Половые продукты
выводятся через половые отверстия.
Современные Cyclostomata делятся на два подкласса: миксино-
вых Myxinoidea, и миноговых — Petromyzontia (рис. 37).
Подкласс и отряд 1. Миксины (Myxinoidea)
Спиппого плавника нет или оп зачаточен. Питуитарный вырост
. открывается, в глотку. Рот окружен четырьмя парами усиков. Жа-
оерпые мешки открываются прямо в глотку. Жаберная коробка
значительно редуцирована. Глаза дегенеративные, скрыты под
кожей, лишены хрусталика п глазных мышц. Полукружный капал
один. Язычный аппарат очень сильно развит, язык вооружен зу-
бами на верхней стенке рта имеется один зуб. Дури в осевом скелете
имеются лишь в хвостовой части тела. Мозг без явственных полу-
шарий и мозжечка. Яйца крупны, и сегментация их неполная.
К отряду относятся два семейства:
Сем. 1. М икс иновые (Му xinida е)— не более чем с 6,парами
жаберных мешков, открывающихся одним общим отверстием по бокам тела.
Род Myxine с видами М. glutinosa — у западных и восточных берегов север-
ной Атлантики, М. garmani у берегов Японии, М. olivacea — в Патагонии
и М. capensis — у берегов мыса Доброй надежды. Наиболее известен пер-
вый вид. Держится у берегов с илистым дном на глубине от 30 до 90 м, до-
ходя до 600 и даже до 900 м. Животное обычно, когда не ищет пищи, зары-
вается в ил в течение дня, выставив только передний кончик тела. Плавает
легко. По образу питания миксины являются ложными паразитами. При-
сосавшись к рыбе, они при помощи своего «языка», действующего как раш-
пиль, въедаются в тело жертвы, пожирая все кроме чешуи и костей. Обычно
они нападают на тресковых рыб, на осетров, въедаются даже в акул. Нападают
особенно на рыб больных, ослабевших, пойманных на крючки или в сети,
а также поедают мертвых рыб. Полагали, что они являются гермафродитами,
причем каждая особь сперва функционирует, как самец, а затем — как самка,
но это не доказано.
Для размножения, повидимому, мигрируют в более глубокие воды. В те-
чение лета приближаются к берегам, зимой уходят дальше от берега — в более
глубокие воды,
Сем. Bdellostomatidae. Жаберных мешков от 6 до 14 пар,
каждый со своим собственным отверстием.
Это семейство, называемое также Polistotremidae, представлено родами:
Bdellostoma (или Polistotrema) с 10—12 жаберными мешками — у тихоокеан-
ских берегов Америки, от Аляски до Чили. Род Heptatrema с 6—7 жабер-
ными отверстиями добыт у берегов Южной Африки и Новой Зеландии. Наи-
более известный вид Bdellostoma stouti — единственный вид отряда, раз-
витие которого известно. По образу жизни и способу питания Bdellostoma
весьма похожа на миксин: въедается в тело даже очень больших рыб, поедая
все, кроме костей и кожи. В Калифорнии наносит большие опустошения
среди уловов сетями.
Подкласс и отряд 2. Миноги (Petromyzontia)
Спиппой плавник хорошо развит. Питуитарный вырост пе от-
крывается в глотку, а оканчивается слепо. Жаберпые мешки от-
крываются в обособленный нижний отдел глотки. Каждый жабер-
ный мешок открывается наружу самостоятельно. Жаберная ко-
робка развита. Рот без усиков, ротовая воронка и язык усажены
многочисленными роговыми зубами. Полушария имеются, мозжечок
также. Полукружных каналов два. Глаза пе дегенерируют. Яйца
мелки, сегментация полная, неравномерная. Развитие происходит
с метаморфозом.
Сюда относится одно семейство — миноговые (Pet-
romyzontidae) с несколькими родами и видами, большей
частью морскими, причем все эти виды, повидимому, входят в реки
для икрометания.
Род минога (Petromyzon) характерен для северного полу-
шария. Также в северном полушарии распространены роды: Ichtyomyzon,
Bathymyzon, Entosphenus, Caspiomyzon, Lampetra. В южном полушарии
встречаются роды Geotria — в реках Чили, Австралии и Новой Зеландии,
Mordacia — вдоль берегов Чили и Тасмании (рис. 38).
Petromyzon marinus — морская минога, до метра длиной; у берегов
Европы, Северной Америки, Западной Африки и Средиземья. Весной миноги
64
приближаются к берегам и входят в реки, где они мечут икру. Развитие миног
происходит с, превращением.
II индо личинки морская минога существует год или два, затем подвер-
tiico ii метаморфозу и мигрирует в море. Здесь молодые миноги совершают
। i iinion.io миграции к берегам весной. Взрослые могут мигрировать довольно
д||'|гио от берега. В настоящее время различные сооружения в устьях рек
и нн рнлнение вод значительно сократили количество морских миног.
К Pclromyzon близки происходящие от этого рода роды Ichtyomyzon
in Великих озер и Верхней Миссури в Северной Америке и Caspiomyzon из
Каспийского моря. Caspiomyzon wagneri входит для икрометания в Куру,
Терек, Волгу и Урал. Эти
формы мигрируют из озер в
реки в целях икрометания и
проходят стадию Ammo-
COClOS.'
Наилучше изучен род
Lam petra. L. fluviatilis,
речная минога, найдена в
Гренландии, обычна в Евро-
пе, в Северной Азии и в Япо-
нии. Южной границей ее
распространения являются
северные реки Средиземья.
М иноги входят в реки для
икрометания в несметном
множестве и служат здесь
легкой добычей. В конце
твоей миграции они разби-
ваются на группы от 2 до
20 и копают ямки для икры.
В виде личинки минога
остается 3—4 года. Ручь-
евая минога, L. pla-
iieri, распространена сходно
с, речной миногой: она обыч-
на в реках Европы, Север-
ной Азии и Японии, найде-
на также в бассейнах Чер-
ного и Каспийского морей.
Личинки живут в стадии
Ammocoetes 3—4 года, а
патом в течение несколь-
Рис. 38. Распространение миног.
Распространение морских миног показано черточ-
ками, речных — точками. В Евразии — Lampetra
fluviatilis и L. planeri, в Аляске —L. аигеа, в Ка-
лифорнии Ь. cibaria, в Сев. Америке — L. wilder!
и Ichthyomyzon, в Тихом океане — Entosphenus, в
Атлантическом — у Европы — Petromyzon, у бере-
гов Америки—Bathymyzon, в Каспии—Caspiomyzon.
к их недель превращаются
по взрослую форму.
Миноги являются предметом промысла во время их хода в реки. Здесь
их ловят особыми ловушками, которые обычно ставятся у дна, а иногда и
выше. Миноги очень жирны, идут в пищу в поджаренном или в маринован-
ном виде. Специальные миножпые промыслы расположены близ Волжской
дельты. Использование миног пока еще недостаточное.
В ископаемом состоянии Cyclostomata неизвестны. Одно время
считали за примитивное круглоротое найденную в нижних слоях
древнего красного песчаника форму Palaeospondylus gunni. Од-
ii.iij) это' предположение не подтвердилось, и некоторые даже счи-
’i.iiiiT Palaeospondylus за личинку примитивной костистой рыбы.
I’ д ругой стороны, за остатки зубов Cyclostomata некоторые
принимают так называемые «конодонты», находимые в слоях
пт нижнего силура до каменноугольных. Но это также не
./!,«нсизано.
Iйинкаров и Стапчивский
65
Класс 2. Панцирные (Ostracodermi)
К этому классу относятся ископаемые позвоночные/ обычно
рассматривавшиеся как рыбы и носящие название Ostracodermi —
кожнопанцырные. Эти существа из пресных водоемов верхнесилу-
рийского и пижнедевоиского времени имели тело, покрытое пап-
цырем из костных пластинок, и чрезвычайно своеобразный внеш-
ний вид. Родство этих древнейших из известных позвоночных
с круглоротыми подозревалось давно по отсутствию челюстей и пар-
ных плавников, по точно установить их систематическое положение
не удавалось до последнего времени. Детальное изучение шлифов
позволило установить их ближайшее родство с круглоротыми (Cyclo-
stomata). Кроме отсутствия челюстей, удалось установить 6—15 жа-
берных отверстий, указывающих на соответственное количество жа-
берных мешков, и обнаружить последние, наличие одного лишь
носового отверстия, присутствие теменного глаза, десять голов-
ных нервов, как у миног, два полукружных канала в слуховом
аппарате, мозг, севершенпо напоминающий мозг миноги, и т. д.
Мы должны считать их ближайшими родственниками современных
круглоротых, происходящих от одних с ними предков, но приспо-
собившихся к донному образу жизни.
К этому классу относятся четыре подкласса своеобразных форм,
дальнейшее изучение которых прольет свет па раннюю историю
позвоночных животных.
Подкласс и отряд 1. Бесщитковыв (ЛпаярМа)
Наиболее примитивные из ископаемых позвоночных. Внутрен-
ний скелет не окостеневал, вероятно, был представлен только хор-
дой. Жаберных отверстий было от 6 до 15. Имелся пинеальный орган
позади органа обоняния.
Рис. 39. Реконструкция Birkenia elegans.
(Из Абеля.)
Сем. Birkeniidae. Единственный вид Birkenia elegans (рис. 39)
имела удлиненное тело, хвост с неравными лопастями. Тело было покрыто
длинными узкими костными пластинками, расположенными в несколько
рядов. На голове группами располагались мелкие шипики, на спине, кото-
рую раньше принимали за брюшную сторону, — несколько крупных шипов;
найдены в верхнем силуре Шотландии. Сем. basaniidae, basanius problema-
ticus похоже на предыдущую форму — из тех же слоев. В верхнем силуре
или нижнем девоне Норвегии найдены роды Pterolepis и Rhyncholepis, от-
носимые к сем. Pterolepidae. Все эти формы жили в пресной воде.
Подкласс и отряд 2. Костнощитковые (Osteostrad, или
Cephalaspidomorphi)
Тело широкое, уплощенное в дорзо-вентральном направлении.
Рот, окруженный кожными окостенениями, помещался па брюшной
Рис. 40. Cephalaspis Murchisoni. Вид сверху.
(По Циттелю, из А. Н. Северцова.)
сторопс, так же как и жаберпые отверстия — в количестве 10 пар.
Глаза помещались па верху головы — близко один от другого.
Между глазами находился пинеальпый
орган. Голова и передняя часть туло-
вища были покрыты одним цельным
костным щитом; задняя — костными че-
тпуями. Под головным щитом поме- 3
щался хрящевой череп. Позади голо-
вы, повидимому, были придатки, по-
добные плавникам, хотя скелета их не
сохранилось.
Костнощитковые были формами при-
донными.
К отряду относятся: сем. Cephalas-
р i d а е с видами Cephalaspis lyelli из древ-
него красного песчаника Шотландии и С. mur-
chisoni из соответственных слоев Англии.
Головной щит у Cephalaspis (рис. 40) назади
оканчивался двумя крыловидными отростками,
поверхность щита была покрыта бугорчатой
скульптурой. К ' тому же отряду относятся
сем. Ateleaspidae с родом Atelaspis из верх-
него силура Шотландии и сем. Tremataspidae
с родом Tremataspis из верхнего силура ост-
рова Эзеля и Kiaeraspis (рис. 41).
Рис. 41. Горизонтальный
шлиф через вентральную
часть головы Kiaeraspis, по-
казывающий вероятное по-
ложение жаберных мешков.
1 — наружные жаберные отвер-
стия, 2—межжаберные пере-
городки, 3 — жаберный мешок,
4 — пищевод. (По Stensio, из
А. Н. Северцова.)
5*
67
Подкласс и отряд 3. Разнощитковые (Heterostract или
Pteraspidomorphi )
У примитивных форм этого отряда еще пет панцырпых пластинок
или щитов. Тело их покрыто конусообразными полыми шипиками,
Рис. 42. Схематическая реконструкция Lanar-
kia spinosa.
(По Traquair, из Goodrich.)
напоминающими так называемые плакоидпые зубы акул (см. ниже).
У более специализоваппых форм из этих зубчиков возникают кост-
Рис. 43. Реконструкция Drepanaspis. Вид
(Л) сверху и (В) снизу.
(По Traquair, из Goodrich.)
иые пластинки, образую-
щие мозаикоподобпый пап-
цырь, и в дальнейшем сли-
вающиеся в большие кост-
ные щиты. Скелет этот со-
стоит из дентина (зубной
ткани). Хвостовой плав-
ник с неравными лопа-
стями.
Сем. Coelolepidae
наиболее примитивное в отря-
де. Роды: Thelodus из верхне-
го силура и девона Европы,
Lanarkia (рис. 42) из верхнего
силура Шотландии.
Сем. Drepanaspi-
d а е, составляющее переход
к панцырным Pteraspidae.
Здесь, на ряду с четырехуголь-
ными бугорчатыми чешуями,
образовавшимися из шипов Co-
elolepidae, имеются широкие
костные щиты в головном и передне-туловищном отделе. Панцырь и на спин-
ной и на брюшной стороне. Drepanaspis dcmimdenensis из нижнего девона
Европы достигал 56 см (рис. 43).
Рис. 44^Реконструкция Pleraspis crouchi.
(S. Woodward, из Абеля.)
Сем. Pteraspidae (рис. 44). Передняя часть тела, как у Drepa-
naspis заключена в панцырь, задняя покрыта чешуями. Передний конец
68
к in iii.iTHiiyT в длинное узкое рыло (rostrum). На спине — подвижной шип.
1'ц । rh’i.ispis из верхнего силура Англии, Европы, Шпицбергена, Сев. Аме-
। in и и из нижнего девона Англии, Шотландии и Европы, образует много-
•11И .ценные виды.
Рис. 45. Реконструкция Pterichthys mulleri.
(По Traquair, из Абеля.)
Подкласс и отряд 4. Крылопанцырные (Antiarchi, или
PtericlitTiyoinorphi)1
У этой группы костный панцирь развит еще сильнее. Папцырь
на голове и туловище состоит из пескольких пластинок. По бокам
тела позади головы имелись особые «боко-
вые органы», состоявшие из отдельных
члеников. Органы эти не являлись пар-
ными конечностями, служившими в каче-
стве весел, но, по всей вероятности, игра-
ли роль аппаратов для схватывания, по-
добных таковым у богомолки (рис. 4G).
Уплощенное тело и расположение глаз на-
верху друг возле друга говорят за то, что
Pterychthyomorplii были донными живот-
ными, быть может, зарывались задним
концом тела в ил. Мощно развитый кост-
ный папцырь мог служить защитой от
крупных хищных членистоногих, населяв-
Рис. 46. Реконструкция
Aslcrolcpis maxima.
(Из Абеля.) 1—глаз, 2 —
боковой орган.
ших в то время воды.
Сем. Asterolepidae. Роды: Astero-
lepis (рис. 46) из верхнего девона Шотландии
и Прибалтики, Pterichthys из девона Шотландии
н Европы, Bothriolcpis из тех же слоев Шотландии и Канады.
ПРОИСХОЖДЕНИЕ И РАННЯЯ ЭВОЛЮЦИЯ ЧЕРЕПНЫХ (CRANIATA)
На основе вышеизложенного мы можем нарисовать пути эво-
люции низших черепных позвоночных — Agnathostomata. Кругло-
ротые и папцырные находятся в несомненном родстве, хотя и зна-
1 Систематическое положение этой группы еще не ясно. Некоторые
помещают их среди рыб., сближая с Arthrodira.
69
чительпо отличаются друг от друга. Они должны были произойти
от общих предков.
Этих предполагаемых предков предложено называть Entobran-
chiata — внутренножаберные. Сами они произошли
от первично-черепных Protocraniata, а те в свою очередь — от
Acrania primitiva, давших начало также и современным Acrania
Branchiostomatidae.
Рис. 47. Схематическое изображение скелета, централь-
ной нервной системы и высших органов чувств гипо-
тетических Protocraniata.
1 — хорда, 2 — верхние дуги, 3 — центральная нервная си-
стема, 4 —жаберные щели, 5 —жаберные дуги, 6 — рот, 7 —
основание черепа, 8—слуховая капсула, 9—глаз, 10—носо-
вые ямки, 11 — гипофиза.
О том, как должны были выглядеть Protocraniata, мы можем
судить на основе анализа строения и развития впутреппожаберных
и наружиожаберпых: они должны были обладать чертами строе-
ния, свойственными тем и другим. '
Protocraniata мы можем представить себе (рис. 47) как живот-
ных с голой кожей, состоявшей, однако, уже из многослойного
эпидермиса и тонкой cutis, лишенных еще парных конечностей,
без зубов, со скелетом из хорды и примитивных верхних дуг,
с большим числом перасчлепепных жаберных дуг, ряд которых
начинался непосредственно позади ограниченного ротовым коль-
цом рта, с черепом, прикрывавшим головной мозг снизу и с боков.
Мускулатура была вполне сегментирована. Жаберпые щели, ве-
роятно, еще не имели жаберных лепестков. Сердце существовало
и было разделено на предсердие и желудочек. Мозг уже был раз-
делен на типичные для позвоночных отделы. Глаза были построе-
ны, как у современных позвоночных; в слуховом органе было два
полукружных канала, обонятельных органов было два. По срав-
нению с бесчерепными первичночерепные, таким образом, в основ-
ном отличались более развитой мускулатурой, более развитым
скелетом, что, несомненно, стоит в связи с более энергичными дви-
жениями этих животных, а затем они отличаются гораздо более раз-
витыми и уже сравнительно совершенно устроенными органами
чувств. Активность в движениях и развитие органов чувств опре-
деленно указывают на наличие у первичпочерепных уже предпо-
сылок к активному способу питания, что и отразилось па прогрес-
сивной эволюции потомков этих форм, приобретших в активном
70
питании преимущества в борьбе за существование и обширные
возможности дальнейших адаптаций. От Protocraniata произошла
группа Entobranchiata (рис. 48), которые имели уже большое коли-
чество энтодермальных жаберных мешков и расположенную по-
верхностно хрящевую жаберную коробку. Потомки Entobranchiata
развивались в двух направлениях. У одних сохранился кольцевой
с.колет рта, но в пем развились роговые зубы и образовался сосущий
Рис. 48. Схематическое изображение переднего конца тела
гипотетических Entobranchiata с энтодермальными жаберны-
ми мешками и жаберной решеткой.
1—хорда, 2 — центральная нервная система, 3 —скелет головы
(parachordalia), 4 —слуховая капсула, 5 — носовая капсула, 6—ги-
пофиза, 7 — рот, 8 — жаберные мешки и щели, 9 — жаберные дуги,
10 — наджаберные комиссуры, 11—поджаберные комиссуры, 12 —
подхордовые комиссуры, 13 — брюшные комиссуры, 14 — трабекулы.
(По А. Н. Северцову.)
рот. От них произошли свободно плавающие, сосущие миноги
и миксины. Другие потомки Entobranchiata перешли к донному
образу жизни и к иному способу активного питания, чем кругло-
ротые: у них образовались как приспособление к донному образу
жизни костный защитный папцырь в коже и ротовой скелет.
Таковы паши представления о происхождении и развитии низ-
ших позвопочпых. Они являются предположением, основанным
па таком большом количестве сравнительноанатомических и срав-
пительпоэмбриологических данных, подтверждаемых, хотя и сла-
быми, свидетельствами палеонтологии, что мы можем считать эту
Гипотезу весьма близкой к истине.
ЧАСТЬ ВТОРАЯ
. РЫБЫ
ПОДОТДЕЛ 2. ЧЕЛЮСТНОРОТЫЕ (GNATHOSTOMATA)
Все остальные позвоночные: рыбы, амфибии, рептилии, птицы
и млекопитающие в отличие от рассмотренных выше круглоротых
и панцирных имеют снабженный челюстями рот — особенность,
имевшую громадное значение в эволюции. Челюсти образовались
из Третьей пары висцеральных дуг протокраниат, первоначальное
значение которых — поддерживание передней дыхательной части
кишечного канала. Для активно питавшихся рыбообразных пред-
ков настоящих рыб приобретение хватательного ротового аппарата
имело, конечно, исключительное значение: опо значительно повы-
сило жизнедеятельность в связи с большой трансформацией энер-
гии и чрезвычайно увеличило возможность дальнейших адаптаций
(приспособлений). Раз появившись, челюсти сохраняются у всех
позвоночных животных, специализируясь соответственно разно-
образнейшим приспособлениям к - способу добывания и роду
пищи.
Челюстноротые обладают также рядом других особенностей
। прогрессивного характера: они обладают парными конечностями
। (отсутствие их у некоторых челюстноротых — вторичное явление),
опи имеют более дифференцированный головной мозг, три полу-
кружных канала в слуховом лабиринте, два носовых отверстия, а
у рыб, дышащих жабрами, жабры эктодермального происхож-
дения и расчлененные жаберные дуги. Челюстноротые распадаются
па пять классов: рыб, амфибий, рептилий, птиц и млекопитающих.
Класс 3. Рыбы (Pisces)
4. ХАРАКТЕРИСТИКА И ОБЗОР КЛАССА
Рыбы могут быть охарактеризованы следующим образом. Пре-
сноводные или морские Gnathostomata
с непостоянной температурой тела, всю
жизнь дышащие жабрами. Органом дви-
жения служит мощный мускулистый хвост,
а дополнительными органами движения
являются парные и непарные плавники,
поддерживаемые собственными скелет-
ными частями. Сердце (за исключением Dipnoi)
72
<• одпнм предсердием. В коже по бокам
г ел а своеобразные органы чувств — орга-
ны боковой линии, связанные с жизнью
и поде. Подобные же органы имеются
н и а голов е.
Рыбы являются высшей ступенью в эволюции сравнительно
<• круглоротыми. Это выражается и в общей форме тела, более со-
nepiucHHOfl, более приспособленной it движению в воде; и в разви-
тии в коже защитного скелета в виде различного рода чешуи;
и в развитии внутреннего скелета, построенного, кроме хорды,
пи хряща и костй; в более дифференцированной мускулатуре, в на-
личии парных плавников, в более развитой нервной системе и т. д.
Лз всех организмов, живущих в воде, рыбы являются наиболее
приспособленными к движению в этой среде. Это обстоятельство
дало рыбам возможность завладеть водами суши и моря, расселиться
всюду и эволюционировать в течение всей геологической истории
земли, начиная- с силура и до наших дней, и конкурировать с дру-
гими обитателями воды в борьбе за обладание водной ареной жизни.
В настоящее время рыбы играют большую роль в круговороте ве-
ществ в природе и в воде, в частности. По числу видов рыбы усту-
пают только птицам, их насчитывают около 12 000 видов. Играя
значительную роль в жизпи природы, рыбы являются весьма важ-
ным объектом и в жизни, и экономике человека. Многие рыбы яв-
ляются важными источниками питания человека.
Однако, ограничиваясь водной средой, рыбы тем самым имеют
ограниченные возможности для эволюции: они приспособляются
лишь к различным водпым местообитаниям. Для дальнейшей эво-
люции позвопочпых был необходим выход из воды па сушу. Уже
у рыб мы видим начало развития тех приспособлений, которые были
необходимы для выхода на сушу как со стороны органов движения,
так и дыхания и кровообращения. Будучи ограничены в пределах
своей эволюции водной средой, оставаясь чисто водными животны-
ми, рыбы чрезвычайно дифференцировались в доступных им преде-
лах и, прогрессивно эволюционируя, представляют в своем разно-
образии, с одной стороны, различные ступени эволюции, с другой —
приспособления к различным условиям обитания. На оспове раз-
личий в организации класс рыб подразделяется следующим об-
разом.
Осповпое деление рыб — па хрящевых (Chondricbthyes)
и па костных рыб (Osteichtbycs). Эти деления представ-
ляют собой эволюционные ступени: хрящевой скелет является
более примитивным и в филогенезе рыб предшествует костному.
К хрящевым относятся акулы (Selachoidei) и происходящие
от них скаты (Batoidei), а также цельпоголовые (Но-
locephali) — древние и примитивные формы, и вымершие
подклассы Acantbodii, Cladoselacbii и Arthrodira; некоторые из
пих сближают хрящевых с вышестоящими костными ры-
бам и (Osteichthyes) . Последние представлены подклассом двоя-
кодышащих (Dipnoi), намечающих тот путь, которым рыбы
73
вышли па сушу, и конечиоротыми (Teleostomi). Из последних
наиболее примитивными и близкими к хрящевым рыбам являются
так называемые хрящевые ганоиды (Chondrosteoidei),
куда относятся осетровые рыбы (Acipenseridae) и лопатоносые
(Polyodontidae). Chondrosteoidei противополагаются рыбам к ост-
н ы м (Holosteoidei). Последние распадаются па две группы: одна —
teleost ei
Elasmobranchii
Holosteoidei
primitive
Chonodrosteoidei
Proiognathostomata
-mi
'Ectobranchiata
Entobranchlato.
Eranchiostomatidae
Protocraniata
Дcrania primitive.
Chondrichthyes
selacholdei
Osteichthyes
primitive
Схема филогенетических отношений у рыб
(Из А. И. Северцова)
ШтреЛа
Dipneusta
Petromyzontes
Holosteoidei..
acjdnopterygu.
'Holosteoidei.
crossoptergqli
Myxinoidei
^finaspida
кистеперые (Holosteoidei crossopterygii), представлена гл. обр.
вымершими формами, которые послужили исходным пунктом для
развития современных многоперых (Polypterini), двоякодышащих
(Dipnoi) и наземных позвоночных: четвероногих (Tetra-
poda); другая группа — лучеперые (Holosteoidei acti-
noplpryi'ii), происшедшие от одних предков с кистеперыми, дала
•..а'п> сопремеппым костным ганоидам (Holostei) —
ii'i । hi Lcpidosteus и Amia и костистым рыбам (Teleostei).
||<>«• ir.'i.iiiie чрезвычайно многочисленны. Три четверти числа их
ihiiiH'T и море, одна четверть — в пресной воде. Костистые распа-
'hiinn n на отряды, некоторые из них объединяются в более круп-
111.10 ГруПНЫ.
II. СТРОЕНИЕ РЫБ
. L Форма тела и движение
Форма тела рыб стоит па высшей ступени эволюции, в сравне-
нии с круглоротыми, а тем более с бесчерепными, так как является
гораздо более приспособленной для движения в воде.
Идеальной формой тела рыбы можно считать форму сельдевой
акулы (Lanina cornubica) (рис. 49). К пей приближается лосось
Рис. 49. Сельдевая акула (Lamna cornubica).
(Salmosalar), треска (Gadus morrhua) или сельдь (Clupea harengus).
Тело их веретенообразпо или торпедообразпо и несколько сжато
с боков. Рыба похожа па двойной клип.
Хвост оканчивается широким и плоским хвостовым плавником.
Все контуры тела мягкие, линии контура постепенно сходят на-
пет, и нигде пе имеется резкого разделения тела, которое могло бы
служить препятствием при движении в воде. Гораздо более проч-
ный, чем у низших Craniata, внутренний скелет и сильные, более
дифференцированные мускулы служат мощным средством для при-
ведения тела в движение.
Главными органами движения являются хвост и хвостовой плав-
ник. Последний служит той поверхностью сопротивления, которая
необходима для толчка вперед. Действует он наподобие кормового
весла. При сгибании, хвоста поверхность сопротивления умень-
шается благодаря сближению лучей плавника и косому его поло-
жению. При разгибании хвостовой плавник расширяется и стано-
вится в нормальное вертикальное положение. Благодаря этому
поверхность сопротивления увеличивается. Вдобавок плавник уда-
ряет теперь с большей силой. Кроме того, двигаясь вперед, рыба
75
производит вокруг себя всасывающее движение воды. Это движение
представляет лишь слабое сопротивление сгибательному движению
хвоста, так как направление обоих движений совпадает, но вы-
прямляясь, хвост встречает всю силу сопротивления течения,а, сле-
довательно, его толкающее вперед действие значительно возра-
стает.
Хвостовой плавник может быть построен симметрично и несим-
метрично. В первом случае он называется изобат и чес к им.
Если у плавника верхняя лопасть больше и длиннее, такой плав-
ник будет эпибатическим. Плавник с большей нижней
лопастью называется гипобатическим. Удары эпи-
батического плавника вызывают движение рыбы головой вниз,
удары гипобатического плавника создают, наоборот, движение рыбы
головой вверх, в результате чего рыба может даже выскочить из
воды, как летучая рыба, например, морская ласточка (Exocoetus).
Выскакивать из воды могут и рыбы с иным устройством хвостового
плавника, по тогда этому способствует также положение грудных
плавников.
Наличие парпых плавников также является большим шагом
вперед в сравнении с Acrania и Cyclostomata. Парные плавники
прежде всего служат для удержания тела рыбы в нормальном для
нее положении. Так как тело рыбы толще спереди, чем сзади,
и сверху шире, чем снизу, то центр тяжести должен лежать ближе
к голове и к спиппой стороне, чем к хвосту и брюху. Благодаря
этому рыба не имеет устойчивого равновесия, и от перевертывания
вверх брюхом через голову ее удерживают поверхности сопротив-
ления, на первом месте — грудные плавники. Последние служат
рулем глубины: при поднятии рыбы в воде кверху плавники уста-
наливаются плоской поверхностью таким образом, что передний
их край лежит выше; при опускании, наоборот, передний край
плавника опускается. При удалении парных плавников па обеих
сторонах рыба перевертывается брюхом кверху. Кроме того, удары
одного плавника спереди назад, в то время как другой остается при-
жатым к телу, поворачивают рыбу в другую сторону. Акула, схва-
тывая добычу, перевертывается брюхом кверху при помощи уда-
ров грудного плавника па одной стороне.
Парпых плавников у рыб обычно две пары: грудная и брюшная;
но иногда задняя пара (у угря — Anguilla, пилы-рыбы — Pristis),
а изредка и обе пары исчезают. Меняется и их положение: часто
брюшные плавники перемещаются вперед под грудные (у макрели —
Scomber), или же вперед от последних на горло (у трески — G-adus).
Кроме парпых и хвостового непарного плавников, у рыб имеются
пепарпйе спиппой и анальный плавники. Последний помещается
между заднепроходным отверстием и хвостовым плавником. Спип-
пой плавник может быть один или их бывает несколько. Эти не-
парные плавники дают рыбе устойчивость и твердость во время
движения, действуя, как скользящий киль яхты, поднимаясь
и опускаясь, когда нужно Удаление этих плавников вызывает
качание и зигзагообразные движения рыб. В исключительных слу-
76
’h'uix попарные плавники играют активную роль в плавании. Так,
у морского конька (Hippocampus) хвост служит органом хватания,
и по движения; последнее
<’ппгрп1ается при помо-
IIIи волнообразных дви-
jhoiiин спинного плавни-
ки, что становится воз-
можным благодаря осо-
бому устройству сплете-
нии между лучами и
осевым скелетом‘И осо-
бому расположению мы-
шечных пучков. Лучи
плавников могут быть
жесткими, перасчлепеп-
пыми и мягкими, рас-
члененными. При дви-
жении рыба обычно пред-
ставляет две кривизны.
У длиниотелых ибыстро-
плавающих рыб, каковы, например, угри (Anguilla), их может
быть и больше, по меньше двух никогда пе бывает; кривизны
всегда расположены по-
парно и дополняют одна
другую (рис. 50).
Форма тела рыб зна-
чительно вариирует, со-
ответствуя характеру
окружающей среды (об
этом см.«Экология рыб»).
Размеры рыб бывают
тоже весьма различны.
Mistichtys luzonensis
представитель семейства
колбпевых (Gobiidae) па
Филиппинских остро-
вах, является самым
маленьким позвоночным
в мире, его длина 12—
14 мм, в то время как
принадлежащий к тому
же семейству Eleotris
marmorata с Борнео и
Суматры имеет размер
в 57 см. -Самая крупйая
из рыб—акула Cetorhi-
IIns (Selache) maximus,
Рис. 50. Диаграмма движений угревых.
Большая стрелка обозначает направление движе-
ния. Короткие стрелки указывают направление
давления при движении изгибов назад.
Рис. 51. Челюсть гигантской ископаемой аку-
лы, реставрированная по зубам, находящимся
в Американском музее естественной истории.
(По Lucas.)
живущая в субарктических водах Атлантического и Тихого океа-
нов, достигает 15 м. Акула Rhinodon из тропических морей дости-
77
гает 20 м. Некоторые ископаемые акулы, насколько можно судить
об их величине по величине зубов, достигали размеров, в срав-
нении с которыми Seiache maxima кажется карликом (рис. 61,
стр. 77). Из костистых рыб тунец (Thynnus thynnus) достигает
почти 3 м, меч-рыба (Xiphias gladius) — почти 4, a Arapaima
Южной Америки превосходит эту длину. Водящийся у берегов
Северной Европы, в Средиземпом море и в Индийском океане
«сельдяной король» (Regalecus) достигает при тонком лентообраз-
ном теле более чем 7 м. Способность достигать больших размеров
также может являться признаком, дающим преимущества в
борьбе за существование, а потому эволюционно-прогрессивным.
Рыбы в этом отношении составляют высшую ступень в сравне-
нии с Acrania и Cyclostomata.
2. Кожа рыб. Железы
Общие покровы рыб значительно сложнее, нежели покровы
круглоротых, а тем более Acrania. Эпидермис многослойный,
па поверхности тонкий слой кутикулы. Ороговения нормально не
бывает. Лишь в период размножения у представителей семейства
карповых (Cyprinidae) и лососевых (Salmonidae) в разных местах
тела наблюдаются ороговения, называемые «жемчужными органа-
ми». Характерно для эпидермиса рыб богатство его одноклеточными
железами. Благодаря этому кожа рыб всегда слизистая. Железы
двоякого рода: слизистые и зерпистые, или белковые, так называе-
мые клетки Лейдига. Соедипительпоткаппая часть кожи
развита хорошо, верхний слой ее богат клеточными элементами,
но беден волокнами, нижний беден клетками, но состоит из трех
взаимно перпендикулярных слоев соединительнотканных волокон,
соединенных в пучки.
Кожа рыб бывает весьма разнообразно окрашена. В некоторых
случаях окраска имеет то или ипое биологическое зпачепие, так
что развитие се у рыб также имеет прогрессивно-эволюционное
значение. Окраска рыб обусловливается тремя причинами: нали-
чием в коже пигмента, отражением и иридировапием благодаря
оптической интерференции. Морфологической основой окраски
является наличие во внешнем и внутреннем слоях кожи специаль-
ных клеток, содержащих пигмепт — хроматофоров и от-
ражающих свет иридоцитов. Хроматофоры сильно вет-
висты и происходят из клеток соединительной ткани. Пигмент
в пих бывает красный, оранжевый и желтый, все виды из группы
липохромов, или черный — меланин. Остальные цвета
окраски получаются благодаря комбинации различно окрашенных
хроматофоров: зеленая окраска — от смешения желтого и черного,
бурая—'от смешения красного и черного, и т. д. Возможно, что
иногда имеется голубой пигмепт. Мелапип является продуктом об-
мена веществ, отбросом, который используется в разных местах
тела. Также и гуанин, из которого состоят иридоциты. Ири-
доциты, имеющие вид полигональных или округлых телец, не-
прозрачны и обладают большой отражающей способностью. В за-
78
ппсимости от того, как отражается от них свет, они кажутся белыми
и.пи ярко серебристыми. Кроме кожи, кристаллы гуанина встре-
чаются в радужной оболочке. Гуанин и меланин, а также другие
пигменты, откладываются иногда не только в коже, но и в других
местах. Большее или меньшее отложение пигмента в различных ме-
стах тела стоит в связи с освещением: чем сильнее освещение, тем
ярче обычно бывают окрашены рыбы. Там, где окраска более темная
(например, па спине), откладывается больше мелапипа, па боках,
окрашенных в серебристый топ, откладывается больше иридо-
цитов, брюхо остается нередко белым. Иридация создается иридо-
цитамп внешнего -слоя кожи. Иридоциты, отлагающиеся у некото-
рых рыб па чешуе (папример, у уклейкп, Alburnus lucidus), служат
для изготовления искусственного жемчуга. Сокращение и экспан-
зия хроматофоров происходят под влиянием нервной системы
(об этом см. «Экология рыб»).
В некоторых случаях одноклеточные железы образуют скопле-
ния, папример, при основании шипов, колючек и т. д. у так назы-
ваемых ядовитых рыб. Но многоклеточных желез у рыб никогда
пе бывает.
3. Наружный скелет
з ' г
Рис. 52. Вид кожных окостенений
Scyllium с поверхности.
1—зубчик, 2 — асповная пластинка >
3 — канал, пронизывающий послед-
нюю, 4 — перекрещивающиеся волок-
на cutis.
Кожа рыб может быть голой, как кожа круглоротых и тогда
в ней обычно имеется большое количество одноклеточных желез.
Чаще же в колее развиваются окостенения, носящие название
чешуи. Она бывает весьма раз-
нообразна по виду и по степеип
развития. Иногда чешуи, сливаясь,
образуют папцырь. По происхож-
дению и способу развития чешуя $
также бывает различна. Различа-
ют чешуи п л а к о и д п ы е, га-
ноидные и к о с т п ы е, ци-
клоидные и к т е п о и д п ы ej
Плакоидпые чешуи (рис. 52)
свойственны сростножаберпым. Ко-
лса последних кажется гладкой,
соли вести рукой от головы к
хвосту, по сильно шероховата, ес-
ли вести рукой от хвоста к го-
лове. Это происходит оттого, что
колса усеяна ромбическими пла-
стинками, несущими по середине
ш и инк с острием, направленным
ннзад («шагрень»). Эти пластинки
кожных з у б о в, или плакоидных чешуй. Название
кожных зубов дано им потому, что по своему строению и по исто-
рии своего развития опи соответствуют зубам ротовой полости.
Эуоы всех позвоночных возникли исторически из плакоидных
’нчпуй. Это — органы гомологичные. Подобно зубам по-
с шипиками носят название
79
лости рта плакоидпая чешуя имеет внутри полость, заполненную
соединительной тканью с сосудами (пульпарная полость). Сама
чешуя состоит из плотного вещества — дентина, пронизан-
ного топкими разветвленными зубпыми канальцами, в которые
входят тонкие отростки клеток пульпы. Снаружи зубчик одет
гомогенным блестящим слоем эмали. Развитие плакоидпой
чешуи происходит таким образом (рис. 53). Сосочек кожи (cutis)
врастает в эпидермис. Клетки основного (мальпигиева) слоя эпи-
дермиса, одевающие этот сосочек, становятся более высокими и
образуют так называемый «эмалевый орган». Клетки
сосочка — одонтобласты — выделяют па своей внешней
1
Рис. 53. Закладка плакоидной чешуи у Scymnus.
1 — сосочек cutis, 2 — эмалевый орган, 3 — эмаль, 4 — эпидер-
мис, 5 — одонтобласты, 6 —дентин. (Из Ihle, Versluys etc.)
поверхности дептип, сам сосочек остается пульпой зубной полости,
а эмалевый орган на своей нижней поверхности выделяет слой эма-
ли. У основания сосочка образуется пластинка чешуи, пе имеющая
строения дентина. Пластинка чешуи и зубчик закладываются от-
дельно И потом срастаются.
Плакоидные чешуи могут чрезвычайно вариировать как по ве-
личине, так и по форме, причем вариируют и пластинка, и сам зуб-
чик. У скатов рода Raja они очень крупны и разбросаны по-телу
сравнительно редко, образуя шипы. У большой акулы Selache
maxima па жаберных дугах имеется сито из длинных до 25 см плот-
ных волосков, служащих для процеживания идущей через жабры
воды. Волоски этого сита представляют не что иное как плакоидные
чешуи. У пилы-рыбы (Pristis) длинный вырост рыла, состоящий из
пропитанного известью хряща, усажен по бокам треугольными
крепкими шипами — плакоидпыми зубами. Плакоидпая чешуя
кое-где. на теле имеется и у целыюголовых—Holocephali. У рыб,
стоящих в системе выше, она заменяется другими видами чешуи,
по кое-где еще сохраняется в виде остатков прежнего покрова.
Так, у панцырпой щуки (Lepidosteus) плакоидные зубы заклады-
ваются и прирастают к заднему краю развивающейся ганоидной
чешуи. Встречаются опи и у Polypterus. И даже у так называемых
панцырпых сомов (Loricaria, Callichthys) развиваются нормальным
80
Рис. 54. Снинорог прямоугольный (Balistes
rectangulus).
(Из Никольского.)
плакоидные чешуи, сидящие также по заднему краю кост-
нЧ1 чешуи этих рыб. Такое появление плакоидных чешуй у выс-
ших рыб является одним из свидетельств происхождения этих рыб
• •г акулонодобных предков.
। Следующим типом чешуи является чешуя ганоидная.
! и ит вид чешуи был широко распространен у вымерших рыб палео-
к hi из отряда Crossopterygii и у Palaeoniscoidei. В настоящее время
ганоидная чешуя имеется в семействах Polypteridae и Lepidostei-
<1ае. Ганоидные чешуи имеют ромбическую форму, толсты,
расположены правильными косыми рядами и сочленяются между
гобой своими краями. Снаружи они покрыты блестящим и очень
плотным слоем так на-
зываемого г а п о и п а,
па поминающего эмаль.
У рода Anna опи имеют
округлую форму, тонки
it налегают друг па дру-
га, своими краями. Не-
смотря на полпое внеш- •
псе сходство ганоина с
змалыо, по происхожде-
нию своему это совсем
другое образование сое-
динительнотканного, ме-
зодермального, ине эпи-
телиального происхож-
дения. Развивается че-
шуя па границе верхнего рыхлого и пижпсго волокнистого слоя
с.utis благодаря деятельности особых клеток —остеобла-
с т о в, образующих кость. Плакоидиая же чешуя образуется
па границе эпидермиса и cutis. Таким образом, современная га-
ноидная чешуя является образованием, как будто бы отличным
<»т плакоидной чешуи. Однако, если мы прибегнем к палеонтоло-
гии и изучим иа шлифах ганоидную чешую ископаемых рыб, мы
найдем ясное свидетельство в пользу того, что первоначальной
формой ганоидной чешуи была чешуя «космоидпая», образовав-
шаяся из слияния плакоидных зубов, базальные пластинки которых
образовали нижнюю часть чешуя, а зубчики, слившись, дали на-
чало дептиноподобному «космиповому» слою и эмалсподобному
гавоину. От такой «космоидной» чешуи произошла уже настоя-
щая галоидная.
Чешуи циклоидные характерны для огромного боль-
шинства костистых рыб. Они имеют вид тонких округлых или элип-
тпческих костных пластинок, помещающихся передним краем
и особом меточке кожи, а задним налегающих иа передний край
нозадилежащей чешуи. Поверхность их часто бывает покрыта тон-
кий скульптурой (так называемые склериты). Если задний
краб чешуи гладкий, чешуя называется циклоидной, если
«•и зазубренный — чешуя ктепоидпая. Иногда чешуи при-
К KniHicanuB и Станчмнский 81
нимают совершенно своеобразный вид костных пластинок (в сем.
Agonidac, TrigJidae, Balistidae) (рис. 54), в которых тело рыбы
заключено как в папцырь, или длинных игл, шипов (в сем. Tetro-
dontidae, Diodontidae). У некоторых костистых рыб чешуя отсут-
ствует вовсе (например, в сем. Siluridae).
Концентрические кольца па поверхности чешуи являются
показателями возраста рыбы (об этом см. в главе о росте).
Происхождение костной чешуи из ганоидной не подлежит сом-
нению. Ромбические чешуи, сочлененные друг с другом, представ-
ляют известные невыгоды при плавании, оказывая сопротивление
при сгибании тела. Поэтому ромбические чешуи, распространенные
у архаических форм, заменяются циклоидными, налегающими
друг на друга, дающими рыбе больше подвижности. Смена ромби-
ческой ганоидной чешуи циклоидной наблюдается не только в эво-
люции рыб, но и у одной и той же рыбы: на туловище — ромбиче-
ские чешуи, на более подвижном хвосте — циклоидные.
Костные щиты осетровых, так называемые «жучки», хотя бле-
стят как эмаль, последней не покрыты, построены из настоя-
щей кости и развиваются за счет одной cutis.
4. Мышцы
Главная масса мышц тела (соматических) у рыб представлепа
двумя большими мускулами, лежащими по бокам тела, и разде-
ленными миосептами иа сегменты, миомеры. Миосспты и здесь не
вертикальные, а зигзагообразные. По у рыб мы уже видим раздс-
U
Рис. 55. Мышцы окуня. (По Cuvier et Valenciennes из Butschli.)
1 — глубокие мышцы спинного плавника, 2 — поверхностные мышцы ту-
ловища, 3 — мышцы хвостового плавника, 4—мышцы анального плавни-
ка, 5 — ребра, 6 — остистые отростки позвонков, 7 — приводящая мышца
нижней челюсти, 8—мышца, поднимающая небную дугу, 9—мышца от
коракоидной кости к подъязычной, 10—мышца, отводящая плавники,
11 —мышца, отводящая брюшные плавники.
лепие миотомов на спинную, или дорзальную, и
брюшную, или вентральную, части благодаря про-
дольной, горизонтально лежащей волокнистой перегородке, иду-
щей от позвоночника к коже (рис. 55). Кроме того, первоначальное
82
иl<<> io'ir.iioo направление, мускульных волокон у рыб в области
|>1>|о\,( начинает переходить в косое. Первоначально цельный
। ши туловищной мускулатуры начинает распадаться на слои,
ним, чаются косой наружный и косой внутренний мускулы. Из верх-
...... нижнего края туловищной мускулатуры дифференцируются
।онанг пучки мышц, поднимающих и опускающих плавниковые
пчи. Уже у миноги было намечено разделение мускулатуры в об-
1п< тп глотки на наджаберную и поджаберную мускулатуру. Здесь
и гпилн с развитием челюстно-жаберного аппарата гипобронхиаль-
паи (поджаберная) мускулатура значительно сложнее. Муску-
латура. конечностей развивается в связи с миотомами и иннерви-
руется, как последние, спинномозговыми нервами. Сложнее ста-
новится и висцеральная мускулатура. Общий сжиматель глотки
пи. constrictor communis) еще имеется, но развиваются отдельные
мышцы в связи с челюстями. У Chondrichthyes мускулатура по-
строена проще, нежели у Osteichtyes.
5. Внутренний скелет
Позвоночник. Внутренний скелет является одной из важней-
ших сторон организации, давшей возможность позвоночным жи-
вотным занять господствующее положение в борьбе за существо-
вание. Он служит каркасом, поддерживающим тело, дающим опору
конечностям, служит для прикрепления мышц, для защиты дру-
гих, более нежных органов. Поэтому в эволюции позвоночных
эволюция скелета играла большую роль. Скелет, если он окосте-
невает, хорошо сохраняется в ископаемом состоянии, тем самым
давая возможность применять палеонтологический метод к восста-
новлению хода филогенетического развития. Наконец, имея тес-
ное отношение к целому ряду органов: к мускулам, нервной системе,
к. кровеносным сосудам и пищеварительному каналу, органам чувств
и т. д., скелет является наиболее удобной системой для применения
в вопросах филогенеза сравнительпоапатомического метода. На
эволюции скелета особенно ясна адаптивная эволюция классов,
отрядов и т. д.
У ланцетника мы видели в качестве скелета только хорду с ок-
ружающей ее перихордальпой соединительной тканью. Движения
амфиоксуса совершаются при помощи боковых изгибов тела, глав-
ным образом хвостового отдела. В связи с этим находится сегмен-
тация мышц и нервов. Дальнейшим шагом в развитии аппарата
движения, в связи с появлением хряща и кости, должно было
явиться расчленение и осевого скелета. Оно намечено уже у кругло-
ротых парными верхними дугами. У рыб осевой скелет эволюциони-
рует дальше, развивается позвоночник из хряща или кости, при-
чем хрящевые или костные части развиваются там, где наиболее
выражено напряжение и где мышцы требуют прикрепления. В раз-
витии позвоночника участвуют хорда, ее оболочка и главным об-
разом перихордальпая соединительная ткань. Последняя возникает,
как мы видели, из мезодермы, из сомитов. От внутренней
6* 83
Диаграмма соединительнотканной си-
туловище Craniata, показывающая
Рис. 56.
стемы в
отношение осевого скелета к поперечным и
продольным септам (по Goodrich.)
1 — стенка спланхноцеля, 2 — верхние, 3 — нижпие
краниальные дуги, 4 — перерезанная стенка тела,
5 — верхняя и 6 — нижняя каудальные дуги, 7 —
«верхнее» ребро, 8 — кишка, подвешенная в цело-
мической полости, 9 — поперечная септа, 10 —
мезентерий, 11 — горизонтальные септы, 12 —
нервная трубка, 13 —срединная спинная септа,
14 — срединная брюшная септа, 15 — оболочки хор-
ды, 16 — нижнее, или плевральное, ребро, ‘"
остистый отросток.
17 —
3
Рис. 57. Поперечный разрез молодого Am-
phioxus, иллюстрирующий происхождение
мезодермы.
1—эктодерма, 2 — кишечная полость, 3—«моз-
говая пластинка, 4 — мезодерма, 4Х—спинная
мезодерма, 4а—брюшная мезодерма (боновая
пластинка). (По Hatschek из Goodrich.)
степки основания сомита
вырастает складка, на-
зываемая склеротомом,
проникающая между
миомером и хордой. По-
лость склеротома делит
его па две части — кра-
ниальную (ближайшую
к голове) и каудальную
(ближе к хвосту). Эта
двойная сегментация на
сомиты и полусклерото-
мы спинной мезодермы
(рис. 56 и 57) продол-
жается и в область голо-
вы. Главная масса сое-
динительной ткани,иду-
щая па построение осе-
вого скелета, происхо-
дит из склеротомов. Па
пересечении миосепт с
соединительно - тканной
обкладкой нервной си-
стемы и сосудов развива-
ются невральные (верхние)
и гемальные (нижние) ду-
ги, вокруг хорды разви-
ваются тела позвонков. Не-
вральные и гемальные дуги
сходятся концами наверху
и внизу и ограничивают
нервный и гемальный ка-
налы. Между этими дугами
появляются еще иптерне-
вральпые и интергемаль-
пые дуги. Таким образом
па каждый сегмент прихо-
дится две пары дуг. На
этой стадии, притом пе во
всем теле и пе вполне вы-
раженной, останавливает-
ся развитие осевого скеле-
та у миног (рис. 26). У рыб
дуги развиваются более
полно и более сильно, верх-
ние и нижние дуги могут
сливаться между собой своими основаниями, обрастая хорду:
волокнистая оболочка хорды может подвергаться охрящевеиию,
и таким образом получаются тела позвонков. Наконец, хрящ
84
может замещаться костью, и таким образом получаются костные
позвонки. У более примитивных рыб, подобно миногам, существует
первичная д и п л о с п о и д и л и я, т. е. на каждый
сегмент приходится две пары дуг: краниальная пара и каудальная.
В дальнейшем мы видим, с одной стороны, слияние дуг, причем
сливаться могут дуги одного сегмента или задняя дуга одного сег-
мента с краниальной позади лежащего, или сливаются три дуги
вместе — две одпого сегмента и одна другого. Иногда одна пара
дуг в сегменте может исчезать.
Из такого рода комбинаций складывается все разнообразие
в строении позвонка. Хорда у рыб пе исчезает. Закладывается
и развивается опа так же, как у миног. Так же образуются и ее
Рис. 58. Lamna cornubica.
Л —участок позвоночного столба туловища, частью разре-
занного вдоль, В — поперечный разрез через центр позвонка.
1—центр, 2— известковое кольцо, 3 — отверстие для спин-
ного корешка, 4 — отверстие дли брюшного корешка, 5 —
верхняя каудальная (basidorsale), 6 — нижняя каудальная
дуга (basiventrain), 7 — верхние и 8 —нижние краниальные
дуги (interealaria, interdorsalia и Intravcntralia), 9 —связка,
10 — спинномозговой канал, 11—хорда, 12 — радиальные
объизвествлепии, 13 — межпозвоночная связка. (По Goodrich.)
оболочки. Развитие позвонков может сузить ее значение, сократить
степень ее развития, но никогда хорда пе исчезает совсем, сохра-
няясь между позвонками.
В осевом скелете Elasmobranch ii дуги закладываются так же,
как у Cyclostomata, — с той разницей, что задние, или каудальные,
дуги развиты сильное. Кроме того, здесь закладываются и нижние
дуги под самой хордой. В волокнистую оболочку хорды проникают
клетки перихордальпои соединительной тканц, в оболочке происхо-
дит охрящсвспис и развивается таким образом тело позвонка
в виде хрящевого кольца в задней половине сегмента. Верхние и
нижние каудальные дуги разрастаются вокруг хорды, образуя
наружный слой тола позвонка. Краниальные дуги остаются сво-
бодными, дклипиваясь между соседними позвонками. Их называют
вставочными дугами (interealaria) (рис. 58). От позвонков
отходят боковые, или поперечные, отростки, служащие для
прикрепления ребер. Образующие нозвоно!£ дуги называют еще:
верхние — basidorsalia, пижпие — basiventralia, а вставочные —
interdorsalia и interventralia. Тела позвонков у элазмобранхий, как
85
пет еще и следов.
и у всех остальных рыб за одним исключением, — двояко-
вогнут ые, или амфицельные. В центре их хорда
сдавливается хрящом, в промежутках между позвонками остается
сильно развитой. В телах позвонков может откладываться известь
или в виде цилиндра, или нескольких, как бы вставленных друг
в друга, колец, или в виде радиальных пластинок, отходящих от
центрального кольца. Кости во внутреннем скелете элазмобранхий
~ позвоночнике различные элементы могут
сливаться или выпадать вовсе.
Сходно с элазмобранхиями построен
осевой скелет у цельноголовых (Holoce-
phali) (рис. 59) с той разницей, что здесь
более развиты краниальные дуги, во-пер-
вых, а во-вторых, хотя клетки перихор-
дальной соединительной ткани и проника-
ют в оболочку хорды, они здесь не охря-
щевевают. Тел позвонков пе образуется,
но в оболочке хорды откладывается коль-
цами известь, четыре или пять колец па
каждый сегмент. Таким образом, позво-
ночник химер еще примитивнее, нежели у
акул. Но эта примитивность, видимо, вто-
ричная, объяснимая некоторой дегенера-
цией у глубинных форм, пользующихся
при плавании больше парными плавни-
ками, чем хвостовым, вследствие его не-
доразвития.
Примитивно и сходно с элазмобранхи-
ями, особенно палеозойскими, строение
позвоночника и у двоякодышащих (Dipnoi).
Хорда сохраняется всю жизнь, тела поз-
вонков отсутствуют, хотя клетки мезодер-
мы и проникают в оболочки хорды. Раз-
виты каудальные дуги, основания кото-
рых сжимают хорду. В хвосте цератода
(Ceratodus) имеются хорошо развитые кра-
ниальные. дуги (in terdorsal i а), в других
местах тела они сливаются с основаниями
широких каудальных дуг. То же и с пиж-
пими дугами. Верхние и нижние дуги не
сходятся своими основаниями, и хордовые
оболочки по бокам остаются незакрытыми.
Основания дуг остаются хрящевыми, ио тонкие невральные (верх-
ние) и гемальные (пижние) дуги окостеневают. Весьма вероятно,
что у современных Dipnoi позвоночник несколько дегенерировап.
С некоторыми отличиями сходно с Dipnoi строение осевого
скелета у хрящевых гапоидов (Chondrosteoidei) (рис. 60).
Здесь также сохраняется хорда и нет еще тел позвонков, по кра-
ниальные дуги лучше развиты, чем у Dipnoi, хотя каудальные
86
7
Рис. 59. Chimaera mon-
struosa. (По Ilesse из
Goodrich.)
А — боковой вид позвонков,
В — поперечный разрез поз-
вонка. 1—известковое коль-
цо, 2— центр, 3 — отвер-
стие для спинного кореш-
ка, 4 — отверстие для брю-
шного корешка, 5 —верх-
няя краниальная дуга, 6 —
верхняя каудальная дуга,
7 — нижняя дуга, 8 — объ-
извествлеиные кольца, 9 —
хорда, 10 — мозговой ка-
пал, И — верхняя дуга.
играют главную роль. И здесь невральные дуги в верхней части
л замыкающие их остистые отростки, а также гемальные остистые
отростки окостеневают. В этом отношении здесь имеется прогресс
в сравнении с селахиями, в остальном
же позвоночник у осетровых стоит
ниже, чем у последних.
У Holosteoidei (Holostei, Crossop-
terygii, Teleostei) позвонки уже впол-
не окостеневают, имеются тела ам-
фицельной формы, за исключением
наицырной щуки (Lepidosteus), у ко-
торой позвонки опистоцельные, т. е.
выпуклые впереди и вогнутые назади.
Первичная диплоспондилия, т. е.
двойственность элементов в каждом
сегменте, у Holosteoidei утрачивается,
по следы ее, указывающие па родство
высших рыб с примитивными хряще-
Рис. 60. Боковой вид предхво-
стовых позвонков осетра (Aci-
penser sturio). (по Bridge)
1 — хорда, 2*—оболочка хорды,
3 — спинномозговой канал, 4 —
краниальная дуга (interdorsale),
5 — каудальная дуга (basidorsale),
6— поперечный отросток, 7— ниж-
няя каудальная дуга (basiventra-
1е), 8 — нижняя краниальная ду-
га (interventrale), 9 —канал для
аорты, 10 — остистый отросток,
11 — ребро.
выми, встречаются у некоторых пред-
ставителей. Например, у представи-
теля Holostei — Amia calva основа-
ния дуг в хвостовой области являются
хрящевыми, имеются и верхние и
нижние каудальные и краниальные
дуги (basidorsalia и intcrdorsalia, Ьа-
siventralia и interventralia), а в зад-
нем отделе хвостовой области па каж-
дый сегмент приходится еще два тела
позвопка (рис. 61). Подобный тип был широко распространен у
ископаемых рыб.отряда Amioidei.
Рис. 61. Amia calva, хрящевая стадия. Вид с левой сто-
роны на передний (Л) и задний хвостовой (В) отделы
позвоночника (по Goodrich.)
1—центр, 2-—верхняя краниальная дуга (interdorsale), 3 —
верхняя каудальная (basidorsale), 4—нижняя каудальная дуга
(basiventrale), 5 — передний полупозвонок, 6 — задний полупо-
вр.онок, 7 — нижняя краниальная дуга (Interventrale), 8 — меж-
сегментальный сосуд, 9 *— отверстие для брюшного корешка,
10 — отверстие для спинного корешка.
,87
Следы парности дуг встречаются и у костистых рыб (Teleostei),
но выражено это здесь гораздо слабее. Здесь при развитии хрящевая
стадия опускается, и кость развивается непосредственно, минуя
хрящ. Обычно позвопок окостеневает как целое, по у примитивных
Teleostei (Cypriniformes, Esociformes, Clupeiformes) верхние и ниж-
ние дуги могут окостеневать раздельно. Таким образом, мы видим,
что общая тенденция развития позвоночника — это тенденция
к созданию прочного костпого позвонка из скелетогепного слоя ме-
зодермы.
Типичный позвонок состоит из тела позвонка, вогнутого спереди
и сзади, пронизанного топким отверстием для прохождения хорды.
Верхние дуги охватывают спинномозговой капал; срастаясь навер-
Рис. 62. А —боковой вид двух туловищных
позвонков Esox lucius, В—вертикальный раз-
рез туловищного позвонка, С—позвонок из
передней туловищной области, D— из задней
туловищной, Е—из хвостовой области Thyn-
nus thynnus.
ху, они образуют ости-
стый отросток. У осно-
вания дуг отходят не-
большие сочленовные от-
ростки для соединения
позвонков друг с дру-
гом. В области тулови-
ща от тела позвонка от-
ходят поперечные отро-
стки, соединяющиеся с
ребрами; в хвостовом
отделе эти отростки,
п ред ста в л я тощие ниж-
ние дуги, загибаются
вниз и срастаются меж-
ду собой при помощи
нижнего остистого отро-
стка, образуя гема,ль-
иый канал, где ирохо-
1 —центр, 2 — нижняя каудальная дуга' (basivent-
rale), 3—верхняя краниальная, 4 — нижняя кра-
ниальная дуга, 5 — эпиплевральнос ребро, 6 —
эпиневральное ребро, 7 — продольная связка, 8 —
левая невральная дуга, (верхняя каудальная), 9 —
ее хрящ (краниальная дуга, interdorsale?), 10 —
хорда, 11 — плевральное ребро, 1 2 — спинномоз-
говой нанал. (Из Goodrich.)
дят сосуды. С позвонка-
ми соединяются ребра.
Последние бывают двоя-
кого рода: одни лежат
в горизонтальной мио-
сснте, разделяющей
спинную и брюшную мускулатуру, — это так называемые в е р х-
н и е ребра; другие охватывают полость тела — н и яс н и е
ребра. Первые закладываются в горизонтальной миосепте,
вторые — в поперечных; они отличаются еще направлением роста.
Тонкие межмускульпые, или «мясные», косточки лежат у кости-
стых рыб в миосептах верхней части туловищной мускулатуры.
У различных рыб ребра развиты различно (рис. 62).
Череп. Сегментация головы. Мы видели, что предки современ-
ных Craniata имели явственно выраженную сегментацию во всем
теле. Эта первичная сегмеитироваппость имеется и у ланцетника.
Она выражена и в туловищном, и хвостовом отделах круглоротых
и рыб. Но в голове уже у круглоротых мы пе видим ее при первом
«8
взглн/ш. Скелет головы, череп не обнаруживает той сегментации,
что ясно выражена в первичной диплоспондилии позвоночника.
Сегментация мускулатуры также не выражена в голове, и нервы
ro.iioin.i иные, нежели в туловище. У рыб череп достигает еще боль-
шего развития, и сегментация совсем неясна. Таким образом можно
думаti>, что голова построена по иному типу, нежели туловище,
что она. пе сегментирована или сегментация выражена только в жа-
огрпых дугах. Однако тщательное исследование головы сравни-
тельно анатомическим, а главным образом эмбриологическим мето-
дом доказало, что голова Craniata является построенной из ряда
сегментов, совершенно подобных сегментам туловища, и что сег-
ментация эта первоначально простиралась до переднего конца тела,
как у Amphioxus. С переходом к активному питанию, с развитием
более совершенного жаберного аппарата и с появлепием сердца
у Protocraniata движения последних стали более энергичными; на
переднем конце тела стал развиваться головной мозг, развились
органы чувств; становилось полезным укрепление переднего конца
тела; понадобилась защита важных органов, здесь находящихся.
Эту роль выполняет развивающийся тут скелет. Материалом для
него послужила та же перихордальная соединительная ткань,
которая в туловищном отделе дала парные верхние дуги, а затем
и позвоночник. Эта перихордальная ткань в туловище происходит
из склеротомов первичных позвонков, иначе говоря, из спинной
сегментированной мезодермы. Эта мезодерма продолжается па ран-
них стадиях развития и в область головы. И здесь опа также сегмен-
тирована, состоит из сомитов, а каждый сомит из миотома, двой-
ного склеротома и пристенной части. Различие от туловища заклю-
чается в том, что здесь склеротомы не образуют парных дуг, дипло-
снопдилия выражена здесь в самой двойственности склеротомов,
последние же очень рано сливаются между собой, образуя основание
будущего черепа так называемые парахордальные х р я-
щи (parachordalia). Так как подвижности, как в позвоночнике,
здесь пет, то мышцы являются ненужными, часть миотомов исчезает,
мпотомы трех сегментов дают начало мышцам, двигающим глаз.
В туловище каждому метамеру соответствует пара спинномозго-
вых нервов, каждый нерв со спинным, смешанным по функции ко-
решком, и брюшным—двигательным. Последний снабжает туловищ-
ную мускулатуру, происходящую из миотома. В спинном кореш-
ке есть чувствующие волокна двоякого рода и двигательные к мыш-
цам внутренностей (висцеральным). Корешки нервов у Amphioxus
раздельны, у Cyclostomata тоже, у рыб и высших позвоночных они
соединяются в туловище в один общий смешанный нерв. В голове
же они раздельны, существуют как отдельные нервы, одни из кото-
рых смешанные, другие — двигательные. Следовательно, в голове
сохраняется более примитивное состояние, чем в туловище. Таким,
образом, область головы первоначально была сегментирована,
голова образовалась в результате слияния передних сегментов
первоначально однородной серии сегментов, в результате чего перво-
начальная сегментация исчезла. Тщательное изучение развития
89
головы у эмбрионов показало, что в состав головы вошло 11 сег-
ментов (рис. 63). Позднее туловищные сегменты с верхними дугами
врастали в череп, сзади.образуя его затылочный отдел у разных по-
звопочпых в различном числе, так что задняя граница головы не
у всех позвоночных тождественна, — она непостоянна. Границей
головы в настоящем смысле слова является десятая пара нервов
(nervus vagus). У круглоротых последний и составляет границу го-
ловы, у других позвоночных граница черепа лежит позади vagus,
так как различное число сегментов вошло здесь в череп.
Рис. 63. Реконструкция зародыша акулы Pristiurus
melanostomus, изображающая сагиттальный разрез
через голову зародыша, видимый с внутренней
стороны.
1 — мозговая трубка, 2 —кишечник (оба органа вскрыты),
3 —слуховая капсула, 4 — миотом, 5 — полость сомита,
6—склеротом, 7 — боковые пластинки сомитов, 8 — спи-
ракулярный жаберный мешок, 9, и 9а — первый и второй
жаберные мешки, 11м, П8, Ш, V, VII, VIII —первый,
второй и т. д. сомиты. (По Матвееву.)
Тазвитие черепа. Хрящевой череп (Chondrocranium). Эволюция
черепа подобна эволюции позвоночника. У пизших форм рыб череп
хрящевой, у высших хрящ заменяется костью. У зародышей выс-
ших рыб он закладывается первоначально в виде мезенхиматозных,
а затем хрящевых элементов (рис. 64), и чем выше та или иная
группа рыб в системе, тем меньше остается в черепе хряща.
^Начало развитию черепа кладет появление в мезенхиме по бокам
переднего конца хорды происходящих из склеротомов продолго-
ватых хрящевых пластинок — околохордовых хря-
щей (parachordalia). Впереди от пих развивается еще одна пара
хрящей, называемая перекладинами черепа (tra-
beculae cranii). По бокам головы закладывается еще пара хрящей —
боковые хрящи. В соседстве с передним отделом парахор-
далий развиваются вокруг органа слуха слуховые кап-
су л ы, а вокруг органов обопяпия— обонятельные кап-
сулы, связанные обычно с передним концом трабекул. В задней
части парахордалий видны явственные следы вхождения в задний
отдел черепа некоторого числа дуг позвоночника. Впереди от хорды,
между парахордальными хрящами и трабекулами, помещается под-
мозговая железка, или гипофиза. (hypophysis cerebri).
90
1 {дальнейшем парахордальные хрящи срастаются с задними кон-
цами трабекул,срастаются между собой, обрастая хорду; срастаются
между собой и передние копцы трабекул. В результате полу-
чается хрящевая плас-
тинка, подстилающая
мозг и являющаяся
дном черепа. Впереди
хорды в этой пластинке
имеется, иногда всю
жизнь, отверстие — п е-
редпяя фонта-
пель основания
черепа (fenestra Ъа-
sicranialis anterior). Упо-
мянутые выше капсулы
очень рано сливаются с
основанием черепа, так
же как и боковые хря-
щи.Хрящевое основание'
черепа разрастается
вверх, образуя боковые
степки черепа. А затем
последние срастаются
между собой наверху,
Рис. 64. Диаграмма, иллюстрирующая разви-
тие черепа Craniata.
А — более ранняя, В — более поздняя стадии. 1 —
слуховой пузырен, 2 —глазной пузырек, 3 — носо-
вой мешон, 4 — хорда, 5 —• позвоночный склеротом,
6 — парахордальная область, 7 — трабекулы чере-
па, 8 —носовая капсула, 9 — глазная капсула,
10 — слуховая капсула, 11 — затылочный сегмент,
или склеротом, 12 — передняя основная фоптанель.
(По Goodrich.)
тканью, у высших закрыто костями.
Рис. 65. Реконструкция головы зародыша
Acanthias.
1 —боковой хрящ, 2 — трабекула, F3 — пара-
хордальный хрящ, 4 — челюстная дуга, 5 —
гиоидная дуга, 6 — жаберные дуги, 7 — слу-
ховая капсула, 8, 9, 10, 11 —5-й, 7-й, 9-й,
10-й черепные нервы, 12 — спинномозговой
ганглий. (По Северцову из Goodrich.)
пад головным мозгом.
Так образуется крыша черепа, которая обычно бывает неполной;
остающееся в крыше отверстие бывает затянуто у низших рыб
(хрящевых) соединительной
Под черепом, независи-
мо от пего, в .мезодерме
боковых пластинок, закла-
дываются висцераль-
ные дуги (рис. 65), раз-
деленные висцеральными
щелями. С сомитами головы
висцеральные дуги не сов-
падают. Дуги охватывают
передний отдел пищевари-
тельного канала и распола-
гаются между жаберными
щелями. Сперва они лежат
подчерепным скелетом, по-
том сдвигаются благодаря
неравномерному росту на-
зад в область туловища.
Передняя пара является наиболее развитой, служит поддержкой
для стенок рта и называется челюстной дугой. Вторая,
тоже сильно развитая, называется гиоидной, или подъ-
язычной, а далее назад лежат обычно пять ж а б е р п ы х
91
Рис. 66. Средний продольный разрез
через neurocranium Heptanchus.
II, III, IV, V и т. д. — отверстия для
черепных нервов. 1, 2, 3 и т. д.—отвер-
стия затылочных окципитальных нервов,
пронизывающие стопку neocranium. (Из
Ihle и др.)
пом — palaeocranium, а часть
дуг. Впереди челюстной дуги, точнее по сторонам ее, лежат еще
иногда хрящевые элементы, называемые губными хрящами,
два с каждой стороны в верхней и одип в пижней половине.
Часть черепа, заключающая в себе головной мозг, называется
осевым черепом (neurocranium), часть, развивающаяся
вокруг переднего отдела пи-
щеварительного канала,—в и с-
ц е р а л ь п ы м черепом
(splanchnocranium). В neuro-
cranium различают затылочный,
слуховой, глазничный и обо-
нятельный отделы. Ту часть
черепа, которая лежит вокруг
хорды и развивается из пара-
хордальных хрящей, называют
хордальным черепом.
Часть черепа, которая лежит
впереди от десятой пары нервов,
называют древним ч е р е-
затылочную, развивающуюся из
позднее вошедших в череп позвонков, — и о в ы м ч е р с п о м—
neocranium (рис. 66).
Верхняя половина челюстной дуги носит название небно-
квадратного хряща— palatoquadratum, нижняя, играю-
з
Рис. 67. Боковой вид черепа акулы (Scyllium canicula).
1 — ростральные хрящи, 2 — обонятельная капсула, 3 — орбита, k —
слуховая капсула, 5 — небноквадратный хрящ, 6 — меккелев хрящ, 7 —
губные хрящи, 8 — гиомандибулярный хрящ, 9 —гиоидный, 10 —жа-
берные дуги, 11 — жаберные лучи, 12 —так называемые наружные
жаберные дуги (extrabranehlaUa.) (Из Видерсгсйма по Паркеру.)
щая у хрящевых рыб роль нижней челюсти, — меккелева
хряща (cartilago Meckeli). Гиоидная дуга и жаберные дуги
распадаются на четыре отдела: pharyngo-, epi-, cerato- и hypohyale,
или branchialia. Дуги одной стороны соединяются внизу с дугами
другой при помощи непарных элементов, называемых basihyale
и basibranchialia, или copulae. В гиоидной дуге обычно два сред-
92
пне отдела бывают сильно развиты и называются: верхний —
г it о м а п д и б у л я р
п д л ы м (hyoideum)
(рис,. 67).
X рящсвой череп в
целом носит пазва-
11 не—chondrocranium.
Число жабер-
ных дуг бывает
различно. У низших
акул их бывает до
семи, обычно же их
пять. Закладываются
они независимо от че-
репа, по потом всту-
пают с ним во вто-
ричную связь при
помощи связок или
непосредственно прн-
членяясь. Особенно
тесно связан бывает с
черепом верхний ко-
нец 11 у от and i bu 1 are.
Il e б и о к в ад pa т-
н ы й х р я щ при-
Рис. 68. Диаграммы: А п D — амфистилический
черен (Ileptanchus)' С и В— гиостилический
череп (Scyllium), Л и С—вид сбоку, D и В —
вид сзади
крепляется к черепу
различно у разных
рыб. У большинства
1 — палатокпадратпый хрящ, 2 —меккелев хрящ,
3—гиомандибулярный, 4--гиоидный хрящи, 5—брыз-
гальце (spiracuhnn). (По Goodrich.)
рыб (акул, костистых, галоидов) гиомандибулярный хрящ крепкой
связкой соединен с задней частью иебиоквадратпого и меккелева
хрящей и служит для прикре-
пления челюстей к черепу,
являясь подвеском. Такой
череп называется г и о с т и-
л и ч е с к и м (рис. 68). В
других, более редких слу-
чаях (у примитивных акул,
как Notidanus, Heptanchus)
пебпоквадратпый хрящ соеди-
няется с черепом непосред-
ственно, а кроме того, к зад-
Рис. 69. Callorhynchus antarcticus. Ске-
лет головы. Пример аутостилического
черепа.
1 — палатокпадратпый хрящ, *'2 — меккелев
хрящ, 3 — гиомандибулярный и 4—гиоид-
ный хрящи. (По Goodrich.)
нему краю челюстей связками
прочно прикрепляется гно-
ма пдибулярпый хрящ. Такой
череп с двойной связью челю-
<• гей с черепом пЬсит пазва-
пin' а м ф и с т и л и ч е с к о г о. У цельпоголовых (Holocephali),
а также у двоякодышащих (Dipnoi) пебпоквадратпый хрящ сра-
стается только с основанием черепа, и гиомандибулярный хрящ
93
не принимает никакого участия в подвешивании челюстной дуги.
Такой череп называется аутостилическим. Этот тип
развивается вторично в связи с более прочной и крепкой поддерж-
кой для массивных зубов (рис. 69).
Между висцеральными дугами находятся жаберные щели. Пер-
вая жаберная щель, находящаяся между челюстной и подъязычной
(гиоидной) дугой, называется брызгальцем.
Типы окостенений черепа. В хрящевом состоянии череп нахо-
дится у хрящевых рыб (Chondrichthyes). У Osteichthyea
в черепе, как и в позвоночнике, появляются окостенения. Сперва
окостенения черепа только дополняют chondrocranium, но у выс-
ших рыб кость совершенно вытесняет хрящ, и тогда вместо chon-
drocranium мы видим костный череп, или osteocranium. Замена
хряща костью является весьма прогрессивным явлением. Кость
прочнее хряща, ее сопротивляемость давлению в 7 раз больше,
нежели у хряща. Поэтому костные скелетные части при меньшем
объеме дают тот же результат. Особенно большое значение имеет
это для наземных животных, но и для рыб не лишено значения.
У рыб, имеющих костный скелет, закладывается он в виде хряща.
Кости, которые участвуют в образовании костного черепа,
двоякого рода: покровные, или кожные окостенения,
и замещающие их хрящевые. Первые называются
еще вторичными, а вторые — первичными. Такое название не-
правильно, так как покровные кости появляются в процессе фи-
логенеза впервые, раньше замещающих, и с этой исторической точки
зрения правильнее было бы их называть первичными, а появляю-
щиеся позднее, у высших рыб, замещающие окостенения — назы-
вать вторичными. Покровные кости развиваются в соединительной
ткани кожи независимо от хряща, па который они только налегают,
отделенные от пего прослойкой соединительной ткани. Но посте-
пенно кости вступают с хрящом в более тесную связь. Первый шаг
к этому заключается в том, что кость развивается в том слое соеди-
нительной ткани, который окружает хрящевые части, в над-
хрящнице (perichondrium), и теспо налегает па хрящ. Такое
окостенение носит название перихондрального. Сле-
дующий этап развития заключается во врастании кости в хрящ:
хрящ растворяется, соединительная ткань врастает в образующиеся
пространства и там отлагает кость. Это так называемое эндо-
хондральное окостенение: оно замещает собой хрящ.
Кость является прсформироваппой хрящом.
Каждая кость в скелете имеет свою историю. Раз возникши,
опа подвергается затем различным изменениям, принимает ту или
иную форму, вступает в связь с другими костями, но пока опа су-
ществует, ее можно сравнивать с тем скелетным элементом предков,
из которого опа возникла. Тождественность, или г около гия,
костей устанавливается по соотношению их с другими костями,
по отношению к выходящим из черепа нервам и сосудам, к так на-
зываемым слизистым каналам, а также по отношению к частям
хрящевого черепа chondrocranium, на которых кость развивается.
94
Зтим методом можпо проследить историю каждой кости в процессе
филогенеза. При этом изучение оптогепетического развития и срав-
нение е ископаемыми формами весьма помогают установлению го-
мологий. У хрящевых рыб окостенение представлено лишь плакоид-
1И.1МП чешуями, которые простираются и на голову. Сходные зуб-
чики покрывают голову более примитивных Ostracodermi. У Ostei-
rhlhyes костный покров головы первоначально имел огромное
сходство с чешуями туловища.
’I «л и у и и пластипки на голове
примитивных Osteichthyes обра-
зовывали первопачальпо сплош-
ной покров. Отверстия остава-
лись для глаз, ноздрей, рта,
для непарного или теменного
глаза, для жаберных отверстий.
Путем разрастания или путем
слияния пластинки станови-
лись крупнее и плотнее соеди-
нялись друг с другом. В ре-
зультате получились определен-
ные окостенения, судьбу кото-
рых можпо проследить у всех
рыб и даже у высших позво-
ночных.
Покровные окостенения в
черепе рыб. На рис. 70 виден
общий план покровных окосте-
нений гипотетической прими-
тивной рыбы. То, что мы видим
у различных представителей со-
временных и ископаемых Oste-
ichthyes, является видоизмсне-
Рис. 70. Диаграмма основного плана
расположения покровных костей
черепа у рыб. Вид сверху па череп
примитивного представителя костных
рыб (Osteichthyes).
1 — ethmoideum, 2 —- nasale, 3 — frontale,
4—parietale, 5 — interfrontale, 6—inter-
nasale, 7 — supraoccipitale dermale, 8—
pracmaxillare, 9—maxlllarc, 10 —prae-
frontalc, it — lacrinialc, t2 —postfrontale,
13 — postorbitale, 14 — jngalc, 15 — squa-
mosum, 16— praeopereulum, 17 —oper-
culum, 18 — postparietale, 19. — tabulare,
(По Goodrich.)
пнем этого плана с теми или
иными особенностями. Крыша черепа образована парными
носовыми костям и (nasalia), лобными (frontalia)
и теменными (parictalia). Глазница окружена предлоб-
ной (praef гоп tale), заднем о б п о й (postfrontalc), слезной
(lacrimal е), з а д п е г л а з и и ч и о й (postorbitale) и скудо-
11 ° ft (jugale) костями. II р е д ч е л ю с т и ы е (praemaxillaria)
и челюстные (maxillaria), несущие обычно зубы, укреп-
ляют верхнюю челюсть. Задний край затылочной области у неко-
торых ископаемых форм был образован пепарпой средипной
з а т ы л о ч и о й (dermsupraoccipitalc) и парными позади-
т е м и пным и (postparietalia и tabularia). По бокам теменной
области над местом. занятым челюстными мышцами, располагаются
щечные» кости, — чешуйчатая (squamosum) и квадрат-
и о - <• к. у л о в а я (quadrato-jugale). Последняя кость прикры-
вала, заднюю, квадратную, область небноквадратного хряща.
В нижней челюсти имеется зубная кость (dentale) с зу-
95
Рис. 71. Череп лосося (Salmo salar), разрезан-
ный вдоль.
1 — basioccipitale, 2 — exoccipitale, 3 —supraoccipitale,
4—baslsphenoideum, 5—prooticum, 6—alisphenoideum,
7 — orbitosphenoidcum, 8 — parasphenoideum, 9 — vo-
mer, 1 0 — praemaxilla, 11—maxilla, 12 — palatinum,
13 — pterygoideum, 14 — mesopterigoidcum, 15 — me-
tapterigoideum, 16 — hyomandibularc, 17 — symplec-
ticum, 18 — stylohyalc, 19 — quadratum, 20 —dentale,
21 — меккелев хрящ, 22 — articulate, ,23 — angulare.
(По Goodrich.)
лес постоянными, нежели окостенения
они развились в качестве поддержки
бами; большая угловая (angulare) прикрывает область сочле-
нения нижней челюсти, простираясь и назад, с внутренней сторо-
ны — н а д у г л о в а я (supraangulare), spleniale, postspleniale,
п р (д с о ч л с п о в п а я (praearticularc), сочленовная (articu-
lare). Над hyomandibularc находится п р е д к р ы ш е ч и а я
к ость (praeoperculum), несколько костей поддерживают складку,
прикрывавшую жаберные щели: крышечная (operculum),
подкрышечная (suboperculum) и межкрышечная
(interoperculum), а также боковые gularia. Непарные срединные
gularia заполняют у некоторых рыб между ними промежуток.
Иногда встречается
еще — р остраль-
пая кость (rost-
ral е), между межче-
люстными и носовы-
ми—м ежи ос о вая
(internasalc), между
nasalia — между-
л о б п а я (intcrfron-
tale). Конечно, в раз-
ных отрядах рыб мы
видим различные ук-
лонения от этой пол-
ной схемы покров-
ных окостенений кры-
ши и боков черепа.
Кроме перечислен-
ных покровных око-
стенений, последние
имеются и па небе, и
па нижней стороне
черепа. Эти окосте-
нения являются бо-
свода. Первоначально
для зубов. Впослед-
ствии зубы могли и исчезнуть. Подстилая дно черепа снизу, ио
средней линии лежит парасфеноид (parasphcnoidcum).
Впереди парасфеноида лежит парный, а в других случаях непар-
ный сошник (vomer). Непарный происходит, видимо, из пар-
ного. На переднем конце palatoquadratum (рис. 71) развивается
кожное окостепепие, песущее обычно много зубов — к о ж и о е
пебпое окостенение (dermpalatinum). Кзади от него
па том же пебноквадратном хряще развиваются три к р ы л о в и д-
н ы х кости (pterygoidea). Из них две передних — endopterygoi-
deum и ectopterygoideum — являются покровными и часто несут
зубы. Третья — metapterygoideum — является преформирован-
пой хрящом.
Замещающие хрящ окостенения в черепе рыб. Среди Osteich-
thyes хрящевой череп, chondrocranium, пе у всех исчезает в одипа-
96
icoimli степени. У низших форм, как у Chondrosteoidei, он сохра-
нился еще полностью, у многих Teleostei, например, у карпа (Су-
pri пня earpio), от него не остается ничего: хрящ замещается костью,
•л 1/1 о хондральными окостенениями. Параллельно с этим уменьшает-
ся количество покровных окостенений. Последние были наиболее
p.TiiiiiTij у низших, древнейших представителей класса, эндохонд-
ральные же — в черепе высших представителей Teleostei.’ Иногда,
впрочем (у Dipnoi), происходила редукция эндохондральных око-
гтг|к‘пий у высших представителей подкласса и частичная реста-
врация chondrocraniuni.
Рис. 72. Продольный разрез черепа сазана (Cyprinus carpio).
1 — basiocclpitale, 2 — exoccipitale, 3 — supraoccipitale, 4 — prooticum, 5 — epio-
tlcum, 6 — alisphenoideum, 7 — orbithosphenoideum, 8 — parasphenoideum, 9 —
vomer, 10 — mesethmoldeum, 11 — rostrale, 12 — maxilla, 13 — praemaxilla, 14 —
dentale, 15 — articulare, 16 — quadratum, 17—pterygoidea, 18—urohyale, 19 —
иижнеглоточные вубы, 20—жаберные дуги, 21—radii branchiostegii. (По Good-
rich.)
В развитом osteocranium (рис. 71 и 72) мы имеем следующие за-
мещающие кости. В затылочном отделе основная затылочная
(hasioccipitale) окружает хорду, у взрослых особей здесь исчезаю-
щую, и на своей задней поверхности имеет вогнутость, как у позвон-
ков; с позвоночником связана посредством связки, без сочленения.
Две боковые затылочные (exoccipitalia, или occipi-
la lia latetaHa) охватывают затылочное отверстие, пронизаны рядом
отверстий для вошедших в череп с позвонками спинномозговых
нервов и ограничивают сзади отверстие для десятой пары нервов
(в. vagus), определяющей заднюю границу palaeocranium. Над за-
тылочным отверстием лежит верхняя затылочная
к петь (supraoccipitale). В слуховой области имеется пять око-
стенений’ наиболее постоянной является передняя слу-
ховая кость (prooticum), пронизанная отверстием для ли-
цевого нерва (n. facialis) и каналом для вены. Prooticum защищает
Киипшров л Стшталский 97
передний полукружный канал. Позади лежит зад неслухо-
вая (opisthoticum), весьма непостоянная и различно развитая.
Верхняя часть слуховой области занята двумя костями: к л и-
нослуховой (sphenoticum) и крылослуховой (ptero-
ticum). В верхне-заднем углу слуховой области, над задним полу-
кружным каналом, лежит верхняя слуховая кость
(epioticum).
Впереди от prootica в основании черепа лежит основная
клиновидная кость (basisphenoideum). Эта кость уже
не принадлежит хордальному черепу, а развивается на трабекулах.
Иногда эта кость отсутствует или сливается с парасфеноидом.
С основной клиновидной соединены парные к р ы л о - к л и н о-
видные (alisphenoidea), лежащие между отверстиями для зри-
тельных нервов и передними слуховыми. Эти кости участвуют в об-
разовании боковой стенки черепа. Впереди от отверстия для зри-
тельного нерва развиваются парные глазнично-клипо-
видные кости (orbitosphenoidea), сливающиеся затем у
позднейших рыб в межглазничпую перегородку. Впереди глазниц
на так называемом предглазничном хряще развивается боковая
этмоидная кость (ethmoideum laterale). На rostrum за-
кладывается средний этмоид (mesethmoideum), распро-
страняясь па носовые хрящи и носовую перегородку.
В областипебноквадратного хряща впереди развивается неб-
ная кость (palatinum), в середине упомянутая заднекрыло-
видная metapterygoideum, и пазади, в месте сочленения с нижней
челюстью — квадратная (quadratum). Упомянутая выше
покровная небная кость срастается с хрящевой palatinum, и тогда
небная кость оказывается смешанной. Такими же смешанными ко-
стями являются sphenoticum, в образовании которой принимает
участие покровная postfrontale, и pteroticum, образующаяся
с участием squamosum. В задней части меккелева хряща раз-
вивается замещающая кость — сочленовная (articulare).
В подъязычной дуге мы видим кости, соответствующие хрящевым
частям: hyomandibularc в верхнем отделе дуги причленяется к пиж-
нему краю pteroticum (squamosum). От нижнего конца hyoman-
dibulare отходит к квадратной кости symplecticum, а небольшая
кость interhyale служит для соединения hyomandibularc с hyoi-
deum — пижпим отделом дуги. Впизу имеется еще hypohyale.
В жаберных дугах мы видим pharyngo-, epi-, cerato- и hypobranchi-
alia, связанные впизу посредством костпых copulae.
Эволюция черепа в разных отрядах рыб. Череп рыб является
хорошим примером эволюции органа. Общее направление эволю-
ции черепа — постепенная замена хрящевого черепа костным.
Сростножаберпые (Elasmobranchii) имеют череп хря-
щевой. Chondrocranium образует вокруг головного мозга сплошную
хрящевую коробку (рис. G6). У большинства акул впереди от че-
репа отходит вырост — rostrum, различия в величине и форме кото-
рого составляют отличия семейств. Рот никогда пе бывает па конце,
а как он, так и носовые отверстия находятся на нижней стороне
98
головы. Череп широкий, платибазальный, т. е. трабекулы у за-
родыша широко расставлены, и мозг у взрослых животных зани-
мает в черепе и глазничную область. В череп сзади вошел ряд по-
звонков, имеются отверстия для затылочных нервов. Обычно череп
гиостилический (рис. 68, с), и hyomandibulare служит подвеском.
У родов Hexanchus и Heptanchus, примитивных акул из Notidani,
череп амфистилический (рис. 68, А), также и у вымерших Асап-
thodii. По бокам рта имеются губные хрящи. Небноквадратные
хрящи сходятся своими передними концами под черепом. Жабер-
ных дуг нормально бывает пять, у представителей Notidani —
Hexanchus и Chlamydoselachus их шесть, а у Heptanchus — семь.
У скатов верхние концы жаберных дуг сливаются с позвоночным
столбом. На наружной стороне жаберных дуг сидят тонкие хря-
щики— жаберные лучи.
Рис. 73. Скелет черепа осетра (Acipenser) по уда-
лении частей наружного скелета.
1 —rostrum, 2—parasphenoideum, 3— позвонки, вошедшие
в состав черепа, 4 — palatoquadratum, 5 — quadratum,
6 — mandibula, 7 — hyomandibulare, 8 — symplecticum,
9 — interhyale, 10 — hyoideum, I, II, III — жаберные дуги.
(Из Видерсгейма по Goodrich.)
У це льпог о л о в ы х (Holocephali, рис. 69) череп также
хрящевой, chondrocranium полный и замкнутый. Характерной
особенностью черепа является полное слияние пебпоквадратного
хряща с черепом, череп автостилический.
Это находится в связи с тем, что Holocephali питаются ракооб-
разными и иглокожими, имеющими твердый панцырь, который опи
должны раздавливать. В соответствии с этим измепепы и зубы.
Губные хрящи развиты сильно.
Из рыб с настоящим ртом (Teleostomi) наиболее примитивен
и более всего приближается к элазмобрапхиям череп Chondrosteoi-
dei. Череп представлен хорошо развитым chondrocranium. Rostrum
развит необычайно сильно. Сзади череп продолжается далеко на-
зад, незаметно переходя в позвоночник, так как в пего входит це-
лый ряд туловищных сегментов (рис. 73). Последнее связано с тем,
что Chondrosteoidei пользуются rostrum для взрывания ила на
дне. Это требует укрепления черепа.
У Chondrostei уже имеются покровпые окостенения. Сильно
развитый parasphenoid поддерживает весь череп снизу, имеется
сошник (vomer), который сдвинут вперед к rostrum. Небноквад-
ратные хрящи сходятся передними концами и пе соединяются самп
7* 99
с черепом, и весь челюстной аппарат, развитый слабо, соединен
с черепом посредством подвеска (hyoman^ibulare). Окостенения
развиваются также в челюсти, в гиоидной и жаберной дугах, не
вытесняя хряща. Сверху череп покрыт многочисленными покровны-
ми костями. Они залегают в коже весьма поверхностно, и назади
переходят в туловищные щитки. Впереди, па rostrum, кожные око-
стенения мелки и не могут еще быть сравниваемы с покровными
костями других Osteichthyes.
Череп Holosteoidei представляет в сравнении с Chondrosteoi-
dei значительный шаг вперед. У современных Cfossopterygii (Ро-
lypterus) chondrocranium сохраняется, но не полностью: свод его
замкнут лишь в этмоидной и затылочной областях, образуя по-
середине большую фонтанель. Имеется фонтанель и в основании
черепа. Однако в черепе имеются уже замещающие хрящ окосте-
нения. Вокруг затылочного отверстия развивается одно затылоч-
ное окостенение. В слуховой области имеются opisthotica, сливаю-
щиеся с epiotica, маленькие prootica и sphenotica. В глазничной
области дно и боковые стенки черепа образованы своеобразным око-
стенением — sphenethmoideum, занимающим место alisphenoidea
и orbitosphenoidea.
Покровные окостенения у Polypterus' лежат еще поверхностно
и назади переходят в чешуи туловища. Основное отличие от по-
кровных окостенений высших рыб (Actinopterygii) заключается
в том, что некоторые элементы сливаются между собой, другие
представлены большим количеством маленьких дополнительных
пластинок.
'Под нижней челюстью на горле находятся большие срединные
горловые пластинки (jugularia).
О замещающих костях ископаемых представителей Crossop-
terygii мы знаем еще очень мало. Покровные кости в этой группе
сходны были в общем с тем, что мы видим у Polypterus, по более
многочисленны и образовывали сплошной папцырь вокруг головы.
У Holostei (Amia и Lepidostcus) строение черепа подни-
мается па еще большую высоту и приближается к строению черепов
у низших Teleostei. Ghondocranium редуцирован гораздо сильнее;
количество замещающих окостенений значительно больше ..Покров-
ные окостенения у Amia в основном уже сходны с таковыми кости-
стых рыб; главнейшие отличия заключаются в отсутствии неко-
торых костей (нет, например, supraoccipitale) и в существовании
некоторых дополнительных костей.
В сверхотряде Teleostei (рис. 71 и 72) мы видим
наивысшее развитие osteocranium. Общий план его, однако, вы-
водится из такового у Holostei. У наиболее примитивных предста-
вителей chondrocranium еще сохраняется в значительной степени
(например,у Salmo), покровные кости обособлены и легко снимаются
с chondrocranium. Но у высших представителей (Gadiformes, Рег-
ciformes и др.) кость совершенно замещает хрящ, и покровные око-
стенения врастают в хрящ и срастаются с замещающими костями
настолько, что лишь с трудом могут быть отделены от него. Осо-
100
бенпостью черепа Teleostei является слияние в некоторых случаях
pharyngo-branchialia, образующих верхнеглоточные ко-
сти. Против них снизу располагаются части недоразвитой пятой
дуги — так называемые нижнеглоточные кости. На
жаберных дужках сидят иногда очень сильно развитые жаберные
тычинки, образующие сито для процеживания микроорганизмов.
Особняком стоит череп двоякодышащих (Dipnoi).
У древнейших ископаемых представителей этого подкласса
(например, у Dipterus) chondrocranium был, видимо, в значительной
А в
Рис. 74. Ccralodus forsleri.
А —вид черепа сверху. В —вид черепа снизу; слева—нижняя
челюсть удалена и жабры разрезаны поперек. 1—ethinoideum, 2 —
задняя срединная н.тастйпна (затылочпая), 3 — боковая пластинка,
4 — pteroticum, 5 — auborbitaie, 6 — operculum, 7 — небный зуб, 8 —
palatoptesygoidcum, 9 — parasphenoidcuni. (По Goodrich.)
мере замещен костями; покровные кости были многочисленны
и лежали сперва поверхностно; некоторые из них сливались в щиты.
У позднейших форм количество костей начинает убывать. Череп
современных Dipnoi имеет хорошо развитый chondrocranium
(рис. 74). Череп платибазальиый. Хрящевые окостенения у Dipnoi
тоже редуцированы: единственными замещающими костями яв-
ляются боковые з а т ы л очи ы е (exoccipitalia). Покров-
ные окостенения тоже развиты неполно. На крыше черепа имеются
две попарные кости: пер о д и я я срединная и за д-
п я я средника я и четыре парных. Сравнение пх с покров-
ными костями Teleostei затруднительно. Жаберпая крышка поддер-
живается посредством operculum и inter operculum. Подъязычная
и жаберпая дужки остаются хрящевыми. В череп вошли три ту-
ловищных позвонка. Из них задний еще несет развитые ребра,
называемые «головными ребрами». Наиболее резкая особенность,
101
М/Л
Рис. 75. Видоизменения хвостового плавника
и отношения эндоскелетных лучей (radialia)
к осевому скелету.
А — Протоцеркальный и дифицернальный хвост
(Dipnoi), В — Гетероцеркальный (Selachii), С — из-
мененный дифицернальный (Coelacanthini), В—гете-
роцеркальный (Chondrostei), В —гомоцеркальный
тип (Teleostei), F — скрытый гетероцеркальный
тип (Amiotdei). (По Goodrich.)
отличающая Dipnoi от других Osteichthyes, — это полная а в то-
ст и л и я черепа. Небноквадратный хрящ прочно сливается с че-
репом впереди и пазади. Hyomandibulare недоразвито или отсут-
ствует Развитие автостилии вызвано здесь, повидимому, развитием
специфических для Dipnoi больших и сложных пебпых зубов. Зубы
эти, видимо, унаследованы от очень далеких предков. Подобные
ясе зубы сидят па нижпей челюсти. Последняя очень прочна. Denta-
le у ископаемых форм развита хорошо, у Neoceratodus опа лишена
зубов и развита слабо, у Lepidosiren и Protopterus отсутствует
вовсе. Зубы несет spleniale. Меккелев хрящ не окостеневает.
Жаберные дуги в числе пяти у современных Dipnoi развиты слабо,
у Lepidosiren и Protop-
terus опи не расчленены
на отделы. Copulae пет.
Таким образом, череп
Dipnoi отличается боль-
шим своеобразием.
Скелет конечностей.
Непарные плавники.
Как указано было вы-
ше, по средпей линии
тела у рыб имеются н е-
парные плавни-
к и. Различают спин-
ной плавник,
хвостовой плав-
ник и анальный,
или подхвосто-
вой. Спинной плавник
может быть один, или их
может быть несколько.
Все плавники имеют поддерживающие их скелетные части. Лишь
так называемый жировой плавник, развивающийся
у некоторых рыб (Salmonidae, Siluridae) па спипе, позади настоя-
щего спинного плавника, пе имеет скелета. Скелетные части плав-
ников сочленяются с особыми поддерживателями плавников, кото-
рые другим, проксимальным (ближайшим к оси тела) концом сидят
на остистых отростках позвонков. Скелетные части самих плавни-
ков называются птеригиофорами, или лучами
(radialia). Лучи эти у Teleostei могут быть сплошными, жесткими
и колючими (у колючеперых, Acanthopterygii из Teleostci), или
же опи состоят из члеников, дихотомически разветвляющихся
наверху, а потому мягких (прежние мягкопсрые Anacantliini). У
низших, хрящевых рыб они построены пе из кости, а из хряща
и роговых нитей.
Форма плавников весьма сильно варнируёт. Особое значение
имеют форма и строение скелета хвостового плавника (рис. 75).
У эмбрионов всех рыб (а также и у Cyclostomata) хвостовой плав-
пик является вполне симметричным, верхняя и нижняя лопасти
102
мы наблюдаем
Г:
Рис. 76. Поперечный
разрез через спинной
плавник акулы (Scyl-
lium).
1 — radii, 2 — ceratotri-
chia, 3 — плакОидные
зубы. (Из Goodrich.)
одинаковы, и осевой скелет идет прямо. Это — протоцер-
ка л ь и ы й плавник. У эмбрионов па более поздней стадии ске-
летная ось загибается кверху, и брюшная лопасть увеличивается
в размерах. Получается плавник гетероцеркальный.
Мы видим такой плавник также во взрослом состоянии у Elasmobran-
chii, Holocephali и Chondrosteoidei. Тут филогенез повторяется
в онтогенезе. У Holosteoidei брюшная лопасть продолжает увели-
чиваться, пижние остистые отростки выпячиваются, и обе лопасти
хвоста становятся одинаковыми. Хвост, не будучи по внутреннему
строению симметричным, становится таковым по внешнему виду
Это хвост гомоцеркальный. У Diponi
хвостовой плавник по внешности протоцер-
кальпый. На самом же деле здесь имеется
вторичное сходство формы. У древпейших
Dipnoi хвост был гетероцеркальным. Посте-
пенно в течение филогенетического развития
осевой скелет начинает отклоняться вниз,
спинная лопасть плавника развивается до
величины брюшной, и плавник становится
вторично симметричным, или д иф и ц ер-
кал ь п ы м. Плавники Dipnoi могут слу-
жить примером этого типа.
Материал, из которого построен скелет
плавников, различен в различных отрядах
рыб. У Elasmobranchii лучи (radii) хряще-
вые, расчлененные па три части. В самую
плавниковую складку они входят недалеко,
и складка поддерживается главным образом
особыми гибкими топкими лучами — ccrato-
trichia из вещества, подобного рогу,—выделяе-
мыми клетками соединительной ткани (рис.76).
Поддерживающие плавник radii первоначаль-
но являются в числе двух па сегмент, соот-
ветствуя первичной диплоспопдилии, у взрослых же форм их боль-
ше. Radii нередко сливаются между собой, образуя хрящевые пла-
стинки (рис. 77), делающие основание плавника более прочным.
Это бывает особенно в тех случаях, когда перед плавниками
имеются шипы (Acanthias, например).
У Osteichthyes этот скелет плавников претерпевает изменения.
У Chondrostci хрящевые radii развиты хорошо, ceratotrichia села-
хпй заменяются костными лучами (lepidotrichia), пред-
ставляющими видоизмененные кожные чешуи. У Holostei, а еще
более у Teleostei костные лучи становятся более развитыми, и ra-
ti ii сокращаются в своем развитии, пе заходя в плавниковую склад-
ку. Ceratotrichia имеются, но в зачаточном виде. У Dipnoi скелет
непарных плавников представлен так называемыми camptotrichia,
которые имеют волокнистое строение и слегка объизвествлепы
у современных Dipnoi и были костными у ископаемых. Снаружи
они покрыты еще чешуями. Происхождение их неясно.
103
Для понимания непарных плавников, а также и парных, о ко-
торых речь будет дальше, много дает рассмотрение развития плав-
ников у эмбриона. Плавники закладываются в виде кожной склад-
ки, идущей по спине до самого заднего конца тела, где она встре-
чается с другой такой же складкой, идущей от анального отвер-
стия. Получается один непрерывный плавник, идущий по спине,
огибающий задний конец тела и продолжающийся вперед до аналь-
ного отверстия, а иногда и вперед от него. Впоследствии обычно
продолжают развиваться лишь отдельные участки этой непарной
складки. В складку врастают мускульные почки от миотомов ту-
ловища, по одной от миотома каждой стороны, т. е. метамерно.
Рис. 77. Скелет переднего спинного плавника Selachii.
Л— Zygaena malleus, В — Ileptanchus cinereus, С — Acanthias. Обозна-
чения те же. (По Fiihrbringer.)
Почки эти закладываются нередко пе только в области развиваю-
щегося плавника, по па протяжении всей складки, подвергаясь
потом в промежуточных местах редукции. Впоследствии эти му-
скульные почки отделяются от своего миотома и становятся муску-
лами плавника. Мезенхима между мускульными почками сгущается,
и в пей закладываются отдельные скелетные элементы в виде пало-
чек, подергающихся затем охрящевонию, по одной между каждой
парой мускульных почек. Таким образом, закладываются хряще-
вые radii. К каждой почке идет веточка от того нерва, которым
иннервируется данный миотом (рис. 78).
Из истории развития непарных плавников мы видим, что
непарные плавники рыб — спинной или спинные, хвостовой и
анальпый — являются частями одной непарной складки, сходной
с той, которая идет по спиппой и огибая задний конец, по брюш-
ной стороне тела до атриопора у ланцетника. Строение непарных
104
плавников метамерпое, соответствующее общей метамерности
туловища.
История развития непарных плавников проливает свет на исто-
рию возникновения парных плавников.
Парные плавники, нх пояса, происхождение и эволюция. У
Cyclostomata парных конечностей нет. У рыб имеются грудные
и брюшные плавники, соединяющиеся с плече-
вым и тазовым поясом. Наиболее примитивен плечевой
пояс у сростножаберпых (Elasmobranchii) и цельноголовых (Ho-
locephali). Он представлен с каждой стороны хрящевой дугой, за-
Рис. 78. Scyllium canicula, непарные и парные плавники у взрослой формы
(Л и В) и у эмбриона (С), у В — плавники развернуты.
1—грудной плавник, 2 —брюшпой плавник, 3 — спинномозговые нервы, 4— мус-
кульные почки в спинных и 5 — в брюшных плавниках, а также в промежутке
между ними. (По Goodrich.)
ключеппой в мускулатуре стенки тела непосредственно позади по-
следней жаберной дуги. Обе половины пояса соединены между собой
внизу и свободны наверху, так что плечевой пояс имеет вид непол-
ного сверху обруча. На заднем краю каждой дуги имеется выступ,
служащий местом для сочленения с плавником. Часть дуги, на-
ходящаяся кверху от этого места, носит название лопатки
(pars scapularis), а книзу — коракоида (pars coracoidalis).
.V ископаемого представителя элазмобранхий — Pleuracanthus по-
ловины плечевого пояса остаются раздельными, не сливаясь между
собой. У ископаемой же примитивной Cladoselache (рис. 79) пижняя
часть плечевого пояса состояла из отдельных частей, невидимому,
radii. У скатов (Batoidei) в связи с очень сильным развитием груд-
ных плавников плечевой пояс тоже развит сильно и прикрепляется
105-
верхними концами дуг к черепу. В связи с этим передние позвонки
сливаются в одно целое. У эмбриона Osteichthyes также заклады-
вается сперва хрящевой пояс, первичный плечевой
пояс, как у акул. Но затем в нем появляются окостенения.
Подобно тому как в истории черепа, сперва появляются покровные
окостенения, азатем и замещающие, и здесь в процессе филогенеза
кость постепенно вытесняет хрящ.
Рис. 79. Реконструкция скелета Cladoselache.
1—часть плечевого пояса, состоящая из не вполпе слившихся
базальных частей radii. (По Dean.)
У Chondrostei первичный хрящевой пояс представлен еще пол-
но, но снаружи на него налегают покровные кости, «вторич-
ный плечевой пояс»; их две: нижняя носит название
ключицы (clavicula), верхняя называется cleithrum. У Dipnoi
также сохраняется хрящевой плечевой пояс из двух половин,
соединяющихся внизу и прикрываемых clavicula и cleithrum сна-
ружи. У Teleostomi обычно имеется в первичном поясе уже
Рис. 80 Плечевой пояс и грудной плавник Polyp-
terus bichir.
1—scapula, 2—coracoideum, 3 —- clavicula, 4 —cleithrum,
5 — supraclavicula, 6 — postciavicula, 7 — posttcmporale, 8 —
metapterygium, 9—propterygium, 10 — mesopterygium, 11 —
radialia, 12 — lepidotrichia. (По Goodrich.)
два замещающих окостенения (рис. 80): верхнее — лопатка
(scapula) и нижнее — коракоид (coracoideum). Между ними
лежит сочленовная поверхность. Вторичный пояс, представленный
clavicula, cleithrum, supraclavicula и другими дополнительными
частями у более примитивных форм, лежит поверхностно и покрыт
скульптурой. Развивается плечевой пояс в связи со скелетом груд-
ных плавников (см. ниже).
406
Рис. 81. Скелет брюшного плавника
лопатоноса (Polyodon folium). Перед-
ний край плавника — справа; прок-
симальные части первоначально раз-
деленных radialia сливаются, обра-
зуя basipterygium.
Тазовый пояс у рыб развит слабо, так как и значение брюшных
iiJianiiiiicoB, местом прикрепления мускулатуры которых оп служит,
дли движения невелико. В связь с осевым скелетом тазовый пояс
in- вступает. У ископаемой Cladoselache он и у взрослой формы
.uiiiiib слабо дифференцирован от основного отдела лучей плавника
и явственно сегментирован. У современных селахий эта сегмен-
тация исчезает. Тазовый пояс представлен здесь непарной пла-
стинкой, связывающей правый и левый брюшной плавники, не-
редко с отростком (processus iliacus) пад местом прикрепления плав-
ников. У цельпоголовых (Holpcephali) тазовый пояс всегда парный,
и processus iliacus настолько велик, что пояс становится подобным
плечевому поясу и подобно последнему состоит из двух частей:
верхней — pars iliaca и пижней — pars ischiopubica. Такое сильное
развитие таза стоит в связи с
сильным развитием брюшных
плавников, служащих здесь ор-
ганами движения, вместо слабо
развитого вытянутого в нить
хвоста.
У Chondrosteoidei тазовый
пояс (рис. 81) лишь очепь слабо
окостеневает, оставаясь хряще-
вым. У молодых особей пояс
явственно сегментирован (см.
Polyodon folium), по у взрослых
форм сегменты сливаются, и по-
лучается цельная парная пла-
стинка, сходная с поясом Holosteoidei. Тазовый пояс Chondrosteoi-
dei является указанием па первоначальное происхождение пояса
путем слияния radii. У Holostei и Teleostei пояс вполне окосте-
невает. У двоякодышащих (Dipnoi) тазовая пластинка непарная,
как у элазмобрапхий.
Скелет самих парных плавников по материалу подобен скелету
непарных. У низших, хрящевых рыб оп построен из хрящевых эле-
ментов (radii), подкрепленных роговидными (ceratortichia); у
Osteichthyes хрящевые лучи плавников сокращаются в числе и сте-
пени развития и заменяются костными лучами (lepidotrichia).
Роговые лучи также редуцируются.
Расположение хрящевых элементов в плавниках весьма разно-
образно (рис. 82). У ископаемой примитивной рыбы Cladoselache
скелет грудного и скелет брюшного плавников, лежащих гори-
зонтально, с широким основанием (рис. 79), представлен рядом
тонких более или менее параллельных перасчлепенных хрящевых
radialia. Плавник напоминает спиппой. У современных элазмобран-
\iiii проксимальные radialia обычно сливаются в два или три хря-
щевых элемента, называемых основными (basalia): pro-, meso-
п metapterygium. К ним снаружи примыкают лучи (radii). Наиболь-
шее значение имеет последний. Такой плавник называется уни-
сериальным, однорядным. У ископаемой рыбы
407
камеппоугольпого периода Pleuracanthug грудной плавник другого
типа: здесь имеется расчленеппая ось, спереди и сзади которой сидят
хрящевые лучи (radii). Такой плавник носит название д в у р я д-
ц о г о, бисериального, или архиптеригия (рис. 83).
I
Рис. 82. Грудные плавники различных рыб:
А—акула Acanthias vulgaris, В—скат Raja, С—Chimaera,
D— Acipenser, Е — Amia calva, F — Ledidosteus, О —Po-
lypterus, H—<Salmo. 1—propterygium, 2—mesopterygium,
3 — metapterygium, 4 — radii. (Иа Гегенбаура до
Boulanger.)
У Chondrosteoidei скелет грудного плавника состоит из немногих
хрящевых лучей, причлепяющихся впереди к поясу, а назади к слабо
развитому metapterygium. Radii развиты слабо, и плавник поддер-
живается костными, вторичными по происхождению лучами. У II о-
Рис. 83. Реставрация Pleuracanthus ducheni. (Из Паркера и Гас-
веля, по Dean.)
lostei (Amia, Lepidosteus) radii в большей или меньшей степени
окостеневают, сидят пепосредствепно па плечевом поясе, и хряще-
вой скелет плавника представлен лишь несколькими маленькими
хрящиками, лежащими кнаружи от окостеневших radii. И у Holo-
108
Л ci и Teleostei главную роль в плавнике играют вторичные костные
iyчп (lepidortichia). У Dipnoi скелет грудного плавника, как и
ipioiiiiioro, хрящевой и построен по тому ясе типу, как у Pleura-
•nnt.lius: к длинной членистой оси прикрепляются спереди и сзади
гонкие расчлененные хрящевые лучи, идущие далеко в лопасть
i.ii.iiiiiiiKa, т. е. здесь имеется бисериальпый архи-
н т <• р и г и й (рис. 84).
.У к и с т е п е р ы х (Crossopterygii) грудной плавник имеет
ппд лопасти и является уппсериальным, касс у элазмо-
>раихии. В нем имеются pro-, meso- и metapterygium, к которым
прпчленяются частично окостеневающие radialia. У ископаемых
кпетеперых были, повидимому, оба типа плав-
никового скелета: унисериальпый и
5 и сериальный.
Скелет брюшных плавников в основном схо-
ден с таковым грудных, по устроен несколько
проще.
Крои схождение парных конечностей. У
Cyclostomata мы пе видим парных конечностей,
пот их и у Acrania. Является вопрос, откуда
получились плавники. Сравпительноанатомичес-
кое изучение плавников, подкрепляемое неко-
торыми палеонтологическими даппыми, по глав-
ным образом эмбриологическое изучение раппих
стадий- развития парных плавников проливает
яркий свет, па эту страницу эволюции низших
позвоночных: появление парных конечностей.
Парные плавники закладываются у зароды-
ша селахии в виде продольной складки эпидер-
миса по бокам тела (см. рис. 78), как непар-
ные — в виде такой же складки па спинной и
брюшпой сторонах тела. В эти складки про-
никают мезенхима и мускульные почки, выро-
сты нижпего конца туловищных миотомов. Они
Рис. 84. Левый
грудной плавник
Neoceratodus.
1 — роговые лучи,
2 — хрящевые radia-
lia, 3 — членики оси.
(Из Видерсгейма.)
образуют мускулы плавника. Меясду спинными и брюшными муску-
лами плавниковой складки закладывается мезепхиматозпая пла-
стинка, в которой вскоре дифференцируются лучи, свободные ди-
стально, а проксимальными концами сливающиеся в пластинку,
которая растет кверху и книзу, образуя зачаток плечевого пояса.
Нервы, идущие от спинного мозга к мышцам копечпости, прони-
зывают плечевой пояс, Число нервов соответствует числу миотомов,
вошедших своими почками в плавниковую складку. Таким образом,
мы видим здесь совершенно тот же способ образования, как у пе-
карных плавников. Тот же тип наблюдается и у Chondrostei и у Ho-
losteoidei. С возрастом начинается сокращение длины основания
плавников, их концентрация, а хрящевые элементы скелета рас-
падаются на ряды — radiit
Такое развитие дает основание думать, что парные плавники, как
п непарные, образовались в процессе филогенеза из непрерывных
109
боковых кожных складок, подобных непарной плавпиковой складке
и метаплевральным складкам ланцетника. Боковые складки слу-
жили для увеличения брюшной поверхности тела. Подобные склад-
ки, слабо развитые, наблюдаются иногда и у современных акул.
В этих складках первоначально пе было скелета. Когда они уве-
личились до более значительных размеров, полезным новоприоб-
ретением явился скелет. Скелет первоначально был в виде ряда пе-
расчлененных лучей, как у зародыша акулы. Развились мышцы,
как выросты миотомов. Сперва складка была длинная и непрерыв-
ная, но так как функция руля глубипы выпадала главным образом
па передний и задний края ее, то средняя часть, потеряв значение,
редуцировалась; образовались грудные и брюшные плавники.
Из мезенхиматозной закладки в проксимальной части развился
плечевой и тазовый пояс. Слияние лучей дало metapterygium.
Построенная на изучении истории развития плавников у эмбрио-
нов теория происхождения парных конечностей из боковой складки
подтверждается и сравнительпоапатомическимп фактами и фактами
палеонтологии. Мы видели уже, что у молодых экземпляров Chon-
drosteoidei видна еще сегментация основных хрящей, образование
их через слияние radii. Она сохрапяется в тазовом поясе с ка-
фир и и х а (Scaphirhynchus), следы ее видны у примитивной акулы
Chlamydoselachus (из Notidani). У примитивной ископаемой Cla-
doselache плавники расположены так, как они располагаются у за-
родыша современных акул. Все подобные факты делают теорию
боковой складки весьма убедительной.
Первоначальные плавники имели широкое основание и лежали
горизонтально. Плавники были унисериальпыми В течение даль-
нейшего хода эволюционного развития рыб в связи с дальнейшим
развитием функции парпых плавников как рулей глубипы и орга-
нов балансирования началось отделение заднего края плавников от
туловища. У селахий это отделение пошло недалеко, у Dipnoi по-
лучилась длиппая лопасть. Вследствие этого ось плавника стала
развиваться в центре плавниковой лопасти. Задний край плавни-
ковой лопасти получил свой скелет в виде radialia, причлепяю-
щихся к оси сзади. Получился бисериальпый архиптеригпй
(рис. 85). У современных рыб мы видим оба типа: уписериальный
и бисериальный. Общая эволюция шла в том же направлении,
как и эволюция других частей скелета: сокращение первичного
хрящевого скелета п развитие вторичного костного, — а также
в изменении горизонтального положения основания плавников
в вертикальное у Teleostei.
Другая теория происхождения парпых плавников от жаберпых
дуг (пояса) и их лучей (сами конечности) в настоящее время совер-
шенно оставлена. Опа пе согласуется пи с тем, что у более древних
рыб (акулы) плавники лежат горизонтально, пи с тем, что муску-
латура их туловищная, происходит из миотомов, а нс висцеральная,
происходящая из боковой пластинки, пц с тем, что пе один нерв —
ветвь п. vagus, как в жаберных дугах, а целый ряд спинномозговых
нервов иннервирует мышцы плавников. Противоречат теории жа-
лю
берной дуги и такие факты, как вырастание поясов конечностей
из закладки скелета конечностей и то, что наиболее примитивна
пе бисериальная форма плавника (этого требует теория жаберной
дуги), а моносериальная и положение плечевого пояса кнаружи
от мускулатуры, а не внутрь от нее, как у жаберных дуг.
Рис. 85. Диаграмма, показывающая результат кон-
центрации скелета и нервов в парном плавнике.
А, В, С ведет к типу селахий, А, В, Т) — к типу Dipnoi.
1П 1а и т. д. — нервы, 2 —спинной мозг, 3 —radialia,
4 —- плавниковая складка. (По Goodrich.)
На примере вопроса о конечностях мы видим блестящее приме-
нение сравнительноэмбриологического, сравнительноапатомиче-
ского и палеонтологического методов в решении вопросов об эволю-
ции позвоночных.
6. Органы дыхания рыб
Жаберные щели, жабры и крышечный аппарат. Органы дыха-
ния рыб тесно связаны со скелетом, стоят в ближайшем отношении
к висцеральным дугам и крышечному аппарату. Органами дыхания
служат жабры, функционирующие всю жизнь. Редко встречается
у рыб иное дыхание, и в связи с ним развиваются тогда особые
органы дыхания. В отношении развития органов дыхания рыбы стоят
па, более высокой ступени, чем Cyclostomata, а тем более — Amphio-
xns. У последнего нет жабр, есть лишь жаберные щели, выстлан-
ные ресничным эпителием. И функция их там пе только дыхатель-
ная, по реснички эпителия вызывают ток воды, несущий пищу
через рот в глотку. У Cyclostomata слизистая оболочка жаберных
мешков сложена в складки, увеличивающие дыхательную поверх-
ность. У рыб жаберные листки представляют поверхность гораздо
• «о чыную, что увеличивает интенсивность кислородного обмена и по-
। i.imaeT общую энергию организма, давая возможность более-
<н.|стро двигаться со всеми вытекающими отсюда последствиями
в борьбе за существование.
ш
Жаберные щели образуются у рыб как парные выросты энто-
дермальпой стенки глотки, навстречу которым растут впячивания
эктодермы. Те и другие выпячивания встречаются, соединяются,
и образуются жаберные щели. Число их больше у низших представи-
телей класса, чем у высших. У Elasmobranchii их бывает от 5 до 7,
причем 7 щелей имеются у Heptanchus, 6 — у Hexanchus и СЫашу-
doselachus, представителей примитивной группыNotidani. У осталь-
ных селахий (Selachii) жаберпых щелей 5. У Holocephali всего
4 жаберпых щели. У костистых рыб (Teleostei) число жаберных дуг
с жабрами 4. У пятой их нет, а иногда имеет место редукция и
4-й и 3-й жабры.
Первая жаберная щель находится между подъязычной дугой
и первой жаберной. Впереди жаберпых щелей, обычно позади
глаза,, у элазмобранхий находится еще отверстие, называемое
брызгальцем (spiraculum), ведущее в глотку. Это отверстие
лежит между пебноквадратным хрящом (челюстной дугой) и hyoman-
dibulare (гиоидной дугой), и у зародышей элазмобранхий па стенках
ее сидят, как и в остальных щелях, жаберные листочки. Таким
образом, щель эта является пе чем ипым, как первой жаберной
щелью. Spiraculum у Osteichthyes обычно исчезает, но сохраняется
у некоторых форм: у осетров (Acipenser). лопатоносов (Polyo-
don) и у Polypterus, в эмбриопальпом состоянии встречается и у не-
которых Teleostei (Salmo). У Holostei (Amia и Lepidosteus), хотя
и в закрытом состоянии, spiraculum встречается и у взрослых форм.
У Chondrostei в spiraculum, как и у элазмобранхий, на передней
степке имеются жаберные листочки, называемые «ложно® а-
б р о й» (pseudobranchia), так как спабжается опа не венозпой,
а артериальной кровью. Так как у современных высших рыб зад-
ние щели (дуги) подвергаются иногда редукции, у Cyclostomata
число жаберных отверстий доходит до 14, у Ostracodermi число их
также до 16, число жаберпых щелей у примитивных акул доходит
до 7, — то нужно думать, что число жаберпых щелей у предков
рыб было больше, чем у современных рыб. Но так как пи у одной
современной или ископаемой рыбы их не имеется более 7, то мояспо
думать, что число их у древнейших рыб было 7 или 8, включая
spiraculum. Большое число жаберпых щелей у Amphioxus—явле-
ние вторичное, приспособление для усиления тока воды через
глотку.
Жаберные щели разделяются межжаберными перегородками —
септами, поддерживаемыми жаберными дугами с впутреппей сто-
роны. У элазмобранхий септы хорошо развиты и выстлапы с обеих
сторон глоточным эпителием, сложенным в складки. Но высти-
лают жаберпые складки межжаберные перегородки не до конца.
Свободный край перегородки срастается с кожей, благодаря чему
жаберные щели могут прикрываться краем предыдущей щели.
На каждой септе находятся две «полужабры», вместе
образующие целую жабру. На гиоидной дуге сидит одна
полужабра на ее задпей стороне, т е. в первой жаберной щели.
В последней жаберной щели имеется полужабра лишь на передней
412
А 8 С О Е
Рис. 86. Схема жабр.
А — Selachii, В — Chimaera, С.1— Acipen-
ser, D, Е — Teleostei. 1 — жаберная дуга»
2 — межжаберные перегородки, 3 — жа-
берные лепестки. (По Bridge, из Ihle,
Versluys, etc.)
стороне,’т. е. па 4-й дуге, 5-я без жаберных листков. В брызгаль-
це, как сказано, имеется «ложножабра».
У целыюголовых (Holocephali) межжаберные перегородки ко-
роче, чем у селахии, благодаря чему жаберные листки слегка вы-
ступают за внешние края септ. Этот процесс редукции септ идет
да ю.пн; у Teleostomi (рис. 86); септы еще короче у Acipenser,
и 1Ч11.С короче у Teleostei. У последних перегородка может и вовсе
исчезать, и тогда жаберных мешков или полостей, открывающихся
наружу особыми отверстиями, уже пе образуется, и на каждой
стороне тела имеется одна жаберная полость, в которую торчат
гребневидные жабры, полость, прикрытая снаружи костями ж а-
б е р н о й крышки (operculum, suboperculum, interoperculum)
и натянутой па лучах подъязыч-
ной кости (radii branchiostegi)
мягкой перепонкой (membrana
branchiostegalis) Из этой жа-
берной полости имеется только
одни выход для воды. Края жа-
берной крышки оторочены упру-
гой мембраной.
Развитие жаберной крышки
налицо уже у Holocephali в ви-
де кожистой складки, идущей
от наружной поверхности гио-
идной дуги и прикрывающей все
таборные щели. Костная жабер-
ная крышка Holosteoidei, до-
стигающая своего максималь-
ного развития у костистых рыб, описана выше. С развитием жа-
берной крышки происходит редукция передней полужабры па
гиоидной дуге, и развивается полужабра па внутренней стороне
крышки, где она носит название оперкулярной жабры,
я паяющейся также ложножаброй.
Жаберный аппарат Dipnoi (Neoceratodus) весьма напоми-
нает таковой у Holocephali: межжаберпые перегородки хорошо
развиты, и жаберпые листки почти все сидят па передней и зад-
ней их стенках, причем полужабры передней и задней степок двух
перегородок соединяются, обрастая щель сверху и снизу. У Рго-
lopterus и Lepidosiren вследствие возрастания значения легких
н.гблюдается соответственная редукция жабр. Имеется жаберная
крышка.
На вогнутой стороне жаберных дуг у большинства рыб сидят
и дна ряда так называемые «жаберпые тычинки», пред-
<• । амиющие выросты слизистой оболочки или костные образова-
нии. ()бычно тычинки переднего ряда одной дуги приходятся между
П.1Ч и и ками заднего ряда вперсдилежащей дуги. Благодаря этому
.li.iui piii.ie тычинки являются ситом, препятствующим пропикно-
...... и жаберпые щели и засорению жабр твердыми частицами,
||||||а.1ан|щими в глотку с водой в процессе дыхания и питапия.
Кiiiiii(ii|)oii и Стапчиискай
113
У некоторых рыб эти тычинки необычайно сильно развиты и много-
численны. Это бывает у тех рыб, которые питаются мелкими орга-
низмами планктона. У Seiache maxima, как указано было выше, они
представлены волосовидными плакоидными зубами.
Жаберные лепестки служат местом, где происходит газообмен.
Два сосуда проходят по наружной выпуклой стороне каждой дуги.
Один сосуд несет кровь от сердца. Хотя кровь эта редуцированная,
бедная кислородом, сосуд является жаберной артерией
(arteria branchialis), от которой в каждый лепесток отходит веточка.
Последняя распадается в лепестке на еще более мелкие артерии и да-
лее на капилляры, распределяющиеся в слизистой оболочке ле-
пестка. Артериальные капилляры переходят в венозные, собираю-
щиеся в мелкие вепы, а последние собираются в вену, идущую па
другой стороне лепестка,
чем артериальная веточка.
Эта вена впадает в ж а-
берпую вепу (vena
branchialis), идущую также
по наружной стороне жа-
берной дуги и несущей
кверху, в парную аорту
кровь артериальную.
Наружные жабры. В
добавление к внутренним
или настоящим жабрам
Рис. 87. Личинка Gymnarchus niloticus
10 дней.
(По Кегг из Goodrich.)
у некоторых рыб развиваются еще наружные жабры. По-
следние бывают двух родов: настоящие наружные
жабры и наружные жаберные нити. У эм-
брионов элазмобранхий встречаются именно последние. Они раз-
виваются в виде тонких нитевидных выростов внутренних жаберных
лепестков задних полужабр и па гиоидной и челюстной дугах.
В каждой нити имеется сосудистое сплетение, между приводящим
и отводящим сосудами, с редуцированной и оксидированной кровью.
Жаберные нити имеются у эмбрионов акул, по вылуплении из яйца
они исчезают. Служат они, вероятно, пе только для дыхания, по
и для поглощения питательного материала из белковой жидкости
внутри яйца, в которой они плавают. Подобные внешние жаберные
нити найдены были и у некоторых личинок Tcleostomi (Chondro-
stei, некоторых Teleostei, папример, Gymnarchus из сем. Mormy-
ridae(pnc. 87).
Настоящие наружные жабры встречаются у эмбрионов и у ли-
чинок Dipnoi (Protopterus и Lepidosiren), у Polypterus и даль-
ше — у амфибий. Они развиваются независимо от внутренних
жабр, иногда еще раньше, чем образуются жаберные щели и жабер-
ная крышка как выросты наружной, покрытой эктодермой поверх-
ности жаберных перегородок. Наружные жабры снабжаются кровью
из аортальных дуг. У личинок Polypterus, Calamoichthys (Cross о-
pterigii) (рис. 88) имеется всего одна пара наружных жабр в виде
больших перисторазветвленпых образований, поддерживаемых ко-
114
роткпм и хрящевым выростом гиоидной дуги. У личинок Protopte-
гня и Lepidosiren (рис. 89) наружных жабр четыре пары. Protop-
l.priiM и во взрослом состоянии сохраняет три задние пары жабр,
хоти в редуцироваппом виде.
Р.швнтие у личинок двоякодышащих и у кистеперых наружных
ливр, свойственных и личинкам амфибий, является весьма заме-
•мтг |ц,пым фактом,
\ 1м1:п.|вающим на род-
ггпп этих рыб с по-
« II’,I,ними.
Механизм дыха-
н и а у рыб и его эво-
люция. Механизм ды-
\.1Н\ИЬПЫХ движений рис gg Передняя часть молодого Polypterus.
V рЫО (рИС. 90) ЯВЛЯеТ- i __ наружная жабра-
с я весьма сложным
и неодинаковым у всех рыб. Наиболее сложным и совершенным
является он у костистых рыб (Teleostei), более простым и несовер-
шенным у акул (Selachii). Остальные Teleostomi, кроме Dipnoi,
которые стоят'особняком, занимают промежуточные ступени меж-
ду этими двумя отрядами. Обычно механизм этот представляют
в виде схемы нагнетательного насоса. По этой
схеме полостью на-
соса являются жа-
оерная иоколожабер-
ная полости, а жа-
оерпые дуги с их му-
скулатурой, у выс-
ших рыб ижаберпая
нрьппка, служат для
расширения и сжима-
ния этой полости.
К, роме того, в этом на-
сосе имеются клапа-
ны. У костистых рыб
т а. । и (ми кл ап анами
Рис. 89. Стадии развития Lepidosiren paradoxa.
1 — наружные жабры, 2 — цементный орган.
являются, с одной стороны, клапан жаберной крышки, с другой —
особый клапан во рту (рис. 90, схема и А2). При расширении жа-
берной полости клапан жаберной крышки прижимается к телу
благодаря давлению воды. Таким образом, жаберная полость ока-
пывается открытой только со стороны рта, давление в ней умень-
шено, и вода устремляется в рот, открывая клапан внутрь. Когда
же жаберноротовая полость пачнет сжиматься, клапан во рту
но л, влиянием давления воды изнутри закроется, а клапан жабер-
|цш крышки откроется, и вода из жаберной полости потечет
‘irpi :i жаберные щели наружу, омывая жаберные листки.
11а самом деле процесс дыхания у Teleostei гораздо сложнее.
' 1деш., кроме нагнетательного насоса, действует еще и играет глав-
ную роль в с а с ы в а ю щ и й насос. В работе его прини-
115
мают участие не только крышечный аппарат, по п жаберпая «ре-
шетка» пз жаберных лепестков. Дыхание может происходить и без
участия ротового клапапа, и при открытом рте, и использование
дыхательной поверхности жаберных лепестков при этом гораздо
полнее и совершеннее. У костистых рыб, как мы видели, межжабер-
ные перегородки редуцированы, гребенчатые жабры сидят свободно
па наружной стороне
дуг, вдаваясь
в
л,
з
4
з
С,
У рыб.
Рис. 90. Механизм дыхания
Ах и Аа —обычно принимаемая схема. В, и Ва —схе-
ма работы жаберной крышки и решетки. Си Cj и С,—
различные фазы работы всасывательной жаберной
крышки. 1 — ротовая и жаберная полость, 2 — около-
жаберная полость, 3—мягкий край жаберной крыш-
ки. (По Воскобойникову).
околожаберпую полость.
Кроме того, в лепест-
ках пмеется свой ске-
лет в виде тонких, со-
стоящих из одного ря-
да хрящевых клеток,
нитей. Хрящ этот, так
называемый клеточ-
ный, весьма топкий
и упругий. У живой
дышащей рыбы жа-
берные лепестки двух
соседних дуг прижаты
друг к другу краями,
и благодаря этому
выход из жаберных
щелей паружу в око-
ложаберную полость
все время закрыт. Во-
да при дыхании долж-
на проходить из соб-
ственно жаберной по-
л ости (п ро дол жени е
ротовой) в околожа-
берную (ограничен-
ную крышкой и шеш-
brana branchioste-
galis) через тончай-
шую решетку, обра-
зованную жаберными
лепестками, и их
складками, вторич-
ными лепестками, уст-
роенным так, что
опи вполне прилегают друг к другу. Жаберная крышка сво-
им мягким эластическим краем прилегает к телу и может
скользить по нем, пе отрываясь, при отхождении крышеч-
ного аппарата от тела (рис. 90, схема В± и В2). В последнем
случае давление в околожаберной полости становится меньше
давления в жаберпо-ротовой полости, вода входит через рот и
устремляется через решетку из жаберпых лепестков в околожабер-
ную полость. При этом достигается максимальное использование
кислорода, так как при просасывапии воды через решетку каждая
116
капля се соприкасается с тончайшей сетью капилляров. Мускула-
тура перегородок используется при этом для продвижения крови
через капилляры перегородок. В следующей стадии процесса
крышка и брапхиостегальпый аппарат прижимаются к телу, дав-
лен не в околожаберпой полости возрастает, и вода механически,
встречая в этом направлении меньше сопротивления, отодвигает
краевой клапан и выходит наружу.
Исследования показали, что у костных рыб работают, глядя по
обстоятельствам, оба насоса; при этом всасывающий насос играет
главную роль, весьма дифференциро-
ван, сложен и является, очевидно
позднейшим приобретением класса.
Наоборот, характерные части нагне-
тательного насоса обнаруживают ре-
дукцию: и ротовой клапан и муску-
латура для движения жаберных дуг.
И работает он почти незаметно. У
сслахий мы имеем также оба типа
насоса, но развит больше насос наг-
нетательный, так как насасывающий
аппарат еще несовершенен. Здесь еще
нет общей околожаберной полости,
жаберпые щели открываются паружу,
причем каждая жаберпая щель при-
крывается складкой кояси (рис. 91),
прирастающей к периферическому
концу впередилеясащей межжаберной
септы. Наружные концы каждой по-
лужабры у живой рыбы отогнуты,
плотно соприкасаются с таковыми
позади леясащей полужабры. Таким
образом, получается ряд околоясабер-
пых полостей, отдельных для каждой
жаберной щели, так как смыкание
концов полуясабр делит жаберную
щель па две части, из которых собст-
венно жаберная щель является все
время замкнутой, а наружная, бо-
ковая или околожаберная замкнута
Рис. 91. Схема строения жабер-
ных листков акулы и их отно-
шения к жаберным перегород-
кам.
1 — жаберная дуга, 2 — канал,
3 — жаберная перегородка, 4 —
пучки межжаберного мускула в
поперечном разрезе. (По Воско-
бойникову.)
при процессе всасывания воды и открывается при выталкива-
пин ее наружу. Вода из собственно жаберной щели прохо-
дит в соответственную околожаберпую через жаберные лист-
ки. Вторичные лепестки сидят не но всей поверхности первичного
листка, по на части, обращенной к ясаберной щели. Благодаря этому
вдоль септы получается канал, через который, прососавшись через
решетку ясаберных листков, и проходит вода из жаберный щели
в околоясаберную. Таким образом отличие от Teleostei заключается
в существовании межжаберных септ. Из рисунков видны сходство
и отличия механизма дыхания у Teleostei и Selachii (рис. 92).
117
Сравнением дыхательного аппарата у различных групп рыб
можно наметить те этапы, которыми шло развитие от типа нагне-
тательного, преобладающего у селахий, к насасывающему у кости-
стых рыб. Развитие это, входить в описание которого здесь нет воз-
можности, показывает, что для развития того или иного признака
па известном этапе развития необходима предпосылка па предыду-
щем этапе. Вся эволюция дыхательного аппарата шла в направле-
нии: 1) освобождения лепестков от межжаберных перегородок;
2) развития в них самостоятельного скелета для поддержания ре-
шетки; 3) использования мускулатуры перегородок для продвиже-
Рис. 92. Схема строения и действия жаберного аппарата акулы и костистой
рыбы.
1—боновые жаберные полости, 2—полость рта и жаберная, 3 —клапан метамера
гиоидной дуги, 4—клапаны жаберных метамер, 5 — жаберные лучи, 6 —мускулы.
(По Воскобойникову).
пия крови через тончайшую сеть капилляров решетки; 4) развития
насасывающего аппарата: жаберной крышки, гиостилии, брапхио-
стегальпого аппарата. Лишь Dipnoi стоят особняком от этого раз-
вития. У них вовсе нет скелета, поддерживающего лепестки, пет
и развитого аппарата решетки^ у них пе образовалось того пасоса,
который развился у других костных рыб (Osteichthyes). Выть мо-
жет, это стоит в связи с развитием у них легочного типа дыхания
при помощи плавательного пузыря.
Из сказанного о дыхательном механизме рыб ясно, что здесь мы
имеем высшую ступень в сравнении с Cyclostomata, у которых в жа-
берных мешках окисление далеко пе так обеспечено, как у рыб с их
жаберной решеткой из лепестков.
Добавочные органы и способы дыхания; плавательный пу-
зырь. Те или иные добавочные органы и способы дыхания развивают-
118
Рис. 93. Голова Anabas scandens с до-
бавочным органом дыхания.
1, 2 —жабры. (По Ihle и др.)
ел у рыб как приспособление к особым условиям жизни. Некоторые
из них важны как предпосылка для выхода рыб на сушу, что некогда
имело место при появлении класса амфибий. Таким способом дыха-
нии является кожное дыхание, которое, видимо, существует у неко-
торых рыб. У рода Periophthalmus хвостовой плавник ъчепъ богат
сосудами и служит для дыхания, когда эта рыбка наполовину выле-
зает из воды. В других случаях приспособление для дыхания на
воздухе заключается в специальном изменении жаберпой полости;
у ЛнаЪаз scandens (рис. 93) имеется так называемый лабиринтовый
орган, представленный волпицтыми костными пластинками, си-
дящими в жаберной полости на первой жаберной дуге. Жабры у
зтой рыбки развиты слабо и играют подчиненную роль. Слизистая
оболочка лабиринтового ор-
гана служит для дыхания,
снабжаясь венозной кровью.
I [аличие органа позволяет
этой и другим рыбкам из. так
называемых «лабиринтовых»
вылезать из бедной кислоро-
дом воды тропических морей
иа сушу и пребывать здесь
некоторое время. В семействе
сомовых (Siluridae) у рода
Clarias добавочные органы ды-
хания представлены ветвистыми и в высокой мере богатыми
сосудистой сетью выростами одной или двух задних жабер-
пых дуг, находящимися в жаберпой полости. У Saccobranchus
добавочным органом дыхания являются длинные слепые выросты
жаберпой полости, простирающиеся назад от спинной части первой
жаберпой щели до хвоста, вдоль позвоночного столба. И здесь
стенки мешков сосудистые и снабжаются венозной кровью от при-
носящих жаберпых артерий.
Некоторые из Teleostei заглатывают воздух, который выходит
затем через апальпое отверстие. Слизистая оболочка кишки весьма
богата сосудами, и здесь происходит газообмен с кровью. Кишечное
дыхание встречается у вьюна (Misgurnus fossilis), у некоторых прес-
новодных видов, сомовых, из так называемых панцирных сомов
(Callichtys, Doras, Loricaria, Plccostomus). От этого приспособле-
ния заглатывать воздух для дыхания один шаг до заглатывания
воздуха в так называемый плавательный пузырь, который у неко-
торых рыб выполняет, кроме гидростатической, еще и дыхательные
функции.
Плавательный пузырь имеется у всех Osteichthyes, кроме неко-
торых костистых рыб, у которых он, очевидно, исчез вторично.
У хрящевых рыб (Chondrichthyes) его пет. Насколько известно из
палеонтологии, явился он давно: уже в отряде Coel асан thini из
(-rossoptcrygii, живших с каменноугольного времени до мелового,
и мелся плавательный пузырь, стенки которого окостеневали, а внут-
ри имелись перегородки, как у двоякодышащих рыб. Как форма,
119
характер, так и местоположение плавательного пузыря весьма ва-
риируют у различных рыб. Обычно оп лежит па спинной стороне,
над кишечником и под позвоночным столбом. С передним отделом
кишечника он может быть связан посредством канала, узкого
и длинного (у части Teleostei) или короткого и широкого (у Holo-
stci), или же связи с пищеводом может и пе быть (у другой части
Teleostei). Подразделение Teleostei на открытопузырпых (Physo-
stomi) и «закрытопузырных» (Physoclysti), однако, пе основательно,
так как и у тех, и у других закладывается пузырь всегда в связи
с кишечником, и признак этот пепостоянеп. Форма весьма измен-
чива, даже у очень близких систематически рыб опа может быть
различной. Иногда пузырь очень длинный, в другом случае корот-
кий и широкий (у Lepidosteus, Amia, Gymnarchus), иногда он
дает от себя добавочные выросты. Такая изменчивость формы стоит
в Связи с изменчивостью формы тела у рыб, изменением поло-
жения центра тяжести. Иногда плавательный пузырь состоит из
двух позади друг друга лежащих половин.
С внутренней стороны пузырь выстлан эпителием, который может
быть плоским или цилиндрическим; стенки построены из соеди-
нительной ткани, содержащей в себе эластические волокна и мы-
шечные. Иногда в стенках пузыря наблюдаются окостенения, на-
пример, у упомянутых вымерших Coelacanthini, из современных —
у сомовых (Saccobranchus, например) и карповых (Diplo-
physa). Стенки пузыря обильно снабжаются кровью, обычно вет-
вями аорты, приносящими артериальную, окисленную кровь.
Местами капилляры образуют на внутренней поверхности пузыря
сплетения, называемые «чудесными сплетениями» (rete mirabile).
Эти сплетения видны на внутренней стенке в виде «красных
тяжей», или «красных тел». На верхней стенке пузыря
у рыб, пе имеющих связи пузыря с пищеводом, развивается особое
образование — «овал»: местное резко ограниченное утопчепие стен-
ки пузыря, богато снабженное сосудами и окруженное круговым
мускулом (sphincter), который, сокращаясь, может изолировать
овал от полости пузыря.
Иннервируется пузырь десятой парой головных нервов, кото-
рой иннервируются и жаберпые щели. В пузыре содержатся кисло-
род, азот и следы углекислоты. Относительное количество каждого
газа очень различно у разных рыб, по всегда оно сильно отличается
от того, что наблюдается в воздухе. Газы эти происходят пе из воз-
духа, который и при наличии соединения с пищеводом (ductus pneu-
maticus) в большинстве случаев попасть в пузырь из кишечника не
может (узость ductus), а выделяются из крови, особенно в красных
телах и в овале. Лишь при дыхании воздухом у некоторых рыб по-
следний попадает в пузырь.
Функция плавательного пузыря у современных рыб — прежде
всего гидростатическая. Когда рыба находится па некоторой глу-
бине, ее удельный вес благодаря накоплению газов в пузыре до
известного предела оказывается равным удельному весу воды.
Рыба в это время пе имеет тенденции пи подниматься, пи опускаться.
120
a
3
способа действия ве-
Рис. 94. Схема
беровских косточек у Cyprynidae.
а — при сильном давлении в пузыре, Ь —
при слабом. 1 — плавательный пузырь,
2 — рычаг, 3 — регулятор, 4 — связна.
Когда пузырь расширяется, объем тела рыбы увеличивается,,
а удельный вес становится меньше: рыба поднимается кверху по
закону физики. При опускании рыбы в глубину происходит обрат-
ное: благодаря уменьшению объема рыбы удельный вес ее возра-
стает, и рыба опускается вниз. Это положение подтверждается
ясно экспериментом. Так как изменения пузыря ограничены извест-
ным пределом, то каждая рыба держится па определенной глубине.
У рыб, держащихся у дпа, пузыря обычно пет. Так, его нет у взрос-
лых (но есть у личинок) камбал (Pleuronectidae). Он отсутствует
у некоторых видов семейства макрелевых (Scombridae) и морских
собачек (Blennidae), имеется у
других. Иногда у одного вида
он есть, а у другого нет.
Для выполнения гидростати-
ческой функции пузырь у неко-
торых рыб может иметь особые
приспособления, связываясь со
слуховым лабиринтом. Этасвязь
может служить и для передачи
звуковых колебаний с поверх-
ности тела на орган слуха. У
тресковых (Gadidae) отходящие
от пузыря впереди мешки упи-
раются в затянутое топкой пе-
репонкой отверстие слуховой
области; в семействах карповых
(Cyprinidae), харациповых (Cha-
racinidae), сомовых (Siluridae),
у Gymnarchus (из Mormyridae)
имеется особый сложно устро-
енный «веберов аппарат», слу-
жащий для связи плавательного пузыря с лабириптом.
При посредстве трех косточек, происходящих из трех передних
позвонков и действующих как система рычагов, изменение давле-
ния в пузыре передастся па перилимфу, окружающую лабиринт,
причем в самом лабиринте существуют соответственные изменения
(рис. 94). Высказывалось мнение, что веберов аппарат служит
для усиления вибраций и для проведения их к уху. У тех рыб, кото-
рые, как Zeus, Dactylopterus, Trigla, производят звуки, мускуль-
ные волокна в стейках пузыря хорошо развиты. В связи с пузырем
у некоторых сомовых (Siluridae) развивается особый производящий
звуки орган.
Вудучи гидростатическим органом у высших Osteihthycs, пла-
вательный пузырь выполняет у более примитивных рыб функции
дыхания (рис. 95): у Amia, Lcpidosteus (Holostei), Gymnarchus
(Mormyridae), Erythrina(Characinidae). Плавательный пузырь Le-
piilosteus более или менее разделен вертикальной соединительно-
тканной перегородкой на две части. От лее отходят боковые пере-
кладины, и вся поверхность оказывается подразделенной на ряд
121
камер. Пузырь широкий и короткий, и таким же протоком соеди-
няется со спиппой стороной глотки, куда открывается щелью, ок-
руженной сложной системой мускулов, расширителей и сжима-
телей, иннервируемых и. vagus и принадлежащих к системе мышц
жаберного апарата. Lepidosteus часто поднимается па поверхность
и заглатывает воздух. Повидимому, жаберное дыхание не обеспе-
чивает организма кислородом, и этот недостаток компенсируется
плавательным пузырем. Сходное, но более простое устройство
имеет пузырь у другого костного гапоида— Amia, у которой он
Рис. 95. Внутреннее строение плавательного
пузыря Lepidosteus.
(По Шимкевичу из Ihlc и др.)
снабжается кровью пе спиппой аорты, а от шестой наджаберной
артерии подобно легким двоякодышащих (Dipnoi), а вена идет
прямо к сердцу через печеночную вену. У американской Erythrina
пузырь функционирует двояким образом: передний его отдел как
гидростатический орган, задний — как дыхательный. Это связано
с образом жизни этой рыбы, которой приходится переживать в сухое
время недостаток воды, когда она и заглатывает воздух. Подобно
упомянутым рыбам вид рода Umbra (сем. Umbridae, Esociformcs),
живущий в стоячих водах Венгрии и США, в сухое время зары-
вается в ил и дышит пузырем, стенки которого, хотя и гладкие,
по снабжены большим количеством «rete mirabile». У Polypterus
(Crossopterygii) плавательный пузырь парный, открывается на
брюшпой стороне пищевода щелью с развитой мускулатурой.
Стенки его ячеек не имеют, по образуют 20—30 продольных скла-
док, снабжаясь кровью из 6-й наджаберной артерии. Пузырная
вепа через печеночную вену впадает в сердце. Положение па брюш-
ной стороне и строение протока говорят за то, что это не гидроста-
тический орган, а орган дыхания, хотя еще мало совершенный.
122
Функцию дыхания выполняет плавательный пузырь у двояко-
дышащих (Dipnoi). Его можно назвать уже легким. У Рго-
topterus и Lepidosiren последние являются парными, имеют па
внутренней поверхности многочисленные камеры, разделяющиеся
на маленькие альвеолы и пузырьки. По внешнему виду, по строе-
нию и по кровоснабжению (у Neoceratodus и Lepidosiren — из
наджаберных артерий), по тому, что кровь из них возвращается
прямо в сердце, легкие двоякодышащих имеют огромное сходство
с легкими наземных, четвероногих. У Neoceratodus легкие непарные,
разбиты септами на ячейки, как у Lepidosteus. Neoceratodus в су-
хое время, когда вода загнивает в водоемах, где он живет постоянно,
поднимается па поверхность и заглатывает воздух, по ив остальное
время года пузырь или легкое служит вспомогательным способом
дыхания вследствие того, что у Dipnoi не развился тот сложный
насасывающий аппарат и решетка из жаберпых лепестков, о кото-
рых говорилось выше. Это в еще большей мере относится к
«двулегочным» Protopterus и Lepidosiren, которые в сухое время
года зарываются в ил, где могут дышать только при помощи
легких. )
Легкие и плавательный пузырь развились филогенетически,
повидимому, из одного источника. Между ними нет существенной
разницы, существует ряд переходов между несвязанным с кишеч-
ником пузырем закрыто пузырных (Physoclisti), чисто гидростати-
ческим органом, и легкими Dipnoi. Различия в кровоснабжении
сводятся па-нет наличием переходов. Непарность пузыря также пе
решает вопроса в отрицательную сторону, ибо у Neoceratodus лег-
кое непарное, а следы первоначальной парности имеются в пузыре
в виде двусторонней иннервации и в других признаках. Наличие
у (Lepidosteus, Amia и других рыб пузыря, используемого для
дыхания, также является связующим звеном.
Предки рыб были, вероятно, прибрежными животными. Сами
рыбы возникли в пресной воде, как тип, приспособительный к те-
кучей среде. Жабры сперва были еще несовершенны. Кроме того,
они могли быть недостаточными в мутной воде рек, пе обеспечивали
достаточно энергичного обмена, необходимого при движении в бы-
строй воде рек. У пих развился добавочный орган из заднего пар-
ного выпячивания глотки, совершенно гомологичного жаберным
выпячиваниям. Этот орган был парный и походил более всего па
плавательный пузырь Polypterus. Заглатывание воздуха, наблю-
дающееся и теперь у некоторых рыб, было предпосылкой для раз-
вития этого органа. Этот добавочный орган дыхания развился
далее в двух направлениях: у тех форм, у которых развился хорошо
жаберный аппарат дыхапия, орган этот превратился в плаватель-
ный пузырь, гидростатический орган, а у форм с недоразвитым аппа-
ратом жаберного дыхапия в соответственных условиях разви-
тей легкие.
Дыхание рыб. Дыхапиеу всех организмов заключается в погло-
щении кислорода и отдаче углекислоты. Эволюция дыхания, не-
сем неппо, важнейшего момента в жизни животных, заключается
а 23
в развитии различных приспособлений для повышения количества
поглощаемого кислорода и отдаваемой углекислоты. На примере
эволюции механизма дыхания у рыб мы видели, что целый ряд мор-
фологических изменений ведет к этому. Появление легочного дыха-
ния является одним из важнейших моментов в эволюции дыхания.
Оно делало животных независимыми от водной среды. Жабры
высыхали бы па воздухе, легкие — пет. Кроме того, дыхапие лег-
кими связано с изменением сердца и с более интенсивным обменом
веществ. Поэтому позвоночные с легкими могли выйти па сушу. Но
по существу процесс дыхания остался тот же: наземные животные
являются водными, поскольку дело идет об их дыхании, ибо и опи
дышат через жидкость, покрывающую альвеолярный эпителий.
В этой жидкости растворяются газы, которыми дышат паземные
животные, как растворяются газы в воде, омывающей жаберпые
листки. И в том и в другом случае напряжение газа определяется
парциальным давлением газа в атмосфере, соприкасающейся с этими
жидкостями. Таким образом, физиология газообмена через мем-
браны дыхательных органов у водных и наземных животных одна
и та же, так как скорость диффузии через мембраны в том и другом
случаях зависит от парциального папряжепия газов, растворенных
в жидкости по обе стороны мембраны. Однако устройство органов
дыхапия у водных животных (жабр) и наземных (легких) существен-
но отлично. Легочные пузырьки, альвеолы представляют закрытую
систему, жаберные листки — открытую, свободно омываемую про-
ходящей через решетку водой. Парциальное давление газов в воде,
омывающей жабры, зависит от состояния внешней среды; у рыб пет
средства изменять напряжение газов, окружающих их жаберные
листки. У паземпых парциальное давление кислорода в альвеолах
в любой момент определяется, с одной стороны, барометрическим
давлением, с другой — скоростью и глубиной вдоха и выдоха;
парциальное же давление углекислоты в альвеолярном воздухе
определяется скоростью образования углекислоты в теле и
глубиной вдоха и выдоха, но не барометрическим давлением,
ибо углекислота в воздухе при всяком давлении находится в незна-
чительном количестве. Таким образом, паземные животные, ды-
шащие легкими, могут в известных пределах регулировать пар-
циальное давление О2 и СО2 во внешней среде (воздух альвеол)
скоростью и глубиной дыхапия; рыбы этого делать не могут, и ре-
гулирование соотношений между парциальным давлением О2, СО2
и кровью выпадает па долю последней. Рыбы живут в воде с разным
содержанием кислорода и углекислоты, и способность устанавли-
вать определенное соотношение между газами воды и газами крови
безусловно имеет очень большое биологическое значение. Как на
один пример такого регулирования можно указать па то, что при
уменьшении напряжения кислорода в воде и при возрастании па-
пряжепия углекислоты число красных кровяных телец в крови
рыб возрастает, чем достигается компенсация: рыба приспособ-
ляется к условиям существования и избегает удушения. Такое же
явление видим мы и у млекопитающих. Дыхапие рыб отличается ог
124
ды\.’|11пп млекопитающих своей малой энергией. Так, например,
Ullin. (Tinea vulgaris) потребляет кислорода в 50 000 раз меньше,
чем человек.
7. Органы кровообращения у рыб
Сердце. Рыбы, как и Cyclostomata, имеют (рис. 96) сердце,
jiiijiiiющееся особо развитой частью продольного брюшного сосуда.
: :.|дача его: насасывать венозную кровь, приносимую вепами из
различных частей тела, и проталкивать эту венозную кровь вперед
и вверх к жабрам. Сердце рыб таким образом является веноз-
ным сердцем. Соответственно со своей функцией сердце
располагается пепосредствеппо позади жабр и впереди от того ме-
черные, с артериальной — штрихованные.
1 — венозный синус (sinus venosus), 2 —предсердие (atrium), 3 — желудочек (vent-
riculus), 4 — брюшная аорта, 5 — приводящие жаберные сосуды, 6 — отводящие
жаберные сосуды, 7—сонная артерия, 8—-спинная аорта, 9 — подключичная ар-
терия, 10 —кишечная артерия, 11 —брюшинная артерия, 12 — семенниковая
артерия, 13 — почечная артерия, 14 — подвзДошТгая артерия,15 —хвостовая арте-
рия, 16 — хвостовая вена, 17 —боковая вена, 18—подключичная вена, 19—воротная
вена печени, 20 — печень (из нее идет к сердцу короткая печеночная вена), 21 — пе-
редняя кардинальная вена, 22 —задняя кардинальная вена, 23 — проток Кювье,
24 —почка, 25 —семенник, 26 —кишка, 27 — жаберная щель. (По Паркеру из
Goodrich.)
ста, где вепы, приносящие кровь из разных мест тела, впадают
в брюшной сосуд. Помещается сердце в особой полости, тйк назы-
ваемой околосердечной, которая у селахий и у Chondro-
steoidei соединяется еще с общей полостью тела, частью которой
опа является.
Сердце рыб состоит из двух главных отделов: предсердия
(atrium) и желудочка (ventriculus). Впереди желудочка
лежит еще так называемый артериальный конус
(conus arteriosus) или ясе луковица аорты (bulbus aortae), а позади
предсердия — венозный сипус (sinus venosus). Все эти
четыре отдела зародыша рыб, как и у Ammocoetes, расположены
в одну линию, по затем образуется изгиб, причем предсердие с ве-
нозным синусом располагается наверху, а яселудочск и bulbus cor-
dis — внизу. В венозный синус впадают вепы, идущие от печени
125
(venae hepaticae), и так называемые кювьеровы протоки (ductus
Cuvieri), слагающиеся справа и слева из яремных вен
(venae jugulares) икардинальных (venae cardinales). Синус
открывается в предсердие отверстием, защищенным двумя кла-
панами. Клапаны имеются и в отверстии, ведущем из тонкостен-
ного предсердия в мускулистый желудочек (атрио-вептрикулярный
А В С
Рис. 97. Схема сердца у Selachii (Л),
Amia (В) и Teleostei (С).
Видны сокращение конуса — 1 и развитие
bill bus arteriosus —2—3 предсердье, 4 — же-
лудочек. (По Видерсгейму.)
клапан). Степки последнего
образованы из сильных мус-
кульных перекладин, вдаю-
щихся в полость желудочка.
Впереди желудочек изливает
кровь через конус или луко-
вицу в ствол брюшной аорты,
который лежит уже вне око-
лосердечной полости. Конус
является по существу частью
желудочка. Стенки его мус-
кулисты, причем мышечная
ткань здесь та ясе, что в же-
луд очке, вместе с которым
конус и сокращается.Вкопусе
находятся продольные ряды
полулунных кармаповидпых
клапанов, направленных от-
крытым концом вперед, благодаря чему кровь может итти в
нем только вперед, так как наполненные кровью карманы—кла-
папы закрывают просвет канала (рис. 97).
Артериальный копус (conus arteriosus), имеется
у селахий, у хрящевых ганоидов, Polypterus и Lepidosteus. Но
Рис. 98. Схема жаберного кровообращения у селахий.
1 — сердце, 2 —брюшная аорта, 3 — приводящие жаберные
артерии (artcriae branchialea), 4 — отводящие жаберные артерии
(artenae epibranchiales), 5—корни аорты, 6 —аорта, 7 — брыз-
гальце (spiraculum), 8 — сонная артерия (a. carotis).
(По Ihle и др.)
у костистых рыб за исключением редких случаев (например, у Clu-
peidae) conus имеет тенденцию к исчезновению и замещается несо-
кратимым вздутием без клапанов, так называемой луковицей аорты
(Amia занимает промежуточное положение, имея и bulbus и
conus). Степки bulbus состоят главным образом из эластических
126
ihijkikoii. От conus у Teleostei остаются лишь следы: узкая мускуль-
n.ni полоска с одним рядом клапанов. Сердце Teleostei представ-
лист правшою степень специализации и не ведет к структуре
сор/ща высших позвоночных, которая выводится скорее из струк-
туры с-грдца низших представителей класса. Сердце Dipnoi будет
рассмотрено ниже, когда мы рассмотрим артериальную и венозную
системы рыб.
Артериальная система (рис. 98). Отходящий от сердца брюш-
н <>и сосуд — arteria ventralis, брюшная аорта идет вперед
под жаберным аппаратом, отдавая от себя к жаберпым дугам
носовые сосуды, приносящие жаберные ар те-
р и и (arteriae branchiales). Число их первоначально G, по затем
число жаберных артерий сокращается до 5. У последней жаберпой
дуги пет жабр, а потому и артерия здесь не развивается, припося-
IIi,iie жаберпые артерии существуют па подъязычной дуге и па 4 жа-
берных.
Приносящие жаберпые артерии распадаются в жаберных ли-
сточках ца капиллярную сеть, последняя же собирается в каждой
дуге в в ы н о с я щ у ю, или эпибранхиальиую, артерию.
11ад глоткой эпибранхиальпью артерии собираются па каждой сто-
роне в один ствол, последние же соединяются в спинную
аорту — aorta dorsalis, идущую назад под позвоноч-
ным столбом до самого заднего конца тела, и отдающую по дороге
ветви к различным частям тела: к парным плавникам идут под-
ключичные артерии — arteriae subclaviae, к печени
и желудку — arteria coeliaca, к кишечнику и поджслудочной
.железе — мезентериальная, б р ы ж е е ч п а я артерия,
к селезенке — селезеночная, к почкам — почечная,
к тазу — подвздошная — arteria iliaca. Первая принося-
щая жаберпая артерия пе развивается, исчезает. Благодаря этому
соответствующая arteria epibranchialis теряет связь с брюшпой
аортой. Она соединяется со тзторой эпибрапхиалыюй артерией,
идущей над подъязычной дугой, и снабжает сниракулярпую жабру
оксидированной кровью, продвигаясь вперед в голову в виде п а-
ружной со и н о и а р т е р и и (arteria carotis externa).
Продолжение вперед парных спинных аорт дает внутренние
сонные артерии (arteriae carotides internae). Эти послед-
ние соединяются между собой в черепе, замыкая кольцо — cir-
culus cephalicus. Сонные артерии снабжают головной мозг окислен-
ной кровью. По этой же схеме построена кровеносная система и у
других рыб, кроме акул. Но так как у Teleostei пи на гиоидной,
пи па челюстной дуге пет жабр, то недоразвитию подвергается
1-я и 2-я артериальные дуги и остаются всего 4.
Своеобразные отличия видим мы в системе артериальных дуг *
у Dipnoi благодаря развитию здесь легочного дыхания. Здесь
развиваются легочные артерии (arteriae pulmonales),
несущие к легким кровь, богатую углекислотой, и легочные
вены (venae pulmonales), по которым кровь (артериальная) идет
от легких к сердцу. Легочные вены являются новообразованием,
127
легочная же артерия есть ветвь шестой эпибранхиальпой артерии.
Это оказывает большое влияние па строение сердца (см. ниже).
У Protopterus существуют 3 пары наружных жабр. Опи (рис.99)
снабжаются венозной кровью через 4-ю, 5-ю, 6-ю приносящие
артерии, дающие веточки к этим жабрам. Оксидированная кровь
возвращается в выносящие,
ю
Рис. 99. Жаберное кровообращение у
Protopterus.
1 — сердце, 2 — брюшная аорта, 3 — принося-
щие жаберные артерии, 4 — эпибранхиальный
сосуд, 5—эпибранхиальная область артериаль-
ной дуги первой жаберной щели, 6 — принося-
щие ветви к наружным жабрам, 7 — наруж-
ные жабры, 8 — выносящие артерии жаберных
дуг, 9 —сонная артерия, 10—левая ветвь
спинной аорты, 11—a. coeliaca, 12 —легоч-
ная артерия. (По Паркеру из Goodrich.)
эпибрапхиальные артерии,
откуда поступает в аорту
и в легочнуй) артерию.
Кроме того, мы видим
у Protopterus, что 3-я и
4-я жаберные дуги вслед-
ствие редукции соответст-
венных жабр пе распада-
ются па капилляры, пе
делятся па приносящую и
выносящую части, а явля-
ются непрерывными, па-
поминая то, что имеется
у амфибий.
У Neoceratodus (рис.
100) этого пет, так как у
него соответственные жаб-
ры сохраняются.
Плавательный пузырь
рыб снабжается, как пра-
вило, кровью от спинной аорты через arteria coeliaca; однако у
Amia он снабжается через артериальные ветви, отходящие от 6-й
пары наджаберных артерий, у Gymnarchus оп снабжается па левой
па правой — от arteria
coeliaca. Также и у
стороне от 5-й и 6-й наджаберных дуг,
Рис. 100. Схема кровообращения в жаберной
области Neoceratodus.
1 —сердце, 2 — легочная артерия, 3 — легочная вена,
4 — протоки Кювье, I—V — жаберные щели. (По Ihle
и др.)
Polypterus пузырь
снабжает 6-я пара
наджаберпых арте-
рий. Таким образом
уже у рыб имеются
предпосылки в стро-
ении кровеносной си-
стемы к развитию ле-
гочного дыхания.
Венозная система.
Венозная система рыб
построена по общему
плану с Cyclostomata. Яремные вепы (venae jugulares) пли
передние кардинальные (v. cardinales anteriores), и два
венозных ствола из органов туловища и хвоста — задние
кардинальные вены (v. cardinales posteriores).
Из хвоста кровь идет по непарной хвостовой вене,
расположенной под позвоночным столбом в канале, образованном.
128
затем в гп о-
в кардиналь-
уже
воротную
Рис.
венозной системы
селахии.
1—брюшная аорта,
2 — хвостовая вена,
3 — почечные вены,
4 — задняя карди-
нальная вена, 5 —
боковая вена (v. la-
teralis), 6— семенни-
ковая вена, 7 — во-
ротная вена, 8—пе-
чень, 9 — подклю-
чичная вена, 10—
передняя кардиналь-
ная вена, 11 — под-
вздошная вена. (По
Bridge из Ihle.)
101. Схема
нижними, или гемальными, дугами позвонков. В туловище хво-
стовая вена делится на две ветви, идущие к почкам — воротные
вены почки (v. portae renales). В последних веточки вен
распадаются на сеть капилляров, собирающуюся
чечные вепы (venae renales), впадающие
ные вены. Таким образом, у рыб мы видим
систему почек. Такая же воротная система
имеется в печени; вены, идущие от кишечного
канала, распадаются в печени на капилляры
(воротная вена печени, v. portae hepa-
ticae), собирающиеся затем в печеночную
вену (vena hepatica) (рис. 96). Печеночная
вепа вливается в венозный синус. Кардинальная
и яремная вены каждой стороны сливаются
перед впадением в последний в так называе-
мые кювьеровы протоки (ductus Cuvieri)
(рис. 101). В кювьеровы протоки впадают так-
же имеющиеся у рыб боковые вены
(venae laterales), несущие кровь из задних ко-
нечностей и из кожи хвоста и туловища, сливаю-
щиеся перед этим с подключичными
венами (venae subcl avaie).
В разных классах рыб существуют различ-
ные уклонения от этой схемы, ив венозной
системе Dipnoi мы видим, на ряду с при-
митивными чертами, такие, которые являются
переходом к состоянию, наблюдаемому у взрос-
лых наземных, воздухом дышащих, позвоночных
(рис. 102). Прежде всего парпые кардинальные
вены заменяются непарной задней полой
в е п о й (vena cava posterior). Эта вена у
Dipnoi, развивающаяся за счет правой карди-
нальной вепы, берет иа себя функцию карди-
нальных вен. Через нее непосредственно в синус
идет кровь и из почек. Затем у Dipnoi впервые
появляется непарпая брюшпая вепа (vena
abdominalis), образующаяся путем частичного
слияния боковых вен и открывающаяся прямо
в правый кювьеров проток. Эту вену мы встре-
чаем потом у амфибий. Интересно, что веноз-
ная система Dipnoi ближе к таковой селахий, чем к венозной
системе Teleostei.
Особенного внимания заслуживает сердце Dipnoi. Здесь на-
чинается тот ряд развития сердца наземных позвоночных, который
заканчивается четырехкамерным сердцем птиц и млекопитающих,
с полным разделением сердца на правую и левую половины, и кро-
пи — на артериальную и венозную, что, конечно, способствует го-
раздо более энергичному обмену веществ в организме. У Neocera-
todus сердце построено (рис. 103) по тому же принципу, как и
З Кашкаров и Стан чин ский 129
Рис. 102. Схема веноз-
ной системы Dipnoi.
1 — сердце, 2 — хвостовая
вена, 3 —почка, 4 —зад-
няя полая вена, 5—-под-
вздошная вена, 6—брюшная
вена, 7—задняя кардиналь-
ная вена, 8—печеночная ве-
на, 9 — воротная вена пе-
чени, 10 — легочная вена,
11 —печень, 12 — подклю-
чичная вена, 13—передняя
кардинальная вена, 14 —
проток Кювье. (По Good-
rich.)
у прочих рыб. Однако па спиппой стороне
предсердия и желудочка находится про-
дольная складка, не доходящая до брюш-
ной стороны этих полостей и потому
разделяющая их па правую и левую поло-
вины неполно. Венозный синус открывает-
ся в предсердие пе прямо сзади, а несколь-
ко справа от средней линии, так что
более широким отверстием открывается
в правое предсердие и меньшим в левое.
В левую половину предсердия откры-
ваются сросшиеся вместе легочные в е-
п ы (venae pulmonales). Таким образом,
в правое предсердие попадает венозная
кровь, в левое немного венозной и
артериальная, окисленная из легочных
вен. Так как при сокращении сердечной
мышцы перегородка прижимается к ниж-
ней стенке сердца, получается полное
разделение в это время венозной от арте-
риальной крови. Длиппый мускулистый
артериальный конус имеет у Dipnoi, как
выше сказано, многочислепные клапаны,
расположенные в 8 поперечных рядов.
Рис. 103. Схема сердца Neo-
ceratodus со спинной стороны.
1 — венозный синус, 2 — проток
Кювье, 3 —правое предсердие, 4
— левое предсердие, 5—желудо-
чек, 6 — спинная перегородка,
7 — артериальный конус, 8 — спи-
ральная складка. (По Ihle и др.)
Клапаны 6 задних рядов, находя-
щиеся по срединной липии брюшной
стороны, соприкасаются друг с дру-
гом, образуя продольную «спираль-
ную складку». Сам конус спирально
скручеп. Поэтому впереди эта спи-
ральная складка из сагиталыюго по-
ложения становится в горизонталь-
ное, фронтальное. Перегородка в же-
лудочке и спиральная в конусе почти
соприкасаются. Благодаря этому в
правую и верхнюю части конуса течет
кровь преимущественно венозная,
в левую — преимущественно арте-
риальная. В верхней части конуса,
конечно, происходит еще некоторое
смешение крови, так как спиральная
складка нс доходит доверху. Но в
момент сокращения конуса половины
последнего опять разделяются впол-
ие. Кровь из правой половины пред-
сердия попадает таким образом через
спиппую часть конуса в 5-ю и 6-ю
arteriae epibranchiales, отходящие от
верхней части конуса. Наиболее ве-
130
позпая кровь идет таким образом к легким через a. pulmonales.
Наиболее оксидированная кровь из вситралыюго отдела конуса
поступает в соппые артерии и в спинную аорту. Так бывает
тогда, когда жабры не функционируют; если же они функцио-
нируют, то во всех эпибрапхиальных артериях течет окислен-
ная в жабрах кровь, попадая и в легкое, которое не работает.
Таким образом, лучшее окисление в организме имеет место в
период нахождения рыбы в воде. Легочное дыхание «выручает
в беде», когда жабры пе могут функционировать. В это время рыба
ведет мепее активную жизнь. Но пе следует забывать, что жабер-
пое дыхание стоит у Dipnoi пе па высокой ступени и развитие лег-
кого является добавочным способом дыхапия.
У Ceratodes — одно легкое (Monopneumones). У имеющих два
легких Protoptcrus и Lcpidosiren (Dipneumoncs) строение сердца
совершеннее, и артериальная кровь почти целиком отделена от
венозной. Достигается это тем, что в конусе спиральная складка
вполне отделяет спинную и брюшную половины конуса. Кроме того,
вепозпый сипус открывается целиком в правое предсердие, и
левое получает только оксидированную кровь. Благодаря этому
в 5-ю и 6-ю жаберпые артерии попадает только вепозная, в 3-ю и
4-ю—: только артериальная кровь. У Lcpidosiren предсердие вполне
разделено па две половины.
По характеру кровообращения Dipnoi могли бы существовать
па суше. Однако к наземному образу жизни они пе перешли. Ne-
oceratodus пользуется легкими при усыхании или загрязнении во-
доемов, в которых он живет, Protoptcrus и Lepidosircn — во время
«летней спячки» в сухое время года, когда они лежат, зарывшись
в ил. Таким образом, Dipnoi — животные в полном смысле слова
двоякодышащие. Сердце Dipnoi не толькц «жаберное сердце», как
у рыб, но и «легочное сердце». У наземных — только последнее.
Следует отмстить, что у Dipnoi, как предпосылка и необходимое
условие легочного дыхапия, носовые ямки открываются в рото-
вую полость, образуя задние ноздри, или choanae.
Кровь рыб имеет, если можпо так выразиться, эмбриональный
характер. Красные кровяные шарики обычно плоски, имеют оваль-
ную форму, двояковогнуты, окрашены гемоглобином и имеют ядро.
У Dipnoi они чрезвычайно крупны, чем Dipnoi еще раз напоминают
амфибий. Лейкоциты вариируют по числу даже у одного н того же
вида в разных физиологических условиях. Особенно многочислен-
ны они у Dipnoi. Из зернистых лейкоцитов у рыб имеются только
• специальные гранулоциты». У некоторых рыб в них пет зернисто-
сти. Эозинофилы встречаются лишь в виде редкого исключения.
8. Органы пищеварения рыб
Органы пищеварения у рыб, как и у остальных позвоночных,
представляют мускулистую трубку, выстланную внутри эпителием
н предназначенную для принятия, проведения, переваривания и
всасывания переваренной пищи. Начинается пищеварительный
* 131
канал ротовой щелью и полостью, выстланной эктодермическим
эпителием (stomodaeum), оканчивается анальным отверстием
(anus), выстланным тоже эктодермическим эпителием (proctodae-
um). Остальная часть пищеварительного капала происходит из
энтодермы и носит название mesenteron. Лежит пищеварительный
капал в полости тела в ее брюшной части. (В другой — околосердеч-
ной— лежит сердце.) Брюшная часть полости тела выстлана внут-
ри брюшиной (peritoneum). Последняя состоит из наружного
соединительнотканного слоя и внутреннего эндотелиального. Брю-
шина одевает все почти органы и подвешивает их к стенкам тела:
кишечник подвешен на брыжейке — mesenterium, представ-
ляющей двойную складку брюшины. Эволюция кишечника заклю-
чается в том, что простая, почти недифференцированная прямая
трубка дифференцируется на ряд отделов с различным строением
и различными функциями. Строение пищеварительного канала
находится в соответствии с родом пищи животного.
В отличие от круглоротых рот рыб представляет поперечную
щель, ограниченную челюстями. Рот может находиться внизу головы
(у акул, например) или на конце ее, как у большинства рыб, или
нижняя челюсть бывает сильнее развита, и рот оказывается тогда
находящимся на верхней стороне рыла. Рот ведет в ротовую по-
лость. Последняя в большинстве случаев бывает снабжена зубами. За
ротовой полостью следует пронизанная жаберными щелями глотка,
в которой также могут находиться зубы. Последние развиваются
филогенетически из плакоидных зубов, которые образуются в по-
кровах, дающих путем впячивания начало stomodaeum. Если зу-
бов нет у некоторых взрослых рыб, то нередко они обнаруживаются у
эмбрионов (например, у осетровых), так что беззубость является вто-
ричным состоянием. У селахий зубы сидят поперечными рядами в ок-
ружности ротовой щели,па меккелевом хряще и на palatoquadratum.
У Holostei они встречаются на всех костях ротовой полости: на ргае-
maxiHae, на palatini, реже на maxillae, па vomer, parasphenoideum,
pterygoidea, на glossohyale, на жаберных дугах. Укреплены зубы
во рту различно. Наиболее примитивные зубы элазмобранхий про-
сто сидят в слизистой оболочке десен, соединенные с челюстями во-
локнистой соединительной тканью. Иногда (у Polyodon) и этого
соединения с челюстями пет. У Teleostei, имеющих подвижные зубы,
последние прикреплены к костям челюстей волокнистой или эласти-
ческой связкой (у щуки — Esox, у морского чорта—Lophius). Лишь
редко (у Characinidae) зубы сидят в ячейках. Почти у всех рыб зубы
сменяются новыми, т. е. рыбы полифиодонтны. По форме,
величине и строению зубы весьма вариируют в связи с различиями
в образе жизни и характере пищи. Наиболее примитивный тип,
встречающийся у некоторых элазмобранхий и многих Teleostei,—
простые конические зубчики. Из этого простого конуса получаются
зубы режущие, пилообразные, зазубренные, иногда очень сложной
формы, и зубы плоские, перетирающие, расположенные наподобие
мостовой (например, у скатов). Зубы могут сливаться между собой,
образуя «жерновые» пластинки (у акул сем. Heterodontidae)
132
(рис. 104), или пластинки режущие (у Diodon, Tetrodon, Scarus).
11иогдау рыб имеется гетеродонтия — различные зубы. Так,
у Heterodontus (Cestracion) передние зубы служат для схватывания
и имеют коническую форму, зад-
ние имеют уплощенную форму и
являются давящими зубами.
Гетеродонтия встречается и у не-
которых Teleostei (Serranidae и
др.). У многих Teleostei зубов
пет вовсе (Cyprinidae, Syngna-
thidae и др.).У некоторых Teleos-
tei зубы развиваются на жабер-
ных дугах, плотно с ними сра-
стаясь. Эти так называемые «гло-
точные зубы» бывают обычно
развиты при недостатке разви-
тия зубов на челюстях. Иногда
они развиты только на cerato-
branchialia последней жаберной
Рис. 104. Верхняя челюсть Hetero-
dontus (Cestacion) philippi.
дуги («нижнеглоточные зубы»),
а в паре с ними работает мозолистое роговое утолщение на верхней
стороне глотки под basioccipitale, так называемый «ж е р н о в о к».
4
Рис. 105. Продольный разрез самца акулы Scyllium.
1 — сердце, 2 — глотна с ж;) верными щелями, 3 — пищевод, 4 —
входная часть желудка (cardium), 5 — выходная часть желудка
(pylorus), 6 — печень, 7—селезенка, 8—поджелудочная железа,
9 —спиральный клапан кишки, 10-правая доля печени, 11 —
кишка, 12 —клоака, 13 —семенник, 14—придаток семенника,
15 —головной мозг, 16 — позвоночник, 17 — спинной мозг. (По
Паркеру из Boulenger.)
Иногда же имеются зубы и на pharyngobranchialia— так "назы-
iiiie.Mi.ic «верхние глоточные зубы».
Мускулистого языка у рыб нет. Зачаток его есть у Elasmobran-
cliii, Dipnoi nCrossopterygii, имеющих в нем мускульные волокна.
Обычно же так называемый «язык» рыб представляет не более
133
как вдающуюся в полость рта entologlossum, одетую слизистой
оболочкой.
Слюнных желез у рыб пе имеется, но диастатический фермент во
рту выделяется, видимо, клетками эпителия последнего. Так как
вода все время омывает ротовую полость, то слюнные железы были
бы здесь излишними.
За глоткой следует пищевод, обычно короткий,, широкий и не-
резко отграниченный от желудка (рис. 105, стр. 133). Мускулатура
его, одпако, поперечнополосатая, и эпителий не цилиндрический,
как в желудке. Наличие цилиндрического эпителия и пищевари-
тельных желез характерно для желудка. Форма его бывает или в
виде колепчатой трубки со входом (cardium) па одном конце и
выходом (pylorus) на другом или в виде слепого мешка, если вход
и выход паходятся па близком расстоянии друг от друга.
Рис. 106. Продольный разрез самца Teleostei (Salmo fario).
1 — сердце, 2 — глотна с жаберными щелями, 3 — пищевод, 4 —желудок, 5—двенад-
цатиперстная кишка, 6 — пилорические придатки, 7 — кишка, 8 — семенник, 9 —
плавательный пузырь, 10 —почки, 11 — позвоночник, 12 —головной мозг. (По
Паркеру и Гасвелю из Boulenger.)
Кишечник у рыб еще очень мало дифференцирован. У Holocephali
оп короток и отделов в пом еще нет. В кишке имеется слабо раз-
витый «спиральный клапан», делающий пе более 8 оборотов. У села-
хий. имеются уже более явственные отделы, в желудке различимы
вход и выход (pylorus), спиральный клапап в заднем отделе кишки
образует до 40 оборотов. Самый задний отдел образован так называе-
мой «клоакой» (cloaca), куда открываются также мочеполовые про-
токи. Кишечник Dipnoi сходен с таковым Holocephali. Наоборот,
кишечник Holostei и Crossopterygii сходен с кишечником селахий,
по в развитии спирального клапана имеется стремление к редукции
и появляются так называемые пилорические придатки
(appendices pyloricae)(pnc.l06). Спиральный клапан имеется у Ро-
lypterus, у Lepidosteus же и у Amia он едва развит. У костистых
рыб Teleostei наблюдаются наибольшая дифференцировка и наиболь-
шее разнообразие в строении пищеварительного канала, в целом
все еще весьма простого.Число пилорических придатков колеблется
134
желудка. Протеаза со-
з
Рис. 107. Поперечный разрез через часть стенки
кишки, комбинированный из того, что наблю-
дается у высших и низших позвоночных.
I*—брюшина (peritoneum), 2—продольный слой мышц,
3 — поперечный слой мышц, 4 — подслизистая (submu-
cosa), 5 —сосочки высших позвоночных, 6 —лим-
фатический фолликул, 7 — лейкоциты, 8—кишечный
эпителий, 9 — одна из либеркюновых желез высших
позвоночных, 10 — лимфатический сосуд. (По Видерс-
гейму из Boulenger.)
зд<‘сь от 2—3 до 200 (у Scomber). О функции пилорических придатков
мы знаем очень мало; повидимому, это не выделительный орган,
а. резорбирующий.
У сельди (Clupea) пилорические придатки содержат фермент
протеазу, возможно попадающий туда из —
храпнет свое дейст-
вие и в солевых рас-
творах. Это имеет зна-
чение призасоле сель-
ди, так как при уда-
лении внутренностей
пилорические при-
датки обычно оста-
ются в брюшпой по-
лости.
Анальное отвер-
стие находится обыч-
но далеко позади.
Лишь у рыб, у кото-
рых брюшные плав-
ники расположены
далеко впереди, на
горле (jugulares) и
анальное отверстие
переносится далеко
вперед. Лежит anus
всегда впереди отвер-
стия урогенитальной
(мочеполовой) систе-
мы. У рыб всегда имеется печепь (hepar), обычно и желчный пузырь.
Поджелудочная железа (pancreas) имеется пе всегда. Опа имеет
примитивное гроздевидное строение, открываясь в кишку позади
pylorus несколькими или одним каналом.
Физиологическая дифференцировка отделов пищеварительного
капала невелика, пепсин вырабатывается видимо и в желудке, и в
кишке; в пилорических придатках кроме слизи встречается и сок
поджелудочной железы; печепь представляет физиологически hepa-
topancreas, т. е. соединяет функцию печени и поджелудочной железы.
Гистологическое строение кишечного канала видно из рисунка
(рис. 107).
9. Органы выделения и размножения (мочеполовая
система) рыб
Почки рыб в виде двух длинных железистых масс лежат по бокам
позвоночного столба вне брюшной полости, т. е. забрюшинпо. Пер-
ричпые почечпые капалы, или вольфовы каналы, служат
выводными протоками. Функционирует у рыб всегда п е р в и ч-
д а я дочка, или mesonephros, но у зародышей имеется головная
435
ю
почка — pronephros. Лишь у немногих Teleostei (Fierasfer, Dacty-
lopterus, Zoarces, Lepadogaster и др.) pronephros не исчезает вполне
до половой зрелости. Выводные (первичнопочечные) протоки у хря-
щевых рыб на конце
расширяются и образу-
ют на каждой стороне
мочевой синус, на об-
щем сосочке открыва-
ющийся в клоаку. У
Osteichthyes, выводные
протоки назади слива-
ются между собой, об-
разуют вздутие, функ-
ционирующее как моче-
вой пузырь, но его не
следует смешивать с
настоящим мочевым пу-
зырем, развивающимся
у наземных из нижней
части конца кишечного
канала. Открываются
протоки или вместе с
половыми отверстиями
или (чаще) на особом
сосочке позади послед-
них.
Характер развития
почек неодинаков в
классе рыб (рис. 108);
у элазмобрапхий pro-
nephros редуцируется,
функционирует meso-
nephros, причем у мно-
гих акул сохраняются
еще nephrosostomata,
воропки, открывающие-
ся в полость тела. Ли-
чинки Dipnoi имеют
хорошо развитую и
функционирующую го-
ловную почку; первич-
ные почки вытянуты в длину, их выводные протоки открываются в
клоаку. Хорошо развитая головная почка—pronephros—имеется
также и у личинок Polypterus (Cr ossopterygii). У Chondrostei Holostei
также имеется на ранних стадиях развития pronephros, канальцы
которого имеют воронку, капсулу и железистую часть. Как у Poly-
pterus, так и у «ганоидов» (Chondrostei, Holostei) имеется первичный
мочевой капал, открывающийся в sinus. И у Teleostei, как сказано,
pronephros имеется в зародышевом состоянии, а у некоторых выше-
136
Рис. 108. Основные модификации выделитель-
ной и половой системы круглоротых и рыб.
A— pronephros и ее проток у зародыша, В— почки
и генитальные поры у Petromyzon, показан зача-
точный pronephros, как у Myxine, С и В —мочепо-
ловые органы самца и самки элазмобрапхий, Е —
самец или самка Teleostei или самец Lepidosteus,
F —самка Polypterus, Amia, Acipenser и Osrnerus.
1 — клоака, 2 —кишка, 3 — анальное отверстие,
4 — абдоминальные поры, 5 —нефростом, 6 —pro-
nephros, головная почка, б!—зачаточная prone-
phros, 7 — архинефричсский проток, 8 — первичная
почка mesonephros, 9 — семенник, 10 —мальпи-
гиево тельце, 11 — метанефрический проток, 12 —
мезонефрический проток, 13 -г семенной пузырек,
14—семенной мешок, 15 —яичник, 16—гонада,
17 — экскреторная часть mesonephros, 18—гениталь-
ная часть mesonephros, 19 — зачаточный meso-
nephros, 20 — перитонеальный канал, 21—гени-
тальная пора, 22—половой проток. (По Boulcnger).
указанных форм и во взрослом состоянии. Mesonephros рыб с ко-
стным скелетом никогда не имеет нефростомов.
Половые органы представлены у самцов семенниками
(testes), у самок — яичниками (ovarii). Развиваются половые
органы из определенного участка эпителия, выстилающего полость
тела и расположенного справа и слева от позвоночного столба.
Как сами гонады, так и их выводные протоки построены по-разному
в разных классах рыб. Есть основания думать, что первоначально
сегментальные трубочки служили для выведения половых продуктов
из полости тела. Эту функцию они еще иногда частично выполняют
у некоторых позвоночных, и мы видим тесную связь выделительных
и половых органов и их протоков.
У селахий (рис. 108, С и Д) семенники лежат на уровне переднего
конца mesonephros. Передний и средний отделы последней совершен-
но теряют здесь свою выделительную функцию, выделяют лишь жид-
кость, в которой движутся сперматозоиды. Семенник соединяется
с передним концом mesonephros при помощи выносящих трубочек,
vasa efferentia, соединяющихся в одну сильно извитую трубочку
epididymis. Мезонефрический проток утрачивает здесь свою функ-
цию и служит для проведения семени к клоаке, образуя перед впа-
дением в последнюю расширение — семенной пузырек,
vesicula seminalis. Он служит для накопления семени. Задняя
часть почки функционирует как выделительный орган. Трубочки
этой части соединяются в функционирующий мочеточник.
Мюллеров проток существует у самцов селахий в виде зачатка.
У самок селахий передняя часть почки также педоразвита, а каналы
задней части соединяются в мочеточник, отдельный от первичного
канала. Яичник имеет вид длинного лопастного органа. Созревшие
яйца попадают в брюшную полость, а оттуда в яйцеводы. Яйце-
воды, образованные срастанием мюллерова капала с воронками
канальцев предпочки, длинны и соединены впереди, открываясь
в полость тела общим отверстием. Другой конец впадает в клоаку.
Яйцеводы состоят из нескольких отделов. В переднем отделе яйце-
водов яйцо, здесь и оплодотворяемое, одевается значительным коли-
чеством белка. В следующем кзади отделе особые железы выделяют
хитиновую скорлупу. В самом заднем отделе яйцеводов имеется рас-
ширение, в котором у живородящих акул происходит развитие за-
родышей. Отдел этот часто называют маткой. Оплодотворение у акул
внутреннее, и для совокупления служат видоизмепеппые передние
и учи брюшных плавников — птеригоподии (pterygopodium).
<>ни скручены в трубку, переднее отверстие которой помещается
у клоаки. Через птеригоподии вводится семя самца в клоаку.
I (ельпоголовые (Holocephali) в основном сходны по организации
мочеполовой системы с селахиями. То, что мы наблюдаем у элазмо-
ьр.шхий, весьма напоминает строение мочеполовой системы у
ПМф||б||Й.
Другой тип этой системы мы видим у-Teleostei. Dipnoi, Chondro-
id с i и Holostei представляют переходное состояние от элазмобрап-
х нН к костистым рыбам. У Dipnoi яичники парные, и яйца выводятся
137
через длинные яйцеводы с воронкой на конце, сходно с селахиями.
Самцы также обнаруживают сходство с последними: семя выво-
дится через посредство mesonephros, хотя пе через передний, а че-
рез задний его отдел. Семенник тоже в соответствии с этим разделен
на переднюю активную часть с семенными трубочками и на заднюю,
которая имеет трубчатое строение и служит для соединения семен-
ника с мезонефрическим протоком. Дальнейшая редукция этих
трубочек приводит к обособлению семяпровода от мочеточника, что
мы видим у костистых рыб. У самок Crossopterygii (Polypterus),
у Chondrostei, у Amia и из костистых у Osmerus (рис. 108, F) яйце-
воды открываются воронкой в полость тела, по они пе столь длинны,
как у элазмобрапхий, укорочены и теспо прилежат к яичникам.
У самцов Chondrostei, Amia и Lepidosteus соединение семенников
с почками примитивное, как у селахий. У Teleostei и у самок,
и у самцов наблюдаются иные отношения: семенники имеют
свои собственные выводные протоки — семяпроводы. Яич-
ники у них и у Lepidosteus имеют мешковидную форму, и стенки
их непосредственно продолжаются в короткие каналы, открываю-
щиеся па мочеполовом сосочке. Яйца выводятся пе через полость
тела, а через эти каналы. Лишь у немногих, притом более прими-
тивных, форм костистых рыб (у лососевых, Salmonidae, у угревых—
Muraenidae, у Mormyridae и из карповых у Cobitis) выводпого ка-
нала яичников нет, яйца выпадают в полость тела и выводятся
наружу через особые отверстия — pori abdominales, брюшные
поры.
У Salmonidae брюшные поры рудиментарны, а яйца выводятся
через особое пепарпое отверстие — половую пору, porus genitalis.
Сравнивая различные формы половых протоков у рыб, мы при-
ходим к следующему представлению об их развитии. У предков
костистых рыб должны были быть нормальные мюллеровы каналы,
т. е. открывающиеся в полость тела воронкой, как у селахий и
«гаиоидов», хотя у последних они несколько редуцированы. Затем
каналы эти исчезли. Функцию выведения яиц стала выполнять
полость тела двумя различными способами. У Salmonidae задняя
часть целотслия сделалась непарной половой воропкой или парной,
но сливающейся в одно отверстие: гспиталыюе или, половое, от-
верстие. Яйца, выпадающие по созревании в полость тела, выводятся
через это отверстие. В другом случае яичники из лептовидпой
формы приняли форму мешка, полость которого является частью
полости тела. Задние пе дающие яиц отделы этих мешков соедини-
лись вместе, образовался выводной проток, открывающийся при
помощи полового отверстия.
Примитивное состояние мужского полового аппарата мы видим
также у элазмобрапхий. Через уменьшение связи семенника с поч-
кой, что наблюдается у Polypterus, развитие пришло к тому состоя-
нию, которое мы наблюдаем у костистых рыб. Половая система
амфибий должна была развиться из примитивного состояния,
сходного с тем, что имеет место у селахий.
У рыб полы раздельны, но у некоторых костистых рыб (Teleostei)
138
гермафродитизм является вполне нормальным явлением. Он наблю-
дался у родов Serranus, Chrysophris как обычное явление и в виде
исключения у карповых, тресковых и сельдей, а также у ерша.
10. Нервная система и органы чувств у рыб
Нервная система рыб организована выше, чем таковая у
Acrania и Cyclostomata, но все еще весьма примитивна. Основные
отделы ее те же, что у круглоротых; развитие нервной системы у
зародыша происходит, как у последних, из впячивания эктодермы
на верхней стороне тела. И здесь первоначально образуются три
мозговых пузыря: передний, средний и задний мозг из передней
Рис. 409. Головной мозг акулы Scyllium canicula.
А — со спины, В — с брюшной стороны. 1 — обонятельная
доля, 2 —полушария, 3 — промежуточный мозг, 4 — сред-
ний мозг (зрительные доли), 5 — мозжечок, 6 — продолговатый
мозг, 7—четвертый желудочек, 8 — питуитарная железа (гипо-
физа), 9 —эпифиза, II, IV, V, VI, VII, VIII, IX и X—че-
репные нервы. (По Видерсгейму из Boulenger.)
части нервной трубки и спинной мозг — из остальной. Последний
обычно тянется через спинномозговой капал, пе образуя шейного
и пояспичного утолщений, как бывает у наземных позвоночных.
1)то попятно, так как парные плавники пе несут еще той работы,
которую песут конечности наземных.
'Головной мозг в различных отрядах обнаруживает весьма боль-
шое разнообразие, по в общем можно выделить три типа строения
головного мозга: тип элазмобранхий, тип костистых рыб и тип двоя-
кодышащих. Во всех этих трех типах он мал и занимает незначи-
тельную часть черепной полости, заполненной жировой рыхлой
тканью.
У элазмобранхий, особенно у акул передний мозг (рис. 109)
значительно развит, от него отходят большие обонятельные доли,
139.
Рис. 110. Схематический срединный раз-
рез через мозг:
А —селахии (ската), В —зародыша форели.
1 — обонятельная доля, 2 — полушария, 21 —
полосатое тело, 3 —эпифиза, 4 —зрительные
доли, 5 — мозжечок, 6 — гипофиза, 7 — воронка
(infundibulum), 8 — продолговатый мозг. (По
Edinger.)
и в крыше переднего мозга есть нервная ткань (рис. 110, Л). Имеется
поверхностное деление переднего мозга на правую и левую половины,
но разделения на два полушария еще нет, так как полость переднего
мозга, или желудочек, остается неразделенной. Обонятельные доли
сидят иногда па ножках (tractus olfactorius). От промежуточного
мозга отходит кверху эпифиза, надмозговой придаток, -а с ниж-
ней стороны — так называемая воронка (infundibulum) и
подмозговой придаток (hypophysis). Хорошо развит
средний мозг, разделенный на две доли — lobi optici.
Мозжечок (cerebellum) сильно развит, прикрывает собой
впереди лежащий средний мозг и позади лежащий продолговатый
и разделен на доли про-
дольной и поперечными бо-
роздками. Продолго-
ватый мозг (medulla
oblongata) хорошо развит.
| На верхней стороне его
• имеется так называемая
ромбоидальная ям-
ка, в полость которой
вдаются вздутия, соответ-
ствующие началу V пары
нервов и X (n. trigeminus и
n. vagus). У электрического
ската (Torpedo) на дне ром-
боидальной ямки находят-
ся очень большие вздутия
с гигантскими нервными
клетками — так называе-
мые электрические
лопасти.
Головной мозг цельно-
головых (Holocephali) в
основном повторяет мозг
селахий, но кроме некото-
рых особенностей формы обнаруживает несколько более высокую
дифференцировку мозговых полушарий.
Головной мозг ганоидов и костистых рыб (Teleostei) (рис. 110, В)
весьма вариирует по форме и развитию отдельных частей. Его осо-
бенностью, отличающей от мозга селахий, является то, что передний
мозг представлен у них непарным пузырем, верхняя стенка которого
очень тонка и образована не нервной тканью, а эпителиальной
(pallium): па брюшной стороне имеются два парных утолщения —
так называемые полосатые тела (corpora striata). В проме-
жуточном мозге у костистых сильнее, нежели у селахий, развиты
нижние доли его. Самыми крупными отделами головного мозга ко-
стистых рыб являются средний мозг и задний. Мозжечок большого
размера закрывает собой продолговатый мозг и вдается в область
среднего мозга в виде складчатой пластинки.
140
Головпой мозг двоякодышащих (Dipnoi) отличается вполне пар-
ными хорошо развитыми полушариями переднего мозга, содержащи-
ми нервную ткань. В этом отношении Dipnoi приближаются к амфи-
оннм. Средний мозг развит слабо. Мозжечок, как у амбифий, в виде
\ ив ого поперечного мостика. Таким образом, и в отношении раз-
ит пн нервной системы Dipnoi представляют сходство с наземными.
Рис. 111 А и В. Схема иннервации соматических и висцеральных
мускулов.
А. Поперечный разрез в области туловища с иннервацией соматиче-
ских мускулов и мускулов кишечника. 1 — спинной мозг, 2 —верхний
рог спинного мозга, 3 — боковой рог, 4 — смешанный спинной корешок
с ганглием, 5 — висцерально-моторные волокна (на правой стороне ри-
сунка не изображены), 6 — соматически моторные волокна» 7 — висце-
рально-моторные волокна, 8 — чувствующая ветвь к коже, 9 — сомати-
чески моторный брюшной корешок, отходящий от брюшного рога
спинного мозга, 10 —хорда, 11 — соматические мускулы, 12 —висце-
ральная ветвь с ганглием n. sympathicus, 13 — чувствительная ветвь
к коже, 14 — кишечная мускулатура, 15 — чувствительная ветвь к коже.
В. Поперечный разрез через жаберную область с иннервацией висце-
ральных мускулов. Висцерально-моторные нервные волокна черные,
соматические моторные — штрихованы поперек, чувствующие — пункти-
рованы. 1 — головной мозг, 2 — смешанный спинной корешок с ган-
глием, 3 — соматический моторный нерв, 4 — соматический мускул,
5—чувствующая ветвь к коже, 6— хорда, 7 — висцерально-моторный
нерв, 8 — висцеральные мускулы, 9 — жаберная щель, 10 —просвет
глотки, И—висцеральные мускулы, 12 — вентральные соматические
мускулы, 13 — скелет, 14 — полость тела, 15 — висцерально-моторные
нервы.
Спинномозговые нервы отходят от мозга двумя корешками: спин-
ным, или «чувствующим», и брюшным, или «двигательным». На са-
мом деле «чувствующий» корешок — смешанный и содержит в себе
ic;ik чувствующие, так и двигательные волокна. Отходят корешки
правой и левой стороны симметрично, причем спинной корешок
и брюшной соединяются друг с другом в один смешанный нерв.Спин-
ной корешок, как и у всех вышестоящих позвоночных,имеет состоя-
щее из нервных клеток (ганглиозное) утолщение, образующее спин-
номозговой узел. Брюшной корешок дает чисто двигательные
ветви (рис. 111) к мускулатуре, происходящей из миотомов, т. е.
к t.’ik называемой туловищной (соматической) мускулатуре и цроис-
ходящей от нее мускулатуре плавников. Спинной корешок делится
441
на волокна чувствующие, идущие к коже и кожным органам чувств
(соматичсски-чувствующие), волокна чувствующие, но идущие
к внутренним органам (висцерально-чувствующие), и на волокна
двигательные, идущие к висцеральным мышцам, происходящим
из боковой пластинки (висцерально-двигательные).
Головных нервов 10 пар (рис. 112). Они соответствуют отдель-
ным корешкам спинномозговых нервов: одни из них соответствуют
спинным смешанным, другие—брюшным двигательным. Двига-
тельными являются: III, IV и VIпары, идущие к мышцам, движущим
глазное яблоко; V, VII, VIII, IX и X пары соответствуют спинным
корешкам. Обонятельные и зрительные нервы нервами в собствеп-
Рис. 112. Схема черепных нервов и их компонентов
у костистой рыбы.
П, Ш и т. д. — че-
репные нервы и их
ветви, а — сомати-
чески чувствующая
ветвь, Ь — нерв бо-
ковой линии, с—вис-
церально чувствую-
щая ветвь, (1 — сома-
тически двигатель-
ная, е — висцераль-
но двигательная,
f — симпатическая
система (по Hedrick
из Ihle).
ном смысле слова пе являются — это выросты самого мозга. Таким
образом, нервы головы обнаруживают в этом отношении более
примитивные черты, чем спинномозговые, напоминая отношения
у ланцетника и у круглоротых.
У рыб мы впервые видим развитую симпатическую систему,
зачаточную у Cyclostomata и отсутствующую у ланцетника. Она
представлена двумя рядами ганглиев, лежащих справа и слева
от позвоночника, и отходящими от них волокнами. Волокна эти
иннервируют внутренние органы, снабжая их двигательными (к ки-
шечнику, сердцу, сосудам) и чувствительными волокнами и обра-
зуя периферические сплетения. У Teleostei и у Dipnoi гапглии сое-
диняются между собой продольными комиссурами, образуя справа
и слева от позвоночника так называемый пограничный
тяж. Симпатическая нервная система стоит в связи со спинно-
мозговыми нервами и частью с головными.
Органы чувств у рыб весьма разнообразны и стоят по развитию
значительно выше таковых у Cyclostomata.
Органами обоняния у всех рыб являются носовые
ямки, лишь у Dipnoi соединяющиеся с ротовой полостью. В отличие
от Cyclostomata обонятельный орган у рыб всегда парный и состоит
142
Рис. ИЗ. Диаграмма отношений капала боковой
линии к чешуям па туловище окуня Perea flu-
viatilis. Продольный разрез.
1 — передний край чешуи, 2 — ипидермпс, 3 — канал
боковой линии, 4 — отверстие капала, 5 — орган чувств
в канале, 6—нерв к органу чувств, 7 — нерв боковой
линии. (По Goodrich.)
из двух ямок, выстланных обонятельным эпителием. У селахий они
помещаются на нижней поверхности головы, у Osteichthyes—на
верхней стороне головы. В большинстве случаев посовал ямка
имеет два наружных отверстия, иногда простых, иногда же с при-
поднятыми в виде трубочек краями. Переднее отверстие служит для
вхождения, заднее — для выхода воды. У акул, химер и скатов
ноздри широки и закрываются клапанами при помощи мускулов.
Органами вкуса у рыб являются так называемые
«концевые почки», или вкусовые, состоящие из чувствующих и
лежащих между ними опорных клеток. Снабжаются эти почки от
пары нервов (n. glossopharyngeus). Вкусовые почки расположены
у рыб па самых различных местах тела: па усиках, па губах, в по-
лости рта, глотки и па жаберных дугах, па некоторых плавниковых
лучах и даже могут быть рассеяны по всей поверхности тела. Но
п в последнем случае
они иннервируются
частями лицевого, 1
языкоглоточного и
блуждащего нервов.
Орган ося-
зания представлен
(юязательными тел ь -
цами, сосредоточен-
ными главным обра-
зом на рыле, в уси-
ках, на передних лу-
чах плавников (у
тригл) и т. д.
Особенно харак-
терны для рыб специализовапные кожные органы, называемые орга-
нами боковой л и и и и. Сами органы представляют утолще-
ния эктодермы, состоящие из двоякого рода клеток: центральные
имеют грушевидную форму и несут па свободном заостренном конце
кутикулярную щетинку; периферические—опорные — идут через
весь орган от поверхности до основания. Грушевидные, чувстви-
тельные клетки связаны с нервными волокнами, опорные облекают
чувствительные наподобие чехла. Эти органы имеются уже у
(’yclostomata, но там опи лежат па поверхности тела. Также и
у рыб в раннем возрасте опи лежат поверхностно, но у взрослых
рыб погружаются в ямки, желобки или каналы. У более примитив-
ных акул, как Chlamydoselachus опи лежат в открытых желобах,
у прочих селахий, а также у Osteichthyes «органы боковой
'пиши» лежат в замкнутых каналах, в большей или^меныпей мерс
нагруженных в кожу (рис. 113). Основной капал — это канал
о<»1ник)Н линии, открывается через некоторые промежутки отвер-
ri'iiiiMii наружу. У Osteichthyes канал этот проходит в чешуях
н нпкровных костях, распространяясь па голову, где от пего отхо-
дит ветви, проходящие над глазом, под глазом и вдоль нижней
чг'погтн. Эти так называемые «слизистые каналы»(рис. 114) отделя-
143
ют и слизь, так как на внутренней стенке их имеются бокаловидные
слизистые клетки. Органы боковой линии иннервируются n. lateralis,
боковым первом, ветвью n. vagus. Можно считать установленным,
Рис. 114. Голова рыбы сбоку; показа-
но распределение органов боковой ли-
нии (черные) и снабжающих их нервов.
(По Goodrich.)
что органы боковой линии
имеют значение органов «ме-
ханического чувства», посред-
ством которого рыба узнает о
направлении и силе токов в
воде и о своем приближении к
твердым предметам. Это имеет
огромное биологическое зна-
чение, так как без этих орга-
нов рыба не могла бы ориен-
тироваться в мутной воде и
находить путь.
Имеется у рыб и способ-
ность воспринимать температурные отличия. Каждый вид рыб
имеет свой температурный оптимум.
'Орган слуха рыб (рис. 115) представлен только внутрен-
Рис. 115. Слуховой орган
рыб и круглоротых. А —
типичной рыбы. В — мик-
сины.
1— utrlculus, 2 — sacculus, 3 —
lagaena, улитка, 4 —ductus
endolymphaticus, 5n 5a, 5,—
полукружные каналы, 6n 63,
6B—ампулы. (По Ретциусу из
Boulengcr.)
ним ухом, перепончатым лабиринтом.
Последний у Holostei и Teleostei частич-
но помещается в боковых стенках чере-
па, значительной же частью помещается
на внутренней поверхности последнего.
Состоит он у рыб уже из трех частей:
верхней — utriculns с тремя полукруж-
ными канальцами, и нижней — sacculus
с маленьким зачатком улитки высших
позвоночных (lagaena). Полукружные
каналы расположены в трех взаимно
перпендикулярных плоскостях. Полу-
кружные каналы при основании имеют
пебольшое расширение — ампулу,
внутри которой находятся окончания
слухового нерва (n. acusticus). Другие
первпые окончания находятся в utriculus
и sacculus. Там же находятся о т о л и-
т ы, камешкоподобные образования из
углекислой извести. У селахий от пе-
репончатого лабиринта отходит пере-
пончатый же капал — ductus endolym-
phaticus, проходящий через крышу че-
репа под кожу головы, где образует,
открывающееся на поверхности головы
расширение. У других рыб канал этот
слепой.
Глаза рыб построены по общему
для всех позвоночных типу, но обнаруживают некоторые отличия
(рис. 116). Прежде всего, как у всех водных животных, р о г о в и-
144
ц a (cornea) у них почти плоская. Хрусталик отличается своей
шарообразной формой. Это обусловливается тем, что рыба живет
в воде, и хрусталик сам по себе лишь немного плотнее жидкости,
в которой рыба живет. Затем радужная оболочка (iris)
у рыб неподвижна, так что зрачок у них не может суживаться и
расширяться. Аккомодации у них, подоб-
но аккомодации в глазу наземных, не
наблюдается, а происходит она при помо-
щи так называемого серповидного
отростка (processus falciformis), иду-
щего от задней стенки главной камеры,
от сосудистой оболочки (cho-
ri oidea) к хрусталику и кончающегося
здесь расширением — колоколом Гал-
лера (campanula Halleri). Сокращение
серповидного отростка оттягивает хруста-
лик назад, и таким образом происходит
приспособление к зрении} на расстояние
10—12 м, тогда как обычно глаз рыб ак-
комодирован для зрения па 1 м, т. е. рыбы
по природе своей близоруки. В склеротике
глаза или в наружной стейке глазного яб-
лока имеются у рыб хрящевые или кост-
Рис. 116. Вертикальный
разрез через глаз Salmo
fario.
1 — роговица, 2 — радужи-
на, 3 — серповидный отро-
сток, 4 — хрусталик, 5 —
колокол Галлера, 6 — сет-
чатка, 7—зрительный нерв.
(Из Boulenger.)
ные пластинки. Характерна для глаза рыб
так называемая серебристая оболочка (argentea), за-
ключающая в себе иризирующие кристаллы гуаниновой извести.
Находится серебристая оболочка между склерой и сосудистой обо-
лочкой. Металлический блеск глаз рыб и
способность некоторых светиться в темноте
обусловливаются наличием этой оболочки.
Войн у рыб зачаточпы. У некоторых акул
имеется в переднем углу глаза третье веко,
так называемая мигательная пе-
репонка — membrana nicticans. Фор-
ма, величина и расположение глаз весьма
вариируют. У глубинных рыб глаза могут
вовсе исчезать. У глубинно-пелагических
встречаются так называемые телеско-
пические глаза (рис. 117).
Теменного глаза, который мы видели
у (iyclostomata и Ostracodermi, Antiacrchi,
а также у некоторых ископаемых Crossop-
l erygii (Osteolepidoti), у современных рыб
Рис. 117. Телескопиче-
ские глаза глубоковод-
ных рыб.
ни имеется. Лишь у некоторых Teleostei имеется намек па него.
Функция головного мозга у рыб. Головной мозг рыб еще далеко
иг достиг того значения, которое он имеет у высших позвоночных.
Поэтому удаление переднего мозга у рыб пе оказывает никакого
влияния иа их движения и на их отношения к внешнему миру. Рыбы
так жп продолжают двигаться, относиться к пище, также ее схваты-
10 I(tin кпроп и Станчииский
145
вают, ведут себя по отношению к другим рыбам, как и нормальные.
Их трудно отличить от последних. Другое дело, если удаляется
средний мозг. Тогда наблюдается ослабление движений, рыба ло-
жится на бок, произвольные движения у нее прекращаются, рыба
реагирует лишь рефлекторно на внешние раздражения. Удалепие
среднего мозга вызывает также слепоту и расстройство координа-
ции локомоторных движений.
III. РАЗВИТИЕ РЫБ
Изучение развития в различных отрядах рыб, казалось, могло бы
пролить нам дополнительный свет на филогенетические между ними
отношения. Однако онтогенез не воспроизводит полностью филоге-
неза, многое в эмбриональном развитии является приспособлением
развивающегося организма к временным
условиям развития (ц е н о г е н е з); мно-
гое, что было в филогенезе, в онтогенезе
исчезло; многое смещено во времени появ-
ления (гетерохрония); многое сме-
щено в пространстве (гетеротопия).
Но все же остается еще многое, по чему мы
могли бы судить о ходе эволюционного
процесса в прошлом Явления развития
рыб еще настолько неполно известны и
настолько спутаны, что дают нам для вос-
становления процесса филогенеза очень
мало. Нередко заведомо генетически близ-
кие группы дают весьма различные кар-
тины развития. Так, например, в преде-
лах одного подкласса Teleostomi существу-
ют большие различия в развитии, чем
между развитием рептилий и млекопитаю-
щих.
Яйца рыб богаты желтком, более или
менее обильным в различных подклассах
и отрядах. В зависимости от количества
Рис. 118. Яйцевая кап-
сула Heterodontus (Се-
stracion) galeatus.
(По Waite из Bridge.)
желтка протекают процессы развития в этих группах.
Яйца акул и скатов отличаются крупной величиной и очень
богаты желтком (телолецитальпые яйца). Если они откладываются
наружу, то бывают снабжены твердой роговидной скорлупой.
Форма их большей частью четырехугольная или иная с длинными
закрученными питями па узлах. Нити эти служат для прикрепле-
ния яиц. Как форма, так и характер оболочек сильно вариируют
у различных элазмобрапхий (рис. 118). Благодаря этому сегмента-
ция неполная, крайне'неравномерная, ограничивается зародыше-
вым диском. Лишь у самых примитивных акул дробление полное,
хотя и неравномерное (Cestracion).
В результате сегментации получается дисковидная бластодерма
(рис. 119), состоящая из поверхностного слоя, в котором клетки
лежат в одцн ряд, и более глубокого — многослойного. Между
446
клетками последнего слоя появляется сегментационная полость.
Гаструляция (рис. 120) и образование первичной кишки происходят
путем образования складки— втягивания на одном из концов за-
родышевого диска. Яйца, точнее оболочки их, у Holocephali также
очень крупны, до 25 см длиной, и одеты волосовидными придат-
ками. О ранних стадиях развития Holocephali ничего не известно.
Сегментация яиц у
Chondrostei (Acipenser)
и Holostei (Amia, Lepi-
dosteus), вероятно, и у
Crossopterygii (Polypte-
rus) имеет характер так
называемой проме-
жуточной сег-
ментации. В яйцах
этих рыб желтка мало
(яйца мезолециталь-
ные), но все же больше,
нежели у Cyclostomata)
Рис. 119. Зародышевый диск акулы (Scyllium
canicula).
1 — сегментационная полость, 2 — эктобласт, 3 —
энтобласт.
или Dipnoi (см. дальше), так что в этом
отношении имеется известное приближение к элазмобранхйям.
У Amia calva (рис. 121), например, уже имеется различие между
верхней частью яйца и нижней. Деление здесь почти полное, но
неравномерное. В результате получаются мелкие шары дробления—
микромер ы—наверху и крупные — макромер ы—внизу.
Яйца Teleostei опять богаты желтком, отличаясь таким образом
от ближайших родственников—Holostei—и сближаясь с селахия-
ми. Яйца Teleostei,’
как правило, очепь
мелки (у сельдей они
имеют около 1 мм в
диаметре), круглы и .
одетыпрозрачпойобо- •
лочкой — zona radia-
ta, снабженной отвер-
стием для проника-
ния спермиев. Заро-
дышевый диск очепь
мал. Он может быть
и па верхнем полюсе
яйца и на нижнем (в
Ь
Рис. 120. Сагиттальный разрез через молодую
гаструлу Pristiurus.
1—первичная кишка, 2 —губа бластопора, 3 —сег-
ментационная полость, 4 — мероциты.
случае присутствия
масляного пузырька-поплавка у пелагических яиц на верхнем
полюсе). Дробление частичное, неравномерное. Отличие от элаз-
мобранхий в том, что вследствие меньшего количества желтка
образующийся из бластодермы зародыш относительно крупнее,
чем у селахий. Как и у последних, бластодерма обрастает желтком.
Ранние стадии развития Dipnoi сближают этих рыб, с одной
стороны, с Cyclostomata (Petromyzon), с другой — с амфибиями.
Яйца их одеты нежной оболочкой, сильно разбухающей в воде.
ю*
147
Само яйцо на верхнем полюсе пигментировано. Сегментация полная
и неравномерная. Гаструляция происходит весьма сходно с тем,
что мы наблюдаем у амфибий. Сходство с последними наблюдается
в позднейшие моменты развития, когда уже образуется за-
родыш.
Образование центральной нервной системы, хорды, рта, ануса,
мезодермы, выделительных органов происходит у рыб в основном
Л в
Рис. 121. Сегментация яйца Amia calva.
А, В, С — последовательные стадии, D —вид с верх-
него полюса. 1— микромеры, 2 —макромеры.
так же, как у круг-
лоротых или у лап-
цетника. Особенности
этого развития у рыб
стоят в связи с боль-
шим количеством
желтка в их яйцах. У
многих костистых рыб
из яйца получается
сперва не рыба мень-
шего размера, а ли-
чинка, отличающаяся
от взрослой рыбы фор-
мой тела, и нередко
наличием так назы-
ваемых провизорных
органов, впоследст-
вии исчезающих. В
зависимости от коли-
чества желтка у ли-
чинок более илй менее
долгое времяЛсохра-
няется на бркЖэ жел-
точный мешок. После
исчезновения желтка тело личинки или приобретает характерные
признаки тела взрослой рыбы, если желтка было много (лосось,
Salmo), или личинка еще некоторое время продолжает оставаться
личинкой, если желтка было мало (камбалы, Pleuronectidae).
IV. КЛАССИФИКАЦИЯ РЫБ
Система какой-либо группы животных должна отображать
путь ее филогенетического развития. О последнем мы судим па ос-
новании всей совокупности фактов, которые нам дает сравнительное
изучение организации животных, фактов, доставляемых историей
развития и палеонтологией. Знания наши в этих трех областях —
анатомии, эмбриологии и палеонтологии — с каждым годом увели-
чиваются. Поэтому система животных все время изменяется, мы
все более и более приближаемся к созданию действительно естест-
венной системы, являющейся «родословным деревом». Это положе-
ние ясно видно на системе рыб, на их классификации.
Основоположником современной классификации рыб является
148
шведский ихтиолог XVIII века А рте ди. Он разделил рыб на три
группы, взяв за основу разделения лучи плавников: Chondropte-
rygii — с хрящевыми лучами, Malacopterygii — с мягкими кост-
ными и Acanthopterygii — с жесткими лучами. В первой половине
XIX века сделан был большой шаг вперед. Кювье и Вален-
сией н в 22-томном труде «Естественная история рыб» положили
в основу деления рыб наличие хряща или кости в скелете и разде-
лили рыб на костных и хрящеперых (Chondropterygii). Это уже
соответствует современному делению рыб на две ступени—Osteich-
thyes и Chondrichthyes. Однако в раздел Chondropterygii у этих авто-
ров отнесены, кроме хрящевых рыб, и осетровые (Chondrostei),
и круглоротые (Cyclostomata).
Эпоху в развитии классификации рыб составили в первой поло-
вине XIX века работы Агассицаоб ископаемых рыбах. Он поло-
жил в основу деления наружный скелет, наиболее доступную для
изучения у ископаемых рыб систему, и разделил рыб на Pla-
coidei с плакоидной чешуей, Ganoidei — с ганоидной, Cycloidei—
с циклоидной и Ctenoidei — с ктеноидной. Тут неправильно воз-
ведение в один ранг Placoidei, с одной стороны, Cycloidei и Cte-
noidei — с другой. Две последних группы гораздо ближе между
собой, чем каждая из них с Placoidei или Ganoidei. Кроме того,
Аг асе иц неправильно составил и группу Ganoidei, объединив
под этим названием Polypterus, Acipenser, Ostracion (из костистых
рыб — Teleostei) и Lepidosiren (из Dipnoi).
Большую ясность в систему рыб внес знаменитый анатом Ио-
ганнес Мюллер, давший капитальную работу о строении и
системе ганоидсв. Он соединил Cycloidei с Ctenoidei, назвав их вместе
Teleostei, и отделил гапоидов от Dipnoi и панцирных от Teleostei.
Сделал он это па основе пе одного признака, а па основе изучения
анатомии рыб. В дальнейшем много цепных данных для построепия
системы рыб дали Гексли, Коп, Циттель, Г ёпт ер, дав-
ший детальную классификацию современных рыб, Траквайр,
Смит-Вудвард, Вуленжер, Иордан, Тэт Ригэн и
другие. Значительную ясность в систему рыб внесли исследова-
ния русских ихтиологов и анатомов: Северцова, Верга и
других. Все более детально изучалась организация различных
групп рыб, и в основу построепия системы классов брали все
больше и больше признаков. Мы уже можем говорить о филогене-
тической системе рыб. Но, несомненно, что с расширением наших
знаний о современных и ископаемых формах система рыб будет
от,е перестраиваться.
СТУПЕНЬ I. ХРЯЩЕВЫЕ РЫБЫ (CHONDRICHTHYES)
Объединяемые под этим названием рыбы являются наиболее
низко организованными среди рыб и сохраняют в большей или
мош,шей мере примитивные признаки: нормально внутренний
сколот хрящевой, наружный скелет из плакоидных чешуй. В пар-
ных п попарных плавниках имеются caratotrichia.
149
Подкласс 1. Акантоди/и (Acanthodii)
Остатки этих рыб известны из наиболее древних слоев, содержа-
щих остатки рыб, а именно из верхнесилурийских (рис. 122).
В старых классификациях их помещали среди «ганоидов», другие
считали их промежуточным звеном между ганоидами (Chondrostei
и Holostei) и элазмобранхиями, третьи соединяли их с последними.
Самое замечательное в их структуре, в общем акулоподобной, —
это то, что все тело у них было покрыто маленькими, тесно прилегаю-
Рис. 122. Реставрация Diplacanthus
striatus. Слева — чешуя Acanthodes
gracilis, справа — чешуя боковой
линии Protacanthodes pinnatus,
(Из Goodrich.)
щими друг к другу чешуями,
по своему строению резко отли-
чающимися от плакоидных че-
шуй акул, но в некоторых от-
ношениях напоминающих га-
ноидные чешуи. Замечательно
также то, что первичная верх-
няя челюсть palatoquadratum и
нижняя построены каждая из
двух, а не из^Дного хряща и
усилены тканью, подобною ко-
сти. Таким образом, челюстная
дуга, оказывается, построена у
них, подобно жаберным дугам,
из четырех отделов. Внутрен-
ний скелет исключительно хря-
щевой, хорда, видимо, сохранялась всю жизнь. Череп был амфи-
стилический. Костеподобная ткань имелась также в сильных ши-
пах, сидевших впереди плавников. Лучи последних были рого-
вые. Крайне интересным является то обстоятельство, что у
некоторых представителей подкласса на брюшной стороне сбоку
находится 6—7 таких шипов.Передние и задние из них соответствуют
грудным и брюшным плавникам, в наличии остальных видят неко-
торое доказательство существования некогда на месте парных плав-
ников боковой складки. '
Более древпим является семейство Diplacanthidae'c двумя
спинными плавниками.. У рода Climatius из девона имелись только
что упомянутые шипы по бокам брюшной стороны тела. Сем.
Acanthodidae имеет только один спинной плавник. Род
Acanthodes—с нижнего девопа до пермской эпохи.
Подкласс 2. Кладосвлахии (Clad о selachii)
К этому подклассу принадлежат некоторые наиболее раппие
ископаемые хрящевые рыбы. Изолированные зубы их известны
из девонских и каменноугольных слоев. По общему виду Clado-
selache напоминает Chlamydoselachus (см. ниже) за исключением
того, что ось в хвостовом плавнике резко загнута кверху и рот
находится почти на конце тела. Жаберных щелей 7. Позвонков еще
не было, не было и копулятивных органов (птеригоподий). Череп
150
пмфпстилический. Тело было покрыто зубчиками, гистологическое
строение которых, однако, неизвестно. Наиболее интересной чертой
является крайне примитивное строение парных плавников, имею-
щих широкое основание и не обнаруживающих никаких следов
слияния лучей. В связи с этим и пояса конечностей только едва на-
мочены. Повидимому, это очень древнее и очень примитивное се-
мейство (7 жаберных щелей, амфистилия, парные плавники), вы-
работавшее некоторые специальные черты после того, как оно ответ-
вилось от общего ствола с селахиями.
Сем. Cl ados el ach idae с родом Cladoselache. (См. рис. 79).
Подкласс 3. Сростножаберные ( Elasmobranchit)
Полужабры прирастают к септам. Рот никогда не бывает на кон-
це. Ноздри снизу головы. Имеется rostrum. Небноквадратные хрящи
сходятся под черепом снизу. Хвост гетероцеркальный Число
жаберных щелей б—7. Хорбшо развитые плавники; задние снаб-
жены копулятивными придатками. Глазные нервы нормально
перекрещиваются. В кишечнике имеется спиральный клапан.
В сердце хорошо развит артериальный конус. Яйца выводятся
через особые яйцеводы, открывающиеся в брюшную полость во-
ронкой: мезонефрические протоки (рис. 108, с) в тесной связи с се-
менниками. Имеется клоака. Подкласс представлен двумя совре-
менными сверхотрядами: поперечноротых (Plagiostomi),
куда относятся акулы и скаты, и цельноголовых (Holo-
cephali), а также одним вымершим сверхотрядом—Pluracanthodii.
Сверхотряд 1. Поперечпоротые (Plagiostomi)
Рот, в виде широкой поперечной или полулунной щели, нахо-
дится на брюшной стороне. Небноквадратные хрящи не прирастают
к черепу. Как верхнечелюстная дуга, так и нижнечелюстная уса-
жены многочисленными зубами. Жаберные щели не прикрыты об-
щей складкой. Глаза с веками. Врызгальце всегда есть. Позвоноч-
ник из амфицельных позвонков. Парные плавники унисериальны.
Отряд 1. Акулы (Selacholdol)
Тело вытянутое, с сильным развитым хвостом. Парные плавники
умеренной величины, с головой не сливаются. Жаберные щели по бо-
кам тела. Зубы острые, конусообразные, иногда с зазубринами или
выступами по бокам.
Акулы в настоящее время весьма богато представлены в морях,
при надлежит к процветающей группе рыб. Зубы и шипы акул встре-
чаются уже в палеозое, цельные экземпляры известны с нижней юры.
Сем. Плащеносные акулы (Chlamydoselachi-
<1 а <•) и сем. гребнезубых акул (Notidanidae) представляют наиболее
примитивную группу современных акул, характеризуемую амфистили-
•юскпм или гиостилическим черепом, 6 или 7 жаберными щелями, с одним
длинным, слабо концентрированным спинным плавником и большим аналь-
ным. Хвост лишь слегка гетероцеркальный. Единственный представитель
норного семейства — Chlamydoselachus anguineus (рис. 123) встречается
451
у берегов Японии, в глубинах, и у берегов Норвегии. Остатки NotidanuS
известны из юры, Chlamydoselachus — из плиоцена. У остальных акул число
жаберных щелей не превосходит 5. Сюда принадлежит несколько семейств.
Рис. 123. Chlamydoselachus anguineus.
(Из Boulenger.)
Сем. Разнозубые (Heterodontidae) акулы. В мезозое это
были наиболее распространенные акулы. Они известны из палеозойских
(каменноугольных) и девонских отложений. В настоящее время представлены
одним родом — Heterodontus (Cestracion) (рис. 124) с несколькими видами —
Н. philippi — у берегов Австралии. Весьма примитивная форма.
Рис. 124. Акула порта Джексон, Heterodontus
(Cestracion) philippi.
(Из Boulenger.)
Сем. Н о к о т н и ц ы (Scyllidae). Сюда принадлежат акулы с двумя
небольшими спинными плавниками без шипов и небольшим заднепроходным,
тоже без игол. Хвост мало загнут кверху, перепонки нет. Зубы маленькие,
без боковых острий. Scyllium canicula.
Сем.
наличием
Настоящие акулы (Carchariidae) характеризуется
мигательной перепонки. Два спинных плавника. К семейству
принадлежат 20 родов и около
60 видов, распространенных во
всех морях. Carchariasglaucus,
голубая акула, до 4 м, пела-
гическая акула Атлантическо-
го и Тихого океанов, питается
рыбами. Zygaena malleus, мо-
лоток-рыба, имеет на голове
боковые выросты, на конце ко-
торых находятся глаза. Сюда
же принадлежит некрупный
род Mustelus.
Сем. Дельфиновые
(Lamnidae) представлено самыми крупными акулами. Lamna
(рис. 125) в водах Атлантического океана и Средиземного моря
4 м и является, опасной для человека. Одной из крупнейших
акулы
cornubica
достигает
акул является Carcharodon rondeletti, пелагическая акула с пилообразными
зубами, распространенная во всех тропических и субтропических морях от
Средиземного до Австралии и Новой Зеландии. Размеры ее доходят до 12 м.
Близок по размерам другой представитель семейства Cetorhinus (Seiache)
maximus пелагическая акула арктических морей, доходящая на юг до Сре-
диземья и Калифорнии. Питается мелкими пелагическими рыбами и беспоз-
152
воночными. В связи с таким питанием у нее существуют упомянутые выше
«жаберные тычинки» из видоизмененных плакоидных зубов. К этому семей-
ству принадлежит самая крупная акула Rhinodon typicus, достигающая
20 м, хотя совершенно безвредная.
Сем. Колючеперых акул (Spin, acidae), спинных плавни-
ков два, каждый с иглой впереди. Анального плавника нет. По бокам рыла с
каждой стороны рта продолговатая глубокая ямка. Без мигательной перепон-
ки. Acanthias vulgaris, морская собака, небольшая, до метра
длиной, акула, наиболее распространенная в умеренном поясе северного и
южного полушарий. Часто встречается в Черном море. Живородяща.
Laemargus borealis, гренландская акула арктических морей, спускающаяся
однако к югу до Мыса Доброй Надежды, хищна, атакует даже китов.
Сем. Пилоносы (Pristiophoridae) характеризуется
длинным плоским rostrum с парой длинных усов на нижней его поверхности.
Вдоль краев rostrum ряд больших зубовидных кожных образований. Груд-
ные плавники широки. Зубы малы. Несколько видов рода Pristiophorus
ограничены в своем распространении австралийскими и японскими берегами.
Почти все акулы являются прекрасными пловцами. Живут они все в мо-
рях, но некоторые виды могут входить в реки и долго в них находиться. Жи-
вут в море в самых разнообразных условиях: в открытом море, на больших
глубинах, и у берегов. Обычно — хищники, но некоторые питаются мелкими
беспозвоночными, икрой рыб и маленькими рыбами несмотря на свою огром-
ную величину.Роль их в экономии природы и круговорота веществ,конечно,
очень велика, так как они должны поглощать огромное количество пищи.
Оплодотворение у акул внутреннее, и большинство акул является живо-
родящими. Яйца, откладываемые некоторыми акулами наружу, бывают оде-
ты толстой и крепкой скорлупой, иногда имеют на концах нити, которыми при-
крепляются ко дну или к растениям. У живородящих акул имеется своеобраз-
ное изменение в яйцеводах, где развиваются зародыши, напоминающие
«плаценту» млекопитающих. Развитие, как вне матери, так и внутри послед-
ней, идет очень медленно. Образ жизни мальков акул неизвестен.
С экономической стороны акулы имеют пока еще мало значения. Их
используют для добывания жира. Мясо едят в Западной Европе, в Китае,
Японии и Корее. У нас его используют на Севере. Мясо вполне пригодно
в пищу. Идет в дело еще кожа под названием «шагрени». Кожа морской со-
баки (Acanthias), употребляется для полировки мягких сортов дерева.
Много акул на Дальнем Востоке, по там эти рыбы пе используются.
Отряд 2. Скаты (Bate id el)
Скаты происходят от акул, и между этими двумя подотрядами
существует целый ряд переходов. Тело скатов более или мепее
сплюснуто в направлении сверху вниз (дорзовептральпо). Труд-
ные плавники развиты чрезвычайно сильно и срастаются с боками
тела и головы. Хвост топкий, редуцированный, менее гетероцер-
кальный, чем у акул. Апальпого плавника пет, спинные развиты
слабо. Жаберные отверстия паходятся па брюшной стороне тела,
так же как и рот и носовые отверстия. Брызгальце больших
размеров остается па спиппой сторопе. Век пет.
Сем. Морские ангелы (Squatinidae) (рис. 126) вклю-
чает в себя формы, составляющие переход к акулам и нередко относимые
к последним. Тело, уплощенное сверху, большие грудные плавники сильно
сдвинуты наперед, но не срастаются с головой. Жаберные отверстия нахо-
дятся частью снизу,частью с боков.Хвостовой плавник маленький, как у ска-
тов; анального плавника нет. Скелет сохраняет признаки скелета акул.
Rostrum нет. Единственный род с одним видом — морской ангел, Squa-
tina squatina, в тропических и умеренных морях обоих полушарий Атлан-
тического и Тихого океанов. По образу жизни более близка к скатам.
153
В остальных семействах скатов грудные плавники сливаются впереди с
боковыми сторонами головы, и жаберные отверстия лежат совершенно на
брюшной стороне. Rostrum у
многих хорошо развит.
Сем. Пилы-рыбы
(Pristidae). Так же как
предыдущее семейство состав-
ляет переход от акул, больше
приближаясь к настоящим ска-
там. Грудные плавники хотя
прирастают к голове, не про-
стираются далеко.
Виды рода Pristis, пилы-
рыбы, распространенного в
„ ,г „ тропических и субтропических
Рис. 126. Морской ангел (Squatina squa- морях, достигают до 6 м. Име-
^па)- ют очень длинное rostrum (до
(Из Boulenger.) 2 м), боковые края которого
усажены серией треугольных
плоских зубов. Ударами этой пилы-рыбы пользуются как оружием при до-
бывании пищи. Как индийские виды, так и американские, доходящие до
Рис. 127. Пила-рыба.
(Из Boulenger)
Мексиканского залива, входят в реки. Ископаемые известны из меловых
слоев. Pristis $
Рис. 128. Электрический
скат Torpedo со вскрытыми
электрическими органами.
1—электрические органы, 2 —
жаберные щели, 3 —n. vagus.
(Ив Goodrich.)
154
antiquorum (рис. 127) встречается в Средиземном морей в Ат-
лантическом океане, доходит до берегов Ан-
глии. Достигает 6 м длины.
Сем. Рохли (Rhinobatidae)
имеет более широкие грудные плавники, бо-
лее придвинувшиеся вперед, но еще не имеет
вида настоящего ската. С юры.
Сем. Настоящие скаты (Ra-
ji d а е,) типичные обитатели дна, имеют те-
ло широкое благодаря сильно разросшимся
вперед и назад грудным плавникам. Хвост
тонкий и почти без плавника и резко отде-
лен от «диска» тела. Кожа покрыта крупными
или мелкими шипами (плакоидные зубы). Яй-
ца в кожистой капсуле с четырьмя выростами
по углам. Семейство представлено 4 родами и
почти 40 видами. Представители рода Raja
обитают в умеренных морях, главным образом
северного полушария, распространяясь далеко
на север. Окрашенные сверху покровитель-
ственно под цвет дна, обычно, песчаного или
каменистого, скаты рода Raja лежат обычно
на дне неподвижно. При приближении ма-
леньких рыб, ракообразных и других подоб-
ных животных, служащих им добычей, скат
быстрым* движением накрывает животное сво-
им телом, а затем пожирает. Имеется половой
диморфизм, выраженный в окраске, в зубах, в расположении шипов наспин-
ной стороне, в наличии у самцов копулятивных органов. Raja radiata —
в северных морях СССР, Raja clavata — в южных.
Сем. Электрические скаты (Torpedinidae). Отли-
чается отсутствием rostrum, круглой формой тела и развитием электрических
органов. Хвост короткий и
толстый, кожа гладкая—
без зубчиков (рис. 128).
Электрические органы ле-
жат между грудными плав-
никами и головой. Обита-
тели теплых морей — до
Средиземного включитель-
но. Наиболее обычный и
наиболее специализирован-
ный вид—Torpedo ocellata.
Род известен с эоцена.
Сем. X востоколы
(Trigonidae). Грудные
плавники срастаются впе-
реди рыла, заменяя собой
rostrum; хвост очень тон-
кий, жгутовидный. Спин-
ных плавников нет, а вме-
сто них одна или две зазу-
бренных иглы. Наиболее
известен род Trygon с видом
Т. pastinaca — м орской кот. Встречается и в Черном и в Азовском
морях. Своим стилетом может наносить опасные раны. Из печени морского
кота в Азовском море вытапливают жир.
Сем. Орляки (Myliobatidae) (рис. 129). Как и в предыду-
щем семействе, грудные плавники срастаются впереди головы, прерываясь в
области последней и появляясь опять впереди в виде головных плавничков.
Голова выдается над диском, благодаря чему глаза и брызгальце лежат по
бокам. Хвост длинный, нитевидный, с длинной иглой на спинной стороне.
Зубы плоские, в виде мостовой, кожа гладкая. Ceratoptera vampyrus, жи-
вущий у берегов Центральной Америки, достигает 500 кг веса, при ширине
диска в 6 м. Myliobatis aquila встречается в Атлантическом океане и Среди-
земном море.
Скаты употребляются в пищу в Западной Европе, в Японии и в Америке.
У нас пока использование скатов только начинается. Мясо их вкусно,
но, как мало плодовитые рыбы, они не могут быть объектами значительного
промысла.
Сверхотряд 2. Цельпоголовые (Holocephali)
Очень древняя и примитивная группа морских рыб. Она из-
вестна с девонского времени в виде отдельных шипов — «ихтиодо-
рулитов» и зубных пластинок. Особенно богато была представлена
эта группа в мезозое. До нашего времени дожили только немногие
представители.
'По общей организации напоминают акул, отличаясь некоторыми
существенными признаками. Скелет хрящевой, как у акул, по тел
позвонков не образуется и вместо них имеются объизвествленные
кольца в волокнистой оболочке хорды, число которых больше, чем
число сегментов. В черепе palatoquadratum и hyomandibulare слиты
друг с другом и с черепом, который является автостилическим.
455
В верхней челюсти находятся четыре, а в нижней два больших зуба.
Жаберные щели прикрыты кожндй складкой, благодаря чему полу-
чается общая жаберная полость, подобная полости у Teleostomi, но
между жаберными щелями имеются септы, вследствие чего жаберный
дыхательный аппарат стоит не на большой высоте. Врызгалец нет.
Глаза не имеют век. Совокупительные органы есть. Клоаки нет.
Имеется rostrum. Спинной плавник с шипами. Весьма интересно,
что недавно ставшие известными ископаемые формы сростноголовых
Рис. 130. Химера (Chimaera monstrosa).
(Из Boulenger.)
обладали костными пла-
стинками.
Из 4 семейств наиболее
известно сем. Ghimaeridae
с родами Chimaera и сем.
Callorhynchidaec родом
Gallorhynchus. У первого ros-
trum короткое, у второго вы-
тянутое и загнутое крючком,
Chimaera monstrosa—глубоко-
водная рыба (рис. 130) дер-
жащаяся на глубине до 1 000 м,
водится у европейских бере-
гов. Доходит почти до 4 м
длиной. Яйца откладываются, повидимому, на дне моря и встречаются
очень редко. Наибольшего расцвета химеры достигли в меловое и ранне-
третичное время. Как можно судить по строению зубов, химеровые пита-
ются ракообразными, червями и иглокожими рыбами.
Промыслового значения они не имеют. Ценен жир из печени химеры как
медицинское средство.
Сверхотряд 3. Плеуракаптодии (Pleuracanthodii)
Сверхотряд представлеп вымершими элазмобрапхиями очень
обобщенного типа, соединявшими в себе признаки нескольких групп.
Наиболее характерной особенностью является скелет грудных
плавников, построенных по типу бисериальпого архиптеригия.
У рода Pleura canthus хвост был дифицернальный, длинный спинной
плавник доходил до хвостового. У нисериальные брюшные плавщики
давали опору копулятивным органам. Череп был амфистилический,
жаберпые дуги хорошо развиты, хорда сохранялась всю жизнь,
тел позвонков не было. Плечевой пояс примитивный. В плавни-
ках были роговые лучи.
Pleuracanthodii интересны тем, что являются группой «обоб-
щенной», совмещающей в себе признаки акул (череп, дуги, копу-
лятивные органы), Dipnoi (архиптеригий грудных плавников) и бо-
лее примитивных форм (неконцентрированный спинной плавник,
строение плечевого пояса из двух дуг, отсутствие тел позвонков).
От такой формы, как Pleuracanthodes, могли произойти и акулы,
и Dipnoi, и Crossopterygii. Однако некоторые черты, как утеря кож-
ного скелета, сильно развитой шип на затылке, являются призна-
ками специализации. Pleuracanthodii появляются в девоне и выми-
рают в перми,
156
Подкласс 4. Артродиры ( Coccosteomorphi, Arthrodira)
Палеозойские рыбы девонской эпохи, защищенные тяжелым пан-
цирем, достигавшие иногда очень большой величины, были ши-
роко распространены, известны из Европы и Северной Америки.
Систематическое положение их до последнего времени было недо-
статочно ясным. Последние тщательные исследования, позволившие
Рис. 131. Реставрация Coccosteue decipiens.
(По S. Woodward иа Boulenger.)
разобраться и во внутреннем строении этих рыб, показывают, что
они стоят ближе всего к селахиям. Об этом свидетельствует строение
черепа, органов чувств, головного мозга и кровеносных сосудов.
Но, с другой стороны, Coccosteomorph! обладали уже костной тка-
нью, из которой был построен панцырь, хотя во внутреннем ске-
лете хорда оставалась постоянной и тел позвонков не имелось.
Костный панцырь был построен весьма сложно. Имелись костные
челюсти, и расположение костей в головном щите уже напоминало
Teleostomi (рис. 131).
Сем. Coccosteidae. Наиболее известен вид Coccosteus decipiens.
Девон Европы.
СТУПЕНЬ 2. КОСТНЫЕ РЫБЫ (OSTEICHTHYES)
Основным отличием от хрящевых рыб являются бблыпая или
мепыпая редукция хрящевого внутреннего скелета и замена его
настоящей костыо. Хотя плакоидпые чешуи в некоторых случаях
сохраняются, по на месте их и под ними развиваются особые кост-
ные чешуи. В плавниках роговые лучи (ceratotrichia) сменяются
костными лучами (lepidotrichia), развивающимися из чешуи.
Подкласс 5. Двоякодышащие (Dipnoi)
Группа рыб, обладающая, па ряду с примитивными признаками,
сближающими их с Chondrichthyes, рядом признаков высокой орга-
низации, что сближает их с наземными позвоночными. У современ-
ных представителей скелет в значительной мере хрящевой. Хорда
сохраняется в течение всей жизни. В кишечнике имеется спираль-
ный клапан, в сердце — сократимый артериальный конус; есть
клоака. Это — примитивные признаки. Однако брызгальца пет,
лучи плавников не роговые, череп автостилический, зубов на че-
люстях нет, а имеются своеобразные «жерновые зубы» на небно-
крыловидных костях и Hasplenialia. Есть легкие, и кровообращение
легочное. Скелет парных плавников представляет бисериальный
архиптеригий. Имеются хоаны. Хвост дифицеркальный. Чешуя
циклоидная.
157
В настоящее время Dipnoi представлены тремя родами, принад-
лежащими к двум разным семействам. В геологическом прошлом
они были представлены несколькими семействами, игравшими зна-
чительную роль в фауне девонской и каменноугольной эпох. Формы
эти были более подвижными, на что указывают гетероцеркальность
и два сниипых плавника. Черепные кости были многочисленными.
Сем. Phaneropleuridae с дифицеркальным или слегка гете-
роцеркальным хвостом. Спинных плавников два или один. Род Р h а п е-
ropleuron из верхнедевонских отложений (рис. 132).
Сем. Uronemidae. Хвост дифицеркальный. Спинной плавник
один. Черепные кости многочисленны. Зубы не слиты в пластиночные. Че-
шуя циклоидная. Род Uronemus из каменноугольных отложений.
Рис. 132.
Phaneropleuron andersonl. (Из Boulenger,
по Traquair и Dollo.)
Сем. Dipteridae (рис. 133). Хвост гетероцеркальный. Два спин-
ных плавника и один анальный. Зубы уже слились в пластинки. Голова
покрыта многочисленными окостенениями с своеобразной примитивной
структурой, сближающей их с Osteolepidoti. Dipterus из девонских слоев
Европы и Северной Америки. По ряду признаков это наиболее примитив-
ные из Dipnoi; в других они обнаруживают специализацию.
Рис. 133. Реставрация
Dipterus vallenciennensis
Сем. Цератодов ы е (Ceratodidae) (рис/134). Как и у всех
современных Dipnoi, хвостовой плавник дифицеркальный. Спинной плавник
один, непрерывный. Черепные кости сильно редуцированы в числе. Зубы
в виде пластинок. Легкое одно. У личинок нет наружных жабр и цементного
органа. Внутренние жабры развиты. Род Ceratodus из триаса и юры Европы,
Северной Америки, Африки, Азии, Австралии и меловых слоев Африки и Па-
тагонии. Баррамунда (Neoceratodus), с одним видом N. forsteri из Квин-
сленда в Австралии. Достигает 2 м длины. Живет в мелких водоемах с или-
стым дном и богатой растительностью, которою, как и находящимися на
ней ракообразными моллюсками и червями, питается. Во время засушливого
периода не впадает в спячку, а когда вода в водоеме портится, то вдобавок
к жаберному дыханию дышет легкими, поднимаясь на поверхность воды и за-
глатывая воздух, причем выпускает ранее заглотанный воздух. Рыба мед-
лительная, малоподвижная, обладает вкусным мясом. Яйца откладывает
между растениями. Разные стадии развития сходны с таковыми амфибий.
Сем. Lepidosirenidae представлено двумя родами: Protopterusв
средней части Африканского материка и Lepidosiren в бассейне реки Амазонки.
Имеют парные легкие (Dipneumones, двулегочные). У личинок имеются наруж-
ные жабры и цементный орган. В засушливое время впадают в летнюю спячку,
158
инкапсулируясь в иле и находясь в этом состоянии около 6 месяцев. В окру-
жающей «спящую» рыбу капсуле имеется отверстие как раз против рта рыбы.
Икра откладывается в гнезде, устраиваемом в иле. Protoptcrus представлен
3 видами. Наиболее известен Р. annectens. Питается лягушками, червями,
ракообразными, насекомыми, рыбами. Достигает 180 см. Lepidosiren рага-
doxa — в бассейне Амазонки (Южная Америка).
Рис. 134. Современные Dipnoi;
ДNeoceratodus forsterl, В — Protopterus annectens, (7—Lepidosiren paradoxa.
(Из Boulenger.)
Подкласс в. Рыбы с совершенным или настоящим
ртом (Teleostomi)
Плакоидных чешуй пет или опи играют совершенно подчинен-
ную роль, а под ними, в слоях cutis, развиваются большие ромби-
ческие, ганоидные или круглые циклоидные и ктеноидпые чешуи.
Последние могут исчезать, по это явление вторичное. На голове
эти чешуи, сливаясь между собой, частично исчезая, образуют по-
кровпые кости черепа, большей частью парные. Вокруг первичных
хрящевых челюстей развивается костный челюстпой аппарат,
несущий зубы. Появляются и вытесняют хрящ в скелете замещаю-
щие окостенения. Череп гиостилический. Обычно имеются парные
горловые пластинки. Позвонки и плечевой пояс большей частью
костные. Хвостовой плавник чаще всего гомоцеркальный, иног-
да гетероцеркальный или вторично дифицеркальный. Парные
плавники не архиптеригиального типа, со слабо развитыми га-
dialia и костными lepidotrichia. Как правило, имеется плавательный
пузырь. Если его пет, его исчезновение вторичное. Жаберпые от-
верстия открываются наружу не по отдельности, а прикрыты кры-
шечным аппаратом, сидящим на гиоидной дуге, открываясь в общую
полость. Жабры свободные. Число жаберных щелей не более 5.
Клоаки нет. Подкласс может быть разбит на два раздела, рано ра-
зошедшиеся в эволюции от общих акулоподобных предков; пред-
ставители одного раздела — хрящекостные или хрящевые ганоиды
159
(Chondrosteoidci) дали начало Chondrostei и представляют слепую
ветвь ствола рыб, другой раздел — Holosteoidei — развивался
прогрессивно. Его эволюция привела к современным костистым
рыбам (Teleostei), к костным ганоидам (вымершие Palaeoniscoidei,
современные Lepidosteoidei и Amioidei). Все эти группы рыб
объединяются в раздел лучеперых (Holosteoidei actinopterygii).
С другой стороны, к Holosteoidei принадлежат Holosteoidei cros-
sopterygii, давшие начало четвероногим наземным животным.
РАЗДЕЛ А. ХРЯЩЕВЫЕ ГАНОИДЫ (CHONDROSTEOIDEI)
Внутренний скелет и череп остаются хрящевыми. Хорда сохра-
няется всю жизнь. Тел позвонков не образуется. Голова покрыта
кожными окостенениями, среди которых уже намечаются основные
виды костей, встречающиеся у высших рыб. Зубов нет или они очень
мелки. Тело или голое, или покрыто рядом костных бляшек, на-
зываемых «жучками» и не имеющих ганоина. Дыхательный механизм
еще не совершенен. В крышке имеется только operculum. Лучей
жаберной перепонки или совсем пет или опи развиты слабо и в не-
большом числе. Хвостовой плавник гетероцеркальный с заходя-
щими на него чешуями — fulcra.
Отряд Осетрообразные (Chondrostei или Acipenseriformos)
Сем. Осетровые (Acipenseridae). Длинное заостренное рыло с
маленьким выдвижным ртом, лишенным зубов, на брюшной стороне. У за-
родышей зубы имеются. Возле рта находится две пары усиков. Тело вытя-
нутое, покрытое пятью продольными рядами «жучек», между которыми на-
ходятся выше боковой линии мелкие костяные пластинки, шипики и бугорки.
К осетровым принадлежат важные промысловые рыбы, называемые
«красной рыбой». Большинство является рыбами «проходными», живут в море,
а для метания икры входят в реки, проводя в последних нередко очень дол-
гий срок. Соленая вода губительна для икры осетровых. Лишь немногие
осетровые, как стерлядь, являются пресноводными. Выметав икру, рыбы
уходят обратно в море. Икра мелкая, липкая. Нерест весной. Плодовитость
велика. Белуга весом 1120 кг дает более 300 кг икры. Пищей осетровым
служат рыбы (рыбой питаются белуга и калуга) или (для большинства осет-
ровых) моллюски, личинки насекомых, черви и т. д. Промысел осетровых более
всего развит в Волжско-Каспийском районе, где добывается свыше 25 млн. кг;
затем в сибирских реках, в Черноморско-Азовском районе, в Архангельском
и в Амурском бассейнах. Всего добывается . до 28 600 000 кг рыбы и до
1 млн. кг икры, расценивающейся весьма высоко.
Род Осетры (Acipenser). Брызгальце есть, жаберные перепонки
прирастают к межжаберному пространству, не образуя складки. Стерлядь,
(A. rhutenus), пресноводная рыба в реках Че рноморско-Азовского
и Каспийского бассейнов, а также в бассейне Ледовитого океана от Оби
и Колымы. В бассейн Белого моря проникла около ста лет назад по каналам
и сильно размножилась в Северной Двине и ее притоках. Русский осетр
(A. giildenstadti) живет в бассейнах Черного и Каспийского мо-
рей. Шип (A. nudiventrls), в бассейнах Черного и Азовского, Каспий-
ского и Аральского морей. Совершает большие нерестовые миграции
ив последнего по Сыр-Дарье до Ферганы. Немецкий осетр (A. sturio,) распро-
странен по берегам Европы к югу от Нордкапа и у восточных берегов Сев.
Америки. Ив Финского залива входит в Неву. Встречается и в сев.-западной
части Черного моря. Сибирский осетр (A. baeri,) — в реках Сибири от Оби
до Колымы, в бассейне озера Байкал. Особый вид осетра. A. schrencki, —
в бассейне реки Амура. В северной части Тихого океана по западному и вос-
160
точному побережью — свой осетр — сахалинский или морской осетр (А. те-
dirostris). Севрюга (A. stellatus) (рис. 135) водится в Каспийском, Черном
и Азовском морях и впадающих в них реках, проникая в Средиземное мо-
ре до Адриатического. В С. Америке есть еще свои осетры. Род Acipenser
известен с эоцена.
Род Белуга (Huso), с двумя видами: Белуга (Huso huso), распростра-
ненным в Черном, Каспийском и Адриатическом морях, и Калуга (Н. dauricus)
в бассейне реки Амура Оба вида дости -дют веса 1000 кг.
Рис. А35. Севрюга (Acipenser stellatus).
(Из Boulenger.)
Род Лопатонос (Pseudoscaphirhynchus) имеет широкое
л< патообразное рыло, spiraculum не развито. Хвост вытянут в конце в нить.
Три вида живут в водах Средней Азии. Р. kaufmanni — в Аму-Дарье, Р.
fedlschenkoi — в Сыр-Дарье. К этому роду очень близок род Scaphirhynchus
с видом Sc. plathyrhynchus, живущий в реке Миссисипи и ее притоках. Такое
прерывистое распространение говорит о большой древности родов. Псевдо-
скафиринхи вкусны, но промыслового значения не имеют.
Сем. Многозубые — Polyodontidae. Рыло вытянуто в
длинный лопатообразный отросток, широкий рот имеет в обеих челюстях ма-
ленькие зубы. Усиков нет. Polydon (Spatularia) folium — в бассейне
Миссисипи. Psephurus gladius — в Китае, в реках Янг-тсе-Кианг и в Хоанхо.
Из меловых отложений Англии и из эоцена Америки известны близкие виды.
РАЗДЕЛЕ. КИСТЕПЕРЫЕ (HOLOSTEOIDEI CROSSOPTERYGII)
В настоящее время раздел представлен недавно найденной ры-
бой Latimeria из отряда Coelacanthini и отрядом Polypte-
rini с двумя родами: Polypterus и Calamoichthys. Polypte-
rini были отнесены к разделу Crossopterygii давно. В последнее
время некоторые ихтиологи выделяют их из этой группы и ставят
или как особый раздел, наряду с Crossopterygii и Actinoptcrygii,
или прямо относят к последним, ставя рядом с Chondrosteoidei.
В палеозое же, отчасти в мезозое отряд был представлен необычайно
богато. Грудные плавники в виде лопасти. У одних ископаемых
опа была короткая и, вероятно, с уписериальпым скелетом, у дру-
гих — заостренная, повидимому, бисериалыюго типа. Чешуи были
ромбические или циклоидные по форме, по «ганоидные» по струк-
туре, со слоем эмалепсдобпого ганоина. Хвост гетероцеркальный
или дифицеркальный. Осевой скелет у разных представителей
группы или просто в виде хорды или с кольцеобразными телами
или с настоящими костными амфицельными позвонками. Жабер-
пая крышка имела пе только operculum, по и suboperculum. Однако
брапхиостегальпый аппарат еще пе был развит, его место зани-
мали широкие горловые пластинки (jugularia) Cros-
sopterygii занимают среди Osteichthyes центральное положение.
Они обнаруживают некоторую, хотя далекую связь с элазмобрап-
хиями и гораздо более тесную — с Holostei и Teleostei
От них же, повидимому, произошли и наземные позвоночные.
11 Каввсаров и Ставчивский 161
Отряд 1. Остеолепиды (Osteolepldoti)
Этот отряд процветал с середины девонского времени (древний
красный песчаник) до конца каменноугольного. Одно семейство
дожило до верхнего мела.
Рис. 136. Реставрация Osteolepis macrolepidota.
(По Traquair из Boulenger.)
Сем. Osteolepidae (рис. 136) имело вытянутую форму, хвост гетеро-
церкальный. Чешуи ромбические. Позвонки кольцевые. Пинеальное отверстие.
Osteolepis macrolepidotus из девона Шотландии.
Рис. 137. Реставрация Holoptychius flemingi.
(По Traquair из Boulenger.)
Сем. Holoptychiidae. Грудные плавники в этом семействе удлинен-
ные и острые. Чешуя крупная, округленная, нос толстым слоем ганоина. Пи-
неального отверстия нет. Зубы крупные, эмаль их образует много входя-
щих складок. Позвонки, повидимому, не окостеневали, и хорда сохранялась
в первоначальном виде. Holoptychius из девона Европы и Америки (рис. 137).
Рис. 138а. Реставрация Undina gulo.
(До S. Woodward из Boulenger.)
Рис. 1386. Рыба рода Latimeria. Найдена
живой у берегов Южной Африки.
162
Отряд 2. Целакаптины (Coelacanthini)
Отряд существовал с девона до мела. Хорда сохранялась всю
жизнь, тел позвонков не было. Хвост дифицернальный из трех
лопастей. Дуги и ребра окостеневали. Череп также окостеневал.
Совсем недавно у берегов Африки поймана крупная рыба,
которая должна быть отнесена к Holosteoidei Crossopterygii и уди-
вительным образом Сходна с родом Undina. Рыбе этой дали название
Latimcria.
Сем. Undinidae. Род Undina из юрских слоев Англии (рис. 138а). Род
Latimeria. Найдена живой у берегов Южной Африки. Крупная рыба. Око-
ло полутора метров длиною (рис. 1386).
Отряд 3. Многоперы (Polypterini)
Как указано выше, сюда принадлежат два ныне живущие рода
кистеперых. Многими опи выделяются из последних и относятся
к Actinopterygii. Грудные плавники у них с хорошо выраженной
Рис. 139. Polypterus bicliir (Л), Calamoichthys calabaricus (В).
(Из Goodrich.)
лопастью. Хорда исчезает и заменяется амфицельными костными
позвонками. К примитивным признакам принадлежат: spiraculum,
развитой артериальный копус и спиральный клапан. Очень ха-
рактерно наличие двойного пузыря, отходящего с брюшной сто-
роны пищевода и ячеистого па внутренней сторопе. Чешуи ромбиче-
ские со слоем гапопна и маленькими зубчиками па поверхности
или па свободном крае. Пипсальпого отверстия пет. К отряду от-
носятся:
Сем. Многой ср ы е — (Polypteridae) (рис. 139) с двумя
родами Polypterus и Calamoichtys. Оба рода африканские, первый с 10 видами,
второй с одним; распространены в реках, текущих в Атлантический океан,
ив Верхнем Ниле. Polypterus bichir держится в ямках и углублениях Нила,
где он живет. Наиболее активен ночью, пользуясь нередко для передвижения
лопастными грудными плавниками. Однако «дыхательный» пузырь Polyp-
terus еще не совершенен. Последний может оставаться вне воды не более
11* ' 163
3—4 часов, да и то при условии, если он покрыт мокрой травой. Замечательна
личинка Polypterus: она имеет наружные жабры (рис. 140), достигающие
половины длины тела и отходящие от края жаберной крышки, спинной плав-
ник не разбит на отдельные плавнички и хвост гетероцеркальный. По своему
питанию Polypterus — хищник. Наружные жабры имеются и у молоди Ca-
lamoichthys.
Рис. 140. Личинка Polypterus senegalus с наруж-
ными жабрами.
(По Budgett пз Boulenger.)
РАЗДЕЛ 0. ЛУЧЕПЕРЫЕ (HOLOSTEOIDEI ACTINOPTERTGII)
Парные плавники не лопастные, radialia более или менее зача-
точны, не выходят за пределы тела, и плавники поддерживаются
почти целиком костными лучами — dermotrichia. Хвостовой плав-
ник может быть гетероцеркальным, гомоцеркальным или гефиро-
церкальпым (с редукцией оси назади и удлинением верхних и ниж-
них дуг в хвосте), но никогда не бывает настоящим дифицеркальным.
У более примитивных представителей чешуи ромбические и ганоид-
ные, у других они циклоидные по форме, но с тонким слоем ганоина,
а у высших представителей группы — просто костные, циклоидные
или ктеноидные, или их нет, или они образуют папцырь на теле
рыбы. К Actinopterygii относятся следующие сверхотряды:
Palaeoniscoidei, Amioidei, Lepidosteoidei и Teleostei.
Сверхотряд 1. Палсописки (Palaeoniscoidei)
Сюда принадлежат вымершие рыбы, относимые многими к Chon-
drostei и считающиеся за предков Acipenscridae. К такому взгляду
приводят данные палеонтологии, так как по внешним признакам
между Palaeoniscoidei и Acipenscridae существуют переходы.
Однако,по данным морфологическим, Acipenseridae являются скорее
примитивными формами. Palaeoniscoidei представлены уже в ниж-
нем девоне, рядом с древнейшими Crossopterygii. Они процветали
в триасе и в юре, но постепенно уменьшались в числе и отсутствуют
уже в мелу. Более древние семейства отряда являются наименее
специализовапными — характеризуются чешуей особого типа,
называемого палеонискоидным.
К сем. Palaeoniscidae относятся рыбы с удлиненным телом и
ромбической или циклоидной ганоидной чешуей. Представителем может
служить Palaeoniscus.
К сем. Platysomidae с более высоким, сжатым с боков «леще-
видным» телом принадлежали рыбы более специализированные, вероятно,
обитатели прибрежной зоны.
164
Три последующих сверхотряда стоят на более высокой сту-
пени развития, нежели предыдущие. Скелет в этих группах око-
стеневает весьма полно. Хвост гомоцеркальный или еще более
измененной формы. Лучи парных плавников еще более укорочены,
чем у Chondrosteoidei. Jugularia исчезают. Спиральный клапан в
кишке еще сохраняется в редуцированном состоянии у низших
представителей—Amia и Lepidosteus, но у костистых рыб (Teleos-
tei) обычно полностью исчезает. То же следует сказать и об
артериальном конусе в сердце. Хотя у более примитивных форм
еще имеется ганоидная чешуя, последняя сменяется у высших
форм, Teleostei, настоящей костной, циклоидной или ктеноидпой.
Сверхотряд 2. Амиевые (Amloldel)
Кроме современной Amia calva, отряд включает ряд ископае-
мых, вымерших семейств. У Amia позвонки хорошо развиты, но
у низших представителей отряда хорда существовала еще в боль-
Рис. 141. Dapediue politus.
(По S. Woodward из Goodrich.)
шом объеме. И у тех форм, которые имели или имеют развитые поз-
вонки, последние — хотя бы в хвостовой области — построены
из чередующихся полуколец (остатки первичной диплоспопдилип).
Череп менее специализован, чем у двух последующих отрядов:
chondrocranium еще в значительной мере сохранен, по большинство
типичных костей черепа уже развито. У Amia conus arteriosus
еще имеется, но уже развит хорошо bulbus arteriosus. Плаватель-
ный пузырь у Amia с легочными артериями, разделен септами на
ячейки.
Возникнув в пермскую эпоху, Amioidei в изобилии жили в триасе,
но максимум их развития, когда они были господствующими рыбами,
падает на юрскую эпоху. К отряду относится несколько семейств,
развившихся в различных направлениях. Некоторые из пих явля-
ются боковыми ветвями и пе дали потомства, другие являются сту-
пенью в развитии костистых рыб (Teleostei). Назовем некоторые.
Сем. Eugnathidae. Тело длинное. Чешуи на теле и кости на
голове «ганоидные» (толстые у одних и тонкие, почти «ганоидные», у других).
Хорошо развитые fulcra. Тел позвонков или не было или они представлены
отдельными полукольцами.
165
Сом. Semionotidae представлено формами с более высоким,
сжатым с боков телом и с более длинными спинными и анальными плавни-
ками. Па всех плавниках имелись fulcra. Костные пластинки на голове еще
с «ганоидным» характером. Осевой скелет примитивен. Ряд форм, идущих
Рис. 142. Mesodon macropterus.
(По S. Woodward из Goodrich.)
от перми к триасу, показывает все возрастающую специализацию в направ-
лении от вытянутой формы к высокой (рис. 141).
Последняя была еще более выражена в семействе Pycnodonti-
d а е, представителем которого может служить вид Mesodon macropterus
из верхней юры Баварии. В этом семействе" fulcra отсутствовали (рис. 142).
Рис. 143. Amia calva, наружный вид и скелет.
(По Goodrich.)
Сем. A m i i d а е представлено одним живущим родом Amia и несколь-
кими ископаемыми. Тело вытянуто, веретенообразно. Спинной плавник из
мягких лучей очень длинный. Чешуи тонкие, циклоидные, налегающие
одна на другую черепицеобразно, как у Teleostei. Ганоин образует лишь
поверхностную орнаментацию чешуи. Fulcra отсутствуют. Амфицельные
166
позвонки развиты в туловище, но в хвостовом отделе существует еще пер-
вичная диплоспондилия: полупозвонки образуют чередующиеся кольца.
Плавательный пузырь ячеистый. Слепых отростков в кишечнике нет.
Amia calva (рис. 143) живет в озерах и реках центральной и южной части
Сев. Америки до Великих озер включительно (Гурон, Эри). Известна с эоцена.
Рис. 144. Коротконосая панцырная щука, Lepidosteus
platystomus.
(Из Dean по Goode.)
Сверхотряд 3. Каймановые рыбы (Lepidosteoideae)
Тело удлиненное. Ромбические ганоидные чешуи, покрывающие тело,
усажены на поверхности плакоидпыми зубиками и прочно между собой со-
членяются. Слой ганоина имеется и па черепных костях. Непарные плав-
ники с fulcra. Верхняя и нижняя челюсти вытянуты в длинное рыло. Позвон-
ки окостеневают, тела опистоцельные (единственный случай среди рыб).
Дентин в основании зубов складчатый. Имеются слепые выросты кишечника.
Плавательный пузырь ячеистый.
Сем. Lepidosteidae с одним родом Lepidosteus (рис. 144) и не-
сколькими видами, живущими в пресных водах Сев. Америки.
«Ганоидная щука» (Lepidosteus osseus) — наиболее известный
вид, ведущий, как и другие виды, хищный образ жизни. Живет в глубоких
местах рек и озер, постоянно поднимается па поверхность, где заглатывает
воздух и выпускает его через жаберные щели, повидимому, после того как он
пройдет плавательный пузырь.
Личинка Lepidosteus, снабженная большим желточным мешком, при-
крепляется предротовым диском к предметам и некоторое время остается
неподвижной, пока пе рассосется желток1. В эоценовое и миоценовое время
Lepidosteus жил в Европе. В Америке этот род живет с эоцена.
Сверхотряд 4. Костистые рыбы (Teleostei)
К сверхотряду Teleostei принадлежит подавляющее число ныне
живущих видов рыб: 11 500 из 12 000. Teleostei характеризуются
следующими признаками. Ганоииовый слой в чешуе утрачивается,
хотя наиболее древние примитивные формы еще имеют ганоидную
чешую. Обычно чешуя округлая и черспицсобразпая, налегающая
одна па другую. Позвоночник окостеневает, и позвонки имеют амфи-
цельную форму. Хвост гомоцеркальный или гефироцеркальный.
Лучи в парных плавниках еще более зачаточны, чем в предыдущих
отрядах. В затылочной области черепа имеется верхпезатылочная
кость (supra occipitale). В пижпей челюсти утрачиваются некоторые
кости, и остаются лишь dentale, angulare и articulare. Срединные
jugularia утрачиваются. Сердце без артериального конуса с раз-
витым bulbus arteriosus. Спирального клапана в кишечпике пет
(у низших представителей последние три признака еще сохра-
няются). Хиазмы зрительных нервов пет.
Из всех Actinopterygii костистые рыбы являются наиболее
молодой группой, появляясь в небольшом числе в верхнем три-
167
асе и Липп» с верхпемелового времени начиная преобладать над дру-
гими Teleostomi. Низшие представители отряда связывают его
с Amioidci.
Классификация костистых рыб очепь сложна и трудна и по-
стоянно изменяется в зависимости от наших успехов в изучении
организации и палеонтологии этого отряда. Ниже будут охаракте-
ризованы только главнейшие из подотрядов, имеющие филогенети-
ческое или хозяйствепное значение. Как раз на долю костистых
рыб выпадает большая роль в хозяйственном отношении.
Рис. 445. Распространение семейства Characinidae.
Отряд 1. Тонкочешуйные (Leptolepiformes)
Связывает низших Teleostomi с костистыми. Вымершая группа,
жившая в мезозое. У них еще сохранялся топкий слой гапоина
на чешуе, которая была уже значительно тоньше, чем чешуя пре-
дыдущих отрядов. Fulcra уже исчезли.
Сем. Leptolepidae имело ряд примитивных признаков и во
внутреннем скелете. Хорда не вполне сжата позвонками и проходит через
последние, дуги не сливаются с телами. Род Leptolepis — из верхнейюры
Баварии.
Отряд 2. Карпообразные (Cypriniformes)
В этот отряд отпосят несколько семейств, обладающих рядом
общих признаков, по весьма различных по внешнему виду. К общим
признакам относятся: наличие плавательного пузыря (за немногими
исключениями), соединяющегося с пищеварительным трактом;
наличие в плечевом поясе так называемого мезокоракоида, своеоб-
разный «Веберов» аппарат между пузырем и слуховым аппаратом.
Сем. Харациновые (Characinidae). К харациновым
чринадлежит обширная группа (число видов 500) рыб, обитающих в прес-
1ых водах Африки и Центральной и Южной Америки (рис. 145). По своей
организации являются «обобщенным» типом, но на ряду с этим образуют ряд
типов, приспособившихся к различным условиям: одни из них являются
травоядными и похожи на карповых, другие — хищники, подобные лососям
л щукам Род Hydrocyon (рис. 146), в переводе — водяная собака, достигаю-
цая более метра длины, является представителем хищных харацинов. У
растительноядных форм зубы очень слабы. У рода Erythrinus пузырь разделен
на гидростатическую переднюю и дыхательную заднюю часть.
Сем. Голые угри (Grymnotidae). Рыбы, внешне похожие
на угрей, но не имеющие с ними ничего общего по происхождению: они яв-
ляются деградированными, сильно измененными харацинидами. Семейство
небогато родами и видами (около 30 видов) и ограничено в своем распростра-
нении Центральной и Южной Америкой. Наилучше изученным видом
является электрический угорь (Grymnotus electricus) из бассейнов рек
Амазонки и Ориноко. Длина его достигает более 2 м. Живет он в болотах
и мелких частях рек. Электрический разряд, который он может давать,
настолько силен, что может сбить с ног крупный скот при переправах через
реки.
Рис. 146. Hydrocyon goliath («водяная собака»).
(По Boulenger.)
Сем. Карповые (Cyprinidae). Большинство рыб наших прес-
ных вод принадлежит к этом семейству. Рот у всех беззубый. В глотке хо-
рошо развиты нижнеглоточные кости с одним, двумя или тремя рядами зу-
бов, пару с которыми составляет роговая подушка на верхней стороне глот-
ки — так называемый «жерновок». Глоточные зубы являются одним из важ-
нейших диагностических признаков, служащих для распознавания родов
и видов. Распространены карповые главным образом в северном умеренном
поясе.
Большинство карповых питается растительной и мелкой животной пищей.
Некоторые являются хищниками. Карповые распадаются на 4 подсемейства.
Подсем. Catostominae распространено в водах Сев. Америки,
Китая и Северной Сибири. Catostomus catostomus, «конек», водится и в Се-
веро-восточной Сибири и в Аляске. Распространение Catostominae говорит
о бывшей связи Восточной Азии и Северной Америки.
Подсем. Вьюновые (Cobit. idinae) распространено в Европе
и в Азии. Вьюн (Misgurnus fossilis) с 10 усиками около рта; щиповка (Cobitia
taenia) из Европы и Азии, с 6 усиками; в Азии Nemacheilus barbatula.
Подсем. Homalopterinae распространено в горных водоемах
Китая, Индии и Малайского архипелага.
Подсем. Карповые (Су prin in ас), очень богатое видами под-
семейство (не менее 1000 видов, принадлежащих к 200 родам), распространено
в пресных водах Старого Света и Сев. Америки. 50 родов живет в СССР. Мно-
гие карповые имеют большое хозяйственное значение для человека и играют
большую роль в экономии природы. Систематическими признаками являются
прежде всего глоточные зубы и число чешуй боковой линии. Род Cyprinus
представлен видом С. carpio, карп, сазан. Распространен в водах бассейнов
Черного, Каспийского и Аральского морей, а также в бассейне Тихого океана
к югу от Амура. Не так давно он заселил Балхаш и искусственно разведен
в Сев. Америке. Чрезвычайно выносливая рыба, переносящая и пресную и со-
лоноватую воду, хорошо переносящая перемены климата, а потому легко
акклиматизируемая там, где вода — хотя летом — хорошо прогревается;
крайне нетребовательна па пищу, можно сказать всеядна. Спермии сазана
дольше спермиев многих других рыб живут в воде, что способствует увели-
чению потомства. Количество яиц велико — от 300 000 до 700 000—у европей-
ского. Благодаря такой плодовитости, нетребовательности к пище и выносли-
169
вости к окружающим условиям (он живет в занимаемой им области в самых
различных условиях) сазан, несмотря на массовую гибель икры и молоди
успешно конкурирует с другими видами; служит предметом серьезного про-
мысла, причем промысел дикого карпа все более заменяется разведением
культурного карпа, представленного рядом рас. Род Carassius с видом карась
(C.carassius) встречается в ряде разновидностей, населяет стоячие воды Европы
и Азии, заходя в Европе в Англию и Скандинавию, Италию и Сицилию. Любит
тину и ил. Имеет рыночное значение. Золотая рыбка (Carassius auratus)
является искусственно выведенной разновидностью карася. Род Rutilus
Рис. 147. Усач каспийский (Barbus brachycephalus caspius).
(Ио Солдатову.)
с видом плотва (R. rutilus) образует большое количество подвидов. Рас-
пространена в водах Средней и Северной Европы, в Западной Сибири и Тур-
кестане, в морях Азовском и Черном, где ее называют таранью, в Кас-
пийском, нося там название воблы, и в Аральском море. Из морей при-
ходит для икрометания в реки. Осенью совершает миграции в море в поисках
мест для залегания на зиму. Плотва имеет большое промысловое значение.
На Каспийском море вобла является одной из важнейших промысловых рыб.
Род усач (Barbus) (рис. 147), самый богатый видами род карповых (до 300 ви-
Рис. 148. Сом обыкновенный (Silurus glanis).
(По Seeley из Goodrich.)
дов), распространенный в умеренной и тропической частях Старого Света.
Мясо усачей, водящихся у пас,в бассейнах Балтийского, Черного, Азовского,
Каспийского и Аральского морей, в Амуре и Уссури, очень ценится, от-
носится к «красной рыбе» (осетровые). Особенно ценны балыки. Из других
представителей карповых можно назвать голавля (Leuciscus cephalus), язя
(Leuciscus idus), красноперку (Scardinius ery thro phthal mus), жереха (Aspius
aspius), подуста (Chondrostoma nasus), уклейка (Alburjius alburnus), шемаю
(Alburnus chalcoides), леща (Abramis brama), чехонь (Pelecus cullratus),
пескаря (Gobio fluviatilis), линя (Tinea tinea) и горчака (Rhodeus amarus).
Последняя рыбка замечательна тем, что в пору икрометания у самки
вырастает длинный трубчатый яйцеклад, которым она откладывает икру
170
в жаберную полость моллюска Anodonta. К перечисленным родам надо доба-
вить еще маринку (Schizothorax) и османа (Dipthychus). Эти роды
населяют горные реки и озера Средней и Центральной Азии, а также бассейн
Инда, Ганга, Брамапутры. Эти рыбы экологически заменяют собой лососевых
в реках Средней и Центральной Азии. Икра считается ядовитой.
Рис. 449. Полунанцырный сом (Synodontis decorus).
(По Boulenger.)
Сем. Сомовые (Siluridae). Сомовые являются, строго говоря,
систематической единицей,более крупной, нежели семейство, включая в себя
крайне разнообразные формы, одни из которых имеют тело голое, другие оде-
ты костяным панцырем, причем чешуи или щитки, его составляющие, нередко
несут на своей поверхности плакоидные зубчики. На челюстных костях всегда
Рис. 150. Clarias anguillaris.
(По Vallenciennes из Boulenger.)
есть усы. Часто есть жировой плавник. Эта обширная группа включает в себя
около 1000 видов, распространенных по всему свету, но главным образом
в тропических странах. Немногие виды являются морскими. Семейство рас-
падается на ряд подсемейств.
Рис. 151. Callithys littoralis.
(По Boulenger.)
Подсем. Silurinae. Сом обыкновенный (Silurus glanis). Почти
или совсем без спинного плавника и с длинным анальным. Эта наи-
крупнейшая из пресноводных европейских рыб достигает 300 кг веса и
5 м в длину (рис. 148). Из рек заходит в Каспийское и Черное моря, также и в
Аральское. Имеет промысловое значение. Род Saccobranchus имеет меш-
ковидные удлинения жаберной полости, живот в Ост-Индии и на Цей-
лоне.
171
Подсем. Doradinae: короткий спинной плавник, имеется жировой, аналь-
ный короткий. Род Synodontis (рис. 149), имеет на голове и за нею панцырь,
остальное тело голос; распространен в Африке. Род Parexostoma, индийский
сомик, небольшого размера, живущий в горных речках в Гималаях и про-
никающий в Среднюю Азию.
Подсем. Clariinae. Спинной и анальный плавники очень длин-
ные, доходят до хвоста. Род Clarias (рис. 150), с покрытой панцырем
головой, живет в Африке. Ведет донный образ жизни, в связи с чем стоит
удлинение спинного и анального плавников.
Подсем. Malapterurinae. Без спинного плавника, есть большой
жировой, анальный короткий. Электрический сом (Malapterurus electricus)
живет в Ниле.
Рис. 152 и 153. Acestra gladius, подсем. Loricariinae.
(По Boulenger.)
Подсем. Callichthyinae. И спинной и жировой плавники
короткие. Тело заключено в панцирь из больших чешуй с плакоидными
зубчиками по заднему краю, плавники с иглами. Род Callichthys (рис. 151),
представители его строят искусные гнезда для икры.
Подсем. Bagrinae. Короткий спинной плавник и удлиненный жиро-
вой. Arius живет в Азии, Африке, Америке, Австралии. Яйца представители
рода носят в ротовой полости. Виды родов Pseudobagrus и Liocassis,
«касатки», в Амуре, Китае, Японии, Корее — мелкие рыбки. Массами
встречаясь в Амуре и Уссури, досаждают рыбакам. Могут быть использованы
на консервы.
Подсем. Loricariinae (рис. 152). Тело заключено в панцырь с
плакоидными зубчиками. Тело вытянутое, рот снизу, живут в горных реках
Центральной и "Южной Америки на большой высоте. 200 видов.
Отряд 3. Сельдеобразные (Clupelfonnes)
Более или меиее искусственная группа, объединяющая ряд
низкооргапизоваппых семейств, характеризующихся рядом при-
митивных признаков в скелете, большим числом позвонков, соеди-
нением плавательного пузыря с пищеводом, циклоидными чешуями
и отсутствием жестких лучей в плавниках. К Clupeiformes относится
более 20 семейств. Из пих отметим следующие.
Сем. Elopidae. Еще сохраняется средняя горловая пластинка.
Elops в тропических морях, маленькая рыбка. Megalops, М. atlanticus,
«тарпон», крупная (до 180 см) рыба, являющаяся предметом спорта у юго-
восточных берегов Сев. Америки, Зап. Индии и Бразилии. Замечателен своей
силой и прыжками из воды.
Сем. Albulidae. Очень древнее семейство, подобно предыдущему,
ведущее начало с мелового периода. Albula с одним видом A. conorhynchus
сохраняет еще артериальный конус в сердце. Личинка чрезвычайно напоми-
нает личинку угрей («Leptocephalus»).
Сем. М о\ my г i d а е. Африканское семейство, обладающее, кроме
примитивных, еще рядом признаков специализации. Замечательно вариирует
172
форма рыла, видимо приспособительно к способу питания и роду пиши,
вариирует в пределах одного рода. Роды: Mormyrus, Gnathonemus и Gymnar-
chus, личинка которого имеет наружные жабры, живут, как и остальные 7 ро-
дов, в пресных водах Африки к северу от тропика Козерога.
Сем. Osteoglossidae, древнее, известно с эоцена, близко к Albu-
lidae и имеет замечательное распространение, указывающее на их древность
(рис. 154). Род Arapaima из Гвианы и Бразилии достигает 4.5 м длины. Не-
tero+is — в Африке и Scleropages — в Австралии.
Сем. Сельдевые (Clupeidae). Тело, сжатое с боков, покрыто
легко спадающей, гладкой и крупной чешуей, брюшко с закругленным или
зазубренным килем из чешуек. Зубов нет или они малы. Боковой линии нет.
Живут в большинстве случаев в море, но есть и пресноводные формы. Мор-
ские формы ходят большими стаями — «косяками», питаясь главным образом
мелкими ракообразными и моллюсками. Имеют огромное промысловое значе-
ние. Наиболее известны следующие виды: североатлантическая сельдь,
Рис. 154. Распространение Osteoglossidae.
(По Boulenger.)
(Clupeaharengus) живет в Баренцевом море, в Белом, в Атлантическом океане
до берегов Испании на юге, на восточном побережье и до Нью-Йорка на западе,
далее — в Беринговом, Охотском и в сев. ч. Японского моря, образуя много-
численные разновидности. Каспийская и черноморская сельдь (Caspialosa)
распадается на ряд видов и подвидов. Род Alosa близок к предыдущему.
A. sapidissima живет по атлантическому побережью Сев. Америки, перенесена
была в Тихий океан, где прекрасно акклиматизировалась. Род Harengula
представлен в Каспийском и Черном морях видами Н. delicatula и Н. grim-
mi, каспийской килькой. Настоящие кильки и шпроты представлены у бе-
регов Европы родом Spratella с видом Spratella sprattus. Сардинки, Sardinella
вид (S. pilchardus) живут в Средиземном море, в Атлантическом океане
от южных берегов Ирландии до острова Мадейры. Имеются особые виды
сельдей в наших дальневосточных водах, например, «иваси» (Sardinella
melanosticta). Как сказано выше, большая часть сельдей — морские, но есть
среди них проходные и такие, что держатся в солоноватых водах. Большин-
ство сельдей совершает миграции в целях икрометания. Из яиц получаются
личинки, отличающиеся от взрослых рыб и подвергающиеся метаморфозу.
Питаются беспозвоночными и планктоном. Расы одного вида нередко отли-
чаются друг от друга характером пищи, временем размножения, темпом ро-
ста и т. д. Морфологические отличия нередко можно установить лишь биомет-
рическим методом. Сельди являются одной из самых важных промысловых
рыб во всем мировом хозяйстве. Громадное значение имеет промысел сель-
дей и у нас.
Сом. Анчоусовые (Engraulidae). К этому семейству,
близкому к предыдущему, принадлежит мелкая рыбка — анчоус, называе-
мый в Черном море хамсой (Engraulisengrassicholus). Эта рыбка, как и сельдь,
173
пелагическая, совершающая миграции. Последние происходят под влиянием
изменений температуры воды. Летом хамса держится в мелководной северной
части Черного и Азовского морей, зимой уходит к югу в более глубокую
воду, вплоть до Босфора. Весной идет обратно. Будучи малоценной рыбой по
своим качествам, хамса имеет значение вследствие необычайного ее обилия.
Сем. Лососевые (Salmonidae). К лососевым принадлежат
более крупные рыбы, чем к сельдевым. Все лососевые имеют позади спинного
плавника маленький жировой плавник. Плавательный пузырь простой.
Плотная мелкая чешуя. Большая часть в прибрежных морях и в пресных
водах умеренного и полярного поясов северного полушария. В геологическом
прошлом некоторые виды, связанные тесно с севером, попали в Аральское,
Каспийское и Черное моря и до сих пор еще сохранились в небольшом коли-
честве. Лососевая икра, искусственно оплодотворенная, может переносить
(со льдом) далекую пересылку; благодаря этому и другим особенностям
Рис. 455. Распространение Salmonidae.
(По Boulenger.)
икры, а также высоким вкусовым качествам, лососи являются объектом ис-
кусственного разведения (рис. 455). Лосось, семга (Salmo, вид S. salar)
распространен у европейского побережья Ледовитого океана, в Белом
море, вдоль берегов Норвегии и к югу до берегов Португалии, в Балтийском
море, также у атлантических берегов Сев. Америки. Кумжа, таймень
(S. trutta) образует проходные формы, речные и озерные. Распростра-
нение сходно с распространением лосося. В Аральском море особый
подвид — S. trutta aralensis, в Каспийском — S. trutta labrax, весом до
50 кг. В озерах образуются особые местные формы всегда меньшего размера —
S. trutta lacustris. Еще меньшего размера нёнаслсдственпые формы — морфы,
называемые форелями, образует таймень в мелких речках и ручьях —
S. trutta morpha far io. Род Восточный лосось (Oncorhynchus) заменяет
род Salmo в водах Тихого океана. Образует ряд видов, живущих как в
водах Атлантического, так и североамериканского побережья. Наиболее
известна у пас кета (О. keta). Этот род образует в озерах карликовые
формы. Лососи, в особенности дальневосточный, служат предметом промысла
большого значения. Сиги (Coregonus). Обширный род с большим количеством
видов, среди которых есть и проходные, и озерные и речные. Ряпушка (Co-
regonus albula) — в морях и озерах Северной Европы. Омуль (С. autu-
mnalis) —в Сибири, от Мезени до Колымы. Проходной сиг (С. lavaretus) — в
бассейне Балтийского моря. Проходные сиги совершают миграции, подобные
миграциям лососей, уходя в реки далеко от устья. Нерестуют осенью.
Пищей сигов являются как донные беспозвоночные: моллюски, личинки насе-
474
комых, так и планктон. Некоторые едят и мелкую рыбу. Вместе с планкто-
ном некоторые сиги совершают вертикальные миграции. Мясо сигов очень
высоко ценится. Озерные сиги могут служить для субституции (заме-
ны) малоценных пород рыб. Род Хариус (Thymallus) похож на сигов,
Рис. 156. Хариус, Thjmallus thymallus.
(По Солдатову.)
но отличается очень длинным спинным плавником. Т. thymallus —
европейский вид (рис. 156). Т. arclicus — сибирский вид. Держатся хариусы
в чистой холодной воде северных рек и в больших озерах. Цо вкусовым
качествам превосходны, но промыслового значения не имеют, а лйшь местное.
Белорыбица (Stonodus S. leucichthys) — в бассейне Каспийского моря,
и S. 1. nelma — в Сибири. Превосходного вкусового достоинства, но про-
Рис. 157. Malacosteus indicus.
(По Гейтеру из Boulenger.)
мысел пока еще имеет малое значение. Разводится искусственно. Osmerus
некоторыми выделяется в особое семейство. Морская корюшка (Osmerus
eperlanus) — в бассейне Балтийского и Немецкого морей. Входит в Неву
для метания икры. Снеток, озерная корюшка (Osmerus eperlanus spirinchus).
Карликовая форма, происходящая от морской, живет в озерах Прибалтий-
ского края. Есть подвиды и в бассейне Ледовитого океана. Мойва (Mallo-
tus, М. villosus) служит наживкой на крючковой снасти при ловле трески.
Сем. Иглоротых (Stomiatidae). Специализованные, глубоковод-
ные рыбы с органами свечения. Роды Chauliodus, Argyropelecus, Stomias,
Malacosteus (рис. 157).
Отряд 4. Щукообразные (EsocIformes)
У предыдущего отряда в плечевом поясе имелся так называемый
мезокоракоид. Здесь его пет. Веберова аппарата нет. Брюшные
плавники па брюхе. Настоящих шипов в плавниках нет. Плава-
тельный пузырь обычно соединен с пищеводом. Щукообразпые
связывают Clupeiformes с некоторыми высшими отрядами. Сюда
принадлежит несколько семейств, из которых назовем:
Сем. У м б р о в ы е (U m b г i d а е) с представителем Umbra umbra из
илистых водоемов бассейна Дуная, Прута и Днестра.
175
Сем. Далиевые (Dalliidae), единственный представитель которого
'маленькая (20 см) Dallia pectoralis живет в промерзающих зимой мелких
речках, озерах и болотах Чукотского полуострова и Аляски, впадая зимой в
а н аб 11 g ти чес кое состояние.
Рис. 1а8. Распространение сем. Gaiaxiidae.
(По Boulenger.)
Сем. G-alaxiidae, с необычайно интересным географическим распро-
странением — в реках Южной Африки, Австралии, Новой Зеландии и юга
Южной Америки (рис. 158).
Сем. Щу новые (Esocidae). R последнему небольшому семейству
принадлежат хищные пресноводные рыбы, в том числе и всем известная
Рис. 159. Распространение сем. щуковых (Esocidae).
(По Boulenger.)
щука. (Esox). Е. lucius живет в Европе, Сибири и Сев. Америке. В.последней
и в Амуре живут еще другие виды. Остатки щуки известны с эоцена. Щука
является одним из общеизвестных объектов промысла, имея широкое рас-
пространение в Северной Америке, Европе и Азии (рис. 159).
Отряд 5. Анчоу сообразные (Scopellformcs)
Сем. Светящиеся анчоусы (Scopelidae), пред-
ставлено более чем 100 видами, большей частью пелагическими
176
и глубоководными рыбами. Известно и много ископаемых начиная
с меловой эпохи.
Виды рода Scopelus, небольшие рыбки, живут почти во всех морях и от-
личаются присутствием светящихся органов — фотофоров — по бокам тела,
на голове. У рода Ipnops, живущего на большой глубине, глаз нет, а их
место занято органом свечения, назначение которого, надо думать, привле-
кать мелкие организмы в пасть этой рыбы, так как без органов зрения и
без органов осязания иной способ добычи пищи для нее был бы крайне за-
труднен.
Отряд 6. Ммкрокарпы (Amblyopsiformos, Mlcrocyprlni)
Включает в себя два семейства — Amblyopsidae и С у р-
rinodontidae, зубастых карпов.
К последнему принадлежат около 200 видов маленьких и очень ма-
леньких рыбок, живущих в пресной или в солоноватой воде, отличающихся
по внешности плоской головой и выдвижным ртом, большими чешуями и от-
сутствием боковой линии. Большинство зубастых карпов живет в Америке,
Рис. 460. Распространение зубастых карпов (Cyprinodontidae).
(По Boulenger.)
Африке, Южной Европе и Южной Азии, а также в Японии (рис. 160). Боль-
шинство видов живородящие, и у многих видов анальный плавник самца
изменен в копулятивный орган, при помощи которого происходит внутрен-
нее оплодотворение. Многие представители этого семейства содержатся в ак-
вариумах и служат хорошими объектами для экспериментов: Fundulus,
Gambusia, Poccilia, Girard inus. У рода Anableps выпяченные кверху глаза
разделены темной полоской па верхнюю и нижнюю половинки. Рыбка обыч-
но плавает таким образом, что верхняя половина глаз торчит наружу.
Отряд 7. Угренодобпые (Augiillliformes)
Представители этого отряда характеризуются, кроме ряда ана-
томических признаков, удлиненным, закругленным или лентовид-
пым телом с большим количеством позвонков. Парные плавники
имеют тенденцию исчезнуть. Плавательный пузырь есть. Чешуи
рудиментарны или вовсе отсутствуют. Развитие с превращением.
Сюда относятся несколько семейств.
32 Кашкаров и Станчицокий 377
Сем. Saccopharyngidae представлено крайне причудливыми
глубоководными рыбами, являющимися деградированным типом угрей.
Скелет объизвествляется слабо, слабо развиты мускулы. Рот огромный, глотка
и желудок сильно растянуты (рис. 161).
Сем. Настоящие угри (Anguillidae). Известно 150 ви-
дов, из которых 50 являются глубоководными, встречаясь до глубины 4300.
Цикл жизни и миграции обыкновенного угря (Anguilla anguilla) и американ-
ского (A. americana) описаны ниже. Первый вид распространен по берегам
Европы, начиная от Белого моря, где изредка встречается, до Черного.
Мясо угря очень высокого качества, богато жиром. Молодь, которую ловят
в огромном количестве, может быть выращиваема в изолированных
водоемах.
Рис. 161. Мешкоглот (Saccopharynx ampullaceus).
(По Hunter из Boulenger.)
Морской угорь (Conger conger) распространен в Атлантическом, Индий-
ском и Тихом океанах. Из Средиземного моря изредка проникает в Черное
море.
Сем. Муреновых (Muraenidae). Представителем является
обыкновенная мурена (Muraena helena) живущая в Средиземном море.
Мясо ее очень ценилось в древности. Есть сведения, что римляне откармли-
вали их рабами.
Отряд 8. Колюшкообразпые (Gasterosteiformes)
Этот отряд включает ряд семейств, из которых паиболее изве-
стны: сем. Колюшковые (Gasterosteidae) и Морские иглы (Syn-
gnathidae).Последнее семейство отличается от всех других рыб тем,
что жаберные листочки у них пе узкие, длинные, по короткие и ши-
Рис. 162. Колюшка трехиглая (Gasterosteus
aculeatus). *
(По Good из Boulenger.)
рокие, напоминающие перистый лист. Тело покрыто тонкими ко-
стными пластинками. Рыло обычно цытяпуто в более или менее
длинную трубку, па конце которой находится маленький беззубый
рот.
Виды большей частью морские,некоторые живут в солоноватой воде пред-
устьевых пространств. Органом движения, видимо, прежде всего служит спин-
ной плавник. Род Морская игла (Syngnathus); обыкновенная игла рыба
(S. acus); морской конек (Hippocampus), с цепким хвостом без плавника. Пла-
178
вает в вертикальном положении, хвостом цепляется за подводные растения.
Н. antiquorum — в Атлантическом океане, Средиземном и Черном морях.
Сем. Колюшки (Gasterosteidae). Рыло не вытянуто
в трубку или лишь слегка вытянуто. Перед спинным плавником ряд изоли-
рованных шипов. GasterosteuS с 2—3 колючками перед спинным плавником.
Каждый брюшной плавник — из одной- крепкой иглы и более короткого
гибкого луча. По бокам голого тела большие плоские щитки. Колюшка трех-
иглая. (G. aculeatus). Водится в солоноватых и пресных водах почти всей
Рис. 163, Распространении uasierosteus.
(По Миг.)
Европы (рис. 462). Колюшки известны своими заботами о потомстве, для
вывода которого из икры строят гнездо или на дне (Gasterosteus), или вися-
чее между растениями (Pygosteus). Все семейство ограничено северным полу-
шарием, проникая на севере до Гренландии и Берингова пролива, на юге —
до Алжира, Средней Азии и Калифорнии (рис. 163). Встречаясь в больших
массах, приносят вред, поедая икру более цепных рыб.
Отряд 9. Макрелещукообразные (Scoinbresociformes)
Представлен рыбами умеренных и тропических морей, большей
частью обитателями открытого моря.
Рис. 164. Сарган (Belune annulata).
(По Cuvier и Vallenciennes из Boulenger.)
Сем. Макрелещуки (Scombresocidae). Род Морская щука,
( Габоне) с видом В. belone, имеет челюсти, вытянутые в виде клюва, усажен-
ного зубами (рис. 164). У Hemiramphus вытянута только нижняя челюсть.
Макрелещука (Scombresox) имеет такое же вытянутое рыло и несколько
179
маленьких плавничков позади спинного и анального, как у Scombridae. Все
представители семейства, построенные по стреловидному типу, имеют при-
вычку выскакивать из воды. Этот инстинкт достигает максимального развития
у летучей рыбы, Exocetus, о полете которой сказано ниже.
Отряд 10. Кефалеобразные (Muglliformes)
Включает в себя несколько семейств, характеризующихся удли-
ненной формой, продвижением вперед брюшпых плавников, двумя
спинными плавниками, причем первый имеет немногочисленные
жесткие шипы. Чешуя циклоидная и, как правило, заходит на
голову. Боковой линии нет или опа неявственна.
Сем. Кефалевые (Mugilidae). Довольно обширное семей-
ство, включающее в себя более 100 видов, живущих в морях, солоноватых и
пресных водах умеренного и тропического поясов. Род Mugil представлен
Рис. 165. Распространение сем. Anabantidae.
(По Boulenger.)
в водах Союза 6 видами рыб средней величины (от 25 до 80 см), держащихся
обычно большими стаями и очень ценимых за вкусное мясо: кефаль, лобан
(М. cephalus), остроносик (М. chelo), сингиль (М. auratus) — в Атлантиче-
ском, Черном, и Азовском морях; М. hacmotochilus — в дальневосточных
водах. Питаются все планктоном. Кефаль имеет промысловое значение.
Сем. Атерин ковые (Atherinidae)c родом Atherina — ма-
ленькие рыбки, ценимые за вкусное мясо, в Черном, Азовском, Каспийском
и Средиземном морях и в Атлантическом океане.
Сем. Лабиринтовые (Labyrinthidae, Anabanthidae).
Чешуя ктеноидная. Замечательные рыбки пресных вод жаркого пояса, имею-
щие описанные выше придаточные полости над жаберной полостью. В этой
придаточной полости находится «лабиринт» — вырост жаберных дуг в виде
свернутых тончайших костных пластинок, покрытых богатой сосудами сли-
зистой оболочкой. Лабиринт дает рыбкам возможность долгое время нахо-
диться вне воды. Anabas scandens в Ост-Индии и на Малайском архипелаге
передвигается на суше при помощи шипов на жаберных крышках, на брюшпых
и на анальном плавниках. Другие виды живут в Африке (рис. 165).
Сем. Змееголовые (Ophiocephalidae) отличается тем,
что крупные чешуи покрывают голову. Также имеют дополнительную над-
жаберную дыхательную полость, имеют совершенно сходное распростра-
нение с предыдущим семейством (рис. 166). Один представитель встречается
в бассейне Амура.
Сем. Хиазмодонтовые (Chiasmodontidae) заклю-
чает хищных глубоководных рыб, сходных с атеринками по строению скеле-
та. Вид Chiasmodon niger добыт в Северной Атлантике в глубине -2590 м
(рис. 167).
180
Сем. Osphromenidae, во многом сходное с Labyrinthidae,
распространено в Юго-восточной Азии и в Африке. Многие известны как аква-
риумные рыбки: петушки (Betta), гурами (Osphromenus), макроподы
(Macropodus viridi-auratus).
Рис. 166. Распространении сем. Ophiocephalidae.
(По Boulenger.)
Группа отрядов Колючеперые (Acanthopterygti)
Наиболее обширная группа во всем классе рыб. Характери-
зуется наличием в спинном и в анальном плавниках нерасчленен-
ных жестких и колющих лучей, хотя из этого правила имеются
и исключения. Брюшные плавники всегда сдвинуты далеко вперед,
располагаясь за редкими исключениями между грудными или па
Рис. 167. Хиазмод черный (Chiasmodus niger).
(По Гептеру из Boulenger.)
горле. Таз обычно прикреплен к плечевому поясу. Плавательный
пузырь обычно, кроме некоторых более примитивных представи-
телей группы, пе имеет .связи с пищеводом. Крышечпый аппарат
хорошо развит, жаберпая щель широкая. Черепные кости погру-
жены в кожу глубоко. Появляются колючеперые только в меловую
эпоху, являясь наиболее совсршеппыми представителями Teleostei.
Отряд 11. Beryclformes
С центральным семейством Berycidae — наиболее древним
семейством среди Acanthopterygii. Архаическими чертами являются
481
большое количество лучей в брюшных плавниках и соединение
пузыря с пищеводом у некоторых родов.
Сем. Berycidae было очень многочисленно в меловое время и яв-
ляется, видимо, предшественником других Acanthopterygii. 13 родов и 70
видов живут в настоящее время главным образом в глубинах морей почти
всего земного шара. Вегух splendens (рис. 168).
Рис. 168. Вегух splendens.
(По Good и Bean из Boulenger.)
Отряд 12. Солнечники (Zelformes>
Отряд этот с семейством солнечники (Zeidae) состав-
ляет переход к следующему разделу, с симметричными древними
формами, видимо, давшими начало несимметричным камбаловым.
Солнечник (Zeus faber) называемый «сельдяным королем», «рыбой
св. Петра», ценится за свое мясо. Достигает он размеров до 1 м и живет в юж-
ной части Атлантического океана и в Средиземном море, откуда проникает
в Черное море. Zeidae, имеющие еще, как у Berycidae, нерасчлененные лучи
в плавниках, связываются с камбаловыми через семейство Amphistidae,
у которого лучи эти утрачены, а тело уже имеет форму тела камбаловых.
Отряд 13. Камбалообразные (Rhomb 1 formes)
Сюда относятся рыбы с сжатым с боков, несимметричным телом.
Одна сторона тела обращена кверху и окрашена, другая, ниж-
няя — бесцветна. Оба глаза находятся па верхней стороне, у одних
форм — на правой, у других — па левой. Непарные плавники без
жестких лучей, очень длинные. Обитатели морей; некоторые виды
заходят и в реки. Молодь построена симметрично (рис. 169). Раз-
вивающаяся впоследствии песимметрия затрагивает и скелет го-
ловы.
Сем. камбаловые (Pleuronectidae) заключает в себе
около 500 видов, большей частью морских. Лишь некоторые формы держатся
в пресных водах Южной Америки и Малайского архипелага. Распространены
они широко — от полярного круга до южных берегов южного полушария.
Все придонные формы, некоторые глубинные. Икра — пелагическая. Стекло-
видно прозрачные личинки ведут также пелагический образ жизни. Питаются
камбаловые различными беспозвоночными и рыбами. Весьма плодовиты: у Во-
thus maximus число икринок исчисляется от 1 000 000 до 2 000 000. Весьма
важное в промысловом отношении семейство; в изобилии вылавливается
у берегов Англии, Голландии, Франции. В СССР их добывают пока еще
в незначительном количестве, хотя запасы камбалы у нас значительны.
Pleuronectes — глаза на правой стороне, рот узкий; морская камбала
182
(P. platessa) — Белое море, Мурман, в Атлантике — до Бискайского за-
лива. Палтус (Hippoglossus hippoglossus)— глаза на правой стороне.
Рот большой. Распространен по обоим берегам северной части Атлантиче-
ского и Тихого океанов. Bothus — глаза на левой стороне, тело высокое,
рот широкий. В. maximus, «палтус», «пильвина большая», камбала, — в
Средиземном море и у западных берегов Европы. В Азовском и Черном мо-
Рис. 169. Три последовательные стадии развития Pleu-
ronectes platessa, стадии А и В еще симметричны.
(По Goodrich.)
рях — В. maeoticus. Solea — глаз,а па правой стороне (Морской язык, соль).
S. nasuta — в Средиземном море и у берегов Западной Европы. S. vulgaris —
в Средиземном море и у западноевропейских берегов.
Отряд 14. Окунеобразные (Perclformes)
Этот отряд и все последующие отряды колючеперых имеют
общий признак: в брюшпых плавниках у них одип перасчлепелпый
колючий и не более пяти членистых мягких лучей. На этом осно-
вании все эти разделы объединяются под именем Percoidei. Эта
группа произошла, видимо, от Berycifornies. Появляются в мело-
вую эпоху.
Сем. Окуневые (Р е г с i d а с). Брюшпые плавники на груди, чешуя
ктепоидная, жаберпая крышка с шипиками или зазубрена. Конические зубы
па многих костях рта; большей частью пресноводные, хищные рыбы, но не-
которые живут в море. Род Рогса — окунь (Р. fluviatilis) — во всей Европе,
исключая Испанию, в Сибири до р. Лены, в Балхаше. В бассейне Амура его
нет. Балхашский окунь (Р. schronki). Судак (Luciopcrca), обыкновенный
судак (L. lucioperca) живот в бассейнах Балтийского, Черного, Азовского,
Каспийского и Аральского морей. Морской судак (L. marina) —
в северо-западной части Черного моря и в Каспийском. Судаки являются
весьма ценной промысловой рыбой благодаря высокому качеству мяса.
Ерши (Acerina). A. cernua — во всей Северной и Средней Европе и Сибири.
Сем. Морские окуни (Serranidae) очень сходно с пре-
дыдущим семейством, но отличается большим числом колючих лучей в аналь-
ном плавнике (3 -вместо 1—2). Serranus, S. scriba и S. cabrilla (рис. 170)
в Черном и Средиземном морях замечателен своим гермафродитизмом, но
183
промыслового значения не имеет. Его имеют некоторые другие роды се-
мейства, живущие в дальневосточных морях.
Сем. Морские лещи (S р а г i d а е). Имеют сжатое с боков
тело и характерные зубы: они сидят только на челюстях, спереди — большие
остроконечные или долотообразные, по бокам же — тупые, жерновые, чрез-
вычайно твердые. Около 200
видов у берегов морей почти
всего света. Некоторые тра-
воядные, большинство—хищ-
ники. Sargus, Морской карась
(S. annularis) — в Атлантиче-
ском океане, в Средиземном и
Черном морях. Там же распро-
Wk странен и род Pagel]us.
уйк Сем. Барбульковые
(М u 1 1 i d а е). Близки к
Рис. 170. Морской окунь (Serranus предыдущему семейству. Мел-
cabrilla). кие рыбы, живущие в море ив
(По Boulenger.) солоноватой воде, питающиеся
беспозвоночными и разлагаю-
щимися органическими веществами. Около 50 видов, почти во всех тропиче-
ских морях, доходят до Северной Европы. Один род Mullus, султанка,
барбулька (М. barbatus), живет у берегов Атлантического океана, в Среди-
земном и Черном морях, где держится большими стаями. Ценится [за ее
вкусовые качества.
Сем. Сциеновые (Sciaenidae). Также близко к Serrani-
dae. В семействе около 150 видов, большей частью морских. Некоторые дости-
гают крупных размеров (до 2 м) и имеют вкусное мясо. Есть представители
в Черном море.
Отряд 15. Чешуеперые (Chaetodontifonnes)
Сюда относятся многочисленные рыбки тропических вод, многие
из которых имеют причудливую форму и очень яркую окраску.
Сем. Щетинозубые (Chaetodontidae). Чешуи покры-
вают голову и непарные плавники почти до верхушки. Небольшие рыбки,
обычно очень ярко окрашенные. Многочисленны в тропических морях,
где держатся возле коралловых рифов. Около 200 видов. (Брызгун Toxotes
jaculator) — в Ост-Индии и Полинезии. Охотится за насекомыми, пуская в них
струю воды.
Сем. Хирурги (Acanthuridae) (80 видов). Составляют
переход к следующему отряду — Род Acanthurus.
Отряд 16. Сростпочелюстпые (Plectognathi)
Чрезвычайно своеобразные рыбы, главным образом тропиче-
ского пояса. Верхнечелюстные и челюстные кости срастаются у них
неподвижно между собой и с зубами. Скелет нередко окостеневает
неполно или построен из совершенно своеобразной кости. Кожа
имеет своеобразный костный скелет, состоящий из шипов, щитков,
игл. Иногда все тело заключено в папцырь. Жаберная щель очень
узкая. Брюшных плавников пет или они заменены иглами. Целый
ряд особенностей во внутреннем строении указывает на дегенера-
цию некоторых форм.
А. ТВЕРДОКОЖИ Е (Sclerodermi)
Сем. Triacanthidae. Межчелюстные кости сливаются с maxilla.
Зубы раздельные, конические или резцоподобные. Мелкие чешуи в виде
184
чешуями. Около 100
Тело покрыто неналегающими друг на
чешуями. Около 100 видов. Один вид»
шипиков. Морские рыбы из Индийского океана и западной части Тихого.
Род Triacanthus (рис. 171).
Сем. Спинороги (Balistidae). Praemaxillae прочно сливаются
с maxillae. Зубы долотоподобные. ”
друга подвижными ромбовидными
Balistes capriscus, случайно
доходит до южных берегов
Англии. Виды рода Balistes
(рис. 172) имеют сильные зубы,
которыми они могут раскусы-
вать кораллы и раковины мол-
люсков, мягкие части которых
съедают.
Сем. Кузовки (О s t-
raciontidae) Praemaxil-
]ae неподвижны, зубы подобны
резцам. Тело заключено в
панцырь из совершенно свое-
образных, по строению шести-
угольных, прилегающих друг
к другу чешуй. 20 видов и три
рода. Род Ostracion (рис. 173)
известен с верхнего эоцена.
Для движения служат здесь
спинной и анальный плавники,
так как тело заключено в
панцырь. Грудные плавники
помогают поддерживать ток
воды через жабры, что иначе
было бы невозможно вслед-
ствие фиксированности жабер-
ной крышки. О. quadricornis —
в тропических морях Атлан-
тики.
Рис. 171. Triacanthus brevirostris.
(По Day из Goodrich.)
’Рис. 172. Спинорог (Balistes steDaris). .
(По Day из Boulenger.)
Рис. 173. Кузовок четырехрогий
(Ostracion quadricornis).
(По Boulenger.)
Род Spheroides па Дальнем Востоке.
В. СКАЛОЗУБЫ Е
(G у in п о d о n t е s)
Сем. Четырохзубые
(иглобрюхи) (Т е t г о-
д! on t i d a e). Челюсти обра-
зуют род клюва. Клюв раз-
делен посредине швом. Око-
ло 60 видов. Род Tetrodon. Т.
hispidus — в Красном море и
в Индийском океане. Мясо ядо-
вито. Т. fahaka—в реке Ниле.
Сем. Двузубые (D iod on t id ас). Клюв без шва посредине.
Мясо также ядовито. D. maculatus (рис. 174), еж-рыба, — в Атлантическом,
Индийском и Тихом океанах. Как и Tetrodon, имеет на брюшной стороне
глотки обширный мешок, в который набирает воздух, вследствие чего тело
сильно надувается, перевертывается — и рыба плавает кверху брюхом.
Сем. М о 1 i d а е. Туловище укорочено, хвостового плавника нет,
спинной и анальный сходятся назади между собой, симулируя хвостовой
плавник. Личинки снабжены иглами и непохожи вовсе на взрослых. Встре-
чается в открытом море и в больших глубинах. Mola mola достигает почти'
тонны веса (рис. 175).
Отряд 17. Сростноглоточпые (Pomacentrlformes)
Этот отряд обнимает формы, у которых нижпеглоточные кости
срастаются в одно целое (Pharyngognathi).
18S
Сем. Pomacentridae. Роскошно окрашенные рыбы тропиче-
ских морей с ктеноидными чешуями. Похожи на Chaetodontidae по форме
чела и образу жизни. Насчитывается около 150 видов.
Сем. Cichlidae — пресноводные рыбки или же из солоноватой
лоды, хорошо известные любителям аквариумов. У некоторых хорошо выра-
жен половой диморфизм во время периода раз-
множения. Забота о потомстве выражается в
том, что самка прячет яйца и молодь в рот
или в глотку. Обитатели жарких или теплых
Рис. 175. Луна-рыба
(Molamola).
(По Goode из Boulenger.)
Рис. 174. Еж-рыба (Diodon
maculatus).
(По Gunter из Boulenger.)
стран: Африки, Мадагаскара, Сирии, Индии и Цейлона, Центральной и
Южной Америки. Известно 45 родов со 150 видами из Африки, 140 из
Америки и 3 из Индии и с Цейлона. Наиболее известны роды: Tilapia
(Chromis) (рис. 176), Paratilapia, Geophagus. Наиболее многочисленны (как
роды, так и виды) в озере Танга-
найка, составляющие большую
часть ихтиофауны последнего
(рис. 177).
Рис. 176. Tilapia dardcnni из озера
Тангапайка.
(По Boulenger.)
Отряд 18. Губапообразпыо
(Labriformes)
Известно около 400 ви-
дов. Большей частью рос-
кошпо окрашеппые рыбки,
обитающие у берегов всех
тропических и умеренных
морей.
Сем. Г у б а н о в ы е (L a b г i d а е). Крепкие острые зубы на челюстях
и конические или бугорчатые глоточные. Наиболее известны роды: Labrus
с видом L. prosostictes, рябчик — в Черном море; Crenilabrus pavo — в Среди-
земном и Черном морях.
Сем. С к а ров ые (Scaridae). Очепь похожи на предыдущее
семейство, так же ярко окрашены, но зубы как в верхней, так и нижней че-
люстях срастаются между собой, образуя клюв. На нижнеглоточных костях
зубы срастаются в одну пластинку, на верхпеглоточных — в две. Этот клюв
служит им для обгрызания кораллов, а также для соскабливания водорослей
•со скал. S. cretensis — в Средиземном море.
Отряд 19. Песчапкообразные (Amodytlformes)
Маленькие рыбки с удлиненным телом, с мелкой циклоидной
чешуей, с длинным анальным и спинным плавником, имеющие
довольно неопределенное положение в системе.
186
Сем. Песчанки (Ammodytidae) живут в морях: Средиземном
Черном, Балтийском, на Мурмане, у берегов Скандинавии и Европы, в Охот-
ском и Японском морях. Держатся на песке вблизи берегов, уходя зимой
в более глубокие части моря. Встречаются в огромном количестве, являясь
пищей лососей, птиц, и служат наживкой для ловли трески (A. tobianus
на Мурмане).
Отряд 20. Драконообразпые (Trachlniformes)
Сем. Trachi ni dae. Брюшные плавники на горле, жаберные крыш-
ки с колючими шипами. Морской дракон (Trachinus draco), живет в
Атлантическом океане, Средиземном и Черном морях. Передними колючими
лучами спинного плавника наносит болезненные уколы. Такие же уколы
причиняются и шипами на жаберной крышечке.
Що Boulenger.)
Сем. Звездочеты (U г а п о s с о р i d а е). Похожи на предыдущее
семейство. Глаза наверху укороченной головы. Звездочет (Uranoscopus sca-
ber.) Под языком находится мясистая, способная выдвигаться нить. Зако-
павшись в песок, откуда видны только глаза и ротовая щель, звездочет
этой нитью подманивает свою добычу — мелких рыбешек, принимающих
нить за червя. Живет в Средиземном и Черном морях.
Отряд 21. Колбпесбразпые (('Obllfornies)
Этот отряд представлен приблизительно 600 видами, большей
частью морскими. Большая часть колбиеГг — рыбки мелкие. Самый
крупный род, выделяемый некоторыми в особое семейство, — Eleot-
ris marmorata, имеет длину почти до 1 м, самый маленький —
Mistichtliys luzonensis с Филиппинских островов — длиной всего
12—1.4 мм. Это самое маленькое существо изо всех позвопочпых.
У многих бычков брюшные плавники срастаются в воропковидпый
диск, при помощи которого рыбы эти могут присасываться к камням
и растениям.
Сем. Колб новые (Ст о bi i dae). Промысловое значение бычков не
очень велико, но все возрастает вследствие уменьшения количества рыбы
других более ценных пород и возрастания опросов на рыбу. После нереста
в Азовском море наблюдается массовая гибель бычков. Пищей их являются
придонные беспозвоночные, сами они являются пищей хищных рыб, а также
187
(в Каспии) и тюленя. Род Gobius представлен многими видами и в фауне
наших морей: Черного, Азовского и Каспийского. G. ophiocephalus, бычок
травяной,—в Черном, Азовском, иногда в Средиземном море.
Отряд 22. Прилппа.юобразпые (Echenelformes)
Одно семейство, прилипаловые(Есйепе1йае). Любопыт-
ная группа Acanthopterygii, у которой передний спинной плавник
Рис. 178. Присасывательный диск
прилипалы.
(По lordann и Evermann из Goodrich.)
превращен в присоску в виде
овального диска (рис. 178), состоя-
щего из поперечных пластипок.
Диск этот помещается па голове.
Семейство немногочисленное, всего
около 10 видов, распределенных
по всем тропическим и теплым мо-
рям. Хищники, питающиеся ры-
бами п присасывающиеся при по-
мощи диска к различным плаваю-
щим предметам: кораблям, лодкам,
акулам, китам, черепахам. Для ловли последних прилипал при-
меняют у восточных берегов Африки.
