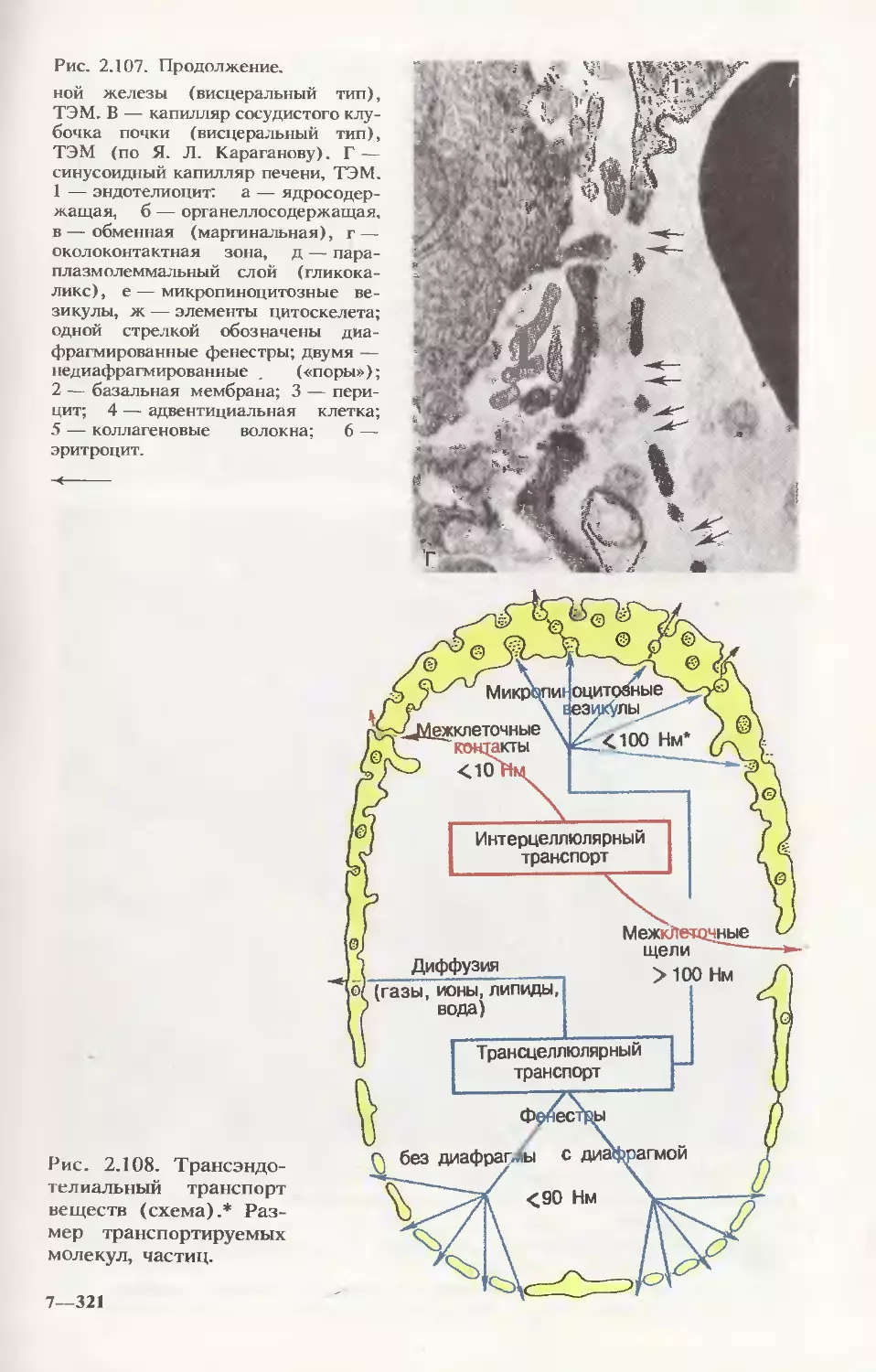
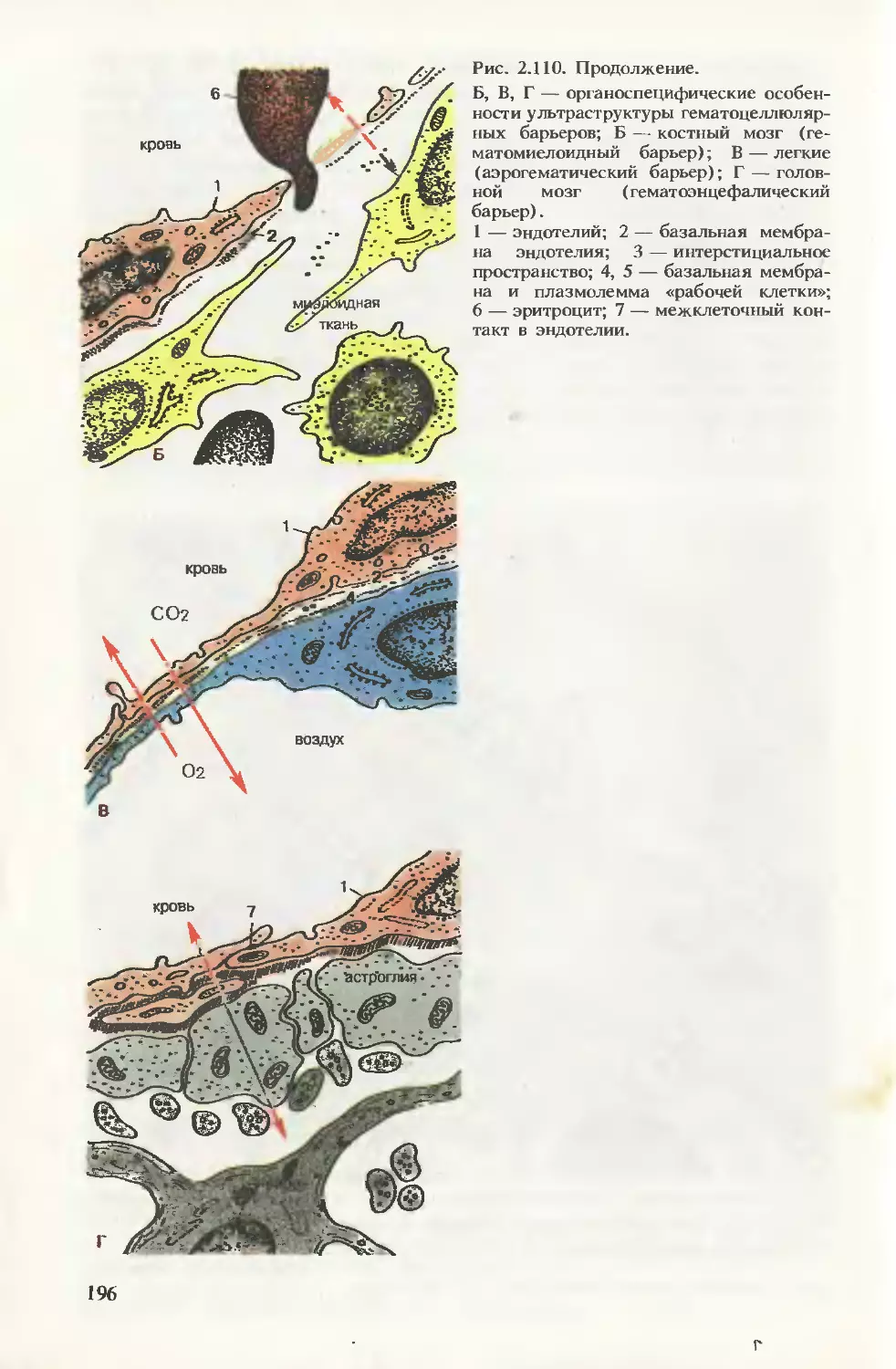
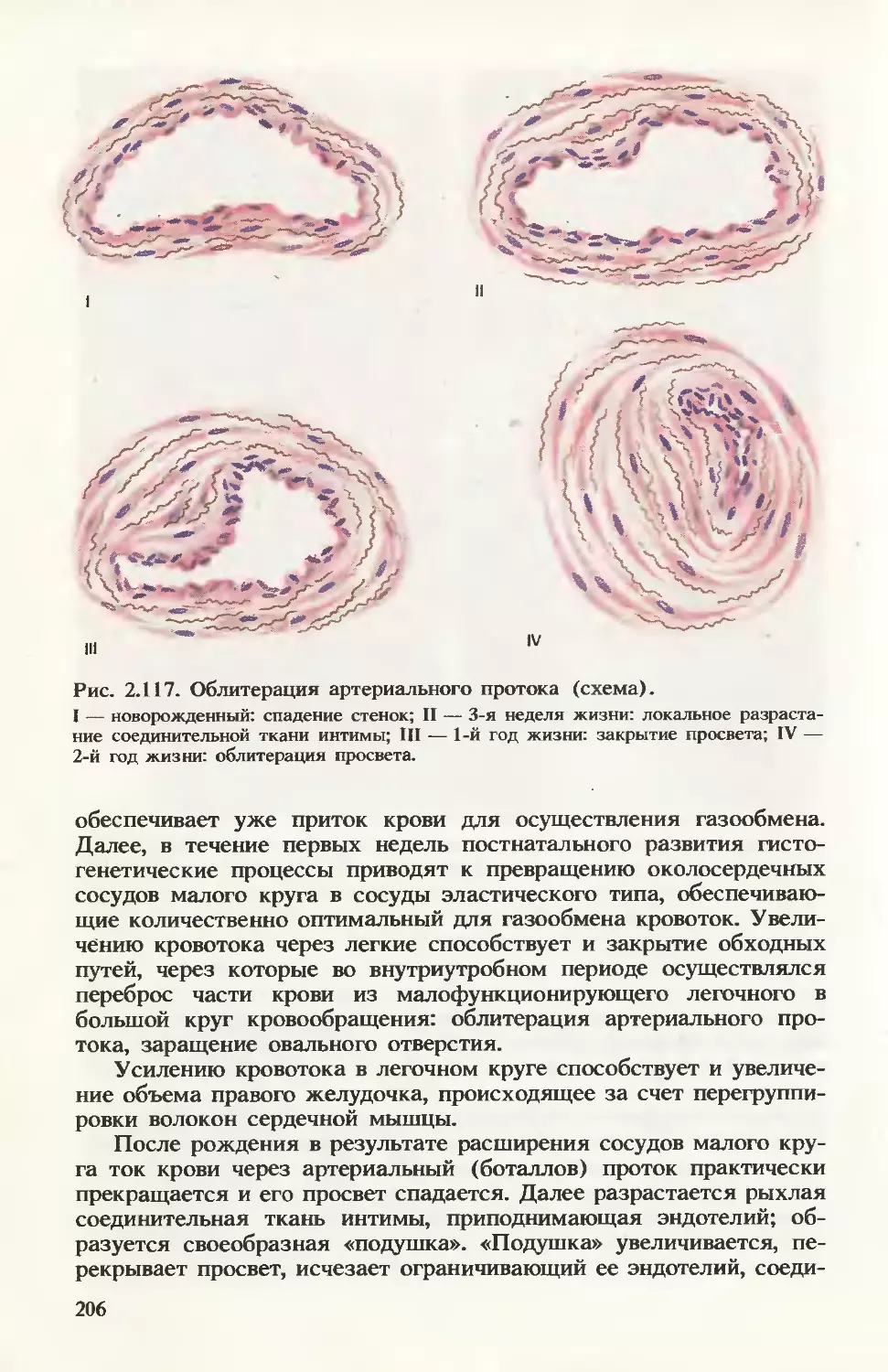
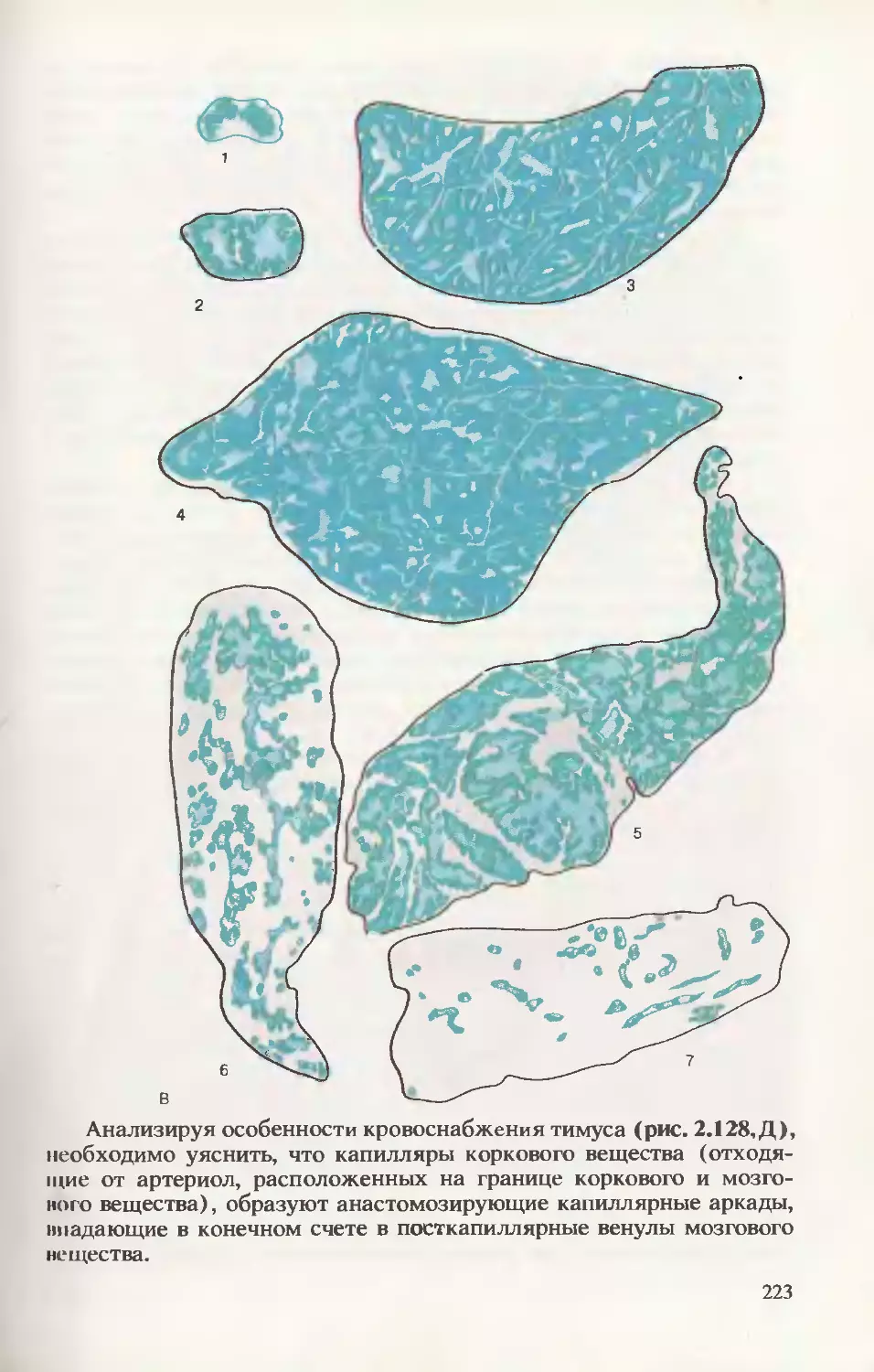
/






























































































































































































































































































































































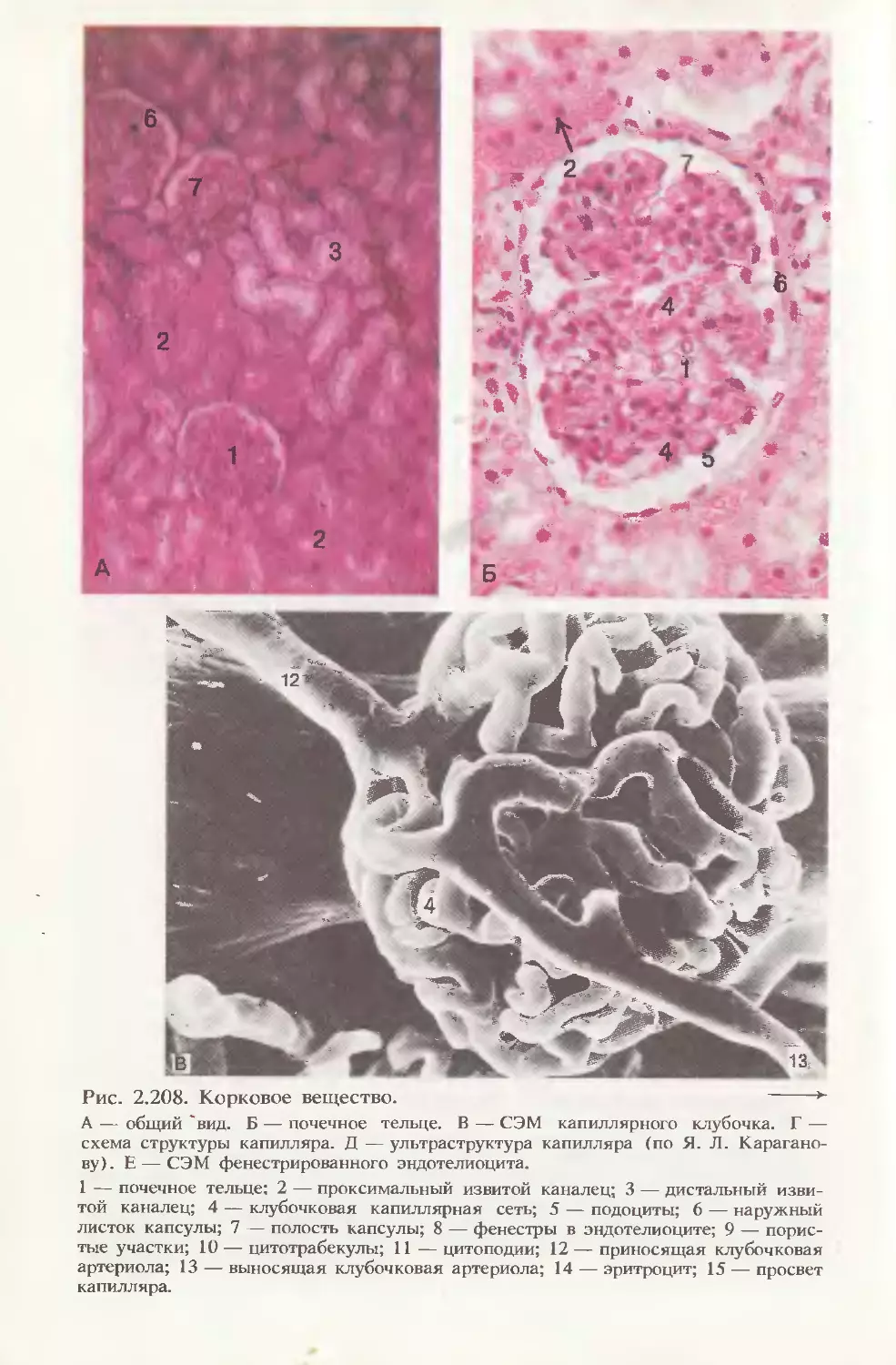





























































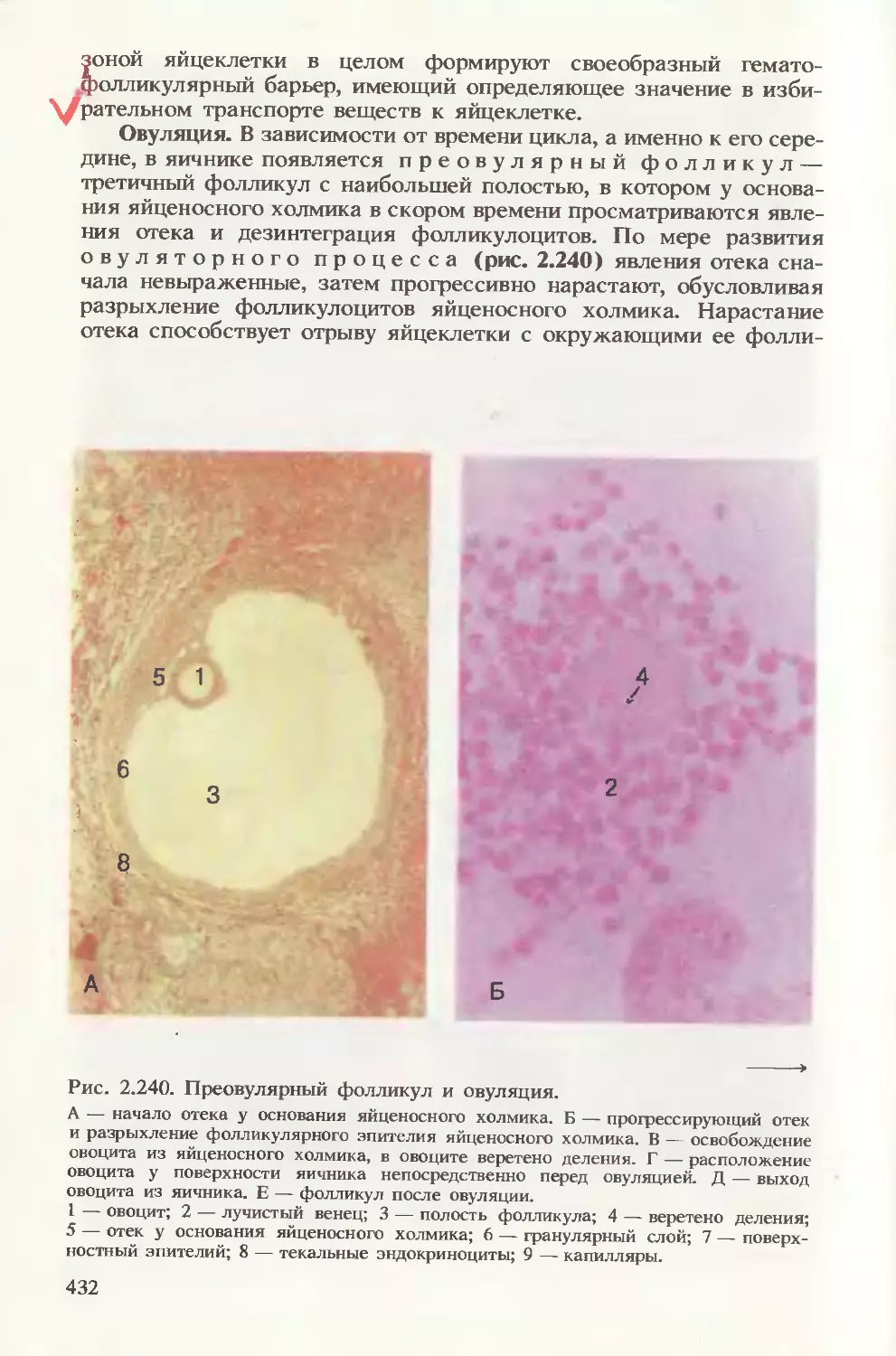



























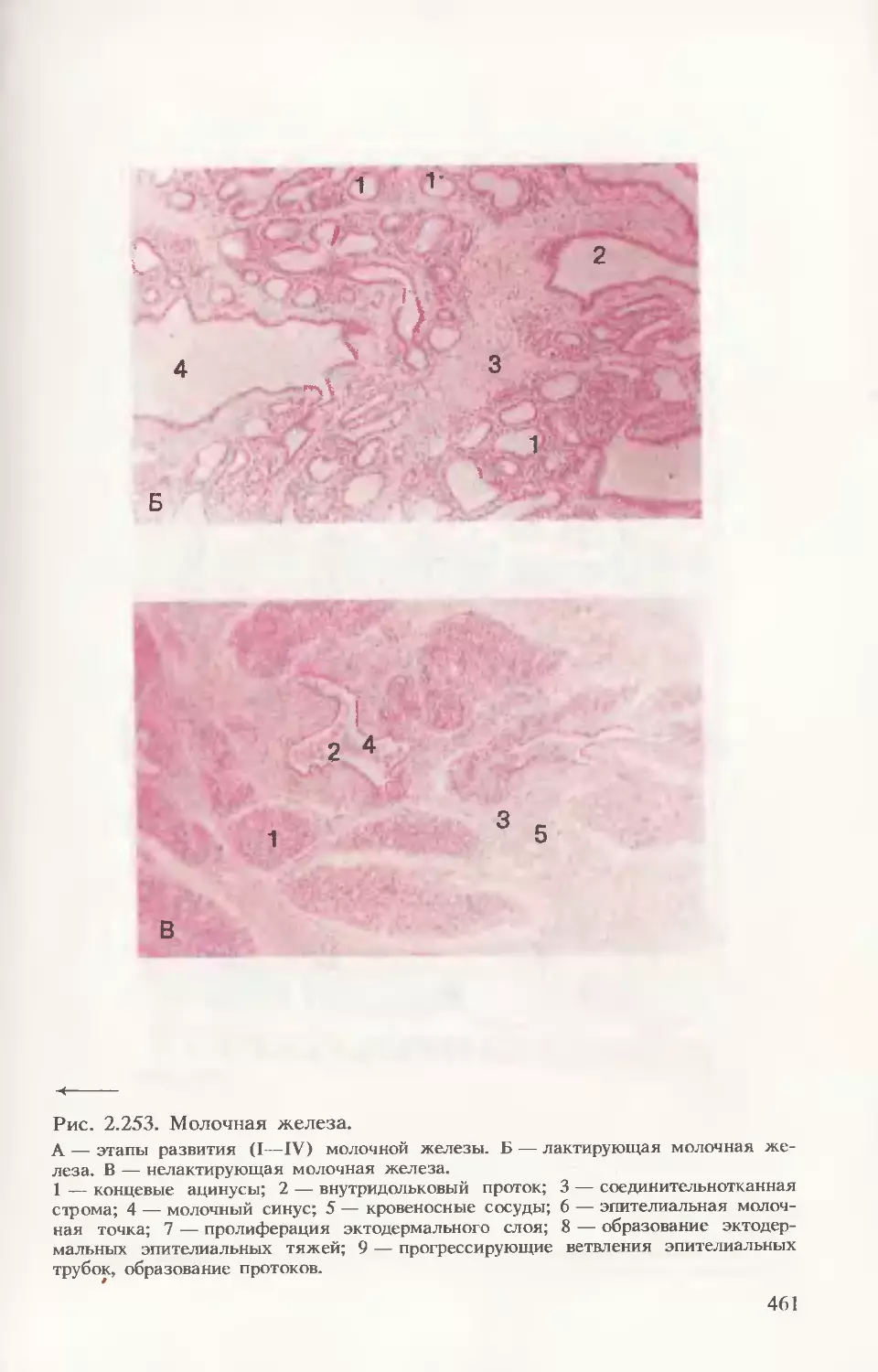














































































Автор: Волкова О.В. Елецкий Ю.К.
Теги: биология физиология гистология цитология учебное пособие эмбриология
Год: 1996




Текст
гистология,
цитология
И ЭМБРИОЛОГИЯ
АТЛАС
Под редакцией
О. В. ВОЛКОВОЙ, Ю. К. ЕЛЕЦКОГО
Рекомендовано Управлением учебных заведений
Министерства здравоохранения и медицинской промышленности
Российской Федерации в качестве учебного пособия
для студентов медицинских вузов
---, . , ....
Ь JJIHOTECI
2Г€ ^йосковскггч »
I X Лг Т .*>14
Москва «Медицина» 1996
Предисловие
Атлас является своеобразным руководством для самостоятель-
ной творческой работы студентов по идентификации конкретных
объектов и их структурных компонентов на микроскопическом и
субмикроскопическом уровне. Эффективная и плодотворная работа
с атласом предполагает исходное знание общих структурно-функ-
циональных основ изучаемого объекта на всех уровнях его орга-
низации — субклеточном, клеточном, тканевом, органном, а также
закономерности развития и возрастные особенности.
Так, анализируя строение клетки, необходимо четко представ-
лять, благодаря какой именно особенности структурной органи-
зации каждый из ее элементов выполняет конкретную функцию,
совокупность которых обеспечивают свойства клетки как элемен-
тарной единицы живого организма. Изучая строение тканей,
важно усвоить, какие специализированные структурные компо-
ненты клеток и их производных обеспечивают функциональные и
биологические свойства каждой группы тканей и их конкретных
разновидностей. При анализе органов и образуемых ими систем
следует определить их тканевый состав, выделить ведущую ткань,
обеспечивающую выполнение специфической функции, и ткани,
обеспечивающие вспомогательные функции — трофическую, за-
щитную и формообразовательную. Материал атласа дает представ-
ление об организации структурно-функциональной единицы орга-
на, особенностях его иннервации и васкуляризации.
Разделы эмбриологии (сравнительной и человека) требуют
предварительного знания динамики морфофункциональных пере-
строек в процессе исторического и индивидуального развития кле-
ток, тканей, органа и организма.
Достижению поставленной цели способствуют, во-первых, сис-
тематизированное изложение и расположение иллюстративного
материала и, во-вторых, тесно взаимосвязанный с ним текстовой
материал. Там, где необходимо, дается обобщающая схема или
рисунок, изображающий общий план организации, топографию и
масштаб составных элементов органа, затем рисунок или микро-
фотография с препарата, носящий обзорный характер и отража-
ющий картину, видимую при малом увеличении микроскопа, далее
представлены фрагменты, освещающие тонкое строение наиболее
3
важных структур. Помимо иллюстраций, отражающих светоонги
ческую микроскопическую характеристику изучаемых объектов, и
атласе приведены фотографии, полученные путем трансмиссион
ной электронной микроскопии структур (ТЭМ), позволяющие
представить и понять ультраструктурное обеспечение специфи
ческих функций. Дан также материал по результатам сканирую
щей электронной микроскопии (СЭМ), демонстрирующий прост-
ранственное изображение изучаемых структур, а также обобщи -
ющие схемы и рисунки.
В целях систематизации иллюстративного материала рисунки
объединены в блоки с общими подрисуночными подписями.
Поскольку большинство гистологических препаратов окрашено
гематоксилином-эозином, в подрисуночных подписях указано
в основном окрашивание специальными методами.
цитология
БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
КЛЕТОЧНАЯ ОБОЛОЧКА
ЦИТОПЛАЗМА
ЯДРО
Цитология — наука о структуре и функции клетки. Современ-
ная многоплановость направлений в их изучении послужила осно-
ванием для возникновения по существу новой науки — биологии
клетки. Клетка — элементарная структурно-функциональная и ге-
нетическая единица организма, составляющая основу его жизне-
деятельности и обладающая всеми признаками живого: раздражи-
мостью, возбудимостью, сократимостью, обменом веществ и энер-
гии, хранением генетической информации и передачей ее в ряду
поколений (способностью к размножению).
В данном разделе будут кратко изложены сведения о принци-
пах структурно-функциональной организации эукариотической жи-
вотной клетки. В составе последней выделяют три относительно
самостоятельных отдела: оболочку (поверхностный аппарат),
цитоплазму и ядро (ядерный аппарат).
Большинство структур клетки состоит из биологических
мембран. Мембраны — тончайшие пластичные образования, разли-
чимые только при электронной микроскопии; они не имеют сво-
бодных концов и не образуются заново, а строятся на предсуще-
ствующих мембранах. Основные химические компоненты мемб-
ран — липиды и белки, некоторые мембраны содержат угле-
воды.
Принцип структурной организации всех мембран един (рис. 1.1, А).
Они имеют липидную основу, представленную двумя слоями соот-
ветствующим образом ориентированных молекул липидов. Липид-
ный бислой выполняет две основные функции: барьерную (для
ионов и водорастворимых молекул) и матричную (для размещения
различных белков). Мембранные белки «плавают» в бислое липи-
дов. Основные функции белков — энзиматическая, рецепторная,
транспортная и ряд других.
1.1. Клеточная оболочка (поверхностный аппарат)
и цитоскелет
Поверхностный аппарат осуществляет регуляцию взаимоотноше-
ний клетки с окружающей средой, активно участвует в межкле-
точных взаимодействиях. Клеточная оболочка выполняет следу-
ющие основные функции: барьерную, транспортную и рецептор-
ную. В ее составе различают три компонента: плазматическую
мембрану, надмембранный и подмембранный слои.
Плазматическая мембрана (плазмолемма) в отличие от внутри-
клеточных мембран утолщена и богата холестерином, гликолипи-
дами, гликопротеинами (рис. 1.1, Б). Для белковых компонентов
плазмолеммы характерно присутствие различных в функциональ-
ном отношении белков. Они находятся в разных пространствен-
ных взаимоотношениях с молекулами липидного бислоя, что отра-
жается на прочности их связей (см. рис. 1.1).
Надмембранный слой (гликокаликс) имеет нежную фибрил-
лярную структуру. В его состав входят периферические белки и
।
6
4
Рис. 1.1. Строение биологической мембраны (А) и клеточной обо-
лочки (Б).
1 — молекула липида; 2 — липидный бислой; 3 — интегральные белки; 4 — пери-
ферические белки; 5 — полуинтегральные белки; 6 — гликокаликс; 7 — субмем-
бранный слой; 8 — актиновые микрофиламенты; 9 — микротрубочки; 10 — проме-
жуточные филаменты; 11 — углеводные молекулы гликопротеинов и гликолипидов.
7
1
A
Рис. 1.2. Микротрубочки и фила-
менты.
А — схема. Б — микротрубочки в клет-
ках эпителия кишечника (ТЭМ, препа-
рат К. А. Зуфарова). В — промежуточ-
ные филаменты (метод иммунофлю-
оресценции, препарат С. М. Светлич-
ной). Г — промежуточные филаменты
(СЭМ). 1 — актиновая микрофиламен-
та; 2 — микротрубочка; 3 — промежу-
точная филамента.
8
углеводные компоненты гликолипидов и гликопротеинов плазмо-
леммы, частично интегральных белков (см. рис. 1.1, Б), а так-
же различные соединения, адсорбированные клеточной поверх-
ностью. Существенно, что в разных клетках организма интенсив-
ность развития надмембранного слоя, его химический состав, а
также функции неодинаковы. Гликокаликс играет важную роль во
взаимоотношениях клеток с окружающей средой и друг с другом,
участвуя в процессах рецепции, адсорбции, в ферментном рас-
щеплении различных соединений и во многих других про-
цессах.
Подмембранный слой, представленный узким участком цито-
плазмы, примыкает к плазмолемме с внутренней стороны. Гиало-
плазма в этой области более вязкая и практически не содержит
органелл. Здесь сосредоточены структурные элементы опорно-
сократительного аппарата клетки — цитоскелета.
Цитоскелет сформирован тремя основными компонентами,
состоящими из разных белков: микротрубочками, микрофиламен-
тами, промежуточными филаментами (рис. 1.2).
Микротрубочки — полые цилиндры, стенка которых состоит из
фибрилл, сформированных молекулами глобулярного белка тубу-
лина. Микрофиламенты — тонкие нити; различают миозиновые и
актиновые микрофиламенты. Микротрубочки и микрофиламен-
ты в основном обеспечивают двигательные процессы в клетке,
а также участвуют в выполнении опорной функции. Промежу-
точные филаменты состоят из белковых нитей, обладающих
высокой прочностью и стабильностью; для их белкового состава
характерна тканевая специфичность. Промежуточные филаменты
выполняют в клетке только опорную функцию.
9
В последнее время выделяют четвертый компонент цитоскеле-
та — систему так называемых тонких филаментов, которая обес-
печивает взаимосвязь основных компонентов цитоскелета;—^Тон-
кие» филаменты пронизывают всю цитоплазму клетки, формируя
особую микротрабекулярную решетку, и, возможно, участвуют в
передаче сигнала от поверхности клетки к генетическому аппа-
рату ядра.
Цитоскелет обеспечивает тургор клетки, поддержание и изме-
нение ее формы, перемещение в пространстве, а также двигатель-
ные процессы внутри клетки.
Описанные три части клеточной оболочки (плазмолемма, над-
и подмембранные слои) взаимосвязаны и работают как единая
система, реализуя различные сложноорганизованные функции:
рецепции, транспорта веществ, образования межклеточных кон-
тактов, передачу сигналов от поверхности в глубь клетки. Через
клеточную оболочку осуществляется высокоизбирательный тран-
спорт веществ, идущий в двух направлениях: в клетку и из нее.
Проницаемость плазмолеммы для воды и ионов обеспечивает
система особых трансмембранных каналов, стенка которых обра-
зована белками.
Другой способ переноса веществ — транспорт в мембранной
«упаковке» — предназначен для поступления и выделения макро-
молекул и различного размера частиц. В зависимости от размера
частиц и механизмов, обеспечивающих поступление веществ в клет-
ку (эндоцитоз), выделяют следующие разновидности этого про-
цесса: фагоцитоз (рис. 1.3), макропиноцитоз и микропиноцитоз,
а также эндоцитоз, опосредованный рецепторами. Последний, в
отличие от других, обусловливает высокую специфичность поступ-
ления веществ. Выделение макромолекул и структур из клетки
называется экзоцитозом.
В некоторых специализированных клетках возникает необхо-
димость морфофункциональной специализации различных отделов
клеточной оболочки, что приводит к образованию соответству-
ющих постоянных структур (рис. 1.4). Так, в связи с интенсифи-
кацией транспортных процессов в определенных участках оболоч-
ки формируются многочисленные выросты, увеличивающие пло-
щадь клеточной поверхности, — микроворсинки и базальные инва-
гинации. Для перемещения образований, расположенных вне
клетки (вблизи ее клеточной оболочки), на последней появляются
реснички (см. главу 2, рис. 2.2, Г).
Сложные изменения в строении клеточной оболочки наблю-
даются при построении постоянных специализированных меж-
клеточных контактов (см. рис. 1.4). По функциональ-
ному значению эти контакты можно разделить на изолирующие,
механические и химические.
Примером контактов изолирующего типа является плотный
контакт (зона слипания). В этом случае изоляция межклеточной
щели от окружающего клетку пространства осуществляется путем
«слипания» глобул интегральных белков мембран соседних клеток.
10
Рис. 1.3. Фагоцитоз (ТЭМ, препарат Л. М. Лихачевой).
1 — цитоплазма фагоцитирующей клетки; 2 — фагоцитированный субстрат (эри-
троцит) .
К механическим контактам относятся сравнительно просто
устроенные образования — пальцевидные соединения (боковые
инвагинации) и более сложно организованные — десмосомы.
В первом случае осуществляется взаимодействие только наружных
слоев клеточной оболочки (гликокаликса), а в области десмосомы
между мембранами соседних клеток из компонентов гликокаликса
формируется центральная пластинка, которая связана с мембрана-
ми контактирующих клеток системой поперечных фибрилл.
Со стороны подмембранного слоя десмосома укреплена при помо-
щи компонентов цитоскелета. В зависимости от протяженности
и особенностей строения различают точечные и опоясывающие
десмосомы, располагающиеся в виде полосок на всем протяжении
контактирующих поверхностей соседних клеток.
К химическим контактам относится щелевой контакт, или
нексус. В области нексуса межклеточное пространство становится
очень узким (щелевидным) и может быть выявлено только при
специальной обработке ультратонких срезов. В составе контакти-
рующих мембран, в области нексуса симметрично расположены
интегральные белки, связанные между собой. При участии белко-
11
1
3
Рис. 1.4. Специализация клеточной обо-
лочки (частично по А. А. Заварзину,
А. Д. Харазовой, 1991).
А — схема. Б — верхушечная и боковая по-
верхности клеток кишечного эпителия (ТЭМ,
препарат К. А. Зуфарова). В — базальные ин-
вагинации плазмолеммы (ТЭМ). I — ресничка;
II — микроворсинки; III — плотный контакт;
IV — десмосома; V — полудесмосома; VI — бо-
ковые инвагинации; VII — щелевой контакт;
VIII — базальные инвагинации.
1 — плазмолемма; 2 — глобулы интегральных
белков; 3 — микрофиламенты; 4 — тонофиб-
риллы; 5 — коннексоны; 6 — митохондрии.
вых глобул формируются межцитоплазматические каналы, по
которым возможен транспорт низкомолекулярных веществ и ионов
из одной клетки в другую.
1.2. Цитоплазма
Цитоплазма состоит из различных структур (органеллы, включе-
ния, компоненты цитоскелета) и основной цитоплазмы (гиало-
плазмы), где осуществляются реакции промежуточного обмена.
По современным представлениям, гиалоплазма весьма неоднород-
ный компартмент клетки; около мембранных и немембранных
структур цитоплазмы формируется особая субсистема — цито-
золь. Его состав в разных органеллах неодинаков.
Основная функция цитоплазмы — обеспечение обмена веществ
и энергии. Для достижения высокой эффективности различных
метаболических процессов вся территория цитоплазмы разделена
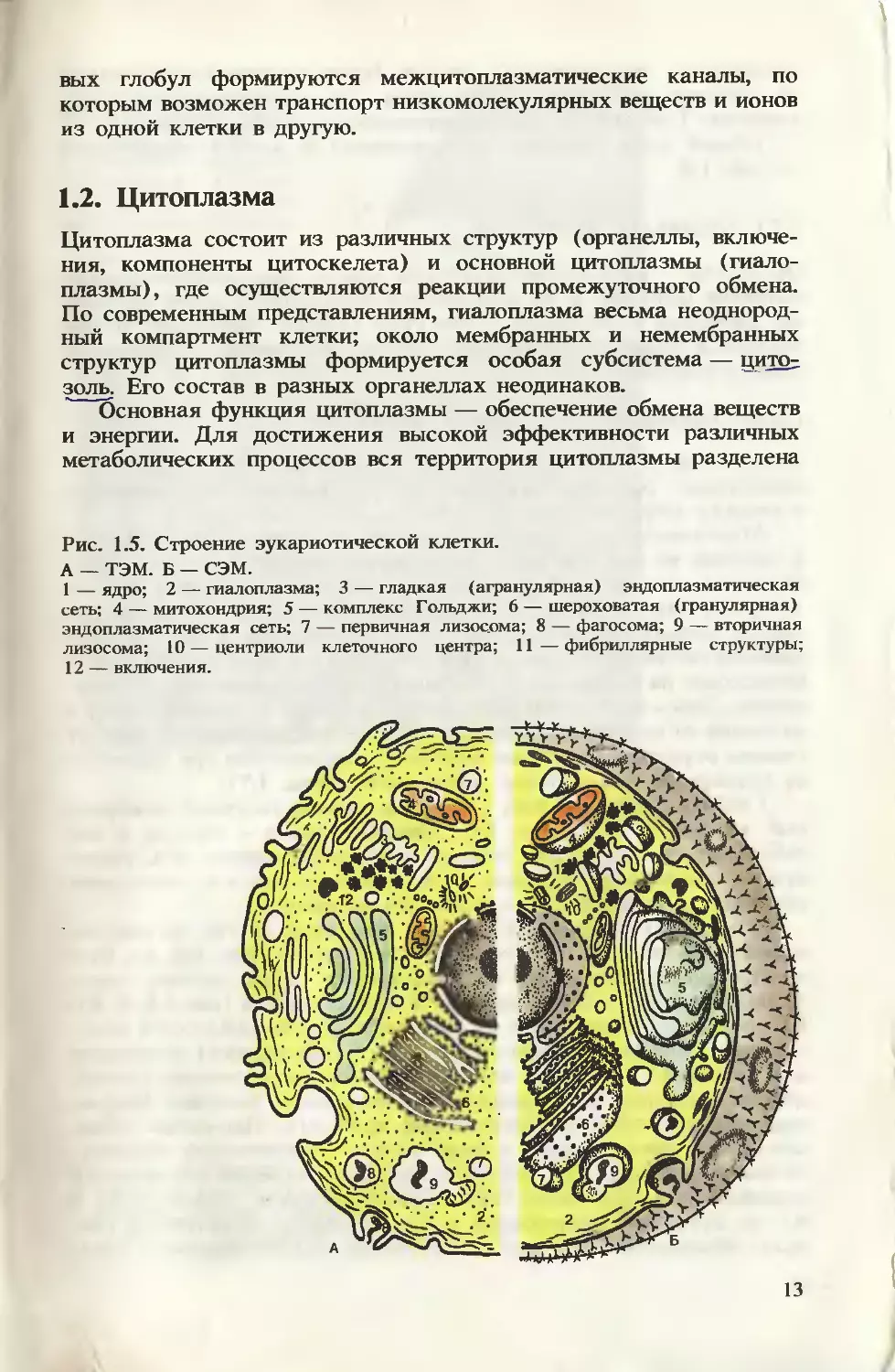
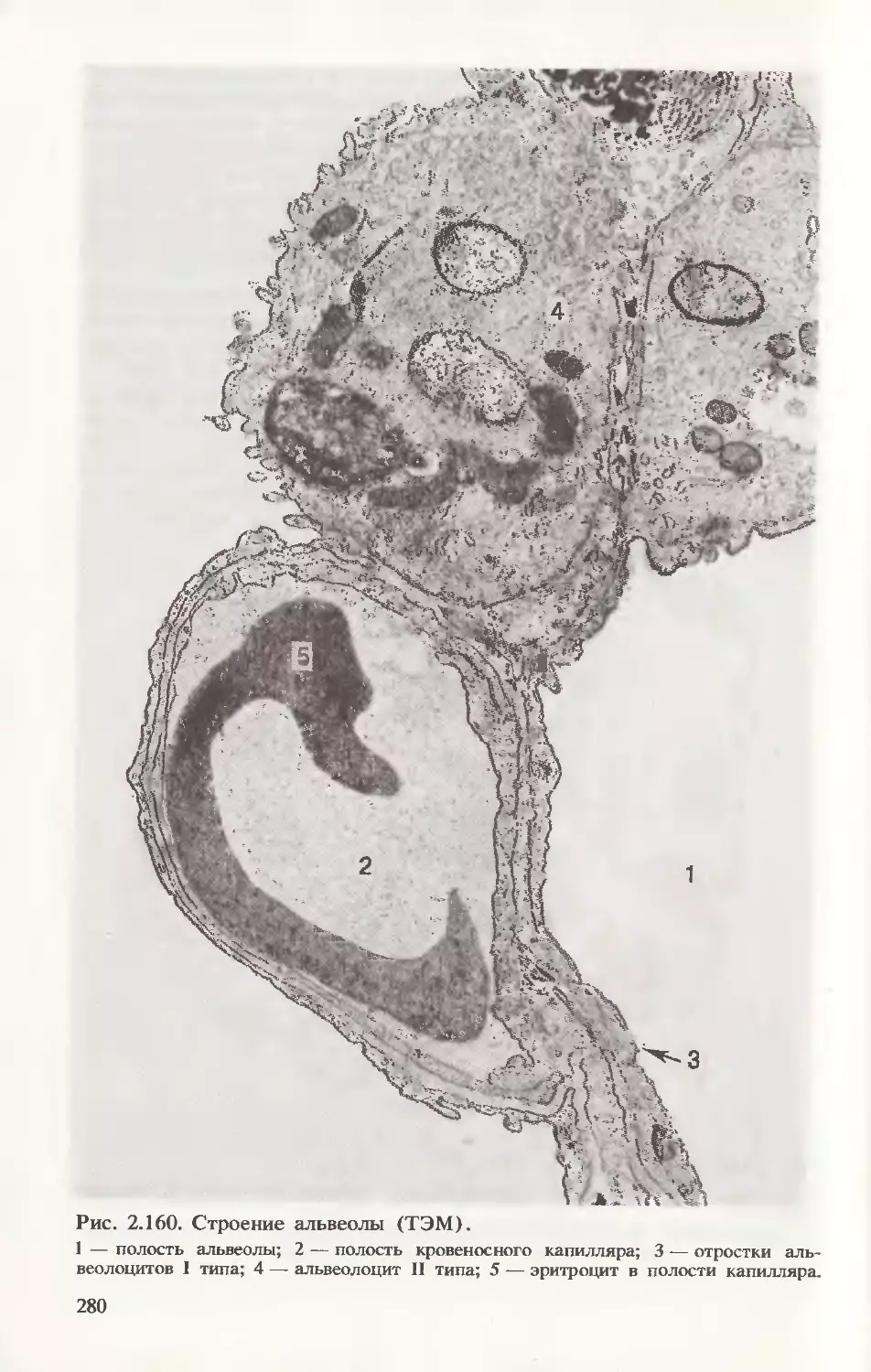
Рис. 1.5. Строение эукариотической клетки.
А — ТЭМ. Б — СЭМ.
1 — ядро; 2 — гиалоплазма; 3 — гладкая (агранулярная) эндоплазматическая
сеть; 4 — митохондрия; 5 — комплекс Гольджи; 6 — шероховатая (гранулярная)
эндоплазматическая сеть; 7 — первичная лизосома; 8 — фагосома; 9 — вторичная
лизосома; 10 — центриоли клеточного центра; 11 — фибриллярные структуры;
12 — включения.
13
мембранами на замкнутые отсеки (компартменты), основными
из которых являются гиалоплазма, эндоплазматическая сеть,
комплекс Гольджи, лизосомы, пероксисомы и митохондрии.
Общий план строения эукариотической клетки представлен
на рис. 1.5.
1.2.1. Органеллы цитоплазмы
Органеллы — постоянные структуры клетки, выполняющие опре-
деленные функции. В их образовании принимают участие мембра-
ны, рибосомы и фибриллы. Далее будут описаны органеллы, встре-
чающиеся практически во всех клетках организма, — органеллы
общего значения.
1.2.1.1. Органеллы, состоящие из мембран
Эндоплазматическая сеть (ЭПС). Существует две морфофунк-
циональные разновидности ЭПС — шероховатая (гранулярная)
и гладкая (агранулярная).
Шероховатая ЭПС (рис. 1.6) состоит из мембранных канальцев
и цистерн, на наружной стороне которых фиксированы рибосомы.
Основная функция этой органеллы — синтез различных белков
(секреторных, мембранных, специальных белков для некоторых ор-
ганелл, например лизосом, митохондрий), участие в посттрансля-
ционной обработке белков. Кроме шероховатой ЭПС, синтез белков
происходит на рибосомах (полисомах), свободно лежащих в гиало-
плазме. Здесь в основном синтезируются белки, не нуждающиеся в
изоляции от основной цитоплазмы. От количества рибосом зависит
степень окраски цитоплазмы основными красителями при подготов-
ке препаратов для световой микроскопии (рис. 1.7).
Гладкая ЭПС (см. рис. 1.5) представлена системой мембран-
ных канальцев и везикул. Ее главная функция — участие в ме-
таболизме углеводов. Как гладкая, так и шероховатая ЭПС участ-
вуют в метаболизме липидов, а также в изоляции и в накоплении
синтезируемых продуктов в полости канальцев.
Комплекс Гольджи (КГ), как и некоторые другие органеллы,
может быть выявлен на светооптическом уровне (рис. 1.8, А). При
электронной микроскопии он представляет собой систему упло-
щенных цистерн, трубочек, вакуолей и мелких везикул (рис. 1.8, Б, В).
КГ отчетливо поляризован по вертикали. Это выражается в нали-
чии двух полюсов: со стороны выпуклого (цис-полюса) поступают
вещества в КГ в составе везикул; со стороны вогнутого (транс-
полюса) «уходят» из КГ вещества в мембранной упаковке (напри-
мер, отшнуровываются секреторные гранулы). Цис-полюс обра-
щен, как правило, к ядру, а транс-полюс — к клеточной оболочке.
Основные функции КГ: участие в процессах секреции (химическая
модификация первичного секреторного продукта, пришедшего в
КГ из ЭПС, его конденсация и формирование секреторных гра-
нул); образование лизосом; «сортировка» синтезированного клет-
14
Рис. 1.6. Шероховатая
(। ранулярная) эндо-
плазматическая сеть.
Л — схема. Б — ТЭМ.
I полость цистерны;
2 рибосома: а — боль-
шая субъединица рибосо-
мы; б — малая субъедини-
ца рибосомы; 3 — инфор-
мационная РНК; 4 — син-
ечируемая молекула бел-
ка; 5 — молекула белка в
>росвете цистерны.
Рис. 1.7. Цитоплазма (окраска основными
и кислыми красителями — гематоксилином
и эозином; рисунок с препарата).
I клетка с базофильной цитоплазмой; 2 —
клюка с оксифильной цитоплазмой.
15
s * vx **•*
Рис. 1.8. Комплекс Гольджи.
А — окраска — импрегнация солями
серебра. Б — схема. В — ТЭМ (пре-
парат Т. Б. Айзенштадт).
1 — цистерны; 2 — транспортные
везикулы; 3 — секреторные гранулы;
4 — лизосома.
16
кой белка (секреторного, лизосомального и т. д.); синтез полиса-
харидов; обновление клеточной оболочки.
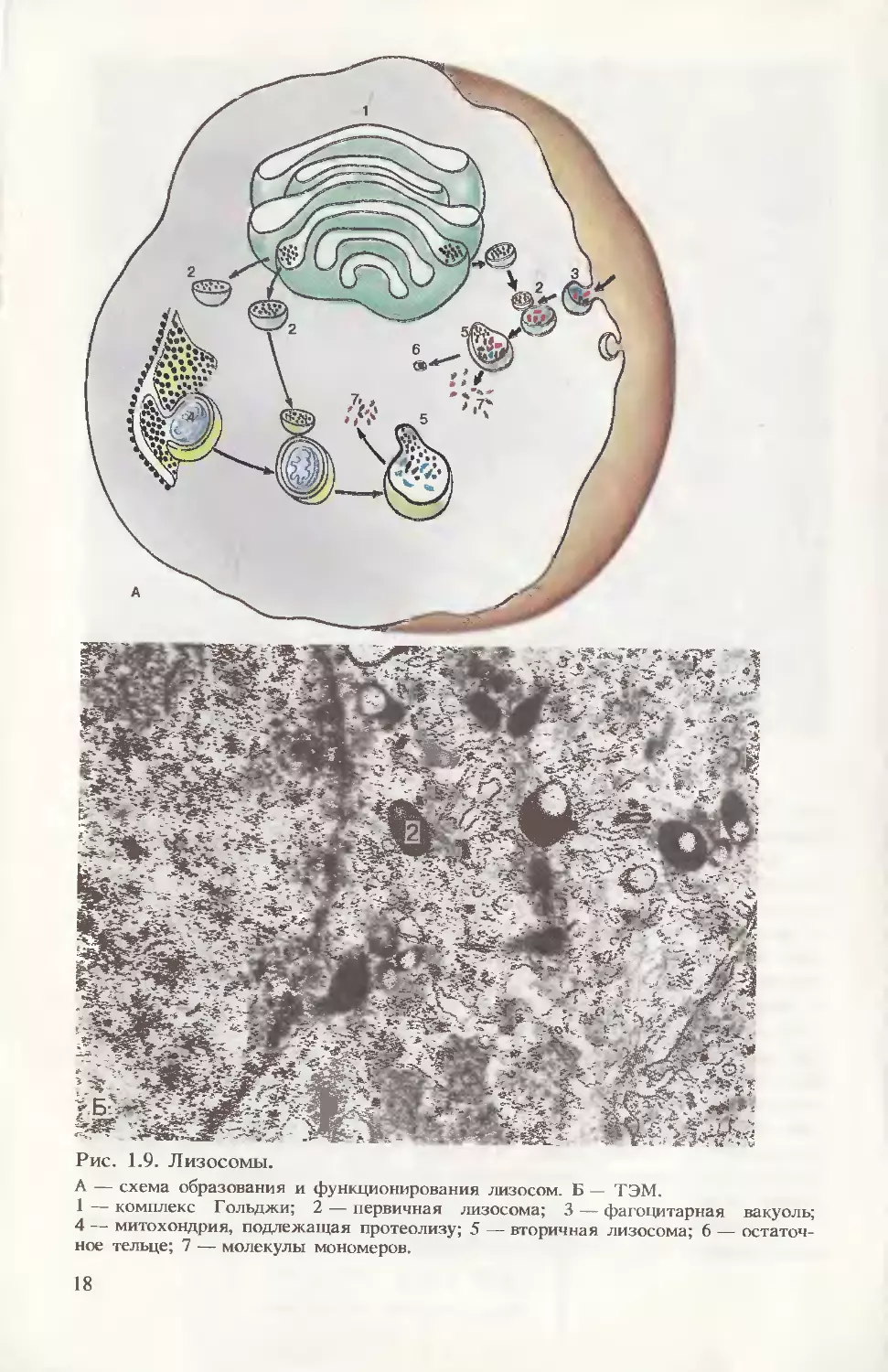
Лизосомы при ТЭМ имеют вид пузырьков, ограниченных
мембраной (рис. 1.9). В состав их плотного гомогенного содер-
жимого (матрикса) входят гидролитические ферменты, способные
к расщеплению различных биополимеров до мономеров. Описан-
ные структуры называются первичными лизосомами, их ферменты
инактивированы. После слияния содержимого первичных лизосом
с вакуолями, образовавшимися в процессе эндоцитоза, формиру-
ется вторичная лизосома (гетеролизосома). В ее полости происхо-
дят активация ферментов и лизис поступивших в клетку веществ.
По такому же принципу может происходить гидролиз внутрикле-
точных структур; при этом вторичная лизосома именуется ауто-
лизосомой (см. рис. 1.9, А). В случае неполного расщепления
компонентов, подлежащих гидролизу, образуются так называемые
остаточные тельца, содержимое которых выводится из клетки
путем экзоцитоза. ____________ , „„ ......
Пероксисома пред тавляет^;^<^»й-.^у^ырек, {ограниченный
мембраной; в его соста, е может*вЙяшгятйё‘Я'‘криста;£тическая (или
2-г« ГА о ск о век :г$
Рис. 1.9. Лизосомы.
А — схема образования и функционирования лизосом. Б — ТЭМ.
1 — комплекс Гольджи; 2 — первичная лизосома; 3 — фагоцитарная вакуоль;
4 — митохондрия, подлежащая протеолизу; 5 — вторичная лизосома; 6 — остаточ-
ное тельце; 7 — молекулы мономеров.
18
Рис. 1.10. Пероксисома (указана
стрелкой), ТЭМ.
аморфная) сердцевина (рис. 1.10). Содержащиеся в пероксисомах
ферменты участвуют в окислении субстратов с образованием перок-
сида водорода, который тут же используется для окисления дру-
гих, в том числе вредных для организма, веществ (детоксицирую-
щая функция). Избыток пероксида водорода разрушается также
ферментативным путем.
Митохондрии в отличие от других органелл состоят из двух
мембран (наружной и внутренней), разделенных узкой щелью, и
матрикса (рис. 1.11). Внутренняя мембрана имеет выросты (кри-
сты) в форме пластин, в некоторых специализированных клет-
ках в форме трубок. Внутренняя мембрана богата ферментами
и обладает избирательноиТтриЦицаемостью. Наружная^мембрана
гладкая, бедна ферментами. Матрикс содержит ДНК, РНК, амино-
кислоты, белки и другие вещества. В его составе есть рибосомы,
встречаются конкреции, состоящие из солей кальция. Главная
функция митохондрий — обеспечение энергетического обмена
(здесь образуются макроэргические соединения). Именно поэтому
митохондрии, как правило, располагаются в тех участках клет-
ки, где активно используется энергия. Кроме того, митохондрии
участвуют в водно-солевом, а также (в некоторых клетках) в
липидном обмене; выполняют белоксинтезирующую функцию.
При этом надо подчеркнуть^ что большая часть митохондри-
альных белков синтезируется в гранулярной эндоплазматической
сети.
19
Рис. 1.11. Митохондрии с пластинчатыми (А, Б) и трубчатыми (В, Г)
кристами.
1 — наружная мембрана; 2 — межмембранное пространство; 3 — внутренняя мем-
брана; 4 — кристы; 5 — матрикс.
20
1.2.1.2. Органеллы, состоящие из фибрилл
Клеточный центр (рис. 1.12) — органелла, образованная микро-
трубочками. Главным элементом клеточного центра являются
центриоли. Микротрубочки в составе центриоли располагаются по
периметру полого цилиндра в виде триплетов. В период интерфа-
1ы в клетке содержатся две центриоли, расположенные под углом
друг к другу (рис. 1.12, А). Основная функция этой органеллы —
формирование полюсов митотического деления.
1.2.2. Включения
Включения — непостоянные структуры цитоплазмы, наличие или
отсутствие которых связано с особенностями клеточного метабо-
лизма. Они могут состоять из химических веществ различной
природы: липидов (рис. 1.13), углеводов (рис. 1.14), белков, вита-
минов и др. Содержимое включений используется клеткой по-
разному; например, как источник энергии и питательных веществ
(трофические включения) включения могут временно накапливать-
ся в клетке для последующего их выделения и использования в
интересах организма (секреторные включения). В виде включений
в цитоплазме могут быть витамины, пигменты и другие вещества.
Рис. 1.12. Центриоли клеточ-
ного центра.
А — ТЭМ; Б — фрагмент (боль-
шое увеличение) центриоли.
1 — продольный срез; 2 — по-
перечный срез: а — трипле-
ты микротрубочек (препарат
Т. Б. Айзенштадт).
21
Рис. 1.13. Жировые включения.
А — окраска Суданом черным. Б —
ТЭМ.
1 — жировые включения.
22
Рис. 1.14. Включения гликогена.
А — ШИК-реакция. Б — ТЭМ.
1 — включения гликогена.
23
1.3. Ядро (ядерный аппарат)
Функциональное значение ядерного аппарата определяется осо-
быми свойствами молекулы ДНК — ее способностью к реплика-
ции (удвоению) и транскрипции. Эти свойства ДНК обеспечивают
основные функции ядра: хранение и передача в ряду поколений
клеток наследственной информации и регуляция клеточного ме-
таболизма (ядро — метаболический центр клетки). Ядро представ-
лено следующими компонентами: ядерной оболочкой, хроматином,
ядрышком, кариоплазмой (рис. 1.15, А, Б). При световой микро-
скопии и соответствующей окраске выявляется базофилия ядра
(см. рис. 1.15, А).
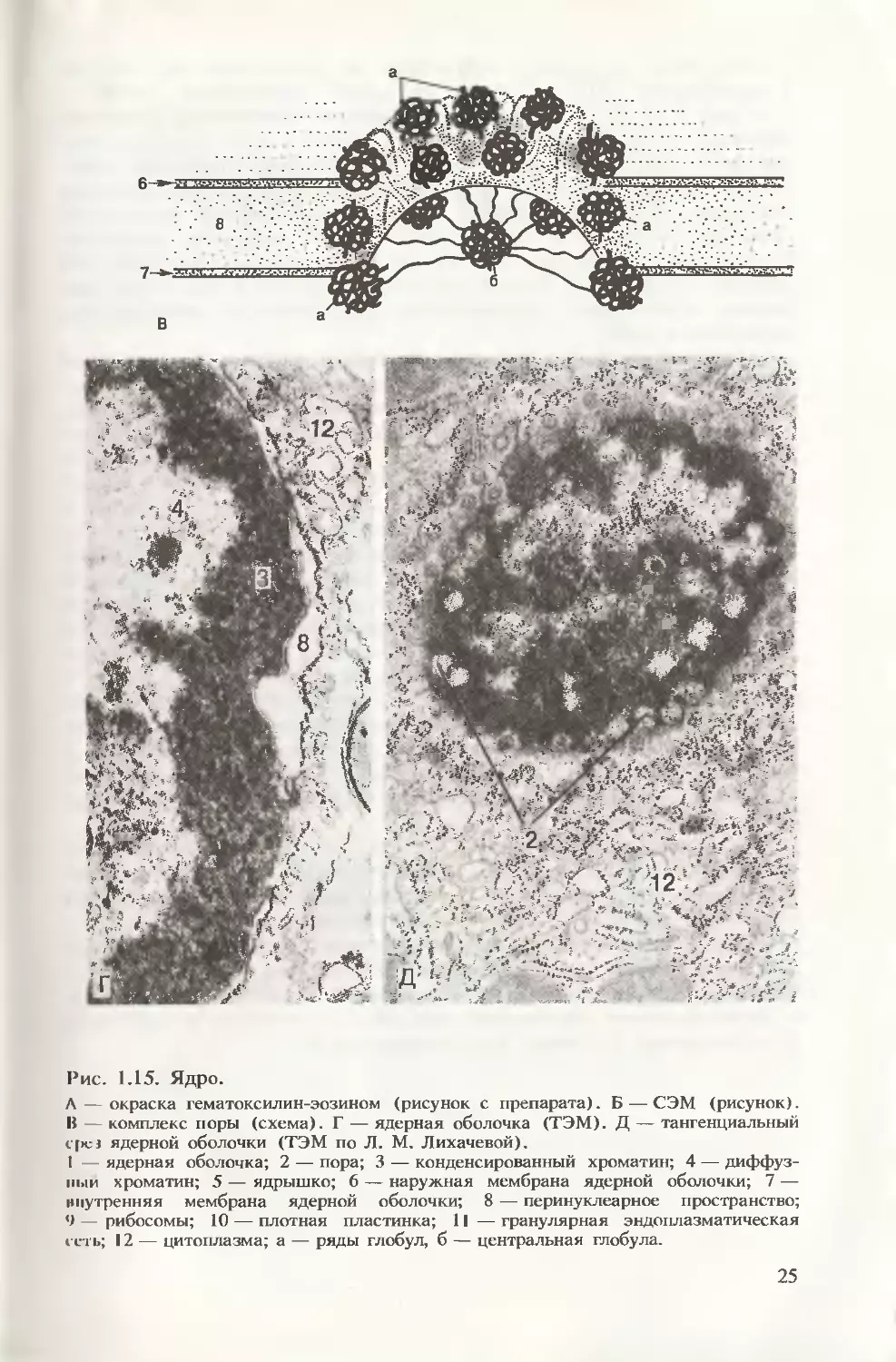
Ядерная оболочка отделяет содержимое ядра от цитоплазмы.
Это обеспечивает пространственное разделение процессов тран-
скрипции и трансляции и создает условия для процессинга. Основ-
ные функции ядерной оболочки: разграничительная и барьерная,
регуляция обмена веществ между ядром и цитоплазмой, участие
в синтезе белка и в обновлении цитоплазматических мембран.
В составе ядерной оболочки (рис. 1.15, Б—Д) выделяют следую-
щие структуры: две мембраны (наружную и внутреннюю), разде-
ленные перинуклеарным пространством, и поровый комплекс.
24
Рис. 1.15. Ядро.
Л — окраска гематоксилин-эозином (рисунок с препарата). Б — СЭМ (рисунок).
В — комплекс поры (схема). Г — ядерная оболочка (ТЭМ). Д — тангенциальный
с[>ез ядерной оболочки (ТЭМ по Л. М. Лихачевой).
1 — ядерная оболочка; 2 — пора; 3 — конденсированный хроматин; 4 — диффуз-
ный хроматин; 5 — ядрышко; 6 — наружная мембрана ядерной оболочки; 7 —
внутренняя мембрана ядерной оболочки; 8 — перинуклеарное пространство;
9 рибосомы; 10 — плотная пластинка; 11 — гранулярная эндоплазматическая
сеть; 12 — цитоплазма; а — ряды глобул, б — центральная глобула.
25
Наружная мембрана структурно и функционально связана
с мембранами ЭПС и имеет на своей поверхности рибосомы
(см. рис. 1.15, Б, Г). Внутренняя мембрана имеет связь с плотной
пластинкой. Последняя представляет собой густую сеть белковых
фибрилл, соединяющихся с аналогичными фибриллярными обра-
зованиями, лежащими по всей территории кариоплазмы. Плотная
пластинка и фибриллярная система кариоплазмы выполняют
опорную функцию, а также участвуют в регуляции процессов
репликации и транскрипции. Кроме того, плотная пластинка при
помощи специальных белков структурно связана с участками хро-
мосом, обеспечивая упорядочение расположения интерфазных
хромосом в ядре.
Поровый комплекс представлен отверстием (порой) в ядерной
оболочке, заполненной тремя кольцами белковых глобул, свя-
занных с центральной глобулой системой фибрилл (см. рис.
1.15, Б, Г). Через поровый комплекс осуществляется транспорт
макромолекул из ядра в цитоплазму и в обратном направлении.
Хроматин — совокупность интерфазных хромосом. Его основ-
ные химические компоненты — ДНК, белки (основные и кислые),
РНК. По морфофункциональной характеристике выделяют декон-
денсированный (эухроматин) и конденсированный (гетерохрома-
тин). Деконденсированный хроматин содержит транскрибируемую
ДНК, а конденсированный представлен в основном участками
ДНК, на которых не происходит процесс транскрипции. При ТЭМ
участки гетерохроматина видны отчетливо, а участки, занятые
эухроматином, трудно различимы (очень светлые) (см. рис. 1.15, Б, Г).
Ядрышко (см. рис. 1.15, Б) является местом образования
субъединиц рибосом. Здесь происходит сложный процесс синтеза
рибосомной РНК, ее созревание и формирование комплекса с
белками. При ТЭМ в составе ядрышка различают ядрышковый
хроматин, фибриллярные и гранулярные структуры, состоящие из
рибонуклеопротеидов.
Кариоплазма представляет собой относительно гомогенное
образование, имеющее сложный химический состав. Кариоплазма
формирует соответствующее микроокружение для структур ядра
и активно участвует в их функционировании.
В заключение следует обратить внимание на то, что описанные
основные отделы клетки (оболочка, цитоплазма и ядерный аппа-
рат) функционируют как единая целостная система и способны
к разнообразным изменениям при дифференцировке и адаптации
к меняющимся условиям жизнедеятельности.
гистология
2.1. Эпителиальные ткани
Из множества классификаций эпителиальных тканей наиболее
предпочтительной является классификация, делящая всю группу
по морфофункциональным признакам на две подгруппы — покров-
ный и железистый эпителий.
2.1.1. Покровный эпителий
Ведущим признаком для выделения данных подгрупп эпителиев
является их пограничность между внутренней средой организма
и ее окружением. Это нашло свое отражение в общем принципе
28
Bl
Рис. 2.1. Однослойный эпите-
лий.
А — плоский эпителий (мезотелий саль-
ника): I — схема, II — препарат, III —
СЭМ эпителиоцита (по Я. Л. Карагано-
ву, А. А. Миронову и В. А. Миронову).
Б — кубический эпителий (почечные
канальцы). В — призматический микро-
ворсинчатый эпителий (тонкая кишка):
I кишечная ворсинка, II — эпителий,
III — ТЭМ апикальной части микровор-
сипчатых эпителиоцитов.
I эпителиоцит; 2 — базальная мембрана; 3 — соединительная ткань; 4 — крове-
носный сосуд; 5 — бокаловидный экзокриноцит; а — ядро, б — микроворсинка,
в межклеточные границы, г — плотное соединение, д — простое соединение,
е десмосома, ж — полудесмосома, и — щеточная каемка.
29
их структурной организации. Качественные же особенности фак-
торов окружения обусловили появление определенных морфологи-
ческих различий.
2.1.1.1. Однослойный эпителий
Однослойный эпителий включает три разновидности: плоский,
кубический и призматический.
Однослойный плоский эпителий представляет
сплошной клеточный пласт, расположенный на базальной мембра-
не, отделяющей его от подлежащей соединительной ткани, содер-
жащей кровеносные сосуды. Постоянное существование сплош-
ного пласта обеспечивается межклеточными соединениями, рас-
положенными в определенном порядке: ближе к поверхности —
плотное (изолирующее от внешней среды), затем простое, а под
ним десмосома — механическое. Прикрепление же к базальной
мембране обеспечивает полудесмосома. На апикальной поверх-
ности эпителиоцитов находятся неупорядоченно расположенные
микроворсинки, увеличивающие--ее длшцадьГ Некоторые~~~Клеткй
могут~имёть два^ТГ иногда и три ядра (рис. 2.1, А).
Однослойный кубический эпителий отличается
от плоского не только (и не столько) формой клеток, но и боль-
Рис. 2.2. Многорядный реснитчатый эпителий (трахея).
А — схема. Б — препарат. В — СЭМ: I — эпителиоциты, II — поверхность слизи-
стой оболочки (по Л. К. Романовой).
1 — реснитчатый эпителиоцит (а — реснички); 2 — бокаловидный экзокриноцит
(б — слизистый секрет); 3 — вставочный эпителиоцит; 4 — базальный эпителио-
цит; 5— эндокриноцит; 6 — базальная мембрана; 7 — соединительная ткань;
8 — кровеносный сосуд. Г — ТЭМ ресничек: I — поперечный, II — продольный
срезы. 1 — внешняя клеточная мембрана; 2 — цитоплазма; 3 — периферический
дуплет микротрубочек; 4 — центральные микротрубочки; 5 — базальное тельце; 6 —
цитоплазма верхушечной части клетки.
30
шим разнообразием их апикальных (щеточная каемка и реснички)
и базальных (инвагинации плазмолеммы) поверхностей, количе-
ством и распределением органелл, а также появлением в его соста-
ве отдельных секреторных клеток (см. канальцы почек, конечные
бронхиолы легких). Это определяется органной спецификой. Меж-
клеточные соединения сходны с плоским (рис. 2.1, Б).
Однослойный призматический эпителий вы-
стилает внутреннюю поверхность большинства полых органов
(•желудка, кишечника, жёлчного пузыря, крупных протокд1Гпечени
и поджелудочной железы, матки и маточных труб, части семявы
носящих путей), что обусловило и большее морфофункциональное
разнообразие его клеток (микроворсинчатые, реснитчатые и беска-
емчатые эпителиоциты, разные типы секреторных клеток).
На рис. 2.1, В представлена одна из его разновидностей —
микроворсинчатый эпителий тонкой кишки, покрывающий поверх-
ность ее ворсинок в виде сплошного однослойного пласта призма-
тических клеток — эпителиоцитов. Среди последних расположены
единичные более светлые клетки бокаловидной формы (бокало-
видные экзокриноциты).
Однослойный многорядный реснитчатый
эпителий выстилает полости, в которых происходит переме-
щение слизи и расположенных на ее поверхности частиц в опре-
деленном направлении (например, воздухоносные пути). Он имеет
более сложный клеточный состав самого пласта. Образующие его
клетки имеют разную форму,
величину, но при этом все со-
храняют связь с базальной мем-
браной — признак однослойно-
сти (рис. 2.2, А).
Особенность строения рес-
нитчатых эпителиоцитов — на-
личие на апикальном полюсе
ресничек и высокое расположе-
ние округлых ядер. Различные
32
уровни локализации ядер и
формируют феномен многоряд-
пости данного эпителия в гисто-
логических препаратах (рис.
2.2, Б).
Общий вид реснитчатого
шителиоцита, бокаловидного
эк юкриноцита, вставочного эпи-
iглиоцита и апикальные по-
верхности первых двух пред-
ел.nvieHbi на рис. 2.2, В, а уль-
граструктурная организация ре-
сничек — на рис. 2.2, Г.
1‘ш . 2.3. Однослойный многоряд-
IIыи плоский эпителий.
Л неороговевающий эпителий. Б —
opoi овевающий эпителий. В—ТЭМ
дпмосомы. Г — ТЭМ фрагментов цито-
II in 1мы эпителиоцитов (по В. И. Сем-
кину)' I — базальный, II — шиповатый,
111 чешуйчатый (зернистый) и че-
шунчатый (роговой) слои, IV — фраг-
М1-П1 чешуйки.
1 ба шльная мембрана; 2 — базаль-
III in слой; 3 — шиповатый слой; 4 —
поверхностный слой (а), зернистый
• |оп(б); 5 — блестящий слой: 6 —
pin оной слой; 7—выводной проток
по.... железы; 8 — соединительная
1КИН.; *>—кровеносный сосуд; 10 —
пшофиламенты; 11 —тонофибриллы;
I (ранула кератогиалина; 13 — ке-
рпгпнопые фибриллы; 14 — полудесмо-
. ими; 15 — шипики; 16 — десмосома.
I Ш
33
2.1.1.2. Многослойный эпителий
В многослойном плоском неороговевающем
эпителии выделяют три слоя: базальный, шиповатый и по-
верхностный (плоский). Первый образован одним рядом призма-
тических эпителиоцитов, второй — более широкой зоной полиго-
нальных клеток, а третий — уплощенными эпителиоцитами
(рис. 2.3, А).
Многослойный плоский оро г о вев аю щ и й
эпителий имеет самое сложное строение, поскольку, покрывая
поверхность тела, он испытывает прямое воздействие физико-хи-
мических факторов окружающей среды. Уже при обзорном рас-
смотрении (рис. 2.3, Б) обращает на себя внимание резкое отли-
чие его общего вида от всех ранее изученных эпителиев; высокая
степень извилистости базальной мембраны, значительно большая
толщина, неоднородность строения и окрашивания эпителиального
пласта. При рассмотрении со стороны базальной мембраны можно
убедиться, что эта зона имеет строение, сходное с многослойным
плоским неороговевающим эпителием — четко дифференцируются
базальный и шиповатый слои. Сравнительная ультраструктурная
характеристика организации их цитоплазмы и межклеточных сое-
динений (рис. 2.3, В, Г, I) отчетливо демонстрирует способ соедине-
ния базального эпителиоцита с базальной мембраной (с помощью
полудесмосом) и шиповатых эпителиоцитов между собой (с по-
мощью десмосом), а также нарастание в цитоплазме последних
тонофибриллярных структур. Все это обеспечивает прочность и
целостность пласта. Остальные же (ранее не встречавшиеся слои)
отражают этапы специфической дифференцировки, сопровождаю-
щейся вертикальной миграцией эпителиоцитов шиповатого слоя.
Так, накопление гранул белка кератогиалина и сопутствующее
уплощение клеток приводят к образованию резко выделяющегося
зернистого слоя. Дальнейшие структурно-химические перестройки
(меняющие тинкториальные и оптические свойства цитоплаз-
мы) — к формированию лежащего над ним (не всегда отчетливо
различимого) блестящего слоя. Верхний роговой слой образован
роговыми чешуйками — безъядерными, заполненными кератино-
выми фибриллами, уплощенными тельцами — корнеоцитами
(см. рис. 2.3, В, Г, IV). Его толщина и плотность в разных участках
кожи сильно варьируют в зависимости от величины и продолжи-
тельности механических воздействий (наибольшая степень ги-
пертрофии — кожная мозоль). Среди базальных и шиповатых
эпителиоцитов находятся и «пришлые» клетки: меланоцит, внутри-
эпителиальный макрофаг (клетка Лангерганса), тактильный эпи-
телиоцит (клетка Меркеля). В толще эпителия могут быть видны
вертикально расположенные округлые светлые полости — попереч-
ные срезы протока потовой железы.
/ Переходный эпителий является разновидностью
многослойного неороговевающего. Его отличтельйая морфологиче-
ская особенность — изменение толщины эпителиального пласта и
34
a
Рис. 2.4. Переходный эпителий (мочевой пузырь).
Л эпителий при нерастянутой стенке. Б — эпителий при растянутой стенке.
I поверхностный слой (а — эпителиоцит); 2 — промежуточный слой; 3 — ба-
|(1лып|й слой; 4 — базальная мембрана; 5 — соединительная ткань; 6 — крове-
носный сосуд.
формы эпителиоцитов поверхностного слоя в зависимости от
состояния стенки органа, которую он выстилает. Так, в нерастяну-
гом состоянии обращает на себя внимание отсутствие феномена
постепенного уплощения эпителиоцитов (характерного для всех
других разновидностей многослойных эпителиев) и наличие в по-
верхностном слое клеток грушевидной (или округлой) формы
(рис. 2.4,А). В растянутом же состоянии (рис. 2.4,Б) эпите-
।сильный пласт утончен, уменьшено число клеточных слоев и уп-
лощены эпителиоциты покровного слоя (эпителий совершил «пе-
реход» в другое состояние).
2.1.2. Железистый эпителий
Железистый эпителий является производным покровного эпите-
лия, специализирующимся на выработке и выделении (в окружа-
ющую его среду) секреторного материала, необходимого для осу-
ществления метаболических процессов и их регуляции в организ-
ме. Это нашло отражение в многообразии разновидностей цитоло-
гической дифференцировки его клеток (гландулоцитов), направ-
ленной на модификацию и гипертрофию систем метаболизма (син-
ieia, транспорта и аккумулирования секрета).
На рис. 2.5 представлены общий план строения многоклеточ-
ных экзокринных (А) и эндокринных (Б) желез. На рис. 2.6 —
морфологические различия гландулоцитов и концевых отделов
35
2*
Рис. 2.5. Многоклеточные железы.
А — строение экзокринных желез (схема). Б — строение эндокринных желез
(схема); I — простые, II — сложные.
Железы: 1 — трубчатая неразветвленная; 2 — трубчатая разветвленная; 3 — аль-
веолярная неразветвленная; 4 — альвеолярная разветвленная; 5 — альвеолярно-
трубчатая; 6 — альвеолярная; 7 — трабекулярная; 8 — диффузная; 9 — фоллику-
лярная; а — эпителий, б — выводной проток, в — концевой отдел, г — гландуло-
цит, д — гемокапилляр.
Рис. 2.6. Концевые отделы желез.
1 — серозной (белковой), II — слизистой (мукоз-
ной), III—смешанной (серомукозной).
I сероцит, 2 — мукоцит, 3 — серозное полулу-
ние, 4 — просвет ацинуса.
Экзокринных желез в зависимости от физико-химических свойств
вырабатываемого секрета; на рис. 2.7 — структурная характери-
стика типов секреций.
2.1.2.1. Основные разновидности строения и локализация
акзокринных желез
Одноклеточные железы находятся в составе однослой-
ных эпителиев (рис. 2.8). Исключение составляет желудок, где
они образуют сплошной железистый покров.
Простые многоклеточные железы расположены
в подэпителиальной соединительной ткани. На рис. 2.9, А, I пред-
ставлена простая неразветвленная трубчатая железа, а на
рис. 2.9, А, II — простая альвеолярная разветвленная железа. Об-
ращают на себя внимание различия в клеточном составе секретор-
ных отделов. В трубчатой — один слой железистых клеток, тогда
как в альвеолах вместо гландулоцитов — клетки на разных ста-
диях жизненного цикла (специфично для сальной железы, секрет
которой образуют разрушенные клетки — голокринный тип секре-
ции).
37
I II III IV
Рис. 2.7. Типы секреции.
А—схема. Б — ТЭМ; I — мерокринный тип, II — микроанокринный тип, III —
макроапокринный тип, IV — голокринный тип (II, III по Е. А. Шубниковой).
