/


























































































































































































































































































































































































































































































































































































Автор: Смит К.
Теги: физиология сравнительная физиология антропология биология нейробиология
ISBN: 978-5-94774-194-0
Год: 2013




Текст
Biology
of Sensory Systems
С. и. M. Smith
Vision Sciences, Aston University,
Birmingham, UK
JOHN WILEY & SONS, LTD
Chichester • New York • Weinheim • Brisbane • Singapore • Toronto
АДАПТИВНЫЕ И ИНТЕЛЛЕКТУАЛЬНЫЕ СИСТЕМЫ
К. Смит
Биология
сенсорных систем
Перевод с английского
доктора биологических наук Ю. Б. Шмуклера
под редакцией
кандидата биологических наук О. Ю. Орлова
Москва
БИНОМ. Лаборатория знаний
УДК 612
ББК 28.706
С50
Смит К. Ю. М.
С50 Биология сенсорных систем / К. Ю. М. Смит; Пер. с
англ. — М.: БИНОМ. Лаборатория знаний, 2013. — 583 с.:
ил. — (Интеллектуальные и адаптивные системы)
ISBN 978-5-94774-194-0 (русск.)
ISBN 471-890901 (англ.)
Книга посвящена строению и работе органов чувств человека и жи-
вотных. Помимо нейробиологических рассматриваются эволюционные,
молекулярные, гистологические, анатомические, а также философские
аспекты. Показано глубокое единство животного мира.
Изучение сенсорных систем животных позволит освободиться от
антропоцентризма и «увидеть» окружающий мир иными органами чувств.
Приведенная в книге библиография и ссылки на интернет-сайты
охватывают период от эпохи Аристотеля до начала нашего века.
Для учащихся старших классов и студентов, изучающих биологию и
поведение животных, а также специалистов, занимающихся моделирова-
нием сложного поведения или интеллекта.
УДК 612
ББК 28.706
По вопросам приобретения обращаться:
«БИНОМ. Лаборатория знаний»
Телефон: (499) 157-52-72,
e-mail: binom@Lbz.ru
http://www.Lbz.ru
ISBN 978-5-94774-194-0 (русск.)
ISBN 471-890901 (англ.)
© 2000 by John Wiley & Sons, Ltd,
Baffins Lane, Chichester,
West Sussex PO19 IUD, England
All rights reserved.
Authorized translation from
the English language edition
published by John Wiley & Sons, Ltd.
© БИНОМ. Лаборатория знаний,
2005
ПРЕДИСЛОВИЕ РЕДАКТОРА
РУССКОГО ПЕРЕВОДА
Так случилось, что знакомство с автором и его книгой состоялось еще до вы-
хода этого произведения в свет, и впечатление было весьма благоприятным.
Когда же профессор Крис Смит любезно прислал книгу, сложилось твердое
убеждение, что она заслуживает того, чтобы быть переведенной и изданной
у нас. Написанная в форме учебного пособия для студентов университетов,
она кроме данных по строению и работе органов чувств человека и животных
содержит вопросы для проверки того, насколько прочитанное усвоено. По
каждому большому разделу приведена литература и дан обзор главных источ-
ников, что может заметно облегчить их поиск в библиотеке и Интернете. Все,
кого интересует биология и поведение животных, будь то студент, старше-
классник или научный работник, найдут эту книгу полезной; она также ока-
жет неоценимую помощь преподавателю для ведения спецкурса по сенсорной
физиологии. Инженер и психолог, имеющие склонность к техническому и
математическому моделированию сложного поведения или интеллекта, также
прочтут ее с пользой.
Вместе с тем, это отнюдь не справочник, ограниченный всего лишь изло-
жением фактических данных; это авторская книга — она несет отпечаток его
собственного исследовательского интереса к ряду фундаментальных проблем,
от главы к главе вновь и вновь встающих за фактолоТией, касающейся того или
иного органа чувств. Эти проблемы, связывающие сенсорику с иными само-
стоятельными областями биологии, обогащают книгу и заслуживают того,
чтобы специально привлечь к ним внимание.
Прежде всего это обращения к временной шкале, относящейся к тому
периоду примерно в полмиллиарда лет, на протяжении которого протекала
эволюция многоклеточных животных. Можно вспомнить, что изначально ис-
точником эволюционных представлений служил преимущественно материал,
поставляемый макроморфологией — прежде всего сравнительной анатоми-
ей животных. Многообразие живых форм и скелетов ископаемых, особенно
позвоночных, бесчисленные вариации в пределах одного плана строения
(«плана творения») — вот что давало пищу для объяснительных построений,
подразумевающих генеалогические взаимоотношения разных ветвей «эволю-
ционного древа». Но с развитием молекулярной биологии и успехами в гено-
мике возник новый источник независимых данных об эволюции — эволюции
того, из чего строятся организмы, и тех генетических текстов, на основе кото-
рых все они строятся. Этот материал несет отпечатки событий, с известной
точностью даже поддающихся датировке. Вот тема, которая постоянно звучит
6 Предисловие редактора русского перевода
в книге благодаря интересу к тому, что дает «сравнительная морфология моле-
кул» — например, образующих мембранные каналы.
Тема эволюции перекликается с оценкой степени ^^ршенства органа в
отношении требований к нему. Приспособительная эволюция подчас упро-
щенно понимается как процесс, гарантированно ведущий к предельному со-
вершенству органа. Мы склонны повсюду видеть целесообразность и ставить
вопрос «зачем?» там, где для этого нет оснований. Есть множество примеров
того, что наблюдаемое является чем-то приемлемым, но отнюдь не идеаль-
ным. Так, у рыб, в том числе и наших далеких предков, местоположение серд-
ца — голова, а не туловище. У нас сердце лежит в грудной клетке, и его иннер-
вацию было бы лучше обеспечить нервом, защищенным шейными позвонка-
ми и покидающим эту защиту уже на уровне груди, а не черепным (как у всех
позвоночных) блуждающим нервом. Каковы бы ни были исторические пред-
посылки того, что глаза позвоночных имеют инвертированную сетчатку (в от-
личие от глаз головоногих и от медиальных глаз самих позвоночных, например
рептилий), не надо искать скрытый приспособительный смысл в том, что свет
на пути к рецепторам вынужден проходить сквозь нервные слои сетчатки.
То же касается адаптивности продукта эволюции, когда речь идет о пре-
дельно малых структурах — молекулах. На этом уровне «морфологии» типич-
но, что чем меньше конструкция, тем больше выражена дискретность возмож-
ных вариантов. В итог$ некоторые свойства того, что закреплено отбором,
оказываются всего лишь попутным следствием жестких требований к иному,
существенному параметру. Так, лишь немногие молекулярные конструкции
пригодны для нужд фотосинтеза или переноса кислорода кровью к тканям.
Следует ли искать целесообразность в красной окраске крови позвоночных
или в зеленом цвете листьев растений? Глубоководным и ночным животным
было бы полезно иметь зрительный пигмент, поглощающий любые доступные
кванты света, т. е. имеющий черный цвет; но все они довольствуются пурпур-
ными родопсинами просто потому, что среди трансмембранных белков нуж-
ного типа — зрительных пигментов — нет черного.
Эволюция глаза позвоночных показывает и такие примеры, когда по тем
или иным причинам удачное решение, найденное предками, оказыватся утра-
ченным их более «прогрессивными» потомками. Именно так обстоит дело
с механизмом аккомодации млекопитающих. Вместо надежного способа
активного сжатия эластичного хрусталика по линии экватора (что ведет к уве-
личению его выпуклости при смотрении вблизь), — выработанного еще реп-
тилиями и сохраненного птицами, — млекопитающие вынуждены полагаться
на его упругость, которая и увеличивает его выпуклость в ответ на ослабление
натяжения хрусталиковой сумки. Этот нелепый способ аккомодации — источ-
ник возрастных проблем с очками для большей части человечества; причина
же его кроется в истории становления млекопитающих, прошедших длитель-
ный период скрытного образа жизни мелких роющихся животных типа земле-
роек, — период, достаточно длительный для того, чтобы был утрачен ряд черт
совершенного глаза, свойственного рептильным предкам млекопитающих.
В результате их потомкам пришлось изобретать заново способ аккомодации, и
найденное решение оказалось не лучшим. Надо помнить, что мы привычно
льстим себе, мимоходом вспоминая нечто на тему «от низшего к высшему»,
Предисловие редактора русского перевода 7
о «братьях наших меньших», и думая о самих себе как о венце творения. Мозг,
речь и рука человека, безусловно, уникально высоко развиты; но это никак не
касается его органов чувств. Вот что имеет в виду Крис Смит, говоря о нашем
антропоцентризме.
Другая сквозная тема — огромная дистанция между рецепцией и восприя-
тием. Самый детальный анализ молекулярной биофизики органа слуха не да-
ет ни малейшего понятия о том, как воспринимается устная речь и где в ней за-
копан «смысл» слов. Когда от работы глаза, органа слуха и т. д. мы переходим
к зрению, к слуховому, тактильному и т. п. восприятию окружающей реально-
сти, — мы постоянно имеем дело с чем-то глубоко отличным от «детектирова-
ния стимула».
Наше восприятие «предметно-ориентировано», нацелено на узнавание
предметов по их предметным свойствам, при этом достойно внимания то, на-
сколько воспринимаемое нами отличается от исходного сенсорного материа-
ла. Всякий без труда отличит наощупь мокрую тряпку от сухой, не подозревая
о том, что у нас нет кожных рецепторов влажности, и что это как бы «первич-
ное» ощущение есть интеграция того, что дают температурные и тактильные
рецепторы. Нам представляется чем-то первичным направление на источник
звука — но наша способность чувствовать это есть результат точнейшего сопо-
ставления того, что «детектируют» два уха, и эта процедура даже не доступна
самонаблюдению. Мы легко узнаем предметы по их окраске — например, от-
личаем ярко освещенный черный предмет от белого, лежащего в тени, даже
если белый при этом отражает света меньше, чем черный; мы «непосредствен-
но» видим цвета этих предметов, не подозревая, какая сложнейшая работа вы-
полняется при этом процедурами зрительной константности в зрительных
центрах мозга. «Световое ощущение не есть еще зрение. Световое ощущение
только тогда становится зрением, когда нам удается при его помощи распозна-
вать внешние предметы; следовательно, зрение заключается в понимании све-
товых ощущений» — говорил Гельмгольц. При этом на пути к предметному
восприятию окружения мы игнорируем огромный поток сенсорной информа-
ции — того, что уже «детектировано». «Как мало мы склонны, при ежедневном
практическом употреблении органов чувств, задумываться над той ролью, ко-
торую они при этом играют; как нас исключительно интересует лишь то из
ощущений, что нам доставляет сведения об отношениях внешне" мира, и как
мало мы обращаем внимания на те ощущения, которые к этому не приуроче-
ны» (Гельмгольц).
Здесь мы вплотную подходим к деликатной теме, которую Смит обознача-
ет как «трудные философские вопросы». Философские потому, что наши чув-
ства (способы восприятия) и мышление были издревле предметом внимания
человека, наивных мыслителей древности, натурфилософов недавнего про-
шлого, философов и психологов современности. Трудные потому, что их об-
суждение сталкивает нас — при первом контакте неожиданно — с двумя нераз-
решимыми проблемами (к чему не всякий готов), и обе имеют прямое отноше-
ние к работе наших органов чувств.
Первая из них звучит вообще как полная нелепость и заключается в том,
что нет никаких способов доказать, т. е. обосновать логически, что столь при-
вычный нам окружающий мир — знакомый нам зрительно, на слух и на-
8 Предисловие редактора русского перевода
ощупь — существует реально, а не только в нашем, скажем, воображении. Да-
же такой яростный полемист и защитник материализма, как автор «Материа-
лизма и эмпириокритицизма», вынужден был признать, что никакими довода-
ми, силлогизмами или рассуждениями невозможно пробить брешь в системе
построений последовательного солипсиста: если я упрямо утверждаю, что все
мое окружение, со всеми плюсами и минусами, включая моих оппонентов и их
высказывания, виртуально, — то выхода из этого тупика нет!
Философские построения в этой области по сути дела есть система выска-
зываний, и этой системе подобает удовлетворять требованиям логики, гаран-
тирующей от явных ошибок. Но как быть, если краеугольный камень постро-
ений — тезис об объективности окружающего мира — не поддается логически
безупречному обоснованию? Если позволить себе принять очевидное, без дока-
зательств, то есть «на веру», то где критерии того, что можно и чего нельзя при-
нимать на веру (без логического обоснования) в дальнейшем? Тот факт, что
в квантовой физике тоже есть как бы похожие трудности — принцип неопре-
деленности (его упоминает Смит), а в математике тоже обнаруживается некая
неполнота аксиоматики, выявленная Гёделем, — не может служить утешени-
ем: это не снимает драматизма положения того, кто одновременно хочет быть
и безупречно корректным в своих высказываниях, и видит неразрешимость
этой проблемы.
Другая проблема звучит не так нелепо, но тоже содержит одну непреодоли-
мую неприятность: нет абсолютно никаких способов непосредственно срав-
нивать наши ощущения, которые суть предмет нашего субъективного мирово-
сприятия. Конечно, каждый склонен полагать, что наши ощущения красного,
соленого и пр. одинаковы, и я тоже верю, что это так; но что же это за наука,
если мы вынуждены прибегать к вере? Действительно, сравниваем ли мы на-
ши ощущения (или слова!), даже когда произносим одинаковые слова, глядя,
скажем, на один и тот же красный объект? А если мы не можем их даже срав-
нивать, то как можно говорить, что они правильно отражают реальность?
Последовательный анализ этой проблемы потребовал бы слишком долгого
обсуждения, здесь не вполне уместного, и мы ограничимся только важным вы-
водом и иллюстрацией к нему. Вывод состоит в том, что сенсорика и вся сис-
тема восприятия служат задаче построения в мозгу модели внешнего мира, и
модель эта в чем-то похожа на комбинацию привычного нам глобуса с поли-
тической картой мира: изображенные на нем пространственные отношения
кардинально отличны от способа его раскраски. Первые отражают свойства,
обязательные для нужд построения действий, и каким бы способом они ни бы-
ли пблучены — зрительно ли, наощупь, с помощью вибрисс, или же на слух
(посредством эхолокации), — все они должны быть между собой в согласии.
Все они должны удовлетворять непреложным императивам нашего трехмер-
ного мира. Совсем другое дело — как раскрашена карта, на которой требуется
различать разные страны и видеть общность принадлежности чьих-то разоб-
щенных территорий. Нам привычно видеть все, принадлежащее Англии, — зе-
леным, Франции — лиловым; но это есть чистая условность, и неуместен во-
прос: «Правильно ли мы делаем, изображая английское — зеленым», и т.'д.
В известном смысле, субъективно знакомая каждому палитра его ощущений
цвета, запахов и вкусов также условна, и ее информативность определяется
Предисловие редактора русского перевода 9
тем, что от чего она позволяет нам отличить. «Видеть — значит различать».
В этом смысл концепции символов (или иероглифов), которой придерживал-
ся Гельмгольц.
Приведенная параллель с глобусом позволяет, быть может, ближе понять
еще одну сквозную тему в книге Смита: то, что другие существа, с иным неже-
ли у нас сенсорным вооружением, обитают в иных, отличных от нашего, сен-
сорных мирах, которые нам, зрительно-ориентирующимся приматам, трудно
себе даже представить. Что значит — быть летучей мышью и «видеть» с помо-
щью эхолокации?
Представим себе, что нужно определить размер и форму предмета на-
ощупь, не глядя на него, например ощупывая чем-то вроде палочки. Решив эту
задачу таким способом, мы вряд ли сможем описать результат в терминах сво-
их тактильных ощущений. Мы объективизируем ощущения, и нужный нам ре-
зультат является сложной интеграцией первичных сенсорных данных, порой
недоступных нашему вниманию и запоминанию. Можно допустить, что лету-
чая мышь также игнорирует акустическую природу своего локационного сиг-
нала и всего сенсорного «сырья», из которого для нее складывется целостная
картина обязательных пространственных отношений — неподвижных препят-
ствий, открытого пространства, мелких порхающих объектов (ее добычи),
и вся эта картина, предназначенная для ориентации в полете, в некотором
смысле не так сильно отлична от привычной нам, получаемой совсем иным
способом — зрительно.
♦ * ♦
Не такая простая задача — охватить круг вопросов «от ионных каналов до
Декарта», т. е. от элементарных процессов на молекулярном уровне, ультра-
структуры и нейрологии в той или иной модальности, до целостного восприя-
тия внешнего мира и места органов чувств в теории познания. Независимо
от достоинств книги К. Смита, осуществить ее перевод и издание в нынешних
условиях тоже непросто. Большой удачей явилось знакомство с издательством
«БИНОМ. Лаборатория знаний», взявшим на себя заботы и риск такого пред-
приятия, и наконец то, что перевод книги был поручен издательством опыт-
ному переводчику и квалифицированному биологу, доктору биологических
наук Ю. Б. Шмуклеру. Его внимательное и ответственное отношение к делу
отчасти отражено в его многочисленных примечаниях переводчика. Вряд ли
кто-либо, тем более редактор перевода, мог бы быть экспертом во всех разде-
лах столь обширной области биологии, как сенорные системы. Редактору по-
могли своими консультациями его коллеги, специалисты по сенсорике:
Н. Г. Бибиков, Б. Д. Васильев, В. И. Говардовский, А. В. Минор, Е. М. Макси-
мова, И. Н. Пигарев, Г. И. Рожкова и А. Я. Супин, — которым он приносит
свою глубокую признательность.
О. Ю. Орлов
ПРЕДИСЛОВИЕ К РУССКОМУ ИЗДАНИЮ
В первую очередь я хотел бы поблагодарить редактора русского издания Оле-
га Орлова, инициировавшего перевод моей книги, переводчика Юрия Шмук-
лера и всех, кто осуществил это предприятие. Знакомству с профессором Ор-
ловым послужил мой запрос на разрешение воспроизвести иллюстрацию
(из книги Ярбуса), которая в моей книге стала рисунком 23.2. Я встретил с
его стороны живой интерес — оказалось, что нас обоих глубоко интересует
сочетание молекулярного и эволюционного подходов в изучении сенсорных
систем, и мы оба чувствуем, как важно не погрязнуть в деталях и видеть
«картину в целом» глазами философа. Я надеюсь, что это издание покажет
читателям в России и во всем русскоговорящем мире интерес такого подхода.
Конечно, мне доставляет большое удовольствие и делает честь, что моя ра-
бота переведена на язык Ивана Павлова, Александра Лурия, Льва Выготского
и многих других выдающихся российских нейробиологов. Я надеюсь, она по-
знакомит русского читателя — студентов и научных работников — с большим
массивом англоязычных публикаций за последние 60 лет, для этого я снабдил
каждую главу книги большой библиографией. С пришествием Интернета и
эры электронных публикаций стало много легче знакомиться с журнальными
статьями. Хотя киберпространство почти исключительно англоязычно, но
возможно, развитие средств электронного перевода даст иноязычным студен-
там больший доступ ко многим электронным литературным источникам.
Исследования сенсорных систем показывают глубокое единство животно-
го мира. Хотя на первый взгляд биосфера кажется населенной почти бесчис-
ленным количеством разнообразных видов, при более внимательном изуче-
нии оказывается, что все они построены на основе общих молекулярных и
биохимических механизмов. Конец XX — начало XXI веков — это героичес-
кая эпоха в изучении генома. Были расшифрованы геномы множества важ-
ных организмов, включая, конечно, Homo sapiens. Потрясающе, что варианты
тех же генов, что у низших форм, таких как круглый червь Caenorhabditis ele-
gans, работают и в нас самих. Нигде это так не проявляется, как в генах, кон-
тролирующих молекулярную структуру и физиологию органов чувств. На-
пример, похоже, что гены, ответственные за механорецепцию у С. elegans (ге-
ны тес) родственны генам, активным в одной из наиболее сложных сенсор-
ных систем человека — внутреннем ухе (см. гл. 7 и 8). Сходным образом ген
Рах-6участвует в раннем развитии глаз во всем животном царстве — от дрозо-
филы до человека и головоногих (гл. 14). В своей основе животный мир един;
возникновение и эволюция всех основных биологических процессов и струк-
Предисловие к русскому изданию 11
тур могут быть прослежены до их общих предковых форм, затерянных в глу-
бинах времени.
Примеры потрясающего внутреннего единства не должны заслонять для
нас огромного разнообразия организмов и их сенсорных систем, которые раз-
вились за более чем полмиллиарда лет эволюции. Царство животных включа-
ет более тридцати групп, или типов, с различным планом строения (гл. 4), и
каждый из них развивался независимо в течение сотен миллионов лет. Соот-
ветственно, и их органы чувств эволюционировали под контролем отличаю-
щихся условий окружающей среды и образа жизни. Удивительны примеры
так называемой конвергентной эволюции, когда к весьма схожим решениям
приходили исходно очень далекие виды (сравните черты сходства и различия
глаз хордовых и белемноидов). С другой стороны, в животном царстве наблю-
дается огромное разнообразие органов чувств, нередко дающих очень похо-
жие результаты при большом различии структур (сравните глаза и слуховые
органы насекомых и позвоночных).
Такие огромные различия сенсорных органов и систем должны, в конце
концов, привлечь внимание философов, занимающихся эпистемологией. Что
значит «быть летучей мышью»? Так называлась знаменитая и оказавшая на
многих влияние статья философа Томаса Нагеля. Действительно, что?* Что
значит — воспринимать мир посредством какой-то из чуждых нам сенсорных
систем, рассматриваемых в этой книге? Этот вопрос не слишком глубоко за-
рыт на многих страницах этой книги, а в главах 22 и 23 рассмотрен напрямую.
Животные существуют в сенсорных мирах, очень отличных от привычного
нам. «Если бы лев умел говорить, — замечает философ Людвиг Витген-
штейн, — мы бы не сумели его понять». Насколько же труднее представить
себе мир в восприятии существ, эволюционно еще более далеких от нас! На-
деюсь, одним из результатов изучения сенсорных систем животных станет ос-
вобождение от узкого антропоцентризма, это заставит включить воображе-
ние, чтобы увидеть мир иными органами чувств, почувствовать свежесть и
новизну и заново ощутить своеобразие нашего собственного восприятия.