Отряд 23. Скорпенообразпые Паицырнощекие (Scorpeniformes
Sclcroparei)
Весьма естественное собрание семейств, происшедшее, видимо,
от Serranidae. Более примитивные представители папцырпощеких
имеют с последними много общего. Происходя от примитивных
окуневых различные
семейства этого под-
отряда дивергирова-
ли(расходились, эво-
люционировали) в
различных направле-
ниях . Характерным
признаком раздела
является сильное раз-
растание одной из
подглазничных ко-
стей, вследствие чего
область щек бывает
Рис. 179. Бородавчатка страшная (Synanceia ЭТОЙ КОСТЬЮ закрыта,
horrida). Сем. scorpaenj_
(По Никольскому.) \ d а е заключает в себе
\ около 30 родов с 250 ви-
дами. Обитатели всех морей, где избирают места с каменистым грунтом.
Рыбы оседлые, странствований не совершают. Съедобны, но некоторые
ядовиты. Род Sebastes. Вид Морской окунь (S. norvegicus) обитает в
северной части Атлантического океана. Держится на глубине не меньше
150 м, обычно 200—300 м; живородящ. Мясо весьма ценится. Род Scorpaena
характеризуется большой головой с сильными шипами и нитевидными
выростами, имеющимися и на туловище. Уколы шипов болезненны.
188
Скорпида, морской ерш (S. porcus) живет в Атлантическом океане, Сре-
диземном и Черном морях. Бородавчатка страшная (Synanceia horrida)
(рис. 179), очень ядовита.
Сем. Голомянковые (Comephoridae). Крайне ин-
тересное семейство. В нем всего 4 рода: один распространен от Аляски до
Калифорнии, другой живет в глубоких частях озер Мичигана и Онтарио,
два — Cottocomephorus и Comephorus в озере Байкал, причем последний
род — в самых больших глубинах этого озера. Представители этого рода
бесцветны, имеют большие глаза, скелет их тонок и мягок, как бывает у
глубинных рыб. Рыба эта живородяща и гибнет после родов.
Сем. Бычки -подкаменщики (Cottidae). Семейство
представлено 220 видами в различных морях. Наибольшее количество живет
в северных частях Тихого океана. Лишь немногие проникают в пресные
воды. Виды рода Cottus принадлежат главным образом к последним. Голова
у них широкая, плоская, тело голое, цилиндрическое, грудные плавники
очень большие. Бычок-подкаменщик (С. gobio) — в пресных водах Средней
и Восточной Европы, Западной Сибири. В Сибири другой вид С. sibiricus;
С. scorpio — в части Ледовитого океана и в Белом, Балтийском и Немец-
ком морях.
Сем. Круглопоровые (Cyclopteridae). Брюшные
плавники, если есть, сливаются в присасывательный диск, жаберная щель
очень мала. Тело короткое, неуклюжее, покрытое толстой кожей, гладкой
или с костяными буграми, представляющими измененную чешую. Мед-
ленно движущиеся рыбы, придерживающиеся дна и питающиеся мелкими
животными и растениями. Около 50 видов из северных арктических и антар-
ктических морей. Liparis liparis живет в наших северных водах. Род Cyclop-
terus — также в северных морях обоих полушарий. Пинагор (С. lumpus)
достигает 50 см. Самец роет в песке ямку, самка откладывает туда икру.
Самец стережет последнюю и охраняет молодь, которая присасывается к его
телу своими присосками.
Сем. Тр игловые (Triglidae). У представителей этого семей-
ства голова как бы заключена в панцырь, благодаря главным образом раз-
растанию подглазничных костей, широких и соединяющихся с praeoperculum-
Два или три передних луча грудных плавников способны к самостоятель-
ному движению и служат органами осязания и движения по дну. Грудные
плавники обычно очень большие и ярко окрашены. Два спинных плавника.
Распространены во всех морях жарких и умеренных областей. 5 родов и 40 ви-
дов. Род Trigla. Много видов в Средиземном море. Мясо съедобно, но не об-
ладает высокими качествами. Морская ласточка (Т. h i.r u n-
d о) — в Атлантическом океане, в Средиземном и Черном морях.
Сем. Л и с и ч к о в ы о (Agon i d ае). Похожи па тригл тем, что
голова заключена в костный панцырь, тело покрыто костными пластинками,
но плавник на спине один. Мелкие рыбки, обитающие у берегов Северной
Атлантики и Тихого океана. Есть глубинные формы. Известно около 40 видов.
Вид Agonus cataphractus — в Сев. Атлантике.
Сем. Летучие петухи (Dactylopteridae). Обык-
новенный летучий петух (Dactylopterus) имеет очень длинные грудные
плавники, с помощью которых может перелетать некоторое пространство над
поверхностью воды. Живет в уморенных и тропических частях Атлантиче-
ского океана.
Отряд 24. Морские собачки (Bleiinlformes, Jugulares)
Все рыбы этого раздела имеют брюшные плавники на горле.
Сем. Морские собачки (В 1 е n n i d а е). Тело удлиненное,
более или менее цилиндрическое, голое и притом очень скользкое или же
покрытое мелкой чешуей. Спинной плавник обычно во всю длину спины,
передняя часть его — из гибких игл, задняя из расчлененных, иногда весь
плавник из игл. Заднепроходный плавник также длинный. Семейство очень
обширное, заключает в себе более 400 видов и 80 родов. Обитатели прибреж-
189
вых частей умеренных и тропических морей. Некоторые виды живородящи.
Виды рода Blcnnius живут у европейских и африканских берегов Атлан-
тического океана, в Средиземном и Черных морях. Род Anarrhichas (рис. 180),
отличающийся сильно развитыми зубами и называемый потому «зубаткой»,
представлен в северной половине Атлантики и у нас вдоль Мурманского
берега и в Белом море видом Морской волк, полосатой или синей зубаткой
(Anarrhichas lupus), кроме мяса, дающим кожу.
Рис. 180. Зубатка (Anarrhichas lupus)4
(По Никольскому.)
Сем. Fierasferidae. Fierasfer живет в полости водных легких
у голотурий.
Сем. Бельдюговые (Zoarcidae является, видимо, от-
прыском семейства Blennidae, широко распространено, многие представи-
тели живут в больших глубинах. Известно около 130 видов. Род Zoarces
с видом Z. vivipara является живородящим (рис. 181).
Рис. 181. Бельдюга живородящая (Zoarces viviparus).
(По Никольскому.)
Отряд 25. Ру копер ые (Pedlculati)
Рыбы этого отряда отличаются плавниками удлиненной руко-
образной формы, коленчатыми и расположенными далеко позади
черепа. Они могут служить для ползания по морскому дну. Жабер-
ная перепонка простирается далеко назад, оставляя лишь малень-
кое круглое жаберное отверстие.
Сем. Морские черти (Lophiidae). Типичные хищники, держа-
щиеся у дна моря. Наиболее известен род Lophius с видом морской чорт,
рыболов (L. piscatorius). Достигает 11/2 м длины. Первый луч спинного плав-
ника превращен в особый придаток, с помощью которого он заманивает
маленьких рыбок, зарывшись в песок и выставив только широкую пасть.
Живет в Атлантическом океане и Средиземном море, откуда заходит в Черное.
Сем. Ceratiidae представлено 25 родами глубинных рыб (до 500 м);
некоторые из них обладают органами свечения. Семейство это, видимо,
происходит от Lophiidae. Кости их отличаются необычайной легкостью и тон-
костью строения. Род Melanocetus с видом М. johnstoni. Имеет светящийся
орган на конце придатка на голове. Живет, видимо, подобно Lophius. В родах
Ceratias и Edriolychnus самцы очень маленькие и у последнего рода срастаются
190
с самкой в области внутренней стороны жаберной крышки, питаясь ее со-
ками (рис. 482).
Сем. Antennariidae. Известно около 40 видов. Представители
рода Antennarius ведут хищный образ жизни, живя среди кораллов, которым,
«подражают» и цветом и придатками на теле. A. marmoratus — в Саргассовом
море, среди водорослей которого строит замечательное гнездо из шелковистых,
нитей.
Рис. 182. Edriolychnus schmidti.
Наверху — самка с паразитом-самцом (внизу), приросшим к
внутренней поверхности жаберной крышки. Натуральная
величина самки — 6 см. Внизу — самец отдельно (сильно увели-
чен). Натуральная величина самца—1.5 см. (По Бергу, из
Солдатова).
Отряд 26. Макролеобразные (Scombrlformes)
Рыбы этого раздела произошли видимо от Perciformes, с кото
рыми тесно связапьт. Рыбы большей частью удлиненной веретено*
видной формы, обычно с мелкой циклоидной чешуей. Характерен
сильно развитый хвостовой стебель с сильно раздвоенным хвостовым
плавником. На хвостовом стебле часто имеются боковые кили.
Спиппой плавник обычно длинный. Задняя часть его нередко раз-
бита па ряд маленьких плавничков. Хищные морские рыбы. Многие
являются пелагическими и широко распространенными.
Сем. Каранговые (Саг angid ае). Сюда принадлежат не-
большие рыбы, живущие в теплых морях большими массами и являющиеся
промысловыми рыбами. Ставридка (Trachurus trachurus) — в Атлантическом
океане у западных и восточных берегов, а также в Балтийском, Средиземном
и Черном морях. Молодь этого вида держится стайками возле медуз, под
колоколом которых укрывается, будучи потревожена.
Сем. Макрелевые (Scombridae). Типичнейшее семейство
отряда, обладающее характернейшими его признаками. Стройные морские
пелагические рыбы, держащиеся обычно большими стаями вблизи поверх-
491
ности. Многие являются важными промысловыми рыбами. Род Scomber
с видом S. scomber (рис. 183), скумбрия, макрель (20—40 видов) — в
Атлантическом океане, Средиземном и Черном морях. В последнем совер-
шает правильные миграции вдоль берегов. Важная промысловая рыба. Более
крупным представителем является распространенная там же и более крупная
(70 см) пеламида (Sarda sarda) питающаяся макрелью. Огромного роста
(до 3 м) достигает тунец (Thynnus thynnus), живущий в Атлантическом
океане и Средиземном море, где держится стаями, охотясь за макрелью и
сельдями. Изредка заходит в Черное море.
Сем. Парусники (Histiophoridae). Крупные океани-
ческие рыбы с rostrum, вытянутым в виде шпаги. Быстрые пловцы. Плаванию
помогает еще высокий спинной плавник служащий парусом.
Рис. 183. Макрель (Scomber scomber).
(По Никольскому.)
Сем. Меченосы (Xiphiidae). Отличается от предыдущего
семейства, в общем будучи весьма сходным с ним отсутствием у взрослых
зубов, полным отсутствием чешуи и брюшных плавников и еще некоторыми
признаками. Верхняя челюсть вытянута в очень длинный мечеобразный отро-
сток. Меч-рыба (Xi phi as gladius) распространена почти по всем морям, обычна
в Атлантическом океане, особенно в Средиземном море, заходит иногда и в
Черное. Достигает 6 м длины.
Отряд 27. Трескообразные, мягкоперые (Cadlformes, AnaeanthIni)
В отличие, от Acanthopterygii у этого отряда плавники с мягкими
расчлепеипыми лучами. Брюшные плавпики или под грудными
или впереди от них. Хвостовой плавпик совершенно симметричный.
Плавательный пузырь, если существует, пе имеет сообщения с пище-
водом. Большинство видов морские, лишь немногие пресноводпы.
Рис. 184. Треска (Gadus callarias).
(Из Boulenger.)
Сем. Долгохвосты (М acrurid ас). К этому семейству
принадлежит около 40 видов, часто имеющих широкое распространение. Глу-
боководные рыбы, встречающиеся до глубины 4500 м. О глубоководности
семейства свидетельствует форма тела его представителей, вытянутое, по-
степенно суживающееся к хвосту; последний заостряется в нить и лишен
хвостового плавника. Рыло заострено, рот на нижней стороне. На подбородке
усик, развивающийся у донных форм.
192
Сем. Тресковые (Gadidae) (рис. 184). Чешуя нателе очень
мелкая, кожа слизистая, голова очень большая, на спине два или три плав-
ника, также и два заднепроходных. Брюшные плавники на горле. На под-
бородке часто имеется усик, и именно у донных форм. Известно около 120 ви-
дов, которые все являются хищниками и распространены главным образом
в умеренных и холодных морях северного полушария. Некоторые приспособ-
лены к жизни на больших глубинах. Имеют большое промысловое значение на
севере. Род Gadus с основными видами: треска (Gadus callarias) — в Север-
ном и Атлантическом океанах; пикша (Gadus aeglefinus) там же. Сайда
(Pollachius virens), навага (Eleginus navaga) в Белом море и далее на
восток; также еще некоторые являются обитателями морей, тогда как
налим (Lota lota) живет в пресных водах Средней и Северной Европы,
Сибири. Главная масса трески, которая приходит к нам на Мурман, приходит
сюда из норвежских вод для целей питания, уходя затем обратно. В Барен-
цовом море есть, впрочем, и местные расы трески, не совершающие далеких
миграций. Миграции трески описаны ниже.
V. ФИЛОГЕНЕТИЧЕСКОЕ РАЗВИТИЕ РЫБ
Система должна служить выражением естественных связей
различных групп животных, явившихся результатом эволюцион-
ного процесса. Система должна отображать филогенез, историю
развития животного мира. Мы уже видели, что знания наши об
этом развитии получаются тремя методами: сравпительиоанатоми-
ческим, эмбриологическим и палеонтологическим. Ни один из этих
методов сам по себе не является универсальным в познании фило-
генетического развития. Однако на основе всех трех методов мы
все ближе и ближе приходим к правильному представлению о ходе
филогенетического процесса. Вспомогательным и очень полезным
может быть экологический апализ форм, выясняющий, в какой
обстановке опи могли возникнуть, какие условия существования
привели к развитию того или иного комплекса признаков.
Относительность каждого метода в отдельности ясна из того,
что онтогенетическое развитие, папример, вовсе не является пол-
ным и неизменным повторением филогенетического процесса,
а обнаруживает массу различного рода уклонений от последнего.
Палеонтологические свидетельства также неполны, а очень часто
и крайне недостаточны вообще, так как необходимы особо благо-
приятные условия для того, чтобы животные сохранились в иско-
паемом состоянии. В особенности же палеонтология пе дает нам
исчерпывающих ответов потому, что как раз наиболее интересные
для построепия филогенеза, наиболее древние примитивные формы
сохраняются хуже всего и менее всего нам известны. Во всяком
случае, несмотря на трудности, мы на основе всех четырех указан-
ных методов имеем более или менее ясное представление о фило-
генетическом развитии класса рыб, что и выражается в приведен-
ной выше системе рыб. Система эта в будущем подвергнется, конечно,
дальнейшей перестройке, когда мы получим новые данпые о строении
различных групп рыб, об их развитии и об ископаемых видах.
Выше было сказано о происхождении Protocraniata. Protocra-
niata дали начало примитивным формам с эптодермальными жаб-
рами (Entobranchiata), от которых произошли современные Cyclosto-
mata и вымершие Ostracodermi, и примитивным формам с экто-
ft з Кашкаров и Станчивокий ft 93
дермальными жабрами (Ectobranchiata). От последних в дальней-
шем произошли рыбы. Но каковы были первичные рыбы, как и по-
чему опи возникли? Для ответа на эти вопросы нам недостает
палеонтологических данных.
На основании сравнительноанатомических и эмбриологических
данных мы можем сказать, что у этих гипотетических эктобранхиат
(рис. 185) хрящевые жаберные дуг» были расчленены на 4 части
и ряд этих дуг достигал рта. На наружной стороне жаберных дуг
возникли сперва эктодермальные жаберные пити, а затем жаберные
листки, что подняло этих животных па высшую ступень (усиление
Рис. 185. Схематическое изображение строения пе-
реднего конца тела гипотетических Ectobranchiata.
(по Северцеву)
дыхательной функции и обмена веществ, повышение энергии) иным
способом, нежели Entobranchiata. Затем передние жаберные щели
закрылись, из передних дуг развились челюстная дута, гиоидная
и губные хрящи. Развившиеся па поверхности тела плакоидные
зубчики во рту, па челюстях стали служить другой функции —
превратились в зубы. Таким образом совершился переход к еще
более активному способу питания, притом иному, нежели у Ento-
branchiata. В связи с этим должны были развиться и органы актив-
ного движения. Образовались хвостовой плавник (протоцеркаль-
иый), непарные и парные плавники с хрящевыми лучами.
Но когда и в каких условиях это произошло? Анализ рыбы как
определенной формы жизни, определенного приспособительного
типа, проливает нам некоторый свет па этот вопрос. Сравнивая
рыбу с беспозвоночными, мы видим, что между ними существует
большое различие в степени их динамичности: рыбы — существа
моторного, двигательного типа, тогда как беспозвоночные в по-
давляющем числе существа спокойные, медленно движущиеся,
часто сидячие. Даже наиболее подвижные из водных беспозво-
ночных — головоногие (Cephalopoda) далеко уступают в динамич-
ности рыбам. У Cephalopoda их подвижность развилась в связи
с их хищпым образом жизни. Неподвижность, привязанность к дну
большинства беспозвоночных явилась ответом па жизнь в стоячих
водах, морских и пресных или в медленно текучих. Если мы видим
беспозвоночных (личинок насекомых, например) в быстро текучих
водах, они вырабатывают приспособления к тому, чтобы их пе
унесло потоком, прикрепляются к неподвижным предметам, пря-
чутся и т. д. Позвоночные должны были явиться в результате от-
бора со стороны среды динамичной, текучих наземных вод, Требе-
ле
вавших развития способности к активному движению как средства
избежать гибели. На это указывают и веретенообразная, вытя-
нутая форма тела, и мощная сегментально расчлененная мускула-
тура, и внутренний прочный и гибкий скелет, и развитие плав-
ников. Все эти характерные признаки, свойственные и более при-
митивным рыбам и эмбрионам рыб, являются приспособлениями
к движению посредством боковых изгибов первоначально вытяну-
того тела в движущейся текучей воде, т. е. пресной, наземной.
Эти особенности могли возникнуть и в море, но море не давало сти-
мулов для развития и прогресса этих особенностей в той мере,
как мы видим это у рыб. Мы видим, что переход к придонному
образу жизни, к неподвижности меняет у рыб эту типичную для
них форму на иную, как бы возвращает их к первоначальному
состоянию. Такой переход мы видим у морских рыб и у рыб, жи-
вущих в медленно текущих водах.
Acrania primitiva и их ближайшие родственники являются
прибрежпыми морскими формами. Реки с обилием взвешенных
пищевых частиц, приносимых течением, были местом обитания этих
пассивно питавшихся форм. Надо думать, что Acrania primitiva
обитали и в пресных водах. Одни их потомки оказались в прибреж-
ной золе моря и не прогрессировали, другие, жившие в текучих
водах, развивались в сторону типа рыб.
Рыбы возникли, вероятно, около 400 млн. лет назад. Их появле-
ние совпадает с временем, последовавшим за высочайшим подня-
тием гор па границе кэмбрийской эпохи. Это поднятие изменило
во мпогих местах ландшафт и сделало стоячие или медленно текшие
до тех пор воды быстро текущими. Тут, вероятно, особенно важным
адаптивным свойством была способность к активному движению
или способность удерживаться па дне с помощью присоски. Но
первые позвоночные, возникшие в то время, к сожалению, нам
неизвестны. Лишь в следующую эпоху, начиная с пижпего силура,
мы видим остатки существ, которые некоторыми принимаются
за челюсти хищных апнелид — Polychaeta errantia, другими —
за остатки позвоночных. Это так называемые «конодонты», по
внешней форме очень похожие па роговые зубы Cyclostomata, но
отличные от них по микроскопическому строению. Наиболее древ-
ними позвоночными, известными в ископаемом состоянии, явля-
ются Ostracodermi. Не следует думать, что эти позвоночные явля-
лись предками рыб: это были специализированные представители
бесчелюстных (Agnathostomata), приспособившиеся к жизни на
дне (бентос). Опи известны из верхнего силура и девонских отло-
жений. Потомства не оставили.
Остатки элазмобранхий известны из верхнего силура в виде
плавниковых лучей — так называемых «ихтиодорулитов», правда,
несколько сомнительных. Однако элазмобранхий, несомненно,
происходят от форм более примитивных, более обобщенных. Такой
наименее специализованпой формой является Cladoselache из ниж-
некаменноугольных отложений. Эта акулоподобная форма еще не
имеет признаков специализации элазмобранхий. Скелет у нее был
43* 195
представлен хордой, рот находился на переднем конце, а не внизу,
совокупительных органов не было, парные плавники прикрепля-
лись горизонтально широким основанием, имели вид частей не-
когда непрерывной складки и были снабжены наиболее примитив-
ным скелетом плавников, нам известным. Хвост был гетероцер-
кальный, но внешне — гомоцеркальный. Эта форма в известной
мере является связующим звеном между Chondrichthyes и Osteich-
thyes,c личинкой которых сходна Cladoselache. Из девонских слоев
мы зпаем еще другую группу акулоподобных рыб — Acanthodii.
Рыбы этой группы имели конечный рот, чешуи в виде мелких пла-
коидных пластинок, жаберпые щели были, как у элазмобрапхий.
Ряд шипов от нескольких пар парных плавников говорит о происхо-
ждении последних из боковой складки. Третья группа вымерших
форм, близких к настоящим акулам, но сближающихся и с другими
рыбами, а именно с Dipnoi, представлена известным с каменно-
угольного до конца пермского периода отрядом Pleuracanthodii.
У Pleuracanthus, как у акул, жаберпые щели открывались наружу
по отдельности каждая, челюсти были также хрящевые, у самцов
были совокупительные органы. Но, с другой стороны, кожа была
без плакоидных чешуй, грудные плавники бисериалыюго типа,
а на голове развиты были покровные окостепепия, подобные та-
ковым у Dipnoi. И хвостовой плавник был симметричный, а пе
гетероцеркальный.
Мы видим, таким образом, что в палеозое существовали формы
рыб, близкие к акулам, но с более примитивными чертами, еще пе
получившие специальных признаков последних. Эти «обобщенные»
формы могли дать начало и настоящим акулам, от которых затем
произошли скаты путем приспособлепия к доппому образу жизни,
и костным рыбам — Osteichthyes. Палеонтология дает пам пе до-
статочно данных, что вполне естественно, так как первые рыбы пе
имели костного скелета и имели мало шансов дойти до нас в иско-
паемом виде. Но нельзя сомневаться в том, что примитивные рыбы
существовали гораздо раньше, нежели Pleuracanthus (каменно-
угольная эпоха), Cladoselache (девон и нижпие каменноугольные
слои) и Acanthodii (нижпий девоп); во всяком случае — опи сущест-
вовали в силуре, ибо, кроме пих, в девоне существовали еще много-
численные представители гораздо более высокоорганизованного
класса рыб, а настоящие акулы известны уже с пижпего каменно-
угольного времени. Cladoselache — это отпрыски древнего ствола.
Акулы и скаты представляют группу весьма древппх рыб, сохра-
нивших еще многие черты своих палеозойских предков, приспосо-
бившихся к определенным условиям существования и потому
процветающих по наше время. Но дальше они пе эволюциониро-
вали, так как пе развили никаких признаков, поднимающих их
общую жизненную энергию па более высокую ступень. Главное —
это то, что жаберное дыхание осталось у пих па той же ступепи
развития, па которой оно было у их палеозойских предков: отсут-
ствие жаберной крышки, межжаберные перегородки, несовершен-
ная жаберная решетка.
196
По другому пути пошло развитие Osteichthyes. Эта группа обиль-
но представлена уже в девоне. Условия существования в силуре
были весьма неустойчивы, разрушительны и стимулятивны: под-
водные извержения, сопровождавшиеся выходом ядовитых газов,
отложения пепла, внезапные подъемы и падение уровня вод, бы-
стрый и разрушительный перенос песка, гравия (об этих явлениях
мы можем судить по характеру геологических напластований) —
все это должно было вызывать миграции, ставило рыб в новые
условия существования, вызывало мутационные изменения, уси-
ливало борьбу за существование и действие отбора и способствовало
эволюции одних и гибели других. Все это происходило в пресных
водах, в море рыбы в те времена еще не вышли: они были слишком
несовершенно организованы, чтобы конкурировать с головоногими
моллюсками и другими беспозвоночными хищниками.
В девоне, как сказано, мы видим очень многочисленную ихтио-
фауну. В девонскую эпоху отложились толщи так называемого
древнего красного песчаника, достигающие колоссальной мощ-
ности. Огромные глубокие озера и котловины непостоянного харак-
тера, большие реки, терявшиеся в болотах, нередко, вероятно,
пересыхавших, кишели рыбами. Тут были и Arthrodira (Coccosteus),
представляющие особый подкласс, и примитивные Dipnoi, Crosso-
pterygii и Palaeoniscidae. Arthrodira не оставили потомков, Osteich-
thyes развивались, эволюционировали дальше.
У примитивных Osteichthyes под плакоидной чешуей развились
метамерпо расположенные ромбические чешуи, из которых затем
образовались па голове покровные кости. Вместо примитивных
хрящевых челюстей развились вокруг рта кости, несущие зубы:
praemaxillae, maxillae, dentalia, palatina, pterygoidea. Роговые
лучи плавников заменились костными (lepidotrichia). Но самые
главные изменения произошли в дыхательной системе. Жаберпые
щели прикрылись крышечным аппаратом — operculum, из костных
чешуй этой области развились кости крышечного аппарата, появи-
лись лучи жаберной перепонки, radii membranae branchi os tegii.
Межжаберные перегородки стали редуцироваться, число жаберпых
щелей сократилось, и из задней пары жаберпых выпячиваний обра-
зовался плавательный пузырь, игравший, вероятно, первоначально
роль дополнительного дыхательного органа. Все эти преобразо-
вания поднимали организм па большую функциональную высоту,
давали возможность шире распространяться, изменяться приспо-
собительно к новым условиям.
От этих примитивных Osteichthyes, вероятно, рано ответвились
Chondrosteoidei. Их можно считать неудачным шагом эволюции.
Они сохранили много признаков своих акулоподобпых предков:
хрящевой скелет, rostrum, палатоквадратный симфиз, примитив-
ные плавники, гетероцеркальный хвост. Кость у них только начала
развиваться, жаберный аппарат не достиг совершенства, крышеч-
ная система еще очепь несовершенна, перегородки между жабер-
ными щелями пе вполне редуцировались. Эта группа дотянула до
наших дней возможно благодаря плодовитости и другим адапта-
197
циям, по не процветает. Другая группа— Holosteoidei — развилась
в прогрессивном направлении, скелет ее окостеневал полнее, плав-
ники совершенствовались, хвост становился гомоцеркальным. Раз-
витие их шло в двух направлениях: в направлении нектонных
Actinopterygii и в направлении придонных Dipnoi и Crossopte-
rygii. Последние две группы первоначально были теснее связаны,
и ископаемые Dipnoi (Dipterus и др.) ближе стоят к Crossopterygii,
чем современные. Actinopterygii распадаются на Palaeoniscoidei,
Lepidosteoidei, Amioidei и Teleostei. Эти группы рыб сохранили
примитивную форму плавников с широким основанием. Кроме
того, у них развился на спинной стороне плавательный пузырь.
Развился он из спинного отдела выпячивания задней жаберной
щели. Древнейшие примитивные Crossopterygii развились в совре-
менных Crossopterygii, дали начало двоякодышащим рыбам Dipnoi
и четвероногим позвоночным. У них сильно развилась вырезка
между парными плавниками и телом, основание плавников стало
узким, и плавники приняли лопастную форму. Кроме того, у них
из вентрального выпячивания последней пары жаберных щелей
развились настоящие легкие. Разделение на Holosteoidei Actinopte-
rygii и Holostoidei Crossopterygii произошло рано. Уже в древнем
красном песчанике девонского времени мы видим представителей
и той и другой групп. Древнейший из Palaeoniscoidei жил здесь
рядом с Crossopterygii и древнейшим Dipnoi. Тело этой рыбы было
вытянутое, покрыто мелкой ромбической ганоидпой чешуей, хвост
был гетероцеркальный, покровпые кости па голове лежали поверх-
ностно, представляя просто видоизмененные ганоидные чешуи
в этой области, глаза были окружены костяным кольцом, тела
позвонков и ребра не окостеневали. От Palaeoniscidae ответвились
Platysomidae, жившие в каменноугольное и пермское время. Это бы-
ли рыбы с высоким телом, с ганоидпой чешуей, с короткими челюстя-
ми. Эта ветвь вымерла без потомства. От Palaeoniscidae лее произош-
ли и Holostei. В сем. Caturidae, жившем в юрскую эпоху до мела
включительно, мы видим постепенное окостенение тел позвонков,
замену гетероцеркального хвоста гомоцеркальным, закругление
ганоидной чешуи. Lepidosteoidei еще храпят в своей организации
многие черты своих ископаемых предков: у них еще сохраняется
ромбическая ганоидная чешуя, имеются остатки плакоидных чешуй,
спиральный клапан, conus arteriosus и т. д. У Amioidei таклсе еще
сохраняются эти примитивные черты, ио чешуя уже типично ци-
клоидного, черепицеобразпо налегающего одна па другую харак-
тера. Появляясь в триасе, Amioidei расцветают в юре, давая разно-
образные типы, и затем вымирают, оставляя до нашего времени
одного представителя — Amia calva. Lepidosteus известен только
с эоцена, но не подлежит сомнению, что это представители более
древней группы. По вымирании Amioidei в мелу доминирующим
типом становятся Teleostei. Они завоевывают первенство благо-
даря тому, что у них развивается целый ряд прогрессивных призна-
ков общего характера, поднимающих их па большую высоту, чем
более древние, вымершие формы рыбы и их современные отпрыски.
198
У Teleostei значительно совершенствуется дыхательный жаберный
аппарат; развивается всасывающий насос, совершенствуется жабер-
ная решетка. Повышение интенсивности дыхательного процесса
влечет за собой повышение активности движения. Этому способ-
ствовала замена тяжелого мало гибкого ганоидного папцыря легкой
черепицеобразно расположенной циклоидной чешуей, также и гомо-
церкальный хвост увеличил силу движения. Длинные отростки
позвонков дают лучшую опору мускулам. Гидростатический аппа-
рат — плавательный пузырь именно у Teleostei приобретает свое
максимальное значение. Благодаря этим чертам Teleostei широко
распространяются и в пресных водах и в море, приспособляясь
к самым разнообразным условиям жизни: и к жизни в поверхно-
стных слоях, и в глубинах, и у берегов, к разного рода пище и т. д.,
давая большое разнообразие приспособительных или адаптивных
форм.
Родословное дерево Teleostei еще не достаточно проработано,
их система поэтому еще пе установилась. Весьма вероятно, что
различные разделы произошли от ископаемых гапоидов самостоя-
тельно, так что филогенетическая схема Teleostei представляет не
дерево, а скорее куст, отдельные ветви которого идут самостоя-
тельно от мезозойских форм. Некоторые современные и ряд вы-
мерших групп костистых сохраняют еще черты своих мезозойских
предков. Таковы Leptolepidae, имевшие еще ганоин в тонких ци-
клоидных чешуйках. Из современных Teleostei наиболее примитив-
ными являются Cypriniformes и Clupeiformes, наиболее специали-
зированными — Acanthopterygii. Что касается другого ствола
Holosteoidei — Н. Crossopterygii, то от него произошли и вымершие,
и современные Crossopterygii и Dipnoi, разошедшиеся очень рано.
За происхождение Dipnoi от Crossopterygii говорит палеонтология:
строение древнейших Dipnoi, принадлежащих к семейству Dipte-
ridae и близких к нему, строение черепа, непарных плавников,
гетероцеркальный хвост и т. д. Раппяя специализация, приспособ-
ление к питанию животными с твердыми защитными образова-
ниями выразилось у пих в развитии автостилии. Недостаточно
совершенный жаберпый дыхательный аппарат привел к образо-
ванию легких как дополнительного органа дыхания. Способность
жить в пересыхающих или загнивающих водоемах избавила Dipnoi
от конкуренции с другими рыбами и сохранила обломки этого
некогда более богатого ствола до нашего времени.
Мы видели, что древнейшие рыбы были пресповодпы. В средине
каменноугольной эпохи мы видим первое вторжение пресновод-
ных элазмобрапхий в море: остатки акул начинают встречаться
вместе с морскими беспозвоночными литторальпой зоны. Остальные
рыбы оставались пресноводными («гапоиды», Dipnoi). Морские се-
лахии встречаются однако только до пермской эпохи, а затем они
исчезают. Видимо, выход в море оказался неудачен, первые вы-
ходцы пе выдержали кошеуренции в море.
Пресноводными оставались рыбы до триаса, в нижних отделах
которого уже есть морские рыбы, по лишь немногие хищные элаз-
499
мобрапхии вышли в море. Этот выход в море стал возможен благо-
даря большей легкости и подвижности, а также хищному образу
жизни. Это стало возможным благодаря эволюции самих элазмо-
брапхий, наступанию моря, давлению со стороны рептилий в водах
суши, изобилию пищи в море и роли Cephalopoda и других хищни-
ков в море. За элазмобранхиями последовали в море «ганоиды»,
т. е. древпие Osteichthyes, например, Pycnodontidae. Инвазия
в море продолжалась и в мелу. Dipnoi, Holostei, Chondrostei,
Crossopterygii, многие Teleostei остались в преспой воде.
В нижнем мелу костистые рыбы становятся преобладающими
и сильными антагонистами медуз, головоногих, развивающихся
высших ракообразных.
VI. экология РЫБ
Каждый вид животного является продуктом долгой эволюции,
происходившей под непрерывным контролем окружавшей его
среды. Поэтому, как правило, виды оказываются приспособлен-
ными к условиям окружающей их среды. Однако приспособление
это всегда временно и относительно. Среда не остается постоянной,
она постоянно изменяется. Изменяется и сам организм, папример
с возрастом. И в том, и в другом случае установившаяся в резуль-
тате долгого отбора приспособленность вида перестает быть действи-
тельной, и наступает та или иная реакция со стороны животного.
Если значительные перемены во внешнем мире наступают
внезапно, то организмы, привыкшие к иным условиям жизпи,
часто не выдерживают и погибают. Так, например, резкие перемены
температуры воды в Ёаренцовом море вызывают иногда гибель
мойвы (Mallotus villosus), так что миллионы ее плавают па поверх-
ности воды. Массовая гибель рыбы зимой подо льдом от изме-
нения газового содержания в воде, известная под названием замо-
ров, задух и т. п., служит другим примером зависимости организма
от изменения внешних условий. Яйца и личинки рыб подвергаются
гибели еще скорее.
Если перемена в условиях жизни происходит постепенно, то
могут иметь место три вида реакции: организмы, пе будучи в со-
стоянии измениться и приспособиться, медленно вымирают, или
же происходит переселение животных в другое место, или ясе опи,
изменяясь, приспособляются к новым условиям ясизни.
Приспособление происходит, конечно, путем отбора средой не-
которых из многочисленных геповариаций (наследственных изме-
нений), большинство которых является пе адаптивными, а безраз-
личными или даже вредными. Последние отбором уничтожаются.
Приспособление к определенным условиям существования со-
провождается, как правило (имеются, правда, исключения), ха-
рактерными изменениями формы органов, а иногда и целого ряда
органов. Животное изменяет свой вид, приобретает повый облик,
соответствующий новым условиям существования. Так как эти
характерные изменения формы органов возникают не сразу, а с боль-
200
шой постепенностью, то возможно бывает установить ступени или
степени приспособления.
Между крайними типами, характерными для крайних, резко
отличающихся условий среды, наблюдается обычно целый ряд
переходных типов.
Условия внешнего мира, к которым приспособляются организмы,
бывают: 1) механические (возникающие при плавании, ползании,
зарывании в ил или песок, при питании твердой или мягкой пищей,
при жизни на дне или в открытом море, в полосе прибоя или в тихих
заливах, на быстром течении или в неподвижной воде и т. д.);
2) температурные; 3) световые (жизнь в хорошо освещенных, слабо
освещенных или вовсе лишенных света слоях воды: жизнь в илу,
дневной или ночной образ жизни и т. д.); 4) химические (жизпь
в соленой, солоноватой или пресной воде; химический состав
пищи; дыхание водное или воздушное и т. д.); 5) биологические
(борьба с врагами или добычей, состязания самцов из-за обладания
самками и т. д.).
В общем условия существования могут быть сведены к следую-
щим трем группам факторов: климатическим (темпера-
тура, свет, давление и влажность атмосферы, осадки и т. д.), био-
топическим (физико-химические особенности среды, оби-
тания), биоценотическим (влияние со стороны других
организмов: пища, хищники и другие виды взаимоотношений).
Все эти факторы сказываются на определенных приспособлениях
в форме тела, в размерах, в строении органов, в поведении, и опре-
деляют степень процветания видов, их вымирание и т. д.
Та часть зоологии, которая занимается изучением взаимо-
отношений вида с условиями среды в его местообитании, изучает
приспособления к последним и возникающие со средой противо-
речия, называется экологией. При этом под средой разу-
меется пе только физическая среда, но и другие организмы, с ко-
торыми животное вступает в те или иные взаимоотношения. Иначе
говоря, экология изучает жизпь видов, изучает их деятельность,
поведение, всегда находящееся под влиянием окружающей среды,
и приспособления к условиям последней, а также и противоречия
с ними.
1. Вода как среда для жизни рыб
Соленость и жесткость воды. Стихия, в которой живет рыба, —
вода. Если рыба и выходит из воды, то только временно. Отсюда —
изумительное приспособление всего организма рыбы к водной
среде. Но вода как среда жизпи есть понятие сложное. Ее хими-
ческий состав, темцература, содержание газов оказывают большое
влияние на жизпь рыб.
Вода в природе всегда содержит соли и газы. Содержанием
солей определяется различие между пресной и морской
водой, а на основе отношения к солености воды подразделяют рыб
на две большие группы: пресноводных рыб и м о р-
201.
ск их. Лишь очень немногие представители рыб могут жить
в той и другой воде.
Пресная вода содержит главным образом двууглекислый и сер-
нокислый кальций, хлористый же натрий и углекислый магний —
лишь в виде следов. Соли кальция служат для построения скелета,
входя в организм рыбы с пищей. В зависимости от того, по каким
породам текут воды данного бассейна, они будут более богаты
теми или иными веществами: углекислыми солями Са и Mg, серно-
кислым Са и т. д. Состав воды колеблется также в зависимости
от времени года.
Морская вода богаче солями. Главным образом хлористыми
(89%) — хлористый натрий и магний и сернокислым магнием.
Большее содержание солей обусловливает большую плотность
морской воды (1.024—1.029). Различные моря и океаны в зависи-
мости от силы испарения и от количества втекающей в море пресной
воды имеют различное содержание солей. Средняя соленость откры-
того моря — 35°/00, колеблется от 32 до Зв0/,». В области пассатов,
где постоянно дуют сухие свежие ветры, в течение года испаряется
слой воды до 3 м толщиной, и здесь вода наиболее соленая. В Сар-
гассовом море, например, соленость доходит до З7.б°/оо. В Немец-
ком море, получающем воду прямо из Атлантического океана,
вода имеет до Зб0/,» солености; в Балтийском же море, соединяю-
щемся лишь узкими проливами с Немецким морем, соленость
падает до Т.б0/,» у острова Рюгена, а в северной части Ботниче-
ского залива вода почти пресная. Черное море имеет 16—18°/00 солей;
Красное море и Персидский залив — 4О°/оо и более. В Мертвом
море человек не тонет, так как содержание соли там 217°/м; орга-
ническая жизнь здесь совершенно невозможна.
Значение содержания солей в воде для жизни рыб и рыболов-
ства огромно. Некоторые рыбы придерживаются воды определен-
ной солености. Этим объясняются, например, совершенно различ-
ные уловы сельдей у берегов южной Норвегии и западной Швеции,
в Скагерраке и в других местах, где сельди в один год приходят
к берегу в колоссальном количестве, в другие же годы почти со-
вершенно отсутствуют. Явление это оказалось связанным с парал-
лельно идущими изменениями в распределении солености морской
воды, зависящими от преобладания поверхностного опресненного
«балтийского течения» или более глубокого течения из соленого
Немецкого моря. В свою очередь эти течения зависят от преобла-
дания тех или иных ветров, гопящих воду из Немецкого моря
в Скагеррак или из Балтийского моря.
Яйца многих рыб (камбала, треска) являются планктонными,
плавают на поверхности; последнее зависит от содержания солей
и удельного веса воды. Там, где содержание солей в воде менее 1О°/оо
мы уже не находим яиц, плавающих па поверхности.
В устьях рек пресная вода смешивается с соленой морской,
и получается солоноватая вода с содержанием соли от 5
до 10 %0. Иногда смешение получается равномерное, иногда же
в спокойных устьях, получается слоистость: менее плотная прес-
202
пая вода находится поверх более плотной соленой. Некоторые
виды рыб живут в такой солоноватой воде, будучи мало чувстви-
тельными к смене пресной и соленой воды. Солоноватоводными
следует считать и морских рыб Каспийского моря. Чувствитель-
ность рыб к перемене воды одной солености па другую зависит
от различия в осмотическом давлении их крови по отношению
к внешней среде. Осмотическое давление в крови пресноводных
костистых рыб значительно ниже, нежели у морских. При попа-
дании морских рыб в пресную воду последняя через жабры путем
эндосмоса проникает в ткапи, вызывая смерть рыбы. Вот почему
так называемые проходные рыбы, проводящие часть жизни в море,
а часть в реках, пробыв некоторое время в море, должны перед
вступлением в реки для икрометания пробыть некоторое время
в предустьевом пространстве, в солоноватой воде, дабы приучить
себя к изменению солености воды (лососи, осетры).
Главную роль, как сказано, играет в морской воде хлористый
патрий. Некоторую роль играют еще соли магния, соли кремневой
кислоты и иодистые соли. Серпокислый и углекислый кальций
стоят совершенно па втором плане.
Большое значение имеет для рыб так называемая жесткость
воды, или содержание в пей известковых солей. Последние необхо-
димы для образования костяка, а также (у форели, например) для
развития жизнеспособных половых продуктов. Количество извести
в воде зависит от того, через какие породы протекает вода. По-
этому в рыбоводстве, например, необходимо принимать во вни-
мание, через какие породы протекает источник. Пределы жесткости
для карповых рыб будут от 3 до 8 немецких градусов (за градус,
или единицу жесткости, в Германии считают одну часть окиси
кальция, растворенного в виде какой-либо соли в 100 000 частях
воды), для форели — от 8 до 16 градусов.
Большое значение имеет иногда и содержание в воде железа,
а также загрязнение воды различными солями отбросов индустри-
ального производства, попадающих в воду. Токсичность солей,
кроме их химической природы, зависит от их концентрации и тем-
пературы воды.
Кроме неорганических солей в воде как в пресной, так и в мор-
ской находится значительное количество растворенных органиче-
ских веществ. Пресная вода богаче морской органическими веще-
ствами. Чем вода богаче последними (при достаточном наличии
кислорода), тем «питательнее» опа для рыб. Сельские пруды, на-
пример, в которые стекают воды с прилежащих дворов, являются
очень удобными для разведения сазанов. Но это только до известного
предела. Если органических примесей в воде слишком много, обра-
зуется сероводород, и рыбы отравляются. Неосторожное обращение
с водоемами и их чрезмерное загрязнение часто ведут к этому.
Вопрос об органических веществах, растворенных в воде, пред-
ставляет очень большой интерес в связи с так называемой «теорией
осмотического питания водных организмов». О ней будет сказано
в главе о питании рыб.
203
Очепь важпо присутствие в воде органических кислот, влияю-
щих па кислородный режим и дыхание рыб.
Содержание газов в воде. Огромное значение для жизни рыб
имеет содержание в воде газов и прежде всего кислорода. Содер-
жание последнего в пресной воде подвержено большим колебаниям
и зависит прежде всего от температуры, а в морской воде и от со-
лености: содержание кислорода уменьшается с повышением тем-
пературы и с повышением солености. Северные моря богаче кисло-
родом, чем южные.
Прилагаемая таблица показывает количество кубических санти-
метров кислорода в 1 л воды при различной температуре и соле-
ности, если вода насыщена газом.
Соленость Температура^4^, в градусах С 0°/оо 2О’/оо 35°/оо
количество О2 е 1 см3
0 10.29 9.01 8.03
40 8.02 7.10 6.40
20 6.57 5.88 5.35
30 5.57 4.96 4.50
В спокойной воде всегда растворяется некоторое количество
кислорода путем диффузии. Осадки, волны и зеленые растения
значительно усиливают накопление кислорода в воде.
Рыбы (как и прочие водные организмы) нуждаются в кислороде
для дыхания и поглощают его из раствора в воде, выделяя угле-
кислоту. То же и растения. Но последние, кроме того, ассимили-
руют углерод, поглощая углекислоту и выделяя кислород, вновь
растворяющийся в воде. Этот процесс образования кислорода,
как известно, происходит под влиянием света. Эксперименты по-
казали, что многие высшие водные растения ассимилируют углерод
лучше в желтом свете, чем в голубом или краном. Так обстоит
дело с зелеными водорослями. Поэтому мы находим их вблизи
поверхности, где достаточно желтого света. Красные водоросли
ассимилируют углерод лучше в голубом свете, нежели в желтом,
а потому живут в более глубоких слоях воды. Требуют света и жи-
вут в поверхностных слоях воды и планктонные водоросли. Таким
образом, в море продукция кислорода имеет место в верхних слоях.
Потребление же его идет во всех слоях, где есть жизнь. Чрезмерное
увеличение в воде животных ведет к уменьшению содержания
кислорода, увеличение числа растений при достаточном свете
вызывает увеличение его. Так как продукция кислорода происхо-
дит лишь на свету, то в небольших водоемах можно отметить за
ночь уменьшение содержания кислорода. То же самое происходит
в закрытых водоемах под покровом льда. Чем слой льда толще,
чем энергичнее идут процессы разложения (гниения) па дне, и чем
медленнее происходит смена воды, тем быстрее идет в водоеме
204
обеднение кислородом. Иногда в результате этого и происходит
массовая гибель рыбы в прудах, озерах и т. д. Наоборот, чём бы-
стрее смена воды, чем больше ее движение, тем вода становится
богаче кислородом. Богаты кислородом горные потоки, водные
бассейны с большим притоком воды, где содержание кислорода
может даже увеличиваться с глубиной.
Повышение температуры не только уменьшает содержание кисло-
рода в воде, но одновременно увеличивает потребность рыбы
в кислороде. Так, например, на килограмм веса в сутки:
Форель при 10° потребляет около -100 см3 кислорода
» » 15° » » 220 » »
Сазан » 10° » » 10—20 » »
» » 15° » » 70—80 » »
Подвижная форель требует кислорода гораздо больше, нежели
менее подвижный сазан.
Определение содержания кислорода в воде данного водоема
чрезвычайно важно для практического рыбоводства.
Обеднение кислородом происходит, однако, пе только вслед-
ствие дыхания животных, но также и в результате различных
химических процессов.
Так, может накопляться на дне сероводород, как это наблю-
дается в Черном море, где сероводород отравляет воду уже с 200-мет-
ровой глубины. Наблюдается скопление сероводорода в некоторых
озерах и в норвежских фиордах. В отравленных сероводородом
слоях невозможна никакая жизнь кроме анаэробных бактерий.
Этим отчасти объясняются, например, неудачные попытки разве-
дения угря в Черном море путем посадки его в Дунай.
Углекислота встречается в виде углекислых и двууглекислых
солей и лишь в пезпачительпом количестве в виде свободного газа.
С глубиной содержание углекислоты увеличивается. Азот погло-
щается водой из атмосферы в довольно большом количестве. Воз-
можно, что морские бактерии частью разлагают азотистые соеди-
нения, освобождая азот, частью же связывают свободный азот
в виде различных солей. Содержание азота идет параллельно
с увеличением планктона. А так как ихтиофауна в значительной
мере зависит от планктона, то содержание азота в воде имеет для
нее огромное значение.
Температура воды. Температура воды имеет первостепенное
влияние па жизнь рыб. Каждый вид рыб процветает при опреде-
ленной температуре. Повышение или* понижение ее за определен-
ный предел вызывает приостановку жизненных проявлений и даже
гибель рыбы. Подвижность, интенсивность питания, например, пря-
мо зависят от температуры; также зависит от температуры и время
икрометания. Температура пресных вод средних широт подвержена
большим колебаниям (0—35°). Меняется опа и в зависимости от
глубипы. При этом летом наивысшая температура будет в верхних
слоях, зимой — в нижних. Осенью и весной бывает момент, когда
вся вода сверху донизу имеет одинаковую температуру: 4°. Изме-
205
пения в распределении в воде температур вызывают изменение
в распределении планктона, а в связи с последним — и рыб.
Средняя температура тропических морей 27°, достигает в Индий-
ском океане 28°, в Красном море 34.4, а в Персидском заливе
даже 35.5°. Умеренные части Великого и Атлантического океанов
имеют 20.3 и 20.7°, в полярных морях температура колеблется
около 0°.
Температура морей в одном и том же месте колеблется сравни-
тельно мало. Зато с глубиной она, начиная с 300—400 м, где является
постоянной, постепенно падает до 2° в среднем. Наиболее пизкая
температура найдена была в глубинах — минус 3.3°. В полярных
морях температура повышается с глубиной до 300—400 м, где вода
наиболее теплая, а затем снова понижается до 2 000 м, где равна
приблизительно 0°.
Как сказано, для каждого вида рыб существует своя наиболее
благоприятная температура, колеблющаяся в больших или мень-
ших границах. Различают виды рыб эйритермные и сте-
нотермные: первые выносят широкие колебания температур,
вторые не выносят. К последним принадлежат рыбы полярных
морей, больших глубин и тропических морей.
При отклонении температуры воды от оптимальной (наилучшей)
для данного вида наступает ослабление жизненных функций и далее
смерть. Этим объясняется так называемая зимняя и лет-
няя спячка рыб. У многих карповых рыб, сомов и угрей
при 4—6° наступает своеобразное летаргическое состояние. Рыбы
перестают есть, зарываются в ил и здесь почти не дышат. Точно
так же падает и сердечная деятельность. Так проводят они зиму,
теряя в весе до 5%. С весенним теплом рыбы вновь становятся
деятельными. В состоянии оцепепепия на дне водоемов зимуют
также осетровые рыбы.
Обычно же рыбы впадают в состояние оцепенения при пони-
жении температуры до 4-1°. В литературе имеются указания
на случаи, когда тело рыбы при температуре от—15° до —20° будто
бы превращалось в кусок льда, а затем при осторожном оттаивании
вновь оживало. Относительно вида Dallia pectoralis указывалось,
что она живет в промерзающих зимой мелких речках, озерах и бо-
лотах Чукотского полуострова и Аляски, впадая зимой в анабио-
тическое состояние. Новейшие эксперименты с карасями, золо-
тыми рыбками, карпами, окунями и угрями показали, что обра-
зование льда во внутренних органах рыб всегда убивает их, и что
еживапие обмерзлых рыб возможно лишь в тех случаях, когда лед
образуется лишь в поверхностных слоях тела рыбы и температура
ее тела падает пе ниже, чем до —0.1 —0.3°. Однако возможно,
что у Dallia отбор выработал способность оживать и при более
низкой температуре.
Летняя спячка встречается у тропических рыб при сильном
перегревании воды или при пересыхании водоемов. Некоторые
сомы (Siluridae), лабиринтовые (Labyrinthici), особенно же двояко-
дышащие (Dipnoi) при наступлении сильной жары и высыхании
206
водоемов зарываются в ил, делают из последнего твердые капсулы
и, свернувшись в них, остаются здесь до нового наступления пе-
риода дождей. И среди наших рыб встречаются случаи «оцепенения
от тепла», именно: у линя (Tinea) и у вьюна (Misgurnus fossilis).
Отмечено такое же явление и у окуня (Регса).
В общем можно сказать, что большинство рыб может перено-
сить изменение температуры в широких границах. Необходимо’
лишь, чтобы изменения эти наступали последовательно, постепенно.
Внезапные изменения температуры ведут, как мы видели, за собой
массовую гибель рыбы.
Глубина, давление и освещение в воде. Как известно, каждые
10 м глубины дают увеличение давления па одну атмосферу. Такое
увеличение давления не может не давать себя знать, не отражаться
на организмах. Так как известны глубины до 10 800 м, то давление
здесь доходит более чем до 1 000 атмосфер. Оно достигает сотен
атмосфер уже на средних глубинах. И однако жизнь здесь суще-
ствует. Это объясняется тем, что в тканях живущих здесь животных
заключается много воды, и газы в пузыре находятся под тем же
давлением, что и наружная среда. При вытаскивании глубинных
рыб на поверхность они тотчас же погибаю! при явлениях сильного
раздутия тела.
Большое влияние па местное распространение рыб оказывают
также условия освещения. Проникновение лучей в воду есть не-
обходимое условие для развития в пей растительности и, в част-
ности, фитопланктона. Распространение последнего ограничено
определенной глубиной, а именно оп пе встречается глубже 400 м.
Планктон служит пищей рыбам. Освещение зависит, кроме широты,
на которой находится данный водоем, еще от прозрачности
воды. Если погружать в воду различных морей белые желез-
ные диски определенного диаметра (диски Секки) и сравнивать
между собой глубины, па которых эти диски еще видимы нами,
то окажется, что вода различных морей далеко пе одинаково про-
зрачна и что она кажется тем синее, чем прозрачнее.
Чем больше в воде взвешенной мути, действующей как ширма,
заслоняющая лучи, тем вода зеленее. Муть состоит частью из ми-
неральных частиц: углекислого кальция, глины, кремнезема. Эта
муть оседает в прибрежных водах. Часто же муть состоит из орга-
нических примесей, в особенности из растительного и животного
планктона.
В зависимости от проникновения лучей и развития планктона
вода океанов делится па три зоны: хорошо освещепну ю—
эйфотическую, простирающуюся до 80 м глубины и бога-
тую фитопланктоном; средне освещенную — дисфо-
т и ч е с к у ю, простирающуюся ^т 80 до 400 м, весьма бедную
растительными организмами, главным образом кремневыми водо-
рослями; пе освещенную — афотическую — без
всяких растительных организмов. Все три зоны населены особыми
видами рыб, о чем речь дальше.
207
2. Рыбы и водная среда. Приспособления б условиям обита-
ния, адаптация формы и движения. Типы приспособления
Условия жизни в различных областях пресных вод, в особен-
ности же в море, накладывают резкую печать на обитающих в этих
областях рыб.
Рыбы могут быть разделены на рыб морских, проходных, полу-
проходных, или приустьевых, рыб солоноватой воды и пресновод-
ных. Уже значительные различия в солености имеют значение для
распространения отдельных видов. То же самое и различия в осталь-
ных свойствах воды: температура, освещение, глубина и т. д.
Форель требует иной воды, нежели усач или сазан; линь и карась
держатся и в таких водоемах, где окунь жить не может из-за слиш-
ком теплой и мутной воды; жерех требует чистой текучей воды
с быстрыми перекатами, а щука может держаться и в заросшей
травой стоячей воде. Наши озера в зависимости от условий суще-
ствования в пих могут быть различаемы как судаковые, лещевые,
карасевые и т. д. Внутри более или менее больших озер и рек мы
можем отметить различные зоны: береговую, открытой воды и при-
донную, характеризуемые разными рыбами. Рыбы одной зоны
могут заходить в другую зону, по в каждой зоне преобладает тот
или иной видовой состав. Более всего богата прибрежная зона.
Обилие растительности, следовательно, пищи, делает эту зону
благоприятной для многих рыб; здесь они кормятся, здесь мечут
икру. Распределение рыб по зонам играет большую роль в рыбо-
ловстве. Например, налим (Lota lota) является придонной
рыбой, и его ловят со дна вентерями, но не плавными сетями, ко-
торыми ловят жереха и т. д. Большинство сигов (Coregonus) питается
мелкими планктонными организмами, главным образом ракообраз-
ными. Поэтому их местообитание зависит от передвижения планк-
тона. Зимой они уходят вслед за последним па глубину, весной же
поднимаются на поверхность. В Швейцарии биологами были ука-
заны места, где зимой держатся планктонные ракообразные, и
здесь возник после этого сиговый промысел; на Байкале омуля
(Coregonus migratorius) ловят сетями зимой на глубине 400—600 м.
Более резко выражено разграничение на зоны в море. Море по
условиям жизни, которые оно предоставляет для организмов может
быть разделено па три зоны: 1) литоральную, или бере-
говую; 2) пелагическую, или зону открытого
моря; 3) абиссальную, или глубинную. Так
называемая сублиторальная зона, составляющая пе-
реход от береговой к глубинной, обнаруживает уже все «признаки
последней. Границей их служит глубина в 350 м. Береговая зона
начинается от берега и простирается до вертикальной плоскости,
ограничивающей область глубже 350 м. Зона открытого моря будет
кнаружи от этой плоскости и кверху от другой плоскости, лежа-
щей горизонтально на глубине 350 м Глубинная зона будет нахо-
диться внизу от этой последней (рис. 186).
Для всякой жизни имеет огромное значение свет. Так как вода
208
пропускает лучи солнца слабо, то в воде на известной глубине
создаются условия существования, неблагоприятные для жизни.
По силе освещения различают, как указано выше, три световых
зоны: эйфотическую, дисфотическую и афо-
тическую.
У берегов тесно перемешаны свободно плавающие и придонные
формы. Здесь находится колыбель морских животных, отсюда
возникают неуклюжие обитатели дна и проворные пловцы откры-
того моря. Таким образом, у берегов мы встретим довольно разно-
Рпс. 186. Схема распределения зон моря.
образную смесь типов. Зато условия жизни в открытом море и на
глубинах очень различны, и типы животных, в частности, рыб,
в этих зонах весьма не похожи друг иа друга. Всех животных,
которые живут иа дне моря, мы называем одним именем: бен-
тос. Сюда относятся ползающие ио дну, лежащие па дне, рою-
щиеся формы (и о д в и ж и ы й бентос) и сидячие формы
(сидячий бентос: кораллы, актинии, трубчатые черви и т. д.).
Те организмы, которые могут свободно плавать, мы называем
и е к т о и о м. Третья группа организмов, лишенная или почти
лишенная способности передвигаться активно, цепляющаяся за
водоросли или беспомощно носимая ио ветру или течениями, на-
зывается и л а и к т о и 6 м. Среди рыб мы имеем формы, отно-
сящиеся ко всем трем группам организмов.
'Пелагические рыбы—нектон и планктон. Организмы, живущие
в воде независимо от дна, ие связанные с пим, называются и е л а-
г и ч е с к им и. К этой группе относятся организмы как живущие
иа поверхности моря, так и в более глубоких слоях его; организмы,
активно плавающие (нектон), и организмы, носимые ветром и те-
чениями (планктон). Глубоко живущие пелагические животные
носят название б а т и тт е л а г и ч е с к и х.
Условия жизни в открытом море характеризуются прежде
всего тем, что здесь пет прибоя, и животным пет необходимости
вырабатывать приспособления для удержания на дне. Хищнику
здесь негде прятаться, подстерегая добычу, последней негде укрыть-
ся от хищников. И те и другие должны полагаться главным образом
на собственную быстроту. Большинство рыб открытого моря явля-
ются поэтому отличными пловцами. Это — во-первых; во-вторых,
44 Кашкаров и Стапчинский 209
окраска морской воды, синяя и в проходящем и в падающем свете
оказывает влияние и на окраску пелагических организмов
вообще и рыб в частности.
Приспособления рыб нектона к передвижению бывают различны.
Мы можем различить несколько типов некто и пых рыб.
Во. всех этих типах способность к быстрому плаванию дости-
гается различными способами.
Тип веретенообразный, или т о р п е д о о б р а з-
н ы й. Орган движения — хвостовой отдел тела. Примером этого
Рис. 187. Парусник (Histiophorus gladius),
(из Брэма)
типа могут служить: сельдевая акула (Lamna cornubica), макрель
(Scomber scomber), лосось (Salmo salar), сельдь (Clupea harengus),
треска (Gadus morrhua).
• .Тип лентообразный. Движения происходят при по-
мощи змеевидных движений сжатого с боков, длинного ленто-
образного тела. Большей частью — обитатели довольно больших
глубин. Пример: сельдяной король, или ремень-рыба (Regalecus
banksii).
Тип стрелообразный. Тело вытянуто, рыло за-
острено, крепкие непарные плавники отнесены назад и располо-
жены в виде оперения стрелы, составляя одно целое с хвостовым
плавником. Пример: сарган обыкновенный (Belone belone).
Тип парус ообразный. Рыло вытянуто, непарные
плавники и общий вид как у предыдущего, передний спинной
плавник сильно^ увеличен и может служить парусом. Пример:
парусник (Histiophorus gladius, рис. 187). Сюда же принадлежит
и меч-рыба (Xiphias gladius),
iio
Рис. 188.Меланоцет (Me]anocetus).
(но iijort)
Рыба по существу своему — животное, активно плавающее.'
Поэтому настоящих планктонных форм среди них нет. Мы можем
различить следующие типы приближающихся к планктону
рыб.
Тип игловидный. Активные движения ослаблены, со-
вершаются при помощи быстрых изгибов тела или же — упдули-
рующих движений спинного и анального плавников. Пример: пе-
лагическая игла-рыба (Syngnathus pelagicus) Саргассова моря.
Тип сдавленпо-сдмметричный. Тело высокое.
Спиппой и анальный плавники расположены друг против друга,
высоки. Брюшных плавников большей частью нет. Движения
очепь ограничены. Пример: лупа-рыба (Mola mola). У этой рыбы
отсутствует и хвостовой плавник.
Активных движений он пе про-
изводит, мускулатура в значи-
тельной мере атрофирована.
Тип ш а р о в и д и ы и. Тело
шаровидно. Тело может у неко-
торых рыб надуваться благодаря
заглатыванию воздуха. Пример:
еж-рыба (Diodon) или глубоко-
водный меланоцет (Melanocetus)
(рис. 188).
Среди взрослых рыб настоящих
планктонных форм нет. Но опи
встречаются среди плапктоппых
яиц и личинок рыб, ведущих
планктонный образ жизни. Способность организма держаться
на воде зависит от ряда причин. Прежде всего важен удельный
вес воды. Организм держится па воде, согласно закону Архимеда
в том случае, если его удельный вес пе больше удельного веса
воды. Если же удельный вес больше, то организм топет со ско-
ростью, пропорциональной разнице удельного веса. Скорость
погружения однако пе всегда будет одинакова. (Мелкие песчинки
тонут медленнее крупных камней одного и того же удельного
веса.)
Это явление зависит, с одной стороны, от так называемой вяз-
кости воды, или, внутреннего трепия, с другой •—
от того, что называется поверхностным трением тел.
Чем больше поверхность какого-либо предмета в сравнении с его
объемом, тем поверхностное сопротивление его больше, и он тонет
медленнее. Малый удельный вес и большая вязкость воды проти-
водействуют погружению. Прекрасные примеры такого изменения
представляют, как известно, веслоногие рачки и ра-
диолярии. В яйцах и у личинок рыб мы наблюдаем то же
самое явление.
Пелагические яйца большей частью мелки. Яйца многих пела-
гических рыб снабжены нитевидными выростами, которые метают
им погружаться, например, яйца макрелещуки (Scombresox)
14* 211
(рис. 189). Личинки некоторых рыб, ведущих пелагический образ
жизни, имеют приспособление для удержания на поверхности
воды в виде длинных нитей, выростов и т. д. Таковы пелагические
личинки глубоководной рыбки Trachypterus. Кроме того, эпителий
этих личинок изменен весьма своеобразно: клетки его почти ли-
шены протоплазмы и растянуты до огромных размеров жидкостью,
что, конечно, уменьшая удельный вес, способствует также удер-
жанию личинок па воде.
Еще одно условие влияет на способность организмов держаться
па воде: осмотическое давление, зависящее от температуры и со-
лености. При большом содержании солей в клетке последняя
Рис. 189. Пелагиче-
ское яйцо: макреле-
щуки (Scombresox).
(По Cunningham.)
впитывает воду, и, хотя она становится тя-
же лее, удельный вес ее уменьшается. Попав
в более соленую воду, клетка, наоборот,
уменьшившись в объеме, станет более тя-
желой. Пелагические яйца многих рыб со-
держат до 90% воды. Химический анализ
показал, что в яйцах многих рыб количе-
ство воды уменьшается с развитием личинки.
По мере обеднения водой развивающиеся
личинки опускаются все глубже и, наконец,
садятся на дно. Прозрачность и легкость
личинок трески (Gadus) обусловливаются
наличием обширного подкожного простран-
ства, наполненного водянистой жидкостью и
тянущегося от головы и желточного мешка
до заднего конца тела. Такое же обширное пространство имеется
у личинки угря (Anguilla) между кощей и мускулами. Все эти
приспособления, несомненно, уменьшают вес и препятствуют
погружению. Но и с большим удельным весом организм будет
держаться па воде, если он представляет достаточное поверхно-
стное сопротивление. Это достигается, как сказано, увеличением
объема и изменением формы.
Отложения жира п масла в организме, служа запасом пищи,
в то же время уменьшают его удельный вес. Яйца и молодь многих
рыб обнаруживают это приспособление. Пелагические яйца пе
прилипают к предметам, они плавают свободно; многие из них
заключают в себе большую жировую каплю па поверхности желтка.
Таковы яйца многих тресковых рыб: обыкновенного мепька (Bros-
niius brosme), часто попадающегося па Мурмане; мольвы (Molva
juolva), которая ловится там лее; таковы яйца макрелей (Scomber
scomber) и других рыб.
Всякого рода воздушные пузыри служат той же цели — умень-
шению удельного веса. Сюда относится, конечно, и плавательный
пузырь.
Совершенно по иному типу построены яйца, погружающиеся —
демерсальные, развивающиеся па дпе. Они крупнее, тя-
желее, темны, тогда как пелагические яйца прозрачны. Их обо-
лочка часто является липкой, так что такие яйца прилипают к кам-
212
ням, водорослям и другим предметам или же друг к другу. У не-
которых рыб, как у саргана (Belone belone), яйца также снаб-
жены многочисленными нитевидными выростами, служащими для
прикрепления к водорослям и друг к другу. У корюшки (Osmerus
eperlanus) яйца прикрепляются к камням и скалам при помощи
наружной оболочки яйца, которая отделяется, но пе вполне, от
внутренней мембраны. Прилипают и крупные яйца акул и скатов.
Яйца же некоторых рыб, как, например, лосося (Salmo salar),
крупны, отдельны и пи к чему не прилипают.
Придонные рыбы, или рыбы бентоса. Рыбы, живущие у дна
по близости берегов, так же как и пелагические, представляют
несколько типов приспособления к условиям их жизни. Главные
Рис. 190. Длиннохвост (Macrurus norvegicus).
(по Iljort)
условия их таковы: во-первых, постоянная опасность быть вы-
брошенными прибоем или в бурю па берег. Отсюда возникает
необходимость в выработке способности держаться за дно. Во-вто-
рых, опасность быть разбитым о камни; отсюда необходимость
приобрести панцырь. Рыбы, живущие па илистом дпе и роющиеся
в нем, вырабатывают различные приспособления: одни для рытья
и для движения в илу, а другие — для ловли добычи, зарывшись
в ил. У некоторых рыб существуют приспособления для того,
чтобы прятаться среди водорослей и кораллов, растущих среди бе-
регов и на дпе, у других — для зарывания в песок во время отлива.
Мы различаем следующие типы донных рыб.
Тип уплощенный д о р з о в с и т р а л ь п о. Тело сжа-
то от спинной стороны к брюшпой. Глаза перемещены па верхнюю
сторону. Рыба может теспо прижиматься к дну. Пример: скаты
(Raja, Trygon и Др.), а из костистых рыб — морской чорт (Lophius
piscatorius).
Тип длиннохвоста. Тело сильно вытянуто, наиболее
высокая часть тела — позади головы, постепенно становится тоньше
и оканчивается заострением. Анальный и спинпой плавники обра-
зуют длинную плавниковую оторочку. Тип распространен среди
глубоководных рыб. Пример: длиннохвост (Macrurus norvegicus)
(рис. 190).
Тип сдавлеппо-а симметрический. Тело сдавлено
с боков, окаймлено длинными спинным и анальпым плавниками.
' 215
Глаза на одной стороне тела. В молодости имеют тело сдавленно-
симметричное. Плавательного пузыря нет, держатся на дне. Сюда
принадлежит семейство камбаловых (Pleuronectidae). Пример:
тюрбо (Rhombus maximus).
Тип угреобразный. Тело очень длинное, змеевидное;
парные плавники зачаточны или отсутствуют. Рыбы донные. Дви-
жение по дну создало ту же форму, какую среди рептилий мы видим
у змей. Примерами могут служить угорь (Anguilla anguilla), минога
(Petromyzon fluviatilis).
Тип астеролепиформный. Передняя половина тела
заключена в костный панцырь, сводящий активные движения до
минимума. Тело в разрезе треугольно. Пример: кузовок (Ostracion
cornutus).
Особые условия господствуют па больших глубинах: огромное
давление, абсолютное отсутствие света, пизкая температура (до 2°),
полное спокойствие и отсутствие движений в воде (если не считать
очень медленного движения всей массы воды из арктических морей
к экватору), отсутствие растений. Эти условия накладывают резкую
печать на организацию рыб, создавая особый характер глубинной
фауны. Мускульная система развита у них слабо, кость мягка.
Глаза редуцируются иногда до полного исчезновения. У тех глу-
бинных рыб, у которых глаза сохраняются, сетчатка по отсутствию
колбочек и положению пигмента сходна с глазом ночных живот-
ных. Далее, глубиппые рыбы отличаются большой головой и топким
туловищем, утончающимся к концу (тип длиннохвоста), большим
растяжимым желудком и очень большими зубами во рту (рис. 191).
Глубинные рыбы могут быть разделены на бентосных
и батипелагических рыб. К донным рыбам глубин
относятся представители скатов (сем. Turpodinidae), камбал (сем.
Pleuronectidae), рукоперых (сем. Pcdiculati), папцырпощских (Са-
taphracti), долгохвостов (сем. Macruridae), бельдюг (сом. Zoarcidac),
тресковых (сем. Gadidae) и др. По и среди батипелагических,
и среди береговых рыб встречаются представители названных
семейств. Провести резкую отчетливую границу между глубин-
ными формами и береговыми пе всегда легко. Многие формы встре-
чаются и тут и там. Также и глубина, иа которой попадаются
батппелагические формы, вариирует в широких пределах. Из бати-
пелагических рыб следует упомянуть светящихся анчоусов (Scope-
lidac).
Донные рыбы питаются малоподвижными животными и их
остатками; это по требует затраты сил, и дойные рыбы обычно
держатся большими стаями. Наоборот, рыбы батппелагические
отыскивают с трудом свою пищу и держатся поодиночке.
Большая часть промысловых рыб принадлежит или к литораль-
ной, или к пелагической фаупе. Некоторые тресковые (Gadidae),
кефалевые (Mugilidae), камбалы (Pleuronectidae) принадлежат бере-
говой зопе; тупцы (Thynnus), макрели (Scombridae) и главные
промысловые рыбы — сельдевые (Clupeidae) — принадлежат к фау-
не пелагической.
214
Рис. 191. Chauliodus sioanci vi.nn «драконы глубин» дср>тсп из-за добычи*
(1 [о Бибу)
Конечно, пе псе рыбы принадлежат непременно к. одному из
указанных типов. Многие рыбы лишь приближаются к тому или
иному из них. Ярко выраженным тип строения — результат
приспособления к определенным, строго обособленным условиям
местообитания и движения. Л подобные условия не всегда бывают
хороню выражены. С другой стороны, для того чтобы выработался
тот пли пион тип, нужно долгое время. Рыба, недавно переменив-
шая свое местообитание, может утерять отчасти свой прежний
приспособительный тин, по еще не выработать нового.
215
В пресной воде нет того разнообразия условий жизни, которое
наблюдается в море, однако и среди пресноводных рыб встречается
несколько типов. Например, елец (Leuciscus leuciscus), предпочи-
тающий держаться па более или менее сильпом течении, имеет
тин, приближающийся к веретенообразному. Наоборот, относя-
щийся к тому же семейству карповых (Cyprinidae) лещ (Abramis
brama) или карась (Carassius carassius) — рыбы малоподвижные,
живущие среди водных растений, кореньев и под крутоярами, —
имеют тело неуклюжее, сдавленное с боков, как у рифовых рыб.
Щука (Esox lucius), стремительно кидающийся на добычу хищник,
напоминает стреловидный тип нектопной рыбы; живущий в типе
и иле, пресмыкающийся у дна вьюн (Misgurnus fossilis) имеет более
или менее угревидную форму. Постоянно пресмыкающаяся по дну
стерлядь (Acipenser ruthenus) напоминает тип долгохвоста.
3. Питание и приспособление к роду пищи у рыб
Вопрос о питании рыб имеет большое хозяйственное значение.
Но знания наши в этой области далеко пе достаточны. Мы нахо-
димся в стадии накопления материалов.
Способ питания рыб весьма различен. Одни рыбы питаются
исключительно животной пищей, другие — главным образом ра-
стительной, третьи являются всеядными. Большинство рыб пи-
тается животной пищей и может быть разделено на две группы:
на питающихся крупными животными, главным образом рыбами
же, иногда лягушками, мышами и птицами, и на питающихся
мелкими животными (беспозвоночными: простейшими, червями,
ракообразными, моллюсками, насекомыми и т.д.). К этим последним,
питающимся мелочью, относятся карповые (Cyprinidae), хариусы
(Thymallus), сиги (Coregonus), осетры (Acipenseridae), сельди
(Clupeidae), макрели (Scombridae) и др. К хищникам, питающимся
более крупными животными, принадлежат лососи (Salmonidae),
сомы (Siluridae), палимы (Lota lota), щука (Esox), скаты (Batoidei),
акулы (Selachii), большое количество глубоководных рыб. Среди
рыб, питающихся мелкими животными, можно различить таких,
которые схватывают животных, попадающих в воду из воздуха,
или питаются планктоном, и таких, которые берут пищу па дпе.
Одни питаются планктоном постоянно, планктон составляет их
главную пищу. Таковы некоторые пелагически живущие рыбы
и прежде всего личинки и рыбья молодь. Другие — лишь временно,
в юности, питаются планктоном, а позднее переходят к питанию
донными животными, лишь иногда и за отсутствием другого корма
кормясь Планктоном. К первым относятся: многие сиги (Coregonus),
питающиеся главным образом планктонными ракообразными; ук-
лейки (Alburnus lucidus), которые, кроме падающих на поверх-
ность воды пасекомых, питаются главным образом планктоном
поверхностных слоев; ко вторым — озерная корюшка, или снеток
(Osmerus eperlanus sprinclius), в молодости питающаяся планкто-
216
ном, а позднее ведущая1 хищный образ жизни и истребляющая
' много мелких особей того же вида.
Уже будучи 10 см ростом, снетки питаются преимущественно
снетками же. Лещ (Abramis brama) в первое лето по выходе из
икры кормится преимущественно мелкими рачками, а затем ста-
новится рыбой донной, питаясь главным образом личинками вод-
ных насекомых и моллюсками. Однако при недостатке такого рода
пищи он переходит в зону открытой воды, питаясь здесь планкто-
ном. Конечно, это весьма плохо отражается па его росте. Таким
образом, может получиться даже особая карликовая раса.
Из морских рыб типичными пожирателями планктона являются
сельди. Планктонными организмами питаются и мелкие представи-
тели сельдевых, как сардина (Sardinella pilchardus), шпрот (Spratella
sprattus), а также представители семейства анчоусов (Engraulidae).
Относящаяся к последнему хамса, вылавливаемая у нас в коли-
честве десятков миллионов килограммов, ведет типично пелаги-
ческий образ жизни, совершая значительные передвижения в за-
висимости от температуры воды и связанного с последней планктона..
Количество поедаемого планктона часто необыкновенно велико.
В желудке сига (Coregonus oxyrrhynchus) было насчитано, напри-
мер, 50 000 рачков рода Bosmina. Между распределением планктон-
ных организмов (которое часто зависит от освещения, температуры,
плотности воды) и питающихся планктоном рыб существует опре-
деленное взаимоотношение, имеющее большое значение в рыбо-
ловстве.
Таким образом, зависимость рыб от планктона очень велика.
А если мы примем во внимание, что молодь питается исключи-
тельно планктоном, то станет попятным, что гибель планктона
часто ведет за собой уничтожение всего рыбьего прироста. Прй
этом имеют значение пе только планктонные организмы из мира жи-
вотных (зоопланктон), по и растительный планктон (ф и то-
пл а п к т о и); последний в конечном итоге играет даже важней-
шую роль, так как за его счет существуют планктонные животные.
Из планктонных водорослей наиболее важны диатомовые, йак
ассимиляторы углерода, присутствующие в воде иногда в колос-
сальном количестве. Планктонные водоросли поедаются личинками
рыб, многими беспозвоночными в ранней стадии развития и соста-
вляют главную пищу планктонных ракообразных.
Большинство хищных рыб отыскивает свою пищу у берегов
и па дпе. Многие из них в ранней юности питаются растениями:
голавль (Squalius cephalus), окунь (Perea fluviatilis), судак (Lucio-
perca sandra), и лишь потом переходят па животную пищу, как
личинки насекомых, бокоплавы, водяные ослики, черви, моллюски,
рыбы. В море камбалы (Pleuronectidae) питаются раками и моллю-
сками; скаровые (Scaridae) обгрызают полипы, губки и кораллы,
перетирая их глоточными зубами. Морской волк (Anarrhichas lupus)
питается крабами и моллюсками. Акулы и тупцы (Thynnus) поедают
массу рыб; в частности, тупцы уничтожают главным образом сельдей
и макрелей.
217
Чисто растительноядных рыб чрезвычайно мало. Многие, осо-
бенно молодь, поедают растительный планктон, объедают водо-
росли с камней (подуст — Chondrostoma nasus). Всеядной рыбой
является сазан (Cyprinus).
Для питания рыб имеет огромное значение их жадность к пище,
которая зависит от температуры. Для каждого вида имеется в этом
отношении свой температурный оптимум, выше и ниже которого
пищеварение продолжается лишь в известных пределах. У форели
оптимум находится при 12—14°, при 2° и при. 18° пищеварение
прекращается. Рыбы, поглощая при благоприятных условиях
большое количество пищи, могут долгое время оставаться и без
нее. Карпа можно держать в аквариуме без пищи 1—1% года.
Хорошо питаясь, рыба отлагает в теле много жира, причем пита-
тельность ее мяса поднимается. Наоборот, при голодании рыба
достигает лишь небольшого роста, делаясь при этом очень вялой.
У рыб умеренного и холодного поясов наблюдается годовая
и суточная изменчивость питания, имеющая весьма важное практи-
ческое значение.
Здесь следует упомянуть о теории осмотического питания рыб
Пюттера. В воде имеется, хотя и слабый, раствор различных ве-
ществ, из которого последние могут путем осмоса проникать в орга-
низм и усваиваться последним. Проникновение это может совер-
шаться через стенки кишечника и через жаберные листочки. Поверх-
ность их очень велика. У рыбы длиной в 24.2 см общая поверхность
жаберных листочков равна 261 см2. Опа почти равна поверхности
тела (равна 0.96 последней). Мы видели, что личинки угрей живут
в океане более двух лет, и за все это время они не могут принимать
пищи. А в то же время они развиваются, у пих образуются новые
органы. Во многих случаях при долгом голодании, при миграциях
количество вещества, теряемого рыбой, значительно меньше (в не-
которых случаях всего 8%) того, чем можно было бы покрыть
общую затрату энергии, при этом производимую. Также и зимой
рыба пе питается, а половые продукты в пей созревают. Эти и много
подобных фактов заставляют отнестись к теории Пюттера со вни-
манием.
Род пищи сказывается па организации рыб. Изменения в орга-
низме, зависящие от рода пищи, касаются прежде всего зубов
и челюстей. Зубы служат для схватывания пищи, для удержи-
вания ее, для разрезания и разрывания и для раздавливания.
Зубы рыбы, раздавливающей твердую пищу, как моллюски или
крабы, конечно, будут совсем-иные, нежели зубы того вида рыб,
который проглатывает свою добычу целиком; для отгрызания ко-
раллов нужны совершенно иные зубы, нежели для схватывания
мелких червей, живущих в мягком <иле. Зубы рыб или просто по-
гружены в ткань десен и соединены с челюстями волокнистой
ткапью (акулы, щуки, морские черти) или же — и это гораздо
чаще — зубы прямо срастаются с костями', развивающимися на
челюстях. Лишь очень редко зубы помещаются в лунках (у хара-
циновых — Characinidae). У большинства рыб зубы заменяются
218
новыми, как только они износятся или выпадают. У химер (Chi-
тпаегае) и двоякодышащих (Dipnoi) зубы не выпадают, а потеря
от стирапия зубов возмещается постоянным ростом их снизу. У ко-
стистых рыб новые зубы образуются
между или при основании старых зу-
бов, и смена их происходит менее пра-
вильно.
Наиболее просты зубы, служащие
для удержания добычи. Они имеют вид
острых конусов или иголок, загнутых
назад, мелки и часто * усеивают кость,
на которой сидят, наподобие щетки. Та-
ковы, например, зубы нашего сома
(Silurus glanis), питающегося главным
образом рыбой и лягушками, которых
сом глотает целиком. У более подвиж-
Рис. 492а. Зубы ската
(Myliobatis aquila).
ных хищников зубы гораздо длиннее, сильно загнуты назад,
пасть очепь широка. Пелагический хищник хаулиод (Chauliodus)
может поглотить добычу более крупную, чем он сам. Пасть его
вооружена длиннейшими зубами. Еще более крупную добычу
может заглатывать хиазмод черный (Chiasmodus niger). Из
наших пресноводных хищников крупные загнутые назад зубы
имеет, например, щука (Esox lucius); у пее небольшие межчелю-
стные и небные кости усажены довольно крупными острыми зу-
бами, а еще более крупные, загнутые назад своей вершиной зубы
находятся па нижней челюсти; сошпик же и даже язык покрыты
мелкими зубами.
У самых больших хищников моря, у акул, зубы имеют вид
полудисков с зазубренным краем или острых кинжалов.
Зубы конической формы, выдающиеся среди других зубов
своей величиной, всегда указывают па хищный образ жизни. Зубы,
служащие для раздавливания раков, крабов, моллюсков и т. д.,
становятся широкими и тупыми. Таковы зубы у скатов (рис. 192 а).
Иногда передние конические зубы служат для отдирания раковйн
ото дна, задние широкие — для их раздавливания (акула Cestra-
cion). Из костистых рыб такими двоякого рода зубами обладают
зубатки, папример, зубатка обыкновенная (Anarrhichas lupus),
питающаяся ракообразными и моллюсками.
Кроме настоящих челюстных зубов для раздавливания и пере-
тирания пищи могут служить еще так называемые глоточные,
или ж а б е р н ы е, зубы, развивающиеся в связи с внутренним
краем жаберных дуг, с которыми они обыкновенно крепко сраста-
ются (рис. 192 6). Иногда опи связаны с нижним отделом глоточных
костей, как у карповых рыб (Cyprinidae), и называются нижне-
глоточными; тогда в паре с ними работает мозолистая по-
душка па верхней стороне глотки, построенная из рогового эпи-
телия, так называемый «жерновок», и глоточные зубы, приводимые
в движение сильными мускулами, работают как пара жерновов,
перетирая проходящую между ними пищу. В других случаях,
219
как у губанов (Labridae), имеются верхние и нижние глоточные
зубы, расположенные друг против друга.
Когда, имеются глоточные зубы, то другие зубы или отсутствуют
или играют второстепенную роль, служа для схватывания и удер-
живания добычи. Так, у карповых рыб рот беззубый, ио нижпе-
глоточпые зубы развиты очень сильпо, спабжены широкой жева-
тельной поверхностью и расположены в один или два ряда. Снаб-
женные верхпе- и нижнеглоточными зубами губаны (Labridae),
многие виды которых водятся у нас, в Черном море, питаются
моллюсками и ракообразными.
Рис. 1926. Глоточные зубы и «жерновок»:
-усача (Barbus fluviatilis), Ъ — карпа (Cyprinus carpio), с —подуста (Chond-
rostoma nasus).
У рыб, которые грызут кораллы, зубы принимают форму резцов
и могут затем срастаться в одно целое, образуя режущий клюв,
вроде клюва попугая, па котором даже не видно отдельных зубов,
а края одеты толстым слоем блестящей эмали. Такие зубы мы встре-
тим в семействе сростпочелюстпых (Plectognathi). Подобные лее
зубы, сросшиеся, однако пе в такой сильной степени, имеются
и в семействе скаровых (Scaridae), представители которого пита-
ются кораллами и твердораковипными моллюсками.
Не только зубы, по и челюсти приспособляются к роду пищи
данного животного, так как в захватывании пищи челюсти играют
большую роль. В связи с тем или иным питанием различают сле-
дующие типы челюстей.
Ударный, или мечевидный, тип челюсти.
У некоторых быстро плавающих хищников челюсти силыю
вытянуты вперед, и кости передней части головы срастаются;
иногда верхняя или нижпяя челюсть длиннее; зубы хвататель-
ного типа. Такой мечевидный орган — орудие для быстрого уби-
вания добычи, которая затем подбирается, или же для нападения
на врагов. Такова челюсть у сарганов (Belonidae), являющихся
большими хищниками, у хорошо известной силой своих ударов
220
меча-рыбы (Xiphias gladius), производящей большие опустошения
среди макрелей и сельдей.
Другой тип, пилообразное рыло, встречается
в группе скатов-пилорыб (Pristis) и акул-пилоносов (Pristiophorus).
Здесь вытянуты не челюсти, а рыло, но значение этого образо-
вания то же самое, что и вытянутых челюстей. У пилоноса
длинное плоское рыло усажено с каждой стороны мощными зубами
и служит для нападения иа косяки сельдей и анчоусов, которыми
пилонос питается. Челюстные же зубы мелки, остры и сидят тесно.
(Тип лопатоносов, плуг-рыло, встречается у не-
которых рыб, рыло которых “Вытянуто в широкую лопасть без
зубов. При его помощи рыба роется, копает дно, поднимая ил,
в. котором находятся мелкие организмы, служащие ей пищей.
Сюда относится американский многозуб
(Polyodon folium), туркестанские ло-
патоносы (Pseudoscaphirhynchus) и до
некоторой степени—осетровые.
Трубкообразное рыло. У
многих рыб, как, например, у иглы-
рыбы (Syngnathus), у морского конька
(Hippocampus) и др., рыло вытянуто в
виде длинной беззубой трубки с кла-
паном внутри, играющей роль всасы-
вающей пипетки.
Необычайно развито трубкообразиое
рыло у многих представителей африкан-
ского семейства длиппорылов (Могшу-
ridae) (рис. 193). Этим рылом, па конце рис. 193. голова Gna-
которого находится еще мясистый чувст- thonemus curvirostris
ВИТеЛЬНЫЙ ОТрОСТОК, рыбы ЗОПДИруЮТ (И^ВоиЫп сг\
почву, залезая им под камни или за- 3 0,1 епесг‘
рываясь носом в ил, как это делают среди птиц вальдшнепы,
бекасы (Scolopax, Gallinago). Здесь мы видим прекрасный пример
конвергенции у представителей двух весьма отдаленных
классов.
У многих рыб, папример, у карповых (Cyprinidae), рот может
в ы т я г и в а т ь с я в короткую трубку, когда рыба схватывает
добычу. Это достигается благодаря тому, что челюстные кости
соединены растяжимыми длинными связками. Так как в передней
части рта также имеется клапан, то возможно, что и здесь ротовая
трубка играет роль пипетки. Такие же растяжимые связки делают
возможным необычайное растягивание рта у многих глубоководных
рыб, глотающих добычу более крупную, чем они сами.
У тех рыб, у которых, как у коралловых, зубы срастаются
между собой, челюстные и предчелюстные кости, срастаясь, до-
стигают мощного развития, служа сильным рычагом.
Наконец, надо упомянуть, что у рыб донных рыло большей
частью очень короткое и широкое, как, например,
у сома (Silurus glanis). У некоторых рыб, питающихся со дна,
221
рот находится иа брюшной стороне (стерлядь — Acipenser rhuthe-
nus). Этих примеров достаточно, чтобы показать, что и ротовые
части, как и зубы, подвергаются значительным изменениям в за-
висимости от условий жизни и рода пищи. Подобные приспособле-
ния возникают медленно. У акул, ведущих пелагический образ
жизни, рот находится на брюшной стороне, как у рыб бентоса.
Это доказывает их происхождение от последних. Вообще при рас-
сматривании приспособительных признаков никогда не следует
упускать из виду их историчности: тот или иной орган может суще-
ствовать потому, что он был адаптивным раньше, и полной при-
способленности к новым условиям еще не наступило.
На пищеварительном канале также сказывается характер пи-
тания. Например, так называемые жаберные тычинки,
сидящие па внутрепней стороне жаберных дуг и образующие род
решетчатого фильтра, защищающего, с одной стороны, жабры от
повреждения посторонними телами, с другой стороны — задержи-
вающего в глотке мелких животных, служащих пищей, в своем
развитии зависят от рода пищи. Чем мельче объекты питапия
(планктон), тем совершеннее построен этот решетчатый аппарат.
У хищников, питающихся рыбой, тычипки расставлены очень
редко, т. е. немногочисленны. Живой пример дают каспийские
сельди—род Caspialosa. Как <и все сельди, они животпоядиы,
но по строению отцеживающего аппарата опи могут быть разде-
лены на три группы. Из следующей таблицы видно различие в числе
тычинок жаберного аппарата в связи с характером питания видов
и подвидов этого рода. Чем больше тычинок, тем больше поедается
планктонных ракообразных и меньше рыб, и наоборот. По числу и
строению тычинок можно делать заключение о роде пищи рыбы.
Число тычинок Ракообраз- ные (вообще) в % Copepoda (в частности) Рыба
С. caspia . . . . 110—130 7G.2 46.6 16.2
С. kessleri . . . 60—90 54.4 19.2 26.4
С. brashnikowi . С. saposhnikovi • * 27—37 22.7 4.5 90.9
В кишечном канале должны быть приспособления к различного
рода пище. Можно указать ряд примеров такого приспособления.
Например, у кефали (Mugil), которая питается мелкими расти-
тельными организмами, смешанными с илом, желудок расши-
ряется в мускульный зоб, подобный таковому у кур. Затем можно
указать па связь, существующую между длиной пищеварительного
канала и пищей рыбы, а имеппо: у рыб, которые питаются расти-
тельной пищей, кишечпик. очень длинен и мпогократпо свернут,
тогда как у рыб, питающихся другими рыбами, — относительно
короток. У живущей в Сев. Америке карповой рыбки Campostoma
anomalum, питающейся илом, который, опа, очевидно, пропускает
222
через себя в очень большом количестве, кишечник в 7 раз длиннее*
тела и образует 15 оборотов вокруг плавательного пузыря. Отноше-
ние длины тела к длине кишечника у разных рыб различное.
Положение рта, строение «губ», наличие усиков, других вы-
ростов разного рода также нередко является приспособительным,
признаком.
И характер пищеварительных ферментов у рыб хищных и расти-
тельноядных (мирных) иной: у первых выделяется пепсин (пере-
варивает белок), у вторых — трипсин (способствует усвоению'
растительности).
4. Окраска рыб
Морфологическая сторона окраски рыб описана выше. Здесь
мы разберем экологическое значение окраски вообще и ее при-
способительной значение.
Немногие животные, не исключая насекомых и птиц, могут
соперничать с рыбами по яркости и изменчивости их окраски,
исчезающей у них большей частью со смертью и после помещения
в консервирующую жидкость. Окрашены столь разнообразно
лишь костистые рыбы (Teleostei), у которых имеются
все способы образования окраски в различных комбинациях. По-
лосы, пятна, ленты комбинируются па основном фоне подчас в очень
сложный рисунок.
В окраске рыб, как и других животных, многие видят во всех,
случаях приспособительное явление, являющееся результатом
отбора и дающее животному возможность стать незаметным, укрыть-
ся от врага, подстеречь добычу. Во многих случаях это несомненно
так, ио далеко не всегда. В последнее время все больше появляется
возражений против такого одностороннего истолкования окраски
рыб. Ряд фактов говорит за то, что окраска является физиологи-
ческим результатом, с одной стороны, обмена веществ, с другой —
действия световых лучей. Окраска возникает благодаря этому
взаимодействию и может совершенно пе иметь защитного значения.
Но в тех случаях, когда окраска может быть важна экологически,,
когда окраска дополняется соответственными привычками рыбы,
когдг1 у нее есть враги, от которых надо скрываться (а это не всегда
бывает у тех животных, которых мы считаем покровительственно
окрашенными), тогда окраска становится орудием в борьбе за су-
ществование, подчиняется отбору и делается приспособительным
явлением. Окраска может быть полезной или вредной не сама по
себе, а будучи связанной коррелятивно с каким-либо другим по-
лезным или вредным признаком.
В тропических водах и обмен веществ и свет более интенсивны.
И окраска животных здесь ярче. В более холодных и менее ярко
освещаемых водах севера, а тем более в пещерах или подводных
глубинах окраска значительно мепее яркая, бывает даже черная.
За необходимость света в продукции пигмента в коже рыб-
говорят эксперименты с камбалами, содержавшимися в аквариу-
22&
г_мах, в которых на свет выставлялась нижняя сторона камбалы.
На последней постепенно развился пигмент, обыкновенно же
нижняя сторона тела камбал белая. Опыты делались с молодыми
камбалами. Пигментация развилась та же, что па верхней стороне;
если камбалы держались таким образом долго (1—3 года), то ниж-
няя сторона становилась совершенно так же пигментированной,
как верхняя. Этот эксперимент, однако, не противоречит роли
отбора в выработке защитной окраски, — оп только показывает
материал, из которого благодаря отбору у камбал выработалась
способность реагировать на действие света образовапием пигмента.
Так как эта способность могла выражаться по в одинаковой мере
у разных особей, отбор мог здесь действовать. В результате у кам-
баловых (Pleuronectidae) мы видим резко выраженную измен-
чивую защитную окраску. У многих камбаловых
верхняя поверхность тела окрашепа в различные оттепки бурого
цвета с черными и светлыми пятнами и гармонирует с преобладаю-
щим тоном песчаных отмелей, па которых опи обычно кормятся.
Попав на грунт иной окраски, опи сейчас же меняют свой цвет
па цвет, соответствующий окраске дна. Опыты с перенесением
камбал на грунты, разрисованные наподобие шахматной доски
с квадратами различной величины, дали поразительную картину
приобретения животным такого же рисунка. Весьма важно, что
некоторые рыбы, меняющие в различное время своей жизни место
обитания, приспособляются в своей окраске к новым условиям.
Например, Pleuronectes platessa в летние месяцы держится на чистом
светлом песке и окрашена светло. Весной же, после икрометания,
Р. platessa, изменив окраску, ищет илистого грунта. Тот же самый
выбор местообитания, соответствующего окраске, точнее — по-
явление иной окраски в связи с повым местообитанием, — наблю-
дается и у других рыб.
Рыбы, живущие в прозрачных реках и озерах, а также рыбы
поверхностных слоев моря обладают общим типом окраски: спина
у них окрашена в темный, большей частью синий цвет, а брюшная
сторона — серебристого топа. Принято считать, что темная синяя
окраска спины делает рыб незаметными для воздушных врагов;
пижняя — серебристая — против хищников, держащихся обычно
на большей глубине и могущих заметить рыбу снизу. Некоторые
полагают, что серебристо-блестящая окраска брюха рыб снизу
является невидимой. Согласно одному мнению, лучи, достигающие
поверхности воды снизу под углом 48° (в соленой воде 45°), целиком
отражаются от пес. Положение глаз па голове рыбы таково, что
они могут видеть поверхность воды самое большее под углом в 45°.
Таким образом в глаза,рыб попадают лишь отраженные лучи, и по-
верхность воды представляется рыбе серебристо-блестящей, как
пижняя и боковые стороны их добычи, становящейся по этой при-
чине невидимой. Согласно другому мнению, зеркальная поверх-
ность воды отражает голубоватые, зеленоватые и бурые тона всего
водоема, серебристое брюхо рыб делает то же. Результат получается
тот же, что и в первом случае.
224
;Однако другие исследователи полагают, что вышеизложенное
толкование белой или серебристой окраски брюха неверно; что
ее полезное значение для рыбы ничем не доказано; что рыба не
атакуется снизу и что она должна казаться темной и заметной
снизу. Белый же цвет брюшной стороны, по этому мнению, есть
простое следствие отсутствия ее освещения. Однако видовым при-
знаком признак может стать лишь в том случае, если он прямо или
косвенно полезен биологически. Поэтому упрощенные физические
объяснения вряд ли оправдываются.
У рыб, живущих на дне водоема, верхняя поверхность тела
темная, часто украшенная извилистыми полосами, бблыпими или
меньшими пятнами. Брюшная сторона серая или беловатая. К та-
ким донным рыбам можно отнести палима (Lota lota), пескаря
(Gobio fluviatilis), бычка (Cottus gobio), сома (Siluris glanis), вьюна
(Misgurnus fossilis) — из пресноводных, осетровых (Acipenseridae),
а из чисто морских — морского чорта (Lophius piscatorius), скатов
(Batoidei) и многих других, в особенности камбаловых (Pleuronecti-
dae). У последних мы видим резко выраженную изменчивую
защитную окраску, о которой сказано выше.
Другой вид изменчивости окраски мы видим в тех случаях,
когда рыбы одного и того же вида становятся более темными в глу-
бокой воде с илистым или торфянистым дном (озера) и более свет-
лыми — в мелкой и прозрачной воде. Примером может служить
форель (Salmo trutta morpha fario). Форели из ручьев с гравиевым
или песчаным дном окрашены светлее, нежели те, которые добыты
из мутных потоков. Для такой перемены окраски необходимо
зрение. В этом убеждают нас опыты с перерезкой зрительных
нервов.
Поразительным примером защитной окраски является австра-
лийский вид морского конька — Phyllopteryx eques, у кото-
рого кожа образует многочисленные, длинные, плоские, развет-
вленные нити, окрашенные бурыми и оранжевыми полосами, по-
добно водорослям, среди которых рыбка живет. Многие рыбы,
живущие среди коралловых рифов Индийского и Тихого океанов,
особенно рыбы, относящиеся к семейству Chactodontidac и Роша-
centridae, имеют в высшей степени блестящую и живую окраску,
часто украшены полосами различного цвета.. В обоих названных
семействах одинаковый цветной узор развился самостоятельно.
Даже у посещающих рифы представителей камбаловых, обычно
окрашенных тускло, верхняя поверхность бывает украшена жи-
выми топами и поразительным рисунком.
Окраска может быть пе только защитной, по и помогать хищ-
нику быть незаметным для его добычи. Такова, например, поло-
сатая окраска нашего окуня и щуки и, быть может, судака;
темные вертикальные полосы на теле этих рыб делают их незамет-
ными среди растений, где они ждут добычу. Всвязи с такой окраской
у многих хищников развиваются на теле особые отростки, служащие
для приманки добычи. Таков, папример, морской чорт (Lophius
piscatorius), окрашенный покровительственно и имеющий передний
15 Кашкаров и Станчинский 225
луч спинного плавника измененным в усик, подвижный благодаря
особым мускулам. Движение этого усика вводит в обман мелких
рыбешек, принимающих его за червя и приближающихся, чтобы
исчезнуть в пасти Lophius.
Весьма возможно, что некоторые случаи яркой окраски служат
у рыб в качестве предупреждающей окраски. Такова,
вероятно, блестящая окраска многих сростночелюстных (Plecto-
gnathi). Она связана с наличием колючих шипов, могущих топор-
щиться, и может служить указанием на опасность нападения на
таких рыб. Значение предупреждающей окраски, быть может, имеет
яркая расцветка морского дракона (Trachinus draco), вооружен-
ного ядовитыми шипами на жаберной крышке и большим шипом
на спине. К явлениям приспособительного характера, возможно,
следует отнести и некоторые случаи, полного исчезновения окраски
у рыб. Многие пелагические личинки Teleostei не имеют хроматофор
и являются бесцветными. Их тело прозрачно, а потому с трудом
заметно, так же как с трудом замечается опущенное в воду стекло.
Прозрачность увеличивается благодаря отсутствию в крови гемо-
глобина, как, например, у Leptocephali — личинок угря. Личинки
Onos (сем. Gadidae) имеют в течение пелагического периода своей
жизни серебряную окраску, обусловленную присутствием в коже
иридоцитов. Но, переходя с возрастом к жизни под камнями, опи
теряют серебряный блеск и приобретают темную окраску,
5. Защитные приспособления у рыб. Ядовитые железы, элек-
трические органы, панцыри и некоторые другие
К числу приспособлений, имеющих значение при защите и на-
падении, относятся шипыплавпиковых лучей или жаберной крышки,
которые рыба произвольно может поднимать и опускать. У не-
которых рыб существует даже особое запирающее приспособление.
Однако одни шипы еще не являются надежным орудием защиты,
и даже такие рыбы, как окунь и колюшка, проглатываются угрем
и щукой. Даже морской кот (Trygon pastinaca) с его страшным
орудием — шипом — поедается акулой молотом (Zygaena). Часто
шипы комбинируются с ядовитыми железами, выде-
ляющими сильнейший яд — т о к с а л ь б у м и п. Уколы шипа
в таком случае вызывают заболевание и даже смерть животного.
Простейший вид ядовитого аппарата рыб — это одноклеточные
эпидермические железы в эпидерме шипов плавников и шипов
покровных костей черепа. У некоторых бычков (Cottus), звездочота
(Uranoscopus scaber) железы образованы двумя эпидермическими
карманами, открывающимися в жолоб соответствующего шипа;
на их внутренней поверхности в период переста появляются секре-
торные клетки, позже исчезающие. У морского кота (Trygon pasti-
naca) па хвосте имеется зазубренный шип, возле которого масса
одноклеточных белковых (серозных) и слизистых желез. Настоя-
щих ядовитых желез еще нет. Наиболее совершенные ядовитые
железы мы видиму скорпены (Scorpaena), дракона (Trachinus draco)
226
и марулек (Sebastes). У Trachinus ядовитые органы стоят в связно
шипом жаберной крышки и с пятью или шестью жесткими лучами
переднего спинного плавника. Железы образованы постоянными
и резко дифференцированными от эпидермиса комплексами клеток,
в которых имеется известное уклонение от строения остального
эпидермиса. Не вполне ясно, каким образом яд попадает в рану.
Можно думать, что благодаря тому давлению, которое получается,
если шип вдавливается в мясо. У представителя скорпено-
в ы х (Synanceia) по обе стороны спинного плавника имеется
длинный, наполненный ядом замкнутый мешок, который лопается
лишь при сильном надавливании, причем яд может быть перенесен
Рис. 194. Схема строения электрического органа;
две электрических пластинки в окружающей соеди-
нительной ткани. Поперечный срез.
1 — нервная конечная сеть, 2 —нерв, 3 — электрический
слой, 4 — центральный слой электрической пластинки,
5— сосочки, G— кровеносный сосуд, 7—электрическая
соединительная ткань, 8—окружающая волокнистая
ткань. {Но Ihle и др.).
в рапу при помощи снабженных бороздами тринадцати плавниковых
лучей. Укол этот вызывает сильнейшую боль в течение нескольких
часов.
К рыбам, хорошо защищенным, несомненно, относятся и элек-
трические рыб ы. Электрические орган ы по-
строены весьма однообразно у всех рыб и состоят из ряда слоев
аналогично вольтову столбу. Эти слои одеты соедини-
тельнотканным чехлом и подразделены соединительнотканными
перегородками па расположенные в порядке ряды. К электриче-
ским пластинкам подходит нерв, обильно разветвляясь и образуя
нервный слой органа. Поверхность этого органа увеличивается
посредством образования многочисленных сосочков. Поверхность
последних граничит со студенистой тканью, выполняющей свобод-
15* 227
ное пространство. Весь электрический орган является таким обра-
зом суммой пластинок, чередующихся со студнеобразными слоями,
соответственно меди и цинку вольтова столба (рис. 194).
У всех электрических рыб (кроме Malapterurus) этот орган пред-
ставляет видоизменение мышц. У Malapterurus он происходит из
желез кожи, имеет ряд отличий в строении, по все же построен
по одному типу с органами остальных рыб. Отдельные электри-
ческие разряды происходят под влиянием нервной системы, с ко-
торой соединяется каждая пластинка. Сила разряда всецело за-
висит от величины и состояния рыбы. У электрического угря (Gym-
notus) она достигает 300 вольт. У электрического ската (Torpedo
marmorata) максимальная сила удара равняется 70—80 вольт.
Для силы удара имеет значение количество отдельных пластинок,
действующих как масса соединенных маленьких элементов. Число
пластинок у Torpedo marmorata равняется 600, у Gymnotus—6 000.
Орган иннервируется 350 спинными нервами. Произведя ряд уда-
ров, рыба должна некоторое время иметь отдых и пищу для вос-
становления сил.
Электрический орган рыбы оглушает и даже убивает добычу,
но может служить и для защиты от врагов. Удары электрического
угря сразу убивают рыб и лягушек, которых он затем глотает.
Он может даже свалить человека. Все рыбы с электрическими орга-
нами медлительны, мало подвижны. Здесь потенциальная энергия
превращается не в движение, а в электрическую энергию.
Приспособительный характер имеют, вероятно, и органы
свечения рыб.
Эти органы развиты у некоторых других животных глубин.
У рыб они развиты независимо в четырех совершенно не связанных
между собой группах: в семействах — иглоротов (Stomiatidae),
складчатогрудых (Sternoptychidae), светящихся анчоусов (Scope-
lidae) и морских дьяволов (Ceratidae). Мы имеем здесь пример
конвергенции, т. е. развития одних и тех же особенностей
строения, у совершенно различных форм, повидимому, в связи
с приспособлениями к одинаковым условиям жизни. Этот случай
тем более интересен, что здесь развиваются не какие-нибудь не-
значительные особенности, а органы совершенно особого назна-
чения и очень сложного строения. Однако при микроскопическом
изучении органов свечения в этих различных группах они обна-
руживают различие в строении, как и бывает обычно в явлении
конвергенции.
Последние исследования показали, что определенные и лока-
лизованные органы свечения находятся главным образом у пела-
гических животных. У донных животных береговых отмелей спо-
собность светиться крайне редка. Многие донные животные глубин
светятся, хотя органов свечения у них найдено не было, и их спо-
собность светиться связана с поверхностным эпителием. Органы
свечения найдены только у океанических форм теплой области
и свойственны главным образом рыбам, живущим выше гори-
зонта в 500 и 600 м глубиной.
228
является железа, вырабатывающая
Рис. 195. Фотофоры Cyclothone
снаружи.
Рис. 196. Разрез органа свечения
Chauliodus.
р —пигмент, dr.—железистая часть органа,
U— линза. (По Brauer).
Органы свечения могут находиться на чешуях боковой линии,
на нижней челюсти, на усиках или на усиковидных выростах,
под жаберной крышкой, около глаз, на хвосте или же они могут
быть рассеяны по. телу. Строение их часто очень сложное. Наи-
более примитивным органом
светящуюся слизь. Желези-
стая часть есть самая глав-
ная часть в каждом органе
свечения. Как правило вок-
руг нее развивается слой
черного пигмента. Часто ис-
пускаемый железой свет про-
ходит через хрусталикоподоб-
пое образование. В общем
такие органы свечения на-
поминают по своему строе-
нию глаз. На рис. 195 изо-
бражены фотофоры рыбки
Cyclothone снаружи, а на
рис. 196 — разрез сложного
органа свечения хаулиода
(Chauliodus). Мы видим здесь
слой черного пигмента, реф-
лектор из плоских светлых,
блестящих клеток, желези-
стую часть органа, линзу
или хрусталик и часть эпи-
телия, соответствующую ро-
говице. Форма многих ор-
ганов свечения в точности
соответствует форме прожек-
торов, употребляемых в воен-
ном деле и для освещения
маяков, и таким образом
способствует наилучшему функционированию, наиболее пол-
ному отражению света.
Как показали эксперименты, свечение с трудом вызывается
различными раздражениями, но легко возникает, если поместить
светящуюся рыбу в щелочную воду, сделанную щелочной путем
прибавки водного раствора аммиака.
Даже куски рыбы, умершей 5—6 часов назад, помещенные
в аммиачную воду, давали свечение. Точно так же и пропускание
сильного тока вызывало свечение. У большинства рыб свечение,
видимо, происходит временами произвольно под влиянием нервного
возбуждения.
Свет, испускаемый органами свечения, холодный, т. е. тепла
здесь почти не производится. Точными физическими методами до-
казано, что, например, у светящегося жука (Photinus) 97.5% затра-
цецгюй энергии превращается в световые лучи и лишь маленький
229
остаток — в тепло. В наших искусственных источниках освещения,
наоборот, лишь от 0.4 до 4% энергии превращается в свет.
Большое значение для свечения имеет вода. Высыхание
светящихся органов прекращает свечение. Смачивание фотофоров,
обращенных в порошок, дает свечение. Точно так же необходим
для свечения организмов и кислород. По старому воззре-
нию, свечение даже стоит в прямой связи с дыханием. Однако этому
противоречит свечение вырезанных и даже высушенных фотофоров.
Это отнюдь пе жизненный процесс. Зато выделение железистыми
клетками фотофора способной к свечепию субстанции есть процесс
жизненный. Возможно, что многие органы свечения окажутся
обязанными своим существованием бактериям, внедряющимся
в клетки рыбы.
По одним морфологическим данным трудно составить себе
представление о функции фотофоров и их биологическом значении.
Вероятнее всего, что различные фотофоры у различных рыб играют
неодинаковую экологическую роль. То обстоятельство, что у всех
особей одного и того же вида фотофоры расположены совершенно
одинаковым образом, т. е. число и расположение фотофоров
является видовым признаком, — приводит к мысли, что в темноте
органы свечения заменяют видовую окраску наземных животных
и служат отличительным признаком при отыскании полами друг
друга. Это своего рода брачный наряд. Кроме того, фотофоры па
конце усикообразных придатков могут служить для приманки
добычи. Возможность использования для рыбной ловли фотофора
Anomalops подтверждает предположение. Внезапное вспыхивание
органа свечения может служить отпугивающим, устрашающим
признаком, подобно тому как развертывание плавников служит
у некоторых рыб для этой цели. Предположение о такой функции
органа находит себе подтверждение в том, что у каракатиц, у ко-
торых в освещенной зоне моря имеется чернильный мешок для
защиты от врагов, в глубинах вместо пего развивается светящаяся
железа; в темноте, где чернильное облако было бы невидимо,
выделение светящегося секрета достигает с успехом цели защиты
от врага. Такое защитное значение свечения выделяемой слизи
подтверждается наблюдениями Биба, спускавшегося в глубины
в так называемой батисфере. Наконец, при помощи сильно разви-
тых органов свечения на голове некоторые рыбы могут освещать
себе путь и добычу. Однако все это предположения, проверить
которые пока невозможно.
К защитным приспособлениям следует, конечно, отнести и тот
панцирь, который мы видим у кузовка (Ostracion), у папцырных
сомов (Loricaria, Callichthys) и некоторых других рыб. В особен-
ности развит был папцырь у древпих рыб девонского времепи,
у рыб из «древнего красного песчаника», и у родственных кругло-
ротых панцыриых (Ostracodermi). Надо думать, что панцырь этих
существ служил им защитой от хищников, из которых можно вспом-
нить гигантских ракообразпых. Борьба за жизнь была, вероятно,
очень напряженной в водоемах того времени.
230
6. Размножение рыб
Изучение размножения рыб, помимо теоретического интереса,
представляет большую практическую важность. Такие вопросы,
как вопрос о плодовитости, местах нерестования, времени нере-
стования, о темпе роста и т. д., имеют прямое прикладное
значение.
Почти все рыбы являются раздельнополыми; самка называется
икряником, самец — молочником. Являются гермафродитами отно-
сящиеся к классу круглоротых миксины (Myxine): задняя часть
половой железы является у них семенником, передняя — яичником;
обе части созревают и функционируют неодновременно. Миксина
является сперва самцом, затем самкой. Встречается гермафро-
дитизм й у костных рыб: так, постоянно гермафродитны некоторые
морские окупи (Serranidae). Изредка встречаются гермафродитные
особи трески (Gadus), макрели (Scomber), сельдей (Clupea).
В яйца и семенные клетки превращается эпителий половых
мешков. Ко времени нерестования — откладывания икры—
и семенники и яичники сильно увеличиваются, достигая 25—40%
веса рыбы, в особенности увеличиваются яичники. Яйца рыб весьма
различны по величине и по виду, а также по характеру их оболочек.
Величина яиц зависит от количества находящегося в них желтка,
служащего для питания зародыша. У селяхий яйца крупные,
у Teleostomi — мельче. Наиболее крупными яйцами у Teleostomi
являются тяжелые и тонущие (демерсальные), самыми
мелкими — легкие и плавающие (пелагические). Боль-
шинство пелагических яиц имеет около 1 мм в диаметре, по не-
которые еще меньше: у угря (Anguilla anguilla) опи равны 0.1 мм,
тогда как у палтусов (Hippoglossidac) они достигают 4.2 мм
в диаметре. Демерсальные яйца крупнее. Так, у лосося диаметр
яйца равен 6.0 мм, у некоторых сомовых (Siluroidei) 17—18 мм.
Но и среди пресноводных рыб встречаются виды с мелкими яйцами.
У ерша (Acerina cornua) яйца имеют 0.8—1 мм в диаметре, у палима
(Lota lota)—1.0 мм, у озерного сига (Coregonus maraena) —
3.2—3.8 мм. Наиболее крупны яйца у акул и скатов. В общем,
как сказано, яйца морских рыб значительно мельче яиц пресновод-
ных. Кроме того, они менее разнятся между собой по величине,
так что для отличия их друг от друга приходится обращаться
к таким признакам, как число масляных капель, пигментирование
зародыша и т. д.
Число яиц стоит в связи с величиной яиц (чем опи мельче, тем
их больше, поэтому их больше у пелагических рыб), и с жизнью
соответственного вида рыбы. Если условия для вывода молоди
благоприятны, число яиц меньше. У мольвы (Molva molva) число
яиц бывает от 12 до 28 млн. У трески (Gadus callarias) и угря (An-
guilla. anguilla) число яиц от 3 до 9 млн. У рыб, которые стерегут
свои япца, их бывает обычно лишь несколько сотен, а у живоро-
дящих рыб — еще того меньше. У колюшки (Gasterosteus aculeatus)
число яиц всего 80—100.
234
Семенные клетки рыб очень мелки. Например, у лосося, диа-
метр яиц которого от 5.5 до 6.0 мм, семенные клетки имеют в длину
всего 0.06 мм. Зато число их гораздо больше числа яиц. По выходе
из семяпроводов в воду семенные клетки начинают совершать
энергичные движения, при помощи которых они продвигаются
вперед и достигают яйца. Живучесть спермиев различна у разных
видов рыб. Более живучи спермин рыб, нерестящихся в спокойной
или в стоячей воде. Спермин гибнут, попав в воду, у ручьевой
форели (Salmo trutta morpha fario) через 23 секунды; у лосося
(Salmo salar) — через 45 секунд; у усача (Barbus fluviatilis) —
через 120 секунд; у щуки (Esox lucius) — 3—4 минуты, сазана
(Cyprinus carpio) через 5 минут. Степень живучести спермиев
имеет огромное значение для практики искусственного оплодотво-
рения.
Чрезвычайно важное значение имеет также определение сте-
пени зрелости икры. Для ведения рыбного хозяйства,
в основе которого должен лежать прежде всего учет рыбных бо-
гатств, не безразлично, становится ли рыба способной к икро-
метанию на четвертом, частью даже на третьем году жизни (напри-
мер, вобла), или же зрелость наступает лишь на 12—13-м году
(осетр) или даже на 16-м (белуга). Рыба, размножающаяся медленно,
менее способна выдерживать сильный вылов, не уменьшаясь в числе.
Не менее важен вопрос и о том, сколько раз размножается в году
данный вид рыбы, существует ли один или несколько сезонов
икрометания. Вскрытие особей данного вида в различное
время года и определение степени зрелости половых продуктов
есть один из методов для решения вопроса. Точно так же и для
определения места икрометания этот метод имеет зна-
чение.
Степень зрелости половых продуктов в настоящее время опре-
деляется по особым таблицам или шкалам, где различные стадии,
характеризующие ту или иную степень зрелости половых про-
дуктов, обозначаются цифрами. Эти таблицы вырабатываются
применительно к данному виду рыбы. Например, для атлантиче-
ской сельди (Clupea harengus) и для каспийских сельдей таблицы
несколько иные. У последних различают следующие стадии зре-
лости. I — стадия юношеская (молодые экземпляры,
пе достигшие половой зрелости). Половые органы зачаточны и имеют
вид топких белых шнурков или узких бесцветных или желтоватых
лепт, прижатых к внутренним стенкам тела под позвоночником.
Пол не различим. II — стадия покоя. Половые органы
вполне дифференцировались. Половые продукты еще не развиты.
Сосудов мало. III — стадия развития. Половые органы
заметно увеличились в объеме и достигают х/2—2/з максимальных
размеров. Очень богаты кровеносными сосудами. IV — стадия
созревания. Половые органы достигают максимальных раз-
меров и заполняют всю или почти всю полость тела. V — стадия
икрометания. Половые продукты вполне созрели и сво-
бодно вытекают при самом легком надавливании на брюшко или
23?
даже без давления. VI — стадия вы боя. Половые органы
дряблы, красны, воспалены. Зрелые продукты из них выброшены,
стенки заметно спались.
Эти данные относятся к первому циклу икро-
метания. Дело в том, что у каспийской сельди рода Caspialosa
икрометание происходит не сразу, но разбито на три отдельных
момента, разделенных промежутком в 1—11/2 недели, в течение
которого созревает следующая порция яиц. Яичник у Caspialosa
наполнен яйцами неодинаковой величины и степени зрелости.
Большинство рыб мечет икру. Но есть и такие рыбы, у которых
эмбрионы развиваются в яйцеводах, играющих роль «матки».
Такие рыбы носят название «живородящих». Обычно желточный
мешок зародыша не связан со стенками матки. Но у некоторых
акул (Carcharias и Mustelus) образуется плацента: сосудистые
стенки желточного мешка образуют складки, которым соответ-
ствуют таковые «матки». Из костистых рыб живородность встре-
чается в пяти семействах: у морских собачек — Blenniidae, зуба-
стых карпов — Cyprinodontidac, скорпеп — Scorpaenidae, голо-
мянок — Comephoridae и в семействе Embiotocidae. У упомянутых
рыб яйца оплодотворяются, находясь еще в яичниковом мешке
или в яйцеводе. Эмбрион питается частью за счет желтка, частью
за счет питательной секреции, происходящей из стенок яичника
или эпителиальных стенок яйцевода. Число молоди, развиваю-
щейся таким образом, различно: у Embiotocidae их 40—50, у мор-
ских собачек (Blenniidae) их до 300, а у норвежской марульки
(Sebastes norvegicus) число зародышей достигает 1 000. Способ-
ность к живорождению рассеяна в разных группах рыб совершенно
спорадически. Поэтому трудно приписать возникновение живород-
ности одним и тем же условиям во всех случаях. Повидимому,
она возникает в некотором смысле случайно, но, несомненно,
в связи с возрастанием полового инстинкта у самца, что подтвер-
ждается прямыми наблюдениями. Как только возникло внутреннее
оплодотворение, развитие яиц внутри тела матери уже нетрудно
объяснить.
В связи с половыми железами у рыб, как и у высших форм
позволенных, находятся первичные половые приз-
наки: копулятивные органы скатов и акул, яйцеклад горчака
(Rhodens amarus) и др. Более интересны так называемые вто-
ричные половые п р и з н а к и, не служащие сами для
оплодотворения, по помогающие особи отыскивать особей другого
пола, возбуждать в пих половой инстинкт и служить для отпуги-
вания соперников. К этим признакам относится прежде всего
пестрый, так называемый брачный наряд многих рыб,
затем удлинение плавников, утолщение их краевых лучей, искрив-
вление нижней челюсти вверх, роговые бородавки на коже («жем-
чужные органы» карповых) и т. д. Эти признаки появляются лишь
на время нереста или же остаются на все время, отличая полы.
Брачный наряд возникает, несомненно, под влиянием внутренней
секреции половых желез и состоит прежде всего в появлении яркой
233
насыщенной окраски. Мы встречаем ее в период нерестования
у многих рыб: у колюшки (Gasterosteus), горчака (Rhodens amarus),
гольяна (Phoxinus laevis), лосося (Salmo salar), восточных лососей
(Oncorhynchus) и др. Особенно ярко выражено это явление у ло-
сосей. Обыкновенно серебристая рыба в брачный период становится
то огненно красной с зеленой головкой (нерка — Oncorhynchus
nerka), то яркочерпой с малиновыми поперечными полосами (кета —
Oncorhynchus keta), то у ней вдоль всего тела проходит огненно-
красная полоса (у аме-
риканских форелей). В
то же время появля-
ются сложная пигмепти-
.ровка, рисунок на че-
шуйках и плавниках.
Эти изменения тракто-
вались частью как сред-
ство для взаимного при-
влечения полов. Но не
только в изменении ок-
раски состоит брачный
наряд. У лососей удли-
няются и изгибаются
челюсти, вырастают не-
обыкновенно мощные
изогнутые зубы, поя-
вляется горб на спипе,
изменения признавались
Рис. 197. Череп кета-самца (Oncorhynchus
keta) в брачном наряде.
1 — maxillare, 2 — dentale, 3 — articulare, 4 —
quadratum, 5 —palatinum, 6 —frontale. (По Черна-
вину.)
придающий рыбе страшный вид. Эти
за средства орудия и защиты в борьбе за самок (рис. 197).
Но на ряду с указанными изменениями развивается еще ряд
признаков, захватывающих внутренние органы и часто вовсе
ненужных и даже вредных: глоточные зубы, мягкий огромный
крюк на челюсти, мешающий действию челюстей, и т. д.
Объяснение брачного наряда выбором самками наиболее кра-
сивых самцов можно считать неудачным уже ио одному тому, что
во многих случаях (лососи) самцы пе конкурируют из-за самок,
а самки не выбирают самцов, в особенности у тех рыб, где самцы
наиболее ярко окрашены (Cyprinodontidae). В брачном наряде рыб
мы имеем пе биологическую окраску, а чисто физиологическую,
результат особых гормонов. Самцы вообще ярче самок потому,
что опи деятельнее, подвижнее и обмен веществ у пих энергичнее.
Что касается величины самца и самки, то у большинства кос-
тистых рыб самка больше самца, у некоторых — в 6 раз. Число
самок обычно больше, нежели число самцов, по пе всегда.
Так, па 100 самок приходится самцов: у атлантической сельди
(Clupea harenguO 101, у бычка (Cottus gobioj—188, у вьюна (Misgur-
nus fossilis) — 11, у окупя (Perea fluviatilis) — 2—10. У камбалы
(Pleuronectes platessa) отношение между числами различных полов
меняется с возрастом. Сначала число самцов относится к числу,
Самок, как 6:4. С возрастом происходит сдвиг в пользу самок.
234
У совсем взрослых камбал известны лишь одни самки. Причина
не вполне ясна. И у других рыб, имеющих плавучую икру, самки
преобладают. Всплывая через слой воды, заполненный молоками,
икра оплодотворяется.
Размножение в общем, как правило, происходит ежегодно
с момента достижения половой зрелости. Но у миног (Petromyzon-
tidac), угрей (Anguilla), восточного лосося (Oncorhynchus) и сельди
черноспинки (Caspialosa kessleri) размножение, по всей видимости,
происходит один раз в жизни, и они, выметав икру, погибают.
Выше говорилось о плодовитости. Она зависит от
целого ряда факторов, прежде всего от вида рыбы. Число икринок
у разных рыб сильно вариирует. Кроме того, чем больше рыба
и чем старше она, тем больше обыкновенно у нее икринок. Плодо-
витость определяется также продолжительностью по-
ловой жизни. Рыба, живущая половой жизнью большее
количество лет, конечно, сможет при прочих равных условиях
дать и большее количество потомства.
Например, атлантическая сельдь (Clupea harengus) имеет боль-
шую продолжительность половой жизни. Атлантическая сельдь
встречается в возрасте и 14 лет. Вся же масса волжской сельди
(Caspialosa volgensis) падает на 3- и 4-летний возраст; пятилетки
являются редкими исключениями. Половая жизнь волжской сельди
продолжается всего три года, так как эта сельдь становится полово-
зрелой лишь в возрасте трех лет, редко двух. А черпоспипка
(С. kessleri) в массе созревает только в возрасте четырех лет, редко —
трех; пятилеток пе встречается вовсе. Таким образом продолжи-
тельность се половой жизни всего один год. Но это обстоятельство
компенсируется, во-первых, тем, что плодовитость разных видов
сельдей не только увеличивается по мере их роста, по при одина-
ковом возрасте бывает тем больше, чем крупнее форма.
Плодовитость увеличиваете^ еще от того, сколько раз
в году мечет данный вид икру. Волжские сельди,
папример, являются одними из наиболее плодовитых рыб благо-
даря тому, что они могут метать икру не в один, а в три приема,
разделенные но только во времени, ио и в пространстве. Это, ко-
нечно, значительно увеличивает шансы в борьбе за существование
и увеличивает плодовитость.
Внешние физические факторы также влияют на плодовитость.
Это очень важно для искусственного рыборазведения. Такими
факторами являются содержание в воде кислорода
и питание. В особенности последнее важно для развития
женских половых желез. Хорошее питание не только ускоряет
наступление половой зрелости, по и увеличивает индивидуальную
продукцию половых клеток. По и здесь должна быть мера. Чрез-
мерное питание ведет к дегенерации половых желез.
По созревании половых продуктов рыбы приступают к икро-
метанию, или нересту. Для этой цели опи отыскивают под-
ходящие места — нерестилища, которые иногда лежат
далеко от места их обитания. Время нереста для каждого вида
235
рыб зависит от ряда условий, и как его наступление, так и про-
должительность колеблются в довольно широких пределах. Обычно
по времени икрометания рыб делят на три категории: нере-
стующих зимой, весной и летом. У большинства
лососевых (Salmonidae), например, время икрометания приходится
па позднюю осень, перед ледоставом (зимний нерест). Благородный
лосось (Salmo salar) начинает у нас нерестовать с последних чисел
августа (молодые особи), кончая в конце октября, а в области
ключей, не покрывающихся льдом, и до конца ноября. Поздней
осенью мечет икру и речная форель (Salmo trutta morpha fario)
и др. Зимой мечут икру сиги (Coregonus). Лишь немногие сиговые,
например, хариусы (Thymallus thymallus), мечут икру весной. Боль-
шая часть карповых (Cyprinidae) мечет икру летом и более поздней
весной. Точно так же и морские рыбы. Так, например, из промы-
словых рыб, водящихся в русских водах, треска (Gadus callarias)
нерестится в финских водах с декабря или января по июнь или
июль, а па Мурмане — с начала января до начала июля, т. е.
опа пе может быть названа пи зимне-, пи весснне-, ни летненере-
стующей. У многих и весьма разнообразных групп проходных рыб
имеются две формы или расы, которые отличаются друг от друга
временами вхождения в реки для нереста. Эти расы называются
сезонпымн. Они обнаруживают аналогию с яровыми и озимыми
расами у хлебов.
Отношения полов во время периода икрометания обычно весьма
беспорядочны, особенно у тех костистых рыб, которые выметы-
вают икру и льют молоки, плавая стаями. Однако самка может
оплодотворяться несколькими самцами (полиандрия), или
самец оплодотворяет несколько самок (полигамия), а не-
которые рыбы, строящие гнездо, дают примеры спаривания одного
самца и одной самки (м о п о г а м и я). Но отношения между
двумя полами являются здесь лишь временными.
В тех случаях, когда несколько самцов следуют за одной сам-
кой, между ними могут происходить схватки, причем соперники
иногда рапят друг друга смертельно. Между самцом и самкой
происходят часто «л ю б о в и ы е и г р ы».
Различают внешнее и внутреннее оплодотворение.
В первом случае самка откладывает яйца в воду, куда и самец
льет сперму. Требуется в общем очень небольшое количество семени
для того, чтобы оплодотворить большое число яиц. Внешнее опло-
дотворение имеет место у всех рыб, кроме селяхий и немногих
костистых рыб. У самцов селяхий для оплодотворения самок служат
копулятивные органы. Копуляция, конечно, имеет
место и у живородящих костистых рыб, где яйца оплодотворяются,
как выше сказало, в яйцеводах или даже в яичниках, и молодь
родится живая.
У некоторых пресноводных рыб для откладки яиц служит то
место, где они обычно находятся; другие выискивают для этого
особые подходящие места, которые бывают у разных рыб весьма
различны. Одпи рыбы выбирают для нереста места мелкие, с теплой
236
водой или места, с определенными видами растений. Например,
лещ (Abranris brama), проводящий зиму в дельте Волги в ямах,
весной поднимается из ям и, скатываясь сначала в предустьевое
пространство, поднимается по рекам в низовые ильмени, где и мечет
икру па водных растениях. Другие, как жерех (Aspius aspius),
откладывают икру па быстром течении. Третьи избирают холодные,
быстрые воды горных речек, как, например, форель (Salmo trutta
inorphafario), хариусы(Thymallus) и др., и зарывают икру в песок;
у лосося, например, икринки падают на дио, у других (окунь —
Perea fluviatilis) яйца связаны в лентообразные массы или кучки,
прикрепляющиеся к подводным растениям и камням. У морских
рыб икра откладывается или в свободную воду (пелагическая)
или на дно (яйца демерсальные). Так как демерсальные яйца мы
встречаем главным образом у пресноводных рыб и у рыб прибреж-
ной зоны, относящихся к более примитивным и древним семействам,
и так как пловучесть яиц, кроме особых приспособлений, о которых
говорилось выше, зависит от плотности воды моря, а плотность
раньше должна была быть меньше, то можно думать, что и пела-
гических яиц раньше не было и демерсальные яйца более древние.
Отсюда можно сделать вывод: рыбы возникли в прес-
ной воде. Развитие пелагических яиц сделало возможным
жизнь и икрометание в открытом море.
Некоторые пелагические яйца, как яйца угря (Anguilla an-
guilla), откладываются па большой глубине (800—ООО м). Дру-
гие — в поверхностных слоях; яйца разных видов — па разной
глубине. Как было указано выше, в большинстве случаев плаваю-
щие яйца имеют меньший удельный вес вследствие наличия в пих
одной или нескольких масляных капель. Число и величина послед-
них облегчают распознавание вида. По распределению пелаги-
ческих яиц, которое определяется путем многочисленных уловов
планктонной сетью, при наличии яиц данного вида в уловах опре-
деляются, между прочим, ареалы икрометания, что
очепь важно как для охраны рыб, так и для рыболовства.
Поимка взрослых нерестующих форм часто очень трудна и тре-
бует много времени. Ловля планктонной сетью плавающих яиц,
наоборот, очень легка и дает достоверное сродство для определения
места икрометания, ибо большинство видов остается некоторое
время на место, где были выброшены яйца. Таким образом, в Нор-
вегии были найдены огромные косяки трески на некоторых отмелях,
где рапее не производилось лова, а затем стали ловить ос миллио-
нами штук.
На горизонтальное распределение пелагических яиц оказывают
сильное влияние морские течения, их направленно и быстрота,
а также температура, продолжительность развития и продолжи-
тельность пелагической жизни. От удельного веса яиц зависит
глубина, на какой яйца будут плавать. Эти данные необходимо
принимать во внимание при определении мест икрометания. В на-
стоящее время роль течений в распространении яиц и молоди тре-
сковых, камбал, угрей и сельдей — главных промысловых рыб
237
моря — деятельно изучается в некоторых местах, особенно в Нор-
вежском море. Значительная работа развернута в советских морях.
Оплодотворенная икра развивается в зависимости от различных
внешних условий и прежде всего от температуры и богатства воды
кислородом. Необходим для развития также определенный солевой
состав. Знание того, как зависит развитие от температуры, имеет
особое практическое значение при искусственном рыборазведении,
где рыборазводчик должен точно знать время вывода молоди.
Время вывода очень колеблется у разных видов: у карповых оно
в среднем равняется неделе, у лососевых, мечущих икру зимой,
2—3 месяцам. Для форели число дней, потребных для развития
икры при данной температуре, помноженное на число, обозначающее
последнюю, является величиной постоянной. Так, например:
При 2° икра развивается
» 5° » »
» 10° »'
в 205 дней (205 X 2 = 410)
» 82 дня (82 х 5 = 410)
» 41 день (41 х 10 = 470)
Точно так же более или менее постоянным является это число
и для других рыб: камбалы (Pleuronectes platessa) и трески (Gadus
callarias). Из яйца сельди развивается молодь в 9 дней при тем-
пературе 12° и в 24 дня при 4°. Можно вычислить, зная это число
и среднюю температуру любого участка моря, время развития
здесь икры и, имея перед собой выловленную икру в известной
стадии развития, сказать, в какое время она была отложена.
При недостатке свободного притока кислорода для дыхания
и при затрудненном удалении выделенной углекислоты икра гиб-
нет и не развивается. Это происходит, например, при промерзании:
у пресноводных при 0°, у морских рыб, живущих в соленой воде,
при более низкой температуре.
Зародыш развивается на питательном желтке и постепенно
обрастает его своими брюшными стенками. Часть желтка остается
для молодой рыбешки как резервный материал в виде желточ-
ного м е ш к а. В некоторых случаях желточный мешок бывает
очень велик. Форма его бывает различна. Рыбешки остаются не-
подвижными, пока желточный мешок по рассосется настолько, что
животные смогут производить собственные движения. У некоторых
рыб отличается весьма существенно молодь по своей организации
от взрослых родительских форм. В таком случае можно говорить
о личинках. Личинок мы видим у миног (Petromyzontidae),
угрей (Anguilla) и камбал (Pleuronectidae). Личинка мипоги —
пескоройка — отличается от мипоги не только по внешним
признакам, но и по внутренней организации. В таком состоянии
личинки живут от 3 до 5 лет, пока не наступает метаморфоза.
Личинки угря долгое время принимались за особый род (Leptoce-
phalus brevirostris), пока пе удалось проследить их превращение
в угря (рис. 198). Молодые камбалы также могут считаться личин-
ками. Они построены вполне симметрично, прозрачны, продолго-
ваты и ведут пелагический образ жизни. Лишь постепенно пере-
238
ходят рыбки к жизни иа дне, тело их уплощается, оба глаза оказы-
ваются па одной стороне.
Молодь многих морских рыб имеет особые приспособительные
образования для удержания иа поверхности воды. Иногда эти обра-
зования имеют филогенетическое значение, ука-
зывая па происхождение данного вида, подобно тому как симметрич-
ное строение личинок камбал указывает на их происхождение от
симметричных пела-
гических форм.
Пелагическая мо- •
лодь подобно планк-
тонным яйцам дер-
жится главный обра-
зом в поверхностных
слоях воды. Она боль-
шей частью прозрач-
на, что, по общерас-
п р о с т р а п о и и о м у
взгляду, защищает
молодь от врагов. О
возрастом развивает-
ся пигментация, и
рыбки опускаются в
более глубокие слои.
Для научно-прак-
тических целей- чрез-
вычайно важно уметь
Рис. 19Я. Личинки угри (Lcploccphali).
(Но lljorl).
различать молодь раз-
ных видов рыб. Она
часто очень сходпа,
поэтому и потребовалось много усилии, чтобы научиться различать
мальков разных рыб. Прилагаемый рисунок. .199 иллюстрирует,
как по распределению пигментных пятен можно отличить друг
от друга совершенно сходных между собой по внешнему виду ли-
чинок трески (Gadus callarias), сайды (Gadus v irons) и G. pollachius.
Остается сказать еще о своеобразных способах з а. щ и т ы с в о-
е г о и о т о м с т в а некоторыми рыбами или, как принято го-
ворить, о «з а б о т а. х о и о т о м с т в е у р ы б».
Обычно рыбы, отложив япца, оставляют их на произвол судьбы.
Некоторые рыбы, как л о с о с ь (Salmo salar), перед откладкой
яиц и их оплодотворением делают в песке и гравии ямку и наблю-
дают, чтобы ею не завладели другие самцы. По как только яйца
отложены и оплодотворены, родители оставляют их. В других
случаях яйца прикрываются песком и гравием. У малоподвижных
рыб, чаще пресноводных, чем морских, дело идет дальше. У В1еп-
nidae яйца, отложенные в расщелину скалы или в ямку на дне,
не оставляются самцом, который их и охраняет.
Дальнейшее усложнение инстинкта состоит в постройке гнезда.
Строит его почти всегда самец. Таков инстинкт у трехиглой ко-
239
люшки (Gasterosteus aculeatus). Вырыв на дне ямку, самец высти-
лает ее мелкой травой, устраивает боковые стенки и свод, склеивая
траву слизью тела. Гнездо имеет вид шара с двумя отверстиями.
После этого самец загоняет в гнездо одну за другой несколько
самок, поливает каждую порцию яиц молоками, а затем в течение
10—14 дней оберегает гнездо, пока вылупившаяся молодь не осво-
бодится от желточного мешка. Во время развития йиц самец про-
изводит при помощи усиленного движения грудными плавниками
обмен воды, способствуя лучшему дыханию. Так же поступает
пинагор (Cyclopterus lumpus),
у которого вылупившаяся мо-
лодь прикрепляется к телу отца
при помощи присосок, и самец
плавает вместе с пими.
Рис. 200. Личинки горчака (Rho-
deus amarus в жабрах Unio).
(Ив oit).
Рис. 199. Личинки трех видов
трески: Gadus callarias, G.
virens, G. pollachius.
(По Schmidt из Hjort).
Дальнейшее усложнение явления заключается в развитии на
теле родителя особых мест для прикрепления яиц. Так, у сомика
(Aspredo) яйца прикрепляются к брюшной стороне тела на стебель-
ках, снабженных кровеносными сосудами, и покрываются при этом
эпителием. Такого же рода приспособления мы наблюдаем у иглы-
рыбы (Syngnathus acus) и у морского конька (Hippocampus). У мор-
ской иглы яйца помещаются па нпжней стороне хвоста, между
двумя складками кожи. У конька эти складки замыкаются в мешок,
имеющий впереди отверстие. Яйца, которыми мешок заполняется,
откладываются несколькими самками в течение нескольких дней.
Потом мешок замыкается, его стенки становятся губчатыми и бо-
гато снабженными кровью. Вышедшая по развитии из мешка мо-
лодь назад не возвращается (рис. 200). У некоторых сомовых
(Siluridae) и в семействе Cichlidae икра развивается во рту одного
из родителей. У Paratilapia из Cichlidae мать водит за собой мо-
лодь, пряча се при опасности в рот. У Chromis pater-familias самец
прячет в рот до 200 эмбрионов.
Замечательное явление в связи с размножением наблюдается
у горчака (Rhodens amarus). Длинным яйцекладом самка откла-
дывает яйца в полости мантии двустворчатого моллюска (Unio или
Anodonta). Здесь яйца оплодотворяются сперматозоидами, про-
240
пикающими с водой через вводной сифон, проходят цикл разви-
тия и выходят, достигнув 10 мм длины (рис. 201).
«Забота о потомстве» встречается опять-таки весьма спорадп-
чески и вариирует у близких видов. Так, один из видов бычков
«заботится» о яйцах, другие не обращают на них внимания; одни
из сомов не делают гнезда, другие — делают; одни берут яйца в рот,
у других они прикрепляются к телу.
Рис. 201. Пинагор (Cyclopterus lumpus) с молодью.
7. Возраст и рост рыб. Продолжительность жизни
Вопросы о возрасте и росте рыб имеют, кроме большого теоре-
тического интереса, громадное практическое значение. Результаты
изучения этих явлений особенно наглядно показывают то значение,
какое имеют научные исследования для хозяйства, в данном случае
для рыболовства. Явления возраста и роста, а также связанный
с этими явлениями вопрос о миграциях (странствованиях)
рыб за последние 25 лет изучаются со все возрастающей энергией
как отдельными учеными, так и специальными научными учре-
ждениями: Международной комиссией по изучению морей, много-
численными научно-промысловыми экспедициями, ихтиологиче-
ски ми лабораториями. О значении для практики рыболовства
исследований возраста и роста можно судить, например, по данным
о возрасте, росте и лове леща (Abramis brama).
Лещ является одной из важнейших промысловых рыб Каспий-
ско-Волжского бассейна. Правильно ли производится его вылов,
16 Кашкаров и Стапчинский * 241
не ведет ли последний к уменьшению запасов леща в море? Для
ответа на этот вопрос необходимо познакомиться с возрастным
составом вылавливаемого леща и с характером роста последнего.
Оказывается, что состав тоневых уловов леща по возрасту в сред-
нем таков (в Каспийском районе):
Возраст . ............1г. 2 г. 3 г. 4 г. 5 лет и старше.
Процентное отношение ... 5.5 67.5 23 3.4 0.3
Иначе говоря, лещ вылавливается
В прилагаемом графике сопоставлена интенсивность прироста
разных поколений
юоо юо
весьма энергично.
с
процентом попадания их в топевых уловах.
Кривая изображает прирост массы тела
леща в процентах к первоначальному сред-
нему весу тела данного поколения, а ве-
личина столбцов показывает процентный
состав неводных уловов в разных возраст-
ных группах. Из графика видно, что воз-
растные группы, дающие Наибольший при-
рост в весе, почти не попадают в улов.
В течение трех кормных периодов лещ
откармливается свободно. Когда же он,
достигнув половой зрелости, устремляется
па места нереста, более двух третей его
(67.5%) попадает в сети. Проскочившая
п 12/&до
Рис. 202. Диаграмма сопоставлений интенсивности прироста тела
разных поколений леща (кривая) с процентом попадания их в
тоневых уловах (столбики).
треть попадает в улов па следующий год. Рыб старше пятилет-
него возраста почти пе имеется среди запасов леща. Эти рыбы,
поедая много пищи и мало увеличиваясь в весе, не оправдывали
бы своего существования. Таким образом, современный лов леща
с выловом форм, закончивших третий сезон питания, в основе
поставлен правильно с точки зрения наиболее продуктивного ис-
пользования корма. Но, с точки зрения коммерческой, имеется
полный расчет перейти к эксплоатации леща на четвертом году
его жизни, ибо, во-первых, прирост в течение четвертого корм-
ного сезона достигает 58% первоначального веса и, во-вторых,
стоимость крупного леща па 30—40% выше, т. е. цена прироста
на четвертом году возрастает па сто с лишпим процентов.
• Изучение темпа роста различных рас одного и того же вида
дает возможность судить как о достоинствах каждой расы, так
и о степени «кормности», или «продуктивности», водоемов, в
которых эти расы живут.
242
Методы определения возраста и темпа роста разработаны в на-
стоящее время хорошо. Первые попытки определения возраста рыб
были сделаны, исходя из их величины У бельдюги живородящей
(Zoarces viviparus) сеголетки имеют размер 8—10 см, молодь
в возрасте 1г/2 лет имеет длину 16—20 см, в возрасте 2% лет—
от 25 до 30 см. Но при помощи этого метода трудно определить
возраст более старых групп; кроме того, один и тот ясе вид растет
иначе в различных условиях.
Возможность определить возраст каждой особи и ее особый
тип роста дает нам другой метод: по кольцевым наслоениям на че-
шуе, костях и отолитах. Выли найдены возрастные кольца вна-
чале на чешуе карпа, а затем на
отолитах палтуса, подобные тем,
которые наблюдаются на стволах
деревьев или на рогах рогатого
скота.
Рост рыбы идет в зависимости
от ее питания. В период ухудше-
ния питания, т. е. зимой, кость
растет слабее, и в кости отлагается
меньше извести. Весной и летом
получается широкое поле нара-
стания, богато пропитанное изве-
стью и резко отделенное прозрач-
ной узкой зимней полоской от
широкого поля нарастания преды-
дущего года. Просчитывая число
Рис. 203. Чешуя сайды (Gadus
virens) в различных возрастах.
зимних колец, мы получаем
число зим, прожитых рыбой. Можпо брать для определения воз-
раста чешую. У других рыб более удобными для определения явля-
ются кости скелета, например, кость плечевого пояса — cleithrum,
жаберная крышка — operculum — или позвонки; наконец, у
третьих — отолиты. Иногда можпо различить годовые слои па
чепгуях, только что вынутых из тела рыб. Этот признак очепь
ценен в прудовом хозяйстве, где валено бывает определить воз-
раст посадочного материала, покупаемого па стороне. Обычно
же приходится, чешую особым образом обрабатывать, чтобы
явственно видеть кольца. Кроме колец зимпих, у некоторых
рыб образуются еще дополнительные кольца, зависящие от пере-
житых неблагоприятных для роста условий.
Дело опыта — уметь отличить эти второстепенные кольца от
годовых. При рассматривании отолитов при падающем свете мы
видим белые кольца, которые соответствуют весеннему периоду
роста, и темные, соответствующие летнему и осеннему. Зимой
отолиты пе растут. Таким образом два кольца— белое и темное —
соответствуют году. Определение возраста по отолитам не всегда
удобно и возможно. Самым падежным, по кропотливым способом
определения возраста рыб является определение его по костям.
В настоящее время мы с точностью можем указать возраст
46*
243
почти всех промысловых рыб: трески (Gadus callarias), пикши
(Gadus acglifinus), сайды (Gadus virens), сельдей, как норвежских
(Clupea harcngus), так и каспийских (Caspialosa), шпрота (Clupea
spratus), камбал (Pleuronectidae), лосося (Salmo salar), осетровых
(Acipcnseridae), карповых (Cyprinidae), как вобла, лещ, сазан и др.
На рис. 203 мы видим ряд чешуй сайды, изображенных пропорцио-
нально величине рыбы. Ясно видно, как количество годовых колец
возрастает с ростом рыбы.
Изучая возраст всех особей в определенном улове, мы можем
получить представление о возрастном составе каждого улова.
Исследуя большое количество уловов, мы можем составить понятие
о возрастном составе дан-
ной породы рыб. На
рис. 204 мы видим резуль-
тат анализа возрастного
состава одного улова сай-
ды. Возрастные группы,
слабо представленные, опу-
щены вовсе. Наиболее мно-
гочисленна группа четы-
рехлеток (IV) со средним
ростом в 54 см. Высота
® кривой пропорциональна
24.
Рис. 204. Возрастной состав косяка сайды, количеству особей данного
возраста в косяке.
Умея определять возраст рыб, мы можем нарисовать историю
их жизни. Оказалось, между прочим, что рост рыбы находится
в большой зависимости от температуры. Удалось установить, что
половая зрелость у многих рыб наступает в разном возрасте в'за-
висимости от местных условий. Например, рост трески идет тем
медленнее, чем севернее, и тем позднее наступает половая зрелость.
О различном росте камбалы (Pleuronectes platessa) из Бал-
тийского и из Немецкого морей говорилось выше.
Таким образом, окружающие условия, местные особенности
среды, температура лета и его продолжительность — все это кладет
определенную печать па организм рыбы и, в частности, па ее
чешую или кость. По чешуе или кости можпо определить историю
роста отдельных ипдивидов, определить место или по крайней мере
условия, в которых опи выросли. Дальше мы увидим, что это дает
ключ к пониманию странствований, или миграций, рыб. Много-
численные исследования показали, что возрастной состав неполово-
зрелых сельдей и косяков мечущей икру сельди значительно раз-
нится из года в год: одни возрасты представлены в косяках обильно,
другие — бедпо. Существуют, следовательно хорошие и плохие
годы, обусловливающие богатство или бедность годичного класса.
Неблагоприятные условия особенно*влияют па более ранние фазы
жизни рыбы. Рыбы, появляющиеся па свет в избытке, гибнут
главным образом па самых ранних стадиях жизни. Поэтому, если
в уловах изобилует какой-либо более взрослый класс сельди, то
244
это поколение будет многочисленно в течение ряда лет. Таким
образом оказывается возможным на основании изучения возрастного
состава косяков в текущем году предсказать состав их (а, следова-
тельно, и хорошие или плохие уловы) в будущем и в последующие
годы. Это, конечно, чрезвычайно важно для регулирования промы-
сла рыбы, особенно там, где она имеет первенствующее значение.
Приводимая таблица дает представление о только что сказан-
ном. Цифры, указывающие процент попадания в уловах наи-
более многочисленного класса, напечатаны курсивом. Видно,
что если какой-либо возраст преобладает в уловах данного года,
то в следующем году будут преобладать особи па один год стар-
ше, т. е. то же самое поколение перейдет и в следующий год в
наибольшем числе. И обратно.
Год Годичные классы;(возраст)
1 г. | 2 г. | 3 г. 1 4 г- 1 I 5 л. | 6 л. | 7 л.
1907 11.5 36.8 61.3 0.4
1908 0.4 51.4 10.3 37.8 — — —
1909 3.1 61.0 13.3 5.0 16.9 0.7 0.2
1910 0.2 50.7 42.0 0.9 1.8 4.5 0.1
По чешуям и костям можно определить не только возраст рыбы,
но также измерить и величину годового прироста рыбы. Кость
с ее годичными наслоениями зарисовывается при помощи рисо-
вального аппарата. Путем наложения соответственного масштаба
измеряется величина этих годичных наслоений. Отношение между
длиной тела и величиной кости, которая берется (например clei-
thrum) для определения возраста и темпа роста рыбы, несколько
неодинаково для рыб разной величины, по изменяется оно законо-
мерно. Длина тела рыбы, разделенная на длину cleithrum, так
называемый знаменатель отношения, есть величина более или
менее постоянная, несколько большая в течение первых двух лет
жизни, чем в последующие. Это видно из следующих цифр:
для сеголеток воблы =16.20
Z (длина тела)
I (длина cleithrum)
» годовиков =15.45
» 2-леток =14.83
» 3 » =14.52
» 4 » =14.48
» 5 » = 14.45
» 6 » = 14.40
Из уравнения т = z мы можем по двум данным находить третье.
V
Ксли мы знаем длину cleithrum и знаменатель отношения для
данной рыбы, то мы можем узнать и длину тела рыбы: L = Ьг.
Нто даст нам возможность по величине наслоений на cleithrum
вычислить рост и прирост каждой рыбы в течение первого, второго
и следующих лет (рис. 205). Данная формула нуждается в поправ-
ках н становится более сложной. По это мы оставим здесь без рас-
245
смотрения, как и технику зарисовки cleithrum и методы опреде-
ления темпа роста по чешуе, тщательно разработанные.
Пользуясь результатами этих методов, мы можем проследить
год за годом прирост веса каждой рыбы в отдельности, что часто
бывает очень важно — папример, при определении продуктив-
ности водоема и для других вопросов (см. выше). Так, устанавли-
вается, папример, отсталость роста воблы в северо-восточной части
Каспия в сравнении с воблой юго-западной, зависящая, видимо,
от характера питания.
Изучение темпа роста дает возможность оценить влияние пере-
лова или недолова на продуктивность бассейна. Человек является
— _______—главным фактором, ведущим к
Z'~ истреблению очень многих ви-
\ ) Д°в животных, в том числе и
Л А ].] ( рыб. Вылов может быть так
W проведен, что водоемы совер-
У / / 2 шепно лишаются рыбного насе-
ления. Но промысел не всегда
Рис. 205. Cleithrum леща. 1а и производит ОДНО опустошение,
т. ц. обозначают годичный прирост. Как в сельском хозяйстве по-
лезно бывает прореживание
растений или в охотничьем — отстрел, так и вылов до известных
пределов может быть полезен. С этой точки зрения, изучение темпа
роста рыбы в данном водоеме в настоящее время и в прежние годы
может дать указание на роль вылова, понижающего конкурен-
цию между отдельными поколениями.
До какого возраста доживает рыба? Относительно рыб на этот
вопрос легче ответить, чем для многих других животных, именно
потому, что па самом организме рыбы, на ее костях записываются
прожитые ею годы. Вообще говоря, рыбы могут жить очень долго.
Это могло быть устаповлепо еще до открытия способа определения
возраста, именно: посредством мечения рыб кольцами с надписью.
Этим путем удалось установить, что карпы и щуки живут более
200 лет. Одна щука была пущена с меткой в Германии в 1230 г.,
а поймана снова в 1497 г., т. е. прожила более 267 лет.
Камбалы (Pleuronectes) из Арктики могут жить, судя по костям,
по крайней мере 60 лет, возможно и дольше. Камбалы же из Не-
мецкого моря редко имеют шансы дожить более чем до 30 лет.
Треска (Gadus callarias) достигает возраста 15 лет, лещ (Abrainis
brama) в Каспийско-Волжском бассейне достигает почти 14 лет;
вобла (Rutilus rutilus caspicus), там же — 8 лет. Aphia minuta
(Gobiidae) и мелкие Scopclidae в Средиземном море живут только
один год.
Что ставит предел жизни рыб? Иногда — ослабление икроме-
танием и далекие миграции (Gadus, Anguilla); иногда, как у неко-
торых лососевых (Oncorhynchus), искривляется рот, делая невоз-
можным питание; иногда, вероятно, наступает старость, как у дру-
гих животных; часто рыба не доживает до естественной смерти
вследствие вылова.
246
8. Изменчивость и наследственность. Образование рас
Материал для эволюции, для выработки новых приспособитель-
ных типов дает изменчивость. Изменчивость дает разнообразные
формы, уклоняющиеся от родительской формы и служащие мате-
риалом для отбора. Отбор происходит со стороны факторов среды;
так, создаются сперва более мелкие систематические единицы,
пока между ними происходит еще скрещивание, потом виды, когда
скрещивание между ними перестает иметь место.
Изменчивость рыб чрезвычайно велика. Мы видим у рыб те же
виды изменчивости, что и у других животных: индивидуальную
ненаследственную изменчивость, или модификации, и на-
следственную — мутации, а если изменение возникает в ре-
зультате скрещивания, то комбинации.
Модификации могут касаться самых разнообразных признаков:
размеров тела, отдельных его частей, числа лучей, числа позвонков,
числа поперечных пятен и т. д. Одни признаки могут быть сосчи-
таны (число позвонков, лучей) и остаются постоянными всю жизнь;
другие могут быть измерены и вариируют с возрастом и полом.
Поэтому при пользовании ими всегда следует соблюдать осторож-
ность и вводить поправки. Так как признаки вариируют, то по
одной особи никак нельзя определить вид или подвид, племя или
расу. Необходимо статистическое изучение. Так, папример, число
чепгуй в боковой линии, играющее столь большую роль в характе-
ристике вида, подвержено довольно широким индивидуальным
колебаниям: • у рыбца (Abramis vimba) число чешуй в боковой
линии от 50 до 60, у густеры (Blicca bjorkna) — от 45 до 49, у жереха
(Aspius тарах) — от 65 до 71, у чехони (Pelecus cultratus) — от 108
до 115 и т. д. Колеблется число лучей в плавниках и число боковых
жучек у осетровых и т. д. Выло бы ошибкой принять число чешуй
у рыбца равным 58, если бы у пас был один экземпляр с таким
числом их. Необходимо в систематической работе считаться с индиви-
дуальной изменчивостью и при изучении се работать с большим
числом особей, беря средние числа. Полученные от измерения
какого-либо признака у ряда особей числа колеблются около не-
которой средней величины, образуя вариационный ряд.
Например, вариационный ряд для числа позвонков у угря (Anguilla
vulgaris) будет таков:
Число поввонков .... 110 111 112 113 114 115 116 117 118 119
Число особей ив 2 725 . . 1 17 89 352 719 865 500 192 36 4
Этот ряд может быть выражен в виде кривой или диа-
грамм ы, где на горизонтали (абсциссе) нанесены размеры
признака (число позвонков), а па перпендикуляре (ординате) —
число особей, у которых данный размер признака встречается в про-
центах к общему числу.
Из приведенного ряда видно, что наиболее частое число позвон-
ков 115. Это число позвонков ближе всего к средней величине.
Характеризовать вид или расу будут, конечно, не отдельные ве-
247
личины признака, а средняя величина, от которой колеблются
индивидуальные изменения.
При анализе вариационного ряда приходится пользоваться
следующими понятиями. Размер признака, или член вариацион-
ного ряда, называется вариантой. Число особей, у которых
данная варианта встречается, называется его частотой.
Та варианта (или класс, если число обозначает размеры при-
знака не у особи, а у группы особей), которая имеет наибольшую
частоту, называется модой (в нашем случае — 115).
Наиболее простой способ вычислить среднюю величину таков.
Вычисляются все уклонения от моды в положительную и отрица-
тельную сторону, сумма всех этих уклонений делится на число
членов ряда, и тогда получается среднее уклонение
(в нашем случае — 0.27). Чтобы получить среднюю вели-
чину признака, следует прибавить к моде это среднее уклонение.
В пашем примере это будет 114.73. Если взять теперь сумму квад-
ратов всех уклонений, умноженных па соответствующую частоту,
разделить это произведение на число членов ряда, вычесть ква-
драт среднего уклонения и извлечь из полученного квадратный
корень, то получим среднее квадратическое укло-
нение ряда. Сумма квадратов уклонений от средней величи-
ны является величиной наименьшей, т. е. она меньше такой же
суммы уклонений от любой другой, не средней величины.
В нашем случае мы будем, таким образом, иметь:
Произведение Произведение
Варианты Частота f Уклонения х частоты на уклонение ж/* квадрата уклонения на частоту
110 1 — 5 — 5 - 25
111 17 > —4 — 68 272
112 89 — 3 — 267 801
113 352 — 2 — 704 1 408
114 719 — 1 — 719 719
115 865 — 0 0 —
116 500 + 1 + 500 500
117 192 + 2 + 384 76»
118 36 + 3 + 108 324
119 4 + '* + 16 64
П ряда) = 2 775 S (*/) = - 755 2 («V) = 4 881
у <xf\
d (среднее уклонение) = ' 11 = — 0.27.
М (средняя величина) = 114.73
Мо (мода) = 115
а (квадратическое уклонение) = ±1/ — —d2= ±1/ —0.07= ±1.30
При помощи среднего квадратического уклонения имеется
возможность определить принадлежность любой особи к соответ-
ствующей расе, когда принадлежность эту нельзя определить обыч-
ным способом. В ы з ы в а ю щая в нас сомнение особь
248
принадлежит к той расе, по отношению
к средним величинам которой сумма квад-
ратов уклонений всех еепризнаков являет-
ся наименьшей. Если перед нами находится сельдь из
Белого моря с числом позвонков 58, то по этому признаку опа
может быть норвежской весенней сельдью, осенней сельдью с ют-
ландской банки или беломорской сельдью. Вычислив сумму квад-
ратических уклонений большого числа признаков этой особи от
средних величин каждой из этих рас мы видим, что она окажется
суммой наименьшей для беломорской сельди, а, следовательно,
к этой расе она и принадлежит.
Примеров изменчивости, подобных приведенному, у рыб можно
пайти сколько угодно. Их изучение методами вариационной ста-
тистики весьма важно, так как, изучая признаки и группы призна-
ков таким образом, возможно отличать местные формы — подвиды
(subspecies), экотипы или расы, которых по отдельным особям
нельзя отличить (трансгрессивная изменчивость).
Изучение «экотипов», групп, па которые распадается вид, групп,
члены которых характеризуются всеми признаками своего вида,
но в то же время обладают общими данной группе признаками
приспособительного к условиям, в которых опи живут, характера,
подводит нас к изучению процесса .видообразования. Среди рыб
мы имеем целый ряд изученных примеров таких экотипов.
Так, папример, в озере Севан так называемая гокчипская форель
(Salmo ischchan) распалась на ряд рас, отличающихся между собой
местом обитания (озеро, река и т. д.), характером питания; местом
и временем нерестования; по морфологически отличимы эти расы
лишь при помощи биометрического метода.
Биометрический метод применяется в ихтиологии в двух слу-
чаях: когда падо установить, существуют ли между теми или дру-
гими рыбами расовые отличия, и когда требуется определить, к ка-
кой из рапсе известных рас относится данная рыба. Как па пример
можно указать па работу по систематике русских корюшек (род
Osmerus). Род этот широко распространен, причем одни виды
живут в морс, другие — в озерах. Род распадается па целый ряд
видов и подвидов. Систематические признаки, па основе которых
строилась их систематика, все трансгрессивны, и пи один из них
пе даст возможности точно отграничить одну форму от другой.
К ним был применен биометрический метод, и выяснилось, что все
корюшки могут быть разбиты па две группы, в пределах каждой
из которых они образуют местные подгруппы, местные формы;
последние более или менее четко характеризуются биометрическими
данными. Биометрическим же анализом удалось установить, что
мурманская сельдь тождественна с норвежской сельдью, и Мурман-
ское побережье является таким образом одним из дальних этапов
описываемых ниже миграций норвежских сельдей на север, воз-
вращающихся затем для икрометания в норвежские воды.
Биометрическим методом можпо уловить отличия у рыб одного
вида, но живущих в различных условиях обитания, т. е. возможно
249
установить начальные стадии видообразования, связанные с раз-
личиями в условиях местообитания, — упомянутые выше «эко-
типы», формы, характерные для данных экологических условий.
Так, папример, изучение биометрическим методом маринки вида
Schizothorax intermedins, нормально живущей в быстрых горных
речках Средней Азии и оказавшейся благодаря геологическим
процессам запертой в глубоком горном озере, показало, что здесь
опа преобразовалась в особую расу, утратившую в некоторой мере
черты обитателя быстрых потоков.
Исследование вариаций, в особенности числа лучей в брюшпых
плавниках, у североамериканской рыбки Leuciscus balteatus по-
казало, что мода различна у рыб, взятых из различных место-
обитаний. Например, у рыбок, пойманных почти на уровне моря,
мода числа лучей была 19; у рыбок, пойманных на высоте
396 м над уровнем моря, ода равнялась 17, а на высоте
179 м— 16.
Исследование вариациопнйм методом ряда признаков у бель-
дюги живородящей (Zoarces viviparus), а именно — суммы позвон-
ков, числа колючек в заднем отделе спинного плавника, числа
темных полос па передней части спинного плавника и числа лучей
грудного плавника, показало, что в направлении от берега в глубь
фиордов заметно сильное уменьшение числа позвонков, числа
колючек спинного плавника и числа пигментных полос. Наоборот,
число лучей грудного плавника слегка увеличилось. Таким образом,
обнаружилось существование явственно и определенно направлен-
ных изменений при проникновении вида в воды защищенные,
бедные солью. Впрочем, нельзя утверждать, что именно различия
в солености вызвали эти морфологические отличия: простое сосу-
ществование двух явлепий пе доказывает еще их причинной связи.
Вообще биометрический метод требует осторожного к себе отно-
шения и учета биологических моментов.
Все подобные явления изменчивости требуют тщательного
изучения для выяснения здесь зависимости между серией
изменений в признаках и вариациями в условиях окружающей
среды. Тогда станут ясны роль наследственности и роль среды
в процессе создания форм.
Указанные примеры представляют явления групповой
изменчивости. Групповая изменчивость может быть наслед-
ственной и пе наследстве п ц ой. Последняя ведет
к образованию так называемых морф. Хорошим примером морфы
могут служить форели.
Потомство лососей, приходящих из моря метать икру в реки,
в том случае, если оно остается жить в пресных водах — в реках,
ручьях или озерах, — превращается в карликовые формы —
форели.
Так, кумжа (Salmo trutta) дает морфы lacustris в озерах Европы
и Кавказа и морфы fario в речках; голец (Salvelinus alpinus) на
Дальнем Востоке дает свою форель — Salvelinus alpinus morpha
pluvialis, и т. д.
250
После перемещения потомства морф в «нормальные» условия
они приобретают нормальный вид: потомство форелей, полу-
чившее возможность выйти в море для окончания своего развития,
превращается в кумжу.
Наследственная групповая изменчивость
ведет к образованию мелких систематических единиц (экотипов,
подвидов), а потом к образованию видов, родов и других высших
систематических единиц.
Встречаются у рыб и мутации. Среди камбал (Pleuro-
nectidae) несимметрично построенных животных, мы встречаем
такие виды, у которых глаза находятся на правой стороне, и таких,
где глаза — на левой стороне. Однако у Pleuronectes flesus, нор-
мально являющейся правосторонней, весьма обыкновенны экзем-
пляры левосторонние, с глазами и окрашенностью на левой стороне.
Положение же внутренних органов и другие признаки совершенно
нормальны. У камбаловых нижняя сторона пе окрашена. Но встре-
чаются случаи частичной или полной окраски нижней стороны.
Также встречаются случаи, когда на верхней стороне имеются
большие или меньшие области неокрашенные. Нормально чешуй-
чатый покров сильнее развит на верхней стороне, но бывают экзем-
пляры о одинаково сильно развитыми чешуями на обеих сторонах.
Объяснить эти случаи извращения нормы действием света, которым
объясняются нормальная окрашенность верхней стороны и не-
окрашенность нижней, — невозможно. Здесь может иметь место
лишь мутация, основанная на изменении зародышевой плазмы,
на изменениях в яйце.
Немало мутаций известно нам среди одомашненных рыб. Правда,
возникновение этих уклонений пе было изучено новейшими мето-
дами, но все обстоятельства говорят, что перед нами — мутации.
Авторитетными специалистами считается, например, что золотые
рыбки (Carassius auratus) явились результатом разведения такой
мутации обыкновенного карася (Carassius carassius); что среди
золотых рыбок так называемые телескопы и вуалехвосты также
являются мутациями, возникшими под влиянием неблагоприятных
условий разведения в аквариумах, папример, вследствие недостатка
кислорода. Так называемые «зеркальные» карпы с чрезвычайно
крупной чешуей, не покрывающей всего тола, и «кожистые» карпы
с голой кожей тоже являются примерами форм, возникших путем
мутаций. Среди линей (Tinea tinea) и вьюнов (Misgurnus fossilis)
иногда попадаются золотые экземпляры, подобные золотой рыбке
по окраске. Эти случаи следует принимать тоже за мутации. Му-
тацией такого же рода является и орфа, имеющая тоже золотую
окраску (Leuciscus idus).
Наконец, среди рыб, как и среди других животных, существует
третий род изменчивости— комбинации, возникающие в ре-
зультите скрещивания различных пород. Мы знаем теперь, что
скрещивание с получением плодовитого потомства происходит даже
между многими различными видами. Среди рыб это самое обыкно-
венное явление. Мы знаем многочисленные случаи образования
251
в природе п о м е с е й, или бастардов, среди осетровых
(Acipenscridae), среди лососевых (Salmonidae), камбал (Pleuro-
nectidae), морских окуней (Serranidae), многих карповых (Cypri-
nidac), окуневых (Percidae). Так, например, окунь (Perea fluvia-
tilis) может быть скрещен с ершом (Acerina cornua), причем резуль-
таты получаются иные в зависимости от того, от какого вида берем
мы самку и от какого вида — самца.
Потомство первого поколения, получающегося при межвидовой
гибридизации, является чрезвычайно разнообразным. В природе
скрещивание различных видов происходит с большой легкостью:
оплодотворение у рыб большей частью наружное, различные виды
соединяются в большие косяки, если время икрометания у них
совпадает. Известны случаи образования межвидовых гибридов.
В высокой степени интересными оказались в промышленном
отношении помеси между расами карпа, которые обнаруживают
пе только плодовитость, но и более быстрый темп роста, бблыпую
мясистость и т. д. Существует несколько различных рас карпа,
и, образуя между ними помеси, можно от той и другой расы полу-
чить лишь ее достоинства, удалив недостатки.
Бастарды ерша и окуня также оказались выше своих родитель-
ских форм в отношении изменчивости, быстроты роста и плодо-
витости.
Возможность бастардирования у рыб является весьма важной
для рыборазведения, для получения новых пород с лучшим э к с-
терьеромит. д.
9. Миграции рыб
Условия существования рыбы по являются чем-то неизменным
и могут меняться самым различным образом. Эти изменения окру-
жающей обстановки в связи с изменениями, происходящими в самом
животном организме в зависимости от возраста, созревания половых
продуктов и т. п., и являются причиной того явления, которое
носит название миграций р ы б и имеет огромное практи-
ческое значение.
Миграции можно разделить па пассивны с, вызываемые,
например, течениями, и па а к т и в и ы е, когда животные пере-
селяются, отыскивая для себя лучшие, более подходящие условия.
Самое большое значение имеют те миграции, которые связаны
с р а з м и о ж е и и е м, с нерестом (нерестовые), и м и-
г рации молоди (личиночные). По, кроме этих миграций,
существуют еще сезонные периодические м и г р а-
ц и и, куда относятся переселения рыбы на места зимовки, из
глубин па поверхность и па мелкие места, а также короткие или
долгие странствования вверх и вниз по реке, — одним словом,
миграции, обусловленные изменениями температуры, отыскиванием
пищи и т. д.
Примитивные рыбы или являются пресноводными или мигри-
руют в пресную воду для размножения (Lampetra, Chondrostei).
252
Акулы имеют демерсальные прикрепляющиеся яйца, и их палеозой-
ские предки были или пресноводными или посещали пресные
воды.
Демерсальные яйца имеются у многих Clupeidae, у лососей
(Salmonidae), у осетровых (Acipenseridae). Пелагические яйца
имеются главным образом у морских рыб. Развитие пелагических
яиц сделало возможными жизпь и икрометание в открытом море.
Личинки (правильнее — молодь) рыб развились также в разных
направлениях. Одни, как личинки тихоокеанского лосося, плывут
сотни миль вниз по реке, чтобы достичь моря; другие, как личинки
лосося атлантического, опускаются на дпо, прячутся под камни,
чтобы пе быть унесенными течением, или зарываются для этого
в песок, прикрепляются и т. д. Третьи не сопротивляются течению,
плывут пассивно и уносятся ипогда очепь далеко от места вывода
из яиц. Таким образом, и личинки могут быть подразделены на
пелагические, переносимые течениями, и па погружающиеся.
Там, где течений пет или где усилия рыбы бороться с ними
увенчиваются успехом, там места икрометания и места кормлепия
молоди почти или вполне совпадают. В большинстве же случаев
такое совпадение бывает редко, оба эти места лежат далеко друг
от друга, и рыбам приходится совершать большие переселения.
Изучение миграций даже главнейших промысловых рыб требует
громадной работы в различных направлениях. Прежде всего не-
обходимо уметь отличить данный вид па каждой стадии его раз-
вития, отличить яйцо, личинку, молодь одного вида от другого,
для чего собираются й изучаются обширные коллекции. Для опре-
деления рас, которые па осиованиии обычных морфологических
методов неразличимы, выработаны особые математические методы,
о которых говорилось выше.
Изучение физических и химических свойств морской воды,
ее температуры и течений также оказывает домощь в изучении
странствования рыб, в особенности важно оно для понимания путей
странствования более молодых классов рьйы, которые плавают
пассивно.
Самое изучение миграций может производиться различными
методами. Весьма точный и верный из них — это «мечение» рыбы,
когда тем или иным способом к пойманной рыбе прикрепляют
металлическое кольцо или пластинку с обозначением места и числа,
когда рыба мечена, и затем рыбу выпускают. Метить воз моле но
лишь более крупные экземпляры, и это было сделано в большом
объеме и с превосходными результатами с камбалами у берегов
Исландии и с лососевыми в Аляске.
В некоторых случаях прекрасным методом изучения миграций
являются ловля планктонных яиц, личинок, молоди и рыб па раз-
ных стадиях их развития и сопоставление всех этих данных. Таким
меточом были изучены в 1901 г. нерестилища трески в северной
Норвегии. Выло сделано большое количество топей во всех на-
правлениях между банками и добыто большое количество яиц.
Считая эти яйца после каждой тони, нашли, что тоня продолжи-
253
телытостыо в 5 минут, сделанная при помощи маленькой шелковой
сетки в 1 м в диаметре, давала тысячи яиц, плававших над этими
банками на глубине 60—80 м, где рыба метала икру. Раз было
найдено нерестилище, оно было использовано и дало не менее
полутора миллиона экземпляров трески. Этим же методом была
выяснена история странствования угрей.
Точно так же можно определять пути странствования, отмечая
различия в величине рыб разных пунктов.
Переселения, связанные с размножением, с отыскиванием места
нереста, бывают раз-
личного типа.
Крайние типы пред-
ставляют миграции угря
и лосося. Первый, дос-
тигая зрелости, скаты-
вается вниз по рекам
и плывет против Гольф-
штрема, чтобы достичь
места икрометания сре-
ди океана; второй вхо-
дит по достижении по-
ловой зрелости в реки
и идет для икрометания
вверх по последним.
Первый тип миграции
носит название к а т а д-
Рис. 206. Места нереста европейского и
американского угрей.
ромальной ми-
грации, второй — ападромальной миграции.
Явления миграций стапут нам более ясными из рассмотрения
отдельных типичных случаев катадромальпой и ападромальной
миграций.
Катадромальпая миграция угрей (Anguilla
anguilla). Вхождение молодых угрей в реки весной и скатывание
в море «серебристых угрей» осенью были известны давно. Лишь
недавно было доказапо, что так называемые т_о н к о г о л о в ы е
(Leptocephali), считавшиеся за отдельный род рыб со многими ви-
дами, являются в действительности личинками угрей, в которых
они превращаются. Местами размножения угрей предполагали
глубины Атлантического океана.
В настоящее время работами норвежских и датских ученых
тайпа угря разгадана, и миграции угрей представляются в таком
свете.
Существуют два вида угрей: европейский угорь — Anguilla
anguilla и американский— Anguilla rostrata, различающиеся между
собой по количеству позвонков: 114 у европейского и 107 у амери-
канского. В конце лета почти половозрелые угри спускаются из
рек и озер в моря — Балтийское, Северное, Средиземное, а отсюда
в океан, к местам икрометания (рис. 206). Последнее удалось уста-
новить лишь совсем недавно. Места икрометания находятся для
254
обоих видов близко друг от друга, частью перекрываясь, в так
называемом Саргассовом море, в западной части Атлантического
океана. Здесь яйца откладываются на большой глубине (800—900 м).
Выметав икру, угри, видимо, погибают, так как к концу нересто-
вания в океане уже не .видно взрослых угрей. В конце зимы или
начале весны из икры выводятся молодые угри в виде листообраз-
ных личинок, называемых тонкоголовыми — Leptocephali. Выра-
стая, они медленно поднимаются в верхние слои воды. Когда они
поднимутся, то дальнейшая история жизни протекает неодинаково
у американского и у европейского угря. Американские Lepto-
cepliali растут гораздо быстрее и уже в течение одного лета
дорастают в среднем до 6—6.6 см, достигая в это время берегов
своей родины. .Зимой происходит метаморфоз в прозрачных «стеклян-
ных» угрей, на вторую весну — вхождение в американские реки.
Европейские Leptocephali растут более медленным темпом
и лишь осенью третьего года вырастают до длины в 7.5 см,
только к этому времени достигая более отдаленных европейских
берегов; зимой имеет место метаморфоз, а лишь на четвертую весну—
вхождение в европейские реки.
К. берегам Европы личинки угря плывут пассивно, увлекаемые
Гольфштромом. Плывет их масса: Шмидт в одну тоню ловил в юж-
ной части Атлантики до 800 особей. Они имеют лентовидное, при-
способление к дрейфу в течении, прозрачное, как вода, тело, сжатое
с боков, маленькую головку и заостренный хвост. В общем форма
их тела напоминает вид лаврового листа; тело их так прозрачно,
что сквозь него виден весь кишечный канал. Прозрачность, столь-
характерная для пелагических животных, происходит от отсут-
ствия окраски, от того, что ткани и кровь лишены пигмента, от
того, что между кожей и мускулами находится обширное простран-
ство, растянутое водянистой жидкостью. Лишь глаза окрашены.
Во время своего пассивного страпствования к берегам личинки
растут таким образом: к концу первого года жизни они достигают
длины в 2.5 см. Такие личипки пе встречаются восточнее 50° за-
падной долготы. На второй год опи вырастают до 6 см и встречаются
между 50 и 20° западной долготы. Лишь па третий год, достигая
континентального подводного плато, отделяющего берега Европы
от глубинной зоны, вырастают опи до 7.5 см. Здесь в течение сле-
дующей зимы происходит превращение личинок в молодых угрей
(рис. 207).
Питаться обычным путем личинки угря, повидимому, не могут,
так как глотка их закрыта эпителиальной перемычкой, в кишечнике
их пе оказывается пищи, т. е. личинки не едят, пока не превра-
тятся в молодых угрей. Превращение начинается с того, что ли-
чинка становится пе такой высокой и длиппой, ио более широкой,
подкожное пространство сокращается, тело становится более круг-
лым, личиночные зубы заменяются настоящими, спинной и задне-
проходной плавники продвигаются вперед, в коже развивается
nni MoiiT, хотя прозрачность теряется пе вполне. Такая личинка
пазыиается стеклянным угрем.
255
В четвертую от рождения весну молодые угри массами входят
ъ реки. Теперь миграция становится активной. В реки угри входят
-большей частью почью (особенно в бурные ночи) и на пути своем
настойчиво преодолевают различные препятствия, обходя их иногда
даже по сырой траве. В некоторые реки опи входят громадными
массами, компактными колоннами и настойчиво стремятся достичь
верховьев рек, потоков, озер и прудов. Особенно интенсивно идет
Рис. 207. Распространение личинки угря и взрослых речных угрей в
Атлантическом океане и по его берегам.
Черная полоса по берегам Европы и Африки отмечает те области, в реки
которых входят прозрачные угри. Заштрихованное пространство, ограни-
ченное линией и?, отмечает область распространения пелагических личинок
речного угря. Кривые ограничивают пространство, где встречаются ли-
чинки меньшей длины (в миллиметрах), чем цифра, стоящая на кривой. Тем-
ная полоса внутри границ 10 мм личинок отмечает область, где были най-
дены личинки в 5—7 мм. (По Joh. Schmidt из П. Шмидта.).
миграция ночью, прерываясь днем и даже в полнолуние. В пресной
воде угорь живет 5—7—10—20 лет, ведя самый хищный образ
жизни, поедая рыбу, икру, личинок насекомых, раков, лягушек,
водяных птиц, водяных крыс и растения. Особенно активны угри
ночью.
Когда приближается время половой зрелости, утри подверга-
ются замечательным изменениям. Они постепенно перестают есть,
нижняя часть тела их становится белее, спина—темнее, больше.
В таком виде угорь называется серебристым угрем.
Осенью «серебристые угри» спускаются вниз по рекам к океану.
Дойдя до моря, угри становятся еще темнее, а глаза у пих больше.
256
Угорь приобретает характер глубоководной рыбы. Невидимому,
угорь размножается раз в жизни и, уйдя из рек в море, никогда
назад не возвращается, а умирает вскоре после икрометания.
Таким образом, все угри, однажды нашедшие свой путь в море,
потеряны для рыболовства навсегда.
У нас угри встречаются в бассейне Балтийского моря.
Таким образом, угри, живущие в пашем Северо-западном крае,
происходят с далекого нерестилища, лежащего в таинственной
пучине Атлантического океана. И туда, в Саргассово море, они
уйдут ио достижении поло-
вой зрелости, чтобы никогда
больше назад не вернуться.
Как могли,возникнуть ми-
грации угря? Повидимому,
объяснение им мы находим в
штеперовской теории смеще-
ния материков.
Личинки угрей плывут к
берегам Европы пассивно, их
несет течение Гольфштрема.
Рис. 208. Атлантический океан с изо-
термами, представляющими распределе-
ние температур воды на глубине 400 м.
Вертикальная штриховка покрывает об-
ласть нереста и икрометания речного угря,
горизонтальная—область размножения аме-
риканского угря. (По Свен Экману из
II. Шмидта.)
Вся организация лептоцефал
является приспособлением к
удержанию па поверхности и
к пассивному перенесению
течением. Будучи пассивным,
движение личинок происхо-
дит не по одной линии, а,
подвергаясь различным случайным влияниям, поток личинок рас-
ходится широким веером, благодаря чему опи оказываются у бере-
гов и Западной Европы, и Северной Африки. Достигпув континен-
тальной ступени (плоскость, определяющая глубины в 1 000 мет-
ров), личинки попадают в область с менее соленой водой. Здесь на-
чинается их превращение, и далее опи движутся уже активно
вверх по течению рек в направлении меньшей солености. Это
попятно.
Но каким образом взрослый угорь находит путь к месту своего
нереста, какие стимулы им управляют? О памяти угря здесь го-
ворить пе приходится. Объяснение, видимо, находится в следующем.
Угорь мечет икру в самой теплой части Атлантического океана,
где температура па глубине 400 м почти круглый год остается
постоянной. Изотермы, т. е. липни, соединяющие места с одина-
конон температурой, располагаются в Атлантическом океане таким
образом (рис. 208), что, если угри при своей миграции в море на-
правляются температурными стимулами, стремясь перейти из более
холодной воды в более теплую, то опи неизбежно должны притти
к мосту своего икрометания, из какого бы пункта побережья Европы
они нн вышли. Что рыбы обладают способностью тонко различать
температурные разницы и руководствоваться ими в своих мигра-
ции.х, не подлежит сомнению. Возможно, что угорь руководится
17 KiiiuKupuu и Станчинский
257
еще изменениями солености, происходящими в Атлантическом
океапе почти параллельно с изотермами.
Согласно теории Вегенера, перед началом третичной эпохи
(рис. 209) Атлантического океана как такового еще не было. Он
был представлен уз-
кой щелью между
континентами Старо-
го и Нового Света. В
эоцене щель раздви-
нулась, образовалось
море, связанное через
область Средиземного
моря с Индийским
океаном, а через Цен-
тральную Америку—
с Тихим. В это время
сюда могли проник-
нуть угри из Тихого
или Индийского океа-
на, где нашли себе
подходящие условия
для нереста па месте
нынешнего Саргассо-
ва моря, откуда со-
вершали миграции к
берегам Европы и
Америки. Но матери-
ки расходились, об-
ласть нерестования и
область пребывания
в стадии угря посте-
пенно удалялась друг
от друга, а вместе с
этим постепенно уд-
линялся и путь ми-
грации угрей.
А н а д р о м а л ь-
п а я м и г р а ц и я
б л аг о род и ого
лосося (Salmo sa-
lar). Странствования семги (или благородного лосося) составляют
полную противоположность катадромальной миграции угря.
Семга является типичной проходной рыбой, приходящей
из моря в реки для нерестования. Входит лосось в реки задолго
до самого икрометания. Последнее бывает осенью и зимой в разных
местностях по-разному: в Шотландии и Норвегии— обычно
в ноябре и декабре, у пас—в сентябре—октябре (осенняя семга).
Бывает еще летний ход.
Уже в реке половые продукты у самца и самки достигают пол-
В
Рис. 209,. Смещение материной согласно вегене-
ровской теории, как объяснение миграции угрей.
258
ной зрелости, и самцы надевают «брачный наряд», становясь «ло-
хом»: рыба становится темнее, красные пятна показываются на
боках и па жаберных крышках, красный цвет появляется и на
брюхе. Верхняя челюсть удлиняется, на нижней вырастает крюк
соединительнотканного происхождения; зубы увеличиваются и вы-
растают новые. Мясо мало-помалу становится безвкусным, менее
жирным и менее питательным. Затем начинается стремительный
ход вверх по реке, по самой быстрине; лосось идет, не зная преград,
перескакивая пороги и водопады высотой до 4 м. Дойдя до места,
где дно состоит из крупного песка или усеяно мелкими камеш-
ками, где вода настолько чиста и течение настолько быстро, что
взмучепный песок не оседает среди отложенной икры, а уносится
далеко вниз, лосось приступает к размножению.
Самцы являются на места нереста раньше самок. Между ними
происходят столкновения из-за обладания нерестилищем. Более
слабые самцы изгоняются. Когда приходят самки, последние при-
ступают к изготовлению нерестовой ямки. Ударами хвоста самка
разбрасывает песок и гальку и делает канавку до 2 м в длину,
1 м в ширипу, иногда до 0.5 м в глубину, обычно же более мелкую.
В эту ямку самка откладывает икру, которую самец, держащийся
возле самки, сейчас ясе оплодотворяет, после чего самка боковыми
двиясениями хвоста забрасывает икру песком или галькой.
После нереста самка оставляет нерестилище, самец же продол-
ясает еще некоторое время держаться около нерестовой ямы, а по-
том часть их (более старые) погибает, остальные в истощенном
и потрепанном виде медленно спускаются обратно в море. Спуск
этот длится месяцами. Быстрее скатываются в море молодые особи
(3—3^-летнего возраста). Во время скатывания в море лососи
большей частью пе питаются, желудки их остаются пустыми.
Откладываемая в количестве от 10 до 40 тысяч икринок, икра
лосося крупна, 6—7 мм в диаметре, немногочисленна, желтовато-
оранжевого цвета и заключена в плотную эластическую оболочку.
Смотря по температуре развитие происходит от 5 до 21 недели,
а на холоду может быть еще более продолжено. Это обстоятельство
дает возможность пересылать икру па далекие расстояния. Из яиц
выходит молодь 2.5—3 см длиной и с огромпым желточным
пузырем.
Личинки остаются лежать пассивно, пока не всосется весь
желток. Бывает это через 6—7 недель. Тогда опи начинают пи-
таться активно личинками пасекомых, мелкими ракообразными
и мало-помалу становятся бойкими, подвижными рыбками, ко-
торых называют «пестрянками» за их пеструю окраску, напоми-
нающую форелей. Зиму пестрянки (английское название «рагг»)
проводят в более глубоких местах, а Па следующую весну начинают
скатываться вниз по течению, к морю. Лишь часть их остается
на мосте. Когда пестрянки доходят до устья реки, окраска их
изменяется: из пестрой становится серебристой, а па спине по-
является синяя окраска; эту стадию англичане называют «smolt»,
что значит синий. «Smolt» мигрирует в море, где он быстро растет
11 * 259
и возвращается в реку в стадии «grilse», проведя в море зиму, т. е.
в возрасте 3—зу2 лет, или даже позднее в стадии «лосося» (salmon).
На севере развитие идет медленнее, до 5—6 лет.
Другим в высшей степени интересным результатом изучения
чешуи лосося явилось позпание того, что процесс размножения
также запечатлевается на чешуе в виде следов образования бахромы,
и чешуя таким образом говорит нам и о том, в какой период жизип
и как часто размножался лосось, и размножался ли оп вообще.
Стада мечущих икру рыб включают в себя небольшое количество
самцов «рагг» и «smolt», достигших зрелости уже в реке, и,обычно,
карликовых самцов «grilse», самок, которым не менее 4 лет отроду,
т. е. проведших по крайней мере две зимы в море, и более взрослых
самцов и самок.
Такова нерестовая миграция лосося. Но лосось вообще каждый
сезоп совершает миграции, пе имеющие ровно ничего общего с раз-
множением, хотя, подобно первым, они анадромальпы и соверша-
ются в том же направлении. Эти сезонные миграции it берегу,
а в некоторых случаях и в реку, ио не так далеко, как для нере-
стования, характеризуются тем, что первыми появляются и исче-
зают молодые лососи. Мечением семги удалось установить, что она
возвращается для нерестования в ту реку, в которой сама развилась.
Впрочем, бывают исключения из этого правила.
К сожалению, о жизни лосося в море мы знаем еще слишком
мало; известно только, что эта рыба ведет хищнический, несомненно,
260
ire чпгнчгг.кий образ жизни па небольшой глубине, питаясь глав-
ным образом летом, преимущественно рыбой, причем очень быстро
откармливается.
Диаграмма (рис. 210) иллюстрирует сказанное выше о миграциях
лососи. Затененная часть диаграммы обозначает жизнь в пресной
иоде, нгзатушеванная часть — жизнь в море.
Миграции восточного лосося (Oncorhynchus).
В северной части Тихого океапа водится другой род лососевых—•
восточный лосось (Oncorhynchus), близко родственный атланти-
ческому лососю и роду Salmo. Существует несколько видов восточ-
ного лосося, жизнь которых наилучше известна в период еже-
годных миграций для размножения. Эти миграции замечательно
интересны: опи совершаются в общем по тому же типу, как ми-
грации лосося обыкновенного, но масштаб их гораздо больше
и представляет некоторые удивительные особенности. К этому
роду относится хорошо известная у нас па рынке кета (Onco-
rhynchus keta), распространенная от Корейского побережья до Ле-
довитого океана.
Побуждаемая наступлением половой зрелости к переселениям,
кета входит в реки в невероятном количестве, служа в это время
предметом лова. Войдя в реку, эти рыбы совершенно не принимают
пищи, а живут за счет запасов жира и белка в теле. Места нереста
находятся часто больше чем за тысячу километров от устья. По-
нятно, что рыба истощается. Приближаясь к месту нерестования,
рыба изменяет свой вид: с моря опа приходит с блестящей светлой
чешуей; здесь опа постепенно темнеет и становится пестрой; мясо ее
бледнеет, теряет свой яркий красный цвет; рыло у самцов вытяги-
вается, и обнаруживаются зубы. Когда же рыба приблизится к ме-
сту икрометания, ее вид становится неузнаваем: прежде серебри-
стая, кета становится темной, почти черной, с черными поперечными
полосами; тело сплющивается с боков, мясо становится белесым,
дряблым; верхняя челюсть вытягивается и загибается крючком
пад нижней; зубы становятся огромными и придают голове рыбы
необычайно хищный вид; вся рыба кажется ослабевшей, истощенной.
Самец и самка совместно роют ямку, самка откладывает туда круп-
ную икру, которую самец оплодотворяет, и оба забрасывают ее
камешками. А затем наступает полный упадок сил, организм окон-
чательно истощается, и начинается гибель рыбы. Невидимому, она
гибнет вся, какого бы возраста и какой бы величины пи была.
Болезнь, ослабевание рыбы начинается тогда, когда рыба
пройдет путь в реке и начнет метать икру. В это время в теле ее
не откладывается пигмента, пе развиваются ткани, пе растут кости.
Наоборот, все изменения говорят за то, что рыба изголодалась,
истощилась, чем-то отравлена: жир в теле исчезает, мышцы дрябнут,
белый пушок паразитического грибка сапролегния покрывает
рыбу, разрушает кожу; па теле появляются ранки и язвы, и часто
кости торчат у рыбы наружу, прободая кожу и мускулы. Рыба
не может сопротивляться течению воды, ее песет вниз, ударяет,
о камни и корни; плавники, и без того уже поврежденные, отламы-
261
ваются... Случается, что оба берега реки, где происходит икро-
метание, бывают густо, непрерывной полосой усеяны трупами
и полумертвыми рыбами (рис. 211). Когда рыбы ослабнут от
икрометания, они делаются легкой и беззащитной добычей выдр,
медведей и других животных, что еще больше увеличивает между
ними смертность. Это явление носит название: «кочевание
до смерти». Если даже допустить, что некоторые особи воз-
Рис. 211. Мертвая горбуша («сненка») на берегах протока реки Мухиной,
на западном берегу Камчатки, в августе 1934 г. (По фото К. И. Панина из
П. Шмидта).
вращаются в море, что мало вероятно, то все же в массе своей
восточный лосось способен к размножению лишь один раз в жизни.
Когда из яиц выведется молодь и станет способной по расса-
сывании желточного пузыря вести самостоятельную жизпь и сво-
бодно плавать, опа начинает скатываться впиз по реке. В реке
молодь питается личинками насекомых — комаров, поденок и др.
Скатившись в море, мальки некоторое время проводят в пред-
устьевых пространствах и в береговой полосе моря, усиленно пи-
таясь мелкими ракообразными и личинками водных насекомых,
а затем уходят в море, где и остаются до того момента, когда ста-
новятся половозрелыми и начинают свои миграции в реки. О жизни
Oncorhynchus в море мы знаем очепь мало. Повидимому, опи ведут
здесь стадную жизнь, подходя летом более близко к берегу и пи-
таясь песчанкой (Ammodytes), сельдью, мелкой треской и т. д.
Таким образом, жизнь тихоокеанского лосося выражается
в чередовании миграции вниз по реке, начинающейся сейчас же
262
по выводе» из икры, и обратной миграции к месту икрометания по
достижении половой зрелости. Прилагаемая диаграмма (рис. 212)
ото ясно показывает. Вся жизнь взрослого лосося на самом деле
проходит в море. Той задержки в реке, которая наблюдается у ло-
соси атлантического, здесь пет.
Существует взгляд, что кета выбирает для нереста ту реку,
г/н' она сама вывелась. Но это обстоятельство объясняется скорее
случаем, чем необходимостью или инстинктом. Инстинкт побу-
Рис. 212. Диаграмма жизни дальневосточного лосося
(Oncorhynchus nerka).
ждаст рыбу искать свежую воду, когда наступает половая зрелость,
заставляющая рыбу двигаться. Рыба идет нс потому, что опа пом-
нит путь, которым опа шла в море, а скорее наоборот: молодь
идет ио пути, по которому пришли родители. Молодь ис выбирает:
опа мигрирует пассивно. По какие ясе условия определяют дви-
жения взрослых? Недавние исследования миграции нерки (Оисо-
rliynchus легка) в Америке показали, во-первых, несомненное влия-
ние направления течения: рыба идет против течения. При выборе
пути в месте соединения двух потоков лосось выбирает поток с более
холодной водой. Меча икру в озере, он опять-таки выбирает более
холодное место Температурный стимул, вероятно, не единствен-
ный, по наиболее заметный. Выбор воды, наиболее холодной летом,
(обилие ключей), спасает отложенную икру от промерзания зимой.
Колебания хода лосося по годам объясняются колебаниями коли-
чества грунтовой воды, режима потока летом, климатических
условий зимой и летом. Неблагоприятные условия могут сильно
повредить миграции.
263
Так как,, как показали опыты с мечепием, в каждую реку входят
главным образом те рыбы, которые в пей вывелись, то уничтожение
их па пути, в предположении, что другие рыбы потом населят
эту реку, ведет к исчезновению лососей в данной реке.
Миграции в море. Странствования атлан-
тических сельдей (Clupea harengus). Сельдь наиболее
Рис. 213. Места обитания разных рас сельдей
Северного моря (обозначение разными кружками).
важная после трески
промысловая рыба.
Поэтому вышеописан-
ные методы были в
широком масштабе
применены к изуче-
нию ее миграций.
Последние соверша-
ются из глубины па
мелкие места, совер-
шаются в значитель-
ной мере против те-
чения в море и пото-
му могут быть отне-
сены к типу анадро-
мальных. Обитающая
в северной части Ат-
лантического океана
с его морями, сельдь
ловится по берегам
Европы весь год; по
для каждой местности
существует время,
когда сельдь появ-
ляется в особенно
большом количестве.
Эти сезоны остаются
постоянными: в од-
ной местности — вес-
ной, в другой—летом,
в третьей — осенью.
Рыбаки давно различали сельдей с отмели в Немецком море, назы-
ваемой Доггербапк, из района Шотландских островов, из Скагер-
рака и с берегов Норвегии (рис. 213). Новейшие методы с приме-
нением вариационной статистики (метод Гейнке) дали возможность
установить действительное существование в Немецком и Норвеж-
ском морях нескольких рас сельдей, отличающихся величиной,
числом позвонков и временем икрометания, а также всей суммой
множества признаков, неуловимых в отдельности.
На чешуе сельди чрезвычайно отчетливо сказывается периодич-
ность роста в виде широких матовых летних и узких прозрачных
зимних колец. Как и в других случаях, на чешуе можно прочесть
всю историю жизни сельди. Если сельдь мечет икру зимой или
264
воспой, то в течепие лета чешуя успевает образоваться, и первая
же зима па ней запечатлевается; если же сельдь мечет икру осенью,
то, так как за зиму чешуя пе образуется, первое зимнее кольца
будит принадлежать второй зиме. Так как различные расыатлан-
Рис. 214. Чешуя различных рас сельдей Северного
моря.
А—норвежская фиордовая, В—норвежская весенняя, С—окагерракская,
D — шотландская сельдь
тической сельди мечут икру в различное время года, так как усло-
вия существования в местах образования рас различны, так как при
миграциях расы эти в различные годы своей жизни бывают в не-
одинаковых условиях, — то па чешуе разных рас будет разная
картина колец. Сравнивая изображения на рис. 214 чешуй раз-
личных рас сельди в возрасте пяти, шести и девяти лет, причем
величина чешуи изображена пропорционально величине рыбы,
265-
tl 13 al is № г~ as .к> да да да да да да да да
Рис. 215. Возрастной состав косяка сельди.
мы видим, что величина их заметно вариирует, и что различия эти
обусловлены различием в условиях роста. Рост норвежской ве-
сенней сельди до пятого или шестого года правилен, рост же фиор-
довой сельди как бы задержан. Это видно, например, из сравнения
шестилетней чешуи той и другой породы. Рост сельдей из Скагер-
рака и шотландской после третьего года становится гораздо мед-
леннее, хотя они вообще крупнее мелкой фиордовой сельди. Цен-
тральная часть чешуи до первого зимнего кольца у весенней нор-
вежской сельди и фиордовой сельди, с одной стороны, и у сельди
из Скагеррака и шотландской, с другой — значительно разнится.
Это объясняется тем, что
первые размножаются зимой
или весной, тогда как сельди
из Скагеррака и шотланд-
ская размножаются летом и
осенью; следовательно, пер-
вое зимнее кольцо на чешуе
образуется у них, когда им
полтора года, а потому че-
шуя значительно крупнее.
По чешуе можпо опреде-
лять, как было указано вы-
ше, возрастной состав кося-
ков. На рис. 215 изображен
проанализированный по че-
шуе возрастной состав трех
косяков весенней
сельди 924 экзем-
пляров. Мы видим,
что одни возрасты
п р едстав лены 66 ль -
шим числом осо-
бей, другие—мень-
шим. Возможность
предсказывания уловов в будущие годы этим путем была указана
выше. Благодаря изучению возрастного состава косяков в раз-
личных пунктах побережья, установлению принадлежности осо-
бей и косяков к той пли иной расе была выяснена история
жизни и миграций атлантической сельди. Опи рисуются нам та-
ким образом.
Переселение сельдей, имеющее целью размножение, совер-
шается в мелкую или сравнительно мелкую воду Это — миграция
анадромальпая. В .зависимости от величины самка кладет
от 20 000 до 40 000 яиц. У маленькой балтийской сельди они всего
1 мм величиной, у сельди из Немецкого моря — до 1.4 мм. Яйца
сельди топут и прикрепляются к зоофитам, раковинам, камням
и т. д. В зависимости от температуры, по приблизительно через две
педели из яиц выходят длинные прозрачные личинки.
Несколько дней, пока не всосется желточный пузырь, личинка
366
Рис. 216. Течение в Северном и Норвежском
морях.
остается у дна, а затем постепенно поднимается кверху и начинает
пелагическу ю жизнь. В течение месяца онавырастает до 1.5—1.8 см.
Пока она ведет планктонный образ жизни, приливные течения
носят ее взад и вперед, а океаническое течение носит ее вдоль
перегон. 1> результате молодь широко распространяется в мелких
110/1.114 но тому направлению, куда идет течение от места икроме-
тании. Сравнивая рисунки 217 и 218, мы видим, что от нерестилища,
Которое Л СИ-СИТ у ЮГО-
.ijiioiiioroGeperaHop-
негнн и обозначено
цифрой V, молодь,
пассивно следуя при-
(11 южному течению
(। ютвь Гольфштрема,
рис. 216), распростра-
няется вдоль берегов
Норвегии к северу.
J эолыпинство перес-
тилищ Северного мо-
ря лежит на западной
стороне, и, следова-
тельно, молодые сель-
ди распространяются
на юг вдоль берегов
Вритании (цифры VI,
VII, VIII, IX). Лишь
на юге течения тако-
вы, что несут личи-
нок сельдей к вос-
току (XII, XI, X).
Личинка растет, по
рост ее в течепие пер-
вого года весьма, раз-
личен, смотря по то-
му, в каком сезоне
происходит икромета-
ние. Весенняя балтийская сельдь достигает 3.6 см в 2—3 ме-
сяца, а осенняя вырастает до той же величины в 7—9 месяцев.
Но па второе лето рост выравнивается.
Наблюдения па разных местах показали, что косяки имеют
наклонность оставаться чистыми и гомогенными.
Распространившись вдоль берега к северу и следуя течению,
не зимовавшая еще группа сельдей уходит зимой в места немного
полое глубокие, а летом выходит в более мелкую воду, т. е. к берегу,
и в то же самое время в направлении, противоположном первому
переселению, т. е. против течепия. И так каждый год, т. е. каждый
следующий год, сельди идут во все более и более глубокие воды,
а летом опять в мелкие и далее — против течепия. Таким образом,
с каждым годом сельдь приближается к месту икрометания, плывя
267
против течения, которое первоначально унесло ее отсюда в стадии
личинок (рис. 217).
Появляется рыба на месте икрометания почти внезапно. Это
показывает, что миграция совершается на некоторой глубине; па
поверх весть сельдь до поры до времени не поднимается. Но, при-
ближаясь к берегу, сельдь приобретает привычку подниматься
ночью па поверхность. Тут во время икрометания и происходит
лов сельди плавными сетями. Сети ставят поперек линии движения
прилива. Косяки сельди плывут всю ночь в поверхностных слоях
воды против течения и за-
путываются в сетях, иног-
да при внезапном подъеме
кверху поднимая па по-
верхность и сети.
Рис. 218 показывает
путь странствований ко-
сяка норвежской сельди,
которая в стадии личинок
была унесена к Лофотен-
ским островам, и распро-
странение истощенной ик-
рометанием «меченной» ры-
бы в Северном море.
Таким образом, мигра-
ции норвежской сельди
происходят по такой схеме.
С места переста большое
количество молоди сно-
сится течениями в область
у Тромзё, которую можно
назвать местом сбора.
Здесь молодь собирается в
косяки, которые начинают
обратное движение к югу.
Сохраняя свой состав, они
одновременно, в зависи-
мости от сезона и дня и
ночи, то выходят па более
мелкую воду, то уходят в глубину. К моменту наступления половой
зрелости, подойдя к месту, где происходит нерест этой расы, они
соединяются со взрослыми косяками этой расы, мечущими икру.
Сельди в косяке обычно бывают одного размера, так как члены
косяка держатся и растут вместе со времени выхода из икры. Но
в Норвежском и Немецком морях, как сказано выше, пе одна раса
сельди. Каждая из них имеет исходную точку в своем месте нереста.
Косяки этих различных рас пе смешиваются между собой в одну
неразличимую массу. Каждая особь песет штамп своей расы.
Разные косяки нерестятся в разных местах в разное время, имеют
различные «пастбища», проходят различную историю жизни.
268
Миграции взрослых сходны с миграциями молоди. Выметав
икру в мелкой воде, косяки проводят несколько месяцев в кормле-
нии, прежде чем возвратиться в глубокую воду, где опи, вероятно,
вновь соединяются для размножения перед следующей миграцией.
IГл миграций каспийских сельдей следует упомянуть миграцию
черноспинки, или так называемого залома (Caspialosa kessleri).
Пта сельдь держится в море, а для метания икры входит в Волгу.
Одиночные рыбы с конца марта, а большие косяки с половины
Рис. 218. Путь миграции косяка норвежской сельди.
апреля входят в реку. Иногда сельдь входит в реку раньше, иногда
позднее, в зависимости от ветра: холодный северный ветер задер-
живает рыбу, теплый южный ускоряет ход сельди. Сначала почти
непрерывно идут громадные косяки; потом они становятся меньше
и меньше, и так проходит несколько педель. В Волгу сельди вхо-
дят, еще пе достигнув половой зрелости, поэтому идут довольно
далеко вверх и льют икру от Куйбышева до Саратова, доходя до
Горького и выше, в Оку. Льют икру, невидимому, не везде в одно
время: и в июне, и в июле. Выметывая икру, сельдь сильно мечется,
выскакивая из воды и кружась как одурелая, почему и названа
бешепкой. Выметав икру, истощенная и исхудалая, сельдь тече-
нием воды сносится обратно к морю, по уже не косяками, а вюди-
ночку или разбитыми группами. Подавляющее большинство после
нереста погибает от истощения и в море уже пе возвращается.
Так как эта рыба мечет икру лишь на 5-м году жизни, то естественно
что этот вид сельди немногочислен.
269
Можно упомянуть еще о миграциях трески (Gadus callarias).
Тресковые принадлежат к самым важным рыбам в промысловом
отношении. Поэтому их миграции были изучены весьма тщательно,
причем главным методом служит статистический подсчет количества
пелагических яиц и пелагической молоди в разных областях моря.
Треска обыкновенная живет в северной части Атлантического
океана и имеет гораздо более широкую область обитания, чем
область икрометания. Главные места последнего находятся у Нью-
фаундленда и у Лофотенских островов в Норвегии. После икро-
метания треска должна распространиться по всей области своего
Рис. 219. Распределение яиц и молоди трески.
обитания. Эта миграция в значительной своей части совершается
путем дрейфа (пассивное следование течению) пелагических яиц
и молоди. Выметывается икра па глубине 20—80 м, над банками.
Личинки приспособлены к дрейфу: легки и прозрачны. Уже яйца,
развивающиеся в срок от одной до двух педель, могут быть унесены
далеко от места икрометания. Дрейф продолжается и тогда, когда
выведется молодь. На рис. 219 изображено передвижение пелаги-
ческих яиц и молоди трески в различные месяцы вблизи Норвегии.
Границы нанесены на основании статистических исследований уло-
вов. Вырастая, молодь постепенно опускается глубже и глубже,
достигая наконец дна. Дрейф яиц и личинок зависит от течений
в море. В результате молодь широко распространяется в области
обитания. Опускание на дно зависит от изменения плотности океа-
нической воды. Таким образом молодь распространяется все шире
и шире. С приближением осени и зимы ее встречает обратная волна
мигрантов. Сеголетки зимуют па небольшой глубине, а следующим
летом снова начинаются миграции, теперь уже сезонные. Перед
перестом начинается новая миграция от берега, треска соединяется
270
п косяки к идет к банкам, где и происходит икромета-
НПО.
Кроме ятой нерестовой миграции, у трески (Gadus callarias)
наблюдается еще кормовая миграция после икрометания
у Лофотенских островов на север и далее в Баренцово море. Мед-
ленно двигаясь в теплой ветви Гольфштрома и усиленно питаясь
мойной, сельдью, песчанкой и ракообразными, треска продви-
гается почти до Новой земли, а с похолоданием спускается вдоль
Перегон Норвегии к югу.
Наконец, важны миграции рыб па места зимовки. У пас наблю-
дается такой ход многих пород в дельте Волги, Урала, Куры и т. д.
Знание путей и времени миграций исключительно важно в хо-
:и1 ветвенном отношении.
10. Роль рыб в биоценозах и в круговороте веществ
Организмы распределены в занимаемом ими пространстве не
беспорядочно. Опи закономерно группируются в объединения,
называемые биоценозами. Биоценозами называются груп-
пировки животных и растений, занимающие участки суши или
части водоемов, характеризуемые особыми, отличными от соседних
участков, условиями существования, т. е. определяемые различными
группировками факторов среды. Оргапизмы, входящие в биоценоз,
11(5 просто существуют рядом.
Оргапизмы находятся между собой в более или менее тесных
взаимоотношениях, прямо или косвенно связаны друг с другом,
('вязь эта существует прежде всего по линии питания. Каждый
биоценоз состоит из ряда «цепей питания»; каждая из них на-
чинается растительными видами, за которыми следуют пита-
ющиеся ими растительноядные; за ними хищные, добычей кото-
рых служат последние, а этими хищниками нередко кормятся дру-
гие хищники. Между многочисленными цепями питания, из которых
слагается биоценоз, имеется в свою очередь та или иная связь, и все
они вместе образуют «цикл, или круг, питания». Таким образом,
каждый вид в биоценозе так или иначе связан с другими. А так
как биоценоз никогда почти пе является изолированным и так или
иначе связан с другими биоценозами, то каждый организм своим
* жизненным оборотом» участвует в общем круговороте веществ
в природе. При этом роль различных организмов далеко пе одина-
кова. Рыбы являются в общем круговороте веществ в природе
чрезвычайно важным звеном благодаря очень сложным и много-
образным отношениям с остальным животным миром, благодаря
споен численности и благодаря тому, что, будучи животными вод-
ными. они являются важным звеном в переработке той колоссаль-
ной энергии, которую накопляют водные растения, многоклеточ-
ные н одноклеточные, недоступные для использования их назем-
ными организмами.
Вода составляет главную часть лика земли, занимая 70.8% ее
площади. В воде сосредоточивается главная масса зеленого веще-
271
ства как в виде зарослей водяных растений и водорослей, так
и в виде растительного планктона из одноклеточных. Зеленое хло-
рофиллоиосное вещество является главным трансформатором све-
товой лучистой солнечной энергии в энергию химическую. Глав-
ная масса водорослей находится па небольшой глубине до 20—60 м,
по встречаются они всюду до 400 м глубины. В сравнении с этой
необъятной, занимающей сотни миллионов квадратных километров
планктонной зеленой сеткой, вес которой должен выражаться
числами 1016—101в тонн (!), неизмеримо мала площадь зеленой
поверхности суши. К этому следует добавить еще Саргассово море,
пловучий лес водорослей, занимающий 100 000 км2, и прибрежные
заросли, равные около х/ю площади океана с мощностью в не-
сколько сот метров.
Растения являются трансформаторами солнечной энергии, источ-
ника всей жизни на земле. Животная жизнь невозможна без ра-
стений. Колоссальная часть солпечпой энергии используется нами
и целым рядом животных благодаря рыбам. Фитопланктон, в ко-
тором главную роль играют диатомовые водоросли, составляет
пищу зоопланктона, поедается бесчисленными беспозвоночными,
среди которых важную роль играют Copepoda, а также молодью
рыб. Зоопланктон поедается рыбами. Таким образом, запасаемая
растениями энергия переводится в мясо последних.
Личинки сельди, например, будучи от 7 до 12 мм размером,
питаются диатомеями, инфузориями, личинками моллюсков, осо-
бенно Gastropoda, копеподами (см. Harpacticidae), сами являясь
добычей медуз, плевробранхий, червей полихет, сагитты. Вырастая,
сельдь питается различными копеподами, теперь уже сама поедает
сагитту. Взрослая сельдь кормится крылоногими моллюсками (Li-
macina), копеподами (Calanus), хвостатыми оболочниками (Oiko-
pleura), ест яйца сельдей же и т. д. Пелагическая скумбрия
(Scomber) питается планктоппыми Crustacea, а после икромета-
ния— рыбьей молодью. Уходя в глубокие воды, опа кормится
шизоподами, амфиподами, Animodytes (песчанкой). Доппые рыбы,
как камбаловые (Pleuronectidae), кормятся донными животными:
червями, офиурами, моллюсками и другими рыбами.
Некоторые рыбы поедают ил или детрит — остатки раститель-
ных и животных организмов, падающих на дно.
Пищей пресноводных рыб являются растения, а также ряд бес-
позвоночных — растительноядных и хищных и их личинок.
Количество низших животных, служащих пищей рыб, опре-
деляет количество последней. Попытки определить «продуктив-
ность моря» делались па основе определения количества планктона,
определяемого по уловам планктонной сетью с определенным диа-
метром в единицу времени, и при помощи уловов донной фауны
особыми аппаратами, берущими пробу с единицы площади дна.
Хотя метод этот встречает большие трудности и данные, им получае-
мые, конечно, приблизительные, по все же, пользуясь им, воз-
можно делать определения запасов рыбы и продуктивности раз-
личных областей моря. .
272
Основной нищей морских животных в северных водах являются
продукты разложения водорослей рода Zostera. Запасы ее оценива-
ете и и H I ООО 000 тонн. Животные, не являющиеся ни пищей чело-
веки, пи рыб и питающиеся непосредственно Zostera, представляют
около !> ооо ООО тонн. Полезные животные, которые способны слу-
жит!. пищей рыб, достигают 1 000 000 тонн. Однако, не весь этот
ми (| иноп то! г п используется промысловыми рыбами. Морские звезды
с ют 200 000 тонн, 500 000 тонн съедают более крупные гастро-
>1,1.1 и ракообразные, часть которых поедается рыбами. Камбалы
съедают 50 000 тонн, давая в год 5 000 тонн пищи для человека.
Треска является менее выгодной рыбой, так как она получает свою
Рис. 220. Некоторые из наиболее важных цепей питания
в биоценозах Каттегата.
(По Martin из Lotka.)
пищу, так сказать, из вторых рук, и каждая тонна из ежегодно
производимых 6 000 топи представляет примерно лишь 1/20 перво-
начально синтезированной органической пищи.
Наиболее важная в пищевом отношении сельдь питается планк-
тоном и поедается треской. Последняя поедает также хищных
раков и больших гастропод (рис. 220).
Из приведенной диаграммы (рис. 220) мы видим, что из перво-
начально огромного количества органического материала произ-
вол,птс,я все меньше и меньше последнего и, в конце концов, срав-
нительно очень небольшое количество пищи человека. Получается
:>т<> потому, что значительная часть материи и связанной с ней
•анергии тратится па жизнедеятельность промежуточных звеньев
между начальным ее запасом (Zostera) и человеком. Было бы вы-
годнее, если бы полезные рыбы питались прямо Zostera.
(IS Кашкарои и Стаичииекий 273
Надо Припять во внимание еще то обстоятельство, что, кроме
видимого планктона, существует еще невидимый нанопланктон
из очепь мелких особей. На литр насчитывается в нем
1 000 000 особей.
Затрачивая на жизнедеятельность значительную часть энергии
полученной с пищей, рыбы часть ее возвращают воде в виде экскре-
ментов, дающих возможность растениям процветать. Сами рыбы
(например, у берегов Перу), поедаемые птицами, превращаются
в ценнейшее удобрение — «гуано».
Таким образом роль рыб в круговороте веществ очень велика.
В биоценозах рыбы прежде всего связаны с теми видами жи-
вотных, которые являются их пищей. Но и сами рыбы служат
пищей многих животных, им угрожают различные враги.
Ряд насекомых — клопов (Ranatra, Notonetca, Naucoris), во-
дяных жуков (Dytiscus, Hydrophilus), личинки стрекоз (Libel-
lula, Aeschna) — нападают на рыб и их молодь. Их поедают ля-
гушки (Rana) из земноводных, ужи (Natrix)— из рептилий.
Из птиц рыбами питаются зимородки (Alcedoj, крачки (Sterna
и др.), чайки (Lari), гагары (Colymbi), все голенастые (цапли—
Ardeae, колпицы — Platalea, аисты — Ciconia и др.), пеликаны
(Pelecani), бакланы (Phalacrocoracidae), орлан белохвост (На-
liaetos albicilla), скопа (Pandion) и многие другие. Количе-
ство поедаемых птицами рыб весьма значительно. Так, малень-
кий зимородок в течение 100 дней уничтожает от 1 000 до
1 200 мальков форелей, в желудке цапли находили одновременно
10—15 рыбок, плотвы и ельцов. Пеликан съедает в день с де-
сяток крупных рыб. Молодой еще баклан съедает зараз трех
больших рыб, которых он с трудом заглатывает. Из млекопитающих
за рыбами охотятся очень многие: водяные куторы (Neomys),
крысы (Epimys), норка (Lutreola), выдра (Lutra), камчатский бобр
(Latax lutris) и др. Огромное количество рыбы поедают ластоногие
(тюлени и др.): так, в одном Балтийском море тюлепи съедают
в год не менее 50 000 тонн рыбы, в Каспийском — 160 000 топп,
еще больше в Белом и Ледовитом океанах; при этом главной пищей
тюленей являются промысловые рыбы: сельди, треска, лососевые.
Наконец, к серьезнейшим врагам рыб следует отнести самих
рыб, причем иногда этими врагами являются рыбы того же вида:
у рыб развит каннибализм — крупные особи поедают мелких.
Но иногда и мелкие рыбы, действуя скопом, поедают крупных;
например, угри нападают на лососей. Вскрытие желудков трески
показало, что они бывают полны песчанками (Ammodytes), сель-
дями и их икрой. Сельдей преследуют макрели (Scomber), тупцы
(Thynnus), дельфины и многие другие. Песчанки поедаются, кроме
трески, скатами, крупными камбалами, лососями и т. д. Очепь
много поедается рыбьей молоди рыбами же и другими животными,
особенно в открытом море. Это — опаснейший момент в жизни рыб.
Также и икре грозит масса опасностей. Сазан, например, в неко-
торых водах Средней Азии поедает икру маринки (Schizothorax),
вытесняя ее; колюшки (Gasterosteus) поедают икру других рыб,
274
форели идут за мечущими икру лососями к гнезду и пожирают их
яйца.
Кроме собственно пищевых взаимоотношений, которыми рыбы
бывают связаны с добычей и с врагами, с целым рядом животных
у них существуют взаимоотношения на почве паразитизма и забо-
леваний.
Против заболеваний рыбы не очень стойки. Нередко простое изме-
нение температуры воды вызывает у них простуду, воспаление жа-
бер, кожи и т. д. Особенно много существует у них заболеваний,
вызываемых внедрением других организмов растительного или
животного происхождения. Большие опустошения вызывают среди
рыб бактериальные заболевания, например, краснуха среди кар-
пов. Большое значение имеют грибные заболевания. Из последних
на первом месте стоит так называемая сапролегпия (sapro-
legnia), играющая огромную роль при искусственном рыбораз-
ведении. Грибки эти могут поражать как яйца, так и молодь и взро-
слых рыб и притом в таком количестве, что сапролегпию долго счи-
тали первичной самостоятельной болезнью. Однако теперь мы знаем,
что она поражает уже заболевших чем-либо рыб или во всяком слу-
чае. ослабленных. Существуют и внутренние паразиты — грибы
из группы Phycomycetes, вызывающие серьезные заболевания
крови и жабер. Есть паразиты и из водорослей.
Грибок Branchyomyces, поселяющийся па жабрах карпа (Cypri-
nus), душит его.
Очень большое значение имеют споровики — Myxosporidia,
вызывающие временами угрожающие эпидемии. Ими вызываются
например, так называемая чума усачей, оспа карпов. Инфузо-
рия Chilodoncyprini наносит огромный ущерб молодому поколению
карповых, паразитируя в коже. Паразитов, живущих в рыбах,
вообще чрезвычайно много как внутренних, так и наружных из чи-
сла червей и ракообразных. Одних трематод, паразитирующих в ры-
бах, более 50 видов, некоторые из них в половозрелом состоянии
живут в чайках, питающихся рыбой.
В кишечнике некоторых, рыб часто в огромном количестве пара-
зитирует Ecliinorhynchus (Acanthocepliali); в глазном яблоке,
вызывая катаракту, — личинка Diplostomum; в мышцах щуки,
окуня, форели, судаков — личинка широкого лептеца (Botrioce-
phalus), начинающего свой цикл в пресноводных Copepoda (Cy-
clops или Diaptomus) и опасного для человека, и т. д. К внешним па-
разитам принадлежат некоторые жгутиковые, простейшие и инфу-
зории. Опи вызывают на коже рыб маленькие беловатые узелки,
разрастающиеся в большие дефекты кожи, ведущие рыбу к гибели.
Далее к внешним паразитам принадлежит ряд ракообразных
из семейств: Lcrnacidae (на карасях, треске и камбале), Lernaeo-
podidae (на окупе), Ergasilidae (па карпах, щуке) и Argulidae
(обычная карповая вошь — Agulus foliaceus, рис. 221). Ряд наруж-
ных паразитов находим мы среди червей из отряда сосальщиков
(Diplozoon paradoxum, Gyrodactylus и др., рис. 221,Б),из пиявок
(Piscicola geometra, рис. 221, А).
18* 275
Мы видели, что у рыб масса врагов среди самых различных
животных.
По вред, приносимый всеми этими животными, незаметен; оп
компенсируется колоссальной плодовитостью рыб, их высоким «би-
отическим потенциалом». Ведь, масса икрышестого поколения одной
только пары осетров равня-
лась бы по величине земпому
шару, если бы осетры размно-
жались беспрепятственно.
Массовая гибель происходит
пе от перечисленных врагов,
а от общих условий, действу-
ющих главным образом па
рыбью молодь. Одной из важ-
нейших причин обеднения
водоемов рыбой является сам
человек. Хищнический лов,
педопускапие рыб к местам
переста, всякого рода соору-
жения в воде и загрязнение
вод — вот те средства, кото-
рыми человек в бесплановом
буржуазном хозяйстве ведет
ихтиофауну к уничтожению,
по крайней мере в пресной
воде. Компенсировать убыль
рыбы могут лишь искусствен-
ное рыборазведение и ряд
других планово проводимых
хозяйственных мероприятий,
о которых будет сказано ни-
же. Одни законодатсльпо-
охранительпые меры пе спа-
сут рыбного населения рек
и озер.
Рис. 221. Паразиты рыб:
А — пиявки (Piscicola), В—черви сосаль-
щики (Diplozoon и Gyrodactylus), С—рако-
образные (Argulus).
11. Географическое распро-
странение рыб
Современное распростра-
нение животных зависит от
многих причин, но в основ-
ном — от с р е д ы, т. е. от
условий существования, необходимых для каждого из пих, и от
истории как самих организмов, так и истории
геологической, истории материков с их внутренними
водами, истории морей и океанов, т. е. от истории среды.
Громадное генеалогическое дерево рыб коренится в слоях глу-
бочайшей древности. Ему примерно 400 миллионов лет. Где-то воз-
276
пик впервые этот ствол, его первые ветви. Они были приспособлены
к определенным условиям существования. Отсюда с ростом всего
ствола расползались его новые и новые ветви по лицу земли, по во-
дам суши и моря, причем рыбы занимали прежде всего такие место-
обитания, к условиям которых опи уже приспособились раньше.
Встречая новые условия при расселении, одни ветви дерева отмирали,
другие сумели приспособиться к ним и сохранялись долгое время,
далее до наших дней; третьи ветви возникали заново, приспособляясь
к новым условиям: дерево разрасталось. Но суша и море пе остава-
лись неизменными. Старая суша исчезала, новая возникала; обшир-
ные материковые массы разбивались на отдельные части; равнины
превращались в горы; проход из одного океана или моря в другие
то открывался, то закрывался. Кроме того, наступали изменения
в климате, теплые воды становились холодными, и, наоборот, из-
менялась соленость воды, менялся целый ряд факторов среды.
Все эти изменения, конечно, отражались самым резким образом
па распространении по лицу земли крупных и мелких ветвей гене-
алогического дерева рыб.
Рыбы обладают широкой приспособляемостью. Поэтому почти
нет места на земле, где бы не было рыб. Они живут в холодных во-
дах северного полушария, доходя до 82°30'сев. широты (Cottus
quadricornis), и в таких же холодных антарктических водах, в теп-
лых тропических морях, в глубипах морей до нескольких тысяч
метров, в пресных и солоноватых озерах, в реках, в быстрых гор-
ных потоках, в холодных и горячих ключах,па равпипах и па огром-
ных высотах.
Во всех этих областях каждый вид приурочен к строго опре-
деленным условиям обитания. При этом одни
виды являются более требовательными к условиям, у других при-
способляемость развита больше. В качестве примера можно при-
вести распределение видов рыб в некоторых бассейнах, изученных
в Сев. Америке, где определенные группировки рыб связаны или
с озерами, или с нижним течением рек, или со средним, или же дер-
жатся только в верховьях рек в зависимости от ничтожных разли-
чий в содержании ионов водорода. Бесчисленные физические, хи-
мические} и биологические факторы служат барьерами, удерживаю-
щими виды в их местообитаниях. И распределение рыб в
небольших районах объясняется прежде всего этими современ-
ными экологическими условиями.
Однако многие виды рыб, отсутствующие в какой-либо области,
прекрасно уживаются и размножаются в пей, будучи перевезены
сюда из другой области, иногда очепь отдаленной, по со сходными
условиями обитания.
Пример мы видим в американской сельди (Clupea sapidissima),
перенесенной на тихоокеанское побережье с атлантического и став-
шей здесь важной промысловой рыбой. Другой пример: сазан (Cypri-
nus carpio), совершенно отсутствовавший в озере Балхаш, а теперь
искусственно туда перенесенный и в полной мере процветающий.
Эти примеры показывают, что естественные границы видов опре-
277
деляются не только подходящими и неподходящими условиями,
по и физическими барьерами для распространения, созданными
историей геологической и историей самого вида, рода и т: д. По-
этому, чтобы понять картину современного распространения рыб,
надо обратиться к геологии, к прошлому земли, и к палеонтологии,
т. е. к. истории генеалогического дерева рыб. Это рассмотрение даст
паи возможность понять распространение больших групп рыб и
ихтиофаун отдельных областей. И обратно: рассмотрение совре-
менного распространения различных групп животных часто дает
нам возможность понять геологическую историю.
Если мы сравним то, что нам известно о генеалогическом дереве
рыб, с тем, как рыбы распространены и где находятся места их не-
рестования, то придем к выводу, что примитивные рыбы, как иско-
паемые, так и дожившие до наших дней, либо являются пресновод-
ными либо совершают миграции в пресные воды. Таковы осетровые,
кистеперые костные гапоиды, двоякодышащие, пресноводные сельди,
костеязычные, лососевые, длипнорылые, карповые, зубастые карпы,
сомовые, харациповые и многие другие. Сельди пе являются прес-
новодными, по держатся в прибрежных водах и имеют демерсаль-
ные яйца: современные селахии, хотя и являются специализиро-
вавшимися морскими формами, но близки к примитивнейшим рыбам;
опи имеют яйца также демерсальные. Ископаемые же формы —
хрящевые акулоподобпые и древпейшие акулы — являются рыбами
пресноводными. Наоборот, группы, более новые филогенетически—
скумбрии, тресковые, камбалы — являются морскими пелагиче-
скими формами. Из тех пыпе живущих родов рыб, которые ведут
свое происхождение с мелового времени или раньше, 320 родов яв-
ляются пресноводными и 155—морскими. Из тех же рыб, которые
возникли после меловой эпохи и в особенности с эоцена и поздпее,
133 рода являются пресноводными и 747 — морскими.
Если мы примем еще во внимание, что ланцетник (Amphioxus),
животное, близкое к предкам рыб, является также прибрежным
существом, то можем притти к выводу, что рыбы возникли
в пресной воде или во всяком случае в прибрежной
полосе. Лишь впоследствии возникли пелагические формы, когда
развились пелагические яйца и сделалось возможным икрометание
в открытом море.
Кроме того, следует отметить, что современное распределение
морей и океанов существовало пе всегда. Например, Атлантиче-
ский океан возник сравнительно недавно, еще недавно па месте
Берингова пролива существовала суша, отделявшая Тихий оксан
от Северного Ледовитого, и т. д. Материки, ныне существующие
самостоятельно, когда-то составляли одно целое, например, Аф-
рика и Южная Америка. Для понимания совремеппого географиче-
ского распространения нельзя поэтому обойти молчанием историю
генеалогического дерева рыб в связи с историей суши, морей и океа-
нов. Лишь в этом случае мы сумеем понять фауны, попять историю
их возникновения, попять, почему в данной стране имеется свой-
ственная ей в настоящее время фауна.
278
Каждая фауна отражает историю занимаемой ею области и со-
ставляющих ее животных. А потому вся суша и все моря могут
быть разбиты на ряд областей и более крупных и более мелких под-
разделений, в зависимости от характера их фаун.
Само собой разумеется, что эти подразделения будут несколько
отличаться в зависимости от того, на основе распространения ка-
ких животных опи получаются, распространение каких классов
положено будет в основу деления. Филогенетическая история птиц
иная, нежели рыб или амфибий; они имеют разную древность, воз-
никли и развивались тогда, когда материки имели уже другое рас-
Рис. 222. Области распространения рыб.
пределепие, пежели в те времена, когда возникли рыбы и начали
эволюционировать, расселяясь ио земле. И способы распростра-
нения у птиц и рыб совсем иные, иная и среда рбитапия. Поэтому
зоогеографическое деление суши по пресноводным рыбам будет
несколько отличаться от того, которое сделано по птицам. Это падо
иметь в виду при чтении глав о географическом распространении
различных классов.1
Обращаясь теперь к рассмотрению современного распределения
пресноводных рыб и рыб внутренних морей и озер, мы видим, что
Австралия, Южная Азия, Африка и Южпая Америка имеют иную
ихтиофауну, пежели Европа, Сибирь и Северная Америка (рис. 222).
13 первых четырех странах, объединяемых вместе под назва-
нием Эогея (Е1— Е4), мы видим следующие группы рыб: д в о я-
1 Можно, конечно, жертвуя многими фактами, составить схему единого
зоогеографического деления и воды и суши, но эту цель авторы не пресле-
довали, считая это нецелесообразным.
279
код i.T in а щ и х (Dipnoi), кистеперых (Crossopterygii), х а-
р а ц и п о в ы х (Characinidae), костеязычных (Osteo-
glossidae), зубастых карпов (Cyprinodontidae), с о м о-
» ы х (Siluridae), Galaxiidae, Cichlidae, Mastacembelidae, Symbran-
cliii, Anabantidae, хромовых (Chroniidae).
В Европе, Сибири и Северной Америке, объединяемых под именем
Лрктогеи (А1—А6), совершенно другой облик ихтиофауны; здесь
живут: миноги (Petromyzontidae), хрящевые ганоиды
(Chondrostei) с осетрами (Acipenseridae) во главе, кост-
ные ганоиды (Holostei: Amia и Lepidosteus), сельди
(Clupeidae), лососевые (Salmonidae), карповые (Cypri-
nidae), щ у к о в ы е (Esocidae), колюшковые (Gasterostei-
dae), налим (Lota), окуневые (Percidae), бычок (Cottus)
и его пресноводные родичи: г оломянка (Comephoridae) и
Cottocomephoridae.
Конечно, Эогея и Арктогея отграничены не рез-
кой линией, а несколько заходят друг за друга; и некоторые
карповые (Cyprinidae), папример, встречаются и в Эогее; немного
представителей Siluridae есть в Арктогее.
Таким образом, Арктогея и Эогея отличаются целым рядом се-
мейств и даже более крупными систематическими единицами. Хотя
примитивные группы, например, осетры, костные га-
ноиды, сельди, лососи, есть и в Арктогее, этих групп больше в Эогее,
притом чисто пресноводных: двоякодышащие, кистеперые, хараци-
новые, костеязычные и другие. Характерно при этом, что ряд групп
обнаруживает прерывистое распространение, встре-
чаясь в Австралии, Африке, Южной Америке или в двух из этих
стран. Лишь некоторые семейства, как, например, некоторые с о-
м о в ы е (Siluridae), распространились в Арктогее, и ареал их рас-
пространения составляет одно целое в обеих областях. Другую кар-
тину дает Арктогея. Здесь за немногими исключениями распре-
деление рыб равномерное, непрерывное, и имеется большое коли-
чество указаний па недавние изменения в распространении неко-
торых рыб. Непрерывно распространены лососевые, щуковые, оку-
невые и т. д.
На основании нынешнего распространения пресноводных (и про-
ходных) рыб Эогея поразделяется па: Африканскую область (Е1),
или Эфиопскую, па Мадагаскарскую (Е2), Индийскую, или Восточ-
ную, с Малайским архипелагом (ЕЗ), Австралийскую (Е4) и Нео-
тропическую (Е5).
Арктогея распадается па Палеарктику, в свою очередь распа-
дающуюся па подобласти: Европейско-Каспийскую подобласть (А2),
Арктическую Азию, или Сибирь (А1), Байкальскую подобласть
(Ala), Мопгольско-Амурскую подобласть (АЗ), Средиземноморскую
подобласть и на Неарктику, распадающуюся на Арктическую Аме-
рику (А4), Западную Североамериканскую, или подобласть Коло-
радо (А5), и Восточную Североамериканскую, или подобласть Мис-
сисипи (Аб). Каждая из областей, конечно, имеет свою ихтиофауну
и свою историю.
280
Пресноводные рыбы Африканской области имеют сходство,
с одной стороны, с рыбами Южной Америки, с другой — с рыбами
Индии. В Африке очень обильно представлено семейство Chara-
cinidae. Семейство Mormyridae целиком ограничено Африканской
областью. Из семейства карповых—Cyprinidae—многие роды име-
ются и в Индии. Также и из сомовых — Siluridae—одни роды, об-
щие с Южной Америкой, другие— с Индией, но некоторые огра-
ничены только Африкой. В Африке имеется один из трех родов
Dipnoi — Protoptcrus, другие — в Южной Америке и в Австралии.
Представлено в Африканской области и сем. костеязычных, Osteo-
glossidae. Семейство Cichlidae, наиболее характерное для Афри-
канской области, представлено и в Южной Америке. Лабиринто-
вые рыбы — Labyrinthidae — представлены здесь и в Южпой Азии.
В Африке живут представители Crossopterygii: Polypterus и Cala-
moichthys, нигде более не встречающиеся.
Восточная, или Индийская, область с Малайским архипелагом,
как сказано, во многом сходна по своей пресноводной ихтиофауне
с Африкой. Здесь очень обильно представлено сем. карповых,
Cyprinidae, более богато, чем где-нибудь еще; также очень много-
численны представленные многими местными родами сомовые — Si-
luridae. Есть один род сем. Cichlidae; лабиринтовые, Labyrinthi-
dae, весьма многочисленны. Сем. Symbranchidae и Osteoglossidae
представлены также в Австралии.
Для Австралийской области, очепь бедной пресноводными ры-
бами, характерны Dipnoi в лице Neoceratodus; имеются здесь нем-
ногие сомовые — Siluridae. Osteoglossidae представлены одним
родом, характерным для этой области, сем. Symbranchidae — двумя
видами. Наиболее здесь многочисленны рыбы сем. Serranidae
из колючеперых (Acanthopterygii), имеющие общий род (Latcs)
с Индийской областью. Чрезвычайно интересно сем. Galaxiidae
(изЕзосИогтез), представленное восемью видами в Новой Зеландии,
несколькими на юге Австралии, несколькими па юге Африки и ря-
дом видов в Чили и Патагонии в Южпой Америке. Живет в Новой.
Зеландии и угорь (Anguilla).
Неотроиичсская область характеризуется в области пресновод-
ной ихтиофауны присутствием третьего рода Dipnoi — Lcpidosiren.
Карповых — Cyprinidae — здесь пот совершенно, и их место зани-
мает общее с Африкой сем. Characinidae. Обильно представлены здесь
и сомовые, Siluridae, многие подсемейства которых только здесь
и встречаются. Характерно для Южной Америки также сем. Gym-
notidac, нигде более пе встречающееся. Имеются здесь представи-
тели и сем. Osteoglossidae. Очень обильно представлено встречаю-
щееся и в Африке сем. зубатых карпов, Cyprinodontidae, представ-
ленное еще па юге Сев. Америки, в Африке, Южной Европе и Индий-
ской области. Многие роды только здесь и встречаются. Имеется одни
вид индийского рода Symbranchus (сем. Galaxiidae). Представлено
здесь, как выше сказано, и сем Cichlidae.
Мадагаскарская область характеризуется главным образом
отрицательно: отсутствием большинства семейств, свойственных
281
другим областям. С Южной Америкой, Африкой и Индией она
сходна по наличию Cyprinodontidae; с теми же областями и с Авст-
ралией — присутствием здесь сем. Kuhliidae (из Acanthopterygii)
и сем. Cichlidae.
Таким образом, целый ряд семейств Teleostei и отряд Dipnoi
объединяют Южную Америку, Африку, Индию, Австралию и Ма-
дагаскар (см. в Отделе систематики карты распространения сем.
Characinidae, Osteoglossidae, Dipnoi, Cyprinodontidae, Galaxiidae,
Cichlidae). Так как семейства эти пресноводны, то опи пе могли рас-
пространиться столь_широко по разным материкам через моря. Со-
вершенно очевидно, что когда-то они населяли одну страну, в ко-
торой находились пункты их теперешнего распространения. Иначе
говоря, когда-то была сухопутная связь Южпой Америки, Африки,
Индии и Австралии с Новой Зеландией (т. е. не было южной части
Атлантического океана). Когда и как указаппая связь осуществля-
лась, показывают нам геология и палеонтология (см. ниже).
Арктогея распадается па две области: палеарктиче-
скую, обнимающую Европу, Северную Африку до Сахары и
Азию за исключением Индийской области, и неарктиче-
скую, представленную Северной Америкой. Та и другая распа-
даются на подобласти.
Палеарктическая область характеризуется отсутствием
Dipnoi, костных ганоидов — Holostei, присутствием «хрящевых
ганоидов» — Chondrostei. Из Teleostei характерны карповые —
Cyprinidae и лососевые—Salmonidae. Сомовые — Siluridae — почти
отсутствуют, представлены лишь несколькими подами. Характерны
также щуковые — Esocidae, хотя представлены опи только двумя
родами — Esox и Umbra. Много видов окуневых — Percidae,
Характерен также пресноводный представитель Gadidae—палим,
Lota, и ряд сельдей — Clupeidae, большая часть которых живет,
однако, в морях.
Неарктическая область очень близка к палеарктической. Здесь
также распространены Salmonidae, Cyprinidae, Esocidae, Percidae,
представленные другими видами и даже нередко родами. Отли-
чается Неарктическая область присутствием в пей Holostei (Amia
и Lepidosteus). Сомов — Siluridae — здесь больше, нежели в Па-
леарктике. В южных частях страны имеются зубастые карпы—
Cyprinodontidae. Из Acanthopterygii для Северной Америки харак-
терно сем. Centrarchidae в реках восточной части.
Таким образом Арктогея резко отличается от Эогеи, и распреде-
ление в ней различных семейств значительно однообразнее, более
непрерывно (см. в Отделе систематики карты распространения
Gasterosteidae, Esocidae, Salmonidae).
Распределение морских рыб определяется теми
же причинами, как и распределение наземных животных. Но в море
гораздо меньше тех непроходимых барьеров, которые разделяют
сушу на определенные участки. Рыбы, приспособившиеся к жизни
в океане, широко в пем распространяются, так как условия явля-
ются сходными на большом протяжении. Кроме того, подразделе-
282
ние океана ца области и участки затруднено тем, что он имеет глу-
бину, а с глубиной условия меняются подобно тому, как они меня-
ются при подъеме на высокие горы. Поэтому, папример, арктиче-
ские полярные рыбы, опускаясь на известную глубину, могут найти
для себя сносные условия и в тропических морях и распространиться
таким образом, в антарктическую область. Это мы и видим на самом
деле: в антарктических водах есть ряд рыб, свойственных арктиче-
ским водам. Выше было сказано, что условия жизни в море весьма
разнообразны в зависимости от глубипы, близости берега и других
факторов. Поэтому было бы совершенно неправильно делить море
на области, пе считаясь с тем, имеем ли мы в виду береговую
фауну, глубинную или пелагическую. Деление на зоогеографиче-
ские области и подобласти может быть проведено для каждой из упо-
мянутых фаун в отдельности. Таким образом, по отношению к фауне
моря мы будет придерживаться следующей схемы.
А. Прибре.жное (литоральное) царство
I. Арктическая циркумполярная область.
II. Атлантическая область.
1. Атлантическая бореальная подобласть.
2. Атлантическая тропическая подобласть.
III. Индийско-тихоокеанская область.
1. Тихоокеанская бореальная подобласть.
2. Тихоокеанская тропическая подобласть.
IV. Антарктическая область.
Б. Глубинное (абиссальное) царство
I. Глубинная (абиссальная) область
В. Пелагическое царство
I. Арктическая область.
1. Арктическая циркумполярная подобласть.
2. Атлантическая бореальная подобласть.
3. Тихоокеанская бореальная подобласть.
II. Индийско-тихоокеанская область.
III. Атлантическая область.
IV. Антарктическая область.
Наиболее разнообразно по условиям жизни прибрежное царство.
Здесь различают различные з о н ы с неодинаковыми физическими
условиями, с различной растительностью, грунтом и с различными
беспозвоночными животными. Меньше разнообразия в пелагиче-
ском царстве. Но и зд<юь существуют различия между богатой рас-
тениями верхней с в с т л о й з о н о й и темной — нижней,
благодаря различиям в физико-географических условиях между
светлой и темной зонами в составе их фауны существуют большие»
отличия. По мерс увеличения глубипы жизнь постепенно замирает.
(’больших глубин, вдали от континентов, были добыты лишь не-
многие рыбы, своеобразно приспособленные к условиям, здесь суще-
ствующим. Фауну больших глубин молено в известной мере сравни-
вать с фауной высоких гор, с той их частью, которая была поднята
вместе; с горообразованием. С древнейших времен сюда попадали
рыбы из береговой области и из верхних слоев моря. Постепенно
283
изменяясь, рыбы приобретали специфический характер, о котором
говорилось выше, и в то же время распространялись все шире
и шире.
Арктическая циркумполярная подоб-
ласть не богата видами береговых рыб. К таковым принадлежит
чстырехрогий бычок (Cottus quadricornis), некоторые виды ликодов:
зеленый ликод (Gymnelis viridis), виды морских собачек: стихей
(Lumpenus maculatus).
Атлантическая бореальная подобласть
богаче формами рыб. Мы видим здесь бычков (Cottidae), лисичек
(Agonidae), пинагоров (Cyclopteridae), морских собачек (Blen-
nidae), тресковых (Gadidae), роды: Molva, Gadus, Onos и др.,
сельдей (Clupeidae), ликодовых (Lycodidae), камбаловых (Pleuro-
nectidae), из акул—гренландскую акулу (Laemargus), и колючепе-
рую акулу (Acanthias), настоящих скатов (Rajidae). В более уме-
ренных водах бореальной подобласти мы видим морских окуней
(Serranidae), рукоперых (Pediculati), из акул появляется морская
собака (Mustelus), из скатов — хвостоколы (Trygon), гпюсы (Tor-
pedo). В Средиземном море примешиваются уже рыбы тропических
морей: из семейств верхоглядов (Uranoscopidae), сфиреп (Sphyrae-
nidae), конских макрелей (Carangidae), морских коньков (Hippo-
cam pidae) и губанов (Labridae).
Тихоокеанская бореальная подобласть
обнаруживает близкое родство с Атлантической, но богаче ее. Здесь
многочисленны бычки (Cottidae), тресковые (Gadidae), камбаловые
(Plueronectidae), лососевые (Salmonidae). В Атлантике нет ни од-
ного семейства, которое пе было бы представлено в Тихом. В пос-
леднем же есть несколько таких семейств, которых пет в Атланти-
ческом: семейство хиров (Hexagrammidac), семейство Embiotoci-
dae и др. Атлантическая бореальная подобласть есть обедненная
тихоокеанская.
Тропические моря имеют много общего и по характеру ихтио-
фауны могут быть сгруппированы в «тропическую зону».
Береговая фауна рыб тропических морей богата как по числу
форм, так и по их разнообразию. Экологические условия жизни
здесь богаче, благоприятнее для жизни: вода теплее, больше света
и т. д. Рыбы тропиков отличаются своей причудливой формой, яр-
кими красками, блеском покровов. Сходство рыб во всей тропической
зоне объясняется сходством условий существования, тем, что тро-
пическая Атлантида заселялась пз Тихого оксана через Панам-
ский канал еще в третичное время, а фауны.Тихого и Индийского
океанов могли сравнительно легко смешиваться. Из наиболее ха-
рактерных семейств назовем: чешуепсрых (Squamipennes), губанов
(Labridae), кузовков (Ostraciontidac), спипорогов (Balistidae) и др.,
щетипозубов (Chaetodontidae), рифовых рыб (Pomaccntridac), отряд
сростночелюстных (Plectognathi). Особенно богаты фауной воды
Индийско-тихоокеанской, или Восточно-ин-
дийской области, .где мощно развиваются коралловые
рифы с их богатейшей фауной рыб.
284
Антарктическая область, как было выше ска-
зано, имеет ряд форм, общих с Арктической. Сюда заходят многие
роды семейства Zoarcidae, род Sebastes, из бычков характерен род
Sclerocottus, из тресковых — род Muraenolepis, встречаются роды
лисичек (Agonus), л и к о д ы (Lycodes) и колючеперые акулы
(Acanthias). Последние формы никогда не встречаются в промежу-
точных теплых морях. Они могли проникпуть сюда только через
глубины с холодной водой. Но в Антарктике есть и свои,
характерные для области рыбы, объединяемые в одно семейство
Nototheniidae, нигде более не живущие.
Пелагических рыб несравненно меньше, нежели береговых.
Особенно мало их в холодных морях. Мы встречаем в арктических
водах, пожалуй, один лишь вид настоящей пелагической рыбы —
гренландскую акулу (Laemargus borealis). В тропических и субтро-
пических водах мы видим опять-таки прежде всего пелагических
акул — настоящих акул (Carcharias), громадных хархародон(СЬаг-
charodon), в более умеренной зоне Атлантики и Тихого океана —
сельдевую акулку (Lamna cornubica). К пелагическим, притом жи-
вущим за счет планктона, акулам принадлежит громадная Cetorhi-
nus (Seiache) maximus в Атлантическом океане. Далее, к пелагиче-
ским рыбам принадлежит богатый видами род летучей рыбы(Ехосое-
tus) из тропических вод Атлантического, Тихого и Индийского
океанов. Атлантические тупцы (Thynnus thynnus и др.), макрели
(Scomber scombrus), меч-рыба (Xiphias gladius) из Атлантического
океана и napycniiK(Histiophorus gladius) из Индийского океана. Из
батипелагических рыб назовем светящихся анчоусов (Scopelidac),
семейство складчатогрудых (Sternoptychidae) с замечательным ро-
дом хаулиод (Chauliodus). Наконец, к пелагическим рыбам следует
отнести и пассивно плавающую иглу-рыбу (Syngnathus pelagicus)
и даже некоторых морских копьков (Hippocampus). Зато молодь
у большого количества рыб является пелагической.
Нарисованная выше картина современного распространения
рыб как пресноводных, так и морских объясняется, помимо эко-
логических условий, историей их возникновения и развития, и уче-
нием Вегенера о смещении континентов. Ряд фактов говорит о том,
что разъединенные ныне материки были раньше связаны. Остатки
рыб, имеющих ныне разорванные ареалы в Эогее, как Dipnoi,
Crossopterygii, найдены в северном полушарии, где опи были ши-
роко распространены. Также Palaeoniscoidei и другие рыбы древ-
него красного песчаника были распространены в Сев. Америке,
в Англии, Европе, Южной Африке, Австралии.
Объяснить этот космополитизм пресноводных рыб можпо лишь та-
ким образом, что раньше материки были связаны, рыбы развивались
в больших реках, бассейны которых занимали не один, а два из сов-
ременных материков. Иначе как бывшей связью Южной Америки и
Африки, существованием большой реки,притоком которой являлась,
г одной стороны, пынепшяя Амазонка, а другой — Нигер и Конго,
нельзя объяснить наличие и в Бразилии, и в Африке харациповых
(Characinidae), поразительное сходство не только ихтиофауны,
285
но и многих групп беспозвоночных в Бразилии и в озере Танга-
найка, в Африке.
Точно так же только связью материков, входящих в Эогею,
можно объяснить нахождение сем. Cichlidae в Южной Америке,
Африке, Индии, а также подобное же распространение зубастых
карпов (Cyprinodontidae), или семейства Galaxiidae в реках Южной
Америки, Австралии, Новой Зеландии.
На непрерывную связь ныне разъединенных материков указы-
вает и распространение так называемых ганоидов.
Нынешние осетровые Polyodon, Pseudoscaphirhynchus и др.—
лишь обломки большого ствола Chondrosteoideae, широко распро-
страненного в юрское время, потомки, сохранившиеся в отдельных
пунктах прежде широкого и непрерывного распространения. Дру-
гим реликтом являются происшедшие от Palaeoniscoidei Amia
и Lepidosteus. Amia в юрскую и меловую эпохи была в Европе,
где сохранялась до нижнего миоцена. Таюке и Lepidosteus был
в Европе. Теперь они сохранились только в Сев. Америке. Кости-
стые рыбы (Teleostei) связаны с Holosteoidei и, быть может, непо-
средственно с Palaeoniscoidei. Связующее между Teleostei и их
«ганоидными» предками звено Leptolepidae имело в меловое время
распространение во всем свете.
На связь в палеозое-мезозое всех континентов между собой ука-
зывают прерывистое распространепие ряда семейств Teleostei
и общность пресноводной ихтиофауны во всей Эогее.
Указанная связь материков осуществлялась пе при помощи «мо-
стов», т. е. перешейков, то погружавшихся в море, то снова подни-
мавшихся. Все материки представляли первоначально одно целое,
покоящееся на пластической магме. Поверхность суши в палеозое
и мезозое была совсем иная, чем теперь; земля была низкая, ровная,
однообразная. По пей медленно текли большие реки, прерывавшиеся
обширными заболоченными озерами и болотами. Были внутренние
моря. Климат был иной, жаркий пояс простирался гораздо дальше
и к северу, и к югу. Целость суши мало нарушалась в палеозое и
мезозое. Наступания и отступания моря имели местный характер.
Опи вызывали смешение пресной и морской воды, последствием ко-
торого была массовая гибель рыб.
Раньше всего целость суши нарушилась в центре и на крайнем
юге, где разделение Южной Америки и Африки закончилось в мелу.
В остальной части опа сохранялась до третичного времени. Отде-
ление Ирландии и Шотландии от Ньюфаундленда и Южной Гренлан-
дии произошло позднее всего, вероятно, в ледниковое время. До на-
чала третичного времени существовало и соединение Южной Индии
с Мадагаскаром. Разрыв и отхождение Австралии от Африки и Юж-
ной Америки произошел еще раньше, вероятно, в лиасе (юра).
Нарисованная геологическая картина выясняет нам и присутст-
вие древних примитивных групп рыб в Эогее, и прерывистое распро-
странепие Dipnoi и Crossopterygii, равно как и позднее появляю-
щихся Characinidae, Cyprinodontidae, Cichlidae; связь Америки
с Австралией объясняет распространепие тех пресноводных форм,
286
которые встречаются и в Америке, и в Австралии. Связь между
Америкой и Африкой и речные системы были таковы, что делали
воможпыми миграции американских форм в Старый Свет.
Распространенные раньше широко и в северном и в южном полу-
шариях древние примитивные формы, как Dipnoi, Crossopterygii
и т. д., были затем вытеснены и исчезли в северной части своего-
прежпего распространения, возникшие же в Эогее формы не смогли
распространиться на север,вАрктогею,таккак вследствие наступив-
ших изменений климата земного шара они не могли найти здесь
подходящих условий для своего существования.
В Арктогее, хотя и происходил ряд перемен, все же суша здесь,
оставалась непрерывной и до педавпего еще времени между Пале-
арктикой и Неарктикой существовала связь. Благодаря этому рыбы
имели в Арктогее больше возможности широко распространиться
и сохранить свои обширные ареалы до наших дней. Например,
щуковые (Esocidae), колюшковые (Gasterosteidae), лососевые (Sal-
monidae) и т. д. Такое широкое распространение объясняется еще
и тем, что некоторые северные семейства являются рыбами пробод-
ными, или же прибрежными, т. е. они могли пользоваться и мор-
скими дорогами для расселения.
Колыбелью морских рыб является, видимо, Тихий океан и вос-
точная часть Индийского. Атлантический океан, позднее образовав-
шийся, заселялся с юго-востока по морским проливам, пересекав-
шим сушу в различные эпохи в направлении от юго-востока Азии
через Европу к Гренландии и Сев. Америке, а также через область
нынешнего Панамского капала.
Последним путем проникли сюда немногие рыбы. Большинство
костистых рыб проникло в европейские и азиатские моря лишь
в меловую эпоху и в эоцене западным путем, образовав здесь пыпе
живущие семейства и роды. Тропическая область Атлантики полу-
чила свое рыбье паселеписиз Тихого оксапа в пачале третичной эры
через Панаму и со стороны Европы. Очень многие роды являются
общими Индийскому океану и Средпеземпоморыо.
С уничтожением непрерывности суши на юге открылась дорога
с юга в северную часть Атлантического океапа. На севере же пос-
ледний был долго закрыт мостов суши менаду северными частями
Евразии и Сев. Америки. Это создавало благоприятные условия
для вторжения в Арктогею с юга главным образом прибрежных
и пелагических рыб. Последние присоединялись к тем, которые
еще задолго до открытия Атлантического океапа проникли сюда
с востока через моря в упомянутые выше морские проливы, к сель-
дям и акулам.
Это пропикповеппе с востока устанавливается на основании
распространения ископаемых и пыпе живущих форм. Тунцы, скум-
брии, рыба-меч пришли сюда этим путем. Многие другие рыбы
пришли с юга. Тресковые могли притти из Тихого океана через Бе-
рингов пролив.
Таким образом, ихтиофауна Арктогеи сложилась из форм, воз-
никших па месте, и из пришельцев из Тихого океана, проникших
287
с востока, частью через Берингов пролив, частью через Панамский
пролив, бывший на месте Панамского перешейка. •
Ледниковый период оказал большое влияние на рыб
Арктогси, так как лед и в Европе и в Америке спускался далеко
к югу. Он оттеснил к югу многих рыб, которые нашли себе приют
в Черпом, Азовском и Каспийском морях, некогда между собой
соединенных. Важным убежищем служил также Амур. То же про-
исходило и в Сев. Америке. Когда же ледник стал отступать, то
часть рыб, загнанная ледником к югу, осталась в той области,
куда она была загнана льдом: белута (Huso huso и Huso dauricus)
осталась в Черном и Каспийском морях и в Амуре, сиги (Core-
gonus) и палья (Salmo salvelinus)—в Англии и в Альпах, туркестан-
ский лосось (Salmo oxianus)— в Аральском море. Оставшись в изо-
лированных озерах, эти формы приспособились к особым условиям
и дали местные расы (племена), подвиды или виды. Они стали прес-
новодными рыбами, в то время как их северные родственники про-
водят часть своего времени в морях, т. е. являются проход-
ными рыбами. Другие рыбы в послеледниковый период ус-
пели широко расселиться. Так, судак (Lucioperca) распростра-
нился на запад, стерлядь (Acipenser ruthenus) — па восток. Пре-
образования моря и суши замкнули в Великих озерах Сев. Америки
мипог, давших здесь новый род Ichthyomyzon.a в Каспийском море—
род Caspiomyzon. Байкал оледенению, видимо, пе подвергался,
так как в нем еще живут роды и виды, которые, надо полагать,
жили здесь в течепие третичной эпохи. Часть видов он получил в
послеледниковое время.
Таким образом, история, приведшая к современному распростра-
нению пресноводных рыб и очерченная здесь лишь несколькими
штрихами, весьма сложна. Конечно, распространяясь, попадая в но-
вую и новую среду обитания, рыбы все время эволюционировали,
изменялись, приспособляясь к новым условиям существования,
давали новые расы, подвиды, виды, роды, семейства.
12. Поведение рыб (высшая нервная деятельность)
Выше была разобрана морфологическая эволюция классов
рыб, эволюция их организации. Нервная деятельность, в высшей
ее форме выявляющаяся в виде поведения и психики, связана не-
разрывно с наличием нервной системы. Последняя в течепие эволю-
ции животного мира прошла путь постепенного усложнения и раз-
вития. Естественно думать, что и поведение также эволюциопиро-
ровало. Каково же поведение рыб? Какова их психика? Ответ па этот
последний вопрос крайне труден, так как психические явления
(ощущения, мысли, чувства, желания) пе доступны объективному
наблюдению даже у других людей.
И какая бы то пи было психология, даже психология человека,
была бы’ невозможна, если бы объективная сторона единого процесса
поведения — деятельность — не была теспо и неразрывно связана
с субъективной стороной—психикой. Чтобы судить о последней,
288
мы должны тщательно изучать поведение, особенности деятельности
животного, и по ним умозаключать о субъективной стороне —
о психике. Трудности, которые встают здесь перед нами, очень ве-
лики, но все же до известной степени, но только до известной, прео-
долимы. Поведение изучается при помощи эксперимепта, кроме
которого необходимо еще принимать во внимание строение и физио-
логию аппарата поведения и психики — нервной системы и орга-
нов чувств.
Рыбы имеют развитую центральную нервную си-
стему, состоящую из головного и спинного мозга. Однако головной
мозг рыб относительно мал и не выполняет черепной полости.
У взрослого палима (Lota lota) вес мозга равняется Vjoo части веса
тела, у щуки (Esox lucius) —Узоо части, а у громадного тунца (Thyn-
nus thynnus) on равен лишь г//з7ооо веса тела. Вместо коры большого
мозга у рыб имеется тонкий эпителиальный слой, «паллиум», не со-
держащий нервных клеток. Лишь у акул имеются зачатки коры
большого мозга. Сильно развиты у рыб обонятельные доли и зри-
тельные бугры, а также мозжечок.
Органом обоняния рыб являются ноздри, представленные
простыми ямками, снабженными нервами от обонятельной доли
мозга. Ямки эти часто снабжены клапанами или края их вытянуты
в виде трубочек. Нельзя с достоверностью сказать, имеют ли рыбы
органы вкуса. На нёбе и в глоточной области у многих рыб сли-
зистая оболочка обильно снабжена нервами, отходящими от лице-
вого нерва.Ветви идут к губам и усикам, а, по Херрику, у некоторых
сомовых окончания этих нервов имеются по всей наружной поверх-
ности тела. Автор этот утверждает, что вкусовые точки па боках
этой рыбы помогают ей находить пищу. Обоняние у рыб, неви-
димому, весьма остро, и рыбы привлекаются к добыче ее за-
пахом, что хорошо известно удильщикам.
Ряд опытов показал, что запах пищи вызывает в рыбе возбужде-
ние, ио пе направляет се движений. Если в аквариуме со Sphcroidcs
maculatus подвешивались два пакета: один с мясом, другой пустой,
то первый бывал укушен 119 раз, второй за то ate время — 18.
Когда путем завязывания трубки ноздрей шелковой питью функция
органа обоняния была устранена, рыбы совсем не обращали вни-
мания па пакет с мясом. Нормальные акулы (Mustelus canis) бы-
стро находили мясо краба, акулы же с закрытыми ноздрями, или
же с перерезанными обонятельными нервами, не обнаружили пи ма-
лейшего интереса к крабам. Золотая рыбка, поедающая личинок
муравья, пе трогает их, когда они памазапы гвоздичным маслом.
Таким образом, рыбы, несомненно, обладают чувством
обоняния, которое у пих локализуется в органе обоняния —
эпптелпи носовых ямок.
'Можно было бы думать, что, хотя рыбы и реагируют на пахучие
стимулы, здесь нельзя говорить об обонянии, так как при обонятель-
ных ощущениях чувствительный эпителий раздражается газами,
в воде же этого пе может быть. Однако было доказано, что и у
воздуходышащих наземных позвоночных пахучие газы действуют
19 Кашкаров и Ставчиаский 289
на обонятельный эпителий, предварительно растворившись в сли-
зи, покрывающей обонятельную поверхность,
Опытами же обнаружена необычайная чувствитель-
ность рыб к кислотности и щелочности. Пе-
ремены состава воды и содержания в ней углекислоты, кислорода,
количества водородных ионов и т. д. вызывают перемены в поведе-
нии рыб. Это обстоятельство важно для понимания миграции рыб.
Орган слуха рыбы состоит только из лабиринта, заключаю-
щего преддверие и обычно три полукружных капала, расширяю-
щихся в ампулы, содержащие один (или более) свободно лежащий
отолит, или слуховой камешек. Ни наружного уха‘, ни барабанной
перепонки, ни евстахиевой трубы у рыб нет. Уже по устройству
органа слуха сомнительно, чтобы рыбы вообще слышали в настоя-
щем смысле слова. Целый ряд экспериментов показал, что к тонам
рыбы чувствительны, а чувствительность к шумам локализована
у них в коже. Известные случаи, когда рыбы приплывали на звон ко-
локольчика, объяснялись тем, что рыбы видели звонящего человека,
и сотрясение земли от шагов подходящего могло передаваться в воду.
Однако опыты с образованием у рыб условного рефлекса «пища —
звук», при условии закрытия производившего звук и кормящего че-
ловека ширмой, говорят все же за то, что звук воспринимается ры-
бами. За это же говорят и те опыты, в которых рыбы реагировали
па звон колокольчика в аквариуме, по переставали реагирова?ь,
если между язычком и колокольчиком была проложена кожа. Опыты,
в которых слуховой перв перерезался, показали, что он необхо-
дим для реакции па вибрации струны. Но это еще не значит, что
рыбы «слышат» так, как мы воспринимаем звуки.
Весьма интересно отношение органа слуха у рыб к органам
чувств боковой линии; функция последних еще пе впол-
не ясна. Совершенно несомненно близкое отношение уха к боковой
линии и пе только морфологически, по и с точки зрения их функций.
Органы боковой линии пе отвечают па раздражения светом, теплом,
соленой водой, пищей, кислородом в воде, углекислотой, гнилой
водой, постоянным течением и звуком. Но рыба, реагирующая
на медленные вибрации — 6 раз в секунду, перестает реагировать,
если перерезать нерв боковой линии. Это приводит некоторых уче-
ных к выводу: раздражение, вызывающее реак-
цию со стороны боковой линии, а именно
медленные неслышные вибрации, имеет ха-
рактер промежуточный между стимулами
для кожи (давление) и стимулами для уха
(частое колебание). Таким образом, ухо есть изменен-
ная часть органов боковой линии, и слух — деликатнейшая
форма осязания.
Несомненно, что лабиринт рыб служит органом равно-
весия. Удаление его ведет к потере равновесия тела.
Глаза рыб по деталям значительно отличаются от глаз на-
земных животных. Роговица лишь очень слабо выпукла. Век пет.
Хрусталик шарообразной формы. Такая крайняя выпуклость его
2 90
является необходимой потому, что хрусталик сам по себе немпого
плотнее жидкости, в которой рыба живет. Затем радужная оболочка
у рыб неподвижна, так что зрачок у пих не может суживаться и рас-
ширяться. У большинства рыб от сосудистой оболочки отходит так
называемый серповидный отросток, идущий к лин-
зе, и образующий здесь расширение, так называемый галле-
ров колокол. Этот отросток представляет мускул, сокраще-
ние которого оттягивает хрусталик назад, что играет роль в акко-
модации рыбьего глаза.
В связи с жизнью в глубинах развиваются рудиментарные, за-
чаточные глаза или же особые телескопические, с парал-
лельно лежащими осями и допускающие бинокулярное зрение.
Нормальная установка глаза рыбы рассчитана пе на дальнее,
а на близкое расстояние. Все рыбы в норме близоруки, глаза их
установлены примерно па один метр расстояния. Аккомодация
к зрению па дальнее расстояние совершается при помощи оттягива-
ния пазад хрусталика посредством серпообразного отростка. При по-
мощи этого приспособления рыбы могут видеть на расстоянии при-
мерно 10—12 м. Аккомодационная сила не одинакова у разных рыб,
по вообще слаба.
Вопрос о том, различают ли рыбы цвета, исследовался прежде
всего методом дрессировки, — устанавливалась ассоциа-
ция цветных предметов с пищей. Рыб, папример, кормили,
давая пищу цветными щипцами. Когда рыба научилась плыть
за пищей к красным щдпцам, когда у пес установилась ассоциация
«красное—пища», тогда пищу (червь) стали давать зелеными щип-
цами. После некоторого периода — переобучения с частыми вначале
ошибками — вместо прежней ассоциации развивалась повая: «зе-
леное — пища».
Другой метод заключался в том, что аквариум освещали раз-
личными цветами и наблюдали, отдают ли рыбы предпочтение
тому или иному освещению. Опыты велись с рыбами, бывшими пред-
варительно па свету или же в темноте. У некоторых авторов ре-
зультат получился положительный: рыбы различают пе только
яркость, по и цвет, когда опи приспособились к зрепию па свету.
То же самое наблюдается у человека. Приспособляясь же к темноте,
рыбий глаз теряет способность различать цвета, подобно томукак
это имеет место у человека.
Другой взгляд был высказан па основании весьма тщательных
опытов и заключается в том, что рыбы ведут себя по отношению
к свету совершенно так ясе, как слепой па цвета человек, так как
относительная яркость различных частей спектра для пих почти
или всецело такова же, как у совершенно слепого на цвета человека
при всякой яркости. У нормального же человека это бывает, когда
его глаз приспособился к темноте, и яркость незначительна. Ате-
рипка (Atherina hepsetus) в опытах па дневном свету искала самого
яркого места. Бассейн с рыбами был освещен наполовину красным,
наполовину голубым светом, причем можно было изменить силу
света обоих цветов. Рыбы предпочитали синее красному. Помещая
49* 291
рыбок в аквариум, освещенный спектром, можно было видеть,
что рыбы собирались в желтозеленой и зеленой части спектра.
Увеличивая яркость других цветов, можно было привлечь туда рыб;
уменьшая яркость — изгнать. Двигая вдоль стенки аквариума
картон с фиолетового конца к красному, можно было гнать влево
и рыбок до известного пункта в красной части спектра, где рыбы
цвета, невидимому, уже не ощущали, так как распределялись
в аквариуме равномерно. То же укорочение спектра слева наблю-
дается и у слепого к цветам человека.
Возможно, что цветное зрение рыбам и не нужно, ибо в глубине
воды цвета спектра не видны благодаря поглощению лучей водой.
Наоборот, различие в яркости очень полезно, а потому способ-
ность различать яркость у рыб развита. Во всяком случае зрение
играет в поведении рыб большую роль.
Осязание развито у рыб хорошо. Многие рыбы имеют особые
органы для ощупывания дна и разыскивания пищи. Таковы, па-
пример, усики и мясистые выросты, которые всегда являются орга-
нами осязания, где бы опи пи развивались, в связи ли с челюстными
органами, или на коже рыла, или па плавниковых лучах. Таковы
усики стерляди, сомов, пескарей, пальцы тригл (Ttrigla gurnar-
dua) и т. д.
Возможно, как указывают некоторые авторы, что органы боковой
линии служат для восприятия, сходного с осязанием, а именно для
восприятия движения воды. Стоящей на месте в аквариуме рыбке
при помощи капиллярной трубки направлялась па боковую линию
струя воды, и рыбка сейчас же ориентировалась в пространстве та-
ким образом, чтобы стоять головой против течения воды.
К изменениям температуры рыбы также очепь чувстви-
тельны. Для каждого вида существует своя наиболее благоприят-
ная температура, одпп рыбы ищут теплой, другие — холодной
воды. Что касается способности рыб чувствовать боль, об этом еще
мало известно. Но те указания, которые есть, говорят за то, что рыбы
чувствуют боль слабо, если вообще чувствуют. Акулу, пожираю-
щую труп кита, можпо повторно багрить за голову, и опа пе поки-
дает своей добычи.
Что касается способности рыб паучаться, образовывать при-
вычку и впредь руководиться этим ощущением, то мы имеем для
ответа па этот вопрос мало данных. Ответа искали в опытах с лаби-
ринтом. Лабирипт применялся чрезвычайно примитивный, простой.
Результат получился положительный. Но поставленные опыты по до-
статочно многочисленны и слитком кратки. Опыты с более сложным
лабиринтом привели к отрицательным результатам. Рыбы но на-
учились находить дорогу в лабирппте и обнаружили такой характер
реакций, который делает мало вероятной способность их паучаться.
В сравнении с млекопитающими и птицами рыба — совершенно
тупое животное, живущее главным образом простыми врожденными
рефлексами и едва способное к научеппю путем опыта. Это связапо
с тем, что у рыб в головпом мозгу отсутствует то, что соответствует
серой коре полушарий большого мозга.
292
VII. ЗНАЧЕНИЕ РЫБ ДЛЯ ЧЕЛОВЕКА
Значительный процент всех продуктов животного происхожде-
ния, потребляемых на территории СССР, дает рыба. Есть области,
где питание рыбой играет исключительно большую роль. Так, на-
пример, народы Северной и Восточной Сибири питаются в значи-
тельной мере рыбой; рыба играет большую роль в диете прибреж-
ного населения бывшей Архангельской губернии. В 1910 г. улов
рыбы в России выражался цифрой 1135 млн. кг по официальным дан-
ным. На самом же деле он достигал 1323 млн. кг. И однако по-
требность в рыбе была настолько велика, что постоянно еще ввозили
значительное количество рыбы.
За последнее пятилетие перед империалистической войной ввоз
был от 331 до 371 млп. кг в год на сумму от 28.4 до 38.6 млн. рублей.
При этом до 288 млн. кг приходилось на соленую сельдь, кото-
рую привозили из Англии, Германии, Норвегии и Швеции. Хотя
в то же время мы вывозили рыбу в количестве от 24 до 46.4 млн. кг
па сумму от 8.4 до 9.2 млн. рублей в год, тем не менее ежегодно пере-
плата за необходимую нам рыбу была от 2Q до 80 млн. рублей.
В 1918—1920 гг. ввоза не было, по с 1921 г. он вновь начался. Однако
теперь даже наибольший привоз в 1924—1925 гг. не превышал 80.5
млп.кг, а в 1927 г. он оценивался всего в 3 594 000 рублей, так как
на рыбные богатства было обращено больше внимания, и добыча
рыбы сильно повысилась. В настоящее время опа значительно воз-
росла.
Таким образом, потребность в рыбе у пас огромная, и рыба яв-
ляется предметом особого внимания со стороны государства. Неко-
торые рыбпые продукты, как, например, икра осетровых имеют боль-
шое значение, являясь предметом вывоза. Другие служат для мас-
сового потребления; к таковым относятся: треска, пикша,
сайда (Gadidae), сельди (Clupeidae), кета (Oncorhyn-
chus), салака (Clupea hareugus nicmbras), снетки (Osmerus
eperlanus spirinchus), вобла (Rutilus rutilus caspius), ч e x о п ь
(Pelecus cultratus), сом (Silurus glanis), сазан (Cyprinus
carpio), судак (Lucioperca lucioperca), л с щ (Abramis brama),
щука (Esox lucius), камбалы (Pleuronectidae), хамса (Eng-
raulis encrassicholus) и др.
Что рыба может заменить собой мясо теплокровных, показы-
вает тот факт что в течение целых поколений люди могут питаться
даже исключительно одной рыбой.
• 'равнение мяса рыб с говядиной показывает ряд преимуществ
в ряд недостатков того и другого. Например, белков в мясе рыб вооб-
ще меньше, чем в мясе вола, но встречаются рыбы и более богатые
ими. Фосфористыми белками, которые имеют преобладающее зна-
чение в питании, рыбье мясо пе беднее говяжьего. Рыбий навар
и желудке вызывает более обильное отделение сока, чем навар мяс-
шin. IЬ (желудочный сок выделяется под влиянием рыбной пищи
в большем количестве, копцептрировапнее и богаче белковым фер-
ментом. По наряду с этими преимуществами, рыбная пища обла-
293
дает и рядом недостатков. Рыбная пища требует при заготовлении
тщательной обработки, в противном же случае продукт теряет всю
свою ценность. Например, разные виды трески вследствие дурного
заготовления были у нас совершенно малоценными с диетической
стороны и даже непригодными. А те же самые рыбы в Норвегии,
Голландии и Сев. Америке перерабывались в сухие, соленые и соле-
повялепые продукты очень высокого качества. В настоящее время
качество наших рыбопродуктов значительно поднялось.
Количество съедобных частей в рыбе колеблется от 46 до 76%.
1. Рыбные промыслы СССР и главнейшие промысловые рыбы
Мы обладаем рядом богатейших в отношении рыбных богатств
водоемов. Даже в довоенное время, при далеко недостаточном вни-
мании к природным богатствам страны, Россия занимала одно
из первых мест в рыболовстве, а именно по ценности рыбного улова
она стояла на втором месте, а по общему количеству улова —
на первом.
Мировой улов рыбы перед войной 1914 г. был 9 508,2 млн. кг,
и добыча в главнейших странах была:
Россия................................. 1 323 млн. кг
Англия (с Шотландией, Ирландией и Уэльсом) . 1 253 » »
Американские соединенные штаты.......... 901 » »
Япония.................................. 737 » »
Норвегия................................ 655 » »
Канада.................................. 416 » »
Франция................................. 358 » »
Послевоенный мировой улов во многих странах понизился.
Однако общая добыча возросла и оценивалась в 1923 г. прибли-
зительно в 800 000 000 долларов. Для отдельных стран оценка
улова такова:
Великобритания ..................... 104830 тыс. долларов
Япония.............................. 89 365 » »
Американские соединенные штаты . . . 85 000 » »
Франция............................. 84 725 » »
Испания............................. 72 200 » »
СССР................................ 50 000 » »
Канада.............................. 49 240 » »
Португалия.......................... 36 875 » »
СССР в связи с гражданской войной занял в 1923 г. шестое
место по ценности улова вместо второго, ио уже в 1927 г. по стои-
мости готового продукта оп вповь перешел па второе место, все
время увеличивая добычу, а одновременно и улучшая качество
продукции. Поисковые и другие исследовательские работы, орга-
низация промысла на коллективных началах, механизация лова,
развитие обрабатывающей промышленности и т. д. чрезвычайно
подняли количественную и качественную сторону пашей рыбной
промышленности и теперь мы по добыче рыбы стоим на первом месте.
У нас выделяются следующие районы рыболовства.
294
Каспийский район, включающий в себя, кроме
Каспийского моря, реки: Волгу, Урал, Терек, и Куру. Этот район
доставляет половину всей рыбы, улавливаемой в пределах СССР,
а именно свыше 690 млн. кг. Рыбные запасы Каспия являются глав-
ной базой нашего промысла. Наиболее старым промыслом здесь
является промысел осетровых — красноловье. Ловятся:
стерлядь, осетр, белуга, севрюга, шип. Из так называемых «части-
ковых» рыб (главным образом карповых, а также окуневых, вклю-
чая сюда также лосося и белорыбицу) ловятся: вобла (Rutilus ru-
tilus caspius), сазан (Cyprinus caprio), судак обыкновенный (Lucio-
perca lucioperca) и морской (Lucioperca marina), лещ (Abramis
brama), com (Silurus glanis), щука (Esox lucius). Это—рыбы, ловимые
в наибольшем количестве. Далее ловится ряд • высокоценных ча-
стиковых рыб: каспийский лосось (Salmo trutta labrax), белорыбица
(Stenodus leucichthys), шемая (Alburnus chalcoides), кутум (Ru-
tilus frisii kutum) и целый ряд менее ценных: густера, или тарань
(Blicca bjoerkna), синец (Abramis ballerus), белоглазка (Abramis
sapa), красноперка (Scardinius erythrophthalmus), чехонь (Pelecus
cultratus) идр. Все большее значение приобретает промысел каспий-
ской миноги (Caspiomyzon wagneri). Несмотря на очень интенсив-
ный, притом механизированный лов частиковой рыбы, запасов
рыбы, как вобла, лещ, сазан, сокращения незаметно.
Наибольшее значение имеет в Каспии лов сельди. Здесь сельдь
иная, нежели в Атлантическом океане или Белом море; опа принад-
лежит к роду пе Harengus, a Caspialosa. Их более 10 видов Наиболее
важны в промысловом отношении следующие: 1) каспийский пуза-
нок (Caspialosa caspia), 2) волжская сельдь (Caspialosa volgensis),
3) черноспинка, бешспка, или залом (Caspialosa kessleri), 4) долгин-
ская, или мапгышлакская, сельдь (Caspialosabrashnikowi), Б) астра-
бадская сельдь (Caspialosa leucoccphala). Лов сельди развился па
Каспии лишь в конце прошлого и пачале нынешнего века, быстро
увеличивался, и увеличение это явилось результатом пе перелова,
а более рациональной и интенсивной эксплоатации.
50-е гг. дали.......................... 53 млп. кг
90-о » ».......................... 423 » »
1912—4915 гг. дали.................... 2G2 » »
Сельдяной лов в Каспии обеспечен па долгие годы, а малые
уловы сельди в некоторые годы объясняются случайными причи-
нами. В настоящее время промыслы па Каспии механизированы
и поставлены па должную высоту.
Особенное значение в Каспийском райопе приобрело изготовле-
ние черной икры, являющейся ценным продуктом для вывоза па
заграничные рынки.
Промыслы находятся главным образом в устьях Волги, имея тен-
денцию передвинуться в море, и в устьях реки Куры. В Астрахани
находится образцово поставленная ихтиологическая станция для
научной разработки вопросов каспийского рыболовства. Другая
станция находится на реке Куре. Третья — в Красноводске.
295
Тихоокеанский, или Дальневосточный,
район охватывает прибрежные области морей Японского, Охот-
ского, Камчатки и Анадырского края, а также и впадающие сюда
реки, на первом месте среди которых стоит Амур. Здесь русское
рыболовство возникло недавно: лишь с 1904—1906 гг. оно сделало
большие успехи и получило государственное значение. За какие-
нибудь 10 лет добыча возросла с 16 до 163 млн. кг в 1913 г., причем
половина улова увозилась в Японию. Главную добычу рыбы в Даль-
невосточном районе составляют проходные лососевые рыбы, при-
надлежащие к роду Oncorhynchus: кета (Oncorhynchus keta), гор-
буша (Oncorhynchus gorbuscha), нерка (Oncorhynchus nerca), кизуч
(Oncorhynchus kisutsch) и чавыча (Oncorhynchus tschawytscha).
Наиболее важными являлись первые два вида. Чрезмерный вылов
лососей во время их хода вверх по рекам для нерестевания уже от-
разился губительно: ход рыбы стал менее правильным, многие реки
и даже целые районы оскудели ею, количество кеты, которая не
только сама служила предметом ввоза в Европейскую Россию,
но и ее икра, сильно уменьшилось.
Из морских промыслов в морях Дальнего Востока
на первом месте стоял промысел тихоокеанской сельди (Clupea
pallasii), вывозившейся в Японию для удобрения.
Кроме того, добывалось небольшое сравнительно количество п а-
в а г и (Gadus navaga), трески (Gadus callarias), скумбрии
и камбал. Перед войной ловля при помощи траулеров как
в Охотском море, так и в океанах обнаружила колоссальные запасы
донных рыб, главным образом тресковых и камбало-
вых (Gadidae и Pleuronectidae), — не меньшие, чем в Атланти-
ческом океане.
Дальневосточному рыболовству принадлежит огромное буду-
щее. Для обслуживания его существует во Владивостоке научно-
промысловая станция.
Реки и озера Европейской части- РСФСР,
дававшие до войны 131 млн. кг рыбы. К этому району относятся
все речные и озерные воды РСФСР.
Особенно важна область больших озер: Ладожского, Онежского,
Чудского, Псковского, Ильменя и Велоозера.
Важнейшей промысловой рыбой здесь являлся снеток, или
озерная корюшка (Osmerus eperlanus sprinchus). Несмотря на край-
не интенсивный вылов, количество снетка не уменьшилось за-
метно. Кроме того, здесь ловятся: окунь (Perea iluviatilis), плотва
(Rutilus rutilus), epm(Acerinacernua), судак (Lucioperca lucioperca),
лещ (Abramis brama), озерный лосось, палия (Salvelinus alpinus),
таймень (Salmo trutta), различные сиги (Coregonus), ряпушка
(Coregonus albula).
Другой, также немаловажный район составляют верхние те-
чения Волги, Дона и Днепра, где добывалось до 50 млп. кг местной
рыбы, в том числе и стерляди (Acipenser ruthenus).
Аральский район, охватывающий, кроме Аральского
моря, реки Сыр-Дарью и Аму-Дарью, а также реки Зеравшащ
296
Чу и озеро Иссык-Куль. Здесь промыслы возникли недавно, всего
50 лет назад, когда по течению реки Сыр-Дарьи царским правитель-
ством были расселены ссыльные казаки-уральцы. Пока не была про-
ведена Оренбург-Ташкентская железная дорога, промысел разви-
вался весьма слабо за отсутствием сбыта. Озеро Иссык-Куль до сих
пор в таком положении. С проведением дороги добыча на промысле
сильно возросла.
Главнейшими промысловыми рыбами Аральского моря являются
сазан и лещ(Сург1пиз carpio и Abramis brama). Кроме того, ловятся
сом (Silurus glanis) и усач аральский (Barbus brachycephalus),
а также шемая (Alburnus chalcoides) и шип (Acipenser nudiven-
tris). Рыба Аральского моря отличается чрезвычайной упитан-
ностью.
На берегу Аральского моря возникли крупные промыслы астра-
ханского типа и восстановлен как речной, так и морской лов.
В настоящее время добыча сильно увеличилась в связи с механиза-
цией лова и его коллективизацией.
В Иссык-Куле добываются сазан, чебак (Leuciscus schmidti),
«селедка иссык-кульская» (Leuciscus bergi) и другие, менее важные
рыбы. Сбыт только местный.
Реки и озера Сибири. Район этот чрезвычайно богат
рыбой, и притом очень ценной (сиговые — род Coregonus; осетро-
вые — Acipenseridae и карповые). Но бездорожье, крайне редкая на-
селенность в наиболее рыбных низовьях и примитивность характера
лова делали здесь промысел рыбы кустарным. Большее значение
имел лов омуля (Coregonus autumnalis) в Байкале.
Район Черноморско-Азовский. Особенно богато
Азовское море благодаря своему мелководью, опреспенности мно-
гими реками и богатству органической жизни. На примере Азовского
моря мы видим результат хищнического лова и изменения режима
впадающих рек, благодаря сооружениям, выражающийся в крайнем
обеднении бассейна. Сто лет назад тут ловилось до 82 млн. кг рыбы,
главным образом осетровых (Acipenseridae), судака (Lucioperca
lucioperca), леща (Abramis brama) и сазана (Cyprinus carpio). Лет
35—40 назад лов пошел па убыль, и в 1913 г. добыто было всего
33.6 млн. кг, да и из пих почти 15 млн. кг приходилось на хамсу,
или анчоуса (Engraulis encrassicholus), недавно ставшую объектом
промысла, а лов осетровых упал до 650 тыс. кг.
Точно так же упал промысел рыбы и па Черном море, которое
и само по себе благодаря глубине у берегов и другим причинам мало
удобно для рыболовства. За последние 35—40 лет перед войной про-
мысел упал здесь с 45 до 13 млн. кг.
Однако запуск рыболовства во время войны и первых лет рево-
люции восстановил рыбные запасы Черноморско-Азовского района
и повысил общий улов здесь до 49 млн. кг в 1925 г., до 56 млн. кг
в 1926 г. и до 65 млн. кг в 1927 г. Лов осетровых возрос до 1.3 млн.
кг, а судак и лещ составляют теперь 65 и 70% общего улова.
Главные рыбы лова в Черноморско-Азовском районе: осетр
(Acipenser guldenstadti), севрюга (Acipenser stellatus), белуга (Huso
297
huso), сельди (Caspialosa pontica, Caspialosa normanni, Caspialosa
tanaica и Caspialosa meotica), морской язык (Solea nasuta), шипова-
тая камбала, глосса (Pleuronectes luscus), скумбрия (Scomber scom-
ber), султанка (Mullus barbatus), кефаль (Mugil auratus и другие
виды), бычки (Cobiidae).
Финский залив. Здесь главнейшим объектом промысла
является сельдь, известная под именем салакушки (Clupea harengus
membras), отличающаяся малыми размерами (от 15 до 23 см) и киль-
ка (Clupea sprattus). Здесь же ловится угорь (Anguilla anguilla).
Наконец, как это ни парадоксально, па последнем месте долго
стоял Ледовит о-Б еломорский район, охватывающий бе-
рега Северного Ледовитого океана с Белым морем и с впадающими
в него реками. Это действительно очень парадоксально, ибо. как
показали научно-промысловые экспедиции, в водах Мурмана, Ка-
нина Носа и острова Колгуева, куда проходит ветвь теплого
Гольфштрома, имеются неистощимые запасы тресковых (Gadidac)
сельдевых (Clupeidae) и камбаловых (Pleuroneclidae) рыб.
Кроме того, в реках этого района в большом количестве ловится
ценная семга (Salmo salar). Причинами слабого развития промысла
раньше являлись: отдаленность, бездорожье, слабая населенность,
слабая техническая оборудоваппость. Данными наших экспедиций
воспользовались до революции иностранцы, главным образом анг-
личане, которые приходили сюда с траулерами. Их лов быстро обо-
гнал русский прибрежный лов. Перед войной 1914 г. иностранный
улов в русских водах Ледовито-Беломорского района равнялся 80%
всего улова этого района. Этим и объясняется издание декрета
(24.Ш 1921 г), запрещающего иностранцам лов ближе 12-миль-
ной полосы вдоль всего северного побережья.
После революции дело с рыболовством в Ледовито-Беломорском
районе, в особенности ясе па Мурмане, приняло совершенно другой
оборот. Здесь появились железная дорога и русские траулеры,
весь рыболовный флот моторизован, промысел технически обо-
рудован, оборудована морская база в Мурманске. Уловы сильно
увеличились.
Однако несмотря па паше богатство рыбой, количество улавлива-
емой рыбы еще не удовлетворяет сильно возросшей потребности на-
селения в рыбе. Кроме того, заготовление рыбы впрок (солка, коп-
чение и консервировка) у нас стояло раньше гораздо ниже, чем
в Европе. В связи с этим в настоящее время проводится коренная
социалистическая реконструкция рыбного хозяйства, рыболовства
и рыбной промышленности па базе индустриализации, организуются
совхозы и коллективные хозяйства рыбаков. В основу хозяйства
положено плановое начало — пятилетпий и перспективные планы,
которые предусматривают пе только увеличение улова па основе ме-
ханизации, рационализации и освоения новых пространств, по и уве-
личение запасов рыбы путем правильно организованного научно
обоснованного отлова, мелиорации рыбных угодий и рыборазведе-
ния. Обращено внимание на качество продукции и широкое исполь-
зование отбросов. За истекшую пятилетку построено много трауле-
298
ров, введены глиссеры/новые способы механического облова, зна-
чительно повысилось качественно и количественно изготовление
консервов и т. д.
2. Охрана рыб и интенсификация рыбного хозяйства
Рыболовство не должно сводиться к массовому истреблению
рыбы. Проводимое нерационально и хищнически, оно скоро ведет
к истощению рыбных запасов. Пример Дальневосточного района,
где до войны быстро уменьшились уловы кеты и горбуши, доста-
точно нагляден. Не менее наглядна история рыбного промысла
и его истощения в Азовском море к началу мировой войны.
Перелов во время пути к месту нереста является наиболее губи-
тельной причиной уменьшения рыбных запасов. Кроме того, самый
характер буржуазного хозяйства, основанного па конкуренции и па
эгоистических стремлениях частных предпринимателей, пе заинте-
ресованных в рыбном хозяйстве как целом, также оказывался гу-
бительным, и опустошение Азовского моря, произведенное перед
мировой войной, служит тому примером. Лов маломерной рыбы
также являлся нерациональным, ибо не давая большой доходности,
в то же время уничтожал рыбу еще до выполнения ею функции раз-
множения. Употребление орудий лова, наносящих вред не пойман-
ной рыбе, также оказывало вредное влияние.
Поэтому во всех странах существует законодательство по охране
рыбы ради сохранения рыбных богатств. Мероприятия касаются вре-
мени лова, места лова, орудий лова и т. д. Некоторые орудия лова,
портящие рыбу, которая таким путем бесполезно пропадает, запре-
щаются. Другие орудия лова допускаются лишь при определенных
ограничениях; например, указываются размеры ячей в сетях, раз-
меры крючков и т. д. Устанавливаются определенные правила
производства топь, расстояний между ними, дабы пебыл совершенно
прегражден путь рыбе, идущей нерестовать. Главное же, быть мо-
жет, это — установление запретных сроков, установление заповед-
ных пространств и минимальных размеров вылавливаемой рыбы.
Однако одними запретами многого пе достигнешь. Рыболовство
должно быть рационализировано, поставлено на научную почву,
подобно тому как ставится рациональное земледелие. Все рыбное
дело должно основываться па учете, па знании тех рыбпых запасов,
которые имеются в море, и па строгом плане, основанном па этом
учете и на всестороннем изучении как самого бассейна, так и насе-
ляющих его животных.
Количество рыбного запаса при современных методах может быть
учитываемо пе только в мелких водоемах, по и в больших озерах
и морях. Конечно, количество штук рыб пе может быть сосчитано.
J |о следить за повышением улова, за падением его, за темпом роста,
т. е. за условиями выкорма рыбы, за возрастным составом уловов,
мы можем. Мы всегда можем иметь наблюдение за состоянием произ-
водительности бассейна. В вышеприведенных данных о леще и
вобле в Каспийском море мы видим этому примеры. Зная запасы,
299
имея возможность предсказывать, как это указано на примере атлан-
тической сельди, уловы будущих лет, мы можем вести более пра-
вильное хозяйство, имея в виду не кратковременную выгоду, а оп-
ределенный план государственного хозяйства. Тогда только мы смо-
жем с уверенностью говорить: «мы берем все, что можем взять,
по сохраняем наши запасы в целости и даже принимаем меры к их
увеличению на будущее время».
Для такого ведения хозяйства водоем должен быть изучен. Нам
должна быть совершенно ясна, ясна до мелочей вся картина жизни
водоема. Мы должны изучить его физические условия жизни; мы
должны знать запасы корма в водоеме — изучить его фаупу, се
видовой состав, распределепие в водоеме по временам года в зави-
симости от климатических условий, от распределения корма (планк-
тона, донной фауны); мы должны знать точно места, периоды и время
размножения видов рыб; должны знать их плодовитость (притом
в разные возрасты), темп роста по возрастам и многое другое. Тогда
мы можем как по часам определять различные моменты рыбного
хозяйства. За научным обследованием, которое должно дать картину
жизни водоема, должны уже следовать практические мероприятия,
касающиеся того, как:
1) повысить уловы в тех местах, где эксплоатацпя уже идет;
2) развить промысел, распространив его па новые объекты;
3) основать промыслы в местах, где промысла еще не существуют;
4) повысить производительность водоема при помощи заселения
его новыми видами рыб или путем массового искусственного рыбо-
разведения;
б) увеличить количество готовых продуктов при помощи введе-
ния более совершенных способов обработки.
Повышать уловы можно лишь тогда, когда есть уверенность,
что это пе поведет к уменьшению запасов. Так, например, сельдяной
лов в Каспийском море можно было без вреда повысить, так как
благодаря громадной плодовитости сельдей количество их заметно
повысилось. С такой же уверенностью можно было повысить улов
судака и леща в Черноморско-Азовском районе, так как эти рыбы
увеличились значительно в числе и качественно.
Осторожнее пришлось отнестись к повышению вылова осетро-
вых в Каспийском море, так как в предшествовавшие революции
годы количество их значительно уменьшилось. Однако запуск в те-
чение ряда лет и мероприятия по искусственному разведению осет-
ровых должны были в течение пескольких лет увеличить запасы
осетровых.
Кроме того, большой вылов может повести к уменьшению кон-
куренции из-за пищи, к увеличению количества последней и более
интенсивному росту. Благодаря этому количество вылавливаемой
рыбы в центнерах может даже возрасти, несмотря на уменьшение
количества штук.
Увеличение лова путем распространения его на новые объекты
все время имеет место на молодых промыслах. Так, на Каспийском
море сначала предметом лова были только осетровые, потом, когда
300
последние уменьшились в числе, перешли к лову судака, леща и са-
запа. Лишь в начале 60-х годов стали ловить сельдь для посола и упо-
требления в пищу. Еще позднее стали ловить воблу. Но и до сих пор
пе все рыбные ресурсы используются па Каспийском море. Некото-
рые рыбы, в других местах уже давно эксплоатируемые серьезным
образом, здесь лишь недавно стали привлекать внимание, несмотря
па огромные запасы их. Таковы бычки и 2—3 вида килек, являю-
щихся настоящими представителями сардинок. Эти кильки (Нагеи-
gula delicatula и Harengula grimmi) находятся здесь в громадном
количестве. Вся северо-западная часть моря представляет сплошное
пастбище взрослой и место выкормки молодой кильки. Несомненно,
что лов этой сельди должен развиться, и товар этот должен пойти
на изготовление консервов.
Количество улова может значительно увеличиться, если будет
произведено обследование в тех районах, где промыслов еще пет
или опи зачаточны. До революции мы недостаточно знали свою
страну. Беглые исследования различных морей, произведенные, не-
которыми экспедициями, обнаружили у пас колоссальные рыбпыё
богатства. Но мало их обнаружить, необходимо их основательно
изучить в фаунистическо-биологическом отношении: образ жизпи
рыб, миграции, время и место икрометания, рост и т. д. Ледовитый
океан, Печорский край, Обская губа, Еписейский район, Байкал,
ряд рек Сибири, Амур, Аральское морс, озера Иссык-Куль, Балхаш
и многие другие потребовали обстоятельного обследования. В на-
стоящее время научное изучение водоемов сильно шагнуло вперед.
Рыборазведение. Возможность искусственного рыбо-
разведения обусловливается возможностью искусственного опло-
дотворения икры рыб. Искусственное оплодотворение производится
таким образом. Путем поглаживания по животу самки с «текучей»
икрой выдавливают в тарелку икру. Затем при помощи такого же
поглаживания самца выдавливают зрелые «молоки» (семя) и сме-
шивают икру и семя без прибавки воды. Через некоторый промежу-
ток времени прибавляют воду, тогда начинается дробление яйца.
Это так называемый «сухой»метод. При «мокром способе» половые
продукты разбавлялись водой. По вследствие малой живучести
спсрмпсв этот способ оказался мало пригодным. В последнее время
советскими учеными пайден способ у пойманных заранее с еще не-
зрелыми половыми продуктами рыб вызывать быстрое их созревание.
Этот способ сильно облегчает работу по искусственному рыбораз-
ведению. Из пресноводных рыб искусственное оплодотворение при-
меняется главным образом у лососевых р ы б (Salmonidae),
применяется еще у леща (Abramis brama), у белорыбицы (Steno-
dus), у осетровых. Оплодотворенная икра выдерживается в особых
аппаратах, сконструированных таким образом, чтобы получились
паивозможпо более естественные условия для вывода. При этом,
конечно, должен быть соблюден целый ряд условий: необходимо
определенное качество воды, температура, содержание кисло-
рода, то или ипое движение ее и т. д. При умелом обращении
молено вывести 76% здоровой молоди.
301
Мальки рыб, еще не всосавшие вполне желточного пузыря, вы-
пускаютсяв естественные водоемы—реки, озера илиморя, в которых
и проходит их дальнейшая жизнь. Часто переносится в водоем
и живая икра в последних стадиях ее развития.
Цели искусственного рыборазведения могут быть различны.
В районах крупного рыболовства оно имеет
целью пополнение и восстановление запасов
наиболее ценных промысловых рыб. И это возможно не только в не-
больших сравнительно и закрытых водоемах, но и, как увидим даль-
ше, в обширных морях и в океане. На Волге производится искус-
ственное разведение белорыбицы (Stenodus leucichthys) и осетровых:
стерляди (Acipenser ruthenus), севрюги (Ac. stellatus) и осетра (Ас.
giildenstadti).
Искусственное оплодотворение икры белорыбицы на Уфимском
заводе чрезвычайно подняло уловы последней на Северном Каспии
и па Нижней Волге. В дальнейшем следует ожидать огромного уве-
личения ходовой белорыбицы. Особенно важны искусственное опло-
дотворение и выпуск молоди осетровых рыб, так как запасы их
сильно уменьшились, а рыбы эти являются весьма ценными, и
можно ожидать очень большого их увеличения от искусственного
рыборазведения.
Чрезвычайно важным является разведение осетровых в Черно-
морско-Азовском бассейне и лососевых — в северных реках и на
Дальнем Востоке.
Громадное значение имеет рыборазведение в районах ма-
лого рыболовства. Здесь главным образом надо иметь
в виду разведение лососевых рыб: лосося и ручьевой форели
(Salmo trutta m. fario), сигов (Coregonus), тайменя (Salmo trulta),
палии (Salvelinus alpinus var. salvclinus), корюшки (Osmcrus eper-
lanus var. spirinchus), ряпушки (Coregonus albula), леща (Abra-
mis brama) и судака (Lucioperca lucioperca). Этими рыбами можно
заселить и такие водоемы, где их до сих пор пе было, по где усло-
вия для их жизни являются подходящими. Так, папример, горное
озеро Иссык-Куль, вероятно, может быть заселспо сигами. В ста-
рой, дореволюционной России было 7 рыбоводных заводов, па кото-
рых разводились осетровые, лососи, сиги и белорыбица. Но работа
этих заводов носила большей частью’ опытный характер. После
революции работа заводов чрезвычайно усилилась. Так, папример,
за 10 лет (с 1915/1916 по 1925/1926) количество выпущенных в круп-
ные водоемы мальков увеличилось в 250 раз. Один из наших круп-
нейших заводов — Волховской — пе имеет себе равного по емко-
сти в Европе. Ленинградским заводом в 1926 г. было выпущено 850
млн. мальков корюшки. Произведен целый ряд удачных пересадок
мальков в новые водоемы.
После войны мальки чудского и ладожского сигов были поса-
жены в различные озера Уральской области, в горное озеро Севан
в Армении, в различные озера Ленинградского, Новгородского и Че-
реповецкого округов, и почти во всех озерах эти рыбы, особенно же
чудской сиг, вполне прижились, начали размножаться и увеличили
302
собой уловы рыбы. В начале 1929 г. с Волховского завода достав-
лено в Японию по просьбе японского правительства по полмил-
лиону икринок чудского и волховского сигов для производства опы-
тов по акклиматизации этих пород сигов.
Наконец, огромное значение имеют рыборазведение в
мелких водоемах искусственного происхождения и и р у д о-
вос рыбоводное хозяйство. Последнее, возникнув
еще в далекие исторические времена в Германии и Австро-Вен-
грии, в настоящее время стоит на научно-обоснованной почве,
основано на знании физиологии питапия разводимых рыб (глав-
ным образом карпа, сазана), па изучении продуктивности бассейна
(бонитировке), па разведении новых, быстро растущих рас и т. д.
Доход от такого рыборазведения очень велик.
Главным объектом рыборазведения в прудах являются не дикий
сазап или карп (Gyprinus), а его культурные расы, т. е. галиций-
ский карп, зеркальный карп, а также форель. О разведении кар-
пов, о карповом хозяйстве, а также о форелевом
существует обширная научная литература.
У нас организационная работа по прудовому рыбоводству нача-
лась только около 1924/1925 г. Карповые хозяйства возникают на Ук-
раине, где их насчитывается уже более 200, а также в Белоруссии,
па Северном Кавказе и даже в Сибири. В Ленинградской области
пробуют восстановить некоторые форелевые хозяйства, бывшие
до войны, а в прудах Никольского рыбоводного района и в других
местах выращивают галицийского карпа.
Для снабжения всех этих хозяйств доброкачественным посадоч-
ным материалом в РСФСР имеются питомники для карпов и форе-
лей. Спрос па карпа со стороны паселеппя очень велик.
Кроме карпов и форелей, можно разводить, в зависимости от ка-
чества водоема, линя (Tinea tinea), щуку (Esox lucius), судака
(Lucioperca lucioperca) и лососевых: ручьевую форель (Salmo trutta
niorpha fario), американскую радужную форель (Salmo irideus),
американскую палию (Salmo fontinalis), палпто (Salvelinus alpi-
nns var. salvelinus), хариуса (Thymallus thymallus), сига проход-
ного (Coregonus lavaretus) и др.
ЧАСТЬ ТРЕТЬЯ
АМФИБИИ, ИЛИ ЗЕМНОВОДНЫЕ
Класс 4. Амфибии, или земноводные (Amphibia)
I. ХАРАКТЕРИСТИКА И ОБЗОР КЛАССА
Основной особенностью амфибий являются их водно-наземный
образ жизни и характер организации: будучи по строению своих пя-
типалых конечностей, по дыханию с помощью легких и по кровооб-
ращению наземными животными, они в личиночном состоянии,
а иногда и во взрослом имеют жаберное дыхание и ведут водный
образ жизни; в воде у них нормально проходит и развитие яиц.
В эволюции — это первые и еще несовершенные наземные позво-
ночные, еще не потерявшие связи с водной стихией.
Вместе с рептилиями, птицами и млекопитающими амфибии со-
ставляют одну группу—ч етвероногих (Tetrapoda), называе-
мую еще Cheiropterygia и противопоставляемую рыбам — Ichthyop-
terygia, как оргапизмам, обладающим конечностями в виде плав-
ников, тогда как четвероногие обладают конечностями, построеп-
пыми по типу руки.
Характеризовать класс амфибий можпо следующим образом. Жи-
вотные с переменной температурой; с кожей, большей частью голой,
богатой железами; конечпости, если есть, построены по типу руки
(хироптеригий); развитие происходит с метаморфозом; дыхание в ли-
чиночном состоянии происходит при помощи жабер, а затем или
при помощи жабер или легких, или же кожи и слизистой оболочки
рта; сочленение с позвоночником осуществляется при помощи двух
сочленовных бугорков; сердце трехкамерное; ногтей на пальцах ист
(за некоторыми исключениями).
С рыбами их сближает ряд существенных признаков, а имеппо:
в сердце имеется артериальный копус (conus arteriosus); артериаль-
ные дуги, хотя часть их исчезает, отходят от брюшпого артериаль-
ного ствола симметрично; сердце сходно с сердцем двоякодышащих;
жаберные дуги еще несут жабры, если пе во взрослом состоянии,
то в личиночном; еще сохраняются, по крайней мере в течение вод-
ного периода жизни, кожные органы чувств системы боковой ли-
нии; у амфибий, как у рыб, нет еще зародышевых оболочек. На ос-
новании последнего признака амфибий вместе с рыбами соединяют
в группу позвоночных, лишенных водной оболочки, или амниона—
A n a m n i а, противопоставляя ее рептилиям, птицам и млекопи-
тающим, объединяемым в группу A m n i о t а. Но от рыб амфибии,
304
кроме преобладания у них легочного дыхания, отличаются еще
парными конечностями типа руки и отсутствием скелета в непарных
плавниках живущих в воде личинок.
Выражением происхождения амфибий от рыбообразных предков
является их местообитание. Это в полном смысле слова живот-
ные земноводные. За редкими исключениями они связаны
с водой, живут в воде или же во всяком случае ищут влажных мест
возле воды. Тропические леса с их жарким, но сырым климатом:
и болотами являются главным местом обитания земноводных.
В период же размножения все амфибии за немногими исключениями
идут в воду, будет ли то река, озеро, болото или вода, скопившаяся
у основания широких листьев, в дуплах и т. д., как это бывает
в девственных лесах тропических частей Южной Америки, Азии
и Африки. И на суше амфибии выбирают места сырые, закрытые,
недоступные лучам солнца, ведут большей частью ночной образ
жизни, а днем, укрываясь под камни, в мох, в трещины и т. д.
В тех случаях, когда амфибии встречаются в сухих пустынных
местностях, опи имеют приспособления, дающие им возможность
избегать неблагоприятных для них условий.
Современные амфибии представлены тремя отрядами. Первый
отряд представлен ползающими формами, подобными тритонам
и саламандрам, имеющими одинаковой длины передние и задние ко-
нечности и длинный хвост. Этот отряд хвостатых— Urodela.
Следующий отряд представлеп формами бесхвостыми, прыгающими,
подобно лягушкам, или имеющими вид жаб, которые хотя и
ползают, но имеют задние конечности более развитые, чем перед-
ние. Это—бесхвостые, Anura.1 Третий отряд представлен
червеобразными, роющимися в земле формами, совершенно
лишенными задних конечностей— безногими— Apoda, или
Gymnophiona.
Перечисленные три отряда амфибий хорошо отграничены один
от другого и пе связаны между собой переходами. Трудно связы-
ваются они в некоторых признаках и с рыбами, и с вышестоящими
наземными позвоночными— рептилиями. Однако связь с теми
и другими легче устанавливается через посредство ископаемого
подкласса амфибий— стегоцефал (Stegocephali), что озна-
чает п о к р ы т о г о л о в ы е.
Стегоцефалы дали начало современным амфибиям и рептилиям.
Их нельзя считать группой, равноценной трем другим группам
амфибий. Их противопоставляют всем последним, вместе взятым,
объединяемым в группу голых а м ф и б и й— Lissamphibia,
как группу п а и ц ы р н ы х амфибий — Phractamphibia. Тогда
Lissamphibia и Stegocephali, или Pliractamphibia, следует расце-
нивать как подклассы, a Urodela, Anura и Apoda— как отряды.
Stegocephali, или Phractamphibia, сами распадаются па ряд отрядов.
1 Urodela по закону приоритета должны называться Caudata,
А п ч г а — S а 1 i е n t i а. Но названия Urodela и Anura яв-
jihiok я более общепринятыми и более логичными, а потому мы их сохраняем.
*Л1 Кшикдров и Станчинский 305
Обычно принято было делить хвостатых амфибий (Urodela)
на два подотряда. В один входили формы, у которых жабры сохра-
нялись всю жизнь—п о с т о я н п о ж а б е р п ы е (Perenni-
branclnata), в другой— формы с отпадающими во взрослом
состоянии ж а б р а м и (Caducibranchiata). В настоящее время
выяснено, что семейства, входившие в эти подотряды, имеют различ-
ное происхождение. Поэтому их возводят в степень подотрядов,
а подотряды Perennibranchiata и Caducibranchiata отпадают
как не естественные. Также и бесхвостые (Anura), делятся па
подотряды.
Удельный вес амфибий в экономике природы сравнительно с дру-
гими классами позвоночных невелик. Число видов амфибий на земле
достигает более 1 300. Из них около 1 150 приходится па бесхвостых,
130 на хвостатых и 43 на безпогих. Значение амфибий для человека
невелико, и интерес их заключается в ипом. Опи стоят в основании
ствола наземных позвоночных, представляют собой древнейших
представителей последних, являются пережитком, обломком далеко-
го прошлого, сохраняя в себе черты последнего. Через ископаемых
представителей стегоцефал опи сближаются и с рыбами и с репти-
лиями. Интерес амфибий в их эволюционном значении, в том, что,
будучи весьма примитивными позвоночными, опи дают весьма инте-
ресный материал для разрешения таких общебиологических вопро-
сов, как регенерация, определение пола и т. д.
II. СТРОЕНИЕ АМФИБИЙ
1. Форма тела и движение
Каждый приспособительный признак возникает из того мате-
риала, который предоставляет в распоряжение отбора сложившийся
исторически орган. Это относится и к форме тела и к способу движе-
ния.
Форма тела стегоцефал, хотя у них уже имелась известная диффе-
ренцировка приспособительного характера, свидетельствует о том,
что стегоцефалы представляли существа, недавно вышедшие па сушу,
недостаточно приспособившиеся к наземной среде. Тело у пих было
еще весьма неуклюжее; конечности несовершенные; опи пе могли под-
держивать тело над землей, и брюхо должно было волочиться по пой.
По форме тела стегоцефалы большой частью походили да тритонов
или саламандр, конечности были короткие, тяжелые и стоялилюпе-
речпо к оси тела. Плечо и бедро располагались почти горизонтально,
предплечье и голень были направлены кнаружи; сочленения в конеч-
ностях были мало подвижны. Для поддержки тела требовалась боль-
шая мускульная сила, что видно из того, как сильно развиты гребни
и выросты на костях конечностей, служивших для прикрепления
мускулов (рис. 223). У некоторых стегоцефал тело было укорочено,
как у современных лягушек (рис. 224), у других оно вытягивалось
в длину, и животные эти приобретали тритоповидпое (рис. 225)
или даже змеевидное тело (Aistopoda).
306
Рис. 223. Реконструкция скелета пермского стегоцефала
(Tremalops milleri) (Л), 13 — левая половина плечевого
пояса и передней конечности Eryops megacephalus (оши-
бочно изображено 5 пальцев, их было 4).
(По Case из Ihlc и др.)
1 — radius, 2 — ulna, 3 — oleocranon, 4 — humerus, 5 — scapula,
6 — cleithrum, 7 — clavicula, 8 — intcrclavicula. (Из Ihle.)
i’nc. 224. Лягвообразный стегоцефал (Mastodontosaurus giganteus).
(Из Ссребровского.)
thМт
Рис. 225. Брапхиозавр
(Protriton), скелет и ре-
конструкция.
(Из Павловой.)
Форма тела современных амфибий, папомииая форму тела раз-
личных стегоцефал, обнаруживает больше приспособлений к усло-
виям их существования и к способу движения.
Безногие амфибии (Ароб.а),или гимнофиопы (Gymnophiona),пред-
ставляли собой небольшую группу червеобразных, роющихся в зем-
ле существ, и форма их тела является приспособлением к такому
образу жизни. У пих вовсе нет конечностей, хвост крайне короток
(рис. 226). Червеобразное голое тело покрыто клейкой жидкостью
и образует многочисленные поперечпые
кольцевидные складки. Роющий образ
жизни отражается и на скелете безногих.
Череп их очепь компактен, образует
сплошную костяную коробку без впадин.
Однако эта компактность черепа, напоми-
нающая вымерших стегоцефал, является
пе древпим, унаследованным от последних
признаком, а результатом разрастания тех
костей, которые есть и у других современ-
ных амфибий, в результате приспособления
к роющему образу жизни. Рот находится
пе на конце головы, так что в пего при
рытье не попадает земля.
Другой тип строения и иной способ пе-
редвижения видим мы у тех амфибий, ко-
торые ведут водный образ жизни; приме-
ром подобного типа может служить аксо-
лотль (рис. 227) или же трит о п .
Тело их вытянуто, имеются две пары пог,
более или менее длинный хвост. Движение
тритона и других подобных форм проис-
ходит при помощи боковых или винто-
образных движений хвоста. Конечности при этом движении пе
играют роли, а направлены назад и остаются приложенными к
телу. У некоторых водных Urodela происходит удлинение тела
до змеевидной формы (папример, у Ainpliiuina); при этом одно-
временно укорачиваются конечности пли передние, или задние,
или те и другие, а иногда и вовсе исчезают. Плавание таких форм
происходит, как у угрей, путем змееобразных изгибов тела.
При движении па суше хвостатые земноводные могут двигаться
пе иначе, как пресмыкаясь, т. о. волоча брюхо по земле. При этом
задние ноги толкают тело вперед, а передние лапы определяют на-
правление движения, все лее тело несколько изгибается змееобразно.
Однако некоторые хвостатые, ведущие наземный образ жизни,могут
бегать быстро, подобно ящерицам.
Иначе движутся в воде и на земле бесхвостые (Аппга^,
представителем которых молеет слулсить лягушка. Тело у пих плот-
ное, более или мепее уплощенное, совершенно лишенное хвоста.
Конечности развиты хорошо, задние большей частью длиннее перед-
них. Их передвижение в воде происходит при помощи сильных толч-
308
Рис. 226. Кольчатая червяга (Siphonops annulatus).
(Из Брэма.)
iltullllllll
Рис. 227. Аксолотль. (Из Брэма.)
ков хорошо развитыми задними ногами, пальцы которых удлинены
и связаны перепонками. Передние же конечности приложены в это
время к телу. Лишь при медленном движении в воде задние ноги
действуют не одновременно, а вытягиваются поочередно, причем
и передние конечности совершают движения, напоминающие ходьбу.
На земле эти амфибии пе-
редвигаются при помощи
прыжков или же, если зад-
ние ноги не очень развиты,
путем поочередного пере-
ставления передних и зад-
них пог (например, жабы).
Многие наземные бес-
хвостые обладают способ-
ностью лазать. В связи с
этим у них па пальцах раз-
вились специальные диски.
Диск очепь богат лимфати-
ческими пространствами, и
в эпителии его находятся
слизистые железы. Иссле-
дование дисков показало,
что прикрепление обуслов-
ливается пе. присасыва-
нием, а молекулярным при-
тяжением двух тесно со-
прикасающихся тел. И чем
меньше между ними воз-
духа, тем присасывание
прочнее. Особые гладкие
мускулы в диске делают
его плоским. Тонкий слой
жидкости между диском и
поверх и остью субстрата,
обладающий липкостью и
вязкостью, усиливает это
притяжение тел. У яван-
ской летучей лягушки
(Rhacopliorus) прикрепле-
ние происходит по только
посредством дисков па
пальцах, по и при помощи кожи брюха, кожных складок па конеч-
ностях и па подбородке. В отличие от других лягушек у этой
лягушки кожа живота тесно срастается с косой мышцей живота
с грудным мускулом, благодаря чему опа может натягиваться.
Некоторые из древесных лягушек имеют особым образом устроен-
ные хватательные конечности. Например у Phyllomedusa burmeis-
teri мы видим налицо все пять пальцев, по 2-й и 3-й укорочены и ре-
дуцируются, тогда как первый, или большой, палец стоит к наиболее
зю
Рис. 228.
1 — задняя конечность Phyllomedusa burmeis-
teri, 2 — левая кисть лемура (Perodlcticus са-
labarensis), 3 — нога лазящей птицы (Momotus
lessoni).