1—гранулярная эндоплазматическая сеть; 2 — комплекс Гольджи; 3 — ми to
хондрия; 4 — секреторный продукт; 5 — отрыв апикальной части с секреюм;
6 — камбиальная клетка; 7 — сальный экзокриноцит (себоцит); 8 — рл tpyiiiu
ющийся экзокриноцит; 9 — плотный контакт; 10 — десмосома; II - восс ы ноилс
ние плазмолеммы; 12 — просвет железы (а — растворение секреторной гранулы),
38
Рис. 2.8. Одноклеточная внутриэпителиальная
железа (тонкая кишка).
Л — препарат (окраска альциановый синий). Б —
I )М.
1 бокаловидный экзокриноцит; 2 — ядро; 3 — слизи-
11 ый секрет; 4 — микроворсинчатый эпителиоцит; 5 —
On шльная мембрана; 6 — соединительная ткань; 7 —
комплекс Гольджи; 8 — микроворсинки.
Сложные многоклеточные железы расположе-
ны преимущественно в подслизистой основе пищеварительного
тракта (рис. 2.9, Б) и воздухоносных путей. Обратите внимание на
обилие концевых отделов (разветвление протока в гистологический
срез попадает редко).
2.2. Кровь
Знание морфологических и возрастных аспектов периферической
крови необходимо врачу любой специальности, так как картина
крови является своеобразным зеркалом состояния различных сис-
тем организма.
Заболевания самой системы крови изучает специальная дис-
циплина — гематология.
1.2.1. Морфологические особенности крови
взрослого человека
Основную массу форменных элементов в препарате-мазке
(рис. 2.10) составляют эритроциты: округлые, безъядерные струк-
39
Рис. 2.9. Многоклеточные
железы.
А — простые: I — неразвет-
вленная трубчатая железа
(эндометрий матки), II —
разветвленная альвеолярная
(сальная железа кожи). Б —
сложная разветвленная аль-
веолярная трубчатая железа
(пищевод).
1 — железа; 2 — гландулоци-
ты; 3 — выводной проток;
4 — альвеолярные концевые
отделы; 5 — базальные экзо-
криноциты (себоциты); 6 —
сальные экзокриноциты (се-
боциты); 7 — разрушающиеся
клетки; 8 — соединительная
ткань; 9 — кровеносный со-
суд; 10 — волос; 11 — покров-
ный эпителий; 12 — миоэпи-
телиоцит.
40
Pm. 2.10. Мазок периферической крови взрослого человека (общий
вид).
I эритроциты; 2 — лимфоциты; 3 — моноцит; 4 — нейтрофильные гранулоци-
111; 5 — эозинофильные гранулоциты; 6 — базофильные гранулоциты; 7 — тром-
б< >1111 гы.
гуры розового цвета. В значительно меньшем количестве встре-
чи ю гея более крупные круглые клетки с сегментированными,
округлыми или бобовидными ядрами сиренево-фиолетовой окрас-
ки лейкоциты. Тромбоциты при малом увеличении видны плохо.
Использование объектива большого • увеличения позволяет
идентифицировать все виды форменных элементов крови по их
м о| урологическим особенностям.
Эритроциты легко диагностируются по отсутствию ядра,
xup.iK герному просветлению в центре, обусловленному спецификой
их формы — двояковогнутого диска, отчетливо определяемой при
и< пользовании метода СЭМ (рис. 2.11). Знание нормальных раз-
меров (7,2 мкм ± 0,5) и формы эритроцитов важно для умения
определять явления анизоцитоза (отклонение от нормальных раз-
41
Рис. 2.11. Эритроцит нормальной
формы (дискоцит), СЭМ (по
Ю. А. Ровенских).
меров в направлении макро- и микроцитов) и пойкилоцитоза
(изменение формы), нередко встречающихся при различных забо-
леваниях крови.
В крови здорового человека содержится небольшое количество
эритроцитов (1—2 %), имеющих синеватый оттенок. За способ-
ность окрашиваться основными и кислыми красителями они полу-
чили название полихроматофилы. При прижизненном окрашива-
нии специальными красителями (ярким крезиловым синим) в ци-
топлазме полихроматофилов выявляются зернисто-нитчатые
структуры (рис. 2.12, А), образующие своеобразную сеть, с чем
связано общепринятое название полихроматофила — ретикулоцит
(предшественник зрелого эритроцита). Зернисто-нитчатые струк-
туры представляют собой остатки органелл, имевшихся в клетках-
предшественниках эритроцитарного ряда (рис. 2.12, Б, В).
Тромбоциты, или кровяные пластинки, в
обычных мазках крови (без принятия специальных мер предосто-
рожности) агрегируют, образуя скопления (рис. 2.13, А). В случае
распластывания отдельных пластинок на поверхности стекла вид-
ны две слабо разграниченные части: наружная (бледно-голубого
цвета) — гиаломер и центральная, содержащая окрашенный ма-
териал в виде зерен-гранул — грануломер.
Знакомство с основными особенностями ультраструктурной ор-
ганизации тромбоцитов позволяет лучше понять их функциональ-
ное значение. Речь идет о системе канальцев, связанных с по-
верхностью (по ним происходит обмен с окружающей средой),
гранулах, содержащих гидролитические ферменты, плотных тель-
цах, содержащих серотонин, кальций, АТФ, АДФ и др., системе
микротрубочек, выполняющих скелетную функцию, микрофила-
ментах, обеспечивающих сократительную функцию (рис. 2.13, Б, В).
Активизация тромбоцитов сопровождается приобретением ими
шаровидной формы (вместо дисковидной) (рис. 2.13, Г) и пере-
мещением гранул в центральную часть пластинок. Основную
функцию тромбоциты осуществляют на различных этапах сверты-
42
Рис. 2.12. Ретикуло-
цп гы.
Л ре 1икулоциты в мазке
периферической крови
(окраска бриллиантовым
крешловым синим). Б —
I >М. В — деталь к «Б».
I митохондрии с пра-
пи п.ио расположенными
крае шми; 2 — митохонд-
рии с признаками дегене-
рации; 3 — элементы ци-
iiuuui гматической сети:
I п «купли; 5 — скопле-
ние рибосом; 6 — зерна
н мшлобина.
1ИИ111И крови, однако их участие в процессе гемостаза наиболее
пырижено в начальном периоде, когда происходит адгезия пласти-
нок к стенке поврежденного сосуда с последующей их агрегацией
(формирование тромбоцитной пробки) (рис. 2.13, Д).
Одной из важнейших функций тромбоцитов в физиологиче-
। к их условиях является ангиотрофическая, нарушение которой
(пцнример, при тромбоцитопении) ведет к повышению сосудистой
проницаемости и возникновению геморрагий.
43
A
Рис. 2.13. Тромбоциты.
А — тромбоциты в мазке перифериче-
ской крови. Б — схема строения тром-
боцита. В — ТЭМ. Г — неактивирован-
ные (отмечены стрелкой) и активиро-
ванные (отмечены двумя стрелками)
тромбоциты, СЭМ. Д — тромбоциты,
прилипшие к стенке аорты в зоне по-
вреждения эндотелиального слоя
(Г, Д — по Ю. А. Ровенских).
1 — микротрубочки; 2 — митохондрии;
3 — а-гранулы; 4 — система плотных
трубочек; 5 — микрофиламенты; 6 —
система канальцев, связанных с по-
верхностью; 7 — гликокаликс; 8 — плот-
ные тельца; 9 — цитоплазматическая
сеть.
44
45
Тромбоциты могут быть и истинными фагоцитами; по мнению
ряда исследователей, удаление инородного материала из крови —
важная функция пластинок. Как и другие форменные элементы
крови, тромбоциты способны фиксировать на поверхности анти-
тела.
Лейкоциты. При изучении лейкоцитов прежде всего сле-
дует отличать гранулярные лейкоциты от агранулярных. Основными
характерными чертами первых являются наличие сегментированно-
го ядра и специфической зернистости. Важно также уметь диагно-
стировать все разновидности зернистых и незернистых форм.
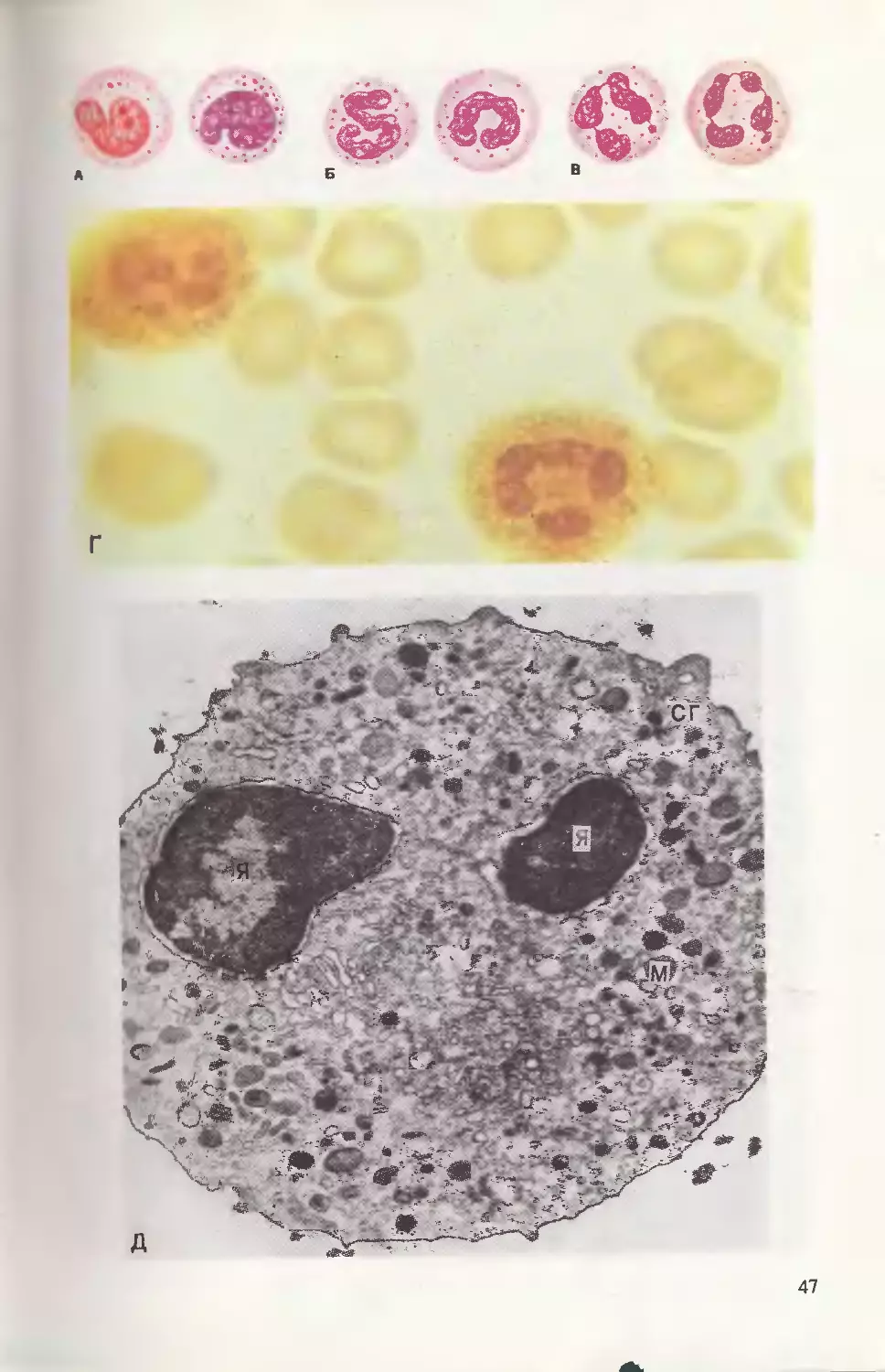
Нейтрофильные гранулоциты. Наиболее часто в
крови взрослого человека встречаются сегментоядерные грануло-
циты (60—65 %); обнаруживаются также палочкоядерные (3—
5 %) и юные формы (с бобовидными ядрами 0—0,5 %). В цито-
плазме этих клеток определяется два вида зернистости: неспеци-
фическая (азурофильная), ранее появляющаяся в процессе гемо-
поэза, и специфическая, имеющая пылевидный характер и при-
дающая цитоплазме лиловый оттенок (рис. 2.14, А, Б, В). Знание
набора ферментов, входящих в состав азурофильных и специфи-
ческих гранул, имеет не только теоретическое значение: обнару-
жение с помощью химических методов некоторых ферментов, в
частности миелопероксидазы, специфичных лишь для нейтрофиль-
ных гранулоцитов, используется в клинических лабораториях в
качестве диагностического теста при дифференцировке различных
заболеваний крови (рис. 2.14, Г). Зернистость зрелых нейтро-
фильных гранулоцитов состоит из 10—30 % пероксидазоположи-
тельных, первичных или азурофильных гранул и 70—90 % специ-
фических вторичных гранул, нелизосомальных пероксидазоотрица-
тельных, т. е. содержащих лизоцим, лактоферрин, фагоцитин,
коллагеназу, щелочную фосфатазу. Ультраструктура зрелого
нейтрофильного гранулоцита продемонстрирована на рис. 2.14, Д.
Основной функцией этого вида лейкоцитов является фагоцитоз,
на первых этапах характеризующийся направленной миграцией
клеток к фагоцитируемому объекту (обычно бактериям). Актив-
ное перемещение в пространстве — наиболее характерный признак
живого нейтрофильного гранулоцита. В последующем цитоплазма-
тические гранулы (сначала специфические, затем — азурофиль-
ные) мигрируют к образовавшейся фагосоме и освобождают в
нее свое содержимое. Стимулирование клеточного дыхания в тече-
ние этого процесса сопровождается продукцией перекиси водоро-
да, причем освобождение активного кислорода из пероксида с
помощью пероксидазы оказывает токсическое действие на различ-
Рис. 2.14. Нейтрофильные гранулоциты.
Нейтрофильные гранулоциты в мазке периферической крови: А — юные, Б —
палочкоядерные, В — сегментоядерные, Г — реакция на миелопероксидазу, Д —
ТЭМ: я — ядро, м — митохондрии, сг — специфические гранулы.
46
47
48
I'm. 2.15. Эозинофильные гранулоциты.
<> шнофильные гранулоциты в мазке периферической крови: А — юные, Б —
ни ючкоядерные, В — сегментоядерные, Г — ТЭМ: я — ядро, сг — специфические
|||||цулы. Д — кристаллическая структура секреторных гранул.
пыс биологические системы: бактерии, грибы, вирусы, микоплазмы
и их токсины. Нелишне заметить, что наряду с лизосомальными
описаны пероксидазосомальные заболевания наследственного и
приобретенного характера.
В зрелых нейтрофильных гранулоцитах описаны особые хрома-
।иноиые тельца («тельца Барра», или «барабанные палочки»),
онрл ювание которых обусловлено наличием неактивной Х-хромо-
гомы. У женщины на 500 нейтрофильных гранулоцитов встре-
ч«1С1ся 6 или более клеток такого типа; обнаружение их имеет
нА ж ное значение при диагностике интерсексуальных состояний.
Эозинофильные гранулоциты (1—5 %) встреча-
пн ся в крови в виде сегментоядерных, палочкоядерных и юных
форм (по аналогии с нейтрофильными гранулоцитами) (рис. 2.15, А,
I», В). Отличительными чертами эозинофильных гранулоцитов по
1 рлшению с нейтрофильными являются более крупные размеры
(диаметр в мазке составляет 12—17 мкм), двудольчатое ядро с
49
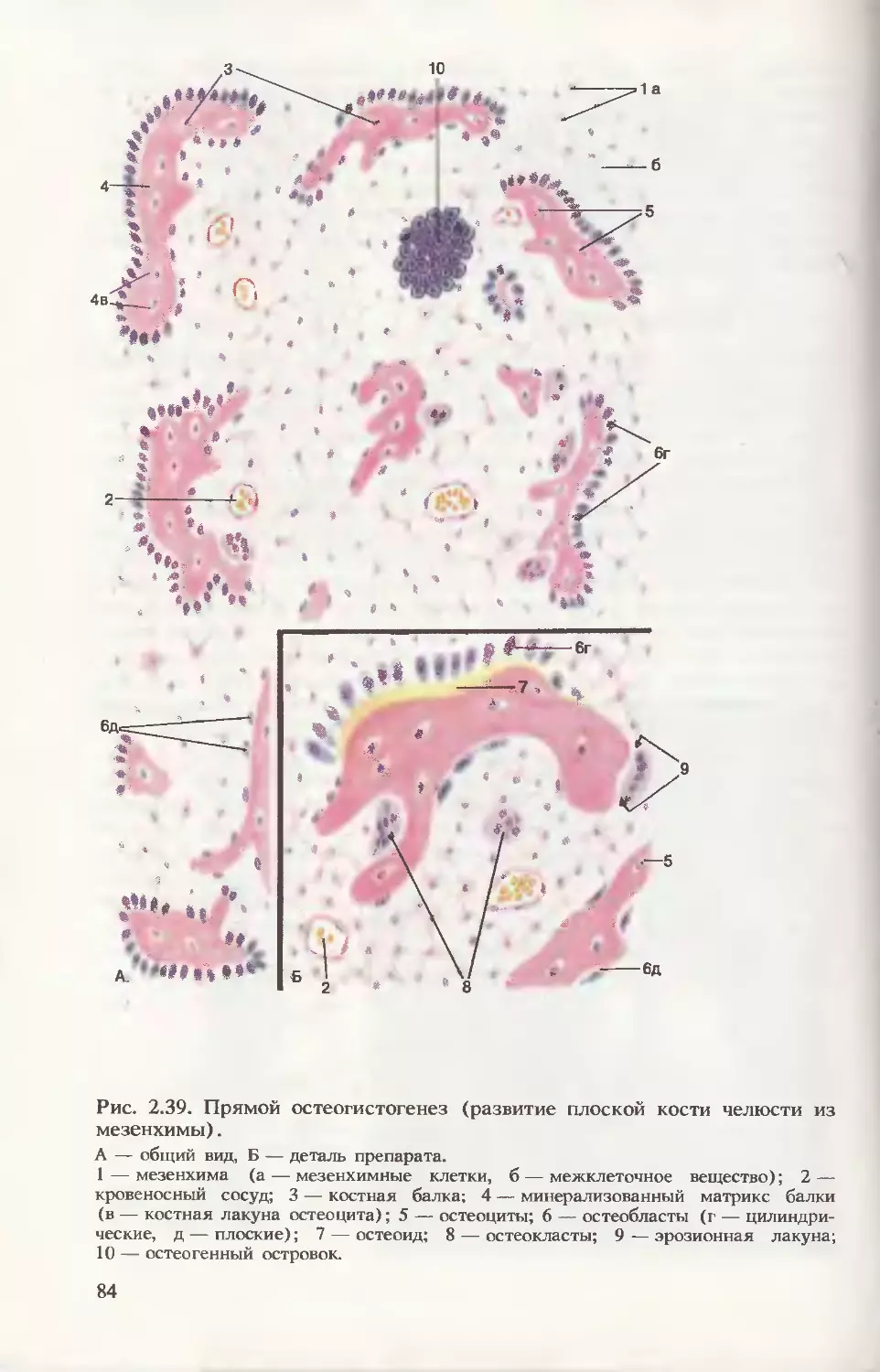
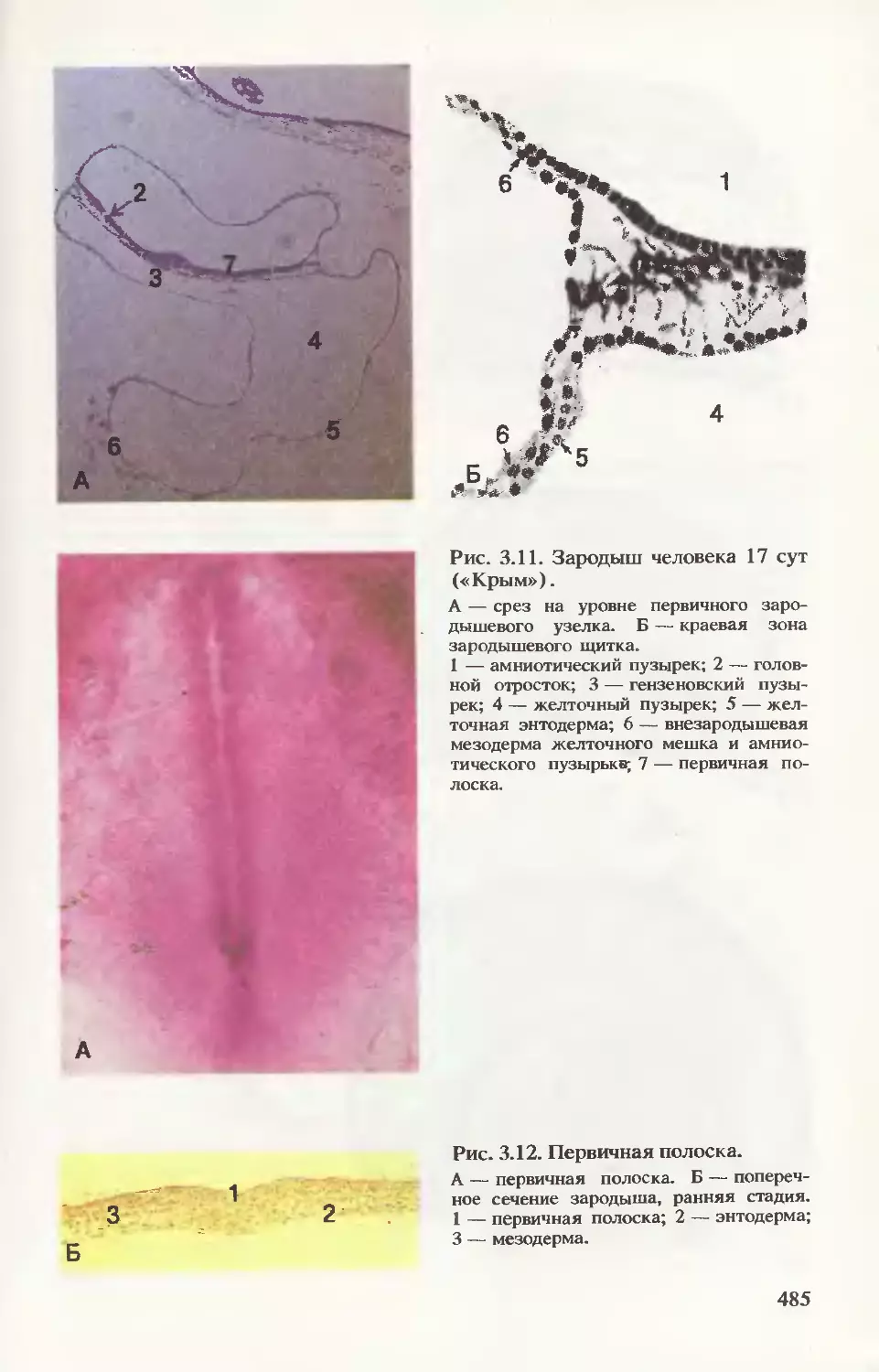
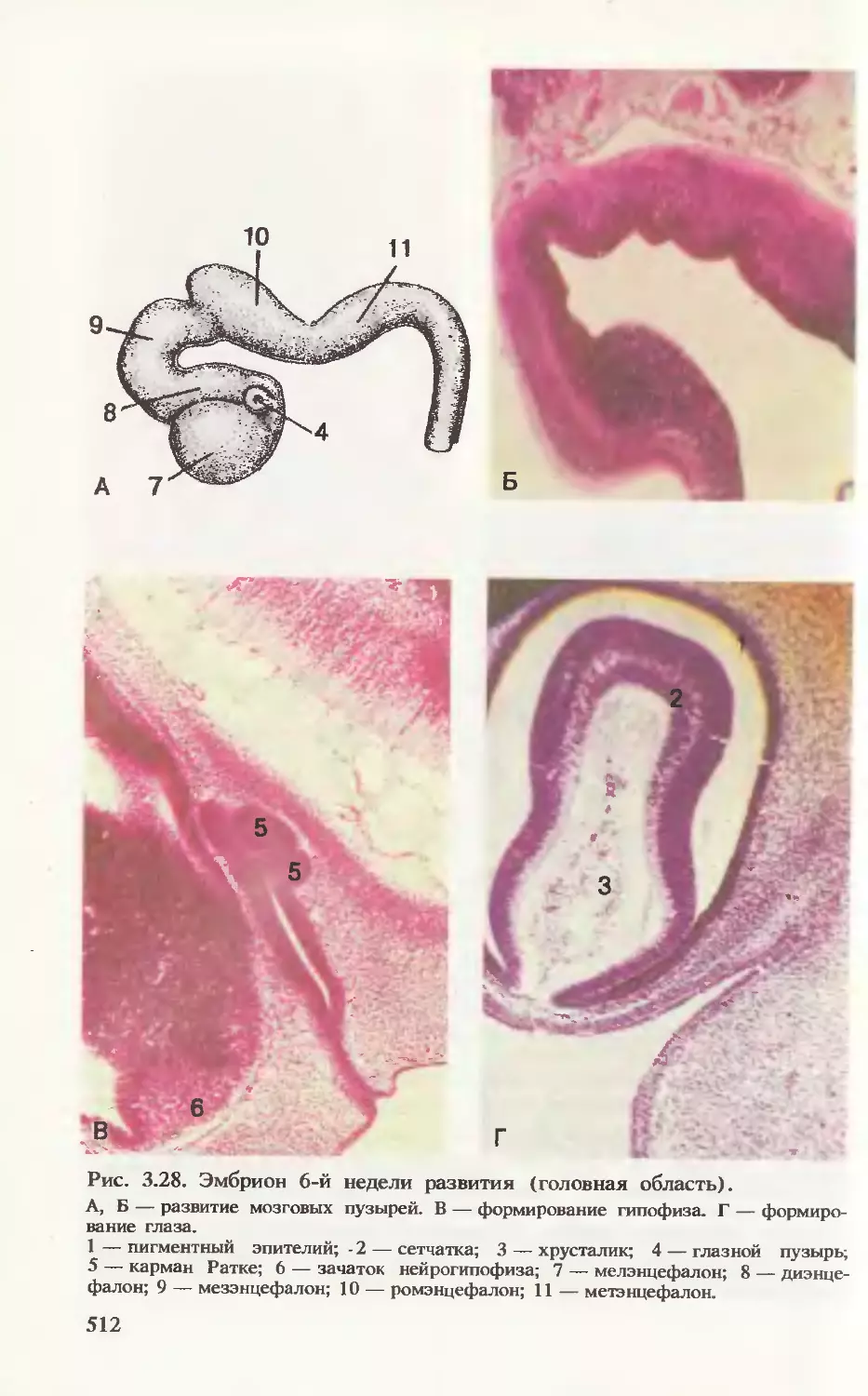
срезах эмбриона (рис. 3.16) видно соотношение развивающихся
частей эмбриона в конце 3-й недели развития.
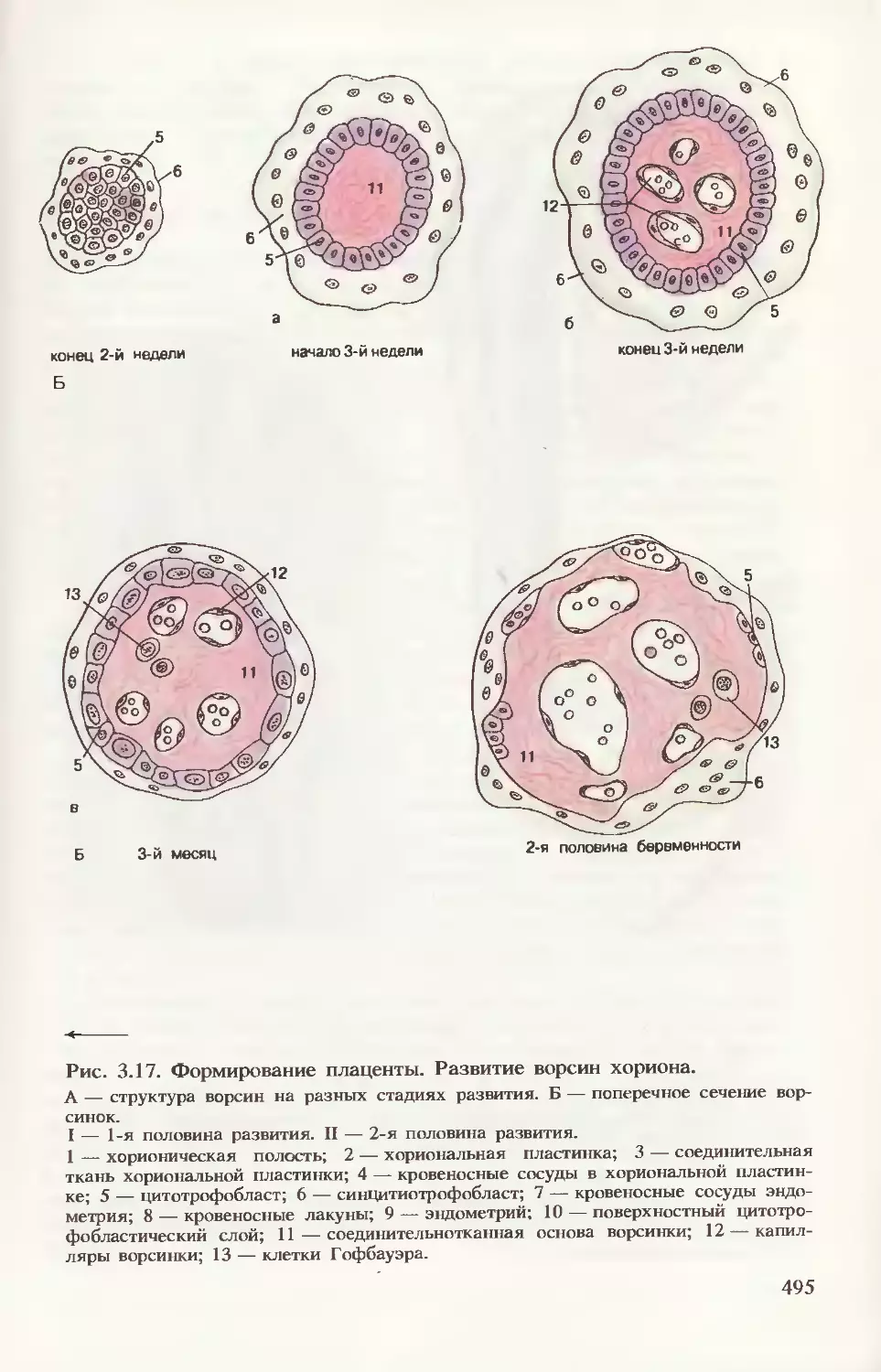
Формирование плаценты. На рис. 3.17 прослеживается процесс
дальнейшего формирования трофобласта. В начале 3-й недели
внезародышевая мезодерма проникает в первичные ворсинки, обра-
зуя их основу, — возникают вторичные ворсинки. К концу недели из
мезодермальных клеток (мезенхимных) дифференцируются кро-
веносные капилляры и клетки крови, формируется артериовеноз-
ная сеть ворсинки. Такие ворсинки называют третичными. Их со-
судистая сеть контактирует с сосудами мезодермальной пластинки
хориона и соединительной ножки, а они в свою очередь с цирку-
лярной кровеносной системой тела эмбриона. Уже к концу 3-й не-
дели развития зародыша эмбриональная кровь циркулирует через
капилляры ворсинок, т. е. четко определяется полное слоистое
строение ворсинки: покровный синцитиотрофобласт, под ним цито-
трофобласт, дифференцированная специфическая соединительно-
тканная строма и кровеносные сосуды.
Для человека (как и для человекообразных обезьян) харак-
терно раннее и мощное развитие трофобласта и внезародышевой
мезодермы, что связано с крайне скудным запасом питательных
веществ в их яйцеклетках и необходимостью быстрого перехода
на контакт с кровеносной системой матери. Ускоренное развитие
ворсинок обеспечивает большую поверхность обмена между
эмбрионом и матерью.
В ворсинках, контактирующих с эндометрием, клетки цитотро-
фобласта пенетрируют через синцитиотрофобласт до тех пор, пока
не достигают тканей эндометрия, где они соединяются с такими
же клетками соседних ворсинок, — образуется тонкий слой (на-
ружный) цитотрофобласта. Этот слой окружает слой синцитио-
трофобласта и «привязывает» хорион к тканям эндометрия.
Рис. 3.16. Соотношение заро-
дышевых и внезародышевых
частей.
1 — гиоидная дуга; 2 — манди-
булярная дуга; 3 — зрительная
плакода; 4 — аллантоис; 5 —
желточный мешок; 6 — стенка
амниона; 7 — сердечный выступ;
8 — полость амниона; 9 — со-
единительная ножка.
493
Рис. 2.16. Базофильные грануло-
циты.
Базофильные гранулоциты в мазке пе-
риферической крови: А — юные и па-
лочкоядерные, Б — сегментоядерные,
В — ТЭМ, Г — схема участия эозино-
фильных и базофильных гранулоцитов в
аллергических реакциях.
1 — дольчатое ядро; 2 — специфические
гранулы.
Эозинофилы
Антиген
(аллерген)
Плазмоцит
Миграция эозинофилов
гистамин, брадикинин,
простагландины, лейкотриены,
в том числе медленно
действующее вещество
анафилаксии
Хемотоксические влияния
lg Е - иммуноглобулин
Рц -рецепторы к lg Е
Е
Переваривание
продуктов дегрануляции,
т.е. ослабление аллергических
реакций
Тканевые базофилы
\ Дегрануляция
Г
Развитие аллергических реакций
(увеличение проницаемости
кровеносных сосудов,
усиление процессов секреции,
возрастание сократительной
активности гладких миоцитов
бронхов, пищеварительного тректа)
50
неплотно упакованным хроматином и самое главное — наличие
и цитоплазме большого количества крупных специфических гранул
красного или оранжевого цвета (оксифильность их объясняется
присутствием основного белка, богатого аргинином). При ультра-
структурном анализе обращает на себя внимание строение специ-
фических гранул, содержащих в своих центральных частях плот-
ные кристаллические тельца, зачастую имеющие форму неправиль-
ных прямоугольников (рис. 2.15, Г, Д). Цитохимический ана-
лиз показал, что вторичные гранулы эозинофильных гранулоцитов
проявляют высокую пероксидазную активность. Поглощение и
деструкция микроорганизмов не являются главной функцией этих
клеток (в отличие от нейтрофильных гранулоцитов); переваривание
иммунных комплексов — их основная задача. Специфическое хе-
мотаксическое действие ряда факторов, проявляющихся в процес-
се развития иммунологических реакций, — основная причина эози-
нофилии при аллергических состояниях и воспалении.
Базофильные гранулоциты, составляющие 0,5—1 % от общего
числа лейкоцитов, характеризуются присутствием в цитоплазме
крупных гранул, обладающих свойством метахромазии, т. е. спо-
собности окрашиваться в тон, отличающийся от цвета красителя.
Базофильная зернистость, густо заполняющая цитоплазму, не
позволяет (на светооптическом уровне) четко различить контуры
сегментированных ядер (рис. 2.16, А, Б). Ультрамикроскопическое
строение клетки представлено на рис. 2.16, В. Большое-содержание
глюкозоаминогликанов (гепарин, гистамин) — характерная черта
базофильных гранулоцитов; практически весь гистамин крови
содержится в специфических гранулах этих клеток. Основная
функция базофильных гранулоцитов — участие в иммунологиче-
ских реакциях немедленного и замедленного типа — в значитель-
ной степени объясняется наличием на их мембранах рецепторов
для иммуноглобулина Е. Взаимодействие антигена с иммуноглобу-
лином Е, связанным с базофильным гранулоцитом, ведет к дегра-
нуляции этих клеток и выделению медиаторов немедленного типа.
Эти теоретические положения, имеющие важное значение для
клиницистов, иллюстрированы на рис. 2.16, Г. Знание структурных
особенностей клеток (в данном случае базофильного грануло-
цита), по нашему мнению, должно сочетаться с пониманием их
функциональной деятельности в курсе гистологии, хотя более по-
дробно эти вопросы будут разбираться при изучении патологиче-
ской физиологии. VV*' I
Среди агранулярных лейкоцитов наиболее много-
численны лимфоциты (20—35 %). В периферической крови взрос-
лого человека в норме встречаются средние и малые лимфоциты
(рис. 2.17, А, Б). Критериями их различия является диаметр клет-
ки (6—8 мкм у малых и более 8 мкм у средних), степень конден-
сации хроматина ядра (значительно большая у малых лимфоци-
тов), ядерно-цитоплазматическое отношение (более высокое у
малых лимфоцитов). Электронномикроскопическое строение ма-
лых лимфоцитов показано на рис. 2.17, В. Подразделение лимфо-
51
цитов на малые и средние (употребляется в клинических лаборп
ториях) не является принципиальным, так как при внешнем мор
фологическом сходстве лимфоидные клетки могут выполнять pa
личные функции в реакциях иммунитета. Речь прежде всего идет
о подразделении на Т- и В-лимфоциты, которые невозможно oi
личить друг от друга на светооптическом уровне, хотя в послед! к>
время появились данные, что для Т-лимфоцитов типична азуро
фильная зернистость. Морфологически Т- и В-лимфоциты диффс
ренцируются лишь при использовании метода сканирующей
электронной микроскопии (рис. 2.17, Г, Д), однако основные pa i
личия заключены в их происхождении, локализации в opraiwx
кроветворения, в продолжительности жизни и роли в осуществле-
нии иммунологических реакций.
Хотелось бы напомнить, что среди клеток, морфологически
идентифицируемых как малые лимфоциты, кроме Т- и В-форм,
могут встречаться стволовые кроветворные клетки (СКК) и нуле
вые клетки, т. е. лимфоциты, не имеющие отличительных маркерон
Т- и В-лимфоцитов.
Немаловажное значение для понимания функции лимфоциты и
имеет тот факт, что они не являются конечными формами дифф»
ренцировки лимфоидного кроветворения: под влиянием специфи
ческих антигенных стимулов они превращаются либо в плазмо-
циты (В-лимфоциты), либо в Т-активированные лимфоциты ( Г
лимфоциты).
В связи со стремительным развитием иммунологии постоянно
пересматриваются и вопросы клеточных механизмов иммунитета,
однако основные этапы развития иммунологических реакций и
роли, выполняемой в процессе их осуществления Т- и В-лимфоци-
тами (наряду с макрофагами), определены и общеизвестны
(рис. 2.18, А).
Наглядной иллюстрацией способности лимфоцитов-киллеров
прикреплять к своей поверхности антигенный материал может
служить реакция образования розеток (прикрепление чужеродных
эритроцитов к лимфоидным клеткам), широко используемая в
клинике для оценки состояния Т-системы иммунитета (рис. 2.18, Б).
Моноциты — самые крупные клетки крови (диаметр 18 -
20 мкм) составляют 5—8 % от общего числа лейкоцитов. Они
характеризуются полиморфизмом ядер (овальные с выемкой,
бобовидные, подковообразные) со средней степенью конденсации
хроматина, ярко-голубой цитоплазмой с мелкими азурофильпы
ми гранулами (рис. 2.19, А).
Рис. 2.17. Лимфоциты.
Лимфоциты в мазке периферической крови: А — средние, Б — малые. В — Г )М
Г—СЭМ: микроотростчатый тип, или В-клетки. Д — СЭМ: «сглаженный» ihii,
или Т-клетки (Г, Д — по Ю. А. Ровенских).
1 — ядро; 2 — ядрышко; 3 — ядерная мембрана; 4 — рибосомы; 5 — митохондрии
52
53
1 мкм
Рис. 2.18. Взаимодействие клеток в иммунном ответе.
А — схема взаимодействия Т- и В-лимфоцитов и макрофагов. Б — «розетка»:
лимфоцит крови человека с эритроцитами барана, СЭМ (по Ю. А. Ровенских).
I — антиген-реактивные Т-лимфоциты с помощью своих рецепторов распознают
чужеродные молекулы антигенов; II—III — комплексы рецепторов Т-лимфоцитов
и антигенов захватываются и перерабатываются макрофагами, имеющими собст-
венные рецепторы; IV — В-лимфоциты, контактируя с макрофагами, с помощью
своих рецепторов получают от них антигенную информацию. Т-лимфоцит-хелпер
выделяет медиатор—«индуктор иммунопоэза» (интерлейкин-2). В результате
них взаимодействий В-лимфоцит трансформируется в антителообразующие
клетки, продуцирующие антитела, инактивирующие антигены.
1 — Т-лимфоциты; М — макрофаги; ИИ — «индуктор иммунопоэза»; АОК — анти-
юлообразующие клетки.
I — рецепторы Т-лимфоцитов; 2 — антиген; 3 — рецепторы В-лимфоцитов; 4 —
рецепторы макрофагов; 5 — антитела.
Рис. 2.19. Моноциты.
А — моноциты в мазке периферической крови. Б — ТЭМ.
55
Для понимания функциональной роли моноцитов следует
помнить, что в крови они циркулируют недолго (в среднем 32 ч), а
затем переходят в ткани и трансформируются там в макрофаги,
продолжительность жизни которых может достичь 100 дней. Зре-
лые макрофаги способны к рециркуляции.
Характерной ультраструктурной чертой моноцитов является
наличие лизосом, обеспечивающих основную функцию этих кле-
ток — способность к фагоцитозу и внутриклеточному переварива-
нию инородных частиц, а также разрушенных клеток собственного
организма (рис. 2.19, Б).
Макрофаги участвуют в начальных этапах иммунологических
реакций, осуществляя захват антигена и выведение его в иммуно-
генной форме на поверхность клетки. Как уже отмечалось при
изучении лимфоцитов, процесс антителообразования возможен
лишь при кооперации трех видов клеток: Т-лимфоцитов, В-лимфо-
цитов и макрофагов, которые часто называют А-клетками за их
адгезивную способность (активно прикрепляются к стеклу и
пластику в культуре тканей).
Одним из навыков, необходимых врачу любой специальности,
является определение лейкоцитарной формулы крови в препарате-
мазке. При качественном изучении мазка крови обращает на себя
внимание то, что на каждый лейкоцит приходится около 1000
эритроцитов, следовательно, лейкоциты легче находить при малом
увеличении (голубые клетки на фоне розовых эритроцитов). Для
подсчета лейкоцитарной формулы крови просматривают не менее
100 лейкоцитов и заполняют 100-клеточную таблицу. В клиниче-
ских лабораториях в этих же целях для большей точности под-
счета просматривают 200 лейкоцитов и более. При подсчете лей-
коцитарной формулы крови начинающему исследователю следует
помнить, что для точного подсчета лучше просматривать весь
мазок, не ограничиваясь какой-либо одной его частью; не следует
также учитывать дегенерирующие лейкоциты и принимать скоп-
ления тромбоцитов за лейкоциты (их легко различить при боль-
шем увеличении).
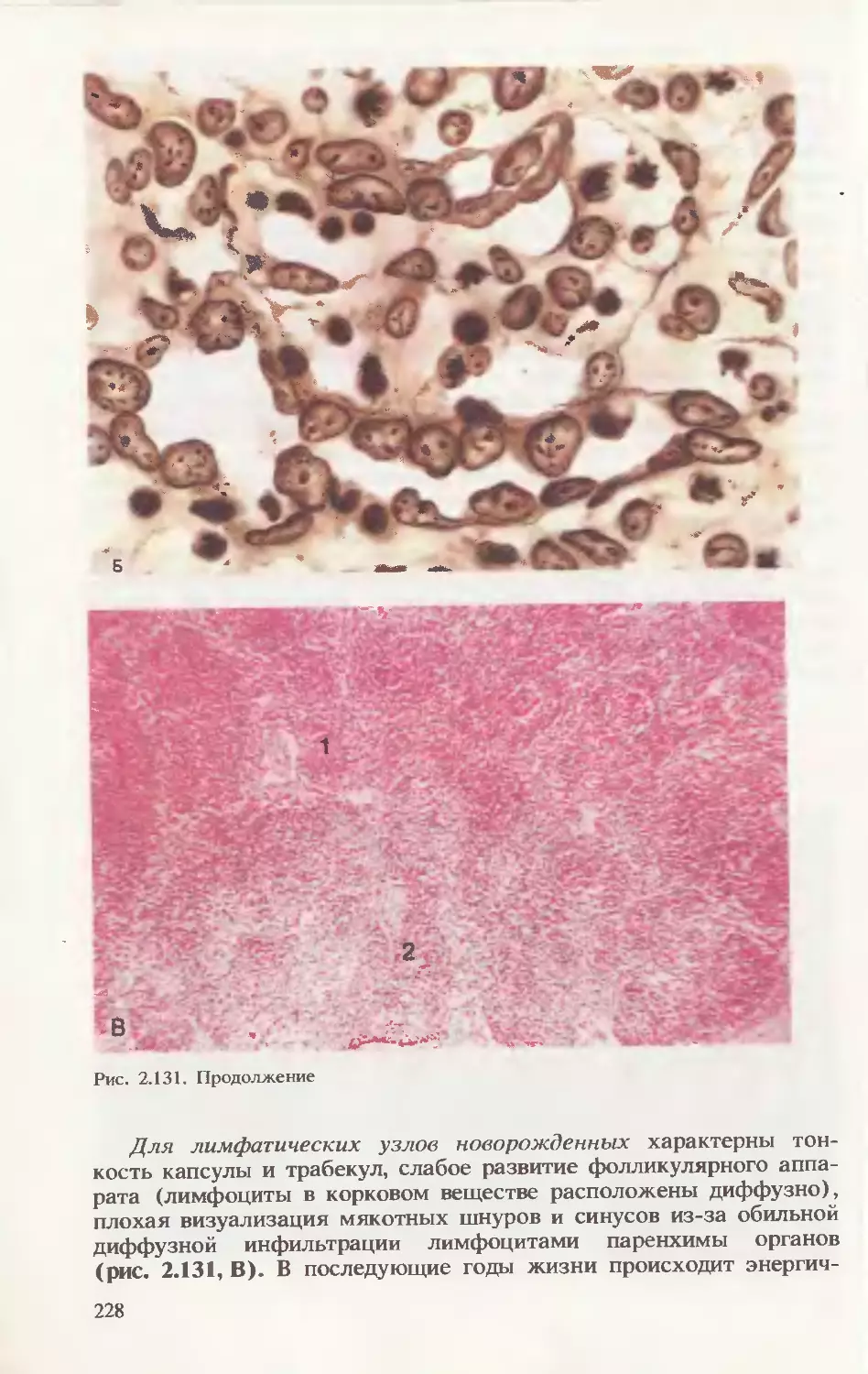
2.2.2. Морфологические особенности крови новорожденного
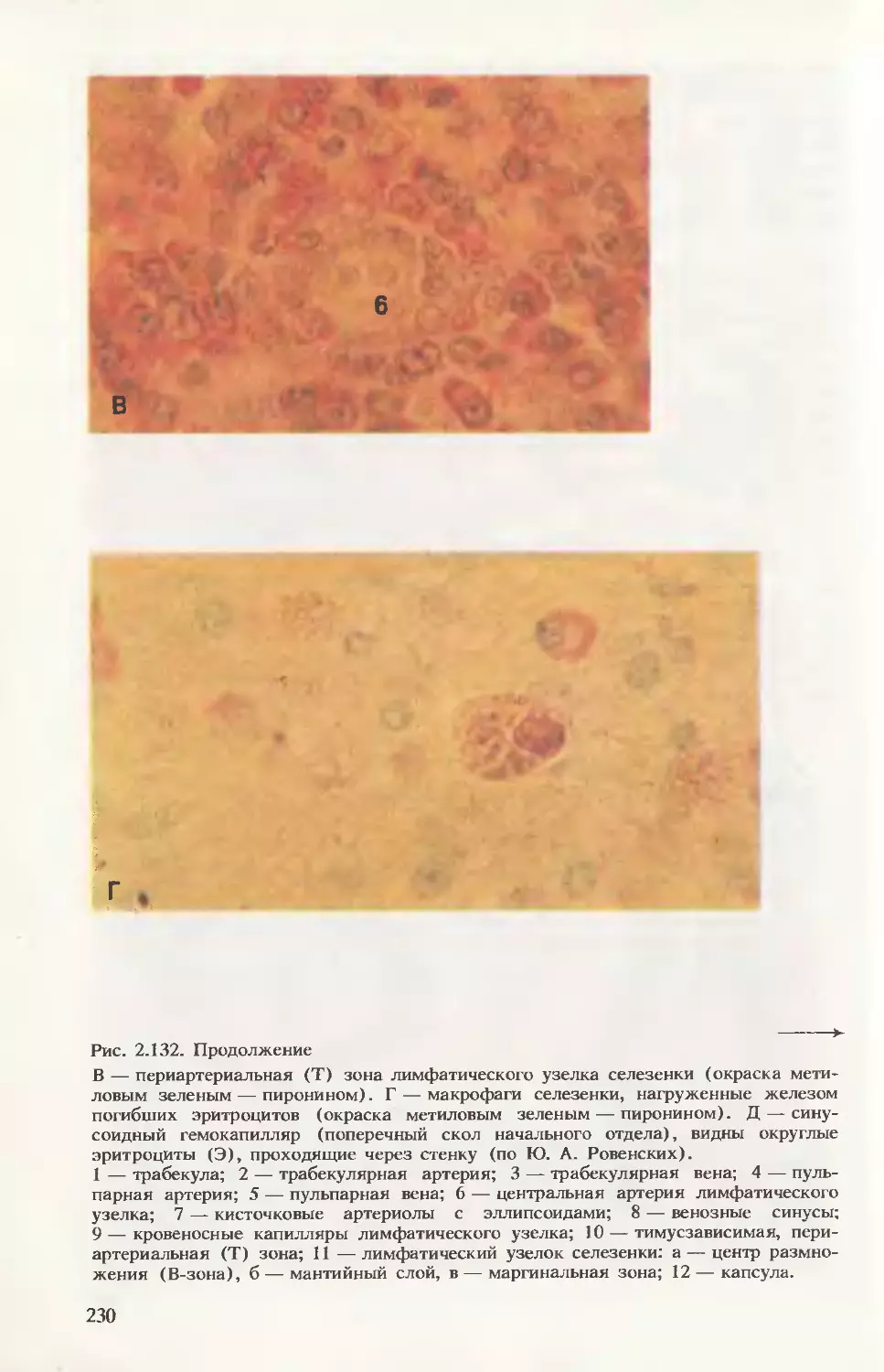
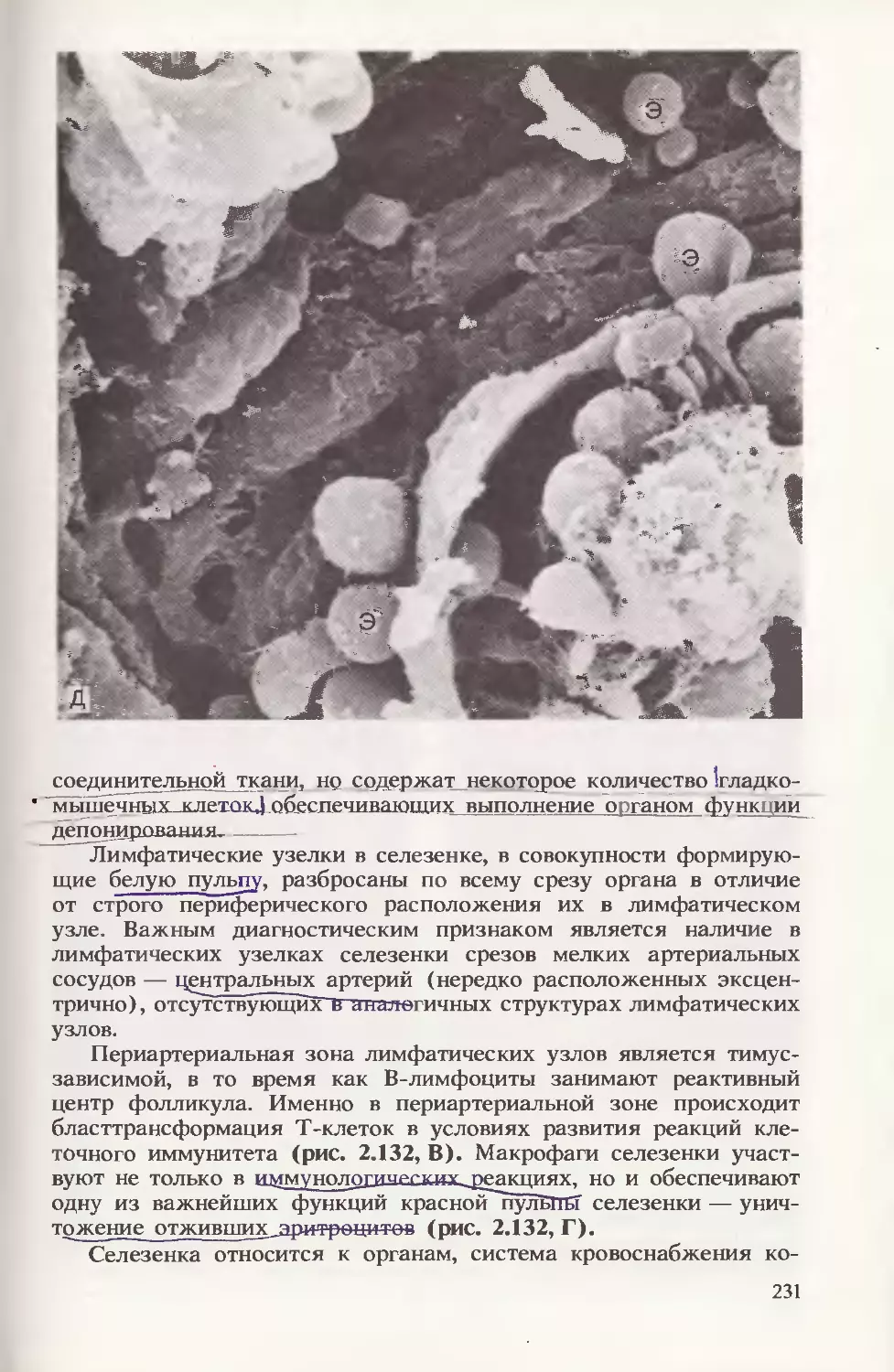
При анализе крови новорожденного (в препарате-мазке) следует
помнить, что форменные элементы практически не отличаются от
таковых у взрослого человека, за исключением того, что в течение
1-й недели постнатальной жизни регистрируется феномен анизо-
цитоза (макроцитоз), увеличено число ретикулоцитов, выявляются
ядросодержащие предшественники эритроцитов, небольшое коли-
чество бластных форм кроветворных клеток (рис. 2.20). В этот
период лейкоцитарная формула аналогична таковой взрослого
человека, однако вскоре число нейтрофильных гранулоцитов сни-
жается, а число лимфоцитов нарастает.
С двухлетнего возраста количество лимфоцитов начинает сни-
56
К Б
I’m. 2.20. Мазок периферической крови новорожденного (общий вид).
Л бластная клетка. Б — полихромазия, анизоцитоз, пойкилоцитоз.
жа гься, а нейтрофилов — нарастать, и приблизительно к 4 годам
количество этих клеток уравнивается, что находит свое отражение
и картине мазка крови детей этого возраста. Обилие лимфоцитов
но сравнению с мазками крови взрослого человека, заметное при
простом просмотре препарата, особенно четко выявляется при
определении лейкоцитарной формулы крови ребенка не старше
4 лет (рис. 2.21).
Для создания четкого представления об особенностях состава
кропи в различные этапы постнатального периода следует руко-
водствоваться рис. 2.22.
2.3. Соединительные ткани
( оединительные ткани осуществляют жизнеобеспечение всех орга-
нон и систем в организме, поскольку наряду с выполнением чисто
механических, опорных функций активно участвуют в важнейших
57
22000
21000
20000
19000
18000
17000
16000
16000
„ 14000
i 1зооо
я 12000
g 11000
I 10000
9000
8000
7000
6000
5000
4000
3000
2000
1000
36 24234567235 369122345678
А Часы Дни Нед. Мес. Годы
Рис. 2.21. Лимфоцитарный профиль крови ребенка.
Моноциты и малые лимфоциты в мазке периферической крови.
1 — лимфоцит; 2 — моноцит; 3 — эритроцит.
Рис. 2.22. Динамика количественных изменений форменных элементов
периферической крови в различные периоды детства.
А — динамика количества лейкоцитов. Б — динамика лейкоцитарной формулы
крови.
сторонах внутреннего обмена: поддержании гомеостаза, механи-
ческой и биологической защите, трофике, депонировании веществ,
транспортных и пластических процессах. Такая специализация
обеспечивает соединительным тканям ведущую роль в реализации
практически всех физиологических и адаптационно-компенса-
торных процессов, поэтому их патология неизбежно влечет за
собой нарушение морфофункционального статуса всего орга-
низма.
58
2.3.1. Собственно соединительная ткань
2.З.1.1. Волокнистая соединительная ткань
2.3.1.1.1. Рыхлая волокнистая ткань
Эта разновидность является наиболее распространенной в орга-
низме, так как участвует в построении всех его органов. Ее гис-
тоструктуру обычно изучают на тотальных препаратах, четко
отражающих составные компоненты ткани и их соотношения
(рис. 2.23, А).
Межклеточное вещество. Его неструктурированный компо-
нент — основное (аморфное) вещество имеет вид
слабо окрашенного гомогенного матрикса. В количественном от-
ношении оно значительно превалирует над волокнистымй компо-
нентами и клетками, поэтому все структурные элементы ткани
располагаются довольно рыхло, на значительном расстоянии друг
от друга.
Коллагеновые волокна определяются как темные,
волнообразно извитые, толстые тяжи, нередко объединенные в
пучки (рис. 2.23, Б). Субмикроскопически в них можно обнару-
жить четкую, периодическую исчерченность, которая впервые
регистрируется на уровне фибрилл (рис. 2.24). Каждый период
исчерченности фибриллы содержит темный и светлый сегменты.
Они отличаются плотностью расположения молекул коллагена и
поэтому контрастируются по-разному.
Эластические волокна в сравнении с коллагеновыми
значительно светлее и тоньше, прямолинейны, могут разветвляться
и анастомозировать между собой, формируя крупноячеистую сеть
(см. рис. 2.23, А, Б).
Ретикулярные волокна в отличие от коллагеновых и
эластических характеризуются аргирофильностью и избирательной
локализацией в организме. Ознакомиться с ними следует при
изучении ретикулярной ткани (см. рис. 2.33).
Клетки. В сложном по составу межклеточном веществе рыхлой
волокнистой соединительной ткани преобладающим является
аморфный компонент. Именно здесь обитают и функционируют
ее многочисленные и разнообразные клетки.
Наиболее постоянными и распространенными клетками явля-
ются фибробласты. Они характеризуются крупными разме-
рами, отростчатой формой, нечеткими (размытыми) границами,
светлой окраской цитоплазмы и ядра (см. рис. 2.23, Б) и способ-
ностью к активному передвижению (рис. 2.25). Особенности
ультраструктурной организации этих клеток (гармоническое раз-
витие гранулярной эндоплазматической сети и комплекса Гольджи)
отражают их главное функциональное назначение — синтез белков
и углеводных компонентов межклеточного вещества (рис. 2.26, А, Б).
Динамику этих сложных процессов можно изучить на примере
59
3
60
Рис. 2.23. Рыхлая волокнистая соединительная ткань
А, Б, В — пленочные препараты (А, Б — окраска железным гематоксилином;
В — окраска трипановым синим).
I — основное вещество; II — коллагеновые волокна; III — эластические волокна;
IV — клетки.
1 — фибробласты, 2 — фиброцит, 3 — макрофаги, 4 — тучные, 5 — плазмоциты,
6 — лейкоциты, 7 — жировая; V — кровеносный сосуд.
Рис. 2.24. Ультраструктура коллагеновых
волокон и фибрилл.
А — СЭМ. Б — ТЭМ: периодическая исчерчен-
ность коллагеновых волокон и фибрилл. В —
схема расположения молекул коллагена в фи-
брилле (А, Б — по Н. П. Омельяненко).
1 — коллагеновая фибрилла: а — светлый участок,
б — темный участок; 2 — молекулы коллагена;
3 — коллагеновое волокно; 4 — поперечный срез
коллагенового волокна.
61
Рис. 2.25. Фибробласт.
А — выявление актиновых микрофиламентов иммунофлюоресцентным методом.
Б — СЭМ: распластывание фибробласта на цилиндрической поверхности.
этапов внутриклеточного синтеза предшественников коллагена и
сборки коллагеновых волокон на поверхности фибробласта
(рис. 2.27).
Фиброцит легко определить по малому размеру, уплощен-
ной веретеновидной форме, более темной окраске цитоплазмы и
ядра, а также высокому ядерно-цитоплазматическому показателю
(см. рис. 2.23, Б). Ультраструктурно он характеризуется слабым
развитием органелл (см. рис. 2.24, В), поэтому в функциональ-
ном отношении является малоактивной клеткой — поддерживает
лишь собственный метаболизм и гомеостаз межклеточного веще-
ства в своем регионе. Таким образом, несмотря на генетическую
общность, фибробласт и фиброцит имеют четкие структурно-
функциональные различия.
Макрофаг. Труднее дифференцировать фибробласт и
макрофаг. Помогут в этом следующие ориентиры: частая локали-
зация макрофагов вблизи мелких сосудов, тенденция к объедине-
нию в обширные группы, мелкий размер этих клеток, непостоян-
62
Рис. 2.26. Ультраструктура клеток фибро-
бластического дифферона.
А — молодой фибробласт. Б — дифференци-
рованный фибробласт. В — фиброцит (ТЭМ).
I — ядро; 2 — свободные рибосомы; 3 — гра-
нулярная эндоплазматическая сеть; 4 — об-
ширная зона комплекса Гольджи; 5 — мито-
хондрии; 6 — коллагеновые волокна (А, В —
по Г. Г. Кругликову).
63
64
ство их формы (от овальной до слабоотростчатой), четко очер-
ченные границы, более насыщенная окраска ядра и цитоплазмы.
И еще один признак — неоднородный «пенистый» вид цитоплазмы
из-за присутствия в ней многочисленных светлых вакуолей
(см. рис. 2.23,Б). При введении в организм витального красителя
макрофаги активно поглощают его, поэтому их вакуоли приобре-
тают цвет фагоцитированной краски (см. рис. 2.23,В). При изуче-
нии ультраструктуры макрофага следует обратить внимание на
длинные пластинчатые отростки, которые придают поверхности
клетки бугристый вид (рис. 2.28). Они служат для поглоще-
ния инородных частиц и одновременно являются аппаратом
передвижения и фиксации к субстрату. Наличие эндосом (пино-
и фагосом), обилие первичных лизосом, присутствие гетерогенных
вторичных лизосом и остаточных телец отражают активность
процессов внутриклеточного пищеварения. По состоянию органелл,
количеству фагоцитированного материала и рельефу клеточной
поверхности можно без труда дифференцировать активную и
неактивную форму функционирования макрофага (см. рис. 2.28, А, Б).
Тучные клетки, или тканевые базофилы.
Так же как и макрофаги, они чаще располагаются группами вбли-
зи сосудов. Форма клеток обычно овально-округлая, а вся цито-
плазма заполнена специфическими базофильными гранулами, ко-
торые частично или полностью маскируют ядро, из-за чего оно
кажется несоразмерно малым (см. рис. 2.23, Б). Эти гранулы об-
ладают метахромазией, которая обусловлена содержанием в них
гепарина. Главными ультраструктурными признаками тучной клет-
ки являются аппарат передвижения — цитоплазматические от-
ростки, центральная локализация ядра, слабое развитие органелл
и обилие гранул, выбухающих на поверхности клетки в виде шаро-
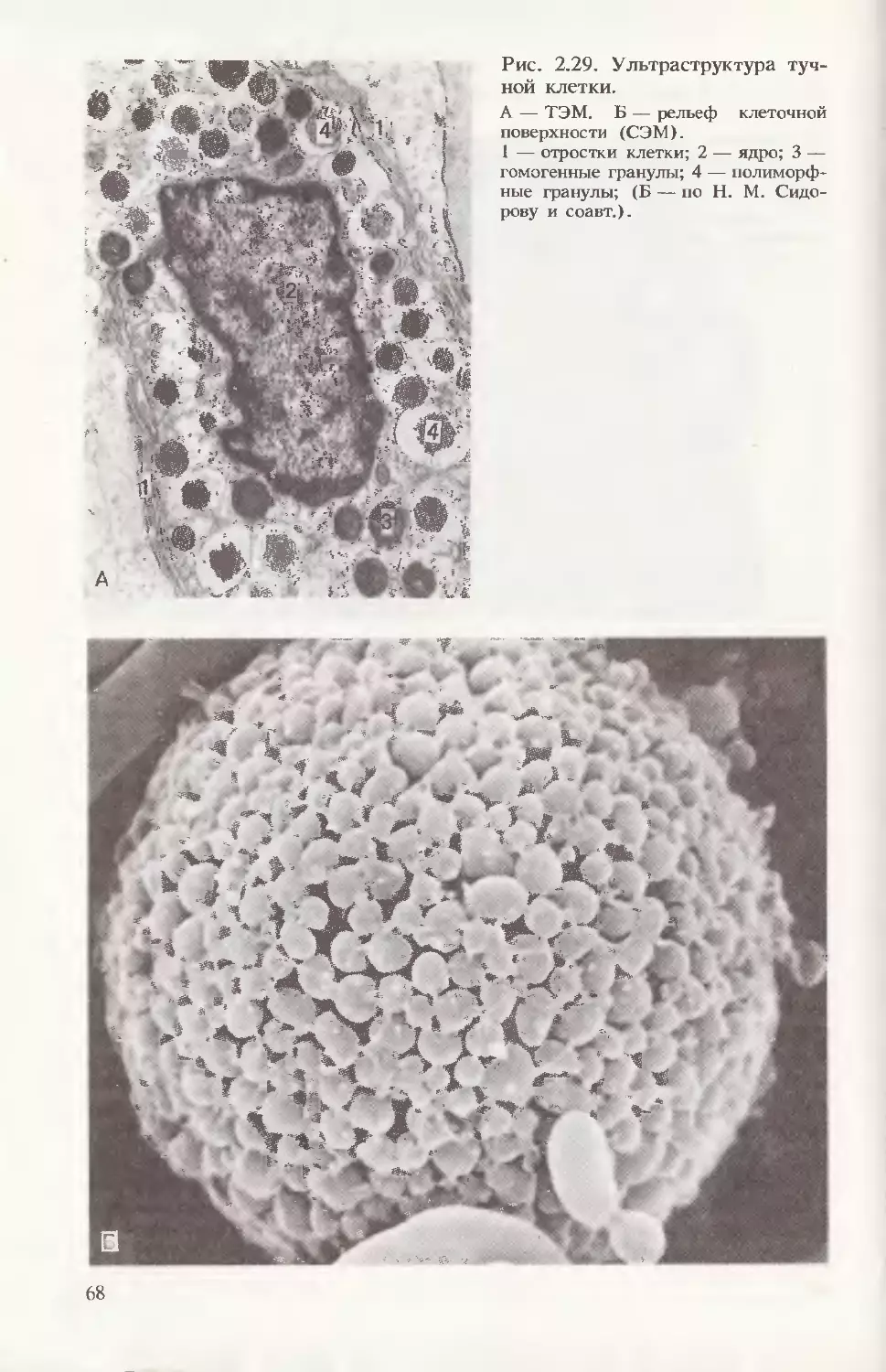
видных структур (рис. 2.29). Следует обратить внимание на то,
что зрелые гранулы темные и гомогенные, тогда как незрелые по-
лиморфны, варьируют по степени плотности и размеру (см.
рис. 2.29,А). Поскольку тучные клетки наряду с гепарином со-
держат в гранулах еще и гистамин, они могут усиливать или ос-
лаблять процессы проницаемости в своем регионе, выступая как
тактические регуляторы местного гомеостаза.
Кроме того, в рыхлой волокнистой соединительной ткани при-
сутствуют единичные жировые клетки, различные виды
лейкоцитов, а также плазматические клетки
(см. рис. 2.23,Б).
Рис. 2.27. Этапы формирования коллагеновых волокон.
А — схема. Б — фрагмент фибробласта с зоной новообразованных фибрилл (ТЭМ).
В — коллагеновые волокна на поверхности фибробласта (СЭМ).
1 — внутриклеточный этап синтеза проколлагена и гликозаминогликанов; 2 —
внеклеточный этап сборки коллагеновых волокон на оболочке клетки; 3 — колла-
геновые фибриллы; 4 — коллагеновые волокна. (Б, В — по Г. Г. Кругликову).
3—321
65
66
Рис. 2.28. Ультраструктура макро-
фага.
А — активная форма. Б — неактивная
форма функционирования макрофага
(ТЭМ). В — поверхность макрофага
(СЭМ).
I — отростки клетки; 2 — ядро; 3 —
первичные лизосомы; 4 — вторичные
лизосомы.
Плазмоциты. Как известно, плазмоциты являются потом-
ками В-лимфоцитов, поэтому их количество прежде всего опре-
деляется состоянием иммунологической защиты в организме. Эти
клетки характеризуются небольшим размером, овальной формой,
эксцентричной локализацией ядра и радиальным расположением
в нем гетерохроматина. Окраска их цитоплазмы неравномерна:
на большем протяжении она насыщенно базофильна, а в около-
ядерной зоне резко просветляется в форме округлой сферы или
«дворика» (рис. 2.30, А). Указанные тинкториальные признаки
плазмоцитов обусловлены своеобразием их ультраструктурной
организации. Так, основную часть цитоплазмы клеток занимает
гранулярная эндоплазматическая сеть, канальцы которой вблизи
дра имеют тенденцию к плотному концентрическому располо-
жению (рис. 2.30, Б, В). В надъядерной же зоне локализован
комплекс Гольджи. Его многочисленные элементы в совокупности
формируют обширную сферу — эквивалент «светлого дворика».
О шадая столь мощным аппаратом биосинтеза, плазмоциты функ-
ционируют как непрерывный конвейер по продукции «экспорт-
ного» белка — секретируют специфические антитела.
Следовательно, в составе рыхлой волокнистой соединительной
1кани наряду с клетками фибробластического ряда, создающими
се межклеточное вещество, широко представлены лейкоциты,
которые заселяют его, поступая из крови. Часть из них специали-
3*
67
Рис. 2.29. Ультраструктура туч-
ной клетки.
А — ТЭМ. Б — рельеф клеточной
поверхности (СЭМ).
1 — отростки клетки; 2 — ядро; 3 —
гомогенные гранулы; 4 — полиморф-
ные гранулы; (Б — по Н. М. Сидо-
рову и соавт.).
68
Рис. 2.30. Плазмоцит.
А — окраска гематоксилин-эозином.
Б — схема. В — ТЭМ.
I — ядро; 2 — радиальный гетерохро-
матин; 3 — концентрическое располо-
жение элементов гранулярной эндоплаз-
матической сети; 4 — обширная зона
комплекса Гольджи; 5 — светлый «дво-
рик»; 6 — митохондрии.
69
Рис. 2.31. Рыхлая волокнистая и
плотная неоформленная волок-
нистая соединительные ткани.
А — общий вид. Б — коллагеновые и
эластические волокна (СЭМ).
I — эпидермис; II — рыхлая волок-
нистая соединительная ткань; III —
плотная волокнистая неоформленная
соединительная ткань.
1 — коллагеновые волокна; 2 — ос-
новное вещество; 3 — клетки (а —
фиброциты); 4 — пучки коллагено-
вых волокон (б — продольный, в —
поперечный, г—косой срезы); 5 —
эластические волокна; 6 — сосуд
(Б — по Н. П. Омельяненко).
зируется в макрофаги, тучные клетки и плазмоциты, участвующие
в регуляции основных свойств этой ткани или обеспечивающие ее
защитные реакции.
Поскольку в дальнейшем Вы будете встречать рыхлую волок-
нистую соединительную ткань не в тотальных препаратах, а в
традиционных срезах, необходимо остановиться на особенностях
ее морфологии в данных случаях. В сравнении с пленочным
препаратом в срезе площадь основного вещества значительно со-
кращается (как следствие обезвоживания в спиртах) и поэтому
волокна располагаются более плотно (рис. 2.31, А), причем видны
только оксифильные коллагеновые волокна, тогда как эластиче-
ские не обнаруживаются, поскольку слабо воспринимают эозин.
Основное вещество также не окрашивается и занятые им участки
выглядят «пустотами». Клеток много, но дифференцировать их
значительно труднее. Таким образом, в срезе существенно изме-
няются количественные и пространственные соотношения компо-
нентов межклеточного вещества и функционирующих в нем клеток.
Заканчивая изучение рыхлой волокнистой соединительной тка-
ни, необходимо суммировать главные черты ее гистофизиологии.
Но-первых, необычайное многообразие клеток и различия в про-
филях их специализации обеспечивают полифункциональные
свойства этой ткани. Во-вторых, принцип организации межкле-
точного вещества создает оптимальные условия для транспортных
процессов и функционирования всех видов клеток и, кроме того,
наделяет ткань в целом формообразующими, опорными и пласти-
ческими свойствами. В-третьих, избирательная периваскулярная
локализация данной ткани позволяет ей формировать единую
трофическую систему с гемокапиллярами, поэтому только она в
организме опосредует гематотканевые взаимодействия.
2.3.1.1.2. Плотная волокнистая ткань
2.3.1.1.2.1. Плотная неоформленная волокнистая
соединительная ткань
Строение этой ткани целесообразно изучить в сравнительном ас-
пекте. Тесное топографическое соседство в дерме кожи сразу двух
соединительнотканных разновидностей — рыхлой волокнистой и
плотной неоформленной тканей — позволяет довольно четко диф-
ференцировать отличительные черты последней (см. рис. 2.31, А, Б).
Прежде всего данная ткань характеризуется отчетливым преоб-
ладанием волокнистых компонентов над основным веществом, а
также скудностью и однотипностью клеточного состава, главным
образом это клетки фиброциты. Коллагеновые волокна для проч-
ности объединены в толстые пучки, имеющие различное направ-
ление: продольное, поперечное и косое. Пучки коллагеновых во-
локон в совокупности с ветвящимися эластическими волокнами
формируют единый механический остов плотной неоформленной
соединительной ткани.
71
23.1.1.2.2. Плотная оформленная волокнистая
соединительная ткань
В организме эта ткань встречается в связках, сухожилиях, фиб-
розных мембранах и т. д. Сухожилие имеет двухкомпонентный тка-
невой состав. Так, рыхлая соединительная ткань формирует перите-
ноний и эндотеноний, организующие плотную оформленную соеди-
нительную ткань в орган — сухожилие. Перитеноний покрывает
его поверхность, а эндотеноний разделяет на сухожильные пучки
второго порядка (рис. 2.32, А, Б). Каждый сухожильный пучок
второго порядка состоит из многочисленных параллельно ориенти-
рованных сухожильных пучков первого порядка, которые разгра-
ничены слоями фиброцитов. Следовательно, волокна и клетки в
сухожилии располагаются упорядоченно и ориентированы строго
вдоль его оси.
Различия в организации волокнистого остова плотных не-
оформленной и оформленной соединительных тканей наглядно
предстают при сравнении двух сканограмм (см. рис. 2.31, Б и
2.32, В).
Таким образом, главную роль в гистофизиологии плотных
соединительных тканей играют уже не клетки и аморфное вещест-
во, а мощный остов волокнистых компонентов, объединенных в
пучки. Сам принцип организации остова и прежде всего ориента-
ция в нем волокон четко адаптированы к тем механическим
нагрузкам, которые предъявляются к конкретной тканевой раз-
новидности.
2.3.2. Специализированные соединительные ткани
2.З.2.1. Ретикулярная соединительная ткань
Обычно ее строение изучают в срезах кроветворных органов,
поскольку она формирует их основу, или паренхиму. Эта ткань
характеризуется рыхлым расположением клеток и имеет вид окси-
фильной сети, образованной отростчатыми ретикулярными клет-
ками (рис. 2.33, А). Представление о взаимоотношениях ретику-
лярных клеток и волокон можно составить с помощью рис. 2.33, Б, В.
2.3.2.2. Белая жировая ткань
Гистохимически участки локализации жировой ткани легко обна-
ружить по ярко-оранжевой окраске (суданофильности) ее клеток
(рис. 2.34, А). Они обычно располагаются обширными группами по
ходу мелких сосудов, имеют шаровидную форму и достаточно круп-
ны, поскольку дифференцируются даже при малом увеличении. Вся
цитоплазма этих клеток заполнена одной большой «каплей» жиро-
вых включений, а ядро оттеснено на самую периферию жировой
клетки (рис. 234, Б).
В жировой ткани подкожной клетчатки отчетливо обнаружи-
72
Б
Рис. 2.32. Плотная оформленная
волокнистая соединительная ткань.
А — поперечный срез сухожилия. Б —
продольный срез сухожилия. В — во-
локнистый остов сухожилия (СЭМ).
1 — перитеноний; 2 — эндотеноний:
3 — сухожильные пучки П порядка;
4 — фиброциты (сухожильные клетки);
5 — сухожильные пучки 1 порядка;
6 — продольные параллельные сухо-
жильные волокна; 7 — поперечные свя-
зующие волокна (А, Б — по И. В. Ал-
мазову; Л. С. Сутулову; В — по
Н. П. Омельяненко).
73
Рис. 2.33. Ретикулярная
ткань лимфатического
узла.
А — общий вид. Б, В — схема
строения.
1 — ретикулярные клетки
(а — отростки, б — ядро, в —
гранулярная цитоплазматиче-
ская сеть, г — комплекс
Гольджи, д — митохондрии);
2 — основное вещество; 3 —
лимфоциты; 4 — ретикуляр-
ные волокна.
74
Рис. 2.34. Белая жировая ткань.
Л. Б — тотальные препараты сальника, окраска Суданом Ш-гематоксилином,
Н. Г — препарат подкожной клетчатки. Д — жировая ткань брыжейки (СЭМ).
I жировая клетка (а — плазмолемма, б — цитоплазма, в — жировые включения,
। пустоты, оставшиеся после удаления жировых включений обезвоживанием,
д ядро); 2 — сосуд; 3 — долька жировой ткани; 4 — соединительнотканная про-
i 1ойка (В, Г — по И. И. Алмазову, Л. С. Сутулову; Д — по А. А. Миронову).
вается дольчатый характер ее строения: дольки различной формы
и величины разграничены соединительнотканными прослойками
(рис. 2.34, В). Липоциты долек настолько плотно прилежат друг
к другу, что между ними не определяется межклеточного вещества.
Сами клетки выглядят пустотелыми (жир экстрагируется при об-
работке материала), поэтому территория дольки имеет ажурный
вид и напоминает пчелиные соты. В тонком оксифильном ободке
периферической зоны цитоплазмы содержится уплощенное ядро
(рис. 2.34, Г). За внешнее сходство эти клетки часто называют
перстневидными. Аналогичное строение липоцитов можно наблю-
дать и на сканограмме (рис. 2.34, Д).
2.3.3. Скелетные ткани
233.1. Хрящевые ткани
2.3.3.1.1. Гиалиновая хрящевая ткань
В большинстве органов эта ткань окружена надхрящницей и в
совокупности с ней формирует самостоятельное анатомическое
образование — гиалиновый хрящ (рис. 2.35, А). Надхрящница
внутрь хряща не проникает, поэтому данная ткань является бессо-
судистой и трофика ее структур обеспечивается за счет диффузии
веществ. В составе надхрящницы различают два слоя: наружный
волокнистый и внутренний хондрогенный, более рыхлый с сосудами
и мелкими веретеновидными хондрогенными клетками и хондробла-
стами. Эти клетки обеспечивают поверхностный аппозиционный
рост хряща, формируя новую хрящевую ткань в дополнение к уже
существующей.
В самой гиалиновой хрящевой ткани различают две зоны.
Под надхрящницей располагается узкая зона молодого хряща
с одиночными уплощенными хондроцитами и гомогенным оксифиль-
ным матриксом.
Вторая зона — зрелого хряща содержит изогенные группы хон-
дроцитов. Ее межклеточное вещество безофильно (см. рис. 2.35, А).
Территориальный матрикс вокруг хрящевых лакун имеет более ин-
тенсивную окраску, чем остальной интертерриториальный матрикс
хрящевой ткани. За счет деления хондроцитов обеих зон происходит
увеличение хрящевой ткани изнутри, или интерстициальный рост.
Поскольку коллагеновые волокна гиалинового хряща при обыч-
ной окраске (гематоксилин-эозин) не выявляются, с помощью
рис. 2.36, А, Б следует изучить особенности их расположения в
различных участках хрящевого матрикса и в стенке хрящевой
лакуны.
2.3.3.1.2. Эластическая хрящевая ткань
Основное отличие этой хрящевой ткани от гиалиновой заключа-
ется в строении межклеточного вещества. При окраске орсеином
(селективным красителем для эластических структур) в его соста-
76
Рис. 2.35. Хрящевая ткань.
А — гиалиновый хрящ ребра. Б — эластиче-
ский хрящ ушной раковины (окраска орсеи-
ном). В — волокнистый хрящ межпозвоноч-
ного диска (А, Б — по И. В. Алмазову,
Л. С. Сутулову).
I — надхрящница; 1 — волокнистый слой. 2 —
хондрогенный слой, 3 — хондрогенные клетки,
4 — хондробласты.
II — зона молодого хряща; 5 — уплощенные
одиночные хондроциты, 6 — хрящевой мат-
рикс.
III — зона зрелого хряща; 7 — изогенная
группа хондроцитов, 8 — хрящевая лакуна,
9 — базофильный хрящевой матрикс (а — тер-
риториальный, б — интертерриториальный),
10 — основное вещество, 11 — эластические
волокна, 12 — пучки коллагеновых волокон,
13 — фиброциты.
77
Рис. 2.36. Клетки и волокна матрикса гиалинового хряща.
А — коллагеновые волокна. Б — хондроцит в хрящевой лакуне (СЭМ) (А, Б — по
Н. П. Омельяненко).
78
ве четко дифференцируются два компонента: гомогенное основное
вещество и густая сеть эластических волокон (см. рис. 2.35, Б).
В хрящевой пластинке эти волокна ориентированы поперечно.
На периферии ее они более тонкие, ветвящиеся, а ближе к центру
заметно утолщаются. В остальном принцип организации эластиче-
ского хряща точно такой же, как и гиалинового: два слоя в над-
хрящнице, аналогичные зоны собственно хрящевой ткани (см.
рис. 2.35, А, Б). Следовательно, эластическая хрящевая ткань в
сравнении с гиалиновой имеет иной волокнистый каркас. Он пред-
ставлен как коллагеновыми, так и эластическими волокнами,
поэтому ткань обладает не только прочностью, но и эластичностью.
2.3.3.1.3. Волокнистая хрящевая ткань
Третья разновидность хрящевых тканей построена совсем иначе,
чем две предыдущие. Межклеточное вещество данного хряща име-
ет самый мощный волокнистый каркас. По своей организации он
точно такой же, как в плотной оформленной соединительной
ткани: коллагеновые пучки ориентированы параллельно друг другу,
их разделяют цепочки сухожильных клеток — фиброцитов (см.
рис. 2.35, В). Однако по мере удаления от сухожилия плотность
расположения волокон меняется и пучки постепенно разрыхля-
ются. При этом между волокнами определяются уже не фибро-
бласты, а хрящевые клетки. Волокнистая хрящевая ткань не толь-
ко по локализации в организме, но и по специфике строения пред-
ставляет собой постепенный переход от сухожилия к гиалиновому
хрящу.