К. Ю. М. Смит
Университет Эстон
Бирмингем
Соединенное Королевство
Сентябрь 2004 г.
С. U. М. Smith
Aston University
Birmingham, UK
September 2004
Посвящается Розмари
ПРЕДИСЛОВИЕ
Каждому человеку свойственно стремление к познанию. Это
видно по тому, какое удовольствие мы испытываем от
собственных ощущений; даже независимо от их пользы, они нам
нравятся сами по себе...
Аристотель, Метафизика, кн. 1, 980а
Эта книга возникла на основе многолетнего опыта преподавания биологии
сенсорных систем в университете. Она отличается от множества других пре-
восходных трудов по этому предмету тем, что, помимо нейробиологических,
охватывает и эволюционные, молекулярно-биологические, а также философ-
ские аспекты. Люди — не единственные существа на поверхности этой плане-
ты, которые обладают сенсорными системами. Мы очень тесно соприкасаем-
ся с другими биологическими формами, населяющими биосферу. Некоторые
из этих форм — другие хордовые, в частности, связаны с нами, пусть и весьма
отдаленно, общим планом строения; другие, такие как моллюски и членисто-
ногие, развивались независимо более полумиллиарда лет. Сравнения между
этими формами часто показывают, как сходные сенсорные проблемы находят
различные, а иногда поразительно схожие решения. В основе всех этих реше-
ний лежат сенсорные клетки и молекулярные механизмы, которые делают их
особенно чувствительными к тем или иным формам энергии, поступающей
из внешней среды. Один из важнейших шагов последних десятилетий в на-
уке — это понимание того, что все эти молекулярные механизмы часто оста-
ются теми же самыми у нематоды и у человека. В основе невообразимого
многообразия сенсорных систем в биосфере лежит их поразительное единст-
во. Наконец, сенсорные системы создают каждому из нас собственный субъ-
ективный мир. Когда мы изучаем сенсорные системы на самих себе или на
животных, с которыми мы делим эту планету, мы не должны терять общего
понимания проблемы, которую ставит перед нами таинственный переход
(если это переход?) между миром молекул, клеток и нервных путей со всеми
их потрясающими тонкостями, к совершенно иным мирам запахов, звуков и
красок, которые мы ощущаем каждый день. Это остается важнейшей фило-
софской проблемой современности, и она будет возникать снова и снова на
страницах этой книги.
Изучение сенсорных систем образует мост между миром биофизики, моле-
кулярной биологии, нейрофизиологии, с одной стороны, и миром высшей
нервной деятельности и психологии — с другой. Я надеюсь, что обзор, сделан-
Предисловие 13
ный на последующих страницах, сможет сформировать основу, на которой бу-
дут строиться конкретные исследования. Предмет науки о сенсорных систе-
мах, как и все другие аспекты современной науки, — безбрежен. Все, что я по-
пытался сделать в последующих главах — это дать основу, рамки последующих
исследований. Чтобы разобраться во всех этих работах, необходимо будет при-
бегнуть к возможностям библиотек и доступных теперь электронных баз дан-
ных, поэтому в книгу включена достаточно полная библиография. Для боль-
шей легкости обращения с литературными ссылками они не включены в текст,
а собраны в конце каждой из шести частей книги и снабжены краткими биб-
лиографическими введениями. Наконец, в конце каждой из частей сгруппи-
рованы вопросы для самоподготовки. Эти вопросы выделены в тексте, после
прочтения главы студенту стоит вернуться к ним, чтобы убедиться, что ответы
на них приходят на ум сами собой. Если — нет, то ссылка на соответствующий
раздел текста поможет освежить память.
Ни одна книга, а уж тем более ни одна книга по такому необозримому
предмету, как биология сенсорных систем, не возникает в совершенном виде,
подобно сияющей доспехами Афине, родившейся из головы Зевса. Все, как
давным-давно сказал Исаак Ньютон, опираются на работы бесчисленных
предшественников и современников. В этой книге литературные ссылки охва-
тывают период с середины 1999 года до эпохи Аристотеля и предшественни-
ков, живших две с половиной тысячи лет назад. Соответственно и я благода-
рен бесчисленным исследователям прошлого и настоящего. В особенности
это касается многих современных исследователей, давших разрешения на вос-
произведение их диаграмм и рисунков, ибо сказано, что одна толковая иллю-
страция стоит тысячи слов, а современные книги и научные статьи изобилуют
высококачественными штриховыми и полутоновыми рисунками. Я чрезвы-
чайно признателен авторам и их издателям за разрешение на воспроизведение
иллюстраций.
В этой краткой главке возможно упомянуть лишь немногих из тех, кто по-
мог мне. Список этих людей в алфавитном порядке (английского алфавита)
включает: проф. Т. Оллисона за самые свежие данные об участках головного
мозга, связанных с чувствительностью лица, проф. Пола Бах-и-Рита за по-
мощь по разделу, касающемуся сенсорного замещения, проф. Джона Э. Брюг-
га за рисунки, показывающие расположение участков слуховой коры у кошек
и обезьян, проф. Алана Дж. Бенсона за улучшенные диаграммы волосковых
клеток улитки, проф. Олега Ю. Орлова за ценную информацию по зритель-
ным системам и зрению, проф. Эдриана Хорриджа за информацию о глазах
членистоногих, проф. Джима Пиклза за сканирующие электронные микрофо-
тографии волосковых клеток улитки. Я хотел бы также поблагодарить двух
анонимных рецензентов чернового варианта текста за компетентные замеча-
ния; там, где возможно, они были учтены. Как обычно, противоречия оста-
лись. Так что все ошибки и недоразумения — на совести автора, а не его кон-
сультантов.
Как указывалось в начале данного предисловия, эта книга возникла из
опыта преподавания биологии, а позднее и оптометрии, студентам универси-
тета Эштон. Список тех, кому я благодарен, был бы неполон без упоминания
их чуткой реакции на мой курс — как всегда, учитель учится не меньше, а ино-
14 Предисловие
гда и больше, чем ученик. В отличие от лабораторий компьютерного обуче-
ния — это двусторонний процесс. Я хотел бы поблагодарить моих универси-
тетских коллег за поддержку и обсуждение книги, а также за то, что они дали
мне время и возможности для выполнения данного проекта. Техническая под-
держка была неоценима, мне очень помог компьютер, информационные тех-
нологии и знания по фотографии. Наконец, я не могу не упомянуть с благо-
дарностью своего издателя, который, как и в случае с моими предыдущими
книгами, справился с тем, чтобы совместить иллюстрации, вопросы, списки
литературы, таблицы, приложения, вставки и оглавление так, чтобы получил-
ся текст, который (надеюсь) найдет признание среди студентов и других чита-
телей в начале нового тысячелетия.
КЮМС
Ноябрь 1999
ПРЕДВАРИТЕЛЬНЫЕ СВЕДЕНИЯ
1. ОСНОВЫ
Происхождение: глубокая древность — РНК и ДНК — «эгоистичные ге-
ны». Аллостерические эффекторы: конформационные изменил — коопе-
ративная аллостерия — протеинкиназы и фосфатазы. Мембраны: липи-
ды — жидко-кристаллическая структура — белки — подвижность. Рецеп-
торные молекулы: семидоменная (7ТМ) конформация — молекулярная
структура — связь с G-белком — десенситизация. Мембранные сигналь-
ные системы: G-белки — структура — биохимия взаимодействий — раз-
личные эффекторы — различные вторичные мессенджеры. Каналы и
воротные механизмы: медиаторные и потенциал-зависимые ионные ка-
налы— Ка+-канал, структура и функция. Заключение: вездесущность мо-
лекулярных элементов
Три с половиной миллиарда лет назад первые предшественники прокариот
возникли в уже тогда древнем океане. Еще до того в первичном бульоне появи-
лись примитивные самовоспроизводящиеся молекулы. Вероятно, наиболее
ранним репликатором была РНК, а не ДНК, поскольку некоторые формы
РНК (рибозимы) обладают ферментативной активностью, а репликация в
этом случае протекает быстрее. Хотя репликация ДНК более эффективна, она
всегда зависима от белковых ферментов, что трудно себе представить в прими-
тивном океане. Следовательно, более сложный процесс репликации ДНК дол-
жен был развиться позднее. Однако, эффективная репликация РНК или ДНК,
a fortiori (в особенности) — последней, зависит от множественности молекул.
Из этого следует, что эти молекулы должны находиться в достаточной близос-
ти друг от друга. Возможно, что все началось с их адсорбции на какой-либо
обычной поверхности, например на глинах. Однако наиболее эффективным
средством удержания взаимодействующих молекул вместе было помещение
их в тонкий мешочек или пузырек.
Простейшие современные прокариоты — это микоплазма. Самое малое из
них имеет диаметр 0,3 мкм и содержит не более 750 различных типов белков, но
даже эти крошечные клетки куда более продвинуты, чем протоклетки, сущест-
вовавшие три с половиной миллиарда лет назад, поскольку они, как и современ-
ные клетки, используют ДНК, а не РНК, как наследственный материал. Так или
иначе, рассматриваем ли мы простейшие из живых клеток или их гипотетиче-
ских предшественников, одно их выделяет — они существуют как отдельности
в окружающей среде. Их внешние мембраны отграничивают внутреннюю сре-
ду от внешней. Биологи с философскими наклонностями прослеживают про-
исхождение индивидуального организма до этого изначального периода.
16 Часть I. Предварительные сведения
Все организмы живут в окружающей среде. Все организмы тем или иным
образом реагируют на окружающую среду. Это то, что отличает их от неоду-
шевленных предметов. Ясно, что поверхностная мембрана, отграничивающая
организм от окружающей среды, должна играть критически важную роль.
Именно здесь развиваются специализации, которые способны детектировать
благоприятные и неблагоприятные изменения. Иными словами, именно здесь
возникает простейшая сенсорная система. Будучи информирован об измене-
ниях в окружающей среде, организм может реагировать на них благоприятным
для себя образом. Благоприятным для чего? В конечном счете, преимущества
могут быть прослежены до уровня физической химии реплицирующихся мо-
лекул. Те из них, которые реплицируются более эффективно, поглощают
больше доступных ресурсов и выживают.
Итак, начнем с начала: рассмотрим элементы, из которых построена сен-
сорная система.
1.1. Аллостерические эффекторы
Учебники биохимии и по молекулярной биологии говорят, что ферментатив-
ные белки имеют сложную трехмерную структуру. Ковалентно связанная пер-
вичная структура, состоящая из одной или более аминокислотной цепочки,
свернута в сложную конформацию, т. н. третичную структуру. Эта структура
стабилизирована многочисленными «слабыми» взаимодействиями: водород-
ными связями, вандерваальсовыми и гидрофобными силами и т. д. Стоит под-
черкнуть, что каждая из этих сил по отдельности слаба. Тогда как энергия еди-
ничной ковалентной связи составляет около 100 ккал/моль (двойные и трой-
ные связи, обладают, соответственно, большими энергиями), водородная
связь характеризуется энергиями всего в 1—5 ккал/моль, а гидрофобные и
очень короткодистантные вандерваальсовы силы — всего около 1 ккал/моль.
Хотя, по сравнению с ковалентными связями, все они очень слабы, однако ча-
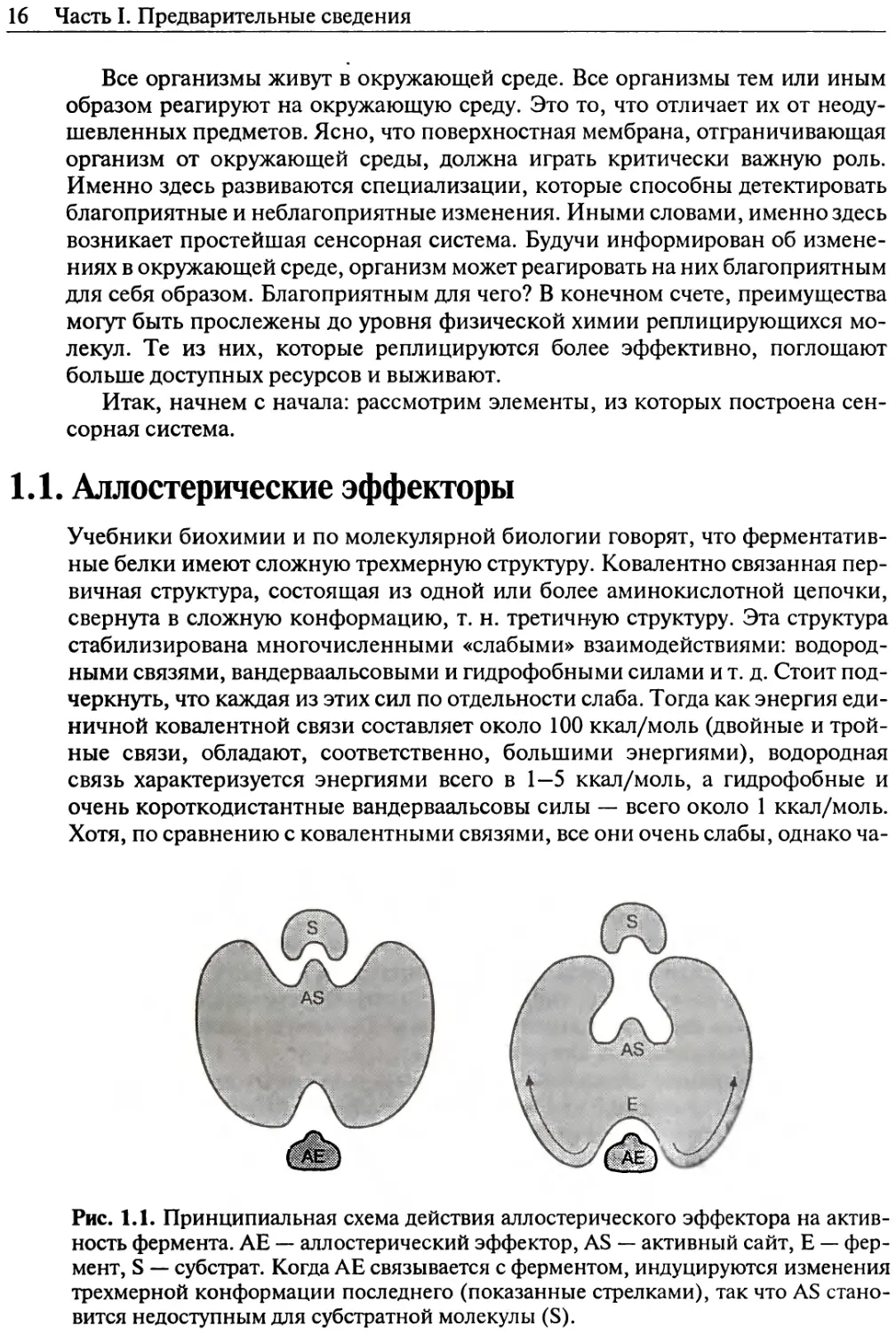
Рис. 1.1. Принципиальная схема действия аллостерического эффектора на актив-
ность фермента. АЕ — аллостерический эффектор, AS — активный сайт, Е — фер-
мент, S — субстрат. Когда АЕ связывается с ферментом, индуцируются изменения
трехмерной конформации последнего (показанные стрелками), так что AS стано-
вится недоступным для субстратной молекулы (S).
1. Основы 17
Рис. 1.2. Белок с боковой цепью (серин, треонин или тирозин), обозначенной на
рисунке «аа». Протеинкиназы фосфорилируют боковую цепь за счет АТФ. Проте-
инфосфатазы затем дефосфорилируют боковую цепь.
сто они чрезвычайно многочисленны. Большое число таких слабых взаимо-
действий и поддерживает сложную конформацию белковой молекулы.
В то же время это означает, что такая пространственная структура фермен-
тативного белка чрезвычайно хрупка и легко ранима. Это также означает, что
такая структура может легко перестраиваться из одной конформации в другую.
Именно эта черта лежит в основе феномена аллостерии. В сущности, это озна-
чает, что когда молекула (или лиганд) связывается с одним из сайтов на по-
верхности белка, это вызывает изменения конформации, которые демаскиру-
ют активные сайты в других участках белка. В известном смысле это может
рассматриваться как наиболее примитивная сенсорная система — белковая
молекула меняет свое поведение в ответ на какой-либо фактор окружающей
среды (рис. 1.1).
Аллостерические переходы играют настолько важную роль, что можно
сказать, что они лежат в основе всей клеточной биологии. Мы еще не раз
столкнемся с этим на последующих страницах. Часто такие аллостерические
переходы встречаются у белков, состоящих из более чем одной субъединицы.
В таких случаях связывание лиганда с аллостерическим сайтом одной из субъ-
единиц вызывает изменения, которые облегчают связывание лигандов с алло-
стерическими сайтами других субъединиц. Такое явление известно как коопе-
ративная аллостерия и может приводить к более существеннным изменениями
поведения аллостерического белка.
Одним из наиболее эффективных факторов, вызывающих аллостерические
переходы, является фосфорилирование. Мы будем сталкиваться с этим меха-
низмом снова и снова при изучении молекулярных основ сенсорных систем,
так что стоит рассмотреть его несколько более’подробно. Реакция фосфорили-
рования катализируется протеинкиназой. Протеинкиназы образуют большое
семейство из нескольких сотен белков, каждый из включает каталитический
домен из 250 аминокислот. Основная реакция состоит в переносе фосфатной
18 Часть I. Предварительные сведения
группы с АТФ на гидроксильную группу боковой цепи аминокислот субстрат-
ного белка. Только три аминокислоты — серин, треонин и тирозин — обладают
гидроксильной группой в боковых цепях, так что только эти аминокислоты и
могут участвовать в реакции, которая схематически представлена на рис. 1.2.
Протеинкиназы, фосфорилирующие субстратные белки (что ведет к алло-
стерическим переходам в последних) сами находятся под аллостерическим
контролем. В этой книге нет необходимости углубляться в биохимические
тонкости процесса, однако стоит упомянуть о других ферментах — протеин-
фосфатазах, которые в цитозоле присутствуют, чтобы ликвидировать послед-
ствия деятельности киназ (рис. 1.2). Эти ферменты устраняют фосфат с суб-
стратного белка, что позволяет последнему возвратиться к исходной конфор-
мации.
1.2. Мембраны
Второй элемент любой сенсорной системы — это биомембраны. Хотя наибо-
лее древние мембраны, сформировавшиеся в первобытные времена, могли
быть построены из аминокислот, все современные биологические мембраны
(или биомембраны) состоят из липидного бислоя с белковыми включениями.
Кроме того, большинство биомембран содержит и углеводы. Липиды образу-
ют матрикс или основу, в которую погружены белки, а углеводы (там, где они
присутствуют) присоединены либо к липидам (гликолипиды) или к белкам
(гликопротеины) (рис. 1.3).
Липиды
На рис. 1.3 видно, что липиды образуют бимолекулярный слой. Они разделя-
ются на три большие группы: фосфолипиды, гликолипиды и стероиды (в част-
ности, холестерин). В этой книге нет необходимости обсуждать детали их
структуры — стандартные учебники по биохимии дают полную информацию.
Важно отметить, что все эти молекулы относятся к амфипатическим, т. е. час-
тично растворимым в воде, а частично — в органических растворителях. Ти-
пичный мембранный липид на одном из концов молекулы несет электроста-
тический заряд и, таким образом, может связываться с водорастворимыми
веществами, а другой конец, ковалентно не связанный с электростатическим
зарядом, чувствует себя, как дома, в органическом растворителе. Поскольку и
внеклеточная, и внутриклеточная среда практически исключительно водные,
липиды, образующие бислойную мембрану, выстраиваются так, что их гидро-
фильные «головы» обращены в водную среду, а гидрофобные «хвосты» — друг
к другу (изолированно как от водной среды снаружи, так и внутри клетки). Не-
которые типичные мембранные липиды показаны на рис. 1.4.
Из того, что было сказано выше, а также рис. 1.3 и 1.4 ясно, что биологиче-
ские мембраны — это весьма тонкие структуры. Составляющие, их липиды
удерживаются гидрофобными силами и случайными электростатическими
взаимодействиями их «головных» групп. Чрезвычайно свободная структура
липидного бислоя означает, что при комнатной температуре индивидуальные
молекулы пребывают в постоянном движении. Действительно, гидрофобные
1. Основы 19
Рис. 1.3. Мембрана быстро заморожена до температуры жидкого азота и вскрыта сколом.
Плоскость скола проходит через середину липидного бислоя. Рисунок показывает, как
белки погружены в мембрану, а также положение углеводных цепей, проецирующихся с
наружной поверхности («strings of sausages»). Воспроизводится по Биргит Сатир «The
final steps of secretion», Scientific American, October 1975, p. 33, с разрешения управляюще-
го имуществом Бунджи Тагава.
жирнокислотные хвосты молекул липидов сравнимы с корзиной змей, пере-
плетенных в постоянном движении. Внутренность мембраны, таким образом,
с любой точки зрения представляет собой органическую жидкость. Мы еще
увидим, что эта чрезвычайная текучесть липидного матрикса биомембран
имеет существенное значение, когда будем рассматривать G-белок-связанную
передачу сигнала.
Не все липидные составляющие биомембран столь же лабильны, как фос-
фолипиды. Холестерин, в частности, — это совершенно другой тип молекулы.
Как показано на рис. 1.4, молекула холестерина состоит из трех составных час-
тей: гидрофильной «головы» — гидроксильной группы, жесткого тарелко-
образного стероидного кольца и гибкого гидрофобного хвоста. Количество
холестерина в мембране значительно варьирует; когда он присутствует, увели-
чивается жесткость мембраны и уменьшается ее текучесть.
Текучесть мембраны определяется, на самом деле, не только количеством
присутствующего в ней холестерина, но и длиной и насыщенностью жирных
кислот, образующих ее основу. Искусственные мембраны, образованные ли-
пидами только одного вида, обладают достаточно резкой характеристикой
20 Часть I. Предварительные сведения
мн,
I
сма
I
сн,
о
I
О"—Р =0
I
о
I I
с=о с=о
I I
сма СМ2
I I
СН2 См,
I I
СН2 Сма
I I
Сна Сна
I I
си, СИ,
сна СНа
I I
СИ. СИ.
Сна сна
I ।
СНа СМ2
I I
Сма CMj
I I
0*2 СНа
I I
сна СМ2
I I
сн3 CMj
I
СНэ
I
СИ.
СН3
I
СМ,
снД
iCHj-N’-CHjI
» । !
\ сн, /
(Г-Р—О
I
________9______________Край липидной
С1_|2___________________фазы мембраны
Н—С—NH-C=O
I I
но-с—н сн2
сн сн,
сн схНг
'сн, ,СНг
' СН2 Липидная
\2 фаза мембраны
СН2 /
СН2 СчНг
СН2 ZCH2
СН2 СН3
СН2
СН2
СН2
сн2
сн2
сн2
СНЭ
Б
® NANA -Gal
I
Glc
CH3
В
сн2
СНэ—СН
Рис. 1.4. Некоторые распространенные липиды мембран. (А) Фосфатилилхолин (леци-
тин); (Б) сфингомиелин; (В) ганглиозид; (Г) холестерин. Пунктирная линия символизи-
рует длинную алифатическую цепь. Gal — галактоза, Glc — глюкоза, NANA — N-ацетил-
нейраминовая кислота.
1. Основы 21
данного фосфолипида — «температурой перехода» от жидкого состояния к ге-
лю. Эта температура варьирует в различных местах естественной биомембра-
ны, в зависимости от количества холестерина и насыщенности фосфолипид-
ных «хвостов». Естественная мембрана может, таким образом, рассматривать-
ся как мозаичная структура с различной степенью текучести.