Рис. 229. Яванская лягушка. (Rhacophorus
pardalis.)
(Иа Boulenger.)
развитому 4-му под углом в 100°. 2-й и 3-й редуцируются, ибо опи
не нужны при образовании хватательной конечности. У некоторых
птиц, а также у лемуров мы видим подобную же картину. Здесь мы
имеем пример конвергенции у лазящих животных (рис. 228).
Другой пример конвергентного развития мы видим в образовании
приспособительных изменепий для планирующего полета у некото-
рых лягушек, именно у представителей рода Rhacophorus (рис. 229).
У большинства видов пальцы соединены перепонкой не более чем
до половины, у немногих опи достигают дисков на концевых фалан-
гах. В этих случаях опи могут пользоваться перепонками как па-
рашютами. Но представления о способности их к полету преувели-
чены, как была преувеличенно вычислена поверхность межпальце-
вых перепонок в 78 см2 вместо 18.8. При полете, когда лягушка по ко-
сой липии спускается с дерева, она растопыривает пальцы, держит
конечности прижатыми'к телу, последнее же надувается вследствие
раздутия сильно развитых легочных мешков. Получается большая
поверхность, служащая для планирования.
Таким образом, у тех из современных амфибий, образ жизни
которых сохранил более древний и примитивный характер,
напоминающий стегоцефал, и форма тела и конечности сохраняют
более древний характер, пригодный лишь для ползания по земле.
Те же амфибии, которые изменили среду своего первоначального
обитания, развивают специальные приспособления как в форме
конечностей, так и в форме тела.
2. Общие покровы амфибий
Ряд особенностей в строении кожи амфибий показывает их род-
ство с рыбами. Покровы амфибий влажны и мягки и не имеют еще
таких специальных особенностей приспособительного характера,
как перо или волос. Мягкость и влажность колеи амфибий стоят
в связи с недостаточно совершенным аппаратом для дыхания, ибо
колса слуясит дополнительным органом последнего. Этот признак
до лисе п был развиться улсе у далеких предков современных амфи-
бий. Это мы и видим па самом деле; улсе у стегоцефал утрачивается
костный колспый папцырь, унаследованный ими от предков рыб,
дольше сохраняясь па брюхе, где он слуясит защитой при ползапье.
Покровы состоят из эпидермиса и к о лс и (cutis).
В эпидермисе еще сохраняются черты, свойственные рыбам: реснич-
ный покров у личинок, сохраняющийся у личинок Лпига до насту-
пления метаморфоза,; ресничный эпителий в органах боковой линии у
Urodela, всю лсизпь проводящих в воде; наличие одноклеточных сли-
зистых лселез у личинок и тех лее водных Urodela. Колса собственно
(cutis) состоит каку рыб из трех взаимно перпендикулярных систем
волокон. У лягушек в колее имеются большие лимфатические полости,
благодаря чему кожа не связана у них с подлежащей мускулатурой.
В колее амфибий, в особенности у тех, которые ведут более наземный
образ жизни (например, исабы), развивается ороговение, предохра-
няющее ниже лелсащие слои кожи как от механических повреждений,
311
так и от высыхания, что связано с переходом к наземному образу
жизни. Ороговение кожи должно, конечно, затруднять кожное дыха-
пие, а потому большее ороговение кожи связано с большим разви-
тием легких (например, у Bufo в сравнении с Rana).
У амфибий наблюдается линька, т. е. периодическое сбрасывание
кожи. Колса сбрасывается как один кусок. В том или ином месте
кожа лопается, и животное выползает из нее и сбрасывает, а
некоторые лягушки и саламандры съедают ее. Линька необходима
амфибиям, ибо они растут до конца своей жизни, и кожа стесняла бы
рост .
На концах пальцев ороговение эпидермиса происходит наиболее
сильно. У некоторых стегоцефал существовали настоящие когти.
Рис. 230. Кожа с железами из паротиды
саламандры. (По Biitschli из Ihle и др.)
1 — ядовитая железа, 2 — слизистая железа, 3 —
секрет, 4 — сосуды, 5 — эпидермис, 6 — corium,
7 — пигментные клетки.
Из современных амфи-
бий они имеются у Хе-
nopus, Hymenochirus и
Onychodactylus.y жабы-
чесночницы (Pelobates)
па задних погах разви-
вается как приспособле-
ние для рытья лопато-
образный вырост.
Боковые органы
чувств, характерные для
рыб, имелись у стего-
цефал, о чем свидетель-
ствуют каналы на череп-
ных костях. Сохраня-
ются опи и у современ-
ных амфибий, а именно
лучше всего у личинок,
у которых опи развиты типичным образом па голове и тремя
продольными рядами идут вдоль тела. С метаморфозом органы эти
или исчезают (у Salamandrinae, у всех Anura, кроме шпорцевой
лягушки Xenopus из Pipidae), или погружаются вглубь, где нахо-
дятся под защитой ороговеватощих опорных клеток. При возвраще-
нии Urodela вводу для размножения органы боковой липни восста-
навливаются.
Кожа амфибий очень богата железами. Одноклеточные железы,
характерные для рыб, еще сохраняются у личинок Apoda и Urodela
и у живущих в воде взрослых Urodela. С другой стороны, здесь
появляются настоящие многоклеточные железы, развившиеся фи-
логенетически, видимо, из скоплений одноклеточных желез, кото-
рые наблюдаются уже у рыб.
Железы амфибий двоякого рода; меньшего размера с л и з и-
с т ы е железы и более крупные серозные, или б е л-
к о в ы е. Первые относятся к группе желез м е з о к р и п и ых,
клетки которых пе разрушаются в процессе секрецпи, вторые —
г о л о к р и и п ы е, клетки которых целиком идут па образование
секрета. Белковые железы образуют бородавчатые возвышения
312
на спинной стороне, спинные валики лягушек, ушные железы (паро-
тиды) у жаб и саламандр. И те и другие железы (рис. 230) одеты
снаружи слоем гладких мышечных волокон. Секрет желез часто бы-
вает ядовит, особенно желез белковых.
Окраска кожи амфибий обусловливается, как и у рыб, присут-
ствием в коже пигмента и рефлектирующих и ридоцит о в.
Пигмент бывает или диффузный или зернистый, располагающийся в
особ।лхклетках — хроматофорах. Диффузныйпигмепт,распре-
деленный в роговом слое эпидермиса, обычно желтый; зернистый
бывает черпого, бурого и красного цвета. Кроме пего, встречаются
белые зерпа гуапипа. Зеленый и голубой цвет окраски некоторых
амфибий является субъективной окраской, обусловленной сме-
щением топов в глазу наблюдателя.
Изучая при небольших увеличениях кожу древесной ля-
гушки, к в а к пт и (Ilyla arborea), мы видим, что при рассмат-
ривании кожи снизу опа кажется черпой вследствие наличия в пей
анастомозирующих и разветвленных черных пигментных клеток,
мелапофоров. Сам эпидермис бесцветен, по там, где свет
проходит сквозь кожу при сократившихся мелапофорах, опа кажется
желтой. Л е й к о ф ор ы, или интерферирующие клетки, содержат
кристаллы гуапипа. К с а и т о ф о р ы содержат золотистожелтый
липохром. Способность мелапофоров изменять свой вид,
то свертываясь в шар, то вытягивая отростки, и обусловливает глав-
ным образом возможность изменения цвета. Точно так же подвижен
желтый пигмент в ксантофорах. Лсйкофоры или интерферирующие
клетки дают серо-голубой, красно-желтый или серебристый блеск.
Совместная игра всех этих элементов создает все виды окраски ам-
фибий. Постоянные черные пятна обусловливаются наличием чер-
ного пигмента. Меланофоры усиливают его действие. Белый цвет
вызывается лейкофорами при отсутствии мелапофоров. При сверты-
вании мелапофоров и при распространении липохрома создается
желтая окраска. Зеленый цвет получается путем взаимодействия
черных и желтых хроматофоров.
Изменения окраски находятся в зависимости от нервной системы.
Кожа амфибий богато снабжена сосудами, служа для дыхания.
У волосатой лягушки (Astylosterniis), имеющей сильно редуцирован-
ные легкие, тело покрыто волосовидными выростами кожи, обильно
снабженными сосудами. Кожа амфибий служит также для восприя-
тия воды и для выделения. В сухом воздухе кожа лягушек и сала-
мандр столт» обильно испаряет, что опи погибают. Имеющие более
развитой роговой слой жабы выживают в тех же условиях значи-
тельно дольше.
3. Наружный скелет
Наличие кожного костного скелета является очень древпей
чертой, возникшей, вероятно, еще у предков Gnatliostomata. Он ши-
роко распространен у рыб. У современных амфибий, как правиле
с редкими исключениями, кожного скелета пет. Зато у стегоцефал
он редко отсутствовал, хотя распространение его ограничивалось,
здз.
большей частью брюшной стороной, продолжаясь и на конечности
(рис. 231). Чешуи стегоцефал имели круглую, овальную или вере-
тенообразную форму, располагаясь косыми рядами на брюхе,
передке черепицеобразно налегая друг па друга, как у рыб. Из этих
рядов чешуи на брюхе возникли, видимо, брюшпые ребра рептилий.
Необходимость развития кожного дыхания и слабо развитые пер-
воначально конечности привели к утрате стегоцефалами панцыря.
Из современных амфибий чешуи в коже развиты у безпогих
(Apoda) (рис. 232). Поверхность тела разделена у них перетяжками
на кольца. Каждое кольцо разде-
лено соединительной ткапыо на-
двое. В переднем отделе находятся
крупные ядовитые железы, в зад-
нем развиваются многочисленные
чешуи. Каждая чешуя является
Рис. 232. Кожа Ichthyophis
glutinosus.
1 — железистый мешок, 2 —
голокринная железа, 3 — ме-
шок шипун, 4—чешуя , 5 —
corlum. (По ППеи др.)
Рис. 231. Брюшной папцырь
Stegocephali.
(По Crcdncr из Циттеля.)
продуктом глубокого слоя кожи, как чешуя рыб, и состоит, подобно
последней, из двух слоев: нижнего волокнистого и верхнего более
объизвествленпого, напоминающего весьма близко поверхностный
слой чешуи чешуйчатпика — Neoceratodus (Dipnoi). Число колец
первоначально соответствует числу сегментов тела. Кожные око-
стенения развиваются также у некоторых Anura: у Ceratophrys
в колее развивается широкий щит, не слитый с позвонками, у 13га-
cliycephalus такой щит сливается с последними.
4. Внутренний скелет
Позвоночник. И в осевом скелете амфибий обнарулшвается ге-
нетическая связь с рыбами. Наиболее ясно это видно в течении раз-
вития амфибий. Хорда зародышей и личинок амфибий очень близка
к рыбьей и по строению, и по развитию. В некоторых случаях еще
обнаруживается та первичная диплоспопдилия, двойственность
314
дуг, которая характерна для низших рыб. Так, у Siredon верхние
хрящевые дуги закладываются двойными, в нижних же имеется
щель, указывающая па их первоначальную двойственность, на со-
став из краниальных и каудальных дуг. У остальных Urodela за-
кладываются только каудальные дуги, а из остатков краниальной
закладывается подобно тому, как это наблюдается у Lepidosteus,
межпозвоночный хрящ, из которого путем образования в нем щели
образуется сочленение. По форме тел позвонки амфибий бывают
амфицельные, о и и с т о ц е л ь п ы е и процельные
(рис. 233), позвоночник хорошо развит, вполне окостеневает,
Рис. 233. Продольный разрез через тола позвонков раз-
личных Urodela.
Л—Ranodon slbirlcus, If— Ambysloina ligrinurn» C—Gyrlnophilus
porpbyricne, J)— Saianiandra perspiclllala.
1 — хорда, 2 — мс)1П1о:шо11оч|цдГ| хрящ, 3 — кость тела позвонка,
4—межпозвоночная связка. (Но Parker.)
хрящ у взрослых встречается редко. Форма тел позвонков со-
ответствует способу движения в различной среде.
У стегоцефал позвонки обнаруживают очень большое сходство
с позвонками древних Teleostomi; пменноу более древних предста-
вителей стегоцефал позвонки состояли из нескольких элементов,
образовавших двояковогнутые тела (рис. 234). В отряде Rha-
cliitomi этих элементов было 4 пары в каждом позвонке: basidorsale,
basiventrale, interdorsale п intervcntrale; у Embolomeri позвонки
имели двойное тело, как в хвостовой части Amia. Позвоночник
из отдельных элементов пе мог, конечно, сохраниться у Tetrapoda,
целиком вышедших на сушу, ибо был для пих слишком слаб. Сег-
ментация, а, следовательно, и первичная диплоспондилия развива-
лись для облегчения сгибания тела при движении в воде. У назем-
ных движение совершается не при помощи изгибов тела, а потому
315
диплоспопдплпя пс только не нужна, но и вредна. Срастание ча-
стей позвонка началом. уже у стегоцефал в отрядах Stereospondyli
(Labyrintliodontia), Branchiosauria (Phyllospondyli). Из позвон-
ков последнего типа развился позвоночник современных амфибий,
развитие шло при этом по двум различным путям у хвостатых и бес-
хвостых. Другой тип образования цельного позвонка, приведшего
Рис. 234. Сверху — рахитомные позвонки Archegosaunis;
снизу — туловищные позвонки Eogyrinus.
А — передние грудные, В —задние туловищные, С—передние
хвостовые, I) и Е — более задние хвостовые.
1—hypocentrum (basiventrale), 2 — neiirocentrum (basidorsalc),
нервная дуга, 3 — pleiirocentniin (inlerdoraale), 4 — hypocenlriim
pleurale (intcrventralc), 5 — передний сочленовный, 6 — задний
сочленовный отросток, 7 — головка ребра, 8 — бугорок ребра.
(По Goodrich.)
к образованию позвонков типа рептилий, мы видим у представи-
телей стегоцефал — Ceraterpetoniorpha, хотя последние и пе были
предками рептилий; сходство здесь конвергентное.
Позвонки амфибий имеют сильно развитые поперечные отростки,
а также передние и задние суставные отростки—п р е- и п о с т з и-
г а п о ф и з ы, соединяющиеся между собой при помощи настоя-
щих сочленений — диартрозов. Ребра у современных амфи-
бий сильно редуцированы, иногда зачаточны или даже вовсе
316
отсутствуют (у Anura), но были более развиты у стегоцефал. У Anura
имеются па месте ребер лишь сильно развитые поперечные отростки.
Более развиты ребра у Apoda и Urodela, по у них они ограничены
только туловищем, у стегоцефал же развивались до половипы хвоста.
На проксимальном конце ребра амфибий вилкообразно раздвоены
и соединяются как с верхней дугой, так и с отростком,отходящим
от тела. Этот тип прикрепления ребер впервые появляется у амфи-
бий, ибо у рыб сочленение имеет место только с нижней дугой,
и повторяется в дальнейшем у высших позвоночных.
сверху.
А—Rana ftsculenla, В— Discoglos-
eus рIctus со свободными ребрами.
1 — поперечные отросли и, 2—реб-
ро, 3—крючковидный отросток,
processus unclnatiis, h — крестцо-
вый позвонок, 5 — копчиковая
кость—уростиль, 6 — иодвздоптап
кость, 7—acetabulum, сочленов-
ная ямка. (Из Ihle.)
з
Рис. 236. Череп молодой Rana
lemporaria после метаморфоза.
Покровные кости иа левой сто-
роне спиты.
1 — cliondrocranium, 2—слуховая кап-
сула, 3 — exoccipitalc, 4 — frontopa-
rietale, 5—iiasule, 6 — maxilla, 7 —
pl.c.rygolfleuni, В — paraquadraliiiu, ‘J —
columella, 10 — qiiadralojugale. (Из
Parker.)
У амфибий наблюдается улсе некоторое расчленение позвоноч-
ника на отделы. Первый позвонок в виде кольца сочленяется с че-
репом при помощи двух сочленовных ямок, соответствующих двум
сочленовным бугоркам па последнем. Второй позвонок— нормаль-
ный. Л т л а с а и э н и с т р о ф е я, характерных для высших
позвоночных, здесь еще пет. Имеется у амфибий к. р е с т ц о в ы й
н о з в о и о к, с которым соединяется тазовой пояс и за которым
следует хвостовой отдел. У Линта последний представлен длинной
палочковидной костью — у р о с т и л е м (рис. 235). Следы сегмеп;
тации у некоторых амфибии (отверстия для первой,зачаточные по-
перечные отростки и зачаточные верхние дуги) показывают, что уро-
стиль образовался из слияния 12 позвонков. У Protobatraclius
из триаса в хвосте были позвонки.
Число позвонков у амфибий вариирует: у Лппга обычно имеется
8 туловищных и 1 крестцовый позвонок, за которым следует
317
уростиль; у Urodela число туловищных позвонков от 14 (Triturus)
до 63 (Amphiuma), хвостовых от 22 (Cryptobranchus) до 36 (Tri-
turus); у Apoda число позвонков от 200 до 300, из которых 25—30
приходится па хвост.
Череп. Череп современных амфибий значительно отличается от
черепа рыб, но легко связывается с последним через посредство чере-
па стегоцефал. Как сказано выше, затылочная часть черепа короткая,
туловищные позвонки
не входят в состав
черепа. Часть черепа,
построенная из тра-
бекул, широка, череп
платибазальпый
как у миног и у древ-
нейших рыб: «гапои-
дов», кистеперых
(Crossopterygii) и дво-
якодышащих. По-
лость череппой короб-
ки простирается да-
леко вперед между
глазницами.
Нёб ноквадратпый
Рис. 237. Пластическая реконструкция choudro-
cranium молодой Rana tcmporaria, 2 см длиной.
1 — подглазничная дуга, 2 — нижние предносовые
хрящи, 3 —• верхние предносовые хрящи, 4 — косые
хрящи, 5—лобиая фонтанель, 6 — теменная фонта-
не ль, 7 — овальное окошко, 8 — сочленовный отрос-
ток quadra turn. (По Parker.)
хрящ прочло соеди-
няется с черепом (автостилический череп). Hyomandibulare пре-
вращается в слуховую косточку—columella auris. Chondrocranium
(рис. 236) у современных амфибий сохраняется еще в значительной
мере; замещающих хрящ костей мало подобно тому, как мы видим
это у двоякодышащих, костных гапоидов и кистепсрых.
Особенностью черепа амфибий в сравнении с рыбами является
образование в слуховой капсуле затянутого перепонкой окошечка—
овального окошечка (fenestra ovalis), в которое упи-
рается columella auris или stapes.
У личинки Anura имеются в черепе еще своеобразные губные
хрящи, верхние пнжпис, возможно гомологичные губным хря-
щам акул. Череп личинки Anura обнаруживает значительное сход-
ство с черепом миноги, однако генетической связи между ними пет
(рис. 237). Сходство это является в том и другом случае приспособле-
нием к образованию сосущего рта.
В гиобрапхиальпом скелете у личинок амфибии закладываются,
кроме подъязычной дуги, еще четыре передних жаберных дуги.
Из пятой развивается скелет гортани. У взрослых гиобранхпальпый
скелет подвергается некоторой редукции.
Таким образом в chondrocranium и висцеральном аппарате выяв-
ляется связь амфибий с рыбами, их предками. *
В черепе современных амфибий имеется мало окостенений как
замещающих, так и покровных. В этом отношении опи значительно
отличаются от рыб. В затылочной области имеются лишь две
боковые затылочные кости (exoccipitalia), каждая
318
с суставным бугорком для сочленения черепа с позвоночником.
В слуховой области, образующей значительные боковые выступы
для соединения нёбноквадратпого отдела, развивается лишь одна
переднеслуховая кость (prooticum) (рис. 238), и лишь.
иногда имеются следы наруж-
ной з а т ы ло ч п о й (epioticum).
В глазничной области у амфибий
бесхвостых развивается характерная
непарная пояспая кость
(sphetlimoideum), охватывающая че-
репную полость: у хвостатых же ам-
фибий в этой области развиваются
парные кости (orbitosphcnoidea), за-
нимающие лишь боковые стопки че-
репа. Обонятельная, этмоидиая об-
ласть остается хрящевой. Из окосте-
нений кожных имеются на крыше че-
репа парные темени ы е (parie-
talia) и лоб и ы е (frontalia) око-
стенения, у Anura сливающиеся в
непарную л о б и о т е м с п и у ю
кость (frontoparietale). Впереди
лобных костей над носовыми кап-
сулами лежат парные и о с о в ы е
кост и (nasaliа), а у хвостатых по
бокам позади последних существуют
еще и р с д л о б и ы е к о с т и (рга-
cfrontalia) (рис. 239). Снизу череп
прикрыт и а р а с фено и д о м (ра-
rasplienoideuni) и с о ш п и к о м (vo-
mer). Последний парный и окружает
з а дине и о с о в ы е о т в е р-
с т и я (choanae). Пёбноквадратный
аппарат у бесхвостых срастается с
черепом назади в слуховой области
и впереди в этмоидальной, в середине
лее- в виде свободной дуги огибает
орбиты. У хвостатых нёбноквадрат-
ный хрящ до этмоидпой области не
доходит, оканчиваясь свободно. Око-
стеневает в пёбноквадратпом хряще
Рис. 238. Дорзальный (у!) и
вентральный (/?) вид черепа
лягушки (Rana oscnlenta). На
рис. В покровные кости с од-
ной стороны удалены.
1—pramaxilla, 2—maxilla, 3 —
nasale, 4 — frontoparietale, 5 —
prooiicinn, 6 — cxocclpitalia, 7 —
plcrygoideuin, 8 — quadra lojugale,
9 — paraquadratuin (squamosum),
1 0 — eUiiiioidcurn, 11 — quadratum,
12 — vomer, 13 — parasphenoideum,
14 — подглазничные дуги. (Из
Parker.)
только небольшой участок — quadra-
turn. У Anura отсюда отходит топкая к в а д р а т и о с к у л о-
в а я кость (quadratojugale), замыкающая дугу, образованную
межчелюстной, верхнечелюстной и квадратпоскуловой костью. На
наружной стороне quadratum развивается покровная кость —
paraquadratum, гомологичная squamosum, а на идущей вперед
дуге — к р ы л о в и д п а я кость (pterygoideum), передним
концом достигающая нёбной кости (palatinum); в виде покров-
319
в
Рис,. 239. Л—черен личинки ак-
солотля. Череп черной сала-
мандры (Salamandra а Ira) свер-
ху (Z?) и снизу (С).
1—ргаетахШа, 2 —maxilla, 3 —
nasale, 4 — praefrontale, 5 — но-
совая капсула, 6 — frontale, 7 —
pterygoidcum, 8 — (на рис. В)—
parietale, 8 —(на рис. О —слу-
ховая капсула и cxoccipil ale,
9 — orbilosphenoidcuin, 10 — рага-
qnadratum (squamosum;, 11—qna-
dratum, 12 —хрящевой ptcrygoi-
deum, 13 — parasphenoideum, 14 —
palatinum. (Из Parker.)
пых окостенений на верхней челюсти
имеются межчелюстные и челюстные
кости (praemaxillaria и maxillaria).
В нижней челюсти на меккелевом
хряще развиваются: зубнаякость
(dentale), кожно сочленовная (goni-
ale) и пластинчатая (spleniale). По-
следняя у Apoda и у постоянно живу-
щих в воде Urodela. Впереди может
окостеневать еще небольшой эле-
мент — mentomandibulare.
Висцеральный скелет современных
амфибий состоит из хорошо развитой
гиоидной дуги, гомологичной гиоид-
ной дуге рыб, и из 4—2 жаберных
дуг. Так как личинки, а также живу-
щие в воде хвостатые дышат жабра-
ми, то у них висцеральный скелет
развит хорошо, весьма наглядно сви-
детельствуя о рыбьих предках амфи-
бий. С метаморфозом от висцераль-
ного скелета сохраняется вполне
лишь первая или подъязычная дуга
и соединяющая обе половинки copula,
идущие па образование скелета силь-
но развитого языка. У хвостатых
остаются после метаморфоза лишь
остатки двух передних дуг, задние
ясе вовсе исчезают. У бесхвостых во
взрослом состоянии имеется лишь
ши роки и подъязычный окостеневаю-
щий хрящ—тело подъязычной кости
(corpus Jiyoideum) н две пары отхо-
дящих от нее «рожков», являющихся,
остатками гиоидной и бранхиальной
дуг (рис. 240). Остатки гиоидной и
3—4 жаберпых дуг сохраняются у
Apoda. Из пятой дуги развивается
гортань.
Череп стегоцефал гораздо больше
походил па черен древних рыб.
Прежде всего в нем было гораздо
больше окостенений. В отряде Ет-
Lolomeri и мелост» 4 затылочных кос-
ти, в слуховой, кроме pfooticum, еще
opistoticum и т. д. С другой стороны,
в черепе примитивных стегоцефал
имелись некоторые черты, сбли-
жающие их с рептилиями: оп был
менее платибазален, чем у современных амфибий, затылочная
область примитивных стегоцефал включала в себя несколько ту-
ловищных сегментов, и через отверстия в боковых затылочных
костях выходили перед-
ние спинномозго-
вые и е р в ы (н. hypo-
glossus), имелось окостене-
ние — epipterygoideum
(= columella cranii), ха-
рактерное для рептилий
(между pterygoideum и
черепом), и transversum
(между первой и maxillare).
Покровных окостенений у
стегоцефал также было
гораздо больше. Кости эти
сходились между собой, бы-
ли широки и образовывали
свод черепа. Columella
auris уже имелась, раз-
Рис. 240. Подъязычная кость Rana escu-
lenta снизу (Л), гиоидная и жаберные ду-
ги у взрослого Iclithyophis glutinosa (R).
1 — тело подъязычной кости, 2 рожки подъ-
язычные, 3 —рожки жаберные. I, II, III —
жаберные дуги.
вившись из освободившегося вследствие развития автостилии
подвеска. Сходство черепа стегоцефала с черепом Crossopterygii
исключительно велико и поэтому убедительно говорит за их род-
ство (рис. 241).
Рис. 241.— Черепная крыша Eustenopteron (А)
девонской рыбы из Crossopterygii, (В)— черепная
крыша Trimororhacliis insignis, стегоцефала.
1— pracmaxiLla, 2 — maxilla, 3—praefrontale, 4—frontalc,
5 — parielale, 6 — postparietalc, 7 —lacrimale, 8 — post-
frontale, 9 — postorbitale, 10 — intertcmporalc, 11—squa-
mosum, 12 — supra temporale, 13 —quadratojugale, 14 — qua-
dra turn, 15 — tabulare, 16 — foramen pineale (теменное
отверстие), 17 — jugale. (Из Ihle и др.)
Осповпая черта черепа стегоцефал — его стегальпость. Покров-
ные кости образуют прочную крышу черепа, спускаются по сторонам
вокруг орбит и через височную область, запятую мускулами челю-
cTeii, п quadratum. Череп сплошь закрыт костями; имеются отвер-
Kuiuicapoo и Ciанчинский 321
стия лишь для наружных ноздрей, для орбит и для теменного глаза.
Грубая скульптура па костях показывает, что кости лежали весьма
поверхностно. На это указывает также наличие каналов боковой
линии, расположенных на тех
же костях, как у рыб (рис.
242). Форма черепа у различных
стегоцефал весьма вариировала.
К современным амфибиям наи-
более приближаются по черепу
Branchiosauria (рис. 243), имев-
шие большие орбиты и значи-
тельно редуцированные эндохон-
дральные окостенения.
Сходство с Crossopterygii об-
наруживают стегоцефалы и в
строении нижней челюсти, бога-
той окостенениями, как у Cros-
sopterygii.
У позднейших форм стегоце-
фал замечается тенденция к ре-
Рис. 242. Реставрация черепной
крыши представителя Archegosauri-
dae из нижнепермских отложений.
Видна грубая скульптура костей.
(Ив Abel.)
У некоторых стегоцефал были
Интересно развитие слухового
дукции как покровных, так и
эндохондральных окостенений,
приводящая (хотя промежуточ-
ных звеньев мы не имеем) к че-
репу современных амфибий,
пайдепы жабры.
аппарата у стегоцефал. У древ-
нейшего представителя Stegocephali Embolomcri columella auris,
4
Рис. 243. Череп Branchiosaurus ambystomus.
А—-со спины, В —с брюшной стороны.
1 — кожное supraoccipltalc-poslparietalc, 2 — tabulare.
(Из Goodrich.)
возникшая из верхнего отдела гиоидной дуги, потерявшего функ-
циональное значение вследствие развития автостилии, просто упи-
рается проксимальным концом в слуховую капсулу. Но уже рано
у стегоцефал появляется овальное окно (fenestra ovalis),
в которое и упирается теперь columella.Вероятно, уже у стегоцефал
322
имелась барабанная перепонка, о чем можпо судить
по форме окружающих барабанную полость частей. Полость же
эта образовалась из щели между челюстной и гиоидпой дугами.
Из современных амфибий наиболее примитивно устройство слухово-
го аппаратау Anura, где columella представлена топкой свободно тор-
чащей в барабанной полости палочкой. У Urodela нет пибарабаппой
перепонки, пи палочковидной columella. У Apoda короткая colu-
mella сочленена с quadratum. Стоит это в связи с подводным и под-
земным образом жизни.
Конечности и их пояса. Конечность типа руки (cheiropterygium)
составляет наиболее характерную черту наземных, в том числе
их от рыб. Развиться конечности этого
и амфибий, отличающую
типа до лисп ы были в
силу того, что па долю
парных конечностей у
наземных животных
выпадает гораздо боль-
шая наг р у зка, п еж ел н
па долю плавников рыб:
парные конечности у
наземных несут па себе
всю тяжесть тела леи пот-
ного и передвигают его.,
чего нс приходится де-
лать леи потным в воде.
Отсюда необходимость в
развитии системы рыча-
гов и удлинении конеч-
ностей. Конечность ла-
Рис. 244. Диаграмма эволюции carpus.
1 — Hcapulororacoid, 2 — плечо, humerus, Л — Sa-
uriptcrus (Crossoptcrygii), В— древние Telrapoda
(гипотетично, па основе личинки Bombina), С —
примитивная амфибии (Eryops). (Но Noble.)
земных—cheiropterygium — развилась из плавника. За это говорит
главным образом эмбриология, показывающая, что наземная ко-
нечность развивается у зародыша из того лее зачатка, из которого
развивается плавник; начальные стадии их развития сходны.
В пользу того ясе взгляда свидетельствует, хотя и слабо, палеонто-
логия: у некоторых ископаемых представителей из Crossopterygyii
(Eustenopteron, ^aurip terns) из верхнего девона мы видим в ске-
лете плавников прнблилссние к типу наземной конечности (рис.
244). Число лучей этой формы плавника, которая иослулшла исход-
ной базой для развития руки, было, видимо, 7. Ото была конеч-
ность, близкая частью к конечности галоидов, частью к таковой
селахий, частью Dipnoi.
Но развиться настоящая конечность наземного типа пе могла
сразу. Опа проделала известную историю, первой известной нам
стадией которой является конечность стегоцефал. Их конечность
состояла уже из тех отделов, которые мы видим у всех наземных.
Последняя состоит из плеча (humerus) в передней и б е д р а
(femur) в задней конечности, за которыми следует предплечье
из локтевой (ulna) и лучевой (radius) или голень из
большой берцовой (tibia) и малой берцовой
2J* 323
(fibula) костей. Наличие в предплечье и голеии двух костей способ-
ствует большей подвижности кисти и стопы следующих за предпле-
чьем и голспыо отделов. Кисть состоит из пескольких косточек, з а-
пястья (carpus), а именно лучепястпой (radiale), п р о-
м е ж у т о ч п о й (intermedium), локтепястпой (ulnare),
одной пли двух центральных (centralia) и пескольких з а-
п я с т п ы х (carpalia), соответствующих числу пальцев. С послед-
ними соединяются лучисто расходящиеся кости пясти (meta-
carpalia), за которыми идут фаланги (phalangae) пальцев.
В задней конечности стопа (metatarsus) состоит из идущих в та-
ком же порядке костей предплюсны: tibiale, intermedium,
fitulare, centraVa, tarsalia, metatarsalia и phalangae.
Рис. 245. Плечевой пояс стегоцефала.
1—scaiMjla, 2—procoracoid, 3—coracoid,
4 — ciavlcula, 5 — cleithrum, 6 — Intercla-
vicula, 7—supraacapulare. (no Jaekel из
Ihle и др.)
Рис. 246. Таз Cacops
aspidephorus сбоку.
1— ilium, 2— ischium,
3—pubis (no Willson из Ihle
и др.)
Конечности стегоцефал были тяжелые, грубо построенные,
неприспособленные к быстрому передвижению. Располагались
опи поперечно к оси тела, плечо и бедро занимали горизонтальное
положение, лапы стояли под углом. Для поддержания тела па неко-
тором расстоянии над землей требовалась большая работа, чем
и объясняется грубая форма конечностей, гребни и выросты па ко-
торых служили местом прикрепления сильной мускулатуры. Это
относится в одинаковой мере пк свободным конечностям и к поясам.
Плечевой пояс стегоцефал очень походил па плечевой пояс при-
митивных Osteichtliyrs, окостеневал пе всегда достаточно и состоял
из лопатки (scapula), к о р а к о и д а, и р о к о р а к о и д а
(procf racoideum), надлопаточного хрящ a (suprascapu-
lar о), клейтрума (cleithrum) п ключиц (claviculae)
(рис. 245). Последние две кости — накладные и налегали па пе-
редний край плечевого первичного пояса. Па средней линии на-
ходилось покровное окостенение — м с ж ключиц a (interclavi-
cula), являющаяся, видимо, расширенной чешуей брюшной серии
и соответствующая падгрудиику (episternum) рептилий.
Тазовый пояс стегоцефал состоял (рис. 246) из хрящевой полу-
дуги с каждой стороны, в которых можно различить следущие око-
стенения: подвздошная кость (os ilem), соединявшаяся
с одним крестцовым ребром; лобковая кость — (os pubis)
224
и седалищная кость (os ischium). Все три кости обра-
зовывали сочленовную ямку (acetabulum). У древнейших
представителей стегоцефал (Eogyrinus, Cricotus) подвздошная кость
соединялась с длинными, мало измененными крестцовыми ребрами
пе при помощи сочленения, а рыхло, с помощью мускулов. Из этого
можно заключить, что первые четвероногие возникли еще в воде.
В свободных конечностях примитивных стегоцефал число эле-
ментов carpus, как сказано, было значительное: radiale, uluare,
intermedium, две centralia, пять дистальных capralia и еще добавоч-
Рис. 247. Л — плечевой пояс Rana natatrix (Firmislcrnia) сверху,
распластанный; /> — плечевой пояс. Cyslignathus (Arcifora) свер-
ху, левая Supraseapiilare удалена; й — плечевой пояс Bombinator
pachypiis, снизу.
Л—1—clavicula, 2—coracoid, 3—scapula, 4—cleithrum, 5—omosternum,
6—acetabulum, 7—suprascapulare, 8 - humerus, 9—sternum, 10—xiphis-
ternum. JJ—1 — cartilage epicoraeold, 2—cartllago procoracoid, 3—cla-
viculn, 4—coracoid, 5—acetabulum, 6—scapula, 7—supraseapiilare,
8—cleithrum, 9—sternum, 1 0— xiphisternum. li— 1— cart, epicoraeold,
2—cartil. procoracoid, 3- clavicula, 4—scapula, 5 - cleithrum, 6—supras-
capula, 7— otostcruiim, 8—sternum. (Л—no Braus, b—no W. Parker,
Ji — no Braus, nee na Ihlc и др.).
пая, так называемая г о р о х о в и д и а я кость (os pisiforme).
У древнейших стегоцефал (Brnbolomeri) было 5 пальцев. У высших
представителей класса число элементов нередко было меньше.
У с о в р е м е и и ы х а м ф и б и й пояса конечностей и сами
шгчпости обнаруживают скорее редукцию, нежели прогрессивное
развптпе по сравнению со стегоцефалами (рис. 247). Плечевой пояс
Hindrl.i посит личиночный характер п состоит с каждой стороны
из \ршHiчюйпластинки, сочленовной ямкой для конечности разде-
ляютсигя па к о р а к ои д н у ю часть и лопаточную. В ко-
paicoippioii части обособляется передний вырост, называемый пр о-
32!>
коракоидом (procoracoideum). Окостенение наблюдается только
возле сочленовной ямки. Коракоидные пластинки налегают друг
па друга, связанные рыхлой соединительной тканью. Позади них
лежит небольшая хрящевая грудина (sternum). У Anura
С
Рис. 248. Тазовый пояс: А—Salamandra maculosa (снизу),
В — Rana esculenta (слева), С — Ascapus (слева).
1—Ilium, 2 — pubis» 3 — ischium, 4 — acetabulum, 5 — epipubis,
6 —postpubis (no Wiedersheim, G-aupp и Nable из IhLe и др.)
окостеневает коракоид, на прокоракоидпом хряще развивается
накладная кость — clavicula. Окостеневает и лопатка, оставляя
Рис. 249. Скелет задней конечности Ranidcns
sibiricus (Л) и Salamandra maculosa {В).
У последней формы па 2 central 1а больше, 1 и 2
tarsalla и тут, и там слипаются.
I-V—пальцы, 1— tarsale postminimi, 2 *—tibtalc,
3 — Intermedium, 4 — fibularc, 5 — проксимальные
centralis, G — дистальные centralia, 7—tarsalla (no
Versluys (В) и Schmalhausen (А) из Ihlc и др.)
нечпостей, пе имеют пи плечевого, пи
наверху, как у Urodela,
надлопаточный хрящ. У
одних Anura обе поло-
вины плечевого пояса
еще налегают друг па
друга, как у Urodela,
у других они срастаются
посредине. Па основа-
нии этого признака без-
хвостых амфибий под-
разделяют па подвпж-
логрудых и пеподвиж-
погрудых (Arcifera и
Firmisternia). У неко-
торых имеются переход-
ные черты между тем
и другим типом. У личи-
нок Anura плечевой по-
яс подвижной.
Apoda, пе имея ко-
тазового пояса. Тазовый
пояс хвостатых и бесхвостых в осповпом повторяет то, что имелось
у стегоцефал. С каждой стороны тела он представлен пластин-
кой. Правая и левая пластипки срастаются между собой по
средней линии. Таз Anura в связи с приспособлением к прыганию
326
имеет очень длинные подвздошные кости, сливающиеся назади с се-
далищно-лобковыми в один вертикальный кружок. Последний не-
сет па себе суставные ямки для задних конечностей. У Urodela под-
вздошные кости узки, соединение их с крестцовыми ребрами рыхлое,
от переднего края таза по средней брюшной линии отходит вперед
вилочкообразно разветвленный надлобковый хрящ (epi-
pubis). Epipubis имеется и у некоторых наиболее примитивных
Anura (Ascaphus, Xenopus) (рис. 248).
Свободная копечпость примитивнее у хвостатых амфибий. При-
митивность эта выражается прежде всего в большей полноте эле-
Рис. 250. Л. Скелет задней левой конечности Rana.
1—tiblale, 2 — fibulare, 3 —ccnlrale, 4—tarsale 2 4-3, 5 — prachallux,
f> — metatarsale 1, 7 — inelatarsale 5, l-V— пальцы. (Ho Howes, из Ihle.)
В . Скелет ноги Bombinator pahypus, очень молодого экземпляра.
1—бедро (femur), 2 —голень (tibia-fibula), 3 —tiblale, 4 — fibulare,
5 — дистальные tarsalla, I—V — пальцы. (По Sleliicr, из Ihle.)
ментов carpus и tarsus (рис. 249), хотя некоторые элементы могут
сливаться или исчезать. У Anura задние конечности изменены для
прыгания: бедро у них очень длинно, кости голени срастаются
между собой, удлиненные fibulare и tibiale образуют третий ры-
чаг конечности, который приобретает тем большую прочность,
что кости эти срастаются между собой своими концами. Многие
элементы tarsus сливаются или исчезают. Пальцы длинны, соеди-
нены перепонкой и служат для плавания (рис. 2Г>0).
Длина задних конечностей у различных Anura значительно ва-
рнирует в зависимости от того, в какой море развита способность
прыгания. Таким образом, конечности современных амфибий строй-
нее, тоньше, длиннее, нежели у тяжелых стегоцефал. Сочленения
у них также развиты гораздо лучше.
5. Мускулатура
Мускулатура амфибий в связи с развитием пятипалых копеч-
iincTrii и переходом к передвижению по земле с их помощью под-
iieprjijci. ряду существенных изменений прогрессивного порядка,
тогда- |«ак висцеральная мускулатура в связи с редукцией жабер-
327
пого аппарата подверглась дегеперации. Особенно сильным изме-
нениям подверглись, конечно, мускулы парных конечностей, тогда
как мускулы хвоста сохранили у хвостатых амфибий свой прими-
тивный характер.
У рыб мускулы туловища лежащие кверху от горизонтальной
миосепты, от головы до хвостового плавника, играют большую роль
при плавании, нежели мускулы, лежащие кпизу от миосепты.
Поэтому и развиты опи гораздо сильнее, масса их значительно
больше. Наземные позвоночные движутся при помощи парных ко-
нечностей, изгибы позвоночника не играют уже прежней роли. Поэ-
тому спинная часть туловищной мускулатуры развита у них значи-
тельно слабее. Мускулы, лежащие книзу от горизонтальной миоссп-
ты, играют у наземных существенную роль, служа для движения ре-
бер и брюшпой стенки в процессе дыхапия и для поддержки внутрен-
ностей. В связи с этим опи подвергаются известпой дифференцировке.
Вполне естественно, что у Urodela, больше времени проводящих
в воде, а иногда и всю жизнь, — еппппая мускулатура, хотя раз-
вита зпачительпо слабее, пежели у рыб, все-таки во многих отно-
шениях напоминает спинную мускулатуру последних п является
наиболее примитивной среди наземных позвоночных.
С правой и левой стороны вдоль спины хвостатых (Urodela)
идет по одному ,мускулу (musculus dorsalis trunci), разделенному
изогнутыми как у рыб мносептами на сегментысоотпетственпо числу
позвонков. В связи с подвижностью головы этот мускул распа-
дается впереди на три порции мускулов, идущих к голове. У Anura
вдоль спины дифференцируются мускулы, непосредственно при-
крепляющиеся к позвоночнику, образующие глубоколежащую меж-
позвоночную мускулатуру. Сппппой мускул туловища в отличие
от Urodela, хотя и остается еще разделенным миосептами па сегмен-
ты, превращается в длинный мускул, так как мпосеиты пе прикреп-
ляются уже здесь к позвонкам; его задача здесь — фиксация головы
поотпошепиюк тазуп копчику. Брюшные мускулы Urodela тоже лег-
ко могут быть выведены из того, что имеет место у рыб; сегментация
мускулатуры выражена здесь совершенно ясно, п мускульные во-
локна идут от миосенты к мпосепте; однако вся мускулатура рас-
падается па четыре слоя, отличающихся направлением мускульных
волокон и местом их прикрепления (рис. 251).
Прежде всего различают наружный и внутренний слои—ко-
сую наружную (musculus obliquus externus) и косую внутреннюю
(musculus obliquus internus) мышцы. Непосредственно под мноссп-
той и па брюшной стороне остаются участки недифференцирован-
ной мускулатуры с продольным направлением волокон. Боковой
и брюшпой прямые мускулы — musculus rectus lateralis и muscu-
lus rectus abdominis. Наружные косые мышцы состоят из волокон,
расположенных спереди и сверху, назад и вниз; волокна внутрен-
них косых идут вниз и вперед. В таком виде мускулы наблю-
даются у личипок, еще. зпачительпо напоминая состояние их у рыб.
С появлением конечностей па внутренней стороне внутренней
косой мышцы образуется слой несколько с иным направлением во-
328
локон — поперечный мускул (nrasculus transversus). Происходит
обособлепие и еще нескольких мышечных образований. Межребер-
ных мышц пе образуется, так как ребра развиты слабо, коротки.
При метаморфозе мускулы претерпевают некоторые изменения, бла-
годаря которым они оказываются менее пригодными для производ-
ства боковых изгибов тела. У Anura в течение развития мышцы,,
служащие для плавания, исчезают.
Рис. 251. Мышцы Siredon pisciformis.
A — сбоку, В — снизу.
1 — спинная мускулатура туловища, 2— in. obliquus cxtcrnus superficial is, 3 — m.
obliquus cxtcrnus profundus, 4 — in. transversus, 5 — subvcrtebralis, 6 — dorsalis sca-
pulae, 7 — latissimus dorsl, 8 — trapezius, 9 — rectus supcrflclalis, 10—depressor man-
dlbulac, 11 — constrictor, 12 — levator arciiuin brancblalium, 13 —(на рис. В) —pec-
toralls, 13 —(на рис. A) — ab ductor inandibnlac, 14 — subcoracoideus. 15 —genio-
hyoldeus, 16 — ceratohyoideus externus, 1 7 — ceratohyoideus internus, 18 — pubo-iscbio-
tibialis, 19 —мышцы плеча, 20—мышцы бедра (пи Biltschli.)
Сложная мускулатура развивается в конечностях в связи со слож-
ными движспияип образующих их частей. Специальные мышцы
связывают конечности с их поясами, другие мышцы располагаются
между частями самих конечностей. Различные мышцы действуют
как антагонисты: сгибатели, разгибатели, приводящие мышцы,
отводящие мышцы.
6. Пищеварительная система амфибий
В системе органов пищеварения также видна связь амфибий с ры-
бами, хотя наземный образ жизни уже здесь накладывает свою
печать иа систему. Рот у Линга, глотающих добычу целиком, очень
широк, также у саламандр. У некоторых амфибий, наоборот, рот
сильно сужен, в связи с особым родом пищи.
(-ходпо с рыбами у амфибий все кости полости рта могут нести
зубы. Обычно опи находятся у Линга па межчелюстных (praemaxil-
laria) и челюстных (maxillaria), а также на сошнике (vomer),
тогда как на нижпейчелюсти (dcntale) п на нёбных костях — редко.
. 32^'
У Pipa и Bufo зубов иет вовсе. Весьма интересно, что у личинок
Anura челюсти одеты роговой оболочкой и имеются роговые зубы.
Рис. 252. Пищеварительная си-
стема самца лягушки.
1 — хоаны, 2— сошниковые зубы, 3—
отверстие евстахиевой трубы, 4 —
жевательная мышца (разрез), 5— гло-
точная часть головного отдела ки-
шечника, 6 — гортанно-трахеальная
камера, 7—левое легкое, 8 — пра-
вое легкое, 9 —желудок, ю —две-
надцатиперстная кишка, 11 —тонкая
кишка, 12 —прямая кишка, 13—кло-
ака, 14 — мочевой пузырь, 15—пра-
вая лопасть печени, 16 — средняя
лопасть печени, 17—левая лопасть
печени, 18 —желчный пузырь, 19 —
желчный проток, 20 — поджелудоч-
ная железа, 21 — селезенка, 22 —
мочеточник, 23 —семенной пузырь.
(По Макушок.)
От рыб амфибии отличаются
наличием мускулистого языка,
У амфибий, живущих в воде, язык
еще мало отличается от языка
рыб; он мал, мало подвижен и мало
мускулист. У некоторых видов оп
пе имеет желез. Зато у Anura и
саламандровых язык развит силь-
но. Но это развитие—явление вто-
ричное, наступающее в онтогенезе
поздно: язык развивается в виде
складки на дне ротовой полости,
охватывающей «первичный язык»,
сперва лишенный желез и обни-
мающий, как у рыб, конец подъ-
язычной кости. Язык обычно очень
богат железами. Иногда (у Sala-
mandrina, у Anura) сидит па осо-
бом стебельке. У лягушек язык
прикреплен передним концом и
может поэтому выбрасываться, обо-
рачиваясь нижней стороной. У ро-
дов Pipa и Xenopus (сем. Pipidae)
язык может вовсе отсутствовать
(Aglossa).
В связи с ротовой полостью
паходятся голосовые меш-
ки — сферические выпячивания
слизистой оболочки основания
ротовой полости. Они могут быть
парными (Rana) и попарными
(Hyla) и служат резонаторами для
усиления звуков, производимых
животным.
У рыб в полости рта находятся
лишь одноклеточные слизистые
железы. Пищеварительные железы
пе могли бы иметь значения, так
как через рот все время проходит
вода, служащая для дыхания.
У амфибий впервые появляются
во рту многоклеточные железы, а
именно одна, выделяющая слизь
для смачивания языка и лежащая между ossa maxillaria —glandula
intermaxillaris, и железы языка; у Anura—еще одна позади хоан.
В глотке личинок и у постоянно живущих в воде Urodela сохра-
няются, как у рыб, жаберпые щели. Слизистая оболочка пищевода
.330
сходна со слизистой рта. Как па примитивный признак можпо ука-
зать па присутствие здесь слизистых клеток и респичатого эпите-
лия. Пищевод переходит в желудок, по форме напоминающий
рыбий. Желудок имеет два слоя мыпщ: наружный продольный и
внутренний из кольцевых волокон. Слизистая оболочка желудка
содержит большое количество трубчатых желез, клетки которых
двоякого рода: слизистые и с зернистой протоплазмой, выделяющие
пепсин и соляную кислоту.
Кишечник еще очепь мало дифференцирован (pifo. 252). Но де-
ление на длинную и топкую среднюю кишку и широкую заднюю
ясно выражено. Топкая кишка образует ряд извилин. Слизистая
ее состоит из бокаловидных и цилиндрических клеток, выполняю-
щих функцию всасывания. Для усиления последнего в слизистой
имеются продольные и поперечные складки, приближающиеся по
строению к ворсинкам высших позвоночных. У Bufo это уже на-
стоящие ворсинки. Задняя кишка, как сказано, переходит в клоаку
с мочевым пузырем па брюшпой стороне.
В связи с кишечником находятся печень (hepar), поджелудочная
железа (pancreas). С печенью связан желчный пузырь. Как кишеч-
ник, так и печень с поджелудочной железой укреплены на месте
при помощи брюшины (peritoneum), образующей брыжейку.
Захватывание пищи производится пастью и лишь редко с по-
мощью передних лапок. Схватыванию у лягушек помогает липкий
язык, выбрасываемый с большой быстротой. Так как он снабжеп
железами с клейким выделением, то добыча — мухи, черви и т. п.—
приклеивается к пему. Заглатыванию добычи помогают глаза,
сильно вдающиеся в ротовую полость, от которой отделены только
слизистой оболочкой. При помощи особого мускула, ретрактора, гла-
за оттягиваются назад и помогают проталкиванию пищи.
П е п с и и и соляная кислота выделяются в желудке. Ими вы-
зывается распадение белков па пептоны. В кишечнике под влия-
нием кислой кашицы, поступающей из желудка, выделяется секре-
тин, попадающий, однако, пе в полость кишки, а в кровь; оп стиму-
лирует к деятельности поджелудочную железу, сок которой, выде-
ляемый в двенадцатиперстную кишку, содержит три фермента:
т р п п с и и, амилазу и липаз у. Под воздействием этих
ферментов происходит переход белков в пептопы, крахмала —
в сахар и расщепление жиров па жирную кислоту и глицерин.
Выделяемый стопками кишки э р е п с и и доводит расщепление
белков до аминокислот. Желчь помогает действию липазы па жиры.
В кишках происходит всасывание продуктов переваривания.
7. Органы дыхания амфибий
У амфибий имеются два рода дыхательпых органов (не считая
кожи): жабры и легкие. Ослабление жаберного дыхапия
и выступание па сцепу легочного дыхапия наблюдается уже у Dip-
noi; изменения в этом направлении наблюдаются у Polypterus,
и Lepidosteus. У амфибий жаберное дыхапие сохраняется прежд(
331
всего у личинок, а затем у тех Urodela, которые всю жизнь проводят
BBofle(Perennibranchiata в прежних системах). Жаберные щели уна-
следованы амфибиями от рыбообразных предков. Жаберные дуги
обнаружены у стегоцефал, у личинок и у некоторых взрослых
(Branchiosauridae). Все современные амфибии в личиночном состоя-
нии дышат жабрами. Нормально у них закладывается б висцераль-
ных мешков и 6-й недоразвитый. Но наружу открываются не все:
жаберных щещй образуется 4 или еще меньше. Иногда щелей значи-
тельно меньше, чем дуг. Наличие щелей и дуг является доказатель-
ством происхождения амфибий от рыб. Внутренние жабры, гомоло-
гичные жабрам рыб, встречаются, однако, только у личинок Anura
в виде коротких выростов покровов па дугах, разделяющих жабер-
пые щели. Мягкой жаберпой крышечкой (operculum), разрастаю-
щейся со стороны гиоидной дуги, жаберная область прикрывается
снаружи. Жаберпые крышечки правой и левой сторон сливаются
между собой с нижней стороны, оставляя у некоторых Anura пар-
ные отверстия, а у большинства — одно непарное на левой стороне
тела.
На раппих стадиях развития у личинок Anura и у всех других
амфибий имеются только наружные жабры, гомологичные, видимо,
наружным жабрам личинок Polypterinin Dipnoi. У Apoda и у Anura
наружные жабры существуют лишь в личиночном периоде, па ран-
них стадиях развития, у Urodela же, вторично вернувшихся к вод-
ной жизпи, они сохраняются всю жизнь. Отсюда название для этих
амфибий — постоянножаберные (Perennibranchiata),
хотя название это, как было сказано, обнимает группы амфибий,
имеющих различное происхождение. Наружные жабры, вероятпо
унаследованы амфибиями от кистеперых рыб.
Легкие амфибии имеют вид длинных цилиндрических мешков
с тонкими стенками (у Urodela) или более коротких (у Anura).
У безногих правое легкое развито гораздо больше левого. Появи-
лись легкие у предков четвероногих еще задолго до выхода па сушу.
Мы видим такие же легкие у двоякодышащих. Появились опи,
очевидно, в качестве дополнительного органа дыхания вследствие
недостаточного развития жаберного дыхания, с одной сторопы, и
возможно неблагоприятных для дыхания условий в пересыхавших
п портившихся водах — с другой. Задняя часть жаберпой полости
развилась у них в дополнительный дыхательный орган. Первона-
чально этот орган, имевший вид двулопастпого мешка, открывав-
шегося па нижней стороне глотки, был несовершенен: степки его
должны были быть топкими, хотя и обильно снабженными кровью,
со слабо развитыми или почти неразвитыми перегородками. Как
все жаберпые выпячивания (щели), оп имел гладкую висцеральную
мускулатуру и иннервировался блуждающим первом.
Легкие амфибий мало изменились в сравнении с этим: у водных
Urodela легкие выполняют скорее роль гидростатического аппарата
и имеют гладкую внутреннюю поверхность; высота организации
их даже ниже, чем у Dipnoi. Нормально же у амфибий внутренняя
поверхность легких является ячеистой благодаря тому, что в по-
332
Рис. 253. Легкое Rana
изнутри.
(по Gaupp.)
лость легкого вдается система перекладин (рис. 253). Весьма ин-
тересно, что чем более наземным является тот или иной вид, тем
более развитыми являются перекладины в легких: у жабы легкое
более ячеисто, чему лягушки. У рода Ascaphus, живущего в горных
ручьях, в воде, богатой кислородом,сильно развито кожное дыхапие,
легкие же, наоборот, маленькие и слабо снабжаются кровью. Ряд
амфибий из подотряда Salamandroidea (Salamandrina, Plethodon,
Spelerpes, Batrachoseps и др.) совершенно утратил легкие, взамен
которого глоточное и кожное дыхапие сильно развилось. .
Легочные мешки в простейшем случае соединяются между собой
впереди, открываясь непосредственно в глотку продольной щелью,
поддерживаемой по сторонам хрящевыми полосками. Эти хряще-
вые полоски при помощи прикрепля-
ющихся к пим мускулов могут рас-
ширять и суживать гортанную щель.
Хрящи эти происходят из послед-
ней жаберпой дуги и в простейшем
виде встречаются у некоторых Uro-
dela. От этих хрящей могут обособ-
ляться хрящики, называемые перст-
невидными хрящами. Их можно
сравнивать с черпаловидными
хрящами (cartilaginea arythenoi-
.dca) высших позвоночных. Некото-
рые Urodela, а также Apoda имеют
довольно длинное дыхательное горло,
поддерживаемое хрящевыми кольцами. У Anura слизистая оболочка
в гортани образует голосовые связки. Гортань имеет сложную
мускулатуру. На дпе или по углам рта находятся резонаторы,
надувающиеся при квакании.
Механизм дыхапия наземных амфибий имеет своим корнем реф-
лексы, наблюдающиеся у рыб и водпых амфибий. Ближе всего
к дыханию рыб дыхапие личинок Anura, имеющих впутреппие жабры,
оперкулярную складку и образованную их срастанием жаберную
полость, открывающуюся паружу одним отверстием. Кроме того,
у личинок амфибий ротовая полость обильно снабжена кровью.
Забирая воду в рот и проталкивая ее путем поднятия челюстей
через ноздри, личинки увеличивают газовый обмен в ротовой по-
лости. Когда же личинки подрастают, опи поднимаются к поверх-
ности, где заглатывают воздух подобно цератоду, и поднятием дпа
ротоглоточной полости проталкивают воздух в легкие. У водных
Urodela наблюдается сходный акт. При опускании дпа ротоглоточ-
ной полости и при закрытых сзади жаберпых отверстиях вода наса-
сывается в ротовую полость через рот или ноздри или через то и дру-
гое вместе. Путем последующего поднятия дпа ротовой полости
при закрытых поздрях вода выталкивается через жаберпые щели.
Благодаря этим движениям слизистая оболочка рта и глотки всту-
пает в соприкосновение с новыми массами воды, а у жабер полу-
чается движение, обновляющее дыхательную среду.
833
У паземных амфибий механизм дыхапия является актом загла-
тывания воздуха благодаря опусканию мускулистого дпа ротовой
полости и проталкиванию его в легкие вследствие поднятия дпа.
Таким образом, дыхание наземных амфибий является актом, осу-
ществляемым по типу нагнетательного насоса, преобладающего
у низших рыб. Непосредственной основой, на которой он разви-
вается, является механизм дыхапия постояпножаберпых амфибий.
Этот последний, наблюдаемый, например, у Necturus, должен был
развиться у отдаленных рыбообразных предков амфибий. Из него
уже развился более сложный тип дыхапия наземных — Anura.
У безлегочных саламандр сильно развит газообмен внутриротовоп
и глоточной полости, совершающийся при помощи частых, до 120—
170 колебаний в минуту диафрагмы рта (у лягушек нх 30).
Вообще следует сказать, что легочное дыхание у амфибий в целом
является вспомогательным способом дыхания. В этом также имеется
указание и па его филогенетическое происхождение.
Дыхание современных амфибий пи в коем случае пе могло быть
источником развития дыхапия высших Tetrapoda (дыхапия при по-
мощи поднятия ребер, расширения грудной клетки и насасывания
таким образом воздуха). Последний тин мог возникнуть, во всяком
случае наметиться у древнейших вымерших амфибий, имевших длин-
ные ребра.
8. Органы кровообращения у амфибий
Дыхание осуществляетсяне только при помощи органов дыхания,
жабер или легких, но и с помощью кровообращения; кровь транс-
портирует кислород, поглощенный ею, и выносит к органам дыхапия
углекислоту. Поэтому переход от жаберного дыхапия к легочному
и к наземной жизни должен был сопровождаться глубокими изме-
нениями в системе кровообращения. Наземный образ жизни ста-
новится возможным при большей энергии обмена веществ. Это до-
стигается соответственными изменениями в дыхательной и сосудистой
системе одновременно. И как развитие легких имело место задолго
до выхода па сушу у рыб, так и изменения в кровообращении по-
звоночных должны были возникнуть одновременно с этим. Эволю-
ция сосудистой системы у наземных заключается в замене симмет-
ричной системы жаберных сосудов, соответствующих по числу жа-
берным щелям, несимметричной системой, состоящей из аорты и ле-
гочной артерии, которую мы наблюдаем у птиц и млекопитающих,
разделением системы кровообращения иа артериальную и венозную
и в соответственных изменениях венозной системы.
Амфибии занимают промежуточное иоложепие между рыбами
и высшими позвоночными.
У них еще сохраняется симметричное расположение артериаль-
ных дуг, по часть жаберных дуг исчезает, развивается система
внешних и внутренних сонных артерий, развиваются легочные,
пока еще симметричные артерии (рис. 254), и происходит, хотя
еще неполное, разделение сердца на артериальную и венозную по-
ловины.
334
В личиночном состоянии артериальная система имеет еще более
рыбий тип (рис. 266). У эмбрионов закладывается обычно шесть
пар артериальных дуг, по первые две почти всегда рано исчезают.
У личинок Urodela пока еще существуют наружные жабры,
от артериального ствола отходят четыре пары приводящих жа-
берных артерий, соответствующие четырем задним дугам рыб, т. е.
3-я — 6-я артериальные дуги. Из них три первые направляются
к жабрам, где распадаются на капиллярную сеть, переходящую
в отводящие жаберпые артерии, идущие к парному корню спинной
аорты. Четвертая, артериальная, дуга прямо переходит в корень
аорты, так как соответственной ждбры пе имеется. От нее идет ветвь
Рис. 254. Диаграмма артериальных дуг различных позвоноч-
ных с брюшной стороны.
А—у эмбриона, В —у рыбы, С—у Urodela.
1—спинная аорта, 2 — брюшная аорта, 3 и 4—первые две эмбрио-
нальные дуги, которые почти всегда исчезают, 5—8 — четыре жабер-
ных дуги, » —легочная артерия. (Из Parker.)
к легким, несущая, следовательно, венозную кровь и являющаяся
как бы продолжением самой четвертой, артериальной, дуги, превра-
щающейся в легочную артерию.
У взрослых Urodela артериальная система остается примерно
в том же положении, ио 1-я из сохраняющихся (3-я) жаберная
артерия обособляется от 2-й (4-й) и дает начало сонным артериям.
2 и 3-я (1 и 5-я) дуги образуют корень аорты, 4-я (G-я) в еще большей
мере становится легочной артерией. У Anura 3-я (5-я) артерия исче-
зает, 2-я (4-я) становится системной дугой аорты, а 4-я (6-я) теряет
связь с аортой и в полной мере становится легочной артерией.
От 1-й (3-й) жаберной артерии отходят вперед к головному мозгу
внутренняя сонная артерия (arteria carotis in-
terna) инару ж пая (carotis externa), идущая к языку. У са-
ламандровых и бесхвостых при основании сонных артерий находится
так называемая «сонная железа», сплетение сосудов (rete mirabile).
Каждая дуга аорты дает подключичную артерию
(arteria subclavia) к передним конечностям; от левой дуги аорты не-
далеко от соединения с правой отходит еще чревная арте-
335
р и я (arteria coeliaca). У Anura от легочной артерии отходит мощ-
ная к о ж п а я артерия (arteria cutanea) к коже.
В венозной системе сравнительно с рыбами имеются существен-
ные изменения, намеченные, однако, подобиоизмепениям артериаль-
ной системы, у Dipnoi. А именно: задние кардинальные вепы, по ко-
Рис. 255. Диаграмма, иллюстрирующая развитие
и судьбу аортальных дуг амфибий, вид слова.
А—личинка Urodela, В—взрослая Urodela, С—Anura.
1, 2, 3, 4, 5, 6 — первичные артериальные дуги, 7 —
предсердие, 8—желудочек, 9 — конус, 10—наружная
сонная артерия, 11 — кровоснабжение наружных
жабер, 12 — боковая спинная аорта, 13 —боталлов
проток, 14 — легочная артерия, 15—легкое, 16 —ле-
гочная вена, 17 — внутренняя сонная артерия, 18 —
общая сонная артерия, 19 —сонная железа, I—V —
жаберные щели. (Из [Goodrich.)
делится па две ветви. Из одной кровь поступает в почки, участвуя
в образовании воротной системы почек (venae por-
tae). Другая вместе с такой же парой другой стороны образует
брюшную вену (vena abdominalis), идущую к печени и
вместе с вепой от внутренностей образующую воротную вепу печени.
Начало редукции задних кардинальных вен мы видели уже у Dip-
noi,так же как и образование нижней полой вепы и брюшной вены.
торым венозная кровь
течет у рыб из моче-
половой системы, а
также из хвостовой
части тела, уступают
свою роль большому
непарному стволу —
нижней или зад-
ней полой вепе
(vena cava inferior).
Эта вена (рис. 256)
впадает в венозный
синус вместе с двумя
верхними по-
лыми венами
(venae cavae superio-
rs), соответствующи-
ми кювьеровым про-
токам рыб. Верхние
полые вепы слага-
ются из я р е м п ы х
(venae jugulares) и
подключичных
в е п (venae subcla-
viae); у Urodela со-
храняются еще па ря-
ду с нижней полой
вепой к а р д ина ль-
и ы е вен ы, назы-
ваемые здесь косы-
м и (vena azygos dex-
tra и vena» azygos si-
nistra) (рис. 257).
Кровь из задних ко-
нечностей идет в под-
вздошную вепу
(vena iliaca), которая
.336
Сердце амфибий состоит, как у рыб, из венозного
синуса, предсердия, желудочка и артери-
ального конуса. Но, как у высших четвероногих, отделы
сердца лежат пе на одной прямой,
а сердце изогнуто, и предсердия
помещаются пад желудочком да-
леко впереди последнего. Основное
же отличие сердца амфибий заклю-
чается в том, что в предсердии
имеется перегородка, разделяю-
щая в известной мере ток крови
Рис. 25G. Схема пснозиой системы
Капа.
1 — задняя полай вена, 2 — передняя
полая вепа, 3 —наружная яремная,
4 — внутренняя яремная, 5—подклю-
чичная вена, G — подвздошная вена,
7 — воротная вепа, 8 — брюшная ве-
па, 9 —бедренная вена, 10—печеноч-
ная вена, 11—легкое, 12 — кишка,
13 —печень. (По Gaupp из Ihlc и др.)
Рис. 257. Схема венозной си-
стемы саламандры.
(По Wiedersheim из Ihle.)
1—сердце, 2—проток Кювье, 3—
наружная яремная вена (v. jugu-
laris externa), 4—внутренняя ярем-
ная иена (v. jugularis interna), 5—
подключичная вена (v. subclavia),
6 — большая ножная вена (v. cu-
tanea magne), 7 — косая вена (v.
azigos—v. cardinal is posterior), 8—
печеночные вепы (v. hepaticae),
9 — печень (hepar), 10 — воротная
вепа (v. portae), И —кишка, 12 —
брюшиан вена (v. abdominalis),
13 — подвздошная вена (v. iliaca),
14 — почка, 15 — хвостовая вена
(v. caudalis).
в сердце. Левое сердце является
артериальным, правое — глш пым
образом венозным: окисленная
кровь по легочным венам прямо
поступает в левое предсердие.
Редуцированная кровь из тела по-
ступает в венозный синус, а отсюда — в правое предсердие, отвер-
стия ясе легочпой вены и венозного синуса находятся на противо-
положных сторонах меясатриальнои перегородки (рис. 258). Между
22 Кашкаров и Стаичинский
337
поступающей сюда артериальной
9
Рис.
258. Продольный разрез через сердце
Rana.
1 — правое предсердие, 2 — левое предсердие,
3 — межатриальная перегородка, 4 — отверстие
венозного синуса в предсердие, 5 — отверстие
легочной вены, 6,7 — правый и левый клапаны
между предсердиями и желудочком, 8 — арте-
риальный нонус, 9 —спиральная складка кону-
са, 10—горизонтальная перегородка, И —
срединная перегородка, 12—клапан между
предсердиями и желудочком. (Из Ihle по G-r.
Кегг.)
синусом и предсердием находятся два клапана, между предсердием
(atrium) и желудочком (ventriculus) — четыре клапана: два боль-
ших п два малых. Желудочек простой, без перегородки. Но в стен-
ках его имеется ряд мускулистых перекладин, мешающих смешению
" и венозной крови. Впереди жНЬгу-
дочка находится длинный,
снабженный поперечнопо-
лосатыми волокнами (т. е.
являющийся частью же-
лудочка) артериаль-
ный конус (conus arte-
riosus), у пачала и конца
имеющий поперечный ряд
клапанов, а внутри спи-
ральной перегородкой раз-
гороженный па верхний
отдел — легочный (cavum
pulmonale) и нижний—ар-
териальный (cavum aorti-
cum). От верхнего отдела
отходят легочные артерии,
от нижнего — системные
дуги аорты и сонные арте-
рии. Благодаря указанно-
му строению сердца, а так-
же порядку сокращения
его частей, артериальная
и венозная кровь пе сме-
шиваются. Из синуса ве-
нозная кровь поступает в
правое предсердие. Когда
последнее сокращается,
артериальный конус рас-
слаблен, а отверстия легоч-
ных артерий широко от-
крыты. Кровь направ-
ляется в последние. Когда
легочные артерии напол-
нятся и давление в них
возрастет, сокращается
конус, спиральный клапан
располагается так,что мешает крови итти в cavum pulmonale, и опа
через cavum aorticum направляется в широко открытые системные
дуги. Сокращение в это время левого предсердия проталкивает
кровь, поступившую через легочные вены, в левую половину желу-
дочка. Здесь кровь лишь немного смешивается с венозной, и в си-
стемные дуги поступает кровь почти артериальная. Под конец же,
когда дуги аорты наполнятся и давление в них возрастет, давле-
ние в желудке достигнет максимума, тогда преодолевается сопротив-
338
ление в сонной железе, и чистая артериальная кровь поступает
по сонным артериям в голову и к мозгу.
Таково строение сердца и распределение крови у Anura. В нем
выражается высшее достижение, имеющееся у амфибий в сравнении
с рыбами. У Urodela и Apoda сердце носит несколько более личиноч-
ный характер, и атриальная перегородка и конус у них построены
проще, так что разделение крови получается не столь полное.
Как в строении артериальной и венозной системы,так и в сердце
амфибий имеется много общего с Dipnoi; у последних аорта уко-
рочена,- как у амфибий, имеется перегородка внутри предсердия,
перегородка в conus arteriosus обусловливает разделение артери-
альной и венозной крови. У двулегочных Dipnoi (Dipneumones)
артериальный конус даже более полно разделен, чем у амфибий.
Кровообращение у последних организовано выше, нежели у Dipnoi
благодаря физиологическим приспособлениям. Сходство системы
кровообращения у Dipnoi и Amphibia говорит за то, что у общего
рыбьего предка тех и других имелась сходная структура сердца,
однако более примитивная, чем у Dipneumones.
У амфибий сильно развита лимфатическая система, состоящая
пз щелевидпых пространств. У бесхвостых обширные лимфатические
пространства имеются под кожей. Во многих местах лимфатической
системы находятся ритмически сокращающиеся так называемые
«лимфатические сердца».
Кровяные тельца амфибий отличаются своей большой величиной
и ядерностью эритроцитов. Эта большая величина, а, следовательно,
малая общая поверхность всей суммы кровяных телец (при большой
величине их число меньше), соответствует малой энергии обмена ве-
ществ у амфибий. В отношении белых кровяных телец амфибии стоят
выше рыб, у которых имеется только один вид зернистых лейко-
цитов — нейтрофилы; у амфибий ясс, как у высших Tetrapoda,
имеются нейтрофилы, базофилы и эозинофилы. Лейкоциты амфи-
бий весьма способны ic фагоцитозу. Красные кровяные шарики ам-
фибий живут дольше, чем у Amniota, что, как и меньшее сродство
их гемоглобина к кислороду, стоит в связи с менее интенсивным
обменом веществ.
9. Мочеполовая система амфибий
Мочеполовая система амфибий напоминает таковую акул.
Постоянной почкой амфибий является mesonephros, или пер-
вичная почка, как и у рыб. Pronephros, или головная
почка, имеется только в эмбриональном состоянии. Наиболее
отчетлива закладка головной почки у Apoda, где она занимает
4—15-й сегменты мезодермы и функционирует довольно долго,
уступая затем место mesonephros и исчезая. Mesonephros развивает-
ся на протяжении 30—100 сегментов. Почки взрослых имеют вид
-длинных, узких лепт, тянущихся от сердца до клоаки и разделенных
друг от друга аортой и нижней полой веной. У эмбрионов Urodela
и Anura pronephros функционирует, но закладывается всего 2—3
22* * 339
канальца головной почки. Ко времени метаморфоза pronephros за-
меняется mesonephros. Почки взрослых Urodela длинны и состоят^
из двух отделов — переднего и заднего. Передний—узкий и нахо-
дится у самцов в связи с половыми железами, задний — более широ-
кий — является почкой собственно. Тот и другой представляют
mesonephros (рис. 259). У Anura этого деления нет. Почечные тру-
Рис. 259. Схема урогенитального
аппарата самца (Л) и самки (В)
Triton taeniatus.
1—семенник, 2 —vas cfferens, выно-
сящий проток, 3 — мюллеров протон,
4 — первичный мочеточник, 5 — соби-
рающие канальцы, 6—хвостовая часть
почки, 7 — краевой канал почки, 8 —
яичник, 9 —воровка яйцевода, 10—
мюллеров проток. (По Wiedcrshelm.)
бочки у амфибий построены уже
весьма сложно, напоминают моче-
вые канальцы Amniota и состоят
из ряда отделов. Как у бесхвостых,
так и у хвостатых на брюшной
стороне почек имеются отверстия в
полость тела, нефростомы. За не-
фростомом идет шейка,’ в расши-
ренную часть которой впячивается
клубочек сосудов, ветвь почечной
артерии, образующая здесь маль-
пигиево тельце (glomeru-
lus). Расширенная часть шейки, в
которую впячивается glomerulus,
называется баумановойкап-
сулой. Образовав ряд изгибов,
мочевые трубочки впадают в моче-
точник, развивающийся из первич-
ного выводного протока почек.
Последний распадается па моче-
точник и па яйцевод (мюллеров
канал). Дистально мочеточники
сближаются между собой и откры-
ваются в клоаку. Все амфибии
имеют обширный тонкостенный м о-
чевой пузырь, отходящий
от брюшной стенки клоаки.
Почки амфибий служат не толь-
ко выделительными органами для
удаления некоторых продуктов об-
мена,по и регуляторным аппаратом,
сохраняющим определенный состав жидкостей тела. Этой функции
помогает мочевой пузырь, через тонкие стенки которого вода может
поступать в ткани, если осмотическое давление в крови нарушено.
Половые железы связаны в большей или меньшей мере с выдели-
тельной системой, составляя одну мочеполовую систему. В общем
эта система у амфибий примыкает к тому, что имеется у селахий.
Семенники (testes) Apoda состоят из многочисленных отдель-
ных частей, соединенных продольным каналом, у Urodela они вы-
тянуты, у Anura — овальны. От семенников отходят выводные ка-
нальцы — vasa efferentia. У Urodela они проходят через передний
отдел почки, имеющий вид дегенерирующего органа, стоящего
па пути к утрате выделительной функции и имеющего главной функ-
340
цией проведение спермы. Почечные канальцы из заднего отдела поч-
ки-открываются в клоаку самостоятельно, а не в остаток первичного
протока, пока служащий здесь только семяпроводом. У большинства
Anura выводные канальцы семенников образуют сеть, открывающую-
ся в выводной канал почки. У Discoglossus и Alytes, частью у Bombina,
семя выводится прямо в мочеточник, минуя почку. Остатки яйцево-
дов, или мюллеровых протоков, обычно имеются у самцов амфибий.
Яичники (ovarium) вытянуты в виде узких пластинок у без-
ногих (Apoda). У хвостатых опи имеют вид полых внутри продолго-
ватых мешков, у Anura перегородками разделены на камеры. Яич-
ники подвешены в полости тела на складке брюшины (mesovarium).
Образованные из мюллеровых протоков яйцеводы открываются
в полость тела воронкообразными отверстиями (ostium abdominale).
Они длинны, извилисты и открываются дистальными концами в клоа-
ку. Яйца, попадая из яичника в полость тела, поступают оттуда
в воронку яйцевода, одеваются, проходя через последний, оболочкой,
взбухающей в воде. В заднем- отделе яйцеводов, называемом м а т-
к о й (uterus), яйца Anura соединяются в комья (у Rana) или в шнур-
ки (у Bufo). У живородящих саламандр в этом отделе проходят
свое развитие зародыши.
У Bufo передняя часть семенника сохраняет характер яичника,
которым первоначально на ранних стадиях развития является
половая железа. Эта часть носит название биддерова ор-
гана. При удалении семенника биддеров орган может развиться
в яичники, зачаток мюллерова капала превратится в яйцевод,
а самец — превратится в самку. Вообще, хотя амфибии раздельпо-
полы и пол определяется рано, можно воздействием внешних факто-
ров направить развитие пола в иную сторону. В связи с половыми
железами (у Anura) или с передним отделом почек (у Urodela) па-
ходятся лопастные, пли лентовидные, органы, содержащие большое
количество жировых отложений — так называемые жировые
тела. Сильнее развиты опи у Apoda. Наибольшего развития
достигают они осенью перед зимовкой, наименьшего — после отклад-
ки яиц. Они, однако, не являются простым запасом жира для зимы,
а стоят в связи с гонадами. Их удаление вызывает дегенерацию по-
ловых желез. Частичная кастрация, обратно, вызывает гипертро-
фию жировых тел.
Клоака у Apoda может выворачиваться и служить совокупитель-
ным органом. Самцы тритонов и саламандр могут охватывать глад-
кую клоаку самки губами своей клоаки, усеянной сосочками,
и производить внутреннее оплодотворение. У Anura оплодотворение
наружное за исключением одного вида (Ascaphus). Спермин Urodela
склеиваются в комочки — сперматофоры, которые захватываются
самкой при помощи клоаки.
10. Нервная система и органы чувств амфибий
Нервная система амфибий стоит еще па низкой ступени разви-
тия. Наиболее близка она с нервной системой двоякодышащих.
Но в пей есть и черты, сближающие ее с нервной системой рептилий.
341
В связи с развитием парных конечностей в спинном мозгу у ам-
фибий имеются плечевое и поясничное вздутия,
или утолщения.
Отделы головного мозга у Anura лежат еще в одной плоскости,
как у рыб, передний мозг лишь слегка прикрывает промежуточный,
как у Dipnoi. Полушария парные, удлиненные (рис. 260), пере-
ходят впереди в обонятельные доли. В каждом полушарии имеется
полость — боковой желудочек. Стенки переднего мозга
построены у амфибий сложнее, нежели у рыб; в них дифференциру-
ются участки с различным строением. Но настоящей коры в верхней
стенке переднего мозга еще нет. Опа появляется только у рептилий.
Рис. 260. Мозг лягушки (Rana temporariaj.
А — сверху, В — снизу, С—сбоку, D—в продольном разрезе. 1 — обонятельные
доли, 2 — полушария, 3 — промежуточный мозг, 4 — пинеальная железа, 5 — зри-
тельная доля, 6— мозжечок, 7 — продолговатый мозг, 8 — хиазма, перекрест зри-
тельных нервов, 9 — питуитарная железа, 10 —• воронка.
(Из Parker по Gaupp и Osborn.)
Проводящих путей от переднего мозга к позадилежащим центрам
у амфибий тоже пока еще пет. Поэтому удаление полушарий
и не вызывает нарушения актов плавания, схватывания добычи,
спаривания. Кора переднего мозга пе дает у амфибий моторных
импульсов.
Промежуточный мозг (diencephalon) лишь слегка прикрывается
спереди полушариями исредпим мозгом сзади. С ним связана верхняя
мозговая железка, или эпифиза (epiphysis); судя по теменному
отверстию в крыше черепа, она была у стегоцефал. Впереди эпифи-
зы отходит еще париетальный орган, или парафиза (paraphysis).
Крыша промежуточного мозга образована сосудистым
сплетением (plexus chorioideus), прикрывающим третий
желудочек, а снизу отходит воронковидное выпячивание —
342
Рис. 261. Вертикальный
разрез органа боковой
линии Siren lacertina,
показывающий нервные
окончания около чувст-
вительных клеток. Пунк-
тиром показаны органы
боковой линии.
(Из Noble.)
с характером место-
нижний мозговой придаток, или гипофиза (hypophysis).
Баковые степки промежуточного мозга образуют утолщения — так
называемые зрительные бугры (thalami optici). Средний
мозг представлен хорошо развитыми округлыми возвышениями —
зрительными долями (lobi optici) или двухол-
мием (corpora bigemina). Зрительные доли являются центром ло-
кализации важных рефлексов: движения передних и задних конеч-
ностей, глазных мышц, охватывания самцом самки. Здесь же лока-
лизуются осязание и рефлекс изменения окраски. Возможно, что
здесь локализуется и центр дыхапия. Таким образом, средний мозг
в известной мере заменяет у амфибий го-
ловной мозг. Желудочек среднего мозга
носит название сильвиева водо-
провода (aqueductus sylvii).
Мозжечок (cerebellum) амфибий
имеет вид узкой поперечной пластинки ме-
жду средним мозгом и продолговатым,
подобно тому, что имеется у Dipnoi. Про-
долговатый мозг (medulla oblon-
gata) представляет непосредственное про-
должение спинного мозга, не имеет сверху
крыщи, образует сверху ромбовидную
ямку (fossa rhomboidalis), или четвер-
тый мозговойжелудочек, при-
крытую задним сосудистым
сплетением (plexus chorioidcus pos-
terior). Этот отдел мозга является центром
многих рефлексов. С мозгом Amniota мозг
амфибий сближается наличием в основа-
нии полушарий утолщений — полоса-
тых тел (corpora striata).
Мозг Urodela носит некоторые личи-
ночные черты и построен более схематично,
нежели мозг Anura. Мозг Apoda характери-
зуется редукцией зрительных центров и
развитием обонятельных, что стоит в связи
обитания и деятельности этих амбифий. Также и головной мозг
различных Anura представляет некоторые отличия, стоящие в связи
с различиями в образе жизни и поведении.
Головные нервы, также и спинномозговые, в основном те же,
что у рыб, с некоторыми отличиями. Число черепных нервов у ам-
фибий десять, последнюю пару черепных нервов образует n. va-
gus. У личинок Anura и у большинства Urodela к этому нерву при-
надлежит и боковой нерв — ramus lateralis, иннервирующий органы
боковой линии. В связи с n. vagus остается и одиннадцатая пара
нервов — п. accessorius, которая у высших позвоночных обособляет-
ся от пего. N. hypoglossus, или двенадцатой пары черепных нер-
вов, cine пет. Ее функцию выполняют свободные спинномозговые
нервы, и и нервирующие мускулы языка.
343
Симпатический ствол (sympaticus) амфибий, лежащий по сторонам
позвоночника, проходит в полость черепа. У Anura этот ствол
Рис. 262. Разрезы обо-
нятельной области
различных Vertebra-
ta, иллюстрирующие
стадии развития якоб-
сонова органа у Uro-
dela (Л—-Я),Apoda (Е)
и Lacerta(F).
образует по сторонам копчиковой кости ряд
ганглиев, указывая па бывшую здесь сегмен-
тацию хвостового отдела.
Органы чу в зтв рыб должны были сильно
измениться с выходом позвоночных на сушу.
Органы боковой линии сохраняются лишь
у личинок и у живущих постоянно в воде
представителей Urodela и у единичных пред-
ставителей Anura. С метаморфозом опи реду-
цируются. Строение их мало чем отличается
от такового у рыб (рис. 261).
Органами осяза щи я являются
свободные нервные окончания в эпидермисе
и специальные осязательные бородавки. Вку-
совые почки ограничены только ро-
товой полостью, где опи сидят па языке, пёбе
и челюстях. Вся поверхность кожи еще сохра-
няет способность воспринимать химические
раздражения, для чего служат свободные
окончания спинных и черепных нервов. Амфи-
бии способны при помощи особых нервных
окончаний воспринимать тепло и хо-
лод. Органы обоняния отлича-
ются от таковых у рыб и сходны с органами
обоняния Dipnoi в том, что имеются не только
наружные, ио и внутренние ноздри, открыва-
ющиеся в ротовую полость. Складки слизи-
стой оболочки или образования, подобные
раковинам (conchae) высших Tctrapoda, па
боковой стороне обонятельной капсулы Uro-
dela, еще лучше развитые у Anura, увеличи-
вают воспринимающую поверхность органа
обоняния. Специальные железы смачивают
эту поверхность, поддерживая чувствующий
эпителий во влажном состоянии. Вполне есте-
ственно, что наибольшего развития обонятель-
ные органы достигают у наземных форм.
Складки слизистой оболочки закрывают хоаны
у личинок Anura и саламандр, особые муску-
лы — у наземных.
У амфибий впервые появляется путем от-
шнуровки части обонятельной полости так
якобсонов орган. (По J г „ .
Symington из Parker.) называемый Я К О б С О Н О В О р Г а П, ГО-
раздо более развитой у рептилий (рис. 262).
Орган иннервируется n. olfactorius. У одних форм он еще связан с
обонятельной полостью, у других (Apoda) отшнуровывается от нее.
Задача органа — оценка пищи, попадающей в рот.
344
Орган слуха у амфибий состоит из лабиринта, или
внутреннего уха, более сложно построенного, чем у рыб, и из
среднего уха с барабанной перепонкой (tympanum)
и слуховой косточкой (columella). Об этом сказа-
но выше в главе о скелете. Среднее ухо возникает из жабер-
ного выпячивания между челюстной и гиоидной дугой, из участка
кожи против первой жаберной щели и из верхнего отдела
гиоидпой дуги, т. е. из тех частей, которые имелись уже у
рыб, но стали ненужными при развитии автостилии и выходе
па сушу. У роющих или водных Anura, а также у всех Urodela
и Apoda нет ни среднего уха, ни барабанной перепонки. Послед-
няя должна была возникнуть у стегоцефал, у которых имелась
columella.
Орган зрения амфибий примитивен и пе имеет тех спе-
циальных приспособлений, которые есть у рыб или у Sauropsida,
рептилий и птиц. Глаза амфибий могут видеть на более далекое
расстояние, чем у рыб. Хрусталик у них уплощен, по крайней мере
спереди, лишь редко будучи круглым. Аккомодация па близкое рас-
стояние происходит путем сокращения ресничного мускула. Осо-
бые мускулы движут линзу вперед. В связи с наземным образом
жизни, с выходом па сушу глаза должны были подвергаться засо-
рению пылью и высыхать. Для защиты глаз у наземных выра-
ботались в с к и и с л е з п ы е железы, которые имеются уже
у амфибий. Веки развиваются у них с метаморфозом. Водные Urodela
век не имеют, и глаза у них лежат иод колеей. У Anura верхнее веко
развито слабо, нижнее хороню и верхняя его половина прозрачна.
У некоторых Anura встречается еще мигательная перепонка (mem-
Ъгапа nicticans). Железы для смачивания глаза есть у всех амфибий,
но развиты они различно. Способность глазного яблока у амфибий
выпячиваться наружу и прятаться в глубину орбиты при помощи
особых мускулов также должна была развиться в связи с перехо-
дом к наземной жизни.
Форма отверстия зрачка служит у амфпбий систематическим
признаком. В сетчатке амфибий уже имеются участки наиболее
острого зрения, где особенно изобильны конусы.
У амфибий нередко встречается дегенерация глаз: у Proteus,
Typlilomoge, Typhlotrion.
III. ДРОБЛЕНИЕ И РАЗВИТИЕ ЯЙЦА У АМФИБИЙ
Яйца амфибий довольно богаты желтком, дробление их полное,
неравномерное (рис. 263). Яйцо лягушки имеет две части: пигменти-
рованную, где сосредоточена главная масса протоплазмы, и белую,
выполненную желтком. Первая обращена кверху. Первая борозда
дробления вертикальная, вторая также вертикальная, но перпен-
дикулярная к первой, третья — горизонтальная, перпендикуляр-
ная к обеим. Уже в скором времени с третьей борозды выявляется
разница между микромерами и макромерами. Деление последних
(белых) идет медленнее, первых — быстрее. Внутри получающейся
345
морулы уже на стадии 8 бластомеров появляется сегментационная
полость, получается бластула. Полость помещается в верхнем полу-
шарии яйца. Верхняя стенка бластулы — более тонкая, чем ниж-
няя. У коупных саламандр, у Necturus, Cryptobranchus, имеющих
большее-количество желтка, дробление приближается к частичному
дроблению. В общем дробление яйца
Рис. 263. Развитие лягушки.
у амфибий происходит по
типу дробления у Dipnoi.
Гаструляция как у Uro-
dela, так и у Anura про-
исходит путем впячивания,
инвагинации, но не нижпе-
го полюса бластулы, а в
так называемой краевой
зоне, па границе микро - и
макромеров. У тритона
(рис. 264) гаструляция от-
личается в основном тем,
что первичная киш-
к a (archenteron) сперва
очепь узкая, и тем, что бла-
стопор долгое время зат-
кнут желточной пробкой.
У лягушки образование
гаструлы сложнее (рис.
265). Кроме первичпоки-
шечной полости во время
впячивапия образуется
«добавочная и о-
л о с т ь», происходящая из
части бластоцеля. Первич-
нокишсчпос впячивапие
прорывается в дополни-
тельную полость, и полу-
чается примитивная
к и ш к а. Здесь мы видим
сходство с тем процессом
образования гаструлы, ко-
наблюдается у Sauropsida — рептилий и птиц. В яйцах
очепь богатых желтком, последний затрудняет ипвагипа-
торый
Anura,
цию, и образование гаструлы происходит путем разрастания верх-
ней губы бластопора, т. е. путем эпибол и и. Так же идет
образование гаструлы у Dipnoi. У Apoda яйца очепь богаты желт-
ком, и дробление никогда не бывает полным в пшкней его половине.
Образование нервной трубки и хорды происходит у Urodela
и у Anura сходно с тем, как это бывает у низших позвоночных.
На спиппой стороне продолговатого зародыша образуется не-
вральная пластинка из высоких клеток эктодермы. Раз-
вивается она спереди назад. Передняя более широкая часть ее об-
разует зачаток головного мозга. В средине невральной пластинки
346
образуется борозда, на краях пластинки развиваются отделяющие
ее от остальной эктодермы нервные валики. Последние сбли-
жаются, срастаются, начиная спереди, и образуется нервная
трубка (рис. 266). Хорда получается путем аналогичного
выпячивания верхней стенки энтодермы, т. е. таким же образом,
как у ланцетника.
Мезодерма у три-
тона получается так-
же сходно с амфиок-
сусом из крыши пер-
вичной кишки (рис.
267). Образование его
начинается сперва
вокруг бластопора и
идет в дальнейшем
справа и слева в па-
Рис. 1264. Срединный раз-
рез через гаструлу три-
тона.
Рис. 265. Четыре средних разреза через гаструлу
лягушки.
1 —- эктобласт, 2 — онтобласт, 3 —- сегментационная
полость, 4— первичная кишка, 5—дополнительная
полость. (Ho Schultze, из Bonnet.)
правлении будущей головы между эктодермой и энтодермой,
сходясь па брюшпой стороне под кишкой. В мезодерме путем
расхождения клеток появляется полость — целом (ши-
зоцсль). Из спинной мезодермы образуются сомиты ил^г первйчные
позвонки. Из вентральной мезодермы образуется висцеральный
и париетальный листки.
Таким образом, развитие идет у Urodela тем ясе путем, что
у ланцетника. У Линга образование среднего листка, или мезобла-
ста, идет не путем образования эктодермальных выпячиваний, но
укороченным путем отщепления от стопки первичной кишки
(рис. 268). Целом возникает путем образования щели в первона-
чально плотной мезодерме. Мезодерма делится па спинпую, состоя-
щую из первичных позвонков, и брюшную несегментированную.
В дальнейшем и у Urodela, и у Anura первичная трубка, разра-
стаясь назад, закрывает бластопор; образуется нервно-кишеч-
ный канал (canalis neurentericus), анальное отверстие на ме-
сте бластопора и новый рот на противоположном конце
(рис. 2G9),
347
Рис. 266,'
Рис. 267*
Рис. 266. Два поперечных разреза через зародыша водяного тритона с
еще открытой нервной бороздой (Л), с закрытой бороздой и отчленяющи-
мися первичными позвонками (Б).
А—1—нервная борозда, 2— нервный валик, 3—хорда, 4—эктоаласт, 5 — парие-
тальный мезобласт, 6 — висцеральный мезобласт, 7 — первичный поз! опок, 8 — це-
лом, 9 — полость кишки, 10 —желточные клетки. Ь’— 1 — нервная пластинка,
2 — хорда, 3 —нервный валик, 4 — париетальный мезобласт, 5 — висцеральный
мезобласт 6 —энтодерма, 7 —эктодерма. (По Hcrtwig из Bonnet.)
Рис. 267. Три поперечных разреза через зародыша тритона. Закладка
хорды, нервного валика и мезодермы.
1 I
Рис. 268. Поперечный разрез через
эмбриона Rana fusca до закладки
нервной борозды.
1 — зачаток хорды, 2 — полость первич-
ной кишки, 3 и 4 —наружный и внут-
ренний слой эктобласта, 5—дополнитель-
ная полость, 6 —мевобласт, 7—энтобласт,
8 — желток, 9 —бластоцель (По Bonnet)
IV. КЛАССИФИКАЦИЯ
АМФИБИЙ
В -настоящее время насчи-
тывается 234 рода и 1300 видов
амфибий. Имеется значительное
число ископаемых форм. Все
амфибии могут быть разделены
на два подкласса, одни из ко-
торых вымер, другой живет и в
настоящее время. Эти подклас-
сы нельзя рассматривать как
равноценные, соподчиненные в
классе. Вымершие формы, не-
сомненно, являются предками
современных, связывают послед-
них с классом рыб. Поэтому
амфибий, как и рыб, следует де-
лить не на подклассы только,
348
to
Рис. 269. A — Сагиттальный разрез через зародыша лягушки. Б — Сагит-
тальный разрез через зародыша жабы, несколько более поздняя стадия.
А — 1 — экто бласт, 2 —нервная трубка, 3 — хорда, 4—canalls neurentericus (нервно-
кишечный канал), 5 —первичный рот, 6—'Остаток желточной пробки, 7 — зачаток
анального отверстия, 8 —мезобласт, 9 — энтодерма, 10—желточные клетки, И — зача-
ток печени, 12 —первичная кишка, 13—ротовая воронка. (По Goette, из Bonnet.)
В —нервная трубка, 2 — полость головного мозга, 3 — эктобласт, 4 — мезобласт, 5 —
хорда, 6 — первичная кишка, 7 — зачаток печени, 8 — анальное впячивание, 9 —
хвостовой отдел кишки, 10 —canalls neurentericus. (По Goette, из Bonnet.)
но па ступени, каждая с одним подклассом. Это дает понятие
о филогенетических взаимоотношениях подклассов, так как пер-
вый подкласс является, несомненно, первой ступенью в эволю-
ции амфибий.
с т у и в и ь I
Подкласс 7. Панцырные, или покрытоголовые амфибии
(Phractamphibia, или Stegocephalia)
Древнейшие наземные четвероногие имели развитой кожпый
скелет, в особенности на голове. Они существовали с каменноуголь-
ной эпохи до начала триаса. Череп, состоявший из многих око-
стенений, был покрыт дермальными костями, оставлявшими отвер-
стия лишь для ноздрей, для орбит и теменного глаза (стегальный
череп). Из многих костей состояла и нижняя челюсть. Зубы си-
дели пе в альвеолах и сменялись путем вырастания зуба рядом.
Позвонки состояли из отдельных элементов или были цельные.
У большинства имелся панцырь па брюшпой стороне. Форма
тела была разнообразна, основная—тритонообразная. Место-
обитанием служили мелкие водоемы. Образ жизни у панцир-
ных был ночной; по роду пищи, они, видимо, являлись хищ-
никами.
Развитие проходило с превращением, личинки вели водный
образ жизни и имели жаберное дыхание. Имелись слизистые ка-
налы на голове. Остатки известны из Европы, Северной и Южной
Азии, из Северной и Южной Америки.
Классификация основывается прежде всего на строении по-
звонков.
349
Отряд 1. Рыбопанцырные (Ichthyostegalia)
Недавно установленный отряд, с рядом признаков, заставляю-
щих признать наибольшее родство с рыбами, именно с Crossop-
terygii. Описаны из девона Гренландии. Ноздри находились на
вентральной стороне черепа.
Отряд 2. Embolomeri
Позвонки из нервной дуги и двух полных амфицельных дисков,
окружавших сдавленную хорду (эмболомерный тип позвонка).
Cricotus — крокодилоподобная форма, из пермских отложений
Тексаса.
Отряд 3. Rhachitomi (Temnospondyll)
Позвонки из 8 самостоятельных элементов, обнимавших хорду
(рахитомный тип позвонка). Archaegosaurus размером
до 1 м, из пермских слоев Германии и каменноугольных Кашмира.
Рис. 270. Реконструкция Cacops aspidophorus из перми Тексаса.
Cacops, из перми Тексаса, с ложновисочной ямой (рис. 270). Dwi-
nasaurus до 1 м длиной, из пермских отложений Северной Двины.
Сохранял жабры во взрослом состоянии.
Отряд 4. Stereospondyli (Labyrinthodonta)
Позвонки цельные из вполне сросшихся между собой частей.
Хорда в позвонках сохраняется лишь иногда. Зубы многосклад-
чатые. Без брюшного панцыря. Триасовые формы. Mastodonsau-
rus (рис. 224) — крупнейший представитель стегоцефал, с чере-
пом до 1.25 м длиной. Metopias, из триаса Вюртемберга (рис. 271).
Отряд 5. Branchlosanria (Phyllospondyli)
Тело позвонка в виде тонкого костного цилиндра из 4 частей,
образованных из basidorsalia и basiventralia, с хордой посредине.
Мелкие саламандрообразные формы. Близки к предкам Urodela
и Anura. Protriton (Branchi osaurus), из пермских слоев Европы
(рис. 225). Pelosaurus, из перми Германии и Франции, длиной 20 см.
350
Отряд 6. Ceraterpetomorpha (Microsauria, Lepospondyli)
Позвопки трубчатые, двояковогнутые, хорда сохранялась,
хвост длинный, форма тела
из верхнекаменноугольных
слоев Ирландии и Англии.
Diplocaulus с очень расши-
ренной треугольной головой
на тонком длинном теле.
Верхнекаменноугольные слои
и пермь Северной Америки
(рис. 272).
Отряд 7. Aistopoda
Форма позвонков, как у
предыдущего отряда, тело
очень длинно, конечности от-
сутствуют. Dolichosoma. Ка-
менноугольные отложения
Ирландии и Богемии. Число
позвонков 150. Череп удли-
нен.
Позвонки образованы по
типу Anmiota, т. е. тела
построены из pleurocentra.
Удлиненное тело и короткий
саламандроподобная. Ceraterpeton
Рис. 271. Скелет Metopias diagnosticus
(из триаса Южной Германии).
хвост, короче половины тела.
Microbrachis, из верхнекамеппоугольных отложений Богемии,
напоминают современную Amphiuma.
СТУПЕНЬ П
Подкласс П. Голые амфибии ( Lissamphibia)
От'ряд 1. Безногие, или червяги (Apoda, Gymnophiona, Coeclilae)
Тело удлиненное, червеобразное, лпшснпое конечностей и раз-
деленное наружными перетяжками па кольца. Кожа богата желе-
зами. У некоторых родов в коже находятся чешуи. Позвонки,
числом 200—300, амфицельны. Между ними сохраняется хорда.
Череп псевдостегальный, приспособлен для рытья в земле. Слухо-
вые органы, как и глаза, зачаточны. Барабанной полости и бара-
банной перепонки пет. Сильно развит обонятельный аппарат.
Левое легкое зачаточно. Почки вытянуты. У самцов имеется ко-
пулятивный орган. Между глазами и ноздрей — щупальце. Яйца
богаты желтком. Распространены в тропическом поясе Южной
Америки, в Африке, в Индии, нет па Мадагаскаре и в Австралии.
Питаются дождевыми червями и живущими в земле насекомыми.
В отряде насчитывается 19 родов с 55 видами.
351
Сем. Цецилиевые (червяги) (Coeciliidae). (Цейлон-
ский рыбо-змей) Ichthyophis glutinosus в Индии, на Цейлоне и на Боль-
ших Зондских островах (рис. 273). Яйца, одетые плотной оболочкой, сцеп-
ляются выростами последней. Самка свертывается вокруг отложенных яиц,
защищая их от слепунов (змей) и безногих ящериц. Личинка имеет перистые
жабры и хвостовой плавник. В воду личинка попадает уже без жабр и остается
здесь довольно долго, а потом переходит к подземной жизни и настолько
к ней приспособляется, что в воде тонет. Кольчатые червяги, (Siphonops).
S. annulatus—в тропической Южной Америке. Земляные червяги (Hypogeo-
phis), в Восточной Африке ина Сейшельских островах (рис. 274).
Отряд 2. Хвостатые амфибии (Urodela, Caudata)
Тело удлиненное, туловище незаметно переходит в сжатый
с боков или округлый хвост. Конечностей две или одна пара. Число
пальцев на них может быть редуцировано. Позвонки амфицельные
Рис. 272. Череп Diplocaulus magnicornis из перми Тексаса.
или опистоцельпые. Ребра есть, раздвоены па проксимальном кон-
це. Верхнечелюстные кости не доходят до квадратной кости. Па-
расфеноид очень широкий, palatinum исчезает или сливается
с vomer. Зубы па пижней челюсти, на верхней и на нёбе, иногда
на парасфеноиде. Язык приросший, слабо развитый. Барабанной
полости и барабанпой перепонки пет. У одних (Proteidae и Sireni-
dae) наружные жабры сохраняются всю жизпь (Perennibranchia-
ta) У других (Caducibranchiata) жабры исчезают, но сохра-
няются жаберные щели (Amphiumidae Derotremata). У третьих
исчезают и жаберные щели.
Распространены Urodela почти исключительно в северном полу-
шарии, главным образом в Северпой Америке. Отдельные виды —
в Центральной и Южной Америке.
Подотряд 4. Lysorophidae (Paterosauria)
Составляют переход от стегоцефал к Urodela, так как сохраняют
в своей организации некоторые черты стегоцефал, например, ко-
сти в затылочном отделе.
352
Сем. Lysorophidae. Lysorophus из пермских отло-
жений Северной Америки соединяет признаки стегоцефал и Uro-
dela. Небольшая постоянножаберная форма, жившая, вероятно,
в иле. Позвонки амфицельны.
Подотряд 2, Hylaeobatrachia
Сем. Hylaeobatrachia. Hylaeobatrachus
из меловых отложений Бельгии. Саламандроподобпая форма. Три
пары жаберных дуг. Andrias scheuchzeri, из миоцена Швейцарии,
Рис. 273. Цейлонский рыбо-змей (Ichthyophis
glutin osus).
(Из Gadow по Saraain.)
Рис. 274. Карта распространения Apoda.
(Из Gadow.)
близкая к следующему подотряду гигантская саламандра (до
1.5 м), описанная как «Homo diluvii testis» («человек — свидетель
потопа»).
Подотряд 3. Cryptobra/nchoidca
Наиболее примитивные Urodela с наименее специализирован-
ным скелетом.
Сем. Азиатские наземные саламандры (Hynobiidae). Си-
бирский четырехпалый тритон (I-Iynobius keyserlingii) — от Урала до Кам-
чатки, в соседних частях Монголии и Манчжурии. Семиреченский тритон
(Ranidens sibiricus) — из Восточного и Южного Семиречья и из западной
части Китайского Туркестана. Обитатель каменистых ручьев. Когтеносные
саламандры (Onychodactylus). Два вида из Хабаровска, Приморской обла-
сти, Кореи и Японии. Оба вида без легких, подобно ручьевым формам
других семейств. Когти помогают держаться за предметы на быстрой воде.
23 Кашкаров и Станчинский 353
Сем. С г у р t о Ъ г a n с h i d а е, без жабр. Дна рода гигантских саламандр,
близких к 11 ynobiidae. Формы полуличиночнью Mcgalobatrachus japonicus —
гигантская саламандра — из Китая и Лионии. Аллеганский скрытожа-
берник (Cryplobranrhus alleganensis) из восточной части США. В ископа-
емом состоянии семейство было распространено шире (рис. 275).
Подотряд 4. Амбистомы (Anibystomattdae)
Происходит от Hynobiidae, но отличается утратой angulare
и выростами от praemaxillaria — назад. Позвонки амфицельны.
Рис. 275. Японская саламандра (Cryptobranchus japonicus).
(Из Gadow.)
Сем. Ambystomidae. Род Ambystoma с И видами широко рас-
пространен в Сев. Америке от Южной Аляски до Мексики. A. tigrinum —
личинки хорошо известны под именем аксолотля. Считался за особый род —
Siredon. В настоящее время считается за неотеническую личинку.
Подотряд 5. Саламандры (Salamandroldea)
Проделывают метаморфоз. На нёбе позади ноздрей имеются
зубы. Наземные и водные формы. Широко распространенный
отряд, возникший от общего корпя с Hynobiidae. Оплодотворение
внутреннее.
Сем. 1. Salamandridae. Позвонки описто цельные, нёбносош-
никовые зубы идут продольно. Triturus (Molge, Triton) (рис. 276) тритоны,
24 вида и много подвидов. Распространены в Восточной Азии, Японии, Север-
ной Африке, Европе и Сев. Америке. Наибольшее разнообразие их в Европе.
Тело стройное, хвост сжат с боков. Весной пишут в воде, где откладывают
яйца на водяных растениях. Весной резкий половой диморфизм. Salamandra с
4 видами, живущими в Европе до Кавказа. Хвост округленный, хвостового
плавника нет, туловище толстое, неуклюжее. Сильно развиты заушные же-
лезы, паротиды (рис. 277). Пятнистая, или огненная, саламандра
(S. salamandra) в Центра льной, Южной и Западной Европе, кроме Британии,
на Корсике, в Алжире, Малой Азии и Сирии. Черная саламандра (S. atra)—
в Альпах на высоте от 600 до 2 500 м. Обе саламандры живородящи, но
огненная саламандра родит детенышей числом от 5 до 50 с длинными наруж-
ными жабрами и с плавниками на конце. Черная родит только двух детены-
шей, прошедших весь метаморфоз в матке матери. '
354
Сем. 2. А м ф и у м ы (Am phiumid а е). Единственный предстА-
витель амфиума угревидная (Amphiuma means). Тело вытянуто, зача-
точные конечности с 2—3 пальцами, по бокам головы постоянно сохраняется
жаберная щель, глава
атрофированы, скрыты
под кожей. До метра
длиной. Полуличиноч-
ная форма, происходя-
щая от саламандр. Живет
в стоячих водах юго-во-
стока США. Самка ох-
раняет яйца, свернув-
шись вокруг них.
Сем. 3. Без легоч-
ные саламандры (Р 1 е-
th od о n ti d а е). Нёб-
ные зубы расположены
поперечными рядами.
Позвонки амфицельные
или опистоцельные.
Хвост круглый или сжа-
тый с боков, иногда за-
кручивающийся и цеп-
кий. Plethodontidae со-
вершенно не имеют лег-
ких, — дыхание — при
помощи кожи и слизи-
стой оболочки глотки.
Число видов более 60,
почти все американские,
лишь пещерная саламан-
дра (Spelerpes fuscus)
живет в Европе. Ответ-
вились от ствола сала-
мандр. Представляют
различные степени спе-
циализации. (Typhlotri-
Рис. 276. Тритон малоазиатский (Triturus
vittatus).
ton) — слепая саламанд-
ра — из пещер Аркан- Рис. 277. Обыкновенная саламандра (Salamandra
заса и Миссури. Рат- maculosa).
бунов колодезный три-
тон (Typhlomolge rath-
buni) из пещер Тексаса, слепая саламандра, всю жизнь остающаяся в личи-
ночном состоянии. Plothodon, 11 видов. 10 из них проводят всю жизнь вдалеке
от воды. Все живут в восточных США. Batrachoseps характеризуется короткими
зачаточными ногами. Распространен в Калифорнии (пещерные саламандры) —
(Spelerpes). Много видов в Северной и Центральной Америке, один — в
Вест-Индии, один — на юге Франции, в Средней и Северной Италии.
Подотряд 6. Протеи (Proteida)
Неотеиическпе личинки неизвестных саламандр выделяются
в особый подотряд, так как отношения их к какой-либо другой
группе Urodela пеяспы. Два рода и два вида, составляющих одно
семейство Proteidae.
Сем. протеев (Proteidae). Proteus anguineus имеет передние и задние
конечности вполне развитыми, но с редуцированным числом пальцев. Глаза
под темной кожей, не функционируют. Живет в подземных водах Крайны
и Далмации. Necturus maculatus имеет глаза функционирующие, так как
23* 355
они прикрыты лишь топкой прозрачной кожей. Кроме трех наружных
жабр —г каждой о,тороны две жаберные щели. Распространен в восточной
половине США и в озерах Канады (рис. 278).
Рис. 278. Американский протей (Necturus
maculatus).
Подотряд 7. Сирены (Meantes, Sirenoidea)
Подотряд представлен также неотеническими личинками с нем-
ногими признаками взрослых. Имеют только передние конечности.
Три пары наружных жабр. Глаза без век, как в предыдущем от-
ряде, по просвечивают сквозь кожу. Происходят от наземных сала-
мандр, возможно от Hynobiidae.
Сем. Sirenidae — S. lacertina, из юго-восточной части
США. Родственные взаимоотношения Urodela см. из схемы (рис. 279).
Отряд 3. Безхвостые амфибии (Anura, Ecaudata)
Бесхвостые являются наиболее многочисленным, наиболее про-
цветающим отрядом амфибий. Их насчитывается около 900 видов.
Тело укорочено, ши-
рокое, число туловищ-
ных позвонков всего
7—9. Позвонки амфи-
цельные, процельные,
опистоцельпые или
представляют комбина-
цию этих форм. Хвост
у пыне живущих от-
сутствует, его место
Рис. 279. Сирена (Siren lacertina).
(Из Gadow.)
занимает позади крестца длинная палочковидная копчиковая
кость (os coccygis). Ребра зачаточны или исчезают вовсе. Зад-
няя конечность удлинена вследствие удлинения astragalus и
calcaneus и пальцев. Глаза — с веками. Зубы есть или отсут-
ствуют. В первом случае они есть только па сошнике или же и
356
на верхней челюсти, на межчелюстной кости, иногда и на ниж-
ней челюсти и на нёбных костях. Язык есть или отсутствует. Раз-
витие всегда с метаморфозом. По образу жизни формы наземные,
древесные, водные и роющие. В ископаемом состоянии известны
мало, что затрудняет естественную классификацию.
t //а суше____
jaiifa- О §
в воде
лица—Q
b
Pldhodon.dc^, Г
^^^^Pletho font I da*
ТурЫоток/е
mtidae
V
lyno
Amphtumidae
Salathai \dridae
Атб/ysft wnidae
,Prole idae
r Siremdae
-----------°
^CryptoOragchi-
Рис. 281. Диаграмма, иллю-
стрирующая филогению
Anura.
(По Noble.)
Рис. 280. Диаграмма, иллюстри-
рующая филогению Urodela.
Толстые черные стрелки показывают
филогенетические отношения. Узкие
горизонтальные представляют онто-
гению различных семейств.
(По Noble.)
Подотряд 1, Protosalienbia
Представлен одним родом Protobatrachus, из триаса Мада-
гаскара. Хвост короткий, но из отдельных позвонков.
Подотряд 2, Aniphicoeltb
Наиболее примитивные из Anura, отпосимые обычно к сем.
Discoglossidae, отличаются от других Anura амфицельными по-
звонками и тем, что хотя хвоста у них нет, еще имеются два муску-
ла, предназначенные для его движения.
Сем. Liopelmidae с двумя родами: Liopelma — живет в Новой
Зеландии, где является единственным представителем амфибий. Род Asca-
phus — в Сев. Америке.
Подотряд 3, Opisthocoela
Позвонки задневогиутые. Имеются свободные ребра. Мускуль*
пая система примитивная.
357
Сем. Р i р i d а о обычно выделяется в особый отряд — Aglossa, безъ-
язычных, примитивно и в родстве с Liopelmidae (рис. 281). Род X е -
п о р u s — шпорцевая лягушка; 5 видов, живут в Африке. Между длин-
ными пальцами задних ног имеется плавательная перепонка, три вну-
тренние пальца снабжены котгями (рис. 282). В верхней челюсти есть
зубы. Имеются органы боковой линии. Род Pipa, с наиболее известным
видом — суринамская жаба (Р. americana) из Гвианы, известная своим ори-
гинальным* способом размножения: яйца развиваются в особых карманах
на спине самки.
Сем. Discoglossidae. Плечевой пояс из свободных несрос-
шихся правой и левой дуг, ребра имеются, крестцовый позвонок свободный,
с двояковыпуклым центром. Жерлянка (Bombina). Паротиды и барабанная
перепонка незаметны. Зрачок треугольный. Язык не обособлен от дна ротовой
полости. Тело плоское. В. bombina обыкновенна в стоячих и медленно теку-
щих водах средней Европы и европейской части Союза до Волги. Жаба-пови-
Рис. 282. Шпорцевая лягушка (Xenopus laevis).
(Из Gadow.)
туха (Alytes). Паротиды и барабанная перепонка явственны. Зрачок верти-
кальный. Лягушка-звонарь (A. obstetricans) — в Испании, Франции, Швей-
царии и Западной Германии. Наземное животное. Живет в холмистых мест-
ностях, до 1500 м над уровнем моря. Название свое получила за мелодичный
голос. Самец, оплодотворяя самку, проталкивает через отложенную массу
яиц сперва одну, а потом другую ногу. Яйца облегают его ногу наподобие
цифры 8, после чего самец уходит со своей ношей в нору, откуда выходит
лишь ночью. Через три недели он идет в ближайшую воду, где из яиц выхо-
дят личинки.
Подотряд 4. Anomocoela
Отряд состоит из одного семейства Pelobatidae, являющегося
промежуточным между Amphicoelan Opistocoela, с одной стороны,
и Procoela, куда Относятся настоящие жабы (Bufonidae), — с дру-
гой. Свободных ребер пет, крестцовые позвонки процельпы, сли-
ваются с копчиком или свободны. Предкрестцовые — или про-
сто процельпы или со свободными межпозвоночными дисками
Сем. жабы - чесно чн и ц bi(Pelobatidae). Сходны с лягушкамг
и с жабами. Живут в северном полушарии, от Мексики через Евразию д<
Филиппин и западных частей Австралийского архипелага. Род чесночниць
(Pelobates); на большом пальце задних ног сидят роговые шпоры, «бойцы»
зрачок вертикальный, кожа гладкая. Обыкновенная чесночница (Р. fuscus
распространена широко в средней Европе, в европейской РСФСР, в За
падной Сибири, проникает на Кавказ. Почти сухопутная форма, идет в вод;
лишь в период икрометания»
358
Подотряд 5. Procoela
Позвонки процельные, двойной сочленовный бугорок для соеди-
нения крестца с копчиковой костью. Ребер нет.
Сем. Palaeo batrachidae, с юры, ископаемые.
Сем. Настоящие жабы (Bufonidae). Жаб, имеющих
зубы, принято было выделять в семейство Cystignathidae, относя его к ля-
гушкам. В настоящее время Cystignathidae соединяются с Bufoninae и дру-
гими подсемействами в семейство Bufonidae. С последними их сближают
подвижной грудной пояс и расширенные крестцовые диапофизы (поперечные
отростки).
Подсем. 1. Австралийские жабы (Criniinae). Пло-
скоголовая рукопалая лягушка (Chiroleptes platycephalus) живет в пу-
стынях Австралии, запасает на засушливый период воду в мочевом пузыре,
в подкожных пространствах и в полости тела, принимая шаровидную форму.
Рис. 283. Ceratophrys cornuta.
Подсем. 2. Ю ж н о а м е р и к а н с к и е ж а б ы (Р s е u d i n а е)—
имеют плотные процельные позвонки, крестцовые диапофизы цилиндриче-
ские или слегка расширенные. Зубы есть. Рогатая лягушка (Ceratophrys)
(рис. 283) — имеет в коже окостенения.
Подсем. 3. Настоящие жабы (Bufoninae). Челюстных зубов
нет, зрачок вертикальный, крестцовые диапофизы расширены, паротиды
хорошо развиты, кожа бородавчатая. Распространение: Африка, главным
образом экваториальная, Южная Азия, лишь один род — Bufo — в Евразии.
Жабы вполне наземные животные. Обыкновенная серая жаба (В. vulgaris)
в Европе, на Кавказе, в умеренной части Азии (Сибирь, Казахстан) на восток
до Великого океана. В Туркестане и Семиречье не найдена. Живет в местах
темных и влажных, выходя оттуда с вечерними сумерками, возвращаясь
до рассвета. В воду идет лишь в период размножения. На зиму зарывается
в землю. Вонючая жаба, или жаба камышевая (В. calamita) в Западной
Европе, доходя на восток до РСФСР. Зеленая жаба (В. viridis) в Север-
ной Европе, Азии и Северной Америке.
Подсем. 4. Свистуны (Leptodactylinae). 35 южно-
и центральноамериканских видов, по внешности относящихся к лягушкам,
по строению — к подвижногрудым жабам. Пятипалый свистун (Leptoda-
ctylus pentadactylus) способен издавать свист, похожий на свист человека,
зовущего собаку. Строит в воде гнездо, похожее на взбитые белки, в котором
и сидит.
Сем. Ко р о тко г ла вы (Вт ас h у с е phalidae). Южноамерикан-
ские бесхвостые амфибии, относившиеся раньше к сем. Ranidae—настоящим ля-
гушкам и сем. Engyetomatidae—- узкоротым лягушкам. По внутренней орга-
359
низации близки к жабам. Позвонки процельные, половины плечевого пояса
более или мопсе полно слиты по средней линии. Разбивается семейство на
три самостоятельных ветви — подсемейства.
Подсем. 1. Rhinodermatinae. У Rinoderma Darwini самец
помещает отложенные самкой яйца в горловой голосовой мешок, где личинки
остаются до конца метаморфоза. Питаются личинки сперва за счет желтка,
а затем, срастаясь со стенкой голосового мешка, за счет соков отца. Оби-
тание — в Чили.
Подсем. 2. Древолазы (Dendrobatinae). Лягушка-кра-
сильщик, или пятнистый древолаз (Dendrobates tinctorius). Выделением кож-
ных желез ее туземцы намазывают стрелы, употребляемые для охоты.
Ид убивает в несколько минут даже крупных животных, как олень, ягуар.
Подсем. 3. Brachycephalinae. У некоторых родов уже
имеется неподвижность грудины. Atelopus — один из наиболее ярко и пестро
окрашенных бесхвостых. Головастики развиваются из яиц в 24 часа. Распро-
странение — Парагвай, Уругвай, Аргентина.
Рис. 284. Молодая сумчатая
лягушка (Nototrema cornu-
tum), с наружными жаб-
Рис. 285. Австралийская квакша
(Нуla coerulea).
(Из G-adow.)
рами.
(Из Брэма.)
Сем. Квакши (Hylidae). Богатое семейство: 16 родов и около 150 ви-
дов. Фаланги на концах пальцев на подобие когтей и присасывательные по-
душки. Большинство — древесные, но есть и водные, и наземные, и роющие
формы. Один вид Hyla arborea—в Палеарктике (в Сибири, Казахстане и Сред-
ней Азии его нет), два близких вида—в Северной Индии и Южном Китае. Все
остальные—либо американские, либо австралийские. Главное местообитание—
влажные тропические леса Южной и Центральной Америки. В Индо-малай-
ском архипелаге их заменяют настоящие лягушки (Ranidae). К сем. квакш
относится и Nototrema — «сумчатая лягушка». У самки на спине имеется от-
крывающаяся назади сумка, в которой и проходят свое развитие отложенные
яйца. Жабры головастиков Nototrema cornutum имеют вид «колоколов»
на двух стебельках (рис. 284).
Представители рода Hyla для размножения уходят в воду. Зимуют также
под водой, закапываясь в ил (рис. 285). Phyllomedusa замечательны спо-
собом размножения, откладыванием яиц между листьями, свисающими над
водой.
Подотряд 6, Diplasiocoela
Крестцовый позвонок спереди выпуклый, сзади с двойным
сочленовным бугорком для os coccygis, восьмой позвонок двояко-
360
вогнутый, передние семь — процельные. Ведренная мускулатура
наиболее специализированная. Космополиты.
Сем. Настоящие лягушки (Ranidae). Амфибии Ста-
рого Света. Лишь один род Rana проникает в Америку. Центром распро-
странения является, видимо, Африка. Несколько подсемейств, из которых
наиболее известно подсемейство Raninae. Водяная, или настоящая, лягуш-
ка — род Rana — включает около 140 видов. Европейские лягушки делятся
на «зеленых» и «бурых». Зеленая лягушка (Rana esculenta) распростра-
нена в Европе, западной Азии и северо-западной Африке. В Средней Азии
распространена озерная лягушка (R. ridibunda); травяная серая, или пло-
скомордая лягушка (R. temporaria), отличаемая темным височным пятном
позади1 глаза, распространена в Европе, в северной и умеренной Азии, до
Японии включительно. В Крыму и в Средней Азии ее нет. Остромордая, или
болотная лягушка (R. terrestris) в восточной половине Европы и в запад-
ной Азии. R. asiatica — в Сибири и Семиречье. Лягушка-бык (R. catesby-
ana = R. mugiens) отличается очень крупными размерами. Родина — США
к востоку от Скалистых гор, к югу до Мексики и к северу до Канады.
Сем. Узкоротые лягушки (Brevicipitidae, Еп-
gystomatidae у других авторов) распространены в обеих Америках,
в Африке, на Мадагаскаре, в южной и восточной Азии и на прилежащих
островах. Два рода, доходят до Квинслэнда в Австралии. Образ жизни —
древесный или роющий. У роющих на ногах имеются специальные лопато-
видные выросты. Breviceps mossambicus имеет короткое тело, короткую го-
лову, узкий рот, длинный приросший язык, большой желудок, короткие
ноги. Пищей являются термиты.
V. ПРОИСХОЖДЕНИЕ АМФИБИЙ
Амфибии являются первыми наземными позвоночными. Проис-
хождение их — величайшее событие в эволюции позвоночных жи-
вотных, выведшее последних из однообразной среды—воды—на
сушу с ее разнообразием условий существования, что обусловило
дальнейшую их эволюцию в условиях наземной среды и привело,
в конце концов, к возникновению рептилий, а затем нтиц и млеко-
питающих. На основе сравнительной анатомии, эмбриологии и па-
леонтологии, хотя свидетельства последней скудны, мы можем себе
представить, как произошел выход позвоночных на сушу.
Мы знаем, что и в настоящее время есть в тропических странах
рыбы, которые на время выходят из воды, проводя на суше довольно
долгое время. Это делается возможным благодаря развитию у них
ряда приспособлений к воздушной среде, дающих им возможность
защититься от высыхания и дышать на воздухе. Так, у выходящих
на сушу Gobiidae наблюдается ороговение кожи, у Blenniidae — об-
разование кутикулы. У них развиваются: кожное дыхание, допол-
нительные оргапы дыхапия в ротовой полости, особые приспособ-
ления для сохранения влажности жаберной полости (Anabas),
своеобразно измененные грудпые плавники, па которые рыбки могут
опираться (Periophtalmus), защитные приспособления в органах
боковой линии и т. д. Но все эти приспособления не такого характера,
чтобы, развиваясь дальше, они могли повести к образованию
наземных форм.
Для выхода рыб на сушу необходимо, чтобы они еще до выхода
приобрели ряд признаков, а именно: жаберное дыхапие должно
361
было уступить место легочному; кроме развития легких должны
были появиться соответственные изменения в системе кровообра-
щения: легочные артерии и вены; в сердце должно было произойти
разделение на артериальную и венозную половины. Для обеспече-
ния возможности легочного дыхания ноздри должны были открыться
в глотку. Должен был развиться особый тип дыхательных движе-
ний. Конечности должпыбыли из плавника превратиться в систему
рычагов для движения по земле, а также должны были развиться
особые рефлексы движения, иные,чем у рыбы. Развитие в связи
с паземной жизнью и изменением питания вызвало развитие более
прочного соединения palatoquadratum с черепом, должна была раз-
виться автостилия. Против высыхания должны были развиться осо-
бые приспособления в коже, прежде всего ороговение. Костный
панцырь, сохранявшийся вначале, должен был исчезнуть как лиш-
няя обуза при слабых еще конечностях. Головной мозг и органы
чувств должны были получить большее развитие. Все или большин-
ство этих признаков должны были развиться более или менее вместе,
одновременно, чтобы могло получиться жизнеспособное существо.
Почти все перечисленные особенности в том или ином виде имеют-
ся уже у Dipnoi, Crossopterygii и частично у некоторых Holostei
(см. главу о рыбах).
Наземные позвоночные могли произойти только от пресновод-
ных рыб, так как вполне понятно, что только у последних могли
возникнуть легкие из выпячиваний жаберного мешка, которые, как
плавательный пузырь, служили одновременно и гидростатическим
органом и дополнительным органом дыхания.
Первое появление стегоцефал относится к каменноугольному
времени. Следы так называемого Thinopus amphibius из верхнего
девопа сомнительны, но во всяком случае совершенно несомненно,
что возникли амфибии именно в это время, быть может даже раньше.
Недавно описанные со Шпицбергена стегоцефалы, выделяемые в от-
ряд Ichthyostegalia, подтверждают эту мысль.
В пресных водах жили тогда кистеперые, двоякодышащие и аку-
лы. К стегоцефалам ближе всего первые, девонские Crosse ptery-
gii, именно сем. Osteolepidoti, происшедшие от одного ствола
с Dipnoi. С Osteolepidoti стегоцефал сближает целый ряд деталей
в строении черепа: теменное отверстие, сложная нижняя челюсть,
большие сошниковые зубы, зубы со складками эмали, кость в склере
глазного яблока. Сходство имелось и в строении плечевого пояса,
и в чешуях, в коже. Наконец, у личинок современного представи-
теля кистеперых, у Polypterus (как у личинок Dipnoi), мы видим
наружные жабры, свойственные личинкам амфибий, вероятно,
и их далеким предкам. Древнейшие из стегоцефал, например,
Eogyrinus из Embolomeri, являлись водными животными, жившими
рядом со своими рыбьими предками. Они были готовы к выходу на
сушу, по пе вполне: таз у них еще пе соединялся с позвоночником;
они проводили жизнь в воде (рис. 286.)
(Толчком к развитию наземной организации, прежде всего дыха-
ния и кровообращения, явилась, вероятно, недостаточность дыха-
362
тельного жаберного аппарата, что мы видим, например, у Dipnoi,
не имеющих ни совершенного насасывающего крышечного аппарата,
ни хорошей жаберной решетки. Вполне естественно, что у них раз-
вился дополнительный дыхательный аппарат: легкие со всеми
необходимыми для дыхания изменениями в других органах. Раз-
витию его способствовали еще условия жизни рыб того времени:
во временно усыхающих водоемах, подобных водоемам современной
Австралии. То же, вероятно, было и у предков Tetrapoda. Конеч-
ности наземного типа развились как приспособление к ползанью по
дну, как мы это видим у личинок Polypterus и Neoceratodus. Пер-
Рис. 286. Eogyrinus atteyi, примитивная эмболомерная амфибия.
Реконструкция скелета.
(Из Watson.)
вые Tetrapoda были еще водными животными и пе вполне заслужи-
вали название земноводных животных. Условия сухого и жаркого
климата в девоне вызывали приспособления к дополнительному
дыханию.
Выйти в это время на сушу первые Tetrapoda не могли. Это
могло осуществиться лишь тогда, когда внешние условия стали
этому выходу благоприятствовать, т. е. во влажном каменноуголь-
ном периоде. Такая форма, как амфибия, совершенно не приспособ-
лена к сухим жарким условиям, особенно первые Tetrapoda не
годились для них. В насыщенных водяными парами лесах каменно-
угольного времени опи, наоборот, должны были найти вполне под-
ходящую среду для выхода из воды. Раз это последнее началось,
отбор должен был работать пад дальнейшим усовершепствовапием
приспособлений к наземной жизни и создатьнастоящих Tetrapoda.
. Легкие первых наземных были несовершенными, папцырь
мешал развитию кожного дыхапия, а потому начал утрачиваться,
дольше всего сохраняясь на брюхе, так как первые наземные Tet-
rapoda, вследствие слабости конечностей, еще волочили брюхо
по земле, и папцырь был здесь полезен. Акт дыхапия наземных,
состоящий в заглатывании воздуха, развился из акта заглатыва-
ния воды, наблюдаемого у Urodela, у которых заглотанная вода
пропускается через жаберные щели, омывая наружные жабры.
Конечности первых Tetrapoda еще пе работали самостоятельно,
а переставлялись при изгибах туловища, так как еще не имели
необходимой иннервации.
Местообитанием взрослых стегоцефал была преспая вода или
суша, или то и другое, как у современных амфибий. Подходящей
средой для них были также болота. Некоторые охотились ночью.
На это указывает, например, нахождение мелких стегоцефал (из
Branchiosauria) в пазухах сигиллярий и лепидодендронов, куда они
363
прятались, видимо, от дневного света. В личиночном состоянии
стегоцефалы дышали при помощи жабр, исчезавших во взрослом
состоянии, по и у них были формы неотенические, сохранявшие
жабры и во взрослом состоянии. Это было, вероятно, в тех слу-
чаях, когда условия на суше вновь становились слишком сухими.
Сильно развитые каналы системы боковой линии служили у ла-
биринтодонтов, очевидно, для смачивания кожи.
Уже рано стегоцефалы начали дифференцироваться в различ-
ных направлениях. Первоначальная форма была тритонообраз-
ная. Дифференцировка шла в двух направлениях: в направлении
удлинения и в направлении укорочения тела. Первое намечено
у представителей группы Aistopoda, напоминающих отсутствием
конечностей Apoda, но не бывших их предками. Укороченное тело
имели лабиринтодонты.
Что стегоцефалы были предками остальных амфибий, в этом
можно не сомневаться. Но они вымерли в пермское время, немногие
дожили до триаса. А затем — до начала третичной эры — ископае-
мые амфибии пам почти неизвестны, и переходных форм от стего-
цефал к современным амфибиям пока еще пе найдено. Отсюда не-
ясность в эволюции современных амфибий.
По общей форме тела к стегоцефалам ближе Urodela. Их древ-
нейшим представителем считали нижпепермского Lysorophus tri-
carinatus. В настоящее время этот вид хорошо изучен, и некоторые
относят его к Stegocephali, так как в черепе у него сохранились
черты стегоцефал. Возможно, что эта жившая в илу форма была
неотенической.
С триаса до эоцена об ископаемых амфибиях не известно, кроме
найденной в нижнемеловых слоях Бельгии амфибии Hylaeobathra-
chus croyi, близкой к Proteidae. В олиго цепе Франции найдена
форма Urodela, названная Megalotriton; в нижнем миоцене уже
жил Triturus. Из верхнего миоцена Швейцарии известна саламанд-
ра, фигурировавшая под названием «грешник — свидетель потопа»
(Andrias scheuchzeri, Cryptobranchidae). Современный тритон
Triturus cristatus известен из дочетвертичных слоев Англии.
Таким образом, палеонтология даст мало данных для построения
филогении современных амфибий. Ес приходится строить па осно-
вании анатомических данных. В настоящее время выводят совре-
менных амфибий от Phyllospondyli (Branchiosauria). От них, вероят-
но, произошли и Anura и Urodela. От Lepospondyli ответвились, ви-
димо, Apoda. Anura известны с нижнего миоцена, где найдено
12 видов рода Palaeobatrachus, близкого к сем. Pipidae (Aglossa);
роды Rana и Bufo известны из верхнего эоцена.
VI. ЭКОЛОГИЯ АМФИБИЙ
1. Приспособление амфибий к условиям среды обитания:
типы — роющий, водный, наземный, древесный
Приспособление к условиям местообитания выражается прежде
всего в форме тела и в структуре некоторых наружных органов.
Мы видели выше, что среда обитания амфибий весьма разпообраз-
364
па. Как. лпчппкп почти всех амфибий, так и многие взрослые формы
леи пут в воде; другие амфибии идут в воду лишь временами, глав-
ным образом в период размножения; третьи становятся совсем
сухопутными формами; четвертые живут на деревьях. Наконец,
есть и такие, которые ведут совершенно подземный образ жизни.
Это различие в среде обитания накладывает печать на органи-
зацию амфибий, представляющих различные приспособительные
типы: водный, полуводный, наземный, древесный, роющий.
Безногие (Apoda) ведут совершенно особый от других амфибий
образ жизни, чисто подземный, роющий. При этом они про-
двигаются вперед при помощи змеевидных изгибов тела, прокла-
дывая себе путь головой. При таком способе движения конечности
пе только пе нужны, — опи мешали бы. Путем долгого отбора опи
подвергались полной редукции. Вместе с ними редуцировались
и пояса конечностей. С другой стороны, при таком способе передви-
жения, который наблюдается у Apoda, большое значение имеет
длина тела: чем оно длиннее, тем сильнее проталкивание тела впе-
ред. Это в свою очередь влияет на строение головы, являющейся
тараном, откуда ее более прочное устройство. В связи с этим число
позвонков у Apoda доходит более чем до 200; большинство позвон-
ков имеет длинные ребра, концы которых при движении упираются
в землю; конечностей пет вовсе, как и поясов; череп очень ком-
пактен, образует сплошную коробку, напоминая череп стегоце-
фал. Одпако оп не стегальный, а псевдостегальный, — сходство
здесь лишь конвергентное: череп образован разрастанием костей,
имеющихся и у других современных амфибий; разрастание яв-
ляется приспособлением к роющему образу жизпи. Приспособ-
лением к последнему является пахождсиие рта не на конце головы,
а снизу, дабы в рот пе попадала земля. Глаза редуцированы, ру-
диментарны, пе функционируют, будучи скрыты под кожей. Бо-
гатство железами и кольчатость кожи служат, видимо, той ясе
цели — облегчению персдвиясепия в земле.
Другой тип строения тела в связи с иной средой обитания мы
видим у водных амфибий. Примером может служить тритон (Tri-
turus). Он имеет вытянутое тело, две пары пог, более или мепее
длинный хвост. Движение тритона в воде происходит при помощи
боковых или винтообразных движений хвоста, имеющего в это
время вертикальный плавник. Конечности при плавании тритона
прижаты к телу и направлены назад.
У некоторых Urodela тело удлинено до змеевидной формы, на-
пример, у Amphiumidae. При этом недоразвиваются передние или
задние конечности. Змеевидная форма развивается у тех Urodela,
которые роются в илу.
Глаза у амфиумы маленькие и без век. Так же и у Sirenidae.
Дегенерировали и скрыты под коясей глаза у протея (Proteus).
Они являются как бы остановившейся стадией развития: роговицы
и передней камеры глаза еще нет, хрусталик зачаточен, в сетчатке
еще пот палочек и колбочек. Здесь мы, однако, видим не наслед-
ственный признак, но недоразвитие наследственного зачатка вслед-
365
лягушка
Рис. 287. Древесная
из Brevicipitidae (Platyhyla
verrucosa).
(Из Noble.)
ствие отсутствия функции, ибо при воспитывании молодых протеев
на свету строение глаз совершенно изменялось: увеличились раз-
меры глаз, образовались роговица и передняя камера глаза, хру-
сталик развился, в ретине появились палочки и колбочки. У ТурЫо-
triton spelaeus из семейства Plethodontidae мы видим обратное:
эта саламандра имеет глаза, пока в личиночном состоянии живет па
свету; позднее, уходя в пещеру, теряет зрение.
На земле тритон передвигается, волоча брюхо по земле. Зад-
ние ноги толкают тело вперед, передние лишь определяют направ-
ление движения, тело изгибается змееобразно. Наземные сала-
мандры имеют тело более стройное и могут бежать более быстро,
подобно ящерицам. Специальные исследования показали, что ре-
флексы ходьбы у Amby stoma вырабо-
тались из ее плавательных реакций.
Первые двигательные нервы, дости-
гающие во время развития до мышц
конечностей, являются ветвями тех
нервов, раздражение которых вызы-
вает сокращение мускулов туловища.
В этом мы имеем еще одно под-
тверждение происхождения амфибий
от рыб.
Иной тип организации и иной тип
движения видим мы у Anura. Тело их
плотное, короткое, лишенное хвоста.
Задние конечности длиннее передних.
В воде они передвигаются при по-
мощи сильных толчков задними но-
гами, пальцы которых удлипепы и
связаны перепонками. Передние
конечности в это время приложены
к телу, как у Urodela. При медленном движении задние ноги вы-
тягиваются поочередно, как при ходьбе. На земле передвижение
происходит скачками или путем поочередного передвижения пе-
редних и задних ног у тех амфибий, у которых задние ноги не длин-
ны (например, жабы).
У тех Anura, которые выработали способность лазать, развился
ряд приспособлений (рис. 287). На пальцах развились специаль-
ные диски, богатые лимфатическими пространствами и слизистыми
железами в эпителии. Прикрепление обусловливается здесь моле-
кулярным притяжением двух тесно соприкасающихся тел, кото-
рое тем сильнее, чем меньше между иими воздуха. Особые гладкие
мускулы делают эти диски плоскими. У яванской лягушки (Rha-
cophorus) прикрепление происходит не только посредством дисков
на пальцах, но и при помощи кожи брюха, кожных складок на ко-
нечностях и на подбородке.
Некоторые из древесных лягушек имеют особым образом
устроенные хватательные конечности. Например, у Phyllomedusa
burmeisteri мы видим такое же изменение в степени развития и в
368
расположении пальцев, как у некоторых птиц и лемуров. Здесь
мы видим пример конвергенции у лазящих животных.
У древесных форм мы видим еще одно приспособление, разви-
вающееся по одному принципу и у рептилий, и у млекопитающих:
способность к планирующему полету. У большинства представи-
телей рода Rhacophorus пальцы соединены перепонкой не более
чем до половины, у немногих они достигают дисков на концевых
фалангах. В этом случае они могут пользоваться перепонками как
парашютами. При полете с дерева лягушка растопыривает пальцы,
держит конечности прижатыми к телу, которое надувается вслед-
ствие раздутия сильно развитых, легочных мешков. Получается
большая поверхность, являющаяся парашютом. Поверхность меж-
пальцевых перепонок у R. pardalis равняется 18.8 см2. R. mala-
baricus может совершать скользящий полет на 10—16 м. Hyla
venulosa может делать прыжки с высоты 40 м.
Среди Anura имеются формы, приспособившиеся к рытью в зем-
ле, куда они зарываются в сухое время. Рытье осуществляется
при помощи задних ног. У чесночницы (Pelobates fuscus) для этого
существует особое приспособление в виде острого лопатообразного
рогового выроста на ступне. У роющих видов родов Scaphiopus
(сем. Pelobatidae) и Pelobates голова тупая, и подкожная соедини-
тельная ткань головы инфильтрирована костной тканью. У дру-
гих роющих видов голова, наоборот, острая и лишена упомянутых
окостенений. Некоторые Urodela, роющиеся в илу, также имеют
острое рыло.
2. Питание амфибий. Приспособление к роду пищи
Во взрослом состоянии амфибии являются хищниками. Опи
питаются червями, улитками, насекомыми, хватая даже пчел.
Некоторые крупные виды лягушек охотятся за другими лягушками
и даже за небольшими теплокровными: так-, рогатая лягушка
(Ceratophrys dorsata) имеет такую пасть, что может проглотить мо-
лодого цыпленка, а крыс и мышей глотает легко. Лягушка-бык
(Rana catesbyana) глотает цыплят, утят, рыбу, змей и себе подоб-
ных. Гигантская саламандра (Megalobatrachus maximus) питается
лягушками и рыбами. Захватывает пищу пастью. Лишь в некоторых
случаях (у лягушки) этому помогают передние лапы. Заглаты-
вание же Происходит при содействии глаз. Глазные яблоки сильно
вдаются в ротовую полость и отделены от нее лишь слизистой обо-
лочкой. Особый мускул оттягивает глаза назад, и таким образом
они способствуют проталкиванию пищи. Когда лягушка, например,
заглатывает схваченную добычу, она всегда закрывает глаза,
которые опускаются в это время в ротовую полость. Так как до-
быча более или менее однообразна, то особых приспособлений
для ее схватывания у амфибий мы почти пе встречаем. Лягушки
могут выбрасывать с большой быстротой язык, прикрепленный
впереди, а не сзади, как это бывает обычно у других позвоночных,
и бьет им как хлопушкой. Так как язык является очень клейким
867
благодаря большому количеству желез, то добыча — мухи, черви
и т. и. — приклеивается к нему (рис. 288). '
Как выбрасывание, так и втягивание языка происходят при
помощи особых мускулов, собственные же мускулы языка дают
ему возможность охватывать добычу. У других амфибий, напри-
мер, у пещерной саламандры
(Spelerpes fuscus), язык сидит на
стебельке, который может втяги-
ваться и сильно вытягиваться
(рис. 289).
У восточноафриканского ко-
роткоголова (Breviceps mossam-
bicus), который, невидимому, пи-
тается термитами, рот очень уз-
кий, язык длинный, приросший,
зубов нет, желудок очень обши-
рен. Здесь мы видим приспособле-
ние того же рода, как и у млеко-
чем заглатывают во множестве мелких
Рис. 289. Язык пещерной саламандры
(Spelerpes fusens):
a — втянутый, Ь— вытянутый. (По Науеп и
Wicdershciin из Ililzhcimer.)
Рис. 288. Голова и язык зеленой питающих, питающихся муравь-
лягушки (Rana escuienta). ями и термитами, например, у му-
(По wemer.) равьеда.
При повышении температуры
прожорливость амфибий возра-
стает, при понижении — падает. Личинки земноводных питаются
пе исключительно животной пищей. В самые первые дни своего
существования опи непрерывно наполняют свой кишечник, при-
животных: инфузорий,
коловраток, ракообраз-
ных, а также диатомо-
вые водоросли. Своими
роговыми челюстями они
скоблят растения и мо-
гут долго существовать
при исключительно рас-
тительной пище. Пере-
ходя же в более взрос-
лое состояние, перед
метаморфозом, они пе-
реходят и к животной
пище. Apoda, роющиеся в земле, питаются мелкими змеями,
именпо слспупами и молодыми щитохвостыми змеями, дож-
девыми червями, а также муравьями, в гнездах которых неред-
ко живут. Urodela способны брать неподвижную добычу. Поэтому
некоторые из пих поедают икру рыб, собственные яйца. Anura,
паоборот, схватывают лишь движущуюся добычу. Древесные ля-
гушки (Hyla) прыгают к добыче.
У амфибий «каннибалов», поедающих себе подобных, разви-
ваются большие «ловчие зубы». Такие же зубы наблюдались у лаби-
риптодоптов. Стегоцефалы питались подобно современным ам-
фибиям.
368
Пища амфибий богата белками, так как оиа состоит из живот-
ных. Излишек, не идущий на пополнение затрат в клетках, от-
кладывается в виде зернышек углевода гликогена главным обра-
зом в печени. Переходя в сахар, он идет на производство работы.
Жир у амфибий откладывается лишь в небольшом количестве,
в «жировых телах», в печени, у Proteus в виде тонкого слоя под
кожей. Это стоит, повидимому, в связи со слабой энергией обмена
веществ у амфибий, животных «холоднокровных». А может быть,
и наоборот: они не стали теплокровными вследствие того, что у пих
не накапливался жир. Во всяком случае это является указанием
на их низкое положение на эволюционном дереве.
Пищеварение происходит в общем типично для позвоночных,
но с некоторыми уклонениями. Так, например, оно начинается
лишь в желудке, так как слюнных желез у амфибий нет. В сек-
рете печени нет веществ, служащих для пищеварения, лишь в жел-
чи содержится липаза, эмульгирующая жиры. Печень — это ме-
сто, где запасается гликоген, образуется мочевина, разрушаются
красные кровяные тельца. Во время зимовок печень служит местом
резервации жира, откуда он быстро поступает в ткани во время
весеннего периода спаривания.
3. Окраска амфибий
Кожа амфибий большей частью окрашена, иногда очень ярко.
У жерлянки (Bombina) кожа брюха окрашена в яркие желтый
и синий или в синий и красный цвета. У огпебрюхого тритона (Tri-
turus pyrrhogaster) брюхо окрашено в интенсивно кармиппо-
краспый цвет. Рогатая лягушка (Ceratophrys cornuta) является
окрашенной в орапжево-краспые, краснобурые и изумрудно-
зеленые тона и т. д. Окраска обусловливается теми же элемен-
тами, что и у рыб: пигментом и рефлектирующими иридоцитами.
Пигмент или диффузио распространен в теле или зернистый. Диф-
фузный пигмент, большей частью желтый, распределен в роговом
слое эпидермиса; зернистый помещается в особых клетках — х р о-
матофорах, способных выпускать амебоидные отростки.
Хроматофоры помещаются в cutis, большей частью в верхнем ее
слое. Сокращение хроматофоров ведет к исчезновению пигмента
на поверхности, вытягивание отростков ведет к его более или менее
равномерному распределению. Пигмент обычно бывает черного,
бурого, желтого и красного цветов. Кроме того, встречаются белые
зерна гуанина и особые красящие вещества с металлическим бле-
ском. Зеленого и голубого пигментов пет, а окраска в эти тона
является «субъективной», обусловленной смешением тонов в глазу
наблюдателя.
Большинство амфибий, особенно обнаруживающие большую
приспособляемость окраски, способны менять свой цвет. Во мно-
гих случаях лягушки, одни быстрее, другие медленнее, могут
менять свой цвет приспособительно к грунту, древесные лягушки—
24 Кашкаров и Станчинский ^69
к цвету листвы. Огненная саламандра, содержимая па желтом
грунте, приобретает преобладание желтого цвета; будучи же содер-
жима па черном грунте, становится почти черной.
Механизм изменения окраски у амфибий в целом тот же, что
у рыб.
Измепение окраски происходит через посредство нервной си-
стемы под ^влиянием внешних или внутренних факторов. Высо-
кая температура, высыхание, повышение интенсивности освещения
вызывают сокращение хроматофоров и посветление животного;
низкая температура, увеличение влажности и затемнение вызы-
вают его потемнение. Если животное ослепить, оно не реагирует
на освещение; тот же результат получается при разрушении дву-
холмия и зрительных бугров. Если раздражать нерв, например,
бедренный, то происходят изменения в окраске бедер. У Rana
pipiens на световую стимуляцию отвечает кожа и непосредственно,
без участия глаз. У квакш (Hylidae) рефлекс изменения окраски
получается и путем осязательных раздражений со стороны паль-
цевых подушек и кожи брюха. На шероховатых листьях квакша
становится темнее, на гладких — зеленой. Темнеет квакша и па
коре деревьев, на листьях становится зеленой даже в совер-
шенной темноте. Тот же рефлекс имеет место и у ослепленных
животных.
Личинки саламандр становятся темными на темпом фоне.
Изменения окраски вызываются также физиологическим со-
стоянием. Так, изменение окраски вызывается расстройством
дыханця, остановкой циркуляции крови, изменением некото-
рых желез внутренней секреции: питуитарной железы и над-
почечников.
Очень интересен вопрос о том, в какой мере окраска амфибий
является приспособительным признаком. Что во многих случаях
она адаптивна — несомненно. Но чаще всего в окраске пе отра-
жаются ни характер местообитания, ни биотические взаимоотноше-
ния. Приспособительный характер имеет окраска Hyla arborea
на листьях и на коре. Покровительственный характер имеет
зеленая окраска наших лягушек. Но нередко лягушки совсем
различной окраски живут бок о бок. Показателем отсутствия
в некоторых случаях защитного значения окраски является
Atelopus varius, лягушка центральной Америки и Колумбии,
отличающаяся чрезвычайным разнообразием и яркостью окраски
верхней стороны тела, причем пи одна окраска не соответствует
обстановке. Яркая окраска ядовитых видов считается «предупреж-
дающей» окраской. Предупреждающей окраской считается чер-
ная с оранжевыми пятнами окраска огненной саламандры (Sa-
lamandra salamandra), окраска жерлянки (Bombina bombina),
которая, будучи потревожена, резко изгибает голову и ноги па
спину и поворачивает ярко окрашенную нижнюю поверхность лап
кверху. Эти амфибии выделяют ядовитые вещества. Но играет ли
здесь окраска приписываемую ей роль, сомнительно. Это следует
проверить наблюдением над экологией видов.
370
4. Защитные приспособления. Регенерация
Амфибии в общем — еще слабо защищенные существа. Число
защитных приспособлений у них весьма невелико, и их организа-
ция еще слишком несовершенна, чтобы широко использоваться
в целях 'защиты и самообороны.
Наиболее важным способом защиты являются выделяе-
мые кожей ядовитые' вещества. Выше, в главе
о морфологии кожи, было указано, что последняя у амфибий
чрезвычайно богата железами, что железы эти многоклеточные,
притом двоякого рода: слизистые. и белковые. Выделенное ими
вещество имеет кислую реакцию и сильный запах. Действие па
организм секрета обоих видов желез различно. Выделение слизи-
стых желез действует как наркотик, яд спинных желез вызывает
судороги. Если в один мешок посадить жерлянок (Bombina)
и амфибий другого вида, то последние могут погибнуть. Особям
того же самого вида яд амфибий невреден; будучи впрыснут в кровь,
яд действует даже на то же самое животное. Яд саламандры, жабы,
даже тритонов, впрыснутый в кровь, убивает млекопитающих,
птиц, рептилий и рыб, если доза соответствует размерам живот-
ного. Мелкие птицы и ящерицы, как правило, гибнут в несколько
минут, более крупные животпые — морские свинки, кролики и со-
баки — в срок менее, чем час. Яд амфибий листолаза (Phyllo-
bates) и древолаза (Dendrobates) употребляется туземцами в Юж-
пой Америке для намазывания наконечников их стрел, в особен-
ности при охоте па обезьян. Действует описываемый яд на сердце
и центральную нервную систему подобно яду наперстянки — ди-
гиталипу, отчасти подобно кураре. Яд амфибий весьма падежпо
предохраняет их от многих покушений: хищные животные или
вовсе пе трогают амфибий или берут их весьма неохотно, так как
яд их при попадании на слизистые оболочки вызывает сильное
жжепие и рвоту.
Другая защитная функция заключается в способности разду-
вать легкие, благодаря чему раздувается все тело, которое ста-
новится более крупным и гладким, так что лягушку трудно бы-
вает взять. Защитными являются и некоторые реакции поведения,
например, так называемый «рефлекс бомбипы», наблюдающийся
у некоторых лягушек: заворачивание головы и яркоокрашенных
копечпостей па спину, как у рода Bombina. Принято думать, что
такого рода движение отвращает нападение змей и птиц.
И у хвостатых и у бесхвостых наблюдается при приближении
опасности своего рода оцепенение, род каталепсии, «мнимая
смерть», обычно продолжающаяся несколько минут. Вряд ли, одпа-
ко, эта реакция имеет защитное значение, так как возникает
она при непосредственном прикосновении, когда’ бывает уже
поздно. Некоторые саламандры, наоборот, принимают «предосте-
регающее» положение или «положение угрозы», помахивая хво-
стом, выделяющим обильный секрет. Большинство амфибий в
случае опасности стремится уйти от последней и скрыться в
24* 371
какое-либо убежище. Движения убегания у амфибий еще весьма
несовершенны.
Некоторые более крупные и сильные амфибии в опасности ведут
себя агрессивно: издают скрипящий звук, раскрывают рот, ку-
саются, как Ceratophrys, Cryptobranchus.
Уисе у наземных саламандр наблюдается, более развитая у реп-
тилий, способность путем автотомии отламывать хвост, за который
животное схвачено. Механизм здесь несколько иной, нежели у яще-
риц: разлом происходит не посредине позвонка, а между позвон-
ками. Как и у ящериц, мускулатура разрывается несколько ближе
к туловищу, нежели кожа, закрывающая^таким образом рану.
К защитным особенностям организма амфибий следует отне-
сти и их способность к регенерации.
Большинство амфибий обладает способностью восстанавливать
утраченные или поврежденные конечности. Чем моложе животное,
тем легче происходит у пего регенерация. Точно так же, как хвост,
регенерируют и наружные жабры. Личинки саламандры нередко
объедают последние друг у друга. И они восстанавливаются.
У хвостатых амфибий хвост и конечности регенерируют даже
во взрослом состоянии.
Если у взрослого аксолотля отрезать кисть, то через четыре
недели получается конический выступ, регенерационная почка.
Через шесть недель выступ становится двойным; через одиннадцать
недель уже различимы три или четыре пальца, а через двенадцать
недель после операции регенерирует и вся кисть. Иногда вместо
четырех пальцев восстанавливается пять. Восстанавливается и
целая конечность, если отрезать ее посреди плечевой кости.
Восстаповление идет из соединительной ткани. У взрослого
тритона (Triturus cristatus) восстанавливаются отрезанные пальцы
в течение пяти или шести педель. Кость и хрящ восстанавли-
ваются позднее. У Proteus утраченная пога целиком восста-
навливается в течение восемнадцати месяцев. У Plethodontidae
восстапавливается утраченный хвост. Личинка Necturus, если
ей отрезать голову, пе погибает. Рапа заживает, животное растет
и дифференцируется в течение 2 месяцев.
Бесхвостые амфибии также способны к регенерации конеч-
ностей, по в значительно мепыпей степени, опа происходит только
у молодых головастиков. Регенерация идет тем быстрее и легче,
чем моложе особь. Если у головастика травяпой лягушки (Rana
temporaria), у которого передние конечности еще скрыты под
кожей, отрезать заднюю копечность посреди бедра, уо через девят-
надцать дпей восстанавливается колено с выступом из двух паль-
цев. В копце концов, восстанавливается и вся конечность. Хвост
головастиков восстапавливается легко и свободно даже в том слу-
чае, если он бывает отрезан незадолго до конца метаморфоза.
Но после последнего бесхвостые амфибии почти всецело утрачи-
вают способность к регенерации, она сохраняется лишь в очень
слабой степени. Способность к регенерации ослабляется и при
переходе на высшую филогенетическую ступень.
372
Весьма интересно, что регенерация, если пе всегда, то в не-
которых случаях, связана с развитием и функцией нервной
системы. Ампутированная конечность взрослого тритона регене-
'рирует лишь при том условии, если при ампутации останутся
неповрежденными волокна симпатической нервной системы. Спо-
собность амфибий к регенерации делает их одним из излюбленных
объектов при изучении этого высоко интересного явления.
Способность к регенерации делает земноводных в высшей сте-
пени живучими. Многие поранения, от которых всякое другое
животное погибло бы, безнаказанно переносятся амфибиями бла-
годаря регенерации. Иногда из убитой змеи выползают совер-
шенно живые жабы с частью переваренными уже задними ногами
и живут.
Способность к регенерации — несомненно высокоадаптивный
признак. Но это не специфическое защитное средство, выработан-
ное отбором. Это общая способность мало дифференцированных тка-
ней, убывающая в филогенезе и онтогенезе. Регенерировать могут
и такие органы и части, которые обычно не подвергаются повреж-
дениям, например, легкие у лягушек. На способность регенера-
ции значительно влияет функция, усиливая процесс.
5. Размножение амфибий
У многих амфибий наблюдается явственно выраженный поло-
вой диморфизм, вторичные половые признаки. У одних эти
различия существуют все время, у других они возникают
лишь в период размножения. Отличия между самцом и самкой
могут выражаться в величипе, причем самец обычно меньше
самки, в наличии «брачной бородавки» па пальцах у самцов,
в яркой окраске, в форме рыла, величине барабанной пере-
понки, в мускулатуре передних конечностей, в развитии в период
спаривания гребней па спипе и на хвосте, в инстинктивном пове-
дении, в строении внутренних органов и т. д.
«Брачная бородавка» у лягушек и жаб развивается на ргае-
pollex. Ее назначение •’-> помогать удержанию самки. В большин-
стве случаев это приспособительный признак. Этого значения не
имеет яркая окраска самцов. Трудпо объяснить ее и теорией по-
лового подбора. У тритона гребенчатого (Triturus cristatus) в пе-
риод спаривания самец становится очепь ярким, па спине выра-
стает зазубренный гребспь, еще более развитый у малоазиатского
тритона (Triturus vittatus). Самец тритона ухаживает за самкой,
но сомнительно, чтобы половые украшения помогали успеху по-
следнего. Яркие пятна па боках появляются у самца слепого
протея (Proteus) в период спаривания, где они никакого значения
иметь не могут. У большинства амфибий в окраске двух полов
различий мало. В некоторых случаях вторичный половой признак
имеет другое приспособительное значение. Так, у южноамери-
канской лягушки — Leptodactylus ocellatus — рыло заострено
и имеет под верхней губой горизонтальный гребень. Получается
373
своего рода лопата, которая используется самцом для рытья ямок
на краю потоков, куда потом заходят самки для откладывания
икры. Волосообразные кожные придатки на боках тела самца во-
лосатой лягушки (Astylosternus robustus) служат для усиления
дыхапия в период спаривания (рис. 290). Легкие у этой лягушки
недоразвиты, а в период спаривания метаболизм усилен.
Голос также является вторичным половым признаком. Голос
появляется среди позвоночных у амфибий впервые. Слух, насколь-
ко возможно судить по устройству слухового аппарата, был уже
у стегоцефал, служа для восприятия опасности. Возможно, что
и голос появился впервые уже у стегоцефал. Особенно известны
Рис. 290. «Волосатая лягушка» (Astylosternus robustus).
(Из Noble.)
своим голосом жерлянка (Bombina), древесные квакши (Hyla),
зеленая жаба (Bufo viridis), лягушки (Rana). Для усиления го-
лоса служат особые парные (у Rana) или непарные (у Hyla) го-
лосовые мешки, представляющие вывороты слизистой оболочки
ротовой полости. Каждый вид лягушек имеет свой голос. Кроме
голоса, служащего вторичным половым признаком, у некоторых
амфибий имеется способность издавать «крик испуга», например,
при нападении змеи.
Urodela большей частью пе имеют голоса.
Кроме голоса у самцов нередко бывают отличия в строении не-
которых органов, например, более крупные легкие, большее чис-
ло кровяных телец (в связи с большей активностью), больший вес
мозга и печени, отличия в сердце.
В общем вторичные половые признаки в большинстве случаев
пе имеют специфического адаптивного значения и обязаны своим
существованием половым гормонам, что доказывается экспери-
ментами с получением половых признаков самца у самки при
пересадке ей мужских семенных желез.
Выбор места для размножения. Амфибии, возникшие в воде,
происходящие от рыб, каждый раз возвращаются в воду в период
размножения. Весь процесс размножения совершается «под зна-
ком воды». Все его особенности у амфибий, включая и так назы-
ваемую заботу о потомстве, проистекают из этого факта. Лишь
374
немногие амфибии откладывают яйца в земле (Amphiuma), на
деревьях (Phyllomedusa, Rhacophorus). Но в этих случаях яйца
всегда защищаются кем-либо из родителей, существует так назы-
ваемая «забота о потомстве». Этот способ кладки яиц не в воде —
явление вторичное, приспособление к особым условиям существо-
вания.
Тем более вторичным явлением, связанным с той же особен-
ностью амфибий — привязанностью к воде и неспособностью раз-
множаться на земле, являются и те виды заботы о потомстве,
когда яйца вынашиваются на спине в коже родителя или в рото-
вой полости, и т. д. Амфибии еще не наземные существа,
вода необходима для их развития. Различные адаптации к жизни на
суше не дают еще им возможности оторваться от воды нацело.
Спаривание и кладка яиц. Половой инстинкт у амфибий очень
силен. В период размножения самцы лягушек на каждое прикос-
новение к груди отвечают крепким охватыванием прикасающегося
предмета, каков бы он ни был, даже неодушевленный. Голос слу-
жит самцам для призыва самок. У_тритона наблюдаются настоя-
щие любовные игры, поднимание и колыхание гребнем и как бы
танцы перед самкой. Этот показатель сильного полового влечения
возбуждает самку. После этого самец приклеивает к какому-
либо предмету сперматофор, который самка захватывает клоакой.
Самец наземной саламандры (Salamandra salamandra) откладывает
светлый сперматофор после продолжительного ухаживания. Самка
захватывает сперматофор, после чего самец оставляет самку.
Совокупление наблюдается лишь у безногих (Apoda), у Urodela
настоящей копуляции никогда не бывает. Самец откладывает
сперматофор, а самка подбирает его клоакой. Поведение полов
во время спаривания весьма разнообразно у различных Urodela:
в одних случаях имеет место продолжительное ухаживание (Tri-
lurus), в других — охватывание самки па короткое время, при-
том па суше, а кладка яиц происходит в воде (Salamandra, Sala-
mandrina, Spelerpes). В одних случаях самец охватывает самку
передними ногами под мышками, в других, имея развитые задние
ноги и цепкий хвост, обнимает самку за поясничную и хвостовую
область.
У Anura охватывание самцом самки — правило. Лишь при
этом условии, если сидящий па спине самки самец с силой сдавли-
вает ее своими передними конечностями, яйца откладываются
в одно время большими порциями. Объятие длится иногда не-
сколько дней. При появлении яиц самец опорожняет па них семя
из семеппых пузырьков. У Anura оплодотворение всегда наружное.
Некоторые Apoda и Urodela являются живородящими. Так,
у пятнистой саламандры (Salamandra salamandra) молодые зароды-
ши разрывают оболочку яйца в тот момент, когда рождаются, и бы-
вают снабжены длинными наружными жабрами; у горной сала-
мандры (Salamandra atra) эмбрионы проходят внутри матки все
свое развитие и метаморфоз. Совершенно готовых зародышей родит
бурая пещерная саламандра (Spelerpes fuscus), а среди безногих —
375
Typhlonectcs compresaicauda и толстокожая червяга (Dermophis
thomensia).
Яйца амфибий одеты, кроме собственной оболочки, еще одной
или двумя студенистыми оболочками, выделяемыми стенками
яйцевода. Наружная оболочка при попадании в воду сильно раз-
бухает. Иногда вся масса отложенной икры одета одной наружной
оболочкой, образуя длинный шнур (у Bufo), иногда же каждое
яйцо одето отдельно наружной оболочкой (у Rana). Оболочки эти
очень хорошо защищают зародыш. Затем они или растворяются,
или зародыш освобождается от них активными движениями. Обо-
лочки дают защиту от механических повреждений, от химиче-
ских воздействий, от поедания птицами. Лишь утки с их широким
клювом могут глотать икру лягушек. Оболочки защищают яйца
и от высыхания. Далее, благодаря сферической форме яиц, обо-
лочки, увеличивая объем последних, оставляют большие проме-
жутки между яйцами, куда проникает вода, необходимая для ды-
хания. Кроме того, прозрачные оболочки действуют как тепличные
стекла, собирая лучистое тепло. Наконец, действуя как собиратель-
ные линзы, они отбрасывают на солнце светлые точки, привлекаю-
щие одноклеточные водоросли, покрывающие икру и способствую-
щие дыханию икринок.
Число икринок и величина их различны у разных амфибий.
Там, где существует забота о потомстве (Alytes, Rhinoderma, Pipa),
яиц откладывается немного, несколько дюжин. У травяной ля-
гушки (Rana temporaria) число яиц 3 000, у зеленой лягушки (Rana
esculenta) число их доходит до 10 000. Тут видна закономерность,
свойственная и другим группам животных.
. Развитие и метаморфоз. Из яйца, пройдя известные стадии раз-
вития, получается личинка с жабрами, впоследствии путем мета-
морфоза превращающаяся во взрослую форму. Развитие и метамор-
фоз в общих чертах протекают у безногих и хвостатых следующим
образом: у них редуцируются жабры, закрываются жаберные щели,
утрачивается жаберпая полость и топкие, служащие для плавания,
края хвоста. Зато у бесхвостых амфибий при метаморфозе происхо-
дит полная реорганизация. Сперва у зародыша имеются голова и ту-
ловище, но хвоста еще нет. Позади налипающего развиваться рта
появляется поперечная складка, развивающаяся в парпый или
непарный присасывательный аппарат, состоя-
щий из большого количества желез, выделяющих вязкий секрет.
При помощи этого секрета личипка прикрепляется к студенистой
оболочке яйца, а затем к водорослям или другим объектам в воде.
Название «присоски» поэтому пе подходит к этому образованию,
ибо здесь имеет место не присасывание, а приклеивание. Положение
присасывательного аппарата па нижпей стороне головы может быть
различно. Он существует очень недолгой исчезает к тому времени,
когда развиваются жабры, хвост и функционирующий рот. Лишь
у немногих форм восточных лягушек этот орган превращается
в брюшной диск, существующий более долгое время (рис. 291).
Рот у ‘личинок бесхвостых снабжен тонкими роговыми зубчиками,
376
заменяющими настоящие зубы. Число их у травяной лягушки
(Rana temporaria) около 640. Каждый зубчик представляет одну
ороговелую клетку, острый конец которой загнут назад и зазубрен.
Одни из этих зубов, окружающие в виде гребня губы, употребляются,
повидимому, для прикрепления или для удержания пищи; дру-
гие, образующие собственно роговой клюв и вооружение челюстей,
действуют подобно терке улиток. У чесночницы (Pelobates fuscus)
число роговых зубов возрастает до 1 000.
Затем, когда у головастиков возникают в виде маленьких бугор-
ков по бокам головы жабры, тело становится овальным, более или
менее шаровидным, и кишечник сильно удлиняется. Открывается
заднепроходное отверстие. Появляются конечности и развиваются
легкие. Головастики начинают подниматься на поверхность, чтобы
1 г в ч
Рис. 291. Четыре стадии развития присасыватель-
ного аппарата (A) Bufo vulgaris. М—рот
(По Thiele из Gadow.)
дышать воздухом. Поздпее жабры дегенерируют, и все органы из-
меняются приспособительно к наземной жизпи. Хвостовой плавник
абсорбируется, роговая обкладка рта и губ сбрасывается, давая
место настоящим зубам, череп изменяется соответственным образом.
Головастики в это время перестают есть, кишечник освобождается от
своего содержимого и перестраивается посредством гистолиза,
сокращаясь до одной шестой части первоначальной длины, так как
прежняя длина, приспособленная к растительной пище, уже пе
нужна. Заднепроходная часть кишки спадается, абсорбируется
и внизу, под корнем хвоста, образуется новый задний проход. Аб-
сорбция хвоста идет более энергично при повышении температуры.
Низкая температура замедляет абсорбцию. Если хвост отрезать, он
регенерирует сейчас же, хотя бы головастик был уже на краю ре-
дукции хвоста. Голодание также ускоряет последние стадии мета-
морфоза. Голодая, организм обращается к собственным запасам
и прежде всего к частям, подлежащим удалению — к хвосту и коже,
прикрывающей передние конечности. Резорбция хвоста производит-
ся при помощи лейкоцитов, которые деятельно поедают распадаю-
щиеся и дегенерирующие ткани хвоста.
Заботы о потомство. Многие земноводные откладывают, подобно
лягушкам, яйца прямо в воду и затем тут же оставляют их. Зато
другие обнаруживают в том или ином виде заботу о потомстве,
охраняют его — то самец, то самка. Забота о потомстве имеет огром-
ный биологический смысл. Она возникает тогда, когда нет подходя-
377
щего места для откладки яиц, пли же в том случае, если в воде
слишком много врагов; если нет воды вследствие слишком большой
сухости местообитания или если вода неподходяща по своим ка-
чествам, папример, вода горных ручьев; если вследствие ежеднев-
ных тропических дождей пе образуется спокойных водоемов или же
образующиеся водоемы быстро высыхают. Ряд приспособлений
в размножении имеет назначение парализовать эти неблагоприятные
условия.
Инстинкт, совершенно неправильно называемый «заботой о по-
томстве», так как общий уровень развития амфибий не позволяет
Рис. 292. Самка червяги (Ichthyophis glutinosus), высиживающая яйца;
яйца и личинки червяги.
(Из Gadow.)
признать у них таких эмоций, как забота, развивался у амфибий
независимо в различных отрядах и семействах. Здесь так же, как
в инстинкте ухаживания, можно наблюдать различные степени его
развития и сложности. Наиболее простую форму этого инстинкта
проявляют Hynobiidae, у которых самец остается на более или ме-
нее продолжительное время возле яиц, оплодотворяя их, причем
он прогоняет самку. У скрытожаберников самец охраняет яйца
до тех пор, пока из них не выведутся личинки. При этом сам он ест
охраняемые им яйца, но, съедая мало, не влияет этим на благо-
получие вида. Истинный характер «заботы о потомстве» ясно виден
из этого примера. Развился инстинкт «заботы» из необходимости по-
коя для прошедших период спаривания и кладки яиц родителей.
Исходя из этого, естественный отбор вырабатывал все более и более
сложные формы инстинкта. У многих амфибий он достигает край-
ней сложности и всегда имеет строго адаптивное значение. У параг-
вайской узкоротой лягушки (Phryniscus nigricans) яйца отклады-
378
Ь а
Рис. 293. Продольный разрез через
переднюю часть ринодермы Дарвина
(Rhinoderma darwini).
(Из Hilzticimer.)
ваются во временные лужи среди травы. Развитие их идет с не-
обычайной быстротой: если в 10 часов утра началась сегментация,
то к 7 часам следующего дня личинки уже вылупились. Стекающая
вода сносит их в более глубокие лужи, для чего форма отдельных
шаров, в виде которых откладывается икра, наиболее удобна. У
чилийской болотной лягушки (Paludicola fuscomaculata) развитие
заканчивается в 24 часа. Такая быстрота развития связана с крат-
ковременностью существования водоемов. Тут еще нет инстинкта
«заботы». Последний мы видим у цейлонского рыбо-змея (Ichthyo-
phis glutinosus). Эта амфибия
воды избегает и тонет в ней,
живет в земле. Опа отклады-
вает яйца в подземной норке,
вырываемой возле воды. Яйца
представляют плотный комок,
так как связаны менаду собой
родом шнурка. Самка обви-
вается вокруг этого комка, и
яйца сохраняются влажными
благодаря кожным выделе-
ниям матери (рис. 292). Это
слизистое выделение идет,
видимо, и для питания за-
родышей, которые, проделав
свое развитие, отправляются
в ближайший ручей. Тот же самый способ заботы о потомстве мы
видим у североамериканской амфиумы (Amphiuma means), и это тем
более замечательно, что вид этот принадлежит к живущим в воде
хвостатым амфибиям. Выход ее па сушу для размножения следует
считать приспособлепием для защиты потомства.
В высыхающих водоемах размножается американский древолаз
(Dendrobates trivittatus). Когда болотце высыхает, головастики при-
сасываются ртом к спине взрослой лягушки в числе 12—18, и в та-
ком виде лягушка ищет нового болотца, более богатого водой. Голо-
вастики длинноногой лягушки с Сейшельских островов (Arthro-
leptis seyschellensis), вылупившись, взбираются па отца или на
мать, сторожащих яйца, и питаются через кожу родителей. Воору-
жение челюстей у них отсутствует. Здесь головастики подвергаются
метаморфозу. Описанные только ^что виды попечения о потомстве
стоят в связи с недостатком спокойных водоемов.
Приспособлепием к неблагоприятным условиям развития яв-
ляется то, что у Rana opisthodon с Соломоновых островов весь мета-
морфоз происходит в яйце.
Совершенно своеобразный способ попечения о потомстве встре-
чаем мы у ринодермы Дарвина (Rhinoderma darwini). Здесь забота
о нем приходится па долю отца. Самец оплодотворяет отложенные
самкой яйца, помещает в рот и проталкивает в горловой мешок.
Сперва развитие идет за счет желтка (рис. 293), но когда послед-
него уже похватает, рост личинок идет за счет питания, получаемого
Рис. 294. Суринамская жаба
(Pipa americana).
(По Wicdersheim.)
от самца. Последний при этом сильно истощается. Его потомство как
бы паразитирует в нем, срастаясь спиной со стенками горлового
мешка и питаясь через кровеносные сосуды родителя. Достигнув
зрелости, лягушки выходят через отверстие горлового мешка и
ползут изо рта паружу.
У сумчатой лягушки (Nototrema) заботу о яйцах берет на себя
самка: яйца развиваются на спине в особой складке кожи. Личин-
ки имеют своеобразные келоколообразные жабры. Самка суринам-
ской жабы (Pipa americana) кладет себе яйца на спину при по-
мощи сильно выпячивающейся клоаки, а самец давлением при
помощи брюха распределяет их в один слой. Каждое яйцо поме-
щается в отдельную ячейку и покрывается роговой крышечкой
(рис. 294). В ячейках личинки и
остаются до полного развития.
Количество их бывает от 40 до
114. Яйца очень богаты желтком.
Перегородки между ямками на
спине очень топки и богаты сосуда-
ми. В позднейших стадиях разви-
тия питание идет еще за счет сту-
денистого слоя белка. Это можно
установить взвешиванием яиц и
выходящих личинок. Сильно раз-
витый хвост служит им органом
дыхапия.
Из наших амфибий заботу о
потомстве мы наблюдаем у жабы-
повитухи(А1у!ез obstetricans). Здесь
самец накручивает себе па ноги
яйцевые шнуры, которые и тас-
кает за собой. Созревшие личинки
выходят в воду. Некоторые ам-
фибии откладывают яйца вне воды,
чем достигается известная защита, так как главные враги находятся,
конечно, в воде. Так, южноамериканская квакша (Phyllomedusa
hypochondralis), найдя себе самца, который усаживается у нее на
спине, взбирается по стеблю растущего у воды растения, схватывает
кончик свисающего пад водой листд и взбирается па пего. Затем,
придерживая задними ногами сближенные края листа вблизи его
копчика, самка откладывает в получившуюся трубку яйца, а самец
их оплодотворяет. Студень, окружающий яйца, склеивает края ли-
ста. Кладка состоит приблизительно из 100 яиц и помещается
в двух листьях. Яйца богаты желтком, студенистая оболочка их
разжижается, и зародыш свободно лежит в кожистой капсуле.
Филломедуза Йерипга откладывает яйца между несколькими
листьями, свисающими над водой. В определенный момент своего
развития личинки падают в воду и здесь подвергаются окончатель-
ному превращению. Упомянутая выше яванская летучая лягушка
(Rhacophorus reinhardtii) приклеивает к листу пенистую массу,
380
внутренняя часть которой разжижается, и молодые головастики
проделывают свое развитие внутри последней (рис. 295). Другой
вид Rhacopliorus делает гнездо в земле, вблизи воды, где в пенистую
массу откладывает яйца. Затем проделывает косой выход к воде.
Через пего выходят личинки наружу.
Квакша-кузнец (Hyla faber) таскает со дна ил и строит из него
кольцевидный вал, который сильно выдается над водой. Наружные
стены его оставляются без внимания, внутренние же аккуратно
сглаживаются. ~
Получается постройка вроде кратера вулкана
(рис. 296). Работу делает одна лишь самка, са-
мец же все время сидит у нее на спине. Работа
идет ночью. Яйца откладываются внутри
вала. Через 4—5 дней вылупляются головас-
тики, продолжающие здесь в течение некото-
рого времени свое развитие.
Рис. 296. Гнездо квакши-кузнеца (Hyla faber).
(По Wicdersheim из Hilzhcimcr.)
Рис. 295. Гнездо
яванской летучей
лягушки (Rhaco-
phorus roinhardtii).
(По Sicdlcckr
из llilzhcimer.)
Неотения. У амфибий мы наблюдаем чрезвычайно интересное
явление, носящее название пеотспии. Оно заключается в том, что
у некоторых амфибий личинки достигают величины взрослых форм
и половой зрелости, сохраняя личипочпые признаки. Неотения
может быть полной, если животные могут размножаться в личи-
ночном состоянии, и неполной, если достигается только определен-
ный рост, по способности к размножению по наступает.
Неполную неотению мы встречаем у Некоторых бесхвостых земно-
водных: у чесночницы (Pelobates), у жерляпки (Bombilia), у ля-
гушки (Rana), у жабы (Bufo) и др. Полная неотения встречается
у хвостатых земноводных. Известны неотепические личинки тритона
(Triturus) и в особенности личинки мексиканской саламандры (Ат-
381
bystoma). Эти личипкп давно были известны под именем jjftredon
axolotl или 8. pisciformc, и их помещали в системе среди постоянно-
жаберных — Perennibranchiata. Возможно, что некоторые из по-
следних являются неотеническими личинками неизвестных
наземных форм.
Значение и происхождение неотении удалось в недавнее время
более или менее выяснить на аксолотле. Уже давно стало известно,
что он может превращаться в амбистому. Давно была также вы-
сказана мысль, что можно принуждать личинок аксолотля про-
делать метаморфоз, ставя его в такие жизненные условия, которые
затрудняли бы употребление жабер и облегчали употребление лег-
ких. Сперва эти опыты не приводили к положительному резуль-
тату, но затем они увенчались некоторым успехом. Была достигиу-
та как задержка в превраще-
•М _____ нии, так и задержка в разви-
тии органов размножения
Q под влиянием изменения та-
--------- ких Факт°РОВг. как темпера-
** тура, пребывание на воздухе
или в воде.
Новый свет был пролит на
явление неотении недавними
\ исследованиями над влиянием
С кормления щитовпдпой же-
лезой. Эти^ опыты показали,
// j I что кормление личипок щи-
/v товидной железой, или тире-
Рис. 297. Изменение аксолотлей под оидином (вытяжка щитовид-
влиянием кормления тиреоидином. ной железы), приостанавли-
(По Кольцову.) вает рост и ускоряет мета-
морфоз; зобпая железа и се
препараты, наоборот, усиливают рост и задерживают метаморфоз.
На основании этих наблюдений были предприняты опыты с корм-
лением личинок аксолотля тиреоидином с целью вызвать у них
метаморфоз. Действие тиреоидина было быстрое и вполне опре-
деленное: молодые аксолотли переставали расти, наружные жабры
сильно уменьшались в размерах, спиппой гребень мало-помалу
исчезал, тело сжималось с боков. Опыты были поставлены со
взрослыми аксолотлями, помещенными в довольно глубоком аква-
риуме, без всякой возможности для пих выбраться па сушу.
Опыты были начаты 28/1. Под влиянием тиреоидина животные
быстрее худели, вес их падал па одну треть первоначального,
кожа становилась гладкой и блестящей, на хвосте появились яркие
белые пятна. К 18/Ш жабры исчезли до размера небольших бугор-
ков, спинной плавник заменился продольной спинной впадиной,
глаза выпятились; получившихся амбистом можно уже было вы-
садить из аквариума в террариум с песком и мхом, и через педелю
жабры исчезли совершенно (рис. 297).
Получилось полное превращение аксолотля ь амбистому под
382
влиянием введения в организм тиреоидина. Повидимому, действие
последнего, т. е. щитовидной железы, у головастиков лягушек
прекращает метаморфоз, образуя неотенические формы. Возмож-
но, что те аксолотли, которые не превращаются в амбистом, обла-
дают недоразвитой щитовидной железой; это, повидимому, особая
раса, так как, на ряду с этими, есть в Мексике другие аксолотли,
нормально превращающиеся в амбистом. Изредка попадают они
и в Европу, давая те случаи самостоятельного обращения в ам-
бистому, которые иногда наблюдались в аквариумах. Но возмож-
но, что какие-либо факторы среды тормозят поступание гормона
щитовидой железы в кровь.
Превращение аксолотлей в амбистому можпо вызвать также со-
держанием их в растворе тиреоидина. Этот заставляет вспомнить
гипотезу Пюттера.
6. Приспособление амфибий к климатическим условиям
существования
Уже распространение амфибий показывает, что они являются
зависимыми от таких факторов среды, как температура и влаж-
ность. Хотя, вообще говоря, амфибии доходят до полярного круга,
а некоторые даже выходят за его пределы, тем не менее главная
масса видов и наибольшая плотность населения амфибий наблю-
даются в странах экваториальных с высокой температурой и боль-
шой влажностью. Леса тропической Америки, Африки и юго-
востока Азии наиболее богаты амфибиями. Здесь они проявляют
и наибольшую активность, что вполне согласуется с эксперимен-
тальными данными, показывающими, что при низкой температуре у
амфибий пе появляется нормальных рефлексов па стимулы среды.
Амфибии, паходящисся в воде, убиваются температурой в 40°
и даже несколько пиже. Но существуют виды, которые переносят
в виде головастиков (в горячих ключах Иеллоустопа) температуру
в 40—41°. Оптимальная температура различна для различных ви-
дов амфибий. Опа обычно соответствует температурным условиям
местообитания данного вида. Так, например, Rana pipiens и
Nocturne наилучше себя чувствуют, видимо, при температуре 18°.
Первый вид при температуре от 5 до 20° проявляет тем более актив-
ности и чаще показывается па поверхность, чем вода теплее. Го-
ловастики лягушки обычно собираются у краев водоемов, где
температура воды выше. С другой стороны, для-бссхвостой амфи-
бии рода Aacaphus, живущей в горах Сев. Америки, оптимум не-
много выше 0°. Наземные амфибии с влажной кожей выносят
высокие температуры лучше, чем водные, понижая свою температу-
ру испарением воды с поверхности кожи. Так, папример, древес-
ная лягушка (Hyla), сидя па листьях, выносит температуру почти
в 50°. Испаряющие меньше влаги жабы избегают солнца, прячутся
от пего.. При высокой температуре и большой сухости у многих
амфибпй наблюдается стадия покоя, так называемая «летняя спяч-
ка», когда амфибия, спрятавшись под камни, листья или закопав-
383
шись в ил, пережидает сухое время, имея крайне пониженный обмен
веществ.
Температура, как указано, повышает чувствительность к раз-
дражениям и быстроту реакции амфибий. Влияет она и на такие
процессы как кладка яиц.
К низким температурам амфибии весьма чувствительны и,
конечно, пе все виды одинаково. Эксперименты с охлаждением ам-
фибий и последующим их оживанием показали, что промерзание
всегда убивает лягушек и жаб. Минимальная температура, под-
вергнувшись действию которой безхвостые амфибии все же ожи-
вают, колеблется от —1.0° до —1.4° (травяная лягушка, Rana
temporaria). После охлаждения Ranapipiens до —1.0° лягушка эта
уже не ожила. Тритоны, обернутые в сухую вату, выдерживают
охлаждение до —1.5° и даже ниже. Молодые лягушки выдер-
живают большее охлаждение, нежели старые.
В связи с песпособностью выживать при низких температурах
амфибии па зиму уходят в места, защищающие их от холода. Так,
зеленая лягушка (Rana ridibunda) и съедобная лягушка (Rana
esculenta) прячутся с осени на дпе прудов и озер, где закапы-
ваются в ил, под упавшие в воду листья. То же делают и те ля-
гушки, которые живут летом на суше (травяная—Rana tempo-
raria и остромордая—R. terrestris). Жабы (Bufo) зарываются в зем-
лю, прячутся под корни деревьев и под камни или в трещины в ка-
менных стенах. В Ташкенте зеленая жаба (Bufo viridis) зимует
часто в подвалах под домами. В степях они зимуют в порах гры-
зунов. Хвостатые амфибии, как тритопы (Triturus), зимуют на
суше, зарываясь в мох или в щелях и норках в земле, под корнями
деревьев, под камнями. То же самое и саламандра (Salamandra
salamandra), иногда по пескольку экземпляров вместе.
Начинается состояние холодового оцепенения у лягушек и жаб
уже при температуре +7, +8°. Животные ищут убежище под кам-
нями или под другими подобными предметами, становятся непод-
вижными, закрывают глаза и перестают реагировать на внешние
воздействия.
Вполне нопятпо, что амфибии, имея голое тело, кожу, богатую
железами, чувствительны к изменениям влажности. Выбор ими
местообитания в широкой мере определяется влажностью. Пони-
жение последней вследствие испарения вызывает у них реакцию
закапывания. У си лспис испарения, благодаря ли повышению тем-
пературы или понижению влажности, или движению воздуха,
вызывает у амфибий защитную реакцию, и они погибают при этом
гораздо скорее, нежели другие животные из сухих местообита-
ний с малой влажностью воздуха. Опять-таки и по отношению к не-
достатку влажности не все амфибии одинаково чувствительны.
Так, папример, жабы с их сухой и более толстой кожей оказы-
ваются мепее чувствительными. Медленное высушивание оказы-
вается менее губительным для амфибий (лягушки, жабы, сала-
мандры), чем быстрое. Лягушка, высушиваемая быстро, гибнет
при потере 15% ее веса, тогда как при медленном высушивании она
384
может потерять в два раза больше. Жабы могут терять до 50% веса,
причем дальнейшая потеря приостанавливается благодаря увели-
чению вязкости крови.
Избегание амфибиями движений днем, максимум деятельности
ночью вызываются в значительной мере тем, что ночью влажность
воздуха бывает больше. Понятно, что дождь оказывает положи-
тельное стимулирующее действие на животных. В периоды засухи
амфибии нередко образуют скопления в более влажных местах.
Скопление, возможно, уменьшает потерю влаги животными, ко-
торые окружаются собственными испарениями. Сухость воздуха
является одним из факторов, определяющих отсутствие амфибий
в пустыне, кроме немногих видов, имеющих для того особые при-
способления.
Указанные выше отношения амфибий к температуре, требование
ими тепла, а также и влажпости воздуха являются отголоском
далекой эпохи, когда амфибии возникли па земле, и являются
опровержением распространенного мнения, что амфибии воз-
никли в пустынных условиях.
Чувствительность к потере влаги, голая, богатая железами
кожа делают понятным действие соли на амфибий. 1% раствор
хлористого натрия убивает и яйца и личинок амфибий. Этим объяс-
няется полное отсутствие амфибий в море, отсутствие их в соленых
озерах; в этом частично лежит и причина отсутствия амфибий в пу-
стынях. Впрочем, в пресных источниках, встречающихся в послед-
них, амфибии иногда живут. Губительно действует иа многих ам-
фибий и известь, растворенная в воде. Однако в некоторых случаях
амфибии встречаются в воде, протекающей в известковых породах,
например, протей (Proteus), живущий в водах известковых
пещер Крайни.
7. Роль амфибий в биоценозах и в круговороте веществ.
Экономическое значение амфибий
Будучи хищниками, питаясь другими животными, амфибии
играют известную, подчас довольно значительную роль в жизни
биоценозов и имеют некоторое значение и для хозяйства человека,
хотя в сравнении с рыбами роль их в этом отношении неизмеримо
меньше уже потому, что численно они несравненно'хуже представ-
лены в природе, гораздо менее подвижны. Будучи зоофагами,
они являются трансформаторами энергии, накопляемой расти-
тельноядными формами.
Наземные Anura, в особенности жабы, поедая массу беспозво-
ночных, в особенности червей и насекомых, являются весьма по-
лезными животными в садах, теплицах, возле парников, в огоро-
дах. Поэтому их надлежит здесь охранять и сюда завозить. Ля-
гушки, в огромном количестве населяющие рисовые поля средней
Азии, Китая, Японии, при их прожорливости, песомнепно должпы
оказывать здесь большое влияние на баланс природы. Они ночью
дополняют ту работу, которую днем производят птицы. С другой
25 Кашкаров и Ставчивский 385
стороны, некоторые амфибии, в особенности зеленые ля-
гушки (Rana csculenta), являются вредителями прудового
хозяйства. Их личинки, головастики, поедают большое количе-
ство корма, который мог бы быть использован рыбами, а взрослые
лягушки поедают и молодь рыб. С другой стороны, сами амфибии
и молодь их являются добычей и пищей многих животных. Они
со всех сторон окружены врагами. Прежде всего от других живот-
ных погибает икра амфибий. Ее поедают сами амфибии, а также
водяные птицы. Водяные клопы, личинки различных других на-
секомых, живущие в воде, водяные жуки, рыбы поедают личинок
амфибий и молодых лягушек. Сами амфибии нередко поедают мо-
лодь или особей другого вида или занимаются каннибализмом.
Головастики одного вида поедают головастиков другого при скоп-
лении их в период усыхания водоема. Одним из злейших врагов
лягушек и головастиков являются змеи. Ужи и змеи заглатывают
целых лягушек, поедают массу их личинок. То же, но в меньшей
мере делают и водяные черепахи, алигаторы. Цапли, аисты, не-
которые совы, вороны охотятся за амфибиями. Некоторые млеко-
питающие также охотятся за амфибиями.
Кроме того, амфибии подвержены инфекции со стороны пара-
зитов, многие из них производят большие опустошения. Таковы
некоторые грибы (Saprolegnia, например), бактерии, среди
которых числится и туберкулезная палочка; но особенно много
паразитов амфибий имеется среди простейших: Trypano-
soma, Opalina, Amoebae, Sporozoa являются самыми обычными
паразитами у хвостатых и бесхвостых амфибий.
Trypanosoma передаются пиявками. По распространению не-
которых паразитов можно судить о миграционных путях амфибий.
Так, род Zelleriella, живущий в различных родах лягушек, встре-
чается лишь в Австралии, в Южной и Центральной Америке,
указывая на давнюю связь этих материков между собой.
Наиболее характерными паразитами амфибий являются Spo-
rozoa. Круглые и плоские черви, нематоды, акаптоцефалы, тре-
матоды, цестоды и т. д. — дополняют список паразитов. Некото-
рые мухи также паразитируют па амфибиях.
Не отказывается от амфибий и человек, причиняющий глав-
ный вред амфибиям осушением болот. Уисе со средних веков ля-
гушачьи лапки стали попадать в Европе па стол под видом «пост-
ного мяса». Теперь их едят как деликатес, и в окрестностях боль-
ших городов количество лягушек даже значительно уменьшилось.
В 1905 г штаты, расположенные по Миссисипи, дали 71 990 кг
лягушачьих лапок. В США в год убивают около 100 млп. лягушек.
В одной только области озера Онейда за 9 месяцев продается ля-
гушачьих лапок на 1.7 млн. долларов при стоимости сотпи лапок
в один доллар, т. е. лягушек здесь было убито 85 000 000. Осо-
бенно ценятся крупные виды Rana pipiens, R. catesbyana, R. pa-
lustris. Для охраны лягушек существуют определенные законы
и сроки охоты. Делались попытки и «лягушкоразведения», но
без особого успеха, так что используются лишь дико живущие
386
лягушки. Существуют особые орудия лова их и правила веде-
ния хозяйства. Так, для успеха лягушачьего хозяйства необхо-
димо, чтобы, кроме болота, для лягушек были и более высокие
места выкорма. Шкурки жаб идут в Японии на выделку тонкой
кожи. Мясо исполинской саламандры (Cryptobranchus japonicus)
считается в Японии очень вкусным. Наконец, амфибии, в особен-
ности лягушки, служат объектом научных работ и обучения.
8. Поведение амфибий
Как мы видели на примере рыб, поведение животного находится
в зависимости от строения его нервной системы и органов чувств,
конечно, и от всей его организации.
Мы видели, что мозг амфибий, и прежде всего его полушария,
стоит па высокой ступени в сравнении с мозгом рыб. Парные полу-
шария построены здесь уже по тому основному плану, по которому
построен передний мозг у высших позвопочпых, хотя наружные
стенки их еще очень тонки и настоящей мозговой коры, как
у высших позвопочпых, здесь еще пет: клетки, соответствующие
пирамидным клеткам высших форм, еще не образуют замкнутого
слоя, как у рептилий и вышестоящих позвоночных.
Органы кожного чувства амфибий сходны с органами боковой
линии рыб. Они встречаются только у ведущих водный образ жизни
амфибий, прежде всего у личинок. У форм, периодически сходя-
щих в воду, например, у тритонов, органы кожного чувства перио-
дически регенерируют и подвергаются редукции. Всюду в коже
имеются осязательные нервные окончания. Вкусовые почки раз-
виты подобно тому, как у всех наземных форм, лишь в ротовой
полости. В органе обоняния имеются наружные и внутренние нозд-
ри. Глаза могут давать отчетливую картину окружающего мира.
Глаз установлен, как и у всех наземных животных, для зрения
вдаль; аккомодация на близкое расстояние происходит при помо-
щи оттягивания линзы ресничным мускулом. Орган слуха устроен
также очень просто. Имеются лишь лабиринт без улитки и сред-
нее ухо со слуховыми косточками. Наружное ухо отсутствует.
Исследования физиологии большого мозга у амфибий показали,
что он еще не играет той роли, которую играек большой мозг
у высших форм.
При осторожном удалении только большого мозга, без повре-
ждения thalamus opticus, животные легко переносят операцию
в течение многих месяцев. Они обнаруживают нормальное по-
ведение: плавают, при поворачивании легко удерживают равно-
весие, ловят мух, погружаются в зимнюю спячку, зарываясь
в землю, прыгая, обходят препятствия, самцы сохраняют сложный
рефлекс обхватывания самки в период спаривания и т. д. Произ-
вольные движения прекращаются в том случае, если повредить
thalamus opticus. Тогда все произвольные движения прекра-
щаются, но рефлекторные, в ответ на внешние раздражения,сохра-
няются. Животное сидит неподвижно, не ловит мух, но зрение
25* 387
у него, повидимому, сохраняется. Нарушения становятся еще
больше, если удалить и средний мозг, lobioptici. После удаления
всего головного мозга, за исключением продолговатого,наблюдается
стремление двигаться вперед: лягушки лишь тогда останавли-
ваются, когда натыкаются иа препятствия.
В соответствии с указанными чертами строения нервной си-
стемы и органов чувств стоит и деятельность, или поведение, ам-
фибий. Ощущения у них развиты равномерно: ни одно чувство
не превалирует. В поведении же преобладают не условные реф-
лексы, не опыт и научение, а врожденные виды поведе-
ния, безусловные рефлексы и инстинкты.
Осязательные ощущения у амфибий, несомненно, имеются.
Помимо окончаний, рассеянных в коже и концентрирующихся
иногда в особых сосочках, осязательным органом является осо-
бое щупальце у Apoda (Gymnophiona), щупальца личинок
шпорцевой лягушки (Xenopus). Так называемое «общее химическое
чувство» свойственно амфибиям в весьма выраженной форме, рас-
пределяясь по всей поверхности тела подобно тому, как это имеет
место у рыб. Растворы хлористого аммония, калия, патрия или
лития, вызывающие у человека вкусовые ощущения, у лягушек вы-
зывают отдергивание лапок. Соответствует ли «химическое чувство»
амфибий чувству вкуса у человека, сказать мы не можем, как не
можем вообще судить о субъективной стороне реакций животных,
о их качественной стороне. Кокаинизированные до исчезновения
боли лягушки все еще реагируют на эти раздражения.
Амфибии чувствительны и к изменениям температуры. Рецеп-
торы для восприятия тепла и холода отличаются как между собой,
так и от рецепторов осязательных, от рецепторов химического
чувства и болевых ощущений. Это установлено методом анесте-
зии при помощи 1% кокаина: через 10 минут исчезают реакции на
прикосновение, через 45 — на тепловые стимулы, тогда как реак-
ции иа химические раздражения (кислотой) и па уколы еще про-
должают сохраняться. Способность к различению тепловых ощу-
щений играет огромную роль в жизни амфибий. Различные виды
амфибий, например, пробуждаются от зимней спячки при различ-
ной температуре. Также охлаждение до известной степени яв-
ляется стимулом, заставляющим амфибий уходить на дно, пря-
таться от света, зарываться в ил.
Обоняние, несомненно, играет известную роль в поведении ам-
фибий. Оно является весьма существенным моментом при оты-
скании пищи, например, тритонами. При этом, па ряду с обонянием
и зрением, играют роль еще «ощущения сотрясения» и зрение.
Если же добыча неподвижна, то «ощущения сотрясения» не играют
роли, и тритоны замечают добычу уже на некотором расстоянии
при помощи обоняния. При перерезке обонятельного нерва или
при закрывания ноздрей коллодием тритоны пищи не находили.
После снятия коллодиальной пленки реакция возобновлялась.
Вынутый из воды тритон судорожными сокращениями глотки
выталкивает изо рта и ноздрей воду и может теперь различать
388
газообразно пахнущие вещества. Даже ослепленные трптопы уже
на расстоянии находят кусочки дождевого червя, комочки ваты,
смоченные соком из червей, льняной мешочек с червями. Пере-
резка обонятельных нервов останавливает реакцию. Таким обра-
зом, тритон может обонять и в воде и на суше.
Органы вкуса, как выше сказано, у амфибий ограничиваются
ртом.
Развитие барабанной перепонки, columella, лабиринта гово-
рит о развитии слуха. Но между хвостатыми и бесхвостыми
имеются в этом отношении различия. У хвостатых орган слуха
близок к тому, что имеется у рыб. То же самое — у личинок бес-
хвостых; у них не найдено никаких реакций на звук. Другое дело
у бесхвостых. За наличие у них слуха говорит уже наличие голоса
у самцов многих лягушек и реакции самки на голос самца. Это
наблюдалось и у жаб, и у лягушек весной: когда самец издает
звук, самка приходит в движение л направляется к поющему
самцу. *
Некоторые эксперименты говорят как будто за то, что лягушки
не слышат, так как не реагируют па звук, ио это потому, что неко-
торые звуки для пих биологически безразличны, не имеют зна-
чения. С другой стороны, наблюдения за лягушками на воле по-
казали, что один только звук — звук шлепанья, вызывает реакцию,
если даже совершенно исключить в опыте зрение. Но ни на какие
другие звуки реакции не было, пока субъект, производящий звук,
оставался невидимым. Звук здесь, видимо, служит сигналом пре-
дупреждения, сигналом же к бегству в воду — зрительный сти-
мул. Когда одна лягушка начинает квакать, другие к пей присоеди-
няются. Замолкают они также одновременно. Веспой реакция ока-
зывалась более сильной. В этом, конечно, есть биологический
смысл. Подражая «пению» квакш, можно заставить их «петь».
Наконец, звук вызывает усиление различных реакций, вызывае-
мых другими стимулами. Звук «настораживает» лягушку.
О зрении амфибий следует прежде всего сказать следующее.
Кожа их, подобно коже многих рыб, чувствительна к свету. Ли-
чинки Rana clamitans, содержавшиеся в темноте и выставленные
затем на свет, начинали быстро плавать. Лягушки и жабы обна-
руживают склонность к движению па свет при высокой температу-
ре, а при понижении ее — к движению от источника £вета. Этот
фототропизм, положительный при высокой температуре и отри-
цательный при низкой, имеет биологический смысл, связан с яв-
лениями пробуждения от зимнего спа весной и с уходом в спячку
осенью.
Что касается зрения собственно, то прежде всего граница спек-
тра у разных видов лягушек, совпадает с таковой человека. Глаз
лягушки настолько совершенен анатомически, что должен да-
вать четкую картину окружающего.
Но опыты этого не подтверждают. По крайне мере в нормаль-
ной обстановке амфибии реагируют только на движение предметов,
но не на их вид. В этом тоже, может быть, имеется известный био-
389
логический смысл: дальше мы увидим, что память у амфибий очень
слаба и пользоваться образом предмета они все равно не смог-
ли бы. Движущийся же предмет в обстановке жизни лягушки —
это всегда почти что-либо съедобное: насекомое, червь и т. п.
Поэтому отбором и выработалась реакция на движение. Вполне
удовлетворительных опытов с реакциями на различную длину
волн, т. е. на цветное зрение, поставлено не было. Но имеющиеся
данные заставляют думать, что амфибии цвета различают.
Выше было указано, что вся почти деятельность амфибий
состоит из рефлексов и инстинктов, т. е. сочетанных, координи-
рованных в цепь рефлексов, при которых стимул вызывает врож-
денную реакцию, эта последняя — другую и т. д., пока не будет
достигнут необходимый результат. Лучше всего виден основной
характер поведения амфибий в акте схватывания добычи. Лягушка
схватывает только движущийся предмет. Неподвижная добыча
(насекомое, например) не является раздражением, вызывающим
акт схватывания. Лягушка умрет с голоду скорее, чем схватит
неподвижную пищу. Обычно движение мухи или червя вызывает
фиксацию взгляда, следующее движение — поворачивание к дви-
жущемуся предмету, затем уже дальнейшее движение последнего
вызывает выбрасывание языка и схватывание. Если муха, при-
шпиленная на булавке, находится перед лягушкой в покое, ля-
гушка не обращает на нее никакого внимания. Лишь только
муха начинает шевелиться, лягушка выбрасывает язык, бьет им
по мухе; но так как последняя прикреплена, то язык возвращается
в рот без мухи. Все же за этим следует акт глотания с закрыва-
нием глаз. Если теперь муха снова шевелится, то следует тот же
самый акт мнимого схватывания и глотания. Весь этот акт есть
цепь безусловных рефлексов, составляющая основу инстинкта.
Питание, ухаживание за самкой, зарывание па зиму в ил или
в землю в сухое время, заботы о потомстве — все это более или
менее сложные инстинкты. К инстинктивному поведению следует
отнести и так называемые миграции переселения и «возвращение
домой», наблюдающееся у многих амфибий. Жабы, например, си-
дящие днем в норах, а ночью выходящие на охоту, причем уда-
ляются от норы довольно далеко, вновь возвращаются в свое убе-
жище. Hyla rosenbergi в Панаме возвращается каждую ночь к по-
строенным ею для обеспечения потомства бассейнам из ила.
Меньше выражен этот инстинкт возвращения домой у саламандр,
но у лягушек и жаб он бывает развит весьма сильно. Были опыты:
лягушки, перенесенные из одного водоема в другой, возвращались
обратно с далекого расстояния.
Ассоциативная память, способность к научению у амфибий
вряд ли выше, чем у рыб, хотя все же амфибии способны паучаться.
При этом наземные научаются быстрее водных. При содержании
жаб в неволе можно убедиться в том, что они научаются узнавать
кормящего, во всяком случае пе персонально. Имеются экспе-
рименты, в которых Rana после 4—7 экспериментов отказывалась
брать мохнатых гусениц, и опыт свой сохраняла в течение 10 дней.
390
Схватывая движущуюся бумагу, лягушка, однако, скоро научается
узнавать ее. Опыты в лабиринте, где лягушка должна была избрать
путь направо или налево от перегородки, дали положительный
результат лишь после 100—120 опытов. После этого лягушка не
ошибалась и выходила из лабиринта немногими короткими прыж-
ками. Окрашивание одной стороны в белый, другой — в красный
цвет ускоряло научение. Если после образования данной привычки
окраска изменялась на обратную, лягушка сбивалась, как будто
ничему не научилась. Научение это чисто физиологического, а не
психологического характера. Видимо вся жизнь амфибий состоит
из рефлексов и инстинктов.
9. Географическое распространение амфибий
Современные амфибии являются обломком некогда широко рас-
пространенной, цветущей ветви животных. Распрострапение ка-
кой-либо группы животных определяется ее возрастом, центром
ее возникновения, способами расселения, доступными данной
группе, ее экологическими требованиями и наличием тех или иных
барьеров на пути ее распространения, а также всеми теми геоло-
гическими, климатическими и прочими изменениями, которые
имели место на протяжении истории данной группы. Понять рас-
пространение какой-либо группы животных легче, если имеются
свидетельства палеонтологии, говорящие о распространении груп-
пы в прошлом. К сожалению, в отношении амфибий палеонтоло-
гия говорит об этом нам мало. Но в отношении некоторых семейств
мы на основании палеонтологических свидетельств знаем о их
некогда гораздо более широком, чем теперь, распространении.
Но и сейчас амфибии распространены достаточно широко;
они встречаются всюду, где есть насекомые в течение достаточно
продолжительного времени, проникают до полярного круга и даже
переходят за него.
Амфибии не могут процветать в районах вечной мерзлоты:
всего два вида Rana живут к северу от полярного круга в Норве-
гии и в РСФСР и одна саламандра живет в арктической Азии.
Большинство амфибий в пределах своего распространения зани-
мают лишь такие местообитания, которые напоминают условия,
в которых когда-то возник этот класс и в которых жили стегоце-
фалы и их предки из класса рыб. Преспая вода и тепло им необхо-
димы. Без воды пресной и по возможности спокойной, стоячей,
пе может жить почти ни одна амфибия. Что касается тепла, то,
хотя есть амфибии, живущие далеко к северу и к югу от экватора,
все же большая часть их живет ближе к последнему. Другим огра-
ничивающим распространение амфибий фактором является ка-
чество воды. Последняя должна быть пресной. Соль является
ядом для амфибий. Даже однопроцентный раствор соли мешает
развитию их личинок. А потому моря, соленые озера и солонова-
тые почвы являются непреодолимыми препятствиями для рас-
пространения амфибий. По морю амфибии могут путешествовать
391
лить на упавших стволах деревьев. Известь также не переносится
амфибиями. Сплошь и рядом в районе известковых пород нет той
или иной амфибии, а рядом они встречаются на песчаниках и
гранитах. В жесткой воде личинки не могут расти правильно. Лишь
протей составляет исключение, живя в известковых пещерах.
Большое значение имеет наличие соответственных мест для
размножения. Руководимые главным образом инстинктами, ам-
фибии в значительной мере являются рабами среды, не будучи
в состоянии выявить инстинкт размножения, если среда отли-
чается от нормальной для данного вида. Те хвостатые или бесхво-
стые амфибии, которые откладывают яйца, например, возле гор-
ных потоков, не уходят далеко от последних и в иной обстановке
размножаться не будут. Кроме того, особенности среды и способ
размножения должны находиться в соответствии: амфибия с мед-
ленным циклом развития не сможет жить там, где водоемы суще-
ствуют периодически и на короткое время. Таких экологи-
ческих барьеров для распространения амфибий может быть
много. Распространение может итти только по «экологиче-
скому руслу».
Горы играют меньшую роль в распространении амфибий, не
являясь непреодолимым препятствием, а скорее содействуя распро-
странению. Так, в некоторых тропических странах фауна земно-
водных па горах такова же, как в более северных областях. Sa-
lamandra atra идет до 2 600 м высоты, Bufo viridis поднимается
еще выше. Само собой разумеется, что древесные формы, вроде
квакш, связаны в своем распространении с лесами и наиболее мно-
гочисленными являются в тропиках; понятно, что пустыни
почти совершенно лишены амфибий. Только в пустынях Австра-
лии найдены амфибии со специальными приспособлениями, даю-
щими им возможность уйти от гибельных для них условий пу-
стыни. В пустынях Средней Азии они встречаются в колодцах
и ключах.
В общем амфибии распространены подобно пресноводным ры-
бам. Это до некоторой степени и попятно, так как амфибии также
связаны с пресной водой. Кроме того, эволюционное дерево ам-
фибий, хотя и моложе эволюционного дерева рыб, но не в такой
мере, как эволюционное дерево птиц или современных млеко-
питающих.
По распространению различных групп амфибий поверхность
земли так же, как и по распространению пресноводных рыб, можно
разделить на северную зону— Арктогею и южную — Эогею.
Северная зона охватывает области Палеарктическую и Неаркти-
ческую; зона южная, или экваториальная, включает в себя обла-
сти Африканскую, или Эфиопскую, Индийскую, или Восточную,
объединяемые в деление афроиндийское и области Южноамери-
канскую, или Неотропическую, и Австралийскую, объединяемые
в деление южноамериканско-австралийское.
Северная зона (Европа, Северная Азия и умеренная Сев. Аме-
рика) характеризуются обилием Urodela и отсутствием Apoda.
392
Южная зона, наоборот, характеризуется присутствием в ней
Apoda и отсутствием Urodela, хотя Apoda встречаются лишь в тро-
пических частях Африки, Азии и Америки, а внетропические части
южной зоны (юг Африки, Австралия, Новая Зеландия, Австра-
лия и юг Америки) имеют из амфибий лишь Anura. Почему-то нет
Apoda на Мадагаскаре и в Папуасии.
Области южной зоны-Эфиопская и Индийская —характери-
зуются, как и вся зона, почти полным отсутствием Urodela; далее,
присутствием Apoda, а из Anura — преобладанием более высоко
организованных семейств с неподвижной грудиной (Firmisternia,
Diplasiocoela). Лишь немногие из более примитивных Anura при-
сутствуют здесь; например, род Xenopus из сем. Pipidae (Opis-
thocoela) — в Африке.
Южноамериканско-австралийское деление южной зоны ха-
рактеризуется, кроме имеющих зубы жаб, объединявшихся рань-
ше в семейство Cystignathidae, еще преобладанием подвижно-
грудых, Arcifera, составляющих здесь 90% Anura. В австралий-
ской области Apoda и Urodela совершенно отсутствуют. Из Fir-
misternia в Австралии только один вид рода Rana. Из островов,
окружающих Австралию, только те населены амфибиями, которые
имеют континентальное происхождение. На океанических остро-
вах амфибий нет. Область Неотропическая, или Южноамерикан-
ская, отличается от Австралийской присутствием Apoda.
Из Urodela здесь есть роды: Spelerpes и Plethodon, распро-
странившиеся сюда из Сев. Америки.
Немногие виды из семейства Ranidae пришли сюда с севера,
как и в Африку и на Мадагаскар, развившись здесь и там под
влиянием жизни в тропическом лесу в особое подсемейство
Dendrobatinae. Имеется в Южпой Америке и представитель груп-
пы Opistocoela — именно род Pipa.
Как в Южной Америке, так и в Австралии представители се-
мейств Bufonidae и Hylidae являются доминирующими. Bufoni-
dae Австралии сходны с африканскими и южноамериканскими. Род
Hyla еще в миоцене находился в Европе. Является ли распростра-
нение Bufonidae и Hylidae указанием па бывшую связь южных
материков или эти семейства возникли на севере и распростра-
нились па юг по существующим ныне материкам? Вероятнее
первое предположение. Оно согласуется с тем, что мы знаем о рас-
пространении рыб в Эогее.
Северная зона, как сказано, является областью распространения
Urodela. Но опа пе однородна. Между Палеарктикой и Неарктикой
существуют значительные отличия и сходство. Между Восточной
Палеарктикой и Неарктикой имеется сходство в присутствии тут
и там семейства Amphiumidae и рода Ambystoma, тогда как под-
семейство Salamandridae характерно для Европы. Сходство Вос-
точной Палеарктики и Неарктики, невидимому, объясняется не-
давней еще сухопутной связью Востока Азии и Сев. Америки.
Связь осуществлялась между Европой и Америкой через Грен-
ландию. Климат здесь был в это в'ремя мягкий и теплый. Нали-
393
чием этой бывшей связи объясняется присутствие в Европе рода
Proteus, в Сев. Америке — рода Necturus, оба одного семейства —
Proteidae. Эта же связь дала возможность одному виду рода Hydro-
mantes из Plethodontidae жить в Калифорнии, другому — в юго-
восточной Франции и северо-западной Италии, третьему — на
острове Сардинии. Связь же восточной Азии и западных штатов
Сев. Америки осуществлялась на месте Берингова пролива. Через
эту связь из Азии проникли Cryptobranchus, Ascaphus, Brevici-
pitidae.
Только.в Палеарктике и на прилежащих островах распростра-
нены Hynobiidae.
Ископаемые стегоцефалы известны из Европы, со Шпицбер-
гена, из Сев. Америки и Южной Африки. Несомненно, что ныне
спорадически распространенные группы были распространены
шире.
ЧАСТЬ ЧЕТВЕРТАЯ
РЕПТИЛИИ, ИЛИ ПРЕСМЫКАЮЩИЕСЯ
Характеристика высших позвоночных (Amniota)
Общий обзор. Пресмыкающиеся, птицы и млекопитающие со-
ставляют группу высших позвоночных животных,
отличающихся развитием зародыша: он снабжен особыми оболоч-
ками — амнион о-ми аллантоисом, откуда эта группа
животных и получила название амниот — Amniota. Осо-
бенность эта возникла в связи с приспособлением к воздушно-на-
земному образу жизни и связана с откладыванием яиц, в которых
развивается зародыш на суше. Уже среди современных амфибий мы
встречаем разнообразные приспособления, направленные к осво-
бождению от воды, к которой амфибии привязаны были развитием
их яиц. У ископаемых амфибий, стегоцефал, эти приспособления
были, вероятно, еще более разнообразными.
Наземно-воздушная среда по сравнению с водной средой пред-
ставляла в камеппоугольпое время громадные преимущества:
она была уже покрыта богатой, хотя и довольно однообразной ра-
стительностью; в ней развивалась своеобразная фауна беспозво-
ночных — черви, моллюски, многоножки, первичпотрахейпые и
низшие насекомые, представлявшие обильную и легко добываемую
пищу, тем более что эти животные еще пе выработали в борьбе за су-
ществование защитных приспособлений от врагов из позвоночных,
которых у пих тогда еще пе было. С другой стороны, па земле со-
вершенно отсутствовали хищники и конкуренты, тогда как в воде
шла обостренпейшая борьба за существование. Врагами амфибий
были там не только разнообразные рыбы, по и сами опи и бесчис-
ленные беспозвоночные, которые питались их икрой. Борьба за су-
ществование в водных бассейнах каменноугольного времени неиз-
бежно приводила прежде всего к отбору всех тех приспособлений,
которые позволяли предкам рептилий дольше держаться на суше
и занимать новые, еще незанятые обширные пространства суши.
Из числа других многочисленных приспособлений к наземно-воз-
душпой жизни, в частности, к развитию без воды, несомненно
чрезвычайно удачным было возникновение особых приспособлений
в яйце и в развитии зародыша сказавшимся в эволюционном процессе
образованием обширнейшей, наиболее высоко организованной груп-
пы высших позвоночных животных. Именно эти приспособления
в яйце и у зародыша дали возможность предкам высших животных
сделаться в полном смысле слова наземно-воздушными животными
и расселиться в самых разнообразных по условиям местах суши.
395
Строение яйца, развитие зародыша и зародышевые оболоч-
ки высших позвоночных. Яйцо рептилий, птиц и яйцекладущих
низших млекопитающих представляет по сравнению с амфибиями ту
замечательную особенность, что зародыш, не проходя стадии личин-
Рис. 298. Схема курино-
го яйца.
1 — известковая скорлупа,
2 — кожистая оболочка, 3—
оболочка желтка (клеточ-
ная), 4—зародышевый диск,
5 — халазы, 6 — воздушная
камера. (По О. Levy.)
ки, проделывает в нем все свое развитие, бу-
дучи снабжен необходимым для этого пита-
тельным материалом в самом яйце (желток)
и в его оболочках (белок) и будучи заклю-
ченным в жидкую среду, заменяющую собой
водную среду амфибий (амниотическая жид-
кость).
Само яйцо отличается огромной ве-
личиной, напоминая этим яйцо селахий.
Оно является меробластическим, так как
содержит большой запас желтка и подвер-
гается дроблению только в бедной желтком
анимальной части, причем дробление
это носит дискоидальный харак-
тер. Яйцо одето тонкой первичной оболоч-
кой и подвергается оплодотворению в верх-
нем отделе яйцевода. Опускаясь по яйце-
воду к выходу, яйцо одевается последо-
вательно белковой оболочкой, выделяемой
стенками яйцевода, и, наконец, у выхода — скорлупой, кожистой
у рептилий и пропитанной известью у птиц. В таком яйце, отло-
женном на земле, в воздушной среде, при соответствующей темпе-
ратуре, зародыш развивается за счет того питательного материала,
который заключается в желтке и белке. У рептилий для развития
яйца оказывается достаточной температура внешней среды, у птиц
(рис. 298) и низших млекопитающих яйца подвергаются согрева-
нию телом родителей.
Рис. 299. Срединный разрез черев бластодерму ящерицы.
1 —эктодерма, 2 — бластоцель, 3 —место начала образования первичной кишки, 4—
желточный слой, 5—дополнительная полость, 6—желточный синцитий. (Из Bonnet.)
Как уже было упомянуто, дробление яйца протекает лишь па апи-
мальном полюсе, где в результате деления клеток образуется заро-
дышевый диск, а под ним—щелевидная сегментацирп-
ная полость, подостланная снизу нераздробившейся частью
желтка и отдельными ядрами и обособившимися вокруг них
клетками — мероцитами (рис. 299).
396
Желток не дробится, оставаясь совершенно пассивным. Будущий
зародыш представлен только зародышевым диском. Благодаря
этому обстоятельству процесс гаструляции крайне затемнен, почти
до неузнаваемости. Он становится понятнее, если сравнить проис-
ходящие здесь процессы с тем, что нам известно уже о гаструляции
и образовании мезодермы у Anura.
У лягушки с самого начала развивался тот участок энтодермы,
который образует стенку первичной кишки, и вслед за этим жел-
точный участок энтодермы, выстилающий дополнительную полость.
У амниот образование впячивания первичпой кишки запаздывает,
а потому сперва образуется выстилающая
дополнительную полость и состоящая из
желточных клеток часть энтодермы. Но
затем, так же как у амфибий, первичная
кишка прорывается в дополнительную по-
лость, и образуется примитивная полость
кишки.
Происходит это таким образом (рис.
299, 300, 301). В задней части среднего
отдела зародышевого диска, т. е. в задней
части так называемого зародышевого щита,
на утолщенной первичной пластин-
к е образуется продольная первичная
бороздка. На переднем конце этой борозд-
ки происходит образование энтодермы
путем инвагинации, — поэтому бороздку
приходится считать соответствующей бла-
стопору амфибий. Впячивание увеличивается, первично-кишеч-
ный канал расширяется и доходит до желточного слоя энтодермы,
отделяющего первично-кишечный капал от дополнительной поло-
сти. Затем происходит соединение этих двух полостей, образует-
ся примитивная кишка.
Мезодерма амниот образуется, как и у амфибий, из энтодермы,
начиная от области первичного рта (рис. 302), в виде полос, идущих
в стороны до края щита. Полость в этих клеточных лентах обра-
зуется позднее путем расхождения клеток (рис. 304).
Под первичной пластинкой закладывается из энтодермы про-
дольный зачаток хорды, по бокам которой лежат мезодермальные
складки. Мезодерма разрастается, распадаясь затем в верхней спин-
ной части на сомит ы, а с брюшпой стороны образуя несегмен-
тироваппые боковые пластинки. Одновременно происхо-
дит впячивание внутрь желобка нервной пластинки, края которой
потом замыкаются, образуя нервную трубку (рис. 303 и 304).
Зародыш, расположенный вдоль средней части зародышевого
диска, теперь начинает формироваться, обособляясь от лежащего
под ним желточного мешка, который, в конце концов, оказывается
подвешенным к зародышу посредством тонкого стебелька, причем все
три зародышевых листка располагаются по его поверхности. Одно-
временно происходит образование зародышевых оболочек (рис. 305).
397
Первой образуется амнион. Зародыш как бы погружается
в желточную массу; пад ним образуется кольцом охватывающая его
складка, образованная всеми зародышевыми листками и лежащей
Рис. 301. Л, В, С—срединный разрез через щит ящерицы
в разных стадиях гаструляции.
1—первичный рот, 2— первичная кишка, 3—желточный слой,
4 — эктобласт.
D — схема поперечного среза Amniota.
1 — зачаток хорды, 2 — полость первичной кишки, 3 —щит,
4 — эктобласт, 5 — дополнительная полость, 6 — мезобласт,
7 — энтобласт.
между ним и мезодермой вторичной полостью. По мере опуска-
ния зародыша в желточную массу и большего замыкания кольце-
вой складки над зародышем полость эта увеличивается, разъеди-
398
няя обращенные друг к другу слои мезодермы. Наконец, кольцевая
складка совершенно замыкается сверху над зародышем, причем экто-
дерма и мезодерма в верхней части складки и внутренней части
в
Рис. 302. А — поперечный разрез области развития головы у Ghrysemys.
В — разрез через область соприкосновения цефалогенной и нотогенной
зоны у Ghrysemys.
А. 1— эктодерма, 2— боковая ограничивающая складка, 3^ зачаток хорды, 4—кишеч-
ный гинобласт, 5—мезобласт, 6—желточный гипобласт.
В. 1— эктодерма, 2 — зачаток хорды, 3 — гипобласт, 4 — мезобласт,’’ 5 — желточный
гипобласт. (Оба рисунка по Brachet.)
Рис. 303. Поперечный разрез через
область развития хорды зародыша
Ghrysemys.
1—нервный желобок, 2—эктодерма, 3—
желточный гипобласт, 4 — хорда, 5 •— ме-
зобласт. (По Brachet.)
Рис. 304. Поперечный разрез через
зародыша Ghrysemys в месте перехо-
да головы в туловище.
1—спинной мозг, 2 —эктодерма, 3 —
амниотическая складка, 4 — первый мезо-
бластический сомит, 5—гипобласт, 6 —
мезобласт. (По Brachet.)
складки соответственно, срастаются, так что образуются две обо-
лочки: одна—внутренняя, с эктодермой, обращенной внутрь, и ме-
зодермой наружу— амнион; другая — наружная, с эктодермой
399
наружу, образует серозную оболочку. Эта последняя по
мере обрастания желтка листками и расщепления мезодермы раз-
растается но желтку все дальше и дальше и, наконец, одевает все
яйцо. Желточный мешок, подвешенный к зародышевому на тонком
стебельке, оказывается внутри серозной оболечки и одет снаружи
мезодермой и эктодермой снутри.
Между стебельком желточного мешка и задней частью зародыша
образуется из стенки вырост энтодермы с мезодермой соответственно
мочевому пузырю зародыша амфибий. Вырост этот (аллантоис)
однако, быстро разрастается под серозной оболочкой, распростра-
няясь и над амнионом, и над желточным мешком. Он не только слу-
жит резервуаром для жидких выделений, но и органом дыхания
зародыша, в связи с чем на его наружной поверхности развивается
Рис. 305. Схема поперечного раврева черев только образующиеся
амниотические складки Sauropsida.
А. 1—зародыш, 2—экзоцель, 3 —эктобласт, 4 — париетальный мезобласт,
5 — висцеральный мезобласт, 6 — желточный листок, 7 — желток, 8 — яй-
цевая оболочка. (По Bonnet.)
В. 1 —экаоцель, 2—париетальный мезобласт, 3 — висцеральный мезобласт,
4—желток, 5—яйцевая оболочка, 6—сходящиеся амниотические складки.
(По Bonnet.)
сеть кровеносных сосудов. В полостях между оболочками находятся
жидкости, а снаружи оболочек над скорлупой находится также
и воздух, которым и дышит зародыш.
Устроенное таким образом яйцо, предоставляя возможность за-
родышу развиваться без воды в жидкостях, заключенных в яичную
скорлупу, явилось тем исключительно широкого значения приспо-
соблением, которое позволило рептилиям быстро распространяться
по всей суше в пермское и последующее мезозойское время и, при-
способляясь затем к различным особеппостям наземной среды оби-
тания, эволюционировать дальше. Мало того, рептилии, как это
мы сейчас увидим, сделавшись типичными наземными обитателями,
затем вторично неоднократно приспосабливались к водной стихии,
образуя здесь на новой основе такие чисто водные формы, как ихтио-
завры или плезиозавры. На основе этого же строения яйца и приоб-
400
ретепия теплокровности и приспособления к полету с помощью кры-
льев, построенных из перьев, развились и эволюционировали дальше
птицы; млекопитающие возникли па этом же основапии: первично
яйцекладущие, они только позднее изменили в длинном эволюцион-
ном процессе этот способ развития на живородность с утробным раз’
витием зародыша за счет питания кровью матери с помощью пла-
центы, которая образуется из аллантоиса, срастающегося с сероз-
ной оболочкой; с помощью многочисленных сосочков, врастающих
в слизистую оболочку матки, аллантоис вместе с серозной оболочкой
вступает в соединение с кровеносной системой матери, которая и до-
ставляет необходимый для питания зародыша материал. Соответ-
ственно с этим яйцо высших млекопитающих теряет свои запасы
питательного вещества, и это, в свою очередь, ведет к некоторым осо-
бенностям дробления яйца и развития зародыша.
Высшие позвоночные животные составляют три класса: класс пре-
смыкающихся, или рептилий — It е р t i 1 i а, класс птиц —
A v e s и класс млекопитающих — M a in in a 1 i а. Родоначаль-
никами двух последних классов являются рептилии.
Класс 5. Рептилии, или пресмыкающиеся (Reptilia)
I. ОБЩАЯ ХАРАКТЕРИСТИКА И ОБЗОР СОВРЕМЕННЫХ И ИСКО-
ПАЕМЫХ ФОРМ КЛАССА РЕПТИЛИЙ
Одной из характернейших особеппостей амфибий, хотя и уте-
рянной некоторыми представителями этого класса, является, как
мы видели, наличие у них четырех пятипалых конечностей,— осо-
бенность, связанная с наземным или полуназемпым образом жизни.
Особенность эта сохраняется и у рептилий, так же как у птиц и мле-
копитающих, хотя и среди рептилий имеются формы, совершенно
утерявшие свои конечности. Конечности типа руки характеризуют
вместе с легочным дыханием лишь первую стадию адаптации к на-
земно-воздушной среде. Яйцо со скорлупой с развивающимся в нем
зародышем, снабженным амниопом и аллантоисом, составляет сле-
дующую стадию перехода к наземно-воздушному образу жизни.
Наличие яйца со скорлупой позволяло рептилиям распростра-
няться по суше вне зависимости от наличия водоемов, бывших ра-
нее столь необходимыми для развития яиц и для личипок. Для широ-
кого распространения по суше и для полного приспособления к на-
земно-воздушному обитанию необходимо было еще освободиться
от кожного дыхания, требующего влажного состояния кожи и влаж-
ной воздушной среды, а это могло быть достигнуто лишц после усо-
вершенствования легочного дыхания и улучшения системы кровооб-
ращения.
Только сухая кожа, защищающая тело от высыхания роговым
покровом из чешуй или щитков, вместе с усовершенствованием
легочного дыхания и кровеносной системы дали возможность реп-
тилиям сделаться настоящими наземпо-воздушпыми обитателями
и эволюционировать дальше, образовав обширный богатый формами
ICniiucapoD и Стаичинский 401
класс высших позвоночных, и дать в процессе эволюции классы
птиц и млекопитающих.
Из других особенностей, характерных для класса рептилий, на-
зовем: наличие одного, а не двух сочленовных бугорков в черепе,
наличие квадратной кости, к которой причленяется сложная нижняя
челюсть, состоящая из сочленовной и ряда покровных костей;
сердце у рептилий или трехкамерное с неполной перегородкой в же-
лудочке или четырехкамерное (у крокодилов); артериальный ствол
совершенно разделен на легочные артерии и дуги аорты, ко-
торых две; функционирующей почкой является вторичная почка
(metanephros), которая снабжена самостоятельным выводным про-
током — мочеточником (ureter), тогда как первичнопочечный про-
ток (вольфов проток) является семяпроводом (vas deferens); яйце-
воды (мюллеровы протоки) в связи с яйцеродностью дифференци-
рованы на отделы, выделяющие белок и кожистую скорлупу.
Рептилии являются родоначальниками высших позвоночных жи-
вотных. На ряду с особенностями, общими с птицами и млекопитаю-
щими, каковыми являются развитие зародыша при наличии амниона
и аллантоиса, начало полного отделения артериальной и венозной
крови в желудочке путем образования перегородки, наличие функ-
ционирующей вторичной почки и отдельных от семяпровода моче-
точников, рептилии обладают рядом свойств, общих им и низшим
позвоночными, отличающих их от птиц и млекопитающих. Такова
прежде всего переменная температура их тела, или их холоднокров-
ность (т. е. температура их тела меняется в зависимости от темпе-
ратуры внешней среды); таковы наличие двух дуг аорты и относи-
тельно примитивное строение легких; таково, наконец, отсутствие
выкармливания и вождения детенышей.
Появившись в конце каменноугольного периода в качестве един-
ственных чисто воздушно-наземных животных, рептилии первона-
чально не имели па суше пи врагов, ни конкурентов. Этим в значи-
тельной мере объясняется быстрая эволюция их в разных направ-
лениях применительно к различным средам обитания, к различным
способам передвижения и к различной пище. Уже в пижнепермское
время жили представители разных групп рептилий — это котило-
завры (Cotylosauria), пеликозавры (Pelycosauria), мезозавры (Meso-
sauria) и дальние предки чешуйчатых (змей и ящериц) — ареосце-
лиды (Areoscelida), причем среди них были пе только наземные,
но также и вторичноводные формы. Однако паивысшего расцвета
достигает класс рептилий в середине и в конце мезозоя — в юре и
в нижнем мелу. В это время он представлен был громадным числом
разнообразнейших форм, из которых мы знаем очепь многие. Одпа-
ко последние составляют лишь незначительную часть рептилий,
живших тогда. Все стихии были заселены ими: вода и суша, от-
крытые пространства и леса; по воздуху летали вместо современных
птиц птерозавры. На ряду с большим разнообразием растительнояд-
ных форм было большое число хищных животных. Одни из них
были мелкие, с современных ящериц, другие достигали громадной
величины, превосходя всех современных нам гигантских млекопи-
402
тающих. Зато в конце мелового периода рептилии начинают быстро
вымирать, и к началу третичной эры они оказываются предста-
вленными лишь ограниченным числом форм, определяющих совре-
менную нам фауну. Их место заняли птицы и млекопитающие.
Правильное представление о рептилиях как о некогда огромном
основном классе воздушно-наземных позвоночных животных может
быть получено, естественно, лишь путем одновременного изучения
как современных, так и ископаемых форм.Однако изучение иско-
паемых рептилий представляет исключительные трудности: реп-
тилии, как мы видели, в основном были животными наземными,
только вторично они сделались водными обитателями. Между тем
среди ископаемых форм преобладают именно водные рептилии,
так как их остатки по сравнению с наземными формами со-
храняются гораздо легче. Это затрудняет построение родословного
дерева рептилий.
Родоначальными рептилиями были котилозавры (Cotylosauria),
обильная формами и довольно разнообразная группа древнейших
рептилий из пермокарбона и пермских отложений, по строению
своего стегального черепа составляющая особую группу (Anapsida),
близких к стегоцефалам форм. Это были или тяжеловесные, непо-
воротливые, медленно двигавшиеся формы — Pareiasauriadae или
более легкие саламандроподобные формы полуводного образа
жизни — Labidosauria или, наконец, похожие на ящериц на-
земные, более подвижные животные, выделенные в отряд Sey-
mouriamorpha, по общему виду, конечностям и черепу близ-
кие к стегоцефалам.
Древнейшие из до сих пор известных рептилий отряда Rhizo-
sauria связывают Cotylosauria с эмболомерными амфибиями —
стегоцефалами.
Очень древними, тоже пермскими, рептилиями являются пели-
козавры (Pely^osauria), дожившие лишь до конца триаса, вероятно,
потомки котилозавров, отличавшиеся от них наличием одной височ-
ной впадины (Synapsida), уже в пермское время эволюционировавшие
в разных направлениях: с одной стороны, опи дали типичных на-
земных растительноядных и хищных разпозубых звероподобных
ящеров с одной скуловой дугой, из которых возникли предки
млекопитающих, подкласс Theromorpha; с другой стороны, от
анапсидных рептилий ведет начало обширнейший сверхотряд
двускуловых (Diapsida). Сюда относится ныпе живущая новозе-
ландская гаттерия (Sphenodon), единственная представительница
подкласса Diaptosauria и древнего триасового отряда (ящерице-
подобных клювоголовых (Rhynchocephalia); сюда же принадлежит
подкласс текодоптов (Thecodontia), — ископаемых чрезвычайно
разнообразных, частью водных, главным же образом назем-
ных, иногда древесных и очень подвижных рептилий, которых делят
на несколько отрядов. От этого подкласса произошли птицы, под-
класс динозавров (Dinosauria), крокодилы (Crocodilia) и летающие
ящеры—птерозавры (Pterosauria). Только крокодилы дожили до на-
шего времени. Динозавры и птерозавры вымерли в конце мелового
26* 403
периода. Наконец, боковая ветвь, отделившаяся, вероятно, от об-
щего ствола двускуловых, привела к подклассу чешуйчатых (Le-
pidosauria), куда, кроме современных ящериц (Lacertilia) и змей
(Opliidia), принадлежит еще отряд ископаемых ареосцелид (Агео-
scclida), близких к предкам ящериц и змей.
От неизвестных предков, возможно близких к водным пелико-
заврам, еще в пермское время произошли, вероятно, водные мезо-
завры (Mesosauria), очень быстро вымершие, а от общего с ними
корня водные морские ихтиозавры (Ichtyosauria), вымершие в мелу.
От неизвестных ближе представителей пермских примитивных
рептилий произошел подкласс плоскозубых (Placodontia) и подкласс
совершенно водных зауроптеригий (Sauropterygia). Первые вы-
мерли в триасе, вторые дожили до верхнего мела.
Наконец, от тех же котилозавров пермской эпохи ответвился
подкласс черепах (Testudinata), отличающийся развитием панцыря
на спине и брюхе. Эта древняя группа рептилий прошла длинную
эволюцию с неоднократными приспособлениями к водной жизни
и дожила до нашего времени в виде наземных и водных, пресновод-
ных и морских черепах.
Как видно из сказанного, только небольшая часть рептилий
дожила до нашего времени — черепахи (Testudinata), ящерицы
(Lacertila) и змеи (Ophidia), составляющие подкласс чешуйчатых
(Lepidosauria), крокодилы (Crocodilia) и клювоголовые (Rhyn-
chocephalia).
II. СТРОЕНИЕ РЕПТИЛИЙ
1. Форма тела рептилий и их движения
Рис. 306. Скелет Diadectes спереди.
1—scapula, 2 — cleithrum, 3 — procoracoid,
4 — interclavicula, 5 — humerus. (По Case из
Ihle и др.)
Ни в одном классе не было такого разнообразия форм и движений,
как в классе рептилий. Исходная форма тела их унаследована была
еще от стегоцефал: это форма
саламандры с туловищем,
волочащимся по земле. Осо-
бенность эта настолько харак-
терна и для современных реп-
тилий, что она запечатлена
в самом их названии — пре-
смыкающиеся. Тяжелое не-
уклюжее тело поддерживалось
широко расставленными ко-
нечностями, перпендикулярно
стоящими к туловищу (рис.
306); впрочем, предплечье и
голень в связи с более совер-
шенным приспособлением к
движению изменили свое пер-
воначальное перпендикуляр-
ное положение к оси тела и получили новые направления: пред-
плечье получило направление вниз и вперед, голень—вниз ина-
404
зад. В связи с таким положением задняя конечность может
подталкивать туловище вперед гораздо более сильно. Все же поло-
жение конечностей по отношению к туловищу остается очень не-
совершенным: требовалась чрезвычайно сильная мускулатура,
чтобы держать туловище на весу и передвигать его. Естественно,
что в эволюционном процессе этот способ передвижения, которым
пользовались пермские котилозавры (Cotylosauria) и пеликозавры
(Pelycosauria), заменился другими, более совершенными. Так, уже
дальнейший поворот конечностей локтевым суставом назад и колен-
ным суставом вперед значительно улучшил движение и дал значи-
тельную экономию сил. У ящериц большую помощь в движении ока-
зывает длинный хвост: животное двигается, не только отталкиваясь
задними ногами, но и одновременно змееобразно изгибая свое тело
и хвост. Дальнейшее развитие этого вида движения, особенно удоб-
ного при жизни среди густой травянистой растительности, приводит к
постепенному укорочению конечностей, становящихся лишь местами
опоры о неровности при движении, и, наконец, к полному их исчез-
новению. Такое исчезновение конечностей встречается в разных
группах ящериц, где оно .возникло совершенно независимо; такое же
исчезновение конечностей привело к характерной форме настоящих
змей.
Движение путем отпихивания двумя задними конечностями од-
новременно привело, как мы видели у амфибий, к прыгающим дви-
жениям. Подобное же приспособление к прыганию мы находим и
у рептилий. Уже многие ящерицы обладают способностью прыгать
с помощью удлиненных задних конечностей. В истории развития
рептилий такой способ передвижения возникал неоднократно.
При этом длинный хвост играл при прыжках, как хвост у тушкан-
чиков или кенгуру, роль руля. Однако прыгающие формы могли
возникнуть и иным путем — из бегавших на задних ногах рептилий,
а эти последнее могли возникнуть как приспособленные к более
быстрому бегу из длинноногих ящерицеобразных животных. Так,
например, прекрасно бегает на задних ногах из современных яще-
риц Chlamydosaurus kingi; бегают на задних ногах некоторые ага-
миды и многие игуаны (ВазШзсизидр.). Среди вымерших форм пе-
редвижение с помощью одних только задних конечностей характерно
было для многих динозавров (Dinosauria). Многие из принад-
лежащих к подклассу животных, которые передвигались на че-
тырех ногах (например, Stegosauridae или Ceratopsidae), произошли,
как то доказывает строение их скелета, из форм, первоначально
передвигавшихся на задних ногах. При передвижении на задних
ногах у разных динозавров, многие из которых достигали громадной
величины, роль хвоста была различна; как показали исследования
отпечатков следов, хвост или служил опорой при медленном пере-
движении с помощью попеременно переставляемых правой и левой
задних ног, или служил опорой при прыжках, или он приподнимался
кверху и служил рулем при поворотах. Различная роль хвоста при
движениях отразилась и па скелете этих животных: те из рептилий,
которые опирались при движениях на хвост, сохранили типичный
405
ние. 307. Таз Allosaurus
из Saurischia.
1 — ilium, 2 — pubis, 3 —
ischium, 4—foramen puboi-
schiadicum, 5 — продыряв-
ленное acetabulum. (Из Ihle
и др.)
для рептилий таз (Theropoda из Saurischia) (рис. 307), тогда как
поднимавшие хвост (Ornithopoda из Ornitischia), имели тазовый
пояс, устроенный сходно с таковым у птиц.
Более быстрое передвижение по земле могло осуществиться у реп-
тилий и иным способом, без прохождения стадии ящерицеобразной
формы. Именно неповоротливые и тяжело-
весные формы из примитивных пеликозав-
ров (Pelycosauria) могли приобрести более
легкий способ движения путем изменения
положения конечностей: последние распо-
лагались все более и более в плоскости,
параллельной сагиттальной плоскости тела,
причем передняя конечность локтевым су-
ставом поворачивалась назад, а задняя
конечность колепным суставом поворачи-
валась вперед. Такой тип строения конеч-
ности мы находим в совершенном виде
лишь у млекопитающих; менее совер-
шенным оп возник независимо в под-
классе звероподобных (Thereinorpha) (рис.
308) и в поклассе динозавров (Dinosau-
ria).
Ящерицеобразное движение, как мы знаем на основании изуче-
ния современных форм, весьма пригодно и для лазания. Хотя дре-
весный образ жизни или лазание по скалам и приводит к отбору
многочисленных и разнообразных приспособлений, которые будут
нами разобраны ниже, однако по самому своему характеру движе-
Рис. 308. Lystrosaurus latirostris из перми Южной Африки.
(Из Abel по Watson.)
ние ящериц, основанное на захвате пальцами неровностей почвы,
уже пригодно и для лазания. Из лазящих по деревьям форм разви-
лись специализированные прыгающие по деревьям формы, привед-
шие к планирующим, вроде летающего дракона, животным с пере-
понками между телом и конечностями и к настоящим летающим пте-
розаврам. У последних полет осуществлялся с помощью крыльев
406
из кожистой перепонки; однако натянута была эта перепонка не ме-
жду четырьмя пальцами, как у летучих мышей, а между телом,
задней конечностью и сильно удлиненным 4-м пальцем (рис. 309).
Приспособление наземных рептилий к водному образу жизни
шло разными путями. К водному образу жизни переходили формы
Рис. 309. Реконструкция Rhamphoi-nynchus gemmingi, из верхней юры
Баварии.
(Из Abel.)
ящерицеобразного типа. Таковы, например, крокодилы (Crocodilia);
такими были предки пермских мезозавров (Mesosauria) и мозозавров
(Mososauria — Pythonomorpha). Все эти формы характеризуются
длинным хвостом, который в большей или меньшей степени превра-
щается в основной орган движения, тогда как передние и задние
конечности превращаются в плавники, служащие главным образом
Рис. 310. Ихтиозавр. Скелет и реконструкция очертаний тела.
(Из Fraas.)
рулями бокового движения и глубипы. Дальнейшее усовершенство-
вание приспособлений к движению в воде приводит к таким совер-
шенным типам водных обитателей, какими являются ихтиозавры
(Ichtyosauria) и мозозавры (Mososauria). Первые (рис. 310) пред-
ставляют собой по форме и движению тот тип, который мы называли
у рыб веретенообразным или торпедообразным типом. Длинный
407
хвост проделал при этом своеобразную эволюцию, пока не превра-
тился в орган, аналогичный хвосту пелагической рыбы. Однако
ихтиозавры, нуждавшиеся в постоянном атмосферном дыхании, в от-
личие от селахий и ганоидных рыб приобрели гйпобатический тип
плавника как основной тип, облегчавший им движение головой вверх
к поверхности воды. Мозозавры, наоборот, двигались,подобно угрям,
путем змеевидных движений своего длинного туловища и длинного
хвоста; хорошо развитые парные конечности в отличие от угрей
позволяли мозозаврам гораздо совершеннее управлять своими дви-
жениями, что дало им возможность
перейти к совершенно пелагиче-
скому образу жизни (рис. 311).
Иной характер движения в во-
де был у перешедших к водному
образу жизни короткохвостых реп-
тилий, происходивших от коротко-
хвостых наземных звероящеров
(Theromorha). Таковы, во-первых,
двуклыковые звероящеры (Dicyno-
dontia), травоядные, обитатели
болотистых побережий и пресно-
водных бассейнов: они плавали с
помощью обеих пар конечностей,
действуя ими как веслами лодки.
Так же плавали более совершен-
ные пловцы — плоскозубые ящеры
(Placodontia), обитатели богатых
моллюсками и иглокожими морских
побережий. Наконец, высшего за-
вершения подобный способ пере-
движения достигает у плезиозавров
(Plesiosauria) из отряда зауропте-
ригий (Sauropterygia). Обе пары
конечностей превращены здесь в
Рис. 311. Мозозавр, реконструк-
ция.
(Из Lucas.)
мощные гребные плавники.
Особняком стоят среди рептилий черепахи со своими панцырями
как защитным от врагов приспособлепием. Возникновение их,
как мы уже отметили выше, относится к глубокой древности. Древ-
нейшие черепахи из верхнего триаса (Triassochelydae) уже обладали
совершенно устроенным панцырем и имели типичный черепаший
вид (рис. 312); они были наземными формами, и папцырьслужил им
надежной защитой от врагов. Между тем возникновение панциря
из роговой чешуи и подлежащих костей никак не может быть объяс-
нено как защитное от врагов приспособление: он уже должен был
существовать как панцырь, чтобы оказаться полезным в защите
от врагов и подвергнуться дальнейшему отбору. Его возникновение
скорее всего может быть объяснено приспособлением к роющему
образу жизни. Плоская форма и расположение передних и задних
конечностей являются безусловно характерными для роющих форм.
408
Наличие твердых роговых пластин на теле защищало тело черепах
от повреждений при закапывании в землю; развившись в качестве
приспособления к роющему образу жизни, роговой панцырь в даль-
нейшем уже мог оказаться полезным в качестве защиты от врагов.
Приспособление к втягиванию головы и шеи, а также конечностей
под панцырь явилось, та-
ким образом, вторичным.
Первоначально наземные
черепахи частично пере-
шли к водно-болотному,
частично к совершенно вод-
ному пелагическому образу
жизни. Наземные черепа-
хи отличаются медленными
движениями. Наличие пан-
цыря, куда втягиваются
конечности, обусловливает
своеобразное положение
последних. Передние ко-
нечности направлены здесь
в своей проксимальной ча-
сти (плечо) вперед; пред-
плечье направлено вниз и
немного наружу; локтевое
сочленение, таким образом,
оказывается тоже совер-
шенно необычным образом
направлено вперед. Что ка-
Рис, 312. Сагарах Triassochelys dux из
триаса Германии.
(По laekel из Abel.)
сается задней конечности, то она сохраняет свое нормальное поло-
жение примитивной наземной конечности. Опираются о землю
черепахи на* ступню. У морских черепах и пресноводных Triony-
chidae конечности превращены в совершенные плавники.
2. Общие покровы рептилий
В отличие от амфибий кожа рептилий суха, лишена желез и одета
роговыми образованиями в виде чсшуй, щитков или папцыря. Та-
кое строение кожи могло возникнуть лишь после потери кожей функ-
ций дыхания, что и было достигнуто, вероятно, уже самыми пер-
выми рептилиями. Первоначальное основное приспособительное зна-
чение такого изменения покрова рептилий состояло в предохранении
кожи от высыхания в связи с переходом к вполне наземному образу
жизни, а затем уже в защите от повреждений в связи с передвиже-
ниями по твердому субстрату. Эпидермис рептилий в связи с этим
дифференцируется па два слоя: на роговой слой (stratum
corneum) имальпигиев слой (str. malpighii). Роговой слой
иногда значительной толщины и в свою очередь распадается на ком-
пцктный наружный слой безъядерных клеток и внутренний с ядрами
409
в клетках, тогда как мальпигиев слой состоит из небольшого числа
(5—10) рядов клеток.
Утолщепие рогового слоя происходит различно. У подвижных
животных, какими являются ящерицы и змеи, утолщение рогового
слоя происходит путем образования или бугоркрв, как это мы видим
у гаттерии, хамелеонов или гекконов, или черепицеобразно рас-
положенных, т. е. налегающих друг на друга своими задними кон-
цами, чешуи, как у ящериц и змей. Между чешуями роговой слой
остается тонким, что не препятствует движению тела. На частях тела,
где при движениях подвижности кожи не требуется, развиваются
плоские роговые щитки или щиты, образующие сплошной панцырь.
Таковы щитки па голове у змей и ящериц, щиты на туловище у кро-
кодилов и, наконец, панцырь черепах. Нередко роговые образования
принимают форму различных выростов — шипов, игл, иногда слож-
ной формы. Будучи первоначально приспособленным к защите от
высыхания, роговой покров постепенно приобретает дополнитель-
ное значение защиты тела от повреждений предметами, с которыми
ему приходится вступать в соприкосновение, в особенности у рою-
щих животных. Эта защита становится еще более полной, когда ро-
говые образования подстилаются развивающимися в коже окосте-
нениями. Таковы костные образования у некоторых ящериц (щитки),
у крокодилов и костные щиты черепах. Такая защита тела от по-
вреждений могла быть полезной также и в качестве защиты от врагов,
от укусов себе подобных и от хищников из рептилий. Путем отбора
мог выработаться, наконец, тип черепахи, покрытой щитом со всех
сторон. Если в настоящее время щиты черепахи и пе являются на-
дежной защитой от хищных зверей и даже хищных птиц, обладаю-
щих рядом приспособлений и прежде всего высокоразвитой высшей
нервной деятельностью, что позволяет им справляться с папцырем
черепах, то не надо забывать, что в то время, когда возникали че-
репахи, врагами этих растительноядных рептилий были неуклю-
жие, малоподвижные хищные рептилии, подобно амфибиям жадно
хватавшие преимущественно двигавшиеся предметы и отступав-
шие перед неподвижными и неуязвимыми черепахами. Вот почему
подобный тип защиты выработался не только у черепах мезозоя, но и
у некоторых других ископаемых рептилий, а позднее у броненосцев
из млекопитающих.
Расположение чешуй и особенно щитков и щитов на теле репти-
лий закономерно; хотя оно и подвержено некоторым индивидуаль-
ным отклонениям, все же в основном оно постоянно для определен-
ных систематических групп, а потому имеет таксономическое зна-
чение. Первоначальное расположение чешуй было, вероятно, сег-
ментальным, причем роговые чешуйки были подостланы костной
чешуей, подобной той, которая встречалась у предков рептилий—
стегоцефал. Такая же костная чешуя была и у некоторых из при-
митивнейших рептилий — котилозавров (Pareiasaurus и др.). Кост-
ные чешуйки сохранились у немногих ящериц (сцинки, веретенница),
у крокодилов и черепах. У других они исчезли. Первоначально и
чисто роговая чешуя имела сегментальное расположение. Впослед-
410
ствии сегментация в связи с различными приспособлениями более
или менее стушевалась или вовсе исчезла вследствие распада на бо-
лее мелкие чешуйки или вследствие их срастания в щитки.
На голове у змей и ящериц различают щитки, имеющие большое
систематическое значение. У змей расположение щитков на голове
сходно с ящерицами. На ряду со срастанием щитков друг с другом,
что имеет место у змей, ведущих подземный образ жизни; нередко
наблюдается и распад щитков на более мелкие. Своеобразным ро-
говым образованием является
ный сросшимися в сплошное
сегментов, которые не сбра-
сываются при линьках, а на-
растают, исполняя роль по-
гремка.
У кожистой черепахи
(Dermochelys coriacea) и у
мягких черепах (Trionycho-
idea) роговые щитки разви-
ваются только в молодости,
а затем исчезают, так что пап-
цырь оказывается покрытым
лишь толстой кожей. Голова
черепах покрыта щитками, а
по краям челюстей щитки сра-
стаются в роговой клюв. На
спине у крокодилов имеются
толстые большие щитки, рас-
погремок гремучей змеи, образован-
кольцо чешуйками двух последних
Рис. 313. Поперечный разрез кожи
Anguis fragilis перед линькой. Можно
видеть три генерации эпидермиса.
положенные сегментально; на
брюхе эти щитки меньше; по
краям туловища и на ногах
щитки расположены непра-
1 —' костяная чешуя, 2 —corium, 3 —маль-
пигиев слой, 4 — промежуточные клетки,
5 —третий роговой слой, 6 — промежуточ-
ные клетки, 7 — второй роговой слой, 8 —
первый роговой слой, 9 — надкожица. (Но
Maurer из Ihle и др.)
вильными рядами и мелки.
Ни у черепах, пи укрокодилов сбрасывания роговых образований
не наблюдается, у змей и ящериц происходит более или менее слож-
ная линька. Она начинается на голове, причем у ящериц и у немно-
гих змей кожа сбрасывается лоскутами, а у всех прочих змей це-
ликом, так что животное выползает из старой кожи, как из мешка.
Линька подготавливается довольно сложным гистологическим про-
цессом, выражающимся в том,, что клетки мальпигиева слоя дают не
одно, а несколько поколений подвергающихся ороговепию клеток,
причем между каждым поколением располагаются так называемые
промежуточные ослизняющиеся клетки. Здесь старый ороговевший
слой отделяется и сбрасывается, а его место занимает более молодой
роговой слой. На разрезе кожи (рис. 313) можно видеть несколько
поколений эпидермиса.
Из роговых образований рептилий необходимо упомянуть когти,
одевающие последние фаланги пальцев; в зависимости от образа
жизни они имеют разное строение: они остры и изогнуты у лазящих
ящериц и плоски у плавающих черепах; у черепах и крокодилов они
411
отрастают, подобно тому как у птиц, по всей поверхности, тогда
как у ящериц отрастание когтя идет только с основания когтя,
как у млекопитающих.
Строение соединительнотканной части кожи сходно с тем, что
мы видели у амфибий; кожа состоит из двух олоев: наружного рых-
лого слоя, образующего сосочки роговых чешуй, и внутреннего,
более толстого слоя из плотной волокнистой соединительной ткани.
Глубже следует слой рыхлой подкожной клетчатки. В соединительно-
тканной части кожи лежат многочисленные пигментные
клетки, обусловливающие цвет кожи. Изредка они встречаются
и в мальпигиевом слое эпидермиса. Меланофоры лежат обычно наи-
более глубоко в плотной соединительной ткани, особенно там, где
она переходит в рыхлую ткань; выше располагаются различно
окрашенные хроматофоры. Среди них преобладают желтые (ксанто-
форы) и красные (эритрофоры). Между ними и меланофорами у не-
которых рептилий лежат наполненные гуаниновыми тельцами
иридирующие гуанофоры.
У хамелеонов имеется чрезвычайно развитая способность менять
свою окраску от беловатой и оранжевой через желтую и зеленую
до пурпуровой и темнокоричневой и черной. Многие ящерицы
обладают способностью также довольно быстро перекрашиваться
в другой цвет. Такая перекраска зависит от сжатия одних и рас-
пространения других хроматофор и имеет по большей части адап-
тивный характер, о чем сказано будет подробно ниже.
Круглоголовки, например (род Phrynocephalus из сем. Agami-
dae) являются более темпыми по утрам, когда холоднее, и светлеют
днем, при повышении температуры.
Кожные железы, как мы уже упоминали, отсутствуют
у современных рептилий совершенно, если не считать встречаю-
щихся у некоторых групп рептилий железоподобных образований
или желез специального назначения. Так, у ящериц в период
размножения имеются на бедрах особые поры, через которые вы-
талкиваются наружу ороговевшие клетки, образующиеся на особом
погруженном в соединительную ткань сосочке; у черепах имеются
особые мускусные железы, расположенные между двумя
щитами; у крокодилов имеются мускусные железы на боках ниж-
ней челюсти. Однако низшие вымершие рептилии — котилозавры
и звероящеры — еще сохраняли, вероятно, кожные железы в зна-
чительном числе на теле, так как иначе трудно понять образование
сальных и потовых желез у млекопитающих, происшедших от зверо-
зубых ящеров (Theriodontia) пермского времени.
3. Наружный, или кожный, скелет
Кожный скелет рептилии частью унаследовали от стегоцефал,
частью он развился у них заново как защитное приспособление.
У стегоцефал, как мы видели, от рыб сохранился костный чешуй-
чатый покров па брюхе в виде круглых или овальных, или веретено-
образных чешуй, расположенных косыми рядами. Эти образования,
412
служившие для защиты брюха при ползании, преобразовались
у рептилий в так называемые брюшные ребра, образова-
ния, и по происхождению и по функции совершенно отличные от
Рис. 314. Скелет Ophiacodon, показывающий положение брюш-
ных ребер.
(По Case па Ihle.)
ребер; они также служат для защиты тела от повреждений при
ползании. Брюшные ребра мы находим у
(рис. 314), у гаттерии, у крокодилов и в
рудиментарном виде у некоторых ящериц.
У черепах па брюхе из этих ясс элементов
и элементов плечевого иояса развился мощ-
ный костный б р ю ш п о й щит — пл а с т-
р о н (plastron), тогда как кожные окосте-
нения на спине, сросшись с остистыми
отростками позвонков и ребрами, образо-
вали спинной щит — карапакс
(сагарах). Вдоль спинного щита тянется ряд
сросшихся с позвонками небольших спин-
ных невральных пластинок (neura-
lia); по бокам от невральных пластинок
располагаются большие сросшиеся с ребрами
реберные или костальные плас-
тинки ‘’(costalia), тогда как края щита
образуют краевые маргинальные
пластинки (marginalia). Самая передняя
непарная краевая, служащая продолжением
невральных пластинок, называется загри-
в о ч н о й (nuchale) (рис. 315). Брюшной
щит образован большой непарной или парной
костью (endoplastron), гомологичной над-
грудиннику (episternum) других рептилий, и
рядом парных костей, из которых передние
(epiplastron) соответствуют ключицам (clavi-
cula); следующие три костных пластинки гомо-
логичны брюшным ребрам и носят названия
liyoplastron, hypoplastron и xiphiplastron.
Сравнительно анатомическое и сравни-
тельно эмбриологическое исследование кост-
ных образований в коже рептилий приводит
к следующим заключениям. Первоначально
многих ископаемых
В ю
Рис. 315. А— сага-
рах Emys orbicularis.
В — plastron Clem-
my? caspica.
1 — neuralia, 2 — costa-
lia, 3— marginalia, 4 —
nuchale, 5 — endoplas-
tron, 6 — eplplastron,
7 — hyoplastron, 8 — me-
soplastron, 9 — hypopla-
stron, 10 — xiphiplast-
orn. (Из Никольского.)
рептилии имели, подобно некоторым рыбам, двоякого рода костные
об|>а:юпания: более глубокие костные чешуйки — панцырь (theca)
413
и более поверхностный слой костных чешуй — надпанцырь
(epitheca). Оба рода чешуй сверху прикрывались рядами роговых че-
шуй. Все чешуи располагались сегментально. От такого первоначаль-
ного расположения чешуй у некоторых ящериц (сцинки, веретенница
и др.) и у крокодилов сохранились поверхностные око-
стенения (epitheca); у крокодилов и у веретенницы каждой
Рис. 316. Гипотети-
ческая исходная фор-
ма черепах.
(По Versluys.)
роговой чешуйке соответствует костная че-
шуя, тогда как у сцинков под роговой чешуй-
кой лежит несколько костных чешуй. К глу-
боким окостенениям принадлежат брюшные
ребра и костный панцырь черепах—кара-
пакс и пластрон. Однако у кожистой че-
репахи (Dermochelys) поверх редуцирован-
ного карапакса, от которого сохранилось
лишь nuchale, располагается верхний пан-
цырь—эпитека, состоящий из большого числа
мелких косточек. У молодой черепахи такая
эпитека отсутствует, зато имеется соответ-
ствующая ей роговая чешуя. Под такой че-
шуей развиваются позднее окостенения, а
сама роговая чешуя исчезает. Прилагаемый
рис. 316 дает представление о гипотетической
исходной форме черепах. Эпитека изображена
черным, тека затушевана пунктиром.
4. Внутренний скелет рептилий
Скелет рептилий соответственно с их пол-
ным переходом к наземной жизни приобре-
тает по сравнению со скелетом их предков,
стегоцефал, целый ряд приспособительных особенностей. Эти при-
способительные особенности охватывают все части скелета.
Позвоночник рептилий приобретает в связи с более совершен-
ным строением позвонков и в связи с дифференциацией их на отделы
и бблыпую прочность и большую подвижность, особенно в шейном
отделе, а иногда и в других отделах.
Форма позвонков низших рептилий, как Cotylosauria и Pely-
cosauria, еще двояковогнутая, амфицельная;при-
чем между позвонками еще сохраняется сдавленная, но непрерыв-
ная хорда; однако у высших рептилий и большинства современных
рептилий позвонки более прочно сочлененные, процельные, и от
хорды пе остается и следа. Лишь у Hatteria и гекконов сохраняются
амфицельные позвонки.
Позвонки рептилии существенно отличаются по своему про-
исхождению от таковых современных амфибий, хотя и те и другие
исходной формой имеют рахитомный тип позвонков стегоцефал..
Позвонок состоял первоначально из четырех кусочков с каждой
стороны, но в то время как формирование тела позвонка современ-
ных амфибий шло за счет гипоцентров и редукции плевроцентров,
развитие тела позвонка рептилий шло, наоборот, зз счет прогрес-
414
пирующего увеличения дорзальных плевроцентров (intercalare)
(рис. 317). Соединение нижних дуг с телом приходится у них интер-
вертебрально; этот процесс, однако, можно проследить еще у стего-
цефал из отряда Gastrocentrophori. Процесс формирования цель-
ного прочного позвонка наземного позвоночного еще не закончился
даже у современных низших рептилий: у гаттерии (Sphenodon)
и у гекконов (Gekkonidae) в некоторых частях тела (шейная об-
ласть, конец туловища *и начала хвоста) между телами позвонков
имеются еще так называемые межцентры (intercentra). К меж-
центрам причленяются ребра, а в хвостовой области — гемальные
дуги. У крокодилов и ящериц межцептры уже прирастают к по-
звонкам, образуя их нижние
отростки. Верхние дуги совер-
шенно срастаются с телом, об-
разуя спинномозговой канал; от
их спинной поверхности отходят
широкие верхние остистые от-
ростки (processus spinosus), слу-
жащие для прикрепления важ-
ных при передвижениях, изги-
бающих туловище спинных
мышц. Прочность позвоночника
достигается развитием сус-
тавных, или сочле-
Рис. 317. Диаграмма, иллюстриру-
ющая расхождение в развитии эле-
ментов позвонков, ведущих от при-
митивных Tetrapoda А: к типич-
ным амфибиям, Р; к типичным
Amniota, Е. D, Е — к туловищному
позвонку добавлен хвостовой.
1 — basiventrale (hypocentrum), 2 — in-
terdorsale (pleumcentrum), 3—-interven-
trale. (Из Goodrich.)
. Причленение ребер происхо-
II ОВНЫ X, отростков
(praezygapophysis и postzyga-
pophysis). Помимо небольших
боковых отростков, на теле у
позвонка рептилий имеются
хорошо развитые попереч-
ные отростки (process!
transversi), отходящие от дуги
дит с помощью развилка, вентральная часть которого (голов-
ка — capitulum) сочленяется с выступом па теле, а дорзальная
часть (бугорок — tuberculum) с концом поперечного отростка.
Такое соединение ребер с позвонками встречается у многих иско-
паемых рептилий и в шейном отделе, и в начале грудной области
(у крокодилов, а также у гаттерии и гекконов). Простое сочленение
ребер с поперечным отростком, встречающееся и широко распро-
страненное у современных рептилий, — явление вторичное.
Подвижность туловища и особенно необходимая при захваты-
вании пищи подвижность головы достигаются у рептилий распа-
дением позвоночника на отделы. Прежде всего выделяется шейная
область, состоящая из различного, иногда большого числа позвон-
ков (у Sauropterygia и некоторых Dinosauria). Наличие шеи чрез-
вычайно облегчает наземным животным добывать себе пищу, позво-
ляя направлять голову независимо от движения туловища в жела-
тельную сторону. Эта подвижность головы увеличивается еще тем,
что ^голова причлепяется к позвоночнику с помощью одного, а не
415
двух, как у амфибий, сочленовцых бугорков, и тем, что первые два
позвонка приобретают особенное строение. Тело первого позвонка,
который здесь получает название атласа (atlas), не срастается
с относящимися к нему дугами, а прикрепляется связками или
даже совершенно срастается с телом второго позвонка, называемого
эпистрофеем (epistropheus), образуя его зубовидный отро-
сток (processus odontoideus). Благодаря такому развитию первый
позвонок, или атлас, оказывается состоящим из одних дуг без тела
и, вследствие срастания дуг с межцентром, представляется устроен-
ным в виде кольца. Так как кольцо это сидит на зубовидном отро-
стке эпистрофея, то получается возможность вращательных дви-
жений головы вместе с атласом. Спинномозговой канал отделен
в атласе соединительной тканью. Необходимо добавить, что между
атласом и черепом раньше был еще один позвонок — проатлас
(proatlas). Тело проатласа прирастает к зубовидному отростку,
а его дуга отмечается иногда в виде самостоятельных костей между
затылочной частью черепа и атласом (у гаттерии, крокодилов
и хамелеонов). Итак, более или менее длинная шея с подвижно
сочлененными позвонками, наличие атласа и эпистрофея и сочле-
нение головы с атласом посредством одного сочленовного бугорка —
все это дает возможность производить головой очень разнообразные
и сложные движения, что, конечно, имело большое значение в борьбе
за существование. В шейном отделе ребра бывают выражены более
или менее сильно; иногда они имеются, как, например, у кроко-
дилов, уже на атласе, обыкновенно же на первых позвонках отсут-
ствуют. У черепах рудиментарные ребра прирастают к позвонкам
шейного отдела. Особенно хорошо развиты ребра последних шей-
ных позвонков. Те позвонки, ребра которых доходят до соединения
с грудиной, относятся к следующему, грудному, отделу. Этот отдел
имеет у рептилий различное число позвонков и в общем характе-
ризуется наличием хорошо развитых самостоятельных ребер, из
которых задние пе доходят до грудины и просто причленяются
к поперечному отростку позвонков. Передние ребра хорошо раз-
виты и состоят из двух половин: из дорзальной — костяной и груд-
ной — хрящевой части, иногда, впрочем, окостеневающей (кроко-
дилы). Вертебральные части ребер у крокодилов и .клювоголовых
снабжены особыми, идущими назад, клювовидными отростками
(processi uncinati), налегающими на расположенные сзади ребра.
Дифференциация грудной области и указанное выше строение ребер
имели исключительно большое значение, так как связаны были
с приобретением более совершенного дыхания, описание которого
будет дано ниже.
Следующая за грудной поясничная область имеется далеко не
у всех рептилий; она характеризуется отсутствием ребер, которые
прирастают к поперечным отросткам, выраженным здесь очень
хорошо. Наличие поясничной области связано с более полной
дифференциацией грудной и брюшной частей туловища и также
является прогрессивной особенностью, существенной прежде всего
при дыхательных движениях. ,
416
Следующий, крестцовый, отдел характеризуется прикреплением
к позвоночнику таза. Подвздошные кости причленяются к попе-
речным отросткам позвонков, с которыми и здесь сливаются руди-
ментарные ребра. Число крестцовых позвонков невелико, у совре-
менных рептилий только два. Хвостовой отдел состоит из чрез-
вычайно различного числа позвонков в зависимости от развития
хвоста. Как уже было упомянуто, в этом отделе у позвонков име-
ются нижние гемальные дуги, которые
в силу развития тел позвонков из плев-
роцентров причленяются интервсртеб-
рально.
Указанные выше отделы позвоноч-
ника вместе с исчезновением конечностей,
когда исчезают и грудина и тазовый пояс
(как это имеет место, например, у змей),
естественно, тоже исчезают. При этом
число позвонков на теле может чрезвы-
чайно увеличиваться (до Б65 у эоценовой
морской змеи — Arcliaeophis).
Грудина у рептилий (рис. 318)
хрящевая и состоит из передней части,
грудины собственно (sternuni) в виде бо-
лее или мепее широкой ромбической пла-
стинки, и более узкой задней (metaster-
num), образовавшейся путем слияния
концов ребер; иногда это слияние осуще-
ствляется частично и у большей части
ребер не идет дальше слияния противо-
положных друг другу ребер каждой сто-
роны (у хамелеонов). К переднему отде-
лу грудины причленяются ребра, иногда
лишь в очень незпачительном числе. У
Рис. 318. Грудина и плече-
вой пояс Lacerta с брюшной
стороны. Хрящ пункти-
рован, кость гладкая.
1 — sternum, 2 — episternum,
3 —clavicula, 4—coracoid,
5 —scapula, 6 — procoracoid.
(По Osborn из Ihle.)
рептилий, лишенных передних конечностей, грудина редуцируется
совершенно. По своему происхождению грудипа рептилий, вероят-
но, гомологична таковой амфибий и возникла независимо от ребер
между концами коракоидов. На грудипу снаружи налегает покров-
ная Т-образная кость — надгрудинник (episternum), про-
исшедшая от соответствующей кости стегоцефал. Поперечная часть
этой кости соединена с ключицами. У форм, лишенных конечностей,
естественно исчезает и эта кость грудины. У черепах она вместе с
ключицами входит в состав брюшного щита, образуя endoplastron.
Череп рептилий в связи с более полным приспособлением к на-
земной жизни и прогрессивным развитием органов чувств
претерпевает ряд существенных изменений.
Во-первых, в связи с большим развитием глаз и соответственно
разрастанием глазпиц череп рептилий становится’ т р о п и б а-
зальный: глазницы настолько сближены, что между ними
остается топкая межглазничная перегородка, которая не окосте-
невает, а остается или хрящевой или перепончатой. В хондро-
27 Кашкаров и Ставчинский
417
краниуме продолжается редукция хряща, идущая значительно
далее, чем у амфибий: вместо цельной хрящевой коробки хондро-
краниум состоит из перекладин, разделенных пространствами,
затянутыми перепонками. Более значительные массы хряща сосре-
доточены в слуховом и обонятельном отделах. В первом, помимо
овального окна у рептилий образовалось еще круг-
лое окно (foramen rotundum), затянутое перепонкой, что
улучшает передачу звуков в перепончатый лабиринт уха.
Окостенение черепа гораздо более полное, чем у амфибий; хрящ
сохраняется незамещенным только в этмоидальной области. Окосте-
нения черепа у наиболее примитивных рептилий (Cotylosauria)
мало отличаются от таковых у эмболомерных стегоцефал; у позд-
нейших представителей класса имеются большие отличия.
Рис. 319. Боковая стенка neurocranium примитив-*
ной рептилии, сбоку.
1 — bassioccipitale, 2 — exoccipitale, 3 — supraoccipitale,
4 —condylus, 5 — prooticum, 6 — opisthoticum, 7 —basis-
phenoideum, 8 — processus basipterygoideus, 9 — parietale,
10 — frontale, 11—nasale, 12 — praemaxilla, 13 —vomer,
14 — pterygoideum (По Ihle.)
В затылочной области (рис. 319) вокруг большого затылочного
отверстия (foramen magnum) окостеневают четыре затылочных кости:
основная затылочная (basioccipitalia), две боко-
вых затылочных (exoccipitale) и верхнезатылоч-
ная (supraoccipitale). Один затылочный бугорок (condylus occi-
pitalis) образован здесь основной и двумя боковыми затылочными
костями, что видно не только по данным эмбрионального развития,
но и у черепах и у низших ископаемых рептилий (Cotylosauria,
Theromorpha). В следующем слуховом отделе окостеневает по три
кости с каждой стороны: переднеслуховая (prooticum),
заднеслуховая (opistlioticum) и верхнеслуховая
(epioticum). Из них верхнеслуховая, одпако, срастается с верхней
затылочной, заднеслуховая тоже обычно срастается с боковой за-
тылочной (у черепах и некоторых ископаемых, например, у плоско-
зубых и ихтиозавров, она остается в виде самостоятельной кости).
В предслуховом, или теменном, отделе в основании черепа окосте-
невает основная клиновидная кость (basisphe-
418 ’
13
Рис. 320. Крыша черепа Sey-
mouria.
1 — parietale, 2 — frontale, 3 — na-
sale, 4 — postparietale, 5— foramen
parietale, 6 — supratemporale, 7 —-
temporale, 8 — postfrontale, 9 —<
praefrontale, 10—squamosum, 11 —
postorbitale, 12 —jugale, 13 —pra-
emaxilla, 14 — maxilla. (По Broil!
из Ihle.)
noideum), характерной особенностью которой является наличие
двух боковых крыловидных отростков (process!
basipterygoidei), к которым причленяются к р ы л о в и д н ы е
кости. Нередко основная клиновидная кость срастается у яще-
риц с основной затылочной и переднеслуховой в одну ком-
плексную кость. Боковые крылоклиповидные кости (alisphe-
noidea) имеются только у крокодилов, у некоторых динозавров,
у Amphisbaenidae из ящериц и немногих других, равно как
и глазничноклиповидные (orbitosphe-
noidea). В обонятельной области заме-
щающих хрящ костей у рептилий не
образуется.
Что касается покровных костей,
то первоначальное их количество со-
ответствовало тому, что имело место
у их предков — стегоцефал: крышка
черепа была стегальной и состояла из
большого числа костей, так же как и
у стегоцефал (рис. 320). Помимо обыч-
ных темени ы х (pariotalia), л о б-
н ы х (frontalia) и носовых (nasa-
lia), имелись еще следующие кости:
позади теменных — заднетемен-
ные (posttemporalia, или postparieta-
lia) обычно в числе четырех; по бокам
от теменных, характеризующихся
здесь тем, что между ними имеется, так
же как и у стегоцефал, теменное отвер-
стие (foramen parietale), лежали над-
височньие кости (supratemporalia), затем височные
(temporalia), позади глазниц — заднелобные (postfrontalia),
а впереди глазниц—п ереднелобные (praefrontalia); в сле-
дующем ряду—ч е ш у й ч а т а я (squamosum), заднеглаз-
ничная (postorbitale), скуловые (jugale), а по бокам черепа
в задней части — квадратноскуловые (quadratojugal е);
спереди череп ограничен межчелюстными (praemaxillare)
и челюстными (maxillare) костями. Такое сплошное, стегаль-
ное, покрытие черепа мы находим лишь у самых низших родо-
начальников рептилий, у котилозавров (Cotylosauria). Дальней-
шая эволюция в связи с развитием жевательных (правильнее
кусательпых) движений, требующих более мощных мышц, с одной
стороны, а с другой стороны — в связи с приобретенной боль-
шой подвижностью головы, при которой существенно полезным
должно было явиться всякое облегчение черепа, привела рептилий
к образованию височных впадин. Этот процесс образования из
сплошного, стегального, черепа у Anapsida черепов с одной (моно-
зигалыптй череп) или двумя затылочными дугами (дизигальный)
шло не по одному, а по нескольким путям.
Во-первых, еще в нижнепермское время из стегального черепа
27* 419
котилозавров образовался монозигальный череп пеликозавров (Ре-
lycosauria) со скуловой дугой, образованной скуловой костью
(jugale) и чешуйчатой (squamosum), и с одной височной впадиной,
ограниченной сверху заднеглазничной (postorbitale) и чешуйчатой
(рис. 321). Этот тип
строения монозигаль-
ного черепа встре-
чается затем у зверо-
яЩеров (Theromorpha)
(рис. 322) и, наконец,
у млекопитающих.
Употребляющееся до
сих пор название та-
кого черепа синап-
сидным (Synapsida) не
совсем удачно, так
Рис. 321. Череп Dimetrodon incisivus. Монози-
гальный (синапсидный) тип черепа Pelycosauria.
1 — jugale, 2 — postorbitale, 3—squamosum, 4—верх-
няя височная яма. (По Broom из Abel.)
как оно исходит из
предположения, что
одна височная впа- •
дина образовалась пу-
тем слияния двух
впадин, что не со-
ответствует действи-
тельности. У пермс-
ких же дейтерозавров
(Deuterosauria) тоже
имеется одна дуга, по-
строенная цесколько
иначе. Она состоит
из чешуйчатой кости,
из заднеглазиичной и
отчасти скуловой; ви-
сочная впадина огра-
ничена сверху заднелоб-
ной и теменной. Так же
устроена скуловая дуга
у плоскозубых (Placo-
dontia), у зауроптеригий
(Sauropterygia) (рис. 323)
и в подклассе Ichthyop-
terygia. И этот монози-
гальный череп относят
к типу Synapsida; так
как он образовался, не-
сомненно, независимо и
инымпутем, его называют
вторым синапсидным
типом черепа. В перм-
Рис. 322. Череп Udenodon (Theromorpha),
монозигальный синапсидный тип
jugale, 2 — postfrontale, 3 —squamosum, 4 —
височная яма.
ское же время из моно-
420
зигального черепа пеликозавров образовался дизигальный диап-
сидный череп, характерный для подкласса диаптозавров (Diapto-
sauria). Здесь две скуловых дуги и две височных впадины. Верхняя
височная впадина соответствует височной впадине дептерозавров,
Рис. 323. Реконструкция черепа Muraenosaurus. Моно-
зигалытый тип 2-го рода.
1 — praemaxilla, 2 — ноздря, 3 —lacrimale, 4 — frontale 4- nasale,
5 — poetfrontale, 6— postorbitale, 7 — jugale, 8 — parietale, 9 —
foramen interp arietale, 10 — squamosum, 11, 12 — angulare, 13 —
orblta, 14 — supraangulare, 15 — complementare, 16 —dentale
в нижнем рисунке, maxilla в верхнем, 17 — maxilla в нижнем
рисунке. (По Andrews из Abel.)
плоскозубых, зауроптеригий; она ограничена сверху postfrontale
и parietal?, а снизу — заднеглазничной (postorbitale) и чешуйчатой
(squamosum), образующими верхнюю скуловую дугу. Последняя
ограничивает нижнюю височную впадину, которая соответствует
таковой пеликозавров и звероящеров (Theromorpha). Такой тип
421
черепа мы встречаем у гаттерии, у многочисленных вымерших
рептилий — текодоптных (Thecodontia), динозавров (Dinosauria),
у крокодилов (Crocodilia) и птерозавров (Pterosauria) (рис. 324).
Четвертый тип крыши черепа мы встречаем у современных
ящериц. У них по своему строению череп очень напоминает дву-
2 л
Рис. 324. Диапсидный тип черепа у
Sphenodon.
1 — frontale, 2— postfrontale, 3 — squamo-
sum, 4 — quadratojugale, 5 —jugale, 6 —-
maxillare, 7 — postfrontale. (По Gadow, из
Abel.)
Рис. 325. Череп черепахи по
типу Emys.
1 — postfrontale, 2 — squamosum,
3 —jugale, 4 —quadratum. (По
Fichs, из Abel.)
Рис. 326. Схематическое изображение
черепа ядовитой змеи. Пунктиром изо-
бражено положение частей челюстного
аппарата при открытой пасти.
1—quadratum, 2 — pterygoideum, 3 —trans-
versum, 4—ядовитый зуб. (Из Biitschli.)
скуловой череп клювоголовых, но без нижней скуловой дуги,
которая должна была бы ограничивать нижнюю впадипу: послед-
няя оказывается снизу свободной, образовавшейся возможно из
двускулового черепа путем прорыва нижней дуги. Однако, как
думают другие, предки дще-
риц — ареосцслиды — имели
лишь верхнюю скуловую ду-
гу и верхнюю височную впа-
дину; в нижней части черепа
образовалась выемка, кото-
рая путем разрастания пре-
вратилась в сходное с ниж-
ней височной ямой образо-
вание.
Наконец, совершенно свое-
образно развивается из сте-
гального черепа монозигаль-
иый череп черепах. Здесь
образовалась ложпая скуло-
вая дуга путем развития вы-
емки в задней части крыши
черепа. Задневисочное отвер-
стие, таким образом, пере-
двигается вперед, образуя ложную скуловую дугу из задне-
лобной (postfrontale) и чешуйчатой (squamosum). Этот процесс
образования задневисочной выемки у черепах молено видеть на
различных стадиях развития у разных представителей Chelonia.
Наиболее примитивным по характеру ложноскуловой дуги яв-
ляется череп Chelonia my das. У ней только намечается развитие
422
задневисочной вырезки; следующая стадия развития представлена
Emys orbicularis (рис. 325) и далее — Testudo oculifera. У Тег-
rapene ornata, наконец, скуловая дуга исчезает совершенно, и
чешуйчатая кость оказывается отделенной от скуловой.
Таким образом, можно установить следующие пять типов строе-
ния черепной коробки: 1) стегальный, или азигальный, череп
(Anapsida); 2) монозигальный череп с нижней височной впадиной
(Synapsida 1-го рода); 3) монозигальпый череп с верхней височной
впадиной (Synapsida 2-го рода); 4) дизигальиый череп с двумя височ-
ными впадинами и двумя скуловыми дугами (Diapsida) и 5) с ложно-
скуловой дугой и задней височной вырезкой черепахи (Testudinata).
Снизу черепа развиваются следующие покровные кости: пара-
сфеноид (parasphenoideum), который срастается с б а з и-
сфеноидом (basisphenoideum), образуя его направленный
вперед отросток (rostrum); в этмоидальной области имеется парный
сошник (vomer), срастающийся у черепах в одну кость.
У некоторых ископаемых и у гаттерии па сошнике имеются зубы.
Что касается в и с ц е р а л ь и о г о скелета, то верхняя
часть челюстной дуги — пёбиоквадратиый хрящ — окостеневает
в заднем углу квадратной костью (quadratum), которая
или прирастает к черепу (моиимостилический череп), или свободно
сочленена с черепом (стрептостилический). Монимостиличезкое по-
ложение квадратной кости характерно для большинства, рептилий;
стрептостилия присуща была некоторым динозаврам и встречается
сейчас у чешуйчатых (Squamata), т. е. у ящериц и змей. У змей эта
подвижность квадратной кости особенно хорошо выражена и имеет
специальное значение, она позволяет особенно широко открывать
пасть (рис. 326).
Вторая кость, которая развивается из нёбноквадратного хряща,
это столбчатая кость, или колонка (epipterygoi-
deum, или columella х), характерная кость черепа рептилий, лежа-
щая на крыловидной кости и упирающаяся своим верхним концом
в теменную. Из покровных костей верхней челюсти, кроме уже упо-
мянутых межчелюстной и верхнечелюстной, имеются крыло-
видные кости (pterygoideum) и нёбные (palatinum)
и у большинства еще поперечные кости (transversum).
У змей эти кости подвижно сочленены таким образом, что при от-
крывании рта ядовитые зубы, сидящие па подвижных межчелю-
стных костях, направляются вперед, как это показано на рис. 326.
Это зависит от того, что поперечные кости являются рычага-
ми, передающими давление от квадратной кости через крыло-
видную и поперечную на челюстную. ~
Подвижность квадратной кости свойственна только змеям
и отчасти ящерицам и связана с особенностями захватывания пищи.
.V черепах, крокодилов и клювоголовых все указанные кости прочло
сое,чинены одна с другой, а крыловидная и нёбная входят в состав
1 Не следует смешивать эту кость, называемую нередко колонкой (co-
lumella cranii), с колонкой уха, или стремечком (columella auris, или stapes).
423
дна черепа. Кроме того, здесь впервые образуется вторичное нёбо.
У амфибий и большей части рептилий верхняя стенка ротовой
полости образована спереди челюстными костями, парасфеноидом
и сошниками, а сзади нёбными и крыловидными. Хоаны открыва-
ются здесь между челюстными костями и сошником. Такое нёбо
можно считать первичным. Его мы находим у ящериц, хотя и у них
намечается уже образование нёбных отростков на челюстных
костях в виде отходящих назад выростов с нижней стороны. У че-
репах и крокодилов эти отростки идут значительно дальше, при-
I
Рис. 327. Череп черепахи (Trionyx) (Л) и кроко-
дила (Alligator) (В).
1 — praemaxillae, 2 —maxillae, 3 — palatini, 4—pterygoi-
Uea, 5 — хоаны, 6 — basisphenoideum. (По Walther (А) и
Hoffmann (В) из Ihlc и др.).
кривая собой первичные хоаны и образуя крышу передней части
ротовой полости, или вторичное твердое нёбо (рис. 327). Сошники,
сросшись в одну кость, образуют перегородку, которая делит
лежащую между передней частью первичного ц£ба и вторичным
нёбом полость на две части — правый и левый носо-
глоточные ходы (ductus nasopharyngeus). Разрастание вто-
ричного твердого нёба назад этим, однако, не ограничивается
и у современных крокодилов достигает максимума своего развития;
вторичные хоаны отодвинуты в заднюю часть нёба, вторичное
твердое нёбо образовано здесь на ряду с нёбными отростками челю-
стных костей также нёбными и крыловидными костями.
Значение образования вторичного твердого нёба двоякое. С од-
ной стороны, чрезвычайно существенным являлось отодвигание
хоан возможно дальше, что значительно облегчало дыхание во
время процесса заглатывания пищи, с другой стороны — чрезвы-
чайное разрастание пасти, являясь приспособлепием к схватыванию
и умерщвлению добычи в связи с сильным развитием жевательных
(кусательпых) мышц, делало переднюю часть черепа очень слабой,
424
Рис. 328. Нижняя челюсть Lacerta agilis.
а — с внутренней, Ъ—с’наружной стороны.
1 — articulare, 2 — dentale, 3 — spleniale, 4 —
angulare, 5 —goniale, 6 — supraangulare, 7 —
согопаге. (По Gaupp.)
тогда как развитие вторичного твердого нёба значительно ее укреп-
ляло.
Нижняя челюсть рептилий является сложной. Из меккелева
хряща развивается сочленовная кость (articulare), с по-
мощью которой нижняя челюсть сочленяется с квадратной костью
верхней челюсти; ее прикрывают уже встречавшиеся у амфибий
зубная кость (dentale), крышечная (spleniale), уг-
ловая (angulare), кожная сочленовная (goniale),
а сверх того, надугло-
вая (supraangulare) и,
наконец, образующая так
называемый венечный
отросток — венеч-
ная кость (согопаге)
(рис. 328).
Что касается гиоидной
дуги, то судьба подвисоч-
ной кости (hyomandibula-
re) нам уже известна —
она превратилась еще у
стегоцефал в колонку
(столбик) уха (colu-
mella auris). Здесь ко-
лонка, однако, окостене-
вает уже двумя окостене-
ниями: наружной (extracolumella) и внутренней, соот-
ветствующей стремечку (stapes). Стремя прилегает к овальному око-
шечку, а наружная колонка — к барабанной перепонке. У гаттерии
и гекконов наружная колонка соединяется еще с квадратной костью
и гиоидным хрящом, указывая путь ее эволюции.
Гиоид вместе с сросшимися непарными частями (copulae) и остат-
ками двух первых жаберных дуг входит в состав подъязычного
аппарата, состоящего из тела и трех пар рожков.
В общем череп рептилий претерпевает ряд существенных изме-
нений прогрессивного порядка, связанных с дальнейшими приспо-
соблениями к наземному существованию, и усовершенствованием
органов чувств, особенно же в связи с приспособлением к захва-
тыванию пищи. Бросаются в глаза маленькая черепная коробка
рептилий и сильно развитая разнообразно устроенная лицевая
часть в связи с различными способами хватания и умерщвления
добычи.
Конечности и их пояса претерпевают у рептилий ещё больше
изменений в связи е чрезвычайным разнообразием тех приспособле-
ний, которые имели место в эволюции рептилий, занимавших разно-
образнейшие среды обитания. Исходной формой их были пяти-
палая конечность и примитивные пояса стегоцефал. Так, у при-
митивных рептилий в плечевом поясе еще имеется много хряща:
надлопаточный (suprascapula), прокоракоидный
(procoracoideum), и эпикора к оидный (epicoracoideum);
425
у них имеется егце покровная кость — к л е й т р у м (cleithrum);
лопатка (scapula) и коракоид (coracoideum) у рептилий
окостеневают как замещающие хрящ окостенения; имеются клю-
чицы (claviculae) и лежащее между ними непарное покровное
А 2 0 с о
Рис. 329. Левая половина плечевого пояса некоторых
рептилий слева:
А — Sphenodon, В — Iguana, С — Ch el one, D — Crocodllus.
1—scapula, 2 — coracoid, 3 —processus acromion, акромиаль-
ный отросток лопатки, 4 и 5 — fossa glenoidalis, сочленовная
ямка (По Goodrich.)
окостенение — падгрудинпик (episternum), соответствую-
щий межключице — (interclavicula) стегоцефал.
У современных рептилий клейтрум (cleithrum) отсут-
ствует, равно как и у большинства ископаемых. Так как коракоиды
причленяются к грудине с помощью эпикоракоидного хряща,
а эпикоракоидпые хрящи обеих сторон, кроме того, соединяются
друг с другом, тогда как ключицы соединяются друг с другом по-
Рис. 330. Таз Anomodon-
tia слева (Moschops са-
pensis).
1 — ilium, 2 — pubis, 3 —
ischium, 4— acetabulum (По
Broom из Ihle.)
средством падгрудипника, а грудина * со-
единяется с некоторым количеством ре-
бер, то плечевой пояс рептилий оказы-
вается очень прочным. Однако в его строе-
нии замечается значительное разнообразие.
Так, у крокодилов ключицы и прокора-
коиды отсутствуют (рис. 329), а у черепах
ключицы входят в состав брюшиого щита,
образуя там переднее боковые пластинки
(epi plastron).
Тазовый пояс рептилий тоже очень
близок к таковому некоторых стегоцефал.
Точно так же п о д в з д о ш и а я (ilium),
седалищная (ischium) и лобко-
в а я' (pubis) кости располагаются вокруг
сочленовной ямки бедра или
вертлужной ямки (acetabulum)
(рис. 330). Позднее обнаруживается между
седалищной и лобковой костями расхожде-
ние: сперва в виде небольшого отверстия, затянутого перепонкой (fe-
nestra ischio-pubica); затем у ящериц, у крокодилов оно значительно
увеличено, а у динозавров лобковые и седалищные кости сильно
развиты, и от лобковой кости отходит направленный назад з а д-
426
Рис. 331. Скелет передней
ноги примитивной рептилии
Ophiacodon.
1—condylus radialis, 2—con-
dylus ulnaris, 3 — foramen en-
tepicondyloideum, 4 — головка
плеча, 5—radius, G — ulna, 7—
radiate, 8 — ulnare, 9 — inter-
medium, 10 — centralia, 11 —
carpalia distalia, 12—metacar-
pales. (По Williston из Ihle.)
нелобковый отросток (processus postpubicus). У че-
репах и крокодилов седалищно-лобковое окно соединяется с ка-
налом для прохождения нерва (canalis obturatorius), образуя
foramen obturatum. Обе половины пояса у рептилий связаны по-
середине хрящом, продолжающимся иногда в направленный вперед
отросток (processus epipubis).
Пятипалая конечность низших стегоцефал с нормальным числом
косточек запястья являлась для рептилий исходной формой и в зна-
чительной мере сохраняет этот харак-
тер у неспециализированных форм. Пле-
чо и бедро у низших примитивных
рептилий лежали еще в горизонталь-
ной плоскости перпендикулярно к оси
тела, однако предплечье и голень уже
приобретали более прочное и эффек-
тивное для движения положение: пред-
плечье направлялось своей дистальной
частью вперед и вниз, голень — назад
и вниз.
В связи с таким несовершенным по-
ложением конечностей, требовавшим
очень сильной мускулатуры для поддер-
жания тела и передвижения по земле,
плечо было устроено у низших репти-
лий сходно с тем, что мы видели у сте-
гоцефал: оно короткое и имеет два
очень сильных расширения для при-
крепления мышц: проксимальное и ди-
стальное (рас. 331). В дистальном рас-
ширении плеча у рептилий имеются два
отверстия. Одно из них лежит наружу
от мыщелка, служащего для причле-
нения лучевой кости (condylus radi-
alis), и продырявливает расширенную
часть плеча, или надмыщелок (epicon-
dylus radialis). Отверстие это называется foramen ectepicondy-
loideum. Другое отверстие — foramen entepicondyloideum — ле-
жит с другой стороны, проходя через надмыщелок локте-
вой кости (cpicondylus ulnaris). Через первое проходит лучевой
нерв (n. radialis), через второе — срединный нерв (n. medianus)
и плечевая артерия (arteria brachialis). Оба отверстия имеют клюво-
головые (Rhynohocephalia); foramen entepicondyloideum имели
котилозавры, пеликозавры, звероящеры и некоторые другие;
ящерицы (Lacertilia) и черепахи (Chelonia) имеют одно отверстие—
foramen ectepicondyloideum. У крокодилов нот этих отверстий.
Головка .плеча у низших рептилий устроена сложно и нередко
разделена на две части гребнем.
У высших рептилий, шагавших и бегавших, плечо соответ-
ственно изменено, оно в большей или меньшей степени направлено
427
назад, длиннее, тоньше и без надмыщелковых отверстий, что стоит
в связи с меньшим развитием мускулатуры, сгибавшей предплечье.
Предплечье у родоначальных рептилий было тоже короткое и на-
правлено косо вниз и вперед. Локтевая и лучевая кости лежали
рядом и были мало подвижны в отношении друг к другу. Локтевой
вырост — processus olecranon — отсутствовал. Малоподвижной бы-
ла также кисть. Она состояла из нормального числа косточек
запястья и пясти, причем нередко имело место сращение carpale
4 и б; из пястных косточек пятая укорочена; число фаланг перво-
начально было, вероятно, 2, 3, 4, 5, 3 (4), но затем в связи с при-
способлением к разным видам движения подверглось сильному
видоизменению.
Что касается задней конечности, то она у низших рептилий
также очень близка к тому, что было у их предков —стегоцефал.
Короткое бедро располагалось горизонтально и перпендикулярно
к телу, имело сильно развитой гребень для прикрепления мускулов.
Большая берцовая кость развита у рептилий больше, как главная
кость, поддерживающая тело и служащая для прикрепления важ-
нейших мышц; она снабжена двумя мыщелками.
Значительные изменения претерпевает ступня рептилий. Только
у самых низших котилозавров в проксимальной ее части имеются
три косточки. У остальных рептилий их только две: таран-
ная (astragalus) и пяточная (calcaneus). Относительно их
гомологизации мнения расходятся; невидимому, верна та точка
зрения, по которой таранная кость (astragalus) соответствует
intermedium, пяточная (calcaneus) — fibulare. Из центральных
сохраняется одна косточка. Четвертая и пятая дистальные
предплюсневые сливаются; число фаланг на пальцах ноги 2, 3,4,5,3.
В связи с приспособлением к различным средам обитания и со-
ответственно к разным способам движения у рептилий мы встре-
чаем ряд существенных изменений в скелете конечностей. При
приспособлении к более совершенным движениям на суше конеч-
ности меняют, как мы видим, свое положение: передняя конеч-
ность все больше и больше поворачивается локтем назад, а задняя
коленкой вперед. Одновременно плечо и голень становятся длиннее
и тоньше. В предплечье radius и ulna соответственно с поворотом
локтя перекрещиваются, так что в локте radius оказывается сна-
ружи, а в кистевом суставе снаружи — локтевая кость. В запястье
исчезает intermedium, и из центральных остается лишь одна ко-
сточка. Между проксимальным и дистальным рядом костей обра-
зуется интертарзальный сустав.
В связи с переходом к передвижению па двух ногах у дино-
завров имел место ряд изменений как в передней, так и в задней
конечностях. Передние конечности становились очень короткими;
запястье часто состояло из плоских косточек; в кисти четвертый
и пятый пальцы рудиментарны или исчезали, остальные три пальца
были снабжены когтями.
Наоборот, задние конечности у динозавров очень мощные;
бедро с хорошо развитым четвертым вертлугом в связи с сильным
428
развитием чрезвычайно важных при двуногом хождении мускулов,
отводящих бедро назад; большая берцовая кость развита гораздо
сильнее, чем малая; в связи с полным развитием интертарзального
сочленения обе берцовые кости вместе с таранной и пяточной
костями становятся одним целым, срастаясь друг с другом в боль-
шей или меньшей степени, образуя голеноплюсневую
кость (tibiotarsus). В связи с тем, что основное давление при
пятипалой конечности приходится на средний палец, последний
приобретал при беге особое значение, тогда как
крайние — первый и четвертый — постепенно те-
ряли свое значение. Нога становилась среднеос-
ной — мезаксиальной, с тенденцией превращения
в трехпалую конечность (рис. 332). Сперва исче-
зал окончательно пятый палец, тогда как первый
или делался рудиментарным или противополагался
остальным. Последнее связано, однако, с приспо-
соблением к древесному образу жизни. Число фа-
ланг 2, 3, 4, б, 0, при третьем самом длинном
пальце. Ноги в общем приобретали в большей или
меньшей степени сходное с птицами строение. При
этом у динозавров, бегавших и ходивших с подня-
тым хвостом, подвергся аналогичному преобразо-
ванию и таз: именно ischium было сильно оттянуто
назад, а от pubis назад отходил длинный отросток
(postpubis). При вторичном переходе от двуногого
передвижения к четвероногому, что в процессе
эволюции рептилий имело место не менее двух
раз (у динозавров и крокодилов), особенности,
полученные при приспособлении к двуногому пе-
редвижению, так или иначе сохранялись, перед-
ние конечности становились естественно в связи
с их короткостью пальцеходящими, пальцы их
короткими, с малым количеством фаланг: 2, 1,
1, О. У крокодилов передние конечности отли-
чаются отсутствием olecranon на локтевой кости; radiale и ulnare
удлинены; intermedium отсутствует, имеется лишь три карпаль-
ных косточки. Три первых пальца развиты сильнее и снабжены
Рис. 332. Ске-
лет правой ноги
Ornithomimus.
1— tibia, 2— Hbu-
la, 3 —calcane-
us, 4 —astragalus,
5a__j — metatarsa-
lia. (По Lambe из
Ihle.)
когтями; при передвижениях крокодилы опираются на пальцы.
Задняя конечность крокодилов — четырехпалая, с хорошо раз-
витым межтарзальным сочленением. При быстром движении кро-
кодилы приподнимаются на ногах довольно высоко и отталкиваются
ногами поочередно.
Лазание по деревьям у рептилий нередко приводит к специали-
зации конечностей, служащих для обхватывания ветвей, с про-
тивоположением одного или двух пальцев остальным. Первое мы
встречаем на задних конечностях у ископаемых древесных Pseudo-
suchia и у древесных динозавров, второе — па передних и задних
конечностях у хамелеонов. В связи с этим меняется расположение
пястных и плюсневых косточек.
429
У многих рептилии ящорицеобразного типа мы наблюдаем обра-
зование эволюционных ветвей, имеющих тенденцию к полному
исчезновению конечностей. При этом конечности исчезают не сразу,
а путем постепенного укорочения.
Своеобразный, хотя и примитивный, характер имеет скелет
конечностей у наземных черепах в связи с наличностью панцыря.
Так, локтевой сустав повернут несколько вперед, плечо сильно
изогнуто, и предплечье становится отвесно к земле, причем обе
кости предплечья сохраняют свое параллельное положение. Число
костей запястья нормально, но из двух закладывающихся централь-
Рис. 333. Левая передняя конеч-
ность черепахи Dermochelys, без
плеча.
1—radius; 2—ulna; 3—radiale; 4—ulnare;
5—intermedium; 6— pisiforme; 7—ccnt-
rale; 8—carpale 1; 9—carpale 4-J-5;
10— пятый палец; 11— кожа.
(По VOlker из Ihle и др.).
Рис. 334. А — передний плавник
Eurypterygius extremus, В—пе-
редний плавник Meriamia zitteli;
триас Калифорнии.
(Из Abel.)
ных косточек остается одна, так как другая обычно сливается с ra-
diale. Имеется гороховидная косточка (os pisiforme).
В задней конечности, кроме типичных для рептилий тараппой
и пяточной костей, имеется четыре тарзальных косточки; единствен-
ная центральная косточка срастается с таранной. Интертарзальпый
сустав развит слабо. Число фаланг на пальцах—2, 3, 3, 3, 3 (2).
В связи с приспособлением к водному образу жизпи у черепах,
как и у многих других водных рептилий, можно проследить па-
раллельное и аналогичное изменение конечностей и превращение
их в плавники.
Они сводятся к следующему: проксимальные части конечности
укорачиваются; наоборот, дистальные части конечности удлиня-
ются или за счет удлинения метатарзальных косточек и фаланг
430
(черепахи, мезозавры, плезиозавры) (рис. 333) или за счет увели-
чения числа фаланг (ихтиозавры и плезиозавры); подвижность
сочленений в конечностях, за исключением, конечно, плечевого
и бедренного сочленений, исчезает; все кости, особенно в средней
части конечности, получают вид плоских косточек, прочно
соединенных друг с другом непосредственно или с помощью хряща,
так что вся конечность становится действующей подобно цельному
веслу (рис. 334). В дистальной части плавников фаланги пальцев
оказываются или широко расставленными в связи с тем, что между
ними натягивается плавательная перепонка (у черепах, мезозавров
и др.), или, наоборот, пальцы сближены друг с другом и даже
прочно соединены (у плезиозавров и ихтиозавров). Число пальцев
па конечностях в одних случаях уменьшается (некоторые ихтио-
Рис. '335. Скелет Pteranodon; правые передняя и задняя
конечности не изображены.
1—лопарка (scapula),2 —сросшиеся грудные позвонки, 3 — крестец
(sacrum), 4 — подвздошная кость (ilium), 5 —- предлобковая кость
(os praepubis), 6 —плечо (humerus), 7 —коракоид (coracoid)*,
8 — киль грудины, 9 — грудина (sternum), 10 — 4-я пястная кость
(metacarpale), 11 —пястные кости и пальцы 1, 2, 3; 12 — палец 4-й.
(По Eaton из Ihle)
завры), и плавники становятся узкими и длинными; в других
случаях, наоборот, число пальцев увеличивается, и плавники
становятся широкими.
Своеобразные изменения претерпевают конечности у летающих
птерозавров. Здесь передняя конечность превращена была в крыло.
Летательная перепонка патяиута была между туловищем и сильпо
удлиненным четвертым пальцем с его удлиненной метакарпальной
косточкой. Три первых пальца были свободны и снабжены ког-
тями. Пятый палец отсутствовал (рис. 335).
Интересен своим строением таз птерозавров, в котором, помимо
рудиментарной лобковой кости, ограничивающей вертлужную впа-
дину (foramen obturatorium) и совершенно слившейся с подвздошной
и седалищной в одну кость, имелась причлепяющаяся к лобковой
кости , предлобковая кость, сходпая с таковой у крокодилов.
Приспособлением к полету такое устройство таза объяснено быть
431
ие может, п наиболее вероятным предположением является до-
пущение, что предками птерозавров, равно как и крокодилов,
были животные, двигавшиеся на задних конечностях подобно
бегавшим на задних ногах динозаврам и птицам.
5. Мускулатура рептилий
Мускулатура рептилий представляет дальнейшую ступень ос-
ложнения в сравнении с мускулатурой амфибий. С полным перехо-
дом рептилий к наземной жизни, с исчезновением жабер и жаберных
дуг, с одной стороны, и с развитием большей подвижности — с дру-
гой, как висцеральная, так и туловищная мускулатура должны были
подвергнуться серьезным изменениям. Вместе с превращением
жаберных дут в часть подъязычного хряща и в хрящи гортани
висцеральные мускулы, снабжаемые от n. vagus должны были
измениться и подвергнуться частично редукции. Зато та группа
иннервируемых n. vagus мышц, которая носит название системы
m. trapezius, в связи с передвижением рептилий по суше, сильно
развилась и специализировалась. Эта система намечена уже у се-
лахий, у которых от спинной фасции и миосепт спинных мускулов
отходит мускул к верхней части плечевого пояса и нередко к верх-
ней части последней жаберной дуги, а в виде исключения — ко
всем жаберным дугам. У амфибий эта группа мышц развита лучше.
У рептилий увеличивается подвижность плечевого пояса, а также
необходимость иногда фиксировать последний. В связи с этим
усиливается развитие группы musculus trapezius. Иннервируется
здесь этот мускул добавочным нервом (n. accessorius XI), ставшим
самостоятельной ветвью блуждающего нерва.
Из мускулатуры гиоидной дуги, принадлежащей к числу мышц,
иннервирующихся лицевым нервом (n. facialis VII), у рептилий
имеются две мышцы: сжиматель шеи (m. sphincter colli) и мускул,
опускающий челюсть (m. depressor mandibulae). Мышцы челюстной
дуги, иннервирующиеся пятой парой нервов (n. trigeminus V),
у рептилий будут: происшедшие из сжимателя — челюстно-
подъязычная (m. mylohyoideus) и происшедшая из т. ad-
ductor mandibulae рыб — височная мышца (т. tempo-
ralis), уже встречающаяся у амфибий; однако в связи с большим
развитием жевательных движений здесь прибавляется начинаю-
щаяся от крыловидной кости крыловидная мышца
(m. pterygoideus). У змей и ящериц в связи с подвижностью квад-
ратной кости и крыловидной кости имеются еще сохранившиеся
от рыб и низших амфибий мускулы, двигающие крыловидную
кость — mm. protractorcs и levatores pterygoidei.
Кожные мышцы у рептилий еще не получили развития. У гат-
терии брюшные ребра соединены с мускулатурой посредством
миокомм; такая же связь кожи с миокоммами имеется у ящериц.
У змей имеется особая мускулатура, выпрямляющая брюшные
щитки при ползании; мышцы эти прикрепляются к концам ребер.
В соматической мускулатуре происходят у рептилий даль-
нейшие изменения в связи с развитием движений по земле и диф-
432
ференциацией туловища на отделы. Эпиаксиальные мускулы туло-
вища дифференцируются на три ряда мышц (рис. 336). Вдоль всего
позвоночника тянется система поперечно-остистых
мышц (mm. transverso-spinales), снаружи от них — система
длинных мышц (m. longissimus) и вдоль ребер — система
подвздошн о-p еберных мышц (m. ileocostalis). Пер-
вые две системы служат для выпрямления позвоночника, тогда
как подвздошные реберпые мышцы служат для боковых змее-
образных искривлений тела. Соответственно они получают особенно
сильное развитие у змей и безногих и коротконогих ящериц.
Миокоммы в этих системах иногда еще хорошо заметны или частич-
но исчезают, и система дифференцируется на ряд продольных мышц
или на слои.
На шее в связи с ее обособлением дифференцируется из этих
систем своя мускулатура, служащая для поднимания головы,
боковых движений шеи и т. д.
Рис. 336. Спинные мускулы ящерицы (Lacerta mu-
ralis) па поперечном разрезе через туловище.
1 — позвонок, 2 —mm. transverso-spinales, 3 —m. longissi-
mus, 4—mm. ileocostales, 5 — гипаксиальпые мускулы. (Но
Vallois из Ihle.)
*
На хвосте, наоборот, сохраняется примитивное метамерное
расположение мыпщ с хорошо выраженными миокоммами.
Что касается гипаксиальпой мускулатуры, то у рептилий
в связи с измененной дыхательной деятельностью с помощью ребер
развивается мускулатура ребер. Так как у примитивных рептилий
ребра были развиты вдоль всего туловища, то и реберная муску-
латура была соответственно развита по всему туловищу, как у гат-
терии. Всего здесь имеется шесть слоев мышц. Два наруж-
ных (m. obliquus externus superficialis и obliquus externus pro-
fundus) и два внутренних (m. obliquus internus и m. trans-
versus) соответствуют таковым амфибий; между ребрами же здесь
появляются два слоя новых мышц: меж реберные внеш-
ние и внутренние (m. intercostalis externus и internus)
(рис,. 337).
На брюхе располагается система прямых мышц (m. rectus)
в виде одной или двух мышц, тянущихся вдоль брюшной стороны и
разделенных на сегменты миокоммами. У Sphenodon прямая мышца
тянется до плечевого пояса, покрывая грудину; у высших рептилий
28 Каш каров и С гаачинекий 433
из области плечевого пояса прямая мышца живота оттеснена раз-
вившимися мышцами конечностей.
На шее развиваются и удлиняются в связи с появлением шеи
мышцы, уже имевшиеся у амфибий: m. sterno-hyoideus и m. genio-
hyoideus.
Рис. 337. Внешняя мускулатура крокодила.
1 — m. obliquus externus superficialis, 2 —т. obliquus externus profundus, 3 —m.
obliquus internus, 4 — m. transversus, 5— m. intercostalis externus, 6—m. intercostalis
internus, 7 — m. rectus. (Из Biitschli.)
В связи с более совершенными движениями по суше и вторичным
приспособлением конечностей к передвижению в воде сильно раз-
вивается и видоизменяется мускулатура конечностей.
6. Органы пищеварения рептилий
В связи с развитием большей подвижности и активности в за-
хватывании добычи ротовая полость рептилий под-
вергается ряду важных изменений в процессе эволюции этого
класса. Во-первых, весьма существенным является ее преобра-
зование в связи с развитием вторичного нёба, опо отделяет носо-
глоточный ход от вторичной полости рта, чрезвычайно облегчая
дыхание при заглатывании пищи, и делает переднюю вытянутую
часть морды более прочпой; во-вторых, значительной эволюции
подвергаются зубы; далее усложняются и специализируются рото-
вые железы и отчасти язык, и, наконец, ротовая полость в целом
специализируется приспособительно к различного рода пище.
Эволюция твердого пёба пами была дана при рассмотрении
скелета рептилий. Что же касается зубов, то первоначально опи
сохраняют примитивное строение и расположение, характерное
для амфибий, т. е. они мелки, имеют простую коническую форму
и сидят пе только па челюстных и межчелюстных костях в верхней
и па зубных в пижпей челюсти, но и на нёбных и крыловидных
костях и на сошниках; у современных рептилий на сошниках сидит
по одному зубу у гаттерии, и па нёбных и крыловидпых костях
зубы имеются у некоторых ящериц и змей. Однако в процессе эво-
люции класса замечается значительная специализация в строении
зубов. Их расположение сосредоточивается на челюстях. Здесь
они первоначально прирастают к внутренней поверхности челюсти,
434
образуя наименее прочную зубную систему, — это п л евро-
до н т н ы е зубы; более прочным типом расположения зубов
будут акродонтные зубы, сидящие соответственно на верх-
ней или нижней поверхности челюстей; наиболее же совершенный
тип положения зубов будет тот, когда зубы сидят в ячейках (аль-
веолах) — текодонтные зубы: такие зубы мы встречаем
у крокодилов и у некоторых ископаемых зверозубых ящеров (Тйе-
riodontia). Далее первоначально одинаковые зубы (гомодонтная
система) уже у рептилий начинают дифференцироваться: у иско-
паемых зверозубых ящеров можно различить уже несколько кате-
горий зубов — передние, соответствующие резцам млекопитающих,
высокие конические клыки и бугорчатые задние — коренные (г е т е-
родоптпая система). У ядовитых змей дифференцируются ядо-
витые зубы, отличающиеся пе только величиной, но и тем, что опи
снабжены бороздой или каналом, по которому яд стекает в рану.
- Замена зубов происходит постоянно по мере снашивания, т. е.
у рептилий существует п о л и ф и о д о и т и я. У многих pen-
тплий происходит своеобразная редукция зубов. У черепах зубы
исчезают совершенно; их заменяет роговой клюв — чехол, покры-
вающий верхнюю и нижнюю челюсти.
Что касается желез во рту рептилий, то сверх уже имеющейся
у амфибий межчелюстной железы (gl. intermaxil-
laris) и нёбных (gl. palatinae) у них имеются подъязыч-
ные железы, (gl. sublinguales) и верхние и нижние
губные железы (gl. labiales superiores и inferiores). Именно
эти верхпегубпые железы и становятся у змей ядовитыми. Опи
находятся в связи с ядовитыми зубами, через желобок или капал
которых происходит излияние ядовитого содержимого в рапу,
благодаря сокращению жевательной мышцы.
. Язык ‘ рептилий по сравнению с амфибиями отличается прежде
всего более сложным строением и происхождением. Он развивается
из двух последовательно расположенных зачатков — пе только
из лежащих впереди гиоидной дуги, по и из зачатка, лежащего
позади, в области второй и третьей жаберной дуг; от слияния
этих зачатков и двух боковых и получается язык рептилий. В связи
с таким более сложным развитием иннервация языка тоже более
сложная: помимо подъязычного и языкоглоточного нерва, он
иннервируется еще тройничным первом. Скелет языка пред-
ставлен подъязычным аппаратом, описанным выше; его непарная
часть — тело (basihyale) — имеет длиппый отросток (ent»glossum).
Длина и подвижность языка очепь различны. У ящериц и змей
язык длинный и подвижный, втягивающийся в особое влагалище;
у хамелеона он очень длиппый, клейкий на конце, где имеются
специальные железы, выделяющие клейкое вещество.
Общая форма головы, шеи и в особенности морды, очертания
ротовой щели, строение зубов, железы ротовой полости, язык —ч
все это части тела, па которых приспособление к роду пищи и спо-
собу ее захватывания сказывается особенно сильно, и у рептилий
они в связи с многообразием приспособлений уже весьма различны
аз* 435
по сравнению с тем, что имеет место у амфибий. Однако и здесь
приспособления не достигают того разнообразия и высокой специа-
лизации, какое мы встречаем у млекопитающих. Приспособления
рептилий к роду нищи касаются прежде всего способа захватывания
растительной или животпой пищи; эволюция рептилий шла по пути
выработки разнообразия формы и величины пасти, величины
и числа зубов, по пути увеличения мощности мускулов, длины
и подвижности головы.
В пропорции частей головы рептилий, особенно ископаемых,
поражает разнообразие в величине всех указанных выше особен-
ностей при неизменно маленькой черепной коробке, в которой
помещался головной мозг. Эволюция рептилий шла по пути усо-
вершенствования конечностей и челюстей, а не по пути развития
инстинктов и привычек, связанных с отысканием пищи, как это
имело место у птиц и млекопитающих. Все же по сравнению с не-
поворотливыми и неуклюжими амфибиями с их примитивным
способом добывания пищи рептилии стали па более высокую,
качественно новую ступень. Что касается характера пищи, то
представляет интерес существование среди них, особенно же сре-
ди ископаемых, большого числа растительноядных форм. Среди
амфибий только личинки и, быть может, некоторые стегоцефалы
были растительноядными; среди рептилий растительноядными
являются из современных многие черепахи, а из ископаемых —
Theromorpha, Placodontia, Dinosauria и др.
Что касается собственно пищеварительного аппарата, то оп
остается па примитивной стадии развития, характерной и для ам-
фибий. Желудок простой, и только у крокодилов намечается де-
ление желудка на железистую кардинальную и мускульную пило-
рическую части; средняя кишка относительно короткая, задняя
кишка отделена слизистой складкой; имеется слепой выступ на
ее границе. Кишечный капал открывается в клоаку.
7. Органы дыхания рептилий
Рептилии как настоящие воздушно-паземиые обитатели совер-
шенно потеряли жаберное дыхание; личинок, обитающих в воде,
опи не имеют, а зародыш, развивающийся в яйце, дышит с помощью
аллантоиса. С другой стороны, у рептилий отсутствует и кожное
дыхание; как настоящие воздушно-наземные обитатели рептилии
приобрели приспособления, защищающие от высыхания кожи в виде
роговых образований, одевающих кожу. Единственным органом
дыхания рептилий являются легкие. Естественно, что у рептилий
мы вправе ожидать более совершенного строения легких и более
совершенного дыхания, а с другой стороны — полной редукции,
связанной с дыханием, жаберного аппарата — жаберных выростов,
жаберпых щелей, жаберных дуг и жаберных сосудов. Действи-
тельно, жаберный аппарат редуцировался совершенно, а из не-
которых его зачатков образовались другие органы. Так, у реп-
тилий из первого жаберного мешка развивается барабанная по-
436
лость; из следующих развиваются зобная же-
леза и околощитовидные железы и, наконец,
жаберные тельца, входящие в систему щито-
видной железы. Что касается жаберпых дуг,
то судьба их описана была выше: три первых
жаберпых дуги превращаются в рожки подъ-
язычного аппарата, а остальные принимают
участие в образовании хрящей гортани.