2.3.3.2. Пластинчатая костная ткань
Поскольку из этой ткани построено большинство плоских и труб-
чатых костей скелета, целесообразно изучить ее строение в составе
диафиза трубчатой кости (рис. 2.37). Пластинчатая костная ткань
в отличие от хрящевой обильно васкуляризована. От надкостницы
(периоста) и эндоста в вещество кости проникают прослойки
соединительной ткани. В них проходят многочисленные сосуды
и нервы, а также содержатся остеогенные клетки, остеобласты
и костные макрофаги — остеокласты. Особенности субмикроско-
пического строения остеобластов и остеокластов (рис. 2.38, А, Б)
свидетельствуют о их специализации для выполнения противо-
положных функций — секреции и разрушения межклеточного
вещества (костного матрикса). Эти клетки обеспечивают пласти-
ческие свойства кости — способность изменять, перестраивать
внутреннюю структуру в течение жизни.
Элементарной единицей построения этой костной ткани являет-
ся костная пластинка. Между костными пластинками в костных
лакунах располагаются тела остеоцитов (рис. 2.38, В, Г), а их
многочисленные отростки лежат в костных канальцах, где контак-
79
80
Рис. 2.37. Пластинчатая костная ткань диафиза трубчатой кости.
А — общий план строения. Б — клеточный состав внутреннего слоя надкостницы.
В — структурные компоненты остеона (схемы). Г — препарат (декальцинирован-
ный срез, окраска по методу Шморля). Д — остеоны. Е — вставочные пластинки
(СЭМ).
I — периост (а — волокнистый слой, б — остеогенный слой, в — остеогенные
клетки, г — остеобласты, д — остеокласты); 2 — кровеносные сосуды; 3 — нервы;
4 — эндост; 5 — слой наружных окружающих пластинок; 6 — прободающие кол-
лагеновые волокна; 7 — прободающий канал; 8 — остеонный слой; 9 — остеон
(е — центральный канал, ж — система концентрических пластинок); 10 — вставоч-
ная пластинка; 11—слой внутренних окружающих пластинок; 12 — остеоциты
(А, Б, В — по Ю. И. Афанасьеву, Д, Е — по Ю. И. Денисову-Никольскому).
тируют с отростками соседних клеток. Лакунарно-канальцевая
система связана с капиллярами кости. По ней циркулирует тка-
невая жидкость, обеспечивающая трофику остеоцитов, поскольку
минерализованное межклеточное вещество костных пластинок
исключает диффузию веществ. В функциональном отношении
остеоциты — малоактивные клетки (органеллы развиты слабо —
см. рис. 2.38, В), основное их значение — поддержание метабо-
лизма в окружающих костных пластинках.
В соответствии с порядком расположения костных пластинок
в диафизе различают три слоя. Первый — наружных общих плас-
тинок. В него из надкостницы проникают прободающие каналы и
81
10
82
волокна (см. рис. 2.37, А, Г). Средний и самый обширный — осте-
онный слой состоит из остеонов (с питающими их сосудами) и
лежащих между ними вставочных пластинок (см. рис. 2.37, В,
Г, Д, Е). Третий — слой внутренних общих пластинок, он форми-
рует сложный рельеф внутренней поверхности кости (см.
рис. 2.37, А, Г). Принцип строения костных пластинок и особен-
ности их архитектоники в кости наделяют ее необычной проч-
ностью, поэтому в механическом отношении кость наиболее совер-
шенна в организме.
2.З.З.2.1. Остеогистогенез
Прямой остеогистогенез. Так называется способ развития костной
ткани, например плоской кости челюсти, непосредственно из ме-
зенхимы. В этом процессе различают четыре стадии. Рассмотрим
одну из них, третью по последовательности — минерализации
(кальцинации) межклеточного вещества новообразованной кости.
Участок остеогистогенеза представлен разветвленной системой
костных балок (перекладин) причудливой конфигурации. Они
окружены мезенхимой, отростчатые клетки которой формируют
рыхлый сетчатый остов. Мезенхимные клетки имеют очень круп-
ное светлое ядро, небольшой объем цитоплазмы и обладают высо-
кой м 1че кой активностью. Между клетками находится го-
могенное неокрашенное межклеточное вещество, в котором опре-
деляются многочисленные кровеносные сосуды. Их легко диффе-
ренцировать по скоплению эритроцитов в просвете (рис. 2.39, А).
В минерализованном гомогенном оксифильном межклеточном
веществе костньщДаадвк замурованы остеоциты. Они располагают-
ся в крстных лг кунах а их отростки — в канальцах ретикулофиб-
РОЗНЙЙ (грубоволокнистой) эмбриональной костной ткани. Однако
канальцевая система, так же как коллагеновые волокна матрикса
этой ткани, в препарате не выявляются. По периферии каждая
балка окружена прерывистыми рядами остеобластов. В тех участ-
ках, где вокруг балки группируются только призматические остео-
бласты с интенсивно базофильной цитоплазмой, происходит
аппозиционный рост этой балки. В таких случаях между остео-
бластами и матриксом балки определяется узкая слабо окрашен-
ная полоска остеоида — некальцинированного межклеточного вд-
Рис. 2.38. Ультраструктура клеток костной ткани.
А — остеобласт. Б — остеокласт (схемы). Остеоцит: В — схема; Г — СЭМ.
1 — стенка костной лакуны; 2 — костный каналец; 3 — отросток остеоцита; 4 —
ядро; 5 — гранулярная эндоплазматическая сеть; 6 — митохондрии; 7 — комплекс
Гольджи; 8 — остеоид; 9 — костный матрикс; 10 — ядра остеокласта; 11—дно
эрозионной лакуны.
1 — гофрированная каемка (а — складки плазмолеммы, б — микроворсинки);
II — светлая зона (микротрубочки и микрофиламенты); III — вакуолярная зона
(в — вакуоли, г — везикулы); IV — базальная зона (Г — по Ю. И. Денисову-Ни-
кольскому) .
83
Рис. 2.39. Прямой остеогистогенез (развитие плоской кости челюсти из
мезенхимы).
А — общий вид, Б — деталь препарата.
1 — мезенхима (а — мезенхимные клетки, б — межклеточное вещество); 2 —
кровеносный сосуд; 3 — костная балка; 4 — минерализованный матрикс балки
(в — костная лакуна остеоцита); 5 — остеоциты; 6 — остеобласты (г — цилиндри-
ческие, д — плоские); 7 — остеоид; 8 — остеокласты; 9 — эрозионная лакуна;
10 — остеогенный островок.
84
щества, синтезированного этими клетками (рис. 2.39, Б). Если же
процессы новообразования кости ослабляются или прекращаются,
то остеобласты приобретают резко уплощенную форму или отсут-
ствуют совсем. В таких участках обычно наблюдается разруше-
ние — резорбция костного матрикса, поэтому вокруг них кон-
центрируются остеокласты — многоядерные макрофаги костной
ткани. В их цитоплазме можно обнаружить от 2 до 10 ядер и
более. В зоне соприкосновения остеокласта с костной балкой
определяется полость разрушенного костного матрикса — эрози-
онная лакуна.
Непрямой остеогистогенез. Это способ развития костной ткани
из мезенхимы через стадию хряща. Так в организме развивается
большинство костей скелета.
Вначале из .мезенхимы образуется хрящевая модель будущей
трубчатой кости. Она состоит из ддаадиново£о хряща, покрытого
надхрящницей. Процесс остеогистогенеза начинается в области
диафиза.ТТрИ"этом скелетогенные клетки надхрящницы диффе-
ренцируются в сторону остеобластов, котррые между -надхрящни-
цей и хрящом, т. е. перихондрально, образуют ретикулофиброз-
ную костную ткань, затем она перестраивается в пластинчатую.
По топографии эту кость называют перихондральной, поскольку
она в виде ажурной манжетки окружает диафиз хряща (рис. 2.40, А).
В дальнейшем надхрящница преобразуется в надкостницу, поэтому
трофика диафизарного хряща нарушается и он подвергается обыз-
вествлению. В минерализованный хрящ из надкостницы через от-
верстия перихойдральной кости врастает ^остеобластическая поч-
ка — кровеносные сосуды вместе с мезенхимными клетками,
остеобластами и остеокластами. Так возникает диафизарный центр
окостенения: элементы остеобластической почки служат источни-
ком образования костной ткани внутри разрушенного диафизар-
ного хряща. По топографии эта кость называется эндохондраль-
ной. Пери- и эндохондральная кости представлены разветвленной
сетью костных балок с замурованными, дстеоцитами, по периферии
их окружают остеобласты, а в местах резорбции встречаются
остеокласты. Отличить балки эндохондральной кости можно, во-
первых, по их топографии, во-вторых, по гетерогенности их мат-
рикса — в нем определяются базофильные метахроматические
участки разрушенного хряща (рис. 2.40, Б). Межклеточное же
вещество балок перихондральной кости равномерно оксифильное
и гомогенное.
Вскоре новообразованная эндохондральная кость в самом
центре диафиза начинает разрушаться qgде.ркаастамн.— возникает
костномозговая полость, формируется строма костного мозга и
начинаются процессы кроветворения.
Распространение фронта диафизарной оссификации в длину
приводит к четкой зональности в строении эпифизарного гиалино-
вого хряща. В его составе можно дифференцировать четыре зоны
(см. рис. 2.40,А). На границе с диафизом располагается зона
резорбции. Она характеризуется разрушением минерализованного
85
Рис. 2.40. Непрямой остеогистогенез (развитие трубчатой кости на месте
хряща).
А — общий вид. Б — деталь препарата (А, Б — по И. В. Алмазову, Л. С. Сутулову).
1 — надхрящница; 2 — зона неизмененного хряща; 3 — зона пролиферации: 4 —
зона гипертрофии; 5 — зона резорбции; 6 — периост; 7 — перихондральная кост-
ная ткань; 8 — эндохондральная костная ткань; 9 — первичная костномозговая
полость; 10 — остеоциты; 11—остеобласты; 12 — остеокласт; 13 — базофильные
остатки хряща; 14 — эрозионная лакуна.
86
хряща и замещением его костной тканью. За ней следует зона
гипертрофии, содержащая вакуолизированные и гипертрофирован-
ные хондроциты. Ее сменяет зона пролиферации, где колонки
уплощенных хондроцитов располагаются как столбики монет.
Самой дистальной и обширной является зона неизмененного хряща,
окруженная надхрящницей. В ее составе клетки равномерно рас-
пределены в слабо-базофильном хрящевом матриксе.
Таким образом, костная ткань может развиваться двумя спо-
собами: непосредственно из мезенхимы или опосредованно через
стадию залоснённой ранее хрящевой модели. Однако, как в первом,
так и во втором случае процесс остеогистогенеза всегда происхо-
дит в тех участках мезенхимы, где есть кровеносные сосуды, Вас-
куляризащнГ-создает условия~длй быстрой минерализации матрик-
са костных балок по мере их образования, поэтому костная ткань
в отличие от хрящевой лишена способности к интерстициальному
росту. Следует усвоить, что единственным механизмом роста любой
костной структуры являетсдиДлпозиционный, т. е. способ наложе-
ния новой костной ткани на уже имеющуюся поверхность.
2.4. Мышечные ткани
Мышечные ткани специализированы главным образом для сокра-
тительных процессов в организме. Они приводят в движение рыча-
ги скелета, обеспечивают ритмическую деятельность миокарда и
гемоциркуляцию в сосудах, активно участвуют в регуляции пе-
ристальтики, функционировании сфинктеров и поддерживают нор-
мальный тонус сосудов и полых внутренних органов.
2.4.1. Поперечнополосатая мышечная ткань
2.4.1.1. Скелетная мышечная ткань
Структурной единицей данной ткани является мышечное волокно.
Кроме того, в ее состав входят соединительнотканные компоненты.
Они представлены эпимизием, перимизием и эндомизием, в кото-
рых проходят сосуды и нервы. Мышечные и соединительнотканные
элементы в совокупности формируют орган — скелетную мышцу.
Ее строение характеризуется продольным и, как правило, парал-
лельным расположением мышечных волокон (рис. 2.41, А), по-
этому направленность производимых ими движений всегда совпа-
дает с ориентацией мышцы. В других органах, например языке,
мышечные волокна и их пучки располагаются в различных направ-
лениях: продольном, косом и поперечном (рис. 2.41, Б), что обес-
печивает этому органу диапазон движений, необходимый для вы-
полнения сложных актов артикуляции и механической обработки
пищи.
Светооптически в продольном срезе волокна можно изучить
симпластический принцип его организации. Это протяженный
оксифильный цитоплазматический тяж, на периферии которого,
87
Рис. 2.41. Скелетная поперечнопо-
лосатая мышечная ткань.
А — строение мышцы как органа. Б —
мышечная ткань языка.
1 — эпимизий; 2 — перимизий; 3 — эн-
домизий: а — сосуды, б — нервы: 4 —
поперечный срез мышечных волокон;
5 — продольный срез мышечных воло-
кон; 6 — сарколемма; 7 — цитоплазма;
8 — ядра волокна; 9 — ядра фиброци-
тов.
88
вблизи его оболочки — сарколеммы, располагаются многочислен-
ные овальные ядра. Они крупные и светлые, их следует дифферен-
цировать от более темных и мелких ядер фиброцитов, лежащих
за пределами волокна в эндомизии.
Субмикроскопическое изучение дает более широкую информа-
цию о структурах мышечного волокна. В частности, в составе его
сарколеммы четко определяются три компонента: плазмолемма
миосимпласта, базальная мембрана и заключенные между ними
камбиальные клетки — миосателлитоциты (рис. 2.42, А, Б). Следо-
вательно, волокно представляет собой не истинный симпласт, а
клеточно-симпластический комплекс, окруженный общей базаль-
ной мембраной.
Важным морфологическим признаком расслабленного мышеч-
ного волокна (а точнее, его симпласта) является наличие в цито-
плазме периодической исчерченности — четкой поперечной (регу-
лярное чередование темных и светлых тонких полосок) и менее
четкой продольной (рис. 2.43, А). Исчерченность обусловлена
присутствием в цитоплазме многочисленных миофибрилл: про-
дольная — параллельной ориентацией миофибрилл вдоль оси во-
Рис. 2.42. Структурные компоненты сарколеммы мышечного волокна.
А — схема. Б — поперечный срез волокна (ТЭМ).
1 — базальная мембрана; 2 — миосателлитоцит; 3 — плазмолемма миосимпласта;
4 — поперечный срез миофибрилл; 5 — митохондрии; 6 — гемокапилляр.
89
1
2
3 5
4
Рис. 2.43. Поперечная и продольная исчерченность цитоплазмы мышеч-
ного волокна.
А, Б — схемы. В — ТЭМ (по Л. П. Сидельниковой).
1 — эндомизий; 2 — ядра фиброцитов; 3 — плазмолемма; 4 — ядра волокна; 5 —
поперечная исчерченность цитоплазмы; 6 — продольная исчерченность цитоплазмы;
7 — диск А: а — светлая зона, б — линия М; 8 — диск I (в — линия Z); 9 — по-
перечный срез миофибрилл.
90
локна, а поперечная — особенностями их морфохимической орга-
низации (рис. 2.43, Б, В).
Подробную информацию о строении миофибриллы, составе и
ультратопографии ее миофиламентов в дисках I и А, химической
природе и структуре этих миофиламентов можно получить с по-
мощью рис. 2.44. Необходимо усвоить, что изотропный эффект
диска I (и его более светлая окраска в препарате) обеспечивается
однородным составом миофиламентов — он сформирован только
из актиновых нитей (рис. 2.44, А, Б). В диск же А входят разные
миофиламенты, как актиновые, так и миозиновые, поэтому он
обладает анизотропными свойствами и имеет более темную окрас-
ку в препарате (рис. 2.44, Б, В, Г). Чтобы понять механизм сокра-
щения, особое внимание следует уделить саркомерному принципу
организации миофибриллы. Саркомер — это ее элементарная
сократимая единица. В его пределах миофиламенты вступают во
взаимодействие: поперечные мостики толстых нитей мгновенно
сдвигают тонкие нити в глубину диска А (рис. 2.45, А—Е). Сле-
довательно, при сокращении длина самих миофиламентов не изме-
няется, просто происходит их «скольжение» друг относительно
друга. Эти события приводят к укорочению площади каждого сар-
комера, всех миофибрилл и волокна в целом.
Регуляция сократительных процессов в скелетной мышце осу-
ществляется на уровне ее структурно-функциональной единицы —
миона (рис. 2.46, А, Б). Возбуждение, инициируемое в эффектор-
ном нервно-мышечном окончании миона, по Т-системе (рис. 2.47, А)
мгновенно достигает триад (рис. 2.47, Б, В, Г). Здесь волна деполя-
ризации с поперечных трубочек распространяется на терминаль-
ные цистерны. В ответ из них в цитоплазму (четко на границе
I и А дисков каждого саркомера) активно поступают ионы каль-
ция, и они сокращаются. При прекращении нервных импульсов
кальций перекачивается обратно в терминальные цистерны, и по-
этому мышца расслабляется.
Метаболизм волокна обеспечивается его трофическим аппара-
том: многочисленными ядрами, органеллами общего значения и
включениями. Из органелл наиболее развиты митохондрии, они
сконцентрированы между миофибриллами или вблизи ядер (см.
рис. 2.43, В).
Заключая общую характеристику скелетной мышечной ткани,
следует подчеркнуть, что все волокна в ее составе метамерны за
счет многократного чередования комбинаций сократительного,
трофического, опорного аппаратов и триад. Поэтому повторяю-
щиеся микрорайоны волокна функционируют на всем его протя-
жении очень синхронно. Метамерность мышечных волокон во
многом обусловлена особенностями их гистогенеза.
2.4.1.1.1. Гистогенез скелетной мышечной ткани
Составить представление об этом процессе и морфологических
критериях каждой из четырех его стадий поможет рис. 2.48. Важ-
91
Рис. 2.44. Строение миофибриллы.
А — состав и структура тонкого мио-
филамента. Б — порядок расположения
миофиламентов в миофибрилле. В —
состав и структура толстого миофила-
мента. Г — принцип «работы» попереч-
ного мостика (схемы).
1 — тонкий миофиламент: а — мономер
актина, б — тропомиозин, в — тропо-
нин; 2 — толстый миофиламент: г —
шесть рядов поперечных мостиков —
головки молекул миозина, д — хвосто-
вые участки молекул миозина, е — свет-
лая зона.
Z M z
%
Рис. 2.45. Саркомер миофибриллы.
Стадия расслабления: А — схема, Б —
ТЭМ. Стадия сокращения: В — схема,
Г — ТЭМ. Д, Е — гексагональная упа-
ковка тонких и толстых миофиламен-
тов в расслабленном (Д) и сокращен-
ном (Е) саркомерах (схемы).
92
Рис. 2.45. Продолжение
(Б, Г — по Л. П. Сидельниковой).
93
Рис. 2.46. Структурные компоненты миона.
А — схема. Б — гемокапилляр миона (СЭМ).
I — эндомизий; 1 — коллагеновые волокна; 2 — ядра фиброцитов; 3 — гемокапил-
ляр; 4 — эффекторное нервно-мышечное окончание — моторная бляшка. II —
мышечное волокно: 5 — базальная мембрана; 6 — миосателлитоцит; 7 — миосим-
пласт: а — плазмолемма, б — ядро, в — миофибриллы (Б — по Я. Л. Караганову).
Рис. 2.47. Компоненты триад миосимпласта.
А — Т-система. Б — агранулярная эндоплазматическая сеть. В, Г — триада (В —
схема, Г — ТЭМ по Л. П. Сидельниковой).
1 — плазмолемма миосимпласта; 2 — митохондрии; 3 — миофибриллы; а — по-
перечные трубочки, б — «ячейки» Т-системы, в — терминальные цистерны, г —
трубчатые элементы, д — сетчатый элемент.
94
95
Рис. 2.48. Схема гистогенеза скелетной мышечной ткани.
А — миобластическая стадия. Б — миосимпластическая стадия. В — стадия мы-
шечных трубочек. Г — стадия формирования дефинитивных мышечных волокон.
1 — эпителиоподобные клетки миотома; 2 — промиобласты; 3 — миобласты; 4 —
квантальный, или дифференцирующий, митоз; 5 — слияние миобластов и начало
синтеза миофибрилл; 6 — образование миосимпласта; а — плазмолемма, б — ба-
зальная мембрана, в — миосателлитоцит, г — ядра миосимпласта, д — миофи-
бриллы.
ним моментом гистогенеза, определяющим переход от клеточной
(рис. 2.48, А) к симпластической (рис. 2.48, Б) стадии, является
дифференцирующий (квантальный) митоз. По его завершении
блокируется репродукция миобластов, они приобретают способ-
ность к слиянию в короткие симпласты, а в их цитоплазме появ-
ляются первые миофибриллы. Однако часть миобластов не всту-
96
пает в симпластическую дифференцировку и дает начало миоса-
теллитоцитам (рис. 2.48, В), поэтому зрелое мышечное волокно
(рис. 2.48, Г) формируется как клеточно-симпластическая единица.
2.4.1.2. Сердечная мышечная ткань
В организме эта ткань формирует мышечную оболочку только
одного органа — сердца. Мышечные элементы миокарда представ-
лены узкими сетевидными волокноподобными структурами, кото-
рые разграничены эндомизием и перимизием (рис. 2.49, А).
Структурной единицей данной ткани является клетка — кардио-
миоцит. Он имеет вытянутую прямоугольную форму и соединяется
с соседними клетками бок в бок с помощью вставочных дисков.
При этом формируется длинная цепочка клеток, очень напомина-
ющая мышечное волокно, поэтому его называют функциональным.
Функциональные волокна ориентированы в миокарде в различных
направлениях: продольно, косо, поперечно. В продольных срезах
можно определить вставочные диски — поперечные оксифильные
полоски, разделяющие каждое волокно на многочисленные сег-
менты, т. е. составляющие его клетки (рис. 2.49, Б). Кардиомио-
циты могут контактировать между собой и с помощью анастомо-
зов — нерегулярных ответвлений от клеток одного функциональ-
ного волокна к клеткам окружающих его волокон. В центре каж-
дого кардиомиоцита располагается одно, реже два ядра, а в пери-
ферических участках его цитоплазмы видна поперечная и про-
дольная исчерченность.
Ориентируясь на рис. 2.50, следует изучить отличительные
признаки субмикроскопического строения желудочковых кардио-
миоцитов. Особое внимание надо обратить на принцип организа-
ции мио-миоцитарных контактов — вставочных дисков и анасто-
мозов (наличие в их составе точечных десмосом, нексусов и опоя-
сывающих десмосом — зон фиксации миофибрилл к плазмолем-
ме). С помощью этих контактов кардиомиоциты желудочков и
предсердий объединяются в структурно-функциональные еди-
ницы — обширные клеточные комплексы, которые благодаря мно-
гочисленности нексусов сокращаются как единое целое.
Важно подчеркнуть, что в миокарде, кроме чисто сократитель-
ных клеток, имеются проводящие кардиомиоциты. Светооптически
они характеризуются крупным размером, овально-круглой формой,
эксцентричной локализацией ядра и светлой цитоплазмой, прак-
тически лишенной исчерченности (см. рис. 2.49, Б). О слабой сок-
ратимости свидетельствуют и особенности их ультраструктурной
организации — малочисленность и ветвящийся тип строения
миофибрилл, низкое содержание митохондрий, отсутствие триад
(рис. 2.51). Более упрощенную структуру имеют их вставочные
диски, а анастомозы отсутствуют. Указанные морфологические
признаки послужили основанием для другого названия этих
клеток — атипические кардиомиоциты. В совокупности они фор-
мируют проводящую систему миокарда, благодаря которой сердеч-
4—321
97
Рис. 2.49. Сердечная поперечнополосатая мышечная ткань.
А — общий вид. Б — деталь препарата.
1 — эндомизий; 2 — перимизий; 3 — вставочный диск; 4 — анастомоз; 5 — кардио-
миоцит; а — продольный, б — поперечные срезы; 6 — ядро кардиомиоцита;
7 — проводящие кардиомиоциты; 8 — миофибриллы (А, Б — по В. Г. Елисееву,
. И. Афанасьеву, Е. Ф. Котовскому).
S 5
Рис. 2.50. Ультраструктура типических
кар ди омиоцитов.
А — желудочковый кардиомиоцит (схема). Б,
В — вставочный диск (схема, ТЭМ).
1 — миолемма: а — базальная мембрана; б —
плазмолемма; 2 — ядро; 3 — трофический ап-
парат; 4 — миофибриллы; 5 — митохондрии;
6 — вставочный диск; в — точечная десмосома,
г — опоясывающая десмосома, д — нексус;
7 — анастомоз.
пая ткань (единственная из мышечных) обладает ритмическим
характером сократительной деятельности. Интенсивность и часто-
та этого ритма контролируются нервными импульсами, , поэтому
кдрдиомиоциты, особенно проводящие, иннервированы очень
обильно.
99
Рис. 2.51. Ультраструктура
проводящих кардиомиоци-
тов.
А — схема. Б — ТЭМ (клетка
синусно-предсердного узла).
В — клетка предсердно-желу-
дочковой проводящей системы
(схема).
1 — миолемма; 2 — ядро; 3 —
миофибриллы; 4 — мелкие
митохондрии; 5 — гликоген;
6 — пучки филаментов; 7 —
вставочный диск.
100
Рис. 2.52. Гистогенез
сердечной мышечной
ткани (схема).
А, Б, В — объяснение в
тексте (по П. П. Румян-
цеву, И. Л. Ерохиной).
Прекардиальная мезодерма
t
Миоциты стенки сердечной трубки
2.4.1.2.1. Гистогенез сердечной мышечной ткани
Согласно схеме (рис. 2.52), направленность этого процесса такова:
вначале из мезодермальных клеток дифференцируются кардио-
миобласты (рис. 2.52, А). Затем они преобразуются в кардиомио-
циты (рис. 2.52, Б), что сопровождается появлением миофибрилл
и возрастанием их числа. Параллельно совершенствуется структу-
ра вставочных дисков, триад, трофического аппарата и форми-
руются дефинитивные кардиомиоциты (рис. 2.52, В). Процессы
репродукции и миофибриллогенеза в этих клетках не находятся в
столь конкурентных отношениях, как в миобластах (см. рис. 2.48).
Так, несмотря на увеличение количества миофибрилл, в кардио-
миоцитах не прекращаются митозы. Более того, они сохраняются
даже на постнатальных этапах развития (см. рис. 2.52, В). Однако
митоз обычно не завершается (эндорепродукция), поэтому обра-
зуются полиплоидные (одно- и двуядерные) кардиомиоциты.
101
Рис. 2.53. Гладкая мышечная ткань.
А — окраска гематоксилин-эозином. Б — ультраструктура гладкого миоцита. В —
зона мио-миоцитарного контакта (схемы).
I — продольный и II — поперечный слои гладких миоцитов; III — эндомизий;
IV — перимизий.
1 — гладкий миоцит; 2 — ядро; 3 — эластические волокна; 4 — миолемма; 5 —
клеточный центр; 6 — митохондрии; 7 — плотные тельца; 8 — миофиламенты;
9 — мио-миоцитарный контакт: а — базальная мембрана, б — плазмолемма, в —
десмосома, г — нексус, д — кавеолы, е — везикулы.
102
2.4.2. Гладкая мышечная ткань
Эта ткань, так же как сердечная, имеет клеточный тип строения.
Ее структурной единицей является гладкий миоцит. Он, как пра-
вило (встречаются и отростчатые миоциты), имеет веретеновид-
ную форму и заостренные концы. В расширенной зоне его цито-
плазмы находится палочковидное ядро. В стенке полых внутрен-
них органов и сосудов гладкие миоциты объединяются в слои,
чаще всего продольный и циркулярный (рис. 2.53, А). В их составе
мышечные элементы разграничены эндомизием и перимизием.
Причем в поперечном сечении миоциты выглядят иначе, чем в
продольном. В этом случае они представлены множеством округ-
лых оксифильных структур. Если срез проходит через периферию
гладкого миоцита, то такая структура будет безъядерной и очень
небольшой по размеру. В случае прохождения среза через сере-
дину клетки диаметр структуры значительно больше, а в центре
ее всегда определяется круглое ядро.
При субмикроскопическом изучении можно определить, что
каждый миоцит (кроме зон контакта с соседними клетками)
окружен базальной мембраной (рис. 2.53, Б, В). В нее вплетается
густая сеть эластических волокон, наделяющих клетку упругостью.
Компоненты этих волокон синтезируются в трофическом аппарате
миоцита, сконцентрированном возле полюсов его ядра (см.
рис. 2.53, Б). Периферическая часть цитоплазмы клетки занята
сократительным и везикулярным аппаратом. Сократительный ап-
парат характеризуется отсутствием миофибрилл, наличием на
плазмолемме и в цитоплазме миоцита хаотично расположенных
плотных телец, к которым фиксируются актиновые миофиламенты
и промежуточные филаменты (рис. 2.54). Везикулярный аппарат
депонирует экзо- и эндогенный кальций и представлен обилием
кавеол и везикул вблизи плазмолеммы, особенно в зонах мио-
миоцитарных контактов (см. рис. 2.53, В). Многочисленность нексу-
сов в этих контактах обеспечивает синхронность функционирова-
ния в гладкой мышечной ткани ее структурно-функциональных
единиц — мио-миоцитарных комплексов.
При сокращении гладкий миоцит укорачивается и утолщается,
что сопровождается деформацией его поверхности и штопорооб-
разным скручиванием ядра (см. рис. 2.54). Характер сокраще-
ния — тонический (волна сокращения медленно и плавно распро-
страняется по длиннику клеток). Он, очевидно, обусловлен отсут-
ствием миофибрилл и триад, иным химическим составом миофила-
ментов и их организацией в гладком миоците, а также принципа-
ми иннервации этой ткани и регуляции сократительного акта в ее
клетках.
Завершая знакомство с гистофизиологией мышечных тканей,
необходимо отметить, что их конкретные разновидности наряду с
общими признаками имеют и много отличительных черт в строе-
нии (табл. 2.1). Подобная специализация в пределах одной ткане-
вой группы позволяет им выполнять все разнообразие сократитель-
ных актов, которое необходимо для жизнедеятельности организма.
103
Таблица 2.1. Сравнительная характеристика видов мышечной ткани
Морфофункцио- нальные критерии Вид мышечной ткани
поперечнополосатая ткань гладкая ткань
скелетная сердечная
Структурная еди- Мышечное волок- Клетка — кардио- Клетка — гладкий
ница но (клеточно-симп- миоцит миоцит
Структурно-функ- ластическая еди- ница) Мион Клеточный комп- Клеточный комп-
циональная еди- леке леке
ница Локализация ядра На периферии В центре кардио- В центре гладкого
миосимпласта миоцита миоцита
Вид сократитель- Миофибриллы Миофибриллы Миофиламенты
него аппарата (длинные) (короткие)
Локализация сок- В центре миосимп- На периферии кар- На периферии глад-
ратительного аппа- ласта диомиоцита кого миоцита
рата Особенности Наличие камбия — Г етерогенность Отсутствие миофиб-
строения миосателлитоцитов клеточного состава рилл
Особенности Метамерность три- ткани (наличие сократительных, мышечно-секре- торных и проводя- щих кардиоцитов) Непостоянство Отсутствие триад
строения ад, сократительно- триад
го, опорного и тро- фического аппара- тов волокна Высокое содержа- Максимальное со- Низкое содержание
ние митохондрий держание мито- митохондрий
хондрий Наличие вставоч- ных дисков и анас- томозов Наличие полипло- М ногочисленность нексусов в зонах контакта гладких миоцитов Наличие плотных
идных кардиомио- цитов Обильная иннерва- телец и везикуляр- ного аппарата Способность к де-
Характер сокраще- Тетанический про- ция Ритмический лению гладких мио- цитов Тонический непро-
НИЯ извольный непроизвольный извольный
Несократительные Участие в термо- Синтез атриопеп- Продукция эласти-
функции регуляции и угле- тидов ческих волокон
Источник иннерва- водном обмене Соматическая Вегетативная нерв- Вегетативная нерв-
ЦИИ нервная система ная система ная система
104
Рис. 2.54. Сократитель-
ный и опорный аппара-
ты гладкого миоцита.
А — фаза расслабления.
Б — фаза сокращения
клетки (схемы).
1 — плотные тельца; 2 —
промежуточные филамен-
ты; 3 — актиновые миофи-
ламенты; 4 — миозиновые
миофиламенты; 5 — ядро.
2.5. Нервная ткань
Нервная ткань обеспечивает восприятие раздражений, трансфор-
мацию их в нервный импульс и проведение импульса по цепи
нейронов к рабочим структурам. В основе выполнения этих и неко-
торых других сложных функций лежит деятельность нейронов.
Нервная ткань состоит из клеток двух популяций, связанных
между собой, — нервных и глиальных, которые формируют еди-
ную морфофункциональную систему.
2.5.1. Нервная клетка
Нервная клетка состоит из тела (перикариона) и отростков
(рис. 2.55). В физиологических условиях нейроны не делятся.
Интенсивность метаболизма в нейроне чрезвычайно высока,
поэтому в нем хорошо развиты органеллы, обеспечивающие про-
цессы обмена веществ и энергии (см. рис. 2.55, А,,). Для нервной
клетки характерен высокий уровень синтеза белка, который, в основ-
ном, происходит в перикарионе. Поэтому в нем содержится
много рибосом в составе гранулярной эндоплазматической сети
и свободно лежащих полисом. Морфофункциональная орга-
низация белоксинтезирующего аппарата нейрона имеет свои
особенности. Структуры, обеспечивающие этот процесс в перика-
105
рионе, получили название субстанции Ниссля (тигроидное, или
базофильное, вещество). Используя специальные методы окраши-
вания срезов, можно при световой микроскопии рассмотреть, что
изучаемое образование имеет вид базофильных глыбок, располо-
женных в перикарионе и в основании дендритов. На рис. 2.56, А
изображены глыбки Ниссля в мотонейронах спинного мозга. При
ТЭМ субстанции Ниссля (рис. 2.56, Б) видно, что в ее состав
входят участки гранулярной цитоплазматической сети и скопления
свободно лежащих полисом. Субстанция Ниссля активно реаги-
рует на изменение режима работы нейрона, гипоксию и другие
неблагоприятные факторы.
Для нейрона характерно большое количество митохондрий, в
которых идет процесс образования макроэргов в присутствии
кислорода. В местах наиболее интенсивной работы клетки их ко-
личество особенно велико (см. рис. 2.55, А, II): в области аксон-
ного холмика, где у большинства клеток происходит генерация
нервного импульса; в месте передачи импульса — в синапсах; в
чувствительных нервных окончаниях, где энергия раздражителя
Рис. 2.55. Нериная клетка (нейрон).
А — общий план строения: I — импрегнация солями серебра, II — схема ультра-
структурной организации. Б — ТЭМ (по Т. Г. Варнавской). В — СЭМ (по А. С. Ха-
ланскому, И. В. Викторову).
1 — ядро; 2 — тело (перикарион); 3 — дендрит; 4 — узел ветвления дендрита;
5 — дендритический шипик; 6 — аксон (нейрит); 7 — аксонный холмик; 8 — кол-
латераль аксона; 9 — гранулярная цитоплазматическая сеть; 10 — свободно лежа-
щие полисомы; 11—агранулярная цитоплазматическая сеть; 12 — включения
гликогена; 13 — митохондрии; 14 — комплекс Гольджи; 15 — лизосома; 16 — ней-
ротубули; 17 — филаменты (нейрофиламенты); 18 — везикулы; 19 — синапс.
106
Рис. 2.56. Базофильное вещество.
А — препарат: глыбки базофильного
вещества в мотонейроне (окраска толуи-
диновым синим). Б — ультраструктура
базофильного вещества, ТЭМ (по Алтай
Готовый).
1 — глыбки базофильного вещества;
2 — гранулярная цитоплазматическая
сеть; 3 — свободно лежащие полисомы.
преобразуется в нервный импульс. Резервным (но в ряде случаев
весьма существенным для деятельности нервной клетки) источни-
ком энергии является гликоген; при его расщеплении, сопровож-
дающемся выделением энергии, не используется кислород. Неко-
торые нейроны, в частности нервные клетки коры больших полу-
108
Рис. 2.57. Нейрофибриллы.
А — мультиполярный нейрон. Б — фиб-
риллярные структуры в составе нейрита,
ТЭМ (по Л. В. Васько). В — микротру-
бочки в составе дендрита, ТЭМ: Ва —
продольный срез (по 3. И. Журавле-
вой), Вб — поперечный срез (по
Н. С. Косицыну).
1 — перикарион; 2 — отросток; 3 —ней-
рофибриллы; 4 — ядро; 5 — микротру-
бочки; 6 — фибриллярные структуры.
шарий не содержат запасов гликогена, что делает их чрезвычайно
чувствительными к недостатку кислорода.
Важной особенностью нейрона является наличие нейрофибрилл,
локализующихся и в теле, и в отростках клетки (рис. 2.57). Они
выполняют опорную и сократительную функции, а также участ-
вуют в транспорте веществ и структур. Особенно важна деятель-
ность нейрофибрилл в отростках, по которым идет перемещение
веществ в двух направлениях. При окрашивании препаратов соля-
ми тяжелых металлов можно рассмотреть нейрофибриллы с ис-
пользованием световой микроскопии (рис. 2.57, А). ТЭМ позволя-
ет дифференцировать в составе нейрофибрилл нейротубулы (мик-
ротрубочки), нейрофиламенты (рис. 2.57, Б, В).
Отростки нейрона представлены дендритами и нейритом (ак-
109
3
1
Рис. 2.58. Морфологи-
ческая классификация
нейронов по числу от-
ростков (А) и по фор-
ме клеточных тел (Б).
1 — биполярный; 2 —
псевдоуниполярный; 3 —
мультиполярный; 4 — ок-
руглый; 5 — веретенооб-
разный; 6 — звездчатый;
7 — пирамидный; 8 — гру-
шевидный; а — дендрит,
б — аксон.
соном). Дендриты несут импульс к телу клетки, а аксон передает
его от одного нейрона к другому либо к рабочему органу. Эти два
типа отростков имеют четкие различия, которые могут быть вы-
явлены при помощи специальных методов световой и электронной
микроскопии. Используя рис. 2.55, А, можно проследить неко-
торые из них. Так, дендриты, как правило, короче аксона, они
интенсивно ветвятся; по ходу дендритов могут быть особые места
расширения (узлы ветвления), а также короткие выросты —
«шипики». В основании дендритов и в узлах ветвлений находится
субстанция Ниссля. Аксон не ветвится, он может давать единич-
ные боковые коллатерали; в месте отхождения аксона от пери-
кариона выделяется территория, лишенная рибосом, — аксонный
холмик.
НО
Классификация нервных клеток. Существует несколько подхо-
дов к классификации нейронов. Наиболее распространенные мор-
фологические классификации основываются на количествё отрост-
ков или на форме клеточных тел (рис. 2.58). В функциональных
классификациях используются различные подходы: по локализации
клеток в составе рефлекторной дуги (чувствительные, ассоциатив-
ные, двигательные); по характеру выделяемого ими медиатора
(холинергические, адренергические и т. д.); по электрофизиоло-
гическим свойствам (возбуждающие и тормозные). Существует
особая популяция нервных клеток, выполняющих секреторную
функцию. О них пойдет речь в разделе «Эндокринная система».
2.5.2. Нейроглия
Нейроглия обеспечивает опорную, разграничительную, трофиче-
скую, секреторную, защитную функции; участвует в регуляции
скорости проведения нервного импульса по нервным волокнам. Раз-
личают макро- и микроглию. Все разновидности макроглии являют-
ся нейроэктодермальными производными, а микроглия имеет моно-
цитарное происхождение и обладает фагоцитарной активностью.
Макроглия представлена астроцитами, эпендимоцитами и олиго-
дендроглиоцитами.
Астроциты (рис. 2.59) — клетки отростчатой формы. Они
входят в состав органов центральной нервной системы и участвуют
в образовании гематоэнцефалического барьера. Кроме того, они
встречаются в составе рецепторов.
Эпендимоциты выстилают полости желудочков мозга и
спинномозгового канала. Они имеют кубическую форму и отлича-
ются большим количеством микроворсинок и ресничек на той час-
ти клеточной поверхности, которая обращена в соответствующую
полость (рис. 2.60). Эпендимоциты принимают участие в образо-
вании и циркуляции спинномозговой жидкости.
Олигодендроглиоциты (рис. 2.61) — мелкие клетки
с небольшим числом отростков, что отчетливо видно при СЭМ
(рис. 2.61, А). Они входят в состав органов центральной и перифе-
рической нервной системы. Одной из главных функций олигоденд-
роглиоцитов является образование миелина. Особой разновид-
ностью этого вида макроглии являются перинейроналъные сател-
литы (рис. 2.61, В) — клетки, окружающие тело нейрона.
2.5.3. Нервные волокна
Нервные волокна обеспечивают проведение нервных импульсов.
В состав волокна входят отростки нейрона и глия. Отростки нерв-
ных клеток в составе волокна называются осевыми цилиндрами.
Их глиальное сопровождение обеспечивается олигодендроглиоци-
тами; в составе волокон периферической нервной системы они
называются леммоцитами, или шванновскими клетками.
111
Рис. 2.59. Астроциты.
А —
(по
А. С. Халанскому и И. В. Викторову).
1 — отростки астроцита.
общий вид, окраска солями серебра
Л. И. Фалину). Б — СЭМ (по
Рис. 2.60. Эпендимоциты, СЭМ
(по А. С. Халанскому).
112
Рис. 2.61. Олигодендроглиоциты.
А — СЭМ (по А. С. Халанскому и И. В. Викторову). Б — ТЭМ (по В. В. Семченко).
В — полутонкий срез (по Т. Г. Варнавской).
1 — олигодендроцит; 2 — астроцит; 3 — клетка-сателлит; 4 — кровеносный сосуд;
5 — ядро олигодендроцита; 6 — цитоплазма олигодендроцита; 7 — тело нейрона.
На основании морфологических и функциональных особенно-
стей различают безмиелиновые и миелиновые волокна. На рис. 2.62
представлен процесс развития периферических нервных волокон.
Принципиальные отличия в характере взаимодействия отростков
нервных клеток и леммоцитов в процессе развития безмиелиновых
и миелиновых волокон отчетливо видны на упрощенной схеме
ранних этапов развития волокон (рис. 2.62, А). Обратите внимание
на то, что процесс развития безмиелинового волокна (рис. 2.62, А, I)
сравнительно прост: несколько отростков нейронов (будущих осе-
вых цилиндров) погружаются в шванновскую клетку (леммоцит),
прогибая ее плазмолемму с образованием так называемого мезак-
сона — дубликатуры плазматической мембраны леммоцита. Каж-
дый осевой цилиндр, охваченный оболочкой леммоцита, лежит как
бы в желобке. Щелевидное пространство между плазматической
оболочкой шванновской клетки и оболочкой осевого цилиндра за-
полнено тканевой жидкостью, что важно для передачи нервного им-
пульса. Безмиелиновые волокна называют волокнами «кабельного
типа», учитывая то, что каждая шванновская клетка (а их по длине
волокна множество) окружает целую группу осевых цилиндров.
Снаружи леммоциты сопровождаются базальной пластинкой.
Процесс развития миелиновых волокон отражает вторая поло-
вина схемы (рис. 2.62, А, II). Анализируя ее, можно проследить,
как только один отросток нервной клетки погружается в цитоплазму
леммоцита, образуя при этом мезаксон. При дальнейшем развитии
мезаксон удлиняется и концентрически наслаивается на осевой
цилиндр; считают, что это происходит в результате вращательных
движений леммоцита. Многочисленные слои мезаксона, окру-
жающие осевой цилиндр, представляют собой миелин. В хими-
ческом отношении миелин — это комплекс липидов (холестерина,
фосфолипидов, гликолипидов) и белков с преобладанием содер-
жания липидов. В процессе образования миелина цитоплазма и
ядро леммоцита оттесняются на периферию волокна, образуя
неврилемму. Снаружи шванновские клетки в составе волокна со-
провождаются базальной пластинкой.
Современные электронно-микроскопические исследования по-
зволили выявить многие детали процесса развития миелиновых
нервных волокон. С некоторыми из них Вы можете ознакомиться
Рис. 2.62. Развитие нервных волокон.
А — упрощенная схема развития волокон: I — развитие безмиелинового волокна,
II — развитие миелинового волокна. Б — схема формирования миелина (по
О. С. Сотникову): а — поперечный срез, б — продольный срез. В — ультраструктура
развивающихся миелиновых волокон: асинхронизм образования миелиновой обо-
лочки в разных волокнах (по Л. В. Васько). Г — ультраструктура развивающе-
гося безмиелинового волокна.
1 — цитоплазма леммоцита; 2 — ядро леммоцита; 3 — отросток нервной клетки —
осевой цилиндр; 4 — оболочка леммоцита (плазмолемма); 5 — мезаксон; 6 — не-
врилемма; 7 — миелиновая оболочка; 8 — базальная пластинка; 9 — промежуточ-
ные линии; 10 — главные линии.
114
Рис. 2.63. Миелиновое нервное волокно.
А — расщепленный нерв, продольно ориентированные нервные волокна (окраска
осмиевой кислотой). Б — поперечный срез нервных волокон (окраска осмиевой
кислотой). В — ультраструктура миелинового волокна, поперечный срез, ТЭМ
(по А. А. Клищеву). Г — насечки миелина по ходу волокна: I — фазово-контраст-
на микроскопия (по О. С. Сотникову), II — ТЭМ (по А. С. Сотникову, О. Е. Хон-
ны). Д — узловой перехват (перехват Ранвье) (по О. С. Сотникову, О. Е. Хой-
ны): I — фазово-контрастная микроскопия, II — ТЭМ.
1 — осевой цилиндр; 2 — миелиновая оболочка; 3 — насечки миелина; 4 — узло-
вой перехват.
117
по рис. 2.62, Б. В отличие от предыдущей упрощенной схемы (см.
рис. 2.62, А, II) здесь плазматическая мембрана леммоцита пред-
ставлена не одной линией, а тремя; эти линии соответствуют липид-
ным и белковым компонентам мембраны. Обратите внимание на
взаимодействие элементов плазмолеммы леммоцита при формиро-
вании миелина: при вращении вокруг осевого цилиндра наружные
стороны плазмолеммы шванновской клетки сливаются в одну истон-
ченную линию (промежуточную); внутренние стороны плазмолеммы
леммоцита тоже сливаются, но образуют при этом довольно плот-
ную главную линию. Территория между главными линиями состоит
из мембранных липидов. При электронной микроскопии структур-
ные компоненты развивающегося миелинового волокна отчетливо
различимы (рис. 2.62, В); главные и промежуточные линии при та-
ком увеличении не выявляются.
Миелинизация нервных волокон в организме протекает асин-
хронно; на рис. 2.62, В, демонстрирующем этот процесс, вцдно, что
у одних волокон миелиновая оболочка представлена лишь несколь-
кими слоями мембран, а у других — она уже довольно толстая.
Процесс миелинизации длительный, он не завершается к рождению.
Имеются существенные отличия и в темпах миелинизации опреде-
ленных систем нервных проводников. Описанный процесс образо-
вания миелина лучше изучен для волокон периферической нервной
системы; в принципе он схож с таковым в центральной нервной
системе, однако в последней имеется ряд особенностей, и в целом
процесс миелинизации в этом отделе более сложен и недостаточно
исследован.
Нарушение образования миелиновых оболочек является причи-
ной многих серьезных заболеваний нервной системы. Миелин чув-
ствителен к различным неблагоприятным факторам (ишемия, ги-
пертермия, инфекции и т. д.), действие которых может привести к
его расслоению и распаду. Так называемая демиелинизация волокон
приводит к нарушению проведения нервных импульсов.
Ультраструктуру развивающегося безмиелинового нервного во-
локна можно рассмотреть на рис. 2.62, Г.
При изучении морфофункциональной организации нервных во-
локон применяют различные методы световой и электронной микро-
скопии. Так, при окраске миелиновых волокон осмиевой кислотой
довольно отчетливо видны их основные компоненты (рис. 2.63, А, Б).
Использование ТЭМ и фазово-контрастного микроскопа дает
возможность проанализировать детали структурной организации
нервного волокна. При изучении соответствующих снимков
(рис. 2.63, В—Д) можно подробно рассмотреть строение осевого
цилиндра, миелина и леммоцитов. В местах контакта шванновских
клеток виден участок, лишенный миелина, где осевой цилиндр ока-
зывается «оголенным». Это место называется узловым перехватом
(перехват Ранвье); здесь аксолемма контактирует с тканевой жид-
костью. Наличие узлового перехвата имеет важное значение в саль-
таторной передаче нервных импульсов. Очень демонстративна об-
ласть перехвата при наблюдении его в фазово-контрастный микро-
118
Рис. 2.64. Безмиелиновое нервное
волокно.
А — общий вид, продольно ориентиро-
ванные волокна (рисунок с препарата,
окраска гематоксилин-эозином). Б —
поперечный срез, ТЭМ (по Л. В. Васько).
1 — осевые цилиндры; 2 — ядро леммо-
цита.
скоп. В составе миелиновой оболочки имеются косо ориентирован-
ные светлые щели — насечки Шмидта—Лантермана (рис. 2.63, Г, I);
при ТЭМ (рис. 2.63, Г, II) установлено, что в этих местах оста-
ются участки цитоплазмы леммоцита, т. е. мембраны, формиру-
ющие миелин, лежат не плотно.
Безмиелиновые нервные волокна существенно отличаются от
вышеописанных миелиновых как по светооптической (рис. 2.64, А),
так и по ультраструктурной характеристикам (рис. 2.64, Б).
2.5.4. Нервные окончания
Различают чувствительные (афферентные) и двигательные (эф-
ферентные) нервные окончания. Первые (рецепторы) обеспечи-
вают восприятие раздражения и преобразование энергии раздра-
жителя в нервный импульс; вторые — выполняют функцию пере-
дачи нервного импульса на рабочий орган. В этом разделе дают-
ся принципы строения нервных окончаний и более подробная
структура лишь некоторых из них. Основным компонентом ре-
цепторов является дендрит афферентного (чувствительного) ней-
рона. По строению различают два типа чувствительных нервных
окончаний — свободные и несвободные.
На рис. 2.65, А изображено свободное нервное
119
Рис. 2.65. Чувствительные нервные
окончания (по В. Т. Елисееву,
Ю. И. Афанасьеву, Е. Ф. Котов-
скому).
А — свободное нервное окончание в ко
же (импрегнация солями серебра). Б
несвободное инкапсулированное нервное
окончание (тельце Фатера — Пачини) и
поджелудочной железе (окраска ivm.i
токсилин-эозином): I — рис. с пренарл
та, II — схема.
120
Рис. 2.65. Продолжение.
а — импрегнация солями серебра, б —
окраска гематоксилин-эозином.
1 — эпителий; 2 — соединительная
ткань; 3 — структурные компоненты
железы; 4 — миелиновое нервное волок-
но; 5 — концевые терминали дендрита;
б — кровеносный сосуд; 7 — внутренняя
колба; 8 — наружная колба; 9 — кап-
сула.
121
окончание. Обратите внимание на его характерную особен-
ность: терминали дендрита располагаются непосредственно между
клетками иннервируемой ткани. Свободные рецепторы не обладают
строгой специфичностью восприятия раздражений. Их характер-
ная локализация — эпителиальная ткань.
В образовании несвободных рецепторов принима-
ют участие разветвления афферентных нервных волокон и глиаль-
ные клетки. Кроме того, такие нервные окончания могут сопро-
вождаться более или менее тонким соединительнотканным окру-
жением в виде капсулы. В зависимости от наличия последней
различают инкапсулированные и неинкапсулированные несвобод-
ные нервные окончания. В качестве примера на рис. 2.65, Б де-
монстрируется тельце Фатера — Пачини.
Несвободные нервные окончания в отличие от свободных более
или менее строго специализированы на восприятие определенных
раздражителей.
Двигательные нервные окончания образуются аксонами мото-
Рис. 2.66. Двигательное нервное
окончание на скелетной мышце
(нейромышечный синапс).
А — общий план строения (по
Е. А. Шубниковой). Б — схема ультра-
структурной организации; деталь к А.
1 — аксон мотонейрона; 2 — миелин;
3 — мышечное волокно; 4 — пальцевид-
ные инвагинации сарколеммы; 5 — ми-
тохондрии мышечного волокна; б — си-
наптическая щель; 7 — синаптические
пузырьки; 8 — базальная пластинка
мышечного волокна; 9 — базальная
пластинка нервного волокна; 10 — ба-
зальная пластинка в синаптической
щели; 11 — леммоцит; 12 — ядра мы-
шечного волокна; 13 — миофибрилла.
122
нейронов на рабочих органах. На рис. 2.66 изображено двигатель-
ное нервное окончание на скелетной мышце. При его рассмот-
рении видно, как миелиновое волокно подходит к поверхности
мышечного волокна и в месте их контакта формируется так назы-
ваемое нервно-мышечное окончание. Электронная микроскопия
позволяет выявить детали строения (рис. 2.66, Б) и установить,
что принцип его морфофункциональной организации аналогичен
межнейрональному синапсу. В связи с этим существует термин
«аксомышечный синапс» для обозначения двигательного окончания
на скелетной мышце.
Анализируя схемы (см. рис. 2.66) можно увидеть, что оконча-
ние аксона лежит в углублении, выстланном оболочкой мышеч-
ного волокна — сарколеммой; между поверхностью аксона и сар-
колеммой сохраняется щелевидное пространство — синаптическая
щель. Обратите внимание на то, что базальная пластинка нервного
волокна переходит в базальную пластинку мышечного волокна и
заходит в синаптическую щель (надо отметить, что в межнейро-
нальных синапсах между пре- и постсинаптическими мембранами
базальная пластинка отсутствует). Сарколемма формирует много-
численные складки, что приводит к резкому локальному увеличе-
нию ее поверхности; полости складок имеют сообщение с синап-
тической щелью. В области моторной бляшки волокно скелетной
мышцы не имеет характерной исчерченности, а в саркоплазме
располагаются ядра и много митохондрий. Окончание аксона со-
держит скопление синаптических пузырьков и обилие митохонд-
рий. По аналогии с межнейрональным синапсом аксолемму в об-
ласти моторной бляшки следует рассматривать в качестве преси-
наптической мембраны, а сарколемму — в качестве постсинаптиче-
ской мембраны. В момент передачи импульса медиатор поступает
в межсинаптическую щель и взаимодействует с рецепторами
сарколеммы.
2.5.5. Межнейрональные синапсы
Синапс — это специализированный контакт между нервными клет-
ками. Воспринимать информацию нейрон может в любом отделе
своей поверхности. В зависимости от того, какие структурные ком-
поненты нейронов взаимодействуют в области синапса, последние
классифицируют на аксодендритические, аксосоматические, аксоак-
сональные.
В межнейрональном синапсе нервный импульс может переда-
ваться двумя способами — химическим и электрическим. В по-
следнем импульс проходит быстро, поскольку контактирующие
мембраны в области синапса сближены настолько, что волна депо-
ляризации беспрепятственно переходит от одной мембраны к дру-
гой. Электрические синапсы у млекопитающих встречаются крайне
редко.
В химических синапсах нервный импульс передается при помо-
123
Рис. 2.67. Межнейрональный химический синапс.
А — общий план строения (схема). Б — структура синапсов в различных функцио-
нальных состояниях (I — схема, II — ТЭМ по Н. С. Косицыну): а — активное
состояние, б — неактивное состояние. В — аксосоматический синапс, ТЭМ (по
С. С. Степанову). Г — синаптические окончания на различных участках дендрита,
ТЭМ (по В. В. Семченко): в—аксодендритическнй синапс, г — аксошипиковый
синапс.
1 — пресинаптический полюс; 2 — синаптическая щель; 3 — постсинаптический
полюс; 4 — аксолемма; 5 — пресинаптическая мембрана; 6 — синаптические пу-
зырьки с медиатором; 7 — молекулы медиатора; 8 — постсинаптическая мембрана;
9 — рецепторы к медиатору; 10 — фибриллярные структуры; 11 — митохондрии;
12 — фрагменты цитоплазматической сети. 13 — уплотнение цитоплазмы вблизи
пре- и постсинаптических мембран; 14 — электронноплотные нити; 15 — электрон-
но-плотная аморфная масса; 16 — базальная пластинка; 17 — перикарион; 18 —
аксон; 19 — дендрит; 20 — дендритический шипик; 21 — активная зона синапса.
124
щи химических посредников — медиаторов; их роль могут выпол-
нять различные вещества (ацетилхолин, норадреналин, дофамин,
нейропептиды и др.). Принцип ультраструктурной организации
химического синапса представлен на рис. 2.67, А. В описываемой
структуре различают три отдела: пресинаптический полюс, синап-
тическую щель и постсинаптический полюс. Пресинапти-
125
ч е с к и й полюс образован окончанием аксона клетки, лере
дающей импульс. В цитоплазме аксона на территории пресинап-
тического полюса содержатся синаптические пузырьки с медиато-
ром, окруженные мембраной, и различные органеллы нейрона; как
правило, здесь много митохондрий. Постсинаптический
полюс представлен постсинаптической мембраной и прилега-
ющим к мембране участком цитоплазмы. Постсинаптическая
мембрана имеет рецепторы к медиаторам. Синаптическая
щель — пространство, ограниченное пресинаптической и пост-
синаптической мембранами.
В непосредственной близости к пресинаптической и постси-
наптической мембранам, а также в синаптической щели выявляют-
ся электронно-плотные образования. Они формируют единую сис-
тему, обеспечивающую прочность синаптического контакта. Суще-
ствует также вполне обоснованное предположение и о другом
функциональном назначении этих структур, в частности, что мак-
ромолекулярные компоненты описываемых электронно-плотных
образований участвуют в проведении нервного импульса в синапсе.
При очень большом увеличении электронного микроскопа можно
обнаружить, что возбужденные и невозбужденные синапсы выгля-
дят по-разному. В активном синапсе (рис. 2.67, Б1, а, БП, а) в си-
наптической щели формируются электронно-плотные нити, ориен-
тированные вертикально, перпендикулярно мембранам, ограничи-
вающим синаптическую щель. Возможно, эти нити обеспечивают
направленный доступ медиатора к постсинаптической мембране.
В неактивном синапсе (рис. 2.67, Б1, б, БП, б) макромолекулы ве-
щества синаптической щели образуют аморфную массу, обладаю-
щую изоляционными свойствами.
Как уже отмечалось, общий принцип структурной организации
синапсов един. В этом можно убедиться, сравнив аксодендритиче-
ский и аксосоматический синапсы на электронных микрофотогра-
фиях (рис. 2.67, В, Г).
2.6. Нервная система
Нервная система выполняет функцию интеграции, обеспечивая
связь составных компонентов организма в единое целое. Под конт-
ролем нервной системы находятся также взаимодействие орга-
низма с внешней средой, обменные процессы в иннервируемых
структурах, сложная и многогранная психическая деятельность
человека. Основным морфологическим субстратом органов этой
системы является нервная ткань.
С анатомической точки зрения в нервной системе выделяют
периферический и центральный отделы. С физиологических пози-
ций нервную систему делят на соматическую и вегетативную; в
последней выделяют симпатический и парасимпатический отделы.
В основе деятельности нервной системы лежат рефлексы. Прин-
цип строения рефлекторных дуг представлен на рис. 2.68. Анали-
126
Рис. 2.68. Виды рефлекторных дуг.
А — соматическая рефлекторная дуга. Б — вегетативные рефлекторные дуги.
I — простая рефлекторная дуга; II — сложная рефлекторная дуга; III — симпа-
тическая рефлекторная дуга; IV — парасимпатическая рефлекторная дуга (рефлек-
торная дуга блуждающего нерва); V — местная (метасимпатическая) рефлектор-
ная дуга.
1 — рецептор; 2 — спинномозговой ганглий; 3 — чувствительный нейрон; 4 —
синапс; 5 — ассоциативный нейрон; 6 — мотонейрон; 7 — вегетативный ганглий
симпатического ствола; 8 — вегетативный ганглий нервного сплетения; 9 — чувст-
вительный ганглий по ходу блуждающего нерва; 10 — продолговатый мозг;
11 — периферический нерв; 12 — двигательное нервное окончание; 13 — скелетная
мышца; 14 — гладкомышечные клетки; 15 — стенка внутреннего органа; 16 — эпи
телий; 17 — спинной мозг.
зируя схемы, можно найти все составные компоненты (от рецеп-
тора до двигательного нервного окончания на рабочем органе) в
составе каждой рефлекторной дуги.
2.6.1. Периферическая нервная система
Этот отдел нервной системы представлен периферическими нерва-
ми, нервными узлами (ганглиями) и нервными окончаниями.
127
Рис. 2.69. Периферический нерв.
А — схема структурной организации.
Б — препарат (окраска гематоксилин-
эозином).
1 — эпиневрий; 2 — периневрий; 3 —
эндоневрий; 4 — кровеносный сосуд;
5 — нервные волокна: а — осевой ци-
линдр.
128
Периферические нервы состоят из нервных волокон
и соединительной ткани, содержащей кровеносные и лимфати-
ческие сосуды. Воспользовавшись схемами (см. рис. 2.68), можно
убедиться, что периферические нервы по составу являются сме-
шанными, поскольку они содержат чувствительные и двигательные
волокна. Схема (рис. 2.69, А) отражает общий план строения
периферического нерва и вполне соответствует характеристике ор-
гана на светооптическом уровне (рис. 2.69, Б). Снаружи нерв на
всем протяжении окружен соединительнотканной «муфтой» —
эпиневрием. Пучки нервных волокон опоясываются периневрием,
также сформированным соединительной тканью. В процессе при-
готовления гистологического среза с последующей его окраской
гематоксилином и эозином (см. рис. 2.69, Б) нерв, взятый для
исследования, контактирует со спиртом, что приводит к растворе-
нию и вымыванию липидного компонента миелина. Поэтому, рас-
сматривая отдельно миелиновое волокно при большом увеличении,
можно четко увидеть в его составе осевой цилиндр, окруженный
пустым пространством — территорией, соответствующей располо-
жению миелина. Это светлое (неокрашенное) пространство сна-
ружи ограничено бледно-розовым ободком, который формируется
узкой полоской цитоплазмы и плазмолеммы леммоцита; здесь же
может располагаться и ядро шванновской клетки, если оно попало
в плоскость среза. Каждое нервное волокно сопровождается
тонкой прослойкой соединительной ткани — эндоневрием.
Ганглии периферической нервной систе-
м ы. Спинномозговой ганглий (чувствительный нервный узел)
(рис. 2.70, А, Б) — это небольшой по размерам орган, состоя-
щий из чувствительных нейронов, глии и соединительной тка-
ни с сосудами, формирующих строму узла и капсулу? Основная
масса нервных клеток расположена на периферии ганглия, а его
центральная часть в основном занята нервными волокнами.
Дендриты нервных клеток ганглиявыходят за пределы органа и
в составе периферических нервов достигают иннервируемых тканей,
где формируют рецептор. Комплекс нейритов нервных клеток, обра—
зуя задний корешок, входит в серое вещество спинного мозга. Тела
нейронов опоясываются слоем уплощенных глиальных клеток. Глио-
циты, плотно контактируя друг с другом, формируют непрерывный
слой в виде капсулы вокруг тел нервных клеток. В связи с такой
пространственной ориентацией глиальные клетки спинномозгового
узла называют капсулярными, мантийными или нейросателлитами;
они являются разновидностью олигодендроглии.
Информацию о геометрии нервных клеток чувствительного
нервного узла можно получить, используя особый метод окраски
срезов солями серебра (рис. 2.70, В). Этот метод позволяет рас-
смотреть отростки псевдоуниполярных нейронов, а также нервные
волокна в центре органа.
Вегетативные ганглии в зависимости от их расположения делят
на паравертебральные, превертебральные и интрамуральные. Пер-
5—321
129
9
8*
A
вне две локализации характерны для симпатической нервной
системы; последняя — для парасимпатической.
Вегетативные ганглии состоят из нервной ткани (мульти-
полярных нейронов и глии) и соединительной, формирующей
строму и капсулу органа. В большинстве симпатических вегета-
тивных ганглиев выделяется особая популяция мелких, интен-
сивно флюоресцирующих (МИФ) клеток, содержащих катехо-
ламины. Предполагают, что они участвуют в модуляции синапти-
ческой передачи в ганглиях, в обеспечении эндокринных и пара-
кринных функций.
Принцип морфофункциональной организации вегетативных
ганглиев един, но вместе с тем есть ряд особенностей, характерных
для ганглиев различной локализации (рис. 2.71). Эти различия в
первую очередь касаются пространственной ориентации, размеров
и функции нервных клеток, а также их медиаторных характе-
ристик. Например, большинство нейронов симпатических ганглиев
содержит катехоламины; основная масса нервных клеток, входя-
щих в состав ганглиев парасимпатического отдела, является хо-
линергическими; в интрамуральных ганглиях выявляются холинер-
гические, адренергические, пуринергические и пептидергические
нейроны.
2.6.2. Центральная нервная система
Этот отдел нервной системы представлен спинным и головным
мозгом, в составе которых выделяют серое и белое вещество.
Основными структурными компонентами серого вещества являют-
ся нейроны, нервные волокна (главным образом их начальные и
конечные отделы) и глия, представленная астроцитами, олигден-
дроглиоцитами и микроглией (глиальными макрофагами). Белое
вещество в основном образовано нервными волокнами (преиму-
щественно миелиновыми). Органы ЦНС богаты кровеносными
сосудами.
Рис. 2.70. Спинномозговой (спинальный) ганглий.
А — схема структурной организации. Б — препарат (окраска гематоксилин-эози-
ном). В — препарат (импрегнация солями серебра).
1 — капсула; 2 — кровеносный капилляр; 3 — нервная клетка: а) — цитоплазма,
б) — ядро; 4 — псевдоуниполярный нейрон (видны отростки нейрона); 5 — глия —
нейросателлиты; 6 — нервные волокна; 7 — передний корешок; 8 — периферический
нерв; 9 — задний корешок.
5*
131
Рис. 2.71. Вегетативные
нервные ганглии.
А — общий план строения
ганглия солнечного сплете-
ния: I — схема, II — ТЭМ
нейроглио^апиллярного комп-
лекса (по Т. Г. Варнавской).
Б — препарат: ганглий солнеч-
ного сплетения. В — интраму-
ральный ганглнй в мышечной
оболочке стенки кишки: а —
окраска гематоксилин-эози-
ном, б — импрегнация соля-
ми серебра. 1 — капсула; 2 —
нервные клетки; 3 — перика-
рион; 4 — отросток нейрона;
5 — клетки глии; 6 — крове-
носный сосуд; 7 — гладкомы-
шечные клетки.
132
д
2.6.2.1. Спинной мозг
Общий план строения органа дан на рис. 2.72, А. В центральной
части поперечного среза спинного мозга расположено темное се-
рое вещество, напоминающее по форме букву «Н»; его окружает
светлоокрашенное белое вещество. Нервные клетки серого вещест-
ва мультиполярные, наиболее крупные из них находятся в перед-
них рогах — это мотонейроны, посылающие свои отростки к ске-
летным мышцам. Более мелкие клетки передних рогов и других
отделов серого вещества являются ассоциативными. Нейроны
располагаются группами, образуя так называемые ядра спинного
мозга.
Белое вещество состоит из двух симметричных половин.
Оно окружено мягкой мозговой оболочкой, плотно прилегаю-
щей к веществу мозга; эта оболочка состоит из рыхлой волокни-
стой соединительной ткани, в которой содержатся кровеносные
сосуды. В веществе мозга находятся мельчайшие кровеносные
сосуды — капилляры.
На поперечном срезе спинного мозга, обработанном солями
серебра, можно обнаружить большинство из вышеописанных
структурных компонентов спинного мозга (см. рис. 2.72, А), рас-
смотреть нейроны в составе серого вещества (рис. 2.72, Б) и
нервные волокна в составе белого вещества (рис. 2.72, В). Эти
волокна представляют собой проводящие пути спинного мозга.
Некоторые структуры спинного мозга лучше видны на срезах,
окрашенных гематоксилином и эозином (рис. 2.72, Г, Д), чем на
импрегнированных препаратах. Это замечание относится, в част-
ности, к сосудам мягкой мозговой оболочки и к клеткам эпендим-
ной глии, выстилающей полость спинномозгового канала.
2.6.2.1.1. Развитие спинного мозга
Процесс развития спинного мозга представлен на рис. 2.73, А. Он
начинается у человека в конце 2-й недели эмбрионального разви-
тия с формирования в дорсальной части эктодермы нервной
пластинки. В результате интенсивного клеточного деления и нерав-
Рис. 2.72. Спинной мозг.
А — общий план строения (рисунок с препарата; импрегнация солями серебра).
Б — препарат: нейроны передних рогов (импрегнация солями серебра). В —
препарат: белое вещество спинного мозга (импрегнация солями серебра). Г —
препарат: центральный канал (окраска гематоксилин-эознном). Д — препарат:
оболочки мозга (окраска гематоксилин-эозином).
1 — задний рог; 2 — передний рог: а — мотонейроны переднего рога; 3 — боковые
рога; 4 — серая спайка; 5 — полость центрального канала; 6 — эпендимная глия;
7 — передняя срединная вырезка; 8 — задняя перегородка; 9 — волокна заднего
корешка; 10 — волокна переднего корешка; 11—задние столбы; 12—-передние
столбы; 13 — боковые столбы; 14 — нервные волокна; 15 — мягкая мозговая обо-
лочка; 16 — кровеносный сосуд мягкой мозговой оболочки.
135
Рис. 2.73. Развитие спинного
мозга (поперечный срез заро-
дыша) .
А — развитие спинного мозга (схе-
ма по А. Г. Кнорре): I — стадия
образования нервнойиластинки; II —
стадия образования нервного желоб-
ка и нервных валиков; III — стадия
образования нервной трубки и ган-
глиозной пластинки; IV — начало
дифференцировки спинного мозга и
формирования спинномозговых ганг-
лиев. Б — нервный желобок и нерв-
ные валики. В — зародыш на стадии
трех зародышевых листков: нервная
трубка. Г — начало дифференциров-
ки спинного мозга. Д — дифферен-
цировка спинного мозга (Б—Д —
препараты, окраска гематоксилин-
эознном).
1 — нервный желобок; 2 — нервный
валик; 3 — кожная эктодерма; 4 —
хорда; 5 — мезодерма; 6 — ганглиоз-
ная пластинка; 7 — нервная трубка;
8 — мезенхима; 9 — зачаток спинно-
го мозга; 10 — зачаток спинномозго-
вого ганглия; 11 — задний корешок
спинного мозга; 12 — передний ко-
решок спинного мозга; 13 — сме-
шанный периферический нерв; 14 —
эпендимный слой; 15 — плащевой
слой; 16 — краевая вуаль; 17 — це-
лом; 18 — аорта; 19 — закладка поз-
вонков; 20 — энтодерма.
номерного роста края нервной пластинки постепенно приподни-
маются и образуют утолщения — нервные валики. Нервная плас-
тинка постепенно прогибается, углубляется в сторону зародыша,
и края валиков смыкаются, образуя трубку. Нервная трубка тя-
нется от головного до каудального конца развивающегося заро-
дыша и является основным источником образования спинного
и головного мозга. Смыкание валиков начинается в конце 3=й не-
дели развития и проходит неравномерно по длине нервной трубки.
Клетки нервной трубки дифференцируются в трех направле-
ниях: в нейробласты — будущие ыойрпны; в глиобласты (спон-
гиобласты) — будущие глиальные клетки (астроциты и олигоденд-
роциты) ; в эпендимоблас.ты— будущие эпендимные клетки.
По мере размножения и миграции клеток в толще нервной~тру бки
(от центра к периферии) в ней постепенно формируются три слоя:
эпендимный, плащевой и краевая вуаль. Клетки эпендимного слоя
на ранних этапах развития выполняют опорную функцию за счет
длинных отростков, идущих от базальной части клеток к наружной
поверхности нервной трубки; позднее эта функция утрачивается.
136
Материал нервных валиков обосабливается в виде полоски и
лежит с обеих сторон от нервной трубки, образуя нервный гребень
(или ганглиозную пластинку). Клетки нервного гребня распада-
ются на цепочку узелков — предшественников спинномозговых
ганглиев.
В развитии спинного мозга, кроме нейроэктодермы, принимают
участие мезенхима, которая является источником образования
сосудов и соединительнотканных оболочек, а также моноциты, из
которых развиваются клетки микроглии, выполняющей защитную
функцию ЦНС.
На серии снимков, выполненных с препаратов, иллюстриру-
ющих развитие спинного мозга, можно найти структуры, соответ-
ствующие определенным этапам этого процесса.
На рис. 2.73, Б представлены нервный желобок и нервные
валики; на рис. 2.73, В — нервная трубка; на рис. 2.73, Г — диф-
ференцировка, пролиферация и миграция клеток нервной трубки.
На рис. 2.73, Д уже отчетливо различимы три слоя в толще нерв-
ной трубки: внутренний (эпендимный), представленный одним
слоем клеток; средний — плащевой, самый широкий слой, в кото-
ром намечаются контуры будущего серого вещества (в этом слое
находятся нейробласты, дифференцирующиеся в нейроны, и глио-
бласты, дифференцирующиеся в астроциты и олигодендроциты);
наружный — краевая вуаль, содержащий отростки нейробластов и
глиобласты. Обратите внимание на то, что полость центрального
канала гораздо уже по сравнению с предыдущей стадией (см. рис.
2.73,В). По обеим сторонам нервной трубки располагаются оваль-
ные структуры — зачатки спинномозговых ганглиев; в дифферен-
цирующихся ганглиях будущие нейроны располагаются довольно
плотно. В этот период развития происходит закладка позвонков.
2.6.2.2. Мозжечок
Этот орган играет важную роль в регуляции равновесия и коор-
динации движений.
Общий план строения мозжечка можно проана-
лизировать по срезам органа, окрашенным гематоксилин-эози-
ном (рис. 2.74, А).
Рассматривая срез мозжечка, импрегнированный солями се-
ребра, невооруженным глазом можно увидеть на его поверхности
множество борозд и извилин. При светооптической микроскопии
(рис. 2.74, Б) отчетливо различимы белое вещество и покрываю-
щее его серое вещество — кора мозжечка. Основным структурным
компонентом белого вещества являются нервные волокна, а серо-
го — нервные клетки и глия. В глубине белого вещества имеются
ядра — локальные компактные образования, состоящие из нейро-
нов и глии. Кора мозжечка состоит из трех слоев — молекуляр-
ного, ганглиозного и зернистого.
Наружный (молекулярный) слой представлен
мультиполярными ассоциативными нейронами разной величины
138
Рис. 2.74. Кора мозжечка.
А — препарат: общий вид (окраска ге-
матоксилин-эозином). Б — препарат
(импрегнация солями серебра): I — об-
щий вид. II — ганглиозные клетки
(клетки Пуркинье).
1 — сосуд мягкой мозговой оболочки;
2 — молекулярный слой; 3 — ганглиоз-
ный слой; 4 — дендриты ганглиозных
клеток; 5 — зернистый слой; 6 — волок-
на, оплетающие тела ганглиозных кле-
ток; 7 — белое вещество.
□К*
Б,II
139
корзинчатой и звездчатой форм и богат нервными волокнами.
Средний (ганглиозный) слой сформирован крупными
ассоциативными нервными клетками с телами грушевидной формы
(клетки Пуркинье); они расположены в один ряд. Глубокий
(зернистый) слой коры состоит из большого количества
мелких ассоциативных нейронов; основную их массу составляют
клетки-зерна, нейриты которых направляются в молекулярный
слой коры и там Т-образно делятся на две ветви, идущие вдоль
извилины мозжечка. Кору покрывает мягкая мозговая оболочка.
В кору мозжечка по системе восходящих путей приходят воз-
буждающие нервные импульсы; при этом лазящие волокна пере-
дают возбуждение непосредственно на клетки Пуркинье, а мохо-
видные (мшистые) — на клетки-зерна. Поступившая информация
обрабатывается нейронами коры; важную роль в этом процессе
играют ассоциативные (сочетанные) связи, существующие между
нервными клетками разных слоев коры мозжечка. Благодаря этим
связям все импульсы «стекаются» к ганглиозным клеткам, по-
скольку только их нейриты формируют систему нисходящих про-
водящих путей, выносящих импульсы за пределы коры. Импульсы,
выходящие из коры мозжечка, являются тормозными.
2.6.2.3. Кора больших полушарий
Кора больших полушарий построена сложно как в структурном, так
и в функциональном отношении. Она ответственна за анализ и син-
тез приходящей информации. Толщина серого вещества, состав-
ляющего кору, около 3 мм, однако эта величина варьирует от
1,5 до 5 мм в различных отделах мозга. Поверхность коры име-
ет множество борозд и извилин, значительно увеличивающих
площадь серого вещества. Основные структурные компоненты
коры — нейроны, глия, нервные волокна и сосуды. Количество
нервных клеток составляет около 14 млрд; они образуют много-
численные связи между собой и с нижележащими отделами нерв-
ной системы.
Для формирования сложных межнейрональных взаимодейст-
вий в коре особое значение имеет геометрия ее нейронов; в первую
очередь это относится к организации дендритов. Для нейронов
ЦНС (и особенно для нервных клеток коры большого мозга) ха-
рактерны очень большая поверхность дендритов (она формируется
за счет многократных дихотомических делений), наличие специ-
альных структур — дендритических шипиков и особых расширен-
ных площадок (или узлов) в зонах деления дендритов. В составе
шипиков корковых нейронов может присутствовать так называе-
мый шипиковый аппарат — комплекс канальцев, ограниченных
мембраной (см. рис. 2.55, А, II). Шипиковый аппарат представляет
собой филогенетически самое молодое образование в нервной
системе и обнаружен только у высших животных. Характерной
особенностью шипиков является обязательное наличие на их по-
140
верхности синапсов и отсутствие в цитоплазме нейротубулей и
нейрофиламентов. Значение шипиков и шипикового аппарата для
деятельности мозга окончательно не выяснено, однако есть вполне
обоснованное предположение об их участии в обеспечении меж-
нейрональных контактов путем увеличения площади синапсов.
Дендритические шипики и их аппарат в онтогенезе созревают
значительно позже других структур. Эти образования очень чув-
ствительны к действию неблагоприятных факторов (алкоголь,
гипоксия и др.).
Другая вышеуказанная особенность дендритов — формирова-
ние специальных расширенных площадок (узлов) в области деле-
ния отростка (см. рис. 2.55, А, II). Как видно на схеме ультра-
структуры нейрона, в этом участке содержится довольно много
митохондрий, элементы гранулярной цитоплазматической сети,
группы свободно лежащих рибосом; установлено, что здесь про-
исходит процесс белкового синтеза. Описываемая зона резко от-
личается от цитоплазмы общего дендритического ствола и по
ультраструктуре похожа на перикарион. Функциональная роль
этих образований изучена недостаточно. Считают, что они выпол-
няют трофическую функцию и, возможно, участвуют в обработке
приходящих нервных импульсов.
Нейроны в составе коры больших полушарий располагаются
нерезко ограниченными слоями. Степень развития слоев и детали
их структуры неодинаковы в разных областях коры, выполняю-
щих различные функции. Общий план строения коры показан
на рис. 2.75, А. Обратите внимание на то, что все нервные клетки
коры мультиполярны, а по размерам и форме тела они существен-
но отличаются друг от друга. Наиболее представительна популя-
ция пирамидных нейронов.
Полиморфизм нейронов отчетливо виден на срезах коры боль-
шого мозга, окрашенных солями серебра (рис. 2.75, Б, I). Особен-
но демонстративны на таких препаратах нейроны ганглиозного
слоя — клетки Беца (рис. 2.75, Б, II). Их треугольное тело обра-
щено вершиной к наружному отделу коры, а основание — к бе-
лому веществу. От апикальной части тела клетки отходит толстый
верхушечный дендрит, достигающий молекулярного слоя; имеются
также боковые дендриты. Аксоны больших пирамидных клеток
выходят за пределы коры в белое вещество, направляются к ядрам
головного и спинного мозга, формируя пирамидный I'plnTT
На электронной микрофотографии (рис. 2.75, В) представлено
ядро и перикарион клетки Беца с отходящим от него апикальным
дендритом. Территория рядом с нейроном занята так называемым
нейрипИлем,~в состав которого входит множество переплетающих-
ся отростков нервных и глиальных клеток (рис. 2.75, Д).
Для общеморфологического исследования структуры коры
используются срезы, окрашенные гематоксилин-эозином
(рис. 2.75,Г). При их анализе можно рассмотреть тела нейронов
и заметить их послойное расположение. Как правило, хорошо
различимы сосуды в мягкой мозговой оболочке.
141
Рис. 2.75. Кора больших полушарий.
А — архитектоника коры: I — схема
(по С. А. Саркисову), II — колонки
нейронов (по Н. С. Косицыну). Б —
импрегнация солями серебра: I — об-
щий вид, II — деталь к I. пирамидный
нейрон. В — ультраструктура пирамид-
ного нейрона и нейропиль, ТЭМ
(по В. В. Семченко). Г — препарат
(окраска гематоксилин-эозином). Д —
густое переплетение отростков нервных
и глиальных клеток, СЭМ (по А. С. Ха-
ламскому и И. В. Викторову).
1 — сосуд мягкой мозговой оболочки;
2— молекулярный слой; 3 — наружный,
зергтстый слой; 4 — пирамидный слой;
5 -— внутренний зернйст&й—Слой;' б~—
ганглиозный слойт~7"—слоййюдиморф-^
ных^клеток; К"— ТТГиропиль; 9 — нерщ’
ное волокбщй ТО — переплетающиеся
отростки нервных и глиальных клеток;
11 — ядро нейрона; 12 — перикарион;
13 — основание дендрита; 14 — колонка
корковых нейронов.
«
143
Л«>
Рис. 2.75. Продолжение.
Важной особенностью нейронной организации коры является
формирование колонок. В состав колонок, располагающихся пер-
пендикулярно поверхности коры, входят нейроны всех ее слоев
(рис. 2.75, А, II). Колонки характеризуются тонкой функциональ-
ной специализацией; имеется структурная и функциональная
связь между вертикальными колонками различных отделов коры.
Кора большого мозга имеет интенсивное кровоснабжение.
Важное значение в регуляции обмена веществ между нервными
клетками и сосудами имеет глия (в основном астроциты), входя-
щая в состав так называемого гематоэнцефалического барьера.
Изменения в структуре стенки сосудов, либо другие изменения в
комплексе сосуд — нейрон — глия приводят к нарушению жизне-
деятельности коры.
2.7. Органы чувств
Являясь периферическим отделом анализаторов, органы чувств
осуществляют специфическую рецепцию сигналов, поступающих
из внешней и внутренней среды организма.
145
При большом разнообразии строения органов зрения, равно-
весия, слуха, вкуса и обоняния все они обладают принципиальной
общностью структурной организации. Их «рабочая» часть состоит
из двух функционально различных видов клеток — сенсорных
(непосредственно воспринимающих действие раздражителя) и
вспомогательных (выполняющих опорно-трофическую и защит-
ную функции). Все сенсорные клетки в апикальной части снаб-
жены микроворсинками или ресничками.
2.7.1. Орган зрения
Изучение строения глаза лучше начинать с основных этапов его
развития, так как это позволит составить более четкое представ-
ление о структурно-функциональной организации его главных
частей (рис. 2.76).