Белки
В «лоскутное одеяло» биомембраны погружены белки. Хотя гликолипиды (та-
кие, как молекулы клеточной адгезии) очень важны для клеточного распозна-
вания, все же наиболее важные характеристики биомембран обусловлены не
липидами, в белками. Количество белка в мембранах варьирует от 20% (мие-
лин) до прибл. 75% массы (внутренняя мембрана митохондрий). Большинство
мембран содержит по массе около 50% белка.
Большинство белков (как показано на рис. 1.4), погружено в мембрану.
Они «плавают», как айсберги в переменчивом «море» фосфолипидов, или,
если посмотреть на это иначе, образуют мозаику в жидком фосфолипидном
матриксе. По этим причинам такая концепция структуры биомембран назы-
вается «жидкостно-мозаичной» моделью. В некоторых случаях белки прони-
зывают весь бислой и соприкасаются как с внутриклеточным, так и внекле-
точным пространствами. В других случаях белки присоединены к мембране
цепью жирной кислоты, фосфолипидом или пренильной группой. В этих
случаях сам белок расположен в цитозоле. Некоторые из этих вариантов при-
соединения белков показаны на рис 1.5. Мы еще увидим впоследствии, что
мембранно связанные белки образуют базовые элементы всех сенсорных ре-
цепторов.
Рис. 1.5. Некоторые пути, которыми белки ассоциируются с мембраной. Цилинд-
ры внутри мембраны символизируют а-спирали. (А) Одиночная а-спираль прохо-
дит через мембрану; (Б) многочисленные а-спирали проходят через мембрану;
(В) белок соединен с цитоплазматической частью бислоя жирнокислотной цепью
или пренильной группой (важный пример такого типа связи — G-белки); (Г) бе-
лок, погруженный в мембрану, нековалентно связан с другим белком в цитозоле.
22 Часть I. Предварительные сведения
Трансмембранные белки построены таким образом, что гидрофобные до-
мены погружены в мембрану, а гидрофильные обращены в водные — внутри-
клеточный и(или) внеклеточный компартменты (см. рис. 1.5) По сравнению с
глобулярными белками водного цитозоля внутримембранные домены мемб-
ранных белков в известном смысле перевернуты: их гидрофобные аминокис-
лотные остатки обращены кнаружи, а гидрофильные загнуты внутрь. Это
обеспечивает способность белка удерживаться в мембране. Очень часто, как
это показано на рис. 1.5, внутримембранные домены состоят из а-спиральных
сегментов. Опять-таки, огромное большинство аминокислотных остатков, об-
разующих а-спирали, гидрофобны.
Исследования включения ферментативных белков в искусственный ли-
пидный бислой показывают, что активность таких белков кондиционируется
их липидным окружением. Характеристики бислоя, такие как длина жирно-
кислотной цепи, степень их насыщенности и природа липидных «голов», вли-
яют на биологическую активность фермента. Точно так же, как на водорас-
творимые ферменты влияют характеристики водного окружения (pH, концен-
трация солей и т. д.), так ферменты, погруженные в слой липидов, зависят от
конкретной структуры последних.
Подвижность белков
Мы уже сравнивали мембранные белки с айсбергами, плавающими в липид-
ном море. Поэтому неудивительно, что многие из них обладают значительной
латеральной подвижностью. В следующем разделе мы увидим, что эта по-
движность с большим эффектом была использована в формировании сиг-
нальных систем, основанных на перемещениях белков в плоскости мембра-
ны. Коэффициент диффузии белков составляет от прибл. 10“9 см2/с для зри-
тельных пигментов наружного сегмента палочек до прибл. 10“11 см2/с для
других белков. В первом случае белок проходит около 0,1 мкм в секунду, а во
втором — 0,001 мкм/с. Существует множество причин для таких больших раз-
личий подвижности. Во-первых, это может происходить вследствие различий
липидного состава мембран, который, как указывалось выше, влияет не ее те-
кучесть. Белок может быть также частью большого комплекса с другими бел-
ками, который оказывается слишком большим, чтобы легко перемещаться.
Подвижность может быть затруднена не принадлежащими собственно мем-
бране структурами, такими как щелевые и тесные контакты, а также десмо-
сомы и т. д. Последнее по порядку, но далеко не последнее по важности —
возможность того, что мембранные белки заякорены за элементы примем-
бранного цитоскелета.
1.3. Рецепторные молекулы
На границе между организмом и окружающей средой располагаются рецеп-
торные молекулы. Не все органы чувств обращены такими молекулами во
внешнюю среду, однако две важные экстероцепторные системы организованы
именно так — хеморецепторы и фоторецепторы. Даже сенсорные системы, ко-
торые не полагаются на рецепторные молекулы в детектировании изменений
1. Основы 23
окружающей среды, механо- и терморецептивная, тем не менее используют
(как мы увидим ниже) мембранные белки, хотя и несколько иным образом.
Хемо- и фоторецепторные клетки имеют особую белковую организацию,
сходную с той, которая обнаружена в рецепторных молекулах, реагирующих
на нейромедиаторы в множестве синапсов. Полипептидные цепи таких рецеп-
торов семь раз проходят через мембрану (рис. 1.6). Из-за такой организации —
с семью трансмембранными сегментами, рецепторы именуются семидоменны-
ми (или 7ТМ), а иногда и змеевидными. Многие, но далеко не все семидомен-
ные рецепторы эволюционно связаны. Они принадлежат огромному суперсе-
мейству белков — согласно расчетам до 2% генома может быть занято кодиро-
ванием этих вездесущих рецепторов.
Схематическое изображение на рис. 1.6 А показывает, как семь трансмем-
бранных сегментов расположены в мембране. Оно также показывает наличие
больших внутри- и внеклеточных доменов. Рис. 1.6 Б показывает, что в реаль-
ности семь трансмембранных сегментов образуют как бы пилоны полой ко-
лонны, ориентированные подобно лепесткам диафрагмы объектива или клеп-
кам в бочке.
Семидоменные рецепторы не только имеют общую архитектуру, но и мем-
бранно-связанные средства усиления сигнала. Этот механизм (и мы увидим
это в следующем разделе) основывается на латеральной подвижности белков,
в данном случае — G-белков, в биомембранах и на том обстоятельстве, что ли-
пидный бислой удерживает эти белки в тесной близости, так что они не могут
Рис. 1.6. Архитектура семидоменного рецептора. (А) схематическое изображение:
семь трансмембранных сегментов обозначены цилиндрами и цифрами от 1 до 7.
N-конец расположен внеклеточно, к нему обычно присоединены углеводные ос-
татки (т. е. он гликозилирован), что обозначено знаками «Y». Внеклеточные петли
обозначены е-1, е-2, е-3, они также могут быть гликозилированы. Внутриклеточ-
ные петли i-1, i-2, i-З представляют собой места распознавания для специфических
G-белков. Темными точками обозначены места фосфорилирования для протеин-
киназ, а крестиками — сайты, на которые воздействуют специфические десенсити-
зирующие протеинкиназы. (Б) Трехмерная конформация рецептора в мембране.
24 Часть I. Предварительные сведения
диффундировать в цитозоль. Тонкий структурно-функциональный анализ се-
мидоменных рецепторов показал, что первая, вторая и третья цитоплазмати-
ческая петли (i-1, i-2 и i-З), а также карбоксильный конец (рис. 1.6 А) критич-
ны для связывания с G-белком, при этом третья петля особенно важна в рас-
познавании специфических G-белков. Кроме того, показано, что рецептор,
подвергнутый длительному воздействию лиганда, значительно уменьшает
свою чувствительность — это явление известно как десенситизация. С ним мы
также неоднократно столкнемся на последующих страницах. Десенситизация
происходит вследствие активности специфических протеинкиназ (см. раз-
дел 1.1), фосфорилирующих гидроксильные группы серинового, треониново-
го и тирозинового остатков карбоксильного конца рецептора. Эти остатки
обозначены крестиками на рис. 1.6 А. Помимо того сериновые, треониновые и
тирозиновые остатки подвергаются воздействию неспецифических протеин-
киназ (обозначены кружками). Фосфорилирование, как было показано в раз-
деле 1.1, меняет трехмерную конформацию рецептора. Чувствительность вос-
станавливается при дефосфорилировании одним из многих фосфатазных фер-
ментов, распространенных в цитозоле.
1.4. Мембранные сигнальные системы
Внешние мембраны клеток, разграничивающие внешнюю среду и цитозоль,
развили множество биохимических механизмов, которые преобразуют внеш-
ние стимулы в передаваемые в цитозоль сигналы.
1.4.1. G-белки
Когда семидоменная рецепторная молекула, локализованная в мембране сен-
сорной клетки, активируется какими-то изменениями во внешней среде, она
претерпевает конформационные изменения. Последние детектируются G-бел-
ками, связанными с мембраной, которые, в свою очередь, активируют эффек-
торные молекулы в мембране. Часто это приводит к выделению вторичных
мессенджеров в цитозоль. Этот процесс схематически показан на рис. 1.7.
G-белки, участвующие в передаче сигнала, являются членами еще одного
большого надсемейства белков, в данном случае — гуанин-связывающих бел-
ков. G-белки — это прецизионные регуляторы, включающие или выключаю-
щие активность других молекул. Все G-белки «включаются» при связывании
с ГТФ и «выключаются» гидролизом ГТФ до ГДФ. Этот гидролиз катализиру-
ется ГТФ-азной активностью самих G-белков. Процесс этот сравнительно
медленный, протекающий в течение секунд — десятков секунд.
G-белки биологических мембран имеют гетеротримерную структуру. Они
состоят из большой а-субъединицы (около 45 килодальтон — кДа), а также
меньших р- и у-субъединиц (рис. 1.8). а-субъединица обладает ГТФ-азной
активностью, в неактивной («выключенной») форме она связывает молекулу
ГДФ на активном сайте. Субъединицы р и у связаны между собой, и в физио-
логических условиях не могут быть диссоциированы. В неактивном состоя-
нии р-у-комплекс непрочно связан с а-субъединицей. у-субъединица связана
с цитоплазматическим листком биологической мембраны геранил-геранило-
1. Основы 25
Рис. 1.7. Схема G-белок сиг-
нальной системы. S — стимул;
R — мембранный рецептор;
Е— эффектор (фермент, ион-
ный канал и т. д.); М — вторич-
ный мессенджер.
Внеклеточное
Цитозоль м
вой цепью (20 атомов углерода в цепи), близкой по структуре к холестерину,
а-субъединица также связана с мембраной жирной кислотой с длиной цепи
в 14 атомов углерода (миристоевая кислота). Такие связи обеспечивают то, что
комплекс G-белка удерживается в плоскости мембраны, но в то же время спо-
собен легко двигаться в этой плоскости. Легко себе представить, как весь ком-
плекс G-белка с присоединенным ГДФ перемещается в плоскости мембраны
под действием тепловых сил.
а-субъединицы чрезвычайно вариабельны. Например те, что обнаружены
в обонятельной системе, существенно отличаются от таковых в зрительной
системе. 0- и у-субъединицы менее гетерогенны, хотя известно множество
различных их типов. Такая молекулярная гетерогенность дает возможность
организовать гибкую высокоадаптивную сигнальную систему. Мы обсудим ее
детали по мере того, как будем сталкиваться с ней в конкретных случаях в сле-
дующих главах.
Когда комплекс G-белка, перемещающийся в мембране, входит в контакт
с активированным семидоменным рецептором, это приводит к освобождению
ГДФ. Вследствие избытка в цитозоле ГТФ диффузия последнего обеспечивает
занятие освободившегося сайта. После того, как это происходит, тримерный
комплекс G-белка разделяется. 0- и у-субъединицы, остающиеся соединенны-
ми между собой, диссоциируют от а-субъединицы, далее эти части комплекса
двигаются своими отдельными путями, а-субъединица с присоединенным с
ней ГТФ способна взаимодействовать с «эффектором» в мембране — фермен-
тами, такими, как аденилатциклаза, или, возможно, ионными каналами.
Фермент может активироваться или ингибироваться, а ионный канал — от-
крываться или закрываться. Конкретные примеры будут рассмотрены в после-
дующих главах. Взаимодействие с эффектором, однако, длится до тех пор, по-
Внеклеточное
пространство
Рис. 1.8. Конформация гетеро-
тримерного мембранно-свя-
занного G-белка, а-субъеди-
ница изображена с полостью,
символизирующей сайт связы-
вания ГДФ или ГТФ.
Мембрана
Цитозоль
26 Часть 1. Предварительныесведения
Рис. 1.9. Сигнальная система G-белка в биологической мембране. (А) фаза покоя;
(Б) лиганд присоединяется и активирует рецептор; (В) G-белок сталкивается с активи-
рованным рецептором, диссоциирует, и а-субъединица теряет ГДФ; (Г) а-субъединица
активируется, присоединяя ГТФ; (Д) а-субъедница присоединяется к эффектору и ак-
тивирует его, эффектор, в свою очередь, катализирует синтез вторичного мессенджера;
(Е) дефосфорилирование ГТФ деактивирует а-субъединицу, которая отсоединяется от
эффектора, готовая начать весь цикл сначала. Пунктирное окрашивание рецептора на
рисунке символизирует активацию; Е — эффектор; L — лиганд; R — рецептор.
ка а-субъединица, являющаяся ГТФ-азой, удерживает ГТФ. Так что, очень
вскоре присоединенный ГТФ гидролизуется до ГДФ. Когда это происходит,
а-субъединица снова меняет свою конформацию и теряет способность акти-
вировать эффектор. После этого а-ГДФ взаимодействует с р-у-комплексом
и снова образует тримерный комплекс, завершая, таким образом, цикл
(рис. 1.9). До недавнего времени полагали, что р-у-комплекс не играет серьез-
ной роли в мембранной сигнализации. Теперь это ставится под сомнение —
существуют данные о том, что этот комплекс может играть независимую роль.
Возможно, что он способен ингибировать активность свободной а-субъеди-
ницы или независимо влиять на мембранные эффекторы.
1.4.2. Эффекторы и вторичные мессенджеры
Существует целый ряд классов эффекторных молекул: циклазные ферменты,
фосфолипазы, фосфодиэстеразы, мембранные каналы. Мы будем сталкивать-
ся с этими эффекторами на следующих страницах этой книги. Соответствен-
но, существуют и различные типы вторичных мессенджеров: цАМФ, цГМФ,
инозитолтрифосфат (ИФ3), диацилглицерин (ДАГ) и вездесущий ион Са2+.
В данном разделе будут рассмотрены только два типа эффекторов — аденилат-
1. Основы 27
циклазы (АЦ) и фосфолипаза С-р (ФИФ2-фосфолипаза), которые продуциру-
ют важные вторичные мессенджеры. Мы также рассмотрим роль фосфодиэс-
тераз и мембранных каналов, а также других вторичных мессенджеров по ме-
ре необходимости в дальнейших частях этой книги.
Аденилатциклазы
Эти ферменты катализируют формирование цАМФ, вездесущего и, возмож-
но, самого важного вторичного мессенджера в клетках животных. Наиболее
существенная роль цАМФ состоит в активации цАМФ-зависимых протеинки-
наз (ПК). Будучи активирован, этот мультимерный фермент фосфорилирует
(при участии АТФ) тот или иной из множества биологически активных бел-
ков, представленных в клетке — ферментативных, рецепторных и канальных
белков, ядерных гистонов, факторов транскрипции и т. д. Фосфорилируются,
как правило, сериновые, треониновые или тирозиновые остатки, что приво-
дит либо к ингибированию (ср. упомянутую выше десенситизацию G-белок-
связанных рецепторов), либо активации белка. Последующее дефосфорили-
рование, восстанавливающее исходное состояние, обеспечивается одним из
множества фосфатазных ферментов цитозоля.
Молекулярно-биологическими методами показано существование в клет-
ках млекопитающих по крайней мере шести различных аденилатциклаз. Все
они имеют молекулярную массу около 120—130 кДа, а исследование их гидро-
фобной части показывает наличие 12 трансмембранных сегментов. Шесть
циклаз различаются по чувствительности к р-у-комплексу G-белков и к Са2+-
связывающему белку кальмодулину. Аденилатциклаза типа 1, например, сти-
мулируется Са2+-кальмодулином и ингибируется p-y-димером, тогда как аде-
нилатциклаза типа 2 не реагирует на первый и стимулируется вторым.
Фосфолипаза С-р (ФЛС-р или ФИФ2-фосфолипаза)
Активация этого важного вторичного эффектора приводит к продукции двух
вторичных мессенджеров: инозитолтрифосфата (ИФ3) и диацилглицерина
(ДАГ). Оба вторичных мессенджера формируются в результате расщепления
фосфолипазой С-р фосфолипида — фосфатидилинозитол 4,5 бифосфата
(ФИФ2), который в основном содержится во внутреннем листке плазматичес-
кой мембраны (рис. 1.10).
Рис. 1.10 показывает рецептор, взаимодействующий с неким внешним сиг-
налом, что приводит через G-белковый механизм к активации погруженной в
мембрану ФЛС-р. Последняя реагирует с ФИФ2, образуя ИФ3 и ДАГ. ИФ3 —
это водорастворимая молекула и потому легко диффундирует в цитозоль.
Здесь он может взаимодействовать с соответствующими рецепторами в мем-
бранах эндоплазматического ретикулума, что приводит к освобождению Са2+.
Эти ионы имеют многочисленные эффекты на клеточную биохимию. В конце
концов ИФ3 инактивируется инозитолтрифосфатазой. С другой стороны,
ДАГ — гидрофобное вещество и потому остается в мембране.
Мы не закончили рассмотрение данной системы, поскольку еще не рассмо-
трели функцию ДАГ. Он взаимодействует с мембранно-связанной протеинки-
назой — протеинкиназой С (ПКС), эта реакция является Са2+-зависимой. Сле-
28 Часть I. Предварительные сведения
Рис. 1.10. Путь формирования вторичного мессенджера, включающий ФЛС-р.
Активация рецептора ведет через G-белок-связывающую систему к активации
ФЛС-р. Последняя расщепляет мембранный липид фосфатидилинозитол-дифос-
фат на ИФ3 и ДАГ. ИФ3 диффундирует в цитозоль; ДАГ остается в мембране. Бо-
лее полное рассмотрение — в тексте.
довательно, когда концентрация Са2+ в цитозоле повышается (эффект, который
мы видели в случае ИФ3), ДАГ активирует ПКС. Такая активация также требу-
ет участия фосфолипида — фосфатидилсерина, который присутствует во внут-
реннем листке мембраны. Активированная ПКС может активировать белки,
вызывающие специфические биохимические ответы. В нейронах продемонст-
рирован целый ряд таких реакций, в том числе синтез и секреция нейромедиа-
торов, изменение чувствительности рецепторов и функции цитоскелета.
Из изложенного выше ясно, что G-белковая система обеспечивает чрезвы-
чайно гибкий способ трансформаци внешнего сигнала во вторичный мессен-
джер, который может диффундировать в цитозоль. Вторичных мессенджеров
достаточно много (в зависимости от эффекторного фермента), однако наибо-
лее распространенным является циклический АМФ (цАМФ). Альтенативным
вариантом, как упоминалось выше, может быть воздействие а-субъединицы
на работу мембранного канала, что в свою очередь может изменять электриче-
скую полярность мембраны.
1.5. Каналы и воротные механизмы
Как показано выше, биомембрана состоит из белков, погруженных в липид-
ный бислой, эффективно препятствующий проникновению гидрофильных
веществ через мембрану. Погруженные в бислой белки часто формируют гид-
рофильные каналы, через которые могут проходить неорганические ионы и
другие водорастворимые вещества. Некоторые из этих каналов, т. н. «каналы
утечки», позволяют ионам, например ионам калия, перемещаться по градиен-
ту концентрации в клетку или из нее; другие играют более активную роль и
действуют как воротный механизм, контролирующий ионные потоки.
Мы встретим множество примеров таких каналов и воротных механизмов на
последующих страницах. Они разделяются на две основные категории: активи-
руемые лигандам и потому называемые медиаторными ионными каналами (МК)
и управляемые потенциалом на мембране (потенциал-зависимые каналы, ПЗК).
1. Основы 29
1.5.1. Медиаторные каналы (МК)
Существует множество МК. Наиболее интенсивно исследуемым является нико-
тиновый ацетилхолиновый рецептор (н-АХР). Он представляет собой большой
(268 кДа) пентамерный белок, погруженный в мембрану. Пентамер объединяет
две (по 461 аминокислоте) а-субъединицы, одну (493 аминокислоты) р-субъ-
единицу, одну — (506 аминокислот) у-субъединицу и одну (522 аминокислоты)
6-субъединицу. Каждая из субъединиц образует 4 трансмембранных сегмента
(рис. 1.11 А), а все вместе 5 субъединиц компактно собраны вокруг центральной
ионной поры (рис. 1.11 Б). Когда лиганд, в данном случае ацетилхолин (АХ),
присоединяется к сайтам связывания двух а-субъединиц, канал открывается и
одновалентные катионы двигаются по электрохимическому градиенту.
Известно много других типов МК, они активируются различными медиа-
торами (серотонином, глицином, у-аминомасляный кислотой — ГАМК и т. д.)
и все эти основные типы МК подразделяются на множество подтипов. Что ка-
сается сенсорных систем, наиболее важные МК, обнаруженные в обонятель-
ных и фоторецепторных клетках, чувствительны к циклическим нуклеотидам
(ЦНВ). Структура ЦНВ-воротных каналов будет описана в главе 13 (раздел
13.2.3). В отличие от н-АХР-каналов, белок субъединицы образует 6 трансмем-
бранных сегментов, а целый канал состоит из четырех субъединиц.
1.5.2. Потенциал-зависимые ионные каналы (ПЗК)
Существует также множество типов ПЗК. Все они активируются изменениями
мембранного потенциала и различаются по типу ионов, движение которых ре-
гулируют. Таким образом, есть огромное множество потенциал-зависимых
К+-каналов, а также различные типы С1“-, Са2+- и Ка+-каналов. В данном
Рис. 1.11. (А) Схематическое изображение а-субъединицы н-АХ-рецептора. Четыре спи-
рали, представленные цилиндрами, пересекают мембрану. И С-, и N-концы локализова-
ны внеклеточно. (Б) Сечение рецептора. Пентамерная структура целого рецептора свер-
ху. Предполагается, что вторая спираль каждой субъединицы образует поверхность поры.
30 Часть I. Предварительные сведения
Внеклеточное
пространство
Рис. 1.12. Схема расположения Ка+-канала в мембране. Четыре домена помечены
I, II, III, IV. В реальности четыре домена формируют трехмерный агрегат с цент-
ральным каналом. Дальнейшие объяснения —• в тексте.
разделе мы ограничимся только рассмотрением потенциал-чувствительного
Ма+-канала. Именно этот канал ответствен за восходящую фазу потенциала
действия и является, таким образом, определяющим элементом возбудимых
тканей, таких, как нервы и поперечно-полосатая мускулатура.
В следующих главах мы увидим, что потенциал покоя на мембране боль-
шинства клеток составляет около 50—60 мВ (заряд внутриклеточной среды от-
рицательный по отношению к внеклеточной). Казалось бы — немного. Одна-
ко, следует помнить, что биологические мембраны очень тонки — не более
6—7 нм, так что разность потенциалов, на самом деле, очень значительна.
Градиент потенциала в 60 мВ на 6 нанометрах составляет 105 В/см. Потенци-
ал-зависимые белки в этом интенсивном электрическом поле ведут себя
очень специфически. Любые изменения градиента потенциала воздействуют
на конформацию ПЗК и, соответственно, открытое или закрытое состояние
ионного канала.