Легкие рептилий постепенно приобретают
гораздо более сложное строение по сравнению
с амфибиями. У гаттерии они еще имеют большое
сходство с легкими амфибий: стенки легкого
имеют лишь более глубокие ячеи, однако уже
имеется пара коротких бронхов, которые входят
сбоку легкого, образуя впереди от бронхов
небольшие верхушки. У более высоко органи-
зованных форм (ящерицы) ячеи увеличиваются
еще больше, образуя глубоко уходящие внутрь
легкого перекладины, которые делят легкое иа
большее или меньшее число отделов. У высших
форм (черепахи, крокодилы) этот процесс
развития внутренних перегородок идет еще
дальше, перегородками заполняется большая
часть легкого — за исключением той средней
части, которая является продолжением брон-
ха; этот канал выстлан мерцательным эпи-
телием и является внутренним дыхательным
путем. Наконец, путем дальнейшего разви-
тия того же процесса, все большего увеличе-
ния перегородок в первоначальных ячеях
образуются перегородки второго порядка,
которые также заполняют в ячеях все про-
странство за исключением средней части,
становящейся продолжением бронха — брон-
хами второго порядка (рис. 338). Таким обра-
зом, все легкое оказывается разделенным на
большое количество ячей, или камер, к кото-
рым ведут внутренние разветвления бронхов.
В стопках внутренних бронхов у черепах и
крокодилов закладываются хрящи, которые
поддерживают его просвет. У ящериц и змей
задние части легкого иногда остаются нераз-
деленными или слабо разделены иа ячеи, а
у некоторых ящериц, у гекконов и хаме-
леонов легкие имеют своим продолжением
тонкие и длинные выросты — л е г о ч и ы е
м е in к и (рис. 339). В связи с удлинением
тела у некоторых рептилий и соответствен-
ным уменьшением ширины тела наблюдается
Рис. 338, Схема
строения легких
рептилий.
1 — внелегочный
бронх, 2 — внутри-
легочный бронх, 3—
главная септа с си-
дящими на ней вто-
ричными септами,
4—вторичный бронх.
(По Vialleton, из
Ihle.)
мешки на легких ха
мелеона.
(По Wiedersheim.)
43
развитие асимметрии. Положение двух рядом лежащих легких, пра-
вого и левого, в топком, вытянутом в длину теле змеи становится
затруднительным, и они располагаются асимметрично: одно легкое
становится короче, другое, наоборот, длиннее. Этот процесс неравно-
мерного развития легких может закончиться редукцией одного
легкого, как это имеет место у змей.
Трахея у рептилий развита очень хорошо и поддерживается
в постоянном просвете многочисленными хрящевыми полными
или неполными кольцами; в задней части трахея делится на пару
бронхов, также поддерживаемых хрящевыми кольцами.
Что касается гортани (larynx), то устройство ее у рептилий
мало подвинулось вперед в сравнении с амфибиями. Передняя
пара хрящей — черпаловидные хрящи (cartilago агу-
tenoidea) — развита хорошо, задние хрящи представлены непар-
ным перстневидным хрящом (с. cricoidea), назади
незамкнутым, образующим таким образом неполное кольцо, по
щитовидного хряща (с. thyroidea) еще пет.
У гекконов и хамелеонов имеется голосовой аппарат, пред-
ставленный парой голосовых складок.
Дыхательный процесс рептилий решительно отличается otf
дыхательных движений амфибий. Вместо заглатывапия воздуха
у рептилий совершается втягивание воздуха в легкие и выталки-
вание его наружу путем периодического расширения и сжатия
грудной полости, что достигается движением ребер, производимым
с помощью брюшпых и реберных мышц. Это гораздо более совер-
шенный тип дыхания, ведущий к более энергичному обмену ве*
ществ и к развитию большей энергии.
Отсюда следует, что родоначальниками рептилий могли быть
только стегоцефалы с хорошо развитыми ребрами. Только более
энергичное дыхание в связи с более совершенным кровообращением
могло компенсировать рептилиям потерю ими кожного дыхапия
и дать возможность эволюционировать дальше.
8. Органы кровообращения рептилий
Передвижение по земле требует гораздо большей затраты
энергии, нежели передвижение в воде, поэтому эволюция рептилий
в качестве воздушно-наземных обитателей возможна была только
при условии усовершенствования их дыхательных процессов и свя-
занного с дыханием кровообращения. В органах кровообращения
рептилий действительно имеется по сравнению с амфибиями ряд
более полных приспособлений как в сердце, так и в артериальной
и венозных-системах.
Сердце большинства рептилий хотя и остается еще трехкамер-
пым, по зато разделение крови на артериальную и венозную у них
осуществляется гораздо более совершенным путем, именно тем,
что в предсердии перегородка полная, и тем, что и в желудочке
имеется поднимающаяся с брюшной стороны перегородка. Она
438
Рис. 340 Схема серд-
ца рептилий.
1 — перегородка между
желудочками, 2 — пра-
вое предсердие, 3 — ле-
вое предсердие. 4 —же
лудочек (По Ihle.)
доходит до спинной стороны только у крокодилов, превращая
тем самым трехкамерное сердце в четырехкамерное; у остальных
рептилий желудочек оказывается разделенным неполно. Тем не
менее при сокращении такого трехкамерпого
сердца перегородка замыкается на спинной
стороне желудочка, отделяя правую веноз-
ную часть от левой артериальной (рис. 340).
Венозный сипус, свойственный амфибиям,
здесь входит в состав предсердия и только
у гаттерии различим снаружи, тогда как
артериальный конус редуцируется и от него
остаются собствепио только клапаны.
Артериальная система. Весьма существен-
ным усовершенствованием является разделе-
ние общего артериального ствола па три са-
мостоятельных ствола. От правой венозной
половины отходит ствол легочных артерий,
который делится на правую и левую арте-
рии; от левой артериальной половины же-
лудочка отходит правая дуга аорты, от ко-
торой по выходе из сердца ответвляются
сонные и подключичные артерии; левая дуга аорты начинается
самостоятельным корнем от правой половины желудочка, но в не-
посредственной близости от перегородки, так что получает сме-
Рис. 341. Схема сердца и
артерий у черепах.
1 — аорта, 2 —дуга аорты, 3 —
arteria coeliaca, 4 — легочная
артерия. (По G-oodrich.)
4 4
Рис. 342. Сердце и дуги аорты крокодила
(Л) и птицы (В).
1 и Г —сонные артерии, 2 и 2'-—правая и
левая дуги аорты, 3 и 3'— легочные артерии,
4 и 4'— правое и левое предсердие, 5 и 5'—<
правый и левый желудочек, 6 — нисходящая
аорта, 7 —сосуды, отходящие к кишечнику.
(По Boas из Шмальгаузена.)
танцую кровь; обогнув с левой стороны сердце, она тонким сосу-
дом соединяется с нисходящим стволом аорты, ответвив предва-
рительно брыжеечную артерию (a. coeliaca). Таким путем легочные
439
артерии, с одной сторопы, правая дуга — с другой, получают
кровь соответственно венозную и артериальную, и только правая
дуга аорты несет смешанную кровь (рис. 341). Отсюда уже один
шаг к полному отделению правой и левой половин путем редукции
левой дуги аорты, что и имеет место в дальнейшей эволюции однако
лишь у птиц (рис. 342).
Нужно считать, что описанное выше строение сердца и распо-
ложение отходящих от пего артериальных стволов, характерное
для современных рептилий, является вторичным, и этому состоя-
нию предшествовала стадия, когда корепь аорты разделился па
ствол легочных артерий, отходящий от правой половины желудочка,
и корень системной аорты, отходящий от левой половины и не-
сущий обе дуги (правую и левую) аорты. Что такая стадия свой-
ственна была рептилиям, ясно из того, что только из такой стадии
может быть выведена та ветвь рептилий, от которой произошли
млекопитающие, так как у этих последних в противоположность
птицам имеется лишь левая дуга аорты. Очевидно, млекопитающие
произошли от рептилий, у которых редукции подверглась не левая,
как у современных рептилий, а правая дуга аорты. Эмбриональное
развитие современных рептилий точно так же указывает па суще-
ствование стадии, па которой ствол легочных артерий был уже
отделен, а ствол системных артерий оставался общим.
Из других особенностей артериальной системы рептилий, ука-
зывающих на ее переходный промежуточный характер, необходимо
отметить, что у ящериц еще сохраняется проток между сонными
артериями и дугами аорты, соответствующий третьей аортальной
дуге рыб, у черепах сохраняется боталлов проток между дугами
аорты и легочными артериями, т. е. между четвертой и шестой
жаберными артериями.
Венозная система. Переходный характер кровепоспой системы
рептилий сказывается также и па венах: начавшаяся у них редук-
ция воротной системы почек не идет до конца. Хвостовая вена
соединяется с подвздошными венами, которые дают общую с пей
ветку в почки — воротную вену почек (v. portae
renales); из почек кровь поступает в нижнюю полую вену. Таков
путь только части крови, идущей от хвоста. Большая часть крови
с задней части тела поступает в брюшные вепы, парные у кроко-
дилов и сливающиеся в один ствол у остальных рептилий; брюшные
вепы впадают в воротную вену печени (рис. 343).
В связи с таким движением крови по вепам происходит редук-
ция задних кардинальных вен, которые превращаются в незначи-
тельные позвоночные вены. Соответственно происходит и редукция
кювьеровых протоков, превращающихся в общие стволы, образо-
вавшиеся от соединения яремных веп с подключичными.
9. Мочеполовая система рептилий
Выделительная система рептилий по сравнению с таковой ам-
фибий также отличается существенными особенностями. Во-первых,
у рептилий функционирующим органом выделения служит уже
440
вторичная почка (metanephros). Эта последняя разви-
вается в задней части полости тела, позади зачатков первичпой
почки, из общей с ней надпочечной ткани, подобно тому как раз-
виваются вторичные пер-
вичнопочечные каналь-
цы, группами, без пра-
вильного метамерного
расположения. Каналь-
цы эти пе имеют воро-
нок (нефростомов) и на-
чинаются сразу мальпи-
гиевым тельцем. Одно-
временно с этим из зад-
ней части вольфова про-
тока вырастает канал,
направляющийся к ме-
сту закладки вторичной
почки, который после
соединения с вторично-
почечными канальцами
превращается в моче-
точник (ureter); сам
вольфов проток превра-
щается в семяпро-
вод (vas deferens). Со-
ответственно с разви-
тием вторичной почки
происходит редукция
первичной ^почки, кото-
рая, равно* как и пред-
почка, становится в
своей выделительной ча-
сти эмбриональкыморга-
ном. Предпочка у заро-
дыша хотя и заклады-
вается, по развита слабо;
Рис. 343. Полу схематическое изображение ве-
нозной системы ящериц с брюшной стороны.
1 — хвостовая вена, 2 —воротная вена почек, 3 —
почечная артерия, 4 — нижняя полая вена, 5 — ле-
ван подключичная вена, 6 — печень, к которой
от кишечника подходит воротная вена, 7 — пече-
ночная вена, 8 — косая вена, vena azygos, 9 —под-
вздошная вена, 10 —аорта, 11 — брюшная вена,
12 — кишечник. (Из В. ПаНсг по llochstetter.)
первичная почка у заро-
дыша и отчасти даже у
молодых животных сей-
час ясе после вылупле-
ния (ящерицы) функцио-
нирует в качестве выде-
лительного органа, по
затем редуцируется в
своей выделительной части совершепио. Задний отдел его постиг
название желтых тел (paradidymis), передний половой отдел
первичной почки превращается у самцов в придаток с е-
м е п п и к a (epididymis). Вторичные почки взрослых рептилий
лежат в области таза и представляют собой многодольчатые тела;
441.
мочевые канальцы впадают в мочевые протоки, которые затем впа-
дают в мочеточник (рис. 344).
Половые органы самца и самки, т. е. семенники и яичники,
закладываются, как и у амфибпй, па спинной стороне полости тела
вдоль брыжейки, между пей
и почками, в виде эпите-
лиальных генитальных скла-
док; эти складки разрастают-
ся, врастают в соединитель-
ную ткань и обособляются
в виде подвешенных к бры-
жейке тел, состоящих из
округлых фолликулов, содер-
жащих в себе эпителиальные
клетки, окруженные соеди-
нительнотканной стромой.
У самцов эти фолликулы
вытягиваются затем в семен-
ные трубочки. С ними в
связь вступают вырастающие
из половой части первичной
почки тяжи из эпителия, пре-
вращающиеся в с е м я в bi-
ll о с я
Tes-
в
Рис. 344. Половой аппарат черепахи
tudo.
А. 1 — семенник, 2 — семенной канал, 3 — почка, 4 —
кишка, 5 —мочевой пузырь, 6 —половой орган (penis).
В. 1 — яичник, 2 — фаллопиева труба, 3 — яйцевод, 4—
клоака, 5 — брюшина. (По В. Haller.)
Оплодо творение у рептилий внутреннее,
юте я совокупительные органы.
442
щ и e канальцы
(vasa efferentia), по
которым идет вывод-
ной путь из семенни-
ков в вольфов капал.
У самок фолли-
кулы сохраняются,
причем каждый фол-
ликул содержит в
себе одно яйцо, окру-
женное многочислен-
ными питающими его
клеточками. Отще-
пившийся от первич-
нопочечного протока
мюллеров капал об-
разует яйцевод с во-
ронкой па переднем
конце. Яйцевод диф-
ференцируется па от-
делы в связи с тем,
что яйцо одевается
белковой и кожи-
стой оболочками,
причем у самцов име-
10. Нервная система и органы чувств рептилий
В спинном мозгу у рептилий в связи с дальнейшим развитием
парных конечностей еще больше развиваются плечевое и крест-
цовое утолщения. Особенного развития достигает утолщение спин-
ного мозга в крестцовой области у динозав-
ров, где диаметр спинного мозга крестца
превышал головной до 10 раз.
Головной мозг (рис. 345) рептилий по
сравнению с амфибиями обнаруживает ряд
прогрессивных особенностей. Так, полу-
шария переднего мозга гораздо сложнее
развиты и образуют кору (cortex) из
серого вещества. Однако опа развита все же
значительно слабее, чем у нтиц, ие говоря
уже о млекопитающих. Основная масса
переднего мозга все же представлена здесь
полосатыми телами (corpora
striata). Обонятельные доли (lobi olfacto-
rii) по большей части состоят из луко-
виц (bulbus olfactorius) и стебель-
ков (pedunculus olfactorius). Промежуточ-
ный мозг благодаря развитию полушарий
прикрыт ими, и ’сверху видны э п и ф и-
з ы (epiphysis) и теменной орган
(paraphysis), развитой у гаттерий, яще-
риц и многих ископаемых форм очень
хорошо; снизу промежуточный мозг отме-
чается воронкой (infundibulum) с по-
задилежащей мозговой железкой — г и -
п о ф и з о й (hypophysis). Строение темен-
ного органа соответствует строению глаза:
сверху имеется прозрачная утолщенная
степка — линза, а задняя, ей противопо-
ложная, часть пузырька, соответствует
ретине с чувствующими и пигментными
клетками (рис. 34G). Нервные волокна
чувствующих клеток объединяются в нерв,
соединяющий теменной глаз с мозгом.
У гаттерий и ящериц и у многих ископае-
мых форм (котилозавры, пеликозавры и др.) теменной глаз поме-
щался в теменном отверстии теменной кости. Средний
мозг (mesencephalon) развит хорошо и явственно заметен сна-
ружи в виде двух зрительных долей или двухолмия (lota
optici, corpora bigemina). Мозжечок развит значительно, имеет
уже выраженную кору, но все же мал по сравнению с птицами
и млекопитающими.
Из особенностей рептилий в отношении органов чувств нужно
отметить полное исчезновение боковой линии. Ее отсутствие у вод-
443
Рис. 345. Мозг черепахи
(Emys europaea); справа
полушария открыты.
1 — обонятельные доли, 2—
верхняя и нижняя часть
полушария, 3 — полосатое
тело, 4 — эпифиза, 5 — зри-
тельные доли, 6 — мозже-
чок, 7 — продолговатый
мозг, III, V, VII, IX, X,
XI, XII—черепные нервы.
(По В. Haller.)
ных рептилий показывает, что опи развились из настоящих на-
земных рептилий, уже успевших совершенно утратить этот орган
чувств, свойственный рыбам и водным амфибиям.
Из осязательных органов чувств рептилий заслуживают вни-
мания так называемые осязательные волоски, которые распола-
гаются па чешуе у многих ящериц (гекконы и агамы). Это пеболь^
шие волоски, образовавшиеся из ороговевших клеток эпидермиса,
которые сидят па клетках основного слоя, превратившихся в чув^
ствующие клетки. Под ними имеется соедипителыюткаппый сосочек,
в который входят первы и кровеносные сосуды.
Органы равновесия и слуха рептилий характеризуютсянекоторым
усложнением лабиринта, именно путем увеличения выроста улитки,
1 — хрусталик, 2 — зрительные клетки, 3 — пигментные клет-
ки, 4 — теменной нерв, 5 — стекловидное тело, 6 — соедини-
тельная ткань. (Но Новикову.)
которая у крокодилов представляет уже значительный капал (са*
nalis cochlearis), и одновременного усложнения в пей слухового
аппарата. Имеется к р у г л о е око ш е ч к о (fenestrarotunda).
В среднем ухе у рептилий, как уже было сказано выше, имеется
единственная косточка — колонка (columella auris).
Дыхательная область носа. В носу у рептилий наблюдается
дальнейшая дифференциация дыхательной и обонятельной областей.
Дыхательная область, лежащая пиже, делится па преддверие
(vestibulum), открывающееся наружу ноздрями, выстланное пло-
ским эпителием, средний отдел, выстланный цилиндри-
ческим мерцательным эпителием (pars respiratoria), и вцутрен-
н и й посоглоточпый ход (ductus nasopliaryngeus), от-
крывающийся внутрь хоапами, которые, как мы видели, отодви-
гаются по мере развития верхнего нёба все дальше и дальше назад.
Орган зрения. Наиболее развитыми органами чувств у реп-
тилий являются глаза. Как у форм вполне наземных, мы видим
у них хорошо развитые веки. У змей верхнее и нижнее веки сра-
444
щены между собой и прозрачны над глазами. Они защищают по-
движное глазное яблоко спереди. Между яблоком и прозрачными
веками находится слезная жидкость, стекающая в посовую полость
через широкий капал во внутреннем углу глаза. У других рептилий
веки пе срастаются между собой, и более развито нижнее веко,
которое может прикрывать все глазное яблоко. У большинства
ящериц, у крокодилов и черепах имеется еще «мигательная пере-
понка», надвигающаяся па глаз из переднего угла глаза. У хаме-
леонов веко кольцеобразно. Зрачок имеет у рептилий вид или
вертикальной щели или (редко) горизонтальной, или он круглый.
Интересные особенности в сетчатке глаза имеются у черепах: жел-
тые маслянистые капли па концах элементов сетчатки. Капли эти
должны задерживать, не пропускать лучей правой части спектра.
III. КЛАССИФИКАЦИЯ РЕПТИЛИЙ (REPTILIA)
Построить естественную систему рептилий, т. е. такую, которая
бы выражала их филогенетическую историю, выражала бы суще-
ствующие и существовавшие в природе группировки форм, весьма
затруднительно. Филогенетическое дерево можно построить лишь
на основании учета совокупности признаков всех представителей
класса как современных, так и вымерших. Последние известны
нам уже в очень большом числе, по те представители вымерших
груцп рептилий, которых мы знаем, составляют лишь небольшую
часть некогда существовавших. При этом в ископаемом состоянии
сохранились главным образом рептилии водные, для сохранения
остатков которых были более благоприятные условия. В виду
такой отрывочности имеющихся у пас данных о прошлом класса
строить естественную систему, разобраться в генетических взаимо-
отношениях групп весьма трудно. Этим объясняется то, что система
рептилий весьма неустойчива, все время меняется, различна у раз-
ных авторов. По мере накопления новых фактических данных, —
а опи накопляются быстрым темпом, система перестраивается,
все более приближаясь к естественной, выражающей филогенети-
ческие взаимоотношения. Относительно некоторых групп уже
теперь мнения различных авторов сходятся; есть группы, зани-
мающие в системе определенное положение. Другие группы пере-
ходят из одного места системы в другое. Пет у различных иссле-
дователей согласия и относительно систематических категорий,
к которым следует относить ту или иную группу рептилий. Группа,
которая одним исследователем считается за подкласс, другим
признается за отряд, третьим — за сверхотряд. Это необходимо
иметь в виду при изучении системы рептилий. .
Хотя корни ствола рептилий нам еще не известны, тем пе мепее
молено сказать, что ствол этот коренится в ископаемых земновод-
ных. Кроме того, от ствола рептилий отчленились, с одной стороны/
птицы, с другой — млекопитающие. Из этого видно, что строение
класса рептилий, взятого в целом, весьма разнообразно, а потому
дать характеристику класса, включая в нее и современных реп-
445
тилий и ископаемых, весьма затруднительно. Рептилии произошли
от земноводных (видимо от стегоцефал). Последние имели развитие
с метаморфозом. Следовательно, должны были существовать не-
когда животные, уже приобретшие некоторые признаки рептилий,
по еще проходившие развитие с превращением. Рептилии дали
начало птицам. Следовательно, существовали рептилии с разви-
тым уже, в той или иной степени оперением и другими признаками
птиц. Точно так же мы должны допустить, что волосяной покров,
отличающий млекопитающих, получил первое свое развитие уже
у рептилий, имевших и другие черты сходства с млекопитающими,
например, двойной затылочный бугорок. Накопец, ко многим
ископаемым рептилиям совершенно пеприложимо самое название
класса «пресмыкающиеся»: так называемые птерозавры (Ptero-
sauria) вели воздушный образ жизни; ихтиозавры (Ichtyosauria)
были обитателями морей и великолепными пловцами; многие
наземные рептилии передвигались на одних задних ногах, будучи
быстрыми бегунами, как, например, аллозавры (Allosaurus).
Дать характеристику всего класса затруднительно еще потому,
что мы не зпаем организации внутренних органов ископаемых
рептилий. Поэтому в дальнейшем мы пе даем характеристики всего
класса рептилий. Современные ясе рептилии могут быть охаракте-
ризованы следующим образом: рептилии являются
животными пойкилотермными, с непостоян-
ной температурой тела; дышат воздухом
при помощи легких, жабры совершенно от-
сутствуют — даже в течение эмбриональной
жизни; кожа отличается отсутствием в пей
желез, покрыта чешуей, папцырем или го-
лая; кладут яйца или яйцеживородящи;
яйца меробластические, оплодотворение-
внутреннее; развитие проходит без мета-
морфоза; череп сочленяется с позвоночни-
ком при помощи одного затылочного бу-
горка.
. Класс Пресмыкающиеся (Reptilia)
Труп па Anapsida или Prosauria
Вымершие рептилии, с черепом, построенным по типу черепа
стегоцефал,. без височных дуг. Повидимому, полифилетическая
группа, содержащая представителей наиболее примитивных реп-
тилий каменноугольной пермской и триасовой эпох. Некоторых
представителей почти невозможно отделить от стегоцефал.
1. Подкласс Котелкочерепные (Cotylosauria)
Древнейшие рептилии из пермо-карбопа, пермских отложений
и триаса. Череп «стегальный» с совершенно замкнутой крышей,
без височных дуг. Кости черепа у большинства с грубой скульпту-
446
рои, как у стегоцефал. Позвонки амфицельные. Конечности пяти-
палые, большей частью с короткими и толстыми фалангами. Пле-
чевая кость нередко с расширенными верхними и нижними кон-
цами, как у стегоцефал. Тяжелые плечевой и тазовый пояса напо-
минают таковые стегоцефал (сохраняются cleithra), но образованы
сросшимися костями, имеют уже вид пояса рептилий. В черепе
было пииеальпое отверстие.
С эволюционной точки зрения, чрезвычайно интересно то, что
некоторых ископаемых лишь с известной натяжкой можно отне-
сти к низшим рептилиям: настоль-
ко близки опи к стегоцефалам,
что некоторые авторы и относят
их к последним. Другие авторы,
наоборот,- некоторых стегоцефал
относят к рептилиям. Таковыми
являются представителп отрядов
Rhizosauria и Seymou-
riamorpha. Мы помещаем их
среди рептилий, так как, сохраняя
в основном характер стегоцефал,
они обладают уже признаками,
свойственными рептилиям. Опи
являются, таким образом, морфо-
логически промежуточными звень-
ями, будучи и геологически наи-
более древними. Некоторые изве-
стны уже из каменноугольных от-
ложений.
Отряд 4. Rhizosauria
Рис. 347. Seymouriabailorensis, из
Сюда принадлежат древнейшие из перми Тексаса.
известных рептилий с весьца примитив- (По WilUston из Abel.)
ной организацией. Хорда сохранялась
всю жизнь. Гемальные дуги были еще
мало редуцированы, напоминая в этом отношении позвонки стегоцефал.
Череп небольшой, с вытянутыми назад углами, напоминающий череп Micro-
sauria, относимых одними к амфибиям, другими — к примитивным репти-
лиям. Eosauravus — из каменноугольной формации Сев. Америки.
Отряд 2. Seymouriamorpha
Одно семейство — Seymouridae, наземные, вероятно, насекомоядные
рептилии, длиной около 0.5 м. Представитель семейства рода Seymouria из
перми Тексаса (рис. 347) является одним из примитивнейших из доселе
известных пресмыкающихся. Череп треугольный с париетальным отвер-
стием. Scymouria, обнаруживающая большое сходство со стегоцефалами, в то же
пр<’ми очень близка к отряДу Cotylosauria, стоя наиболее близко к роду
babidosaurus. На Северной Двине была найдена близкая к Seymo-
nria форма, являющаяся ее подродом и названная Kotlassia. Длина ее была
4 25 см.
447
Отряд 3. Cotylosauria
It этому отряду прежде всего относится характеристика под-
класса, данная выше. В сравнении с двумя предыдущими отряда-
ми представители отряда Cotylosauria обнаруживают несколько
высшую специализацию. Приспособление к различной пище на-
чинает вызывать у пих дифференциацию зубов и модификацию кры-
ши черепа благодаря развитию мускулов.
Подотряд Парейазавры (Pareiasau/ria)
Подотряд Парейазавры (Pareiasauria) представлен тремя
семействами.
Сем. Diadectidae. Крепко сложенные животные, от 1 до 3 м
длиной, с мощно, развитым плечевым поясом и крепкими, расширенными на
конце ребрами, крышеобразно налегавшими друга на друга. Передние ко-
нечности очень сильно развитые, пятипалые. Эти рептилии вели образ жизни
броненосцев, были по крайней мере частично роющими животными. Кости
черепной крыши сильно скульптурированы. Diadectes phaseolinus (рис. 306)
Рис. 348 a. Pareiasaurus karpinskii. Пермо-карбон, Северная
Двина.
(По Павловой)
из пермских конгломератов Тексаса и Новой Мексики. Животное медленно
передвигавшееся, наземное, травоядное, рывшее, вероятно, норы для укры-
тия.
Сем. Pareiasauridae. Довольно крупные (до 3 м) рептилии,
тяжелого сложения, с короткими конечностями. Род Pareiasaurus известен
из пермских же отложений на Северной Двине. Р. karpinskii (рис. 348 а)
достигал в длину 2 м 45 см, череп его — 48 см. Род Elginia сходен с Pareia-
saurus, но черен покрыт шипами, особенно в задней части, весьма напоминая
этим голову современной ящерицы из Agamidae—Moloch и голову рода Phry-
nosoma из Iguanidае, у которых, однако, шипы роговые. Образ жизни, ве-
роятно, был, как у названных ящериц.
Подотряд Labidosauria
Отряд обнимает три семейства, из которых наиболее известны роды
Labidosaurus (сем. Labidosauridae) и Limnoscelis (сем. Limnoscelidae).
Labidosaurus, из пермских отложений Тексаса. Небольшое животное
(70 см). Характерны передние зубы в межчелюстных костях, загнутые, как
и последние, вниз (рис. 348 6). Зубы в челюстях— в один ряд. Ловя добычу,
должен был поднимать голову кверху,чтобы ударить передними зубами.
Жил в прибрежной полосе, спускаясь в воду. Limnoscelis paludis, из пермо-
448
карбона Новой Мексики, был значительно крупнее: м. Череп похож на
череп Labidosaurus. Судя по коротким широким конечностям, вел земновод-
ный образ жизни.
Группа Synapslda 1-го типа
Группа Synapsida с монозигальным черепом обпимает два под-
класса: Theromorpha и Pelycosauria, представители которого имели
одну пару височных дуг и, следовательно, одну пару височных
ям (кроме Ophiacodontidae), напоминая в этом отношении мле-
копитающих. Пермские
и триасовые формы. От
примитивных Tlieromor-
pha произошли, видимо,
млекопитающие. Оба
подкласса сипапсид об-
наруживают сходство в
низших представителях,
а высшие их представи-
тели отличаются значи-
тельно.
2. Подкласс.
Pelycosauria
Пермские и триасо-
вые рептилии, главным
Рис. 348 б. Череп Labidosaurus hamatus. Из
перми Тексаса.
1 — parietale, 2 — squamosum, 3 — postorbitale, 4 —
postfrontale, 5—lacrimale, 6 — nasale, 7 —jugale,
8 — quadrato-jugale, 9 — supramaxillare, 10 —adlacri-
male, 11—носовое отверстие, 12 — praemaxillare,
13 —орбита. (По Williston из Abel.)
образом североамериканские, размером от 70 см до 1.5 м. В отли-
чие от Cotylosauria имели не сплошной «стегальный» череп, а с
одной парой височных ям, т. е. череп «зигальный». Височная яма
ограничена сверху посредством postorbitale и squamosum, снизу—
посредством jugale и squamosum, т. е. опа соответствует ниж-
ней височной яме современной гаттерии.
На челюстях сидели*круппые зубы, па нёбе — мелкие. Имелось
отверстие для темепного органа. У многих представителей под-
класса наблюдается па костях черепа скульптура. Некоторые
группы подкласса имели нормальные спинные отростки позвон-
ков, другие — сильно удлиненные и представляли весьма при-
чудливых животных (они вымерли нацело, пе оставив потомков
(‘роди более молодых рептилий).
Отряд 1. Шлемочерепные (Pelycosauria)
Специализированные формы с сильно развитыми остистыми отростками.
X шцные наземные рептилии, достигавшие 1—3 м длины. На образ жизни ука-
зывают острые, сжатые с боков зубы. Наблюдается некоторая дифференци-
ровка зубов; они неодинаковой величины, и между зубами па praemaxillare
и maxiHare имеется свободный от зубов промежуток (диастемма). Череп
yuhiHi и высокий, откуда и название отряда.
Г‘»д Dimetrodon (рис. 349), из пермских отложений Сев. Америки. Ости-
<vi'i4o отрос тки в 25 раз выше тел позвонков и должны были придавать живот-
ному письма страшный вид. Была ли между отростками кожистая перепон-
ки пнипвестпо, мнения расходятся.
29
KiMUiiupou и Станчинекий
449
Отряд 2. Edapliosanria
Как у предыдущего семейства, остистые отростки необычайно длинны,,
в АО—20 раз выше* тел позвонков. Остистые отростки позвонков с попереч-
ными отростками. Сем. Edaphosauridae, с двумя родами: Naosaurus, ив
пермских отложений Германии и Сев. Америки, и Edaphosaurus, очень к
нему близкий, из пермских слоев Тексаса (рис. 350).
3. Подкласс. Звероподобные (Theromorpha, Therapsida)
К этому подклассу принадлежат рептилии также с одной парой
височных ям и одной скуловой дугой, процветавшие в пермскую
и триасовую эпохи. Подкласс замечателен тем, что одип из состав-
Рис. 349. Череп и нижняя челюсть Dimetrodon
incisivus, из перми Тексаса.
(По Broom из Abel.)
ляющих его отрядов Theriodontia развивался в направлении, во
многом сходном с классом млекопитающих, с которым у пего на-
столько большое сходство, что вполне естественна мысль
о происхождении млекопитающих от этого отряда.
На костях черепа Therapsida пе наблюдается скульптуры,
как у Cotylosauria или Pelycosauria. Височная яма ограничена
снизу jugale и squamosum. Во многих чертах строение скелета
450
Рис. 350. Реконструкция Edaphosaurus cruciger, из перми
Тексаса.
(По Сазе из Abe].)
Therapsida сближается с таковым у американских Pelycosauna, но,
как указано выше, эти два подкласса кульминируют в различных
группах. Подкласс распа-
дается па несколько отрядов.
Отряд 1. Зверозубые (Therlo-
dontla)
Большей частью хищные
рептилии, зубы, череп и стро-
ение тела которых носят,
с одной стороны, явпые следы
приспособления к способу
питания, с другой—обнару-
живают огромное сходство
с млекопитающими. Зубы
дифференцированы н£ слу-
жившие для схватывания, для
разрывания и для раздробле-
ния добычи аналогично рез-
цам, клыкам и коренным
млекопитающих. «Коренные»
у более примитивных форм
с одним острием, у более вы-
соко специализированных —
с тремя. В связи с характе-
ром строения и функцией
зубов происходит и изме-
нение пижпей челюсти: зуб-
ная кость (dentale) полу-
чает преобладающее разви-
тие, у tree развивается венеч-
ni.iii отросток (processus со-
rniioidns), arliculare, angulare
Рис. 351. Череп и нижняя челюсть
Theriodontia Scylacosaurus sclateri (Л)
и Gynognathus crateronotus (В) из перми
(А) и триаса (В) Южной Африки. (Пер-
вый из ранних Therocephalia, второй —
из Gynodontia).
1 — praemaxillare, 2 — sept omaxill are, 3 —
maxillare, 4 — nasale, 5 — frontale, 6 — lacry-
male, 7 — adlacrymale, 8 — postfrontale, 9—
postorbitale, 10 —нижняя височная яма (В)
и parietale (А), И—jugale, 12—squamosum,
13 — quadratum, 14 — dentale, 15 — angulare,
16 — supraangulare, 17—articulare (По Broom
и Watson из Abel.)
451
и supraangulare постепенно редуцируются и теряют связь с
dentale (рис. 351). Зубы в альвеолах. У более поздних
представителей отряда имеется сформированное вторичное
нёбо.
У триасовых представителей имелось два сочленовных затылоч-
ных бугорка. Quadratum сильно редуцировано. Имелся наружный
слуховой проход, как у млекопитающих. Сходство с последними
имелось и в разделении позвоночника па отделы, в форме плече-
вого пояса, плеча, в наличии локтевого выроста у Ulna, в строе-
нии запястья, в одинаковом с млекопитающими числе фаланг па
передних и задних конечностях, в построении крестца более чем
из двух позвонков, в строении таза, в сосудистой системе черепа,
в смене зубов всего один раз.
Отряд Theriodontia распадается на несколько под-
отрядов: из них отметим следующие.
Подотряд Звероголовые (Tfoerocephalla)
Относительно мелкие, главным образом хищные формы. Род Scylaco-
saurus (рис. 351 Л).
Подотряд Цинодонты (Cynodontia)
К этому отряду принадлежит группа высших Therapsida, в ряде призна-
ков обнаруживающих более сходства с млекопитающими, нежели до сих пор
рассмотренные группы отряда Theriodontia. Резцы и клыки подвергались
Рис. 352. Нижняя часть
черепа Diademodon browni,
из триаса Южной Африки.
(По Broom, из Abel.)
смене, как у млекопитающих. Ложнокорен-
ные зубы многих Gynodontia сходны с та-
ковыми хищных млекопитающих, имея боль-
шой главный конус и маленькие передний и
задний. Как у млекопитающих, коренные пе
подвергались смене. Слезная кость и слезное
отверстие были, как у млекопитающих. Те-
менные кости образуют между височными яма-
ми гребень, как у I'herocephalia. Пинеальное
отверстие всегда развито хорошо у более ран-
них Gynodontia, ио у более поздних часто ма-
лое. Скуловая дуга как у млекопитающих.
Род Gynognathus, из триаса Южной Афри-
ки. С. crateronotus (рис. 351, В) достигал 2 м
длины. По общему облику это животное пред-
ставляло что-то сроднее между рептилией и
млекопитающим. Судя по ничтожному объему
мозговой капсулы, животное это должно было
быть медлительным и неповоротливым, мед-
ленно выслеживавшим добычу. Пищей пита-
лось, невидимому, живой, как и современные
рептилии. Род Diademodon был растительно-
ядным, как показывает строение зубов
(рис. 352). У него коренные зубы были снаб-
жены бугорками, по четыре и по пяти на боль-
ших зубах и два на последнем маленьком. Это
зубы типа млекопитающих. Род Permocynodon
ив пермских слоев Северной Двины, настолько млекопитающеподобен,
что его уже трудно назвать настоящей рептилией.
452
Отряд 2. Дицинодонты (DIcynodontla, Anomodontia)
Значительное число сходных с млекопитающими рептилий, живших
в Южной Африке в верхнепермское и триасовое время, относится к этому от-
ряду. Естественная группа с весьма замечательной специализацией. Резцов
нет, обычно (кроме примитивных форм) нет совершенно и коренных. В не-
которых случаях утрачиваются и клыки (рис. 353), обычно сильно развитые.
Praemaxillae всегда соединены и образуют покрытый рогом черепахообраз-
ный клюв.
Размерами аномодонты были от величины небольшой крысы до величины
тапира. Большей частью это были тяжелые животные с крупной головой.
V»
Рис. 353. Череп Oudenodon gracilis (Dicynodon-
tia) из перми Южной Африки.
(По Broom из Abel.) Обозначения как па рис. 351.
Некоторые более длинны и стройны. По способу питания эти рептилии были
травоядными и служили добычей хищных Therocephalia, Gorgonopsida
и других Therapsida. 9
Как показывает изучение сочленовных поверхностей и положения раз-
меров и формы костей передних и задних конечностей единственного извест-
ного почти полностью скелета Lyslrosaurus latirostris (рис. 308), Dicynodon-
tia были обитателями болот и спокойных, тихо текущих вод. Нищей их были
водные растения. Клыки служили орудием защиты. Род Dycinodon,?H3 перми
и триаса Южной. Африки, а также из пермских слоев Северной Двины.
Отряд 3. Dlnocephalla
Группа рептилий, появляющихся в отложепиях Карру одно-
временно с примитивными Theriodontia, и вымирающая в пермскую
эпоху. Череп короткий и высокий, сжатый. У более специали-,
зировапных форм кости черепа пепормальпо толсты, что стоит
в связи с сильным развитием гипофизы. У более примитивных
Dinocephalia этого нет. Зубы развиты слабо. У некоторых под
височной ямой находится еще другая, маленькая. Более прими-
тивные Dinocephalia являлись хищными, питались исключитель-
но мясом. Так, у Archaeosuchus, из средпепермских слоев Карру,
453
клыки с лезвием па задней стороне, коренные являются маленькими,
короткими конусами. У рода Rhopalodon, из пермских слоев Баш-
кирии, клыки большие, коренные сжатые, острые. Так же и у рода
Deuterosaurus, найденного там же. У более специализированных
форм — зубы однородные, не хищного типа, указывающие на тра-
воядный образ жизни.
Отряд 4. Ictidosauria
К этому отряду некоторыми относятся рептилии, происходя-
щие из триасовых отложепий Южной Африки, сравнительно мало
еще известные, но являющиеся, по Broom, наиболее близкими
к млекопитающим из перечисленных выше отрядов. К Ictidosauria
относится Karoomys, являющийся самым мелким представителем
Therapsida из доселе известных. Сходство с млекопитающим
столь велико, что к последним эту рептилию и отнесли, когда опа
была пайдена. Размер dentale у Karroomys всего 1.8 см. Сюда же
относят некоторые и род Dromatherium, относимый другими к низ-
шим млекопитающих (Protodonta).
Группа Synapsida 2-го типа
Рептилии, имеющие одну височную яму, по ограниченную иначе,
нежели у Synapsida 1-го типа и млекопитающих: опа ограни-
чена снизу или postfrontalia, postorbitalia и squamosum (у Meso-
sauria, Placodontia, Sauropterygia) или посредством postfrontalia
и supratemporalia (у Ichthyosauria).
4. Подкласс. Hxmuonmepuzuu(Ichthyopterygia)
Отряд 1. Мезозавры (Mososaurla)
В отряде одно семейство, Mesosauridae, с родами: Stereoster-
num, из пермских отложепий Южной Америки и Южпой Африки
(рис. 354), и Mesosaurus, пайдеппый там же, где и предыдущий
род, и, кроме того,в юго-западпой Африке. Вытянутые четвероно-
гие рептилии, приспособленные к плаванию, с длинным рылом,
отодвинутыми назад ноздрями, большими глазами. Зубы сидели
в альвеолах, были длинны, заострены и слегка загнуты. Пояса
плоские снизу. Конечности превращены в плавники паподобие
Sauropterygia, по превращение пе пошло столь далеко; они топки
и сравнительно длинны, так что отделы конечностей видны были
снаружи, перепонка связывала пальцы. Позвонки амфицельны.
Имелись брюшные ребра. Длиппый хвост в переднем отделе имел
длинные поперечные отростки, в средней — длинные остистые
и гемальпые отростки, что указывает на сжатый с боков хвост,
служивший органом плавания. Оба рода были обитателями прес-
ных вод, передвигаясь в них, подобно тритонам и крокодилам, так
как задние ноги были зпачительпо сильнее развиты, чем передние.
454
Отряд 2. Ихтиозавры, рыбоящеры
(Ichthyosauria)
Наиболее приспособленные к жизни
в море рептилии, у которых приспособ-
ление выражено не только в общей
форме тела, но и в деталях, благодаря
чему мы имеем в пих яркий пример
конвергентного развития, сближающего
их с рыбами и китообразными. Форма
тела веретенообразная, рыло вытянутое,
большие глаза, с костным кольцом в
склере, шея отсутствует, ноздри да-
леко назади, непосредственно впереди
глаз, позадиглазпичная часть черепа
короткая, невысокая. Височная яма
ограничена снизу посредством postfron-
talia и supratemporalia (рис. 355).
Имеется пинеальное отверстие. Заты-
лочный бугорок один. Зубы, сидев-
шие в общем жолобе или в альвеолах,
многочисленные, острые, покрытые тол-
стым слоем эмали, образующей продоль-
ные складки, благодаря чему зубы па
поперечном разреЗе имеют сходство с
зубами лабиринтодонтов из стегоцефал.
Амфицельные позвонки многочисленны,
их более 150, причем большая часть их
приходится па хвост. Последний с дву-
мя лопастями, причем конец позво-
ночника направляется в нижнюю ло-
Рис. 354. bleroosternum
tumidum (отр. Mesosauria),
из перми Южной Америки.
(По Gregor из Abel.)
пасть (рис. 356). Дуги пе срастаются
с телами. Ребра с двуйя головками, длинные, особенно в средней
части туловища. Грудины ист, имеются брюшные ребра из
Рис. 355. Реконструкция черепа Stenopterygius acutirostris, ихтиозавра
из верхнего лиаса Южной Германии.
1— praemaxillare; 2 — maxillare; 3 —nasale; 4 — носовое отверстие; 5 — adlacrymale;
6—lacrymale; 7—frontale; 8—parietale; 9—postfrontale; 10—squamosum; 11—sup-
ratemporale; 12—postorbitale; 13—quadrato-jugalc; 14—jugale; 15—dentale; 16—sple-
niale; 17—angulare; 18 — теменное отверстие; 19—верхняя височная яма.
455
отдельных частей. Тая значительно редуцирован: маленькая
ilium не соединяется с туловищем, pubes и ischia не образуют
симфиза. Конечности укорочены, имеют большое число излишних
фаланг п пальцев и превращены в плавники. На спине
кожный плавник. Тело голое.
Рис. 356. Филогенетическое раз-
витие хвостового плавника у
ихтиозавров.
Сем. Mixosauridae. Зубы
сидят в альвеолах. Первый и второй
шейные позвонки не срастались. В
строении скелета имеются черты, ука-
зывающие на близость к наземным фор-
мам.
Более древние, среднетриасовые
ихтиозавры.
Сем. Ichthyosauri d а е.
Более поздние ихтиозавры, из юры и
мела. Зубы сидят в общем жолобе, а у
более поздних форм, с переходом на
пищу, которая заглатывалась целиком
и являлась более мелкой и пассивной,
исчезают вовсе. Хвост двулопастный,
конечности стали короче и более при-
способленными, как плавники. Род
Eurypterygius ( = Ichthyosaurus, группа
многопалых ихтиозавров) был широко
распространен с нижнего лиаса до верх-
него мела. Плавники широкие, с мно-
гими пальцами (рис. 334). Ряд видов. Род
Stenopterygius (Ichthyosaurus), из лиаса
Германии. Плавники с редуцированным
редко нормальным числом пальцев, хотя
могут быть и очень большими и широ-
кими благодаря ширине элементов. К
этому роду принадлежат наиболее из-
вестные виды: St. quadriscissus, St.
tenuirostris, St. acutirostris и др.
(рис. 357).
А — Mixosaurus, из триаса Шпицбер-
гена, В — Ichthyosaurus (Stenoptery-
gius) quadriscissus, молодая стадия,из
верхнего лиаса Южной Германии,
С — тот же вид, взрослый экземпляр,
1) — (нижний рисунок) — Ichthyosau-
rus trigonus, верхняя юра Баварии.
(Из Abel, по Fraas.)
Об образе жизни ихтиозавров
можно судить уже по их внеш-
нему виду, по их организации, о
которой мы имеем весьма полное
представление, так как в юрских
сланцах Южной Германии остатки
ихтиозавров находятся в огромном
количестве и в прекрасной со-
хранности. Ихтиозавры—один из наилучше известных ископае-
мых. От многих сохранилось содержимое желудков, содержимое
полости тела, отпечатки кожи, копролиты. Пищей ихтиозавров
служили рыбы, белемпиты и другие головопогие моллюски, как
это видно из содержимого их желудков, нередко сохранившегося
в виде прекрасных отпечатков, и из анализа копролитов («ока-
меневшего кала»), полных остатков костей и ганоидных чешуй рыб
и остатков головоногих. На копролитах видны следы спиральной
складки кишки. У некоторых экземпляров внутри тела находятся
456
молодые экземпляры ихтиозавров же. Большей частью — это
проглоченные особи, в некоторых же случаях — эмбрионы. В свя-
зи с совершенно водным образом жизни ихтиозавры были живо-
родящи. На землю выходить ихтиозавры были, конечно, неспособны.
Размеры их тела колебались от 1 до 12 м. Череп самого крупного
Рис. 357. Ихтиозавр рода Stenopterygius.
(Из Fraas)
из известных ихтиозавров достигал в длину значительно более
метра. Местообитанием ихтиозавров было главным образом от-
крытое море.
5. Подкласс. Зауроптеригии (Sauropterygia)
Морские рептилии, происходящие от паземпых форм, характе-
ризуются длиппой шеей, конечноелями, измененными в плавники,
узкой плоской головой с удлиненным рылом, отодвинутыми назад
ноздрями, небольшими глазами, укороченным хвостом, пе играв-
шим роли в передвижении животного, которое осуществлялось
при помощи парных конечностей. Одна височная яма ограничи-
валась снизу при помощи postfrontale и squamosum. Имелось
пинеальное отверстие. Зубы простые, круглые в сечении, с про-
дольными гребнями. Позвонки слегка амфицельные и пролизан-
ные хордой у триасовых родов, вполне плотные и плоские у боль-
шинства плезиозавров. Дуги прирастают к телам. Ребра длинные,
сочленяются только с телами позвонков. Число шейных позвон-
ков колеблется, доходя до 30—40. Крестцовые позвонки раздель-
ны. Грудипы пет, по есть хорошо развитые брюшные ребра. Ко-
нечности представляют значительные различия у разных групп
плезиозавров, позволяя проследить их эволюцию от наземного
типа, по всегда опи пятипалы.
Отряд Зауроптеригии (Sauropterygia)
К отряду принадлежит несколько подотрядов. Одни из пих
(трахелозавры, нотозавры) представляют начальные стадии при-
способления к воде, при которой конечности не редуцируются,
457
а служат в качестве весел. Плечо и бедро еще относительно тонки,
хвост также относительно длинен. Оба подотряда — из триаса
Германии. Наиболее специализованы плезиозавры.
Подотряд Плезиозавры (Pleslosaurla)
Пе будучи потомками предыдущих подотрядов, представляют
дальнейшую ступень приспособления к жизни в воде. Однако
Рис. 358. Thaumatosaurus victor, пле-
зиозавр 3.44 м длины, из верхнего
лиаса Южной Германии.
(По Fraas.)
плезиозавры не были жите-
лями открытого моря, как
ихтиозавры, а держались у
берегов. Конечности превра-
тились в длинные узкие плав-
ники, причем передние и
задние примерно одинаковой
величины. Туловище корот-
кое, так же как и хвост.
Плечевой и тазовый пояса па
брюшной стороне сильно рас-
ширены и вместе с палочко-
образными, расположенными
в несколько слоев брюшными
ребрами образуют своего ро-
да брюшной панцырь, функ-
ционально играющий ту же
роль, что брюшной щит чере-
пах (рис. 358). Череп с раз-
витыми височными ямами и
большими топкими хвата-
тельными зубами, сидевшими
в глубоких альвеолах в одип
ряд. Обе пары конечностей
построены одинаково. У более
древних триасовых и нижне-
юрских представителей плечо
и бедро относительно сильны
и длинны, кости пред-
плечья и голени примерно в
два раза короче, но еще резко
отличаются от carpaliа и tar-
salia. У более молодых ро-
дов, из юры и мела, предплечье и голень сильно укорочены и пре-
вращены в плоские косточки, нелегко отличимые от таковых car-
pus и tarsus. Позвонки платицельпые, большая часть их прихо-
дится па шею (у рода Elasmosaurus — 76 шейных позвонков).
В хвосте около 40 позвонков.
Распространены в триасе, юре, мелу Европы и Америки.
Причина вымирания неясна. Плезиозавры не были исключительно
свободно плавающими существами, подобными ихтиозаврам или
458
китам. Как показывает наличие брюшного панцыря, опи вели
жизнь, подобную таковой тюленей, лежа на плоских песчаных отме-
лях в свободное от охоты время. Возможно, что лежали большими
стадами. В море пх врагами были крупные мозозавры. Как и у
современных тюленей, в трупах плезиозавров находились «га-
стролиты», заглотанные камни. Тело было голо.
Рис. 359. Реконструкция черепа Muraenosaurus.
1—nasalc, 2 — ноздри, 3 — lacrymale, 4 — frontalc, 5 — postfron-
tale, 6—postorbitale, 7 — jugale, 8—parietale, 9 — пинеальное
отверстие, 10 — squamosum, It— quadratum, 12 —- articulare,
13 —angulare, 14—supraangulare, 15 — complcmentare, 16 —den-
tale, 17 — maxillare. (Ho Andrews из Abel.)
Плезиозавры были представлены двумя различными типами: один —
с длинной шеей, маленькой головой и стройным телом—сем. Elasmosauridae,
другой—с короткой шеей, большой головой и мощным туловищем — сем.
Pliosauridae. К Первому семейству относятся: род Plesiosaurus с многими
видами, от нижнего триаса до нижнего мела, главным образом в Европе.
Из одной Англии известно более 30 видов. Размеры тела от 2 до 5 м. Род
Elasmosaurus. Наиболее длинношеяя форма, с коротким хвостом, из верх-
459
немеловых отложений Сон. Америки, 13 м длиной, причем длина шеи от-
носится к длине тела, как 23 : 9. Ко второму семейству относятся роды:
род Pliosaurus, найденный в Англии, гигантской величины: череп дости-
гал почти 1.5 м в длину и более 1.5 м в ширину, вся длина животного
Эми род Muraenosaurus (рис. 359).
6. Подкласс Лластинчатозубые (Hlacodontia)
Рис. 360. Череп Placodus из
среднего триаса Баварии.
1 — praemaxilla, 2 —vomer, 3 —
maxilla, 4 — palatinum, 5 — ptery-
goideum, 6 — quadratum, 7—squa-
mosum. (Ho Broili из Abel.)
Своеобразная группа рептилий, приспособившаяся к жизни
в море и к питанию моллюсками. Небольшого размера. Самый
крупный череп имеет размеры около 25 см. В черепе большие
орбиты, одна височпая яма, соответ-
ствующая верхней яме диапсид.
Нижняя граница ямы образована
postfrontale, postorbitale и supratem-
porale. Большое пинеальное отвер-
стие. Хоапы соединены вместе впе-
реди palatina. Широкое костное пёбо
образовано palatina и pterygoidea.
Весьма замечательны зубы. У Р1а-
codus па praemaxillae две или три
пары тупых конических или ложкооб-
разных зубов (резцы), па maxillae—
4 пары широких и плоских зубов,
служивших для раздавливания пищи,
и три пары еще больших, совершенно
плоских зубов на пёбе. На нижней
челюсти три пары таких же зубов,
как на верхней. Эти зубы указы-
вают (рис. 360) па то, что пищей
Placodontia служили моллюски с
твердыми раковинами, что подтверж-
дается нахождением их остатков в
раковинном известняке, чисто мор-
ских отложениях, полных моллюс-
ками. Позвонки слегка амфицельиы.
Грудная коробка Placochelys из очепь крепких ребер. Имелись
и хорошо развитые брюшные ребра. В коже был па спиипой сто-
роне папцырь из отдельных окостенений. Пояса и конечности
пока известны лишь у одного скелета. Таз плоский, что говорит
за плоскую брюшную сторону, подобно плезиозаврам. Конеч-
ности пе известны, по, ввиду общего сходства с водяными че-
репахами, следует думать, что конечности Placodontia представ-
ляли плавники, подобные плавникам Chelonia.
7. Подкласс Черепахи (Testudinata, Chelonia)
Черепахи представляют подкласс, специализированный чрез-
вычайно односторонне и сохраняющий древние черты организа-
ции. Наиболее характерной чертой подкласса является наличие
460
туловищного папцыря. Последний состоит из костных элементов,
образующих на спине спинной щит — сагарах, па нижней сто-
роне брюшной щит — plastron. Свод черепа является полным,
ио не соответствующим тому, что есть у Cotylosauria (рис. 361),
или имеет одну яму,
являющуюся задней
височной вырезкой, рас-
пространившейся впе-
ред и пе соответствую-
щей верхним или ниж-
ним височным ямам
других рептилий. Име-
ются вторичное пёбо,
одна скуловая дуга и
тройной затылочный бу-
горок. Челюсти лишены
зубов и покрыты рого-
вым клювом. Quadra-
tum прочно фиксиро-
вано между костями че-
репа. Конечности пяти-
палы, с примитивной
пальцевой формулой,
наземного типа или пре-
вращены в ласты. Ребра
с одной головкой, крест-
цовых позвонков всего
два. Копуляторпый ор-
ган непарный, клоа-
кальная щель продол-
говатая. Размножаются
яйцами. Известны с
триаса, ио возникли,
Рис. 361. Череп Emys orbicularis {А) и Che-
Ionia mydas (В).
1 — max Шаге, 2 — praefrontale, 3 — frontalc, 4 —
parictale, 5 — postfrontale, 6 — jugale, 7 — squa-
mosum, 8 — quadrato-jugale, 9 — quadratum. (По
Rabi из Abel.)
видимо, с перми, дости-
гают максимального развития к концу мезозойской эры и в начале
третичного времени. Количество семейств и родов в настоящее
время сильно редуцировано, по число видов достигает 232. Рас-
пространены черепахи по всему миру, кроме холодных стран.
Отряд Chelonla
Подотряд A'mphichelydia
Голова и шея пе втягивались под панцырь. Иа шейных позвон-
ках пет или имеются лишь слабые поперечные отростки.
Сем. Trias sochelydae. Род Triassochelys. Из верхнего триаса
Германии. Известен один вид Т. dux, один только экземпляр. Череп массив-
ный, «стегальный», с задней височной дырой. По краям нижней и верхней
челюсти спрятанные в альвеолах сидят зачаточные зубы.
461
Подотряд Боном ейные черепахи (Pleurodira)
Шейные позвонки с сильно развитыми поперечными отростками.
Благодаря этому шея не втягивается вместе с головой в щит, а из-
гибается в сторону и помещается между спинпым и брюшным
панцирями. Таз неподвижно срастается как с брюшным, так и со
спинным папцырем.
Современные бокошейные черепахи распространены в Африке,
па Мадагаскаре, в Бразилии и в Австралии. Ископаемые формы —
наземные или водные, современные—все обитатели пресной
воды, которую или вовсе не оставляют или оставляют лишь на
время. Конечности у них превращены в плавники.
Сем. Пеломедузы (Pelomedusidae). Сюда принадлежит 3 ро-
да с 44 видами, живущими в тропической Южной Африке, на Мадагаскаре и
в Южной Америке, в ручьях и реках. Род Pelomedusa живет на Мадага-
скаре и во всей Африке к югу от Сахары, а также на Синайском полуост-
рове. В Абиссинии в течение сухих сезонов спит. Род Podocnemis с ря-
дом видов в Южной Америке, главным образом в бассейне Амазонки и на
Мадагаскаре. Вида Р. expansa, no-местному «аррау», представляется весь-
ма ценным с экономической точки зрения, так как яйца его, откладываемые
на суше, собирают в огромном количестве (десятки миллионов), главным обра-
зом для добывания жира. Яйца откладываются в ямку глубиной около полу-
метра и засыпаются песком. Живет в разливах лесных рек, уходя в послед-
ние в засушливое время. Пища состоит главным образом из падающих в во-
ду с деревьев фруктов.
Сем. Змеиношейные черепахи (Chelydidae). Шея
сгибается под край щита, но остается частично выставленной. Выйный
щиток есть. 42 пар краевых щитков. 40 родов с 27 видами живут в Южной
Америке, Австралии и на Новой Гвинее. Род С h е 1 у s с одним видом—
бахромчатая черепаха (Ch. fimbriata). Передние и задние конечности с пере-
понками. Достигает почти метра в длину. Питается рыбами, лягушками, го-
ловастиками.1) Род Hydromedusa. Реки Южной Америки. Длина щита около
20 см; 2 вида.
Подотряд Скрытоголовые черепахи (Cryptodirа)
Сюда относятся сухопутные, пресноводные и морские черепахи,
шейные позвонки которых или вовсе лишены поперечных отрост-
ков, или опи очепь слабо развиты. Благодаря этому шея может,
изгибаясь S-образпо в вертикальной плоскости, втягиваться
прямо назад под щит. Таз пе сливается со спинпым и брюшным
папцырем. Копечпости сухопутные или плавательные.
Сем. Каймановые черепахи (Chelydridae). Два
рода, каждый с одним видом, в Америке. Пальцы пятипалых конечностей со-
единены перепонками. Род Chelydra. Кусающаяся черепаха (Ch. serpentina).
Достигает в длину 4 м. Распространена от озер Канады к востоку от Скали-
стых гор через Соединенные Штаты и Центральную Америку до Эквадора.
Живет в стоячих или медленно текущих водах. Большой хищник. Пи-
тается рыбами, рептилиями и всякой другой живностью, попадающей
вводу. Иногда оставляет воду. Съедобна, особенно молодые особи, мясо же
старых пахнет мускусом. Яйца съедобны. С этой черепахой сходен род гри-
фовая черепаха (Macroclemmys, рис. 362) — самая крупная из всех пресно-
водных черепах, живущая в южной части США, достигающая 4.4 м в длину.
Клювом своим действует, как гарпуном, вонзая его в тело добычи, которую
затем поджимает под себя и съедает. Яйца несколько самок откладывают
вместе в одну яму — до 60—70 яиц.
462
Сем. Наземные черепахи (Testudinidae). Обшир-
ное семейство, распространенное по всему свету за исключением Авст-
ралии и прилежащих индо-малайских стран. К семейству принадлежат
как вполне наземные, так и совершенно водные черепахи; многие являются
земноводными. У наземных форм щит более выпуклый, короткие пальцы
без перепонок, формы травоядные. У водных форм щит более плоский, паль-
цы ног соединены перепонками, ногти длиннее и тоньше, животные плотояд-
ные. Представлено семейство 25 родами и более чем 140 видами, распреде-
ляющимися между двумя подсемействами.
Подсемейство пресноводных черепах (Emydinae) включает в себя главным
образом водные формы, подсемейство наземных черепах (Testudininae) заклю-
чает в себе только наземных черепах. Пресноводные черепахи обитают в ре-
ках с медленным течением,
Рис. 362. Грифовая черепаха. (Macroclemmys
temminchii)
(Из Gadow.)
в озерах, прудах, некоторые
в солоноватой воде моря.
Они являются изумитель-
ными пловцами. Хищники,
приносящие нередко суще-
ственный ущерб рыбному
и птичьему населению
водоемов. Зимой зарываются
в землю. Яйца отклады-
вают в землю или песок.
Род Болотная черепаха
(Emys). Голова покрыта
мягкой кожей. Ноги по-
крыты крупной чешуей.
Пальцы с развитыми пла-
вательными перепонками.
Хвост у молодых длин-
ный, затем становится от-
носительно более коротким.
Европейская болотная или
речная черепаха (Е. orbi-
cularis, рис. 363), длиной
32 см. Родина — Южная Европа и восточная часть Средней Европы.
Живот в низовьях Днепра, Днестра, Дона, Волги и Урала, в Крыму, на
Кавказе. В Африке встречается к северу от гор Атлас, встречается в Сред-
ней Азии, в Южной Туркмении и в низовьях Сыр-Дарьи, в Персии, Курди-
стане и Малой Азии. Самая северная из черепах. Раньше встречалась даже в
Скандинавии и Англии. Держится в стоячих водах или в медленно теку-
щих реках. Дном выходит па сушу погреться. Активна ночью. Зимой зары-
вается в ил. Питается червями, водяными насекомыми, лягушками, трито-
нами и их личинками, нападая и на рыб. Спаривание происходит на суше.
Яиц откладывается 13—15 в ямки, которые самка роет в относительно твер-
дой почве, смачивая ее из мочевого пузыря. Отве^тие самка сверлит при
помощи хвоста, расширяя его затем задними ногами. Затем, отложив яйца,
черепаха засыпает их землей, придавливает ногами и задней частью брюшно-
го панцыря. Отложенные в конце мая яйца развиваются примерно 3 меся-
ца, когда из них вылупляются детеныши. Род Clemmys, собственно пре-
сноводная или водяная черепаха. Вид чисто арктический. В Европе жи-
вут: каспийская черепаха (Clemmys caspica) и мавританская водяная че-
репаха (Cl. leprosa) Несколько видов живет в Америке. Каспийская чере-
паха распространена от восточной половины Средиземноморья до Западной
Персии.
На(тоя цне паземные черепахи подсемейства Testudininae имеют, как
сказано, короткие пальцы, лишенные плавательных перепонок. Кожа пе-
редних конечностей покрыта сильно развитыми роговыми чешуями, иногда
с кожными окостенениями. Plastron прочно соединен швом с сагарах, обычно
очень выпуклым. Вершина головы покрыта щитками. Хвост короткий. В на-
стоящее время—лишь несколько родов, группирующихся вокруг центрального
463
рода Tesludo. Последний является космополитичным в более теплых странах
уMepciinoii зоны н в тропических областях. Нет его лишь в Австралии и на Ма-
лайских островах.Туркестанская степная черепаха (Testudo ЬогзНеИ1)водится
но всей (вредней Азии и в Южном Казахстане. Обитает в пустынях, главным об-
разом в глинистой, или пустыне эфемеров, в степях, заходит в предгорья. Пи-
тается растительной пищей, а потому цикл ее жизни связан с циклом жизни ра-
стительности. Появляясь в массовом количестве весной, черепаха эта исчезает,
когда наступает вследствие летней засухи перерыв вегетации, зарываясь в это
время в землю. За короткий период активной жизни (март — июнь) успевает
размножаться. Яйца откладываются в землю. Своей роющей деятельностью
вредит арыкам, железнодорожным насыпям. Съедобна как сама, так и ее
Рис. 363. Европейская водяная черепаха (Emys orbicularis}.
(Из Gadow.)
яйца. В период максимального выхода черепах весной область их распро-
странения посещается крупными хищниками — грифами, которым черепаха
служит в это время пищей.
Гигантские наземные черепахи отличаются от описанных главным обра-
зом величиной. Наиболее крупной из этих черепах является Testudo atlas
(Colossochelys), найденная в поздних миоценовых или ранних плиоценовых
слоях Сиваликских холмов у подножья Гималаев. В третичное время ги-
гантские черепахи жили в Европе. Черепаха эта немного не достигала 2 м.
В настоящее время гигантские черепахи встречаются в двух удаленных пунк-
тах: на Галапагосовых островах, но имени испанского названия черепах
и получивших свое название, и в западной части Индийского океана, на
островах Маскаренских, Коморских, Сейшельских и некоторых других.
Еще недавно вид гигантской наземной черепахи существовал и на Мадага-
скаре. Изолированное существование черепах в этих условиях, невозможность
межвидового скрещивания привели к тому, что каждая группа островов, а на
Галапагосах почти каждый остров имел свою особую форму гигантской чере-
пахи. Ни на одном из указанных островов не было мелких видов черепах.
Параллельно с увеличением размеров утоньшался костный панцырь. Воз-
можно, что на этих островах, где черепахи были единственными крупными
животными, в панцыре не было надобности, и отбор не поддерживал этот
признак. Попасть на указанные острова черепахи могли только вследствие
того, что эти острова являлись частью некогда существовавшего здесь мате-
рика. На оставшихся континентальных частях последнего гигантские чере-
пахи вымерли, на островах вследствие отсутствия или слабой здесь борьбы
за существование они сохранились. Теперь, с приходом человека, эти черепахи
подверглись нападению ради вкусного мяса. На большинстве островов они
были почти уничтожены.
464
рынках. В Закавказье встречается
Рис. 364. Сагарах Toxochelys из
верхнего мела Канзаса.
(По Wieland из Abel.)
Из видов рода Testudo упомянем еще европейский вид — греческую че-
репаху (Testudo greaca), обитающую в сухих местностях, поросших кустар-
ником, в странах, окаймляющих берега Средиземного^моря с севера и с во-
стока. Мясо ее съедобно и продается на
еще распространенная в Северо-западной
Африке, в части Сирии в Малой Азии,
Месопотамии и Персии кавказская или
мавританская черепаха (Testudo ibera).
Сем. Морские черепахи
(Chelonidae). Высоко специализирован-
ная группа скрытошейных, приспосо-
бившихся к жизни в море. Передние и
задние конечности превращены в плав-
ники, причем передние более развиты,
чем задние. Metacarpalia и metatarsalia
и большинство фаланг удлинены, и
между ними не образуется сочленений.
Кроме того, элементы эти уплощены и
кисти расширены благодаря широкой
os pisiforme. Распадаются на два подсе-
мейства.
Поде. Cheloninae. Панцырь,
покрытый роговыми пластинками, пло-
ский. Между спинным и брюшным
щитами имеется ряд щитков, называе-
мых субмаргинальными. Острые рого-
вые челюсти загибаются на конце
крючком. Современные виды, объеди-
няемые в два рода, являются вполне
морскими животными, встречающимися
на сотни километров от берега. Пло-
тоядны, питаются главным образом моллюсками и ракообразными. Размно-
жаются на суше, откладывая многочисленные (до 100) яйца в вырытую зад-
ними ногами яму. Суповая или зеленая черепаха (Chelonc mvdas), живет в мо-
Рис. 365. Archcion ischyros, сверху (Л) и снизу (/У).
(Но Wieland из Abel.)
рях жаркого и подтропического пояса. Питается преимущественно водорос-
лями и вообще морскими растениями (главным образом Zostera), но не брез-
гует и животной пищей. Яйца откладывают на песчаных берегах или остро-
вах вдалеке от человека. Мясо и яйца съедобны. Иногда мясо бывает весьма
ядовито. Настоящая каретта (Chelone imbricata), напоминает суповую чере-
30 Каш каров и Станчинский 46а
паху, но несколько меньше. Встре-
чается в тропических и субтропи-
ческих морях Старого и Нового Све-
та. Также съедобна. Роговые плас-
тинки каретты легко свариваются в
массу и идут на изготовление раз-
личных изделий.
Подсем. Кожистых чере-
пах (Dermochelyinae) заключает
в себе своеобразных морских чере-
пах. Сагарах состоит из многочис-
ленных маленьких полигональных
пластинок и покрыт кожей без рого-
вых щитков. Конечности превращены
в весла. Шея не втягивается. В
настоящее время живет один только
род и вид этого подсемейства, но в
ископаемом состоянии мы знаем ряд
представителей его, проливающих
свет на эволюцию кожистых черепах.
Род Toxochelys, из нижнего мела
Германии и верхнемеловых отложе-
ний Сев. Америки (рис. 364). Боковые
фонтане л и открыты. На средних
Рис. 366. Protosphargis veronensis.
1 — epiplastron, 2 — ребра, 3 — hyoplas-
tron, 4 — hypoplastron, 5— xiphiplastron,
6 — marginalia. (Из Abel.)
Рис. 367. Кожистая черепаха (Dermochelys
coriacea), скелет передней конечности.
(По Vblker из Abel.)
пластинках панцыря кост-
ные бугорки, остатки эпи-
телиального кожного по-
крова. Род Archelon, из
верхнего мела Сев. Амери-
ки. Гигантский вид A. Ischy-
ros (рис. 365), достигал
4.25 м в длину при длине
черепа в 1 м. Редукция пан-
цыря как на спинной, так
и на брюшной стороне ушла
гораздо дальше: фонтане л и
занимают большую часть
сагарах. Род Protosphargis,
из верхнего мела Италии.
Редукция ушла еще дальше:
костальные (реберные) пла-
стинки утратили связь с
краевыми пластинками,
костный plastron редуци-
ровался до степени кольца
из 4 элементов (рис. 366).
У перечисленных форм,
живших, несомненно, в от-
крытом море, мы видим шаг
за шагом развивающееся
приспособление к жизни в
море. Редукция панцыря
соответствует редукции ске-
лета у всех позвоночных
обитателей открытого моря;
она служит для облегчения
веса животного. Род Der-
mochelys — Кожистая чере-
паха. (D. coriacea). Самая
крупная из всех ныне жи-
вущих черепах, с колос-
сальными передними плав-
466
никами (рис. 367). Спинной щит состоит не из роговых пластинок, за-
крывающих фонтанели в костном панцыре, но из маленьких, мозаично
расположенных тонких кожных окостенений. Также и брюшной щит ив
многочисленных разбросанных в коже костных бугорков. Под этим щитом
на брюшной стороне находится костное кольцо, на спинной — костная
выйная пластинка. Последние соответствуют костному панцырю меловых
морских черепах. У третичных черепах рода Psephophorus мы видим под
мозаикообразпым костным панцырем рудименты первоначального костного
панцыря меловых морских черепах. Этот мозаичный панцырь возник как
приспособление при возврате к прибрежной жизни.
Подотряд Мягкие черепаоси (Trion/ychoidea,)
Панцырь плоский, овальной формы или почти круглый и
покрыт гладкой и нежной кожей вместо роговых щитов, чем эти
черепахи отличаются от всех остальных. На ногах плавательные
перепонки. Голова с хоботом, па конце которого помещаются
поздри. Поперечные отростки шейных позвонков слабо развиты,
почему голова и шея могут втягиваться под панцырь в виде
S-образного изгиба. Хвост короткий. Появляются Trionychoidea
в верхнем мелу Северной Америки. В третичное время жили
в Европе, Африке, Ост-Индии и Сев. Америке. Известно 80 иско-
паемых видов рода Trionyx. Современные формы—жители прес-
ной воды и отсутствуют лишь в Австралии и Южной Америке,
где они известны, одпако, в ископаемом виде.
Сем. Trionychidae. Род Trionyx известен с верхнего мела. Т. ferox —
наиболее известная мягкокожая черепаха. Живет в США. Длина щита 45 см.
Большой хищник. Укусы этой черепахи очень опасны. Проводит большую
часть времени в воде, где прячется под стволами упавших деревьев, под кам-
нями и т. д. Иногда выходит наружу и зарывается в ил, оставляя отверстие
для дыхания. Весной самка откладывает яйца (до 60) на песчаных местах
вдоль берега. Мясо чрезвычайно вкусно, почему эти черепахи являются объ-
ектами охоты. Распространен вид в Сев. Америке широко. Другие виды трио-
никсов имеются в Африке, в Индии, Индо-Китае, в Китае, от Амура до
острова Тимора, в Монголии, в Японии*. Род Amyda встречается у нас на
Дальнем Востоке.
Группа Diapsida
Рептилии с дизигальпым типом черепа. Две височпых дуги
ограничивают две височные ямы. У некоторых представителей
(Lacertilia) нижняя височная дуга исчезла. Diapsida в лице Rhyn-
chocephalia появляются, как и Synapsida и Anapsida, вероятно,
в пермское время. Однако на основе сравнительпоанатомических
данных можно полагать, что апапсидпое состояние черепа яв-
ляется наиболее древним и примитивным.
Группа Diapsida обнимает древнюю группу клювоголовых
(Rhyncliocephalia и другие близкие подотряды), называемую не-
которыми Prosauria, предъящерами, далее — чешуйчатых (Lepi-
.dosauria), архозавров (Archosauria или Thecodontia) динозавров
(Dinosauria) и птерозавров (Pterosauria). Все эти группы могут
считаться подклассами.
30* 467
8. Подкласс JDiaptosauria
Формы с двумя явственными височными ямами и двумя височ-
ными дугами.
Отряд Клювоголовые (Bhynchocephalia)
В настоящее время представлен одной только так называемой
новозеландской ящерицей. Особого расцвета отряд не достигал
никогда и особой роли в филогенезе других групп рептилий также
Рис. 3G8. Sphenodon punctalum (Hatteria).
(Из Никольского.)
не играл. Общий вид ящерицеобразный. Позвонки большей
частью амфицельные с остатками хорды. Обычно хорошо развитые
intercentra. Грудные ребра с одной головкой. Имеются брюшные
ребра. В черепе характерны неподвижное quadratum и наличие
двух пар височных ям и дуг. Зубы акродонтные. Грудина хряще-
вая. Конечности пятипалые, у некоторых форм приспособленные
к плаванию. Кожа с роговыми чешуйками. С триаса до современ-
ности.
Сем. Sphenodontidae. Род Sphenodon (Hatteria)
очень примитивная форма, сходная с нижнепермской Palaeohatteria. Это
468
в настоящем смысле слова живое ископаемое. Размер тела до 75 см (рис. 368).
Из наружных признаков в глаза бросается гребень из треугольных чешуй
на затылке, вдоль спины и на хвосте. На голове и туловище — мелкие зер-
нистые чешуйки. Крупные чешуи лишь на брюшной стороне. У туземцев
зовется «туатера». В настоящее время живет только местами на островах
возле северного острова Новой Зеландии. В остальных местах истреблена.
Днем большей частью спит, ча-
сами может лежать в воде без
дыхания. Питается животной
пищей, весьма разнообразной,
главным образом ночью. Яйца
откладывает в песок, где они
развиваются под влиянием солнца
около года. В движениях ленива
и скоро устает. Роет себе на бе-
регу норы, в которых живет вмес-
те с буревестниками, птенцов и
яиц которых гаттерия не трогает,
но защищает нору отвсякого дру-
гого животного.
9. Подкласс Чешуйчатые
( Lepidosauria )
Наиболее богато пред-
ставленная в современности
группа, характеризуемая сле-
дующими признаками. Тело
покрыто чешуей, с длинным
хвостом, иногда змеевидное.
Позвонки процельные, редко
амфицельные, ребра с одной
головкой. Quadratum под-
вижное, вторичного нёба нет.
Крестец из двух позвонков
или отсутствует. Конечности
пятипалые, иногда рудимен-
тарные’ или вовсе отсутст-
вуют.
Рис. 369. Реконструкция черепа Агае-
oscclis gracilis, из перми Тексаса.
1 — praemaxillare, 2 — maxiHare, 3 — na-
sale, 4 — iacrirnale, 5 —frontale, 6 —parie-
tale, 7 — postfrontale, 8 — postorbitale, 9 —
squamosum, 10—supratcmporale, 11 —fora-
men pineale, 12 — dentale, 13 — supraangu-
lare, 14 — angulare. (По Williston из Abel.)
О т p я д 1. Areo.scelhla
Com. Arcoscelidae. Род Arcoscelis — древнейший представитель
Lepidosauria, найденный в перми Тексаса. Размер тела 80 см. Общий вид —
ящеричный. Длинный хвост, хорошо развитые пятипалые конечности. Уд-
линенный 4-й палец на передних и задних конечностях, а также строение
тела говорят за то, что эта рептилия могла лазать по деревьям. От ящериц
отличается тем, что имеется одна височная ямка, соответствующая верхней
ямке Sphcnodon. Нижняя височная ямка вторично закрыта тонкой костной
пластинкой от squamosum (рис. 369). Благодаря этому подвижность черепа
(стрептостилия), характеризующая череп ящериц, здесь отсутствует. Сбли-
жаясь с ящерицами, эта форма сближается и с древнейшими Rhynchoce-
phalia.
Сверхотряд Sqnamata
К сверхотряду относятся ящерицы и змеи. До сих пор процвета-
ющая группа, известная лишь с триаса (род Paliguana). Впрочем,
469
в юре и в мелу лишь очень мало остатков чешуйчатых. Наиболее
характерно строение черепа. Последний пронизан рядом дыр,
представляя в итоге систему перекладин, в особенности у змей.
Такое устройство дает элементам черепа большую подвижность.
Зубы плевродонтныеилиакродонтные. В конечностях никогда не
бывает более 5 пальцев, даже у водных форм, за одним исключе-
нием (Mosasauria). Чешуи покрывают и череп.
Отряд 2. Ящерицевые (Lacertilla)
Тело покрыто чешуей, с длинным хвостом, ипогда змеевидное.
Череп большей частью с палочковидным столбиком черепа (colu-
mella cranii) и подвижным quadratum. Нижняя височная дуга
всегда отсутствует, верхняя — иногда. Межглазничная перего-
родка пе окостеневает. Ветви нижней челюсти соединены впереди
швом. Зубы акродонтные или плевродонтпые. Грудина почти всег-
да есть. Конечности пятипалы, иногда отсутствуют (Anguis,
Ophisaurus). Тело в большинстве случаев покрыто роговыми че-
шуями. Позвонки процельны (у Geckonidae они амфицельны),
верхние дуги плотно срастаются с телами позвонков. Зубовидный
отросток прочно соединен с эпистрофеем. В крестце — 2 позвонка.
Ребра одноглавы. Грудина отсутствует лишь у некоторых Ampliis-
baenidae, ключица и Т-образная episternum отсутствуют лишь
у хамелеонов и некоторых безногих форм. Foramen entepicondy-
loideum отсутствует. Таз у безногих сильно редуцируется, но
никогда не исчезает полностью. Обитают в жарких и умеренных
странах, известно более 1 800 видов. В ископаемом состоянии из-
вестно немного форм. Из триаса Южной Африки известен род
Paliguana* (Iguanidae). Представитель Anguidae известен из верх-
ней юры. Мало остатков известно из меловых отложений (сем.
Tejidae), больше — из нижнего эоцепа Сев. Америки (сем. Не-
lodermatidae, Anguinidae, Iguanidae). Еще больше — из верх-
него эоцена и олигоцена.
Подотряд Ящерицы собственно (Lacertae)
Сем. Агамовые (Agamidae). Обширное семейство, заключаю-
щее в себе 30 родов и более 200 видов. Окостенений в коже нет. Форма тола
весьма разнообразна — плоская у наземных и сжатая с боков у древесных
форм. Большинство родов и видов насекомоядны, некоторые агамы едят и
растительную пищу. Род Uromastix питается исключительно плодами и
растениями. Обитатели Старого Света, его теплых частей. Нет их на Мада-
гаскаре. В Австралии, Индии и на Малайском архипелаге живет большин-
ство; мало их в Африке. Род Draco — «летающий дракон». Живет па остро-
вах Малайского архипелага. Обладает способностью, захватив внутрь себя
большое количество воздуха, становиться благодаря длинным ребрам подо-
бием плоского балона. Это дает ему возможность совершать планирующие по-
леты. Род Agama; треугольная голова, более или менее примитивное туловище,
длинный круглый хвост, на глотке у самцов небольшой мешок, гребня на спине
нет или, он развит слабо. Степная агама, A. sanguinolenta, обитает в равнинах
низовьев Поволжья, Средней Азии и Южного Казахстана до подножья Тар-
багатая и Алатау, заходя на Северный Кавказ и в Закавказье. Кавказский
стеллион (A. caucasia), живет в восточной части Закавказья, в Туркменистане,
470
, в горах Копет-Даг и в Северной Персии. Род Phrynocephalus; типичные
для полупустынь и пустынь Средней и Центральной Азии представители
агам; отличаются короткой и толстой головой, коротким, плоским туловищем
и скрытыми ушами; мешка на горле нет. Ушастая круглоголовка (Phr. mys-
taceus) — обитатель песков. Такырная круглоголовка (Phr. helioscopus).
Обе круглоголовки обладают удивительно сходной с окружающей обстанов-
кой окраской: первая — с песком, вторая — с глинистой почвой. Спасаясь,
могут быстро зарываться в песок, как бы тонут в нем. Род плащеносная яще-
рица (Chlamydosaurus). Своеобразная ящерица Квинслэнда и вообще Се-
верной и Северо-западной Австралии. Отличается зубчатым на краях капю-
шоном — кожным выростом с каждой стороны шеи. Оба выроста срастаются
на глотке. При испуге расправляет воротник и бежит, причем может бежать
Рис. 370. Летающий дракон (Draco volitans).
(Из Gadow.)
на двух задних ногах (рис. 371). Род Шипохвост (Uromastix). Крупные не-
уклюжие агамы, живущие в пустынях Северной Африки и Южной Азии.
Наиболее характерное их отличие — плоский толстый хвост, покрытый ши-
пами (рис. 372). Шипохвосты растительноядны (в Алжире едят финики,
ягоды, траву, цветы) и съедобны. Род Молох (Moloch), — колючая ящерица,
«колючий чорт». Обитатель песков Южной и Западной Австралии. Страшная
по виду, но безобидная ящерица. Шипы ее слабы и опасности не представ-
ляют (рис. 373). .
Сем. Игуан (Iguanidae). Дифференцировки зубов, в известной
мере имеющейся у агам, здесь нет. Кожных окостенений нет.Распространены
главным образом в Америке, где заменяют агам, а также на Мадагаскаре и
островах Фиджи. Большинство — насекомоядны, некоторые Iguana, Basi-
liscus — растительноядны. Живут в различных условиях, иные при опас-
ности идут в воду. Некоторые виды .съедобны. Один вид, живущий на Гала-
пагосовых островах, является морским животным, питается водорослями.
Род Basiliscus замечателен высоким и способным напрягаться гребнем на спи-
не и на хвосте у самцов (рис. 374). Несколько видов в Центральной Америке
и в странах, прилежащих к ней с юга. Род Iguana, тело сжато с боков и по-
крыто мелкими чешуями. На конце головы они крупны. На спине высокий
471
гребет, из отдельных плоских и мягких шипов. Подобный же гребень на гор-
ловом мешки. Два вида в тропической Америке, абсолютно растительноядны.
Род Phrynosoma (рис. 375), жабовидная ящерица, «рогатая жаба». Сходна с мо-
лохом и с Phrynocephalus. Плоское широкое тело, без гребня. На голове боль-
шие шипы. Около дюжины видов в западной половине Соединенных Штатов
и в Центральной Америке. Все виды — живородящи.
Рис. 371. Плащеносная ящерица (Chlamydosaurus).
(По Saville Kent из Hilzheimer.)
Сем. Поясохвостые ящерицы (Zonuridae). Неболь-
шое семейство, представленное 4 родами с дюжиной видов в южной и тро-
пической Африке и на Мадагаскаре. С окостенениями в коже, по крайней
мере на черепе, где они покрывают височную яму. Род Zonurus по внешнему
виду напоминает агам. Тело покрыто поперечными рядами расположенных
в поперечные ряды щитовидных чешуй (рис. 376а).
Рис. 372. Шипохвост (Uromastix acanthinurus).
Из Gadow.)
Сем. Беретеницевые (Anguidae). Передняя часть языка
может втягиваться в заднюю часть. Кожные окостенения имеются на теле
и на голове, где покрывают верхнюю височную яму. Конечности большой
частью редуцированы. Хвост длинный, ломкий, легко регенерирует. Живут
в Америке, Европе и Индии. Плотоядны. Некоторые живородящи. Род желто-
пузик (Ophisaurus). По бокам тела глубокая продольная борозда от области
затылка до хвоста. Конечности редуцированы лишь от задних остается
пара тонких незаметных отростков. Похож на змею, но отличается легко но
вполне развитым векам. Тело покрыто ромбическими чешуями, расположен-
472
ными правильными рядами. Живет в Венгрии, Далмации, Греции, Малой
"Азии, Сирии, Персии, Закавказье, Средней Азии и в Нижнем Поволжье
(рис. 3766). Род Anguis с одним видом веретеница ломкая, или медяница
(Anguis fragilis). Также безногая ящерица, водящаяся почти по всей Европе
и в Европейской части СССР, кроме северных губерний и Крыма. OrOphisau-
rus отличается отсутствием боковых складок и полной редукцией конечно-
стей. Живородяща; молодые змейки выходят из яиц немедленно после их
откладки. Главная пища— дождевые черви и улитки.
Рис. 373. Молох (Moloch horridus).
(Из Gadow.)
Сем. Ядозубы (Helodermatidae). Плевродонтны. Нижние-
зубы с желобком спереди и сзади; имеются ядовитые железы. Язык разд-
военный. Кожа сверху зернистая с поверхности, с многочисленными непра-
вильными костными бугорками. Низ покрыт плоскими чешуями. Единствен-
ный род Heloderma с двумя видами — на юге Сев. Америки. Животное тол-
стое, неуклюжее, сплюснутое, с толстым хвостом. Ночное животное, питаю-
щееся мелкими животными: червями, многоножками, лягушками и яйцами
больших ящериц. Яд вызывает при укусе тяжелые симптомы, изредка смерть.
даже человека.
Рис. 374. Василиск (Basilibcus amuricanus).
(Из G-adow.)
Сем. А м е р и к а п с к и е вараны (Т е j i d а е). Зубы плотные*
почти акродонтные. Язык длинный, глубоко раздвоенный, с сосочками. Око-
стенений в коже нет. Конечности иногда редуцированы. Большое семейство,
включает около 40 родов и более 100 видов. Некоторые формы древесны, дру-
гие наземны, обитатели жарких песчаных равнин, третьи, наконец, живут
под поверхностью земли, превращаясь в червеобразных, почти лишенных
конечностей и слепых животных. Живут по всей Южной Америке, на Вест-
47а
'Индских островах, в Центральной Америке и в южных частях США. Наи-
более известен род Tupinambis (рис. 377а), напоминает по внешности варана,
достигает почти метра в длину, причем на хвост приходится 3/б. Яйца часто
откладывает в термитники. Мясо очень ценится.
Сем. Настоящие ящерицы (Lacertidae). Плевродонт-
ные ящерицы Старого Света, без окостенений в коже тела, но с таковыми в
Рис. 375. .Жабовидная ящерица (Phrynosoma
cornutum).»
(Из Gadow.)
области верхневисочных ям. Конечности всегда хорошо развиты, пятипалы.
На голове крупные щитки и мелкие чешуи, большая часть которых с кожными
окостенениями. Чешуи спинной поверхности весьма вариируют по величине
от больших чешуй до мелких зернышек. На брюшной стороне чешуи крупнее
и часто расположены поперечными или косыми рядами. Хвост длинный и лом-
кий. 20 родов и около 100 видов распространены в Европе, Азии, Африке за
Рис. 376а. Поясохвостая ящерица (Zonurus giganteus).
(Из Gadow.)
исключением Мадагаскара; отсутствуют в Австралийской области. Наиболее
многочисленны в Африке. Северная граница распространения совпадаете гра-
ницей вечной мерзлоты. Плотоядны, питаются главным образом насекомыми,
червями и улитками, а также (более крупные) другими ящерицами. Род
Lacerta — центральный род с 20 видами— в Европе, Северной и Западной
Азии и в Африке к северу от экватора. Живородящая ящерица (L. vivipara)
живет в Северной и Средней Европе и во всей Сибири до Сахалина включи-
474
тельно. Заходит за полярный круг, на юге в Азии идет до реки Лепсы в Се-
миречье, не проникая в Туркестан. Родит живых детенышей. Глазчатая яще-
рица (L. ocellata) распространена на Пиренейском полуострове, в Южной
Франции и по северо-западному побережью Италии, в Алжире и Тунисе к
северу от Сахары. Имеет по бокам спины крупные синие глазки. Зеленая яще-
Рис. 3766. Желтопузик (Ophisaurus a pus).
(Из Никольского.)
рица (L. viridis) живет в’ средиземноморских странах, в южной части евро-
пейской части Союза, кроме Крыма, на Кавказе, в Персии, в Малой Азии,
в Сирии и Палестине. Прыткая ящерица (L. agilis), на восток доходит до
Саянского хребта, на юг — до Семиречья, многочисленна в Крыму. Как
и предыдущий вид, образует многочисленные разновидности. Род степные яще-
рицы, или ящурки (Eremias), живут в Средней и Центральной Азии, а так-
же в Африке. Держатся главным образом в пустынях. Похожи на представи-
Рпс. 377а. Американский варан (Tupinambis
nigropunctatus).
(Из Gadow.)
телей рода Lacerta, отличаясь расположением чешуй и щитков. Ящурка
быстрая (Е. velox), обитательница пустынь. Разноцветная ящурка (Е. arguta)
более свойственна полупустыне. Некоторые имеют отороченные роговыми
зубчиками пальцы, живут в Средней Азии и в Южной Африке. Ящурка
полосатая (Е. scripta) и сетчатая (Е. grammica).
475
Сом. С ц и п к о в ы о (S с i п с i d а е). Плевродонтны. На голове и на теле
сильно развиты ножные окостенения. Язык короткий, очень слабо выемча-
тый. Нижнее веко часто с прозрачным кружком посредине. На голове круп-
ные щитки, на теле черепицеобразная чешуя, напоминающая рыбью. Рас-
пространено семейство во всех частях света, от экватора до умеренного
пояса; многочисленны в Австралии, Африке, Ост-Индии и на островах Ве-
ликого океана. 25 родов и около 375 видов. Большинство обитает в песке,
под поверхностью которого могут двигаться, выходя на поверхность ночью.
Род гологлаз (Ablepharus) — несколько видов в пустынях и горах
Средней Азии. Род Scincus с видом сцинк аптечный (S. officinalis) живет в
Сахаре и пустынях, окружающих Красное море. Род длинноногие сцинки
(Eumeces) водятся в Северной Африке, в Закавказье. Персии, Белуджи-
стане и в Средней Азии, во всей южной половине. Род Seps (Chalcides) с
видом трехпалый сепс (S. tridactylus) имеет редуцированные, крошечные
ножки и напоминает веретеницу (Anguis).
Рис. 3776. Пятнистая двуходка (Amphisbaena
fuliginosa).
(Из Брима.)
Подотряд Амфисбеповые ( Amphlsbaenica)
Сем. Двуходков ые (Amphisbaenidae). Червеобразные ящерицы,
ведущие подземный, роющий образ жизни. Кожа без чешуи с многочислен-
ными кольцами, каждое из которых разделено на многочисленные мелкие
квадратики, являющиеся следами чешуй. Последние ограничиваются голо-
вой. Глаза и ушные отверстия скрыты. Конечности, за исключением рода
Chiroles, имеющего четырехпалые передние конечности, редуцированы. Пояса
также—лишь в виде следов. Хвост очень короток. 12 родов и 66 видов. Род
Хирот (Chirotes) в Мексике и Калифорнии. Род Amphisbaena, с 27 видами—
в тропической Америке и Африке, три вида в Малой Азии. Вполне подзем-
ные животные, роющиеся подобно червям, часто в гнездах муравьев (рис.
3776).
Подотряд Хамелеоны (Chamaeleontes)
Сем. Chamaeleontidae, род Chamaeleo — акродоптные
рептилии Старого Света со сжатым с боков телом, хватательным хвостом
и хорошо развитыми конечностями, пальцы которых образуют своеобразную
клешню. Африканское семейство, заходящее в Индию, Малую Азию, Пале-
стину и Аравию. Пятьдесят видов, из которых половина живет на Мадага-
скаре. Ряд своеобразных особенностей заставляет отделить хамелеонов от
ящериц и выделить в особый подотряд (выдвижной язык, равный в вытянутом
виде половине длины тела, ряд особенностей в строении черепа (отсутствие
columella cranii и др.), отсутствие claviculae и interclaviculae, кожа, покрытая
не чешуями, а гранулами и т. д. Род Chamaeleo. Обыкновенный хамелеон (Ch,
vulgaris) живет в Северной Африке, проникает в Андалузию. Глаза покрыты,
сросшимися в виде колпачка веками, по середине которых имеется отвер-
стие для прохождения света. Глаза движутся независимо друг от друга,
476
что дает хамелеону возможность хорошо высматривать добычу. Наличие
добавочных мешков в легких позволяет хамелеону надуваться. Язык
выбрасывается на 10 см. Окраска тела способна меняться в известных пре-
делах. Яйца откладывает в землю, зарьвая их при этом. Питается мухами,
кузнечиками и другими насекомыми,
пауками и т. п. (рис. 378).
Подотряд Гекконы (Geckones)
Позвонки амфицельные, хорда
долго сохраняется, череп без ко-
стных височных дуг, ключицы
расширены; теменные кости раз-
дельные; глаза за немногими
исключениями покрыты прозрач-
ной кожей, без подвижных век;
язык мясистый и широкий, слегка
вырезанный спереди и способный
выдвигаться. Кожа покрыта зерни-
стыми бугорками, иногда содержа-
щими кожные окостенения или
объизвествления, в особенности на
голове, где они сливаются иногда
с костями черепа. Нижняя сторо-
на обычно покрыта мелкими че-
шуями. У большинства пальцы
снабжены присасывательными об-
разованиями, дающими им воз-
можность лазать по абсолютно
гладким и вертикальным поверхно-
стям. Хвост у большинства лом-
кий и легко регенерирует. Все—
насекомоядны, некоторые одоле-
вают и более крупную добычу. За немногими исключениями —
ночные формы. Распространены космополитически в более теп-
лых зонах, особенно многочисленны во всех странах между тро-
пиками и на островах, даже в Новой Зеландии. Около 50 родов
и 270 видов.
Сем. Це п к о п а лыс или гекконы ( G е с Г< о n i d а е). Род
Terafosr incus с видом спнпковый геккон (Т. scincus) обитает в песчаных пу-
стынях Средней и Центральной Азии и в Персии, где ведет ночной образ жиз-
ни, днем укрываясь в таллинах почвы и под корнями кустарников. Пальцы
без нрисасывател] вых дисков, но с роювой оторочкой. Тело покрыто чешуей,
напоминающей рыбью. Па хвосте ряд больших поперечных ногтеобразных че-
шуй, которыми при закручивании и раскручивании хвоста геккон производит
звук, подобный стрекотанию кузнечика. Это, видимо, половой признак
(рис. 379).
Род Tarentola. Стенной геккон (Т. mauritanica) (рис. 380), с бугорками
па спине и с развитыми присосками на конечностях. Живет в странах, рас-
положенных вокруг Средиземного моря, особенно часто — в Испании, на
островах близ Италии и в Северной Африке. Род голопалые гекконы (Gym-
nodactylus) без присосок на пальцах. Каспийский геккон (G. caspius, рис. 381)
распространен в районе Каспийского моря, в Персии и Туркестане. Серый го-
477
Рис. 378. Хамелеоны (Chamaeleon
vulgaris и Chamaeleon pumilis)
(направо в углу).
(Из Gadow.)
лопалый геккон (G. russovi), — в Средней Азии. Род геккончик (Alsophylax)
A. pipiens — в пустынях Средней Азии. Род гребнепалый геккон (Gros-
sobamon) с гребешками на пальцах, живет в Средней Азии, Персии и Афгани-
стане. Род Eublepharus. Представители этого рода, выделяемого иногда
Рис. 379. Сцинковый геккон (Teratoscincus scincus).
(Из Никольского.)
Рис. 380. Стенной геккон (Tarentola mauritanica).
в особое подсемейство, отличаются от настоящих гекконов процельными по-
звонками и слиянием двух parietalia в одну кость. Веки не редуцированы.
Немного видов в Индии, Западной Африке и Центральной Америке. Род
Uroplates представлен немногими видами на Мадагаскаре., У рода Ptycho-
zoon пальцы с присосками и связаны перепонками; по бокам головы, туло-
вища и хвоста кожистые расширения. Живет на Малайских островах и Ма-
лакке. Складки кожи служат парашютами.
478
Подотряд Гидрдзавровые (Hydrosauria)
Более или менее приспособленные к жизни в воде формы,,
иногда чисто водные, иногда наземные, но сходящие в воду.
Сем. Варанов (Varanidae). Ящерицы Старого Света с длинным
глубоко раздвоенным и высовывающимся языком, с длинной шеей, мелкими,
рядом располагающимися чешуйками и бугорками на спинной стороне и рас-
положенными в ряды четырехугольными чешуями
на брюхе. Длинный хвост часто сжат с боков.
Из верхнего эоцена Франции известен род Palaeo-
varanus, из плейстоцена Австралии — гигант-
ский (10 м) род Megalania. Современный род
Varanus содержит около 30 видов, живущих в
Африке, Южной Азии и Австралии. Нет их на
Мадагаскаре. Род этот, называемый также мони-
тором, заключает в себе виды, ведущие полу-
водный образ жизни, другие виды — обитатели
сухих и жарских пустынь, третьи — живут в
заросших деревьями местах. Все они хищники,
хватают все, что могут одолеть. Некоторые виды,
достигают в длину 2 м. V. niloticus — во всей
Африке, V. griseus (рис. 382) — в Средней Азии,
V. salvator — в Юго-восточной Азии.
Сем. Долихозавры (Dolichosa-
u г i d а е). Водные рептилии с маленькой голо-
вой, длинной шеей (13 позвонков), длинным (ив-
Рис. 381. Каспийский Рис. 382. Серый варан (Varanus griseus).
геккон (Gymnodacty- (Из Gadow.)
lus caspius).
(Из Никольского.) 60 позвонков) плавательным хвостом. Конечности
редуцированы. Конечности — для плавания, пе-
редние вполовину короче задних. Зубы плевродонтные. Род Dolichosaurus —
верхний мел Англии.
Сем. AiirHflO3aBpH(Aigialosauridae). Обитатели бере-
гов, с большим черепом, напоминающим череп следующей группы. Шея ко-
роткая, из 7 позвонков, тело короткое. Передние и задние конечности реду-
цированы лишь слегка и обнаруживают уже приспособления к водному обра-
зу жизни. Занимают промежуточное положение между Varanidae и Mosasau-
ridae. Известны из нижнего мела. Достигали 1—2L/2 м. Род Aigialosaurus
найден в Далмации.
Сем. Mosasauridae (Pythonomorpha). Крупные, длинные, оде-
тые чешуей морские рептилии с конечностями, превращенными в плавники.
Позвонки процельны. Крестцовый позвонок только один. Височная впадина
одна — верхняя. Большое подвижное quadratum. В непарном parietale —
большое пинеальное отверстие. Глаза со склеротическим кольцом. Ветви
479
нижней челюсти были соединены связкой. Кроме того, каждая ветвь имела
посредине подвижное сочленение элементов (рис. 383). Большие конические
:вубы сидели на челюстях и pterygoidea, срастаясь с костями. Пояса были раз-
виты, но таз у более поздних форм не связан с позвоночником. Кости ко-
нечностей очень коротки, 5 тонких пальцев, иногда с лишними фалангами
(рис. 384). Череп очень сходен с черепом Varanus. Некоторые из мозазавров
(Mosasaurus) были обитателями поверхностных слоев воды (Plioplatecarpus).
Органом движения являлся хвост. Характер зубов указывает, что это были
опасные хищники. Жили в верхнемеловую эпоху в морях Новой Зеландии,
Рис. 383. Правая половина нижней челюсти Ту-
losaurus dyspelor.
1 — сочленение посредине. (По Пиепе из Abel.)
Рис. 384. Реконструкция скелета Tylosaurus dyspuior из верхнего мела
Канзаса.
(По Osborn из Abel.)
Северной и Южной Америки и Европы. Род Mosasaurus — из мела Голлан-
дии, Северной Германии, Франции, Италии, Сев. Америки и Новой Зелан-
дии. Некоторые виды достигали 12 м. Род Platecarpus — из мела Сев. Аме-
рики и, быть может, Франции. Размер до 5.6 м.
Отряд 3. Змеи (Ophhlia, Serpentes)
Так как змеи являются лишь специализованпой ветвью La-
certilia, отделившейся в меловое время, то некоторыми зоологами
змеи и ящерицы соединяются даже в один отряд и рассматри-
ваются как подотряды. Тело покрыто чешуей, сильно вытянуто
в длину. Конечностей и плечевого пояса нет. Parietale без темен-
ного отверстия. Позвонки процельные, с дополнительными сочле-
новными приспособлениями (zygosphena и zygantra). Передние —
с сильными нижними (гемальными) отростками. Ни височных дуг,
ни columella cranii пет. Quadratum, pterygoideum,’ maxillare
и palatium очень подвижны и рыхло соединены с черепом. Ргае-
480
maxillae—очень малы; ветви нижней челюсти связаны связкой.
Череп, в сравнении с ящеричным, окостеневает солиднее. Зубы
могут находиться па верхней челюсти, межчелюстных пёбных
и птеригоидных костях и dentale. У ядовитых змей передние
челюстные зубы — с желобком или внутренним каналом. Глаза
скрыты под кожей, без век. Движение совершается путем боковых
изгибов позвоночника, а также при помощи ребер, нижние концы
которых могут сдвигаться вперед и назад.
Большая часть змей является наземными обитателями, живу-
щими преимущественно в более теплых зонах. Различают около
400 родов и 1800 видов. Меловые остатки очень скудны или сом-
нительны. Подлинные остатки известны с эоцена. О происхождении
змей они говорят мало.
Сем. Palaeophidae, вымершие формы из эоцена, частью, ги-
гантские, свыше Им длиной. Тело, судя по позвонкам, было сжато с боков,
что говорит за то, что это были водные животные, именно морские. По строе-
нию позвонков уклоняются от современных змей. Найдены и в Европе, и в
Сев. Америке. Род Palaeophis — из нижнего эоцена Англии.
Сем. Archaeophidae. Известен один вид Archaeophis proavus пз
эоцена Италии с заостренным и вытянутым рылом и огромным числом по-
звонков (565). Тело также сжато с боков. Видимо, обитатели моря.
Сем. Слепуны (Т у р h 1 о р i d а е). Роющиеся под землей змеи,
покрытые сверху и снизу однообразной циклоидной чешуей, причем чешуйки
черепицеобразно налегают друг на друга. Зубы находятся только на верхней
челюсти, по 2—5 с каждой стороны. Тело червеобразное, голова не ограни-
чена от туловища, хвост короткий, так что заднепроходное отверстие нахо-
дится почти на конце тела. Глаза очень небольшие, иногда скрытые под кож-
ными щитками. Кости черепа плотно между собой соединяются. Конечности
редуцированы, имеются лишь следы таза в виде одной косточки справа и слева.
Живут в земле, питаясь червями, личинками, насекомыми. Размер 70 см
в длину. Широко распространены во всех тропических и субтропических стра-
нах, кроме Новой Зеландии (нет их в Сев. Америке), встречаясь и на уеди-
ненных островах. Typhlops vermicularis живет на Балканском полуострове
и в юго-восточной Азии. Кроме этого вида, имеется еще 125 видов слепунов,
заключенных в 3 рода.
Сем. Л о ж н о н о г и е (Boidao). Обычно крупные змеи со следами таза
и задних конечное*,тей, видимых снаружи в виде когтевидных придатков по
сторонам анального отверстия. Чешуи верхней части тела гладкие и обычно
мелкие, на брюхе образуют один ряд, на хвосте снизу два ряда. Quadratum
связано со squamosum, имеющей вид длинной горизонтальной кости, рыхло
связанной с боковой стороной черепа, чем достигается большая подвижность
нижней челюсти, способность ее широко раскрываться. Зубы сидят на den-
talc, pterygoidea, palatina, maxillaria, у Pythoninae и па praemaxillaria.
Распространены 60—70 видов Boidae во всех тропических и субтропических
странах, включая и острова, кроме Новой Зеландии. Большинство — лес-
ные формы, лазящие по деревьям при помощи хватательного хвоста. Некото-
рые связаны с водой, некоторые — с песком (Егух). Все — хищники, пи-
таются теплокровными, которых душат, обвив кольцами своего тела, раз-
давливают и затем заглатывают. У некоторых наблюдается забота о потом-
стве:
Подссм. 1. П и т о н о в ы е (Ру thoninae). 20 видов распростра-
нены в Палеотропической и Австралийской областях, один вид — в Южной
Мексике.
Род Python — основной род. Тигровый питоц (Р. molurus) распро-
странен в Индии (рис. 385), на Цейлоне и в Индо-Китае. Достигает 8.5 м.
Питается мелкими млекопитающими, на человека нападает лишь случайно.
В Африке живут другие виды питонов, достигающие 4.25 м длины.
31 К&шкаров и Стаичжиокий
Подсем. 2. Уд а в о в ы е (В о i п а е). Род песчаный удав (Егух). Жи-
вет в пустынях и сухих степях Восточной Европы, Северной Африки, Перед
ней и Средней Азии, отыскивая добычу не только на поверхности земли, но
и в песке, в котором роется и движется весьма быстро. Добычу величиной до
крупного суслика включительно душит и заглатывает, как более крупные
представители удавов в жарких странах. Размер змеи до 75 см. Шкурка
очень красивой раскраски идет на поделки. Род Воа. Удав обыкновенный
(В. constrictor). Область распространения — тропическая Южная Америка,
где придерживается сухих лесистых или покрытых кустарниками мест. Не-
редко влезает на деревья, но никогда не идет в воду. Животное ночное.
Длина более 3 м. Род водяные удавы (Eunectes). Анаконда (Е. murinus).
Отличается от Воа тем, что у нее лишь передняя часть головы покрыта щит-
ками. Самая крупная из американских змей, достигает более 8 м длины, жи-
Рис. 385. Питон (Python molurus).
(Из Gadow.)
вет большей частью в воде, в которой может оставаться долго. Ловит рыбу,
а на суше — водосвинок, агути и других подобных животных, а также птиц.
7Кивородяща, как и предыдущий вид.
Сем. В альков а ты е змеи, сперташки (II у si id ас).
Занимают промежуточное место между удавами и щитохвостыми, напоми-
ная по внешности и слепунов. Чешуи мягкие и мелкие на спине и боках,
крупные на брюшной стороне. Хвост очепь короткий и тупой. Голова
очепь маленькая. Подобно удавам имеют зачаточный таз и зачатки задних
конечностей. Живородящи. Ведут частью подземный образ жизни, питаясь
червями, насекомыми и маленькими Typhlopidae. Живут (С> видов) в тро-
пических странах Южной Америки, Цейлона, Малайского архипелага и
Индо-Китая. Коралловая свсрташка (Ilysia scytalc), кораллово-красного
цвета с многочисленными черными кольцами. Живет в Южной Америке.
Сем. Щитохвостые (Uropeltidae). Небольшие змеи,
ведущие роющий образ жизни на Цейлоне п в Южной Индии, с коротким
цилиндрическим телом и очепь коротким хвостом, па конце которого нахо-
дится своеобразный щит. Красиво окрашенные змейки.
482
Сем. Ужовые (Colubridae). Обширное семейство, включающее»
около 9/10 всех современных видов. Голова ясно отграничена от туловища
и покрыта правильными щитками. Обе челюсти с зубами. Остатков таза и зад-
них конечностей нет. Семейство космополитично, представлено формами на-
земными, древесными и водными. Многочисленные представители семейства
могут быть разделены на три параллельных ряда.
Рис. 386. Полоз (Zamenis mucosus).
(Из Gadow.)
Ряд. 1. Гладкозубые змеи (Л glyph а). Все зубы одинако-
вы, плотны, без жолобов на поверхности и без каналов. Все относящиеся
сюда змеи безвредны.
Подсем. Настоящие ужи (Colubrinae). Продолговатая
голова более или менее ясно отделена от тела. Хвост вытянут в острие. На
верхних и нижних челюстях зубы сидят во всю их длину. Чешуи черепице-
образные, гладкие или снабженные килем. На брюшной стороне — большие
щитки. К подсемейству относится большин-
ство змей, около 1 000 видов, не достига-
ющих, за редкими исключениями, величины
более 2 м. Большинство яйцеродящи, но
некоторые (Coronella) живородящи. Кос-
мополиты, ограниченные в своем распро-
странении па север вечной мерзлотой, де-
лающей невозможной перезимовку. Род
водяные ужи (Natrix). Желтоухий уж
(N. natrix) распространен в Европе, Алжи-
ре, в Западной и Центральной Азии,
в европейской части Союза — всюду кро-
Рис. 387. Полоз Карелина
(Zamenis karelini).
(Из Никольского.)
ме крайнего севера. В Сибирь идет до
Забайкалья, есть па Кавказе и в Закавказье, в Средней Азии — лишь по
побережью Каспийского моря. Кормится лягушками, жабами, ест яще-
риц, мелкую рыбу и крупных жуков. Держится у воды. Водяной уж (N.
tesselata). Отличается от первого незначительными признаками. Водится в
Средней, южной и юго-восточной Европе, в юго-западной, в средней и цен-
тральной Азии. Пресноводная форма. Поедает мальков рыб, ест лягушек. Род
медянка (Coronella). Плоская и короткая голова слабо отграничена от туло-
вища. Зрачок круглый. С. austriaca. Живет во всей Европе до 62.5° северной
широты, на Кавказе, в Сирии. Добычу свою душит, обвиваясь вокруг нее
три раза. Яйцеживородяща. Род Полозы (Zamenis). Два задних верхнечелю-
стных зуба часто несколько крупнее и отделены от остальных беззубым
промежутком — диастеммой. Чешуи гладки, с ямкой у вершины. Глаза боль-
31*
483
шие с круглым зрачком. 30 видов рода распространены во всей Палеаркти-
ке. Z. mucosus живет на острове Яве, Малайском полуострове, Цейлоне, в
Индии, Бирме, Южном Китае, Сиаме, Афганистане, в Туркмении. Выделяет-
ся из других змей острой спиной благодаря выпячиванию остистых отростков
(рис. 386). Ряд полозов живет в пределах СССР. Из них Z. rhodorhachis,
распространенный в Египте, Аравии, Сирии, Персии, Белуджистане, запад-
ных Гималаях, в Туркменистане и в Дарвазе; Z. karelini (рис. 387). Кроме
стран Ирана, распространен еще в Туркестане (Средней Азии и Южном
Казахстане). Z. ravergieri распространен так же. Род Elaphe. Узорчатый по-
лоз (Е. dione) распространен от долины Волги по всей умеренной Азии до
Амурского края и Японии. Род древесные ужи (Dendrophis). Имеют длин-
ное, тонкое, сжатое с боков, тело. Болес других приспособлены к жизни
на деревьях. Распространены в тропических странах обоих полушарий
(рис/ 388). Род Lytorhynchus. Литоринх афганский (L. ridgewayi) имеет
длинную заостренную морду,
Рис. 388. Древесная змея (Dendrophys
punctulatus).
(Из Gadow.)
повидимому приспособление для рытья.
Живет в Афганистане, найден
в Туркменистане. Род лазящие
ужи (Coluber). Состоит из мно-
гих видов. Название лазящих
присвоено им не совсем по
праву, так как они лазят не
лучше других ужей. Живут
они в большей части Европы,
Азии, северной и тропической
Америки. Сильные змеи, пи-
тающиеся главным образом
млекопитающими и птицами.
Держатся большей частью на
кустарниках и деревьях. Мно-
гие хорошо плавают. Эеку ла-
нова змея (С. aesculapii) до-
стигает 1.9 м длины, живет в
южной, отчасти и в средней
Европе до Каспийского моря
и в Закавказье. Близка к дре-
весным змеям по способ пости
лазить.
Подсем. Глоточно-
зубые (R h а с h i о d о п-
t i п а е). Сюда относятся ужо-
вые с немногими зубами на
задних частях верхнечелюст-
ных костей, небных и нижнечелюстных. Как бы в виде компенсации недостатка
зубов у них необычайно развиты нижние остистые отростки шейных позвон-
ков, направленные вперед и пронизывающие пищевод. Отростки эти служат
для раздавливания птичьих яиц, составляющих главную пищу этих змей.
Dasypeltis scabra — жительница тропической и южной Африки.
Ряд 2. 3 а д н е б о р о з д ч а т ы е (О р i s t h о g 1 у p h а). Змеи
этого ряда имеют один или более задних максиллярных зубов с желобком
на передней стороне. Зубы эти связаны с ядовитой железой, хотя и слабо
развитой. Этих змей следует считать подозрительными в смысле ядовитости.
Некоторые таковыми, несомненно, являются. Этот род гораздо беднее пре-
дыдущего представителями. Из представителей трех относящихся сюда подсе-
мейств назовем род Taphrometopon. Стрела-змея (Т. linoolatum) обитатель-
ница глинистых и песчаных пустынь Средней Азии, Афганистана, Персии
и Монголии. Длина доходит почти до 1 м, тело стройное, топкое, ползает
змея с необыкновенной быстротой, гоняясь за ящерицами, являющимися
ее добычей. Лазит по кустам. По середине ряда верхнечелюстных зубов имеется
один очень длинный, выдающийся над другими. На нижней челюсти самый
длинный зуб находится спереди. Быть может благодаря наличию этих зубов
местное население считает стрелу-змею ядовитой, повидимому, напрасно.
484
Рис. 389. Очковая змея (Naja tripu-
dians).
(Из Gadow.)
Ряд. 3. Переднебороздчатые (Proteroglypha). Передние
верхнечелюстные зубы с глубоким желобком; последний, бывает столь глу-
боким, что ауб кажется полым. Позади ядоносных зубов сидят зубы плот-
ные. Относящиеся к этому ряду змеи крайне ядовиты. Распространены они
в областях Австралийской, Палеотропической и Неотропической, нет их на
Мадагаскаре и в Новой Зеландии. Из коренной области своего распростра-
нения они идут к северу, в более теплые части Северной Америки и в Пале-
арктическую область, встречаясь и в Северной Африке и в Юго-западной
Азии. Распадаются на два подсемейства: Elapinae и Hydrophinae.
Подсем. Аспидов ые, или ядовитые, ужи (Elapi-
nae). Тело и хвост цилиндрические, хвост короткий, заостренный. В под-
семействе почти 150 видов. Большинство австралийских змей принадлежит
к этому семейству. Род кобра (Naja). Одна из самых опасных ядовитых
змей, укусы которой смертельны, обладает способностью сильно раздувать
шею благодаря подвижным длинным шейным ребрам. Приподнимая в
раздраженном состоянии переднюю
часть тела, змея раздувает шею.
У очковой змеи (N. naja), живущей
в Ост-Индии, Индо-Китае и на Малай-
ском архипелаге, на спинной сторо-
не шеи находится темный рисунок,
напоминающий очки (рис. 389). У
N. n. oxi ana этого рисунка на шее
нет. Этот подвид живет в Средней
Азии до Самарканда, в Южной Азии
до Китая и Малайского архипелага.
Род Elaps — чрезвычайно красивая
американская змея, в окраске кото-
рой красное и черное чередуются.
Один из наиболее красивых видов,
коралловый аспид (Е. coraHinus),
живет в лесах тропической Южной
Америки и Малых Антильских остро-
вов.
Подсем. Морские змеи
(Hydrophinae). Тело и в
особенности хвост сильно сжаты с
боков. Около 50 видов распростра-
нены в Индийском и Тихом океанах,
живут почти исключительно в море,
лишь редко заходя в реки или вы-
ходя на сушу. Размеры тела достигают 1.5—2 м. Очень ядовиты. Живо-
родящи. Распространены в Индийском и Тихом океанах. Особенно часто
встречаются между берегами Китая и Австралии. Питаются рыбой. Род
Hydrus, или Pelamis, в лице единственного представителя вида Н. pla-
turus, водящегося в Индийском океане и в тропических и субтропиче-
ских частях Великого океана, найден у берегов Восточной Сибири, зане-
сенный сюда, видимо, случайно.
Сем. Г а д ю к о в ы е, или виперовые (Vi р е г i d а е). Толстое
тело. Голова треугольная или сердцевидная, ясно отграниченная, покрыта
сверху гладкими чешуями или маленькими щитками. Верхняя челюсть
короткая, очень подвижная и может выдвигаться вперед вместе с большими
ядовитыми зубами, кроме которых (исключая замещающих) нет других.
Зубы полые. Зрачок вертикальный. При утрате зуба на его месте встают дру-
гие. За исключением одного рода, гадюковые живородящи. Они включают
паземные, древесные, полу водные и роющие типы. Семейство космополи-
тично, встречаются и в Старом и Новом Свете, кроме Мадагаскара. Делится
на два подсемейства: Viperinae и Crotalinae.
Подсем. Гадюки (Viperinae). Между глазом и носовой ямкой
пот ямки. Жители Старого Света. Нет их только на Мадагаскаре. Северная
граница определяется вечной мерзлотой. Род настоящая гадюка (Vipera),
485
Гадюка обыкновенная, или козюлька (V. berus). Почти всегда на спине имеет-
ся темная полоса, составленная из ромбов, соприкасающихся углами. Окра-
ска весьма вариирует, самец всегда светлее самки. Зрачок имеет вид косой,
продольной щели. В Европе — преимущественно в северной, в средней —
попадается в горах, иногда встречается и в горах южной, а также в горах
Кавказа и во всей Сибири. В Европейской части СССР — от берегов Ледови-
того океана до черноморских и прикаспийских степей, где ее заменяет
степная гадюка (Vipera renardi), идущая от Черноморья через Киргизские сте-
пи до Алтая в Туркестан. Обе гадюки ведут ночной образ жизни. Степная днем
укрывается в норах грызунов. Питается мелкими грызунами, ящерицами
и, вероятно, насекомыми. Еж, хорек и луни являются ее врагами. В пределах
СССР живут еще другие гадюки. В юго-западной Европе и в Закавказье
живет гадюка-носорог, или песчаная гадюка (Vipera ammodytes), отличаю-
щаяся конусообразным наростом на конце носа, торчащим кверху. Очень
ядовита; укус ее вызывает смерть. Гюрза (V. lebetinus) в Закавказье, Персии,
Туркменистане. Род рогатые гадюки (Cerastes, рис. 390) отличается сразу
по двум рожкам, располагающимся над глазами, хотя этот признак имеется
не у всех видов. Рогатая гадюка — обитатель пустыни, с окраской грунта
которой сходна ее окраска. Водится во всей Северной Африке, кроме Марок-
ко, в Аравии и в Судане. Ядовита. В Персии, близ советской границы, най-
ден особый род ложных рогатых гадюк (Pseudocerastes).
Рис. 390. Рогатые гадюки (Cerastes cornutus и Vipera ammodytes).
(Из Gadow.)
Подсем. Гремучники, или ямкоголовые змеи (Сго-
Нп ае). Между глазами и носовым отверстием с каждой стороны находится
глубокая ямка. Представители этого семейства крупнее и сильнее гадюк,
и ядовитость их сильнее. Распространены они в Америке и в Азии. Род
щитомордник (Ancistrodon) встречается между Волгой и Уралом, в Сред-
ней Азии и в Монголии и дальше па Восток, а также в Америке. Не имеет
погремушки. В Союзе живет несколько видов. Палласов щитомордник
(A. halys) от нижнего течения Волги и Закавказья до верхнего течения Ени-
сея. A. halys intermedius распространен в Восточной Сибири до Семиречья
на западе, в Монголии и в Восточном Китае. Укус щитомордника нередко
бывает смертельным. В Сев. Америке до Флориды и Тексаса живет A. pis-
civorus. Род гремучая змея (Crotalus). На конце хвоста находится трещотка,
состоящая из ряда вложенных одно в другое роговых колец, являющихся
видоизменением концевых чешуй хвоста. Одна из самых страшных и ядо-
витых змей. Распространена в Северной и Южной Америке. Сг. horridus —
в США.
10, Подкласс Луночнозубые (Thecodonntia)
Сюда относятся несколько отрядов, из которых только один —
Crocodilia — дожил до наших дней; остальные вымерли. Имея
корпи, вероятно, еще в пермской эпохе, Thccodontia достигают
большого развития в триасовую эпоху в сухих областях того вре-
мени.
Отряд 1. Первокрокодилы (Eosuclila)
Представлен семейством Younginidae и родом Youngina. Череп
этой формы обнаруживает весьма примитивные черты, напоминаю-
486
щие череп Pelycosauria. В пем имеется tabulare, пинеальное от-
верстие. Но, с другой стороны, в нем нет предглазничного отвер-
стия и имеются две височных дуги.
Отряд 2. Пар асу хин (Parasuchia)
Триасовые рептилии, распадающиеся па две резко отличных
группы: подотряд Erythrosuchia (Pelycosimia), с высоким коротким
черепом й носовыми отверстиями на конце рыла, и подотряд Phy-
tosauria, с чрезвычайно длинным рылом и носовыми отверстиями,
отодвинутыми назад. Помимо этого различия, являющегося у вто-
рой группы приспособлением к водному образу жизни, в осталь-
ном у обоих подотрядов большое соответствие в строении. Длинно-
рылых Phytosauria следует рассматривать как происшедших от
короткорылых Erythrosuchia.
Подотряд Vhytosciiuria,
Крупные рептилии с длинным рылом, образованным почти
исключительно за счет praemaxillac. Череп с грубой скульпту-
рой. Шея, спина и хвост, иногда и брюхо были покрыты кост-
ными пластинками. Фитозавры весьма походили па крокодилов
как по форме, так и по образу жизни. Название фитозавры —
«растительные ящерицы»—неудачно, ибо рептилии эти были, не-
сомненно, хищниками. Вымерли до наступления мелового вре-
мени. Центральный род — Phytosaurus (Belodon). Череп более
метра длиной. На спине — могучий панцырь. Верхний триас
Южной Германии, триас Повой Мексики и других мест США.
Несмотря па свое внешнее сходство с крокодилами, Parasu-
chia представляют целый ряд отличий в строении черепа и не
являются предками последних.
Отряд 3. Ложные крокодилы (Pseudosuchla)
Очепь важпая в филогенетическом отношении и богатая группа
рептилий триасовой эпохи, живших главным образом в Европе.
Мелкие формы, приспособленные к жизни па суше. Морда корот-
Рис. 391. Aetosaurus ferratus, реконструкция.
(По Marsh из Abel.)
кая. Носовые отверстия па конце морды между praemaxillaria
и nasalia. Большие глаза сбоку. Передние конечности короче
задних. На спипе папцырь из длинных пластинок, расположенных
в два ряда. Образ жизни ложных крокодилов был весьма различен,
как это можно судить по форме тела и по скелету конечностей.
487
Пссвдосухии являются исключительно важной в филогенети-
ческом отношении группой рептилий, давшей начало динозаврам,
птерозаврам, крокодилам и птицам. Они стояли в близком род-
стве с Parasuchia, отошедшими от одного с ними корня.
Сем. Aetosauridae. Род Aetosaurus из верхнего триаса Вюртем-
берга. Имел треугольную голову с острой короткой мордой. Кости черепа
гладкие. Зубы в глубоких альвеолах. На спине и на хвосте прочные по-
перечные цластинки. Хвост длинный. Найдена плита со скоплением этих
рептилий, внешне более похожих на ящерицу, чем на крокодила (24 осо-
би) (рис. 391). Жили и на суше и в воде.
Сем. Ornithosuchidae из триаса Южной Африки. Особенно замеча-
тельно строение таза, напоминающего строение такового у Ornithopoda
(Dinosauria). Род Euparkeia из триаса Капской колонии. Род Ornitho-
suchus — ив среднего триаса Шотландии. Оба рода являлись назем-
ными.
Сем. Scleromochlidae. Род Scleromochlu s—ив среднего три-
аса Шотландии. Маленькая рептилия с необычайно развитыми задними
конечностями, что дает основание думать, что эта рептилия была
прыгающей.
Отряд 4. Настоящие крокодилы (Crocodlliа)
Тело вытянутое, с длинным хвостом большей частью значи-
тельной или средней величины. Носовые отверстия лежат па перед-
нем конце морды и слиты вместе. Внутренние ноздри (хоаны) сое-
динены вместе и .значительно отодвинуты назад. Имеется образо-
ванное посредством maxillaria, palatina иногда и pterygoidea,
сливающимися посредине, вторичное (твердое) нёбо. Непарное
parietale без пинеального отверстия. Две пары височных ям. Quad-
ratum неподвижное. Зубы в ячейках (текодоптные). Глаза боль-
шей частью без окостенений в склере. Имеются брюшные ребра.
Ключицы нет. Pubis не участвует в образовании сочленеиовпой
ямки. Пальцы четырехпалых задних конечностей соединены пла-
вательной перепонкой. Тело покрыто роговыми щитками, под
которыми паходятся костные пластинки. Среди современных реп-
тилий крокодилы — наиболее высокоорганизованные формы. В на-
стоящее время живет всего около 25 видов, относящихся к одному
из 3 подотрядов, именно к Eusuchia— «настоящим крокодилам».
Кроме этого подотряда, к крокодилам относятся Mesosuschia —
«средние крокодилы» и Talattosuchia — «морские крокодилы».
Подотряд «Почти крокодилы», «Средние крокодилы»
(Mesoauchla,)
Включает в себя много разнообразных форм, живших с нижнеюрского до
начала третичного времени. Отличительным признаком их являлись прими-
тивные позвонки амфицельного типа. Внутренние ноздри открываются еще
не столь далеко назади, как у Eusuchia. Панцырь их развит полнее. Более
древние представители подотряда, семейство Teleosauridae, имели длинное
узкое рыло, многочисленные зубы. К концу юрского времени появляются
широкорылые формы, большей частью небольшой величины. Те и другие
продолжали жить еще в меловое время.
488
Подотряд Морение крокодилы (Talattosuchia)
Отличался весьма сильно от предыдущего и следующего подотрядов каш
по строению, так и по образу жизни. Представители его существовали на земле*
недолго и известны в числе немногих форм только из позднеюрских отложе-
ний Европы. Наружность их несет отпечаток среды и образа жизни. Происхо*
дя от речных форм типа современного гавиала, они постепенно приспособи-
лись к добыванию пищи в море и стали вполне водными животными. Кожа их
утеряла скелет, ставший бесполезным, в склере развилось костное кольцо,
отсутствующее у других крокодилов, зубы стали хватательными, тело удли-
нилось, спинные и брюшные отростки хвостовых позвонков увеличились,
в размерах и стали служить для поддержки вертикального плавника, перед-
ние конечности превратились в несовершенный еще плавник. Задние ноги
в плавники не обратились. «Морские крокодилы» не выработали способности*
живорождения и должны были выходить па берега, а потому не могли вьь-
держать конкуренции с ихтиозаврами и плезиозаврами.
Рис. 392. Черен Geosaurus suevicus (Mesosuchia). (Из белой юры
Швабии.)
1 — praemaxilla, 2 — maxilla, 3 —nasale, 4— adlacrymale, 5 — lacrimale,
6 — sclera, 7 — jugate, 8 — postfrontale, 9— parietale, 10 — quadratum
на нижнем и frontale на верхнем рисунке, 11 — quadratojugale, 12 —dentale,
13 — supraangulare, 14 — angulare, 15 — articulare, 16 — нижняя височная
. яма. (Uo Fraas из Abel.)
Сем. Metriorhynchidao. Род Mctriorhynchus, из юры северной
Франции и Англии и Патагонии, представлен более широкорылыми и узко-
рылыми формами.
Сем. Geosa uridae. Род Geosaurus — из верхней юры Южной
Германии, Франции, Англии (рис. 392).
Подотряд Eusiichia,, или Crocodtlicb vera
Голова широкая и плоская, тело плоское, узкое, хвост длин-
ный и служит главным образом при плавании, хотя пальцы задних
пог более или менее связаны перепонкой. Конечности наземного
489>
типа, и животные могут легко передвигаться по суше. Тело по-
крыто роговыми пластипками, подстилаемыми кожными окосте-
нениями. Есть формы с широким и с узким рылом. Появляются
в верхпемеловое время. В третичное время жили крокодилы, до-
стигавшие 12.5 м. Распространены в настоящее время в Южной
Азии, в более теплых частях Северной и Южной Америки, Африки
и па Мадагаскаре. Еще шире были распространены в третичное
время. Крокодилы живут в реках и озерах, выходя и на сушу.
В воде хорошо плавают при помощи хвоста, сжатого и снабжен-
ного для увеличения поверхности сопротивления еще зубчатым
гребнем на спинной стороне. Более неуклюжи они па суше, по ко-
торой все же могут передвигаться довольно сносно. На сушу вы-
ходят, чтобы погреться па солнце, а также для кладки яиц. По-
следние снабжены твердой скорлупой.
Сем. Крокодилы собственно (Crocodilidae). Ры-
ло довольно длинное. Род (Crocodilus, С. niloticus) нильский крокодил.
В древности встречался во всем Ниле, доходя и до его дельты, те-
перь становится редкостью вследствие истребления его человеком. Это
так называемый «левиафан» древних. Кроме Нила, распространен еще
в Судане и вообще по всей Африке, как по берегам ее, так и вну-
три страны, а также на Мадагаскаре, Сейшельских и Каморских остро-
вах. Для отдыха выбирает песчаные отмели, где лежит иногда большими
группами. Сильный и опасный хищник, опасный и для человека. Охотится
на суше лишь ночью. Яйца откладывает в песок, в глубокую ямку, которую
затем засыпает. Детеныши, только что вышедшие из яйца, уходят в воду.
Мускусные железы, развивающиеся у крокодила в период размножения,
представляют ценное медицинское средство.
Узкорылый крокодил (С. cataphractus), с более длинным и узким рылом,
живет в Западной Африке. С. americanus обитает на Вест-Индских
островах, во Флориде, в центральной и в более жарких частях Южной
Америки.
Сем. Аллигаторовые (Alligatoridae). Голова с относительно
коротким и широким рылом. Три рода. Род кайман (Caiman), представлен
в тропической части Южной Америки видом С. trigonalus, достигающим
всего 1.4 м длины. Род lacare. Черный очковый кайман (I. nigra) главным
образом в реке Амазонке, достигает 6 м длины. Род аллигатор (Alligator),
представлен одним видом в южной части США, A. missisipionsis, достигаю-
щим 5 м длины.
Сем. Gavialidae, отличается узким и очень длинным рылом.
Представлено двумя видами, относимыми нередко к различным родам: Gavia-
lis gangeticus — области реки Ганга в Индии, и G. (или Tomistoma) schle-
geli — на Борнео, Малашю, Яве и Суматре. Достигает 4 с лишним метров
(рис. 393).
В геологическое время крокодилы были распространены гораздо шире,
были находимы и в Европе.
11. Подкласс. Динозавры (Dinosauria)
К этому подклассу отпосили весьма различных паземпых пре-
смыкающихся, имевших длинную шею, длиппый хвост, две ску-
ловые дуги и две височные ямы, две предглазпичные ямы, меньшие
передние и более крупные задние конечности, не имевших пине-
ального отверстия, за редкими исключениями. Их считали за есте-
ственную, хорошо отграниченную группу, жившую с триаса
490
до нижнего мела. В настоящее время на основе различий в ске-
лете, главным образом в строении таза, группа эта разбита на два
отряда: хищных Saurischia и травоядных Ornitschia. Но так как
название динозавры очень упрочилось в науке, с другой стороны,
эти с самого начала разошедшиеся группы все ясе имеют один ко-
рень, здесь оставлено объединяющее для двух подотрядов назва-
ние — Dinosauria («чудовищные ящеры»).
Отряд 1. Ящеротазовые (Saurischia)
Таз трехлучевой, ossa pubica направлены, вперед и срастаются
внизу по средней линии. Череп небольшой с предглазпичным от-
Рис. 393. Гавиал (Gavialis gangeticus).
(Из Никольского.)
верстием. Голова под прямым углом к шее. Зубы, сидящие спереди
и по краю челюстей, сжаты или пожевидны, с зазубренными
краями, или в виде ложки или карандаша. Нижняя челюсть без
предзубпой кости (praedentalae). Окостенения сухожилия вдоль
позвоночника нет. В этом отряде много гигантских форм, обязан-
ных своим развитием увеличению гипофизы мозга, т. е. это фор-
мы акромегалические, как припято говорить в патологической
анатомии. С нижнего триаса до конца мезозоя. Следы этих дино-
завров известны уже из пермских отложений, в особенности же мно-
гочисленны из триаса Европы, Сев. Америки и Южной Африки.
Отряд распадается на четыре подотряда.
491
Подотряд ( Pach/ypodosauria)
Группа динозавров, давшая, видимо, начало подотрядам Sauropoda,
Theropoda и ограниченная триасом. Без расширения ossis pubis.
Подотряд (Coelurosauria)
Двуногие динозавры, являвшиеся, видимо, обитателями сте-
пей, где прыгали на задних ногах, подобно кенгуру. Легко по-
Рис. 394. Пра-
вая задняя нога
Ornithomi m u s
altus (Compso-
gnathidae) из
верхнего мела
Канады.
(Из Abel.)
строенные, с кавернозными позвонками и костя-
ми конечностей. Задние поги тонкие, metatarsa-
lia — удлиненные, но не сливавшиеся между со-
бой. Хищные по образу жизни.
Сем. Compsognathidae. Род Compsogna-
thus, из верхней юры Баварии. 75 см длиной, с длинной
шеей, длинной голенью, длинными и тонкими Metatarsalia,
на конечностях по три функционировавших пальца. Хвост
длинный. Род Ornithomimus. Череп маленький с больши-
ми орбитами, длинные тонкие шейные позвонки. Длинные
и тонкие конечности; передние с тремя пальцами и длин-
ными слегка отогнутыми когтями. Голени длиннее бедра.
Pubis с расширением у основания. Metatarsalia длинные
и прижатые друг к другу, пальцы и ногти короткие (рис.
394). Длинный хвост. Сходен с ним род Struthiomimus.
Подотряд Звероногие (Theropoda Megalosauria)
Хищпые динозавры умеренной или крупной
величины, с острыми зазубренными зубами. Рн-
Семейство
bis с расширением у основания. Голень короче
бедра.
Сем. Ceratosauridae. Род Ceratosarus, из
верхней юры Сев. Америки. На носовых костях тре-
угольный гребень (рог) и два костных бугра перед глаз-
ницами. В нижней челюсти отверстие. Передние конечно-
сти и пальцы редуцированные. Род Allosaurus (рис. 395)
из верхней юры Сев. Америки. Редукция пальцев ушла
еще дальше. Длина тела 7—10 м.
Megalosauridae. Род Megalosaurus, гигантские дино-
завры, из юрских и меловых слоев Европы, а также Индии, Австралии, Ма-
дагаскара и Южной Америки. Размером был со слона.
Семейство Thyrannosauridae. Род Thyrannosaurus — почти
такой же гигант, как предыдущий род.
Подотряд Ящероногие (Sauropoda)
Растительпоядпые, ведшие земноводный образ жизни рептилии.
Голова маленькая с одной или реже с двумя предглазничпыми
дырами. На челюсти шпателеобразпые или палочковидные зубы.
Большинство позвонков перед крестцом опистоцельны, позади ясе
лежащие или амфицельные или плоские, редко процельпые. По-
звонки обычно с полостями. В области крестца певральпый капал
сильно расширен. В этом месте помещалось поясничное вздутие
мозга. Копечпости массивные, вероятно, без костномозговой поло-
492
ста. Передние конечности обычно короче, но иногда равны зад-
ним или длиннее их. Полустопоходящие пятипалые формы,
по с редуцированными фалангами на внешней стороне, с массив-
ными костями. Жили с юры до конца меловой эпохи. Четверо-
ногость их вторичная. Тело относительно короткое с очень длин-
ной шеей и хвостом. Вероятно, самые крупные животные из когда-
либо живших.
Сем. Диплодоков ые (Diplodocidae). Род Diplo-
d о с u s. Длина 24 м. Тело удлиненное. Шея из 15 позвонков, длинная,
тонкая. В хвосте 75 позвонков. Наиболее быть может популярная форма из
этой группы динозавров. В отличие от большинства динозавров имел пи-
Рис. 395. Лллозавр (Allosaurus), пожирающий труп.
неальное отверстие. Череп похож на лошадиный. Зубы сидели по нескольку
в одной ячейке и, стираясь, постепенно заменяли друг друга сзади. Форма
их была цилиндрическая с несколько сжатой короткой коронной, с длинными
корнями. Ноги тяжелые, пятипалые (рис. 396).
Судя по строению зубов и тола, Pacliypodosauria и в особенности Thero-
poda являлись хищниками, питавшимися за счет многочисленных тогда ра-
стительноядных, а иногда, быть может, и своих же собратьев. Coclurosauria
не могли быть такими же хищниками: они были менее мощны, пальцы у них
нередко были очень длинны и топки, когти крайне загнуты. Конечно, и они
являлись плотоядными, но питались ли опи рыбой, птицей или наземными
формами рептилий же — нельзя сказать.
Sauropoda были земноводны. Кроме того, они были растительно-
ядны. Об этом говорит форма зубов, не зазубренных, расположенных в пе-
редней части челюстей и или шпателевидных, или палочковидных, быстро
сменявшихся. На концах они обычно стерты. Такие зубы не могли быть
у хищников. Между ребрами некоторых экземпляров находятся отполирован-
ные камни (гастролиты), говорящие эа то, что здесь имелся мощный муску-
листый желудок, в котором перетиралась растительная пища. Обычно встре-
чающийся на передней конечности мощный ноготь мог служить для отрывания
водяных растений. Пищей служили, быть может, водоросли, подобные Chara,
или внутренность стволов цикадовых.
493
Отряд 2. Ппшцетановые (Ornitischia, Orthopoda, Praedentata)
Таз четырохлучевой, pubis с двумя ветвями: направленной впе-
ред praepubis и топкой postpubis, располагающейся параллельно
с седалищной костью. По образу питания — травоядные, с зу-
бами тина коренных, тесно расположенными в верхнечелюстных
и нижнечелюстных костях (за исключением двух форм, у которых
praemaxillae лишены зубов). В симфизе двух половин нижней
челюсти находится лишенная зубов кость — praedentale. Пред-
глазпичпая яма редуциро-
вана. Передняя беззубая
часть челюстей одета ко-
жистым или роговым клю-
вом. Брюшных ребер нет.
Сухожилия вдоль позво-
ночника окостеневают.
Позвонки плотные, кости
конечностей полые или
плотные. Па льцеходящи е
или полустопоходящие, со
сдавленными когтями.
Иногда сильно развитый
кожный скелет. Жили с
верхнего триаса до самого
верхнего мела. Распадают-
ся на три подотряда.
Подотряд Птиценоггсе
(Ornithopoda Ignanodon-
tia)
Двуногие прямо ходя-
щие динозавры с головой
Рис. 396. Диплодок (Diplodocus carnegii)
из верхней юры Сев. Америки.
умеренного размера, по-
ставленной под прямым
углом к шее. Praemaxillae,
как правило, без зубов; зубы па maxillae и dentalia с режущим
зазубренным краем. Позвонки туловища опистоцельные. Передние
конечности короче задних; пальцеходящие формы со сдавленными
копытообразными когтями. Postpubis длинная и тонкая. Кожного
скелета, как правило, пет. В отличие от Theropoda, насколько
можно судить по отпечаткам пог, иногда касались передними по-
тами земли.
Сем. Iguanodontidae. Praemaxillae беззубы. Род Iguanodon
(рис. 397) из вельдских (нижнемеловых) слоев Бельгии и Англии. В Бельгии
найдено целое кладбище — 29 особей. В высоту животные достигали 5 м,
в длину — 10 м. Огромные животные эти передвигались на двух задних
трехпалых ногах; устройство передних конечностей было таково, что совер-
шенно не допускало ходьбы или бега на четырех ногах. Реконструкция этого
животного изображает его подобным гигантской ощипанной бегающей ку-
494
рице. На хвост игуанодоны опирались, обгладывая листья с древовидных ра-
стений, вовремя ходьбы и бега они хвостом балансировали (рис. 398).
Сем. Trachodontidae, отличается от предыдущего семейства,
четырьмя пальцами на передних конечностях вместо 5, сжатым с боков плава-
тельным хвостом, перепонками на пальцах и утиным клювом. Панцыря не-
было, кожа была покрыта мелкими пятиугольными чешуями, не налегавши-
ми одна на другую. Зубы сидели многочисленными рядами, от 45 до 60 вер-
Рис. 397. Череп Iguanodon bernissartensis из нижнего мела’
Бельгии.
1 — praemaxillare, 2 —maxillare, 3 —nasalla, 4 — supraorbitalia, 5 —adla-
crimale, 6 — frontale, 7 — postfrontale, 8 — postorbitale, 9 — parietale,
10 — squamosum, 11 — левое, quadratum, Ila —правое quadratum, 12 —
quadratojugale, 13 —jugale, 14—нижняя височная яма, 15 — praedentale,..
16 —dentale, 17 —орбита. (По Dollo из Abel.)
Рис. 398. Реконструкция картины жизни Iguanodon bernissar-
tensis: быстрый бег. (Из Abel.)
тикальных и 40—14 горизонтальных. Такие зубы были в высшей степени'
пригодны (числом около 2 000) для перетирания жесткой растительной пищи
(рис. 399). Род Trachodon — верхний мел Сев. Америки. Известны не-
только скелет, но и мумия.
Подотряд Панцыркые динозавры (Stegosauria)
Чстырехпогие, покрытые папцырем дипозавры, с малым чере-
пом, с беззубой передней частью рта, с функционирующими зубами,
расположенными в один ряд. Большие расположенные впереди
495»-
ноздри, широким тяжело вооруженный панцырь. Позвонки
амфицельные или плоские, весьма массивные. Задние ноги длиннее
передних; полустопоходящие формы с 3—5 пальцами и копыто-
образными когтями. Сильный кожный панцырь, явившийся причи-
ной вторичного перехода от двуногого к четвероногому положе-
нию тела. С нижней юры до конца мелового периода, в Европе
и в Сев. Америке.
Сом. Stegosauridae. Род Scelidosaurus. Панцырь дифференцирован
не сильно. Длина — 4 м. Нижняя юра Англии. Род Stegosaurus — весьма
Рис. 399. Trachodon annectens из верхнего мола Сев. Америки.
причудливое животное с маленьким удлиненным черепом, расширением
в крестцовой области, в двадцать раз превосходящим полость мозга. Задние
ноги длиннее передних. На спине два ряда прямо стоящих костных пласти-
нок, на хвосте — длинные костные шипы в два ряда (рис. 400). Верхняя юра
Северной Америки.
Подотряд Рогатые динозавры. (Ceratopsta)
Рогатые динозавры с огромным черепом, по длине равнявшимся
одной трети длины всего животного. Рога в числе трех помеща-
лись на посу и над глазами. Затылок и сросшиеся 4 шейных по-
звонка покрывались костным воротником, образованным squa-
mosum и dermo-supraoccipitalia. Впереди лишенные зубов ргае-
maxillae — непарный ростральный элемент, образовавший вме-
сте с соответствующей predentale нижней челюсти черепаховидный
клюв. Зубы, как ни у каких других рептилий, с раздвоенпымикор-
496
нями. Позвонки и кости конечностей плотные. Передние конеч-
ности короче задних. Когти копытоподобные, кожа покрыта тол-
стой мозаикоподобной чешуей. Род Triceratops (рис.401), из верхне-
го мела Сев. Америки, с носовым рогом и двумя длинными, направ-
Рис. 400. Stegosaurus ungulatus из верхней юры Сев. Америки.
(По Abel.)
Рис. 401. Реконструкции Triceratops.
(По Night.)
ленными вперед рогами над глазницами. Род Protoceratops с менее
развитым воротником и рогами. Найден в Монголии так же, как
его яйца (рис. 402).
Строение зубов Ornithischia совершенно определенно указы-
вает, что пища их была растительной. Некоторые из них были все-
ядны. Чертами, указывающими па растительпоядпость, являются
последовательная смена зубов в верхней челюсти и в dentale;
32 Кашкаров и СтанчнпскиИ 497
отсутствие за пемпогими исключениями зубов в передней части
рта, наличие режущего клюва у одних и утинонодобного клюва
у других, жевательный характер зубов в задних частях рта. Ра-
стителыгоядность еще не выражена у низших представителей
Ornitliopoda. Iguanodontidae, имевшие хватательный клюв и пе-
редние конечности, сильное зубное вооружение, являлись живот-
ными, объедавшими ветви. Зубы Trachodontidae указывают на пи-
тание их жесткой растительной пищей, главным образом хво-
щами, которые они отрывали своим клювом. Stegosauria имели
маленькие зубы, игравшие сравнительно малую роль. Строение
симфиза нижней челюсти говорит за то, что Stegosaurus имел хва-
тательный язык. Роговыекрая челюстей по функции замещали зубы.
Рис. 402. Яйца Protoceratops, найденные в Монголии.
(По Andrews.)
Стегозавры обитали в низких, более или менее болотистых ме-
стах. Triceratops, представитель рогатых динозавров, являлся,
видимо, существом, кормившимся растительностью кустарной,
как кормятся современные носороги.
Динозавры имели самое широкое распространение, почти ио
всему миру, обнаруживая самые разнообразные формы приспо-
собления.
12. Подкласс Крылатые ящеры (Pterosauria)
Рептилии, приспособившиеся к жизпи в воздухе и приобрет-
шие специальные для этого приспособления. Кости топки и полы,
кожа голая, передние конечности превращены в крылья путем
удлинения последнего пальца. Череп заостренный, зубы хвата-
тельные. Большая предглазпичпая и маленькая височная яма.
Большие глаза окружены костным кольцом. Длинным пальцем
является 4-й. 1-й, 2-й, 3-й имеют нормальное строение и снабжены
498
когтями. В тазу имеется сходная с тем, что есть у крокодилов,
кость praepubis. Подкласс представлен главным образом в юре,
из мела известно очень мало форм.
Отряд 1. Трибелезодоиты Trlbelesodontla
Сем. Tribelesodontidae с родом Tribelesodon — примитив-
ная длиннохвостая форма. Палец крыла очень короткий, наоборот, задняя
конечность очень длинная, грудина узкая, таз большой, широкий praepu-
bis. В нижней чрлюсти направленные зазубринами вверх зубы, чего нет ни
у каких других рептилий. Крылья короткие, служившие не для свободного
полета, а в качестве парашюта.Форма — несомненно примитивная.
Отряд 2. Длиннохвостые птерозавры Rhainphorliyncholdea
Длинный тонкий хвост, оканчивающийся ромбической руле-
вой пластинкой, играл при полете большую роль. Он был совер-
шенно плотный, сухожильный, причем сухожилия окостеневали.
Крылья, длинные и узкие, нс складывались. В черепе небольшое
отделенное от носовых отверстий, предглазиичпое отверстие.
Сем. Dimorphod out id ае. Род Dimorphodon из триаса и ниж-
ней юры Англии. Большая голова, короткая шея и туловище, две височных
дуги. Глаза окружены костным кольцом. Конечности тоньше, на задних ко-
нечностях 5-й палец отставлен в сторону.
Сем. Rhamphorhynchidae. Род Rhamphorhynchus
из верхней юры Южной Германии. Хвост такой же длинный, как у Dimor-
phodon, с рулевой пластинкой на конце, но череп более длинный, ящеричного
типа, с длинными коническими зубами в верхней и нижней челюстях. Костное
кольцо в склере также существовало. Летательная перепонка оканчивалась,
как показывает ископаемая находка, в начальной части хвоста и склады-
валась, видимо, продольными складками.
Отряд 3. Короткохвостые птерозавры Pterodaclyloldea
Хвост совершенно укорочен, а потому при полете пе играл ни-
какой роли. Крылья более широкие, чем в предыдущем отряде.
4-й палец складывался вместе с крыльями. Глазницы не были впол-
не отделены от носовых отверстий.
Сем. Р t о г о d а с t у 1 i d а с. Род Pterodactyl us. Передняя
часть черепа удлинена. Орбита пе вполне отделена от преорбитальной ямы.
У некоторых видов намечается редукция зубов. Летательный палец склады-
вался. Грудина относительно мала. Пятый палец задних ног рудиментарный.
В верхней юре Южной Германии и Франции. Найден также и в Англии
(рис. 403).
Сем. Pteranodontidae — самое оригинальное семейство из
птерозавров. Череп с длинным топким затылочным гребнем. Scapula причле-
пялась к слившимся 8 позвонкам. Длинное вытянутое рыло было лишено зу-
бов. Череп равнялся но длине туловищу с вытянутыми задними ногами.
Глаза со склеротическим кольцом. Симфиз нижней челюсти очень прочный.
С черепом пижняя челюсть сочленялась сходно с тем, что имеется у пеликана,
у которого задние концы черепа могут раздвигаться, открывая мешок нахо-
дящийся под челюстями. Такой мешок, видимо, был у и рода Pteranodon.
Тазовые кости у этого рода срастались с 10 крестцовыми позвонками напо-
добие таза птиц. Pteranodon являлся самым крупным из когда-либо живших
летающих существ и достигал 8 м в размахе крыльев (рис. 411).
Rhamphorhynchoidea и Pterodactyloidea представляли два различных
приспособительных к полету типа. На это указывает вся их организация,
32* 499
главным образом средних конечностей. Полет представителей первого отря-
да должен был быть спокойным, равномерным, скользящим, уверенным поле-
том. Полет птеродактилей должен был быть тяжелым, неуверенным гребным
полотом. У Rhamphorhynchoidea длинное узкое крыло имело слегка изогну-
тый передний край и заостренную форму. Между плечом и предплечьем
спереди натягивалась складка — propatagium. Летательная перепонка про-
стиралась и от задних конечностей к хвосту. У Pterodactyloidea крылья были
короткие, широкие. На хвост перепонка не могла распространяться, ибо
и хвоста не было. Тело птерозавров было голо, без чешуй, кожа в склад-
ках. Пищей их являлась рыба, которую они добывали из воды подобно
Рис. 403. Реконструкция Pterodactylus spectabilis ив верх-
ней юры Баварии.
(По Abel.)
чайкам. Населяли они, видимо, коралловые острова, где подобно летучим
мышам висели на деревьях, когда отдыхали. Были они дневными или ноч-
ными животными,—неизвестно. Сами нередко попадали в пищу плезиозав-
рам. Вымерли, не оставив потомства.
IV. ФИЛОГЕНЕТИЧЕСКОЕ РАЗВИТИЕ РЕПТИЛИЙ
Изменения среды в эпоху появления рептилий, отношение
рептилий к амфибиям, адаптация к жизни на суше. Происхожде-
ние земноводных приурочено к девону. Рептилии происходят от
стегоцефал.
Низшие рептилии, как было указано выше, с трудом отли-
чаются от последних, и некоторыми исследователями относятся
к стегоцефалам. Таковы, папример, Rliizosauria, которые по
строению позвоночника отпосятся к эмболомерпым стегоце-
фалам, а по строению конечностей являются рептилиями. Такой ясе
переходный тип меяеду двумя классами представляет и Seymouria.
Последняя известна из пермских отложепий, Rliizosauria— из
пижпепермских. Нуясно думать, что возникли рептилии уясе в кон-
це каменноугольного времени. Чем их появление и развитие
было вызвано?
Появление земноводных было подготовлено развитием в де-
вонское время двоякодышащих рыб (предков Dipnoi и Crossopte-
rygii). В каменноугольное время опи вышли па сушу при усло-
виях, благоприятных для выхода па сушу ишвотпых с весьма еще
500 *
несовершенным легочным дыханием. Появление рептилий было
подготовлено развитием среди стегоцефал форм, более наземных.
Толчком к развитию вполне наземных позвоночных — рептилий —
явилось изменение условий существования при переходе от ка-
менноугольной эпохи к пермской.
В конце каменноугольной эры имели место большие поднятия
земной коры, повлекшие за собой развитие более сухого климата,
а затем захолодапия; к началу триаса сухость достигла еще боль-
шего развития. Условия жизпи улсе с конца каменноугольной
эпохи стали изменяться в неблагоприятную для земноводных сто-
рону. Обширные каменноугольные леса с их болотами и озерами
стали сильно сокращаться. Сырой оранжерейный климат стал
сменяться на сухой, иногда далее пустынный, под влиянием кото-
рого откладывались гипсы, каменная соль и т. д. Там, где сохра-
нялись еще прежние условия, сохранялись и стегоцефалы, до-
жившие до триаса. Некоторые из земноводных под влиянием
усыхания среды вторично приспособились к жизни в воде, стали
вторичноводпымн, подобно современным Urodela, как, например,
северодвинский Dwinasaurus. По для большинства возврат в воду
оказался невозможным. Ставка в борьбе за существование была
поставлена па приспособление к новым условиям, на приспособ-
ление к существованию в сухом климате. Роговой покров, под-
стилавшийся вначале еще костными образованиями, утрата кожных
желез предохраняли их от высыхания. Более совершенные легкие
и соответственное изменение кровообращения дали возможность
существовать без жаберного дыхапия, оставить воду и прибрежные
сырые местообитания. Но главное, что дало возможность сделать
это, было радикальное изменение в строении яйца, богатого
желтком и снабженного оболочками, благодаря которым разви-
вающийся в яйце зародыш может дышать воздухом.
Более стройное, нежели у стегоцефал, тело, более совершенные
конечности, полное освобождение от водной среды, развитие лег-
ких и соответствующие изменения в кровообращении вывели вновь
возникший класс на широкий простор, и он начал быстро разви-
ваться.
Начавшаяся, вероятно, еще в каменноугольную эпоху диффе-
ренцировка рептилий достигает значительных размеров в перм-
ское время, и еще больших в триасовое. Не встретив па суше кон-
курентов, так как млекопитающих еще пе было, рептилии начали
быстро расселяться, приспособляясь к различным местообита-
ниям па суше, приобретая различные форму и образ жизпи. Не-
которые вторично возвратились к жизпи в воде, и у них развился
целый ряд приспособлений для существования в этой среде. На-
конец, третьи, пе имея конкурентов в воздухе, приспособились
к жизни в этой стихии. У тех рептилий, которые приспособлялись
к жизпи на суше, развивались иногда приспособления, аналогич-
ные тем, которые отличают млекопитающих (у Theriodontia);
у водных рептилий развивались черты поразительного сходства
с рыбами (у Ichtyosauria) и китообразными; у воздушных рентн-
ый
лий (Ptcrosaiiria) выработались многие черты, свойственные пти-
цам. Рептилии долго властвовали па суше, па воде и в воздухе.
Затем часто по непонятным причинам начали вымирать, и их
место заняли от них ясе ведущие свое начало птицы и млекопитаю-
щие. Некоторые группы рептилий вымерли еще раньше, так как
одностороннее развитие приводило их в тупик эволюции, и они
по могли следовать за изменениями среды. Вообще же эволюция
рептилий, как и эволюция других групп позвопочпых животных,
происходила под контролем среды, следовала за се изменениями.
При этом, под средой следует понимать пе только внешнюю фи-
зическую среду, но и — для каждой группы — изменения окру-
жающего растительного и животного мира. Эволюция отдельных
Рис. 404. Родословное дерево рептилий.
форм шла в связи с эволюцией окружающего комплекса. Это
можно проследить па истории отдельных ветвей родословного
дерева рептилий.
В настоящее время сохранились лишь некоторые древние ветви
последнего, как черепахи (подкласс Testundinata), крокодилы (от-
ряд Crocodilia), гаттерия (подотряд Sphenodontia) и другие более
новые ветви: ящерицы (отряд Lacertilia) и змеи (отряд Ophi-
dia), приспособившиеся к особым условиям существования, выра-
ботавшие ряд особенностей, дающих им возможность пе бояться
конкурентов.
Филогенез отдельных групп рептилий (рис. 404). В пижпеперм-
ских слоях нам встречаются уже представители подклассов Coty-
losauria, Pelycosauria, Ichthyopterygia и отряда Araeoscelida; из
них 56% приходится па Synapsida, 44% — па Anapsida и 1% — па
остальных. Предки рептилий должны были иметь цельный, сте-
гальный череп, как у стегоцефал, т. е. принадлежать к Anapsida.
502
К этим предкам наиболее близки Cotylosauria с замкпутым,
стегальным черепом. По черепу опи вполне примыкают к стего-
цефалам. Различие в строении позвопков говорит за то, что,
происходя от стегоцефал, опи очень рано, вероятно еще в камен-
ноугольное время, пошли в своем развитии собственным путем.
Среди Cotylosauria были формы роющие (сем. Diadectidae), на-
земные (сем. Pareiasauridae) и земноводные (сем. Labidosauridae
и сем. Limnoscelidae). Cotylosauria, вероятно, дали начало ряду
других ветвей рептилий.
Pelycosauria появляются одновременно с Cotylosau-
ria, и наиболее примитивные из пих весьма сходны с Cotylosau-
ria, от которых или от форм, близких к которым, они, видимо,
и произошли. Высшие формы обнаруживают значительную спе-
циализацию, давая иногда весьма причудливые формы. Интересно,
что у пих, как и у представителей следующего подкласса — The-
romorpha, в строении черепа появляются черты черепа млеко-
питающих. Здесь мы имеем дело с параллельным развитием
у форм, генетически близких друг к другу. Pelycosauria вымерли
нацело, не оставив потомков среди более молодых рептилий.
Подкласс Theromorpha (Therapsida) является одной
из наиболее богатых как формами, так и потомками групп реп-
тилий. Он дал начало млекопитающим, процветал в холодную перм-
скую эпоху и в нижнем триасе. Наиболее примитивные предста-
вители в высшей степепи близки к наиболее примитивным пред-
ставителям Pelycosauria. От общего корпя с Pelycosauria видимо
и происходит Theromorpha.
Эволюция подкласса прослежена довольно хорошо благодаря
феноменально большому количеству ископаемых представителей
его в пустыне Карру в Южной Африке, где, по подсчетам, скрыто
в земле до 1000 ископаемых па квадратную милю (около 3 км2).
Центральным отрядом подкласса является отряд Theriodontia,
процветавший в пермское время и в триасе. Этот отряд привел к раз-
витию млекопитающих. К этому отряду примыкают отряды Dino-
cephalia и Dicynodontia (Anomodontia). Ступени, которые в под-
классе Theromorpha привели к млекопитающим, связаны с изме-
нением мест обитания и рода пищи, с переходом от мало подвиж-
ных форм ко все более активным. Прослеживая смепу различных
тероморф в последовательных слоях Карру, мы видим, что большие
медленно двигавшиеся формы вымирали вместе с теми, па кого они
охотились. Менее спсциализовапные мелкие формы, более пла-
стичные, приспособлялись к новым условиям. Наиболее млеко-
питающеподобпыми являются Ictidosauria — почти млекопитаю-
щие, а также Gynodontia, папример Permocynodon. Питались они
очень активными ящерицеподобпыми Eosuchia. Весьма вероятно,
что затруднения в добывании и поедании последних заставляли
их питаться насекомыми и были причиной развития особенностей,
приведших к развитию млекопитающих типа Monotremata. Dicyno-
dontia (Anomodontia) и Dinocephalia вымерли без потомков. В этих
отрядах мы видим приспособления к различному образу жизни.
503
Предки Ichtyopterygia нам неизвестны. Mesosau-
ria и Ichthyosauria происходят, повидимому, от общего корня,
но разошлись, приспособляясь к жизни в воде различными путями.
Эволюция ихтиозавров в общем шла в направлении развития все
более совершенного торпедообразного типа обитателя моря. Ин-
тересны изменения зубов в зависимости от развития окружающего
комплекса и изменения пищи. Древнейшие ихтиозавры обладают
мощно развитыми хватательными зубами. В верхней юре увели-
чение в морях мягкотелых головоногих моллюсков привело к из-
менению питания ихтиозавров, развитию «теутофагии» и атрофии
зубов, так как пища глоталась целиком.
История Sauropterygia в настоящее время более
ясна. Различные подотряды этого подкласса — Trachelosauria,
Nothosauria и Plesiosauria—происходят от одного корпя, от на-
земных предков, вероятно, близких к общим предкам примитивных
Theriodontia и примитивных Pelycosauria. Различие строения го-
ловы, зубов, шеи у плезиозавров являются результатом различия
в пище: одни с более короткой шеей являлись охотниками за бы-
стро двигавшейся рыбой, другие с длинной шеей (у Elasmosaurus
было 76 шейных позвонков) питались бентосом, доставая его по-
добно некоторым водяным птицам. Нотозавры, пе будучи ‘ пред-
ками плезиозавров, показывают, однако, каким образом могли
последние развиться.
Подкласс Placodontia представляет одностороппе спе-
циализированную, приспособившуюся к водной жизни и питапию
твердой пищей ветвь рептилий, ответвившуюся от каких-то прими-
тивных форм. Потомства он не оставил,
Представители подкласса Testudinata известны с
триаса. Это — замкнутая группа, ведущая свое начало, вероятно,
от пермских Cotylosauria,. типа Diadectidae, бывших роющими
животными. Древнейшие черепахи были пе водными, а роющими
существами. Панцырь развился у них как приспособление к этому
образу жизпи подобно тому, как у броненосцев. Древнейшие пред-
ставители, как Triassochclys, были наземными формами. Из
остальных черепах скрытоптсйпыс — Cryptodir а, а из
них Testudinidae являются наиболее примитивными
из живущих. В конце юры от черепах, живших в болотах, озерах
и реках, происходят морские формы, представленные ныне такими
формами, как Dermochelys. Переход совершился, конечно, не сра-
зу. Таким образом, семейство Chclonidac является специализиро-
ванной боковой ветвью основного наземного ствола черепах. Так же
и Trionychoidea происходят от наземных черепах. Бокошейиыс
черепахи Pleurodira также очепь рано ответвились от Cryptodira.
Подкласс Diaptosauria, представленный в настоящее
время только гаттерией в Новой Зеландии, представляет собой
примитивную лилию, рано отделившуюся от ствола Cotylosauria.
Их нельзя ставить в основу чуть пе всех линий рептилий, как это
делалось неоднократно. Эта группа никогда пе имела большого
значения в истории развития класса рептилий.
504
О происхождении подкласса Lepidosauria известно'
очень мало. Можно думать, что оп возник рано, где-то от общего
ствола с Diaptosauria. На древность их указывает Araeoscelis,
найденный в пермских слоях, хотя считать эту форму прямым
предком остальных Lepidosauria нельзя. Опа является, повиди-
мому, боковой ветвью, ответвившейся от самого основания ствола
Lepidosauria. В верхнем мелу ствол Lepidosauria делится на две
ветви — Lacertilia и Ophidia.
Весьма интересным является неоднократное ответвление от
ствола Lepidosauria форм, приспособлявшихся к жизни в море и
приобретавших при этом высокую специализацию: из них Doli-
chosauria — до третичной эры. Мезозавры были очень крупными
формами.
Остальные три подкласса — Thecodontia, D i п о-
sauria и Pterosauria — развиваются как параллель-
ные ветви от ствола, общего с пачалом Diaptosauria и Lepidosauria
(группа Diapsida). В этой группе рептилий берут начало птицы.
Древнейший представитель Thecodontia, род Youngina, об-
наруживает примитивные черты, напоминающие череп Pelyco-
sauria. Отряд Parasuchia, триасовых рептилий, жил лишь короткое
время — в течение триаса и не дал потомков. Наоборот, отряд
Pseudosuchia представляет собой чрезвычайно важную в филоге-
нетическом отношении группу рептилий. Происходят псевдосухии
от одного корня с парасухиями. Но опи очень рано разошлись,
приспособившись: Parasuchia — к жизни в воде и приобретая
внешне сходство с крокодилами, Pseudosuchia — к жизни на
суше. Последние являются родоначальниками как крокодилов,
так и динозавров и птерозавров. Птицы также ведут свое проис-
хождение от псевдосухии. Расхождение псевдосухии и парасухий
произошло пе позднее пермского времени, динозавры отчленились
ранее триаса, птицы — в юре или раньше. По образу жизни раз-
личные псевдосухии значительно отличались друг от друга. Одни
вели наземный образ жизни, другие полуводный, третьи лазали
по деревьям; были даже прыгающие формы с сильпо развитыми
задними конечностями, как Sclcroinochlus.
Крокодилы, происходящие от псевдосухии, повидимому, ответ-
вились от них уже в триасе и наибольшего расцвета достигали
в юрское и меловое время. Mcsosuchia и Talattosuchia вымерли
без потомков. Eusuchia, появившиеся в мелу, живут и по сие
время.
От неизвестных предков типа пссвдосухий ведут свое начало
и представители подкласса Dinosauria. Динозавры распадаются на
две группы — Saurischia и Ornithischia. Эти две группы, видимо,
развивались параллельно, после того как опи ответвились от не-
известного предка из Pseudosuchia. Возможно, по менее вероятно,
что опи ведут начало не от одной родопачальпой группы. Ornith-
ischia обнаруживают родственные отношения с Pterosauria
и Aves, чего пет у Saurischia, более близких с крокодилами. Ди-
нозавры обнаруживают большое разнообразие строения формы и
505
величины. Эволюция их в значительной мере связана с изменения-
ми среды.
Из Saurischia наиболее примитивен триасовый подотряд
Pachydosauria. От пего возникли подотряды Sauropoda и Thero-
poda, вымершие в меловом периоде. От одного основания с Pachy-
dosauria возник и подотряд Coelurosauria, вымерший также в мелу.
Отряд Ornitischia возник в триасе от одного корпя с птицами и пте-
розаврами. Корнем этим являлись псевдосухии, именно Ornitho-
suchidae. Уже в триасе этот отряд распадается па подотряды —
Ornithopoda, Stegosauria и Ceratopsida. Последние бтветвляются
от основания ствола Stegosauria.
Как указано выше, появление тех или иных форм динозавров
связывается с условиями существования. Так, в сухости климата
триаса и в связанном с этим развитий пустынных равпип следует
искать причину развития двупогости динозавров. Влажный кли-
мат и изобилие стоячих вод создали тип Sauropoda. Огромная ве-
личина этих рептилий предшествовала переходу к земноводпому
образу жизпи, который в некоторой мерс разрешил вопрос, свя-
занный с увеличением веса тела, а также вопрос о пище. Чрезмер-
ное же расходование энергии, связанное с необходимостью добы-
вать много пищи, привело Sauropoda к гибели. Меловые Tracho-
"dontidae также приспособлялись к полуводному образу жизпи,
но они более плавали, нежели ходили. Внезапное исчезновение
с лица земли динозавров в конце мелового времени является загад-
кой, еще не вполпе разрешенной, но во всяком случае связанной
с высокими горными поднятиями в это время.
Подкласс Pterosauria имеет, подобно динозаврам, ди-
филетическое развитие. Уже древнейшие представители подкласса
прошли различные пути развития от псевдосухии, и между двумя
главными отрядами — Rhamphorhynchoidea и Pterodactyloi-
dea — пет переходов. В одпой группе из парашютного полета раз-
вился парящий полет, в другой — гребной тип полета. Возникнув
в триасе, птерозавры господствовали в юре, когда археоптерикс
только начинал порхать. Вымерли птерозавры в верхпемеловое
время.
V. экология РЕПТИЛИЙ
1. Приспособления к условиям обитания. Типы: наземный,
подземный, древесный, водный
Современные рептилии — лишь обломок огромного филогене-
тического дерева, процветавшего в конце палеозоя и в мезозое.
Тогда опи господствовали па земле, в воде и воздухе, занимая все
среды обитания, причем у них вырабатывались приспособления
к ним разнообразными путями. В некоторых случаях приспособ-
ленность рептилий, как ископаемых, так и современных, к условиям
обитания является изумительной.
Приспособления к наземному и подземному образу жизни.
Возникли рептилии как тип, прогрессивно приспособлявшийся
506
к жизни на суше. Но первые рептилии были еще весьма несовер-
шенно приспособлены к движению и жизни па земле. Пока у них
пе было конкурентов, они могли господствовать на суше, но по-
том конкуренция различных форм рептилий между собой, напря-
женная борьба за существование обусловили появление более со-
вершенного типа паземпого существа. Первые рептилии из под-
класса Cotylosauria, например Seymouria, имели еще настолько
большое сходство со стегоцефалами, что некоторыми учеными они
и относились к последним. Неуклюжее тритонообразное тело на
коротких массивных ногах, короткий хвост с ребрами, массивная
голова характерны для животного, которое еще очень плохо дви-
галось по земле. Так же мало приспособленными наземными живот-
ными, жившими па берегах и спускавшимися в воду, были Labi-
dosaurus и Pareiasaurus. Одпако длинный путь развития привел
к созданию весьма совершенных наземных форм. Примером по-
следних могут служить ящерицы из родов Agama, Lacerta или
Eremias. Эти ящерицы ведут дпсвпой образ жизни. Каждому ви-
девшему их известна быстрота, с которой передвигаются они по
земле, убегая от опасности или гоняясь друг за другом в период
спаривания. Длинное тонкое тело на стройных ногах, с длинным
тонким хвостом сильно отличает этих животных от их палеозой-
ских предков и совершенно не оправдывает названия «пресмы-
кающиеся».
При очень быстром беге становится необходимым уменьшение
поверхности трения. Одновременно стопа, направленная наружу
и назад, должна повернуться внутрь и вперед. Колено благодаря
этому приближается к туловищу, и тело более опирается па внут-
реннюю часть стопы, чем па наружную. Благодаря всему этому
утрачивается крайний наружный, т. е. пятый палец.
Из современных рептилий способность к быстрому бегу наблю-
дается у относящейся к семейству Agamidae австралийской плаще-
носной ящерицы— Chlamydosaurus. Мы видим у нее способность
пробегать небольшие расстояния только на задних ногах, держа
передние в несколько опущенном состоянии и балансируя хвостом.
Так убегает эта ящерица от преследования. Кроме Chlamydo-
saurus, эта способность встречается лишь у немногих современных
ящериц, по была, как мы видим, весьма распространена у мпогих
ископаемых, передвигавшихся па задних погах, подобно кенгуру.
Как мы видели из систематического обзора, среди них были не
только мелкие формы, как Compsognathidac, по и весьма крупные,
как Allosaurus (6—7 м длины) и Iguanodon (10 м длиной и 5 м в вы-
шину). В скелете этих двуногих динозавров мы видим ряд при-
знаков, сближающих их с птицами, по являющихся, видимо,
результатом приспособления к одинаковым условиям передвиже-
ния. Конечности весьма сходны с птичьими; в особенности сходны
с последними опи у Ornithomimus, хищного динозавра из меловых
отложений Сев. Америки. Сходство это и в наличии tibio-tarsus
и в устройстве metatarsus. Os ischium у птиценогих динозавров
длинна и направлена назад, как у птиц, и параллельно ей распо-
507
лагалась os postpubis, длиппый отросток pubis, аналогично тому
как располагается os pubis у птиц. Весь таз, соединявшийся о
V—VI крестцовыми позвонками, сходен с птичьим. Если бы одгп.
мускулатуру на скелет такого динозавра, то последний выглядел бы
подобно бегающей птице. Мы знаем, что двуногость связана с пали
чием сухих равнин. Подобные условия должны были создать и Си-
гающих динозавров.
Жизнь в пустыне, в частности, в песчапой пустыне, вызывает
еще целый ряд приспособлений у населяющих ее рептилий. Наи-
более важными являются приспособления, мешающие погру-
жению в песок. Это достигается, например, расширением самого
Рис. 405. Ступни задних конечностей Phrynocephalus mystaceus,
Eremias grammica, Callisaurus scoparius (слева направо).
(Из Buxton.)
тела животного, превращающегося в округлый диск (Phrynosoma);
это же менее, но все же выражено у круглоголовки (Phrynoce-
phalus).
Преимущественно же для предохранения от проваливания жи-
вотного в песок (и для рытья) служат конечности. Опи превра-
щаются в своего рода лыжи с широкой поверхностью (рис. 405).
Пальцы, отороченные как бы щетками, развиваются совершен-
но самостоятельно в ряде семейств: Agamidae, Lacertidae, Igua-
nidae, Geckonidae. Это является указанием на условия жизни
в песке. Особенно развита роговая щетка на пальцах у Phrynoce-
phalus и Eremias. Даже гекконы, живущие в пустыне, своеобразно
приспособляются к бегу по песку. У одних присасывательные пла-
стинки на концах пальцев исчезают, у других (Ptyodactylus), на-
оборот, расширяются в широкие пластинки, играющие опять-
таки роль лыж; у третьих, как у Palmatogecko rangei, живущего
в песчаных пустынях юго-западной Африки, между пальцами на-
тянута как бы плавательная перепонка, и, пе зная его настоящего
местообитания, можно подумать о приспособлении к плаванию.
Здесь же перепонки служат для закапывания в землю.
В качестве приспособления к жизни и движению в густой траве,
повидимому, развился тип рептилий, лишенных копечностей, т. е.
змеевидный. Длинная «змеиная» форма тела встречается уже у яще-
риц. Специфическим признаком являются утрата копечностей и уве-
личение числа позвонков.
Вслед за конечностями исчезают и пояса конечностей. Среди
ящериц мы видим формы, у которых хотя и есть передние и зад-
ние конечности, но они очепь малы, слабо развиты, число пальцев
уменьшается; например, у гологлаза — Ablepharus lineatus —
508
•J и 3 пальца, или у халцида трехпалого — Chalcides tridactylus 3
У других имеются лишь задние конечности, иногда лишь в ’
выступа, как у желтопузика, — Ophisaurus apus; у третьих имеютвя
лишь передние конечности; у четвертых нет ни тех, ни ДруГих
(перетенпица — Anguis, двуходка — Amphisbaena). У некоторЬ1Х
змей передние конечности отсутствуют, задние представлены Незна-
чительными рудиментами таза и конечностей. У Python sebae
африканского питона, эти рудименты имеют коготь, у других
Нас рудименты эти выражены менее яспо. Имеются их следы у ко.
ралловой змеи (Ilysia scytale) и у других представителей Семей-
ства Ilysiidae, также и у роющих слепунов — Typhlopi(jae
У всех остальных змей передние и задние конечности исчезают
новее.
Движение совершается при помощи изгибов тела в горизонталь-
ной плоскости в виде следующих друг за другом букв S. При Таком
движении конечности становятся излишними.
Весьма интересно, что такое исчезновение конечностей разви-
лось независимо в разных семействах. Оно является, песомце11но
«приспособлением» к определенным условиям — к жизпи
нпю в густой траве, в ветвях, в лесном хламе или, как у рц^__
приспособлением к жизни па илистом дне. ’
Приспособлением к жизпи па суше является и роговой покров
Ото подтверждается тем обстоятельством, что ихтиозавры, 1гле1
янозавры и другие вымершие обитатели моря из среды рептцЛий
были голы.
Многие рептилии зарываются в землю. Одни зарываюТся
сами, другие копают поры лишь для зарывания своих яиц. у не_
которых рептилий эта привычка к рытью привела, в кс^нце коццов
к совершенно подземному образу жизни. Такой образ жизцц да
и вообще способность к рытью связаны с определенными измене-
ниями в строении тела. У некоторых, например, у аптечного сцин-
ка (Scincus officinalis), который роется при помощи конечностей
мы видим превращение их в настоящую лопату для рытья, цпи
помощи которой он с необыкновенной быстротой может зарываТь^я
в песок.Такое же, примерно, изменение видим мы у некоторых пес-
чаных ящериц, которые имеют удлиненные пальцы и рогоВые
щетки на последних.
Обычно же рептилии
помощи головы. Голова в таком случае приобретает форму
к липа, слегка вогнутого C^pvy мптпгтттогп pnvwnTL --
Рот, как у роющих представителей всех классов, смещается
нижнюю сторону, чтобы при рытье я него не попадали песок
земля. У некоторых представителей роющихся в песке рептилий
।..... —-----------—------------—----------------------
ком с острым краем. Мы видим это у живущего под землей
роются пе при помощи лап, а ЦрИ
сверху, могущего служить лопатой
— - — {
на
или
голова снабжена впереди большим ростральным (рыльцевым) 1цИт.
ком с острым краем. Мы видим это у живущего под землей сле-
пуна (Typhlops vermicularis), у пустынного удава (Егух jacuiug\'
у роющих змей Цейлона и Южной Индии из семейства Uropeitjl
<1ае, у литоринха афганского (Lytorhynchus ridgewai), найден-
ного в Закаспийской области и отличающегося удлиненной и за_
509
Рис. 406. Голова литоринха (Li-
torhynchus) — А; голова и задний
конец тела змеи Uropeltis—В.
(Из Hllzheimer.)
остренной мордой, выдающейся над ротовой щелью в виде хобота,
У Uropeltis для рытья имеется еще хвостовой щиток, сильно уве-
личенный, служащий для упора заднего конца тела при рытье
(рис. 406).
Параллельно с приспособлением головы для рытья идет умень-
шение ее ширины. Так как подземноживущие рептилии питаются
мелкими животными, большей частью червями, то им нет надобно-
сти в широко раскрывающемся рте и в’сильпо развитых зубах.
, Как и всегда у роющих форм, ведущих подземный образ жизни,
тело становится равномерно вальковатым, голова мало отделяется
от туловища, конечности начинают редуцироваться, а в крайних
типичных случаях даже совершенно исчезают; так, например,
х (Amphisbaenidae) следы пле-
чевого и тазового пояса скрыты
под кожей.
Покровы становятся у под-
земноживущих форм более одно-
родными, большие щитки на
брюшной стороне исчезают, а у
амфисбен тело покрыто вместо
чешуи твердой кожей, разделен-
ной поперечными бороздами и
продольными углублениями.
Для защиты органов чувств
от песка или земли мы видим у
роющих форм различные при-
способления.. У афганского ли-
торипха ноздри закрываются особым клапаном. В более слабой
степени это приспособление развито также у степного удава
(Егух), у полозов (Zamenis) и некоторых других. У жабовидной
ящерицы (Phrynosoma cornutum), зарывающейся на ночь в целях
самозащиты в песок, ноздри на это время всецело закрываются
клапанами.
Наружное ухо или совсем исчезает, как у круглоголовок (Phry-
nocephalus) и змей, или прикрыто выступающими чешуями (у ро-
дов Agama и Eumeces), или очень уменьшено, как у сцинка
(Scincus).
Глаза у роющих змей по большей части малы, а у слепунов (Ту-
phlopidae) и двуходок (Amphisbaenidae) совершенно скрыты под
кожей, просвечивая сквозь нее в виде маленьких темных пятнышек.
Век нет, так как верхнее и пижпее веки срастаются, становясь
прозрачными, и дают надежную защиту глазу. Такие приспособ-
ления мы встречаем у змей и некоторых ящериц. Самое устрой-
ство глаза при этом весьма примитивно.
Зарывание в землю является приспособлением главным обра-
зом для жизни в пустыне. Этим способом одни, как степной удав
(Егух jaculus), подстерегают добычу, другие , как круглоголовки
(Phrynocephalus), укрываются, от врагов, зарываясь с необычайной'
быстротой на глазах у преследователя при помощи боковых дви-
510
жений туловища. В песке же, где всегда достаточно влажно, от-
кладывают пустынные рептилии свои яйца. Расширенное тело ме-
шает зарыванию в песок. Но у ящериц с расширенным телом зары-
вание осуществляется при помощи особого приспособления.
У круглоголовки на боках тела имеется складка кожи, снабженная
выступающими чешуями. Плотно прижавшись к земле, круглого-
ловка может при помощи особой мускулатуры этих складок забра-
сывать себе на спину песок и в буквальном смысле утопать в по-
следнем. Подобное же приспособление имеют живущая в пустыне
Северной Африки рогатая гадюка
(Cerastes cornutus), эфа (Echis carina-
tus) и некоторые другие. У неко-
торых песчаных ящериц расширяет-
ся пе туловище, а хвост.
Рис. 407. Lyriocephalus scu-
tatus (Aganridae).
(Из Gadow.)
Рис. 408. Древесная змея (Zaocys
carin atus).
(Из Gadow.)
Приспособления к жизни на деревьях и к полету. Многие пре-
смыкающиеся ведут древесный образ жизни, лазая по ветвям,
другие же лазят по стенам и скалам, иногда совершенно отвесным.
Этот образ жизни опять-таки накладывает печать на строение жи-
вотного.
Древесный образ жизни в наиболее типичных случаях, напри-
мер, в семействах игуан (Iguaaidae) и хамелеонов (Chamaeleonti-
dae), создает совершенно другую форму тела, пежели жизпь на
земле: тело сжато с боков, спина же и брюхо образуют как бы
гребень. Такую же форму в большей или меньшей мере имеет хвост.
Эта форма тела в соединении с соответственной окраской придает
животным значительное сходство с листом или с обломанной ветвью.
Такая сжатая форма развилась независимо в разных группах. Так,
например, в семействе агам (Agamidae) мы находим такие формы,
как Lyriocephalus scutatus (рис. 407) с острова Цейлона или
Й 511
Gonyocephalus chamaeleontinus с островов Малайского архипелага,
совершенно сходные по внешней форме тела, хвоста и головы
и по окраске, но сразу отличающиеся от хамелеона по целому
ряду признаков строения. Это сходство есть пример конвергентного
развития под влиянием одинакового древесного образа жизни.
Такую же форму тела имеют американские древесные игуаны
(Iguanidae) и многие древесные змеи (рис. 408). Все лазящие формы
отличаются длинными пальцами и острыми сильными ногтями,
серповидно изогнутыми. Это хорошо видно на стопе двух видов яще-
рицы тейю (Tupinambis), из которых один живет на земле, а другой
лазит.
У других видов пальцы расширены и снабжены снизу листовид-
ными поперечными складками, которые позволяют им держаться
две
Рис. 409. Конечности трех видов гекконов с нижней
стороны:
(Из Hilzheimer.)
даже на нижней стороне ветви или скал. Такие хватательные орга-
ны развиваются независимо в двух семействах: у игуан (Iguani-
dae) и у гекконов (Geckonidae) (рис. 409) Представители этих
семейств, живущие в пустыне, лишены такого аппарата. Все при-
сасывательные приспособления построены по принципу образова-
ния безвоздушного пространства, благодаря складкам при прижи-
мании поги к поверхности получается целый ряд безвоздушных
камер.
Благодаря этим приспособлениям некоторые виды бегают по
самым гладким скалам и деревьям, даже по обращенной вниз сто-
роне стекла, с такой же быстротой, как другие по земле. Заме-
чательное приспособление для лазания мы видим у одного геккона
с Малайских островов и полуострова Малакки, у Ptychozoon
homalocephalum. У пего по бокам тела, головы, ног и хвоста
имеется кожистое расширение, служащее пе для планирования,
по присасывательным аппаратом (рис. 410). Для планирования слу-
жат оторочки на хвосте, ногах и между пальцами.
Геккоп Lygodactylus picturatus имеет присоски па пижней
стороне хвоста и может, присосавшись ими, висеть вниз головой,
освободив конечности.
Гекконы, лазящие по стволам деревьев, бывают иногда окра-
шены покровительственно под цвет коры, и тело их уплощено
и может прижиматься к дереву. Таков мадагаскарский лишайни-
ковый геккон (Uroplates fimbriatus licheni), совершенно не отли-
чимый от покрытой лишайниками коры дерева.
512
Копальский серый геккон (Gymnodactylus russowi copalensis),
живущий в саксауловых зарослях, окрашен в тон мертвого сак-
саула, под которым он держится днем, ведя активный образ жиз-
ни ночью.
Совершенно другой характер приспособления для лазания по
ветвям мы видим на конечностях всем известного хамелеона
(Chamaeleon). Конечности его тонки, длинны, и два пальца всегда
противополагаются трем другим. На пе-
редних конечностях первые три пальца
противополагаются четвертому [и пятому,
на ноге первые два — остальным трем.
Получаются своего рода щипцы, которые
служат для зажатия ветвей. Конечности
у хамелеона не отстоят в стороны, а опу-
щены вертикально, так что тело может
высоко подниматься над ветвями. Лазанию
помогает у хамелеона еще хвост, кото-
рый может закручиваться вокруг ветвей.
Хвост очень прочен, пе отламывается, а
будучи насильственно отломан, —не реге-
нерирует.
Способность вращать каждым глазом в
отдельности также является приспособле-
нием хамелеона к жизни на деревьях,
где иногда приходится, сидя на веточке,
фиксировать одним глазом добычу, а дру-
гим отыскивать путь.
Лазящие рептилии известны нам и из
числа ископаемых. У динозавра Hypsilo-
phodon большой палец задних ног про-
тивополагался остальным пальцам; жи-
вотное это, несомненно, было древесным.
В связи с древесным образом жизни
развиваются й приспособления, для скользящего полета. Рептилий
со способностью к скользящему полету мы встречаем главным
образом в лесах юго-востока Азии. Весьма любопытно, что здесь
же встречаются и планирующие лягушки, и снабженные приспо-
соблениями для планирования млекопитающие.
► Выше уже был упомянут геккон (Ptychozoon homalocephalum)
с Зондских островов, у которого вдоль боков, головы, туловища
и хвоста имеются кожистые оторочки, служащие парашютом.
Перепонки между фалангами пальцев увеличивают поверхность,
сопротивления при падении. Такие же приспособления Мы встре-
чаем еще у другого геккона — Mimetozoon с Малакки, развив-
шиеся совершенно независимо от первого. Как и всякое другое
приспособление, приспособление для скользящего полета разви-
вается постепенно. Целый ряд переходных форм связывает ле-
тающих гекконов с нормально устроенными. У одних уплощен
хвост, у других на нижней стороне выступают большие чешуи, у
33 Кашкаров и Станчинский 513
третьих пальцы или вполне соединены перепонкой или отчасти.
Все эти формы, иногда встречающиеся вместе, показывают,
что скользящий полет мог развиться постепенно.
Наилучше развит аппарат для планирования у знаменитого
летающего дракона (Draco volans), принадлежащего к семейству
Agamidae.
У этого животного ребра, лежащие позади грудной клетки,
чрезвычайно длинны, не соединены на брюшной стороне, подвижно
сочленены с позвонками и при помощи межреберных мышц могут
складываться или принимать перпендикулярное к оси Тела поло-
жение. Ребра поддерживают служащую при планировании складку
(рис. 411 и 370). Пользуясь
своей способностью к полету,
летающий дракон ловит насеко-
мых. Наблюдается способность
к планирующему полету и у
некоторых змей, например, у
древесной змеи (Dendrophis).
Эти змеи могут бросаться вниз
на землю с кокосовых пальм,с
высоты в 15—20 м, плавно
скользить в воздухе и под углом,
не производя шума, опускаться
на землю. Это достигается тем,
что ребра у змеи растопырива-
ются, брюхо втягивается, че-
шуи по бокам его сильно вы-
ступают. Змея в таком виде
похожа на расщепленную вдоль
бамбуковую палку, что увели-
чивает сопротивление воздуха
и заставляет змею не падать, а
Рис. 411. Скелет Draco fimbriatus, планировать.
летающего дракона. Чрезвычайно развиты были
(Из вокег.) приспособления для полета у
вымерших летающих ящеров
(Pterosauria). Здесь целый ряд признаков являлся общим с пти-
цами, а именно: кости были пневматичны, как у птиц,череп
стоял под прямым углом к шее, был заострен впереди, как у
птиц, кости его срастались. Но крыло птерозавров не имело ни-
чего общего с крылом птиц. Было развито четыре пальца. Первые
три были малы и служили, невидимому, для того, чтобы подвеши-
ваться на деревьях за ветви. Четвертый палец был очень длинен
и служил в качестве опоры летательной перепонки.
Развилась у рептилий способность к активному полету из спо-
собности к полету планирующему, а этот в свою очередь развился
на основе способности лазать у древесных рептилий, как мы это
наблюдаем у амфибий и у млекопитающих и как, по всей вероят-
ности, было при развитии класса птиц из лазящих рептилий. Орга-
514
пизация летающих рептилий рассмотрена
выше (в главе о систематике).
Приспособления для жизни вводе. Мно-
гие рептилии не только способны плавать,
но имеют для водной жизни особые приспо-
собления. Почти все пресмыкающиеся могут
плавать, хотя при продолжительном пребы-
вании на воде ящерицы тонут, наземные же
черепахи сразу погружаются на дно водо-
ема. Самое плавание совершается при по-
мощи различных органов. У одних хвост
сжат с боков и служит органом движения.
У таких рептилий, движущихся как бы при
помощи кормового весла или винта, и тело
обыкновенно сжато с боков, а часто есть и
спинной плавник, шея короткая, а рыло
сально вытянуто и заострено.
Примером может служить крокодил или
морская игуана с Галапагосовых островов,
Amblyrhynchus cristatus (рис. 412), отлично
плавающая.При плавании последняя, подобно
тритону, прижимает конечности к телу и дви-
жется при помощи боковых движений удли-
ненного и сильно уплощенного с боков хво-
ста. Еще более выражен этот способ движе-
ния у крокодилов, имеющих к тому же
плавательные перепонки между пальцами
задних ног, а крайней степеии выражения
достигает у морских змей, например, у двух-
цветной пеламиды (Hydrus platurus), имею-
щей вид плоской ленты.
Движение при помощи изгибов тела яв-
ляется у рептилий медленным и обычно не
продолжается долго. От крокодилоподобных
(по форме тела) предков произошли рептилии
иного приспособительного типа, способные
к быстрому, устремленному и продолжи-
тельному плаванию. Это тот тип, который мы
встречаем у ихтиозавров. Ихтиозавры имели
вполне рыбообразный вид не только по
форме тела, но и по наличию спинного и
хвостового плавника, по присутствию плав-
ников грудных и брюшных. Однако позво-
ночник у них был загнут в нижнюю лопасть
хвоста, а не в верхнюю, как у рыб (см.
рис. 356).
Другие рептилии (черепахи) плавают
при помощи широких веслообразных ног
(речные черепахи) или ластов (морские чере-
зз*
Рис. 412. Amblyrhynchus cristatus в положении плавания.
(По Abel из Bdker.)
515
пахи). У- таких форм тело не вынятуто, маленькая голова сидит
на длинной подвижной шее, хвост круглый.
К этому типу принадлежат и ископаемые плезиозавры,
двигавшиеся также при помощи парных ластов. У них для управ-
ления движениями служила длинная шея и сильный, хотя не длин-
ный, хвост; у морских черепах рулем служат задние ноги, органом
движения— передние.
Таким образом у рептилий можно установить следующие типы
приспособления к плаванию.
А- Двигательный орган движения на-
ходится на заднем конце тела (принцип кор-
мового весла).
I. Ящерицеобразный (тритоновидный) приспособительный вид.
Задние конечности принимают участие в движении и развиты луч-
ше, чем часто сильно уменьшенные передние конечности. Тело вы-
тянуто, хвост сильно сжат с боков. Крокодилы и многие ископаемые.
II. Веретенообразный приспособительный тип. Парные конеч-
ности не принимают больше участия в движении. Задние всегда
меньше передних, рудиментарны или даже отсутствуют, передние
же служат органами управления и балансирования. Тело веретено-
образно. Орган движения вертикален. Ихтиозавры.
В. Органы движения по бокам тела
(принцип весельной лодки). <
III. Черепахообразпый приспособительный тип. Обе конеч-
ности принимают участие в движении. Тело сжато. Ящерицы.
Черепахи. Плезиозавры.
IV. Тип мозазавра. Хвост длинный, оба плавника сильно раз-
виты, или широкие или узкие. Питономорфы (рис. 310).
Плавательные приспособления, как и другие, развиваются по-
степенно, и путь, которым нынешние водные черепахи произошли
от сухопутных предков, можно проследить даже на ныне живу-
щих формах: постепенное увеличение плавательной перепонки
и расширение конечностей.
Приспособлениями к водному образу жизни являются, далее,
смещение ноздрей на верхнюю сторону головы, закрытие их при
помощи пещеристой ткани, так что, с одной стороны, дыхатель-
ные пути закрываются от проникающей воды, с другой — в них
задерживается воздух. При дыхании закрывающая ткапь сжи-
мается при помощи особого мускула. Также и глаза поднимаются
кверху и выдаются над уровнем головы, особенно у тех форм, кото-
рые, плавая у поверхности воды, высовывают оттуда лишь глаза.
Приспособляясь к водному образу жизни, рептилии утра-
чивают кожный панцырь. Так, у царетты (Chelone imbricata)
спинной щит сильно редуцирован. Эта редукция панцыря у вод-
ных рептилий наблюдается и у ископаемых форм: у черепах,
морских крокодилов, плезиозавров, большинства питономорфов
и. у ихтиозавров. Здесь мы видим интересную параллель с потерей
волосяного покрова у китов. Потеря панцыря произошла не сразу:
вначале крупные чешуи превратились в мелкие, как, например,
516
у морских змей из подсемейства Hydrophinae, затем в этом чешуй-
чатом одеянии появились пробелы, которые, все увеличиваясь,
привели, в конце концов, к полной потере панцыря.
Приспособление к водному образу жизни у рептилий резко
сказывается и на скелете конечностей, приобретающих характер
плавников. Изменение идет постепенно, и в конце получается плот-
ное весло из коротких и многочисленных костных элементов
(рис. 413).
У черепах, которые подолгу могут оставаться под водой, раз-
вивается особое приспособление для дыхапия в воде, а именно
в глотке развиваются богатые кровью сосочки, служащие для по-
глощения кислорода. Благодаря этому животное может оставаться
Рис. 413. Конечности рептилий: наземной Uromastix
(Л), водных Platecarpus (В) и Ichthyosaurus (С).
(Из Hilzhelmer.)
под водой до десяти часов, все время набирая воду в рот и вытал-
кивая ее обратно. Такое же приспособление находим мы и у мор-
ских змей. Упомянутая выше морская игуапа более часа может
оставаться под водой. У многих черепах для дыхания слу-
жат еще сильно васкуляризованные стенки анальных мешков.
У большинства змей имеется гладкостенный воздушный резер-
вуар, позволяющий им часами оставаться под водой. У крокоди-
лов для этой цели служит мешковидное расширение легкого.
2. Питание рептилий и приспособления к роду пищи
По характеру питания рептилии, как современные, так и иско-
паемые, обнаруживают большие отличия. Иногда в пределах одной
группы, даже у близких форм, питание бывает различно. Преоб-
ладает у современных рептилий пища животная. Даже те пресмы-
кающиеся, которые питаются растительной пищей, не брезгают
пищей животной. Все крокодилы являются исключительно хищ-
ными животными, нападая на крупных животных и даже на чело-
века. Главной их пищей являются рыбы. Змеи нападают также
517
главным образом на позвоночных животных; при этом одни ку-
сают добычу, другие ее давят (удавы Bcidae), ломают кости своими
кольцами и уже потом заглатывают иногда даже очепь крупных
животных. Однако целые семейства змей питаются червями и на-
секомыми, некоторые—моллюсками. Почти все рептилии гло-
тают добычу целиком. Лишь немногие, а именно черепахи и кро-
кодилы, в состоянии откусывать куски, подобно тем пресмыкаю-
щимся, главную пищу которых составляют растения. Среди чере-
пах наземные формы питаются растениями, водяные — мясом. Но
и многие представители семейства Testudinidae охотно едят мясо.
Из ящериц растительноядные формы находятся среди агам (Aga-
midae) и игуан (Iguanidae), но и они не отказываются от живот-
ной пищи. Вообще же ящерицы питаются мелкими млекопитаю-
щими, птицами, другими ящерицами, моллюсками, насекомыми,
червями, амфибиями и рыбами. С другой стороны, ящерицы пус-
тынь, по крайней мере в определенные периоды, поедают зеленую
растительность. Возможно, что делают они это главным образом
ради той воды, которая содержится в свежей зелени.
То или иное питание вызывает развитие приспособительных
признаков. Прежде всего реагируют на изменение пищи зубы.
По характеру зубов мы судим о характере пищи и о способе ее
захватывания у ископаемых рептилий. Так, мы встречаем' чистых
хищников, нападавших на других животных, как, например,
хищные динозавры; растительноядных, питавшихся водорослями
(Trachodon); формы, питавшиеся моллюсками, которых они раз-
давливали своими плоскими зубами (Placodontia); питавшиеся
рыбой (Ichthyosauria, Plesiosauria, Pterosauria). Некоторые пте-
розавры ловили рыбу так, как это делают чайки.
Изменение окружающей среды, изменение окружающего мира
животных и растений сопровождалось и вызывало (путем отбора)
изменение характера пищи, что в свою очередь вызывало изменение
зубов. Например, у древнейших ихтиозавров имелись ловчие
зубы, а у верхнеюрских наблюдается редукция зубов. Причина
(контролирующая) лежала в переходе ихтиозавров от пита-
ния рыбой (ихтиофагия) к питанию головоногими моллюсками
(теутофагия). Об этом говорит содержимое желудков.Переход же
к теутофагии объясняется увеличением в юре числа головоногих
моллюсков.
Кроме зубов, изменялись в связи с изменением рода пищи и че-
люсти. У водных рептилий, питающихся мелкими организмами,
поднимаемыми со дна, нижпие челюсти удлиняются, расширяются
на переднем конце и усажены косо кнаружи расположенными зу-
бами. Такая челюсть была у некоторых плезиозавров (Thauma-
tosaurus, Plesiosaurus). Приспособлением к роду пищи является
и ложкообразная форма «клюва» у динозавра Trachodon, питав-
шегося подобно уткам. Длинные клювоподобные челюсти Ptera-
nodon (рис. 414) являются приспособлением для ловли рыбы,
которой эти летающие ящеры питались.
То или иное питание вызывает развитие и других приспособи-
518
тельных признаков. Прежде всего, у растительноядных форм тон-
кая кишка значительно длиннее, чем у плотоядных. У них обычно
развита также и слепая кишка, отсутствующая у насекомоядных.
Там, где толстая кишка коротка, может быть и слепая кишка.
Таким образом, увеличивается всасывающая поверхность бла-
годаря образованию слепого мешка, в котором долго могут на-
ходиться трудно перевариваемые части, например, ноги насекомых.
Сильное развитие этой части кишечника при растительной пище
есть явление приспособительное, так как здесь части растений
разлагаются при помощи ферментов и бактерий, даже такие
вещества, как целлюлоза.
Специальным приспособлением к способу питания является
язык хамелеона. Сидя на ветке и заметив насекомое, он с необы-
Рис. 414. Реконструкция картины жизни Pteranodon ingens
из верхнего мела Канзаса.
чайной быстротой выбрасывает длинный, до 15 см, язык, смочен-
ный на конце клейкой жидкостью, и схватывает насекомое. В со-
стоянии покоя язык втянут в глотку, в случае же надобности выбра-
сывается в виде эластического стержня. В движение он приводится
особыми мышцами. Задняя часть языка в покое образует длинную
складчатую оболочку, в стенке которой лежит так называемый
подъязычный мускул — musculus hypoglossus. Если теперь при
помощи подбородочно-язычного мускула (m. geniohyoideus) и
мускула диафрагмы рта (m. mylohyoideus) подъязычная костЦ
быстро высовывается вперед, то язык также быстро выбрасывается
вперед и тащит за собой оболочку. Быстрому выбрасыванию языка
способствует еще усиленный и быстрый приток крови в кровенос-
ные сосуды языка. Втягивается язык с такой же быстротой с по-
мощью подъязычно-язычного мускула (m. hyoglossus). В момент
выбрасывания благодаря давлению мускулов выдавливается со-
держимое слизистых желез. Специальный мускул на конце языка
делает его плотным.
Другие, не менее оригинальные приспособления к роду пищи
наблюдаем мы у яйцеядных змей (Elachistodon и Dasipeltis).
519
Зубов у этих змей почти нет, они стали рудиментарны. Но для
раздавления яиц у них существует следующее приспособление:
нижние отростки передних позвонков прорастают сквозь верх-
нюю стенку глотки, выступая в просвет последней; эти отростки
и раздавливают заглотанное яйцо.
Характер пищи стоит у змей в связи с числом чешуй. Абсолют-
ная и относительная растяжимость тела больше у тех, которые
имеют большее число чешуй. Характер поедаемой пищи и способ
ее заглатывания связаны, 1аким образом, с числом чешуй.
Характер питания рептилий естественно зависит от характера
обитаемой ими среды. Водпые рептилии (современные) являются
почти исключительно ихтиофагами, гораздо реже опи питаются
беспозвоночными (обитатели маленьких водоемов). Обитатели
сырых болотистых мест питаются лягушками. Живущие в земле
(Typhlops, например), в общем питаются мелкими животными:
насекомыми, многоножками, червями. Но некоторые из них спе-
циализируются также йа питании змеями (Cylindrophis), другие
не брезгуют и теплокровными (Xenopeltis из Boidae)..
Многие древесные змеи кормятся яйцами и птенцами птиц
(змеи родов Ptyas, Elaphe, Boiga и др.). В некоторых странах
имеется значительное количество лазящих ящериц, например,
гекконов. Этими ящерицами питаются лазящие по деревьям змеи.
В Китае, где в домах живет большой геккон (Gecko gecko), живет
красивый древесный уж — Chrysopelea ornata. Мухи привлекают
гекконов, гекконы — эту змею, способную к скользящему полету.
Более разнообразна пища у форм, живущих на земле. Среди них
есть и растительноядные (черепахи, игуаны) и плотоядные, пое-
дающие мелких млекопитающих, птиц, их яйца, лягушек, змей,
ящериц, насекомых.
Находят свою добычу рептилии различным способом. Питон
(Python), индийская плетевидная змея (Dryophis), гадюка (Vipera)
и некоторые другие лежат спокойно, пока голод пе станет чувстви-
тельно заметным, а затем внезапно с большой быстротой кидаются
на добычу, попавшую в поле досягаемости. Будучи голодными,
все змеи отправляются на поиски пищи. Сперва находят добычу
при помощп «химического чувства», языка и якобсонова органа.
Открытое таким образом животное преследуется в дальнейшем
уже при помощи зрения. При отыскивании пищи обследуются
все уголки. Некоторые змеи ищут добычу на большом простран-
стве, другие, в особенности те, которые охотятся ночью за мелкими
грызунами, придерживаются местообитания последних. Пресле-
дование добычи у многих бывает весьма энергичным. Иногда змеи
делают прыжок сверху вниз на добычу (Chrysopelea).
Неядовитые (Aglypha) проглатывают добычу живой. Elaphe,
Python, l?ryx, Boiga имеют длинное тело и длинный хвост, кото-
рыми охватывают добычу и душат. Сопротивление животного лишь
усиливает рефлекс схватывания. Opistoglypha схватывают добычу
и запускают в нее зубы, пока яд пе подействует. Elapidae и Vipe-
ridae проглатывают лишь мертвую добычу, умертвив ее ударом
520
верхних ядовитых зубов, наносимым с поразительной быстротой.
Все рептилии, кроме тех, что обитают в пустыне или в воде,
пьют воду, лакая ее или черпая нижпей челюстью. Пустынные
обходятся без воды, получая ее с пищей.
Чем выше температура, тем больше едят рептилии. Некоторые
питаются периодически и,- наевшись досыта, могут затем меся-
цами не принимать пищи (например, удавы, черепахи).
3. Окраска рептилий
По разнообразию окраски рептилии не уступают другим клас-
сам. Окраска их обыкновенно похожа на окраску той среды, в ко-
торой данный вид обитает. Древесные рептилии (хамелеон, змеи)
большей частью — зеленые. Среди же обитателей пустынь мы
совершенно пе видим рептилий зеленого цвета. Наоборот, желтый,
буроватый и красноватый цвета являются обычными среди песча-
ных форм. Те рептилии, которые живут в болотах, имеют илисто-
зеленую окраску, морские змеи — сине-зеленую или сине-серую,
а змеи, живущие на известняках, — светло-серую; те, которые
живут среди сухой травы, часто бывают полосатыми. Мозаичная
окраска питона (Python reticulatus), яркая в музеях, делает
его незаметным среди ярких солнечных бликов в его родном место-
обитании, в лесах Малайских островов. Окраска многих ящериц
может быть в виде рисунка: продольных полос, пятен, поперечных
полос. Наблюдения над изменением рисунка у европейских и аме-
риканских ящериц показывают, что первоначальной стадией яв-
ляется полосатость; второй стадией является распадение полос на
пятна; эти пятна могут затем сливаться в поперечные полосы
или исчезнуть вовсе, делая окраску одноцветной. У птиц и млеко-
питающих мы видим такую же последовательность в смене окра-
ски. Повидимому, полосатость есть первичная исходная форма
окраски, однотонная же или пятнистая окраска пустынных или
древесных форм — явление приспособительное.
Многие ящерицы и некоторые змеи обнаруживают значительные
различия окраски в связи с полом и возрастом. Иногда рисунок
и окраска взрослой формы совершенно непохожи на окраску мо-
лодых особей. Большие колебания в окраске бывают и независимо
от пола и возраста. В некоторых случаях окраска особей одного
и того же вида значительно вариирует в зависимости от грунта,
на котором они находятся.
Некоторые рептилии могут изменять свою окраску и изменять
ее произвольно как под влиянием изменений в окружающей
среде, так и под влиянием внутренних состояний: голода, возбуж-
дения. Наиболее известны такие изменения у хамелеона (Chamae-
leon vulgaris). В мальпигиевом слое эпидермиса хамелеона нахо-
дятся иризирующие клетки с очень тонкой волнистой исчерчен-
ностью на поверхности. В волокнистом же слое кожи имеется
большое число маленьких, тесно лежащих клеток, наполненных
сильно преломляющими свет кристаллами гуанина. Диффузное
521
отражение прямого света от этих клеток создает белый цвет.
Клетки, которые лежат ближе к поверхности, заполнены масля-
нистыми каплями желтого цвета. Глубже лежат большие хромато-
форы, большей частью с зернами черновато-коричневого или крас-
новатого пигмента, которые движутся в разветвленных отростках
хроматофор. Когда эти отростки сокращаются, пигмент соби-
рается в основной части хроматофора, и кожа кажется желтой или
белой. Когда весь пигмент сдвигается к поверхности волокнистого
слоя кожи, животное выглядит темным, иногда черным. В проме-
жуточном состоянии цвет изменяется в зеленый благодаря дифрак-
ции лучей в желтоватом внешнем слое и в тонко исчерченных ири-
зирующих клетках мальпигиева слоя. Те части, в которых хрома-
тофоры не посылают пигмента, кажутся желтыми пятнами.
Кроме хамелеона, обладают способностью изменять свой цвет
и другие ящерицы. Например, самцы степной агамы (Agama
sanguinolenta при поимке получают яркосинюю стальную окраску
брюшной стороны тела. Такую же способность к изменению окра-
ски, как у хамелеона, приспособительно к изменениям мы ви-
дим у анолиса (Anolis carolinensis) из семейства игуан (Iguani-
dae). Эксперименты с этой ящерицей установили' что быстрота
изменения окраски увеличивается по мере упражнения. Кроме
того, движения пигмента к поверхности кожи вызываются пониже-
нием температуры, при повышении же ее пигмент уходит глубже.
Свет действует иногда таким же образом, иногда же — обратно.
Часто окраска рептилий носит характер покровительственный,
защитный, например зеленая окраска тех рептилий, которые жи-
вут среди листвы, и эта окраска приобретается независимо в раз-
ных группах и в разных странах: в Южной Америке, в Австралии,
в Африке. Точно так же принято считать приспособительной,
именно покровительственной, и окраску пустынных рептилий
(как и других животных). Полагают, что она помогает животному
укрываться от врагов.
Однако против такого толкования пустынной окраски многое
можно возразить, так как эта теория не приложима к животным,
которые охотятся или за которыми охотятся ночью; она не прило-
жима к тем животным, за которыми никто не охотится, и к тем,
которые сами не охотятся; она не приложима к тем случаям, когда
бледная окраска простирается на брюхо и подошвы ног. Она
совершенно пе приложима ни к подземным животным, ни к чер-
ным. Конечно, окраска служит средством защиты при некоторых
обстоятельствах и для некоторых животных, делая их незамет-
ными. Но эта теория покрывает лишь часть фактов. Должна суще-
ствовать какая-то другая причина, притом характера не биоцено-
логического, а физического, одна во всех пустынях и для всех
животных: слишком она универсальна. Вопрос может быть раз-
решен только экспериментально и основательным изучением всей
проблемы заново.
Много фактов, несогласных с теорией покровительственной
окраски, встречаем мы и среди рептилий: например, окраска зе-
522
леной ящерицы (Lacerta viridis) яркозеленая, между тем как
в Далмации она живет на местах совсем не зеленых, на скалах,
чаще всего на стенах. .
«Покровительственно» окрашенная ушастая круглоголовая
(Phrynocephalus mystaceus), обнаруживая в окраске тела порази-
тельное сходство с песком, ведет себя так, что ее видно в жаркое
время дня лучше, чем какую-либо другую ящерицу: она встает
высоко на четырех ногах на гребне бархана и крутит хвостом.
Виды круглоголовок в каменистой пустыне не имеют врагов, от
которых надо укрываться, в то же время весьма быстро меняют
свою окраску в зависимости от того, на каком грунте они оказы-
ваются: на светлом светлые пятнышки становятся больше, темные
сокращаются; на черном, обожженном солнцем грунте— наоборот.
Но те же изменения окраски происходят и под влиянием измене-
ний температуры: при более низкой температуре окраска темнее,
при высокой — светлее. А изменения температуры связаны с осве-
щением. Возможно, что окраска играет роль в регулировании тем-
пературы тела (см. ниже).
Некоторые ящерицы и змеи окрашены необыкновенно ярко:
в черные, желтые, синие и красные цвета. Некоторые из этих яще-
риц ядовиты: ядозуб (Heloderma horridum), коралловый аспид
(Elaps corallinus) и др. Эту окраску опять-таки считали за приспо-
собительную, предостерегающую, извещающую врага, что добыча
опасна и что к ней не следует приближаться. Но и это толкование
не совпадает с действительностью. Там же, где живет коралловый
аспид, живет ряд совершенно безобидных змей того же цвета, что
и аспид, и существует еще целый ряд подобных примеров. Эти
случаи объяснялись мимикрией: подражанием неядовитых змей
ядовитым по окраске, дающим возможность избежать нападения.
Но враги, поедающие ядовитых змей, поедают их независимо от их
окраски. Вернее говорить здесь о конвергентном развитии, о раз-
витии одинаковых черт под влиянием одинаковых условий. Сход-
ная с коралловым аспидом, Coronella распространена не только
там, где есть Elaps, но и в других странах, за сотни миль от этих
мест, где нет никакой пользы от сходства с аспидом. Да и там, где
эти змеи живут обе, они редко встречаются бок о бок.
Обычная для всех позвоночных смена эпидермиса и его произ-
водных происходит у рептилий путем сбрасывания его целиком
зараз или несколькими кусками. Сбрасывается при этом и рого-
вая оболочка глаза. Сбрасывание эпидермиса связано с ростом.
Молодые змеи Liopeltis, например, первый раз сбрасывают кожу
на 8—10-й день, становясь, при этом ярче. Большинство ящериц
линяет долго, целыми месяцами. Некоторые ящерицы съедают
свою сброшенную кожу подобно тому, как это делают амфибии.
4. Приспособления для защиты и нападения
Приспособительными изменениями следует считать и те, кото-
рые служат в качестве оружия для защиты и нападения.
623
У рептилий много врагов, и даже у тех, которые являются
ядовитыми. Например, еж и свинья едят змей без вреда для себя.
Из птиц также некоторые истребляют змей: секретарь, кариама,
орел-змееяд. И среди самих рептилий находятся такие, которые
поедают себе подобных.
Средства защиты у рептилий различны. Некоторые защи-
щаются, сильно кусаясь, или нападают, пользуясь когтями, уда-
рами хвоста, например, вараны (Varanus griseus), кайманы (Caiman).
Панцырь как ныне живущих (крокодилы, черепахи), так и иско-
паемых рептилий (некоторые динозавры, котилозавры, пластинчато-
зубые) является, конечно, средством защиты. Панцырь стего-
завра, скелидозавра и других родов стегозавров имел защитное
значение. У стегозавра костные пластинки, покрывавшие спину,
переходили на хвост, на конце которого имели форму шести пар
острых костных шипов, функционировавших как оружие защиты.
Формы эти были травоядные, и панцырь служил им защитой
против хищных динозавров. Остистые отростки позвонков у Ре-
lycosaurus также служили орудием защиты, как и у Naosaurus.
Той же цели служили рога на голове у пеликозавров, точно так же
и мощные, иногда поддерживаемые костной основой, шипы (как
у Phrynosoma). Служат ли для этой цели колоссально развитые
шипы австралийского молоха (Moloch horridus), неизвестно,
так как они оказываются необычайно гигроскопичными и впиты-
вают в себя воду, как пропускная бумага. Быть может, они и слу-
жат для накапливания воды. Особенно развиты были всевозмож-
ные выросты, шипы и рога у вымерших рептилий.
Защитным же средством является и способность к автотомии,
к отламыванию хвоста у ящерицы, когда, схваченная врагом, она
энергичными боковыми движениями отламывает себе хвост, при-
чем разрыв происходит посредине позвонка, состоящего как бы
из двух половиц. Этому способствует и устройство мускулатуры,
которая не тянется вдоль всего хвоста, а состоит как бы из отдель-
ных конусов, вставленных друг в друга. Движение отламывания
есть рефлекс на болевое раздражение, и никакой намеренности,
произвольности в этом акте нет.
Вместо отломанного хвоста может отрасти (регенерировать)
новый, и уже такой хвост, имеющий цельные позвонки, не может
отламываться путем автотомии. Если хвост только поранить,
то в месте поранения вырастает новый хвост. Таким образом
могут возникнуть дву- и много хвостые ящерицы. В тех случаях,
когда хвост имеет для ящерицы какое-либо особое значение, слу-
жит, например, хватательным органом, он не обладает способ-
ностью автотомии.
К защитным приспособлениям следует отнести и способность
надуваться и таким образом увеличивать объем своего тела, на-
блюдаемую, например, у хамелеонов; у них от легкого идут во все
тело слепые выросты, и таким образом все тело может надуваться.
В добавление к этому животное поднимается па задние ноги
и шипит. Так же шипят, выпуская воздух, варан, многие змеи
524
и т. д. У ушастой круглоголовки по бокам головы оттопыриваются
широкие, розовые с внутренней стороны складки, пасть разе-
вается и кажется благодаря складкам еще шире, а круглоголовка
в добавление сильно шипит и подпрыгивает навстречу врагу. Все
это создает устрашающий вид. Прыгание навстречу врагу вообще
является одним из видов устрашения у рептилий. Это особенно
часто наблюдается у змей. Способность очковой змеи (Naja naja)
раздувать шею принято считать устрашающим признаком: на ней
имеется резкий рисунок в виде очков.
Особенно интересно устрашающее положение, принимаемое
австралийской плащеносной ящерицей (Chlamydosaurus kingi).
Имеющийся у нее на шее воротник зазубрен на краях и поддер-
живается особыми хрящевыми палочками. Будучи испугана,
ящерица расправляет воротник и бежит, стараясь скрыться на де-
реве. Бежит она на двух ногах. Застигнутая врасплох, она при-
жимает заднюю часть тела к земле, поднимает переднюю, показы-
вает зубы, а иногда переходит и в наступление, кусая все, за что
можно уцепиться.
Защитным свойством черепахи (Clemmys leprosa), а также не-
которых других является способность выделять паховыми желе-
зами вонючее вещество и привычка некоторых ящериц и змей
выбрасывать вонючую жидкость из клоаки.
5. Размножение рептилий
Рептилии раздельнополы и могут отличаться вторичными поло-
выми признаками. Часто эти различия незначительны или даже*
вовсе отсутствуют. Иногда самцы лишь крупнее самок, как бы-
вает обычно у видов рода Lacerta, за исключением живородящей
ящерицы (Lacerta vivipara), где самка крупнее, что связано с раз-
витием детенышей в теле. То же самое у живородящих змей. У сам-
цов многих видов ящериц, змей и черепах хвост вздут при основании.
Это зависит от того, что здесь находится в свернутом состоянии
двойной совокупительный члеп. У черепах полы можно отли-
чить по виду грудного щита — у самцов он вогнут, тогда как у са-
мок он плоский или выпуклый. В других случаях смысл вторич-
ных половых различий неясен; например, различие в числе щит-
ков у многих змей, большая узость шеи. У самок многих змей
имеются килеватые гребни на спинных чешуях в период половой
жизни. Этот признак служит для раздражения самки во время
спаривания.
Непонятен смысл различия в окраске, наблюдаемого у неко-
торых рептилий. Например, у самцов черепахи Cistudo глаза
красные, у самок — бурые. У кровососа (Calotes versicolor) цвет
тела самца может меняться по произволу. Иногда он бывает весь
блестяще красного цвета с черными пятнами; в некоторых случаях
цвет кожи меняется только на голове, в других — на всем теле,
не исключая хвоста. Самка же остается одноцветной. Впрочем,
не всегда яркая окраска связана с полом. Далее полы могут от-
525
личаться наличием многочисленных выростов на голове, напри-
мер, у некоторых видов хамелеона, где у самца развиваются рога,
подобные рогам носорога. Сюда же относятся гребни на затылке
и на спине у многих игуан. Так называемые бедренные поры у сам-
цов ящериц более сильно развиты. У некоторых рептилий самцы
отличаются еще голосом, и не исключена возможность, что он слу-
жит для привлечения самок. У гекконов и аллигаторов голос
определенно служит для привлечения самок в период размно-
жения.
Соотношение числа особей обоих полов не всегда одинаково.
У змей оно бывает 1:1, 1:3, 1:4, 4 : 11. Иногда у одного
и того же вида соотношение полов различно в различных районах.
Так, китайская форма Dinodon septentrionale во всем ареале оби-
тания дает цифры 3 :13, а в Южной Азии — 0:8. Меньшее коли-
чество самцов происходит видимо от большей смертности послед-
них.
С другой стороны, иногда наблюдается временное преобла-
дание самцов. Так, в Средней Азии в конце лета (август) встре-
чаются только самцы змей, самки же отсутствуют и неизвестно,
где в это время они находятся.
В период спаривания часто происходит борьба между самцами.
Крокодилы свирепо преследуют в это время друг друга и вступают
в драки. То же самое наблюдается у ящериц; некоторые из них
во время драки меняют окраску. Змеи в период размножения со-
бираются в большом числе, свиваются в клубки и шипяТ до тех
пор, пока не соединятся с самкой, после чего половое возбуждение
проходит.
У рептилий в период спаривания часто наблюдаются любовные
игры. Самцы ящерицы кусают самок, чтобы понудить их к спа-
риванию; то же наблюдалось иногда и у черепах, где самцы издают
свистящий тон, ползают за самкой, кусают ее за голову и тол-
кают до тех пор, пока та не придет в возбуждение. После спарива-
ния возбуждение исчезает, и особи расходятся. Лишь редко на-
блюдается продолжительное пребывание самца и самки вместе.
У черепахи Testudo polyphemus наблюдалась жизнь парами в но-
рах. Известно еще несколько подобных примеров,
Большинство рептилий откладывает яйца, другие родят живых
детенышей. Яйца одеты твердой скорлупой или похожей на пер-
гамент. Первое состояние, типичное для черепах, является, по-
видимому, более примитивным. Среди ящериц яйца в скорлупе
встречаются лишь у гекконов. Но отвердевает у них скорлупа лишь
постепенно, воспринимая углекислоту из воздуха. У змей яйца
со скорлупой уже не встречаются. У большинства змей яйца при
откладке клейкие и приклеиваются к предметам, с которыми со-
прикасаются. Они вариируют нередко как по величине, так и по
форме.
Число яиц от 2 до 150. Как количество яиц, так и способ, раз-
множения обнаруживают признаки приспособления к условиям
внешнего мира и ими обусловливается. Наибольшая (до 400 в год)
526
продукция яиц наблюдается у морских черепах. Обусловливается
это, видимо, тем, что молодые черепашки мало приспособлены
первое время к жизни в воде: они плавают, но не погружаются,
выбрасываются на берег и служат добычей рыб и птиц. Чрезмер-
ное увеличение веса тела от такой массы яиц и слишком большая
потребность в питательном материале избегаются благодаря отло-
жению яиц партиями и утратой скорлупы. Очень малое количество
яиц можно также рассматривать как приспособительное явление.
Черепахи, размножающиеся в Китае в культурных областях,
где животные, враги их, почти всецело отсутствуют, откладывают
минимальное количество яиц (2). Гекконы откладывают яиц боль-
ше на юге Азии, чем на севере, что можно поставить в связь с на-
личием их врага древесной змеи — Chrisopelea ornata. Лазящие
формы кладут яиц меньше, чем наземные.
Откладываются яйца или в специально вырытые норы, или
среди мха и листвы, во влажных теплых местах. Змея США Dio-
dophys punctatus откладывает яйца в самых различных местах:
в грязи, выкинутой из каменоломни, в трухе под гнилым деревом,
в пне, под камнем, в ходах муравьев с трухой — вообще во влаж-
ных, но не мокрых местах, подставленных солнцу. В 95% случаев
это ямка в земле. Обычно самка оставляет яйца на произвол
судьбы. Лишь некоторые змеи и крокодилы ведут себя иначе;
некоторые американские формы последних устраивают для своих
яиц настоящие гнезда во влажных местах. Гнездо это состоит из
слоя растений, на которые откладываются яйца, снова затем по-
крываемые растениями. Благодаря гниению растений в гнезде
возникает столь высокая температура, что гнездо дымится. Это
тепло служит для скорейшего развития молоди.
На Мадагаскаре самка крокодила стережет гнездо до тех пор,
пока молодь не разовьется. Самка будто бы узнает это по особому
звуку, который молодые крокодилы производят в яйце, помогает
им откапываться из песка и сейчас же ведет их к воде. Так же
ведет себя самка каймана. Африканские крокодилы, невидимому,
не охраняют своих гнезд. Североамериканские аллигаторы за-
кладывают гнездо вблизи места обитания самки, которая бро-
сается на всех, кто к ней приближается, и таким путем охраняет
гнездо. Тут мы имеем интересный ряд постепенного усложнения
инстинкта, показывающий, каким образом он мог возникнуть.
Как было указано в систематической части, динозавры отклады-
вали яйца гнездами.
Самки некоторых крупных змей (питонов, например), отложив
яйца, ложатся на них таким образом, что образуют над ними пло-
ский свод, внутри которого температура оказывается выше окру-
жающей на 10—12° по Цельсию, что способствует развитию яиц.
Так же поступает щитомордник Ancistrodon. В это время самка
не принимает ни воды, ни пищи и защищает яйца от всякого пося-
гательства на них. В этом можно видеть и заботу о потомстве.
Ящерица, называемая тейю (Tupinambis teguixin), раскапывает
гнезда термитов и туда откладывает свои яйца. Выводящиеся из
527
последних молодые ящерицы сразу находят себе корм в виде
термитов.
Многие пресмыкающиеся производят на свет живых детены-
шей, являются живородящими. Детеныши прорывают
тонкие оболочки яиц еще в теле матери или сейчас же после рож-
дения. Правильнее называть их не живородящими, а яйце-
живородящими. Не известны яйцеживородящие формы
лишь у крокодилов и черепах. Этот вид размножения мы встре-
чаем у многих ящериц, особенно у живущих в горах, у многих
исполинских американских змей, морских змей, гадюк, у различ-
ных ужей и прочих. Лишь у немногих пресмыкающихся мы встре-
чаем и настоящую живородность, когда питание зародыша в теле
матери происходит при помощи кровеносных сосудов желточного
мешка, вступающих в связь с сосудами того отдела яйцеводов,
который играет роль матки. Живородящими были и ископаемые
ихтиозавры (Ichtyosauria). Эта особенность стояла у них, как у мор-
ских змей, в связи с переходом к пелагической жизни в море.
По строению конечностей ихтиозавров можно судить, что ихтио-
завры никогда не выходили на берег, но вели жизнь, подобную
жизни современных китов.
Первоначальной формой размножения у рептилий является
размножение путем откладки яиц. От него ряд переходов ведет
к яйцеживородности и далее — к живородности. Переход этот
облегчается у рептилий тем обстоятельством, что при наступлении
неблагоприятных для откладывания яиц обстоятельств последняя
может затягиваться на недели и яйцо оставаться в теле. Условия,
при которых живородность возникла у рептилий как приспо-
собление к ним, могут быть названы следующие: а) чисто водная
жизнь (Hydrophis); у видов крупных яйцеродящих родов (Natrix,
Elaphe), переходящих к жизни в воде (Natrix annularis, Elaphe
rufodorsata), развивается живородность; б) распространение в хо-
лодных областях, на холодных границах их вертикального и гори-
зонтального ареала, где ночные температуры слишком низки для
яиц и требуют защиты в материнском теле (Phrynocephalus из
центральноазиатских высокогорных областей, Lacerta vivipara,
Vipera berus, Ancistrodon в Азии). Живущий в Тибете на высоте
4 200 м, но у горячих источников, вид Natrix откладывает яйца;
в) жизнь' под землей (Scincus officinalis, Echis carinata, Vipera
ammodytes); *г) жизнь на деревьях и кустарниках (Dryophis,
Boiga). Если какая-либо группа рептилий с древнейших геологи-
ческих времен живет в условиях, вызывающих живородность, то
последняя свойственна всем членам группы. Так, у Scincidae,
сцинковых, живородность — признак всей группы.
Продолжительность инкубации у рептилий различная. За-
висит она от условий, в которых яйца развиваются; в среднем
у змей от 2 до 3 месяцев. Вылупление из яйца может длиться
много часов, до суток. Не все яйца вылупляются одновременно.
Для выхода змей из всех яиц кладки требуется 2—3 дня.
Выйдя из яйца или родившись живыми, рептилии растут бы-
528
стро, но достигают половой зрелости очень медленно: например,
китайский уж (Natrix piscator) на 4-м году, древесная змея
(Dryophis)—в конце 2-го года, самцы питона—в конце 3-го
и самки — 4-го года. Зато рептилии достигают глубокой старо-
сти. Относительно черепах, например, известны случаи, когда они
выживали в неволе до 54 лет. Гигантская черепаха (Testudo su-
nieri) достигла в неволе возраста в 150 лет. Известны случаи, когда
черепахи доживали до 250 лет. Большого возраста достигают
и крокодилы. Болезням пресмыкающиеся, повидимому, мало
подвержены, хотя в крови рептилий нередко встречаются па-
разиты из типа простейших (Protozoa). Надо полагать,
что в естественной обстановке большинство рептилий умирает
не от старости и болезпей, а насильственной смертью или от
каких-либо внешних неблагоприятных причин. Впрочем, суще-
ствует подозрение, что ящерицы являются носителями и передат-
чиками некоторых болезпей (например, лейшманиоза).
6. Отношение рептилий к климатическим факторам
Рептилии возникли в эпоху, когда благодаря поднятиям зем-
ной коры наступил сухой климат. Изменения, которые отличают
рептилий как класс от амфибий, указывают на приспособление
к сухому климату. Как сказано было выше, яйцо рептилий яв-
ляется самым существенным из этих приспособлений. Эту чер-
ту — приспособленность к сухому климату — рептилии сохра-
няют и по сие время.
Местообитания рептилий весьма различны. Про них можно
сказать лишь одно: в большей своей массе это животные наземные
и любящие сухость. Болота, арыки, тихие речки также служат
местом обитания немногих пресмыкающихся. Есть немногие реп-
тилии, связанные с морем (черепахи и змеи). Все же остальные
живут на суше, выбирая здесь большей частью места сухие. Не-
которые виды живут в лесах, влезая на деревья; большинство же
предпочитает пустынные бесплодные места, песчаные, глинистые
или каменистые. В противоположность большинству амфибий
рептилии — любители солнца, легко переносят летом высокую тем-
пературу. К холоду, наоборот, они чрезвычайно чувствительны.
Жизнедеятельность их при пониженной температуре ослабевает,
подвижность и аппетит уменьшаются. Зимой в холодном или уме-
ренном климате рептилии впадают в спячку. И чем холоднее
страна, тем дольше длится соп и тем оп крепче. При этом они
зарываются в землю, в ил (водяные) и залегают в спячку не в оди-
ночку, а по две или по нескольку особей вместе. Гремучие змеи
собираются осенью па зимовку в особых логовах, например, в тре-
щинах скал, где свертываются в большие шары. Собираются их
сотни. Выходят из логова весной. В спячке змеи лежат с закрытыми
глазами, не производя заметных дыхательных движений, со-
здавая впечатление мертвых. Отогретые, они приходят в себя.
34 Каш каров и Станчинский 529
Однако обмен веществ, характеризующий жизнь, не прекращается
вполне в спящих зимой рептилиях, он только сильно понижается.
Запасом питательного материала на это время и на первое время
после пробуждения, когда еще нет для них достаточно пищи,
служат жировые массы, располагающиеся в области таза. Край-
няя жара и главным образом сухость и отсутствие воды в жарких
странах, так же как холод, действуют на пресмыкающихся.
В сухое время года они впадают в летнюю спячку. Необыкновенно
многочисленные весной в степях и пустынях Туркестана черепахи
зарываются глубоко в землю и засыпают до следующей весны,
лишь только равнины начнут выгорать. Аллигаторы Сев. Америки
и крокодилы Южной залегают в одно время, одни — под влия-
нием холода, другие — под влияниет жары и сухости, лишающей
их пищи. Засыпают и кайманы и африканские крокодилы, зары-
ваясь по мере усыхания водоемов в мягкий ил. Засыпают, свер-
нувшись кольцом, и южноамериканские анаконды (Eunectes muri-
nus), причем они также зарываются на это время на некоторую
глубину в ил.
В огромном числе случаев, однако, рептилии прекрасно пере-
носят самый сухой климат. Для этой цели у них имеется целый
ряд приспособлений. Прежде всего сильно понижено испарение
благодаря отсутствию кожных желез. Далее, ими, подобно птицам,
выделяется не мочевина, а мочевая кислота, что требует меньшего
расхода воды. Поэтому они могут жить в совершенно безводных
пустынях, довольствуясь той водой, которая поступает в них вме-
сте с пищей. У некоторых черепах с Галапагосовых островов, кото-
рые должны совершать длинные путешествия за пресной водой
внутрь острова, мочевой пузырь и перикардий служат для на-
копления воды.
В связи с высыханием местообитания у некоторых рептилий
наблюдаются переселения. Так, например, европейская водяная
черепаха (Emys orbicularis) совершает такие переселения.
В пустынях Туркестана рептилии не впадают в летнюю спячку,
несмотря на сильную сухость. Но они живут здесь строго по часам,
выходя наружу и обнаруживая деятельность лишь по достижении
воздухом определенной температуры. Одпако чрезмерно высокой
температуры и рептилии, подобно всем другим животным, не пере-
носят. В пустынях они различными способами избегают ее дей-
ствия. Если же выставить пустынную ящерицу Eremias на солнце
на почве, то при нагреве последней свыше 50° ящерица неминуемо
погибает в срок от 11/3 до 4х/2 минут. Избегание губительного пе-
регрева достигается разными способами. Некоторые ящерицы,
например, агамы (Agama sanguinolenta) или (реже) ящурки (Ere-
mias velox), залезают в полдень при максимуме жары на кусты,
где температура значительно (на 20° и больше) ниже, чем на земле.
Другие, как ушастая круглоголовка (Phrynocephalus mystaceus),
в полдень забираются на вершины барханов, где стоят, высоко
поднявшись на ногах. Здесь температура также ниже на несколько
градусов. Ящерицы пустыни, где почва накаливается до 65е
530
и больше, уклоняются от перегрева, забегая в тепь, совершая
очень быстрые перебежки по освещенным участкам, забиваясь
в траву, где температура значительно ниже, закапываются в зем-
лю. Благодаря отношению к высокой температуре у них создается
суточный цикл жизни. Рано утром их пет. Появляясь, когда солн-
це в известной мере нагреет землю; они уходят снова в порки или
под кусты растительности в наиболее жаркое время дня. У жи-
вущих на каменистом или на глинистом грунте круглоголовок
(Phrynocephalus), у Uroniastix и других ящериц.температура тела
регулируется работой хроматофоров и быстрым дыханием. Благо-
даря этому колебания температуры ящерицы мепее колебаний тем-
пературы среды. Некоторые ящерицы, как сцинки (Teratoscin-
cus), избегают высоких температур, ведя ночной образ жизни. Орга-
низм изолирует себя от температурных условий среды при помощи
автоматических реакций, выработанных долгим отбором в процессе
борьбы за существование.
Будучи животными с непостоянной температурой тела, реп-
тилии не переносят охлаждения ниже некоторой не слишком низ-
кой температуры. Так, черепахи выдерживают охлаждение до
—8°, но погибают при температуре —9°. Этим определяются среди
ряда других причин границы их распространения. Сильные по-
нижения температуры ночью также могут, повидимому, служить
ограничивающим распространение фактором. Понижения тем-
пературы ночью в пустыне вызывают ослабление энергии движе-
ния, переходящее в оцепенение. Различные приспособления к ха-
рактеру местообитания — наземному, подземному, древесному,
водному — рассмотрены выше.
7. Биоценотические отношения рептилий
О роли рептилий в биоценозах и вообще в экономии природы
немногое можно сказать, так как под этим углом зрения рептилий
изучали мало. Во всяком случае роль эта не слишком велика вслед-
ствие их относительной малочисленности, малых размеров и хо-
лоднокровности. Лишь в некоторых странах и биотопах числен-
ность рептилий значительно возрастает, и роль их становится
также значительной, хотя числовых данных, касающихся этой
роли, мы пе имеем. Так, в пустынях рептилии нередко являются
столь многочисленными, что превосходят всех других позвоноч-
ных; очень многочисленны они в тропиках, а также местами
в море. В этих случаях опи не могут пе играть роли в хозяйстве
природы, составляя одно из звеньев в круговороте веществ. В да-
лекие геологические эпохи, когда развивавшийся класс рептилий
в лице своих многочисленных отрядов не встречал сопротивле-
ния распространению своего господства, роль птиц и в особенности
млекопитающих была значительно больше.
Роль какой-либо группы или какого-либо вида в сообществах
и в экономии всей природы определяется их численностью
и местом в цикле питания, т. е. тем, что данный вид ест, сколько
34* 531
ест и кто его ест. Взаимоотношения иного порядка, порядка взаимо-
помощи, играют значительно, меньшую роль.
Пища и роль рептилий в различных средах обитания. Пища
рептилий, живущих в морях, может быть или растительной или
животной. Морские черепахи (Chelonidae) питаются ракообраз-
ными, моллюсками и другими беспозвоночными, а один вид (Che-
lone mydas) питается преимущественно морскими растениями,
в особенности Zostera, не брезгуя и животной пищей. Имея в виду
большое количество этих черепах в некоторых местах, следует
признать, что роль их в соответственных биоценозах не мала, но
мы о ней слишком мало знаем. Исключительно животной пищей
питается каретта (Ch. imbricata). Пеламиды (Pelamis) из морских
змей (Hydrophinae) питаются главным образом головоногими мол-
люсками. Другие морские змеи питаются преимущественно ры-
бами. Ужи в Каспийском море истребляют массу бычков. Пресные
и солоноватые воды тропической, субтропической и южпой части
умеренной зоны дают приют рептилиям, которые питаются рыбой,
лягушками и другими водными позвоночными, нападая и на водя-
ную птицу (представители каймановых черепах, Chelydridae),
рыбами, раками и моллюсками (кожистая черепаха, Dermoche-
lys; мягкие черепахи, Trionychidae). Крокодилы, во многих местах
тропиков являющиеся довольно многочисленными, питаются ры-
бой, раками, моллюсками, схватывают птиц, млекопитающих.
Змеи едят рыб, реже лягушек, вероятно, также беспозвоночных.
Подземные, роющиеся рептилии получают свою пищу под зем-
лей, причем пища эта состоит из червей и насекомых.
Рептилии, обитающие на земле, питаются самой разнообраз-
ной пищей. Одни, как, например, сухопутные черепахи (Testu-
dinidae), питаются растениями. Принимая во внимание огромное
количество последних в некоторых местах, например, в степях Сред-
ней Азии весной, надо думать, что роль их в круговороте веществ
в данном районе пе слишком мала. Растительная пища составляет
значительную часть диэты игуан (Iguanidae), поедающих также
и насекомых. Крупные змеи, как питоны (Python), нападают па
крупных позвоночных, менее крупные змеи кормятся мелкими
млекопитающими, птицами, яйцами птиц, амфибиями, реп-
тилиями, насекомыми, пауками и другими беспозвоночными.
Некоторую —• и быть может не маленькую — роль в круговороте
веществ играют опи там, где их много, как, папример, в песчаных
пустынях. Здесь они должны играть роль в накоплении в почве
органического вещества.
Точно так же играют известную роль в биоценозах и те реп-
тилии, которые, как, например, древесные змеи (Chrysopelea
и др.) питаются насекомыми, яйцами птиц и птенцами.
К сожалению, мы пе имеем данных ни о количестве репти-
лий в разных группировках, ни о количестве поедаемой ими
пищи.
Сожительства рептилий с другими животными немногочислен-
ны. Так, новозеландская «ящерица» (Sphenodon), или Hatte-
532
ria живет в одних норах с буревестниками, крокодил допускает
птичку Trochilus копаться у него в зубах, выковыривая оттуда
застрявшие между зубами остатки пищи. Известно о сожитель-
стве в одной норе луговых собачек (Cynomys) и гремучей змеи,
впрочем, не вполне невинном, ибо молодых Cynomys змея
съедает.
В то же время у ррцтилий немало врагов. Главным из них
является, повидимому, человек^ уничтожающий их прежде всего
изменениями ландшафта при помощи своих культурных воздей-
ствий на природу, охотящийся за многими видами ради их мяса,
яиц, шкурок, роговых щитков и других целей. За змеями охотятся
многие млекопитающие. Домашние кошки убивают даже кобру.
В желудках диких кошек (Felis viverrina) находили даже остатки
питона. Вероятно, большинство виверровых поедает рептилий.
Это во всяком случае делают мунго, охотящиеся за змеями груп-
пами. Молодых змей уничтожает насекомоядное млекопитающее
Pachyura swinhoei. Даже мыши иногда нападают на змей. Олени,
козы, свиньи, коровы нередко нападают па змей и топчут их.
Интересен способ, которым еж поедает змей, например, гадюку.
Он схватывает ее за хвост и сейчас же свертывается. Змея не может
укусить его, так как он защищен иглами. Еж же постепенно втяги-
вает и съедает змею.
Змей и ящериц уничтожают многие хищные и другие — как
крупные, так и мелкие птицы. Особенно известны в этом отноше-
нии секретарь (Secretarius) в Африке, далее орлы белохвостые,
змееяд, некоторые луни, коршуны, совы, вороны, сорокопуты,
цапли, кваквы и др.
Из рептилий ест животных своего же класса варан (Varanus).
Крупные ящерицы поедают мелких, многие змеи едят ящериц.
Даже сколопендры с успехом нападают иногда на змей.
Одно значение рептилий в экономии природы следует отме-
тить — истребление ими грызунов и насекомых.
Совершенно другую роль играли рептилии в мезозойскую эру,
когда в морях кишели ихтиозавры, питавшиеся рыбой и голово-
ногими, плезиозавры, лотозавры-и другие водные рептилии; па
суше двигались колоссальные травоядные, разнообразные хищ-
ные динозавры, а воздух населяли птерозавры. Тогда на долю
рептилий выпадала более важная роль в «цепях питания», связы-
вающих между собой различных животных и растения. Рептилии
тогда завершали эти цепи, являясь наиболее мощными превра-
тителями энергии.
8. Значение рептилий для человека
Значение рептилий для человека невелико в сравнении с ры-
бами и высшими классами. Но все же ряд рептилий используется
человеком. С другой стороны, многие рептилии, являясь ядови-
тыми, имеют для него немаловажное значение, особенно в некото-
рых странах.
533
Съедобными являются некоторые черепахи, местами едят
также и черепашьи яйца. Исполинские слоновые черепахи были
истреблены в значительной мере ради мяса, так как служили
чрезвычайно удобным для плавающих в океане парусных кораблей
запасом провианта. Они могли долго жить на судне и давали
40—100 кг прекрасного, свежего и вкусного мяса. Их едят и сей-
час там, где они еще сохранились. Греческая черепаха (Testudo
graeca) постоянно выносится на рынки Италии; из нее приготов-
ляют суп, считающийся лакомством. Близкая к ней туркестанская
степная черепаха (Testudo horsfieldi) также вполне съедобна;
суп из нее напоминает куриный. Зеленая, или суповая, черепаха
(Chelone mydas), очень крупное животное (до 450 кг); она вполне
съедобна, так же как и каретта (Ch. imbricata). Съедобны также
и яйца черепах или же они служат для вытапливания жира. На
яйца некоторых черепах, папример, «аррау» (Podocnemis ехрап-
на) из бокошейных черепах, производится настоящий организован-
ный промысел.
Употребляется в пищу мясо некоторых крупных ящериц..
Имеют значение некоторые рептилии и в качестве второсте-
пенных видов сырья. Из кожи крокодилов, крупных ящериц,
как вараны и ящерицы тейю, а также из кожи многих змей, в осо-
бенности удавов, и многих более мелких выделывают различные
изделия: дамскую обувь, портфели, портсигары, галстуки, отделку
для платьев. Благодаря своей прочности и красивому рисунку кожа
этих рептилий и изделия из нее очень ценятся. Ради кожи многие
виды местами сильно преследуются, притом чаще без учета дипамики
запасов этих животных, что грозит их истреблением. Из роговых
щитков тех черепах, у которых роговая масса от горячей воды
разбухает и при давлении «сваривается», изготовляют различные
предметы повседневной жизни и различные художественные изде-
лия. Из варанов добывают ценный жир, идущий в кожной про-
мышленности. Правильное использование рептилий как сырья
требует знания их экологии, учета количества, времени размно-
жения, величины приплода и т. д. Это мы, до сих пор знаем очень
мало.
Ядовитые змеи. Гораздо большим является вред, приносимый
ядовитыми змеями. Правда, страх перед ними несколько преуве-
личен, по крайней мере в умеренных странах, но все же в неко-
торых странах, ежегодно умирает от укуса змей большое число
людей.
Так, папример, в 1911 г. в Индии (по официальным данным) по-
гибло от укуса змей 24 312 человек и 10 534 голов скота; в Бра-
зилии за год в среднем бывает укушено змеями 24 000 человек
с 4800 случаями со смертельным исходом (смертность уменьшают
впрыскивания сыворотки).
Змеи ядовиты далеко не все, хотя различие между ядовитыми
и неядовитыми змеями вовсе не принципиальное, ибо и у неядо-
витых есть весьма ядовитая слюна. Из всего количества змей —
2 300 видов — около 250 являются ядовитыми. Распространены
534
ядовитые змеи преимущественно в тропических странах: в Европе
8 видов ядовитых змей, в Африке — около 75, в Азии и на Малай-
ском архипелаге — 165, в Америке, преимущественно в Южной —
91, в Австралии — 80.
Ядовитыми называют тех, у которых имеются «ядовитые зубы»,
расположенные так, что при укусе яд попадает в рану. Выше
было указано деление змей на гладкозубых (Aglypha), задпеборозд-
4aTHX(Opisthoglypha), у которых зубы с бороздкой сидят па зад-
нем конце верхней челюсти, и переднебороздчатых (Proterogly-
pha), у которых ядовитыми (бороздчатыми) зубами являются перед-
ние зубы. Лишь эти последние являются настоящими ядовитыми
змеями, так как при укусе ими бороздчатый зуб или полый делает
глубокую рапу (зубы достигают 2.5 см в длину), и яд изливается
в глубину.
Желез в голове у змей несколько, но лишь одна, лежащая
ниже и позади глаза (паротида), является «ядовитой»; гомологи
ее есть и у неядовитых змей. Выделение яда происходит при по-
мощи сокращения височного мускула (m. temporalis).
Ядовитые змеи встречаются в разных семействах и подсемей-
ствах (сем. Colubridae с подсем. Elapinae и Hydrophiinae и сем.
Viperidae с подсем. Viperinae и Crotalinae). К сожалению, нет
такого признака, по которому всякую ядовитую змею можно было
бы отличить от неядовитой. Часто признаком ядовитости является
широкая голова — широкая вследствие развития ядовитых желез.
Но это пе всегда. Многие неядовитые змеи имеют также широкую,
плоскую голову, а ядовитые, наоборот (например, очковая змея,
Naja tripudians), имеют голову узкую.
Даже наличие ядовитых зубов не абсолютно характерно Труб-
чатые зубы связаны переходами с бороздчатыми, а последние
с гладкими. Также и физиологическое действие секрета ротовых
желез не принципиально отлично у тех и других: безвредные для
человека змеи с бороздчатыми зубами убивают укусом мышь,
ящерицу, воробья; верхние губные железы безвредного ужа (Na-
trix natrix), морфологически гомологичные ядовитым железам
гадюки (Vipera), выделяют секрет, .убивающий морскую свинку.
Яд стрелы-змеи (Taphrometopon из Opisthoglypha), которую турк-
мены считают безопасной, а казахи боятся, будто бы убивает мел-
ких ящериц. Вовсе не парадокс говорить о яде неядовитых змей.
Слюна каждой змеи более или менее ядовита.
Змеиный яд представляет собой соломенно-или бледножелтую
жидкость с кислой реакцией, содержащую около~'ЗО% твердых
веществ. Он безвкусен. Лишь у кобры — неприятно горек. В высу-
шенном виде сохраняется неопределенно долго и легко растворяется
в воде.
В составе яда находятся различные токсические (отравляющие)
вещества, действующие на различные ткани_ и органы. Содержа-
щийся в нем нейротоксин возбуждает нервные клетки; геморрагии
яда действует па внутреннюю оболочку сосудов — эндотелий,
на фибрин крови и мускулы действует протеолитическая часть
535
яда. Под влиянием тромбазы яда освобождается фибринфермепт,
вызывающий свертывание крови. Вообще состав яда змей очень
сложен.
Яд легко разрушается веществами, разлагающими протеиды,
например, азотнокислым серебром, перекисью марганца, хлорной
известью, хлором, бромом, иодом. Количество выделяемого яда
у разных змей различно. У гадюки обыкновенной (Vipera be-
rus) — 30 мг, у песчаной гадюки-носорога (V. ammodytes) —
65 мг, у кобры (Naja tripudians) до 220 мг, у гремучей змеи (Crotalus
durissus) — 300—370 мг.
Действие яда зависит от количества яда и от места внесения
его, а также от вида змеи. После укуса яд поступает в подкож-
ную соединительную ткань и затем всасывается при помощи лимфы
и кровеносных сосудов. Будучи введен непосредственно в кровь,
вызывает действие немедленно. Через слизистую оболочку рта
или пищеварительного канала не всасывается, если только здесь
нет раны. Действие яда различное на теплокровных и холодно-
кровных животных. На человека яд действует, вызывая, с одной
стороны, местные явления, с другой — общие явления отравления.
То и другое действие различно у разных видов змей.
После укуса гадюкой (Vipera) вокруг раны тотчас по-
является краснота и опухание. Чувствуется сильная местная
боль. Опухоль расплывается по всей укушенной конечности и даже
по телу. Опухоль спадает на 4—7-й день. На месте укуса может
произойти отмирание тканей, их распадение и образование язв.
На ряду с этими местными явлениями наблюдаются явления общие:
сонливость, упадок сил или возбуждение, тошнота, рвота; пульс
становится частым и слабым; тело холодеет, пострадавшему нехва-
тает воздуха, в моче появляется кровь. Слабость может оста-
ваться 2—3 месяца. Иногда получается слепота. В тяжелых слу-
чаях через промежуток от 12 часов до 7—8 дней может наступить
смерть. При сильном местном поражении больные очень долго
мучаются с язвами.
При укусе некоторыми тропическими змеями местное действие
бывает во много раз сильнее, и распад тканей (гангрена) может
привести к тому, что отваливается нога, или ткани распадаются
до самой кости.
При укусе коброй (Naja), наоборот, местные явления незна-
чительны. Хотя окружность раны вздувается, резкой боли пет.
Зато очень сильны общие явления отравления: похолодание уку-
шенной части, усталость, непреодолимая сопливость; позднее за-
темняется сознание, дыхание становится тяжелым, деятельность
сердца падает, сильно выделяется слюна, и через 27 часов насту-
пает смерть. Если отравление было не сильно, то через 1—2 дня
человек поправляется.
Сходная картина наблюдается и при укусе гремучей змеей
(Crotalus): местные явления слабы, гангрены никогда не бывает,
по яд сильнейшим образом действует на нервную систему, вызывая
нарушения чувствительности, параличи, слепоту и смерть от па-
536
ралича дыхания. Если отравление былопе слишком сильное и про-
текает не столь быстро, то наблюдается воспаление разных органов:
печени, почек, мозга.
Иная картина — при укусе Elaps, где местной реакции вовсе
не бывает.
Яд одной и той же змеи действует различно в разное время.
Яд кобры, например, в 10 раз сильнее после линьки, чем до линьки,.
сильнее после приема пиши, нежели
у голодных. Зависит сила действия яда
и от времени года, а также и от места
укуса. Чем больше в этом месте сосу-
дов, тем сильнее и скорее действие. Дей-
ствие бывает молниеносным, если яд по-
пал в большую вену. Через здоровую
кожу и слизистые оболочки яд пе про-
никает.
Когда яд попадает в организм пе
в очень большой дозе, организм с ним
успешно справляется: поднимается тем-
пература тела, наблюдается усиленная
деятельность белых кровяных шариков
(гиперлейкоцитоз). При смертельных до-
зах эта защитная мобилизация белых кро-
вяных телец, наоборот, падает (гипо-
лейкоцитоз), падает и температура тела.
Выше было указано, что и неядови-
тые змеи имеют ядовитую слюну. Яд
некоторых «неядовитых» африканских
заднебороздчатых змей (Opislhoglyplia),
будучи впрыснут, убивал бабуина, ко-
шек, даже лошадь. Наблюдались и боль,
Рис. 415. Расположение
зубов у Aglypha (Л), Opi-
slhogly pha (В) и Protero-
glypba (С).
(Ив Павловского.)
и отек, и некроз, и нарушение дыхания. По укусы их неопасны
(по крайней мере не всегда опасны), ибо ядовитые зубы сидят
у них далеко назади. Чтобы змея была на деле ядовитой, надо,
чтобы наличие ядовитой железы сопровождалось еще наличием,
зубов, которыми змея могла бы наносить раны, и по которым яд
проникал бы в тело жертвы. Между ядовитыми и неядовитыми
змеями существует ряд переходов. Мы видим в них как бы ряд.
ступеней эволюции.
При укусе ядовитой змеей необходимо: 1) пометать распро-
странению яда из раны в организм; 2) удалить яд, разрушить или
нейтрализовать его; 3) лечить поврежденные органы. Поэтому
прежде всего необходимо наложить лигатуру (перевязку) выше
места укуса, туго закрутить перевязку при помощи палочки;
затем сделать надрез в рапе, высосать из нее яд, если на губах
и во рту нет ссадин. Массировать конечность книзу, к ране, не-
сколько раз. Посыпать рану порошком хлорноватистокислого
кальция. Лучше всего ампутировать палец, если его укусила
опасная змея. Продержав лигатуру полчаса, снять ее, дать крови
637
свободно разойтись и снова наложить перевязку. Внутрь прини-
мают небольшие дозы алкоголя, еще лучше аммиака или стрих-
нина. Хорошо действует подкожное впрыскивание стрихнина.
Лишь одно лечение действительно вполне: это лечение сыворот-
кой — впрыскивание сыворотки животных, которые были имму-
низированы повторным впрыскиванием небольших доз змеиного
яда. К сожалению, это не всегда доступно. В тропических странах,
где много ядовитых змей и часты случаи смерти от их укуса, устраи-
ваются специальные институты для изготовления змеиной сыво-
ротки. Такие институты имеются в Индии (два), в Японии, в Аф-
рике (порт Елизавет), в Алжире (институт Пастера), в Австралии
(Сидней), в Северной Америке (Филадельфия), в Южной Америке
(в Сан-Паоло-Бутантан, Вуэнос Айрес), в Лилле (Пастеровский
институт). Наиболее знаменит институт в Сан-Паоло-Бутантан.
В институтах имеются «змеиные сады», где отгороженные от внеш-
него мира стеной и глубокой цементированной канавой с водой,
живут, прячась от солнца, в особых домиках змеи, от которых берут
яд. Яд берут у змеи, держа ее за шею щипцами, раскрыв ей пасть
и давая кусать маленький стеклянный сосуд. Яд высушивают,
и в растворенном виде малыми дозами впрыскивают лошадям.
У последних в крови получаются антитоксины, развивается
активный иммунитет. Сыворотку этих животных впрыскивают
укушенным змеями, вызывая у них пассивный иммунитет. Это
лучшее средство помощи, действующее, как специфическое
средство.
Яд змей, несомненно, защищает их от других животных. Однако
некоторые млекопитающие, птицы и рептилии от природы иммунны
против змеиного яда и поедают ядовитых змей. Так, например,
еж поедает гадюк, то же некоторые представители куниц (Muste-
lidae), лисица, кабан. Поедает змей и домашняя свипья; это живот-
ное в долине реки Миссисипи дрессируют на гремучих змей. Им-
мунна к яду гадюк и кошка. Славятся своими способностями уни-
чтожать ядовитых змей ихневмон (Herpestes ichneumon) в Египте
и мунго (Н. griseus) в Индии. Поедают ядовитых змей многие
птицы: кондор (Sarcorhamphus gryphus), орел змееяд (Circaetus
gallicus), канюк (Buteo), вороны (Corvus), секретарь (Serpentarius
secretarius), китайская сова (Ketupa). Из змей Lachesis mutus,
Naja, Elaps едят других ядовитых и неядовитых змей. Особенно
известна как пожирательница ядовитых змей «муссурана» — не-
ядовитая Oxyrhopus cloelia из Coluhridae, достигающая 1.5—2.35 м.
Яд даже гремучей змеи на нее не действует. Оца схватывает по-
следнюю за спину, добирается, перебирая зубами, До головы,
ломает добыче шейные позвонки, повреждает мозг и заглатывает.
Борьба длится иногда до 1.5 часа. Поедает она змей, которые
в 1.5 раза больше нее.
Не все животные, поедающие ядовитых имей, иммунны к их
яду. В крови домашней свиньи нет никаких антитоксинов. Она
не боится змей, ибо защищена толстым слоем плотного жира, и яд
не распространяется по организму. У других животных имеется
538
иммунитет в крови. Например, сыворотка крови ежа удвояет со-
противляемость кролика яду гадюки. Но доза яда гадюк в 40 раз
более сильная, чем та, что нужна для умерщвления кролика,
убивает и ежа.
В крови ядовитых змей имеется антитоксин против их соб-
ственного яда. Сыворотка ядовитых змей действует как энергичный
антифермент против яда той же змеи. Но сильной дозой и впрыски-
вая яд в соответственном месте, можно убить каждую змею ее же
ядом. Например, гадюка гибнет от собственного яда в количестве
100—200 мг под кожу или 2—4 мг в мозг. Другие холоднокровные
животные восприимчивы к яду змей в различной степени. Эффект
его общий и местный. Главный местный эффект — это выход из
сосудов красных кровяных шариков (геморрагия). У некоторых
это происходит не только в месте поражения. У укушенных змеей
рыб кровь из жабер окрашивает воду. У акулы кровоизлияние
происходит внутри черепа. Чаще это бывает в коже. Нервная
система поражается нейротоксином. Рыбы теряют равновесие,
первоначально придя в сильное возбуждение. Сильпо действует
яд на черепах. Саранча гибнет лишь от больших доз. На игло-
кожих влияние еще меньше.
Ядовитых змей человек преследует, стремясь избавиться от
них.
Из других рептилий, кроме змей, ядовита только одна ящерица
ядозуб (Heloderma horridum), водящаяся в1 Мексике и соседних
частях США. У нее зубы нижней челюсти загнуты крючком, утол-
щены при основании и некоторые из пих снабжены бороздкой,
по которой стекает яд слюнных желез, гомологичных неядовитым
нижне-челюстным железам змей. Специальных мускулов для вы-
давливания яда у Heloderma пет. Сухой «секрет желез сохраняет
свои свойства год. О силе действия яда ядозуба мнения различны.
После укуса ядозубом наблюдалась боль, вздутие и покрасне-
ние места укуса. Одновременно наступали общие явления: через
5 минут — пот, бледность лица, головная боль, а через час — обмо-
рок с копвульсиями. Последствия пе оставляли укушенного пять
месяцев.
Собака, которой было впрыснуто 5 — 6 капель свежего яда,
сдохла через 15—18 минут. На холоднокровных животных действие
слабее.
9. Поведение рептилий (высшая нервная деятельность)
Поведение рептилий сложнее, нежели амфибий, их нервная
деятельность находится на более высоком уровне. Это стоит
в связи с более высокой ступенью развития у них головного мозга.
Полушария здесь развиты сильнее. В передней их части хорошо
развиты обонятельные доли. В своде полушарий (pallium) уже
намечено разделение бороздой па верхний отдел, являющийся
центром высших способностей, и нижний, служащий для восприя-
539
2
1
тия обонятельных раздражений. Сильно развито полосатое тело.
В деталях строения полушарий рептилий уже намечаются, хотя
еще слабо, особенности полушарий млекопитающих. Помимо уве-
личения объема имеется явно слоистая кора. Средний мозг (зри-
тельные доли, lobi optici) развит у рептилий слабее, нежели у рыб
и амфибий, у которых он являлся зрительным центром. Здесь
волокна зрительного нерва частич-
но идут уже к зрительным буг-
рам — thalami optici, значительно
развитому ганглию в основании
полушарий. Отсюда волокна идут
к коре полушарий, как (в боль-
шей мере!) это имеет место у мле-
копитающих (рис. 416).
Удаление переднего мозга, да-
же боковых его частей только
вызывает у ящерицы потерю спо-
собности принимать пищу.
Вкусовые органы расположены
у многих ящериц и черепах в
ряд, параллельный зубам, у ве-
ретенницы (Anguis fragilis) — па
кончике языка, у Lacerta — на
поперечных складках. Они не
отсутствуют и у змей, и слабо
развиты у крокодилов. Орган обо-
няния снабжен особым дополни-
тельным якобсоновым органом.
Улитка в органе слуха достигает
большого развития только у кро-
кодилов. Орган зрения является,
невидимому, главным рецептором
у рептилий, в особенности при рас-
познавании добычи. В сетчатке
глаза у рептилий преобладают колбочки, а у черепах и ящериц —
исключительно одни колбочки. Последние снабжены особыми «жи-
ровыми каплями», т. е. каплями жидкого жира, окрашенного липо-
хромами в желто-красный цвет, поглощающий голубую часть
спектра. У многих рептилий имеется, кроме парных, еще непар-
ный теменной глаз, связанный с мозгом. Он сильно
вариирует в строении, состоит из преломляющей части, из сетчатки
и из пигментного слоя. Последний устанавливается различно в за-
висимости от освещения, подобно тому как это имеет место в пар-
ных глазах. Функция этого рудиментарного глаза неясна, хотя
какую-то функцию он, повидимому, судя по движению пигмента,
имеет.
Рептилии гораздо более активны, нежели амфибии. Как мы
видели, большинство их является наземными формами, притом
хищными. Исключением являются малоподвижные наземные чере-
540
Рис. 416. Схема прохождения
волокон opticus.
I —глаз, 2 — tectum opticum, 3— pal-
liium, 4—talamus opticus. (Ho Mones-
cow из Ihle.)
пахи. В связи с таким образом жизни рептилии должны более
быстро реагировать на изменяющиеся обстоятельства. Этому и
служит лучшее, чем у амфибий, развитие головного мозга и органов
чувств.
Относительно деятельности органов ощущения мы знаем еще
недостаточно. Но все же можно сказать, что опи имеют большее
значение в жизни рептилий, нежели у амфибий. Черепаха, как
и змея, чует свою добычу или пищу на расстоянии. Черепаха
выбирает свою пищу из смеси разных сортов; змея преследует
убежавшего грызуна по следу, руководясь, видимо, обонянием.
Осязание у ящериц и змей сосредоточено на языке. Их раздвоен-
ный язык, называемый иногда неправильно «жалом», постоянно
высовывается наружу, как бы совершая нащупывающие движения.
На нем сосредоточено много осязательных нервных окончаний.
Змеи могут при помощи языка воспринимать твердые тела на рас-
стоянии, не придя еще с ними в соприкосновение. Несомненно
чувствительны рептилии и к температурным воздействиям. По на-
блюдениям в природе и по экспериментам мы знаем, что змеи,
ящерицы, крокодилы выбирают места с одними температурами
и избегают других.
Слух у рептилий развит плохо. Черепахи па звукпе реагируют.
О слухе гремучей змеи и щитомордника (Ancistrodon) мы знаем
лишь, что он рудиментарен, и вряд ли гремучая змея слышит
в пашем смысле этого слова. Вряд ли музыка играет какую-нибудь
роль в тех представлениях, когда «заклинатели змей» «очаровы-
вают» последних при помощи музыки. Ящерицы, видимо, слы-
шат. По крайней мере их можно дрессировать на звук, а в природе
ящерица реагирует на звуки, производимые ее добычей. Вообще
следует заметить, что животное может не реагировать на звуки
йо потому, что оно вообще не слышит, а потому, что эти звуки
чужды ему в его обычной жизни, на звуки же, которые имеют для
животного жизненное значение, оно будет реагировать.
Относительно зрения черепах мы знаем, что правая часть
спектра у них сильно укорочена благодаря окрашенным каплям
в колбочках. Глаз черепах, приспособляясь к темноте, становится
более чувствительным. Такое животное схватывает (в эксперименте)
движущийся кусок мяса, невидимый человеческим глазом. Приспо-
собляемость глаза к темноте в монохроматическом свете одинакова
у черепахи и у человека, смотрящего через оранжевое стекло.
Относительно чувствительности к длине волны с рептилиями не
было поставлено опытов, которые дали бы удовлетворительные
результаты. Опыты с различением черного—белого, поставленные
с черепахой Chrysemys marginata, дали положительные результаты.
Опыты с различением той же черепахой формы и величины тела
их не дали. Неподвижную добычу могут распознавать лишь гекконы
(на 8—10 см). Другие, например, агамиды, сразу теряют из вида
бабочку, которую они преследовали, если она, сложив крылья,
сядет. Змеи видят движущегося человека на расстоянии 5 м.
Что некоторые, по крайней мере, рептилии так или иначе,
541
по цветности или по яркости различают цвета, видно из того, что
многие из них способны изменять свою окраску при изменении
цвета окружающей среды, как, например, знаменитый этой спо-
собностью хамелеон.
Способность к научению у рептилий еще очень невелика, но
все же она имеется. Ящерицы, змеи и крокодилы обнаруживают
иногда хорошее знание окрестностей своего местообитания. Они
способны к запоминанию некоторых обстоятельств, к образованию
условных рефлексов. Крокодилы, папример, содержимые в неволе,
приходили в возбуждение каждый раз, как приносили доску,
на которой им разрезали мясо. Черепахи подходили, когда ставили
им сосуд с питьем. Когда змеям давали кусать железную палку
или глиняный черепок, опи скоро переставали кусать пе только
их, но и все похожие по виду предметы.
Опыты с простым лабиринтом показали, что черепаха может
научиться паходить дорогу в очень простом лабиринте, по опыты,
эти не достаточно убедительны; дрессировка варапа удается с боль-
шим трудом, причем никогда нельзя быть уверенным, что оп пе
забудет выученного. В общем умственные способности рептилий
невелики, но все же опи выше таковых у амфибий. При этом те
рептилии, которые ведут охотничий образ жизни, обнаруживают
больше способностей, чем те, которые питаются червями.
В основном поведение рептилий базируется еще па безуслов-
ных рефлексах и инстинктах. Последние у рептилий тоже не до-
стигают большой сложности. Некоторые виды обнаруживают
«заботу о потомстве». Но, как мы видели, эта «забота» представляет
инстинкты еще весьма простые, например, у крокодила, охраняю-
щего гнездо. Эволюция рептилий шла, видимо, по линии выработки
морфологических адаптаций, средств адаптивной защиты и напа-
дения на добычу. Этими свойствами должны были обладать в вы-
сокой мере многие ископаемые рептилии, как, например, дино-
завры.
10» Географическое распространение рептилий
Ни один большой отряд современных пресмыкающихся не
является характерным для какой-либо области. Это потому, что
различные отряды и подотряды рептилий распространились по
земному шару очень широко еще в те отдаленные времена, когда
различные, ныне разъединенные, континенты были соединены.
Поэтому те деления суши, которые установлены по млекопитаю-
щим и птицам, здесь не годятся. Но более мелкие систематические
единицы, семейства и роды часто имеют свое определенное и огра-
ниченное распространепие, которое удобно рассмотреть не по
странам, а отдельно, по систематическим единицам.
Вымершие подклассы и отряды рептилий имели очень широкое
распространение. Остатки котилозавров были найдены
в Европе, в Сев. Америке, в Южной Африке. Пеликозавры —
в Сев. Америке и в Европе. Тероморфы — в Южной Африке
542
и на Северной Двине. Мезозавры — в Южной Америке,
Южной Африке, па Шпицбергене, в Калифорнии. Сауропте-
р и г и и — в Европе, Америке и Индии. Черепахи — по
всему миру. Диаптозаурии — в Сев. Америке, Южпой
Африке, Англии, Новой Зеландии. Текодонты — в Сев.
Америке, Индии, Европе, Африке и Южпой Америке. Дино-
завры — в Европе, Сев. Америке, Южной Африке, в Азии.
Все эти нахождения относятся к периоду от пермской эпохи и до
меловой. Как наземные, так и пресноводные рептилии имели самое
широкое распрострапепие. Современные рептилии представляют
лишь жалкие обломки огромного ствола, уцелевшие в разных
местах земного шара.
Рис. 417. Карта распространения гекконов (Geckonidae).
(Из Gadow, с изменениями.)
О всех пресмыкающихся можно сказать, что они более, чем
все остальные классы позвоночных, убывают к полюсам, с одной
стороны, и по мере поднятия вверх над уровнем моря — с другой.
Все рептилии — любители тепла, а потому они представлены
гораздо большим числом форм и особей и крупнее в жарких стра-
нах, нежели в умеренных и холодных. Так, например, на острове
Ява 122 вида рептилий, па Суматре — 127, на Филиппинах — 204,
в Китае к северу от Янцзе — 25, в Германии — 12, в Манчжурии—6.
За полярный круг переходят лишь очень немногие виды. Наи-
большей высотой, на которую поднимаются рептилии в горах,
является высота в 5400 м, на которой в Тибете еще встречаются
круглоголовки (Phrynocephalus). По мере приближения к экватору
не только увеличивается число форм и особей, но рептилии ста-
новятся крупнее, красивее окрашены и более жизнедеятельны.
Влажность не играет в распространении рептилий той роли, какую
она играет в распространении амфибий; например, рептилии могут
существовать и в безводной пустыне, обходясь совершенно беа
воды.
54а
ние. 418. Карта распространения агам (Aga-
midae).
(Из Gadow, с изменениями )
Обратимся к рассмотрению географического распространения
в пределах отдельных отрядов. Начнем с ящериц. Вероятно, наи-
более древпим семейством из современных ящериц являются гек-
коны (Geckonidae). И
они являются почти
космополитами, отсут-
ствуя лишь в холодных
и умеренных областях.
Даже на океанических
островах, как, напри-
мер, Сандвичевы остро-
ва, мы находим гекко-
нов (рис. 417). Такое
широкое распростране-
ние возможно, помимо
истории происхождения
материков в прошлом,
благодаря выносливости
гекконов по отношению
к недостатку пищи и
благодаря возможности
распространения па пла-
вающих деревьях. Кос-
мополитично также се-
мейство сцинковых (Scin-
cidae), хотя опи и не
так широко распростра-
нены по островам океа-
на. Их главное распро-
странение в Австралии.
Агамы (Agamidae) (рис.
418), варапы (Varanidae)
(рис.419), ящерицы в со-
бственном смысле слова
(Lacertidae) (рис. 420) и
хамелеоны (Chamaeleon-
tidae) (рис. 421) ограни-
чены только Старым Све-
том. Наиболее широко
распространены первые
два семейства, встречаю-
щееся всюду в Старом
Свете, кроме Мадагаска-
ра и Новой Зеландиих
Lacertidae встречают-
ся в Палеарктической,
Индийской и Эфиоп-
Рис. 4АЭ. Карга распространения ва-
ранов (Varanidae).
(Из Gadow, с изменениями.)
Рис. 420. Карта распространения настоящих
ящериц (Lacertidae).
(Из Gadow, с изменениями.)
ской областях и также отсутствуют на Мадагаскаре. Хамелеоны
свойственны главным образом Африке и Мадагаскару, где живет
544
половина известных (50) видов, остальные — на африканском
материке. Один вид, именно обычный хамелеон, живет на севере
Африки и в Андалузии, два—в Южной Аравии и Палестине, один—
в Южной Индии и на Цейлоне. Но и па островах Индийского океана,
Сейшельских и св. Маврикия,
имеются свои виды хамелеона. Это,
быть может, указывает, что прежде
они существовали на той суше,
которая соединяла Южную Индию
и Южную Африку. Наоборот, се-
мейство игуан (Iguanidae) — спе-
циально американское, если по
считать двух родов на Мадагаска-
ре и одного на островах Фиджи и
островах Товарищества. В Аме-
рике, в Вест-Индии, Африки кроме
Мадагаскара и средиземноморских
стран, распространены двуходко-
вые (Amphisbaenidae) (рис. 4 22).
Это весьма странно при современ-
ном распределении материков,
Рис. 421. Карта распространения
хамелеонов (Chameleontidae).
(Из Gadow, с изменениями.)
если принять во впима-
особеппо
ние, что эти ящерицы живут в земле и ведут роющий образ жиз-
ни подобно дождевым червям.
Веретеппицы (Anguidae) встречаются в Северной и Южной
Америке, в Европе,
Африки и в Ипдии, за
Гангом.
Змеи, как и ящери-
цы, являются молодой
ветвью рептилий. Весь
отряд в целом космопо-
литичен, и Новая Зе-
ландия является един-
ственным большим ост-
ровом (кроме полярных),
не имеющим змей. Осно-
ву современного рас-
пространения. более
древних семейств надо
искать в миоценовом пе-
риоде. Наиболее древни-
ми семействами, еще со-
в средиземноморских частях Северной
Рис. 422. Карта распространения двуходко-
вых (Amphisbaenidae).
ХраНИВШИМИ СЛеДЫ зад- (Из Gadow, с изменениями.)
пих конечностей и таза,
являются роющие змеи семейства слепунов (Typhlopidae), щито-
хвостов (Uropeltidae) и других, а также удавы (Boidae). Поэтому
нет ничего удивительного, что эти группы имеют широкое распро-
странение, кроме щитохвостов, ограниченных Южпой Индией
и Цейлоном. Из распространенных почти космополитически во-
35 Кашкаров и Станчинский
545
круг тропиков Boidae одно подсемейство — питопы (Pythoninac) —
ограничено почти исключительно Старым Светом, включая и Австра-
лию; другое подсемейство — боа (Boinae) — характерно главным
образом для тропической Америки, родины анаконды (Eunectes
murinus). Относящийся к Boinae степной удав (Егух) живет
Рис. 423. Карта распространения гадюк (Vipe
rinae).
(Из Gadow, с изменениями.)
Рис. 424. Карта распространения гремучников (С го tai in ае).
(Из Gadow, с изменениями.)
в Северной Африке, Греции и юго-западной и Средней Азии. Colu-
bridae, ужовые, большей частью являются космополитами. Аспи-
ды (Elapinae) распространены в тропических и подтропических
частях Старого Света, кроме Австралии. Морские змеи (Hydro-
phinae) распространены от Персидского залива до Центральной
Америки. Гадюковые (Viperidae) (рис. 423) почти космополитичны,
отсутствуя на Мадагаскаре и в Австралии, причем настоящие га-
546
дюки (Viperinae) являются обитателями Старого Света, гремучники
(Crotalinae) представлены в тропической Азии, Северной и Юж-
ной Америке (рис. 424).
Все главные современные группы черепах и многие черепахи,
вымершие в прошедшие времена, встречались в тех же странах
вместе. Поэтому сделать какие-либо выводы общего характера из
Рис. 425. Географическое распространение бокошейных
черепах (Pleurodira).
(Из Gadow, с изменениями.)
Рис. 426. Географическое распространение скрытошейных черепах (Стур-
todira).
(Из Gadow.)
их распространения трудно. Кожистые черепахи (Derm ochelinae
широко распространены по всем тропическим морям, но всюду
редки. Из панцирных бокошейпые (Pleurodira) ограничены рас-
пространением в южном полушарии (рис. 425), причем семейство
Pelomedusidae свойственно Африке, Мадагаскару и Южной Аме-
рике, семейство же Chelydidae живет в 1Ожной Америке и Австра-
лии. Из скрытошейных (Cryptodira) (рис. 426) семейство Chelydri-
dae является американским, но в миоцепе было представлено
в Европе, наземные же черепахи (Testudinidae) космополитичны,
35* 547
не встречаясь только в Австралии. Что касается мягких черепах
(Trionychidae), то они обитают в Африке, тропической Азии и во-
сточной части Сев. Америки. Их нет в Южной Америке и Австра-
лии. Сем. Chelydidae ограничено Австралией и восточной частью
Южной Америки (рис. 427).
Crionychidde Chelydidae
Рис. 427. Географическое распространение Trionychidae и Chely-
didae.
(Из Gadow, с изменениями.)
Рис. 428. Географическое распространение крокодилов
(Crocodilia).
(Из Gadow.)
Географическое распространение крокодилов становится по-
пятным, если принять во внимание распространение ископаемых
форм. А именно и аллигаторы, и крокодилы, и гавиалы в третичную
эру сущестговали в Европе. Единственный вид аллигатора, живу-
щий в Китае в настоящее время, является последним свидетель-
ством о прежнем их кругоарктическом распространении. Теперь
крокодилы, взятые в целом, обитают между тропиками. Крокодилы
(Crocodilidae) собственно обитают в Палеотропической области
вместе с гавиалами (Gavialidae)—в Восточной подобласти, алли-
гаторы же (Alligatoridae) и кайманы и небольшое количество кроко-
548
дилов живут в Америке (рис. 428). Подкласс Rhynchocephalia
имеет очень узкое распространение, встречаясь только в Новой
Зеландии. В ископаемом состоянии известны почти всюду.
В распространении рептилий мы видим также подтверждение
тому, что до начала третичной эры материки Старого и Нового
Света составляли один материк. Иначе трудно понять распростра-
нение двуходковых (Amphisbenidae), хамелеонов, сухопутных ги-
гантских черепах и др. Отсутствие рептилий в северных частях
суши, кроме их холоднокровности, стоит в связи с тем, что в лед-
никовое время они были вытеснены с севера наступившим похоло-
данием.
Распространение сухопутных, вообще наземных и ископаемых
рептилий также говорит в пользу теории плавания континентов.
Лишь таким образом можно понять широкое, по всему почти свету
распространение динозавров, котилозавров, мезозавров и других
подклассов.
ЧАСТЬ ПЯТАЯ
ПТИЦЫ
Класс 6. Птицы (Aves)
I. ХАРАКТЕРИСТИКА КЛАССА ПТИЦ
Основными свойствами птиц, сделавшими их обширным классом
позвоночных животных, следует считать их теплокровность, со-
храняющий тепло тела покров из перьев, способность летать с по-
мощью крыльев, летательная поверхность которых образована
перьями, и способность свободно передвигаться по деревьям и земле
только с помощью специализированных задних конечностей.
Теплокровность уже в умеренном климате давала птицам гро-
мадные преимущества по сравнению с холоднокровными рептилиями,
впадавшими в депрессию и оцепепепие при попижениях темпера-
туры; полет давал возможность неизмеримо шире использовать
пространства по сравнению с нелетающими животными, а пре-
красно приспособленные к передвижению по земле и по деревьям
задние конечности давали громадные преимущества птицам по
сравнению с летавшими птерозаврами, пе обладавшими к тому
же, вероятно, теплокровностью.
Птицы составляют вместе с пресмыкающимися и млекопитаю-
щими группу амниот (Amniota), так как па ряду с другими
признаками (почки, сердце, улитка уха) в эмбриональном разви-
тии птиц появляются особые зародышевые оболочки — ампион и
аллантоис.
По своей организации птицы очень близки к пресмыкающимся
и могут быть объединены с ними в группу ящерообраз-
ных — Sauropsida — и противопоставлены млекопитающим: как
и у рептилий, кожа у них очень бедна железами, причем молочные
железы отсутствуют, и покрыта ороговелыми придатками — перья-
ми; череп соединяется с позвоночником одним непарным сочленов-
ным бугорком; пижняя челюсть сочленяется с черепом посредством
хорошо развитой квадратной кости; имеется клоака.
От рептилий и млекопитающих птицы отличаются тем, что тело
их покрыто перьями, передние конечности превращены в крылья
с летательной поверхностью из перьев, задние конечности хорошо
развиты и служат для передвижения по земле или деревьям; малая
берцовая кость редуцируется, прирастает к большой берцовой,
сюда же прирастают проксимальные косточки предплюсны, а пу-
тем срастания трех плюсневых костей с дистальными косточками
предплюсны образуется цевка. Передняя часть черепа вытянута
550
вперед в клюв, который у современных птиц лишен зубов и покрыт
роговым чехлом; крестец состоит из нескольких позвопков; сердце
с совершенной перегородкой желудочка, четырехкамерное, имеется
только одна правая дуга аорты; мочевого пузыря нет. Правый
яичник и яйцевод редуцированы. Температура тела постоянная;
самки несут яйца.
В общем птиц можно кратко определить как теплокров-
ных Sauropsida, тело которых покрыто
перьями и которые приспособлены к пере-
движению по воздуху с помощью передних
к oj3 ечностей, превратившихся в крылья
с летательной поверхностью из перьев,
тогда как задние конечности хорошо при-
способлены для передвижения по землей
деревьям.
п. СТРОЕНИЕ птиц
1. Покровы
Кожа птиц характеризуется топким эпидермисом и собственно
кожей (cutis), состоящей из плотного, богатого соединительно-
тканными волокнами наружного слоя — собственно кожи (corium)
и рыхлой подкожной клетчатки, в которой образуются жировые
отложения, у некоторых птиц, например, у водоплавающих, до-
стигающие большого развития.
Характерными кожными образованиями птиц являются перья.
Типичное, или контурное, перо (plumae) со-
стоит из ствола (rhachis) и опахала (pogonium), состоя-
щего и^ бородок первого (rami) и второго по-
рядка (radii) (рис. 429). Эти последние, сцепляясь между собой
тонкими крючочками (рис. 430), образуют прочную, эластичную
и в то же время очень легкую пластинку, которая оказывает весьма
сильное сопротивление воздуху. Нижняя часть ствола представляет
полый роговой цилиндр и называется очином (calamus).
Конец очина помещается в мешочке кожи, и на его основании
имеется отверстие, через которое во время развития в перо прони-
кают с сосочком кожи кровеносные сосуды, питающие перо. У раз-
витого пера в полости очина можно найти роговые колпачки, от-
мершие и ороговевшие остатки сосочка кожи.
Так устроены типичные перья птиц, придающие птице ее кон-
туры и называемые контурными. Это наиболее сложно
устроенные перья. На крыле и хвосте эти перья особенно сильно
развиты^ служат плоскостью опоры о воздух. В том случае, если
бородки пера длинны, мягки и не сцеплены друг с другом, а ствол
развит нормально, то оно называется пуховым пером
(semiplumae) (рис. 431, 4 и 5). Если же ствол короткий, а бородки
длинные и свободные, то получается настоящий пух (plu-
mulae). Длинный и тонкий ствол с зачаточными бородками или вовсе
551
Рис. 429. Строе-
ние пера птицы.
1 — очин (cala-
mus), 2—стержень
(rhachls), 3—по-
бочный стержень
(hyporhachis), 4—
опахало (pogoni-
um) (ориг.)
Рис. 430. Схематическое изобра-
жение строения бородок:
1 — первого порядка (ramus), 2 — вто-
рого порядка (radius). На последних
расположены крючочки (radiolus) (3).
(По Бючлп.)
Рис. 431. Различные видоизменения пера.
1 — эмбриональное перо, 2 —нитевидное перо, 3 — перо с сильно развитым
побочным стержнем (эму Dromaeus), 4 и 5 — пуховидные перья (ориг.)
552
без пих образует нитевидное перо (filoplumae) (рис. 395,2);
если такой ствол более короткий и толстый, получается щ е-
т и н к а.
У многих птиц контурные перья снабжены побочным стерж-
нем, отходящим от очина (hyporhachis). У эму и казуаров побоч-
ный стержень развит особенно сильно (рис. 395, 3).
Наконец,, есть еще одна категория перьев или пуха — п уд-
ре т к и (pulviplumae); бородки пера здесь крошатся по мере роста
и в виде пудры пересыпают перья, как это мы видим у цапель,
попугаев и у некоторых других птиц.
Рис. 432. Нтерилии (1) и аптерии (2) петуха. (По Нитцшу.)
Между всеми этими категориями перьев существуют много-
численные переходы.
Так называемое- эмбриональное перо, или э м-
бриональный пух, составляет первоначальное одеяние
вылупившихся птенцов neossoptile, или neoptile (рис. 431, 1).
Контурные перья за немногими исключениями (пингвины,
паламедеи, страусовые птицы) располагаются участками — так
называемыми птерилиями, между которыми остаются места,
лишеппые перьев, — аптерии (рис. 432). Так как у зароды-
шей страуса и киви имеются уже птерилии и аптерии, нужно ду-
мать, что предки пелетающих страусовых птиц имели нормальное
для всех птиц расположение перьев по птерилиям и лишь вторично
приобрели их равномерное расположение. Самое расположение
перьев по птерилиям можно поставить в связь со способностью
летать, ибо расположение перьев участками представляет удоб-
ство при сокращении мускулов во время полета.
Пух у разных групп птиц располагается неодинаково. Развитие
553
пуха связано с тем или иным образом жизни, и у водоплавающих
обычно он развит особенно сильно.
К кожным мешочкам крупных перьев прикрепляются гладкие
мышцы, которые приводят их в движение. Перья, располагаясь
определенным образом, и одевая тело птицы, придают ей харак-
терный контур. Для облегчения описания определенные участки
тела птиц имеют свои названия. Они даны на рис. 433.
Рис. 433. Номенклатура отдельных частей тела и оперения птицы.
1 — подклювье, 2 — надклювье, 3 — конек клюва, 4 — ноздри, 5 — уздеч-
ка, 6 —лоб, 7 — темя, 8 — затылок, 9 — шея, 10 — спина, 11 —большие
кроющие крыла, 12 — надхвостье, 13 — второстепенные маховые перья,
14 — первостепенные маховые, 15 — верхние кроющие хвоста, 16 — ру-
левые, 17 — нижние кроющие хвоста, 18 — кроющче маховых перьев,
19 — подхвостье, 20 — гузка, 21—плюсна, 22 — задний палец, 23 — го-
лень, 24 — бок брюха, 25 — бок груди, 26 — крылышко, 27—средние
кроющие крыла, 28 — плечевые, 29 — наружный палец, 30 — средний
палец, 31 — внутренний палец, 32 — брюхо, 33 —грудь, 34 — малые
кроющие крыла, 35 — кистевой сгиб крыла, 36 —воб, 37 — шея, 38 —бок
шеи, 39 —щеки, 40 —бока головы, 41 подбородок, (Ориг.)
Онтогенетическое развитие перьев указывает на их генетиче-
скую связь с чешуйчатыми образованиями рептилий. Подобно
этим последним зачаток пера представляет бугорок мезодермаль-
ных клеток кожи, который, будучи покрыт снаружи эпидермисом,
по мере роста направляется своей вершиной назад, тогда как осно-
вание бугорка постепенно спускается в кожу, образуя мешочек,
или влагалище пера. Позднее в эпидермисе образуются утолщения,
которые ороговевают и превращаются в бородки пера после того,
как чехлик (одевающий перо наружный слой эпидермиса) спадает.
Мезодермальный бугорок превращается в сосочек пера, обильно
снабженный кровеносными сосудами, питающими перо (рис. 434).
554
Развитие контурного пера отличается лишь сложностью строе-
ния утолщений роговой части, которые идут косо по отношению
в продольному главному утолщению—стержню. При расщепле-
нии этих утолщений опи превращаются в бородки обоих опахал
(рис. 434).
На генетическую связь перьев и чешуи указывает также факт
наличия роговых чешуй там, где у птиц отсутствуют перья. Так,
у громадного большинства птиц пальцы и большая часть плюсны
одеты роговыми щитками, или чешуйками; у некоторых групп
птиц роговыми щитками одета и нижняя часть голени.
Рис. 434. Развитие пуха и пера (схема).
1 — эпидермис, 2 —собственно ножа (corium), 3 — закладка лучей
пуха, 4 —сосочек (pulpa), 5—лучи пуха (radii), 6 —стержень кон-
турного пера, 7 — бородка первого порядка (ramus), 8 — бородка
второго порядка (radius). (Из Бючли.)
Последние фаланги пальцев одеты роговыми чехликами —
когтями, клюв тоже одет роговыми образованиями различной
формы, что связано с характером и способом добывания пищи.
Характерно для птиц отсутствие кожных желез. Единственная
кожная железа у птиц — копчиковая железа. Она помещается
пад хвостом у громадного большинства птиц (ее нет у бескилевых,
некоторых попугаев и некоторых голубей) и выделяет маслянистый
секрет, которым птица смазывает себе перья с помощью клюва.
Особенно развита копчиковая железа у водоплавающих птиц,
и обильно смазанные жирным ее секретом перья этих птиц не сма-
чиваются водой.
2. Скелет птиц
Скелет птиц по своему строению чрезвычайно близок к скелету
рептилий. Характерные особенности скелета птиц главным обра-
зом связаны с теми преобразованиями, которые вызваны были
приспособлением к полету.
555
Рис. 435. Скелет гуся.
1—второй шейный позвонон (epistro-
pheus), 2— акрокоракоидальный отросток
вороновой кости (proc, acrocoracoideus),
3 —ключица (clavicula), 4 —воронова
кость (coracoideum), 5 — клювовидные
отростки ребер (proc, uncinati), 6 — киль
грудной кости (carina), 7—чашечка (pa-
tella); 8 — коленный отросток большой
берцовой кости (proc. cnemialis,rotularis),
9 — сочленовная головна бедра с малой
берцовой костью (epicondylus fibularis),
10 — большая берцовая кость (tibia), 11 —
второй палец (digitus II), 12 —третий па-
лец (digitus III), 13 —четвертый палец
(digitus IV), 14—первый палец (digitus I)
15—плюсневая косточка первого паль-
ца (metatarsale I), 16 —цевка (tarsus me-
tatarsus), 17— fibulare и 18 —tibiale,
сросшиеся с большой берцовой в 19—tibio-
tarsus, 20—лобковая (pubis), 21—седа-
лищная (ischium), 22 —малая берцовая
(fibula), 23 —малый вертлуг (trochanter
minor), 24 — хвост (cauda), 25—копчикова
кость ( os coccygis), 26—сложный крестец
(ileosynsacrum), 27—большой вертлуг
(trochanter major), 28—лопатка (scapula),
29—плечевая (humerus), 30 — дельтовид-
ный гребень (crista deltoidea), 31 — локте-
вая (ulna), 32 —лучевая (radius), 33 —
третий палец (digitus III), 34—первая
(phal. II), 35 —вторая фаланга второго
пальца (phal. II), 36 — метакарпальные
третьего (met. Ill), и 37 — второго (met.
II),38—неправильно наз. ulnare=metacar-
pale V, 39 —первый палец (digit. I),
4 0—carpometacarpus, 41—metacar. I,
42 —radiale (=radiale+centrale), 43—пер-
вый шейный позвонок (atlas).
В отличие от других летаю-
щих позвоночных (летучих мы-
шей, ископаемых Pterosauria)
крыло птиц имеет несущую по-
верхность из перьев. Преобра-
зованию передней конечности
в крыло очевидно предшество-
вало освобождение передней ко-
нечности от опоры тела при пе-
редвижении по земле или по
деревьям. Уже предки птиц,
еще не умевшие летать, долж-
ны были иметь характерные
особенности двуногих животных
с коротким туловищем, в кото-
ром все внутренние органы бы-
ли передвинуты назад, вслед-
ствие чего положение тела, опи-
рающегося на задние конечно-
сти, становилось более устойчи-
вым. Этим объясняются адап-
тивные свойства своеобразно
измененных крестцовых позвон-
ков и таза (рис.*435).
Сами задние конечности,
являющиеся исключительной
опорой тела при передвижении
по деревьям и земле, постепенно
удлинялись и приобрели, в кон-
це концов, характерное строение
птичьей ноги. Кисть видоизме-
нилась, преобразовавшись в опо-
ру для самых важных при поле-
те перьев —маховых.
При такой специализации
задних и передних конечностей
основным хватательным органом
является рот. Лицевые части
скелета черепа вытягиваются
^вперед и одеваются роговым
чехлом, так что получается
клюв весьма разнообразной фор-
мы и строения в связи с раз-
нообразием принимаемой пти-
цами пищи. Полное слияние
большинства костей черепа
птиц придает особенную проч-
ность хватательному аппарату
птицы.
556
Туловище птиц представлено в скелете прочной костяной кор-
зинкой, дающей возможность сохранять устойчивость как при
опоре на крылья при полете, так и при передвижениях по земле.
Приспособление к полету выразилось у птиц также в ппевма-
тичпости костей: вместо костного мозга кости птиц выполнены в
большей или меньшей степени воздухоносными полостями. Особенно
это выражено у хорошо летающих птиц (буревестники); у нелетаю-
щих страусообразных' пневматичпы только некоторые кости
черепа.
В связи с указанными общими преобразованиями у птиц мы
наблюдаем ряд характерных особенностей в строении отдельных
частей скелета.
Позвоночник птиц состоит из различного числа позвонков
(от 39 до 63) и весьма своеобразно специализирован по отделам.
Шейный отдел в связи с хватательными функциями головы обла-
дает значительной длиной и чрезвычайно подвижен. Тела отно-
сительно длинных позвонков, число которых колеблется от 9 до 25,
сочленяются седловидными (вогнуто-выпуклыми) поверхностями,
образуя так называемые гетероцельпые позвонки, что
позволяет изгибать шею в разных направлениях, сохраняя прочное
соединение между позвонками. Вследствие срастания зачаточных
ребер с позвонками поперечные отростки оказываются продыря-
вленными: через отверстия в них проходят позвоночные артерии.
Первый позвонок — атлас (atlas) сочленяется с одним затылоч-
ным бугорком черепа, что дает возможность голове совершать
вращательпые движения. Эта способность к вращательным движе-
ниям у птиц увеличивается еще вследствие прирастапия тела атласа
ко второму позвонку — эпистрофею (epistropheus) в виде его
зубовидного отростка (processus odontoideus). Благодаря этому
атлас может вращаться, как па оси, на зубовидном отростке
эпистрофея. Задние шейные позвонки спабжепы свободными реб-
рами.
Спипные, или грудные, позвонки (в числе от 3 до 10) мепее по-
движны, иногда срастаются и спабжепы ребрами, доходящими до
грудной кости; ребра состоят из двух отделов, вполне окостенев-
ших, — грудного и позвоночного, находящихся под углом, на-
правленным назад; опи подвижно сочленены друг с другом, с по-
звонками и с грудной костью и несут па прилегающем к позвонкам
отделе особые отростки — клювовидные отростки (processi unci-
nati). Такое строение грудной корзины имеет большое значение
при дыхании: с помощью соответствующей мускулатуры угол, под
которым отделы ребер находятся друг к другу, может увеличи-
ваться, и грудина, отдаляясь от позвоночника, увеличивает этим
полость грудной корзины, что обусловливает наполнение воздухом
легких и воздушных мешков.
Следующие отделы позвоночника у птиц образуют сложный
крестец (synsacrum), состоящий из различного числа (от 10
до 22) позвонков, сросшихся между собой и с подвздошной костью.
Среди позвонков сложного крестца обращают на себя внимание
557
два позвонка, которые выделяются своими крестцовыми ребрами.
Они соответствуют крестцовым позвонкам современных рептилий
и потому носят название истинных крестцовых по-
звонков. Впереди них в сложном крестце лежат позвонки
поясничной области и даже иногда еще и грудные позвонки, а по-
зади — позвонки хвостовой области. Число свободных хвостовых
позвонков у настоящих птиц невелико (6—9), а последние хвостовые
позвонки (4—7) срастаются между собой в так называемый п и го-
ст и л ь, или копчик (pygostyl или os coccygis) — пластинку,
к которой прикрепляются рулевые перья, играющие большую роль
при полете.
Грудина у птиц состоит из длинной пластинки с возвышаю-
щимся вдоль длины высоким гребнем, или килем (crista
sterni, carina). Сильное его развитие, как и всей грудной кости,
стоит в связи с наличностью мощных грудных мускулов, приво-
дящих в движение крылья. У нелетающих птиц киль бывает недо-
развит или вовсе отсутствует, как то имеет место у большинства
«древних птиц» (Palaeognathae), выделяемых по этому признаку в
группу бескилевых птиц (Ratitae). 1
Череп птиц в связи с чрезвычайным развитием глаз, наиболее
важными органами чувств при полете, характеризуется большими
глазницами, отодвинувшими мозговую коробку назад и обусло-
вившими тонкую межглазничную перегородку, а образование
клюва как органа хватания привело к удлинению вперед лицевой
части черепа. Характерно, далее, полное срастание большей части
костей черепа у взрослых птиц, что тоже стоит в связи с развитием
клюва, требующим большой прочности от скелета, служащего
основанием для клюва.
Границы отдельных костей можпо видеть только у очень мо-
лодых птиц и птенцов. Вокруг затылочного отверстия располага-
ются обычные четыре кости (basioccipitale, две exoccipitale или occi-
pitale laterale с боков и supraoccipitale сверху). Имеется, как
и у рептилий, один сочленовный бугорок, сильно сдвинутый вниз
на основание черепа.
Три ушных кости (prooticum, opistoticum и epioticum) сра-
стаются между собой и с затылочными костями в одно целое.
В межглазничной области снизу у зародыша закладываются
основная и передняя клиновидные кости
(basisphenoideum и praesphenoideum), которые рано срастаются
в одну кость; к ним спереди и снизу прирастает покровная кость
парасфеноид (parasphenoideum), образующая спереди так
называемый клюв клиновидной кости (rostrum). Для системати-
ческих подразделений птиц имеет значение наличие или отсутствие
боковых отростков клиновидной кости — так называемых крыло-
видных отростков (processi ptqrygoidei или. basipterygoidei). По
бокам, образуя пижнюю часть передней степки мозговой коробки,
лежат крыло-клиновидные кости (alisphenoidea),
1 От слова ratis — бескилевая лодка.
558
а между глазницами—г лазничпо-клиновпдные (orbi-
tosphenoidea).
В обонятельной области имеется лежащая посредине обо-
нятельная кость (mesethmoideum). Снизу здесь налегает
покровная кость — сошник (vomer), образующийся путем
слияния двух костей в одну.
Сверху и с боков череп прикрывается покровным костями —
теменными (parietale), лобными (frontale) и чешуй-
чатыми (squamosum). Спереди лежат две п о с о в ы х ко-
сти (nasale), а по бокам перед глазницами — слезные (lacry-
Рис. 436. Схематизированный рисунок черепа птенца хищной
птицы (канюк малый, Buteo buteo vulpinus).
1 — затылочное отверстие (foramen magnum), 2 —основная затылочная
(basioccipitale), 3 — боковые затылочные (exoccipitale), 4 — верхняя за-
тылочная (supraoccipitale), 5 — слуховое отверстие (foramen auditivum),
6 —основная клиновидная (basisphenoideum), 7 — крылоклиновидная (ali-
sphenoideum), 8 — парасфеноид (parasphenoideum), 9 —средняя обоня-
тельная (mesethmoideum), 10 —сошник (vomer), 11 — теменная (parie-
tale;, 12 —лобная (frontale), 13 — слезная (Lacrymale), 14 — носовая (na-
sale), 15 — квадратная (quadratum), 16 — квадратно-скуловая (quadrato-
jugaie), 17 — крыловидная (pterygoideum), 18 —нёбная (palatinum), 19 —
челюстная (maxillare), 20 — межчелюстная (praemaxillare), 21—чешуй-
чатая (squamosum). (Из Бючли по препарату Станчинского.)
male). Строение носовых костей имеет большое систематическое
значение. Различают: голоринальные носовые кости
с закругленным краем, обращенным к носовому отверстию, и ш и-
зоринальпые с узким в виде расщепа краем.
Висцеральный скелет у птиц представлен более
или мепее сросшейся с черепом верхней частью челюстной дуги.
Так, спереди, образуя клюв, срастаются друг с другом и с черепом
межчелюстные кости (praemaxilla); позади них — не-
большие в е р х н е ч е л ю с т-n ы е кости (maxilla), от которых
отходят участвующие снизу в образовании нёба нёбные отростки
(processi palatini); еще дальше кзади лежат с каждой стороны
тоненькие палочкообразные квадратноскуловые кости
559
Рис. 437. Дромэогнатическое
нёбо эму.
1 —квадратная кость (quadratum),
2 — клюв клиновидной кости (ros-
trum parasphenoideum), 3 — кры-
ловидная (pterygoideum), 4 —нёб-
ная (palatinum), 5 —сошник (vo-
mer), 6 — нёбный отросток (pro-
cessus palatinus) челюстной кости
(maxillare), 7 — межчелюстная
кость (praemaxillare). Крыловид-
ные кости и сошник заштрихова-
ны. (Ориг).
вых, трогопов, козодоев и
(quadratojugalе). Последние причленяются к подвижной (стрепто-
стилия) квадратной кости (quadratum).
Нёбо птиц образовано тонкими палочкообразными косточка-
ми и подвижно. Квадратные кости каждой стороны сочле-
няются с крыловидным и (pterygoideum) костями, а эти
последние обычно сочленяются снёбными (palatinum). Местами
соединения они прилегают к клюву основной клиновидной кости
и во время движения челюстями скользят по его поверхности,
тогда как подталкиваемая сзади верх-
няя челюсть поднимает надклювье
вверх.
По взаимному расположению ко-
сточек, образующих нёбо, различают
следующие четыре типа черепов у
птиц:
1. Дромэогнатический
череп (рис. 437) характеризуется
широким сошпиком (5), который спе-
реди соединяется с нёбными отро-
стками челюстных костей (6), а сзади
с нёбными и крыловидными костями,
тогда как нёбные кости (palatinum)
срастаются с передними концами
крыловидных костей (pterygoideum).
Такой череп свойствен бескилевым
птицам (Ratitae) и тинаму (Tinami).
2. Шизогнатическое иё-
б о (рис. 438) отличается тем, что
нёбные, кости (palatinum) здесь сочле-
няются сзади с крыловидными, а спе-
реди соединяются с межчелюстными
(praemaxillae) и челюстными (maxil-
lae) костями. Нёбные отростки че-
\люстпых костей прилегают к нёбным
спереди и снутри, не соединяясь друг
с другом, тогда как сошник остается
свободным и оканчивается спереди
острием. Такой череп характерен для
очень многих птиц— гагароподобных,
куриных, журавлиных, пастушко-
колибри.
3. Деомогнатический череп (рис. 439) характе-
ризуется тем, что пёбные кости снизу сочленяются с крыловид-
ными, а спереди срастаются с челюстными и частично с их нёбными
отростками, которые всегда соединены или друг с другом своими
внутренними краями, или с костным основанием обонятельной
полости. Сошник, если он имеется, сзади соединен с основной
клиновидной костью и задними концами нёбных костей, спереди
же он соединяется с нёбными отростками челюстных костей. Такой
560
тип черепа свойствен отрядам сверхотрядов гусиных (Anseriformes),
дневных, хищных птиц (Falconiformes), аистообразных (Ciconiifor-
mes), попугаям (Psittaci) и сизоворонкообразиым (Coraciiformes).
~4. Эгитогнатическим (рис. 440) называют такое нёбо,
где нёбные кости сзади широкие, с выдающимися задними углами,
сочленяются с крыловидными костями, а спереди — прирастают
к челюстным костям. Сошник спереди оканчивается широким
свободным, большей частью вильчатым концом. Если смотреть
Рис. 438. Шизогнатическое
нёбо дрофы.
Обозначения, как на рис. 437.
Крыловидные ности и сошник
заштрихованы. (Ориг).
Рис. 440. Эгитогнатиче-
ское нёбо вороны.
Обозначения, как на рис.
437. Крыловидные кости и
сошник заштрихованы.
(Ориг.)
Рис. 439. Десмогна-
тическое нёбо утки.
Обозначения, как на
рис. 4 37. К рыло вид-
ные кости и сошнин
заштрихованы (Ориг).
на него снизу, то над ним своими внутренними концами выдаются,
но не соединяются друг с другом топкие нёбные отростки челюстных
костей. Этот тип характеризует воробьиных птиц (Passeriformes).
Нижняя челюсть птиц образуется с каждой сто-
роны из замещающей меккелев хрящ кости — сочленовной
(articulare) и покровных: зубной (dentale), пластин-
чатой (operculare), угловой (angulare), надугловой
(supraangulare) и венечной (complementare). Все эти око-
стенения очень рано сливаются друг с другом и с костями противо-
положной стороны, образуя у взрослой птицы одну нижнечелю-
стную кость.
Подъязычный аппарат, или подъязычная кость (оз
36 Кашкаров и Станчиисвий 561
hyoideum), образован остатками подъязычной и первой жаберной
дуг. Первая представлена непарной косточкой (copula I = tasi-
hyale), к которой спереди примыкает парная или непарная кость
(os entoglossum), соответствующая, очевидно, нижним кусочкам
гиоидной дуги (hypohyale). Сзади прирастают базальные члены
первой и второй жаберной дуг, тогда как боковые члены первой
жаберной дуги образуют длинные задние рога.
Из сказанного видно, что по своему общему строению череп
птиц очень сходен с черепом рептилий, особенно псевдозухий.
Исчезновение височных дуг и иное расположение черепных и висце-
ральных костей обусловлены с одной стороны сильным развитием
глазных яблок и увеличением объема головного мозга, с другой
стороны — образованием клюва.
Конечности претерпели в связи с приспособлением к полету осо-
бенно сильные видоизменения. Пояс передних конечностей в связи
с тем, что он становится у птиц опорой для крыла, приобретает
особенную прочность и своеобразное устройство (рис. 399).
Он образован тремя костями, сходящимися своими прокси-
мальными концами, где причленяется плечевая кость крыла. Ло-
патка (scapula), соответствующая таковой рептилий, становится
длинной, направленной назад параллельно позвоночнику сабле-
видной костью, лежащей на ребрах, и у хороших летунов заходит
назад до последних ребер; спереди она снабжена у большинства
птиц акромиальным отростком (acromion). Вороновы кости, или
коракоиды (coracoideum), очень хорошо развиты и плотно при-
членяются к грудине в углублениях по обеим сторонам киля.
Иногда они соприкасаются своими внутренними концами друг
с другом и даже срастаются (у фрегатов и гоацина) или налегают
друг на друга (цапли и бананоеды). На внутренней стороне верхних
отделов коракоидов имеются прокоракоидные отростки (processus
procoracoidalis), гомологичные прокоракоидам рептилий. У ки-
левых птиц верхние концы коракоид образуют акрокоракоидаль-
ные отростки (processus acrocoracoideus).
Ключицы (clavicula) срастаются друг с другом, образуя «ви-
лочку» (furcula), которая придает не только прочность, но и не-
обходимую упругость поясу передних конечностей.
У некоторых птиц, в связи с потерей способности летать клю-
чицы редуцируются (казуары, совиный попугай и др.) или даже
вовсе исчезают (киви, страусы, нанду и пастушковые куропатки —
Mesites).
В самой конечности происходят важные перемены в кисти
в связи с превращением ее в крыло. Плечо такой же длины, как
и предплечье, сочленяется с помощью элипсовидной поверхности
с проксимальными концами лопатки и коракоидов, которые обра-
зуют вместе сочленовную ямку (fossa glenoidalis).
Проксимальный конец плеча сильно расширен и снабжен двумя
буграми и сверху гребнем для прикрепления мускулов, поднимаю-
щих и опускающих, крыло (crista deltoidea).
Около среднего бугорка располагается ямка, на дне которой
562
имеется одно или несколько отверстий, через которые в полость
плеча проходят воздухоносные пути.
Лучевая и локтевая кость лежат в плоскости крыла и, так как
к последней, расположенной внутрь, прикрепляются маховые перья,
она более развита, чем лучевая.
Отдел запястья редуцируется до двух косточек — ulnare (=meta-
carpale V) и radiale (radiale + centrale), дистальные косточки
запястья трех сохранившихся пальцев прирастают к пястным;
пястные кости второго и третьего пальцев срастаются друг с другом
концами, а короткая пястная кость первого пальца прирастает по
всей длине к таковой же второго пальца. Образовавшаяся одна слож-
ная костьбудет carpometacarpus. Число фаланг, для первого пальца—
одна или две, для второго — две или три и для третьего — одна.
У зародыша еще видпо расположение элементов запястья в два
ряда. В проксимальном ряду закладываются следующие элементы
этого ряда: radiale и centrale radiale, которые сливаются друг с
другом, centrale ulnare и ulnare. В следующем ряду видна заклад-
ка четырех карпальных элементов, слившихся вместе, которые
затем прирастают к метакарпальным соответствующих паль-
цев. Метакарпальная кость пятого пальца закладывается само-
стоятельно и образует отдельную косточку, неправильно называе-
мую ulnare.
Передние конечности ограничены в движении: части их могут
двигаться только в одном направлении, именно в плоскости крыла,
складывая и расправляя крыло. Благодаря этому при взмахе
крыло двигается все целиком и обладает при этом большой устой-
чивостью. При расправлении крыло никогда не может быть вы-
тянуто совершенно, и его части остаются соединенными всегда
под некоторым углом. Это достигается при помощи особых эла-
стичных перепонок (patagium) между грудной клеткой и плечом
и между плечом и предплечьем. Последнее к тому же обладает
на наружном крае особым эластическим сухожилием мускула,
который короче, чем сумма длины плеча и предплечья. Это сухо-
жилие прикрепляется, перегнувшись через сочленовную головку
плеча, к основанию лучевой и локтевой кости и, натягиваясь при
разгибании предплечья, пассивно разгибает и выпрямляет кисть,
что тоже играет важную роль при полете.
В поясе задних конечностей в связи с опорой тела при ходьбе
и лазании только лишь на задние конечности произошли следующие
изменения. Путем значительного перемещения тяжелых внутрен-
ностей назад за точку прикрепления ног к тазу при наклонном
положении туловища птицы достигается большее равновесие между
длинной передней и короткой задней частью тела.
Такое отодвигание назад внутренностей имело следствием раз-
витие задней части седалищной кости, сильное удлинение под-
вздошной кости и направление лобковой кости не вперед от верт-
лужной ямы, а назад. Необходимость поддерживать тяжелые
Внутренности, свешивающиеся вниз спереди и сзади от точки опоры
на ноги, требовало особенной прочности горизонтально расположен-
36* 563
пого позвоночника. Эта прочность позвоночника была достигнута,
во-первых, срастанием крестцовых, поясничных и грудных позвон-
ков; во-вторых, срастанием с первыми седалищной и подвздошной
костей, в результате чего образуется одна прочная позвоночно-
тазовая кость в виде вогнутой длинной пластины, па которой под-
вешены внутренности. Сходное преобразование таза с некоторыми
отличиями мы видим и у двуногих динозавров из группы орнито-
по д о в (Orni thopoda).
Высокой специализации достигают у птиц задние конечности,
эволюция которых шла по пути приспособления к древесному
образу жизни. В первую очередь в связи с древесным образом жизни
преобразовалась ступня, где образовались длинные пальцы, причем
первый палец стал противополагаться остальным, что позволяло
птицам обхватывать ветки. Как лишний, пятый палец редуциро-
вался. Передвижение прыжками и лазанье по ветвям деревьев
вызвало дальнейшие приспособления в ногах.
Сильного развития достигает большая берцовая кость (tibia),
в то время как малая берцовая кость (fibula) становится зачаточ-
ной и прирастает к большой берцовой; а так как проксимальные
косточки предплюсны tibiale и fibulare срастаются между собой
и прирастают к большой берцовой кости, то последняя увели-
чивается еще больше, так что в отличие от tibia других классов
их называют здесь ti bi о tarsus. Дистальные части предплюсны
редуцируются в числе, срастаются и прирастают к плюсневым
костям, которые сливаются в длинную косточку, образуя цевку
(tarsometatarsus). Плюсневая косточка первого пальца, если он не
редуцирован, сохраняется и причленяется с задней стороны цевки.
Обычно имеются четыре пальца. Первый противополагается осталь-
ным трем; часто число пальцев редуцируется до трех, а у афри-
канского страуса — до двух. Первый палец состоит из двух фа-
ланг, второй — из трех, третий — из четырех, а четвертый — из
пяти фаланг. Этот последний нередко вместе с первым противо-
полагается двум остальным.
3. Мышечная система
В связи со сложностью выполняемых движений при полете,
при передвижениях с помощью ног и при работах клювом у птиц
наблюдается усложнение соответствующих отделов мышечной
системы.
На голове наибольшего развития достигают мускулы, откры-
вающие и закрывающие клюв, т. е. опускающие и поднимающие
подклювье (нижнюю челюсть). Открывают клюв мышцы (в числе
2—3), опускающие нижпюю челюсть (m. depressores mandibulae,
или m. digastricus), прикрепляющиеся позади точки сочленения
ее с квадратной. Этими же мускулами поднимается кверху над-
клювье, так как квадратная кость при этом передает давление
вперед на крыловидные и нёбные, на скуловые и верхнечелюстные
кости.
564
Мышцы, поднимающие подклювье, т. е. приводящие нижнюю
челюсть (m. adductores mandibulae), у птиц распадаются на ле-
жащие глубже височные мускулы (в числе до 4 — m. temporales)
и на лежащие снаружи жевательные мускулы (m. masseteres);
с внутренней стороны лежат крыловидные мышцы (m. pterygoidei).
Хорошо развита у птиц мускулатура подъязычного аппарата.
Одни мускулы втягивают язык внутрь (m. stylohyoideus, т. thy-
reohyoideus, т. mylohyoides posterior, т. ceratoglossus), другие
выдвигают язык вперед (т. geniohyoideus, т. genioglossus); при
неодновременном сокращении этих мускулов по обеим сторонам
эти мускулы в разной комбинации их сокращений могут обусловли-
вать очень сложные движения языком, как то мы наблюдаем,
например, у дятлов и попугаев.
Мускулатура шеи имеет у птиц чрезвычайное развитие, что
при подвижности позвонков и длине шейного отдела стоит в свя-
зи со сложностью движений, выполняемых головой. Помимо мышц,
соединяющих следующие друг за другом позвонки шеи (ш.
interspinales, m. intertransversarii и transversospinales), и мышц,
идущих вдоль всей спины до головы (m. iliocostales и m. longissi-
mus dorsi), обычно весьма усложненных, имеется еще ряд систем
мышц, выпрямляющих (система m. spinalis), сгибающих (m. longus
colli anticus), поворачивающих шею и голову (m. coinplexus, ш.
rectus capitis posticus, major и minor) и др.
Хорошо развита у птиц мускулатура ребер и живота в связи
с характером дыхательных движений: одни из них увеличивают
объем грудной клетки (mm. levatores costarum и mm. intercostales),
другие уменьшают (m. obliquus abdominis). Серия мускулов живота
играет роль брюшного пресса (m. transversoabdominalis и т. rectus
abdominis).
Довольно сложна и хорошо развита мускулатура хвоста, где
имеются мускулы, поднимающие и поворачивающие хвост (m. le-
vatores coccygis), опускающие хвост (m. depressores coccygis) и рас-
правляющие или складывающие рулевые перья (т. pubi-coccygeus).