Основными эмбриональными источниками развития г л а-
з а являются глазной пузырек (выпячивание стенки переднего
мозгового пузыря) и эктодерма головной части зародыша. Первый
путем инвагинации превращается в глазной бокал, соединенный с
мозговым пузырем глазным стебельком. Из внутренней его стенки
развивается нервный слой сетчатки, из наружного — пигментный
слой, а из края бокала — миоциты радужки и ресничного тела.
Эктодерма дифференцируется в хрусталиковую плакоду, которая
вначале путем инвагинации и отделения превращается в хруста-
ликовый пузырек, дифференцирующийся затем в хрусталик, а
расположенная над ним эктодерма — в роговицу. Все же осталь-
ные компоненты глаза развиваются из мезенхимы.
Схема (рис. 2.77, А) отражает общий план структурной органи-
зации глаза. Обратите внимание на то, что его основу составляют
три оболочки (склера, сосудистая и сетчатая); вместе со своими
производными они и формируют все функциональные аппараты
глаза: аккомодационный (ресничное тело, радужка), диоптричес-
кий (роговица, хрусталик, стекловидное тело) и рецепторный
(сетчатка).
Первые два расположены в передней части глаза. Здесь же
находятся структуры, обеспечивающие регуляцию оттока водяни-
стой влаги из передней камеры глаза (рис. 2.77, Б).
Ресничное (цилиарное) тело — продолжение со-
судистой оболочки в направлении хрусталика. Его основу состав-
ляют пучки гладких миоцитов, формирующие ресничную мышцу,
а ресничный поясок соединяет его отростки с хрусталиком (при
обычной окраске почти не выявляется). Радужка выглядит как
вырост ресничного тела, расположенный над передней поверх-
ностью хрусталика. Именно морфофункциональные изменения
этого комплекса структур глаза ведут к нарушению аккомодации
(близорукость, дальнозоркость).
Венозный синус склеры (шлеммов канал)
выглядит в виде полости, расположенной в склере на уровне рес-
ничного тела, а гребенчатая связка представлена пучками волокон
146
Рис. 2.76. Развитие глаза.
А — схема, иллюстрирующая начальные этапы (первые недели). Б — препараты
последующих стадий: I — эмбрион 7 нед; II — плод 2 мес.
1 — эктодерма; 2 — хрусталиковая плакода; 3 — глазной пузырек; 4 — мозговой
пузырь; 5 — мезенхима; 6 — начало инвагинации стенки глазного пузырька; 7 —
глазной стебелек; 8 — хрусталиковый пузырек; 9 — внутренняя и 10 — наружная
стенки глазного бокала; 11—роговица; 12 — хрусталик; 13 — стекловидное тело;
14 — сетчатка: а — нервный слой, б — пигментный эпителий; 15 — радужка; 16 —
передняя камера глаза; 17 — ресничный поясок; 18 — сосудистая оболочка.
147
Рис. 2.77. Глаз.
А — схема строения глазного ябло-
ка. Б — угол глаза (передняя часть).
1 — роговица; 2 — хрусталик; 3 —
передняя камера; 4 — радужка; 5 —
задняя камера; 6 — ресничное тело;
7 — ресничный поясок; 8 — склера;
9 — сосудистая оболочка; 10 — сет-
чатка; 11 — решетчатая пластинка;
12 — зрительный нерв; 13 — цент-
ральные сосуды сетчатки; 14 —
мозговые оболочки, окружающие
нерв; 15 — стекловидное тело; 16 —
конъюнктива; а — пятно сетчатки,
б — диск зрительного нерва, в — от-
росток ресничного тела, г — реснич-
ная мышца, д — венозный синус
склеры, е — гребенчатая связка.
соединительной ткани в углу между роговицей и радужкой. Вместе
они составляют дренажную систему глаза, обеспечивающую отток
водянистой влаги, поступающей в переднюю камеру (обозначено
стрелками). Именно структурно-функциональные нарушения в
этой системе приводят к повышению внутриглазного давления
(одна из главных причин развития глаукомы).
При более детальном рассмотрении строения радужки
(рис. 2.78, А) обращает на себя внимание то, что в ее соединитель-
но тканной основе (условно подразделяемой на слои — наружный
148
Рис. 2.78. Радужка.
А — общий вид (по В. Г. Елисееву
с изменениями). Б — СЭМ наруж-
ного пограничного слоя (по
Н. Е. Думбровой).
1 — наружный пограничный слой;
2 — сосудистый слой; 3 — внутрен-
ний пограничный слой; 4 — задний
пигментный слой; 5 — передняя ка-
мера глаза; 6 — фибробласт; 7 —
пигментоцит; 8 — гладкий миоцит;
9 — кровеносный сосуд.
Рис. 2.79. Хрусталик.
1 — капсула; 2 — эпителий; 3 — волок-
на; 4 — ядра волокон; 5 — ядро хру-
сталика.
149
пограничный, сосудистый, внутренний пограничный), помимо фиб-
робластов, находятся пигментоциты (количество их варьирует у
разных особей), гладкие миоциты и много кровеносных сосудов.
Имеются существенные отличия и в строении внутренней и наруж-
ной поверхности радужки. Первая покрыта двухслойным пигмент-
ным эпителием, наружная же не имеет сплошного эпителиального
покрытия (рис. 2.78, Б).
При рассмотрении строения хрусталика обратите внима-
ние на отсутствие кровеносных сосудов и на наличие эпителия
только на передней половине (рис. 2.79). Это обусловлено тем,
что смещение его клеток по мере превращения в хрусталиковые
волокна сопровождается гибелью ядер.
Строение роговицы отличается выраженной слоистостью
(рис. 2.80, А). Ее наружная поверхность покрыта многослойным
плоским эпителием (передний эпителий), под ним расположена
гомогенная светлая полоска (передняя пограничная пластинка),
ниже следует самый широкий волокнистый слой (собственное
вещество роговицы). Затем вновь кажущаяся бесструктурной
полоска — задняя пограничная (десцеметова) пластинка. Ее ниж-
няя поверхность покрыта плоским эпителием из полигональных
клеток, напоминающих по строениею мезотелий, — задний эпите-
лий. На рис. 2.80, Б представлена СЭМ поверхности переднего
эпителия, а на рис. 2.80, В — СЭМ перпендикулярного среза соб-
ственного вещества, состоящего из извитых пластин, образован-
ных коллагеновыми волокнами, пространства между которыми
при жизни заполнены основным веществом, определяющим опти-
ческие свойства роговицы. Обратите особое внимание на то, что
роговица не содержит кровеносных сосудов. Именно поэтому она
не отторгает трансплантат, полученный от донора (не развивается
иммунный ответ).
Наиболее сложно устроена задняя стенка глаза. Она
состоит из трех оболочек — фиброзной (склера) (I), широкой,
довольно плотной зоны волокнистой соединительной ткани; сосу-
дистой (II) — лежащего под ней темно-коричневого слоя и внут-
ренней (сенсорной) — сетчатки (III), имеющей вид чередующихся
горизонтальных зернистых и светлых слоев (рис. 2.81, А). Изуче-
ние последней заслуживает особого внимания, поскольку именно
сетчатка является главным функциональным компонентом глаза.
Чтобы лучше разобраться в ее микроскопическом строении, вна-
чале необходимо рассмотреть схему состава и расположения ее
структурных компонентов (рис. 2.81, Б), а затем сопоставить их
с рис. 2.81, В.
Сетчатка состоит из пигментного слоя, представленного
специализированным нейральным эпителием, и нервной части. Пиг-
ментный слой относится к вспомогательным ее компонентам —
обеспечивает трофику и защиту периферических отростков нейро-
сенсорных клеток, их дифференцировку в эмбриогенезе, выработ-
ку ретиналя (компонент родопсина) и ряда других функций, а
также является одним из основных элементов гематоофтальми-
150
Рис. 2.80. Роговица глаза.
А — общий вид. Б — СЭМ пе-
реднего эпителия. В — СЭМ
собственного вещества (Б — по
Я. Л. Караганову, А. А. Миро-
нову и В. А. Миронову, В —
по Н. П. Омельяненко).
1 — передний эпителий; 2 — пе-
редняя пограничная пластинка;
3 — собственное вещество; 4 —
задняя пограничная пластинка;
5 — задний эпителий (на Б
стрелка — микроворсинки и мар-
гинальные складки в местах
соединения клеток); 6 — марги-
нальные складки.
5
А
151
Рис. 2.81. Строение задней
r'<y»LV
стенки глаза.
А — общий вид препарата: I —
склера, II — сосудистая оболоч-
ка, III — сетчатка. Б — схема
клеточного строения сетчатки и
ее слоев. В — срез сетчатки:
I — пигментный, II — фотосен-
сорный, III — наружный погра-
ничный, IV — наружный ядер-
ный, V — наружный сетчатый,
VI — внутренний ядерный, VII —
внутренний сетчатый, VIII — ган-
глионарный, IX — нервных воло-
кон, X — внутренний погранич-
ный.
152
Рис. 2.81. Продолжение.
Г —ТЭМ пигментоцита. Д — фрагмент Г (по Н. Е. Думбровой).
1 — пигментоцит; 2 — палочковая и 3 — колбочковая нейросенсорные клетки;
4 — горизонтальный нейрон; 5 — биполярный нейрон; 6 — радиальный глиоцит;
7 — амакринный нейрон; 8 — мультиполярный нейрон; 9 — базальная пластинка;
10 — базальные инвагинации плазмолеммы: а — палочки, б — колбочки, в —
отросток пигментоцита, г — синаптический комплекс, д — комплекс Г ольджи,
е — гранулы пигмента, ж — фагоцитоз поврежденных наружных сегментов.
ческого барьера. На рис. 2.81, Б, Г представлена ультраструктурная
организация пигментоцита, осуществляющего фагоцитоз повреж-
денных фрагментов наружных сегментов фоторецепторных клеток.
В его цитоплазме хорошо видны гранулы пигмента.
В процессе изучения нервной части необходимо хорошо усво-
ить, что, будучи частью головного мозга, «вынесенной на перифе-
рию», она также состоит из нейронов и глии. В ее составе можно
выделить две группы нейронов. Первая обеспечивает рецепцию и
проведение светового сигнала в мозг (нейросенсорные, биполярные
и мультиполярные нейроны); вторая регулирует их функцию в пре-
делах сетчатки (горизонтальные, амакринные и интерплексиформ-
ные нейроны). Глиальные же элементы представлены в основном
специализированными глиоцитами, выполняющими не только
опорно-трофическую функцию, но и являющимися вторым компо-
нентом гематоофтальмического барьера. Их участие в формиро-
вании последнего обеспечивается благодаря наличию плотных си-
153
наптических контактов между своими периферическими отростка-
ми и внутренними сегментами нейросенсорных клеток. Поскольку
эти контакты в световом микроскопе определяются в виде сплош-
ной полоски, данное образование было названо наружной погра-
ничной мембраной. Между ней и пигментным эпителием создается
оптимальная микросреда для палочек и колбочек. Центральные
же отростки радиальных глиоцитов, прикрепляясь к истинной
базальной мембране, участвуют в формировании внутренней по-
граничной мембраны, ограничивающей сетчатку от стекловидного
тела. Заканчивая рассмотрение схемы, обратите внимание на то,
что наружные сегменты фоторецепторных клеток, будучи окруже-
ны отростками пигментоцитов, не соединены с ними, в силу чего
при некоторых обстоятельствах нервный слой может отделяться
от пигментного, вызывая заболевание, именуемое отслойкой сет-
чатки.
На основании вышеизложенного в сенсорной оболочке выде-
ляют следующие слои: пигментный (I) — узкая полоска пигменто-
цитов, в цитоплазме которых можно различить зерна пигмента.
Фотосенсорный слой (II) — лежащая ниже светлая безъядерная
зона, состоящая из вертикальных удлиненных структур — палочек
Рис. 2.82. Схема ультраструктурной
организации нейросенсорных кле-
ток.
А — палочки. Б — колбочки. В — ТЭМ
их фрагментов (III — по Н. Е. Думбро-
вой; I, II, VI — по К. Койчеву, Е. Ива-
нову) .
I — наружный сегмент, II — связующий
отдел, III — внутренний сегмент, IV —
тело, V — центральный отросток, VI —
зона синапса.
1 — плазмолемма: а — ее инвагинации;
2 — диски; 3 — реснички; 4 — мито-
хондрии; 5 — комплекс Гольджи; 6 —
зернистая ЭПС; 7 — ядро; 8 — синапс:
б — пресинаптическая, в — постсинап-
тическая части; 9 — микроворсинки ра-
диального глиоцита; 10 — синаптиче-
ский комплекс между глиоцитом и
внутренним сегментом.
154
и конусовидных колбочек периферических отростков нейросен-
сорных клеток. Наружный пограничный слой (III) описан выше
(синаптические комплексы). Наружный ядерный слой (IV) —
зона густорасположенных гиперхромных ядер (тела нейросенсор-
ных клеток). Наружный сетчатый слой (V) —нежноволокнистая
безъядерная полоска (центральные отростки нейросенсорных
клеток и дендриты биполярных нейронов). Внутренний ядерный
слой (VI) — зона ядер, окрашенных несколько светлее и распо-
ложенных менее плотно (тела биполярных, горизонтальных, ама-
кринных и интерплексиформных нейронов, а также радиальных
глиоцитов). Обращает на себя внимание то, что общее их коли-
чество меньше, чем в наружном зернистом слое. Внутренний сет-
чатый (VII) — нежноволокнистая светлая зона (аксоны биполяр-
ных и дендриты нижележащих мультиполярных нейронов). Гангли-
озный (VIII) — более крупные и светлые клетки (тела мультипо-
лярных нейронов). Ширина этой зоны и количество клеток еще
меньше, чем у предыдущих ядерных слоев вследствие конверген-
ции аксонов (см. рис. 2.81, Б) нескольких клеток вышележащего
слоя к одной в нижележащем. Поэтому число волокон в составе
оптического нерва примерно в 100 раз меньше, чем количество
нейросенсорных клеток в сетчатке. Слой нервных волокон (IX) —
аксоны мультиполярных нейронов. Внутренний пограничный
слой (X), отграничивающий сетчатку от стекловидного тела (опи-
санный ранее комплекс окончаний центральных отростков глио-
цитов с базальной мембраной).
Нейросенсорные клетки (палочковая и колбочковая) имеют
сходный общий план строения. Их периферические отростки (па-
лочки и колбочки) состоят из трех частей: наружной (I),
содержащей мембранные диски, и внутренней (III) с хорошо раз-
витым синтетическим аппаратом и расположенным между ними
связующим отделом (II), содержащим реснички (рис. 2.82, А, Б).
Главные же структурные отличия заключаются в размере и форме
наружных сегментов их мембранных дисков и более сложном
строении окончаний аксонов колбочковых клеток. На электроно-
граммах, демонстрирующих основные фрагменты палочковых кле-
ток (рис. 2.82, В), обратите внимание на глубокие инвагинации
плазмолеммы у основания наружного сегмента (начало формиро-
вания мембранных дисков), взаимоотношения отростков и микро-
ворсинок радиальных глиоцитов с внутренними сегментами.
Сетчатка глаза имеет две зоны с особой структурно-функцио-
нальной характеристикой. Первая — центральная ямка сетчатки
глаза, или зона наилучшего видения (рис. 2.83, А), представляет
небольшое углубление в месте проекции центральной оптической
оси. Ганглионарный слой здесь отсутствует (как бы раздвинут в
стороны), внутренний и наружный ядерные слои резко истончены,
так как в наружном расположены только колбочковые нейросен-
сорные клетки, а во внутреннем — соответствующее им количест-
во биполярных нейронов. Такое строение создает наиболее опти-
мальные условия для прохождения светового импульса к рецеп-
156
Рис. 2.83. Сетчатка глаза.
А — пятно. Б — диск зри-
тельного нерва.
I — склера, II — сосудистая
оболочка, III — сетчатка.
1 — пигментный слой; 2 —
фотосенсорный слой; 3 — на-
ружный ядерный слой; 4 —
наружный сетчатый слой; 5 —
внутренний ядерный слой;
6 — внутренний сетчатый
слой; 7 — ганглионарный
слой; 8 — слой нервных воло-
кон; 9 — пятно сетчатки;
10 — диск зрительного нерва;
11 — центральные вена и ар-
терия; 12 — решетчатая пла-
стинка; 13 — зрительный
нерв; 14 — рыхлая волокни-
стая соединительная ткань.
торам, а высокая концентрация колбочек, обладающих большей
(в сравнении с палочками) остротой зрения, обеспечивает наи-
лучшие условия видения объекта. Вторая зона — место формиро-
вания и выхода зрительного нерва, именуемое «диском зритель-
ного нерва», или слепым пятном (рис. 2.83, Б), расположена в
центре сетчатки. Здесь собираются аксоны всех мультиполярных
нейронов ганглионарного слоя. Обратите внимание на наличие в
толще нервного ствола кровеносных сосудов, формирующих со-
судистую систему сетчатки, которая вместе с глиоцитами образует
второй гематоофтальмический барьер сетчатки (подобный гемато-
энцефалическому). Эти сосуды чутко реагируют на многие пато-
логические процессы в организме. Изменения их «рисунка», выяв-
ляемые при осмотре глазного дна, нередко бывают первым симпто-
мом заболевания, не связанного с патологией глаза (диабет, опу-
холь головного мозга и др.).
2.7.2. Преддверно-улитковый орган (орган слуха и равновесия)
Развитие перепончатого лабиринта внутреннего уха (рис. 2.84)
начинается с закладки слухового пузырька путем погружения
157
Рис. 2.84. Развитие внутреннего уха
у человека (схема).
А — образование слуховых плакод (ста-
дия 9 сомитов). Б — появление слухо-
вой ямки (стадия 16 сомитов). В —
формирование слуховых пузырьков
(стадия 30 сомитов). Г — общий вид
развивающегося перепончатого лаби-
ринта: I — зародыш 11 мм, II — заро-
дыш 20 мм (по Пэттену). Д — правый
перепончатый лабиринт сформирован-
ного внутреннего уха (по И. В. Алма-
зову, Л. С. Сутулову).
1 — эктодерма; 2 — плакода; 3 — мезо-
дерма; 4 — дорсальная аорта; 5 —
глотка; 6 — слуховая ямка; 7 — мозго-
вой пузырь; 8 — вентральная аорта;
9 — слуховой пузырек; 10 — закладка
преддверия; 11—улитка; 12 — сфери-
ческий мешочек; 13 — эллиптический
мешочек; 14 — верхний полукружный
продольный проток; 15 — внешний по-
лукружный проток (горизонтальный);
16 — задний полукружный проток
(фронтальный); 17 — межмешочковый
проток; 18 — эндолимфатический про-
ток (срезан); 19 — артерии; 20 — вены.
158
участка эктодермы — плакоды, который в дальнейшем дифферен-
цируется в трех направлениях — формирование преддверия (орган
равновесия), улитки (орган слуха) и объединяющий их эндолим-
фатический проток. Схема окончательного строения всего пере-
пончатого лабиринта дана на рис. 2.84, Д.
Таким образом, сенсорные и вспомогательные клетки всего
органа — специализированные эпителиальные клетки — производ-
ные определенного участка эктодермы. Из окружающей же мезен-
химы формируются его соединительнотканная оболочка и хряще-
вая бластема, которая позднее замещается костной тканью (кост-
ный лабиринт).
2.7.2.1. Орган слуха
Орган слуха расположен в улитковом протоке, находящемся меж-
ду вестибулярной (сверху) и тимпанальной (внизу) лестницей
перепончатого лабиринта улитки, заполненной перилимфой
(рис. 2.85, А, Б). Верхнюю стенку протока составляет вестибуляр-
ная мембрана, наружную — сосудистая полоска. Необычность
составляющего ее многорядного эпителия — наличие в нем гемо-
капилляров, участвующих в выработке эндолимфы и трофике спи-
рального органа, — рис. 2.85, Д. Внутренняя стенка образована
лимбом спиральной пластинки, нижняя — базальной пластинкой.
Последняя с наружной стороны прикреплена к выступу спираль-
ной связки, а с внутренней — к клиновидному выступу стержня
улитки — спиральной костной пластинке, в основании которой
расположено скопление более крупных округлых клеток (тела
нейронов спирального ганглия). Именно на базилярной пластинке
и расположен основной рецепторный аппарат органа слуха —
спиральный (кортиев) орган, над которым лежит гомогенная по-
лоска — покровная мембрана. Детали строения спирального ор-
гана представлены на рис. 2.85, В. В центре над базилярной пла-
стинкой расположена полость — внутренний туннель; в нем про-
ходят нервные волокна. Его стенка образована высокими стоя-
щими наклонно наружными и внутренними столбовыми эпителио-
цитами. По обе стороны от них находятся главные компоненты
органа: поддерживающие эпителиоциты (наружные и внутренние,
лежащие на базилярной пластинке) и расположенные на их
апикальной поверхности соответствующие наружные и внутренние
волосковые сенсорные эпителиоциты (в препарате их можно раз-
личить лишь по верхнему ряду ядер). Рис. 2.85,Г,демонстрирует
вид поверхности органа. Количественное различие рядов и формы
расположения волосков наружных и внутренних волосковых сен-
сорных клеток связано с дифференцировкой восприятия интен-
сивности звукового сигнала. На рис. 2.86, А, Б представлены
ультраструктурная организация волоскового сенсорного эпителио-
цита, его взаимоотношения с поддерживающими эпителиоцитами
и нервными элементами. Фрагменты их апикальной и базальной
частей можно видеть на рис. 2.86, В, Г.
159
Рис. 2.85. Орган слуха.
А — аксиальный разрез улитки (по В. Г. Елисееву). Б — разрез через улитковый
160
4
5
лабиринт. В — схема спирального органа (по А. Хэм, Д. Кормак, 1983). Г — СЭМ
его поверхности (по К. Койчеву. Е. Иванову). I — волосковые клетки, II — под-
держивающие клетки. Д — сосудистая полоска.
1 — костная стенка улитки; 2 — вестибулярная лестница; 3 — тимпанальная' (ба-
рабанная) лестница; 4 — улитковый проток; 5 — вестибулярная мембрана; 6 —
сосудистая полоска: а — эпителиоциты, б — кровеносный капилляр; 7 — спираль-
ная связка; 8 — лимб спиральной пластинки; 9 — базиллярная пластинка; 10 —
стержень улитки; 11 —спиральная костная пластинка; 12 — спиральный ганглий;
13 — спиральный (кортиев) орган; 14 — покровная мембрана; 15 — внутренний
туннель; 16 — наружные и 17 — внутренние сиолбовые эпителиоциты; 18 — наруж-
ные и 19 — внутренние поддерживающие эпителиоциты; 20 — наружные и 21 —
внутренние волосковые сенсорные эпителиоци^ь
6—321
Рис. 2.86. Ультраструктурная орга-
низация наружного волоскового
сенсорного эпителиоцита.
А — схема. Б — ТЭМ волосковых сен-
сорных эпителиоцитов. В — СЭМ апи-
кальной и Г — ТЭМ базальной их ча-
стей (по К. Койчеву, Е. Иванову).
I — волосковая клетка, II — поддержи-
вающая клетка.
1 — плотный контакт; 2 — кутикула;
3 — волоски; 4 — отростки поддержи-
вающей клетки; 5 — микрофибриллы;
6 — эндоплазматическая сеть; 7 — ми-
тохондрии; 8 — ядро; 9 — стенка «ни-
ши» в поддерживающей клетке; 10 —
эфферентное и 11 — афферентное
нервные волокна.
162
2.7.2.2. Орган равновесия
Орган равновесия образован двумя мешочками — элиптическим
и ампулярным, расположенными в вестибулярной части перепон-
чатого лабиринта. В месте сближения полостей в их стенках име-
ются утолщенные участки разной конфигурации. В глиптическом
мешочке — в форме горизонтального утолщения — пятно, а в ам-
пулярном мешочке в виде складки — ампулярный гребешок
(рис. 2.87, А). Их структурная организация более наглядно пока-
зана на схемах (рис. 2.87, Б). В составе пятна глиптического ме-
шочка, как и в спиральном органе, различают два типа клеток —
поддерживающие эпителиоциты (расположенные на базальной
мембране) и лежащие между ними волосковые сенсорные клетки,
волоски которых расположены в студенистом веществе, содержа-
щем на поверхности зернистость (статоконии). Аналогичный
состав клеток и их расположение имеет и ампулярный гребешок,
только вместо мембраны статоконии на апикальной поверхности
расположен желатинозный купол, лишенный статоконий. К волос-
ковым сенсорным клеткам обоих образований (пятна и мешочка)
подходят нервные волокна.
Ультраструктурная организация волосковых сенсорных эпите-
лиоцитов дана на рис. 2.88. Обратите внимание на наличие двух
типов сенсорных клеток — грушевидных и столбчатых эпителио-
цитов и различие в топографии нервных окончаний.
6*
163
Рис. 2.87. Орган равновесия.
А — поперечный срез через пятно эллиптического мешочка преддверия и ампулы
полукружного канала (по И. В. Алмазову и Л. С. Сутулову). Б — схема их
строения (по Кольмеру): I — пятно, II — ампулярный гребешок.
1 — костная стенка и 2 — полость эллиптического мешочка; 3 — ампулярная по-
лость; 4 — эпителий; 5 — поддерживающие эпителиоциты; 6 — волосковые сенсор-
ные эпителиоциты: а — волоски; 7 — мембрана статоконий; 8 — статоконии;
9 — желатинозный купол; 10 — ганглий преддверного нерва; 11 — нервные волокна;
12 — нервные окончания; 13 — выстилающий эпителий.
164
8
«J CD
165
Рис. 2.88. Ультрамикроскопическое
строение клеток пятна эллиптиче-
ского мешочка.
А — схема (по Я. А. Винникову). Б —
СЭМ апикальной части волосковой сен-
сорной клетки (по К. Койчеву).
I — грушевидный волосковый сенсор-
ный эпителиоцит; II — столбчатый во-
лосковый сенсорный эпителиоцит.
1 — киноцилия; 2 — стереоцилия; 3 —
кутикула; 4 — поддерживающий эпите-
лиоцит; 5 — чашевидное нервное окон-
чание; 6 — эфферентное нервное окон-
чание; 7 — афферентное нервное окон-
чание; 8 — миелиновое нервное волокно.
166
2.7.3. Орган вкуса
Как и у органа слуха и равновесия, поддерживающие и сенсорные
реснитчатые клетки являются производными эктодермы.
Орган (вкусовая почка) располагается в эпителии боковых
поверхностей сосочков языка (кроме нитевидных). Наибольшее их
количество встречается в листовидных сосочках (рис. 2.89, А).
Вкусовая почка сохраняет общий принцип строения органов ре-
цепции. Вкусовой сенсорный эпителиоцит окружен поддерживаю-
щими эпителиоцитами, а его апикальная поверхность снабжена
микроворсинками (рис. 2.89, Б). Последние погружены в вещество,
способствующее взаимодействию молекул вкусовых компонентов
пищи с рецепторами мембран микроворсинок. В нижней части
органа расположен базальный эпителиоцит (камбиальная клетка),
не сообщающийся с внешней средой. К вкусовой почке подходит
афферентное нервное волокно, образующее синапсы с поверх-
ностью вкусовых сенсорных эпителиоцитов. Ультраструктура вку-
совой почки представлена на рис. 2.89, В.
Рис. 2.89. Орган вкуса.
А — листовидные сосочки языка. Б —
вкусовая почка. В — схема ультраструк-
турной организации вкусовой почки.
1 — эпителий; 2 — соединительная
ткань; 3 — вкусовая почка; 4 — эпите-
лиоциты вкусовой почки; 5 — вкусовая
пора; 6 — микроворсинки; 7 — вкусовой
сенсорный эпителиоцит; 8 — поддержи-
вающий эпителиоцит; 9 — базальный
эпителиоцит; 10 — нервное волокно;
11—синаптическая зона; 12 — десмо-
сома; 13 — базальная мембрана; 14 —
гемокапилляры: а — мукопротеиды, б —
ЭПС, в — комплекс Гольджи, г — се-
креторные гранулы.
167
5
В
Рис. 2.89. Продолжение.
2.7.4. Орган обоняния
По источникам развития он наиболее сложный из органов чувств,
так как его обонятельные нейросенсорные клетки имеют нейраль-
ное происхождение, а поддерживающие и базальные являются
производными эктодермального эпителия.
Орган расположен в обонятельной области слизистой оболочки
носа в виде клеточного пласта, названного обонятельным эпите-
лием (рис. 2.90, А). Обычно клеточный состав эпителия определить
в гистологическом препарате не удается. Обратите внимание на
сохранение общего принципа организации органов рецепции (осо-
бенно с органом вкуса). Как следует из рис. 2.90, Б, обонятельные
нейросенсорные клетки находятся в окружении поддерживающих
эпителиоцитов. Базальные же эпителиоциты (камбиальные клетки,
обновляющие эпителиоциты) и малодифференцированные нейро-
ны (обновляющие нейросенсорные клетки) расположены в нижней
зоне и не сообщаются с внешней средой.
168
1
/
A 2
Рис. 2.90. Орган обоняния.
А — слизистая оболочка
(обонятельная). Б — схема
ультраструктурной организа-
ции обонятельного эпителия.
В — ТЭМ обонятельной бу-
лавы (по К. Койчеву, М. Кок-
левой, Д. Лазареву): I — обо-
нятельный эпителий; II — соб-
ственная пластинка слизистой
оболочки.
1 — реснички; 2 — концевые
отделы обонятельной железы;
3 — кровеносный сосуд; 4 —
поддерживающий эпителио-
цит; 5 — обонятельная нейро-
сенсорная клетка; 6 — базаль-
ный эпителиоцит; 7 — мало-
дифференцированный нейрон;
8 — базальная мембрана: а —
дендрит, б — луковица денд-
рита, в — реснички, г —
микроворсинки поддерживаю-
щего эпителиоцита; д — аксон.
169
2.8. Сердечно-сосудистая система
Сердечно-сосудистая система — одна из интегрирующих систем,
играющая важную роль в поддержании гомеостаза организма. Она
способствует реализации функций нервной и эндокринной систем,
а также органов иммунологической защиты. Заболевания сердца и
сосудов широко распространены, и их профилактика и лечение —
важнейшая задача медицины.
Сердечно-сосудистая система представляет собой замкнутую
систему полых трубок. Структура сосудов сильно варьи-
рует и в каждом конкретном случае соответствует их положению
в круге кровообращения и особенностям функции. Вместе с тем
можно говорить об общем плане строения стенки сосуда. На схе-
ме, представленной на рис. 2.91, видны три основные оболочки
стенки сосуда: внутренняя (I), средняя (II), наружная (III), раз-
нообразие тканевого состава оболочек. При этом практически все
разнообразие тканевых компонентов сосудов состоит из производ-
ных мезенхимы.
В принципе слоистую трубку, только значительно более сложно
построенную, представляет собой и сердце. Прежде всего следует
отметить соответствие структуры эндокарда общему плану строе-
ния сосуда. Это единство основано на общности эмбрионального
источника: сосуды и эндокард развиваются из мезенхимы. Мио-
кард и эпикард развиваются из другого источника — миоэпи-
кардиальной пластинки и соответствий в стенке сосуда не
имеют.
2.8.1. Сердце
Для понимания строения сердца целесообразно ознакомиться со
схемой (рис. 2.92). При изучении структуры сердца
взрослого легко определяются три оболочки стенки сердца:
внутренняя — эндокард, средняя — миокард, наружная — эпи-
кард.
Эндокард со стороны полости сердца выстлан непрерывным
слоем эндотелия. Под эндотелием определяется базальная мембра-
на, глубже — субэндотелиальный слой, представленный элемента-
ми рыхлой соединительной ткани, затем мышечно-эластический
слой, состоящий из волокон, преимущественно эластических и
гладких миоцитов. К миокарду прилегает наружный соединитель-
нотканный слой эндокарда, соответствующий наружной оболочке
сосуда с большим количеством кровеносных сосудов, нервов,
жировых клеток, образующих местами значительные скоп-
ления.
Миокард представляет собой сложную многотканевую оболоч-
ку. Основной тканевый компонент миокарда — типическая, или
сократительная, сердечная мышца. Состоит из поперечноисчер-
ченных кардиомиоцитов, состыкованных друг с другом по типу
«конец в конец» и образующих систему анастомозирующих воло-
170
кон. Особенности архитектоники миокарда особенно наглядны в
сканирующем электронном микроскопе (рис. 2.93). При окраске
железным гематоксилином и гематоксилин-эозином ясно опреде-
ляются вставочные диски. На электронограммах (см. рис. 2.92)
хорошо видно, что они представляют собой границу двух кардио-
миоцитов. Кардиомиоциты не представляют собой однообразную
клеточную популяцию, а имеются некоторые отличия: кардиомио-
циты предсердий меньше, чем желудочковые, у них менее развита
Т-тубулярная система.
Преимущественно в правом предсердии открыта особая попу-
ляция клеток — предсердные секреторные (эндокринные) кардио-
миоциты (рис. 2.94), морфофункциональная характеристика ко-
торых (см. гл. 2.4) весьма близка к параметрам пептидсекретор-
ных клеток так называемой диффузной эндокринной системы.
Значение эндокринного аппарата трудно переоценить: модифици-
руя уровень давления крови и объем циркулирующей жидкости,
сердце создает более благоприятные условия для своей работы.
< . f Таким образом, за правым предсердием следует признать целый
ряд важных функций: сократительную, эндокринную, функцию
ритмоводителя.
В сердце выделяют и атипическую («проводящую») мышечную
систему. Микроанатомия проводящей системы сердца отражена
на схеме (рис. 2.95). В синоатриальном узле, располо-
женном практически в стенке верхней полой вены в области веноз-
/ < ного синуса, находятся кардиомиоциты — водители ритма. По
У ' сравнению с типическими кардиомиоцитами это клетки более
узкие, содержат меньшее количество миофибрилл. Их цитолемма
обладает способностью к спонтанной ритмической поляризации
и деполяризации. Ритмоводители окружены значительным коли-
чеством соединительной ткани, богатой коллагеновыми и эласти-
ческими волокнами; узел имеет обильную васкуляризацию и ин-
нервацию. От клеток-ритмоводителей синоатриального узла воз-
буждение распространяется к следующему коллектору атипичес-
ких кардиомиоцитов — атриовентрикулярному узлу.
Клетки этого узла выполняют функцию проведения возбуждения
и его преобразования (замедление ритма), но в некоторых усло-
виях принимают на себя функцию ритмоводителя. Состоящие из
проводящих клеток волокна Пуркинье можно наблюдать
на следующем специальном препарате. Чаще всего волокна прово-
дящей системы расположены субэндокардиально: образованные
атипическими миоцитами волокна Пуркинье примыкают к эн-
докарду.
Клетки по сравнению с сократительными кардиомиоцитами
большие. Ядра расположены эксцентрично, окраска цитоплазмы
более бледная и неравномерная. Неравномерность окраски объяс-
няется неупорядоченным ходом миофибрилл, что подтверждается
электронно-микроскопически (рис. 2.96).
Миокард — многотканевая система. Соединительная ткань
сердца, «прослаивающая» и окутывающая мышечные волокна,
171
Рис. 2.91. Строение кровеносного
сосуда (схема по Ю. И. Афанась-
еву).
I — внутренняя оболочка; II — средняя
оболочка; III — наружная оболочка.
1 — эндотелий; 2 — базальная мембра-
на; 3 — субэндотелий; 4 — внутренняя
эластическая мембрана; 5 — гладкие
миоциты; 6 — эластический аппарат;
7 — наружная эластическая мембрана;
8 — волокнистая соединительная ткань;
9 — сосуды сосудов.
Рис. 2.92. Стенка сердца взрослого
человека.
I — эндокард, II — миокард, III — эпи-
кард.
1 — эндотелиоцит; 2 — эластические во-
локна; 3 — гладкий миоцит; 4 — миофи-
бриллы; 5 — вставочные диски; 6 —
кардиомиоциты; 7 — волокна Пуркинье;
8 — мезотелиоцит; 9 — липоцит.
Рис. 2.93. Миокард.
А — строение кардиомиоцита (схема). Б — вставочный диск (по Б. В. Втюрину).
В — архитектоника мышечных волокон. Г — архитектоника кровеносных сосудов.
1 — контакт кардиомиоцитов;' 2 — ядро; 3 — миофибриллы; 4 — митохондрии;
5 — комплекс Гольджи.
172
Рис. 2.94. Предсердие.
А — секреторный (эндокринный) кар-
диомиоцит. Б — деталь к А (по
И. А. Червовой).
1 — секреторные гранулы; 2 — ядро;
3 — миофиламенты; 4 — митохондрии.
Рис. 2.95. Проводящая система
сердца (схема).
1 — синусный узел; 2 — атриовентрику-
лярный узел; 3 — ножки пучка Гиса;
4 — волокна Пуркинье.
174
1
2
3
Рис. 2.96. Атипические мышечные
клетки (сердце быка).
А — атипические кардиомиоциты. Б —
ТЭМ (по И. А. Червовой).
1 — эндокард; 2 — волокна Пуркинье;
3 — миокард; 4 — миофибриллы; 5 —
митохондрия.
богата ретикулярными элемен-
тами, особенно у детей. На их
основе при некоторых заболева-
ниях возникают экстрамедул-
лярные очаги кроветворения.
На срезах миокарда можно ви-
деть большое количество сосу-
дов: различного калибра арте-
рии, лимфатические сосуды,
многочисленные гемокапилля-
ры, а также нервные элементы,
принадлежащие к интра- и эк-
стракардиальному иннервацион-
ному аппарату.
Эпикард по структуре соот-
ветствует другим серозным обо-
лочкам. Со стороны полости
сердечной сорочки эпикард по-
крыт мезотелием. Под базаль-
ной мембраной определяется
рыхлая соединительная ткань,
богатая коллагеновыми и эла-
стическими волокнами, крове-
носными сосудами, жировыми
клетками, местами образующи-
ми значительные скопления —
прослойки жировой ткани.
2.8.2. Сосуды
До изучения структуры различ-
ных кровеносных сосудов следу-
ет вновь рассмотреть общий план
строения сосуда, представленный
176
3
Рис. 2.97. Артерия мышечного типа.
А — окраска гематоксилин-эозином. Б — окраска альдегид-фуксином.
1 — внутренняя оболочка; 2 — средняя оболочка; 3 — наружная оболочка.
на схеме (см. рис. 2.91). По составу средней оболочки выделяют
сосуды (артерии и вены) мышечного, мышечно-эластического и
эластического типа. На рис. 2.97 представлена артерия мышечного
типа. Хорошо различимы три оболочки сосуда: внутренняя, сред-
няя и наружная. Внутренний слой интимы представлен плоскими
эндотелиальными клетками. Наиболее четко видны их ядра. Внут-
ренний край у артерии неровный, эндотелиоциты видны как на
поверхности, так и в глубине складок. Рельеф внутренней поверх-
ности сосуда представлен на электронной сканограмме (рис. 2.98).
За эндотелием расположен субэндотелиальный слой, состоящий
из соединительной ткани. Последний слой внутренней оболочки —
внутренняя эластическая мембрана, имеющая также извитой вид.
Эластические мембраны хорошо выявляются специальными мето-
177
Рис. 2.98. Границы эндотелиальных клеток интимы коронарной ар-
терии (СЭМ).
диками (см. рис. 2.97). Средняя оболочка артерии мышечного
типа построена из нескольких слоев гладких миоцитов, располо-
женных преимущественно циркулярно (именно из-за посмертного
сокращения миоцитов внутренняя оболочка извита). В мышечных
артериях крупного калибра на границе средней и наружной оболо-
чек присутствует наружная эластическая мембрана. Покрываю-
щая сосуд наружная оболочка состоит из рыхлой волокнистой
соединительной ткани, богатой коллагеновыми и эластическими
волокнами.
Отличительные особенности строения вен мышечного типа по-
казаны на рис. 2.99. При сохранении принципа трехслойного
строения общая толщина стенки вены значительно меньше тол-
щины средней оболочки артерии (это один из ориентиров при
идентификации типа сосуда). Внутренняя оболочка не образует
складок из-за преимущественно продольного расположения глад-
ких миоцитов в средней оболочке. Внутренняя и наружная эла-
стические мембраны у вен не сформированы, и, как правило, на
178
3
Рис. 2.99. Вена мышечного типа.
1 — внутренняя оболочка; 2 — средняя оболочка; 3 — наружная оболочка.
Рис. 2.100. Сосудисто-нервный пучок.
1 — артерия мышечного типа; 2 — вена мышечного типа; 3 — нерв.
их месте наблюдается лишь некоторая концентрация эластичес-
ких волокон.
Рис. 2.100, представляющий сосудисто-нервный пучок, позво-
ляет сравнить строение артерий и вен, предварительно идентифи-
цировав сосуды среди других структур.
179
Рис. 2.101. Аорта (артерия эласти-
ческого типа).
А — общий вид. Б — окончатые мем-
браны средней оболочки (СЭМ).
1 — внутренняя оболочка; 2 — средняя
оболочка; 3 — окончатые мембраны;
4 — наружная оболочка.
180
Стенка аорты (рис. 2.101) отличается прежде всего значитель-
ной толщиной, главным образом за счет средней оболочки. Изме-_
нение тканевого состава средней оболочки артерий (замещение
•тттадКихПмиоцитов, эластическими волокнами) отражается на мор-
Трблогической картине всей стенки артерий. За внутренней ободоч-
кой, образованной эндотелием_и умеренно базофильным субэндо-
телиальным слоем,) распола; ается ширбкая~^рёдняя ~ оболочка"
'внутренняя эластическая мемб^ана^заполненная довольно густым
сплетением эл5стйчёских*“вЬлок< н „Основу средней оболочки со-
ставлятот*МТГПГо4исленные (несколько десяткошряддв1_<<окончатые»
мембраны, среди которых видны удлиненные ядра гладких мирци-
тов. Гладкие миоциты сосудов такого типа не могущцри„сокраще-
ний существенно изменить диаметр просвета сосуда, а являются
лишь Дополнительным элементом эластичности'ГР'овньш внутрен-
ний рельеф изобилие эластически? волокон”наряд/ с другими осо-
бёнИОстям1Г'( большой npocfcet неспособность к спадению) прида-
ют аорте и другим сосудам эластического типа способность ока-
зывать минимальное гемодинамическое сопротивление при распре-
делении больших потоков крови. Это позволяет с функциональной
стороны определять сосуды эластического типа как сосуды распре-
деления, емкостные1. Наружная оболочка содержит большое коли-
чество эластических волокон. В ней легко можно видеть питаю-
щие сосуды, нервные стволики. Обилие эластических, элементов
стенке-аортьПгаиболее демонстративно npif "использовании спе-
"цйальных методов их окраски.
'Строение верхней и нижней полых вен весьма своеобразно и
определяется гемодинамическими и гидростатическими условиями,
связанными с прямохождением.
В структуре верхней полой вены (рис. 2.102, А) хорошо опреде-
ляются внутренняя, средняя и наружная оболочки; обращает вни-
мание слабое развитие мышечных элементов в средней оболочке.
При рассмотрении структуры нижней полой вены (рис. 2.102, Б)
возникает определенная сложность в идентификации сосудистых
оболочек, что связано с изменением привычной топографии глад-
комышечных элементов: мощный мышечный аппарат стенки со-
средоточен в наружной оболочке, толщина которой значительно
превышает толщину внутренней и средней оболочек вместе взя-
тых. В адвентиции гладкие миоциты формируют продольно ориен-
тированные пучки, пронизанные прослойками соединительной тка-
ни, в изобилии содержащей кровеносные и лимфатические сосуды,
нервы. Такая архитектоника стенки является оптимальной для
продвижения большой массы крови к сердцу против направления
силы тяжести.
Система лимфотока начинается «корнями» — начальными лим-
фатическими капиллярами (см. ниже). Крупные лимфатические
сосуды по общему плану строения полностью соответствуют и по
Последняя характеристика относится главным образом к венам.
181
Рис. 2.102. Крупные вены.
А — верхняя полая вена. Б — нижняя полая вена.
1 — внутренняя оболочка; 2 — средняя оболочка; 3 — наружная оболочка.
структуре стенки близки к кровеносным, особенно венам, но систе-
ма клапанов в них более развита.
На рис. 2.103 представлена стенка экстраорганного лимфати-
ческого сосуда, состоящая из трех оболочек: внутренней (I), сред-
ней (II), наружной (III). Однако границы между ними, особенно
между средней и наружной оболочкой, определяются нередко с
трудом. Гладкие миоциты часто разделены большим количеством
коллагеновых аргирофильных волокон, ориентированных главным
образом в виде пологой спирали (в положении, близком к про-
дольному). В некоторых сосудах гладкие миоциты локализуются
не только в средней оболочке, а лежат в несколько слоев в субэн-
дотелиальном слое, могут определяться и в адвентиции. Выделение
среди магистральных сосудов лимфатических сосудов безмышеч-
ного типа (так же, как и в отношении соответствующих вен)
нуждается в оговорке. В ряде лимфатических сосудов миоциты
располагаются по длиннику неравномерно, и в полость среза могут
попадать именно безмышечные участки.
182
Рис. 2.103. Магистральный лимфа-
тический сосуд мышечного типа.
1 — внутренняя оболочка; 2 — средняя
оболочка; 3 — наружная оболочка; 4 —
створка клапана.
2.8.3. Микроциркуляторное русло
Микроциркуляторное, или микрососудистое, русло — система кро-
веносных сосудов небольшого диаметра (как правило, до 100 мкм),
соединяющих артериальные и венозные коллекторы. На ее уровне
регулируется оптимальный режим кровоснабжения органа (и даже
его отдельных микрорайонов), осуществляется гематотканевый
обмен, обеспечивается поддержание внутриорганного гемостаза.
Этот микрососудистый комплекс формируют пять основных типов
микрососудов: артериолы, прекапилляры (прекапиллярные арте-
риолы), капилляры, посткапилляры (посткапиллярные венулы),
венулы; вспомогательную роль играют артериоловенулярные ана-
стомозы. В последние годы понятие «микроциркуляторное русло»
расширено благодаря включению в эту систему лимфатических
капилляров и посткапилляров.
Каким же образом при однотипном ансамбле микрососудов
достигается многообразие, неповторимость условий васкуляриза-
ции различных в морфофункциональном отношении органов?
183
Рис. 2.104. Органоспецифические особенности пространственной органи-
зации микроциркуляторного русла (коррозионные препараты, СЭМ).
А — надпочечник (по Л. И. Полянской). Б — сальник (по В. А. Миронову).
1 — артериола; 2 — прекапилляр; 3 — кровеносные капилляры.
Базисным моментом, помимо органной специфики строения самих
микрососудов (преимущественно гемокапилляров и венул), служит
строгая индивидуальность пространственной организации микро-
циркуляторного русла, генетически закрепленная в соответствии со
структурой и функцией кровоснабжаемого органа. Диапазон вариа-
ций внутриорганной ангиоархитектоники демонстрирует рис. 2.104.
Органоспецифичность строения микрососудистого русла заложена
в композиции его отдельных блоков: в сосудистых ассоциациях
многих органов выделены повторяющиеся сегменты (модули) с
характерной для того или иного органа структурой. Такие комп-
лексы, включающие все звенья микроциркуляторного русла, обла-
дают относительной автономией и квалифицируются как его
структурные единицы. Наиболее наглядно принцип -модульной
конструкции прослеживается в плоскостных препаратах (серозные
оболочки и др.) с двухмерной ориентацией сосудистой сети
(рис. 2.105, А). Формальной границей такого модуля полигональ-
184
ной формы служат анастомозирующие артериолы и сопровож-
дающие их венулы; каждая артериола отдает внутрь модуля не-
сколько прекапилляров, дающих начало капиллярной сети, из
которой кровь по посткапиллярам отводится в венулы. Множест-
венность каналов притока и оттока крови является определенным
гарантом стабильного кровообеспечения тканевого микрорайона.
Важно отметить включение в модуль лимфатических микрососу-
дов, функционирование которых направлено на поддержание ба-
ланса жидкости и белка в тканях. Исключение в этом плане со-
ставляют лишь немногие органы: мозг, селезенка, хрящ, некоторые
отделы глазного яблока, где лимфатические сосуды отсутствуют.
Микрососудистый модуль является удобной моделью для рас-
крытия понятия «система микроциркуляции», более широкого, не-
жели «микроциркуляторное русло». Система микроциркуляции
объединяет каналы транспорта биологических жидкостей — кро-
веносное микроциркуляторное русло, лцмфоносные пути и интер-
стициальное пространство (неклеточный компонент соединитель-
ной ткани, окружающей микрососуды и клетки). Постоянный
перенос воды, ионов, макромолекул между тремя жидкостными
«отсеками» (компартментами) — кровью, интерстициальной (тка-
185
Рис. 2.105. Микроциркуляторное
русло.
А — структурно-функциональная еди-
ница (модуль) микроциркуляторного
русла брюшины (по В. В. Куприянову,
импрегнация солями свинца, фоторе-
конструкция). Б — артериола, венула,
капилляры в мягкой мозговой оболочке
(пленочный препарат).
1 — артериола; 2 — прекапиллярная ар-
териола; 3 — кровеносный капилляр;
4 — посткапиллярная венула; 5 — вену-
ла; 6 — артериоловенулярный анасто-
моз; 7 — лимфатический капилляр; 8 —
лимфатический посткапилляр; 9 — ар-
терия.
186
невой) жидкостью и лимфой — составляет суть процесса микро-
циркуляции и лежит в основе поддержания гомеостаза и собствен-
но жизнеобеспечения тканей и органов.
Тонкостенность микрососудов позволяет идентифицировать их
типы даже при обзоре тотальных (пленочных) препаратов
(см. рис. 2.105). Микрососуды наиболее крупного диаметра —
артериолы и венулы — дифференцировать достаточно просто. Ар-
териолы, как правило, имеют меньший диаметр и более плотную
стенку с характерной «исчерченностью»; последняя обусловлена
визуальным пересечением ядер циркулярно ориентированных мио-
цитов с ядрами эндотелиальных клеток (рис. 2.106, А, Б). Венулы
мягкой мозговой оболочки, впадая в безмышечные вены, не содер-
жат в своей стенке гладких миоцитов. Отсюда — более тонкая
стенка, четко различим просвет сосуда, заполненный эритроцита-
ми, описанная «исчерченность» отсутствует; ядра эндотелиоцитов
в венулах имеют более округлую форму, нежели в артериолах.
Прекапилляры, или прекапиллярные артериолы, отходят от арте-
риолы и еще содержат в своей оболочке разрозненные гладкие
миоциты. Посткапилляры, или посткапиллярные венулы, впадают
в собирательные венулы; строение стенки посткапилляров близко
к строению стенки капилляров, но от последних они отличаются
большим диаметром. Между пре- и посткапиллярами локализуется
сеть гемокапилляров — самых тонкостенных микрососудов с уз-
ким просветом.
Артериолы и венулы могут соединиться напрямую, минуя ка-
пиллярную сеть, посредством артериоловенулярных анастомозов
(АВА). Их задача как механизма регуляции гемодинамики свя-
зана с оперативным перераспределением потоков крови в тканевом
микрорайоне.
Особенности строения стенки различных типов микрососудов
ярко проявляет электронная микроскопия.
Артериолы. В стенке артериолы еще сохраняются три оболочки,
типичные для артерий. Однако в артериоле средняя оболочка
представлена одним слоем миоцитов, внутренняя эластическая
мембрана развита слабее, наружная оболочка менее выраже-
на (рис. 2.106, В, Г). Мышечная оболочка является основной
рабочей структурой артериолярной стенки, так как сокраще-
ние миоцитов служит ведущим механизмом изменения про^
света микрососуда и как следствие регуляции кровотока. Син-
хронность сократительной активности отдельных гладкомышечных
комплексов обеспечивают мио-миоцитарные контакты — нексусы.
Тонус артериол находится под двойным нейрогуморальным кон-
тролем. К стенке артериол подходят нервные проводники, гумо-
ральные же стимулы поступают к миоцитам как через адвенти-
циальную оболочку, так и с током крови. Одним из путей передачи
этой информации, вероятно, служат миоэндотелиальные щелевые
контакты, возникающие благодаря перфорации базальных и внут-
ренней эластической мембран клеточными выростами. Прека-
пиллярную артериолу отличает меньший диаметр, отсут-
187
Рис. 2.106. Строение стенки артериолы.
А, Б — ориентация эндотелиоцитов (одной стрелкой обозначено ядро эндотелио-
цита) и гладких миоцитов (двумя стрелками обозначено ядро миоцита) в стенке
артериолы (по Я. Л. Караганову и соавт., импрегнация серебром). В — фрагмент
стенки артериолы (ТЭМ). Г — артериола (вазоконстрикция, ТЭМ). Д — прекапил-
лярная артериола, ТЭМ (по Т. А. Алимову).
1 — эндотелий; 2 — внутренняя эластическая мембрана; 3 — мышечная оболочка;
4 — мио-миоцитарный контакт; 5 — миоэндотелиальный контакт; 6 — наружная
оболочка; 7 — нервное волокно.
188
ствие эластических мембран и прерывистость монослоя гладких
миоцитов (рис. 2.106, Д). Сплошное мышечное «кольцо» восста-
навливается только в местах отхождения капилляров, формируя
прекапиллярный сфинктер. Эта особенность структуры и «стратеги-
ческое» положение прекапилляров как каналов входа в капилляр-
ное русло (см. рис. 2.105, А) определяет их кардиальное значение
в распределении потоков крови между отдельными участками ка-
пиллярной сети.
Кровеносные капилляры. Функция этих микрососудов — непо-
средственное участие в процессах гематотканевого обмена —
нашла отражение в структуре их стенки (Рис. 2.107). Стенка
капилляра состоит лишь из двух слоев: эндотелия и базаль-
ной мембраны с заключенными в ней перицитами; наружная обо-
лочка отчетливо не выявляется. Для эндотелиоцитов капилляра
характерна функциональная специализация отдельных областей
клетки, значительная протяженность периферической (обменной)
зоны (рис. 2.107, А).
Различают три основных типа капилляров: соматичес-
кий с непрерывными эндотелием и базальной мембраной
(см. рис. 2.107, А); висцеральный с фенестрированным эн-
дотелием и непрерывной базальной мембраной (рис. 2.107, Б, В) и
синусоидный с прерывистыми эндотелием и базальной мем-
браной (рис. 2.107, Г). Соматические капилляры локализуются в
мышцах, сердце, коже, коре головного мозга; висцеральные —
в слизистой оболочке пищеварительного тракта, пищеварительных
и эндокринных железах, почке; синусоидные, которые, помимо
структуры, отличаются максимальной шириной просвета, — только
в печени, костном мозге и селезенке. Тип строения кровеносных
капилляров достаточно объективно отражает активность гемато-
тканевых отношений в органах и тканях.
Представление о путях трансэндотелиального переноса ве-
ществ, транспортных свойствах их структурных эквивалентов
(микропиноцитозные везикулы, фенестры, межклеточные соедине-
ния) облегчает оценку функциональных параметров обменных
микрососудов (рис. 2.108). Значительные успехи в этом направле-
нии достигнуты с помощью метода «замораживание — скалыва-
ние», который позволяет получить информацию о пространствен-
ных (объемных, топологических и др.) характеристиках тран-
спортных систем эндотелия (рис. 2.109). Следует иметь в виду, что
на органном уровне интенсивность обмена определяется не только
структурой стенки капилляров (и прежде всего эндотелия), но и
геометрией капиллярного русла (плотностью распределения ка-
пилляров, характером их взаимосвязи и др.).
Итогом изучения строения кровеносных капилляров является
формирование представления о принципах организации гистоге-
матических, или гематоцеллюлярных, барьеров — своеобразного
форпоста на границе между кровью и «рабочими» клетками орга-
на. Схема (рис. 2.110) иллюстрирует наиболее распространенный
вариант структуры гематоцеллюлярного барьера (рис. 2.110, А),
190
Рис. 2.107. Ультраструктура кро-
веносных капилляров.
А — капилляр соматического типа
(схема). Б — капилляр поджелудоч-
191
Рис. 2.107. Продолжение.
ной железы (висцеральный тип),
ТЭМ. В — капилляр сосудистого клу-
бочка почки (висцеральный тип),
ТЭМ (по Я. Л. Караганову). Г —
синусоидный капилляр печени, ТЭМ.
1 — эндотелиоцит: а — ядросодер-
жащая, б — органеллосодержащая,
в — обменная (маргинальная), г —
околоконтактная зона, д — пара-
плазмолеммальный слой (гликока-
ликс), е — микропиноцитозные ве-
зикулы, ж — элементы цитоскелета;
одной стрелкой обозначены диа-
фрагмированные фенестры; двумя —
недиафрагмированные . («поры»);
2 — базальная мембрана; 3 — пери-
цит; 4 — адвентициальная клетка;
5 — коллагеновые волокна; б —
эритроцит.
Рис. 2.108. Трансэндо-
телиальный транспорт
веществ (схема).* Раз-
мер транспортируемых
молекул, частиц.
7—321
Рис. 2.109. Микропиноцитозные везикулы и фенестры в эндотелии
кровеносных капилляров. Метод «замораживание — скалывание» (ТЭМ
по Г. А. Алимову).
А — скол эндотелия соматического капилляра. Б — скол базальной плазмолеммы
эндотелия перитубулярного капилляра почки.
1 — просвет капилляра; 2 — эндотелий; 3 — микропиноцитозные везикулы, «сво-
бодные» и «связанные» с плазмолеммой эндотелиоцита; 4 — группы фенестр.
а также ее специфику в некоторых органах (рис. 2.110, Б, В, Г).
Ясно, что стенка капилляра является важнейшим, но не един-
ственным компонентом барьера. Более того, при необходимости
повышения изоляционных свойств его композиция может услож-
няться за счет паравазального «футляра» из стромальных клеток
кровоснабжаемого органа (см. рис. 2.110, Г). В противном случае,
(«облегчение обмена») составные компоненты барьера редуцируют-
ся. Так становится прерывистой или совсем исчезает базальная мем-
брана перфорированного эндотелия в синусоидных капиллярах
(см. рис. 2.110, Б), сливаются базальные мембраны капилляров и
респираторных эпителиоцитов в зонах интенсивного газообмена
(см. рис. 2.117, В). Важно усвоить, что морфогенез любого органа
всегда сопряжен с изменением характера гематотканевых отно-
шений, в основе которых лежит органоспецифическая дифферен-
цировка структуры гематоцеллюлярных барьеров.
Рис. 2.110. Гематоцеллюлярные барьеры (схема).
Л — общий принцип строения.
195
7'
Рис. 2.110. Продолжение.
Б, В, Г — органоспецифические особен-
ности ультраструктуры гематоцеллюляр-
ных барьеров; Б — костный мозг (ге-
матом иелоидный барьер); В — легкие
(аэрогематический барьер); Г — голов-
ной мозг (гематоэнцефалический
барьер).
1 — эндотелий; 2 — базальная мембра-
на эндотелия; 3 — интерстициальное
пространство; 4, 5 — базальная мембра-
на и плазмолемма «рабочей клетки»;
6 — эритроцит; 7 — межклеточный кон-
такт в эндотелии.
196
г
Рис. 2.111. Венула
(ТЭМ).
Оболочки: I — внутренняя;
II — средняя; III — на-
ружная.
1 — эндотелиоцит; 2 — ба-
зальная мембрана; 3 —
гладкие миоциты средней
оболочки; 4 — коллагено-
вые волокна; 5 — нервное
волокно.
Посткапилляры, по современным представлениям, также участ-
вуют в обменных процессах, что объясняет сходство их структуры
со структурой кровеносных капилляров. Однако посткапилляры
имеют больший диаметр и более густое перицитарное окружение,
уже имеется адвентициальная оболочка.
В венулах, как правило, восстанавливается четкая трехслой-
ность строения стенки (рис. 2.111). В большей мере это касается
венул достаточно высоких порядков, в частности венул мышечного
типа. В мелких (собирательных) венулах, а также в венулах, впа-
дающих в безмышечные вены, граница между средней и наружной
оболочками сглажена. Мышечную венулу от артериолы отличают
197
Рис. 2.112. Лимфатические микрососуды.
А — лимфатический капилляр; тотальный препарат брюшины (импрегнация солями
серебра). Б, В — ультраструктура стенки лимфатических микрососудов (схема).
Г — фрагмент стенки лимфатического посткапилляра (ТЭМ, по А. В. Королю).
1 — лимфатический капилляр; 2 — лимфатический посткапилляр; 3 — клапан;
4 — эндотелиоцит; 5 — стройные филаменты; 6 — коллагеновые фибриллы; 7 —
кровеносный посткапилляр; 8 — мышечная венула.
198
более округлая форма ядер эндотелиоцитов, отсутствие эласти-
ческих мембран. Нередко наблюдаются прерывистость мо-
нослоя гладких миоцитов в средней оболочке, а в интиме —
клапаны.
Лимфатические микрососуды (капилляры, посткапилляры).
В отличие от кровеносной истоки лимфатической системы лежат
в органах и тканях. Истоками лимфатической системы явля-
ются лимфатические капилляры — слепые широкие туннели в
интерстиции, выстланные крупноклеточным эндотелием (рис.
2.112, А). Их диаметр в несколько, иногда в десятки раз пре-
вышает диаметр гемокапилляров.
Обращает на себя внимание идентичность строения лим-
фатических капилляров и посткапилляров — переход одного
сосуда в другой знаменует лишь появление клапанов (рис.
2.112, Б, В).
Структурные особенности лимфатических капилляров — мень-
шая, нежели в кровеносных, толщина эндотелия, отсутствие
базальной мембраны и перицитов, наличие стропных, или якор-
ных, филаментов, фиксирующих положение микросбсуда в
соединительной ткани, большая подвижность межклеточных
контактов (рис. 2.112, Г, Д)—отвечают ведущей функции лим-
фатических капилляров — резорбции воды, электролитов, макро-
молекул белков и липидов (дренаж интерстициального про-
странства) .
199
2.8.4. Развитие сердца и сосудов
Рассматривая развитие сердца и сосудов, обратим внимание на
ряд особенно важных общих моментов.
Являясь жизненно важной интегративной системой, кровенос-
ная система закладывается в эмбриогенезе рано и рано же вклю-
чается в функцию. Первые кровеносные сосуды появляются в ме-
зодерме стенки желточного мешка, где возникают очаги проли-
ферации мезенхимы, в результате чего образуются так называе-
мые кровяные островки — скопления мезенхимных клеток. Коли-
чество кровяных островков увеличивается, они растут, вытяги-
ваются и, анастомозируя, образуют сеть. Тем временем в них
параллельно идут процессы гемангиогенеза: формируются прими-
тивные сосуды, содержащие первые клетки крови. Описанные
процессы совпадают по времени с началом сегментации мезодермы
(2-я неделя развития). Происходящее в мезодерме желточного
мешка обозначается как процессы гемангиогенеза В отличие от
этого начинающиеся чуть позже процессы закладки сосудов в
теле зародыша (ангиогенез) не сопровождаются процессом кро-
ветворения.
Необходимо отметить, что все возникающие в эмбриогенезе
сосуды первоначально имеют структуру тонких эндотелиальных
трубочек — примитивных капилляров, предназначенных для прове-
дения крови и трансмурального обмена.
На рис. 2.113 представлены начало процесса сегментации мезо-
дермы, появление кровяных островков и происходящие в них
процессы гемангиогенеза. На периферии островков формируются
примитивные (эмбриональные) эндотелиоциты, образующие стен-
ку сосуда, в центре возникают элементы крови — примитивные
эритробласты и эритроциты.
Положение о том, что любой первичный сосуд представляет
собой эндотелиальную трубку, иллюстрирует магистральный сосуд
зародыша человека, представленный на рис. 2.114, А.
В ходе дальнейшего гистогенеза магистрального сосуда в окру-
жающей эндотелиальную трубку мезенхиме обнаруживаются мио-
бласты, идет формирование гладких миоцитов, коллагеновых и
эластических волокон (рис. 2.114,6).
Закладка сердца в эмбриогенезе начинается с выселения из
мезодермы тяжей мезенхимных клеток, образующих билатерально
симметричные «сердечные линии», которые быстро превращаются
в эндотелиальные трубки, являющиеся закладками эндокарда
(рис. 2.115). Эндокард, собственно, закладывается из мезенхимы,
как любой сосуд, и это объясняет возникающее впоследствии
сходство общего плана строения этих структур. Огибающие эндо-
телиальные трубки части висцеральных листков спланхноплевры —
миоэпикардиальные пластинки — представляют собой закладку
миокарда и эпикарда. Билатеральные закладки сердца сближаются
и объединяются, образуя простое трубчатое инвертированное серд-
це, весьма напоминающее сердце рыб: желудочковые отделы зани-
200
Рис. 2.113. Зародыш курицы (сегментация мезодермы).
А — общий вид. Б, В — гистогенетические процессы в кровяных островках.
1 — осевой комплекс; 2 — сомиты; 3 — кровяные островки.
201
Рис. 2.114. Развитие сосудов.
А — магистральный сосуд зародыша человека (4—5 нед, обозначен стрелками).
Б — брюшная аорта человека (7—8 нед).
1 — эндотелий; 2 — гладкие миоциты.
мают головное положение, предсердные — каудальное. Вследствие
неравномерного развития разных частей трубки, ее роста в малом
пространстве средостения начинаются изгибы сердца, приводящие
в конечном счете к формированию четырехкамерного сердца.
В эмбриогенезе человека стенка сердца рано приобретает зна-
чительную толщину, особенно в области желудочков: сердце во
внутриутробном периоде выполняет очень большую работу. Очень
мощное развитие получают правые отделы сердца: сосуды легочно-
го круга дают в эмбриогенезе большое гемодинамическое сопро-
тивление. Рост осуществляется главным образом за счет размно-
жения кардиомиобластов и кардиомиоцитов. Эти клетки имеют
небольшую длину, расположены плотно друг к другу. Соедини-
тельной ткани, кровеносных сосудов практически нет. Трофика
столь мощной стенки осуществляется благодаря тому, что значи-
тельная ее часть (внутренняя) как бы «разбита» на трабекулы,
202
6 12 6
Рис. 2.115. Закладка сердца человека (схема по Штралю, Гису и Борну).
А — парные закладки сердца. Б — сближение закладок. В — слияние закладок в
одну непарную закладку.
1 — эктодерма; 2 — энтодерма; 3 — париетальный листок мезодермы; 4 — висце-
ральный листок мезодермы; 5 — хорда; 6 — нервная пластинка; 7 — сомит; 8 —
вторичная полость тела; 9—эндотелиальная закладка сердца (парная); 10—
нервная трубка; 11 — миоэпикардиальная пластинка; 12 — нисходящая аорта (пар-
ная); 13 — образующаяся головная кишка; 14 — головная кишка; 15 — дорсаль-
ная брыжейка сердца; 16 — полость сердца; 17 — эпикард; 18 — эндокард; 19 —
миокард; 20 — перикард; 21 — перикардиальная полость; 22 — редуцирующаяся
вентральная брыжейка сердца; 23 — нервная трубка. Г, Д, Е — три стадии
развития внешней формы сердца: 1 — артериальный проток (конус); 2 — колено
артериального отдела; 3 — венозный отдел; 4 — венозный синус; 5 — ушковый
канал; 6 — ушки сердца; 7 — правый желудочек; 8 — левый желудочек. Ж — разрез
сердца зародыша на стадии формирования перегородок: 1 — левое предсердие;
2 — правое предсердие; 3 — левый желудочек; 4 — правый желудочек; 5,6 — кла-
паны вен; 7 — перегородка предсердий; 8 — овальное отверстие; 9 — атриовен-
трикулярное отверстие; 10 — перегородка желудочков.
203
1
2
3
Рис. 2.116. Развитие сердца человека.
А, Б — конец 1-го — начало 2-го месяца развития: 1 — трабекулярный миокард;
2 — компактный миокард; 3 — закладка атриовентрикулярного клапана. В —
сердце плода человека (7 мес). Г— миокард новорожденного.
омываемые со всех сторон кровью. В стенке сердца различают
наружный компактный и внутренний трабекулярный миокард
(рис. 2.116, А, Б). В области перехода желудочков в предсердия
обнаруживаются большие разрастания мезенхимы, значительно
выпячивающие эндотелий в полость сердца. Это так называемые
эндокардиальные подушки, являющиеся закладками клапанов.
На 7-м месяце внутриутробного развития хорошо определяют-
ся три оболочки стенки сердца. В миокарде значительно укрупня-
ются кардиомиоциты, формируется характерная структура анасто-
мозирующих волокон сердечной мышцы, заметным компонентом
миокарда становится рыхлая соединителная ткань со значитель-
ным количеством кровеносных сосудов (рис. 2.116, В).
204
В миокарде новорожденного (рис. 2.116, Г) обращает на себя
внимание дальнейшее значительное увеличение размеров кардио-
миоцитов. Митозы в мышечных клетках отсутствуют: способность
к размножению у них оказывается репрессированной и дальней-
шее развитие осуществляется за счет роста кардиомиоцитов. Уве-
личивается и количество соединительной ткани и кровеносных
сосудов, о чем можно судить по большим расстояниям между
мышечными тяжами и по увеличению количества темных поли-
морфных ядер между ними. При сравнении с миокардом взрос-
лого необходимо отметить большую плотность ядер кардиомио-
цитов в поле зрения (клетки мельче), меньшее количество соеди-
нительной ткани и сосудов.
В постнатальном периоде в сердечно-сосудистой системе в
весьма сжатые сроки происходят широкомасштабные изменения,
связанные прежде всего с включением легких в функцию газо-
обмена. Во внутриутробном периоде сосуды малого круга кровооб-
ращения развиваются как сосуды мышечного типа, как резистив-
ные. Благодаря этому к легким поступает относительно небольшое
количество крови. После рождения эти сосуды расширяются, что
205
«5*
Рис. 2.117. Облитерация артериального протока (схема).
I — новорожденный: спадение стенок; II — 3-я неделя жизни: локальное разраста-
ние соединительной ткани интимы; III — 1-й год жизни: закрытие просвета; IV —
2-й год жизни: облитерация просвета.
обеспечивает уже приток крови для осуществления газообмена.
Далее, в течение первых недель постнатального развития гисто-
генетические процессы приводят к превращению околосердечных
сосудов малого круга в сосуды эластического типа, обеспечиваю-
щие количественно оптимальный для газообмена кровоток. Увели-
чению кровотока через легкие способствует и закрытие обходных
путей, через которые во внутриутробном периоде осуществлялся
переброс части крови из малофункционирующего легочного в
большой круг кровообращения: облитерация артериального про-
тока, заращение овального отверстия.
Усилению кровотока в легочном круге способствует и увеличе-
ние объема правого желудочка, происходящее за счет перегруппи-
ровки волокон сердечной мышцы.
После рождения в результате расширения сосудов малого кру-
га ток крови через артериальный (боталлов) проток практически
прекращается и его просвет спадается. Далее разрастается рыхлая
соединительная ткань интимы, приподнимающая эндотелий; об-
разуется своеобразная «подушка». «Подушка» увеличивается, пе-
рекрывает просвет, исчезает ограничивающий ее эндотелий, соеди-
206
Новорожденный ребенок
Ребенок 2 мес
Рис. 2.118. Перестройка мышечных
волокон в стенке сердца после
рождения (объяснения в тексте).
Рис. 2.119. Стенка сердца ребенка
4 лет.
207
нительная ткань становится более грубой, и происходит облите-
рация (заращение) протока (рис. 2.117).
Как отмечалось выше, в это же время увеличивается полость
правого желудочка за счет истончения его стенок. Этот процесс
нередко обозначали ранее как «физиологическую атрофию» право-
го сердца. Установлено, однако, что истончение стенки правого
желудочка и соответствующее увеличение объема правого желу-
дочка происходят за счет перегруппировки мышечных волокон,
без заметной потери мышечных клеток и, следовательно, без
уменьшения сократительной способности (рис. 2.118).
В постнатальном периоде, как уже упоминалось, кардиомиоци-
ты не размножаются. Развитие и рост сердца осуществляются
главным образом за счет синтеза специфических белков и соот-
ветственно роста кардиомиоцитов, развития соединительной ткани,
сосудов (рис. 2.119).
2.9. Органы кроветворения
Количественные и качественные параметры периферической крови
являются результатом сбалансированного протекания двух процес-
сов: кроветворения и кроворазрушения. Большой интерес представ-
ляют вопросы, связанные с гистофизиологией органов иммуните-
та, ибо от их адекватного функционирования зависит успешное
протекание реакций защиты организма от живых организмов и
веществ, несущих на себе признаки чужеродного генома. Умение
диагностировать основные структурные элементы кроветворных
органов необходимо не только для формирования теоретических
представлений; в современной клинике врачу нередко приходится
иметь дело с результатами биопсийных исследований костного
мозга, селезенки, лимфатических узлов.
Приступая к изучению структурно-функциональных особен-
ностей органов кроветворения, необходимо уяснить основные эта-
пы гемопоэза в онтогенезе, их сроки, принципиальные характе-
ристики (табл. 2.2).
Первые клетки крови появляются вне тела зародыша, в ме-
зенхиме стенки желточного мешка эмбриона человека в возрасте
13 дней в виде эритроидных клеток, дифференцирующихся по
мегалобластическому типу.
Начиная со 2-го месяца внутриутробного развития функцию
кроветворения берет на себя печень. В сосудах печени продолжа-
ется эндоваскулярный мегалобластический эритропоэз, впослед-
ствии сменяющийся экстраваскулярным эритропоэзом нормоблас-
тического типа. В печени впервые возникают гранулоциты и
мегакариоциты; в последнее время получены данные, что на-
чиная с 9-й недели эмбрионального развития в печени появляются
В-лимфоциты. Небольшое количество очагов кроветворения сохра-
няется в печени новорожденного, особенно в случае недоношен-
ности. В последующем гемопоэз перемещается в селезенку, лим-
фатические узлы и костный мозг.
208
Таблица 2.2. Развитие гемопоэтической системы человека
Локализация кроветворения Период эмбриогенеза (недели)
Желточный мешок 3—4-я
Печень, начало кроветворения 5—6-я
Появление селезеночного рудимента Конец 6-й
Эритропоэз в селезенке (начало) Конец 12-й
Лимфопоэз в селезенке (начало) 20-я
Появление гемопоэтических очагов в кост- ном мозге 13—14-я
Появление эпителиального тимусного ру- димента Коней 5-й
Появление больших лимфоцитов в тимусе 9—20-я
Лимфопоэз в лимфатических узлах 16—17-я
Появление циркулирующих малых лим- фоцитов 17-я
2.9.1. Костный мозг
Для оценки костномозгового кроветворения используют мазок
костного мозга, полученный путем прижизненного пунктирования
грудины. Именно на таких препаратах в клинических лаборато-
риях подсчитывают миелограмму — соотношение различных кле-
точных элементов. Для диагностики кроветворных клеток удобно
воспользоваться схемой гемопоэза (рис. 2.120).
Элементы ранних стадий развития гемопоэза (клетки I—III
класса, или клетки предшественники)', стволовые кроветворные
клетки— (СКА) (рис. 2.121, А, Б), полустволовые клетки и уни-
потентные клетки-предшественники невозможно отличить друг от
друга, применяя морфологические и цитохимические методы ана-
лиза. По внешнему виду все названные клетки напоминают малый
лимфоцит.
Клетки следующего IV класса — бластные элементы — харак-
теризуются значительными размерами (Д = 16 мкм, т. е. в 2 раза
больше, чем в предшествующих формах), крупным ядром с неж-
ным рисунком хроматина и отчетливо видными ядрышками, не-
большим, слабо базофильным ободком цитоплазмы (рис. 2.121, В).
На уровне привычного морфологического анализа невозможно
сказать, в каком направлении (эритроцитарном, гранулоцитарном
и т. д.) развиваются клетки-бласты; это можно сделать лишь при
применении цитохимических методов исследования.
209
2
Рис. 2.120. Схема постэмбрионального гемопоэза (И. Л. Чертков,
А. И. Воробьев, 1973).
1 — стволовая кроветворная клетка; 2 — полипотентная клетка-предшественница
лимфопоэза; 3 — полипотентная клетка-предшественница миелопоэза; 4 — клетка-
предшественница В-лимфоцитов; 5 — клетка-предшественница Т-лимфоцитов; 6 —
лейкопоэтинчувствительная клетка; 7 — эритропоэтинчувствительная клетка; 8 —
тромбопоэтинчувствительная клетка; 9 — плазмобласт; 10 — проплазмоцит; 11 —
плазмоцит; 12 — лимфобласт; 13 — пролимфоцит; 14 — В-лимфоцит; 15 — лимфо-
бласт; 16 — пролимфоцит; 17 — Т-лимфоцит; 18 — монобласт; 19 — промоноцит;
20 — моноцит; 21 — миелобласт; 22 — базофильный промиелоцит; 23 — базофиль-
ный миелоцит; 24 — базофильный метамиелоцит; 25 — палочкоядерный базофиль-
ный гранулоцит; 26 — сегментоядерный базофильный гранулоцит; 27 — нейтро-
фильный промиелоцит; 28 — нейтрофильный миелоцит; 29 — нейтрофильный мета-
миелоцит; 30 — нейтрофильный палочкоядерный гранулоцит; 31 — нейтрофильный
сегментоядерный гранулоцит; 32 — эозинофильный промиелоцит; 33 — эозинофиль-
ный миелоцит; 34 — эозинофильный метамиелоцит; 35 — эозинофильный па-
лочкоядерный гранулоцит; 36 — эозинофильный сегментоядерный гранулоцит;
37 — эритробласт; 38 — пронормоцит; 39 — нормоцит базофильный; 40 — нормоцит
полихроматофильный; 41 — нормоцит оксифильный; 42 — ретикулоцит, 43 — эри-
троцит; 44 — мегакариобласт; 45 — промегакариоцит; 46 — мегакариоцит; 47 —
тромбоцит.
210
1
Рис. 2.122. Эритропоэз. Выброс
ядра (схема, ТЭМ).
в
Рис. 2.121. Характеристика стволо-
вой кроветворной клетки.
А — колонии гемопоэтических клеток,
развившихся из донорских стволовых
клеток у летально облученных реципи-
ентов. Б — улвграструктура стволовой
кроветворной клетки. В — морфологи-
чески неидентифицируемые бласты в
пунктате костного мозга.
1 — ядро; 2 — цитоплазма.
212
При диагностике элементов эритропоэза следует помнить, что
их дифференцировка сопровождается уменьшением размеров кле-
ток, огрубением хроматина ядра с последующим «выталкиванием»
его из клетки (рис. 2.122), изменением окраски цитоплазмы: вна-
чале базофильной (обусловленной синтезом белковой части гемо-
глобина), затем полихроматофильной (синтез белка сочетается
с накоплением готового гемоглобина) и оксифильной (синтез ге-
моглобина окончен). При изучении конечных форм эритропоэза
следует иметь в виду, что при обычной окраске ретикулоцит не-
возможно отличить от эритроцита; сделать это можно лишь при
использовании специальных методов (см. раздел «Кровь»),
Среди элементов тромбоцитарного ряда легко определяется
мегакариоцит — гигантская клетка костного мозга (Д = 50—
80 мкм) с крупным многодольчатым ядром и скоплением азуро-
фильной зернистости в цитоплазме (рис. 2.123, А). Для изучения
процесса образования тромбоцитов путем их отшнуровки по так
называемым демаркационным линиям (каналам агранулярной
цитоплазматической сети) можно использовать рис. 2.123,'Б.
При идентификации элементов гранулоцитопоэза следует
учесть, что по мере созревания клеток происходят уменьшение их
Рис. 2.123. Отшнуровка тромбоци-
тов от мегакариоцита.
А — на мазке костного мозга. Б — схе-
ма, ТЭМ.
1 — плотные тельца; 2 — гранулы; 3 —
митохондрии; 4 — агранулярная цито-
плазматическая сеть, представляющая
собой демаркационные линии отделения
тромбоцитов; 5 — многодольчатое ядро;
6 — отделяющиеся тромбоциты.
213
Рис. 2.124. Красный костный мозг.
А — схема. Б — окраска гематоксилин-эозином. В — деталь к «Б». Г — СЭМ
макрофага и ретикулоцита (по Ю. А. Ровенских).
1 — жировые клетки; 2 — островки кроветворных клеток; 3 — мегакариоцит;
4 — артерия; 5 — стромальная клетка; 6 — кровеносный синусоидальный капилляр;
7 — макрофаг; 8 — оксифильный нормоцит; 9 — ретикулоцит; 10 — поры.
размеров, изменение формы ядер (приобретение ядром сегменти-
рованного вида у зрелых форм), снижение базофилии цитоплаз-
мы, появление неспецифической (азурофильной) зернистости на
стадии промиелоцита и специфической (нейтрофильной, окси-
фильной и базофильной зернистости) на стадии миелоцита.
Среди предшественников моноцитов морфологически можно
выявить лишь монобласты и промоноциты, характеризующиеся
большими, нежели у зрелых клеток, размерами, базофилией цито-
плазмы, высоким ядерно-цитоплазматическим соотношением, от-
сутствием азурофильной зернистости цитоплазмы (появляется
на стадии промоноцита), изменением формы ядер.
Среди элементов лимфопоэза в мазке костного мозга можно
увидеть клетки с морфологией лимфобластов, пролимфоцйтов, ма-
лых лимфоцитов. При изучении лимфоидного ряда необходимо
четко уяснить, что морфология (особенно на светооптическом
уровне) не является основным диагностическим тестом, характе-
ризующим роль, выполняемую данным элементом в обеспечении
их основной функции — участие в развитии иммунологических
215
реакций. Так, клетки с морфологией лимфобластов и пролимфо-
цитов могут быть промежуточными элементами как антигеннеза-
висимого, так антигензависимого этапа дифференцировки, а среди
клеток, морфологически идентифицируемых как малый лимфоцит,
могут находиться такие разнообразные элементы, как клетки-
предшественники I—III класса любого гемопоэтического ряда,
конечные формы антигеннезависимого этапа дифференцировки
Т- и В-линий, иммунные Т-лимфоциты, 0-лимфоциты.
216
Рис. 2.125. Кровеносный синусоид-
ный капилляр красного костного
мозга.
А — окраска гематоксилин-эозином.
Б — СЭМ (по Ю. А. Ровенских). В —
выход оксифильного нормоцита из крас-
ного костного мозга (ТЭМ).
1 — синусоидный гемокапилляр; 2 —
мигрирующие эритроциты и лимфоци-
ты; 3 — жировые клетки; 4 — гемопо-
этическая ткань; 5 — оксифильный
нормоцит; 6 — эндотелиальные клетки.
В поле зрения могут встретиться и клетки плазмоцитарного
ряда (0,9 %), вероятно, попадающие в костный мозг через крово-
ток из периферических лимфоидных органов. Количество этих
элементов может быть увеличено при некоторых (В-лимфопроли-
феративных) заболеваниях системы крови: плазмоцитоме, плазмо-
клеточном остром лейкозе. В процессе антигензависимого этапа
дифференцировки В- и Т-лимфоцитов наряду с эффекторами
(плазмоциты, Т-киллеры, Т-хелперы, Т-супрессоры) образуются
В- и Т-клетки памяти.