Структура Ка+-канала была объектом интенсивных исследований и к на-
стоящему времени хорошо известна (схематически показана на рис. 1.12).
Этот массивный полипептид (1820 аминокислот), как показано на рис. 1.12,
состоит из четырех последовательных гомологичных друг другу доменов, каж-
дый из которых образует шесть трансмембранных спиралей. Четвертая из них
(S4) в каждом домене содержит множество положительно заряженных амино-
кислотных остатков (особенно, аргинина и лизина), что и образует, как пола-
гают, «сенсор потенциала», чувствительный к изменениям потенциала на
мембране. Предполагается, что между пятой (S5) и шестой (S6) трансмембран-
ной спиралями в каждом домене полипептидной цепи формируется «шпиль-
ка» (Н5), входящая в мембрану. Если представить трехмерную форму белка
как полого цилиндра, «шпилька» выстилает пору и обеспечивает ионную се-
лективность. Наконец, внутриклеточный сегмент полипептида между гомоло-
гичными доменами III и IV ответствен за инактивацию канала.
Физиология Na+-канала также глубоко исследована. Показано, что, когда
разность потенциала на мембране падает ниже определенного порога, канал
открывается на прибл. 1 мс и пропускает ток порядка 2 пА. По прошествии
1 мс времени открывания канал закрывается и не откроется больше, пока мем-
брана остается деполяризованной. Инактивация, как говорилось выше, связа-
на с сегментом полипептида между доменами III и IV, блокирующим канал.
1. Основы 31
Рис. 1.13. Цикл изменения конформации натриевого канала. (А) В покоящейся
мембране натриевый канал закрыт. Активирующий воротный механизм (AG) за-
крыт, инактивирующий (IG) — открыт. (Б) Когда мембрана деполяризуется, изме-
нение потенциала детектируется «сенсором потенциала», и активирующий меха-
низм открывается. Ионы натрия двигаются по электрохимическому градиенту.
Они перепрыгивают с одного сайта в канале на другой (как показано на рисунке)
и, таким образом, «двигаются колонной». (В) Через 1 мс инактивирующий меха-
низм закрывается. (Г) Когда мембрана возвращается к уровню потенциала покоя,
активирующий механизм закрывается, а инактивирующий — открывается снова.
Таким образом, канал существует в трех основных конформациях: закрытой,
открытой и инактивированной. Этот цикл показан на рис. 1.13.
Точное время открывания и закрытия канала точно непредсказуемо, иными
словами — это стохастический процесс. Кроме того, каждый участок возбуди-
мой мембраны обладает большой популяцией Ка+-каналов, и порог открыва-
ния каждого из них несколько варьирует. По мере того, как возбудимая мем-
брана деполяризуется, открывается все больше №+-каналов. Входящий поток
ионов Na+ деполяризует и, в конце концов, даже реполяризует мембрану, обра-
зуя то, что электрофизиологи называют потенциалом действия (спайк). Био-
физика потенциала действия будет рассмотрена в следующей главе (раздел 2.6).
1.5. Заключение
В этой главе мы построили фундамент для дальнейшей работы. Рассмотрен-
ные здесь элементы будут возникать в дальнейшем снова и снова при обсуж-
дении специализированных и часто очень сложных сенсорных систем, воз-
никших в эволюции царства животных. В частности, будет показано, что био-
химия биомембран, рецепторов и G-белковых систем связаны с механизмом,
контролирующим полярность мембран сенсорных клеток и окончаний. Это
именно та тема, которой посвящена следующая глава.
2. МЕМБРАНА И ПОТЕНЦИАЛ ДЕЙСТВИЯ
Ионы и вода: распределение в мембране. Потенциал покоя (Кт): измере-
ние — гигантский аксон — уравнение Нернста — уравнение Голдмана —
примеры. Электротонический потенциал и кабельное проведение: локаль-
ные токи — электротоническое проведение — рецепторный потенциал —
генераторный потенциал. Сенсорная адаптация: быстрая и медленная
адаптация — биофизические механизмы. Потенциал действия: рефрак-
терный период устанавливает предел частоты — миелинизация — ско-
рость распространения — классы аксонов. Заключение: вездесущность
молекулярных и биофизических компонентов — редукционизм — про-
блема соотношения мозга и мышления
Как было показано в гл. 1 ’ с самого начала липидная биомембрана является гра-
ницей между первичными клетками и окружающей средой. Поскольку ныне су-
ществующие клетки эукариот содержат множество внутренних мембран (мито-
хондриальные, мембраны эндоплазматического ретикулума, лизосом и т. д.),
стоит выделить внешнюю мембрану как плазматическую. Мы видели выше, что
белковые элементы плазматической мембраны эволюционировали в направле-
нии детектирования изменений во внешней среде и (в некоторых случаях) сигна-
лизации об этих изменениях через мембранно-связанный G-белок. Плазматиче-
ская мембрана также развила средства контроля движения различных веществ в
клетку и из нее. Следует помнить, что основой современных биомембран являет-
ся липидный бислой. Гидрофильные вещества не могут (по определению) диф-
фундировать через гидрофобный барьер липидного бислоя. Здесь не место для
рассмотрения множества механизмов, которые сформированы клеткой, чтобы
обойти это препятствие, однако, один из них должен быть рассмотрен здесь в не-
которых деталях. Это — механизм или, скорее, группа механизмов, которые кон-
тролируют движение малых неорганических ионов через мембрану.
Неорганические ионы, конечно, водорастворимы. Электростатический за-
ряд позволяет им входить в водную структуру и смешиваться с ней (рис. 2.1).
С другой стороны, он препятствует их смешиванию с органической фазой, ко-
торая в данном случае представлена липидным бислоем. Чтобы пропустить их
внутрь, необходимы гидрофильные «поры» в биомембране. Множество видов
таких пор возникло с тех пор, как первые прокариоты возникли три с полови-
ной миллиарда лет назад. Во всех случаях — это сложные белковые структуры,
погруженные в мембрану. Как мы увидим в последующих главах, таких кана-
лов много и типы их различны. В целом, каналы очень специфичны в отноше-
нии того, какие ионы они пропускают.
2. Мембрана и потенциал действия 33
Рис. 2.1. Ионы в водной структуре. Молекулы воды электрически поляризованы.
Поскольку кислород оттягивает на себя электрон от водорода, локальный отри-
цательный заряд связан с этим концом молекулы. Равный ему и противополож-
ный по знаку заряд связан с другим концом. Вследствие этого молекуля воды
стремятся образовать структуру вокруг положительно или отрицательно заря-
женных ионов, таких как С1_ или Na+, изолируя их друг от друга в растворе.
Сам факт, что ионы — это заряженные частицы, означает что их распреде-
ление у мембраны может быть выражено в электрических величинах. Еще со
времен Гальвани и Вольта в конце восемнадцатого столетия известно, что
функционирование нервной системы связано с электрическими явлениями.
Однако только к середине XX века, когда были развиты электрические (в осо-
бенности, электронные) технологии, а также адекватные биологические пре-
параты (гигантский аксон) удалось добиться истинного понимания «животно-
го электричества».
В данной главе мы рассмотрим в первую очередь происхождение потенциала
покоя ( Ут ), который существует на плазматической мембране, и покажем, как
он меняется, когда рецепторные клетки подвергаются воздействию стимула и
формируют т. н. «рецепторные потенциалы». Затем мы рассмотрим пассивные
свойства электрических токов — т. н. кабельное или элекгротоническое проведе-
ние, которое играет важную роль во всех сенсорных и нейросенсорных клетках.
Наконец, мы кратко рассмотрим природу рецепторного и генераторного потен-
циалов, сенсорной адаптации и важного феномена потенциала действия.
2.1. Измерение потенциала покоя
Уже говорилось, что потенциал покоя формируется на всех плазматических
мембранах. Большинство клеток, в особенности нейронов и сенсорных кле-
ток, крайне малы. Хотя сейчас методы исследования их электрических харак-
теристик уже развиты, очень долго не удавалось провести точные измерения.
Осознание того факта, что огромная трубчатая структура с диаметром
500—600 мкм у кальмара Loligo представляет собой гигантский аксон, имело
огромное значение для электрофизиологов. Наконец-то они получили воз-
можность ввести тонкие стеклянные микропипетки, заполненные электро-
литом, в аксон и измерить электрическую полярность мембраны непосредст-
венно (рис. 2.2). Оказалось возможным и выдавить аксоплазму, как пасту из
тюбика, и подвергнуть ее химическому анализу (табл. 2.1). Большинство пио-
2 Биология сенсорных систем
34 Часть I. Предварительные сведения
Рис. 2.2. На схеме показан стеклянный
микроэлектрод, заполненный раствором
КС1, введенный в гигантский аксон. Элек-
трический контур проходит через вольт-
метр на пластину электрода на внешней
поверхности аксона.
нерских работ, которые установили физические основы потенциалов покоя и
действия были выполнены на этом общепринятом препарате.
Используя условия, показанные на рис. 2.2, было обнаружено, что внутри-
клеточный электрод регистрирует на мембране падение потенциала на прибл.
50 мВ. Этот потенциал был определен как потенциал покоя, Кт. Что же вызы-
вает формирование разности потенциалов на мембране?
2.2. Ионные основы потенциала покоя
Мы уже отмечали, что был определен ионный состав аксоплазмы (табл. 2.1).
Известно также, что существуют уравнения физической химии, которые свя-
зывают электрический потенциал на мембране с распределением ионов, спо-
собных проникать через нее. Наиболее известно уравнение Нернста, которое
может быть записано в форме:
(2.1)
RT [1]0
И =------In —
1 ZXF [1],
В уравнении 2.1 Kj — потенциал на мембране (в вольтах, хотя обычно это ве-
личины порядка милливольт), возникающий вследствие распределения иона I;
R — газовая константа; Т — температура по Кельвину; F— число Фарадея (ве-
личина электрического заряда на моль одновалентного иона); Zx — валент-
ность иона (+1 для Na+ и К+; -1 для СГ); In — натуральный логарифм; [1]о и
[I]i — концентрации иона I по внешнюю и внутреннюю стороны мембраны,
соответственно. Важно отметить, что одной из посылок, из которой получено
уравнение Нернста, является то, что мембрана полностью проницаема для
рассматриваемого иона. При выполнении этого условия и известных равно-
весных концентрациях иона на мембране можно предсказать равновесный
электрический потенциал.
2. Мембрана и потенциал действия 35
Таблица 2.1. Концентрации ионов внутри и во внешней среде некоторых из
обсуждаемых клеток
Ион Внутриклеточная концентрация Внеклеточная концентрация
7. Гигантский аксон кальмара мМ/кг Н2О ' мМ/кг Н2О
К+ 400 20
Na+ 50 440
Са2+ 0,4 10
Mg2+ 10 54
сг 100 560
Органические ионы =385 —
2. Мышечные клетки млекопитающих мМ мМ
К+ 155 4
Na+ 12 145
Mg2+ 30 1-2
Са2+ 1-2 2,5-5
Cl- 4 (только 10 4 в свободной форме) 120
Органические ионы =150 —
3. Моторный нейрон кошки мМ мМ
К+ 150 5,5
Na+ 15 150
СГ 9 125
Проверим уравнение заменой некоторых значений концентраций К+ по
обе стороны мембраны. Во-первых, если предположить, что концентрации К+
по обе стороны мембраны равны, тогда подстановка их в уравнение Нернста
обращает часть под логарифмом в единицу. Поскольку логарифм 1 — ноль, то
уравнение дает нулевой потенциал на мембране. Это, собственно, то, что и
наблюдается. Когда клетка погибает, мембрана утрачивает целостность, а ее
насосные механизмы исчезают. Ионы движутся по своим градиентами кон-
центраций до тех пор, пока их концентрации по обе стороны мембраны не
уравниваются, а потенциал на ней не исчезает.
Теперь проверим уравнение подстановкой значений концентраций [К+]о
(около 5,5 мМ) и [К+][ (около 150 мМ), показанных в табл. 2.1. Тогда
или
RT 5,5
Vy.------In -----
k ZlF 150
= -0,027 In 0,036
= - 0,089 В
= — 89 мВ
36 Часть I. Предварительные сведения
Эта величина известна как нернстовский калиевый потенциал Ик или потен-
циал реверсии для калия. Смысл последнего термина состоит в том, что, когда
на мембране удерживается такой градиент потенциала, отсутствует какой-ли-
бо результирующий поток этого иона через мембрану. Измерение реального
потенциала покоя Ит на клеточной мембране обычно дает значения между
—50 мВ и —75 мВ. Кк очевидно больше этих величин, однако совпадает по зна-
ку и порядку величины. Если, однако, подставить в уравнение Нернста зна-
чения концентраций других существенных ионов (СГ, Na+ и т. д.) получаю-
щиеся значения (ИС1, KNa и т. д.) отличаются от Ит очень сильно. Особенно это
заметно, если подставить значения концентраций для Na+.
Причины таких отклонений понять нетрудно. Клеточные мембраны —- это
очень сложные структуры. Их проницаемость для разных ионов резко разли-
чается и, как было отмечено выше, уравнение Нернста работает только в слу-
чае ионов, для которых мембрана полностью проницаема. Известно, однако,
что и ионы натрия, и ионы хлора имеют очень маленькие коэффициенты про-
ницаемости через покоящиеся биомембраны.
Далее, важно, что Ит зависит не от трансмембранного распределения како-
го-то одного вида ионов, а от распределения всех ионов. Таким образом, для
более полного понимания природы потенциала покоя следует обобщить урав-
нение Нернста. Оно должно учитывать различную проницаемость мембраны
для разных ионов и то, что существенен не один тип ионов, а много.
Уравнение, о котором идет речь, выведено Дэвидом Голдманом и впослед-
ствии стало известно как уравнение Голдмана. Его иногда называют и «уравне-
нием постоянного поля», поскольку предполагается, что электрическое поле на
мембране (градиент электрического потенциала Кт) неизменно, что, конечно,
очень большое допущение. Тем не менее, уравнение Голдмана дает полезное
первое приближение оценки биофизической ситуации на мембране. Оно запи-
сывается следующим образом:
V=1LX ^[К+]0+Рка^а+]0+Рс1[СГ];
. F П PK[K+]i+PNa[Na+]i+Pcl[Cn0
где Р — константа проницаемости иона, квадратные скобки обозначают, как
это принято, концентрации иона с внутренней (обозначены «i») и внешней
(обозначены «о») стороны мембраны, a R, Ти F— их обычные значения.
Отметьте, что, если внешние концентрации катионов К+ и Na+ помещены
в числитель уравнения, то внешняя концентрация аниона СГ — в знамена-
тель.
Проведем еще несколько опытов. Во-первых, если мы примем константы
проницаемости для Na+ и СГ равными нулю, то уравнение вырождается в
уравнение Нернста для калия. Сходным образом, если принять Рк = РС1 = 0,
уравнение сводится к уравнению Нернста для Na+ и дает HNa равным потенци-
алу на мембране (величина проницаемости для Na+ удалена).
Липидный бислой плазматической мембраны, как это было показано в на-
чале данной главы, полностью непроницаем для неорганических ионов. Они
перемещаются через мембрану по каналам, образованным мембранными бел-
ками. Многие из этих т. н. «каналов утечки» еще не охарактеризованы пол-
2. Мембрана и потенциал действия 37
ностью. Проницаемость плазматической мембраны для гидрофильных ионов
зависит от этих каналов. «Пропускная способность» этих каналов у разных
клеток варьирует. Нейроглиальные клетки, например, вероятно, более прони-
цаемы для К+, чем нейроны.
Большинство сенсорных клеток, однако, сходны с нейронами по их боль-
шей проницаемости к К+, чем к С” и Na+:
РК >:> ^Na
Можно привести некоторые данные по сравнительной проницаемости, по-
лученные из измерения потоков радиоизотопов ионов через плазматические
мембраны:
Рк = 10“7 см/с
РС1 = 10”8 см/с
= 10“8 см/с
Подставим эти константы проницаемости и соответствующие концентра-
ции ионов (табл. 2.1 — мотонейрон кошки) в уравнение Голдмана:
!Z П ПОТ , Х 10-7 I5’5!) + Х 10'8 11501) + О Х 10-8 19D
Vm = 0,027 In----------------------s-------------;-----
(lx IO"7 [150]) +(1 x 10"8 [15]) +(1 x 1(Г8 [125])
(55 х IO"8) +(150 х IO"8) + (9x IO"8)
= 0,027 In----------z-----------------------
(1500 x 10"8) +(15 x 10"8) +(125 x IO"8)
= -0,055 В = -55 мВ
Это значение Km действительно очень близко к значению потенциала покоя
у мотонейронов кошки, наблюдаемого при микроэлектродной регистрации.
Теперь посмотрим, что происходит, если повысить калиевую проницае-
мость на порядок. Если подставить в уравнение величину Рк = 1 х 10“6 см/с,
все остальные параметры оставив неизменными, получим
Ит = —83 мВ
Уже было отмечено, что некоторые глиальные клетки существенно более
проницаемы для К+, чем мембраны нейронов. Потому-то Ит на мембранах
глиальных клеток заметно выше, чем потенциал покоя мембран нейронов.
В сетчатке, как мы увидим в гл. 16, крупные глиальные клетки — мюллеров-
ские клетки, имеют Ит от —70 до —90 мВ. Эта большая, чем обычно, К+-про-
ницаемость, вероятно, имеет существенное значение для устранения избытка
К+, образующегося в клетках сетчатки в ответ на освещение. Избыток К+ за-
тем растворяется в жидкости стекловидного тела. Напротив, если повысить в
уравнении Голдмана константу проницаемости для Na+, можно предсказать
значительное понижение Ит. Мы столкнемся с важным случаем такого изме-
нения концентрации, когда будем рассматривать биофизику палочек сетчатки
в глдве 16.
38 Часть I. Предварительные сведения
В конце данной главы стоит отметить, что значительно легче измерять от-
носительные, а не абсолютные проницаемости для ионов, поэтому уравнение
Голдмана часто записывают в несколько ином виде:
RT [К]о + />[Nal + с[С1]=
V=-----In —---------------—— (2.3)
F [K]j + *[Na]. + c[Cl]o
где
„ b=P^/PK
И
C = PC\/PK
Поскольку ион хлора играет меньшую роль во многих нейрофизиологических
функциях, уравнение иногда упрощают еще больше:
RT , [К] + ^[NaJi
----In-----------------
р [K]i + 6[Na]i
(2.4)
Мы еще, однако, увидим, что хотя ион хлора несущественен в многих областях
нейрофизиологии, он играет ключевую роль в гиперполяризации в тормозных
синапсах и в других случаях. В таких и подобных случаях важно использовать
полную форму уравнения Голдмана.
2.3. Электротонические потенциалы
и кабельное проведение
Рассмотрим рис. 2.3. Невозбудимая мембрана слабо деполяризована. Это сле-
дует особо подчеркнуть — на невозбудимой мембране нет риска вызвать фор-
мирование потенциала действия. Подобраны условия для детектирования
очень маленьких, т. н. локальных токов. Внутренняя среда клетки, по сути де-
ла, — ионный раствор, так же как и внеклеточная жидкость. Из этого следует,
что они пропускают электрический ток. Если деполяризовать мембрану в точ-
ке «х», установится некоторая разность потенциалов между ней и точкой «у»,
находящейся от нее на некотором расстоянии. Расстояние, конечно, неболь-
шое — редко больше, чем 1 микрон. Тем не менее, на этой дистанции малые
электрические токи протекают до тех пор, пока не будет устранена разность
потенциалов. Следовательно, и в точке «у» мембрана будет слегка деполяризо-
вана. Такие небольшие потенциалы известны как электротонические, а ло-
кальные токи — как электротонические токи или кабельное проведение.
Амплитуда электротонических потенциалов по крайней мере на порядок, а
иногда и на два или более порядков, меньше, чем потенциала действия. Тем не
менее, как ни малы электротонические потенциалы, их эффекты могут быть
весьма значительны. Чтобы убедиться в этом, достаточно лишь вспомнить о
чрезвычайной чувствительности потенциал-зависимых ионных каналов, та-
ких как Ка+-каналы, рассмотренных в гл. 1. Если локальные токи, распрост-
раняющиеся из области, где мембрана деполяризована, достигнут участка,
2. Мембрана и потенциал действия 39
Рис. 2.3. Электротоническое
проведение. Ток входит в точке
«х» и приводит к формирова-
нию потенциала на мембране
Их. Отводящий электрод в точ-
ке «у» измеряет электротони-
ческий потенциал. Локальные
токи показаны стрелками. Для
упрощения рисунка токи из
внешней среды не показаны.
Генератор тока
Аксоплазма
изобилующего Ка+-каналами, возникающая деполяризация может запустить
потенциал действия, передающийся в мозг, что само по себе может иметь не-
предсказуемые последствия.
2.4. Рецепторный и генераторный потенциалы
Рецепторный и генераторный потенциалы — это частные случаи электротони-
ческих потенциалов. Когда рецепторная (сенсорная) клетка, например меха-
ночувствительная волосковая или вкусовая, подвергается воздействию со-
ответствующего стимула, реализуется более или менее сложный набор собы-
тий (который будет рассмотрен в деталях в последующих главах), ведущих
к изменениям электрической полярности участка их мембраны. Это явление
именуется рецепторным потенциалом. В большинстве случаев рецепторные по-
тенциалы (как и в случае, рассмотренном в разделе 2.3) — это деполяризация,
в других, однако, в частности в палочках и колбочках сетчатки, — это гиперпо-
ляризация. Так или иначе, результат — одни и тот же — возникают токи меж-
ду подвергающимся воздействию участком мембраны и другими участками
мембраны рецепторной клетки (рис. 2.4). В общем случае, изменения электри-
ческой полярности (увеличение ее или уменьшение) влияет на выделение
медиатора на подлежащий сенсорный нейрон.
Не все сенсорные системы развили специализированные сенсорные клет-
ки. Обонятельные и некоторые механорецептивные системы построены на
нейросенсорных клетках. В таких случаях функции детектирования соответст-
вующих факторов внешней среды и передачи информации в мозг совмещают-
ся в одной клетке (рис. 2.5). Электрофизиологические феномены при этом
аналогичны только что описанным. Когда чувствительные окончания нейро-
сенсорной клетки подвергаются воздействию стимула, ряд биохимических
процессов приводит к изменению электрического потенциала (в случае ней-
росенсорных клеток — это всегда деполяризация). Механизмом локальных то-
ков деполяризация распространяется в область мембраны, изобилующую по-
тенциал-зависимыми Ка+-каналами. Если деполяризация достаточно велика,
Ка+-каналы открываются, в результате чего генерируется потенциал дейст-
вия, который без декремента передается в центральную нервную систему. По-
скольку первоначальная деполяризация происходит не в специальной рецеп-
40 Часть I. Предварительные сведения
Рис. 2.4. Рисунок показывает локаль-
ные токи, распространяющиеся от де-
поляризованного участка мембраны
(отмечен черным) рецепторной клетки.
В общем случае это ведет к деполяриза-
ции клетки, а это путем сложного био-
химического процесса к выделению
медиаторного вещества.
Рис. 2.5. Деполяризация окончаний
нейросенсорной клетки ведет к фор-
мированию локальных токов, которые
инициируют потенциал действия.
торной клетке, она часто именуется генераторным потенциалом. Многие, од-
нако, оба варианта называют рецепторными потенциалами.