Однако более всего развиты у птиц мышцы конечностей. Мышцы
крыла можно разделить на несколько групп. Во-первых, имеются
мускулы, приводящие в движение пояс передних конечностей:
лопатку, коракоиды и ключицу, или — правильнее — удерживаю-
щие пояс конечностей от движения при полете. Из них одни тянут
лопатку к позвоночному столбу (mm. rhomboideus), другие идут
от ребер к лопатке (mm. serratores), оттягивая лопатку в противо-
положном направлении, третьи идут от передней части грудины
к коракоиду (m. sterno-coracoideus), оттягивая его вниз и назад.
Вторая группа мышц приводит в движение плечевую кость. Здесь
мы имеем группу мышц, прикрепляющихся к плечу сверху: ши-
рокий спинной мускул (m. latissimus dorsi), который идет от ости-
стых отростков спинных позвонков к спипной стороне проксималь-
ной части плеча; он отводит плечо назад. Дельтовидные мускулы
(ш. deltoideus major и m. deltoideus minor) отходят от пояса пе-
редней конечности к плечу — двигают крыло вверх и вперед..
56&
Имеется ряд других мышц, идущих от пояса передней конечности
к плечу, которые модифицируют движения при поднимании крыла
вверх. Исключительного развития достигают у птиц мышцы, опу-
скающие крыло. Здесь самыми большими среди мышц птиц явля-
ются грудные мышцы (m. pectorales). Они лежат на грудине, при-
крепляясь к ней и к ее килю. При полете они опускают крыло
книзу. Являясь мышцами, от деятельности которых зависит бы-
строта полета, они естественно достигают наибольшего своего раз-
вития у хороших летунов, составляя до половины веса всей птицы
(в среднем веса тела).
Предплечье приводится в движение девятью мускулами, причем
одни из них, например, трехглавая мышца (m. triceps), разгибают,
а другие, например, двуглавая мышца (m. biceps), сгибают крыло.
Имеются мускулы, сгибающие (flexores) и разгибающие (extensores)
кисть.
Также своеобразна мышца, начинающаяся от плеча, прикре-
пляющаяся к основанию маховых перьев (mm. rectores remigum),
поворачивающая их при полете.
Что касается задних конечностей, то там насчитывается до
35 мышц. На голени располагается главный разгибатель пальцев
(т. extensor digitorum communis) и несколько сгибателей пальцев
(mm. flexores perforantes digitorum profundi и другие); мелкие
сгибатели и разгибатели пальцев и приводящие мышцы пальцев
расположены на цевке. У некоторых птиц имеется особый тонкий
бедренный мускул (m. ambiens), который идет от лобковой кости
и снабжен длинным сухожилием. Это последнее проходит в борозде
через надколенную чашечку, частично прикрепляясь к передней
поверхности голени; частично этот мускул соединен с сухожилием
глубокого сгибателя пальцев (m. flexor digitorum profundus);
этот последний соединен с голенью при помощи связок, так что
он не может соскочить и, пройдя через пяточный сустав, распадается
по числу пальцев на отдельные сухожилия. При сгибании ноги
в колене происходит натяжение всего аппарата, и пальцы сгиба-
ются сами собой. Хотя это приспособление чрезвычайно важно
для садящихся на деревья птиц, особенно во время сна, однако
у таких типичных древесных обитателей, какими являются во-
робьиные, m. ambiens отсутствует.
4. Органы пищеварения
Пищеварительный канал птиц характеризуется прежде всего
особенностями ротовой полости,' которая является здесь полостью
клюва. Она выстлана во внутренней части слизистой оболочкой,
а спереди одета роговым чехлом. Форма клюва соответствует ха-
рактеру принимаемой птицей пищи и весьма разнообразна. Длинное
щелевидное углубление на нёбе ведет к хоанам, другое отверстие
у корня языка — в трахею. Зубов у ныне живущих форм нет, но
у ископаемых форм — у юрских первоптиц (Archaeornithes) и у ме-
ловых — ихтиоринра (Ichthyornis) и гесперорниса (Hesperornis) —
566
рот был усажен зубами. Язык птиц вариирует в форме и строении
в зависимости от пищи. По большей части он жесткий и заострен-
ный с ороговевшей кожей на переднем конце; у дятлов язык уса-
жен острыми щетинками и может далеко выбрасываться вперед
(рис. 441), у попугаев, наоборот, язык толстый и мягкий, у страу-
сов он рудиментарен. Слюнные железы у птиц развиты весьма
различно и могут даже отсутствовать (козодои — Caprimulgi);
у стрижей и дятлов, наоборот, они хорошо развиты. Обычно —
это железы нёба и подчелюстные. Пищевод у многих птиц
имеет расширение — зоб, который у хищных, куриных обра-
зует боковой карман, а у голубей раздвоен и в период выкармли-
Рис. 441. Схема аппарата для выдвигания явыка
у дятла.
(Из Хильцхеймера.)
вания птенцов выделяет жирное беловатое вещество (молочко),
которое отрыгивается г. рот птенцам. Пищевод ведет в желези-
стый желудок (proventriculus), богатый пищеварительными
железами, а этот последний переходит в мускульный же-
лудок (ventriculus), который у птиц, питающихся раститель-
ной пищей, в особенности у тех, которые питаются зернами (кури-
ные, голуби), бывает очень хорошо развит, снабжен сильными
мускулами и выстлан внутри твердой оболочкой, кутикулой,
получающейся в результате затвердения секрета особых желез
желудка. Мускульный желудок служит для перетирания пищи
с помощью заглотанных камешков и песка. У плотоядных птиц
мускульный желудок, наоборот, слабо развит (хищные птицы
Accipitres). Выход из желудка (pylorus) помещается рядом с входом
в пего и у некоторых птиц (гагары, цапли, веслоногие) ведет в обо-
собившийся пилорический желудок.
Средний отдел кишечника: тонкая кишка у птиц длин-
ная, так что образует сложные петли, характеризующие отдельные
систематические группы. Толстая кишка короткая, ясно обособлена
от тонкой. На границе между ними у громадного большинства
дтиц имеются (отсутствуют у зимородка, удода, страуса, некоторых
567
попугаев и др.) большей частью два (у цапель — один) различно
развитых слепых отростка — слепые кишки (coecum).
Толстая кишка открывается в клоаку, на спинной стороне ко-
торой у многих птиц имеется
Рис. 442. Кишечный канал голубя.
1 — пищевод, 2 — яоб, 3 — железистый
желудок, 4 мускульный желудок,
5 — двенадцатиперстная кишка, 6 — пе-
чень, 7 — печеночные протоки, 8 — под-
желудочная железа, 9 — выводные про-
токи поджелудочной железы, 10 —тон-
кая кишка, 11—две слепых кишки,
12 — клоака. (Из Штреземана.)
большой длиной, проходит в
численные анастомозирующие
парабронхи, от которых отходят тончайшие воздушные
ветвящиеся мешочки. Вокруг последних разветвляются капил-
слепой мешок — так называемая
фабрициева сумка (bursa
Fabrici i). Клоака запирается
мускульным кольцом (sphincter
cloacae).
Печень птиц двулопастная,
желчный пузырь (vesica
fellea) отсутствует только у
некоторых птиц (голуби, стра-
усы, нанду, многие попугаи).
Печеночный проток (ductus he-
paticus) берет начало в обеих
лопастях, тогда как печеночно-
желчный проток (ductus hepa-
tico-cysticus) начинается в пер-
вой доле печени и ведет в желч-
ный пузырь; последний одна-
ко иногда отсутствует (страус,
нанду, голуби, попугаи, ко-
либри); пузырный проток (du-
ctus cysticus) впадает обыкновен-
но в двенадцатиперстную кишку
отдельно от печеночного про-
тока, не образуя общего желч-
ного протока (ductus choledo-
chus).
Поджелудочная железа (pan-
creas) лежит в петле двенад-
цатиперстной кишки и имеет
два протока.
5. Органы- дыхания птиц
Весьма интересно устроены
у птиц, опять-таки в связи , с
летанием, органы дыхания.
Воздух через носовые отвер-
стия, хоаны и через трахею
(trachea), которая у всех птиц
отличается благодаря длине шеи
бронхи, затем оттуда — в много-
между собой тонкие каналы —
ляры кровеносных сосудов (рис. 443). Легкие представляют, таким
образом, губчатую массу с очень обширной площадью воздушных
568
капилляров. Часть бронхов продолжается за пределами легких
и переходит в так называемые воздушные мешки,
которые располагаются под кожей, между внутренностными орга-
Рис. 443. Два парабронха гуся, в левом воздушные
полости инъицированы.
(Из Хильцхеймера.)
нами, мускулами и заходят даже в полости трубчатых костей
(рис. 444). Отчасти легочные мешки содействуют сохранению тепла
птицы и уменьшают ее удельный вес. Но главная их роль — при
дыхании. Воздух наполняет возду-
шные мешки, пройдя через легкие,
где происходит окисление гемогло-
бина: из воздушных мешков воздух
выходит вновь через легкие, чем
обусловливают более полное исполь-
зование кислорода. .
Полнота окислительных процессов
тела достигается также большим
совершенством кровеносной системы,
а именно полным отделением арте-
риальной крови от венозной. Тем-
пература тела птиц благодаря этим
приспособлениям выше температуры
тела млекопитающих.
Характерно для птиц наличие
двух гортаней: верхней г о р-
т а и и (larynx), устроенной так же,
как у рептилий, в основном из
Рис. 444. Воздушные мешки
голубя (изображены черным
цветом).
перстневидного хряща <Из Штреземана.)
(cartilago cricoidea) и двух черпало-
видных хрящей (cartilago arytenoidea), и нижней гортани
(syrinx), лйкащей в месте разветвления трахеи на два бронха.
Верхняя гортань не имеет голосовых связок, и голосовым аппа-
ратом слуясит только сиринкс.
6. Кровеносная система
В связи с полетом и теплокровностью, требующими большой
траты энергии, у птиц кровеносная система устроена более совер-
шенно, чем у рептилий
(рис. 445).
Сердце птиц четы-
рехкамерное, с полной
перегородкой между желу-
дочками; оно относительно
очень велико, причем пра-
вый желудочек имеет по
сравнению с левым более
тонкие стенки и охватывает
этот последний. Венозный
синус, так же как и у
рептилий, входит в состав
правого предсердия. Пра-
вый атриовентрикулярный
перепончатый клапан, ха-
рактерный для рептилий,
редуцируется и заменяется
здесь мускульным клапа-
ном, развившимся по срав-
нению с крокодилами, у
которых он впервые отме-
чается. Перепончатый ле-
Рис. 445. Сердце лебедя с венами (темные)
ц артериями (светлые); венозная часть
сердца — темная, артериальная—светлая.
1—яремные вены (v. jugularis), 2 —плечевые
вепы (V. brachialis), 3 —верхняя полая вена
правая и 4—левая (v. cava superior), 5 —
правое предсердие, 6 — правый желудочек, 7 —
левая легочная артерия (a. pulmonalis), 8 —
дуга аорты (arcus aortae), 9 — безыменные арте-
рии (a. anonymae), 10 —сонная артерия (а. ca-
rotis), 11 — пищеводная артерия (a. oesophagea),
12 — позвоночная артерия (a. vertebralis), 13 —
подключичная артерия (a. sublavia), 14—плече-
вая артерия (a. brachialis), 15 — нисходящая
аорта (aorta desecendens), 16 —кишечная арте-
рия (a. coeliaca), 17 — брыжеечная артерия (а.
mesenteriaca). (Из Штреземана по Гадову.)
вый атриовентрикулярный
клапан трехстворчатый. От
правого желудочка отхо-
дит общая легочная
артерия (a. pulmona-
lis), которая идет налево и
делится на правую и левую
артерии. Левая дуга
аорты исчезает
совершенно, и вся
артериальная кровь напра-
вляется по правой дуге.
У большинства птиц от
дуги аорты начинаются две безыменных артерии (а.
anonymae), которые делятся затем на сонную артерию
(a. carotis) и большую подключичную (a. subclavia);
последняя снабжает кровью мощные грудные мышцы. Однако
есть птицы, у которых не две, а только одна сонная артерия (а.
carotis primaria), идущая обыкновенно с левой стороны (Passeres,
многие Coraciiformes, Rhea и др., только у фламинго — Phoenicop-
terus, — она идет с правой стороны).
От нисходящей аорты (a. descendens) далее кзади отходят
$70
кишечная (a. coeliaca), брыжеечная (a. mesenterica), почечная
(a. renalis), седалищные (a. ischiadicae) и более мелкие артерии —
межреберные, крестцовые и др.
Весьма характерно строение венозной системы птиц.
Кровь поступает в правое предсердие из передней части тела по
правой и левой верхним полым венам (v. cava supe-
rior), собираясь в них по яремным венам (v. jugulares)
из головы, по подключичным венам (v. subclavia)
из крыльев и грудных мышц и по позвоночным венам
со спины (v. vertebrales). Последние представляют, повидимому,
остатки кювьеровых протоков.
Из задней половины тела кровь поступает в нижнюю
полую вену (v. cava inferior), которая непосредственно вли-
вается в правое предсердие. Она образуется путем слияния двух
подвздошных вен (v. iliacac), которые в свою очередь возникают
с каждой стороны от слияния бедренной вены, несущей кровь от
ноги (v. femoralis), с проходящей через почку почти без ответвлений
в нее веной (v. hypogastrica), образовавшейся путем разветвления
хвостовой вены (v. caudalis). Воротная система почек,
таким образом, сохраняет у птиц лишь следы своего прежнего
существования. В месте разветвления хвостовой вены от нее отхо-
дит третья — копчико-брыжеечная вена (v. coccygo-
mesenterica), которая вместе с рядом других вен, идущих от вну-
тренностей брюха, образует воротную вену печени (v.
postae hepatis ). Из печени же кровь несколькими печеноч-
ными венами (v. hepaticae) вливается в нижнюю полую вену
вместе с брюшной веной (v. abdominalis). Из легких кровь по двум
сливающимся в общий ствол легочным венам (v. pulmo-
nales) вливается в левое предсердие.
Птицы обладают лимфатической системой сосудов и же-
лез. Последние лежат на нижней части шеи и в грудной полости. Два
грудных лимфатических сосуда впадают в верхнюю полую вену,
другие — в седалищные и другие вены хвостовой области. У не-
которых птиц (гуси, лебеди, страусы, казуары и некоторые другие)
найдено в хвостовой области лимфатическое сердце, которое гонит
лимфу в вены почек.
Из желез внутренней секреции необходимо отметить гипофизу
(hypophysis), надпочечники (glandulae suprarenales), ле-
жащие у переднего края почек, разную степень развития селе-
зенки (lien) (у страусовых имеются даже добавочные селезенки),
парную щитовидную железу (gl. thyreoidea), лежащую
близ сердца и начала бронхов, и зобную железу (gl.
thymus), хорошо различимую у молодых птиц.
7. Мочеполовая система птиц
Почки у птиц, как у всех амниот, являются по своему
происхождению вторичными почками (metanephros). Они
лежат в области сложного крестца, плотно прилегая к тазу,
57<
и состоят обычно в каждой половине из трех долей. От почек отхо-
дят мочеточники, открывающиеся на спинной стороне клоаки.
Мочевого пузыря у птиц не бывает.
Половой аппарат самцов (рис. 446) представлен у птиц
двумя семенниками (testiculi) в виде овальных белых
в
Рис. 446. Мочеполовой
аппарат самца голубя.
1 — прямая кишка, 2, 2а,
2в — передняя, средняя и
задняя лопасти правой поч-
ки, 3—мочеточник, 4 —
семенник, 5—семяпроводы,
6—отверстие мочеточника в
клоаку, 7—отверстие семя-
провода в клоаку. (По
Резелеру и Латпрехту из
Ше и др.)
Рис. 447. Половой аппарат
курицы.
1т—яичник, 2 —пузырьки с
растущими в них яйцами,
3 —лопнувший пузырек, 4 —
воронка яйцевода, 5— яйце-
вод, 6 —мезентерий, его под-
держивающий, 7—задняя часть
яйцевода с яйцом (вскрыта),
8 — отверстие яйцевода в кло-
аку, 9—отверстия мочеточни-
ков в клоаку. (По Тихоми-
рову.)
тел, лежащих над передней долей почки у ее переднего края, и двумя
семяпроводами., открывающимися в клоаку рядом с мочеточниками.
У страусовых и у гусиных (Anseres) имеется совокупитель-
ный аппарат (penis) — толстый выступ на брюшной стен-
ке клоаки, выпячивающийся из клоаки во время совокупления.
У самок (рис. 447) имеются лишь левый яичник (ova-
rium) и левый яйцевод (только у некоторых хищных птиц встре-
чается рудиментарный правый яичник). Лежит яичник на краю
верхней доли левой почки. В период, когда птица несется, он до-
572
стигает громадного развития, в прочее же время он сокращается
до небольшой продолговатой пузырчатой пластинки желтоватого
цвета. Яйцевод вблизи яичника открывается в полость тела широкой
воронкой; в его переднем отделе желток яйца окружается белком
(собственно яйцевод-—oviductus), далее в отделе, называемом
маткой (uterus), яйцо одевается скорлупой.
Оплодотворение у птиц внутреннее и происходит в самом верх-
нем отделе яйцевода. В это время яйцо, освободившееся из фолли-
Рис. 448. Схематическое изобра-
жение желтка зрелого куриного
яйца из яйцевода.
1 —зародышевый пузырек (ядро), 2 —
образовательный желток, 3—«белый*
питательный желток, 4 — желтый
питательный желток, 5 — латебра.
Рис. 449. Схематическое изображе-
ние развития зародышевых оболочек
куриного зародыша.
1—зародыш, 2—-амнион, 3 —серозная
оболочка, 4—желточный мешок, 5—жел-
ток, 6 — аллантоис, 7 —эктодерма, 8—
мезодерма, 9 — энтодерма.
кула яичника и попавшее в воронку яйцевода, представлено желт-
ком с «зародышевым пузырьком» и подстилающим его «образова-
тельным желтком», т. е. ядро с окружающей его протоплазмой
с малым количеством питательного желтка. Последний распреде-
ляется в желтке неравномерно, образуя две зоны: «белый желток»
по периферии, который подстилал «образовательный желток»,
опускается к центру яйца, образуя так называемую латебру
(latebra), и «желтый желток», расположенный внутри кругом
спускающейся в него латебры (рис. 448). Снаружи яйцо одето
клеточной оболочкой — «желточной оболочкой» — (membrana
vitellina).
К моменту выхода из фолликула яйцо уже образовало первое
направляющее тельце, тогда как второе направляющее тельце вы-
деляется по входе яйца в яйцевод. Этим заканчивается «созревание»
яйца и оно готово к оплодотворению. Так как яйца птиц «гетеро-
зиготны», т. е. незрелое яйцо — яйцеклетка (оогонии и ооциты
первого порядка) содержат в себе лишь по одной половой Х-хромо-
соме, то во время первого созревания, сущность которого заклю-
чается в редукции хромосом, образуются яйцеклетки (ооциты вто-
рого порядка) двух родов: одни содержат Х-хромосому, другие ее
573
не содержат. Наличием или отсутствием Х-хромосомы, как извест-
но, определяется пол образующегося из яйца организма, так как
у птиц самцы гомозиготны, т. е. содержат во всех сперматозоидах
половые Х-хромосомы.
Сперматозоиды птиц у большей части отрядов птиц сходны с та-
ковыми рептилий. У отряда воробьиных, Passeres, они, однако,
существенно отличаются более сложным строением и длиной (длина
хвостовой части спермин зяблика достигает 259 pt).
После оплодотворения происходит дробление яйца, а затем
формирование зародыша и образование зародышевых оболочек —
амниона, серозной оболочки и аллантоиса (рис. 449).
S. Нервная система и органы чувств
В связи с большим совершенством органов чувств, сложным
поведением и развитием многочисленных инстинктов у птиц зна-
чительно усложняется строение головного мозга. Строение же
Рис. 450. Головной мозг утки сбоку.
1 — полушария переднего мозга, 2 — височные
лопасти, 3 — обонятельные луковицы, 4 — про-
межуточный мозг, 5—эпифиза, 6 —гипофиза,
7 —зрительные доли среднего мозга, 8—моз-
жечок, 9 —боковые выступы (flocculi) моз-
жечка. I—XII головные нервы. (По Плате.)
спинного мозга мало чем
отличается от такового
рептилий.
Так же как и у репти-
лий, у птиц имеются утол-
щения спинного мозга в
области нервов плечевого
и поясничного сплетений.
При этом в сложном
крестце центральный канал
расширяется настолько,
что образует ромбическую
полость (sinus rhomboi-
deus). Подобное же расши-
рение крестцовой области
спинного мозга имелось у
некоторых ископаемых реп-
тилий (Dinosauria).
Головной мозг (рис. 450, 451) птиц характеризуется сильным
развитием полушарий мозга, что происходит, однако, главным
образом благодаря разрастанию полосатых тел (corpora
striata), а не крыши (pallium), как это имеет место у млеко-
питающих. В связи со слабым развитием органов обоняния обоня-
тельные доли очень малы. Изгибы мозга у птиц выражены очень
резко. Промежуточный мозг сверху закрыт полушариями и мозжеч-
ком; совершенно между ними скрывается зачаточная м о з г о в ая
железка (glandula pinealis). Снизу ясно видны зритель-
ные бугры (thalami optici), прикрытые перекрестом громад-
ных зрительных нервов (chiasma) и позади них хорошо развитой
гипофизой (hypophysis). Благодаря сильному развитию моз-
жечка и полушарий зрительные доли среднего мозга оттеснены на
бока. Мозжечок прикрывает большую часть изогнутого продолго-
574
ватого мозга, посредине снабжен поперечными бороздами (червя-
чок), а по бокам — небольшими выступами (flocculi). На сагитталь-
ном разрезе явно обнаруживаются радиальные углубления серого
вещества (дерево жизни — arbor vitae).
Периферическая нервная система не пред-
ставляет существенных отличий по сравнению с тем, что мы видели
у рептилий.
Кожные органы чувств у птиц представлены осязательными
тельцами, разбросанными по телу у оснований контурных перьев.
Их скопления находятся па клюве у куликов и на восковице у
хищных птиц.
Рис. 451. Схематический парасагиттальный разрез через
головной мозг утки.
1 — полушария переднего мозга, 2 — полосатое тело (corpus (striatum),
3, 4, 6—дифференцированные части полосатого тела, 5 — обонятельная
луковица (bulbus olfactorius), 7 — обонятельная лопасть (lobus olfac-
torius), 9 — передняя комиссура (comissura anterior), 10—воронка (infun-
dibulum), И—гипофиза (hypophysis), 12 —варолиев мост и про-
долговатый мозг (medulla oblongata), 13 —IV желудочек, 14 —III
желудочек, 15 —мозжечок (cerebellum), 16—средний мозг, 17—задняя
комиссура (comissura posterior), 18—сильвиев водопровод (aquaeductus
Sylvii), 19 —эпифиза (epiphysis), 20 — переднее сосудистое сплетение
(plexus chorioideus anterior). (По Плате, изменено.)
Орган слуха и равновесия у птиц устроен так же, как и у реп-
тилий, но канал улитки (canalis cochlearia) развит здесь вышей отде-
ляется от мешочка (sacculus) перехватом (canalis reunions). Узкие
евстахиевы трубы открываются в глотке одним отверстием. Они
отходят от барабанной полости среднего уха, которая сообщается
с воздухоносными полостями черепных костей. К барабанной пере-
понке прилегает единственная слуховая-косточка — колонка
уха (columella auris). Другим своим концом она упирается через
овальное окно (fenestra ovalis) в слуховую камеру. По-
добно тому как у рептилий, птицы обладают в стенке слуховой кап-
сулы еще другим отверстием — так называемым круглым
окном (fenestra rotunda). Перепонка этого окна отделяет бара-
банную полость oj паралимфатического пространства костного ла-
биринта. Этим достигается более совершенная передача звуковых
колебаний на внутреннее ухо. У птиц собственно наружного уха
575
нет; только у сов имеется особая кожистая складка, прикрывающая
обширную полость, соответствующую наружному уху.
Глаза птиц устроены весьма совершенно, что обусловливает их
острое зрение. Во-первых, они очень велики, хотя и мало подвиж-
ны, они не шаровидной, а чечевицеобразной формы и имеют слабо
развитую мускулатуру. Характерной особенностью глаза птиц
является наличие особого выроста — так называемого гребе-
шка (pecten), который в виде складчатой пластинки, богатой сосу-
дами, выпячивается в стекловидное тело глаза у места вхождения
в глаз зрительного нерва.
Аккомодация глаз достигается не только передвижением хру-
сталика, но и изменением его кривизны под давлением ресничатого
тела (corpus ciliare).
Глаз птиц снабжен мигательной перепонкой
(membrana nicticans), которая приводится в движение особым муску-
лом (ш. nicticans). Имеются верхние и нижние веки (palpebrae),
большая подвижность которых обусловливается наличием обособ-
ленных мышц, открывающих глаз (m. levator palpebrae superioris
и т. depressor palpebrae inferioris), тогда как закрытие глаза произ-
водится действием кольцевой мышцы (m. orbicularis oculi).
Орган обоняния птиц развит слабо. По сравнению с рептилиями
носовая полость все же усложнена выростами. Помимо средней че-
люстной раковипы, уже имеющейся у рептилий, здесь имеются еще
две: одна раковина развивается в преддверии, без обонятельного
эпителия, другая—над челюстной. Эта последняя (nasoturbinale)
является единственной обонятельной раковиной птиц.
Ноздри лежат обычно в задней части клюва и нередко прикрыты
перьями. Носовая железа сообщается простым каналом с носовой
полостью. Якобсонов орган редуцирован п намечается только у за-
родыша.
III. ПРОИСХОЖДЕНИЕ ПТИЦ
Птицы настолько близки по своей организации к пресмыкаю-
щимся, что происхождение их от этих последних не вызывает сом-
нений, однако путь эволюции птиц благодаря отсутствию прямых
палеонтологических данных, остается до сих пор еще не вполне
ясным.
Из ископаемых рептилий наиболее близкой к птицам группой
является отряд псевдосухий (Pseudosuchia) из подкласса текодопт-
ных (Thecodonta). С птицами псевдосухий сближает целый ряд при-
знаков. Череп псевдосухий очень похож на череп юрских перво-
птиц, но имел неподвижную квадратную кость; число крестцовых
позвонков 2—3; задние конечности развиты были сильнее перед-
них и у некоторых являлись главным органом передвижения по
земле, тогда как передние конечности могли служить для схваты-
вания веток и обычно имели типичное пятипалое строение, у Orni-
thosuchus было всего лишь три пальца. Несомненно, что древней-
шие предки птиц были четвероногими, однако, прежде чем передние
конечности превратились в крылья, они должны были освободиться
576
Рис. 452. Археорнис (Archaeornis siemensi)
из литографских сланцев Баварии.
(Из Steinnrann-DOderleln.)
от поддержания тела при движениях по земле или деревьям, и эту
обязанность должны были взять на себя исключительно задние ко-
нечности. Одпако более близкими предками птиц могли быть только
такие двуногие рептилии, которые имели па погах пе менее 4 паль-
цев, из которых первый палец был хорошо развит и мог противо-
полагаться трехМ осталь-
ным. А такими живот-
ными могли быть только
лазящие древес-
ные п р е с м ы к а ю-
щ и е с я.
На это и на дальней-
ший путь эволюции птиц
указывает прежде всего
то обстоятельство, что
все известные как ныне
живущие, так и ископа-
емые птицы, если имеют
меньшее число пальцев
на ногах, то исключи-
тельно благодаря вто-
ричному приспособле-
нию для передвижения
по земле или в воде
вследствие перемены дре-
весного образа жизни на
другой.
Наиболее древней
группой ископаемых
птиц являются пер в о-
птицы (Archaeorni-
thes), известные лишь
по двум экземплярам
двух разных родов, най-
денных в литографских
сланцах Баварии, отно-
сящихся к верхпеюр-
ским отложениям мезо-
зоя (рис. 452). Это были
средней величины пти-
цы, ростом с сороку, соединявшие в себе удивительным обра-
зом признаки птиц и рептилий. Так, мы видим у них длинный
хвост из 18—21 свободного позвонка, как у ящериц; тела шейных
позвонков были пли двояковогнутое или с гладкими сочленовными
поверхностями, а не седлообразными, как у современных птиц.
Ребра — как у пресмыкающихся, с одной сочленовной головкой
без клювовидных отростков, характерных для птиц, причехм раз-"
виты они не только в грудной области, но и в поясничной.
Грудная кость, к сожалению, известна только по одному перед-
37 Кашкаров и Станчинский 577
нему отделу одного из экземпляров. Череп в общем носит птичий
характер с большой черепной коробкой и свободной квадратной
костью; передняя его часть удлинена, и челюсти снабжены
зубами.
Пояс передних конечностей устроен, каку птиц: саблевидные ло-
патки и сросшиеся в дужку ключицы. Вороповы кости, как пока-
зали новейшие исследования, различно устроены у обоих экзем-
пляров: у хранящегося в Британском музее в Лондоне археоптерик-
са (Archaeopteryx) каракоиды типа бескилевых птиц, тогда как у на-
ходящегося в Берлинском музее археорниса (Archaeornis) они,
как у килевых. Однако сама передняя конечность очень странного
типа: ее с таким же правом можно назвать крылом, как и ногой.
Плечо и предплечье так же сходны с птичьими, как и с таковыми
рептилий: кисть с тремя свободно двигающимися пальцами, снаб-
женными когтями. В то же время пояс задних конечностей и сами
задние конечности имели такое же строение как у птиц, но остава-
лись на более низкой стадии развития. Тазовый пояс далеко не был
так полно сросшимся с позвоночником; вместо 10—22 позвонков
здесь с подвздошной костью соединены только 4 позвонка; под-
вздошные кости короткие и не соединяются друг с другом. Седалищ-
ные кости, как у современных птиц, не образуют симфиза, что, ве-
роятно, стоит в связи с откладыванием крупных яиц, одетых твер-
дой скорлупой. Лобковые кости, как и у птиц, направлены назад,
по у археоптерикса, так же, как и у псевдорухий, образуют сим-
физ. В предплюсне имелись две косточки: свободное fibularc и срос-
шиеся друге другом tibiale п intermedium. В плюсне три длинных
не сросшихся metacarpalia, IT, III и IV, лежащие в одной
плоскости.
Тело первоптиц было покрыто перьями, за исключением шеи
и головы. На крыле было расположено десять длинных первостепен-
ных маховых перьев и большее число второстепенных маховых.
И те и другие были прикрыты так называемыми кроющими перьями
крыла. Длинный, как у ящерицы, хвост был усажен двумя рядами
длинных перьев (рулевых), расположенных по бокам.
По строению крыла и хвоста первоптиц можно заключить, что
онц не обладали способностью активно летать. Эти своеобразные
птицы могли лишь перепархивать с одного места на другое. Строе-
ние задних конечностей, где задний палец противополагается трем
остальным, говорит за то, что птицы вели древесный образ жизни.
Они жили в лесах, где перепархивали с ветки на ветку, с дерева па
дерево, распустив свой длинный хвост и махая крыльями, упот-
ребляя в то же время свободные пальцы крыла, чтобы цепляться
за ветки.
Первоптицы показывают нам, как из рептилий могли возник-
нуть птицы. Птицы, несомненно, имели древесных предков. За это
говорит строение их передних и задних конечностей. Такими опи
могли сделаться лишь в результате приспособления к лазанию.
Но как возникло первое оперение, как получилась стадия, которую
представляют первоптицы, мы достоверно не знаем. Возможно, как
578
думают некоторые (Heilmann, 1926 ), что тело лазящих рептилий
было покрыто относительно большими чешуями, особенно боль-
шими па конечностях и хвосте, что увеличивало поверхность тела
при прыгании с ветки на ветку. А затем эти чешуи, удлиняясь и
расщепляясь, превратились в перья. Но нам кажется более вероят-
ным, что первые перья имели вид Нитевидных перьев или щетинок
и служили вначале для сохранения температуры тела в связи с при-
обретенной птицами теплокровностью, а потом приобрели вид пу-
ховидных (рис. 453) и только позднее — строение коЪтурных
перьев, сперва на крыльях
п па хвосте, а быть может,
и па ногах, если принять,
как думают некоторые
(Beebe, 1915), существова-
ние переходной «четырех-
крылой» стадии.
Но когда перья уже
были приобретены, тогда,
махая крыльями и рас-
правляя перья хвоста, ро-
доначальники птиц могли
значительно легче дер-
жаться па воздухе при
прыжках, в то время как
свободные пальцы с ког-
тями на крыльях давали 2
возможность хвататься за
ветки деревьев. Таковы
были первоптицы. Только
дальнейшее усовершенство-
вание крыла превратило
птиц в настоящих власте-
Рис. 453. Гипотетический одетьЩ дзухом
предок птиц.
(Ориг.)
липов воздуха, и оно шло рука об руку с уничтожением свобод-
ных пальцев и превращением кисти в своеобразную костяную
опору для маховых перьев. Что эволюция птиц шла таким обра-
зом, свидетельствует также онтогенетическое развитие современных
птиц и такие формы, как птенцы г о а ц и н a (Opisthocomus),
которые пользуются для лазания передними конечностями с
пальцами, снабженными изогнутыми когтями, причем внутренний
и второй пальцы вполне подвижны, а внутренний даже способен
противополагаться.
Дальнейшая эволюция птиц шла путем все большего и большего
завоевания воздуха вследствие усовершенствования крыла, в ко-
тором три сохранившихся пальца частью редуцируются, частью
срастаются; одновремепно с укорочением хвоста и превращением
его в копчиковую кость (os. coccygis) развивается грудина, к которой
прикрепляются мощные мускулы, приводящие в движение
крыло.
Только после совершенного приспособления птиц к полету на-
^37* 579
чалось вторичное приспособление птиц к жизни в других условиях
со вторичной утратой способности летать. Еще позднее происходит
видоизменение переднего отдела черепа в роговой клюв и уничтоже-
ние зубов. Птицы мелового периода, как видно по имеющимся в не-
большом числе остаткам, уже приспособились к разным средам оби-
тания, по все еще сохраняют у большинства форм настоящие зубы на
челюстях.
Итак^ предками первоптиц следует считать ископаемых репти-
лий, псевдосухий (Pseudosuchia), от которых, кроме птиц,
произошли динозавры, птерозавры и крокодилы. Pseudosuchia,
давшие начало первоптицам, должны были вести древесный образ
жизни. Обладающие большим сходством с птицами динозавры
рано отщепились от общего с ними ствола, вернувшись к назем-
ному образу жизпи.
IV. КЛАССИФИКАЦИЯ ПТИЦ
Всех птиц, как современных, так и ископаемых, можпо подразде-
лить на два подкласса: древних птиц, или первоптиц — Archaeor-
nithes и новых птиц — Neornithes.
Подкласс If Первоптицы ( Arc7iaeornithes9 Saururae)
К этому подклассу отпосят ископаемых юрских птпц, которые
характеризуются: амфицельными шейными позвонками, длинным
хвостом с большим числом мелких позвонков (20—21), па котором
рулевые перья сидели двумя продольными рядами; тремя свобод-
ными пальцами с когтями па крыле; кольцом склеротики из косто-
чек вокруг глаза. Известно всего лишь два экземпляра из юрских
литографских сланцев Баварии, два рода.
Археоптерикс (Archaeopteryx macrura) из литографских сланцев
Золенгофена, хранится в Лондоне, в Британском музее. Отличается от археор-
ниса узким по середине и широким на концах коракоидом, как у бескилевых
птиц, лобковыми ’ костями с симфизом и некоторыми другими призна-
ками.
Археорнис (Archaeornis simensii) из юрских сланцов Эйхштедта,
хранится в Берлинском музее естествознания. Отличается нормальным
для килевых птиц коракоидом и отсутствием симфиза лобковых костей.
Подкласс II. Новые 9 или настоящие птицы
(N eornithes)
Характеризуется гетероцельными шейными позвонками (за
исключением ихтиорниса — Ichthyornis, — у которого позвонки
амфицельпы), коротким хвостом (не более 13 позвонков) и типич-
ным для птиц строением кисти крыла.
Делятся настоящие птицы на два отдела: на древненёб-
ных — Palaeognathae и новонёбных — Neognathae.
580
ОТДЕЛ I. ПАТЕОГНАТИЧЕСКИЕ (ДРЕВ НЕ НЁБНЫЕ) ПТИЦЫ
(PALAEOtiNATHAE) *
К, этому отделу относят наиболее примитивных птиц, которые
характеризуются так называемым дромэогнатическим
пОГюм, т. е. таким, где нёбные кости тесно срастаются, не образуя
(•оч.'К’пепия с крыловидными костями и где широкий сошник сзади
соединяется с клювом основной клиновидной кости, а спереди —
е нёбными отростками челюстных костей. Сюда относятся, во-пер-
иых, давно потерявшие способность летать, так называемые бес-
килевые птицы из ныне живущих: эму и казуары, страусы и нанду
л ископаемые: моа и рокки. Надо думать, что потеря способности
летать развивалась у этих птиц параллельно по крайней мере в трех
группах — у австралийских, африканских и американских бески-
левых птиц, так как каждая из этих групп характеризуется доста-
точно резко и потому заслуживает выделения в самостоятельные
отряды. К этому ж? отделу относят южноамериканских тинам,
или скрытохвостов (Tinami), куроподобных птиц, не утерявших
способности летать. Однако тинамы хотя и являются очень древ-
ней группой птиц, близкой к бескилевым птицам, но не связаны
с ними непосредственным родством, почему их целесообразнее вы-
делить в особый подотдел.
ПОДОТДЕЛ 1. БЕСКИЛЕВЫЕ (RATI ТАЕ)
Как уже сказано было выше, для всех относящихся сюда птиц
характерна давняя вторичная утеря способности летать, что при-
вело к целому ряду особенностей конвергентного порядка. Слабое
развитие грудных мышц привело к изменению грудины, лишив-
шейся киля, укоротившейся и ставшей плоской; произошло сраста-
ние вороновых костей с грудиной, угол между этими костями уве-
личился; произошла редукция ключиц до их полного исчезновения
(за исключением эму); оперение распределилось равномерно, так
что исчезли птерилии и аптерии; изменилось строение перьев,
ставших пухлявыми, исчезла копчикова железа и т. д. Все эти осо-
бенности могли возникнуть в разных группах самостоятельно
как последствие утери способности летать. Все же наличие других
общих особенностей указывает на древний характер всех бески-
левых и на их связь с основанием общего ствола развития птиц.
Помимо строения костного нёба, укажем на примитивное строение
хрящевых колец трахеи и наличие penis.
Отряд 1. Страусы (Struthlonos)
Самые крупные из ныне живущих птиц; характеризуются дву-
палой конечностью (3-й, 4-й пальцы); перья без побочного ствола,
крыло и хвост с крупными перьями. Область распространения —
Африка, Аравия, Месопотамия. В плиоцене были распространены
в южной Европе и в Азии. Единственное семейство.
681
Рис. 454. Эму (Dromacus novae hollandiae) в акклиматизационном
парке Аскания-Нова.
(Фот. И. Озерова.)
Рис. 455. Нанду (Rhea), самец на гнезде в акклиматизационном
парке Аскания-Нова.
(Фот. II. Озерова.)
Сем. Страусы (Struthionidae). Единственный pdflStruthio ивидб.
camelus с шестью подвидами. Ради красивых перьев хвоста и крыльев страусов
раньше добывали в Африке. С конца прошлого столетия страуса разводят
в полудомашнем состоянии в Южной и Северной Африке. Он успешно аккли-
матизируется у нас в Союзе в заповеднике Аскания-Нова. »
582
Отряд 2. Казуары (CasuarH)
Крупные нелетающие страусоподобные птицы; ноги трехпалые;
перья узкие, ланцетовидные с побочным стволом, так же хорошо
развитым, как и само главное перо; перья хвоста и крыльев, ко-
торые настолько малы, что не видны среди оперения, такие же, как
и иа теле. В своем распространении ограничены Австралийской об-
ластью. Два семейства.
Сем. Эму (Dromaeidae). Широкий’’ плоский клюв, голова покры-
та волосовидными перьями, без шлема. Распространен в Австралии и Тасма-
нии. Единственный род эму (Dromaeus) распадается на два вида. Воз-
можна акклиматизация этой птицы у нас на юге (рис. 454). Эму Кенгуровых
островов недавно вымер.
Сем. Казуаровые (Casuariidae). С коническим слегка
загнутым клювом, на голой голове «шлем». Единственный род (casuarius)
с 5 видами и большим числом подвидов обитает в. Новой Гвинее, северо-во-
сточной Австралии, на островах Ару, Церам и некоторых других.
Отряд 3. Нанду (Rheae)
Крупные страусообразные птицы с широкими, как у страу-
сов перьями, по без больших перьев на хвосте и крыльях и с
трехпалыми ногами. Распространены в степях Южной Америки,
в. средней и южной Бразилии, в Боливии, Аргентине и Патагонии.
Единственное семейство.
Сем. Нандовые (R h е i d а е). Единственный род Rhea с двумя
видами. На. родине нанду промышляют ради мяса, перьев и яиц. У нас в
Союзе нанду акклиматизируется в зоопарке Аскания-Нова (рис. 455). Rhea
americana в степях Бразилии и Аргентины, Rhea pennata — Патагойия.
Отряд 4. Киви или бескрылы (Apreryges)
этому отряду относятся современные новозеландскре неболь-
шие (величиной с крупных куриных) нелетающие с очень редуци-
рованным крылом и волосовидным оперением бескрылы, или киви
и недавно вымершие плейстоценовые большие и даже гигантских
размеров птицы м о а. Общими особенностями, объединяющими
современных и ископаемых новозеландских бескилевых являются:
сильное развитие обонятельной полости, небольшие орбиты, сла-
бая, почти квадратная грудина и некоторые другие признаки.
Кроме тре$ хорошо развитых пальцев, имеется зачаточный пер-
вый палец. Два семейства.
Сем. Бескрыловые (Apterygidae). Современные ново-
зеландские киви образуют один род Apteryx, представленный тремя видами.
Ведут ночной образ жизни; у них хорошо развито чувство обоняния, кото-
рым они и пользуются главным образом при добывании пищи. Соответствен-
но глаза у них, в противоположность прочим ночным птицам, развиты слабо.
Наиболее известный вид Apteryx australis (рис. 456).
Сом. М о a (Dinornithidae). Сюда относятся совсем недавно
вымершие страусоподобные*’птицы Новой Зеландии, среди которых в на-
стоящее время установлено 22 вида. Это были крупные и гигантские птицы;
583
ив них самые мелкие были в один метр высотой, тогда как наиболее крупный
вид достигал 3.30 м. Они питались, повидимому, растительной пищей.
Гигантский моа (Dinornis giganteus) — плейстоцен
северного острова Новой Зеландии.
Отряд 5. Мадагаскарские страусы, или рокки (Aepyornlthes)
Ископаемые плиоценовые и плейстоценовые громадные страусооб-
разные птицы с очень широкой грудиной совершенно зачаточными
крыльями и более
Рис. 456. Киви (Apteryx mantelli)
(Из Гаане.)
короткими, чем у моа,
пальцами ног. Извест-
но несколько видов,
из которых самый
крупный достигал
почти 5 м высоты.
Громадные яйца это-
го вида весом в 9 кг
равнялись пяти стра-
усовым яйцам. Един-
ственное семейство.
Сем. Мадага-
скарские страу-
сы (Aepyorni-
t h i d a e). Гигантский
роки (Aepyornis maxi-
mus).
ПОДОТДЕЛ 2. СКРЫТО-
ХВОСТЫ (CRYPTURI)
Отличаются от бес-
килевых наличием
грудины с развитым
как у прочих птиц,
килем, и несколько
отличным строением
палеогнатического нё-
ба, а также отсутст-
вием всех вышеука-
занных особенностей,
связанных с потерей
способности летать.
Отряд 6. Типами или скрытохвосты (Tlnami)
В отличие от всех перечисленных выше отрядов тинамы не по-
теряли способности летать и имеют хорошо развитый киль грудины.
По внешнему виду мелкие формы напоминают куропаток, крупные
же производят впечатление маленьких страусов.
584
Сем. Тинамовые (Tinamidae) (9 родов и около 50 видов) рас-
пространено в Южной Америке, заменяя там наших куропаток и перепе-
лов; обитают преимущественно в открытых пространствах. Самцы у боль-
шинства видов меньше самок и выводят птенцов, Tinamus solitarius в
Южной Бразилии.
ОТДЕЛ 2. НЕОГНАТИЧЕСКЙЕ (НОВОНЁБНЫЕ) ПТИЦЫ (NE0GX4THAE) i
Сюда относятся все остальные птицы, имеющие различно устроен-
ное нёбо черепа, но не дромэогнатического типа. В этом отделе
32 отряда, которые могут быть объединены в четыре группы соот-
ветственно их взаимному родству и положению в системе.
Группа!. Гагароподобные (Colymbomorphae)
Сюда относятся водные, преимущественно морские птицы с ши-
зогнатическим нёбом и голорпнальными носовыми костями; иско-
паемые гесперорнисы (Hesperornithes), гагары (Со-
lymbi), пингвины (Sphenisci) и .буревестники (Procellariae).
Гагароподобные птицы являются, несомненно, самой древней
группой новонёбных птиц.
Отряд 7. Гесперорнисы (Hesperornithes)
Ископаемые водоплавающие, утерявшие способность летать,
крупные, величиной с пеликана, птицы мелового периода с зубами,
сидящими в борозде. Позвонки
гетероцельные; тазовые кости не
сросшиеся. Грудина без киля.
Единственный род Hesperornis
из верхнего мела Канзаса Сев. Аме-
рики.
Отряд 8. Гагары (Colymbl)
Водоплавающие хорошо ны-
ряющие выводковые птицы с
ногами, далеко посаженными
назад, почему прп передвижек
нии по земле держат тело более
или^мепее вертикально. Клюв
прямой и острый. Плюсна силь-
но сжата с боков (рис. 457).
Базиптеригоидные отростки от-
сутствуют.
Два подотряда; оба имеют
промысловое значение, так как
шкурки идут на выделку меха
для шапок и муфт.
Рис. 457. Гагары (Colymbi).
*1—гагара чернозобая (Urlnator arctl-
cus), 2 —клюв чернозобой гагары, 3 и
4—ее нога, плюсна сбоку и спереди, 5—
нога ушастой поганки (Colymbus nigri-
collls). (Ориг.)
585
Подотряд 1. Гагары (Urinatores)
Три пальца соединены перепонкой.
Сом. Гаг а ровые (Urinatoridae); один род, 4 вида распростра-
нены в северных частях Европы, Азии и Сев. Америки. У нас три вида, из
них наиболее широко—распространен—гагара полосатая—Urinatdr arcticus.
Подотряд 2. Поганки (Col/утЫ)
Каждый палец снабжен лопастной перепонкой. Единственное
космополитическое семейство.
Сем. Пог анковые (Colymbidae) —18 видов, относящихся к И
родам. В нашей фауне 5 видов, из которых наибольшее' промысловое зна-
чение имеет большая поганка (Colymbus cristatus).
Отряд 9. Пингвины (Splienlscl)
Пелагические нелетающие, хорошо ныряющие птенцовые птицы,
с опистоцельными позвонками, отличающиеся от всех птиц тем,
что на крыле у пих вместо маховых перьев расположены че-
шуевидные перышки, причем крыло служит ластом (плавником)
Рис. 458. Пингвины (Sphe-
nisci).
1 — южноафрйканский пинг-
вин (Spheniscus demerseus), 2—
крыло пингвина. (Ориг.)
при пырянии. Ноги отодвинуты назад,
и при движении по земле пингвины
держат туловище вертикально. Насушу
выходят только на время вывода птенцов.
Единственное семейство. (Рис. 458).
Сем. Пингвиновые (S ph е n i-
s с i d а о) принадлежит южному полушарию,
преимущественно его полярным областям; 6
родов с 17 видами. Очкастый пингвин (Sphe-
niscus demersus) из Южной Африки.
Отряд 10. Буревестники (Procellarll)
Пелагические, преимущественно оке-
анические птенцовые прекрасно летаю-
щие, парящие и плавающие птицы: зад-
ний палец зачаточный или отсутствует;
от всех других отрядов птиц отличается
тем, что ноздри расположены на концах
трубочек (рис. 459).
Два семейства с более чем 20 родами и более чем 100 видами, распростра-
ненными по морям всего света.
Сем. Б у ре в естн и ков ые (Procellariidae). Качурки
(Oceanodroma) — маленькие птички, величиной со скворца. Вилохвостая
качурка (О., furcata), в Тихом океане. Буревестники (Puffinus) средней
величины и крупные птицы, из пих Р. puffinus атлантический вид, встре-
чается в Черном море. Глупыш (Fulmarus) — единственный вид этого рода.
[F. glacialis] принадлежит Арктике, где на отвесных скалах островов обра-
зуют обширные гнездовые колонии.
Сем. Альбатросов ые (Diomedeidae), крупные птицы,
распространенные преимущественно в океанах южного полушария. Стран-
ствующий альбатрос [Diomedea exulans].
586
Рис. 459. Буревестники (Procellariae).
1 — качурка (Procellaria pelagica), 2 — клюв ее, 3 — ее нога.
(Ориг.)
Группа II. Листо-
подобные (Р о largo-
m о г р h а о)
Вторая группа отря
дов, характеризующая-
ся десмогнатическим че-
репом, соответствует
более высокой ступени
развития неогнат, хо-
тя еще представляет
ствол очень древнего
происхождения. Сюда
относятся ископаемые
ихтиорнисы (Ich-
thyornithes), весло-
ногие (Steganopo-
des), цапли (Ardeae),
аисты — (Ciconiij,
фламинго (Phoeni-
copteri), паламедеи
(Palamedeae), гуси
(Anseres) идневные
хищные п т иц ы
(Accipitltes).
Рис. 460. Веслоногие (Steganopodes).
1 — баклан большой (Phalacrocorax carbo), 2 — его
клюв, 3 — его нога, 4 — голова и шея анхинги
(Anchinga anchinga), 5 — голова кудрявого пели-
♦ кана (Pelecanus crispus.) (Ориг.)
Отряд 11. Пхтиорписы
(lehthyorulthos)
Ископаемые, хорошо
летавшие, небольшие,ве-
личиной с чайку птицы мелового периода с зубами, лежащими в аль-
веолах, и с амфицельными позвонками. Известно несколько родов и
видов из верхиемеловых отложений Канзаса Сев. Америки.
587
Отряд 12. Веслоногие (Steganopodies)
Сюда относятся водоплавающие, хорошо летающие птен-
цовое птицы с клювом, одетым сложным чехлом, с нозд-
рями в виде узких щелей, с короткими ногами, на которых все
четыре пальца соедш епы плавательными перепонками. К этому
отряду относятся пять семейств (рис. 460).
Сем. Бакланов ые ( Р h а 1 а сг о г а с i d а е) отличаются силь-
ным крючковатым клювом, маленьким горловым мешком, длинным наружным
паяцем и полными плавательными перепонками на ногах. (Рис. 460, 1—3)
Два рода: Бакланы (Phalacrocorax) с 29 видами во всех частях света,
из них в фауне СССР 4 вида, гнездящихся по морям; питаются рыбами.
Змеептицы (Andringa); 3 вида на пресных водах Африки, Индии, Юж-
ной Америки и Австралии. (Рис. 460, 4).
Сем. Пеликановые (Pelecani-
d а е) крупные, с громадным клювом и обшир-
ным чгорловым мешком птицы. (Рис. 460, 5).
Единственный род (Pelecanus) с 8 видами. У нас
на юге два вида, имеющие некоторое промысловое
значение, так как их добывают ради перьев. Розо-
вый пеликан (Р. onocrotalus) и кудрявый пеликан
(Р. crispus).
Три остальных семейства:
Сем. олушевые (Sulidae), 5 видов;
Сем. фаэтоновые (Ph a St on id ае)
с тремя видами;
Сем. фрегатовые (Fregatidae)—
океанические, преимущественно тропические
птицы, 5 видов.
Рис. 461. Цапли(Ardeae).
1 — голова серой цапли
(Ardea cinerea), 2—ее нога,
3 — коготь среднего пальца
с наружной стороны, 4 —
летящая серая цапля.
(Ориг.)
Отряд 13. Цапли (Ardeae)
Птенцовые длинноногие голенастые пти-
цы с хорошо развитым задним пальцем
без плавательных перепонок на ногах, с
прямым острым длинным клювом без крюч-
ка на конце и без восковицы. Пух распо-
лагается только на аптериях, причем име-
ются боковые аптерии. Внутренний край
среднего пальца гребенчатый. Три семей-
ства, из которых в пашей фауне имеется
только одно (рис. 461). Около 67 видов.
Сем. UanfleBHe(Ardeidae). В нашей фауне следующие ро-
ды: цапли (Ardea), серая и рыжая цапли (A. cinerea и A. purpurea). Белые
цапли (Herodias), большая (Н. alba) и малая, (Н. garzetta), распространен-
ные на юге, представляют предмет промысла ради особых украшающих
перьев, так называемых эгреток (рис. 462). Вследствие беспощадного
истребления белые цапли были почти окончательно уничтожены не только
на юге Европейской части бывшей России, но и на Кавказе; в .настоя-
щее время, благодаря принятым мерам охраны, белые цапли вновь раз-
множились. К этому же семейству относятся: чепура (Ardeola), кваква
(Nyctiaorax), малая выпь или волчок (Ardetta minuta), очень вредный
истребитель яиц и птенцов других птиц, большая выпь (Botaurus) и неко-
торые другие.
Сем. Сумеречные цапли (Scopidae)n сем. К и т о г о-
ловые цапли (Balenicepidae) распространены в Африке.
588
Рис. 462. Белая цапля (Herodias alba).
Отряд 14. Аисты (Clcoulae)
Это тоже птенцовые длинноногие голенастые птицы без плава-
тельных перепонок на ногах с хорошо развитым задним пальцем,
с длинным клювом без крючка и без восковицы, но с пухом, сидя-
щим по аптериям и птерилиям, без боковых аптерий и с гладким
Рис. 463. Аисты (Ciconii).
1 — клюв белого аиста (Ciconia
ciconia), 2 — клюв каравайки (Р1е-
gadis falcinellus), 3 — клюв колпи-
ка (Platalea leucorodia), 4 — нога
белого аиста. (Ориг.)
Рис. 464. Фламинго (Phoe-
nicopteri).
1 —розовый фламинго (Phoeni-
copterus roseus), 2 — его клюв,
3 — его нога. (Ориг.)
589
внутренним краем среднего когтя; сирипкс без мышц и голосового
аппарата. Три семейства, все в нашей фауне, число видов — 47
(рис. 463).
Сем. 1. Аистовые (Ciconiidae) — с прямым заостренным клю-
вом; у нас белый и черный аисты (Ciconia ciconia и С. nigra) (рис. 463, 1).
Сем. 2. К о л п и к о в ы е (Plataleidae)c широким ложкооб-
разным клювом, у нас один вид (Platalea leucorodia) (рис. 463, 3).
Сем. 3. Иб’исовые (I b i d i d a e), каравайка [Ibis (Plegadis)
falcinellus] с изогнутым клювом (рис. 463, 2).
Отряд 15. Фламинго (Phoenlcopterl)
Длинноногие голенастые выводкоьыа птицы, характеризующиеся
своеобразным клювом, изогнутым коленом; носовые кости голори-
нальные, ноздри сквозные, клюв с рого-
выми иластипками, ноги с тремя паль-
цами, соединенными плавательной пере-
понкой и с недоразвитым задним паль-
цем, который прикреплен выше перед-
них; одно семейство с 6 видами (рис. 464).
Сем. Фламинговые (Phoenicop-
teridae) в нашей фауне один род и вид —
розовый фламинго (Phoenicopterus roseus)
(рис. 464).
medeae).
1 — голова рогатой паламедеи
(Palamedea cornuta), 2 — ее
крыло, 3 — ее нога. (Ориг.)
Отряд 16. Паламедеи (Palainedeae)
Своеобразные довольно крупные вы-
водковые птицы с голенастыми длин-
нопалыми погами, без плавательных пе-
репонок, с хорошо развитым задним
пальцем, с крючковатым куриным клю-
вом, одетым восковицей, со шпорами
на крыльях, и — что отличает паламе-
дей от всех остальных настоящих птиц
(Neornithes) —без клювовидпых отрост-
ков (prosessi uncinati) ребер ^рис. 465).
Единственное семейство Паламеде-
е в ы е (Palamedeidae) с тремя вида-
ми, распространенное в Ю>кной Америке.
' Рогатая паламедея (Palamedcn cornuta).
Рис. 465. Паламедеи (Pala-
Отряд 17. Гуси (Aiiseres)
«0
Как правило, хорошо летающие и
ныряющие выводковые водоплавающие
птицы с клювом, усажеппым по краям
роговыми пластинками или зубцами, с ноготком на конце и
с щелевидными ноздрями; поги с тремя пальцами, соединенными
плавательной перепонкой; задний палец развит слабо и прикреп-
590
лги выше остальных; имеется совокупительный аппарат (penis)
(рис. 466). 160 видов образуют четыре семейства; представлен-
ные у нас имеют большое промысловое и сельскохозяйственное
значение.
Рис. 466. Гуси (Anseres).
1 — клюв лебедя кликуна (Cygnus cyg-
nus), 2 — клюв серого гуся (Anser anser),
3 — клюв гуменника Melanonyx, 4—клюв
утки кряквы (Anas platyrhyncha), 5 —
клюв большого крохаля (Mergus mergan-
ser), 6—клюв гаги (Somateria molassima),
7 — нога утки (Anas platyrhyncha), 8 —
нога нырка (Fuligula fuliguta). (Ориг.)
Сем. 1. Лебединые (Cygnidae) характеризуются длинной
узкой шеей, короткой плюсной, голой уздечкой и узким ноготком клюва
(рис. 466, 1). Несколько видов, имеющих промысловое значение. Разводятся
и полу домашнем состоянии в качестве декоративной птицы на прудах и озе-
рах. Лебедь кликун (Cygnus cygnus) распространен на севере: лебедь шипун
(<;. olor) —на юге Союза; черный лебедь (С. atratus)— Австралия; черношеий
лебедь (С. mel an ocory phus) — Южная Америка.
Сем. 2. Гусиные (A n s е г i d а е). Шея средней длины, длинная
плюсна, широкий ноготок на клюве, края которого усажены зубцами
(рис. 466,2), длинные крылья. Несколько видов; все имеют промысловое
значение. Гусь (Anser) — клюв и ноги светлые. Серый гусь (Anser anser) —
родоначальник наших домашних гусей; гумеши^к (Melanonyx) — клюв
двуцветный черный с желтым или красным. Виды, распространенные в тун-
драх, имеют большое промысловое значение. Казарки (Branta) — клюв чер-
ный, несколько видов их у нас на севере; белый гусь (Chen). Сухонос
(Cygnopsis), вероятный родоначальник китайского домашнего гуся. Ряд
других родов в разных частях света.
Сем. 3. У т и п ы о (A n a I i d а о). Характеризуется короткой шеей
и плюсной, широким клювом с, пластинками. Различают 4 подсемейства.
Подсем. 1. У т к и с о б ст-
в е п п о (А и a I i п а с). Перепон-
ка заднего пальца зачаточная, плюс-
на относительно короткая. Сюда от-
носится большое число родов: утка
(Anas), кряква (A. platyrhincha—
родоначальница наших домашних
уток; несколько других видов.
Широконоска (Spatula) с лопатооб-
разным расширенным клювом, ши-
лохвость (Dafila) с удлиненными ру-
лями у самцов. Серая утка (Chaule-
lasmus), чирок (Querquedula) — не-
сколько видов, свиязь (Матеса) и др.
Все являются охотничье промыс-
ловыми птицами.
Промышляют уток весной на
«подсадную утку» — самку дворовой
утки, которая во время спаривания
привлекает селезней; молодых про-
мышляют с собаками. На зимовках
в Закавказье ловили уток сетями.
Там, где уток много, существовал
ранее также промысел по сбору
утиных яиц.
Подсем. 2. Нырковые
(F u 1 i g u 1 i n a e). Сюда относится
большое число родов и видов. Среди
них промысловое значение имеют
гаги (Somateria), пух из гнезд ко-
торых очень дорого ценится; чернети
(Nyroca), гоголи (Bucephala) и др. Особенно важное хозяйственное значение
имеют гаги, которых эксплоатируют ради очень ценного пуха. Они распростра-
нены на севере. По берегам Мурмана, на островах Белого моря, Новой земле,
Вайгаче, Колгуеве живет гага обыкновенная (Somateria molissima); отчасти
севернее и дальше на восток по сибирскому побережью до Берингова моря
591
гага-гребенушна (S. spectabilis), вместе co стеллеровой гагой (S. stelleri);
последняя распространена к востоку от Таймыра и на Мурмане. Гнездятся
гаги колониями. Самка выстилает свое гнездо пухом и откладывает от
5 до 8 яиц.
Неразумный промысел пуха и яиц, сопровождаемый избиением самих
птиц, привел почти к полному уничтожению богатых колоний гаг на Мур-
манском берегу. Правильный промысел, однако, дает чрезвычайно ценные
результаты. Колонии гаг старательно оберегают от хищников. Пух
и даже яйца отбирают, прежде, чем самка села на яйца, после чего самка вновь
откладывает яйца, заменяя пух водорослями или выдирая пух у самца.
Вторично гнездо гаги уже не трогают. При таком уходе гаги размножаются
быстро и даже привыкают к людям.
Недавно на Мурманском побережье организован гагачий заповедник
«Семь островов».
Подсем. 3. Савков ые (Erismaturinae) с длинным ступен-
чатым хвостом. Один вид: Савка белоголовая или синеклювая утка (Eris-
matura leucocephala).
Подсем. 4. Огаревые (переходное к гусям) (Tadornin ае): гал-
газ или пеганка (Tadorna tadorna), так же как и гаги имеет промысловое
значение ради пуха. Огарь (Casarca) — несколько видов.
Сем. 4. Крохалевые (Mergidae). Характеризуются относи-
тельно короткой шеей и плюсной и узким клювом, ноготок которого обра-
зует на конце клюва крючок, а края клюва усажены зубцами. Несколько
родов и видов. Крохаль большой (Merganser merganser). Луток (Mergus
albellus).
К этому же отряду может быть отнесено весьма своеобразное семейство,
объединяющее ископаемых нижнеэоценовых птиц гасторнисов (Gas-
tornithes). Это были гигантские птицы, утерявшие способность летать.
Отряд 18. Дневные хищные птицы (Acclpltres)
Это птепцовые дневные птицы с «хищным» крючкообразно за-
гнутым верхним клювом, с ясно выраженной восковицей на нем
и хищными ногами с длинными изогнутыми когтями на длинных
пальцах. Носовые кости голоринальпые. Отряд распадается на
три подотряда (рис. 467).
Подотряд 1. Грифы Нового Света (Cathartae)
Перья без побочного ствола, крыловидные отростки развиты хо-
рошо, копчиковая железа голая, сиринкс без мышц. Задний палец
на одном уровне с передними. Питаются падалью. Распространены
в Южной и Сев. Америке.
Сем. Американские грифы (Cathartidae). Кондор (Sarcor-
hamphus gryphus). Гриф —индейка (Cathartes aura).
Подотряд 2. Секретари (Serpentarii)
Перья без побочного ствола, крыловидные отростки хорошо раз-
виты, задний палец причленяется выше передних. Копчиковая
железа оперена, сиринкс с мышцами. Единственное семейство.
592
Сем. Секретаревые (S е г р е n t а г i i d а е); род и вид —
< гьрстарь (Scrpentarius serpentarius) — Африка.
Подотряд 3, Ястреба (Асе ip it res)
Рис. 467. Дневные хищные птицы (Acci-
pitres).
1 — голова королевского грифа (Cathartes
papa), 2 — голова стервятника (Neophoron
perenopterus), 3 — голова орла (Aquila chry-
saetus), 4—голова сокола (Falco peregrinus),
5 — нога скопы (Pandion haliaetus), 6 —
нога ястреба-тетеревятника (Astur palumbar-
tus), 7 — нога орла (Aquila chrysaetus), 8 —
часть плюсны осоеда (Pernis apivorus).
(Ориг.)
Перья с побочным стволом, крыловидные отростки основной
кости развиты слабо; сиринкс с мышцами. Задний палец причле-
пистей иа одном уровне с остальными. Копчиковая железа оперена.
< 'юдл относятся два семейства
с |оо видами.
Сом. 1. Соколиные
(I ’ а I с о n i d а е). Граница опе-
рении на боках клюва не вы-
latvi’oi вперед в виде острого угла.
Мелкие сокола: пустельги (Tin-
пн пси Ins tinnunculus и др.) и
кончики (Erythropus vespertinus
и др.) очень полезны в сельском
хоиийстпе, как истребители мел-
ких грызунов, тогда как дорбиик
(Lilliofalco аона1он) и чеглок
(llypotrlorchl* NUblnileo) пита-
ются птицами. Крупные сокола:
гашиш (Falco) и кречет (llloro-
fiilro) пита юте л крупной птицей,
обычнодобыпаи го* на лету. Jle-
ецолько других родов со. многими
нидами. Четыре рода с голой обла-
стью глаз и уздечкой —из Амери-
ки, Африки и Индии—выделяются
и подсемейство каракар (Polybo-
rlnae).
Сом. 2. О р л и н ы е (A qu i-
I I (I но). Многочисленные роды
крупной передней величины хищ-
ных: скопа (Pandion), питается
рыбой, змееяд (Circaetus ferox)
питается гадами; орлы (Aquila
clirynaelos, clanga, orientalis, he-
lia(’a) и др. крупные виды, пи-
таются млекопитающими и пти-
цами). Морской орел (Haliadtus
.ilhicilla) питается рыбой. Ястреб — тетеревятник (Astur palumbarius) и
иг треб-перепелятник (Accipiter nisus) — два чрезвычайно вредных хищ-
ника, из которых первый питается крупными птицами, второй — мел-
кими. Сарычи, или канюки (Buteo buteo и др.) и луни полевые, степ-
ные и луговые (Circus cyaneus, С. macrurus и С. pygargus) являются весь-
ма полезными птицами, истребителями мелких и средних грызунов. На-
оьорот, болотный лунь (С. aeruginosus) — вредитель, истребляющий водопла-
пающих птиц и куликов на болотах и водоемах. Грифы (Vultur, Gyps, Neoph-
ron)— стервятники, образуют группу, аналогичную грифам Нового Света,
распространены в Старом Свете. Бородач (Gypaetos barbatus)— обитатель
гор, питается мелкими копытными.
Группа III. Куроподобные (А 1 е с t о г о m о г р h а е)
Относящиеся сюда отряды характеризуются шизогнатическим
небом, но представляют собой более молодой ствол эволюции'птиц
ЗЯ Knniicnpou и Станчинский 593
по сравнению с гагароподобными и притом приспособившийся к са-
мым разнообразным средам обитания. Всего сюда относят двенад-
цать отрядов.
Отряд 19. Куры (Galli)
Наземные выводковые птицы с сильными погами бегательного и
роющего типа с хорошо развитыми когтями, с оперенным, по край-
ней мере спереди, голенопяточпым суставом, с коротким, слегка
Рис. 468. Куры (Galli).
1 — голова банкивского петуха (Gallus bankiva), 2 — голова фазана (РЬа-
sianus colchicus), 3 — голова тетерера (Lyrurus tetrix), 4 — нога 'сорной
курицы (Megapodius), 5 — нога банкивского петуха (G. bankiva), 6 — нога
тетерева (L. tetrix), 7 — нога трехперстки (Turnix sylvatica).
(Ориг.)
изогнутым клювом, прикрытым у основания мягкой кожей; носо-
вые кости голоринальные; грудина с высоким килем и двумя глу-
бокими вырезками; хорошо развит зоб; крыло короткое. 5 семейств
(рис. 468).
•
Два первых семейства характеризуются длинными ногами с задним
пальцем, прикрепленным на одном уровне с передними.
Сем. Сорные, или большеногие куры (Megapodi-
d а е) замечательны тем, что не высиживают сами свои яйца, а пользуются
для вывода птенцов теплотой, получающейся в результате тления раститель-
ных остатков. Несколько родов, объединяющих около 19 видов, распро-
страненных в Австралии, Новой Гвинеи, на Филиппинских, Молуккских,
Никобарских и некоторых других островах. Megapodius reinhardti
Австралия.
594
Сем. К р.а к с о в ы е (С г а с i d а е), крупные птицы с крючковатыми
ногтями на пальцах. Обитают в лесах Центральной и Южной Америки. Около
40 видов. Гонко (Сгах alector).
У трех следующих семейств задний палец прикреплен выше передних.
Особенно многочисленно видами (рис. 468).
Сом. Фазановые (Phasianidae). Включает 220 видов.
Плюсна в большей своей части не оперена, нижняя часть клюва по краям
6(”<i зазубрин. Сюда относятся несколько родов куропаток, из которых
у пас имеет промысловое значение широко распространенная серая ку-
роиатка (Perdix perdix); горные куропатки (Caccabis), турачи (Franco-
linns) и крупные горные индейки (Tetraogallus), представленные у
пас па Кавказе и в Средней Азии несколькими видами промысловых
птиц; сюда же относятся роды более мелких птиц — перепелов, из кото-
рых у нас всем известен перепел обыкновенный (Coturnix coturnix). Далее
следуют многочисленные и разнообразные обитатели Азии — фазаны,
из которых пользуется широким распространением хорошо акклиматизиро-
вавшийся кавказский фазан (Phasianus colchicus)„ тан же как и близкие
ему виды из Средней Азии и Манчжурии, Китая и Японии, легко образу-
ющие между собой плодовитые помеси: павлины (Pavo cristatus), аргусы
(Argusianus) и др.
К семейству фазановых относятся также куры (Gallus) вместе с диким
предком наших домашних кур — банкивской курицей (Gallus bankiva) из
Индии; цесарки (Numida), представленные в Африке многими видами с ро-
доначальницей наших домашних цесарок (Numida meleagris); распространен-
ные в Сев. Америке индейки (Meleagris), откуда вывезена и наша домашняя
индейка (Mel. gallopavo).
Из семейства фазанов (Phasianidae) наибольшее промысловое и главным
образом охотничье значение имеет серая куропатка (Perdix perdix). Это
птица слепей и лесостепи. Вместе с расчисткой леса она проникла довольно
далеко в область тайгщ. Так как серая куропатка легко мирится с культурой
человека, то в будущем в СССР она будет, несомненно, основной дичью, по-
добно тому, что мы видим на Западе. В охотничьих хозяйствах куропаток
разводят специально и во время глубоких снегов и гололедиц подкармли-
вают.
Фазаны (Phasianus) на Западе, а отчасти и у нас специально разво-
дятся ради спортивной охоты. И хотя фазаны — южные птицы, вывезенные
когда-то давно с Кавказа в Европу, они отлично акклиматизировались по-
всюду в Западной Европе; в условиях же средней части СССР нуж-
даются в подкормке зимой.
Перепел (Coturnix coturnix) представляет предмет спортивной охоты
и любителями перепелиного «боя» (т. е. пения) содержится в клетках. Рас-
пространены были в степи и лесостепях и с вырубкой лесов и разработ-
кой полей распространились далеко на север.
Сем. Зубатые куропатки (Odontophoridae), —
названо так потому, что нижняя челюсть по внешнему краю зазубрена. Сюда
относятся распространенные в Америке мелкие и средней величины (с куро-
патку) птицы, калифорнийская куропатка (Lophortyx californicus)..
Сем. Тетеревиные (Т е t г а о n i d а е) — объединяет виды
с оперенной плюсной. Сюда относятся самые важные наши промысло-
вые птицы («боровая птица»), а именно: рябчики (Tetrastes bonasa)
тетерев (Lyrurus tetrix) и кавказский тетерев (L. mlokosiewiczi), глу-
харь (Tetrao urogallus), белая куропатка (Lagopus lagopus), тундря-
ная (L. mutus) и др.; в Сев. Америке еще около 10 видов степных
тетеревов — Tympanuchus и др.
На первом месте по промысловому значению нужно поставить рябчика
(Tetrastes bonasa). Он живет в лесах и распространен по всей лесной зоне
СССР. Промышленники рябчиков добывают преимущественно самоловным»
приборами (петлями), реже бьют их из ружья. Промысел начинается с осени.
Промышленник расставляет на своей охотничьей тропе до 1000 силков.
Осенью же и весной добывают рябчиков «на пищик», подражая голосу самца
или самки.
38 ♦
595
Второе место среди промысловых птиц принадлежит тетереву (Lyrurus
tetrix), тоже очень широко распространенному в лесной и лесостепной зонах.
Держатся они по опушкам, полянам, вырубкам. Промышленники добывают
их петлями, сетями (шатрами) или с помощью ружья на чучела и на токах.
Чрезвычайно распространена спортивная охота на молодых тетеревов во
второй половине лета с охотничьей собакой.
Глухарь (Tetrao urogallus, рис. 469) занимает видное место среди промыс-
Рис. 469. Токующий глухарь (Tetrao
urogallus) (из Гааке).
ловых птиц и добывается как
самодействующими приборами,
так и с помощью ружья.
Глухарь во время второй по-
ловины своей песни делается со-
вершенно глухим и даже не слы-
шит выстрела из ружья. Охота
заключается в том, что под песню
к нему «подскакивают» на ружей-
ный выстрел. Глухарь обитает в
больших, преимущественно хвой-
ных лесах.
Белые куропатки (Lagopus
lagopus) населяют тундру и мо-
ховые болота зоны тайги. Явля-
ются преимущественно промыс-
ловой птицей. Ловят их петлями
и добывают с помощью ружья
(рис. 470).
Отряд 20. Гоацпны
(Opistocoml)
По своему строению близки
к курам, отличаясь от послед-
них своеобразной грудиной
с одной неглубокой вырезкой
и приплюснутым спереди ки-
лем, а также тем, что кора-
коиды срастаются с грудиной
и с ключицами, тогда как эти
последние своей общей ди-
стальной частью прирастают
к килю грудины. Имеют сложно устроенный пищевод с большим
зобом, в котором подготовляются к перевариванию твердые листья
некоторых деревьев тропических лесов. Птенцы на двух пальцах
крыла имеют когти.
Сем. Гоациновые (О pistoc omidae). Единственный вид
обитает по заливаемым тропическим лесам в Южной Америке—гоацнн (Opisto-
comus hoazin), похож на кракса, близок по многим признакам к курам.
Отряд 21. Стереорнксы (Stereornithes)
Ископаемые из эоцена Европы и Сев. Америки гигантские птицы
с недоразвитыми крыльями и массивным черепом с узким, высоким
и с крючком на конце клювом, близкие по организации к журавлям,
выделяются в особый отряд.
596
Отряд 22. Трехперстки (Ttirnlces)
Сюда откосятся небольшие наземные, похожие на перепелов
111.1НОД1СОНЫС птицы, отличающиеся от всех куриных пшзориналь-
ними носовыми костями, редукцией заднего пальца, одной глубо-
кой вырезкой на грудине и отсутствием зоба. От журавлей трех-
перстки отличаются наличием базиптеригоидных отростков. Как
привило, самки крупнее и ярче окрашены, чем самцы, которые вы-
сиживают яйца и водят птенцов. Два семейства. Тропики и субтро-
пики Старого Света.
Сем. Трехперстковые (Turnicidae). 13 видов без заднего
пальца. Африка, Южная Азия и Мадагаскар — Turnix sylvatica в Юж-
ной Европе и . Северной
Африке.
Сем: Pedionomi-
d а е с слабо развитым пер-
вым пальцем. Австралия.
Единственный вид Pediono-
mus torquatus.
Отряд 23. Журавли
(Grnes)
Голенастые крупные
птицы с задним паль-
цем, слабо развитым, и
прикрепленным выше
остальных, без базипте-
ригоидных отростков в
черепе, с трахейно-брон-
Рис. 470. Белая куропатка (Lagopus lagopus)
в летнем пере.
хиальным сиринксом.
Отряд объединяет не-
сколько уклоняющихся
форм, которые нередко выделяют даже в отдельные отряды; мы
выделяем их в подотряды.
Подотряд 7. Мезнты (Mesttes)
Небольшие, величиной с дрозда, наземные птицы с короткими
ногами, шиз орина льными носовыми костями, длиппым хвостом и ко-
роткими слабыми крыльями; ключицы отсутствуют. Одио семейство.
Сем. Мезитовые, или пастушков ые куропатки
(Mesitid ае) отличаются недоразвитием ключиц. Два рода с тремя видами
на острове Мадагаскаре.
Подотряд 2. Солнечные цапли (Eurypygae)
Похожие на мелких цапель с относительно длинными плюснами
птицы тропической Америки, отличающиеся сквозными ноздрями,
шизорипальными носовыми костями, слабо развитым боковым
стержнем перьев, голой кобчиковой железой и наличием пудреток.
597
Птицы вылупляются одетые пухом и выкармливаются родителями.
Вилочка имеется.
Сем. Солнечные цапли (Е и г у р у g i d а е). Единствен-
ный род с двумя видами в тропической Америке. Eurypyga helias—тропическая
Южная Америка.
Подотряд 3. Кагу (Rhino cheti)
Птица, похожая папочных цапель, величиной с курицу. Остров
Новая Каледопия. По наличию голой кобчиковой железы, пудреток
Рис. 471. Журавли, Grues; дрофы Otides и пастушки Ralli.
1 — голова серого журавля (Grus grus), 2 — его нога. Пастушки
(Ralli), 3 — голова пастушка (Rallus aquaticus), 4 — его нога, 5 —
голова лысухи (Fulica atra), 6 — ее нога. Дрофы (Otides), 7 — голова
стрепета (Microtis tetrax), 8 — его нога. (Ориг.)
и по тому, как выкармливает птенцов, сходна с солнечными цап-
лями, но ноздри не сквозные; носовые кости шизоринальные. Клю-
чицы имеются.
Сем. К а г у в ы е (Rhinochetidae). Единственный вид острова
Новой Каледонии — Rhinochetus jubatus.
Подотряд 4. Кариамы (Cariamae)
Крупные, величиной с дрофу, птицы с ключицами, с шизориналь-
ными носовыми костями, несквозпыми ноздрями, неоперенной коп-
чиковой железой и десмогнатическим черепом. Пудреток пет.
598
Единственное семейство с двумя видами, относимыми к двум родам.
Стени и полупустыни Южной Америки.
Сем. Кариамовые (Cariamidae). Бразильская кариама —
Cariama cristata.
Подотряд 5. Журавли (Grues)
Ноздри сквозные, носовые кости или шизо,- или голоринальные
(сем. трубачей). Вилочка нормальная, пудретки отсутствуют, зад-
ний палец прикреплен выше передних. (Рис. 471).
Сем. Журавлиные (Gruidae) объединяет журавлей (Grus
grus), красавок или степных журавлей (Anthropoides virgo), венценосных
журавлей (Balearica) и др.; имеют некоторое промысловое значение.
Сем. Трубачи (Psophiidae), птицы величиной с крупную ку-
рицу, с. коротким загнутым клювом (3 вида из тропической Южной Аме-
рики).
Сем. Арамовые (Aramidae), птицы более крупные, с длинным
узким клювом, как у пастушков; один род в Южной и Центральной Аме-
рике— Aramus scolopaceus.
Отряд 24. Дрофы (Otldes)
Выводковые голенастые крупные и средней величины птицы
открытых сухих пространств; с сильными трехпалыми бегатель-
ными ногами (без заднего пальца); с относительно коротким, слегка
изогнутым клювом, с голоринальпыми носовыми костями, без коп-
чиковой железы и без аптерий на боках шеи. Распространены в Ста-
ром Свете, 24 вида. Единственное семейство.
Сем. Дрофиные (Otididae) содержит несколько родов. У нас
Два вида степей имеют промысловое значение: дрофа (Otis tarda) и стрепет
(Microtis tetrax).
Отряд 25. Пастушки (Ralll)
Мелкие и средней величины голенастые птицы топкого, сплюсну-
того с боков телосложения, приспособленного для продвижения сре-
ди густой травянистой болотной растительности; болотные и водно-
болотные птицы с бегательными или плавательными ногами, причем
в последнем случае они образованы пе перепонкой, соединяющей
пальцы, а отдельными лопастями каждого пальца; ноздри сквоз-
ные, носовые кости голоринальные, базиптеригоидные отростки
отсутствуют: копчиковая железа имеется и оперена, аптерии по
бокам шеи развиты. Космополиты. Два семейства; каждое принадле-
жит особому подотряду. 50 родов, около 130 видов.
Подотряд 1. Пастушки (Ralli)
Типичные для отряда птицы с вилочкой, не доходящей до
грудины.
Сем. Пастушков ые (R а 1 1 i d а е). Эти выводковые и типич-
ные для отряда космополитические виды могут быть разбиты на два под-
семейства:
Подсем. П асту ш ковые (R а 11 i п а е), куда из встречающихся у нас,
599
кроме пастушка (Rallus aqualicus, рис. 471, 3 и 4) еще относятся коростель
(Сгех сгех) и болотные курочки (Porzana porzana, Р. pusilia) и др.; замеча-
тельны утерявшие способность летать новозеландские пастушки (Ocydromus).
Подсем. Камышницевые (Gallinulinae), характеризующиеся
лобным щитом над клювом (рис. 474,5 и 4): камышницы (Gallinula chlo-
ropus), синие султанские курицы (Porphyrio), лысухи (I?ulica atra) с лопаст-
ными перепонками на пальцах. Все пастушки являются дичью и некоторые
из них имеют промысловое значение.
Подотряд 2. Лапчатоноги ( Hellornlthes)
Хорошо плавающие и пыряющие птицы с лапчатыми перепонками
на пальцах ног, с вилочкой, срастающейся с грудипой.
Сем. Лапчатоногие (Heliornithidae) по одному роду
и виду в Африке, Индии с Суматрой и Южной Америке. Podica senegalensis
Африка.
Отряд 26. Ржанки (Charadrii)
Сюда относятся выводковые голенастые, по большей части
длинноногие и длиппоклювые, мелкие илп средней величины птицы—
Рис. 472. Кулики (Charadrii).
t — голова дупеля (G-allinago media), 2 — его нога,3 — голова кроншнепа (Nu-
menius arquatis), 4 — его нога, 5 — голова степной тиркушки (G-lareola melanopteT
га), 6 — ее нога, 7 — голова авдотки (Oedicnemus oedicnemus), ее нога. (Ориг.)
обитатели болот и побережий, и лишь в виде исключения вторична
приспособившиеся к жизни в степях или лесах; сошник развит
хорошо, спереди заостренный, коракоиды разделены, сиринкс
трахейно-бронхиальный, копчикова железа оперена. Космополиты.
Около 180 видов (рис. 472).
К этому отряду мы относим уклоняющихся как по организации,
так и по образу жизни южноамериканских зобастых бекасов
и якан в качестве подотрядов.
600
Подотряд 1. Кулики ( Llnticolae)
(’ и । hi jilik.im п отряда, без развитого зоба и лобного щита па клюве.-
। ‘i’m I. Ржанковые (Charadriidae) — со щелевидными нс-
Г1.НМ HH.IMII ноздрями, лежащими в ямках и ограниченными спереди кожи-
। и>н ।и11и• ।к>и।;ой. 6 подсемейств.
Нидгсм. I. Зуйковые (Charadriinae)—с короткой плюсной
и коротким клювом, несколько утолщенным перед концом. Ржанки — Cha-
i idrins apriearius и Ch. fulvus— обитатели тундр; зуйки: галстушник (Aegi-
.ililii hiaticula); малый
( \cgial i I is dubius), морской
( \og. alcxandrinus) и др.
’Гу.'Н’с (Squatarola squata-
rola), чибис (Vanellus, vanel-
lus), кречотка (Chaettusia
gregaria).
Подсем. 2. К p и в к о-
в ы е (Haematopodi-
11 а е) с приподнятым
вверх клювом: кулик-сорока
(Н aemato pus os tr alegus.)
Подсем. 3. X о д y-
л о ч н ивовые (Н i-
inantopodinae) с очень
длинными ногами (плюсна
в I1/2 раза длиннее сред-
него пальца); шилоклювкп
(Recurvirostra avocetta) с
загнутым вверх клювом
и ходулочник (Himantopus
himan topus).
Подсем. 4. Плавун-
ч и к о в ы е (Phalaro-
р i п а е) с лопастными
перепонками на пальцах —
плавунчик (Phaloropus 1о-
batus) и др.
Подсем. 5. Улито-
в ые ( Tot an in ае).
Многочисленные роды и ви-
ды: песочники (Limonites
minutus, L. temmincki),
чернозобик (Pelidna alpina),
краснозобик (Amylochilus
ferruginous) и др., песчанка Рис. 473. Вальдшнеп (Scolopax rusticola).
(Calidris arenaria), трав-
ники (Totanus totanus),
улиты (Glottis nebularius), черныш (Helodromas ochropus), турухтаны
Philomachus pugnax), перевозчик (Actitis hypoleucos), веретенник (Liniosa
limosa), кроншнепы (Numenius arquatus) и др.
Подсем. 6. Вальдшнепиные (Scolopacina е)—отличаются
большими глазами, отодвинутыми далеко назад. Наиболее ценная дичь.
Вальдшнеп (Scolopax rusticola) (рис. 473), на которого охота «на тяге»
псом известна; бекасы (Gallinago gallinago), дупель (G. media), гарш-
неп (Limnocryptes gallinula), спортивная охота на которых в большом
почете.
Сем. 2. Т и р к у ш к о в ы е (G 1 а г е о 1 i d а е) (рис. 472,5) — отли-
чаются иссквозными округлыми ноздрями, лежащими в ямках, и тем, что
короткий клюв спереди слегка загнут вниз. Тиркушка (Glareola melanoptera)
и др.; бегунок (Cursorius gallicus) и другие роды и виды.
601
Сем. 3. Авдотковые (Oedicnemidae) (рис. 472, 7), отли-
ваются сквозными ноздрями, не лежащими в ямках, и слегка утолщенным
к концу клювом. Обитатели степей и полупустынь. Авдотка (Oedicnemus
oedicnemus).
Сем. 4. Лопастноклювые ржанки (Chionidae); на корот-
ком клюве чешуйки, прикрывающие носовые ямки. 5 видов. Фалкландские
и Кергуленские острова.
Подотряд 2. Зобастые бекасы (Thiпосогг)
Похожие на перепелов, но длиннокрылые зерноядные птицы
с большим зобом, коротким, как у куриных, клювом и недоразви-
тым задним пальцем. Единственное семейство.
Рис. 474. Чайки (Lari) и чистики (Alcae).
1—голова поморника (Stercorarius parasiticus), 2—голова- чайки сизой (Larus canurs),
3 — ее нога, 4 — клюв крачки (Sterna fluviatilis), 5 — ее нога. Чистики (Alcae),
6 — голова чистика (Cephus), 7 — его нога, 8 — голова гагарки (Alca torda). (Ориг.)
Сем. Зобастые бекасы (Thinocoridae) южноамерикан-
ские обитатели гор. Attagis malouinus Огненная Земля и Фалкандские
острова.
Подотряд 3. Яканы (Jacanae)
Отличаются наличием лобного щита и чрезвычайно длинными
пальцами, снабженными длинными когтями, причем задний палец
хорошо развит и тоже с длинным когтем. Такое строение ног позво-
ляет якапам бегать по плавающим па воде растениям. Одно се-
мейство, объединяющее 6 видов, распространенных в тропиках
всех частей света.
Сем. Я кановы е (Parridae). Jacana jacana — тропическая
Америка.
Отряд 27. Чайки (Lari)
Выводковые водоплавающие, голенастые, прекрасно летающие
птицы с длинными крыльями и короткими ногами, с плаватель-
ными перепонками; со щелевидными шизорипальпыми ноздрями
без базиптеригоидпых отростков, с хорошо развитым сошником,
с соединяющимися коракоидами и оперенной копчиковой железой.
4 семейства с более чем 140 видами (рис. 474).
602
Сем. 1. Помории к ов ы е (S t е г с о г а г i i d а е) — отличаются
сложным роговым чехлом оканчивающегося крючком клюва; являются
наиболее хищными среди чаек. Четыре вида в Арктике и Антарктике.
Поморник длиннохвостый (Stercorarius parasiticus) и др.
Сем. 2. Чайков bie(Laridae) — около 35 видов; имеют клюв, оде-
тый простым чехлом, слегка утолщающийся перед концом. Чайка сизая (La-
ruscanus), морская (L. marinus), хохотун (L. cachinnans), чайка клуша (L. fus-
cus), голубок (L. gelastes) и др.; рыболовы (с черной головой в брачном наряде);
рыболов обыкновенный (Chroicocephalus ridibunadus), малый (Ch. minutus)
черноголовый мартын (Ch. melanocephalus), трехпалая чайка (Rissa
tridactyla) и др.
Сем. 3. К ра чк ов ые (S tern id а е), 42 вида с длинным сужи-
вающимся клювом, одетым простым чехлом. Крачки: речная (Sterna hirundo),
пестроносая (St. cantiaca) малая (St. minuta), чеграва (St. caspia); крачка
черная (Hydrochelidon nigra) и др.
Сем. Водорезов ые (Rhyncho pidae). 3 вида в Африке,
Америке и Индии; имеют своеобразный клюв: короткое надклювье и выдаю-
щийся вперед нижний клюв. Птицы летают над водой, опустив нижний
клюв в воду.
Отряд 28. Чистики (Alcae)
Выводковые, или птенцовые, морские, плохо летающие, но зато
прекрасно ныряющие водоплавающие птицы с ногами, посаженны-
ми, как у гагар, пазади, но с нормальной и не сжатой с боков, как
у гагар, плюсной; с острым клювом, нередко очень сжатым с боков,
и шизорипальными носовыми костями с хорошо развитым сошником
и без базиптеригоидных отростков. Коракоиды разъединены, си-
ринкс трахейно-бронхиальный. 22 вида: обитатели северных, преи-
мущественно полярных морей, где образуют обширные колонии
гнездящихся птиц — птичьи базары; играют важную роль в жизни
полярпых стран, составляя главную пищу для ездовых собак, а ме-
стами и для людей.
Сем. Ч и с т и к о в ы е (А 1 с i d а е); гагарка (Alca torda), кайра (Uria
troille), чистик (Cepphus grylle), топорик (Fratercula arctica) и др.
Отряд 29. Рябки (Pterocletes)
Небольшие выводковые, очень быстро летающие птицы открытых
пространств с коротким клювом, с очепь короткими, иногда трех-
палыми ногами и оперенной плюсной, с длинными и острыми
крыльями. Носовые кости шизоринальные, сошник зачаточный,
базиптеригоидные отростки имеются, коракоиды разделены, коп-
чиковая железа голая.
Сем. Рябков ые (Pteroclidae). Единственное семейство
с 3 родами и 14 видами в Африке, Мадагаскаре и Южной Азии. Рябок (Ptero-
cles arenarius) и др. Попытка или саджа (Syrrhaptes paradoxus).
Отряд 30. Голуби (Columbae)
Птенцовые, в основном древесные и хорошо летающие птицы,
обычно мелкие или достигающие средней, только иногда значитель-
ной, величины; с характерным, прямым и обычно тонким клювом,
603
одетым у осповапия мягкой кожей, которая за немногими исклю-
чениями прикрывает ноздри; ноги с хорошо развитым задним паль-
цем, сидящим па одном уровне с остальными; носовые кости шизо-
ринальпые, сошпик — рудиментарный, базиптеригоидные отростки
обыкновенно имеются, коракоиды разъединены, копчиковая железа
голая, хорошо развитый зоб разделен на два кармана. Детей вы-
кармливают «молоч-
ком» — отделением
зобных желез. Свыше
300 видов, распрост-
раненных преимуще-
ственно в тропиках,
особенно в индий-
ской и австралийской
областях. Два подот-
ряда (рис. 475, 1 и 2).
Подотряд 1. Дронты
(Didi)
Крупные нелета-
ющие птицы Маска-
репских островов.
Были в XVII столе-
Рис. 475. Голуби (Columbae) и рябки
(Pterocletes).
1 — голова сизого голубя (Columba 11 via), 2 — его
нога. Ряб ни (Pterocletes), 3 — голова саджи (Syrrhap-
tes paradoxus), 4 — ее нога сбоку, 5 — снизу. (Ориг.)
тии истреблены.
Сем. Дронтов ые (Raphidae) — два вида: один из них дронт
(Raphus cucullatus или Dibus ineptus) жил на острове Маврикия, а дру-
гой— пустынник (Pezophus solitarius) — на острове Бурбон.
Подотряд 2. Голуби (Columbae)
Распадаются на четыре семейства.
Сем. 1. Крючкоклювые голуби (Didunculidae)
представлены единственным видом—небольшим голубем острова Самоа,
отличающимся от всех голубей своим высоким клювом, загнутым крючком.
(Didunculus strigirostris).
Сем. 2. Наземные голуби (Geotrygonidae) отли-
чаются относительно длинными ногами, так как плюсна значительно длиннее
среднего пальца. Обитают в тропиках. Особенно замечательны среди них
венценосные голуби (Goura) из Новой Гвинеи и соседних островов.
Сем. 3. Плодоядные голуби (С arpohagidae) с короткими,
оперенными более чем наполовину ногами, отличаются яркой окраской, оби-
тают в тропической Америке, Африке, юго-восточной Азии, Австралийской
области и Новой Зеландии.
Сем. 4. Настоящие голуби (С olumbidae) распространено по
всем частям света.
У нас, кроме полудомашнего сизого голубя (Columba livia), родиной
из Южной Азии, родоначальника домашних голубей встречаются еще сле-
дующие: витютень (С. palumbus), клинтух (С. oenas), горлица (Turtur
turtur) и др.
Группа IV. Сизоворонкоподобные (Coraciimorphae)
Эта группа объединяет птенцовых, преимущественно мел-
ких, реже средней величины древесных птиц, характеризующихся
604
древесными зигодактильными или оборотнопалыми
ногами при ,/!«*<• м<платизме или шизогпатизме, а также нормальными
ногами, <нн.1’П1о при эгитогпатизме.
Отряд 31. Кукушки (Cuculi)
JljH iHu uhic птицы с лазательными ногами, у которых 2-й и 3-й
‘ iiin ni l гд.1 направлены вперед, а 1-й и 4-й или направлены назад
I II на лги. может отгибаться назад; череп десмогпатический,
Ппп||птгр|11’о||диые отростки отсутствуют, носовые кости голори-
нл ii|.ппн•, имеются 5-е второстепенное маховое и 10 первостепенных
млчоных. Два подотряда (рис. 476, 1, 2 и 3).
Рис. 476. Кукушки (Cuculi), попугаи (Psittaci) и дятлы (Pici).
К у к ушки (Cuculi): 1 — голова кукушки (Cuculus canorus), 2 — голова бананоеда
(Musophaga violacea), 3 — его нога.
Попугаи (Psittaci): 4 — голова серого попугая (Psittacus erithacus), 5—его
нога.
Дятлы (Pici): 6 — голова пестрого дятла (Dendrocopus major), 7 — его пога.
(Ориг.)
Подотряд 1. Кукушки (Cuculi)
Характеризуются настоящей зигодактилией (1-й и 4-й пальцы
обращены назад), голой копчиковой железой и разъединенными ко-
ракоидами. Два семейства (рис. 476, 1), 43 рода приблизительно
с 200 видами.
Сем. 1. Кукушковые (Cuculidae)— с 10-рулевыми перьями
в хвосте; часть из них строит гнезда и высиживает, часть является птица-
ми-паразитами; около 200 видов распространены по всему свету. Кукушки
(Cuculus canorus) и др.; пятнистые кукушки (Coccystes glandarius) и многие
другие роды и виды; земляные кукушки; (Geococcyges) из Африки, Ма-
дагаскара и Индии; шпорцевые кукушки (Centropous) с 40 видами из раз-
ных частей Старого Света и др.
Сем. 2. Личинкоедовые (Crotophagidae) отличаются
8 рулевыми; 2 рода с 4 видами из Средней и Южной Америки. Ани (Croto-
phaga ani).
Подотряд 2. Бапаноеды (Musophague)
Имеют 4-й палец, способный противополагаться передним; коп-
чиковая железа оперена, коракоиды перекрещиваются. Обитают
605
в лесах Африки в количестве около 40 яркоокрашенных видов»
Одно семейство (рис. 476, 2 и 3).
Сем. Бананоедовые (Musophagidae) (рис. 476, 2 и 3).
Турако (Turacus cristatus) величиной с фазана, зеленого цвета с красными
крыльями.
Отряд 32. Попугаи (Psittacl)
Древесные с типичной лазательной ногой (1-й и 4-й пальцы
противоположны 2-му и 3-му) и большим высоким крючковатым
клювом, снабженным восковицей и подвижно сочлененным с че-
репом; нёбо десмогнатическое, базиптеригоидные отростки отсут-
ствуют; носовые кости голорииальпые; коракоиды пе соединяются,
ключицы нередко отсутствуют; пятое второстепенное маховое от-
сутствует. Очень хорошо очерченная древпяя группа птиц, рас-
пространенная в тропиках и субтропиках по всему свету, за
исключением Палеарктики 8 семейств, которые могут быть разде-
лены на две группы (рис. 476, 4 и 5).
I. Гладкоклювые попугаи из Австралийской
и Новозеландской областей. Около 60 видов.
Сем. 1. Несторов ые (Nestoridae) два вида новозеландских
попугаев величиной с ворону отличаются длинным крючком своего клюва;
кса (Nestor notabilis).
Сем. 2. Лориевые (Loridae). И родов с полсотней видов как
средней величины, так и мелких попугаев; распространены в Австралии,
Новой Гвинее, на Полинезийских и Малайских островах; отличаются щеткой
роговых сосочков на конце языка.
Сем. 3. Карликовые попугаи (Cyclopsittaci). Мел-
кие, величиной с воробья попугайчики из Австралии, Новой Гвинеи и сосед-
них островов.
11. Бороздчато клювы е попугаи. 60 родов, о' б ъ е-
диняющих 230 с лишним видов.
Сем. 4. Какаду (С а с a t u i d а е). Как средних, так и крупных
размеров попугаи; отличаются подклювьем более широким, чем надклювье;
обитают главным образом в Австралии, также в Новой Гвинее и на сосед-
них островах до Целебеса. Очень распространены в качестве комнатных птиц.
Сем. 5. Совиные попугаи (Stringopidae). Своеобразные
новозеландские большие попугаи, утерявшие способность летать и ведущие
ночной образ жизни. Единственный вид какапо (Stringops habroptilus.)
Сем. 6. Широкохвостые попугаи (Platycercidae).
Средней величины и небольшие, чрезвычайно пестро окрашенные попугаи,
обитающие в количестве около 80 видов в Австралийской области до Целе-
беса включительно; в противоположность распространенным одинаково с ними
какаду, это птицы открытых пространств, умеющие бегать по земле.
Сем. 7. Настоящие попугаи (Psittacidae) распа-
дается на три подсемейства: восковоклювые попугаи (Palae-
ornithinae) 120 преимущественно индийско-папуасские, отчасти африканские
виды; тупохвостые попугаи (Psittacinae) распространены в Аф-
рике и в Южной Америке: клинохвостые попугаи (Conuri-
пае) принадлежат исключительно Америке, где они распространены от 40°
северной широты до 50° южной широты.
Отряд 33. Совы (Strlges)
Ночные хищные птицы с оборотнопалой ногой (3-й палец может
отгибаться назад), с хищным/ крючковатым клювом, одетым вос-
606
ковицей, хищными ногами с крючковатыми когтями и мягким
оперением.
Нёбо шизогнатическое, базиптеригоидные отростки имеются,
сопшпк хорошо развит, коракоиды перекрещиваются, копчиковая
железа голая, 5-е второстепенное маховое отсутствует. Только одно>
семейство с 400 видами из разных частей света.
Рис. 477. Совы (Striges), козодои (Caprimulgi)
и стрижи (Cypseli).
Совы (Striges): 1 — голова уральской совы (Syrnium
uralense), 2 — ее нога, 3—голова ушастой совы (Asio otus).
Козодои (Caprimulgi): 4—голова козодоя (Caprimulgus
europaeus). С т р и ж и (Cypseli): 5—голова стрижа (Apus
apus), 6 — его нога. (Ориг.)
Сем. Совиные (Strigidae) разделяются на несколько под-
семейств.
Поде. Голоногие совы (Ketupinae) — 10 африканских и
южноазиатских видов.
Поде. Ушастые совы (Buboninae) около 150 видов; филины
(Bubo bubo) и другие самые крупные из сов виды; ушастая сова (Asio otus)
болотная (A. accipitrinus), сплюшка (Scops scops) и многие другие.
Поде. Дневные совы (Nycteinae), белая сова (Nyctea nyctea)
и ястребиная сова (Surnia ulula) — северные кругополярные виды.
Поде. Пенсы тевые (Syrniinae) — свыше 100 видов, у нас:
сыч — домовый (Athene noctua), сыч воробьиный (Glaucidium passerinum),
неясыть обыкновенная (Syrnium aluco), уральская (S. uralense) и лапланд-
ская — (S. lapponicum).
Поде. Сипуховые (gtriginae), сипуха (Strix flammea).
Отряд 34. Козодои (Caprimulgi)
Ночные широкоротые птицы со слабыми небольшими ногами, у
которых из трех передних пальцев средний значительно длиннее
двух других; оперепие этих птиц мягкое, крылья острые и длинные.
Нёбо шизогнатическое или десмогнатическое, носовые кости голори-
нальпые, коракоиды разъединены, копчиковая железа голая; 10 пер-
востепенных маховых, 5-е второстепенное маховое отсутствует.
Сем. Гуахаровые (Steathornitidae) своеобразные гуа-
харо (Steathornis caripensis) — из Южной Америки — крупные плодоядные
607
штицы, названные жировыми птицами за обилие.у птенцов жира, употребляе-
мого в пищу.
Сем. Козодоев ые (Caprimulgidae) объединяет около
100 видов, распространенных преимущественно в теплых странах, особен-
но в тропиках. У нас полуночник (Caprimulgus europaeus).
Сем. Совиные козодои (Podargidae) объединяет не-
сколько крупных козодоев, отличающихся между прочим десмогнатическим
лёбом; распространено в Австралийской области и на Малайском архипелаге.
Отряд 35. Стрижи (Cypseli)
Дневные, прекрасно летающие птицы со слабыми ногами, длин-
ными острыми крыльями и различно устроенным клювом. В че-
репе носовые кости голорпнальные, нёбо устроено различно, по
сошпик имеется всегда и пет базиптерпгоидпых отростков; коракои-
ды разъединены, копчиковая железа голая. 10 первостепенных ма-
ховых. Два подотряда.
Подотряд 1. Стрижи (Cypseli)
Короткий плоский и широкий клюв. Ловят насекомых на лету.
Сем. Стрижевые (Cypselidae). Около 100 видов; распро-
странены по всему свету. У нас стриж (Apus apus}; на Кавказе и в Средней
Азии также горный стриж (A. melba).
Подотряд 2. Колибри (Tpochili)
Самые мелкие из всех птиц; замечательны тем, что питаются некта-
ром цветов и насекомыми, находящимися в цветах; со стрижами их
-сближает помимо анатомических признаков неутомимость в полете,
так как колибри собирают нектар, останавливаясь в воздухе, быстро
махая крыльями. Отличаются устройством своего языка, вытяну-
того в двойную трубочку и одетого роговым покровом. Свыше 500
видов преимущественно Южной и Центральной Америки. Несколько
видов заходят в Сев. Америку.
Сем. Колибревые (Trochilidae) является единственным се-
мейством.
Отряд 36. Сизоворонки (Coraciae)
Сюда относится большое число разнообразных, по большей части
небольших древесных птиц, с очепь малым числом общих призна-
ков: кроме древесного образа жизпи, беспомощных и голых птен-
цов, отсутствия пуха па птерилиях, отсутствия базиптеригоидных
отростков и рудиментарности сошника, голоринальпых носовых
костей и пепрободенных ноздрей, для них нет более общих при-
знаков. Характерное для большинства десмогнатическое устройство
нёба заменено у трогонов шизогнатическим, а сростнопалость
ног отсутствует у многих сизоворонок и у удодов. 8 семейств, из
которых большинство можно считать в качестве отдельных под-
отрядов (рис. 478).
Сем. 1. Сизоворонковые (Coraciidae) — яркоокрашен-
ные птицы с преобладанием синих и зеленых цветов, свыше 30 видов, рас-
пространенных во всех частях света; у нас сизоворонка (Coracias garrulus)
(рис. 481, 1 и 2).
608
Ом. 2. Момотовые (М о m о t i d а е) — также ярко окрашенные
птицы из тропической, Средней и Южной Америки с зубчатыми краями клюва,
некоторые с так называемым ракетообразным хвостом. На Вест-Индских
островах эндемичен уклоняющийся род маленьких птичек — Todus.
Сем. 3. Удодовые (U р u р i d а е) —отличаются нормальной ногой
при десмогнатизме. В Африке и в Азии; в Европе один вид удода (Upupa
epops) (рис. 481, 3 и 4).
Сем. 4. Зимород новые (Alcedinidae). Коротконогие,
большеголовые птицы, с короткой шеей, большим клювом и ярким оперением.
Больше 100 видов, преимущественно в тропиках. У нас только один вид —
зимородок (Alcedo ispida) (рис. 479, 5 и 4, 482).
Рис. 478. Сизоворонки (Coraciae).
1—голова ^сизоворонки (Coracias garrulus), 2 — ее нога, 3 — голова удода
(Upupa epops), 4 — его нога, 5 — голова зимородка (Alcedo ispida), 6 —
его нога, 7 — голова щурки (Merops apiaster).
Сем. 5. Щурковые (Mero’pidae) — отличаются тонким тело-
сложением, тонким изогнутым клювом; очень ярко окрашенные тропиче-
ские птицы из Африки, Мадагаскара, Индии и Австралии с Папуасией. У нас
щурка золотистая (Merops apiaster) (рис. 478, 7) и др.
Сем. 6. Птиц ы-- носороги (Bucerotidae) — замеча-
тельны своим громадным клювом, изогнутым вниз и снабженным сверху вы-
ростом. В тропиках Африки, Азии и Индо-Малайских островов (рис. 480).
Сем. 7. Мышиные птицы (Col i id а е). Длиннохвостые мелкие
птички с четырьмя пальцами, направленными вперед. Несколько африканских
ПИДОВ.
Сем. 8. Трогоны (Trogonidae) — отличаются шизогнатиче-
ским нёбом и гетеродактилией: 1-й и 2-й пальцы обращены назад, а вперед —
3 Й и 4-й; величиной с кукушку яркие птицы с длинным хвостом из Центр,
и Южн. Америки, Восточной и Эфиопской обл.
Отряд 37. Дятлы (Пс1)
11 этот отряд объединяются небольшие птицы с типичными лаза-
тгпьными ногами, причем первый и четвертый пальцы обращены
назад, а второй прирос по крайней мере одним члеником к третьему
39 1 (пипсароо и Станчинский
пальцу. Клюв разной формы с ноздрями непрободенными, с нёбом
шизогнатическим у дятлов, десмогнатическим у жакмар и перцея-
дов и эгитогнатическим у бородатой и медоведов; носовые кости
голоринальные, базиптеригоидных отростков пет, коракоиды раз-
делены, копчиковая железа почти всегда оперена. 4 семейства.
Рис. 479. Зимородок (Alcedo ispida).
(Из Гаане.)
Сем. 1. Дятловые (Picidae) с длинным вытяжным языком
и долотовидным клювом. Около 500 видов, распространены в Европе, Азии,
Африке и Америке, но отсутствуют в Австралийской области и на Мада-
гаскаре. Уклоняется несколько род вертишейка (lynx torquilla) серая, не ла-
зящая по стволам птичка. Зеленый дятел (Picus viridis) и седой (Р. canus);
большой пестрый дятел (Dryobates major), средний (D. medius), белоспин-
ный (D. leuconotos), малый (D. minor), трехпалый дятел (Picoides tridacty-
lus), черный дятел (Dryocopus martius).
Сем. 2. Перцеяды или ту к ан о вы е (Rh a m ph a s t i d а е).
Замечательны своим длинным и массивным клювом, как у птиц-носорогов,
но без рога. Распространены в Центральной и Южной Америке.
610
Сом. 3. Во р о д а т к о в ы е (Capitonidae) названы так за сильно раз-
витые в области клюва щетинки; около 150 видов обитает в тропической
Азии, Индии и Америке.
Сем. 4. Жакмаровые (Galbulidae) южноамериканские
тропические птицы с яркой металлической окраской и длинным клювом.
Сем. 5. Me д ов е д овые (In die a t orid а е) около 20 видов из
тропической Азии и Африки.
Отряд 38. Воробьиные (Passeres)
К этому отряду относится громадное количество, свыше 6000 ви-
дов преимущественно мелких птиц. Все они характеризуются нор-
мальной древесной ногой, эги-
тогнатическим нёбом, отсутст-
вием базиптеригоидных отрост-
ков, голоринальными носовыми
костями, не соединяющимися
коракоидами, и голой копчико-
вой железой. Всех воробьиных
птиц делят на два подотряда.
Подотряд 1. Кричащие воробъи-
ные (Clavnatores)
Характеризуется устройством
голосового аппарата, имеющего
по одной паре мускулов (или по
2—3 мускула), которые прикре-
пляются или к середине полу-
колец сиринкса или ко всей их
широте (анизомиодный тип).
9 семейств свыше 1000 видов,
из которых громадное большин-
ство принадлежит Америке, за-
меняя там певчих птиц Старого
Света.
Сем. 1. Рогоклювов ые Рис. 480. Гомрай (Buceros bicornis).
(Е и г у 1 aemidae) являются как (Из Гаане.)
бы переходной формой между сизо-
воронками и воробьиными; почти ограничены Индийской областью,
только один вид в Африке. Один уклоняющийся род на Мадагаскаре.
Сем. 2. IlHTTOBbie(Pittidae)c длинной плюсной, одетой спереди
и сзади одним щитком; в Африке, Индии, Австралии, Австромалайских
островах и Новой Зеландии.
Сом. 3. Тиранов bie(Tyrannidae). Обширное семейство аме-
риканских птиц с плюсной, одетой спереди и с внутренней стороны многими
щитками; распространены от Аляски и Лабрадора до Огненной Земли, от-
чисти заменяя в Америке наших мухоловок, славок и каменок. Свыше
!»00 ппдов.
Сем. 4. Котинговые (Cotingidae). Это неотропическое
семейство объединяет весьма различных по величине (от величины вороны до
величины крапивника) и форме тела птиц, отличающихся общим устройством
сиринкса и тем, что плюсна спереди одета щитками, а сзади — мелкими
чешуйками. Многие из них отличаются ярким оперением, украшениями и
громким металлическим голосом.
39* , 611
Сем. 5. Древолазов ые (Dendrocolaptidae). Плюсна
одета щитками снутри так, что только небольшая полоса с внутренней сторо-
ны остается голой; форма клюва различна. Большая часть форм живет в тро-
пических лесах и обладает способностью лазить по стволам, подобно нашим
пищухам или поползням, но некоторые походят на дроздов, а некоторые
живут в открытых пространствах, напоминая каменок и даже жаворонков.
Центральная и Южная Америка.
Сем. 6. Муравьиные — (Formicariidae). Птицы тропических
лесов Южной Америки; имеют плюсну, одетую спереди щитками, а по
бокам — рядами щитков.
Подотряд 2. Певчие воробьиные (Oscines)
Отличаются от кричащих воробьиных устройством своего голо-
сового аппарата: 5—7 пар мышц сиринкса прикрепляются не на сере-
дине, а по бокам хрящевых полуколец (акромиоидный тип сиринк
Рис. 481. Воробьиные певчие (Passer es).
1 — голова жаворонка (Alauda arvensis), 2 — его нога, 3 — голова пополз-
ня (Sitta епгораеа), 4 — его нога, 5 — голова ласточки (Chelidon rustica)
сбоку, 6 — сверху, 7 — нога ласточки. (Ориг.)
са); плюсна одета спереди щитками, которые иногда срастаются
в один сплошной щиток, по бокам продольными щитками, нередко
распадающимися внизу па отдельные щиточки (рис. 481). Около
5000 видов, распространенных по всему земному шару. Обилие
форм и громадная изменчивость приспособительных признаков и
сложная эволюция относящихся сюда филогенетических ветвей,
дававших в разных местах у филогенетически разных групп одина-
ковые приспособления, а отсюда обилие конвергенций, — все это
чрезвычайно усложняет классификацию подотряда и делает крайне
затруднительной характеристику семейств и подсемейств. Всего
различают следующие 28 семейств.
Сем. 1. Лирохвостовые (М enuridae) отличаются непол-
ным голосовым аппаратом, состоящим только из 3 пар мускулов, и замеча-
тельным лирообразным хвостом самцов из 16 рулевых. Крупные птицы вели-
чиной с фазана. 3 вида в Австралии. Лирохвост. (Menura superba).
Сем. 2. Вороновые (Corvidae) — объединяют самых круп-
ных из певчих птиц, отличающихся сильным, слегка изогнутым или прямым
клювом с ноздрями, прикрытыми (за немногими исключениями) щетинкооб-
612
рапными перьями. Распространены по всему свету. У нас: ворон (Corvus
corax), ворона (С. cornix), грач (Corvus frugilegus), галка (Coloeus mone-
dnla), сорока (Pica pica), сойки (Garrulus glandarius), кедровка (Nucifraga
enryocotactes), саксаульная сойка (Podoces panderi) и др.
Сем. 3. Райские птицы (Paradiseidae). Характерные
птицы Австралийской области, замечательные своим ярким оперением и пла-
стическими украшениями; около 100 видов. Особенно замечательны роды
Paradisea, Pteridophora, Parotia, беседковые (птицы Ptilonorhynchus, Aelu-
roedus и др.).
Сем. 4. Д р о н г о в ы е (D i с г u г i d а е). Близкие к вороновым,
обыкновенно черного цвета птицы, величиной с дрозда. Распространены
в Африке, Индии, на Папуасских островах и в Северной Австралии. Один
вид на Мадагаскаре.
Рис. 482. Воробьиные певчие (Oscines Passeres).
Головы: 1 — иволги (Oriolus oriolus), 2 — щегла (Carduelis carduelis), 3 —
пгленушки (Chloris chloris), 4 — нлеста (Loxia curvirostra), 5 — овсянни
(Emberiza citrinella), 6 — сорокопута (Lanius minor), 7 — трясогузки бе-
лой (Motacillalba), 8 — пищухи (Certhia tamiliaris), 9 — славки садовой
(Sylvia borin), 10 —мухоловки серой (Muscicapa striata). (Ориг.)
Сем. 5. Иволговые (Oriolidae). Примерно такого же рас-
пространения, как дронго, ярко окрашенные в желтый, зеленый или красный
с черным цвета. У нас иволга (Oriolus oriolus).
Сем. 6. Скворцовые (Sturnidae). Около 200 видов, пре-
имущественно в тропических частях Старого света, в Америке отсутствуют.
У пас — обыкновенный скворец (Sturnus vulgaris), розовый скворец (Pastor
roseus) и др.
Сем. 7. Касси ковые (Icteridae) — заменяют в Америке
скворцов Старого света.
Сем. 8. Ткачи ковые (Ploceidae). Обширное семейство мел-
ких птичек, охватывающее свыше 600 видов птиц, очень сходных с вьюрками,
<»т которых они отличаются наличием 10 маховых. Распространены преиму-
щественно в Африке, меньшая часть обитает в Индии на Мадагаскаре, в Ав-
стралии и Полинезии. Из птиц нашей фауны к этому семейству относятся;
домашний воробей (Passer domesticus), полевой (Р. montanus), испанский
(Р. Iiispaniolensis). ч
Сом. 9. Вьюрковые (Fringillidae). Мелкие пташки с кони-
ческим клювом и 9 первостепенными маховыми; разделены на 150 родов;
распространены повсеместно за исключением Австралийской области. У нас
613
Рис. 483. Лирохвост (Menura superba).
(Из Гааке.)
дубонос (Coccothraustes coccothraustes), зеленушка (Chioris chloris). клесты
(Loxia curvirostra) и др., снегирь (Pyrrhula pyrrhula), чечевица (Eryth-
rina erythrina), щур (Pinicola enucleator), длиннохвостый снегирь
(Uragus sibiricus), чиж (Spinus spinus), щегол (Carduelis carduelis), коноп-
лянка (Linaria cannabina), чечотка (L. linaria), горная чечотка (L. flavirost-
ris), зяблик (Fringilla coelebs), юрок (F. montifringilla).
Сем. 10. Овсянковые (Emberizidae). Близки к вьюркам, отли-
чаясь от них строением своего клюва. Овсянка (Emberiza citrinella), садовая
(Е. hortulana), черноголовая
(Е. melanocephala), дубров-
ник (Е. aureola), желтая (Е.
luteola), просянка (Miliaria Ca-
landra), болотная овсянка
(Суnchramusschoeniclus) и др.,
подорожник (Plectrophenax
nivalis).
Сем. 11. Танагров ы.е
(Т a n a g г i d а е). Около 700
видов распространены в Аме-
рике, преимущественно в тро-
пической, представляют как бы
переход от вьюрков к славкам.
Сем. ^.Жаворонко-
вые (Alaudidae) — от-
личаются от всех воробьиных
тем, что плюсна одета по сторо-
нам рядами щитов. 225 видов,
главным образом Евразии и
Африки; в Австралии — только
один род, а в Америке только
рогатые жаворонки. Рогатые
жаворонки (Eremophila alpest-
ris), полевой жаворонок (Alauda
arvensis), хохлатый (Galerida
cristata), лесной (Lullula arbo-
rea), малый (Alaudula minor),
короткопалый (Calandrella
brachydactyla), степной (Mela-
nocorypha calandra), (M. si-
birica), черный (M. yeltonen-
sis).
Сем. 13. T p я с о г у з-
ковые (Motacilli-
d а е). Тоненькие длинные
птички с тонким клювом.
Космополиты, 110 видов. Пе-
стрые трясогузки; белая (Мо-
tacillaalba), горная (М. melanope); желтые (Budytes flava, В. boarula)
черноголовая (В. melanocephala), желтоголовая (В, citreola) и др.; конь-
ки: лесной (Anthus trivialis), луговой (A. pratensis), полевой (А. сат-
pestris), красногрудый (A. cervinus).
Сем. 14. Медососы (М е 1 i р h a g i d а е). Обладают расщеплен-
ным на конце языком, приспособленным для высасывания меда из цветов
и вылавливания со дна цветов насекомых; с 10 маховыми первого порядка.
Около 280 видов ограничены Австралийской областью.
Сем. 15. Белоглазки (Z о ste.ro pi dae). Медососы с 9 ма-
ховыми и с белым кругом около глаз у типичных видов. Около 160 ви-
дов, похожих на пеночек, в тропических странах Старого Света.
Сем. 16. Нектарницевые (Nectariniidae). Маленькие
птички, близкие к медососам, от которых отличаются строением языка.
Очень напоминают колибри, которых заменяют в тропической Африке, на
614
Мадагаскаре, в Индийской области и на Австромалайских островах до
Северной Австралии, где найден лишь один вид.
Сем. 17. Сахарные птицы (Dacnididae). Похожи на
медососов строением языка, от которых отличаются между прочим 9 махо-
выми; 100 видов из Южной и Средней Америки и Среднеамериканских
островов.
Сем. 18. Цветососы (Dicaeidae). Очень близки к сахар-
Рис. 484. Райская птица (Paradisea
apoda).
(Из ГаакеД
ным птицам, но имеют совсем другое распространение: они принадлежат
Старому Свету, где распро-
странены в Индийской и Ав-
стралийской областях; на Га-
вайских и Зондских островах
особое подсемейство.
Сем. 19. Пищуховые
(С е г t h i i d a e). Лазающие
но стволам мелкие птицы с
длинным и тонким загнутым
вниз клювом; около 30 ви-
дов, распространенных пре-
имущественно в северном по-
лушарии. У нас: пищуха (Сег-
thia familiaris), стенолаз
(Tichodroma muraria).
Сем. 20. Поползне-
вые (S i t t i d a e). Также
лазящие по стволам птицы,
по с более массивным прямым
пли даже вздернутым вверх
клювом. Широко распростра-
ненное семейство, отсутству-
ющее в Южной Америке, на
Мадагаскаре и в южной поло-
вине Африки. Свыше 60 видов.
У нас поползень (Sitta euro-
раеа).
Сем. 21. С и н и ц е в ы е
(Р а г i d а е). Мелкие, пре-
красно лазнщиелд’ицы с силь-
ными ногами, Ф коротким и
прямым клювом. Около 300
пидов, распространены по все-
му свету. Три подсемейства.
Синицы с ноздрями, при-
крытыми щетинками (Parinае).
У нас синица большая (Parus
major), черная (Periparus ater),
лазоревки (Cyanistes coeruleus
и C. cyaneus), хохлатая
(Lophophanes cristatus), гаич-
ки (Poecile palustris), пухляк (Poecile borealis), длиннохвостая (Aegithalos
< iiinlalus), усатая (Panurus biarmicus), ремезы (Anthoscopus pendulinus)
п др.
Два уклоняющихся подсемейства. Одно в Азии — попугаеобразные
। пиццы (Paradoxornithinae), другое в Америке — комароловы (Poliop-
1111 шт).
Сем. 22. С в и р и с т е л е в ы е (Ampelidae) 4 вида в Северной
Кироне, Северной Азии и Сев. Америке; отличаются пластинчатыми придат-
ками на второстепенных маховых. Свиристель (Bombycilla garrula).
I ’.ем. 23. Сорокопутовые (L a n i i d а е) характеризуются
< ильным нагнутым крючком клювом с зубцом перед вершиной. Свыше 450 ви-
дон, рвиделипных на несколько подсемейств. Распространены в Старом Свете,
615
Рис. 485. Нектарница (Cinnyris fal-
kensteini).
(Из Брэма.)
только немногие в Америке. У нас: сорокопут большой (Lanius excubitor),
чернолобый (L. minor) и др.; жулан (Enneoctonus collurio).
Сем. 24. Мухоловковые (Muscicapidae) отличаются ши-
роким клювом, усаженным по бокам длинными щетинками и крючочком
на конце клюва, ловят летающих насекомых. Обширное семейство с 750 ви-
дами принадлежит Старому Свету, где распространено преимущественно в тро-
пиках. У нас: мухоловка серая (Muscicapa striata), пеструшка (Ficedula atri-
capilla), белошейка (F. collaris) малая (Siphia parva) и др.
Сем. 25. Славковые (Sylviidae). Самое обширное из всех семейств,
охватывает около 2500 видов, распространенных по всему земному шару.
Вследствие большого разнообразия
видов, входящих в состав семейст-
ва, и разнообразия их приспособ-
лений дать характеристику семей-
ства чрезвычайно трудно. В общем
это — мелкие птички, отличающиеся
своим прекрасным пением, с 10 пер-
востепенными маховыми, причем
первое длиннее половины второго или
очень короткое, короче кроющих
крыла. Различают 8 подсемейств, из
которых пять принадлежат нашей
фауне.
Подсем. 1. Тималиевые
(Timal i i n ae). Это—самое при-
митивное из семейств; принадлежит
исключительно тропикам и субтропи-
кам восточного полушария. Харак-
теризуется коротким, закругленным
крылом с 12 рулевыми перьями.
Подсем. 2. Славк и-п о р т-
нихи (Cisticolinae) сход-
ные с камышевками птицы, отлича-
ющиеся круглым крылом. Распро-
странены одинаково с предыдущим
подсемейством.
Подсем. 3. Корольковые
(Regulinae) объединяют не-
большую группу маленьких птичек.
У нас королек (Regulus regulus)
и др.
Подсем. 4. Славковые (S у 1-
v i i n a e) имеют в хвосте 12
рулевых;, первое маховое короче кро-
ющих. Принадлежат восточному по-
лушарию, где распространены до
Полярной области. У нас большое число видов, из них назовем пе-
ночек: весничка (Phylloscopus trochilus), желтобровка (Phylloscopus sibi-
latrix), теньковка (Phylloscopus collybita), малиновка (Hippolais ic-
tgrina), пересмешник (Hippolais caligata); камышевки отличаются более
тонким телосложением и более длинным клювом: болотная (Acrocephalus
palustris), тростниковая (A. streperus), дроздовидная (A. arundinaceus),
барсучок (Calamodus schoenobaenus), водяная (С. aquaticus), сверчки (Lo-
custella naevia, L. fluviatilis, L. luscinioides), славка садовая (Sylvia
borin), ястребиная (S. nisoria), серая (S. communis), черноголовая (S. atrica-
pilla), завирушка (S. curruca) и др.
Подсем. 5. Пересмешники (М i m i п а е). Американские птицы
сходные с дроздами.
Подсем. 6. Дроздовые (Turdinae). Сюда относятся самые
крупные из славковых. Многие характеризуются пятнистостью оперения;
распространены в Южной и Северной Америке, в Африке, а также в Европе
616
и \ ши. У нас дрозды: певчий (Turdus philomelus), белобровый (Т. mu-
п|< нм), рябинник (Т. pilaris), серый (Т. viscivorus), черные дрозды (Merula
HK'rula), белозобый дрозд (М. atrogularis); каменные дрозды: пестрый (Моп-
Ih'olu saxalilis), синий (М. solitarius) и др.
Подсем. 7. Чеканов ые (Saxicolinae). Более мелкие, чем
1роады, птицы с длинным хвостом; обитатели открытых пространств. У нас:
снменки (Oenanthe oenanthe), плешанка (Saxicola . pleschanka), луговые
(S.ixicola rubetra) и др.
Подсем. 8. Соловьиные (Erithacinae)c острым крылом
и длинным хвостом, преимущественно держащиеся около земли птицы,
нас: соловей (Luscinia luscinia), западный соловей (L. megarhynchos),
юрника (Erithacus rubecula), горехвостка (Phoenicurus phoenicurus), вара-
кушка (Cyanecula svecica) и др.
Сем. 26. Крапивниковые (Troglodytidae) объединяют
мелких и средней величины пташек с тонким клювом, мягким оперением
н короткими рулевыми. Около 250 видов, главным образом в Южной Америке,
и большом числе в Сев. Америке, частично в Южной Азии и два рода в Пале-
арктике.
Подсем. 1. Крапивниковые (Troglodytinae). У нас
один вид — крапивник (Troglodytes troglodytes).
Подсем. 2. Оляп новые (Cinclinae). Приспособлены к бе-
ганью по дну ручьев и речек с быстротекущей прозрачной водой. У нас
о л я п к а ' (С i п с 1 u s с i п с 1 u s) и др.
Сем. 27. Завирушковые (Асе е n tor i d а е) отличаются
широким у основания и узким на конце клювом с ноздрями, прикрытыми
перепонкой. Завирушка лесная (Prunella modularis) и др.
Сем. 28. Ласточковые (Hirundinidae) легко характе-
ризуются широким клювом, очень длинными крыльями и очень маленькими
ножками и тем, что ловят насекомых налету; около 125 видов распространены
но всему свету. У нас: ласточка деревенская, или касатка (Hirundo rus-
lira), ласточка городская (Delichon urbica),; ласточка береговая (Ripa-
ria riparia) и др.
V. ЭВОЛЮЦИЯ ПТИЦ В СВЯЗИ С ПРИСПОСОБЛЕНИЕМ
К ^ЛИЧНЫМ СРЕДАМ ОБИТАНИЯ
Завоевав воздух, птицы получили широкую возможность при-
способляться к самым разнообразным условиям существования, поч-
ти не встречая конкурентов.
Жившие в мезозое с триаса и до конца мелового периода пте-
розавры, или летающие ящеры (Pterosauria) не могли быть
серьезными конкурентами птицам вследствие их меньшей при-
способленности к полету и меньшей пластичности их организа-
ции. Птерозавры, вероятно, не были теплокровными животными,
какими являются птицы, так как не имели покрова из перьев. Их
крыло являлось весьма нежным и несовершенным летательным
л и паратом, на что указывает и относительно слабая грудная кость,
правда, снабженная, как и у птиц, килем; задняя конечность не
и ри обрела той специализации, какую мы видиму птиц, и птерозавры
для передвижения по земле и деревьям должны были опираться
па все четыре конечности, что не позволяло совершенствоваться
крылу. Птерозавры достигли большой специализации, но она по-
шла, так сказать, по тому пути, который не давал возможности даль-
нейшего совершенствования, поэтому они бесследно вымирают
к концу мелового периода, когда более совершенно организован-
617
ные птицы развиваются в самых различных направлениях и через
воздушную среду завоевывают другие среды обитания. Вымерли
и все формы переходных стадий эволюции от рептилий к птицам,
как менее приспособленные по сравнению с этими последними.
Колыбелью птиц, как мы видели, был лес. В лесу начали образо-
вываться лазящие по деревьям ящеры, в лесу развились переход-
ные ящероптицы, там же появились и первоптицы. И дальше лес
оставался осповной стихией птиц; в нем происходила дальнейшая
эволюция класса от самых примитивных форм до наиболее высо-
коорганизованных современных птяц, как воробьиные (Passeres).
Именно лесные формы птиц разных этапов эволюции давали вы-
ходцев в другие среды обитания: в открытые пространства, в болота
и воды. Там, в борьбе за существование, путем длительного отбора
они приобретали особые приспособления к условиям существова-
ния, специализировались соответствующим образом и теряли затем
всякую связь с лесом. Это приспособление к новым условиям оби-
тания отражалось прежде всего на строении ног: нога, приспособ-
ленная для лазания по деревьям, теряла свое характерное строе-
ние с охватывающими ветку пальцами, по мере приспособления
к передвижению в других средах. При передвижении по земле
задний палец редуцировался и, наконец, пропадал совершенно,
остальные пальцы делались более короткими, и нога приспособля-
лась к бегу по земле. У болотных и береговых форм ноги
становились длинными «бродными»; у водоплавающих они связы-
вались плавательными перепонками, приспособляясь к передвиже-
нию в воде. Соответственно видоизменялась, конечно, и вся орга-
низация птицы. Эти приспособления к другим средам обитания
могли настолько специализировать птиц, что возвращение их
обратно к древесному образу жизни становилось уже невоз-
можным.
Такое переселение лесных древесных форм птиц в другие среды
обитания мы встречаем в разных группах птиц, возникновение кото-
рых должно быть отнесено к разным моментам эволюции. Несом-
ненно, что значительное число таких групп, особенно древних,
мелового и начала третичного периода, вымерло, не оставив даже
следов в современной фауне, тогда как ископаемые остатки их не
найдены. Таким образом, многие из боковых ветвей эволюции
птиц остаются нам неизвестными.
Отсутствие прямых свидетельств палеонтологии не да$т нам воз-
можности судить с полной достоверностью обо всем ходе эволюции,
но систематика птиц, в настоящее время достаточно полно разрабо-
танная сравнительпоанатомическим методом, позволяет установить
главнейшие этапы этой эволюции в связи с приспособлениями к раз-
ным средам обитания.
Такими основными средами обитания птиц являются: 1) леса
и кустарники (деревянистая растительность), 2) открытые простран-
ства (луга, степи, пустыни), 3) болота и 4) воды. В каждой среде
обитания мы находим группы разного происхождения д различной
древности. Если они приспособлены более или менее к одинаковым
618
WL* <>п> Кр ; г>ространгтпва Болота Воп-н.ые r»pf
Летание на о в о ho и Плана нм? и нг.д^иуг
Чайки (Lari)
Чистики (Alcae)
Си iceгр: нкопгообные
Кулики (Limicolae)
Рябки ( Pterocletes)
Куроподобнъге)А lector omorphae)
Дрофы (Otides)
Куры (Galli/К
Журавли (Grues)
' Пастушки (Ralli)
Триперстки | Жура1влеобр<мпъм
(Turmcesp^( Gruifor^sj
Голуби (Colum Ъае)
Гоацм/къг
Opistocomi
Дневные
хищники
(Accipitres)
Дневные хищники
(Accipitres
p^^uaxcLaufdeu
/ (Palamedei)
Дневнъъе хищники
( Accipitres )
Дневные хищники
(Accipitres)
Трубконосъге
(Pro cel lariiformes)
—~~'~~7Г)лалгинго (Phoenicopteri
Цаиш (Ardeae) \Д 1———1
Аисты (Ciconia^
Аистпообразнъге
( Ciconiiformes)
Pclaryomorphcic Аистоподобные
Настоящие птицы —|.
(He од______________*
Птицы
( Neornithes)
Первоптицъъ
Archaeorndthes
4
Muytponm и цъъ_^
(№>rpetorni thes)
Древние птплщъъ
(Paia воgnat,hue) |
( Tinamiformes. St ru-
tion (form es, Aptery-
угГогтп es,Rheiform es)
ГагаропоЪобнъ ъе
( (P)lyinbomx)rphae)
* Гуси (Anseres)
Веслоногие
( Sfeganopodcs)
Гагар ообразнъге
( Colymbitormes)
Пингвинообразн ые
( SpKenisiformes)
(Hesperomiihes)
Рис. 48G. Эволюция птиц в связи с приспособлением к различным средам.
условиям, то они образуют аналогичные ряды система-
тических групп.
Черты организации, дававшие птицам возможность при рассе-
лении в новые среды обитания вытеснять’ конкурентов и рассе-
ляться, приспособляясь к разнообразным условиям существования
внутри новой среды, были двоякого рода.
Свойства, позволяющие животным приспособляться к раз-
нообразным условиям существования среды, мы называем
эйритоадапт явными. Таковыми в классе птиц явля-
ются теплокровность и совершенный полет; быстрый и лег-
кий полет у голубей, рябков, чаек и ржанок; хищный
образ жизни дневных хищников; ночной образ жизни сов
и т. д.
Наоборот, приспособления специального характера к сравни-
тельно ограниченным условиям существования, как, папример,
приспособления дятлов к лазанию по деревьям, стрижей и ласточек
к добыванию пищи налету и т. п., удерживают эти группы в узких
рамках приспособлений. Такие группы могут давать приспособле-
ния к еще более узким и еще более специальным условиям суще-
ствования, тем самым в силу изменения всей организации в эволюции
не выходят за пределы данной среды обитания. Признаки
узкоприспособительного характера носят название стеноадап-
тивных.
Эволюция птиц в общем шла обоими путями. Мы видим, что
в разное время возникают в классе птиц эйритоадаптивные ветви,
отдельные этапы развития, дающие возможность такой группе на но-
вом основании приспособиться к разным средам обитания и разно-
образным условиям внутри среды. И в то же время внутри таких
групп развивается большее или меньшее количество ветвей, кото-
рые ограничивают свое развитие узкими рамками специальных
условий существования.
Схема на странице 619 (рис. 486) дает представление о вероят-
ном ходе эволюции птиц в связи с их приспособлениями к основ-
ным средам обитания.
Первоптицы, как мы видели, были лесными птицами.
Следующие за ними предки всех современных нам птиц, родона-
чальные Neornithes, были тоже лесными птицами, но они очень
рано дали выходцев в открытые пространства, которые превратились
в типичных жителей степей и полупустынь. Из них отряд южно-
американские т и н а м у (Tinarni) сохранил способность летать,
тогда как остальные отряды утеряли эту способность совершенно:
таковы казуары (Casuarii), страусы (Struthiones), киви
(Apteryges) и нанду (Rheae), также ископаемые моа (Dinornithes)
и рокки (Aepyornithes).
Родоначальные лесные формы этой группы древних птиц
(Palaeognathae) остались неизвестными, но их потомки, развиваясь
дальше в лесах, дали, очевидно, еще в меловой период выходцев
во все среды обитания, образовавших сообразно с четырьмя кате-
гориями основных сред обитания четыре родоначальных группы
620 ;
соответственно основным систематическим подразделениям н а-
стоящих птиц (Neognathae).
К водному образу жизни приспособились гагароподоб-
и ы е (Colymbomorpbae), представителем которых в меловых отло-
жениях является утративший способность летать весьма специа-
лизированный гесперорнис (Hesperornis). В связи с двумя
основными способами добывания пищи в воде — путем ныряния
и плавания в воде и летания над водой — эта группа дала ныряю-
щих пингвинов (Sphenisci), гагар (Colymbi) и летающих и парящих
буревестников (Procellarii).
К жизни среди болот приспособилась большая часть отрядов
из группы аистоподобных (Pelargomorphae), тогда как
коренная их ветвь, обитавшая в лесу, в связи с приобретением
эйритоадаптивных признаков дневных хищников дала свой анало-
гичный ряд в разные стихии — сверх отряд дневные хищники
(Falconiformes).
Типичными обитателями болот являются отряды: цапли (Ardeae
аисты (Ciconiae) и фламинго (Phoenicopteri). Последние, од-
нако, скорее являются обитателями отмелей, а не болот. Отно-
сящийся к этой же группе отрядов, отряд веслоногих (Steganopodes)
дал аналогичный ряд водных ныряющих птиц. Следующий сверх-
отряд этой же группы дал через формы открытых мест, ка-
кими являются паламедеи (Palamedei), типичных водных обита-
телей отряда гусей (Anseres),— в общем четвертый аналогичный
ряд этой стихии.
К жизни преимущественно среди открытых стаций приспосо-
бились центральные отряды группы куроподобных (А1е-
ctoromorphae). Таковы куры (Galli), трехперстки (Turnices), дрофы
(Otidies). $
Древесный образ жизни сохранили гоацин (Opisthocomi)- и
голуби (Columbae), последние благодаря приобретенной эйритоадап-
тивпой способности к быстрому полету дали ряд аналогичных групп
в другие стихии. Так, рябки (Pterocletes) приспособились
к жизни в открытых местах, пустынях, кулики (Limicolae) —
болотах, чайки (Lari) и чистики (Alcae) — в воде, примени-
тельно к разному -способу добывания пищи: первые — на лету,
а вторые — нырянием. Последнее приспособление делало уже
ненужным быстрый полет, и мы видим, как в этой группе посте-
пенно теряется эта способность вплоть до полной потери умения
чстать, как то было у вымершего гигантского чистика (А1са
impennis).
Преимущественно лесными остались отряды группы сизо-
и о р о п к о п о д о б н ы х (Coraciimorphae). В своих более мелких
подразделениях они путем эйритоадаптивных приспособлений да-
Н.11П многочисленные аналогичные ряды, наиболее интересными из
которых являются совы (Striges), приспособившиеся к ночному об-
разу жизни, стрижи (Cypseli) и козодои (Caprimulgi), приспособив-
шиеся к добыванию пищи в воздухе, первые — днем, а вторые —
ночью; колибри (Trochili) и др.
621
VI. ПОЛЕТ ПТИЦ
Различаются несколько видов полета птиц: 1) планирующий,
или скользящий, полет, когда птица летит с более или менее рас-
простертыми крыльями, не двигая ими, или опускаясь вниз с вы-
соты, сохраняя или увеличивая скорость за счет высоты, или сохра-
няя высоту и даже поднимаясь вверх, но теряя скорость; 2) паря-
щий, или парусный, полет, когда птица летает, не двигая
крыльями, сохраняя и высоту и скорость полета или даже увеличи-
вая их за счет силы движения воздуха; 3) гребной или про-
пеллирующий полет (обычный вид полета), когда птица машет
крыльями и тем осуществляет опору о воздух и поступательное дви-
жение. Этот последний вид полета имеет ряд разновидностей, из ко-
торых особого внимания заслуживает
пульсирующ ий полет,
когда
Рис. 487. Чертеж распределения давления
на плоскости, двигающейся в воздухе.
вибрационный или
птица с помощью чрезвы-
чайно быстрых движений
крыла или висит в возду-
хе, или двигается и притом
может не только верти-
кально подниматься вверх,
но даже двигаться назад.
Теория полета птиц в
настоящее время разрабо-
тана в связи с успехами
воздухоплавания очень
подробно. В общем летя-
щая птица подчиняется
законам движения пласти-
нок в воздухе. Теория дви-
жения таких пластинок и
многочисленные эксперименты установили следующее:
1. Если пластинка двигается в воздухе под некоторым углом
а (угол атаки) к оси движения (рис. 487), то давление встречного
воздуха на пластинку R будет направлено почти перпендикулярно
к ней, разлагаясь на подъемную силу Р и лобовое сопротивление Q.
Подъемная сила и лобовое сопротивление возрастают прямо про-
порционально площади пластипок и квадрату скорости движения.
Для получения больших скоростей выгоднее малый угол накло-
на пластинки, а при большой скорости увеличение угла а до из-
вестных пределов ведет к увеличению подъемной силы.
Центр давления (точка приложения давления на пластинку снизу,
точка опоры о воздух ) тем ближе передвигается к переднему краю
пластинки, чем быстрее она двигается.
2. В продолговатых пластинках давление зависит от положения
пластинки, а именно — пластинки, поставленные длинной своей
стороной перпендикулярно направлению движения, получают боль-
шее давление снизу, а потому более выгодны для полета.
3. Вогнутые пластинки дают большую подъемную силу, чем пло-
ские, причем:
622
а) направление равнодействующей наклонено вперед, вследствие
чего такая пластинка сохраняет свою подъемную силу не только
при горизонтальной, но даже при несколько наклонной вперед
хорде;
б) если передняя сторона такой пластинки утолщена, то она
пе только не увеличивает лобовое сопротивление, а, наоборот, ока-
зывает благоприятное действие па подъемную силу и лобовое сопро-
тивление (рис. 488), тогда как такое же утолщение задней стороны
очень неблагоприятно;
в) лучшие изгибы кривизны дают 7ю—V15 стрелы прогиба;
г) для устойчивости оказывается полезным отгиб задней
части пластинки несколько п
Р ___ О
вверх.
4. Устойчивость дви- f
жущейся пластинки дости- /
гается: /
а) расположением цент- /
I >а тяжести ниже плоскости /
приложения равнодейству-
Ютой аэродинамических ' Q
сил совпадают с центром --------------------—
тяжести; Рис. 488. Профиль крыла: линия АВ на-
зывается хордой крыла; угол а, под ко-
торым движется крыло, — угол атаки;
В — сила давления.
б) наличностью позади
главной несущей плоскости
еще дополнительной плос-
кости так называемого стабилизатора.
В общем горизонтальный полет определенной скорости летя-
щего объекта с крыльями (птицы или самолеты) определяется сле-
дующими формулами:1
1) р =
2) Q=F=CxpSV\
где Р — подъемная сила, G — вес, Q — лобовое сопротивление,
/*' — сила тяги, р — плотность воздуха, 8 — площадь несущей
поверхности, К—скорость движения, Су и Сх коэффициенты про-
порциональностей (подъемной силы и лобовых сопротивлений)
в основном зависящие от качеств несущей поверхности (формы
крыльев) и угла атаки.
Плоскость опоры о воздух у птиц представлена крыльями и хво-
стом. Крылья птиц удовлетворяют именно всем вышеуказанным
требованиям; они вытянуты в направлении, перпендикулярном по-
'П’ту, представляют пластинки, выгнутые вверх с утолщенным
передним краем и выпрямленной задней частью. Последняя эла-
стична и может отгибаться вверх. Хвост выполняет роль стаби-
лизатора.
1 Формулы эти имеют в виду равновесие летящих объектов, имеющих
ужо определенную скорость движения.
623
Общая форма тела с острым клювом, маленькой головой и плотно
прилегающим к телу оперением представляет наименьшее сопро-
тивление воздуху.
Центр тяжести у птиц лежит значительно ниже плоскости опоры,
что достигается высоким положением крыла и тем, что все тяжелые
органы птицы — пищеварительные органы и грудные мускулы—
находятся снизу, а легкие и воздушные мешки лежат выше. Такое
положение центра тяжести придает летательному аппарату птицы
большую устойчивость. Самая плоскость опоры о воздух, т. е.
крылья и хвост, может по произволу легко уменьшаться, что дости-
гается большим или меньшим расправлением крыльеви хвоста. Та-
ким образом, птица может менять отношение площади крыльев к весу
своего тела. Между тем, чем больше плоскость опоры о воздух, тем
большее сопротивление испытывает падающая вертикально вниз
горизонтальная плоскость. Если же такая плоскость двигается
вперед, то она встречает сопротивление воздуха, которое увеличи-
вается пропорционально квадратам скорости и прямо пропорцио-
нально плоскости сечения, проведенной под прямым углом к на-
правлению движения.
Таким образом чем быстрее летит птица, тем легче ей держаться
в воздухе. Но так как летящей птице приходится преодолевать
также сопротивление воздуха в направлении движения, то есте-
ственно, что птица при движении вперед постепенно теряет раз
полученную скорость; вместе с уменьшением скорости будет умень-
шаться сопротивление воздуха снизу, и птица принуждена будет
опускаться. Чтобы не опускаться, птица должна вновь приобрести
скорость, соответственно увеличив маханием крыльев силу тяги.
При движении плоскости играет большую роль тот угол, кото-
рый она образует с осью движения, угол атаки. От величины этого
угла, как мы видели, зависит сила, поднимающая птицу вверх,
а также лобовое сопротивление.
Изменений этого угла достигается перемещением центра тяжести.
При увеличении быстроты полета птица должна переносить центр
тяжести все более вперед, чтобы центр опоры совпал с центром
тяжести.
Скользящий, или планирующий, полет
возможен только при негоризонтальном полете, именно когда он
направлен косо книзу. Движущей силой в этом'случае является
сила тяжести птицы. Отношение площади крыльев к весу птицы
обусловливает при определенной скорости и при определенном по-
ложении крыла по отношению к встречному течению воздуха (угол
атаки) угол планирования, т. е. угол направления полета к горизон-
тальной плоскости. Чем больше площадь крыла, чем меньше вес пти-
цы и чем быстрее полет, тем меньше может быть угол планирования.
Уменьшая площадь крыльев, птица может достигнуть большей
скорости скольжения и может эту быстроту движения использовать
для подъема вновь на известную высоту.
Управляют птицы своим скользящим полетом разными способа-
ми. Во-первых, птица легко может увеличивать и уменьшать пло-
624
щадь плоскости опоры о воздух, расправляя или складывая крылья
и хвост; во-вторых, она может перемещать центр опоры о воздух
по отношению к центру тяжести двояким образом: или, сохраняя
положение центра тяжести, изменять положение центра опоры, сги-
бая крылья, распуская хвост ит. д., или переносить центр тяжести,
вытягивая шею вперед или втягивая ее назад; последнее, впрочем,
имеет значение лишь у птиц с длинной шеей. Это перемещение цент-
ра опоры в отношении центра тяжести может вести к измене-
нию угла атаки крыла, т. е. угла плоскости крыла к встречному
движению воздуха, а вместе с тем и к изменению угла плани-
рования.
У птиц разной величины площадь крыльев изменяется пропорцио-
нально квадрату, а вес птицы изменяется пропорционально кубу.
Отсюда и из приведенных выше формул видно, что с увеличе-
нием птицы втрое площадь крыла увеличивается в 9 раз, а вес
птицы увеличится в 27 раз. Следовательно, птица должна увели-
чить подъемную силу тоже в 27 раз. Так как несущая поверхность
крыла при этом увеличится вч 9 раз, то для сохранения высоты
необходимо увеличение скорости в р/3 раз. При этом лобовое
сопротивление увеличится тоже в 27 раз, и птица должна со-
ответственно произвести большую работу, чтобы увеличить ско-
рость bj/З раз.
Из этого следует, что птицы крупные тратят гораздо больше энер-
гии, чем мелкие птицы. Этим, повидимому, кладется известный
предел увеличению размеров птиц вообще. Вероятно, поэтому
к рупные птицы часто пользуются именно парящим, а не гребным
полетом.
Птицы, быстро спускающиеся вниз, складывают свои крылья и
распускают хвост; центр опоры о воздух передвигается значительно
назад по сравнению с центром тяжести, и планирующая плоскость
нагибается передней стороной вниз. Наоборот, когда птица хочет
выравнять свой планирующий полет или подняться кверху, она
распускает крылья и передвигает их вперед: центр опоры о воздух
становится впереди центра тяжести, и у планирующей плоскости
передняя сторона поднимается вверх; тот же эффект достигается
опусканием хвоста вниз. Поворот вправо и влево достигается или
сгибанием соответствующего крыла, или поворачиванием головы,
или вытягиванием в соответствующую сторону шеи, или поворотом
распущенного хвоста в противоположную сторону.
Гребной, или пропеллирующий, полет. При
этом типе сохраняются условия скользящего полета; к ним присое-
диняется еще поступательная сила — сила тяги, которая достигается
взмахами крыльев. Чем больше скорость движения птицы, тем легче
।ггп це держаться в воздухе; поэтому попятно, что на получение имен-
но начальной скорости птице приходится затрачивать больше всего
энергии. Встречный ветер в этом отношении оказывает птице некото-
рую помощь, так как при определенной силе и определенном отноше-
нии площади крыльев к весу птицы ветер может поднять птицу вверх
40 Кашкаров Станчинекий 625
подобно воздушному змею. Поэтому все птицы при ветре под-
нимаются на крылья, становясь против ветра. В других случаях
первоначальная скорость достигается разбегом или прыжками,
после чего уже птица поднимается вверх. Или, наконец, птице
приходится делать частые и сильные взмахи крыльями, чтобы по-
лучить начальную скорость. При этом крылья иногда ударяются
концами одно о другое за спиной, производя у разных видов птиц
характерный звук при подъеме. Затрата энергии при подъеме так
велика, что птицы, которым приходится несколько раз кряду под-
ниматься и спускаться, легко обессиливают. Есть птицы, которые
вовсе не могут подняться с горизонтальной поверхности и начальную
скорость, необходимую для полета, получают, падая вниз с вы-
соких предметов, как, например, стрижи.
Для поднятия птицы на воздух необходимо, чтобы оба крыла
могли захватить определенную по отношению к весу тела массу
воздуха. Отсюда понятно, что птицы с небольшими короткими
крыльями машут крыльями гораздо чаще, чем птицы с большими
крыльями. Так, воробей при полете делает 13 взмахов в секун-
ду, утка — 9, ворона — 3—4, аист — 2 и пеликан — 1 х/н-
На поднятие крыла птицы вообще затрачивают меньше времени,
так что в среднем время, потребное на поднимание, относится ко
времени опускания, как 2:3.
Как видно из прилагаемой моментальной фотографии (рис. 489),
во время полета птица двигает крыло не только вниз, но и вперед.
При поднимании крыла сопротивление воздуха уменьшается тем,
что крыло сверху выпукло, что при поднятии оно несколько согнуто
в сгибе и направлено задней частью книзу, и, наконец, тем, что
маховые, благодаря большей ширине внутренних опахал, расхо-
дятся, пропуская воздух.
Получив достаточную поступательную скорость полета, птица
уже не нуждается в затрате большой энергии, ей необходимо лишь
сохранить приобретенную скорость; так как сопротивление воздуха
движущейся поверхности снизу возрастает, как мы видели пропор-
ционально квадрату скорости движения, то птица при быстром
полете при прочих равных условиях будет подниматься вверх. От-
сюда быстро летящие птицы и реже машут крыльями и уменьшают
поверхность плоскости опоры, складывая крылья и хвост. Так,
галка, например, при подъеме делает около пяти взмахов в секун-
ду, а разлетевшись — только три взмаха.
Многие мелкие пташки, сделав несколько быстрых взмахов
крыльями и поднявшись на некоторую высоту, затем складывают
крылья, летят несколько секунд без взмахов, немного опускаются
при этом, затем вновь машут крыльями, поднимаясь на первоначаль-
ную высоту, вновь перестают махать и т. д. Здесь, благодаря незна-
чительному весу и более благоприятному соотношению между ве-
сом и площадью крыльев, мелкие птицы при гребном полете, махая
крыльями, очень быстро увеличивают подъемную силу.
'Парящий полет птиц, или парение, изучено
теперь очень хорошо благодаря разработанной теории летательных
626
машин без двигатель-
ных аппаратов—пла-
неров. Птица вовсе не
двигает крыльями, а
между тем не только
сохраняет скорость,
но может поднимать-
ся вверх. Такой по-
лет мыслим в том
случае, когда птица
использует горизон-
тальные или, еще луч-
ше, негоризонталь-
ные воздушные тече-
ния или неоднород-
ность воздушных те-
чений в горизонталь-
ном и вертикальном
направлениях. Раз-
личные воздушные
вихри, завихрения,
пульсация воздуха
(так называемая тур-
булентность воздуха)
обусловлены разно-
го рода причинами:
например, неравно-
мерным нагреванием
разных частей земли,
благодаря чему со-
гретый воздух подни-
мается вверх, а хо-
лодный опускается
вниз; препятствиями,
которые встречают
на своем пути воз-
душные течения в
виде гор, лесов, волн
к т. п. Для ис-
пользования воздуш-
ных течений необхо-
димы некоторые ус-
ловия. Во-первых,
и । ч »б ходима известная
абсолютная и отпо-
ен те л i > пая величина
врыльев. Хорошо па-
|иицпх птиц, которые
могутпаритьдаже при
40*'
Рис. 489. Кинематограмма летящего какаду.
(Из Хильцхеймера.)
$27
малейших, как будто бы незаметных движениях воздуха, мы нахо-
дим именно среди крупных видов птиц с мощными крыльями. Та-
ковы грифы Старого и Нового Света, жители преимущественно
гор, где, как мы видим, турбулентность воздуха должна быть осо-
бенно сильна. Далее, хорошо парят орлы и другие крупные хищ-
ники, аисты, вороны, чайки, буревестники, пеликаны и многие
другие. Все это или крупные птицы с мощными крыльями, или
птицы мелкие и средней величины с очень длинными крыльями.
Во-вторых, необходимо некоторое определенное строение крыль-
ев, именно: они должны быть достаточно длинны по сравнению с ши-
риной, прицем здесь мы встречаем два типа крыльев. Первый тип
мы находим у материковых птиц: крылья относительно широки, и
первостепенные маховые могут широко растопыриваться. У мор-
ских птиц — буревестников и чаек — крылья очень длинны, узки
и остры. В обоих случаях птицы управляют своим полетом одинако-
во, каждую минуту меняя сообразно с потребностью величину по-
верхности крыльев: первые — путем большего или меньшего рас-
топыривания маховых и распусканием хвоста, вторые — сгибанием
крыла.
Интересно, что парение в большинстве случаев сопровождается
кружением. При этом птица описывает большие или меньшие круги
над одним и тем же местом, то поднимаясь, то опускаясь без малей-
шего движения крыльями.
При сильном ветре птица позволяет относить себя назад, дер-
жась в направлении, встречном ветру, постепенно поднимаясь
и уменьшая быстроту движения. Достигнув наивысшей точки,
птица круто повертывает назад и, постепенно опускаясь, приобре-
тает значительную скорость, чтобы, описав дугу, стать опять против
ветра и вновь подниматься в исходном месте.
Пульсирующий полет свойственен многим мелким
птичкам: королькам, пеночкам, мухоловкам, но большой специали-
зации он достигает у питающихся нектаром цветов птиц у пектар-
ниц (Nectariniidae) Старого Света и в особенности у колибри (Trochi-
]idae). Подлетев к цветку, колибри быстро махая крыльями, по-
добно бабочкам-бражникам, повисают в воздухе перед цветком,
пока не высосут из цветка нектар. При этом они делают до 50 взма-
хов в ^секунду, так что движение крыльев заметить невозможно,
а кажется, что птичка окружена туманным ореолом. Колибри при
этом могут не только вертикально подниматься в воздух, но и един-
ственные из птиц обладают способностью двигаться в воздухе задом.
С пульсирующим полетом сходен висячий, или трепе-
щущий, полет, свойственный многим хищникам — пустельге,
мохноногим канюкам, а также крачкам и некоторым другим птицам.
Быстро махая крыльями, птицы повисают в воздухе над одним ме-
стом, высматривая добычу. Жаворонки тоже обладают способ-
ностью во время пенья висеть в воздухе.
Быстрота полета довольна различна у разных видов, как и у од-
ной и той же особи, — в зависимости от условий полета. Для поч-
товых голубей установлена быстрота в 1000—1500 м в минуту.
628
Наиболее быстро летающей птицей считается стриж рода Chae-
tura—2400 м в минуту; другие птицы имеют значительно меньшую
быстроту полета, а именно: почтовые голуби — 1320 м (и до 1950 м)
в минуту, скворцы — 1230 м, зимородок— 960 м, зяблик — 870, во-
рона — 840 м в минуту. В общем нужно заметить, что приведенные
цифры дают среднюю нормальную скорость, тогда как временами
скорость полета может быть значительно больше, например, у дого-
няющего добычу сокола или стремящейся избегнуть опасности
вороны. Так, было установлено, что баклан, преследуемый аэро-
планом, пролетел 15 к»| со скоростью 105 км в час, тогда как сред-
няя скорость его полета около 70 км в час.
В общем мелкие птицы развивают большую скорость полета,
хотя каждый удар крыльев большой птицы дает в полете больший
эффект. Так, среди отряда гусей чирок-свистунок весом 330 г летит
со скоростью 1980 м в минуту, кряква весом 1250 г — со скоростью
1740 м в минуту, а серый гусь весом 3500 г со скоростью 1170 м
в минуту.
Продолжительность полета без отдыха у птиц поразительна.
Так, например, стрижи за исключением короткого отдыха ночью
в длинные летние дни носятся в воздухе почти целые сутки.
Некоторые кулики во время перелета пролетают пространство в
3000 км без отдыха.
VII. пища птиц
Возможность приспособлений к разного рода пище, к ее оты-
скиванию, захватыванию и перевариванию зависит прежде всего
от особенностей организации тех родоначальных форм, от которых
развилась данная группа животных.
Как форма челюстей, так и характер зубов совершенно опреде-
ленно говорят в пользу предположения, что пищу родоначальников
птиц составляли различного рода животные. Вероятно, подобно
разным современным ящерицам они питались разнообразной живот-
ной пищей: насекомыми, их личинками, амфибиями и многочислен-
ными и разнообразными в то время рептилиями. Конусообразные
ост ,ые зубы служили исключительно для удерживания во рту схва-
тываемых животных. Каким образом, однако, произошло видоизме-
нение челюстей в настоящий клюв, покрытый роговым чехлом, и от-
чего произошла потеря зубов, мы, к сожалению, не имеем возмож-
ности судить с достоверностью за отсутствием палеонтологических
данных. Возможно, что потеря зубов стояла в связи с развитием ро-
гового клюва с острыми краями и загнутой крючком верхней че-
люстью. Роговой клюв делал излишним присутствие зубов такого
примитивного характера, когда они служили только лишь для за-
хватывания и удержания добычи.
Получив своеобразное строение органа хватания, одетый ро-
говым чехлом, клюв птиц после потери зубов мог в дальнейшем спе-
циализироваться в различных направлениях, но не мог дать
приспособлений для пережевывания пищи. Эту последнюю функ-
629
цию выполняет у растительноядных птиц мускулистый желудок,
который все же слишком слаб, чтобы заменить зубы жующих ртом
млекопитающих. Клювы птиц являют удивительное разнообразие
в связи с разнообразием принимаемой птицами пищи (рис. 490).
Большая часть птиц питается животной пищей, является з о-
о ф а г а м и; растительноядные — фитофаги — представляют
значительное меньшинство. Среди последних многие, на ряду с ра-
стительной пищей, употребляют пищу животную, причем некоторые,
Рис. 490. Клювы птиц:
1 — сокола балобана, 2 — стервятника, 3 — болотной совы, 4 — шилохвости, 5 —
белолобой казарни, 6 — крохаля, 7 — шилоклювки, 8 — кроншнепа, 9 — бекаса,
10 — выпи малой, И — чайки, 12 — пеликана, 13 —колпицы. (Из Мензбира.)
являясь неразборчивыми в пище, могут быть причислены к группе
всеядных (эврифагия). Как среди растительноядных, так и среди
животноядных птиц можно различить таких, которые специализи-
ровались к принятию определенного сорта пищи — монофагов
(стенофагия) и таких, которые питаются разнообразной пищей
(полифагия).
Среди зоофагов хищными птицами мы называем таких, кото-
рые питаются преимущественно теплокровными позвоночными —
птицами и млекопитающими. Сюда относятся, во-первых, много-
численпые представители отряда нормальных дневных хищных
630
птиц (Acci ])i Ires), обладающих специальными приспособлениями
для добывания пищи в виде хищных ног с длинными острыми крюч-
кообразными когтями и крючкообразной верхней половиной клюва
(рис. 498). Когтями хищные птицы схватывают и умерщвляют до-
бычу, а клюв служит для разрывания на части тела животных.
Приспособление хищных птиц к роду пищи шло в разных на-
правлениях. Во-первых, происходила специализация в отношении
объектов добычи. В то время как одни из хищников птиц специа-
лизировались на добывании млекопитающих, другие специализиро-
вались на ловле птиц, третьи — на добывании рептилий и амфи-
бии, четвертые — на добывании рыб из воды. Так выработались
в процессе эволюции формы питающихся млекопитающими орлов
п сарычей, с широкими крыльями, относительно коротким хвостом,
способностью парить или висеть в воздухе, с ногами, пригодными
для передвижения по земле, с короткими пальцами и длинной плюс-
ной; формы соколов и ястребов, питающихся птицами, которых они
ловят на лету. Из них первые—птицы преимущественно открытых
пространств—обладают, благодаря длинным и узким крыльям,
чрезвц^айно быстрым полетом; короткая плюсна и длинные пальцы
их пог не пригодны к передвижению по земле, но удобны для схва-
тывания летящей птицы. У ястребов крылья относительно короткие,
по длинный хвост, что дает им возможность ловить птиц в лесу, на
быстром полете лавировать между сучьями и стволами. Питаю-
щиеся гадами хищники обладают длинными ногами: таковы
орлы — змееяды, или карачуны (Circaetus), у которых плю-
сна снабжена настоящим панцырем из щитков, защищающих
от укуса змей. Замечательны по сходству своей организации питаю-
щиеся змеями африканский секретарь (Serpentarius serpentarius)
и южноамериканская кариама (Cariama cristata). Относимые на
основании анатомических признаков к совершенно различным груп-
пам птиц, они очень напоминают друг друга внешним видом: имеют
длинные ноги, защищенные от укусов, загнутый крючковатый клюв,
защищенную перьями шею и хохол на голове. Обе птицы одинаково
обращаются со змеями, которых растаптывают ногами. Здесь мы
имеем пример экологического викариата, с одной
стороны, и конвергенции — с другой (рис. 491).
У других хищных мы видим приспособление к ловле рыб в воде,
особенно ярко выраженное у космополитических скоп (Pandion).
(’копа имеет четвертый палец, отворачивающийся назад, когти
острые и длинные, в поперечном разрезе суживающиеся к нижнему
крало, лапа снизу покрыта многочисленными иглообразными рого-
выми образованиями.
Сходное со скопами строение когтей имеют питающиеся рыбой
рыбные орлы (Polioaetus), распространенные в Южной Азии.
На ряду с дифференциацией по роду пищи шла дифференциация
\iini.liих птиц применительно к величине добычи. Крупные хищ-
ники ягнятник, орлы, гарпии, орланы — питаются крупными
млекопитающими, от зайца до серны и средних обезьян; более мел-
кие, как сарычи, луни, балобаны, питаются сусликами, хомяками
631
и т. д.; еще более мелкие питаются мышевидными 'грызунами; то же
относится и к питающимся птицами хищникам: крупных птиц бьют
ястреба-тетеревятники, соколы-сапсаны и кречеты, мелких —
ястреба-перепелятники и соколы-дербники.
Мелкие хищные птицы ловят также насекомых. Так, например,
наш кобчик (Erythropus vespertinus), чеглок (Hypotriorcbis виЬ-
Рис. 491. Африканский секретарь (Serpentarius serpentarius).
(Из Брэма.)
buteo) охотно питаются крупными насекомыми, ловя их налету,
особенно же обитающие в юго-восточной Азии соколы-крошки
(Microhierax).
У нашего осоеда (Pernis) приспособление к питанию перепон-
чатокрылыми выразилось, менаду прочим, в оперении уздечки, ко-
торая покрыта чешуйчатыми жесткими перьями, защищающими го-
лову от жала перепончатокрылых.
Совы, по своей организации относящиеся к далекой от настоящих
хищных птиц группе сизоворонковых птид (Coraeiimor-
phae), по образу жизпи являются хищниками, приспособившимися
к ночному образу жизпи. У них мы имеем сходную с дневными хищ-
никами форму клюва и когтей; однако, у сов четвертый палец отво-
рачивается назад, так что они могут охватывать животных когтями
632
пли с трех сторон, или двумя когтями спереди и двумя сзади. И у сов,
так же как и дневных хищников, дифференпиация шла в разных,
направлениях: по пути приспособления к разному роду пищи —
к ловле птиц, млекопитающих, крупных и мелких, или амфибий,
рептилий и рыб, а также к разным средам обитания, причем образо-
вывались приспособления, сходные с таковыми дневных хищных
птиц. Так, например, добывающая рыб сова-рыболов (Ketupa)
имеет поразительное сходство с ногой скопы.
Отчасти хищниками являются поморники (Stercorarius) из от-
ряда чаек (Lari), сорокопуты (Lanius) из отряда воробьиных. И те,
и другие имеют загнутый крючком клюв.
Что касается птиц, питающихся рептилиями и амфибиями, то,
помимо указанных выше секретаря, кариамы, змееяда, сюда отно-
сятся аисты, цапли, многие из крупных зимородков и мел-
ких сов.
Рыбами питается большое количество птид из разнообразных от-
рядов. Отряды: гагары (Colymbi), пингвины (Sphenisci), веслоногие
(Steganopodes), чайки (Lari), чистики (Alcae) целиком или большей
частью являются рыболовами. Цапли (Ardeae), крохали (Merginae)
и некоторые нырки из гусиных (Anseres), указанные выше скопы
и рыбные орлы из хищных, зимородки (Alcedinidae) из сизово-
ронковых (Coraciae) питаются или исключительно рыбой, или преи-
мущественно таковой. При этом мы встречаем весьма разнообраз-
ные приспособления для ловли рыб.
Гагары, пингвины, веслоногие, крохали и чистики ловят рыб,
ныряя за ними, причем пингвины и некоторые чистики во время
пыряния двигаются под водой с помощью крыльев. Чайки, хищные
птицы, питающиеся рыбой, бросаются с высоты в воду, высматри-
вают добычу, летая над водой, в то время как зимородки бросаются
за рыбой в воду, высмотрев предварительно ее, сидя где-нибудь
над водой.
Цапли караулят рыбу, стоя в воде, и хватают добычу подобно
змеиным птицам, или анхингам (Plotus), быстро выбрасывая вперед
благодаря длинной шее свою голову с длинным и острым клювом.
Для удерживания пойманной рыбы у многих из рыбоядных
птиц имеется крючок на конце клюва, Аа у крохалей (Mergus) по
краям клюва имеются роговые зубцы.
У пеликанов (Pelecanus) на нижней челюсти подвешен мешок,
и котором сохраняется пойманная рыба.
Немалое количество птиц питается трупами других животных,
падалью. Таковы, во-первых, грифы (стервятники) Старого (из
Л(। ui 1 idae) и Нового Света (Cathartae), во-вторых, крупные виды нор-
мальных дневных хищников, как, например, орлы, орланы, подор-
лики и коршуны, вороны, вороны из воробьиных и буревестники,
или трубконосы (Procellarii), для которых падаль не является,
пдплкп, исключительной пищей.
В спнзи с переходом на питание падалью у грифов Старого иНо-
iHini Спета изменилось строение пальцев и особенно когтей, так как
ипп перестали здесь играть роль органа умерщвления' добычи.
633
Далее падалью и другими отбросами питается аист марабу
(Leptoptilus), представленный африканским и южноамериканским
видами. Типичные представители видов, питающихся падалью,
имеют как приспособление в большей или меньшей степени оголен-
ные части головы и шеи. У некоторых голой или слабо оперенной
оказывается лишь часть головы, у других — вся голова и даже
шея. Таковы марабу, королевский гриф и кондор Америки, сипы
и другие азиатские грифы. Это помогает птицам забираться головой
в глубь трупа, не боясь запачкать перья. Грифы разыскивают падаль
с помощью удивительно острого зрения, поднимаясь чрезвычайно
высоко над землей. Большинство питающихся падалью птиц спо-
собно одновременно съедать громадное количество пищи и затем
подолгу оставаться без всякой пищи.
Чрезвычайно большое количество птиц питается мелкими жи-
вотными из различного рода беспозвоночных.
Водными беспозвоночными питаются буревестники (Procella-
rii), чайки, утки, нырки, фламинго, ибисы и мелкие кулики.
Буревестники вылавливают различных беспозвоночных на по-
верхности воды, неутомимо летая над волнами; чайки ныряют за
пищей в воду, бросаясь с воздуха, тогда как утки и нырки ныряют
за беспозвоночными, плавая по поверхности. У утиных имеются
для добывания особые приспособления в клюве. Внутренняя по-
верхность верхнего и нижнего клюва снабжена по краям особыми
пластинками; вместе с бахромчатыми краями языка они составляют
особый цедильный аппарат, свободно пропускающий воду, но удер-
живающий все съедобное во рту. Этому процеживанию помогает
обилие нервных осязательных окончаний на пластинках. Такое же
строение имеет массивный согнутый коленом клюв фламинго.
Эта птица, по своей организации близкая к аистам, обладает длин-
ными ногами и длинной шеей и питается так же, как и утки, т. е.
пропуская через'клюв, как через сито, воду с илом. У колпицы
(Platalea) клюв ложкообразно расширен, у ибисов (Ibis) и многих
куликов — загнут серпообразно, в большинстве случаев вниз
(кроншнепы, серпоклювы, краснозобики и др.), а у шилоклювки
(Recurvirostra) — вверх. У кривоклювого песочника клюв загнут
вправо. Клювы многих куликов достигают высокой специализации
в связи с родом их пищи. Таков, например, клюв кулика — сороки
(Haematopus ostralegus), питающегося преимущественно двуствор-
чатыми моллюсками (Mytilus, Ostrea и др.).
Многочисленные кулики добывают себе пищу (разного рода
беспозвоночных) из земли с помощью своего клюва, который у се-
мейства вальдшнепов (Scolopacidae) особенно обильно снабжен
осязательными окончаниями на поверхности, что дает птице воз-
можность разбираться в попадающихся в земле предметах.
Громадное большинство мелких пташек (Passeres) питается
насекомыми и их личинками (рис. 492). Как общее правило, клюв
насекомоядных птиц тонкий и узкий, с крючком на конце и иногда
с зазубринкой по краям. У птиц, которые добывают себе насекомых
налету, клюв очень широкий, плоский, с широким разрезом. Таков
634
он у ночных насекомоядных — козодоев, и у дневных — стрижей и
ласточек. У мухоловок, которые схватывают налету насекомых,
по пе гоняются,а только бросаются за ними с ветки дерева, клюв
приближается по строению к клюву ласточек.
У птиц, добывающих себе насекомых на деревьях, имеются спе-
циальные приспособления к лазанию: цепкие лапы, как, например,
Рис. 492. Клювы птиц:
1 — черного дрозда, 2 — дубоноса, 3 — клеста, 4 — козо-
доя, 5 — ржанки, 6 — пищухи, 7 — овсянки, 8 — голубя,
9 — снегиря, 10 — поганки, 11 — дятла и 12 — перепела.
(Из Мензбира.)
V синиц, поползней, дятлов, пищух; жесткий хвост, дающий воз-
можность упираться в дерево, и различные формы клюва специаль-
ною назначения. Так, пищухи и стенолазы (Tichodroma) имеют
очин, топкий недлинный клюв, с помощью которого они достают
ннюкомых из щелей; нектарки, маленькие прекрасные птички
('ыриго Спета, и колибри Нового Света, добывающие насекомых из
'ifiH’ion. откуда они пьют также пектар, имеют, кроме узких и длин-
1и.1\ h'lionoii весьма разнообразной формы, еще длинный растяжимый
и илк А’ дятлов, добывающих насекомых, живущих в древесине,
помимо очень крепкого нестрого клюва, исполняющего роль долота,
635
имеется длинный язык, способный далеко выбрасываться вперед
благодаря длинным, загибающимся на темя рожкам, снабженный
на конце роговым острием с боковыми зазубринками.
Птиц, питающихся растительной пищей, вообще говоря, меньше,
чем зоофагов. Большое количество мелких пташек из отряда во-
робьиных питается семенами растений — это зерноядные птицы.
Сюда относятся вьюрковые птицы (Fringillidae), ткачи (Ploteidae),
жаворонки (Alaudidae) и вороновые птицы (Corvidae). Клюв этих
птиц толстый, конический, приспособленный для шелушения или
разгрызания семян. Особенно толстый клюв у дубоноса (Coccothraus-
tes), который может раздроблять даже косточки (костянки) плодов
(вишен, слив). У клеста (Loxia) верхний клюв перекрещивается
с нижним, что дает ему возможность легко вышелушивать шишки
хвойных деревьев. Ореховки и сойки могут раздроблять ореховую
шелуху.
Семенами по преимуществу питаются птицы из отряда куриных
(Galli), трехперстки (Turnices), тинаму (Tinami).
Что касается голубей (Columbae), то среди них мы видим и таких,
которые питаются зернами, и таких, которые питаются мягкими
плодами.
Мягкими плодами питаются попугаи (Psittaci), туканы (Rhampha-
stidae), птицы-носороги (Bucerotidae), бананоеды (Musophagidae).
У попугаев большой массивный клюв, похожий на клюв хищ-
ной птицы, подвижно сочленяется с черепом, что делает его при-
годным к объеданию мякоти плодов. У южноамериканских ту-
канов (Rhamphastidae) клюв очень длинный и толстый, но легкий
благодаря наличности спонгиозной (губчатой) костной ткани. Края
клюва зазубрены пилообразно. Это дает возможность разрезать ко-
жистую оболочку плодов, а длинный клюв позволяет им отрывать
плоды с тонких веток деревьев, до которых добраться иначе не было
бы возможности. Птицы-носороги экологически заменяют туканов
в тропических лесах Старого Света.
VIII. РАЗМНОЖЕНИЕ ПТИЦ
Вторичные половые признаки птиц. Помимо органов воспроиз-
ведения, полы птиц отличаются рядом других особенностей, и ни
в одном классе животных нет такого разнообразия в развитии вто-
ричных половых признаков, как у птиц. Опо выражается в различ-
ных размерах самца и самки, в окраске, в форме перьев, в наличии
разного рода придатков, в порядке линьки, в способности издавать
различные звуки, в пении, в различного рода движениях, в осо-
бенностях образа жизни и связанных с этим инстинктах.
Как общее правило, самцы крупнее самок. Исключение из этого
правила представляют дневные хищные птицы, большинство
куликов, трехперстки, тинаму и казуары; самки некоторых видов
отряда трехперсток (Turnices) не только крупнее самцов, но и
ярче окрашены. К подобным особенностям самок этих птиц присоеди-
няются и другие черты, характерные для самцов других видов.
Так^ у трехперсток самки, а не самцы токуют и дерутся во время то-
636
ков, а самцы, в противоположность прочим птицам, высиживают
япца и воспитывают птенцов. Подобно трехперсткам более яркую
окраску имеют самки нашего куличка-плавунчика (Phalaropus
lobatus). И здесь самец принимает на себя высиживание яиц.
Примеров более яркой окраски самцов можно привести бес-
численное множество даже среди нашей фауны. Достаточно ука-
зать на окраску петухов по сравнению с курами, индюков и ин-
дюшек, селезней и уток, павлинов, фазанов, тетеревов и т. д.
Очень часто яркое оперение самцов сопровождается развитием осо-
бой формы «украшающих» перьев или других дридатков. Таковы
Рис. 493. Турухтаны (Philomachus pugnax) самцы во время драки.
удивительные перья павлина, фазанов,хвост нашего обыкновенного
тетерева, перья на шее турухтанов (рис. 493), поразительные укра-
шения райских птиц, многих колибри, эгретки белых цапель
и многих других; к таким же «украшениям» должны быть от-
несены голые придатки па голове и шее индюка и других птиц.
У очень многих птиц яркая окраска самцов (а иногда и самок)
развивается только в период спаривания, после чего птица линяет
и надевает более скромный наряд (самцы турухтанов, селезни
уток); у таких самцов имеется тогда одна лишняя по сравнению
г. самками дополнительная линька. К тому же в период ухажива-
ния ярко окрашенные и украшенные самцы имеют обыкновение
производить своеобразные телодвижения, надуваться, ерошить
перья, как бы выставляя эти украшения напоказ.
I {прочем, особые телодвижения, игры, ерошепие перьев в период
сваривания свойственны не только тем самцам, которые имеют
ирное оперение, по и тем, у которых самцы очень скромно окрашены
и попсе не отличаются от самок, как, например, у воробьев.
Ко вторичным половым признакам относятся также те инстинк-
ты, которыми обладает только один какой-либо пол. Они обычно
637
связаны с размножением. Таковы все инстинкты самцов, служащие
для обеспечения оплодотворения самки. Так как оплодотворение
у птиц может быть достигнуто только при условии одновременной
готовности к этому как самца так и самки, то у птиц черезвычайного
развития достигают различного рода акты, значение которых за-
ключается в том, чтобы вызвать соответствующие психофизиоло-
гические рефлексы, обеспечивающие оплодотворение. Как правило,
именно самцы обладают этими инстинктами, обусловливающими
деятельность, направленную на половое возбуждение самки.
Пение, брачные крики, брачные полеты, ерошенье перьев, пози-
рование, игры, пляски, токование, гнездостроение, поднесение
корма и т. д., иногда очень сходная деятельность самцов птиц в брач-
ный период являются необходимой рефлекторной подготовкой
половой деятельности. У некоторых видов птиц, однако, актив-
ная роль этой деятельности принадлежит самкам: таковы трехперст-
ки (Turnices), золотой кулик (Rostratula), плавунчик (Phalaropus).
Токование. Игры, пляски и драки в брачный период спари-
вания, часто сопровождаемые особого рода звуками, называются
токованием.
Наиболее типичен ток у п о л и г а м о в, т. е. у тех птиц, у ко-
торых самцы спариваются в брачный период нес одной, а с несколь-
кими самками, более или менее случайно.
У таких птиц в определенное время суток в определенных ме-
стах собираются самцы, своеобразными движениями и характер-
ными звуками привлекают самок, ожесточенно дерутся между собой,
чтобы затем вместе с прилетевшими на ток самками образовать па
короткий срок пары, а на следующий день в том же самом месте и в
то же самое время вновь повторяют ту же игру (рис. 493). Тетерева
токуют ранним утром па открытых местах, собираясь на токовище
там, где их еще много, в количестве до сотни штук. Распустив веером
хвост, опусти^ крылья, пригнув к земле и вытянув шею, они издают
своеобразное «бормотание» или шипящие звуки — «чуфыкание»,
хлопают крыльями и вступают друг с другом в драку. Глухари
токуют в лесах по утренним зорям, издавая короткую трель, во
время второй половины которой запрокидывают голову кверху,
становясь совершенно глухими. Вальдшнепы токуют вечерами,
летая над лесом, «хоркая» и «циркая». Это — так называемая
тяга вальдшнепов. Бекасы токуют в воздухе, поднимаясь на
значительную высоту, время от времени стремительно опускаясь
вниз, издавая своим хвостом дребезжащие звуки, похожие на бле-
яние барана. Дупеля токуют по болотам, собираясь большими об-
ществами. Кукование кукушек тоже является своего рода током.
Токование в период спаривания, сопровождаемое играми, тело-
движениями и пепием, встречается также у многих м о н о г а м ов.
Такой ток кончается образованием пар, которые и приступают
к гнездованию.
Однако самцы многих видов уже после образования пар про-
должают воспроизводить движения, издавать крики и пение
и драться друг с другом даже тогда, когда самки уже сядут на яйца.
638
Так бывает, например, у белых куропаток (Lagopus lagopus),
у стрепетов (Microtis tetrax) и у других.
Что касается брачных звуков, то они свойственны самцам боль-
шинства видов птиц. Простые брачные крики, вроде тех, что из-
даются многими утками, нырками, куликами и др., переходят в более
мелодичное воркование голубей и, наконец, в пение, которое столь
характерно для отряда воробьиных (Passeres). Птицы обла-
дают особым голосовым аппаратом в нижней части трахеи, там, где
она ветвится на двое бронхов. Здесь образуется сложно построен-
ная нижняя гортань, или syrinx (рис.
494).Наиболее сложно устроен голосовой
аппарат певчих птиц; совсем отсутствует
он у аистов и американских грифов, ко-
торые, таким образом, вовсе лишены
голоса. Между этими крайностями
имеются многочисленные переходы.
В общем в зависимости от развития
той или иной части голосового аппарата
различают т р а х е й и у ю (древола-
зы), бро нхиальную (кукушки
и совы) и трахейно-бронхи-
альную гортани (большая
часть птиц). Из осложнений голосо-
вого аппарата можно назвать расши-
рение трахеи у утиных или удли-
нение до такой степени, что у некото-
рых птиц она образует петли, врастая
8
у журавля и лебедя-кликуна в гребень
грудины, а у райской птицы (Baryta)
складываясь в спиральную петлю и
окостеневая. В связи с этим лебеди и
журавли обладают особенностью произ-
водить громкие трубные звуки, а самец
Рис. 494. Нижняя гортань
(syrinx) птиц.
1 и 2—трахейно-бронхиальная
система, гортань ворона: 1 —
вид сбоку, 2 —вид со спинной
стороны, 3 — трахейная гор-
тань (схема), 4—бронхиальная
гортань (схема). (Из Мензбира.)
Baryta издает звуки, сходные со звоном колокола. Наиболее разви-
тым голосовым аппаратом обладают певчие птицы (Oscines).
Пение птиц представляет большое разнообразие, причем
в одних группах близкие виды очень сходны между собой по пению,
в других, наоборот, близкие, виды резко отличаются в этом отно-
шении. Многие птицы владеют способностью звукоподражания
и вплетают в свою песнь звуки и строфы из песни других птиц или
звуки, воспроизводимые другими животными. Поющие птицы,
несомненно, получают от пения большое удовлетворение и тратят
на него много времени. Для большинства птиц время пения огра-
ничено в году временем воспроизведения: начинается незадолго до
начала спаривания и оканчивается с выводом птенцов и связано та-
ким образом с размножением. Самки по большей части вовсе не
поют и имеют редуцированный голосовой аппарат. В редких слу-
чаях исключения из этого правила пение самок значительно проще
и слабее.
639
Наконец, есть птицы, которые производят звуки не голосом.
Так, лишенные голоса аисты издают громкое щелканье клювом,
куриные и голуби хлопают крыльями, а дятлы воспроизводят клю-
вом барабанную трель о дерево. Эти звуки могут заменять песню
и воспроизводятся тогда самцами в период размножения.
Чрезвычайно интересными являются удивительные сооруже-
ния некоторых австралийских птиц из подсемейства шалашников
(Ptilonorhynchidae), связанные с периодом воспроизведения, —
так называемые «беседки» и «сады». Беседки строятся на земле из
прутьев и стеблей, основания которых представляют пол, а верши-
ны сходятся сводом, образуя как бы коридор. Такой шалаш укра-
шается раковинками, красивыми камнями, белыми костями и яр-
кими перьями попугаев. Сады устраивают птицы того же семейства
в Новой Гвинее. Помимо беседки, которая строится из прутьев в виде
шалаша у подножия какого-либо дерева и устилается мхом, около
беседки устраивается площадка, которая также устилается зеленым
мхом, а на нем раскладываются яркие цветы и плоды растений.
Вся такая площадка производит впечатление миниатюрного са-
дика, который поддерживается в образцовом порядке: трава
тщательно выпалывается, а увядшие цветы и плоды заменяются
новыми.
Спаривание птиц. Брачные звуки и пение птиц, помимо полового
возбуждения, имеют еще другое значение. Заняв определенный
гнездовый участок, самцы звуками дают знать другим самцам того же
вида о том, что участок занят, обусловливая тем более равномер-
ное распределение по стациям гнездящихся пар; своими криками
опи предупреждают самок, которые у многих перелетных птиц
прилетают позднее, о том, что участок для гнездования выбран
и дают им возможность распределиться между самцами. Самцы
ревниво охраняют занятый участок от вторжения других сам-
цов, а самки, выбраЪ самца, не допускают в участок других самок.
В результате токов у полигамов самки оплодотворяются, кла-
дут яйца и приступают к высиживанию яиц. Самцы заканчивают
токи и живут одиноко или небольшими обществами, совершенно не
принимая участия в заботах о потомстве. У моногамов же обра-
зуются пары, причем самцы в разной степени принимают участие
в заботах о потомстве.
Высиживает яйца обычно одна самка; у многих птиц самец по-
могает высиживать яйца, обычно в то время, когда самка кормится,
и у очень немногих птиц высиживание яиц целиком или главным
образом падает на долю самца. Это мы видим у нанду (Rhea),
золотого кулика (Rostratula) и у птиц нашей фауны: у пла-
вунчика (Phalaropus lobatus), лапландского веретенника (Li-
mosa lapponica) и глупой сивки (Eudromias morinellus).
У громадного большинства птиц пары образуются лишь на
время вывода птенцов, и после окончания забот о потомстве пары
расходятся, а перед выводом птенцов на следующий год заклю-
чаются новые брачные союзы в новых комбинациях. Однако у не-
которых видов пары образуются на более продолжительный период,
640
иногда быть может на всю жизнь. Так, многие хищники, особенно
крупные, образовав пару, живут совместно в продолжение мно-
гих лет.
Гнезда птиц. Почти все птицы устраивают гнезда для высижи-
вания птенцов; очень многие птицы выкармливают в этих же гнез-
дах своих птенцов. Однако существуют птицы, которые вовсе не
устраивают гнезд и даже не высиживают яиц.
Гнездостроение, несомненно, явилось приспособлением для
высиживания яиц. Дальнейшее усложнение гнезд обусловлено было
сперва приспособлением к сохранению яиц и высиживающих их
Рис. 495. Самка африканского страуса садится
на яйца.
(Фото Н. Озерова.)
самок или самцов, а затей приспособлением к сохранению выкар-
мливаемых в гнезде беспомощных птенцов. Период высиживания
ниц и особенно выкармливания птенцов в гнезде является одним
пи самых узких мест в экологии птиц, так как яйца и птенцы, если
инн пе скрыты и не помещаются в защищенных или недоступных
'мн врагов местах, могут легко поедаться рептилиями, млекопитаю-
щими и некоторыми птицами. Эволюция гнездостроепия шла в раз-
ных систематических группах по разным путям в связи с особен-
ностями других приспособлений: к средам обитания, к пище и
в связи с особенностями местообитаний. Наиболее совершенно
устроенные гнезда мы встречаему мелких птиц, особенно у воробь-
иных, которые, как мы видели, являются в то же время наиболее
молодой и богатой видами группой.
Тогда как высиживание яиц является характерной особенностью
птиц, у рептилий мы высиживания в собственном смысле не встре-
I KiuiiKHpou и Стапчинский 641
чаем; нужно считать, что эта особенность явилась у птиц новооб-
разованием и связана с приобретенной птицами теплокровностью
и потерей чешуйчатого покрова, который заменился перьями
и пухом. А так как у птиц забота о потомстве выражается не только
в высиживании яиц, но и в воспитании птенцов, а у очень многих
птиц в продолжительном выкармливании птенцов в гнезде, то
можно было бы думать, что и эти особенности являются благо-
приобретенными и что те группы птиц, у которых птенцы вылуп-
ляются из яиц более развитыми, у которых отсутствует гнездо-
строение и яйца не высиживаются, должны считаться наиболее при-
митивными. Однако бли-
жайшее знакомство с
этим вопросом показы-
вает, что у современных
птиц не сохранились ^эти
примитивные черты, и
случаи отсутствия гнезд
и высиживания в боль-
шинстве случаев пужно
рассматривать как вто-
ричную утрату суще-
ствовавшей ранее спо-
собности.
Наиболее оригиналь-
ны по сравнению сдруги-
ми птицами в отношении
высиживания так назы-
ваемые сорные куры
Рис. 496. Гагарка (Alca torda). (Megapodidae), распро-
страненные в Австралии
ина островах'Тихого океана. Свои яйца они зарывают в песок или в
кучи гнилых растительных веществ и не высиживают их. У птиц, ко-
торыекладут свои яйца впесок на доступных солнцу открытых местах,
зародыши в яйцах развиваются за счет теплоты солнца; у птиц, от-
кладывающих яйца в кучи гниющих растительных веществ, —
за счет температуры гниения. Любопытно, что температура гние-
ния по времени и интенсивности как раз соответствует потребно-
сти, между тем как постройка куч пачинается задолго до кладки яиц.
Птенцы мегапод выклевываются весьма развитыми и способны-
ми самостоятельно двигаться и брать пищу, а у некоторых сорных
кур покрыты перьями и способны почти тотчас же летать.
Полное отсутствие гпезд у других птиц, несомнеппо, является
вторичным приспособлением. Прямо на землю кладут свои яйца
чистики (Cepphus), кайры (Uria) и гагарки (Alca) (рис. 496), гнездя-
щиеся по выступам прибрежных скал; крачка (Sterna hirundo) и
песочники откладывают яйца между гальками, козодои (Capri-
mulgus) — прямо на голой почве лесов. Интересно, что яйца пер-
вых двух настолько сходны по цвету с гальками, что заметить
их чрезвычайно трудно, и высиживающие яйца птички оставляют
642
нч при приближении малейшей опасности; у козодоя, наоборот,
полые с буроватыми пестринками яйца настолько резко отличаются
от ветоши почвы леса, что издали бросались бы в глаза, если бы
и х пе прикрывала птица, чрезвычайно сходно окрашенная с окру-
жающей обстановкой. И в противоположность песочникам козодой
сидит на яйцах очень крепко.
Многие кулики (Limicolae) кладут свои яйца в ямки, как на-
пример, чибис, бекас. Рытье ямок на обрывах легко могло перейти
к рытью норок, как, напри-
мер, делают береговые ласточ-
ки (Riparia riparia), щурки
(Merops), красные утки и
пеганки (Tadorna tadorna),
зимородок (Alcedo) и очень
многие другие. С другой сто-
роны, от таких птиц, кото-
рые несут свои яйца на ка-
кой-либо подстилке, нарочито
принесенной для кладки яиц,
существуют многочисленные
переходы к настоящим гнез-
д остро ениям.
Олуши (Sula bassana), мно-
гие чайки (Lari), некоторые
кулики (Limicolae), многие
гуси (Anseres) и куры (Galli)
делают для яиц весьма при-
митивную подстилку из мор-
ской травы, листьев, стеб-
лей трав, собственного пу-
ха, мха и т. п. Коньки
(Anthus), трясогузки (Mota-
ci 11а), жаворонки (Alaudidae)
и многие другие птицы, гнез-
дящиеся на земле, вьют из
стеблей трав гнезда разной
степени сложности. Крупные
птицы пользуются для этого
пе только стеблями, ио ве-
Рис. 497. Гнезда саланганы (Collocalia
sp.J.
точками и сучьями кустарников. Если подобные гнезда поме-
щаются не на земле, а на горизонтальных сучьях деревьев
пли в корнях кустов, то переход к гнездящимся на ветвях пред-
ставляется само собой понятным. Таковы примитивные гнезда
дневных хищников (Accipitres), голубей (Columbae) и многих
других. Постройка таких гнезд может все более и более ослож-
няться, пока не достигает удивительного совершенства гнезд мно-
гочисленных воробьиных. От гнездящихся в норах, расщелинах
скал и в развилинах деревьев естественным кажется переход к пти-
цам, делающим гнезда в дуплах. Таких птиц очень много. Назо-
41“
643
йем для примера дятлов (Pici), сов (Striges), поползней (Sit-
tidae), синиц (Paridae). Дятлы сами выдалбливают себе дуп-
ла, другие птицы довольствуются готовыми дуплами. У наших
поползней (Sitta) мы встречаем привычку обмазывать глиной от-
верстие дупла, чтобы тем уменьшить его диаметр, а у своеобраз-
ных птиц-носорогов (Buceros), относящихся к отряду сизоворонок
(Coraciae), мы находим удивительный обычай замуровывать
особым похожим на резину веществом сидящую в дупле на яйцах
самку, так что остается лишь небольшое отверстие, через которое
самка может просунуть только свой клюв, чтобы получить пищу от
самца.
От птиц, которые выполняют те или иные лепные работы при
приспособлении гнезд, можно перейти к таким гнездостроителям,
у которых гнезда лепятся из различных материалов. Ласточки
склеивают грязь застывающей слюной; съедобные гнезда саланган
(из отряда стрижей — Cypseli) вылепляются из затвердеваю-
щей в студень слюны, к которой бывают примешаны морские
водоросли (рис. 497). Гватемальский стриж (Panyptila sancti
hieronymi) подвешивает свое удивительное лепное гнездо на
тонкие ветки деревьев, свисающие над водой, употребляя во-
локна, которые склеивает слюной; гнездо висит как бы на веревке;
входное отверстие внизу ведет в расширение верхней части, где
помещается собственно гнездо. Такое гнездо можпо считать вер-
хом совершенства в смысле безопасности.
Часто гнезда удивительны по прочности и изяществу. Прекрасно
свиты и изящно украшены с целью скрыть гнездо от внимания гнезда
наших зябликов и щеглов; маленькие крапивники вьют искусные
шаровидные гнезда из мха; длиннохвостые синицы сооружают
также шарообразные гнезда из мха, шерсти, паутины, волос и
перьев. Ремез свивает из растительного пуха мягкое войлокооб-
разиое висячее гнездо в виде реторты.
Особенно интересны колониальные гнезда африканского об-
щественного ткача. Большая колония птичек (до 300) совместно
сооружает в виде зонта навес, под которым помещаются висячие
гнезда отдельных парочек.
Нельзя не упомянуть также про пловучие гнезда некоторых птиц.
Такие гнезда устраиваются лысухами (Fulica atra) и поганками
(Podiceps). Первые прикрепляют свои пловучие гнезда к камы-
шу, последние вьют их на открытой воде из разных плавающих
растительных веществ.
Некоторые виды вовсе не строят гнезд, а занимают готовые
гнезда других видов птиц. Большинство птиц каждый раз строит
новые гнезда, но некоторые виды селятся в старых гнездах из года
в год (орлы, аисты, цапли, ласточки).
В постройке гнезд принимают участие или оба пола или постройка
производится одной лишь самкой, а самец или вовсе не участвует
или приносит материал. Можно считать установленным, что птицы
руководствуются при постройке гнезд врожденными инстинктами.
Инстинкты, связанные с гнездостроением, чрезвычайно интересны
644
и иногда удивительно сложны. У многих птиц в период гнездова-
ния большого развития достигает общественность, которая ведет
к колониальному гнездованию.
Яйца птиц. Подобно громадному большинству рептилий птицы
являются яйцекладущими животными. Оплодотворенное яйцо, снаб-
женное большим количеством питательного желтка и тонкой обо-
лочкой (membrana vitellina), в яйцеводе одевается белком, рас-
положенным тремя слоями: во-первых, халазообразующей оболоч-
кой, переходящей по обеим сторонам в шнуровидные, закруженные
спирально халазы, служащие осью, на которой подвешен в белке
желток. Благодаря халазам желток всегда ориентирован вверх
своей более легкой частью, в которой происходит дробление яйца.
Далее, яйцо одевается тонким слоем жидкого белка и толстым слоем
плотного. В конце яйцевода яйцо одевается тонкой двойной пере-
пончатой прозрачной оболочкой, между листками которой распо-
ложена воздушная камера. В матке белок яйца, во-первых, снаб-
жается значительным количеством воды (до 50—60%), а, во-вто-
рых, яйцо одевается скорлупой.
Скорлупа состоит главным образом из углекислого кальция,
и (.‘значительного количества фосфорнокислого кальция и органи-
ческого вещества. Скорлупа снабжена многочисленными порами,
через которые происходит свободно газообмен, имеет различную
толщину и различное строение. Иногда она совершенно гладкая,
как бы полированная (Tinami); у других она, наоборот, зер-
нистая, шероховатая. У пеликанов, фламинго, личинкоедов
собственно скорлупа покрывается сверху еще губчатой известковой
массой.
Толщина скорлупы яйца, повидимому, стоит в связи со способом
откладки яиц. У птиц, гнездящихся в дуплах и вообще в закрытых
местах, скорлупа очень тонкая и хрупкая; наоборот, она толстая
и прочная у птиц, не строящих гнезд, а откладывающих яйца прямо
па землю, особенно на камни.
У очень многих птиц яйца покрыты снаружи тонкой кожицей,
снабженной порами. Она у разных видов имеет различную толщину.
< Особенно мало она развита у водоплавающих птиц и еще слабее
у птиц, гнездящихся в дуплах; предполагают, что назначение этой
кожицы — защищать зародыш от сырости.
Чрезвычайное разнообразие представляет окраска яиц. Пиг-
менты, обусловливающие окраску, откладываются обыкновенно
г птк,ладкой скорлупы и могут даже лежать так глубоко, что сна-
ружи незаметны вовсе. Пигменты могут быть сведены к двум ос-
ипни ым типам: голубому (ооциан) или желтому (оохлорин) и красно-
му рому (оопорфирин), которые в зависимости от интенсивности
н расположения в скорлупе, обусловливают все бесконечное раз-
нообразие окраски яиц. Существуют, однако, яйца, совершенно
'iHiiienиые пигмента. Тогда они кажутся белыми, а при тонкой
скорлупе — нежнорозовыми благодаря просвечиванию содержи-
мого. Таковы яйца дятлов, стрижей, колибри, мышиных птиц, сов,
•и у рок, зимородков, попугаев и почти всех голубей.
645
Окрашенные яйца бывают или одноцветные без пестрин, осо-
бенно часто голубые или зеленовато-голубые, или покрыты пе-
стринами в виде мелких и крупных точек, пятен, мазков или линий.
Глубже лежат более бледные, на поверхности — более яркие цвета.
Несомненно, каждый вид птиц характеризуется определенной
окраской яиц, но иногда окраска сильно вариирует у различных
особей того же вида, как, папример, у колониально гнездящихся
кайр (Uria troille), у кукушек. В общем каждая самка обладает
своими особенностями, которые передаются по наследству. Однако
существуют иногда очень значительные вариации в окраске яиц
одних и тех ясе самок.
Несомненно, что окраска яиц—признак, приобретенный пти-
цами, так как рептилии несут яйца без всякой окраски. Возможно,
что окраска яиц выработалась как защитное средство: с одной сто-
роны, как покровительственная окраска, когда яйца по своему
цвету подходят к окружающей обстановке и оказываются поэтому
мало заметными, с другой стороны, — как защита от солнечных
лучей. В этом отношении белая окраска яиц у птиц, гнездящихся
в закрытых помещениях, и темная у гнездящихся открыто — вполне
отвечала бы такому предположению. Однако белые и пестрые яйца
встречаются как у тех, так и у других.
Белая окраска яиц встречается: иногда у птиц, гпездящихся
открыто, но которые, оставляя гнезда, прикрывают яйца или соб-
ственным пухом (Anseres), или растениями; или у птиц, ко-
торые очень крепко сидят на яйцах и слетают с них очепь редко;
или затем у птиц достаточно крупных и сильных.
Пестрая окраска яиц у птиц, гпездящихся в дуплах, может
быть объяснена тоже как сохранившееся приспособлепие птпц,
ранее гнездившихся в открытых гнездах (например, галка).
Во многих случаях покровительственная окраска яиц выражена
чрезвычайно сильно/Достаточно привести в качестве примеров яйца
пигалиц, бекасов и куликов, по окраске гармонирующих с мши-
стой поверхностью почвы; песочников, морских ласточек, которые
кладут свои яйца прямо на песок с разбросанными между ними
гальками, — яйца как нельзя более походят на гальки; яйца
тетеревов, глухарей и белых куропаток гармонируют по цвету
с прошлогодней ветошью; яйца лысухи, окрашенные под цвет
камышей, и т. д.
Окраска япц для одних групп птиц является удивительно по-
стоянной, тогда как в других группах даже близкие виды птиц
несут разно окрашеппые яйца.
Величина яиц только до известной степени стоит в коррелятив-
ной зависимости от величины птицы этого вида. Весьма сильно
сказывается па величине яиц то состояние, в котором вылупляются
из яиц птенцы. У птиц выводковых яйца относительно зпачительпо
крупнее, чем у птенцовых. Особенно малы по сравнению с величи-
ной птицы яйца кукушки. Это вполне соответствует инстинкту ку-
кушки откладывать свои яйца в гнезда различных мелких
пташек.
646
Форма лиц для систематических групп птиц иногда весьма
\.iрактерпа, хотя и вариирует у одного и того же индивидуума.
В общем по форме можно различить четыре типа яиц: округ-
। у ю (совы, зимородки), овальную (козодои, колибри, ряб-
ки, дневные хищники, голуби и др.), удлиненно-оваль-
ную (гагары, веслоногие) и грушевидную (ржанки, чи-
онг.ы, кулики и чистики). Форма яиц имеет иногда определенное
значение. При грушевидной форме четыре яйца лежат крестооб-
разно своими острыми концами к центру и тем занимают мини-
мал г.ное пространство; у чистиков, которые кладут яйца на утесах
без всякого гнезда, грушевидная форма не дает возможности
япцам при ветре или толчках упасть, так как такое яйцо не катит-
ся, а только вертится на месте.
Количество яиц в кладке колеблется от 1 до 22. Одно яйцо в клад-
ке характерно для отряда буревестников (Procellarii), для боль-
шей части чистиков (Alcae), для наиболее крупных хищников
(Accipitres), для некоторых голубей (Columbae), чаек (Lari), неко-
торых стрижей (Cypseli) и козодоев (Caprimulgidae).
Два яйца откладывают киви (Apteryx), пингвины (Sphenisi),
гагары (Colymbi)/ журавли (Grues), прочие чистики (Alcae),
большая часть голубей (Columbae), козодоев (Caprimulgi), стрижей
(Cypseli) и колибри (Trochili).
Три яйца характерны для рябков (Pterocletes) и большей части
чаек (Lari).
Четыре яйца в кладке имеют трехперстки (Turnices) и боль-
шая часть куликов (Limicolae).
В общем, как правило, можно считать, что птицы, питающиеся
позвоночными животными, откладывают наименьшее число яиц
в сравнении с птицами, питающимися растительной пищей; вывод-
ковые птицы откладывают наибольшее число яиц. У нанду (Rhea)
несколько самок откладывают в одно гнездо до 40 яиц; высиживает
пх один самец.
Большинство птиц выводят один раз в год, но некоторые — два
и даже три раза в год.
Продолжительность ^высиживания* весьма различна и отчасти
зависит от абсолютной величины яйца. В общем продолжительность
колеблется от 10 до 14 дней у мелких пташек, от 15 до 26 — у птиц
средней величины, от 26 до 40 — у крупных птиц, до 55 — у кондора
п до 58 дней — у эму, 60 дней у буревестника (Diomedea). У ку-
риных, водоплавающих и хищников средняя продолжительность
высиживания в общем около 24—28 дней.
Птенцы. Различают птиц выводковых и птенцовых.
У выводковых птиц птенцы выклевываются из яиц довольно раз-
витыми. Тело у них покрыто пухом, а у некоторых сорных кур
(Megapodidae) — даже настоящими перьями; обсохнув, они могут
бегать или плавать и следовать за матерью; опи умеют клевать корм
ji быстро научаются его отыскивать. Примером выводковых птиц
могут служить все наши домашние птицы: куры, утки, гуси, индейки
л др. (за исключением голубя). Выводковыми являются псе палеог-
647
Окрашеппые яйца бывают или одноцветные без пестрин, осо-
бенно часто голубые или зеленовато-голубые, или покрыты пе-
стринами в виде мелких и крупных точек, пятен, мазков или линий.
Глубже лежат более бледные, на поверхности — более яркие цвета.
Несомненно, каждый вид птиц характеризуется определенной
окраской яиц, но иногда окраска сильно вариирует у различных
особей того же вида, как, например, у колониально гнездящихся
кайр (Uria troille), у кукушек. В общем каждая самка обладает
своими особенностями, которые передаются по наследству. Однако
существуют иногда очень значительные вариации в окраске яиц
одних и тех же самок.
Несомненно, что окраска яиц — признак, приобретенный пти-
цами, так как рептилии несут яйца без всякой окраски. Возможно,
что окраска яиц выработалась как защитное средство: с одной сто-
роны, как покровительственная окраска, когда яйца по своему
цвету подходят к окружающей обстановке и оказываются поэтому
мало заметными, с другой стороны, — как защита от солнечных
лучей. В этом отношении белая окраска яиц у птиц, гпездящихся
в закрытых помещениях, и темная у гнездящихся открыто — вполне
отвечала бы такому предположению. Однако белые и пестрые яйца
встречаются как у тех, так и у других.
Белая окраска яиц встречается: иногда у птиц, гпездящихся
открыто, но которые, оставляя гнезда, прикрывают яйца или соб-
ственным пухом (Anseres), или растениями; или у птиц, ко-
торые очень крепко сидят на яйцах и слетают с них очепь редко;
или затем у птиц достаточно крупных и сильных.
Пестрая окраска яиц у птиц, гнездящихся в дуплах, может
быть объяснена тоже как сохранившееся приспособление птиц,
ранее гнездившихся в открытых гнездах (например, галка).
Во многих случаях покровительственная окраска яиц выражена
чрезвычайно сильно. Достаточно привести в качестве примеров яйца
пигалиц, бекасов и куликов, по окраске гармонирующих с мши-
стой поверхностью почвы; песочников, морских ласточек, которые
кладут свои яйца прямо на песок с разбросанными между ними
гальками, — яйца как нельзя более походят на гальки; яйца
тетеревов, глухарей и белых куропаток гармонируют по цвету
с прошлогодней ветошью; яйца лысухи, окрашенные под цвет
камышей, и т. д.
Окраска яиц для одних групп птиц является удивительно по-
стоянной, тогда как в других группах даже близкие виды птиц
несут разно окрашеппые яйца.
Величина яиц только до известной степени стоит в коррелятив-
ной зависимости от величины птицы этого вида. Весьма сильно
сказывается на величине яиц то состояние, в котором вылупляются
из яиц птенцы. У птиц выводковых яйца относительно значительно
крупнее, чем у птенцовых. Особенно малы по сравпепию с величи-
ной птицы яйца кукушки. Это вполне соответствует инстинкту ку-
кушки откладывать свои яйца в гнезда различных мелких
пташек.
646
яиц для систематических групп птиц иногда весьма
'нIиhTcpii;i,, хотя и вариирует у одного и того же индивидуума.
Н «нянем но форме можно различить четыре типа яиц: округ-
i\ in (emu,I, зимородки), овальную (козодои, колибри, ряб-
ки. дневные хищники, голуби и др.), удлиненно-оваль-
ном (гагары, веслоногие) и грушевидную (ржанки, чи-
oiiei.i, кулики и чистики). Форма яиц имеет иногда определенное
ni.’iченпо. При грушевидной форме четыре яйца лежат крестооб-
р.гпю своими острыми концами к центру и тем занимают мини-
м.|'н.ное пространство; у чистиков, которые кладут яйца на утесах
hei всякого гнезда, грушевидная форма не дает возможности
лицам при ветре или толчках упасть, так как такое яйцо не катит-
<•11, а. только вертится на месте.
Количество яиц в кладке колеблется от 1 до 22. Одно яйцо в клад-
ке характерно для отряда буревестников (Procellarii), для боль-
шой части чистиков (Alcae), для наиболее крупных хищников
(Aeei pitres), для некоторых голубей (Columbae), чаек (Lari), неко-
торых стрижей (Cypseli) и козодоев (Caprimulgidae).
Два яйца откладывают киви (Apteryx), пингвины (Sphenisi),
гагары (Colymbi),’ журавли (Grues), прочие чистики (Alcae),
большая часть голубей (Columbae), козодоев (Caprimulgi), стрижей
(Cypseli) и колибри (Trochili).
Три яйца характерны для рябков (Pterocletes) и большей части
чаек (Lari).
Четыре яйца в кладке имеют трехперстки (Turnices) и боль-
шая часть куликов (Limicolae).
В общем, как правило, можно считать, что птицы, питающиеся
позвоночными животными, откладывают наименьшее число яиц
в сравнении с птицами, питающимися растительной пищей; вывод-
ковые птицы откладывают наибольшее число яиц. У нанду (Rhea)
несколько самок откладывают в одно гнездо до 40 яиц; высиживает
п х один самец.
Большинство птиц выводят один раз в год, но некоторые — два
и даже три раза в год.
Продолжительность высиживания* весьма различна и отчасти
зависит от абсолютной величины яйца. В общем продолжительность
колеблется от 10 до 14 дней у мелких пташек, от 15 до 26 — у птиц
средней величины, от 26 до 40 — у крупных птиц, до 55 — у кондора
и до 58 дней — у эму, 60 дпей у буревестника (Diomedea). У ку-
риных, водоплавающих и хищников средняя продолжительность
высиживания в общем около 24—28 дней.
Птенцы. Различают птиц выводковых и птенцовых.
У выводковых птиц птенцы выклевываются из яиц довольно раз-
витыми. Тело у них покрыто пухом, а у некоторых сорных кур
(Megapodidae) — даже настоящими перьями; обсохнув, они могут
бегать или плавать и следовать за матерью; они умеют клевать корм
п быстро научаются его отыскивать. Примером выводковых птиц
могут служить все наши домашние птицы: куры, утки, гуси, индейки
и др. (за исключением голубя). Выводковыми являются все палеог-
647
Окрашенные яйца бывают или одноцветные без пестрин, осо-
бенно часто голубые или зеленовато-голубые, или покрыты пе-
стринами в виде мелких и крупных точек, пятен, мазков или линий.
Глубже лежат более бледпые, на поверхности — более яркие цвета.
Несомненно, каждый вид птиц характеризуется определенной
окраской яиц, но иногда окраска сильно вариирует у различных
особей того же вида, как, например, у колониально гнездящихся
кайр (Uria troille), у кукушек. В общем каждая самка обладает
своими особенностями, которые передаются по наследству. Однако
существуют иногда очень значительные вариации в окраске яиц
одних и тех же самок.
Несомненно, что окраска яиц — признак, приобретенный пти-
цами, так как рептилии несут яйца без всякой окраски. Возможно,
что окраска яиц выработалась как защитное средство: с одной сто-
роны, как покровительственная окраска, когда яйца по своему
цвету подходят к окружающей обстановке и оказываются поэтому
мало заметными, с другой стороны, — как защита от солнечных
лучей. В этом отношении белая окраска яиц у птиц, гнездящихся
в закрытых помещениях, и темная у гнездящихся открыто — вполне
отвечала бы такому предположению. Однако белые и пестрые яйца
встречаются как у тех, так и у других.
Белая окраска яиц встречается: иногда у птиц, гнездящихся
открыто, но которые, оставляя гнезда, прикрывают яйца или соб-
ственным пухом (Anseres), или растениями; или у птиц, ко-
торые очень крепко сидят на яйцах и слетают с них очепь редко;
или затем у птиц достаточно крупных и сильных.
Пестрая окраска яиц у птиц, гнездящихся в дуплах, может
быть объяснена тоже как сохранившееся приспособление птиц,
ранее гнездившихся в открытых гнездах (например, галка).
Во многих случаях покровительственная окраска яиц выражена
чрезвычайно сильно. Достаточно привести в качестве примеров яйца
пигалиц, бекасов и куликов, по окраске гармонирующих с мши-
стой поверхностью почвы; песочников, морских ласточек, которые
кладут свои яйца прямо на песок с разбросанными между ними
гальками, — яйца как нельзя более походят на гальки; яйца
тетеревов, глухарей и белых куропаток гармонируют по цвету
с прошлогодней ветошью; яйца лысухи, окрашенные под цвет
камышей, и т. д.
Окраска яиц для одних групп птиц является удивительно по-
стоянной, тогда как в других группах даже близкие виды птиц
несут разно окрашенные яйца.
Величина яиц только до известной степени стоит в коррелятив-
ной зависимости от величины птицы этого вида. Весьма сильно
сказывается на величине яиц то состояние, в котором вылупляются
из яиц птенцы. У птиц выводковых яйца относительно значительно
крупнее, чем у птепцовых. Особенно малы по сравнению с величи-
ной птицы яйца кукушки. Это вполне соответствует инстинкту ку-
кушки откладывать свои яйца в гнезда различных мелких
пташек.
646
•I'и। mi.। ниц для систематических групп птиц иногда весьма
'tip нптрпа, хотя и вариирует у одного и того же индивидуума.
Н ।чипом по форме можно различить четыре типа яиц: округ-
1\ (совы, зимородки), овальную (козодои, колибри, ряб-
hii. диопп ые хищники, голуби и др.), уд линенно-оваль-
|ц ю (гагары, веслоногие) и грушевидную (ржанки, чи-
"ik'i.i, кулики и чистики). Форма яиц имеет иногда определенное
iii.'i’юнпо. При грушевидной форме четыре яйца лежат крестооб-
Iи ню своими острыми концами к центру и тем занимают мини-
ма ’п.1100 пространство; у чистиков, которые кладут яйца на утесах
oo'i цепкого гнезда, грушевидная форма не дает возможности
пипам при ветре или толчках упасть, так как такое яйцо не катит-
еп, а. только вертится па месте.
Количество яиц в кладке колеблется от 1 до 22. Одно яйцо в клад-
вс характерно для отряда буревестников (Procellarii), для боль-
шой части чистиков (Alcae), для наиболее крупных хищников
I Acci pitres), для некоторых голубей (Columbae), чаек (Lari), неко-
торых стрижей (Cypseli) и козодоев (Caprimulgidae).
Два яйца откладывают киви (Apteryx), пингвины (Sphenisi),
гагары (Colymbi), журавли (Grues), прочие чистики (Alcae),
(миыпая часть голубей (Columbae), козодоев (Caprimulgi), стрижей
(Cypscli) и колибри (Trochili).
Три яйца характерны для рябков (Pterocletes) и большей части
чаек (Lari).
Четыре яйца в кладке имеют трехперстки (Turnices) и боль-
шая часть куликов (Limicolae).
В общем, как правило, можно считать, что птицы, питающиеся
позвоночными животными, откладывают наименьшее число яиц
в сравнении с птицами, питающимися растительной пищей; вывод-
ковые птицы откладывают наибольшее число яиц. У напду (Rhea)
несколько самок откладывают в одно гнездо до 40 яиц; высиживает
их один самец.
Большинство птиц выводят один раз в год, но некоторые — два
и даже три раза в год.
Продолжительность высиживания* весьма различна и отчасти
зависит от абсолютной величины яйца. В общем продолжительность
колеблется от 10 до 14 дней у мелких пташек, от 15 до 26 — у птиц
средней величины, от 26 до 40 — у крупных птиц, до 55 — у кондора
и до 58 дней — у эму, 60 дней у буревестника (Diomedea). У ку-
риных, водоплавающих и хищников средняя продолжительность
высиживания в общем около 24—28 дней.
Птенцы. Различают птиц выводковых и птенцовых.
У выводковых птиц птенцы выклевываются из яиц довольно раз-
витыми. Тело у них покрыто пухом, а у некоторых сорных кур
(М egapodidae) — даже настоящими перьями; обсохнув, они могут
бегать или плавать и следовать за матерью; опи умеют клевать корм
и быстро научаются его отыскивать. Примером выводковых птиц
могут служить все наши домашние птицы: куры, утки, гуси, индейки
и др. (за исключением голубя). Выводковыми являются все палеог-
647
паты (Palaeognathae) и следующие отряды неогнат (Neognathae):
гагары (Urinatores), поганки (Colymbi), гуси (Anseres), куры (Galli),
пастушки (Ralli), журавли (Grues), дрофы (Otides), кулики (Limi-
colae), чайки (Lari), рябки (Pterocletes).
У многих выводковых птиц птенцы остаются некоторое время
в гнезде, куда родители прийосят им корм (чистики, чайки); по-
ганки, пастушки, журавли, дрофы, трехперстки и некото-
рые. 498. Птенец гоацина и его
нрыло.
рые другие подносят корм
птенцам в клюве, куры и не-
которые кулики подзывают
птенцов к себе, указывая
им на корм клювом; у других
выводковых птиц птенцы с
первого дня добывают себе
корм сами.
У птенцовых птиц
птенцы выходят из яиц бес-
помощными, недоразвитыми
и должны выкармливаться
родителями более или ме-
нее продолжительное время.
Птенцы этой категории птиц
выходят иногда На свет даже
слепыми и совершенно голы-
ми. Наиболее беспомощными
выклевываются птенцы в сле-
дующих отрядах и подотря-
дах: у воробьиных (Passeres),
дятлов (Pici), удодов (Upupi-
dae), кукушек (Cuculi), у стри-
жей (Cypseli), сизоворонок
(Coraciae), у зимородков (А1-
cedinidae) и веслоногих (Ste-
ganopodes).
В зависимости от харак-
тера одеяния птенцов птиц
этой категории можно разбить
на несколько групп.
К первой группе относятся такие, у которых птенцы родятся оде-
тыми в эмбриональное оперение; таковы, например, дневные хищ-
ники (Accipitres), голуби (Columbae), совы (Striges) и козо-
дои (Caprimulgidae). К этой же группе близки птицы, у ко-
торых птенцы хотя и родятся голыми, но скоро получают одея-
ние из эмбриональных перьев, а именно: веслоногие (Stegano-
podes), аисты (Ciconiae), цапли (Ardeae), грифы Нового Света
(Cathartae). У воробьиных (Passeres) и у удодов (Upupidae)
птенцы рождаются голыми, с редкими эмбриональными пе-
рышками; у кукушек (Cuculi), попугаев (Psittaci), стрижей (Cyp-
seli), дятлов (Pici) и сизоворонок (Coraciae) птенцы вы-
648
клевываются совершенно голыми и не надевают наряда из пер-
вичных перьев вовсе, а прямо одеваются настоящими перьями.
Птенцы своеобразной южноамериканской птицы гоацина (Opis-
thocomus hoazin), выделяемого в особый отряд (Opisthocomi)
выклевываются зрячими, почти голыми, свободно лазят по
веткам, но выкармливаются родителями (рис. 498). Лазят они,
цепляясь за ветки не только ногами и клювом, но также и когтя-
ми, которые развиваются на первых двух пальцах крыла, напо-
миная этим ископаемого археорниса.
Выкармливание птенцов производится обыкновенно и самцом
и самкой, иногда одной самкой и в виде исключения лишь одним
самцом. У журавлей (Grues), которые имеют двух птенцов,один
птенец выкармливается самцом, другой — самкой. У птиц — носо-
рогов (Buceros), на долю самца выпадает не только выкармли-
вание единственного птенца, но и кормление сидящей в затворниче-
стве матери. Иногда между самцом и самкой существует из-
вестное разделение труда при кормлении. Так, у дневных
хищников (Accipitres) самец только приносит пищу, а самка
терзает ее и кормит птенцов. У орлов и аистов птенцы сами
хватают принесенную пищу; у_других,. у_. веслоногих (Steganopo-
<les) и пингвинов (Sphenisci) родители берут голову птенцов в
(•вой рот, чайКйХВ'ап) и некоторые аисты (Ciconiae) отрыгивают
пищу перед ^птенцами, большинство же птиц кладет пищу в
открытый рот птенцов.
Пища птенцов в общем соответствует пище родителей, но очень
многие растительноядные птицы выкармливают птенцов, особенно
первое время, животной пищей; зерноядные пташки выкармливают
детей преимущественно насекомыми. Голуби выкармливают птен-
цов особой творожистой массой, которая отделяется железами
зоба, — так называемым молочком. Количество корма, приноси-
мого птенцам, у разных видов неодинаково, и родители кормят
птенцов через различные промежутки времени. В то время как
у некоторых птиц (Phaeton) птенцы получают корм лишь
один раз в сутки, у других—кормление производится очень
часто.
С помощью особого аппарата-самописца было зарегистрировано,
что пара родителей синицы за день присаживались к гнезду для
кормления от 350 до 390 раз, пара поползней — 370—380 раз, пара
горихвосток — 220—240 раз.
Помимо кормления, родители проявляют и другие виды забот.
’Гак, у некоторых птиц, гнездящихся в дуплах, родители во избе-
жа и не загрязнения убирают помет птенцов. У птенцов в свою оче-
редь имеются приспособления к этому: у них экскременты одеты
ToiiKoii оболочкой, так что легко могут быть взяты полностью клю-
нем ‘ родителей; затем опи обладают рефлексом немедленно после
ни пучения корма испражняться, принимая при этом удобную для
гхнитывания экскрементов позу.
У хищных и некоторых других птиц птенцы с силой выпрыски-
вают испражнения, чем избегается загрязнение гнезда.
649
Забота о потомстве у птиц выражается также в предохранении
птенцов от неблагоприятных изменений погоды, от холода, дождя,
солнца, в предупреждении об опасности и в защите птенцов от
врагов.
Во время дождей, холодов, а у некоторых птиц по ночам, роди-
тели, большей частью самка, прикрывают птенцов своим телом.
При приближающейся опасности многие виды птиц издают особые
крики и стараются разными способами отвлечь внимание врагов
от гнезда на себя и отвести последних в противоположную сторону
от гнезда. Иногда па тревожные крики родителей слетаются особи
того ясс вида, гнездящиеся по соседству, чтобы общими усилиями
отвлечь или прогнать врага. Нередко па эти крики слетаются особи
других видов.
Окреппув и оперившись, птенцы покидают гнездо, и тогда ро-
дители водят птенцов так же, как это делают с самого начала вывод-
ковые птицы. Этот период вождения птенцов кончается обычно
тогда, когда птенцы вполне оперились и научились сами избегать
опасности и самостоятельно отыскивать пищу.
У хищных птиц родители находятся при детях до тех пор,
пока дети не научатся сами ловить себе добычу. Для обучения этому
искусству самец и самка ловят добычу и не умерщвляют ее, а вы-
пускают па глазах у детей живой, чтобы те могли ее сами поймать.
Когда птенцы окончательно подрастут, молодые и старые птицы
у одних видов разбиваются и живут поодиночке, папример, у хищ-
ников; у других видов молодые живут выводком, а старые живут
поодиночке; у третьих молодые собираются в стаи; у четвертых и ро-
дители, и молодые остаются вместе или образуют стадо; наконец,
некоторые старые птицы образуют особое от молодых стадо.
Обычно в это время происходят более или менее отдаленное
кочевье и перелеты чтиц, и начинается линька молодых и старых
птиц.
Птицы-паразиты. Совершенно особняком в деле воспитания
птенцов стоят некоторые виды птиц, которые пользуются для вы-
сиживания яиц и воспитания птенцов услугами других птиц. Наша
обыкновенная кукушка в этом отношении является наиболее со-
вершенно приспособленным видом.
Кукушка откладывает свои яйца в гнезда других видов, вы-
кармливающих своих птенцов насекомыми. В общем насчитывается
более 150 видов птиц, которых могут считать приемными родите-
лями кукушки, но наиболее частями приемными родителями можпо
считать пеночек, коньков, трясогузок, зорянок, славок и камы-
шевок.
В связи с таким паразитизмом стоит целый ряд особенностей в по-
вадках взрослой кукушки и ее птенцов. Во-первых, у кукушек мы
наблюдаем чрезвычайно продолжительный период откладывания
яиц; свои несколько яиц самка кукушки кладет в продолжение меся-
ца с лишним приблизительно через 6—8 дней одно после другого.
Во-вторых, яйца кукушек чрезвычайно мелки по сравнению с ве-
личиной птицы, что гармонирует с привычкой откладывать яйца
650
и гнезда мелких пташек, и отличаются удивительным разнообразием
своей окраски, что опять-таки соответствует паразитизму, так как
замечено, что яйца, откладываемые кукушкой, соответствуют по
окраске яйцам того вида, в гнездо которого откладывается яйцо.
11-третьих, кукушка кладет в каждое гнездо только по одному яйцу
и по откладывает его непосредственно в гнездо, а приносит в клюве.
11-четвертых, яйца кукушки развиваются очень быстро. В пятых,
птенцы кукушки обладают инстинктом в самые первые дни своего
существования выбрасывать из гнезда все яйца, из которых еще пе
111 »i л упились птенцы,
и всех птенцов, чтобы
(ютаться единствен-
п ым обитателем гнез-
да и тем сосредото-
ч и ть исключительно
па себя внимание
приемных родителей
(рис. 499).
Как могли вырабо-
таться эти удивитель-
п ые свойства у куку-
шек, остается нераз-
гаданным. Подойти к
разрешению вопроса
<>6 эволюции этого
явления можно, лишь
сопоставляя наблю-
дающиеся у некото-
рых видов кукушек
и У других птиц бо-
лею* простые случаи
аналогичного характера. Так, есть виды кукушек, которые несут
яйца нормальной величины, откладывают их в гнезда других видов
одинаковой с ними величины, пе по одному, а по нескольку, причем
и'гепцы их не выкидывают яйца и птенцов приемных родителей (род
(Joccystes), а желтоклювые кукушки Сев. Америки (Coccyzus) строят
собственные гнезда, но откладывают в нем яйца через большие про-
межутки времени.
Птицами-паразитами являются также американские желтуш-
ники (Molotus), у которых разные виды обладают разной степенью
развития паразитизма.
IX. ЛИНЬКА птиц
От рептилий, которые периодически сбрасывают свой роговой
покров, птицы унаследовали линьку, т. е. периодическую смену,
перьев, а у некоторых птиц — и других роговых образований кожи:
щитков пог, когтей, шпор и даже рогового одеяния клюва.
Являясь омертвелыми придатками кожи, окончательно отрос-
шие перья изнашиваются под влиянием различных климатических
651
Рис. 499. Птенец -кукушки, выбрасывающий
яйцо из гнезда.
факторов и в силу механического вытрепывапия, а потому должны
заменяться новыми.
Громадному большинству птиц свойственна ежегодно одна
полная линька, которая совершается в послебрачный период;
только у страусов и у крупных хищников перья сменяются пос-
тепенно в течение всего года, а у последних маховые и рулевые
в тот год, когда птицы не гнездятся.
Кроме послебрачной линьки, очень распространена у птиц
вторая линька — предбрачная. Полной предбрачная линька
бывает только у очень немногих птиц (гагары, чайки, кулики, жу-
равли, коростели и др.), обыкновенно она ограничивается лишь
сменой мелких перьев, а иногда сводится преимущественно к появ-
лению новых «украшающих» перьев.
У некоторых куриных (например, белая куропатка) и у рябков
имеется еще третья дополнительная линька, ограничи-
вающаяся мелкими перьями головы и шеи.
Очень часто предбрачная линька бывает только у самцов (на-
пример, у селезней), тогда как у самок этого же вида она отсутствует
совершенно.
У молодых птиц имеются линьки, свойственные только им, при-
чем они могут быть полными и неполными.
Самая линька идет у разных видов неодинаково, но всегда зако-
номерно в определенном порядке и на правой и левой сторонах одно-
временно и симметрично. В громадном большинстве случаев линька
идет постепенно и захватывает одновременно лишь такое количе-
ство перьев, какое позволяет птице вести нормальный образ жизни.
Только у немногих, именно у некоторых уток, у гусей,
лебедей, чистиков, фламинго, у змеиных птиц и некоторых пастуш-
ков маховые выпадают сразу целыми сериями, так что итнцы
лишаются возможности летать до тех пор, пока новые перья не
отрастут.
Обычно линька идет на крыле одновременно в трех местах: на
кисти, начиная с самого внутреннего махового первого порядка и за-
канчиваясь наружными маховыми, а на предплечье смена маховых
второго порядка идет одновременно с проксимального и дисталь-
ного концов.
У пингвинов (Sphenisci) и казуаров (Casuarii) линька отли-
чается от всех прочих птиц тем, что сосочки новых перьев врастают
в душку старых, при своем росте выталкивают их так, что старые
перья некоторое время держатся па концах новых и затем лишь
отпадают или, как у пингвинов, срываются клювом и ногами. У не-
которых хорошо летающих птиц линька маховых идет постепенно,
когда мелкое оперение уже сменилось, и иногда продолжается
целый год.
Возрастные и сезонные изменения оперения. От линек зависят
иногда очень сложные возрастные изменения в нарядах и сезонные
и брачные наряды птиц, которые в связи с обнашиванием пера
дают характерную для каждого вида последовательность смены
нарядов.
652
। .1 мi.im первым нарядом нужно считать тот наряд,
hninpi.ni мы видим у вылупившихся птенцов и который обусловли-
h.irrni одеянием из эмбриональных перьев (neoptiles).
Второй наряд — это гнездовой наряд, т. е. первое одея-
... ни настоящих перьев, которое сменяет «пуховое» (эмбриональ-
ное; одеяние. Эти перья отличаются мягкостью и рыхлостью (те-
eopiiles) и поэтому легко отличимы от последующих нарядов из
окончательных (дефинитивных) перьев (teleoptiles).
11 этом паряде молодые птицы, если самцы и самки окрашены оди-
ii.'iicoiio, иногда похожи на родителей (вороны, галки, сороки),
иногда резко от них отличаются (у соловьев, у мухоловок птенцы
имеют пятнистую окраску). При неодинаковой окраске самцов
п самок молодые большей частью бывают похожи на самок (напри-
мер, у зябликов, у домашнего воробья), иногда отличаются и от
г.чмцов и от самок (например, у соловьев и мухоловок, у которых
птицы пятнисты, или у луней, где молодые птенцы иногда совсем
не похожи на родителей). В редких случаях молодые птенцы похожи
на самцов.
Некоторые птицы начинают сменять этот наряд, как только по-
кинут гнездо, другие сменяют его перед отлетом, третьи, как, на-
пример, ласточки, сменяют его только на зимовках.
Эта линька (вторая по порядку) носит название юноше-
с к о й, бывает или полной со сменой маховых и рулевых (напри-
мер, у скворцов) или неполной, когда не сменяются не только круп-
ные перья, как у большинства, но и некоторые мелкие.
Третий наряд, в который облекаются птицы после юноше-
ской липьки, может быть весьма различным. В тех случаях, когда
самцы и самки одинаково окрашены, молодые птенцы становятся
похожими на родителей уже в этом третьем наряде, который ста-
новится, таким образом, окончательным.
Однако у очень многих птиц существуют значительные отличия
этого наряда. Он носится птицами или в продолжение целого года,
до следующей послебрачпой линьки, или заменяется через полгода
с.л('дующим, четвертым нарядом, который возникает
п.'1и в результате предбрачной липьки, или в результате снашивания
пера.
Этот четвертый (он же первый брачный) наряд в случае, если
он образовался в результате частичной липьки, может быть очень
сложным и состоять из перьев второго, третьего и четвертого наря-
дов. Только осенью происходит полная смена пера, и птица оде-
вается тогда в пятый наряд, который у большинства птиц,
нс имеющих предбрачной линьки, будет окончательным.
У птиц же с брачной линькой, или перекраской, ч е т в е р-
т ы й или шестой наряды, т. е. или первый, или второй брач-
ные наряды, будут окончательными брачными нарядами, после
чего птицы уже ежегодно их надевают.
< )дпако птицам с сезонной перекраской, которую мы встречаем,
например, у белых куропаток, свойственны еще дополнительные
липьки. Так, молодые птицы, надев юношеский (третий) наряд,
653
имеют перед зимой еще частичную линьку с заменой темного пера
на белое. Такую же дополнительную линьку имеют и взрослые пти-
цы, так как они вместе с послебрачной полной линькой одеваются
не в сплошь белое перо, а только па нижней стороне получают белое
оперение; лишь после дополнительной линьки зимой они становятся
сплошь белыми.
У уток селезни линяют весной, меняя свое брачное, отличное
от самок яркое одеяние на скромный наряд, похожий на наряд са-
мок. Этот летний наряд, благодаря линьке осенью и снашиванию
пера, превращается зимой в брачный наряд.
Явления липьки вообще протекают у птиц болезненно. В это
время они наиболее подвержены заболеваниям, легче делаются
добычей хищников, особенно у тех водоплавающих, которые, как
мы видели, теряют способность летать. Естественно, что во время
линяния птицы ведут поэтому очепь скрытый образ жизни, пря-
чась в зарослях.
Обыкновенно линька совершается до отлета, но многие виды
летят, не перелиняв окончательно, тогда как другие виды, па-
пример, ласточки, линяют только на зимовках.
X. ОКРАСКА ПТИЦ
Окраска птиц зависит главным образом от окраски пера. Кожа
птиц, за исключением голых частей тела, которые иногда приобре-
тают особую яркую окраску, бывает слабо окрашена или вовсе
не окрашена. Окраска перьев зависит от пигмента, но также от ми-
кроструктуры пера.
Что касается пигментов, то у птиц различают две группы:
меланины и липохромы. Меланины — это зернистые пигменты от
желтокоричневого до черного цвета, причем зерна черных и темно-
коричневых пигментов имеют форму палочек, носят название эуме-
ланппа, а желтокоричпевые пигменты в виде крупных зернышек
называются фэомеланином.
Липохромы обычно растворены в жиру, диффузно реже в виде
пятен с неясным контуром. Это — многочисленные пигменты крас-
ного, желтого, зелено-голубого или фиолетового цвета.
Природа этих пигментов недостаточно выяснена.
Различают три красных пигмента: 1) зооэритрин, самый рас-
пространенный из них, обусловливающий красную, розовую и бу-
рую окраску большинства птиц, 2) зоорубин, найденный в опере-
нии райских птиц, и 3) турацин, красный пигмент перьев бананоедов
(Musophagidae).
Желтый пигмент — зооксантин, или зоофульцин, обусловли-
вающий желтую окраску, а вместе с красным — оранжевую ок-
раску птиц.
Наконец, имеется еще зеленый пигмент — тураковердин, встре-
чающийся только в зеленых перьях бананоедов.
Столь распространенные у птиц синие и фиолетовые цвета перьев
объясняются соединением разной пигментации, а также сложной
654
г грун ту poii пера. При проходящем свете окраска таких перьев
оур;|}|, так как в таком случае сказывается лишь действие пигмента;
такого же цвета будут казаться синие, голубые и фиолетовые перья,
«•«•ли их подвергнуть механической обработке, которая разрушила
ih.i структуру пера. Последняя представлена лежащей поверх глу-
боких пигментных клеток роговой оболочкой, под которой распо-
лагаются многоугольные призматические клетки, преломляющие
спет. Оту окраску, поскольку опа обусловливается не только пиг-
ментом, ио и строением пера, можно назвать структурной
объективной окраской. *
Другое дело субъективная структурная ок-
раска перьев — та блестящая металлическая окраска, котора^
отливает различными цветами в зависимости от положения птицы
по отношению к источнику света и наблюдателю. Эта окраска обус-
ловлена дифракцией света, благодаря отражению света от гладкой
поверхности, или интерференцией, обусловливаемой тончайшими
пластинками, расположенными на верхней стороне пера.
Металлическая окраска довольно распространена среди птиц.
Всем известны металлические узоры перьев павлинов, фазанов,
петухов, «зеркальца» крыла уток, но особенного развития металли-
ческая окраска достигает у удивительных тропических семейств
Старого Света — райских птиц (Paradiseidae), нектарок (Nectari-
niidae) и у американского семейства колибри (Trochilidae), которые
относятся к отряду стрижей (Cypseli).
Вообще окраска птиц до чрезвычайности разнообразна и выра-
жается не только в разнообразии цветов, по и в сложности и разно-
образии рисунков.
Обычно особенно яркой окраской отличаются самцы, тогда как
самки окрашены в тусклые серые тона, имеют так называемую
«покровительственную» окраску. Однако имеются виды, у которых
п самцы и самки окрашены одинаково, и тут встречаются виды
п с яркой и со скромной покровительственной окраской.
По тому значению, которое имеет окраска у птиц, различают:
1) брачную окраску, 2) покровительственную окраску, 3) мимити-
рующую окраску, 4) предупреждающую окраску, 5) распознава-
тельную окраску.
Под брачной окраской разумеют ту, по большей
части более яркую, окраску, которая, как мы видели, нередко воз-
никает как результат предбрачной линьки. Часто она бывает при-
сущей и самцам, и самкам, как, папример, у гагар (Urinatores), по-
ганок (Colymbi), у чаек (Lari) и др., но по большей части опа
бывает свойственна только самцам и принадлежит, таким образом,
ко вторичпым половым признакам самцов.
Иногда чрезвычайно яркая окраска самцов сопровождается
удивительно сложными рисунками, часто также особым развитием
украшающих перьев или других придатков кожи (павлины,
фазаны, куры и др.). Объяснявшая раньше такую окраску
теория полового отбора встречает, однако, ряд серьезных затруд-
нений.
655
Яркая окраска самцов и целый ряд других украшающих особен-
ностей объясняются как коррелятивно возникшие и, являясь
нередко вредными для особей, допускаются естественным отбором
в значительных и резких проявлениях лишь по отношению к самцам.
Может быть, часть вторичных половых признаков возникла как
приспособление для более легкого нахождения и распознавания
особей противоположного вида среди близких и сходных видов.
Тогда их развитие одновременно обусловливается и половым и есте-
ственным отбором.
Распознавательная окраска может иметь и
другое значение. Для молодых птиц она облегчает нахождение роди-
телей, особенно матери, которая водит птенцов. Такой смысл,
быть может, имеет белое подхвостье у водяной курочки (Gallinula
chroropus), которая имеет обыкновение держать хвост вертикаль-
но, так что следующим за матерью птенцам белый цвет служит путе-
водным сигналом.
Для птиц, образующих стаи, особые отметины, отличающие птиц
данного вида от особей близкого вида, облегчают образование стай,
примером чего могут служить яркие «зеркальца» на крыльях у раз-
ных видов утиных.
Что же касается покровительственной окраски, мимикрии, пре-
дупреждающей окраски или отпугивающей окраски, то они имеют
защитное значение, и о пих будет речь в следующей главе.
XI. ПРИСПОСОБЛЕНИЕ ПТИЦ В ЦЕЛЯХ ЗАЩИТЫ
Сравнительно небольшое число видов птиц обладает вооружением
для защиты от врагов. Гораздо чаще приспособления в целях защиты
выражаются у птиц в иаличпи соответствующей окраски, которая
делает их незаметными в окружающей обстановке, когда они пря-
чутся.
В редких случаях такая окраска сопровождается пластическими
особенностями, делающими такое сходство с окружающей обста-
новкой еще более разительным; в очень редких случаях мы встречаем
подражательное сходство, или мимикрию.
Сюда же нужно отнести те особенности окраски и другие осо-
бенности, которые служат птицам для отпугивания врагов. Помимо
свойственных всем птицам инстинктов самосохранения, все пере-
численные выше особенности сопровождаются особыми инстинк-
тами, дающими возможность использовать эти особенности.
Рассмотрим вначале защитные вооружения,
встречающиеся у птиц. Обыкновенно для целей защиты служат клюв,
реже когти ног. Наилучше в этом отношении защищены хищные
птицы, которые при защите пускают в дело не столько клюв, сколь-
ко когти; цапли, аисты, журавли и другие птицы бьют длинным
и острым клювом. Некоторые крупные птицы, в особенности гуси
и лебеди, защищаются сильными ударами крыльев.
Из специальных органов защиты на первом месте нужно поста-
вить так называемые шпоры — роговые острые выросты на цевке
656
V куриных птиц, служащие здесь главным образом, если не исклю-
•iirrr. ii.iio, самцам при их боях друг с другом. У шпорцевых чибисов,
шпорцевых пастушков и паламедей имеются такие шпоры на
крыльях.
К. специальным органам защиты нужно также отнести длинные
перья на воротнике у турухтанов, выполняющие роль щита при
драпах; длинные ноги у секретаря и кариамы, защищающие этих
эмггядиых птиц от укусов змей; чешуйчатое оперение головы
у осоедов, защищающее их от укусов перепончатокрылых, и некото-
рые другие.
Самым распространенным защитным приспособлением является
защитная о к р а с к а у птиц.
Чрезвычайно распространена среди птиц окраска, соответствую-
щая общему тону окружающей среды. Виды, живущие среди зеле-
ных ветвей и трав, имеют нередко зеленую или зеленоватую окра-
ску. Достаточно упомянуть в виде примеров среди птиц нашей фауны
зеленушку (Chloris chloris), пеночек (Phylloscopus), камышевок
(Acrocephalus), желтых трясогузок (Budytes), зеленых дятлов
(Picus). Виды, обитающие в пустыне, имеют желтую окраску, как,
например, рябки (Pterocletes), авдотки (Oedicnemus), коньки (Ап-
llius). Виды, скрывающиеся на земле среди прошлогодней ветоши,
листьев и стеблей, земли и трав, имеют желтобурую окраску в пе-
стринах и чрезвычайно подходят к окружающей среде, как бекасы
п дупель (Gallinago), вальдшнеп (Scolopax rusticola), тетерки и мо-
лодые тетерева (Tetrao tetrix), серые куропатки (Perdix), белые куро-
патки (Lagopus) летом и очень многие другие птицы. Белые и тундро-
пые куропатки зимой линяют в белое перо и па снегу становятся
мало заметными. Все эти птицы в случае опасности остаются на
месте без движения и тем становятся незаметными. В некоторых слу-
чаях сходство с окружающей обстановкой увеличивается тем, что
птицы принимают соответствующие позы, походя па окружающие
неодушевленные предметы. Так, козодои, цвет которых походит
на кору дерева, садятся параллельно горизонтальным веткам и при-
падают к ним, авдотки ложатся на землю, вытягивая шею, напоминая
но форме камни или глыбы глины; выпи вытягиваются вертикаль-
но, получая благодаря этому сходство с камышами.
Подражательное сходство, или мимикрия
(миметизм), когда один вид сходен с другим видом совсем другого
семейства или отряда и извлекает из этого определенную выгоду,
столь распространенное у насекомых, находит свое выражение и у
птиц, хотя и далеко не с такой очевидностью. Примером мимикрии
среди птиц нашей фауны может служить обыкновенная кукушка
((Jnculus canorus), которая по своей окраске, по полету и форме
крыльев и хвоста сильно напоминает ястреба-перепелятника. Такое
с ходство дает кукушке то преимущество, что она своим видом ястреб-
ка пугает мелких птиц и через это получает возможность положить
сноп яйца в их гнезда.
Другого характера подражательное сходство в том случае,
когда хищная птица походит на птицу безвредную и таким образом,
*2 ICuuiKupou и Станчинский 657
получает возможность менее пугать добычу, которой кормится.
Так, например, дымчатый коршун (Elanus) походит на чаек (Larus).
Защитное приспособление может иногда выражаться в противо-
положном покровительственной окраске направлении, когда жи-
вотное как раз выделяется из окружающей среды. Это так называе-
мая предупреждающая окраска. Обладатели такой окраски, осо-
бенно распространенной среди насекомых, имеют защитные свой-
ства, предохраняющие их от опасности. Такова, например, спо-
собность насекомых выделять ядовитые или сильно пахнущие во-
Рис. 500. Птичий базар.
нючие вещества.
Быть может, заметная пе-
страя окраска наших удодов
(Upupa epops) играет такую
же роль. Они в период гнез-
дования выделяют вонючий
секрет своей копчиковой же-
лезой, и потому ими брезгают
хищники.
У вертишейки (Yunx) под-
ражательное сходство выра-
жается в том, что во время
опасности она вытягивается
на земле, поднимает свою
голову и вращает головой
так, что производит впечатле-
ние отнюдь не птицы, а ско-
рее змеи.
Это отчасти имитация, от-
части угрожающая поза. Сре-
ди птиц мы чрезвычайно ча-
сто встречаемся с принятием
определенных поз угрозы,
чтобы испугать врага. Птицы
ерошат перья, поднимают
крылья, открывают клюв или щелкают им, производят другие отпу-
гивающие звуки.
Так поступают многие мелкие птички, так поступают и крупные
птицы, например, совы, лебеди и др.
Некоторые птицы «притворяются» в момент опасности мертвыми.
Это мы наблюдаем, например, у больших синиц (Parus major).
К числу защитных приспособлений могут быть отнесены также
общественная жизнь птиц и взаимная защита
у необщественных видов, иногда у особей, относящихся к раз-
ным видам. На первом месте необходимо поставить гнездовые коло-
нии птиц.
Образование таких колоний иногда может быть вызвано малым
количеством мест, подходящих для гнездования, что вынуждает
птиц гнездиться в непосредственной близости друг от друга. Таковы
колонии морских птиц — так называемые птичьи горы или базары
658
по побережью или островам морей, где в непосредственной близости
гнездятся тысячи чаек, чистиков, кайр и других морских птиц
(рис. 500), или гнездовые колонии буревестников, пингвинов и др.
В других случаях общественность обусловливает взаимную за-
щиту, как, например, в колониях грачей, чибисов, чаек.
При приближении к такой гнездовой колонии врага первый
заприметивший его член колонии дает об этом знать характерным
криком. Это сейчас же вызывает общий переполох, и навстречу
опасности бросаются соседи, чтобы общими усилиями отогнать
врага или по крайней мере отвлечь его.
Очень многие птицы, не гнездящиеся колониями, собираются
осенью в стаи и проводят в стадной жизни и осень и зиму; перелет-
ные птицы стаями перелетают на юг, ведут общественную жизнь на
зимовках и стаями возвращаются. Такие стаи, между прочим, ока-
зывают иногда взаимную защиту и, что бывает гораздо чаще, пре-
дупреждают об опасности.
Впрочем, многие птицы издают крики предосторожности неза-
висимо от стадного образа жизни, причем крики предосторожности
понимаются не только особями того же самого вида, но и другими
видами.
Нередко в таких случаях на крик об опасности слетаются особи
того же или разных видов и общими силами отгоняют врагов.
XII. ЗАВИСИМОСТЬ ЖИЗНИ ПТИЦ ОТ УСЛОВИЙ СРЕДЫ
1. Климатические факторы существования итиц
Влияние температуры. Птицы, являясь теплокровными живот-
ными, гораздо менее зависят от изменений температуры, чем живот-
ные холоднокровные. Этим объясняется то, что на земном шаре нет
таких мест, в которых при наличности достаточного количества
нищи не жили бы птицы даже при самых низких температурах
полярных стран и высоких температурах знойных пустынь.
Для предохранения тела от охлаждения у птиц имеется ряд
приспособлений. Для этой цели служит плохо проводящий тепло
покров из перьев и пуха. Перья сами по себе являются плохим
проводником тепла; таким же плохим проводником является воз-
дух, который находится между перьями. Чем чаще сидят перья, чем
они длиннее, тем лучшую защиту от охлаждения представляет
оперение. Еще лучше сохраняет тейло пух. Обильный пуховой по-
кров вместе с густым и плотным оперением является в этом отно-
шении самым надежным покровом.
Так как в защите от широких колебаний температуры нуждаются
главным образом птицы, обитающие в полярных странах и странах
е континентальным климатом, а также водные птицы вообще и холод-
ных стран в особенности, то естественно, что у этих птиц мы на-
ходим и обильный пуховой покров и густое плотное оперение. Та-
ковы из водоплавающих знаменитые своим пухом гаги (Somateria),
лебеди (Cygnus) и вообще гусиные (Anseres) севера. Удивитель-
42“ 659
но густые и плотные перья гагар (Urinator) и поганок (Colymbus),
служащие вместе со шкуркой предметом особого промысла и иду-
щие даже па приготовление костюмов, являются другим прекрас-
ным примером. У наземных птиц, остающихся на зиму в холодных
странах, после осенней линьки вырастает более длинное и пухля-
вое перо.
Чем меньше животное, тем больше тепла излучает его тело,
согласно так называемому правилу Бергмана, по сравнению с теплом
им вырабатываемым. И дей-
ствительно у мелких птиц за-
щита тепла от охлаждения через
излучение достигает наибольше-
го совершенства. Особенно пу-
шистым и длинным оперением
отличаются наши самые мелкие
зимующие птицы, как синицы,
пищухи и корольки.
Особенно подвержены охла-
ждению голые части тела, на-
пример, у марабу (Leptoptilus)
(рис. 501), и птиц с голыми
частями как грифы, страусы и
др., мы встречаем только в теп-
лых или умеренно теплых стра-
нах.
Далее для защиты тела от
охлаждения служит слой
подкожного жира, ко-
торый опять-таки развивается
преимущественно у водоплаваю-
щих птиц холодных стран: гагар
(Colymbi), чистиков (Alcae), гаг
(Somateria), пингвинов (Sphe-
nisci) и др.
Важным условием для со-
Рис. 501. Марабу (Leptoptilus cru- хранения определенной темпе-
menifer). ратуры тела при низкой темпера-
туре среды служат энергичные
движения,при которых освобождающаяся теплота согревает тело.Все
зимующие в холодных странах птицы отличаются в это время уси-
ленной деятельностью. Так как энергичная деятельность обусловли-
вается сильными окислительными процессами, то необходимо по-
стоянное возобновление сгорающих органических веществ тела,
следовательно, нужны и обильная пища, и энергичный газообмен.
Все это требует усиленной деятельности органов кровообращения.
В пределах вида соответственно этому популяции более холодных
стран обладают сердцем большего объема. Сидящая спокойно
в полутемноте канарейка вырабатывает в час при 22.3° 319 калорий,
тогда как при 14° — 992 калории. Чем холоднее среда, тем больше
660
отдает тепла птица, тем больше пищи опа должна принимать, что-
бы восполнить сгорающие в теле резервные вещества; и чем меньше
птица, тем больше, согласно правилу Бергмана, требуется пищи.
Для некоторых птиц понижение температуры среды при одно-
временном уменьшении дня и продолжительности кормежки в те-
чение суток могут вызвать невозможность восполнения пищей рас-
ходуемых на поддержание температуры тела веществ. Птицы даже
при обильном корме будут в таком случае голодать, худеть и гиб-
нуть.
Само собой разумеется, что не все птицы одинаково стойки как
к высоким и низким температурам, так и к колебаниям температур.
И среди птиц мы можем различать виды, которые могут жить при
широких колебаниях температуры — эйритермические,
и такие виды, которые приспособлены лишь к определенным, срав-
нительно узким ее колебаниям, — виды стенотермиче-
ские. К первым относятся во первых, все остающиеся у нас на
зиму птицы, как, например, вороны, галки, сойки, клесты, сини-
цы, корольки, дятлы и др. Во-вторых, сюда относятся птицы, оби-
тающие в странах с ярко выраженным континентальным климатом,
как нагорья центральной Азии или внутренние части Сахары, где
суточные колебания темйературы достигают наибольшей амплитуды
(в Тибете, например, до 55°), таковы — рябки, саджи, горные индей-
ки, горные вьюрки и др. В третьих, к эйритермическим видам
можем отнести птиц, поднимающихся во время полета на большие
высоты с низкой температурой, как многие крупные хищники,
например, грифы, орлы.
К стенотермическим формам относятся птицы тропических стран
с ровным приморским климатом и перелетные птицы умеренного
пояса.
Как средство против перегревания млекопитающие отделяют
нот, который, испаряясь, охлаждает тело. Птицы, как мы видели,
н(5 имеют подобных желез, и роль охлаждающего аппарата берут
па себя органы дыхания, испаряющие воду.
Благодаря указанным выше приспособлениям птицы могут пере-
носить колебания температуры, как довольно высокие, так и низкие.
При прочих равных условиях растительный мир тем богаче,
чем выше температура года и чем ровнее она в продолжение года;
богатый растительный мир обусловливает богатство мира насеко-
мых и других наземных беспозвоночных, а это в свою очередь по-
зволяет развиться более богатой фаупе птиц. Таковы области с
приморским тропическим климатом. Насколько разнообразен пти-
чп11 мир тропических стран, показывают следующие цифры: из об-
пито числа свыше 10000 видов птиц для всей палеарктической об-
ihh’tii насчитывается около 1000 видов, тогда как почти такое же
число пидов принадлежит тропической Амазонке; в Европе мы имеем
около зоо видов оседлых и перелетных, тогда как на острове Борнео
и ч вдвое больше: около 600 видов. Многие из крупных система-
тических подразделений ограничены исключительно тропическими
ГТ| hl.ll.IM п.
661
Наоборот, страны с холодным, континентальным климатом, с рез-
кими колебаниями температуры бедны флорой и фауной вообще
и птицами, в частности.
Температурные максимумы и минимумы гораздо сильнее ска-
зываются на жизни птиц, чем средние месячные и годовые. При
этом играют роль не столько абсолютные величины температуры,
сколько относительные их отклонения от нормы, время этих откло-
нений в году и продолжительность отклонений.
Очень низкий минимум может заморозить яйца птиц, птенцов,
или даже взрослых птиц. Но и небольшой мороз весной или в начале
лета может иметь большое влияние на фауну птиц, в то время,
как гораздо больший мороз зимой не окажет на фауну никакого
влияния.
Продолжительность отклонений температуры очень важна.
Резкие уклонения от нормы могут иметь иногда катастрофический
характер. Так, поздние весенние морозы, ранние осенние заморозки,
неожиданные морозы в местах зимовки, во время перелета и т. д.
могут обусловливать гибель птиц массами.
Суточные изменения температуры, особенно сильные в конти-
нентальных странах с сухим климатом, переносятся обитающи-
ми там птицами без всякого ущерба, между тем как даже меньшие
колебания в других местах сказываются на птицах катастрофи-
чески.
Влияние света. В то время как для растений свет является ос-
новным фактором, ибо без энергии солнечных лучей зеленые расте-
ния не могут ассимилировать углерод, т. е. не могут питаться, для
животных свет не является безусловно необходимым фактором. Од-
нако влияние света на животный мир чрезвычайно велико.
По условиям проявления жизнедеятельности птицы разделяются
на ночных и дневных. Громадное большинство птиц —
дневные птицы. Днем они отыскивают пищу, проявляют различного
рода деятельность, а ночью находятся в состоянии покоя. Ночные
птицы, наоборот, отдыхают днем, а ночью находятся в деятельном
состоянии. Однако есть еще группа птиц, которая занимает проме-
жуточное положение: их деятельность проявляется утром и вече-
ром. Это сумеречные птицы. В связи с тем или иным обра-
зом жизни птицы имеют соответствующие приспособления. Днев-
ные птицы при ориентировке пользуются главным образом зре-
нием.
Ночными птицами являются совы (Striges), совиный попугай
Австралии (Stringops), многие цапли (Ardeae), многие буревест-
ники (Procellarii) и отчасти козодои (Caprimulgidae). У сов, наи-
более совершенных ночных птиц, мы видим специальные приспо-
собления для ночного образа жизни. Так, глаза у них относи-
тельно больше, с сильно вытянутой осью, так что они имеют верх-
нюю часть в форме цилиндра. Сетчатка их также имеет свои осо-
бенности (рис. 502).
Ночным птицам, помимо глаз, приходится в темноте ориенти-
роваться при помощи других органов. У сов чрезвычайно развито
662
наружное ухо, образованное находящейся позади слухового отвер-
стия кожной складкой, снабженной мускулатурой; некоторые ноч-
ные буревестники (Fulmarus, Pogodroma) и киви (Apteryx) обла-
дают хорошо развитым органом обоняния.
Третье приспособление к ночному образу жизни — это осо-
бенная мягкость оперения, чрезвычайно выраженная у сов.
Мягкое перо делает полет сов неслышным, что дает возможность
полнее использовать свой орган слуха и бесшумно подлетать
к добыче.
Четвертое приспособление ночных птиц — цвет оперения. Оно
всегда выдержано в бурых и серых тонах и потому стушевывается
в темноте. Исключение из этого правила составляет белый цвет по-
лярной совы (Nyctea nivea). Но эта птица — обитательница дале-
кого севера, летом живет в условиях незаходящего солнца, беспре-
рывного летнего дня и боль-
шую часть года проводит сре-
ди снега, с которым цвет ее
гармонирует.
Сумеречных птиц очень
много в разных отрядах. Пре-
1 i мущественно сумеречными
птицам могут считаться ко-
зодои (Caprimulgidae), сре-
ди которых много и ночных
птиц.
Суточная жизнь птиц скла-
дывается для каждого вида
вполне определенным обра-
зом. Периоды деятельности
сменяются периодами покоя.
()собенно резки такие периоды
в странах тропических и под-
Рис. 502. Разрез через глаз совы.
(Из Мензбира.)
тропических, где ночь также
продолжительна, как и день, и где переход от дня к ночи совер-
.шлется быстро. В умеренных поясах эти переходы имеют еще до-
статочно явно выраженный характер, особенно под осень, тогда
как дальше на север они все больше и больше стушевываются,
пока, наконец, в условиях летнего сплошного дня почти пропадают
совершенно. Там нет ночных животных, и северные совы, как по-
ли риал сова (Nictea nivea), ястребиная сова (Surnia ulula) и воробьи-
iii.hi сычик (Glaucidium passerinum) проявляют свою деятельность
при свете незаходящего солнца.
Период деятельности птиц выражается главным образом в оты-
скивании пищи; многие птицы после кормежки передвигаются
и поисках воды. Иногда птицы передвигаются вцелях защиты от хо-
лода пли жары, или от других неблагоприятных метеорологических
uii.ueiiiiii. Весной деятельность выражается, между прочим, в явле-
ниях, связанных с воспроизведением, а у перелетных птиц дважды
и год имеют место явления перелета.
663
В общем суточная жизнь птиц может быть сведена к следующей
схеме:
Дневные птицы Ночные птицы
Утро 1. Просыпание 2. Поиски пищи, перелеты на жировку 3. Жировка Возвращение с ночной охоты (перелет на дневной отдых)
День 1 1. Передвижение на отдых 2. Иногда перелет на водопой 3. Дневной отдых Период покоя
Вечер 1. Поиски пищи, перелеты на жировку 2. Жировка 3. Поиски ночлега, перелеты на ночлег Вылет (перелет) на ночную охоту
Ночь 1. Период покоя Ночная охота
Просыпаются птицы пе все сразу, а по видам: одни раньше, дру-
гие — позже. В общем можно сказать, что насекомоядные птицы
просыпаются раньше, их период деятельности более оживленный,
и они позднее успокаиваются па ночь по сравнению с птицами зер-
ноядными. Пищу одни виды птиц отыскивают непосредственно вбли-
зи того места, где отдыхают, другие облетают более или мепее обшир-
ный охотничий район, иногда с замечательной правильностью появ-
ляясь в определенный час па определенном месте, третьи улетают кор-
миться на значительное расстояние от места отдыха (цапли, аисты,
утки, гуси). На водопой летают далеко не все птицы. Такие перелеты
наблюдаются у гусей (на зимовках), у тиркушек и рябков и др.
Изменения в суточной жизпи птиц по временам года очень зна-
чительны, особенно у птиц, обитающих в умеренном и холодном поя-
сах. По мере сокращения дня к концу лета дневной отдых, посте-
пенно сокращаясь, наконец, пропадает совершенно, особенно у птиц
насекомоядных. Но и зерноядные птицы едва успевают насобирать
достаточное количество пищи, чтобы наесться досыта.
Еще более меняется образ жизпи птиц в период воспроизведения.
Птицы бывают настолько поглощены заботами воспроизведения,
что тратят очень мало времени на отыскивание пищи. Самцы больше
всего времени уделяют в это время пению, взаимным дракам и уха-
живанию за самками. Некоторые птицы в это время поют чуть пе
круглые сутки, почти совершенно пе отдыхая.
Но особенно меняется образ жизпи птиц во время перелетов.
Очень многие, даже чисто дневные птицы летят по ночам. Многие
так спешат во время перелетов, что почти не останавливаются на
кормежку, а лишь делают остановку па краткосрочный отдых.
Влияние влаги. Вода имеет большое значение в жизпи птиц:
во-первых, вода — это среда обитания и среда добывания пищи
в виде водоемов и в виде болот; во-вторых, вода необходима для
обмена веществ как вещество, принимаемое внутрь организма;
664
в-третьих, вода— фактор климатический в виде гидрометеоров:
влажности, испарения, облачности и осадков.
По своему отношению к влаге всех птиц можпо разделить на
три категории: гидрофилов (водолюбивых), г и г р о ф и-
л о в (влаголюбивых) и ксерофилов (сухолюбивых). По
строению своих покровов птицы отлично защищены от высыхания
и поэтому могут выносить большую сухость воздуха без вреда
для себя. Однако в отношении стаций и пищи они приспособлены
к разным степеням влажности климата.
К гидрофилам относятся птицы водных бассейнов и их берегов
и птицы топких болот, например, кулики; они добывают себе пищу
в воде, питаясь гидрофильными беспозвоночными — червями, мол-
люсками и амфибиями. Гигрофильными птицами является боль-
шинство видов — обитатели влажных лесов, лугов, моховых и тра-
вяных болот. Ксерофильными птицами можпо считать тех птиц, ко-
торые живут в степях, полупустынях и прежде всего птиц пустынь.
Влажность климата влияет на состояние водных бассейнов и поч-
венных вод, а тем самым косвенно — на жизнь растений и животных;
с другой стороны, влияние климатической влажности сказывается
па жизпи животных непосредственно.
В связи с уменьшением тепла к полюсам и увеличением холодных
зимних периодов меняется аггрегатное состояние влаги. С известных
широт на известное время в году вода замерзает, а осадки выпадают
в виде снега и инея, образуя снежный покров. Это обстоятельство
имеет весьма важное значение в жизни животных. Замерзание водое-
мов лишает водоплавающих и болотных птиц их стаций, и опи при-
нуждены отлетать на это время туда, где вода не замерзает. Замер-
зание почвы делает ее недоступной для птиц, которые добывают
пищу в земле, так как снежный покров скрывает от птиц спрятав-
шихся в земле животных и рассеянные по земле семена.
Отсюда границы мест зимовок многих птиц стоят в непосред-
ственной связи с границами замерзания воды зимой и наличностью
снежного покрова.
В общем к колебаниям в пределах нормы в распределении влаги
во времени и пространстве птицы каждой местности так или иначе
приспособились. Однако совсем другое дело, когда мы встречаемся
с изредка, пе ежегодно повторяющимися уклонениями от нормы по
временам года. Эти уклонения важны не только своими абсолютными
величинами, но и своей продолжительностью и тем временем в году,
когда они происходят.
Снегопады зимой в местах, где обычно они не бывают и где зи-
мует большое число птиц, добывающих себе пищу в почве или на
почве; глубокий снежный покров там, где он бывает обычно мелкий;
обильный проливной дождь весной, вызывающий чрезмерное сма-
чивание почвы или разливы рек; обложной продолжительный дождь
в это же время или летом; крупный и сильный град, поздний снег
весной и ранний снег осенью; продолжительное ненастье осенью
во время перелетов — все это может сказываться весьма губитель-
но ini фауне птиц; вторичный выход из берегов рек весной и в на-
665
чале лета, когда птицы гнездятся, иногда вызывает сплошное уни-
чтожение выводков птиц, гнездящихся в приречпой пойме на земле
или в кустах.
Влияние атмосферного давления. Давление воздуха уменьшается
с поднятием пад уровнем моря, так что птицы, летая высоко и затем
опускаясь, попадают в слои воздуха с разным давлением и поэтому
должны иметь соответствующие приспособления, чтобы переносить
эту быструю смену. Таким приспособлением являются воздушные
мешки, которые распределяются между внутренностями и заходят
даже в трубчатые костил
Ветер. Изменение давления на поверхности земли вызывает ветры.
Господствующие ветры в определенной местности в определенное
время года в связи с распределением морей и материков обусловли-
вают погоду, которая играет чрезвычайно важную роль в жизни птиц.
Разная степень нагревания воды и суши и различных частей
суши в зависимости от1 топографии местности, от почвы и раститель-
ного покрова вызывает различные местные токи воздуха, как гори-
зонтальные, так и вертикальные. Эти токи играют весьма важную
роль во время полета птиц и именно при парении. Чтобы во-время
пользоваться этими токами, птица должна обладать способностью
чувствовать их па расстоянии, до известной степени их предвидеть.
Таким тонким барометром, вероятно, являются те же воздушные
мешки. Впрочем, далеко не все птицы в этом отношении одинаковы.
Наблюдения показывают, что в то время как не умеющие парить
птицы, вроде галки, часто не могут справиться с воздушными то-
ками, хорошо парящие птицы, как чайки, пользуются ими с боль-
шим совершенством. Далее, птицы обладают способностью до из-
вестной степени предчувствовать перемену погоды и нередко еще
до наступления ненастья, бури или во время непастья перед на-
ступлением хорошей погоды дают знать своим Поведением о пред-
стоящей перемене погоды/
Ветер сам по себе играет роль, помимо влияния на полет, только
в своих крайних проявлениях. Сильный ветер раскачивает деревья,
сваливает гнезда, вываливает яйца и птенцов из гнезд, препят-
ствует полету во время перелетов, иногда заносит летящих птиц
в сторону от пролетных путей, а, застигая птиц в открытом море,
иногда является причиной гибели целых стай.
Ветер зимой при сухом снеге обусловливает метели, сдувая
с возвышенных мест снеговой покров, облегчает зимующим птицам
возможность находить пищу.
2. Сезонные явления в жизни птиц в зависимости от клима-
тических факторов
На земном шаре только полоса экваториальных климатов харак-
теризуется отсутствием сезонной периодичности в ходе метеоро-
логических элементов, иными словами однообразной изо дня в день
погодой. Это—полоса тропических лесных дебрей с ежедневными
дождями, с колебанием месячных температур в пределах 25—28° и
666
суточных температур пе более как на 8—10°. Оптимальные жизнен-
ные условия и их равномерность круглый год обусловливают чрезвы-
чайное разнообразие и богатство жизни. Годичная цикличность в жиз-
ни растений и животных отсутствует. Вместо сезонных циклов здесь
господствуют видовые и индивидуальные циклы, обусловливающие
в течение всего года полноту тонуса жизни природы. В этих условиях
разные виды и разные индивидуумы птиц гнездятся, выводят птен-
цов и линяют в разные календарные сроки, в зависимости от воз-
раста, наступления половой зрелости, продолжительности гнездо-
вого периода, сношенности пера и т. д.
В тех же тропических странах, однако, на континентах по обеим
сторонам экватора, в области действия тропических муссонов рас-
полагаются климаты с более или менее ясно выраженным периодом
засухи. Здесь более благоприятные условия жизни чередуются с ме-
нее благоприятными, причем, по мере удаления от экваторов эти
различия становятся все сильнее и сильнее и в области континен-
тальных пассатов становятся настолько сильно выраженными, что
ведут к образованию сухих областей—пустынь и полупустынь с ко-
роткими периодами дождей.
Постепенные изменения равномерных благоприятных условий
жизни птиц в продолжение года в сторону чередования благоприят-
ных условий с неблагоприятными, обусловленными периодом засу-
хи, вызвали у птиц целый ряд приспособлений, приведших к раз-
витию сезонных явлений жизни.
Еще дальше от экватора в полосе умеренных климатов к не-
благоприятным климатическим условиям прибавляется перроди-
ческое понижение температуры, которая, падая ниже 0°, обуслов-
ливает наступление зимних периодов.
Смена по временам года обусловливает годичные циклы в жизни
птиц. Наступление неблагоприятных условий, во-впервых, ведет
к тому, что птицы гнездятся и выводят птенцов только в наиболее
благоприятное время года, а во-вторых, приводит к кочевкам или
перелетам.
Отношение к месту обитания. По характеру пребывания в
данной местности различают птиц оседлых, т. е. таких,
которые круглый год живут в данной местности. К оседлым же
обыкновенно относят птиц кочующих, так как они, хотя не
остаются на местах, в которых выводили птенцов, но все же не по-
кидают на зиму данную область.
Те птицы, которые на зиму улетают в другие местности, чтобы
возвратиться лишь весной, называются перелетными пти-
цами. Эти последние в данной местности могут появляться только на
лето для вывода птенцов, — это летующие птицы; если же
они в данной местности бывают только на пролетах, тогда — это
пролетные птицы; далее опи могут появляться в данной мест-
ности только на зиму — это зимующие птицы.
Очепь часто особи одного и того же виД для данной местности
могут быть и летующими и пролетными, пролетными и зимующими,
зимующими и оседлыми. Например, большинство прилетающих
667
к нам на лето видов в то же самое время пролетает дальше на север
(зяблики, утки, вальдшнепы, бекасы и т. д.); у очень многих зимую-
щих видов часть особей пролетает южнее, например, чечетки, сне-
гири, подорожники, и очень часто зимующие виды встречаются на
зимовках с оседлыми особями того же вида: таковы у нас вороны,
галки, а южнее — вальдшнепы, черные дрозды и др.
Жизнь птиц в тропических странах в сухое время года. В са-
ваннах, расположенных к югу и северу от полосы экваториальных
климатов тропических лесов — дебрей, в тропических и субтро-
пических степях, полупустынях и пустынях северного и южного
полушарий в периоды засухи жизпь замирает: прекращается веге-
тация растений, беспозвоночные,амфибии и большая часть рептилий
впадают в анабиоз, мелкие млекопитающие в значительной части
впадают в спячку, и условия жизни птиц становятся неблагоприят-
ными. Поэтому в этот период птицы пе гнездятся и частью или от-
кочевывают или улетают к северу или к югу, соответственно тому,
где в это время условия являются благоприятными.
Жизнь птиц зимой. По мере движения с юга от тропиков к се-
веру количество видов птиц, остающихся на зиму, все уменьшается.
Это стоит в прямой зависимости от климатических условий зимы.
Из неблагоприятных для жизни птиц условий зимы наиболее
характерны следующие.
Понижение температуры ниже 0° обусловливает
замерзание воды, а следовательно, и почвы. Замерзание воды ли-
шает возможности сохранить свое местопребывание птиц водопла-
вающих и болотпых. Такие птицы, очевидпо, могут существовать
только там, где нормально температура не падает настолько низко,
чтобы болота и водоемы замерзали. В тех случаях, когда такое
замерзание происходит в местах нормальных зимовок птиц водо-
плавающих и куликов, птицы сперва скопляются там, где налич-
ность ключей задерживает замерзание водоемов и болот, а в случае
более или менее полного замерзания птицы или вынуждены отлетать
дальше к югу или, проголодав более или менее продолжительное
время, гибнут, если погода не меняется.
Понижение температуры до 0° и пиже прекращает вегетацию
растений, и таким образом те птицы, которые питаются за счет
растений, оказываются в условиях, при которых они вынуждены
откочевывать или отлетать. Таковы гуси, питающиеся травой.
Понижение температуры вместе с прекращением вегетации ра-
стений ведет за собой прекращение жизпенной деятельности ряда
животных,, представляющих пищу для птиц. Большая часть насе-
комых, моллюски, черви, амфибии, рептилии и часть млекопитаю-
щих впадают в анабиоз или спячку, прячутся, и птицы, питаю-
щиеся за счет этих животных, не могут оставаться на зиму.
Вторым важным фактором зимней жизни птиц является снеж-
ный покров. Для птиц, которые питаются семенами расте-
ний, даже при замерзании почвы в отсутствии спежного покрова
остается возможность питаться, собирая те семепа, которые оста-
лись па самих растениях, и те, которые рассеялись на почве. Точно
668
так же насекомоядные птицы могут выискивать спрятавшихся
и траве и в почве насекомых. Уже незначительный снежный покров
ставит таких птиц в более тяжелые условия, так как для отыски-
вания пищи птице приходится разрывать снег. Условия становятся
еще тяжелее, если снежный покров толще, а глубокие снега не толь-
ко делают почву недоступной для сбора семян, но закрывают и са-
мые стебли трав с семенами.
Поэтому глубина снежного покрова является важным фактором
для растительноядных и зерноядных птиц. Так, серые куропатки
которые зимой питаются зернами и травой — зеленью озимых хле-
бов, при неглубоком снеге добираются до земли, разрывая снег
ногами; при более глубоких снегах они пользуются услугами зай-
цев, которые, конечно, легче добираются до зеленей, при очень же
глубоких снегах они сильно страдают и гибнут от голода массами.
Третьим важным фактором зимней жизни птиц являются в е т-
р ы, действие которых выражается главным образом в связи со
с.нежным покровом. Сильные ветры при сухом и рыхлом снежном
покрове обусловливают метели (низовые), сдувают снег с возвы-
шенных мест, обнажая почву. Снежные зимы без оттепелей, но
с сильными ветрами для зимующих птиц являются, таким образом,
гораздо более благоприятными, чем такие же зимы без ветров с ров-
ным покровом из снега.
Четвертым и очень важным фактором для жизни зимних птиц
являются оттепели или повышение температуры зимой выше
о°. Оттепели обусловливают образование ледяной корки поверх
почвы, поверх снега (наст), что делает даже при неглубоких снегах
недоступными для пользования семена, разбросанные на почве,
а при глубоких снегах даже па стеблях трав.
Если оттепель сопровождается снегопадом или дождем, вслед
за которым следует мороз, то образуется так называемая г о л о-
л е д и ц а. Коркой льда покрываются не только почва и снег, но
травы, кусты и деревья. При этом птицы не могут пользоваться не
только семенами почвы, но даже семенами, упавшими с деревьев,
семенами и почками кустов, деревьев, спрятавшимися в деревьях
насекомыми и т. д. Гололедицы зимой являются самым страшным
для зимующих птиц фактором.
Таким образом, мы видим, что наиболее благоприятными зи-
мами являются малоснежные зимы с частыми ветрами, без оттепе-
лей и гололедиц. Наиболее неблагоприятными будут зимы с глу-
бокими снегами, без ветров, с частыми оттепелями и гололедицами.
Па севере могут оставаться на зиму только очень немногие виды
птиц, а именно: из насекомоядных только те, которые обладают
специальными приспособлениями для добывания зимующих и
спрятавшихся в деревьях насекомых. Таковы пищухи, синицы, не-
которые дятлы; синицы и дятлы питаются также семенами. Из ра-
стительноядных птиц — лишь те, которые питаются хвоей, почками
деревьев и кустов, семенами деревьев и кустов (рябчики, белые
куропатки, тетерева, глухари, чечетки, клесты и др.). Ягод слишком
мало па севере, и их запас удерживает птиц, питающихся ими, на
669
время, пока все ягоды пе будут съедены (дрозды-рябинники, сви-
ристели и др.) (рис. 503). Из хищных птиц остаются лишь те, кото-
рые питаются птицами, белками, зайцами (ястреба-тетеревятни-
ки, лапландская сова). Южнее количество птиц, остающихся
на зиму, увеличивается все более и более. Вороны и галки, отле-
тающие с севера, оседло живут в средней части РСФСР. Сюда при-
Рис. 50*3. Свиристель (Ampelis garrulus).
(Из Гаане.)
летают с севера чечетки,
снегири, подорожники. Еще
южнее, где покров снега
меньше, количество зиму-
ющих птиц увеличивается
за счет зерноядных птиц,
за счет хищников, питаю-
щихся мелкими грызунами,
а там, где морозы редки,
начинаются зимовки водо-
плавающих, куликов и на-
секомоядных птиц.
Уже на юге европейской
части Союза зимует много
зерноядных птиц (зяблики,
зеленушки, коноплянки и
щеглы), водоплавающие
(утки, крохали, гагары),
хищники, питающиеся
грызунами (сычи, совы,
мохноногие канюки).
Еще больше птиц остает-
ся на зиму на побережье
Черного и Каспийского
морей (утки, нырки, лебе-
ди, гуси, вальдшнепы, бе-
касы, дупеля, жаворонки
и т. д.).
В Средней Азии, Закав-
казье, в Малой Азии мы
встречаемся уже с чрезвы-
чайно богатой оседлой фау-
ной птиц и обильной фауной зимующих птиц Севера.
Северная Африка, долина Нила и различные части Средней
и Южной Африки, Индии, острова Индийского океана и Австра-
лия служат тоже местом зимовок многочисленных видов Севера.
Для Америки место зимовок тянется от умеренных частей Се-
верной Америки через Центральную Америку до Южной Америки.
Нередко ареалы зимовок видов занимают большую площадь,
чем ареалы гнездований, например, у белого аиста, у деревенской
ласточки и др. У иных видов, наоборот, ареал зимнего пребывания
бывает очепь ограничен. Так, краснозобая козарка (Rufibranta
ruficollis), гнездящаяся в тундрах Западной Сибири, зимует в юго-
670
западном углу Каспийского моря. Малая мухоловка (Siphia parva)
и розовый скворец (Pastor roseus) из обширных областей своего
гнездового распространения собираются на зиму в Индию на срав-
нительно очень ограниченном пространстве. Зимовки популяций
одного и того же вида, относящихся нередко к разным подвидам,
если они занимают на гнездовье более северные области, оказы-
ваются лежащими далее к югу. Обычно на зимовках птицы держатся
сходных экологических условий с их местами зимовок, но иногда
опи и на пролетах и во время зимовок меняют стации и живут в иных
условиях. Так, например, малый и большой веретенники (Limosa
limosa, L. rufa) гнездятся в болотах, а на зимовках живут первый
по берегам морей, второй — по берегам пресных водоемов.
За исключением хищников, которые никогда не образуют стай,
и дятлов, которые по большой части ведут одиночный образ жизни,
зимние птицы живут большими стаями или стайками, или неболь-
шими обществами.
Общественность зимних птиц стоит в связи с кочевками.
Очень часто у одного и того же вида южные популяции живут
оседло, севернее — являются кочующими, а еще севернее — пере-
летными. Таковы галки, вороны, чижи, щеглы, зеленушки и многие
другие.
Перелет птиц. Кочевки птиц, которые обусловливаются пря-
мым недостатком корма в результате неблагоприятных условий зимы,
непосредственно связываются с перелетами.
В самом простейшем виде перелет птиц представляет лишь более
или мепее удлиненную кочевку, которая не выходит даже за пре-
делы области, запятой гнездящимися или оседлыми особями того же
вида. Необходимость в таких перелетах объясняется просто недо-
статком корма зимой, который исчерпывается постепенно, вслед-
ствие чего особи, занимающие более северные части области рас-
пространения, принуждены откочевывать все дальше и дальше,
пока не попадут в более южные части, где условия зимы настолько
благоприятны, что дают возможность провести зиму одновременно
и оседлым и зимующим особям того же самого вида. Таковы пере-
леты у галок, ворон, грачей, снегирей, московок и др. Область
гнездования у них смежна с областью зимовок, где часть особей
того же вида живет оседло.
В случае удлинения той части области распространения вида,
из которой особи этого вида зимой бывают вынуждены отлетать
к югу, мы получаем уже более удлиненный перелетный путь, ко-
торый только этим и будет отличаться от предыдущей категории
и связывается с ней рядом переходов. Таковы перелеты овсянок
(Emberiza citrinella), зеленушек (Chloris chloris), зорянок (Eritha-
cus rubecula) и др.
Такой перелет мог возникнуть или путем расселения вида к северу
из области, где он жил оседло, в область, откуда ему приходи-
лось откочевывать зимой, причем по мере расселения на север ко-
чевки все более и более удлинялись; или перелет мог возникнуть
в результате постепенного изменения климата в более северных
671
частях области распространения в худшую сторону, пока возник-
шие как следствие ухудшающихся условий кочевки путем их удли-
нения пе превратились в перелеты.
К этой же категории простых перелетов можно отнести и сле-
дующую категорию, когда все особи из области своего гнездования
отлетают на зиму в смежную с ней область зимовок. Примером таких
видов могут служить чечетки (Linarialinaria), подорожники (Plectro-
phenax nivalis) и др.
В только что нами рассмотренных случаях область распростра-
нения видов (гнездования и зимовок) представляет непрерывное це-
лое, и такие виды можпо назвать моноареальными.
У других видов область зимовки разобщена от области гнездо-
ваний территорией, по которой особи данного вида пролетают и где
опи, следовательно, бывают лишь в качестве пролетных птиц; это
виды с двумя ареалами распространения, гнездования и зимовки,
или диареальныс виды. Таково большинство перелетных
птиц.
Такое положение могло произойти в том случае, когда на пути
следования с зимовок к северу условия местности, в которой вид гнез-
дился, изменились настолько, что вытеснили отсюда гнездящихся
особей. Тут возможны два случая. Во-первых, когда гнездящиеся
особи вытеснены из области, лежащей по соседству к местам зимо-
вок, — тогда ареал летнего и зимнего пребывания разобщен со-
вершенно, или, во-вторых, когда гнездящиеся особи вытеснены из
области, более удаленной от зимовок, — тогда разорванным оказы-
вается ареал гнездования, и вид имеет две гнездовых области, свя-
занные между собой местностью, в которой данный вид бывает толь-
ко на пролете.
Ко второй группе перелетов этой категории можно отнести
хотя бы вальдшнепа (Scolopax rusticola), который гнездится всюду
в лесной области и на Кавказе и не гнездится в степях юга России
и Украины.
К первой группе относится громадное большинство перелетных
видов. Если мы допустим, что неблагоприятные условия действо-
вали продолжительное время, что область их воздействия распро-
странялась одновременно и на гнездовую область, оттесняя ее
все дальше к северу,' и на область зимовок, оттесняя ее дальше
к югу, то разобщение этих ареалов могло получиться весьма и весьма
значительное.
Так, из простых кочевок путем естественного расселения
видов при неменявшемся климате с одной стороны, и путем
переселений под воздействием менявшихся климатических и иных
условий местности — с другой стороны, возникли в длительном
историческом процессе эволюции перелеты птиц.
Дифференциация в тоТтографии местности в третичной эре в связи
с изменениями климатов и растительности, что имело место еще
в конце миоцена и в йлиоцене, не могла не обусловить сложно-
сти явлений в расселении видов птиц, а следовательно, и их пере-
летов.
672
Уже в плиоцене, когда климатические пояса располагались как
современные, большая часть птиц севера должна была сделаться
постепенно из кочующих птиц перелетными.
Когда наступил ледниковый период, эти перелеты должны были
все более и более осложняться. А если принять во внимание, что
великое оледенение по крайней мере дважды прерывалось межлед-
никовыми эпохами, то та сложность перелетов, которую мы наблю-
даем у современных птиц становится попятной. Наступление ледни-
ков с севера должно было вытеснять птиц из гнездовых областей все
дальше и дальше к югу, к западу или востоку. Перелетные птицы в
местах зимовок должны были в силу этого скопляться в больших ко-
личествах, а это должно было вызвать обостренную между ними кон-
куренцию, заставляя их откочевывать все дальше и дальше к югу.
Так могли сложиться перелеты из южных частей Европы и Малой
Азии в Африку, из Туркестана и Китая в Индостан и Индокитай.
А потом, когда ледник стал отступать обратно на север, распро-
странившиеся вслед за ним виды птиц продолжали свои перелеты
ла зиму туда, куда опи уже привыкли летать раньше.
Изменения, которые претерпевала природа в послеледниковое
время, усложнили распространение видов, сделали ареалы многих
из них прерывчатыми; усложнились вместе с этим и пути переле-
тов птиц.
Таким образом возникли перелеты у большинства видов. В це-
лом направление перелетов или перелетные пути выражают то
направление, по которому шло расселение вида из мест прежних
гнездований во время наибольшего оледенения до мест их совре-
менного гнездования, а для многих видов и дальше: до прежних
мест зимовок, которые и по сие время остаются местами зимовок.
Эти пролетные пути мы можем назвать путями первичными.
Так как виды расселялись в северные части Евразии и Сев. Аме-
рики не только с юга, юго-запада и юго-востока, по и с востока и за-
пада, то и направление первичпых пролетных путей должно иметь
соответствующий характер.
Так мы и видим на самом деле. Для западноевропейских видов
наиболее характерно направление полета на юго-запад. Так дол-
жно было итти в общем расселение видов птиц в послеледниковое
громя. Однайо есть много видов, которые летят па юг и на вос-
ток почти в противоположных направлениях.
В некоторых случаях первичные пролетные пути идут так,
что своим направлением указывают на имевшие место ранее иные
экологические условия. Так, папример, гнездящаяся на Лабрадо-
ре длиннохвостая крачка (Sterna macrura) летит к своим зимов-
кам в Антарктике по побережью Америки и пересекает Атланти-
Hoehiiii океан около 51° северной широты, указывая на имевшее
место ранее соединение Сев. Америки с Европой, откуда и шло
первоначальное заселение Лабрадора.
Однако нзложеная нами схема не объясняет направления про-
летных путей некоторых видов. Например, исландский песочник
(('aiiulus canutus) (рис. 504) и камнешарка (Arenaria interpres),—
luiiuKiipou и Станчинский 673
птицы, гнездящиеся на крайнем севере, зимуют в Австралии и
Новой Зелапдии, а амурский кобчик (Erythropus amurensis)
гнездится в Восточной Сибири, а зимует в Южной Африке. Оче-
видно, эти перелеты не могли слджиться ни во время леднико-
вого периода, ни в плиоцене, так как и Австралия с Новой Гвинеей
и тем более Новая Зеландия в это время были отделены уже от Азии
(ангарского материка) и Европы (палеарктического материка). Ве-
роятно, перелеты этих птиц, равно как и других птиц, зимующих
в южном полушарии (ласточки, белый аист, козодой и др.), сложи-
лись иначе, раньше.
Наконец, у некоторых видов первичные пролетные пути, повто-
ряющие пути расселения вида, могли подвергнуться постепенным
Рис. 504. Исландский песочник (Camitus canutusj,
(Из Мензбира.)
изменениям в сторону их сокращения и превратились во вторич-
ные пути пролета. Такие пролетные пути могли измениться
настолько, что не выражают уже расселения вида. В некоторых слу-
чаях, как это точно установлено методами кольцевания для особей
обыкновенного рыболова, чайки (Chroicocephalus ridibundus), гнез-
дящиеся в южной Германии и Чехо-Словакии, во время осеннего
перелета птиц летят не на юг, как это можно было бы ожидать
а почти на север: часть их летит на юго-запад к Адриатическому морю
по долинам Дуная и Савы, через Триеит или через Боденское
и Женевское озера, по Роне и ее ущелью и далее по побережью Сре-
диземного моря; другая часть их летит долинами Эльбы и Рейна
на северо-запад, т. е. почти в противоположном направлении
(рис. 505).
~ Вторичное изменение пролетных путей происходит у некоторых
видов на наших глазах. Так, например, рогатый жаворонок (Еге-
674
11lopliila alpestns), заселивший c 1837 г. северную Скандинавию, стал
появляться впервые на зимовках в Англии, где ранее вовсе не
встречался.
Бывают случаи, когда пролетные пути весной и осенью не сов-
падают, как то, например, доказано методом кольцевания для
ряда птиц.
Один из этих путей несомненно является вторичным.
Таким образом, для перелетных птиц, как мы видели, могут быть
5 казаны определенные пролетные пути. Это, конечно, пе пути в пря-
Рис. ,505. Карта пролетных путей чайки (Chroicocephalus
ridibundus).
(Из Lucanus.)
мом смысле, не дороги, но более или менее широкий район, по ко-
торому двигаются перелетные птицы. В одной и той же области
отн пути для разных видов могут итти в разных, иногда в почти
противоположных направлениях. Так, из средней части РСФСР
дубровник (Emberiza aureola) и чечевица (Erythrina erythrina)
'ютит почти па восток, в то время как вальдшнепы (Scolopax rus-
IicoI.t.) и ряд других видов летят на юго-запад (рис. 505).
Но и особи одного и того же вида из разйых мест летят особыми
пролетными путями. Аисты (Ciconia ciconia) из Западной Европы,
западнее Везера, летят в Южную Африку через Гибралтар, Сахару
и норхоиья Конго; аисты из области восточнее Везера летят туда же
черен Малую Азию, Палестину и дальше вдоль долины Нила.
675
Западный путь следует считать вторичным путем.
Даже особи одного и того же вида из одной и той же местности
могут лететь разными путями; так, например, чайки (обыкновен-
ный рыболов — Chroicocephalus ridibundus) имеют три пролетных
пути; (рис. 505); вальдшнепы из северо-западной области СССР летят
или вдоль побережья Балтийского моря через Шотландию и далее
вдоль побережья Северного моря, или через Восточную Пруссию,
Баварию, Южную Францию и Испанию в Марокко или, наконец,
через Каринтию, Истрию и Сицилию в Тунис (рис. 506).
Рис. 506. Карта пролетных путей вальдшнепа (Scolopax rus-
ticola).
(Из Lucanus.)
Для многих видов птиц пролетные пути совпадают с берегами
морей, с долинами больших рек, расположением озер, опушками
лесов и другими подходящими для видов экологическими усло-
виями.
Время отлета и прилета для видов весьма различно, различны
также и причины, побуждающие птиц лететь. Тут можно установить
три категории птиц. Некоторые птицы начинают отлетать очень
рано, когда еще нет никаких признаков наступающей осепи и лет-
няя жизпь других животных в полном разгаре. Очень рано отле-
тают стрижи (Cypselus apus), многие кулики: веретенники (Li-
mosa limosa), турухтаны (Philomachus pugnax), дупеля (Gallinago
m^dia) и др.; иволги (Oriolus oriolus), сизоворонки (Coracias garru-
676
lus) и некоторые другие отлетают из средней европейской части
СССР в начале августа; несколько позднее — ласточки (Hirundo
urbica, Chelidon rustica, Riparia riparia) и многие насекомоядные.
У этих рано отлетающих птиц отлет (равно как и прилет) происхо-
дит без всякой видимой зависимости от изменения во внешних
условиях среды, почти с календарной точностью. Очевидно, побу-
дительной причиной к отлету (а весной к прилету) следует здесь
считать пробуждение врожденной потребности к перелету или пере-
летного инстинкта, который активируется какими-то неизвестными
еще нам периодически действующими внутренними возбудите-
лями. Возможно, что таким возбудителем являются инкреты поло-
вых желез птиц, периодическая деятельность которых так всеобща
у животных. Поставленные для решения этого вопроса эксперимен-
ты, однако, пе дали определенных результатов. Экспериментально
установлено, что из впешпих факторов на пробуждение перелетпого
инстинкта оказывает влияние изменение в продолжительности
освещения. Инстинкт у рано отлетающих птиц так неудержимо
сильно действует, что другой даже такой сильный инстинкт, каким
является у птиц материнский инстинкт, заглушается. Бывают слу-
чаи, что наступает время отлета, а птенцы у запоздалых выводков
еще пе успели окрепнуть, родители их бросают на произвол судьбы
и пускаются в путь.
Это — первая категория перелетных птиц. Зимовки этих птиц
лежат далеко, пролетные пути у них длинные, и их перелеты, сло-
жившиеся очепь давно, путем естественного отбора, наследственно
закрепились настолько, что обусловливаются в настоящее время
внутренними причинами: пробуждением инстинкта независимо от
ухудшения условий жизни.
Ко второй категории перелетных птиц можно отнести таких,
которые начинаю? отлетать, когда в окружающей природе уже
происходят заметные изменения: понижается температура, умень-
шается количество пищи и т. д., ио еще пе настолько, чтобы вид
находился в условиях неблагоприятных. Очевидно, у этих птиц
необходимость перелета обусловливается тоже пробуждением ин-
стинкта, но пробуждепие его, по крайней мере осенью, связы-
вается с внешними факторами. К этой категории могут быть отне-
сены гуси, журавли, очень многие насекомоядные птицы и боль-
шинство перелетных зерноядных птиц.
Наконец, к третьей категории можно отнести тех перелет-
ных птиц, которые остаются на родине до тех пор, пока не
наступит такая осенняя погода, что птицам становится трудно
жить: количество пищи резко уменьшается, почва и водоемы за-
мерзают и т. д. Это — поздно отлетающие птицы. Их отлет и прилет
стоят в прямой связи с общим состоянием наступающей осени или
весны. У этих птиц отлет идет медленно, напоминая кочевки, а зи-
мовье находится сравнительно близко. Сюда относятся многие виды
уток, нырков, грачи, вороны, жаворонки, бекасы и др.
Отмечают, что птицы обычно возвращаются на свои прежние
места гнездования т. е. на свою родину, улетают на свои места
677
зимовок и на пролетах держатся определенных пролетных путей.
Кольцевание птиц дало много фактов, подтверждающих это поло-
жение. Имеются, однако, и исключения из этого общего правила.
Так опытами кольцевания доказано, что утки (кряква, шилохвость
и некоторые другие) часто пе попадают на родину, а оказываются
в местах очень далеких от родины.
Чтобы найти свою родину, птицы, очевидно, должны обладать
способностью ориентироваться в пространстве. Можно думать, что
птицы ориентируются с помощью памяти, острота которой у птиц
экспериментально доказана. Осенью молодые птицы отправляются
па зимовки под руководством старых опытных птиц, которые пе
раз проделывали перелеты и которые хорошо знают дорогу. Долины
больших рек, берега морей, большие озера, границы пустынь,
острова, по которым, как мы видели, проходят пролетные пути, облег-
чают птицам нахождение дороги. То обстоятельство, что мно-
гие птицы летят ночью, как будто не противоречит этому, так как
воды, отражая звездное небо, обозначаются ночью звездными доро-
гами, по которым птицы могут ориентироваться очень легко. Тот
факт, что во время туманов птицы легко сбиваются с пути, только
подтверждает такое предположение. И, действительно, у многих
птиц, как, например, у гусей, лебедей, журавлей, уток, нырков,
мы видим пе только образование стай, но и строгий порядок в строе
этих стай с вожаком стаи во главе. Мы видим, что у этих птиц моло-
дые отправляются в путь одновременно со старыми птицами в од-
ной, общей с пими стае.
Однако есть птицы, которые отправляются в путь в одиночку,
папример, кукушка (Cuculus canorus), удод (Upupa epops) и др.;
молодые птицы этих видов, в первый раз отправляясь на зимовки,
безошибочно находят туда дорогу, двигаясь всегда в соответствую-
щем направлении. У некоторых видов молодые птицы образуют
стаи отдельно от взрослых и иногда первыми отправляются на зи-
мовки. Так, у скворцов (Sturnu^ vulgaris) стаи молодых птиц от-
летают задолго до начала отлета старых и направляются к местам
зимовок так же уверенно, как и старые птицы. Кроме того, очень
многие чисто дневные птицы совершают свои перелеты ночью
притом часто в темные ночи, когда небо закрыто облаками и тучами;
летят они при этом нередко через море или сушу, вдали от долин
больших рек. Есть виды, которые зимуют на островах среди океа-
нов и так же легко находят к ним дорогу, как вооруженные кар-
тами и компасом мореплаватели.
Для последней категории птиц объяснение способности нахо-
дить места зимовок осенью и свою родину весной по памяти не-
пригодно. Инстинкт побуждает птицу лететь и притом лететь не-
пременно в определенном направлении. Какой механизм управ-
ляет при этом птицей, остается пока неизвестным. Мы знаем только,
что перелетный инстинкт обусловливается внутренними причинами,
которые у части птиц стоят в связи с внешними изменениями при-
роды; мы знаем, что этот инстинкт действует с большой силой, так
что птица не может ему противостоять,^ что он действует в продол-
678
жепие сравнительно короткого времени, чтобы па время, до весны,
заглохнуть. Перелетные птицы в неволе, когда наступает время
отлета, обнаруживают беспокойство, бьются в клетке, стремятся
лететь, издают призывные крики, но как только время пролета
кончается, они снова совершенно успокаиваются. Запоздавшие
особи, часто не долетев до места обычных зимовок, останавливаются
па полпути и часто весьма страдают от тяжелых условий зимы
и даже гибнут, по не летят дальше, так как время пролета кончи-
лось и перелетный инстинкт перестал действовать побуждающим
образом.
Что касается высоты перелета, то прежние предположения
о больших высотах в 4 000—5 000 м совершенно опровергнуты
новейшими наблюдениями с помощью летательных аппаратов. Ока-
залось, что только как исключение птицы поднимаются на высоту
более 1000 м. Нормально птицы летят на высоте всего лишь 100—
500 М.
Быстрота перелета тоже в значительной степени переоценива-
лась прежними авторами. Точные наблюдения последнего времени
установили, что па осеннем .перелете птицы вообще двигаются го-
раздо медленнее, чем па весеннем. В среднем для некоторых птиц
получены определенные числа. Так, аист летит со скорость^ около
200—250 км в сутки, вальдшнепы — около 400—500 км в сутки.
Один из наиболее длинных перелетов в 15 000 км плавунчик (Cry-
mophilus fulicarius) совершает в 47 дней, т. е. в среднем делает по
300 км в сутки.
Некоторые птицы прилетают громадные пространства без вся-
кого отдыха. Это относится к тем птицам, которые летят через
океаны. Так па Гавайских островах зимуют кроншнепы (Numenius
lhaitiensis) и ржанка (Charadrius dominions fulvus). Оба вида гнез-
дятся в тундрах Сев. Америки. Они должны проделывать без от-
дыха путь в 3300 км.
Условия весенней, летней и осенней жизни птиц. Весен-
няя ж и з п ь птиц выражается прежде всего у перелетных птиц
пролетом и прилетом, у кочующих птиц — прекращением кочевок
и возвращением на обычные места гнездования. После этого птицы
приступают к размножению. У полигамов оно начинается токова-
нием, у мопогамов — образованием пар. У этих последних пары
образуются нередко еще на зимовке. Далее следуют гнездостроение,
откладка и высиживание яиц, вывод и выкармливание птенцов.
Только у клестов (Loxia) (рис. 507) период размножения падает и на
зиму. Собственно весенний период заканчивается выводом птенцов.
Выкармливание и вождение птенцов происходят уже в летний пе-
риод.
Весенпий прилет происходит в определенной закономерной по-
следовательности. Для средней европейской части РСФСР первыми
появляются грачи, вначале марта, затем около 20-го прилетают жа-
воронки, за ними скворцы, потом коноплянки, зеленушки, зяблики,
трясогузки, чибисы, утки и т. д. Это первый период весеннего при-
лета. Вскрытием рек открывается второй период, когда пролетает
679
большинство водоплавающих птиц и куликов, происходит валовой
прилет зерноядных птиц начинается прилет насекомоядных. Он
продолжается до распускания лиственных деревьев. В это время
начинается массовое появление насекомоядных птиц, прилетают
кукушки и ласточки и закапчивается прилет и пролет водоплаваю-
щих птиц и куликов. Четвертый и последний период начинается
с момепта установления
ровной и теплой погоды. В
этот период заканчивается
прилет и пролет большин-
ства насекомоядных птиц.
Появляются поздно про-
летающие птицы: перепел-
ки, коростели, стрижи, си-
зоворопки, чечевицы, зеле-
ные пеночки и др.
Если на птицах первого
и второго периодов приле-
та в высокой степени ска-
зывается общее состояние
погоды, и прилет и пролет
их стоят в прямой зави-
симости от поступательно-
го хода весны, птицы треть-
его и особенно четвертого
периодов прилетают почти
с календарной точностью.
Это обычно совпадает с
установлением ровной ве-
сенней погоды.
Различного рода небла-
гоприятные климатические
факторы поэтому гораздо
сильнее сказываются на
птицах, прилетающих в
первые периоды весны. Но,
с другой стороны, они ока-
зывают иа этих птиц го-
Рис. 507. Клест еловик (Loxia curvi ro-
stra).
раздо менее губительное влияние, чем на птиц, поздно при-
летающих. Дело в том, что птицы, которые прилетают рано, руко-
водствуются состоянием погоды и в случае наступления неблаго-
приятной погоды нередко отправляются в обратный путь. Птицы,
прилетающие поздпее, этого пе делают и в случае наступления моро-
зов или выпадения снега в середине или конце мая гибнут массами.
Не менее губительно отзывается неблагоприятная погода весной
па яйцах и птенцах.
Внезапное понижение температуры до 0° и ниже вызывает за-
мерзание яиц и гибель птенцов от холода и голода. Выпадение снегов
имеет следствием еще большую бескормицу. Продолжительное пена-
680
гтье обусловливает подмокание яиц. Вызванный длительными
дождями вторичный разлив рек часто уничтожает весной и летом
гнезда и целые выводки птенцов.
Условия летней жизпи птиц уже были отчасти рас-
смотрены нами выше. Выкармливание птенцов, вождение их у вывод-
ковых и следующая за этим послебрачная линька птиц исчерпывают
содержание летней жизни. Неблагоприятные условия погоды ска-
зываются здесь главным образом продолжительным ненастьем, так
как понижение температуры пиже нормы и до 0° бывает в это время
очень редко. Помимо гибели птенцов, которые мокнут и просту-
жаются, продолжительное ненастье вызывает голод у насекомояд-
ных птиц, особенно у птиц, кормящихся летающими насекомыми;
обложные дожди во время линьки, затрудняя добывание пищи,
делают их одновременно легко подверженными заболеваниям.
Засуха, наоборот, ставит в благоприятные условия насекомо-
ядных птиц, по вредно отражается на болотных птицах, так как
болота пересыхают, и птицам приходится откочевывать в поисках-
подходящих местообитаний. В случае длительной засухи травяни-
стая растительность выгорает, урожай семян падает, и зерноядные*
птицы тоже попадают в тяжелые условия бескормицы.
Осенняя жизнь птиц выражается в образовании стай,
кочевках и отлете. Отлет идет в общем у большинства видов за
исключением первой категории перелетных птиц гораздо медлен-
нее, чем прилет.
И тут мы наблюдаем строгую последовательность в порядке. Мы
пи дели, что отлет начинается уже в июле, когда летят молодые,
скворцы, затем улиты (Limosa limosa), турухтаны (Philomachus),
в начале августа — стрижи (Cypselus apus) и крачки (Hydroche-
lidon), позднее — иволги (Oriolus oriolus), сизоворонки (Сога-
cias garrula), удоды (Upupa epops) и др., за ними летят прочие
насекомоядные и дупеля (Gallinago media); в сентябре заканчивается
отлет насекомоядных, происходит отлет зерноядных и начинается
отлет водоплавающих.
Неблагоприятными климатическими факторами в этот период
жизпи птиц могут быть: рано наступившие холода с замерзанием
почвы, водоемов и болот, ранний снегопад, продолжительное нена-
стье, сильные и продолжительные встречные ветры и, наконец, бури.
Рано наступившая осень или зима могут застать перелетных
птиц врасплох. Они еще не приготовились к отлету, перелетный
инстинкт еще не успел заговорить с достаточной силой, птицы пе
трогаются в путь и, попадая в тяжелые условия, гибнут. В пути пе-
релетные птицы могут сильно страдать от ненастья, от сильного
встречного ветра, а бури губят их или заносят иногда далеко в чуж-
дые страны, где опи гибпут в борьбе со стихией.
3. Экотопические факторы
Изучая современных нам птиц в отношении их распределения по
средам обитания, мы видим, что птицы приспособились к самым
разнообразным условиям существования—к годному образу жизни,
68 Ь
к жизни в болотах, в открытых, лишенных деревянистой раститель-
ности местах, будь то равнины или скалы, к жизни в лесах и ку-
старных зарослях, не дав, однако, совершенно подземных, живу-
щих в земле форм. Последнее попятно самой собой, так как слишком
уже неподходящей для летающих форм оказывается подземная
среда и слишком специализировалось крыло, чтобы оно могло пре-
вратиться в роющую конечность, неудобен для этого покров из
перьев и слишком специализированы задние конечности. Лишь
небольшое число видов роет норки для вывода птенцов или для
обеспечения безопасности на время покоя. Ко всем остальным сре-
дам обитания, как то мы видели, птицы приспособились уже на
заре своей эволюции.
Воздух как среда обитания птиц. Хотя воздух не является сре-
дой обитания в полном смысле слова, так как все животные при-
нуждены бывают для отдыха и для размножения опускаться на
землю, однако для некоторых птиц именно воздух нужно считать
основной средой обитания, так как эти птицы питаются исключи-
тельно в воздухе летающими животными. Таковы стрижи (Сур-
seli), ласточки (Hirundinidae), таковы отчасти козодои (Capri-
mulgi), тиркушки (Glareolidae), крачки (Sterninae), таковы неко-
торые сокола, которые бьют птиц па лету например, сокол cancan
(Falco peregrinus).
Ограниченность летающих в воздухе животных (главным обра-
зом насекомых), служащих пищей для птиц, обусловливает и малое
количество форм, приспособившихся к исключительному добыва-
нию пищи в воздухе. Между тем большая часть птиц пользуется
воздухом как средой передвижения при добывании пищи из других
сред обитания.
Использование воздуха одновременно и как среды передвиже-
ния и как среды добывания пищи требует особой специализации.
Конкурентами птицам в этом использовании воздуха в пастоящее
время являются летучие мыши из млекопитающих, а в мезозое,
быть может, являлись птерозавры.
Птицы, проводящие большую часть своей суточной жизни в по-
лете, обладают наиболее совершенным летательным аппаратом,
который совершенствовался в двух разных направлениях. У птиц
мелких и средних мы видим чрезвычайное удлинение крыла, ко-
торое бывает заострено к вершине, как у стрижей, ласточек,
тиркушек, крачек, соколов. Птицы с такими крыльями обладают
чрезвычайно быстрым полетом, которым очень легко управляют,
делая быстрые и неожиданные повороты в разных направле-
ниях. Хвост у этих птиц по большей части глубоко вырезанный,
или вильчатый, что, очевидно, тоже помогает воздушным эволю-
циям.
У более крупных видов летательный аппарат приспособляется
к парению, причем у морских форм крыло бывает относительно
длинное и узкое (буревестники,- чайки), а у сухопутных форм оно
шире и короче (хищные птицы), что стоит в связи с особенностями
парения над волнами или сушей.
<682
Большая часть птиц, добывающих пищу в воздухе, являет-
ся или жителями скал и обрывов или древесной раститель-
ности.
Стрижи и ласточки вторично приспособились к гнездованию
в постройках человека, стены которых заменяют собой отвесные
склоны скал и берегов.
Вода как среда обитания птиц. Вода, как и воздух, не может
быть исключительной средой обитания птиц. В период кладки яиц
и высиживаяния яиц водные птицы вынуждены выходить на сушу,
где и происходят кладка яиц и вывод птенцов.
Использующие воду как среду обитания и добычи пищи птицы
обнаруживают разнообразные приспособления. Возможность ис-
пользовать для передвижения как крылья, так и ноги дает два на-
правления в приспособлениях. Первая группа птиц, летая над водой,
отыскивает пищу, находящуюся в воде или на воде, и, бросаясь
затем в воду, хватает ее. У птиц этого рода, естественно, разви-
ваются крылья, птицы приобретают все более и более мощный по-
лет, приобретают способность парить и достигают возможности по-
долгу, целыми днями, носиться над водой, превращаясь в воздуш-
ных океанических обитателей. Наиболее совершенным типом птиц
такого рода являются, несомненно, буревестники, или трубконосы
(Procellarii). Сюда относятся и крупные виды, до 3 м в разма-
хе крыльев, как, например, альбатросы (Diomedea) или гигантский
буревестник (Ossifraga gigantea) и средней величины птицы и даже
мелкие виды вроде качурок (Procellaria), величиной со скворца.
Все относящиеся сюда птицы обладают чрезвычайно длинными
крыльями и весьма совершенной способностью парить. Опи неуто-
мимо носятся над открытым морем, одинаково свободно как в бурю,
так и в относительно тихую погоду. В случае надобности птицы
опускаются па воду и могут плавать па воде.
Совершенно аналогичный ряд приспособлений выработался
у птиц другого отдела, аистоподобпых, только в отряде веслоногих
(Steganopodes), —таковы океанические фаэтоны (Phaetonidae),
олуши (Sulidae) и фрегаты (Fregatidae). Третий аналогичный ряд
приспособлений мы находим в отряде чаек (Lari).
Наконец, дневные хищные птицы со своими хищными лапами
н крючкообразным носом, получив возможность схватывать добычу
когтями и разрывать ее на части острым клювом, тоже дали формы,
приспособившиеся к добыванию пищи в воде; таковы: скопа (Pan-
fl ion haliaCtus), рыбные орлы (Polioaetus) и морские орланы (Tha-
lassaetus).
Вторая группа птиц использует крылья для передвижения в воде
при пырянии. Пингвины (Sphenisci). и..цистики (Alcae) действу-
ют в воде . крыльями "как веслами; при этом крыло пингвинов
достигло в этом отношении высокой специализации: перья крыла
превратились в чешуевидные образования, крыло получило ха-
рактерный изгиб, и птицы совершенно утратили способность ле-
тит!.. Вымерший недавно чистик (Alca impennis) тоже, аналогично
пингвинам, утратил способность летать.
683
Остальные водные птицы при плавании и нырянии пользуются
ногами, для чего пальцы их срастаются перепонками. У большин-
ства водоплавающих три передних пальца соединены перепон-
ками, а у веслоногих (Steganopodes) все четыре пальца соединены
перепонкой; у поганок (Colymbi), лысух (Fulica), куличка-пла-
вунчика (Phalaropus) пальцы снабжены лопастными перепонками.
Впрочем, неплохо плавающая водяная курочка (Gallinula chlo-
ropus) вовсе не имеет перепонок па ногах.
Болота как среда обитания птиц. Болота представляют своеоб-
разную среду обитания птиц, хотя и связываются незаметными пе-
реходами, с одной стороны, с водными стациями, с другой — с на-
земными.
Характерными экологическими особенностями болот являются:
обилие воды, вязкость почвы, богатство и доступность фау-
ны беспозвоночных и амфибий. Растительность болот может
быть довольно различной, и па основании ее могут быть уста-
новлены три типа’ стаций: 1) тростниковые болота,
2) осоковые или луговые болота и 3) моховые
болота.
Экологические особенности болот близки к таковым неболо-
тистых берегов водоемов, почему ряд видов птиц является общим
как для болот, так и для берегов водоемов.
Приспособления, которые мы встречаем у болотных птиц, обу-
словливаются упомянутыми экологическими особенностями. Как
и в других средах, так и здесь мы видим два способа приспособле-
ния к добыванию пищи в болотах. Часть нтиц пользуется главным
образом крыльями при движении, другие — ногами. В связи
с наличностью мелких вод и вязкостью почвы у последних ноги
длинные, и голепопяточпос сочленение пе оперено (голенастые),
передние пальцы пог длинные, часто связанные перепонками, что
позволяет противостоять вязкости почвы; иногда длина пальцев
увеличивается за счет когтей.
Наличие вод в болотах и переход от болот к водным ста-
циям дает возможность селиться в болотах ряду водоплавающих
видов.
Типичными обитателями болот являются кулики (Limicolae),
затем цапли (Ardeae), аисты (Ciconiae), пастушки (Ralli) и жу-
равли (Grues) Далее в болотах мы встречаем мпогих утиных,
некоторых дневных хищных, сов, чаек и воробьиных.
Наиболее частыми комбинациями болотных стаций с другими
являются, конечно, водно-болотные констации, к которым приспо-
соблены очень многие водоплавающие из утиных и чаек, многие
аисты, цапли, водяные курочки, лысухи и др.
Очень часты сочетания болот с зарослями кустарников, дающие
болотно-кустарные и водо-болотно-кустарные коистации, причем
из кустарниковых при этом наиболее обыкновенной бывает лоза.
Нередко сюда присоединяются деревья.
Открытые пространства как среда обитания птиц. Очень многие
птицы приспособились к жизпи среди открытых, лишеппых
684
деревянистой растительности пространств. Таковы луга, степи,
пустыни. Общие для открытых стаций экологические особенности:
твердость почвы, отсутствие воды, сухость воздуха, значительная
суточная амплитуда колебания температуры, отсутствие древесной
растительности, отсутствие защиты от ветров и бури, но зато отсут-
ствие препятствий для передвижения по земле и воздуху, отсут-
ствие удобных мест для защиты от врагов, обилие семян, насеко^
мых, рептилий и грызунов.
В зависимости от степени развития травянистой растительности
можно различить две категории открытых стаций: 1) стации с бед-
ной растительностью (пески, полупустыни, пустыни) и 2) стации
Рис. 508. Попытка, или саджа (Syrrhaples paradoxus).
(Из Брэма.)
с богатой растительностью (луга, степи). Между обеими катего-
риями, конечно, есть переходы.
Как и среди древесных обитателей, мы встречаем здесь два на-
правления в развитии приспособлений. С одной стороны, существуют
птицы, которые все больше и больше приспособляются к передви-
жению по земле в поисках за пищей и пользуются ногами, чтобы
спасаться от врагов, почти пе пользуясь крыльями; с другой сто-
роны, наоборот, есть птицы, которые пользуются при передвиже-
ниях главным образом крыльями совершенно почти не пользуясь
ногами. Первое направление в приспособлении повело к полной
потере способности летать и к редукции крыла; одновременно
сильно развиваются и специализируются ноги: для бега полезно
уменьшение поверхности соприкосновения с землей, что достигается
укорочением пальцев и уничтожением заднего пальца, который,
очевидно, лишь мешает при беге.
685
Второе направление в эволюции ведет к усовершенствованию
крыла в связи с более совершенным полетом; ноги при этом могут
сохранять свое нормальное развитие или даже до известной сте-
пени редуцироваться.