При изучении структуры костного мозга на гистологическом
срезе (рис. 2.124, А, Б, В) обращает на себя внимание соотноше-
ние между участками, занятыми кроветворными элементами и
стромальными клетками, в том числе жировыми. Функциональную
связь стромальных и кроветворных клеток наглядно иллюстрирует
рис. 2.124, г, где представлен своеобразный «островок» из макро-
фага и эритробластов, при контактировании которых происходит
передача молекулярного железа (освобожденного из неполноцен-
ных эритроцитов) для построения геминовой части гемоглобина.
Особое внимание следует уделить изучению микроциркулятор-
ного русла органа. Идентификация кровеносных синусоидных
капилляров или синусов (рис. 2.125, А) затруднена тем, что их
просвет занят форменными элементами крови, а строма между
ними — большим количеством гемопоэтических клеток. Объемная
картина строения синусоида представлена на рис. 2.125, Б. Учи-
тывая то, что одной из важнейших функций синусоидных капил-
ляров является обеспечение перехода форменных элементов в
кровоток, следует подчеркнуть наличие своеобразного устройст-
ва — пор в эндотелиоцитах, пропускающих в циркуляцию лишь
зрелые формы (рис. 2.125, В).
217
Рис. 2.126. Красный костный мозг (плод 7 мес, окраска гематоксилин-
эозином).
1 — костные перекладины; 2 — синусоидный гемокапилляр; 3 — островки крове-
творных клеток.
Рис. 2.127. Красный костный мозг новорожденного (окраска гематок-
силин-эозином).
1 — островки кроветворных клеток; 2 — мегакариоцит.
Единственной кроветворной клеткой, легко диагностируемой
при малом увеличении, является мегакариоцит. При большом уве-
личении на препарате можно идентифицировать также миелоциты
(характеризующиеся специфической зернистостью и бобовидным
ядром), элементы эритропоэза, зрелые форменные элементы кро-
ви, «бластные» формы.
С 7 мес антенатальной жизни костный мозг становится
основным центром миелоидного кроветворения: среди многочис-
ленных островков костеобразования регистрируются экстраваску-
218
лярные очаги гемопоэза. Основную массу среди всех форм гемо-
поэза в этот период составляют эритропоэтические элементы
(рис. 2.126).
У новорожденного костномозговые полости заполнены актив-
ным (красным) костным мозгом, а жировая ткань (в отличие от
костного мозга взрослого) представлена лишь отдельными липо-
цитами или их небольшими скоплениями. Наряду с эритроидным,
миелоидным и мегакариоцитарным ростком в этот период встре-
чаются и лимфоидные клетки (рис. 2.127).
2.9.2. Вилочковая железа (тимус)
Изучение структурной организации тимуса следует начинать с пе-
риода новорожденное™, так как максимальная функциональная
активность органа, находящая свое отражение в его строении,
наблюдается в эмбриональном, неонатальном и раннем постна-
тальном периоде. При изучении строения тимуса (рис. 2.128, А, Б)
3
4
2
Б
Рис. 2.128. Тимус новорожденного.
Л окраска гематоксилин-эозином. Б — слоистое эпителиальное тельце (по
К). Л. Ровенских).
219
Рис. 2.128. Продолжение.
В — СЭМ: клеточные элементы и ретикулярные волокна. Г — контакт Т-лимфоцитов
с ретикулоэпителиальной клеткой (в центре) тимуса, (ТЭМ). Д — схема крово-
снабжения дольки тимуса. Е — ультраструктура гемотимического барьера.
1 — междольковая соединительная ткань; 2 — корковое вещество; 3 — мозговое
вещество; 4 — слоистое эпителиальное тельце; 5 — лимфоциты; 6 — ретикулоэпи-
телиальная клетка; 7 — просвет гемокапилляра; 8 — эндотелиоцит; 9 — перикапил-
лярное пространство.
221
Рис. 2.129. Возрастные изменения тимуса.
А — закладка тимуса (эмбрион, конец 2-го месяца). Б — развитие и инволюция
тимуса (первые два рисунка отражают вид органа, последующие — вид одной
доли). В — тимус взрослого человека (гематоксилин-эозин).
1 — эмбрион 34,9 мм; 2 — эмбрион 119 мм; 3 — новорожденный; 4 — ребенок
7 лет; 5 — мужчина 20 лет; 6 — мужчина 30 лет; 7 — мужчина 66 лет.
обращает на себя внимание выраженная дольчатость и подразде-
ление каждой дольки на корковое (темное) и мозговое (бледное)
вещество, содержащее меньшее количество лимфоидных клеток.
Септы, являющиеся продолжением соединительнотканной капсу-
лы, не полностью разделяют ткань тимуса на дольки: в централь-
ной части каждой из них ткань тимуса непрерывна.
Эпителиальную основу органа легче рассмотреть в мозговом
веществе (в связи с меньшим количеством лимфоцитов): речь
идет об отростчатых эрителиоцитах и слоистых эпителиальных
тельцах (или тельцах Гассаля), образованных концентрически
наслоенными клеточными элементами. Во избежание идентифика-
ции этих структур как мелких кровеносных сосудов можно ис-
пользовать объектив большого увеличения (рис. 2.128, В). Изучая
структурные особенности эпителиального компонента тимуса, сле-
дует учесть, что он выполняет не только опорную функцию, но и
обеспечивает создание специфического микроокружения путем
выделения гуморальных факторов, обусловливающих дифферен-
цировку лимфоидных клеток в направлении Т-лимфоцитов
(рис. 2.128, Г).
222
Анализируя особенности кровоснабжения тимуса (рис. 2.128, Д),
необходимо уяснить, что капилляры коркового вещества (отходя-
щие от артериол, расположенных на границе коркового и мозго-
вого вещества), образуют анастомозирующие капиллярные аркады,
впадающие в конечном счете в посткапиллярные венулы мозгового
вещества.
223
Характерно, что капилляры коркового вещества (с периваску-
лярным пространством с расположенными в нем макрофагами)
окружены непрерывным рядом эпителиальных клеток (рис.
2.128, Е), что обеспечивает образование гематотимусного барь-
ера. Последний предохраняет корковое вещество органа от попа-
дания чужеродных антигенов, особенно в эмбриональном пе-
риоде. V _
Для тимуса взрослого человека характерны признаки инволю-
ции органа, выражающиеся в уменьшении количества лимфоцитов
(особенно в корковом веществе), обильном разрастании соедини-
тельной ткани, возрастании количества жировых клеток, редукции
мозгового вещества (но в меньшей степени, чем коркового), при-
обретении тельцами Гассаля гомогенной структуры. Подчеркивая
гистологически идентифицируемую инволюцию тимуса, не следует
забывать, что и в этот период тимус не полностью теряет функ-
циональную активность, что особенно наглядно проявляется в
условиях иммунологического конфликта.
Характеризуя особенности эмбрионального развития тимуса,
следует обратить внимание на то, что закладка органа на первом
месяце эмбриональной жизни представлена эцителиальными поч-
ками на вентральной части третьего и дорсальной Части четвер-
того эмбриональньРгжаберщ^х-карманов (рис. 2.1297АГ.По совре-
менным данным, парная закладка тимуса происходит из III экто-
дермального жаберного кармана, а энтодерма не принимает участия
в органогенезе тимуса. Впоследствии эпителиальные почки увеличи-
ваются в размерах и сближаются своими концами (но без полного
слияния правой и левой закладок). Первое появление лимфоцитов
отмечено в органе с 7,5 нед развития.
С возрастом тимус постепенно атрофируется: абсолютная мас-
са органа увеличивается до периода половой зрелости, а затем
снижается; относительная же масса органа снижается с момента
рождения (рис. 2.129, Б).
2.9.3. Лимфатический узел
Основные черты структурной организации лимфатического узла
представлены на рис. 2.130, А. Снаружи—димфатический узел по-
крыт соединительнотканной капсулой с обилием липоцитов, от
которой отходят тонкие_перегородки — трабекулы. Непосредст-
венно под капсулой расположено корковое вещество, представлен-
"ноё скоплениями лимфатических узелков, а еще глубже по направ-
лению к воротам органа — мозговое вещество, в котором лимфо-
циты располагаются диффузно или в составе мякотных шнуров.
Строму" органа^ составляет ретикулярная ткань, структуру ко-
торой лучше рассмотретьГв"мозгдвом веществе.
Основная функция лимфатических узлов — участие в иммуно-
логических реакциях — находит свое отражение в / структурной
организации органа: антигены, проникающие через приносящие
лимфатические сосуды, вначале захватываются макрофагами па-
224
1
Рис— 2.130. Лимфатический узел.
А — общий вид. Б—В- и Т-зоны лимфатического узла (гематоксилин-эозин).
В — накопление краски в фагоцитирующих клетках лимфатического узла.
К—321
225
Рис. 2.130. Продолжение.
Г — клетки плазмоцитарного ряда (ок-
раска метиловым зеленым — нирони-
ном). Д — миграция лимфоцитов через
посткапиллярную венулу (схема). Е —
СЭМ (по Ю. А. Ровенских).
1 — капсула; 2 — рыхлая соединитель-
ная ткань; 3 — трабекулы; 4 — рыхлая
соединительная ткань в области ворот;
5 — артерии и 6 — вены в области во-
рот; 7 — краевой синус; 8 — промежу-
точный синус; 9 — центральный синус;
10 — лимфатические узелки; 11 — моз-
говые тяжи; 12 — центр размножения;
13 — тимусзависимая (паракортикаль-
ная), или Т-зона; 14 — бурсазависимая,
или В-зона; 15 — эндотелиальные клет-
ки; 16 — лимфоциты.
226
ракортикальных (Т-зон), так называемыми интердигитирующими
клетками, затем в реакцию включаются макрофаги герминативных
центров лимфатических узелков (В-зон) — дендритные клетки
(рис. 2.130, Б). Таким образом, макрофагальная реакция — пер-
вый этап в развитии иммунологического ответа (рис. 2.130, В).
В реакциях клеточного иммунитета конечным этапом является
образование Т-киллеров, вызывающих лизис клеток-мишеней, диа-
гностировать которые морфологическими способами не представ-
ляется возможным. В противоположность этому конечными фор-
мами В-лимфоцитов (в реакциях гуморального иммунитета) яв-
ляются плазматические клетки, обладающие характерной струк-
турой и обычно скапливающиеся в мякотных шнурах лимфатиче-
ских узлов (рис. 2.130, Г).
Наряду с описанными выше процессами — усиление макрофа-
гальной активности, увеличение размеров реактивных центров,
плазматизация — в условиях антигенной стимуляции увеличивает-
ся клеточность органа, стираются границы между корковым и моз-
говым веществом, возрастает миграция лимфоцитов в кровоток
через посткапиллярные венулы (рис. 2.130, Д, Е).
Особенности развития лимфатических узлов в эмбриональном
периоде состоят в том, что они закладываются и дифференциру-
ются неодновременно. Первые закладки появляются на втором
месяце, большая часть — во второй половине внутриутробной жиз-
ни. Основные особенности гистогенетических процессов в лимфа-
тических узлах представлены на рис. 2.131, А, Б.
А
Рис. 2.131. Развитие лимфатических узлов.
А — начало формирования краевого сплетения и паренхимы по периферии петель
кровеносных сосудов (эмбрион 9 нед). Б — начальные этапы формирования си-
нусов в виде эндотелиальных трубочек, эмбрион 16 нед. В — лимфатический узел
новорожденного.
I — корковое вещество; 2 — мозговое вещество.
227
Рис. 2.131. Продолжение
Для лимфатических узлов новорожденных характерны тон-
кость капсулы и трабекул, слабое развитие фолликулярного аппа-
рата (лимфоциты в корковом веществе расположены диффузно),
плохая визуализация мякотных шнуров и синусов из-за обильной
диффузной инфильтрации лимфоцитами паренхимы органов
(рис. 2.131, В). В последующие годы жизни происходит энергич-
228
ное развитие лимфатичес-
кой ткани, новообразование
фолликулов, окончательное
формирование трабекул, но
только к 12 годам структура
лимфатических узлов при-
ближается к дефинитивной.
2.9.4. Селезенка
Ознакомление с рис. 2.132,
А, Б позволит понять микро-
скопическое строение орга-
на. Капсула, покрытая сероз- 6
ной оболочкой, и отходящие в-
от нее трабекулы образовав _ б-
ны не только элементами 11
6
Рис. 2.132. Селезенка.
А — схема строения. Б — схема
кровоснабжения (штрихами обо-
значены стромальные клетки, тре-
угольниками — макрофаги, круж-
ками и точками — лимфоциты). А
229
Г .
Рис. 2.132. Продолжение
В — периартериальная (Т) зона лимфатического узелка селезенки (окраска мети-
ловым зеленым — пиронином). Г — макрофаги селезенки, нагруженные железом
погибших эритроцитов (окраска метиловым зеленым — пиронином). Д — сину-
соидный гемокапилляр (поперечный скол начального отдела), видны округлые
эритроциты (Э), проходящие через стенку (по Ю. А. Ровенских).
1 — трабекула; 2 — трабекулярная артерия; 3 — трабекулярная вена; 4 — пуль-
парная артерия; 5 — пульпарная вена; 6 — центральная артерия лимфатического
узелка; 7 — кисточковые артериолы с эллипсоидами; 8 — венозные синусы;
9 — кровеносные капилляры лимфатического узелка; 10 — тимусзависимая, пери-
артериальная (Т) зона; 11 —лимфатический узелок селезенки: а—центр размно-
жения (В-зона), б — мантийный слой, в — маргинальная зона; 12 — капсула.
230
соединительной ткани, hq содержат_некоторое количество 1гладко-
' ~ мышечных^клетокЗобеспечиваюших выполнение органом функций
депонирования,_______
Лимфатические узелки в селезенке, в совокупности формирую-
щие белую пульпу, разбросаны по всему срезу органа в отличие
от строго периферического расположения их в лимфатическом
узле. Важным диагностическим признаком является наличие в
лимфатических узелках селезенки срезов мелких артериальных
сосудов — центральных артерий (нередко расположенных эксцен-
трично), отсутствующих" в аналогичных структурах лимфатических
узлов.
Периартериальная зона лимфатических узлов является тимус-
зависимой, в то время как В-лимфоциты занимают реактивный
центр фолликула. Именно в периартериальной зоне происходит
бласттрансформация Т-клеток в условиях развития реакций кле-
точного иммунитета (рис. 2.132, В). Макрофаги селезенки участ-
вуют не только в иммунологическиу^реакциях, но и обеспечивают
одну из важнейших функций красноипульГТБГ селезенки — унич-
тржение отживших ^эритроцитов (рис. 2.132, Г).
Селезенка относится к органам, система кровоснабжения ко-
231
Рис. 2.133. Развитие селезенки.
А — закладка селезенки (эмбрион конца 2-го месяца). Б — селезенка плода 7 мес.
В — селезенка новорожденного.
232
торых неразрывно связана с их структурно-функциональными
особенностями (рис. 2.132, Д).
Развитие селезенки. Закладка органа происходит на 5—6-й
неделе антенатального развития и представлена плотным скопле-
нием мезенхимальных клеток внутри развивающегося большого
сальника (рис. 2.133, А).
В течение 2—4-го месяца внутриутробного развития идет ак-
тивный гистогенез сосудов селезенки, а первые гемопоэтические
очаги (миелоидные) появляются в начале 5—6-го месяца эмб-
риогенеза.
На 7-м месяце внутриутробного развития возрастает интен-
сивность лимфопоэза, однако оформленные лимфатические узелки
еще не диагностируются и лимфоидные клетки располагаются
диффузно (рис. 2.133, Б).
У новорожденного гистогенез селезенки далеко не завершен
(рис. 2.133, В). Капсула и трабекулы тонкие (гладкомышечный
компонент их развит слабо). Трабекулярные артерии и вены диф-
ференцируются с трудом, в красной пульпе (экстраваскулярно)
выражена миелопоэтическая активность. Белая пульпа развита
слабо; нередко можно наблюдать не лимфатические узелки, а их
закладки — периартериальные лимфоидные муфты.
2.10. Эндокринная система
Значение эндокринной системы как интегрирующей системы орга-
низма состоит в координации различных метаболических процес-
сов на уровне целостного организма, т. е. в поддержании его го-
меостаза. Эндокринопатии, связанные с дисфункцией той или иной
железы, занимают значительное место в ряду важнейших заболе-
ваний человека, в том числе болезней детского возраста, в генезе
аномалий развития.
2.10.1. Центральные звенья эндокринной системы
I Нейросекреторные ядра гипоталамуса. Общие принципы органи-
зации нейросекреторных образований мозга можно проиллюстри-
ровать на примере крупноклеточных ядер переднего гипоталамуса.
На схеме, показывающей топографию супраоптического и пара-
вентрикулярного ядер (рис. 2.134, А), можно видеть центрально
расположенную полость III желудочка (I), выстланную низкими
кубическими клетками эпендимы. По обе стороны от него в виде
«крыльев бабочки» располагаются комплексы мультиполярных
нейронов с базофильной секреторной зернистостью — паравентри-
кулярные ядра (рис. 2.134, Б). Перпендикулярно к просвету
III желудочка, в базальной части мозга, непосредственно под моз-
говой оболочкой виден срез перекрестка зрительных нервов (или
зрительного тракта при большой глубине среза) — рис. 2.134, В.
Супраоптические ядра — комплексы более крупных нейросекре-
торных клеток с базофильной зернистостью — лежат непосред-
233
R
Рис. 2.134. Нейросекреторные (крупноклеточные) ядра гипоталамуса.
А — схема топографии ядер. Б, В — препарат (окраска альдегид-фуксином-метиле-
новым синим).
1 — полость 111 желудочка; 2 — оптический тракт; 3 — супраоптическое ядро; 4 —
паравентрикулярное ядро.
Рис. 2.135. Ультраструктурная организация нейросекреторной клетки
гипоталамуса (схема).
1 — ядро; 2 — перикарион; 3 — цитоплазматическая сеть; 4 — пластинчатый ком-
плекс; 5 — лизосома; 6 — секреторные гранулы; 7 — нейрофиламенты; 8 — крове-
носный капилляр; 9 — аксон; 10 — дендриты; 11—аксосоматический синапс;
12 — аксодендритический синапс.
ственно над зрительным-перекрестом по его боковым сторонам.
Они лучше васкуляризованы, чем окружающая ткань мозга. Об-
щие черты организации и специфика клеточного состава супра-
оптического и паравентрикулярного ядер'заключаются в том, что
первое образовано в основном крупными мультиполярными ней-
росекреторными клетками, во втором — наряду с ними широко
11редставленапоПуляция мелких аД|Юнергичеекихн£Йросекреторных
клеток. Отметим, что отростки крупных нейроцитов паравентрику-
235
Рис. 2.136. Развитие ги-
поталамической обла-
сти. Фронтальный срез
мозга эмбриона чело-
века 2 мес (по И. Н. Бо-
голеповой).
1 — хиазма; 2 — паравен-
трикулярное ядро; 3 — су-
праоптическое ядро.
лярного ядра контактируют со стенкой Ш желудочка, что допускает
возможность поступления нейросекрета в спинномозговую жид-
кость. При комбинированной окраске препарата в клетках круп-
ноклеточных ядер одновременно выявляются секреторная зер-
нистость и хроматофильная субстанция. Соответственно нейро-
секреторные клетки совмещают морфологические характеристики
нейронов (мультиполярность, присутствие субстанции Ниссля)
и гландулоцитов (признаки секреции).
«Двойственность» строения нейросекреторных клеток четко
проявляется при ТЭМ (рис. 2.135) и выражается в активном раз-
витии синтетического аппарата в перикйрионе клетки, присутствии
секреторных гранул и нейрофибрилл. Наличие в цитоплазме аксо-
нов этих клеток (см. также «Нейрогипофиз») как нейросекреторных
гранул, так и типичных синаптических везикул указывает на па-
раллельный синтез нейрогормонов и нейромедиаторов (пептидхо-
линергические клетки). Генетическая и функциональная связь
секреторных нейроцитов с нервной системой проявляется также
присутствием в области перикариона нейро-нейрональных синап-
сов, т. е. включением нейросекреторных клеток в рефлекторные дуги.
Принцип цитоархитектоники мелкоклеточных ядер гипотала-
муса близок к описанному, однако здесь клетки отличаются мень-
шими размерами и длиной отростков, преобладанием структурных
черт типичных нейроцитов и связью с адренергическими нейронами
(пептидадренергические клетки).
Существуют различия и в процессе развития гипоталамических
ядер. Миграция и пролиферация нейробластов приводят к выде-
лению у 2-месячного зародыша клеточных коопераций, топогра-
236
Рис. 2.137. Гипофиз кошки.
А — связь гипофиза с гипоталамусом
(по В. Г. Елисееву, Ю. И. Афанасьеву,
Е. Ф. Котовскому). Б — строение адено-
и нейрогипофиза. Окраска гематоксили-
ном и эозином.
1 — воронка мозга; 2 — туберальная
часть гипофиза; 3 — передняя доля;
4 — промежуточная доля; 5 — задняя
доля (нейрогипофиз); 6 — остаток кар-
мана Ратке.
фически идентифицируемых как крупноклеточные ядра гипота-
ламуса (рис. 2.136). Закладка большинства мелкоклеточных ядер
определяется лишь у 3—4-месячных плодов. Дифференцировка
секреторных нейроцитов переднего гипоталамуса отчетливо прояв-
ляется у 4—5-месячных плодов, в элементах мелкоклеточных ядер
она регистрируется значительно позже (к 6-му месяцу развития).
Гипофиз. Фронтальный срединный срез через межуточный
мозг и гипофиз кошки (рис. 2.137) раскрывает микроанатомию
гипофиза и топографоанатомические характеристики гипоталамо-
гипофизарных связей. При этом отчетливо выявляются различия
формы, размеров и окраски трех основных частей гипофиза: наи-
237
Рис. 2.138. Развитие гипофиза (схема).
1 — ротовая полость; 2 — полость желудочка мозга; 3 — карман Ратке; 4 — ди-
вертикул промежуточного мозга; 5 — язык; 6 — эпителий ротовой полости; 7 —
мезенхима; 8 — кровеносные сосуды; 9 — передняя стенка кармана Ратке (перед-
няя доля гипофиза); 10 — задняя стенка кармана Ратке (промежуточная доля
гипофиза); 11—задняя доля гипофиза; 12 — туберальная часть; 13 — эпендима;
14 — твердая мозговая оболочка; 15 — мягкая мозговая оболочка; 16 — клино-
видная кость; 17 — гипофизарная ножка.
более крупной, интенсивно окрашенной передней доли, слабо окси-
фильной округлой задней доли (нейрогипофиз) и промежуточной
части аденогипофиза, расположенной между первыми и охва-
тывающей «полукольцом» нейрогипофиз. Щель, разделяющая пе-
реднюю и промежуточную части аденогипофиза, является рудимен-
том полости гипофизарного кармана (см. ниже). Такие топогра-
фические отношения промежуточной и задней долей характерны
для некоторых видов млекопитающих, но не для человека. Основой
анатомической связи гипофиза и гипоталамуса служит как гипо-
физарная ножка, воронка III желудочка мозга, так и тесный контакт
туберальной части аденогипофиза с нижней поверхностью медиаль-
ной эминенции (область локализации первичной капиллярной
сети).
238
Схема эмбрионального развития органа объясняет различия
структуры адено- и нейрогипофиза и характера их связи с гипота-
ламусом (рис. 2.138).
Гипофиз человека отличает более компактное расположе-
ние его долей, слабое развитие промежуточной части адено-
гипофиза. Адено- и нейрогипофиз легко идентифицировать
при малом увеличении микроскопа благодаря выраженным раз-
личиям структуры и интенсивности окрашивания ткани его долей
(рис. 2.139, А).
Гистологическое строение паренхимы передней части
аденогипофиза (рис. 2.139, Б) может служить иллюстра-
цией принципа организации большинства эндокринных желез. Это
ветвящиеся тяжи (трабекулы) эпителиальных клеток, пронизан-
ные широкими капиллярами. Паренхима разделена прослойками
рыхлой волокнистой соединительной ткани на отдельные клеточ-
ные комплексы — аденомеры, включающие две разновидности
эндокриноцитов: хромофобные (около 60 %) и хромофильные
эндокриноциты (аденоциты).
Хромофобные аденоциты отличаются небольшими размерами,
нечеткими контурами, крупным (относительно величины клетки)
ядром и слабой окраской цитоплазмы, лишенной секреторных вклю-
чений. Хромофилъные аденоциты располагаются по периферии
трабекул и содержат в цитоплазме интенсивно окрашивающиеся
секреторные гранулы. По характеру окраски гранул хромофильные
аденоциты подразделяются на базофильные и_ ацидофильные
(оксифильные). Ацидофилы (соматотропоциты*й маммотр'опоци-
ты) имеют несколько меньшие размеры, но более многочисленны.
В группе базофилов можно различить два клеточных типа, представ-
ленных гонадотропоцитами и тиреотропоцитами. Гонадотропоциты
определяются по округлой форме, более слабой базофилии цито-
плазмы, эксцентричному расположению ядра, присутствию в центре
клетки просветленного участка (макулы), соответствующего лока-
лизации пластинчатого комплекса. Тиротропоциты отличаются не-
правильной, угловатой формой и более высокой базофилией цито-
плазмы. Популяция базофилов хорошо маркируется при использо-
вании ШИК-метода (выявление углеводных групп), так как гор-
моны этих клеток являются гликопротеидами. Для идентификации
типов ацидофильных аденоцитов на светооптическом уровне пока
не существует надежных критериев. Это относится и к кортикотро-
поцитам, которые крайне трудно выделить из хромофобной попу-
ляции аденоцитов.
При электронно-микроскопическом анализе идентификация
аденоцитов основывается на цитоспецифических особенностях
их формы, внутриклеточной организации ультраструктуры и разме-
ров секреторных гранул (рис. 2.140). Клетки, вырабатывающие
белковые гормоны (сомато- и маммотропоциты), содержат очень
крупные секреторные гранулы (400—700 нм) и хорошо выраженный
аппарат белкового синтеза. В клетках, продуцирующих гормоны
гликопротеидной природы (гонадотропоциты, тиреотропоциты),
239
240
Рис. 2.139. Гипофиз человека (окраска по методу Маллори).
А — схема: AI — передняя доля, АП — промежуточная доля, АШ — задняя доля.
Б — передняя доля (аденогипофиз). В — промежуточная часть аденогипофиза.
Г — задняя доля (нейрогипофиз).
1 — ножка гипофиза; 2 — капсула; 3 — оксифильные аденоциты; 4 — базофиль-
ные аденоциты; 5 — хромофобные (главные) аденоциты; 6 — кровеносные ка-
пилляры; 7 — интермедиоциты; 8 — псевдофолликул; 9 — питуициты; 10 — нервные
волокна.
241
Рис. 2.140. Ультраструктура хро-
мофильных аденоцитов передней
доли гипофиза.
А — схема. Б, В, Г — ТЭМ.
1 — тиротропоцит; 2 — соматотропо-
цит; 3 — лактотропоцит; 4 — гонадо-
тропоцит; 5 — кортикотропоцит; 6 —
гранулярная цитоплазматическая сеть;
7 — пластинчатый комплекс; 8 — мито-
хондрия; 9 — секреторные гранулы; 10 —
базальная мембрана аденоцита; 11 —
базальная мембрана гемокапилляра;
12 — фенестры в эндотелии капилляра
(указано стрелкой); 13 — эритроцит.
242
243
гранулы мельче (150—180 нм), гранулярная цитоплазматическая
сеть развита менее активно. Следует отметить, что каждый эндо-
криноцит тесно контактирует с капилляром висцерального типа.
Промежуточная часть аденогипофиза чело-
века (см. рис. 2.140, В) представлена короткими плотными тяжа-
ми довольно крупных клеток, разделенных прослойками волокнис-
той ткани с проходящими в ней микрососудами. Характерная
структура — псевдофолликулы, которые являются результатом
накопления секрета в межклеточных пространствах.
Нейрогипофиз (см. рис. 2.140, Г) отличает от аденоги-
пофиза более слабое окрашивание ткани, скудность клеточных
элементов — питуицитов, обилие нервных волокон. При окраске
препарата альдегид-фуксином выявляются паравазальные нако-
пительные тельца с базофильным нейросекреторным веществом —
терминали аксонов нейросекреторных клеток гипоталамуса
(рис. 2.141, Б).
Принцип ультраструктурной организации нейрогипофиза отра-
жен на схеме (рис. 2.141, Б). Примечательно, что терминали ак-
сонов содержат секреторные гранулы и синаптические везикулы
и граничат с истонченными маргинальными зонами эндотелио-
цитов капилляров (рис. 2.141, В).
Особенности функции нейросекреторных и нейрогемальных
образований мозга (ядра гипоталамуса, нейрогипофиз) отразились
в своеобразии структуры гематоцеллюлярных барьеров: в отличие
Рис. 2.141. Взаимоотношение тер-
миналей аксонов нейросекреторных
клеток гипоталамуса с питуицитами
и кровеносными капиллярами в
задней доле гипофиза (нейрогипо-
физе) .
А — контакт терминалей аксонов с ка-
пилляром; окраска альдегидфуксином.
Б — ТЭМ (рисунок). В — ТЭМ. I —
аксоны нейросекреторных клеток; II —
питуициты; III — кровеносный капил-
ляр.
1 — питуицит; 2 — митохондрии; 3 —
нейрофибриллы; 4 — липидная гра-
нула; 5 — лизосома; 6 — секреторные
гранулы; 7 — синаптические везикулы;
8 — базальная мембрана; 9 — коллаге-
новые фибриллы; 10 — фенестры в эндо-
телии кровеносного капилляра; 11 —
эндотелий; 12 — накопительное тельце.
244
МП
8
246
от других областей мозга здесь локализуются капилляры висцераль-
ного типа, отсутствует паравазальный глиальный «футляр» (по-
стоянные компоненты гематоэнцефалического барьера).
____Эпифиз так же, как нейрогипофиз^ является производным ней-
'_роэктодермы. Однако от нейрогипофиза его морфологически от-
личают присутствие (параллельно с глиоцитами) специфических
секреторных клеток — пинеалоцитов^Для эпифиза (рис. 2.142)
характерно дольчатоЕ строениеГопределяемое отходящими от
капсулы соединительнотканными перегородками, обилие кровенос-
ных капилляров, типичное для эндокринных органов. Присутствие
в паренхимеТсбнкрементов, так называемого мозгового песк , свиде-
тельствует о возрастной инволюции эпифиза.
Рис. 2.142. Эпифиз человека.
А — общий вид. Б — структура паренхимы при импрегнации азотнокислым се-
ребром.
1 — капсула; 2 — междольковая соединительная ткань; 3 — кровеносные капилля-
ры; 4 — пинеалоциты; 5 — глиальные клетки; 6 — мозговой песок.
В, Г — ТЭМ пинеалоцитов.
1 — ядро пинеалоцита; 2 — митохондрия; 3 — секреторные гранулы; 4 — просвет
капилляра; 5 — перикапиллярное пространство; 6 — отросток пинеалоцита.
247
В эпифизе идентифицируются два основных типа клеточных
элементов. Пинеалоциты представлены более широко и, как пра-
вило, образуют клеточные комплексы. Это довольно крупные клет-
ки с округлыми крупными ядрами, светлой цитоплазмой и длин-
ными отростками. Глиоциты также имеют отростчатую форму, но
отличаются меньшими размерами, более базофильной цитоплаз-
мой и уплощенными гиперхромными (темными) ядрами; они рас-
сеяны по всей паренхиме, часто составляют группы из 2—3 кле-
ток. Пространства между телами пинеалоцитов и глиоцитов за-
няты их отростками и кровеносными микрососудами.
В ультраструктуре пинеалоцитов сочетаются черты, общие для
всех гландулоцитов и типичные только для производных нейро-
эктодермы. О железистой функции пинеалоцитов говорит сравни-
тельно активное развитие органелл синтеза (рис. 2.142, В) и при-
сутствие мелких секреторных гранул, концентрирующихся преиму-
щественно в булавовидных расширениях клеточных отростков, про-
стирающихся в перикапиллярные пространства (рис. 2.142, Г).
Форма и обилие в цитоплазме микротрубочек и микрофиламентов
придают пинеалоцитам черты строения нейросекреторных клеток.
В популяции пинеалоцитов могут быть выделены «светлые» и «тем-
ные» клетки, которые по аналогии с другими железами, вероятно,
отражают вариации их функционального состояния. В «темных» пи-
неалоцитах органеллы развиты слабее, нежели в светлых, цито-
плазма более осмиофильна.
Ультраструктура кровеносных капилляров эпифиза свидетель-
ствует об активности транспортных процессов и отвечает законо-
мерностям строения гемокапилляров в эндокринных железах
(капилляры висцерального типа).
2.10.2. Периферические звенья эндокринной системы
Щитовидная железа — эндокринный орган, функциональная мор-
фология которого по ряду параметров отличается от других эндо-
кринных желез. Это связано с тем, что закладываясь в эмбриоге-
незе подобно железам внешней секреции, она и в дефинитивном
состоянии сохраняет некоторые присущие им черты. Так, струк-
турно-функциональные единицы щитовидной железы (фолликулы)
по своей конструкции напоминают концевые отделы экзокринных
желез, секреция тироглобулина (1-й этап гормонопродукции)
Рис. 2.143. Щитовидная железа.
А — общий вид. Б — стенка фолликула. В — коррозионный препарат микрососудов,
СЭМ (по В. А. Миронову, А. А. Миронову, Ю. В. Погорельскому).
1 — капсула; 2 — соединительнотканные междольковые перегородки; 3 — долька;
4 — междольковые кровеносные сосуды; 5 — кровеносный капилляр; 6 — фолли-
кул; 7 — коллоид; 8 — резорбционные вакуоли; 9 — фолликулярные тироциты;
10 — парафолликулярные тироциты; 11 —межфолликулярный островок; 12 — око-
лощитовидная железа.
248
Б
249
Рис. 2.144. Особенности строения
фолликулов при различных функ-
циональных состояниях щитовид-
ной железы.
А — препарат. Б (ТЭМ) — гипофунк-
ция. В — препарат. Г (ТЭМ) — гипер-
функция.
1 — просвет фолликула; 2 — фоллику-
лярный тироцит; 3 — гранулярная цито-
плазматическая сеть.
250
251
252
Рис. 2.145. Фрагмент стенки фолликула щитовидной железы.
А — верхушечная поверхность тироцитов, СЭМ (препарат В. А. Миронова,
А. А. Миронова, Ю. В. Погорельского). Б — ТЭМ: 1 —полость фолликула (кол-
лоид) ; 2 — ядро тироцита; 3 — гранулярная цитоплазматическая сеть; 4 — комп-
лекс Гольджи: 5 — митохондрия; 6 — секреторные гранулы; 7 — лизосома; 8 —
интрацеллюлярная капля коллоида; 9 — десмосома; 10 — базальная мембрана
фолликулярного эпителия; 11 — фибробласт.
В — схема биосинтеза гормонов в фолликулярных тироцитах (по Б. В. Алешину с
изменениями): 1 —поступление йодидов в клетку: 2 — поступление тирозина и
других аминокислот; 3 — синтез белка на рибосомах гранулярной цитоплазмати-
ческой сети; 4 — транспорт секрета из цитоплазматической сети в зону комплекса
Гольджи; 5 — гликозилирование белка и транспорт гликопротеида в составе секре-
торных гранул к апикальной плазмолемме клетки; 6 — экзоцитоз секреторного
продукта (тироглобулина) в просвет фолликула; 7 — окисление ионов йода в ато-
марный йод под влиянием фермента пероксидазы; 8—12 — стадии йодирования
тироглобулинов с образованием йодтиронинов и йодтирозинов; 13—фагоцитоз
иодированного тироглобулина; 14—слияние капель фагоцитированного коллоида
с лизосомой; 15 — протеолиз тироглобулина ферментами лизосом; 16 — отщепле-
ние тироксина и трийодтиронина; 17, 18 — поступление гормонов в кровоток; 19 —
интрафолликулярный протеолиз коллоида на поверхности тироцита, пиноцитоз и
интрацеллюлярный транспорт тиреоидных гормонов (дополнительный вариант
гормонопоэза).
253
происходит в просвет фолликула, т. е. через апикальную плазмо-
лемму тироцитов, что несвойственно эндокринным клеткам. Но
изучение препаратов щитовидной железы позволяет выявить
морфологические признаки органа с инкреторной функцией:
отсутствие выводных протоков, обильная васкуляризация парен-
химы. -
Щитовидная железа человека (рис. 2.143, А, Б) имеет дольча-
тое строение: от плотной соединительнотканной капсулы, покры-
вающей железу, отходят прослойки рыхлой волокнистой соедини-
тельной ткани — междольковые перегородки, разделяющие парен-
химу на дольки. По междольковым перегородкам проходят маги-
стральные сосуды — междольковые артерии и вены.
Паренхима долек преимущественно представлена образовани-
ями округлой формы — фолликулами с оксифильным содержи-
мым — коллоидом. Между фолликулами локализуются компакт-
ные скопления тиреоидных клеток — островки интерфолликуляр-
ного эпителия (косые срезы фолликулов). О высоком уровне васку-
ляризации фолликулов и ангиоархитектонике дольки позволяет
судить рис. 2.143, В.
Фолликулы выстланы однослойным кубическим эпителием
и заполнены гомогенным коллоидом с резорбционными вакуо-
лями — результат разжижения секрета. Нередко наблюдаются
некоторое «сморщивание» и отслойка коллоида от эпителия,
что является артефактом фиксации. Коллоид, как правило, окси-
филен, но на начальных этапах своего образования может прояв-
лять легкую базофилию.
В стенке фолликула идентифицируются 2 типа клеток, лежа-
щих на базальной мембране: фолликулярные эндокриноциты (ти-
роциты) — основной компонент эпителиальной выстилки фолли-
кула и немногочисленные парафолликулярные эндокриноциты
(кальцитониноциты), которые одиночно, реже группами из 2—
3 клеток, располагаются между тироцитами.
Гистологически можно объективно оценить уровень функцио-
нальной активности органа. Основными критериями служат:
размеры и форма фолликулов, высота тироцитов, состояние кол-
лоида (плотный или разжиженный — «пенистый» с обилием резорб-
ционных вакуолей), митотическая активность клеток. Различия
структуры щитовидной железы в условиях гипо- и гиперфункции
иллюстрирует рис. 2.144.
При анализе ультраструктурной организации фолликула обра-
щает на себя внимание кубическая форма клеток, соединен-
ных специализированными контактами в сплошной монослой, по-
лярность их строения, умеренная осмиофилия коллоида (рис.
2.145). Обилие микроворсинок на апикальной плазмолемме тиро-
цитов является выражением ее высокой транспортной активности
(см. рис. 2.145, А). Ультраструктурные характеристики фоллику-
лярных тироцитов типичны для клеток, активно синтезирующих
секреторные белки (тироглобулин — гликопротеид); примеча-
тельны присутствие лизосом и неоднородность популяции секре-
254
Рис. 2.146. Парафолликулярные тироциты.
А — импрегнация кальцитониноцитов азотнокислым серебром. Б — топография
клеток в стенке фолликула (ТЭМ, схема). В — ТЭМ парафолликулярной клетки.
I — парафолликулярный тироцит; 2 — гранулярная цитоплазматическая сеть; 3 —
комплекс Гольджи; 4 — митохондрия; 5 — лизосома; 6 — секреторные гранулы;
7 — фолликулярный тироцит; 8 — десмосома; 9 — кровеносный капилляр.
торных включений (мелкие секреторные гранулы и более осмио-
фильные интрацеллюлярные «капли» коллоида) (рис. 2.145, Б).
Функциональная специализация органелл и особенности секретор-
ного процесса в тироцитах представлены на рис. 2.145, В.
Кальцитониноциты легко выявляются при импрегна-
ции серебром благодаря аргирофилии их цитоплазмы, свойственной
элементам диффузной эндокринной системы (APUD-система),
производным нейроэктодермы: клетки четко маркируются как в
составе стенки фолликулов, так и в межфолликулярных пространст-
вах (рис. 2.146, А). В препаратах, окрашенных гематоксилином и
эозином, кальцитониноциты отличают более крупные размеры,
светлая цитоплазма и особенность топографии в стенке фоллику-
ла — они не соприкасаются с коллоидом, будучи отделены от про-
света фолликула аркообразным соединением тироцитов (рис.
2.146,Б, см. рис. 2.143, Б).
Схема (рис. 2.146, В) ультраструктуры парафолликулярных
клеток дает четкие ориентиры для их идентификации на субмикро-
скопическом уровне. Обращает на себя внимание отсутствие кон-
такта апикальной плазмолеммы кальцитониноцита с просветом
фолликула, его округлая форма, слабая «поляризация» цитоплазмы,
обилие в ней мелких секреторных гранул с осмиофильной «сердце-
виной» (еще один морфологический признак принадлежности пара-
фолликулярных клеток к APUD-системе), а также более слабое,
нежели в фолликулярном тироците, развитие гранулярной цитоплаз-
матической сети. Сравнение
ультраструктуры фолликуляр-
ного тироцита (см. рис.
2.145, А) и кальцитониноцита
(см. рис. 2.146, В) убеждает в
достаточной простоте их иден-
тификации при ТЭМ. Рис. 2.147
иллюстрирует механизмы эф-
фекта кальцитонина (гипокаль-
циемия) .
Развитие щитовидной же-
лезы. Зачаток железы появля-
ется на дне глотки между I и
II парой глоточных карманов в
экскреции Са2* в почках,
торможение всасывания
Са24 в кишечнике
f
Нормализация содержания Са2* в крови
Рис. 2.147. Механизмы снижения
кальцитонином содержания кальция
в крови (схема).
256
Рис. 2.148. Закладка щитовидной
и околощитовидной желез (схема).
А — вид сбоку. Б — вид спереди.
I, II, III, IV — глоточные карманы.
1 — трахея; 2 — пищевод; 3 — зачаток
щитовидной железы; 4 — зачатки око-
лощитовидных желез; 5 — карман Рат-
ке; 6 — нервная трубка; 7 — эктодерма;
8 — зачаток тимуса.
конце 1-го месяца внутриутробного развития (рис. 2.148). Клеточ-
ные массы, врастая в область шеи, в течение некоторого времени
сохраняют связь со стенкой глотки в виде щитоязычного протока,
который затем редуцируется. До 9—10-й недели железа представ-
лена в основном плотными эпителиальными тяжами, но к концу
12-й недели ее паренхима приобретает фолликулярное строение.
Общее представление об этапах становления функциональной
морфологии органа дает сравнительный анализ строения щитовид-
ной железы 15-недельного плода и щитовидной железы новорож-
денного (рис. 2.149, А, Б).
Околощитовидные железы (у человека их, как правило, четы-
ре), прилежат к задней поверхности долей щитовидной железы,
будучи покрыты вместе с последней глубокой фасцией шеи. Эта
топографическая связь объясняет частое присутствие фрагментов
щитовидной железы в гистологических препаратах околощитовид-
ной и наоборот (см. рис. 2.144).
Паренхима околощитовидной железы человека (рис. 2.150, А)
представлена тяжами эпителиальных клеток, пронизанных крове-
носными капиллярами. Характерно, что отходящие от тонкой кап-
сулы соединительнотканные прослойки имеют различную протя-
женность и не обеспечивают полного разграничения паренхимы
на железистые дольки. В этих прослойках проходят кровеносные
сосуды и нервы, часто обнаруживаются комплексы жировых кле-
ток. Популяция паратироцитов неоднородна. Она представлена
главными и оксифильными паратироцитами.
Главные паратироциты — основной клеточный тип
эпителиальных тяжей — характеризуются небольшими размерами.
Выделено две их разновидности: «темные» клетки со слабо базо-
фильной цитоплазмой и «светлые», цитоплазма которых практи-
257
9—321
Рис. 2.149. Щитовидная железа плода 15 нед (А, ШИК-реакция) и ново-
рожденного (Б, окраску гематоксилином и эозином).
Рис. 2.150. Околощитовидная железа.
А — общий вид. Б — ультраструктура паратироцитов (схема). В, Г — возрастные
особенности клеточного состава паренхимы (схема): В — ребенок до 6 лет; Г —
взрослый.
1 — капсула; 2 — главные клетки; 3 — оксифильные клетки; 4 — кровеносные
сосуды; 5 — соединительная ткань; 6 — митохондрия; 7 — цитоплазматическая
сеть; 8—комплекс Гольджи; 9 — липидные включения; 10 — гликоген; 11 —
секреторные гранулы; 12 — базальная мембрана; 13 — гемокапилляр; 14 — фибро-
бласт.
258
чески не окрашена. Оксифильные паратироциты
отличаются от главных большими размерами и выраженной эози-
нофилией цитоплазмы. Они обычно образуют небольшие клеточ-
ные скопления на периферии железы.
Более ярко различие строения двух типов паратироцитов
проявляется при ТЭМ (рис. 2.150, Б). Ультраструктура г л а в-
9*
259
260
Рис. 2.151. Надпочечник.
А — схема строения. Строение корково-
го (Б) и мозгового (В) вещества
(окраска железным 1ематоксилином).
Г — липиды в пучковой зоне (окраска
Суданом).
I — корковое вещество; II — мозговое
вещество.
1 — капсула; 2 — клубочковая зона;
3 — пучковая зона; 4 — сетчатая зона;
5 — кровеносные сосуды; 6 — централь-
ная вена.
ных паратироцитов типична для железистых клеток, про-
дуцирующих белки: активное развитие цитоплазматической сети,
пластинчатого комплекса, присутствие секреторных гранул. Обилие
гликогена в цитоплазме «светлых» главных паратироцитов объясня-
ет ее негативное окрашивание на гистологических препаратах.
Своеобразие субмикроскопической картины оксифильных парати-
роцитов состоит в обилии митохондрий и липидных включений,
отсутствии секреторных гранул. Функция оксифильных паратирео-
цитов неясна; они дифференцируются только в постнатальном пе-
риоде. Отсутствие этих клеток — главный отличительный признак
гистоструктуры околощитовидных желез детей первых лет жизни
(см. рис. 2.150, В, Г).
Надпочечник человека и млекопитающих — еще один (наряду
с гипофизом) пример объединения в единый орган эндокринных
желез различного происхождения, строения и функции, представ-
ленных здесь корковым и мозговым веществом.
Надпочечник (рис. 2.151, А, Б) покрыт толстой соединительно-
тканной капсулой, в которой постоянно обнаруживаются крове-
носные сосуды и жировые клетки.
Корковое вещество надпочечника (рис.
2.151, В)—единственная эндокринная железа с «послойной»
функциональной специализацией паренхимы. Его зоны легко иден-
тифицировать, так как конфигурация эпителиальных тяжей доста-
точно точно соответствует названию зон.
Непосредственно под капсулой в клубочковой зоне
эпителий формирует округлые образования, лежащие, как правило,
одним слоем. В подлежащей наиболее широкой пучковой
зоне секреторные клетки сгруппированы в прямые тяжи («пуч-
261
ки») толщиной в 1—2 клетки, ориентированные перпендикулярно
поверхности органа. В сравнительно узкой сетчатой зоне,
локализующейся на границе с мозговым веществом, архитектоника
идущих в различных направлениях анастомозирующих эпители-
альных тяжей напоминает сеть. Клетки клубочковой зоны близки
по форме к цилиндрическому эпителию, их цитоплазма более ба-
зофильна, чем у клеток других зон, ядра более мелкие и темные.
Клетки пучковой зоны отличаются полигональной формой, круп-
ными ядрами, сильно вакуолизированной цитоплазмой (вакуоли-
зация цитоплазмы — результат действия спиртовой проводки на
липидные включения). Максимальную насыщенность клеток пуч-
ковой зоны липидными включениями убедительно демонстрирует
рис. 2.151, Г. Клетки сетчатой зоны близки по форме клеткам
пучковой, но уступают им в размере и содержании липидов.
В паренхиме коркового вещества обращает на себя внимание
обилие кровеносных капилляров с широким просветом, проходя-
щих в узких прослойках рыхлой волокнистой соединительной
ткани между клеточными тяжами. О высоком уровне развития
капиллярной сети и соответствии ангиоархитектоники ходу эпи-
телиальных тяжей дает представление рис. 2.152.
На границе с мозговым веществом гемокапилляры коркового
вещества формируют сеть обменных микрососудов большего диа-
метра (венозные синусоиды), которые затем собираются в централь-
ную вену мозгового вещества. Эта особенность пространственной
организации русла определяет поступление насыщенной кортико-
стероидами и другими биологически активными веществами крови
в мозговое вещество и является морфологической основой функцио-
нальной связи между ним и корковой зоной. /
Обращаясь к строению мозгового вещества надпо-
чечника, прежде всего отметим более рыхлое (по сравне-
нию с корковым) расположение секреторных элементов, зани-
мающих пространства вокруг кровеносных сосудов. В отличие от
коркового вещества здесь обнаруживаются срезы магистральных
сосудов, в частности центральной вены, зияющей благодаря оби-
лию в ее стенке продольно расположенных гладких миоцитов.
Мозговые эндокриноциты — крупные клетки овальной фор-
мы — содержат в цитоплазме зернистость. Идентифицировать
светлые (эпинефроциты) и темные (норэпинефроциты) эндокри-
ноциты в препаратах трудно. Эта задача упрощается при обработ-
ке срезов окислителями (в частности, бихроматом калия): нор-
адреналин является более сильным восстановителем, и поэтому
цитоплазма норэпинефроцитов окрашивается более интенсивно.
При анализе электронных микрофотографий
клеток коркового и мозгового вещества прослеживаются параллели
между особенностями их ультраструктуры и химической природой
продуцируемых гормонов.
Ультраструктура клеток пучковой и сетчатой зон коркового ве-
щества (рис. 2.153) характерна для всех эндокриноцитов, специали-
зированных на продукции стероидных гормонов. Для этих клеток
262
с)
5
с
Рис. 2.152. Архитектоника паренхимы и сосудов надпочечника (СЭМ).
А — скол ткани. Б — коррозионный препарат сосудов (по В. А. Миронову и соавт.).
1 — капсула; 2 — корковое вещество; 3 — мозговое вещество; 4 — кровеносные
сосуды.
263
Рис. 2.153. Ультраструктура клеток пучковой зоны (А) и мозгового ве-
щества (Б) надпочечника, ТЭМ.
1 — ядро; 2 — липидная гранула; 3 — митохондрия; 4 — гладкая цитоплазмати-
ческая сеть; 5 — секреторные гранулы.
264
типичны обилие липидных включений, особая тубуло-везикулярная
структура крист многочисленных митохондрий и активное раз-
витие агранулярной цитоплазматической сети. Отметим, что в
стероидопродуцирующих клетках предшественники гормонов син-
тезируются в митохондриях и процесс секреции лишен этапа
образования секреторных гранул. Отличительные черты ультра-
структуры эндокриноцитов клубочковой зоны (менее активное
развитие агранулярной цитоплазматической сети, митохондрии с
пластинчатыми кристами, относительная скудность липидных
включений) служат отражением специфики биосинтеза минерало-
кортикоидов: предшественники альдостерона, в частности корти-
костерон поступает в эндокриноциты из пучковой зоны.
В мозговых эндокриноцитах обилие секреторных гранул соче-
тается с относительной скудностью органелл, в частности элемен-
тов гранулярной цитоплазматической сети. Это объяснимо, так
как катехоламины являются продуктом последовательного пре-
вращения одной аминокислоты (тирозина), не требующим
активного развития аппарата белкового синтеза. Ультраструк-
тура двух типов хромаффиноцитов отличается лишь строением
секреторных гранул: в эпинефроцитах они содержат плотный
зернистый продукт, в норэпинефроцитах — гомогенный. Как
производных нейроэктодермы хромаффиноциты характеризует
тесный синаптоподобный контакт с нервными терминалями. Такие
нервно-тканевые отношения нетипичны для эндокринных желез
эпителиальной природы.
Схема динамики возрастной морфологии надпочечника (рис.
2.154; 2.155, А) иллюстрирует особенности гистогенеза отделов
органа в анте- и постнатальном периоде, активное преобразование
его гистоструктуры в течение первого года жизни ребенка.
У эмбриона 7-й недели развития надпочечник лежит дорсаль-
но под диафрагмой над окончательной почкой (рис. 2.155, Б).
Отмечается раннее разделение его паренхимы на 2 зоны: узкую
периферическую, построенную из нескольких рядов мелких базо-
фильных клеток (зачаток постоянной коры) и широкую централь-
ную, содержащую крупные клетки с оксифильной цитоплазмой
(фетальная кора). Мозговое вещество отсутствует; иногда в этот
период в субкапсулярной зоне удается увидеть небольшие темные
скопления мелких клеток (симпатогонии) — «мозговые шары».
В надпочечнике плода 6 мес постоянная кора развита слабо
(рис. 2.155, В). Основной объем органа занят фетальной корой.
По всему корковому веществу рассеяны «мозговые шары», скопле-
ния клеток, выделяющиеся более темной окраской; часть «мозго-
вых шаров» достигает центра органа, формируя мозговое вещество.
Надпочечник новорожденного характеризует незавершенность
процесса развития и активность морфогенетических перестрой-
ки (рис. 2.155, Г). В фетальной коре наблюдаются признаки
инволюции (гибель клеток, резкое расширение кровеносных сосу-
дов, кровоизлияния и др.). Эти процессы как бы маркируют
область фетальной коры, благодаря чему она легко определяется
265
Рис. 2.154. Основные этапы морфогенеза надпочечника (схема по О. В. Волковой, М. И. Пе-
карскому) .
Рис. 2.155. Структура надпочечника в процессе развития.
А — надпочечник эмбриона 7 нед. Б — надпочечник плода 6 мес. В — надпочечник
новорожденного.
1 — надпочечник; II — почка.
1 — постоянная кора; 2 — фетальная кора; 3 — мозговые шары.
267
на препарате. В постоянной коре, занимающей субкапсуляр-
ную зону, еще отсутствует четкое зональное подразделение па-
ренхимы. Центральное мозговое вещество занимает значительно
меньший объем, чем у взрослого, продолжается миграция «моз-
говых шаров».
Гормональное обеспечение организма не ограничивается толь-
ко системой желез внутренней секреции. Эндокринные клетки,
обнаруженные во многих органах (диффузная эндокринная сис-
тема), осуществляют как местную (паракринную), так и дистант-
ную модуляцию биологических процессов (см. гл. 2.8, 2.13, 2.14).
2.11. Дыхательная система
Из многочисленных функций органов дыхательной системы —
газообменной, метаболической, синтетический и др. — первая
обеспечивает адекватный потребностям клеточного метаболизма
обмен кислорода и двуокиси углерода между атмосферным возду-
хом и кровью. Знание конкретного морфологического субстрата,
обеспечивающего эту и другие функции дыхательной системы,
создает предпосылки для правильного понимания физиологии ды-
хания, а представления об особенностях ее строения в различные
возрастные периоды — базис для правильной диагностики и ле-
чения патологии дыхательной системы у взрослых и у детей.
В дыхательной системе принято различать воздухоносные пути
и газообменные участки. В последних и происходит собственно
обмен Тазами между кровью и воздухом, так как там эти две среды
наименее разделены.
2.11.1. Воздухоносные пути
Воздухоносные пути — система древовидно ветвящихся трубок,
начинающаяся с трахеи. У человека она имеет 16 генераций дихо-
томически ветвящихся бронхов, заканчивающихся терминальной
бронхиолой.
К воздухоносным путям относятся также носовая полость,
глотка, гортань, а в ряде случаев и ротовая полость.
Основными функциональными задачами воздухоносных путей
являются, помимо проведения воздуха, его очищение от пыли,
согревание и увлажнение. При анализе препаратов воздухоносных
путей можно увидеть морфологические субстраты, обеспечиваю-
щие выполнение каждой из указанных функций. С функциями
верхних отделов воздухоносных путей тесно связаны способность
определять запахи (обоняние) и звукообразование.
Трахея. Строение верхних отделов воздухоносных путей мож-
но изучить на примере трахеи. На срезе трахеи (рис. 2.156, А)
различают четыре оболочки. С_л изистая оболочка т дэа-
х е и (рис. 2.156, Б) выстлана псевдомногослойным (многоряд-
ным) столбчатым реснитчатым эпителием. Клеточный состав это-
го эпителия схематически изображен на рис. 2.156, В.
268
Рис. 2.156. Трахея.
А — поперечный срез. Б — структура стенки. В — клетки эпителия верхних отде-
лов воздухоносных путей. Г — реснитчатая клетка. Д — поверхность эпителиаль-
ной выстилки слизистой оболочки трахеи.
1 — слизистая оболочка; 2 — подслизистая основа; 3 — фиброзно-хрящевая обо-
лочка; 4 — адвентициальная оболочка; 5 — однослойный многорядный мерцатель-
ный эпителий; 6 — собственная пластинка слизистой оболочки; 7 — концевые
отделы желез; 8 — кровеносный сосуд; 9 — реснитчатый эпителиоцит (РК); 10 —
бокаловидная клетка (БК); 11 — высокая базальная клетка; 12 — низкая базаль-
ная клетка.
269
Под относительно ровной базальной мембраной эпителия рас-
полагается собственная пластинка слизистой оболочки, представ-
ленная рыхлой волокнистой соединительной тканью. Особенностью
этого слоя является обилие продольно ориентированных пучков
эластических волокон, легко обнаруживаемых с большим увели-
чением микроскопа в виде блестящих розовых точек.
Другой особенностью собственной пластинки является нали-
чие в ней венозных сплетений, располагающихся близко от по-
верхности и предназначенных для согревания вдыхаемого воз-
духа.
В трахее человека, а также в главных бронхах слизистая обо-
270
лочка без выраженных границ переходит в п о д с л изистую
основу, представленную рыхлой волокнистой соединительной
'тканью. На препаратах, окрашенных гематоксилином-эозином,
практически невозможно определить границу между собственной
пластинкой слизистой оболочки и подслизистой оболочкой. Если
же применить красители, выявляющие элементы эластической
ткани, то в собственной пластинке слизистой можно увидеть ее
слой, не обнаруживаемый в подслизистой. Отличительным при-
знаком трахеи человека является наличие в ее подслизистой осно-
ве большого скопления концевых отделов смешанных белково-
слизистых желез. Назначение этих желез — выработка и доставка
слизи на поверхность воздухоносных путей. Обладая адгезивными
свойствами, а также определенным бактерицидным действием,
слизь играет важную роль в мукоцилиарном механизме очищения
вдыхаемого воздуха.
Подслизистая оболочка трахеи переходит в плотную со-
единительную ткань надхрящницы, которая по-
крывает незамкнутые на дорсальной поверхности трахеи ее хря-
щевые кольца. Свободные концы незамкнутых хрящевых колец
соединены пучками миоцитов. В промежутках между кольцами
по длине органа расположена плотная неоформленная соедини-
тельная ткань. Все это вместе с гиалиновым хрящом составляет
фибр о зн о-хрящеву K)j оболочку трахеи. _____________
Надхрящница с наружной поверхности хряща переходит в
рыхлую неоформленную соединительную
ткань ад в е тгт'И циальной оболочки, связывающей
трахею с пищеводом и сосудисто-нервными пучками.
Изучению строения центрального органа системы дыхания —
легкого — необходимо предпослать краткую характеристику внут-
рилегочных воздухоносных путей. При этом необходимо иметь
в виду, что применяемая гистологами классификация бронхиаль-
ного дерева (главные -> крупные -> средние -*• мелкие —► терми-
нальные) является условной, достаточной для изучения гистоло-
гического строения легкого, но не применяемой в практической
медицине. Клиницисты (терапевты, фтизиатры, хирурги) исполь-
зуют другую классификацию, основанную на представлении о
бронхо-легочном сегменте. С этой классификацией студенты зна-
комятся в курсе анатомии и в клинике.
2.11.2. Легкое
Разбор строения легкого человека целесообразно начать с обзор-
ной схемы бронхиального дерева (рис. 2.157), где прежде всего
необходимо научиться дифференцировать бронхи в соответствии
с приведенной выше классификацией.
Бронхи крупного калибра характеризуются, по-
мимо размеров, наличием всех четырех оболочек, складчатостью
слизистой, выраженностью мышечного слоя слизистой, наличием
пластин гиалинового хряща и «пакетов» концевых отделов желез.
272
Рис. 2.157. Бронхиальное дерево.
А — схема (по В. Г. Елисееву и соавт.). Б — крупный бронх. В — средний бронх.
Г — мелкий бронх. Д — деталь к Г.
Т — трахея; ГБ — главный бронх; КБ — крупные бронхи (внелегочные и внутри-
легочные — долевые и сегментарные); СБ — средние бронхи (субсегментарные);
МБ — мелкие бронхи и бронхиолы (внутридольковые бронхи); ТБ — терминаль-
ная бронхиола; Ац — ацинус; Аб — дыхательная (респираторная) бронхиола;
Ах — альвеолярный ход; Ам — альвеолярные мешочки; Эо — эпителий однорядный
реснитчатый кубический, с — собственная пластинка слизистой оболочки, м — мы-
шечная пластинка слизистой оболочки; А — альвеола; Ма — межальвеолярная
перегородка.
1 — просвет бронха; 2 — многорядный мерцательный эпителий; 3 — собственная
пластинка слизистой оболочки; 4 — мышечная пластинка; 5 — концевые отделы
желез; 6 — хрящ; 7 — кровеносный сосуд; СК — секреторная клетка; РК — рес-
нитчатая клетка.
Рис. 2.157. Продолжение.
276
Рис. 2.158. Терминальная и респи-
раторная бронхиолы.
А — препарат. Б — поверхность терми-
нальной бронхиолы (СЭМ). В — рес-
пираторная бронхиола (СЭМ). Г —
поверхность дистальной части респи-
раторной бронхиолы (СЭМ).
1 — терминальная бронхиола; 2 — рес-
пираторная бронхиола; 3 — альвеолы;
СК — секреторные клетки в различных
фазах секреторного цикла; РК — рес-
нитчатые клетки; ЩК — щеточные
клетки.
277
Бронхи среднего калибра имеют также складчатую
внешнюю поверхность, еще большее относительное содержание
гладких мышц, островки гиалинового хряща, между которыми
располагаются концевые отделы слизисто-белковых желез.
Бронхи мелкого калибра отличаются резкой склад-
чатостью слизистой оболочки, максимальным развитием мышеч-
ной ткани, отсутствием хряща и желез. Отсутствие хрящевого
каркаса в сочетании с хорошо развитой гладкой мышечной тканью,
расположенной циркулярно, является морфологической основой
полного перекрытия просвета, возникающего при различного рода
спастических процессах. Последнее лежит в основе приступов
удушья, возникающих, например, при бронхиальной астме.
Рис. 2.158 демонстрирует пространственные взаимоотношения
между респираторной и терминальной бронхиолой и альвеоляр-
ными ходами в легком человека. Широкие темные полоски на ри-
сунке — кубический эпителий. Более детальное изображение аци-
нуса легкого дано на рис. 2.159. Здесь показаны все компоненты
ацинуса, начиная с респираторной бронхиолы.
Ключевой схемой, позволяющей правильно понять принцип
278
строения легкого в целом и, в частности, его респираторных от-
делов, является схема строения структурно-функциональной еди-
ницы легкого — а ц и н у с а. .. На схеме ацинуса широкая линия
обозначает однослойный кубический,"“или недыхательный, эпите-
лий (терминальная бронхйола,'участки респираторных бронхиол).
Тонкие—линии-хубпзначакУГ^плоскйЙ^т. ie. дыхательный эпителий. .
Обратите внимание на принцип дихотомического деления воздухе-,
носнйх'Тг респираторных отделов, строго- соблюдающийся почти
на всем их протяжении.
Перед~идентификацией участков респираторного отдела легко-
го целесообразно рассмотреть общий вид респираторного отдела
с помощью сканирующего электронного микроскопа (см. рис.
2.158).
Обнаружение и идентификация терминальной брон-
Рис. 2.159. Ацинус легкого.
А — схема, общий вид. Б, В — микроструктура ацинуса при малом и большом
увеличении.
1 — терминальная бронхиола; 2 — респираторная бронхиола; 3 — альвеолярные
ходы; 4 — альвеолы; 5 — реснитчатый эпителий; 6 — кубический эпителий; 7 —
полость альвеолы; 8 — капилляры.
279
Рис. 2.160. Строение альвеолы (ТЭМ).
1 — полость альвеолы; 2 — полость кровеносного капилляра; 3 — отростки аль-
веолоцитов I типа; 4 — альвеолоцит II типа; 5 — эритроцит в полости капилляра.
280
х и о л ы основывается на тонкости стенки этого участка возду-
хоносных путей и на наличии покрывающего ее однослойного
(простого) кубического эпителия. В составе этого эпителия, по-
мимо мерцательных клеток (реснитчатых эпителиоцитов) (см.
рис. 2.158, Б), располагается значительное количество секретор-
ных клеток (клеток Клара), значение секреторной активности
которых пока не установлено. Они не имеют ресничек, апикальная
поверхность часто куполообразно выпукла. В терминальной брон-
хиоле нет бокаловидных клеток.
Респираторные бронхиолы у человека и большин-
ства млекопитающих относительно короткие, в силу чего их обна-
ружение затруднено. Характерной чертой этого типа бронхиол
является чередование на их поверхности однослойного кубиче^
ского и плоского эпителия, так как~стейка этих- бронхиолсодер-
жит альвеольь_В проксимальной части эпителий респираторных
'броюЖотГсбдё^ит'рёгйитйатыёТйетЮГТГ’клётки'Клара. По мере ~
прбдЕИжёнйя~ТГ*глубь~ацинуса реснитчатые клетки исчезают, но
появляются «щеточные пневмоциты» (brush-клетки). Эти клетки
содержат на апикальной поверхности крупные и широкие микро-
ворсинки, которым приписывают сенсорные функции. Они встре-
чаются, правда, более редко, и в воздухоносных путях.
Большую часть среза легкого занимают срезы наполненных
воздухом мешочков — альвеол (рис. 2.160). Именно через их
стенки происходит диффузия газов между кровью и воздухом.
Альвеолы представляют собой полигональной формы образования,
открытые только со стороны выхода из легкого. Совокупность
альвеол, расположенных по ходу воздуха, образует альвеолярный
ход. У места окончания альвеолярного хода скопление альвеол,
называемое альвеолярным мешочком, имеет общий выход.
Полость альвеолы выстлана на большем своем протяжении
(95 % поверхности) плоскими клетками, называемыми респи-
раторными дыхательными альвеолоцитами
(респираторные альвеолярные клетки, или альвеолоциты I типа)
(рис. 2.161). На электронограмме такой клетки видны длинные
тонкие отростки, которыми покрыта большая часть полости аль-
веолы. Цитоплазма в отростках настолько тонка, что при световой
микроскопии неразличима. Отражением процесса газообмена через
наиболее тонкие участки респираторных альвеолярных клеток
являются микропиноцитозные пузырьки в их цитоплазме.
Другой разновидностью клеток, формирующих внутреннюю
стенку альвеолы, являются секреторные альвеоляр-
ные клетки (большие альвеолоциты, или альвеолоциты II ти-
па), строение которых представлено на рис. 2.162. Альвеолоциты
II типа называют также септальными клетками, так как они рас-
полагаются главным образом в септальных соединениях, где часто
выбухают в просвет альвеолы. Количество альвеолоцитов II типа
примерно равно количеству альвеолоцитов I типа, но они занимают
приблизительно 5 % поверхности альвеолы. Характерными чер-
тами этих клеток являются их кубическая или полигональная
281
Рис. 2.161. Респираторная альвео-
лярная клетка (альвеолоцит
I типа).
А — схема строения. Б — ТЭМ.
1 — тело альвеолоцита; 2 — отросток;
3 — цитоплазма эндотелиоцита капил-
ляра; 4 — ядро альвеолоцита.
282
Рис. 2.162. Секреторная альвеолярная клетка (альвеолоцит II типа).
А — схема строения. Б — ТЭМ. В — выход ламеллярного тельца из альвеолоцита
II типа на поверхность альвеолы (ТЭМ). Г — схема образования сурфактанта.
1 — ядро альвеолоцита II типа; 2 — ламеллярные пластинчатые тельца; 3 — мик-
роворсинки.
283
Рис. 2.162. Продолжение.
форма, а также наличие в цитоплазме осмиофильных телец, со-
держащих материал поверхностно-активной бесструктурной вы-
стилки альвеол — сурфактанта. Выделение сурфактанта на по-
верхность альвеолы происходит путем секреции и является важ-
нейшим процессом, обеспечивающим нормальные физиологиче-
ские функции легких. Доказательства способности альвеолоцита
II типа к секреции получены с помощью сканирующего электрон-
ного микроскопа. Наплывы на апикальной поверхности альвеоло-
цита II типа являются, очевидно, зонами сурфактанта. В экспери-
ментах на животных показано, что альвеолоциты II типа обеспе-
чивают репарацию альвеолярного эпителия после его повреждения.
Третьей клеточной формой, входящей в состав стенки альвео-
лы, являются альвеолярные макрофагоциты (рис.
2.163), часто называемые «пылевыми клетками».
На электронограммах альвеолярных макрофагов можно видеть
осмиофильные пластинчатые тельца, что свидетельствует о том,
что по крайней мере часть макрофагоцитов легкого происходит
из альвеолоцитов II типа (предшественниками других, как пола-
гают, являются моноциты крови). На электронограммах макро-
284
Рис. 2.163. Альвеолярный макрофагоцит (ТЭМ, препарат Г. Г. Кругликова).
фаги альвеол выглядят не связанными непосредственно со стенкой
альвеолы и имеют морфологические признаки фагоцитов — оби-
лие лизосом, фагоцитированных частиц и др.
В респираторных отделах легкого очищение вдыхаемого воз-
духа от пыли и бактерий происходит главным образом путем
фагоцитоза (заметим при этом, что в норме полость альвеол
стерильна).
Альвеолярные макрофаги способны фагоцитировать и эритро-
циты, если они появляются в полости альвеол, что имеет место,
например, при сердечной недостаточности. В этом случае макро-
фаги наполняются железосодержащим пигментом — гемосидери-
ном, возникающим при разрушении гемоглобина. В патологии они
называются клетками сердечной недостаточности.
Комплексное представление о взаимоотношении альвеолы с
капиллярным руслом дает схема, представленная на рис. 2.164.
Как видно из этой схемы, расправленная альвеола большей частью
своей поверхности контактирует с кровеносными капиллярами.
Строение сети кровеносных капилляров межальвеолярных пере-
городок представлено на коррозионном препарате.
Совокупность структур, через которые в физиологических ус-
ловиях проходят газы на пути из воздуха в кровь и обратно, носит
285
название аэрогематического барьера. Его схематическое изобра-
жение и электронная микрофотография представлены на рис.
2.165. Толщина аэрогематического барьера у человека составляет
всего 0,1—0,2 мкм. Даже очень незначительное его увеличение,
например при отеке или воспалении, может привести к снижению
проницаемости и грубым нарушениям газообмена.
Между соседними альвеолами в легком человека обнаружены
небольшие отверстия диаметром 7—9 мкм, называемые порами
Кона (рис. 2.166). Они расположены в пространствах между ка-
пиллярами альвеолярной стенки. Предполагают, что они могут
служить для коллатерального воздухообмена при спадении аль-
веол (ателектаз) и невозможности их аэрации вследствие заку-
порки бронха.
Даже при самом глубоком выдохе стенки альвеол полностью
не спадаются вследствие наличия эластической тяги легких. На
рис. 2.167 представлены избирательно выкрашенные эластические
волокна и их пучки, формирующие эластический каркас.
Межальвеолярные перегородки в легком человека содержат в
своем составе и рыхлую волокнистую соединительную ткань со
значительным количеством коллагеновых волокон. При заболева-
ниях, сопровождающихся деструкцией легочной ткани, участки
коллагеновых волокон могут попадать в отделяемое (мокроту).
Основные этапы развития легкого представлены на рис. 2.168,1.
Источником развития легкого у человека являются энтодерма пе-
редней (глоточной) кишки и мезенхима. Очень рано (длина эмб-
риона 3 мм) вырост передней кишки делится на две несимметрич-
ные почки: левую и правую, из которых впоследствии образуются
легкие. Существенно отметить, что дифференцировка ткани легко-
го идет снаружи кнутри, т. е. от бронхов к альвеолам.
Во внутриутробном (функционально не активном) периоде
развития легкого человека принято различать три четко дифферен-
цируемые во времени и морфологически стадии. Первая из них —
железистая (рис. 2.168, II, III). Препарат легкого на этой стадии
характеризуется картиной крупных бронхов на фоне недифферен-
цированной мезенхимы, что делает легкое в этот период действи-
тельно похожим на экзокринную железу.
Вторая стадия развития, завершающаяся формированием всего
бронхиального дерева, называется каналикулярной, а третья, на-
чало которой у человека приходится на середину 6-го месяца внут-
Рис. 2.164. Взаимоотношения альвеолы с кровеносными капиллярами
(по Е. Ф. Котовскому).
А — схема. Б — наливка капилляров кармином. В — СЭМ капиллярного русла
вокруг альвеолы (коррозионный препарат, стрелками показаны сколы поперечных
сечений капилляров).
1 — полость альвеолы; 2 — альвеолоцит I типа; 3 — базальная мембрана альвеоло-
цита; 4 — эндотелиоцит кровеносного капилляра; 5 — базальная мембрана эндо-
телия; 6 — клетки и волокна соединительной ткани в межальвеолярных перегород-
ках; 7 — эластические волокна.
287
«л*.
Рис. 2.165. Аэрогематический барь-
ер (по В. А. Шахламову).
А — схема. Б — ТЭМ.
1 — ядро эндотелиоцита; 2 — просвет
капилляра; 3 — эритроцит; 4 — цито-
плазма эндотелиоцита; 5 — цитоплазма
альвеолоцита I типа; 6 — базальные
мембраны эндотелиоцита и альвеолоци-
та; 7 — аэрогематический барьер; 8 —
просвет альвеолы.
288
Рис. 2.166. Пора Кона.
1 — капилляры; 2 — пора; 3 — сурфак-
тант.
Рис. 2.167. Эластический каркас
альвеол (окраска орсеином).
1 — альвеолы; 2 — эластический каркас.
10—321
289
06Z
Рис. 2.168. Основные этапы развития легкого.
I — схема ранних этапов развития легкого человека. [I — легкое эмбриона чело-
века 6-й недели развития (железистая стадия). III — легкое плода 3-го месяца
развития. IV —- легкое новорожденного. V — легкое мертворожденного (недышав-
шее легкое).
А — эмбрион длиной 4 мм; Б — эмбрион длиной 6 мм; В — эмбрион длиной 7 мм:
1 —- трахея; 2 — первичные бронхи; Г — эмбрион длиной 8,5 мм: 1 — трахея; 2 —
правый ствол бронха; Д — эмбрион длиной 10 мм: 1 — трахея; 2 — левый ствол
бронха; Е — эмбрион длиной 20 мм: 1 — бифуркация трахеи; 2 — верхняя доля;
3 — левый бронх; 4 — мезенхимная закладка стромы легкого; 5, 8 — нижняя доля;
6 — легочная вена; 7 — сердечный бронх; 9 — закладка висцеральной плевры;
10 — средняя доля; 11 — правый бронх; 12 — верхняя доля; 13 — верхушечный
бронх (по Б. М. Пэттену); 14 — закладывающиеся бронхи; 15 — нерасправленные
альвеолы.
риутробного развития, — альвеолярной. В этой связи полезно по-
мнить, что жизнеспособность недоношенного ребенка во многом
определяется степенью развития альвеол.
Легкое доношенного новорожденного ребенка характеризуется
наличием в его нижних (придиафрагмальных) отделах незапол-
ненных воздухом и нерасправленных групп альвеол (рис. 2.168, IV).
Легкое мертворожденного ребенка отличается полной нерас-
правленностью альвеол (рис. 2.168, V).
10*
291
2.12. Кожа и ее производные
Анатомо-топографи«еское положение кожи как внешнего покрова
тела определяют ее значение в поддержании гомеостаза и защите
организма от внешних воздействий.
В состав кожи входят различные виды и типы тканей — мцщхь
слойный ороговевающий и железистый эпителии, рыхлая волокни-
стая соединительная ткань, плотная неоформленная соединитель-
наяткань, а также многочисленные нервные окончания; Опреде-
ленные закономерности их внутрикожной топографии, функцио-
нальная специализация различных слоев и структур кожи обеспе-
чивают многообразие ее функций (защитная, экскреторная,
кроведепонирующая, теплообменная), а также функцию органа
чувств и др. В функциональной морфологии производных кожи —
волоса, потовых и сальных желез прослеживаются как общие чер-
ты, так и выраженные различия (отражающие полипотентность
кожной эктодермы).
Регионарные особенности в строении внешнего покрова служат
основанием для выделения двух типов кожи — толстой и тонкой.
Толстая кожа покрывает ладони и подошвы, тонкая — остальные
части тела. При общем плане строения эти разновидности кожи
отличаются некоторой спецификой морфологии.
Развитие кожи из эктодермы и мезенхимы проявляется в двух-
слойное™ ее строения (рис. 2.169, А). Четко выделяются наруж-
ный пласт многослойного плоского ороговевающегр эпителия —
эпидермис й подлежащая соединительнотканная основа — дерма,
или собственно кожа, которая без резкой границы переходит в
подкожную клетчатку, или гиподерму. Дерма представлена двумя
разновидностями собственно соединительной ткани, формирую-
щими соответственно ее сосочковый и сетчатый слои. Подлежащий
эпидермису более узкий сосочковый слой выполнен ^ыхлой, а сет-
чатый -—плотной волокнистой неоформленной соединительной
тканью. Тесная взаимосвязь эпидермиса и сосочкового слоя, оби-
лие в последнем кровеносных микрососудов обеспечивают трофику
эпидермиса. Примечательно, что наружный рельеф эпидермиса
повторяет контуры соединительнотканных сосочков, которым при-
сущ строго индивидуальный характер. Этот феномен положен в
Рис. 2.169. Кожа ладонной поверхности пальца («толстая кожа»).
А — схема. Б — потовые железы. В — осязательное тельце.
I — эпидермис; II — собственно кожа (дерма); III — подкожная жировая клет-
чатка.
1 — роговой слой; 2 — блестящий слой; 3 — зернистый слой; 4 — шиповатый слой;
5 — базальный слой; 6 — базальная мембрана; 7 — сосочковый слой; 8 — сетча-
тый слой; 9 — выводной проток потовой железы; 10 — концевой отдел мерокри-
новой потовой железы: а — железистые клетки; б — миоэпителий; 11 — пластин-
чатое тельце; 12 — осязательное тельце; 13 — кровеносные капилляры; 14 — арте-
риола.
292
Oi (Л
293
основу дактилоскопии — метода идентификации личности по от-
печаткам пальцев в криминалистической и судебно-медицинской
практике. Дактилоскопия оказывается полезной и при диагностике
у новорожденных генетических заболеваний, связанных с хромо-
сомными аномалиями. Так, у преобладающего большинства детей
с тяжелой врожденной патологией — синдромом Дауна — дакти-
лоскопически обнаруживаются комбинации «узоров», никогда не
встречающиеся у здоровых людей. Сосочковый слой пронизан
протоками потовых желез (рис. 2.169, Б), в субэпителиальной
зоне часто обнаруживают осязательные тельца (рис. 2.169, В).
Обращает внимание отсутствие четкой границы между сосоч-
ковым и сетчатым слоем. Она лишь намечена срезами более круп-
ных, нежели в сосочковом слое, микрососудов — артериол и венул,
входящих в подсосочковое сосудистое сплетение. В сетчатом слое
дермы мощные различно ориентированные пучки коллагеновых
волокон как бы формируют широкопетлистую сеть, ячейки кото-
рой часто заполнены жировой тканью. Такая конструкция придает
коже прочность, способность противостоять разнонаправленным
механическим воздействиям. Вместе с тем архитектоника и состав
этой сети (в частности, содержание эластических волокон) варь-
ируют в различных областях кожного покрова адекватно к испы-
тываемым кожей нагрузкам. Среди пучков волокон часто обнару-
живают крупные пластинчатые тельца — тельца Фатера—Пачини,
которые легко идентифицировать благодаря своеобразной кон-
центрической структуре. Примечательна различная топография
рецепторов тактильной чувствительности (осязательные тельца) и
механорецепторов (пластинчатые тельца), представляющих в коже
два вида инкапсулированных окончаний. Другие элементы обшир-
ного иннервационного аппарата кожи в препаратах, окрашенных
гематоксилином и эозином, выявить трудно. Для этой пели исполь-
зуют специальные нейрогистологические методы окраски (импрег-
нация солями тяжелых металлов).
Для сосудистого русла сетчатого слоя характерна меньшая
(по сравнению с сосочковым слоем) плотность расположения
гемокапилляров, локализующихся в основном вблизи кожных же-
лез и волосяных фолликулов; здесь проходят крупные магистраль-
ные сосуды, артерии и вены, связывающие поверхностное и глу-
бокое сосудистые сплетения.
Рис. 2.170 помогает понять принцип организации системы кро-
воснабжения кожи и функциональную обоснованность кожной
ангиоархитектоники. Так, система двух мощных сосудистых спле-
тений обеспечивает функцию кожи как депо крови. Поверхностное
расположение обширного микроциркуляторного бассейна сосочко-
вого слоя с многочисленными артериоловенулярными анастомо-
зами создает основу ее теплообменной функции.
Плавный переход сетчатого слоя в гиподерму, с которой он
связан отдельными пучками коллагеновых волокон, придает коже
некоторую подвижность относительно подлежащих тканей, что
обеспечивает ей дополнительную прочность.
294
Рис. 2.170. Строение и кровоснабжение тонкой кожи (схема по В. Е. Соко-
лову с соавт. с изменениями).
1 — эпидермис; II — сосочковый слой дермы; III — сетчатый слой дермы; IV —
подкожная клетчатка.
1 — концевой отдел мерокриновой потовой железы; 2 — выводной проток меро-
криновой потовой железы; 3 — корень волоса; 4 — концевой отдел апокриновой
потовой железы; 5 — выводной проток апокриновой потовой железы; 6 — сальная
железа; 7 — мышца, поднимающая волос; 8 — пучки коллагеновых волокон; 9 —
жировая ткань; 10 — пластинчатое нервное тельце (Фатера — Пачини); 11 — сво-
бодные нервные окончания; 12 — осязательное тельце (Мейснера); 13 — нервное
волокно; 14 — артерии и 15 — вены глубокого сосудистого сплетения; 16 — арте-
риолы и 17 — венулы поверхностного сосудистого сплетения; 18 — кровеносные
капилляры.
Эпидермис толстой кожи состоит из нескольких десятков плас- .
тов клеток, которые объединяются _д~,5 основных слоев (рис.
2.Т71, А). Эпидермис тонкой кожи.отличает отсутствие блестяще-
го^еньшаяТплтЦ1ТТ13П11иповатого_.и„роговох11_СЛ оев,.(рис. 2.171, Б).
Кромё~того, в поверхностном рельефе эпидермиса тонкой кожи
практически отсутствуют бороздки и гребешки.
Анализируя строение эпидермиса, важно понять, что его много-
слойность связана с постоянным передвижением клеток из базаль-
ного слоя в поверхностные. По мере этого продвижения клетка
295
Рис. 2.171. Эпидермис «толстой» и
«тонкой» кожи.
А — кожа пальца («толстая» кожа).
Б — кожа лица («тонкая» кожа).
I — роговой слой; 2 — блестящий слой;
3 — зернистый слой; 4 — шиповатый
слой; 5 — базальный слой; 6 — базаль-
ная мембрана; 7 — сосочковый слой
дермы.