Амплитуда генераторных и рецепторных потенциалов зависит от величины
стимула — между потенциалом и интенсивностью стимула существует прак-
тически прямая пропорциональная зависимость (рис. 2.6). Из-за того, что ло-
кальные токи должны быть достаточно значительными по величине, чтобы
запустить выделение медиатора (рис. 2.4) или активировать хотя бы часть по-
пуляции потенциал-зависимых №+-каналов до порогового уровня, запуск
потенциала действия в сенсорном нерве наблюдается только, когда рецептор-
ный или генераторный потенциал достигают определенной амплитуда. Ины-
ми словами, потенциал действия не генерируется до тех пор, пока стимул не
достигнет критической величины (Sc на рис. 2.6).
стимула
Рис. 2.6. Соотношение интен-
сивности стимула и величины
рецепторного (генераторного)
потенциала. График показы-
вает первичные ответы на сти-
мулы разной интенсивности.
Как указывается в тексте,
адаптация снижает амплитуду
рецепторного (генераторного)
потенциала при большой про-
должительности стимула.
2. Мембрана и потенциал действия 41
2.5. Сенсорная адаптация
Все сенсорные системы демонстрируют адаптацию. Это означает, что ответ
(каким бы они ни был) на постоянный стимул с течением времени уменьшает-
ся. В сенсорных системах многоклеточных это выражается в том, что частота
импульсации в волокне сенсорного нерва со временем снижается. В таких си-
стемах присутствует несколько форм адаптации. Обычно различают две край-
ние формы — быструю и медленную адаптацию (рис. 2.7). В первом случае при
включении стимула сначала наблюдается быстрый залп активности в сенсор-
ном волокне, которая быстро уменьшается еще до того, как стимул будет вы-
ключен. Выключение стимула также сопровождается залпом активности. Во
втором случае также наблюдается залп активности при включении стимула,
которая, хотя и уменьшается со временем, никогда не достигает нуля. Актив-
ность остается на некотором уровне плато до выключения стимула, когда она
переходит на исходный уровень. В обоих случаях именно частота первого зал-
па характеризует интенсивность стимула.
Биологические причины сенсорной адаптации многочисленны и разнооб-
разны. Мы встретим многие из этих вариантов на последующих страницах.
Например, в некоторых случаях бактериальной хемочувствительности это
происходит в результате метилирования белков «переноса рецептора». В мно-
гих животных системах, в которых рецепторные молекулы связаны с системой
G-белков, это — результат инактивации рецепторной молекулы путем дефос-
форилирования (см. раздел 1.4.2). Присутствие Са2+-каналов и кальций-зави-
симых К+-каналов (КСа) наряду с 1Ча+-каналами, в которых зарождается по-
тенциал действия в сенсорных нервных окончаниях, также очень существенно.
Когда мембрана деполяризуется в ответ на локальные токи, приходящие от
стимулированных окончаний, потенциал-зависимые Са2+-каналы открыва-
ются, и поток Са2+ устремляется по градиенту концентрации в нейрон. Повы-
шение внутриклеточного уровня Са2+ внутри клетки открывает Са2+-зависи-
мые К+-каналы и избыток К+ также двигается по градиенту концентрации во-
вне клетки. Другими словами, мембрана становится необычно проницаемой
для К+. Если подставить эту увеличенную константу проницаемости для К+
в уравнение Голдмана, то оно предскажет гиперполяризацию мембраны. В та-
ких условиях локальным токам значительно труднее открыть потенциал-зави-
симые Ка+-каналы и инициировать потенциал действия. Таким образом, ско-
А Быстрая адаптация
Б Медленная адаптация
Рис. 2.7. Сенсорная адап-
тация. (А) Быстро адапти-
рующееся волокно; (Б)
медленно адаптирующее-
ся волокно.
Включение Выключение
Стимул
Включение Выключение
Стимул
42 Часть I. Предварительные сведения
Рис. 2.8. Один из механизмов, вызывающих адаптацию сенсорных окончаний. Де-
поляризация вызывает открытие Са2+-воротного механизма; ионы Са2+ входят в
цитоплазму и воздействуют на Са2+-зависимые К+-каналы, что ведет к увеличе-
нию мембранной полярности, вследствие чего снижается вероятность инициации
потенциала действия.
рость импульсации снижается, в чем и проявляется сенсорная адаптация. Эта
последовательность событий схематически отражена на рис. 2.8.
Биофизическая и молекулярно-биологическая ситуация в сенсорных
окончаниях животных несомненно чрезвычайно сложна. Данный раздел кос-
нулся проблемы адаптации лишь в общих чертах, в дальнейших главах мы рас-
смотрим ее конкретные случаи.
2.6. Потенциал действия
Данная книга — не место, где следует излагать все известное о биофизике по-
тенциалов действия, более уместно это в учебниках по нейрофизиологии. Од-
нако, поскольку органы чувств животных передают информацию в аналитиче-
ский аппарат центральной нервной системы в форме потенциалов действия
в сенсорных нервных волокнах, их следует описать хотя бы в общих чертах.
Мы уже отмечали выше (и в гл. 1), что когда мембранное окружение попу-
ляции Ка+-канальных белков деполяризовано, каналы с наиболее низким по-
рогом открываются, и Na+ начинает по градиенту концентрации поступать
в аксон. Этот процесс имеет эффект положительной обратной связи. По мере
того, как Na+ во все большем количестве входит в клетку, мембрана деполяри-
зуется еще больше, вследствие чего открываются соседние более высокопоро-
говые Ка+-каналы. Поток Na+ увеличивается, мембрана деполяризуется и так
далее. Мембрана достигает нулевого потенциала и реполяризуется (отрица-
тельный заряд снаружи, положительный — внутри) до достижения нернстов-
ского потенциала для Na+ (KNa). Вследствие эффекта положительной обратной
связи реверсия потенциала происходит очень быстро. В большинстве случаев
KNa достигается в течение 0,5 мс, затем воротный механизм для Na+ начинает
закрываться.
Очень вскоре после открытия Ка+-каналов (<0,5 мс) срабатывает другой
воротный механизм — открывается калиевый. Это позволяет ионам К+ поки-
нуть аксон, а, поскольку Ка+-механизм закрывается, мембрана возвращается
к исходной полярности, несколько превышает ее и приближается к нернстов-
скому К+-потенциалу, Кк. Активность АТФ-зависимого Na+/K+-Hacoca в
мембране возвращает ее к нормальному уровню потенциала покоя (Vm). Все
2. Мембрана и потенциалдействия43
Рис. 2.9. Основные ионные
каналы и характеристики
проведения, ответственные
за формирование потенци-
ала действия. (A) Na+- и
К+-проведение показано
кривыми, помеченными
gNa и gK (левая ордината
градуирована в мСи/см2).
Пунктирная линия показы-
вает потенциал на мембра-
не, образующийся за счет
этих проводимостей. Гори-
зонтальные пунктирные
линии вверху и внизу ри-
сунка показывают KNa и
Кк, соответственно. (Из
Hodgkin and Huxley, 1952,
Journal of Physiology, 117,
500—544; воспроизводится
с разрешения Физиологи-
ческого Общества). (Б) Ось
времени сильно растянута
по сравнению с (А). Верх-
нюю часть рисунка (Б) сле-
дует сравнивать с пунктир-
ной линией на рисунке (А).
Нижняя часть рисунка (Б)
показывает потоки Na+ и
К+, которые возникают
вследствие открывания и
закрывания Na+- и К+-ка-
налов аксональной мемб-
раны.
эти события и результирующие изменения мембранной полярности показаны
на рис. 2.9.
Ну, теперь все в порядке, однако смысл потенциала действия (нервного
импульса) заключается в его распространении. Как это драматическое измене-
ние мембранной полярности распространяется? И снова ответ заключается в
локальных токах. Мы уже видели в разделе 2.3, что деполяризация небольшо-
го участка мембраны ведет к возникновению малых электрических токов, рас-
пространяющихся отсюда и деполяризующих следующие участки мембраны
на небольшом расстоянии. То же самое происходит и когда мембрана резко
меняет полярность при прохождении потенциала действия, описанного выше.
Локальные токи, деполяризуя соседний участок мембраны, так же открывают
в нем Ка+-каналы и запускают последовательность событий, ответственную
за формирование потенциала действия, описанную выше. Можно сказать, что
потенциал действия похож на огонь, бегущий по дорожке из пороха. Каждый
44 Часть I. Предварительные сведения
Таблица 2.2. Некоторые характеристики афферентных волокон млекопитающих
Мышечные нервы Кожные нервы, диаметр (мкм) Диаметр волокна (мкм) Скорость проведения (м/с)
Миелинизированные
крупные I 13-20 80-120
малые II Ар 6-12 35-75
самые малые III AS 1-5 5-30
Немиелинизированные IV С 0,2-1,5 0,5-2
По Kandel et al., 1991
активированный участок запускает последующий, оставляя за собой дорожку
из пепла. В случае нервного волокна инактивация, символизируемая пеплом,
представляет собой гиперполяризацию мембраны вслед за прошедшим потен-
циалом действия (рис. 2.9) и, что более существенно, то, что Ка+-воротный
механизм остается в закрытом состоянии в течение прибл.1,5 мс после их по-
тенциал-зависимого открывания. Это состояние мембраны именуется ре-
фрактерностью.
Рефрактерный период длительностью 1,5—2 мс имеет существенное значе-
ние для сенсорной сигнализации. Он ограничивает частоту, с которой импуль-
сы могут следовать по сенсорному нервному волокну. Такая длительность
означает, что сенсорное волокно в состоянии проводить импульсы с частотой
не выше 500 Гц. Это важно, например, в детектировании частоты звука и час-
тотной дискриминации в слуховой системе (см. гл. 8).
В то время, как рефрактерный период ограничивает частоту, с которой им-
пульсы могут проводиться по нервному волокну, диаметр последнего (при
прочих равных) определяет скорость проведения (табл. 2.2). Чем больше диа-
метр, тем дальше по аксоплазме распространяются локальные токи и на боль-
шем расстоянии они могут открыть Ка+-воротный механизм. Поэтому у мно-
гих беспозвоночных, например, кольчатых червей и головоногих моллюсков,
развились гигантские волокна, обеспечивающие быстрое проведение, необхо-
димое для реакции избегания. Позвоночные развили иной механизм увеличе-
ния скорости проведения — миелинизацию.
Миелин состоит из нескольких слоев мембран шванновских клеток, обви-
вающихся вокруг аксон, и обеспечивает очень эффективную электрическую
изоляцию. Последняя предупреждает утечку внутриклеточного компонента
локального тока из аксоплазмы, «удерживая» его внутри волокна, и обеспечи-
вает его распространение. У беспозвоночных такого специфического и эффек-
тивного механизма повышения скорости проведения не развилось. Однако
когда мы будем рассматривать функцию некоторых сенсилл насекомых
в гл. 12, мы увидим, что существуют альтернативные средства изоляции ло-
кальных токов, обеспечивающие эффективное электротоническое проведение
в длинных внешних сегментах хемочувствительных клеток.
Табл. 2.2 показывает скорость проведения импульсов в афферентных во-
локнах млекопитающих. Афферентные волокна мышц состоят из четырех
2. Мембрана и потенциал действия 45
перекрывающихся классов (I—IV), чья скорость проведения составляет от
120 м/с в наиболее крупных миелинизированных волокнах (диаметр ок. 20 мкм)
до прибл. 0,5 м/с в самых тонких немиелинизированных (диаметр ок. 0,5 мкм)
В сенсорных нервах кожи отсутствуют наиболее быстрые волокна класса I
(или Аос). Кожные нервы, таким образом, подразделяются на три группы Ар,
А6 и С, что соответствует группам II, III и IV мышечных афферентов.
2.7. Заключение
В гл. 1 мы рассмотрели некоторые вездесущие молекулярные элементы, ответ-
ственные за активность сенсорных клеток. В данной главе мы увидели, как эти
молекулярные элементы реализуются в равным образом вездесущих биофизи-
ческих характеристиках сенсорных клеток и волокон. Мы снова убедились в
той центральной роли, которую играют биомембраны. Мы увидели, как био-
физика ионных потоков через мембрану порождает потенциал покоя Кт. Мы
отметили, как электротоническое (кабельное) проведение позволяет распро-
страняться на малые расстояния по мембране небольшим изменениям Кт и
что такое проведение ответственно за формирование генераторного и рецеп-
торного потенциалов. Хотя Ит кажется небольшим, всего —60 мВ, благодаря
тому, что мембрана так тонка (ок. 7 нм), градиент потенциала чрезвычайно
высок — ок. 105 В/см. Ясно, что какой-либо потенциал-чувствительный бе-
лок, погруженный в такую мембрану, испытывает действие мощного электри-
ческого поля. И конечно это касается таких белков, как Са2+- и Ка+-каналь-
ные. Любые изменения электрической полярности мембраны оказывают зна-
чительный эффект на конформацию этих каналов. Мы отметили важность
этих конформационных изменений в сенсорной адаптации и в проведении
потенциалов действия.
В следующей главе мы сделаем небольшой шаг назад и рассмотрим некото-
рые глобальные черты сенсорных систем. Мы снова столкнемся с множеством
сходств. Надо постоянно помнить, что все их свойства могут быть прослежены
вглубь до уровня молекулярных механизмов и их биофизических эффектов,
рассмотренных в двух первых главах. Действительно, по мере дальнейшего из-
ложения мы увидим, что, несмотря на многочисленные различающиеся дета-
ли, нет разрыва между основными биохимическими и биофизическими прин-
ципами и феноменами более высокого уровня в зрительной, слуховой и иной
сенсорной коре, рассматриваемыми в последующих главах. Это «первый зво-
ночек» главной нерешенной дилеммы науки о чувствах: как могут физика и
химия, лежащие в основе функционирования мозга, быть связаны с миром
цветов, запахов и звуков, образующих ткань нашей субъективной жизни? Ко-
нечно, это лишь еще один путь постановки проблемы соотношения мозга и
мышления. Она всегда в неявном виде присутствует в науке о чувствах и,
вследствие этого, будет возникать на последующих страницах снова и снова.
Хотя попытки ее решения часто полагают предметом другой науки — филосо-
фии, студентам не следует думать, что эта проблема несущественна и о ней
можно просто забыть.
3. ОБЩИЕ ЧЕРТЫ СЕНСОРНЫХ СИСТЕМ
Психофизика и поведение против сенсорной физиологии: будущее объеди-
нение? Классификация чувств: по адекватным стимулам — по внешней
или внутренней локализации. Модальность: меченные линии — экспери-
менты по «переключению» проводящих путей. Интенсивность: частотная
модуляция — порог и психометрия — соотношение сигнал/шум — разли-
чение минимальных отличий и закон Вебера—Фехнера — психофизиче-
ский закон Стивенса. Адаптация. Рецептивные поля (РП): определение —
организация — вариативность по площади. Картирование: неизоморф-
ные сенсорные карты — «проблемы связывания». Иерархическая и парал-
лельная организация. Выделение отличительных характеристик и пусковые
стимулы: иерархии — «понтификальные клетки». Заключение: сенсорные
системы и «гонка вооружений» хищник—жертва
В первых двух главах этой книги мы рассмотрели некоторые молекулярные и
биофизические элементы, образующие сенсорные системы. В этой главе мы
рассмотрим глобальные черты, общие для различных сенсорных систем. Од-
нако, прежде, чем мы займемся этим, стоит провести разграничение между
двумя подходами к изучению сенсорных систем.
Первый подход сводится к установлению корреляций между физическим
стимулом и ответной реакцией животного (и человека). Сенсорная система
рассматривается как «черный ящик». Исследователь анализирует (часто —
в мельчайших деталях), что может, а чего не может сделать «черный ящик»,
что он может или не может детектировать или дискриминировать. При этом
исследователь, однако, не лезет внутрь ящика, чтобы разобраться, какие шес-
теренки, колесики и проводки важны для того, чтобы он работал. В случае изу-
чения человека экспериментатор пытается коррелировать физический стимул
со словесным ответом испытуемого, выражающего его или ее субъективную
реакцию. Такие исследования по очевидным причинам именуются психофизи-
ческими. В случае животного психофизический подход более сложен. Приходит-
ся устанавливать корреляции между физическим стимулом и каким-либо аспек-
том поведения животного. Поведенческие реакции могут быть весьма утончен-
ными, животное можно натренировать давать ответ на стимул (кондициониро-
вать), сам стимул можно варьировать по интенсивности, чтобы изучить, на каком
уровне он может детектироваться. Можно отметить, что многие и делают, что
вербальный ответ испытуемого — тоже не более, чем форма поведения. И снова,
как и всегда при изучении сенсорных систем, мы оказываемся в непосредствен-
ной близости от проблемы соотношения мозга и мышления. Все же, оставляя
3. Общие черты сенсорных систем 47
в стороне все эти сложности, можно сказать, что намного проще коррелировать
стимул и ответ у подготовленного человека, чем у других животных. Действи-
тельно, психофизические исследования человека принесли некоторые наиболее
точные определения того, что центральная нервная система может, а чего — нет.
Второй подход состоит в том, что исследователь пытается залезть внутрь чер-
ного ящика психофизики. Внимание здесь сфокусировано на физиологии сен-
сорной системы, включая, в случае млекопитающих, сенсорную кору, а не на
поведенческом ответе. Реакция этой системы на адекватные стимулы исследует-
ся на различных уровнях — от молекулярного и субклеточного до уровня сен-
сорных анализаторов в мозге. Исследователь может изучать и ответы индивиду-
альных сенсорных клеток на стимул или — на противоположном конце спект-
ра — регистрировать физиологические ответы коры мозга. Вся надежда на то,
что эти два подхода — сенсорно-физиологический и психофизическо-поведен-
ческий — в конце концов объединившись, принесут всестороннее понимание
того пути, которым человек и другие животные реагируют на внешнюю среду.
3.1. Классификация чувств
Все организмы живут в среде, полной изменений, полной событий. Какие-то
из этих изменений благоприятны для организма, какие-то неблагоприятны.
Некоторые происходят независимо от организма, а некоторые являются пло-
дом деятельности самого организма. Для выживания организму необходима
максимальная информация о том, что происходит, поэтому он развил целый
арсенал сенсорных датчиков. Как всегда, работу лучше всего делают специа-
листы. Сенсорные детекторы специализированы для того, чтобы отвечать на
определенный тип изменений окружающей среды. Как мы увидим, они стано-
вятся чрезвычайно чувствительными к специфическому типу стимула, во мно-
гих случаях достигая физического предела. Основные изменения в окружаю-
щей среде (тех, что касаются большинства организмов) могут быть сгруппиро-
ваны в большие категории механических, химических, электромагнитных и
термических. Соответственно, мы можем классифицировать основные сен-
сорные системы как механорецептивную, хеморецептивную и фоторецептивную.
В отличие от этих трех основных классов рецепторов, терморецепторы, хотя и
так же вездесущие, за исключением некоторых зачатков у змей, не развили
сложного чувствительного органа. Помимо этих четырех классов рецепторов
у некоторых животных развились электрорецепторы, чувствительные к элект-
рическим полям и магниторецепторы, известные у прокариот и подозреваемые
у некоторых животных. Тип стимула, на который настроен специфический
рецептор называется адекватным стимулом.
Классификация сенсорных рецепторов в соответствии с типом энергии из
окружающей среды, к которой они наиболее чувствительны, — важный и оче-
видный путь группировки чувствительных органов. Именно такой класси-
фикации мы придерживаемся в этой книге. Альтернативная классификация
построена не на типе детектируемой энергии, а на том, направлены ли чувст-
вительные окончания вовнутрь организма, контролируя события внутренней
среды организма, либо вовне — во внешнюю среду. Эта классификация — на
интеро- и экстерорецепторы. Тогда как фоторецепторы являются исключитель-
Часть I. Предварительные сведения
Информация 3.1
ГЕРМАН ФОН ГЕЛЬМГОЛЬЦ
Герман Людвиг фон Гельмгольц родился в Потсдаме 31 августа 1821 года. Старший
из четырех детей. У отца унаследовал глубокий интерес к музыке и живописи, а у
матери — уравновешенный характер, который немало пригодился ему в жизни. Его
отец активно участвовал в германском романтическом движении в начале 19-го ве-
ка. Сын Фихте Иммануил Герман Фихте был частым гостем в доме Гельмгольца,
где идеалистическая философия, литература и наука были постоянными темами
обсуждения. Неудивительно, что философские аспекты науки, особенно сенсор-
ной физиологии, занимали Гельмгольца всю его научную карьеру.
В школе юный Гельмгольц не выделялся, но рано проявил интерес к физике.
Отец его, не имея средств, чтобы послать сына в университет, отправил его изучать
медицину, где можно было получить финансовую поддержку от государства. В ре-
зультате Герман фон Гельмгольц в 1837 году получил пятилетний грант на обучение
в Королевском медико-хирургическом институте Фридриха-Вильгельма в Берлине.
Находясь в Берлине, он посещал курсы в университете, в том числе курс физиоло-
гии у Йоханнеса Мюллера. Он воспользовался возможностью написать диссерта-
цию под руководством Мюллера. Работая в университете, Гельмгольц столкнулся
со студентами Эрнстом Брюке и Эмилем Дюбуа-Реймоном, а также Карлом Люд-
вигом. Вместе они основали «школу 1847 года» по физиологии, где избегали не-
физических терминов и «виталистических» сил.
После получения степени доктора медицины в 1842 году, он поступил в армию
в качестве хирурга. При этом он не порывал связей с академическими кругами и
в 1848 году получил место профессора физиологии в Кёнигсберге. Именно здесь он
изобрел офтальмоскоп (1851), измерил скорость проведения в седалищном нерве ля-
гушки, составившую около 90 футов в секунду, намного медленнее, чем считалось
согласно предшествующим исследованиям (1850). В 1855 году с помощью Александ-
ра фон Гумбольдта он получил место профессора в Бонне, и именно здесь он начал
писать и публиковать свой потрясающий основы трехтомный труд по физиологиче-
ской оптике Handbuch der physiologischen Optik (т. 1, 1856). Его механистический под-
ход к сенсорной физиологии весьма огорчил отца с его идеалистическими воззрени-
ями, и возникшая между ними холодность остановила сына в продолжении публи-
кации. Его слава, однако, продолжала расти, и в 1858 году ему была предложена ка-
федра, которую он и принял, в научном центре Германии — Гейдельберге. Здесь он
продолжал интересоваться акустикой, что привело к публикации Die Lehre von den
Tonenpfindungen alsphysiofische Grundlage fur die Theorie der Musik (1863). В этом трак-
тате он ввел т. н. теорию места в частотной дискриминации (см. гл. 8, раздел 8.3.2).
В 1858 году умирает отец, и это устраняет препятствие к публикации остав-
шихся томов его работы по физиологической оптике. Заключительный том
(т. 3) Handbuch был опубликован в 1867 году. Три тома этого учебника содержали
массу новых данных по цветному зрению, по аккомодации хрусталика (Гельмгольц
изобрел офтальмометр, который позволил измерить кривизну хрусталика) и по-
следовательным образам. В частности, он опирался на концепцию своего учителя
Йоханнеса Мюллера о «специфических нервных энергиях». Согласно ей различные
3. Общие черты сенсорных систем 49
Продолжение
сенсорные нервы специализированы к реакции на специфические типы энергии из
внешней среды (стимулы) — т. е. это та самая концепция, которая, как мы видели,
стала основой современного понимания в форме «адекватных стимулов».
Гельмгольц использовал концепцию «специфических нервных энергий» в своей
трихроматической теории цветного зрения и теории места в частотной дискримина-
ции. Он также очень много думал о соотношении между тем, что в нашем сенсорном
восприятии обусловлено опытом, а что — врожденными качествами. Гельмгольц,
в отличие от натурфилософов, придерживался строго эмпирического подхода и на-
стаивал на том, что наши знания о внешнем мире основываются на длительном
опыте — мало что или совсем ничего дается от рождения (см гл. 23).