Примером птиц, приспособившихся к бегу по земле, может слу-
жить своеобразная группа южноамериканских птиц тинаму (Tina-
mi), по виду напоминающих наших куропаток. Опи великолепно
бегают по земле, пользуясь крыльями лишь в случае явпой опас-
ности, причем летят стремительно, но могут пролетать лишь корот-
кое пространство. Ноги у них сильные и специализировались для
бега. Задний палец редуцирован.
Таковы же многие из наших куриных (Galli). Трехперстки
(Turnicies) дают подобную же группу птиц, приспособившихся к
передвижению по земле, у которых редукция заднего пальца доведена
до полной его потери. Наиболее же резким уклонением в этом отно-
шении является группа бескилевых птиц (Ratitae). К этой группе
относятся африканские страусы (Struthiones), южноамериканские
панду (Rhea), австралийские эму (Dromaeus) и новозеландские
киви (Apteryx). Приспособление ног для бега выражено у этих
птиц весьма сильно, особенно у африканских страусов, где реду-
цировался четвертый палец и укоротился третий.
В качестве примеров птиц, приспособившихся к жизни среди
открытых пространств, но усовершенствовавших для передвиже-
ния не ноги, а крылья, мы можем привести, во-первых, рябков
(Pterocletes), во-вторых, тиркушек (Glaredidae),напоминающих свои-
ми легким и быстрым полетом ласточек, к тому же последние при-
способились к добыванию пищи — насекомых — на лету. И у тир-
кушек ноги относительно слабо развиты, а задний палец очень мал.
К этой же группе могут быть отнесены многие дневные хищпые
птицы и некоторые совы.
Для добывания пищи из твердой почвы птицы могут прибегать
к клюву и лапам. Клюв таких птиц в противоположность клюву
куликов должен быть короткий и достаточно прочный, что мы видим
у кур, трехперсток, тинаму. Ноги для разрывания земли должны
быть снабжены короткими и сильными когтцми, что мы и видим
опять-таки у кур.
Отсутствие деревянистой растительности делает условия само-
сохранения птиц более тяжелыми, а потому защитная окраска у птиц
открытых пространств достигает особенного развития. Меньшая
обеспеченность яиц и птенцов от хищных млекопитающих и реп-
тилий вызывает необходимость в компенсации или большими клад-
ками или дву- и даже троекратным выводом птенцов; отсутствие
дупел вынуждает птиц пользоваться норами для того, чтобы пря-
тать гнезда и яйца, а иногда, чтобы и самим прятаться.
Отсутствие воды в пустынях обусловливает у птиц необхо-
димость перелетов на водопой, ипогда за многие километры; или
же у них развивается способность обходиться без воды. Резкие
суточные колебания температуры и отсутствие защиты от ветров
вызывают образование более густого оперения.
686
Древесная растительность как среда обитания птиц. Древесная?
растительность в течение всей эволюции птиц и в настоящее-
время является основной стихией этого класса. Благодаря
умению летать, благодаря строению ног птицы могли исполь-
зовать для обитания все горизонты деревянистой раститель-
ности как в отношении пищи, так и в отношении убежища для
гнезда.
У птиц с древесным образом жизни нога устроена всегда таким
образом, что свободные пальцы противополагаются друг другу,
так что получается возможность охватывать с двух сторон ветки.
Обычно хорошо развитый первый палец противополагается трем
остальным. Так устроена нога у большинства птиц, такой она была
у археоптерикса, и такую ногу нужно считать исходной для всех
других видоизменений ног. У некоторых птиц передние пальцы
срастаются при основании ног, причем сила давления падает пре-
имущественно на третий и четвертый пальцы; второй палец может
в таком случае укоротиться и даже стать совершенно рудиментар-
ным. Еще большая сила при охватывании получается тогда, когда
двум передним противополагаются два задних пальца. Иногда
один из передних пальцев, именно четвертый, по произволу или
обращен вперед или отворачивается назад, в помощь первому пальцу.
Иногда четвертый палец окончательно обращен назад, так что два
пальца обращены вперед, а два назад, например, у попугаев и дят-
лов. Иногда даже первый палец редуцируется, как, папример,
у трехпалых дятлов (Picoides).
У птиц, лазящих по вертикально стоящим стволам деревьев,
когти пальцев сильно загнуты и остры, что дает возможность птице
прочно держаться за поверхность коры. При достаточной силе
мускулатуры некоторые птицы могут, цепляясь когтями, лазать по
дереву в разных направлениях и вверх и вниз головой, как, напри-
мер, наши поползни (Sittidae). В других случаях птицы при
лазании по стволам помогают себе хвостом, перья которого
делаются особенно прочными, жесткими. Так лазят дятлы (Pici),
пищухи (Certhiidae). Хвост служит здесь отчасти опорой, не
дающей телу сползать вниз по стволу, отчасти для удержания
тела в равновесии при откидывании передней части тела. Сте-
нолаз красиокрылка (Tichodroma muraria) помогает себе при ла-
зании крыльями, а попугаи пользуются одновременно и клювом
и пальцами ног, хватаясь попеременно то клювом, то ногами за
ветки и подтягивая затем тело. Вообще у птиц, лазящих по стволам,
ноги укорочены, тогда как птицы, которые приспособились лазать
по веткам, имеют ноги более длинные, особенно те, которые при-
способились к лазанию по вертикально стоящим веткам, густым
зарослям кустарников. У этих последних тело обыкновенно более
удлинено и голова нередко заострена.
Многие птицы приспособлены лазать по веткам, подвешиваясь
к ним снизу. Тут необходимы особенно крепкие и цепкие ноги и
большая ловкость движений. Из наших птиц этой способностью
в совершенстве владеют синицы (Paridae); такой способностью об-
687
ладают и другие птицы, папример, некоторые из семейства выор-^
новых (Fringillidae).
Как и в других средах, обитатели древесной растительности спе-
циализировались в двух направлениях. Большинство приспособи-
лось к передвижению в лесу с помощью ног, т. е. к лазанию в целях
добывания пищи, по некоторая часть, конечно, меньшая, приспосо-
билась к передвижению в целях добывания пищи с помощью крыльев.
К таким, по преимуществу летающим в поисках пищи, мы должны
отнести голубей (Columbae)—птиц с мощным полетом и слабыми
ногами, стрижей (Cypseli), которые могут только подвешиваться
к веткам своей маленькой ножкой, родственных им колибри (Тго-
chili), также козодоев (Caprimulgi), некоторых воробьиных (неко-
торые ласточки, мухоловки) и многих дневных хищных птиц
и сов.
Наконец, немалое число видов в лесах приспособилось к жизни
на земле, используя почву леса как среду обитапия, частью вторич-
но перекочевав в лес из открытых стаций, как, папример, нелетаю-
щие казуары (Casuarii), некоторые пастушки (RaHi) и куры (Galli),
частью спустившись на землю с ветвей и превратившись в ти-
пичных наземных обитателей, например, некоторые голуби и
попугаи.
Экологические особенности закрытых стаций: более или менее
высокие деревянистые растения; густая чаща листвы или хвои,
в которой легко скрываться и прятать гнездо и птенцов; сучья, на
которых можно строить гнезда, где птенцы и яйца недоступны бе-
гающим по земле млекопитающим и рептилиям; обилие семян, ягод,
плодов и насекомых; стволы, в которых живут и прячутся насеко-
мые и в которых могут быть сделаньГдупла для гнезд или имеются
естественные дупла; наличность защиты от ветра и бурь; более
ровные температура и влажность.
4. Биоценотические факторы
Взаимоотношения между птицами и растениями. Зависимость
птиц от растительности чрезвычайно велика. Растения в виде опре-
деленных растительных сообществ представляют прежде всего
определенную среду обитания, в которой птицы передвигаются,
добывают пищу, прячутся от врагов и воспроизводят потомство.
Для очень многих видов птиц растения непосредственно яв-
ляются пищей: семена, плоды, почки, молодые побеги, пыльца,
нектар цветов и даже листья могут служить пищей для птиц.
Далее, растения служат пищей для животных, преимущественно
насекомых, которыми в свою очередь питаются птицы.
Наконец, растения доставляют убежища для гнезд и материал
для постройки гнезд и убежище для взрослых животных.
Б свою очередь птицы служат весьма важным фактором в рас-
пространении семян растений. Во-первых, объедая мясистые части
многих ягод и других мясистых плодов, птицы выплевывают семена
и тем самым их рассеивают, иногда вдали от материнского растения;
.688
no-вторых, проглатывая мясистые плоды вместе с семенами, птицы
переваривают только мясистые части, а семена выходят из пищева-
рительного канала, нисколько не теряя своей всхожести; в-третьих,
многие птицы, питаясь семенами, часть семян съедают и перевари-
па ют, другую часть рассеивают, расклевывая плоды или не пере-
паривая их. Некоторые из птиц устраивают склады семян на слу-
чаи бескормицы (кедровка, сойка); наконец, семена с прицепками,
цепляясь за оперение птиц, переносятся ими на другое место,
а иногда семена приклеиваются к ножкам птиц вместе с илом.
Колибри Нового Света, нектарки и щетиноязычные попугаи
Старого Света, питаясь медом растений (нектаром) или посещая
цветы для ловли приютившихся там насекомых, содействуют, по-
добно насекомым, перекрестному опылению цветов. Цветы многих
тропических растений обладают в этом направлении специальными
11 риспособлениями (орнитофилия).
Большое влияние на жизнь растений оказывают насекомоядные
птицы тем, что поедают великое множество насекомых, питающихся
растениями, защищая тем самым растения.
Такую же роль играют хищные птицы, питаясь грызунами, кото-
рые истребляют растения. Есть, однако, птицы, которые питаются
насекомыми, полезными для растений, например, пчелами, шмелями
и др. (щурки, осоеды и др.).
Другие птицы, наоборот, вредят растениям, питаясь почками,
молодыми побегами или обламывая на постройку гнезд ветви (грачи),
продалбливая кору и древесину деревьев (дятлы) и т. д.
Уже из сказанного видно, в каких сложных взаимоотношениях
находятся птицы и растения. Эта сложность в значительной степени
увеличивается, если изучать те взаимоотношения, которые возни-
кают между животными при использовании условий, предоставлен-
ных им растениями.
Взаимоотношения между птицами и другими животными.Взаи-
моотношения между птицами и зооценозом существенно отличаются
в зависимости от того, обусловливаются ли они взаимодействием
особей того же вида или других видов птиц или других животных.
Эти отношения между птицами одного какого-либо вида и другими
видами животных выражаются: 1) в том, что одни виды являются
пищей для других видов животных (отношения между хищни-
ком и добычей), 2) в явлениях синойкии,
или такого сожительства, когда один вид живет в большей или
меньшей степени за счет другого, не причиняя ему вреда; 3) в яв-
лениях п а р а з и т и з м а; 4) в явлениях оказывания слу-
чайных услуг видами, обитающими в биоценозе, не связанными
непосредственным сожительством; 5) в явлениях с о р е fi-
ll о в а н и я, или конкуренции, из-за пищи, из-за ме-
ста 11 пр.
Взаимоотношения между добычей и зоо-
ф а г а м и чрезвычайно сложны.
Во-первых, жизнь зоофага теснейшим образом связана с налич-
ностью тех животных, которыми он питается. Если животные, кото-
‘V* Кашкаров и Станчянокий §89
рыми питается данный вид, исчезает, вид принужден переходить на
Другую пищу или переселяться в другую местность или вымирать.
Всякое улучшение в условиях существования животных, состав-
ляющих пищу, благоприятно отражается на жизни соответствую-
щих зоофагов, обусловливая их численное увеличение. Размножив-
шиеся зоофаги могут повести тогда к уменьшению пищи, что вызо-
вет гибель их самих, а это снова благоприятно отразится на уве-
личении пищи оставшихся в живых и т. д.
Сложность этих явлений обусловливается еще тем, что существует
обычно несколько степеней зависимости в отношении пищи, идущей
по разным направлениям. Есть виды хищных птиц, которые пи-
таются другими птицами. Эти последние могут быть или фитофагами
или зоофагами. В первом случае, зависимость хищников связывается
с благоприятными условиями в развитии соответствующих расте-
ний. Во втором случае, мы имеем зависимость от размножения дру-
гих животных, составляющих пищу зоофага. Эти другие животные,
по большей части насекомые, могут быть в свою очередь или зоофа-
гами или фитофагами. Всякое улучшение жизненных условий расте-
ний, увеличивая количество и разнообразие питающихся растения-
ми насекомых, птиц и грызунов, ведет к улучшению положения хищ-
ников, и наоборот.
С другой стороны, влияние хищников на добычу не только отри-
цательного характера, но, как выяснено непосредственными на-
блюдениями, оказывается в известных пределах благотворным.
Во-первых, хищники не дают тем животным, которыми они питаются,
выходить в своем размножении за известные пределы и тем избежать
вредных последствий перенаселения; во-вторых, в добычу хищникам
достаются прежде всего или больные или более заметные или ме-
нее ловкие животные. Таким образом, хищники содействуют отбору
и препятствуют тем вырождению вида.
Среди птиц можно найти примеры синойкии, т. е. поль-
зования совместным жилищем разных видов.
В Новой Зелапдии буревестники (Procellaria и Puffinus) поль-
зуются вырытыми гаттерией норами. Некоторые мелкие птицы из
воробьиных (Oscines), например, воробьи (Passer) и скворцы
(Sturnus), устраивают гнезда в гнездах крупных хищников (орлов
или аистов).
Другой вид подобного сожительства мы встречаем, когда мелкие
птицы поселяются в колониях других, более крупных видов. Так,
скворцы часто селятся вместе с грачами; грачи селятся вместе с ко-
лониями цапель. Тут более слабые птицы прячутся от врагов под
прикрытие более сильных. Скворцы нередко сопровождают в конце
лета стаи грачей, с которыми кочуют.
Примерами такого вида совместной жизни, когда один вид деры
жится подле другого, чтобы пользоваться защитой или извлекать
другую выгоду, может служить также постоянное сопутствование
ласточек, а иногда скворцов и галок пасущимся в поле стадам
крупных млекопитающих, домашних и диких. Крупные млеко-
питающие привлекают к себе многих двукрылых, которых лас-
690
точки ловят па лету или ловят, подобно скворцам тех насекомых,
которых вспугивают двигающиеся животные.
Страусы обыкновенно сопровождают стада антилоп, достигая
этим большей безопасности.
Спутник крокодила Hoplopterus armatus забирается в открытую
пасть крокодила, чтобы добывать завязшую в зубах пищу. Стервят-
ники и коршуны сопровождают иногда крупных хищников из мле-
копитающих, чтобы воспользоваться остатками от их пищи.
Серые куропатки зимой при глубоком снеге пользуются услугами
зайцев, которые разрывают снег, чтобы добраться до травы.
К явлениям этого же порядка, когда из совместной деятельности
извлекается выгода обоими видами, можно отнести любопытную
совместную рыбную ловлю бакланов и пеликанов. Образовав сме-
шанную стаю, птицы располагаются в море дугой и начинают дви-
гаться к берегу, загоняя рыбу к отмели, причем пеликаны бьют кры-
льями по воде, а бакланы ныряют. Когда концы дуги упрутся в бе-
рег и рыба будет отрезана от моря, птицы начинают ее вылавливать.
Случайное сотрудничество в целях охраны и
защиты выражается среди птиц прежде всего в том, что многие
птицы при виде опасности издают особые предупреждающие крики
(ласточки, дрозды, вороны, чибисы и т. д.). Эти крики понимаются
не только особями того же вида, но и другими членами биоценоза.
Птицы замолкают и прячутся.
Некоторые птицы спешат на крик опасности, и нередко на об-
щего врага набрасываются особи разных видов.
Примеры нахлебничества, или к о м е н са-
дизма, мы встречаем у птиц, преимущественно приспособив-
шихся к человеку, а также и к другим млекопитающим. В послед-
нем случае птицы пользуются экскрементами, в которых отыскивают
личинки мух и жуков, непереварившиеся зерна или паразитов.
В свою очередь птицы имеют нахлебников. В грязном от скопив-
шихся экскрементов и вонючем гнезде удода выводятся личинки
мух. Старыми гнездами пользуются различные насекомые, между
прочим, муравьи. В гнездах хищников нередко бывают разные
трупоеды из насекомых.
Что касается паразитизма, то птицы-паразиты отчасти
описаны уже, когда говорилось о гнездовании у кукушек и желтуш-
ников. Паразитами в отношении гнезд являются воробьи, которые
изгоняют из гнезд городских ласточек, иногда выбрасывая из них
птенцов или яйца. Многие хищники не строят гнезд, а отбивают их
у других птиц, менее слабых.
Поморники (Stercorarius) отбивают пойманную добычу у чаек.
Птицы в свою очередь являются хозяевами многочисленных па-
разитов. Временными паразитами птиц являются различные колю-
щие насекомые из двукрылых, комары, доошки.
Внешними паразитами (эктопаразитами) являются различные виды
из насекомых: пухоеды (Mallophaga), вши (Ornithobius, Goniocotes)
и многочисленные виды клещей. Пухоеды и пероеды питаются
отделяющимися ороговевшими чешуйками кожи и перьями.
№ 691
Из клещей упомянем: чесоточных клещей (Dermaryctes), кото-
рые делают ходы в коже, кожеедов (Symbiotes), которые живут
на поверхности кожи; клещей, которые живут в мешочках перьев
(Harprirhynchus, Syringophilus и др.). Интересны мухи кровососки
(Hippoboscidae), обладающие специальными приспособлениями для
жизни между перьями на теле птиц.
Внутренние паразиты очень многочисленны, и нет таких органов
птиц, в которых бы не было паразитов. В коже птиц встречаются
трематоды (Collyriclum), в подкожной клетчатке — филярия
(Hamatospiculum), в глазу—нематоды (Thelasia) и трематоды
(Philophtalmus), в крови — филярии и простейшие (Тгура-
nosomatidae, Leucocytozoon, Halteridium и др.), в дыхательных
путях — трематоды (Cyclocoeliidae), в мочевых путях — тре-
матоды (Eucotyle, Renicola и др.), в кишечном канале — мно-
гочисленные черви из трематод, ленточных глист и нематод
и простейшие.
Многие паразиты являются опасными и вызывают болезни и
смерть птиц.
Явления соревнования, или конкуренции,
среди птиц могут быть чрезвычайно сложны. Наиболее частым и
простым видом такого соревнования являются соревнования из-за
мест гнездования и из-за пищи.
В первом случае, конкуренция возникает тогда, когда пищи для
вида в какой-либо стации достаточно, но нехватает мест для гнездо-
вания. Обыкновенно птицы, за исключением общественно гнездя-
щихся, занимают определенный район, в пределы которого не допу-
скаются другие птицы. Общественно гнездящиеся виды часто тоже
не допускают птиц других видов селиться вблизи своей колонии.
Чем крупнее птица, тем больший район она занимает. Между птй-
цами нередко возникает соревнование за гнездовой район, причем
птицы вступают в драку, и сильнейший выгоняет из пределов райо-
на слабого. При гнездовании в дуплах у птиц, которые пользуются
готовыми дуплами, идет упорная борьба из-за них: при недостатке
деревьев, как то бывает в степи, каждое дерево может сделаться аре-
ной борьбы из-за места для гнезда; то же нужно сказать про тех хищ-
ников, которые не строят собственных гнезд.
Так как летом пищи для большинства видов вполне достаточно,
то конкуренция из-за гнезд принимает чрезвычайно острый харак-
тер, особенно у гнездящихся в дуплах.
Устройство искусственных насаждений и развешивание специ-
альных гнездилищ (например, дуплянок) показывают, что при на-
личности удобных мест гнездования количество гнездящихся птиц
может увеличиться на данной площади во много раз.
Конкуренция из-за пищи возникает тогда, когда количественно
фауна питающихся одинаковой пищей итиц достигает такого раз-
мера, что ее запасов нехватает для прокормления населения. В на-
ших условиях при отсутствии удобных для гнездования мест такая
конкуренция летом в период гнездования у большинства птиц не
дает себя чувствовать.
692
Больше всего недостаток пищи сказывается на хищниках. Мно-
гие из них имеют свой определенный охотничий район, в который ста-
раются не допускать других хищников. Такие районы мы наблюдаем
11 У Других птиц, например, у дятлов.
Конкуренция из-за пищи сказывается сильно осенью и зимой
Особенно заметна она у тех птиц, которые питаются ягодами. При
урожае ягод некоторые птицы (например, дрозды-рябинники —
Tiirdus pilaris) задерживаются осенью очень долго, пока ягоды
не будут объедены; конкурентами им являются прилетающие
с севера свиристели (Ampelis garrulus) и снегири (Pyrrhula
pyrrhula).
Сказывается такая конкуренция и среди питающихся семенами
деревьев особенно при неурожае семян.
Взаимоотношения между особями одного и того же вида. Такие,
взаимоотношения вызываются прежде всего раздельнополостью
птиц. В отношении численного соотношения полов необходимо от-
метить, что у птиц количество самцов, как правило, преобладает
над количеством самок. Во всяком случае замечено, что в случае
гибели самца самка сейчас же находит себе другого. Замечено также,
что у большинства птиц наблюдаются в период воспроизведения хо-
лостые самцы и очень редко холостые самки.
Таким образом, между самцами должна существовать известная
конкуренция из-за самок.
В период спаривания самцы, которые поют, имеют более яркую
окраску и токуют, подвергаются большей опасности по сравнению
с самками, но зато во время насиживания и в период вождения птен-
цов большей опасности подвергаются самки. В общем для вида вы-
годнее, чтобы самцов было больше, чтобы все самки были использо-
ваны для воспроизведения. Недостаток самцов, иногда даже у по-
лигамов, ведет нередко к тому, что часть самок остается неоплодот-
ворецной. Далее важно, чтобы и самки и самцы были распределены
па территории равномерно. Так как большинство птиц образует
смешанные стаи самцов и самок, то спаривание у них происходит
еще на зимовках, как у некоторых наших оседлых птиц: ворон,
juhok, сорок и др., или ранней весной, как у синиц, дятлов и др.
хИ этих птиц равномерное распределение самок и самцов может
быть легко достигнуто. Другие птицы образуют отдельно стаи сам-
цов и самок, например, зяблики. У них, как и у целого ряда дру-
гих птиц, спаривание происходит перед самым гнездованием. Пе-
ние и другие брачные звуки, издаваемые самцами, имеют при этом
большое значение: самки легко находят самцов, и пары равномерно
распределяются по территории.
Конкуренция из-за пищи между особями одного и того же вида
обусловливается или недостатком пищи в силу неблагоприятных
климатических явлений или перенаселением.
II тут, в случае, если на одной и той же территории живет не-
сколько видов, приспособленных к одинаковой пище, конкурен-
ция тем сильнее, чем больше видов с одинаковой пищей и чем боль-
шее число' особей каждого вида живет вместе.
693
Этим объясняется тот любопытный факт, что количество особей
вида, населяющего местность, обратно пропорционально количеству
видов даппой местности: чем большее число видов составляет насе-
ление, тем меньшим числом особей представлен каждый вид.
Недостаток подходящих для гнездования мест может повести
к ложному общественному колониальному гнездованию.
Такая общественность случайна, с настоящей общественностью
имеет мало общего, хотя может быть некоторые виды стали обще-
ственными именно в силу такого первоначально случайного совме-
стного гнездования.
Недостаток пищи и ее скопление в определенных местах тоже
могут повести к общественности.
Однако общественность может возникнуть и как результат вож-
дения птенцов.
У некоторых птиц общественность достигает высокой степени
развития: образование стай с вожаком, строгий строй стай на пе-
релетах, способность понимать друг друга, совместная защита от
врагов, самопожертвование при опасностях, угрожающих колонии.
Нередко образуются стаи смешанного характера из особей двух
йболее видов. Кочующие зимой стайки синиц состоят из нескольких
видов птиц: пищух,0 поползней, а иногда корольков и дятлов.
XIII. ЭКОНОМИЧЕСКОЕ ЗНАЧЕНИЕ ПТИЦ
Экономическое значение птиц вырая^ется в том вреде и той
пользе, которые они приносят человеку ц его хозяйству. Во-пер-
вых, человек использует птиц в качестве домашних животных,
во-вторых, многие дикие птицы являются объектом охоты и промы-
сла; в третьих, многие дикие птицы приносят пользу человеку и его
хозяйству, истребляя вредных животных: грызунов, насекомых,
вредителей сельского хозяйства и переносчиков инфекций; нако-
нец, польза птиц выражается в их эстетическом значении, когда че-
ловек получает удовольствие от созерцания красивых птиц или слу-
шая их пение.
Что касается вреда, приносимого птицами, то он выражается
в том, что немногие виды истребляют домашних и промысловых жи-
вотных и некоторые птицы вредят хозяйству, уничтожая посевы
или полезных для хозяйства животных.
1. Домашние птицы
Домашней птицей мы называем тех птиц, которые приручены че-
ловеком и успешно разводятся в домашнем состоянии с целью полу-
чения мяса, яиц, перьев и пуха. Подбирая по желательным призна-
кам производителей и воспитывая их в соответствующих (улучшен-
ных) условиях кормления и содержания, человек в сильной мере
влиял на изменчивость домашних животных и путем селекции
и скрещивания вывел ряд новых, весьма ценных и разнообразных по
складу и производительности пород домашней птицы,
694
В качестве домашней птицы разводятся: куры, гуси, утки,
индейки, фазаны, цесарки, павлины, лебеди и страусы, из кото-
рых в СССР наибольшее хозяйственное значение имеют куры, гуси
и утки.
Большое хозяйственное значение птицеводства определяется
прежде всего ценностью получаемых от птицы продуктов. Так,
яйца птиц отличаются исключительной питательностью, легко пере-
париваются, употребляются в пищу даже в сыром виде, входят
в состав разнообразных кушаний и употребляются для технических
надобностей; мясо птиц очень питательно, особенно белое мясо,
легко переваримо; пух и перо применяются для подушек, одеял,
перин, теплых вещей и украшений; птичий помет (гуано) является
первосортным удобрением, превосходя конский нагоз по качеству
в 3—4 раза. Не меньшее значение имеют особенные условия этой
отрасли хозяйства по сравнению с другими видами животноводства.
Вследствие большой яйценоскости, быстрого роста и созревания
птица может быть очень быстро размножена, особенно если приме-
нять искусственную инкубацию; она очепь быстро становится при-
годной для использования; будучи очень неприхотливой на корм,
довольствуется малым количеством корма, при содержании на
свободе сама себе добывает корм, причем истребляет вредных для
сельского хозяйства насекомых; с помощью птицы могут быть ис-
пользованы в качестве кормов многие отбросы сельского хозяй-
ства и промышленности, могут быть использованы зерна, оставшиеся
в поле после уборки; птица требует очень ограниченного места для
разведения, портативна и довольствуется самыми простыми и де-
шевыми постройками и т. д. Наконец, благодаря разнообразию по*
род и видов домашней птицы, птицеводством можно заниматься по-
всеместно, начиная от крайнего севера до крайнего юга.
У нас, в СССР, и в наиболее передовых европейских странах, в
(ЯНА и в Канаде птицеводство играет в хозяйстве весьма видную
роль. В этих странах достижения по птицеводству за последнее
время очень большие: имеются крупные хозяйства, разводящие
десятки тысяч птиц, широко поставлена опытная и научно-исследо-
вательская работа по птицеводству.
В дореволюционное время и в восстановительный период раз-
вития советского хозяйства промысловое птицеводство у нас явля-
лось почти исключительно крестьянским. 4
Будучи доступно самому мелкому хозяйству, птицеводство,
однако, как и другие отрасли сельского хозяйства, только при ус-
ловиях крупного хозяйства может использовать все новейшие до-
стижения науки и техники и быть особенно выгодным. Только круп-
ному хозяйству доступны большие инкубатории, только большие
хозяйства могут быть специализированы в том или ином направле-
нии, только крупные хозяйства могут создать специальные сево-
обороты, приспособленные для обеспечения птицы кормами и вы-
гулами, только крупные" хозяйства могут обеспечить производство
специально подготовленным техническим персоналом и высококва-
лифицированными специалистами. Таким образом, колхозное
695
и совхозное хозяйство у наев СССР создает чрезвычайно благопри-
ятные условия для развития птицеводства, гораздо более бла
гоприятные, чем в передовых в отношении птицеводства США
и Канаде.
Птицеводство в СССР существует во всех областях, но далеко
не везде оно имеет характер промыслового птицеводства. Первое
место по птицеводству занимает Украина, затем Воронежская и
Курская области, Центрально-промышленная область, Северный
Кавказ, Средневолжская область, Сибирь и, наконец, Урал. Осталь-
ные области занимают второстепенное значение.
К домашним птицам относятся: куры, индейки, цесарки, утки
и гуси.
Рис. 509. Банкивская курица (Gallus bankiva).
(Из Брэма.)
Куры. Со времени исследования Ч. Дарвина является обще-
признанным, что все многочисленные породы домашних кур проис-
ходят от дикой банкивской курицы (Gallus bankiva, рис. 609),
которая и сейчас обитает в диком состоянии в Индии, Бирме и на
Малайском архипелаге. По мнению Ч. Давенпорта, в образовании
пород домашних кур, помимо банкивской курицы, давшей начало
легким яйценоским породам, принимал участие другой неизвест-
ный предок, давший начало малайским курам «азиль», от которой
произошли тяжелые азиатские породы — кохинхина, брама, ланг-
шан. Одомашнение курицы относится к глубокой древности. В
Индии они были одомашнены по крайней мере за 1200 лет до нашего
летоисчисления, но, быть может, даже значительно раньше, так
как о курах имеется упоминание в Ведах, составление которых про-
исходило за 1 V2—2 тысячи лет до нашей эры, а китайский бог-
дыхан Фу-Ши, живший в четвертом веке до нашей эры, уже издавал
указы, касающиеся разведения птиц.
696
Так как происхождение пород кур в большинстве остается невыяснен-
ным, то нет до сих пор установленной генетической классификации их.
В общем, согласно Гиги, их можно разделить на три группы.
1. Гомеосоматические породы кур, несомненно
происшедшие от банкивской курицы. Сюда относятся итальянские, или
легхорны, минорка, испанские, андалузские и др., происшедшие от заве-
зенных в Европу во времена Аристофана или Александра Македонского
(IV—V в. до нашей эры) индийских кур.
2. Гетеросоматические породы — кохинхина, брама —
отличаются тяжелым телосложением, короткими крыльями, свислым хво-
стом, коротким и толстым клювом, гороховидным низким гребнем. По мне-
нию Давенпорта, эти породы происходят от малайской породы «азиль».
Рис. 510. Плимут-роки.
3. Смешанная группа пород (миксосоматическая) полу-
чена в результате скрещивания пород вышеописанных двух групп. Сюда
относится большинство так называемых общепользовательных и мясных
пород.
Из всей массы пород остановимся лишь на самых важных, имеющих
значение в нашем Союзе.
1. Итальянские куры, или л етхоркы. Получили свое
первое название от места своего происхождения, второе получили от США.
'Гам, а позднее в Англии легхорны дали замечательно яйценоских кур с про-
изводительностью в 130—200 яиц. Среди белых легхорнов получены были
даже линии с яйценоскостью в 300—315 яиц. Скороспелы. Уже через 4—5 ме-
сяцев курочки начинают нестись, петушки в возрасте 21/2 месяцев пригодны
л ли откорма. Самая широко распространенная порода, составляющая в США
но % всего поголовья.
2. Минорка занимает второе место по распространенности. Про-
исходит от испанских кур с острова Минорка. Улучшена в Англии. Средняя
год,оная носкость 130—180. Скороспела. Курочки начинают нестись 472—5
месяцев.
3. Р о д-а й л е.н д красные, выведены в США, в штате Род-Айленд, из
местных кур путем скрещивания их с завезенными ив Индии шанхайскими
и крнснобурыми малайскими. Это крупные, крепкие, выносливые, ноские
и мясные куры типичного общепользовательного типа. Как общепользова-
К'лшый тип импортируется в СССР.
697
4. Ви а н доты выведены в США в шестидесятых годах прошлого
столетия; общепользовательная порода.
5. П л и м у т-р о к. Выведены в Америке в результате разнообраз-
ных скрещиваний. Являются прекрасной общепользовательной породой
(рис. 510, стр. 697).
Из мясных пород назовем английскую породу доркинг с прекрас-
ными качествами мяса; французскую породу л а-ф л е ш с крайним раз-
витием мясных свойств и прекрасного качества мясом и значительной нос-
костью (до 120 яиц в год); китайскую породу л а н г ш а н, улучшенную
в Англии, кохинхин, брама. Интересны специализированные
Рис. 511. Индейка бронзовая.
бойцовые куры, многочисленные декоративные породы кур (бен-
тамки, фениксы с длинными хвостами, бесхвостые, курчавые, шелковистые
и т. д.), показывающие, какие удивительные результаты можно получить
путе^м скрещиваний и отбора.
Что касается наших местных пород, то на территории СССР их имеется
большое количество, указывающее на их общее происхождение, отличное
от средиземноморских (итальянских, испанских и др.), и на близость, с одной
стороны, к польским и германским, с другой — к западноазиатским. Назовем
распространенных в Воронежской области и на Украине ушанок с хо-
рошими хозяйственными качествами, орловских кур, похожих на
ушанок, и юрловских голосистых кур, названных так потому, что
выведены в селе Юрлове Ливенского района Воронежской области люби-
телями петушиного пения; интересны распространенные в южных районах
СССР голошейные, или семиградские куры, довольно скороспелые,
яйценоские, средней величины куры с голой шеей. Надо заметить, что хозяй-
ственные качества местных кур СССР недостаточно изучены.
Индейки одомашнены были в Мексике в глубокой древности.
Полагают, что в образовании домашних пород индеек принимали
участие три формы дикой индейки (Meleagris gallopavo): мексикан-
ская, североамериканская из восточных и центральных штатов и
гондурская. В Европу завезена индейка в 1530 г.
698
Имеется довольно большое количество пород, из которых назовем
•. । и >н;и »цу ю индейку (рис. 511), которая имеет блестящее бронзовое'оперение,
. пегом самцов до 10.4 кг и самок до 8.3 кг. В СССР разводятся в незначи-
।елыюм числе еще так’ называемые белые русские индейки. Наши индейки,
\отп и уступают иностранным по величине, но по плодовитости и выносли-
Н1И-ТП их превосходят. Индейки начинают нестись в возрасте 9 месяцев, дают
/ю 15 -40 яиц в год, великолепно высиживают и воспитывают цыплят.
Цыплята требуют внимательного
\ кода и совершенно не выносят
< ырости.
Цесарки (Numidia ше-
le.igris) (рис- 512) одомашнены
на несколько столетий до
нашей эры в Африке, где они
и диком состоянии встреча-
ются и сейчас. Разводятся
ради очень вкусного мяса и
лиц, которых несут до 100
штук в год. Очепь неприхот-
ли вы на корм и летом не тре-
пу ют ухода, сами себе добы-
вают корм. Существует не-
сколько пород цесарок.
Утки. Все породы до-
машних уток произошли от
дикой утки кряквы (Anas
platyrhyncha), причем при-
ручение утки происходило в
Рис. 512. Цесарка.
разных местах в разное вре-
мя; в Китае раньше, чем в Европе, где она была приручена во
времена древнего Рима. Несмотря на значительную древность
одомашнения утка не богата породами. Хозяйственное значение
имеют следующие породы.
Руанские утки, выведенные во Франции, по оперению не отли-
чаются от дикой утки; интересны своей крупной величиной до 4—5.5 кг
несом, особенно вкусным мясом и значительной носкостью.
Пекинские утки выведены были в Китае, откуда вывезены были
в Америку и там улучшены. Они характерны приподнятым положением
туловища и вздернутым хвостом, величиной, скороспелостью, отличным мя-
сом и значительной носкостью.
Индийские бегуны выведены были в Индии и улучшены
и Англии. Они имеют очень характерное гагарообразное положение туло-
вища вследствие того, что ноги значительно отодвинуты назад, что очень
облегчает их передвижение по земле. Это небольшие утки замечательны
। впей яйценоскостью — до 200 яиц в год — и качеством своих яиц, по вкусу
Ь.ИПЗКИХ к куриным.
X а к и-к е м п б е л л —новая, недавно выведенная в Англии порода
уки; цвета хаки; она по своему складу похожа на бегунов, от скрещивания
г которыми и получена эта очень ценная, яйценоская и в тоже время мясная,
порода уток.
Мускусные у т к и, или шипуны, составляют отдельный
род угон (Cairina moschata), видиком состоянии живущий в Южной Аме-
рико — в Бразилии и Парагвае; обладает мускусным запахом кожи и жира.
Самец значительно больше самки.
' 6*
2. Промысловые птицы
Использование в хозяйственном отношении дикой фауны птиц
производится путем промысловой охоты и разведением диких жи-
вотных с промышленной целью. Кроме того, охота как спорт может
послужить источником побочного использования сельскохозяйствен-
ных угодий.
В общем, дикие птицы используются на мясо или ради шкуры,
перьев и пуха, или, наконец, производится сбор птичьих яиц.
По тому значению, которое они имеют в качестве объектов про-
мысла или спортивной охоты, птиц можно разбить на две группы:
на чисто промысловых и спортивно-охотничьих. Не все имеют одина-
Рис. 515. Поганка большая (Colymbus
cristatus). (Из Гааке).
ковое значение вообще,
и один и тот же вид
имеет разное значение
в разных районах.
Отряд Гагары (Colym-
bi). У нас па севере во-
дятся три вида гагар
(Urinator), из которых
чернозобая гагара (U.
arcticus) доходит до сред-
ней части РСФСР. Дру-
гой подотряд этого отря-
да с лопастными пере-
понками на ногах — по-
ганки (Colymbus)—пред-
ставлен в СССР 5 видами, из которых самая крупная хохла-
тая, или большая, поганка (С. cristatus) (рис. 515) распространена
наиболее широко. Добываются эти птицы ради шкурВк, кото-
рые идут на разного рода отделки дамских нарядов.
Отряд Веслоногие (Steganopodes). Сюда относятся пеликаны,
или птицы-бабы (бабуры) (Pelecanus), которые распространены в юж-
ной подобласти Палеарктики. Пеликаны добываются ради перьев.
Отряд Цапли (Ardeae). Белые цапли (Herodias), большая
(Н. alba) и малая (Н. gerzetta), являются предметом ценного промысла
ради особых украшающих перьев — так называемых эгреток, 400 г
которых до войны стоили на месте 300 руб., были почти оконча-
тельно уничтожены. На юге европейской части СССР и на Кавказе,
в настоящее время в ряде заповедников — Астраханском, Черно-
морских, Кизил-Агачском и др.—размножились.
Отряд Гуси (Anseres) содержит целый ряд весьма ценных про-
мысловых и охотничьих птиц.
S Лебеди (Cygnidae) представлены 3 видами, из которых лебедь-
кликун (Cygnus cygnus) был раньше широко распространен ра тер-
ритории Союза ССР, но в настоящее время истреблен и в значи-
тельном количестве гнездится лишь в тундрах и в самой северной
части лесной области и на юге. Зимует у нас на юге Средней Азии
и на Каспийском и Черном морях. Ценная охотничья птица.
702
I1 у с n (Anseridae) у пас представлены родами серых гусей
(Anser), куда кроме серого гуся (A. anser) относят белолобых ка-
зарок. Серый гусь был широко распространен по СССР, но сейчас
почти истреблен всюду за исключением юга европейской части СССР
и севера; белолобые казарки, так же как гуменники (Melanonyx),
и числе нескольких видов, и другие роды казарок (Branta и др.)
распространены на севере в тундре. Там существует специальный
промысел линяющих гусей, когда они вследствие потери маховых
неспособны летать. Их окружают облавой и загоняют в сети. Мясо
гусей заготовляется на зиму впрок, одновременно собирая пух.
Охота на гусей практикуется во время перелетов и на зимовках.
Утки (Anatidae) распадаются на подсемейство благородных
уток (Anatinae) и нырковых (Fuligulinae). К первым относятся
кряквы (Anas platyrhyncha) и два вида чирков (Querquedula quer-
quedula и Q. сгесса), очень широко распространенные; представ-
ляют предмет охоты и промысла. Реже встречаются шилохвость
(Dafila acuta) и широконоска (Spatula clypeata), а свиязь (Матеса
penelope) принадлежит северу, тогда как серая утка (Chaulelasmus
strep era) — югу.
К семейству нырковых (Fuligulinae) относятся многочи-
сленные утки, которые прекрасно ныряют, добывая пищу со дна.
Особенно важное хозяйственное значение имеют гаги, которых
оксплоатируют ради очень ценного пуха. Они распространены на
севере по побережью морей. По берегам Мурмана, на островах
Белого моря, Новой земле, Вайгаче, Колгуеве живет гага обыкно-
венная (Somateria molissima); отчасти севернее и дальше на восток
по сибирскому побережью до Берингова моря — гага-гребенушка
(S. spectabilis); стеллеровагага(8. stelleri) распространена к востоку
от Таймыра и на Мурмане.
Гнездятся гаги колониями. Самка выстилает свое гнездо пухом
и откладывает от 5 до 8 яиц.
Неразумный промысел пуха и яиц, который сопровождался
избиением самих птиц, привел почти к полному уничтожению бо-
гатых колоний гаг на Мурманском берегу. В настоящее время
там организуется правильный промысел, которой должен дать
чрезвычайно ценные результаты. Колонии гаг старательно обере-
гаются от хищников. Пух и даже яйца отбираются, прежде чем
самка села на яйца, после чего самка вновь откладывает яйца,
заменяя пух водорослями или выдирая пух у самца. Вторичное
гнездо гаги уже .не трогают. При внимательном уходе гаги
разможаются быстро и даже привыкают к людям. На Мурман-
ском берегу организован специальный птичий заповедник «Семь
островов», где охраняются колонии гаг.
Отряд Куры (Galli). Этот отряд содержит наиболее ценных про-
мысловых и охотничьих птиц, особенно из семейства тетеревов
(Telraonidae)—«боровая птица».
Па первом месте по промысловому значению нужно поставить
рябчика (Tetrastes bonasa).
Второе место среди боровой дичи принадлежит тетереву
703
(Lyrurus tetrix), тоже очень широко распространенному в южной
части тайги, в зоне смешанных лесов и в лесостепи. Глухарь
(Tetrao urogallus) занимает видное место среди промысловых жи-
вотных и добывается как самодействующими приборами, так и с по-
мощью ружья.
Из семейства фазанов (Phasianidae) наибольшее промысловое и
главным образом охотничье значение имеет серая куропатка (Perdix
perdix).
Фазаны (Phasianus) на Западе, а отчасти и у нас специально
разводятся ради спортивной охоты.
Перепел (Coturnix coturnix) представляет предмет спортив-
ной охоты и содержится в клетках.
Горные куропатки (Caccabis), горные индейки
(Tetraogallus) и т у р а ч и (Francolinus) относятся к охотничьим
птицам и промыслового значения почти не имеют.
Отряд журавлей (Grues). Сюда помимо журавлей (Grus grus),
распространенных очень широко по степям и болотам, однако не
представляющих ни охотничьего, ни промыслового значения вслед-
ствие плохого достоинства мяса и осторожности птицы, относятся
птицы степей: журавль-красавка (Anthropoides virgo), дрофа (Otis
tarda) и стрепет (Microtis tetrax). Дрофа крупная, очень вкусная,
но редкая и осторожная птица степей. Стрепет меньше, добывается
легче при охоте с собакой, но очень требователен в смысле стаций
и плохо мирится с распашкой степи, поэтому местами исчез совер-
шенно.
Пастушки (Ralli) — отчасти брлотные, отчасти водные
птицы, имеют малое промысловое и охотничье значение.
Чайки (Lari) добываются промышленниками ради шкурок
и перьев. Там, где эти птицы массами гнездятся, особенно на так
называемых «птичьих базарах» вместе с различными видами отряда
чистиков (Alcae), местные жители собирают яйца всех этих птиц.
Кулики (Limicolae) включают несколько видов очень цен-
ных объектов спортивной охоты, но почти не имеют промыслового
значения. На ряду с большим числом видов, которые бывают лишь
случайной добычей охотника, сюда относятся такие виды, как
вальдшнеп (Scolopax rusticola, рис. 481), бекасы (Gallinago galli-
nago), дупель (G. media), гаршнеп (Limnocryptes gallinula) —
объекты спортивной охоты.
Промысловая охота на птиц далеко не имеет того значения, как
промысел пушных зверей. Все же промысловые птицы кормят не
только промышленников (рябчик, тетерев и др.), но и население се-
вера (гуси), они дают ценное вкусное мясо, пух и перья. Как объект
охоты и промысла птицы, конечно, имеют большое экономическое
значение. Однако при бесплановом, нерегламентированном истреб-
лении охотничьи и промысловые птицы чрезвычайно быстро уничто-
жаются.
В целях сохранения дичи правительством устанавливаются оп-
ределенные правила охоты, которые имеют целью: 1) охранить птиц
в период брачной жизни: во время спаривания, кладки яиц и вы-
704
вода птетщов, 2) ограничить время охоты вообще, 3) ограничить
контингент охотников путем разрешения охоты только лицам, по-
лучившим специальный охотничий билет.
Наивысшей формой правильной постановки охоты будет органи-
зация правильного охотничьего хозяйства с таким расчетом, чтобы
дичь ие только пе уничтожалась, по, наоборот, размножалась. По-
мимо мер законодательных, регламентации охоты и наблюдения за
исполнением законов охоты, правильная организация охотничьего
хозяйства должна иметь в виду охрану дичи от вредных влияний
среды, а именно: от уничтожения хищниками, собаками, кошками;
подкормку дичи во время неблагоприятных климатических явлений;
организацию заказников, т. е. участков, в которых на время запре-
щается охота на тот или иной вид дичи или всякая охота; регламен-
тацию отстрела только определенного количества особей каждого
вида и, наконец, разведение новых видов дичи.
3. Полезные в хозяйстве птицы
Всем хорошо известно, какой громадный вред приносят нашему
лесному и сельскому хозяйству насекомые. В иные годы некоторые
из насекомых размножаются в такой массе, что выедают поля,
огороды, листву фруктовых деревьев, плоды и ягоды, листву или
хвою лесных насаждений буквально начисто.
Насекомые вредят не только лесам, полям, огородам и садам,
они вредят домашним животным тем, что сосут их кровь, беспокоят,
паразитируют в их теле, вызывая болезни, и тем, что разносят за-
разные болезни.
Насекомые приносят такой вред потому, что опи обладают чрез-
вычайной плодовитостью. Вычислено, что обыкновенная комнатная
муха, самка которой откладывает по 120 яиц и за лето может дать
7 поколений, к концу лета, если бы потомство ее сохранилось,
дала бы 5 598 8G0 ООО 000 мух. Недаром было высказано когда-то
остроумное замечание, что потомство одной мясной лгу хи может ско-
рее съесть слона, чем лев.
С вредпыми насекомыми ведется неустанная борьба. Затрачива-
ются значительные средства и силы па уничтожение вредителей раз-
иыми методами: проводятся в жизпь мероприятия, препятствующие
быстрому размножению вредителей; имеется специальная служба
* защиты растений» с многочисленным штатом, наблюдательными
пунктами, станциями и научно-исследовательскими институтами.
В этой борьбе с вредителями-насекомыми человек в птицах,
преимущественно в воробьиных, имеет чрезвычайно ценных союз-
ников. Опи массами истребляют насекомых во всякое время года.
Летом целая армия прилетевших насекомоядных птичек — мухо-
ловок, трясогузок, коньков, славок, пеночек, камышевок, дроздов,
соловьев, горихвосток, крапивников, ласточек, стрижей, куку-
шек, козодоев и других неустанно занята уничтожением насеко-
мых. Зерноядные птицы, как, например, разные вьюрки, зяблики,
Кошкарои и Станчинский 705
Рис. 516. Про-
дольный разрез
через дуплянку.
коноплянки, овсянки и другие выкармливают своих птенцов на-
секомыми и сами в значительной мере питаются ими. Подсчитано,
что одна ласточка за лето съедает до миллиона летающих насекомых;
выводок маленьких корольков за год уничтожает до 10 миллионов*
Зимой, над уничтожением насекомых трудятся зимние птицы:
дятлы, пищухи, поползни, синицы. Выло установлено, что за зиму
до 70—80% гусениц бабочки-боярышницы, зимующих в листовых
паутиновых гнездах, уничтожается синицами. Синицы — лазоревка
и московка — съедают за день такое количество насекомых, что их
вес равен 30% веса самих птиц. Отсюда ясно, что чем больше гнез-
дится птиц в наших садах, парках, огородах, тем надежнее будет за-
щита их от насекомых. Опыт показал, что соответствующими меро-
приятиями можно привлечь полезных нам птиц
на гнездование в гораздо большем количестве,
чем их гпездилось бы, если бы не принимать
соответствующих мер. Так, например, в одном
из парков в Германии площадью около 5 га
около города Мюльгаузена соответствующими
мероприятиями и постановкой искусственных
гнезд было привлечено так много птиц, что в
иных местах приходилось на 11 м2 по одному
гнезду. И вот в годы, когда в районе Мюльгау-
зепа (в Западной Пруссии) от нашествия насеко-
мых леса и сады подверглись опустошению, рощи
и сады окрестностей упомянутого парка стояли
зелеными, не тронуты^ вредителями. Птицы
отстояли их от нашествия этих врагов.
При нашем социалистическом строительстве,
когда природные условия нашей родины меня-
ются с поразительной быстротой, когда вместо
пустырей, залежей, неосвоенной целины, мало-
ценных зарослей кустов и т. п. пустопорожних
мест, возникают плодородные поля, цветущие
сады й огороды, когда города и деревни при-
обретают культурный вид и озеленяются парками, садами и буль-
варами, перед нами встает задача, чтобы в этих условиях макси-
мально содействовать привлечению к нам полезных птиц и созданию
для них благоприятных условий для размножения. Они не только
будут представлять надежную защиту от вредных насекомых, но
и оживлять природу своим пением, ярким оперением и веселой
деятельностью.
Как уже было сказано выше, летом наиболее трудным для птиц
моментом в жизни является недостаток подходящих мест для гнездо-
вания. Птицы в этом отношении, как мы видели, очень требователь-
ны. Гнездо должно быть хорошо защищено, спрятано, укрыто. Боль-
шое количество птиц строит свои гнезда в дуплах. Отсутствие
естественных дупел не позволяет гнездиться этим птицам. Есте-
ственные дупла можно заменить искусственными. Специальными
исследованиями выяснено, какой величины и как должны быть при-
зов
готовлены для разных видов полезных птиц искусственные гнезда,
чтобы удовлетворить всем их требованиям. А среди наших птиц
есть такие очень требовательные в этом отношении птицы, как, на-
пример, дятлы. Наоборот, серая мухоловка совсем малотребователь-
ная птица.
Лучшими искусственными гнездами являются «дуплянки»
(ряс. 516). Сделанная по всем правилам дуплянка должна иметь та-
кой внешний и внутренний вид, как это показано на нашем рисунке.
Белыми линиями показаны контуры выдолбленной сверху полости
внутри обрубка дерева или круглого полена, которое сверху при-
крыто наклонной деревянной крышечкой. Дно должно быть не тонь-
ше 6—8 см; стенки тоже по возможности такие же. Леток должен на-
ходиться на расстоянии не меньше 6 см. В общем при изготовлении
дуплянок нужно руководствоваться следующей таблицей:
Для птиц Глубина (в см) Диаметр (в см) Диаметр летка (в см)
меньший больший
мелких .... средних . . . крупных . . . 15—17 23—25 30—32 7.5—8.5 9—10 12—13 9—10 12—13 1.6—18 3 5 8—9
Вез специальных инструментов выдолбить с одной стороны та-
кую дуплянку трудно. Можно упростить работу тем, что, расколов
1’иг 517. Полудуплянка Рис. 518. Скворешницы.
дли ri'pi.ix мухоловок и
Гн’.1||,1\ трясогузок.
полено пополам, выдолбить дупло в каждой половине отдельно,
а патом их сколотить или связать, но аккуратно так,чтобы не было
що io,II, лучше на пазу. ,
45* 707
Дуплянки делаются стоячие или лежачие. Последние рекомен-
дуются для больших птиц, как голуби-клиптухи, сизоворонки,
галки, удоды и совы.
Для серых мухоловок и белых трясогузок делают полудуплянки,
как показано на рисунке (рис. 517).
Не так охотно птицы занимают более простые, сколоченные из
досок «скворешницы». Формы им придают разнообразные. Вид стан-
дартных скворешниц показан на прилагаемом рисунке (рис. 518), а
из следующей таблицы видны размеры (в см), которые должны иметь
скворешницы для разных птиц:
Для птиц | Длина Ширина Высота Диаметр летна
мелких . * . . 10—12 11—12 25—27 3
средних . . . 15—18 15—18 30—32 5
крупных . . . 22—24 22—24 42—52 8—9
На дно скворешницы следует насыпать опилок в смеси с торфом
или трухи гнилушек.
Очень важно правильно повесить дуплянки и скворешницы. Во-
первых, для птиц не безразлична высота от земли, па которой поме-
щается гнездо. Многие птицы в этом отношении очень требовательны.
Для синиц лучшая высота будет в 2—2.5 м; мухоловки и горихво-
стки любят гнездиться повыше, до 4 м; поползни и дятлы предпо-
читают высоту от 4 до 7 м; удоды, наоборот, и трясогузки селятся
на небольшой высоте до 2 м.
Скворешницы и дупляпки должны быть прочно привязаны так,
чтобы они пе шатались. Они должны стоять или отвесно, или слегка
быть наклоненными в сторону летка, чтобы косой дождь пе заливал
гнезда. Не следует ставить скворешницы на солнечном припеке и на
очень видном месте. Первые скворешницы следует ставить в лесу
по опушкам и па полянах, а затем постепенно ставить их и в глубине
леса.
Лучшее время расстановки скворешниц осень или зима, чтобы
зимние птицы успели их осмотреть за зиму, и весенние после при-
лета застали их на месте.
Кроме искусственных гнезд, большое значение для привлечения
птиц имеют посадки кустов и деревьев и специальная их подрезка,
чтобы вызвать тем особенно густое ветвление. Даже если придер-
живаться простого правила не вырубать без надобности лишние
деревья и кустарники, сохраняя их всюду, где это возможно без
ущерба для хозяйства, и то это будет иметь большое значение для
привлечения птиц. Затем, очень полезно заменять деревянные из-
городи живыми изгородями из колючих кустарников или молодых
елей, которые все время подрезаются на определенной высоте.
Далее, полезно всюду, где это позволяют хозяйственные цели и
декоративные требования, рассаживать густые заросли кустарни-
708
кип — спиреи, жимолости, боярышника, шиповника, бузины, си-
рени и т. п.
Деревья и кустарники хорошо подрезать так, чтобы они выше
линии отреза образовывали густую мутовку ветвей.
Для привлечения зимних птиц чрезвычайно полезна их зимняя
подкормка. Зима, как мы видели, самая тяжелая пора для зимую-
щих птиЦ. Если их подкармливать, то они держатся всю зиму на
одном месте и весной расселяются по окрестностям.
Подкармливание производится на специально отведенном месте.
Оно расчищается от снега или там ставится специальный столик,
на который небольшими порциями ежедневно разбрасывают корм:
семена конопли, проса, овса, подсолнуха; .крошки хлеба, толченые
сухари, кусочки сала и мяса, сушеные ягоды рябины, бузины,
боярышника и др. Чтобы семена высыпались не сразу, устраивают
специальные кормушки вроде маленьких закромов.
Рис. 519. Скворешница
с защитой от кошек.
Рис. 520. Защита ствола
дерева с гнездами птиц от
кошек с помощью при-
вязанных к стволу сучьев.
Но важно не только привлечь птиц; важно, привлекши их, ох-
ранить от врагов. Злейшим врагом птиц являются кошки. Во-пер-
вых, надо защитить от кошек гнезда птиц. Для этого скворешницы
и дупляпки делаются так, чтобы кошка не могла достать птенцов
лапой (рис. 519). Во-вторых, к стволу привязывают сучья так,
чтобы кошка, добравшись до них, пе могла взобраться выше (рис.
520). В-третьих, кошек просто подвергают уничтожению капканами
или особыми ловушками. Это — ящик более узкий в начале и рас-
ширенный в конце, в котором па оси качается доска. Более длинный,
а потому и более тяжелый конец обращен ко входу. Противополож-
ная сторона затянута проволочной сеткой. Кошка, забравшись
< 709
в ящик, опускает доску своей тяжестью и не может выбраться
обратно, так как этому мешает привешенная на шарнире к доске
палочка, которая выпрямляется, когда поднимается от тяжести
кошки доска.
Большие опустошения в наших полях и огородах производят все-
возможные грызуны — мыши, полевки, суслики, хомяки и некото-
рые другие. Временами полевки и некоторые виды мышей размно-
жаются в таком множестве, что производят сплошные опустошения
в полях и огородах.
В борьбе с сусликами и всевозможными мышевидными грызу-
нами помогают нам наши хищные птицы и совы.
Так, по произведенным подсчетам, наш обыкновенный канюк,
или сарыч, за лето съедает не менее тысячи мышей, а пустельга не
мепее полутысячи. Но в годы, обильные мышами, птицы их истреб-
ляют в неизмеримо большем количестве. В желудке сарычей их на-
ходили до 17 штук зараз, у ушастых сов — до 12, а у пустельги —
до 9 штук. Сусликов и мышевидных уничтожают степные орлы, подор-
лики, луни, канюки, мелкие соколы — пустельги и кобчики,
а также большинство сов — неясыти, ушастые совы, болотные
совы, синухи, сычи‘домашние, мохноногие и воробьиные.
Эти птицы подлежат тоже нашим заботам. Мелкие соколы
охотно селятся в дуплах, почти все совы тоже жители дупел.
Нельзя не отметить и санитарного значения птиц. У нас грачи,
вороны, галки и коршуны очищают свалочные места вблизи горо-
дов. Здесь они выбирают все доступное или съедобное, что, валяясь
на свалках, иначе подвергалось бы гниению и отравляло воздух.
4. Вредные в хозяйстве птицы
Таких птиц немного, по все же имеются и вредные птицы. Так,
воробьи, домашние, полевые, а в Средней Азии испанские воробьи
приносят большой вред зерновым полям, садам и огородам.
Они вредны еще и тем, что вытесняют из гнезд полезных птиц,
гнездящихся в дуплах, и ласточек. Воробьи оклевывают се-
мена хлебов, конопли, гороха, поедают ягоды, вишни, черешни,
урюк и др.
Питаются хлебами также грачи, вороны, галки, голуби, жаво-
ронки. Скворцы и дрозды склевывают ягоды в садах. Дубоносы пи-
таются косточками вишен. Этим они приносят несомненный вред.
Однако в другое время большинство этих птиц приносит пользу, а от
вреда, приносимого этими птицами, нетрудно избавиться. Поэтому,
прежде чем огульпо зачислить этих птиц в стаи вредителей, следует
внимательно разобраться в пользе и вреде, который приносят эти
птицы в конкретной обстановке, и уже тогда приступать к их уничто-
жению. Так, в больших городах грачи и вороны приносят больше
пользы, тогда как в полях, в степи опи бывают вредны. Грачи,
кроме того, очень вредят деревьям тем, что ломают сучья и,
селясь колониями, портят сады, парки и другие искусственные
посадки.
710
Безусловно вредны хищники — ястреб-тетеревятник, ястреб-
игреислятпик, камышевый лунь, сокол-сапсан, соколик-дербник и
1П.1111» малая (волчок), так как во множестве уничтожают птиц,
в том числе крупные из них — дичь, а также таскают домашних
кур. Сокол-чеглок, хотя и поедает много насекомых, но главным об-
разом полезных стрекоз, и питается все же преимущественно мел-
кими птицами, а потому в общем вреден, а не полезен.
Пчеловодству вредят щурки, так как уничтожают пчел. Как
мы видим, список вредных птиц невелик. Борьбу с ними вести
легче, чем с вредными насекомыми и млекопитающими. Однако,
борьба с воробьями вовсе нелегка: приходится не только стрелять
их, разорять гнезда, но и отравлять ядами, и все же они настолько
быстро размножаются, что уничтожить их трудно.
ЧАСТЬ ШЕСТАЯ
МЛЕКОПИТАЮЩИЕ
Класс 7. Млекопитающие (Mammalia)
I. ХАРАКТЕРИСТИКА КЛАССА МЛЕКОПИТАЮЩИХ
Основными особенностями млекопитаю-
щих как класса являются их теплокров-
ность, наличие волосяного покрова и спо-
собность к вынашиванию детенышей в утро-
бе матери или в сумке и выкармливание мо-
локом. Именно эти особенности и дали млекопитающим возмож-
ность, одержав победу в жизненной борьбе над рептилиями, при-
способиться к самым разнообразным условиям существования и
превратиться в богатый формами, процветающий обширный класс
позвоночных.
Теплокровность давала млекопитающим возможность приспособ-
ляться к разнообразным климатическим неблагоприятным услови-
ям, а вынашивание и выкармливание молоком детенышей — со-
хранять потомство при самых различных приспособлениях к среде.
В общем млекопитающие характеризуются следующими при-
знаками: тело их покрыто волосами (у голых млекопитающих по
крайней мере во время утробной жизни), кожа богата трубча-
тыми и гроздевидными железами, снабжена мускулами; всегда
имеются молочные железы для выкармливания детенышей; череп
имеет два сочленовных бугорка, с помощью которых он сочле-
няется с видоизмененным в атлас первым позвонком; этот последний
вместе с черепом может вращаться на зубовидном, отростке, т. е.
на теле первого позвонка, сросшемся с телом видоизмененного
второго позвонка. Тела позвонков с плоскими хрящевыми эпи-
физами (во всяком случае хвостовые позвонки), эпифизами же
снабжены на концах и длинные кости конечностей. Нижняя
челюсть с каждой стороны состоит лишь из одной, несущей зубы,
кости (dentale), которая непосредственно сочленяется при помощи
выпуклой сочленовной поверхности с чешуйчатой костью (squamo-
sum). Барабанная перепонка натянута на барабанные кости (tym-
panicum). Имеются три слуховые косточки: стремя (stapes), на-
ковальня (incus), молоточек (malleus), имеется наружное ухо.
Скуловая дуга одна и состоит из отростков чешуйчатой и че-
люстной кости. Зубы сидят только на челюстной (maxillare), меж-
712
«|'"1 нir.Tiioii (praemaxillare) и нижнечелюстной костях (dentale), при-
чем имеется пе больше двух генераций зубов.
Красные кровяные тела безъядерны, округлы и лишь как исклю-
чение овальны. Сердце четырехкамерное, имеется лишь одна дуга
in|рты, которая загибается налево. Гортань снабжена щитовидным
хрящом и прикрывается надгортанным; мускульно-сухожильная
/шафрагма разделяет грудную и брюпшую полости. В мозгу осо-
бенно сильно развиты полушария, связанные мозолистым телом
и породней комиссурой. Характерно развитие обонятельных долей
мозга, которые состоят из многочисленных нитей обонятельных нер-
вов п проходят в носовую полость через многочисленные отверстия
решетчатой кости. По рождении функционирует лишь вторичная
ночка, всегда имеется мочевой пузырь; сперма, большей частью,
и моча выводятся через совокупительный аппарат; по способу
размножения млекопитающие животные живородящие или яйце-
кладущие.
Указанные выше особенности характеризуют млекопитающих
как хорошо отграниченный класс. Однако среди современных мле-
копитающих существует небольшая группа форм, которые отли-
чаются довольно резко от прочих млекопитающих тем, что имеют
клоаку, песут яйца ине имеют настоящих молочных желез, а также
рядом других особенностей, приближающих их к рептилиям.
Ото — однопроходные (Monotremata), входящие в состав
подкласса первозвери (Prototheria) и составляющие особую
ветвь млекопитающих, отошедшую от общего ствола развития тогда,
когда млекопитающие не приобрели еще способности вынашивать
детенышей. В другой подкласс млекопитающих—низшие
звери (Metatheria) входят сумчатые (Marsupialia), которые
вынашивают детенышей в особой сумке, имеющейся у самок на
брюшной стороне. Остальные млекопитающие — подкласс в ы с-
1п и е звери (Eutheria), называемые также одноутроб-
ными (Monodelphia) и плацентарными (Placentalia) —
вынашивают детенышей в матке (причем оболочка зародыша обра-
зует так называемую плаценту).
В настоящее время известно свыше 6 000 современных и иско-
паемых видов, которые приспособлены к самым разнообразным ус-
ловиям существования.
II. СТРОЕНИЕ МЛЕКОПИТАЮЩИХ
' 1. Покровы
Покровы млекопитающих характеризуются целым рядом осо-
бенностей.
Кожа. Кожа состоит из многослойного эпидермиса (epi-
dermis) и соединительной ткани (cutis). Эпидермис в свою очередь
состоит из двух слоев: из ороговевшего мертвого наружного слоя —
роговой слой (stratum corneum) и из слоя живых размножающихся
клеток — мукозный слой (stratum mucosum, или malpighii). Наруж-
ный слой снашивается, стирается и возобновляется за счет размно-
жения клеток мукозного (мальпигиева) слоя. Ороговение наружного
713.
слоя является сложным химическим процессом. Линька всего эпи-
дермиса заменяется у млекопитающих частичной сменой его и
только у некоторых животных она имеет место в эмбриональ-
ном состоянии. В глубоких частях эпидермиса имеется пигмент, по
большей части темный, в виде мельчайших зернышек, расположенных
между или внутри клеток (рис. 521).
Из эпидермиса развиваются волосы, железы, а также различного
рода роговые образования, о которых будет сказано ниже.
Рис. 521. Разрез через
кожу губы гиппопотама.
1 — роговой слой (stratum
corneum), 2—мукозный сл^и
(str. Malplghii), 3 — сосоч-
ки соединительнотканного
слоя кожи, 4 — кровенос-
ные сосуды, 5 — собственно
кожа (corium). (Из Вебера).
Под эпидермисом лежит соединитель-
нотканный слой кожи, который заходит в
эпителий тонкими сосочками. Толщина эпи-
дермального слоя различна у разных мле-
копитающих и у одного и того же млеко-
питающего на разных частях тела. Там,
где эпидермис достигает сильного разви-
тия, сосочки с кровеносными сосудами
заходят в него глубоко, что связано с пи-
танием эпидермиса, который не имеет своих
сосудов.
Кожа собственно, или соедини-
тельнотканная часть кожи (cutis, corium), со-
стоит у млекопитающих из плотной волокни-
стой соединительной ткапи (stratum reticu-
lare), которая постепенно переходит в под-
кожную клетчатку (stratum subcutaneum)—
рыхлую соединительную ткань, прилегаю-
щую к лежащим под кожей органам, мус-
кулам, костям и пр.
Кожа содержит многочисленные эла-
стические соединительнотканные волокна,
которые в разных местах тела и у разных
животных развиты в разной степени, при-
давая своим присутствием коже большую
прочность ибблыпую грубость. Многочислен-
ные кровеносные сосуды и нервы образуют
ответвления, заходящие в сосочки наружно-
го слоя, подстилающего эпидермис; пигмент-
ные клетки, хроматофоры, встречаются в соединительной ткани обыч-
но в самом ограниченном количестве. Толщина кожи у разных жи-
вотных и у одного и того же животного на разных частях тела очень
различна. То же нужно сказать и про подкожную клетчатку, которая
у некоторых животных характеризуется способностью откладывать
жир, образуя под кожей мощный слой сала (panniculus adiposus),
как то встречается у свиней и у китообразных; у некоторых млеко-
питающих такое накопление жировых запасов приурочено к опре-
деленным местам тела — хвост жирнохвостых овец, горб верблюда
или зебу и т. п.
Из эпидермальных образований у млекопитающих наиболее
характерным является волосяной покров, тогда как чешуйчатый
714
iinh’iMHi, столь характерный для рептилий, здесь играет лишь вто-
|HirTpiir.iiную роль и встречается лишь у очень ограниченного числа
форм. Несомненно, однако, что у предков млекопитающих, у репти-
'IIIи, чешуя была развита нормально и была вытеснена волосами
шин. постепенно, в связи с приобретением млекопитающими тепло-
кровности. Сравнительноанатомические данные учат нас, что че-
шуя и волосы встречаются одновременно, закономерно чередуясь
в своем расположении; что даже у животных, не имеющих чешуи,
первоначальное расположение волос такое, как
будто они чередуются с чешуей; что у многих жи-
вотных в тех местах, где сохранился чешуйчатый
покров, он тоже сочетается с более или менее ред-
ким волосяпым покровом. Все это указывает, что
волосы и чешуя у родоначальников млекопитающих
существовали одновременно и лишь в дальнейшие
эволюции млекопитающих волосы вытеснили че-
шую совершенно. Только у ящеров и у броненосцев
чешуя развита как основной покров. У ящеров
(Tholidota) — это черепицеобразно налегающие
друг на друга, обращенные назад роговые плас-
тинки значительной величины, между которыми
у азиатских видов располагаются одиночные ще-
тинообразные волосы, в эмбриональном своем раз-
витии закладывающиеся на несколько более позд-
Рис. 522. За-
кладка волоса у
новорожденной
мыши.
1— роговой слой,
2 — эпидермичес-
кий сосочек.
(Из Бючли).
них стадиях, чем чешуя, что указывает также на
филогенетически более позднее их появление. У броненосцев
(l)asypodidae) чешуя подстилается костяными пластинками, обра-
зуя папцырь. У зародышей и у молодых броненосцев мелкие
чешуйки сопровождаются волосами; позднее они сливаются в
белес крупные пластинки, налегая на кости, а волосы в большей
или меньшей степени, а иногда и совсем, исчезают.
Хорошо выражен чешуйчатый покров на хвосте у бобра, у мно-
гих сумчатых, насекомоядных и грызунов; далее, чешуйчатый
покров встречается на конечностях, на морде, на хвосте целого
рида других млекопитающих. Чешуя представляет сильно утол-
щенный ороговевший участок эпидермиса, сидящий на плоском
сосочке соединительной ткани.
Волосы, подобно чешуе, являются также эпидермическим обра-
нопапием, но соединительная ткань играет здесь гораздо меньшую
ро и., только постольку, поскольку это необходимо для укрепле-
нии и для питания волоса. Так, развитие волоса в отличие от тако-
вого чешуи или пера начинается образованием эпидермического со-
точка, который путем деятельного размножения образующих его
кчегок глубоко врастает в соединительнотканную часть кожи
(рис. г>22). Только поздпее, когда входящие в состав сосочка клетки
начинают дифференцироваться на концентрически расположенные
цн 11111дрнческие слои, в нижнюю расширенную часть волоса, в его
чукоппцу, вдается соединительнотканный сосочек с кровеносными
сосудами и нервами.
715
Рис. 523. Схема строения волоса,
поперечный и продольный разрезы.
1 — наружное, 2—внутреннее влагалище
волоса, 3 — кожица (кутикула) волоса,
4 — корковый слой, 5 — сердцевина во-
лоса, 6 — луковица, 7 — соединительно-
тканный сосочек, 8 — роговой и мальпи-
гиев слой эпидермиса, 9 — соединитель-
нотканная часть кожи (corium), 10 —
сальная, 11 — потовая железа.
(Из Бючли.)
В результате концентрической дифференцировки слоев и роста
осевой части эпидермического зачатка волоса образуются волос
с луковицей, сидящей на соединительнотканном сосочке, и сумка
волоса с двуслойным влагалищем и слоем соединительной ткани,
окружающим его снаружи. Луковица состоит из недифференцирован-
ных эпидермических клеток, размножение которых обусловливает
рост волоса. Стержень волоса состоит из сердцевины, построенной из
сплющенных в поперечном направлении клеток, из коркового слоя
с волокнистыми клетками и наружного — кожицы (кутикулы) из
плоских клеток, по большей
части черепицеобразпо нале-
гающих друг на друга (рис.
523). Корень волоса, т. е. си-
дящая в сумке часть, прилегает
к внутреннему влагалищу во-
лоса, т. е. к слою эпидерми-
ческих клеток, которые очень
рано ороговевают, и молодой
волос пробивается наружу.
Внутреннее влагалище не дости-
гает . до верхней части сумки
и своей наружной стенкой при-
легает к наружному влагали-
щу, которое является пе чем
иным* как продолжением маль-
пигиева слоя эпидермиса, спус-
тившегося в сумку волоса.
В сумку волоса выше края
внутреннего влагалища откры-
ваются протоки сальных же-
лез, секрет которых смазывает
волос. К корню волоса под-
ходят пучки гладких мышц
(mm. arrectores pilorum), при-
поднимающих волосы, и нервные
окончания.
Как показали новейшие
исследования, волосы возни-
кают из того же зачатка, из которого развиваются и чешуи, и
перья. Однако в последнем случае изменения в развитии наступают
значительно позднее, развитие же волоса идет по новому пути,
отличному от пути развития чешуи, уже с самой ранней стадии
закладки. Таким образом филогенетически и волос в конце кон-
цов связан с чешуей.
Можно различить несколько родов волос в волосяном покрове
млекопитающих. Во-первых, обычно имеются более длинные и же-
сткие волосы — ость (pili), которые образуют шерсть; во-вто-
рых, более короткие и мягкие волосы — подшерсток (1апа).
Эти последние встречаются преимущественно у обитателей холодного
□16
к я и мата и у водпых животных. Нередко отсутствуют совершенно.
Нндонзменением шерсти, или ости, являются щетина у свиней
и и г л ы у ежей, дикобразов и др. Третий вид волос —осяза-
т <1 л ь п ы с волос ы, или вибриссы (vibrissae). Это осо-
бенно длинные волосы, расположенные преимущественно на голове,
гпабжеппые венозным синусом, поперечнополосатыми мышцами
и многочисленными нервными окончаниями. На отдельных местах
тела развиваются волосы специального значения, например, ки-
сточки па ушах, грива на шее, хвост, борода и т. д.
Большинство волос подвержено смене, которая у животных хо-
лодных и умеренных климатических поясов носит сезонный харак-
тер. Перед выпадением луковица волоса ороговевает, волос отде-
ляется от сумки и выпадает; в то же время в сумке из нижней части
наружного влагалища образуется зачаток нового волоса путем вра-
стания в пего сосочка соединительной ткани.
Рис. 524. Продольный разрез через конечные фаланги пальцев:
А — исходная стадия,' В — ноготь, С — коготь, D — копыто.
1 —подушечка пальца, 2 — подошвенная роговая пластинка, 3 — ногте-
вая пластинка, 4 — ногтевой валин, 5 — конечная фаланга. (Из Вебера.)
Целый ряд групп среди млекопитающих лишен волосяного по-
крова. Однако всегда удается доказать, что отсутствие представляет
вторичное явление. Во время утробной жизни у всех млекопитающих
развивается сплошной волосяной покров.
Когти, ногти и копыта представляют видоизмененные и сильно
утолщенные роговые части эпидермиса, прикрывающие фаланги
пальцев. Они произошли от когтей, которые имели их предки — реп-
тилии. Исходной формой является коготь, сходный с таковым у
черепах или крокодилов. Здесь мы имеем с верхней (тыльной)
стороны роговую пластинку, которая прикрывает конец .последней
фаланги сверху и с боков; с нижней стороны эта пластинка, по-
строенная из плотного рогового вещества, переходит в более мягкую
п легче стирающуюся подошвенную пластинку. Когти такого рода,
(нагодаря неравномерному стиранию, всегда острые, встречаются у
очнонроходных, у сумчатых, насекомоядных, хищных, грызунов.
Нижняя часть пальца под ногтем образует здесь подушечку
на и ьца, так как подошвенная пластинка здесь не идет так же далеко
назад, как когтевая пластинка с верхней стороны (рис. 524», С).
II о г о т ь представляет видоизменение когтя; он стал значительно
более плоским, и подошвенная пластинка значительно редуциро-
вались, в связи с чем сильно увеличились подушечки на концах
717
Рис. 525. Развитие рогов оленя.
1— первоначальное развитие рога нан
отростка лобной кости, 2 — рог голый
с развившимся в нем синусом в ре-
зультате разрушения костной ткани,
За — пенёк рога с отвалившимся ро-
гом ЗЬ, 4 — процесс зарастания пень-
ка кожей, 5 — образование венчика
в пеньке, обросшем кожей, 6-разви-
тие рога на второй год, 7 — очистив-
шийся от кожи рог с двумя отрост-
ками — вилка. (Из Вебера).
пальцев. Ногти заменяют когти у приматов (Primates). Копыта
являются тоже видоизменением когтя, развившегося в другом
направлении. Здесь когтевая пластинка видоизменилась в наруж-
ное копыто, подошвенная пластинка образовала подошву копыта,
а подушечка пальца .превратилась в так называемую «стрелку
копыта» (рис. 549, D).
На нёбе у млекопитающих тоже иногда имеет место образование
роговых пластинок. Это нёбные пластинки. У беззубых
китов из них образуются большие
расщепленные пластинки — «кито-
вый ус».
Что касается рогов млекопитаю-
щих животных, которые развива-
ются на голове, то в их образова-
нии по большей части принимает
участие также и кость.
Рог носорога представ-
ляет чисто эпидермальное обра-
зование, где чрезвычайно длинные
волосовидные роговые нити объе-
диняются в конусообразный рог,
а многочисленные, тоже очень
длинные, сосочки эпидермиса глу-
боко заходят в соединительноткан-
ную кожу. Один или два таких рога
сидят па носовых костях. У иско-
паемого Elasmotherium па кости
имелось возвышение, служившее
для поддержания громадного рога.
Рога полорогих жи-
вотных являются уже пе только
роговыми образованиями: роговые
чехлы помещаются здесь па более
или менее значительных костяных
стержнях, отходящих от лобных
костей. История онтогенетиче-
ского развития показывает, что
костные стержни рогов не явля-
ются выростами лобных костей,
а являются результатом роста самостоятельных кожных око-
стенений (os cornu), которые очень рано срастаются с лоб-
ными костями. Только у вилорога (североамерикапская
антилопа — Antilocarpa) происходит периодическое сбрасыва-
ние роговых чехлов; у всех остальных полорогих животных они
сохраняются в течение всей жизни и продолжают медленно
отрастать.
У жирафа (Giraffidae) кожные окостенения срастаются с лобно-
теменными выростами кости и бывают покрыты кожей, одетой
волосами.
718
12
Рога оленей в отличие от только что описанных форм яв-
ппются плотными, чисто костными образованиями и притом перио-
дически возобновляемыми. Они свойственны только самцам, за един-
отиенным исключением северных оленей, где они имеются у обоих
iHHion. Отрастающие рога покрыты волосами, мягки, только позд-
ней окостеневают, причем эпидермис с волосами отмирает и сбрасы-
вается, оголяя пигментированную кость. После прекращения по-
'iniioli деятельности рога оленей отпадают вследствие разрушения
костной ткани у основания рога. Остается лишь так называемый-
«пенек». Пенёк зарастает кожей,
н эта последняя продуцирует по-
in.iii рог (рис. 525).
Как видно из изложенного, не-
смотря на различие всех перечис-
ленных типов рогов, рога оленей
н полорогих связаны друг с дру-
гом переходами и эволюционно мо-
гут быть выведены из рога поло-
рогих.
Кожные железы у млекопитаю-
щих, в противоположность птицам
н пресмыкающимся, очень мно-
гочисленны. Этим млекопитающие
напоминают амфибий, и, хотя
кожные железы млекопитающих
но своему строению значитель-
но отличаются от желез амфибий,
однако они произошли, несом-
ненно, от последних. Наиболее
похожи на многоклеточные же-
лезы амфибий простые труб-
ч а т ы е потовые железы. Эти
железы, так же как и у амфи-
бий, состоят из одного слоя же-
лезистых клеток и одеты сна-
ружи слоем эпидермальных глад-
ких мускульных клеток; секрет
этих желез — пот—выделяется
клетками (рис. 526). Трубча-
тки1. железы подвергаются значительным видоизменениям: тру-
бочка может делаться длинной, скрученной клубочком, или она
ветвится, расширяясь по концам веточек.
В других случаях мускульная обкладка исчезает; выделяемое
железами вещество получается в результате перерождения и
отмирания самих клеток, которые располагаются в несколько
(‘.лоев. Такими именно железами являются альвеоляр-
п ы е, или гроздевидные, сальные железы млеко-
питающих, составляющие другую группу желез. Основным продук-
том выделения трубчатых желез является пот, служащий для сма-
Рис. 526. Схема разреза через
кожу млекопитающего.
1 — роговой слой (stratum corneum),
2 — мальпигиев слой (str.’ Malpighii),
3 — гроздевидная железа, 4 — труб-
чатая железа, 5 — сосочек волоса»
6 — волос, 7 •- влагалище волоса, 8 —
эпителий волосяной сумки, 9 - во-
локнистая сумка волоса 10 — мускул
волоса (m. arrector pill,) 11 — жиро-
вое скопление, 12 — сосочек кожи.
(Из Вебера.)
чивания кожи, для охлаждения тела путем испарения воды и для
выделения некоторых продуктов обмена веществ; у лошадей пот
содержит белок (пепа), у бегемота он содержит красный пигмент,
так же как у красного кенгуру, где он, высыхая в порошок, при-
дает красную окраску шерсти. У водпых животных (киты, сирены)
потовые железы отсутствуют вовсе, а у грызунов опи располагаются
только на подошвах. Вопреки распространенному мнению потовые
железы имеются также и у кошек и у собак, хотя и пе многочислен-
ны. Обыкновенно выводное отверстие потовых желез открывается
в сумку волоса выше отверстий сальных желез. Последние в числе
двух или большем открываются в сумку, выделяя жирное вещество,
основное значение которого смазывать жиром волосы, что предо-
храняет их от высыхания и снашивания. Запах, издаваемый этими
железами, играет большую роль в распознавании особей вида и
пола, в половом возбуждении.
Однако существуют специальные пахучие железы, более слож-
но построенные, по большей части из потовых желез, реже из саль-
ных, а иногда из тех и других одновременно и открывающиеся
в особые мешки. Иногда такие железы выполняют и некоторые
другие специальные функции. Таковы анальные ж!елезы мно-
гих мелких хищников, выделяющие резко вонючий секрет, который
может иногда выпрыскиваться наружу, как, например, у вонючек
(Mephitis, Conepatus) и др.; перинеальные (промежно-
стные) железы, расположенные между анальным и половым отвер-
стием у виверр (Viverra, Genetta) и др.; мускусные железы,
открывающиеся у самца кабарги {Moschus moschifcrus) в препу-
циум пениса, у выхухоли (Desmana moschata). У многих грызунов
в препуциальный мешок самцов и, соответственно, у основания
клитора самок открываются две железы, причем у бобра (Castor
fiber) именно этими большими железами вырабатывается возбуж-
дающий секрет — «бобровая струя»; у основания хвоста у олепей,
по бокам туловища у землероек, па спине у пекари и даманов имеют-
ся аналогично устроенные пахучие железы.
Из сложных желез специального назначения следует указать еще
слезные мешки многих жвачных (олепи, антилопы, овцы),
челюстную железу некоторых антилоп (x^ntilope maxwelli) и носо-
вые железы у большей части млекопитающих, выделяющие жидкий
секрет, поддерживающий голый пос во влажном состоянии. На-
конец, у очепь многих копытных па конечностях развиваются слож-
ные железы, служащие для смазывания трущихся друг о друга
поверхностей. Такими железами являются прежде всего копыт-
ные железы многих жвачных, смазывающие прилегающие
друг к другу поверхности двух пальцев. Однако в виду того, что
эти железы встречаются у некоторых двукопытных лишь на задпих
конечностях, а у крупного рогатого скота и у благородного оленя
они не встречаются вовсе, есть основание предполагать, что железы
эти имеют также и другое значение, являясь пахучими железами,
оставляющими запах на следу. У самцов однопроходных есть на
задних конечностях особая шпорцевая железа, открывающаяся
720
u.'i конце роговой шпоры, лежащей в пяточной области. Значение
.mil железы все еще остается неясным, и хотя впрыскивание ее
гекрота кролику оказалось смертельным, надо думать, что железа
на не является ядовитой, а каким-то образом связана с половыми
процессами, так как именно в период половой деятельности она
Рис. 527. Самка ехидны (Echidna hystrix) с сумкой.
(Пе Гаане.)
становится заметно больше. Кожные железы млекопитающих фи-
ло генетически стоят в несомненной связи с кожными железами
амфибий. Те древнейшие ископаемые рептилии, от которых про-
изошли млекопитающие и которые сами произошли от стегоце-
фал, должны были иметь кожные железы в отличие от современ-
ных рептилий, которые их утратили.
Самыми характерными кожными железами млекопитающих яв-
ляются молочные железы. Несмотря на их гроздевидпое
строение, можно считать доказанным, что опи развились из трубча-
М Кишкнрог и Станчинский ^21
тых потовых желез. Наиболее примитивное строение имеют они у
однопроходных (Monotremata). Самка ехидны (рис. 527) имеет на зад-
ней части брюха кожную складку, разрастающуюся в период раз-
множения в сумку, в которой вынашивается сперва яйцо, а потом
детеныш. По бокам этой сумки лежат ветвящиеся трубчатые мо-
лочные железы, одетые снаружи мускульными клетками и собран-
ные в пучки, по одному с каждой стороны. Железы эти открываются
в сумки волосков и собраны на небольшом поле. У утконоса, ко-
торый не имеет сумки и, повидимому, высиживает яйцо в гнезде,
Рис. 528. ' Вскрытая сумка кенгуру с детенышем.
(Из Брэма.)
молочные железы открываются непосредственно на нижней поверх-
ности брюха. Соски у однопроходных отсутствуют, и молоко тягучей
консистенции слизывается детенышами прямо с поверхности волос.
У самок сумчатых (Marsupialia) для вынашивания детенышей,
которые рождаются очень несовершенными, имеется на брюшной
стороне сумка. Сумка эта образуется из двух продольных складок
кожи, которые затем срастаются, образуя сумку у большинства
с передним, реже с задним отверстием. Только у наиболее прими-
тивных сумчатых, у сумчатых крыс (Didelphyidae) сумка остается
неполной в виде двух складок в течение всей жизни, а у некото-
рых даже отсутствует совершенно. Однако у сумчатого муравьеда
(Myrmecobius), у которого сумка отсутствует, наличие рудимента
722
лпткула, соответствующего сфинктеру сумки, указывает, что отсут-
ггниг су мин здесь явление вторичное. Открываются молочные
железы и сумке на конце сосков, которые сидят в особом кар-
M/HI'inко. Отот последний во время кормления выворачивается соском
нлружу и превращается в длинный сосок, плотно сидящий у детены-
ша по рту и даже срастающийся с эпителием рта; детеныш первое
прем и беспомощно висит на соске, тогда как мать с помощью осо-
1'нц’о мускула впрыскивает ему в рот молоко (рис. 528).
Число сосков у сумчатых от 2 до 22, причем они сидят иногда
и 2, а иногда в 4 ряда.
У плацентарных млекопитающих положение молочных желез
различно, что стоит в связи с их образом жизни. На груди они рас-
полагаются у обезьян, у летучих мышей, у сирен и некоторых не-
полнозубых, на брюхе — у большинства млекопитающих, в обла-
сти наха—у непарно-и парнокопытных и китов. Число сосков
обычно стоит в связи с плодовитостью вида и соответствует макси-
мальному числу7 одновременно рождаемых детенышей; максималь-
ное число в 22 соска у мадагаскарского насекомоядного тенрека
(Centetes ecaudatus), минимальное — 2 — у человека, обезьян, ло-
шадей, овец, коз; слонов. Что касается строения сосков, то у пла-
центарных можно различить два типа сосков. Один тип, соответ-
ствующий соскам сумчатых, т. е. вывернутому наружу карману, —
:н’о так называемые истинные сосцы; у них выводные протоки желез
открываются отдельными отверстиями на вершине сосца. Они встре-
чаются у полуобезьян, обезьян и человека. Второй тип — ложные
госцы образуются за счет разрастания кожи валика и иногда имеют
кнутри соска общий выводной проток, в который открываются
мелкие выводные протоки желез, что характерно для жвачных
копытных (рис. 529).
Можно себе представить, что процесс возникновения и развития
молочных желез протекал следующим образом. Предки млекопита-
ющих были, как известно, пойкилотермными рептилиями, не утра-
тившими еще своих многочисленных желез и жившими, вероятно,
и условиях достаточно влажного климата; они были яйцекладущими
животными. Приобретенная затем гомотермия или теплокровность
развилась одновременно с возникновением волос. Теплокровность
сделала необходимым высиживание яиц, что являлось для млеко-
питающих чрезвычайно опасным, ибо привязывало их к месту и под-
вергало большому риску жизнь потомства. Отсюда чрезвычайно
полезной должна была оказаться приобретенная возможность пере-
носить в случае опасности яйцо с помощью складок на брюхе и с по-
мощью так называемых сумчатых костей, столь харак-
терных для низших млекопитающих. Впоследствии способность
перетаскивать яйца заменилась более совершенной способностью
вынашивать яйца в сумке, где они, будучи согреты телом, могли
быстрее и безопаснее развиваться. Так появилась сумчатость.
Молочные железы возникали в сумке в связи с обильным отделе-
нием пота, когда из яйца выходил детеныш. Постепенно выработа-
лась способность детеныша питаться отделением потовых желез,
4ь‘ 723
сперва как влагой, а потом, с увеличением в поту питательных ве-
ществ (жира, белка) как универсальным питательным веществом.
На этой стадии остановились в своем развитии однопроходные.
Так как молочные железы имеются у обоих полов, у всех млекопита-
ющих, а сумчатые кости характерны также для обоих полов одно-
проходных и сумчатых, вероятно, оба пола первоначально вынаши-
вали яйца и детенышей и выкармливали их молоком. Только позд-
нее, когда яйцо стало развиваться в утробе матери и особенно когда
стали рождаться живые детеныши, что могло иметь место только
у самок, вынашивание и выкармливание детенышей пало исклю-
чительно на долю самки.
Рис. 529. Схема строения сосков разных млекопитающих:
1а —соски сумчатых животных до периода сосания, 1b—
в период сосания молока, 2 — соски приматов, 3 — соски хищных,
4 — соски жвачных. (По Бючли.)
Выкармливание детенышей молоком, когда они в связи с раз-
витием сосков приобрели способность сосать, постепенно привело
к сокращению эмбрионального периода, а это в свою очередь сде-
лало возможным развитие яйца в утробе матери и вынашивание
совершенно беспомощных детенышей в сумке. Это — стадия со-
временных сумчатых. Последующее возникновение и развитие пла-
центы дало возможность рождать все более и более совершенных де-
тенышей и привело в связи с вторичным удлинением эмбрионального
периода к полному исчезновению сделавшейся ненужной сумки.
Таким образом, по нашему мнению, однопроходные, сумчатые и пла-
центарные млекопитающие составляют последовательно следующие
одна за другой ступени эволюции, а не три ствола эволюции,
независимо развившиеся из общего корня, как обычно принято
думать.
724
2. Скелет
()дпоП из особенностей скелета млекопитающих является боль-
шие развитие хряща у молодых животных и продолжительное сохра-
нен не его в постэмбриональный период развития. Это объясняется
особенностями роста хряща и кости в связи с ростом самого живот-
ного. Именно хрящ растет всей своей массой (интерстициально), рост
совершается быстро и пропорции частей при этом могут сохраняться,
тогда как кость растет медленнее и только с поверхности, так что,
появившиеся части скелета остаются всегда на прежнем расстоя-
нии и сохранение пропорции частей при росте кости оказывается
6
Рис. 530. Схема эндохондрального и перихондрального окостенения
в длинных костях млекопитающих.
Горизонтальной штриховкой обозначены эндохондральные окостенения,
вертикальной — перихондральные окостенения, черным — компактная
костная масса, белым по концам кости — хрящ, внутри — полость кост-
ного мозга.
1 — закладка перихондрального и эндохондрального окостенений диа-
физы в хряще, 2 — средняя часть, т. е. диафиза, окостенела изнутри (эн-
дохондрально), снаружи (перихондрально) концы или эпифизы остаются
еще хрящевыми, 3 — первая закладка эндохондрального окостенения в
верхней эпифизе, 4 — эндохондральное окостенение в обеих эпифизах,
появление внутримозговой полости и компактных костных масс в диа-
физе, 5 — в связи с дальнейшим окостенением остаются хрящевыми лишь
участки дальнейшего роста кости между эпифизами и диафизой, 6 — в
связи с исчезновением хряща между эпифизами и диафизой рост костей
прекратился, окончательно сформировалась полость костного мозга. (По
Дювалю из Иле и др.)
невозможным. Вот почему молодые животные всегда содержат мно-
го хряща и особенно в таких местах, где сильно осуществляется
рост. Так, длинные кости конечностей всегда состоят из средней
части — диафизы, которая окостеневает раньше всего, и кон-
цов, которые долго сохраняются хрящевыми — эпифиз. Око-
стенение диафизы начинается прежде всего с периферии — перихонд-
рально, т. е. за счет клеток, происходящих из соединительной ткани,
одевающей хрящ, — из перихондрия; затем к нему присоединяется
внутреннее окостенение хряща — эндохондральное окостенение;
только позднее, когда в диафизах появляются путем распадения
кости полости костного мозга, эпифизы начинают окостеневать эндо-
хондрально; между диафизой и эпифизами, однако, долго остается
хрящ, с помощью которого осуществляется рост кости в длину
с сохранением пропорций (рис. 530).
725
Позвоночник млекопитающих характеризуется в отличие от
Sauropsida отсутствием свободных сочленений между позвонками,
за исключением первого и второго позвонков. Между телами поз-
вонков залегают межпозвоночные эластические соединительно-
тканные диски. В общем это обусловливает меньшую гибкость поз-
воночника, что стоит в связи с более специализированным движе-
нием млекопитающих с помощью конечностей.
Окостенение тел позвонков происходит при посредстве хряще-
вых эпифиз. Тела имеют или плоские поверхности или слабо выпук-
лые, у большинства современных копытных слегка опистоцельные.
На позвонках хорошо развиты, особенно в грудной области, ости-
стые отростки, а также поперечные отростки, которые сидят на
дугах; у основания последних имеются сочленовные отростки.
Рис. 531. Скелет собаки, врисованный в контуры животного.
1—7 — шейные позвонки, 3 — 13 — грудные позвонки, 1 — 7 — пояснич-
ные позвонки, 1 — 12 —ребра, I — лопатка, II — плечо, III—лучевая,
IV — локтевая, V — запястье, VI — пястье, VII — пальцы, VIII — под-
вздошная, IX — седалищная, X — лобковая, XI — бедренная, XII —ча-
шечка, X III — большая берцовая, XIV — малая берцовая, XV — пред-
плюсна, XVI — плюсна, XVII — пальцы. (Из Вебера.)
Число шейных позвонков, как правило, 7 (рис. 531), независимо
от длины шеи, так что длина шеи, в противоположность птицам, за-
висит исключительно от длины позвонков. Только у ламантина
(Manatus) и у одного вида ленивца (Choloepus hoffmani) 6 шейных
позвонков, тогда как у трехпалых ленивцев (Bradypus) их 9. Почти
исчезнувшая в связи с водным образом жизни шея китообразных
все же содержит 7 плоских, частично сливающихся позвонков, тогда
как в шее жирафа эти же 7 позвонков очень длинные. Первые 2 шей-
ных позвонка видоизменены в атлас и эпистрофей, соединены на-
стоящими суставами, причем у однопроходных зубовидный отро-
сток функционирует еще в виде особой кости, а пе сросся, как
у прочих млекопитающих, с телом эпистрофея. Рудиментарные реб-
726
1> । ni«iiin*n области срастаются с позвонками, вследствие чего обра-
ц iiirini продырявленные поперечные отростки. Частичное или пол-
П' о’ срас, такие шейных позвонков имеет место у китообразных; у туш-
I' । и'in ii'uii срастаются все шейные позвонки за исключением 7-го,
) сумчатого крота (Notoryctes) сливаются от 2-го до 6-го, у крота—
"I :! го до 4-го.
11пело грудных позвонков, которые характеризуют-
| >| прпчлепением к ним ребер, вариирует от 9 [у одного китооб-
разного (Hyperoodon) и броненосца (Tatusia)] до 25 [у ленивца
О'holoepus)], но обыкновенно их бы- ,
Рис. 532. Развитие грудины челове-
ческого зародыша.
1 — ключица, 2 — episternum, 3 — заклад-
ка грудины в виде двух полос, 4 — ме-
чевидный отросток, I—-X — ребра. (Из
Вебера по Руге.)
Рис. 533. Грудина
лемура.
1 — ключица, 2 — руко-
ятка грудины, 3 — мече-
видный отросток, 4 —
ребра. (По Боасу.)
хорошо развиты отростки: остистые (proc, spinosus), поперечные,
или диапофизы (proc, transversus), сочленовные, или зига-
пофизы (proc, articularis); ребра причленяются головками к телу
одного или двух соседних позвонков и бугорками к поперечным
отросткам. Они состоят из двух частей: костяного ребра — про-
ксимального и хрящевого — дистального. Только ребра передних
грудных позвонков доходят до грудины, задние ребра оканчиваются
свободно (ложные ребра). Поясничных позвонков, которые
.характеризуются отсутствием ребер, вошедших здесь в состав
поперечных отростков, у млекопитающих от 2 до 9, обычно же
их 6—7.
Крестцовые позвонки млекопитающих — это те позвон-
ки, которые, сросшись между собой, сочленяются с подвздошной
костью. Таких позвонков 1—10, но только 1—2 из них являются
истинными крестцовыми позвонками, т. е. такими, у которых сочле-
нение с тазом происходит посредством самостоятельных окостене-
ний, соответствующих крестцовым ребрам; остальные крестцовые
позвонки являются ложными, т. е. приросшими к крестцу хвосто-
выми позвонками.
727
Число позвонков, входящих вхвостовой отдел, вариирует
чрезвычайно: от 3 (летучие мыши, гиббон) до 49 [ящер (Manis ma-
crura)].
Грудина (sternum) млекопитающих развивается из слияния
брюшных концов ребер, которые затем образуют одну непарную
пластинку, состоящую из ряда отделов (рис. 532).
Соединенный с ключицами (если они имеются) и с первой парой
ребер расширенный передний отдел грудины носит название ру-
коятки (praesternum, manubrium); далее следует отдел, назы-
ваемый телом (mesosternum, corpus), нередко распадающийся
на ряд косточек, последовательно расположенных между следую-
щими друг за другом парами ребер; еще дальше назад лежит так
называемый мечевидный отросток (xiphisternum). Да-
леко не все ребра находятся в связи с грудиной, более или менее
значительное число ложных ребер оканчивается свободно
(рис. 533).
У клоачных млекопитающих (Monotremata), в связи с наличием
отсутствующих у всех прочих млекопитающих коракоидов, впере-
ди от рукоятки грудины закладывается хрящ предгрудин-
н и к (prosternum), находящийся в связи с коракоидами, кото-
рый затем вытесняется развивающейся костью — н а д г р у-
динником (episternum), что сближает клоачных с репти-
лиями.
Череп млекопитающих целым*рядом особенностей отличается от
черепа рептилий. Однако эти особенности появились не сразу, а пу-
тем постепенного изменения, с которым можно познакомиться, изу-
чая черепа ископаемых звероящеров (Theromorpha), особенно
из отряда зверозубых (Theriodontia). Череп млекопитающих —
череп тропибазальный, но с укороченной межглазничной ча-
стью, благодаря чему он выглядит как платибазальный, с боль-
шой черепной коробкой, с ясно выраженной тенденцией к сра-
станию отдельных костей между собой.
О расположении костей в черепе млекопитающих можно судить
по прилагаемой схеме (рис. 534). й
На боковых затылочных костях, наружу от сочленовных бугор-
ков, обыкновенно имеется затылочный отросток (proc,
paroccipitalis), особенно хорошо развитый у грызунов и у многих
копытных. Впереди от сочленовных бугорков обычно имеется от-
верстие для выхода наружу подъязычного (XII) нерва (for. condy-
loideum anterius).
В хряще слуховой капсулы закладывается несколько окостене-
ний, которые, однако, очень рано сливаются в две кости: впереди
лежащую — каменистую (petrosum) и лежащую снаружи
и сзади — сосцевидную (mastoideum); обе эти кости, сра-
стаясь вместе, дают каменисто-сосцевидную, или просто каме-
нистую кость (petromastoideum, petrosum или perioticum),
заключающую внутренний слуховой лабиринт. Perioticum граничит
в задней части с основной и боковой затылочными костями, а в пе-
редней части с клиновидной и крыло-клиновидными костями; так
728
if.iic между этими костями и каменистой костью проходят сосуды
и нервы, то здесь остаются значительные щели: сзади — заднее
рваное отверстие (for. laceruin posterius), или яремное отверстие
(Гог. jugulare) для прохождения языкоглоточного (IX) нерва,
нетвей блуждающего (X) нерва и для добавочного (XI) нерва,
Рис. 534. Схема строения черепа млекопитающего.
Замещающие кости заштрихованы.
1—межтеменная (interparietale), 2 — ламбдовидный шов (sutura lambdoidea), з — те-
менная (parietale), 4 — венечный шов (sutura coronal is), 5 — позадиглазничный отросток
(proc, postorbitalis), 6 — лобная (frontale), 7—предглазнидный отросток (ргос.ргаеог-
bitalis), 8 —носовая (nasale), 9 — межчелюстная (ргаетахШаге), 10 —сошник (vomer
(=»parasphenoideum), 11 — предглазничное отв. (for. infraorbitalis), 12 —челюстная
(maxillare), 13 — нёбная (palatinum), 14 — скуловая (jugale), 15 —слезная (lacrymale),
16 — глазнично-клиновидная (orbltosphenoideum), 17 — передняя клиновидная (ргае-
sphenoideum), 18 — крыловидная (pterygoideum, 19 — крылоклиновидная (allsphenoi-
deum), 20 — засочленовный отросток (processus postglenoidalis), 21 — тимпанальный
пузырь (bulla tympani), 22 — барабанная (tympanlcum), 23—чешуйчатая (squamosum),
24 — сосцевидная (mostoideum), 25 — основная затылочная (basioccipitale), 26— со-
членовный отросток (condylus), 27—боковая затылочная (exoccipitale), 28—верхняя
затылочная (supraoccipitale), 29 — затылочный отросток (processus paroccipital is),
30 —переднее рваное отв. (foramen lacerum anterius), 31—скуловой отр. (processus
zygomaticus), 32 — основная клиновидная (basisphenoidale), 33 — круглое отверстие
(foramen rotundum (V2), 34 — клино-глазничное отв. (foramen sphenorbitale (III,
IV, Vi, VI), 35 — глазничное отв. (foramen opticum), 36 —слезное отв. (foramen
lacrymale), 37 — обонятельная (ethmoideum).
а также для внутренней яремной вены, а спереди — переднее
рваное отверстие (for. lacerum anterius), через которое проходит в
череп внутренняя сонная артерия. Иногда связь каменистой кости
с черепом настолько слаба, что она легко может быть отделена
вместе с приросшей к ней барабанной костью или даже при
мацерации черепа совершенно выпадает, как это имеет место у
китообразных.
729
Снаружи ушной области образуются две покровных кости. Одна
из них, происхождение которой до сих пор остается еще не достаточ-
но выясненным, развивается в области наружного слухового про-
хода в виде полукружной небольшой косточки. Это — бара-
банная кость (tympanicum), которая, вероятно, является
гомологом угловой кости (angulare) нижней челюсти рептилий,
перешедшей в околоушную область вместе с другими костями ниж-
ней челюсти в связи с переменами, происшедшими в челюстном со-
членении. Располагаясь первоначально в виде полукружной или
кольцевой кости в области слухового прохода, она нередко раз-
растается и образует, обычно вместе с каменистой костью и иногда
с чешуйчатой, наружный слуховой проход (meatus auditorium
externus). На барабанной кости бывает натянута барабанная
перепонка. Глубже внутрь барабанная кость может принимать
участие также в образовании стенок среднего уха. Нередко бара-
банная кость на нижней стороне слуховой области вздувается в
тонкостенный пузырь (bulla ossea).
Вторая кость кожного происхождения, принимающая участие
в образовании скелета ушной области, — это чешуйчатая кость
(squamosum); она получила у млекопитающих исключительное раз-
витие и значение в связи с причленением к ней нижней челюсти. У
обезьян и человека все эти кости ушной области срастаются вместе,
образуя сложную височную кость (temporale); чешуйчатая кость
здесь составляет лишь часть, именно »чешую (squama temporalis)
височной кости, тогда как барабанная кость—барабанную часть
(pars tympanicum), а собственно околоушные хрящевые кости об-
разуют части каменистую (pars petrosa) и сосцевидную (pars ma-
stoidea). Степень развития чешуйчатой кости и участие ее в образо-
вании стенок мозговой коробки и слухового прохода весьма различ-
ны, но всегда чешуйчатая кость имеет сочленовную поверхность (са-
vitas glenoidea) для причленения нижней челюсти. Таким образом,
у млекопитающих в отличие от всех остальных позвоночных имеется
совершенно особенное оригинальное причленение нижней челюсти:
не к квадратной кости, а к чешуйчатой кости черепа. Сочленовная
поверхность для нижней челюсти лежит на основании идущего от
чешуи наружу и вперед скулового отростка (proc, zygomaticus).
У однопроходных над сочленовной поверхностью он пронизан ви-
сочным каналом (can. temporalis), особенно хорошо выраженным
у утконоса. Позади сочленовной ямки имеется отросток (proc,
postglenoideus).
Спереди от слухового отдела основание черепа образовано о с-
новной клиновидной костью (basisphenoidale), на
внутренней поверхности которой имеется более или менее зна-
чительное углубление — так называемое турецкое седло (sella
turtica), в котором помещается гипофиза. По бокам от основ-
ной клиновидной кости дежат, образуя нижние части передней
стенки черепной коробки, крыло-клиновидные кости (ali-
sphenoidea), обычно срастающиеся с ней в одну кость, образуя боль-
шие крылья (alae magnae) этой кости. Крыло-клиновидные кости
730
i iin'iiu4ii.i двумя отверстиями: овальным (for. ovale) для прохожде-
нии третьей ветви тройничного нерва (V3) и круглым (for. rotun-
diiini для прохождения второй ветви того же нерва (72). Впрочем,
iiepiuui нередко сливается с передним рваным отверстием (for.
Immruiu anteris), а второе часто сливается с клино-глазничным
11 гнсрстием (for. sphenorbitale), ограниченным одновременно как
в рыло-клиновидной костью, так и лежащей впереди от нее глазнич-
1н> клиновидной костью. Через это отверстие проходят нервы глазо-
пп1гательный (oculomotorius III), блоковидный (trochlearis IV),
Н1ТК1ДЯ1ЦИЙ (abducens VI) и первая ветка тройничного нерва (trige-
minus Vj).
Черепная крышка здесь прикрыта теменными костя-
ми (parietale), которые нередко срастаются друг с другом (у од-
нопроходных, некоторых сумчатых и копытных); между ними и
порхней затылочной костью закладываются еще две кости, сливаю-
щиеся в одну меж теменную кость (interparietale), ко-
торая в некоторых отрядах сохраняется в виде самостоятель-
ной кости (китообразные, сумчатые, даманы, трубкозуб, шерсто-
крыл и некоторые грызуны), тогда как в других она срастается
пли с теменными (у некоторых грызунов, сирен и жвачных),
или с верхней затылочной костью (у хищных, однокопытных,
приматов).
Впереди основной клиновидной кости лежит передняя
клиновидная кость (praesphenoideum) с углублением
для хиазмы глазничных нервов; по бокам от нее, обычно срастаясь
с ней, лежат глазничн о-к линовидные кости (orbi-
tosphenoidea) с отверстием для прохождения глазничных нервов
(for. opticum), называемые в случае срастания с передней и основ-
ной клиновидными костями малыми крыльями (alae parvae). Как
уже сказано выше, между обеими крыловидными костями с каждой
стороны имеются щелевидные отверстия, через которые проходят
нерЬы глазных мышц и первая ветка тройничного нерва (for.
sphenorbitale).
Сверху этому ряду костей соответствуют покровные лобные
кости (frontale) — обычно хорошо развитые парные кости, иногда
срастающиеся между собой и несущие у полорогих стержни, а у оле-
ней — пеньки для рогов. Покрывая черепную коробку, лобные кости
ограничивают сверху и с внутренней стороны глазницы, образуя
нередко отростки: предглазничный (proc, praeorbitalis и позади-
глазничный (proc, postorbitalis). Иногда вокруг глаз образуется
вследствие срастания скуловой кости (jugale) с лобной
костью окологлазничное кольцо (некоторые насекомоядные и хищ-
ные, жвачные, однокопытные, полуобезьяны). У обезьян процесс
разъединения глазной впадины от височной идет так далеко, что
между глазницей и височной впадиной образуется костяная
стенка.
В передней стенке глазницы лежит небольшая покровная
слезная косточка (lacrymale), продырявленная отверстием
слезного канала (for. lacrymale). *
731
Рис. 535. Схематическое
изображение расположе-
ния носовых раковин на
поперечном разрезе че-
рез носовую полость.
I—V—endoturblnalla, Ь—$—
ectoturbinalia. (Из Бючли.)
В носовой области закладывается в хрящевой носовой капсуле
несколько окостенений. Во-первых, вдоль хрящевой носовой пере-
городки развивается посредине костная перегородка (lamina perpen-
dicularis), соответствующая средней обонятельной кости
(mesethmoideum); во-вторых, в задней стенке носовой капсулы
и в передней стенке мозговой полости развиваются две боковых пла-
стинки, продырявленные многочисленными [только у утконоса
эти отверстия заменены одним (for. olfactorium)] отверстиями для
прохождения обонятельного нерва — решетчатые пластинки (lamina
cribrosa), соответствующие боковым этмоидальным костям (ethmoi-
dalia lateralia). От носовой перегородки окостенение распростра-
няется с дорзальной стороны по бокам, идет по боковым стенкам
носовых капсул и заканчивается на дне ее, образуя со сторон много-
численные выступы внутрь носовой поло-
сти. Это обонятельные раковины (ethmo-
turbinalia), образующие вместе с боковыми
пластинками и с полостями ячеистого строе-
ния иногда очень сложный носовой лаби-
ринт. Расположение раковин видно из прила-
гаемого схематического рисунка (рис. 535).
От покровной носовой ко-
сти (nasale), покрывающей сверху эту
область, свешиваются внутрь носовой по-
лости, тоже по одной с каждой стороны,
раковины (nasoturbinalia). Снизу на носо-
вую перегородку налегает непарная кость,
считавшаяся раньше за с о ш н и к (vomer).
Однако исследования последнего времени
показали, что, кроме этой непарной покров-
ной кости, в основании черепа развиваются
еще две парных кости, названные п р е д-
сошником (praevomer); а так как у
рептилий сошник имеется в виде парной
кости, а непарной костью в этой области
был парасфеноид, то гомологом этой
последней кости рептилий можно считать именно сошник млеко-
питающих, а предсошники считать сошниками. Спереди стенками
носовой полости являются челюстные кости (maxillare), на
которых образуются раковины, врастающие в носовую полость;
это — челюстные раковины (maxilloturbinale).
Лицевая сторона черепа заканчивается спереди межчелю-
стными костями (praemaxillare). Эта кость у антропоморф-
ных обезьян и у человека очень рано срастается с челюстной ко-
стью, а у рукокрылых редуцирована до небольшой косточки или
даже отсутствует вовсе. Своим вертикальным отростком межчелю-
стная кость ограничивает спереди носовое отверстие и заходит меж-
ду носовой костью и челюстной далеко назад, иногда до лобной ко-
сти. Снизу межчелюстные кости принимают участие в образовании
твердого нёба. Они ограничивают спереди и с боков на твердом нёбе
732
п«,|н’тпл пёбно-посового (степсонова) канала (for. incisiva), соеди-
II июпюго ротовую полость с якобсоновым органом. Лежащие кнару-
жи пт итих нёбно-носовых отверстий отростки межчелюстных костей
|/1к.||адынаются для каждой кости независимо, вероятно являются
1ом().||огами парного сошника рептилий (praevomer) и у утконоса
сохраняются в виде самостоятельных костей. На межчелюстной
кости располагаются резцы.
Челюстные кости (maxillare), как уже было сказано, ограничи-
вают дно носовой полости, образуя твердое нёбо горизонтальными
отростками. G внутренней стороны носовой полости к боковым стен-
кам се, образованным челюстными костями, прирастает нижняя но-
совая раковина (maxilloturbinae), закладывающаяся в качестве
г а мостоятельной кости.
Рис. 536. Череп дельфина.
1 — боковая затылочная (exoccipitale), 2 — condylus occiplta"
lis, 3 — теменная (parietale), 4—верхи, затылочная (supraoc"
cipitale), 5 — лобная (frontale), 6 — носовая (nasale), 7 — носо-
вое отверстие, 8 — межчелюстная (praemaxilla), 9 — челюст-
ная (maxilla), 10 —нёДная (palatinum), 11 — крыловидная
(pterygoideum), 12 — скуловая (jugale), 13 — лобная (frontale),
14—барабанная (tympanicum), 15 — чешуйчатая (squamosum)-
(Из Вебера.)
Продолжением твердого нёба назад служат горизонтальные
отростки нёбных костей (palatinum), ограничивающих в ка-
честве боковых стенок внутренний носовой проход. Они стоят вер-
тикально, и к ним сзади примыкают также вертикально стоящие
крыловидные кости(pterygoideum), у человекасрастающиеся с клино-
видной костью, образуя ее крыловидные отростки (proc, pterygoidei).
Значительные видоизменения претерпевает передний отдел че-
репа в связи с переходом к водному образу жизни у китообразных,
сирен и в связи с развитием хобота у слонов. /
У китообразных (рис. 536) ноздри отодвинуты далеко назад в
межглазпичную область и ведут в вертикальный канал, соответ-
ственно открывающийся хоанами в полость рта; носовые кости ле-
жат позади ноздрей, налегая на лобные кости, и почти прилегают
к межтеменной кости; также и челюстные кости налегают на лобные
кости, заходя почти до теменных костей, сильно сдвинутых на бока
межтеменной костью.
733
У сирен имеет место такое же перемещение назад ноздрей, но не
так далеко; носовые кости здесь еще больше редуцированы и могут
даже отсутствовать (рис. 537).
У слонов с непомерным развитием бивней и мощных коренных
зубов и в связи с наличием хобота сильно видоизменяется нормаль-
ное расположение костей: ноздри сильно отодвинуты назад, в связи
с этим носовой канал принимает почти перпендикулярное положение;
лобные кости очень коротки, межчелюстные кости очень велики и за-
кости сдвинуты под основание че-
ходят далеко назад; челюстные
Рис. 537. Череп ламантина (Manatus
latirostris) с брюшной стороны.
1 — basioccipitale, 2 — exoccipitale, 3 — pet-
rosum, 4 — tympanicum, 5— basisphenoideum,
6 — alisphenoideum, 7 — squamosum, 8 —
jugale, 9 —palatinum, 10 —maxilla, 11 —
praemaxilla. (Из Вебера.)
репа; весь череп делается
очёнь массивным с чрезвы-
чайно развитой ппевматично-
стыо костей (рис. 538).
Рис. 538. Череп слона (Elephas
maximus), в продольном разрезе.
1 — мозговая полость черепа, 2 —
exoccipitale с сочленовным бугорком,
3 — supraoccipitale, 4 — parietale, 5 —
frontale (в этих костях видны мно-
гочисленные полости, наполненные
воздухом), 6 — mesethmoideum, 7 —
носовой проход, 8 — praemaxillare,
9 —maxillare, 10 —первый корен-
ной зуб (Мг), 11 — второй корен-
ной зуб (М3). (По Циттелю из Ве-
бера.)
Нижняя челюсть млекопитаю-
щих в отличие от всех остальных
позвоночных развивается с каж-
дой стороны только лишь из одной
зубной кости (dentale); обе половины или остаются соеди-
ненными друг с другом посредством хряща, шва, пли же срастаются
между собой (однокопытные, слон, рукокрылые, обезьяны). Каж-
дая половина состоит из горизонтального тела (corpus), на котором
сидят зубы, и восходящей части с отростками: передним венеч-
ным отростком (proc, coronoideus), лежащим позади пего сочленов-
ным отростком (proc, articularis) и у низших млекопитающих иду-
734
Рис. 539. Череп опоссума (Didelphys marsu-
pialis).
1 — exoccipitale, 2 — supraoccipitale, 3 — tympa-
niettv, 4 —squamosum, 5 — parietale, 6 — frontale,
7 —lacrymale, 8—nasale, 9 — praemaxillare, 10 —
maxillare, 11—jugale, 12 —proc, paroccipitalls,
13 —proc, coronoideus, 14 —proc, angularis.
Шим назад угловым отростком (proc, angularis); у большинства сум-
ч.ггых и у отдельных грызунов этот отросток превращается в про-
пирающуюся внутрь пластинку (рис. 539).
Такое строение нижней челюсти и своеобразное ее причле-
игние к черепу приводят нас к вопросу относительно происхо-
ждения этих особенностей у млекопитающих. История индиви-
дуального развития и палеонтологические данные дают ответ на
этот вопрос. У эмбрионов млекопитающих закладываются нёбно-
кнадратпый и меккелев хрящи. Однако окостенение нёбноквад-
раткого хряща происходит вне связи с сочленением нижней челю-
сти и не приводит к образованию квадратной кости, как у рептилий,
л даст новую слуховую косточку — наковальню (incus),
тогда как меккелев хрящ, окостеневая в своей задней части, дает
не сочленовную кость (articulare), как у прочих челюстноротных,
а третью слуховую ко-
сточку—м олоточек
(malleus). Первая слу-
ховая косточка, с т р е-
м я (stapes), соответ-
ствует столбику амфи-1
они, рептилий и птиц ।
н, как мы уже знаем,
я вляется видоизменен -
н ым подвеском (hyoman-
dibulare), т. е. прокси-
мальным отделом подъ-
язычной дуги. Так же
как и столбик (columella
auris), стремя прикры-
вает овальное окно слу-
хового лабиринта, и про-
дырявлено, как у неко-
торых рептилий, отвер-
стием, через которое у
некоторых низших мле-
копитающих проходит лицевая артерия (а. facialis) (рис. 540 и 541).
Таким образом, у млекопитающих квадратная кость превра-
тилась в наковальню, сочленовная — в молоточек; а так как к
молоточку прирастает еще небольшая покровная косточка, обра-
зующая средний его отросток, то полагают, что этот последний
соответствует покровной сочленовной кости (goniale) рептилий.
Лежащая по соседству с этой последней косточкой /Покровная
угловая кость рептилий также переходит в ушную область, стано-
вясь барабанной костью (tympanicum). Указанные изменения
видны на прилагаемой схеме (рис. 541).
Палеонтологические данные подтверждают возможность ука-
занных выше превращений. Среди зверозубых звероящеров (The-
riodontia) встречаются такие, у которых зубная кость получает
особенно большое развитие, достигая сзади места сочленения
735
нижней челюсти с квадратной кистью. В то же время чешуйчатая
кость спускается по квадратной кости до этого же сочленения.
Зубная и чешуйчатая кости, таким образом, все более и более
сближаются.
Рис. 540. Нижняя челюсть зародыша кролика со стреме-
нем, наковальней, молоточком и барабанной костью.
1 — меккелев хрящ, 2 — молоточек (malleus), 3 — наковальня
(incus), 4 —стремя (stapes), 5 —зубная (dentale), 6 — goniale, 7 —
барабанная (tympanicum). (По Гаупу из Иле, Кампен и др.)
Наконец, у некоторых зверозубых зубная и чешуйчатая кости
принимают участие в сочленении наряду с сочленовной и квад-
Рис. 541. Схема, иллюстрирую-
щая изменения висцерального
аппарата у млекопитающих по
сравнению с рептилиями в свя-
зи с развитием слуховых ко-
сточек. А — рептилии, В —
млекопитающие.
А. 1 — dentale, 2 — angulare, 3 —.
goniale (articulare не заштрихо-
ван), 4 — quadratum, 5 — stapes,
6 — cornu hyale. »
B. 1 — dentale, 2 — tympanicum,
3 — proseseus anterior mallei (mal-
leus не заштрихован), 4 — incus
5 — stapes, 6 — cornu hyale.
(По Гаупу из Иле, Кампен и др.)
ратной костями. *
Что касается подъязычного аппа-
рата млекопитающих, то он состоит
из тела, видоизмененной базальной
части (copula) гиоидной дуги, из пе-
редних рожков (cornua anteriora), об-
разованных нижним отделом гиоид-
ной дуги, и задних рожков (cornua
posteriora), преобразованных из пер-
вой жаберной дуги. Передние рож-
ки состоят из нескольких окостенений
(до четырех); обращенный к черепу
отрезок нередко прирастает к каме-
нистой кости, образуя шиловидный
отросток (proc, styloideus).
Вторая и третья жаберные дуги
дали начало образованию щитовидно-
го хряща (cartilago thyreoidea.)
Пояса конечностей. Плечевой
пояс млекопитающих характе-
ризуется прежде всего редукцией
вороновых костей (procoracoi-
deum, epicoracoideum, coracoideum),
тенденцией к исчезновению ключиц
(clavicula) и хорошо развитым греб-
нем лопатки (spina scapulae),
продолжающимся в акромиаль-
ный отросток (acromion).
Однако у однопроходных плечевой
пояс имеет большое сходство с репти-
736
'in и мн. Л опатка снабжена акромиальным выступом, к которому при-
чгглет спереди ключица, асзади к лопаткепричленяютсяширокие и
к• цнiTKiie вороновы кости (coracoideum), идущие к грудине. Спереди
пг них имеются в виде самостоятельных костей широкие прокора-
в• «иды (procoracoideum). Ключицы(clavicula)медиально соединяются
г надгрудциком (episternum) (рис. 542). У прочих млекопитающих
коракоиды закладываются в виде двух окостенений, но теряют
<’||>1зь с грудиной и прирастают к лопатке, образуя коракоидный
огросток (proc, coracoideus); прокоракоиды исчезают совершенно
(рис. 543), ключицы исчезают у некоторых хищных и копытных в свя-
III с приспособлением к быстрому бегу, а у китообразных и сирен
н с.вязи с превращением передних копечностей в плавники.
Рис. 543. Плечевой пояс зародыша и детены-
ша сумчатого — кузу (Trichosurus).
вина.
I — лопатка (scapula), 2 —
коракоиды (coracoideum),
3 — прокоракоиды (procora-
coideum), 4 — ключица (cla-
vicula), 5— надгрудник (epi-
al, ernum), 6—грудина (ster-
Unum). (По Веберу.)
Рис. 542. Плечевой пояс
утконоса, правая поло-
ся,- во-первых, наличие
1 — лопатка, 2 —коракоид, 3- прокоракоид, 4 —
ключица, 5 —грудина, 6—ребро. (По Бруму из
Иле, Кампен и др.)
Тазовый пояс млекопитающих
имеет при закладке типичное расположе-
ние элементов; особенностью его явля?т-
дополпительной кости в вертлужной ямке
(acetabulare), во-вторых, сращение всех этих костей с каждой сто-
роны в одну безыменную кость (innominatum) и сра-
щение между собой лобковой и седалищной костей, так что
образуется замкнутое со всех сторон отверстие между этими костями
(for. obturatum)(pnc. 544). У однопроходных и сумчатых к лобко-
вой кости снизу причлепяется особая кость для поддержания сумки
(marsupiale).
Скелет конечностей у исходных форм млекопитающих обу-
словлен их приспособлением к быстрым движениям — бегу при
помощи четырех конечностей, которые располагаются под туло-
вищем, так что последнее не висит между конечностями, как
у большинства рептилий, а непосредственно на них опирается.
Способность к бегу стоит в связи с иным положением конечностей
4 7 Кашкаров и Ставчввсквй
737
у млекопитающих. Именно у них конечности ориентированы так,
что локтевые суставы передних конечностей направлены назад,
а коленные сочленения задних — вперед. При движениях перед-
ние конечности выбрасываются вперед и притягивают туловище
сгибанием кистевого и локтевого суставов; задние конечности
отталкивают туловище вперед путем выпрямления ноги в колен-
ном суставе.
По количеству входящих в состав конечностей костей и по
их расположению скелет конечностей родоначальных форм млекопи-
тающих имеет очень типичное строе-
ние примитивной пятипалой конечно-
сти и подвергся большим изменениям
в пределах класса лишь в процессе
приспособления к различного рода
движениям в разных условиях среды.
В передней конечности
характерно наличие в дистальном
отделе плечевой кости отверстия (for.
entepicondylare), наличие проксималь-
ного выступа локтевой кости (olecra-
non), наличие гороховидной косточки
(pisiforme) и слияние четвертой и
пятой дистальпых карпальных косто-
чек (carpalia distalia 4+5 = hama-
tum, или uncinatum) и централь-
ной проксимальной с интермедиаль-
пой (centrale dist. + intermedium =
lunatum, или semilunare). В связи
с тем, что нередко употребляется
номенклатура косточек запястья мле-
копитающих, принятая в анатомии
человека, приводим таблицу этих названий в схеме, указы-
вающей расположение костей.
Рис. 544. Тазовый пояс кроли-
ка (Oryctolagus cuniculus) с ле-
вой стороны.
1 — подвздошная (ileum), 2 — се-
далищная (Ischium), 3 — лобковая
(pubis), 4 — вертлужная ямка (ace-
tabulum), 5 — for. obturatum, 6—
os acetabulare. (По Краузе из Иле,
Кампен и др.)
Pisiforme — гороховидная
Intermedium + centrale- Ulnare-
lunatum (semilunare) triquetrum (cuneiforme)
полулунная трехгранная
Radiale-
naviculare (scaphoideum)
ладьеобразная
Centrale — центральная
Carpale distale 1 Carpale distale 2 Carpale distale 3 Carpale distale
multangulum multangulum mi- capitatum (mag- 4-}-5hamatum
majus (trapezium) nus (trapezoideum) num) (uncinatum)
большая много- малая много- головчатая крючковатая
угольная угольная
Jletacarpale 1 Metacarpale 2 Metacarpale 3 Metacarpale 4 Metacarpale 5
I II III IV V
2 фаланги 3 фаланги 3 фаланги 3 фаланги 3 фаланги
Задняя конечность млекопитающих характеризуется
прежде всего тем, что повернута коленным сочленением впереди
738
Педро имеет хорошо развитые на проксимальной части отростки —
пгртлуги в числе 2—3 (trochanter major, minor и tertius); в голени
полыпеберцовая кость развита сильнее, имеется костная колен-
ная чашечка (patella), защищающая коленный сустав. В пред-
плюсне имеются только две проксимальных кости, хорошо
разлитые: внутренняя — т а р а н н а я (talus=astragalus), образо-
вавшаяся от слияния промежуточной и центральной проксималь-
ной (intermedium + centrale proximate), и наружная и я т о ч -
и а. я (calcaneus), соответствующая фибулярной косточке пред-
плюсны (fibulare). Из косточек среднего ряда центральная дисталь-
на я (centrale distale) превратилась в ладьеобразную (naviculare=
Hcaplioideum). Тибиальная косточка или редуцировалась или
приросла к этой последней.
Схема расположения косточек предплюсны и плюсны и сино-
нимика названий таковы:
I и I ermedium 4- centrale proximate =
talus (astragalus) таранная Fibulare =
Centrale distale= calcaneus — пяточная
naviculare (scaphoideum) ладьеобразная
Tarsal e 1 =
cuneiforme 1 —
клиновидная 1
Tarsale 2 =
cuneiforme 2 —
клиновидная 2
Tarsale 3 =
cuneiforme 3 —
клиновидная 3
Tarsale 4+ 5 =
cuboideum —
кубовидная
Metatarsale 1
I
2 фаланги
Metatarsale 2
II
3 фаланги
Metatarsale 3
III
3 фаланги
Metatarsale 4
IV
'3 фаланги
Metatarsale 5
V
8 фаланги
В связи с приспособлением к передвижению в различных средах
конечности млекопитающих подверглись весьма существенным
изменениям, причем эти изменения особенно хорошо можно про-
следить на скелете. К сожалению, палеонтологических данных
о строении конечностей родоначальников млекопитающих мы не
имеем. Несомненно, что у первых млекопитающих были мало
специализированные конечности описанного выше типа. Такие
именно конечности имеют из современных нам млекопитающих
наиболее примитивные сумчатые, например, опоссумы — сумча-
тые крысы (Didelphys), и насекомоядные, ведущие древесный
образ жизни. Однако было бы неправильно делать отсюда вывод,
что родоначальниками млекопитающих непременно были жители
лесов и притом лазавшие по деревьям формы. К тому же копечность
опоссума, хотя и служит для передвижения по сучкам деревьев,
является отнюдь не типичной специализированной древесной
конечностью, а сохраняет свой универсальный пятипалый тип.
Такие конечности могли лучше всего сохраниться у животных,
по только бегавших по земле, но и взбиравшихся на деревья, а
также у животных, лазавших по горам. Специализация конечностей
млекопитающих шла одновременно с дифференциацией отрядов и
других более мелких систематических единиц, причем имели место
конвергенции.
47? 739
. Особенно глубоким видоизменениям подвергаются конечности
при приспособлениях к полету, к плаванию в воде, к передвиже-
нию под землей; медленные движения по земле и древесный образ
жизни пе стоят в связи с глубокой специализацией.
У летучих мышей (Chiroptera) передняя конечность превра-
щена в крыло, причем летательная перепонка натягивается между
четырьмя (2—5) сильно удлиненными пальцами, лишенными когтей;
первый палец короткий и имеет на конце сильно развитый коготь;
этот палец не играет никакой роли при летании, служит для лаза-
ния; в запястье ладьеобразная, полулунная и центральная кос-
точки срастаются в одпу; предплечье удлинено и состоит из хорошо
развитой лучевой кости, тогда как локтевая кость редуцирована
и прирастает к лучевой. Лета-
тельная перепонка захватывает
Рис. 546. Передняя конечность си-
рены (В) и кита (Л).
Н — плечевая кость, В — лучевая, U —
локтевая, I — V — пальцы, с — centrale,
и — ulnare, I — intermedium. (По Абелю.)
Рис. 545. Скелет летучей мыши (Pte-
ropus edulis), врисованныйв контуры
тела.
1 — ключица (clavicula), 2 — лопатка (sca-
pula), 8 — плечевая кость (humerus), 4 —
лучевая (radius), 5 — локтевая (ulna),
6 — запястье (carpus), 7 — пястные кости
(metacarpus), 8— бедренная кость (femur),
9 — большеберцовая (tibia), 10—мало-
берцовая (fibula), И — предплюсна (tar-
sus), 12—плюсна (metatarsus), I — V—
пальцы по порядку, Chp — chiropatagium,
Pip — plaglopatagium, Prp — propatagium,
Urp — uropatagium. (Из Вебера.)
и задние ноги. Между послед-
ними она развита у рукокры-
лых различно (рис. 545).
У плавающих в воде млекопи-
тающих конечности превращают-
ся в плавники подобным же образом, как это мы уже видели у репти-
лий. Особенно сильно изменились при этом конечности у китов
и сирен. В противоположность крылу передняя конечность зна-
740
•iiiTr.ii 1,110 укорачивается в своей проксимальной части. У китов
ы>гти плеча и предплечья превратились в короткие и широкие
п.ппстиики, дающие место для прикрепления мощной мускулатуре.
Her отделы конечности неподвижно соединены друг с другом,
образуя прочное весло. При этом число фаланг может быть силь-
но увеличено. Те же изменения мы видели у рептилий. Увели-
Рис. 547. Скелет дюгони, врисованный в контуры животного; в скелете
черным врисованы головной и спинной мозг.
(Из Вебера, по Декслеру^и Егеру.)
черне числа фаланг зависит от того, что диафизы фаланг не сра-
стаются с эпифизами и окостеневают из отдельных центров. Такое
отдельное окостенение стоит в связи с замедленным процессом око-
стенения у этих животных. Замедленность процесса окостенения
ведет к тому, что большинство костей в местах соединения их
остается хрящевыми.
Плавники сирен (Sirenia) имеют с таковыми китообразных
лишь внешнее сходство: плечо и предплечье между собой сочлене-
ны, так что могут сгибаться; имеются почти все косточки запястья
и все пять пальцев с малоизмепенным числом фаланг (рис. 646).
Задние конечности у современных китов и сирен редуци-
рованы настолько, что от них остаются лишь остатки таза
(рис. 547 и 548).
Рис. 5у48. Скелет кита (Balaena mysticetus).
(Йа Вебера, по Эшрихту и Рейнгардту.)
Другой тип приспособления конечностей к передвижению в
воде мы видим у ластоногих (Pinnipedia). Здесь хвост почти исче-
зает, а задние ноги передвигаются назад, играя роль двигатель-
ного органа. Три семейства этого отряда представляют по степени
развития приспособленности к передвижению в воде как бы три сту-
пени (не генетически). У моржей и у ушастых тюленей (Otariidae)
задние конечности еще могут подгибаться вперед под туловище,
741
но у моржей в устройстве конечностей больше примитивности,
они более коротки и подвижны. У тюленей (Phocidae) задние ноги
уже не могут подгибаться под тело вперед, однако на пальцах
ног (ласта) еще сохраняются когти.
У роющих животных конечности подвергаются также весьма
сильным видоизменениям: особенно сильно изменились конечности
сумчатого крота (Notoryctes typhlops). У него на лопатке имеется
два гребня, как и у многих роющих животных, что стоит в связи
с сильным развитием трехглавой мышцы плеча; имеется ключица,
как у всех роющих млекопитающих. На плечевой кости имеется
очень сильный гребень, разветвляющийся вилкообразно и слу-
жащий для прикрепления сильных (необходимых при рытье) дель-
товидной и грудной мышц; сильно развит внутренний мыщелок
плеча благодаря сильному развитию мускулов сгибателей лок-
тевой кости (proc, olecranon), служащий местом прикрепления
трехглавой мышцы; запястье своеобразно изменено, становясь
очень прочным; 1-й и 2-й пальцы отодвинуты на ладонную сторону,
и для рытья служат лишь 3-й и 4-й, снабженные огромными ког-
тями, особенно 3-й; число фаланг у пальцев очень мало: 2 и 1.
5-е metacarpale с широким плоским когтем, что увеличивает рою-
щую поверхность; в плоскости ладони находится сесамовидная
косточка, к которой прикрепляется мускул, сгибающий паль-
цы, играющий большую роль при рытье; это увеличивает проч-
ность запястья; как и у других роющих форм, симфиз лоб-
ковых костей очень короток; седалищная кость срастается с попе-
речными отростками обоих последних крестцовых позвонков; на
бедре, колене и большеберцовой кости сильно развиты гребни
и выросты — места прикрепления мускулов; большеберцовая
и малоберцовая кости (tibia и fibula) прочно срастаются, как
у златокрота (Chrysochloris); подошва отворочена наружу, как и
у Chrysochloris; 5-й палец загпут на подошвенную поверхность.
5-й ноготь па ступне широк и лопатообразен, 4-й длинный и серпо-
видный, 3-й и 2-й становятся короче и шире; на грудной кости
сильно развитой гребепь, как у златокрота, слепыша (Spalax) и
крота (Talpa) (рис. 549).
Сильное развитие гребней на костях для прикрепления мускулов,
укорочение carpus, прочное соединение carpalia, редукция одних
фаланг и усиление других, играющих роль при рытье, — все эти
признаки свойственны роющим млекопитающим вообще. Но в
зависимости от того, в каком грунте животное роется, будут наблю-
даться различия. При рытье в мягком грунте (папример, у крота —
Talpa) (рис. 550) кисть сильно расширяется, она действует как
лопата и имеет наклонность к развитию 6-го пальца. Передняя
конечность крота сравнима с лопатой, служащей для рытья в мяг-
ком грунте. Наоборот, у тех форм, которые роются в твердом грун-
те, кисть становится уже, в рытье принимают участие лишь не-
многие пальцы, большею частью 3-й и 4-й, развивающиеся сильнее
других, как у Notoryctes и Chrysochloris. Эта кисть сравнима с
заступом для рытья в твердом грунте.
742
То же видим мы у млекопитающих, разрывающих твердые
постройки термитов, например, у муравьеда (Myrmecophagus),
а Ъ с d
Рис. 549. Передняя (а и Ь) и задняя (с и d) конечности сумчатого крота
(Notoryctes typhlops); а и d — с внутренней, b и с — с наружной стороны.
Обозначения в передней конечности: гс — radiale, uc — ulnare, тес — метакарпальные
кости, mt — карпальная, 1—5 — пальцы; в задней: tm — большой вертлуг, ast — та-
ранная, cat — пяточная, nav — ладьеобразная, met — метатарзальные косточки, mes—
тарзальная П. (Из Абеля по Карльсону.)
у тамапдуа (Tamandua). Эти животные ходят, опираясь на наруж-
ный край ступней, подогнув когти.
Приспособления млекопитающих к быстрому передвижению
по земле вызвали весьма существенные изменения в конечностях.
Передвижение по земле,
которое было свойственно
примитивным млекопитаю-
щим, осуществлялось пу-
тем опоры на всю кисть
и ступню; исходные фор-
мы млекопитающих были
первично стопоходящими
или плантиградными, т. е.
опирались при ходьбе на
всю ступню, как, напри-
мер, медведи. Но такое
строение допускает лишь
относительно медленное хо-
ждение в сравнении с бы-
строногими антилопами или
лошадьми, или прыгаю-
щими зайцами, кенгу-
ру, тушканчиками. В те-
чение филогенетического
Рис. 550. Правая передняя конечность кро-
та (Talpa europaea).
II — плечевая, R —лучевая, U —локтевая кость,
г — малая лучепястпая или ладьеобразная, 1 —
иптермедиальпая или полулунная, и — локте-
пястная или трехгранная, pi — гороховидная,
се — центральная, си са, с3 — карпальные ди-
стальные, I—V — пальцы. (Из Абеля.)
развития мы видим постепенное приподнятие над землей за-
пястья и предплюсны (carpus и tarsus,) за которыми затем
743
последовали пясть и плюсна, но фаланги еще касались земли цели-
ком. В конце концов, выпрямились и фаланги, передние и задние
конечности стали опираться на концы пальцев. Этапы этой эволю-
ции мы можем теперь проследить шаг за шагом не только у копыт-
ных, но и у хищных млекопитающих, приобретших способность >
к быстрому бегу. По мере выпрямления конечностей крайние пальцы
перестают соприкасаться с землей, перестают функционировать
и подвергаются атрофии. Получается меньшая поверхность со-
прикосновения с землей. Изменения в дистальной части конечно-
стей влекли за собой изменения в проксимальной: carpalia и tar-
salia располагаются все более плотно друг около друга, ulna и fibula
редуцируются, конечности начинают совершать движения лишь
в сагиттальной плоскости, срединные metapodia сливаются в одну
кость.
Рис. 551. Филогенетическое развитие передней конечности лошади. Северо-
американский ряд.
а — Eohippus pernix, Ъ — Orohippus agilis, с — Mesohippus celer, d — Mlohippus anceps,
e — Hypohippus equinus, f — Neohipparion Whitneyi, g — Protohippus pernix.
(Из Абеля.)
Два пути развития ведут к образованию конечностей современ-
ных копытных. В одном ряду ось конечности проходит в середине
третьего пальца и последний развивается наиболее сильно, осталь-
ные же становятся рудиментарными. Это — непарноко-
пытные (Perissodactyla), мезаксиальные (Mesaxonia).
У других ось проходит между третьим и четвертым пальцами,
развивающимися равномерно: парнокопытные (Artiodac-
tyla), параксиальные (Paraxonia). Боковые пальцы
редуцируются. Развитие непарнокопытных нагляднее всего видно
на филогенетическом развитии конечностей лошади (рис. 561).
Развитие парнокопытного состояния обусловлено было, между
прочим, тем, что ему предшествовала редукция первого паль-
ца, так что при переходе от стопохождения он почти не при-
744
intM.i.n никакого участия, тогда как у предков однокопытных
|i.'i:iiiiithc пятого пальца первоначально было нормальным.
Интересно, что и парнокопытпость и непарнокопытность воз-
никали в эволюционном процессе неоднократно и в некоторых слу-
чаях приводили к весьма специализированному типу, например,
\ южноамериканских Proterotheriidae (вымерших).
('охраняющиеся пальцы одеты копытами, которые могут диффе-
ренцироваться дальше в зависимости от образа жизни и место-
обитания. Так, у козлов, держащихся на скалах, копыта круты
н узки и снабжены острым краем; у северного оленя они очень
д линны, широко раздвоены и служат своего рода лыжами, не даю-
щими животному проваливаться в снег и болота.
Одновременно с пальцами при развитии хождения на паль-
цах, происходит изменение запястья (carpus) и предплюсны (tarsus).
Оно идет различно у парнокопытных и непарнокопытных в связи
с иным распределением силы тяжести. Наконец, в связи с пере-
ходом к пальцехождеиию мало-помалу уменьшается значение
локтевой кости (ulna) как опоры для плеча: она уменьшается
или сливается с лучевой (radius). Точно так же редуцируется и,
наконец, исчезает в задних конечностях малоберцовая кость
(fibula).
Значение и характер описанных изменений ясно видны из
сравнения конечности верблюда и слона (рис. 552). У слона кисть
(стопа) остается короткой, а удлинены плечо и предплечье (бедро
и голень). Наоборот, у верблюда удлиняются срастающиеся meta-
podia; плечо и предплечье (а также бедро и голень) остаются корот-
кими; Благодаря выпадению ряда костей происходит упрощение
скелета конечности, последняя становится более легкой. Редук-
ция пальцев ведет к меньшему трению, уменьшается опасность
повреждения, разрыва связок. Срастание metapodia создает боль-
шую прочность. Конечность слона гораздо тяжелее. Обилие костей
в кисти требует для получения устойчивости соединения всех
пальцев общей кожей. Конечность слона — это колонна боль-
шой прочности, но лишенная той легкости, которую имеет конеч-
ность верблюда.
Другим способом быстрого передвижения по земле является
прыгание. Это приспособление возникало неоднократно в исто-
рии млекопитающих и притом в самых разнообразных филоге-
нетических стволах: в ряду сумчатых — у кенгуру (Macropus),
среди насекомоядных — у прыгунчиков (Macroscelidae), и, наконец,
в разных рядах развития грызунов: сем. зайцы (Leporidae) из
Duplicidentata; в ряду дикобразоподобных (Hystricomorpha) раз-
вилась группа долгоногов (Pedetinae); в ряду мышеобразных
(Myomorpha) мы имеем австралийского тушканчика (Conilurus),
тушканчиков Старого Света (Jaculus, Alactaga), американских
прыгунчиков (Zapus) и мешетчатых прыгунов (Dipodomys).
Развитие способности к прыганию связано с удлинением задней
конечности, а именно костей плюсны (metatarsus). Кроме того,
при этом происходит потеря липших крайних пальцев, и кости
745
предплюсны соединяются между собой очень прочно. Примером
могут служить ноги земляного зайца (Alactaga) и настоящих туш-
канчиков (Dipus).
Передние ноги у Alactaga малы и коротки. Земляной заяц поль-
зуется ими при рытье, но не касается ими земли при прыгании.
Прыгание осуществляется лишь при помощи задних ног и даже
Рис. 552. Передние (а и Ь) и задние (с и d) конечности верблюда и слона.
Передние конечности:
Sc — лопатка (scapula), Н — плечевая кость (humerus), U — локтевая к. (ulna), R —луче-
вая к. (radius), С—запястье (carpus), Мс—пястье (metacarpus), Ph—фаланги пальцев.
Задние конечности:
В —таз (os innominatum), F —бедреная к. (femur), Т —большая берцовая к. (tibia),
Fib— малаяберцовая к. (fibula), Tars — предплюсна (tarsus), Mt —плюсна (metatarsus),
Ph —фаланги пальцев.
(Из Абеля.)
без помощи хвоста. Последний касается земли лишь при спокой-
ном положении, когда животное опирается на хвост. Скелет ноги
имеет ту особенность, что плюсневые кости 2-го, 3-го и 4-го паль-
цев сливаются в одну кость. Концы плюсневых костей внизу не
слиты, и срединная (3-й палец) выступает дальше вперед. Из соеди-
746
ценных с ними фаланг опять-таки средние наиболее длинны. 1-й
и b-й пальцы рудиментарны й не касаются земли. Оба эти пальца
имеют одинаковую длину благодаря тому, что плюсневая кость
большого пальца длиннее, нежели 5-го. Первая имеет 2, вторая—
з фаланги.
У Dipus специализация ушла еще дальше: 1-й палец исчез
|>ез следа, от 5-го сохраняется лишь рудимент. Средний палец
вдвое слабее, нежели два крайние, и лишь немного длиннее их.
Пальцы параллельны. Средний палец редуцируется потому, что
его функцию берут на себя 2-й и 4-й, на концы которых прихо-
дится упор при сидении и прыгании. В дальнейшей эволюции 3-й
палец должен исчезнуть: мы имеем у Dipus незаконченный тип
приспособления.
3. Мускулатура
Особенности мускулатуры млекопитающих по сравнению с
рептилиями обусловливаются характером производимых ими дви-
жений. Все движения млекопитающих гораздо сложнее и со-
вершеннее в смысле соответствия функции задачам ее, нежели
движения рептилий. Сложнее действие челюстного аппарата, слож-
нее движения головы и шеи, сложнее движения аппарата дыха-
пия, движения туловища, движения конечностей, в особенности
в связи с быстрым движением по земле и с древесным образом
жизни.
Поэтому мускулатура млекопитающих, как соматическая, так
и висцеральная, гораздо сложнее и дифференцированнее муску-
латуры рептилий, хотя от последней она происходит филогене-
тически, и источник развития каждой мышцы млекопитающих
может быть найден у рептилий.
На голове некоторому изменению подверглась висцеральная
мускулатура. Особенного развития достигают у млекопитающих
жевательные мышцы. У хищных и насекомоядных сильно
развиты только височные мышцы (m. temporalis),
цак служащие для простого приведения нижней челюсти. При
сильном своем развитии они вызывают образование на черепе,
в теменной и затылочной областях гребней (crista sagit-
talis и crista occipitalis). У грызунов и у копытных в связи с энергич-
ным пережевыванием пищи большего развития достигает собственно
жевательная мышца (m. masseter), которая не только
поднимает (приводит) нижнюю челюсть, но и толкает ее сзади
наперед; мышца эта начинается от скуловой дуги и прикрепляется
к углу нижней челюсти (рис. 553).
Жевательные боковые движения совершаются с помощью двух
крыловидных мышц с каждой стороны — внутрен-
ней (m. pterygoideus externus), начинающейся от крыловидного
углубления крыловидной кости (отростка) и идущей к внутрен-
ней поверхности угла нижней челюсти, и наружной (m. pte-
rygoideus externus), начинающейся сбоку от только что упомянутой
и идущей к сочленовному отростку нижней челюсти.
747
м
а т е л я шеи
553. Фронталь-
разреэ через че-
Рис.
ный
реп человека в височ-
ной области.
Мышцы: 1 — височная
мышца (musculua tempo-
ralis) , 2 — жевательная
мышца (m. masseter), 3—
крыловидная внутрен-
няя (m. pterygoldeus in-
ternus), 4 — крыловид-
ная внутренняя (m. pte-
rygoideus externus), по-
следняя в поперечном
разрезе. Черным изоб-
ражены кссти: 5—'Ску-
ловая дуга,6 — нижняя
челюсть. (Из Вебера по
Меркелю.)
Эти мышцы иннервируются тройничным нервом (V). Опускается
нижняя челюсть помимо тяжести самой кости еще деятельностью
подбородочно-подъязычной мышцы (ш. mylohyoi-
deus), отделившейся от межчелюстной мышцы, и двубрюш-
ной мышцы (m. digastricus), также иннервируемых тройнич-
ным нервом (V) и отчасти принадлежащих системе лицевого
нерва (VII).
Из мышц системы лицевого нерва (VII) на шее из общего с ж и-
(m. sphincter colli) обособляются подкож-
ная мышца шеи (platysma myoides) и
ряд мышц, образующих сложную систему
подкожной мускулатуры лица, или
м и м и ч е с к у ю мускулатуру обезьян и
человека. Таковы мышцы наружного слу-
хового прохода, мышцы глазной щели, мыш-
цы ротовой щели, мышцы носа и некоторые
Другие.
Мышцы системы языкоглоточного (IX) и
блуждающего нерва (X), т. е. мускулы жа-
берного аппарата, сохраняются лишь в виде
мышц подъязычного и глотательного аппа-
ратов; зато большого развития достигают
трапециевидные мышцы. Передняя часть этих
мышц делается самостоятельной грудино-
к л ю ч и" ч н о - с о с ц е в и д н о й мышцей
(m. sternocleidomastoideus), иннервируемой
дополнительным (XI) нервом; собственно
трапециевидная мышца (m. tra-
pezius) идет от головы и позвоночного стол-
ба к акромиальному отростку и гребпю ло-
патки; иннервируется n. accessorius (XI).
Из соматических мышц головы у млекопи-
тающих сохраняются в том же виде, как и у
нижестоящих форм, прямые мышцы глаза
и н и ж н я я косая, тогда как верх-
няя косая мышца (m. obliquus supe-
rior, IV) своим прикреплением уходит в
глубь глазницы и перекидывается через блок
у края глазницы; имеется мышца, втягиваю-
щая глазное яблоко (m. retractor bullbi, VI),
а от верхней прямой мышцы обособляется
мышца, поднимающая верхнее веко (m. levator
palpebrae superioris, III).
Соматические мышцы подъязычного аппа-
рата ничем существенным не отличаются у
млекопитающих: имеются, кроме груди-
о й мышцы (m. sternohyoideus), л о п а т о ч-
но-подъязычн
но-поДъязычной мышцы (m. omohyoideus) еще несколько
мышц языка — п одбородочн о-я зычная мышца(т. genio-
748
tflossus, XII) для выдвигания и подъязычно-язычная
мышца (m. hyoglossus, XII) для втягивания языка.
Соматические мышцы туловища в своей дорзальной (эпакси-
.гп.ной) части ограничиваются областью позвоночника, испол-
ни н задачу сгибания позвоночника и его фиксации в определенном
положении. Их имеется много, но они могут быть объединены в сле-
дующие системы: мышцы, соединяющие остистые отростки (mm.
int.erspinales), мышцы, протягивающиеся между поперечными отро-
стками (mm. intertransversales), мышцы, протягивающиеся между
поперечными и остистыми отростками (mm. transversospinales);
от поперечных отростков к ребрам идут поперечнореберные
мышцы (mm. transversocostales), вдоль спины тянется система длин-
ной мышцы спины (m. longissimus dorsi); к голове от позвонков
спины идут прямые и косые мышцы головы (mm. recti
capitis и obliqui capitis). В задней части туловища от подвздошной
кости к ребрам идут подвздошно-реберные мышцы
(il cocos tales) и, наконец, хвостовая система мышц.
Вентральные мышцы (гипаксиальпые) туловища образуют на
ребрах внутренние и наружные межреберные
и зубчатые мышцы (mm.intercostales interni и extend и m. ser-
rati); они являются продолжением косых брюшных мышц
(mm. obliqui interni и extern!), и еще дальше к заду — квадрат-
ной поясничной (m. quadratus lumborum), тогда как их
продолжением вперед является группа лестничных мышц
(mm. scaleni); на брюшной стороне располагаются посередине п р я-
м ы е мышцы живота (mm. recti abdominis) а по бокам от
внутренних поверхностей ребер идут поперечные мышцы
живота (mm. transversi abdominis). У сумчатых имеется еще
пирамидальная мышца (m. pyramidalis), отходящая от
сумчатых костей и идущая вперед до грудины.
Чрезвычайно характерной для млекопитающих является муску-
листая перегородка, отделяющая грудную полость от брюшной,
развитие которой связано с приобретением энергичных и очень
совершенных дыхательных движений, а это в свою очередь ^мело
большое значение для приобретения теплокровности и общего по-
вышения жизнеспособности.
Что касается мускулатуры передних конечностей, то, помимо
упомянутых уже висцеральных мышц — трапециевидной и груди-
но-ключично-сосцевидной, — ее приводит в движение около 60
тчышц. Хорошо развиты в плечевом поясе ш и ро к и е мышцы
спины (m. latissimus dorsi), возникает из дельтовидной
мышцы (m. deltoideus) круглая малая мышца (т. teres
minor), обособляются из грудной большая и малая мыш-
цы (т. pectoralis major и minor). Мышцы свободной конечности,
расположенные со спинной стороны, являются разгибателями
(extensores) и вращающими конечность наружу (supinatores), а
лежащие с брюшной стороны являются сгибателями (flexores) и
поворачивающими кисть внутрь (pronatores). Из разгибателей упо-
мянем трехглавую мышцу (m. triceps brachii), из сгибателей—
749
двуглавую мышцу плеч (m. biceps brachii), которая также
является супинатором, и плечевую мышцу (m. brachialis);
иа предплечье располагаются сгибатели и разгибатели кисти, паль-
цев и пронаторы и супинаторы предплечья.
Так как тазовый пояс у млекопитающих срастается с позвоноч-
ником, то у них, естественно, отсутствует мускулатура, приводя-
щая в движение тазовые кости, однако имеются мускулы туловища,
которые, как мы видели, прикрепляются к тазу. Из мускулов, при-
водящих в движение бедро, упомянем широкий мускул бедра
(m. vastus), лежащий спереди, прикрепляющийся к чашечке и за-
тем прикрепляющийся к больше-берцовой кости; позади распола-
гается группа ягодичных мышц (mm. glutei). Из мышц
бедра упомянем двуглавую мышцу бедра (m. biceps femoris),
полусухожильную мышцу (m. semitendinosus), полу-
перепончатую мышцу (m. semimembranosus); с внутрен-
ней стороны помещаются приводящие голень мышцы: тон-
кая (m. gracilis), гребешковая (т. pectineus) и порт-
няжная (т. sartorius). На голени располагаются сгибатели и
разгибатели плюсны и пальцев; упомянем очень важную икро-
ножную мышцу (m. gastrocnemius), оканчивающуюся так
называемым ахилловым сухожилием.
Само собой понятно, что развитие одних и редукция других
мышц стоят в связи с различного рода приспособлениями в движе-
ниях.
4. Органы пищеварения
Органы пищеварения млекопитающих подверглись в процессе
эволюции весьма большим изменениям в связи с приспособления-
ми к чрезвычайно разнообразной пище.
Уже на характере ротовой щели отражается приспособление к
захватыванию той пищи, которой питается данное млекопитающее;
например, пасть летучих мышей, ловящих насекомых на лету, не-
обыкновенно широка. Здесь мы имеем полную параллель с устрой-
ством рта у птиц: ласточек, стрижей и козодоев, ловящих насекомых
на лету. Язык летучих мышей снабжен большими роговыми шипа-
ми, служащими для удержания попавших в рот насекомых.
Колоссально развита пасть у китов; благодаря узости своей
глотки они могут глотать лишь мелких животных, не крупнее сель-
ди, а вследствие своих громадных размеров должны поглощать их
в огромном количестве. У зубатых китов конические зубы являются
еще весьма несовершенным ситом, напоминающим сито на клюве
уток; у беззубых китов, питающихся планктоном, роль сита выпол-
няет столь известный китовый ус — роговые пластинки, развиваю-
щиеся из слизистой оболочки рта и соответствующие нёбным склад-
кам других млекопитающих.
Полную противоположность представляет рот и язык питающихся
муравьями и термитами млекопитающих: ехидны, муравьеда, ящера.
У них сильно удлиняются обе челюсти: будучи до самого перед-
него конца одеты кожей, они образуют длинную трубу с маленьким
750 •
u nil']и’'г|щм впереди, через которое высовывается червеобразный
hi и к. У новорожденных млекопитающих также имеются изменения
приспособительного к питанию характера. А именно мы видим у
мо'и।дых млекопитающих сильное укорочение челюстей, что связа-
..... актом сосания. У сумчатых детеныш родится слабым и беспо-
м> и иным н присасывается к соску. Сосок во рту разбухает, края
|и.| срастаются, и детеныш таким образом крепко повисает на соске.
JI, hi того чтобы разбухший сосок умещался во рту, на верхней сто-
liiiiH1 языка детеныша имеется углубление; эпителий стенок рта сра-
ГГЗ.|'ТСН с соском.
Приспособление к роду пищи выражается, далее, в строении
IV". У молотоголового крылана (Hypsignathus monstrosus) рот
ограничен очепь широкими отвислыми губами, замечательно при-
способленными к тому, чтобы высасывать сочное содержимое фиг
и других мягких плодов, служащих им пищей. У многих форм губы,
соответственным образом изменяясь, играют роль при щипании
ip.'iin.i и обрывании ветвей; например, губы очепь подвижны у
л ни;идей; при помощи подвижных и мясистых губи языка коровы
и и ил ют траву; у жирафы, которая питается ветвями, губы подвижны,
и заострены, являясь хватательным органом. Длинные губы верб-
iiii,i.a играют важную роль при захватывании пищи.
Коли вместе с губой удлиняется и нос, то развивается хобот,
но лее короткий у тапиров и предков современных слонов и более
। । пи цый у последних. С развитием такого органа, как хобот, связа-
но развитие соответственно сильной мускулатуры, с чем в свою оче-
pe.li, связано развитие черепа, построенного из пористых костей
hale места прикрепления мускулатуры хобота. В некоторых случаях
разрастается преддверие, т. е. полость между губами и зубами,
uieitoii и зубами; оно выстлано бывает не настоящей слизистой обо-
лочкой, а более тонким эпителием кожи и снабжено иногда приспо-
соблениями к механическому удержанию или обработке пищи, а
также чувствующими окончаниями. К преддверию рта относятся
также защечные мешки, встречающиеся у отдельных сумчатых,
очень многих грызунов иу большинства обезьян Нового Света, слу-
жащие для временного сохранения или переноса запасов пищи.
Иногда они достигают чрезвычайной длины; например, у хо-
ти ко в они простираются до груди и втягиваются внутрь с помощью
особого мускула.
У мешотчатых крыс (Geomyidae) защечные мешки являются
л о ж н ы м и мешка м и, так как они не являются продолже-
нием преддверия рта, а являются карманами наружной кожи.
Рот ограничен сверху твердым нёбом, на котором развиваются
расположенные поперек нёбные пластинки, нередко
ороговевающие, у сирен и китов сильно разрастающиеся (китовый
ус); закапчивается твердое нёбо свешивающимся вниз мягким нёбом.
В задней части ротовой полости от дна ее отходит мускулистый
язык, чрезвычайно богато снабженный железами и сосочками раз-
ной величины (вальковатые, нитевидные, грибовидные по поверх-
ности и листовидные у основания). Величина и форма языка весьма
751
различны. Он очень подвижен у жвачных и играет важную роль
при захватывании пищи. Особенно длинный и тонкий язык у питаю-
щихся муравьями и термитами животных: он здесь смачивается лип-
ким выделением подъязычных и подчелюстных желез, благодаря
чему к нему прилипают муравьи и термиты, и снабжен необыкно-
венно сильно развитой мускулатурой. Такой же язык мы встречаем
у замечательного сумчатого пяткохода (Tarsipes rostratus), име-
ющего длинную острую морду и весьма растяжимый червеобразный
язык. Пяткоход питается насекомыми цветов. В поисках за ними он
засовывает в цветы свой длинный язык, слегка зазубренный по
краям. Такое устройство языка встречается и у некоторых птиц,
питающихся подобным же образом.
У целого ряда млекопитающих сильно ороговевший язык играет
за отсутствием редуцированных зубов роль перетирающего пищу
органа, а именно: у ехидны, утконоса, плодоядных крупных летучих
мышей (Macrochiroptera).
Подвижной язык медведей (Ursus) служит для схватывания насе-
комых, срывания травы и ягод и добывания меда; язык ленивца
(Bradypus), питающегося листьями, ложкообразно складывающийся
язык собаки, которым последняя подбрасывает воду в рот, также
являются приспособлением в связи с родом пищи.
Ротовые железы у млекопитающих хорошо развиты и спе-
циализированы. Многочисленны мелкие железы: язычные (glandu-
lae linguales), пёбпые (gl. palatinae), щечные (gl. buccales), губные
(gl. labiales), у копытных сверх того молярные железы (gl. mola-
res), тогда как у собак и зайцев — глазничные железы (gl. orbita-
les), выделяющие слизь. Ряд крупных желез носит общее назва-
ние слюнных желез. Различают с каждой стороны слизистую—
подъязычную (gl. sublingualis), лежащую в углу между ветвями
нижней челюсти, и заднеязычную (gl. retro-lingualis); смешанную
слизисто-серозную подчелюстную (gl. submaxillaris), лежащую с вну-
тренней стороны и позади нижней челюсти, открывающуюся вар-
тоновым протоком под языком позади резцов, и чисто серозную
околоушную (gl. parotis). Эта последняя имеет длинный выводной
стенонов проток, открывающийся у коренных зубов верхней
челюсти (рис. 554).
Сильное развитие и специализация ротовых желез млекопи-
тающих стоят в связи с большим разнообразием их пищи, и яв-
ляется одним из условий хорошего усвоения последней, что
повышает общую активность этого класса.
Зубы. Эволюция млекопитающих шла как по пути приспособ-
ления к различным средам обитания, так и по пути приспособления
к разного рода пище, причем оба эти пути имели место иногда в
одной и той же систематической группе: или при жизни в одной среде
шла дифференциация по путям приспособления к разнообразной пи-
ще, или при одинаковой пище животные приспособлялись к разным
средам. При приспособлении к разного рода пище большое значение
имели зубы. Зубы млекопитающих, как правило, дифиодонтны,
т. е. являются в двух генерациях (молочные зубы и постоянные),
752
Рис.
554. Железы головы собаки.
1 — околоушная железа (glandula parotis), 2 — подче-
люстная (gl. submaxillaris), 3 и 3' — подъязычная (gl.
sublingualis), 4 — вартонов проток подчелюстной же-
лезы (ductus Whartonianus), 5 — бартолияиев проток
подъязычной железы (ductus Bartholin!), 6 — глазнич-
ная железа (gl. orbitalis). (По Эленбергу и Бауму из
Вебера.)
и гетеро доп.тны, т. е. состоят из различно построенных
компонентов: резцов, сидящих на межчелюстной кости, к л ы-
к о в, расположенных позади резцов, и коренных. Последние,
и зависимости от того, имеют они молочную генерацию или нет,
называются ложнокорепными или коренными. Последние расположе-
ны в заднем конце зубного ряда и не имеют молочной геперации.
Зубы сидят всегда в альвеолах, т. е. тек о д о н т н ы, на меж-
челюстных и челюстных костях. Зуб, как известно, состоит из корон-
ки и корня. Эти зубы имеют ограниченный рост. Но есть еще зубы,
лишенные корней, притом совсем или отчасти, и лишенные эмали.
Эти зубы имеют неограниченный рост. Эмаль в таком случае или
сидит на одной сторо-
не зуба (резцы грызу-
нов) или на конце зу-
ба в виде шапочки,
скоро стираясь (бивни
слонов).
В связи с измене-
нием пищи изменя-
лись и зубы: их строе-
ние, размеры и число.
Исходным числом для
зубов млекопитающих^
считается 60. Эти
шестьдесят зубов рас-
пределены были по
следующей формуле
(для каждой полови-
ны челюстного аппа-
рата):
т 1-2-3-4-5 р £
1 1-2-3-4-5 b 1
pi-2-3-4 м1-2.3-4-5
1 1-2-3-4 М 1-2-3.4-5
1.2-3.4.5
I—Incisivi — резцы, С—Canini — клыки, Р—Praemolares — лож-
нокоренные, М—Molares — коренные; числа, стоящие над чертой,
показывают зубы или их число в верхней челюсти, числа, стоящие
под чертой, — зубы нижней челюсти.
В разных группах млекопитающих число зубов менялось, вы-
падали те или иные зубы; эти изменения всегда могут быть выражены
в формуле.
В большинстве случаев постоянным зубам предшествуют зубы
молочные. При этом число зубов, подвергающихся смене, может
быть различно. Смены может и не быть, например, у крысы. Но это
уже явление вторичной редукции.
Исходное число молочных зубов 32, а именно:
тл 12 3 1 pi 12 3 4
IdT-2T CdT Pdl 2 3 4
(d — decidous — выпадающий).
48 Кашкаров я Станчинский
753
При смене зубов зуб заменяется другим, вырастающим под ним.
Глубокие различия в строении зубов возникают при изменении
рода пищи. Поэтому зубы являются лучшим диагностическим при-
знаком для распознавания форм млекопитающих.
Исходной формой для млекопитающих являются рептилии. У
последних зубы конические с одной вершиной — гаплодонт-
ная стадия. Они стоят в верхнем и нижнем ряду, чередуясь
(рис. 555,1). Эти простые зубы усложняются у примитивных триасо-
вых млекопитающих благодаря тому, что к каждому главном конусу
(protoconus) прибавляются еще два добавочных конуса — передний
и задний: в верхней челюсти — paraconus и metaconus, а в нижней —
Рис. 555. Эволюция зубов млекопитающих.
1 — гаплодонтный зуб рептилий, 2 — протодонтпый зуб Dromatherium, 3 — трикопо-
донтный зуб Microdonodon, 4 — тригонодонтный, или тритуберкулярный, зуб Spa-,
lacotherium, 5 — туберкулосекториальный зуб Amphitherium, 6 - зубы Miacis сбоку,
7 — сверху, 8 — тритуберкулярпые коренные в верхней (С) и нижней (А) челюсти.
(По Осборну из Вебера.)
paraconid и metaconid. Таким образом, возникает первая прото-
донтная стадия (рис. 555,2), затем с увеличением добавоч-
ных конусов следующая — вторая триконодонтная ста-
дия (рис. 555,3). Так как верхний ряд зубов и нижний располага-
лись не вертикально один под другим, а нижний несколько кнутри
от верхнего, то произошло смещение конусов : добавочные бугорки
верхнего ряда :— paraconus и metaconus — сдвинулись кнаружи,
бугорки зубов нижнего — paraconid и metaconid — кнутри. Такие
трехбугорчатые, или тритуберкулярпые, зубы
(рис. 555,3) господствуют у млекопитающих с мела по эоцен. Из
современных млекопитающих они сохраняются частично и лишь
у немногих насекомоядных, — например, у златокрота (Chysochlo-
ris), у двуутробки (Didelphys) и у лемуров (Lemuridae).
Такая форма зубов пригодна, главным образом, для размель-
чения насекомых с твердым хитиновым панцырем. Ее дальнейшим
754
усовершенствованием является соединение бугорков режущими греб-
пями, так что получается форма треугольника (trigon, trigonid).
Устроенные таким образом зубы называют тригонодонт-
п ы ми (рис. 556d).
Дальнейшее развитие форм зуба заключается в развитии у ниж-
него зуба придатка или пятки (talon, talonid) сперва с одним бугор-
ком (hypoconus, hypoconid). Такую форму мы встречаем в хищ-
ных зубах у многих хищников, например, у виверр (Viverridae).
С
л VAVAV
vava '
Рис. 5Ь6. Схема эволюции зубов.
а — гаплодонтная стадия, Ь — протодонтпая стадия, с — трикоподонтная
стадия, d — тригонодонтная стадия, в — переход зубов нижней челюсти
к туберкулосекториальной стадии, f — в нижней челюсти туберкуло-сек-
ториальные зубы, на талониде гипоконид, в верхней — тригонодонтные»
д — на пятке нижней челюсти образуются дополнительные бугорки, Л — Ъ
верхней и нижней челюсти туберкуло-секториальные зубы, i — переход
к четырехбугорчатым зубам, j — четырехбугорчатые зубы. (По Осборну.)
Ото бугорчато-режущие зубы — туберкуло-секториаль-
ные (секодонтные). Пятка становится больше и может нести
еще добавочные бугорки. Вслед за нижними коренными развитие
пятки наблюдается и у верхних зубов. Затем острия становятся срав-
нительно маленькими и округленными бугорками, крона теряет
свою треугольную форму и становится более или менее квадратной
с четырьмя или шестью бугорками. Из такой основной формы раз-
виваются коренные зубы большинства травоядных животных, хищ-
ных и приматов, причем развитие зубов того или иного типа проис-
ходит в связи с тем или иным способом питания. (См. схему, рис. ББ6).
48* 755
У большинства хищных коренные зубы построены по принципу
ломающих ножниц. Но путь к разрешению проблемы таких нож-
ниц весьма различен и неодинаков у чисто плотоядных форм, как
например, у куниц (Mustelidae) и более всеядных, как, например,
медведей (Ursidae).
Роль таких ломающих ножниц выполняют преимущественно
два зуба: расположенный впереди последний ложнокоренной зуб
верхней челюсти и лежащий соответственно несколько позади ко-
ренной зуб нижней челюсти Это так называемые хищные зубы.
У собаки в передней половине челюсти зубы верхнего и ниж-
него рядов чередуются, образуя ветви ножниц; в задней они при-
ходятся друг против друга, действуя как части пресса. Передняя
часть зубного ряда служит, таким образом, для разрыва мяса,
задняя — для раздавливания растительной пищи. Их расположе-
ние соответствует механическим принципам. У питающегося исклю-
чительно мясом красного волка (Суоп) коренные зубы развиты слабо,
и последний коренной в нижней челюсти даже вовсе утерян; пятка
нижнего хищного зуба также утеряна. Наоборот, южноамерикан-
ский гривистый волк (Canis jubatus), охотно поедающий сахарный
тростник и другие подобные растения, имеет очень широкие корен-
ные зубы.
У бурых медведей, в питании которых растительная пища играет
большую роль, коренные зубы развиты сильно, хищный зуб раз-
вит незначительно; остальные ложнокоренные развиты очень
слабо. У более хищного белого медведя коренные зубы развиты
меньше. У целого ряда млекопитающих, питающихся растительной
пищей, мы видим усиление перетирающих пищу коренных зубов.
Другое дело хищники, питающиеся, главным образом, мясом,
папример, кошки и гиены (Felidae, Hyaenidae). Служащие для пере-
тирания растительной пищи зубы исчезают у них в нижней челюсти
совершенно. Исчезает и пятка (talon) хищного зуба. Лишь в верх-
ней челюсти сохраняется рудиментарный, негодный к употребле-
нию коренной зуб. Наоборот, хищный зуб верхней челюсти стано-
вится длинным узким клином, как и впереди него лежащий ложно-
коренной; то же и в нижней челюсти. Благодаря этому у кошек обе
челюсти действуют, как половины ножниц; у гиен, которые дробят
кости, зубы несколько массивнее.
К млекопитающим, питающимся животной пищей, относится и
семейство куниц (Mustelidae). Идя по пути приспособления в этом
направлении, они утеряли два верхних и нижний коренные, но, дойдя
до этой стадии, многие из них вновь стали приспособляться ко все
более и более растительной пище. Для пережевывания раститель-
ной пищи они должны были использовать последний из оставшихся
верхних коренных и пятку нижнего хищного зуба. Особенно далеко
пошли в этом отношении барсуки (Meles).
У выдр (Lutra), питающихся исключительно животной пищей,
мы видим однако, значительное расширение верхнего хищного зуба
и уплощение всех бугорков. Такая особенность, говорящая как
бы о растительной пище, связана с тем, что выдра, кроме рыбы,
756
сст моллюсков и раков, а эти животные с твердой раковиной раздав-
ливаются как раз там, где развивается наибольшая сила: назади,
вблизи сочленения нижней челюсти. У капской выдры (Lutra сареп-
sis), живущей преимущественно за счет моллюсков, эта особенность
выработалась в борьбе за существование еще сильнее.
Таким образом, уже
в одном отряде хищ-
ных животных, плото-
ядных по преимуще-
ству, мы имеем большое
разнообразие в строе-
нии зубов, стоящее в свя-
зи с теми или иными при-
способлениями к пище.
На основе такого при-
способления происходи-
ла и эволюция хищных
зубов. У насекомояд-
ных, являющихся родо-
начальниками хищных,
третий коренной зуб
верхней челюсти лежит
так далеко назади, что
в этом направлении он
уже не может увеличи-
ваться. Увеличению его
в переднем направлении
мешают другие зубы. Ме-
жду тем, для того чтобы
ряды зубов могли рабо-
тать, как ножницы, необ-
ходимо, чтобы самый
сильный зуб лежал в че-
люсти возможно дальше
назад. Поэтому давящие
ножницы не могли осу-
ществиться при помощи
заднего коренного. Это
могдо быть осуществлено
Рис. 557. Три различных пути развития зу-
бов хищных млекопитающих:
(Pseudocreodi) а — Hyaenodon, b—Oxyaena, с —
Eucroedi: — волк (Canis lupus). (По Абелю.)
путем увеличения второ-
го верхнего коренного,
а для этого должен был
исчезнуть задний коренной. Еще лучше достигается то же приспособ-
ление с помощью первого коренного, развивающегося за счет 2-го
и 3-го. То же и в нижней челюсти. Но так как в нижней челюсти зубы
стоят на один зуб впереди по сравнению с верхней челюстью, то при
увеличении в пижпей челюсти 2-го коренного в верхней существенно
должен был увеличиваться 1-й коренной; при увеличении в нижней
челюсти 1-го коренного в верхней увеличивается задний (4-й) лож-
757
покоренной. Таким образом, развитие хищных зубов могло итти
тремя путями:1
М- М-; М - М-5-; Р- Мт.
о а 1
До настоящего времени дожили лишь те хищники, у которых
эволюция шла последним путем, остальные вымерли (рис. 557).
Еще большего разнообразия в связи с пищей достигают зубы
копытных. Такого разнообразия нет ни в одном другом отряде, —
и оно проявляется не только в различной форме и в относительной
величине зубов, но и в деталях строения коронки. Происходя от
разных плотоядных предков, копытные прошли стадию всеядности
и перешли к питанию растительной пищей. На зубах ясно отражена
эта эволюция. Сперва коренные зубы имели низкие кроны с немно-
гими тупыми бугорками на жующей поверхности. В связи с пере-
ходом к питанию мягкой и сочной или наоборот, более твердой,
соломообразной, растительной пищей коренные зубы стали изме-
няться в различных направлениях. Мы различаем следующие типы
в строении зубов копытных (рис. 558).
a bed е f
Рис. 558. Различные типы коренных зубов копытных.
а — бунодонтный, Ь—селенодонтный, с — лофодонтный типы; d — буноселенодонт-
ный, е — лофобунодонтный и/*—лофоселенодонтный смешанные типы.
1. Бунодонтный тип. Зубы с бугорками. Сперва бугор-
ков немного, затем их становится больше. Исходной для других
формой являются зубы с немногими бугорками. Такие зубы мы
встречаем у гиппопотама, у свиней, у которых мы видим зубы с 4
бугорками и зубы очень сложной формы со многими добавочными и
промежуточными бугорками. Такие же зубы имеются у приматов:
это зубы всеядных.
2. Лофодонтный тип. Зубы с поперечными или косо
стоящими гребнями, происходящими от слияния бугорков. Здесь
опять такие зубы могут быть с 2—3 (олиголофодонтные зубы) или
со многими гребнями (полилофодонтные). Первые мы встречаем у
носорогов, вторые — у слонов. Эволюция зубов последних просле-
жена хорошо и стоит в связи с переходом к питанию более грубой
пищей, как, например, ветками кустов и деревьев.
3. Селенодонтныйтип. Бугорки принимают форму бук-
вы V, так что мы имеем два V-образных наружных бугорка и, как
правило, два таких же внутренних, постепенно принимающих по-
лулунную форму. Такие зубы мы встречаем у оленей.
1 Цифра над чертой означает порядковый номер зуба верхней челюсти,
стоящая под чертой — нижней.
758
Кроме указанных трех главных типов, имеется еще целый ряд
форм переходных, встречаемых частью у ископаемых, частью у со-
временных форм: буноселенодонтный — с бугорками и
полулуниями, лофобунодонтный — с бугорками и греб-
нями, лофоселенодонтный—с гребнями и полулу-
ниями.
Все эти типы сводятся, в конечном счете, к четырехбугорчатым
зубам плотоядных. Кроме изменения в строении поверхности кроны,
переход от мягкой пищи к более твердой сопровождается еще изме-
нением низкой кроны в высокую. Этот переход от «брахиодонтии»
к «гипселодонтии» совершается независимо и параллельно в раз-
личных группах.
Рассмотрим вкратце связь той или иной формы зубов копытных
с родом пищи. Мягкой растительной пищей питается гиппопотам.
Его пищу составляет прежде всего лотос, а затем ряд других водя-
ных растений, как папирус. Жует гиппопотам медленно и долго.
Благодаря этому пища не требует особой спо-
собности к сопротивлению со стороны зубов,
и кроны коренных зубов снабжены 4 бугор-
ками, стирающимися при жевании. Кроме
коренных зубов, гиппопотам имеет 2 резца, 1
клык и 4 ложнокоренных в каждой челюсти:
Цс1рАм|.
Совсем иные зубы видим мы у лошади
(рис. 559). У нее имеется три резца, клыки
очень малы, первый ложнокоренной находит-
ся на пути к исчезновению. Форма остальных
трех ложнокорепных и коренных в каждой
челюсти одна и та же, кроме последнего ко-
ренного:
4
Рис. 559. Стадия раз-
вития гипселодонтии
у лошадей.
а — брахидонтный ко-
ренной зуб Anchithe-
rium, Ъ — гипселодонт-
ный зуб Hippotherium,
с — призматический зуб
лошади. (По Годри, из
Прежде всего следует' отметить очень высокую крону и рудимен-
тарные корни. Бугорки видны на кронах лишь не работавших еще
зубов и с наступлением жевания исчезают. Поверхность кроны ста-
новится тогда очень сложной, покрытой складками эмали (стенки
стертых бугорков), между которыми находится цемент. Зубы лошади
настолько отличаются, что с первого взгляда трудно предполо-
жить, что они произошли от зубов, сходных с зубами гиппопотама.
Однако палеонтология ясно свидетельствует нам о том, что копытные
с высокими зубными кронами произошли от форм с низкими кронами.
Особенности зубов лошадей связаны с питанием твердыми злаками,
что стоит в связи с их жизнью в степях — коренном местообитании
лошадей. Приспособления зубов к пережевыванию жестких злаков
совершенно иные, нежели приспособления к пережевыванию
мягких болотных растений.
759
Elephas 1/д
Современ. период
Плейстоцен
Stegodon Va
Нижний плиоцен
Верхний миоцен
Средний миоцен
Нижний миоцен
Верхний плиоцен
Elephas
I Mastodon
' Gomphotherium
Gomphotherium
Верхний олигоцен
Нижний олигоцен^
Верхний эоцен
Средний эоцен
Нижний эоцен
Рис. 560. Эволюция хоботных.
Направо черепа, налево зубы — именно, последний нижний коренной дуб. (Иа
Абеля.)
760
Еще большее сопротивление оказывают зубы при пережевыва-
нии такой грубой пищи, как ветви растений. Тогда развиваются
такие сложные зубы, как у слонов, историю которых нам прекрас-
но очерчивает палеонтология. У эоценовых предков хоботных зубы
были бунодонтные (рис. 560). В связи с переходом к питанию ветвя-
ми и жесткими листьями бугорки начинают соединяться в гребни,
ясно выраженные уже у верхнеплиоценовых и нижнеплиоценовых
форм, например, у гомфотерия (Gomphotherium).
Счастливая находка обнаружила, что гомфотерии питались вет-
вями и жесткими листьями, которые они не могли пережевывать
обычным образом, а должны были перетирать движением челюсти
спереди назад. При такой работе челюсти необходимо было приобре-
тение поперечных гребней. У более позднего верхнеплиоценового
рода Stegodon количество этих гребней увеличивается. У молодых
слонов на гребнях еще имеются бугорки, но потом они стираются,
и получаются гладкие поперечные гребни. Увеличение их числа и
выполнение поперечных углублений цементом приводят, наконец,
к зубам настоящих слонов, представляющим великолепную перети-
рающую поверхность.
Африканский слон кормится преимущественно ветвями деревьев,
до 4—Б см толщиной и 10—12 м длиной, которые он поедает вместе
с древесиной. В равнинах он выкапывает из земли сочные корни.
Наоборот, индийский слон предпочитает злаки, которыми преиму-
щественно и питается, лишь случайно употребляя в пищу ветви с
листьями. Соответственно с таким различием в пище у обоих слонов
различаются и зубы. У африканского слона поперечных дуг мало,
но они широки, и сильные эмалевые петли выдаются над цементом.
Гребни на зубах индийского слона более многочисленны, узки, и их
эмаль слабо выдается над цементом. Это еще лучше выражено у ис-
копаемого мамонта. Современный индийский слон, ближайший
родственник мамонта, имеет более грубую зубную поверхность и
косо поставленные эмалевые петли, так как питается мягкими,
сочными растениями, вроде сахарного тростника.
Увеличение перетирающей поверхности зуба и удлинение его
новели, в конце концов, к укорочению зубного ряда, к уменьшению
числа зубов и к замене их в горизонтальном направлении: у слонов
из шести коренных зубов в употреблении находится всего лишь
один, по мере изнашивания заменяющийся позади лежащим зубом.
Интересно, что по общему характеру коренные зубы у слонов в
высокой мере напоминают зубы водосвинки (Hydrochoerus capy-
bara) — самого крупного из современных грызунов, питающегося
корой молодых деревьев.
Параллельно с изменением коренных зубов у слонов в течение
их эволюции изменялись приспособительно к пище челюсти, и
бивни.
Совершенно своеобразны изменения зубов в связи с характером
пищи у различных грызунов (Rodentia). Специализация зубов у
грызунов касается их формы, положения и строения и стоит в связи
с их способом питания (рис. 561).
761
Первой особенностью, бросающейся в глаза, является необычное
развитие верхних и нижних резцов, сточенных впереди наподобие
стамески. Сточены и верхние резцы и нижние. Очевидно нижние
резцы так же действуют стачивающим образом на верхние, как верх-
ние на нижние. Это возможно лишь при том условии, если нижняя
челюсть совершает движения, параллельные оси черепа, спереди
назад и обратно. Сочленовная головка нижней челюсти представляет
цилиндрический валик, помещающийся в параллельном оси черепа
углублении височной кости. Резцы, сточенные стамескообразно,
могут отгрызать части растений, в то же время горизонтальным
движением челюстей коренные зубы перетирают пищу, состоящую
из твердых растительных частей.
Однако перетирание ведет к изнашиванию коренных зубов. Как
приспособление против этого жующая поверхность последних при-
Рис. 561. Череп мешотчатой крысы (Geomys).
(Ив Вебера.)
обрела многочисленные
складки эмали, и зубы
увеличились в продоль-
ном направлении. Склад-
ки лежат поперечно, как
у слонов, тогда как у
копытных, где имеет ме-
сто движение челюстей
справа налево, складки
эмали (гребни) лежат
продольно, что опять-та-
ки является соответст-
вующей адаптацией.
Так же как в ряду ко- •
пытных, мы видим у сло-
нов и у грызунов, кроме
изменения жующей по- .
верхности кроны, еще и
другого рода изменения.
Исходная, бунодонтная,
форма зубов сопровож-
дается развитием корня и низкой кроны, покрытой толстой эмалью:
зубы брахиодонтны. Однако такая крона быстро стирается.
Это стирание компенсируется тем, что зуб растет постоянно или по
крайней мере очепь долгое время. Отсутствие корня или позднее
его образование позволяет это. Зуб становится гичселодонт-
н ы м. В конце концов, корень исчезает вовсе, и зуб становится
призматическим. Соответственное этим отдельные бугорки становят-
ся колонками, между которыми лежит цемент, изнашивающийся
быстрее эмали. Благодаря этому получается очень неровная поверх-
ность кроны с многочисленными петлями эмали. Так как эти петли
сильно вариируют, то они имеют большое значение в система-
тике.
Почти во всех группах позвоночных мы встречаем формы, которые
по тем или иным причинам утеряли зубы. Причины этой утраты
762
могут быть весьма различны. Среди млекопитающих мы видим
редукцию зубов у китов, у сирен и у тюленей.
Киты приспособились к водной среде. Понятно, что в Воде жр,
ванне затруднительно, если не невозможно. Поэтому даже у зубатых
китов, еще сохраняющих зубы, весь жевательный аппарат с муску-
лами и нижней челюстью подвергается редукции. Зубы становятся
простыми коническими и служат только для схватывания ц удепжа-
пия добычи. Свойственная млекопитающим дифференцировка на
резцы, клыки и т. д. исчезает за ненадобностью: зубы становятся
гомодонтными; число зубов значительно увеличивается (до
250), так что зубы напоминают зубы рептилий, особенно ихтиозав-
ров. Таковы зубы у китообразных, питающихся рыбой; у китов"
пищу которых составляют моллюски, зубы подвергаются иекотовой
редукции, достигающей наибольшего размера в подсемействе Zinhii-
пае. Палеонтологическая история последних хорошо изучена и
поэтому можно проследить путь, по которому шла редукция зубов
в этой группе. у
Исчезновение зубов у Ziphiidae стоит в связи с тем, что эти ки-
тообразные питаются преимущественно мягкотелыми головоногими
которых они схватывают беззубыми, но твердыми краями
и глотают целиком.
У одного из представителей этих китообразных, Hyperoodon го-
stratus, на нёбе имеются поперечно расположенные ороговевшие
полоски эпителия. Эти роговые полоски есть первое указание
па то, как возникли роговые «усы» у беззубых китов (Mystacoceti)
Другим указанием на путь возникновения «усов» является то
что у некоторых Ziphiinae ветви нижней челюсти сильно выдаются
сбоку из-за ветвей верхней, как у Mystacoceti. Последние пеово-
начально имели большое количество (свыше 200) зубов; Это fCH0
из того , что у эмбрионов беззубых китов имеются рудименты зу-
бов. Потеря зубов, твердые края челюстей, нёбные гребни и высту-
пание ветвей нижней челюсти у Ziphiinae указывают на то что v
предков беззубых китов питанию планктоном предшествовало пита-
ние головоногими моллюсками. Современные беззубые киты питаются
исключительно планктонными организмами, главным обпазом
мелкими ракообразными, которых в кишечнике одного кита полоса-
тика (Balaenoptera) нашли 200 литров. Иногда пищей слузкат''мел1
кие рыбы, иногда крылоногие моллюски. Этих животных беззубы^
киты захватывают, бороздя море с открытой пастью, причем мел-
кие животные задерживаются на усах. При закрывании рта и ппи
поднимании языка к нёбу попавшие в рот организмы попадают
в глотку. При таком образе питания зубы становятся совершенно
излишними.
Редукцию зубов мы видим в отряде сирен (Sirenia) Знаме-
нитая стеллерова морская корова (Rhytina gigas), уничтожен-
ная уже почти полтора столетия назад, была совершенно лишена
зубов. Это были крупные животные, которые паслись в мелком
море, кормясь водорослями. Для перетирания этой совершенно
мягкой пищи им служили шершавые роговые пластинки на нёбе
. 763
и между ветвями нижней челюсти. На тот путь, каким возникла эта
специализация, указывают зубы другого представителя сирен —
дюгони (Halicore). Роговые пластинки есть и здесь, но сохраня-
ются и зубы, а именно: кроме бивней, у самца еще 6 коренных в каж-
дой челюсти в виде штифтообразных зубов с открытым корнем. Эти
шесть зубов являются молочными зубами, не сменяются настоящими
и остаются до полного изнашивания. У древнетретичных сирен
три передних молочных коренных зуба еще сменялись окончатель-
ными ложнокоренными.
Приспособлением к роду пищи, а именно к питанию мягкими
насекомыми, червями, отчасти и моллюсками, следует считать
исчезновение зубов и замену их роговой обкладкой челюсти
у однопроходных (Monotremata), у которых рыло служит для
рытья. Доказательством вторичного исчезновения зубов служит
наличие двух коренных зубов в верхней и трех в нижней челюсти
у молодых утконосов (Ornithorhynchus).
Питание насекомыми вызвало редукцию зубов и у ящеров
(Manis) и муравьедов (Myremecophaga), у которых зубы или недо-
развиты и представляют лишенные эмали призмы, или же отсут-
ствуют вовсе.
Кишечный канал. Глотка и пищевод млекопитающих не пред-
ставляют каких-либо интересных особенностей; желудок же,
наоборот, подвергается ряду изменений в связи с характером при-
нимаемой пищи.
В простейшем виде он представляет расположенный поперек
и мешкообразно расширенный кзади участок пищеварительной
трубки. Уже в самой простой своей форме он дифференцируется в
зависимости ст характера покровов на две части: область пищевода,
покрытую снутри многослойным эпителием, как и пищевод, и лишен-
ную желез, I прочую часть, одетую однослойным эпителием и
снабженную железами; эти железы устроены различно: одни имеют,
кроме цилиндоических, «главных» клеток, еще и округлые обкла-
дочные клеткх, другие состоят исключительно из цилиндрических
клеток. Таком клетки области пилоруса. Расположение желез
в желудке видно из прилагаемых схематических рисунков (рис. 662).
Строение желудка изменяется в связи с родом пищи. Он прост
у плотоядных и может быть чрезвычайно сложен у травоядных.
Наиболее сложен желудок жвачных (рис. 563). Он состоит из
4 отделов: 1) рубец (rumen), внутренняя поверхность кото-
рого покрыта твердыми бородавками; 2) сетка (reticulum),
степки которой разделены на альвеолы, подобно сети; 3) к н и ж-
к a (psalter.um, omasus), на внутренней стенке которой находятся
продольные сктадки подобно листам книги. Во всех этих трех
отделах железн отсутствуют, и, следовательно, эти отделы при-
надлежат к пицеводной части пищеварительного тракта; эпителий
их многослойный и частью ороговелый. Собственно желудок состав-
ляет 4-й отдел: железистый желудок, сычу г (abomasus).
Функции отделов таковы. Рубец является мацерационным и бро-
дильным желудюм. Кормовые массы под влиянием слюны, тепла и
764 *
деятельности бактерий подвергаются брожению (молочнокислому,
маслянокислому и уксуснокислому); крахмал частично перевари-
вается под влиянием слюны. Из рубца, благодаря перистальти-
Рис. 562. Схематическое изображение расположения от-
делов в желудках:
А — человека, В — собаки, С — крысы, D — хомяка, Е — лошади,
F — жвачного.
Область пищевода с многослойным эпителием отмечена горизон-
тальными штрихами, область кардиальных желез — косыми
штрихами, область донных желез — пунктиром, область пило-
рических — крестиками. (Из Шмальгаузена.)
ческим движениям, пища поступает в сетку, откуда путем отрыги-
вания — снова в рот, где размельчается и основательно смачивает-
ся слюной. Получившая-
ся таким образом киселе-
образная масса стекает по
желобку, края которого
тесно смыкаются, в книж-
ку и сычуг. Отрыгивание
есть акт произвольный и
происходит при помощи
сокращения диафрагмы и
сокращения рубца под
влиянием воздействия нер-
вной системы. Остальная
часть акта жвачки рефлек-
торная. Выгода от наличия
жвачки у травоядных, ко-
торые питаются трудно пе-
ревариваемой пищей, оче-
видна. В желудке жвачных
живет огромное количест-
во бродильных бактерий
Рис. 563. Сложный желудок овцы.
а — пищевод, Ь — рубец (rumen), g — сетка (re-
ticulum), i — книжка (psalterium) и h — сычуг
(abomasus). (По Краусу и Отто.)
и инфузорий, которые,
быть может служат, тоже
пищей для своих хозяев.
Ту или иную степень сложности желудка мы видим у целого
ряда других млекопитающих.
765
У китов, пе жующих пиши, передний отдел желудка предста-
вляет лишенное желез расширение пищевода с сильно развитой
мускулатурой, выстлан ороговевшим эпителием и служит для ма-
церации пищи. Желудок с толстыми стенками служит для пере-
тирания пищи у беззубых муравьедов. У ящеров прилежащая к
пищеводу пасть желудка выстлана ороговевшим эпителием. Му-
равьи и термиты перетираются здесь при помощи песка и маленьких
камешков. В средней части желудка пища пропитывается пищева-
рительными соками, а в пилорической части снова мы видим пере-
тирающее приспособление с роговыми зубчиками и сильной муску-
латурой. Здесь опять-таки наблюдается компенсирование недо-
статка зубов.
Кишечный канал млекопитающих очень разнообразен в смы-
сле соотношений длины между отделами. Соотношение между
длиной кишечного канала и тела животного колеблется от 2:1 у
некоторых рукокрылых и до 32:1 у некоторых китообразных; у
корг вы соотношение 20:1, у лошади 12:Г, в общем же, как правило,
и у млекопитающих в связи с животной пищей кишечный канал
бывает короче, с связи с растительной — длиннее.
Двенадцатиперстная кишка (duodenum) содержит особые брунне-
ровы железы; задняя кишка подразделяется на толстую (colon) и
прямую (rectum); слепая кишка (coecum) по большей части одна,
в очень разной степени развитая, или зачаточная или достигает ко-
лоссальных размеров; она заменяет у нежвачных травоядных слож-
ный желудок жвачных, так как здесь, как и в толстой кишке,
происходит бактериальное брожение клетчатки. Дистальный конец,
слепой кишки при ее редукции превращается в рудимент —
червеобразный отросток (appendix).
У клоачных прямая кишка открывается в клоаку, отсутствую-
щую у прочих млекопитающих.
Печень (hepar) имеет три больших выводных протока, которые
соединяются в общий печеночный (ductus hepaticus); положе-
ние желчного пузыря (vesica fellea) различно; такое положение
желчного пузыря, когда его выводной проток (ductus cysticus)
открывается в печеночный проток, становящийся желчным протоком
(ductus choledochus), кроме человека, встречается редко. Обычно
желчный пузырь сидит на одном из печеночных протоков.
Органы дыхания
Гортань млекопитающих в отношении входящих в ее со-
став хрящей представляет дальнейшие изменения по сравнению
с тем, что имело место у рептилий — предков млекопитающих.
У однопроходных гортань соединена с copula подъязычного
аппарата, задние рожки которого, соответствующие 1-й жабер-
ной дуге, входят в состав гортани. Непосредственно позади распола-
гаются на гортани пять хрящей: один непарный, соответствую-
щий копуле жаберных дуг, и два парных, соответствующие 2-й
и 3-й жаберным дугам.
765
У прочих млекопитающих подъязычный аппарат вместе с зад-
ними рожками отделен от гортани, а хрящевые элементы 2-й и
3-й жаберных дуг сливаются, образуя черпаловидный
хрящ.
Такое строение гортани живородящих млекопитающих уста-
навливается эмбриональными стадиями. Кроме щитовидного хряща,
для млекопитающих характерно присутствие спереди надгор-
танника (epiglottis). Голосовые связки располагаются в виде
складок слизистой оболочки между щитовидным хрящом и чер-
паловидными; связки могут натягиваться, меняя звук, про-
изводимый воздухом, проходящим мимо в легкие и обратно.
Между гортанными хрящами или выше над ними помещают-
ся у некоторых животных мешкообразные выросты — резо-
наторы, свойственные многим обезья-
нам, парнокопытным, хищным и. некото-
рым другим. Таковы Морганиевы желу-
дочки обезьян, особенно развитые у ре-
вунов, являющиеся частью резонаторами,
частью защитой от ушибов. Соответству-
ющие мешки у китов служат резервуаром
воздуха.
Трахея делится на двое бронхов,
которые в свою очередь делятся мно-
гократно, оканчиваясь на конце тон-
чайших трубочек — бронхиол — конеч-
ными пузырьками с ячеистыми стенка-
ми — альвеолами. Бронхиоли хря-
щей не имеют.
Легкое млекопитающего устроено
гораздо совершеннее, чем у рептилий, хотя
и является завершением уже начавшегося
у рептилий процесса последовательного
ветвления бронхов. Очень совершенным по
сравнению с таковым рептилий является
дыхательный процесс; особенно важным
органом дыхательного процесса является
диафрагма (рис. 564).
Воздух всасывается в легкое и выжи-
мается из него периодически, во-первых,пу-
темизмепения объема грудной полости вслед-
ствие движения ребер при помощи ребер-
ной мускулатуры, описанной выше, и,
Рис. 564. Схематический
фронтальный разрез че-
рез грудную полость
млекопитающего.
1 — трахея, 2 — ветвящиеся
бронхи, 3 — париетальный
листок плевры, 4 — легоч-
ный листок плевры, 5 —
легкое, 6 — диафрагма, 7 —
нижняя полая вена, 8 —
аорта, 9 — желудок, 10 —
брюшная полость. (Из Ви-
дерсгейма.)
во-вторых, вследствие последовательного сокращения или расслаб-
ления диафрагмы, куполообразно вдающейся в грудную полость.
Усовершенствование дыхания вместе с приспособлениями в
кровеносной системе и наличие волосяного покрова дали воз-
можность предкам млекопитающих приобрести теплокров-
ность.
767
6. Кровеносная система
менее развитой правую
№
Кровеносная система млекопитающих достигает такого же
высокого развития, как и кровеносная система птиц, но путь
развития особенностей кровеносной системы млекопитающих совер-
шенно самостоятельный. Дело в том, что у млекопитающих имеется
только одна и притом левая дуга аорты. Так как у птиц
имеется одна правая дуга аорты, а у современных рептилий левая
дуга развита слабее (обе подключичные артерии отходят от пра-
вой аорты), т. е., следовательно, уже имеет тенденцию к сокраще-
нию, то, очевидно, предки млекопитающих — териодонтные реп-
тилии — имели в противоположность современным рептилиям
дугу аорты.
Сердце разделено полной перегород-
кой, в которой могут иногда (у копытных)
появляться дажеокостенения. Левая дуга
аорты отходит от левого, легочные арте-
рии— от правого желудочка. Венозный
синус слился с правым предсердием, пра-
вый атриовентрикулярный клапан —
трехстворчатый (valvula tricuspida-
Нз);левый—двуст ворчатый (val-
vula bicuspidalis) у живородящих снаб-
жен сухожильными нитями к мускуль-
ным выступам (mm. papillares). В отхо-
дящих от сердца артериях (аорта и ле-
гочная) обычные полулунные клапаны.
Главные артериальные стволы отхо-
дят от аорты различно. Подключичные
артерии отходят самостоятельно или
с сонными артериями, или сонные арте-
рии отходят общим стволом (рис. 565).
Из других особенностей артериальной
системы млекопитающих, которая сход-
на в остальном с тем, что имеет место
у рептилий, укажем, что подвздошная
артерия делится на подвздошную (a.iliaca
interna) и бедренную (a. femoralis).
В венозной системе для млекопитаю-
щих характерна глубокая ассиметрия.
Кроме характерной и для других на-
земных позвоночных животных зад-
ней полой вены (V. cava posterior),
(верхняя у человека,) полая вена (v. cava
anterior), получившаяся путем развития безыменной вены соеди-
няющей левую полую вену с правой (v. innominata); безыменная вена
у млекопитающих является новообразованием.
Остатки кардиальных вен, так называемые здесь непарное
правая (V. azygos) и левая (v. hemiazygos), редуцируются так, что
768
Рис. 565. Схематическое
изображение сердца и ар-
терий млекопитающих.
1 — правое предсердие, 2 —
трехстворчатый клапан, 3 —
правый желудочек, 4 — полу-
лунный клапан, 5 — легочная
артерия, 6 — левое предсердие,
7 — двустворчатый клапан, 8 —
левый желудочек, 9 — аорта,
10 — левая подключичная, 12—
правая подключичная, 13 — об-
щий проток сонных артерий
(truncus caroticus), 14 — общая
сонная артерия (carotis com-
munis), 15 — внешняя сонная,
16 — внутренняя сонная. Эм-
бриональные дуги отмечены
пунктиром. (По Гудричу.)
имеется передняя
одна из них, по большей части правая (v. azy^os) получает к
и с другой половины тела по поперечной Bei3ej лежащей далеко
позади кювьеровых протоков, после чего о<статок карДиальпой
ноны превращается в короткую непарную ве^у
Правая непарная вена впадает в передпюкэ цолую в р
.ко вену впадает через левую анонимную вену (v anonvm-i?w г™ J
ный лимфатический сосуд. ' ' у ' и глав
Кровеносные тела по большей части округл^ и всегда безъядеп-
ны. Температура крови постоянная (млекопитгиОщие__‘г 0 м ей 0_
термн ы е животные), колеблется между 3g и ^qo у gXI^Hbj
_____" ___ .___ __4 9
температура колеблется в связи с па-
дением температуры воздуха весьма
значительно, обыкновенно она 28°,
но, колеблясь в пределах 10°, па-
дает иногда от 35 до 5°.
Рис.
56;
ппчрчнь3, Схема расположения
почсчнь^ канальцев у млеко.
питающих.
той канаанова капсУла, 2 — изви-
bulus соглЛ?+ первого порядна (tu-
Генле, £tortus primus), 3 — петля
рого пор, извитый каналец вто-
канал, вндка’ 5 “ собирательный
papillaris?- канал сосочка (ductus
мозговая5'* К корковая и М. —
MUd,UBdH части почки. (По Эбнеру.)
7. Органы выделения
Почки млекопитающих, как и
у других амниот, вторичные
11 очки (metanephros); первич! i ая
почка является эмбриональным орга-
ном, и только у однопроходных и у
сумчатых она функционирует в моло-
дом возрасте, а затем редуцируется.
Почки представляют компактные
гола бобовидной формы, состоящие из
наружного коркового зернистого слоя
н внутренней радиально исчерченной
сердцевины. В корковом слое распо-
лагаются мальпигиев ы тела
и извитые канальцы, во-втором—
прямые канальцы (рис. 566);
последние группами открываются па
конце сосочков, опускающихся в по-
чечную лоханку (pelvis)—ворон-
кообразное начало мочеточника (ure-
ter), расположенное на внутренней
вогнутой части почки. Группа пря-
мых канальцев, объединенных общим
местом окончания, составляет так
называемые пирамиды, между
которыми проникает корковое веще-
ство (рис. 567).
Иногда каждой пирамиде соответствует и наружное деление
почки, которое таким образом распадается ш отдельные дольки
(renicoli), что особенно сильно выражено у китообразных (до 100
дэлек почки), также у Pinnipedia, у некоторых Carnivora и yArtio-
49 Кашка ров и Стапчинокий
769
моча выводится
через короткое
Рис. 5G7. Продольный
разрез через почку
кролика.
1 — корковый слой, 2—
мозговое вещество, 3 —
пирамиды, 4 — сосочек,
5 — лоханка, 6 — моче-
точник. (По Фохту и
Юнгу.)
У однопроходных мочеточники открываются в мочеполовой
синус (sinus urogenitalis) независимо от мочевого пузыря, так что,
моча попадает в мочевой пузырь через синус. У всех остальных
мочеточники открываются непосредственно в пузырь; из пузыря
самок через остатки мочеполового синуса —
щверие влагалища, тогда как у самца синус
вытягивается в длиный мочевой канал, от-
крывающийся на конце копулятивного органа
(рис. 568).
8. Органы воспроизведения
Яйца млекопитающих значительно отли-
чаются по подклассам. У однопроходных яйца
крупные (2.5 —4.0 мм) и содержат большое
количество питательного желтка, являются
меробластическими и имеют сверх желточной
вторичной оболочки (zona pellucida) еще тре-
тичные оболочки.
Сюда относится белковая оболочка —
proalbumen, которая, попав в яйцевод, пре-
вращается от впитывания жидкости в белок;
в яйцеводе же как результат выделения ма-
точных желез образуется вокруг яйца пер-
гаментообразная скорлупа из кератина с из-
вестью. В матке яйцо под влиянием усвоения маточной жид-
кости увеличивается, достигая у ехидны диаметра в 16.6—13 мм.
У сумчатых яйца значительно меньше (0.3 мм) и одеты белковой обо-
лочкой. Совсем маленькие они у плацентарных (у собаки 0.18 мм,
у мыши 0.06 мм), голобластические и не имеют белковой оболочки.
Яичники (ovarium) парные и одинаковой величины за
исключением утконоса, у которого левый яичник развит силь-
нее; у однопроходных опи гроздевидные, как и у некоторых
сумчатых, тогда как у плацентарных они компактнее, продолго-
ваты, часто бобовидной формы.
Яйцеводы у однопроходных разделены на всем протя-
жении, образуют фаллопиевы трубы, открывающиеся в полость
тела, а на противоположном конце открываются в мочеполовой
синус каждый самостоятельно; последний открывается в клоаку
(рис. 568, А). У сумчатых мы наблюдаем тенденцию к срастанию на
некотором расстоянии от синуса правого и левого яйцеводов, что
приводит к двум кольцеобразно сросшимся влагалищам; от перед-
ней стороны этого кольца отходит по направлению к синусу сле-
пой мешок; если последний прорывается в синус, получается так
называемое третье влагалище. Оставшаяся часть яйцеводов обра-
зует матку (uterus) и собственно яйцеводы с фаллопиевыми тру-
бами (рис. 568, В и С).
У плацентарных млекопитающих из прилегающих к зачаточному
мочеполовому синусу частей мюллеровых протоков развивается
770
непарное влагалище. Из остальных частей мюллеровых протоков
или образуются две матки, каждая с самостоятельным отверстием—
двойная матка (uterus duplex) большинства грызунов,
трубкозуба, отдельных рукокрылых и некоторых хищных, или
две матки с общим отверстием — двураздельная матка,
(uterus Ыpartitus) хищных, свиней, некоторых грызунов, или
сращение идет дальше, так что образуется, как мы встречаем у
китообразных, копытных, насекомоядных и у большинства руко-
* Рис. 568. Схема эволюции яйцеводов и матки у млекопитающих.
А — клоачное, В — низшее сумчатое (опоссум), С — высшее сум-
чатое, D — плацентарное млекопитающее с двойной, Е — с двуро-
гой, F — с простой маткой.
cl — клоака, od — яйцеводы, г — прямая кишка, аир — мочеполовой синус,
иг — мочеточник, ut — матка, vag — влагалище, и. и. — мочевой пузырь.
(Из Шмальгаузена.)
крылых и у полуобезьян, двурогая матка (uterus bicor-
nis), и, наконец, при полном сращении получается простая
матка (uterus simplex) некоторых рукокрылых, обезьян и
человека (рис. 568, Ь’).
Сперматозоиды млекопитающих очень мелкие и просто
устроены; они определяют пол, так как одни из них содержат,
а другие пе содержат Х-хромосому.
Семенники (testes) имеют яйцевидную форму, компактны
и соединяются с длинным, сильно извилистым в начале, семяпро-
водом и р так называемым придатком (epididymis), происшед-
шим из первичной почки и состоящим из многочисленных изви-
тых топких канальцев, которые открываются в эту сильно изви-
листую часть семяпровода (canalis epididymis). Последний переходит
49* 771
в ровный прямой семяпровод (vas deferens), который впадает
в мочеполовой сипус, вытянутый в мочеполовой капал (ca-
nalis urogenitalis, или urethra); последний продолжается в копу-
лятивный орган (penis) и открывается на его конце иногда
суженной частью — семяизвергательным каналом (ductus ejacu-
latorius.) :
В связи с семяпроводами находятся особые железы—семен-
ные пузырьки (gl. vesiculares); в связи с мочеполовым
каналом развита предстательная железа (gl. pros-
tata), отсутствующая у китообразных, у однопроходных и сум-
чатых, на месте которой развиты железы мочеполового канала
(gl. urethrales), встречающиеся и на ряду с первой. Почти у всех
млекопитающих имеются также куперовы железы (gl. Couperi).
Значение секрета этих желез в том, что в нем разбавляется
сперма; кроме того, у грызунов после совокупления из пего обра-
зуется во влагалище пробка, которая не дает сперме выливаться
наружу. У летучих мышей нашего климата такой пробкой сперма
задерживается в матке в течение всей зимы, так как оплодотво-
рение яйца происходит лишь весной следующего года.
У клоачных, у слонов, китообразных, сирен, у некоторых
насекомоядных и неполнозубых семенники в течение всей жизни
остаются в брюшной полости тела, у всех остальных они спу-
скаются наружу, помещаясь в особом мешке — мошонке (scrotum).
Полость мошонки — полость тела и выстлана брюшиной; она или
соединяется с полостью брюха большим отверстием, через которое
семенники периодически спускаются в мошонку и втягиваются
обратно особым мускулом (m. cremaster), или это отверстие зара-
стает, семенники постоянно остаются в мошонке.
Внутреннее оплодотворение яйца в матке самки осуществляется
с помощью хорошо развитых копулятивных органов.
У клоачпых половой член (penis) самца помещается в особом
влагалище клоаки, откуда он может выдвигаться наружу. По
нему проходит канал, соединяющийся с мочеполовым синусом.
Когда отверстие мочеполового синуса в клоаку закрывается, сперма
вытекает по каналу члена.
В связи с исчезновением клоаки у живородящих млекопитаю-
щих мочеполовой синус срастается с каналом, образуя мочепо-
ловой канал, а половой член оказывается наружи. Помещаясь
первоначально около заднепроходного отверстия и будучи повер-
нут головкой назад (у сумчатых), он постепенно перемещается
на брюхо. У сумчатых, соответственно двойному влагалищу, penis
раздвоен спереди. Копулятивный орган снабжен боковыми пеще-
ристыми телами и пещеристым телом па конце, образующим
головку. У сумчатых, рукокрылых, грызунов, китообразных,
хищных, ластоногих, полуобезьяп и некоторых обезьян в поло-
вом члене развивается кость (os penis). Влагалище копулятивного
органа образует вокруг головки особую кожную складку' (ргае-
putium), куда нередко открываются особые кожп/с препудиатть-
пые железы.
772
недоразвитыми конечностя-
Но характеру размножения млекопитающие делятся на три
группы сообразно трем подклассам.
Однопроходные (Monotremata) в отношении размноже-
пин очень близки к рептилиям и сохраняют унаследованные от
них примитивные особенности, — опи кладут яйца.
Яйца высиживаются утконосом в гнезде, а ехидной вынаши-
ваются в особой выводковой сумке, которая ко времени откладки
ипца развивается на брюхе самки. Тут же вынашивается вылу-
пившийся детеныш, который питается молоком млечных желез.
Яги последние в отличие от млечных желез других млекопита-
ющих не имеют сосцов.
Сумчатые (Marsupialia) живородящи, но рождают детены-
iiH'ii малоразвитых, голых, слепых, с
ми. Детеныши донашиваются в осо-
ihhI сумке, расположенной на брюхе
мятери, в которой находятся сосцы
молочных желез. Детенышей мать по-
мещает в сумку так, что довольно
продолжительное время, пока не pa-
in вьются органы чувств и конечно-
мн, детеныши висят на сосках, при-
чем молоко впрыскивается в рот
г помощью особого мускула.
Высшие м л ек о п и та ю-
щ и о (Eutheria) рождают детенышей
\ же более развитых, во всяком слу-
чае настолько развитых, что опи мо-
। \ г самостоятельно сосать и имеют
полое или мепее развитые копечнс-
гги, могущие производить движения.
У гребное развитие зародыша более
ш>'пню и происходит в матке, при-
чем питание плода осуществляется
через посредство плаценты, или дет-
। него места, откуда эта группа и получила название плацен-
г ,1 р и и х млекопитающих (Placentalia) (рис. 569). Аллан-
гипс, врастая между серозной оболочкой и амнионом и плотно
нрн кч’ая к серозной оболочке, образует на большем или мень-
шем пространстве двойную оболочку, или хорион. На поверхпо-
। гн хориона образуются сосочки, которые врастают в слизистую
пГш'|очиу матки, где ветвятся, так что между кровеносными систе-
MIMH м.чтери и зародыша устанавливается тесная связь. Область
ыhell связи тела матки и хориона и носит название плацент ы.
При рождении сосочки или выходят более или мепее свободно
н » ге 1.1 матки (неотпадающая плацента,) или настолько плотно
• Р К Г.1ЮТСЯ со слизистой оболочкой матки, что при рождении часть
»• п4'пядает вместе с хориопом (отпадающая).
Нернед беременности млекопитающих различный. У сумчатых,
р•• шщющпх неразвитых детенышей, беременность продолжается
773
Рис. 569. Схема эмбриональ-
ных оболочек у плацентарных
млекопитающих.
1 — зародыш, 2 — матка, 3 — ам-
нион, 4 — желточный мешок, 5 —
аллантоис, 6 — плацента.
лишь незначительную часть всего времени, необходимого для раз-
вития детенышей, которые развиваются дальше в сумке ма-
тери.
В общем, чем больше животное, тем меньшее число детенышей
оно рождает, и чем более развитыми производятся на свет дете-
ныши, тем продолжительнее период беременности. Так, у мыши
беременность продолжается всегда лишь 21 день, тогда как у сйопа
до 628 дней; у морской свипки, рождающей способных бегать
детей, беременность продолжается 63 дня, а у кролика, рождаю-
щего беспомощных детей — 23 дня; у коров 280 дней, тогда как
у свиней, рождающих по многу детенышей сразу, — всего лишь
120 дней.
Вторичные половые признаки далеко не дости-
гают у млекопитающих такого развития, как у птиц.
Они выражаются прежде всего в росте. Как правило, самцы
больше самок; особенно это различие выражено у морских живот-
ных, у которых самцы нередко, как, например, у кашалотов
(Physeter), у сивуча (Eumetopias stelleri), превосходят самок вдвое
и даже больше. У котика (Callorhinus ursinus) самец тяжелее самки
в 4 — 5 раз.
Кроме величины, самцы от самок отличаются пропорциями
отдельных частей тела, большей развитостью мускулатуры, а
потому и большей силой, что видно хотя бы на наших домашних
животных.
Далее самцы нередко в противоположность самкам имеют
особые органы нападения — оружие для боев. Достаточно указать
на рога' у оленей, которые развиваются только у самцов, причем
как раз ко времени течки самок. Только у северного оленя самки
тоже имеют рога. У антилоп можно проследить постепенные пере-
ходы от видов, у которых самцы и самки имеют одинаковые рога,
к таким, у которых самки вовсе лишены рогов. У коз (Сарга) и
овец (Ovis) рога самок обычно развиты гораздо слабее, иногда
зачаточны или вовсе отсутствуют. Неизвестно ни одного вида
млекопитающих, у которых рога были бы только у самок или
были развиты у них сильнее, чем у самцов.
Хищные животные, у которых зубы и когти являются орудиями
нападения на добычу, не имеют специальных орудий для поедин-
ков самцов. У других животных, однако, зубы могут играть роль
вторичных половых признаков как орудий борьбы. У многих
копытных такую роль играют клыки. Так, у самца кабарги (Мо-
schus), которые составляют подсемейство оленей, нет рогов, по
зато большого развития достигают клыки. У самцов верблюдов,
некоторых антилоп и у лошадей развиваются в верхней челюсти
клыки, которые у самок слабо развиты или отсутствуют. У сам-
цов нарвалов один из клыков достигает исключительного развития.
У некоторых видов слонов бивни развиваются только у самцов
(индийский слои), а там, где они присущи и самкам, у этих послед-
них они развиты слабее. У самцов ехидны и утконоса на ноге раз-
вивается шпора с особой железой.
774
На ряду с органами борьбы и нападепия у самцов развиваются
и органы защиты. Таково, вероятно, назначение длинных волос
на голове и шее, которые развиваются преимуптр.р,твртттто у самцов,
как грива у льва, у зубров, некоторых антидОп, многих баранов^
как борода козлов, бакенбарды у рыси и пр.
В качестве вторичных половых признаков могут быть также
и железы, которые нередко выделяют особью сильно пахнущие
вещества. Так, самцы козлов и в меньшей степени баранов издают
особые запахи, или вовсе несвойственные сайкам или свойствен"
пые им в слабой степени. Особенно сильны^ запах издают они
в период половой деятельности. Таковы мускусные железы самца
кабарги, некоторых оленей и антилоп. Некоторые самцы в период
течки откладывают сильно пахнущий жир, например, северо-
американские кроты (Condylura).
Цветовые отличия между полами, которые так развиты у птиц,
у млекопитающих встречаются редко. Таковы., например, отличия
у кенгуру (Macropus rufus), у которых самец Красноватого, а самки
голубовато-серого цвета. Иногда, как папример, у некоторых
обезьян, голые места самцов гораздо ярче, че^ у самок (мандрил).
Ко вторичным половым признакам нужно отнести также голос,
который может быть различным у самцов и у (;амок или отличается
у первых большей силой. При этом различные приспособления,
вроде резонаторов, оказываются у самцов сильнее развитыми,
чем у самок, и присущи только им. Так, у олеНей, антилоп, у мно-
гих обезьян имеются также резонаторы па го^ле. Особенного раз-
вития и сложности они достигают у обезьян рода ревунов (Mycetes).
Если орудия борьбы, защиты и нападения могли выработаться
путем отбора в борьбе из-за самок, то происхождение других вто-
ричных половых признаков у млекопитающих остается так же
мало выясненным, как и цветовые и голосовью различия у птиц.
При кастрации признаки эти обычно исчезают и, таким образом,
стоят в связи с гормональной деятельностью половых желез. Ско-
рее всего здесь просто структурная корреляция.
К числу вторичных половых признаков нужно отнести также
некоторые инстинкты и нравы, которые иногда резко отличаются
у обоих полов; они обычно сглаживаются ПрИ кастрации.
9. Железы внутренней секреции
Не останавливаясь здесь на значении д роли чрезвычайно
важных органов внутренней секреции, как на проблеме чисто
физиологической, мы ограничимся только кратким морфологи-
ческим описанием этих органов у млекопитающих.
‘Щитовидная железа (gl. thyreoidea) состоит у мле-
копитающих из двух лопастей и помещается :за брюшной стороне
в области нижней части гортани; иногда она простирается назад
вдоль трахеи, иногда распадается на две железы, которые связаны
лишь соединительнотканной перемычкой istnUs (некоторые Маг-
supialia, Chiroptera, Rodentia, Lemuroidea).
775
Щитовидная железа закладывается у зародышей в виде непар-
ного вентрального выроста энтодермы и первоначально представ
вляет сложную ветвящуюся трубчатую железу с выводным про-
током, который, однако, скоро исчезает, и железа становится эндо-
кринной, т. е. выделяющей свой инкрет в кровеносные сосуды.
Врастающая соединительная ткань разделяет ветвящиеся трубочки
на две доли, правую и левую, образуя перешеек, а затем на от-
дельные пузырьки — фолликулы с характерным гомогенным со-
держимым, так называемым коллоидом.
Гормон щитовидной железы — тироксин — оказывает большое
влияние па общий обмен веществ. Другой гормон щитовидной
железы — ди-иод-тирозин является антагонистом тироксина в его
влиянии на обмен веществ.
Эпителиальные тельца, или околощито-
видные железы (gl. parathyreoidea), образующиеся из
эпителия внутренней дорзальной стороны 3-го и 4-го жаберных
мешков, занимают разное положение, будучи отодвинуты к спинной
стороне развивающейся щитовидной железой. Гормон околощито-
видных желез повышает содержание кальция в крови. Удаление
этих желез приводит к тяжелой тетании. Эпителиальное тельце
2-го жаберного мешка превращается в железку сонной ар-
терии.
Развивающиеся из 5-й пары жаберпых мешков ультимо-
бранхиальпые тельца (gl. ultimo-branchialis) рас-
полагаются в непосредственной близости от щитовидной железы;
иногда они вовсе отсутствуют.
Зобная, или вилочковая, железа (thymus) раз-
вивается у млекопитающих главным образом из 3-го отчасти 4-го
жаберного мешка. Эпителиальный зачаток железы окружается
и прорастает соединительной тканью с нервами и сосудами, раз-
деляя железу на многочисленные дольки. Зобная железа сильно
развита у молодых животных и растет до половой зрелости, после
чего она уменьшается. Ее инкрет — тимокресцин — влияет на раз-
витие, содействуя преодолеванию противоречий развития.
Гипофиза (hypophysis cerebri, или gl. pituitaria). Перед-
няя доля образуется, как известно, частью из дна третьего желу-
дочка мозга, частью из крыши ротовой полости (мешочек Рат-
ке), а от нижней стенки промежуточного мозга развивается зад-
няя доля гипофизы, которая остается соединенной с промежуточ-
ным мозгом мозговой воронкой (infundibulum). На границе между
передней и задней долями гипофизы находится промежуточный отдел
железы (pars intermedia). Гипофиза является одним из чрезвычайно
важных органов внутренней секреции и выделяет, по новейшим
данным, большое число гормонов (16—22), в том числе такие важные
гормоны, как гормон роста, гормон, ускоряющий половое соз-
ревание (пролан А и В), гормон, стимулирующий разрастание
щитовидной железы; ряд гормонов, влияющих на другие железы
наружной и внутренней секреции; лактогенный гормон, или про-
лактин, возбуждающий деятельность молочных желез; гормон,
776
вызывающий сон; гормон, повышающий кровяное давление; гор-
мои, повышающий тонус гладкой мускулатуры и др.
Эпифиза (gl. pinealis) у млекопитающих, как и у прочих
позвоночных, развивается из крыши промежуточного мозга и обра-
зует плотный железистый шишковидный орган. Гормоны этой
железы оказывают тормозящее влияние на половую деятельность
и па развитие половых органов.
Надпочечники, или околопочечные тела,
млекопитающих состоят из внутреннего мозгового вещества,
образовавшегося из эктодермы, как производное симпатической
нервной системы и из коркового вещества, образовавшегося из
мезодермы.
Мозговое вещество вырабатывает гормон — адреналин, дей-
ствующий на все органы, иннервируемые симпатической нервной
системой и поддерживающий тонус самой симпатической нервной
системы. Корковое вещество выделяет гормон кортин, вызываю-
щий расширение кровеносных сосудов и понижение кровяного
давления.
Гормональная деятельность присуща также половым ж е-
л е з а м. Они вырабатывают гормоны, определяющие развитие’
вторичных половых признаков и половых инстинктов. Кастрация
приводит к угасанию половых инстинктов и к обратному разви-
тию вторичных половых признаков.
Кастрация и последующая трансплантация половых желез
противоположного пола вызывают у подопытных животных обра-
зование ряда вторичных половых признаков противоположного
пола.
В плаценте беременных животных вырабатываются гормоны,
стимулирующие развитие молочных желез.
Наконец, в моче беременных самок млекопитающих открыт выше
упомянутый сильно стимулирующий развитие половых органов
гормон.
10. Нервная система
По развитию и степени дифференцировки нервной системы
млекопитающие стоят выше всех классов позвоночных, хотя в.
низших подклассах и отрядах млекопитающих мозг еще далеко-
пе достигает той высоты, которую мы видим у высших.
Спинной мозг млекопитающих характеризуется общим-
его укорочением и редукцией хвостового отдела. В связи с этим,
нервы, идущие от конца спинного мозга к задним частям тела,
проходя совместно по спинномозговому каналу, образуют так
называемый конский хвост (cauda equina). У человека
собственно спинной мозг кончается уже между первым и вторым
поясничными позвонками, тогда как у летучих мышей он окан-
чивается даже на девятом грудном позвонке. Однако редукция
заднего отдела спинного мозга не идет до конца, так как вместо
спинного мозга остается так называемая конечная нить (filum
terminale), т. е. тонкий рудимент, тянущийся до хвоста.
777
Головной мозг млекопитающих отличается весьма
существенно от головного мозга прочих позвоночных животных,
причем особенно характерно развитие у млекопитающих полуша-
рий переднего мозга. Однако принципиальное различие головного
мозга млекопитающих от мозга других позвоночных животных за-
ключается пе в величине полушарий, а в качественно ином их раз-
витии. Кора мозга получила здесь особенно сильное развитие,
что наметилось у рептилий лишь в виде небольших лобных отделов
первичной мантии. Уже у однопроходных и сумчатых разрастание
этой части — так называемой новой мантии (neopallium)
пошло настолько далеко, что лежащие сзади и кверху участки
первичной мантии оттесняются внутрь медиальной стенки полуша-
рий, где они образуют складки, превращаясь в так называемый
гиппокамп, или аммонов рог (hippocampus, s. cornu
Ammonis). Разрастание новой мантии у высших млекопитающих
прогрессирует все дальше и дальше, вследствие чего происходит
все большее и большее увеличение полушарий, а затем и уве-
личение их поверхностей путем возникновения складок коры мозга.
Впрочем, образование борозд и извилин па головном мозгу
нельзя считать находящимся только в зависимости от высоты
организации и нервной одаренности той или иной группы мле-
копитающих, так как установлено, что образование извилин стоит,
между прочим, в связи с величиной животных. Крупные и мел-
кие виды одного и того же отряда довольно резко в этом отноше-
нии отличаются, причем у крупных видов извилины бывают хо-
рошо развиты, у мелких же видов, наоборот, мозг остается глад-
ким. Приведем несколько примеров: маленькая луговая собачка
(Cynomys) имеет гладкие полушария, тогда как крупная
водосвинка (Hydrochoerus) имеет извилистые полушария, хотя
оба относятся к отряду грызунов; из парнокопытных упомянем
маленьких оленьков (Tragulidae) и крупных оленей или крупный
рогатый скот; из полуобезьян: долгопята (Tarsius)n индри (Indris).
Заметим, что у таких крупных млекопитающих, как киты, слоны,
извилины хорошо развиты.
Полушария головного мозга, однако, у всех млекопитающих раз-
деляются на отделы по крайней мере тремя бороздами: уже у низших
млекопитающих на нижней стороне полушарий проходит продоль-
ная ринальная бороздка (fovea rhinalis), отделяющая обонятельную
часть, так называемую грушевидную (lobus pyriformis), от мантии;
другая борозда отделяет в задней части снизу и медиально изви-
лину, образованную гиппокампом (fisusra hippocampi); третья рас-
полагается спереди, отделяя передний отдел обонятельной области,
обонятельный бугорок (tuberculum olfactorium) (рис. 570).
У высших млекопитающих дифференциация отделов коры голов-
ного мозга идет значительно дальше.
Изменения, происшедшие в развитии вторичной мантии в коре
головного мозга, обусловили изменения и во внутреннем строепии
мозга, в частности, в конфигурации боковых желудочков, в поло-
жении и строении верхней передней комиссуры, или комиссуры мап-
778
тии. Обособляются отдельные участки выросшего вместе с ростом
полушарий желудочка в виде узких щелей, получивших название
рогов: передние рога (cornu anterius), полость которых рас-
полагается над полосатыми телами, нижние рога (согни
inferius), в полость которых вдается гиппокамп, и задние рога
(cornu posterius).
В области промежуточного мозга у млекопитаю-
щих имеется небольшая эпифиза (epiphysis), которая, однако,
у неполнозубых и сирен отсутствует.
Зрительные бугры (thalami
optici) хорошо развиты; вследствие
разрастания соединяющей их мягкой
комиссуры (commissura mollis) иног-
да, как, например, у человека, III же-
лудочек оказывается разделенным на
верхний и нижний отделы. Позади
воронки лежит непарное или у при-
матов парное вздутие — так называе-
мое сосцевидное тело (corpus
mammilare); гипофиза всегда имеется,
по не бывает сильно развита.
В среднем мозгу, который
здесь крайне мал и содержит узкий
сильвиев водопровод (aque-
ductus Sylvii), обращают на себя
внимание четыре вздутия на крыше
(corpora quadrigemina), образующие
четверохолмие.
Очень хорошо развит м о з ж е -
чок, который дифференцирован на
среднюю часть (vermis), соответ-
ствующую мозжечку прочих позвоноч-
ных, и две боковых части — п олу-
ша р и я — с придатками (flocculi).
Полушария и их придатки являются
новообразованием, так же как и силь-
Рис. 570. Мозг утконоса снизу.
1 — обонятельные луковицы (bul-
bils olfactorius), 2 — грушевидные
доли, 3 — обонятельная бороздка
(fovea rhinalis), 4 — обонятельный
бугорок (tuberc. ol factorium), 5 —
конечная пластинка (lamina ter-
minalis), 6 — серый бугор (tuberc.
cinereum), 7 — сосковидные тела
(corpora mammilaria), 8 — мост,
9— придатки (flocculi), I—XII —
головные нервы. (Из Вебера по
Элиоту.)
ное развитие поперечных волокон, соединяющих друг с другом
оба полушария; располагаясь снизу продолговатого мозга волокна
эти образуют характерный для млекопитающих варолиев мост
(pons Varolii). IV желудочек слегка заходит в область мозжечка.
По бокам крыши этого желудочка идут в мозжечок волокна,
образующие задние ножки мозжечка, или веревоч-
ные тела (corpora restiformia); снизу валики продольных во-
локон образуют пирамиды. У приматов и у некоторых других
млекопитающих кнаружи от пирамид лежат еще небольшие утол-
щения — оливы.
Из особенностей, периферической нервной системы млекопитаю-
щих необходимо указать на полное обособление от блуждающего
нерва (X) добавочного нерва (XI) (n. accessorius). Он является чи-
779
I—XII.
571.
нервы кошки:
Головные
ophtalmicus, , Va — ramus maxillaris,
Рис.
— ramus . х__________, _____ ___________,
V3 — ramus mandibularis, 1 — nervus infraorbitalis,
2 — верхняя прямая мышца глаза, 3 — боковая,
4 — нижняя, 5 — косая мышца глаза, 6 — n. denta-
1 is, 7 — n. lingualie, 8 — chorda tympani, 9 — gang-
lion gasseri, 10 —ganglion vagi, 11 — n. laryngeus.
(По Миварту, из Вебера.)
сто висцерально двигательным нервом и иннервирует трапециевид-
ную и грудино-ключично-сосцевидную мышцы.
К комплексу головных нервов у млекопитающих прибавляется
еще XII — подъязычный нерв (n. hypoglossus); этот последний со-
стоит из соматических
двигательных волокон и
иннервирует мускулату-
ру языка.
Относительно распре-
деления по голове вет-
вей головных нервов да-
ет представление рис.
571.
11. Органы чувств .
Органы осязания у
млекопитающих разви-
ты хорошо, хотя и не-
равномерно. Что касает-
ся отдельных осязатель-
ных клетокили ихгрупн,
объединенных в тела
М е р к е л я, то они на-
блюдаются лишь па ме-
стах, лишенных волос;
более сложно устроенные
осязательные органы из клеток, окруженных соединительной ткапыо,
с ветвящимся среди них первом —. тельца Мейснера — распола-
гаются в соединительной ткапи под кожей; такие чувствующие ор-
ганы развиты на мякишах пальцев передних и задних конечностей
у лазящих животных. Очень тонкими осязательными органами яв-
ляются у летучих мышей летательные перепонки, наружные уши
и носовые выросты, позволяющие им осязать малейшее сотрясение
воздуха о предметы, производимое ими во время полета; конец рыла
свиньи, конец хобота слона тоже очень восприимчивы к осязатель-
ным раздражениям вследствие обилия в коже осязательных телец.
Кроме того свободные нервные окончания в эпителии также служат
для восприятия осязательных раздражений. На ряду с этим опи
воспринимают также и температурные раздражения.
Однако главными органами осязания у млекопитающих являются
волосы. Мы уже видели, что во влагалищах корней волос всегда
имеются нервные окончания, и каждый волос является, таким об-
разом, органом осязания. Одпако имеются и специализированные
осязательные волосы, или в и б р и с ы (pili). Такие
волосы отличаются своей длиной, прочностью, ранним, появлением
в эмбриональном развитии и дольше удерживаются после потерн
волосяного покрова. Волосы эти сидят глубоко в коже, окру-
женные кровеносным синусом и многочисленными нервными развет-
влениями по поверхности корня волоса; они расположены ио опреде-
780
лепным местам, в особенности же на голове. Различают pili
labiates, сидящие па губах, pili submentales — па подбородке,
pili supraorbitales — над глазами, pili angulares — по углам
рта и pili zygomatici на — щеках или в области скуловой дуги.
Такие волосы разбросаны также по телу; особенно большую роль
опи играют у млекопитающих, пользующихся норами; наконец,
имеются вибрисы и на конечностях, в области запястья.
Органы равновесия и слуха у млекопитающих развиты очепь
хорошо, и именно в этом классе орган слуха достигает своего наи-
высшего развития, в то время как орган равновесия сохраняет приб-
лизительно тот же уровень, на котором он был уже у рептилий.
В слуховом аппарате произошли во всех частях весьма существен-
ные измепепия. В наружном ухе достигает значительной длины
наружный слуховой проход и развивается наружная слу-
ховая раковина (concha), которая приводится в движение
специальной мускулатурой и служит для собирания звуков. Опа
редуцируется, а затем и исчезает совсем в связи с приспособлением
к водному или подземному образу жизпи. В среднем ухе, как мы
Рис. 572. Схема органа слуха млекопитающих. Вид
спереди и сбоку, правое ухо. I — наружное, II —
среднее, III —внутреннее ухо.
1 — барабанная перепонка, 2 — молоточек, 3 — наковальня,
4 — стремячко и musculus stapedius, 5 — tensor tympani, 6
и 7—chorda tympani и н. facialis, 8 — слуховой нерв, 9 —
овальное окошечко (f. ovale), 10 — круглое окошечко (f. го-
tundum), 11 — перилимфатический канал, 12 — эндолимфа-
тический канал, 13 — желудочек, 14 — мешочек, 15 — улит-
ка, 1 — барабанная лестница, 17 — евстахиева труба. (Из Ве-
бера.)
уже видели, усложняется и совершенствуется передаточный аппа-
рат путем присоединения к столбику, или стремени уха (columella
auris, stapes), еще двух косточек — наковальни (incus =
781
quadratum) и м о л о т о ч к a (malleus=articulare+goniale). Сотря-
сение барабанной перепонки звуками передается по косточкам пере-
понке овального окна. Имеется мускул (m. tensor tympani), натя-
гивающий барабанную перепонку и прикрепляющийся к ручке мо-
лоточка, тогда как мышца стремени (m. stapedius) натягивает пере-
понку овального окошка. Барабанная полость, т. е. полость сред-
него уха, заходит в ограничивающие ее кости (mastoideum, tympa-
nicum и squamosum) (рис. 672).
Однако особенной сложности достигает строение внутреннего
уха. Именно здесь разрастается в улитку (cochlea) выступ круглого
мешочка (sacculus), причем канал улитки соединяется с полостью
круглого мешочка (sacculus) посредством тонкого соединительного
канала (canalis reuni ens); в свою очередь мешочек соединяется с
овальным мешочком, или желудочком (utriculus), посредством эндо-
лимфатического канала (ductus endolymphaticus). Перепончатый
лабиринт, окруженный перилимфатической жидкостью, л ежи г
в костном лабиринте, заключенном в каменистой кости (os petrosum).
В костном лабиринте имеются два отверстия в среднее ухо, из кото-
рых верхнее овальное отверстие прикрыто стременем. Нижнее круг-
лое отверстие затянуто барабанной перепонкой (membrana tympani
secundaria).
Слуховой нерв (n. acusticus, VIII) дает три ветви, подходя глав-
ной своей массой (n. cochlearis)
к улитке, где располагается
очень сложно устроенный слу-
ховой аппарат — кортиев
орган. Дно перепончатой улит-
ки натянуто между внутренней
и наружной стенками костной
улитки, причем с внутренней
стороны в улитку заходят нерв-
ные волокна от лежащего вдоль
улитки спирального ганглия
(ganglion spirale). Наружная
сторона основной перепонки
(membrana basilaris) (так назы-
вается дно улитки) прикреп-
ляется к наружной стенке спи-
ральной связкой (ligamentum
spirale); поперек перепонки на-
тянуты многочисленные соеди-
нительнотканные нити; над пе-
репонкой помещается восприни-
мающий звуковые раздражения
кортиев орган, состоящий из чувствующих клеток, соединенных
с нервными волокнами, и поддерживающих клеток разного строе-
ния (рис. 573).
Глаза у млекопитающих характеризуются немногими особен-
ностями, прежде всего отрицательными: полным отсутствием око-
782
Рис.‘573. Кортиев орган.
1—основная перепонка, 2 — спираль-
ная пластинка, 3 — чувствующие клетки,
4 — слуховой нерв с ганглием, 5 — клет-
ки столбов, 6 — средняя лестница (канал
улитки), 7 —лестница преддверия, 8 —
барабанная лестница, (Из Иле, Кампен
и др. по Гессе.)
стенепий в склеротике и отсутствием гребешка или сходного с ним
органа; иногда имеется отражательное зеркальце (tapetum), боль-
шей частью три века, т. е. имеется мигательная перепонка (у чело-
века и обезьян зачаточная); верхнее и нижнее веко снабжены рес-
ницами. Хорошо развита слезная железа; слеза стекает по слезному
каналу в носовую полость (ductus naso-lacrymalis). Нередко
имеется еще гардерова железа в области верхнего века. У основания
век иногда лежат видоизмененные сальные железы — мейбомовы
железы.
Форма глазного яблока довольно различна. Хрусталик у водных
животных по форме приближается к форме шара. Зрачок иногда бы-
вает в виде щели.
У сумчатого крота (Notoryctes typhlops) нет вовсе глаз; у
других отдельных подземных обитателей они нередко скры-
ты под кожей (златокроты и слепыши) или не функционируют
(крот).
Органы обоняния у млекопитающих, подобно органам слухаг
достигают наибольшего совершенства, хотя они редуцируются у вод-
ных животных.
Обонятельная часть носовой полости, имеющей, как мы уже виде-
ли при рассмотрении скелета, многочисленные раковины, сосредото-
чивается в задней части в области раковин решетчатой кости (ethmo-
turbinalia). Раковины эти возникают на боковых стенках (lamina
lateralis) этмоидальной кости. В простейшем виде они образуют
нередко ветвящиеся, закрученные внутрь пластинки; в более слож-
ных случаях между ними образуются дополнительные раковины,
между которыми в свою очередь могут возникнуть дополнительные
раковины. Тогда первоначально образовавшиеся, как наиболее
внутрь лежащие, получают название внутренних раковин (endotur-
binalia) (рис. 535, стр. 732). У животных с сильно развитым обо-
нянием число раковин больше, они длиннее и имеют более слож-
ную форму. Над обонятельными раковинами и внизу от них по-
мещается носовая раковина (nasoturbinale), а спереди и снизу
раковина челюстной кости (maxilloturbinale). Якобсонов орган
у млекопитающих вполне обособлен и отсутствует лишь у неко-
торых групп (обезьян).
Для млекопитающих характерен наружный мясистый нос, кото-
рый у многих форм вырастает в хобот со сложной мускулатурой.
Органы вкуса весьма сильно развиты у млекопитающих. Вку-
совые почки помещаются прежде всего па языке, на так назы-
ваемых грибовидных сосочках (papillae fungiformes), на
их свободной поверхности, затем па окруженных валом
сосочках (papillae circumvallatae), где имеется приспособление
для удержания растворенных веществ в непосредственной близости
от вкусовых почек, расположенных по бокам. сосочка на уровне
ямки, окружающей сосочек; наконец, вкусовые почки поме-
щаются по сторонам* задней части языка, где они сидят
на расположенном по одному с каждой стороны листовидном
сосочке.
783
III. ПРОИСХОЖДЕНИЕ МЛЕКОПИТАЮЩИХ
Перечисленные выше особенности млекопитающих вполне опре-
деленно указывают на то, что предками млекопитающих могли быть
только рептилии и притом такие, которые обладали некоторыми осо-
бенностями, отсутствующими у современных рептилий. Так,
у всех известных нам современных рептилий, как бы далеко опи
пи стояли друг от друга в систематическом отношении, развивается
сильнее правая дуга аорты, которая выходит из левого желудочка,
тогда как левая дуга выходит из правого, между тем у млекопи-
тающих имеется только левая дуга аорты, которая выходит из левого
желудочка; у всех современных рептилий кожа утеряла многочис-
ленные железы, которые были свойственны их предкам — амфибиям,
сохранив лишь самое ограниченное их число в виде желез специаль-
ного значения; кожа покрыта ороговевшими производными эпидер-
миса — чешуей, тогда как млекопитающие имеют кожу, богатую
железами, и одеты покровом из волос, происхождение которых,
согласно данным эмбриологических исследований, можпо допустить
лишь из ранних стадий развития чешуй. Череп рептилий имеет один
затылочный бугор, тогда как у млекопитающих он имеет два затылоч-
ных бугорка как у амфибий. Как у этих последних, у млекопитающих
число фаланг пальцев 2,3,3,3,3, и отсутствует падгрудинпик. Строе-
ние среднего уха с тремя слуховыми косточками, из которых нако-
вальня и молоточек преобразовались первая из квадратной, а второй
из сочленовной косточки. Сочленение нижней челюсти, состоящей
из зубной кости (dentale), с чешуйчатой (squamosum), совершенно
отлично от того, что мы встречаем у современных и громадного
большинства ископаемых рептилий.
Из сказанного следует, что млекопитающие могли произойти
только от таких рептилий, которые еще не успели приобрести ука-
занные признаки, отличающие их от млекопитающих, и стояли
ближе к своим предкам — амфибиям. Таковыми, повидимому,
являются пермские рептилии из отряда зверозубых (Theriodontia)
подкласса звероподобных (Theromorpha).
Большинство этих рептилий вели хищный образ жизни и обна-
руживали поразительное сходство с млекопитающими в строении
зубов, в общей форме черепа и в общей форме тела. Зубы у них диф-
ференцировались па резцы, клыки и коренные и помещались в аль-
веолах, подвергаясь однократной смене. Квадратная кость сильно
редуцировалась, как и элементы нижней челюсти, за исключением
зубной (dentale) которая, вследствие прогрессирующего развития зу-
бов, увеличивалась, доходя до скулового сочленения. В черепе была
одна скуловая дуга, состоявшая, как и у млекопитающих, изотро-
стка чешуйчатой кости (squamosum) и скуловой (jugale); имелся
длинный слуховой проход; затылочный бугорок у некоторых форм
был двойной. Сходство было и в делении позвоночника на отделы,
и в строении плечевого пояса, таза и конечностей.
Рептилии эти бегали на высоких ногах, построенных и ориен-
тированных в отношении туловища так же, как и ноги млекони-
тающих; туловище, было высоко поднято, на ногах, которые имели
сходное с ними строение кисти и ступни с числом фаланг 2, 3, 3, 3, 3.
У териодонтов имелся olecranon, лопатка была с гребнем и
акромиальным отростком, а коракоиды имели уже тенденцию
к редукции.
В то же время большинство известных нам представителей зве-
розубых ящеров, на ряду с этими признаками, обладали рядом спе-
циализированных особенностей. Прямыми предками млекопита-
ющих, следовательно, были более примитивные Theriodontia. Они,
следует думать, были близки к амфибиям по особенностям своей
кожи, снабженной многочисленными железами, лежащими между
чешуями, где имелись не специализированные зачатки чешуй,
которые послужили материалом для развития волосяного покрова;
у них очевидно левая дуга аорты была развита сильнее правой и отхо-
дила от левого желудочка. Такие териодонты могли быть только среди
примитивных форм, близких к котилозаврам, а, следовательно,
среди пермских териодонтов. У нас имеются все основания предпо-
лагать, что это были мелкие животные, так как первые млекопита-
ющие были именно мелкими формами и притом вели полудревес-
пый образ жизни, потому что наиболее примитивные пятипатые
конечности современных млекопитающих — это копечности опос-
сума (Didelphys), ведущего полудревесный образ жизни, при кото-
ром пятипалая конечность может сохранить свое наиболее универ-
сальное строение. В последнее время палеонтология обогатилась
новыми данными, подтверждающими эти предположения. В 1927 г.
описан был (Сушкиным) териодонт Permocynodon, а в 1929 г. три-
асовые териодонты из Южной Африки Ictidosauria (Брумом),
которые стояли чрезвычайно близко к млекопитающим и были не-
большой величины.
Мало этого, у нас есть основание думать, что родиной млекопи-
тающих были части суши, где климатические условия могли содей-
ствовать развитию теплокровности, и что родоначальные мелкие те-
риодонтные звероящеры были энергичными, проворными формами,
так как энергичные окислительные процессы в связи с движениями
при большем, чем у рептилий, совершенстве кровеносных органов
и органов дыхания являются необходимыми предпосылками для
приобретения теплокровности. В самом деле, при отсутствии
теплокровности всякое понижение температуры вызывало неизбеж-
ную депрессию и даже окоченение холоднокровных животных;
отсюда огромное преимущество первых теплокровных животных
в борьбе за существование. Теплокровность связана с развитием
волос и потовых желез как терморегуляторов.
Повышенная жизнедеятельность и теплокровность, требовавшие
большой траты энергии, могли появиться только с усилением обмена
веществ и более обильным питанием. Это в свою очередь не могло
пе быть связано с прогрессивным развитием зубов и органов чувств,
в особенности слуха и обоняния. Все эти приобретенные пред-
ками млекопитающих свойства могли развиться только в связи
с прогрессивным развитием центральной нервной системы, что
^0 Каш каре в к Станчшский 785
выразилось в усложнении строения мозга и развитии neopal-
lium.
Однако этого все же, повидимому, было недостаточно для того,
чтобы быстро эволюционировать в жизненной борьбе.
Первозвери Prototheria — первый этап эволюции, характери-
зующийся теплокровностью при волосяном покрове и яйценоскостью
при наличии клоаки. Нам кажется особенно вероятным это пред-
положение потому, что млекопитающих удерживало что-то от.бы-
строй эволюции в первое время вторичной эры. Приобретенная теп-
локровность при яйценоскости требовала высиживания яиц, а это
представляло большие затруднения, если принять во внимание оби-
лие всевозможных хищных рептилий. Только сумчатость, а затем
живородпость в связи с сумкой и, наконец, плацентарное вынашива-
ние детенышей в матке дали ряд последовательных этапов эволю-
ции, приведших, в копце копцов, к пышпому расцвету класса мле-
копитающих в самом конце мелового периода. Этот последний этан
совершился, как то показали недавние открытия американских па-
леонтологов в Монголии, повидимому, в Азии, где, вероятно, хра-
нятся в необъятных просторах пустынь, льдов и лесов самые ин-
тересные страницы первобытной истории класса млекопитающих.
IV. КЛАССИФИКАЦИЯ МЛЕКОПИТАЮЩИХ
' Подразделение на подклассы, отряды и более мелкие система-
тические единицы современных млекопитающих не представляет
трудностей, так как промежуточные формы хорошо дифференциро-
ванных систематических групп вымерли. Совсем другое, когда при-
ходится иметь дело с ископаемыми млекопитающими, куда входят
многочисленные переходные формы. Трудности увеличиваются
здесь еще потому, что от ископаемых форм ^сохранились только ске-
леты, а от низших, наиболее интересных и важных в систематиче-
ском отношении мезозойских млекопитающих, по большей части
сохранились лишь черепа или всего лишь нижние челюсти. А так
как основные подразделения млекопитающих на подклассы осно-
ваны на особенностях, связанных с воспроизведением и со строе-
нием органов воспроизведения (яйценоскость, клоака, сумка, пла-
цепта, строение молочных желез), о состоянии которых у ископа-
емых животных мы судить не можем, то отнесение ископаемых,
особенно мезозойских млекопитающих, к тому или ипому подклассу
является чрезвычайно затруднительным. Приходится довольство-
ваться теоретическими соображениями, косвенно освещающими этот
вопрос. Мы полагаем, что эволюционный путь млекопитающих,
прошел через следующие основные этапы развития: 1) стадия яй-'
ценоскости без инкубации в сумке, 2) стадия инкубации в сумке
яйца, 3) стадия инкубации яйца в сумке и выкармливания и вына-
шивания в ней детенышей, 4) стадия живородности с последующим .
выкармливанием и вынашиванием детеныша в сумке или стадия j
живородности и плацептарности без последующего вынашивания j
754
i>, еумке. Среди современных млекопитающих мы имеем уцелевшими
стадии 2 и 4. Если бы и 3-я стадия уцелела, мы отнесли бы ее
к первому подклассу однопроходных, так как яйценоскость и свя-
занное с ним строение половых выводных путей мы считали бы
определяющими. Об отношении ископаемых млекопитающих к этим
стадиям, к сожалению, мы ничего не знаем, однако мы знаем, что
млекопитающие прошли несколько последовательных стадий в раз-
витии коренных зубов. Эти стадии таковы: 1) стадия протодоптная,
2) стадия триконодонтная, 3) стадия тритуберкулярная и затем
стадии последующей дифференциации зубов в связи с приспособле-
ниями к использованию различной пищи. У нас нет данных считать,
что стадии развития воспроизведения соответствуют стадиям раз-
вития зубов, по совершенно несомненно, что низшие стадии разви-
тия должны были совпадать: яйцекладущие однопроходные без
инкубации в сумке должны были обладать протодонтной стадией
зубов. Вот почему протодонтпых млекопитающих мы объединяем
в одип подкласс с однопроходными. Весьма вероятно, что млекопи-
тающие с трикоподоитпыми зубами должны быть отпесепы сюда же.
Только потому, что наличие у пих углового отростка пижней челю-
сти напоминает сумчатых, мы объединяем их в один подкласс с этими
последними.
Подкласс 1. Первозвери (Pvototheria)
Осповпой особенностью этого подкласса являются унаследован-
ные от рептилий яйценоскость и наличие клоаки при характерной
для млекопитающих теплокровности и волосяном покрове и целом
ряде других особенностей класса. Молочные железы трубчатые и не
имеют еще сосков, а открываются многочисленными отверстиями
па железистом поле; имеется или редуцирована ипкубаторная сумка,
в которой вынашиваются яйца.
В современной фауне представлены отрядом однопроходных —
Monotrernata. Из ископаемых форм по указанным выше соображе-
ниям мы отпосим сюда первично-
зубых — Protodonta.
Сюда же мы относим мезозой-
ских Multituberculata, так как по
строению их зубы похожи на вре-
менные зубы утконоса.
Рис. 574. Нижняя челюсть Dro-
Отряд!. Псрвичпозубые (Protodonta) math (^вебера'Г^'
Мезозойские млекопитающие с
наиболее просто устроенными зубами, в которых низкие допол-
нительные бугорки располагаются в одной плоскости с главным
бугорком; известны только по сохранившимся нижним челюстям
из верхнего триаса.
' Дроматерий (Dromatherium) (рис. 574) из верхнего триаса Сев. Аме-
рики. Мигроконодон (Microconodon) и др.
50* 787
Отряд 2. Однопроходные (Monotremata)
Зубы отсутствуют или встречаются только в молодости; перед-
ний отдел морды превращен в клюв, одетый роговым чехлом; в пле-
чевом поясе имеются прокоракоиды и коракоиды; в тазовом —
сумчатые кости.
Ограничены в своем распространении Австралией, Тасманией и Новой
Гвинеей.
Сем. Ехидновые (Echidnidae). Кроме волосяного покрова,
одеты иглами; узкий изогнутый клюв и червеобразный язык; у самок
имеется выводковая сумка. Два рода, Echidna и Zaglossus. (Echidna aculeata,
Zaglossus bruyni (рис. 575).
Сем. Утконосовые (Ornithorynchidae). Клюв, как
у утки, широкий, сумка отсутствует; ведут водный образ жизни. Яйца вы-
сиживают в гнезде. Один род и вид Ornithorynchus paradoxus (рис. 576).
Отряд 3. Многобугорчатые (Multituberculate, Allotheria)
Мезозойские небольшие и мелкие млекопитающие со своеобраз-
ным устройством зубов: коренные зубы с 2—3 продольными рядами
бугорков, тогда как Р4 отличается большой высотой и режущей
Рис. 575. Черноиглая ехидна (Zaglossus bruyni).
(Из Брэма.)
поверхностью. Появились в триасе и вымерли в эоцене; распро-
странены были в Европе и Сев. Америке.
Сем. Plagiaulacidae. Верхняя юра Европы и Сев. Америки, Р1а-
giaulax, Новая Мексика, эоцен, величиной с бобра.
Подкласс II. Низшие звери (Metatheria)
Сюда, кроме хорошо характеризующихся современных сум-
чатых животных (Marsupialia), мы относим отряд трех-
конусозубых (Triconodonta). Основная характеризующая
подкласс особенность этих животных — рождение недоразвитых
детенышей, которые развиваются в сумке. Есть сумчатые кости
(в случае отсутствия сумки наличие последних указывает на редук-
788
цию бывшей ранее сумки). Нижняя челюсть снабжена обычно бол
нгим, направленным внутрь угловым отростком (proc, angulari
твердое нёбо с отвер- Б
ггиями.
Отряд 4. Трехконусо-
зубые (Triconodonta)
В этот отряд мы
включаем пе только
тех мезозойских мле-
копитающих, у кото-
рых коренные зубы
имеют по три сидящих
в одной плоскости ко-
нуса — триконодопт-
ные зубы, но и тех, у
которых дополнитель-
ные конусы распола-
гаются внутрь в ниж-
ней и наружу в верх-
ней челюстях, т. е.
трехбугорчатые—три-
туберкулярные зубы,
а также тех, у кото-
рых появляется поза-
ди пятка — тритубер-
кулярпо - векториаль-
ные зубы; первона-
чальное число зубов:
1ср|.
Принадлежность
этих животных к сум-
чатым очень условна,
так как опи занимают
но строению зубов пе- А?
реходное место между
сумчатыми и плацен-
тарными и так как
нижняя челюсть не у *
всех них имеет угло-
вой отросток, направ-
ленный внутрь. Три
подотряда.
Рис. 576. Утконос (Ornithorhynchus anatimun
(Из Гаане.) '•
Рис. 577. Коренные зубы TpexKOHyconvfit.v л
Amphitherium, В - Tinodon, С —- Triconodon~
(Из Вебера.)
конусозубые (в узком смысле) it ,;
пис. S77. Cl Arnnhilcustes 1
Подотряд 1. Трех „ ------------------- _---------- „„„
и о d о п t a) Triconodon (рис. 577, С), Amphilestes, Phascolotherium ~ £
789
верхнеюрских отложений Англии; Priacodon, Phascolodon и др. —из верхней
юры Сев. Америки.
Подотряд 2. Симметрозубые (Symmetrodonta). Три-
туберкулярные зубы без пятки Spalacotherium, Peralestes и др. — из верх-
ней юры Англии; Tinodon — из верхней юры Сев. Америки (рис. 577 В).
Подотряд 3. Трехбугорчатые [Trituberculata
(Pantotheria)] — с тритуберкулярно-секториальными зубами (Ат-
phitherium) и др. — из верхней юры Англии (рис. 577 A); Phascolestes,
Stylodon и др. — из верхней юры Сев. Америки.
Отряд 5. Сумчатые (Marsupialia)
Помимо особенностей, указанных в характеристике подкласса,
следует упомянуть наличие у самок двойного влагалища и двой-
ной матки, у самцов penis обычно на конце раздвоен; детеныши
развиваются в матке обычно без плаценты, но иногда имеется
аллонтоидная или желточная плацента. Зубы за исключением
Рг, Рг имеют лишь одну генерацию; число ложнокоренных зубов
колеблется между тремя и одним. Коренных зубов обычно бывает
четыре. Коракоиды редуцируются до отростка лопатки (proc, сога-
coideus). В мозгу пет мозолистого тела. Мошонка у самцов впереди
пениса, семяпроводы не перекрещиваются с уретрами. Клоака
редуцирована.
230 видов распространены в настоящее время в Австралии с Тас-
манией и Новой Гвинеей; немногие виды заходят на Новую Брита-
нию, Ару, Кей, Сангир, Целебес и др.; семейство двуутробок (Didel-
phyidae) и ценолест (Caenolestidae) обитают в Южной Америке,
откуда первые заходят в Сев. Америку. В конце юрского периода
и в меловое время, сумчатые были распространены по всему земному
шару и вымерли в Арктогее в третичное время, дожив в Европе до
начала миоцена и до конца олигоцена в Сев. Америке. Три подотряда.
Подотряд 1, Многорезцовые (Polyprotodontia)
Сюда относятся насекомоядные, хищные или — реже — всеяд-
г т5 4 3
ные сумчатые с большим числом резцов: I— или — редко —; пиж-
т: О О
ние средние резцы пе удлинены; древесные или наземные, иногда
водные и подземные животные.
Сем. 1. Двуутробковые или сумчатые крысы (Didelphyi-
d а е). Американские насекомоядные или всеядные сумчатые, наиболее
примитивные из современных. Сумчатые крысы, или опоссумы (Didelphys) —
древесные животные, заменяющие в Южной Америке насекомоядных живот-
ных, например, тупай Восточной области; Peramys — сумчатая землеройка;
Chironectes — плавающая сумчатая крыса (вернее, экологически сумчатая
выхухоль).
Сем. 2. Сумчатые куницы (Dasyuridae). Насекомоядные
f4 3
мелкие животные, от которых произошли более крупные хищники. 1у, — ,
Австралия и восточная часть Индо-Австралийских островов. Сумчатые земле-
ройки (Phascologale); сумчатый дьявол (Sarcophilus) — довольно крупный
хищник Тасмании (рис. 578).
790
Сем. 3. Сумчатые волки (Thylacinidae). Единствен-
ный род и вид — сумчатый волк (Thylacinus cynocephalus), крупный хищник
Тасмании.
Сем. 4. Сумчатые мурашееды (Myrmecobiidae).
Единственный род и вид, распространенный в Австралии, — Myrmecobius
fasciatus — питается муравьями; замечателен между прочим и тем, что самка
нс имеет сумки и детеныши висят на сосках, спрятанные в пушистом меху
брюха.
Сем. 5. Сумчатые кроты (N о t о г у с t i d а с). Единствен-
ный род и вид — Notoryctes typhlops в Южной Австралии.
Рис. 578. Сумчатый дьявол (Sarcophilus sataninus).
(Из Брэма.)
Сем. 6. Сумчатые барсуки (Peramelidae). Роющие
насекомоядные или всеядные сумчатые, величиной с кролика, Австралия
и Новая Гвинея. Сумчатый барсук — Perameles.
Подотряд 2. Ценолестовые (Caenolestoldea)
Занимают промежуточное место между многорезцовыми и дву-
резцовыми сумчатыми. Число резцов, как у многорезцовых боль-
ше двух в нижнеи челюсти: I посредине резцы удлинены, как
у двурезцовых; пятипалая конечность. Самки без сумки, с одной
парой сосков. Небольшие ночные животные, обитающие в Южной
Америке в Андах, Боливии и Венецуэлле. 2 рода с 3 видами.
Ближайшими родственниками этих видов являются ископаемые
южноамериканские сумчатые из семейства Epanortnidae.
Сем. 7. Ценолестовые (Caenolestidae). Род Caenolestes; С.
fuliginosus.
Подотряд 3, Двурезцовые ( Dlprotodontla)
Растительноядные — травоядные или плодоядные, реже насеко-
моядные и всеядные сумчатые с характерным числом (2) удлиненных
• 3
резцов в нижней челюсти, I —; М в связи с растительной пищей
буподонтны, лофодонтны или селенодонтны; пальцы 2-й и 3-й более
или мепее укорочены, 4-й имеет тенденцию к увеличению, 1-й про-
тивополагается у первичнодревесных, исчезает у наземных. Хорошо
развитая сумка, максимум 4 соска. В Австралии, Тасмании и Новой
Гвинее.
791
Сем. 8. Сумчатые медведи (Phascolarctidae). Это
семейство объединяет лазающих сумчатых медведей, куда относится хорошо
известный коала (Phascolarctus cinereus), и роющих типичных сумчатых гры-
зунов — вомбатов (Phascolomys), представленных в Австралии несколькими
видами.
Сем. 9. Лазающие сумчатые (Р h а 1 a n g е г i d а е). Сюда
относятся главным образом древесные растительноядные лазающие живот-
ные, величиной колеблющиеся между величиной кошки и мыши; ноги пяти-
палые с противополагающимся первым пальцем. Кускус (Phalanger) со сход-
ным с опоссумами строением ног; наиболее крупные широко распростра-
ненные виды. Сумчатые белки (Acrobates) с летательной перепонкой (рис. 579)
и Gymnobelideus без летательной перепонки, сумчатые летяги (Petaurus),
род белкообразных сумчатых с хорошо развитой летательной перепонкой;
сумчатые сони (Distoechurus) без летательной перепонки, мелкие лазающие
зверьки, пяткоход (Tarsipes) и др.
Сем. 10. Кенгуровые (Macropodidae). Прыгающие,
с удлиненным плюсневым отделом, растительноядные, преимущественно
травоядные животные. Кускусные кенгуру (Hypsiprimnodon), величиной
с крысу, недавно приспособившиеся к жизни на земле, а потому имеющие
в себе ряд признаков, свойственных древесным кускусам. Кенгуровые
крысы (Potorous) и др. мелкие, величиной с крысу и кролика, кенгуру.
Настоящие кенгуру (Macropus, Halmaturus и др.) — высоко специализиро-
ванные к быстрому бегу прыжками животные, начиная от величины кролика
до величины в рост человека (рис. 580). Горные кенгуру (Petrogale), древес-
ные кенгуру (Dendrolagus), вторично приспособившиеся к жизни на де-
ревьях кенгуру.
Подкласс III. Высшие звери (Eutheria)
Этот подкласс в противоположность первым двум можпо назвать
одноутробными животными—Monodelfia—в отличие от двуутробок,
так как при отсутствии сумки эти животные вынашивают детены-
шей только в матке. Их называли также плацентарными животны-
ми — Placentalia, что, однако, неправильно, так как плацента пе
является признаком, характеризующим только высших зверей:
плацента встречается, как то мы видели выше, уже у некоторых
сумчатых животных, да и сумка у многих сумчатых уже исчезла.
Тем не менее именно развитие плаценты в связи с рождением более
совершенных детенышей, могущих сосать самостоятельно молоко
матери, является основным свойством высших млекопитающих,
тем повььм качеством, которое позволило млекопитающим эволю-
ционировать дальше, приспособляясь к самым разнообразным усло-
виям обитапия. Матка только как исключение двойная с призна-
ками исчезнувшей разделенности влагалища;, penis простой; семя-
проводы перекрещиваются с мочеточниками; твердое нёбо имеет из
отверстий только for. incisivi. В нижней челюсти, если имеется раз-
витой угловой отросток (proc, angularis), он не бывает загнут внутрь.
Этот подкласс может быть разделен по характеру приспособле-
ний и по генетическому родству в следующие сверхотряды:
I. Насекомоядные (Insectivora), объединяющие насе-
комоядных в узком смысле (Insectivora s. str.), летучих мышей (Chi-
roptera), шерстокрылов (Dermoptera) и ископаемых Tillodontia.
II. Неполнозубые (Edentata) с отрядами: ящеры (Pholidota)
и собственно неполнозубые (Xenarthra). III. Грызуны (Roden-
792
Рис. 579. Сумчатая соня (Acrobates pygmaeus).
(Из Брэма.)
7ЭЗ
Рис. 580. Красный кенгуру (Halma-
turus rufus).
(Ив Гаане.)
tia). IV. Хищники (Carnivora) с отрядами: креодонты (Creodonta),
хищные в узком смысле (Fissipedia), ластоногие (Pinnipedia), ки-
тообразные (Cetacea). V. Копытные (Ungulata) с отрядами:
жиряки (Hyracoidea), хоботные (Proboscidea), эмбритоподы (Еш-
brithopoda), сирены (Sirenia), нотунгуляты (Notungulata), пиро-
терии (Pyrotheria), двукопытные (Artiodactyla), однокопытные
(Perissodactyla). VI. Трубкозубы (Tubulidentata) и VII. При-
маты (Primates) с отрядами: полуобезьяны (Prosimiae) и обезья-
ны (Simiae).
Отряд 6. Насекомоядные (Insectivora)
Наиболее примитивные из высших зверей, появившиеся еще в кон-
це мезозоя, из которых развились все прочие сверхотряды и отря-
ды. Это по большей части мелкие плотоядные, преимущественно
насекомоядные млекопитающие. Первоначальная зубная формула
3 14 3 2
I С р , Р —, М —; резцы могут быть редуцированы до I - , клыки
очень часто похожи или на резцы, или па ложнокоренпые; эти пос-
ледние и коренные примитивны — триконодоптны или тритуберку-
ло-секториальпы, с тенденцией к квадритуберкулярности. Мозго-
вая полость маленькая. Матка раздельная и двурогая. По большей
части ночные животные, ведущие скрытый образ жизпи, стопохо-
дящие и полустопоходящие, обычно пятипалые с когтями, нередко
приспособленные к подземному или водному и изредка к древесному
образу жизни.
Два мезозойских семейства, недавно описанные из верхнемеловых
отложений Монголии, представляют интерес, потому что в них обособились
уже два типа: тип насекомоядных — сем. Zalambdalestidae
и тип низших хищных — Deltatheriidae. Такой же архаический
переходный характер к древним хищникам (Creodonta) имеет эоценовое
семейство Pantolestidae, несколько специализированное вероятно водным
образом жизни. Другое эоценовое семейство Plesiadapidae, наоборот,
является переходным к приматам, соединяя насекомоядных с полуобезья-
нами.
Сем. Тупайевые (Tupaiidae). Напоминающие белок, дневные,
j.9 1 2-3*4 1.2-3
древесные, с зубной формулой I Су, у М^ , распространен-
ные начиная с северо-восточного Индокитая через Малайский архипелаг
до Филиппинских островов. Тана (Tupaia tana). 1
Сем. Прыгунчиковые (Macroscelidae). Мелкие, напоми-
нающие тушканчиков, прыгающие, полустопоходящие, насекомоядные,
с длинным хвостом и узкой мордочкой, вытянутой в хоботок. Берцовые
кости срастаются, плюсневые кости удлинены, зубная формула: 1 —-—,
о
С у, Ру, Му. Распространены в Африке. Обыкновенный прыгунчик
(Macroscelides typus) Южная Африка.
Сем. Златокротовые (Chrysochlorid-ae). Ведут подобно
кротам подземный образ жизни, похожи на кротов, но со специализирован-
1 Некоторыми авторами сем. тупай относится к приматам, а именно
они составляют низший отряд последних, переход от насекомоядных к полу-
обезьянам. Тупайи совмещают в себе черты тех и других.
794
Рис. 581.. Златокрот (Chrysochloris aurea).
(Из Брэма./
ными передними конечностями по-
добно сумчатым кротам (Notoryc-
tes); имеют v золотистозеленый ме-
таллический блеск шерсти. Зубная
3 1 3 3
формула I —, С j, Р у, М у.
Распространены в Южной и Цент-
ральной Африке; златокрот—Chry-
sochloris aurea (рис. 581).
Сем. Тенрековые (Сеп-
1 е t i d а е). Ограниченное Мада-
гаскаром семейство, чрезвычайно
интересное тем, что среди него
в связи с малым количеством на
острове других млекопитающие
развились формы, аналогичные
хищным, мелким наземным рою-
щим или водным насекомоядным;
зубная формула, как у златокро-
та. Иглистый тенрек (Ericulus)
похож на ежа; некоторые тенреки
(Centetes) заменяют мелких хищ-
ных; животные из рода Oryzoryc-
tes роются, как кроты; род Micro-
gale заменяет на Мадагаскаре
землероек, тогда как род Lim-
nogale приспособился к жизни
в воде. Тенрек (Centetes ecauda-
tus).
Сем. Выдровые зем-
леройки (Potamogali-
<1 а е). Зубная формула, как у
златокрота; ключица в связи с
водным образом жизни исчезла.
Два рода. Один эндемичен для
Мадагаскара (Geogale), другой
Рис. 582. Выдровая землеройка (Pota-
mogale velbx).
(Из Брэма.)
795
для Африки — Potamogale. Единственный вид этого рода — западноафрикан-
ская водная выдровая землеройка, величиной с норку, с мощным плоским
хвостом, приводящим в движение в воде тело (рис. 582). Potamogale
velox.
Сем. Щелезубовые (Solenodontidae). Близкие к тенрекам
насекомоядные, обитающие на Больших Антильских островах. Единствен-
ный род — Solenodon. Щелезуб кубский (S. cubanus), остров Куба.
Сем. Ежовые (Erinaceidae). С полной и прочной скуловой ду-
гой. Настоящие ежи (Erinaceinae). Одеты иглами, распространены в тропи-
ческих и умеренных частях Старого Света. В пределах СССР обыкновенные
ежи (Erinaceus): Е. europaeus с коротком ухом и грубым мехом и нор-
мальными иглами; длинноухие ежи (Hemiechinus), длинные уши и мягкий
мех низа: Н. auritus.; длинноиглый еж (Paraechinus); крысиные ежи
(Gymnurinae) без игл, распространены в Индокитае и на Малайском архи-
пелаге.
Сем. Землеройковые (Soricidae). Мелкие мышевидные жи-
вотные с острой, вытянутой в хоботок мордочкой и коротким бархатистым
мехом. Скуловые дуги неполные; барабанная кость кольцеобразная, не со-
3 । 3_2 ' 3
единена с черепом. Зубная формула I -—-, С г—5, Р ——, М — .
х—1 U—1 1 о
Подсем. Бурозубки (Soricinae) с бурокрасными окончаниями
всех зубов: землеройка-бурозубка (Sorex aranaeus), малая бурозубка (S. minu-
tus), кутора, водяная землеройка (Neomys fodiens).
Подсем. Белозубки
(Crocidurinae)c белыми
оконечностями зубов; бело-
зубка белобрюхая (Crocidura
leucodon) и др.
Сем. Кротовые (Та1-
р i d а е). Приспособленные к
подземной жизни животные с
коротким и бархатистым мехом
и коротким хвостом; скуловые
дуги полные; барабанные кости
срастаются с черепом, образуя
bulla tympani. В пределах
СССР обыкновенный крот (Tal-
pa europea), слепой крот (Т.
соеса) на Кавказе, дальнево-
сточный крот (Mogera robusta).
Приносит некоторый вред унич-
тожением земляных червей,
улучшающих свойства почвы,
и своей роющей деятельностью
на огородах, в питомниках и
т. п.; является промысловым
животным, так как шкурки
крота идут на мех (кротовый
мех).
Сем. Выхухолевые
(D е s m a n i d а е). По призна-
кам близки к кротам, водные
обитатели с широким, сжатым
Рис. 583. Выхухоль (Desmana moschata).
(Из Брэма.)
с боков и покрытым чешуей
хвостом и вытянутой в хобот мордой. У нас выхухоль русская (Des-
mana moschata), в бассейнах рек BoJirn, Урала и Дона. Ценный пушной зве-
рек, сильно истребленный и находящийся в настоящее время под специальной
охраной закона (рис. 583). Организованы специальные выхухолевые запо-
ведники на р. Оке, на р. Клязьме и на р. Хопер.
796
Рис. 584. Ухо ночни-
цы (Myotis myotis).
1 —козелок (tragus), 2—
antitragus. (Из Вебера.)
Отряд 7. Рукокрылые (Chiroptera)
В результате приспособления насекомоядных животных к по-
лету образовался в длинном эволюционном процессе отряд летучих
мышей, или рукокрылых. Крыло вырабатывалось, очевидно, по-
степенно, путем изменения первоначальной летательной перепонки,
натянутой между передней и задней конечностями, наподобие той,
какая имеется у шерстокрылов. Надо думать, что уже в меловой
период появились первые рукокрылые, так как все третичные иско-
паемые находки говорят нам о наличии уже
высокоспециализированных рукокрылых.
Летательная перепонка образована голой
кожей, снабженной многочисленными чувст-
вующими волосками; в ней мы различаем
передний отдел между головой, шеей и перед-
ней конечностью (propatagium), средний, со-
стоящий из медиальной части между тулови-
щем и передней задней конечностями (plagio-
patagium), дистальной части, натянутой меж-
ду длинными метакарпальными костями и фа-
лангами 2-го, 3-го, 4-го и 5-го пальцев (chiro-
patagium) и задней между хвостом, ногой и
задней частью туловища (uropatagium). —
(рис. 545) На поге, на пяточной кости, сидит
костный тонкий придаток га п о р ц а, к кото-
рой прикрепляется летательная перепонка.
Хорошо развито наружное ухо, в котором у основания ушной рако-
вины обыкновенно бывает направленная внутрь лопасть — козелок
(tragus) (рис. 584). Зубы построены по типу зубов насекомоядных. Чис-
ло резцовредуцируется до двух, и, однако, как в верхней, так и в ниж-
ней челюсти всегда имеются клыки; ложпокоренные зубы одновер-
шинны, коренные четырех-, пятибугорчаты, с острыми режущими
краями в форме W. Ведут ночной или сумеречный образ жизни;
день проводят, подвесившись когтями задних пог головой вниз.
Два подотряда с большим числом видов — около 1000.
Подотряд 1. Плодоядные летучие мыши, или летучие собаки
( Macrochiroptera)
Крупные, до 1.5 м в размахе крыльев летучие мыши с тупобугор-
чатыми зубами, приспособленными для питания плодами; являются
хотя и специализированными, но более примитивными формами.
Около 250 видов образуют одно семейство.
Сем. Крылановые (Р tero podid ае). Тропические и суб-
тропические страны Старого Света. Летучая собака или калонг — Pteropus
celano — самый крупный вид, длиной 40 см. Острова Ява, Суматра, Банд
и Тимор. .
Подотряд 2. Насекомоядные летучие мыши ( Microchiroptera)
В отличие от предыдущих эти летучие мыши имеют острозубча-
тые зубы, так как сохранили основной характер питания своих
предков, но специализировались по добыванию насекомых на лету.
797
Сосцы (1 пара) расположены на груди; один-два детеныша при-
сасываются к сосцу и держатся, зацепившись за мать, во время по-
лета. Зимнее время летучие мыши проводят в спячке или, подобно
птицам, отлетают к югу. Спаривание летучих мышей происходит
осенью, по сперма в матке сохраняется, не оплодотворяя яйца,
до весны. Уничтожая большое количество насекомых, среди кото-
рых большая часть ночных являются очень вредными, летучие мыши
приносят большую пользу; особенно полезны они истреблением
вредителей древесных пород. Распространены по всему земному
шару, образуя 12 семейств, из которых мы остановимся только па
некоторых.
Сем. Подковоносовые — (Rhinolophidae). Свыше 100 видов
этого семейства распространено в тропических странах Старого Света; от-
личаются наличием листообразных придатков на носу; межчелюстные кости
редуцированы, и верхние резцы расположены на хрящевых отростках нёба,
,1
I —, не более. У нас в Союзе подковонос большой (Rhinolophus ferrum equi-
num) и малый подковонос (Rh. hipposideros) на юге: в Крыму, на Кав-
казе, в Туркестане и Узбекистане.
Сем. Листоносовые (Phyllostomidae). Названы так
потому, что имеют листовидный нарост на носу; резцы расположены нор-
мально. Приурочены исключительно к Америке и главным образом н Южной
Америке, где двумя своими тропическими подсемействами соответственно
заменяют два подсемейства плодоядных летучих мышей Старого Света,
а третье подсемейство заменяет подковоносов.
Сем. Кровососовые (Desmodontidae.) Характеризуются
острыми и длинными верхними клыками и наружными резцами, служа-
щими для прокалывания кожи теплокровных животных, кровью которых
они питаются. Тропическая Америка до Ю. Мексики.
Сем. Гладконосые (Vespertilionidae). Без ко-
жистых наростов на носу и с хорошо развитым козелком. Около 40 видов
этого семейства встречается в СССР. Ночницы (Myotis) — несколько видов
крупных, средних и мелких с узким и заостренным козелком; нетопыри
(Pipistrellus) — несколько мелких видов с булавовидным козелком; ве-
черницы (Nyctalus) средней величины или крупные виды с булавовидным
козелком; кожаны (Eptesicus, Vespertilio и др.), ушаны (Plecotus) и
некоторые другие.
Сем. Свободнохвостые (Emboelomuridae) со
свободной межчелюстной костью и со свободным хвостом, т. е. летательная
перепонка не приросла к хвосту. Тропические страны Старого и Нового Света.
Сем. Молоссовые (Molossidae). Характеризуются, по-
мимо сплюснутой спереди морды, широкими задними ногами с хорошо раз-
витой малоберцовой костью и тем, что перепонка может собираться в складки,
оставляя конечности свободными. Это дает им возможность не только про-
водить во время полета сложные повороты, но и быстро передвигаться по
земле. Тропики и субтропики Старого и Нового Света.
Отряд 6. Шерстокрылы (Dermoptera)
Шерстокрылые произошли от низших насекомоядных, с которыми
они имеют много общего. Они легко характеризуются наличием
прекрасно развитой летательной перепонки, идущей от головы
к пальцам передних и задних конечностей и к концу длинного хво-
ста, причем эта перепонка покрыта волосами; это лазящие по де-
2 1
ревьям растительноядные животные, с зубной формулой I—, С ->
о 1
798
Р^-, М~; конечности пятипалые; две пары сосков на груди; рож-
& о
дают одного детеныша. Всего лишь два вида.
Сем. Шерстокрыловые (Galeopithecidae). Един-
ственный род (разделяемый некоторыми на два рода) шерстокрылы, или
летучие маки (Galeopitheus), с двумя видами, распространенными в юго-
Рис. 585. Летучий маки (Galaeopithecus volans)’.
(Из Брэма.)
восточной Азии: один вид (G. temmincki) — от южного Китая через Мл-
лчНекий полуостров до Малайского архипелага, другой живет на Филиппи-
нах (рис. 585).
Отряд 9. Тиллодонты (Tillodontia)
В нижнем и среднем эоцене Сев. Америки были распространены довольно
крупны»» животные, с зубной формулой, очень напоминающей грызунов,
причем можно наблюдать три последовательных ступени такого приближения.
799
2 13 3
У нижнеэоценового Esthonyx зубная формула: I Су, Ру, Му, при-
13 2
чем редуцируются: I—, ly, I— и I—, тогда как I — и I— увеличивают-
ся, С тоже имеют тенденцию к исчезновению. У среднеэоценового Tillo-
tberium (рис. 586) пара больших резцов без корня в каждой челюсти, как
у грызунов; клыки зачаточны, между резцами и коренными зубами обра-
зуется диастема. Несмотря на сходство зубов тиллодонтов с грызунами их
надо считать ближе стоящими
к древним хищным Creodonta
и в особенности к насекомо-
ядным, от которых они ответ-
вились, приспосабливаясь к
растительной пище подобно
грызунам.
Отряд 10. Ящеры (Pholidota)
Ящеры резко отличают-
ся от всех млекопитающих
тем, что тело, их и длинный
хвост покрыты крупными
роговыми чешуйками, ца-
Рис. 586. Череп Tillotherium fodiens. летающими Друг па Дру-
(Из Вебера по Маршу.) га подобно Чешуйкам еЛО-
вой шишки; между че-
шуйками сидят редкие волосы; конечности пятипалые, стопоходя-
щие, снабженные когтями; в плечевом поясе отсутствуют ключицы;
зубы отсутствуют совершенно, язык длиппый, червеобразный, при-
способленный к питанию муравьями, желудок специализирован
к перетиранию пищи и снабжен роговым покровом и роговыми зу-
бами.
Происхождение и систематическое положение ящеров остаются
до сих пор еще мало выясненными; повидимому, они ответвились
очень рано от насекомоядных в виде общего ствола с американскими
неполнозубыми; палеонтологических данных, уясняющих происхож-
дение и эволюцию ящеров, мы пе имеем.
Сем. Панголинов ые (Manidae). Несколько видов этого
семейства распространено в Африке и в Южной Азии, Индостане с Цейло-
ном, Индокитае и прилегающих островах, объединены в последнее время
в несколько родов. Большинство азиатских видов ведет древесный образ
жизни (Paramanis javanica); некоторые африканские виды живут в степях:
степной ящер (Smulsia temmincki), панголин (Manis pentadactyla) в Индо-
стане, на Цейлоне и др.
Отряд 11. Неполнозубые (американские) (Xenarthra)
Этот отряд объединяет исключительно американских, но мало
сходных по внешнему виду животных, а именно броненосцев, ле-
нивцев, муравьедов из современных нам и целый ряд ископаемых
форм, как глиптодонт, мегатерий, милодон. Все эти формы,однако,
связаны общим происхождением и рядом общих им признаков:
несмотря на приспособление к разным условиям существования:
к жизни в лесах, па деревьях — ленивцы и отчасти муравьеды;
к роющему образу жизни — броненосцы; к питанию муравьями и
800
термитами — муравьеды и отчасти броненосцы; к питанию насекомы-
ми, другими беспозвоночными и отчасти растительной пищей — бро-
неносцы и к питанию листьями и плодами — ленивцы. Общим для
всех является дополнительное сочленение между последним грудным
и первым поясничным позвонками с помощью добавочных сочленов-
ных отростков. По особенностям скелета муравьеды и ленивцы
соединяются с ископаемыми тихоходами, тогда как эти последние
через глиптодонтов связываются с броненосцами, происходя от
пижпеэоценовых Palaeanodonta.
Отряд делится на 2 подотряда в зависимости от наличия или от-
сутствия панцыря.
Подотряд 1. Панцырные неполнозубые (Hicanodonta)
Одеты кожным панцырем из костных пластинок, прикрытых сна-
ружи роговыми пластинками эпидермального происхождения. По
7
мепыпей мере-у постоянных коренных зубов. Южная и Централь-
ная Америка до Мексики.
Сем. Броненосцевые (Dasypodidae). Объединяет совре-
менных и ископаемых броненосцев, характеризующихся нормальной ску-
ловой дугой без спускающегося вниз отростка, наличностью ключицы и*
белее или менее нормальной нижней челюстью (рис. 587).
Рис. 587. Трехпоясный броненосец (Tolypeutes conurus).
(Из Брэма.)
Ведут ночной образ жизни, на день прячутся в норах и обладают спо-
собностью легко и быстро зарываться в землю.
Три подсемейства:
Подсем. Броненосцевые (D а в у р о di n а е). С панцырем,
в котором костным пластинкам соответствуют роговые. Девятипоясной бро-
неносец — Dasypus novemcinctus.
51 Кашкаров и Станчинский 804
Подсем. Армадиллов ые (Eu ph г act i п а е). Костной пла-
стинке соответствуют одна большая и несколько мелких роговых. Гигантский
броненосец — Priodonteg giganteus.
Подсем. Щ итонос цов ые (Ch 1 a my d о ph о г i п а е). Мелкие
животные с панцырем из полос щитков сверху и вертикальным щитом по-
зади. Аргентинский щитоносец — Chlamydophorus truncatus.
Сем. Глиптодонтов ые (Glyptodontidae). Огромные
травоядные миоценовые и плиоценовые вымершие животные с массивным
высоким и неподвижным панцырем, короткой и высокой головой, массивной
нижней челюстью и характерным, обращенным вниз скуловым отростком.
Подотряд 2. Беспанцырные неполнозубые (Anicanodonta)
Без панцыря или лишь с его зачатками. Зубы редуцированы;
наибольшее их количество Q или их нет вовсе. Скуловая дуга
неполная. Сюда относятся муравьеды, ленивцы и ископаемые
тихоходы.
Сем. Тихоходовые (Gravigrada). Ископаемые гигантские
травоядные животные. Гигантский мегатерий (Megatherium) дожил до четвер-
тичных отложений Южной Америки; милодон (Mylodon) имел наружный
скелет из костяных пластинок.
Сем. Муравьедов ые (Myrmecophagida е). Имеют длин-
ное узкое рыло, совершенно лишены зубов и при помощи очень длинного и
подвижного языка питаются муравьями и термитами, жилища которых рас-
капывают приспособленными для копания передними конечностями с остры-
ми и длинными когтями. Несколько видов, среди которых есть крупные и
мелкие, наземные и лазящие по деревьям.
Муравьед (Myrmecophaga jubata), от Гватемалы до Парагвая; двупалый
муравьед (Cyclops didactylus), тропические леса Южной Америки, ведет
древесный образ жизни.
Сем. Лен и в цов ые (Bradypodidae). С коротким черепом,
неполной скуловой дугой, рудиментарным хвостом и конечностями, приспо-
собленными для подвешивания к ветвям: с длинными 2—3 сросшимися паль-
цами, с очень длинными загнутыми когтями. Питаются листьями и плодами.
Распространены в Южной Америке. Двупалый ленивец (Choloepus dida-
ctylus), трехпалый ленивец (Bradypus torquatus).
Отряд 12. Грызуны (Rodentia)
Наиболее многочисленный своими представителями отряд гры-
зунов охватывает около 3000 видов современных животных, т. е.
более трети всех известных видов млекопитающих, в наше время
населяющих земной шар. Грызуны распространены по суше всего
земного шара, будучи приспособлены к самым разнообразным био-
топам. Они живут в тундрах, лесах, степях, лугах и пустынях; они
населяют болота, пресные воды и горы; опи являются древесными
обитателями, обитателями открытых пространств, обладая, быстрым
бегом, или приспособлены к быстрому прыганию или роются
под землей подобно кроту. В то же самое время грызуны наиболее
многочисленны по количеству особей. Будучи приспособленными
к многообразным условиям среды, являясь экологически универ-
сальными по общей своей организации, грызуны имеют чрезвычайно
большое количество особенностей в связи с приспособлениями к раз-
нообразным условиям. Ив то же самое время, несмотря на колос-
сальное разнообразие своих форм, грызуны в основных чертах со-
802
храняют удивительное постоянство, что позволяет всегда безошибоч-
но и легко определить принадлежность к отряду его представителей.
. Таким характерным для отряда признаком является прежде
всего строение их зубов — именно строение их резцов.
В нижней челюсти всегда только два резца, в верхней челюсти
обычно тоже два, но у небольшой группы грызунов (семейства зай-
цев и пищух) в верхней челюсти позади двух больших резцов сидит
пара маленьких дополнительных — подотряд двупарпорезцовых
(Duplicidentata). Резцы развиты необычайно и, пе имея корней,
растут в течение всей жизпи, постоянно снашиваясь с вершины.
Так как эмалевый слой располагается только па наружной поверх-
ности резцов или здесь более мощно развит, то, вследствие беспре-
рывного, но неравномерного снашивания
режущей поверхности при работе зуба
(эмаль как более прочная часть стирает-
ся медленнее), резцы все время остают-
ся острыми. Долотообразные резцы, дне-
ствующие как стамески, несомненно,
являются основной особенностью грызу-
нов. Остальные признаки являются со-
провождающими эту особенность: силь-
ное развитие межчелюстных костей и
нижней челюсти связано с большими раз-
мерами резцов и их укреплением. Отсут-
ствие задних резцов, клыков и недораз-
витие передних ложнокоренных зубов
обусловливает наличие между резцами
и коренными зубами промежутка, ли-
шенного зубов, — диастемы (diastema).
Образование последней является след-
ствием приспособления резцов для
грызущей деятельности, так как зубы,
расположенные непосредственно позади
резцов, только препятствовали бы ра-
боте резцов; коренные зубы при расти-
тельноядности грызунов служат для
перетирания стеблей, корней, зерен, дре-
III
Рис. 588.
, IV V
Коренные зубы
грызунов.
1 — верхние, а — нижние корен-
ные зубы, I — полевки обык-
новенной (Microtua arvalis),
II-—бобра (Castor fiber), III —
крысы (Rattus norvegicus),
IV — тушканчика (Dipus), V —
морской свинки (Cavia). (Из
Вебера.)
вссины и т.п.; их поверхность соответственно бугорчатая, плоская
или снабжена эмалевыми петлями; грызение и перетирание требуют
приспособлений и в суставах и в мускулатуре челюстей: суставная
ямка челюсти грызунов продолговатая, сочленовная поверхность
нижней челюсти поперечно вальковатая, что дает возможность про-
изводить продольные движения нижней челюстью; соответственно
особо сильного развития достигает жевательная мышца (m. masseter),
передняя порция которой проходит через подглазничное отверстие
(for. infraorbitale) (рис. 689).
Соответственно удлинению межчелюстных костей, доходящих
до лобных, сильно удлиняется вся носовая область. Отсюда сильное
развитие носовых костей. Характерна для грызунов еще одна осо-
803
бенность, стоящая в связи с растительноядпостью: при значительной
длине кишечника особенно развита слепая кишка, функциониру-
ющая в качестве бродильного чана.
Рис. 589. Жевательные мышцы грызуна — агути (Da-
syprocta aguti). (По Веберу.)
1 — носовая (nasale), 2 — межчелюстная (praemaxillare), 3—*
челюстная (maxillare)» 4—слезная (lacrymale), 5—скуловая
(jugale), 6—скуловой отросток (processus zygomaticus), 7 —
барабанная(1утрап1сит), 8—processus paroccipitalis, 9—лобная
(frontale), 10 —теменная (parietale), 11 — верхи, затылочная
(supraoccipitale), 12 — чешуйчатая (squamosum), 13 — нижн.
челюсть (mandibula), 14 — жевательная мышца (musculus
masseter), 15 — волокна последней, проходящие через canalis
inf raorbi tails.
Рис. 590. Paramus delicatus.
(Из Мэтью.)
Эти именно особенности грызунов, их совершенное приспособ-
ление к грызущей деятельности, возникшей как известная специа-
лизация при растительпоядности, явилась в то же время особен-
ностью широко приспособительной — эйритоадаптивной, позволив-
шей обладателям гры-
зущего типа рта при-
спосабливаться к са-
мым разнообразным
условиям среды, при-
чем одновременно с
экотопической спе-
циализацией шла спе-
циализация в отноше-
нии пищи.
,Уже древнейшие
из грызунов, извест-
ные нам из нижнего
эоцепа, обладают ос-
новными особенностя-
ми отряда, как то вид-
но на рис. 590, где изображен череп самого древнего из известных
нам грызунов, Paramys. Как и когда произошло ответвление грызу-
нов от общего ствола млекопитающих, палеонтологические данные
ничего определенного не говорят, и наиболее вероятным предположе-
нием остается происхождение грызунов от меловых насекомоядных.
804
В своей дальнейшей эволюции грызуны уже очень рано разде-
лились на два ствола: на более примитивных двупарнорезцовых —
Duplicidentata и на более совершенно специализированных парно
резцовых — Simplicidentata. Обе ветки имеют значение подотрядов.
Подотряд 1. Двупарнореацовые (DupUcidentata9 Lagontorpha).
Позади двух основных резцов в верхней челюсти лежит пара
небольших дополнительных резцов с молочными предшественни-
ками; формула резцов I—; резцы со всех сторон одеты эмалью,
которая на задней поверхности развита слабо. Общее число корен-
^65
ных зубов -— причем число ложнокоренных постоянно, а имен-
234 123
по: Р '3'4, тогда как число коренных у зайцев М 123, а у пищух
„ 1-2 • -п
М -y-x-z. В противоположность парнорезцовым, у двупарнорезцовых
сохраняется характерное для прочих млекопитающих поперечное
движение челюстей во время жевания, а в связи с этим сочленовная
ямка более широкая, допускающая боковые движения челюсти, а из
жевательных мышц m. masseter не достигает такого развития, как
у других грызунов; подглазничное отверстие слабо развито, зато
гораздо сильнее, чем у других грызунов, развита височная мышца
(m. temporalis). Все эти признаки указывают па примитивность
этого подотряда. Два семейства.
Сем. Пищуховые (Ochotonidae). Небольшие, величиной
несколько больше серой крысы, но без хвоста и с короткими ушами, голар-
ктические, главным образом высокогорные животные, замечательные тем, что
делают себе запасы из сена (откуда их другое название — сеноставцы).
2 0 3 2
Зубная формула I —, С Р —, М —.
1 U Z о У
У нас в Союзе распространена в степях северного Казахстана и южного
Заволжья малая пищуха (Ochotona pusilia), даурская пищуха (Och. dau-
rica) — в Забайкалье, несколько видов в горах. Приносит незначительный
вред полям. Ископаемые предки пищух — олигоценовые и миоценовые
Titanomys Европы.
Сем. Зайцев ые (Leporidae). Довольно крупные, с удлинен-
ными задними ногами и длинными ушами животные, которые были широко
распространены по всему свету за исключением Мадагаскара и Австралии,
2 0 3
куда завезены были человеком кролики. Зубная формула: I —, С —, Р —,
10 2
3
м з-
Предками зайцев являются олигоценовые Palaeolagus Сев. Америки.
К этому семейству относятся зайцы (Lepus), около 100 видов. У нас в Союзе
беляк (L. timidus) распространен в зоне тундры, тайги и лиственных лесов;
зимой линяет в белый цвет за исключением ушей; русак (L. europaeus) —
обитатель степей, лесостепи и отчасти зоны лиственных и смешанных ле-
сов, отчасти белеет на зиму; в южных частях СССР, в Азии толай (L. tolai),
который зимой вовсе не меняет цвета; на Дальнем Востоке темный манчжур-
скнй залц (L. mandschuricus) и др. Очень плодовиты, дают два-три помета
805
по 3—5 зайчат; размножение зайцев удерживается большим количеством
врагов из хищников млекопитающих" и птиц. Вредят плодовым садам,
питомникам, но имеют большое промысловое значение, доставляя мех
и мясо.
Кролики (Oryctolagus) отличаются от зайцев более короткими задними
конечностями и тем, что живут в норах, которые себе выкапывают в земле.
Дикий кролик (О. cuniculus) родиной из Северной Африки и Южной Европы,
с помощью человека широко распространился в других странах. В Австралии
размножился до громадных размеров, сделавшись злостным вредителем
полей. Разводится в качестве домашнего животного, образуя многочислен-
ные породы.
Подотряд 2. Парнорезцовые (Slmplicidentata)
В каждой половине челюсти лишь по одному резцу; ложпокорсп-
пые зубы имеют тенденцию к полному исчезновению.
По характеру изменений, развившихся в процессе эволюции,
этот подотряд можпо разбить па ряд групп большего объема, чем
семейство.
Надсем. Гаплодонтные (Haplodontoidea)c одним северо-
американским семейством.
Сем. Гаплодонтовые (Н а р 1 о d on t i dae), величиной с бай-
бака. Роющие животные; интересны тем, что у них число коренных зубов
2 3
Р у, М- совпадает с примитивным жующим аппаратом, т. е. относительно
слабой жевательной мышцей и относительно сильной височной мышцей и
соответствующей сочленовной поверхностью; очень близкими к ним оказа-
лись наиболее древние из известных нам ископаемых грызунов: уже упомя-
нутая эоценовая Paramys, дошедшая до олигоцена, где в североамериканской
фауне ее сопровождало несколько родов (Sciuravus, Ischyromys и др.); из
этой же группы грызунов можно вывести и следующую группу семейств,
так как представленные несколькими североамериканскими и европейскими
олигоценовыми родами близкие к гаплодонтовым через Prosciurus приводят
к белкам.
Надсем. Белковидные (Sciuroidea).
Сем. Беличьи (Sciuridae) охватывает белок, летяг, бурун-
дуков, земляных белок, сусликов и сурков, которые выделяются в отдельное
подсемейство; все они характеризуются хорошо развитыми заглазничными
10 2 3
отростками и зубной формулой: I у, С —, Р у, М у, причем коренные зубы
с короткими кронами и длинными корнями.Древесными обитателями являются
прежде всего белки, распространенные по всему свету, за исключением Ма-
дагаскара и Австралийской области, в количестве почти 200 видов, соста-
вляющих несколько родов. Все они приспособлены к лазанью по деревьям,
прыганию с ветки на ветку и имеют пушистый мех, длинные уши и хвост.
У нас в Союзе два вида белок: обыкновенная (Sciurus vulgaris) и закавказ-
ская (S. anomalus). Первая распространена по лесам всего Союза, отсут-
ствует лишь в лесах Северного Кавказа и Крыма; она образует многочислен-
ные подвиды, отличающиеся окраской и качеством меха, которые более или
менее совпадают с сортами беличьих мехов — так называемыми «кряжами».
Промысловое значение белки в СССР очень велико. Бурундук (Eutamias) —
лесной зверек с полосатой спинкой, распространенный по всем лесам на
восток от северо-востока Европейской части Союза; местами является вре-
дителем зерновых культур; имеет некоторое промысловое значение как
пушной зверек. Байбаки, или сурки (Marmota) (рис. 591), в Сибири на-
зываемые тарабаганчиками, обитатели открытых пространств; в европей-
ской части Союза почти истребленный байбак (М. bobac), в Забайкалье
(М. sibirica), алтайский горный (М. baibacina), камчатский (М. camtschatica)
и др. Имеют промысловое значение, зерновым хлебам не вредят. Суслики
(род Citellus) представлены в СССР 14 видами, из них некоторые виды яв-
806
ляются весьма вредными животными полеводства, другие являются опасными
разносчиками чумы, третьи могут быть полезными в качестве пушных зверь-
ков. На крайнем западе Украины распространен европейский суслик (Citellus
citellus), на правобережной Украине, в Воронежской и Курской областях и
к западу от Волги — крапчатый суслик (С. suslicus), на левобережной Укра-
ине, на Северном Кавказе и в Нижневолжском крае, к западу.от Волги, за
Волгой — рыжий суслик (С. rufescens), краснощекий суслик (С. erythro-
genys) и др.
Наибольшее промысловое значение имеет желтый суслик, или песчаник
(С. fulvus), из Казахстана и среднеазиатских республик. В Сев. Америке
род луговые собачки (Cynomys).
Летяги (Pteromys, Sciuropterus) нередко выделяются в особое семей-
ство — Pteromyidae, так как отличаются наличием летательной
перепонки. Летяга (Sciuropterus russicus) (рис. 592) водится в СССР.
Выделяется в отдельное семейство группа африканских крупных назем-
ных белок с центральным родом Xerus — Xeridae.
Рис. 591. Байбак (Marmota bobac).
(Из Брэма.)
Надсем. Бобровидные (Castoroide а).
Сем. Бобровые (Castoridae). Довольно крупные водные
обитатели с плоским хвостом, покрытым чешуей, и задними конечностями,
снабженными плавательными перепонками. Зубы I -у, С-^-, Р у-, М . Живут
в норах или специальных сооружениях из перегрызенных обрезков дерева—
так называемых «хатках». У нас речной бобер (Castor fiber) почти совершенно
истребленный, благодаря охране вновь стал размножаться. Очень ценен своим
мехом, так что возможна организация специальных промысловых хозяйств.
Успешно разводится у нас в Воронежском бобровом заповеднике, откуда
перевезен в ряд других заповедников. В Сев. Америке (С. canadensis)
(рис. 593). В олигоцене и миоцене бобры были представлены большим числом
родов.
Надсем. Мешотчатые крысы — Geomyoidea.
Сем. Мешотчатые крысы (Geomyidae). По зубной фор-
муле сходны с бобрами, имеют наружные защечные мешки, обитают главным
образом в Сев. Америке, отчасти в Центральной и Южной. Некоторые
ведут подземный образ жизни подобно кротам. Гофер (Geomys bursarius)
в долине Миссисипи.
Надсем. Аномальные (Anomaluroidea).
807
Рис. 592. Летяга (Sciuropterus russicus).
(Из Гаане.)
Рис. 593. Речной бобер (Castor fiber)c
(Из Гаане.)
808
Сем. Аномальные белки (Anomaluridae). Исключи-
1
только африканское семейство древесных белкообразных животных с Р -р
М
3 ’
некоторые виды имеют хорошо развитую летательную перепонку
(Anomalurus).
Сем. Длинноногов ые (Pedetidae). Южноафриканские,
похожие на тушканчиков длинноноги (Pedetes) очень близки к аномальным
белкам и связываются с ними ископаемыми Issiodoromys.
Надсем. Соневидные (М у о х о i de а).
Сем. Соневые (Myoxidae) объединяет небольших зверьков,
по одним признакам — пушистому хвосту и древесному образу жизни — на-
поминающих мелких белок, по другим — острой мордочке, голым ушам —
напоминающих мышей или крыс. Коренные зубы сонь Р —, М —. В связи с пе-
1 о
реходом частью на питание плодами и насекомыми, иногда и мелкими позво-
ночными, у сонь в отличие от остальных грызунов отсутствует функциони-
Рис. 594. Кафрский долгоног (Pedetes caffer).
(Из Гааке.)
рующая слепая кишка. У нас в Союзе распространена лесная соня (Dryomys
nitedula), садовая соня (Eliomys quercinus); наиболее крупная из сонь —
полчек (Grlis glis) и мушловка, самая мелкая из сонь (Muscardinus avel-
lanarius), распространены по лиственным лесам, преимущественно европейской
части. Делают запасы, впадают в продолжительную спячку.
Надсем. Тушканчиковидные (Dipodoidea).
Сем. Ту шкан чиков ые (Dipodidae). К этому семейству
относятся две группы грызунов, но внешнему виду мало сходных друг с дру-
гом, собственно тушканчики (Dipodinae) и мышовки (Sicistinae), выделяемые
в особые подсемейства, связанные друг с другом переходами; коренных
13 1
зубов Р —, М —, причем Р —имеет тенденцию к исчезновению. У нас мы-
V о
шовка Нордмана (Sicista nordmani) распространена в степях, мышовка горная
,(S. montana) — в лесах и горах. Тушканчики представлены разными ста-
диями последовательной специализации этой группы к прыганию. У рода
Sapus Сев. Америки, еще очень близкого к мышовкам, шейные позвонки
нормальные, все пять метатарзальных костей самостоятельны, хотя и удли-
нены приспособительно к прыганию, 1-й и 5-й пальцы хорошо развиты.
809
Рис. 595. Хомяк (Cricetus cricetus).
(Из Гаане.)
У рода Alactaga (земляные зайцы) три средних метатарзальных кости сильно
удлинены и срастаются, тогда как 1-я и 5-я сильно укорочены. Сюда отно-
сится самый крупный тушканчик (A. jacalus); он распространен в степях на
восток до Алтая, отчасти вредит посевам и является переносчиком чумы.
У рода Dipus 1-й и 5-й пальцы исчезают вместе с их метатарзальными ко-
стями — мохноногий тушканчик (Dipus sagitta) живет на Северном Кавказе
и по бахчам Средней Азии, емуранчик (Scirtopoda teium) распространен
там же и на песках нижнего Днепра.
Надсем. Мышевидные (Myoidea).
Сем. Слепышевые (Spalacidae). У слепыша (Spalax) целый ряд
особенностей, связанных с подземным образом жизни: зачаточные глаза спрй-
таны под кожей, широкая ло-
патообразная голова, ушных
раковин нет, нос имеет орого-
вевшую кожу, роющие ко-
нечности. Зубная формула
I с| Р р м|. Наиболее
распространен у нас обыкно-
венный в черноземной части
слепыш (S. microphtalmus),
вредящий полям тем, что по-
едает корни и клубни и вы-
брасывает кучки земли на по-
верхность, затрудняя уборку
урожая машинами.
Сем. Netomyidae —
единственные мадагаскарские
грызуны, близкие к слепышам.
Сем. Мышиные (М и-
r i d а е). Обширнейшее семей-
ство с космополитическим рас-
пространением, распадается на
12 семейств. У них МГ более
длинный, чем последующие.
Повидимому это семейство
не составляет естественной
группы.
Подсем. Хомяки (С ri-
ce t i в а е) характеризуются
двумя продольными рядами бу-
горков на жевательной поверх-
ности коренных зубов и за-
щечными мешками. Обыкновен-
ный хомяк (Cricetus cricetus)
* является во многих местах
серьезным вредителем полей,
хотя и получил в последнее
время значение в качестве пуш-
ного животного; средние хомя-
ки (Mesocricetus) вредят полям в Закавказье, хомячки (Cricetulus), не-
сколько видов маленьких хомяков СССР не имеют большого значения как
вредители сельскохозяйственных / культур.
, Подсем. Полевки (Microtinae) имеют коренные зубы с треуголь-
ными петлями (рис. 588) и по большей части с беспрерывным ростом, тупую
морду и короткий хвост. В СССР около 50 видов, среди которых много злост-
ных вредителей сельского хозяйства. Упомянем немногих: водяная крыса
(Arvicola amphibius) — самая крупная из полевок, обитающая как в воде,
так и далеко от воды, иногда вредящая садам и огородам; обыкновенная
полевка (Microtus arvalis) — один из самых распространенных и опасных
вредителей полей СССР; полевка лесная (Microtus agrestis), неудачно назван-
ию
пая по-латыни пашенной, так как обитает в лесах и болотах, подобно эко-
номке (М. oeconomus); в степях серьезным вредителем являются обще-
ственная полевка (М. socialis) и степная пеструшка (Lagurus lagurus) и др.
В лесных районах распространены рыжие полевки (Evotomys). Очень свое-
образны слепушонки (Ellobius), роющие полевки с выдающимися резцами
и зачаточными ушными раковинами. Для тундр характерны пеструшки,
пли лемминги (Lemmus lemmus). В Сев. Америке большое промысловое зна-
чение имеет ондатра (Fiber zibeticus) — крупная водная крыса с очень цен-
ным мехом; в настоящее время акклиматизирована в Западной Европе и
у нас.
Подсем. Ц окори (Myospalacinae) являются полевками, при-
способившимися к подземному образу жизни подобно слепушонкам. Три
вида в пределах азиатской части СССР.
Подсем. Мыши (Ми г i п а е) вместе с крысами составляют особое
подсемейство с очень большим числом видов (около 400); они имеют на ко-
ренных зубах бугорки, расположенные в три продольных ряда, и длинный
хвост с короткими редкими волосками и роговыми чешуйками. Серая крыса,
или пасюк (Rattus norvegicus), черная крыса (Rattus rattus), туркестанская
крыса (R. turkestanicus) — главные вредители складов, амбаров, продо-
вольственных магазинов, вредители птицеводства и кролиководства, пере-
носчики чумы, трихины и других паразитов и инфекционных болезней.
Па юге Узбекистана и Туркмении живет и приносит большой вред полям
и амбарам пластинчатая крыса (Nesokia indica). Из мышей (Mus) назовем
домашнюю (М. musculus); лесную мышь (М. silvaticus) и желтогорлую мышь
(М. flavicollis) выделяют в отдельный род так же, как и мышь полевую
(М. agrarius) и малютку (М. minutus).
Подаем. Песчанки (Gerbilli ща е) — отличаются от мышей
и крыс пушистым хвостом и коренными зубами с плоской жевательной по-
верхностью. Полуденная песчанка (Meriones meridianus) — характерный
обитатель песков, пустынь Ср. Азии, большая песчанка (Rhombomys opimus)
Средней Азии, местами вредит полям.
Сем. Австралийские мыши (Hydromyidae). Состав-
ляют особое семейство отчасти водных (Hydromys), отчасти наземных
обитателей (Xeromys и др.) с доведенной до крайности редукцией зубов:
т '1 „ 0 п 0 .. 2 А
I —, С Р —, М —. Австралия и прилежащие острова.
Надсем. Дикобразовидные (Hystricoidea).
Сем. Дикобразовые (Hystricidae). Характеризуется
наличием игл на спине и четырьмя коренными зубами в каждой половине
челюсти. Распространены в теплых странах Старого Света за исключением
Мадагаскарской, Австралийской и Новозеландской областей.
У нас дикобраз (Hystrix hirsutirostris) — встречается в Закавказье
и Средней Азии.
Сем. Иглошерстовые (Erethizontidae). Лазящие по
деревьям американские дикобразы с длинными когтями и хватательным
хвостом. Erethizon представлен многими видами, от Канады до Мексики;
Coendu — с цепким хвостом, из тропической Южной и Центральной Америки
(рис. 596).
Сем. Свинковые (Caviidae) принадлежит исключительно
Южной и Центральной Америке и объединяет наиболее крупных грызунов, на-
чавших приспособляться к быстрому бегу подобно копытным животным, с от-
носительно высокими конечностями, снабженными копытообразными когтями.
Морские свинки (Cavia) представлены многими видами, из них домашняя
морская свинка (С. porcellus); водосвинка (Hydrochoerus) — самый крупный
из грызунов — живет по рекам и озерам лесной зоны; мара (Dolichotis ра-
tagoniea) — быстро бегающий грызун пустынь и полупустынь Патагонии;
агути (Dasyprocta) с 12 видами и пака (Coelogenys) из лесов тропической
Южной Америки являются основными охотничьими животными Бразилии.
Сем. Ill ин шил ловы е (Chinchillidae.)K этому семейству отно-
сятся обитатели горных степей Южной Америки, высоко ценимые пуш-
ные зверьки: шиншилла (Chinchilla) и вискаша (Viscacia).
ВИ
Рис. 59G. Древесный дикобраз (Coendu prehensilis).
(Из Гаане.)
Сем. Осьмизубые (Octodontidae). Очень интересна отно-
сящаяся к этому южноамериканскому семейству нутрия, или бобровая
крыса (Myopotamus coypus), — очень ценный пушной зверек, распростра-
ненный по Южной Америке и акклиматизируемый в настоящее время
в Европе и у нас (рис. 597).
Рис. 597. Нутрия (Myopotamus coypus).
(Аснания Нова, фото Озерова). \
812
Отряд 13. Креодонты (Creodonta)
Еще в конце мезозоя в меловое время ответвились от насекомо-
ядных низшие всеядные и хищные животные, перешедшие от пита-
ния насекомыми к питанию позвоночными животными. Это были
креодонты (Creodonta), мелкие или средней величины животные
с короткими и полустопоходящими конечностями. Сделавшись все-
ядными хищниками, креодонты дали несколько ветвей развития,
приспосабливаясь к разным условиям обитания. Очень рано от
них ответвились китообразные — Cetacea, позднее первичноко-
пытные (Protungulata), тогда как их продолжением явились
настоящие хищные (Carnivora fissipedia) с ответвившимися от
них ластоногими (Pinnipedia). Еще среди насекомоядных мы
познакомились с семейством Deltatheriidae из верхнего мела Мон-
голии с характерными особенностями в строении зубов, напоми-
нающими креодонтов. Близкие к ним палеоценовые и нижнеоце-
новые мелкие хищники Oxyclaenidae из Сев. Америки и Европы
вместе с крупными всеядными хищниками, напоминающими медведя,
3 14
объединяются в подотряд Acreodi, с зубной системой I—, С—, Р—,
3
М— с режущими коренными, но без обособления хищных зубов. Из
<5
родоначальной группы примитивных креодонтов эволюция пошла
в двух направлениях: в сторону специализации в направлении
усовершенствования хищного образа жизни и в сторону отхода от
хищного питания — к большей растительноядности и к быстрому
бегу среди открытых пространств. Первое направление привело,
в конце концов, к современным хищным, второе дало ту-
пую, быть может, параллельную, эоценовую ветку примитивным
парнокопытным. У начала ствола настоящих хищников получает
развитие подотряд так называемых Pseudocreodi, причем у семейства
2
качестве хищных зубов развиваются М-и М —,
о
Hyaenodontidae в
а у семейства Oxyaenidae таковыми являются М — и М —. Этот
подотряд Pseudocreodi противополагается подотряду Eucreodi,
в котором уже в эоцене в семействе Miacidae выявились два под-
семейства, ставших родоначальниками двух основных групп со-
временных нам хищников: подсемейство Viverravinae дало семейства
виверр (Viverridae), гиен (Hyaenidae) и кошек (Felidae), а под-
семейство Miacinae дало семейства куниц (Mustelidae), медведей
(Ursidae), енотов (Procyonidae) и собак (Canidae) (рис. 598).
Отряд 14. Хищные в узком смысле или наземные хищные (Carnivora
s. str., или Carnivora fissipedia)
Отряд хищных возник и развился в связи с приспособлением
к питанию позвоночными животными, сперва, вероятно, рептилиями
и амфибиями, а потом птицами и млекопитающими. Основными ор-
813
ганами, подвергшимися изменениям кроме зубов, преобразования
которых здесь были особенно характерными, явились конечности,
в связи с приспособлением к разным средам обитапия и к ловле и схва-
тыванию добычи, и оргапы чувств, важные при отыскании добычи.
3 3 1
Резцов обычно I— , только как исключение —; Сротличаются осо-
бенно сильным развитием и служат для удержания и умерщвления
добычи; коренные зубы тритуберкуло-секториальные, обычно с ре-
жущими краями, реже бугорчатые с четырьмя или большим числом
бугорков; при этом передние Р и задние М имеют тенденцию к исчез-
4
новению. Р—и М — образуют хищные зубы. Сочленение нижней
челюсти в глубокой сочленовной ямке. На конечностях б-й палец
иногда зачаточный или даже отсутствует; обонятельный аппарат
Carnivora fissipedia
Рис. 598. Схема эволюции хищных (Carnivora).
(По Абелю.)
с 5—7 складками на четырех внутренних раковинах. Наконец,
у хищных в связи со сложной высшей нервной деятельностью, необ-
ходимой при добывании пищи, хороро развиты полушария голов-
ного мозга. Две ветви развития — виверровые и медведевые.