утрачивает способность делиться и специализируется на продук-
ции рогового вещества. Таким образом^ базальный эпителиоцит
и роговая чешуйка соответственно начальная и конечная ста
дии жизненного дикла_одной_клетки. Рис. 2.172 позволяет про-
следить^ как~при этом постепенно изменяется форма и ультра-
структура клеток, происходит дестабилизация межклеточных
связей за счет редукции десмосом. Важно отметить закономерное
присутствие в составе эпидермального пласта элементов неэкто-
дермального происхождения — пигментные клетки (меланоциты)
и внутриэпидермальных макрофагов (клеток Лангерганса). Эти
клетки не связаны с эпителиоцитами десмосомами, что пред-
полагает возможность их миграции. Очевидно, что особенности
биологии эпителиоцитов, включение в состав эпидермиса клеточ-
ных элементов с различной функциональной специализацией яв-
ляются морфологической основой широкого диапазона защитной
функции кожи. В частности, кооперация лимфоцитов и макрофа-
гов (интра- и экстраэпидермальная) обеспечивает реакции мест-
ного иммунитета. В базальном слое эпидермиса встречаются ося-
зательные эпителиоциты, входящие в состав свободных нервных
окончаний.
296
Важная роль в реализации защитной, экскреторной и термо-
регулирующей функций кожи принадлежит производным эпидер-
миса — кожным железам.
Потовые железы кожи представлены двумя разновидностя-
ми — э к к р ин овы М-И—-й а п окриновыми п о т о в ы м и
ж е л одз~а_м_и, отличающимися” топбграфйёй~концевых отделов,
протяженностью и локализацией устья выводных протоков, а так-
же степенью представительства в кожном покрове. Более широко
распространены эккриновые (мерокриновые) потовые-железы, кон-
севБге~тгтделы-~которых располагаются к. глубине_дермы,„часто_=^
^_гащодермёДсмГ^йсГ27169). Обратившись к схеме строения кожи
(см. рисГ2Л 70), легко объяснить регистрируемое в гистологических
препаратах обилие срезов концевого отдела и начальных сегментов
выводного протока каждой потовой железы. Причина этого явле-
ния — своеобразие конструкции секреторного отдела (трубочка,
свернутая в клубочек) этих простых неразветвленных желез.
Схема иллюстрирует также особенности расположения (более
поверхностное) и микроанатомии (более короткий выводной про-
ток, впадающий в волосяную воронку) апокриновых потовых же-
лез, имеющих ограниченную область локали ации'~Тподмцшечные _
впадины, бблдсти заднего прохода ^и_.лобка, вокруг соска модемной ,,
железы) Г~Апокринодыс_ потовые железы отличаются от меро-
криновь!Х ^"большим диаметром _концёвых отделов., В тех же
случаях, когДа они открываются непосредственно на поверхность
кожи, минуя волосяной фолликул, внутриэпидермальную часть их
выводных протоков отличает (по сравнению с мерокриновыми
железами; см. ниже) более прямой ход и сохранность стенки;
потовая пора не формируется.
Рассматривая строение потовых желез, следует подчеркнуть,
что принцип многослойности эпителия как признак его эктодер-
мального происхождения постоянен на всем протяжении железы
(исключение составляет лишь внутриэпидермальный отдел вывод-
ных протоков мерокриновых желез). Эти протоки, выстланные
двуслойным кубическим эпителием, имеют вид"тонкйх интенсивно
окрашенньцГ'Трубочек, которые^ проходя через всю толщу дермы,
штопорообразно извиваясь, прободают эпидермис в виде канала,
лишенного собственной стенки, и открываются в потовую пору.
В концевых отделах потовых желез, образованных однослойным
цилиндрическим железистым эпителием, двуслойность стенки со-
храняется благодаря присутствию веретеновидных миоэпителио-
цитов, лежащих на базальной мембране и облегчающих своими
сокращениями экструзию секрета.
Волос и сальные железы. Для изучения строения других при-
датков кожи — волоса и сальных желез — используют препараты
тонкой кожи (рис. 2.173). Отметим, что, помимо особенностей
структуры эпидермиса, от толстой кожи ее отличает более мощное
развитие сетчатого слоя, где располагаются корни волос и кожные
железы.
В волосе различают две части: выступающий над поверх-
297
298
Рис. 2.172. Субмикроскопическая организация эпидермиса.
А — схема. Б — ТЭМ (по Л. Д. Степановой).
I — базальный слой; II — шиповатый слой; III — зернистый слой; IV — блестящий
и роговой слои.
1 — базальная мембрана; 2 — базальный эпидермоцит; 3 — меланофороцит; 4 —
внутриэпидермальный макрофаг; 5 — полудесмосомы; 6 — десмосомы; 7 — тоно-
фибриллы; 8 — промеланосома; 9 — меланосома; 10 — лизосома; 11 — межклеточ-
ная щель; 12 — гранула кератогиалина; 13 — кровеносный капилляр сосочкового
слоя дермы.
299
11
11
Рис. 2.173. Кожа волосистой части головы («тонкая» кожа).
А — общий вид. Б — фрагмент волосяной луковицы (продольное сечение). В —
сальная железа. Г, Д — ультраструктура клеток потовой (Г) и сальной (Д) желез
(по Л. Д. Степановой).
I — эпидермис; II — дерма; III — подкожная клетчатка.
1 — выводной проток потовой железы; 2 — концевой отдел мерокриновой потовой
железы; 3 — сальная железа: а — ростковый слой, б — сальные экзокриноциты.
в — гибнущие клетки (голокриновая секреция), г — выводной проток; 4 — мышца,
поднимающая волос; 5 — корень волоса; 6 — внутреннее эпителиальное корневое
влагалище; 7 — наружное эпителиальное корневое влагалище; 8 — волосяная сум-
ка; 9 — волосяная луковица; 10 — волосяной сосочек; 11 — стержень волоса; 12 —
волосяная колба; 13 — липидная гранула; 14—миоэпителиоцит.
300
301
Рис. 2.173. Продолжение.
Рис. 2.174. Строение и иннервация волоса.
А, Б — поперечное сечение корня волоса: А — общий вид; Б — СЭМ (по
Н. П. Омельяненко, Л. Д. Жеребцову). В — рельеф поверхности кутикулы корня
волоса (СЭМ). Г — стержень волоса в зоне выхода на поверхность (СЭМ). Д —
иннервация волосяной луковицы.
I — поперечное сечение корня волоса на уровне нижней трети; II — поперечное
сечение корня волоса на уровне сальной железы.
1 — кутикула; 2 — корковое вещество; 3 — мозговое вещество; 4 — внутреннее
эпителиальное корневое влагалище: а — внутренний (гранулосодержащий) слой,
б — наружный (бледный) слой; 5 — наружное эпителиальное корневое влагалище;
6 — волосяная сумка; 7 — сальная железа; 8 — проток и 9 — концевой отдел по-
товой железы; 10 — жировая ткань.
302
II
303
Рис. 2.174. Продолжение.
304
ностью кожи стержень и глубоко погруженный в дерму корень.
Стержень волоса состоит из коркового вещества и кутикулы. Строе-
ние корня зависит от вида волос: в длинных и щетинистых волосах
он образован корковым, мозговым веществом и кутикулой, в пушко-
вых — только корковым и кутикулой. На продольных срезах кор-
ня волоса легко определяются элементы его оболочки — волося-
ного фолликула (внутреннее и наружное эпителиальные влагали-
ща, соединительнотканная волосяная сумка), а также волосяная
луковица и волосяной сосочек. При идентификации отдельных ком-
понентов корня волоса удобно пользоваться представлением о нем
как своеобразной дупликатуре эпидермиса (ороговевающими сло-
ями внутрь, ростковыми — наружу). Отметим, что пролифери-
рующие эпителиальные клетки, ответственные за рост волоса, со-
средоточены в волосяной луковице, и выпадение волос связано
с ороговением матрицы луковицы, превращением последней в так
называемую волосяную колбу, которая постепенно выталкивается
на поверхность.
Детали строения и иннервации волоса лучше видны на его
поперечных (тангенциальных) срезах (рис. 2.174). Ориентируясь
на уровень среза, можно четко проследить постепенное изменение
структуры корня и его оболочек: редукцию мозгового вещества воло-
са, ороговение кутикулярных клеток, исчезновение внутреннего
эпителиального влагалища на уровне сальной железы и др. Ме-
тод СЭМ позволяет увидеть поверхностный рельеф волоса, кути-
кула которого построена из черепицеобразных роговых чешуек.
Волосяные фолликулы обильно иннервированы, максимальная
концентрация нервных окончаний обнаруживается в наиболее
активной «ростковой» зоне волоса — волосяной луковице.
Анализируя строение корня волоса, нельзя не обратить внима-
ния на тесную взаимосвязь волосяного фолликула и сальной
железы, открывающейся в волосяную воронку (см. рис. 2.173).
Эта простая альвеолярная разветвленная железа с лишенным про-
света концевым отделом иногда расценивается как участок много-
слойного эпителия с особым типом обмена, ведущим к гибели желе-
зистых клеток (голокриновая секреция). Действительно, схемы
жизненного цикла эпидермальных эпителиоцитов и сальных экзо-
криноцитов достаточно близки: пролиферация клеток (ростковый
слой эпидермиса — ростковые клетки железы), продвижение кле-
ток в поверхностные слои и приобретение ими способности к син-
тезу специфических веществ (кератиноциты — сальные клетки
железы) и, наконец, гибель и отторжение клеток. Однако в отли-
чие от эпидермиса в случае сальной железы мы имеем пример
экзокриновой (голокриновой) секреции с утилизацией организмом
материала погибших клеток.
Развитие кожи. Некоторые моменты гистогенеза кожи и ее
производных отражены на рис. 2.175. Хорошо видна сочетанная
дифференцировка производных эктодермы (эпидермис) и мезен-
химы (дерма), изменение характера эпидермо-дермальных взаи-
мосвязей. Обращает внимание диффузная протрузия всех слоев
305
Рис. 2.175. Развитие кожи, волоса, потовых желез (по Е. А. Загорученко).
А — стадия дифференцировки эпидермиса человека. Б — развитие потовых желез
(плод 19 нед). В — зачаток волоса (плод 17 нед).
306
дермы инвагинациями эпидермиса при формировании потовых же-
лез, представительство всех эпителиальных слоев в зачатке волоса.
2.13. Пищеварительная система
Пищеварительная система — единый комплекс органов, обеспечи-
вающий поступление в организм всех питательных веществ, необ-
ходимых для его жизнедеятельности. Болезни органов пищеваре-
ния нередко являются причиной возникновения других заболева-
ний — гиповитаминозов, диабета, болезней крови и др. Изучение
общей морфофункциональной характеристики пищеварительной
трубки, тканевого состава и функций оболочек, микроскопическо-
го и субмикроскопического строения органов системы и особен-
ностей их строения, связанных с выполнением функций, способ-
ствует пониманию физиологии пищеварения, причин и законо-
мерностей патологических процессов.
2.13.1. Передний отдел
2.13.1.1. Зуб
По строению зуб значительно отличается от остальных органов,
имеющих общие принципы структурной организации. На рис.
2.176, А можно рассмотреть части зуба: коронку, шейку и корень
и его тканевые компоненты. Эмаль определяется как наружный
слой коронки, дентин составляет основу всего органа, цемент по-
крывает дентин в области корня и шейки. В центре рисунка видны
полость коронки зуба и канал корня зуба. На рис. 2.176, Б следует
обратить внимание на исчерченность эмали, которая является
результатом расположения пучков эмалевых призм, наличие па-
раллельных полосок через весь дентин (дентинные трубочки),
в которых размещаются отростки одонтобластов, а также двух
зон в цементе — клеточной и бесклеточной.
Развитие зуба. Самая ранняя стадия (стадия закладки зубной
пластинки) представлена на рис. 2.177, А, на котором виден мно-
гослойный эпителий и врастающий от него в подлежащую мезен-
химу эпителиальный тяж (источник формирования эмали) —
зубная пластинка. Обращает на себя внимание начало концентра-
ции клеток мезенхимы вокруг пластинки (источник всех осталь-
ных компонентов зуба). На следующей стадии (дифференцировка
зубного зачатка) (рис. 2.177, Б) формируется эмалевый орган,
по форме напоминающий двухстенный бокал — результат диффе-
ренцировки зубной пластинки и окружающей мезенхимы. При
этом можно отметить, что его шейка еще соединена с базальным
слоем эпителия ротовой полости. Компоненты эмалевого органа
составляют: наружный эмалевый эпителий — выгнутая часть стен-
ки; внутренний эмалевый эпителий — вогнутая часть стенки; пуль-
па органа — пространство между эпителиями со скоплением от-
ростчатых клеток. Кроме этого, видны зубной сосочек — «чаша
307
Рис. 2.176. Зуб.
А — продольный шлиф однокорневого зуба. Б — СЭМ шлифа эмали (по
Л. С. Строчковой).
I — коронка зуба; II — шейка зуба; П1 — корень зуба.
1 — эмаль: а — эмалевые призмы; 2 — дентин: б — дентинные канальцы; 3 — це-
мент: в — бесклеточный, г — клеточный; 4 — полость коронки зуба; 5 — канал
корня зуба.
бокала», заполненная недифференцированными клетками и содер-
жащая кровеносные сосуды,’ зубной мешочек — несколько слоев
уплощенных клеток, покрывающих эмалевый орган снаружи. Во-
круг зачатка зуба расположена мезенхима с оксифильно окрашен-
ными островками развивающейся костной ткани — закладка че-
люсти. На поздней стадии развития зуба в составе его зачатка
(рис. 2.178, А) можно рассмотреть следующие компоненты: эмаль-
продуцирующие клетки — амелобласты, расположенные снаружи
в один строго упорядоченный сплошной ряд призматические клет-
ки; эмаль — ярко окрашенная гомогенная зона под амелобласта-
ми; дентин — следующий также бесклеточный, но более широкий
и отличающийся по окраске слой; дентинопродуцирующие клетки
308
Рис. 2.177. Стадии развития зуба (разрез головы зародыша).
А — закладка зубной пластинки. Б — дифференцировка зубного зачатка.
1 — эпителий ротовой полости; 2 — мезенхима; 3 — зубная пластинка; 4 — кро-
веносный сосуд; 5 — эмалевый орган; 6 — шейка эмалевого органа; 7 — наружный
и 8 — внутренний эмалевый эпителий; 9 — пульпа эмалевого органа; 10 — зубной
сосочек; 11 — зубной мешочек; 12 — костный островок.
одонтобласты — отростчатые клетки, образующие зону под ден-
тином. Вся внутренняя часть — зубной сосочек — рыхлая волок-
нистая соединительная ткань, формирующая пульпу органа. Во-
круг зачатка могут встречаться остатки наружного эмалевого эпи-
телия и пульпы эмалевого органа (дающей кутикулу эмали), а
иногда и эпителия зубной пластинки. Следует отметить, что на
данной стадии происходит инверсия ядер амелобластов (рис.
2.178, Б), что отражает общую картину смены базальных и апи-
кальных полюсов клетки, в результате которой апикальный полюс
стал обращен не в сторону пульпы эмалевого органа, а в сторону
зубного сосочка. На рис. 2.179, А, В, Г представлена ультраструк-
турная организация одонтобласта, на рис. 2.179, Б — амелобласта.
309
Рис. 2.178. Гистогенез зуба.
А — общий вид. Б — фрагмент.
1 — амелобласты; 2 — эмаль; 3 — дентин; 4 — одонтобласты; 5 — зубной сосочек;
6—8 — остатки эмалевого органа; 9 — костный островок.
Рис. 2.179. Ультраструктурная организация элементов зуба.
А — одонтобласт. Б — амелобласт. В — ТЭМ отростка. Г — ТЭМ тела одонто-
бласта (по Н. Л. Яковлевой).
I — дентин, II — предентин.
1 — зона минерализации; 2 — отросток; 3 — секреторные гранулы; 4 — десмосомы;
5 — комплекс Гольджи; 6 — гранулярная эндоплазматическая сеть; 7 — мито-
хондрии; 8 — ядро; 9 — коллагеновые волокна.
310
311
Рис. 2.179. Продолжение.
2.13.1.2. Язык
Изучение строения языка целесообразно начать с рассмотрения
общего плана его структурно-тканевой организации (рис. 2.180).
Рельеф слизистой оболочки и размеры площади, занимаемой со-
единительной тканью на верхней и нижней поверхностях, суще-
ственно отличаются, а основная масса органа представлена попе-
речнополосатой скелетной мышцей, волокна которой расположе-
Рис. 2.180. Язык.
А — схема строения (продольный разрез): I — стенка, II—нижняя поверхность,
III — корень языка.
Сосочки языка: Б — нитевидный, В — грибовидный, Г — листовидный, Д — жело-
боватый. Е — СЭМ желобоватого сосочка (по В. А. Шахламову).
1 — эпителий слизистой оболочки; 2 — собственная пластинка слизистой оболоч-
ки; 3 — подслизистая основа.
Сосочки: 4 — нитевидные; 5 — грибовидные; 6 — листовидные; 7 — желобоватые:
а — первичные, б — вторичные.
Пучки скелетных мышц: 8 — вертикальные; 9 — горизонтальные; 10 — поперечные.
Железы: 11 — мукозные; 12 — серозные; 13 — язычная миндалина; 14 — скопле-
ние липоцитов; 15 — кровеносные сосуды; 16 — вкусовые почки; 17—валик со-
сочка; 18 — желобок сосочка; 19 — лимфатический узелок.
312
11 12
7
13
313
Рис. 2.180. Продолжение.
ны в трех взаимно перпендикулярных направлениях. Между ними
находятся железы и скопления липоцитов. Характерная особен-
ность верхней поверхности языка — наличие разнообразных по
форме выростов — сосочков. Нижняя же поверхность гладкая,
соединительнотканная прослойка значительно шире за счет нали-
чия подслизистой основы. Изучение сосочков языка обычно про-
изводят на разных участках языка из-за различной их локализа-
ции. Основной же принцип их строения сходен — это вырост со-
единительной ткани собственной пластинки слизистой оболочки,
покрытый многослойным плоским неороговевающим или частично
ороговевающим (нитевидные сосочки) эпителием. На рис. 2.180, А
представлена слизистая оболочка верхней поверхности языка.
При этом обращает на себя внимание, что ее собственная плас-
тинка, образующая основу сосочка, непосредственно сращена с
мышцей языка и снабжена слюнными железами. Нитевидный со-
сочек (рис. 2.180, Б) отличается конической формой, а грибовид-
ный (рис. 2.180, В) — более широкой верхней частью, придающей
ему сходство с грибом (в толще его эпителия могут встречаться
вкусовые почки). Листовидные сосочки располагаются группами,
314
в эпителии их боковых поверхностей много вкусовых почек (рис.
2.180, Г). Под сосочками и глубже залегают слюнные железы,
серозный секрет которых промывает межсосочковые щели. Же-
лобоватые сосочки имеют крупные размеры (видны невооружен-
ным глазом) и более сложную форму (рис. 2.180, Д). При этом
хорошо прослеживается собственно сосочек, окружающий его ва-
лик и расположенный между ними желобок. В эпителии сосочка
и валика, образующем стенку желобка, видны вкусовые почки.
В соединительной ткани сосочка могут встречаться лимфатические
узелки. На рис. 2.180, Е представлена СЭМ желобоватого сосочка.
2.13.1.3. Слюнные железы
Помимо множества слюнных желез, расположенных в слизистой
оболочке органов ротовой полости, в последнюю выделяют свой
секрет три пары крупных желез, являющихся также производны-
ми эктодермального эпителия ротовой полости. Они оформлены
в виде самостоятельных органов, обладающих общим принципом
структурной организации. Для лучшего понимания особенностей
данных желез ознакомьтесь с рис. 2.181, А, из которого следует,
что они имеют дольчатое строение, состоят из концевых отделов
и систем выводных протоков, отличающихся по строению и лока-
лизации. Большую часть дольки околоушной ж ел е з_ы
составляют ацинусы — базофильно окрашенные, _округлые и
овальные образования без выраженного просвета в центре. Среди
них находятся трубчатые структуры — протоки. Наиболее узкие
из них, определяемые по тесно расположенным концентрически
темным округлым ядрам выстилающих эпителиоцитов, — вставоч-
ные, более широкие, выстланные призматическими эпителиоци-
тами— исчерченные (рис. 2.181, Б). Ультраструктурной основой,
определяющей феномен их исчерченности, является сильно раз-
витая система инвагинаций базальной плазматической мембраны,
включающая значительное количество митохондрий (см. рис.
2.182, В). В составе дольки располагается также внутридольковый
проток, характеризующийся большими размерами и двухслойным
эпителием. К структурам, расположенным в междольковой пере-
городке, относятся междольковый проток с эпителиальной вы-
стилкой из двух и более слоев (признак эктодермального проис-
хождения) и кровеносные сосуды.
При изучении морфологии сероцитов важно обратить внимание
на форму и расположение ядер секреторных клеток и уровень
плотности цитоплазмы. Ультраструктурная организация клетки
представлена на рис. 2.182, А, Б. Изучая строение ацинуса, вста-
вочного и исчерченного протоков, необходимо обратить внимание
на плотно примыкающие снаружи удлиненные ядра миоэпителио-
цитов. При знакомстве с микроскопическим строением подче-
люстной железы (рис. 2.181, В) отмечаются более частая
встречаемость срезов исчерченных протоков (следствие большей
их длины и степени ветвления), неравномерность окраски и боль-
315
co
Рис. 2.181. Слюнные
железы.
А — основные компоненты
желез: I — концевой от-
дел; 11 — протоки. Б —
околоушная железа. В —
подчелюстная железа.
Г — развитие околоушной
железы: I — железа но-
ворожденного; II — желе-
за ребенка 7 лет (ШИК-
реакция).
Концевые отделы: 1 —
серозный; 2 — мукозный;
3 — смешанный; 4 — аци-
нус; 5 — сероцит; 6 —
миоэпителиоцит; 7 — му-
коцит; 8 — серозное по-
лулуние; протоки: 9
вставочный; 10 — исчер-
ченный; 11 — внутридоль-
ковый; 12 — междолько-
вый; 13 — долька; 14 —
междольковая перегород-
ка; 15 — артерия.
317
Рис. 2.182. Слюнные железы, ТЭМ.
А — сероцит (по Е. А. Шубниковой).
Б — мукоцит. В — базальная зона клетки
исчерченного протока.
1 — ядро; 2 — митохондрия; 3 — секретор-
ный продукт; 4 — базальная мембрана;
5 — инвагинация базального участка плаз-
молеммы; 6 — десмосома.
318
ший размер ацинусов. В остальном же строение сходно с около-
ушной железой. Кроме того, можно заметить, что у большинства
ацинусов основная центральная часть занята светлоокрашенными
гландулоцитами со сплюснутыми ядрами, оттесненными к их ба-
зальной части (признаки, характерные для мукоцитов). Их окру-
жают в виде полулуния клетки меньших размеров, окрашенные
базофильно и с округлыми ядрами, — серозные полулуния. Как
и в околоушной железе, ацинусы, вставочный и исчерченный про-
токи «оплетены» миоэпителиоцитами.
Развитие слюнных желез представлено на рис. 2.181, Г. За-
кладка подчелюстных желез происходит на 6-й неделе, а около-
ушных — на 8-й неделе эмбриогенеза. В процессе развития из
эпителиальных тяжей первоначально формируются выводные про-
токи, а затем концевые отделы. К моменту рождения (рис.
2.181, Г, I) в железах ►Сформированы все структурные компоненты
органа, но заметны малочисленность ацинусов и обилие междоль-
ковой соединительной ткани с междольковыми выводными про-
токами. Дальнейший рост и структурно-функциональная диффе-
ренцировка слюнных желез продолжаются в течение нескольких
лет постнатальной жизни, о чем свидетельствует значительное
количество слизеобразующих клеток в околоушной железе ребен-
ка 7 лет (рис. 2.181, Г, II).
319
Рис. 2.183. Миндалина.
1 — многослойный плоский неороговевающий эпителий; 2 — крипта; 3 — собст-
венная пластинка слизистой оболочки; 4 — лимфатические узелки; 5 — гладкие
миоциты; 6 — подслизистая основа; 7 — слюнные железы; 8 — кровеносный сосуд.
2.13.1.4. Миндалины
Данный орган представляет собой складку слизистой оболочки
(крипту), покрытую многослойным плоским эпителием (местами
инфильтрированным лимфоцитами). Под эпителием залегает во-
локнистая соединительная ткань — собственная пластинка и рас-
положенные в ней скопления лимфатических узелков — главный
элемент органа. В миндалине, как и в языке, нет четкого разделе-
ния между слизистой оболочкой и подслизистой основой. В под-
слизистой основе располагаются слюнные железы, а также можно
определить кровеносные сосуды (рис. 2.183).
2.13.1.5. Общий план строения пищеварительного тракта
Поскольку все органы пищеварительной системы в эмбриогенезе
формируются из пищеварительной трубки, а последняя в постна-
тальном онтогенезе представляет морфофункциональную основу
320
Рис. 2.184. Строение пищеварительной трубки (схема).
I — слизистая оболочка; II — подслизистая основа; III — мышечная оболочка;
IV — наружная оболочка.
А — пищевод. Б — желудок. В — тонкая кишка. Г — толстая кишка.
1 — эпителий; 2 — собственная пластинка слизистой оболочки; 3 — мышечная
пластинка слизистой оболочки; 4 — желудочные ямочки; 5 — кишечные ворсинки;
6— кишечные железы (крипты); 7 — одноклеточные внутриэпителиальные же-
лезы; 8 — желудочные железы; 9 — железы в подслизистой основе; 10 — лимфа-
тические узелки; 11 — кровеносные сосуды; 12 — лимфатические сосуды; 13 —
подслизистое нервное сплетение; 14 — межмышечно-кишечное нервное сплетение;
15 — подсерозное нервное сплетение; 16 — внекишечный проток железы; 17 —
серозная оболочка (а — брыжейка); 18 — адвентициальная оболочка.
всей системы, необходимо начать изучение темы с усвоения зако-
номерностей общего плана ее тканевой организации и наличия
органных различий (рис. 2.184).
Оболочки стенки пищеварительного канала различаются по
тканевому составу и сложности строения. Особенно это относится
к слизистой оболочке, являющейся ведущей в обеспечении пище-
варения. Она образована эпителием, собственной и мышечной
пластинками и более разнообразна по структурной организации.
Так эпителий может быть многослойным или однослойным. По-
гружаясь в стенку пищеварительной трубки на различную глуби-
11—321
321
ну, он формирует ямки, железы и крипты, а вместе с выростами
собственной пластинки — ворсинки. Покровный эпителий слизистой
оболочки желудка состоит полностью из одноклеточных желез,
а в кишечнике содержит отдельные гландулоциты. Наличие в соб-
ственной пластинке лимфатических узелков подчеркивает преоб-
ладание в ее структуре ретикулярной ткани (участие в иммунной
защите организма). Мышечная пластинка слизистой оболочки
представлена двумя (в желудке — тремя) слоями гладких миоци-
тов. Подслизистая основа, располагающаяся между слизистой и
мышечной оболочками и содержащая большое количество рыхлой
волокнистой соединительной ткани, обеспечивает подвижность
слизистой оболочки. Мышечная оболочка состоит из двух (в же-
лудке — из трех) слоев гладких миоцитов (исключение составляет
верхняя треть пищевода, где данная оболочка представлена ске-
летной мышцей). Серозная оболочка, состоящая из мезотелия
и тонкого слоя волокнистой соединительной ткани, покрывает
лишь органы, находящиеся в брюшной полости, способствуя их
взаимному скольжению.
Изображение кровеносных и лимфатических сосудов, нервных
ганглиев и обилия желез на всех фрагментах схемы подчеркивает
важную их роль в обеспечении оптимального функционирования,
а также отражает преимущественную локализацию перечисленных
структур.
2.13.1.6. Пищевод
Изучение пищевода целесообразно начать с рассмотрения компо-
нентов слизистой оболочки (рис. 2.185, А). Внутренняя поверх-
ность органа покрыта многослойным плоским неороговевающим
эпителием. Под ним расположена зона рыхлой волокнистой тка-
ни — собственная пластинка, и подстилающие ее пучки гладких
миоцитов — мышечная пластинка слизистой оболочки. Следующая
оболочка — подслизистая основа — выглядит как более широкая
зона рыхлой волокнистой ткани, в которой расположены концевые
отделы собственных желез — серозных и слизистых (рис.
2.185, Б). Мышечная оболочка прослеживается хорошо в виде
Рис. 2.185. Пищевод.
А — средний отдел. Б — фрагмент (поперечный срез). В—развитие пищевода
(по Л. И. Фалину): а — эмбрион человека 8 нед; б— эмбрион человека 16 нед.
Г — переход пищевода в желудок (по И. В. Алмазову, Л. С. Сутулову): I — сли-
зистая оболочка; II — подслизистая основа; III — мышечная оболочка; IV: а — ад-
вентициальная, б — серозная оболочка.
1 — многослойный плоский неороговевающий эпителий; 2 — собственная пластин-
ка; 3 — мышечная пластинка; 4 — собственные железы пищевода; 5 — проток
железы; 6 — внутренний и 7 — наружный слои мышечной оболочки; 8 крове-
носный сосуд; 9 — однослойный призматический железистый эпителий; 10 — же-
лудочные ямочки; 11 — кардиальные железы; 12 — мышечно-кишечный нервный
узел.
322
11*
323
Рис. 2.185. Продолжение.
двух довольно широких слоев мышечной ткани, срезанной попе-
речно и продольно. Тип ткани зависит от уровня органа, на кото-
ром сделан срез (в верхней трети поперечнополосатая, на осталь-
ном протяжении — гладкая), а структурная характеристика сло-
ев — от направления среза по отношению к длинной оси органа.
Поперечный внутренний (циркулярный) слой представлен про-
дольным сечением миоцитов (или мышечных волокон), а наруж-
ный (продольный) — срезанными поперечно. Адвентиция выгля-
дит как тонкая оболочка из волокнистой ткани, покрывающая
орган снаружи.
Развитие пищевода представлено на рис. 2.185, В. Пищевод
закладывается в виде короткой трубки из отдела передней кишки,
лежащего каудальнее трахеального желобка. На ранней стадии
(1*/2 мес) полость закладки органа покрыта высоким цилиндри-
ческим эпителием, мезенхимная оболочка развита слабо. На более
поздней стадии (4 мес) выстилка органа представлена многоряд-
324
4
2
11
ным мерцательным эпителием, который затем преобразуется в
многослойный неороговевающий. Под эпителием залегают собст-
венная и мышечная пластинки слизистой оболочки.
При изучении перехода пищевода в желудок (рис. 2.185, Г)
обращает на себя внимание резкое (скачкообразное) изменение
структурной организации слизистой оболочки пищеварительного
канала. Так, гладкая поверхность многослойного плоского неоро-
говевающего эпителия пищевода в желудке заменяется однослой-
нь!м_^1ризматическим эпителием, образующим^углубления — же-
лудочные^ ямочки^ В собственной пластинке слизйстой оболочки
пищевода появляются железы, сходные по строению с таковыми
в собственной пластинке стенки желудка — кардиальные железы
пищевода. Заметно увеличивается толщина собственной пластинки
слизистой оболочки желудка, в которой располагаются ее карди-
альные железы, а в месте перехода появляется дополнительный
слой пучков гладких миоцитов в мышечной пластинке. Подсли-
зистая основа заметных различий не имеет, так как и здесь могут
еще встречаться концевые отделы слизистых желез. Мышечная
325
оболочка в области перехода более толстая (место образования
сфинктера). Снаружи весь участок стенки покрыт серозной обо-
лочкой.
2.13.2. Средний отдел
2.13.2.1. Желудок
Анатомическое подразделение желудка на отделы представлено
на схеме (рис. 2.186).
Поскольку функции и строение стенки желудка в разных его
отделах отличаются (особенно по составу желез), обычно изучают
два отдела — дно желудка и его пилорический отдел. .Слизистая
оболочка дна желудк а покрыта эпителием, усеяна ямочка-
ми, в которые впадают вертикально ориентированные трубчатые
клеточные тяжи — собственные железы, занимающиедщактически
цсю собственную пластинку, лёжащую на трехслойной мышечной
гГластйнке. Расположенная под ней волокнистая зона — подсли-
з стая основа, за которой следует более компактная широкая, рав-
номерно окрашенная мышечная оболочка (также трехслойная),
между слоями которой находится мышечно-желудочный нервный
узел (рис. 2.187, А, Д). Серозная оболочка всех органов полости
выглядит однотипно. Более детально строение слизистой оболочки
желудка можно рассмотреть на рис. 2.187, Б. По сюжетам, представ-
ленным на рис. 2.187, Б, Г, можно убедиться в секреторной природе
всех клеток покрывающего ее однослойного цилиндрического эпи-
телия (продуцируют мукопротеины — протектор слизистой от хи-
мических и механических повреждений). На рис. 2.187, В представ-
Рис. 2.186. Отделы желудка
(схема).
1 — пищевод; 2 — кардия; 3 — дно; 4 —
тело; 5 — преддверие; 6 — привратник;
7 — двенадцатиперстная кишка.
326
Рис. 2.187. Желудок, дно.
А — общий вид. Б — гликопротеиды в поверхностных эпителиоцитах (ШИК-реак-
ция). В — СЭМ поверхности слизистой оболочки (по В. А. Шахламову). Г — ТЭМ
поверхностного эпителиоцита (по Я. Р. Мацюку). Д — мышечно-желудочный
нервный узел (импрегнация серебром).
I — слизистая оболочка; II — подслизистая основа; III — мышечная оболочка;
IV — серозная оболочка.
1 — эпителий; 2 — желудочная ямочка; 3 — собственная железа.
Пластинки слизистой оболочки: 4 — собственная; 5 — мышечная, 6 — кровенос-
ный сосуд. Слои мышечной оболочки: 7 — косой, 8 — циркулярный, 9 — продоль-
ный, 11р — мышечно-желудочный нервный узел, 11—шеечные мукоциты, 12 —
лавные экзокриноциты. 13 — париетальные экзокриноциты, 14 — капилляр; а —
эндоплазматическая сеть, б — митохондрии, в — секреторные гранулы, г — плот-
ный контакт, j — нейрон.
Рнс. 2.187. Продолжение.
лена СЭМ слизистой оболочки дна желудка, на которой хорошо
видны поверхность эпителия и желудочные ямочки с открывающи-
мися в них железами. В собственной железе при обычной окраске
можно дифференцировать только париетальные и главные экзокри-
ноциты. Первые по оксифильной окраске, более крупным размерам
и округлой форме (преобладают в области шейки и тела железы),
вторые — меньших размеров, окрашены базофильно и составляют
большинство клеток железы. Под дном железы залегает мышечная
пластинка.
Для более углубленного изучения строения собственных желез
целесообразно вначале рассмотреть схему (рис. 2.188, AI) с ука-
занием типов гландулоцитов и их топографии в железе, затем на
рис. 2.188ГАП — обзорную ТЭМ, демонстрирующую взаимоотно-
шения гландулоцитов в железе, а на рис. 2.188, Б—Д — ТЭМ их
ультраструктурной организации. При этом необходимо обратить
внимание на существенные различия в их строении, связанные с
функциональной принадлежностью. В главном экзокриноците от-
четливо выражены все ультрастр^ктурныё признаки гландуло-^
1 цита,“интенсивно’~зарабатывающего’ белковый секрет. В шеечном
Й8
мукоците и эндокриноците органеллы белкового синтеза менее
развиты и другой характер гранул. Наибольшей спецификой стрр-
~7ения обладает -париетальный экзокрйноцит. Поскольку это" основ-
ной продуцент соляной кислоты — клетка лишена комплекса
Гольджи, практически отсутствует эндоплазматическая сеть, но
зато много внутриклеточных канальцев (для выведения соляной
329
Рис. 2.188.
330
331
Рис. 2.188. Собственная железа желудка.
AI, АН — строение собственной железы желудка: 1 — желудочная ямочка, 2 —
перешеек, 3 — шейка, 4 — главная часть (тело и дно), 5 — поверхностный эпи-
телиоцит, 6 — недифференцированный эпителиоцит (митоз), 7 — париетальный
экзокриноцит, 8 — шеечный мукоцит, 9 — главный экзокриноцит, 10 — желудоч-
но-кишечный эндокриноцит.
Ультраструктурная организация различных типов гландулоцитов: Б1, БП — глав-
ный экзокриноцит; BI, ВП — париетальный экзокриноцит; Г1, ГН — шеечный му-
коцит; Д1, ДП — желудочно-кишечный эндокриноцит; а — эндоплазматическая
сеть, б — комплекс Гольджи, в — митохондрии, г — секреторные гранулы, д —
внутриклеточный каналец, е — тубуловезикулы, ж — границы клетки.
кислоты) и митохондрий (процесс образования кислоты — энер-
гоемкий).
Основной отличительной чертой в строении слизистой оболоч-
ки пилорического отдела желудка являются более
широкие иглубокие желудочные ямочки. Сами железы выглядят
как косо и поперечно срезанные трубочки (следствие разветвлен-
ности их концевых отделов), большего диаметра с выраженным
просветом. Расположены они значительно реже, благодаря чему
отчетливо видна ткань собственной пластинки (рис. 2.189). При
этом клетки выглядят однотипно — в подавляющем их большин-
стве цитоплазма окрашена светло и однородно, а ядра деформи-
рованы и оттеснены к базальной мембране. Однако следует по-
мнить, что как по количеству, так и по качеству вырабатываемых
гормонов эндокриноцитов здесь больше, чем в собственных же-
лезах.
332
Рис. 2.189. Желудок (пило-
рический отдел).
I — слизистая, II — подслизис-
тая основа, III — мышечная обо-
лочка, IV — серозная оболочка.
1 — эпителий; 2 — желудочная
ямочка; 3 — собственная плас-
тинка; 4 — железы; 5 — мы-
шечная пластинка.
Рис. 2.190. Развитие желудка
человека в области дна.
А — эмбрион 8 нед. Б — эмбри-
он 24 нед.
1 — эпителий; 2 — закладка же-
лудочных ямочек; 3 — мезенхи-
ма; 4 — мышечная оболочка; 5 —
собственные железы; 6 — мы-
шечная пластинка; 7 — подсли-
зистая основа; 8 — дифференци-
рующаяся соединительная ткань.
333
Развитие желудка. Желудок закладывается в виде веретено-
образного расширения передней кишки. На ранней стадии (8 нед)
он покрыт ложномногорядным эпителием, который, врастая в под-
лежащую мезенхиму, образует закладки желудочных ямочек; мы-
шечная пластинка еще отсутствует, по периферии органа идет
формирование мышечной оболочки (рис. 2.190, А). На более позд-
ней стадии (24 нед) в составе слизистой оболочки желудка уже
отчетливо прослеживаются все ее структурные компоненты —
ямки, железы, собственная и мышечная пластинки. Выявляемая
в клеточных элементах желез активность лактатдегидрогеназы
(одного из ключевых биоэнергетических ферментов) свидетель-
ствует о достаточно высоком уровне их дифференцировки
(рис. 2.190, Б).
2.13.2.2. Тонкая кишка
Иное функциональное назначение органа (химическая переработ-
ка и всасывание пищевых веществ) обусловливает значительную
перестройку его слизистой оболочки. Резкое изменение ее рель-
334
Рис. 2.191. Тонкая кишка.
А — схема строения стенки. Б — СЭМ поверхности слизистой оболочки (по
В. А. Шахламову). В — поперечный срез стенки. Г — кишечная ворсинка. Д —
СЭМ кровеносного русла кишечной ворсинки (коррозионный препарат по А. А. Ми-
ронову и А. В. Миронову). Е — ТЭМ эпителия кишечной ворсинки (по В. А. Шах-
ламову). Ж — бокаловидный экзокриноцит (гистологический препарат, ШИК-
реакция). 3 — СЭМ поверхности слизистой над сгруппированным лимфатическим
узелком (по Ю. А. Гайдару). И — схема иммунной системы кишечника.
1 — слизистая оболочка; II — подслизистая основа; III — мышечная оболочка;
IV — серозная оболочка.
1 — однослойный призматический каемчатый эпителий; 2 — собственная пластин-
ка слизистой оболочки; 3 — кишечная ворсинка; 4 — кишечная железа (крипта);
5 -— лимфатический узелок; 6 — мышечная пластинка слизистой оболочки; 7 —
циркулярный и 8 — продольный слои мышечной оболочки; 9 — кровеносный сосуд;
10 — бокаловидный экзокриноцит; 11 —лимфоцит; 12 — купол; 13 — волокнистые
и клеточные компоненты соединительной ткани; 14 — миоцит; 15 — кровеносные и
16—лимфатические микрососуды кишечной ворсинки; 17 — регионарный лимфа-
тический узел: 18 — грудной лимфатический проток; 19 — общее кровеносное
русло; 20 — плазмоцит; 21 — М-клетка; 22 — сенсибилизированный лимфоцит;
a -L мукоидный секрет, б — микроворсинки, в — антиген.
335
336
Рис. 2.191. Продолжение.
I
337
ефа наглядно можно представить, сравнивая СЭМ поверхностей
слизистых оболочек желудка (см. рис. 2.187, В) и тонкой кишки
(рис. 2.191, Б). Цоследняя образована пальцевидными выпячи-
ваниямш_слизцстой ободочки^кишечными ворсинками^ _значи-
телыю увели чивающими гьтощадь контакта пищевых веществ (нут-
риентов)£ п<>верхностьк)_слизис10Й—оболбчки (рис. 2.191, А, Б).
На рис. 2.1917Вможно видеть ее оболочки и рассмотреть особен-
ности их строения. В слизистой оболочке пальцевидные выпячи-
вания эпителия и собственной пластинки образуют кишечные вор-
синки, а погружения эпителия до уровня мышечной пластинки
формируют кищечдашкрипты (в собстве11ной пластинке могут
встречаться лимфатическиеузелки). Следует отметить, что мы-
шечная пластинка и мышечная оболочка тонкой кишки в отличие
от таковых желудка состоят из двух слоев. При более подробном
изучении строения ворсинки можно убедиться, что ее стенку со-
ставляет однослойный призматический эпителий, состоящий из
микроворсинчатых эпителиоцитов и содержащий бокаловидные
экзокриноциты (рис. 2.191, Г). В собственной пластинке непосред-
ственно под эпителием залегает сильно развитая капиллярная
сеть (см. СЭМ коррозионного препарата верхушки кишечной вор-
синки — рис. 2.191, Д), а также лимфатические сосуды, между
которыми находятся волокнистые и клеточные структуры соеди-
нительной ткани и гладкие миоциты. Электронно-микроскопиче-
ское изображение участка эпителия ворсинки представлено на
схеме (см. рис. 2.192, А) и ТЭМ (см. рис. 2.192, Б и 2.191, Е). На
этих рисунках видно, что микроворсинчатые эпителиоциты и бо-
каловидные экзокриноциты лежат на тонкой базальной мембране,
причем первые имеют призматическую форму (с ядром в середи-
не клетки), вторые — форму бокала (с ядром в базальном полю-
се). Апикальная плазматическая мембрана микроворсинчатых
эпителиоцитов образует многочисленные пальцеобразные вырос-
ты — микроворсинки. В цитоплазме этих клеток можно видеть
митохондрии и другие органеллы. В апикальном полюсе бокало-
видных экзокриноцитов наблюдается скопление гранул мукоид-
ного секрета, который можно наблюдать в окрашенном виде на
рис. 2.191, Ж (фрагменты рис. 2.191,3, И см. далее в статье «Ап-
парат иммунной защиты»),
В пространстве между клетками эпителия находится межэпи-
телиальный лимфоцит. Непосредственно под базальной мембраной
эпителия залегают кровеносные микрососуды. Поскольку именно
микроворсинчатый эпителиоцит — основная клетка эпителия,
обеспечивающая главные функции тонкой кишки, рассмотрим
схему его ультраструктурной организации и межклеточные взаи-
моотношения (рис. 2.192, А). При этом особое внимание следует
обратить на специализацию апикального отдела. Микроворсинки,
образующие щеточную кайму, не только увеличивают всасываю-
щую поверхность клетки, но и, будучи покрыты развитым глико-
каликсом, обеспечивают мембранное пищеварение, сопряженное
со всасыванием, а фибриллярный компонент служит не только
338
Рис. 2.192. Ультраструктурная
организация эпителиоцитов.
А — схема. Б — ТЭМ апикальной
части контактирующих клеток (по
К. А. Зуфарову). В — ТЭМ слоя
слизистых наложений (по И. А. Мо-
розову) .
1 — микроворсинка: а — гликока-
ликс, б — фибриллярные структуры,
в — терминальная сеть; 2 — плот-
ное соединение; 3 — зона прилежа-
ния; 4 — десмосома; 5 — пиноци-
тозные везикулы; 6 — лизосомы;
7 — фаголизосома; 8 — хиломикро-
ны; 9 — эндоплазматическая сеть;
10 — митохондрия; 11 — комплекс
Гольджи; 12 — отросток энтероци-
та; 13 — базальная мембрана; 14 —
мембранные везикулы.
каркасом, но и, взаимодей-
ствуя с филаментами терми-
нальной сети, способствует их
сокращению. Исследования-
ми последних лет показано,
что эпителий кишечных вор-
синок покрыт мощным слоем
слизистых наложений, в со-
став которого входят глико-
протеиды (продуцируемые
бокаловидными экзокриноци-
тами), кишечные и панкреа-
тические ферменты, субстрат-
связывающие белки. Ультра-
структура слоя слизистых на-
ложений характеризуется
значительной гетероген-
ностью: в нем встречаются
фрагменты слущенных кле-
ток, мембранные везикулы
(отпочковавшиеся от микро-
ворсинок) , лимфоидные клет-
ки, пищевые частицы, бакте-
рии (рис. 2.192, В). Функцио-
нальное значение данного
слоя заключается в том, что в
нем осуществляется опреде-
ленный этап химической об-
работки пищевых веществ,
занимающий промежуточное
положение между полостным
и пристеночным пищеварени-
339
Рис. 2.192. Продолжение.
ем (слизистое пищеварение). Сложная система межклеточных свя-
зей в апикальном отделе в свою очередь обеспечивает герметизацию
пласта (плотное соединение) и его механическую прочность (зона
прилежания и десмосомы). Отсутствие же таковых на остальных
участках боковых поверхностей способствует выделению из клет-
340
7
Рис. 2.193. Кишечная железа (крипта).
А — схема строения. Б — препарат. В — препарат (дно железы, гистохимическая
реакция на цинк). Г — ТЭМ участка дна железы (по А. Юлдашеву). Д — ТЭМ
эндокриноцита открытого типа (по К. А. Зуфарову).
I — основание кишечной ворсинки; П — кишечная железа.
1 — микроворсинчатый эпителиоцит; 2 — бокаловидный экзокриноцит; 3 — эндо-
криноцит закрытого типа; 4 — эндокриноцит открытого типа; 5 — недифференци-
рованный (делящийся) эпителиоцит; 6 — экзокриноциты с ацидофильными гра-
нулами; 7 — соединительная ткань: а — секреторные гранулы, б — комплекс
Гольджи, в — митохондрии, г — просвет железы.
341
ки части продуктов через межклеточные щели. Базальные отрост-
ки, проникающие через базальную мембрану, облегчают миграцию
энтероцитов к вершине ворсинки. Крипта в отличие от ворсинки
имеет более сложный клеточный состав (рис. 2.193, А). При обыч-
ной окраске препарата видны только два типа клеток — микро-
ворсинчатые эпителиоциты и бокаловидные экзокриноциты
(рис. 2.193, Б). С остальными типами клеток, выявленными с по-
мощью специальных методов, можно ознакомиться на схеме (см.
рис. 2.193, А). Некоторые из указанных в ней клеточных форм
представлены на электронно-микроскопических изображениях
крипты (рис. 2.193, Б, Г), в частности недифференцированные
эпителиоциты, экзокриноциты с ацидофильной зернистостью, эн-
Рис. 2.194. Двенадцатиперстная кишка.
I — слизистая оболочка; II — подслизистая основа; III — мышечная оболочка.
1 — кишечная ворсинка; 2 — однослойный призматический микроворсинчатый
эпителий; 3 — собственная пластинка слизистой оболочки; 4 — кишечная железа;
5 — мышечная пластинка слизистой оболочки; 6 — дуоденальные железы.
342
Рис. 2.195. Развитие тонкой кишки человека.
А — эмбрион 5—6 нед. Б — эмбрион 7—8 нед (ШИК-реакция).
1 — просвет кишки; 2 — эпителий; 3 — мезенхима; 4 — циркулярный и 5 — про-
дольный слои мышечной оболочки; 6 — начало формирования кишечных ворсинок.
343
докринные клетки. Последние по своей общей морфологической
характеристике сходны с таковыми в желудке, но большинство
их отличается по гормональному профилю, а следовательно, и по
ультраструктуре гранул. На рис. 2.193, Д можно видеть эндокри-
ноцит открытого типа (контактирует с содержимым полости киш-
ки). Принципиально новый тип клеток, встречающихся группами
на дне крипт только тонкой кишки, — экзокриноциты с ацидо-
фильными гранулами. На рис. 2.193, В представлена микрофото-
графия дна крипты с такими гландулоцитами, содержащими в
гранулах цинк-положительный материал (одна из функций —
экскреция цинка из организма). Расположенные рядом безкаем-
чатые клетки — недифференцированные эпителиоциты на разных
стадиях дифференцировки. Последние по мере миграции в направ-
лении ворсинки дифференцируются, давая все типы зрелых кле-
ток (аналогичны шеечным клеткам желез желудка, но месторас-
положение иное).
Главным отличительным морфологическим признаком две-
надцатиперстной кишки является наличие дуоденаль-
ных желез, лежащих в подслизистой основе (рис. 2.194). Следует
обратить внимание на меньшее количество бокаловидных экзокри-
ноцитов в эпителии ворсинок.
Развитие тонкой кишки. Первоначально кишка закладывается
как эпителиальная трубка, которая позже одевается висцеральным
листком спланхноплевры, за счет которого формируется мезен-
химная оболочка и мезотелий. На стадии 5—6 нед (рис. 2.195, А)
можно уже различить эпителиальную выстилку, за ней зону рас-
положения мезенхимы, окруженную обозначившейся мышечной
оболочкой, дифференцированной на циркулярный и продольный
слои. Обращает на себя внимание значительная толщина эпите-
лия, существенно суживающего просвет трубки и нарушение в нем
рядности ядер (результат опережающего рост трубки размноже-
ния клеток эпителия). На стадии 7—8 нед (рис. 2.195, Б) уже
четко обозначается начало формирования кишечных ворсинок
путем врастания мезенхимы, в результате чего просвет трубки
приобретает складчатый характер. Мышечная оболочка заметных
изменений не претерпевает. Окончательного формирования сли-
зистой оболочки на данной стадии еще не происходит.
2.13.2.3. Толстая кишка
Отличительные особенности строения стенки толстой кишки в пер-
вую очередь касаются слизистой оболочки (рис. 2.196, А, Б). Ее
рельеф формируют исключительно кишечные крипты. В эпителии
значительно возрастает количество бокаловидных экзокриноцитов
и уменьшается число микроворсинчатых эпителиоцитов. Лимфа-
тические узелки расположены не только в собственной пластин-
ке, лимфоциты инфильтрируют дно крипт и «прорываются» час-
тично сквозь мышечную пластинку в подслизистую основу. В на-
344
Рис. 2.196. Толстая кишка.
А — схема. Б — препарат. В — раз-
витие толстой кишки.
I — слизистая оболочка; П — под-
слизистая основа; III — мышечная
оболочка; IV — серозная оболочка.
1 — эпителий: а — микроворсинча-
тый эпителиоцит, б — бокаловидный
экзокриноцит; 2 — собственная
пластинка слизистой оболочки; 3 —
кишечная крипта; 4 — мышечная
пластинка слизистой оболочки; 5 —
лимфатический узелок; 6 — крове-
носный сосуд; 7 — просвет кишки;
8 — закладка крипты; 9 — мезен-
хима.
345
ружном слое мышечной оболочки образуются вдавления и он при-
обретает складчатый характер.
Развитие толстой кишки эмбриона человека 9—10 нед пред-
ставлено на рис. 2.196, В. Уже видно формирование крипт и двух-
слойной мышечной оболочки. Однако мышечная пластинка сли-
зистой оболочки еще не формируется и все пространство от эпи-
телия до мышечной оболочки занято мезенхимой.
При ознакомлении с микроскопическим строением червеобраз-
ного отростка бросается в глаза обилие лимфатических узелков,
преимущественно крупных размеров, некоторые из которых могут
занимать значительные территории (рис. 2.197, А, Б). Из-за этого
и отсутствия четко оформленной мышечной пластинки слизистой
оболочки дифференцировать последнюю от подслизистой основы
практически не удается. Лимфатические узелки нарушают также
регулярное расположение кишечных крипт. Необходимо отметить,
что в отличие от толстой кишки в эпителии червеобразного от-
ростка преобладают микроворсинчатые эпителиоциты, а мышеч-
ная оболочка сходна по строению с таковой у тонкой кишки. Оби-
лие лимфоидной ткани в червеобразном отростке некоторые ис-
следователи связывают с возможностью выполнения им роли
центра созревания В-лимфоцитов.
Развитие червеобразного отростка. У новорожденного лимфо-
цитов в соединительной ткани отростка немного и расположены
они диффузно. В эпителии и криптах много бокаловидных экзо-
криноцитов. Остальные компоненты стенки в основном уже сфор-
мированы (рис. 2.197, Ва). После рождения лимфоидный аппарат
активно формируется. На рис. 2.197, Вб червеобразный отросток
ребенка 2 лет. Обращает на себя внимание значительное развитие
лимфатических узелков.
Аппарат иммунной защиты. Важной морфофункциональной
особенностью кишечника является наличие у него сильно разви-
того аппарата иммунной защиты, что связано с присутствием в
пище разнообразных чужеродных веществ, антигенов или ксено-
биотиков. Основными его структурными элементами являются
межэпителиальные лимфоциты, лимфоциты собственной пластин-
ки слизистой оболочки (располагающиеся диффузно или органи-
зованные в лимфоидные фолликулы), лимфатические узлы. На
СЭМ (см. рис. 2.191, 3) представлен участок слизистой оболочки
тощей кишки в виде купола, окруженного кишечными ворсинками,
расположенного над лимфатическими узелками, куда и мигрируют
Рис. 2.197. Червеобразный отросток.
А — схема. Б — препарат. В — развитие червеобразного отростка человека: Ва —
новорожденный, Вб — ребенок двух лет.
1 — эпителий; 2 — кишечная крипта; 3 — собственная пластинка слизистой обо-
лочки; 4 — лимфатические узелки; 5 — подслизистая основа; 6 — циркулярный
и 7 — продольный слои мышечной оболочки; 8 — серозная оболочка; 9 — крове-
носный сосуд.
346
347
внутриэпителиальные лимфоциты после антигенной стимуляции,
«запуская» иммунный ответ. С принципиальной схемой функцио-
нирования иммунной системы кишечника можно ознакомиться
на рис. 2.191, И.
2.13.3. Пищеварительные железы
2.13.3.1. Поджелудочная железа
>терией (надежный критерий
Гкапрбтока). При более
Несмотря на смешанный экзо-эндокринный характер, орган по
своей общей структурной организации имеет сходство с околоуш-
ной железой, поскольку основу составляет экзокринная часть,
вырабатывающая белковый секрет. .Последняя имеет дольчатое
строение. Долька включает_4? себя ацинусы, вставочные _и_внутри-
дольковые протоки. Панкреатические островки выглядят как ком-
пактаиё^бледноокрашенные образования, состоящие из различных
клеток (рис. 2.198, А). В соединительнотканных перегородках
между дольками располагаются кровеносные сосуды и междоль-
ковые протоки. Важно отметить, что ^независимо отдиаметра про-
токи всегда выстланы однослойным кубическим или призматиче-
ским :шителиегуГ2Д признак энтбдермального происхождения).
1Г~стенке рудных протоков встречаются гладкие—миоциты, что
придает им внешнее^~СХодсТвб- С—др- ; “ ‘
дифференцировки — эпителиальная выс
подробном рассмотрении строения ацинуса обращает на себя вни-
мание различие в окраске апикальной и базальной частей экзо-
кринных панкреацитов. Первая выглядит светлой и содержит окси-
фильную зернистость (зимогенные гранулы секрета), вторая —
темной (окрашенная базофильно-зернистая эндоплазматическая
сеть) (рис. 2.198, Б). Обращает на себя внимание, что соотноше-
ние этих зон в ацинусах и даже отдельных клетках неодинаково
(результат различного их функционального состояния).^В цент-
раддной части ацинуса часто встречаются одиночные ядра светло-
окрашенных клеток — центроацинозныё кледжц: Экзокринным
па^креацита1^~ггрису1ци^основ11Ь1е признаки белоксинтезирующих
Рис. 2.198. Поджелудочная железа.
А — общий вид. Б — фрагмент к А (препарат). В — СЭМ ацинуса (по А. А. Ми-
ронову и В. А. Миронову). Г — ТЭМ экзокринного панкреацита (по К. А. Зуфа-
рову) . Д — ТЭМ центроацинозного эпителиоцита. Е — ТЭМ нервно-сосудистого
обеспечения ацинуса.
1 — долька; 2 — концевой отдел; 3 — панкреатический островок; 4 — вставочный
проток; 5 — междольковая перегородка; 6 — междольковый проток; 7 — кровенос-
ные сосуды; 8 — центроацинозный эпителиоцит; 9 — экзокринный панкреацит;
10 — ядро; 11 — зимогенные гранулы; 12 — гранулярная эндоплазматическая сеть;
13 — митохондрия; 14 — комплекс Гольджи; 15 — межклеточное соединение; 16 —
просвет ацинуса; 17 — гемокапилляр: п — просвет, э — эндотелиоцит; 18 — ба-
зальная мембрана; 19 — безмиелиновое нервное волокно: а — отросток леммоцита,
б — «оголенный» участок аксона, в — скопление пресинаптических везикул.
348
1
349
350
Рис. 2.198. Продолжение.
351
Рис. 2.199. Панкреатический ост-
ровок.
А — схема строения. Б — препарат (ок-
раска альдегид-фуксином), бета-инсу-
лоциты сиреневые. В — кровоснабжение
островка (инъекция тушь-желатина,
препарат И. А. Михайлюка). Г — ТЭМ
ацино-островковой клетки типа А. Д —
ТЭМ разных типов панкреатических
инсулоцитов: I — альфа-эндокриноцит,
II — бета-эндокриноцит, III — дельта-
эндокриноцит, IV — ПП-эндокриноцит.
1 — панкреатические эндокриноциты;
2 — гемокапилляры островка; 3 — бета-
эндокриноциты; 4 — другие эндокрино-
циты; 5 — экзокринный панкреацит;
6 — зимогенные гранулы; 7 — альфа-
гранулы; 8 — прекапиллярная артерио-
ла; 9 — капилляры, окружающие аци-
нусы.
352
CD
Рис. 2.199. Продолжение.
гландулоцитов: выраженность зернистой эндоплазматической сети,
комплекса 1 ольджи, зимогенные гранулы (рис. 2.198, В, Г). Суще-
ственное структурное отличие экзокринного отдела паренхимы
поджелудочной железы от слюнных желез — отсутствие миоэпи-
телиальных клеток (признак эктодермального происхождения),
исчерченности протоков и наличие центроацинозных клеток
(рис. 2.198, Д). Нервно-сосудистое обеспечение ацинуса представ-
лено на рис. 2.198, Е.
Изучение строения панкреатического островка целесообразно
начать с рассмотрения его схематического изображения (рис.
2.199, А) и гистологического препарата (рис. 2.199, Б), из которого
следует, что панкреатические эндокриноциты располагаются не-
упорядоченно (диффузный тип строения). Между ними находятся
полости — синусоидные гемокапилляры, общая архитектоника
которых в островке представлена на рис. 2.199, В. Помимо бета-
эндокриноцитов, имеющих характерную темно-фиолетовую окрас-
ку и занимающих большую часть площади островка, все другие
типы клеток расположены в основном по его периферии. Вообще
же типы панкреатических эндокриноцитов удается четко иденти-
фицировать иммуноцитохимически или по ультраструктурной ха-
рактеристике секреторных гранул (рис. 2.199, Г). В составе ост-
ровков имеется особый вид клеток, получивших название ацино-
354
Рис. 2.200. Развитие поджелудочной железы человека.
А — эмбрион 8 нед. Б — плод 17 нед (общий вид).
1 — закладка железы; 2 — эпителиальные трубочки; 3 — мезенхима; 4 — кишечная
трубка; 5 — панкреатический островок; 6 — ацинус; 7 — проток; 8 — соединитель-
ная ткань; 9 — кровеносный сосуд.
12*
355
островковых (смешанных), характеризующихся одновременным
содержанием зимогенных (крупных) и эндокринных (мелких)
гранул (рис. 2.199, Д). При этом последние подразделяют на со-
ответствующие типы по характеру эндокринных гранул (на рис.
показан тип А). Функция этих клеток окончательно не установ-
лена.
Развитие поджелудочной железы. Зачаток железы появляется
в конце 3-й недели эмбриогенеза в виде двух выростов эпители-
альной (энтодермальной) выстилки развивающейся двенадцати-
перстной кишки, из которых на 3-м месяце дифференцируются
экзокринный и эндокринный отделы органа. На 8-й неделе в зачатке
железы наблюдается формирование эпителиальных трубочек, об-
разование ацинусов и панкреатических островков еще не началось
(рис. 2.200, А). Через 17 нед в органе можно увидеть дифферен-
цирующиеся ацинусы и панкреатические островки. Отмечается
также обилие соединительной ткани, в которой видны протоки,
дольчатость железы не выражена (рис. 2.200, Б). С помощью гис-
тохимической реакции удается выявить секреторный продукт
экзокринных панкреацитов ацинуса, содержащий гликопротеиды,
что свидетельствует об их незрелости.
2.13.3.2. Печень
Этот орган занимает особое положение среди всех желез (не
только пищеварительных). Размещаясь на «стратегических» путях
оттока крови от непарных органов брюшной полости и нижней
полой вены и пропуская ее через себя, печень выполняет не толь-
ко (и даже не столько) железистую функцию, но и множество
других (метаболическую, защитную, депонирующую и др.). Это
находит свое отражение и в особой организации долек органа,
основу архитектоники которых определяет конструкция сосудис-
того русла. Поэтому вначале целесообразно ознакомиться с общей
схемой его организации и классической дольки в целом по схеме,
представленной на рис. 2.201, А. При этом необходимо обратить
внимание на следующие особенности: углы долек формируются
сосудисто-протоковым комплексом1 — печеночной триадой, от
каждого компонента которой отходят горизонтальные ветви (уча-
ствующие одновременно в образовании периметра трех смежных
долек) — вокругдольковые сосуды и протоки. Ответвления вокруг-
дольковых вен и артерий, вступая в дольку, образуют сложную,
анастомозирующую сеть синусоидных сосудов (сходящихся в
центральной зоне), между которыми расположены разнообразные
по форме и количеству гепатоцитов печеночные пластинки.
В связи *с тем что в печени человека слабо развита междоль-
ковая соединительная ткань и дольки отчетливо не выявляются,
1 Сосудисто-протоковый комплекс включает в себя желчный проток, артерию, вену
и лимфатический сосуд; ввиду того что последний выявляется редко, весь комп-
лекс традиционно именуют триадой.
356
Рис. 2.201. Печень.
А — строение классической дольки и ее сосудистого аппарата (схема). Б — пре-
парат (печень свиньи). В — печень человека: I — общий вид дольки, II—триада.
Г — СЭМ дольки: I — печеночная пластинка, II — внутридольковые сосуды (кор-
розионные препараты по А. А. Миронову и В. А. Миронову).
1 — долевая вена; 2 — сегментарная вена; 3 — междольковые сосуды и желчный
проток; 4 — вокругдольковые сосуды и желчный проток; 5 — триада; 6 — синусо-
идные сосуды; 7 — центральная вена; 8 — поддольковая вена; 9 — печеночная
пластинка; 10—-желчный каналец; 11 — междольковая соединительная ткань:
а — вена, б — артерия, в — лимфатический сосуд, г — желчный проток (сплош-
ными стрелками обозначено направление кровотока, пунктирными — ток желчи
и лимфы).
357
7
5
Bl 10 6
358
Рис. 2.201. Продолжение.
359
Рис. 2.201. Продолжение.
представляется целесообразным начать изучение органа с печени
свиньи (рис. 2.201, Б). Здесь контуры долек четко определяются
расположением «триад» и находящимися между ними тяжами
соединительной ткани. В печени же человека (рис. 2.201, BI) гра-
ницы долек идентифицируются только по триадам, которые хо-
рошо выявляются по сочетанию выраженного просвета междоль-
ковой вены с лежащей рядом узкой трубкой (выстланной темными
круглыми ядрами) — междольковым желчным протоком и арте-
рией, а иногда и лимфатическим сосудом. Наряду с этими струк-
турами видны центральная вена (круглая или овальная полость,
расположенная в центре дольки), печеночные пластинки (клеточ-
ные тяжи, идущие от периферии к центру) и синусоидные сосуды
(щели между печеночными пластинками). Наглядно пространст-
венная организация и взаимоотношения между перечисленными
выше структурами представлены на СЭМ (рис. 2.201, Г), демон-
стрирующей разнообразие их архитектоники. При более подроб-
ном рассмотрении строения триады обращают на себя внимание
значительные различия диаметров артерии и вены (связанные со
спецификой кровоснабжения органа), а также однослойность эпи-
телия, выстилающего междольковый желчный проток (признак
эктодермального происхождения) (рис. 2.201, BIT).
360
Детальнее сведения о тонком строении печеночных пластинок
и синусоидных сосудов, обеспечивающих уникальную полифунк-
циональность органа, представлены на рис. 2.202.
У гепатоцитов хорощо_выражены практически все органеллы.
Их поверхность неоднозначна. Одни участки снаб^енымикрс вор-
синками и обращены к синусоидным сосудам^от которых _их от-
деляет перисинусоидальное пространство (пространство Диссе),
содержащёе ртдельные ретикулярные волокна, липоциты и Pit-
клетки; другие — примыкают к рядом лежащим гепатоцитам и
путем симметричных инвагинаций определенных участков плазмо-
лемм образуют желчные канальцы (рис. 2.202, А—В). «Запаиваю-
щие» их плотные контакты, а также десмосомы обеспечивают
физическую целостность пластинок и препятствуют попаданию
желчи в межклеточные щели и синусоидные сосуды. В перисину-
соидальном пространстве (вклинившись между гепатоцитами),
расположены перисинусоидальные липоциты (накапливающие
липиды и растворимые в них вещества и обладающие способ-
ностью к фибриллогенезу) (рис. 2.202, Г), функции которых изу-
чены недостаточно. Стенка синусоидных сосудов по своему строе-
нию не имеет аналогов среди гемокапилляров, так как состоит из
Рис. 2.202. Печень.
А — ультраструктурная организация гепатоцитов, их взаимосвязей с синусоидными
сосудами (схема). Б — ТЭМ гепатоцита. В — СЭМ скола участка: I — печеночная
пластинка, II — синусоидный сосуд (по А. А. Миронову и В. А. Миронову). Г —
ТЭМ участка сосуд — гепатоцит (по Л. М. Лихачевой). Д — клетка Купфера.
1 — гепатоцит; 2 — эндотелиоцит; 3 — клетка Купфера; 4 — перисинусоидальное
пространство; 5 — ретикулярное волокно; 6 — желчный каналец; 7 — плотные кон-
такты; 8 — десмосома; 9—перисинусоидальный липоцит; 10 — эритроцит; 11 —
Pit-клетка: а — ядро, б — митохондрия, в — эндоплазматическая сеть, г — комп-
лекс Гольджи, д — лизосома, е — включение гликогена, ж — микроворсинки гепа-
тоцита, з — фенестры и поры, и — липидные включения.
361
Рис. 2.202. Продолжение.
362
363
двух генетически различных типов клеток —федестриров
эндотелиоцитов и звездч^х^^лакрофа ов — .клеток Купфера и
лишена на большемгтротяжйпн! базальной мембраны (рис.
2.202, Д). Именно такая конструкция максимально способствует
активному обмену веществ между кровью и гепатоцитами и ее
«очистке» от чужеродных частиц.
Особый интерес представляет рассмотрение ультраструктурной
организации эндотелиоцитов и звездчатых макрофагов и их про-
странственных взаимоотношений. Эндотелиоциты, за исключени-
ем выступающего в просвет сосуда ядросодержащего участка,
представлены плоской удлиненной периферической зоной, снаб-
женной многочисленными фенестрами, размеры которых значи-
тельно варьируют (см. рис. 2.202, Г). Форма же клеток Купфера
и рельеф поверхности чрезвычайно разнообразны в зависимости
от локализации в дольке и функционального состояния. Различен
и характер их взаимоотношений с эндотелиоцитами: в одних слу-
чаях они как бы лежат на поверхности эндотелиоцитов, в дру-
гих — большая часть тела расположена между ними (см. рис.
2.202, Д). В последние годы выделяют особый комплекс структур,
играющий ключевую роль в обеспечении функционирования ор-
гана, в который входят клетки синусоидного сосуда, Pit-клетки,
содержимое перисинусоидального пространства с расположенны-
ми в нем лйпоцитами (перисинусоидальная функциональная еди-
ница и синусоидальный полюс гепатоцита). Поскольку представ-
ление о классической дольке не соответствует общепринятой
структуре экзокринных желез и не в состоянии объяснить ряд
аспектов гистофизиологии и патологии печени, существуют и дру-
гие подходы к определению структурно-функциональных единиц
органа. В соответствии с одним из них выделяют так называемую
портальную дольку. В ее основу положена внешнесекре-
торная деятельность органа. Трабекулы рассматриваются как
трубчатые периферические секреторные отделы, а междольковые
желчные пути (в центре) — начало протоковой системы (рис.
2.203, А). Согласно другому подходу, структурно-функциональной
единицей печени является печеночный ацинус. В основу
положены особенности сосудистой архитектоники дольки, приво-
дящие к неравномерности (количественного и качественного)
кровоснабжения различных ее зон. Диаметр вокругдольковых со-
судов в соответствии с их отдалением от триады уменьшается,
так как уменьшается количество крови. По направлению к цент-
ральной вене характер сосудов меняется качественно. Это дало
основание определить контуры дольки в виде ромба, длинная ось
которого соединяет центральные вены двух долек, и наметить
условно три зоны по степени интенсивности их кровоснабжения
(рис. 2.203, Б).
Развитие печени. Печень закладывается в виде желобообраз-
ного выпячивания утолщенного эпителия кишечной трубки, перед-
няя часть которого, разрастаясь, формирует паренхиму органа,
а задняя — желчный пузырь. Развитие печени имеет свои особен-
364
Рис. 2.203. Печень.
А — портальная долька (треугольни-
ком обозначен контур дольки). Б —
печеночный ацинус (ромбом обозначен
контур ацинуса).
1 — классические дольки; 2 — централь-
ные вены; 3 — триада (стрелками от-
мечено направление тока желчи); 4 —
вокругдольковые кровеносные сосуды:
а — зона оптимального, б — среднего,
в — наихудшего кровоснабжения (стрел-
ками отмечено направление кровотока).
Рис. 2.204. Развитие печени.
А — печень плода человека 5 мес. Б —
печень новорожденного.
1 — очаг кроветворения; 2 — комплексы
гепатоцитов.
365
ности, связанные с ее кроветворной функцией в период эмбрио-
нального развития организма. На рис. 2.204, А представлена пе-
чень плода человека 5 мес. Обращают на себя внимание много-
численные очаги кроветворения среди скоплений гепатоцитов,
еще не сформированных в пластинки. К моменту рождения очаги
кроветворения исчезают или встречаются единичные мелкие (рис.
2.204, Б). Однако пластинчатое строение паренхимы еще не вы-
ражено.
2.13.4. Желчный пузырь
Характерной особенностью структурной организации желчного
пузыря является наличие слизистой оболочки, которая благодаря
обилию анастомозирующих между собой складок не имеет строгой
упорядоченности (рис. 2.205, А, Б). Обращает на себя внимание
также отсутствие мышечной пластинки и подслизистой основы.
Мышечная оболочка тонкая и не имеет четкой слоистости. Хоро-
шо развита адвентиция, участки которой, обращенные в брюш-
ную полость, покрыты мезотелием.
366
2
1
Рис. 2.205. Желчный пузырь.
А — СЭМ поверхности слизистой оболочки желчного пузыря (по В. А. Шахламо-
ву). Б — строение стенки (схема).
_ I — слизистая оболочка; II — мышечная оболочка; III — адвентициальная оболочка.
1 — эпителий; 2 — собственная пластинка; 3 — складки слизистой оболочки; 4 —
лимфатический узелок; 5 — кровеносный сосуд; 6 — липоциты; 7 — мезотелий.
2.14. Мочевыделительная система
Знание гистофизиологии мочевыделительной системы важно для
понимания сущности физиологических процессов образования
мочи, генеза дисфункции почек, анализа множественной патоло-
гии выделительной системы. В педиатрии знание особенностей
гистоструктуры почки новорожденного и детей раннего возраста,
правильная оценка степени недифференцированности этого органа
и несовершенства его функции позволяют правильно назначать
лекарственную терапию и рациональное кормление.
2.14.1. Почка
Взаимоотношения нефронов и их составных
частей, сосудистого и других тканевых компонентов почки по-
казаны на схемах (рис. 2.206 и 2.207), где видно следующее:
а) особенности в системе кровоснабжения (наличие капиллярных
клубочков, расположенных между двумя артериолами — «чудесная
сеть»; разность диаметров артериол, приносящих и выносящих кровь
367
Рис. 2.206. Строение почки (схема).
А — топография кровеносных сосудов. Б — структура и топография корковых и
околомозговых нефронов. В — гистологическое строение.
I — капсула; II — корковое вещество; III — мозговое вещество.
1 — корковый нефрон; 2 — околомозговой нефрон; 3, 4 — дуговые артерия и вена;
5, 6 — прямые артерия и вена; 7, 8 — артерии, вены, капилляры мозгового вещест-
ва; 9 — приносящая клубочковая артериола; 10 — сосудистый клубочек; 11 —
выносящая клубочковая артериола; 12 — перитубулярная капиллярная сеть; 13 —
звездчатые вены; 14 — почечное тельце; 15 — проксимальный извитой каналец;
16 — проксимальный прямой каналец; 17 — нисходящая часть петли; 18 — вос-
ходящая часть петли; 19 — дистальный извитой каналец; 20 — связующий отдел;
21 — собирательная трубочка.
368
Рис. 2.207. Сосудистая система почки.
А — взаимоотношения сосудистой системы и компонентов почки (схема). Б —
инъекция сосудов коркового вещества.
1 — капиллярная сеть клубочка; 2 — приносящая клубочковая артериола; 3 —
выносящая клубочковая артериола; 4 — перитубулярная капиллярная сеть; 5 —
просвет капсулы почечного тельца; 6 — внутренняя стенка капсулы; 7 — наружная
стенка капсулы; 8 — проксимальный извитой каналец; 9 — проксимальный прямой
каналец; 10 — нисходящая часть петли; 11—восходящая часть петли; 12 —
дистальный извитой каналец; 13 — связующий отдел; 14 — собирательная трубочка;
15 — дуговая артерия; 16 — дуговая вена; 17 — прямая артерия; 18 — прямая вена.
369
Рис. 2.208. Корковое вещество. -----*-
А — общий "вид. Б — почечное тельце. В — СЭМ капиллярного клубочка. Г —
схема структуры капилляра. Д — ультраструктура капилляра (по Я. Л. Карагано-
ву). Е — СЭМ фенестрированного эндотелиоцита.
1 — почечное тельце: 2 — проксимальный извитой каналец; 3 — дистальный изви-
той каналец; 4 — клубочковая капиллярная сеть; 5 — подоциты; 6 — наружный
листок капсулы; 7 — полость капсулы; 8 — фенестры в эндотелиоците; 9 — порис-
тые участки; 10— цитотрабекулы; 11 — цитоподии; 12— приносящая клубочковая
артериола; 13 — выносящая клубочковая артериола; 14 — эритроцит; 15 — просвет
капилляра.
Рис. 2.208. Продолжение.
из клубочков — обеспечение высокого гидрационного давления в
клубочке); б) особенности в системе кровоснабжения, связанные
с реабсорбцией. В первую очередь обращает внимание разное топо-
графическое положение корковых и околомозговых (юкстамедул-
лярных) нефронов. В корковой части расположены в основном по-
чечные тельца, проксимальные и дистальные извитые канальцы, а
в мозговом веществе — петли нефронов и прямые собирательные
трубочки. Корковые нефроны почти полностью локализованы в
корковом веществе либо в составе мозговых лучей. При этом часть
поверхностных нефронов не имеет тонкого отдела петли. У около-
мозговых нефронов почечные тельца расположены более глубоко —
в юкстамедуллярной зоне. Они имеют длинные нисходящие и вос-
ходящие отделы петли. Особенно характерен тонкий сегмент, кото-
рый пенетрирует глубоко во внутреннюю зону мозгового слоя.
При окраске по Маллори при небольшом увеличении четко
идентифицируются составные компоненты органа: на поверхности
среза — соединительнотканная капсула, снаружи от нее — брю-
шина и затем последовательно вглубь — корковое и мозговое ве-
щество. Корковое вещество (рис. 2.208) определяется
как более темное, состоящее из почечных телец и в основном про-
ксимальных и дистальных частей нефрона. Мозговое веще-
ство окрашено светлее, более однородно, так как содержит сре-
зы канальцев петель нефрона и собирательных трубочек. Хорошо
372
видно, что на границе с корковым мозговое вещество разделено
на пирамиды, между которыми располагаются колонки коркового
вещества, в то время как мозговое вещество проникает в корковое
в виде тонких мозговых лучей.
Почечные тельца состоят из капиллярного клубочка,
заключенного между приносящей и выносящей артериолами. Ка-
пилляры анастомозируют между собой. Характерна фенестрирован-
ность эндотелиоцитов капилляра, фенестры диафрагмированные и
недиафрагмированные (поры). Эндотелиоциты подстилает базаль-
ная трехслойная мембрана. Капилляры плотно охвачены эпите-
лиоцитами внутреннего листка капсулы — подоцитами (рис. 2.209).
Наружный листок капсулы, выстланный низким эпителием, огра-
ничивает полость капсулы тельца. Подоциты, контактирующие со
стенкой капилляров, на светооптическом уровне определяются с
трудом — как очень плоские, неправильной формы клетки. Тонкая
структура подоцитов представлена на электронограмме и скано-
Рис. 2.209. Подоциты почечного тельца.
А — СЭМ. Б — ТЭМ. В — контакт отростков подоцита с базальной мембраной
(Деталь к Б) (по А. В. Миронову и Я. Л. Караганову).
1 — подоцит; 2 — цитотрабекула подоцита; 3 — цитопедикулы подоцита; 4 — щель
между цитопедикулами; 5 — диафрагма; 6 — базальная мембрана; 7 — перифери-
ческая часть фенестрированного эндотелиоцита; 8 — просвет капилляра.
373
Рис. 2.209. Продолжение.
грамме (см. рис. 2.209), где видны их многочисленные отростки —
цитотрабекулы и цитоподии, благодаря которым эти клетки кон-
тактируют с базальной мембраной. Цитоподии между собой не
контактируют, а отделяются фильтрационными щелями, затянуты-
ми тонкой диафрагмой. Указанные морфологические особенности
капилляров и подоцитов способствуют процессу фильтрации мочи;
ведущую роль играет базальная мембрана, обеспечивающая избира-
тельную проницаемость почечного фильтра. Весь комплекс струк-
турных компонентов фильтрационного барьера почечного тельца
виден на рис. 2.210.
Проксимальные извитые канальцы в корковом
веществе занимают максимальную площадь. Эти канальцы распо-
лагаются вокруг почечных телец (рис. 2.211). От других каналь-
цев их можно отдифференцировать по более темной окраске, не-
большому неправильной формы просвету. Образующие их стенку
высокие эпителиоциты характеризуются мутной цитоплазмой, со-
держащей большое количество разнообразных включений, нечет-
костью межклеточных границ, наличием щеточной каемки на апи-
кальной поверхности. Эта каемка представляет собой комплекс
многочисленных микроворсинок, резко увеличивающих реабсор-
бирующую поверхность эпителия этих канальцев (по аналогии
с микроворсинками энтероцитов). В базальной части эпителиоци-
тов видна «исчерченность», которая связана с множественными
инвагинациями базальной плазматической мембраны и особым,
чаще параллельным, расположением митохондрий, концентрирую-
щихся в этой области. Эти особенности базальной части клетки
обеспечивают оптимальную возможность транспорта ряда веществ
из канальцев в кровяное русло, а обилие митохондрий свидетель-
ствует, что этот процесс идет с большой затратой энергии.
Дистальные извитые канальцы в сравнении с
проксимальными более короткие, поэтому общее их количество
на срезах почки значительно меньше, чем проксимальных. Они
выстланы кубическим эпителием, имеют светлую цитоплазму,
меньший диаметр и хорошо выраженный правильной формы про-
свет. Обращает внимание, что в одних из составляющих их эпи-
телиоцитов щеточная каемка может быть, в других — отсутство-
вать. В базальной части этих клеток имеются впячивания базаль-
ной мембраны, длинные митохондрии.
Мозговое вещество (рис. 2.212) состоит в основном из петель
нефронов и собирательных трубочек. Тонкие канальцы
нисходящей части петли определяются по плоскому
эпителию, с выступающими в просвет канальца ядросодержащими
зонами эпителиоцитов. Бедность органеллами цитоплазмы эпите-
лиоцитов нисходящей части является морфологическим свидетель-
ством пассивности протекающих здесь процессов. В канальцах
восходящей части петли эпителиоциты значительно
выше, кубической или цилиндрической формы, с более сложной
внутриклеточной организацией и обилием митохондрий в базаль-
ной части.
375
376
Рис. 2.210. Структура фильтрационного барьера почечного тельца.
А — общий вид капилляра, окруженного подоцитами. Б — ТЭМ компонентов барь-
ера (по Я. Л. Караганову).
1 — просвет капилляра; 2 — фенестрированный эндотелиоцит; 3 — перифериче-
ская часть фенестрированного эндотелиоцита; 4 — трехслойная мембрана; 5 —
подоцит; 6 — цитоподии подоцита; 7 — щели между цитоподиями; 8 — цитоплазма
подоцита; 9 — ядро подоцита; 10 — просвет капсулы клубочка; 11 — кубический
эпителий париетального листка капсулы.
377
Рис. 2.211. Извитые канальцы.
А — строение проксимального каналь-
ца (схема). Б — ультраструктура про-
ксимального канальца. В — апикальная
часть каемчатого эпителиоцита. Г —
СЭМ стенки проксимального канальца.
Д — базальная часть каемчатого эпи-
телиоцита. Е — схема строения дис-
тального канальца. Ж — ультраструкту-
ра дистального извитого канальца (по
К. А. Зуфарову).
1 —- просвет канальца: 2 — ядра каем-
чатых эпителиоцитов; 3 — митохонд-
рии; 4 — щеточная кайма; 5 -— базаль-
ная мембрана; 6 — базальная исчер-
ченность; 7 — капилляр; СБ — складки
базальной мембраны.
378
379
Рис. 2.211. Продолжение.