Работы Гельмгольца по сенсорной физиологии, таким образом, привели его к
гносеологии, которая обсуждалась когда-то в доме его отца. Он пришел к убежде-
нию, что сенсорная физиология в какой-то степени продолжает и поддерживает гно-
сеологию Канта. Иными словами, он полагал, что органы чувств, в соответствии с за-
коном Мюллера о специфических типах нервной энергии, обусловливают субъек-
тивную картину мира. Он, однако, критически оценивал воззрения Канта. В середи-
не 60-х годов XIX века Гельмгольц показал, что аксиомы евклидовой геометрии весь-
ма далеки от априорных синтетических положений, как полагал Кант, а фактически
построены на визуальном опыте, т. е. построены a posteriori, а не a priori.
Работы Гельмгольца отнюдь не ограничивались сенсорной физиологией. Он
внес вклад в несколько областей физики, особенно энергетики, физической хи-
мии, электродинамики и гидродинамики. Он был одним из последних, кто оказал-
ся способен на плодотворную деятельность во всех естественных науках. Им двига-
ло то, что он воспринял в доме отца в Потсдаме и от его друзей — пламенная страсть
найти великие единые принципы, лежащие в основе естественного мира и связы-
вающие его. Когда он умер в Шарлоттенбурге 8-го сентября 1894 года, научный мир
уже стал слишком велик и многосложен, чтобы один человек смог сделать фунда-
ментальный вклад более, чем в одной области науки. Гельмгольц был последним,
кто мог бы получить Нобелевские премии и по физике, и в области медицины и
физиологии, и по химии.
Литература
Cahan, D., ed., 1993, Hermann von Helmholz and the Foundation of Nineteenth-Century
Science, Berkely: University of California Press.
Turner, R. S., 1972, «Helmholz, Hermann von» in Dictionary of Scientific Biography,
С. C. Gillispie, ed., New York, Scribner’s.
но экстерорецепторами (не существует существенных электромагнитных по-
лей внутри организма, которые требуют контроля), остальные три модальнос-
ти чувств могут быть связаны как с экстеро-, так и интерорецепторами. Ин-
терорецепторы, хотя они и не развитых столь же впечатляюще, как многие
экстерорецепторы, тем не менее также важны. У высших животных, особенно
млекопитающих и птиц, контроль внутренней среды важен для гомеостатиче-
ского механизма, от которого зависит сама их жизнь.
50 Часть I. Предварительные сведения
3.2. Модальность
Мы отмечали в гл. 2 (раздел 2.6), что биофизика нервного импульса идентич-
на во всех нервных волокнах. Эволюция, единожды «найдя» эффективный ме-
ханизм передачи информации, придерживалась его, лишь едва усовершенст-
вуя в геологических масштабах времени. Это означает, что информация из
всех сенсорных органов трансформируется в идентичные сигнала в централь-
ную нервную систему (ЦНС). Из этого следует, что ЦНС не имеет иного пути
распознать, о чем говорят поступающие в нее по нервным волокнам импуль-
сы — звуке, свете, температуре или запахе, — кроме как по тому, какие имен-
но волокна активированы. Имеются данные, что, если переключить эти «ме-
ченные линии» хирургически — направив слуховые волокна в зрительную ко-
ру, а зрительные волокна — слуховую, то мы услышим молнию, как гром, и
увидим гром, как молнию. Это, естественно, ставит перед нами потрясающие
философские вопросы. Поскольку зрительная и слуховая кора на клеточном
уровне существенно не различаются (еще менее — на биофизическом), то по-
чему активность в одном участке коры вызывает ощущение звука, а в другом —
совершенно иное ощущение — зрения? Еще интереснее и, может быть важнее
для ответа на философские вопросы то, что если хирургическое «переключе-
ние» выполнено точно на эмбрионах млекопитающих, то слуховая кора разви-
вает некоторые характеристики зрительной и наоборот. Мы отсылаем студен-
тов к библиографии в конце этой части, где они смогут познакомиться с этой
восхитительной работой.
3.3. Интенсивность
События во внешнем мире различаются не только по типу, но и по интенсив-
ности. Грубо говоря, интенсивность стимула кодируется частотой потенциа-
лов действия в волокне сенсорного нерва. То есть ЦНС необходимо «разо-
браться», какой именно сенсорный нерв активирован (что определяет сенсор-
ную модальность) и оценить частоту импульсации в нем (которая определяет
интенсивность модальности), чтобы получить адекватную информацию о со-
бытиях во внешней и внутренней среде.
Наиболее слабые стимулы, которые организм способен детектировать на-
зываются пороговыми (сенсорный порог). Организму (или в случае человека —
субъекту) предъявляется серия стимулов возрастающей интенсивности и вы-
ясняется (выработкой рефлекса или словесным ответом в случае человека),
когда стимул детектируется впервые. Порог определяется как стимул, детекти-
руемый в половину случаев предъявления (рис. 3.1). Кривая, представленная
на рис. 3.1, известна как психометрическая функция или кривая.
Сенсорные пороги не постоянны. Они зависят от множества факторов, в
особенности от утомления, ситуации, опыта и т. д. Это подчеркивает тонкость
физиологии мозга, здесь задействованы многочисленные петли прямых и об-
ратных связей. Мы сможем увидеть, когда будем рассматривать боль (в гл. 21),
что наша чувствительность к этой пренеприятной сенсорной модальности
варьирует очень сильно. В разгар событий мы можем и не почувствовать по-
вреждения, и только потом будем страдать от боли. Сдвиг психометрической
3. Общие черты сенсорных систем 51
Рис. 3.1. Психометрическая кри-
вая. Порог определен как интен-
сивность стимула, при которой
половина реакций правильна.
Ордината дана в условных еди-
ницах. Кривая сдвигается впра-
во или влево в зависимости от
обстоятельств.
кривой вправо встречается и при родах. Болевой порог варьирует и в зависи-
мости от типа культуры. То, что неприемлемо для человека одной культуры —
в порядке вещей для другой.
Возвращаясь к физиологии, лежащей в основе психофизики, следует, в
первую очередь, отметить, что даже в отсутствие стимула по сенсорным волок-
нам потенциалы действия все же передаются, но с малой частотой. Это — ак-
тивность покоя. Она может варьировать (в разных сенсорных волокнах) от
1-2 имп./с до более, чем 50 имп./с. Когда подается адекватный стимул доста-
точной интенсивности, частота импульсации возрастает. Ясно, что для ЦНС
очень важно отличать сигнал, свидетельствующий о наличии адекватного сти-
мула малой интенсивности, от случайных флуктуаций. То есть, иными слова-
ми, отличать сигналы от шума.
И активность покоя, и разряд, возникающий на предъявление адекватного
стимула, варьируют около некоторых средних. Например, первая из них име-
ет частоту 20 ± 5 Гц, а реакция на стимул — 25 ± 6 Гц. Ясно, что эти величины
в крайних значениях перекрываются, т. е. ЦНС «не может быть уверена» в том,
был стимул или нет. Иногда реакция будет верной, иногда — ошибочной. Эта
ситуация приблизительно схожа с сенсорным порогом, определенным нами
выше. По мере увеличения интенсивности стимула, сигнал все более и более
удаляется от уровня шума. При 30 ± 7 Гц «перекрытие» с активностью покоя
становится меньше, а при 35 ± 8 Гц уже нет сомнений, что стимул действи-
тельно присутствует.
Отметим, однако, что в приведенном гипотетическом примере разброс ча-
стот вокруг среднего значения растет с увеличением интенсивности стимула.
Это означает, что с ростом интенсивности стимула амплитуда физических из-
менений стимула, способного создать минимально различимое отличие стимулов
(МРО) тоже растет. Это соотношение было подмечено Вебером в 1834 году и
известно как закон Вебера:
Д\|/ = к х \|/
52 Часть I. Предварительные сведения
где Д\р — МРО, — субъективное восприятие стимула, а к — константа. Наи-
более часто упоминаемый пример действия закона Вебера связан с восприяти-
ем массы. Легко различить массы в 1 и 1,5 кг, но очень трудно, если не невоз-
можно, — 25 и 25,5 кг.
Позднее в XIX столетии (1860 г.) Фехнер расширил закон Вебера, дав соот-
ношение между субъективно воспринимаемыми пороговым и надпороговым
стимулами. Это соотношение стало известно как закон Вебера—Фехнера:
V = A: log ф/ф0
где — субъективная величина стимула по сравнению с пороговой, ф — физи-
ческая величина стимула, ф0 — величина порогового стимула, а к — костанта.
Почти через столетие, в 50-е годы XX века Стэнли Стивенс исследовал боль-
шое количество сенсорных модальностей и показал, что «психофизический
Б
Амплитуда стимула (условные единицы)
Амплитуда стимула (условные
единицы, логарифмическая шкала)
Рис. 3.2. Психофизические соотношения. (А) Когда субъективно воспринимаемая
величина стимула наносится на график против амплитуды стимула в линейных ко-
ординатах, линии часто изгибаются вверх или вниз. (Б) Когда кривая наносится
в логарифмических координатах, образуются прямые линии, угол наклона кото-
рых зависит от степенного показателя п. По Stevens, 1961.
3. Общие черты сенсорных систем 53
закон» следует несколько более сложному соотношению, лучше всего выража-
емому степенной функцией:
у = Л(Ф - Фо)"
или
log V = log к + п log (ф - ф0)
то есть,
log \|/ = К + п log (ф - ф0)
Показатель степени п варьирует от одной модальности к другой: п « 1 для вос-
приятия длины, п » 0,4 для восприятия яркости, п » 3,5 — для электрического
шока. Некоторые из этих соотношений между стимулом и ответом показаны
на рис. 3.2. Их гораздо легче сравнивать, когда они представлены в логариф-
мической форме, поскольку это прямые.
Хотя в предыдущем изложении мы рассматривали случай, когда импульса-
ция, ответственная за субъективное восприятие, проводится по одиночному
сенсорному волокну, — это большая редкость, если вообще возможно. На са-
мом деле все естественным образом воспринимаемые надпороговые стимулы
вовлекают больше, чем одно волокно, и чем выше интенсивность стимула, тем
больше сенсорных волокон участвуют в проведении.
3.4. Адаптация
Мы видели в главе 2 (раздел 2.5), что все сенсорные волокна демонстрируют
адаптацию. Здесь нет необходимости распространяться на эту тему больше.
Тем не менее, стоит отметить, что в некоторых случаях адаптация может сни-
жать уровень импульсации до уровня, ниже порогового. Субъективно мы мо-
жем перестать ощущать стимул. Только при включении и(или) выключении
стимула поток импульсации возрастает настолько существенно, чтобы вызвать
осознанное ощущение. Всем нам случалось испытывать, что только когда не-
который постоянный стимул внезапно исчезает, мы осознаем, что он был.
3.5. Рецептивные поля
Сенсорные нервные окончания, как правило, чувствительны к стимуляции
малых участков своего окружения. Эти участки называются рецептивными по-
лями (РП) сенсорного волокна. Во многих случаях, как мы увидим в последу-
ющих главах этой книги, рецептивные поля имеют свою внутреннюю органи-
зацию. Часто стимуляция одной из частей РП ведет к возбуждению сенсорно-
го нейрона, в другой — к торможению.
Рецептивные поля существенно разнятся и по площади. В тех участках сен-
сорной поверхности, где важно, чтобы стимул был точно локализован, РП
малы. Напротив, там, где локализация стимула не так важна, РП больше. Это
хорошо показано опытом Фехнера по изучению порога в двух точках. Некото-
рые части тела — кончики пальцев и губы — способны различать два близко
расположенных стимула (укола), тогда как другие (тыльная сторона ладони,
спина) — только значительно дальше разнесенные стимулы. В некоторых слу-
54 Часть I. Предварительные сведения
чаях есть и связь между чувствительностью и точностью локализации ощуще-
ния. Если только строго определенное количество энергии взаимодействует
с сенсорной поверхностью, как это происходит, например, при постоянном
освещении сетчатки, то большее рецептивное поле способно получить больше
энергии, чем меньшее. При прочих равных оно обладает большей чувстви-
тельностью. Но по некоторым причинам оно неспособно обеспечить точную
информацию о том, какая именно часть сетчатки стимулирована.
Хотя сенсорные поверхности, такие как сетчатка или кожа, могут рассмат-
риваться как мозаика РП, не следует думать, что РП имеют ясно очерченные
границы, отделяющие их друг от друга, подобно кусочкам стекла, составляю-
щим декоративную мозаику классического или восточного типа. На самом де-
ле, РП в сенсорной поверхности образуют широкие перекрытия и, более того,
особенно в сетчатке, варьируют в размерах в зависимости от физиологических
условий. Это — одно из проявлений динамического характера сенсорных
систем, который накладывается на сравнительно «жесткую» сеть нейронов и
сенсорных клеток протекающими в них активными биохимическими и моле-
кулярно-биологическими процессами. Это очень резко отличает их от неиз-
меняющихся неорганических компонентов нашей, основанной на кремнии,
электронной технологии.
3.6. Карты сенсорных поверхностей
Сенсорные окончания и их рецептивные поля часто (но не всегда) организо-
ваны в двумерные структуры — карты. Очевидным образом это так в случае
сетчатки и кожных рецепторов прикосновения. Менее очевидно это примени-
тельно к базилярной мембране улитки. Пространственные связи этих структур
поддерживаются ЦНС. У людей, как мы увидим в последующих главах, в со-
матосенсорной коре постцентральной извилины имеется карта поверхности
тела, карта сетчатки в первичной зрительной коре затылочной доли, и карта
базилярной мембраны в первичной слуховой коре височной доли. Эти карты
называются, соответственно, соматотопической, ретинотопической и тонотопи-
ческой.
Не следует думать, однако, что все эти карты изоморфны сенсорным по-
верхностям. Во всех случаях те части сенсорной поверхности, которые имеют
большее биологическое значение, где требуется большая точность дискрими-
нации сенсорной информации, занимают непропорционально большее место
в коре. Это не-изоморфное картирование частично отражает тот упомянутый
выше факт, что части сенсорных поверхностей, имеющие большее биологиче-
ское значение, характеризуются меньшими по размеру рецептивными полями
(или большим числом сенсорных волокон на единицу поверхности). Это так-
же связано с большей сложностью нейрональных связей в таких участках ко-
ры. Хотя неизоморфное картирование характерно для всех карт сенсорных
поверхностей в ЦНС, вероятно наиболее широко известно это для соматосен-
сорной коры. Сенсорные «гомункулусы» (рис. 7.15) с их огромными губами,
.гигантскими кистями рук, на которых выделяется еще и большой палец, и на-
против — с крошечными ножками и ступнями, уже почти превратились
в икону.
3. Общие черты сенсорных систем 55
В некоторых сенсорных системах, особенно в зрительной системе прима-
тов, карта из первичного сенсорного экрана проецируется далее во вторич-
ный, третичный и более высокие уровни. Однако в целом, эти более высокие
уровни становятся топографически все менее и менее точными. Одновремен-
но, несмотря на эту неточность, высшие уровни обработки сигнала могут од-
новременно обеспечивать точную карту других свойств стимула. Например, в
зрительной системе приматов (макак), наиболее изученной среди высокораз-
витых сенсорных систем, картируются скорее 4 цвета, чем пространственное
расположение на сенсорной поверхности. Длины волн света, на которые реа-
гируют клетки V4, определенным образом меняются от одной клетки к другой,
соседней. Сходным образом, в зрительном поле 5, которое связано с восприя-
тием движения зрительного стимула, обнаружено, что направление движения,
на которое реагируют соседние клетки, меняется регулярным образом.
Но почему карты? Почему вообще сенсорные системы развили карты сен-
сорных поверхностей организма? Краткий ответ на этот вопрос — этого не
знает никто, но спекуляций на эту тему множество. Мы вернемся к этому во-
просу в гл. 22 (раздел 22.4). Возможно, дело в том, что сенсорные волокна, ко-
торые в норме срабатывают одновременно, стремятся и пространственно
сгруппироваться вместе. В пользу такого предположения есть, как мы увидим,
существенное свидетельство из области биологии развития. Понятно, что во-
локна из соседних участков сенсорной поверхности (например, сетчатки) с
большей вероятностью срабатывают одновременно. Есть также аргумент, свя-
занный с экономией организации. Высказывается мнение, что сенсорный
анализ, обработка сигнала, более эффективна, если схожие части сенсорной
картины, как топографически, так и функционально содержатся близко друг
от друга. В отличие от компьютеров, основанных на кремнии и меди, мозг не
может работать со скоростью света. Скорость обмена информацией в немие-
линизированных коротких аксонах и дендритах серого вещества на несколько
порядков ниже, чем у электрического проведения в компьютере. Если же сиг-
налу требуется пройти большие расстояния (сантиметры, а не миллиметры),
то связность «целого» объекта перцепции рискует оказаться невозможной.
Здесь мы оказываемся в окрестностях проблемы связывания — того, каким об-
разом получается, что наше восприятие целостно и связно, хотя мозг в высшей
степени гетерогенен и построен из модулей. Решения проблемы связывания у
нас нет. Не знаем мы и ответа на наше «почему?» о сенсорных картах. Нельзя
исключать, что оба эти вопроса — аспекты одной и той же проблемы. Когда
удастся достичь нового понимания в одном из них, это может пролить свет и
на второй.
3.7. Иерархическая и параллельная организация
Из предыдущего раздела могло сложиться впечатление, что сенсорные систе-
мы организованы исключительно иерархическим образом. Могло бы пока-
заться, что сенсорные поверхности дают проекцию в первичную сенсорную
кору, где сохраняются топографические взаимоотношения, та дает свои даль-
нейшие проекции, несколько утрачивая топографическую точность, те пере-
дают информацию в следующую в ином участке коры, и так далее. Хотя это и
56 Часть I. Предварительные сведения
правда, но не вся. Мы увидим, когда будем рассматривать основные сенсор-
ные системы, особенно зрительную систему приматов, что существуют много-
численные проекции сенсорной информации в мозг. То есть это не один эк-
ран, проецирующийся в более высокие уровни по иерархическому принципу,
а несколько параллельных экранов, взаимосвязанных и взаимодействующих
между собой. К тому же, иерархическая последовательность карт связана и в
центрипетальном, и центрифугальном направлениях. Организация сенсорно-
го мозга позволяет и параллельную, и последовательную обработку информа-
ции, и лучше всего описывается термином Хофштадтера как «усложненная
иерархия».
3.8. Выделение отличительных характеристик
и пусковые стимулы
В заключение еще одна общая черта сенсорных систем должна быть упомяну-
та. Мы увидим по ходе дальнейшего изложения, что в высокоразвитых систе-
мах, таких как слуховая и зрительная, в ходе обработки в мозге в восходящих
уровнях выделяются специфические черты полученной информации. Эти чер-
ты связаны с теми аспектами окружающего мира, которые имеют первостепен-
ное значение для животного. Лишь небольшое количество клеток в слуховой
коре отвечают на продолжительные чистые тоны, которые так эффективны
в стимуляции базилярной мембраны улитки. Напротив, щелчки и потрескива-
ния, издаваемые хищником, подкрадывающимся через подлесок, или частот-
но-модулированные тоны, повышающиеся или понижающиеся по высоте,
оказываются значительно более эффективными. Мы увидим в гл. 9, что эти
черты широко используются в высокоспециализированных участках коры че-
ловека (в формировании звуков речи) и летучих мышей (в эхолокации). Сход-
ным образом, в зрительной системе яркие пятна света или длительное нефо-
кусированное освещение, эффективно стимулирующие сетчатку, вызывают в
клетках зрительной коры лишь небольшой эффект. Напротив, контуры объек-
та вызывают выраженный эффект в первичной зрительной коре. С повышени-
ем уровня переработки сигнала ответ запускают все более содержательные
черты объекта. Например, в высших уровнях у макак (нижне-теменная кора)
есть клетки, отвечающие исключительно на характерные черты обезьяньих
морд (в фас и в профиль) (гл. 17, раздел 17.4).
Эти специфические черты сенсорного мира, которые имеют в жизни жи-
вотного важное значение, называются триггерными стимулами. Этой категории
стимулов посвящено большое количество исследований. Сначала популярной
стала концепция иерархии уровней выделения специфических черт. Предпо-
лагалось, что выделяются еще более специфические, чем обезьяньи морды,
черты визуальной картины — в случае человека вплоть до распознавания лица
бабушки. Признание возможности существования клеток распознавания лица
бабушки или понтификальных клеток приобрело широкое хождение, одна-
ко, не исключено, что это все же следует принимать как reductio ad absurdum
(упрощение до абсурда) или, по крайней мере, с большой осторожностью.
Хватит ли клеток в коре, чтобы разобраться со всеми бабушкиными прическа-
3. Общие черты сенсорных систем 57
ми, бижутерией, шляпками и приступами дурного настроения? Что же, отбро-
сить прочие специфические детали визуального восприятия? А что, если клет-
ка, ответственная за бабушку, по каким-то причинам повредится или заболе-
ет, мы бабушку узнавать перестанем? К тому же, осложняются и философские
проблемы. Можно ли всерьез предполагать, что активность одной-единствен-
ной клетки преобразуется в зрительный образ бабушки? Кажется все же более
вероятным, как мы увидим в гл. 17, что распознавание бабушки зависит от ак-
тивности популяции клеток, распределенной в зрительных участках коры и
других участках мозга, включая (что важно) эмоциональные центры лимби-
ческой системы. Вероятность этого возрастает, если признать упомянутую в
предыдущем разделе роль параллельной обработки информации в зрительной
системе. Таким образом, идея об однонаправленном потоке информации, пе-
редаваемой в иерархически высшие центры мозга, очевидно, устарела.
3.9. Заключение
«Эволюция — халтурщица», — сказал Франсуа Жакоб. Но в случае сенсорных
систем она употребила биофизику и молекулярную биологию, рассмотренные
в гл. 1 и 2, компоновала их в течение многих миллионов лет в удивительные и
чувствительные устройства для обнаружения событий в окружающей среде.
Эволюция, может быть и халтурщица, но халтурщица, движимая страхом
смерти. Сенсорные системы чрезвычайно важны для борьбы за существо-
вание, в том, что некоторые называют «эволюционной гонкой вооружений».
Как и в технологической гонке вооружений, с которой мы знакомы, важна
информация. Битва за Британию и битва за Атлантику были выиграны (и про-
играны) благодаря радарам и взлому радиокодов. Сенсорные системы подверг-
лись всем безжалостным испытаниям естественного отбора, но если вернуть-
ся к метафоре Жакоба, у эволюции не было возможности отбросить лишнее и
начать все заново. Халтурщица работала с тем, что получилось. Потому и сен-
сорные системы большинства высокоразвитых животных сохраняют следы
организации далеких предков. Несмотря на это, они развились до такого уров-
ня чувствительности, который во многих случаях недоступен для человечес-
кой технологии.