Сем. Вив е*р ров ые (Viverridae). Наиболее примитивные
мелкие и средней величины хищники, происшедшие из эоценового подсемей-
ства креодонтных хищников (Viverravinae). Коренные зубы в самом начале
редукции
П 1 2 3 4 .Л 2 п >2 3 4 1 2
Р 1 2 3 4 ’ М 1 2 ИЛИ Р -2 3 4 ’ М 1 2'
Два подсемейства, отличающиеся втяжными — виверры (Viverrinae) и не-
втяжными когтями — мангусты (Herpestinae). Виверры (Viverra), представ-
лены несколькими видами, из них циветты (V. civetta и V. zibetha) замеча-
тельны своими промежностными железами, выделяющими цибет. Несколько
видов виверр с Мадагаскара, представляющих там всех хищников, объединяют
в несколько родов и даже в три подсемейства, что стоит в связи с эволюцией их
814
в разных направлениях параллельно прочим хищникам. Мангусты (Herpestes)
представлены в Африке и Восточной области многими видами, по большей
части копающими норы: фараонова мышь (Herpestes ichneumon), мунго
(Mungo mungo) и др.
Сем. Г и е н о в ы е (Н у а е n i d а е). Небольшая группа быстро
бегающих собакообразных животных с очень сильными специализирован-
ными зубами, в которых редукция коренных доведена до М у,тогда как
п*234 й ™
Р 2 3 , причем М — становится малым, а хищные Р£ и Mi очень вели-
ки и служат для разгрызания самых прочных и толстых костей, недоступных
другим хищным животным. Ноги пальцеходящие, с 4 пальцами на передних
и задних или с 5-м пальцем на задних конечностях. Происходят от преды-
дущего семейства, с которым их связывает верхнемиоценовый Ictitherium
Европы.
Полосатая гиена (Hyaena hyaena) из Северной Африки и Южной Азии.
Пятнистая гиена (Crocotta crocuta) из Центральной и Южной Африки. Обыкно-
венная в пещерах Европы в конце третичного и в начало четвертичного пе-
риода С. spelaea близка к последней.
Рис. 599. Ирбис (Felis uncia). (Из Сатуйина)
Сем. Кошачьи (Felidae). Хорошо ограничиваются среди совре-
менных хищных круглой головой в связи с укороченными челюстями, в кото-
, т 3 п * tj 3 шт *
рых коренные зубы редуцируются до одного; I —, С —, Р —, М —; очень раз-
О 1 X 1
виты хищные зубы; передние конечности с 5, задние с 4 пальцами, по
большей части со втяжными когтями. Семейство кошек ответвилось от
виверр еще в нижнем олигоцене, причем эволюционировало в двух различ-
ных направлениях: первое привело к современным кошкам, причем шло
постепенное уменьшение большого верхнего клыка, увеличение нижнего
и развитие хищных зубов от Dinictis через Nimravus к Felis; второе напра-
вление путем прогрессивного увеличения верхних клыков привело к сабле-
зубым тиграм — Machairodus Европы и Азии и Smilodon Америки. Эта ветка
вымерла в четвертичное время. Современные кошки — прыгающие формы:
лев (Leo 1ео) доходил в виде пещерного льва (L. spelaea) в историческое
время в южную и среднюю Европу; тигр (Tigris tigris) встречается у нас в
Союзе в Закавказье, в Средней Азии и Уссурийском крае; леопард (Pantera
pardiis) —с таким же распространением; ирбис (Felis uncia) в горах Средней
Азии; в Америке ягуар (F. опса); дикий кот (Felis sylvestris) — в лесах
юго-яападпой Украины и Кавказа; камышовый кот (F. chaus), нубийская
кошка (F. maniculata) —предок домашней; рысь (Lynx lynx) в лесах средней
и северной Европы, Сибири, Кавказа и др.; в Америке — пума (Puma
815
concolor). Ив быстро бегающих форм интересен гепард (Acinonyx jubatus),
дрессируется в Индии в качестве охотничьего животного.
Сем. Собачьи (Gan id ае). Вместе с остальными семействами
хищных происходят от сем. Miacidae, креодонтов, которые очень рано,
выделив ветку, ведущую через олигоценового Amphicyon к медведям, в даль-
нейшем своем развитии дали через олигоценовых же Cynodictis, Nothocion,
Cynodesmus собак, а через Temnocyon ведут к красным волкам (Cyoninae).
Семейство это эволюционировало в сторону приспособления к быстрому
бегу, что сказалось в удлиненных пальцеходящих ногах, с 4—5 передними
и 4 задними пальцами. Зубы от примитивной формулы I-|-, С Р у, М
2
по большей части эволюционировали в сторону сокращения до М j.
Волк (Canis lupus) представлен рядом видов и подвидов как в Европе
и Азии, так и в Сев. Америке, вреден истреблением домашнего скота и как
разносчик бешенства; в юго-западной Азии шакал (С. aureus). Домашняя
собака (С. familiaris) произошла от какого-то средней величины волка из
юго-восточной Азии, вероятно, от вымершего С. ferus; ее старейшим домаш-
ним потомком была неолитическая С. poutiatini. В прериях Сев. Америки
кайот (Canis latrans).
Лисица (Vulpes vulpes) — ценный пушной зверь и истребитель грызу-
нов — образует ряд подвидов и цветовых мутаций, разводится на зоофермах.
Питаясь разнообразной животной пищей, лисицы приносят вред уничто-
жением охотничьих и промысловых птиц и зверей (зайцев) и в то же
время приносят большую пользу истреблением мелких грызунов — вре-
дителей сельского хозяйства. Лисицы довольно быстро размножаются,
ежегодно приносят от 3 до 12 лисят. Живут лисицы в глубоких норах.
Степная лисица, или корсак (V. corsac) — в степи; ценные пушные
зверьки Севера — песцы (Alopex lagopus). Красные волки (Cyoninae) в со-
ставе трех родов Африки и Южной Азии составляют особое подсемейство.
В Приамурье и Уссурийском крае енотовидная собака (Nyctereutes amu-
rensis).
Сем. Медведев bie (Ursidae). В противоположность вол-
кам имеют тяжелое сложение и не приспособлены к быстрому бегу, имеют
пятипалые стопоходящие ноги; приспособительно к всеядному образу пи-
тания медведи имеют бугорчатые коренные зубы и слабо развитые хищные
зубы. Развитие медведей прослежено довольно подробно от современных
медведей до мелких величиной с волка форм рода Ursavus из миоцена Европы.
Бурый медведь (Ursus arctos) и близкие виды живут в лесах Европы, Азии
и Сев. Америки (гризли). В четвертичное время в Европе жил пещерный
медведь (U. spelaeus), черные медведи (Euarctos) — в Сев. Америке, барибал
(Е. americanus) в Восточной Азии (Е. japonicus), тибетский медведь (Тге-
marctos thibetanus) и др. Белый медведь (Talassarctos maritimus), круго-
полярный обитатель Северного полярного мо^ря, имеет промысловое зна-
чение. Особняком стоят бамбуковые медведи (Ailuropus).
Сем. Енотовые (Рг осу on id ае). Еноты связываются с мед-
ведями общим корнем происхождения и ведут свое начало или от североаме-
риканского олигоценового (Phlaocyon) или от европейского (Pachycynodon).
Панда (Ailurus fulgens) из Гималаев; еноты (Procyon lutor) и др., а также
цепкохвостый кинкажу (Cercoleptes) и носухи (Nasua) из Америки.
Сем. Куницевые (Mustelidae). Подразделяется на три под-
семейства: выдровые (Lutrinae), приспособившиеся к водному образу жизни,
всеядные барсуковые (Melinae) и собственно куницевые или хорьковые (Muste-
Ипае). Для всех них характерна редукция задних коренных М у или у. Ко-
нечности пятипалые.
Подсем. Куницевые (Mustelinae). Хорьки (Muetela) у нас в
Союзе представлены несколькими видами: обыкновенный хорек (М. putorius)—
в лесной зоне, степной хорек (М. ewersmani), колонок. (М. sibirica) имеют
промысловое значение как пушные звери и полезны как истребители вредных
грызунов. Норка (Lutreola lutreola) ведет водный образ жизни, имеет ценный
816
мех*; горностай (Ictis erminea) и ласка (I. nivalis) — истребители грызунов;
пушные мелкие зверьки, зимой одевающие белый мех.
Куница лесная (Martes martes) (рис. 600) и белодушна (М. foina) —
очень ценные пушные звери, распространенные в Европе; соболь (М. zibel-
lina) ценнейший пушной зверек, распространен в лесах Сибири. Вследствие
векового преследования сохранился только кое-где на Урале и на Алтае,
в Саянах, в Забайкалье, на Камчатке, в Приморском, Дальневосточном
Рис. 600. Куница (Martes martes).
(Из Гаане.)
крае. В настоящее время соболь находится под специальной охраной закона
и успешно размножается в целом ряде заповедников.
В самое последнее время удалось заставить соболей размножаться
в неволе.
Соболь питается преимущественно грызунами (белками, бурундуками,
пищухами, мышами), птицами, а также растительной пищей, ягодами, орехами.
Перевязка (Vormela peregusna) распространена в степях Европы. Ро-
сомаха (Gulo gulo) — в лесах Европы и G. luscus в Сев. Америке; медоеды
(Mellivora).
Подсем. Барсуковые (Melini) отличаются неуклюжим телом и
грубой шерстью и всеядностью. В СССР живет барсук обыкновенный (Meles
52 Кашкаров и Станчинсний 817
meles), и др., в Америке его заменяет другой род, Taxidea; сюда же относятся
американские вонючки (Mephitis), доставляющие ценный скунцевый мех.
Подсем. Выдровые (Lutrinae) представлены в СССР речной выд-
рой (Lutra lutra), с ценным мехом, и крупным морским «бобром», правильнее
морской выдрой, или каланом (Latax lutris), который раньше в большом
количестве жил в северной части Тихого океана, у берегов Камчатки,
Курильских и Командорских островов. Вследствие жестокого преследования
повсеместно близок к полному исчезновению. В виду этого промысел камчат-
ских бобров находится под контролем.
Отряд 16. Ластоногие (Pinnipedia)
Хотя и не подлежит никакому сомнению, что ластоногие прои-
зошли от наземных хищников, однако палеонтологические сви-
детельства не дают прямых указаний на то, когда это произошло и
Рис. 601. Морской котик (Arctocephalus ursinus).
(Из Гаане.)
какие именно наземные хищники были непосредственными предше-
ственниками ластоногих. Мы знаем только, что моржи и ушастые
тюлени имели в миоцене общих предков и что в это время они были
отделены уже от хищных. Всего вероятнее, что общий корень от-
ряда опускается до эоценовых креодоптных хищников. Приспособ-
ление ластоногих к водному образу жизни очень полное, но пе
настолько, как у китообразных, и в разной степени выражено в пре-
делах отряда.
Характеристика отряда вкратце может быть сведена к следую-
щему. Водные хищные млекопитающие с пятипалыми конечностями,
318
превращенными вплавники (ласты); хвоста нет, и з.адпие конечности
обращенные назад, функционально заменяют хвостовой плавник,
являясь главным органом движения в воде. Характерны в связи
< водным образом жизпи упрощенность зубов и отсутствие хищного
зуба. Три семейства.
Сем. Ушастые тюлени (Otariidae). Менее других
изменились в связи с водным образом жизни; имеются наружные уши, задние
конечности могут подгибаться под туловище при движении по суше.
Котик (Arctocephalus ursinus) (рис. 601) обитает в северных водах
Тихого океана: один подвид—на Командорских островах и на Тюленьем, вто-
рой—на острове Прибылова. На Командорских островах организовано пра-
вильное промысловое хозяйство, и убой котиков производится по опреде-
ленному плану.
Сивуч (Eumetopias jubatus), из Тихого океана, также имеет очень
ценную шкуру. Морской лев (Eumetopias californianus), в Калифорнии.
Сем. Моржевые (Odobenidae). Характеризуются огром-
ными клыками верхней челюсти, отсутствием наружного уха и подгибаю-
щимися вперед задними конечностями. У нас в Полярном море два подвида
моржей (Odobenus rosmarus и О. obesus) имеют промысловое значение
(рис. 602). Моржи дают мясо, жир, шкуру, клыки. Вследствие преследования
моржи быстро уменьшаются в числе.
52*
819
Сем. Тюленевые (Р h oci dae). Задние конечности не под-
гибаются под туловище. У нас в СССР лысун (Phoca groenlandica), нерпа
(Phoca foetida), каспийский тюлень (Ph. caspica), сибирский тюлень (Ph.
sibirica) на о. Байкале, морской заяц (Erignatus barbatus) и др.
Отряд 16. Китообразные (Cetacea)
В настоящее время китообразные представлены двумя подотря-
дами — беззубыми (Mystacoceti) и зубастыми китами (Odontoceti).
Это целиком водные, почти исключительно морские животные с ви-
доизмененным в горизонтальный широкий плавник мускулистым
Рис. 603. Лысун (Phoca groenlandica).
хвостом; волосяной покров исчез, так что от него сохранились лищь
следы на голове; отсутствуют потовые и сальные железы; передние
конечности видоизменены в плавник, причем все обычные сочлене-
ния костей в конечности за исключением плечевого исчезают; клю-
чица отсутствует; задние конечности исчезли, и от них сохраняется
лишь рудимент тазового пояса. Лицевая часть черепа вытянута
далеко вперед, носовые отверстия, наоборот, отодвинуты на темя —
особенности, связанные с водным образом , жизни и способом
захватывания пищи; потеря функции жевания связана со стро-
ением и сочленением нижней челюсти: опа узкая, длинная, без
восходящей ветки и с сочленением, лежащим на уровне нижней
челюсти.
Такое совершенное приспособление первично наземных четверо-
ногих животных к водной жизни, связанное с коренной рекон-
струкцией организации, могло произойти только в результате дли-
тельной эволюции. К сожалению, нам остается неизвестной ранняя
история развития китообразных. Самые ранние ископаемые остатки
относятся к началу среднего эоцена, они составляют особый под-
отряд.
820
Подотряд 1. Древние киты (Archaeoceti)
В окрестностях Каира, в среднеэоцеповых отложениях, найден
череп замечательного китообразного, который определенно указы-
вает, что путь эволюции китообразных идет от низших хищных жи-
вотных. Этот «первокит» — Protocetus atavus имеет череп хищника
с чрезвычайно вытянутой мордой, как это можно видеть из прила-
гаемого рисунка (рис. 604).
Зубная система еще сохраняет типичные черты низших хищных:
1^-, С—, Р—, М-^-, однако зубы упрощены, более однородны и рас-
о 1 4 о
пределены более равномерно по ротовой щели; носовые отверстия
еще расположены в передней половине длинной морды, носовые
кости нормальны, на темени сохранен характерный гребень.
Рис. 604. Череп Protocetus artavus.
(Из Вебера.)
Protocetus очень близок к довольно многочисленной группе иско-
паемых эоценовых китов, составляющих семейство Zeuglodonti-
dae вместе с немногими другими, относящимися к родоначальной
группе всех китообразных, к ископаемому подотряду древних
китов (Archaeoceti).
Так как все известные нам представители этой группы являются
уже вполне водными животными с передними конечностями, пре-
вратившимися в плавники, и редуцированными задними конечностя-
ми, то нужно думать, что ответвление китообразных от хищных
произошло по крайней мере в самом начале эоцена; при этом опи
должны были пройти стадию водных животных с малоизмененными
передними и еще не успевшими исчезнуть задними конечностями
и удлиненной мордой хищного типа.
Ископаемый эоценовый Apterodon из сем. Hyaenodontidae
отряда Creodonta, быть может, указывает нам, какой была эта
переходная ступень.
Archaeoceti дали, с одной стороны, через олигоценового Patrio-
cetus и Agriocetus настоящих беззубых китов. (Mystaco-
ceti), история развития которых хорошо прослежена и начинается
с нижнего миоцена; с другой стороны, через олигоценовое и особен-
но миоценовое семейство Squalodontidae древние киты эволюцио-
нировали до современных зубастых китов (Odontoceti).
821
Подотряд 2. Беззубые ниты (Mystacoceti)
Характеризуются отсутствием зубов во рту, где вдоль костного
нёба располагаются два ряда роговых пластинок, образующих так
называемый кптовый ус; имеют два носовых отверстия. Сюда отно-
сятся самые крупные киты.
Подотряд содержит три семейства.
Сем. Гладкие киты (В а 1 а е n i d а е). В водах, омывающих
берега Союза, встречаются: гренландский кит (Balaena mysticetus), до 15 м
длины (раньше добывались киты до 20 м длины), и японский кит (В. japonica)
Тихого океана до 12 м длины.
Сем. Полосатиков ые (Bal.aenopteridae). В наших
морях встречаются: синий кит (Balaenoptera musculus), самый крупный из
современных китов, достигающий до 30 м длины (обычно 25—27 м), более
мелкий мойвенный полосатик, или финваль (В. physalus), малый полосатик
(В. rostrata).
Сем. Серые киты (Rhachianectidae); Rhachianectes
glaucus — серый кит Тихого океана.
Китобойный промысел в прежней России почти совершенно отсутство-
вал, в последнее время, в связи с освоением Арктики и с введением спе-
циально оборудованного механизированного флота, особенно на Дальнем
Востоке, имеет все шайсы на успех.
Рис. 605. Касатка (Orca orca).
(Из Бихнера.)
Подотряд ^3. Зубастые ниты (Odontocett)
Отличаются наличием зубов, которых то очень много, то они пред-
ставлены единично; имеют одну ноздрю. Три семейства.
822
Сем. 1. Речные дельфины (Platanistidae).
Небольшая группа из трех видов из Индии и Южной Америки, ранее была
представлена многочисленными вымершими видами; иния (Inia amasonica)
|мжи Амазонки, сусук (Platanista gangetica) реки Ганга.
Сем. 2. Кашалотов ые (Physeleridae).
Имеют в верхней челюсти рудиментарные зубы. Кашалот (Physpter
< al,odon) тропических и субтропических морей величиной до 18 м, добы-
вается ради ворвани, спермацета и амбры. Изредка доходит до Камчатки.
Кашалоты дают до 24 кг жира и, кроме того, клея до 1600 кг, спер-
мацет и благовонную амбру.
Питающиеся моллюсками кашалоты выделяются в особое подсемейство—
Ziphiinae.
Сем. 3. Белуховые (Delphinapteridae).
Белуха (Delphinapterus leucus) в Ледовитом океане и Белом море со-
ставляет объект промысла; нарвал (Monodon monoceros) замечателен тем,
что у самца в верхней челюсти с левой стороны сильно вырастает зуб, соот-
ветствующий клыку, образуя бивень.
Сем. 4. Дельфиновые (Del phinid ае). Сюда относятся с хорошо
развитыми зубами касатки(Огса orca)(рис. 605),они у нас водятся в Ледовитом
океане, Охотском и Беринговом морях; в Черном море многочисленны дель-
фины (Delphinus del phis), морские свиньи (Phocaena relicta).
Дельфинов в северных и в Черном море добывают пока исключительно
ради подкожного жира. Каждый дельфин дает от 8 до 16 кг жира.
Сверхотряд. Копытные (Ungulata)
Следующие отряды, несмотря па большое разнообразие в об-
разе жизни, приспособлениях, величине, генетически оказыва-
ются связанными друг с другом и притом все прошли стадию при-
способления к быстрому передвижению на пальцеходящей конеч-
ности. Исходя из этой стадии, они затем разветвились в самых
разнообразных направлениях, дав в основном животных, быстро
бегающих, обитателей открытых пространств, опирающихся или
на четыре и два средних пальца, или па три и один средних пальца,
причем когти видоизменяются в копыта.
Все копытные происходят от первичных хищных животных —
Creodonta. Они образуют несколько ветвей, из которых основными
будут:
1) группа полукопытных (Subungulata) с двумя отря-
дами наземных животных: жиряки (Hyracoidea) и хобот-
н ы е (Proboscidea), с отрядом водных—с и р с а ы (Sirenia) и
ископаемым — эмбритоподы (Embrithopoua);
2) группа парнокопытных (Artiodactyla, Paraxonia)
с подотрядами;
3) . группа м е з о а к спальных с одним отрядом (Perisso-
dactyla, Mesaxonia) с подотрядами первичпокопытные
(Protungulata), непарнокопытные (Perissodactyla) сли-
топтернами (Litopterna) и
4) группа своеобразных южноамериканских
копытных (Notungulata).
Отряд 17. Даманы, или жиряки (Hyracoidea)
Даманы, или жиряки, составляют небольшую группу живот-
ных, обитающую в настоящее время в Африке, Аравии, Сирии
823
и Палестине, а в ископаемом состоянии найденную также в Гре-
ции; это небольшие, лазящие по скалам и деревьям растительно-
ядные животные, у которых па передних ногах 4 пальца с самым
длинным 3-м, а на задних — 3, тоже с осью, проходящей через
3-й палец. Пальцы одеты плоскими погтеобразными ’ когтями,
которые на конце пальца переходят в подошвенную ороговевшую
кожу, как в копытах. Несмотря на такое строение ног, указываю-
щее обычно на приспособление к пальцехождению, даманы явля-
ются стопоходящими, причем нога здесь снабжена присасыва-
тельной, лазательной подошвой. Стопохождение, таким образом,
у даманов является, несомненно, вторичным признаком и связано
с приспособлением к лазанию по скалам, а затем уже и по деревьям.
Даманы имеют характерно устроенные зубы. В молочной генера-
з
ции полное число зубов, обычное для низших хищников: Id-g-,
О
Cd|, Pd|.
4 4
В постоянном составе зубов Id- заменяется постоянно расту-
щим зубом, наподобие грызунов; внизу 2 резца, первые 3 ложно-
коренные могут выпасть, причем они заменяются появляющимися
коренными зубами.
Происхождение даманов относится к началу олигоцена или,
что более вероятно, к эоцену. Уже в начале олигоцена име-
ются остатки вымершего более примитивного семейства Saghathe-
riidae с большим числом разной величины животных, приспособлен-
ных к питанию разнообразной растительной пищей. Впрочем,
только наиболее крупные достигали величины льва. Зубная фор-
мула их 1у, Су, Ру, М-|-. По ряду признаков даманы близки
к низшим хоботным (Probospidea).
Сем. Дама новые (Procaviidae). Это единственное семейство
отряда даманов, или жиряков, состоит из нескольких родов с неболь-
шим числом видов. Род Procavia имеет виды, обитающие в горах, жиряк
капский (Р. capensis) в степях, род Dendrohyrax лазит по деревьям.
Отряд 18. Хоботные (Proboscidea)
Современные хоботные представлены всего лишь тремя ви-
дами слонов. Они характеризуются своей массивностью, тол-
стыми высокими ногами с пятью пальцами, одетыми копы-
тами; нос и губа вытянуты в хобот, в связи с чем своеоб-
разно изменен череп, где носовые отверстия сильно сдвинуты
назад и кости сильно пневматизированы: в зубной системе,
I -ц, т. е. верхние резцы, превращены в бивни, отсутствуют
клыки С ложнокорейные молочные РЬ yg-g > которые
постепенно заменяются громадными лофодонтными коренными, ко-
,т 3
ТОрЫХ М -д.
824
В своем происхождении хоботные связываются с даманами
в общий корень, так как наиболее древние из них близки к при-
митивным даманам.
Сем. Меритериевые (Moeritheriidae) (рис. 606). Эти
животные из верхнего эоцена и нижнего
миоцена, представляют наиболее древних и
наиболее примитивных хоботных, величиной
с тапира, с небольшим хоботом; зубы по
3 13 3 2
формуле I -5-» С —, Р М , причем I —
и U и о
И
превращены уже в маленькие бивни;
Рис. 606. Череп меритерия
(Moeritherium lyonsi).
(Из Абеля.)
конечности у них сходны с таковыми других
хоботных.
Дальнейшая эволюция хоботных пошла
в двух направлениях. Одна ветка дала вы-
мершее семейство динотериев (Dinotheriidae).
Другая ветвь привела к развитию семейства мастодонтов.
Сем. Динотериевые (Dinotheriidae). Эти крупные
и гигантские животные с короткой шеей, хоботом и нижними резцами,
превращенными в бивни, напоминали слонов, от которых однако резко
отличались строением черепа; последний обладал длинными носовыми
костями, маленькими глазными и огромными скуловыми впадинами.
Европа, от среднего миоцена до верхнего плиоцена. Dinotherium gi-
ganteum. (рис. 607).
Сем. Мастодонтовые (Mastodontidae). Это семей-
ство характеризуется наличием как
верхних, так и нижних бивней и по-
степенным усложнением строения ко-
ренных зубов. Начинается семейство
мастодонтов с нижнего олигоцена ро-
дом палеомастодон (Palaeomastodon),
с небольшими бивнями и билофодонт-
ными коренными. Можно проследить
всю постепенную эволюцию семейства
мастодонтов, ставших очень разнооб-
разными и многочисленными в миоце-
не и плиоцене Старого Света, откуда
они перекочевали в Америку. Здесь
они распространились до Южной
Америки в конце плиоцена, тогда как
дольше всего сохранились в Север-
ной Америке в плейстоцене. Масто-
донты в одной своей линии через
Zygolophodon дали следующее се-
мейство (рис. 560, стр. 760).
Сем. Слоновые (Elephan-
t i d a e). Представлено в миоцене
и плиоцене Южной Азии еще близ-
Рис. 607. Динотерий (Dinotherium
giganteum).
(Реконструкция Абеля, из Вебера.)
ким к мастодонтам родом Stegodon,
с-коренными зубами, с 6—12 поперечными гребнями. В дальнейшем они
ведут, с одной стороны, к современному роду африканских слонов (Loxo-
don), с простыми гребнями коренных зубов, с другой стороны — к до-
вольно большому числу плиоценовых видов рода индийских слонов (Elephas).
Начавшись с вида Е. planifrons восточной Индии, слоны перекочевали
в Европу, где дали полуденного слона (Е. meridionalis), из которого про-
изошли, с одной стороны, лесной слон (Е. antiquus) ледникового времени,
с другой стороны—стенной слон(Е. trogonotherii) с последующим ледни-
ковым тундряным мамонтом (Е. primigenius); из этой группы произошли
825
и американские плейстоценовые слоны, а также мелкие слоны среди-
земноморских островов.
В настоящее время представители рода Elephas — индийский слон
(Е. maximus) из юго-восточной Азии отчасти является домашним животным;
другой вид (Е. sumatrensis) — на Суматре.
Отряд 19. Эмбритоподы (Embrithopoda)
Весьма своеобразный единственный ископаемый представитель
этого отряда арзиноте-
рий Циттеля (Arsino-
therium Zitteli) найден
в нижнеэоценовых отло-
.жениях Египта. Будучи
величиной с носорога,
он имеет на носовых ко-
стях два громадны^
в ыр оста, образующих
двойной рог. По строе-
нию скелета он ближе
всего к хоботным и дама-
нам.
Рис. 608. Три последовательных стадии эво-
люции мастодонтов Европы.
А —- Bunolophodon angustidens (миоцен), В—В. 1оп-
girostris (нижний плиоцен), С—В. arvernensis (сред-
ний и верхний плиоцен). (По Абелю из Вебера.)
ч Отряд 20. Сирены
(Sirenia)
Это чисто водные,
морские или пресно-
водные животные, кото-
рых раньше сближали
. с китообразными в ка-
честве травоядных ки-
тов; при изучении их
эволюционного развития
они оказались приспо-
собившейся к водной
жизни ветвью общего
ствола развития копыт-
ных животных. В связи
с водным образом жизни
они имеют вытянутое в
длину грузное тело с
длинным хвостом, снаб-
женным горизонтальным
плавником; волосяной
покров редуцирован, со-
храняются чувствующие
волосы на морде; перед-
ние конечности пре-
вращены в плавники, причем в отличие отГкитов сохраняется
локтевое сочленение и частично сочленение кисти; задние конеч-
ности редуцируются до полного исчезновения самой конечно-
сти (иногда сохраняются остатки бедра) и сохранения лишь
S26
o< i'.itkoii тазового пояса. Сирены — травоядные животные. Их
иуоы нерпоначально сохраняют особенности зубов низших копыт-
пi.iх животных, но имеют тенденцию к полному исчезновению,
к.щ то было у стеллеровой морской коровы (Rhytina stelleri); желу-
док довольно сложный в связи с растительной пищей.
Эволюционная ветвь сирен начинается со среднего эоцена.
I!<iнболсс древняя и примитивная форма э о т е р и й (Eotherium
всрд ptiacum). Хотя это животное и было уже типичным водно-
мирским животным, однако оно имело еще довольно хорошо раз-
витую заднюю конечность, о чем можно, судить по тазу с хорошо
развитой вертлужной ямкой; зубы имели характерную для при-
митивных хищных формулу: 1-|, С~, Р-^-, М-|- ; череп по своему
характеру очень близок к черепу низших копытных. Ряд последо-
вательных стадий развития: эосирен (Eosiien) верхнего эоцена и
гллитериум (Halitherium) приводят к современным дюгоням
сем. Halicoridae и недавно уничтоженным Rhytinidae. Что касается
ламантинов, то они отделились от общего ствола еще в олйгоце-
нг; Manatherium из олигоцена Бельгии.
Сем. Ламантиновые (Manatidae) с функционирующими
коренными зубами, которые постепенно сменяются горизонтальной сменой
(до 20 зубов в последовательно сменяющемся ряду); хвостовой плавник
лопатообразный. Ламантин (Manatus) — два вида по берегам и рекам Се-
верной Бразилии и берегам Антильских островов, один вид на западном бе-
регу Африки.
Сем. Дюгоневые (Halicoridae). У самцов бивни; 5—6 ко-
ренных зубов в каждой стороне верхней и нижней челюсти, без корня; хво-
стовой плавник полулунный. Три вида рода Halicore по берегам Индии,
в Красном море и по берегам Австралии и Новой Гвинеи.
Сем. Морские коровы (Rhytinidae). Единственный
вид — стеллерова морская корова (Rhytina stelleri) — был открыт у берегов
Камчатки и Командорских островов в 1741 г. Стеллером. Уже 100 лет спустя
была уничтожена моряками. Последний экземпляр был отмечен в 1854 г.
Это были крупные до 8 м стадные животные, питавшиеся морскими водорос-
лями.
О т р я д 21. Парнокопытные (Artiodactyla)
Парнокопытные составляют самую обширную ветвь современ-
ных копытных животных. Она естественно распадается на две
основных группы: на небольшую группу всеядных животных или
пежвачных, отличающуюся буподонтными зубами — подотряд
Neobunodontia, и группу, обильную формами животных
чисто травоядных, обладающих жвачкой (жвачные — Ruminantia)
и селенодонтными зубами—подотряд Selenodontia. Помимо
этих двух, довольно резко отличающихся групп, в ископаемом со-
стоянии известно очень много форм из эоцена и олигоцена, которые
занимают промежуточное место между ними, не являясь в то же
время предками ни тех, пи других, а потому объединяются в само-
стоятельные параллельные группы. Мы объединяем их в один
подотряд — буноселенодонтных (BunosQlenodontia).
Происхождение парнокопытных остается недостаточно выяс-
ненным. Мы знаем только, что в своем происхождении они непос-
827
родственно не связаны с непарнокопытными, которые произошли
от первичнокопытных (Protungulata); мы знаем, что прошлое парно-
копытных коренится где-то среди иижнеэоценовых предков хищ-
ников, в отряде креодонтов (Creodonta), но не тех, которые имели
хорошо развитый 1-й палец, а тех, у которых 1-й палец был уже
значительно редуцирован.
Наиболее характерной особенностью этого отряда служит так
называемое параксиальное строение конечностей, при котором
равнодействующая веса тела для каждой конечности проходит
между 3-м и 4-м пальцами, давая каждому из них равномерную
нагрузку, при этом 1-й палец всегда отсутствует, тогда как 2-й
и б-й имеют тенденцию в эволюционном ряду к постепенной редук-
ции. Эта редукция боковых пальцев у той группы животных,
которые привели к процветающим в наше время формам, сопро-
вождалась расширением проксимальной части двух средних мета-
подий, что привело к более прочному сочленению их с запястьем
и предплюсной. У животных тех боковых ответвлений ствола
эволюции, которые вымерли к миоцену, такого расширения про-
ксимальных частей метаподий не было и редукция боковых паль-
цев не отражалась на запястье и предплюсне.
Чрезвычайно существенны изменения, происходившие в строе-
нии зубов в процессе эволюции парнокопытных. Исходная фор-
мула и строение зубов соответствовали таковым низших креодон-
тов (Acreodi): зубы с корнями образуют замкнутый ряд 1-|-, С^-,
О 1
4 з
Р у, М-g-, коренные зубы с низкими коронками с 3—6 бугорками.
Преобразованию прежде всего подверглись коренные зубы в связи
с переходом на растительную пишу. Они становятся четырех-
бугорчатыми, бунодонтными, а затем селенодонтными. Как пока-
зали подробные палеонтологические исследования, это преобра-
зование шло по трем разным направлениям, одно из которых
привело к зубам современных парнокопытных.
Дальнейшие видоизменения зубов парнокопытных связаны
с адаптацией к более соломистой пище. Вырабатывается приспо-
собление для обрывания травы и листьев с помощью языка и ниж-
них резцов, что ведет к полному исчезновению верхних резцов
и превращению нижнего клыка в резцеподобный зуб.
Трава, являясь пищей, содержащей большое количество клет-
чатки, принадлежит к объемистым кормам. Животные травояд-
ные должны иметь приспособления к одновременному пережевы-
ванию больших объемов травы, что стоит в связи с чрезвычай-
ным усилением жевательных мышц, увеличением в длину ротовой
полости, увеличением слюнных желез и изменением бунодонтных
зубов в селенодонтные; лицевая йасть черепа удлиняемая, и между
резцами и сходным с пим клыком и коренными зубами в нижней
челюсти образуется диастема; в верхней челюсти вслед за редук-
цией исчезает клык, который сохраняется лишь у немногих
жвачных в качестве органа защиты. Для защиты же в основном
828
приспособлены рога. Коренные зубы делаются высокими (гипсело-
доптными), призматическими, со стирающейся коронкой, с поздно
замыкающимся корнем или даже вовсе без корня, т. е. с постоянным
ростом. Одновременно с приспособлениями, связанными с обры-
ванием и перетиранием пищи, должны были соответственно при-
способляться органы переваривания пищи, и у ныне живущих
многочисленных травоядных парнокопытных имеется весьма совер-
шенное приспособление к перевариванию пищи в сложном желуд-
ке, соединенное со вторичным пережевыванием отрыгиваемой в рот
Рис. 609. Бегемот (Hippopotamus
amphibius).
(Из Гааке.)
жвачки. Несомненно, что в высоком приспособлении зубной сис-
темы и в жвачке мы имеем эйритоадаптивное приспособление,
позволившее животным эволюционировать в различных направле-
ниях, используя разнообразные среды для жизни и разнообраз-
ные ^равы для пищи. Три подотряда: бунрдонтные (Neobuno-
dontia), селенодонтные (Selenodontia) и ископаемые — буноселе-
иодоитпые (Bunoselenodontia).
» Подотряд 1. Бунодонтные (Neobunodontia)
Из современных нам млекопитающих сюда относятся свиньи
и бегемоты, составляющие два отдельных семейства.
Это всеядные неуклюжие ясивотные на низких ногах с четырьмя
пальцами и песросшимися метакарпальными костями, с вытянутым
829
черепом и вытянутыми назад орбитами, с бунодоптными зубами,
происшедшими из полу селенодонтных зубов; желудок различной
сложности; характерна способность откладывать подкожный слой
сала.
Сем. Гиппопотамов ые (Hyppopotamidae), или бе-
гемоты, хорошо плавающие и ныряющие толстые неуклюжие животные,
в настоящее время представленные двумя родами бегемота: Hippopotamus —
крупный бегемот с тремя видами, распространенный по Северной, Восточной
и Южной Африке, и мелкий род бегемота—Choeropsis—из Западной Африки,
интересный тем, что его ископаемые остатки известны из плейстоцена Мада-
гаскара, Кипра, Крита и Мальты.
Сем. Свиньи (Suidae). Характеризуются клиновидным черепом
в связи с развитием роющего рыла и хорошо развитыми клыками, из которых
верхние загнуты наверх. Род свинья, или кабан (Sus), представлен целым
рядом видов; из них упомянем родоначальников домашней свиньи — евро-
пейско-азиатского кабана (S. scrofa) и восточноазиатских свиней (S. vitta-
tus); бабирусса (Babirussa babirussa) с очень длинными загнутыми вверх
и назад верхними клыками, прободающими во время роста верхнюю губу,
распространена на Целебесе и Молуккских островах; бородавочник (Phaco-
choerus) — несколько видов Южной и тропической Африки. -
В особое подсемейство выделяются южноамериканские пекари (Dicotyli-
пае), обитатели лесов Южной и Центральной Америки; имеют на спине
пахучую железу.
Свиней производят от нижнеэоценового Propalaeochoerus и через мио-
ценовых Palaeochoerus и Hyotherium приводят к верхнеолигоценовым Sus
palaeochoerus, S. antiquus и S. major.
Подотряд 2. Лунчатозубые (Selenodontia)
Самая богатая видами группа копытных, обязанная своим про-
цветанием многочисленным приспособлениям эйритоадаптивного
порядка: 1) к питанию самой разнообразной растительной пищей—
от сочпой травь! листьев деревьев до сухих колючих пустынных ра-
стений и молодых побегов деревянистой растительности; 2) к бы-
строму бегу с помощью приспособленных соответственно двупалых
конечностей; 3) ко вторичному пережевыванию заглотанной в боль-
ших объемах пищи; 4) к защите от нападений путем развития ро-
гов.
В отличие от бугорчатозубых они покрыты густой шерстью,
имеют замкнутую сзади орбиту и специализированные зубы, при-
чем верхние резцы обычно исчезают, нижние клыки становятся
резцоподобными, а коренные—лунчатыми. Метаподии 3-й и 4-й
срастаются, 5-й — в разной степени редукции.
Этот подотряд может служить чрезвычайно поучительным при-
мером того, как приобретенные эйритоадаптивные свойства могут
привести к перерастанию группы низшего порядка в группу выс-
шего систематического порядка: указанные выше эйритоадаптив-
ные свойства привели к разнообразнейшим приспособлениям сте-
ноадаптивпого порядка и широкому распространению, так что луп-
чатозубые могут быть подразделены на большое число семейств,
в свою очередь объединенных в более общие группы. Можно уста-
новить, не считая ископаемых, по крайней мере три надсемейства:
мо золеногих (Tylopoda), оленьков (Traguloidea) и
жвачных (Ресога) в узком понимании.
830
Надсем. I. Мозоленогие (Ту lo pod а)
Ом. Верблюд о’вые (Gamelidae) отличается от всех прочих пар-
нокопытных своеобразным строением своих ног, два пальца которых не имеют
копыт и опираются тремя фалангами на подушкообразное эластическое
утолщение кожи, лежащее на толстой ороговелой подошве; спереди и сна-
ружи имеется плоский роговой ноготь (рис. 610). Боковые пальцы исчезли
совершенно: метаподии срослись в бегательные кости, причем дистальные
концы их несколько расходятся.
Расходящиеся в стороны пальцы, их опора на мозолевидную эластиче-
скую подошву являются приспособлением к обитанию в песчаных пусты-
нях, позволяющим беспрепятственно и легко двигаться по сыпучему рас-
каленному песку. Стеноадаптивное приспособление, явившееся эйритоадап-
тивным в пределах пустынной среды и биотопов. Таким же приспособле-
нием является наличие горба, который служит резервуаром для жира, что
повволяет этим животным подолгу оставаться без пищи и воды в знойных
пустынях. В верхней челюсти имеются клык и резец с каждой стороны.
Происхождение верблюдов прослежено до верхнего эоцена. Родиной их
।шляется Сев. Америка. Наиболее примитивной формой является эоцено-
вый Protylopus — маленький, величиной с зайца, предок верблюдов с еще
полной зубной системой I ~ , С~ , Р М , но с уже редуцированными
<5 1 Ч О
боковыми пальцами: к нему примыкает олигоценовый Poebrotherium, вели-
чиной с овцу, далее следуют миоценовые Protolabis
и Procamelus и, наконец, плиоценовые верблюды рода
Carnelus. Из Сев. Америки они распространяются
в Старый Свет и Южную Америку. Интересна бо-
ковая ветка миоценовых Alticamelus, которая была
родом параллельно развившимся наподобие жираф
с длинными конечностями и длинной шеей.
Два рода единственного семейства Camelidae:
верблюд (Carnelus) и лама (Auchenia). Верблюд дву-
горбый (Carnelus bactrianus) — единственный дикий
вид верблюдов в наше время. Центральная Азия.
()домашнены. Одногорбый верблюд (С. dromedarius)—
только в домашнем состоянии. Два диких вида ламы:
гуанако (A. huanachus) и викунья (A. vicugna)—
в горах Южной Америки. Одомашнены: лама (А.
glama) и альпака (A. pacos).
Надсем. 2. Оленьки (Traguloidea)
Сем. Оленьков ые (Tragulidae). По-
добно верблюдам во многом отличаются от прочих
парнокопытных, сохраняя многие примитивные чер-
ты строения и представляя боковую ветвь жвачных
Рис. 610. Продольный
разрез через палец
передней конечности
ламы Auchenia glama.
животных, ответвившуюся от общего ствола разви-
тия парнокопытных по меньшей мере с олигоцена.
Сюда относятся самые мелкие из копытных, причем
самые крупные из них не превосходят величины
косули. Характеризуются наличием боковых пальцев
и хорошо развитыми клыками, особенно у самцов,
i—пястная (metacarpa-
le), 2, 3, 4—три фалан-
ги, 5 — роговая ногте-
вая пластинка, 6— рого-
вая подошвенная пла-
стинка, 7—эластическая
подушка. (Из Вебера.)
। де О1*и служат для защиты. Желудок из трех отде-
лов, так как нет книжки (psalterium). Два рода; кроме того, одно иско-
вой моо семейство (Gelocidae).
Оленек (Tragulus)—несколько видов в юго-восточной Азии и прилегающих
островах. Другой род Hyaemoschus в Западной Африке.
Падсем. 3. Настоящие жвачные (Ресога)
Наиболее высокоорганизованные парнокопытные — олени, быки, козы,
овцы, антилопы и жирафы — составляют три семейства: олени (Cervidae),
быки (Bovidae) и жирафы (Giraffidae) со следующими общими признаками:
831
Зоновые пальцы представлены лишь рудиментами, желудок состоит из
четырех отделов, на голове обыкновенно имеются рога или только у самцов
или у самцов и у самок.
Сем. Оленевые (Cervidae) объединяет стройных животных
на высоких ногах, с отпадающими ежегодно, обычно ветвистыми рогами
у самцов; только у северного оленя (Rangifer) рога имеются и у самок, а у
самца кабарги (Moschus и Hyelaphus) рога отсутствуют вовсе, но вместо
них орудием защиты и нападения у самцов являются сильно развитые клыки;
Форма рогов оленей имеет большое систематическое значение, так как
в процессе эволюции шло прогрессирующее увеличение и ветвление рогов.
В связи с этим проведена была гомологизация отростков оленьих рогов,
которая видна из прилагаемого рисунка (рис. 611). Побочные копыта име-
ются (за исключением мунтджака); в черепе два слезных отверстия; коренные
зубы с низкими коронками — брахиодонтные.
ft)
Рис. 611. Эволюция рогов оленей и семь стадий развития ро-
гов у благородного оленя.
Рога: 1—Mazama и 2 — Hippocamelus (оленчики), 3 — Axis, 4 — Rusa,
5—Sika (пятнистые олени), 6 — Daina (лань), 7 — Alces (лось), 8— Сарге-
olus (косуля), 9 —Rangifer (северный олень), 10—> семь стадий развития
рогов у благородного оленя. Заштрихованы средние отростки, черным
отмечены глазничные отростки, пунктированы задние отростки.
(Из Вебера.)
Подсем. Кабарговые (RJoschinae). Кабарга (Moschus moschi-
ferus) — единственный вид и род, отличается отсутствием рогов у самцов
и развитием взамен их клыков, а также одним слезным каналом. Горы
Сибири, Центральной и Восточной Азии.
Подсем. Оленевые (Cervinae) делятся на две группы.
А. Первая группа (Telemetacarpalia) — метакарпальные боковые кости
редуцированы в своей проксимальной части.
Косули (Capreolus capreolus) — в Европе; другие виды в Азии (С. ру-
gargus, С. bedfordi).
Лось (Alces alces) — самый крупный из оленей, типичный обитатель
лесов севера Европы и Азии: образует несколько подвидов. Нет лося в Крыму,
на Кавказе, в Туркестане, на Камчатке и на Сахалине. В лесах Европейской
части СССР сейчас сделался большой редкостью и, вероятно, скоро исчез
бы совершенно, так как промышленники и население преследую г лосей ради
мяса и шкуры. В виду повсеместного исчезновения в Европейской части
СССР охота на лосей запрещена совершенно, а в остальной части Союза
ограничена ноябрем и декабрем. Организованы специальные лесные запо-
ведники: Мордовский, Центрально-лесной, Бузулукский бор, Куйбышев-
ский, в которых лоси успешно размножаются.
Северный олень (Rangifer tarandus) незаменимое домашнее животное
Севера. Дикий северный олень занимает своим распространением всю зону
тундр и почти всю полосу тайги до южных горных хребтов и Великого океана.
832
(hi и образуют на этой обширной территории несколько подвидов.
I'ни.ню дикие олени были очень многочисленны, и жизнь охотничьих на-
poioii Крайнего Севера главным образом обеспечивалась охотой на них,
преимущественно во время их перекочевок из тундры в леса и обратно.
< »гл ичается присутствием рогов у самок.
Рис. 612. Крымские благородные олени и маралы в акклиматизационном
парке Аскания-Нова.
(Фот. Н. Озерова.)
В Америке несколько уклоняющихся родов оленей.
. В. Вторая группа (Plesiometacarpalia) — боковые метакарпалии сохра
няются в проксимальной части. Благородный олень (Cervus elaphus) — охот
ничье животное Западной Евро-
пы; кое-где встречается в лесах
Украины, тогда как в лесах Кры-
ма и Кавказа встречаются его
подвиды (рис. 612).
В южной Сибири и на Даль-
нем Востоке распространены ма-
ралы и изюбри, подвиды северо-
американского оленя-вапити (С.са-
nadensis). Пятнистые олени (Pseu-
daxis hortulorum и Р. sica, рис.
673) на Дальнем Востоке являют-
ся наиболее ценными пантовыми
оленями; в настоящее время ши-
роко акклиматизируются в систе-
ме заповедников РСФСР; (Axis)
в Индии; мунтджак (Muntjacks)
из Южной Азии и островов. Лань
(Dama dama, рис. 613) в четвертич-
ный период широко распростра-
ненная в Европе, в настоящее вре-
мя в диком состоянии живет лишь
в Средиземноморье, разводится в
парках Европы.
Олени в ископаемом состоянии
известны со времени нижнего мио-
*>3 Каш каров и Станчинский
Рис. 613. Лань (Dama dama).
(Аскания-Нова, фот. Озерова.)
833
цена Европы: Amphitragulus — наиболее примитивная форма родоначальных
оленей; затем Dremotherium, обе формы близки и к примитивным полорогим
и выделяются в отдельное сем. Cervulidae. Paleomeryx ив среднего и верх-
него миоцена является уже настоящим оленем; замечательные гигантские,
вероятно, степные, олени плейстоцена (Cervus megaceros) и гигантские степ-
ные лоси (Alces latifrons).
Все олени промышляются ради мяса и шкуры, а на маралов, изюбров
и пятнистых охотятся главным образом ради еще неокостеневших рогов,
которые в высушенном виде дают весьма ценный в Китае товар «п а н т ы»,
имеющие в китайской медицине чрезвычайное распространение. В пантах
содержится сильно возбуждающее вещество — пантокрин.
Рис. 614. Антилопа канна (Taurotragus oryx) в акклиматизационном парке
Аскания-Нова.
(Фот. Н. Озерова.)
В виду ценности пантов оленей содержат в загородях, ежегодно от-
пиливают в нужное время рога.
Благородный олень и марал при надлежащей охране быстро размно-
жаются в лесах, отведенных под правильно организованные охотничьи
хозяйства. В виду имевшего место повсеместного их уменьшения прово-
дится организация соответствующих заказников и разведение их в заповед-
никах.
Сем. Полорогие (Bovidae) — охватывает чрезвычайно много-
численных и разнообразных антилоп, серн, козлов, овец, мускусных быков,
буйволов и настоящих быков. Характерная особенность их — роговые рога,
сидящие на выростах лобных костей; верхние клыки отсутствуют. Свыше
150 видов. Семейство полорогих в настоящее время разбито на 13 подсе-
мейств, причем 8 подсемейств представлены антилопами. Последних мы
рассматриваем как одно подсемейство.
Подсем. А н т и л о п о в ы е (А n t i 1 о р i п а е) — (свыше 100 видов)
распространены по степям и саваннам Старого Света. В пределах СССР встре-
чаются: сайга (Saiga tatarica), (рис. 683) до половины XIX века жив-
шая по степям на запад до Днепра, в настоящее время сохранилась в Калмыц-
ких степях, в Казахстане и Средней Азии; джейран (Gazella subgutturosa) в
Закавказье и Средней Азии, дзерен (G. gutturosa) в Монголии (рис. 641).
834
В Африке! своеобразные бубалы (Bubalus), гну (Connochaetes)
(рис. 615)—похожи на рогатых лошадей, крупные сернобыки (Oryx), канны,
или оленебыки (Taurotragus oryx), лошадиные антилопы (Hippotragus) и др.
Из азиатских видов упомянем индийскую нильгау (Boselaphus tragocamelus),
гарну (Antilope cervicapra) (рис. 614).
Подсем. Серновые (Rupicaprinae), Горные антилопы или
серны с рожками, загнутыми назад, выделяются в особое подсемейство,
которое ванимает среднее место между антилопами и козами. Серна (Rupi-
capra rupicapra) — у нас на Кавказе и в Западной Европе, на хребтах
Приамурья — горал (Nemorhaedus caudatus), в Америке — снежная коза
(Oreamnos americanus).
Подсем. Козо-барановые (Caprinae или С а р г о v i-
пае); у нас в Союзе обитают: сибирский горный козел (Capra sibirica)—
в Тянь-Шане, Алтае и Саянах; безоаровый козел (Capra aegargus)
(рис. 616) — в Закавказье и в горах Туркмении; горные туры (Capra
Рис. 615. Антилопы голубые гну (Connochaetes taurinus), и самка антилопы
сайги (Saiga tatarica) в акклиматизационном парке Аскания-Нова.
f (Фото Озерова.)
caucasica) — на Кавказе, в Альпах— козерог (Capra ibex). Домашних коз
производят из трех центров доместикации: от дикой ископаемой козы Европы
(С. prisca), от беэоарового козла (С. aegargus, рис. 616) Западной Азии
и от винторогого козла (С. falconeri)(pnc. 664) Кашмира и Гималаев.
Из диких баранов многие виды принадлежат фауне Союза: азиатский
муфлон (Ovis orientalis) из южной Туркмении, качкар (Ovis poloi) с подви-
дами в Тянь-Шане и на Памире, аргали (Ovis ammon) на Алтае и Саянах,
( нежный баран (Ovis nivicola) по горам Восточной Сибири.
В Европе раньше был широко распространен муфлон (Ovis muusimon),
сейчас ограниченный горами Корсики и Сардинии. Домашних овец (Ovis
arics) Производят не только от муфлона, от которого произошли некоторые
примитивные породы овец, но и от других овец, прежде всего степных овец,
распространенных в Передней Азии (О. vignei), а также упомянутых азиат-
ского муфлона и аргали.
• Чрезвычайно интересные североамериканские овцебыки, или мускусные
быки (Ovibos moschatus), ранее с кругополярным распространением, являются
боковой веткой ствола развития парнокопытных, занимающей промежуточ-
ное место между овцами и быками (рис. 617).
Подсем. Быки (В о v i п а е) объединяет быков с широкой голой на
конце мордой, загнутыми наружу рогами и коренными зубами с высо-
53* 835
кими призматическими коронками и добавочными столбиками эмали между
луночками. Буйволы (Bubalus) представлены обыкновенным южноазиатским
буйволом, частью диким, частью одомашненным^В. bubalus, рис. 618)', с боль-
шим числом рас, маленьким аноа (В. depressicoFnis) с острова Целебес и
африканским буйволом (В. caffer). Обыкновенный буйвол в качестве домаш-
него животного распространен также в Северной Африке и Южной Европе,
а у нас на Кавказе. Род быков в узком смысле (Bos и Bibos) представлен
Рис. 616. Безоаровый козел (Capra aogargus).
(Из Гааке.)
в настоящее время немногими дикими видами: гаял (В. frontalis) с очень
толстыми рогами и гаур (В. gaurus) в Индии; бантенг (В. banteng) —Сума-
тра, Ява, Борнео; все три вида легко приручаются (рис. 619 и 620).
Предками многочисленных пород домашнего крупного рогатого скота
(Bos taurus) были дикие виды животных: тур (Bos primigenius), последний
живой экземпляр которого пал в 1627 г. в Польше, и близкий к туру чет-
вертичный азиатский бык номадов (Bos nomadicus), происшедший, вероятно,
из плиоценового индийского В. planifrons. Наконец для происхождения за-
падноевропейского скота вводят еще одну форму более мелкого короткорогого
быка, предки которого встречались в четвертичное время (В. brachyceros).
Род бизон (Bison) в Сев. Америке представлен бизоном (В. ameri-
canus), в Европе — зубром (В. bonasus). Последний был представлен двумя
подвидами: беловежским и кавказским. Кавказский зубр окончательно ис-
треблен в период интервенции, а от беловежского зубра, которые в эна-
836
Рис. 617. Мускусный быц (Ovibos moschatus).
(Ия Гаане.)
Рис. 618. Домашний буйвол (Bubalus bubalus).
(Асканин Нова, фот. Озерова.)
837
Рис. 619. Еантенг (Bibos banteng).
(Аснания-Нова.)
Рис. 620. Гаял (Bibos frontalis).
чительном числе жили до империалистической войны в Беловежской пуще
в западной Белоруссии (бывш. Гродненской губ.) сохранилось в разных
странах на 1/1 1938 г. всего лишь 9-7 чистокровных особей. В настоящее
время восстановление зубра в СССР проводится в заповедниках (Аскания—
Нова, Крымский и Кавказский).
838
Сем. Жирафовые
(Giraffidae). Два рода
итого семейства принадлежат
исключительно Африке. Эти
очень высокие, с длинной шеей,
животные приспособились к
питанию эа счет листьев древес-
ной растительности. Передние
ноги длиннее задних. На го-
лове, на границе лобных и те-
менных костей, небольшие кост-
ные рожки, одетые кожей (рис.
621). Окапи (Okapia johnsto-
ni) —в лесах Центральной Аф-
рики; жираф (Giraffa Camelo-
pardalis и др.) — в саваннах
Центральной и Восточной Аф-
рики. Интересно, что в плио-
цене различные роды и виды
жираф населяли Грецию, ост-
ров Самос, Бессарабию, Пер-
сию, С. Индию и Китай. Пред-
ками жираф были близкие к
оленям безрогие мелкие живот-
ные с нормальной шеей и нор-
мальными ногами.
Очень близки к жирафам
по своему происхождению ис-
копаемые сиватерии (Si-
vatherium, Bramatherium и др.)
Рис. 621. Жираф (Qiraffa camelopardalis);
;Иэ Брэма.)
с громадными костными выро-
стами для рогов между лобной
и теменной костями — из ниж-
него плиоцена Индии.
Кроме перечисленных выше семейств в исков шшргтмп
еще большое количество форм парнокопытных?™™„^^’’следующие
три семейства очень близки к жвачным, почему “3 "° I приводим
Сем. HypertraguHdae. Небольшие H3”
Европы и главным образом из миоцена Сев. АмеР„„„ r 1О93г найдены от-
носящиеся сюда формы из эоцена Монголии, при • которые обна-
руживают замечательные признаки, связывающ/ семейства жвач-
ных, так что можно считать их даже предками » Р животных. Таков,
например, Archaeomeryx, имеющий уже слитными i ладьевидную
косточки, тогда как в передней конечности совери^У0™ « У ивн« дое^ы
четыре пальца; локтевая и Малоберцовая кости редуцированы, так же
Рис. 622. Реставрация ископаемых
tomeryx) и Oreodontidae (
(Из Осборна.)
В —e{tagulidae (А — Lep-
Ireodon).
839
как и верхние резцы. Эти животные были величиной с кролика. У некоторых
на черепе были рога в виде одной или двух пар; у других были хорошо раз-
виты клыки (Blastomeryx, Leptomeryx) (рис. 622, А).
Сем. Oreodontidae. Высокоспециализированное североамери-
канское семейство, появившееся, вероятно, из Азии в эоцене, достигшее
своего наибольшего развития в миоцене и исчезнувшее в плиоцене. Известно
около 100 видов небольших, величиной с зайца или свинью видов, питав-
шихся растительной пищей и живших, вероятно, среди степей и пустынь.
Отличается большим черепом с примитивной зубной формулой и 5—4-па-
лыми низкими конечностями (Oreodon) (рис. 622, В).
Сем. Xiphodontidae. Отличается высокими конечностями, на-
поминает оленей. Из верхнего олигоцена Европы.
Подотряд 3. Бугорчато-лунчатозубые (Bunoselenodontia)
Сюда мы относим целый ряд групп, относящихся, вероятно,
пе менее как к пяти семействам парнокопытных животных, появив-
шихся в эоцене и затем вымерших в разное время миоцена и плио-
цена. Отметим некоторых: Anthracotheriidae из олигоцена и эоцена
Южной Азии, Сев. Америки и Европы— крупные, то похожие на
свиней, то на бегемотов болотные формы; Anoplotheriidae — вели-
чиной с тапира, из эоцена и олигоцена Европы; Dichobunidae и
Elotheriidae с бунодонтными зубами, с характерным развитием
гипоконуса (НуросопЦега), Caenotheriidae с такими же зубами,
образованными с помощью протоконулуса.
Отряд 22. Южно-американские копытные (Notungulata)
Южная Америка, отделенная в течение большей части третич-
ного времени от остальных материков, была, подобно Австралии,
местом совершенно своеобразной эволюции млекопитающих жи-
вотных. Однако в связи с тем, что в Южной Америке были иные
условия существования и в ней были открытые пространства от пу-
стынь до прекрасных луговых степей, с одной стороны и, с другой
стороны, в связи с тем, что первоначальный состав фаупы был значи-
тельно иной, чем в Австралии (имелось уже достаточно мощное ядро
высших плацентарных зверей), эволюция южноамериканских мле-
копитающих пошла по совершенно своеобразному пути, отличному
как от развития млекопитающих Австралии, так и от эволюции их
в Старом Свете и Сев. Америке. В связи с указанными выше особен-
ностями среды чрезвычайного развития достигли здесь травоядные
копытные, притом такие, которые в значительной степени заменяли
здесь пе только копытных Старого Света, по отчасти грызунов, по-
лучивших здесь своебразпое развитие. Как теперь начинает вы-
ясняться, предки многочисленных копытных Южной Америки про-
исходят из Азии и являются очень древней группой, от которой
произошли даже такие древние примитивные формы, как первично-
копытные. Они ответвились от очень древних креодоптных млеко-
питающих еще, вероятно, на самой заре их развития, т. е. в конце
мелового периода. Счастливое стечение обстоятельств, а именно то,
что па южноамериканский континент почему-то не попали предки
настоящих хищных и не образовались в эволюционном процессе
840
свои хищники, имело тоже немаловажное значение для эво-
люции.
Было бы очень интересно и поучительно для понимания эволю-
ционного процесса проследить, каким образом шла эволюция
млекопитающих этой группы животных в связи с приспособлением
к разным условиям обитания, однако это работа будущего. Мы
видим здесь только большое разнообразие групп, связанное
с большим разнообразием направлений в приспособлениях, а
в связи с этим большое разнообразие
' как в строении конечностей, так и
в строении зубов.
Подотряд I. Типотерии (Typotheria) '
Мелкие, величиной с кролика и
не больше свиньи, своеобразные жи-
вотные; похожие на даманов или
грызунов, или низших копытных. Рис. 623. Череп Protypotherium,
Зубы растительноядного типа, пер- (Из Вебера,)
воначально брахиодонтного, без диастемы, затем становятся похо-
жими на зубы грызунов, с постоянным ростом, Protypotherium
(рис. 623). Пятипалые конечности переходят в четырехпалые. Не-
сколько семейств. С эоцена до плейстоцена.
Подотряд 2. Токсодонты (Toxodontia)
С олигоцена до плейстоцена в Южной Америке жили крупные
формы, величиной с носорогов или бегемотов, — копытные живот-
ные с тяжелым складом, с толстыми трехпалыми ногами и громад-
ным черепом. Коренные зубы селенолофодонтного типа, у высших
форм с постоянным ростом (Toxodon) (рис. 624).
Рис. 624. Скелет Toxodon platensis.
(Из Вебера.)
Подотряд 3. Астрапотерии (Astrapotherioidea)
Весьма своеобразный отряд с небольшим числом форм, отличаю-
щихся отсутствием верхних резцов, длинными верхними клыками и
841
запоминающими носорогов коренными зубами; имели хобот и были
величиной от свиньи до крупного носорога. С олигоцена до миоцена.
Подотряд 4. Пиротерии ( Pyrotheria)
Пожалуй, это самые удивительные из южноамериканских иско-
паемых животных; они имели укороченный череп, длинный хобот,
коренные зубы, как у динотерия, и бивнеобразпые резцы; пятипалые
конечности, невидимому, роющего типа.
Отряд 23. Непарнокопытные (Mesaxonia)
Относящиеся сюда животные все связаны между собой общно-
стью происхождения из общего родоначального ствола и тем, что
эволюция всех относящихся сюда форм шла по пути перехода от
пятипалой конечности к конечности трехпалой и, наконец, у наи-
более специализированных форм— однопалой.
Родоначальной группой всех непарнокопытных являются при-
митивные палеоценовые и эоценовые первичнокопытные Сев. Аме-
рики.
Подотряд 1. Первичнокопытные ( Protungulata)
Эти животные по своим признакам и по внешнему виду очень близ-
ки к низшим креодонтам, но образуют ряд переходов к непарноко-
пытным животным; они имеют пятипалые конечности, череп по об-
щему очертанию напоминает череп насекомоядных или происшед-
3 14
ших от них низших креодонтов. Зубная формула 1у, Су, Ру,
,,з
Му ; ложнокоренпые принимают постепенно вид настоящих корен-
ных, которые из трех- и пятибугорчатых становятся шестибугор-
чатыми. Несколько семейств, из которых особенно интересно:
Рио. 625. Реконструкция фенакодуса (Phenacodus primaevus).
(Из Вебера.)
сем. Фенакодонтовые (Phenacodontidae) (рис. 625).
Небольшие и средней величины животные, от зайца до рыси, с вытянутой
в длину головой, с плоским лбом и небольшим хоботом, с небольшими клы-
ками, трехбугорчатыми ложнокоренными зубами, с шестибугорчатыми верх-
ними и пятибугорчатыми нижними коренными; в переднем поясе ключиц
нет, конечности пятипалые, пальцеходящие, с копытами, причем средний па-
лец наиболее развит; с длинным хвостом. Палеоцен и нижний эоцен Сев.
Америки. Phenacodus считался раньше родоначальником лошадей. В на-
стоящее время считается наиболее близкой формой к предкам хорошо изу-
ченного ряда лошадей Tetraclaenodon.
842
Подотряд 2. Непарнокопытные (Perissodactyla)
Этот подотряд охватывает три эволюционных ветви непарнопа-
лых животных, от цветущего состояния которых в среднетретичное
время сохранились лишь жалкие остатки: тапиры, носороги и ло-
шади; к ним необходимо добавить еще многочисленных громадных
и грузных титанотериев (Titanotheriidae) олигоцена Сев. Америки.
Все эти группы, столь различные, сходятся в общем своем проис-
хождении, и их родоначальники в эоцене имели еще очень много
общего между собой. Зубная система была еще непрерывной, зубы
были брахиодонтные, конечности были четырех- или трехпалые.
Разделение на эти четыре группы намечается уже в эоцене.
Сем. Лошадиные (Equidae). Обзор непарнокопытных удобнее
всего начать с лошадей, так как эта группа является в отношении своего
развития лучше всего изученной; особенно исследования за последние 20 лет
дали чрезвычайно богатый материал для восстановления родословной этой
группы, так что мы с большой достоверностью можем говорить сейчас о про-
исхождении и эволюции лошадей.
Как мы уже указывали, лошади берут свое начало в семействе фена-
кодонтов ых (Phenacodontidae) подотряда первичнокопытных из палео-
цена Сев. Америки (Tetraclaenodon). Следующая стадия эволюции относится
уже к группе лошадей и дает две ветви: одна развивается в Европе, образуя
шесть параллельных рядов форм с нижнего эоцена до нижнего олигоцена,
когда эта ветвь окончательно вымирает; другая, наоборот, прогрессивно
развивается в Сев. Америке, посылая оттуда не менее трех раз выходцев
в Старый Свет. Первую европейскую эволюционную тупую ветку лошадей
объединяют в подсемейство древних лошадей (Palaeohippi-
п ае); вторая ветвь, приведшая к современной лошади, составляет семейство
настоящих лошадей (Equinaе). Мы ограничимся, рассмотре-
нием эволюционных рядов лишь этого последнего семейства, пользуясь
прилагаемой ниже таблицей (см. стр.844).
Развитие приведенного в таблице ряда лошадей сопровождалось изме-
нением всей организации от близких к низшим хищным, небольших пяти-
палых первичнокопытных — к современным лошадям. Это преобразование
особенно хорошо шаг за шагом может быть прослежено в изменении строения
зубов и пальцев конечностей. У Eohippus и Hyracotherium передняя конеч-
ность с четырьмя пальцами, задняя — с тремя; Р3 и Р4 по форме и числу
бугорков приближаются к коренным, которые имеют 6 бугорков. У Ого-
hippus и Epihippus имеется дальнейшее приближение ложнокоренных зубов
к коренным, у которых бугорки удлиняются и сливаются нижними концами.
Mesohippus имеет уже 6 зубов одинакового строения, тогда как Р1 остается
конусовидным; бугорки приобретают форму, близкую к луночкам, средние
и внутренние вытягиваются; хвост становится короче; конечности трехпалые,
причем все пальцы достигают земли. Mesohippus дает две ветви: одна при-
водит к европейскому Anchitherium, который вымирает в верхнем миоцене;
другая ветка дает родоначальника европейских и американских лошадей —
средне- и верхнемиоценового Merychippus. Происходит значительное услож-
нение коренных зубов путем превращения внешних бугорков в слившиеся
друг с другом лунки и усложнения расположения эмали, что уже напоми-
нает зубы лошади; одновременно происходит превращение брахиодонтного
типа зубов в гипселодонтный. Изменения эти имели крупное экологическое
значение: зубы становились, таким образом, способными перетирать более
твердую растительную пищу, а не только сочную и мягкую траву и листья.
Трехпалые конечности характеризуются значительным укорочением боковых
пальцев. Новейшие исследования позволяют вновь вернуться к прежнему
представлению, что предком современной лошади был европейский род
с промежуточной формой, представленной гиппарионом (Hipparion), тогда
как американский род лошадей вымирает в плейстоцене. За это время, однако,
после соединения Северной Америки с Южной, в плиоцене североамерикан-
843
Европа
Северная Америка
Наше время Equus А > Equus
Плейстоцен Equus > Equus
Т Neohippus
Equus Pliohippus
Плиоцен А 1 А 1
Hipparion Protohippus
А | А i
Mery chippus
Миоцен Anchitherium t
А Para hip pus
Олиго цен * Miohippus
— A 1 Mesohippus
А
Palaeohippidae Epihippus , A ।
Эоцен Pachynolophus и др. Orohippus ' 1
Protohippus
Hyracotherium T >Eohippus
ские лошади успели перекочевать туда и дать там несколько родов, тоже
вымерших в плейстоцене. Вместо них уже в плейстоцене перебираются туда
новые формы ив Сев. Америки (Neohippus). Древнейшая настоящая лошадь
Европы (Equus stenonis) известна из’верхнего плиоцена средиземноморских
стран.
В ледниковое и послеледниковое время в Европе были следующие дикие
лошади: степные или лесостепные крупные тяжелого склада лошади (Equus
germanicus), тундряная и лесные лошади (Е. abeli, mosbachiensis и др.) и
мелкие степные: тарпан (Е. gmelini) и лошадь Пржевальского (Е. przewalskii)
(рис. 626). Лесные и степные тяжелые лошади вымерли, оставив после себя
след в крови домашних западноевропейских пород лошади. Тарпан был ис-
треблен в наших степях недавно, всего лишь в 1876 г.; лошадь Пржевальского
обитает в степях Монголии. К семейству лошадей мы относим: полу ослов —
кианга (Е. kiang) из Центральной Азии, джигитая (Е. hemionus) из Средней
Азии, кулана (Е. onager) в юго-западной Азии; ослов (Asinus), из которых
африканский осел (A. africanus) считается предком домашнего осла (A. asi-
nus); зебры, полосатые африканские лошади (Hippotigris и Dolichohippus)
(рис. 627), тоже относятся к этому семейству.
Сем. Т а п и р о в ы е (Т а р i г i d а е). Из современных нам живот-
ных сюда относятся лишь два рода: южноамериканские (Tapirella) с несколь-
кими видами (Т. terrestris и др.) и более крупный южноазиатский (Tapirus indi-
cus), от Тенассерима до Суматры и Борнео. Они имеют передние ноги с че-
тырьмя, а задние с тремя пальцами, пальцеходящи, зубы имеют формулу
1Ср Рр м|-, причем коренные с двумя поперечными гребнями. Имеется
небольшой хобот. Происходят от общего ствола с носорогами, были очень
многочисленны и широко распространены в Европе и Сев. Америке, в мио-
цене и плиоцене.
844
Рис. 626. Джунгарская дикая лошадь (Equus przewalskii).
(Аскания-Нова, фот, Озерова.)
К тапирам близко ископаемое семейство — лофиодонтовые (Lo-
phiodontidae) из эоцена Европы и Северной Америки.
Сем. Носороговые (Rhinocerotidae). В настоящее время
это семейство представлено небольшим числом родов и видов, тогда как
Рис. 627. Зебра квагга Чапмана (Hippotigris quagga
chapmani).
(Аксания-Нова, фот. Озерова.)
в ископаемом состоянии известно весьма большое число форм, то объединяемых
в одно подсемейство с носорогами, то выделяемых в разные подсемейства.
У современных представителей собственно носорогов (Rhinocerotinae)
все конечности трехпалые (тогда как у их предков передние имели по 4 паль-
ца), причем опираются на землю мозолистой подушкой, на которую
налегают не только фаланги пальцев, но и метаподии. Зубы: I сЯ, Р^,м|- ;
а и тк О
на носу один или два рога; тело одето чрезвычайно толстой кожей. Жи-
вотные приспособлены к тому, чтобы своей тяжестью беспрепятственно
845
прокладывать себе дорогу через колючие кустарники и густые заросли тро-
пического леса. В Африке двурогие носороги с гладкой кожей (Diceros
bicornis) и белый (Ceratotherium simus). В юго-восточной Азии три вида
со складчатой кожей и одним или двумя рогами (Rhinoceros unicornis,
R. sumatrensis и др.).
В четвертичное время в Европе были распространены волосатый носо-
рог (Tichorhinus antiquitatis) и носорог Мерка (Rhinoceros Mercki).
Из многочисленных плиоценовых и миоценовых носорогов мы упомянем
лишь гигантских эласмотериев (Elasmotherium), которые оби-
тали в Средиземноморье и в самое последнее плиоценовое время.
Сем. Титанотериевые (Titanotheriidae). Массивные
североамериканские животные, достигавшие величины слонов, появившиеся
в эоцене и вымершие в начале миоцена, с костными выростами впереди орбит,
громадных размеров.
Подотряд 3. Литоптерны (IMopterna)
Эта группа является самостоятельной, своеобразной ветвью
непарнокопытных, развившейся в Южной Америке из перекоче-
вавших туда североамериканских первичнокопытных. Она появ-
ляется в эоцепе, достигает наибольшего расцвета в миоцене Пата-
гонии и исчезает лишь в плейстоцене. Чрезвычайно интересным
является эволюционный путь этих животных, который представ-
ляет удивительный параллелизм с ходом развития лошадей.
Сем. Proterotheriidae. Начиная от Notodiaphorus с тремя
функционирующими пальцами ног через Diadiaphorus с укороченными
боковыми пальцами, специализация конечностей приводит к Thoatherium,
у которого разви!1 и функционирует один третий палец, одетый копытом;
конечность становится очень похожей на лошадиную. Существенным отли-
чием от этих последних является сериальное расположение карпальных
косточек.
Сем. Macraucheniidae с эоцена просуществовало до плейсто-
цена и представлено было стройными животными с рядом весьма усовершен-
ствовавшихся зубов; приспособлены они были к жизни в болотах.
Подотряд 4.Ам6липоды ( Amblypoda)
Вымершие палеоценовые и эоценовые весьма своеобразные ко-
пытные Сев. Америки, заменявшие в то время современных носоро-
гов или слонов. Это были большие тяжеловесные животные с тол-
стыми пятипалыми конечностями; начали они свое развитие с мел-
ких животных, которые едва отличаются от описанных выше пер-
вичнокопытных (род Pantolambda). Наибольшей специализации
к пережевыванию жесткой пищи достигли среднеэоценовые дино-
церы (Dinoceros).
Подотряд 5. Анцилоподы ( Ancylopoda)
Небольшая группа ископаемых копытных, появившаяся в эоцепе
Южной Америки, по затем в олигоцене и миоцене встречавшаяся
в Сев. Америке, в Европе и Азии. При сокращенном числе паль-
цев они опирались на наружную часть согнутой кисти подобно
неполнозубым, пальцы были приспособлены к роющей деятель-
ности.
Коренные зубы с буполофодонтной коронкой.
846
Or ряд 24. Трубкозубые (Tubulidentata)
В отдельный отряд приходится выделять единственный род аф-
риканских трубкозубов (Orycteropus), которых раньше соединяли-
с неполнозубыми вследствие характерной для них редукции зубов.
Исследования же последнего времени показали, что трубкозубов-
нужно считать близкими к копытным животным; они произошли,
вероятно, от первичнокопытных животных.
Это пятипалые, полустопоходящие копытные, насекомоядные
животные с полной молочной Id |-, Cd^-, Pd-^- и редуцированной
3 16
окончательной зубной формулой, без резцов, клыков и передних лож-
нокоренных зубов; 4—5 коренных зубов без корней с постоянным
ростом без эмали, но с отделением цемента, более прочного, чем ден-
тин. Зубы построены из многочисленных шестигранных призм»,
откуда животные и получили свое пазвапие.
Рис. 628. Трубкозуб эфиопский (Orycteropus aethiopicus).
(Из Брэма.)
Сем. Трубкозубые (О гу ctero pi d а е). Единственный род..
Трубкозуб капский (Orycteropus afer), распространенный через всю
Южную Африку до Сахары;! несколько других видов и подвидов в Африке
(рис. 628).
Сверхотряд. Приматы (Primates)
Полуобезьяны и обезьяны составляют общий ствол развития
млекопитающих, очень рано, еще в палеоцене, ответвившийся от
насекомоядных. Уже в самых древних третичных отложениях в па-
леоцене мы находим целый ряд ископаемых форм, объединяемых
в особое семейство плезиадаписовых (Plesiadapidae), близкое к со-
временным мадагаскарским руконожкам (Chiromyinae) из полу-
обезьян, очень примитивное, близкое к насекомоядным, но не-
сколько специализированное по строению своих зубов. Другое очень
845 -
древнее эоценовое семейство, адаписовые (Adapidae) (рис. 629) яв-
ляется также очень близким к насекомоядным и в некоторых отноше-
ниях (зубы) менее специализированным; оно представляет основной
родоначальный корень полуобезьян (Lemuroidea). Уже в нижнем эо-
цене жили формы, в которых мы узнаем представителей другого под-
отряда полуобезьян—долгопятов (Tarsioidea). Немного позднее, но
все еще в эоцене, мы находим вместе с полуобезьянами и долгопятами
также предков отряда обезьян
Нового и Старого Света. Все
приматы оказываются, таким
образом, связанными общим
корнем своего происхождения
из насекомоядных и, несмотря
па длинный эволюционный
путь, во многих отношениях
сохраняют до сих пор при-
митивные черты своего строе-
ния. Их эволюция в основном
связана с древесным образом
жизни, что отразилось на
строении конечностей (боль-
шой палец или противопола-
гается другим или по крайней
мере отгибается; он всегда
бывает одет погтем, а не ког-
тем). Первоначально будучи
Рис. 629. Череп Ada'pis parisiensis из
верхнего эоцена Франции.
(Из Вебера.)
насекомоядными, приматы эволюционируют в сторону плодояд-
ности, делаются более или менее растительноядными или всеяд-
ными, но никогда не делаются хищниками. Исходная зубная фор-
мула:
12 ’ ° ррр м У’ редуцируются до 1^-, С Р М
или
_ 1 _ о „ 1 .. з , т 2 л 1 „2 /з \ . - з(2)
11 ’ С 0 ’Р О’ М 3’ обычно 1 2 ’ ° 1 ’ Р2 \3 / ’ м 3(2) ‘
Из органов чувств характерно сильное развитие глаз, смещение их на
переднюю сторону, что приводит к стереоскопическому зрению, а
также приспособление многих видов к ночному образу жизни. На-
оборот, орган обоняния находится па разных стадиях редукции.
Чрезвычайно важным для понимания эволюции этой группы яв-
ляется факт прогрессивного развития головного мозга. Приматы
делятся на два отряда.
Отряд 25. Полуобезьяны (Prosimiae)
У полуобезьян, которые, как мы видели, принадлежат к древ-
нейшей группе современных млекопитающих, к тому же мало изме-
нившейся за длинный период их существования, сохранилось очень
много признаков, общих с насекомоядными. От обезьян полуобезь-
848
Рис. 630. .Лемур катта (Lemur catta).
(Из Гаане.)
япы отличаются вытянутой лицевой частью черепа и малой черепной
коробкой; глазница у многих форм не отделен^ от височной впадины,
по меньшей мере второй палец снабжен когтем. Это небольшие или
средней величины, очень часто ночные, Животные. Они в наше
время распространены главным образом на Мадагаскаре, где соста-
вляют чуть ли не половину всего населения млекопитающих; не-
большое число видов свойственно тропической Африке и Южной Азии
с прилегающими островами. В палеоцене и эоцене примитивные
обезьяны были широко рас-
пространены в Европе и Сев.
Америке. Два подотряда.
Подотряд!. Лемуры (Lemurог-
dea)
Древесные ночные или су-
меречные животные, частью
питающиеся насекомыми, ча-
стью плодами или всеядные;
они одеты густой пушистой
шерстью и имеют по большей
части длинный хвост; второй
палец ноги всегда с когтем.
Два семейства.
Сем. Лемуров ые (L emu-
rid а е) разделяется на четыре под-
семейства мадагаскарских живот-
ных с большим количеством преи-
мущественно ночных видов (рис.
630).
Из них особенно интересны
руконожки (Chiromyinae), напо-
минающие своими постоянно ра-
стущими, как у грызунов, резцами;
наиболее крупные из лемуров —
индри (Indrisinae).
См. Лориевые (Lori-
s i d а е) объединяет африканских
и индийских лемуров; бесхвостые
лори—ночные маленькие, стройные полуобезьяна тлитгтлт, ттл»
£ Nyc,‘“b“"
Подотряд 2. Долгопяты (T<ir8ioidea)
Этот подотряд представлен в паше врэмя единственным родом
удивительных ночных, резко отличающихся по своему виду живот-
П1ЫХ. Это маленькие животные с длинными прыгательными задними
ногами, у которых сильно удлинились плюсневые кости и длинный
снабженный кисточкой хвост; пальцы снабжены присасывательными
и <>ду точками, а большая круглая голова имеет громадные глаза и
^аленькие голые ушки. Долгопяты — обитатели Малайского ар-
хипелага. F
Чрезвычайно интересно, что долгопяты были представлены боль-
шим числом родов в Европе и Сев. Америку в древнетретичное время
М Наши и г ов и Станчинский ,
и играли некогда большую роль в фауне того времени и сравни-
тельно мало изменились с тех пор.
Сем. Долгопятовые (Tarsiidae).
> Долгопяты (пугало) (Tarsius tarsius) живут на Суматре, Борнео, Яве
и др. (рис. 631).
Отряд 26. Обезьяны (Simiae)
Обезьяны представляют высшую ступень приспособления к дре-
весному образу жизни, к питанию преимущественно древесными пло-
хие. 631. Долгопят (Tarsius tar-
sius).
(Из Гааке.)
дами и стереоскопическому зрению;
одновременно с этим они характе-
ризуются наиболее сильно раз-
витыми полушариями головного
мозга.
Древесный образ жизни наибо-
лее сильно выражен в строении ко-
нечностей, причем большой палец
отделен и противополагается про-
чим. На ногах человека в связи с
вторичным приспособлениемк стопо-
хождению в выпрямленном состоя-
нии большой палец утерял свою
подвижность и отклоняется только
в детском возрасте. Пальцы одеты
ногтями, но у сем. игрунковых (На-
palidae) все пальцы за исключе-
нием большого, имеют ногти ког-
теобразного вида. Зубная система
всеядного, или правильнее, плодо-
ядного типа, так как долотообраз-
ные резцы, в числе четырех, сидят
сплошным рядом (откусывание пло-
дов), конусообразные клыки возвы-
шаются над остальными зубами (уде-
рживание во рту плодов), коренные
зубы приспособлены для пережевы-
вания мягкой растительной пищи, с
низкими коронками и четырьмя ту-
пыми бугорками (у игрунковых и не-
которых ревунов с тремя). У некото-
рых гиббонов вторично сильно развит клык как орган защиты и напа-
О Т2 П1 -П 2 (3) ™ 3 (2) V
дения. Зубная формула I—, С—, Р М ; Хорошо развитые
а 1 4 (о) О (2)
глаза, в противоположность полуобезьянам, за редким исключени-
ем приспособлены к дневному свету, сидят спереди в больших,
совершенно замкнутых орбитах.
Появляются первые обезьяны в Новом Свете лишь в миоцене Юж-
ной Америки, где Homunculus patagonicus имеет уже все особенно-
сти обезьян Нового Света; однако происхождение обезьян следует
350
отнести к эоцену Сев. Америки, где среди полуобезьян семейства
Adapidae род Notarctos является вероятным их предком. Обезьяны
Старого Света имеют своих древ-
нейших представителей в олиго-
цене Египта. Так, парапи-
тек (Parapithecus fraasi), более
близок к человекообразным
обезьянам, чем к собакоголовым,
а а п и д и й (Apidium phio-
mense) (рис. 632) — ближайший
предок этих последних. Очень
интересна находка в этих же
отложениях нижней челюсти
обезьяны, которая уже очень
близка к таковой человекооб-
разных обезьян: и р о п л и о-
п и т е к (Propliopithecus hae-
ckeli). Родину обезьян Старого
Ряс. 632. Кусок челюсти Apidium
phiomense.
А — сверху, Ах — с наружной стороны,
А2 —с внутренней стороны. (Из Вебера.)
эпы (Франция) найдены остатки
и т е к a (Pliopithecus antiquus)
Света надо искать в Азии.
Дальнейшая эволюция про-
слежена, начиная с миоцена, хо-
рошо только для человекообраз-
ных обезьян. Так, в миоцене Е]
несомненного гиббона — и л и о
(рис. 633), который, повидимому, был
редок; в среднем миоцене Франции,
Египта и Южной Азии найдены ос-
татки нескольких видов дриопи-
тека (Dryopithecus) обезьяны, из
которых одни имеют признаки зу-
бов, сходные с гориллой, другие—с
шимпанзе; есть основания думать, что
именно дриопитеки являлись предка-
ми шимпанзе через ископаемую евро-
пейскую шимпанзе (Antropopithecus-
Pan vetus). Несколько позднее в
верхнем миоцене восточной Индии (си-
валикские отложения) найден верх-
ний коренной зуб, по которому мож-
но все же было установить, что это
зуб человекообразной обезьяны, про-
си м и и (Prosimia), близкой к оранг-
утану (Simia), который в тех же си-
вал и кских отложениях найдеп в верх-
нем плиоцепе; в этих же отложениях
найдшпл остатки особенно интересной
обезьяны — с ивапитека (Siva-
pithecus), которая по своим зубам
уже во многих отношениях оказы-
в лесах Европы не так уж
Рис. 633. Куски верхней челю-
сти А — плиопитека (Pliopi-
thecus antiquus) и В—гиббона
(Hylobates 1аг). (Из Вебера.)
54*
851
А
вается приближающейся к человеку. Наконец, в нижнем плиоцене
тех же отложений найдена верхняя челюсть с зубами, близкая к
таковой современной гориллы, но отличающаяся от нее рядои
особенностей; это челюсть палеопитека (Palaeopithecus),
вероятно, предка гориллы.
Наиболее известной и наиболее изученной является находка в са-1
мых нижних четвертичных отложениях Явы в 1881 г. первых остат-
ков, которые не могут быть отнесены
ни к обезьянам, ни к человеку. Это
был уже один из прямоходящих пред-
ков людей (сем. Hominidae), прямо-
ходящая человекообразная обезьяна—
питекантроп (Pithecanthropus
errectus). Наконец, уже совсем недав-
но наука обогатилась новыми данны-
ми о происхождении человека, так
как в Китае, в районе Пекина, в
1928 г. найдены были остатки живот-
ного уже несомненно гораздо более
близкого к человеку, чем питекант-
роп. Его выделяют в особый род
синантроп (Sinanthropus). От-
ложения принадлежат раннему чет-
вертичному времени. К этому же вре-
мени относится знаменитая гейдель-
бергская нижняя челюсть, принадле-
жащая очень древнему человеческо-
му виду—Homo heidelbergensis. Этот
и следующие этапы эволюции принад-
лежат уже человеку, который, бу-
дучи общественным, приобрел речь
и, взяв в руки орудия, перестал пас-
сивно приспособляться к природе, а
начал приспособлять природу к
себе.
Громадное значение при этом в
эволюционном процессе человекооб-
разных обезьян при переходе их к
человеку сыграло освобождение рук,
вызванное прямым хождением по зем-
ле. Это хождение было связано и с
соответствующим преобразованием
всей организации.
Современных обезьян мы делим на два подотряда в соответствии
с особенностями их организации, обусловленной их самостоя-
тельной эволюцией: на обезьян Нового Света, или широконосых
обезьян (Platyrrhina), и обезьян Старого Света, или узконосых обезь-
ян (Catarrhina).
\t тг
Рис. 634. Нижние челюсти.
А — шимпанзе, В — пильтдаунско-
го человека (Eoanthropus dawsoni),
С — гейдельбергского человека,
D — современного человека.
852
Подотряд 1, Широконосые обезьяны (Platy rrhina).
Отличаются широкой носовой перегородкой, вследствие чего
ноздри оказываются обращенными наружу; имеют длинный, иногда
хватательный, хвост; имеется третий ложнокоренной зуб Р'-, по-
О
этому формулазубной системы:
I- С— Р— м^ з
*2 ’ С1 ’ Р3 ’ М2—3*
Два семейства.
Сем. 1. И г ру н к ов ы е и
(Н а р а 1 i d а е). Отличаются ког-
теобразными ногтями за исклю-
чением большого пальца, пуши-
стым хвостом и зубной формулой:
]?_ qL р§_ Эд —
2 ’ С1 ’ Р3 ’ М2 *
Несколько родов в тропичес-
ких лесах Южной Америки; мел-
кие пушистые обезьяны, напо-
минающие своими движениями
белок (рис. 635).
Сем. 2. Цепкохвостые
(С е b i d а е). Названы так пото-
му, что большинство имеет цеп-
кий хвост. Зубы имеют формулу
>2 U „3 „3
I—, Су, Ру, Му ; распростране-
ны в Южной и Центральной Аме-
рике. Одно семейство, распадаю-
щееся на пять подсемейств.
Ночные обезьяны (Nyctipite-
cinae), составляют группу пуши-
стых кошкоподобных обезьян,
хвост не цепкий. Ревуны (Мусе-
tinae) (рис. 636), самые крупные из
американских обезьян, замечатель-
ны своими резонирующими аппа-
ратами в гортани. Паукообразные
обезьяны (Atelinae) с длинным
и цепким хвостом Jрис. 642) и др.
Рис. 635. Игрунка львиная (Midas го-
salia).
(Из Гааке.)
Подотряд 2. Узконосые
обезьяны ( Catarrh/ina)
Обезьяны Старого Света
отличаются от обезьян Нового Света большей величиной и тем,
про у них никогда не бывает хватательного хвоста, а также зуб-
2 12 3
пой формулой I—, С-, Р—, М—. Большой палец передней конеч-’
А 1 о
пости только у Colobus рудиментарный. Три семейства.
Л
Сем. 3. Собакоголовые (С ercopithecidae). Большая
группа обезьян, передвигающихся по земле с помощью четырех конечностей.
853
Лицо более или менее голое, равно как и седалищные мозоли. По большей
части имеются защечные мешки. Много видов, объединяемых в 13 родов;
широко распространены по Старому Свету, отсутствуют только в Австра-
лийской области й Северной подобласти Палеарктики. Два подсемей-
ства.
Поде. Мартышковые (Ccrcopithecinae) объединяют проворных
обезьян с передними и задними конечностями одинаковой длины: макаки (Ма-
cacus), бесхвостый макак (М. sylvanus)—единственная обезьяна Европы
(Гибралтар), Туниса, Алжира и Марокко; павианы (Papio) и мандрилы
(Mandrillus) (рис. 637) встречаются
в Африке — преимущественно жи-
Рис. 636. Рыжий ревун (Mycetes seni-
culus).
(Из Гаане.)
тел и скал; мартышки (Cercopite-
cus) — многочисленные виды в
Африке.
Поде. Тонкотелые обе-
зьяны (Colobinae) имеют
более длинные задние конечности
и отличаются малой подвижно-
стью. Носатая обезьяна (Nasalis)
с острова Борнео, священная
обезьяна (Pithecus entellus) и
др. из Индии, Р. schistaceus под-
нимаются в горах до 2000 м вы-
соты.
Сем. 4. Гиббонов ые (Ну-
lobatidae). Типичные обитате-
ли деревьер с очень длинными ру-
ками, небольшими седалищными
мозолями; охотно спускаются на
землю, где передвигаются в вер-
тикальном положении; не имеют
хвоста; одеты густой шерстью.
Род Hylobates распространен в
юго-восточной Индии, на остро-
вах Суматра, Ява, Борнео (рис.
638). Один вид (Hylobates haina-
nus) на острове Формоза. Дру-
гой род с единственным видом
Symphalangus syndactylus встре-
чается исключительно на остро-
ве Суматра.
Сем. 5. Человекообраз-
ные (An tr о рот о г ph а е). Эти
крупные бесхвостые обезьяны, от-
личают ся отсутствием седалищных
мозолей, тем, что при передви-
жении по земле держатся прямо,
опираясь на согнутые пальцы рук,
которые значительно длиннее ног.
Лучевая кость имеет возможность
полной пронации и супинации.
Три рода. Из них на островах
Борнео и Суматра оранг-утан(Б1ш!а
satyrus рис. 639) и другие виды и
подвиды. С 12 ребрами. Горилла
(Gorilla) — Африка — самая круп-
ная из обезьян, представлена
несколькими видами, имеет 13 ребер. Шимпанзе (Antropopithecus) — также
несколько видов (рис. 640).
Сем. Люди (Hominidae) с одним видом Homo sapiens — человек
разумный и с рядом ископаемых форм.
854
855
Рис. 637. Мандрил (Mandrillus sphinx).
(Из Гаане.)
Рис. 638. Гиббон (Hylobatcs 1аг).
(Из Гаане.)
Рис. 639. Оранг-утаи (Simia satyrus).
(Из Гааке.)
Рис. 640. Шимпанзе (Anthropopithecus niger.)
V. ЭВОЛЮЦИЯ МЛЕКОПИТАЮЩИХ В СВЯЗИ С ПРИСПОСОБЛЕНИЕМ
К СРЕДЕ
Резюмируя кратко все то, что сказано нами относительно про-
исхождения и эволюции отдельных систематических групп млеко-
питающих, мы можем построить общую схему их эволюции в связи
с приспособлением к среде.
Первый период в эволюции млекопитающих — это период яй-
ценоскости при наличии сумки, а может быть, и при ее отсутствии
еще, но при наличии первичных зубов и дальнейшей их Дифференциа-
ции. Млекопитающие должны были вести скрытый образ жизни.
Это были мелкие, полудревесные животные; они не могли эволюцио-
нировать, распространяясь и приспособляясь к новым условиям
существования, потому что яйценоскость при теплокровности в свя-
зи с необходимостью высиживания яиц сильно ограничивала воз-
можности их приспособления и эволюции.
Только сумчатость, появившаяся, вероятно, в конце триаса
и развившаяся в юре, и живородность позволили значительно шире
эволюционировать, приспособляясь к разным средам обитания. По-
лудревесные примитивные сумчатые дали еще в юрское время жи-
вотных, приспособлявшихся к использованию различной пищи и раз-
личных сред обитания. Достаточно указать насекомоядных Tri-
conodonta, растительноядных Tritylodontidae, Plagiaulacidae, Ро-
lymastodontidae, у которых уже имелась довольно высокая специа-
лизация, указывающая на различные приспособления к раститель-
ноядности. Однако и сумчатые животные не дали, да, вероятно,
и не могли дать вследствие их недостаточной эйритоадаптивности
форм, способных к дальнейшей эволюции в условиях обостренной
борьбы за существование при наличии богатого видами класса реп-
тилий. Только в условиях отделившихся материков — Австралии
и отчасти Южной Америки — сумчатые могли дать разнообразные
филогенетические ответвления в разные среды до двум основным
разделам: по способу питания — животноядных (Polyprotodontia)
и растительноядных (Diprotodontia). Мы уже познакомились с этой
в высшей степени интересной, параллельной высшим зверям, эво-
люцией в систематической части. Богатство и разнообразие ав-
стралийской фауны сумчатых, вероятно, может быть объяснено
островным характером Австралии, отсутствием богатой фауны хищ-
ных рептилий и отсутствием высших плацентарных за исключением
прёдставителей летучих мышей и мышиных.
Что касается высших зверей, то их эволюция в связи с приспо-
соблением шла по следующей схеме. Получив широкоадаптивные
(эиритоадаптивные) особенности — вынашивание детенышей в матке
и выкармливание их, мелкие плацентарные млекопитающие, вед-
шие скрытый полудревесный образ жизни, начинают прежде всего
дифференцироваться в отношении пищи. Из насекомоядно-плотояд-
ных. они дифференцируются: на насекомоядных преимущественно,
родоначальников сверхотряда Insectivora; на преимущественно пло-
тоядных — креодонтных Acreodi, родоначальников Carnivora; па
857
растительноядных грызущего типа — грызуны (Rodentia); на всеяд-
нотравоядных — предков первичнокопытных, предков двукопыт-
ных и предков полукопытных (Ungulata). Эти приспособления
к пище оказались настолько эйритоадаптивными, настолько ве-
дущими, что дальнейшая эволюция млекопитающих пошла уже по
пути приспособления насекомоядных, хищных, грызунов и траво-
ядных к разным средам обитания.
Представители различных отрядов млекопитающих, становясь
лазающими формами, приобретают складки кожи по бокам тела.
В группе насекомоядных это приводит, в конце концов, к разви-
тию летающих млекопитающих — летучих мышей — отряд Chirop-
tera, напоминающих по принципу устройства летательного аппа-
рата птерозавров. От травоядных млекопитающих произошли вод-
ные сирены (Sirenia), от хищпых тоже водные — сперва китообраз-
ные (Cetacea) и позднее ластоногие (Pinnipedia). Дальнейшая диффе-
ренциация идет уже различно: в одних случаях, эволюция идет по
пути большей специализации к средам обитания (например, у насе-
комоядных, у неполнозубых, у копытных); в других случаях, эво-
люция идет по пути дальнейшей специализации в отношении пищи —
летучие мыши, наземные хищные, многие группы копытных. Так,,
в чередовании разных видов приспособления к среде складывалась
'Систематическая единица высшего и низшего порядка, образуя
многочисленные параллельные ряды, особенно усложненные от-
делением и вновь соединением материков. Эйритоадаптивные при-
способления приводили к образованию крупных систематических
'единиц; степоадаптивная эволюция давала преимущественно специа-
лизированные формы мелких систематических единиц.
VI. ЗАВИСИМОСТЬ МЛЕКОПИТАЮЩИХ ОТ УСЛОВИЙ СРЕДЫ.
ЭКОЛОГИЯ МЛЕКОПИТАЮЩИХ
1. Приспособление к условиям обитания среды как субстрата
(приспособление к биотопическим условиям)
Первые родоначальники млекопитающих были, по всей вероят-
ности, небольшими насекомоядными или всеядными формами с про-
стыми зубами, с короткими ногами, снабженными когтями. Образ
жизпи их был наземно-древесный. Постепенно распространяясь,
занимая все новые и новые среды обитания (новые биотопы)
млекопитающие изменялись, дифференцировались в различных
направлениях. Одни из них приспособлялись к жизни в земле,
ведя полуподземпый или же совершенно подзем-
н ы й образ жизпи, становились роющими животными; другие
приспособлялись к жизпи в воде и становились земноводными
или же всецело водными формами, причем можно уста-
новить ряд переходов от плавающих наземных, иначе земновод-
ных форм до пелагических морских млекопитающих;1 третьи
занимали открытые пространства суши: степь, пу-
стыню и т. д., становясь ходящими, бегающими или прыгающими
млекопитающими; четвертые приспособлялись к жизни в лесу,став
.858
древесными, лазящими животными; наконец, некоторые
приспособлялись к жизни в воздухе, становясь летающими
животными.
Иногда, присное обившись к одной среде, млекопитающие пере-
ходили к иному образу жизни, и у них развивались новые приспо-
собления. Но тут имеются определенные закономерности. К воздуш-
ному образу жизни млекопитающие перешли, видимо, пе от назем-
ной жизни, а от древесной. Древесные формы иногда становились
снова наземными и бегающими (многие сумчатые Австралии и дру-
гие из плацентарных). Но мы не знаем пи одного примера, чтобы
ставшее водным млекопитающее снова перешло к наземной жизпи.
Также никогда животное резко выраженного роющего или бегаю-
щего типа не превращалось в медленно двигающийся тип ходящего
животного.
Таким образом, эволюцию млекопитающих в связи с приспособ-
лением к различным средам обитания можно изобразить следую-
щим образом (см. схему):
Роющие формы Летающие формы
Древесные формы
Коротконогие, стопоходящие,
пятипалые, коготные
Ходящие или наземно-древес-
ные формы
Плавающие земноводные
формы
Бегающие пальцеходящие
Чисто водные
Когтеходящие, или копытные
Одновременно с приспособлением к субстрату, где животное дви-
жется, строит гнездо, добывает пищу (биотопическая адаптация),
млекопитающие приспособлялись к разного рода пище (трофоце-
нотическая адаптация).
Эти приспособления шли самостоятельно, и характер зубов не
совпадает пепремепно с характером конечностей. Например, ра-
стительноядными могут быть и бегающие (копытные), плавающие
(манат, дюгонь) и древесные (ленивец) формы.
Млекопитающие открытых мест обитания. Млекопитающие,
имеющие своим местопребыванием открытые места — луга, степи,
пустыни, — встречаются во многих отрядах: среди насекомоядных,
хищных, грызунов, сумчатых; копытные по самому существу —
животные открытых мест и вместе с грызунами составляют главную
массу их населения.
Млекопитающие открытых мест обладают особыми характерными
приспособлениями. Для животных, живущих в открытой местности,
во-первых, весьма важно иметь возможность издалека узнать опас-
859
ность или увидеть добычу или пищу: во-вторых, весьма, важно иметь
быстрые ноги, чтобы убегать при опасности или догонять добычу,
с одной стороны, и быстро передвигаться в поисках корма и воды—
с другой, или уметь укрыться в надежное убежище— в нору.
Наконец, важно иметь малую потребность в воде. Для животных,
служащих добычей хищникам, очень важно быть мало заметными.
Такого рода приспособления и являются характерными для млеко-
питающих открытых мест: саванн, лугов, степей, полупустынь и
пустынь.
Млекопитающие открытых мест благодаря открытым далям при-
обрели в борьбе -за существование прекрасное зрение. У жирафы
(Giraffa Camelopardalis) получается очень широкий кругозор еще
благодаря тому, что передняя часть тела приподнята в сравнении
с задней. У родственного ей окапи (Okapia johnstoni), жителя леса,
эта особенность далеко не так выражена. Эту особенность, хотя
в меньшей степени, мы видим у целого ряда степных млекопитающих;
взять хотя бы бубалов (Bubalus) или гну (Connochaetes), лошадиных
антилоп (Hippotragus), живущих в открытых местностях, поросших
низким кустарником. У всех у них передние ноги длиннее задних
и шея вытянута вверх.
Другие млекопитающие, а именно многие грызуны, для обо-
зрения местности встают па задние ноги и садятся на хвост, «стано-
вятся столбиком». Мы видим это у зайцев (Lepus), байбака (Маг-
motta), у сусликов (Citellus), у кенгуру (Macropus) и др.
Способность к быстрому передвижению достигается двумя пу-
тями: бегом или прыганием. Приспособление к бегу выражается
в переходе от стопохождения к пальцехождению, в потере ключицы,
в удлинении метаподиалыюй части конечности, в развитии копыт,
в сокращении числа пальцев до двух или одного, в слиянии запяст-
ных и предплюсневых костей и соответствующих изменений муску-
латуры. В морфологической части при описании скелета конечностей
и в систематической части при описании копытных млекопитающих
нами указаны были те эволюционные пути, по которым шло в разных
группах млекопитающих и в разное время приспособление к бы-
строму бегу. Среди современных млекопитающих наиболее совершен-
ным бегом обладают из непарнокопытных лошади, полуослы, ослы
и зебры, а из парнокопытных— антилопы и олени. Хищники откры-
тых мест, догоняющие добычу, имеют соответственное приспособле-
ние конечностей. Таковы быстро бегающие волки и собаки из сем.
Canidae, гепард из сем. Felidae.
Прыгание как приспособление к быстрому передвижению харак-
теризуется удлинением задних конечностей, особенно из метаподи-
альной части, и укорачиванием передних конечностей. Передние ко-
нечности служат сперва лишь опорой о землю при беге, причем зад-
ние конечности выносятся иногда вперед по сравнению с передними.
Так, у зайцев характерное расположение следа зависит от того,
что задние конечности выносятся вперед по бокам расположенных
друг за другом передних конечностей. При дальнейшей специали-
зации передние конечности при передвижении уже не принимают
860
участия, как, например, у тушканчиков, они служат лишь для
схватывания пищи или для рытья. Хвост, наоборот, удлиняется
и служит точкой опоры при стоянии и рулем для быстрых пово-
ротов при прыжках.
Способность к прыганию развивается конвергентно в разных
отрядах: сумчатые — кенгуру, насекомоядные — прыгунчики и
целый ряд прыгающих грызунов. Из хищных прыжками передви-
гаются многие кошки. Те обитатели открытых мест, которые не обла-
дают быстрым бегом и имеют небольшую величину, прячутся в норы.
Эта способность рыть норы, чтобы прятаться от врагов, может при-
вести при дальнейших приспособлениях к настоящему подземному
образу жизни.
В качестве убежища прячутся в норы мыши, суслики, хомяки,
сурки, тушканчики и многие другие грызуны из ведущих надзем-
ный образ жизни. Многие полевки переходят к подземному пита-
нию; таковы обыкновенная полевка (Microtus arvalis), обществен-
ная полевка (М. socialis) и другие полевки. Кавказская прометеева
мышь (Prometheomys schaposchnikovi), слепушонка (Ellobius) яв-
ляются уже типичными подземными обитателями, нормально пи-
тающимися подземными частями растений.
Помимо сЦособности укрываться или убегать, млекопитающие
открытых мест защищены обыкновенно также покровительственной
окраской; достаточно указать на окраску джейрана (Antilope sub-
gutturosa) (рис. 641), желтого и тонкопалогосуслика (Citellus fulvus
и Spermophilopsis leptodactylus), песчанок (Gerbillus) и др.
Млекопитающие открытых мест имеют еще одну особенность:
они, как правило, очень мало пьют, довольствуясь той влагой,
которая заключена в их пище — растениях. Сцособность верблю-
дов выносить жажду общеизвестна. Они могут пить воду соленую и
горькую. Долго могут не пить многие антилопы, жирафы. Неко-
торые млекопитающие не пьют вовсе. Не пьет вовсе карликовая
газель (Gazella arabica) с островов Красного моря; быть может,
и другие газели тоже не пьют. Желтый суслик (Citellus fulvus)
не пьет даже тогда, когда есть вода; также не пьют и некоторые дру-
гие суслики, например, тонкопалый (Spermophilopsis leptodacty-
lus), песчанки (Gerbillus, Rhombomys).
Млекопитающие роющие и подземные. Среди млекопитающих
мы находим не только очень большое число роющих форм, но также
наиболее далеко ушедшую специализацию конечностей приспосо-
бительно к роющему образу жизни.
Наибольшее число роющих млекопитающих представлено гры-
зунами. Среди последних роющие формы есть во всех почти груп-
пах. И хотя они часто не связаны между собой близким родством
и живут в разных частях света, образ жизни накладывает на них
одинаковую печать. Роющими среди грызунов являются: сурки
(Marmotta), суслики (Citellus, Spermophilopsis), бурундуки (Та-
mias) и многие другие среди группы Sciuroidea; среди Myoidea:
мно'гие крысы (Murinae) и полевки (Microtinae), слепушонки (Ello-
bius), песчанки (Gerbillus, Rhombomys), хомяки (Cricetinae),
861
пеструшки (Lagurus), слепыши (Spalax), тушканчики (Dipus,
Alactaga); капский долгоног (Pedetes); дикобразы Старого Света
(Hystrix) из группы Hystricomorpha; южноамериканские туко-
туко (Ctenomys), вискаши (Viscacia) из сем. Chinchillidae и, нако-
нец, из группы Lagomorpha: кролики (Oryctolagus cuniculus),
пищухи (Ochotona). Эти грызуны живут часто огромными коло-
ниями, и тогда вся почва так ими изрыта, что бывает опасно ехать
верхом, так как лошадь все время проваливается в норы. Такую
картину можно наблюдать и в Азии (пищухи в Монголии, песчанки
в Средней Азии) и в Африке (долгоног), и в Патагонии (Ctenomys).
г
Рис. 641. Джейран (Gazella subgutturosa).
(Из Бихнера.)
Однако немалое количество роющих форм имеется и в других от-
рядах млекопитающих. Среди сумчатых в Австралии необычайно
ловко роет, живя под землей, сумчатый крот (Notoryctes); броне-
носцы (Dasypodidae) из неполнозубых; капский трубкозуб (Orycte-
ropus) и степной ящер (Manis temminckii). Африканский боро-
давочник (Phacochoerus) расширяет норы трубкозуба в африкан-
ских степях. Роют и некоторые хищные: барсуки (Meles), лисица,
фенек (Megalotis zerda) в африканских пустынях и Cerdocyon
brasiliensis — в патагонских пустынях и др.
Способность рыть норы развилась, невидимому, у млекопитаю-
щих как приспособление для защиты-с^-Фрагов и от холода и вы-
сокой температуры. Можно отметить различные ступени в разви-
862
тии этого приспособления, начиная от простого соскребания земли
до настоящих нор с различными камерами: для запасов, для вы-
вода детей и т. д. Роющих млекопитающих можно разделить на
две группы: на роющих временно и выходящих наружу, и на под-
земных, т. е. все время пребывающих под землей. В первом случае
изменения в теле животного будут невелики. Примером могут
служить те изменения, которым подвергалась организация кро-
лика, происходящего, несомненно, от зайца. Череп кролика более
сжат с боков; углубление для глаз, область, где помещаются орган
обоняния и дыхательные пути, более сжаты. Зато место прикрепле-
ния для жевательных и для затылочных мускулов более обширно.
Такое более плотное и прочное строение черепа кролика в сравне-
нии с черепом зайца стоит в прямой связи с тем обстоятельством,
что кролик, скрываясь от опасности, бросается головой в нору:
узкая головка, прочно построенная, в таком случае более подходит.
Сильные челюсти и зубы служат кролику для рытья под землей,
для перегрызания корней.
Различие между кроликом и зайцем существует и в строении
конечностей. У зайца плечи гораздо длиннее, предплечья тоньше и
представлены главным образом локтевыми костями, тогда как луче-
вая развита гораздо слабее и отодвинута назад. У кролика обе кости
предплечья почти одинаково сильно развиты и лежат в одной
плоскости. Несколько дальше, чем у кролика, пошли изменения
у пищухи (Ochotona). Но и это животное большую часть времени
проводит снаружи, и изменения организации потому не слишком
велики.
Изменения организации у роющих форм прежде всего касаются
передних конечностей, по затем захватывают и череп, и органы
чувств, и позвоночник, и окраску.
Примером роющего животного, ставшего совершенно подземным
и в высокой степени специализировавшимся, может служить сум-
чатый крот (Notoryctes typhlops). Среди сумчатых это единствен-
ное животное, ведущее вполне подземный образ жизни. Скелет
этого животного сильно уклонился ст того, что мы видим у других
сумчатых; зато во многих признаках мы видим у этого животного
совпадение, конвергентное развитие с роющими формами из пла-
центарных животных, особенно с южноафриканским роющим на-
секомоядным златокротом (Chrysocliloris aureus), кротом, слепы-
шом и другими подземными млекопитающими: 1) шерсть короткая,
густая и шелковистая; 2) глаза рудиментарны, зачаточны: их ре-
дукция связана с их бесполезностью; 3) заглазпичный отросток
в черепе (proc, postortitalis), дающий нормально развитому глазу
защиту, отсутствует; 4) боковые затылочные отростки (proc, ра-
roccipitales) отсутствуют; обычно эти отростки по бокам затылоч-
ных сочленений служат для прикрепления мускулов, поворачи-
вающих голову в сторону; при подземном образе жизни эти
движения отсутствуют; 5) сочленовные бугорки черепа, которые
у сумчатых лежат более снизу, смотрят назад, так что череп и
позвоночник оказываются лежащими на одной оси, что весьма
863*
помогает при рытье; 6) черепные кости срастаются очень рано,
благодаря чему череп получает большую прочность; 7) череп имеет
коническую форму легче проникающую в землю; 8) передние но-
совые отверстия расположены на нижней стороне заостренного
рыла; благодаря этому земля, песок и т. д. не попадают в ноздри;
9) обе половины нижпей челюсти прочно срастаются, что увеличи-
вает прочность действующего, как клин, черепа; 10) на лопатке
имеются гребни, что стоит в связи с сильным развитием трехглавой
мышцы плеча; 11) имеется ключица; 12) па плечевой кости имеется
сильный гребень — место прикрепления роющих мышц — дель-
товидной (m. deltoideus) и грудной (m. pectoralis); 13) на плече
сильно развит внутренний прицепок в связи с сильным развитием
мускулов сгибателей; 14) сильно развит proc, olecranon локтевой
кости — место прикрепления трехглавой мышцы (m. triceps);
15) ряд изменений в дистальной части конечности: укорочение
пястья и пальцев, более сильное развитие средних пальцев, проч-
ное соединение карпальных косточек; 16) у подземных млекопитаю-
щих наружное ухо отсутствует; хвост редуцирован.
Но в зависимости от того, в каком грунте животное роется,
будут наблюдаться различия. При рытье в мягком грунте (на-
пример, у крота — Talpa) кисть сильно расширяется, опа дейст-
вует, как лопата и имеет наклонность к развитию шестого
пальца (см. стр. 742).
Передняя конечность крота сравнима с ложкой для выскабли-
вания, которую берут за рукоятку вблизи ее самой, работая при
помощи короткого рычага. Наоборот, у тех форм, которые роются
в твердом грунте, кисть становится уже, в рытье принимают уча-
стие лишь немногие пальцы, большей частью 3-й и 4-й, разви-
вающиеся за счет других, как у Notoryctes и Chrysochloris. То же
мы видим у млекопитающих, разрывающих твердые постройки
термитов, например, у муравьеда (Myrmecophaga), у тамандуа
(Tamandua). Чтобы когти при хождении не тупились, эти живот-
ные ходят, опираясь на наружный край ступней и подогпув когти.
У слепушонки (Ellobius) для рытья служат резцы, сильно вы-
дающиеся вперед. Ими этот зверек действует как мотыгой, отку^1
санную же землю выбрасывает из норы, пятясь задними ногами.
Чтобы земля не попадала в рот, волосистая часть верхней и ниж-
ней губы загибается между резцами и коренными, причем остается
лишь очень маленькое ротовое отверстие, закрываемое волосами,
У тех млекопитающих, которые выбрасывают землю головой,
изменяется сама голова. Так, у крота (Talpa) хобот поддерживается
специальным хрящом (os praenasale), достаточно, впрочем, гиб-
ким, чтобы хоботок мог действовать как орган осязания. У No-
toryctes на конце носа -находится твердый роговой щит; у Chryso-
chloris на переднем конце морды имеется широкая голая кожная
поверхность, оканчивающаяся клиновидным, уплощенным в го-
ризонтально^/ направлении краем. В расположении костей черепа
у форм, роющих головой, имеются уклонения; например, затылоч-
ная часть располагается косо вперед, а не вертикально.
864
Тело роющих млекопитающих в разрезе вальковато, округло
вместо того, чтобы быть, сжатым с боков, как у других млекопи-
тающих, и одинаковой толщины спереди и сзади. Такое тело у крота,
у сумчатого крота, у златокрота, у слепца, у слепушонки; такую же
форму имеет тело у тех мелких хищпых, которые, не роя сами,
охотятся за роющими, залезая в поры: у перевязки, хорька, гор-
ностая, у ласки и др.
Лесные млекопитающие. Далеко не все млекопитающие, ко-
торых мы встречаем в лесу, могут быть названы лесными живот-
ными по своему происхождению. Многие из них приспособились
к жизни в лесу вторично, перейдя в него из открытых про-
странств, из саванн или редких кустарников, перенеся туда чуж-
дые лесу особенности своей организации. Таковы, например, олени
(Cervus), косули (Capreolus), лоси (Alces), кабаны (Sus), росомахи
(Gulo), медведи (Ursus), барсуки (Meles), бобры (Castor) и др.
Что олень и лось вовсе не типичные лесные животные, видно
уже из того, что это копытные животные, в своем происхожде-
нии и эволюции связанные с открытыми биотопами, к тому же
огромные рога их сильно затрудняют движение в лесу; кабапы
населяют не только леса, но и камыши; медведи встречаются в
безлесных горах; росомаха — в альпийской зоне; барсуки —
обитатели также степей и пустынь; бобры обитают по рекам
в луговых поймах, если имеются кустарники по берегам.
Леса от тропических дебрей до редколесья и заросли кустов,
несмотря на чрезвычайное разнообразие в высоте, густоте и
составе растительности обладают некоторыми общими чертами,
позволяющими их рассматривать как естественную группу «за-
крытых» биотопов.
Все же заросли кустарников по биотопическим условиям
сильно отличаются от лесов. Кустарники поднимаются па не-
значительную высоту и не имеют голых стволов, а обычно обра-
зуют густую сеть топких веток. Они связываются по своим био-
топическим особенностям с травянистыми открытыми простран-
ствами, где в некоторых случаях травы не уступают им в росте
и где полукустарники очень сходны с кустами своими ветвле-
ниями. Для крупных и тяжелых млекопитающих, как носороги
и слоны, кустарники представляют недостаточное препятствие
движениям. Над кустами имеется свободное пространство, и вы-
сокие млекопитающие могут выше кустов обозревать окрестности.
Другое дело леса. Здесь имеется особая экологическая ни-
ша— стволы деревьев с дуплами и толстые сучья, на которых
может жить своя фауна млекопитающих и по которым они могут
лазать. Движения по земле в лесу для крупных животных стеснены
и деревья являются серьезным препятствием для бега. В лесу
имеется более или менее выраженная многоярусность, причем
нередко кустарники входят в состав леса, образуя особую нишу.
Из лесов целесообразно выделить в качестве отдельной группы
биотопов те тропические леса, где благодаря исключительно
благоприятным климатическим условиям развивается такая бо-
55 Кашка ров и Станчинский 865
гатая растительность, что древостой переплетенный лианами,
воздушными корнями и усаженный громадным количеством эпи-
фитов, образует сплошную зеленую массу — дебри (гилеи), по
которым можно только лазать. Для свободных движений по
земле пет места, полет между стволами, лианами и листьями
эпифитов так же затруднен, как и в густых кронах деревьев.
Основная масса млекопитающих обитателей лесных тропических
дебрей — лазающие формы.
Изменения, являющиеся приспособлением для лазания, ка-
саются главным образом конечностей и хвоста. Если же при этом
животное приобретает более или менее вертикальное положение
тела, то изменяется и форма тела. Это изменение мы видим, напри-
мер, на грудной клетке антропоморфных обезьян, у которых она,
вместо обычной для ходящих млекопитающих сжатой с боков
формы, приобретает форму, сжатую спереди назад.
Среди низших позвоночных, амфибий и рептилий мы видели
немало примеров лазания при помощи особых приспособлений
присасывательного характера на нижней стороне фаланг и т. д.
У млекопитающих такие присасывательные органы развиваются
редко и весьма несовершенны. У даманов, или жиряков (Hyra-
coidea) для присасывания служат голые подошвы, которые
представляют несколько эластических мякишей, разделенных
глубокими бороздами. Даманы — обитатели скалистых местно-
стей, и древесные даманы вторично приспособились к лесу; такое
устройство подошв помогает им держаться на скалах и деревьях.
У маки домового (Tarsius spectrum) мякиши на концах пальцев
увеличены в виде присасывательных подушечек. Наилучше раз-
виты органы присасывания у летучих мышей из рода Thyroptera
из Южной Америки и Mysopoda с Мадагаскара, которых можно
назвать «цепкокрылыми» и «лицконогими» летучими мышами. У
Tyroptera и Mysopoda подошва ноги совершенно превращена в
круглый присосок с углубленной внутренней поверхностью, на-
поминающей присоски на щупальцах осьминога.
Гораздо большее число форм среди млекопитающих обладает
способностью лазать или прицепляться при помощи острых когтей.
По коре деревьев могут взбираться кошки, куницы, медведи,
белки, летучие мыши и т. д.
Превращение конечностей в своего рода щипцы для лазания
по ветвям встречается у некоторых млекопитающих, главным
образом у лемуров и сумчатых. У лемура потто (Perodicticus potto)
большой палец передней конечности и оба наружные пальца (4-й
и 5-й) лежат па одной оси, 3-й же палец развит весьма слабо, а 2-й
существует лишь в виде маленького выступа, лишенного ногтя.
У близко родственного лемуру лори (Stenops tardigradus) указа-
тельный палец несколько больше’. Устройство руки у потто чрез-
вычайно напоминает таковое у амфибии (Phyllomedusa). У сумча-
того медведя, или «коалу» (Phascolarctos), пятипалые конечности
приспособлены для хва^ния, На передних ногах оба внутренних
пальца могут быть противопоставлены трем остальным, на задних
866
ногах толстый большой палец также может противополагаться,
остальные же пальцы вооружены острыми длинными и изогну-
тыми когтями. Подобное тому, что имеется у хамелеона, а именно
«клещевидное» устройство передней ноги, мы видим у кольцехво-
стых кускусов (Phseudochirus). У них большой и указательный
пальцы противополагаются трем остальным.
Третий способ приспособления к лазящему образу жизни пред-
ставляет развитие из конечпостей в некотором роде крючка, на
котором животное подвешивается, вися па ветвях вниз головой.
Такой способ лазания мы видим у ленивцев (Bradypus), у малого
муравьеда (Cyclopes didactylus). Этот способ лазания ведет за собой
Рис. 642. Паукообразная обезьяна (Ateles paniscus).
(Из Гаане.)
ряд изменений; например, свисание шерсти вниз с брюха к спине
у лвнивцев и ряд изменений в скелете конечностей. Когти у этих
животных сильно изогнуты (предки висящих неполнозубых были
роющими животными, у которых уже редуцировались боковые
пальцы и развились сильные когти).
Четвертый способ передвижения на деревьях заключается в том,
что животное перебирается с ветви на ветвь, раскачиваясь. У та-
ких животных большой и 5-й пальцы обычно исчезают. Этот тип
приспособления — дальнейшее развитие того, что мы видели у под-
вешивающихся млекопитающих. Мы встречаем такое лазание
у некоторых обезьян. У них большие пальцы руки и ноги реду-
цируются, а у паукообразных обезьян (Ateles) (рис. 642) большой
палец руки исчезает вовсе. Кроме того, при таком способе лазания
55* 867
передние конечности, как плечо, так и предплечье и пальцы,
сильно удлиняются. Часто развивается синдактилия, т. е.
пальцы одеваются общей кожей. В той или иной мере мы видим
это у лемура Lichanotus и одного вида гиббонов (Hylobates
syndactylus).
У видов шимпанзе (Anthropopithecus), орапга (Simia satyrus)
и гориллы (Gorilla gorilla) также наблюдается приспособление к та-
кому способу лазания по деревьям, что ведет к некоторой редук-
ции большого пальца. В этом отношении у человека пет такой спе-
циализации. У него большой палец руки развит сильпо. Отсюда
следует, что его предки двигались не так, как гиббоны или шим-
панзе, а медленно лазали, пользуясь рукой как щипцами;
при этом приспособление к такому лазанию пе заходило у пего
далеко, а было на той ступени, па которой оно находится у сумча-
той крысы. Люди (Hominidae) представляют древнюю ветвь
ствола антропоморфных, никогда не бывшую строго приспособлен-
ной к лазанию по ветвям, а скорее к схватыванию и пригиба-
нию ‘веток низкорослых деревьев и кустарников и обрыванию
плодов.
Пятый способ лазания — с помощью цепкого хвоста. У реп-
тилий (у хамелеона, например) мы видим в качестве орудия
лазания цепкий хвост. Такой хвост мы встречаем и у млеко-
питающих, главным образом среди обезьян и сумчатых в лесных
дебрях Южной Америки и Австралии. В Америке у малого муравьеда
(Cyclopes didactylus), у обезьян: цепких (Ateles), ревунов (Му-
cetes), сапажу (Cebus) и некоторых других, у сумчатых у опос-
сума (Didelphis marsupialis). В Австралии цепкий хвост раз-
вивается у ряда сумчатых и одного грызуна из мышиных (Chiru-
romys). В развитии цепкого хвоста можно найти ряд ступеней
постепенного совершенствования и перехода от простого хвоста,
служащего опорой, к хвосту, голому снизу и снабженному по-
перечными бороздами. Такой хвост может служить хвататель-
ным органом — «пятой рукой».
Особенно богаты формами млекопитающие тропических ле-
сов, где господствующими среди них являются обезьяны. Боль-
шинство обезьян прекрасно приспособлены к жизпи на де-
ревьях, где питаются главным образом плодами. К древесным
же тропическим формам принадлежат и полуобезьяны (Prosimiae).
Оба эти отряда сверхотряда приматов в своем происхождении
и эволюции принадлежат тропическим лесным дебрям, откуда они
давали выходцев в другие биотопы. Из. других отрядов совершенно
древесными животными являются ленивцы (Bradypus), малые
муравьеды (Cyclopes) из неполнозубых. Многие летучие мыши,
главным образом летучие собаки (Megachiroptera), пользуются
деревьями, для дневного пребывания. Последнц^ питаются пло-
дами деревьев. .
Многие обитатели тропического леса, живя и двигаясь глав-
ным образом на земле, в силу указанных выше особенностей
лесных дебрей способны, однако, взбираться и на деревья, где
868
охотятся за добычей или укрываются па почь. Так, например,
горилла (Gorilla) слишком велика и тяжела, чтобы жить па де-
ревьях, по самка гориллы и дети почуют на деревьях. Ягуар (Felis
опса), леопард (Felis pardus), посухи (Nasua rufa) едят других,
более мелких млекопитающих, лазая по деревьям, но отлично
двигаются и по земле.
Из млекопитающих, держащихся в тропическом лесу па земле
и не лазящих, одни прилагают себе путь среди густой чащи и под-
леска, действуя своей тяжестью, — таковы африканские слоны
(Elephas africanus), носороги (Rhinoceros), буйволы (Buffalus),
тапиры (Tapirus), свиньи (Sus); другие, более мелкие, пробираются
между кустами и деревьями по’длеска, — таковы, например, са-
мые мелкие из копытных: 04effiKn"(Tragulidae) в малайских и аф-
риканских лесах.
Наиболее типичными лесными животными умеренной зоны яв-
ляются: белка (Sciurus) и летяга (Sciuropterus volans), соня (Elio-
mys), мышловка (Muscardinus), полчек (Glis glis), лесная куница
(Martes martes), рысь (Lynx lynx), дикая кошка (Felis sylvestris),
соболь (Martes zibellina) и др.
Летающие млекопитающие. Летающие млекопитающие, не-
сомненно, развились среди лесных форм и возникли путем при-
способления к прыганию с ветки на ветку, с дерева па дерево. И хотя
летающие млекопитающие составляют особый отряд рукокры-
лых (Chiroptera), совершенно специально измененных живот-
ных, однако среди млекопитающих других отрядов мы встречаем
формы как бы переходные к летающим, обладающие способностью
к планирующему полету. Это достигается у млекопитающих раз-
витием кожных складок, натянутых между конечностями па боках
и между головой и передними конечностями, между хвостом и зад-
ними конечностями. Складка между головой, шеей и передними
конечностями называется propatagium; между туловищем и обеими
парами конечностей складка носит название plagiopatagium, между
задними ногами и хвостом — uropatagium и, наконец, между
пальцами — chiropatagium (см. рис. 545, стр. 740). В некото-
рых случаях увеличение поверхности сопротивления парашюта
достигается путем сильного удлинения волос на краю кожпой
складки.
В развитии складок наблюдается некоторая определенная
закономерность, а именно прежде всего возникает plagiopatagium.
Если отвлечься совершенно от филогенетических взаимоотношений
и иметь в виду только степень сложности в развитии, то можно
составить следующий ряд с постепенным усложнением в развитии
способности планировать.
На первом месте в начале ряда следует поставить белку (Sciurus
vulgaris). Этот зверек во время прыжка широко расставляет ко-
нечности. Патагиума еще нет, по на боках тела имеются длинные
волосы, которые при прыжке отодвигаются в стороны. При пере-
прыгивапии помогает хвост. Опыты показали, что белки, у которых
отрезали хвост, не могли прыгать так хорошо и далеко.
869
Следующую ступень специализации представляет лемур си-
фака (Propithecus). Эта полуобезьяна прыгает с высокого места
(с ветви косо вниз), подняв руки высоко над головой. Подоб-
ное приспособление мы видим у целого ряда лемуров. Боковые
складки по бокам тела еще весьма слабы, при прыжке плотные
и длинные боковые волосы расправляются. Длинный пушистый
хвост служит при прыжке рулем.
У обезьяны, носящей имя сатана (Pithecia satanas), пред-
ставителя семейства Cebidae, кожные складки имеются на туло-
вище, на передних и задних конечностях. Небольшая складка
на передней стороне рук — первый намек на propatagium.
У австралийской сумчатой сони (Acrobates pygmaeus) plagiopa-
tagium тянется между локтем, туловищем и коленом и усажен
по краю длинными волосами. (Рис. 579, стр. 793).
У сумчатой белки (Petauroides volans) летательпая перепонка
на передних ногах доходит до локтя, а на задних — до основания
большого пальца. Этот зверек великолепно прыгает на значитель-
ные расстояния и в полном смысле слова перелетает с одной ветви
на другую, с вершины дерева на вершину.
У сахарной белки [тоже из сумчатых (Petaurus sciureus)] pla-
giopatagium очень широк и натянут между пятым пальцем перед-
ней и основанием большого пальца задней поги. На переднем краю
передней конечности имеется хорошо развитый propatagium. Хвост
длинный и пушистый. Прыгает зверек великолепно и может, пры-
гая с высоты 10 м, достичь другого дерева, удаленного на 20—30 м.
Еще дальше ушло развитие летательного аппарата у летяг
из Rodentia. У одпого рода тагуапов (Pteromys) летательная пе-
репонка начинается от передних ног, тянется вдоль боков туло-
вища, прикрепляется к задним ногам и отсюда распространяется
в виде узкой складки кожи к хвосту. Большой пушистый хвост
является при прыгапии мощным рулем. У обыкновенной летяги
(Sciuropterus volans) uropatagium очень мал; наоборот, хвост чрез-
вычайно расширен. (Рис. 592, стр. 808).
У африканских летяг из рода Anomalurus (Rodentia) мы
имеем два замечательных приспособления к древесной жизни
и планирующему полету. На нижней стороне основания хвоста
имеется два ряда роговых чешуй, упирающихся при лазании в кору
деревьев и облегчающих таким образом влезание. Увеличение pla-
giopatagium* происходит благодаря хрящевой палочке, отходящей
от верхнего конца локтевой кости до края парашютной складки,
как у Pteromys и Sciuropterus. ‘
Наконец, у летучего маки (Galaeopithecus volans) летательная
перепонка начинается иа шее, соединяется с передними, ногами,
охватывает их вплоть до кисти, далее переходит к задИи погам
и отсюда направляется к копчику хвоста. (Рис. 585, стр. 799).
Таким образом в этой перепонке лежат все части тела, кроме
головы. Летучий маки с высоты 12 м спускается па дерево, отстоя-
щее на 55 м.
Таким образом приспособления к планирующему полету раз-
870
виваются у представителей самых различных отрядов и семейств мле-
копитающих. Рассмотренный ряд указывает нам путь, которым воз-
никала эта специализация. Переход к одинаковому образу жизни
вызывал и одинаковые приспособления. При этом процессе эволюции
приспособление не явилось внезапно, а для пего использовались
уже существующие черты организации. Однако лишь в одном отря-
де— рукокрылых — развилась способность к настоящему полету.
Рукокрылые — животные воздушной среды. Здесь они движутся,
здесь ловят свою добычу, здесь спариваются. Летательная пере-
понка развилась у них иначе, нежели у рассмотренных до сих пор
форм. Подобно тому как это было у ископаемых летающих ящеров
(Pterosauria) и как это имеет место у птиц, летательный аппарат
несут у них передние конечности. Повидимому, именно такая кон-
струкция необходима для осуществления настоящего полета.
Происхождение рукокрылых от насекомоядных несомненно.
Мы можем с уверенностью утверждать, что предки рукокрылых
имели приспособления для планирующего полета подобно описан-
ным формам, но мы, к сожалению, не знаем переходных форм,
которые вели бы к образованию рукокрылых.
Переход к воздушному образу жизни вызвал целый ряд спе-
циальных приспособлений. Тело приобретает большую прочность,
кости черепа срастаются, как у птиц. Во время полета животное
усиленно дышит, и у рукокрылых сильно расширяется грудная
клетка, увеличивая тем полость для легких; грудная кость сильно
отходит от позвоночника, и позвоночник в этой области искрив-
ляется дугой вверх. Мощные трудные мускулы, приводящие в дви-
жение крылья, вызывают на месте своего прикрепления развитие
гребля или киля па грудной кости, как у птиц. Чтобы дать грудной
кости больше опоры, ребра сливаются друг с другом, с грудиной
и со спинными позвонками. Постепенное развитие этого явления
можно наблюдать в различных семействах рукокрылых. Прочное
соединение плечевого пояса с осевым скелетом достигается при
помощи сильной ключицы. Лопатка сильно развита. • Передние
конечности удлинены, особенно предплечье и пальцы. Удлинение
последних достигается путем удлинения отдельных элементов,
главным образом пястных костей и вторых фаланг. Наоборот,
ногтевые фаланги укорочены, сохраняясь лишь на втором пальце
у плодоядных и на большом пальце передних ног у всех рукокры-
лых. Этот палец не играет никакой роли при летании, служит для
лазания, не удлинен и несет острый коготь. Летательная пере-
понка захватывает и задние ноги. Между последними она развита
у рукокрылых различно. О присасывательных дисках па ступне
у некоторых видов говорилось выше. Они помогают животным
подвешиваться.
Жизнь в воздухе вызвала у летучих мышей развитие приспо-
собления к тому, чтобы детеныши после рождения не падали па
землю. Перед родами самка подвешивается пе при помощи задних,
как обыкновенно, а с помощью передних конечностей и подгибает
хвост с хвостовым отделом летательной перепонки к брюху.
Благодаря этому получается нечто вроде мешка или чашки,
куда и падает новорожденный детеныш. Последний крепко цеп-
ляется за грудь матери, когда последняя перекусит пуповину.
Водные млекопитающие. Приспособлепие к жизни в водной
среде встречается среди млекопитающих весьма часто. Приспо-
соблепие это может быть или полным или частичным: животное
может или всецело перейти к жизни в воде или же только частично,
ведя двоякий образ жизни: и на суше, и на воде. В последней мле-
копитающее может находить себе убежище от врагов или от жары,
добывая себе пищу на суше; таков, например, гиппопотам, или
бегемот (Hippopotamus amphibius). Или же, живя на суше, жи-
вотное идет в воду для добывания себе пищи, как охотящаяся
за рыбой выдра (Lutra). В том и другом случаях животное по об-
разу жизни является земноводным. Водным млеко-
питающее можно назвать лишь в том случае, если оно всю свою
жизнь проводит в воде, выходя, быть может, па сушу лишь для
размножения, как мы видим это у морского котика (Callorhinus
ursinus). Чисто водные млекопитающие должны были, конечно,
пройти стадию земноводных млекопитающих, прежде чем стать
чисто водными, хотя, история их далеко не всегда нам ясна.
Если считать не только чисто водных млекопитающих, но и
земноводных, то мы найдем приспособлепие к воде среди целого
ряда отрядов. Киты, сирены и ластоногие являются по существу
водными животными. В той или иной мере водными животными
являются из однопроходных: утконос (Ornithorhynchus paradoxus),
из отряда сумчатых плавун (Chironectes minimus), из грызунов:
мускусная крыса (Fiber zibethicus), бобер (Castor fiber), болотный
бобер, или нутрия (Myopotamus coypu), водосвинка (Hidroochoerus
capybara), три рода австралийских мышей (сем. Hydromyidae)
и др.; из отряда насекомоядных: кутора водяная - (Crossopus
fodiens), выхухоль (Myogale mas’chata), тибетская водяная земле-
ройка (Nectogale elegans), выдровая землеройка (Potamogale
velox), длиннохвостые тенреки (Limnogale); из хищных: норки (Lut-
reola lutreola), выдра (Lutra lutra), камчатский бобер (Latax lut-
ris); среди кШйтных собственно водных, кроме гиппопотама, нет,
но ряд форм обитает в болотистых местностях, по болотистым бе-
регам: тапиры (Tapirus), свиньи (Suidae), водяной козел (Cobus
cobus), буйвол (Buffalus). Из ископаемых копытных водно-болот-
ными было семейство Macrauchenidae, из отряда Litopterna.
Частичный или полный переход к водному образу жизни при-
водит к ряду приспособлений. Эти приспособления обусловли-
ваются двумя моментами: животное должно уметь плавать, т. е.
держаться и двигаться в воде, с одной стороны, и встречать наивоз-
можно меньше препятствий в воде — с другой.
Здесь мы наблюдаем очень интересное явление. Приспособле-
ния, которые мы видим у плавающих млекопитающих, оказыва-
ются чрезвычайно сходными с приспособлениями жизни в воде,
наблюдаемыми у плавающих рептилий. Сходство в общей форме
тела между крайними членами приспособительных к воде рядов
872
у тех и других между ихтиозаврами и китообразными настолько,
велико, что заставляло некоторых говорить об их родстве. Само
собой разумеется, что даже поверхностное рассмотрение внутрен-
него строения сразу обнаруживает между ними огромное разли-
чие: здесь имеет место лишь конвергенция признаков.
Тело и тех, и других становится вальковатым и в крайних слу-
чаях (у дельфлиов) даже рыбообразным. Становится маленьким
и даже исчезает наружное ухо. Шея укорачивается, становится
малоподвижной благодаря укорочению шейных позвонков и их
срастанию (у китов). Волосы делаются короткими и шелковистыми,
смазанными кожпым жиром, благодаря чему они не намокают
(у бобра, выдры, тюленей). Иногда волосы совсем исчезают (у ки-
тов и сирен), сохраняясь в той или иной степени у зародыша во
время утробной жизни. Мощный слой жира имеет значение как
защита от охлаждения, он имеет еще и другое значение: умень-
шает удельный вес тела и ослабляет давление при погружении
животного в глубину. Наибольшего развития достигает этот жи-
ровой слой у китов. У крайних членов приспособительного ряда,
у некоторых китов, развился спинной плавник, а именно у тех,
которые являются быстроплавающими пелагическими формами.
В отличпе от спинного плавника рыб этот плавник не поддержи-
вается скелетными частями, так же как и хвостовой плавник китов
и сирен.
Выше, в главе о рептилиях, было указано на существование
двух способов продвижения тела в воде: при помощи ног наподо-
бие весельной лодки и при помощи хвоста, действующего наподо-
бие пароходного винта. Оба способа мы находим у водных млеко-
питающих (у чисто водных и земноводных).
Изменения ног у водных млекопитающих идут в следующих
направлениях. Ноги становятся короткими, задние ноги разви-
ваются сильнее передних и отодвигаются назад, пальцы стано-
вятся широко расставленными, между ними развиваются пере-
понки. Все эти признаки бывают развиты в различной степени
в зависимости от степепи приспособления к водному образу жизни.
У водяной куторы и других плавающих, насекомоядных на
подошве и пальцах задних пог имеются длинные жесткие волосы,
прилегающие к ногам, когда животные находятся на суше; эти
волосы, оттопыриваясь, увеличивают поверхность стопы во время
плавания. У длиннохвостого тепрека и выхухоли пальцы, кроме
того, соединены перепонкой. Эти перепонки имеются в слабо раз-
витом состоянии у целого ряда наземных млекопитающих (у со-
бак, куниц), у водных же они значительно увеличиваются в раз-
мере, соединяя, например, у выдры, пальцы почти целиком. Пе-
репонка прежде всего и в большей степени развивается на задних
ногах, которые обычно бывают и развиты сильнее, чем передние
поги, например, у камчатского бобра (Latax lutris).
У ряда плавающих млекопитающих упор при плавании сосре-
доточивается главным образом па 4-м пальце, который становится
благодаря этому длиннее всех, например, у выхухоли (Myogale
873
moschata), у водяной куторы (Neomys fodiens). У земноводных
млекопитающих, плавающих при помощи лап, хвост играет роль
руля. Обычно он горизонтально плоский, как, папример, у бобра.
Лишь у насекомоядных (выхухоль, длиннохвостый тенрек, выдро-
вая землеройка) хвост образует вертикальную плоскость.
Еще дальше идут приспособления к водному образу жизпи
у сирен и китов. Организация их представляет массу конвергент-
ных признаков. У тех и других тело имеет веретенообразную
форму, шея отсутствует, задние конечности редуцированы. Раз-
растания кожи хвоста в стороны образуют •горизонтально распо-
ложенный хвостовой плавник. Передние конечности превратились
в плавники, задние конечности редуцируются, волосяной покров
редуцирован, и зато сильно развит жировой слой.
На ряду с редукцией задних конечностей идет редукция таза.
Весьма интересно, что у китообразных редукция таза протекает
теми же самыми путями, какими она протекала у третичных и совре-
менных сирен, отличаясь лишь в деталях. Общий же характер
редукции совершенно тот же, несмотря на различное происхож-
дение китов и сирен.
Несмотря па сильную конвергенцию во внешних в анатоми-
ческих признаках между сиренами и китами существует большое
различие. У первых приспособление зашло не столь далеко, как
у вторых. Плечо и предплечье у них хотя и укоротились, еще
относительно длинны и подвижны. Кисть хотя и стала уже
плавником, еще мало изменилась.
Многие скелетные части у китов и сиреп соединяются между
собой очень рыхло. Например, скелет грудной клетки становится
очень растяжимым, что имеет большое значение при захватывании
большого количества воздуха перед погружением кита в глубину.
Рубчатость костей уменьшает удельный вес. Этому способствует
и чрезвычайное богатство костей жиром.
Череп большинства водных млекопитающих, как и других
водных позвопочпых, сплющен в вертикальном направлении,
становится низким и широким. Это видно уже на черепе' выдры,
но еще больше у тюленя. Межглазпичная часть черепа очень су-
жена, так как глаза у шЖсдвигаются кверху. Надглазничные
отростки исчезают. Сдвигание глаза, а также носовых отверстий
кверху наблюдается уже у бегемота, проводящего день в погружен-
ном в воду положении так что над поверхностью воды выдаются
только глаза и ноздри. В этом отношении голова его напоминает
голову крокодила.
Дальше всего ушли в этом приспособлении киты. У них носо-
вые кости редуцированы до состояния маленьких косточек, ле-
жащих позади носа, в углублении лобных костей. Таким образом,
ноздри оказываются лежащими па самом верхнем пункте головы
и при поднятии из воды первые появляются над поверхностью
последней.
Как ш у крокодила, у китов воздухоносные пути защищены
ют проникновения воды. Твердое нёбо вытянуто назад при помощи
874
сближающихся посредине крыловидпых костей. В то же время
гортань вытягивается благодаря удлинению щитовидного хряща
в трубку и открывается не в ротовую полость, а в носовые каналы,
которые при помощи особых мускулов в мягком нёбе могут совер-
шенно изолироваться от глотки. У зубастых китов (Odontoceti)
носовые ходы соединяются наверху в одну общую носовую полость,
открывающуюся наружу «брызгалом». Сложная система клапа-
нообразных складок надежно закрывает носовое отверстие. Та-
ким же образом закрываются носовые отверстия и у других вод-
ных млекопитающих: у ластопогих, бобра, выдровой землеройки.
Закрываются они благодаря эластичности стенок, открываются
при помощи особых мускулов.
Органы обоняния у водных млекопитающих становятся ру-
диментарными, обонятельный эпителий исчезает, обонятельные
части мозга редуцируются, а у дельфинов обонятельные доли и
обонятельный нерв исчезают вовсе.
Ухо также подвергается изменениям. Наружное ухо, как ука-
зано выше, исчезает; мускулы, которые служили для приведения
его в движение у сухопутных предков водных млекопитающих,
у последних служат для замыкания наружного слухового про-
хода. У тех животных, которые сохранили наружное ухо, для его
закрывания служат другие приспособления. У бобра очень ко-
роткие ушные раковины просто закрываются назад и плотно при-
жимаются. У выдры наружный слуховой проход закрывается осо-
бой кожной складкой. У водяной куторы для этого служат две
кожных складки внутри ушной раковины.
Так как на глубине большое давление воды, то bullae osseae
черепа становятся у тюленей и в особенности у китов толсто-
стенными.
Изменяется у водных млекопитающих и глаз. У тюленей
и китов, как и у рыб, роговица становится плоской, а хруста-
лик — шаровидным. Такой глаз мало способен к нормальной
аккомодации. Последняя обеспечивается у тюленей и китов
развитием особого меридионально лежащего мускульного пучка,
сокращение которого влечет линзу вперед, устанавливая глаз
для зрения на близкое расстояние. Глаз водных млекопитающих,
испытывающих при погружении в воду значительное давление,
обычно имеет те или иные приспособления для защиты от послед-
него. Склера у тюленей и китов очень толста и тверда, глазные
мускулы очень сильно развиты, сильнее, чем это нужно для дви-
жения глазного яблока; глазной нерв защищен толстым слоем
«дивной сети» (сосудов), образующих эластическую трубку.
Слезная железа редуцируется за ненадобностью, так как глаз
пепрерьгопо омывается водой. Зато большего развития достигает
так называемая гардерова железа, выделяющая жирный секрет,
защищающий глаз от вредного влияния морской воды. Секрет
этой железы образует у ламантина (Manatus) перед глазом столь
надежный желеобразный слой, что даже прикосновение к рого-
вице не вызывает с ее стороны рефлекса.
875
Изменяются у китов и внутренние органы. Исчезновение зу-
бов, развитие так называемого китового уса описано выше. О ра-
стяжимости легких также было уже сказано. Внутреннее устрой-
ство легких с плотными стенками легочной паренхимы, с хрящом
вокруг мельчайших бронхов (у дельфинов) приспособлено к захва-
тыванию большого количества воздуха и большому внутреннему
давлению. Такое устройство дает возможность китам и сиренам
долго дышать захваченным воздухом. Обычно дельфины остаются
под водой до 3 минут, а кит (Balaenoptera) может оставаться
под водой 8—12 часов.
Приспособлением к водной жизни является и устройство груд-
ной железы у китов. Соски спрятаны у них в кожном кармане.
Большая железа имеет резервуар, в котором скопляется молоко.
Присосавшемуся детенышу молоко это впрыскивается в рот при
помощи особого мускула, так как сосать в воде детеныши не могут.
2. Приспособления к климатическим особенностям среды
Приспособления к температурным условиям. Млекопитающие —
животные гомотермные,. с постоянной температурой тела. Это
делает их в общем животными эйритермическими,
способными переносить значительные колебания температуры,
которые в полярных странах доходят до 60 и 70°. Выравни-
вание температуры тела достигается у млекопитающих путем
усиленной продукции тепла и уменьшения его отдачи благо-
даря сжатию периферических сосудов при понижении темпера-
туры воздуха или усилепия отдачи тепла, благодаря расширению
периферических сосудов и усиленной деятельности потовых желез
при повышении окружающей температуры.
Кроме того, более или менее независимыми от температурных
колебаний млекопитающих делает их волосяной покров.
Сильное развитие волосяного покрова как защитного средства
наблюдаем мы у животных холодных стран и высоких нагорий. Та-
кое значение имеет густая и длинная шерсть яка (Poephagus grun-
niens) (рис. 643), поднимающегося до высоты 5 200 м, густой мех
рыжего хомяка (Cricetulusfulv^fe), горных полевок (Alticola), памир-
ского зайца (Lepus pamirensis) и др. Из северных животных
образцом животного, защищенного от холода густыми волосами, яв-
ляются северный олень (Rangifer tarandus)n мускусный бык (Ovidos
nioschatus) (рис. 617), а также представители полярных трызупов:
лемминг (Myodes), полярный заяц, песец (Alopex alopex). Шерсть
полярных животных отличается своей длиной, шелковистостью
и густым подшерстком. У мелких млекопитающих ойа покрывает
все тело, даже ступни. Длинная шерсть на хвосте песца служит
ему как бы покрывалом, когда он свертывается в клубок во время
сна. Волосы северного оленя заключают в себе воздух, а потому
являются еще более плохим проводником тепла.
Волосяной покров мелких животных, которые рок?тся под спе-
том и всегда могут укрыться там от ветра, бывает мяЬок и шелко-
876
вист; наоборот, пе могущие укрыться от ветра более крупные жи-
вотные имеют шерсть более жесткую, грубую.
Так как у крупных животных относительно к объему поверх-
ность тела меньше, чем у мелких, то мелкие животные выра-
батывают по сравнению с крупными меньше тепла, а излучают
его сравнительно много; поэтому они нуждаются в более теплом
мехе. Так, у северного оленя длина каждого волоска составляет
лишь небольшую часть диаметра тела, а у лемминга, живущего
с оленем в одних и тех же местах, длина волоса равна диаметру
тела или даже превосходит его. Таким же приспособлением
к защите от охлаждения может служить увеличение размеров
Рис. 643. Як (Phoephagus grunniens).
(Из Гаане.)
животных, обитающих в холодных странах, по сравнению с
особями того же или близкого вида из более теплых стран.
Это правило, установленное Бергманом, подтверждается ина мле-
копитающих. Так, например, с юга на. север увеличиваются раз-
меры зайцев — русаков и беляков, медведей и др.
В качестве приспособления к изменениям температуры в тече-
ние года следует рассматривать и периодическую смену волося-
ного покрова. Смена постоянная, непрерывная не имеет приспо-
собительного характера к изменению температуры по сезонам
и является более древним свойством, унаследованным от далеких
предков. Сезонная линька связана с приспособлением к сезонным
изменениям температуры. Зимняя* шерсть бывает более густой
и длинной, летняя — более короткой и редкой. Приурочивается
линька в холодных и умеренных странах к осени и весне или же
877
только к началу осени. Смена шерсти происходит иногда очень
быстро. Так, например, мускусные быки уже в течение несколь-
ких дней после выпадения первого снега одеваются в зимнюю
шерсть, так же быстро сменяя ее весной.
Запасаемый на зиму под кожей жир имеет также и защитное
от холода значение.
Зимняя и летняя спячка являются приспособлением к небла-
гоприятным климатическим сезонным сменам. Она наступает, од-
нако, не под непосредственным влиянием изменения температуры,
а вследствие педостатка пищи или воды, наступающего благодаря
изменениям температуры.
Зимняя и летняя спячка — глубоко отличное от обычного сна
явление. Летняя спячка зависит от недостатка воды. Примером
может служить желтый суслик в Туркестане (Citellus fulvus),
бодрствующий З1^ месяца в году, а остальное время предающийся
спячке. Засыпает он, уходя глубоко в пору в конце мая, когда
окружающая растительность пустыни совершенно выгорает, ста-
новясь сухой. В лёссовой пустыне, где живет этот суслик, нет воды,
и свежая растительность является единственным источником воды,
необходимой для нормального обмена веществ. Такой же летней
спячке подвергается американский колумбийский суслик (Citellus
columbianus).
На Мадагаскаре впадает в летнюю спячку тенрек (Centetes
ecaudatus), мелкие лемуры (Otolemur, Chirogale).
Физиологические явления, сопровождающие летпюю спячку,
повидимому, одинаковы с теми, что наблюдаются при зимней
спячке.
Что зимняя спячка связана с недостатком пищи, вызванным
низкой температурой, ясно из состава залегающих в спячку
животных. Засыпают насекомоядные (Insectivora) и летучие
мыши (Chiroptera), питающиеся насекомыми, которых зимой
они достать не могут. Засыпают суслики, тушканчики, сурки,
питающиеся степными и альпийскими травами. Те же пред-
ставители грызунов, которые запасают на зиму пищу, не засы-
пают, например, полевки (Microtus). Также не засыпают крот
(Talpa) и землеройки (Soricidae), относящиеся к насекомоядным:
крот живет под землей и может добывать себе пищу в течение всей
зимы, землеройки же вообще хищничают, нападают на мышей
и тоже не остаются без пищи. Из хищных млекопитающих засы-
пают медведь (Ursus) и барсук (Meles); первый питается главным
образом растительной пищей, которую не может добыть зимой,
второй же — смешанной пищей, причем из животных он ест глав-
ным образом низших, недоступных зимой. При этом впадают
в спячку медведи и барсуки, обитающие в более холодных обла-
стях; кавказские и закавказские медведи и барсуки в спячку не
впадают.
Продолжительность спячки зависит от продолжительности пе-
риода недостатка пищи. На севере и в высокогорной области она
продолжительнее, чем па юге и внизу. Упомянутый выше Citellus
878
columbianus па 10 дней спит меньше на южных склонах хол-
мов, чем на северных, где инсоляция меньше и вегетация запаз-
дывает.
Животные, делающие на зиму запасы, спят с перерывами. Дру-
гие животные, как летучие мыши, запасов не делают, спят не про-
сыпаясь, даже не выделяя мочи, что делают, например, сурки»
просыпающиеся для выделения мочи или кала.
Для спячки выбираются места укрытые, защищенные от хо-
лода: норы, дупла деревьев, берлоги. Животпое свертывается при
этом в шар, чем достигается уменьшение поверхности тела, излу-
чающей тепло. Летучие мыши обертывают свое тело крыльями,
как плащом, что ведет к той же цели. #
Обмен веществ у спящих животных замедляется, дыхание ста-
новится медленным. Именно это ослабление обмена веществ является
причиной понижения температуры у спящих зиму животных.
У суслика было обнаружено понижение до 2°, у летучих мышей
до 7—7.2°.
Температура спящих зиму млекопитающих находится в зави-
симости от окружающей среды. Так, у сурка при внешней темпе-
ратуре 10.5°, 6.2°, 5.5°, 10.8°, 8.25° температура животного 10.6°,
5.4°, 5.9°, 12.1°, 8.25°.
Таким образом, получается сходство с пойкилотермными, так
называемыми холоднокровными животными.
Перед спячкой млекопитающие накапливают много жира. Сон,
повидимому, наступает лишь тогда, когда накопится достаточное
количество жира. В неволе спящие зиму животные не засыпают,
если они пе накопили достаточно жира; наоборот, очень жирные
животные засыпают уже летом. В этой зависимости засыпания
от накопления жира, несомненно заключается большая приспособ-
ленность, так как без этого засыпающее животное было бы обре-
чено на голодную смерть, ибо во время спячки жир служит мате-
риалом для поддержания жизни. Просыпающиеся весной живот-
ные бывают очень худы.
Физиологическая причина, заставляющая спящих зиму живот-
ных по наступлении холодов залегать в спячку, еще не выяснена.
Повидимому, такой причиной является непосредственное влияние
низкой температуры в связи с известным состоянием организма
к этому времени. Выть может, и накопление жира играет здесь
известную роль.
К спящим зиму и спящим лето животным относятся: летучие
мыши, ежи, тенрек, енот, барсук, соня, хомяк, некоторые мыши,
тушканчики, сурки, белки, суслики, бурундуки, летяги, неко-
торые лемуры, ехидна и утконос. Причислить медведей к подвер-
женным зимней спячке животным не совсем легко. Они ложатся
в берлогу и спят гораздо больше, чем летом, потребляя в это время
собственный жир, но все же это не настоящая спячка, и они очень
легко от нее пробуждаются. Во время этой так- называемой спячки
у медведей в феврале сходит кожа с подошв, почему они и сосут
лапы. Спячка помогает им переждать неблагоприятное время.
872
Что это не настоящая спячка, показывает п то обстоятельство,
что медведица в январе мечет детенышей. Самцы й холостые самки
белых медведей бодрствуют всю зиму. Беременные же самки с осени
накапливают много жира и дают снегу занести себя, причем для
дыхания остается небольшое отверстие. Здесь самка мечет дете-
нышей, кормя их молоком за счет накопившегося жира до того
момента, пока медвежата не подрастут.
На ряду с рассмотренными выше пассивными приспособле-
ниями к неблагоприятным температурным условиям, млеко-
питающие приобрели в борьбе за существование целый ряд а к-
тивпых приспособлений, обусловленных большой
подвижностью этих животных. Сюда относятся передвижения,
а также зарывание в землю, копание нор, укрывание под
камнями и деревьями. В норах и под камнями температура
гораздо ровнее, мягче, не испытывает тех резких колебаний, кото-
рые испытывает температура воздуха. В Тибете, например, на
высоте 5 200 м температура под камнем дает в течение суток коле-
бание 7°, тогда как рядом на воздухе она колеблется в пределах
24°. В норах температура еще ровнее. Пеструшки, или норвежские
леммипги, прогрызают в слое мхов и лишаев, которыми покрыта
тундра, ходы, проходящие под кочками и под корнями низкорос-
лой полярной березы. Здесь они и проводят день, выходя через
отдушины в снегу. Роют ходы полевки (Microtinae), сурки
(Marmota), обитатели альпийской зоны, и другие. В жарких
пустынях большинство млекопитающих укрывается в порах
днем и проявляют свою деятельность вечером, утром и почью.
И это в одинаковой степени относится к представителям разных
•систематических групп: насекомоядных, грызунов, хищных и др.
Запасание корма па зиму имеет также защитно-приспособитель-
ный характер к неблагоприятным изменениям температурных
условий по сезонам.
Такую заготовку корма мы видим у пищух (Ochotona) в аль-
пийской зоне, которые на зиму не засыпают, а делают запасы корма,
складывая под камни, где находятся их норы, стожки сепа. Делает
запасы и песец, когда имеет пищу в изобилии.
Еочевки млекопитающих. Одним из средств, при помощи ко-
торых млекопитающие избавляются от тяжелого времени в связи
с колебанием температуры, являются перекочевки. Но,
надо сказать, что здесь температура и вообще климат действуют
обычно не непосредственно, а тем, что вызывают недостаток пищи.
Такие перекочевки млекопитающих особенно заметны в горах:
как только там выпадут снега и температура упадет ниже извест-
ных границ, ряд высокогорных животных спускается вниз, где
меньше снега, теплее и легче добыть пищу. Горные козлы (Capra
sibirica), серны (Rupicapra), косули (Capreolus), олени (Cervus); барс
ирбис (Leopardus uncia), медведи (Ursus)n др. спускаются в ниже-
расположенные места. Весной с таянием снегов эти животные по-
степенно поднимаются все выше в горы.
В равнинах такие сезонные перекочевки имеют место или там,
*80
где выпадают глубокие снега, или там, где бывают более суровые
зимние условия. Так, северные олени из тундры откочевывают
в полосу криволесья; из районов с более глубоким снежным по-
кровом откочевывают в менее снежные районы; куланы и анти-
лопы дзераны уходят из высоких холодных полупустынь в более
северные, по мепее холодные равнины.
Переселение млекопитающих животных наблюдается и в со-
вершенно иных условиях, например, во внутренней и Южной
Африке, где огромные стада жвачных животных, зебр и слонов,
периодически передвигаются через огромные пространства по на-
правлению то к северу, то к югу, в зависимости от чередующихся
периодов дождей и засухи.
Зависимость от света. Среди млекопитающих можно разли-
чить дневных, ночных и сумеречных. Зависимость здесь стоит
в связи с добыванием пищи и избегапием врагов и конкурен-
тов, а отчасти и от неблагоприятных климатических условий.
Среди ночных млекопитающих опять-таки есть такие, что ведут
более или менее исключительно ночную жизнь, — глаза у них
приспособились к ночной жизни. Другие же млекопитающие,
охотясь преимущественно ночью или в сумерках, отлично видят
и днем.
Примером животного чисто дневного могут служить сурки или
суслики. Ночью они спят. Другой пример дневного животного
и суточной периодичности представляет нам горный козел (Capra
sibirica). Ночью стадо спит под прикрытием скал. С раннего утра
оно просыпается и начинает медленно взбираться вверх по скло-
нам ущелья, пощипывая траву. К восходу солнца стадо добирается
до гребня. Покормившись, козлы начинают спускаться вниз к во-
допою. Напившись, снова поднимаются кверху и располагаются
на охдых. Затем снова спускаются к водопою и, напившись, под-
нимаются немного вверх, располагаясь под скалами на ночлег.
Пример ночного животного дает нам дикобраз (Hystrix hirsu-
tirostris и другие виды). День он проводит в норе, вырытой им,
или заняв для этого расселину в горе, ночью же выходит для оты-
скивания пищи. Австралийский утконос (Ornithorhynchus) яв-
ляется примером животного сумеречного.
Приспособлепие к ночному образу жизни сказывается, в ряде
приспособлений, во-первых, в сильном увеличении глаза, что стоит
в связи с получением на сетчатку возможно большего количества
световых лучей. Это требует способности к особо сильному со-
кращению со стороны зрачка, который может иногда сокращаться
до размеров точки, как, например, у кошки. Такое чрезмерное
увеличение глаз мы наблюдаем в сильной степени у ведущего
ночной образ жизни долгопята или маки домового (Tarsius tarsius
(рис. 631). У этого животного и у ночных животных вообще
наблюдается расположение глаз с почти параллельными осями,
со 1 [ровождаемое укорочением челюстей.
Очень часто у ночных животных развиваются огромные уши,
свидетельствующие о сильном развитии слуха. Мы видим такие
56 Кашкаров и Стаичинский 881
уши у ряда лемуров: ушастого маки, или галаго (Galago galago)
(рис. 644), ай-ай, или руконожки (Chiromys madagascarenis),
у упомянутого выше долгопята (Tarsius). Сильно развиты уши
у маленькой степной лисицы ф е н е к a (Megalotis zerda), а также
у летучих мышей, большинство которых летает в сумерки и в ран-
ние часы ночи, пользуясь, повидимому, главным образом слухом
для отыскания своей добычи — насекомых. Голые уши летучих
мышей, богатые нервными окончаниями, служат также и органами
осязания па ряду с летательной перепонкой и кожистыми выро-
стами на голове.
Окраска. Несомненно, в зависимости от света находится
окраска млекопитающих. Обычно кожа у них бывает пе окра-
шена. Пигмепт сосредоточивается в волосах. На ряду с пигментом
окраска зависит и от структуры поверхности волоса и от содер-
жания в нем воздуха.
Мы еще очень мало зна-
ем о действительных при-
чинах появления той или
иной окраски, что принуж-
дает нас остановиться преи-
мущественно на фактах ок-
раски, не вдаваясь каждый
раз в их объяснение.
Окраска млекопитаю-
щих редко бывает одно-
цветной. Волосы большей
частью состоят йз разпо-
_ цветных колец, из совме-
Рис. 644. Ушастый маки, или галаго (Ga- СТНОГО действия которых И
Iago galago). получается или смешанный
(Из врэма.) цвет или рисунок. Есть ос-
нование думать, что наибо-
лее примитивной окраской является продольнополосатая, путем
разрыва полос переходящая в пятнистую. Последняя же путем
слияния пятен переходит в поперечнополосатую.
Наиболее интересными являются, конечно, случаи покрови-
тельственной окраски, очень частые и у млекопитающих. По по-
вода7 нее можно лишь повторить то, что было выше сказано о покро-
вительственной окраске других классов. Если речь идет об одно-
образной, малозаметной окраске, то с покровительственным зна-
чением ее можно согласиться.
Но, когда речь идет о покровительственной якобы окраске
зебры или тигра, дело меняется. По распространенном}7 мнению,
полосы на их теле имитируют тень, падающую на пих. Но
среди зарослей на них падали бы на самом деле эти тепи. Зачем
тогда рисунок, и зачем зебре или тигру вечно носить за собой даже
на открытом месте, на солнце, изображение этих теней? Оно де-
лает их сразу заметными. Далее, зебры пасутся всегда вместе с
антилопами, жирафами, которые окрашены совершенно иначе.
882
Наконец, тигр живет пе только в джунглях, где его волосатость
могла бы подражать тепям, — он живет и в. снегах Сибири, имея
ту же полосатую окраску. Многие млекопитающие, имеющие
якобы покровительственную пустынную окраску, не могут ею
пользоваться, ибо являются животными ночными, днем же спят,
свернувшись в поре, например, тушканчики (Alactaga, Dipus).
Сумчатый крот (Notoryctes) живет под землей, а окрашен в песча-
ный цвет. Наконец, самое представление об окраске животных,
как о чем-то однообразном, весьма схематично: окраска большин-
ства видов сильно вариирует.
В полярных странах большинство животных бело, но и здесь
трудно говорить всегда с уверенностью о покровительственной
окраске, ибо мы сразу натолкнемся па ряд противоречий. Почему,
папример, белый медведь (Ursus maritimus) бел и летом, когда
белая окраска бросается в глаза?
Нисколько пе сомневаясь в существовании и большом эколо-
гическом значении покровительственной окраски, мы считаем
необходимым предостеречь от упрощенных объяснений в этом во-
просе и от излишней схематизации.
Помимо покровительственной окраски, выражающейся в об-
щем тоне окраски или в рисунках, особенно замечательна способ-
ность многих млекопитающих менять летнюю серую и бурую покро-
вительственную окраску па белую покровительственную окраску
зимой. Такую сезонную перекраску мы встречаем из северных
млекопитающих: у зайца-беляка, песца, горностая, ласки. Такая
окраска может иметь пе только покровительственное значение,
но и является более теплой, так как содержит в волосе воздух.
Вообще цвет окраски может служить защитой от излучения,
от вредного воздействия лучей солнца или, наоборот, для боль-
шего поглощения лучей определенного значения.
Зависимость от влаги. Влажность воздуха, несомненно, яв-
ляется одним из существенных факторов, определяющих жизнь
млекопитающих и их распространение. Влажность влияет, дей-
ствуя на животных и непосредственно и косвенно. Хорошо из-
вестно, что в сыром климате развиваются различные болезни,
вызываемые животными организмами: малярия, лейшманиоз, пиро-
плазмоз и т. п. Эти болезни могут ограничивать млекопитающих
в их распространении. Чрезмерная влажность воздуха является
гибельной для многих животных степей и пустынь. Примером
является верблюд, который не может жить долго в слишком
влажном климате.
Целый ряд млекопитающих, подобно верблюду, оказывается
приспособленным к чрезвычайной сухости воздуха и довольствуется
минимальным количеством влаги. Это,— млекопитающие пустынь.
Примером может служить упомянутый желтый суслик (Citellus
fulvus), для которого единственным источником воды является
свежая растительность. С уменьшением содержания воды в расте-
ниях суслик становится вялым и скоро засыпает. Другие члены
того же биоценоза, как песчанка Эверсмана (Gerbillus eversmanni),
56* 883
живущая в тех же условиях, довольствуется водой, заключающейся
в семенах растений, и пе засыпает по окончании вегетации.
Наоборот, другие млекопитающие требуют для себя влаги,
без которой быстро умирают. Недостаток пресной воды часто бы-
вает причиной массовой гибели животных. Такая массовая гибель
млекопитающих наблюдается в пампасах Южной Америки во время
засухи и описана была Дарвином. Засухи часто являются причи-
ной довольно правильных переселений. Антилопы и другие ко-
пытные в Африке по высыхании ручьев и рек переселяются на
сотни километров туда, где нет такой засухи.
В виду сказанного большие пояса пустынь являются весьма
важными препятствиями на пути расселения влаголюбивых жи-
вотных. Сахара в Африке образует для целых групп животпых
границу (граница между палеарктической и эфиоп-
ской областями).
Но во всяком случае температура оказывает большее влияние,
нежели влажность. Следующий пример наглядно это показывает.
При сравнении влажной восточной и ксерофитной (сухой) западной
частей Сонорской подобласти в Сев. Америке мы видим, что сухая
часть отличается от влажной наличием в ней 1 семейства и 34 ро-
дов птиц и млекопитающих, которых нет в последней. Влажная
часть в свою очередь имеет 1 семейство и 11 родов, которых нет
в сухой. Общими для той и другой части являются 25 семейств
с 58 родами. В то же время вся Сонорская подобласть, располо-
женная широтно, отличается от расположенной широтно же Не-
арктической или Канадской подобласти 18 семействами и 141
родом, Неарктическая же подобласть от Сонорской—9 семействам^
и 70 родами. Общими же той и другой оказываются всего 26 родов.
Прямое влияние осадков сказывается, например, па
сусликах.. Ливневые дожди при определенной интенсивности за-
ливают норы сусликов и нередко являются причиной их массовой
гибели, особенно если после дождя бывает холодная ночь. Глубо-
кие снега и запосы, особенно гололедица, очень опасны для многих
животпых, не впадающих в спячку: зайцев, мышей. ;
Косвенное влияние осадков на животных через растительность
еще больше. За годы 1929—1933 заповедная степь в Аскания-Нова
резко меняла свои летние аспекты благодаря преимущественному
развитию разных представителей растительности, что сейчас же
отражалось на мире насекомых, а также и на позвоночных.
Недостаток влаги вызывает иногда правильные сезонные пе-
рекочевки млекопитающих. Так, в центральной и южной Африке,
в областях, где ежегодно происходит смена сухого периода пе-
риодом дождей, наблюдается массовое передвижение антилоп,
зебр и слонов с юга на север и с севера на юг.
3. Биоценотические факторы существования млекопитающих
Млекопитающие подобно птицам находятся в тесной зависи-
мости от окружающего мира живых существ, входя в состав биоце-
нозов. Однако в редких случаях они, являются непременными чле-
384
пами только этих биоценозов, так как в большинстве случаев живут
пе за счет одной или двух трофических систем (рядов), а за счет
гораздо большего числа растительных или животных организмов,
входящих в состав двух, а иногда и большего числа биоценозов,
образующих ландшафтные единицы большего порядка. Так, сус-
лики, например, живя на юге Украины в плакорной степи, а не
па подах, где их заливала бы вода, живут за счет большого числа
растений, главным образом за счет тонконога (Роа bulbosa), и охот-
но питаются растительностью пода. Вто же времязаяц, лисица, сайга
обитают во всем комплексе биоценозов степи, живя за счет еще
большего разнообразия животных и растений. Основным биоцено-
тическим фактором является, конечно, пища.
Пища млекопитающих. Приспособление млекопитающих
к разного рода пище. Млекопитающих подобно, другим позвоноч-
ным животным, можпо разделить по роду их пищи па растительно-
ядных, или фитофагов, и плотоядных, или зоофагов.
Однако в связи с тем, что среди них имеется большое количество
видов, питающихся одновременно как растительной, так и животной
пищей, может быть выделена группа всеядных животных, или
пантофагов (omnivora). В свою очередь млекопитающие,
питающиеся животной пищей, могут быть разделены па две группы:
па хищных, питающихся позвоночными, преимущественно
теплокровными (carnivora), и насекомоядных, питаю-
щихся беспозвоночными животными, преимущественно насеко-
мыми (insectivora).
Подобно другим животным, среди млекопитающих встречается
и питающиеся разнообразной пищей: эйритофаги, или
полифаги, и, наоборот, приспособленные к однообразной
пище стенофаги, или олигофаги и монофаги.
Мы уже видели выше, какое большое значение в эволюции мле-
копитающих имело приспособление их к различного рода пище.
В связи с дифференциацией по роду и способам добывания пищи
применительно к различным средам обитания возникли и эволю-
ционировали основные группы млекопитающих.
Первоначальной пищей млекопитающих, как уже указано
-выше, были мелкие беспозвоночные животные: насекомые, мол-
люски, черви, а также, вероятно, и мелкие амфибии и реп-
тилии. Современные одпопроходные, некоторые сумчатые, боль-
шинство насекомоядных и мелкие летучие мыши сохранили до
нашего времени этот первоначальный способ питания, приспосо-
бившись к разнообразным способам добывания пищи в разных
средах.
0 поверхности земли, из верхних слоев ее, добывают себе бес-
позвоночных многие сумчатые: землеройки, ежи, тенреки, неко-
торые броненосцы; в земле: сумчатый крот, златокрот, крот, рою-
щие- тенреки и некоторые другие; в воде: утконос из однопроход-
ных; плавуны из сумчатых; водяные тенреки, водяные землеройки,
выхухоли и другие из насекомоядных; из воздуха — летучие мыши,
на деревьях — лемуры. Это по большей части полифаги.
885
Специализированными в отношении питания насекомыми — оли-
гофами — нужно считать млекопитающих, питающихся муравьями
и термита™. Таковы ехидна, муравьеды, трубкозуб, ящеры, сум-
чатый муравьед (Myrmecobius). Морда у них вытянута в длинную
трубку с маленьким ротовым отверстием, через которое высовы-
вается длинный червеобразный язык. Он смачивается липким выде-
лением подъязычных и подчелюстных желез, благодаря чему к нему
прилипают муравьи и термиты, и спабжен чрезвычайно развитой
мускулатурой. Высокой специализации в отношении питания
насекомыми достиг сумчатый пяткоход (Tarsipes rostratus). Он
питается насекомыми, сидящими в цветах. Туда он засовывает
свой длинный язык, слегка зазубренный ио краям, и достает
им насекомых.
Путем перехода к питанию мягкими и сочны™ побегами и пло-
дами возникли первые растительноядные животные как из сумча-
тых и насекомоядных (вместе с летучи™ мышами), так и из хищ-
ных, и из полуобезьян, и неполнозубых. Первоначально это были
малоспециализированные формы смешанного питания (omnivora),
затем опи приобрели в большей или мепыпей степени специаль-
ные приспособления для питания разного рода растительной
пищей.
Такими малоспециализированными ^всеядными формами явля-
ются некоторые многорезцовые сумчатые, часть насекомоядных,
некоторые виверры, медведи, носухи, барсуки, медоеды из хищ-
ных, большинство полуобезьян и многие обезьяны. Обычным
приспособлением к растительиоядпости являются сильное развитие
губ, долотообразное строение резцов, укорочение клыков и бу-
нодоптный характер коренных зубов, удлинение кишечного канала
и развитие слепой кишки.
Особенности эти получают ,свое полное выражение у таких
высокоспециализированных млекопитающих, как двурезцовые сум-
чатые, грызуны и могочисленные отряды сверхотряда копытных
(Ungulata).
Травами, главным образом зерновыми злака™, питаются по-
лорогие, олени, оленьки, верблюды, тапиры, носороги, лошади,
кенгуру и ™огие грызуны: зайцы, пищухи, суслики, сурки, по-
левки, песчанки, шиншиллы, полукопытные.
У парнокопытных характерным приспособлением к травоядной
пище являются редукция верхних резцов, видоизменение нижних
клыков и превращение коренных зубов в гипселодонтные и селе-
нодонтные зубы, высокая подвижность губ и языка и усложнение
желудка. У лошадей специализация зубов, как мы видели шла в
ином направлении: режущему ряду пижних резцов соответствует
ряд верхних; коренные зубы получают сложное строение от скла-
док эмали, образующихся вследствие стирания бугорков, и имеют
высокие коронки (гипселодонтны); в общем зубы здесь приспособ-
лены к пережевыванию более жестких, сухих трав — степных зла-
ков соответственно основному местообитанию лошадей. Вместо
сложного желудка жвачных, в котором происходят бродильные
8 86 '
процессы, важные для переваривания клетчатки, у лошадей бро-
жение происходит в толстой кишке.
У грызунов, зубная система которых получила типичное свое
строение в связи с приспособлением к питанию твердой расти-
тельной пищей, коренные зубы в разной степени совершенства
приспособлены к перетиранию или мягкой и сочной травы или
сухих стеблей злаков и зерен. Из квадратных они становятся все
более и более удлиненными и получают все более и более
сложное строение путем образования складок эмали. Броже-
ние клетчатки происходит у грызунов в сильно развитой слепой
кишке.
Листья деревьев составляют пищу жирафов, обладающих спе-
циальным приспособлением^для их срывания в виде чрезвычайно
длинной шеи, длинных передних конечностей и специализирован-
ного языка и губ; листьями и молодыми побегами деревьев питаются
многие олени, отчасти медведи, тогда как слоны способны переже-
вывать более толстые ветки с листьями, питаясь также злаками и
корнями. Соответственно с таким питанием у слонов имеются хобот,
приспособленный к обрыванию веток, бивни, приспособленные
к разрыванию земли, и чрезвычайно большие полилофодонтные
зубы, приспособленные к перетиранию такой твердой пищи, какой
являются ветви и корни деревьев и кустарников. Древесиной и
корой питаются из грызунов бобры с высокоспециализироцанными
резцами и водосвинки (Hydrochoerus capybara) с лофодонтными
коренными зубами, напоминающими зубы слона. Корнями, клуб-
нями и луковицами растений питаются многочисленные роющие
грызуны: слепыши (Spalax), цокоры (Myospalacinae), водяные
крысы (Arvicola), слепушонки (Ellobius) й многие другие.
Плодами травянистых растений и упавшими с деревьев плодами,
ягодами, зернами и семенами питаются очень многие животные,
преимущественно грызуны; бурундуки, хомяки, полевки, мыши,
песчанки, долгоноги, дикообразы, пластинчатозубые крысы и др.,
но также и многие хищники: медведи, барсуки, медоеды, а из ко-
пытных — свиньи.
Плодами, растущими на деревьях, питаются белки, летяги,
летучие собаки (Megachiroptera), многие полуобезьяны и обезьяны,
а также и некоторые хищники (медведи). У летучих мышей, пита-
ющихся мягкими и сочными плодами, растущими на деревьях,
имеются большие мясистые губы, приспособленные к обсасыванию
плодов.
Что касается водных животпых, то растительноядным является
отряд сирен (Sirenia). Этот отряд возник путем приспособления
к видному образу жизни растительноядных (травоядных) примитив-
ных копытных животных. Питаются они сочными мягкими водя-
ными растениями. В связи с такой пищей мы видим у сирен тенден-
цию к редукции зубов, описанную нами выше, приведшую к полной
их потере у стеллеровой морской коровы (Rhytina stelleri); вместо
зубов для перетирания мягкой пищи у нее служат шершавые рого-
вые пластинки на нёбе и между ветвями нижней челюсти.
887
Плотоядные хищные звери возникли путем перехода к хищному
образу жизни мелких насекомоядных зверьков. Таковы еще сум-
чатые крысы (Didelphys) и сумчатые кроты (Notoryctes), сумчатые
барсуки (Peramelidae) и в особенности сумчатые куницы; сумчатые
волки и т. п. из семейства Dasyuridae становятся уже настоящими
хищниками, приспособившимися к питанию птицами и мле-
копитающими, мелкими и более крупными. Ископаемые креодонты
(Creodonta) возникли также из насекомоядных животных, а из них,
как уже нам известно из вышеизложенного, развились настоящие
наземные хищники (Carnivora fissipedia) и два отряда водных хищ-
ников: китообразные (Cetacea) и ластоногие (Pinnipedia).
Если добывание пищи у растительноядных и насекомоядных жи-
вотных сопряжено с трудностями лишь в случаях ее недостатка, то
добывание добычи хищниками всегда сопряжено с большими труд-
ностями: необходимо не только отыскать добычу, но и поймать ее
и умертвить. При этом добываемое животное — птицы и млекопи-
тающие — обладают рядом приспособлений, позволяющих им из-
бегать опасности быть пойманными хищником: они или обладают
быстрым бегом или полетом, или умеют скрыться, или обладают
органами защиты, могущими оказаться опасными для врагов:
рогами, иглами, зубами, когтями и т. п.
Итак, хищники должны обладать приспособлениями, чтобы оты-
скать добычу. Большая часть кошек (Felidae) отыскивает добычу
преимущественно с помощью острого зрения; собаки (Canidae) оты-
скивают добычу с помощью чутья, острого обоняния; многие хищни-
ки, особенно ночью, пользуются слухом (кошки, собаки, куницы).
Ловля добычи осуществляется различно. Большинство хищников
старается подкрасться к добыче как можно ближе, а затем или бро-
сается на нее прыжками (кошки) или бежит вдогонку в расчете пой-
мать ее благодаря более быстрому бегу или большей выносливости.
Другие подкарауливают добычу на звериных тропах, на водопоях
или на кормежке и набрасываются на нее из-за засады. Третьи ищут
добычу в ее же собственных убежищах, в норах, дуплах, гнездах.
Соответственно способу добывания пищи вырабатываются у хищ-
ных млекопитающих и соответствующие приспособления. Покрови-
тельственная окраска делает хищника мало заметным. Таковы бу-
рые, серые, ржавые тона лесных животных (лисица, волк, куница,
соболь, рысь), пятнистый рисунок их меха (рысь, пантера, ягуар),
смена бурого наряда на белый зимой у песца, ласки, горностая
Формы тела соответствуют способу добывания добычи: компактная
гибкая форма кошки, тигра, льва и других прыгающих на жертву
кошек прямо противоположна стройным высоконогим и длинноте-
лым формам гепарда (кошки, догоняющей добычу при быстром
беге) или поджарой и длинноногой форме тела волков, догоняющих
добычу: и те, и другие совсем не похожи на тонких, длиппотелых
коротконогих изящных горностаев, ласок, хорьков, мангуст,
ихневмонов и других хищников, добывающих себе пищу из нор,
Ягуар и отчасти куница ловят добычу на деревьях.
Для умерщвления добычи обыкновенно употребляются зубы.
888
Длинные и острые клыки играют эту роль у хищников. Особенно
достопримечательны в этом отношении высокоспециализированные
клыки вымерших саблезубых кошек (Machairodus), служившие,
вероятно, для умерщвления крупных толстокожих животных:
носорогов и слонов. При схватывании зубами добычи многие хищ-
ники стараются прокусить наиболее жизненно важные места: гор-
ностай и ласки прокусывают сонные артерии на горле, за горло хва-
тают зубами собаки, волки, лисицы, рыси и др. Кошки и рыси схва-
тывают добычу выдвижными когтями передних лап; львы, тигры,
медведи ударами передних лап валят и убивают добычу. Рыси,
тигры, львы и другие крупные хищники при нападении на крупных
копытных, вооруженных рогами, стараются прыгнуть животному
иа спину. Это делали и саблезубые тцгры.
Для разрывания мяса и дробления костей служат коренные зубы,
в особенности хищпые зубы, описанные выше.
Среди грызупов имеет место вторичный переход к плотоядному
образу жизпи; так, например, крысы охотно поедают других жи-
вотных, в том числе и себе подобных, водяная крыса (Arvicola)
нападает далее па уток, тогда как соня (Eliomys quercinus) поедает
мелких птиц, птенцов, яйца и мелких млекопитающих. Явление
каннибализма, отмеченное только что для крыс, встречается у вол-
ков во время зимних голодовок, у соболей, кротов и у хомяков,
а также у некоторых летучих мышей (подковоносов и ушанов).
Типичными некрофагами (трупоедами) являются гиены, облада-
ющие особенно сильными хищными зубами, позволяющими им
разгрызать такие кости, которые для других хищников остаются
недоступными; для отыскания трупов они обладают острым чутьем
и неутомимостью при передвижениях. Некрофагами в значительной
мере являются шакалы, песцы; волки зимой охотно идут на падаль,
иногда и медведи; дохлых животных едят крысы, многие другие мы-
шиные, еж и даже свиньи.
Копрофагия среди млекопитающих — редкое явление. Шакалы,
гиены, многие мышевидные и свиньи поедают экскременты других
млекопитающих. В некоторых случаях, как известно, например,
у кроликов, хомяка, бобра, животные поедают свои собственный
экскременты в случае недостаточного переваривания в них пищи.
Водные хищники питаются или рыбой (выдра, морской бобер,
порка, водяные землеройки), или беспозвоночными: раками, мол-
люсками (также выдра, норка, енот; из насекомоядных: выхухоль,
водяная кутора и др.). Белый медведь питается тюленями и-мор-
жами. Ластоногие питаются рыбой, морскими беспозвоночйымн,
а также морскими растениями. Китообразные зубастые и беззубые
по характеру пищи резко отличаются друг от друга, в то время кок
зубастые являются хищниками, питающимися или рыбой (дельфи-
ны, белуга и др.), или млекопитающими, как, например, касатка,
которая питается тюленями, проглатывая мелкие виды (нерпу
и молодых тюленей целиком, или нападает на крупных беззубых
китов, отрывая от них мясо кусками. Беззубые киты питаются мел-
кими планктонными организмами.
Плотоядные хищные звери возникли путем перехода к хищному
образу жизни мелких насекомоядных зверьков. Таковы еще сум-
чатые крысы (Didelphys) и сумчатые кроты (Notoryctes), сумчатые
барсуки (Peramelidae) и в особенности сумчатые куницы; сумчатые
волки и т. п. из семейства Dasyuridae становятся уже настоящими
хищниками, приспособившимися к питанию птицами и мле-
копитающими, мелкими и более крупными. Ископаемые креодонты
(Creodonta) возникли также из насекомоядных животных, а из пих,
как уже нам известно из вышеизложенного, развились настоящие
наземные хищники (Carnivora fissipedia) и два отряда водных хищ-
ников: китообразные (Cetacea) и ластоногие (Pinnipedia).
Если добывание пищи у растительноядных и насекомоядных жи-
вотных сопряжено с трудностями лишь в случаях ее недостатка, то
добывание добычи хищниками всегда сопряжено с бблыиими труд-
ностями: необходимо не только отыскать добычу, по и поймать ее
и умертвить. При этом добываемое животное — птицы и млекопи-
тающие — обладают рядом приспособлений, позволяющих им из-
бегать опасности быть пойманными хищником: они или обладают
быстрым бегом или полетом, или умеют скрыться, или обладают
органами защиты, могущими оказаться опасными для врагов:
рогами, иглами, зубами, когтями и т. п.
Итак, хищники должны обладать приспособлениями, чтобы оты-
скать добычу. Большая часть кошек (Felidae) отыскивает добычу
преимущественно с помощью острого зрения; собаки (Canidae) оты-
скивают добычу с помощью чутья, острого обоняния; многие хищни-
ки, особенно ночью, пользуются слухом (кошки, собаки, куницы).
Ловля добычи осуществляется различно. Большинство хищников
старается подкрасться к добыче как можно ближе, а затем или бро-
сается на нее прыжками (кошки) или бежит вдогонку в расчете пой-
мать ее благодаря более быстрому бегу или большей выносливости.
Другие подкарауливают добычу на звериных тропах, на водопоях
или на кормежке и набрасываются на нее из-за засады. Третьи ищут
добычу в ее же собственных убежищах, в норах, дуплах, гнездах.
Соответственно способу добывания пищи вырабатываются у хищ-
ных млекопитающих и соответствующие приспособления. Покрови-
тельственная окраска делает, хищника мало заметным. Таковы бу-
рые, серые, ржавые тона лесных животных (лисица, волк, куница,
соболь, рысь), пятнистый рисунок их меха (рысь, пантера, ягуар),
смена бурого наряда на белый зимой у песца, ласки, горностая
Формы тела соответствуют способу добывания добычи: компактная
гибкая форма копией, тигра, льва и других прыгающих на жертву
кошек прямо противоположна стройным высоконогим и длинноте-
лым формам ,гепарда (кошки, догоняющей добычу при быстром
беге) или поджарой и длинноногой форме тела волков, догоняющих
добычу: и те, и другие совсем не похожи на тонких, длиннотелых
коротконогих изящных горностаев, ласок, хорьков, мангуст,
ихневмонов и других хищников, добывающих себе пищу из нор,
Ягуар и отчасти куница ловят добычу па деревьях.
Для умерщвления добычи обыкновенно употребляются зубы.
888
Длинные и острые клыки играют эту роль у хищников. Особенно
достопримечательны в этом отношении высокоспециализированные
клыки вымерших саблезубых кошек (Machairodus), служившие,
вероятно, для умерщвления крупных толстокожих животных:
носорогов и слонов. При схватывании зубами добычи многие хищ-
ники стараются прокусить наиболее жизненно важные места: гор-
ностай и ласки прокусывают сонпые артерии на горле, за горло хва-
тают зубами собаки, волки, лисицы, рыси и др. Кошки и рыси схва-
тывают добычу выдвижными когтями передних лап; львы, тигры,
медведи ударами передних лап валят и убивают добычу. Рыси,
тигры, львы и другие крупные хищники при нападении на крупных
копытных, вооруженных рогами, стараются прыгнуть животному
па спину. Это делали и саблезубые тцгры.
Для разрывания мяса и дробления костей служат коренные зубы,
в особенности хищные зубы, описанные выше.
Среди грызунов имеет место вторичный переход к плотоядному
образу жизни; так, папример, крысы охотно поедают других жи-
вотных, в том числе и себе подобных, водяная крыса (Arvicola)
нападает даже па уток, тогда как сопя (Eliomys quercinus) поедает
мелких птиц, птенцов, яйца и мелких млекопитающих. Явление
каннибализма, отмеченное только что для крыс, встречается у вол-
ков во время зимних голодовок, у соболей, кротов и у хомяков,
а также у некоторых летучих мышей (подковоносов и ушанов).
Типичными пекрофагами (трупоедами) являются гиены, облада-
ющие особенно сильными хищными зубами, позволяющими им
разгрызать такие кости, которые для других хищников остаются
недоступными; для отыскания трупов они обладают острым чутьем
и неутомимостью при передвижениях. Некрофагами в значительной
мерё являются шакалы, песцы; волки зимой охотно идут на падаль,
иногда и медведи; дохлых животных едят крысы, многие другие мы-
шиные, еж и даже свиньи.
Копрофагия среди млекопитающих — редкое явление. Шакалы,
гиены, многие мышевидные и свиньи поедают экскременты других
млекопитающих. В некоторых случаях, как известно, например,
у кроликов, хомяка, бобра, животные поедают свои собственные
экскременты в случае недостаточного переваривания в них пищи.
Водные хищники питаются или рыбрй (выдра, морской бобер,
порка, водяные землеройки), или беспозвоночными: раками, мол-
люсками (также выдра, норка, енот; из насекомоядных: выхухоль,
водяная кутора и др.). Белый медведь питается тюленями и мор-
жами. Ластоногие питаются рыбой, морскими беспозвоночными,
а также морскими растениями. Китообразные зубастые и беззубые
по характеру пищи резко отличаются друг от друга, в то время как
зубастые являются хищниками, питающимися или рыбой (дельфи-
ны, белуга и др.), или млекопитающими, как, например, касатка,
которая питается тюленями, проглатывая мелкие виды (нерпу
и молодых тюленей целиком, или нападает на крупных беззубых
китов, отрывая от пих мясо кусками. Беззубые киты питаются мел-
кими планктонными организмами.
Зависимость млекопитающих от растений. Зависимость эта
очень велика. Наибольшее количество млекопитающих приходится
на долю растительноядных. Многие виды связаны с определенными
растениями, питаясь только их листьями или плодами.
В пустынных местностях растения служат млекопитающим пе
только пищевым материалом, но являются для них единственным
источником воды.
Наконец, для многих млекопитающих растения являются сре-
дой, в которой они движутся, и убежищем, где они прячутся от
врагов, отдыхают, размножаются. Уничтожение растительности
ведет за собой исчезновение млекопитающих. В своем географиче-
ском распространении млекопитающие чрезвычайно зависят, прямо
или косвенно от растительности. Стоит посмотреть на карту распре-
деления по земной поверхности многих млекопитающих, чтобы
сразу бросилась в глаза зависимость их от распределения расти-
тельности. Если взять распространение рода Alactaga из тушкан-
чиков," то оно совпадает с распространением полынных и ковыльных
степей; распространение желтого суслика (Citellus fulvus) в средней
Азии строго ограничено распространением осоково-злаковых и
злаково-осоковых растительных сообществ. Таких примеров можно
найти очень много. Взаимоотношения между растительностью и
животными могут быть чрезвычайно сложными. Остановимся на
немногих примерах.
Растения пустыни можно разделить на три биологических груп-
пы: однолетние, многолетние, пережидающие неблагоприятное
время года под землей, и многолетние, существующие на поверх-
ности земли в течение всех сезонов.
Для млекопитающих пустыни растения первой группы имеют
мало значения; впрочем, иногда зависимость от них выявляется
резко. Интересное наблюдение было сделано над сумчатым Phas-
cologale cristicaudata в Центральной Австралии: в конце благо-
приятного сезона, вслед за периодом дождей и пышным развитием
вегетации, этот грызун весил на 40—60% больше, чем в неблаго-
приятный сезон.
Гораздо более тесная связь существует между млекопитающими
пустыни и растениями второй группы, накопляющими питатель-
ный материал в луковицах, клубнях и других подземных образова-
ниях и вегетирующими надземно лишь короткое время. Обилие
мелких грызунов в некоторых пустынях объясняется тем, что они
питаются этими луковицами и другими подземными частями. В норе
слепца в Египте было найдено 68 луковиц растений; главная пища
слепушонки (Ellobius) — луковицы тюльпанов.
Распространение малого суслика (Citellus pygmaeus) в целин-
ной степи стоит в теснейшей зависимости от тонконога (Роа bul-
bosa), клубеньками которого он питается. Так как тонконог посе-
ляется на участках степи, выбитых выпасом, то и количество
сусликов в степи увеличивается по мере увеличения сбоя.
Наиболее тесная связь существует с растениями третьей группы.
Эти растения доставляют пищу и воду животным даже в самые не-
890
благоприятные периоды года. Примером могут служить в Турке-
стане большие песчанки (Rhombomys opimus), обитатели совершен-
но безводных мест. Кроме осоки (Carex physodes), они питаются
ветвями саксаула, которые обгрызают по концам. Саксаул служит
для них среди лета единственным источником воды. Другой при-
мер — уже упомянутый выше желтый суслик (Citellus fulvus),
живущий всецело за счет членов осоково-злакового сообщества,
главным образом осок и малькольмии. Когда последние в середине
лета высыхают (относятся скорее ко второй группе растений), сус-
лик, лишаясь единственного источника воды, залегает в спячку.
Карликовая песчанка (Dipodillus dasyurus) в Месопотамии живет
на солончаках за счет небольшого кустарника Suaeda, растущего
в пустыне на пятнах солончака. Питаясь этим растением и насеко-
мыми, находимыми па его ветвях, песчанка находит себе между
его корнями убежище и защиту от лисиц и шакалов. Последние,
а также заяц (Lepus connori) представляют с указанной песчанкой
всех млекопитающих, населяющих солончаки этих пустынь, и при-
надлежат к быстро бегающим формам, которые могут бегать далеко
от места обитания в поисках воды и пищи. За счет многолетников
пустынь Австралии — сочных солянок — живут овцы, за счет как-
тусов они живут в сухих частях южных штатов.
Но растения тоже зависят от млекопитающих. Упомянутые
выше большие песчанки, обгрызая ветви саксаула, вызывают уси-
ленное разрастание ветвей и, кроме того, разрыхляют вокруг почву.
Благодаря этому уже издали по особенно пышному разрастанию
кроны видно место колоний песчанок. Байбаки (Marmota bobac)
значительно изменяют состав растительности возле их колоний.
Разрыхленная почва, выброшенная из их нор, дает приют расте-
ниям, пе встречающимся нигде больше в степи. Распространение
семян растений млекопитающими общеизвестно.
Имеются веские данные для того, чтобы считать копытных и гры-
зунов тайим элементом степи, без которого степной растительный
покров резко изменяется.
Многочисленные растения пустыни имеют семена с колючками
и шипами, которые пристают к шерсти лисиц, газелей, тушканчи-
ков. Однако, млекопитающие часто наносят растительности огром-
ный вред.
В западной Азии возле стоянок бедуинов нельзя найти ни одного
не испорченного домашними животными растения. В пустынях
Америки кладбища, недоступные животным, являются местом оби-
тания многочисленных растений, нигде больше не встречающихся.
Белки в условиях еловой тайги, живущие почти исключи-
тельно за счет семян, ели в годы, когда ель плодоносит, чрезвы-
чайно размножаются, в годы же, когда ель совершенно или
почти пе плодоносит, они принуждены откочевывать в другие
районы в поисках пищи и при этом иногда массами гибнут.
В условиях елово-широколиственных или широколиственных
лесов белки лучше обеспечены более разнообразным кормом
(желудями, орехами, ягодами и т. п.) и поэтому живут там оседло.
891
От урожая семяп ели в значительной мере зависит в тайге
численность рыжей полевки (Evotomys glareolus).
Взаимоотношения млекопитающих с другими животными
биоценоза
Взаимоотношения между животными биоценоза бывают иногда
весьма тесны и обычно сложны. Между плотоядными млекопитаю-
щими и их добычей существует известная пропорциональность.
Так, папример, лисица питается разными мелкими грызунами. Она
во множестве поедает полевок, ест песчанок, сусликов. Увеличение
в благоприятные годы числа этих грызунов влечет за собой
и увеличение лисиц. Наоборот, наблюдались случаи, когда истреб-
ление последних вызывало в ближайшее же время значительное
увеличение грызунов, создавало «мышиную опасность». Хорошо
известно, что в годы «мышиной опасности» появляется огромное
количество хорьков, ласок, лис, хищных птиц. В лесах Евразии
и Сев. Америки за счет зайца-беляка живут рыси. Уменьшение по-
головья беляка ведет к уменьшению поголовья рыси и наоборот.
В Сев. Америке статистика числа поступивших шкур зайцев (Le-
pus americanus) и рыси (Lynx canadensis) показала поразительное
совпадение в увеличении числа вторых с увеличением числа
первых. Истребление в Лабрадоре дикого олепя (Rangifer cari-
bou) повело за собой исчезновение зависящего от него волка. Барс
(Felis unica) всегда встречается в одном сообществе с горными
козлами (Capra sibirica) и горными индейками (Tetraogallus). Зимой
снег заставляет козлов и индеек спускаться ниже. За ними
следует барс. На каменистых осыпях гор Туркестана под боль-
шими камнями устраивают свои гнезда пищухи (Ochotona). Здесь
же держатся в большом количестве горные полевки подрода Alti-
cola. Те и другие составляют добычу ласок (Mustela nivalis) и
ферганского горпостая (Mustela ferghanae), входя с ним в один
биоценоз. В него же вхбдят еще ушастая сова.(Asio otus) и неясыть
(Syrnium), обитающие здесь же, на арчевых деревьях.
Ни полного истребления, ни длительного чрезмерного размноже-
ния грызунов не происходит, а имеется всегда колеблющееся в не-
которых пределах числовое соотношение между хищниками и их
добычей. Чрезмерное развитие грызунов должно неизбежно вызы-
вать чрезмерное развитие хищников. Опи истребляют грызунов и тем
самым сами ставят пределы своему дальнейшему размножению.
Изучение животных сообществ и биоценотических факторов
существования млекопитающих едва начато. Понимание их чрезвы-
чайно важно для учета промысловых животных и их защиты. Пред-
ставление о сложности взаимоотношений между животными в био-
ценозе дает диаграмма, изображающая взаимоотношения между
животными прерии в умеренной части Америки. Круги и эллипсы
в диаграмме заключают животных, служащих пищей одним и тем же
хищникам, и группы, питающиеся одинаковой пищей. Стрелки ведут
от животных, служащих пищей, к тем, чьей добычей они являются
(рйс. 645).
892
Волки, например, уничтожают бизона; если почему-либо волки
возрастают в числе, число бизонов становится меньше. Большое
уничтожение мышей размножившимися волками вызовет, в конце
концов, уменьшение числа волков благодаря недостатку больших
животпых зимой и мышей летом. Это даст возможность бизонам
и мышам восстановить или даже превысить свое прежнее число.
Уменьшение мышей может заставить койота, или лугового волка
(Canis latrans), поедать больше сусликов (Spermopliilus). Это,
несомненно, окажет влияние па растительную часть биоценоза.
Все члены биоценоза различными узами связаны друг с другом;
эти связи, иногда очепь сложны. Знание этих отношений большое
практическое значение .в деле правильной постановки охотничьего
хозяйства, в деле борьбы с вредителями и т. д. Иногда самым
действительным приемом в борьбе с вредителями является введе-
ние в сообщество нового члена, враждебного вредителю. Иногда
животное, считающееся вредным, оказывается и полезным,
так как уничтожает других, более опасных вредителей. Так, в
Индии местами тигр, истребляя чрезвычайно многочисленных
кабанов и оленей, портящих чпосевы, является желательным
гостем.
Конкуренция из-за пищи может чрезвычайно обостряться в тех
случаях, когда, вследствие усиленного размножения вида, проис-
ходит «перенаселение», т. е. число индивидуумов вида становится
так велико, что пищи оказывается недостаточно. Тогда наступает
голод, вызывающий массовую гибель млекопитающих или их пере-
селение, которое может возникать периодически. В таком случае
животные не возвращаются обратно, а обычно тоже погибают.
Примером могут служитьпереселениямышей и полевок в годы «мы-
шиной папасти». Грызуны эти иногда размножаются в таком коли-
честве, что местность, ими обитаемая, не может их прокормить, и они
вынуждены переселяться в соседние районы, двигаясь несметными
полчищами, уничтожая все на своем пути, поедая слабых и боль-
ных из своей среды. За ними обычно движутся многочисленные хищ-
ники. Такие передвижения известны также для водяных крыс,
и особенно для белок и других грызунов. Следом за грызунами идут
хищники: росомахи (Gul о borealis), волки (Canis lupus), лисы
(Vulpes vulpes), песцы (Vulpes lagopus), соболи (Mustela zibellina),
горностаи (Mustela ^erminea). Особенно грандиозны бывают мас-
совые передвижения лемминга на севере, когда в несметном коли-
честве эти грызуны, сопровождаемые песцами, лисами, белыми со-
вами и другими хищниками, идут то к югу, то к северу, то в других
направлениях.
В других случаях передвижения млекопитающих зависят не от
недостатка пищи, а от сезонного развития жалящих насекомых.
Таковы передвижения северного оленя на севере. Летом, когда
в лесу и тундре появляется огромное количество комаров, оводов,
слепней, олени огромными стадами начинают двигаться к северу,
на берег моря, где дующий с моря ветер отгоняет насекомых.Осенью
же, с исчезновением комара и захолоданием, олени движутся к югу.
894
Взаимоотношения между самцами и самками и отношение
к детенышам
Характер размножения, как мы видим, явился у млекопитающих
определяющим их эволюцию моментом: на трех основных этапах
эволюции размножение у них было различное.
В связи с различным способом воспроизведения меняется соот-
ветственно и отношение между самцами и самками и особенно отно-
шение матерей к детенышам.
В общем взаимоотношения между самцами и самками и дете-
нышами посят обычно сезонный характер, так как у громадного
большинства млекопитающих половая жизнь протекает в связи
с сезонами. При этом чем резче годовые сезонные изменения кли-
мата, тем больше размножение бывает приурочено к сезонам. Яв-
ление это объясняется тем, что период рождения детенышей и их
воспитания является наиболее тяжелым и опасным для матери
и самих детенышей. Как приспособление к сезонным сменам выра-
боталось у зверей рождение детенышей в такое время, когда они
могли бы быть выкормлены с наибольшим успехом. В зависимости
от продолжительности беременности, таким образом, отодвигается
момент оплодотворения. Отсюда и спаривание у большинства зве-
рей становится тоже сезонным и приурочивается к соответствен-
ным срокам. Этот период носит название течки, так как у самок он
сопровождается особыми выделениями из половых органов. К этому
же времени приурочена бывает деятельность семенпиков самцов.
Одновременно и у самцов и у самок (у разных видов различно)
проявляется усиленная деятельность особых желез, выделяющих
пахучие вещества, помогающие полам отыскивать друг друга или
действующие возбуждающим образом.
Период спаривания, или течки, сопровождается нередко особыми
играми: самцы гоняются за самками (почему у некоторых зверей
этот период называется «гоном», или «гоньбой»), издают звуки, ревут
(олени), причем самцы сражаются друг с другом из-за самок. Эта
борьба из-за самок иногда носит характер поединка насмерть.
И среди млекопитающих можно различить мопогамов и полига-
мов. Моногамов мало: таковы большинство обезьян, некоторые гры-
зуны (например, бобры, сурки), отдельные хищники, в общем не-
большое число форм. Громадное большинство млекопитающих по-
лигамно. При этом одни из них являются полигамными в собствен-
ном смысле, т. е. отдельные самцы являются обладателями целого
стада самок, как это, например, имеет место у лошадей и у многих
парнокопытных. Таких видов тоже немного. Это стадные животные.
У громадного большинства самцы и самки не образуют семей и жи-
вут до периода спаривания отдельно, нередко даже враждебно от-
носясь друг к другу (многие хищные, насекомоядные, многие гры-
зуны). В зависимости от того, какая связь между самцом и самкой,
создаются в значительной степени взаимоотношения между самцами.
В связи с этим у самцов выработались в результате различные вто-
ричные половые признаки, описанные выше.
895
Так, при случайном спаривании самцы и самки, живущие разъ-
единенно, нуждаются в приспособлениях, чтобы найти друг друга.
Это достигается тем, что в этот период животные делаются гораздо
подвижнее, дают много следов и потому легче могут быть найдены;
во-вторых, этот период сопровождается выделением половых
органов и специальных пахучих желез; в-третьих, животные из-
дают различные звуки, производят шум, привлекая этим внимание
самок,
У млекопитающих, образующих семьи, можно различать семьи
сезонные, как например, у волка, когда ой остается при самке от
спаривания до осеннего образования стай и течки, а потом драка
между самцами решает вопрос, кому достанется самка па следую-
щий сезон; или семьи бывают более или менее постоянные, как у мно-
гих обезьян, где, однако, другой самец может силой отбить себе
самку. Здесь приспособления и инстинкты у самца вырабатываются
и применительно к заботам о потомстве и к его защите.
Наконец, у стадных полигамов только сильные самцы дер*
жатся с самками, более слабые держатся в стороне или образуют
небольшие стада и пасутся отдельно от самок. Эти последние
пасутся с молодыми и с одним старым самцом. Здесь выработались
приспособления у самца для защиты стада и для его вождения,
так как самец является обычно и вожаком стада.
Детеныши однопроходных вылупляются из яиц, причем утконос
высиживает яйца в гнезде подобно птицам, а ехидна вынашивает
яйца в сумке, которая имеется на брюхе самки. Детеныши их за
неимением сосков не сосут молока, а должны слизывать выделение
молочных желез.
У сумчатых детеныши рождаются настолько несовершенно раз-
витыми, что долгое время донашиваются в сумке, где они висят
некоторое время на сосках матери, причем длинный сосок заглаты-
вается ими до желудка, тогда как края рта срастаются вокруг соска.
Дыхание совершается исключительно через нос с помощью гортани,
которая подымается до внутренних носовых отверстий. Молоко
впрыскивается матерью в рот детенышам при помощи особого мус-
кула (рис. 528).
У остальных млекопитающих детеныши рождаются более со-
вершенными, во всяком случае настолько, что могут сами сосать
молоко.
У некоторых детеныши голые, слепые и не способны к передвиже-
нию, у других опи настолько развиты, что почти сразу могут следо-
вать за матерью.
К первой группе нужно отнести насекомоядных, летучих мышей,
отчасти американских неполнозубых, грызунов, хищных, полу-
обезьян и обезьян. Ко второй — копытных.
Забота о потомстве свойственна в громадном большинстве слу-
чаев только самкам, и только у некоторых насекомоядных, грызу-
нов, хищных и обезьян заботу о детях разделяют также самцы.
Самки в большинстве случаев чрезвычайно привязаны к детям
и до того момента, пока они не научатся сами добывать себе пищу,
8%
проявляют по отношению к детям разнообразные заботы: охраняют
и защищают их от врагов, прячут в случае опасности, переносят
в случае надобности на другое место, облизывают, поедают их экс-
кременты, выискивают насекомых и т. д.
Однако как только молодые достигли известной самостоятель-
ности, всякие заботы о детях у матери быстро исчезают, и она, если
не ведет общественный образ жизни, старается от них избавиться
и начинает отгонять от себя.
Для детенышей многие млекопитающие устраивают гнезда или
логовища. Чаще всего у животных открытых местообитаний бывают
специально для этой цели вырытые в земле норы. Так поступают
однопроходные, многие сумчатые, большей частью неполнозубые,
насекомоядные и грызуны, некоторые хищники, например, барсуки
Рис. 646. Хатка бобра (Castor fiber).
(По фот. А. В. Федюшина.)
и лисицы. Другие пользуются готовыми норами, расселинами скал,
выворотами ит. п.: выдры, норки, хорьки и многие другие. Лесные
млекопитающие устраиваются в дуплах деревьев, как, например,
сопи, древесные хищники, вроде куниц, и другие. Четвертые строят
гнезда или устраиваются прямо на земле, как ежи, или на траве, как
малая мышь (Micromys), или на деревьях, как, например, белка,
некоторые обезьяны.
Сложные постройки устраивает речной бобер (Castor fiber), ко-
торый в некоторых случаях, именно когда берега водоемов недо-
статочно высоки (рис. 646), устраивает над норой деревянную по-
стройку из перегрызенных па обрубки деревьев, которую затем
обмазывает илом и грязью. Если вода в проточном водоеме падает
ниже необходимого уровня, бобры устраивают плотины из подгры-
зенных деревьев, обрубков, хвороста, водных растений и грязи.
57 Капшаров и Станчипсвий 897
VII. ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ МЛЕКОПИТАЮЩИХ
У млекопитающих мозг достигает наивысшего развития. Только
в этом классе позвоночных мы встречаем полушария мозга, снаб-
женные извилинами, наиболее многочисленными у обезьян и чело-
века. В соответствии с этим и чувственная жизнь млекопитающих
является наиболее сложной, реакции поведения наиболее совершен-
ными. Однако не все млекопитающие стоят в этом отношении на
одном уровне. В пределах класса можно установить восходящий
ряд увеличения и усложнения головного мозга, а параллельно
с этим и усложнение поведения. В мозгу у представителей различ-
ных отрядов развивается больше то тот, то другой отдел: обонятель-
ный, слуховой, теменной, двигательный, лобный. Как известно из
физиологии, эти отделы имеют различную функцию. Поэтому пове-
дение 'и психические особенности млекопитающих различных от-
рядов весьма не одинаковы. Так, например, у собаки хорошо развит
обонятельный отдел мозга. В связи с этим обоняние играет у нее
особенно большую роль. У кошки этот отдел развит гораздо
слабее — кошка пользуется преимущественно зрением. Особенно
слабо развиты обонятельный отдел и обоняние у ластоногих, без-
зубых китов, приматов.
Однако можно говорить и о чувственной жизни млекопитающих
в целом, противополагая ее чувственной жизни других классов и
отвлекаясь от различий в пределах самого класса млекопитающих.
Обоняние у большинства млекопитающих развито очень
хорошо. Опыты и наблюдения показали, что собаки, например,
не только весьма чувствительны к запахам, по обладают способ-
ностью из суммы запахов узнавать отдельные. Пользуясь обонянием,
многие животные выслеживают добычу (волки); горные козлы или
бараны издалека, пользуясь ветром, по запаху узнают присутствие
человека; дикие свиньи по запаху узнают о присутствии под землей
съедобных корней и т. д.
Слух у млекопитающих также играет большую роль. Слух
изучался у собаки методом условных рефлексов. Уже различие
в четверть тона давало при этом положительный эффект. Зашивание
глаз не влияло на успех опыта, разрушение улитки в слуховом ор-
гане разрушало и слуховую привычку (условный рефлекс). Развит
слух и у кошек. Лошади выучиваются по тому или иному звуковому
сигналу производить те или иные эволюции и т. д. Многие млеко-
питающие по слуху находят пищу, другие пользуются слухом,
чтобы избегнуть опасности.
Зрение млекопитающих подвергалось многочисленным и
точным исследованиям. Согласия здесь не достигнуто по вопросу
о цветном зрении. Одни авторы находят, что некоторые млекопи-
тающие (кошки) различают цвета не на основе длины волны, а на
основе их яркости. По этому взгляду кошки слепы на цвета, вйдят
все в одном цвете, ио различают степень яркости. Другие' исследо-
ватели признают у млекопитающих (обезьян) способность раз-
личать и цвета как таковые. Во всяком случае на основе-ли цвет-
898
пости, на основе ли яркости, но цвета млекопитающие хорошо раз»
личают. Различают иногда самые тонкие оттенки; шимпанзе нау»
чается выбирать среди комбинаций несколько цветов. Многие мле-
копитающие различают также простые рисунки.
В к/у с у млекопитающих развит хорошо. Хорошо развит^
и кожная чувствительность (осязание), а также так назы-
ваемая органическая и кинестетическая чув-
ствительность. Для осязания па теле млекопитающих развива-
ются особые волосы, вибриссы. Высоко специальные, чувстви-
тельные окончания существуют в коже и мускулах. Ряд экспе-
риментов показал, что многие» млекопитающие при образовании
двигательных привычек руководятся мышечным чувством, в го-
раздо большей «мере, нежели зрением или обонянием. Крысы,
выучившиеся находить себе дорогу в лабиринте, сбивались и дела-
ли неправильные повороты, если лабиринт удлинялся или укора-
чивался, а именно — продолжали итти вперед и упирались в стенку,
если лабиринт укорачивался, и делали преждевременные повороты,
если лабиринт удлинялся. Одним словом, органы ощущений у мле-
копитающих развиты очень хорошо, и млекопитающие получают из
внешнего мира большое количество впечатлений, которые ассоци-
ируются; таким образом, создается привычка чувственная (при-
вычка к определенного рода стимулам) или двигательная (привычка
к определенного рода движениям).
В способности к образованию привычек млеко-
питающие превосходят всех остальных позвоночных.
У большинства исследованных в условиях эксперимента млеко-
питающих привычка образуется весьма быстро: у крыс, собак, ко-
шек и других она вырабатывается в совершенстве в течение 3—4
десятков опытов. В сущности эти привычки образуются почти с пер-
вого раза, а потом идет уже лишь усовершенствование в выполнении
действия. Такая быстрота в научении, конечно, играет большущ
роль в жизни млекопитающих, является для них могучим
оружием в борьбе за существование, помогая быстро избегать
всякой опасности, с которой животное однажды столкнулось,
находить быстро пути к выходу из положения. Такая способ-
ность является более мощным оружием, нежели физические ору-
дия защиты.
Можно считать установленным, что некоторые млекопитающие
способны не только реагировать на действующие стимулы, но и На
стимулй, переставшие действовать, на стимулы,от которых остался
только след в мозгу; многие млекопитающие способны, таким обра-
зом, руководиться, действовать под влиянием «образа», «представле-
ния». Так, например, собака после некоторых попыток переплыть
реку, по пе желая этого делать, например, из-за холодной воды,
опрометью бросается к мосту, находящемуся далеко и отсюда
видному, чтобы перейти реку по мосту, по которому она раньше
переходила реку. Или, например, перед собакой, находящейся
в комнате, выбрасывают из окна лакомый кусок. Упавшего куска
собаке не видно. Но она, взволнованно повертясь у окна, кидается
57* 899
к двери па противоположной стороне комнаты и бежит кругом вниз
к выброшенному куску.
Особенного развития достигает мозг у человекообразных обезь-
ян. И психические особенности оранга или шимпанзе оказываются
очень высокими, приближающимися к таковым человека. Шимпанзе
способны пользоваться орудием для достижения цели. Они в со-
стоянии сами приготовить себе это орудие, например, сломать прут
и оборвать на нем листья, чтобы воспользоваться им для продвига-
ния лакомого кускаг лежащего за решеткой. Они оказались способ-
ными собрать в одно место и поставить друг на друга 3 ящика,
чтобы достать висящие под потолком бананы. Делали они это со-
обща, причем одна об^ьяна влезала на ящик, другая поддерживала,
остальные располагались вокруг в ожидании. Если ящики были
с песком или камнями, шимпанзе опорожняли их, а затем тащили
на место.
Однако между человеком и шимпанзе все же существует раз-
личие в том отношении, что только человек способен к членораз-
дельной речи и к образованию общих понятий, необходимых для ло-
гического мышления. Поэтому той сложности, которую имеют поступ-
ки человека, они не достигают даже у человекообразных обезьян.
Высокоразвитая высшая нервная деятельность, способность
к быстрому образованию привычек, т. е. к быстрому научению из
личного опыта, является для млекопитающих могучим оружием
в борьбе за жизнь.
VIII. ЭКОНОМИЧЕСКОЕ ЗНАЧЕНИЕ МЛЕКОПИТАЮЩИХ
Экономическое значение птиц и млекопитающих, как и прочих
животных, выражается в пользе и вреде, которые приносят они
человеку и его хозяйству.
Форма использования млекопитающих чрезвычайно разнооб-
разна. Во-первых, они доставляют человеку пищу в виде мяса,
жиров и молока; во-вторых, их шкуры, меха и шерсть идут на
изготовление одежды и других хозяйственных предметов, в-третьих,
из многих млекопитающих добываются вещества технического
применения и лекарственные вещества, далее млекопитающие до-
ставляют человеку рабочую силу в виде упряжпых, вьючных
и верховых рабочих животных, и, наконец, опи являются пред-
метом спорта и эстетического наслаждения. Кроме того польза от
млекопитающих может косвенно выражаться в том, что они истреб-
ляют вредных нам животных или являются пищей для полезных.
Вред, приносимый млекопитающими, выражается прежде всего
в той опасности, которую представляют некоторые из них непо-
средственно жизни’человека; во-вторых, многие из них являются
разносчиками опасных для человека болезней; в-третьих, они’
могут быть опасны прямо или через паразитов для домашних жи-
вотных или диких животных, которых эксплоатирует человек;
в-четвертых, млекопитающие вредят тем растениям, которые раз-
водятся в хозяйственных целях человеком, или запасам хозяй-
900
стненпых продуктов и предметам домашнего обихода. Многие из
млекопитающих являются одновременно и полезными и вред-
ными, значение других меняется в зависимости от экологических
условий среды, от географического положения местности или от
высоты уровня сельскохозяйственной культуры. Изучение жи-
вотных вообще и млекопитающих, в частности, все более и более
приводит к той идее, что безусловно вредных млекопитающих
очень мало и что с ростом культуры все большее и большее число
животпых находит и будет находить то или иное применение
в жизни человека. Отсюда вытекает необходимость сохранения
но возможности всех за исключением немногих безусловно вред-
ных представителей' Класса, ибо раз исчезнувшие с лица земли
животные уже пе. могут быть вновь воспроизведены, между тем как
возможности использования диких и даже в том числе некоторых
вредных животных в будущем представляйся бесконечными.
По характеру использования всех млекопитающих можно раз-
делить на домашних и диких.
1. Домашние млекопитающие, их история и экология
Домашними мы называем тех млекопитающих, которые, будучи
приручены человеком, разводятся им ради тех или иных целей,
живут под попечением человека и размножаются в прирученном
(домашнем) состоянии под его контролем. Сюда относятся прежде
всего те млекопитающие, которые приручены были в глубокой
древности, подверглись длительному воздействию человека как
со стороны искусственного содержания, так и отбора, образовав
значительное число пород сообразно с темп требованиями, которые
предъявлял к ним человек. Таковы собаки, крупный
рогатый скот, овцы, свиньи, лошади, кро-
лики..
Другая группа домашних млекопитающих хотя и размножается
под непосредственным контролем человека, не образовала еще
пород, так как не подверглась еще значительному отбору. Сюда
относятся северные ‘олени, яки, верблюды, бу й-
во л.ы и некоторые другие.
Третья группа млекопитающих представлена многочисленными
видами, которые с различными целями приручены человеком
сравнительно недавно и хотя размножаются в неволе, но еще не
могут быть названы домашними, так как не имеют массового раз-
ведения. Сюда относятся разные разводимые с промысловыми
целями пушные звери, некоторые охотничьи звери, а также млеко-
питающие, которые шока разводятся лишу, ради нужд зоологиче-
ских садов и зверинцев. Таковы маралы, ла ни, лисицы,
песцы, антилопы, зебры и мгюгие другие. В отличие
от домашних животных их можно назвать животными приручен-
ными.
Все эти группы представляют лестницу разных ступеней при-
ручения и одомашнения, связывающую диких животных с настоя-
901
гцими домашними животными, которая наглядно показывает, как
постепенно человек распространял свое могущественное влияние
на диких животных, подчинял их себе и видоизменял сообразно
со своими целями.
Приручепие и одомашнение началось в- глубокой древности,
на заре человеческой культуры, и продолжается по сие время.
Детальное исследование вопроса о происхождении домашних
животных показало, что одомашнение производилось разными
народами и в разное время и шло в известной закономерности.
Всегда материалом для приручепия и одомашнения служили те
дикие -животные, которые водились в данной местности; самое
приручение разных видов имело известную последовательность.
Восстановление истории различных пород, их происхождения
от диких предков, места и времени их возникновения представляет
большие трудности.% Однако, пользуясь данными всестороннего
изучения ныне живущих пород каждого вида домашних живот-
ных, сравнивая их с дико живущими видами, прослеживая путем
изучения ископаемых остатков историю изменения пород домаш-
них животных в историческое и доисторическое время, пользуясь
далее методом скрещивания и данными археологии и этнографии,
удается подойти к выяснению даже - очець трудных вопросов
прохождения наиболее древних домашних пород.
Особенно много дали для выяснения вопросов происхождения
пород домашних животных сравнительно недавние раскопки в Аиау,
близ Ашхабада в Туркменистане.
Собака. Домашняя собака представлена чрезвычайно много-
численными породами, которые иногда очень резко отличаются
друг от друга. Достаточно указать па крупных догов, сеп-бернаров,
быстроногих поджарых борзых, своеобразных бульдогов, мопсов,
кривоногих такс, голых египетских собак и маленьких болонок
и японских' собачек. Все эти породы, однако, оказывается сравни-
тельно недавно выведены человеком в определенными целями. По
своему значению их можно разделить на Собак охотничьих,
сторожевых и комнатных.
Произошли все породу домашних собак от нескольких диких,
собак (род Canis).
Собака была первым домашним животным первобытного чело-
века, занимавшегося охотой и ^рыбной ловлей. На стоянках чело-
века ранцего неолитического периода Европы мы находим остатки
этого первого домашнего животного. Что собака была домашним
животным, т. е., что она жила с человеком, размножаясь в при-
рученном состоянии, свидетельствуют следующие данные. Скелеты
собаки находятся в местах обитания неолитического человека
(человека «нового каменного века», употреблявшего шлифованные
камни) со скелетами других животных, которых человек добывал
охотой в целях пропитания. Но в то время как черепа и кости,
содержащие мозг, у диких животных всегда оказываются раз-
дробленными (для извлечения из них мозга), черепа и кости собак
всегда целы. Самый скелет собаки носит следы одомашнения, так
902
как кости оказываются более гладкими’по сравнению с таковыми
диких животных; наконец на костях диких животных, употребляв-
шихся в пищу неолитическим человеком, нередко можно видеть
следы зубов собаки, из чего ясно следует, что и тогда собака жила
за счет остатков от стола человека. Это были средней величины
собаки со стоячими ушами и повислым непушистым хвостом, низ-
ким лбом и средней длины мордой. Наиболее древняя неолити-
ческого времени, собака, найденная в России, получила название
собаки Путятина (Canis poutiatini); к ней близка более
поздняя торфяная собака (Canis palustris) свайных построек.
Рис. 647. Шпиц
(Из Брэма.)
Детальное изучение торфяной собаки показало, что опа близка
к современной породе небольших домашних собак со стоячими
ушами, именно так называемым шпицам ненцев и лопарей.
Это небольшие сторожевые и отчасти ездовые собаки; их потомки —
комнатные шпицы белого, или черного цвета, с пушистым загну-
тым крючком хвостом (рис. 647).
От типичных шпицев произошли позднее пинчеры, терье-
ры, китайские и японские комнатные собачки.
К собаке Путятина очень близки австралийский динго,
вероятно уже одичавший в Австралии (рис. 678), а также полудикие
восточноазиатские собаки парна, похожие на динго и на Шпица.
903
Новейшие исследования даксг основания предполагать, что эти
древние собаки произошли от небольших вымерших волков юго-
восточной Европы (Canis ferns).
Дальнейшая эволюция торфяной собаки приводит к Canis
intermedins конца неолита и бронзового века. Эта собака средней
величины дала разные породы гончих, а также принимала участие
в образовании пород легавых собак.
Крупная собака бронзового века (Canis matris optimae) может
быть названа бронзовой овчаркой, так как примитивные
европейские овчарки сходны с ней по типу.
Рис. 648. Украинская овчарка
(Аскания-Нова, фот. Озерова.)
Овчарки представлены в настоящее время рядом пород. Больше других
по своему внешнему виду’напоминает своего дикого преДка немецкая
овчарка. От овчарок произошла английская колли, от них же
ведут свою родословную и пудели. Значение овчарок для первобытных
пастушеских племен было так велико, что повело к обоготворению их на ряду
с огнем.
От скрещивания бронзовой овчарки с северными волками получились
многочисленные породы крупных пастушеских собак, в том числе и у к р а-
инские овчарки (рис. 648), а также английские собаки (сен-бер-
нары) и доги, потомками которых являются различные боевые собаки
(медвежьи собаки, боксеры, бульдоги и др.). *
Вероятно от другого дикого волка (Canis lupaster) берут начало древне-
египетские борзые собаки:- стройные, высоконогие со стоячими ушами»
короткой шерстью й первоначально с длинным опущенным хвостом. Отли-
чаясь быстрым бегом, они-служили для травли зверей. От них произошли,
с одной стороны, более поздние египетские борзые с загнутым крючком
хвостом, с другой стороны—разные породы европейских борзых й, наконец,
таксы — маленькие собаки на коротких ‘кривых ногах, как нельзя лучше
приспособленных для лазания в норах.
Есть данные предполагать, что и легавые собаки выведены были
204
уже в Египте. Ив многочисленных легавых собак мы упомянем н е м е ц-
них легавых, сетеров и пойнтеров (рис. 649). <
Северные крупные борзые возникли от скрещивания южных борзых с се-
верными волками. Ирландский волкодав, шотландская
оленья собака относятся к числу их потомков. Наконец большое ко-
личество крови северных волков имеется у полярных собак и лаек.
Лайки — незаменимые помощники северных промышленников. Они упо-
требляются там одновременно в качестве упряжных животных. Не исклю-
чено участие в образовании домашней собаки шакала.
Рис. 649. Пойнтеры на стойке.
Домашняя кошка. Домашняя кошка разводится с целью уни-
чтожения крыс и мышей и как комнатное животное. Домашние
кошки и в особенности одичавшие являются вредителями птице-
водства и уничтожают массу диких птиц и их птенцов, а также зай-
чат, но в то же время истребляют также мелких полевых грызунов.
Приручение кошки произошло в .Африке. Ее родоначальником
является нубийская буланая кошка (Felis mani-
culata), распространенная в Нубии, Судане, Абиссинии, Цен-
тральной Африке и Палестине. Судя по изображениям па египет-
ских памятниках, домашняя кошка была приручена более чем
6000 лет назад и почиталась в древнем'Египте животным священ-
ным в связи с той ролью, которую играла она в истреблении мелких
грызунов, уничтожавших зерновые хлеба и запасы египетских
хлебопашцев. В Европу кошка завезена была поздно.
Крупный рогатый скот.. К крупному рогатому скоту относятся
представители подсемейства быков (Bovinae), семейства п о-
дорогих животных.
©05
В домашнем состоянии живут многочисленные породы евро-
пейских быков. Предками этих пород домашних быков были
дикие быки. Из них только один известен нам не только по
ископаемым остаткам, но и по письменному свидетельству лиц,
видевших его. Это — тур (Bos primigenius). Он был широко рас-
пространен еще в исторические времена по лесостепи и степи,
и самец, по свидетельству Герберштейна, имел черную окраску
с белой полосой вдоль спины. Последний тур (самка) был убит
в 1627 г. Имея широкое распространение, тур образовал ряд ме-
стных рас (рис. 650). От тура произошли степные породы домаш-
него скота, шотландский скот и скот североевропейских низменно-
стей: голландский, остфрисландский, нормандский, фламандский,
Рис. 650. Тур (Bos primigenius).
(С древней картины, из Абеля.)
а также и широколобый скот Германии, Австрии, симменталы
и др.', скот южных низменностей получился однако путем скре-
щивания с короткорогим лесным скотом (В. brachyceros). Этот
последний отличался от тура небольшим ростом, более корот-
кими и слабыми костными отростками ротов и рядом других
признаков. От этого «лесного тура» произошли и примитивный
скот свайнйх построек и многочисленные породы рогатого скота
Европы. Таковы культурные породы альпийского скота: швицкий,
альгаузский, обертальский и др., туземный скот Центральной
и значительной части Восточной Европы, в том числе местный скот
значительной части СССР.
Что касается восточных пород крупного рогатого скота — кал-
мыцкого, киргизского скота и др., сходных с ним местных, а также
многочисленных пород горбатого, скота или зебу, — то они могут
быть выведены от дикого индийского быка времен четвертичного
периода Bos nomadicus. Черепа этой формы, с одной стороны,
приближаются к Bos primigenius, с другой — к роду бантенга,
S06
гаяла, гаура — к роду Bibos. Эти последиие являются тоже родо-
начальниками своих пород: от бантенга (Bibos banteng) произошел
балийский скот (о. Вали), от гаяла ( Bibos frontalis) и гаура
(Bibos gaurus) выведены домашние формы в западной и северо-
восточной Индии.
Породы крупного рогатого скота по продуктивности могут быть
разделены иа примитивные, или п р о с т.ы е, т. е. такие,
которые не подвергались еще племенной работе, и культу р-
н ы е, или заводские, т. е. такие, над которыми уже велась
племенная работа, приведшая к выделению более продуктивных
популяций. Мы остаповимся только на немногих культурных и тех
из примитивных пород, над которыми в виду их перспективности
началась заводская племенная работа.
Рис. 651. Голландская корова.,
По основному направлению продуктивности все породы могут
быть разделены на молочный скот, скот мясо-молочный, мясо-
рабочий, мясной и рабочий.
Молочный скот. Из иностранных пород мы назовем лишь немногие породы.
Голландская порода (рис. 651), выведенная в окрестностях Амстер-
дама. В СССР принят к разведению в ряде районов в Северном ' крае, в
Западносибирском, Восточносибирском, на Дальнем Востоке и вокруг
крупных городов.
Фрисландская порода южной Голландии.
Джерсейская порода выведена - на острове Джерсей, близ берега
Франции в Английском канале и принадлежит к мелким молочным поро-
дам, дающим молоко с высоким содержанием жира.
Ангельнская порода выведена на полуострове этого названия
и Шлеявиг-Голштинии.
Из русских пород к голландским близка холмогорская порода,
которая получилась путем улучшения местного скота привозными голланд-
ским и производителями. Разводятся преимущественно на нижнем течении
С<чн»р|юй Двины. Признана плановой породой для всего Северного края,
дли большой части Ленинградской области, части Московской области и
Горьковского края.
Нрославская порода выведена крестьянами б. Ярославской гу-
бернии путем улучшенного содержания и отбора из некультурной вели-
907
корусской старой породы, или первичнолесного скота. Ярос-
лавская порода признана плановой для части Московской, Ленинградской,
Ивановской и некоторых других областей.
На Юге Европейской части СССР, особенно на Украине в Харьков-
щине и Мелитопольщине, образовалась из вывезенного немецкими коло-
нистами скота очень интересная и ценная порода красного молочного
скота, которая известна под названием красного немецкого
(рис. 652). Признана плановой для степной Украины и Западной Сибири.
Белоголовая красная полесская очень близка к красно-
немецкой, так как произошла путем скрещивания местного скота с выве-
зенным на Украину голландским и немецким. Продуктивность при одина-
ковом росте с предыдущей несколько ниже.
Рис. 652. Корова красной немецкой породы.
Горбатовский скот выведен в б. Горбатовском у. б. Нижегород-
ской губ. путем метизации местного приокского скота со скотом, вывезенным
ив Тироля. В Горьковском крае и Ивановской промышленной области.
Тагильская порода выведена в районе Нижнего Тагила путем скре-
щивания местного скота с холмогорской и голландской породами.
В Уральской области и Башкирской республике.
Сибирский скот отличается удивительной приспособленностью
к климату, покрывается на зиму длинной шерстью, вымя покрыто воло-
сами; дает жирное молоко. Плановая порода для значительной части
Сибири.
Кавказский скот является чисто местной породой, чрезвы-
чайно приспособленной к условиям горной жизни; очень мелкий с живым
весом коров 160 кг. 7
Мясо-молочный скот. Ш в и ц к а я порода выведена в Швейцарии
в кантоне Швиц из местного горного скота без всякого участия других пород.
Плановая порода для Смоленской обл., части Московской обл., для Северного
Кавказа и др. .
Симментальская порода в одинаковой степени может считаться
как молочной, так и рабочей и мясной. Разводится во многих местах Швей-
царии, Германии, Австрии, Польши и СССР.
Плановая порода для ЦЧО, для Средней и Нижней Волги, для части
Сибири, Смоленской области, Горьковского края и Украины (рис. 653).
908
Бесту ж*е’в с к а я порода. Выведена была путем скрещивания
местного скота с шортгорнской,1 симментальской и голландской породой в на-
чале XIX века в Ульяновском районе (б. Симбирская губ.). Плановая по-
рода для Среднего Поволжья, Татарской и Башкирской АССР.
Рис. 653. Бык симментальской породы.
Мясной скот. Шортгорнская порода. Благодаря своей ско-
роспелости, быстроте откладывания жира, тонкости костей и качества мяса
стоит настолько высоко, что в.настоящее время разводится по всему свету.
Выведена она в графстве Дюргэймском из ввезенного в Англию голландского
скота. Ввозится в СССР для разведения в чистом виде и для скрещивания
с местным скотом.
Рис. 654. Корова сероукраинской породы
(Аскания-Нова, фот. Озерова.)
909
Герефордская порода выведена также в Англии, причем
по своим мясным качествам и скороспелости она не уступает шортгорнам,
отличаясь большей выносливостью, но меньшей молочностью; выведена из
местного скота путем отбора в середине XIX столетия. В последнее время
ввозится в СССР.
Мясо-рабочий скот. Калмыцкая порода является азиатской
породой, сильно отличается от европейских пород, напоминая зебу Индии;
распространена в юго-восточной части Европейской СССР, начиная от Кур-
ской области до Казакстана и Туркменистана.
. Одна из лучших пород в СССР по весу и качеству мяса. Плановая по-
рода для юго-востока.
Киргизская порода является лучшей мясной породой в
СССР, так как обладает при менйшем росте и весе большей молочностью
й лучшего качества мясом по сравнению с калмыцким скотом. Очень ценна
своей выносливостью и неприхотливостью.. Путем метизации с мясными
породами возможно получить исключительно ценных жйвотных.
Рис. 655. Индейский зебу, бык
(Аснания-Нова.)
Рабочий скот. Сероукраинский скот является одной из
примитивнейших степных пород, особенно близко стоящей к дикому туру.
Очень позднеспелый, так как телята обычно воспитывались под матками,
а матки почти круглый год ходят на подножном корму в степи. Прекрасный
рабочий скот (рис. 654).
Зебу (Bos indicus) представляет вероятно особый вид, происшедший
от дикого Bos nomadicus Индии. В виде многочисленных пород распростра-
нен в Южной Азии от Малой Азии и Аравии до Японских островов и в' Аф-
рике. Замечателен своей приспособленностью к очень тощим пастбищам,
своей невосприимчивостью к пироплазмозу и прекрасными мясными и молоч-
ными качествами при большом содержании жира в молоке (до 6%).
Очень интересных результатов можно ожидать от гибридизации его
с культурными породами (рис. 655).
Буй в_оJiJ^iibalu&--bubahis)-OTmriaeTCff OT крупного рогатого
скотатбм' что^дзещ>-хорошО"яереносит”сырейижаркий климат
и прекрйПЖб^приспособлен к жизни в болотистых местностях;
Дикие яки (Poephagus grunniens) обитают по* горам и
910
плоскогорьям Центральной Азии. Там же живут и их домашние
прямые потомки. *
Яки — типичные обитатели горных стран с суровым конти-
нентальным климатом. Они имеют приземистее тело, крепкие
прямые ноги, широко раздвоенные копыта и одеты чрезвычайно
густой шерстью черного цвета. Используются в качестве верховых
и вьючных животных, дают хорошее молоко и прекрасную шерсть.
Приручение яка произошло в глубокой древности.
Мелкий-рогатый скот. Домашние овцы. Овцы, доволь-
ствуясь тощими и сухими пастбищами, хорошо переносят холод
и жару и значительные передвижения в поисках корма и водопоев,
а потому являются незаменимыми животными при экстенсивных хо-
зяйствах с большими пространствами незапаханных сухих и тощих
земель.
У нас в Союзе в таких местах, как Казахстан, Узбекистан, Турк-
менистан с полупустынями и пустынями, юго-восток Европейской
части СССР, Кавказ, Крым, овцеводство будет долго играть очень
важную роль в сельском хозяйстве. Но и в условиях интенсивного
хозяйства овцеводство играет и будет играть большую роль, так
как, во-первых, многие породы овец дают чрезвычайно ценную
шерсть, овчину и смушек, будучи очень продуктивны, используя
в то же время незначительное количество кормов на пастбищах,
довольствуясь тем, что остается от крупного рогатого скота.
Все породы овец произошли не меньше как от трех видов ди-
ких баранов: европейского и малоазиатского муфлона, аркала и ар-
гали; вероятно, также влияние на образование домашних пород
североафриканского гривистого барана.
От живущего иа юге Европы на островах Средиземного моря
и Малой Азии муфлона (Ovis musimon) произошло несколько
пород с слаборазвитыми рогами и коротким хвостом, как например
русские короткохвостые овцы, вересковые и др., а также безрогие
молочные овцы Европы. Приручение муфлонов по имеющимся
данным началось не более 4000 лет назад, между тем как уже
в торфяных отложениях неолита мы имеем остатки домашних
овец. Есть основания думать, что предками торфяной овцы были
малоазиатские муфлоны, если только торфяная овца не средне-
азиатского происхождения. По данным раскопок в Анау прируче-
ние овец произошло в Средней Азии около 9000 лет назад, причем
материалом для приручения был степной баран, или
а р к а л - (Ovis arkal, рис. 656). On и сейчас живет в полупусты-
нях и .степях Туркменистана большими стадами до 100 и больше
голов. Можно проследить, как из аркала получились породы,
близкпе к торфяным овцам Европы.
Потомки торфяных овец сейчас разводятся в Швейцарии — это
небольшого роста горные овцы с маленькими торчащими вверх
рогами у баранов.
Третьим видом диких баранов, от которых произошел ряд
пород овец, отличающихся своими сильно изогнутыми наружу
рогами, являются аргали (Ovis ammon), крупные животные
911
темносерой масти с мощными завитыми наружу рогами — бараны,
живущие в горах Центрально!» Азии. '
Сказать с уверенностью, каким породам домашних овец дал
начало тот пли иной из перечисленных диких видов, пока* еще
трудно. Но три центра образования овец уже ясны.
Рис. 656. Аркал (Ovis arkal).
(Из Бихнера.)
Наконец гривистому барану
Северной Африки (Ammotragus
tragelaphus) нужно приписать
образование некоторых африкан-
ских домашних пород с длин-
ными рогами и гривой на шее.
Длительным содержанием в
певоле, продолжительным отбо-
ром и скрещиванием домашние
овцы в большой степени изме-
нены по сравнению с дикими
предками, образовав громадное
количество пород, из которых
нами упомянуты будут глав-
нейшие.
По характеру направлений
породы овец могут быть разде-
лены на шерстных овец, овчин-
ных, мясных, 'смушковых и мо-
лочных, а также ряд комби-
нированных пород, например
мясо-шерстных или молочно-смушковых.
Шерстные овцы. К чисто шерстным овцам, дающим очень ценную тонко-
рунную шерсть, относятся разные типы мериносов.
Многочисленные породы мериносовых овец вывезены из разводившихся
еще до нашей эры в Средиземноморской области овец, отличавшихся своими
сильно закрученными рогами и особенностью своей шерсти без ости с тон-
кими, правильно извитыми волосками пуха, пригодной для изготовления
самых тонких шерстяных тканей. До XVIII в. мериносы разводились почти
исключительно в Испании, откуда с XVIII в. распространяются по всей
Европе. Вывезенные из Испании мериносы быстро благодаря отбору обра-
зуют ряд пород, которые обыкновенно различаются по трем типам: э л е к т о-
р а л ь — первый выведенный в Саксонии тип мериносов с тонкой шерстью
и нежным сложением; негретти с большим развитием складок кожи,
с сильным и’грубым сложением, развитым костяком и грубой шерстью и
камвольная мериносовая овца.
Последний тип представлен породами: без складок—с уасонеи ша-
т и л ь о н е, складчатым — рамбулье и русской — маваевски-
м и камвольными овцами.
Рамбулье прекрасно акклиматизировались в степях УССР. Они обла-
дают помимо прекрасной шерсти хорошими мясными формами и большой
продуктивностью шерсти. Рекордный настриг барана в Аскании-Нова —
16 кг. Однако рамбулье не скороспелы (рис. 657).
Прекосы выведены во Франции с целью придания мериносам боль-
шей скороспелости путем скрещивания с лейстеровскими английскими длинно-
шерстными овцами. В настоящее время с успехом ввозятся в СССР (рис. 658).
Цигайские овцы распространены у нас на юге, главным обра-
зом в Крыму. Обладают хорошей шерстью, хорошими мясными качествами
и неплохими овчинами.
912
Рис. 657. Мериносовая овца породы рамбулье, 2-х лет.
(Аскания-Нова.)
Овчинные овцы. Сюда относятся овцы северной части Европы и Запад-
ной Сибири, из которых особенно славится романовская порода,
выведенная по берегу р. Волги в Ярославской обл. Благодаря высоким
качествам овчины сероголубого цвета, многоплодности и большой мо-
лочности является очень ценной плановой породой, идущей для улучшения
местных овец при овчинном направлении овцеводства (рис. 659).
Мясо-шерстные овцы. Интенсификация овцеводства в таких странах, как
Англия, привела к необходимости комбинированного направления, чтобы
получать с овец не только максимум шерсти, но и максимум мяса. Благо-
даря соответствующей селекции выведены были короткошерстные скоро-
Рис. 658. Баран прекос, 2-х лет.
(Аскания-Нова.)
58 Кашкаров и Станчинсний
913
Рис. 659. Баран романовской породы
(Аснания-Нова.)
спелые овцы саусдаунской породы с- прекрасными мясными ка-
чествами; от них же получены гемпширские овцы, белые с темно-
коричневой мордой и ногами, очень скороспелые. Ввозятся в СССР, но тре-
бовательны к кормам и уходу, так что необходима их специальная акклима-
тизация (рис. 660). К этой же группе овец относятся шропшир ы.
Из длинношерстных английских овец назовем очень крупных с богатым
настригом и очень скороспелых линкольнских овец. Произошли
Рис. 660. Баран гемпширской породы
(Аскания-Нова.)
914
Рис. 661. Баран линкольнской породы
(Аскания-Нова.)
от скрещивания местной овцы восточной Англии с лейстерской по-
родой. Линкольны ввозятся в СССР, где однако требуют специальной аккли-
матизации (рис. 661).
Р о м н и - м а р ш несколько меньше линкольнов, но хорошо откар-
мливаются, так же скороспелы и дают прекрасную баранину.
Из наших мясошерстных пород овец назовем длиннохвостых
жирнохвостых овец, куда относятся волошские овцы,
довольно крупные с длинными хвостами овцы, и азиатского происхождения
Рис. 662. Овца каракульской породы
(Аскания-Нова.)
58*
915
длиннохвостых тощехвостых овец, широко распространен-
ных по территории СССР, где они являются в большинстве случаев универ-
сальными овцами; дают мясо, шерсть, овчину и * смушку; они образуют
ряд отродий. Из них назовем серую сокольскую овцу и черную
решетиловскую, являющихся уже смушково-молочными овцами.
Лучшими овцами этого направления является бухарская каракуль-
ская овца (рис. 662), дающая ценную каракульчу (шкурка
недоношенного ягненка), каракуль (шкурка новорожденного ягненка)
и молоко. Путем метизации с каракульской овцой проводится плановое улуч-
шение местных южнорусских и украинских овец.
Наконец. к мясо-сальным породам овец относятся курдючные овцы.
Курдючные овцы отличаются своими жировыми отложениями по
бокам хвоста и на крестце — «курдюком», в числе нескольких пород распро-
странены в Африке и в Азии и у нас в Средней Азии и на юге Европейской
части СССР. (Рис. 663) Курдючные овцы отличаются крепкими ногами, даю-
Рис. 663. Баран гиссарской породы
(Аскания-Нова.)
щими им возможность далеко перекочевывать в поисках за кормом; их курдюк
служит в качестве кладовой питательных веществ на время зимних голодовок,
когда овцам приходится довольствоваться скудной сухой травой или добы-
вать себе корм из-под снега. Достоинства шерсти низки, но как мясо-саль-
ные овцы они очень ценны. Гиссарское отродье достигает 200 кг
веса при 80 кг чистого жира.
Домашние козы. Экономическое значение коз, которые раз-
водятся человеком с глубокой древности, стоит в связи главдым
образом с нетребовательностью их к содержанию и пище и вы-
носливостью к климату. Коза дает много хорошего, богатого са-
харом молока, мясо козлят употребляется в пищу и близко по своим
качествам к мясу ягнят, кожа высоко ценится при выделке, а длинно-
шерстные козы дают хорошую шерсть. Доставляя все это при ни-
чтожной затрате труда и капитала, коза совершенно справедливо
называлась «коровой бедняка». Довольствуясь самой грубой и скуд-
916
ной пищей: листьями и молодыми побегами древесной раститель-
ности и сорными травами, коза может жить в условиях, при ко-
торых не могут жить другие домашние животные, а, будучи гор-
ным животным, позволяет использовать крутые склоны гор. Благо-
Рис. 664. Винторогий козел (Capra falconneri).
(Из Бихнера.)
даря .козе человек мог поселиться даже в таких суровых условиях
климата, какими являются вершины высоких гор.
Родина домашних коз — Азия. Хотя в Европе живут дикие
козы, именно козероги (Capra ibex), но можно считать уста-
новленным, что домашние козы произошли не от них, а от распро-
страненного в Юго-западной Азии и на Кавказе б е з о а р о-
917
в о г о козла (Capra aegargus) (рис. 616) и от европейского
вымершего недавно (в четвертичное время) галицийского
козла (Capra prisca). Домашняя коза новокаменного века,
быть может подобно другим торфяным домашним животным, при-
ведена была перекочевавшими с востока первобытными племе-
нами.
Другим, диким родоначальником домашней козы является
марку р, винторогий козел (Capra f alconneri) (рис. 664),
житель Гималайских гор. Он отличается завитыми рогами и длин-
ной шерстью на передней части тела. От него произошли длинно-
шерстные породы коз, дающих ценные шкуры и шерсть, ангор-
ские и кашмирские козы.
Рис. 665. Кабан (Sus scrofa).
Домашние свиньи. Приручение свиньи произошло очень давно,
так как в раскопках, относящихся к позднему неолиту, уже встре-
чаются скелеты домашней свиньи. Правда, свинья появилась много
позднее собаки, крупного рогатого скота, овцы и козы.
» Родоначальником пород домашних свиней нужно считать не
только европейского кабана с его подвидами (Sus scrofa) (рис. 665),
но и азиатских свиней (S. vittatus с подвидами S. leu-
comystax), а также недавно установленный вид: средиземно-
морскую свинью (S. mediterraneus) промежуточного харак-
тера между европейским и азиатским типами.
Самая древняя европейская домашняя свинья — свайных по-
строек, «торфяная свинья» неолитического века — оказывается
разного происхождения. На юге Европы (в Швейцарии) была
приручена средиземноморская свинья. Эта же порода свиней была
918
распространена в древней Греции и Риме. Остатками этого древ-
него корня являются различные «романские» породы свиней, к ко-
торым, однако, была примешана кровь восточной свиньи. Восточные
свиньи приручены были в глубокой древности, они отличаются
коротким и высоким черепом, расходящимися кпереди рядами
коренных зубов, легко откармливаются, но нежны и неплодовиты.
Дикий европейский кабан был приручен позднее, но все же
является «торфяной свиньей» каменного века. От него произошли
многочисленные малокультурные «простые» свиньи. Будучи менее
плодовиты и не так производительны, европейские породы отли-
чаются зато невзыскательностью к содержанию и климату и дают
более высокого качества мясо и сало. Различают породы больших
длинноухих европейских свиней от малых короткоухих.
Рис. 666. Матка крупной йоркширской породы
(Аскания-Нова.)
Так как недостатки азиатских свиней покрываются достоинствами евро-
пейских свиней, то следовало ожидать, что путем умелого скрещивания этих
двух пород можно получить породы, которые при плодовитости, невзыска-
тельности к климату и содержанию, высокой доброкачественности мяса и
сала европейских свиней отличались бы в то же самое время скороспелостью
и хорошей способностью к откорму китайской свиньи. Путем такого скрещи-
вания и выведены в действительности почти все породы культурных свиней,
из которых особенно славятся английские свиньи. Среди этих последних
различают: белые породы свиней — мелких, средних и крупных, среди
которых наибольшей известностью пользуются иоркш иры (рис. 666)
и пестрые породы, мелкие и средние; среди последних наиболее распро-
страненной является черная беркширкская порода.
Домашние лошади. Задолго до приручения дикие лошади были
излюбленным охотничьим животным первобытного человека. При-
ручение лошади произошло позднее других домашних животных,
причем первоначально она играла роль убойного животного, за-
тем стала употребляться на войне и охоте и, наконец, как рабочее
животное. Сообразно с такой последовательностью назначения
919
лошади мы видим, что большинство выведенных в прежнее время
пород лошадей отвечает именно назначению верховой, военной
и охотничьей лошади, и только в последнее время стали выво-
диться культурные породы рабочей лошади — упряжной, сельско-
хозяйственной рабочей и ; ’ ^довозов. В зависимости от того,
насколько сильно сказало' Лошадях влияние отбора человека
в связи с определенными г ' лми, можно различать породы лошадей:
естественные, простые и культурные.
Что касается особенностей происхождения, то на современных
породах оно сказалось в существовании двух типов, отличающихся
между прочим по строению черепа. Первый несомненно более
древний тип домашней лошади — восточный, характери-
зуется. сильным развитием лобной, мозговой части черепа, выдаю-
щимися глазницами и вогнутостью профиля головы; лошади этого
типа мельче. Западный тип — это более крупные лошади с
узким и коротким лбом, менее выдающимися глазницами и выпук-
лым профилем.
Первый тип представлен многочисленными породами Востока
и мелкими породами Запада; второй — более крупными породами
преимущественно Западной Европы.
Домашняя лошадь в Европе появляется поздно — в бронзовый
век, когда налицо были уже другие домашние животные, причем
появляется сразу, неожиданно, в виде лошади восточного типа.
На Востоке лошадь, судя по неоднократно упоминавшимся раскоп-
кам в Анау, была приручена 6000—7000 лет назад. Из Азии, нужно
думать, домашняя лошадь восточного типа проникла в Европу
и послужила здесь поводом к приручению бывших еще много-
численными тогда диких лошадей. В то время в Европе жило не-
сколько видов (или подвидов) лошади. Одни из них были близки
к домашним крупным породам западного типа (Equus robustus).
Это были: Е. germanicus из лёсса Северной Германии — степная
или лесостепная лошадь, Е. abeli — тундряная и лесная ло-
шадь; Е. mosbachensis, Е. sequanius и др. — лесные лошади, все
четвертичного времени. Другие жили в степях восточной Евро-
пы и были близки восточному типу. Первые вымерли бесследно,
оставив следы в упомянутых западных породах домашних живот-
ных, вторые еще сравнительно недавно были широко распростра-
нены в степях СССР и Азии, а в настоящее время представлены
лишь дикой джунгарской лошадью, открытой Пржевальским.
Дикие степные лошади или тарпаны юга СССР (Equus
gmelini) отличались от лошади Пржевальского (Equus przewalski)
мышастой мастью с черным ремнем на спине и более коротким,
но более широким черепом. О пих сохранились многочисленные
описания, относящиеся к XVIII в.; что же касается последних
тарпанов, пойманных в середине XIX в., то они представляют
собой или одичавших лошадей или сохранившихся естественных
ублюдков домашней лошади и тарпанов.
Потомками тарпана Антониус считает некультурных лошадей
Восточной Галиции, Балканского полуострова, отчасти Литвы
92.1
и Польши и юго-западных районов СССР; сюда же относятся пер-
сидские и арабские лошади.
Адамец считает, что приручение тарпана произведено было
в южноукраинских и южнорусских степях скифами, откуда коне-
водство перешло к хеттидам (арменоидам) в Западную Азию.
В начале второго тысячелетия до нашей эры коневодство перешло
в Вавилон, из Вавилона в Египет; только в начале нашей эры арабы
сделались коневодами.
Лошадь Пржевальского — мелкая лошадь светлобуро-желтой
или светлокоричневой масти с темным ремнем вдоль спины, без
чолки и со стоячей гривой и с длинными жесткими волосами по
бокам нижней челюсти. В настоящее время распространена исклю-
чительно в джунгарской пустыне Центральной Азии.
Приручается дикая лошадь Пржевальского с большим трудом,
даже рожденная и воспитанная в неволе. Единственный в мире
питомник этих вымирающих лошадей—в Аскания-Нова на Украине.
Во времена палеолита лошадь Пржевальского была распро-
странена гораздо шире и доходила до Франции и Испании (вероятно
в виде несколько отличающихся экологически рас или экоти-
пов). Одомашнена была эта лошадь монголами когда-то очепь
давно; потомки их — монгольские лошади, киргизские, манчжур-
ские и отчасти китайские.
Что касается времени приручения западной лесной лошади,
которая отличалась большими размерами и более грубым складом,
то данных об этом пока нет никаких.
К лошадям западного типа относятся многочисленные породы,
в том числе тяжеловозы, па образовании которых сказался позднее
отбор периода рыцарства, когда от верховой лошади требовались
большая сила и мощное сложение, чтобы выдержать всадника с его
тяжелыми боевыми доспехами.
Из пород западного типа назовем: бельгийскую, или фла-
мандскую, породу, к которой относятся крупные тяжеловозы и более
мелкие горные лошади сельскохозяйственного склада — ардены и др.
Из бельгийских тяжеловозов лучшей лошадью считается б р а б а н-
с о н, тяжелая, рослая (170 см), сильная и выносливая лошадь. У нас она
признана плановой улучшающей лошадью в Курской и Воронежской обла-
стях, по Волге, в Московской и Смоленской областях.
Першерон — наиболее известная древняя порода французских тя-
желовозов, происхождение которой относится еще к крестовым походам.
У нас эта порода тоже признана улучшающей, но не имеет такого распро-
странения. Нельзя не упомянуть английскую клейдесдальскую
лошадь как прекрасную породу лошадей-тяжеловозов, весьма распростра-
ненную в Европе, особенно в юго-западной Шотландии (рис. 667).
К восточному типу должны быть отнесены древние породы лошадей,
выведенных в Ассирии, Египте, в Греции, и следующие породы верховых
лошадей: знаменитая арабская лошадь, игравшая такую важную роль
в коннозаводстве, туркменская, карабахская, турецкая
и знаменитая английская скаковая; далее к этому типу нужно
отнести мелких лошадей Европы, как пони, эстляндский клеп-
пер, вятка и об пинка; сюда же относятся степные породы ло-
шадей так называемого монгольского типа: киргизская, калмыц-
кая, башкирская, донская, забайкальская и неко-
торые другие.
921
Рис. 667. Жеребец клейдесдальской породы.
Все лошади восточного типа за исключением английской могут считаться
естественными породами, так как выведены путем бессознательного отбора
в сьязи с природными условиями и укладом жизни народов. С одной сто-
роны, это типичные верховые лошади воинственных восточных племен и ско-
товодов номадов, с другой — это сильные лошади сельскохозяйственного
значения, как вятка и клеппер.
Родоначальниками скаковой английской лошади являются вывезенные
из Аравии три прекрасных жеребца, от скрещивания которых с английскими
кобылами местной породы при примеси крови других верховых лошадей
путем длительного систематического отбора на скачках получился тип лучшей
к мире верховой лошади (рис. 668).
Рис. 668. Английская скаковая лошадь.
S22
Кроме указанных выше «типичных» пород лошадей еще есть довольно
много лошадей, которые произошли от скрещивания лошадей восточного и
западного типов.
Ив них особенно интересны русские рысаки, выведенное
в конце XVIII в. Орловым-Чесменским путем соединения лучших качеств
легких арабских и английских лошадей с силой и массивностью лошадей
низменностей Запада — датской и голландской (рис. 669).
Американский рысак выведен недавно и происходит
от английского жеребца, вывезенного в Америку в 4788 г.
Г ё н т е р — охотничья лошадь Англии, выведенная от скрещивания
местной породы с английской чистокровной.
Англо норманны представляют рысаков, получившихся от скре-
щивания чистокровных английских лошадей с местной тяжелой породой.
Рис. 669. Орловский рысак.
У нас в СССР в настоящее время поставлена большая работа по улуч-
шению качеств нашей лошади. Сохранение в чистоте, размножение лучших
лошадей и усовершенствование пород и создание нового племенного ядра
для дальнейшего улучшения всего конского поголовья СССР возложены на
Коневодтрест.
Для покрытия возможно большего числа кобыл лучшими жеребцами
широко применяется искусственное осеменение (до 250 маток на одного же-
ребца).
Домашний осел. Осел был приручен раньше лошади и служил
в качестве рабочего животного, главным образом вьючного. Не-
сомненным его родоначальником является дикий африкан-
ский нубийский осел (Asinus africanus), и первые
сведения о нем как о домашнем животном, относящиеся еще ко
времени за 6000—7000 лет до нашего времени, мы имеем именно
из Египта. Из Египта домашний осел проникает в Палестину.
Еще во времена Гомера он хорошо известен древним грекам. Из
Греции осел распространился по Европе. Возможно, что кроме
африканского осла в образовании домашнего осла принимал уча-
стие дикий сомалийский осел (Asinus somalensis), более крупный
и сильный; от него, по Антониусу, произошли массайский
осел и м а с к а т.
923
Очень давно употребляются в качестве рабочих упряжных и
вьючных животных, отличающихся большими достоинствами, помеси
между лошадью и ослом. От осла и кобылы получаются так
называемые мулы, а от жеребца и ослицы получаются л о-
ш а к и.
Очень интересные опыты по выведению новых пород поставлены
в настоящее время в Аскания-Нова, Полученные там еще до рево-
люции зеброиды (помеси зебры и домашней лошади) показали боль-
шую силу и выносливость, равно как помеси домашних лошадей
с лошадью Пржевальского.
Верблюды, лама и альпака. Две породы домашних верблюдов:
одногорбый, или дромадер, и двугорбый при-
надлежат зоне пустынь и являются их типичными обитателями.
Рис. 670. Двугорбые и одногорбые верблюды.
(Аскания-Нова.)
Одногорбый имеет более широкое распространение от Африки
до Индии, двугорбый принадлежит Азии (рис. 670). Оба верблюда
скрещиваются друг с другом, образуя вполне плодовитые помеси.
В настоящее время разводятся преимущественно метисы. Вся
организация верблюдов приспособлена к сухому климату пустынь
и пескам, и только ему обязан человек своим существованием
среди безводных песчаных пространств.
Верблюд употребляется для сельскохозяйственных работ и как
упряжное, вьючное и верховое животное. Приручены верблюды
были по всей вероятности в Азии, где Пржевальским в пустыне
Гоби найдены были двугорбые их родоначальники. Верблюд ест
вдвое меньше вола и гораздо меньше лошади, подолгу может оста-
ваться без питья, не боится жары и холода. Помимо работ, выпол-
няемых верблюдом, он полезен также своей шерстью, которая
снимается раз в год, причем дает от 7.5 до 10 кг; верблюжье молоко
употребляется в пищу непосредственно или идет на приготовление
молочных продуктов, между прочим «чала» (верблюжий кумыс),
верблюжье мясо употребляется в пищу.
‘24
Южноамериканские представители семейства верблюдов (Саше-
lidae, подотряд Tylopoda) дали двух домашних животных: ламу
и альпака. Первое является вьючным животным, тогда как
второе дает ценную шерсть. Так как оба животных встречаются
только в домашнем состоянии в области горных плоскогорий Юж-
ной Америки, в то время как там же обитают дикие близкие им виды
гуанако (Auchenia huanacus) и викунья (Auchenia vi-
cugna), предполагают, что лама произошла от гуанако, а альпака —
или от викупьи или от скрещивания ламы с викуньей. Альпака
и лама дают метисов. Возможна их акклиматизация в СССР.
Северный олень. Северный олень для жизни человека на севере
так же необходим, как верблюд для жителей пустынь. Будучи
приспособлен к жизни в тундрах и тайге, он содержится круглый
год на подножном корму. Весь уход за ним состоит только в охране
стад и в передвижениях сообразно времени года в поисках удоб-
ных кормпьгх мест. Между тем олени дают жителям севера мясо,
молоко, разнообразную одежду, материал для 'жилища и освеще-
ния и служат упряжным, вьючным и верховым животным.
Выпорки выкидышей оленя — «пешка» — идут на пригото-
вление шапок и детской одежды; из шкуры «пеплюя» — 3—6-ме-
сячного теленка — приготовляются «малица» (рубашка), дохи и
парки; шкуры взрослых оленей идут на приготовление постелей,
покрышки, обивку и т. и. Сапоги приготовляются из «комоса» —
шкуры оленьих ног; сухожилья и перепонки кишок используются
для сшивания; из рогов добывается клей, из костей получается
жир; сало употребляется для освещения.
Оленеводством занимаются в СССР лопари, ненцы, остяки, юраки,
долганы, коряки, чукчи, ламуты, тунгусы, юкагиры, якуты, коми
и русские. Наибольшее число домашних оленей сосредоточивается
в б. Архангельской и Тобольской губерниях, в каждой около
У2 млн. голов.
Домашний олень произошел от живущего там в полосе тундры и
отчасти тайги дикого северного оленя (Rangifer tarandus)
и отличается от последнего разве только изменчивостью цвета масти.
Кролики. Многочисленные породы домашних кроликов выве-
дены путем отбора человеком от общего дикого предка — евро-
пейского кролика (Oryctolagus cuniculus), распростра-
ненного в Средиземноморской области. Разведение кроликов на-
чалось, как предполагают, в Испании. Благодаря своей громадной
плодовитости при чрезвычайной дешевизне и простоте содержания
и при малой затрате капитала кролики служат прекрасным объек-
том мелкого животноводства. Они дают довольно вкусное мясо,
пух и мех. Самка дает в год до 48 штук молодых, которые уже
с 8-месячного возраста могут плодиться. Если считать приплод
вдвое меньше, и то каждая самка может дать в год до 80 кг мяса
и 20 кроличьих шкурок.
При выводе одних из многочисленных пород домашних кроли-
ков имелось в виду увеличение веса и улучшение мясных достоинств,
как например при разведении фламандского кролика,
925
доставляющего 5—6 кг мяса; при выводе других — улучшение
качества шерсти или ее цвета, как например у серебристого
кролика; выводя третьих, имели целью увеличение плодо-
витости — китайский кролик; наконец много пород
кроликов выведено исключительно со спортивными целями и не
имеет никакого хозяйственного значения.
Необходимо упомянуть еще об охотничьем кролике, который
разводится с охотничьими целями на свободе. Благодаря своей
плодовитости охотничьи кролики так быстро размножаются, что
быстро делаются опасными вредителями сельского хозяйства.
Во многих странах так размножились одичавшие кролики, заве-
зенные туда европейцами. Еще за 100 лет до нашей эры жители
римских колоний на Балеарских островах жаловались в Рим на
бедствия, причиняемые их посевам одичавшими кроликами. В Ав-
стралии в Новом Южном Валисе привезенные туда кролики раз-
множились так быстро, что сделались буквально бедствием для
населения. С ними пришлось вести упорную и продолжительную
борьбу. Из этой колонии за один только год было вывезено более
15 млн. шкурок кроликов. На маленьком островке Порто-Санто
одичавшие с 1418 г. кролики образовали совершенно особенную
мелкую расу своеобразной окраски — рыжий верх и белое брюхо.
2. Промысловые и охотничьи млекопитающие
На обширной территории СССР с его необъятными лесами и
тундрами на севере Европы и Сибири и величественными гор-
ными странами на юге промысел диких животных имеет большое
значение в экономике страны. Особенно большое значение имеет
Рис. 671. Ондатра (Fiber zibethicus).
(Из Брэма.)
926
промысел млекопитающих на пушнину. Еще дореволюционная
Россия занимала второе после СИГА место в мировом экспорте
пушнипы, доставляя на мировой рынок свыше 20% этого това-
ра. В настоящее время СССР по экспорту пушнины занимает
первое место и дает на мировой рынок около 30% всей пушни-
ны; второе место занимают США (около 20%), третье — Канада
(около 15%).
В 1928—1929 гг. стоимость пушнины, заготовленной в СССР,
определялась в 16 миллионов руб.
О количестве добываемого зверя дает представление следующая
таблица (по данным за тот же год):
Общая
т_ заготовка
Наименование вверя по СССР
(в штуках)
Барс......................... 118
Барсук.................... 64 807
Белка.................. 13 067 873
Бурундук................ 1407 143
Волк...................... 26 200
Выдра...................... 7 544
Горностай................ 945 278
Енотовидная собака . . . 784
Заяц-беляк............. 6 176 665
Заяц-русак.............. 2 464 346
Заяц-песчаник........... 1 869 768
Заяц земляной............. 26 475
Колонок.................. 432 557
Котик...................... 4 000
Корсак.................... 9113
Кошка дикая............... 17 393
Крот ................... 2 784 396
Крыса водяная........... 2 758 204
Куница.................... 34 862
Кроме того добыто морского
том океане:
Гренландского тюленя .
Моржа................
Морского зайца ....
Белухи ..............
Белого медведя ....
Общая
тт заготовка
Наименование зверя п0 СССР
(в штуках)
Ласка.................... 105 579
Летяга..........• . . . . 31 383
Лисица................... 320 647
Медведь.................... 1 672
Норка..................... 59 781
Песец..................... 53 694
Перевязка .............. 1264
Росомаха.................... 738
Рысь ...................... 4 729
Соболь.................... 13 716
Сурок . . . ........... 576 320’
Суслик ................. 9 487 009
Суслик-песчаник .... 3,143 319
Харза................... ' 122
Хомяк................... 5 741 174
Хорь светлый............1181 569
Хорь темный.............. 411 389
Шакал..................... 13 433
зверя в Белом море и Ледови-
1929 г. 1930 г.
. . . 95 000 ,165 000
... 116 390
... 74 84
... 75 40
... 18 125
Из млекопитающих охотничье и промысловое значение имеют
следующие виды, встречающиеся в СССР.
Отряд насекомоядные (Insectivora). Из этого отряда
исключительно большое промысловое значение у нас имеет в ы-
х у х о л ь (Desmana moschata) именно потому, что, обладая оиень
цепным мехом, она живет только в СССР.
Это водное животное, величиной несколько больше крысы
с выдающимся вперед хоботообразным концом морды и густым
теплым мехом. Живет в реках и озерах бассейна Волги и Дона,
питается различными беспозвоночными и рыбой. Из-за ценного
меха выхухоль, которая раньше была очень многочисленна, под-
927
верглась истреблению и была близка к полному исчезновению.
Чтобы предотвратить это, добыча выхухоли совершенно воспре-
щена законом; в 1935 г. организованы специальные выхухолевые
заповедники: Окский на р. Пре (приток Оки), Клязьминский и
Хоперский.
Под названием выхухоли в продажу поступает мех северо-
американского водяного грызупа опдатры (Fiber zibethicus),
которая с успехом акклиматизируется у нас в Союзе (рис. 671).
В настоящее время приобрели промысловое значение также
кроты (Talpa), шкурки которых употребляются на меха. Про-
мысел кротов производится главным образом в Европейской части
СССР.
Отряд грызуны (Rodentia). Из грызунов промысло-
вое значение имеют белки, зайцы, сурки и бобры, достав-
ляя ценную пушнину. Зайцы дают вкусное питательное мясо
и являются охотничьим зверем. Вредители сельского хозяйства
хомяки и суслики дают шкурки, которые употребляются для
выделки и поступают в продажу в возрастающем количестве.
Наконец грызуны служат пищей для многочисленных видов цен-
ных хищных животных, а речной бобер дает употребляю-
щуюся в медицине «бобровую струю».
Зайцы (Lepus) распространены по всей стране и образуют
несколько видов и подвидов.
Всю зону тайги и отчасти тундры занимает своим распростра-
нением заяц-беляк (L. timidus), образующий несколько под-
видов. Зимой беляк становится совершенно белым за исключе-
нием черных копчиков ушей. Другой заяц — русак (L. europeus)
занимает большую часть лесной области и всю зону степей
Европейской части СССР. В Туркестане, в степях Западной Сибири,
в Забайкалье* в Приамурье и в Закавказье живут другие • виды
зайцев.
Зайцы плодятся два-три раза в год, принося каждый раз от 3
до 5 зайчат. Первый помет в марте, а последний— в сентябре.
При таком размножении зайцы быстро увеличились бы в числе,
если бы у них не было большого числа врагов из хищников-млеко-
питающих и птиц.
Заячьи шкурки занимают па пушном рынке по количеству по-
ступающего товара второе место, немалое количество их идет на
изготовление меховой одежды населения прямо от охотников,
значительное количество вместе со шкурами продается на рынках
как дичь.
Местами, где их много, зайцы вредят плодовым садам, особенно
питомникам и молодым лесным посадкам.
Белки (Sciurus vulgaris), образуя несколько подвидов, оби-
тают во всей лесной зоне за исключением Камчатки; нет их в Крыму
и на Кавказе. В Закавказье живет другой вид белки (S. anomalus).
Мех белки, одетой в зимний серый наряд, очень теплый, ноский
и легкий, почему имеет всегда большой спрос на рынке. В СССР
белка — главный промысловый йверек. В зависимости от цвета
928
и качества шерсти различается несколько сортов беличьего меха
различной ценности так наз. «кряжи». Особенно ценятся во-
сточносибирские и дальневосточные белки с темным мехом*
Вред, приносимый белками, может быть заметным только там,
где их очень много, и при условии специального хозяйства. Обу-
словливается он тем, что белка питается семенами хвойных
деревьев, орехами, жолудями, плодами и ягодами, а также
яйцами и птенцами различных птиц.
Сурки (Marmota) в числе нескольких видов обитают в степ-
ной зоне и в безлесных поясах горных стран. Это довольно крупные
зверьки, живущие в норах, около которых они насыпают холмики
земли — так называемые байбаковины или сурчины.
Европейский степной сурок, или байбак (М. bobak), еще
недавно был очень широко распространен по всей степной полосе,
в настоящее время почти совершенно исчез вследствие распашки
целинных степей и сохраняется в заповедниках.
Другие виды сурков, по-сибирски тарабаганов, обитают в
степных предгорьях, на безлесных склонах и на плоскогорьях
Средней Азии и Сибири. Шкурки сурков дают довольно хороший
мех. К концу лета, перед тем как впасть в спячку, сурки накопляют
большие запасы жира, который отличается тем, что не сгущается
даже па морозе. Сурков промышляют ради меха, жира; местами
мясо их употребляется в пищу. В то же время сурки опасны тем,
что являются переносчиками чумы.
Суслики (Citellus) в противоположность суркам прекрасно
уживаются с распашкой степей и подобно суркам живут в норах.
Размножаясь в большом количестве, приносят большой вред хлебам,
так что с ними приходится вести ожесточенную борьбу. Шкурки
сусликов в последнее время идут на выделку мехов, особенно
крупных восточных видов и среднеазиатского желтого суслика.
Подобно суркам суслики могут распространять чуму.
Речной бобер (Castor fiber) был некогда широко распро-
странен по всей лесостепной и южной части таежной зоны и на
ряду с соболем и куницей был главным промысловым животным.
К концу прошлого столетия бобры были почти повсеместно истреб-
лены; небольшое число бобров сохранилось лишь в Полесье, на
Кавказе, в Тобольской губ. и кое-где в Северной Монголии.
Забытый промышленниками бобер в начале этого столетия
местами заметно размножился и успел значительно расширить свою
область распространения и встречается не только в Белоруссии,
но и на Украине и в ЦЧО. Охота на бобров совершенно запрещена?
законом, и правительством организованы бобровые заповедники.
Из них наибольшее значение имеет Воронежский заповедник,
где за короткий срок его существования (с 1927 г.), за 12 лет,
бобры со 150 особей увеличились в числе почти в 10 раз. В
Белорусском заповеднике бобров сейчас более 800. Из Воронеж-
ского заповедника бобры завезены в ряд других заповедников,
где они успешно размножаются. В Канаде в специальном запо-
веднике бобры настолько размноашлись, что правильный отстрел
59 Кашнарои и Стиичинский 929
их и продажа на сторону производителей даю? значительный
доход.
Помимо цепного меха речной бобер доставляет «бобровую струю»,
употребляющуюся в медицине, — экстракт особой придаточной
железы, встречающейся у обоих полов. Бобры живут колониями
в норах высоких берегов рек и озер, возводят особые постройки —
«хатки» из отрезков деревьев, которые подгрызают и валят; на
ручьях и речках возводят «плотины», чтобы поднять упавшую воду
выше уровня выходов из нор.
Питаются корой деревьев, явором, тростником и водорослями.
Размножаются довольно медленно, принося в год 1—3 детенышей.
Отряд хищных (Carnivora). Значение хищных живот-
ных в жизни человека и природы чрезвычайно велико. С од-
ной стороны, среди крупных хищников мы имеем действитель-
ных врагов самого человека и его домашних животпых, как
тигры, барсы, волки, медведи; с другой стороны — наиболее цен-
ную пушнину доставляют именно хищники (соболь, куница, ли-
сица, выдра, енот, норка, скунс, горностай и др.); хищники же
являются злейшими врагами вредителей сельского хозяйства,
грызунов (лисицы, хорьки, горностаи, ласки и др.). Таким обра-
зом, чтобы оценить значение хищников для человека, необходимо,
анализировать значение каждого вида в отдельности и притом в кон-
кретных условиях хозяйства каждой местности.
Сем. кошачьих (Felidae). Сюда относятся крупные
и средней величины хищники, питающиеся исключительно пти-
цами и млекопитающими. Из крупных видов в СССР живут:
тигры (Tigris tigris), несколько видов с подвидами живут в За-
кавказье, Средней Азии, Уссурийском крае, Манчжурии и Мон-
голии; пантеры, или барсы (Leopardus pardus), образуют местные
подвиды на Кавказе, в Закавказье, в Манчжурии и Уссурийском
крае; ирбис (Leopardus uncia) обитает в Средней Азии. Несмотря
на свою редкость эти виды все еще приносят ощутительный вред,
истребляя крупную дичь из млекопитающих, нападая на стада,
и иногда являются опасными самому человеку. Хотя меха этих
кошек и ценятся высоко как ковры благодаря своей редкости,
животные эти не имеют промыслового значения, а являются лишь
объектом спорта и разводятся в зоологических садах.
Рысь (Lynx lynx) в нескольких подвидах распространена
по всей лесной зоне и па Кавказе. Истребляет большое количество
крупных птиц, имеющих промысловое значение, и млекопитаю-
щих — от зайцев до косуль и молодых лосей и оленей. Имеет не-
которое промысловое значение, так как добывается ежегодно
в довольно большом количестве и доставляет довольно высоко
расцениваемый мех. В Закавказье живет небольшая красивая
парделевая рысь (Lynx pardellus), питающаяся преиму-
щественно фазанами, турачами и другими птицами.
Кошки собственно (Felis) представлены лесными ко-
тами (F. sy Ives tris) в Закавказье. Истребляют мелких птичек
и пернатую дичь.
930
Камышевый кот (Catolynx chaus) обитает в тростни-
ковых зарослях бассейнов Каспийского и Аральского морей и при-
носит большой ущерб гнездящимся там фазанам и водоплавающей
птице. С т е п н ые к о т ы:1 в Средней и Восточной Азии м а-
н у л (Trichaelurus manul), в Туркестане — длиннохво-
стый кот (Felis caudata), в Закаспийской области — кара-
кал (Caracal caracal), хотя и питаются также птицами, в то же
время истребляют весьма усердно различного рода грызунов.
Все эти кошки дают пушпипу, однако в таком незначительном ко-
личестве, что серьезного промыслового значения не имеют.
Сем. куниц (Mustelidae). Это семейство дает наиболее
цепных промысловых пушных зверей. Из видов куниц лес-
ная куница (Martes inartes) распространена в лесах Европы,
Крыма и Кавказа, тогда, как б е л о д у ш к а, или каменная
куница (Martes foina), живет в открытых местностях — в сте-
пях и в горах, особенно в Крыму, на Кавказе и в Средней
Азии.
Хотя обе куницы, особенно лесная куница, усиленно истреб-
ляют птиц и охотничьих млекопитающих (белок, зайцев и даже
молодых косуль), однако они доставляют настолько ценный мех,
что подобно соболю не могут считаться вредными и подлежат
охране.
В Приамурье и Уссурийском крае живет самая крупная из ку-
ниц — х а р з a (Martes flavigula), не имеющая однако промыс-
лового значения, так как мех ее относительно малоценен.
Соболь (Martes zibelina) был некогда распространен широко
в таежной зоне Европейской части СССР и Сибирй. Однако благо-
даря вековому преследованию из-за ценнейшего меха в настоящее
время сохранился в значительном количестве только на Северном
Урале, па Алтае, в Саянах, в Якутии, в Забайкалье, на Камчатке,
в Приморском Дальневосточном крае. Ценность меха соболя в
очень больших размерах колеблется тз зависимости от густоты, неж-
ности оттенков и окраски. Наиболее ценные соболи добываются в
Прибайкалье (баргузинские), по правым притокам Лены (витимские)
и по Амуру (зейские) и в Якутии. Исчезновение соболей вызвало
необходимость принятия срочных мер к его сохранению. В 1913 г.
охота была запрещена на 3 года. Было приступлено к организации
соболиных заповедников в Саянах и Забайкалье. Однако только
после Октябрьской революции сохранение соболя и увеличение
о го поголовья проводится планомерно и успешно в заповедниках:
II ечерско-Ылычском, КондОгСосвенском, Алтайском, Саяпском,
Сихотэ-Алинском, Баргузинском и Кроноцком (на Камчатке).
В самое последнее время удалось заставить соболей размно-
жаться в неволе, что обеспечивает развитие соболеводства в QCCP.
Хорьки — обыкновенный (Putorius putorius), степной хо-
рек (Put. eversmanni) и колонок (Kolonocus sibiricus) —
служат предметом значительного хорькового промысла. Пер-
вый широко распространен в степной и лесной зонах Европей-
ской части СССР; второй обитает в степной зоне Европейской
59* 931
части СССР и Западной Сибири; третий распространен по Южной
Сибири от Уральского хребта до берегов Великого океана за исклю-
чением западного степного района, Средней Азии, Камчатки и Са-
халина. В Забайкалье и на Алтае встречается небольшой хорек —
сусленик (Kolonocus alpinus). В степях юга Европейской части
СССР, Крыма, Кавказа и Средней Азии живет пестрый хорек —
перевязка (Vormela sarmatica).
Хорьки живут в норах, питаются главным образом грызунами и,
доставляя прочную и довольно ценную пушнину, должны считаться
ценными зверями. В редких случаях хорьки вредят птицеводству.
Из хвостов хорьков, особенно же колонков, изготовляются аква-
рельные кисточки.
Горностаи (Mustela erminea) и ласки (М. nivalis)
распространены несколькими подвидами почти по всему СССР и
питаются преимущественно мелкими грызунами. Оба к зиме наде-
вают белоснежный наряд. Горностаи большей величины, имеют
черный кончик хвоста и добываются как пушные зверьки из-за кра-
сивого меха значительной ценности.
Росомаха (Gulo gulo), обитательница таежной зоны,
раньше была распространена в Европейской части СССР гораздо
шире. В данный момент обитает только в северо-восточной ее части
и по всей Сибири. Своим истреблением промысловых зверей и птиц,
а также уничтожением пойманной промышленниками в ловушки до-
бычи наносит вред охотничьим промыслам; доставляемый же ею
мех груб и малоценен.
Барсуки представлены несколькими видами. Обыкно-
венный барсук (Meles meles) принадлежит Европе и Кавказу; в
Киргизских степях, в Юго-западной Сибири живет песчаный
барсук (М. arcnarius), в Восточной РСФСР — амурский
барсук (М. amurensis). Барсуки роют норы, в которых живут
летом и проводят в спячке зиму. Питаются не только животной,
но и растительной пищей. Мех малоценный, шкура идет на обив-
ку, из волос готовятся кисточки. Мясо барсуков употребляется
в пищу.
Норка (Lutreola lutreola) является водным животным и
живет по берегам водоемов. Распространена во всей Европей-
ской части СССР (за исключением Крыма) и на Кавказе. Питается
рыбой, земноводными, птицами и зверьками. Дает ценный краси-
вый мех.
Выдра (Lutra lutra) — типичная жительница вод распро-
странена по всему Союзу. Питается главным образом рыбой, ра-
ками, иногда добывает водоплавающих птиц. Дает ценный и проч-
ный мех.
Камчатский бобер, или калан (Latax lutris) — до-
вольно крупный морской зверь, раныпев большом количестве живший
в северной части Тихого океана у берегов Камчатки, Курильских и
Командорских островов. Вследствие очень ценного меха был объектом
морского промысла и благодаря жестокому преследованию повсеме-
стно близок к полному исчезновению. В наибольшем числе ветре-
932
чается около Командорских островов, где является для жителей
источником дохода. В виду этого промысел камчатских бобров на-
ходится под контролем, устанавливающим размеры добычи. В по-
следние годы отстрел морских бобров запрещен вовсе. Питаются
бобры рыбой, моллюсками, кишечнополостными и иглокожими.
Главное Управление по заповедникам в настоящее время прово-
дит опыт акклиматизации этого ценного зверя на Мурманском
побережье.
Сем. собачьих (Canidae). Волк (Canis lupus), об-
разуя несколько подвидов, распространен по всей стране и
всюду является врагом животноводства, опасным и для жизни че-
ловека, главным образом потому, что является разносчиком бе-
шенства. Количество волков особенно велико в малокультурных
местах с массовым животноводством — в полосе тундр и степей.
В лесной зоне количество волков увеличивается по мере увеличения
населения до известного предела, после чего оно вновь падает. Это
объясняется тем, что в малонаселенной тайге для волков мало пищи,
а в местностях с густым населением для волков нет удобных мест
для вывода детей и кроме того с волками ведется жестокая борьба.
Вред, приносимый волком, обусловливается еще тем, что он ис-
требляет диких цепных копытных животпых и препятствует ак-
климатизации последних.
По совокупности приносимого волками вреда они подлежат пла-
новому возможно более быстрому и полному уничтожению, в том
числе и в заповедниках.
Лисица (Vulpes vulpes) распространена в СССР повсемест-
но, во всех зонах, образуя большое число подвидов.
Отличаясь цветом и качеством меха, эти подвиды имеют раз-
личную ценность, соответствуя разным кряжам лисьего меха. Осо-
бенно ценны лисицы лесных зон и горных местностей. Помимо геогра-
фических подвидов лисицы дают цветные расы от белого до буро-
черного, известные в торговле под особыми названиями: огневка,
крестовка, сиводушка, белодушка и черно-
бурая. Последняя. вариация, встречаясь спорадически глав-
ным образом на севере, представляет очень цепную и дорогую расу?
В виду этого чернобурых лисиц разводят с промышленными
целями на лисьих фермах, причем здесь они подвергаются отбору;
из мутации удалось вывести весьма ценных серебристо-черных
лисиц. '
По тому значению, которое имеет лисица как промысловое
животное, и характеру вреда и пользы, приносимых лисицей, ей
следует считать в общем ценным, подлежащим охране видом,
особенно в районах земледельческих.
В степях юго-восточной Европейской части СССР, в Казахстане
и дальше в степях Сибири до Забайкалья живет другой вид лисицы—
корсак (Megalotis corsac), имеющий менее ценный мех.
Песец (Alopex lagopus) обитает в тундрах Европы и Азии, па
островах Ледовитого океана, и другой вид (Alopex beringiensis) —
па Командорских острорах. Песцы меняют к зиме свой летний жел-
933
товатобурый мех на чисто белый. Встречается раса так называе-
мых голубых песцов, надевающих вместо чисто белого го-
лубоватый пепельно-серый мех. Как цветовая мутация голубые
песцы встречаются в известном проценте среди белых повсюду, но
к востоку — в большем проценте; на о. Медном живут только голу-
бые, на о. Беринга процент белых очень невелик.
Песцы живут в норах и питаются разной животной пищей п
между прочим отбросами моря. Из-за ценного меха песцов усиленно
промышляют, причем не только взрослых, но и молодых, которые
с момента рождения последовательно называются, копанцем, нор-
пиком, крестоватиком, синяком и недопеском.
Шакал (Canis aureus) живет на Кавказе и в Ср. Азии.
Питается различной животной и отчасти растительной пищей, па-
далью и отбросами; иногда нападает на домашних птиц и скот. Мех
шакала грубый и потому дешевый.
Енотовидная собака (Nyctereutes amurensis) живет
в Приамурье и тоже служит объектом промысла, хотя и дает мало-
ценный грубый мех. Питается разной животной пищей, особенно
рыбой. В Уссурийском крае живет другой подвид. В последнее
времц проведены многочисленные опыты по акклиматизации
енотовидной собаки в разных местах СССР, давшие пока про-
тиворечивые результаты.
Сем. медведей (Ursidae). В нашей фауне живут три
вида медведей, относящиеся к двум родам.
Бурый медведь (Ursus arctos), рапсе широко распростра-
ненный по всей лесной, кактаежпой зоне, так и зоне смешанных ле-
сов, и на Кавказе, образует несколько подвидов. В пределах Европей-
ской части СССР бурый медведь становится все более и более редким,
скрываясь в обширных лесах малонаселенных районов. Медведи
питаются смешанной пищей и главным образом растительной. Раньше
там, где их было много, они очень вредили пчеловодству, особенно
бортневому; вредят также посевам яровых хлебов, особенно овсам и
кукурузе и нередко нападают на домашний скот. Медведи не являют-
ся промысловыми животными в полном смысле, так как добываются
случайно, хотя шкура медведя ценится довольно дорого и идет на
выделку ковров. Мясо медведя съедобно, а сало идет для приготов-
ления медикаментов. Зимой медведи повсеместно, за исключением
Кавказа, впадают в спячку.
Белый медведь (Talassarctos maritimus) обитает в
прибрежной полосе Ледовитого океана и на островах, являясь мор-
ским океаническим видом. Из-за ценной шкуры, съедобного мяса
и жира белый медведь служит предметом промысла дальнего се-
вера. Питается морскими млекопитающими и рыбой.
Огряд ластоногих (Pinnipedia). Из 31 вида, представля-
ющих этот отряд, у наших берегов встречается около половины—
14 вилов; из них мы остановимся на следующих имеющих очепь
большое промысловое значение.
Моржи (Trichechus rosmarus) представлены в СССР двумя под-
видами: атлантическим, распространенным в Европе и на
934
большом протяжении берегов Сибири, и тихоокеанским,
заменяющим атлантический подвид в восточной части Ледовитого
океана и,в северной части Берингова моря. Последний имеет осо-
бенно большое промысловое значение в жизни жителей восточного
побережья Ледовитого океана. Моржи дают мясо, жир, шкуру,
клыки. Благодаря преследованию моржи быстро уменьшаются
в числе.
Тюлени представлены несколькими родами и видами. Из них
особенное промысловое значение имеют: лысун (Phoca groen-
landica) (рис. 603), нерпа (Phoca foetida), сибирский
тюлень (Phoca sibirica), каспийский тюлень (Phoca
caspica), серый тюлень (Halichoerus gripus), морской
заяц (Erignathus barbatus). Каспийский тюлень обитает в Кас-
пийском море; сибирский — живет на Байкале, два подвида
нерпы — на озерах Сейма и Ладожском, остальные виды принад-
лежат Северному Ледовитому океану, Белому и Балтийскому
морям.
Для жителей полярных побережий тюлени являются черезвы-
чайно ценными в жизпи животными, так как они дают жир, кожи
и мясо. Жир тюленя служит главным образом в качестве горючего
и смазочного материала. Кожи идут на обивки.
Центром промысла тюленей служат: горло Белого моря, Мур-
манский берег, Канип, Зимний берег, острова Новой Земли, Вай-
гач, Каспийское море и Байкал. Пищей тюленей служит главным
образом рыба; однако мнение, что тюлени вредят рыболовству,
преувеличено.
Морские котики (Callbrrhinus ursinus), образуя два
подвида, обитают в северных водах Тихого океана и во время раз-
множения с мая по октябрь держатся на островах: первый подвид—
на Командорских островах и па острове Тюленьем, второй — на
островах Прибылова.
Будучи довольно крупным жийотпым (секач-самец достигает
2 м, матка —1У4 м), котик дает чрезвычайно ценный мех. Раньше ко-
тики были очень многочисленны, и промысел их давал большой до-
ход государству. Однако вследствие*чрезвычайного их истребления
число котиков стало уменьшаться, и опи были близки к полному ис-
чезновению. Поэтому между Англией, Северо-американскими со-
единенными штатами, Россией и Японией была заключена конвен-
ция — бой котиков был приостановлен на 5 лет (с 1911 г.), и затем
на- оспове специальных исследований зоологов организовано
правильное промысловое хозяйство, и убой котиков производится
по определенному плану. У нас такое хозяйство организовано
па Командорских островах.
Си в у ч (Eumetopias stelleri) относится подобно котику к пуш-
ным зверям, но и^еет плохой редкий подшерсток и добывается пре-
имущественно ради кожи. Он живет в северной части Тихого оке-
ана к югу до берегов Японии. Подобно котикам летом держится на
лежбищах.
Благодаря истреблению сивуч стал очень редок и, видимо,
935
скоро совершенно исчезнет, если не будут приняты решительные
меры к его сохранению.
Отряд непарнокопытные (Perissodactyla).
Охотничьим животным этого отряда у нас можно шщтать
только полуосла—к улана, или джигетая (Asians hemio-
nus), распространенного в степях от Урала до Алтая и в Монго-
лии. Добывается ради мяса и шкуры и близок к окончательному
истреблению.
Отряд парнокопытные (Artiodactyla). Из группы
нежвачных парнокопытных (Nonruminantia) только семейство
свиней (Suidae) представлено в нашей фауне кабаном
(Sus), который в числе двух видов и нескольких подвидов распро-
странен в южной части Союза. Он живет сейчас в Белорусском и
Украинском полесье, на Кавказе, по берегам Каспийского моря,
в Средней Азии и по южным хребтам до Великого океана и к югу
от них.
На крайнем востоке распространены подвиды японского
кабана (Sus leucomystax), в остальной части — подвиды обык-
новенного европейского кабана (Sus scrota). Охотятся,
за кабаном ради мяса, жира и щетины. Местами кабаны приносят
вред сельскохозяйственным культурам — картофельным, рисовым,
кукурузным полям и бахчам.
Из группы жвачных (Ruminantia) у нас распространены
семейства оленей и полорогих.
Сем. оленей (Cervidae) представлено очень важными охот-
ничьими и промысловыми животными.
Северный олень (Rangifer tarandus) занимает своим
распространением всю зону тундр с некоторыми островами Ледо-
витого океана и почти всю полосу тайги до южных горных хребтов
и Великого океана с Сахалином. В Европейской части СССР,
в большой части таежной полосы северные олени истреблены.
Раньше, когда дикие олени были очень многочисленны, охотничьи
народы крайнего севера Сибири жили в значительной мере
охотой на них, во время их перекочевок из тундры в леса
и обратно. Теперь прежних массовых перекочевок оленей не
бывает, и олений промысел заменяется все больше и больше* оле-
неводством.
Лось (Alces alces) принадлежит всей полосе тайги и
смешанных лесов. Сравнительно недавно был еще многочис-
ленен в лесах Европейской части СССР, сейчас сделался большой
редкостью и находится под специальной охраной закона и раз-
множается во многих лесных заповедниках; охота на лосей за-
прещена совершенно, а в остальной части Союза ограничена ноябрем
и декабрем.
Два вида косули, живут у нас: европейская (Сар-
reolus capreolus), принадлежащая лесам Западной Европы и запада
Европейской части СССР, и сибирская косуля (Capreo-
lus pygargus), распространенная по крайней мере тремя подвидами
по всей лесной зоне Сибири, Ср. Азии и Кавказа. Европейская
936
косуля в пределах Европейской части Союза, будучи до войны до-
вольно многочисленной в охотничьих хозяйствах и казенных лесах
Западной России, Украины и Белоруссии, в период гражданской
войны была истреблена настолько, что стала редкой, почему и
находится под покровительством закона, который совершенно запре-
щает охоту на нее. В настоящее время после организации запо-
ведников, при правильной постановке охотничьего хозяйства, при
подкормке в зимнее время, косули стали размножаться и в недале-
ком будущем станут важным объектом охоты. Добывают косуль
ради мяса и шкур.
Рио. 672. Пятнистый олень (Pseudaxis hortulorum).
(Аскания-Нова, фот. Озерова.) *
Олень (Cervus) у нас подставлен несколькими видами.
Европейский благородный олень (С. elaphus), охра-
няемый в охотничьих хозяйствах Западной Европы, был распро-
странен и у нас в некоторых крупных охотничьих хозяйствах. В на-
стоящее время содержится в Воронежском заповеднике; особый
подвид разводится в Крыму Под охраной закона в Крымском за-
поведнике. На Кавказе обитает восточный подвид — кавказ-
ский олень (С. elaphus maral). От горного Туркестана на
восток через Китай до Тихого океана по южным сибирским
горам рапространены подвиды американского вапити
(Cervus canadensis), марал (С. canadensis asiaticus) на за-
паде до Саянских гор и и з ю б р (С. canadensis luedorfi) на
восток отсюда. В Уссурийском крае обитает пятнистый
олепь (С. hortulorum) и манчжурский олень (С. sika
mantschuricus). В Бухаре живет бухарский олень (С. bact-
rianus).
Все олени промышляются ради мяса и шкуры, а на маралов,
изюбров и пятнистых охотятся главным образом ради еще неоко-
937
стеневших рогов, которые в высушенном виде дают весьма ценный
в Китае товар «п а н т ы», из которых готовят лекарства, име-
ющие в китайской медицине чрезвычайное распространение. Особен-
но ценятся панты пятнистого оленя. В последнее время панты
приобрели значение в мировой медицине. Из паптов извлекается
целебное вещество пантокрин, поднимающее общую жизнедеятель-
ность.
В виду ценности пантов промышлепники начали ловить живых
оленей и содержать их в загородях, ежегодно отпиливая в нужное
время рога, чем было положено начало промышленному мара-
ловодству, или пантовому хозяйству, получившему у пас на
юге Сибири и особенно на Дальнем Востоке значительное разви-
тие. Оленей содержат или в одомашненном виде в оленниках или
в парках.
Благородный и пятнистые олени и маралы при некотором
надзоре и надлежащей охране размножаются в лесах, отведенных под
правильно организованные охотничьи хозяйства и в заповед-
никах. В последнее время Гл. Управлением по заповедни-
кам успешно проводится акклиматизация пятнистых оленей
в заповедниках на Кавказе, в лесостепной полосе и южном
Урале.
Кабарга (Moschus moschiferus) живет в горах тайги от Ал-
тая до Великого океана. Самец кабарги рогов не имеет, но имеет удли-
ненные клыки. Этот маленький олень служит предметом промы-
сла ради мяса, шкуры и главным образом «кабарговой струи» —
маслянистого выделения особой железы, которая находится у сам-
цов позади пупка. Струя служит для изготовления мускуса.
Сем. полорогие (Bovidae-Cavicornia). Бараны (Ovis)
представлены в пределах Союза целым рядом видов. На Кав-
казе в южной его части живет муфлон Г м е л*и н а (О. orien-
talis orientalis); дальше на восток между Каспийским и Аральским
морями живут архары: на Усть-Урале (О. orientalis агсаг)
и в Копет-Даге (О. orientalis cycloceros); еще дальше па восток,
в пределах Туркестана до Иртыша на восток, обитают подвиды
к а ч к а р а (О. poloi); за Иртышом на восток Забайкалья жи-
вет аргали (О. ammon); еще далее на восток и севернее по го-
рам ^Восточной Сибири живут подвиды северных баранов
(О. nivicola). Барапы — обитатели степных плоскогорий и гор.
Охотятся за ними ради мяса и шкур, но в виду трудности охоты
промыслового значения бараны не имеют.
Козлы (Capra) представлены горными козлами и турами.
Сибирский горный козел, или т э к э (С. sibirica),
распространен по всей горной части Южной Сибири от Тянь-
Шаня до Байкала. Держится в скалистой безлесной части гор.
Безозровый козел (С. aegargus) принадлежит Кавказу,
ц Персии; винторогий козел (С. falconneri) обитает в горах
южной Бухары.
Туры представлены видами, живущими на Кавказе:
дагестанским (Capra cylindricornis) (рис. 673) и к а в к а з-
938
с к и м (С. caucasica). Они принадлежат высокогорным лугам и
держатся стадами. Козлы и'туры служат предметом охоты ради
мяса, кожи и рогов. Охраняются туры в Кавказском заповеднике.
Серна (Rupicapra rupicapra) у нас представлена кавказ-
ским подвидом, распространенным по горам Кавказа в зоне альпий-
ских лугов. '
Рис. 673. Дагестанский тур (Capra cylindricornis).
(Ив Бихнера.)
В Приамурье и горной части Забайкалья живет горная анти-
лопа— горал (Nemorhoedus caudatus). Оба вида промыслового зна-
чения не имеют и являются скорее объектом спортивной охоты,
так как мясо их очень низкого качества, а шкура расценивается
невысоко.
Сайга (Saiga tatarica) некогда была очень широко распро-
странена по всей степной зоне Европейской части СССР и в Киргиз-
ских степях. В настоящее время* встречается как большая редкость
в Калмыцкой степи и еще обыкновенна местами восточнее р. Урала
и Каспийского моря. Охотятся зй сайгой ради мяса, шкуры и имею-
щих целебное значение рогов.
939
Джейран (Gasella suhgutturosa) и д з e p e и (G. guttu-
rosa) принадлежат к группе газелей. Первый вид живет в степях
Закавказья, в Закаспийской области и в Туркестане; второй — в Се-
верной Монголии, откуда заходит в степи Забайкалья и в долины
Аргуни и Онона. Мясо этих антилоп очень вкусно, но в виду труд-
ности охоты за ними они не имеют промыслового значения. Джей-
ран легко приручается и в неволе хорошо плодится.
Отряд китообразные (Cetacea). Из подотряда без-
зубых китов (Mystacoceti) в водах, омывающих берега
Союза, встречается не менее девяти видов, из которых мы остано-
вимся только на следующих: синий кит (Balaenoptera mus-
culus), самый крупный вид современных китов, достигающий 27.5
и 30.5 м длины, встречается как в восточных морях, так и в Баренце-
вом море; гренландский кит (Balaena mysticetus) ве-
личиной до 15.25 м, нередок в Охотском и Беринговом морях и Ле-
довитом океане. В восточных морях нередки еще японский
кит (Balaena japonica) и с е р ы й кит (Rhachianect es glaucus),
а в Баренцовом море — мойвепный полосатик, или
ф и н в а л ь (Balaenoptera physalus), и малый полосатик
(Balaenoptera rostrata).
Из подотряда зубастых китов кашалот (Physe-
ter macrocephalus) изредка заходит до Камчатки; белуха
(Delphinapterus leucas) обыкновенна в Ледовитом океане, спу-
скается в Белое море на западе и до Сахалина па юге. Белуха
имеет наибольшее значение в китобойном промысле Союза, так
как она дает чрезвычайно цеппый жир. Нередки в Ледовитом
океане, Беринговом и Охотском морях касатки (Orca orca)
и нарвалы (Monodon monoceros). В Черном море обыкновенны
дельфины (Delphinus delphis) и морские свинки
(Phocaena phocaena), реже — более крупные^ черные дельфи-
ны рода Tursiops.
Китобойный промысел, в дореволюционное время почти совер-
шенно отсутствовавший, если не считать кустарного способа добы-
вания китов населением Чукотской земли, в последнее время
начал быстро развиваться, особенно на Дальнем Востоке, и имеет
все шансы для успеха. При полной утилизации убитого кита по-
лучается от 7000 до 28 000 руб. дохода. Из подкожного слоя поло-
сатика мойвенного получается до 4000 кг ворвани, а от синего
кита до 12 500 кг. Из туши вытапливается еще до 1600 кг жира;
китового уса получается около 160 кг и более. Остальное мясо идет
на приготовление удобрительного тука (до 5000 кг). Кашалоты
дают до 24 000 кг жира и, кроме того, клея до 1600 кг, спермацет
и благовонную амбру. Дельфинов в Черном море добывают пока
исключительно ради подкожного жира. Каждый дельфин дает от 8
до 16 кг жира.
Неизбежно прогрессирующее повсеместное уменьшение промы-
словых животных в связи с развитием интенсивных видов сельско-
хозяйственного использования территории заставило всюду при-
нимать меры к охранению существующих естественных запасов
940
промысловых животпых, с одной стороны, а с другой стороны, выз-
вало необходимость искусственного увеличения числа ценных пуш-
ных зверей путем их разведения. Охрана объектов промысла однако
при все развивающейся культуре и уменьшении площадей охотни-
чьих угодий, при все возрастающей интенсификации сельского хо-
зяйства пе может остановить без принятия активных мер падение
промысла, а может лишь на более или менее долгий срок задержать
это неизбежное падение. Искусственное же разведение пушных
зверей после весьма удачных попыток, сделанных в Канаде, привело
к быстро прогрессирующей новой отрасли животноводства —
к промышленному разведению пушных зверей в полудомашнем
состоянии.
В целях сохрапепия промысловых зверей от их уничтожения
принимаются следующие мероприятия:
1. Устанавливаются законом в целях ограничения промысла
определенные сроки охоты, чтобы защитить зверя от добычи в то
время, когда он, не вылиняв к зиме, имеет малоценный мех, чтобы ох-
ранить их в период размножения, защитить самок, выкармливаю-
щих детей, и т. п.
2. Регламентируется самый способ, охоты: запрещается и огра-
ничивается отравление зверей, ловли самодействующими прибо-
рами, при которой звери могут быть легко истреблены и при кото-
рой известный и весьма значительный процент пойманных зверей
пропадает для самого охотника.
3. В интересах сохранения и увеличения промысловых и охот-
ничьих зверей организуются правильные охотничьи хозяйства, в ко-
торых количество добываемого зверя определяется наперед в соот-
ветствии с его естественным размножением и проявляется известная
забота о зверях путем уничтожения их врагов или организации под-
кормки, когда звери вынуждены голодать.
4. Организуются заказники, на^оторых на определенное
время запрещаются всякая охота и ловля промысловых и охотни-
чьих животных или заказывается охота только па определенные виды
животных.
Однако принятие одних только ограничительных мероприятий
в надежде на естественное восстановление природных ресурсов
не обещает быстрых желательных результатов. Гораздо важнее
принятие активных мер к восстановлению и обогащению при-
родных ресурсов промысловых животных, что может быть дости-
гнуто:
1) заботой о животных, особенно в тяжелые момепты их суще-
ствования, путем их подкормки, отстрела хищников и т. п.
2) отбором лучших производителей путем отстрела или отлова
отбракованных животных;
3) улучшением хозяйственно-полезных особенностей животных
путем подсадки специально для этой цели лучших производителей;
4) обогащение местной фауны путем акклиматизации новых бо-
лее ценных форм промысловых животных вместо менее ценных и
вселение (ингабитация) их сверх имеющихся уже видов;
941
5) изменение природных условий существования промысловых
животных применительно к созданию наилучших (оптимальных)
условий для размножения ценных животных;
6) замена менее цепных биоценозов другими более ценными био-
ценозами;
7) выведение путем гибридизации новых особенно ценных про-
мысловых животных.
Однако при все возрастающем спросе на пушнину такие пра-
вильно организованные промысловые хозяйства в будущем все же
не в состоянии будут покрыть потребностей на пушнину, и возни-
кает необходимость разведения пушных зверей с промысловой
целью. Так, возникло в начале этого столетия новое направле-
ние животноводства —промышленное звероводство. Некоторые
фермеры острова принца Эдуарда в Восточной Канаде начали уси-
ленно заводить промышленное лисоводство. Уже в 1912 г. число
лисьих ферм на острове достигло 200 с общим числом до 1000
лисиц, из которых более половины чернобурых. Кроме острова
лисьи фермы быстро начинают распространяться в других районах
Канады.
Помимо лисиц в настоящее время с успехом разводят песцов,
хорьков, ондатр, нутрий и др.
Широко организуется разведение пушных зверей у нас в Союзе,
где с этой целью учреждены крупные звероводческие совхозы,
главным образом на севере и в Сибири.
3. Вредные млекопитающие
Вредными млекопитающими являются, во-первых, хищники,
опасные для самого человека или уничтожающие его домашних жи-
вотных; во-вторых, грызуны — враги хозяйства человека, и нако-
нец, в-третьих, разные виды млекопитающих — разносители бо-
лезней человека и домашних животных.
Что касается вреда от хищников, то очень немногие виды
представляют непосредственную опасность для человека. Таковы
тигры, львы, пантеры, волки и отчасти медведи. В настоящее
время этот вред имеет серьезное значение только по’ отношению
к немногим странам, где еще сохранились упомянутые выше
хищники.
Тем не менее, по данным конца прошлого столетия (1899 г.),
людей, заеденных хищниками, насчитывалось 1351 человек.
Лошадей и жеребят.....................147 000
Коров и телят........................ 122 484
Верблюдов.............................. 1 605
Северных оленей....................... 17 083
Мелкого домашнего скота.............. 585 173
Домашней птицы.......................561177
Всего убыток, причинённый хозяйству человека, за один только
год измеряется в 5 млн. рублей.
942
Если сравнить вред, приносимый крупными и мелкими хищ-
никами, то оказывается, что убыток
(В %)
от крупных хищников равняется........ 4 433 000 89
» мелких » » ..... 99000 2
» хищных птиц » • . . . . 427 000 9
Другими словами, мелкие хищники приносят относительно ни-
чтожный вред по сравнению с крупными хищниками из млекопитаю-
щих и даже по сравнению с птицами.
Если принять во внимание, что мелкие хищники приносят од-
новременно пользу, уничтожая гораздо более вредных грызунов,
и дают ценный мех, то станет совершенно ясно, что они не только не
заслуживают названия вредных, а, наоборот, в известных случаях
заслуживают охраны и разведения.
Многие грызун ы являются, наоборот, чрезвычайно опас-
ными врагами хозяйства человека. Помимо двух видов крыс —
черной (Rattus rattus) и бурой, или серой (Rattan
norvegicus) — и домашней мыши (Mus musculus), вредя-
щих преимущественно в домах, амбарах, складах, вагонах и ко-
раблях, и п о левых мышей (Apodemus agrarius) к главным
вредителям из мышевидных грызунов относятся еще полоса-
тая пеструшка (Lagurus lagurus), обыкновенная
полевка (Microtus arvalis), общественная п о л е в-
к а (М. socialis) и рыжая полевка (Evotomys gla-
reolus), лесная мышь «(Sylvemus sylvaticus) и др. Кроме
мышевидных грызунов полям иногда приносят колоссальный вред
суслики (Gitellus) и хомяки (Cricetus).
Размножаемость грызунов колоссальна: черная крыса мечет
за раз от 4 до 8 детенышей, бурая — от 6 до 23. Беременность про-
должается всего лишь 21 день; за время^полового периода в 9 меся-
цев самка может произвести потомство 8 раз. Если считать каждый
приплод по 10 крыс, то за год самка бурой крысы может дать до 80
крыс, а так как молодые самки могут воспроизводить уже после &
недель, то станет понятно, как трудно бороться с крысами. В Анг-
лии было высчитано, что если общее количество крыс, определяе-
мое там в 40 млн. особей, уменьшить вдвое, то уже через 7 лет
они достигли бы прежнего количества.
Приносимые грызунами убытки исчисляются миллионами. В не-
которые годы они >гак размножаются, что становятся страшным бед-
ствием для населения. Так, в 1916—1917 гг. в Австралии мыши так
размножились, что уничтожили одних только запасов пшепицы на
10 млп. рублей. В 1906—1908 гг. крысы в Канаде уничтожили на
площади в 180(10 акров посевы люцерны. В 1914 г. в бывшей Рос-
сии в одном только У йанском уезде Киевской губ. было зарегистри-
ровано 100 тыс. га поля, поврежденного полевками. В 1922 г.
в Сибири было повреждено сусликами свыше 966 тыс. га и мы-
шами — свыше 50 тыс. га. В Волжском районе в 1923 г. было
занято сусликами 3.5 млн? га, а на Кавказе в 1022 г. — 2. 67 млн.
га хлебов.
943
Если приблизительно только, с явным преуменьшением, исчис-
лить вред, приносимый грызунами, и перевести его на деньги, то
получаются удивительные результаты.
Исчислен вред, причиненный крысами в Англии. Оказалось, что
40 млн. крыс Англии, съедая в день на ^копейки, дают в год убы-
ток в 150 млн. рублей; при увеличенном расчете убыток от крыс
Англии доходит до 660 млн. рублей. Во Франции убыток от крыс
в 1904 г. исчислялся в 200 млн. франков, в Германии — в 200 млн.
марок, в Соединенных штатах — в 200 млп. долларов.
В СССР убыток от крыс исчисляется в 234 млн. рублей. При-
нимая в среднем доход одного человека в год в 2000 рублей, мы полу-
чаем, что для содержания одних только крыс у нас работает 117 000
людей.
Естественно, что с грызунами человеку приходится вести упор-
ную борьбу. Меры борьбы с грызунами могут быть преду-
предительными и истребительными.
К первым относится, во-первых, соблюдение чистоты и порядка.
Необходимо, чтобы для крыс и мышей не было доступа к помойным
ямам, разного рода отбросам, отхожим местам и т. д., чтобы все
продукты были убраны так, чтобы крысы и мыши не имели к ним до-
ступа. Необходимо далее, чтобы не было мест, в которых мыши и
крысы могли бы гнездиться и прятаться: разрушенных зданий,
груд камней, закоулков и т. п. Во-вторых, необходимо содержа-
ние хищников, уничтожающих грызунов, как кошки и собаки:
таксы, фоксы, пинчеры, которых следует специально натаскивать
для этой цели. Дикие хищники — хорьки и ласки — уни-
чтожают грызунов массами и селятся иногда около жилья человека.
Очень полезны в отношении уничтожения грызунов также лисицы.
Этих животных необходимо охранять, предоставляя им необходи-
мые для размножения условия.
Многие виды хищных птиц питаются исключительно грызу-
нами: сарычи (Buteo), мелкие соколы (Tinunculus,
Erythropus), сычи (Athene), совы- неясыти (Strix aluco),
ушастая сова (Asio otus) и др.
Истребительные меры борьбы могут быть: меха-
нические (различного рода ловушки, капканы, западни и пр.)
и химические. Эти последние заключаются в том, что гры-
зунов отравляют, или давая им яд в пище: углекислый барий, мы-
шьяк, стрихнин, фосфор, или отравляя их газами: сероуглеродом,
сернистым ангидридом, хлором, фосгеном и другими удушливыми
газами.
Наконец чрезвычайно важным оказался метод истребления гры-
зунов с помощью прйвивки грызунам заразного бактериального за-
болевания — крысиного тифа.
4. Роль млекопитающих в распространении заболеваний
Многие домашние млекопитающие — и на первом месте д о-
машние собаки — являются распространителями . страш-
ной болезни — бешенства; из диких животных распространяет
944
бешенство волк. 'Собака кроме бешенства передает человеку па-
разитического червя из ленточных червей (Cestodes) —
эхинококка (Taenia echinococcus); от собак же происходит
заражение эхинококком домашних животных. Собака заражает
другим лентецом (Т. coenurus) — мозговиком —домаш-
них овец. ‘
Через свиней происходит заражение людей солите-
ром (Т. solium), трихинами (Trichina spiralis).
Крупный рогатый скот является промежуточным
хозяином другого солитера (Т. saginata).
Лошади и крупный рогатый скот служат источником заражения
людей сибирской язвой, а лошади, кроме того, заражают
человека с а п о м.
Крысы способствуют распространению трихин среди свиней
и являются вместе с мышами, сусликами и тарабаганами распростра-
нителями чумы. Переносчиками чумной инфекции от крыс к кры-
сам и человеку являются блохи, но человек может заразиться от
больных крыс и другим путем. Инфекционная желтуха, обуслов-
ливаемая спирохетой, передается также через посредство крыс.
Многие грызуны передают чуму и туларемию: сурки, суслики,
мыши, крысы, зайцы, водяные крысы и другие. Целый ряд болез-
ней отчасти человека, как возвратный тиф и энцефалит, и осо-
бенно болезней домашних животных, как то пироплазмозы,
анаплазмозы и спирохетозы, переносятся с помощью клещей,
которые живут за счет диких млекопитающих, являющихся вре-
менными хозяевами клещей. *
5. Влияние человека на изменение фауны млекопитающих
Влияние человека на распространение млекопитающих гро-
мадно. Оно сказывается прежде всего в том, что человек непосред-
ственно уничтожает некоторые виды зверей.
Так, совершенно уничтожена-человеком жившая в Беринговом
море морская корова (Rhytina gigas), принадлежавшая
к отряду сирен (Sirenia), открытая т\ неудачной экспе-
дицией Беринга и описанная зоологом этой экспедиции Стеллером.
Морская корова многочисленными стадами обитала тогда вблизи
Командорских островов, где питалась на мелкой воде морскими ра-
стениями. Это громадное, животное благодаря своему вкусному мясу
и полной беззащитности дало возможность пережить участникам
неудачной экспедиции Беринга вынужденную стоянку па Командор-
ских обтровах. Прошло каких-нибудь 27 лет, и морская корова была
окончательно уничтожена человекам. Предполагают, что в_ 1768 г.
убит последний экземпляр этого чрезвычайно интересного и ценного
животного, которое несомненно имело все шансы сделаться един-
ственным в мире домашним морским скотом. Стада морских коров
всегда держались на определенных местах *и не уходили с них до
тех пор, пока не был убит в безрассудном преследовании послед-
ний экземпляр.