380
381
Рис. 2.212. Мозговое вещество.
А — продольный и поперечный срезы (схема). Б — тонкий каналец нисхо-
дящей части петли нефрона (ТЭМ). В — прямая собирательная трубочка (ТЭМ)
(по К. А. Зуфарову).
1 — капилляр клубочка; 2 — проксимальный каналец; 3 — нисходящая часть петли
нефрона; 4 — собирательная трубочка; 5 — дистальный каналец; 6 — восходящая
часть петли нефрона; 7 — светлые клетки собирательной трубочки; 8 — темные
клетки собирательной трубочки.
382
383
Рис. 2.212. Продолжение.
Г — схема строения противоточно-множительного аппарата почки (в модифика-
ции по Е. Ф. Котовскому).
384
Рис. 2.213. Строение
почечного тельца с юк-
стагломерулярным комп-
лексом.
А — схема строения. Б —
ультраструктура миоидно-
го эндокриноцита.
1 — приносящая клубоч-
ковая артериола; 2 — вы-
носящая клубочковая ар-
териола; 3 — капилляры
сосудистого клубочка; 4 —
эндотелиоциты: 5 — подо-
циты внутреннего листка;
6 — базальная мембрана;
7 — мезангиальные клет-
ки; 8 — полость капсулы
клубочка; 9 — наружный
листок капсулы клубочка;
10 — дистальный каналец
нефрона; 11 — плотное
пятно; 12 — эндокриноци-
ты (юкстагломерулярные
клетки); 13 — юкставаску-
лярные клетки; 14 — стро-
ма почки; 15 — секретор-
ные гранулы в эндокри-
ноците.
13—321
385
Собирательные трубочки составляют основную
часть мозгового вещества. Они выстланы низкими, цилиндрической
формы эпителиоцитами с гиперхромными круглыми ядрами, имеют
светлую цитоплазму. Выявляются отличия в ультраструктуре этих
клеток: более многочисленные из них — светлые клетки, а темные
клетки снабжены более длинными ворсинками. Близость органи-
зации темных клеток собирательных трубочек и обкладочных
клеток желудка отражает их функциональное сходство (выработка
хлоридов, участие трубочек в подкислении мочи).
На рис. 2.213 представлена схема почечного тельца
с прилежащими структурами юкстагломеру-
лярного комплекса. Верхняя часть дистального канальца
(macula densa) структурно отличается от остальной части каналь-
ца, это эллипсоидный диск из высоких клеток. Полярность пла-
стинчатого комплекса этих клеток направлена к миоидным эндо-
криноцитам (в то время как в эпителиоцитах других участков
канальцев — к его полости). Базальная мембрана чрезвычайно
тонка — имеется тонкая связь клеток macula densa с миоидными
эндокриноцитами. В приносящей (и частично выносящей) артерии
у проксимального участка ее вхождения в тельце среди гладко-
мышечных клеток имеются специфические клетки с секреторны-
ми гранулами. Это миоидные эндокриноциты (юкстагломеруляр-
ные клетки). С одной стороны они контактируют с интимой арте-
рии, с другой — с основанием клеток macula densa в стенке дис-
тального канальца. С ними контактируют и экстрагломерулярные
клетки, локализованные у основания клубочка в углу между арте-
риями. Миоидные эндокриноциты имеют слегка базофильную ци-
топлазму, умеренно выраженную гранулярную сеть и хорошо вы-
раженный пластинчатый комплекс. Секреторная природа гранул
установлена — это ренин. Установлена связь между уровнем ренина
и степенью грануляции клеток. Мезангиоциты располагаются по
ходу капилляров и составляют остов капиллярного клубочка.
Развитие почки. Как известно^цочка в эмбриогенезе человека
закладывается в виде трех генерации: Развитие предпочки, пер-
вичной и вторичной почки, взаимоотношения их закладок пока-
заны на рис. 2.214. П р!5 д п о ч к а закладывается в головных
сегментах и не имеет связи с кровеносной системой, она быстро
редуцируется (на 4-й неделе). Первичная почка харак-
теризуется локализацией в области туловищных сегментов и на-
личием непосредственной связи с кровеносной системой, сравни-
тельно длительным существованием. Часть канальцев первичной
почки не редуцируется, эти остающиеся канальцы являются источ-
ником образования выносящих канальцев семенника.
Первичная' почка у эмбриона 4-й недели развития выглядит
как вытянутое параллельно позвоночнику образование. Каудаль-
ные части его представляют собой только самые начальные пре-
образования мезодермы, а более краниальные отражают прогрес-
сирующие стадии: формирование канальцев и клубочков.
У эмбриона 6—7-й недели развития первичная почка иден-
386
Рис. 2.214. Развитие предпочки,
первичной и постоянной почки.
А — схемы (I, II). Б — первичная поч-
ка: эмбрион 4 нед. В — первичная поч-
ка: эмбрион 6 нед.
1 — канальцы пронефроса (дегенери-
рующие) ; 2 — канальцы мезонефроса;
3 — метанефрогенная ткань; 4 — ин-
дифферентная гонада; 5 — клоака; б —
дивертикул; 7 — парамезоиефральные
протоки; 8 — проток мезонефроса; 9 —
яичник; 10 — яичко; 11—функциони-
рующие клубочки и канальцы первич-
ной почки; 12 — легкое; 13 — желудок;
14 — спинномозговые узлы; 15 -— пе-
чень; 16 — спинной мозг.
13*
387
тифицируется как плотное образование, лежащее под медиально
расположенным однородным, еще более плотным тяжем — буду-
щей гонадой. Составные части первичной почки по длиннику име-
ют особенности строения, отражающие этапы ее морфогенеза.
В каудальном конце виден процесс образования канальцев и по-
чечных телец, в центральной части — срезы уже сформированных
структур, в краниальной части — процесс деструкции канальцев
и телец. Эта почка к концу эмбрионального периода (8 нед)
достигает максимального развития, но к середине беременности
редуцируется.
На рис. 2.215 можно видеть развитие вторичной,почки:
уже имеет место врастание дивертикула в нефрогенную ткань, его
ветвление, образование нефрогенной бластемы. Вокруг последу-
ющих ветвлений дивертикула активизируется нефрогенная ткань и
происходит начальное образование клубочков и канальцев и их
последовательная дифференцировка.
Динамика развития почечного тельца показана также на рис.
2.214: капиллярный клубочек только намечается под будущим
внутренним листком капсулы почечного тельца рядом с каналь-
цем нефрона, затем видно, что в клубочке уже увеличивается чис-
ло петель капилляров и клубочек становится очерченной струк-
турой. Процесс прогрессирует — клубочки увеличиваются в раз-
мере; параллельно с увеличением клубочка дифференцируется
внутренний листок капсулы, эпителий которого, становясь плос-
ким, плотно прилежит к эндотелию капилляров. Затем происходит
увеличение полости капсулы и уплощение эпителия ее наружного
листка. На 7-й неделе появляются миоидные эндокриноциты, но
они еще агранулярные (без признаков секреторной активности).
Плотное пятно дифференцируется позже на неделю, а клетки
юкстагломерулярного комплекса только к 12—13 нед (рис. 2.216).
Чтобы лучше понять процесс дифференцировки составных
частей нефрона, следует еще раз последовательно рассмотреть
схему (см. рис. 2.215), на которой виден первый этап — контакт
собирательной трубки с нефрогенной бластемой, затем начало
формирования пузырьковидных канальцевых структур, затем обра-
Рис. 2.215. Развитие постоянной почки.
А — этапы (I—IV) образования и дифференцировки компонентов нефрона (со-
ответственно: вверху — поверхностные, внизу — глубокие слои развивающейся
почки). Б — контакт собирательной трубочки с нефрогенной бластемой (деталь
к А). В — дифференцировка эпителиоцитов проксимального канальца (деталь кА):
а — эмбрион 8 нед, б — плод 6 мес. Га — Гг — последовательные стадии формиро-
вания почечного тельца. Д — эмрион 6 нед. Е — эмбрион 8 нед. Ж — плод 6 мес.
1 — гонада; 2 — первичная почка; 3 — постоянная почка; 4 — надпочечник; 5 —
ветвление дивертикула; 6 — продвинутые стадии развития канальцев и клубочков
нефронов; 7 — незрелые стадии образующихся канальцев и почечных телец; 8 —
наиболее дифференцированная генерация почечных телец; 9 — формирующиеся
почечные тельца; 10— нефрогенная бластема; 11 —собирательная трубочка; 12 —
формирующаяся капсула; 13 — сосудистый клубочек; 14 — внутренний листок кап-
сулы; 15 — наружный листок капсулы; 16 — полость капсулы; 17 — легкие.
388
A
a
390
2
17
10
1
3
391
Рис. 2.216. Эмбриональное развитие юкстагломерулярного комплекса.
А — эмбрион 8 нед. Б — плод 12 нед.
1 — капилляры; 2 — плотное пятно.
392
зование S-образного канальца. Параллельно прослеживается обра-
зование клубочка. На периферии почки располагаются самые не-
зрелые формы клубочков, а по мере приближения к ее центру клу-
бочки и канальцы соответственно уже более дифференцированы.
В почке плода 7 мес достаточно четко прослеживаются ход
образования нескольких генераций нефронов и последовательные
этапы формирования почечных телец и канальцев. Видно, что чем
ближе к периферии почки, тем нефроны менее продвинуты в раз-
витии, а в подкапсульной области даже еще имеются последние
ветвления (ампулы) собирательных трубок. Вокруг этих ветвлений
активизированная ткань — бластема, т. е. в этот период развития
эмбриона еще продолжается образование S-образных канальцев.
Почечные тельца маленькие, плотно расположенные. Нефроны,
локализованные центральнее, более сформированы: уже проявля-
ются четкие отличия проксимального и дистального отделов из-
витых канальцев. Продолжается дифференцировка эпителиоцитов
проксимального канальца. Недифференцированность структуры
клубочков и канальцев соответственно свидетельствует о выражен-
ной недостаточности фильтрационного и реабсорбционного аппа-
рата почки у недоношенных детей.
В почке доношенного новорожденного ребенка
(рис. 2.217) еще имеется дольчатость строения и сохранность
черт эмбрионального строения: тонкий корковый слой, нечетко
сформированные лучи и пирамиды, еще сравнительно короткие
канальцы (вследствие этого плотность клубочков на единицу пло-
щади значительно выше, чем у взрослых). При этом в самых пе-
риферических участках почки у некоторых детей еще может про-
должаться процесс образования клубочков и канальцев. Сосудис-
тые клубочки малы, длина капилляров небольшая, значительная
часть их еще не функционирует (спавшиеся капилляры) и эндо-
телиоциты мало фенестрированы. Подоциты в большинстве почеч-
ных телец еще кубической формы с малым количеством отростков.
Такие особенности строения почечных телец обусловливают в це-
лом малую поверхность фильтрующего аппарата почки, несовер-
шенство фильтрации. Также еще несовершенен и аппарат реаб-
сорбций: отсутствуют четкие дифференциальные признаки между
отделами нефронов, проксимальные канальцы коротки, еще не пол-
ностью дифференцированы. Особенно в поверхностных нефронах
слабо развиты микроворсинки эпителиоцитов, не выражена базаль-
ная исчерченность. Юкстагломерулярный комплекс в этот период
еще в процессе дифференцировки, лишь к концу первого года
жизни ребенка он структурно сформирован.
В почке ребенка двух лет обращает на себя внима-
ние прогрессирование процесса дифференцировки: увеличивается
величина почечных телец, снижается плотность клубочков на еди-
ницу площади, увеличивается длина канальцев нефронов.
В почке ребенка пяти лет уже видно четкое под-
разделение на корковое и мозговое вещество, их соотношение бо-
лее близко к дефинитивному. Увеличивается длина канальцев и
393
5
394
Рис. 2.217. Постнатальное формирование почки.
А — почка недоношенного новорожденного. Б — почка доношенного новорожден-
ного. В — почка ребенка 2 лет. Г — почка ребенка 5 лет.
1 — слой еще образующихся клубочков и канальцев; 2 — слой уже образовавших-
ся клубочков и канальцев; 3 — слой более зрелых почечных телец и канальцев;
4 — подкапсульный бесклубочковый слой.
соответственно расстояние между почечными тельцами; поэтому
число клубочков на единицу площади значительно меньше. Клу-
бочки имеют более крупные размеры, и подоциты всех клубочков
(на любом уровне коркового вещества) становятся более плоски-
ми. Эта морфологическая картина свидетельствует о совершен-
ствовании физиологии процесса фильтрации мочи. Высокая диф-
ференцированность отделов нефрона указывает и на достаточно
высокий уровень процесса реабсорбции в этот возрастной период
ребенка. Однако все эти показатели остаются все еще ниже, чем
у взрослого человека, — полное формирование с оптимальным
функционированием происходит только к 10—12 годам жизни
ребенка и даже позже.
2.14.2. Мочевой пузырь
В мочевом пузыре (рис. 2.218) виден трехкомпонентный состав
его стенки и складчатость слизистой оболочки. Ее выстилает пе-
реходный эпителий, под которым расположена собственная плас-
тинка слизистой оболочки, образованная рыхлой неоформленной
соединительной тканою, с локализованными в ней кровеносными
сосудами. Без резкой границы она переходит в подслизистую обо-
лочку, построенную из того же вида соединительной ткани (что
оправдывает термин «слизисто-подслизистая оболочка»), Мышеч-
395
Рис. 2.218. Мочевой пузырь.
А — общий вид. Б — слизистая оболочка (окраска гематоксилин-эозином). В —
рельеф поверхности слизистой оболочки в опорожненном состоянии (СЭМ).
I — слизистая оболочка; II — подслизистая основа: III — мышечная оболочка;
IV — серозная оболочка.
1 — эпителий; 2 — соединительнотканная основа; 3 — гладкие мышцы; 4 — мезо-
телий.
Рис. 2.219. Эмбриональное развитие мочевого пузыря.
А — эмбрион 8 нед. Б — плод 3 мес.
1 — слизистая оболочка; 2 — мышечная оболочка; 3 — серозная оболочка.
ная оболочка образована гладкомышечной гканью, пучки которой
прослоены рыхлой соединительной тканыд. В оболочке определя-
ются три нерезко разграниченных слоя, различающиеся ориента-
цией миоцитов. Во внутреннем слое мышечные пучки идут пре оль-
но, в среднем — циркулярно "и в наружном — продольно. ТВ самой
наружной оболочке мочев’ого пузыря"’— ентицйальной — в со-
ставляющей ее рыхлой соединительной ткани видны срезы крове-
носных.. сосудов, жировые клетки к интрамуральные ганглии. В дне
мочевого пузыря адвентициальную оболочку покрывает м^з^телий
(серозная оболочка).
Развитие мочевого пузыря. Закладка мочевого пузыря состоит
из эпителиальной выстилки и окружающей мезенхимы. На рис.
2.219 видна прогрессирующая дифференцировка эпителия и со-
единительнотканных структур, начало формирования мышечной
оболочки.
397
Рис. 2.220. Мочеточник.
1 — слизистая оболочка; 2 — подсли-
зистая оболочка; 3 — мышечная обо-
лочка; 4 — серозная оболочка.
3 2
1
41 2 2.14.3. Мочеточник
Мочеточник (рис. 2.220) имеет
сл истое строение. Его звездча-
5 2 - той формы просвет образован
2 складками слизистой оболочки,
внутренняя поверхность которой
выстлана переходным эпители-
ем. По г. ним просматривается
собственная пластинка слизи-
стой оболочки, образованная
рыхл “ неоформленной соеди-
нительной тканью. Характерно,
что слизистая оболочка, так же
как пузыре, без
резких границ переходит в соединительную ткань подслизи-
стой оболочки. В нижней части мочеточника *й' подслизистой
оболочке видны еще и железы. Подстилающая подслизистую
мышечная оболочка образована гладкими мышцами. В верх-
ней трети органа ^рдны два слоя мышц ^внутренний продоль-
ный и наружный циркулярный); в нщ ней трети — три сфор-
мированных слоя (как и в мочевом пузыре). Для соединительной
ткани наружной адвентициальной оболочки характерно обилие
сосудов „и еток.
2.15. Мужская половая система
Мужская половая (репродуктивная) система обеспечивает муж-
скому организму выполнение двух взаимосвязанных функций —
образование половых клеток (сперматозоидов) и выработку муж-
ских половых гормонов, главным из которых является тесто-
сдерон.
При изучении гистофизиологии мужской репродуктивной сис-
темы необходимо помнить, что ее патология может являться при-
чиной бесплодия, врожденных и приобретенных заболеваний по-
томства. Заболевания мужской половой системы составляют пред-
мет быстро развивающейся в последние годы самостоятельной
науки — андрологии.
398
2.15.1. Яичко
Яичко (семенник) — центральный орган всей системы, сочетаю-
щий в себе сложную трубчатую экзокринную железу, в которой
происходит образование сперматозоидов с элементами эндокрин-
ных структур.
На схеме микроанатомических взаимоотно-
шений (рис. 2.221) показано, что яичко окружено толстой фиб- .
ручной оболочкой (белочной), хорошо определяемой и на препа-
ратах, еав1 ^ЯзаНа периферия органа. На заднелатеральной по-
верхности яичка человека располагается утолщение белочной обо- *
лочки, вдающееся в железу, образующее ее средостение. От сре-
достения отходят тонкие фиброзные перегородки — септ^д, раз-
деляющие половую железу человека примерно на 250 долек, каж-
дая из которых имеет пирамидную форму. В своих дистальных
отделах септы имеют поры, обеспечивающие сообщение между
дольками. В каждой дольке располагается от 1 до 4 изддтых се-
менных ^анальц^в — единственного и самого протяженного от-
дела канальцевой системы яичка, в котором происходит процесс
сперматогенеза. Извитые канальцы представляют собой концевой
отдел экзокринной части мужской половой железы, секреторным
продуктом которой являются половые клетки. Извитые канальцы
чаще всего образуют петли, но иногда ветвятся или заканчивают-
ся слепо. На верхушке каждой дольки извитой .семенной каналец
довольно резко переходит в £?Ha&eU< представляющий со-
бой начальный отдел системы выводных протоков железы. На
схеме представлены и последующие этапы выводных протоке».
Рис. 2.221. Микроанатомические
взаимоотношения в половой желе-
зе взрослого мужчины (схема).
1 — белочная оболочка; 2 — сеть яичка;
3 — септа; 4 — извитые семенные ка-
нальцы; 5 — прямые канальцы; 6 — вы-
носящие канальцы; 7 — головка придат-
ка; 8 — проток придатка; 9 — хвост
придатка; 10 — семявыносящий проток;
11 — интерстициальная ткань.
399
Все промежутки между извитыми семенными канальцами за-
полнены интерстициальной тканью. Она обнаруживается на срезах
яичка между срезами извитых канальцев.
2.15.2. Сперматогенез
Процесс сперматогенеза представлен на рис. 2.222.
Современная периодизация этого процесса подразумевает наличие
в нем трех последовательных фаз: сцерматэ.ониальной. содержа-
нием которой является митотическое делениеисдмых ранних кле-
ток сперматогенеза —„сцеркшуогоний (эта фаза часто называется
сперматоцитогенезом), мейотической и формирования, называе-
мой спермиогенезом.
Сперматогонии, представляя собой разнородную в степени
своей дифференцировки популяцию клеток, все являются
иднымд. Делясь только путем митоза, они дают начало генераци-
ЛГТлеток, вступающим в сперматогенез.
Важнейшей с общебиологической точки зрения является вто-
рая, мейотическая фаза сперматогенеза. Мейоз
представляет собой особый процесс клеточного деления, свойст-
венный только половым клеткам.
События и их последовательность, происходящие в ядрах
мейотически делящихся клеток, представлены на рис. 2.223. Они
принципиально одинаковы как в мужском, так и в женском opia
низмах, т. е. в сперматогенезе и в овогенезе.
Напомним, что первому мейотическому дел<•
нию (первому делению созревания) предшествует удвоен и?
сперматогонии
Рис. 2.222. Сперматогенез
(схема).
400
ДНК. Характерной особенностью первого деления мейоза явля-
етсЙ длительная профаза, в ходе которой происходят спаривание
гомологичных хромосом и кроссинговер.
События первого мейотического деления происходят в клетках,
носящих название сдгрмчтудщгод плчяДкд (сперматоци-
тов I). В конце первого деления мейоза происходит цитотомця
(неполная), приводящая к образованию из одного сперматоцита I
двух неразделенных до конца последующих клеточных форм мейо-
тической фазы сперматогенеза — сперматоцитов второго порядка
(сперматоцитов II).
Второе деление мейоза (второе деление созревания,
эквационное деление) осуществляется сразу вслед за телофазой I,
без предшествующей редупликации ДНК. Оно представляет собой
обычное митотическое деление. Сперматиды — клетки, за-
вершающие мейотическую фазу сперматогенеза, представляют
собой небольших размеров округлые клетки, характеризующиеся
генетическим набором пс, т. е. истинно гаплоидные. —
Мейоз
Рис. 2.223. Мейоз в мужских и женских половых клетках животного,
имеющего в кариотипе 2 хромосомы (схема).
401
Фаза формирования, называемая также спермиогенезом, при-
водит к превращению округлой сперматиды в вытянутую высоко-
специализированную клетку — сперматозоид. Наиболее сущест-
венные события этого процесса продемонстрированы на рис. 2.224.
Они заключаются в образовании акросомы, развитии жгутика,
конденсации хровд^тина, изменении формы и размеров ядра, поте-
ре части цитоплазмы. Эти сложные морфологические преобразо-
вания сопровождаются синтезом ряда белковых молекул, свой-
ственных только сперматозоиду.
При высвобождении зрелого сперматозоида из пласта клеток
извитого канальца (процесс называется спермиацией) избыточная
цитоплазма окончательно отделяется, оставаясь ограниченной
мембранной структурой, которая называется резидуальным тель-
цем. В последующем резидуальное тельце подвергается фагоци-
тозу поддерживающими клетками яичка (см. ниже).
А — схема. Б — начальная стадия формирования акросомы (ТЭМ). В — продви-
нутая стадия формирования акросомы.
1 — пластинчатый комплекс; 2 — акросомальный пузырек; 3 — акросома; 4 —
жгутик; 5 — манжетка; 6 — фиброзная оболочка жгутика; 7 — кольцо; 8 — мито-
хондрии; 9 — проксимальная центриоль; 10 — преакросомные гранулы; 11—ядро
сперматиды.
402
403
Время, необходимое для полного превращения сперматогонии
в сперматозоид, у человека составляет 74 сут.
Сперматозоид — конечная клетка сперматогенеза — яв-
ляется во многих отношениях уникальной. Помимо уже упоми-
навшихся генетических характеристик (22 + Х; 22-|-Y), она резко
вытянута и высокоспециализирована по своему длиннику (рис.
2.225). Головка сперматозоида содержит ядро, около двух третей
которого спереди покрыто а^росомой. 1 Гостакросомальная зона
головки сперматозоида является тем участком, цитоплазматическая
мембрана которого при оплодотворении сливается с мембраной
яйцеклетки.
Шейка — самая короткая часть зрелой мужской половой клет-
ки. Она содержит проксимальную центриоль и соединительную
ветвь.
Рис. 2.225. Строение зрелого сперматозоида человека.
А — общий вид. Б — ТЭМ.
1 — головка сперматозоида; 2 — шейка; 3 — хвостовая часть; 4 — акросома; 5 —
постакросомальная зона; 6 — средняя часть хвоста; 7 — главная часть хвоста;
8 — конечная часть.
404
Хвостовая часть сперматозоида наиболее протяженная. На
электронной микрофотографии ее продольного сечения следует
обратить внимание на высокую степень упорядоченности распо-
ложения митохондрий. Концевая часть хвоста сперматозоида со-
держит только аксонему, покрытую цитоплазмой.
Большое практическое значение в андрологической практике
имеет исследование мазка спермы. На таком препарате,
микрофотография которого представлена на рис. 2.226, необхо-
димо убедиться в морфологической дифференцировке этих клеток,
описанной выше. Естественными компонентами спермы человека
являются лимфоцщ^ (в небольшом количестве), а также безъ-
ядерные ча&Ы' РП?гопл43нь|, отделившиеся от сперматид в период
спермиогенеза и не фагоцитированные цругами ^летками. В опре-
деленном количества: встречаются и аномаЖИгеформы сперма-
тозоидов, особенно у лиц пожилого возраста.
После ознакомления с процессом сперматогенеза и строением
зрелого сперматозоида целесообразно перейти к разбору строения
яичка на гистологических препаратах.
На рис. 2.227, А обратите внимание на степень извитости
Рис. 2.226. Мазок -спермы че-
ловека.
1 — сперматозоиды; 2 — лимфоциты;
3 — безъядерные участки цитоплаз-
мы; 4 — спущенные эпителиоциты;
5 — кристаллы спермина.
405
Рис. 2.227. Извитой семенной каналец крысы (СЭМ).
А — вид канальца в продольном сечении. Б — вид канальца в поперечном сечении.
406
канальца, находящегося в дольке яичка. На СЭМ видно, что
каналец снаружи окружен довольно прочной оболочкой, называе-
мой собственной оболочкой извитого семенного канальца. В ее
составе находятся клетки, напоминающие миоциты и называемые
миоидными, а также неклеточдь^ Миоидные клетки извитых
семенных канальцев человека в отличие от хорошо изученных в
этом отношении лабораторных грызунов не обладают контрак-
тильной. активностью. В ряде случаев мужской инфертильности
собственная оболочка извитых канальцев может резко утолщаться,
нарушая трофику клеток, находящихся внутри канальца.
На СЭМ поперечного среза канальца (рис.
2.227, Б) можно видеть тела клеток, составляющих сперматог.ен-
ны эпителий (пласт), а также хвостовые части поздних сперма-
тид и сперматозоидов, сходящиеся к просвету канальца.
2.15.2.1. Клетки сперматогенного пласта
У взрослого человека извитые семенные канальцы выстланы слож-
ным комплексом клеток, не совсем корректно называемым спер-
матогенным «эпителием». Из множества клеточных форм этого
пласта только один тип клеток, а именно поддерживающие клет-
ки (сустентоциты, клетки Сертоли), являются собственно эпите-
лиальными. Эти же клетки являются единственными внутри ка-
нальца соматическими его элементами. Все остальные клеточные
формы, образующие сперматогенный пласт, представляют собой
клетки сперматогенеза, т. е. являются половыми по своей природе.
Соматические клетки сперматогенного пласта — сустентоциты
имеют чрезвычайно сложную трехмерную конфигурацию, не вы-
являющуюся с помощью световой микроскопии. Обладая цилиндри-
ческой формой, большим количеством длинных цитоплазматиче-
ских островков, расходящихся латерально и к просвету канальца,
они расположены, как и любой эпителий, на базальной мембране
и имеют особый тип контактов друг с другом. На рис. 2.228, А,
представляющем взаимоотношения сустентоцитов с клетками
сперматогенного ряда, видно, что цитоплазматические отростки
сустентоцитов, охватывая со всех сторон клетки сперматогенного
пласта, отделяют их друг от друга, создавая своеобразное микро-
окружение для них и заполняя промежутки между половыми
клетками. Вместе с тем клетки сперматогенного ряда остаются
связанными между собой межклеточными мостиками. В своей
совокупности в пределах сперматогенного пласта сустентоциты
не только «поддерживают» половые клетки разных стадий разви-
тия, но и пространственно упорядочивают их расположение. По-
следнее проявляется в четкой закономерности: наиболее ранние
клетки сперматогенеза .(сперматогонии, прелептотенные сперма-
тоциты) располагаются вблизи базальной мембраны, по мере своей
дифференцировки они перемещаются все ближе к апикальному
полюсу сустентоцита, как бы двигаясь вдоль боковых его поверх-
ностей к верхушке.
407
80tz
Сустентоциты обладают характерным ядром. Оно располага-
ется у базальной мембраны, имеет овоидную или треугольную
форму и глубокие инвагинации цитолеммы. На светооптическом
уровне (см. рис. 2.228) ядра сустентоцитов легко определяются
по расположению относительно базальной мембраны, частоте
встречаемости и светлому диспергированному хроматину, на фоне
которого хорошо определяются одно или два ядрышка.
Сустентоциты у взрослого человека относятся к категории
непролиферирующих клеток. Свою митотическую активность они
прекращают, по-видимому, еще внутриутробно, не делясь ни при
каких условиях после пубертации. Вместе с тем это наиболее
стабильная, устойчивая популяция клеток сперматогенного пласта.
Если половые клетки довольно легко и быстро погибают от раз-
личных воздействий (ионизирующая радиация, сперматогенные
яды, аутоиммунные конфликты и др.), то сустентоциты часто
остаются после таких воздействий единственными клетками
внутри извитого канальца.
На рис. 2.228, А показано, что поддерживающие клетки содер-
жат вытянутые митохондрии, ориентированные вдоль длинной
оси клетки. Обычными для цитоплазмы этих клеток являются
липидные капли и хорошо развитая агранулярная сеть в непосред-
ственной близости от развивающейся акросомальной шапки спер-
матид. Значение этой близости неизвестно, но оно рассматрива-
ется в качестве одного из проявлений функции сустентоцита как
«клетки-няньки» для половых.
Сертоли-сертолиевы соединения. Соседствующие друг с другом
сустентоциты соединяются необычным типом соединений — так
называемыми сертоли-сертолиевыми соединениями, не встречаю-
щимися ни в одном другом типе эпителиев. В их образовании
принимают участие наиболее базально расположенные отростки
сустентоцитов. Сертоли-сертолиевы соединения являются чрезвы-
чайно прочными, надежно отграничивающими расположенный
над ними адлюминальный отсек извитого семенного канальца от
проникновения туда многих химических соединений и, наоборот,
препятствующими контакту содержимого адлюминального отсека
с иммунокомпетентными клетками организма (практически все
применявшиеся для изучения гистогематических барьеров вещест-
ва не способны их преодолевать). По современным представлениям
смысл наличия столь прочных разграничивающих каналец струк-
тур заключается в изоляции поздно появляющихся в онтогенезе
Рис. 2.228. Взаимоотношения сустентоцитов с клетками сперматогенного
ряда.
А, Б — схемы.
1 — базальная мембрана; 2 — сперматогонии; 3 — соединительный комплекс меж-
ду двумя соседними сустентоцитами (сертоли-сертолиево соединение); 4—спер-
матоцит I; 5 — соединительный комплекс со сперматидой; 6 — ранняя сперматида;
7 — липиды; 8 — гладкая цитоплазматическая сеть; 9 — микротрубочки; 10 —
поздняя сперматида; 11 — сустентоцит; 12 — миоидные клетки.
409
продвинутых форм сперматогенеза, начиная со сперматоцита I,
вступившего в мейоз. Именно поэтому в базальном отсеке изви-
того канальца, расположенном кнаружи от сертоли-сертолиевых
соединений, располагаются только ранние формы сперматогенеза
(сперматогонии, прелептотенные сперматоциты).
Физиологические и морфологические данные позволили уста-
новить, что сертоли-сертолиевы соединения вместе с оболочкой
извитого семенного канальца в совокупности образуют очень проч-
ный гистогематический барьер, называемый гематотестикулярным
барьером (ГТБ). Он избирательно пропускает молекулы как из
полости канальца, так и в обратном направлении, обеспечивая
тем самым существенную разницу ионного, аминокислотного и
углеводного состава крови (лимфы) и содержимого канальца.
Кроме того, ГТБ препятствует возникновению аутоиммунного
конфликта — реакции собственной иммунной системы на неиз-
вестные ей белки клеток сперматогенного ряда. Нарушение це-
лостности ГТБ в результате травмы яичка или после воздействия
некоторых веществ (после начала пубертации) приводит к ауто-
иммунному процессу, часто полностью уничтожающему спермато-
генный пласт.
Как следует из рис. 2.229, представляющего общий вид
участка яичка, помимо сертоли-сертолиевых соединений,
в образовании ГТБ участвует также собственная оболочка изви-
того семенного канальца. Значение упоминавшихся выше периту-
булярных клеток и неклеточных слоев оболочки в ГТБ неясно.
Однако следует помнить, что перитубулярные миоидные клетки у
человека не имеют сократительной активности, в связи с чем их
значение в физиологии извитого канальца остается загадочным.
На препарате яичка взрослого человека (рис. 2.229, А) боль-
шую часть поля зрения занимают срезы извитых семенных каналь-
цев. Обязательной задачей работы с этим препаратом является
идентификация клеток сперматогенеза и поддерживающих клеток.
Ее следует начать с обнаружения ядер сустентоцитов в самом на-
ружном отсеке канальца по следующим признакам: крупные, редко
расположены, овоидной или треугольной формы, светлый хроматин,
ядрышко на его фоне.
Примерно на уровне ядер поддерживающих клеток распола-
гаются ядра сперматогоний, отличающиеся небольшими размера-
ми, компактным хроматином, часто встречающимися фигурами
митоза.
Следующий ярус ядер в сперматогенном пласте составляют
ядра сперматоцитов I. Это самые крупные ядра пласта, их размер
позволяет их легко отличать от других ядер. Кроме того, для
них характерен специфический рисунок хроматина, соответствую-
щий профазе I мейоза.
На этом же уровне находятся ядра вторичных сперматоцитов.
Но в связи с короткостью их жизненного цикла и циклических
изменений, о которых будет упомянуто позже, они редко встреча-
ются на препаратах.
410
Рис. 2.229. Фрагмент яичка.
А — общий вид. Б — шесть клеточных ассоциаций, составляющих цикл сперма-
тогенного эпителия у человека.
1 — базальная мембрана; 2 — сперматогонии группы А; 3 — сперматогонии груп-
пы В; 4 — поздние сперматиды; 5, 6 — сперматоцит II; 7 — сустентоциты; 8 —
прелептогенТТьТе-спЕрматоциты; 9 и 10 — сперматоцит I.
411
Только что образовавшиеся в результате второго деления со-
зревания ранние сперматиды являются небольшими, округлой
формы клетками. Более поздние сперматиды характеризуются
вытянутой формой, большей степенью конденсации хроматина
и развитием акросомы и жгутика.
В процессе изучения распределения различных генераций
сперматогенных клеток возникло представление об ассоциациях
клеток сперматогенного ряда и о цикле сперматогенного эпителия.
Цикл сперматогенного эпителия определяется как серия измене-
ний в данной зоне сперматогенного пласта между двумя появле-
ниями одной и той же клеточной ассоциации. Эта последователь-
ность изменений наиболее тщательно изучена у крысы. У человека
цикл сперматогенного эпителия включает 6 стадий (рис. 2.229, Б)
и длится примерно 16 сут.
У всех изученных в этом отношении видов, кроме человека,
определенная стадия цикла занимает, как правило, довольно длин-
ный участок извитого семенного канальца. Поэтому на поперечном
срезе чаще всего встречается только одна клеточная ассоциация.
При этом рядом с ней располагается следующая по счету стадия,
а с противоположного конца — предыдущая (рис. 2.230). В изви-
Рис. 2.230. Различия в организации
семенного эпителия у человека
и лабораторных грызунов (схема).
412
Рис. 2.231. Структура интерстициальных клеток яичек человека. Ультра-
структура гландулоцита яичка.
413
том канальце человека одна ассоциация клеток занимает короткий
неправильной формы участок, что и объясняет наличие на срезе
одновременно 2—3 и более ассоциаций.
Для полного завершения развития сперматогонии в зрелый
сперматозоид необходимо 4,6 цикла.
Эндокринный компонент мужской половой железы — интер-
стициальные гландулоциты, или клетки Лейдига,
располагаются в промежутках между извитыми семенными ка-
нальцами (см. рис. 2.229, А; рис. 2.231). Здесь же кровеносные
сосуды образуют перитубулярные сплетения. Интерстициальная
ткань яичка человека представляет собой рыхлую соединительную
ткань, в значительной степени насыщенную внеклеточной жидко-
стью. Помимо гландулоцитов, в ней находятся фибробласты, ма-
крофаги, небольшое количество лаброцитов, а также некоторое
количество недифференцированных клеток мезенхимного проис-
хождения, которые под влиянием гонадотропинов аденогипофиза
способны дифференцироваться в гландулоциты.
У взрослого человека гландулоциты располагаются либо в виде
скоплений возле сосудов (собственно интерстициальные), либо
прижаты к канальцам (перитубулярные гландулоциты). Незави-
симо от топографии эти клетки являются классическими стероид-
продуцирующими элементами, аналогичными по своей морфологии
находящимся в коре надпочечников, желтом теле, интерстици-
альной ткани яичника. В случае близкого расположения к сосудам
они, по-видимому, выступают как истинно эндокринные клетки,
выделяя тестостерон в общий кровоток.
Интерстициальные гландулоциты периваскулярной локализации
имеют неправильную многоугольную форму, особенно в центре
скопления. На периферии скопления могут располагаться и вытя-
нутые, и даже веретеновидной формы клетки.
В случае перитубулярной локализации они поставляют тесто-
стерон прямо в извитой семенной каналец, т. е. паракринно.
Тестостерон необходим сперматогенному пласту прежде всего
для завершения с пермиогенеза.
Одной из особенностей интерстициальных гландулоцитов чело-
века является наличие в них кристаллоидных структур, называе-
мых кристаллами Реинке. Плохо окрашиваясь обычными гистоло-
гическими красителями, они остаются бесцветными на препаратах,
предназначенных для светооптического изучения. На электронных
микрофотографиях они имеют упорядоченную организацию,
особенно при поперечном сечении. Интересно отметить, что кри-
сталлы Рейнке появляются в клетках Лейдига вплоть до глубокой
старости. Значение их остается невыясненным.
Развитие яичка проходит несколько стадий. После закладки
органа в виде утолщения целомического эпителия на поверхности
первичной почки наступает так называемая индифферентная ста-
дия (рис. 2.232, А), называемая так потому, что светооптически в
этот период времени невозможно определить гонадный пол заро-
дыша.
414
Рис. 2.232. Развитие яичка чело-
века.
А — зародыш человека 5 нед. Б — се-
менник зародыша 6 нед. В — яичко
новорожденного ребенка.
I — гонада; II — первичная почка.
1 — поверхностный эпителий; 2 — сре-
зы извитых семенных тяжей (каналь-
цев); 3, 4 — мезенхима (интерстициаль-
ная ткань).
415
На срезе зародыша человека 6—7 нед (рис. 2.232, Б) гонада
обнаруживается в виде тяжа, тесно связанного с первичной
почкой. Снаружи она покрыта цилиндрическим эпителием, парен-
химу образуют мезенхимные клетки. При более детальном изуче-
нии гонады можно различить комплексы, состоящие из погру-
зившихся в нее первичных половых клеток и окружающих их
клеток целомического эпителия. Последние образуют половые
шнуры, являющиеся предшественниками извитых семенных ка-
нальцев.
Следует помнить, что появлению половых шнуров предшеству-
ет дифференцировка в полевой железе первой, так называемой
плодной генерации клеток Лейдига. Без тестостерона, который
они продуцируют в самом начале морфогенеза гонады, невозмож-
но ее развитие по мужскому типу. Примерно к середине внутри-
утробного развития человека начинается постепенная инволюция
плодной генерации гландулоцитов, приводящая к практически
полному их исчезновению вскоре после рождения.
Изменения в развивающейся мужской гонаде зародыша и пло-
да человека после 6—7-й недели очень незначительны. Они заклю-
чаются в постепенном удлинении канальцев, некоторой дифферен-
цировке первичных половых клеток и нарастании общего числа
сустентоцитов.
Яичко новорожденного ребенка (рис. 2.232, В) характеризуется
дольчатостью строения, рыхлым расположением интерстициальной
ткани, которая сильно напоминает мезенхиму и практически не
содержит гландулоцитов, редкостью расположения срезов извитых
семенных канальцев (половых шнуров), большая часть которых
лишена просвета. Из клеточных форм в сперматогенном пласте
в этот период находятся лишь сустентоциты и половые клетки,
занимающие промежуточное положение между первичными поло-
выми и сперматогониями, — так называемые пресперматогонии.
К моменту рождения доношенного нормального мальчика его
половые железы в подавляющем большинстве случаев находятся в
мошонке. Примерно в 2—3 % случаев нормы они опускаются в
мошонку в течение первых двух недель постнатальной жизни.
Отсутствие яичек в мошонке более старших детей, а также взрос-
лых людей носит название крипторхизма (одностороннего или
двустороннего) и является одной из наиболее распространенных
форм аномалий развития мужской половой железы.
Критическим моментом развития всего организма и половой
системы в частности является период полового созревания (пубер-
тация). Ни одна другая система организма не претерпевает столь
серьезной перестройки и одновременно не играет столь большой
роли в пубертации, как половая. В этот период под влиянием
соответствующих гормонов аденогипофиза происходит быстрый
рост яичка, его придатка и добавочных половых желез. При этом
увеличивается длина и диаметр извитых семенных канальцев,
в них появляются продвинутые формы сперматогенеза (10—
12 лет), появляется и функционально дифференцируется интер-
416
стициальная ткань (12—14 лет). К возрасту 15—17 лет спермато-
генез характеризуется наличием своих конечных форм, становясь
в дальнейшем более устойчивым.
Исследования последних лет показали, что процесс спермато-
генеза течет с примерно одинаковой интенсивностью вплоть до
глубокой старости.
2.15.3. Придаток яичка
Значительную роль в гистофизиологии всей мужской репродук-
тивной системы играет придаток яичка (рис. 2.233).
В нем происходит не только накапливание (депонирование) осво-
бодившихся в процессе спермиации сперматозоидов, но и придание
им фертильной способности (физиологического дозревания, капа-
цитации в пределах мужского организма). Сперматозоиды, взя-
тые из сети семенника или непосредственно из извитых семенных
канальцев, не способны оплодотворить яйцеклетку.
В придатке яичка принято выделять головку, тело и хвост.
Рис. 2.233. Срез головки придатка
яичка.
1 — срезы выносящих канальцев; 2 —
срезы канала придатка.
417
14—321
В головке придатка на краю среза обязательно присутствуют сре-
зы выносящих канальцев железы, имеющих ряд характерных черт
выстилающего их эпителия. Необходимо обратить внимание на
крайне неровный внутренний контур этих канальцев, обусловлен-
ный чередованием в эпителии высоких реснитчатых клеток и
низких нереснитчатых. Последние могут содержать гранулы
(лизосомы). Однако значение низких клеток в функционировании
системы не определено.
Высокие цилиндрические клетки снабжены подвижными рес-
ничками, колебательные движения которых позволяют содержи-
мому продвигаться по направлению к каналу придатка.
Тело придатка яичка содержит в себе только канал придатка
общей длиной примерно 6 см (у взрослого человека). Канал при-
датка имеет мощную наружную оболочку, содержащую значитель-
ное количество гладких миоцитов, толщина которых увеличивается
по ходу канала. Эпителий канала придатка состоит из цилиндриче-
ской формы главных клеток, апикальная поверхность которых
снабжена пучком очень длинных неподвижных стереоцилий. Эти
структуры не имеют базальных телец и на электронограммах
оцениваются как очень высокие микроворсинки. На гистологиче-
ских препаратах стереоцилии склеиваются на поверхности каждой
клетки, образуя как бы язычок пламени. Главные клетки
придатка играют большую роль в процессе обратного всасывания
проходящей по придатку жидкости. Показано, что более 90 %
жидкости, покидающей извитые семенные канальцы, абсорбирует-
ся в выносящих канальцах и в канале придатка. Кроме того,
главные клетки обладают секреторной активностью.
Второй тип клеток стенки канала придатка — базальные
клетки — небольшой округлой формы, ядра которых обнаружи-
ваются у базальной мембраны эпителия. Их функция не уста-
новлена.
2.15.4. Предстательная железа
Предстательная железа человека — крупнейшая из добавочных
желез мужского полового аппарата, вырабатывает слегка кислый
секрет, содержащий, помимо прочих компонентов, лимонную ки-
слоту. и кислую фосфатазу. Секрет железы служит для разбавле-
ния содержимого, выходящего из яичка.
В качестве главных функциональных тканей железа содержит
железистую и гладкую мышечную ткань (рис. 2.234). В центре
органа можно обнаружить звездообразный просвет — простатиче-
скую часть уретры, выстланный переходным эпителием. Железы
в составе органа образуют в совокупности разветвленную альвео-
лярно-трубчатую железу. У пожилых людей разрастание желези-
стой ткани, часто перекрывающее уретру, носит название аденомы
предстательной железы.
418
Рис. 2.234. Предстательная железа
человека.
А — центральная зона. Б — перифери-
ческая (подкапсульная часть). В —
структура стенки железы.
1 — уретра; 2 — гладкая мышечная
ткань; 3 — простатические железы; 4 —
эпителий; 5 — простатические камни.
14*
419
2.16. Женская половая система
Изучение морфологических основ овогенеза важно для понимания
патологии гаметогенеза, нарушений репродуктивной функции жен-
щины. Квалифицированное знание гистофизиологии яичников —
процессов роста и созревания фолликулов, овуляции, функциони-
рования эндокринных компонентов — необходимо для анализа
причин дисфункции женской половой сферы. Знание процессов
развития половой системы поможет врачу в выявлении врожден-
ных пороков развития и в оценке гипопластических состояний
женской половой системы.
2.16.1. Яичник
Топографическую локализацию, структуру и взаимоотношения
многочисленных составных компонентов яичника иллюстрирует
рис. 2.235.
Орган покрывает белочная оболочка. Для ее поверхност-
ного эпителия характерно наличие множественных микро-
ворсинок на апикальной части этих клеток (рис. 2.236). Соедини-
тельная основа содержит несколько слоев коллагеновых волокон и
фибробластов, строение и ход которых зависят от активности
развития фолликулов, атретических тел и т. д. Без резких границ
белочная оболочка переходит в строму яичника, содержащую
специфические веретенообразные клетки. Корковое вещество осо-
бенно богато клетками, напоминающими фибробласты, их тяжи
вместе с волокнами образуют своеобразные завихрения. В сравне-
нии с корковым мозговое вещество богаче сосудами. Здесь пред-
ставлены спиральные артерии и обширные венозные сплетения.
В корковом веществе расположены фолликулы на раз-
личн^к^-ста тниые--па-заития (рис. 2.237). Примордиальные фолли-
кулы занимают наиболее периферическое положение. Они состоят
из овоцит§, окруженного одним слоем плоских фргцщкударных
кле^рк. В растущем первичном фолликуле эпителии становится
столбчатым. Во вторичном фолликуле появляется многослойный
эпителий, в третичном появляется полость с жидкостью. У вторич-
нЙВГитфетичных фолликулов вокруг яйцеклетки четко определя-
ется однородная пр.о.З.р.ач на .я зона, которая появляется в
конце стадии первичного фолликула и является продуктом деятель-
ности и фолликулярного эпителия, и.овоцита. Зону окружает слой
фодликулоцитов — гранулярный слой (рис. 2.238). Образующие его
фолликулрциты богаты органеллами синтеза и содержат много-
численные пицоцитознд[£_пузырьки, что является морфологиче-
ским свидетельством их активного участия в трофике овоцита и
продукции фолликулярной жидкости. Многочисленные отростки
этих клеток внедряются в прозрачную зону. У третичных фолли-
кулов (граафовы пузырьки) зона имеет очень характерные перепле-
тения ~фйбрййлярйьи, структур,- с множественными окончатыми
420
Рис. 2.235.
421
6
3
4
5
Рис. 2.235. Яичник, у/
А — схема микроскопического стро-
ения яичника (в модификации по
Б — структура коркового слоя.
1 — поверхностный эпителий; 2 —
белочная оболочка; 3 — соедини-
тельнотканная строма; 4 — примор-
диальные фолликулы; 5 — вторич-
ные фолликулы; 6 — третичные фол-
ликулы; 7 — тека фолликулов; 8 —
атретические фолликулы; 9 — фол-
ликулярная жидкость; 10 — овоцит;
11 — старое желтое тело; 12 — кро-
веносные сосуды; 13 — яйценосный
холмик; 14 — эндокриноциты.
образованиями, через которые на этой стадии осуществляется ак-
тивный контакт эпителиоцитов и овоцита.
Овоцит в третичных фолликулах «оттеснен» к одному полюсу,
окружен слоем фолликулоцитов (лучистым венцом). Такой яйце-
носный холмик на сканограмме виден со стороны полости фолли-
кула. Остальной участок фолликулярного эпителия — грануляр-
ный слой — ограничивает полость фолликула, заполненную жид-
костью.
На последовательных стадиях развития фолликулов и особенно
пузырчатых форм прослеживается постепенное образование во-
круг них с о е дили.д»ях _р_б.0_д Ряе_к: на-
ружуру у унутрениай-теки (рис. 2.239). Внутренняя тека, рас-
положенная под базальной мембраной фолликула, имеет выражен-
ную капиллярную сеть, богата фибробластоподобными клетками и
характерными крупными полигональными, светлыми клетками
Рис. 2.236. Покровный эпителий яичника.
А — СЭМ. Б — микроворсинки на апикальной поверхности эпителиоцитов (де-
таль к А).
422
423
424
Рис. 2.237. Фолликулы.
А — примордиальные фол-
ликулы. Б — ультраструк-
тура примордиального
фолликула (ТЭМ). В —
первичный фолликул. Г —
вторичный фолликул. Д —
третичный фолликул. Е —
общий вид яйценосного
холмика. Ж — СЭМ хол-
мика.
1 — овоцит; 2 — прозрач-
ная зона; 3 — слой фол-
ликулоцитов; 4 — базаль-
ная мембрана; 5 — фолли-
кулярная жидкость; 6 —
лучистый венец; 7 — яйце-
носный холмик; 8 — внут-
ренняя текальная оболоч-
ка; 9 — наружная текаль-
ная оболочка.
425
426
Рис. 2.238. Оболочки овоцита.
А— общий вид слоя фолликулоцитов (СЭМ). Б — поверхность прозрачной зоны
(СЭМ). В — ультраструктура овоцита, фолликулярных эпителиоцитов и их кон-
такт с прозрачной зоной.
I — фолликулоциты и их отростки; 2 — прозрачная зона; 3 — овоцит; «окна» в
прозрачной зоне; 4 — полость фолликула; 5 — гранулярный слой.
427
428
Рис. 2.239.
429
Рис. 2.239. Оболочки фолликула.
А — ультраструктура оболочек. Б — СЭМ. В — функциональные взаимодействия
текальных эндокриноцитов и фолликулоцитов. Г — ультраструктура эндокрино-
цитов.
БМ — базальная мембрана; Т — текальный эндокриноцит; К — капилляр; Я —
ядро; А — артериола; ВТ — внутренняя тека; НТ — наружная тека; Ф — фолли-
кулярные эпителиоциты; ФБ — фибробласт; М — миоидная клетка; ПФ — полость
фолликула; ФСГ — фолликулостимулирующий гормон; ЛГ — лютеинизирующий
гормон; Л — липиды.
тека — эндокриноцитами. Эндокриноциты богаты липидами и
митохондриями, с трубчатыми кристами — свидетельство их сте-
роидопродуцирующей деятельности. Взаимодействие этих клеток и
фолликулоцитов очень важно в процессе образования гормонов
яичника: текапьрые- эидокринопиты.. участвуют в образовании
андроуеыо-и, я фолликуло ты — в последующей ароматизации
андрогенов С образованием,.астрогенов. В сравнении’Т внутренней
наружная оболочка фолликула имеет типичное строение волокни-
стой соединительнотканной оболочки.
Обе оболочки вместе с фолликулярным слоем и прозрачной
430
431
зоной яйцеклетки в целом формируют своеобразный гемато-
фолликулярный барьер, имеющий определяющее значение в изби-
рательном транспорте веществ к яйцеклетке.
Овуляция. В зависимости от времени цикла, а именно к его сере-
дине, в яичнике появляется преовулярный фолликул —
третичный фолликул с наибольшей полостью, в котором у основа-
ния яйценосного холмика в скором времени просматриваются явле-
ния отека и дезинтеграция фолликулоцитов. По мере развития
овуляторного процесса (рис. 2.240) явления отека сна-
чала невыраженные, затем прогрессивно нарастают, обусловливая
разрыхление фолликулоцитов яйценосного холмика. Нарастание
отека способствует отрыву яйцеклетки с окружающими ее фолли-
Рис. 2.240. Преовулярный фолликул и овуляция.
А — начало отека у основания яйценосного холмика. Б — прогрессирующий отек
и разрыхление фолликулярного эпителия яйценосного холмика. В — освобождение
овоцита из яйценосного холмика, в овоците веретено деления. Г — расположение
овоцита у поверхности яичника непосредственно перед овуляцией. Д — выход
овоцита из яичника. Е — фолликул после овуляции.
1 — овоцит; 2 — лучистый венец; 3 — полость фолликула; 4 — веретено деления;
5 — отек у основания яйценосного холмика; 6 — гранулярный слой; 7 — поверх-
ностный эпителий; 8 — текальные эндокриноциты; 9 — капилляры.
432
433
Рис. 2.241. Атретический фолликул.
А — расширение сосудов, дезинтеграция гранулярного слоя, увеличение липидов
в фолликулоцитах. Б — дистрофия овоцита. В — атретическое тело. Г — эндокри-
ноцит, митохондрии с тубулярными кристами.
1 — овоцит; 2 — фолликул; 3 — текоциты; 4 — остатки прозрачной зоны; 5 —
соединительнотканный рубец; 6 — гемокапилляры.
434
435
кулярной и прозрачной оболочками от основания яйценос-
ного холмика, с последующим ее свободным положением в
фолликулярной жидкости. Появление веретена деления соз-
ревания в сводите овуляторного фолликула свидетельствует
о снятии блока мейоза, что обусловлено пиком лютеинизирую-
щего гормона.
Процесс овуляции в физиологических условиях происходит в
каждый менструальный цикл, примерно в его середине. При этом
возникают истончение гранулярного слоя преовуляторного фол-
ликула, направленного к поверхности яичника, отек и последую-
щее истончение поверхностного эпителия над таким фолликулом.
На рис. 2.240, Б, В, Г видны положение яйцеклетки у образую-
щегося отверстия (разрыва) в покровном эпителий, возникновение
веретена деления созревания и выход овоцита из фолликула на
поверхность яичника — овуляция.
Основная часть начавших развитие фолликулов подвергается
атрезии. Для таких фолликулов характерны прогрессирующее
уплотнение яйцеклетки, утолщение прозрачной зоны, регрессия
фолликулярного эпителия. Начальным моментом процесса являет-
ся застой крови в сосудах тека, накопление липидов в фолликуло-
цитах (при параллельном развитии дистрофических процессов в
овоците). Одновременно наблюдаются пролиферация текоцитов
и капилляров. Следствием этого процесса является образование
a и е с к и х тел (рис. 2.241), состоящих из упорядо-
че 1 ь ’ -и^тудиал» на расположенных, активно разросшихся на месте
погибших фолликулоцитов стероидопродуцирующих клеток тека,
прослоенных вросшими сюда капиллярами, т. е. образуется струк-
тура, идентичная эндокринной железе. В центре атретического
тела в зависимости от периода его развития могут быть погибаю-
щий овоцит, либо только остатки уплотненной прозрачной зоны,
либо на конечной стадии развития — сформировавшийся соеди-
нительнотканный рубец.
Развитие желтого тела. На рис. 2.242 прослеживаются эта-
пы развития желтого тела — от стадии расцвета до
образования белого тела. Возникает трансформация клеток
гранулярной зоны (фолликулоцитов) в лютеиновые клетки.
Этот процесс начинается еще до овуляции и активизируется
после нее. При этом происходит активное врастание капилляров
в ранее бессосудистый фолликулярный слой и последующее
прослоение образующихся лютеиновых клеток капиллярами.
Периферическую часть желтого тела составляет слой тека —
лютеоцитов, образованных при врастании сюда активной тека-
ткани вместе с капиллярами. Дифференцируются 2 вида лю-
теоцитов: светлые и темные. При обратном развитии происходит
жировое перерождение лютеоцитов, запустевание капилляров, раз-
растание соединительной ткани и соответственно уменьшение
размеров и изменение формы желтого тела на препарате. В за-
висимости от стадии развития желтого тела в центре его может
быть кровоизлияние (самый ранний период), образование соеди-
436
Рис. 2.242. Желтое тело.
А — период расцвета. Б — период регрессии. В — период белого тела. Г — учас-
ток желтого тела в его периферической части. Д — участок желтого тела в цент-
ральной части. Е — ТЭМ гранулезных лютеоцитов.
1 — гранулезные лютеоциты; 2 — текальные лютеоциты; 3 — кровеносные капил-
ляры; 4 — запустевшие кровеносные капилляры; 5 — гиалиновый материал; 6 —
кровеносные сосуды стромы; 7 — соединительнотканная строма яичника. СЛ —
светлые лютеоциты; ТЛ — темные лютеоциты; ЛГ — липидная гранула; МК —
межклеточные канальцы; Я — ядро.
437
1
5
7
Рис. 2.242. Продолжение.
нительнотканного центра или рубцовой ткани (поздний пе-
риод) .
При сравнении яичников животных и яичников человека обра-
щает внимание более активно развитая соединительнотканная
строма у человека, «скудность» развивающихся фолликулов и
желтых тел.
438
д
439
Рис. 2.242. Продолжение.
440
Развитие яичника. Гонобласты образуются в эпибласте. Затем
их положение в желточной энтодерме идентифицируется при
обработке на гликоген, щелочную фосфатазу. Гонобласты мигри-
руют по кровеносным сосудам и мезенхиме в половые валики —
область на медиальной поверхности первичной почки, состоящую
из целомического эпителиального покрова, подлежащей мезен-
химы и элементов первичной почки. Так происходит закладка
яичника (рис. 2.243). Установление контакта половых клеток
с элементами специфической стромы является ключевым момен-
том гаметогенеза. У эмбриона 5-й недели развития медиальнее
продольно срезанной первичной почки уже различим развиваю-
щийся половой валик — эмбриональная гонада. В клеточном мас-
сиве тканей гонады поселившиеся здесь гонобласты (овогонии)
идентифицируются как более крупные и светлые клетки на фоне
более мелких темных клеток дифференцирующейся мезенхимы.
Динамика начала развития фолликулов, пролиферации и
структурных изменений гоноцитов, развивающихся в половых ва-
ликах, соотношение первичных овогоний и овоцитов в различные
периоды эмбрионального развития показаны на рис. 2.244, А.
Обращает внимание высокая активность процесса деления половых
Рис. 2.243.
441
Рис. 2.243. Развитие яичника.
А — миграция половых клеток из жел-
точного мешка в зачаток гонады (схе-
ма) . Б — начальный этап формирова-
ния полового валика. В — половой ва-
лик на 5-й неделе развития: соотно-
шения полового валика (а) и первич-
ной почки (б), сагиттальный срез.
Г — структурные компоненты полового
валика (деталь к В).
1 — первичные половые клетки; 2 —
желточная энтодерма; 3 — мезенхима:
4 — сосуды; 5 — первичная почка; 6 —
эпителий полового валика; 7 — хорда;
8 — нервная трубка; 9 — капиллярный
клубочек первичной почки; 10 — ка-
нальцы первичной почки.
442
клеток: возникают несколько миллионов овогоний. Часть овогоний
рано прекращает деление и переходит в профазу мейоза — обра-
зуются овоциты. При их окружении эпителием (фолликулярным)
возникает блок профазы мейоза. Другая часть овогоний при этом
еще продолжает процесс митотического деления. Постепенно все
овогонии переходят в профазу мейоза и становятся овоцитами.
У новорожденной девочки остаются только единичные овогонии,
т. е. процесс увеличения половых клеток у женской особи практи-
чески заканчивается еще в эмбриональном периоде.
Схема (рис. 2.444, Б) иллюстрирует взаимоотношения
овоцита с элементами целомического эпителия (внедрение его
клеток в область расположения овоцитов) и сетью яичника —
мезонефральной бластемой (выселение клеток из сети в эту же
область). Участие именно сети в индукции мейоза и последующем
Рис. 2.244. Развитие фолликулов.
А — половые клетки на разных этапах эмбрионального развития яичника. Б —
взаимодействие половых клеток с сетью яичника, элементами стромы и текаль-
ными эндокриноцитами (по Л. Ф. Курило).
Д — овоцит в диплотене; ДТ — овоцит в диктиотене; ДКХ — овоцит в прелепто-
генной деконденсации хромосом; 3 — овоцит в зиготене; КХ — овоцит в прелеп-
тотенной конденсации хромосом; Л — овоцит в лептотене; СИ — овогония в ин-
терфазе; ОМ — овогония в митозе; П — овоциты в пахитене; ПФ — примордиаль-
ный фолликул; ПЧФ — первичный фолликул; ПЭ — поверхностный эпителий;
ТК — тяжи соединительнотканных клеток.
443
Рис. 2.244. Продолжение.
444
Рис. 2.245. Формирование яичника.
А — плод 12 нед. Б — новорожденный ребенок.
1 — половые клетки; 2 — префолликулярные клетки; 3 — гибель развивающихся
фолликулов.
445
формировании эпителиального окружения овоцитов, равно как
участие сети в образовании тека — эндокриноцитов, в настоящее
время признается многими учеными. Окружение овоцитов фолли-
кулярным эпителием создает длительную (на десятки лет) блокаду
мейоза. И только пик лютеинизирующего гормона снимает эту
блокаду в доминантном преовуляторном фолликуле.
В яичнике новорожденно^ девочки (рис. 2.245)
корковый слой насыщен примордиальными фолликулами, их окру-
жает нежная соединительнотканная строма. Имеются единичные
фолликулы, находящиеся на разных стадиях развития (следствие
воздействия гормонального фона матери). В течение короткого
времени происходит гибель этих форм. Для яичника ребен-
к а первых лет жизни характерно индифферентное состояние
гонады — слой примордиальных фолликулов, с малым числом
ранних форм роста фолликулов.
2.16.2. Матка
Строение и взаимоотношение матки с маточ-
ными трубами видны на рис. 2.246. В матке различаются
А — взаимоотношения матки, маточных труб, яичника (схема). Б — общий вид
матки. В —строение матки женщины в репродуктивном периоде в разные фазы
цикла: I — фаза пролиферации, II — фаза ^секреции, III — фаза десквамации.
а — эпителий; б — соединительнотканная основа; в — железы; г — гладкие мышцы;
д — сосуды; е — гемостаз и диапедез элементов крови.
1 — воронка маточной трубы; 2 — ампула маточной трубы; 3 — перешеек маточной
трубы; 4 — маточная часть трубы; (5 — эндометрий;^ — миометрий;^— перимет-
рий; 8 — полость матки; 9 — шейка матки; 10 — влагалище; 11—яичник.
446
7
в
447
3 оболочки: эндометрий, миометрий, периметрий. Эндометрий об-
ращен в полость матки, имеет складчатость, выстлан рднослой-
ным цилиндрическим эпителием, под ним хорошо выражена
соединительнотканная стромгГТгрубчатыми железами, количество
и форма которых варьируют в зависимости от стадии цикла.
Миометрий — мощный слой из гладких мышц, пучки которых рас-
положены продольно, поперечно и косо и пронизаны хорошо раз-
витой сосудистой сетью. В периметрии видны крупные сосуды,
вступающие в матку, интрамуральные ганглии.
В зависимости от фазы цикла имеют место явные отличитель-
ные особенности эндометрия (см. рис. 2.246 и рис. 2.247).
В стадию ранней пролиферации слой сравнительно тонок, еще сла-
бая пролиферация желез, малая васкуляризация. Эпителиальный
покров эндометрия состоит из секреторных и реснитчатых эпите-
лиоцитов. В отличие от этой фазы в последующей фазе пролифе-
рации (середина цикла у женщин) происходит активное разраста-
ние трубчатых желез, обильно васкуляризируется основа слизистой
оболочки, происходит трансформация ее клеточного состава.
Рис. 2.247. Эндометрий.
А — фаза пролиферации. Б — фаза секреции. В — поверхность эпителиального
покрова и устье железы (СЭМ). Г — мерцательные эпителиоциты эндометрия.
1 — эпителий; 2 — соединительная ткань; 3 — гладкие мышцы; 4 — железы; 5 —
реснички; 6, 7 — межклеточные контакты.
448
15—321 ‘
449
A
Рис. 2.248. Развитие матки.
А — динамика формирования матки и
ее широкой связки (I—III). Б— пара-
мезонефральные и мезонефральные ка-
налы эмбриона 2-го месяца развития.
В — образование зачатка матки — слия-
ние парамезонефральных каналов. Г —
матка плода 3-го месяца развития. Д —
матка плода 6 мес. Е — матка ново-
рожденной девочки.
1 — нервная трубка; 2 — хорда; 3 —
кишка; 4 — гонада; 5 — мезенхима; 6 —
канал мезонефроса; 7 — парамезонеф-
ральный канал; 8 — мезонефрос; 9 —
мочеточник; 10 — широкая связка мат-
ки; 11 — полость матки; 12 — эндомет-
рий; 13 — миометрий.
450
15*
451
Рис. 2.248. Продолжение.
Покровный эпителий при этом становится высоким цилиндриче-
ским, активно секреторным. В последующей фазе секреции железы
особенно активно секретируют и принимают вид бухтообразных
расширений, заполненных секретом. Появляются богатые гликоге-
ном и каплями жира многочисленные децидуальные клетки в
соединительнотканной строме, эндометрий утолщен. В следующей
фазе — пременструальной — нарушается кровоснабжение, в стро-
ме эндометрия возникает стаз (застой). Дезинтеграция эндомет-
рия прогрессирует, возникает некроз функционального слоя эндо-
метрия с последующим отторжением и кровотечением.
Развитие матки. В образовании матки (рис. 2.248) принимают
участие дистальные отделы парамезонефральных протоков и
окружающая их мезенхима. В срезе через каудальную часть
эмбриона 2-го месяца развития парамезонефральные каналы за-
нимают центральное положение, латеральнее расположены прото-
ки первичной почки — мезонефральные. (Верхние отделы параме-
зонефральных каналов формируют маточные трубы.) Слияние в
нижних отделах парамезонефральных каналов дает образование
матки, которое заканчивается к концу 3-го месяца развития.
Сначала виден формирующийся эпителиальный канал, вокруг ко-
торого затем возникает активация мезенхимы и ее последующая
дифференцировка в гладкую мускулатуру и соединительную ткань.
452
У новорожденной девочки эндометрий находится в состоянии по-
коя. Мышечный слой еще сравнительно тонкий, васкуляризация
слабая.
2.16.3. Маточная труба
Маточная труба имеет 3 оболочки: слизистую, мышечную, се-
розную. Слизистая оболочка имеет продольные складки,
которые в зависимости от отдела трубы и стадии цикла имеют
выраженные отличия (рис. 2.249), особенно в ампулярной части.
Покровный эпителий также имеет разные количественные соотно-
шения реснитчатых и секреторных клеток в зависимости от отдела
трубы (при специальной окраске на глюкозаминогликаны секре-
торные клетки эпителиального покрова особенно четко иденти-
фицируются). Если реснитчатые клетки способствуют продви-
жению зародыша, то секреторные принимают участие и в его тро-
фике.
Развитие маточных труб. В образовании маточных труб прини-
мают участие верхние отделы парамезонефральных каналов и
окружающая их мезенхима. На рис. 2.250 прослеживается после-
довательный процесс развития трубы в эмбриональный и пост-
натальный периоды: образование и дифференцировка слоев трубы,
формирование сложного характера складок. Соотношение разме-
ров матки и маточных труб в разные возрастные пе-
риоды различно: прогрессирующее смещение соотношений дли-
ны шейки и тела матки, изменение размеров и положения яич-
ника в брюшной полости, изменение размеров и степени извили-
стости маточных труб (рис. 2.251).
2.16.4. Влагалище
Тканевые компоненты стенки влагалища видны на
рис. 2.252. Обращает внимание десквамация поверхностного эпи-
телия. При этом количество, форма, степень ороговения слущи-
вающихся клеток варьируют в зависимости от гормонального фона
в организме, определяемого активностью яичников. Характери-
стика этих показателей может в общих чертах указать на стадию
цикла. Метод анализа цитологического состава содержимого вла-
галища (мазки) используется как метод диагностики.
2.16.5. Молочная железа
В молочной железе (рис. 2.253) четко определяется
дольчатость органа, разделение долек прослойками рыхлой соеди-
нительной и жировой ткани. По этим прослойкам проходят кро-
веносные сосуды и междольковые выводные протоки, образующие
расширения — молочные синусы, заполненные секретом. Внутри-
дольковые выводные протоки — молочные ходы — заканчиваются
концевыми отделами — альвеолами, где происходит процесс се-
453
454
Рис. 2.249. Маточная труба.
А — наружная часть ампулы. Б — истмическая часть трубы. В — интерстициаль-
ная маточная часть. Г — слизистая оболочка трубы в ампулярной части. Д —
слизистая оболочка трубы истмического отдела (ШИК-реакция).
1 — складки слизистой оболочки; 2 — эпителий; 3 — соединительнотканная осно-
ва слизистой оболочки; 4 — мышечная оболочка; 5 — полость трубы; 6 — серозная
оболочка; 7 — секреторные клетки; 8 — кровеносные сосуды.
455
456
Рис. 2.250. Развитие маточной трубы.
Л — плод 20 нед. Б — плод 30 нед. В — ново-
рожденная девочка. Г — пубертатный период.
I — эпителий; 2 — соединительнотканная ос-
нова; 3 — мышечная оболочка; 4 — складки;
5 — полость трубы.
Рис. 2.251. Возрастные изменения размеров и особенности топографии
яичника, матки и маточных труб (в модификации).
А — у новорожденной. Б — в пубертатном периоде. В — в репродуктивном периоде.
Г — в постклимактерическом периоде.
457
Рис. 2.252. Стенка влагалища.
А — общий вид. Б — эпителиальный покров в фазе пролиферации (ШИК-реакция).
1 — слизистая оболочка; 2 — мышечная оболочка; 3 — адвентициальная оболочка;
4 — многослойный эпителий; 5 — собственная пластинка слизистой оболочки;
6 — кровеносные сосуды.
креции. Лактоциты куполоообразной формы заполнены каплями
липидов крупных размеров, вакуолями, белковыми гранулами.
На схеме виден процесс формирования секрета в лактиру то-
щей железе, участие в нем цитоплазматических структур,
формирование и отшнуровка апикальной части. К базальной по-
верхности альвеолоцитов примыкают миоэпителиальные клетки.
В нелактирующей молочной железе паренхима
железы в сравнении с лактирующей резко уменьшена, а основная
масса органа представлена волокнистой соединительной и жиро-
вой тканью. Не развиты концевые отделы, у мелких протоков не
видны просветы.
458
Рис. 2.252. Продолжение.
В — цитология влагалищных мазков: I — 1-й день цикла (ранняя пролифератив-
ная фаза); II — 11 — 14-й дни цикла (овуляторная фаза); III — 21-й день цикла
(лютеиновая фаза); IV — 28-й день цикла (предменструальная фаза): а — про-
межуточные клетки, б — поверхностные (ацидофильные клетки), в — поверхност-
ные базофильные клетки, г — полиморфно-ядерные лейкоциты.
Развитие молочной железы. На рис. 2.253 у эмбриона 6-й не-
дели развития видна эктодермальная закладка молочной железы.
Последующие инвагинации и ветвления эпителиальных тяжей
происходят до 4-го месяца развития, их активное разраста-
ние — к 6-му месяцу развития. К рождению формируются только
главные протоки, в последующем активно накапливается жировая
ткань и при половом созревании происходит активное увеличение
эпителиальных тяжей. Только наступление беременности вызывает
максимальную дифференцировку структурных компонентов мо-
лочной железы, причем ацинусы дифференцируются только в по-
следней трети беременности.
459
460
Рис. 2.253. Молочная железа.
А — этапы развития (I—IV) молочной железы. Б — лактирующая молочная же-
леза. В — нелактирующая молочная железа.
1 — концевые ацинусы; 2 — внутридольковый проток; 3 — соединительнотканная
строма; 4 — молочный синус; 5 — кровеносные сосуды; 6 — эпителиальная молоч-
ная точка; 7 — пролиферация эктодермального слоя; 8 — образование эктодер-
мальных эпителиальных тяжей; 9 — прогрессирующие ветвления эпителиальных
трубок, образование протоков.
461
Рис. 2.253. Продолжение.
Г — схема секреции.
1 — пластинчатый комплекс; 2 — цитоплазматическая сеть; 3 — жировые гранулы;
4 — белковые гранулы; 5 — ядро.
462
ЭМБРИОЛОГИЯ
ОСНОВНЫЕ ЭТАПЫ РАЗВИТИЯ
ЭМБРИОНАЛЬНЫЙ ПЕРИОД
ПЛОДНЫЙ ПЕРИОД
3
Эмбриология человека
Изучение основ эмбриологии человека, знание физиологиче-
ской динамики развития, его особенностей в различные сроки
эмбрионального периода позволяют врачу понять и по возмож-
ности скорригировать патологию развития, а в ряде случаев
даже осуществить ее профилактику. Представление о взаимоотно-
шениях нЯЬда человека и материнского организма дает возмож-
ность оценить весь цикл биологических явлений, сопровождаю-
щих течение беременности, что важно для тактики ведения родов,
прогнозирования здоровья плода. Знание медицинских аспектов
эмбриологии способствует решению таких задач, как лечение,
профилактика врожденных ненаследственных и наследственных
заболеваний, проблемы бесплодия, трансплантации органов и
тканей и др.
Во внутриутробном периоде развития человека разграничивают
эмбриональный (первые 8 нед) и плодный периоды.
Эмбриональный период — самый ответственный в эмбриональ-
ном развитии. В это время начинают свое развитие все эмбрио-
нальные и неэмбриональные структуры. Образовавшиеся три заро-
дышевых листка в течение этого времени дифференцируются в
различные ткани и органы. К концу эмбрионального периода
все главные органные системы не только начинают свое развитие,
но многие из них проявляют определенную функциональную
активность. В развитии человека эмбриональный период является
критическим; он особенно подвержен влиянию материнского орга-
низма и самых различных факторов внешней среды.
Плодный период — период дальнейшего развития гистогенети-
ческих и формообразовательных органных процессов. Их суть
изложена в соответствующих разделах этого атласа, где приво-
дится динамика развития той или иной органной системы.
3.1. Эмбриональный период
3.1.1. Первая неделя развития
В этот период происходит оплодотворение, дробление, начало им-
плантации зародыша.
Оплодотворение у человека моноспермное и в своей динамике
не отличается от других млекопитающих. После овуляции яйце-
клетка (овоцит II) попадает на фимбрии трубы и проходит в ам-
пулярную часть. Она окружена лучистым венцом (рис. 3.1) и
находится в метафазе второго деления созревания.
На рис. 3.2 представлен процесс оплодотворения в динамик^?
Спермий, войдя в половые пути, проходит процесс капацитации,
который длится около 7 ч: гликопротеиновое покрытие и протеины
семенной плазмы удаляются с плазматической мембраны в обла-
464
Рис. 3.1. Овоцит перед оплодотворением.
А — овоцит из фолликула. Б — овоцит с первым полярным тельцем.
465
Рис. 3.2. Оплодотворение у человека.
А — схема (I, II). Б — яйцеклетка в маточной трубе. В, Г — стадии сближения
пронуклеусов (по Петрову). Д — ультраструктура эмбриона на стадии пронуклеу-
сов. Е — стадия пронуклеусов через 16—18 ч после оплодотворения (фазовый
контраст, препарат).
1 —просвет трубы; 2 — стенка трубы; 3 — яйцеклетка (вторичный овоцит); 4 —
прозрачная зона; 5 — лучистый венец; 6 — ламеллярные структуры; 7 — микро-
ворсинки; 8 — сперматозоид в процессе капацитации (а), в процессе акросомной
реакции и выделения ферментов (б), пенетрирует мембрану яйцеклетки, оставляя
плазматическую мембрану (в), проникает в цитоплазму яйцеклетки (г); 9 — жен-
ский пронуклеус; 10 — мужской пронуклеус; 11 —мейотическое веретено деления;
12 — митотическое веретено деления; 13 — хромосомы пронуклеусов; 14 — поляр-
ные тельца; 15 — дегенерирующий хвост спермия; д — образование мужского про-
нуклеуса, дегенерация хвоста сперматозоида, е — слияние пронуклеусов, ж —
зигота, митотическое веретено 1-го деления дробления.
сти акросомы спермия, что позволяет осуществиться затем акро-
сомальной реакции. После капацитации при внедрении спермия
в лучистый венец происходит акросомальная реакция. Наружная
мембрана акросомы во многих участках «сплавляется» с перед-
ними 2/з цитоплазматической мембраны головки спермия. Затем
сплавления мембран разрываются, образуются маленькие пу-
зырьки и множественные отверстия, через которые выходят
энзимы акросомы. Фермент гиалуронидаза инициирует «рас-
сеивание» клеток лучистого венца, а трипсиноподобная субстан-
ция способствует прохождению спермия через прозрачную зону.
Прозрачная зона прикрепляет спермин, что связано с наличием
на ее поверхности рецепторов для сперматозоидов — гликопротеи-
нов (число рецепторов достигает колоссального количества).
Эти гликопротеины индуцируют и акросомальную реакцию. Про-
467
зрачная зона контактирует с акросомальными ферментами, отго-
роженными от остальной части спермия внутренней частью акро-
сомальной мембраны. Ферменты разрушают прозрачную зону, и
один из множества спермиев проходит в перивителлиновое про-
странство — пространство между прозрачной зоной и мембраной
овоцита. На рис. 3.2 видно, что происходит слияние мембраны овоци-
та и сохранившейся после акросомальной реакции части мембраны
на головке спермия. Внутрь цитоплазмы у овоцита человека входят
головка и хвостовая часть спермия, плазматическая мембрана спер-
мия при этом остается за пределами поверхности овоцита. В течение
нескольких секунд изменяются свойства мембраны овоцита и она
становится невосприимчивой к другим спермиям (предотвращается
полиспермия). В овоците возникает кортикальная реакция—
выброс ферментов кортикальных гранул, под действием которых
теряется способность прикреплять спермин — т. е. утрачивается ре-
цепторная активность. Вошедший спермий активирует клеточные и
молекулярные процессы в яйцеклетке, которая немедленно заканчи-
вает второе деление мейоза, образуя второе полярное тельце (ядро
такой яйцеклетки называют женским пронуклеусом). У спермия,
продвигающегося в это время к женскому пронуклеусу, дегенери-
рует хвост. Его ядро вначале более темное, чем женское, затем
вследствие его обводнения становится светлее и эти отличия реду-
цируются (ядро спермия называют мужским пронуклеусом).
Оплодотворение заканчивается потерей ядерных мембран прону-
клеусами, их слиянием и объединением хромосом.
Таким образом, оплодотворение восстанавливает диплоидный
набор хромосом, обеспечивает новые вариации в их структуре.
Результатом оплодотворения является инициация дробления зи-
готы. Без оплодотворения овоцит дегенерирует на 2-е сутки после
овуляции.
Дробление. На рис. 3.3 и рис. 3.4 видно, что характер дробле-
ния у человека в общих чертах сходен с дроблением у других
млекопитающих и особенно с этим же процессом у обезьян: ми-
тотическое деление зиготы на 2 дочерние клетки (бластомеры)
начинается сразу же после оплодотворения и продолжается
примерно 30 ч. Затем происходит серия митозов, нарастает коли-
чество бластомеров (четырехклеточная стадия наступает через
40 ч), прогрессивно формируются все меньшие по размерам
бластомеры. После 3 делений через 3 сут после оплодотворения
образуется морула, состоящая из 12—16 клеток, а к 4-му дню —
поздняя морула.
Этот процесс происходит в маточной трубе, а затем дробле-
ние продолжается в матке до стадии бластоцисты. Полость бласто-
цисты появляется у эмбриона, состоящего из 32 клеток, на
4-е сутки развития. Так же как и у животных, у человека прозрач-
ная зона сохраняется на протяжении всего периода дробления,
исчезая всего за несколько часов до имплантации.
Периферически расположенные светлые мелкие бластомеры
резко увеличиваются в числе — это будущие клетки трофобласта.
468
Б
Рис. 3.3. Дробление зародыша.
А — стадия двух бластомеров. Б —
стадия трех бластомеров. В — стадия
четырех бластомеров. Г — восьмикле-
точная стадия. Д — морула (по
Э. М. Китаеву). Е — стадия восьми
бластомеров (ТЭМ). Ж — стадия ран-
ней бластоцисты (ТЭМ). 3 — структура
бластомеров на стадии бластоцисты.
И — схема миграции и взаимоотноше-
ния дробящегося зародыша в маточной
трубе и матке.
1 — прозрачная зона; 2 — ядро; 3 —
бластомеры; 4 — клетки трофобласта;
5 — клетки эмбриобласта; 6 — полость
бластоцисты; 7 — ядрышки.
469
ж
470
Рис. 3.3. Продолжение.
471
Крупные — более темные бластомеры — занимают центральное
положение (внутренняя клеточная масса), их число значительно
меньшее (в среднем 1 клетка на 10—12 клеток трофобласта).
В этот период динамично изменяются и усложняются межклеточ-
ные связи между бластомерами. В клетках двухклеточного заро-
дыша ядрышко плотное, что свидетельствует об отсутствии соб-
ственных синтезов в бластомерах. Затем со стадии 8 бластомеров
оно постепенно превращается в активное ретикулярное ядрышко.
Это период активации синтезов бластомеров, развития в них гра-
нулярной цитоплазматической сети.
Соотношение внутренней клеточной массы (эмбриобласта) и
трофобласта в бластоцисте ранней и поздней стадии видно на
рис. 3.3.
После свободного положения бластоцисты в матке (2—3 дня)
(см. рис. 3.4) к моменту имплантации на 6—7-е сутки за не-
сколько часов до имплантации происходит сбрасывание прозрач-
ной зоны. Это освобождение имеет своим следствием прекращение
Рис. 3.4. Стадия дробления.
А — зигота в полости маточной трубы (труба свернута в виде улитки по С. Хватову).
Б — зародыш Да стадии двух бластомеров в маточной трубе. В — зародыш на стадии
морулы. Г — бластоциста в полости матки.
1 — зародыш; 2 — полость трубы; 3 — слизистая оболочка трубы; 4 — эпителий
трубы; 5 — бластомеры; 6 — ядра бластомеров; 7 — эпителий матки; 8 — полость
бластоцисты; 9 — эмбриобласт; 10 — трофобласт.
472
473
Рис. 3.5. Зародыш в пре-
дымплаитационный период
и в начале имплантации.
Контакт зародыша (в об-
ласти эмбриобласта) с эпи-
телием эндометрия и нача-
ло имплантации.
А, Б — эмбрион обезьяны (по
Б. М. Пэттену). В — схема им-
плантации у человека на 7-е
сутки.
1 — полость матки; 2 — эндо-
метрий; 3 — эмбриобласт; 4 —
полость бластоцисты; 5 — эпи-
бласт; 6 — гипобласт; 7 — ци-
тотрофобласт; 8 — синцитио-
трофобласт; 9 — эпителий эн-
дометрия; 10 — железы; 11 —
кровеносные сосуды эндомет-
рия; 12 — трофобласт.
474
«дробления» и переход на обычный митоз, что сопровождается
бурным ростом зародыша.
Матка. Имплантация (рис. 3.5). В матке в этот период (к концу
1-й недели развития) имеет место секреторная фаза. Артерии
приобретают спиралеобразный вид, ткань эндометрия богато
васкуляризована, формируются богатые капиллярные сети эндо-
метрия. Железы усиливают секреторную активность.