4. ЭВОЛЮЦИЯ И КЛАССИФИКАЦИЯ
ОРГАНИЗМОВ
Систематика: эволюционная и кладистическая классификации. Шесть
царств: природа типов животных. Одноклеточные: прокариоты (Archae-
bacteria и Eubacteria) — эукариоты (протесты). Многоклеточность: грибы,
растения и животные — Parazoa и Metazoa — беспозвоночные и позво-
ночные. Первичноротые и вторичноротые: спиральное и радиальное дроб-
ление — шизоцель и энтероцель — формирование рта — выраженные
эмбриологические различия отделяют последовательность «кольчатые
черви — членистоногие — моллюски» от последовательности «иглоко-
жие — хордовые» и предполагают независимую эволюцию в течение
более полумиллиарда лет. Классификация многоклеточных Metazoa:
стрекающие (Cnidaria) — плоские черви (Platyhelmintes) — круглые чер-
ви (Nematoda) — моллюски (Mollusca) — кольчатые черви (Annelida) —
членистоногие (Arthropoda) — хордовые (Chordata). Заключение: неко-
торые типы организации лучше других — три основных типа (членисто-
ногие, моллюски, хордовые) оказались самыми успешными, потому они
в основном и фигурируют в этой книге
В данной главе мы предпримем широкий экскурс по миру живого в нашем ис-
следовании сенсорных систем. Не все читатели этой книги хорошо знакомы
с таксономическим и эволюционным положением рассматриваемых организ-
мов. Эта глава направлена на то, чтобы устранить такие пробелы.
4.1. Систематика
Стороннему наблюдателю может показаться удивительным, что биологичес-
кая систематика до сих пор не завершена и не согласована. Даже число круп-
нейших групп (типов), на которые подразделяется царство животных, остает-
ся предметом противоречий. Кроме того, с введением Хеннигом в 50-е годы
XX века не-эволюционной кладистической схемы классификации, системати-
ки оказались в центре горячей полемики. К счастью, нам нет необходимости
вступать на это «минное поле». Наш интерес к положению организмов в клас-
сификации имеет выраженную эволюционную природу. Наш интерес к орга-
нам чувств животных, а не только нашим собственным, обусловлен не просто
тем, что они существуют (хотя и это вполне уважительная причина), но глав-
ным образом тем, что их изучение способно пролить свет на сенсорные систе-
мы человека, а также продемонстрировать альтернативные пути решения тех
же самых проблем. В некоторых случаях — это радикально иные решения (гла-
4. Эволюция и классификация организмов 59
за насекомых и позвоночных), в других — поразительно схожие (глаза голово-
ногих и позвоночных). Понимание эволюционных связей между группами
животных — это основа полного понимания сенсорных систем животных и,
в частности, тех, которые приобрели мы сами.
4.2. Классификация на шесть царств
В этой книге мы будем использовать общепринятую схему классификации ор-
ганизмов на шесть царств: Archaebacteria (архебактерии), Eubacteria (собствен-
но бактерии), Protista (протесты), Plantae (растения), Fungi (грибы) и Animalia
(животные). Внутри этих шести царств организмы подразделяются далее на
большие группы со сходным планом строения, которые называются типами.
Такой принцип классификации, как и можно было ожидать, куда более очеви-
ден на многоклеточных — грибах, растениях и животных, чем в трех царствах
одноклеточных. Действительно, классификация оказывается общепринятой
только для царства животных. В изучении сенсорных систем именно оно зани-
мает доминирующее положение, потому и наше внимание по большей часта
будет сконцентрировано на нем.
4.3. Одноклеточные
Archaebacteria и Eubacteria часто объединяются в единую группу «бактерий»
или, иногда, монер (Moneres — греч., одинокий). Известно свыше 5 000 видов,
представляющих собой маленькие клетки без ядра и органоидов. Поскольку
эти клетки не имеют ясно выраженного ядра (т. е. нуклеиновые кислоты не от-
делены от цитоплазмы ядерной мембраной), они получили наименование про-
кариот. В течение прибл. 2 миллиардов лет (начиная с 3 600 млн лет назад до
1 600 млн лет назад) монеры владели миром. Сенсорные системы бактерий,
конечно, предельно просты. Однако они представляют для нас значительный
интерес, поскольку на них часто можно проследить, по крайней мере на био-
химическом уровне, зачатки тех бесконечно сложных систем, которые разви-
лись у животных.
Спустя 2 млрд лет появились более крупные и более сложные клетки —
эукариоты. Вероятно, они более тесно связаны с архебактериями, чем с собст-
венно бактериями. Все эукариоты имеют выраженное ядро (их генетический
материал сосредоточен за ядерной мембраной) и органоиды, такие как мито-
хондрии, хлоропласты и т. д., локализованные в цитоплазме. Ранние класси-
фикации объединяли автотрофных эукариот с растениями в Prophyta, а гетеро-
трофных — с животными в Protozoa (простейшие). Более современный альтер-
нативный анализ предполагает, что одноклеточные эукариоты схожи друг с
другом значительно больше, чем с многоклеточными животными, растениями
и грибами, и к тому же сама многоклеточность могла возникать несколько раз.
Исходя из этого одноклеточные эукариоты и объединены в отдельное царство
Protista или Protoctista (protos — первичный, kristos — устанавливать, греч.).
Протесты — чрезвычайно разнообразная группа, подразделяющаяся на типы,
которых насчитывается до 30, однако, в этой книге они, в основном, не фигу-
рируют. С нашей точки зрения, самое интересное, возможно, заключается
60 Часть I. Предварительные сведения
Рис. 4.1. (А) (а) Спиральное
и (б) радиальное дробление.
По Caroil, 1988. (Б) Первич-
норотые и вторичноротые,
(а) Бластула; (б) инвагина-
ция при формировании гас-
трулы. Бластоцель в основ-
ном смыкается, и образуется
первичная кишка или архэн-
терон, открытый во внеш-
нюю среду (бластопор). У
первичноротых он в даль-
нейшем формирует рот, а у
вторичноротых возникает
еще одна инвагинация (на
противоположном от блас-
топора конце эмбриона), ко-
торая прорывается в архэн-
терон. Здесь формируется
рот, тогда как по соседству
(рядом с зарастающим блас-
топором) прорывается вто-
ричный анус.
в наличии у них наиболее ранних фоторецепторов — различных типов свето-
чувствительных пятен. Они обнаружены в типе Eustigmatophyta, Chlorophyta и,
особенно, Euglenophyta (Euglena viridis), а также у некоторых представителей
динофлагеллят (Pouchetia sp., Erythropsidinium pavillardii).
4.4. Многоклеточность
Многоклеточность, вероятно, возникла около 1 млрд лет назад. Бйологи вы-
деляют три царства многоклеточных — грибы, растения и животные. Только
последнее представляет интерес для данной работы. Здесь тоже имеется не-
сколько различных методов классификации. В старых работах царство живот-
4. Эволюция и классификация организмов 61
Рис. 4.2. Эволюционные
взаимоотношения между
основными группами ор-
ганизмов. Числами (в млн
лет) показана давность
дивергенций. Псевдоце-
ломические животные
представлены нематодами
(напр. С. elegans). Данные
из Doolittle et al., 1996.
грам-положительные
ных подразделялось на три подцарства: Protozoa (простейшие), Parazoa
(Mesozoa, губки) и Metazoa (истинно многоклеточные). Позже, как мы виде-
ли, Protozoa были объединены с другими одноклеточными формами в царст-
во Protista. Animalia (животные), в свою очередь, подразделяются на два под-
царства — Parazoa и Metazoa. Parazoa далее подразделяются на Porifera (губ-
ки) и Placozoa (с единственным видом Trichoplax adhaerans, размером с боль-
шую амебу). Все остальные животные образуют Metazoa — многоклеточных.
Старая (в высшей степени антропоцентрическая) классификация делит мно-
гоклеточных на позвоночных и беспозвоночных — в зависимости от наличия
или отсутствия позвоночного столба. Такая классификация остается обще-
признанной и общеупотребительной, но необъективной из-за нашего само-
мнения. Позвоночные (если быть более точным — черепные) образуют два
подтипа внутри типа Chordata (хордовые), одного из более чем тридцати
в подцарстве Metazoa.
4.5. Первичноротые и вторичноротые
Если внимательно рассмотреть ранний эмбриогенез целомических многокле-
точных, то мы обнаружим несколько фундаментальных черт, которые предпо-
лагают разделение их на две большие группы форм животных: ветвь кольчатых
червей — членистоногих — моллюсков (КЧМ) и ветвь иглокожих — хордовых
(ИХ) (рис. 4.3). У кольчатых червей — членистоногих — моллюсков первые де-
ления зигот (деления дробления) имеют спиральный характер (рис. 4.1 А(а)).
Слои дочерних клеток (бластомеров) выстраиваются со сдвигом относительно
друг друга. Проспективная судьба клеток таких зародышей детерминирована.
Если одна из них удалена, соответствующая часть организма не сформируется.
Напротив, в линии развития иглокожих — хордовых зигота претерпевает ради-
альное дробление\рис. 4.1 А(б)), а слои клеток расположены строго один над
(под) другим. Судьбы бластомеров на этой стадии строго не детерминированы.
Если один из них удален, остальные способны это компенсировать, так что
формируется нормальный зародыш.
62 Часть I. Предварительные сведения
СЛ
ш
сл
1
£
Q
О
О
Jen
LU
ш
I
§
i Ш
•О
О
I ш
। ш
‘1
•о
•о
JLU
8
DEUTEROSTOMATA * •, J •
X LOPHOTROCHOZOA U
\ \ ECDYOZOA *.
800 млн лет назад ххх<>
PROTOSTOMATA
BILATERALIA
1 млрд лет назад
PROTISTA
ш
LJL-
сс
Рис. 4.3. Эволюционное
древо царства животных.
Всего насчитывается около
30 типов, но на рисунке по-
казаны только те, которые
включают виды, упоминаю-
щиеся на последующих
страницах этой книги. Воз-
никновение царства из про-
тестов произошло около
1 млрд лет назад. Некоторые
важные стадии формиро-
вания плана строения пока-
заны на основном стволе
схемы: билатеральная сим-
метрия (BILATERALIA);
выраженная полость тела
(целом), отделяющая кишку
от мускулатуры (COELO-
MATA); развитие рта из
бластопора (PROSTOMA-
TA— первичноротые) или
на противоположном конце
(DEUTEROSTOMATA -
вторичноротые). Все эти
важные события эволюции
имели место в докембрий-
ский период.
№
К
Q
Z
о
Далее, две эти большие ветви демонстрируют глубокие различия путей
формирования целома (полости тела). В линии КЧМ это происходит в резуль-
тате расщепления мезодермы (шизоцель). В линии ИХ это происходит в ре-
зультате отшнуровки выпячиваний кишки (энтероцель).
Наконец, важное различие — в том, как формируется рот. Сфера из клеток,
образующаяся в результате делений дробления оплодотворенной яйцеклет-
ки — бластула — затем инвагинирует, формируя первичную кишку — архэнте-
рон. Этот процесс называется гаструляцией. В ветви КЧМ отверстие первич-
ной кишки — бластопор — в конце концов развивается в рот взрослой особи.
Животные, у которых рот развивается таким образом, относятся к первичноро-
тым. Напротив, в ветви ИХ рот развивается иначе. В этом случае новое отвер-
4. Эволюция и классификация организмов 63
стие в конце концов возникает на конце, противоположном бластопору, а бла-
стопор преобразуется в анус (рис. 4.1 А(б)). Животные с таким типом развития
относятся к вторичноротым (поскольку рот развивается у них «вторично»). Эти
два термина — первичноротые и вторичноротые — обычно употребляются для
обозначения двух упомянутых выше крупных ветвей развития.
Хотя т. н. «биогенетический закон» Геккеля (онтогенез отражает филоге-
нез), в настоящее время принимается со значительными оговорками, нет со-
мнений, что последовательность событий раннего эмбриогенеза сложилась в
эволюционной истории очень давно. Из этого ясно, что и принципиальные
различия эмбриогенеза первично- и вторичноротых возникли в филогенезе
очень рано. Молекулярно-биологические данные предполагают, что их общий
предок существовал более, чем 670 млн лет назад. Недавние работы отодвину-
ли этот срок на еще более давний срок — порядка 830 млн лет. Таким образом,
эти две ветви в течение более трех четвертей миллиарда лет развиваются неза-
висимо друг от друга. Это делает возможность сравнения органов чувств меж-
ду этими ветвями особенно интересной — нам нет необходимости лететь на
другую планету, чтобы получить возможность сравнения с эволюционными
альтернативами форм жизни, основанных на углероде.
В заключение, накапливаются молекулярно-биологические данные, даю-
щие основания полагать, что первичноротые очень рано разделились на две
крупных группы. Первая из них — Ecdyozoa — включающая, помимо прочих,
нематод и членистоногих и характеризующаяся твердым наружным покровом,
что требует периодических линек (ecdysis) для обеспечения возможности рос-
та. Другая — Lophotrochozoa — жестким покровом не обладает (что, правда,
отрицательно сказывается на их защищенности) и включает плоских и кольча-
тых червей, моллюсков, а также несколько других типов животных.
4.6. Классификация многоклеточных
Теперь сосредоточим внимание на основных группах животного царства. Си-
стематики подразделяют его на типы. Тип, как упоминалось выше, включает
животных, обладающих общим планом строения. Изучение эволюции такого
плана за геологические промежутки времени называется филогенией. Внутри
типов зоологи классифицируют организмы на подтипы, классы, отряды, се-
мейства, роды и, наконец, виды и подвиды. В некоторых случаях зоологи груп-
пируют отряды в надотряды, а семейства — в надсемейства. Таким образом,
например, человек классифицируется как
Царство Animalia (животные)
Тип Chordata (хордовые)
Подтип Gnathostomata (челюстноротые)
Класс Mammalia (млекопитающие)
Отряд Primates (приматы)
Семейство Hominoidea (гоминиды, человекообразные)
Род Ното (человек)
Вид sapiens (разумный)
Подвид sapiens (разумный)
64 Часть I. Предварительные сведения
Все животные по определению вынуждены искать пропитание в перемен-
чивой окружающей среде, вследствие чего все эти многочисленные формы,
сгруппированные в приблизительно 30 типов, развили те или иные органы
чувств. Рис. 4.3 показывает эволюционные взаимоотношения между группами
животных, которые займут важное место на следующих страницах этой книги.
Выделяются среди них членистоногие, моллюски и, конечно, хордовые.
Кратко опишем типы, показанные на рис. 4.3. Cnidaria (стрекающие, ранее
объединявшиеся с Ctenophora в тип Coelenterata — кишечнополостные) —
наиболее просто организованные среди многоклеточных. Их тела не имеют
целомической полости и просто состоят из двух слоев клеток — эктодермы и
эндодермы. У них не развивается истинной центральной нервной системы, но
формируется диффузная нервная сеть. Хорошо известны такие представители
этого типа, как медузы (класс Scyphozoa), гидроиды, такие как гидра Hydra, а
также Obelia и т. д. (класс Hydrozoa), и, наконец, кораллы и актинии (морские
анемоны, класс Anthozoa).
Platyhelmintes (плоские черви) демонстрируют эволюционный прогресс
по сравнению с Cnidaria, поскольку их тела построены из трех слоев клеток:
экто-, мезо- и эндодермы. Полость тела по-прежнему отсутствует, но это уже
билатерально симметричные животные с выраженными передним и задним
концом. Свободно живущие формы развили «лестничную» центральную нерв-
ную систему, в которой на головном конце наблюдается аккумуляция нервных
клеток, где и концентрируются органы чувств, включая пигментированные
глазки. Включают три класса: Turbellaria (свободно живущие хищные плоские
черви, напр. планария), Trematoda (паразитические трематоды печени) и
Cestoda (ленточные черви). Большинство зоологов полагают, что формы,
сходные с современной планарией, представляют собой общего предка, из ко-
торого 600—700 миллионов лет назад и развилось все многообразие истинно
многоклеточных.
Тип Nematoda (обычно — круглые черви) состоит из большого количества
видов (вероятно, около 1 миллиона), из которых наиболее известны эндопара-
зиты, такие как аскарида и трихинелла. Круглые черви имеют зачатки полос -
титела, которая именуется не целомом, а псевдоцеломом, поскольку она об-
разована не мезодермой. Нематоды имеют очень простые дорзальную и вент-
ральную нервные цепочки, объединенные в нервное кольцо вокруг глотки.
Многие нематоды известны благодаря своим выдающимся способностям к
воспроизводству — самки иногда содержат десятки миллионов яйцеклеток и
выделяют их в темпе до 100 000 в день. Благодаря такой чрезвычайно плодови-
тости и очень короткого генеративного периода одна из свободно живущих
почвенных нематод Caenorhabditis elegans широко используется в молекуляр-
но-биологических исследованиях и теперь, наверное, является наиболее изу-
ченным в этом отношении видом. Замечательно, что на уровне молекулярной
биологии обнаружилось удивительное сходство между основными чертами
этого вида и высших животных, включая человека.
Тип Molhisca состоит из большого количества весьма разнообразных
форм — более 100 000 видов. Это мягкотелые несегментированные животные
с истинной полостью тела (целомом). По размерам они могут составлять от не-
скольких миллиметров до более 20 м (как в случае гигантского кальмара
4. Эволюция и классификация организмов 65
Architheutis, крупнейшего из беспозвоночных). Среди моллюсков найдено не-
сколько наиболее интересных и ценных объектов для нейробиологических ис-
следований. Как показано на рис. 4.3, они развивались независимо от хордо-
вых около 700 или 800 миллионов лет, и вполне могут рассматриваться как
продукт отдельного «акта творения».
Моллюски подразделяются на 7 классов. Monoplacophora, включающие
всего один вид с плоской раковиной, обитают в глубоких субтропических и
тропических водах. Aplacophora включают червеобразных соленогастров (бо-
роздчатобрюхих моллюсков), также обитающих в глубинах теплых морей.
Polyplacophora включают обычных обрастающих прибрежные камни хитонов.
Pelycypoda включают, наверное, наиболее известных моллюсков — двухствор-
чатых, таких, как устрицы, мидии и гребешки. В гл. 14 мы увидим, что гребеш-
ки обладают удивительными глазами. Gastropoda (брюхоногие) включают все
формы, которые ползают на собственном брюхе — улитки, береговички и
слизни. У Scaphopoda — конические раковины, они зарываются в ил или пе-
сок. Наконец, наиболее импозантные моллюски относятся к классу
Cephalopoda (головоногие). Они подразделяются на три подкласса: Ammonoi-
dea — аммониты, ныне вымерший, но часто обнаруживаемый в ископаемых
отложениях; Nautiloidea, представленному жемчугоносным Nautilus^ и
Belemnoidea, к которым относятся кальмары и осьминоги. Мы увидим, что эти
удивительные морские хищники, несмотря на более чем полмиллиарда лет не-
зависимой эволюции и принципиально иной план строения тела, нашли пора-
зительное схожие решения в организации и функции фоторецепторов с теми,
которые мы встречаем у позвоночных.
Тип Annelida (кольчатые черви) составляют обычные наземные и водные
(в основном, морские) черви. Как и моллюски, это целомические первично-
ротые, но что сильно их отличает от моллюсков — это выражение сегментиро-
ванные животные. 4 класса: Polychaeta (многощетинковые) — в основном,
морские черви, напр. обычный прибрежный нереис {Nereis). Многощетинко-
вые черви — часто крупные, активные формы с хорошо развитой нервной си-
стемой и органами чувств. Класс Oligochaeta (малощетинковые) представлен
обычным дождевым червем Lumbricus. Класс Hirudinea включает эктопарази-
тических пиявок, напр. Hirudo. Наконец, Myzostomaria объединяет множество
мелких паразитов иглокожих.
Тип Arthropoda (членистоногие) — вероятно, наиболее успешный из всех
типов животного царства. Многие зоологи полагают, что если бы все ныне жи-
вущие виды членистоногих удалось идентифицировать и пересчитать, то их
оказалось бы больше 10 миллионов. Членистоногие населяют все экологичес-
кие ниши и отличаются размерами в диапазоне от микроскопических клещей
до гигантских крабов, больше метра в диаметре (считая с ногами). Наземные
членистоногие, представленные в подавляющем большинстве насекомыми,
преимущественно невелики. Это обусловлено основным принципом их стро-
ения — твердый хитиновый наружный скелет не позволяет организму расти
постоянно. Рост возможен, когда кутикула сбрасывается в ходе линьки, одна-
ко в эти периоды членистоногие, особенно наземные, чрезвычайно уязвимы.
Поэтому такие «мягкотелые» периоды должны быть елико возможно коротки-
ми. Если бы не такая катастрофическая организация индивидуального разви-
3 Биология сенсорных систем
66 Часть I. Предварительные сведения
тия членистоногих, они могли бы достичь чудовищных размеров персонажей
научно-фантастических романов.
Членистоногие, как и кольчатые черви, четко сегментированные целоми-
ческие первичноротые. Они отличаются от аннелид твердым наружным хити-
новым скелетом и членистыми конечностями, которые и дали им название.
Существуют две большие группы — подтипы Mandibulata (жвалоносные или
челюстные) и Chelicerata (хелицеровые). Челюстные имеют три отдела тела
(голова, грудь, брюшко), а у хелицеровых две передние части сливаются в го-
ловогрудь.
Подтип Mandibulata подразделяется на шесть классов. Класс Crustacea (ра-
кообразные) представлен большим числом водных жаберно-дышащих члени-
стоногих. Наиболее значительный отряд этого класса — Decapoda — включает
омаров, раков, крабов и разнообразных креветок. Ракообразные включают
также множество более мелких отрядов, таких как водяные блохи, веслоногие
рачки, усоногие раки и т. д. Второй класс, Diplopoda, объединяет двупарноно-
гих. Класс Chilopoda включает многоножек; класс Pauropoda — формы, схожие
с многоножками, как и класс Symphyla (симфилы). Последний класс подти-
па — Insecta (насекомые) — крупнейший среди членистоногих. Насекомые
присутствуют во всех экологических нишах, за важным исключением — мо-
рей. В большинстве своем — это весьма активные животные, а потому развили
целый ряд чрезвычайно тонко организованных органов чувств. Из-за того, что
насекомые покрыты твердым хитиновым наружным скелетом (кутикулой), их
органы чувств, главным образом, основываются на кутикулярных сенсиллах.
Такая организация сильно отличается от наблюдающейся у мягкотелых форм
(моллюски, позвоночные), а потому изучение сенсорной биологии насекомых
дает неоценимый материал для сравнительных исследований. Мы обсудим
многочисленные примеры этого на последующих страницах.
Второй подтип членистоногих, хелицеровые, получили свое название
вследствие того, что первая пара конечностей у них преобразована в хелицеры,
предназначенные для разрывания и измельчения пищи. 3 класса: Pycnogonida
представлен морскими пауками, обитающими в придонном слое литоральной
зоны; класс Merostomata включает мечехвостов (Limulus), чьи простые состав-
ные глаза — очень важный объект исследований сенсорных физиологов; нако-
нец, большой и важный класс Arachnida с пауками, скорпионами, сенокосца-
ми и различными клещами.
Мы пропустим тип Echinodermata (морские звезды, морские ежи, голоту-
рии и т. д.), несмотря на их удивительно красивую эмбриологию, поскольку у
них нет органов чувств, заслуживающих внимания в этой книге, и перейдем
прямо к тому, что с нашей точки зрения, является последним существенным
типом — Chordata (хордовые). Поскольку этот тип включает и Ното sapiens,
многие могут подумать, что это и самый важный из всех. Наш краткий обзор,
однако, показывает, что это — всего лишь один из многих типов, к тому же не-
большой, насчитывающий всего около 45 000 видов. Тип включает млекопита-
ющих, пресмыкающихся, птиц, земноводных и рыб (т. е. всех позвоночных) и
несколько групп беспозвоночных, объединяемых в протохордовые. Как ука-
зывалось выше, хордовые — вторичноротые и, как это очевидно по строению
позвоночного столба, сегментированные. Для хордовых характерны три черты
4. Эволюция и классификация организмов 67
Рис. 4.4. Вероятные эво-
люционные взаимосвязи
позвоночных. По Caroil,
1968.