60 ГСашкароп и Станчинский 945
Совершенно уничтожен человеком дикий бык, один из родона-
чальников пород наших домашних животных — тур (Bos pri-
migenius). Некогда стада диких быков были широко распространены
по всей лесостепной зоне Европы. Преследование туров ради их
вкусного мяса повело в конце концов к тому, что в 1567 г. был убит
последний тур.
Такая же судьба постигла и диких европейских лошадей — тар-
панов. Вероятно не мене'е двух видов диких лошадей жило в
Европе одновременно с первобытным человеком каменного века.
Дикая лошадь была излюбленным охотничьим зверем в то время.
Об этом свидетельствует громадное количество скелетов лошадей,
которые находятся около стоянок первобытного человека. Объяс-
няется это конечно не тем, что мясо лошадей было особенно вкусным,
а тем, что добывать лошадей было легче, чем других крупных
зверей. Очевидно человек умел их загонять в засады еще тогда,
когда у него не было домашней верховой лошади, которая, конечно,
облегчила охоту за ними. Раньше других был уничтожен более
крупный вид лесной европейской лошади, кровь которой чечет еще
в жилах некоторых пород домашних лошадей.
В степях Европы жила другая форма дикой лошади, собственно
тарпап (Е. gmelini), которую по ее преимущественному распростра-
нению можно было бы назвать южнорусской степной дикой лошадью.
Отдельные стада тарпанов, вероятно уже со значительной примесью
крови одичавших домашних лошадей, еще бродили среди безгра-
ничных просторов степей и по лугам лесостепной области сравни-
тельно совсем недавно. В древней Руси за ними усиленно охотятся,
как то видно из «поучения» Владимира ьТопомаха. Но уже во вре-
мена знаменитых экспедиций Академии паук в середине XVIII в.
они становятся редкими, распространенными еще в обширных неза-
паханных степях юго-востока. Последние тарпаны, более похожие
на домашних лошадей, чем на диких, были пойманы в середине
XIX в.
От диких лошадей сохранилась лишь открытая Пржевальским
джунгарская лошадь (Equus przewalskii), малодоступ-
ных полупустынь западной Монголии. Ранее этот вид был
широко распространен по степям и полустепям Азии и заходил
в Европу.
Зубр (Bison bonasus), будучи раньше широко распространен
в зоне лиственных лесов Европы и лесостепи, истреблен почти со-
вершенно. Зубры содержались в заповедной Беловежской пуще
в б. Гродненской губ., где перед войной их насчитывалось до 700
голов. Во время войны большая часть зубров была уничтожена;
незначительное число их сохранилось в Польше и в Германии.;
Другая колония зубров обитала на Северном Кавказе,где они окон-
чательно были уничтожены во время гражданской войны и интер-
венции. Работа по восстановлению зубра ведется успешно в запо-
веднике Аскания-Нова путем поглотительного скрещивания зубра
с гибридами — зубро-бизонами (рис. 674). В настоящее время часть
зубров из Аксания-Нова перегнана в Крымский государственный
946
заповедник для ингабитации. Намечено восстановление зубра в
Кавказском заповеднике.
Почти совершенно были стерты с лица земли человеком и амери-
канские бизоны (Bison americanus). Еще совсем недавно в се-
редине прошлого столетия сотни тысяч бизонов паслись в прериях
на запад от Миссури. Начавшееся потом в связи с проведением же-
лезных дорог истребление бизонов'ради мяса, языков и шкур по-
вело к тому, что в несколько лет бизоны были почти совершенно
уничтожены. В 1889 году их насчитывалось всего лишь 635 голов.
Если бы не специальная охрана оставшихся бизонов и организа-
ция крупных заповедников, они были бы совершенно уничтожены.
В настоящее время количество бизонов сильно увеличилось, и ©ни
считаются уже тысячами.
Рис. 674. Зубр (Bison bonasus).
(Аснания-Нова.)
Почти совершенно был истреблен бобер речной (Castor
fiber), когда-то одно из главных промысловых животных. Чтобы
предотвратить их полное исчезновение, правительством организо-
ваны бобровые заповедники.
Соболь (Martes zibellina) был широко распространен в севе-
ро-восточной таежной части Европейской части Союза и был так обы-
кновенен в Сибири, что туземцы ходили в собольих мехах и подби-
вали лыжи собольими шкурами, а «бабы били соболей коромыс-
лами». В настоящее время -уничтожение соболей привело к тому,
что пришлось организовать соболиные заповедники.
В Африке почти совершенно уничтожены белые н о с © р © ги
(Rhinoceros simus), насчитывающиеся сейчас всего лишь в коли-
честве 20 экземпляров.
Чрезвычайно быстро, идет уничтожение лосей, косуль, изюб-
ров, маралов, кавказских туров и многих антилоп Африки.
Кроме непосредственного истребления, на распространении мле-*
копитающих сказываются изменения в естественных условиях
среды, которые производятся культурной деятельностью человека.
Человек изменяет естественные стации (биотопы)^ к которым при-
60* 947
способлены те или иные виды. Вырубаются леса, распахиваются
степи, осушаются болота. Вместо естественных биотопов возникают
новые биотопы, которые удобны лишь небольшому числу млекопи-
тающих.
Распахивание степи, связанное с уничтожением естественных
биотопов, повело к исчезновению ранее обыкновенных в южпых
степях байбаков (Marmota bobac); вырубание лесов ведет к
сокращению распространения многочисленных видов таежпой и
лесостепной фауны: медведи, рыси, лоси, косули, куницы, многие
другие хищники и даже белки становятся все более и более ред-
кими.
С Другой стороны, обширные площади засеянных полей дают
прекрасные биотопы для мелких грызупов — хомяков, сусликов,
мышей и полевок. Вместе с постройками человека быстро рассе-
ляются приспособившиеся к жилью человека как к биотопу к р ы с ы
и мыши.
Наконец человек изменяет естественные биоценозы. В этом
отношении его влияние сказывается особенно сильно через по-
средство домашних животных. Стада домашних животных, двигав-
шиеся вместе с кочевниками по степям издревле, должны были
влиять на распространепие крупных видов копытных млекопитаю-
щих: лошадей, куланов, антилоп и т. д. На распространении туров
и зубров сказалось разведение домашнего крупного рогатого скота.
Домашние собаки и кошки уничтожают грызупов, птиц и т. д.
Многие ввезенные человеком в другие страны млекопитающие
одичали и превратились в диких млекопитающих, влияние которых
на фауну местных млекопитающих иногда чрезвычайно велико.
Рост влияния человека и его культуры на природу идет так бы-
стро и с таким прогрессирующим ускорением, что дальнейшая
судьба дикой природы, флоры и фауны, особенно крупных млеко-
питающих, оказывается предопределенной.
Необходимо разумное вмешательство человека в этот процесс
разрушения природы и уничтожения ее богатств, и прежде всего
необходимо принять меры к сохранению тех видов крупных зве-
рей, исчезновения которых можно ждать со дня на день.
Дальнейшее существование дикой фауны, большей части круп-
ных млекопитающих, возможно будет только в специально органи-
зованных резерватах — заповедниках.
Что же касается прочей территории, то забота человека должна
быть направлена к тому, чтобы всемерно содействовать размноже-
нию полезных видов и всячески препятствовать размножению вред-
ных. Самотеку в изменении фауны должна быть противопоставлена
плановая перестройка фауны с заменой бедных биоценозов более
богатыми с видами ^хозяйственно и культурно цепными.
Встает задача о науке совершенно новой, науке о конструиро-
вании биоценозов и фаун, о плановом в государственном и миро-
вом масштабе перераспределении диких животных с той целью,
чтобы максимально использовать их в интересах хозяйства и- куль-
туры. Роль заповедников при этом громадна.
348
ЗАПОВЕДНИКИ СССР
л и <• и гд II и к и являются государственными учреждениями,
। i\ih.iiniiMii для охраны и восстановления природных ресурсов
। |||Г1г|1гг.1\социалистического хозяйства; естественные ландшафты
и .ынпцгдппках сохраняются как примеры естественных при-
родных условий и тем помогают понимать происходящие в природе
под цоздгйствием культуры изменения географической среды
и.) 1ПГГО хозяйства; являясь лабораторией в природе, заповедники
до<|/1ли.1 наметить и разработать пути и методы работ по измене-
нию природного комплекса в интересах человека, его хозяйства и
нулвтуры. .
Заповедники СССР, занимая наиболее интересные и по большей
части чрезвычайно живописные участки природы в разных угол-
ках нашей обширной родины, имеют огромное культурно-прос-
ветительное значение; они являются базой для развития туризма
широких масс трудящихся и подготовки научно-исследователь-
ских кадров в области географических и биологических дисцип-
лин, в особенности по зоологии позвоночных.
В настоящее время в СССР организовано 46 государственных
заповедников союзного и республиканского значения, с общей
площадью свыше 10 млн. гектаров. Из них мы назовем только
те, которые имеют особенно большое значение для сохранения,
восстановления и акклиматизации млекопитающих и птиц.
Заповедники Европейской части СССР
1. Лапландский заповедник. Площадь 200 тыс. га. На-
ходится в западной части Кольского полуострова в полосе
лесотундры. Дикий северный олень, лось, лесная куница, росо-
маха и др.; проводится реакклиматизация речного бобра и аккли-
матизация опдатры.
2. Заповедник Семь островов. У Мурманского побережья.
Гнездовья гаги и «птичьи базары».
3. Печерско-Ы л ычский. 1000 тыса га. Восточноевро-
пейская тайга. Кидас (помесь соболя и куницы), соболь, лесная
куница, колонок, песец, цепные кряжи лисицы, росомаха, север-
ный олень, лось и др.
4. Белорусский. 72 дас. га. В Белоруссии между рр.
Зап. Двина и Березина. В зоне смешанных лесов. Речной бобер,
лось, косуля, кабан, лесная куница, медведь и др.
Б. Центрально-лесной. 34 тыс. га. В Калининской обл.,
на водоразделе Зап. Двины и Волги. Притаежная подзона зоны
смешанных лесов. Лось, косуля, лесная куница, медведь, рысь
и др. Проводится ингабитация бобра.
6. Выхухолевые заповедники: Окский по р. Пре,
41 тыс. га; Клязьминский на р. Клязьме, 14 тыс. га; Хо-
перский па р. Хопер, 17 тыс. га. Служат для сохранения и
восстановления выхухоли. В Окский и Хоперский заповедники
завезены пятнистые олени и бобры.
«49
7. Мордовский. 60 тыс. га. Лесной массив зоны смешан-
ных лесов, вклинившийся в лесостепь. Лось, куница, выхухоль
и др. Произведен запуск речных бобров, сибирской косули и
пятнистого оленя.
8. Воронежский. 30 тыс. га. На рр. Усманка и Ивпица;
в лесостепной полосе. В основном это — бобровый заповедник.
Имеется единственное в РСФСР стадо благородного европейского
оленя.
9. Куйбышевский. Состоит из пяти участков в среднем
Поволжье общей площадью 24 тыс. га; в зоне лесостепи. Лоси.
Завезены пятнистые олени.
10. Ильменский. 41 тыс. га. На Ильменском хребте в
Южн. Урале. Горные леса и многочисленные горные озера. Лоси,
косули; завезены пятнистые олени. В основном минералоги-
ческий заповедник.
11. Башкирский. 56 тыс. га. Горные леса Южн. Урала;
сибирская косуля, лось.
12. Степные заповедники: Центрально-черноземный.
3.7 тыс. га: из трех участков в Курской обл.; Аскания-
Нова в левобережной У крайне; там же акклиматизационный парк.
13. Черноморские заповедники. В низовьях Днепра
несколько участков по берегу Черного моря и некоторые острова.
Места гнездовья водоплавающей птицы и места скопления птиц
на перелетах.
14. Астраханский. 23 тыс. га. В дельте р. Волги. Ка-
бан, разнообразные птицы: пеликаны, колпики, каравайки,
большая и малая белые цапли, фламинго, разнообразные утки;
массовые и колониальные гнездовья, скопление водоплавающей
птицы на пролетах; нерест промысловых рыб.
15. Крымский. 40 тыс. га. В Крымских горах. Крымский
олепь, косуля, акклиматизировавшиеся муфлоны; завезены для
акклиматизации из Аскания-Нова зубро-бизоны.
Заповедники Кавказа
16. Кавказский. 337 тыс. га. Горные леса и альпийские
луга западного Кавказа; кавказский олень, кавказская косуля,
тур Северцова, кавказская серна, кабан, лесной кот, барс,
кавказский медведь, куницы лесная и белодушка, кавказский
тетерев, горная индейка и др.
Намечается восстановление истребленного здесь во время
интервенции зубра.
17. Тебердипский. 62 тыс. га. Горные леса, альпийские
луга и ледники восточной части Зап. Кавказа. Акклиматизация
пятнистого оленя, енотовидной собаки и белки-телеутки.
18. Кизыл-Агаче кий. 120 тыс. га. Занимает 5-километро-
вую полосу вокруг аалива того же наименования на берегу Кас-
пийского моря. Место массовых зимовок водоплавающих птиц
и некоторых куликов и цапель.
950
Заповедники Азиатской части.СССР
19. Кондо-Со свей ск ий. 800 тыс. га. В верховьях рек,
давших название заповеднику, между Уральскими горами и р. Обь.
Западносибирская тайга. Уральский соболь, речной бобер, лось,
северный олень; произведен запуск ондатры и американской норки.
20. Алтайский. 1000 тыс. га. Горы Алтая к северу от
Телецкого озера; горная тайга, горные озера. Марал, северный
олень, кабарга, сибирская косуля, соболь, колонок, горный
сурок — тарабаган и др.
21. Сих отэ-Ал инский. 1500 тыс. га. На хребте того же
наименования в Приморской области ДВК. Смешанные и листвен-
ные уссурийские и южноуссурийские леса с характерной флорой.
Уссурийская фауна Китайско-Гималайской подобласти. Соболь,
харза, манджурский и гималайский медведи, уссурийский тигр,
леопард, енотовидная собака, пятнистый олень, изюбрь, лось,
сибирская косуля, кабарга, горал, восточный кабан и др.; из
птиц — каменный глухарь, дикуша, фазан и много других инте-
ресный видов уссурийской фауны.
22. Варгузинский. 700 тыс. га. На восточном берегу оз.
Байкал. Восточносибирская горная тайга. Варгузинский соболь,
имеющий чрезвычайно ценный темный мех; фауна восточносибир-
ской тайги.
23. Кроноцкий. Свыше 1 000 тыс. га на Камчатке; камчат-
скаЯчшйга. Из млекопитающих особенно ценен камчатский соболь,
медведь, снежный баран и др.
24. Алма-Атинский. 800 тыс. га. В Казахстане. Горы и
предгорья Заалийского Алатау. Горные леса, альпийские луга.
Марал, косуля, архар, горный козел, кабан, дикобраз; улар,
фазаны и др.
25. Ак-Су-Джебаглы. 50 тыс. га. В горах Таласского
Алатау Казахской ССР. Горные леса и альпийские луга. Архар,
горный козел, косуля, кабан, барс, каменная куница и др.
26. Наурзумский. Всего 300 тыс. га в двух участках.
Степная полоса Казахской ССР. Пресные и соленые озера. Степ-
ные млекопитающие, много гнездящихся и пролетных водопла-
вающих птиц.
27. Саянский. 1100 тыс. га.Изюбрь, кабарга, лось, соболь и др.
28. Кандалакшский. 10 тыс. га. Гнездовья гаги, леж-
бища тюленей: гренландского, нерпы, морского зайца.
IX. ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ ПТИЦ И МЛЕКОПИ-
ТАЮЩИХ
Если, как мы видели, жизпь птиц и млекопитающих находится
в самой теслой зависимости от условий окружающей среды, то по-
нятно, что распространение и распределение их будут зависеть
прежде всего от наличия подходящих для жизни условий: подхо-
дящего климата, наличия необходимых биотопов и т. п.
Однака^сследование границ распространения отдельных видов
951
легко убеждает нас в том, что влияние всех этих факторов хотя и
очень велико, но его часто бывает недостаточно для объяснения со-
временного распространения и распределения животных.
Современное распространение животных может быть понято
только в том случае, если изучить распространение животных
в исторической перспективе.
В процессе эволюции, как мы видели, все время возникали но-
вые формы животных, которые, естественно размножаясь, расселя-
лись за пределы своей родины все дальше и дальше, всюду, куда
могли проникнуть и где находили для себя подходящие условия;
они изменялись вновь, давали новые формы, лучше приспособлен-
ные к условиям существования, или приспосабливались к иным из-
менившимся условиям существования; эти новые формы в свою
очередь распространялись, тогда как родоначальные формы, как
менее приспособленные, вымирали в борьбе за существование, если
им не удавалось сохраниться где-либо на отделившемся от материка
уединенном острове.
Так в продолжение бесчисленных веков происходили распростра-
нение, изменение и приспособление к‘ разным условиям существо-
вания различных групп животных, встречавших на своем пути
распространения разного рода преграды в-виде морей, высоких гор
и т. п. На отделившихся от материка островах эволюция могла итти
своим путем, равно как и на тех материках, которые надолго или
окончательно (т. е. до сего времени) теряли связь с другими матери-
ками. Чем раньше произошло такое отделение части суши, будь то ма-
терик или остров, от других материков, тем своеобразнее должна быть
на пих фауна в силу изменений в совершенно изолированных усло-
виях существования и тем более древний характер она должна носить.
Отсюда ясно, что всякая более или менее продолжительная изо-
ляция той или иной территории обусловливала своеобразное изме-
нение фауны ее, и чем раньше происходила и чем продолжительнее
была эта изоляция, тем своеобразнее становилась фауна.
В разъединенных фаунах при одинаковых экологических усло-
виях самостоятельно приспособлялись к одинаковым условиям су-
ществования разные группы животных, образуя параллельные эко-
логические ряды или, как их называют в зоогеографии, викарные
группы (виды, роды, семейства).
При новом соединении разобщенных частей суши фауны должны
были смешиваться, причем приспособленные к одинаковым усло-
виям формы неизбежно должны были вступать между собой в обо-
стренную борьбу за существование, в которой менее приспособлен-
ные гибли, уступая место более приспособленным. В результате
должна была появиться крайне пестрая и сложная картина, в ко-
торой чрезвычайно трудно разобраться.
Однако при детальном изучении отдельных фаун, при сравнении
их друг с другом и с фаунами прежних геологических периодов
удается* выяснить общность развития фаун значительных террито-
рий и проследить их историю.
Те фауны, которые отличаются резкими чертами, т. е. такие,
952
в которых мы находим только им одним свойственных (эндемич-
ных) представителей целых подклассов или отрядов и большого
числа семейств, очевидно давно получили самостоятельное напра-
вление в развитии, были отделены от других фаун и явно носят
в себе черты большей или меньшей древности.
Такие фауны принадлежат высшей единице зоогеографических,
подразделений, которые мы называем’ царством.
Приходится установить три царства: нотогейское, нео-
гейское и арктогейское. Каждое из них развивалось про-
должительное время совершенно изолированно,и эта изоляция нача-
лась с определенного исторического момента глубокой древности.
НОТОГЕЙСКОЕ ЦАРСТВО llllllllllllll Эфиопская область ПТТПТ! Восточная область
НЕОГЕЙСКОЕ« д Мадагаскарская область | | Голарнтичесная область
АРКТОГЕЙСКОЕ ЦАРСТВО
Рис. 675. Карта зоогеографических подразделений.
Царства в свою очередь могут быть подразделены на области,
области подразделяются дальше на отделы, на подобласти и дальше
на провинции и участки.
Этц зоогеографические подразделения устанавливаются, как
мы видели, на основе тех различий, которые зависят от историче-
ских причин. Экологические современные условия тут не играют
роли (см. карту зоогеографических подразделений — рис. 675).
1. Нотогейское царство
Оно состоит, во-первых, из Австралии с Тасманией и Новой Гви-
неей с прилежащими мелкими островами; во-вторых, сюда отно-
сился Новая Зеландия и многочисленные острова, которые в це-
лом носят название Полинезии; в-третьих, сюда по большей части
1 953-
относят острова Молуккские, лежащие на запад от Новой Гвинеи.
•Эти территории имеют свои отличительные черты, причем
особенности Новой Зеландии и Полинезии настолько своеоб-
разны и существенны, что их необходимо выделить в отдельные
области.
Острова же, лежащие к западу от Новой Гвинеи и Молуккских
островов, характеризуются чертами, в которых сказывается их
переходный характер к фауне Азии, причем элементы Азии в них
господствуют. Таким образом нотогейское царство слагается из
трех самостоятельных областей — Новозеландской, Полинезий-
ской и Австралийской; третья же переходная полоса должна быть
выделена в особую подобласть и отнесена скорее к другому
царству — арктогейскому.
Новозеландская область, кроме обоих островов Новой Зелан-
дии, охватывающая также острова Окленд, Кампбел и Чатам,
уже по одному тому заслуживает выделения в самостоятельную
зоогеографическую единицу, что в ней отсутствуют свои млеко-
питающие, за исключением летучих мышей и одного вида крысы
(Rattus exulans maorium). Отсутствие наземных млекопитающих
в этой области — явление первичное, т. е. нужно считать, что их
там и не было никогда, другими словами, что Новая Зеландия
отделилась от материков в то время, когда млекопитающие еще не
успели окончательно выработаться как класс и распространиться,
т. е. еще в меловое время.
Два рода летучих мышей и маорийская крыса проникли в Но-
вую Зеландию вторично. Первые залетели туда, а крыса была за-
везена случайно.
Этому древнему обособлению обязана также оригинальная фауна
птиц Новой Зеландии и прилежащих островов, состоящая из зна-
чительного числа только ей свойственных семейств и даже отрядов
птиц.
Наличие очень древпей группы рептилий (Rynchocephalia)
вполне согласуется с древним отделением Новой Зеландии.
Так, только Новой Зеландии принадлежит отряд бескилевых—
бескрылы или кивы (Apteryges), представленные современным се-
мейством Apterygidae и ископаемым семейством моа — Dinorni-
thidae. В Новой Зеландии эндемичны семейства попугаев —
несторов (Nestoridae), совиных попугаев (Stringopidae) и новозеланд-
ских крапивников (Xenicidae). Имеется значительное число укло-
няющихся родов птиц, как, папример, кривоносый кулик (Апа-
rhynchus), водяной пастушок (Ocydromus) и целый ряд воробьи-
ных (Turnagra, Greadion и др.).
Характерно большое число нелетающих форм с недоразвитыми
крыльями, как киви (Apteryx), — водяной пастушок (Ocydro-
mus), совиный попугай (Stringops), султанские курочки (Notornis),
из которых белая султанская курочка вымерла, и ряд других
вымерших нелетающих птиц: своеобразный гусь (Cnemiornis) и бе-
скилевая гигантская Dinornis, или моа, близкая современным
киви (рис. 676). Своеобразие фауны Новой Зеландии и указанных
S54
островов вполне оправдывает выделение ее в особую фаунистиче-
скую Новозеландскую область.
Ряд общих форм Новой Зеландии и Австралии показывает, что
фауна первой обязана своим происхождением этому ближайшему
материку.
Полинезийская область, куда мы относим все острова, лежа*
щие в Тихом океане между Южной Америкой и Австралией, из-
вестные под сборным названием Полинезии, начиная от острова
Новая Каледония и
Ново-Гибридских ост-
ровов на север, севе-
ро-восток и восток до
Гавайских островов и
островов Товарище-
ства включительно,
имеет то общее с Но-
возеландской обла-
стью, что и ей чуж-
ды наземные млеко-
питающие. Из этого
класса ей свойствен-
ны только летучие мы-
ши, с одним эндемич-
ным родом. Что ка-
сается фауны птиц, то
она состоит частью
из типичных остров-
ных форм, т. е. по-
томков случайно за-
несенных на острова
животных, или из ви-
дов, которые способ-
ны к продолжитель-
ным странствованиям
через моря. Мы от-
носим полинезийскую
фауну к нотогейско-
му царству не только
потому, ЧТО там ОТ- Рис. 676. Моа (Dinornis) (реставр.).
сутствуют наземные
млекопитающие, но и потому, что среди птиц ей присущи формы,
несомненно, очень древнего происхождения, как, например, кагу
(Rhinochetus jubatus) на острове Новая Каледония; кагу принад-
лежит особому подотряду (Rhinocheti) отряда журавлей (Grues) и,
может быть, заслуживает выделения в особый отряд; на острове
Самоа обитает крючкоклювый голубь (Didunculus strigirostris),
несомпенно очепь древнего происхождения птица, выделяемая в
особое семейство (Didunculidae); имеется немалое число уклоня-
ющихся родов птиц, рассеянных по разным островам.
955
На Гавайских островах, которые можно выделить в особую
подобласть, имеется эндемическое семейство воробьиных птиц —
Drepanididae, образовавшееся сравнительно недавно из семейства
вьюрков. Такое же недавнее происхождение имеют многие эндеми-
ческие роды и виды полинезийской фауны, образовавшейся из
выходцев Старого и Нового Света северного полушария.
Австралийская область. Австралия с Тасманией, Новой Гвинеей
с Молуккскими островами и островами Бисмарка (Новая Британия),
Соломоновыми и другими прилегающими мелкими островами харак-
теризуются весьма своеобразным миром млекопитающих и птиц.
Прежде всего этой области принадлежит замечательный отряд одно-
проходных (Monotremata), который выделяют в особый подкласс
млекопитающих (Prototheria), представляющий собой древнейшую
и примитивнейшую группу млекопитающих, ближайшими родствен-
никами которой считают ископаемых первичнозубых (Protodonta).
Однопроходные представлены тремя родами и двумя семействами.
К первому (Ornithorynchidae) относится утконос (Ornithoryn-
chus anatinus), роющее водное животное с вытянутой наподобие
утиного клюва мордой и одетыми перепонкой ногами. Он живет
в Квинсленде, к югу от 18° южной широты, в Новом Южном Ва-
лисе, в Виктории, в Южной Австралии и в Тасмании.
Семейство ехидн (Echidnidae) представлено двумя родами:
пятипалой ехидной (Echidna aculeata), распространенной от
юго-восточной Новой Гвинеи через Австралию до Тасмании, и
трехпалой ехидной (Zaglossus), которая встречается только в
Новой Гвинее.
Так как оба семейства известны лишь из плейстоцена Австра-
лии, то современных однопроходных приходится считать коренными
аборигенами этой области, развившимися, вероятно, еще в мезо-
зое в качестве отдельного ствола.
Другой замечательной особенностью Австралийской области
является пышное развитие там другого отряда млекопитающих —
сумчатых (Marsupialia), которых тоже с полным правом вы-
деляют в особый подкласс — Metatheria.
Современные сумчатые распадаются на три подотряда: на менее
специализированных плотоядных и насекомоядных (Polyprotodon-
tia) и растительноядных (Diprotodontia). Третий подотряд цепо-
лестовые (Caenolestoidea) занимает промежуточное место.
Полипротодонтиые сумчатые распадаются на шесть семейств:
бандикуты, язвицы, илй сумчатые барсуки (Peramelidae), хищ-
ные и насекомоядные сумчатые куницы, или шерстохвосты
(Dasyuridae), сумчатые волки (Thylacinidae), сумчатые кроты
(Notoryctidae), сумчатые мурашееды (Myrmecobiidae) и сумчатые
крысы, или двуутробксйч^ (Dydelphyidae). Последнее семей-
ство принадлежит исключ ^дьно Америке, тогда как пять осталь-
ных ограничены в своем распространении только - Австралийской
областью, где образуют большое количество видов.
Дипротодонтные сумчатые представлены чрезвычайно большим
числом родов и видов разнообразных, преимущественно раститель-
956
поядпых млекопитающих, которых объединяют в три семейства.
Первое семейство (Macropodidae) объединяет кенгуру и близкие
к ним формы, а также кенгуровых крыс; второе семейство содер-
жит сумчатых медведей (Phascolarctidae); третье семейство
(Phalangeridae) объединяет кускусов (Phalanger), кузу (Trichos-
urus) и сумчатых мышей и белок (Petaurus, Petauroides, Acrobates).
Все эти сумчатые живут исключительно в Австралийской об-
ласти. Ценолестовые (Caenolestidae), куда относятся три рода
мелких сумчатых, принадлежит Южной Америке.
Во время плейстоцена фауна сумчатых в Австралии была еще
богаче: особенно поразительно было развитие крупных сумчатых:
гигантских кенгуру, вомбатов, хищных, величиной с леопарда Thy-
lacoleo, исполинских Dyprotodon и др.
Рис. 677. Динго (Canis dingo).
(Из Брэма.)
Что касается других отрядов млекопитающих, то из них в Ав-
стралийской области мы находим немногих грызунов (Rodentia),
все из надсемейства мышевидных (Murioidea) из эндемичного се-
мейства Hydromyidae: водные Hydromys, близкие к нему Xeromys,
роды Coniluru^, Mastacomys; очень крупные Mallomys и др.;
несколько видов широко распространенного космополитического
рода Mus, вероятно, завезенных; из парнокопытных (Artiodactyla)
единственный вид свиней — Sus papuensis; из хищных (Carnivora),
единственный вид собаки — Canis dingo (рис. 677) и, наконец, из ле-
тучих мышей (Chiroptera) — представителей всех семейств, за исклю-
чением американского семейства листоносых (Phyllostomidae).
Нахождение всех этих высших млекопитающих в Австралийской
области стоит в противоречии с остальной фауной зверей области,
состоящей из однопроходных и сумчатых млекопитающих, и за-
ставляет думать, что проникновение в Австралийскую область не-
летающих плацентарных млекопитающих произошло в более позд-
нее время частью, вероятно, вместе с человеком, частью путем-слу-
чайного заселения посредством плавающих в море деревьев; лету-
чие же мыши могли сюда залететь.
957
Ископаемые остатки однопроходных нигде за исключением плей-
стоцена Австралии найдены не были. Что же касается распростра-
нения сумчатых, то в мезозойскую эру менее специализированные
предки полипротодонтных сумчатых были широко распространены
по северному царству (Arctogea) как в Сев. Америке, так и в Ев-
ропе, но вымерли здесь в начале третичной эры. Дольше других
сохраняются формы, близкие к северноамерикапскому опоссу-
му, — в Европе до начала миоцена, а в Сев. Америке до олигопепа.
Очевидно сумчатые проникли в Австралию еще в конце мезозой-
ской эры, тогда, когда не успели дифференцироваться еще другие
отряды высших плацентарных млекопитающих. Другое место, где
сохранились потомки древпих полипротодонтных сумчатых, — это
Южная Америка (рис. 678).
Рис. 678. Карта распространения однопроходных, сумчатых-крыс и прочих
сумчатых.
Дипротодонтные сумчатые^ очевидно, возникли в Австралии.
А так как в Южной Америке мы находим в отложениях Сан-
та-Круц в Патагонии относящихся к нижнему миоцену остатки
близких к ним форм, то приходится думать, что еще во время по
крайней мере эоцена Южная Америка находилавъ в соединении
с Австралией.
Таким образом, фауна млекопитающих нотогеи — это сохра-
нившаяся до наших дней измененд^^рауна конца мезозойской эры,
дополненная позднее немногими Случайными переселенцами. Это
и оправдывает выделение ее в самостоят'ельно^царство.
Из птиц Австралийской области ’характеры эндемичный отряд
(Casuarii), куда относятся бескилевые казуары и эму и целый
958
ряд эндемичных семейств, из которых упомянем: медососов (Meli*
phagidae), ласточковых сорокопутов (ArtamicLae), кукушечьих со*
рокоиутов (Campophagidae), беседковых птиц (Ptilonorhynchidae),
лирохпостов (Menuridae) и др.
Из богатого видами отряда попугаев характерны семейства
лори (Loridac), какаду (Cacatuidae); из голубей, которые в Австра-
лии очень многочисленны, характерно семейство крупных венце-
носных голубей (Gouridae).
Такое своеобразие фауны птиц Австралийской области вполне
подтверждает предположение о древнем мезозойском происхожде-
нии се фауны.
Австралийскую область делят на две подобласти: Австралийскую
и Папуасскую.
В состав Австралийской подобласти входит почти вся
Австралия за исключением лишь самой северной ее части, остров
Тасмания и прилежащие к материку и Тасмании острова. Для нее
эндемичны утконосы (Ornithorynchidae) и большое число родов
и видов сумчатых [мурашеед — (Myrmecolius), сумчатые куни-
цы — Dasyurus, сумчатый волк — (Thylacinus), сумчатый барсук
(Pcrameles), кузу —(Trichosurus), вомбат —(Pascolaretos) и др.].
Из птиц для Австралийской подобласти эндемичны семейства эму
(Dromaeidae), лирохвосты (Menuridae), сем. Atrichornitidae и Stre-
peridae. В общем Австралийская подобласть является наиболее-
типичной частью области, в которой в большей степени сохрани-
лись ее архаические черты и менее сказалось влияние других фаун.
Папуасская подобласть, охватывающая самую северную-
часть Австралии, Новую Гвинею, Молуккские, Соломоновы острова
и острова Бисмарка (Новая Британия), наоборот, подвергались
значительному влиянию фауны арктогеи, а потому ее архаические
черты перекрыты позднейшим заселением территории выходцами из.
Восточной области. Однопроходные представлены здесь ехиДнамиг
причем род Zaglossus эндемичен для подобласти. Сумчатые здесь,
представлены менее полно, причем древесными лазящими формами
в связи с экологическими условиями (лесные тропические дебри).
Из птиц для подобласти характерны казуары (Casuaridae), боль-
шеногие (сорные) куры (Megapodidae), венценосные голуби . (Gou-
ridae), а из попугаев: какаду (Cacatuidae), широкохвостые (Platyce-
riidae) и лориевые (Loridae); из воробьиных—райские птицы (Рага-
diseidae), питтовые (Pittidae) и др.
2. Неогейское царство
После выделения Австралийской, Новозеландской и Полине*
зийской областей в самостоятельное царство самой своеобразной
страной по своей современной и ископаемой фауне млекопитающих
является Южная Америка.
Поэтому ее приходится выделить в особое фаунистическое цар-
ство, состоящее из одной области — Неотропической.
Неотропическая область/ Она охватывает всю Южную Аме-
рику с Фалкландскими, Галапагосскими и Вест-Индскими остро-
• 959'
вами и Центральную Америку до Мексиканского плоскогория па
севере.
Своеобразие неотропической фауны выражается прежде всего
в том, что мы находим здесь представителей сумчатых млекопи-
тающих (Marsupialia).
Эндемичное семейство сумчатые крысы (Didelphyidae) пред-
ставлено несколькими родами, среди которых центральное место
занимает опоссум (Didelphys), с рядом видов, распространенный
до Соединенных штатов. В отложениях Санта-Круц представлено
было еще другое семейство полипротодонтных сумчатых (Micro-
biotheriidae). В этих же отложениях найдены остатки других
сумчатых, которых приходится отнести к семейству сумчатых ку-
ниц (Dasyuridae). Кроме упомянутых полипротодонтных сумчатых,
в тех же отложениях найдены остатки близких к дипротодонтпым
сумчатым, из которых одно семейство (Caenolestidae) представлен о
в настоящее время тремя родами, живущими в Аидах от Экуадора
до Чили, из которых наиболее известен мышеобразный Caenole-
stes fuliginosus; это семейство выделяется в отдельный подотряд
Caenolestoidea.
Другой замечательной особенностью рассматриваемой области
является наличие эндемичного отряда своеобразных плацентарных
млекопитающих, . именно американских неполнозубых (Хепаг-
thra).
Сюда относятся броненосцы, муравьеды и ленивцы — самые
удивительные из всех современных южноамериканских млекопи-
тающих. Родоначальники их происходят из нижнего эоцена Сев.
Америки, где, однако, скоро вымирают. Наибольшего своего раз-
вития неполпозубыс достигают в миоцене, плиоцене и плейстоцене
Южной Америки. Их можно разделить па два больших ствола:
на группу муравьедов и ленивцев — подотряд беспанцырных (Ani-
canodonta) и группу броненосцев—подотряд паи-цырные (Hicano-
donta).
В настоящее время муравьеды, которых следует выделить в от-
дельное семейство (Myrmecophagidae), представлены тремя вида-
ми, которых относят к трем родам, а ленивцы—семейством
(Bradypodidae), с двумя родами: двупалые (Choloepus) и трехпа-
лые ленивцы (Bradypus). Начиная с олиго^ена, миоцена и позднее,
до плейстоцена включительно, были весьма многочисленны пред-
ставители вымершего подотряда тихоходов (Gravigrada). Это были
огромные и массивные травоядные жи^тные, достигавшие вели-
чины слона, с длинным хвостом, н^который опи опирались.
Таковы Megalonyx, Mylodon, Grypothenum.. Этот последний дожил
до появления в Южной Америкс^еловека. Имеются данные счи-
тать, что человек даже охотилс^Да пим и употреблял в пищу
этого гиганта. •
Броненосцы представлены в настоящее время несколькими ро-
дами одетых панцирем средней величины животных, которых
относят к особому семейству (Dasypodidae). Они распространены
в настоящее время до Мексики включительно и известны с древпей-
•JGO
iiiiix третичных отложений. Ископаемые другого семейства —
глиптодонтов (Glyptodonta), вымершего уже в послетретичное
время, известны с олигоцена и были тогда чрезвычайно многочис-
ленны, причем некоторые из них достигали очень крупных раз-
меров (до 5 м длины) и были одеты сверху сплошным толстым пан-
цы рем.
Отряд насекомоядных (Insectivora) совершенно отсутствует
на материке, если не считать двух землероек, которые, вероятно,
недавно проникли в Гватемалу и Коста-Рику из Сев. Америки.
На Вест-Индских островах, однако, имеется отдельное и очень
своеобразное семейство щелезубов (Solenodontidae), близких к ма-
дагаскарскому семейству тепреков (Centetidae).
Наоборот, отряд грызунов (Rodentia) представлен в Южной
Америке очень богато и притом значительным числом эндемичных
семейств и родов.
Из группы белкообразных (Sciuroidea) в Южной Америке пред-
ставлены только белки, перекочевавшие сюда, очевидно, из Сев.
Америки. Из группы мышеобразных (Myoidea) имеются только
космополитическое семейство мышиных (Muridae) и одни род пе-
рекочевавших с севера мешотчатых крыс (Geoinyidae)1.
Группа дикобразных грызунов (Hystricoidea), наоборот, пред-
ставлена в Иеотропической области очень богато. Сюда относятся
четыре эндемических семейства: семейство морских свинок (Cavii-
dae) с настоящими морскими свинками (Cavia), патагонскими
свинками (Dolichotis) и водосвинками (Hydrochoerus); семейство
агутиевых (Dasyproctidae) с агути (Dasyprocta) и пака (Coeloge-
nus) семейство Dinomyidae, с единственным родом, и семейство
шиншилловых (Chinchillidae) с шиншиллой (Chinchilla) и внека-
шей (Viscacia). Семейство иглошерстов (Erethizontidae) представле-
но в Иеотропической области древесными дикобразами (Coend и),
а семейство осьмизубых (Octodontidae ) имеет в Южной Америке
эндемичное подсемейство Octodontinae с несколькими весьма хпрак
терпыми родами, как туко-туко (CtcnoniyH), дегу (Oclodoii)
и др.; из подсемейства Caproinyinae характерны болотные бобры
(Myocastor), ежскрысы (Caproniys). Подотряд двунарнорезцоных
(Duplicidentata) представлен Двумя видами зайцев.
Хищные (Carnivora) гораздо менее характерны,• чем грызуны,
хотя и представлены в настоящее время довольно богато. Интересно
полное отсутствие гиеновых (Hyaenidae) и виверровых (Viver-
ridae) и почти полное отсутствие куницевых (Mustelidae), за исклю
чением выдр (Lutra,) и вонючек (Mephitis). Семейство кошачьих
представлено пумой (Felis 'concolor), ягуаром (F. овса) и рядом
других видов, не характерных в смысле распространения.
Собаки (Canidae) представлены в числе других нехарактерных
видов эндемичным родом кустарнйх собак (Icticyon). Медведи
(Ursidae) имеют своего представителя в виде Ursus ornatns. Кио-
ты (Procyonidae) распространены как в Неотропической области,
так и в Сев. Америке и в Восточной области. Опи, как и другие
представители отряда, проникли в Южную Америку с севера.
61 Кашкаров и Станчинский 961
В высшей степени характерными для фауны рассматриваемого
царства являются копьпные (Ungulata), именно ископаемые фор-
мы их, которые не менее своеобразны, чем рассмотренные выше
неполнозубые.
В настоящее время мы находим в Южной Америке из отряда -
парнокопытных (Artyodactyla) пекари (Dicotyles), подсемейства,
широко распространенного в третичное время в Сев. Америке и
Азии, Dicotylinae; ламу, гуанако и викунью (Lama) из верблю-
довых (Camelidae); оленей из американского рода Cariacus. Из не-,
парнокопытных (Perissodactyla) в области имеются тапиры (Tapirel-
1а), ранее в миоцене и плиоцене весьма широко распространенные
в Сев. Америке, Европе и Азии, г в настоящее время представлен-
ные еще другим родом на Малайских островах н в Индо-Китае.
Лошади (Equus) были введены в Южную Америку человеком
и там одичали. Однако в плиоцене имеются остатки более древних'
лошадей (Hippidium, Onohippidium), которые перебрались сюда
из Сев. Америки.
Чрезвычайно своеобразны ископаемые формы копытных, кото-
рые приходится отнести к трем отрядам: Protungulata; Notungu-
lata с четырьмя подотрядами и Litopterna. Свое происхождение
они ведут от проникших еще в нижнем эоцене из Сев. Америки
первичных копытных (Protungulata), которые развивались здесь
совершенно независимо в разных направлениях частью очень резко
отличающихся от прочих копытных, частью удивительно повторя-
ющих их эволюцию. Подотряд Pyrotheria, вымерший раньше дру-
гих в плиоцене, имел животпых величиной со слона, и по стро-
еншыягрепных зубов напоминал динотериев. Прочие Notungulata
вымерли всего лишь в плейстоцене; частью опи совмещали в себе
признаки копытных и грызунов и были небольшой величины (под-
отряд Typotheria), частью (в подотряде Taxodontia) напоминают
гиппопотамов и носорогов, хоботных и грызунов и были различной
величины — до весьма значительных размеров (крупнее носорогов).
Из эоценовых первичцых копытных Protungulata в эоцене воз-
никли и быстро эволюционировали своеобразные непарнокопыт-
ные Litopterna: семейство трехкопытных болотных Macrauchcnii-
dae и быстробегавших подобно лошадям однокопытных Proterothe-
riidae. Оба семейства вымерли лишь в плейстоцене.
Из вообще очень космополитического отряда рукокрылых (Chi-
roptera) Неотропическая область вс> же имеет характерных для
нее летучих мышей из семейства листоносов (Phyllostomidae),
одно подсемейство которых заменяет экологически подковоно-
сов Старого Света, два других — плодоядных летучих мышей,
тогда как подковоносы и плодоядные летучие мыши там совер-
шенно отсутствуют.
Чтобы покончить с характеристикой млекопитающих рассмат-
риваемого царства, остается сказать про обезьян (Primates).
Полуобезьяны (Prosimiae) совершенно отсутствуют даже в
ископаемом виде, тогда как собственно обезьяны (Simiae) принад-
лежат к особому подотряду широконосых обезьян (Platyrrhina)
962
и представлены двумя эндемичными семействами: когтистыми1
или игрунками (Hapalidae), и цепкохвостыми (Cebidae).
Оценивая изложенные данные о фауне млекопитающих Неотро-
ппческой области с исторической точки зрения, мы должны конста-
тировать некоторую общность ее истории с историей Австралии.
Как и Австралия, Южная Америка очепь рано теряет свою связь
с большей частью суши, па которой под нивелирующим действием
обмена фаунами различных частей с различными экологическими
условиями происходила более быстрая и разносторонняя эволюция
млекопитающих. Благодаря этому разобщению, которое произош-
ло в начале третичной эры (позднее чем в Австралии), Южная
Америка сохраняет примитивных млекопитающих вроде полипро-
тодонтпых сумчатых и вырабатывает целый ряд своеобразных отря-
дов,, подотрядов и семейств, которые после установившейся в плио-
цене связи с Сев. Америкой большей частью быстро вымирают под
наплывом проникнувшей с севера фауны с крупными хищниками
во главе.
Особенности фауны птиц Неотропической области подтверждают
ее архаический характер.
Мы имеем там эндемичный отряй палеогнатических птиц нан-
ду, или американских страусов (Rheae), ближайшие родственники
которых известны в ископаемом состоянии в отложениях Мадага-
скара и Новой Зеландии. Очевидно, что эта очень древняя группа
птиц во вторичную эру была широко распространена в южном
полушарии; второй отряд палеогнат, тоже эндемичный, — это
тинаму (Tinami), курообразные летающие птицы древнего проис-
хождения.
Отряд пингвины (Sphenisci) тоже принадлежит только этой
области вместе с полярной антарктикой и островами. Эндемичны
подотряды солнечные цапли (Eurypygae), близкие к своеобраз-
ной новокаледонской кагу (Rhinochetes), тоже очень древний под-
отряд отряда журавлей (Grues), и подотряд кариамы (Cari-
amae).
Чрезвычайно интересный отряд шпорцевых гусей, или пала-
медей (Palamedeae), близкий к отряду гусей (Anseres); подотряд
американских грифов (Cathartae), отряд гоацинов (Opistocomi)
и подотряд колибри (Trochili) отряда стрижей (Cypseli) — все эти
своеобразные, уклоняющиеся с признаками древнего обособления
группы, принадлежащие или исключительно Южной Америке,
или частично в последнее время проникшие в Сев. Америку,
в достаточной степени подтверждают необходимость выделения ее
не только в особую область, по и в особое царство. Прибавим
к этому перечню отрядов и,подотрядов те семейства, которые
являются эндемичными для Неотропической области. Таковы се-
мейства трубачей (Rsopliidae) из журавлей; Conuridae из попу-
гаев; момотов (Momotidae) и тоди (Todidae) из сизоворонковых;
гуахаро (Steatornithidae), из козодоев; якуар (Galbulidae) и тука-
нов (Rampliastidae) из дятловых; тираннов (Tyrannidae), мопаки-
нов (Pipridae), котипг (Cotingidae), древолазов (Dendrocolaptidae);
61* *963
муравьедов (Formicariidae), американских мухоловок (Conopopha-
gidae), Pteroptochidae и некоторые другие из воробьиных.
Неотропическая область делится на четыре подобласти: 1) Бра-
зильская — включает, кроме Бразилии, Гвиану, Вен’ецуэлу,
Колумбию, Экуадор, Парагвай и части Перу и Боливии к востоку
от Андов; она имеет наиболее характерных представителей области;
2) Чилийская — охватывает Чили, Аргентину, Уругвай,
Патагонию и западные части Перу и Боливии; опа характеризуется
наличием рода лама, очкастого медведя, коипу, вискаши, мелких
броненосцев и некоторых других; 3) Антильская — охва-
тывает Вест-Индские острова (за исключением острова Тринидада)
и, помимо большей бедности фауны, связанной с островным харак-
тером ее, и присутствия североамериканских форм, характери-
зуется наличием эндемичного семейства из отряда насекомоядных
щелезубых (Solenodontidae) и рядом эндемичных родов и видов;
4) Мексиканская — занимает Центральную Америку* от
Панамского перешейка до южной Мексики включительно и характе-
ризуется своим переходным положением между двумя царствами.
3. Арктогейское царство
Вся остальная суша должна быть отнесена к третьему царству.
Различие между ее отдельными частями, как бы они удалены ни
были, всегда значительно меньше, чем с перечисленными выше об-
ластями. Это объясняется тем, что в продолжение третичной эры
развивавшиеся в различных частях ее самостоятельные фауны пе
оставались разобщенными и подвергались нивеллирующему дей-
ствию обмена. Более обостренная борьба за существование обусло-
вила б шиш энергичную эволюцию, благодаря которой вырабаты-
вались более новые и разнообразные формы, вытеснявшие древние
группы млекопитающих. Так, совершенно вымерли еще в мезо-
зойскую эру близкие к однопроходным первичнозубые
(Protodonta), близкие к сумчатым многобугорчат ые
(Multituberculata), вымерли, не оставив до пас потомков, с у м ч а-
т ы е (Marsupialia), вымерли и отдельные отряды — первобытные
представители разных групп: трехбугорчатые (Panto-
tlieria), ти л л о’д онт ы (TiTlodonta), креодонты (Creo-
donta), первичные копытн ьге (Protungulata). Вымерли
многие уклоняющиеся в сторону ветви млекопитающих, вроде
анцилоподов (Ancylopoda) и др., тогда как некоторые из
них, как ящеры (Pholidota), трубкозубы (Tubuliden-
tata), пока еще сохранились в незначительном числе. Вымерли и
уступили место новым многочисленные переходные и уклоняю-
щиеся семейства. ,
Отдельные части обширной территории рассматриваемого фау-
нистического царства, однако, прошли различную историю, и в ме-
стах, более удаленных от арены наиболее обостренной борьбы и бы-
строй эволюции, сохранились некоторые черты более примитивных
и архаических фаун. Это и дает нам возможность говорить о подраз-
делении всего царства на несколько областей.
964
Области эти следующие.
Мадагаскарская область. Мадагаскар с прилежащими остро-
вами: Коморскими, Маскаренскими, Сейшельскими и Амирапскими,
хотя и представляет очепь незначительную территорию и близко
лежит от материка Африки, тем не менее имеет настолько
своеобразную фауну, что заслуживает выделения в особую об-
ласть.
Характерная особенность маммологической фауны Мадагас-
карской области заключается, с одной стороны, в ее отрицательных
чертах: на Мадагаскаре пет таких характерных для материка Аф-
рики млекопитающих, как антилопы, жирафы, носороги, слоны,,
бегемоты, свипьи, большие кошки, павианы, человекообразные
обезьяны; с другой стороны —па Мадагаскаре мы находим громадное
^количество полуобезьян (Prosimiae), как ни в одной из других
стран, хищные млекопитающие представлены семейством виверр
(Viverridae), насекомоядные — своеобразными тенрекаАми (Centeti-
dae), которые здесь дали и водных, и древесных, и роющих насе-
комоядных, а грызуны — эндемичными родами космополитического
семейства мышей (Muridae).
Полуобезьяны имеют па Мадагаскаре не менее 36 ныне живу-
щих видов, которые относятся к 11 родам и двум семействам
(вместе с огромными ископаемыми Megaladapidae — к трем). Кроме
Мадагаскара, полуобезьяны обитают в Африке в количестве двух
родов и в Восточной (Индийской) области — в количестве трех ро-
дов, тогда как в эоцене и нижнем олигоцене Европы и Сев. Аме-
рики полуобезьяны были обыкновенны и вымерли к началу мио-
цена. Виверры (Viverridae) на Мадагаскаре, кроме своеобразной
фоссы (Cryptoprocta), которую вследствие особенной, как у львов,
зубной формулы выделяют в особое подсемейство, представлены
еще родом настоящих виверр (Viverrinae), ближайшие родствен-
ники которых находятся в Восточной области, и четырьмя видами
мангуст (Herpestinae), из которых один мелкий вид (Eupleres)
заслуживает тоже выделения в особое семейство. Большое коли-
чество видов тепреков (Centetidae, 6 родов) указывает па древность
мадагаскарской фауны, так как ближайшие им родственные формы
(щелезубые) принадлежат Вест-Ипдским островам.
Фауна Мадагаскарской области представляет, таким образом^
весьма своеобразную фауну, близкую к эоценовой и олигоцеповой
фауне Голарктической области, которая, очевидно, в то время со-
ставляла коренную фауну Африки. Отделение Мадагаскара
от материка должно было произойти не раньше и не позднее начала
миоцена, когда новая индийская фауна копытных и хищных еще
не успела проникнуть до юга Африки.
Этот эоценовый и олигоценовый характер Мадагаскара и заста-
вляет считать его самостоятельной областью. Остатки вымершего
носорога и наличие одного вида свиней из рода речпых свиней (Ро-
tamochoerus) заставляет считать их случайно пробравшимися на
остров (переплывшими) уже позднее, а землеройку (Crocidura) —
туда завезенной.
965
Из птиц, кроме ископаемых рокки (Aepyornis), составляющих
особый отряд, близкий новозеландским моа 'и американским
нанду, и вымершего дронта (Didus) с островов Маврикия и Бур-
бона, мы паходим особый отряд пастушковых куропаток (Mesites)
и целый ряд эндемичных родов из сизоворонок, славок, мухоло-
вок и др. В то же время на Мадагаскаре отсутствуют такие ха-
рактерные для Африки семейства, как бапаноеды (Musophagidae),
мышиные птицы (Coliidae), птицы-носороги (Bucerotidae) и др.
Эфиопская область. Границы этой области в общем совпадают
с границами Африки за исключением севера, где вся страна к . се-
веру от тропика Рака относится к северпой — Голарктической
области, тогда как лежащая к югу от тропика часть Аравии долж-
на быть отнесена сюда.
Отметим прежде всего некоторые отрицательные черты эфиоп-
ской фаупы, так как они тоже характеризуют отличия ее от 'Восточ-
ной и Голарктической областей.
В Эфиопской области совершенно отсутствует семейство оле-
ней (Cervidae); почти совершенно отсутствует подсемейство коз
и овец (Caprovinae), так как два вида этого семейства принадле-
жат прилежащим к Голарктической области частям. (роды Capra
и Heimtragus); пет в Эфиопской области медведей (Ursidae) и ено-
тов (Procyonidae), настоящих свиней (род Sus), семейства бобров
(Castoridae), полевок (Microtinae), пищух (Ochotonidae), байбаков
(Marmota), сусликов (Citellus) настоящих землероек (Sorex), кро-
тов (Talpidae).
С другой стороны, характерны следующие группы млекопитаю-
щих: человекообразные обезьяны здесь представлены шимпанзе
(Antropopithecus) и гориллой (Gorilla), причем важно знать для
понимания генезиса эфиопской фаупы, что ископаемые остатки
шимпанзе пайдепы в плиоцене Ийдии.
Собакоголовые обезьяны (Cercopithecidae) имеют пять родов,
ограниченных в своем распространении Африкой, причем род
павианов (Cynocephalus) встречается в’ ископаемом виде в плио-
цене и плейстоцене Индии. Йолуобсзьяпы (Prosimine) представле-
ны двумя эндемичными родами^
•Насекомоядные имеют два семейства исключительно африкан-
ских: прыгунчиков (Macroscelididae) и золотых кротов (Chryso-
chloridae). Первое семейство имеет близких родичей в олигоцене
Европы, а златокроты близки к мадагаскарским тенрекам.
Третье семейство насекомоядных — это крупные водные выд-
рообразные землеройки (Potamogalidae), один род которых встре-
чается также па Мадагаскаре.
Из хищных кошки представлены общими с Восточной областью
двумя родами и четырьмя видамп: виверры тоже являются общей
для обеих областей группой, но имеют эндемичные роды в Африке.
Характерен земляной волк (Proteles), выделенный в особое
подсемейство (Protelinae), семейства гиеп (Hyenidae); последние
были широко распространены по Арктогейскому царству еще в
плейстоцене.
966
Из собак характерны два рода: гиеновая собака (Lycaon) и
кафрская лисица (Otocyon); интересно, что из семейства купиц
(Mustelidae) медоеды (Mellivora) представлены также и в Индии.
Из грызупов в Эфиопской области нет настоящих летяг, ко-
торые заменены аномальными (Anomaluridae), настоящие белки
(Sciurus) нехарактерны, а род мелких белок (Nannosciurus) яв-
ляется общим для рассматриваемой области и Малайского архи-
пелага. Далее, в Эфиопской области имеются эндемичные роды
сонь (Myoxidae), мышей (Muridae), слепышей (Spalacidae) и три
рода семейства восьмизубых (Octodontidae), столь характерных
для Неогейского царства.
Особенно же характерны представители сверхотряда копыт-
ных млекопитающих.
Из парнокопытных в Эфиопской области имеются два эндемич-
ных семейства и большое * число эндемичных родов. Семейство
гиппопотамов (Hippopotamidae) принадлежит в настоящее время
исключительно данной области, тогда как в плиоцене и плейсто-
цене было распространено в Европе и Азии, а также было раньше
и на Мадагаскаре; второе эндемичное семейство — жирафы (Giraf-
fidae) — в пижпем плиоцепе было представлено в Евразии. Се-
мейство свиней представлено эндемичным родом бородавочников
(Phacochocrus), и семейство полорогих (Bovidae) хорошо предста-
влено эндемичными родами подсемейства антилоп (Antilopinae),
некоторые из них в ископаемом состоянии найдены в Индии.
Из семейства оленьков (Tragulidae) род водяных оленьков
(Dorcatherium) встречается исключительно в Западной Африке,
ио в миоцене обитал в Европе, а в плиоцепе найден в Индии.
Из непарнокопытных носороги (Rhinocerotidae) представлены
несколькими видами, ближайшие к пим формы были в плиоцене
и плейстоцене, обыкновенны в Европе и Азии; а из семейства
лошадей (Equidae) в Африке мы находим эндемичных полосатых
лошадей: кваг и зебр (Hippotigris).
Очень характерен для Африки с Сирией здесь только встречаю-
щийся отряд дамапов (Hyracoidea).
Слопы (Elephantidae), ранее в плиоцепе и плейстоцене обык-
новенные в Европе н сохранившиеся по сие время в Индии, пред-
ставлены широко распространенным видом ^эндемичного рода
(Loxodon).
Чрезвычайно интересен исключительно африканский своеобраз-
ный трубкозуб (Orycteropus), относящийся к особому отряду
Tubulidentata. Из отряда ящеров (Pholid.ota) Эфиопская область
имеет несколько видов.
Из сказанного о фауне Мадагаскара и Африки видно, как сло-
жилась эфиопская фауна. К сожалению, даппых об ископаемой фа-
уне Африки в третичное время мы почти пе имеем. Во всяком слу-
чае, судя по Мадагаскару, в олигоцеповое время фауна Африки ре-
шительно отличалась от ее теперешней фауны; там отсутствовали
обезьяны тропических лесов и крупные хищники, пе было там и
современного обилия и разнообразия копытных.
967
Только в плиоценовое время, после отделения от Африки Ма-
дагаскара, в Африку проникает сложившаяся в это в^емя в Евра-
зии фауна. При этом более детальное изучение распространения
отдельных форм и сравненпе пыне живущих с ископаемыми плиоце-
новыми формами разных частей Европы и Азии убеждают пас в том,
что движение в Африку плиоценовой евразийской фауны проис-
ходило неодновременно. Сперва в Африку проникает фауна тропиче-
ских лесов с человекообразными обезьянами, а затем уже, после из-
менения климата в сторону большей сухости с образованием сте-
пей, — фаупа антилоп, лошадей, носорогов, слонов и крупных хищ-
ников.
Характерными эфиопскими птицами являются: отряд двупалых
страусов (Strutiones), подотряд секретарей (Serpentarii), отряда
дневных хищных птиц и целый ряд семейств, подсемейств и ро-
дов птиц: цесарки (Numidinae) из куриных, настоящие попугаи
(Psittacini), бапаноеды (Musophagidae), африканские удоды (Irri-
sarinae) из сизоворонковых, птицы-мыши (Colii) из дятловых;
нектарки (Nectariniidae) и ткачики (Ploceidae) из воробьиных
принадлежат преимущественно к этой области.
Фауна Эфиопской области носит, таким образом, ясно выражен-
ный миоценовый и плиоценовый характер. Этим и оправдывается
выделение ее в особую зоогеографическую область.
В Эфиопской области различают три подобласти: западно-
африканскую, восточноафриканскую и южноафри-
канскую, отличающиеся различной степенью воздействия па эту
область пришельцев из Евразии. Самая древняя фаупа—это фауна
южпой' АфрикЯкс златокротами, зсмляпым волком и виверрами.
Восточная, или Индийская область. Занимает юго-восточную
часть Азии, а именно Индостан до альпийской зоны Гималаев пасе-
вере и до горных хребтов южной Персии и Белуджистана на за-
паде, Ипдо-Китай до верхнего течения Меконга и водораздела между
Ян-цзе-киангом и Хуап-хе, а также острова Цейлон, Формозу,
Филиппинские, Суматру, Яву, Борнео и мелкие острова Зондского
архипелага к западу от острова Лембока и Целебеса. Что касается
Целебеса и мелких островов, к западу от о. Тимора, то вопрос
об их припадлежности к той или иной области всецело зависит от
того, какому элементу придавать более важное значение: присут-
ствию ли там сумчатых животных или присутствию высших млеко-
питающих. Мы придерживаемся той точки зрения, что фауна этих
Островов есть фауна Восточной области со значительной примесью
австралийских животных, и потому она должна быть отнесена
к Восточной области и выделена в самостоятельную переходную
подобласть.
В общем фауна Индийской области очень близка к Эфиопской
области, образовалась путем смешения миоценовой и плиоценовой
фауны с местной эоценовой и олигоценовой. Индийская область
сохраняет более чисто плиоценовую фауну со значительной приме-
сью более древних форм, и примесью более молодых плиоценовых
животных, перешедших из Голарктической области.
968
Человекообразные обезьяны (Antropomorphae) представлены
в лице оранг-утана (Simia), в настоящее время распространен-
ного на Суматре и Борнео, но в плиоцене жившего в Индии.
Эндемично семейство гиббонов (Hylobatidae), в миоцене принад-
лежавших Европе.
Собакоголовые обезьяны (Cercopithecidae), представленные
макаками (Macacus), заходящими и в Голарктическую область,
где, равно как и следущий род
тонкотелых обезьян (Semno-
pithecus), встречались в Ев-
ропе в плиоцене; сюда же от-
носятся эндемичные носатые
обезьяны (Nasalis) с острова.
Борнео. Полу обезьяны (Рго-
simiae), кроме лемуров (Nyc-
ticebus и Loris), представле-
ны еще долгопятами (Таг-
siidae).
Отряд шерстокрылов (Der-
moptera) эндемичен: летучий
маки (Geleopithecus) принад-
лежит Малайской подобласти.
Насекомоядные имеют здесь
эндемичное семейство тупай
(Tupajidae, рис. 679), заменяю-
щее здесь семейство хоботных
насекомоядных Африки; ежи
представлены эндемичными
родами ежекрыс (Gymnura и
Halomys), а землеройки —
родом Soriculus.
Отряд хищных представ-
лен здесь большим числом ко-
шек (Felidae) и большим чис-
лом эндемичных родов.виверр
(Viverridae), тогда как соба-
ки области нехарактерно; мед- Рис. 679. Тупайя перохвостая (РШо-
ведиг представлены тремя ви- cercus lowi).
дами, из которых большой ин- (Из Брэма->
дийский й цейлонский мед-
ведь относятся к отдельному роду (Melursus). Очень интересны
распространенные здесь в восточных Гималаях и Бирме панды
(Ailurus), представители семейства енотов (Procyonidae), круп-
ный вид которых найден в плиоцене Англии, тогда как представи-
тели данного семейства принадлежат Новому свету. Из семейства
купиц (Mustelidae) имеются один вид медоедов (Mellivora), сви-
ные барсуки (Arctomyx) и эндемичные роды (Helictis и Mydaus).
1'Из семейства мышей (Muridae) в Восточной области не менее^
одиннадцати родов эндемичны, а род — кустарных крыс (Galun-
969
da)—общий Ипдии и Эфиопской области. Из семейства белок
характерен живущий на острове Борией род длинноухих белок
(Rhithrosciurus), а мелкие белки из рода Nannosciurus представ-
лены несколькими видами; типичные белки (Sciurus) многочислен-
ны в Малайской подобласти, а летяги представлены, кроме голар-
ктического рода (Sciuropterus), еще эндемичным родом (Pteromys).
Семейство дикобразов (Hystricidae) имеет па острове Борнео
эндемичный род (Trichys).
Из сшряда парнокопытных представители' рода Bos Восточной,
области отличаются некоторыми общими признаками; далее для
области характерны короткррогие козы (Hemitragus), антилопы:
пильгау (Boselaphus), гарна (Antilope), четырехрогая (Tetra-
ceros) и некоторые другие. В общем по сравнению с Эфйопской
областью Восточпая область бедпа антилопами в настоящее время,
тогда как в плиоценовое время антилопы были здесь многочисленны
и разнообразны. Настоящие козы и овцы для области нехарактерны,
и встречающиеся в северной части представители принадлежат
Голарктической области.
Отличительной особенностью рассматриваемой области по срав-
нению с предыдущей является многочисленность представителей
семейства олепей (Cervidae). Кроме центрального рода оленей
(Cervus), характерного для Голарктической области, здесь имеется
свой эндемичный род оленей мунтжак (Cervulus), из которого
только один заходит в Тибет.
Из оленьков (Tragulidae), столь обыкновенных в олпгоцене
и миоцепе Европы, в Восточной области имеет широкое распро-
странение род Tragulus, известный здесь с плиоцена.
Свиньи (Suidae) представлены многочисленными видамп; носо-
роги (Rhinocerotidae) имеют три вида, отличающихся общим приз-
наком — присутствием развитых резцов. Интереспо, что пх иско-
паемые остатки находят в плейстоцене вместе с остатками эфиоп-
ских носорогов. Верблюды (Camelidae) в диком состоянии в
области отсутствуют, по в плиоцепе имеются остатки верблюдов
рода Camel us.
Лошади (Equidae) представлены дикими ослами; несколько ви-
.дов известны из плейстоцена Ипдии.
На полуострове Малакка и на островах Суматре и Борнео жи-
вет тапир (Tapirus indicus). Индийский слон (Elephas indicus),
который очень широко распространен по области, близок к иско-
паемому мамопту. В плиоцене и плейстоцене имеются остатки дру-
гих видов слопов.
Отряд ящеров (Pholidota), распространенный также в Эфиоп-
ской области, представлен здесь несколькими видами.
Птицы Восточной области чрезвычайно разнообразны, по пе бо-
гаты эндемичными семействам^. Большого разнообразия достигает
фауна куриных: настоящие куры (Gallus), фазаны, павлины;
из попугаев характерно подсемейство ошейниковых попугаев (Ра-
laeornithidae); многочисленны распространенные и в Африке птицы-
носороги (Bucerotidae); характерны семейства воробьиных: широ-
ко
короты (Eurylaemidae), питты (Pittidae), многие бюльбюль (Рус-
nonotidae) и ряд эндемичных родов.
Итак, Индийская область по составу и происхождению своей
фауны очень близка к Эфиопской области, отличается от последней
наличием более молодых выработавшихся в верхнем плиоцене
форм, присутствием древних со времени миоцена коренных групп,
а также отсутствием еще более древних (олигоценовых, эоценовых)
форм, характерных для Эфиопской области.
Индийская область делится на следующие подобласти: И п-
д п й с к у ю, И н д о - К и т а й с к у ю, Малайскую, Фи-
липпинскую и Целебесскую.
Голарктическая область. Вся остальная часть северной суши
с прилежащими островами относится к одной области, которая не-
смотря па большое протяжение имеет так много общего в своей
фауне, что может быть подразделена дальше лишь на единицы мень-
шего значения. Эта общность фауны легко характеризуется как по-
ложительными, так и отрицательными чертами, что в полном согла-
сии с данными палеонтологии указывает на общность ее истории.
Для Голарктической области, именно для ее обеих частей: для
Сев. Америки и Старого света, характерны следующие общие
им семейства: кротовые (Talpidae), бобровые (Castoridae), туш-
канчиковые (Jaculidae). Землеройковые кроты (Urotrichus) пред-
ставлены двумя близкими видами в Сев. Америке и Японии. Из
хищных эндемичен род рысей (Lynx), распространенный по всей
области; род белых медведей (Talassarctos) свойствен обоим полу-
шариям; бурые медведи* (Ursus) тоже эндемичны и представлены
близкими видами. Также широко распространен род росомах
(Gulo).
Из грызунов семейство летяг (Pteromyidae), хотя и имеет
большое число малайских видов, тем не менее широко распростра-
нено по всей Голарктической области, а суслики (Citellus) и бай-
баки (Marmota) эндемичны и характерны для нее; также распро-
странен в обоих полушариях и бурундук (Tamias).
Целиком голарктическим является подсемейство полевок (Mic-
rotinae). Исключительно голарктическим является также подсе-
мейство мышовок (Sitistinae); американские мышовки представ-
лены в Китае близким родом; эндемично и семейство пищух
(Ochotonidae).
Из копытных в плейстоцене был распространен по всей Голар-
ктической области мускусный бык (Ovibos), который в настоящее
время в Старом Свете вымер; род бизонов представлен в Сев. Аме-
рике бизоном (Bison americanus), а в Европе — зубром (Bison
bonasus).
Чрезвычайно сближает фаупу Сев. Америки (неарктическую)
с фауной голарктической части Евразии (палеарктической) наличие
общих для обеих стран видов. Таковы волк (Canis lupus), ли-
сица (Vulpes vulpes), песец (Alopex lagopus), бурый медведь
(Ursus arctos), белый медведь (Talassarctos maritimus), горно-
стай (Mustela erminea), лось (Alces alces), вапити, или марал
971
(Cervus canadensis), северный олень (Rangifer tarandus) и мно-
гие другие.
Из птиц эндемичным для Голарктики является подотряд га-
гар (Colymbi) и отряд чистики (Alcae) и семейство тетеревиных
(Tetraonidae).
Общими для Голарктики является также большое число родов
и даже видов птиц. Таковы сороки (Pica), вороны (Corvus), кед-
ровка (Nucifraga), сорокопут (Lanins), конек (Anthus), рога-
тый жаворонок (Eremophila), дрозд (Turdus), сипица (Parus),
гаичка (Pfecile), поползень (Sitta), пищуха (Certhia), королек
(Regulus), чиж (Spinus spinus), чечевица (Erythrina) и др.
Так как, однако, между фауной самой северной части Голарк-
тики обоих материков и фаунами более южной части Сев. Аме-
рики с одной стороны и более южной голарктической частью евро-
пейско-азиатского материка, с другой стороны имеются существен-
ные различия, которые особенно резко выражены в южных частях
Голарктики обоих материков, что обусловлено особенностями
исторического хода развития их фаун, то приходится эти части
выделить в фаунистические единицы — подобласти неравноценного
значения. В свете современных данных зоогеографии прежнее
упрощенное деление Голарктики па Неарктику и Палеарктику
должно быть оставлено: история фауны разных частей Голарктики
была различна. Так, более южная часть Сев. Америки к югу от
40° северной широты, в отличие от расположенной к северу, пе
подвергалась воздействию ледникового периода; па пей значительно
слабее также „сказывалось нивелирующее' воздействие северных
соединений Америки с Азией и Европой, и потому развитие ее фауны
шло самобытным путем. С другой стороны, соединение Южной и
Северной Америки, имевшее место в древпетретичное время и осо-
бенно в последнее время в плиоцене, сохранившееся до наших дней,
обусловило вторжение в южные части Сев. Америки неотропических
форм. Южную часть Сев. Америки поэтому следует выделить в осо-
бую Сонорскую подобласть.1 Лежащая с северу от этой
подобласти часть Сев. Америки, за исключением крайнего севе-
ра приполярной части материка, должна быть выделена во вто-
рую подобласть — Канадскую, или Неарктическую в узком
смысле/
В Старом Свете вся Европа, северная Африка, лежащая к северу
от тропика Рака и вся Азия за исключением южной части Аравии
к югу от тропика, которая относится к Эфиопской области, а также
Индокитая и Индостана, относящихся к Восточной области, вхо-
дят в состав так называемой Палеарктики. Ранее ее выделяли в от-
дельную фаунистическую область. Ее. северная часть находилась
с конца плиоцена под сильным воздействием климатических изме-
1 Это влияние фауны Южной Америки на фауну Сонорской подобласти
так велико, что является возможным рассматривать Сонорскую, Неаркти-
ческую, Палеарктическую и Арктическую подобласти как отделы, подчинив
им более низкие подразделения как подобласти. ,
972
пений ледникового периода, тогда как в южных частях влияние
ледника сказывалось значительно слабее. Здесь сохранились в боль-
шей степени плиоценовые элементы фауны, причем в западной по-
ловине южной части Палеарктики на ее фауну большое влияние
оказывала эфиопская фауна Африки, и па востоке — индийская
фауна. Этим обусловливается деление Палеарктической части
Голарктики на три подобласти: Средиземноморскую, или
Южную, Китайско-Гималайскую, или Восточную и
Европейско-Сибирскую, или Северную.
Крайний север приполярной части Голарктики представлен
фауной, которая находилась под наибольшим воздействием клима-
тических влияний ледникового времени, более или менее одинаково
сильно выраженных в обоих полушариях. Эта фауна представляется
единой кругополярной фауной, которая поэтому заслуживает вы-
деления в особую- Арктическую подобласть.
Сонорская подобласть охватывает переходную фауну
Сев. Америки от границы Неогеи, лежащей около тропика Рака
на берегах Тихого и Атлантического океанов и языком выдаю-
щейся к югу по плоскогорью Мексики до границы Канадской
подобласти, лежащей к северу приблизительно до 40° север-
ной широты в США. Фауна характеризуется присутствием таких
форм, которые сближают ее с фауной Южной Америки, как,
например, опоссумы (Didelphyidae), броненосцы (Dasypodidae),
пекари (Dicotyles), носуха (Nasua), еноты (Ргосуон) и др. Энде-
мичными для Сонорской подобласти являются семейство бобровых
белок (Haplodontidae), подсемейство вилорогих антилоп (Antilo-
caprinae) и несколько родов насекомоядных, летучих мышей и
грызунов.
Фаупа птиц Сонорской подобласти носит тоже переходный ха-
рактер, благодаря большому числу вторгшихся в Сев. Америку
неотропических форм.
Таковы славки из сем. Mniotiltidae, тиранны (Tyrannidae),
кассики (Icteridae), попугаи из сем. Conuridae, колибри (Trochi-
lidae) и некоторые другие. Эндемичны подсемейство индеек (Ме-
leagrinae) и целый ряд родов птиц; семейство зубатых куропаток
(Odontophoridae) в основном является сонорским.
Канадская, или Неарктическая, подобласть, помимо не-
которых общих с Сонорским отделом родов и видов имеет не-
сколько эндемичных родов, из которых упомянем болотных земле-
роек (Neosorex), звездорылых кротов (Condylura), ондатру (Fiber
zibethicus), иглошерста (Erethizon), снежную козу (Oreamnos).
В остальном фаупа млекопитающих очень сходна с таковой Па-
леарктики.
Фауна птиц Канадской подобласти тоже очень близка к фаупе
Палеарктического отдела, но имеется немало характерных ро-
дов, как степные тетерева (Tympanuchus, Pediocetes и др.),
своеобразные кукушки (Coccyzus), пересмешники подсемейства
Miminae и семейство синицеобразных крапивников (Chamaeidae)
и др.
W3
Палеарктический отдел Голарктики охватывает гро-
мадную область — всю .Европу, Северную Африку, Аравию до
тропика Рака, Малую, Среднюю и Центральную Азию до Гималаев
и водораздела между Хуан-хе и Ян-цзе-киангом, остров Сахалин,
Японские и Курильские острова.
Для этого отдела характерны: куторы (Neomys), путораки
(Diplomesodon), тибетская кутора (Nectogale), выхухоль (Desma-
па), кроты (Talpa), некоторые виверровые, хомяки (Cricetus),
мушловки (Muscardinus), земляные зайцы (Alactaga), тушканчики
(Dipus), слепыши (Spalax), настоящие мышовки (Sicista), большое
число видов козлов (Capra) и овец (Ovis), кабарга (Moschus),
косули (Capreolus), серпа (Rupicapra), сайга (Saiga) и, наконец,
некоторые макаки (Macacus).
Южная, или Средиземноморская, подобласть
охватывает страны, окружающие Средиземное море, Пиренейский
полуостров, Апеннинский полуостров, Малую Азию, Северную
Африку, Аравию до тропика Рака, Кавказ, южную часть Крыма,
Персию, Афганистан, Белуджистан, Среднюю и Центральную Азию,
включая Монголию и Тибет.
Фауна Средиземноморской, или, правильнее, Южной, подобла-
сти представляет собой реликт плиоценовой фауны, измененной
влиянием ледникового периода; кроме того, здесь сохранились так-
же некоторые древние архаические формы третичной эры.
Из обезьян мы находим здесь бесхвостую мартышку (Macacus
inuus); насекомоядные представлены прыгунчиками (Macroscelidae)
и тибетской куторой (Nectogale); грызуны представлены гунди (Cte-
nodactflus), дикобразом (Hystrix), эндемичными видами байба-
ков (Marmota), су с л идо в (Citellus), пищух (Lagomys), полевок
(Microtinae) и мышей (Murinae).
Из хищпых мы находим крупных кошек: львов (Leo), которые
иедавпо были широко распространены, тигров (Tigris), паптер
(Leopardus pardus), ирбиса (Felis uncia), гепарда (Acinonyx juba-
tus), длиннохвостую кошку (Felis caudata), каракала (Caracal
caracal), манула (Otcolobusmanul), камышевого KOTa(Catolynx), ры-
сей (Lynx pardellus, L. isabellina), купиц (Martes intermedia),
виверр: ихневмона (Herpestes ichneumon) и генетту (Genetta де-
лена); шакала (Canis aureus), полосатую гиену (Hyaena), медве-
дей (Ursus leuconyx, U. lagomyarius).
Из парнокопытных весьма характерны многочисленные виды
козлов (Сарга) и барапов (Ovis), серны (Rupicapra), яки (Poepha-
gus), верблюды (Camelus), олени (Cervus affinis, С. hagenbecki),
антилопы (Gazella).
Из парнокопытных интересны дикая лошадь Пржевальского
(Equus przewalskii) и дикие ослы (Asinus hemionus, A. kiang, А.
onager) (рис. 680). Из характерных птиц мы назовем настоящих
фазанов (Phasianus), тетеревидных фазанов (Tetraophasis), рябков
(Pterocles), попыток (Syrrhaptes), горных индеек (Tetraogallus),
дроф красоток (Houbara), грифов (Vulturidae) и многочисленных
характерных воробьиных, из которых укажем: пустынных соек
074
(Podoces) (рис. 681), горных вьюрков (Chinospiza, Leucosticte).
синичек (Leptopoecile) и др. На фауне Египта в значительной
степени сказывается влияние фауны тропической Африки, а в.
Афганистан проникают многие индийские виды.
Рис. 680. Онагр (Asinus onager).
(Из Гаане.)
Гималайско-китайская, или Восточная,,
подобласть занимает Уссурийский край, Манчжурию, Се-
веро-восточный Китай до водораздела между реками Хуан-хе и Ян-
цзе-киангом, Ганьсу, Гималаи
до Кашмира включительно
и Японские острова. Фауна
этой подобласти носит следы
сильного влияния Индийской
области. Здесь мы находим
обезьян из рода макак (Ма-
cacus, гульманов (Semnopi-
thecus); из хищных имеется
особый род медведей (Ailu-
ropus), тибетский медведь (Ur-
sus tibetanus), красный волк
(Суon), волосатый леопард Рис. 681. Пустынная сойка (Podoces
(Leopardus pardus viHosus), pander!).
енотовидная собака (Nycte- <Из гаане.)
reutes), амурская лесная кош-
ка (Oncoides), мангуста (Herpestes) и др.; из грызунов харак-
терны черный заяц (Caprolagus brachyurus),. гималайский байбак
(Marmota hymalayensis), ряд видов пищух (Lagomys), козлы, и»
975-
которых интересен Capra falconnen—випторогий козел, несколько
видов оленей (Pseudaxis sica, Р. hortulorum, С. cashmirensis), анти-
лопа горал (Nemorchaedus caudatus).
Из птиц эта фауна характеризуется обилием фазанов (ушас-
тые — Crossoptilon, блестящие — Lophophorus, трагопаны — Се-
riornis) и большим числом своеобразных воробьиных (Paradox urus,
Sutora и др.).
Что касается Северной подобласти, то границы ее
иа севере. ограничены тундрой и побережьем Ледовитого океана,
а па юге определяются системами гор Европы и южной Сибири и
морями: Средиземным, Черным, Каспийским и Аральским.
Огромная территория этой подобласти протянулась в широтном
направлении от Атлантического океана до Тихого и на этом протя-
жении дает постепенно и медленно меняющуюся картину с запада
на восток, особенно в ее южных частях, благодаря тем влияниям,
которые сказывались в ее фауне в послеледниковые, ледниковые и
доледниковые эпохи. В общем ее можно разделить па три провин-
ции: Европейскую, Европейско-Сибирскую и Восточносибирскую.
Европейская провинция характеризуется присутствием следую-
щих млекопитающих: крот (Talpa), пиренейская и русская выху-
холи (Desmana), некоторые виды землероек (Sorex alpinus, Neomys
fodiens), типичный еж (Erinaceus europaeus), куница (Martes mar-
tes), мышовки (Sicista loniger), сони (Eliomys quercinus и Muscar-
dinus), полчок (Glis glis), хомяк (Cricetus cricetus), несколько ви-
дов рода Pitymys, некоторые виды мышей рода Apodemus и рода
Mus; некоторые виды сусликов (Citellus citellus, С. suslica), сурок
(Marmota marmota), благородней олень (Cervus elaphus), европей-
ская косуля (Capreolus capreolus), зубр (Bison bonasus).
Из птиц для этой провинции характерны: зеленый дятел
(Picus viridis), хохлатая сипица (Lophophanes cristatus), вьюрок
(Serinus citrinella), некоторые виды славок (Sylvia) и другие
виды.
К Европейско-сибирской провинции мы относим северо-восточ-
ный угол Европы от низовьев Северной Двины и низовьев Волги
и всю Западную Сибирь до Енисея на востоке и Алтая па юге вклю-
чительно.
Здесь характерны: соболь (Martes zibellina), колонок (Putorius
sibiricus), сусленник (Putorius alpinus), красный волк (Cyon alpinus),
алтайский крот (Talpa altaica). Несколько видов полевок, кра-
сяощекий суслик (Citellus erythrogenys), мышовки (Sicista
concolor), сеноставец (Ochotona alpina), марал (Cervus canadensis
asiaticus), сибирская косуля (Capreolus pygargus), кабарга (Moschus
moschiferus), сайга (Saiga tatarica) (рис. 682) и некоторые другие.
• Для этой провинции характерны следующие виды птиц: глухая
кукушка (Cuculus optatus), клест-сосновик (Loxia pityopsittacus)
и белокрылый клест (В. leucoptera), юр (Fringilla montifringilia),
буроголовый пухляк (Poecile cinetus), малая овсянка (Е. pusilia),
дрозд чернозобый (Turdus atrogularis), свиристель (Bombycilla
garrulus).
9 76
Восточносибирская провинция охватывает Восточную Сибирь
с Саянами на западе. Здесь, кроме некоторых упомянутых для
Западной Сибири видов, мы находим целый ряд эндемичных
форм.
Из хищных—камчатский медведь (Ursus beringianus), амурский
барсук (Meles amurensis), могера (Mogera robusta), амурский еж (Еп-
naceus amurensis), суслик эверсманов (Citellus eversmanni), особые
виды пищух, полевок, особые виды байбаков (Marmota kamtscha-
tica), восточносибирский лось, китайский кабан (Sus leucomys-
tax), изюбр (Cervus canadensis luedorfi), кабарга (Moschus mos-
chiferus), снежный баран (Ovis nivicola) и др.
Рис. 682. Сайга (Saiga tatarica).
(Ив Гааге.)
Для Восточной Сибири можно указать много типичных птиц—
дикуша (Falcipennis falcipennis), каменный глухарь (Tetrao
parvirostris), японский свиристель (Bombycilla japonica), дрозды
(Turdus naumanni, T. obscurus и T. sibiricus), чечевица розовая
(Erythrina rosea), овсянки (Emeriza rutila, E. chrysophrys), ne-
ночка-зорничка (Ptylloscopus inornatus) и другие.
Арктическая подобласть занимает всю круглополярную тер-
риторию, т. е. область полярного Северного моря с островами и по-
бережьем материков Евразии и Сев. Америки, а также полосу тун-
дры этих материков. Современная фауна этой подобласти складыва-
лась под непосредственным воздействием климатических изменений
четвертичного периода, а потому является во многих своих чертах
62 Кашкаров и Станчинский 977
самой молодой фауной. Характерными формами этой подобла-
сти являются из млекопитающих, помимо пелагических тюленей,
моржа, белого медведя (Talassarctos) и камчатского бобра (Latax),
обитатели тундры, песцы (Lagopus), лемминги (Lemmus, Dicrosto-
myx), овцебык (Ovibos), в значительной мере северный олень
(Rangifer tarandus).
Из птиц — многие гагары (Urinator adamsi, U. stellatus), гаги
(Somateria, Polysticta), гуси (Melanonyx, Rufibrenta, Chen Bran-
ta), чистики (Uria, Acthia, Brachyramphus, Phaleria) и др., чайки:
белая, полярная и др. (Sterna paradisea, Ebornea pogophila, Хеша
sabini, Larus hyperboreus и др.), белая сова (Nyctea nivea).
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА1
1. Кашкаров и Станчинский, Курс биологии позвоночных, 1934,
(Заменяет экологическую часть настоящего курса).
2. Шмальгаувен, Основы сравнительной анатомии, 1935.
3. Бобринской и Матвеев, Курс зоологии позвоночных, 1938.
(Может служить конспектом.)
4. Паркер, Курс зоологии, т. II, Позвоночные, 1908.
5. Солдатов, Рыбы и рыбный промысел, 1928.
6. Брэм, Жизнь животных, тт. 4, 5, 6, 7, 8, 9, 10, Издательство то-
варищества «Деятель», под ред. Книповича, 1914.
7. Мензбир, Птицы, Издательство «Просвещение», 1904.
8. Мензбир, Миграции птиц, 1934.
9. Мензбир, Очерк истории фауны Европейской части СССР, 1934»
10. Огнев, Звери СССР и прилежащих стран, тт. I, II, III, IV.
11. Промптов, Птицы в природе, 1937.
12; Шмидт, Миграции рыб, 1936.
13. Вебер, Приматы, 1936.
14. Се в ер цо в, Главные направления эволюционного процесса, 1934.
15. Грегори, Эволюция от рыб до человека, 1934.
16. Рей Л ан к е с те р, Вымершие животные.
1 № 1, 2, 3, 4 могут в известной мере частично заменять курс. Осталь-
йые являются пособиями для желающих расширить круг своих знаний.
I. ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Абиссальная вона 208
Авдотка 600, 602, 657
Авдотковые 602
Австралийская область, фауна 956
Агама степная 470
Агамиды 405, 541
Агамовые 470
Агамы 444, 511, 518, 530, 544
Агути 804, 811, 961
Агутиевые 961
Адаписовые 848
Адаптации 31
Азиль 696, 697
Ай-ай 882
Айгилозавры 479
Аист белый 589, 590, 670
— черный 590
Аистовые 590
Аистообразные 561
Аистоподобные 587, 621
Аисты 587, 589, 590, 621, 626, 628, 633,
639, 640, 644, 648, 649, 656, 664, 670,
675, 679, 684, 690
Акантодии 150
Аксолотль 308, 309, 354, 382, 383
«Акула голубая 152
— молот 226
— пелагическая 152
— сельдевая 75, 85, 152, 210
Акулы 73, 77, 80, 90, 92, 109, 115, 117,
133, 137, 151, 213, 216, 217, 218, 219,
222, 231, 233, 253
— гребнезубые 151
— дельфиновые 152
— ископаемые 77, 78
— колючеперые 153
— настоящие 152
— оплодотворение у них 137
— пилоносы 221
— плащеносные 151
— разнозубые 152
Аллигаторовые 490
Аллигаторы 490, 526, 527, 530, 548
Аллозавр 492, 493
Альбатросовые 586
Альбатросы 586, 683
Альпака 831, 925
Амбистомы 354, 383
Амблиподы 846
Амиевые 165
Амниоты 304, 395, 550
Амфибии 33, 50, 72» 304 сл.
— бесхвостые 356
— биологическое значение 306
— голые 351
— дыхательный процесс у них 331 сл.
— жаберное дыхание у них 331, 332
— защитные приспособления у них 371
сл.
— классификация 306, 348 сл.
— обзор класса 305, 306
— панцырные 349 сл.
— паразиты и враги их 385, 386
62*
— питание их 367 с л.
— покрытоголовые 349
— происхождение 361 сл.
— размножение у них 373 сл.
— регенерация конечностей у них 372
— характеристика 304, 305
— хвостатые 352
— число видов на земле 306
— экология 364 сл.
— Якобсонов орган у них 344
Амфиоксус 84, см. также ланцетник
Амфисбеновые 476
Амфиума 355, 379
— угревидная 355
Анадромальная миграция 254
Анаконда 482, 530, 546
Ангельнский молочный скот 907
Английская скаковая лошадь 921, 922
Англонорманны 023
Ани 605
Аннелидная гипотеза происхождения типа
хордовых 48
Аноа 836
Анолис 522
Аномальные 807
— белки 809
Аномодонты 453
Антилоповые 834
Антилопы 691, 718, 720, 743, 774, 775,
831, 860, 861, 881, 882, 884, 939,
940, 947, 948, 965, 967, 968, 970, 974
— вилорогие 973
— горные 835
— Канна 834
— лошадиные 835, 860
Анхинга 587
Анцилоподы 846, 964
Анчоусовые 173
Анчоусообразные 176
Анчоусы 173, 297
— промысловое значение 297
— светящиеся 176, 177
Апидий 851
Аппендикулярии 28
Арабская лошадь 921
Арамовые 599
Аргали 835, 911, 938
Аргусы 595
Ардены 921
Ареосцелиды 402, 404, 422
Арзинотерий Циттеля 826
Аркал 911, 912
Арктогейское царство 964 сл.
Арктогея, ихтиофауна ее 280 сл.
Армадилловые 802
Ароморфозы 32
Аррау 462, 534
Артродиры 157
Архары 938, 951
Археоптерикс 578, 580
Археорнис 577, 578, 580
Архозавры 467
Аспид коралловый 485, 523
979
Аспидовые ужи 485
Аспиды 546
Ассоциативная память у амфибий 390
---у млекопитающих 899
--- у рыб 291
Астрапотерии 841, 842
Асцидии 22 сп., 28
Атеринка 180, 291
Атеринковые 180
Афотическая зона 207, 209
Вабируссы 830
Бабуры 702, см. также пеликаны
Байбаки 806, 807, 860, 891, 929, 948,
974, 975, 977
Баклановые 588
Бакланы 587, 588, 629, 691
Балийский скот 907
Балобаны 630, 631
Бинаноедовые 606
сананоеды 605, 636, 654, 968
Бандикуты 956
Банкивская курица 594, 595, 696
Бантенг 836. 838, 906, 907
Баран гршДстый 911, 912
— снежжи 835, 977
— степной 911
Бараны 775, 938, 974
— дикие 835, 911 сл.
Барбулька 184
Барбульковые 184
Барибал 816
Барсук амурский 977
— обыкновенный 817
Барсуки 756, 817, 862, 865, 878, 879,
886, 887, 897, 932, 977
Барсуковые 816, 817
Барсучок 616
Барсы 892, 930, 950, 951
Бастардирование у рыб 252
Бастарды 252
Батипелагические рыбы 209, 214
Башкирская лошадь 921
Бегемоты 720, 829, 830, 872, 965
Бегунок 601
Бегуны 699
Безногие (амфибии) 305, 308 сл., 351
Безоаровый козел 835, 836, 917, 918, 938
Бекасы 601, 602, 630, 638, 643, 646,
657, 668, 670, 704
Беличьи 806
Белки 806 сл., 866, 869, 879, 887, 891,
894, 897, 928 сл., 948, 961, 967, 970,
973
Белковидные 806
Белоглазки 295, 614
Белодушка 817, 931, 933
Белозубки 796
Белорыбица 295, 301, 302
Белошейка 616
Белуга 161, 295, 297 *
Белуха 823, 940
Белуховые 823
Белый медведь, см. медведь белый
Бельгийская лошадь 921
Бельдюга живородящая 190, 243
Бельдюговые 190
Веляк, см. заяц-беляк
ентос 209, 213 сл.
Бергмана правило 660, 661
Береговая эона 208, 209
Беркширская свинья 919
Беседковые птицы 959
Бескилевые птицы 581, 686
Бескрыловые 583
Бескрылы 583, 954
Беспанцырные неполнозубые 802
Бестужевский мясо-молочный скот 909
Бесхвостые (амфибии) 305, 308
Бесчелюстнце 50 сл.
Бесчерепные 33 сл.
— положение в системе 47
Бесщитковые 66
Бешенка 269, 295
Бизоны 836, 894, 947, 971
Биометрический метод, применение в их-
тиологии 249 сл.
Биотопические условия существования
организмов 201
Биоценотические отношения в прериях
Америки 892, 893
— условия существования амфибий
385 сл.
—------млекопитающих 884 сл.
------- организмов 201
---- — птиц 688 сл.
----— рептилий 531
-------рыб 201 сл., 271 сл.
Биоценозы, определение понятия 271
Боа 482, 546
Бобровая крыса 812, см. также нутрия
♦Бобровая птица» 703
♦Бобровая струя» 720, 928, 930
Бобровидные 807
Бобровые 807, 971
Бобры 715, 720, 807, 808, 865, 872, 873,
874, 875, 887, 889, 895, 897, 928, 929,
930, 947, 949, 950, 951
— болотные 961
Боксеры 904
Болонки 902
Болота как среда обитания птиц 684
Борзые собаки 902, 904
Бородавочник 830, 862, 967
Бородавчатка страшная 188, 189
Бородатковые 611
Бородач 593
Бочонок 30
Брабансон 921
Брама 696, 697, 698
Бранхиозавр 308
♦Брачная бородавка» у лягушек и
жаб 373
Брачная окраска у птиц 655
Брачные звуки у птиц 638 сл.
— игры у млекопитающих 895
---- у птиц 638
---- у рептилий 526
----у рыб 236
Брачный наряд у рыб 233 сл.
Броненосцевые 801
Броненосцы 715, 727, 800, 801, 802» 862»
885, 960, 973
Брызгун 184
Бубалы 835, 860
Бугорчато-лунчатозубые 840
Буйвол африканский 836
— домашний 837
— южноазиатский 836
„Буйволы 836, 869, 872, 910
Бульдоги 902, 904
Бунодонтные 829 сл.
Буноселенодонтные 827, 829
Буревестник гигантский 683
Буревестники 533, 585, 586, 587, 621,
628, 633, 634, 647, 659, 662, 682, 683,
690
— ночные 663
Буревестниковые 586
Бурозубки 796
Бурундуки 806, 861, 879, 887, 971
Бык дикий 906, 946
— номадов 836
Быки 831, 835 сл., 905 сл.
Бычки 187, 188, 225, 226, 234, 241, 298,
301
— подкаменщики 189
— травяные 188
Бюльбюль 971
Вальдшнепиные 601
Вальдшнепы 601, 634, 638, 657, 668»
670, 672, 675, 676, 679, 704
Вальковатые змеи 482
980
Вапити 833, 937, 971
Варакушка 617
Вараны 473, 475, 479, 524, 533, 534, 544
Варианта 248
Вариационный метод 250
— ряд 247
Василиск 471, 473
Вагенера теория смещения материков
257, 258, 285
Венценосные голуби, см. голуби
— журавли, см. журавли
Верблюд двугорбый 831, 924
— одногорбый 831, 924
Верблюдовые 831
Верблюды 745, 746, 751, 774, 831, 861,
883, 886, 924, 974
Веретенники 601, 640, 671, 676
Веретенница 410, 473, 509, 545
Веретеницевые 472
Вертишейка 610, 658
Верхогляды 284
Веслоногие 567, 587, 588, 621, 633, 647,
648, 649, 683, 684, 702
Весничка 616
Ветры, влияние на птиц 666, 669
Вечерницы 798
Виандоты 698
Виверровые 814, 974
Виверры 720, 755, 813, 814, 815, 886,
965, 969, 974
Викарные группы 952
Викунья 831, 925, 962
Вилорог 718
Винторогий козел 835, 917, 918, 938, 976
Виперовые 485
Виснаша 811, 862, 961
Витютень 604
Вкусовые органы млекопитающих 783,
899
— — рептилий 540
----рыб 143, 289
Влажность, влияние ее на амфибий 384,
385
— — — на млекопитающих 883 сл.
-------- на птиц 664 сл.
----— на рептилий 530
Внутренножаоерные 70, 71
Вобла 170, 244, 246, 293, 295
Вода, давление, влияние на рыб 207
—* жесткость 203
•— значение в жизни птиц 664 сл.
----для свечения рыб 230
— как среда для жизни рыб 201 сл.
-------- обитания птиц 683 сл.
— морская 202
— пресная 202
— прозрачность 207
— свойства, значение для распростра-
нения отдельных видов рыб 208
— содержание кислорода, влияние на
жизнь рыб 204, 205
— соленость ее 202
— температура, влияние на жизнь рыб
205 сл.
Водные млекопитающие 872 сл.
Водоплавающие птицы 647, 659, 660,
670, 683, 684
Водорезовые 603
Водоросли Zostera в качестве пищи мор-
ских животных 273
Водосвинка 761, 778, 811, 872, 887
Водяная землеройка 872, 885, 889
— крыса 810, 887, 889, 894
— курочка 656, 684
— собака 168
Водяной козел 872
— пастушок 954
Водяные тенреки 885
Воздух, как среда обитания птиц 682
Воздушные мешки птиц 568
Возраст рыб 241 сл., 245
----методы определения его 243
Возрастные кольца у рыб 243 сл.
Волк гривистый 756
— земляной 966
— красный 756, 816, 975, 976
Волки 757, 816, 860, 888, 889, 892, 894,
896, 933, 942, 945, 971
Волкодав 905
Волосатая лягушка 313, 374
Волосяной покров млекопитающих 715
сл., 876 сл.
Волошские овцы 915
Волчок 588
Вомбаты 792, 957, 959
Вонючки 720, 818, 961
Воробьи 626, 637, 690, 691, 710
— домашние 613, 710
— полевые 613, 710
— испанские 613, 710
Воробьиные 561, 574, 611, 618, 636,
639, 641, 643, 648, 684, 688, 710, 954,
956, 974, 976
— кричащие 611
— певчие 612, 613
-Ворон J13. 639
Ворона 613, 626, 629
Вороновые 612, 613, 636
Вороны 533, 538, 561, 628, 633, 661,
670, 671, 677, 691, 693, 710, 972
Восточная область, фауна 968 сл.
Восьмизубые 967
Вредные млекопитающие 942 сл.
Вторичноротые 48
Вторичные половые признаки у амфибий
373 сл.
-------у млекопитающих 774 сл., 895,
896
--------у птиц 636 сл.
—------у рептилий 525
--------у рыб 233
Вуалехвосты 251
Выводковые птицы 647 сл.
Выделения защитные у птиц 658
----- У рептилий 525
Выделительные органы амфибий 339 сл.
-----круглоротых 56
— — ланцетника 40
-----млекопитающих 769 сл.
— — птиц 571 сл.
-----рептилий 440 сл.
— — рыб 135 сл.
Выдра капская 757
— морская 818
— речная 818
Выдровые 816, 818
— землеройки 795, 796, 872, 874
Выдры 756, 872, 873, 875, 889, 897, 932.
961
Выпи 657
Выпь большая 588
— малая 588, 630, 710
Высшие звери 713, 792 сл.
-----классификация 792, 794
— млекопитающие 773
Выхухолевые 796
Выхухоли 720, 796, 872, 873, 874, 885,
889, 927 сл., 949, 950, 976
Выхухоль русская 796
Вьюн 169, 216, 225, 234, 251
Вьюновые 169
Вьюрки 705, 975, 976
— горные 661
Вьюрковые 613, 614, 636, 688
Вятка 921
Гавиалы 490, 491, 548
Гага гребенушка 592, 703
— обыкновенная 591, 703
— стеллерова 592, 703
Гагара полосатая 586
— чернозобая 585, 702
Гагарка 602, 603, 642
981
Гагаровые 586
Гагароподобные 560, 585, 621
Гагары 567, 585, 586, 621, 633, 647, 648,
652, 655, 660, 670, 702, 972, 978
Гаги 591, 659, 660, 949,- 951, 978
Гадюка настоящая 485
— носорог 486, 536
— обыкновенная 486, 536
— песчаная 486
— рогатая 486, 511
— ложная 486
— степная 486
Гадюки 485, 520, 528, 533, 535, 538, 539
546, 547
Гадюковые 485, 546
Газель карликовая 861
Газы, содержание в воде, значение для
рыб 2Ю4, 205
Гаичка 615, 972
Галаго 882
Галгаз 592
Галитериум 827
Галки 613, 626, 646, 661, 670, 671, 690,
. 693, 710
'ГалстушникЬ 601
£аноиды 140
Гаплодонтовые 806
Гаплодонтные 806
?Гарну 835, 970
Гарпии 631
Гаршнеп 601, 704
Гастр
Гасторнисы 592
Гаттерия 403, 410, 414, 415, 422, 425,
437, 443, 468, 469, 502, 504, 532, 533,
690
Гаур 836, 907
Гаял 836, 838, 907
Гейдельбергский человек 852
Геккон гребнепалый 478
— каспийский 477, 479
— серый 477, 478
— — копальский 513
— стенной 477, 478
— сцинковый 477, 478
Геккончик 478
Гекконы 410, 414, 415, 425, 438, 444,
477, 508, 512» 513, 520, 526, 527,
543, 544
Гемпширские овцы 914
Гейтер 923
Географическое распространение амфи-
бий 391
— — миног 65
----млекопитающих 951 сл.
---- птиц 951 сл.
— — рептилий 542 сл.
----рыб 276 сл.
Гепард 816, 860, 974
Герефордский мясной скот 910
Гермафродитизм у рыб 231
Гесперорнисы 566, 585, 621
Гиббоновые 854
Гиббоны 728, 850, 851, 854, 855, 868,
969
Гидрозавровые 479
Гиена полосатая 815
— пятнистая 815
Гиеновая собака 967
Гиеновые 815
Гиены 756, 813, 889, 966, 974
Гимнофионы 308 сл.
Гиппарион 843
Гиппопотамовые 830
Гиппопотамы 714, 830, 872, 967
Гиссарская овца 916
Гладконосые 798
Глиптодонты 802, 961
Глосса 298
Глоточнозубые 484
Глубинная эона 208; 209
Глубинныё рыбы 214 сл.
982
-----классификация 214
Глупыш 586
Глухари 595, 596, 638, 646, 669, 704,
977
Гнезда птиц 641 сл.
— — искусственные 706, 707
— колониальные 644
— съедобные саланган 644
Гнездовые колонии птиц 658, 659
Гнездостроение у птиц 641, 643 сл.
Гну 835, 860
Гнюсы 284
Гоациновые 596
Гоацины 579, 596, 621, 648, 649, 963
Гоголи 591
Гокко 595 *
Голавль 170, 217
Голарктическая область, фауна 791
Голец 250
Голландский молочный скот 907
Головной мозг амфибий 342 сл.
----- круглоротых 58
— — млекопитающих 778 сл.
— — птиц 574
— — рептилий 443
-----рыб 139 сл., 145
— — — функция его 145
Головохордовые 33 сл.
Гологлаз 476
Голомянки 233
Голомянковые 189
Голос у амфибий 374
— у птиц 569, 639
Голуби 567, 568, 569, 603, 604, 620, 621,
635, 636, 639, 640, 645, 647, 648, 649,
688, 700, 701, 710
— венценосные 604, 959
— клинтухи 708
— крючкоклювые 604, 955
— наземные 604
— настоящие 604
— плодоядные 604
— породы их 701
— почтовые 628, 629
Голубок 603
Голубь горный обыкновенный 701
— сизый 604
Голые амфибии 305, 351
— угри 169
Гольян 234
Гомрай 611
Гомфотерии 761
Горал 835, 939, 951, 976
Горбатовский молочный скот 908
Горбуша 262, 296
Горилла 851 854, 868, 869, 966
Горихвостки 617, 649, 705, 708
Горлица 604
Гормоны у млекопитающих 776, 777
Горностай 817, 865, 883, 888, 889, 894,
932, 971
— ферганский 892
Горчак 170, 233, 234, 240
Гофер 807
Грачи613, 659, 671,677, 679, 689, 690, 710
Гремучая змея, см. змея гремучая
Гремучники 486
Гризли 816
Гриф-индейка 592
Гриф королевский 593, 634
Грифы 628, 634, 660, 661, 974
— американские 592, 639, 963
— Нового Света 592, 648
— стервятники 593, 633
Грызуны 715, 720, 728, 735, 747, 751,
761 сл., 771, 772, 792, 802 сл., 858,
860, 861, 862, 887, 889, 891, 892, 894,
896, 897, 945, 957, 961, 973
— вред, причиняемый ими 942 сл.
— меры борьбы с ними 944
— размножаемость 943
Гуанако 831, 925, 962
Гуахаро 607, 608, 063
Гуахаровые 607, 608
Губановые 186
Губанообразные 186
Губаны 220
Гульманы 975
Гуменник 591, 703
Гунди 974
Гурами 180
Гуси 556, 569, 571, 587, 590 сл., 621,
629, 643, 647, 648, 652, 656, 664,
670, 677, 678, 695, 696, 700, 702, 703,
704, 978
— арзамасские 700
— китайские 700
— породы 700
— тулузские 700
— тульские 700
— холмогорские 700
— эмденские 700
Гусиные 561, 591, 633, 659
Густера 247, 295
Гусь белый 591
— дикий серый 700
---шишковатый 700
— серый 591, 629, 700, 703
Далиевые 176
Дамановые 824
Даманы 720, 823, 824, 866, 967
Двоякодышащие 73, 74, 86, 93, 101,
129, 130, 131, 157 сл., 219
Двузубые 185
Двукопытные 794, 858
Двупарнорезцовые 805, 806
Двурезцовые 791, 792
Двускуловые 403
Двуутробки 754, 790
Двуутробковые 790, 956
Двуходка 509, 510
— пятнистая 476
Двуходковые 476, 510, 545, 549
Дегу 961
Дейтерозавры 420
Дельфиновые 823
Дельфины 733, 823, 873, 876, 940
— речные 823
Демерсальные яйца 212, 253
Дербник 593
Джейран 834, 861, 862, 940
Джерсейсний молочный скот 907
Джигитай 844, 936
Джунгарская дикая лошадь 920, 946,
см. также Пржевальского лошадь
Дзерен 834, 881, 940
Диаптозаурии 543
Диареальные виды птиц 672
Дикая кошка 533, 869
Дикий кот 815
Дикобраз древесный 812
Дикобразовидные 811
Дикобразовые 811
Дикобразы 811, 862, 881, 887, 951, 961,
970, 974
Дикуша 977
Динго 903, 957
Динозавры 403, 405, 406, 419, 422, 423,
426, 428, 429, 467, 488, 490 сл.,
498, 505, 507, 513, 518,524, 527, 543,
549, 580
— панцырные 495
— рогатые 496
Динотериевые 825
Диноцеры 846
Диплодок 494
Диплодоковые 493
Дисфотическая зона 207, 209
Дицинодонты 453
Дичь, охрана ее 705
Длинноноги 809
Длинноноговые 809
Длиннорылы 221
Длиннохвост 213, 214
Длиннохвостая кошка 974
Дневные хищные птицы 561, 587, 592 сл.,
|62О, 621,630, 631, 636, 643, 647, 648,
649, 683, 684, 968
Доги 902, 904
Долгоноги 745, 862, 887
Долгопятовые 850
Долгопяты 778, 848, 849 сл., 881, 882,
969
Долгохвосты 192
Долихозавры 479
Домашние быки 906
— кошки 905 сл.
— лошади 919 сл.
— млекопитающие 901 сл.
— овцы 911 сл.
— олени 925
— ослы 923
— птицы 694 сл. „
— свиньи 918 сл.
— собаки 902 сл., 944, 945
Донные рыбы 213, 214
---типы 213 сл.
Донская лошадь 921
Доркинг 698
«Дракон» глубин 215
— летающий 470, 471, 514
Дракондобразные 187
Древесная растительность как среда
обитания птиц 687
Древненебные птицы 580, 581
Древние птицы 558, см. также перво-
птицы
Древолазовые 612
Древолазы 360, 379, 963
Дриопитек 851
Дрозд белобровый 617
— белозобый 617
— каменный 617
— певчий 617
— пестрый 617
— рябинник 670, 693
— серый 617
— синий 617
— чернозобый 976
— черный 617, 635
Дроздовые 616, 617
Дрозды 617, 691, 705, 710, 972, 977
Дромадер 924
Дроматерий 787
Дронговые 613
Др оптовые 604
Дронты 604, 966
Дрофиные 599
Дрофы 561, 598, 599, 621, 648, 704,
Дубонос 614, 635, 636, 710
Дубровник 614, 675
Дупели 600, 601, 638, 657, 670, 676,
681, 704
Дуплянки 706 сл.
Дыхание рыб 123, 124
Дыхательные органы амфибий 331 сл.
— — млекопитающих 766 сл.
--- птиц 568 сл.
---рептилий 436 сл., 444
---рыб 111 сл.
Дюгоневые 827
Дюгони 741, 764, 859
Дятел белоспинный 610
— зеленый 610, 657, 976
— малый 610
— пестрый 605, 610
— седой 610
— средний 610
трехпалый 610, 687
— черный 610
Дятловые 610
Дятлы 567, 605, 609| 610. 635, 640, 644,
645, 648, 657, 661, 669, 687, 689 сл.,
693, 706, 707, 708, 976
983
Еж-рыба 185, 186, 211
Ежекрысы 961, 969
Ежи 533. 538, 539, 879, 885, 889, 897,
976, 977
— длинноиглые 796
— длинноухие 796
— крысиные 796
— настоящие 796
Ежовые 796
Елец 216
Емуранчик 810
Енотовидная собака 816, 934, 950, 951,
975
Енотовые 816
Еноты 813, 816, 879, 889, 961. 973
Ерш морской 18й
Ерши 231, 252, 496
Ехидна 596, 721, 722, 750, 752, 773, 774,
879, 886, |?6, 959
Ехидно вые 78BL
Жаба вонючая 339
— зеленая 359, 374, 384
— камышевая 359
— обыкновенная серая 359
— повитуха 358, 380
— суринамская 358, 380
— чесночница 358
Жабы 381, 384, 385, 387, 389, 390
— австралийские 359
— настоящие 358, 359
— южно-американские 359
Жабры 111 сл.
— у амфибий 331 сл.
Жаворонки 612, 628, 636, 643, 670, 677,
679, 710, 972
Жаворонковые 614
Жаворонок малый 614
— короткопалый 614
— лесной 614
— полевой 614
— рогатый 614, 674, 675
— степной 614
— хохлатый 614
— черный 614
Жакмаровые 611
Жвачные 752, 765, 827, 830, 831, 831 сл.
Железы внутренней секреции у млеко-
питающих 775 сл.
— кожные у млекопитающих 719 сл.
— млекопитающих 775
Желтобровка 616
Желтопузик 472, 475, 509
Желтушники 651
Желудок млекопитающих 764 сл.
Жерех 170, 208, 237, 247
Жерлянка 358, 374, 381
Живородящие рептилии 528
— рыбы 233
Животноядные 857
Жирафовые 839
Жирафы 718, 751, 831, 839, 860, 861,
882, 887, 965, 967
Жиряки 794, 823, 824, 866
Жулан 616
Журавли 597, 598, 599, 639, 647, 648,
649, 652, 656, 677, 678, 684, 704,
963
— венценосные 599
— степные 599
Журавлиные 560, 599
Журавль красавка 704
— серый 598
Забайкальская лошадь 921
Забота о потомстве у амфибий 377 сл.
---у млекопитающих 896 сл.
--- у птиц 649, 650
— т— у рыб 239 сл.
Завирушка 616, 617
Завирушковые .617
Заднебороздч^тые 484
984
Зайцевые 805, 806
Зайцы 743, 745, 752, 805, 860, 863, 877,
884, 886, 891, 928, 961
Заказники 941
Закрытопуэырные 120
Залом 295
— миграция его 269
Заповедники 948, 949 сл.
Зауроптеригии 404, 408, 420, 421, 457,
543
Защитная окраска, см. покровитель-
ственная окраска
Защитные приспособления у птиц 656 сл.
-----у рыб 226 сл.
Заяц-беляк 805, 883, 892, 928
— манчжурский 805
— памирский 876
— полярный 876
— русак 928
Звездорылые кроты 973
Звездочеты 187
Звероводство промышленное 942
Звероголовые 452
Зверозубые 451, 728, 735, 736, 784
Звероногие 492
Звероподобные 406, 450, 784
Звероящеры 408, 412, 421, 427, 728, 735
Зебра квагга Чапмана 845
Зеброиды 924
Зебры 844, 845, 860, 882, 884. 967
Зебу 910
Зеленушка 613, 614, 657, 670, 671, 679
Землеройка бурозубка 796
— водяная 796
Землеройки 720, 878, 885, 965, 966, 969,
973, 976
Землеройковые 796
— кроты 971
Земляные зайцы 746, 810, 974
Земноводные см. амфибии
Зерноядные птицы 670, 705
Зимородки 567, 609, 610, 629, 633, 643,
645, 647, 648
Зимородновые 609
Златокротовые 794
Златокроты 742, 754, 783, 795, 865,
885, 966
Змееголовые 180
Змеептицы 588
Змееяд 533, 538, 593, 631, 633
Змеи 404, 410, 411, 417, 422, 423, 432,
435, 437, 438, 469, 480 сл., 502, 509,
510, 517, 518, 520, 523, 524, 525, 526,
527, 528, 532, 533, 534 сл., 541, 542,
545
— великоватые 482
— географическое распространение
545 сл.
— гладкоаубые 483, 535
— древесные 484, 511, 512, 514, 520,
527, 529, 532
— ваднебороэдчатые 535
— линька их 411
— морские 417, 485, 515, 517, 528, 532,
546
— переднебороэдчатые 535
— роющие 509, 510
— ядовитые 534 сл.
— яйцеядные 519
— ямкоголовые 486
Змеиная сыворотка 538
Змеиный яд 535 сл.
Змея гремучая 486, 529, 533, 536, 538,
541
—7коралловая 509
— очковая 485, 525, 535
— плетевидная 520
— стрела 484, 535
— эскулапова 484
Золотая рыбка 170, 251
Зоографическое подразделение млемеяи-
тающих 953
Зоопланктон 217, 272 сл.
Зорянка 617, 650, 671
Зрительный аппарат амфибий 345, 389,
---млекопитающих 782, 783, 898
---птиц 576, 662, 663
--- рептилий 444, 445, 540, 541
---рыб 144, 145, 290, 291
Зубатна 190, 219
Зубры 775, 836, 838, 946, 947, 948, 971,
976
Зубы амфибий 329
— грызунов 803
— копытных, типы 758 сл.
— млекопитающих 752 сл.
— рептилий 434, 435
— рыб 132 сл., 218 сл.
Зуйки 601
Зуйковые 601
Зяблики 614, 629, 644, 668, 670, 679,
693, 705
Ибисовые 590
Ибисы 634
Иваси 173
Иволги 613, 676, 681
Иволговые 613
Игла - рыба 178, 211, 221, 240
Иглобрюхие 185
Иглокожие 50
Иглоротые 175
Иглошерстовые 811
Игрунка львиная 853
Игрунки 963
Игрунковые 850, 853
Игуанодоны 494, 495
Игуаны 405, 471, 472, 511, 512, 518, 520,
522, 526, 532, 545
— древесные 512
— морские 515, 517
Изменчивость групповая 250
Изюбры 833, 834, 937, 947, 951, 977
Икра, время потребное для развития 238
— лососевая 174, 259
— осетровая 293, 295
— степень зрелости, значение 232
— черная 295
Икрометание 232, 235
— циклы 233
Индейка бронзовая 698, 699
— дикая 698
Индейки 595, 647, 695, 696, 698, 699,
973
— белые русские 699
— горные 595, 661, 704, 892, 974
Индийская область, фауна 968 сл.
Индри 778, 849
Иния 823
Инстинкт перелетный 677
Йоркширская свинья 919
Ирбис 815, 880, 930, 974
Ихневмоны 538, 888, 974
Ихтиодорулиты 195
Ихтиозавры 404, 407, 431, 455, 456, 504,
515, 516, 518, 528, 533
Ихтиоптеригии 454
Ихтиорнисы 566, 587
Ихтиофагия 518
Кабаны 538, 830, 865, 894, 918, 936, 949,
950 951 977
Кабарга 720, 774, 775, 832, 938, 951,
974, 976, 977
«Кабарговая струя» 938
Кабарговые 832
Кавказский молочный скот 908
Кагу 598, 955, 963
Кагувые 598
Казарки 591, 630, 703
Казуаровые 588
Казуары 571, 581, 583, 620, 636, 652»
688, 958, 959
Кайман 490, 524, 527, 530, 548
Каймановые рыбы 167
— черепахи 462
Кайра 603, 642, 646. 659
Какаду 606, 627, 959
Калан 818, 932
Калмыцкая лошадь 921
Калмыцкий мясо-рабочий скот 910
Калош» 797
Калуга 161
Камбаловые 182, 214, 224, 225, 272, 298
Камбалообразные 182
Камбалы 121, 182, 183, 217, 224, 234,
238, 244, 246, 251, 252, 253, 273, 274,
275, 293, 296, 298
Камвольная мериносовая овца 912
Каменка 617
Каменная куница 931
Камнешарка 673
Камчатский бобер 818, 872, 873, 932»
933, 978
Камышевка болотная 616
— дроздовидная 616
— тростниковая 616
Камышевки 616, 650, 657, 705
Камышовый кот 815, 931, 974
Камышницевые 600
Камышницы 600
Канарейка 660, 701
Каннибализм у рыб 274
Канны 834, 835
Канюк 538, 559, 593, 628 670, 710
Карабахская лошадь 921
Каравайка 589, 590
Каракал 931, 974
Каракар 593
Каракуль 916
Каракульская овца 916
Каракульча 916
Каранговые 191
Караси 170, 208, 216, 275
Карась морской 184
— обыкновенный 251
Карачунь* 631
Каретта 465, 466, 516, 532, 534
Кариамовые 599
Кариамы 598, 599, 631, 633, 657, 963
Карп галицийский 303
Карповая рыбка 222
Карповые 78, 120, 121, 169, 216, 219,
220, 221, 236, 244, 252, 297
Карпообразные 168
Карпы 97, 169, 218, 220, 243, 246, 252,
275, 3 03
Карпы зеркальные 251, 303
— зубастые 177, 233
Касатни 172, 617, 822, 823, 889, 940
Кассики 973
Кассиковые 613
Катадромальная миграция 254
Качкар 835, 938
Качурки 586, 587, 683
Кашалотовые 823
Кашалоты 774, 823, 940
Квагф 967
Кваквы 533, 588
Квакша 360
— австралийская 360
— древесная 374
— кузнец 381
— южноамериканская 380
Кедровка 613, 689, 972
Кенгуровые 792
— крысы 792, 957
Кенгуру 720, 743, 745, 775, 792, 861т
886, 957
— горные 792
— древесные 792
— красный 793
— кускусные 792
$85
Кета 174, 234, 261, 293, 296
— миграции ее 261 сл.
Кефалевые 180
Кефалеобразные 16»
Кефаль 180, 222» 298
Кианг 844
Киви 583, 584, 620, 647, 663, 686, 954
Кидас 949
Кивуч 296
Килька 173, 298, 301
Кинкажу •W*'
Киргизская лошадь 921
Киргизский мясо-рабочий скот 910
Кислород, значение для свечения рыб 230
— потребление рыбами для дыхания
204, 205
Кистеперые 74, 161
Кит гренландский 822, 940
— серый 822, 940
— синий 822, 940
— японский 822, 940
Китайские собачки 903 i
Китайский кролик 926
Китобойный промысел 940
«Китовый ус» 750, 763, 822
Китообразные 726, 727, 729, 733, 766,
769, 771, 772, 794, 813, 820, 858, 874,
888, 889, 940
Киты 720, 723, 740, 741, 750, 763, 766,
778, 872, 873, 874, 875, 876
— беззубые 820, 821, 822, 889, 898,
940
— гладкие 822
— древние 821
— зубастые 820, 821, 822, 875, 889, 940
— промысловые, значение их 940
Кишечнодышащие 14 сл.
Кладоселахии 150
Классификация амфибий 306, 348 сл.
— высших зверей 792, 794
— млекопитающих 786 сл.
— птиц 580 сл.
— рептилий 445 сл.
— рыб 148 сл.
Клейдесдальская лошадь 921, 922
Клеппер 921
Клест белокрылый 976
— еловик 680
— сосновик 976
Клесты 613, 614, 635, 636, 661, 669,
679
Климатические условия существования
амфибий 383 сл.
-------млекопитающих 876
------- организмов 201
-------птиц 659 сл.
-------рептилий 529
Клинтух 604
Клюв птиц 629, 630
---форма 630, 635
Клювоголовые 403, 404, 422, 423, 427,
467, 468
Коала 792, 866
Кобра 485, 533, 536, 537
Кобчик 632, 710
— амурский 674
Когтистые, см. игрунки
Кожа, см. покровы
Кожаны 798
Кожнопанцырные 66
Коса дикая ископаемая 835
— снежная» 835
Козарка краснозобая 670
Козел безоаровый 835, 836, 917, 918,
938
— винторогий 835, 917, 918, 938, 976
— галицийский 918
— горный 880, 881, 892, 951
--- сибирский 835
Козерог 835, 917
Козлы ^45, 775, 938, 974, 975
Козо-барановые .835
Козодоевые 608
Козодои 560, 567, 607, 621, 635, 642,
643, 647, 648, 657, 662, 663, 682, 688,
705
— совиные 608
Козы 533, 723, 774, 831, 973
— ангорские 918
— домашние 835, 916 сл.
— кашмирские 918
— короткорогие 970
Козюлька 486
Койот 816, 894
Колбневые 187
Колбнеобразные 187
Колибревые 608
Колибри 560, 608, 621, 628, 635, 637,
645, 647, 655, 688, 689, 963, 973
Колли 904
Колониальные гнезда 644
Колонок 816, 931, 949, 951, 976
Колпики 589, 590
Колпиковые 590
Колпица 630, 634
Колючеперые 181
Колючий чорт 471
Колюшка 179, 231, 234, 239, 240, 274
— трехиглая 178, 179
Колюшковые 178
Колюшкообравные 178
Комароловы 615
Коменсализм 691
Конвергенция 228, 873, 874
Кондор 538, 634, 647
«Конек» 169
Конечноротые 74
Конечности амфибий 323 сл.
— млекопитающих 737 сл.
— птиц 562 сл.
— рептилий 425 сл., 507, 508, 512 сл.,
517
— рыб 102 сл.
Конодонты 195
Коноплянка 614, 670, 679, 706
Коньки 614, 643, 650, 657, 705, 972
Копрофагия у млекопитающих 889
Копчики 593
Копытни 603, 685, 974
Копытные 728, 747, 752, 758, 771, 774,
794, 823, 858, 859, 891, 896, 964
— южно-американские 840 сл.
Коровы 533, 766, 774
Королек 616, 628, 660, 661, 972
Корольновые 616
Коростель 600, 652, 680
Короткоглавы 359
Корсак. 816, 933
Коршун дымчатый 658
Коршуны 533, 633, 691, 710
Корюшка морская 175
— озерная 175, 216
Корюшки 213, 249, 296, 302
Костистые рыбы 75, 88, 140, 167 сл.
— — классификация 168
Костпощитковые 67
Костные ганоиды 75
— рыбы 73, 74, 157 сл.
Косули 832, 865, 880, 936, 937, 947,
948, 949, 950, 951, 974, 976
Котелкочерепные 446
Котик 774, 818, 819
Котилозавры 402, 403» 404, 405, 410,
412, 418, 419, 427, 428, 443, 524,
f 542, 549
Котинги 963
Котинговые 611
Коты 930, 931, 975
Кохинхина 696, 697, 698
Кочевки млекопитающих 880, 881, 884
— птиц 671 сл.
Кошачьи 815, 930 сл.
Кошка диная 533, 869
— домашняя 533, 905
*986
Кошка лесная 975
— нубийская 815, 905
— саблезубая 889
Кошки 538, 720, 756, 780, 813, 815, 866,
888, 889, 898, 899, 930, 965, 969
Краксовые 595
Крапивники 617, 644, 705, 973
Нрапивниковые 617
Красавки 599
Красная рыба 160, 295
Нраснозобик 601, 634
Красноловье 295
Красноперка 170, 295
Красный волк 756, 816, 975, 976
— молочный скот 908
Крачки 602» 603, 628, 642, 673, 681, 682
Крачковые 603
Креодонтные 857
Креодонты 794, 813, 888, 964
Крестовка 933
Кречет 593, 64
Кречет 593, 632
Кречотка 601
Кривковые 601
Кривоносый кулик 954
Кричащие воробьиные 611
Кровеносная система амфибий 334 сл.
---- асцидий 24
----кишечнодышащих 16
----ланцетника 38
----млекопитающих 768
---- птиц 570
---- рептилий 438 сл.
----рыб 125 сл.
Кровосос 525
Кровососовые 798
Крокодил нильский 490
— узкокрылый 490
Крокодилы 403, 404, 407, 410, 411, 412,
'1415, 419, 422, 423, 424, 426, 427, 429,
;432, 434, 437, 445, 488, 502, 505, 516,
|517, 518, 528, 530, 532, 533, 534, 541,
• |542, 548, 580, 691
•—( африканские 527, 530
— ложные 487
— морские 488, 489
— настоящие 488, 489, 490
— собственно 490
— средние 488
Кролик дикий 806
Кролики 736, 738, 770, 774, 806, 862,
863, 889, 925 сл.
Кроншнепы 600, 601, 630, 634, 679
Крот алтайский 976
— дальневосточный 796
— обыкновенный 796
— североамериканский 775
— слепой 796
Кротовые 796, 971
Кроты 727, 742, 743, 775, 783, 863, 864,
865, 878—885, 889, 928, 976
Крохалевые 592
Крохали 630, 633, 670
Крохаль большой 591, 592
Круглоголовка такырная 471
— ушастая 471, 523, 525, 530
Круглоголовки 412, 510, 511, 523, 531,
543
Круглоперовые 189
Круглоротые 51 сл.
— классификация, распространение и
экология 63
Круговорот веществ, роль амфибий в нем
385 сл.
-------рептилий в нем 531 сл.
-------рыб в нем 271 сл.
Крылан молотоголовый 751
Крылановые 797
Крылопанцырные 69
Крыса мешетчатая 751, 762
— пластинчатая 9
— пластинчатая 811
— серая 811
— туркестанская 811
— черная 811
Крысы 765, 811, 861, 887, 889, 899, 943,
945, 969
Кряквы 591, 629, 678, 703
Кса 606
Кузовок 185, 214, 230
Кузу 737, 957, 959
Кукушка глухая 976
— обыкновенная 657
— паразитизм ее 650 сл.
Кукушки 605. 638, 646, 648, 650, 678,
680, 705, 973
— желтоклювые 651
— земляные 605
— пятнистые 605
— шпорцевые 605
Кукушковые 605
Кулан 844, 881, 936, 948
Кулик золотой 638, 640
Кулик-сорока 601, 634
Кулики 600, 601, 621, 629, 634, 636,
639, 643, 646, 647, 648, 652, 670, 676,
680, 684, 704
Куличок-плавунчик 637, 684
Кумжа 174, 250
Куница лесная 817, 869
Куницевые 816
Куницы 538, 756, 813, 866, 873, 888,
897, 931, 948, 949, 950, 951, 974,
976
Курдючные овцы 916
Куриные 560, 567, 636, 640, 647, 657, 686
Курицы синие султанские 600
Куропатки 595, 966, 973
— белые 595, 596, 597, 639, 646, 652,
653, 657, 669
— горные 595, 704
— зубатые 595
— калифорнийские 595
— мезитовые (пастушковые) 597
— серые 595, 657, 669, 691, 704
— тундровые 595, 657
Куроподобные 593 сл., 621
Курочки болотные 600
Куры 594, 621/643, 647, 648, 688, 695,
696, 703, 790
— андалузские 697
— большеногие, см. куры сорные
— горные 652
— испанские 697
— одомашнение их 696
— орловские 698
— породы их, 696, 697, 698
— семиградские 698
— сорные 594, 642, 647, 959
Кускусы 792» 867, 957
Кутора водяная 796, 872, 873, 874, 889«
974
Кутум 295
«Лабиринт» у рлэ 180
Лабиринтовые 180
Лазающие формы млекопитающих 866
Лазоревки 615
Лайки 905
Лама 831, 925, 96>
Ламантиновые 827
Ламантины 726, 734, 827, 875
Лангшан 696, 698
Ланцетник 33 сл., 83 сл., 88, 89, 278
— развитие 42 сл.
— строение 33 сл.
Лань 832, 833
Лапчатоноги 600
Лапчатоногие 600
Ласки 817, 865, 883, 888, 889, 892, 932
944
Ластоногие 741, 772, 794, 818, 858, 872,
889, 898, 934
987
Ласточка береговая 617, 643
— городская 617, 691
— деревенская 617, 670
— морская 646
Ласточки 612, 635, 644, 677, 680, 682,
683, 688, 690, 691, 705, 706
Ласточковые 617
Ла-флеш 698
Лебеди 570, 571, 591, 639, 652, 656, 658,
659, 670, 678, 695, 701, 702
Лебединые 591
Лебедь-кликун 591, 639, 702
— черношеий 591
— черный 591
— шипун 591
Легавые собаки 904
Легхорны 697
Лейстерсние овцы 912, 915
Лемминги 811, 876, 880, 894, 978
Лемуровые 849
Лемуры 727, 754, 849, 966, 868, 870, 878,
879, 882, 885
Ленивец двупалый 802
— трехпалый 802
Ленивцовые 802
Ленивцы 726, 727, 752, 800, 801, 802,
859, 867, 868, 960
Леопард 815, 869, 975
Лесные млекопитающие 865 сл.
Летательный аппарат птиц 682, 685, 686
Летающие млекопитающие 869 сл.
Летающий дракон, см. дракон летающий
Летучая ‘лягушка 310
— рыба 180
Летучие мыши 723, 728, 740, 750, 752,
772, 777, 780, 792, 797, 798, 858,
866, 868, 871, 878, 879, 882, 885,
887, 889, 896, 954, 962, 973
— петухи 189
— собаки 797, 868, 887
Летучий маки 799, 870
Летяги 807, 808, 869, 870, 879, 887, 970,
971
Лещи 170, 216, 217, 237, 241, 242, 244,
246, 293, 295, 296, 297, 301, 302
— морские 184
Ликодовые 284
Ликоды 284
Линкольнские овцы 914, 915
Линь 170, 208, 251, 303
Линька у амфибий 312
— у млекопитающих 877, 878
— у птиц 651 сл.
— у рептилий 411
Лирохвост 612, 614, 959
Лирохвостовые 612
Лисица степная 816
Лисицы 538, 816, 862, 888, 889, 891, 892,
894, 897, 933, 949, 967, 971
Лисичковые 189
Лисюводство промышленное 942
Листоносовые 798
Листоносы 962
Литоптерны 823, 846
Литоральная зона 208
Литоринх афганский 484, 509, 510
Личинки рыб 238 сл.
— угрей 254 сл.
Личинкоедовые 605
Личинкоеды 645
Личйночна-хордовые 21 сл.
---значение их для эволюционного
учения 30
— — развитие 25 сл.
— — систематический обзор 27 сл.
--- строение 22 сл.
Личиночные 28
Лобан 180
Ложноногие 481
Ложные крокодилы 487
Лопатоносы 112, 161, 221
Лопатоносые 74
988
Лори 849, 866, 959
Лориевые 606, 849
Лососевые 78, 174, 236, 246, 252, 253,
296, 301, 303
Лососи 75, 96, 174, 210, 213, 231, 232,
234, 237, 239, 244, 250, 253, 254, 258
сл., 274, 302
— миграция их 258 сл.
Лосось благородный 236
--- миграции 258 сл.
— восточный 174, 234, 235, 261
--- миграции 261
— каспийский 295,
— озерный 296
Лось 832, 865, 936, 947, 948, 949, 950,
951, 971, 977
Лофиодонтовые 845
Лошади 720, 723, 743, 744, 759, 765, 766,
774, 860, 886, 887, 895, 945, 948,
962, 968
— африканские полосатые 844
— дикие 920
— домашние 919
— древние 843
— лесные 844
— лесостепные 844
— настоящие 743
— породы 920
— степные 844
— типы 920
— тундряные Г844
— дикая, джунгарская 844, 845, 920,
946
— одомашненная 920, 921
— Пржевальского 844, 845, 920, 921,
924, 946, 974
— эволюция 843, 844
Лошадиные 843
Лошаки 924
Луговой волк, см. нойот
Луговые собачки 807
Луна-рыба 185, 186, 211
Луни 533, 593, 631, 710
— болотные 593
— камышевые 710
— луговые 593
— полевые 593
— степные 593
Лупочновубые 486
Лунчатозубые 830
Луток 592
Лучеперые 74, 164
Лысун 820» 935
Лысухи 598, 600, 644, 646, 684
Львы 775, 815, 888, 889, 942, 974
Люди 852, 854, 868
Лягушка болотная 361
Лягушка-бык 361
— водяная 361
— волосатая 313, 374
— длинноногая 379
— древесная 383
Лягушка-звонарь 358
— зеленая 361, 376, 384, 386
Лягушка-красильщик 360
— озерная 361
— остромордая 361, 384
— плоскоголовая рукопалая 359
— плоскомордая 361
— развитие 346
— съедобная 384
— сумчатая 360, 380
— травяная 376, 384
--- серая 361
— узкоротая 361
— чилийская болотная 379
— шпорцевая 358
— яванская 310
---летучая 380, 381
Лягушки 374, 381, 384, 385, 386, 387,
389, 390, 391
— настоящие 361
Мадагаскарская область, фауна 965
Мазаевские камвольные овцы 912
Макаки 854, 969, 974, 975
Маки 849
— домовый 866, 881
— летучий 969
— ушастый 882
Макрелевые 121, 191
Макрелеобразные 191
Макрелещуки 179, 211, 212
Макрелещукообраэные 179
Макрель 76, 192, 210, 212, 216, 231, 274
Макроподы 180
Малиновка 616
Мамонт 825
Манат 859
Мангусты 814, 815, 888, 965, 975
Мандрилы 775., 854, 855
Манул 931, 974
Маорийская крыса 954
Мара 811
Марабу 634, 660
Маралы 833, 834, 937, 947, 951, 971, 976
Маринки 171, 250, 274
Маркур 918
Мартышки 854, 974
Мартышковые 854
Марульки 227, 233
Маскат 923
Массайский осел 923
Мастодонтовые 825
Мастодонты, стадии эволюции 823
Мегатерий 802
Медведевые 816
Медведи 743, 752, 756, 813, 865, 866,
877, 878, 879, 880, 886, 887, 889, 934,
942, 948, 949, 950, 951, 961, 969, 974,
975
Медведь бамбуковый 816
— белый 756, 816, 880, 883, 889, 934,
971, 978
— бурый 756, 816, 934, 971
— камчатский 977
— пещерный 816
— тибетский 816
— черный 816
Медоведовые 611
Медоеды 817, 886, 887, 967, 969
Медососы 614, 959
Медяница 473
Медянка 483
Мезаксиальные 744, 823
Мезиты 597
Мезозавры 402, 404, 407, 431, 454, 505,
543, 549
Мезозойские млекопитающие 787, 788
Меланоцет 211
Менек 212
Мериносы 912
Меритериевые 825
Метаморфоз у амфибий 376 сл.
Меч-рыба 78, 192, 210, 221
Мечение рыбы 253
Меченосы 152
Мешкоглот 178
Мешкожаберные 51
Мешетчатые крысы 807, 961
Миграции рыб 241, 252 сл.
---изучение 253 сл.
Микрокарпы 177
Микроконодон 787
гМиксиновые 64
Динеины 51, 63, 64
— развитие 62
Мил о дон 802
Мимикрия у птиц 657
Минога каспийская 295
— морская 64, 65
— речная 65
— ручьевая 65
Миноги 51 сл., 64, 65, 83, 214, 235, 238
— классификация 64 сл.
— развитие их 60 сл.
— распространение 65
Миноговые 64
Минорка 697
Млекопитающие 33, 50, 72, 401, 712 сл.
— взаимоотношения между самцами и
самками 895 сл.
— — с другими животными биоценоза
892 сл.
— водные 872 сл.
— вредные 942 сл.
— высшая нервная деятельность 898 сл.
— зависимость их от растений 890 сл.
— их роль в распространении заболе-
ваний 944 сл.
— классификация 786 сл.
— кочевки их 880, 881
— лазящие формы 866
— лесные 865 сл.
— летающие 869 сл.
— открытых мест обитания 859 сл.
— отношение к детенышам 895 сл.
— охотничье и промысловое значение
927 сл.
— подземные 861 сл.
— приспособление к температурным ус-
ловиям 876
— продолжительность периода беремен-
ности 774
— роющие 861 сл.
— способность к образованию привычек
899
— строение 713 сл.
— температура крови 769
— характеристика класса 712 сл.
— эволюция 786 сл., 857 сл.
— экономическое значение 900 сл.,
927 сл.
Многобугорчатые 788, 964
Многозуб американский 221
Многозубые 161
Многоперы 163
Многоперые 74, 163
Многорезцовые 790, 791
Моа 581, 583, 584, 620, 954, 955, 966
— гигантский 584
Могера 977
Мозозавры 487, 408, 480, 516
Мозоленогие 830, 831
Мойвенный полосатик, см. полосатик
мойвенный
Молоссовые 798
Молоток-рыба 152
Молох 471, 473, 524
Молочный скот, породы 907
Мольва 212, 231
Момотовые 609
Момоты 963
Монакины 963
«Монитор» 479
Моно ареальные виды птиц 672
Моногамия у птиц 638
Мопсы 902
Море, схема распределения эон 209
Моржевые 819
Моржи 741, 742, 819, 889, 934, 978
Морская вода, состав 202
— корова, см. стеллерова морская корова
Морские ангелы 153, 154
— змеи 485
— иглы 178, 240
— котики 872, 935
— ласточки 189
•— свинки 774, 811, 823, 940, 961
— собачки 121, 153, 189, 233
Морской бобер 889
— волк 190, 217
— дракон 187, 226
— заяц 820, 935
— конек 77, 178, 221, 225, 240
— кот 155,[226
— лев 819
989
Морской чорт 132, 190, 213, 218, 225
— язык 183, 298
Морфы 250
Московки 671
Мохноногие 710
Мочеполовая система, см. выделительные
органы
Мулы 924
/Мунго 533, 538, 815
Мунтжак 833, 970
Муравьед двупалый 802
— малый 867, 868
— сумчатый 722
Муравьедовые 802
Муравьеды 743, 750, 764, 766, 800, 801,
802, 864, 886, 960, 964
Муравьиные 612
Мурашеед 959
Мурена 178
Муреновые 178
Мускулатура, см. мышечная система
Мускусная крыса 872
Мускусные утки 699
Мускусный бык 835, 837, 876, 971
Муссурана 538
Мутации у рыб 247, 251
Муфлон 835, 911, 938, 950
Мухоловка малая 671
— серая 613, 616, 707
Мухоловки 628, 635, 688, 705, 708, 964,
966
Мухоловковые 616
Мушловка 809, 974
Мышевидные 810
Мышечная система амфибий 327 сл.
— круглоротых 51
— млекопитающих 747 сл.
— птиц 564 сл.
— рептилий 432 сл.
— рыб 82 сл.
Мыши 715, 774, 811, 861, 884, 887, 894,
943, 948, 965, 967, 969, 974, 976
Мышиные 810, 961
Мышиные птицы 609, 645 *
Мышловка 869
Мышовка 809, 971, 974, 976
— горная 809
— Нордмана 809
Мышь желтогорлая 811
— домашняя 811
— лесная 811
— малая 897
— малютка 811
— полевая 811
Мягкоперые 192
Мясной скот 909 сл.
Мясо рыбы, сравнительная оценка его
293, 294
Мясо-молочный скот 908, 909
Мясо-рабочий скот 910
Мясо-сальные овцы 916
Мясо-шерстные овцы 913
Навага 193, 296
Налим 193, 208, 216, 225
Нандовые 583
Нанду 581, 582,*583, 620, 640, 647, 686,
816, 963, 966
Нанопланктон 274
Нарвалы 774, 823, 940
Насекомоядные птицы 669, 689, 705,
715, 739, 771, 772, 792, 794 сл., 857,
878, 885, 896, 897
Наследственность у рыб 247 сл.
Нахлебничество у птиц 691
Негретти 912
Некрофагия у млекопитающих
Нектарки 635, 655, 689, 968
Нектарницевые 614
Иектарницы 616, 628
Нектон 209
990
Нектонные рыбы 210
----типы 210 сл.
Немертинная теория происхождения тина
хордовых 49
Неогейское царство 959 сл.
Неогнатические птицы 585
Неогнаты 648
Неотения 381 сл.
Неотропическая область, фауна 959 сл.
Непарнокопытные 723, 744, 823, 842, 843
Неполнозубые 772, 792, 800 сл., 897
Нервная система амфибий 341 сл.
---- асцидий 24
----кишечнодышащих 16
---- круглоротых 58
---- ланцетника 39
----млекопитающих 777 сл.
----птиц 574 сл.
---- рептилий 443 сл.
----рыб 139 сл., 288 сл.
Нерест 231, 235
Нерка 234, 263, 296
— миграция 263
Нерпа 820, 935
Несторовые 606
Нетопыри 798
Неясытевые 607
Неясыти 710, 982, 944
Неясыть лапландская 607
— обыкновенная 607
— уральская 607
Низшие звери 713, 788 сл.
Нильгау 835, 970
Новозеландская область, фауна 954
Новонебные птицы 585
Нокотницы 152
Норки 816, 872, 889, 897, 932, 951
Носорог белый 846, 947
— волосатый 846
— двурогий 846
— Мерка 846
Носороги 718, 845, 846, 869, 886, 965,
967, 968
Носороговые 845
Носухи 816, 869, 886, 973
Нотогейское царство 953 сл.
Нотозавры 457, 504, 533
Нотунгуляты 794
Ночницы 797, 798
Ночные животные 881, 882
Нубийская кошка 815, 905
Нубийский осел 923
Нутрия 812, 872, 942
Нырки 591, 633, 634, 639, 670, 677, 678
Нырковые 591, 703
Обезьяны 723, 734, 748, 751, 772, 783,
794, 850 сл., 867, 868, 886, 887, 895,
896, 897, 962, 963, 965, 969
— носатые 854
— ночные 853
— паукообразные 853, 867
— священные 854
— тонкотелые 854
— узконосые 852, 853 сл.
— человекообразные 851, 900, 968
— широконосые 852, 853
— эволюция 850, 851, 852
Обнинка 921
Оболочники 21 сл.
Обонятельные органы амфибий 344, 388
----млекопитающих 783, 898
---- птиц 576
---- рептилий 540
----рыб 142, 143, 289
Овсянки 613, 614, 635, 671, 706, 976,
977
Овсянковые 614
Овцебыки 835, 978
Овцы 720, 723, 765, 774, 831, 891, 974
— домашние 835, 911
Овцы, породы 912
— степные 835
— торфяные 911
Овчарки 904
Овчинные овцы 913
Огаревые 592
Огарь 592
Огневка 933
Однокопытные 734, 794
Одноноздревые 50
Однопроходные 713, 722, 764, 770, 772,
773, 787, 788, 896, 897, 956
Одноутробные 713, 7-92 сл.
Одомашнение животных 902
— курицы 696
Озера, как водная среда для рыб 208
Окапи 839, 860
Окраска амфибий 369 сл.
— млекопитающих 882, 883
— птиц 654 сл.
— — предупреждающая 658
— птичьих яиц 645, 646
— рептилий 521 сл., 541, 542
— рыб 183 сл.
Окуневые 183, 252
Окунеобразные 183
Окунь 82, 183, 208, 217, 225, 234, 237,
; 252, 275, 296
— Балхашский 183
— морской 183, 184, 188, 231, 252
Оленебыки 834, 835
Оленеводство 925
Оленевые' 832
Олени 533, 719, 720, 774, 775, 778, 831,
860, 865, 880, 886, 887, 894, 895,
936, 937, 950, 970, 974, 976
— акклиматизация 938
— промышленное значение 834
— рога у них 719
— эволюция и стадии развития рогов
у них 832
Оленчики 832
Олень благородный 832, 833, 834, 937,
950, 976
— дикий 892
— домашний 925
— пятнистый 832, 833, 937, 949, 950,
951
— северный 745, 774, 832, 833, 876,
877, 881, 894, 925, 936, 949, 951,
972, 978
Оленьки 778, 830, 831, 869, 886, 967, 970
Оленьковые 831
Оленья собака 905
Олушевые 588
Олуши 643, 683
Оляпка 617
Оляпковые 617
Омуль 174, 208, 297
Онагр 975
Ондатра 811, 926, 928, 942, 949, 951,
973
Онтогенез рыб 146
Оперение птиц 652 сл., 659 сл.
Оплодотворение у амфибий 340, 341
— у птиц 573, 574
— у рептилий 442
— у рыб 137 сл., 231 сл.
-----внешнее и внутреннее 236
-----искусственное 301 сл.
Опоссумы 735, 739, 785, 790, 868, 960,
973
Оранг-утан 851, 854, 856, 868, 900, 969
Органы чувств амфибий 344, 387 сл.
----- круглоротых 59
----- ланцетника 40
-----млекопитающих 780 сл., 898 сл.
-----птиц 575 сл., 662, 663
-----рептилий 443, 540, 541
-----рыб 142 сл., 289 сл.
Ореховки 636
Орланы 631, 633, 683
Орлиные 593
Орлы 593, 628, 631, 633, 644, 649, 66i„
690
— белохвостые 533
— змееяды 533, 538, 593, 631, 633
— морские 593
— рыбные 631, 633, 683
— степные 710
Орляки 155
Орнитоподы 564
Орфа 251
Осадки, влияние на млекопитающих 884
Осел африканский 844
— дикий 970, 974
— домашний 844, 923
— нубийский 923
— сомалийский 923
Ослы 844, 860
Осетр немецкий 160
— русский 160
— сахалинский или морской 161
— сибирский 160
Осетровые 74, 82, 160, 221, 225, 244, 2527
253, 297, 300, 301
Осетрообразные 160
Осетры 87, 99, 112, 160, 295, 297, 302
Осман 171
Осмотическое питание рыб 218
Осоед 593, 632, 689
Остео лепиды 162
Остроносик 180
Осьмизубые 812
Осязательные органы у амфибий 344,
388
----у млекопитающих 780, 781
----у птиц 575
----рептилий 444, 541
----рыб 143, 292
Отравление змеиным ядом 535 сл.
Открытопузырные 120
Охота, регламентация ее 941
Охотничий кролин 926
Охотничьи млекопитающие 926 сл.
Павианы 854, 965, 966
Павлины 595, 637, 655, 695, 701, 970
Пака 811, 961
Паламедеевые 590
Паламедеи 587, 590, 621, 657, 963
Паламедея рогатая 590
Палеогнатические птицы 581
Палеогнаты 647, 648
Палеомастодон 825
Палеониски 164
Палеопитек 852
Палия 296, 302, 303
— американская 303
Палтус 183, 231, 243
Панголин 800
Панголиновые 800
Панды 969
Пантеры 888, 930, 942 974
«Панты» 834, 937
Панцырная щука 80, 167
Панцырнощекие 188
Панцирные 66
— амфибии 305, 349 сл.
— неполнозубые 801
— сомы 80
Паразитизм у птиц 650 сл., 691 сл.
Паразиты и враги амфибий 386
-------рыб 275, 276
Параксиальные 744
Парапитек 851
Парасухии 487, 505
Парделевая рысь 930
Парейазавры 448
Парение птиц 626 сл.
Парна 903
Парнокопытные 723, 744, 823, 827
Парнорезцовые 805, 806 сл.
991
Парусник 192, 210
Пастушки 598, 599, 600, 648, 652, 657,
684, 688, 704
Пастушковые 560, 599, 600
Пасюк 811, см. также крыса серая
Певчие воробьиные 612, 639
Пеганка 592, 643
Пекари 720, 830, 962, 973
Пелагическая вона 208, 209
Пелагические яйца 211, 212, 253
— рыбы 209
Пеламида 192 (Sard a sard а)
— двухцветная 515 (Hydrus Platurus)
Пеламиды 532 (Pelamis)
Пеликан кудрявый 587, 588
— розовый 588
Пеликановые 588
Пеликаны 626, 628, 630, 633, 645, 691,
702
Пеликозавры 402, 403, 405, 406, 421,
427, 443, 542
Пеломедузы 462
Пение птиц 639
Пеночка-ворничка 977
Пеночки 616, 628, 650, 657, 705
— зеленые 680
Первичнозубые 787, 964
Первичнокопытные 813, 823, 842, 858
Первичноротые 48
Первоввери 713, 786, 787 сл.
Первокит 821
Первокрокодилы 486
Первоптицы 566, 577, 580 сл., 620
Перевозчик 601
Перевязка 817, 865, 932
Переднебороздчатые 485
Перелет птиц 671 сл.
--- высота его 679
--- скорость 679
Перелетные птицы 667, 671 сл.
---пути перелета 673 сл.
Перелетный инстинкт птиц 677
Перепел обыкновенный 595
Перепела 595, 635, 680, 704
Пересмешники 616, 973
Перистожаберные 18 сл.
Перцеяды 610
Першерон 921
Перья птиц 551 сл.
Перья птиц, генетическая связь с чешуей
554 сл.
Пескарь 170, 225
Пескоройка.65, 238
Песочник исландский 673, 674
Песочники 601, 634, 646
Пеструшка 616, 811, 862, 880, 943
— степная 811
Пестрянки 259
Песцы 816, 876, 880, 883, 889, 894, 933,
934, 942, 949, 971, 978
— голубые 934
Песчаник 807, см. также суслик жел-
тый
Песчанка (рыба) 187, 234
Песчанка большая (птица) 811, 891
— карликовая 891
— полуденная 811
— Эверсмана 883
Песчанки (птицы) 601, 811, 861, 862,
886, 887, 891, 892
Песчанкообразные 186
Петухи 655
Петушки 180
Пигалицы 646
Пикша 193, 244, 293
Пила-рыба 76, 80, 154
Пилоносы 153
Пильвина 183
Пинагор 189, 240, 241
Пингвин очкастый 586
— южноафриканский 586
Пингвиновые 586
992
Пингвины 585, 586, 621, 633, 647,
649, 652, 659, 660, 683, 963
Пинчеры 903
Пиротерии 794, 842
Питание амфибий 367 сл.
— млекопитающих 885 сл.
— птиц 629 сл.
— рептилий 436, 517 сл., 532
— рыб 216 сл., 272 сл.
Питекантроп 852
Питон африканский 509
— тигровый 481
Питоновые 481
Питономорфы 516
Питоны 481, 482, 520, 521, 527, 529,
532, 533, 546
Питтовые 611, 959
Питты 971
Пищеварительный аппарат амфибий
----ланцетника 36
----млекопитающих 750 сл.
----птиц 566 сл.
---- рептилий 434 сл.
----рыб 131 сл.
Пищуха даурская 805
— малая 805
Пищухи 613, 615, 635, 660, 669, 687.
706, 862, 863, 880, 886, 892, 971, 972
974, 975, 977
Пищуховые 615, 805
Плавательные приспособления у реп-
тилий 516
Плавательный пузырь рыб 118, 119 сл.
-------первоначальная роль его 197
-------генетическая связь с легкими
123
—------гидростатические функции ег©
120 сл.
-------функции его 121 сл.
Плавники 75 сл., 102 сл.
— непарные 102 сл.
----история развития 103 сл.
— парные 105 сл.
— — происхождение 109
— форма 102 сл.
Плавун 872, 885
Плавунчик 601, 638, 640, 679
Плавунчиковые 601
Планктон 209, 216 сл.
— типы рыб, приближающихся к нему
211
Пластинчатозубые 460, 524
Плацентарные 713, 723, 770, 773
Плезиадаписовые 847
Плезиозавры 408, 431, 458, 459, 460,
504, 516, 518, 533
Плеурокантодии 156
Плешанка 617
Плиопитек 851
Плимут-роки 697, 698
Плоскозубые ящеры 404, 408, 420, 421
Плотва 170, 296
Поведение (высшая нервная деятель-
ность) амфибий 387 сл.
----------млекопитающих 898 сл.
---------- рептилий 539 сл.
----------рыб 288 сл.
Поганка большая 586, 702
— /ушастая 585
Поганки 586, 635, 644, 648, 655, 660,
684, 702
Поганковые 586
<1одковоносовые 798
Подковоносы 798, 889
Подорлики 633, 710
Подорожники 614, 670, 672
Подуст 170, 218, 220
Позвоночник амфибий 314 сл.
— млекопитающих 726 сл.
— птиц 557 сл.
— рептилий 414
Позвоночник рыб 83 сл.
Позвоночные 31 сл.
— высшие 395 сл.
Пойнтеры 905
Покровительственная окраска амфибий
370
— млекопитающих 860, 882, 883
--- птиц 657
— — рыб 223 сл.
---хищников 888
Покровы амфибий 311 сл.
— млекопитающих 713 сл.
— птиц 551 сл.
— рептилий 409 сл.
— рыб 78
Покрытоголовые амфибии 305, 349 сл.
Полевка лесная 810, 811
— общественная 811
— обыкновенная 810
— рыжая 811, 892
Полевки 810, 861, 876, 878, 880, 886,
887, 892, 894, 943, 948, 971, 974, 976,
977
Полет птиц 622 сл.
— — быстрота 628, 629
---висячий (трепещущий) 628
— — гребной (пропеллирующий) 625 сл.
---парящий 626
— — скользящий (планирующий) 624
Полинезийская область, фауна 955
Полифиодонтия 435
Половой аппарат амфибий 339 сл.
— — круглоротых 58
— — ланцетника 41
---млекопитающих 770 сл.
---птиц 572 сл.
— — рептилий 442
---рыб 137 сл.
Полоз Карелина 483
Полозы 483, 484, 510
Полорогие 834, 905
Полосатик мойвенный 940
Полосатики 822
Полосатиковые 822
Полудупляпки 707, 708
Полукопытные 823, 858, 886
Полуночник 608
Полуобезьяны 771, 772, 794, 848 сл.,
868, 870 886, 887, 896, 965, 966,
969
— отличие от обезьян 848, 849
Полуослы 844, 860
Полухордовые 13 сл.
Полчек 809, 869, 976
Полярные собаки 905
Поморник длиннохвостый 602, 603
Поморники 633, 691
Поморниковые 603
Пони 921
Поперечноротые 151
Поползневые 615
Поползни 612, 615, 644, 649, 687, 706, 972
Попугаи 561, 567, 568, 605, 606, 636,
645, 648, 687, 688, 689, 954, 959, 968,
970, 973
— бороздчатоклювые 606
— восковоклювые 606
— гладкоклювые 606
— карликовые 606
— клинохвосты <606
— настоящие 606
— серые 605
— совиные 606, 662
— тупохвостые 606
— широкохвостые 606
Попугайчики 606
— волнистые 701
Потто 866
Правило Бергмана 660, 661
Предъящеры 467
Прекосы 912, 9М
Пресмыкающиеся 33, 395 сл., 4 01 сл.
63 Кашка ров и Станчинский
Пресная вода, возникновение рыб в ней
278
— — состав 203
Пржевальского лошадь 844, 845, 920,
921, 924, 946, 974
Привычки, способность к образованию
их у млекопитающих 899 сл.
Придонные рыбы 213 сл.
Прилипаловые 188
Прилипалообразные 188
Прилипалы 188
Приматы 794, 847 сл., 898
Приручение животных 902
Происхождение амфибий 305 сл., 361 сл.
— млекопитающих 784 сл.
— птиц 576 сл.
— рептилий 403 сл.
— рыб 193 сл.
— хордовых 47 сл.
Пролетные пути птиц 673 сл.
Прометеева мышь 861
Промысловые млекопитающие 926 сл.
— птицы 702 сл.
— рептилии 534
— рыбы 294 сл.
Проплиопитек 851
Просимия 851
Просянка 614
Протей 355, 356, 385
Проходные рыбы 203, 288
Прудовое рыбоводное хозяйство 303
Прыгунчик обыкновенный 794
Прыгунчики 745, 861, 966, 974
Прыгунчиковые 794
Прыгуны мешетчатые 745
Псевдоскафиринхи 161
Псевдосухии 487, 488, 505, 506, 576,
580
Птенцовые птицы 647 сл.
Птенцы 647 сл.
Птеродактили 499, 500
Птерозавры 403, 422, 431, 432, 467, 488,
505, 514, 518, 533, 580, 617
— длиннохвостые 499
— короткохвостые 499
Птицеводство 695 сл.
Птиценогие 494
Птицетазовые 494
Птицы 550 сл.
— бескилевые 558, 560
— болотные 668, 684
— взаимоотношения между особями од-
ного и того же вида 693
— — с другими животными 689
— весенняя жизнь их 679, 680
— влияние атмосферного давления 666
— влияние ветров 666
— влияние влаги 664 сл.
— влияние света на их жизнедеятель-
ность 662 сл.
— внешние паразиты у них 691
— внутренние паразиты у них 692
— вода как среда обитания 683 сл.
— воздух как среда обитания 682
— вредные в хозяйстве 710
— выводковые 647 сл.
— годичные циклы в их жизни 667
— дневные 662 сл.
— домашние 694 сл.
— древесная растительность как среда
обитания 687
— жизнь в зимнее время 668 сл.
— жизнь в тропических странах в сухое
время года 668
— защитные приспособления их 656 сл.
— зимующие 667
— конкуренция пз-за пищи 693, 694
— кочующие 667
— летательный аппарат у них 682, Я85,
686
— летняя жизнь 681
— летающие 667
993
Птицы, линька их 651 сл.
— настоящие 580 сл.
— новые 580 сл.
— номенклатура частей тела и оперения
554
— ночные 662 сл.
— общественность у них 658, 659, 694
— оплодотворение 573, 574
— оседлые 667
— осенняя жизнь 681
— открытые пространства как среда
обитания 684 сл.
— перелет их 671 сл.
— перелетные 667, 671 сл.
— перелетный инстинкт у них 677
— полезные в хозяйстве 705
— полет их 622 сл.
— приспособления к ночному образу
жизни 662, 663
— происхождение 576
— пролетные 667
— пролетные пути их 673 сл.
— промысловые 702 сл.
— птенцовые 647 сл,
—сезонные явления в зависимости от
климатических факторов 666 сл.
— спортивно-охотничьи 702
— строение 551
— стенотермические 661
— сумеречные 662 сл.
— суточная жизнь их 664
— характеристика класса 550
— эйритермические 661
— экономическое значение 694 сл.
— экотопические факторы их жизни
681, 682
— явления соревнования среди них 692,
693
Птицы-бабы, см. пеликаны
Птицы-мыши 968
Птицы-носороги 609, 611, 636, 644, 649
Птицы-паразиты 605, 650 сл.
Птичьи базары 658, 659, 704, 949
«Пугало» 850
Пудели 904
Пузанок каспийский 295
Пума 815, 961
Пустельга 593, 628, 710
Пустынник 604
Путораки 974
Пухляк 615, 976
Пуховой покров птиц 659 сл.
Пушнина 927
Пушные звери, добыча 927
Пюттера теория осмотического питания
рыб 218
Пяткоход 752, 792
Рабочий скот 910
Равновесия органы у млекопитающих
781, 782
--- у птиц 575
— — У рептилий 444
--- у рыб 290
Развитие, см. эволюция
Размножение амфибий 373 сл.
— птиц 636 сл.
— рептилий 525 сл.
— рыб 231 сл.
Разнощитковые 68
Райские птицы 613, 637, 639, 654, 655,
959
Рамбулье 912, 913
Растительность, роль ее в жизни млеко-
питающих 890 сл.
— в жизни птиц 687 сл.
Растительноядные птицы 669
— рыбы 218
Ревун рыжий 854
Ревуны 775, 850, 853
Регенерация у амфибий 372, 373
994
— у рептилий 524
Резонаторы 775
Ремез 615, 644
Ремень-рыба 210
Рептилии 33, 50, 72, 395 сл., 401 сл.,
574, 784 сл.
— адаптация к жизни на суще 500 сл.
— враги их 533
движения их 404 сл.
— действие температуры на них 529 сл.
— дыхательный процесс у них 438
— живородящие 528
— защитные приспособления у них
523 сл.
— значение в экономике природы 533
---для человека 533 сл.
— лазящие 512, 513
— летательные приспособления у них
514 сл.
— линька у них 411, 523
— любовные игры у них 526
— обзор современных и ископаемых
форм 401 сл.
— общая характеристика 401 сл.
— окраска их 521 сл.
— отношение к амфибиям 500 сл.
— переселения у них 530
— питание 517 сл.
— плавательные приспособления 516
— приспособления для жизни в воде
515
— I— для защиты и нападения 523 сл.
— •' — к жизни на деревьях и к полету
1511 сл.
---к наземному и подземному образу
жизни 506 сл.
— продолжительность жизни 529
— регенерация у них 524
— роговой слой эпидермиса их 409 сл.
— родословное дерево 502
— сожительство с другими животными
532, 533
— способность к скользящему полету
513, 514
— спячка у них 529 сл.
— съедобные 534
— типы приспособлений к плаванию
516
— умственные способности 542
— филогенетическое развитие 500 сл.
— форма тела их 404 сл.
— яйца их 526 сл.
— яйцеживородящие 528
Решетиловская овца 916
Ржанка 600, 601, 620, 635, 647, 679
— лопастноклювые 602
Ржанковые 601
Ринодерма Дарвина 379
Рога млекопитающих 718 сл.
— оленей, эволюция 832
Рогатая жаба 472
Рогатый скот крупный 905 сл., 945
---— породы 907
--- мелкий 911 сл.
Рогоклювовые 611
Родайленд 697
Рокки 581, 584, 620, 966
Романовская овца. 913, 914
Ромни-марш 915
Росомаха 817, 865, 894, 932, 949,
971
Рохли 154
Роющие млекопитающие 861
Рукокрылые 734, 771, 772, 797, 798,
869 871
Руконожки 847, 849, 882
Рукоперые 190
Русак 805, см. также заяц-русак
Рыба, мировой улов 294
— мясо ее, сравнительная оценка 293,
294
Рыба св. Петра 182
рыбец 247
Рыбное хозяйство 299 сл.^
---интенсификация 301 сл.
— — правильное ведение его 300
Рыбное хозяйство СССР 294 сл.
Рыбоводство прудовое 303
Рыбо-змей цейлонский 352, 353, 379
Рыболов 190, 603, 676
Рыболовство 294 сл., 299 сл.
Рыбопанцерные амфибии 350
Рыборазведение искусственное 301 сл.
Рыбоящеры 455
Рыбы 72 сл.
— батипелагические 209, 214
— в качестве пищи для различных жи-
вотных 274
— влияние давления воды 207
— влияние освещения в воде 207
— возникновение 237, 278
— время появления 195
— генеалогическое дерево 276 сл.
— географическое распространение
276 сл.
— глубинные 214 сл.
— движения их 75 сл., 77 сл.
— долговечность 246
— живородящие 233
— заболеваемость 275
— зависимость от планктона 217
— значение водной среды 208 сл.
— значение для них газов, содержа-
щихся в воде 204, 205
— изменчивость у них 247 сл., 250
— искусственное оплодотворение 302
— классификация 148
— комбинации у них 251, 252
— мероприятия по охране 299 сл.
— миграции 252 сл.
— модификации у них 247
— морские 201, 278
--- распределение 282 сл.
— мутации у них 251
— наследственность 247 сл.
— нектонные типы 210
— нервная деятельность их 139 сл.,
288 сл.
— область распространения 279
— образование рас 247 сл.
— окраска их 223 сл.
---приспособительное значение ее
223 сл.
— онтогенез 146
— оплодотворение у них 137 сл.
— определение величины годового при-
роста 245 сл.
— определение их возраста 243
— органы и механизм дыхания 111 сл.,
115 сл.
— органы пищеварения 131 сл.
---свечения у них 228 сл.
-------приспособительный характер их
228
---чувств у них 142 сл., 289 сл.
— паразиты и враги их 275, 276
— пелагические 209
— питание их 216 сл., 272 сл.
— пищеварение, температурный оптимум
218
— плавательный пузырь у них 119 сл.
— плавники у них 75 сл., 102 сл.
— плодовитость их 235 сл.
— поведение (высшая нервная деятель-
ность) 288 сл.
— половые органы их 137 сл.
— половые признаки 233
---продукты, определение степени
зрелости 232
— пресноводные 201, 278 сл.
--- распределение 279 сл.
— придонные 213 сл.
— приспособление к условиям обитания
208 сл.
63*
— приспособления к передвижению, ти-
пы 210 сл.
— продолжительность половой жиэни
235
— промысловое значение их 294 сл.
— проходные 203, 288
— развитие их 146
— разделение по признаку водной среды
208
— размножение 231 сл.
— растительноядные 218
— сегментация яиц у них 146 сл.
— скрещивание у них 251, 252
— способы дыхания у них 118 сл.
— спячка у них 206
— стенотермные 206
— строение 75 сл.
— темпы роста у них 242 сл.
— теория осмотического питания Пют-
тера 218
— уловы 244 сл.
— филогенез 146, 193 сл.
— форма тела 77 сл.
— характеристика и обзор класса
72 сл.
— эйритермные 206
— экономическое значение 293 сл.
— электрические органы у них 227
— явления возраста и роста 241
— ядовитый аппарат у них 226 сл.
— яйца их 146 сл.
Рысаки 923
Рысь 775, 815, 869, 888, 889, 892, 930,
948, 949, 971, 974
Рябинник 617
Рябки 603, 604, 620, 647, 648, 657, 661,
664, 686, 974
Рябковые 603
Рябчик (рыба) 186
Рябчики 595, 669, 703, 704
Ряпушка 174, 296, 302
Саблезубые кошки 889
Савка белоголовая 592
Савковые 592
Саджа 603, 661, 685
Сайга 834, 835, 939, 974, 976, 977
Сайда 193, 239, 244, 293
Сазан 97, 169, 208, 218, 232, 244, 274,
277, 293, 295, 297, 303
Салака 293
Салакушка 298
Саламандра азиатская наземная 353
— безлегочная 355
— гигантская 353, 354
— горная 375
— когтеносная 353
— мексиканская 381
— наземная 375
— обыкновенная 355
— пещерная 355, 375
— пятнистая (огненная) 354, 375
— слепая 355
— черная 354
— японская 354
Саламандры 353 сл., 384
Саланганы 643, 644
Сальпы 30 сл.
Сапролегния 275
Сапсан 593
Сарган 179, 213, 220
— обыкновенный 210
Сардинки 173, 217
Сарычи 593, 631, 710, 944
Сатана 870
Саусданские овцы 914
Сахарная белка 870
Сахарные птицы 615
Сверташка коралловая 482
Сверташки 482
Сверчки 616
995
Свет, влияние на жизнедеятельность
млекопитающих 881
----------- птиц 662 сл.
—----------рыб 207 сл.
Свечение рыб 228 сл. *
Свинковые 811
Свиные барсуки 969
Свиньи 533, 771, 774, 780, 829, 830,
869, 872, 887, 889, 936, 945, 965,
970
Свинья домашняя 538, 918 сл.
— средиземноморская 918
Свиристелевые 615
Свиристель 615, 670, 693, 976
— японский 977
Свистуны 359
Свиязь 591, 703
Свободнохвостые 798
Северный олень, см. олень северный
Севрюга 161, 295, 297, 302
Секретаревые 593
Секретари 533, 538, 592, 593, 968
Секретарь африканский 631, 632, 633
Селахии, см. акулы
Селенодонтные 829, 830 сл.
Сельдевые 298
— промысловое значение их 173, 295 сл.
Сельдеобразные 172
Сельди 75, 135, 210, 216, 231, 238, 244,
249, 272, 273, 274, 293, 298, 301
— миграция их 264 сл,
Сельдь американская 277
— астрабадская 295
— атлантическая 232, 234, 235
-----миграции 264 сл.
— волжская 235, 295
.— долгинская 295
— иссык-кульская 297
— каспийская 173, 222, 232
— мангышланская 295
— североатлантическая 173
— тихоокеанская 296
— черноморская 173
— черноспинка 235
Сельдяной король 78, 182, 210
Семга 174, 258 сл., 298
Семга, миграции 258 сл.
Сен-бернары 902, 904
Сеноставец 805, 976
Сепс трехпалый 476
Серебристый кролик 926
— угорь 256
Сернобыки 835
Серповые 835
Серны 880, 939, 950, 974
Сероводород, значение для жизни рыб
205
Сероукраинский рабочий скот 910
Серпоклювы 634
Сетеры 905
Сибирский молочный скот 908
Сивапитек 851
Сиватерии 839
Сивка глупая 640
Сиводушка 933
Сивуч 774, 819, 935
Сиг озерный 175, 231
— проходной 174, 303
Сиги 174, 175, 208, 216, 217, 236, 296,
302
Сиговые 297
Сизоворонки 608, 609, 644, 648, 676,
680, €81, 708, 966
Сизоворонковые 608, 632, 633
Сизоворонкообразные 561
Сизоворонкоподобные 604 сл., 621
Симментальский мясо-молочный скот 908
Симметрозубые 790
Синантроп 852
Сингиль 180
Синдактилия 868
Синец 295
996
Синица большая 615
— хохлатая 976
— черная 615
Синицевые 615
Синицы 615, 635, 644, 649, 658, 660, 661,
669, 687, 706, 708, 972
— попугаеобразные 615
Синички 975
Синойкия у птиц 690
Синухи 710
Сипуха 607
Сипуховые 607
Сипы 634
Сирены 356, 720, 723,740, 741, 763, 764,
772, 794, 823, 826 сл., 858, 872,873,
874, 887
Сифака 870
Скадозубые 185
Сяаровые 186, 217, 220
Скаты 73, 80, 153 сл., 213, 216, 219, 225,
231, 233, 274,
— настоящие 154
Скаты-пилорыбы 221
— электрические 154, 155, 228
Скворец обыкновенный 613
— розовый 613, 671
Скворешницы 707 сл.
Скворцовые 613
Скворцы 629, 678, 679, 681, 690
Скелет амфибий 313 сл.
— круглоротых 52
— ланцетника 35
— млекопитающих 725 сл.
— птиц 555 сл.
— рептилий 404 сл., 412 сл.
— рыб 79 сл., 83 сл., 102 сл.
Сколопендры 533
Скопа 593, 631, 633, 683
Скорпена 226, 227, 233
Скорпенообразные 188
Скорпида 189
Скрещивание у рыб 252
Скрытоголовые черепахи 462
Скрытожаберник аллеганский 354
Скрытохвосты 581, 584
Скрытошейные черепахи 547
Скумбрия 192, 272, 296, 298
Славка садовая 613, 616
— серая 616
— черноголовая 616
— ястребиная 616
Славки 650, 705, 966, 973, 976
Славки-портнихи 616
Славковые 616
ЬчСлепуны 481, 509, 510, 545
—“роющие 509
Слепушонки 811, 861, 864, 865, 887,
890
Слепышевые 810
Слепыши 742, 783, 810, 862, 863, 865,
887, 967, 974
Слон африканский 761, 825, 869
— индийский 761, 774, 825, 826, 970
— лесной 825
— полуденный 825
— степной 825
Слоновые 825
Слоны 723, 734, 745, 746, 753, 760 сл.»
772, 774, 778, 780, 824 сл., 884, 887,
965, 967, 968
Слуховой аппарат у амфибий 345, 389
---у млекопитающих 781 сл., 898
— — у птиц 575
---У рептилий 444, 540, 541
---у рыб 144, 290
Смушково-молочные овцы 916
Снегирь 614, 635, 670, 671, 693
— длиннохвостый 614
Снеток 175, 216, 217, 293, 296
Собака домашняя 816, 902 сл.
— Путятина 903
— торфяная 903
Собаки 720, 726, 752, 753, 756, 765, 813,
860, 873, 888, 889, 898, 899, 900, 902
— кустарные 961
Собакоголовые 853
Собачки луговые 533, 778
Собачьи 816, 933
Соболь 817, 869, 888, 889, 894, 931, 947,
949, 951, 976
Сова белая 607, 894, 978
— болотная 607, 630, 710
— китайская 538
— лапландская 670
— полярная 663
Сова-рыболов 633
— уральская 607
— ушастая 607, 710, 892
— ястребиная 607, 633
Совиные 607
Совы 533, 606, 607, 620, 621, 632, 633,
644, 645, 647, 648, 658, 662, 663, 670,
684, 708, 710, 944
— голоногие 607
— дневные 607
Совы-неясыти, см. неясыти
Сойка саксаульная 613
СОйки 613, 636, 661, 689, 974, 975
Сокол-балобан 630
Сокол-сапсан 632, 682, 710
Сокол-чеглок 710
Соколиные 593
Соколы 593, 629, 631, 682, 944
Соколы-дербники 632, 710
Сонолы-крошни 632
Сокольская овца 916
Соли, содержание в воде, значение для
рыб 203
Солнечники 182
Соловьи 617, 705
Соловьиные 617
Соль (рыба) 183
Сом обыкновенный 171
— панцырный 80, 230
— полу панцырный 171
— электрический 172
Сомик 240
— индийский 172
Сомовые 120, 121, 171, 231, 240
Сомы 171, 172, 216, 219, 221, 225, 293,
297
Сони 809, 869, 879, 889, 897, 967, 976
Соневидные 809
Соневые 809
Соня лесная 809
— садовая 809
Сороки 613, 693, 972
Сорокопут большой 616
— чернолобый 616
Сорокопутовые 615, 616
Сорокопуты 533, 613, 633, 959, 972
Спаривание у амфибий 375 сл.
— у млекопитающих 895, 896
— у птиц 640 сл.
Спинная струна 26, 31
— — ланцетника 35, 43
Спинорог прямоугольный 81
Спинороги 185
Сплюшка 607
Спячка у амфибий 384
— у млекопитающих 878 сл.
— у рептилий 529, 530
— у рыб 206
Сростноглоточные 185
Сростножаберные 98, 151
Сростночелюстные 184, 220, 226
Ставридка 191
Стации обитания птиц 685 сл.
Стегозавры 498, 524
Стегоцефал пермский 307
— лягвообразный 307
Стегоцефалы 305, 306 слГ.. 403, 415,
501
Стеклянный угорь 255
Стелигозавр 524
Стеллерова морская корова 763, 827F
887, 945
Стеллион кавказский 470
Стеноадаптивные признаки 620
Стенолаз 615, 635
— нраснокрылка 687
Стенотермические птицы 661
Стенотермные рыбы 206
Стервятник 630, 691
Стереорнисы 596
Стерлядь 160, 216, 222, 295, 296, 302
Стихей 284
Страусы 567, 571, 581 сл., 620, 641, 652,
660, 691, 695, 968
— мадагаскарские 584
Стрела-змея 484, 535
Стрепет 598, 599, 639, 704
Стриж гватемальский 644
— горный 608
Стрижевые 608
Стрижи 567, 607, 608, 621, 626, 629, 635,
645, 647, 648, 655, 676, 680, 681,
682, 683, 686, 688, 705
Суасоне 912
Сублиторальная зона 208
Судак 183, 217, 225, 275, 293, 296, 297,
302, 303
— морской 183, 295
— обыкновенный 183, 295
Султанка 184, 298
Султанские курочки 954
Сумка сумчатых 722
Сумчатая выхухоль 790
— землеройка 790
— лягушка 380
Сумчатость 722 сл., 857
Сумчатые 713, 715, 722, 735, 751, 770,
772, 773, 788, 789, 790, 857, 861,
866, 868, 896, 897, 956, 960, 964
— барсуки 791, 888, 956, 959
— белки 792, 870, 957
— волки 791, 888, 956, 959
— кроты 727, 742, 743, 783, 791, 862,
863, 865, 883, 885, 888, 956
— крысы 722, 790, 888, 956, 960 (см.
также опоссумы)
— куницы 790, 888, 956, 959, 960
— лазающие 792
— летяги 792
— медведи 792, 866, 957
— мурашееды 791, 956
— мыши 957
— сони 792, 793, 870
Сумчатый дьявол 790, 791
— муравьед 722, 886
— пяткоход 886
Сурки 806, 861, 878, 879, 880, 881, 886,-
895, 929, 976
Сусленик 932, 976
Суслик европейский 807
— желтый 807, 878, 883, 890, 891
— колумбийский 878
— крапчатый 807
— краснощекий 807
— малый 890
— рыжий 807
— эверсманов 977
Суслики 806, 807, 860, 879, 881, 884,
886, 892, 929, 943, 971, 974, 976
Сусук 823
Сухонос 591
Сциеновые 184
Сцинк аптечный 476, 509
Сцинки 410, 510, 531
— длинноногие 476
Сцинковые 476, 528, 544
Сыч воробьиный 607
— домашний 710
— домовый 607
Сычи 670, 944
Сычик воробьиный 663
997
Тагильский молочный скот 908
Тагуаны 870
Таймень 174, 296, 302
Таксы 902, 904
Тамдндуа 743, 864
Т5Гйа 794
Танагровые 614
Тапировые 844
Тапиры 751, 844, 845, 869, 872, 886,
962, 970
Тарабаганчики 806
Тарабаганы 929, 951
Тарань 170, 295
Тарпаны 844, 920, 921, 946
Тарпон 172
Твердокожие 184
Тейю 512, 527
Текодонты 403, 543
Текодонтные 422, 576
Телескопические глаза у рыб 145, 291
Телескоп 251
Температурные условия существования
амфибий 383 сл., 388
---------млекопитающих 876 сл., 884
— — — птиц 659 сл., 668
— рептилий 529
— рыб 205 сл.
Тенрек иглистый 795
Тенреки 723, 795, 872, 873, 874, 878,
879, 885, 961, 965
Тенрековые 795
Тецьковка 616
Териодонты 785
Тероморфы 542
Терьеры 903
Тетерев кавказский 595
Тетерева 594, 595, 596, 637, 646, 657,
669, 703, 704, 973
Тетерева степные 595
Тетеревиные 595, 972
Тетерки 657
Теутофагия 518
Течка у млекопитающих 895
Тигры 815, 882, 883, 888, 889, 894, 930,
942, 951, 974
— саблезубые 815
Тиллодонты 799, 964
Тималиевые 616
Тинамовые 585
Тинаму 560, 581, 584, 620, 636, 645,
686, 963
Типотерии 841
Тиранны 963, 973
Тирановые 611
Тиркушка степная 600, 601
Тиркушки 664, 682, 686
Тиркушковые 601
Титанотериевые 846
Тихо ходовые 802
Тихоходы 802, 960
Ткачи 636, 644
Ткачики 968
Ткачиковые 613
Тоди 963
Токование 638
Токсодонты 841
Толай 805
Тонкоголовые 254 сл.
Тонкочешуйные 168
Топорик 603
Торфяная овца 911
— свинья 918
Травники 601
Трагопаны 976
Трахелозавры 457
Треска 75, 76, 192, 193, 210, 212,
231, 236, 238, 239, 240, 244, 246,
253, 270, 271, 273, 274, 275, 293,
296
— миграции 270 сл.
Тресковые 121, 193, 298
Трескообраэные 192
998
Трехбугорчатые 790, 964
Трехконусоэубые 788, 789, 790
Трехперстки 594, 597, 621» 636, 638,
647, 648, 686
Трехперстковые 597
Трибелезодонты 499
Тригловые 189
Триглы 189
Тритон малоазиатский 355
— ратбунов колодезный 355
— семиреченский 353
— сибирский четырехпалый 353
Тритоны 308, 353, 354, 375, 381, 384
Трогоны 560, 609
Тропический климат, богатство фауны
птиц 661
Трубачи 599, 963
Трубкозуб 771, 794, 886, 964, 967
— капский 847, 862
— эфиопский 847
Трубкозубые 847
Трубконосы 633, 683
Трясогузка белая 613, 614
— горная 614
— желтая 614, 657
— желтоголовая 614
— пестрая 614
— черноголовая 614
Трясогузки 643, 650, 679, 705, 708
Трясогузковые 614
Тукановые 610
Туканы 636, 963
Туко-туко 862, 961
Тулес 601
Тунец 78, 192, 217, 274
Тупайевые 794
Тупайи 794, 969
Турако 606
Турачи 595, 704
Турецкая лошадь 921
Туркменская лошадь 921
Турухтаны 601, 637, 657, 676, 681
Туры 835, 836, 906, 938, 939, 946, 947,
948
Тушканчик мохноногий 810
Тушканчики 727, 743, 745, 746, 809,
810, 861, 862, 878, 879, 883, 890,
974
Тушканчиковидные 809
Тушканчиковые 809, 971
Тэкэ 938
Тюленевые 820
Тюлени 742, 763, 873, 874, 875, 889,
935, 951, 978
Тюлень каспийский 820
— сибирский 820
— ушастый 741, 819
Тюрбо 214
Тяжеловозы 921
Угорь американский 178
— европейский, отличие от американ-
ского 254
— морской 178
— обыкновенный 178
— электрический 228
Угреподобные 177
Угри 76, 77, 212, 214, 235, 237, 238, 247,
254 сл., 274, 298
— миграция их 254 сл.
— настоящие 178
Удав водяной 482
— обыкновенный 482
— песчаный 482
— пустынный 509
— степной 510, 546
Удавовые 482
Удавы 482, 518, 521, 534, 545
Удодовые $09
Удоды 567, 609, 648, 58, 678, 681, 691,
708, 968
Уж желтоухий 483
— китайский 529
Ужи 528, 532, 535
— водяные 483
— древесные 484
— лазящие 484
— настоящие 483
— ядовитые (аспидовые) 485
Ужовые 483, 546
Уклейка 79, 170, 216
Улар 951
Улитовые 601
Улиты 601, 681
Уловы рыб 244 сл.
Умбровые 175
Усач 170, 208, 220, 232, 275
— аральский 297
Утиные 591, 639, 684
Утка дикая (кряква) 699
— серая 591, 703
— синеклювая 592
Утки 561, 574, 575, 591, 626, 634, 639,
647, 652, 654, 655, 664, 668, 670, 677,
678, 679, 695, 696, 699, 703
— благородные 703
— красные 643
— мускусные 699
— пекинские 699
— породы 699
— руанские 699
— собственно 591
Утконос 722, 737, 752, 764, 770, 773,
774, 779, 788, 789, 872, 879, 881,
885, 896, 956, 959
Утконосовые 788
Ушанки 698
Ушаны 798, 889
Ушастые совы, см. совы ушастые
Уши у ночных животных 881, 882
Фазан кавказский 594, 595
Фазановые 595
Фазаны 595, 637, 655, 695, 701, 704, 970,
974, 976
Фараонова мышь 815
Фауна млекопитающих, влияние чело-
века на ее изменение 945 сл.
Фаэтоновые 588
Фаэтоны 683
Фенакодонтовые 842, 843
Фенакодус 842
Фенек 862, 882
Филины 607
Филломедуза Иеринга 380
Филогенез рептилий 445, 500 сл.» 502 сл.
— рыб 146, 193 сл.
Финваль 822, 940
Фитозавры 487
Фитопланктон 217, 272 сл.
Фламандская лошадь 921
Фламандский кролик 925
Фламинго 570, 587, 589, 590, 621» 634,
645, 652
— розовый 589, 590
Фламинговые 590
Форель 174, 208, 218, 225, 237, 238, 250,
251, 275, 302, 303
— американская радужная 303
— гокчинская 249
— речная 236
)— ручьевая 303
'Фосса 965
Фотофоры 229, 230
Фрегатовые 588
Фрегаты 683
Фрисландский молочный скот 907
Хаки-Кемпбелл 699
Халцид трехпалый 509
Хамелеон обыкновенный 476
Хамелеоны 410, 412, 417, 429, 435, 438,
476, 477, 511, 513, 519, 521, 522,
526, 542, 544, 545, 549
Хамса 173, 174, 217, 293, 297
Харациновые 121, 168, 218
Харза 931, 951
Хариусы 175, 216, 236, 237, 303
Хаулиод 219, 229
Хвостатые (амфибии) 305, 352
Хвостатые (личиночные) 28
Хвостоколы 155
Хиазмод черный 181, 219
Хиазмодонтовые 180
Химеры 156, 219
Химическое чувство амфибий 388
---рептилий 520
Хироты 476
Хирурги 184
Хищники 774, 794, 813, 885, 888, 889,
896, 930 сл., 968
— вред, причиняемый ими 942 сл.
— значение в жизни человека 930 сл.
— схема эволюции 814
Хищные птицы 567, 647, 650, 656, 670,
671, 682, 689, 710, 892
---дневные, см. дневные хищные
птицы
Хоботные 794, 823, 824 сл.
— эволюция их 760
Ходулочник 601
Ходулочниковые 601
Холмогорский молочный скот 907
Хомяк обыкновенный 810
— рыжий 876
Хомяки 751, 765, 810, 861, 879, 887,
889, 943, 948, 974, 976
Хомячки 810
Хорда 26, 31
— ланцетника 35, 43
Хордовые 13 сл. •
— происхождение 47
Хорек обыкновенный 816
— степной 816
Хорьки 816, 865, 888, 892, 897, 931, 942,
944
Хорьковые 816
Хохотун 603
Хрящевые ганоиды 74, 86, 160
— рыбы 73, 149 сл.
Паплевые 588
Цапли 533, 567, 568, 587, 588, 621, 633,
637,644, 648, 656, 662, 664, 684, 690,
702
— белые 588
— китоголовые 588
— рыжие 588
— серые 588
— солнечные 597, 598, 963
— сумеречные 588
Цветососы 615
Целакантины 163
Цельноголовые 73, 86, 93,99, 113, 155 сл.
Ценогенез 146
Ценолестовые 791
Ценолесты 790
Цепкопалые 477
Цепкохвостые 853, 963
Цератод 86
Цератодовые 158
Цесарки 595, 695, 696, 699, 968
Цепилиевые 352
Циветты 814
Цигайские овцы 912
Цинодонты 452
Цокоры 811, 887
Чавыча 296
Чайка клуша 603
— морская 603
— сизая 602, 603
999
Чайка трехпалая 603
Чайки 602, 603, 620, 621, 628, 630, 633,
634, 643, 647, 648, 649, 652, 655, 658,
659, 674, 675, 676, 682, 683, 684, 691,
704, 978
Чайковые 603
Частиковые рыбы, промысловое зна-
чение их 295
Чебак 297
Чеглок 593, 632
Чеграва 603
Чекановые 617
Человек 723, 748, 765, 771, 854, 900
Человекообразные 854
Челюстноротые 72 сл.
Чепура 588
Червяга 351, 352, 378
— земляная 352
— кольчатая 309, 352
— толстокожая 376
Череп амфибий 318 сл.
— круглоротых 52, 53
— млекопитающих 728 сл.
— птиц 558
— рептилий 417 сл.
— рыб 88 сл.
Черепаха <аррау» 462, 534
— бахромчатая 462
— болотная 463
— водяная европейская 463, 464, 530
— гигантская 464, 529, 549
— греческая 465, 534
— грифовая 462, 463
— зеленая 465, 534
— кавказская 465
— каретта, см. каретта
— каспийская 463
— кожистая 411, 466, 467, 532, 547
— кусающаяся 462
— мавританская 465
----- водяная 463
— речная 463
— степная 464
— суповая 465, 534
— туркестанская степная 534
Черепахи 408, 409, 410, 422, 423, 424,
426, 427, 430, 431, 436, 437, 440, 442,
443, 445, 460 сл., 502, 515, 516, 517,
518, 520, 521, 524, 525, 526, 527, 528,
529, 530, 531, 534, 541, 542, 543
— бокошейные 462, 547
— змеиношейные 462
— исполинские слоновые 534
— каймановые 462, 532
— морские 465, 515, 516, 527, 532
— мягкие 411, 467, 532, 548
— наземные 463, 547
— пресноводные 463
— происхождение 504
— скрытоголовые 462
— скрытошейные 547
— сухопутные 532
Черепные 50 сл.
— происхождение и эволюция 6 9
Чернети 591
Чернобурая лиса 933, 942
Черноголовый мартын 603
Чернозобик 601
Черноспинка 295
— миграция 269
Черный заяц 975
Черныш 601
Чесночница 381
— обыкновенная 358
Четвероногие 74, 304
Четырехзубые 185
Чехонь 170, 247, 293, 295
Чечевица 614, 675, 680, 972
— розовая 977
Чечотка 614, 669, 670, 672
— горная 614
Чешуеперые 184
1000
Чешуйчатые 404, 423, 467, 469
Чешуя 79 сл., 243, 244, 247, 260, 265
— ганоидная 81
— ктеноидная 81
— плакоидная 80
— ромбическая 82
— циклоидная 81
Чибисы 601, 643, 647, 657, 659, 679, 691
Чиж 614, 671, 972
Чирок 591, 703
Чирок-свистунок 629
Чистик гигантский 621
Чистики 602, 603, 621, 633, 642, 647,
648, 652, 659, 660, 683, 704, 972,
978
Чистиковые 603
Шакал 816, 889, 891, 905, 934
Шалашники 640
Шатильоне 912
Швицкий мясо-молочный скот 908
Шемая 170, 295, 297
Шерстные овцы 912
Шерстокрыловые 799
Шерстокрылы 792, 798, 799
Шерстохвосты 956
Шерсть млекопитающих 876 сл.
Шилоклювка 601, 630, 634
Шилохвость 591, 630, 678, 703
Шимпанзе 851, 854, 856, 868, 900, 966
Шиншилла 811, 886, 961
Шиншилловые 811
Шип 295, 297
Шипохвост 471, 472
Шипуны 699
Широконоска 591, 703
Широкороты 970
Шлемочерепные 449
Шортгорнский мясной скот 909
Шпицы 903
Шпроты 173, 217, 244
Шропширы 914
Щеглы 613, 614, 644, 670, 671
Щелезубовые 796
Щелезубы 796, 961
Щетинозубые 184
Щиповка 169
Щитомордник 486, 527, 541
— палласов 486
Щитоносец аргентинский 802
Щитоносцовые 802
Щитохвосты 545
Щитохвостовые 482
Щука 132, 176, 208, 216, 218, 219, 225,
232, 246, 275, 293, 295, 303
— ганоидная 167
— морская 179
— панцирная 80
----коротконосая *167
Щу новые 176
Щукообразные 175
Щур 614
Щурка золотистая 609
Щурки 643, 645, 689, 710
Щурковые 609
Эволюция амфибий 376 сл.
— ланцетника 42 сл.
— личиночно-хордовых 25 сл.
— миног 60 сл. *
— млекопитающих 786 сл., 857 сл
— птиц 577 сл., 617 сл.
----- схема 619
— рыб 146 сл.
Эйритермические птицы 661
Эйритермные рыбы 206
Эйритоадаптивные свойства 620
Эйфотическая зона 207, 209
Экология амфибий 364 сл., 392
Экология круглоротых 63 сл.
— миксин 64
— миног 64 сл.
— млекопитающих 858 сл.
— птиц 659 сл.
— рептилий 506 сл.
— рыб 200 сл.
Экономическое значение амфибий 386,
387
---млекопитающих 927 сл.
— — птиц 694 сл.
--- рептилий 534
---рыб 293 сл.
Экономка 811
Экотопические факторы жизни птиц 681,
682
Элазмобранхий 85, 86, 195 сл., 200
Эласмотерии 846
Элсктораль 912
Электрические рыбы 227 сл.
Электрический угорь 169
Эмбритоподы 794, 823, 826
Эму 560, 581, 582, 583, 647, 686, 958
Эогея, ихтиофауна ее 279 сл.
Эосирен 827
Эотерий 827
Эскулапова змея 484
Эфа 511
Эфиопская область, фауна 966
Юр 976
Юрок 614
Ягнятник 631
Ягуар 815, 869, 888, 961
Яд змеиный 535, 536
Ядовитые змеи 534 сл.
— рыбы 79
— кожные выделения у амфибий 371
Ядовитый аппарат рыб 226 сл.
Ядозубы 473, 523, 539
Язвицы 956
Язык амфибий 330
— птиц 567
— рептилий 435, 519, 541
— рыб 133, 134
Язь 170
Яйца амфибий 345 сл., 375 сл.
— демерсальные 212, 231
— млекопитающих 770
— пелагические 211, 212, 231
— птиц 573, 645 сл.
---высиживание их 641 сл.
— — количество яиц в кладке 647
окраска 645, 646
-----продолжительность высиживания
647
-----форма 646, 647
— рептилий 390 сл., 526 сл.
— рыб 146 сл.
— черепах 534
Якановые 602
Яканы 602
Яки 876, 877, 910, 911, 974
Якмары 963
Якобсонов орган у амфибий 344
-----у млекопитающих 783
— — у птиц 576
----- У рептилий 540
Японские собачки 902, 903
Ярославский молочный скот 907
Ястреб-перепелятник 593, 632, 657, 710
Ястреб-тетеревятник 593, 632, 670, 710
Ястребы 593, 631
Ящерица глазчатая 475
— жабовидная 472, 474, 510
— живородящая 474, 525
— зеленая 475, 523
— плащеносная 471, 472, 507, 525
— прыткая 475
— степная 475
— ядозуб 539
Ящерицевые 470
Ящерицы 404, 405, 410, 411, 412, 415, 419,
422, 423, 424, 426, 427, 432, 433,
435, 437, 440, 441, 443, 444, 469 сл.,
480, 502, 507, 508, 516, 518, 521. 522,
523, 525, 526, 528, 529, 530, 531, 533,
534, 541, 542, 544, 545
— линька их 411
— настоящие 474
— поясохвостые 472
— тейю, см. тейю
Ящероногие 492
Ящерообразные 550
Ящеротазовые 491
Ящеры 715, 728, 750, 764, 766, 792, 800,
886, 964, 967, 970
— зверозубые 412
— крылатые 498 сл.
— степные 800, 862
Ящурна 475, 530
— быстрая 475
— полосатая 475
— разноцветная 475
— сетчатая 475
II. УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ЖИВОТНЫХ
Ablepharus 476
— lineatus 508
Abramis balerus 295
— brama 170, 216, 217, 237, 241, 246,
293, 295, 296, 297, 301, 302
— вара 295
— vimba 247
Acanthias 91, 103, 284, 285
— vulgaris 108, 153
Acanthodes 150
— gracilis 150
Acanthodidae 150
Acanthodii 73, 99, 150, 196
Acanthopterygii 102, 149, 181 сл., 188,
199, 281, 282
Acanthuridae 184
Acanthurus 184
Accent or idae 617
Accipiter nisus 593
Accipitres 567, 587, 592, 593, 631, 643,
647, 648, 649
Acerina 183
— cernua 183, 252, 296
Acestra gladius 172
Acinonyx jubatus 816, 974
Acipenser 99, 108, 112, 113, 147, 149,
160, 161
— baeri 160
— gtildenstadtl 160, 297, 302
— medirostris 161
— nudiventrls 160, 297
Acipenser ruthenus 160, 216, 222, 288,
296, 302
— schrencki 160
— stellatus 161, 297, 302
— sturio 160
Acipenseridae 74, 160, 164, 216, 225,
244, 252, 253, 280, 297
Acipenseriformes 160 сл.
Acrania 32, 33, 45, 46, 47, 48, 50, 109
— primitiva 50, 195
Acreodi 813, 828, 857
Acrobates 792, 957
— pygmaeus 793, 870
Acrocephalus 657
— arundinaceus 616
— palustris ol6
— streperus, 616
Acthia 978
Actihopterygli 100, 161, 198
Actitis hypoleucos 601
Adapidae 848, 851
Adapis parisiensis 848
Adelochordata 13, 24
Aegialitis alexandrinus 601
— dubius 601
— hiaticula 601
Aegithalos caudatus 615
Aeluroedus 613
Aepyornis 966
— maximus 584
Aepyornithes 584, 620
1002
Aepyornithidae 584
Aetosauridae 488
Aetosaurus 488
— terr at us 487
Agama 470, 507, 510
— caucasia 470
— sanguinolenta 470, 522, 530
Agamidae 412, 448, 470, 507, 508, 511
514, 518, 544
Aglossa 358, 364
Aglypha 483, 520, 535, 537
Agnathostomata 33, 50, 63, 69, 195
Agonidae 82, 189, 284
Agonus 285
— cataphractus 189
Agriocetus 821
Aguilla 246
Aigialosauridae 479
Aigialosaurus 479
Ailuropus 816, 975
Ailurus 969
— fulgens 816
Aistopoda 306, 351, 364
Alactaga 745, 746, 810, 862, 883, 890
974
— jacalus 810
Alauda arvensis 612, 614
Alaudidae 614, 636, 643
Alaudula minor 614
Albulidae 172
Alburnus alburnus 170
— chalcoides 170, 295, 297
— lucidus 79, 216
Alca 642
— impennis 621, 683
— torda 602, 603, 642
Alcae 602, 603, 621, 633, 647, 660, 683
704, 972
Alcedinidae 609, 633, 648
Alcedo 274, 643
— ispida 609, 610
Alces 832, 865
— alces 832, 936, 971
— latifrons 834
Alcidae 603
Alectoromorphae 593, 621
Alligator 424, 490
— missislpiensis 490
Alllgatoridae 490, 548
Aliosaurus 406, 446, 492, 493, 507
Allotheria 788
Alopex alopex 876
— beringiensis 933
— lagopus 816, 933, 971
Alosa 173
— sapidissima 173
Alsophylax 478
— pipiens 478
Alticamelus 831
Alticola 876, 892
Alytes 341, 358, 376
— obstetricans 358, 380
Amblypoda 846
Amblyopsidae 177
Amblyopsiformes 177
Amblyrhynchus crist at us 515
Ambystoma 354, 366, 381, 382, 393
— tigrinujn 315, 354
Ambystomatidae 354
Ambystomidae 354
Amia 75, 81, 100, 108, 112, 120, 121, 122,
123, 126, 128, 134, 138, 147, 165, 280,
282, 286
— calva 87, 108, 147, 148, 165, 166, 167,
198
Amildae 166
Amioidei 87, 102, 160, 164, 165 сл.,
198
Ammocoetes 55, 61, 65, 125
Ammodytes 262, 272, 274
Arnmodytidae 187
Ammod у ti formes 186
Ammodytus tobuanus 187
Ammotragus tragelaphus 912
Amniota 304, 339, 340, 343, 351, 395,
415. 550
Ampelidae 615
Am pells garrulus 693
Amphibia 33, 50, 304 сл.
Amphichelydia 461
Amphicoela 357, 358
Amphicyon 816
Amphilestes 789
Amphioxides 46, 47, 48
Amphioxididae 33, 46
Amphioxus 21, 32, 33, 34, 38, 42, 44,
46, 47, 61, 84, 111, 112
Amphirhina 50
Amphisbaena 476, 509
— fuliginosa 476
Amphisbaenica 476
Amphisbaenidae 419, 470, 476, 510, 545,
549
Amphistidae 182
Amphitherium 754, 789, 790
Amphitragulus 834
Amphiuma 308, 318, 351, 375
— means 355, 379
Amphiumidae 352, 355, 365, 393
Amylochilus ferruginae 601
Anabanthidae 180, 280
Anabas 361
— scandens 119, 180
Anableps 177
Anacanthini 102, 192
Anamnia 304
Anapsida 66, 403, 419, 423, 446, 467, 502
Anarhynchus 954
Anarrhichas 190
— lupus 190, 217, 219
Anas 591
— platyrhyncha 591, 699, 703
Anatidae 591, 703
Anatinae 591, 703
Anchinga 588
Anchitherium 759, 843, 844
Ancistrodon 486, 527, 528, 541
— halys 486
-----intermedins 486
— piscivorus 486
Ancylopoda 846, 964
Andri as scheuchzeri 353, 364
Anguidae 470, 472, 545
Anguilla 76, 77, 212, 235, 238, 281
— americana 178
— anguilla 178, 214, 231, 237, 254, 298
— rostrata 254
— vulgaris 247
Anguiliformes 177
Anguillidae 178
Anguinidae 470
Anguis 470, 473, 476, 509
— fragilis 411, 444, 473, 540
Anicanodonta 802, 960
Anodonta 171, 240
Anolis carolinensis 522
Anomalops 230
Anomaluridae 809, 967
Anomaluroidea 807
Anomalurus 809, 870
Anomocoela 358
Anomodontia 426, 453, 503
Anoplotheriidae 840
Anser 591, 703
— anser 591, 700, 703
Anseres 572, 587, 590, 621, 633, 643,
646, 648, 659, 702, 963
Anseridae 591, 703
Anseriformes 561
Antennariidae 191
Anthoscopus pendulinus 615
Anthracotheriidae 840
Anthropoides virgo 599, 704
Anthropomorphae 854, 969
Anthropopithecus 851, 854, 868, 966
— niger 856
Anthus 643, 657, 972
— campestris 614
— cervinus 614
— pratensis 614
— trivialis 614
Antiarchi 69, 145
Antilocaprinae 973
Antilocarpa 718
Antilope 970
— cervicarpa 835
— maxwell! 720
— subgutturosa 861
Antilopinae 834, 967
Anura 305, 306, 308, 312, 314, 317, 318,
319, 323, 326, 327, 329, 330, 332, 333,
334, 335, 336, 339, 340, 341, 342, 343,
344, 345, 346, 347, 350, 356, 357, 364,
366, 367, 368, 375, 385, 393
Aphia minuta 246
Apidium phiomense 851
Apoda 305, 308, 312, 314, 317, 318, 320,
323, 326, 332,333, 339, 340, 341, 343,
344, 345, 346, 351,-353, 364, 365, 368,
375, 388, 392, 393
Apodemus 976
— agrarius 943
Appendicularia 28
Appendiculariidae 28
Apterodon 821
Apteryges 583, 620, 954
Apterygidae 583, 954
Apteryx 663, 686, 954
— australis 583
— manteHi 584
Apus apus 607, 608
— melba 608
Aquila chrysaetas 593
— clanga 593
— heliaca 593
— orientals 593
Aquilidae 593, 633
Aramidae 599
Aranus scolopaceus 599
Arapaima 78
Archaeoceti 821
Archaeomeryx 839
Archaeophis 417
— proavus 481
Archaeopteryx 578
— macrura 580
Archaeornis 578
— simensli 577, 580
Archaeornithes 566, 577, 580
Archaeosuchus 453
Archaeophidae 481
Arch egos auridae 322
Archegosaurus 316, 350
Archelon 466
— ischyros 465, 466
Archosauria 467
100 3
Arcifera 325, 326, 393
Arctocephalus ursinus 818, 819
Arctomyx 969
Ardea cinerea 588
— purpurea 588
Ardeae 274, 587, 588, 621, 633, 648, 662,
684, 702
Ardeidae 588
Ardeola 588
Ardetta minuta 588
Arenaria interpres 673
Areoscellda 402, 404, 469, 502
Areoscelidae 469 f
Areoscelis 469, 505
— gracilis 469
Argusianus 595
Argyropelecus 175
Arius 172
Arsinotherium Zitteli 826
Artemidae 959
Arthrodlra 69, 73, 157, 197
Arthroleptls seyschellensis 379
Artrichornitidae 959
Artiodactyla 744, 769, 794, 823, 827, 936,
OS7 QRo
Arvicoia 887, 889
— amphibius 810
Ascaphus 327, 332, 341, 357, 383
Ascidia 27, 29
— mentula 22
Ascidiae 28
— compositae 29
Ascidiae luciae 29, 31
— salpaeformes 29
— simplices 29
Asinus 844
— africanus 844, 923
— asinus 844
— hemionus 936, 974
— kiang 974
— onager 974, 975
— somalensis 923
Asio accipitrinus 607
— otus 607, 892, 944
Aspius aspius 170, 237
— тарах 247
Aspredo 240
Asterolepidae 69
Asterolepis maxima 69
Astrapotherioidea 841
Astur palumbarius 593
Astylosternus 313
— robustus 374
Asymmetron 33, 46
Atelaspis 67
Ateleaspidae 67
Ateles 867, 868
— panlscus 867
Atelinae 853
Atelopus 360
— varius 370
Athene 944
— noctua 607
Atherina 180
— hepsetus 291
Atherinidae 180
Attagis malouinus 602
Auchenia 831
— glama 831
— huanachus 831, 925
— paces 831
— vicugna 831, 925
Aves 33, 50, 401, 505, 550 сл
Axis 832, 833
Babirussa babirussa 830
Bagrinae 172
Balaena japonica 822, 940
— mysticetus 741, 822, 940
— physalus 822
— rostrata 822
Balaenidae 822
1004 ”
Balaenoptera 763, 876
— musculus 822, 940
— physalus 940
— rostrata 940
Balaenopteridae 822
Balanoglossus 14, 16, 21, 24, 49
Balearica 599
Balenicepidae 588
Balistes 185
— capriscus 185
— rectangulus 81
— stellaris 185
Balistidae 81, 82, 185, 284
Barbus 170
— brachycephalus 297
— — caspius 170
— fluviatilis 220, 232
Baryta 639
Basiliscus 405, 471
— americanus 473
Batoidei 73, 105, 153, 216, 225
Batrachoseps 333, 355
Bathymyzon 64, 65
Bdellostoma 54, 55, 57, 64
— dombeyi 63
— stouti 64
Bdellostomatidae 64
Belodon 487
Belone 179
— annulata 179
— belone 179, 210, 213
Belonidae 220
Berycidae 181, 182
Beryciformes 181, 183
Beryx splendens 182
Betta 181
Bibos 836, 907
— banteng 838, 907
— frontalis 838, 907
— gaurus 907
Birkenia elegans 66
Birkeniidae 66
Bison 836
— americanus 836, 947, 971
— bonasus 836, 946, 947, 971, 976
Blastomeryx 840
Blennidae 121, 189, 239, 284
Blenniformes 189
Blenniidae 233, 361
Blennius 190
Blicca bjorkna 247, 295
Boa 482
— constrictor 482
Boidae 481, 509, 518, 520, 545, 546
Boiga 520, 528
Boinae 482, 546
Bombina 341, 358, 369, 371, 374, 381
— bombina 358, 370
Bombinator pahypus 325, 327
Bombycilla garrula 615, 976, 977
— japonica 977
Bos 836, 970
— brachyceros 836, 906
— indicus 910
— nomadicus 836, 906, 910
— planifrons 836
— primigenius 836, 906, 946
— taurus 836
Boselaphus 970
— trangocamelus 835
Bosmina 217
Botaurus 588
Bothriolepis 69
Bothus 183
— maeoticus 183
— maximus 182, 183
Bovidae 831, 834, 938, 967
Bovinae 835, 905
Brachycephalidae 359
Brachycephalinae 360
Brachycephalus 314
Brachyramphus 978
Bradypodidae 802, 960
Bradypus 726, 752, 867, 868, 960
— torquatus 802
Bramatherlum 839
Branchiosauria 316, 322, 350, 363, 364
Branchiosauridae 332
Branchiosaurus 350
— ambystomus 322
Branchiostoma 33, 46
— lanceolatum 33, 36, 37, 40, 41
Branchiostomatidae 33, 46
Branta 591, 703
Breviceps mossambicus 361, 368
Brevicipitidae 361, 366, 394
Brosmius brosme 212
Bubalus 835, 836, 860
— banteng 836
— bubalus 836, 837, 910
— caffer 836
— depressicornis 836
— frontalis 836
— gaurus 836
Bubo bubo 607
Buboninae 607
Bucephala 591
Buceros 644, 649
— bicornis 611
Bucerotidae 609, 636, 966, 970
Budytes 657
— boarula 614
— citreola 614
— flava 614
— melanocephala 614
Buffalus 869, 872
Bufo 312, 330, 331, 341, 359, 364, 376,
381, 384
— calamita 359
— viridis 359, 374, 384, 392
— vulgaris 359, 377
Bufonidae 358, 359, 393
Bufoninae 359
Bunolophodon angustidens 826
— arvernensis 826
— longirostris 826
Bunoselenodontia 827, 829, 840
Buteo 538, 944
— buteo 593
-----vulpinus 559
Cacatuidae 606, 959
Caccabis 595, 704
Cacops aspidephorus 324, 350
Cadiformes 192
Caducibranchiata 306, 352
Caenolestes 791
— fuliginosus 791, 960
Caenolestidae 790, 791, 957, 960
Caenolestoidea 791, 956, 958, 960
Caenotheriidae 840
Caiman 490, 524
— trigonatus 490
Cairina moschata 699
Calamodus aquaticus 616
— schoenobaenus 616
Calamoichthys 161, 163, 164, 281
— calabaricus 163
Calanus 272
Calandrella brachydactyla 614
Calidris arenaria 601
CaHichthyinae 172
Callichthys 80, 119, 172, 230
— littoralis 171
Callisanrus scoparius 508
Callorhynchidae 156
Callorhynchus 156
— antarcticus 93
— ursinus 774, 872, 935
Calotes versicolor 525
Camelidae 831, 925, 962, 970
Camelus 831, 970, 974
— bactrianus 831
— droniedarius 831
Campophagldae 959
Canidae 813, 816, 860, 888, 933, 961
Canis 902
— aureus 816, 934, 974
— dingo 957
— famHiaris 816
— ferus 816, 904
— intermedius 904
— jubatus 756
— latrans 816, 894
— lupaster 904
— lupus 757, 816, 894, 933, 971
— matris optimae 904
— palustris 903
— poutiatini 816, 903
Canutus canutus 673, 674
Capitonidae 611
Capra 774, 938, 966, 974
— aegargus 835, 836, 918, 938
— cashmirensis 876
— caucaslca 835, 939
— cylindricornis 938, 939
— falconer! 835, 918, 938, 976
— ibex 835, 917
— prisca 835, 918
— sibirica 835, 880, 881, 892, 938
Capreolus 832, 865, 880, 974
— bedfordl 832
— capreolus 832, 936, 976
— pygargus 832, 936, 976
Caprimulgi 567, 607, 621, 682, 688 ,
Caprimulgidae 608, 647, 648, 662,
663
Caprimulgus 642
— europaeus 607, 608
Caprinae 835
Caprolagus brachyurus 975
Capromyinae 961
Capromys 961
Caprovinae 835, 966
Caracal caracal 931, 974
Carangidae 191, 284
Carassius 170
— auratus 170, 251
— carassius 216, 251
Car ch arias 233, 285
— glaucus 152
Carchariidae 152
Carcharodon rondeletti 152
Carduelis carduelis 613, 614
Carex physodes 891
Cariacus 962
Cariama cristata 599, 631
Cariamae 598, 963
Cariamidae 599
Carnivora 769, 794, 813, 857, 885, 930,
957, 961
— fissipedia 813 сл., 888
Carpohagidae 604
Casarca 592
Caspialosa 173, 222, 233, 244, 295
— brashnikowi 222, 295
— caspia 222, 295
— kessleri 222, 235, 295
— leucocephala 295
— meotica 298
— normanni 298
— pontica '298
— saposhnikovi 222
— tanaica 298
— volgensis 235, 295
— caspiomyzon 64, 65, 288
— wagneri 65, 295
Castor 865
— canadensis 807
— fiber 720, 803, 807, 808, 872, 897, 929,
947
Castoridae 807, 966, 971
Casuarii 583, 620, 652, 688, 958
Casuariidae 583, 959
Cataphracti 214
Catarrhina 852, 853 сл.
1005
Ca th artае 592, 633, 648, 963
Cathartes aura 592
Cathartidae 592
Catolynx 974
— chaus 931
Catostominae 169
Caturidae 198
Caudata 352
Caulelasmus 591
Cavia 803, 811, 961
— porcellus 811
Cavicornia 938
Caviidae 811, 961
Cebidae 853, 870, 963
Cebus 868
Centetes ecaudatus 722, 795, 878
Centetidae 795, 961, 965
Centrarchidae 282
Centropous 605
Cephalaspidae 67
Cephalaspidomorphi 67
Cephalaspis Lyelll 67
— Murchison! 67
Cephalochordata 33, 45
Cephalodiscus 14
— dodecalophus 18, 19
Cephalopoda 194, 200
Cepphus 602, 642
— grylle 603
Cerastes 486
— cornutus 486, 511
Ceraterpetomorpha 351
Ceraterpeton 351
Ceratias 190
Ceratidae 228
Ceratiidae 190
Ceratodidae 158
Ceratodus 86, 131, 158
— forsteri 101
Ceratophrys 314, 372
— cornuta 359, 369
— dorsata 367
Ceratopsida 506
Ceratopsidae 405
Ceratopsin 496
Ceratoptera vampyrus 155
Ceratosauridae 492
Ceratosaurus 492
Ceratotherium simus 846
Cercoleptes 816
Cercopithecidae 853, 966, 969
Cercopithecinae 854
Cercopithecus 854
Cerdocyon brasiliensis 862
Ceriornis 976
Certhia 972
— familiaris 613, 615
Certhiidae 615, 687
Cervidae 831, 832, 936, 966, 970
Cervulidae 834
Cervulus 970
Cervus 865, 880, 937, 970
— affinis 974
— bactrianus 937
— canadensis 833, 937, 972
----asiaticus 937, 976
----luedorfi 937, 977
— elaphus 833, 937, 976
— — maral 937
— hagenbecki 974
— hortulorum 937
— sika mandschurikus 937
— megaceros 834
Cestracion 133, 146, 152, 219
Cetacea 794, 813, 820, 858, 888, 940
Cetorhinus (Seiache) maximus 77, 78,
152, 285
Chaetodontidae 184, 225, 284
Chaetodontiformes 184
Chaettusia gregaria 601
Chaetura 629
Chalcidcs 476
1006
— tri dactylus 509
Chamaeidae 973
Chamaeleon 476, 513
— pumilis 477
— vulgaris 476, 477, 521
Chamaeleontes 476
Chamaeleontidae 476, 511, 544, 545
Characinidae 121, 132, 168, 218, 280,
281, 282, 285, 286
Charadrii 600
Charadriidae 601
Charadriinae 601
Charadrius apricarius 601
— dominicus fulvus 679
— fulvus 601
Charcharodon 285
Chaulelasmus strepera 703
Chauliodus 175, 219, 229, 285
— sloanei 215
Cheiropterygia 304
Chelidon rustica 612, 677
Chelone 426
— imbricata 465, 516, 532, 534
— mydas 465, 532, 534
Chelonia 422, 427, 460, 461, 462
— mydas 422, 461
Chelonidae 465, 504, 532
Cheloninae 465
Chelydidae 462, 547, 548
Chelydridae 462, 532, 547
Chelys 462
— fimbriata 462
Chen 591, 978
Chiasmodontidae 180
Chiasmodus niger 180, 181, 219
Chimaera 108, 113, 156
— monstrosa 86, 156
Chimaerae 219
Chimaeridae 156
Chinchilla 811, 961
Chinchillidae 811, 862, 961
Chinospiza 975
Chionidae 602
Chirogale 878
Chiroleptes platycephalus 359
Chiromyinae 847, 849
Chiromys madagascarenis 882
Chironectes 790
— minimus 872
Chiroptera 740, 775, 792, 797, 858, 869,
878, 957, 962
Chirotes 476
Chiruromys 868
Chlamydophorinae 802
Chlamydophorus truncatus 802
Chlamydosaurus 471, 472, 507
— kingi 405
Chlamydoselachidae 151
Chlamydoselachus 99, 110, 112, 143, 150,
152
— anguineus 151, 152
Chloris chloris 613, 614, 657, 671
Choeropsis 830
Choloepus 960
— didactylus-802
— hoffmani 726, 727
Chondrichthyes 73, 83, 94, 119, 149, 196
Chondropterygii 149
Chondrostei 99, 103, 106, 109, 112, 114,
136, 137, 138, 147, 149, 150, 160 сл.,
200, 252, 280, 282
Chondrosteoidae 286
Chondrosteoidei 74, 86, 97, 99, 100, 103,
107, 108, 160 сл., 197
Chondrostoma nasus 170, 218, 220
Chordata 13, 27, 49
Chromidae 280
Chromis pater familias 240
Chrochloris aurae 795
Chroicocephalus melanocephalus 603
— minutus 603
— ridibundus 603, 674, 675, 676
Chrysemys 399
— margtnata 541
Chrysochloridae 794, 966
Chrysochlorls 742, 754, 864
— aureus 863
Chrysopelea 520, 532
— ornata 520, 527
Chrysophrls 139
Cichlidae 187, 240, 280, 281, 282, 286
Ciconia 274
— ciconia 589, 590, 675
— nigra 590
Ciconiae 621, 648, 649, 684
Cicinii 587, 589
Ciconlidae 590
Ciconi iformes 561
Cinclinae 617
(’Indus cinclus 617
Cinnyris falkensteini 616
Ciona 29
Clrcaetus 631
— ferox 52
— gallicus 53 8
Circus aeruginosus 593
— cyaneus 593
— macrurus 593
— pygargus 593
Cisticolinae 616
Cistudo 525
Citellus 800, 860, 929, 943, 966, 971, 974
— citellus 807, 976
— columbianus 878
— crythrogenys 807, 976
— cvcrsmani 977
— fulvus 807, 8G1, 878, 883, 890, 891
— pygmaeus 890
— rufescens 807
— Buslica 807, 976
Cladoselache 105, 106, 107, 110, 150, 151,
195, 196
Cladoselachidae 151
Cladoselachli 73, 150
Clamatores 611
Clarias 119, 172
— anguillaris 171
Clariinae 172
Clavellina 29, 31
Clemmys 463
— caspica 463
— leprosa 463, 525
Clirnatius 150
Clupea 135, 231
— harengus 75, 173, 210, 232, 234, 235,
244, 264
— — membras 293, 298
— pallasil 296
Clupea sapldissima 277
— spratus 244, 298
Clupeidae 126, 173, 214, 216, 253, 280,
282, 284, 293, 298
Clupeiformes 88, 172 сл.» 175, 199
Cnemiornis 954
Coblidae 298
Cobitidinae 169
Cobitis 138
— taenia 169
Cobus cobus 872
Coccosteidae 157
Coccosteomorphi 157
Coccosteus 197
— decipiens 157
Coccothraustes 636
— coccothraustes 614
Coccystes 651
— glandarius 605
Coccyzus 651, 973
Coed line 351
Coeciliidae 352
Coolacanthini 102, 119, 120, 161, 163
Coelogenys 811, 961
Coelolepldae 86
Coelurosauria 492, 493, 506
Coendu 811, 961
— prehensilis 812
Colli 968
Coliidae 609, 96G
Collocalia 643
Colobinae 854
Colobus 853
Coloeus monedula 613
Colossochelys 464
Coluber 484
— aesculapii 484
Colubridae 483, 535, 538, 546
Colubrinae 483
Columba livia 604, 701
— oenax 604
— palumbus 604
Columbae 603, 604, 621, 636, 643, 647,
648, 688
Columbldae 604
Colymbi 274, 585, 586, 621, 633, 648»
660, 684, 702, 972
Colymbidae 586
Colymbomorpliae 585, 621
Colymbus 660, 702
— nigricollis 585
— cristatus 586, 702
Coracias garrulus 676
Comephoridae 189, 280, 283 i
Comephorus 189
Compsognathidae 492, 507
Compsognathus 492
Condylura 775, 973
Conepatus 720
Conger conger 178
Conllurus 745, 957
Connochaetes 835, 860
— taurinus 835
Conopophagldae 964
Conuridae 963, 973
Conurinae 606
Copelata 28
Copepoda 272
Coraciae 608, 609, 644, 648
Coracias garrulus 608, 681
Coracndae 608
Coraciiformes 561, 570
Coraciimorphae 604, 621, 632
Coregonus 174, 208, 216, 236, 288, 296»
297, 302
— albula 174, 296, 302
— automnalis 174, 297
— lavaretus 174, 303
— maraena 231
— migratorlus 208
— oxyrrhynchus 217
Coronella 483, 523
— austriaca 483
Corvidae 612, 636
Corvus 538, 972
— corax 613
— cornix 613
— frugilegus 613
Cotingidae 611, 963
Cottidae 189, 284
Cottocomephorldae 2 80
Cottocomephorus 189
Cottus 189, 226, 280
— gobio 189, 225, 234
— quadricornis 277, 284
— Scorpio 189
— sibiricus 189
Coturnix coturnix 595, 704
Cotylosauria 402, 403, 405, 414, 418, 419,
446, 447, 448, 449, 450, 460, 502,
503, 504, 507
Cracidae 595
Craniata 33, 45, 46, 50, 51, 52, 53, 69
Crax alector 595
Creadion 954
Crenilabrus pavo 186
Creodonta 794, 800, 813, 821, 828, 888,
964
1007
Crex er ex 600
Cricetinae 810, 861
Crlcetulus 810
— fulvus 876
Cricetus 943, 974
— cricetus 810, 976
Cricotus 325, 350
Criniinae 359
Crocidura 965
— leucodon 796
Crocidurinae 796
Crocodilia 403, 404, 407, 422, 486, 488,
502, 548
Crocodilia vera 489, 490
Crocodilidae 490, 548
Crocodilus 426, 490
— catapbractus 490
— niloticus 490
Crocotta crocuta 815
— spelaea 815
Crossopterygii 81, 87, 100, 109, 114, 119,
122, 133, 136, 138, 145, 147, 156, 161,
197, 198, 199, 200, 280, 281, 285, 286,
287, 350, 362, 500
Crossoptilon 976
Crossopus fodiens 872
Crotalinae 485, 486, 535, 546, 547
Crotalus 486
— durissus 536
— horridus 486
Crotophaga anl 605
Crotophagidae 605
Crymophilus fulicarius 679
Cryptobranchidae 354, 364
Cryptobranchoidea 353
Cryptobranchus 318, 346, 372, 394
— alleganensis 354
— japonicus 354, 387
Cryptodira 462, 504, 547
Cryptoprocta 965
Crypturi 584
Ctenodactylus 974
Ctenoidei 149
Ctenomys 862, 961
Cuculi 605, 648
Cuculidae 605
Cuculus canorus 605, 657, 678
— optatus 976 4.
Cursorlus gallicus 601
Cyanecula svecica 617
Cyanistes coeruleus 615
— cyaneus 615
Cycloidei 149
Cyclopes 868
— didactylus 802, 867, 868
Cyclopsittaci 606
Cyclopteridae 189, 284
Cyclopt erus 189
— lumpus 189, 240, 241
Cyclothone 229
Cyclostomata 33, 46, 50, 51, 63, 65, 66.
,85, 102, 105, 109, 111, 112, 118, 128
142, 143, 145, 147, 149, 193, 195
Cygnidae 591, 702
Cygnopsis 591
— cygnoides 700
Cygnus 659
— atratus 591
— cygnus 591, 702
— melanocoryphus 59!
— olor 591
Cylindrophis 520
Cynchramus schoeniclus 614
Cynocephalus 966
Cynodesmus 816
Cynodictis 816
Cynodontia 451, 452, 503
Cynognathus 452
— crateronotus 451, 452
Cynomys 533, 778, 807
Cynthia 29
Cyon 756, 975
4008
— alpinus 976
Cyoninae 816
Cyprinidae 78, 121, 133, 169, 216, 219,
221, 236, 244, 252, 280, 281, 282
Cypriniformes 88, 168, 199
Cyprininae 169
Cyprinodontidae 177, 233, 234, 280, 281,
282, 286
Cyprinus 169, 218, 303
— carassius 170
— carpio 97, 169, 220, 232, 277, 293,
295 297
Cypsell 607, 608, 621, 644, 647, 648, 682,
688, 963
Cypselidae 608 . x
Cypselus apus 676, 681 \
Cystognathidae 359, 393 '
Cystignathus 325
Dachidldae 615
Dactylopteridae 189
Dactylopterus 121, 136, 189
Dafila 591
— acuta 703
Dallia 206
— pectoralls 176, 206
DalUidae 176
Dama 832
— dama 833
Dapedius politus 165
Dasypeltis 519
— scabra 484
Dasypodidae 715, 801, 862, 960, 973
Dasypodinae 801
Dasyprocta 811, 961
— aguti 804
Dasyproctidae 961
Dasyuridae 790, 888, 956, 960
Dasyurus 959
Dasypus novemclnctus 801
Delichon urbica 617
Delphinapteridae 823
Delphinapterus leucus 823, 940
Delphlnidae 823
Delphinus delphis 823, 940
Deltatheriidae 794, 813
Dendrobates 371
— tinctorius 360
— trivittatus 379
Dendrobatinae 360, 393
Dendrocolaptidae 612, 963
Dendrocopus major 605
Dendrolagus 792
Dendrophis 484, 514
— punctulatus 484
Dermochelyinae 466, 547
Dermochelys 414, 430, 466, 504, 532
— coriacea 411, 466
Dermophis thomensis 376
Dermoptera 792, 798, 969
Derotremata 352
Desmana 974, 976
— moschata 720, 796, 927
Desmanidae 796
Desmodontidae 798
Deuterosauria 420
Deuterosaurus 454
Deuterostomia 48, 49
Diadectes 404
— phaseolinus 448
Dladectidae 448, 503, 504 *
Diademodon 452
— browni 452
Diadiaphorus 846
Diapsida 403, 423, 467, 505
Diaptosauria 403, 421, 468, 504, 505
Dibus ineptus 604
Dicaeidae 615
Diceros bicornis 846
Dichobunidae 840
Dicotyles 962, 973
Dicotylinae 830, 962
Dicrostomyx 978
Dicrurldae 613
Dirinmlon 453
Dleynodofitla 408, 453, 503
DhMphyhhic 722, 790, 956, 958, 960, 973
Dhlrlpliys 739, 754, 785, 790, 888, 960
— 111 n rs и p I a 1 Is 735, 868
I) hl I 604
I)i<11111c111hlnc 604, 955
Dldiinriihis strigirostris 604, 955
I thins 966
1)1 nir irmlon 449
inrlHlvus 420, 450
Itliiini phiHlon 499
I)I in।»i।ihodontidae 499
Dlnh-IlH K15 -
|)lni>rr|dialta 453, 503
IU ii«h’(‘ii>s 846
DiihhIiiii scytentrionale 526
lUnimiyhlae 961
liliiiiinla 954, 955
idganteus ori 584
I >I tit>i'n iUles 620
IiiiinrnIthidae 583, 954
IHni'saiiria 403, 405, 406, 415, 422t 436,
467, 488, 4.90, 505, 574
J »lmd heriidae 825
!>>notheriuin giganteum 825
lilmlnn 133,, 211
inaculat us 185, 186
I ihnlnnl hlae 82, 185
Iilodoplivs punctatus 527
Iihimcdea 647, 683
exnlans 586
I ihmn drhlac 586
I и pl acanlh'idae 150
IИplacaiil.lnis striatus 150
I H pl i JtH'orla 360, 393
1 il phan iila 49, 50
I H phH jiiliis 351
niagnh'ornis 352
I il phahichlae 493
I )l plinhirns 493
t'lirnrg11 494
I ik phiinrsodun 974
I >1 pl <iph уча 120
I)I pnciiiiioik'4 131, 158
IMpiiol 7”, 73. 74. 86, 93, 97, 101, 102,
|i)3. HU’., 107, 109, 110, 111, 113, 114,
116, I 18, 122, 123, 127, 129, 130, 131,
I 13. 134, 136, 137, 141, 142, 147, 149,
166. 157 сл., 196, 197, 198, 199, 200,
2IMI, 219. 280, 281, 282, 285, 286, 287,
3.19. 342, 343, 344, 346, 362, 363, 500
I il pixllllus dasyurus 891
111 pi и! I о .ip 809
111 pi id < 11 d (*a 809
I H pnil villus 745
I И pi "I tidiuil I a 857, 956
lilph i hl.ir I 58
Dlplinm HU, 158, 198
v al Iriii'leu urns Is 158
I и pl Ii V’ Iuih 171
I il pi и /4 Г., 747, 803, 810, 862, 883, 974
n nd Hu 8 10
I U n o ’haeihhic 357, 358
111 и < ip I h i ат 341
• pi» 111ч 3 17
I и d ।*i4 IniriiH 79*2
| ltd II IlnpliHSHH 1 4
11.d i> liolil ppiin 844
11. d 11 111<>\ л 11г I a 5(1.5
I на h 111i'i,iиi hhic 479
I )'d h 111mani in 479
I nd 11 Ind p. Hid
pa 1 и*i'ti leu 811
I • -I I "I || iи 3)1
I...... line 17?
Iini'ii I Hl
I ха । il In i him 967
I........ 4 Hi
11iii Ii i lul ни M4 ’
h aiiiidipun и Стапчинский
— volitans 471, 514
Drepanididae 956
Drepanaspidae 68
Drepanaspis demimdenensis 68
Dromaeidae 583, 959
Dromaeus 583, 686
— novae hollandiae 582
Dromatherium 454, 754, 787, 834
— sylvestre 787
Dryobates leuconotos 610
— major 610
— medius 610
— minor 610
Dryocopus martins 610
Dryomys nitedula 809
Dryophis 520, 528, 529*
Dryopithecus 851
Duplicidentata 745, 803, 805, 961
Dwinasaurus 350, 501
Dyprotodon 957
Ebornea pogophila 978
Ecaudata 356
Eeheneidae 188
Echeneifonnes 188 •
Echidna 788 .
— aculeata 788, 956
— hystrix 720
Echidnidae 788, 956
Echinoderinata 50
Echis carinatus 511, 528
Ectobranchiata 194
Edaphosauria 450
Edaphosauridae 450
Edaphosaurus 450
— cruciger 4M
Edentata 792
Edriolychnus 190
— schmidti 191
Elachistodon 519
Elanus 658, 718
Elaphe 484, 520, 528
— dionc 484
— rufodorsata 528
Elapidae 520
Elapinae 485, 535, 546
Elaps 485, 537, 538
— corallinus 485, 523
Elasmobranchii 85, 98, 103, 105, 112, 133,
151
Elasmosauridae 459
Elasmosaurus 458, 459, 504
Elasmotherium 846
Elcginus navaga 193
Eleotris marmorata 77, 187
Elephantidae 825, 967
Elephas 760, 825
— africanus 869
— antiquus 825
— indicus 970
— maxirnus 734, 826
— meridionalis 825
— planifrons 825
— primigenius 825
— sumatrensis 826
— trogonotherii 825
Elginia 448
Eliomys 869
— quercinus 809, 889, 976
Ellobius 811, 861, 864, 887, 890
Elopidae 172
Elotheriidae 840
Emberiza aureola 614, 675
— chrysophrys 977
— citrinella 613, 614, 671
— hortulana 614
— luteola 614
— melanocephala 614
— pusilia 976
— rutila 977
Emberizidae 614
Embiotocidae 233, 284
1009
Emboelomuridae 798
Embolomeri 315, 320, 322, 325, 350
Embrithopoda 794, 823, 826
Emydinae 463
Emys 422, 463
— europaea 443
— orbicularis 413, 423, 461, 463, 464, 530
Engraulidae 173, 217
Engraulis encrassicholus 173, 293, 297
Engystomatidae 359, 361
Enneoctonus collurio 616
Enteropneusta 14
— pterobranchia 14
— vermiformia 14
Entobranchiata 70, 71, 193, 194
Entosphenus 64, 65
Eoanthropus dawsoni 852
Eogyrinus 316, 325, 362
Eohippus 843, 844
Eosauravus 447
Eosiren 827
Eosuchia 486, 503
Eotherium aegiptiacum 827
Epanorthidae 791
Epihippus 843, 844
Epimys 274
Eptesicus 798
Equidae 843, 967, 970
Equiuae 843
Equus 844, 962
— abeli 844, 920
— germanicus 844, 920
— gmelini 844, 920, 946
— hemionus 844
— kiang 844
— mosbachensis 844, 920
• — onager 844
— przewalski 844, 845, 920, 946, 974
— robustus 920
— sequanius 920
— stenonis 844
Eremias 475, 507, 508, 530
— arguta 475
— grammica 475, 508
— scripta 475
— velox 475, 530
Eremophila 972
— alpestris 614, 674, 675
Erethizon 811, 973
Erethizontidae 811, 961
Ericulus 795
Erignatus barbatus 820, 935
Erinaceidae 796
Erinaceinae 796
Erinaceus 796
— amurensis 977
— europaeus 976
Erismatura leucocephala 592
Erismaturinae 592
Erithacinae 617
Erithacus rubecula 617, 671
Eryops megacephalus 307
Erythrina 121, 122, 972
— erythrina 614, 675
— rosea 977
Erythrinus 169
Erythropus 944
— amurensis 674
— vespertinus 593, 631
Erythrosuchia 487
Eryx 481, 482, 510, 520, 546
— jaculus 509, 510
Esocidae 176, 280, 282, 287
Esociformes 88, 122, 275 сл. 281,
Esox 132, 176, 216, 282
— lucius 88, 176, 216, 219, 232, 289, 293,
295, 303
Esthonyx 800
Euarctos 816
— americanus 816
— japonicus 81G
Eublepharus 478
1010
Eucrcodi 757, 813
Eudromias morinellus 640
Eugnathidae 165
Eumetopias californianus 819
— jubatus 819
— stelleri 774, 935
Eumeces 476, 510
Eunectes 482
— murinus 482, 530, 546
Euparkeia 488
Euphractinae 802
Eurylaemidae 611, 971
Eurypterygius 456
— extremus 430
Eurypyga helias 598
Eurypygae 597, 963
Eurypygidae 598
Eustenopteron 321, 323
Eusuchia 488, 489, 490, 505
Eutamias 806
Eutheria 713, 773, 792
Evotomys 811
— glareolus 892, 943
Exocoetus 285
Falcipennis falcipennis 977
Falco 593
— peregrinus 682
Falconidae 593
Falconiformes 561, 621
Felidae 756, 813, 815, 860, 888, 930, 969
Felis 815, 930
— caudata 931, 974
— chaus 815
— concolor 961
— maniculata 815, 905
— onca 815, 869, 961
— pardus 869
— sylvestris 815, 869, 930
— viverrina 533
— uncia 815, 892, 974
Fiber zibethicus 811, 872, 926, 928, 973
Ficedula atricapilla 616
— col laris 616
Fierasfer 136, 190
Fierasferidae 190
Firmisternia 325, 326, 393
Fissipcdia 794
Formicariidae 612, 964
Franeolinus 595, 704
Fratercula arctica 603
Fregatidae 588, 683
Fringilla coelebs 614
— montifringilla 614, 976
Fringillidae 613, 636
Fulica 684
— atra 598, 600, 644
Fuligulinae 591, 703 *
Fulmarus 586, 663
— glacialis 586
Fundulus 177
Gadidae 121, 193, 214, 226, 282, 284, 293,
296, 298
Gadiformes 100
Gadus 76, 193, 212, 231, 246, 284
— aeglifinus 193, 244
— callarias 192, 193, 231, 236, 238, 239,
240, 244i 246, 270, 271, 296
— morrhua 75, 210
— navaga 296
— pollachius 239, 240
— virens 239, 240, 243, 244
Galaginae 849
Galago galago 882
Galamoichtus 114
Galaxiidae 176, 280, 281, 282, 286
Galbulidae 611, 963
Galeopithecidae 799
Galeopithecus 799
— volans 799, 870
— temmincki 799
Galerida cristata 614
Galli 594, 621, 636, 643, 648, 686, 688, 703
Gallinago 221, 657
— gallinago 601, 704
— media 600, 601, 676. 681, 704
Galllnula chloropus 600, 656, 684
Gallinulinae 600
Gallus 595, 970
— bankiva 594, 595, 696
Galunda 969, 970
Gambusia 177
Ganoidel 149
Garrulus glandarius 613
Gasterosteidae 178, 179, 280, 282,
Gasterosleiformes 178
Gasterosteus 234, 274
— aculcetus 178, 179, 231, 240
Gastornithes 592
Gastrocentrophori 415
Gastropoda 272 .
Gavialidae 490, 548
Gavialis gangeticus 490, 491
— schlegeli 490
Gazella 974
— arabica 861
287
— gulturosa 834, 940
— subgutturosa 834, 862, 940
Gecko gecko 520
Geckones 477
Gekkonidae 415, 470, 477, 508, 512, 544
Geleopithecus 969
Gelocidae 831
Genetta 720
— genetta 974
Geococcyges 605
Geogale 795
Geomyidae 751, 807, 961
Geomyoidea 807
Geomys 760
— bursarius 807
Geophagns 186
Geosauridae 4 89
Geos.iurus 489
— suevicus 489
Geotria 64
Geotrygonidae 604
Gerbillinae 811
Gerbillus 861
— eversmanni 883
Giraffa Camelopardalis 839, 860
Giraffidae 718, 831, 839, 967
Girardinus 177
Glaredidae 686
Glared a melanoptera 600, 601
Glaredidae 601, 682
Glaucidtum passerinum 607, 663 >
Glis glis 809, 869, 976
Glossobalanus 14, 15, 16
— minutus 14
Glottis nebularius 601
Giyptodonta 961
Glyptodonti'dae 802
Gnathonemus 173 ’сл.
— curvirostris 221
Gnathostomata 33, 50, 72 сл.
Gobiidae 77, 187, 246, 361
Gobiiformes 187
Gobio fluviatilis 170, 225
Gobius 188
— ophiocephalus 188
Gomphotherium 760, 761
Gonyocephalus chamaeleontinus 512
Gorgonopsida 453
Gorilla 854, 869, 966
— gorilla 868
Goura 604
Gouridae 959
Gravlgrada 802, 960
Grossobamon 478
Grues 597, 598, 599, 648, 649, 68£, 704,
955, 963
Gruidae 599
Grus grus 598, 599, 704
Grypotherium 960
Gulo 865, 971
— borealis 894
— gulo 817, 932
— luscus 817
Gymnarchus 114, 120, 121, 128, 173
— niloticus 114
Gymnells vtridls 284
Gymnobelideus 792
Gymnodactylus 477
— caspius 477, 479
— russovi 478, 513
Gymnodontes 185
Gymnophiona 305, 308, 351, 388
Gymnotidae 169, 281
Gymnotus 228
— electricus 169
Gymnura 969
Gyinnurinae 796
Gypaetos barbatus 593
Gyps 593
Gyrinophilus porphyricus 315
Haematopodinae 601
Haematopus ostralegus 601, 634
Hallaetus albicilla 274, 593
Halichoerus gripus 935
Halicore 763, 827
Halicoridae 827
Halitherium 827
Halmaturus 792
— rufus 793
Halomys 969
Hapalidae 850, 853, 963
Haplodontidae 806, 973
Haplodontoidea 806
Harengula 173
— delicatula 173, 301
— grimmi 173, 301
Harpacticidae 272
Hatteria 468, 532, 533
Heimtragus 966
Helictis 969
Heliornithes 600
Heliornithidae 600
Heloderma 473
— horridum 523, 539
Helodermatidae 470, 473
Helodromas ochropus 601
Hemichordata 13, 18, 35, 44, 49, 50
Hemiechinus 796
— aeritus 796
Hemiramphus 179
Hemitragus 970
Heptanchus 92, 93, 99, 112
Hept a trema 64
Herodias 588, 702
— alba 588, 589, 702
— garzetta 588, 702
Herpestes 815, 975
— griseus 538, 589
— Ichneumon 538, 815, 974
Herpestinae 814, 965
Hesperornithes 585
Hesperornis 566, 585, 621
Heterodontidae 132, 152
Heterodontus (Cestracion) 133, 146, 152
— philippi 133, 152
Heterostraci 68
Hexagrammidae 284
Hexanchus 99, 112
Hicanodonta 801, 960
Hidroochoerus capybara 872
Hierofalco 593
Himantopodinae 601
Himantopus himantopus 601
Hipparion 843, 844
Hippidium 962
Hippocamelus 832
Hippocampidae 284
Hippocampus 77, 178, 221, 240, 285
1011
G4*
Hippocampus antiquorum 178
Hippoglossidae 231
Hippoglossus hippoglossus 183
Hippolais caligata 616
— icterina 616
Hippopotamidae 830, 967
Hippopotamus 830
— amphibius 829, 872
Hippotigris 844, 967
— quagga 845
Hippolragus 835, 860
Hirundinidae 617, 682
Hirundo russica 617
— urbica 677
Histiophoridae 192
Histiophorus gladius 210, 285
Holocephali 73, 80, 86, 93, 99, 103, 105,
107, 112, 113, 134, 137, 147, 151,
155 сл.
Holoptychiidae 162
Holoptychius 162
— flemingi 162
Holostei 75, 87, 100, 103, 107, 108,
112, 120, 121, 132, 134, 136, 137, 144,
147, 150, 161, 198, 200, 280, 282, 362
Holosteoidei 74, 87, 99, 103, 107, 109,
113, 160, 198, 199, 286
— actinopterygii 74, 75, 160, 164 сл., 198
— crossopterygii 74, 160, 161 сл., 198,
199
Homalopterinae 169
Hominidae 852, 854, 868
Homo heidelbergensis 852
— sapiens 854
Homunculus patagonicus 850
Hoploptcrus armatus 691
Houbara 974
Huso 161
— dauricus 161, 288
— huso 161, 288, 297, 298
Hyaernoschus 831
Hyaena 974
— hyaena 815
Hyaenidae 756, 813, 815, 961, 966
Hyaenodon 757
Hyaenodontidae 821
Hydrochelidon 681
— nigra 603
Hydrochoerus 778, 841, 961
— capybara 760, 887
Hydrocyon goliath 169
Hydromantes 394
Hydromedusa 462
Hydromyidae 811, 872, 957
Hydromys 811, 957
Hydrophinae 485, 517, 532, 535, 546
Hydrophis 528
Hydrosauria 479
Hydrus 485
— platurus 485, 515
Hyelaphus 832
Hyla 330, 368, 374, 383, 393
— arborea 313, 360, 370
— coerulea 360
— faber 381
— rosenbergi 390
— venulosa 367
Hylaeobatrachia 353
Hylaeobatrachus 353
— croyi 364
Hylidae 360, 370, 393
Hylobates 854
— hainanus 854
— lar 851, 855
— syndactylus 868
Hylobatidae 854, 969
Hymenochirus 312
Ilynobiidae 353, 354, 356, 378, 394
Hynobius keyserlingii 353
Hyothcrium 830
Hyperoodon 727
— rostratus 763
1012
Hypertragulidae 839
Hypoconifera 840
Hypogeophis 352
Hypohippus equinus 744
Hypotriorchis subbuteo 593, 631
Hypsignathus monstrosus 751
Hypsiloihodon 513
Hypsiprimnodon 792
Hyracoidea 794, 823, 866, 967
Hyracotherium 843, 844
Hystricidae 811, 970
Hystricoidea 811, 961
Hystricomorpha 745, 862
Hystrix 862, 974
— hirsutirostris 811, 881
lacare 490
— nigra 490
Ibididae 590
Ibis 634
— falcinellus 590
Ichthyophis glutinosus 314, 321, 352, 353,
378, 379
Ichthyopterygia 304, 420, 454, 502, 504
Ichthyornis 566, 580
Ichthyornithes 587
Ichthyosauridae 456
Ichthyosaurus 456, 517
— trigonus 456
Ichthyostegalia 350, 362
Ichtyomyzon 64, 65, 288
Ichtyosauria 404, 407, 446, 454, 455, 501,
503, 504, 518, 528
Icteridae 613, 973
Icticyon 961
Ictidosauria 454, 785
Ictis errninea 817
— nivalis 817
Ictitherium 815
Iguana 426, 471
Iguanidae 448, 470, 471, 508, 511, 512,
518, 522, 532, 545
Iguanodon 494, 507
— bernissartensis 495
Iguanodontia 494
Iguanodontidae 494, 498
Ilysia scytale 482, 509
Ilysidae 482, 509
Indicatoridae 611
Indris 778
Indrisinae 849
Inia amasonica 823
Insectivora 792, 794, 857, 878, 885, 927,
961
Ipnops 177
Irrisarinae 968
Ischyromys 806
Issiodoromys 809
Jacana jacana 602
Jacanae 602
Jaculidae 971
Jaculus 745
Jagulares 189
Karoomys 454
Ketupa 538, 633
Ketupinae 607
Kiaeraspis 67
Kolonocus alpinus 932
— sibiricus 931
Kotlassia 447
Kuhliidae 282
Labidosauria 403, 448
Labidosauridae 448, 503
Labidosaurus 447, 448, 507
— hamatus 44 9
'Labridae 186, 220, 284
Labriformps 186
Labrus 186
— prosostictes 186
Labyrinthic! 206
Labyrinlhidae 180, 281
Labyrinthodonta 350
Labyrinthodontia 316-
Lacerta 344, 417, 474, 507, 525, 540
— agilis 425, 475
— muralls 397, 433
— ocellata 475
— viridis 475, 523
— vivipara 474, 525, 528
Lacertae 470
Lacertidae 474, 508, 544
Lacertilia 404, 427, 467, 470, 480, 502,
505
Lachesis mutus 538
Laemargus 284
— borealis 153, 285
Lagomorpha 805, 862
Lagornys 974, 975
Lagopus 657, 978
— lagopus 595, 596, 597, 639
— rnutus 595
Lagurus 862
— lagurus 811, 943
Lama 962
Lainna cornubica 75, 85, 152, 210,
285
Lamnidae 152
Lampetra 64, 65, 252
— aurea 65
— cibaria 65
— fluviatilis 65
— planer! 65
- w 11 <1(4’1 65
Lanark la spinosa 68
1 .пн 11<1 ar 615
I.util us 633, 972
— rxrubitor 616
llllnor 613, 616
l.nrl 274, 602, 621. 633, 647, 648, 649,
6H3, 704
Liu I (I nr. 603
Lin Ihii'inrs 643
Lai ня 05H
( iu liinnans 603
« nun i 6112, 603
Iiim’iih 603
uchinlrH 603
h \ pi’i linlrriiH 978
ni n11 и ।in 003
LuiviKi'd ’.'H, 31
I n « n 1111«I ar 00
I >i in о I и к piiilil ruin I Icus 66
I n|n< V/H
lull In v Z4, HIK, 872, 873, 932
I him
I и inn i In in I, 162, 163
I I Hl III IPI V / H
Il 11111111*1 Mil .
I । ill и i 111 iu« / f14 , К49
I । hi и11.1 «I i*.i J L>, M4H, 840
I in. V/’|
bn hfli
4 |<| I III II Mtn
I । ..Г II «Ilin pniiliM 030, 974
t i IIii«ih V/5
inn In NHII, V.lll
I . | >>l*n i H i I 'III
I । Ы. ин hi Mi4 , 467,460, Г»05
1*1'1 >iiiii ни, ||3, 114, tit». 123, 131,
li' l Ml, ,'П |
-I n । *i... .i Uh, | Ml
I । ) i*i ........ 11.H
I III 11 i<l i. Hi, (01
I I >1 । * । । ih I I Oil. 104, 107, IVH
I । । • । * и • i * но, к/, I ini, | пн 11?,
I " II I 1 •. I 'll, 134, LIM, 14/, 165,•
L - I ......I, '.'li7, 7M0
и lb >
I ' i I * in и i I «I /
I ( Ml........... HlHi
I i । il l t H iHi |, 304
Leptocephali 226, 239, 254, 255
Leptocephalus brevirostris 238
Leptodactylinae 359
Leptodactyhis ocellatus 373
— pentadactylus 359
Leptolepidae 168, 199, 286
Leptolepiformes 168
Leptolepis 168
Leptomeryx 839, 840
Leptopoecile 975
Leptoptilus 634, 660
Lepus 805, 860, 928
— americanus 892
— connori 891
— europaeus 805, 928
— mandschuricus 805
— pamircnsis 876
— tirnidus 805, 928
— tolai 805
Leuciscus balteatus 250
— bergi 297
— cephalus 170
— idus 170, 251
— leuciscus 216
— schmidti 297
Leucosticte 975
Licbanotus 868
Limacina 272
Limax lanceolatus 33
Limlcolae 601, 621, 643, 647, 648, 684
Limnocryptes gallinula 704
Limnogale 795, 872
Limnoscelidae 448, 503
Limnoscelis 448
— paludis 448
Limnonites minutus 601
— teinmincki 601
Limosa lapponica 640
— limosa 601, 671, 676, 681
— rufa 671
Linaria cannabina 614
— flavirostris 614
— linaria 614, 672
Liocassis 172
Liopelma 357
Liopelrnidae 357, 358
Liopeltis 523
Liparis liparis 189
Lissamphibia 305, 351
Lithofalco aesalon 593
Litopterna 823, 846, 872, 962
Litorhynchus 510
Locustella fluviatilis 616
— luscinioides 616
— naevia 616
Lophiidae 190
Lophiodontldae 845
Lophius 132, 190
— piscatorius 190, 213, 22$. 226
Lophophanes cristatus 615, 976
Lophophoras 976
Lorlcaria 80, 119, 230
Lor I car! inae 172
Loridae 606, 959
Loris 969
Lorlsidae 849
Lota 280, 282
— lota 193, 208, 216, 225, 289
Lox I a 636, 67 9
— curvirostra 613, 614, 680
LoxL’i leucoptcra 976
— pHyopslltacus 976
Loxodon 825, 967
Lucloprrca 183, 288
— lurloperrji 183, 293, 295, 290, 207.
30?, 3(13
— iiinrlna 183, 295
— snndra 217
Lnlluln artiorra 614
1 .ii in I <*<11 ar. 704
I nun prniiH ill ih’iiLiI us 284
l.uMclniu lusclnln 617
1013
Luscinia megarhynchos 617
Lutra 274, 756, 872, 961
— capensis 757
— lutra 818, 872, 932
Lutreola 274
— lutreola 816, 872, 932
Lutrinae 816, 818
Lycaon 967
Lycodes 285
Lycodidae 284
Lygodactylus plcturatus 512
Lymnocryptes gallinula 601
Lynx 971
— canadensis 892
— isabellina 974
— lynx 815, 869, 930
— pardellus 930, 974
— tolquilla 610
Lyriocephalus scutatus 511
Lyrurus mlokosiewiczi 595
— tetrix 594, 595, 596, 704
Lysorophidae 352, 353
Lysorophus 353
— tricarinatus 364
Lystrosaurus latirostris 406, 453
Lytorhynchus 484
— rldgewayl 484, 509
Macacus 854, 969, 974, 975
— innus 974
— sylvanus 854
Machairodus 815, 889
Macraucheniidae 846, 872, 962
Macrochiroptera 752, 797
Macroclemmys temmincki 462, 463
Macropodidae 792, 957
Macropodus viridi-auratus 181
Macropus 745, 792, 860
— rufus 775
Macroscelidae 745, 794, 974
Macroscelides typns 794
Macroscelididae 966
Macruridae 192, 214
Macrurus norvegicus 213
Malacopterygii 149
Malacosteus 175
— indicus 175
Malapterurinae 172
Malapterurus 228
— electricus 172
Mallomys 957
Mallotus 175
— villosus 175, 200
Mammalia 33, 50, 401, 712
Manatherium 827
Manalidae 827
Manatus 726, 827, 875
— latirostris 734
Mandrillus 854
— sphinx 855
Manidae 800
Manis 764
— macrura 728
— pentadactyla 800
— temincki 862
Mareca 591
— penelope 703
Marmota 806, 860, 861, 880, 929, 966,
971, 974
— baibacina 806
— bobac 806, 807, 891, 929, 948
— camtschatica 806, 977
— hymaiayensis 975
— marmota 976
— sibirica 806
Marsupialia 713, 722. 773, 775, 788,
790, 956, 958, 960, 964
Martes flavigula 931
— foina 817, 931
— intermedia 974
— martes 817, 869, 931, 976
— zibellina 817, 869, 931, 947, 976
1014
Mastacembelidae 280
Mastacomys 957
Mastodon 760
Mastodontidae 825
Mastodontosaurus 350
— glganteus 307
Mazama 832
Meantes 356
Megachiroptera 868, 887
Megaladapidae 965
Megalania 479
Megalobatrachus japonicus 354
— maximus 367
Megalonyx 960
Megalosauria 492
Megalosauridae 492
Megalosaurus 492
Megalotis corsac 933
— zerda 862, 882
Megalotrion 364
Megapodidae 594, 647, 959
Megapodius 594
— reinhardti 594
Megatherium 802
Mel anocetus 190, 211
— johnstonl 190
Melanocorypha calandra 614
— sibirica 614
— yeltonensis 614
Melanonyx 591, 703, 978
Meleagrinae 973
Meleagris 595
— gallopavo 595, 698
Meles 756, 862, 865, 878
— amurensis 932, 977
— arenarius 932
— meles 817, 818, 932
Melinae 816
Mellni 817
Meliphagidae 614, 959
Mellivora 817, 967, 969
Melopsitacus undulatus 701
Melursus 969
Menura superba 612, 614
Menuridae 612, 959
Mephitis 720, 818, 961
Merganser merganser 592
Mergidae 592
Merginae 633
Mergus 633
— albellus 592
Meriamia zitteli 430
Meriones meridianus 811
Meropidae 609
Merops 643
— apiaster 609
Merula atrogularis 617
— merula 617
Merychippus 843, 844
Mesaxonia 744, 823, 842
Mesites 597, 966
Mesitidae 597
Mesocri cetus 810
Mesodon macropterus 166
Mesohippus 843, 844
— celer 744
Mesosauria 402, 404, 407, 454, 455, 504
Mesosauridae 454
Mesosaurus 454
Mesosuchia 488, 489, 505
Metatheria 713, 788, 956
Metopias 350
— diagnostics 351
Metriofhynchidae 489
Metriorhynchus 489
Miacidae 813, 816
Miacinae 813
Miacis 754
Microbiotheriidae 960
Microbranch is 351
Microchiroptera 797
.Microconodon 787
Mlcrocyprlni 177
M кTutioiiodon 754
Microgale 795
M hToh Ic.rax 632
Mtcroinys 897
Mh-rosaurla 351, 447
Microlinae 810, 861, 880, 966, 971, 974
Microtis tetrax 598, 599, 639, 704
Microtus 878
— agrestls 810
— arvalis 803, 810, 861, 943
— oeconomus 811
— socialis 811, 861, 943
Midas rosalia 853
Miliaria calandra 614
Mirnetozoon 513
Miminae 616
Mlohippus 844
— anceps 744
Misgurnus fossilis 119, 169, 207, 216,
225, 234, 251
Mistichthys luzonensis 77, 187
Mixosauridae 456
Mixosaurus 456
Mnioliltidae 973
Moerithcriidae 825
Moeritherium 760
— lyonsi 825
Mogera robusta 796, 977
Mola mola 185, 186, 211
Molge 354 *
Molgula 29
Molidae 185
Moloch 448, 471
— horridus 473, 524
Molossidae 798
Molotus 651
Mclva 284
— molva 212, 231
Mornotidae 609, 963
Momotus lessoni 310
Monodclphia 713, 7 92
Monodon monoceros 823, 940
Monopneumones 131
Monorhina 33, 50
Monosauria 407 >
Monotremata 503^ 713, 722, 764, 773,
787, 788, 956, 958
Monticola saxatilis 617
— solitarius 617
Mordacia 64
Mormyrid.ae 114, 121, 138, 172, 221, 281
Morniyrus 173
Mosasauria 407, 470
Mosasauridae 479
Mosasaurus 480
Mosch inae 832
Moschus 832, 974 *
— moschiferus 720, 832: 938, 976, 977
Motacilla 643
— alba 613, 614
— melanope 614
Motacillidae 614
Mugil 180, 222
— auratus 180, 298
— cephalus 180 ,
— chelo 180
— haemotochilus 180
Mugilidae 180, 214
Mugilitormes 180
Mnllidae 184
Mullus 184
— batrbaus 184, 298
Multituberculata 787, 788, 964
Mungo mu ng о 815
Munt jacks 833
Muraena helena 178
Muraenidae 138, 178
Muraenolepis 285
Muraenosaurus 421, 459, 460
Muridae 810, 961, 965, 967, 969
Murinae 811, 861, 974
Murioidea 957
Mus 811, >57, 976
—; agrariu: 811
— flavicolis 811
— minutu 811
— muscuhs 811» 943
— silvaticis 811
Muscardims 869, 974, 976
— avelanmrius 809
Muscicapa striata 613, 616
Muscicapicae 616
Musophaga violacea 605
Musophagas 605
Musophagilae 606, 636, 966, 968
Must el a 8 6
— ermine? 894, 932, 971
— ewersmjii 816
— ferghame 892
— nivalis 892, 932
— putoriu. 816
— sibirica 816
— zibellini 894
Mustelidae 538, 756, 813, 816, 931, 961,
967, 96)
Mustelinae 816
Mustelus 152, 233, 284
— canis 239
Mycetes 7r5, 868
— seniculis 854
Mycetinae 853
Mydaus 9(9
Myliobatidae 155
Myliobatis aquilla 155, 219
Mylodon 8)2, 960
Myocastor 961
Myodes 876
Myogale maschata 872, 873, 874
Myoidea 810, 861, 961
Myomorph। 845
Myopotamns coypus 812, 872
Myospalachae 811, 887
Myotis 791
— myotis 7 97
Myoxidae 309, 967
Myoxoidea 809
Myrmecobius 722, 886, 959
— fascia tie 791
Myrmecobiiflae 791, 956
Myrmecoptaga 764, 864
— jubata 802
Myrmecoptagidae 802, 960
Myrmecoptagus 743
Mysopoda 866
Mystacoceti 7d63, 820, 821, 822, 940
Mytilus 634
Myxine 54, 55, 231
— capensis 64
— garman, 64
— ghitinosa 63, 64
— olivacei 64
Myxinoidea 57, 63, 64
Naja 485, 538
— naja 485, 525
----- oxiana 485
— tripudUns 485, 535, 536
Nannosciuius 967, 970
Naosaurus 450, 524
Nasalis 854, 969
Nasua 816, 973
— rufa 869
Natrix 274, 483, $28
— annularis 528
— natrix 483, 535
— piscator 529
— tesselata 483
Nectariniidae 614, 628i 968
Nectogale 974
— elegans 872
Necturus 334, 346, 372, 383, 394
— maculatus 355, 356
Nemachelius barbatula 169
1015
Hippocampus antiquorum 178
Hippoglossidae 231
Hippoglossus hippoglossus 183
Hippolais caligata 616
— icterina 616
Hippopotamidae 830, 967
Hippopotamus 830
— amphibius 829,- 872
Hippotigris 844, 967
— quagga 845
Hippolragus 835, 860
Hirundinidae 617, 682
Hirundo russica 617
— urbica 677
Histiophoridae 192
Histiophorus gladius 210, 285
Holocephali 73, 80, 86, 93, 99, 103, 105,
107, 112, 113, 134, 137, 147, 151,
155 сл.
Holoptychiidae 162
Holoptychius 162
— flerningi 162
Holostei 75, 87, 100, 103, 107, 108,
112, 120, 121, 132, 134, 136, 137, 144,
147, 150, 161, 198, 200, 280, 282, 362
Holosteoidei 74, 87, 99, 103, 107, 109,
113, 160, 198, 199, 286
— actinopterygii 74, 75, 160, 164 сл., 198
— crossopterygii 74, 160, 161 сл., 1,98,
199
Homalopterinae 169
Hominidae 852, 854, 868
Homo heidelbergensis 852
— sapiens 854
Homunculus patagonicus 850
Hoplopterus armatus 691
Houbara 974
Huso 161
— dauricus 161, 288
— huso 161, 288, 297, 298
Hyaemoschus 831
Hyaena 974
— hyaena 815
Hyaenidae 756, 813, 815, 961, 966
Hyaenodon 757
Hyaenodontidae 821
Hydrochelidon 681
— nigra 603
Hydrochoerus 778, 811, 961
— capybara 760, 887
Hydrocyon goliath 169
Hydromantes 394
Hydromedusa 462
Hydromyidae 811, 872, 957
Hydromys 811, 957
Ilydrophinae 485, 517, 532, 535, 546
Hydrophis 528
Hydrosauria 479
Hydrus 485
— platurus 485, 515
Hyclaphus 832
Hyla 330, 368, 374, 383, 393
— arborea 313, 360, 370 •
— coerulea 360
— faber 381
— rosenbergi 390
— venulosa 367
Hylaeobatrachia 353
Hylaeobatrachus 353
— croyi 364
Hylidae 360, 370, 393
Hylobates 854
— hainanus 854
— lar 851, 855
— syndactylus 868
Hylobatidae 854, 969
Hymenochirus 312
Hynobiidae 353, 354, 356, 378, 394
Ilynobius keyserlingii 353
Hyothcrium 830
Ilyperoodon 727
— rostratus 763
1012
Hypertragulidae 839
Hypoconifera 840
Hypogeophis 352
Hypohippus equinus 744
Hypotriorchis subbuteo 593, 631
Hypsignathus monstrosus 751
Hypsiloihodon 513
Hypsiprimnodon 792
Hyracoidea 794, 823, 866, 967
Hyracotherium 843, 844
Hystricidae 811, 970
Hystricoidea 811, 961
Hystricomorpha 745, 862
Hystrix 862, 974
— hirsutirostris 811, 881
lacare 490
— nigra 490
Ibididae 590
Ibis 634
— falcinellus 590
Ichthyophis glutinosus 314, 321, 352, 353,
378, 379
Ichthyopterygia 304, 420, 454, 502, 504
Ichthyornis 566, 580
Ichthyornithes 587
Ichthyosauridae 456
Ichthyosaurus 456, 517
— trigonus 456
Ichthyostegalia 350, 362
Ichtyomyzon 64, 65, 288
Ichtyosauria 404, 407, 446, 454, 455, 501,
503, 504, 518, 528
Icteridae 613, 973
Icticyon 961
Ictidosauria 454, 785
Ictis erminea 817
— nivalis 817
Ictitherium 815
Iguana 426, 471
Iguanidae 448, 470, 471, 508, 511, 512,
518, 522, 532, 545
Iguanodon 494, 507
— bernissartensjs 495
Iguanodontia 494
Iguanodontidae 494, 498
Uysia scytale 482, 509
Ilysidae 482, 509
Indicatoridae 611
Indris 778
Indrisinae 849
Inia amasonica 823
Insectivora 792, 794, 857, 878, 885, 927,
961
Ipnops 177
Irrisarinae 968
Ischyromys 806
Issiodoromys 809
Jacana jacana. 602
Jacanae 602
Jaculidae 971
Jaculus 745
Jagulares 189
Karoomys 454
Ketupa 538, 633
Ketupinae 607
Kiaeraspis 67
Kolonocus alpinus 932
— sibiricus 931
Kotlassia 447
Kuhliidae 282
Labidosauria 403, 448
Labidosauridae 448, 503
Labidosaurus 447, 448, 507
— hamatus 449
Labridae 186, 220, 284
Labriformes 186
Labrus 186
— prosostictes 186
I .nbyrinthlci 206
Labyrlntbidae 180, 281
L.ibyrinthodonta 350
Inibyrlnlbodontia 316
Lacerta 344, 417, 474, 507, 525, 540
— agilis 425, 475
— uiuralls 397, 433
— ocellata 475
— vindis 475, 523
— vivipara 474, 525, 528
Laceitac 470
Laccrtidae 474, 508, 544
Lacertilia 404, 427, 467, 470, 480, 502,
505
Lachesis mutus 538
Lacniargus 284
— borealis 153, 285
Lairomorpha 805, 862
Lagornys 974, 975
Lagopus 657, 978
— lagopus 595, 596, 597, 639
— mutus 595
Lagurus 862
— lagurus 811, 943
Lama 962
Lainna cornubica 75, 85, 152, 210,
' 285
Lamnidae 152
Lampetra 64, 65, 252
— aurea 65
— cibaria 65
— fluviatilis 65
— planeri 65
— wilderi 65
Lanarkia spinosa 68
Laniidae 615
Lanins 633, 972
— excubitor 616
— minor 613, 616
Lari 274, 602, 621. 633, 647, 648, 649,
683, 704
Laridae 603
Lariformes 643
Larus 658
— cachinnans 603
— canus G02, 603
— fuscus 603
— gelastes 603
— hyperboireus 978
— marinus 603
Larvacea 28, 31
Lasaniidae 66
Lasanius probl ematicus 66
Latax 978
— lutris 274, 818, 872, 873, 932
Lates 281
Latirnerla 161, 162, 163
Lein m us 978
— lemmus 811
Lemuridae 754, 849
Lemuroidea 775, 848, 849
Leo Q74
— leo 815
— spelaea 815
Leopardus pardus 930, 974
— — villosus 975
— uncia 880, 930
Lep ad og aster 136
Lcpidosauria 404, 467,469, 505
Lepidosiren 102, 113, 114, 115, 123, 131,
149, 158, 281
— paradoxa 115, 159
Lepidosirenidae 158
Lcpidosleidae 81, 167
Lepidosteoidei 160, 164, 167, 198
Lepidosteus 75, 80, 87, 100, 108, 112,
120, 121, 122, 126, 134, 138, 147, 165,t
167, 198, 280, 282, 286
— osseus 167
— platystomus 167
Leporidae 745, 805
Lepospondyli 351, 364
Leptocephali 226, 239, 254, 255
Leptocephalus brevirostris 238
Leplodactylinae 359
Leptodactylus ocellatus 373
— pentadactylus 359
Leptolepidae 168, 199, 286
Leptolepiformes 168
Leptolepis 168
Leptomeryx 839, 840
Leptopoecile 975
Leptoptilus 634, 660
Lepus 805, 860, 928
— americanus 892
— connori 891
— europaeus 805, 928
— mandschuricus 805
— pamircnsis 876
— timidus 805, 928
— tolai 805
Leuciscus balteatus 250
— bergi 297
— cephalus 170
— idus 170, 251
— leuciscus 216
— schmidti 297
Leu cost icte 975
Lichanotus 868
Limacina 272
Limax lanceolatus 33
Limicolae 601, 621, 643, 647, 648, 684
Limnocryptes gall inula 704
Limnogale 795, 872
Limnoscelidae 448, 503
Limnoscelis 448
— paludis 448
Limnonites minutus 601
— temmincki 601
Limosa lapponica 640
— limosa 601, 671, 676, 681
— ruta 671
Linaria cannabina 614
— flavirostris 614
— linaria 614, 672
Liocassis 172
Liopelma 357
Liopelrnidae 357, 358
Liopeltis 523
Liparis liparis 189
Lissamphibia 305, 351
• Lithofalco aesalon 593
Litopterna 823, 846, 872, 962
Litorhynchus 510
Locustella fluviatilis 616
— luscinioides 616 <
— naevia 616
Lophiidae 190
Lophiodontidae 845
Lophius 132, 100
— piscatorius 190, 213, 22$. 226
Lophophanes cristatus 615, 976
Lophophorus 976
Loricaria 80, 119, 230
Loricariinae 172
Loridae 606, 959
Loris 969
Lorisidae 849
Lota 280, 282
— lota 193, 208, 216, 225, 289
Loxia 636, 679
— curvirostra 613, 614, 680
Loxia leucoptera 976
— pityopsittacus 976
Loxodon 825, 967
Lucioperca 183, 288
— lucioperca 183, 293, 295, 296, 297,
302, 303
— marina 183, 295
— sandra 217
Lullula arborea 614
Lumicolae 704
Lumpenus maculatus 284
Luscinia luscinia 617
1013
Nemorhaedus caudatus 835, 939, 976
Neobunodontla 827, 829, 830
Neoceratodus 102, 109, 113, 123, 128,
129, 130, 158, 281
— foster! 158, 159
Neosorex 973
Neognathae 580, 621, 648
Neohipparion whitneyi 744
Neohjppus 844
Neomys 274, 974
— fodiens 796, 874, 976
Neophron 593
Neornithes 580, 590, 620
Nesokia indica 811
Nestor notabilis 606
Nestoridae 606, 954
Netomyidae 810
Nimravus 815
Nonruminantia 936 •
Not arc'toss 851
Nothocion 816
Nothosauria 504
Notidani 99, 110, 112
Notidanidae 151
Notidanus 93, 152
Notodiaphorus 846
Notornis 954
Notoryctes 727, 795, 862, 864, 883,
888
— typhlops 742, 743, 783, 791, 863
Notoryctidae 791, 956
Nototheniidae 285
Nototrema 380
— cornutum 360
Notungulata 794, 823, 840, 962
Nueifraga 972
— caryocotactes 613
Numenius arquatis 600, 601
— thaitiensis 679
Numida 595
— meleagris 595, 699
Numtdinae 968
Nyetalus 798
Nyctea nivea 663, 978
— nyctea 607
Nycteinae 607
Nyctereutes 975
— amurensis 816, 934
Nycticebus 849, 969
N yeti cor as 588
Nyctipitecinae 853
Nyroca 591
Oceanodroma 586
— furcata 586
Ochotona 862, 863, 880, 892
— alpina 976
— daurica 805
— pusilia 805
Ochotonidae 805, 966, 971
Octodon 961
Octodontidae 812, 961, 967
Ocydrornus 600, 954
Odobenidae 819
Odobenus obesus 819
— rosmarus 819
Odontoceti 820, 821, 822, 875
Odontophoridae 595, 973
Oedicnemus 657
— oedicnemus 600, 602
Oedicnimidae 602
Oenanthe oenanthe 617
Oikopleura 28, 272
Okapia johnstoni 839, 860
Omnivora 885
Oncoides 975
Oncorhynochus 174, 234, 235, 246, 261,
262, 293, 296
— gorbuscha 2 96
— keta 174, 234, 261, 296
— kisutsch 2 96
— nerka 234, 263, 296
4016
Oncorhynochus tschawytscha 296
Onohippidium 962
Onos 226, 284
Onychodactylus 312, 353
Ophiacodon 413, 427
Ophiacodontidae 449
Ophidia 404, 480, 502, 505
Ophiocephalidae 180
Ophisaurus 470, 472, 473
— apus 475, 509
Opisthoglypha 484, 520, 535, 537
Opistocoela 357, 358, 393
Opistocomi 621, 963
Opistocomidae 596
Opistocomus' 579
—’hoazin 596, 649
Orca orca 822, 823, 940
Oreamnos 973
— americanus 835
Oreodon 839, 840
Oreodontidae 839, 840
Oriolidae 613
Oriolus oriolus 613, 676, 681
Ornithomimus 429, 492, 507
— altus 492
Ornithopoda 406, 488, 494, 498, 506, 564
Ornithorynchidae 788, 956, 959
Ornithorynchus 764, 881
— aoatinus 789, 956
— paradoxus 872
Ornitischia 406, 491, 494, 497, 505, 506
Ornithosuchidae 488, 506
Ornithosuchus 488
Orohippus 843, 844
— agilis 744
Ortopoda 494
Orycteropidae 847
Orycteropus 847, 862, 967
— aethiopicus 847
Orycteropus afer 847
Oryctolagus 806
— cuniculus 738, 806, 862, 925
Oryx 835
Oryzoryctes ,795
Oscines 612, 613, 639, 690
Osmerus 175, 249
— eperlanus 175, 213
----sprinchus 175, 216, 293, 296, 302
Osphromenidae 181
Osphromenus 181
Ossifraga gigantea 683
Osteichthyes 73, 83, 94, 95, 96, 100, 102,
103, 106, 107, 112, 118, 119, 121, 143,
149, 157 сл., 161, 196, 197, 200
Ostecglossidae 173, 280, 281, 282
Osieolepidae 162
Osteolepidoti 145, 162, 362
Osteolepis macrolepldota 162
Osteostraci 67
Ostracion 149, 185, 230
— cornutus 214
— quadricornis 185
Ostraciontidae 185, 284
Ostracodermi 33, 46, 50, 51, 66, 95, 112,
145, 193, 195, 230
Ostrea 634
Otariidae 819
Otcolobus manul 974
Otides 598, 599, 621, 648
Otididae 599
Otis tarda 599, 704
Otocyon 967
Otolemur 878
Oudenodon gracilis 453
Ovibos 971, 978
— moschatus 835, 837, 876
Ovis 774, 938, 974
— ammon 835, 911, 938
— aries 835
— arkal 911, 912
— musimon 835, 911
— nivicola 835, 938, 977
Ovis orientalis 835
----- archar 938
-----cycloceros 938
-----orientalis 938
— poloi 835, 938
— vignei 835
Oxyclaenidae 813
Oxygaena 757
Oxyropus cloelia 538
Pachycynodon 816
Pachynolophus 844
Pachypodosauria 49?, 493, 506
Pachyura swinhoei 533
Pagel las 184
Pal aeanodonta 801
Palaeobatrachidae 359
Palaeobatrachus 364
Palaeochoerus 830
Palaeognathae 558, 580, 581, 648
Palaeohatteria 468
Palaeohippidae 844
Palaeohippinae 843
Palaeolagus 805
Palaeomastodon 760, 825
Palaeomeryx 834
Palaeoniscidae 164, 197
Palaeoniscoidei 81, 160, 164, 165, 198, 285,
286
Palaeophidae 481
Palaeophis 481
Palaeopithecus 852
Palaeornithidae 970
Palaeornithinae 606
Palaeospondylus gunni 65
Palaeovaranus 479
Palainedea cornuta 590
Palarnedeae 587, 590, 963
Palamedei 621
Palamedeidae 590
Paliguana 469, 470
Palmatogecko range! 508
Paludicola fuscomaculata 379
Pandion 274, 593, 631
— haliaetus 683
Pantera pardus 815
Pantolambda 846
Pantolestidae 794
Pantotheria 790, 964
Panurus biarmicus 615
Panyptila sancti hieronymi 644
Papio 854
Paradisea 613
Paradiseidae 613, 959
Paradokurus 976
Paraechinus 7 96
Parahippus 844
Paramanis javanica 800
Paramys 804, 806
— deli cat us 804
Parapithecus fraasi 851
Parasuchia 487, 505
Paratilapia 186, 240
Paraxonia 744, 823
Pareiasauria 448
Pareiasauridae 403, 448, 503
Pareiasaurus 410, 448, 507
— karpinskii 448
Parexostoma 172
Paridae 615, 644, 687
Parinae 615
Parioxornithinae 615
Parotia 613
Parridae 602
Parus 972
— major 615, 658
Pascolaretos 959
Passer 690
•— domesticus 613
— hispaniolensis 613
— montanus 613
Passeres 570, 611, 613, 634, 639, 648
Passeriformes 561, 612
Pastor roseus 613, 671
Paterosauria 352
Patriocetus 821
Pavo crist at us 595
Pecora 830, 831
Pedetes 809, 862
— caffer 809
Pedetidae 809
Pedetinae 745
Pediculati 190, 214, 284
Pediocetes 973
Pedionomidae 597
Pedionomus torquatus 597
Pedoces pander! 975
Pelamis 485, 532
Pelargomorphae 587, 621
Pelecani 274
Pelecanidae 588
Pelecanus 588, 633, 702
— crispus 588
— onocrotalus 588
Pelecus cultratus 170, 247, 293, 295
Pelidna alpina 601
Pelobates 312, 358, 367, 381
— fuscus 538, 367, 377
Pelobatidae 358, 367
Pelomedusa 462
Pelomedusidae 462, 547
Pelosaurus 350
Pelycosauria 402, 403, 405, 406, 414, 420,
449, 450, 451, 502, 503, 504, 505
Pelycosaurus 524
Pelycosimia 487
Peralestes 790
Perameles 791, 959
Peramelidae 791, 888, 956
Perarnys 790
Perea 183, 207
— fluviatilis 143, 183, 217, 234, 237, 252.
296
— schrenki 183
Percidae 183, 252, 280, 282
Perciformes 100, 183, 191
Percoidei 183
Perdix 657
— perdix 595, 704
Perennibranchiata 306, 332, 352, 382
Periophtalinus 119
Periparus ater 615
Perissodactyla 744, 794, 823, 843, 936, 962
Permocynodon 452, 503, 785
Pernis 632
Perodicticus calabarensis 310
— potto 866
Pet auroides 957
— volans 870
Petaurus 957
— sciureus 870
Petrogale 792
Petromyzon 64, 65, 147
— fluviatilis 63, 214
— marinus 52, 53, 64
Petromyzontia 63, 64 сл.
Petromyzontidae 64 cp., 235, 238, 280
Pezophus solitarius 604
Phacochoerus 830, 862, 967
Phaeton 649
Phaetonidae 588, 683
Phalacrocoracidae 274, 588
Phalacrocorax 588
Phalanger 792, 957
Phalangeridae 792, 957
Phalaropinae 601
Phalaropus 638, 684
— lobatus 601, 637, 640
Phaleria 978
Phaneropleuron 158
— andersoni. 158
Phaneropleuridae 158
Pharyngognath! 185
Phascolarctidae 792
1017
Phasedarctus 866
— cinereus 792
Phascolestes 790
Phascolodon 790
Phascologale 790
— cristicaudata 890
Phascolamys 792
Phasianidae 595, 704
Phasianus 595, 704, 974
— colchicus 594, 595
Phenacodontidae 842, 843
Phenacodus 842
— primaevus 842
Philomachus 681
— pugnax 601, 637, 676
Phlaocyon 816
Phoca caspica 820, 935
— foetida 820, 935
— groenlandica 820, 935
— sibirica 820, 935
Phocaena phocaena 940
— relicta 823
Phocidae 742, 820
Phocnicirus phoenicirus 617
Phoenicopteri 587, 589, 590, 621
Phoenicopleridae 590
Phoenicopterus 570
— roseus 589, 590
Pholidota 715, 792, 800, 964, 967, 970
Photinus 229
Phoxinus laevis 234
Phractamphibia 305, 349
Phryniscus nigricans 378
Phrynocephalus 412, 471, 472, 508, 510,
528, 531, 543
— helioscopus 471
— mystaceus 471, 508, 523, 530 4»
Phrynosoma 448, 472, 508, 524
— cornutum 474, 510
Phyllobates 371
Phyllomedusa 360, 375, 866
— burmeisteri 310, 366
— hypochondralis 380
Phyllopteryx eques 225
Pbylloscopus 657
— collybita 616
— sibilatrlx 616
— trochilus 616
Phyllospondyli 316, 350, J-64
Phyllostomidae 798, 957, 962
Physeter 774
— catodon 823
— macrocephalus 940
Physeteridae 823
Physoelysti 120, 123
Physostomi 120
Phytosauria 487
Phytosaurus 487
Pica 972
— pica 613
Plci 605, 609, 644, 648, 687
Picidae 610
Picoides 687
— tridactylus 610
Picus 657
— canus 610
— viridis 610, 976
Pinicola enucleator 614
Pinnipedia 741, 769, 794, 813, 818, 858,
888, 934
Pipa 330, 358, 376
— americana 358, 380
Pipidae 312, 358, 364, 393
Pipistrellus 798
Pipridae 963
Pisces 33, 50, 72 сл.
Pithecanthropus errectus 852
Pithecia satanas 870
Pithecus entellus 854
— schistaceus 854
Pittidae 611, 959, 971
Pitymys 976
1018
Placentalia 713, 773, 792
Placochelys 460
Placodontia 404, 408, 420, 436, 454, 460,
504, 518
Placedus 460
Placoidei 149
Plagiaulacidae 788, 857
Plagiostomi 151
Plascolarctidae 957
Plascolotherium 789
Platale.a 274, 634
— leucorodia 589, 590
Plataleidae 590
Platanista gangetica 823
Platanistidae 823
Platecarpus 480, 517
Platycercidae 606
Platyceriidae 959
Platyhyla verrucosa 366
Platyrrbina 852, 853, 962
Platysomidae 164, 198
Pl ecot us 798
Plectognathi 184, 220, 226, 284
Plectrophenax nivalis 614, 672
Plegadis 590
— falcinellus 589
Plesiadapidae 794, 847
Plesiometacarpal»a 833
Plesiosauria 408, 458, 504, 518
Plesiosaurus 459, 518
Plethodon 333, 355, 393
Plethodontidae 355, 366, 372, 393, 394
Pleuracanthodes 156
Pleuracanthodii 156, 196
Pleuracanthus 105, 108, 109, 156, 196
— ducheni 108
Pleurodira 462, 504, 547
Pleuronectes 182, 246
— flesus 251
— luscus 298
— platessa 183, 224, 234, 238, 244
Pleuronectidae 121, 148, 182, 183, 214,
217, 224, 225, 238, g44, 251, 252, 272,
284, 293, 296, 298
Pliohippus 844*
Pliopithecus antiquus 851
Plioplatecarpus 480
Pliosauridae 459
Pliosaurus 460
Ploceidae 613, 968
Ploteidae 636
Plotus 633
Pluracanthodii 151
Poa bulbosa 885, 890
Podargidae 608
Podica senegalensis 600
Podiceps 64 4
Podoces panderi 613
Podocnemis 462
— expansa 534
Pogodroma 663
Poebrotherium 831
Poecile 972
— borealis 615
— cinetus 976
— palustris 615
Poecilia 177
Poephagus 974
— grunniens 876, 877, 910
Polloaetus 631, 683
Polioptilinae 615
Polistotrema 64
Polistotremidae 64
Pollachius virens 193
Polyborinae 593
Polychaeta errantia 195
Polymastodontidae 857
Polyodon 112, 132, 286
— (Spatularia) folium 107, 161, 221
Polyodontidae 74, 161
Polyprotodontia 790, 857, 956
Polypteridae 81, 163
Polypterini 74, 161, 163
Polypterus 80, 100, 108, 112, 114, 115,
122, 123, 126, 134, 136, 138, 147, 149,
161, 163, 164, 281, 362
— bichir 106, 163
— senegalus 164
Polysticta 978
Poinacentridae 186, 225, 284
Pomacen tri formes 185
Porphyrio 600
Porzana porzana 600
— pus ilia 600
Potainochoerus 965
Pot a mog ale 796
— veloj 795, 796, 872
Potamogalidae 795, 966
Potorous 792
Praedcntata 494
Prlacodon 790
Primates 718, 794, 847, 962
priodontes giganteus 802
Prlstiophoridae 153
Pristiophorus 153, 221
Pristidae 154
Pristis 76, 80, 154, 221
— antiquorum 154
Pristiurns 147
— melanostomus 90
Proboscidea 794, 823, 824
Proceliariae 585, 587, 683, 690
Proucliarii 586, 621, 633, 634, 647, 662, 683
Procel lari idae 586
Prochordata 50
Procoela 358, 359
Procyon 973
— Intor 816
Procyonidae 813, 816, 961, 966, 969
Produces 975
Proinctheomys schaposchnikovi 861
Propalaeochoerus 830
Propilheeus 870
Prophopithecus haeckell 851
Prosaiiria 446, 467
Proscinrus 806
J'roHlmla 851
Proahnlae 794, 848, 868, 962, 965, 966, 969
Ptoiacanthodes pinnatus 150
Protelda 355
Pruh-ldae 352, 355, 364, 394
ProlelcH 966
j'rolrllnar 966
Prol<'i iI у plia 485, 535, 537
ProlrrolbrilIdae 745, 846, 962
ProlciiH 345. 365, 369, 372, 373, 385, 394
— anguIncus 355
Protoioit rnilnis 317, 357
Protocnmcliis 831
Prolorrtiis alavus 821
ProtorcratopH 497, 498
Protoci nnlala 50, 70, 71, 193
Protodmita 787, 956, 964
Protoh I ppua 844 -
— pern lx 74 4 "
Protolabls 831
Proloplenm 10?, 113, 114, 115, 123, 131,
158, 159, 281
— annectcns 159
Protosalirnl.ia 357
Protosphargls 466
— veronensls 466
Protostornla 48, 4 9
Protolheria 713, 780, 787, 956
Protrllon 308, 350
Protungulata 813, 823, 828, 842, 962, 964
Protypotherluni 841
Prunella modularls 617
Psephophorus 467
Psephurus gladius 161
Pseud a xis hortulorum 833, 937, 976
— sica 833, 976
Pseudochirus 867
Pseudinae 359
Pseudobagrus 172
Pseudo cerastes 486
Pseudocreodi 757, 813
Pseudoscaphirhynchus 161, 221, ?86
— tedtschenkoi 161
— kaufmanni 161
Pseudosuchia 429, 487, 505, 576, 580
Psittaci 561, 605, 606, 636, 648
Psittacidae 606
Psittacinae 606
Psittacini 968
Psittacus erithacus 605
Psophiidae 599, 963
Pteranodon 431, 499, 518
— ingens 519
Pteranodontidae 499
Pteraspidae 68
Pteraspidomorphi 68
P ter as pis 69
— crouchi 68
Pteromyidae 971
Pterichtyomorphi 69
Pterichtys 69
— mtiller! 69
Pteridophora 613
Pterobranchia 14, 18
Pterolepidae 66
Pterolepis 66 *
Pterocles 974
— arenarius 603
Pterocletes 603, 604, 621, 647, 648, 657,
686
Pterocrtdae 603
Pterodactylidae 499
Pterodactyloidea 499, 500, 506
Pterodactylus 499
— spectabilis 500
Pteromyidae 807
Pleromys 807, 870, 970
Pteropodidae 797
Pteroptochidae 964
Pieropus cel ano 797
— edulis 740
Pterosauria 402, 403, 446, 467, 498, 502,
505, 506, 514, 518, 871
Ptilonorhynchidae 959
Ptilonorhynchus 613
Ptyas 520
Ptychodcra 14, 15
Ptychozoon 478
— homalocephalum 512, 513
Ptylonorhynchidae 640
Ptyiloscopus inornatus 977
Ptyodactylus 508
Puffinus 586, 690
— puffinus 586
Puma concolor 815, 816
Putorius alpinus 976
— eversmanni 931
— putorius 931
— sibiricus 976
Pycnodontidae 166, 200
Pym on о t idae 971
Pygosteus 179
Pyrosoma 29
Pyrosomatidae 29
Pyrotheria 794, 842, 962
Pyrrhuia pyrrhuia 614, 693
Python 481, 520, 532
— molurus 481, 482
— reticulatus 521
— sebae 509
Pythonlnae 481, 546
Pythonomorpha 407, 479
Raja 80, 108, 154, 213
— clavata 155
— radiata 155
Rajidae 154, 284
Ralli 598, 599, 648, 684, 688, 704
Rallidae 599
Rallinae 599
1019
Rallus aquaMcus 598, 600
Ramphastidae 963
Rana 274, 312, 327, 330, 333, 337, 338,
341, 361, 364, 374, 376, 381, 390, 391
— asiatica 361
Rana catesbyana 361, 367, 386
— clamitans 389
— esculenta 317, 319, 321, 361, 368,
376, 384,- 386
— fusca 348
— mugiens 361
— natatrix 325
— opisto
— opistbodon 379
— palustris 386
— pipiens 370, 383, 384, 386
— rlbibunda 361, 384
— teinporaria 317, 318, 342, 361, 376,
377, 384
— terrestris 361, 384
Rangifer 832
— caribou 892
— tarandus 832, 876, 925, 936, 972, 978
Ranidae 359, 361, 393
Ranidens sibiricus 326, 353
Raninae 361
Ranodon sibiricus 315
Raphidae 604
Raphus cuculatns 604
Ratitae 558, 560, 581, 686
Rattus exulans maorium 954
— norvegicus 803, 811, 943
— rattus 811, 943
— turkestanicus 811
Recurvirostra 634
— avocetta 601
Regalecus 78
— banksii 210
Regulinae 616
Regulus 972
— regulus 616
Reptilia 33, 50, 401, 446
Rhabdopleura 14
— normanni 18, 19, 21
Rhachianectes glaucus 822, 940
Rhachianectidae 822
Rbachiodontinae 484
Rhachitorni 315, 350
Rhacophorus 366, 367, 375, 381
— rnalabaricus 367
— pardalis 310, 311, 367
— reinhardtii 380, 381
Rhamphastidae 610, 636
Rhampborhynchidae 49Q
Rhamphorhynehoidea 499, 500, 506
Rhamphorhynchus 499
— gernmingi 407
Rhea 570, 582, 640, 647, 686
— americana 583
— pennata 583
Rheae 583, 620, 963
Rheidae 583
Rhinobatidae 154
Rhinoceros 869
— mercki 846
— simus 947
— sumatrensis 846
— unicornis 846
Rhinocerotidae 845, 967, 970
Rhinocerotinae 845
Rhinochetes 963
Rhinocheti 598, 955
Rhinochetidae 598
Rhinochetus jubatus 598, 955
Rhinoderma 376
— darwini 360, 379
Rhinodermatinae 360
Rhinodon 77
— typicus 153
Rhinolophldae 798
Rhinolophus ferrum equinum 798
— hipposideros 798
1020
Rhithrosciurus 970
Rhizosauria 403, 447, 500
Rhodeus amarus 170, 233, 234, 240
Rhonibiforrnes 182
Rhombomys 861
— opinius 811, 891
Rhombus maximus 214
Rhopalodon 454
Rhynchocephalia 403, 404, 427, 467, 468
469, 549
Rhyncholepis 66
Rhynchopidae 603
Rhytina gigas 763, 945
— stelleri 827, 887
Rhytinidae 827
Riparia riparia 617, 643, 677
Rissa tridactyla 603
Rodentia 760, 775, 792, 794, 802, 858
870, 928, 957, 961
Rostratula 638, 640
Rufibranta 978
— ruficollis 670
Ruminantia 827, 936
Rupbapra 880, 974
— rupicapra 835, 939
Rupicaprinae 835
Rusa 832
Rutilus 170
— frisii kutum 295
— rutilus 170, 296
---caspicus 246, 293, 295
Rynchocephalia 954
Querquedula 591
— crecca 703
— querquedula 703
Saccobranchus 119, 120, 171
Saccopharyngidae 178
Saccopharynx ampullaceus 178
Saghatheriidae 824
Saiga 974
— tatarica 834, 835, 939, 976
Salamandra 354, 375
— atra 320,* 354, 375, 392
— maculosa 326, 355
— perspicillata 315
— salamandra 354, 370, 375, 384
Salamandridae 354, 393
Salamandrina 330, 333, 375
Salamandrinae 312
Salamandroidea 333, 354
Salmo 100, 108, 112, 148, 174, 261
Salmo fario 134, 145
— fonlinalis 303
— irideus 303
— ischchan 249
— oxianus 288
— salar 75, 96, 174, 210, 213, 232, 234
236, 239, 244, 258, 298
— salvelinus 288
— trutta 174, 250, 296, 302
— — aralensis 174
--- labrax 174, 295
— — lacustris 174
— — morph a fario 174, 225, 232, 236
237 302 303
Salmonidae 78, 102, 138, 173, 216, 236
252, 253, 280, 282, 284, 287, 301
Salpa 30
Salvelinus alpinus 250, 296
— — morpha pluvialis 250
— — var. salvelinus 302, 303
Sarcophilus 790
— sataninus 791
Sarcohamphus gryphus 538, 592
Sarda sarda 192
Sardinella 173
— melanostica 173
— pilchardus 173, 217
Sargus 184
— annularis 184
Saurlptcrus .323
Saurisclua '«06, 491, 505, 506
Sauropoda 492, 493, 506
Sauropsida 345, 346, 400
Sanroptorygia 404, 408, 415, 420, 454,
457, 458, 504
Saururae 580
SaMcola pleschanka 617
— rubetra 617
Saxlcoilnae 617
Seaph iopus 367
Scapbirhynchus 110, 161
— pl a th yrh у rich us 161
Scardinius erythrophtalmus 170, 295
Scaridae 186, 217, 220
Scarus 133
— eretensis 186
Seel idosaurus 496
Schizolhorax 171, 274
— intermedins 250
Sclerodermi 184
Sciaenidae 184
Scincidae 476, 528, 544
Scincus 510
— officinalis 476, 509, 528
— tridactylus 476
Scirtopoda teium 810
Sciuravus 806
Sciuridac 806
Sciiiroidea 806, 861, 001
Sciuropterus 807, H70, 970
— riisslrus 807, 808
— Volans 869, 810
Sciiirus 869, 967. 970
— aiiomalim 806, V?M
— vulgaris 806 869, 928
Scler orol I ih
Sclrromoeblldiin 488
Sclcr (iinorhl us 488, 505
Sclri Opili H 188
ol it par I d <ir 634
Srolopnclо не 601
Scolupnx 2 2 I
— rusllrola 601, 657, 672, 675, 676, 704
Scomber 76, 135, 19?, 231, 272, 274, 285
— scomber 19?, 210, 212, 298
Scoiiibrertm lfoi mrs 179
Scornbresox 17 9, ?1t, 21?
Scombrldae 121. 180, 191, 214, 216
Scombrifonnes 191
Scopelidae 176, 214, 228, 240, 285
Scopelilormes 176, 177
Scopelus 177
Scopidae 588
Scops scops 607
Scorpacna 188, 226
— porcus 189
Scorpaenidae 188, 233
Scorpeniformei 188
Scylacosaurus 451, 452
Scvllidae 152
Scyllium 103, 133
— canicula 92. 93, 105, 139, 147, 152
Sebastes 188, 227, 285
— norvegicus 188, 233
Secretarius 533, 538
Seiache maxima 80, 114
Selachii 102,104,112,113,115,117,126,216
Selacg
Selachoidel 73, 151
Selenodontia 827, 829, 830 сл.
Semionotidae 166
Semnopithccus 969, 975
Seps 476
Serinus canarlus 701
Serpentaril 5 9?, 968
Serpentariidae 593
Serpent arius serpent arlua 53R, 593, 631,632
Serpentrs 4SO
Serranldar 133. 183, 188, 231, 252,281, 284
Serranus 139. 183
— cabi'llla 183
— scriba 183
Seymouria 419, 447, 500, 507
— bailorensis 447
Seymouriarnorpha 403, 447
Seymouridae 447
Sferna paradisea 978
Sicisla 974
— concolor 976
—loniger 976
— rnontana 809
— nordinani 809
Slelstinae 809
S i к a 8 3
Siluridae 102, 119, 121, 171, 206, 216.
240, 280, 281, 282
Silurinae 171
Slluroidei 231
Silurus glanis 170, 171, 219, 221, 225,
293, 295, 297
Simla 851, 969
— satyrus 854, 856, 868
Slmiae 794, 850, 962
Simplicidentata 805, 806 сл.
Sinanthropus 852
Slphla parva 616, 671
Siplionops 352
— annulatus 308, 352
SI redon 315, 354
— axolotl 382
— p*sciformls 329, 382
Siren lacert.ina 343, 356
Slrenla 741, 763,794,823,826, 858,887, 945
Sirenldae 352, 356, 365
Slrenoidea 356
Slllstinae 971
Sitta 644, 972
— curopaea 612, 615
Slttidae 615, 644, 687
Slvapilhecus 85t
Sivatherium 839
Smilodon 815
Smuts I a temmlnckl 800
Sofortyx cahfornicus 595
Solea 183
— nasuta 183, 298
— vulgaris 183
Sol enod on 796
— (mb an us 796
Sole.nodontidae 796, 961, 964
Soniateria 591, 659, 660, 978
— rnollssima 591, 703
— spectabilis 592, 703
— «teller! 592, 703
Sorex 966
— alpliius 976
— aranriis 796
— minutiis 796
Soricldae 796, 878
Sorlrlnae 7 96
Sori cuius 969
Spalacidne 810, 967
Spalacolherlum 754, 790
Spalax 742. 810, 86?, 877, 974
— rnicrophtalinus 810
Sparidae 184
Spatula 591
— clvprata 703
Spelerpes 333, 355, 375, 393
— fuscus 355, 368, 375
Spermophilopsis leptodactylus 861
Spermophilus 894
Sphenisci 585, 586, 621, 633, 649, 652,
660, 683, 963
Spheniscidae 586
Sphenisciformes 586
Spheniscus dernerseus 5Я6
Sphenodon 403, 415, 422» 426, 433, 468,
469, 532
— punctatum 468
Sphenodontia 502
Spheroides 185
— maculatus 289
1021
Sphyraenidae 284
Spinacidae 153
Spinus spinus 614, 972
Spratella 173
— sprattus 173, 217
Squalius cephaius 217
Squalodontidae 821
Squamata 423, 469
Squamipennes 284
Squatarola squatarola 601
Squatina squatlna 153, 154
Squatinidae 153
Steathoruis caripensis 607
Steathornitldae 607, 963
Steganopodes 587, 588, 621, 633, 648,
649, 683, 684, 702
Stegocephali 305, 314, 322, 364
Stegocephalia 349
Stegodon 760, 761, 825
Stegosauria 495, 498, 506
Stegosauridae 405, 496
Stegosaurus 496, 498
— ungulatus 497
Stenodus 175, 301
— leiwichthys 175, 295, 302
— — nelma 175
Stenops 849
— tardigradus 866
Stenopterygius 456
— acutirostris 455, 456, 457
— quadriscissus 456
— tennirostris 456
Stercorariidae 603
Stercorarius 633, 691
— parasiticus 602, 603
Stereornithes 596
Stereospond.yli 316, 350
Stereosternum 454
— tumidum 455 >
Sterna 274
— cantiаса 603
— caspia 603
— fluviatilis 602
— hirundo 603, 642
— macrura 673
— minuta 603
Sternidae 603
Sterninae 682
Sternoptychidae 228, 285
Stomiatidae 175, 228
Stomias 175
Streperidae 959
Striges 606, 607, 621, 644, 648, 665
Strigidae 607
Striginae 607
Stringopidae 606, 954
Stringops 662, 954
— habroptilus 606
Strix aluco 944
— flaminea 607
Struthiomimus 492
Stnithiones 581, 620, 686, 968
Struthionidae 582
Sturnidae 613
Sturnus 690
— vulgaris 613, 678
Stylodon 790
Subungulata 823
Suidae 830, 872, 936, 970
Sulla bassana 643
Sulidae 588, 683
Surnia ulula 607, 663
Sus 830, 865, 869, 936, 966
— antiquus 830
— leucomystax 918, 936, 977
— major 830
— mediterraneus 918
— palaeochoerus 830
— papuensis 957
— scrofa 830, 918, 936
— vittatus 830, 918
Sutora 976
1022
Sylvaemus sylvaticus 943
Sylvia 976
— atricapilla 616
— borlu 613, 616
— communis 616
— curruca 616
— nisoria 616
Sylvidae 616
Sylviinae 616
Symbranchidae 281
Symmetrodontia 790
Symbranchii 280
Symbranchus 281
Symphalangus syndactylus 854
Synanceia 227
— borrida 188, 189
Synapsida 423, 449, 450, 467, 502
Syngnathidae 133, 178
Syngnathus 178, 221
— acus 178, 240
— pelagicus 211, 285
Synodontis 172
— decorus 171
Syrrhaptes 974
— paradoxus 603, 604, 685
Syrniinae 607
Syrnium 892
— aluco 607
— lapponicum - 607
— uralense 607
Tadorna tadorna 592, 643
Tadorninae 592
Tai ass a ret os 971, 978
— maritimus 816, 934, 971
Talattosuchia 488, 489, 505
Talpa 742, 864, 878, 928, 974, 976
Talpa altaica 976
— coeca 796
— euporaea 743, 796
Talpidae 796, 966, 971
TarnTindua 743, 864
Tam las 861, 971 *
Tanagridae 614
Tapitrometopon 484, 535
— lineolatum 484
Tapirella 844, 962
Tapiridae 844
Tapirus 869, 872
— indicus 844, 970
— terrestris 844
Tarentola 477
— mauritanica 477, 478
Tarsiidae 850, 969
Tarsioidea 848, 849
Tarsi pes 792
— rostratus 752, 886
Tarslus 778
— spectrum 866
— tarsius 850, 881, 882
Tatusia 727
Taurotragus oryx 835
Taxidea 818
Taxodontia 962
Tejidae 470, 473
Telematacarpalia 832
Teleostei 75, 87, 88, 97, 100, 102.
103, 107, 109, 110, 112, 113, 114, 115,
117, 119, 120, 126, 127, 129, 132, 133,
134, 136, 137, 138, 140, 142, 144, 145,
147, 149, 161, 164, 167, 168, 198, 199,
200, 223, 226, 282, 286
Teleostomi 74, 99, 106, 113, 114, 115,
146, 159 сл., 231
Tmenocyon 816
Temnospondyii 350
Teratoscincus 477, 531
— scincus 477, 478
Terrapene ornata 423
Testudinata 404, 423, 460, 502, 504
Testudinidae 463, 504, 518, 532, 547
Testudinlnae 463
Testudo 442, 464
— atlas 464
— greaca 465, 534
— horsfieldi 464, 534
— ibera 465
— ocidifera 423
— polyphemus 526
— sunierl 529
Tetrabelodon 760
Trirareros 970
Tetrad aenodon 842, 843
Tetnio parvlroslris 977
— tetri x 657
— uriignilus 595, 596, 704
TrtriHigalliiH 595, 704, 892, 974
Trtrnonldiir 595, 703, 972
TrlrimpliaHls 974
ЪЧ! Н|н>1|п74,304,315,334.339,344,303,415
'IrlriMii-.i boniiHa 595, 703
'I el l Dilun 185
hilinkn 185
• lilHphluK 1НГ»
* 1 ell • >d (in 11 (I lie 8*/, I 85
:l liiilliii ch .'ll), 111
‘I hnllhnniirl uh 0H,'l
* 1 liiiuiiliil iiMiiill III hill '
V li I hi 4hit
'I lii’i Hilniillii 403, 422, 407, 486, 505, 576
*1 lie 11ill им Illi
*1 |iri и pi I it и 4t.o, 451, 452, 453, 454, 503
'I In i liuliml lit 412, A.'lb. 4h0, 451, 452,
4bl. hill, hn.'l, 504, 728. 735, 784, 785
mi vl >i’ '"I nUl UM iii'l ill cl I 4hi
'I |ii<i in । । hul Iк 4h 1, 41»2, 453
Hu i Hill'» I'liH 40'1, 400, 407, 418, 420, 421,
4 in 4И>. 4ho, 503, 7','H, 784
*1 In i ii|H'.lii 4011, 4W, 493, 500
............ mi"
............. 007
'I In .Hu i инн I» 4 (I
'I I. I и Hilil >r /01, Vhfl
'Ill 'l l! I IHI I V'lV
। . и * .. । pliiiliiM 791
*1 II I I' I'll И O'l I
I II I nil 11 III I /, у til, 237
и 11< и । HO
Hi . и» »lIим I /h, 238, 303
'll.-I...- 14, VI I. V/4
H'.unu. in нА. toy, 285, 289
III ,i hi । in ‘ uni III He 4 01,'
|i । । iiiiei ‘.nil Il’i 4U2
Ii I" ill i'MIii Il li.
и........ lx О I'i, НИ/
I l О । Ii liiii'i uni li|iilt ill Ih 846
i l II i-i i
|||||||. Hill, 0'10
I 1 ч I ' -I" <0 nut 188
I I H > L i 11 и I'll 1
I 111 • 111111 a i Щ'. 799
I III i О i Inin IHIII
Г ill । ih Ini II
| ) i >। < 111ii 11- 0 | II
i'll nil 1'0 |||I| I.H4, 820, 036,045, 080,963
I I........ Ill'll
I I...... ""I || HI IIIM hNh
th' ।
Hu i | * II el, | ( MUD
..I.. и I . I ,'l|
......... I " I i'll
............ I'H
и....и hIiih hlill
I..................
I । ii 11.. । ii.l.,) Ii4'lt М4П
I ) ' i . • । inn ни nil|
I I hi и \ID
, | I I I I I I! I .1. V t 4
I I ' 111. I'li, vul
• и и । и. • 11 ii V'/M
I I I H | I II
i ..I 'iihi'ii UUl
Tot anus tetanus 601
Toxochelys 465, 466
Toxodon 841
— platensis 841
Toxodontia 841
Toxotes jaculator 184
Trachelosauria 504
Trachinidae 187
TTachiniformes 187
Tracbinus draco 187, 226, 227
Trachodon 495, 518
— annectens 4 96
Trachodontidae 495, 498, 506
Trachurus trachurus 191
Trachypterus 212
Tragulidae 778, 831, 869, 967, 970
Traguloidea 830, 831 „
Tragulus 831, 970
Tremarctos thibetanus 816
Tremataspidae 67
Tremataspis 67
Trematops miller! 307
Tiracanthidae 184
Tri acanthus 185
— brevirostris 185
Trlassochelydae 408, 461
Triassochelys 461, 504
— dux 409, 461
Tribelosodon 499
Tribelosodonta 499
Tribelosodontidae 499
Triceratops 497, 498
TrichaeluruX manul 931
Triehechus rosmarus 934
Trichosurus 737, 957, 959
Tricbys 970
Triconodon 789
Triconodonta 788, 789, 857
Trigla 121, 189
— gurnardus 292
— hirundo 189
Triglidae 82, 189
Trigonidae 155
Trimerorachis Insignis 321
Trionycbidae 409, 467, 532, 548
Trionychoidca 411, 467, 504
Trionyx 424, 467
— ferox 467
Triton 354
— taeniatus 340
Trituberrulata 790
Tri turns 318, 354, 364, 365, 375, 381, 384
— cristatus 364, 372, 373
— pyrrhogaster 369
— vitlatus 355, 373
Tritylodonlidae 857
Trochill 608, 621, 647, 688, 963
Trochilldae 608, 628, 973
Trochilus 533
Troglodytes troglodytes 617
Troglodytidae 617
Troglodytinae 617
Trogonldac 609
Trygon 155, 213, 284
— pnstinaca 155, 226
Tubulidenlata 794, 847; 964, 967
Tunienta 13, 21, 47
Tupaln tana 794
’I’lipaildjic 794, 969
Tu pin am bls 474, 512
— tpgulxin 527
Tiirncus cnstatus 606
Tnrdinne 610
Tuning 972
— и 1.1 ogularls 976
— inusiciis 617
— niiiiinaiinl 977
— оЬчгпгиа 977
— ph I huticlus 617
— pllnrlH 617, 003
— nlblrh’us 977
— VlMdvurns 017
iO2.5
Turnagra 954
Turnices 597, 621, 636, 638, 647, 686
Turnicidae 597
Turnix sylvatica 594, 597
Tursiops 940
Turtur turtur 604
Tylopoda 830, 831 сл., 925
Tylosaurus dyspelor 480
Tympanuchus 595, 973
Typhlomolge 345
— rathbuni 355
Typhlonecles compressicauda 376
Typhlopidae 481, 509, 510, 545
Typhlops 520
— vermicularis 481, 509
Typhlotriton 345, 355
— spelacus 366
Typotheria 841, 962
Typannidae 611, 963, 973
Tyroptera 866
<Jdenodon 420
Umbra 122, 282
— umbra 175
Umbridae 122, 175
Undina 163
— gulo 162
Undinidae 163
Ungulata 794, 823, 858, 962
Unio 240
Upupa epops 609, 658, 678, 681
Upupidae 609, 648
Uragus sibiricus 614
Uranoscopidae 187, 285
Uranoscopus scaber 187, 226
Uria 642, 978
— troille 603, 646
Urinator 660, 702 *
— adamsi 978
— arcticus 586, 702
— stellatus 978
Urinatores 586, 648
Urinatoridae 586
Urochordata 13, 18, 21, 27, 35, 45, 47, 50
Urodela 305, 306, 308, 311, 312, 315,
317, 318, 320, 323, 327, 328, 330, 332,
333, 334, 336, 339, 340, 341, 343, 344,
345, 346, 347. 350, 352, 353, 355,
356, 357, 363, 364, 365,366, 367, 368,
374, 375, 392, 393, 501
Uromastix 470, 471, 517, 531
— acanthinurus 472
Uronemidae 158
Uronemus 158
Uropeltidae 482, 509, 545
Uropeltis 510
Uroplatcs 478
— fimbriatus lichen! 512
Urotrichus 971
Ursavus 816
Ursidae 756, 813, 816, 934, 961, 966
Ursus 752, 865, 878, 880, 971
— arctos 816, 934, 971
— beringianus 977
— lagomyarius 974
— leu cony x 974
— rnaritimus 883
— ornatus 961
— spelaeus 816
— tibetanus 975
Vanellus vanellus 601
Varanidae 479, 544
Varanus 479, 480, 533
— griseus 479, 524
— niloticus 479
— salvator 479
Vertebrate 13, 18, 31
Vespertiiio 798
Vespertilionidae 798
Vipera 485, 486, 520, 535, 536
— ammodytes 486, 528, 536
— berus 486, 528, 536
— lebetinus 486
— renardi 486
Viperidae 485, 520, 535, 546
Viperinae 485, 535, 546, 547
Viscacia 811, 862, 961
Viverra 720, 814
— civetta 814
— zibetha 814
Viverravinae 813 .
Viverridae 755, 813, 814, 961, 965, 969
Viverrinae 814, 965
Vormela peregusna 817
— sarmatica 932
Vulpes corsac 816
— lagopus 894
— vulpes 816, 894, 933, 971
Vultur 593
Vulturidae 974
Xema salini 978
Xenarthra 792, 800, 960
Xenicidae 954 .
Xenopeltis 520
Xenopus 312, 327, 358, 388, 393
— laevis 358
Xeridae 807
Xeromys 811, 957
Xerus 807
Xiphias gladius 78, 192, 210, 221, 285
Xiphiidae 192
Xiphodontidae 840
Youngina 486, 505
Younginidae 486
Yunx 658
Zaglossus 788, 956, 959
— bruyni 788
Zalambdalestridae 794
Zamenis 483, 510
— karclini 483, 484
— mucosus 483, 484
— ravergieri 484
— rhodorhachis 484
Zaocys carinatus 511
Zapus 745, 809
Zeidae 182
Zeiformes 182
Zeuglodontidae 821
Zeus 121
— faber 182
Ziphiidae 763
Ziphiinae 763, 823
Zoarces 136, 190
— viviparus 190, 243, 250
Zoarcidae 190, 214, 285
Zonuridae 472
Zonurus 472
— giganteus 474 *
Zosteropidae 614
Zygaena 226
— -malleus 152
Zygolophodon 825
Редактор издательства 3. И. Берман
Технический редактор О. Н. Персиянинова Корректор Л. Г. Афанасьева
Сдано в набор 5/VIII 1939 г. Подписано к печати 1/VIII 1940 г. Формат 60X92
в J/te. Объем 64 п. л. Учетно-изд. 75.7. В 1 л. л. 46 540 печ. зн. Тираж 15000 экз.
Уполн. Главлита № А-22898. РИСО № 1082. АПИ № 1227. Заказ № 3459
4-я Образцовая типография Огиза РСФСР треста «Полиграфкыига». Москва,
Валовая, 28
ЗАМЕЧЕННЫЕ ОПЕЧАТКИ
Стр, Строка Напечатано Следует По чьем ezine
55 4 и 5 сн. кардиальные кардинальные Abt.
под рис.
30, 9 и 9г кардиальная кардинальная
77 9 сверху сплетении сочленений
156 26 ,, червями и иглоко- червями, иглокожи- Ред.
жими рыбами ми и рыбами
157 4—5 > сн. Есть легкие, и кро- Есть легкие п ле-
вообращение ле- гочное кровообра- Авт.
гочное. щение.
173 16 п вид (S. pilchardus) (вид S. pilchardus) Kopp.
183 18 разделы отряды Abt.
187 3 п опросов спроса Kopp.
192 13 Cadiforines Gadiformes _ n
275 14 СВ. saprolegnia Saprolegnia Ред.
283 3 СН. их ее Авт.
298 5 св. Cobiidac Gobiidae Ред.
324 2 СИ. os ilem os ilium j,
352 9 5? Amphiumidae Amphiumidae,
403 21 св. Pareiasauriidae Pareiasauridae 5,
453 12 СП. Dycinodon Dicynodon A bt.
604 25 Di bus Didus Kopp.
609 7 св. 481 478 Ред.
618 21 55 они пальцы Авт.
658 19 Yunx lynx
686 25 Glared id a e Glared idae Kopp.
704 14 сн. рис. 481 рис. 473 Ред.
718 6 св. рис. 549, D рис. 524
736 4 сн. под proseseus processus Kopp.
рис. 541 действующие
803 17 св. днествующпе ,5
959 20 Pascoiaretos Phascolarctos Abt.
971 14 сн. Sitistinae Sicistinae Abt.
974 16 Otcolobus Octolobus Kopp.
978 4—5 св. Dicrdstomyx Dicrostonyx Abt. -
8 Rufibrcnia Rufibranta
10 Sferna Sterna Kopp.
В указателе (при правильном начертании в тексте книги):
995 6 СП. Саусданские Саусдаунские Состави- тель
999 9 У) Ееиилиевые Цецплпсвыс Kopp.
1992 16 св. A guilla Anguilla Состави- тель
1009 1 СН. Draco Draco
rimbriatus fimbriatus Kopp.
1014 20 св. Megalotrion Megalotriton Состави- тель
1016 12 СИ. Oedicniinidao Oedicuemidae ?>
1017 8 св. Oxygaena Oxyaena
1018 9 Plascolarctidae Phascolarctidae
10 Plascolotheriuia Phascolotherium
1020 13 opisto — Тип.
1021 14 СИ. selaeg •—
1023 23 св. Tiracanthide® Triacanthidae Kopp.
1024 19 •• Typaimidac Tyrannidae
25 СИ. Zalainbdalesteine Zalambdalcstidae