Место имплантации зародыша человека может быть различ-
ным, но значительно чаще это происходит на передней или задней
стенке матки, в верхней ее части. В отличие от других млекопи-
тающих, имплантация у человека погружная (интерстициальная).
Погружение зародыша происходит «плашмя» вперед будущей дор-
сальной стороной тела. При этом трофобласт лизирует эпителий,
соединительную ткань и стенку сосудов. Имеет место и другое
отличие имплантации у человека от других млекопитающих:
довольно быстрое погружение бластоцисты в толщу матки
(за 24 ч бластоциста погружается больше чем наполовину, а через
40 ч — целиком).
3.1.2. Вторая неделя развития
На 2-й неделе развития продолжается имплантация зародыша,
разделяется внутренняя клеточная масса — эмбриобласт на эпи-
бласт и гипобласт, формируются амниотический и желточный
пузырьки, начинаются гаструляция, дифференцировка в трофобла-
сте двух слоев и др. (рис. 3.6).
Эмбриобласт. В этот период происходит трансформация мате-
риала эмбриобласта, его дифференцировка на 2 слоя: гипобласт
и эпибласт. Оба слоя вместе составляют зародышевый диск.
Г ипобласт — мелкие кубоидальные пенистого вида клетки,
граничащие с полостью бластоцисты. Они формируют тонкий
слой, подстилающий эпибласт. К концу 2-й недели в головной ча-
сти гипобласта обнаруживается небольшое утолщение, известное
как прехордальная пластинка (она указывает на место будущего
рта). Эта область высоких клеток, которые тесно связаны с эпи-
бластом. Гипобласт, разрастаясь по внутренней поверхности
трофобласта, формирует тонкую экзоцеломическую мембрану,
которая окаймляет полость, известную как первичный желточный
мешок — экзоцеломическая полость. У человека она не содержит
желтка. К концу 2-й недели развития клетки крыши первичного
желточного мешка, мигрируя и пролиферируя, формируют новую
полость — вторичный (дефинитивный) желточный мешок значи-
тельно меньших размеров, чем первичный. К этому времени по-
лость первичного желточного мешка спадается. Могут оставаться
лишь экзоцеломические пузырьки (см. рис. 3.6).
Эпибласт — прилежащий к трофобласту слой клеток
эмбриобласта, представлен упорядоченно расположенными клет-
ками, напоминающими псевдомногослойный призматический
эпителий. Между клетками эпибласта появляются маленькие
475
476
Рис. 3.6. Дифференцировка эмбриобласта и трофобласта.
А — 9-й день развития. Б — 13-й день развития (в модификации по Langhmann).
В — 3-я неделя развития (в модификации по Keith L. Moore).
1 — эпибласт; 2 — гипобласт; 3 — амниотическая полость; 4 — экзоцеломическая
полость и ее остаток — экзоцеломический пузырек (на Б); 5 — экзоцеломическая
мембрана; 6 — цитотрофобласт; 7 — синцитиотрофобласт; 8 — лакуны в синцитиотро-
фобласте; 9 — внезародышевый целом (полость хориона); 10 — внезародышевая
мезодерма (пластинка хориона); 11 — вторичный желточный мешок; 12 — соедини-
тельная ножка; 13 — крыша вторичного желточного мешка; 14 — соединительная
ткань эндометрия; 15 — железы эндометрия; 16 — кровеносные сосуды эндометрия;
17 — коагулированный фибрин; 18 — эпителий эндометрия.
477
полости, которые, сливаясь, образуют общую полость — будущую
амниотическую полость. Вследствие расхождения клеток крыши
полости в стороны участок трофобласта оказывается временно
ее крышей. Но затем боковые участки разрастающегося эпибласта
образуют направленные вверх складки (амниотическая эктодерма),
которые, сливаясь, формируют полный покров амниотической по-
лости — формируется амниотический пузырек. Осталь-
ная часть эпибласта, составляющая дно образованного амниоти-
ческого пузырька, представляет собой материал всех зародышевых
листков.
В период между 9—14-ми сутками в эпибласте зародыша уже
начинается видимый процесс дифференцировки и перемещения
клеточных элементов, известный как процесс гаструля-
ц и и с образованием первичной полоски. Этот процесс
в деталях прослеживается на рис. 3.7; 3.8; 3.9. Из материала первич-
ной полоски выселяется зародышевая энтодерма. Ее клетки, сдвигая
клетки гипобласта, занимают крышу желточного мешка. В это время
выселяется и внезародышевая мезодерма. Мигрируя, она тонким
слоем окружает амниотический и желточный пузырьки. Кроме того,
она занимает пространство между экзоцеломической мембраной
и трофобластом. Здесь в ее массе появляются лакуны, которые
затем, сливаясь, сформируют общую полость — полость внезаро-
дышевого целома — полость хориона. Внезародышевая мезодерма,
прилегающая к трофобласту, дифференцируется, образуя в будущем
хорионическую пластинку. В области будущей каудальной части
зародыша внезародышевая мезодерма формирует соединитель-
ный стебелек (соединительную ножку) — плотный тяж кле-
ток, связывающий амниотический и желточный пузырьки с трофо-
бластом.
Трофобласт. Динамика преобразований трофобласта в течение
2-й недели развития представлена на рис. 3.7—3.9. Еще раньше —
во время контакта бластоцисты с эндометрием — активно разра-
стающийся трофобласт приобретает четко выраженное двухслой -
ное строение (см. рис. 3.5). Внутренний слой — цитотрофо-
бласт тз^-иденФифицируется по четким границам клеток, в то время
как наружный слой — синцитиотрофобласт представляет собой
синцитий с Ьеспорядочно расположенными 'ядрами. Синцититро-
фобласт вскоре приобретает вид массивных многоядерных прото-
плазматических масс, наиболее развитых в области эмбриобласта.
В цитотрофобласте видны митозы, в синцитиотрофобласте их нет.
Однако масса последнего нарастает чрезвычайно быстро, что по-
зволяет клеткам цитотрофобласта встраиваться в слой синцитио-
трофобласта с потерей их собственных мембран.
В активно развивающемся синцитиотрофобласте появляются
изолированные пространства —- лакуны. К концу 2-й недели разви-
тия с лакунами синцитиотрофобласта затем соединяются расширен-
ные капилляры эндометрия и материнская кровь входит в лакунар-
ную систему. Процесс прогрессирует и затем постепенно устанавли-
вается маточно-плацентарная циркуляция. Разрастающиеся слои
478
Рис. 3.7.
479
Рис. 3.7. Имплантированный эмбрион человека в эндометрии матки (по
М. Д. Пэттену).
А — эмбрирн 9 дней. Б — эмбрион 11 дней. В — общий вид эндометрия и им-
плантированного зародыша 11 дней.
1 — эмбрион; 2 — эпителий эндометрия; 3 — железы эндометрия; 4 — соедини-
тельная строма и сосуды эндометрия; 5 — лакуны; 6 — амниотический пузырек;
7 — желточный пузырек; 8 — эпибласт; 9 — гипобласт; 10 — внезародышевая ме-
зодерма; 11 — цитотрофобласт; 12 — синцитиотрофобласт.
480
Рис. 3.8. Двухнедельный зародыш человека.
А — общий вид. Б — деталь (по А. Г. Кнорре).
1 — внезародышевая полость; 2 — хориальный эпителий; 3 — мезенхима хориона;
4 — эпителий амниона; 5 — амниотическая ножка; 6 — полость амниона; 7 — эпи-
бласт; 8 — первичная энтодерма; 9 — полость желточного мешка; 10 — стенка
желточного мешка.
трофобласта (будущий эпителий хориона) образует выросты —
первинчые ворсинки: их центр занимают клетки цитотрофобласта,
а периферию — синцитиотрофобласта.
Матка. В этот период развития в области нидации строма
эндометрия, окружающая зародыш, отечна, высоковаскуляризова-
на, с бухтообразными, активно секретирующими железами. Клет-
ки эндометрия богаты гликогеном и липидами. Эта реакция
известна как децидуальная реакция.
К 9-му дню бластоциста полностью погружается в строму
эндометрия. Дефект слизистой оболочки и имплантационный
кратер заполняются массой фибрина с примесью свернувшейся
крови и частиц материнских тканей. Полное закрытие дефекта за
счет регенерационных процессов в эпителии и соединительной
ткани занимает около 5 сут, и к 12—13-му дню эпителий эндо-
метрия покрывает полностью дефект стенки матки.
16—321
481
Рис. 3.9. Первичная полоска и миграция материала зародышевых листков
(с конца 2-й до конца 3-й недели развития).
А — дорсальная поверхность зародыша. Б — поперечное сечение зародыша.
1 — первичная полоска; 2 — гензеновский узелок; 3 — прехордальная пластинка;
4 — головной отросток; 5 — мезодермальный зачаток; 6 — энтодермальные клетки;
7 — внезародышевая мезодерма; 8 — эпибласт; 9 — будущая амниотическая экто-
дерма; 10 — гипобласт; 11 — клоакальная мембрана.
3.1.3. Третья неделя развития
В этот период полностью образуются 3 зародышевых листка,
формируются такие важнейшие структуры, как первичная полоска,
хорда, нервная трубка.
Зародыш в этот период превращается из плоской почти круг-
лой структуры в вытянутую с широким головным (вследствие
активного роста головной части нервной трубки) и узким каудаль-
ным концом.
Суть процессов, происходящих на 3-й неделе развития, обоб-
щают рис. 3.10 — рис. 3.15, на которых в динамике виден процесс
482
гаструляции, начавшийся еще на 2-й неделе развития. В ходе
этого процесса из эпибласта образуется первичная полоска —
источник материала энтодермального и мезодермального зароды-
шевых листков и будущая эктодерма.
Образование первичной полоски у человека идентично тако-
вому у других млекопитающих. Сначала в части диска, соответ-
ствующей будущему хвостовому концу зародыша, появляется
16*
483
Рис. 3.10. Период гаструляции.
А, Б, В, Г — образование хордального отростка (с центральным каналом), фор-
мирование мезодермы, начало образования складок (в модификации по Б. М. Пэт-
тену).
1 — первичная полоска; 2 — первичная ямка; 3 — хордальный отросток с цент-
ральным каналом; 4 — эктодерма; 5 — энтодерма; 6 — прехордальная пластинка;
7 — амниобласт; 8 — амниотическая мезодерма; 9 — полость амниона; 10 — нерв-
но-кишечный канал; 11 — мезодерма; 12 — кардиальная область; 13 — энтодерма;
14 — полость желточного мешка; 15 — желточная энтодерма; 16 — желточная
мезодерма; 17 — аллантоис; 18 — соединительная ножка; 19 — пластинка хориона;
20 — ворсинки хориона; 21 — клоакальная мембрана; 22 — передняя кишка; 23 —
задняя кишка; 24 — нервная трубка.
484
1
3
Б
5
Й *'
Za
В<*-!
д W
- L#*5
Б ».,*«>
Л.
Рис. 3.11. Зародыш человека 17 сут
(«Крым»).
А — срез на уровне первичного заро-
дышевого узелка. Б — краевая зона
зародышевого щитка.
1 — амниотический пузырек; 2 — голов-
ной отросток; 3 — гензеновский пузы-
рек; 4 — желточный пузырек; 5 — жел-
точная энтодерма; 6 — внезародышевая
мезодерма желточного мешка и амнио-
тического пузырьке; 7 — первичная по-
лоска.
А
2
Рис. 3.12. Первичная полоска.
А — первичная полоска. Б — попереч-
ное сечение зародыша, ранняя стадия.
1 — первичная полоска; 2 — энтодерма;
3 — мезодерма.
485
486
5
9
15
17
13
14
16
Б
Рис. 3.13. Образование нервной трубки и дифференцировка мезодермы.
AI—AIV — 17—21-й день развития, поперечное сечение зародыша, схема. Б —
окраска гематоксилин-эозином.
1 — мезодерма амниона; 2 — эпителий амниона; 3 — хорда; 4 — эктодерма; 5 —
полость амниона; 6 — нервная пластинка; 7 — нервная ямка; 8 — нервные валики;
9 — нервная трубка; 10 — мезодерма; 11—энтодерма; 12 — сомиты; 13 — ножки
сомитов; 14 — эмбриональный целом; 15 — соматоплевра; 16 — спланхноплевра.
А
Рис. 3.14.
487
Рис. 3.14. Развитие кровеносных сосудов.
А — образование сосудов в мезодерме желточного мешка, хориона и соединитель-
ной ножке. Б — кровяные островки в мезодерме желточного мешка зародыша 17 дней.
В — кровеносные сосуды в теле зародыша и их связь с ворсинками хориона в конце
3-й недели развития (в модификации по Keith L. Moore).
1 — кровеносные сосуды; 2 — кровяные островки; 3 — желточный мешок; 4 —
сердечные трубки; 5 — перикардиальная полость; 6 — амнион; 7 — амниотическая
полость; 8 — ворсинки; 9 — аллантоис; 10 — соединительная ножка; 11 — хори-
альная пластинка; 12 — сосудистое сплетение в желточном мешке; 13 — вителли-
новые артерии; 14 — вителлиновые вены; 15 — умбиликальная вена; 16 — умбили-
кальные артерии; 17 — дорсальная аорта; 18 — дорсальные интерсегментарные
артерии; 19 — дорсальные кардиальные вены; 20 — сосуды третичных ворсинок;
21 — артериовенозная сеть ворсинки.
утолщение эпибласта, которое, распространяясь, формирует плот-
ную линейную структуру, состоящую из колбообразных клеток
(первичная полоска) и лежащую вдоль длинной оси зародыша в
середине дорсальной поверхности эмбрионального диска. На го-
ловном конце первичная полоска заканчивается утолщением из
плотно упакованных клеток — гензеновским первичным узелком.
Он образует приподнятую область, окружающую первичную ямку.
488
Рис. 3.15. Развитие сердца (за-
родыш на 17—21-й день разви-
тия).
1 — эмбриональный целом; 2 — эн-
тодерма; 3 — спланхноплевра; 4 —
ангиогенные клетки; 5 — эктодерма;
6 — нервная ямка; 7 — хорда; 8 —
нервная трубка; 9 — первичная киш-
ка; 10 — мезодерма; 11 — аорта;
12 — эндокардиальные трубки.
489
Как уже указывалось, первыми из области передней части пер-
вичной полоски выселяются клетки будущей зародышевой энто-
дермы. Они включаются в подлежащей гипобласт, перемещая его
клетки кнаружи, к головному концу и латерально.
Уже упоминалось, что еще в конце 2-й недели развития часть
ранней мезодермы, выселяющаяся из области заднего участка пер-
вичной полоски, мигрировала за пределы зародышевого диска —
это внезародышевая мезодерма. Продолжая распространяться,
она примет участие в образовании мезодермального слоя амниоти-
ческого и желточного пузырьков, в формировании хориальной
пластинки, подстилающей цитотрофобласт, и в образовании
соединительной ножки между зародышем и трофобластом.
Несколько позднее инвагинирующие и мигрирующие массив-
ные слои мезодермальных клеток первичной полоски образуют
уже зародышевую мезодерму, материал которой распространяется
к головному концу и в латеральных направлениях. В головном
конце мезодермальные слои охватывают прехордальную пластинку
и, встречаясь впереди этой области, формируют сердечную пла-
стинку (см. рис. 3.13). Инвагинация клеток первичной полоски
и их миграция к головной части зародыша и латерально продол-
жаются всю 3-ю неделю. Уже с 17-го дня развития слой зароды-
шевой мезодермы полностью разделяет верхний слой эпибласта
(будущую эктодерму) и энтодерму, исключая область прехордаль-
ной пластинки и клоакальной мембраны в каудальном конце пер-
вичной полоски. Оба этих участка, как указывалось выше, содер-
жат тесно прилегающие слои клеток эктодермы и энтодермы. Сама
первичная полоска прогрессивно уменьшается в размере и полно-
стью исчезает к концу 4-й недели развития.
В материале эпибласта головной части, расположенного до
первичной ямки, по средней линии зародышевого диска происхо-
дит активная пролиферация клеток. Они перемещаются, и в обла-
сти первичной ямки инвагинируют в свободное пространство
между пластами мезодермы, формируя подобный трубке отросток
(нотохорд, или головной отросток). Этот отросток
«растет» в направлении будущей головной части зародыша, пока
не достигнет прехордальной пластинки. Это материал будущей
хорды. У эмбриона человека хорда является срединной осью и
основанием осевого скелета (позвоночника, ребер, грудины и т. д.),
т. е. это структура, вокруг которой формируется позвоночный столб.
Она дегенерирует и исчезает, когда окружается позвоночными те-
лами. Рудименты хорды в виде пульпозных ядер обнаруживаются
в межпозвоночных дисках.
Вся остальная часть эпибласта является материалом эктодер-
мы. Та ее часть, которая занимает место над инвагинированным
материалом хорды, т. е. по длинной оси зародыша в середине его
дорсальной поверхности, является зачатковым материалом для
нервной системы. Переднее расширение первичной ямки образует
маленький канал (нейрокишечный), который временно связывает
желточный мешок и амниотическую полость.
490
Таким образом, на 3-й неделе развития сформировались 3 заро-
дышевых листка, которые затем дифференцируются асинхронно,
в зависимости от области тела зародыша: в головной части уже
в середине 3-й недели, в каудальных отделах — на неделю позже.
Процесс образования нервной трубки показан на рис. 3.13.
Во время краниального продвижения хорды ее клетки оказывают
воздействие на лежащие над ними клетки эктодермы, которая в
этих участках утолщается, давая начало образованию будущей
нервной пластинки. Нервная пластинка сначала появляется в об-
ласти, близкой к первичному узелку, но по мере роста хорды
к прехордальной пластинке она образуется и в этой области.
На 18-й день нервная пластинка инвагинирует, формируя нервный
желобок с нервными валиками на каждой стороне. Последние
к концу 3-й недели смыкаются и образуют нервную трубку. Обра-
зование и замыкание нервного желобка в нервную трубку начи-
нается в будущей шейной области (на уровне 4-го сомита) и про-
должается в каудальном направлении. При этом в головной части
нервная пластинка имеет большие размеры, быстро разрастается,
но замыкается позднее (передний нейропор). Также позднее за-
мыкается нервная трубка и в каудальной области (задний нейро-
пор).
Эктодермальные клетки, лежащие в месте соединения невраль-
ной и кожной эктодермы, образуют нервный гребень, клетки ко-
торого, мигрируя, распространяются по всему организму, диффе-
ренцируясь в зачатки отделов периферической нервной системы.
Происходящий процесс прогрессирующей дифференцировки
зародышевой мезодермы прослеживается на той же схеме (см.
рис. 3.13). Мезодерма, лежащая по бокам от хордального отростка,
утолщается, расщепляясь в продольном направлении, сегментирует-
ся, образуя симметричные кубоидальные парные структуры (напо-
минающие кирпичики) — сомиты. Обращает внимание, что первые
пары сомитов формируются в головной части рано — уже в по-
следние дни 3-й недели. Затем в сутки в среднем прибавляется по
2—3 сомита. Соответственно возраст зародыша можно определить
по количеству сомитов. В каудальной области остаются несегмен-,
тированные массы (нефрогенная ткань).
От более компактных сомитов отходят тоже сегментирован-
ные, но более тонкие пласты мезодермальных клеток — ножки
сомитов (нефротомы). Нефротомы соединяют сомиты с несегмен-
тированной массой латеральной мезодермы. Это более рыхлые
участки мезодермы (латеральная пластинка). В этой мезодерме
появляется много изолированных пространств, которые, соединя-
ясь, образуют полость — эмбриональный целом. Полость окай-
мляют тонкие слои мезодермальных клеток, заполняющие боковые
и брюшные части зародыша. Полость разделяет латеральную
мезодерму на 2 слоя: а) наружный, «подстилающий» эктодерму
{париетальная мезодерма, соматоплевра) и продолжающийся в
мезодерму амниона; б) внутренний, «подстилающий» энтодерму
{висцеральная мезодерма, спланхноплевра) и продолжающийся в
491
мезодерму желточного мешка. На каждой стороне зародыша
эмбриональный целом в данный период развития еще связан с
внеэмбриональным целомом. В течение 2-го месяца эмбриональ-
ный целом разделяется на 3 полости тела — перикардиальную,
плевральную, перитонеальную.
С первых дней 3-й недели развития (см. рис. 3.10) виден ма-
ленький, пальцеобразный вырост из задней стенки желточного
мешка — аллантоис. В течение этой недели он мало увеличивается
в размере. Аллантоис связан с внеэмбриональным формированием
' кровеносных сосудов и образованием крови (при развитии моче-
вого пузыря он становится урахусом).
Прехордальная пластинка, как указывалось, представляет со-
бой эктодермально-энтодермальную мембрану. На этом этапе
развития ее называют орофарингеальной мембраной (позже она
прорывается, образуя отверстие, связывающее амниотическую
полость и первичную кишку).
В начале 3-й недели во внезародышевой мезодерме желточ-
ного мешка и хориона, а также в соединительной ножке форми-
руются первые кровеносные сосуды (см. рис. 3.14). В теле заро-
дыша они образуются двумя днями позже (раннее формирование
сосудистой системы у человека, вероятно, связано с бедностью
желтка в яйцеклетке и желточном мешке). Мезодермальные
(мезенхимные) клетки — ангиобласты — образуют изолированные
массы, тяжи — кровяные островки, в которых появляются поло-
сти. Клетки, окружающие полость, формируют первичный эндо-
телий, в полости появляются 3 типа клеток: недифференцирован-
ные мезенхимные клетки, гемоцитобласты, первичные эритро-
бласты.
Обращает внимание, что в начале 3-й недели развития жел-
точный мешок крупный. Зародыш на нем распластан. В середине
недели верхняя часть желточного мешка несколько впячивается в
тело зародыша, образуя первичную' кишку. В последующем проис-
ходит формирование передней и задней кишки. В этот период чет-
ко ограничивается клоакальная мембрана, изменяется положение
нейрокишечного канала, который может быть закрытым или в виде
выемки.
В течение 3-й недели развития уже выявляются закладки ряда
систем.
К концу 3-й недели развития возникает зачаток двухкамерного
сердца (см. рис. 3.15) и ряд крупных сосудистых стволов: разви-
ваются сердечные эндокардиальные трубки, которые, сливаясь,
формируют примитивные сердечные трубки. К 21-му дню они
присоединяются к кровеносным сосудам соединительной эмбрио-
нальной ножки, хориона, желточного мешка, формируя примитив-
ную сердечно-сосудистую систему. Это практически первая орган-
ная система, которая в этот период достигает функционального
статуса.
Предпочка (пронефрос), образующаяся на этой неделе разви-
тия, очень быстро редуцируется. На сагиттальном и фронтальном
492
7
494
а
начало 3-й недели
коней 3-й недели
конец 2-й недели
Б
Б 3-й месяц
2-я половина беременности
Рис. 3.17. Формирование плаценты. Развитие ворсин хориона.
А — структура ворсин на разных стадиях развития. Б — поперечное сечение вор-
синок.
I — 1-я половина развития. II — 2-я половина развития.
1 — хорионическая полость; 2 — хориональная пластинка; 3 — соединительная
ткань хориональной пластинки; 4 — кровеносные сосуды в хориональной пластин-
ке; 5 — цитотрофобласт; 6 — синцитиотрофобласт; 7 — кровеносные сосуды эндо-
метрия; 8 — кровеносные лакуны; 9 — эндометрий; 10 — поверхностный цитотро-
фобластический слой; 11 — соединительнотканная основа ворсинки; 12 — капил-
ляры ворсинки; 13 — клетки Гофбауэра.
495
6.
Рис. 3.18. Эмбрион в конце 3-й — начале 4-й недели развития.
А — схема строения (срединный срез). Б — вид с поверхности после вскрытия
хориона и амниона (19 сут) (по Б. М. Пэттену).
1 — передний нейропор; 2 — задний нейропор; 3 — слуховая плакода; 4 — пер-
вый сомит; 5 — желточный мешок; 6 — стенка амниона; 7 — сердце; 8 — нервная
трубка; 9 — хорда; 10 — орофорингеальная мембрана; 11—зачаток щитовидной
железы; 12 — зачаток легкого; 13 — передняя кишка; 14 — зачаток печени; 15 —
клоакальная мембрана; 16 — задняя кишка; 17 — средняя кишка; 18 — аллантоис;
19 — соединительная ножка.
496
К концу 3-й недели хорионическая полость увеличивается в
размере, и эмбрион оказывается связан с трофобластическим
слоем только узкой соединительной ножкой. Рис. 3.18 показывает
соотношение развивающихся частей зародыша на 3-й неделе раз-
вития.
3.1.4. Четвертая неделя развития
В этот период возникают характерные изменения внешней фор-
мы зародыша. Зародыш растет быстро, утраиваясь в размере. Его
форма изменяется от плоской до цилиндрической благодаря обра-
зованию складок. Образуются поперечные складки, направленные
к средней линии эмбрионального диска (рис. 3.19). Быстро ра-
стущие сомиты способствуют сгибанию эмбриона в латеральном
направлении (латеральные складки), и в результате широкая зона
контакта между зародышем и желточным мешком становится
узкой, принимая постепенно вид узкого длинного протока —
желточного стебля. С развитием и ростом мозговых пузырьков
эмбриональный диск начинает выпячиваться в амниотическую
полость. В связи с образованием продольных складок (головной
и хвостовой изгиб) зародышевый щиток становится выпуклым
(С-образный вид) и приподнятым в амниотической полости.
Первичная полоска продолжает еще поставлять мезодермаль-
ный материал до конца недели. Она прогрессивно уменьшается в
размере, сохраняясь лишь в крестцовом отделе, затем исчезает.
В мезодерме продолжается процесс сегментации, что ведет к
появлению к концу 4-й недели 30 и более сомитов. На рис. 3.20
прослеживается дальнейший процесс их дифференцировки: клет-
ки, формирующие вентральную и медиальную стенки сомитов,
становятся полиморфными, выселяются и окружают хорду, обра-
зуя зачаток мезенхимы и позвоночного столба (склеротом).
Материал оставшейся дорсальной стенки сомитов образует новый
слой клеток — миотом. Каждый миотом дифференцируется в
скелетную мускулатуру своего сегмента. Из миотомов выселяются
клетки, распространяющиеся под эктодермой. Образуется дерма-
том, дифференцирующийся в дерму и подкожную ткань (этот
процесс, начавшийся на 4-й неделе, активно продолжается на
5-й неделе).
Продолжается процесс замыкания нервной трубки: к 25-му дню
(20 сомитов) замыкается передний нейропор, а к 27-му дню
(25 сомитов) — задний. Мозговой отдел нервной трубки увели-
чивается в размере и объеме (почти половина нервной трубки—
это будущий головной мозг). Он растет краниальнее орофарин-
геальной мембраны и вскоре свешивается над первичным
сердцем. К концу недели головной мозг образует выраженное
разгибание. Прослеживается начало формирования спинных нер-
вов и ганглиев. Ткань хорды простирается от конца головной
кишки до каудального отдела. Нейрокишечный канал может быть
открытым.
497
498
Рис. 3.19. Образование складок.
А — сагиттальное сечение по средней линии эмбриона на различных стадиях раз-
вития (I—IV; видно краниально-каудальное сгибание зародыша и как его резуль-
тат втягивание энтодермы). Б1— БШ — поперечное сечение.
1 — полость амниона; 2 — сердце; 3 — энтодерма; 4 — передняя кишка; 5 — сред-
няя кишка; 6 — задняя кишка; 7 — аллантоис; 8 — желточный мешок; 9 — пер-
вичная кишка; 10 — прехордальная пластинка (орофарингеальная мембрана);
11—мезодерма; 12 — нервная трубка; 13 — целомическая полость; 14 — соеди-
нительная ножка.
499
Рис. 3.20. Дифференцировка мезодермы.
I—-IV — стадии дифференцировки.
1 — эктодерма; 2 — нервная трубка; 3 — сомиты; 4 — нефротом; 5 — спланхнотом;
6 — целом; 7 — энтодерма; 8 — хорда; 9 — аорта; 10 — мезенхима; 11 — дерматом;
12 — миотом.
500
Сердце уже начинает биться, и сформированная еще к 21-му
дню развития сосудистая сеть ворсинок обеспечивает трофику
зародыша (рис. 3.21). Сердце принимает S-образную конфигу-
рацию, выделяются правые и левые желудочки, дифференциру-
ется венозный синус, возникает межжелудочковая перегородка,
появляются атриовентрикулярные утолщения.
Как указывалось выше, с развитием и ростом головного мозга
эмбриональный диск выпячивается в амниотическую полость и
образует изгиб в краниально-каудальном направлении. В резуль-
тате энтодерма крыши желточного мешка выпячивается в тело
эмбриона — образуется первичная кишка. Передняя кишка закан-
чивается слепо у орофарингеальной мембраны, отделяющей кишку
от амниотической полости. Возникают выросты кишки — зачатки
аденогипофиза, щитовидной и околощитовидной желез, развивает-
ся легочный зачаток и зачаток желудка, дорсальная закладка
поджелудочной железы, закладка печени, которая очень быстро
принимает вид эпителиально-трабекулярной структуры.
Складки в каудальном направлении возникают позже, ее ре-
зультатом являются впячивание аллантоиса в тело эмбриона,
где и формируется клоака. Дистальная часть аллантоиса остается
в соединительной ножке. Хвостовая часть оказывается выше
клоакальной мембраны. Начинается образование производных
эктодермы: в головной области становятся видимыми ее утолще-
ния — ушная (рис. 3.22) и хрусталиковая плакоды. Ушная плако-
да инвагинирует и формирует ушной пузырек. К концу недели
эктодерма и подстилающая ее мезодерма формируют вентральные
и латеральные стенки тела. Дифференцируется граница нефрото-
мов и нефрогенной ткани, развиваются клубочки и канальцы
мезонефроса. Гоноциты мигрируют в каудальную кишку и затем в
медиальную часть над мезонефросом.
В течение 4-й недели происходит и активное развитие в не-
эмбриональных структурах (см. рис. 3.22). В области соединитель-
ной ножки ворсинки хориона достигают наибольшего развития:
они увеличиваются в числе и нарастает сложность их ветвления.
Активно формируется ворсинчатый хорион (детская часть плацен-
ты). Там, где ворсинки лежат не свободно, а соприкасаются с
тканью матки, их цитотрофобластические клетки пенетрируют
слой синцитиотрофобласта, и соединившись, формируют наруж-
ную цитотрофобластическую оболочку (поверхностный цитотрофо-
бласт), покрывающую материнские ткани. Ворсинки лежат в лаку-
нах — обширных пространствах, которые еще в течение 2-й недели
образовались из слияния лакун синцитиотрофобласта. Эти лакуны
ограничены хориальной пластинкой и наружным, прилежащим к
тканям эндометрия слоем цитотрофобласта. Ворсинки омываются
материнской кровью и лимфой. Эта область в первые несколько не-
дель резко расширяется. В материнской ткани (decidua basalis) —
материнской части плаценты — нарастает сосудистая сеть, откры-
вающаяся в межворсинистое пространство, увеличивается число
децидуальных клеток.
501
Рис. 3.21. Развитие сердечно-сосу-
дистой системы.
А — схема кровообращения (по
Б. М. Пэттену). Б — структура сердца
(4-я неделя развития).
I — сердце; 2 — ветви аорты; 3 — пе-
редняя кардиальная вена; 4 — общая
кардиальная вена; 5 — дорсальная аор-
та; 6 — задняя кардиальная вена; 7 —
пупочная вена и артерия; 8 — желточ-
ные вены; 9 — желточные артерии;
10 — хориональная пластинка; 11 —
ворсинки хориона; 12 — внутренняя ка-
ротидная артерия; 13 — синцитиотро-
фобласт; 14 — цитотрофобласт; 15 —
артериовенозная капиллярная сеть вор-
синок; 16 — цитотрофобластические
разрастания на конечных ворсинках.
502
Рис. 3.22.
503
Рис. 3.22. Эмбрион 4-й недели развития.
А — общий вид. Б — срединный срез зародыша 4 мм. В — срединный срез зародыша
6 мм (по Л. И. Фалину). Г — структура пищеварительной трубки. Д — формирование
головного мозга. Е — закладка слухового пузырька. Ж — формирование глаза.
И — развитие печени и первичной почки.
1, 2, 3 — мозговые пузыри; 4 — желудок; 5 — черепно-глоточный карман; 6 — язык;
7 — сердце; 8 — печень; 9 — дорсальная часть поджелудочной железы; 10 — кро-
веносный сосуд; 11 — трахея; 12 — пищевод; 13 — передняя кишка; 14 — желточный
мешок; 15 — первичная почка; 16 — аллантоис; 17 — соединительная ножка; 18 —
амнион; 19 — аорта; 20 — спинной мозг; 21 — хорда; 22 — щитовидная железа;
23 — задняя кишка; 24 — средняя кишка; 25 — орофарингеальная мембрана;
26 — кишечная петля; 27 — сердечный выступ; 28 — 4 пары жаберных дуг; 29 —
инвагинированный глазной пузырек; 30 — вентральная аорта; 31 — дорсальная
аорта; 32 — глотка; 33 — продолговатый мозг; 34 — слуховая ямка; 35 — слуховой
пузырек.
504
505
У зародыша в этот период полость амниотического пузыря
увеличивается, желточного — резко уменьшается. Видны зачатки
верхней, а к концу недели и нижней конечности, резко выбухает
область сердца, к концу недели видны 4 пары жаберных дуг, уш-
ные и хрусталиковые плакоды, носовые ходы.
3.1.5. Пятая неделя развития
Развитие зародыша на 5-й неделе развития представлено на
рис. 3.23—3.26. Форма тела в этот период мало отличается от
таковой 4-й недели. Экстенсивному росту подвергается только
голова, что связано с быстрым развитием мозга. К концу недели
верхняя конечность становится лопатообразной, а нижняя —
ластоподобной, т. е. верхняя опережает развитие нижней. К этому
же сроку заканчивается процесс сегментации мезодермы, обра-
зуется 42—44 пары сомитов. Обращает внимание наличие в кау-
дальном отделе несегментированной мезодермы — нефрогенной
ткани. Прогрессирует развитие спинного и головного мозга, все
черепные нервы и многие спинальные уже образованы, возникает
черепноглоточный карман. Глазная плакода инвагинирует и фор-
мирует линзу. Хорошо видны 2 сердечные трубки, зачатки вторич-
ных бронхов и долей, большую площадь занимают развитые мезо-
нефросы. Начинается развитие метанефроса, виден урогениталь-
ный синус. Прямая кишка изолируется от мочевого пузыря.
Тонкая кишка образует первичную кишечную петлю. К дорсаль-
ной закладке поджелудочной железы добавляется вентральная.
Трахея изолируется от пищевода. Формируются половые валики.
Рис. 3.23. Эмбрион в начале 5-й
недели развития.
А — общий вид. Б — схема строения
(в модификации). В — срединный срез
(по Л. И. Фалину). Г — пищевари-
тельная трубка.
1 — мелэнцефалон; 2 — диэнцефалон;
3 — мезэнцефалон; 4 — метэнцефалон;
5 — миелэнцефалон; 6 — спинной мозг;
7 — пищевод; 8 — легкое; 9 — сердце;
10 — селезенка; 11 — печень; 12 — под-
желудочная железа; 13 — желчный про-
ток; 14 — средняя кишка; 15 — аллан-
тоис; 16 — кишечная петля в пуповине;
17 — дивертикул; 18 — щитовидная же-
леза; 19 — мезонефрос; 20 — язык;
21 — желудок; 22 — мочеполовой синус;
23 — прямая кишка.
506
507
Рис. 3.24. Эмбрион 5-й недели развития (срез на уровне головной части).
А — образование зачатка гипофиза — черепно-глоточный карман. Б — образование
глазного пузырька. В, Г — формирование слухового пузырька (В — схема, Г —
препарат).
1 — черепно-глоточный карман; 2 — язык; 3 — хорда; 4 — хрусталиковая плакода;
5 — мозг; 6 — глазной бокал; 7 — дорсальная аорта; 8 — глотка; 9 — слуховой
пузырек; 10 — вентральная аорта; 11 — четвертый желудочек мозга.
508
Рис. 3.25. Эмбрион 5-й недели раз-
вития. Гистогенез органов (А, Б,
В) (наддиафрагмальный отдел).
1 — желудочки сердца; 2 — предсердия;
3 — сердечный клапан; 4 — печень; 5 —
легкое.
509
Рис. 3.26. Эмбрион 5-й недели развития. Гистогенез органов (А, Б).
1 — желудок; 2 — легкое; 3 — канальцы мезонефроса; 4 — кишечник; 5 — спин-
номозговые узлы; 6 — печень; 7 — мезонефрос; 8 — закладка гонады; 9 — задняя
кишка.
3.1.6. Шестая неделя развития
У зародыша в этот период вырисовывается первичное лицо,
его голова значительно больше по сравнению с туловищем и
более наклонена по отношению к сердечному выросту (такая
позиция является результатом изгиба мозга в цервикальной об-
ласти). И только к концу недели туловище и шея начинают вы-
прямляться. Конечности приобретают отчетливые региональные
отличия, особенно верхние (рис. 3.27 и рис. 3.28). Локтевая и
запястная зоны становятся определимыми; в лопатообразной руке,
на ее периферии, начинают образовываться будущие пальцы
(при этом, как и на 5-й неделе, дифференцировка нижней конеч-
ности по-прежнему несколько отстает от верхней). Сомиты видны
в люмбосакральной зоне вплоть до середины недели. Вокруг
жаберных щелей (между первыми двумя дугами) видны неболь-
шие вздутия — ушные бугорки — отверстия, которые дадут
начало наружному слуховому проходу; вздутия сливаются, образуя
ушную раковину. Глаз становится более заметным, потому что в
клетках сетчатки появляется пигмент. Конец дорсальной хорды
510
16
18
20 23
27
Рис. 3.27. Эмбрион начала 6-й недели развития.
А — общий вид. Б — сагитальный срез. В — схема строения пищеварительной труб-
ки (по А. С. Леонтюк).
1 — глотка; 2 — трахея; 3 —- желудок; 4 — печень; 5 — дорсальная закладка под-
желудочной железы; 6 — хорда; 7 — задняя кишка; 8 — клоака; 9 — аллантоис;
10 — желточный стебелек; 11 — вентральная закладка поджелудочной железы;
12 — карман Ратке; 13 — первичная почка; 14 — корень языка; 15 — пищевод;
16 — глаз; 17 — прямая кишка; 18 — верхняя конечность; 19 — мочеполовой
синус; 20 — сердечно-печеночный выступ; 21 — позвоночник; 22 — кишечник;
23 — пуповина; 24 — мозговые пузыри; 25 — спинной мозг; 26 — сердце; 27 —
нижняя конечность; 28 — аорта; 29 — кишечная петля.
511
Рис. 3.28. Эмбрион б-й недели развития (головная область).
А, Б — развитие мозговых пузырей. В — формирование гипофиза. Г — формиро-
вание глаза.
1 — пигментный эпителий; - 2 — сетчатка; 3 — хрусталик; 4 — глазной пузырь;
5 — карман Ратке; 6 — зачаток нейрогипофиза; 7 — мелэнцефалон; 8 — диэнце-
фалон; 9 — мезэнцефалон; 10 — ромэнцефалон; 11 — метэнцефалон.
512
10
Рис. 3.29. Эмбрион 6-й недели развития. Гистогенез органов (А, Б, В,
Г, Д, Е, Ж, И).
1 — желудочки сердца; 2 — предсердия; 3 — легкое; 4 — бронхи; 5 — пищевод;
6 — аорта; 7 — поджелудочная железа; 8 — вторичная почка; 9 — надпочечник;
ю — печень; 11 —первичная почка; 12 — гонада; 13 — спинальные узлы; 14 —
тонкая кишка; 15 — желудок.
17—321
513
Рис. 3.29. Продолжение.
514
окружается основанием черепа. Специальные ганглии локализуют-
ся в межпозвоночных пространствах. Продолжается развитие
нервной трубки, появляются зачатки моста мозга и мозжечка.
Кишечная петля доходит до пупочной границы. Сливаются дор-
сальная и вентральная закладки поджелудочной железы, желчные
протоки внедряются в печень. В закладке легких возникают
бронхолегочные сегменты. Формируются зачатки грудных желез.
Гонады отделяются от мезонефроса, их структура уже имеет
половые отличия. Подробно гистогенез органов в этот период
изложен выше в каждой главе. Рис. 3,29 позволяет видеть со-
отношение развивающихся органных систем.
3.1.7. Седьмая неделя развития
Сильно изменяются конечности: верхние простираются под серд-
це, четко определяются будущие пальцы. Связь между примитив-
ной кишкой и желточным мешком редуцируется до относительно
малого протока — желточного стебля. Петли кишки входят в вне-
эмбриональный целом в проксимальной части пупочного шнура
(умбиликальная грыжа). Происходит разрыв клоакальной мем-
браны. Продолжается дифференцировка метанефрогенной тка-
ни — увеличиваются в числе почечные тельца. Парамезонефраль-
ные каналы сливаются и достигают урогенитального синуса.
В семенниках видны тестикулярные трубочки — тяжи, в яични-
ках — яйценосные шары. Гистогенез и топография развивающихся
органов представлены на рис. 3.30 и рис. 3.31.
3.1.8. Восьмая неделя развития
В начале последней недели эмбрионального периода пальцы
конечностей короткие и заметно перепончатые, между ними луча-
ми определяются выемки. Еще присутствует хвост, но он уже
имеет вид обломка. Глаза в течение 8-й недели обычно открыты,
но к концу недели веки встречаются друг с другом, закрывая их.
Ушные раковины приобретают свою окончательную форму, но
остаются пока расположенными низко. Появляется черепное со-
судистое сплетение. ,
К концу недели четко определяются отделы конечностей,
удлиняются пальцы рук, появляются отчетливые пальцы и на
ногах, совсем исчезает хвост. Черепное сосудистое сплетение
теперь образует полосы на верхушке черепа. Эмбрион приобретает
явно человеческий вид. Голова более округлая и приподнятая,
еще непропорционально большая, занимает почти '/г всего орга-
низма. Устанавливается область шеи; надбровные дуги становятся
более отчетливыми. Пуповина уменьшается в размерах. Кишечник
все еще находится в проксимальной части пупочного шнура.
Между наружными гениталиями существуют различия, но внешне
это крайне трудно определяется. На рис. 3.32 видно соотношение
развивающихся органных систем.
17
515
Рис. 3.30. Эмбрион
7-й недели развития.
А — общий вид. Б — сре-
динный срез. В — форми-
рование пищеварительной
трубки (по А. С. Леонтюк).
Г — формирование глаза.
1 — спинной мозг; 2 —
оболочки глаза; 3 — мозго-
вые пузыри; 4 — печень;
5 — желудок; 6 — легкое;
7 — сердце; 8 — кишечная
петля; 9 — пищевод; 10 —
глотка; 11 — хрусталик;
12 — сетчатка.
516
Рис. 3.31. Эмбрион 7-й недели развития. Гистогенез органов (А, Б, В, Г,
Д, Е, Ж).
517
Рис. 3.31. Продолжение.
1 — предсердие сердца; 2 — желудочек
сердца; 3 — желудок; 4 — поджелудоч-
ная железа; 5 — легкое; 6 — надпочеч-
ник; 7 — вторичная почка; 8 — спиналь-
ные узлы; 9 — гонада; 10 — кишка:
11—спинной мозг; 12 — позвоночник.
518
Рис. 3.32. Эмбрион 8-й недели развития.
А — общий вид. Б — срединный срез. В, Г, Д, Е, Ж, И — гистогенез органов.
1 — аорта; 2 — сердце; 3 — легкое; 4 — печень; 5 — почка; 6 — надпочечник;
7 — парамезонефральные каналы; 8 — мезонефральные каналы; 9 — мочевой
пузырь; 10 — матка; 11—кишка; 12 — спинной мозг; 13 — позвоночник; 14 —
кишечная петля; 15 — поджелудочная железа; 16 — бронхи; 17 — пищевод; 18 —
прямая кишка.
519
Рис. 3.32. Продолжение.
520
Оценивая процесс развития у человека в течение 5—8 нед
развития, следует отметить, что в отличие от других видов мле-
копитающих период после имплантации характеризуется замед-
ленным развитием.
3.2. Плодный период
3.2.1. Формирование плаценты
Используя рис. 3.33 и рис. 3.34, можно проследить динамику
изменения положения зародыша в матке и формирования его
зрелой плаценты. Ворсинки хориона покрывают поверхность за-
родыша до 8-й недели развития. Затем (рис. 3.35), по мере увели-
чения размеров зародыша, ворсинки, обращенные к поверхности
матки, не развиваются, их кровоснабжение редуцируется. По мере
роста плода decidua capsularis истончается, происходит сближение
с эндометрием противоположной стенки матки (decidua parieta-
lis) и щель между ними прогрессивно сокращается. К 4-му меся-
цу развития ворсинки атрофируются полностью, образуется глад-
кий (лысый) хорион. Decidua capsularis еще больше растягивается,
и в конечном итоге ткань гладкого хориона соприкасается с
decidua parietalis (т. е. полость матки не видна).
Ворсинки области эмбрионального полюса в области соедини-
тельной ножки формируют ветвистый ворсинчатый хорион —
детскую часть плаценты. Здесь ворсинки особенно интенсивно
васкуляризуются и гипертрофируются, увеличиваются в числе и
нарастает сложность их ветвления. Они располагаются в обшир-
ных пространствах, образовавшихся из слияния лакун синцитио-
трофобласта, и ограничены тканями эндометрия (decidua basalis)
и хориальной пластинкой. Ворсинки омываются материнской
кровью и лимфой. Эта область уже в первые несколько недель
резко расширяется. Следует отметить, что в ходе беременности,
особенно от начала 4-го месяца, структура~~торСинок_А1еняется:
клетки цитотрофобласта, покрывающие соединительную основу,
начинают исчезать. Уменьшается и площадь соеди и ельной
ткани, нарастает площадь капилляров. Синцитиотрофобласт и
эндотелии ка ров остаются теми двумя слоями, которые в
основном~разграничивают материнскую циркуляцию и циркуляцию
плода. ' ~ —
** Decidua basalis (материнская часть плаценты) — децидуальная
пластинка в этот период достигает своего максимального раз-
вития: состоит из компактного слоя крупных богатых липидами
и гликогеном клеток, насыщена сосудами и синусоидами, откры-
вающимися в межворсинистое пространство. Видна тесная связь
decidua basalis и хориона: в местах контакта decidua basalis ворсин-
ками хориона клетки их наружного цитотрофобластического слоя
тесно соприкасаются с ее клеточными элементами; эти участки
богаты мукополисахаридным материалом. В плаценте плода
4—5 месяцев и на более поздних этапах развития уже видны
521
522
Рис. 3.33. Дифференцировка эмбриобласта, трофобласта, оболочек
эмбриона человека (А—Е; в модификации по Б. М. Пэттену).
1 — эмбриобласт; 2 — трофобласт; 3 — синцитиотрофобласт; 4 — цитотрофобласт;
5 — эпибласт; 6 — гипобласт; 7 — полость амниона; 8 — желточный мешок; 9 —
внезародышевая полость; 10 — внезародышевая мезодерма; 11 — амниотическая
ножка; 12 — аллантоис; 13 — пуповина; 14 — сосуды пуповины; 15 — первичные
ворсинки; 16 — вторичные ворсинки; ,17 — третичные ворсинки хориона; 18 —
экзоцеломическая мембрана; 19 — экзоцеломическая полость; 20 — атрофия вор-
синок.
Рис. 3.34. Взаимоотношения развивающегося эмбриона человека с эндо-
метрием матки в различные сроки беременности.
А — 2-я неделя. Б — 3-я неделя. В — 4-й месяц.
1 — амниотическая полость; 2 — внезародышевая полость; 3 — желточный мешок;
4 — decidua parietalis; 5 — полость матки; 6 — decidua basalis; 7 — эмбрион; 8 —
decidua capsularis; 9 — влагалище; 10 — амниотическая ножка; 11 — ворсинки хо-
риона; 12 — миометрий; 13 — шейка матки.
523
4
Рис. 3.35. Строение ворсинок хориона в различные сроки беременности.
А — 2-й месяц беременности. Б — 2-я половина беременности. В — ТЭМ ворсинки
на 2-м месяце беременности. Г — ТЭМ ворсинки во 2-й половине беременности.
Д — характер ветвления ворсинок зрелой плаценты (СЭМ) (по А. И. Брусилов-
скому и С. И. Колесникову).
1 — ворсинки; 2 — соединительная ткань; 3 — синцитиотрофобласт; 4 — цитотро-
фобласт; 5 — сосуды ворсинки; 6 — клетки Гофбауэра. <
524
'я
9 8
7
526
Рис. 3.36. Строение плаценты.
А, Б — схемы. В—Е -— плацента в 1-й
половине беременности. Г — плодная
часть. Д — поперечное сечение ворсин-
ки (деталь к Г). .Е — децидуальная
оболочка матки.
1 — терминальные ворсинки хориона;
2 — соединительная ткань ворсинки,
3 — синцитиотрофобласт; 4 — капилля-
ры; 5 — меливорсинчатое пространство;
стволовая ворсинка; 7 — хориальная
пластинка; 8 — соединительная ткань
амниона; 9 — эпителий амниона; 10 —
край амниона; 11 — пупочный канатик;
12 — основа слизистой оболочки матки;
13 — децидуальные клетки; 14 — по-
верхностный цитотрофобластический
слой.
Е
527
сформированные септы в decidua basalis, которые разделяют кро-
вяные лакуны. Так как они не достигают хориальной пластинки,
то связь между компартментами лакун сохраняется. Эти септы
соответственно покрыты прилежащим поверхностным цитотрофо-
бластическим слоем, так что есть слой трофобласта, разделяющий
циркуляцию крови матери и плода. Зрелая плацента имеет до
25 см в диаметре, массу до 500—600 г.
На рис. 3.36 видна зрелая плацента человека второй половины
беременности. Она морфологически четко разделяется на плодную
и материнскую части.
В плодной части амнион виден как тонкий слой, в котором
со стороны амниотической полости различаются однослойный
амниотический эпителий и тонкий слой «подстилающей» мезо-
дермы, без резких границ переходящей в хориальную мезодер-
му — хориальную мезодермальную пластинку. От нее отходят
якорные и стволовые ветвящиеся ворсинки хориона. Они свободно
лежат в лакунах материнской крови и видны в различных сече-
ниях. Рассматривая ворсинки в различных вариациях сечений
(косых, поперечных, продольных), можно отчетливо увидеть их
составные компоненты и доминирование синцитиотрофобластиче-
ского покрова. Только в небольших участках остается цитотрофо-
бласт. На рис. 3.35 и 3.36 можно еще раз увидеть внешний вид
ворсинок и соотношения их тканевых компонентов.
На рис. 3.37 прослеживается ход формирования пуповины,
образование соединительной ножки, соединяющей зародыш с тро-
фобластом, врастание в нее аллантоиса, «подтягивание» к ней
желточного мешка. Прослеживается в динамике «окружение» этих
компонентов стенкой амниотического пузыря по мере увеличения
размеров пузыря и прогрессирующего сближения его стенок с
Рис. 3.37. Пупочный канатик.
1 — артерии; 2 — вена; 3 — студенистая ткань; 4 — остатки желточного мешка.
528
Рис. 3.38. Оболочки эмбриона.
А — эмбрион 5 нед. Б — эмбрион 6 нед.
1 — амниотическая оболочка; 2 — ворсинка хориона; 3 — пупочный канатик; 4 —
соединительнотканная основа хориона; 5 — эпителий и соединительная ткань
амниона; 6 — полость амниона; 7 — желточный мешок.
529
соединительной ножкой. Наружный покров пуповины — эпителий
амниотического пузыря, под ним расположена специфическая
студенистая соединительная ткань, являющаяся основой пупо-
вины, в которой видны артерии, вена, остатки желточного меш-
ка — желточный стебелек, аллантоис. Рис. 3.38 представляет про-
странственное соотношение оболочек плода.
Плодный период в данной главе дан очень кратко, ибо в преды-
дущих главах процесс развития каждой системы и каждого органа
изложен детально.
Здесь же акцент сделан на демонстрацию взаимоотношений
развивающихся органов, их топографические особенности, раз-
витие и особенности плаценты.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Агрегация 43
Адгезия 43
Аденогипофиз, часть передняя 239
— — задняя 245
Аденоциты 239
— хромофильные 239
— — ацидофильные 239
— — базофильные 239
— — оксифильные см. Аденоциты хро-
мофильные ацидофильные
— хромофобные 239
Аксон см. Нейрит
Альвеола (ы) 280, 281
Альвеолоциты большие см. Клетки аль-
веолярные секреторные
— дыхательные см. Клетки альвеоляр-
ные респираторные
— респираторные см. Клетки альвео-
лярные респираторные
— типа I см. Клетки альвеолярные
респираторные
— — II см. Клетки альвеолярные се-
креторные
Амелобласты 308, 309
Анастомозы артериовенулярные 183,
' 187
Анизоцитоз 41
Аорта 180, 181
Аппарат защитный иммунный 346
— поверхностный см. Оболочка кле-
точная
— ядерный см. Ядро
— — поверхностный см. Оболочка
ядерная
— ядра генетический 19
Артерии 177
— типа мышечного 177
— — мышечно-эластического 177
— — эластического 177
Артериола (ы) 183, 187
— выносящая 373
— прекапиллярные см. Прекапилляры
— приносящая 373
Астроциты 111, 112
Аутолизосома 17
Ацинус 279, 348
— печеночный 364
Базофилы тканевые см. Клетки тучные
Балки костные 83
Барьер(ы) аэрогематический 287, 288
— гематоофтальмический 157
— гематоцеллюлярные 195
— фильтрационный почечного тельца
375
— — — — структура 377
— гематотестикулярный 410
— гематоэнцефалический 157
Бластомозы 468
Бластоциста 468
Бляшка моторная 122, 123
Бронхи калибра крупного 272
— — мелкого 278
— — среднего 278
Бронхиола респираторная 277, 281
— и терминальная 277, 279
Вены 177
— типа мышечного 177
— — мышечно-эластического 177
— — эластического 177
Венулы 183, 187
— посткапиллярные см. Посткапилляры
— собирательные 197
Вещество аморфное см. Вещество основ-
ное
— базофильное см. Субстанция Ниссля
— белое 131
531
— межклеточное 59
— надпочечника корковое 261
— — мозговое 262
— основное 59
— почки корковое 372
— — мозговое 372, 375
— серое 131
— тигроидное см. Ниссля субстанция
Включения 21
— гликогена 23
— жировые 22
— секреторные 21
— трофические 21
Влагалище 453
В-лимфоциты 52
Волокна коллагеновые 59
— нервные 111
— — безмиелиновые 114, 119
— — миелиновые 114, 116
— — развитие 114
— Пуркинье см. Пуркинье волокна
— ретикулярные 59
— эластические 59
Волос 297
— иннервация 302
— развитие 306
— строение 302
Ворсинки кишечные 331, 346
Ганглий(и) 128
— вегетативные 129, 132
— — интрамуральные 129
— — паравертебральные 129
— — превертебральные 129
— нервной системы периферической
129
— спинальный см. Ганглий спинно-
мозговой
— спинномозговой 129, 131
Гаструляция 483
Гемокапилляры 187
Гемопоэз 209
Гепатоциты 361
Гетеролизосома см. Лизосома вторичная
Гетерохроматин см. Хроматин конден-
сированный
Гиаломеры 42
Гипобласт 475
Гистология 27
532
Глаз 148
— развитие 146, 147
Гландулоциты интерстициальные 414
Гликокаликс см. Слой надмембранный
Глиоциты 248
— радиальные 154
— специализированные 154
Глюкозаминогликаны 51
Головка придатка яичка 418
— сперматозоида 404
Гольджи комплекс 14, 16
Гранулоциты 42
Гранулоциты базофильные 51
— нейтрофильные 46
— — палочкоядерные 46
— — сегментоядерные 46
— — юные 46
— эозинофильные 49
— — палочкоядерные 49
— — сегментоядерные 49
— — юные 49
Гранулоцитопоэз 213
Гребешок ампулярный 163
Деление мейотическое второе 401
— — первое 400
— митотическое 21
Дендрит(ы) 106, 119
Дентин 307, 308
Дерево бронхиальное 273
Дерма 292
Десмосома 11
Диск зрительного нерва 157
Долька портальная 364
Дробление 468
Дуга (и) рефлекторная (ые) 126
— — автономная см. Дуга рефлектор-
ная местная
— — вегетативные 127
— — виды 127
— — местная 127
— — соматическая 127
Железа (ы) вилочковая 219
— кишечная 341
— многоклеточные простые 37
— — сложные 39
— молочная 453, 461
— — лактирующая 458
— — нелактирующая 458
— — развитие 459
— околощитовидная 256
— пищеварительные 348
— поджелудочная 348
— — развитие 355, 356
— потовые 297
— — апокриновые 297
— — мерокриновые 297
— — развитие 306
— предстательная 418
— сальные 297
— слюнная (ые) 315, 317, 319
— — околоушная 315
— — подчелюстная 315
— — развитие 319
— щитовидная 248
— развитие 256
— экзокринные 37
— — локализация 37
— — одноклеточные 37
— — многоклеточные простые 37
— — — — неразветвленная трубчатая
37
— — — — разветвленная альвеоляр-
ная 37
— — — сложные 39
— — основные разновидности строе-
ния 37
Желудок 326
— дно 326, 327
— отделы 326
— развитие 334
Звенья системы эндокринной перифе-
рические 248
— — — центральные 233
Зернистость базофильная 51
Зигота 468
Зона(ы) сетчатки 156
— — вторая 157
— — первая 156
— коркового вещества надпочечника
клубочковая 261
— — — — пучковая 261
— — — — сетчатая 262
— наилучшего видения см. Ямка сет-
чатки центральная
Зуб 307
— корень 307, 308
— коронка 307, 308
— развитие 307
— шейка 307, 308
Имплантация 475
Инвагинации базальные 10
Канал шлеммов см. Синус склеры ве-
нозный
Канальцы почки извитые 378
— — — дистальные 375
— — — проксимальные 375
— семенные извитые 399
— — прямые 399
Кальцитониноциты см. Эндокриноциты
парафолликулярные
Капилляр(ы) 183
— висцеральный 190
— лимфатические 199
— синусоидный 190
— соматический 190
Кардиомиоцит (ы) 97
— желудочковые 97
— предсердные секреторные 171
— — эндокринные см. Кардиомиоциты
предсердные секреторные
— проводящие 97
Кариоплазма 24, 26
Кишка двенадцатиперстная 342, 344
— тонкая 334, 335
— — развитие 343, 344
— толстая 344, 345
— — развитие 346
Клетка (и) 6
— альвеолярные респираторные 228
— — секреторные 228
— ациноостровковые 354
— жировые 65
— кроветворные стволовые 52
— Купфера см. Макрофаги звездчатые
— Лангерганса см. Макрофаги внутри-
эпидермальные
— Лейдига см. Гландулоциты интер-
стициальные
— мезенхимные 83
— нейросенсорные 156
— — палочковая 156
— — колбочковая 156
— нервные 105
533
— — классификация 111
— плазматические 65
— пласта сперматогенного 407
— — — половые 407
— — — соматические 407
— предшественники 209
— — ряда эритроцитарного 42
— придатка яичка базальные 418
— — — главные 418
— «пылевые» см. Макрофагоциты аль-
веолярные
— септальные см. Клетки альвеолярные
секреторные
— Сертоли см. Сустентоциты
— тучные 65
— — ультраструктура 68
— хондрогенные 76
— шванновская 114
— экстрагломерулярные 386
— эритроидные 201
— эукариотическая 14
— — строение 13
— юкстагломерулярные см. Эндокрино-
циты миоидные
Кожа 292
— производные 292
— развитие 305, 306
— «толстая» 292
— «тонкая» 300
Колбочки 154
Комплекс Гольджи см. Гольджи комп-
лекс
— поровый 24
— юкстагломерулярный 384, 386
Компоненты ядра 24
Кона пора 289
Контакт (ы) липоклеточные 10
— — изолирующего типа 10
— — механические 10
— — химические 11
— — щелевой см. Нексус
— мио-миоцитарные 97, 187
Кора больших полушарий 140, 143
— — — — слой ганглиозный 143
— — — — — зернистый внутренний
143
— — — — — — наружный 143
— — — — — молекулярный 143
— — — — — пирамидный 143
— — — — — полиморфных клеток
143
— надпочечника постоянная 268
— — фетальная 265
— мозжечка 139
— — слой ганглиозный см. Кора моз-
жечка, слой средний
— — — глубокий 140
— — — зернистый см. Кора мозжеч-
ка, слой глубокий
— — — молекулярный см. Кора моз-
жечка, слой наружный
— — — наружный 138
Корень зуба 307
Кортиев орган см. Орган спираль-
ный
Кости скелета плоские 79
— — трубчатые 79
Крипты кишечные 338
Кристаллы Рейнике см. Рейнике кри-
сталлы
Кристы 19
— пластинчатые 20
— трубчатые 20
Кровь 39
— морфологические особенности см.
Особенности крови
— морфологические
Купол желатинозный 163
Купфера клетки см. Макрофаги звезд-
чатые
Лабиринт внутреннего уха костный 159
— — — перепончатый 159
Лакуна (ы) костные 83
— эрозионная 85
Легкое 272
— ребенка новорожденного доношенно-
го 291
— — мертворожденного 291
— этапы развития 287
— Лейдига клетки см. Гландулоциты
интерстициальные
Лейкоциты 46, 65
— агранулярные 51
— гранулярные
Леммоцит см. Клетка шванновская
Лестница перепончатого лабиринта
улитки вестибулярная 159
— — — — тимпанальная 158
534
Лизосома (ы) 17, 19
— вторичная 17
— первичные 17
Лимфоциты 51, 52
— киллеры 52
— межэпителиальные 346
— пластинки собственной 346
Листки зародышевые 482
Мазок спермы 405
Макроглия 111
Макропиноцитоз 10
Макрофаг(и) 52, 56, 62
— внутриэпидермальные 296
— звездчатые 364
Макрофагоциты альвеолярные 284, 285
Матка 446, 481
— развитие 450, 452
Мезаксон 114
Мезангиоциты 386
Меланофороциты 296
Мембрана 6
— оболочки ядерной внутренняя 24, 26
— — — наружная 23, 26
— плазматическая 6
— покровная 159
Мешочек ампулярный 163
— эллиптический 163
Миелин 114
Миелограмма 209
Микроворсинки 10, 338
Микроглии 111
Микропиноцитоз 10
Микрососуды 183
— лимфатические 198, 199
Микротрубочки 9
Микрофиламенты 9
Микроциркуляция 183
Миндалина 320
Миокард 170, 172
Миометрий 448
Мион 91
Миосателлитоциты 89
Миофибриллы 91, 92
Миофиламенты 91
— актиновые 103
— промежуточные 103
Миоцит гладкий 103, 170
Митохондрии 19
Модуль микрососудистый 185
Мозг головной 131
— костный 209
— — красный 215, 218
— спинной 131, 135
— — развитие 125, 136
Мозжечок 138
Моноциты 52, 55
Морула 468
Мочеточник 398
Надкостница 79
Надпочечник 261
Насечки Шмидта—Лантермана
Шмидта—Лантермана насечки
Неврилемма 114
Нейрогипофиз 245
Нейроглия 111
Нейрон (ы) 105, 106
— амакринные 153, 156
— биполярный ПО, 153, 156
— веретенообразный ПО
— горизонтальные 153
— грушевидный ПО
— звездчатый ПО
— интерплексиформные 153, 156
— мультинолярный ПО, 153
— нейросенсорные 153
— округлый ПО
— пирамидный ПО
— псевдоуниполярный ПО
Нейротубулы 109
Нейрофибриллы 109
Нейрофиламенты 109
Нексус 11, 187
Нервы периферические 127
Нефроны 367, 393
Ниссля субстанция 106
Оболочка (и) глаза 146
— — сетчатая см. Сетчатка
— — сосудистая
— клеточная 6
— — специализация 12
— матки 448
— мочеточника 398
— — мышечная 398
— — подслизистая 398
— — серозная 398
см.
535
— — слизистая 398
— овоцита 427
— стенки глаза задней 150
— — — — сетчатая см. Сетчатка
— — — — сенсорная см. Сетчатка
— — — — сосудистая 150
— — — — фиброзная 150
— — канала пищеварительного 321
— — — — адвентициальная см. Обо-
лочка стенки канала пищеваритель-
ного серозная
— — — — мышечная 322
— — — — серозная 322
— — — — слизистая 321
— — сердца 170
— — — внутренняя см. Эндокард
— — — наружная см. Эпикард
— — — средняя см. Миокард
— — сосуда 170
— — — внутренняя 170
— — — наружная 170
— — — средняя 170
— — — лимфатического 182
— — — — внутренняя 182
— — — — наружная 182
— — — — средняя 182
— трахеи адвентициальная 272
— — слизистая 268
— — фиброзно-хрящевая 272
— трубы маточной 453
— — — мышечная 453
— — — серозная 453
— — -— слизистая 453
— фолликула 430
— эмбриона 529
— ядерная 24
— яичка белочная 399
— яичника белочная 420
— — соединительнотканные 422
Овоцит 422
Овуляция 432
Одонтобласт 309
Окончание(я) нервное (ые) 119, 127
— — афферентные см. Окончания
нервные чувствительные
— — двигательные 119
— — несвободные 119, 122
— — — инкапсулированные 122
— — — неинкапсу лированные 122
— — свободные 119
— — чувствительные 119, 120
— — эфферентные см. Окончания
нервные двигательные
Олигодендроциты 111, 113
Оплодотворение 464
Орган (ы) вкуса 166
— зрения 146
— Кортиев см. Орган спиральный
— кроветворения 208
— обоняния 168, 169
— пре дверно-улитковые 157
— равновесия 163, 164
— слуха 159
— — и равновесия см. Орган пред-
дверно-улитковый
— спиральный 159
— чувств 145
Органеллы цитоплазмы 14
— — значения общего 14
— — состоящие из мембран 14
— — — — фибрилл 21
Основа подслизистая тракта пищева-
рительного 39
— — трахеи 272
Особенности морфологические крови
взрослого человека 39
— — — новорожденного 56
Остеобласты 79
Остеогистогенез 83
— непрямой 85, 86
— прямой 83, 84
Остеокласты 79
Остеоциты 79, 81
Островки панкреатические 348, 352
Отросток червеобразный 346
— — развитие 346
Отдел (ы) желез слюнных концевой (ые)
317
— — — — мукозный 317
— — — — серозный 317
— — — — смешанный 317
— желудка 326
— — пилорический 322, 326
— системы пищеварительной передний
307
— — — средний 326
536
Палочки 154
Панкреациты экзокринные 348
Паратироциты главные 257
— оксифильные 258
Печень 356, 357, 365
— развитие 364
Перикарион 105
Периметрий 448
Перимизий 87
Периост см. Надкостница
Перитендон 72
Пероксисома 17, 19
Пинеалоциты 248
Пиносомы 65
Pit-клетки 364
Пищевод 322
— развитие 324
Плазмолемма см. Мембрана плазмати-
ческая
Плазмоциты 67, 69
Плакоза 159
Пластинка (и) костная 79
— кровяные см. Тромбоциты
— печеночные 360
— субмембранная плотная 24
Плацента, формирование 493, 521
Подоциты 373
Пойкилоцитоз 42
Полисомы 14
Полихроматофилы 42
Полоска первичная 483
Полюс деления митотического 21
— постсинаптический 126
— пресинаптический 125
Пора Кона см. Кона пора
Посткапилляры 183, 187, 195
— лимфатические 199
Почка 367
— вкусовая см. Орган вкуса
— первичная 386
— постоянная, развитие 388
— развитие 386
— ребенка двух лет 393
— — новорожденного 393
— — пяти лет 393
Преддверие см. Орган равновесия
Предпочка 386
Прекапилляры 183, 183
Придаток яичка 416
Производные кожи 292
Проток улитковый 159
— эндолимфатический 159
Пузырь желчный 366
— мочевой 395, 396
— — развитие 397
Пузырьки синаптические 123
Пульпа селезенки белая 231
— — красная 231
Пуркинье волокна 171
Пути воздухоносные 268
Пучок сухожильный порядка второго 72
— — — первого 72
Пятно слепое см. Диск зрительного
нерва
— электрического мешочка 163
Радужка 148, 149
Реакция децидуальная 481
Рейнике кристаллы 414
Репликация ДНК 24
Реснички 10
Ретикулоцит 42, 43
Рецепторы см. Окончания нервные чув-
ствительные
Рибосомы 14
Роговица 150, 151
Рост интерстициальный 76
Русло микрососудистое см. Русло мик-
роцирку ляторное
— микроциркуляторное 183, 186
— — коркового вещества надпочечника
262
Саркомер 91, 92
Сарколемма 123
Сателлиты перинейренальные Ill
Селезенка 229, 231
— развитие 233
Семенник см. Яичко
Сердце 170
— развитие 199
Сетчатка 150
— зоны 156
— слой ганглиозный 156
— — внутренний пограничный 156
— — — сетчатый 156
— — — ядерный 156
— — наружный пограничный 156
537
— — — сетчатый 156
— —- — ядерный 156
— — нервных волокон 156
— — пигментный 150, 154
— — фотосенсорный 154
— часть нервная 153
Сеть эндоплазматическая 14
---агранулярная см. Сеть эндоплаз-
матическая гладкая
— — гладкая 14
— — гранулема см. Сеть эндоплазма-
тическая шероховатая
— — шероховатая 14
Синапс (ы) 123, 126
— аксоаксональные 123
— аксодендритические 123, 126
— аксомышечный 123
— аксосоматический 123, 126
— аксошипиковый 126
— дендродендритические 123
— межнейрональные 123
— — химические 123, 124
— — электрические 123
Синус склеры венозный 146
Синцитиотрофобласт 478
Система дыхательная 268
— нервная 126
— — периферическая 127
— — — ганглии 129
— — центральная 131
— мочевыделительная 367
— мышечная атипическая 171
— — проводящие см. Система мышеч-
ная атипическая
— пищеварительная 307
половая женская 420
— — мужская 398
— сердечно-сосудистая 170
— сосудистая почки 369
— эндокринная 233
— — звенья периферические 248
— — — центральные 233
Склера см. Оболочка стенки глаза
задней фиброзная
Слой надмембранный 6
— подмембранный 9
Соединения сертоли-сертолиевы 409
Сосочки языка 312, 314
— — вторичные 312
— — грибовидные 312
— — желобоватые 312
— — листовидные 167, 312
— — нитевидные 167, 312, 314
— — первичные 312
Сосуды 176
— лимфатические 181
— развитие 199, 202
— синусоидные 360
— структура 170
Сперматиды 401
Сперматогенез 400
— фазы 400
Сперматогенный 400
Сперматозоид 402, 404
Сперматоцит порядка второго 401
— — первого 401
Сперматоцитогенез 400
Спермиогенез 400, 402
Стадия закладки зубной пластинки 307
— дифференцировки зубного зачатка
307
— развития легкого альвеолярная 291
— — — железистая 287
— — — каналикулярная 287
Статоконий 163
Стенка глаза задняя 150
— — — строение 152
— сосуда
Субстанция Ниссля см. Ниссля суб-
станция
Суданофильность 72
Сустентоциты 407
Тело(а) атретические 436
— белое 436
— желтое, развитие 436
— придатка яичка 418
— ресничное 146
— стекловидное 146
— цилиарное см. Тело ресничное
Тельце (а) остаточные 17
— осязательные 294
— почечные 373, 386
— резидуальные 402
— Фатера—Пачини см. Фатера—Почи-
ни тельце
Тимус см. Железа вилочковая
538
Тироциты см. Эндокриноциты фолли-
кулярные
Ткань (и) жировая белая 72, 75
— костная пластинчатая 79, 81
— мышечная (ые) 87
— — гладкая 103
— — поперечнополосатая 87
— — скелетная 87
— — — гистогенез 91
— — сердечная 97
— — — гистогенез 101
— нервная 105
— собственно соединительная 59
— соединительная 57
— — волокнистая 59
— — — рыхлая 59, 61
— — — плотная 71
— — — — неоформленная 71
— — — — оформленная 72, 73
— — ретикулярная
— — специализированная (ые) 59, 72
— скелетные 76
— хрящевая 76, 77
— — волокнистая 79
— — гиалиновая 76
— — эластическая 76
— эпителиальные 28
— яичка интерстициальная 414
Т-лимфоциты 52
Трансляция 24
Транскрипция ДНК 24
Тракт пищеварительный, общий план
строения 320
Трахея 268, 269
Триада печеночная 356
Тромбоциты 42, 44
Трофобласт 477
Труба маточная 453, 455
— — развитие 453, 457
Тубулин
Узел (узлы) атриовентрикулярный
171
— лимфатические 224, 346
— нервные 128
— синоатриальный 171
Узелки лимфатические 346
Улитка см. Орган слуха
Фагосомы 65
Фагоцитоз 10
Фаза(ы) сперматогенеза 400
— — мейотическая 400
— — сперматогониальная 400
Фатера—Пачини тельце 122
— — строение 384
Фибробласт(ы) 59, 62
Фиброцит 62
Филаменты промежуточные 9
— «тонкие» 10
Фолликул (ы) железы щитовидной 248
— лимфоидные 346
— яичника 420, 425
— — атретический 434
— — вторичные 420
— — первичные 420
— — преовулярные 432
— — примордиальные 420
— — развитие 441, 443
— — третичные 420
Формула крови лейкоцитарная, опреде-
ление 56
Функции оболочки ядерной 24
— ядра 24
Хроматин 24, 26
— деконденсированный 26
— конденсированный 26
Хромаффиноциты 265
Хрусталик 149, 150
Центр клеточный 21
— окостенения диафизарный 85
Центриоли 21
Цилиндр осевой 114
Цитозоль 13
Цитология 6
Цитоплазма 13
— органеллы 14
Цитоскелет 6, 9
Цитотрофобласт 478
Часть хвостовая сперматозоида 405
Шейка зуба 307
— сперматозоида 404
539
Шлеммов канал см. Синус склеры ве-
нозный
Шмидта—Лантермана насечки 119
Щель синаптическая 12, 126
Экзоцитоз 10
Эмаль 307, 308
Эмбриобласт 472
Эмбриология 463
— основные этапы развития 463
— период плодный 464, 521
— — эмбриональный 464
— человека 464
Эмбрион 504
— оболочки 529
Эндокард 170
Эндокриноциты 239
— липоидные 386
— мозговые 262
— панкреатические 354
— парафолликулярные 254
— фолликулярные 254
Эндомизит 87
Эндометрий 129
Эндоплазматическая сеть (ЭПС) см.
Сеть эндоплазматическая
Эндосомы 65
Эндост 79
Эндотендоний 72
Эндотелиоциты фенестрированные 364
Энтодерма зародышевая 480
Эндоцитоз 10
Эпендимоциты 111, 112
Эпидерма 292
Эпидермис кожи «толстой» 295
— — «тонкий» 295
Эпибласт 475
Эпикард 170
Эпимизий 87
Эпителий задний 150
— железистый 28, 35
— многослойный плоский неорогове-
вающий 34, 150
— — — ороговевающий 34
— однослойный 30
— — кубический 30
— — многорядный реснитчатый 30, 32
— передний.150
— переходный 34
— — плоский 30
— — призматический 32
— покровный 28
— — яичника 422
Эпителиоцит (ы) базальный 167
— железистый 338
— микроворсинчатая 338, 344
— поддерживающие 159, 163
— — внутренние 159
— — наружные 159
— почки вкусовой см. Эпителиоцит
сенсорный вкусовой
— сенсорный(е) вкусовой 167
— — волосковые 159, 163
— — внутренние 159
— — наружные 159
Эпифиз 247
Эритропоэз 212
Эритроциты 41
Эухроматин см. Хроматин деконден-
сированный
Ядро (а) 23, 25
— гипоталамуса нейросекреторные 233,
234
— комплекс поровый 26
— компоненты 24
— мембрана внутренняя 26
— — наружная 26
— функции 24
Ядрышко 26
Язык 312
Яичко 399
— придаток 416
— развитие 414
Яичник 420, 422
— закладка 441, 442
— девочки новорожденной 446
— развитие 441
Ямка сетчатки центральная 156
540
Оглавление
Предисловие................................................... 3
1. Цитология. — Т. К. Дубовая, Н. С. Миловидова.............. 5
1.1. Клеточная оболочка (поверхностный аппарат) и цитоскелет 6
1.2. Цитоплазма........................................... 13
1.2.1. Органеллы цитоплазмы........................... 14
1.2.2. Включения...................................... 21
1.3. Ядро (ядерный аппарат)............................... 24
2. Гистология .............................................. Т1
2.1. Эпителиальные ткани. — Ю. К. Елецкий..................28
2.1.1. Покровный эпителий............................. 28
2.1.2. Железистый эпителий............................ 35
2.2. Кровь. — Е. А. Поскребышева.......................... 39
2.3. Соединительные ткани. — А. Б. Строганова............ 57
2.3.1. Собственно соединительная ткань................ 59
2.3.2. Специализированные соединительные ткани .... 72
2.3.3. Скелетные ткани................................ 76
2.4. Мышечные ткани. — А. Б. Строганова................... 87
2.4.1. Поперечнополосатая мышечная ткань...............87
2.4.2. Гладкая мышечная ткань........................103
2.5. Нервная ткань. — Т. К. Дубовая.......................105
2.5.1. Нервная клетка.................................105
2.5.2. Нейроглия......................................111
2.5.3. Нервные волокна................................111
2.5.4. Нервные окончания..............................119
2.5.5. Межнейрональные синапсы........................123
2.6. Нервная система. — Т. К. Дубовая.....................126
2.6.1. Периферическая нервная система.................127
2.6.2. Центральная нервная система....................131
2.7. Органы чувств. — Ю. К. Елецкий.......................145
2.7.1. Орган зрения...................................146
2.7.2. Преддверно-улитковый орган (органы слуха и равно-
весия) ...............................................157
541
2.7.3. Орган вкуса....................................167
2.7.4. Орган обоняния...................................168
2.8. Сердечно-сосудистая система. — М. И. Пекарский, М. Д. Дон-
скова ......................................................170
2.8.1. Сердце...........................................170
2.8.2. Сосуды...........................................176
2.8.3. Микроциркуляторное русло........................183
2.9. Органы кроветворения. — Е. А. Поскребышева...............208
2.9.1. Костный мозг.....................................209
2.9.2. Вилочковая железа (тимус)....................... 219
2.9.3. Лимфатический узел...............................224
2.9.4. Селезенка........................................229
2.10. Эндокринная система. — М. Д. Донскова, Н. С. Милови-
дова .......................................................233
2.10.1. Центральные звенья эндокринной системы . . . 233
2.10.2. Периферические звенья эндокринной системы . . 248
2.11. Дыхательная система.—С.Б.Тарабрин.......................268
2.11.1. Воздухоносные пути.............................268
2.11.2. Легкое ........................................272
2.12. Кожа и ее производные. — М. Д. Донскова.................292
2.13. Пищеварительная система. — Ю. К. Елецкий, А. Ю. Цибу-
левский.....................................................307
2.13.1. Передний отдел.................................307
2.13.2. Средний отдел..................................326
2.13.3. Пищеварительные железы.........................348
2.13.4. Желчный пузырь.................................366
2.14. Мочевыделительная система. — О. В. Волкова..............367
2.14.1. Почка.........................................367
2.14.2. Мочевой пузырь.................................395
2.14.3. Мочеточник.....................................398
2.15. Мужская половая система. — С. Б. Тарабрин...............398
2.15.1. Яичко.........................................399
2.15.2. Сперматогенез..................................400
2.15.3. Придаток яичка.................................417
2.15.4. Предстательная железа..........................418
2.16. Женская половая система. — О. В. Волкова................420
2.16.1. Яичник.........................................420
2.16.2. Матка..........................................446
2.16.3. Маточная труба.................................453
2.16.4. Влагалище.....................................453
2.16.5. Молочная железа................................453
3. Эмбриология. — О. В. Волкова..............................463
3.1. Эмбриональный период.....................................464
3.1.1. Первая неделя развития...........................464
542
3.1.2. Вторая неделя развития........................475
3.1.3. Третья неделя развития........................482
3.1.4. Четвертая неделя развития..................’ . 497
3.1.5. Пятая неделя развития.........................506
3.1.6. Шестая неделя развития........................510
3.1.7. Седьмая неделя развития.......................515
3.1.8. Восьмая неделя развития.......................515
3.2. Плодный период.......................................521
3.2.1. Формирование плаценты........................521
Предметный указатель.......................................5 1
Учебное пособие
ОЛЬГА ВАСИЛЬЕВНА ВОЛКОВА,
ЮРИЙ КОНСТАНТИНОВИЧ ЕЛЕЦКИЙ,
ТАТЬЯНА КЛЕОНИКОВНА ДУБОВАЯ и др.
Гистология, цитология и эмбриология
Атлас
Зав. редакцией А. К. Владимирова
Научные редакторы Т. Н. Лосева, М. Н. Соловова
Редактор 3. В. Колесникова
Художественный редактор С. М. Лымина
Технические редакторы В. И. Табенская,
Г. Н. Жильцова
Корректор А. К. Баскакова
ИБ № 5785
ЛР № 01 02 15 от 11.03.92. Сдано в набор 03.05.95.
Подписано к печати 22.05.96. Формат бумаги
60Х90’/1б. Бумага офс. № 1. Гарнитура тайме.
Печать офсетная. Усл. печ. л. 34,0. Усл. кр.-отт.
136,0. Уч.-изд. л. 29,22. Тираж 10 000 экз.
Заказ 321. «С» 063.
Ордена Трудового Красного Знамени издательство
«Медицина». 101000, Москва, Петроверигский
пер., 6/8.
ОАО «Ярославский полиграфкомбинат».
150049, г. Ярославль, ул. Свободы, '97.
Напечатано Следует читать
с.7, рис. 1.1 ,Б цифры 4 (верхняя) и 5 Не нужны
с 29. рис 2.1.811 цифра 3 Не нужна
с.31. рис.2.2,ГП цифра 4 е нижней правом углу Ненужна
с.98, рис.2.49.Б две цифры 7 (справа и верхняя Ненужны
с.172, рис 291 Слева — артерия.справа — вена
с 209, табл 22 лимфоцитов в тимусе 9 — 20-я* •„.S-10H.-
c222.pMC.2129 Б в
с222.рмс.2129 в Б
с 266, рис.2 154,А цифры 1 и г 2 — фетальная кора
С 303, рис 2.174.Б цифра 4 ^фре 4 указывает на стержень
с.358. рис.2.201.81 цифра IO од.’ст«н“Ы’“'Т “ "вЧ"’°Ч‘
с ЗМе подписи под рис.2 21S (I-W) (I-VQ
с.396, рис.2 220 цифра S Не нужна
с.403 Б В
С. 403 в
0 447. рис 2.246.Б цифры 1.2.3 Не нужны
'45ггиЧЯ"ИС" "ад •-1-1-Й день...* •...1 -7-й день...*
с,512. рис 3.28,8 цифра е цифра 5 (сверху) Цифра 6
с.516, рис.3.30,Б цифра е цифра 4 цифра? Цифра 4 с отводом влево Цифра 7 с отводом влево
с 516: рис З.ЗО.Г цифра 4 Не нужна
с.527. 10-я строка *..5 — меливорсинчатое...* •...5-межаорсиичетое...-