(присутствующие хотя бы только на определенных стадиях онтогенеза): дор-
зально локализованный полый нервный тяж; расположенный между ним и киш-
кой жесткий стержень — хорда, который у позвоночных трансформирован в
позвоночный столб; присутствие на некоторых стадиях развития жаберных
щелей, соединяющих пищевод с внешней средой.
68 Часть I. Предварительные сведения
Наиболее широко распространенная классификация подразделяет хордо-
вых на четыре подтипа: два подтипа — бесчерепные: это Tunicata (оболочники)
и Cephalochordata (головохордовые), и два подтипа — черепные: Agnatha (бес-
челюстные) и Gnathostomata (челюстноротые). Два последних образуют «по-
звоночных». Взрослые оболочники — это «сидячие» животные, многие из них,
например асцидия Ciona intestinalis, населяют литораль. Только личиночная
стадия, напоминающая головастика, активно подвижна, развивает хорду,
нервный тяж и жаберные щели. Головохордовые представлены ланцетником
Amphioxus lanceolatum, обитающим на песчаном дне морского мелководья. Из-
давна их рассматривают как обобщенный упрощенный образ хордовых —
у них есть дорзальный спинной нервный тяж (спинной мозг), хорда и жабер-
ные щели (в том числе и у взрослых животных). Хотя на головном конце отсут-
ствует расширение спинного мозга, у ланцетника развились некоторые, хотя и
рудиментарные, органы чувств.
Два подтипа черепных включают всех известных позвоночных, чьи веро-
ятные эволюционные взаимосвязи показаны на рис. 4.4. Подтип Agnatha
представлен в основном ископаемыми формами, у которых отсутствуют обе
челюсти и парные конечности. Единственные современные представители
подтипа — миноги и миксины — относятся к классу Cyclostomata (круглоро-
тые). Подтип Gnathostomata, характеризующийся наличием челюстей и пар-
ных конечностей, подразделяется на два надкласса — Pisces (рыбы), куда
входят все рыбы, и Tetrapoda (четвероногие). Рыбы, в свою очередь, подраз-
деляются на два больших класса — Chondrichthyes и Osteichthyes. Класс
Chondrichthyes (или Elasmobranchii) включает всех хрящевых рыб с грубой
дентиновой плакоидной чешуей и представлен акулами, катранами, скатами
(в т. ч. ромбовые скаты, включая электрического Torpedo) и пилой-рыбой.
Класс Osteichtyes включает всех костных рыб; чешуя — циклоидная или кте-
ноидная, в зависимости от формы — гладкой или зазубренной, соответствен-
но. По числу видов и разнообразию форм костистые рыбы намного превос-
ходят хрящевых. Вероятно наиболее продвинутым является отряд Teleostei
(костистые рыбы), куда входят сельдь, форель, лосось, карп, угорь, летучая
рыба и т. д.
Надкласс Tetrapoda подразделяется на четыре больших класса: Amphibia
(земноводные), Reptilia (пресмыкающиеся), Aves (птицы) и Mammalia (млеко-
питающие). Как видно из рис. 4.4, земноводные — наиболее ранний из них по
происхождению. Ископаемые формы могут быть прослежены на почти
400 миллионов лет назад — в девонском периоде. Современные земноводные
относятся к трем разным подклассам: Urodela (хвостатые, напр. тритоны, сала-
мандры и т. д.), Anura (бесхвостые, в т.ч. лягушки и жабы) и Apoda (безногие
слепые закапывающиеся тропические формы).
Для класса Reptilia характерно развитие из яиц, имеющих, в отличие от
земноводных, амниотическую оболочку, которая позволила им адаптировать-
ся к условиям существования на суше. Таким образом, пресмыкающиеся —
это первые полностью наземные позвоночные. Зоологи иногда используют
собирательный термин «амниоты» для всех полностью наземных позвоночных
(пресмыкающихся, птиц и млекопитающих), в отличие от «анамний» (земно-
водные и рыбы), у которых амниотическая оболочка отсутствует, а потому они
4. Эволюция и классификация организмов 69
по необходимости часть жизененного цикла или весь целиком вынуждены
проводить в воде. Известно огромное количество ископаемых отрядов репти-
лий (включая, конечно, ужасных ящеров — динозавров), но только четыре
ныне существующих: Crocodilia (крокодилы), Chelonia (водные и сухопутные
черепахи), Rhynchocephalia (клювоголовые, единственный современный
представитель — гаттерия Spenodon) и Squamata (чешуйчатые — ящерицы, ва-
раны, игуаны, змеи и т. д.). У змей, которые, как предполагается, утратили ко-
нечности за долгий период роющего образа жизни, развились (в некоторых
случаях — повторно) очень интересные органы чувств.
Класс Aves (птицы) иногда рассматривают как «оперенных рептилий». Су-
ществует более 9 000 видов, распространенных во всех частях света и практи-
чески в любых условиях обитания. Как и следовало бы ожидать исходя из ак-
тивного образа жизни этих животных, у птиц развились наиболее сложные
сенсорные системы, известные на этой планете.
Класс Mammalia (млекопитающие) обычно подразделяется на три под-
класса: Prototheria (Monotremata, однопроходные), Allotheria (включает только
вымершие виды), Metatheria (Marsupaliana, сумчатые) и Eutheria (Placentalia,
плацентарные). Подкласс однопроходных включает яйцекладущих млекопи-
тающих Австралазии — утконоса (Ornithorhynchus) и ехидну (Tachyglossus).
Подкласс Metatheria опять-таки в основном представлен австралазийскими
видами, но имеются и северо-, и южноамериканские, он выделяются наличи-
ем наружной сумки для вынашивания детенышей, которые рождаются очень
незрелыми. Подкласс Eutheria включает все современные отряды млекопи-
тающих— Insectivora (насекомоядные — ежи, землеройки, кроты, тупайи,
и т. д.), Chiroptera (рукокрылые — летучие мыши), Rodentia (грызуны — кры-
сы, мыши и т. д.), Lagomorpha (зайцеобразные — кролики, зайцы), Carnivora
(хищные — кошачьи, собачьи, медведи, куньи, тюлени), Cetacea (китообраз-
ные — киты, дельфины, морские свиньи), Perrisodactyla (непарнокопытные —
лошади и т. д.), Artiodactyla (парнокопытные — коровы, свиньи и т. д.) и, ко-
нечно, Primates (приматы), к которым принадлежим и мы, наряду с лемурами,
долгопятами, галаго, обезьянами Старого и Нового Света и человекообразны-
ми обезьянами.
4.7. Заключение
Можно ли на основании такого короткого обзора сделать какие-либо обоб-
щения? Многие полагают, что все многообразие жизненных форм, населяю-
щих ныне нашу планету — результат чистой случайности, и все они могли
быть совершенно иными и по форме, и по функции. Стивен Гулд популяри-
зовал эту точку зрения своими оценками странных и удивительных ископае-
мых, обнаруженных в отложениях Бэрджес Шале и датируемых 525 миллио-
нами лет назад. Другие, особенно Конвей Моррис, опираются на тот факт,
что только небольшое число крупных групп животных успешно дожило до на-
ших дней. Наш обзор показывает, что три таксона, три плана строения до-
стигли потрясающих успехов, это моллюски, членистоногие (особенно насе-
комые) и хордовые. Несомненно, что не было какого-то предопределения
в том, будут ли способны выжить те или иные виды, включая самый для нас
70 Часть I. Предварительные сведения
интересный — Homo sapiens\ однако постоянное действие фактора естествен-
ного отбора выделило те основные планы строения организмов, которые
обеспечили развитие форм, способных просуществовать миллиард лет. На
следующих страницах, где предпочтение из-за нашего эгоцентризма отдано
сенсорным системам млекопитающих, в том числе человека, все же часто бу-
дет проводиться сравнение с моллюсками и членистоногими — двумя други-
ми типами, демонстрирующими решения проблем выживания в течение
1 миллиарда лет на Земле.
МАТЕРИАЛЫ К ЧАСТИ I
Для самоподготовки
Нижеследующие вопросы подготовлены для того, чтобы помочь вам оценить
собственное понимание материалов Части I, а также обратить ваше внимание
на наиболее существенные аспекты изложенного материала.
Глава 1: Основы
1.1. Почему считают, что ДНК не была исходным носителем генетической инфор-
мации?
1.2. Укажите приблизительные даты возникновения наиболее ранних прокариот и
эукариот.
1.3. Чем отличаются первичная, вторичная и третичная структуры белков?
1.4. Объясните, что обозначает термин «аллостерия» и «аллостерический эффек-
тор». Что такое кооперативная аллостерия?
1.5. Что такое протеинкиназы и каким образом они участвуют в аллостерических
переходах? Какую роль играют протеин-фосфатазы?
1.6. Почему существенно, что мембранные липиды являются амфипатическими
молекулами?
1.7. Гидрофильна или гидрофобна внутриклеточная сторона биологической мемб-
раны? Почему?
1.8. Сказано, что биомембраны — это «жидкостно-мозаичная» структура. Что это
означает?
1.9. Какие структурные свойства удерживают белки в биомембране? Опишите
важный структурный элемент, присутствующий у многих рецепторных бел-
ков.
1.10. На какое расстояние и с какой скоростью могут перемещаться белки в био-
мембранах? Почему эта подвижность важна для сенсорных клеток?
1.11. На что похожа трехмерная конформация семидоменного рецептора?
1.12. Каким образом протеинкиназы участвуют в десенситизации рецепторных бел-
KUD?
1.13. Что обозначает буква «G» в термине G-белок?
1.14. Объясните, как G-белки могут быть связаны с таймерными функциями?
1.15. Назовите три типа мембранно-связанных эффекторов?
1.16. Назовите три вторичных мессенджера.
1.17. В нескольких фразах и (желательно) схемой объясните, как основанная на G-
белках система вторичных мессенджеров обеспечивает работу гибкого меха-
низма «стимул — ответ».
1.18. Что означают аббревиатуры ПЗК и МК (LGIC и VGIC)? Приведите примеры.
1.19. Нарисуйте схему конформационного цикла №+-канала.
1.20. Одинаков ли порог открывания №+-каналов, локализованных в одном участ-
ке мембраны? Почему ответ на этот вопрос важен?
Глава 2: Мембрана и потенциал действия
2.1. Какова приблизительная величина Кт? Почему головоногие моллюски так
важны для ее оценки?
2.2. Напишите уравнение Нернста. Определите Vb когда [1+]о = 150 мМ, а [1+], =
15 мМ; когда [1+]о = 5,5 мМ, a [I+]j = 150 мМ; и когда [Г]о = 125 мМ, а [Г^ =
9 мМ.
72 Часть I. Предварительные сведения
2.3. Используя уравнение Голдмана и значения наружных и вйутриклеточных кон-
центраций К+, С1“ и Na+ при Рк, РС1 и PNa, данных в тексте, определите Ит на
мембране гигантского аксона кальмара и мотонейрона кошки.
2.4. Используя те же значения К+, СГ и Na+, что в предыдущем вопросе, найдите зна-
чение Ит мотонейрона кошки, если 1) PNa = 5 х 10“7 см/с; 2) Рк = 5 х 10-6 см/с.
2.5. Используя упрощенную форму уравнения Голдмана (уравнение 2.4) и концен-
трации К+ и Na+, данные в табл. 2.1, определите Кт гигантского аксона каль-
мара.
2.6. С помощью рисунка объясните природу электротонического (кабельного)
проведения.
2.7. В чем разница между рецепторным и генераторным потенциалом?
2.8. Почему, хотя рецепторный потенциал линейно зависит от интенсивности сти-
мула, сенсорные и нейросенсорные клетки реагируют только когда достигает-
ся определенный критический порог стимула (Sc)?
2.9. Определите, что такое «сенсорная адаптация». В чем состоит разница между
«быстрой» и «медленной» адаптацией?
2.10. Объясните роль, которую кальций-зависимые калиевые каналы (Кса-каналы)
играют в некоторых случаях сенсорной адаптации.
2.11. Какое событие происходит первым при развитии потенциала действия?
2.12. Опишите последовательность изменений, которые претерпевает Na+-канал
в ходе потенциала действия.
2.13. Какой ионный механизм обеспечивает реполяризацию мембраны после пика
потенциала действия?
2.14. С чем связана гиперполяризация после потенциала действия и как долго она
длится?
2.15. Какова приблизительная длительность рефрактерного периода? Каково его
физиологическое значение? Если рефрактерный период нервного волокна со-
ставляет 3 мс, то, какова максимальная частота импульсов в нем?
2.16. Как скорость проведения связана с диаметром волокна и почему? Каким обра-
зом объясняется наличие гигантских волокон у многих беспозвоночных?
2.17. Какой альтернативный гигантским волокнам вариант развился у позвоноч-
ных? Какие преимущества дает эта альтернатива по сравнению с гигантскими
волокнами беспозвоночных?
2.18. Влияет ли диаметр волокна на скорость проведения в волокнах позвоночных?
Назовите еще один фактор, влияющий на скорость проведения импульса.
Глава 3: Общие черты сенсорных систем
3.1. В чем различия между двумя основными подходами к изучению сенсорных си-
стем? Приведите примеры.
3.2. Опишите два пути классификации сенсорных рецепторов и объясните, что оз-
начает термин «адекватный стимул».
3.3. Объясните, почему эксперименты по хирургическому переключению сенсор-
ных волокон у эмбрионов хорька ставит интригующие вопросы о взаимоотно-
шениях ощущений в мозге.
3.4. Определите, что такое «сенсорный порог». Меняется ли порог данной модаль-
ности стимула или остается постоянным?
3.5. Скажите, что такое минимально различимое отличие стимулов (МРО). Как
оно связано с интенсивностью стимула?
3.6. Приведите «психофизический закон» Стивенса и примеры стимулов с различ-
ными характеристиками.
3.7. Что такое рецептивные поля (РП)? Объясните, почему существует взаимосвязь
между чувствительностью и локализацией.
3.8. Что понимают под термином «неизоморфный»? Почему сенсорные карты
главным образом не изоморфны?
Материалы к части I 73
3.9. Что Йоханнес Мюллер имел в виду под «специфической нервной энергией»?
Какой термин используется в настоящее время для концепции Мюллера?
3.10. Заглянув также в гл. 22 (раздел 22.4), обдумайте, почему сенсорная информа-
ция, доставленная в мозг, должна быть организована в карту.
3.11. Что такое «проблема связывания»?
3.12. В чем разница между «иерархической» и «параллельной» организацией? Как
построены сенсорные системы — на основе первой, второй или обеих вмес-
те?
3.13. Что такое «триггерный стимул»? Приведите пример для зрительной системы.
3.14. Что такое «клетка, распознающая лицо бабушки»? Верите ли вы в то, что такое
возможно?
3.15. Объясните концепцию эволюционной «гонки вооружений».
Глава 4: Эволюция и классификация организмов
4.1 Перечислите три существенных различия между прокариотами и эукариотами.
Когда приблизительно эукариоты произошли от прокариот?
4.2 Назовите три царства, на которые подразделяются многоклеточные.
4.3. Опишите три различия между первичноротыми и вторичноротыми. Почему
зоологи считают, что это подразделение знаменует собой очень ранние эволю-
ционные различия этих групп животных?
4.4. Что означает термин «филогения»? Объясните, что означает фраза «в онтоге-
незе воспроизводится филогенез».
4.5. Почему подразделение царства животных на позвоночных и беспозвоночных
рассматривается как антропоцентрическое?
4.6. Используя схему классификации в разделе 4.6, классифицируйте 1) макака,
2) домашнюю кошку, 3) дрозофилу, 4) кальмара. Какое из животных согласно
это схеме наиболее близко к человеку?
4.7. Какой из типов включает животных, напоминающих формы, из которых раз-
вились все современные животные?
4.8. К какому типу относится Caenorhabditis elegansl Почему это животное так важ-
но для молекулярной нейробиологии?
4.9. К какому типу относятся улитки, сердцевидки, кальмары и осьминоги?
4.10. Какая особенность членистоногих исключает их рост до больших разме-
ров?
4.11. Назовите два подтипа, на которые подразделяются членистоногие.
4.12. Влияет ли твердый наружный скелет на природу органов чувств, которые раз-
вились у членистоногих? Если да, то как?
4.13. Сколько приблизительно существует видов хордовых? Сколько — моллюсков?
Сколько — членистоногих?
4.14. К какому подтипу относится ланцетник Amphioxus?
4.15. В чем состоят отличия между Chondrichthyes (Elasmobranchii) и Osteichthyes?
Приведите примеры.
4.16. Какие классы позвоночных относятся к анамниям, а какие — к амниотам?
4.17. По каким эволюционным причинам змеи утратили конечности?
4.18. К какому отряду млекопитающих относятся летучие мыши? К какому — мы-
ши и крысы? К какому — макаки и люди?
4.19. Думаете ли вы, что формы животных могли бы существенно отличаться от тех,
которые мы видим сегодня, или вы полагаете, что естественный отбор привел
к ограничению числа морфологий до небольшого количества оптимально ор-
ганизованных? Приведите аргументы.
74 Часть I. Предварительные сведения
Замечания и библиография
Глава 1: Основы
Вступительная часть первого раздела этой главы опирается на «эгоистичные
гены», для популяризации которых так много сделал Ричард Докинс (Richard
Dawkins, 1976, 1995). Детали структуры и функционирования рибозимов даны
у Narliker & Herschlag (1997) и Herschlag (1998). Исчерпывающие сведения по
молекулярной биологии можно найти, например, в Alberts et al. (1994), а об-
суждение структуры мембраны, рецепторных белков и каналов — в Smith
(1996). Вездесущность семидоменных рецепторов подчеркнута Shepherd et al.
(1998), а G-белок связанные сигнальные системы обсуждаются у Birnbaumer
et al. (1990) и Bourne, Sanders & McCormick (1991), тогда как общий обзор
сделан Smith (1995). Shepherd et al. (1998) описывают проект «Мозг челове-
ка», начатый в 1990-е годы и финансируемый рядом агентств США. Проект,
использующий интернет-технологии высокого уровня, направлен на накоп-
ление огромного количества информации по нейрологическим наукам на
различных уровнях — от молекулярного до целого мозга — полученного в ис-
следовательских лабораториях. В этом отношении проект сравним, хотя он и
намного более сложен, с чрезвычайно успешным проектом «Геном человека»
в молекулярной биологии.
Alberts, В., D. Bray, J. Lewis, М. Raff and J. D. Watson, 1994, Molecular Biology of the Cell
(3rd edn), New York: Garland.
Birnbaumer, L. et al., 1990, «Roles of G-proteins in coupling receptors to ion channels», in
Transmembrane Signalling: Intracellular messengers and implications for drug development,
S. R. Nahorski, ed., Chichester: Wiley.
Bourne, H. R., D. A. Sanders and E. McCoemick, 1991, «The GTPase superfamily: con-
served structure and molecular mechanism», Nature, 349, 117-127.
Dawkins, R., 1976, The Selfich Gene, Oxford: Oxford University Press.
Dawkins, R., 1995, River out of Eden, London: Weidenfeld and Nicolson.
Herschlag, D., 1998, «Rybozyme crevices and catalysis», Nature, 395, 548—549.
Narliker, G. J. and D. Herschlag, 1997, «Mechanistic aspects of enzyme catalysis: Lessons
from comparison of RNA and protein enzymes», Annual Review of Biochemistry, 66,
14-59.
Shepherd et al., 1998, «The Human Brain Project: neuroinformatics tools for integrating,
searching and modelling multidisciplinary neuroscientific data», Trends in Neuroscien-
ces, 21, 460-468.
Smith, C. U. M., 1995, «Membrane signalling systems», in Biomembranes: General princi-
ples, A. G. Lee, ed., Greenwich, CT., JAI Press, vol. 1, pp. 245—270.
Smith, C. U. M., 1996, Elements of Molecular Neurobiology (2nd edn), Chichester: Wiley.
Глава 2: Мембрана и потенциал действия
Данные по потенциалу покоя и потенциалу действия на возбудимых мембра-
нах можно найти во всех учебниках по нейрофизиологии. Aidley (1989) дает
более продвинутый подход к вопросу, a Hille (1992) — вероятно, лучший обзор
данных по биофизике и молекулярной биологии ионных каналов. Классичес-
кое изложение данных по электротоническому проведению дано Rail (1977), а
менее математизированное введение в материал можно найти у Shepherd
(1998) и Smith (1996).
Материалы к части I 75
Aidley, D. J., 1989, The Physiology of Excitable Cells (3rd edn), Cambridge: Cambridge
University Press.
Hille, B., 1992, Ionic Channels in Excitable Membranes (2nd edn), Sunderland, MA:
Sinauer.
Rail, W., 1977, «Core conductor theory and cable properties of neurons», in Handbook of
Neurophysiology, Bethesda, MD, American Physiological Society, vol. 1, section 1,
pp. 39-97.
Shepherd, G. M., 1998, The Synaptic Organisation of the Brain (3rd edn), Oxford: Oxford
University Press.
Smith, C. U. M., 1996, Elements of Molecular Neurobiology (2nd edn), Chichester: Wiley.
Глава 3: Общие черты сенсорных систем
Пионерские работы по хирургическому «переключению» сенсорных волокон у
зародышей млекопитающих описаны Metin & Frost (1989) и Sur, Garraghty &
Roe (1989). Stevens (1957) опубликовал классическую статью по соотношениям
физической и субъективной интенсивности. Материалы по «проблеме связы-
вания» можно найти у Crick (1994). Felleman & van Essen (1991) дали плодо-
творный подход к «многослойной иерархии» участков зрительной коры, где
реализуется параллельная и последовательная обработка информации, кото-
рая передается, как «вверх», так и «вниз». Jacob (1977) обсуждает разнообразие
эволюционного процесса.
Crick, F. Н. С., 1994, The Astonishing Hypothesis, London: Simon and Schuster.
Felleman, D. J. and D. van Essen, 1991, «Distributed hierarchical processing in the primate
cerebral cortex», Cerebral Cortex, 1, 1—47.
Hofstadter, D.R., 1979, Godel, Escher, Bach: an eternal golden braid, Hassocks: Harvester
Press.
Jacob, F., 1977, «Evolution and tinkering», Science, 196, 1161—1166.
Metin, C. and D. Frost, 1989, «Visual responses of neurons in somatosensory cortex of ham-
sters with experimentally-induced retinal projections to somatosensory thalamus»,
PNAS(USA),Z6, 357-361.
Stevens, S. S., 1957, «On the psychophysical law», Psychological Review, 64, 153—181.
Sur, M., P. E. Garraghty and A. W. Roe, 1989, «Experimentally-induced visual projections
into auditory thalamus and cortex», Science, 242, 1437—1441.
Глава 4: Эволюция и классификация организмов
Две публикации Hennig (1950, 1966) — это основы метода зоологической клас-
сификации, известной как кладистическая. Студенты, интересующиеся аль-
тернативными системами классификации, могут познакомиться с более об-
щепринятой эволюционной классификацией, почитав Nelson & Platnick
(1984) и книгу Wiley (1981). Различие между Archaebacteria и Eubacteria впер-
вые проведено Woese & Fox(1977), см. также Woese (1981). Archaebacteria в на-
стоящее время по большей части связаны с экстремальными условиями суще-
ствования: горячие источники, анаэробные, обогащенные метаном, условия и
т.д. Возможно, это в какой-то степени схоже с условиями, которые существо-
вали на ранних этапах истории Земли. То есть Archaebacteria могут рассматри-
ваться как реликтовая группа наиболее древних обитателей земного шара. Они
отличаются рядом черт своей биохимии и молекулярной биологии от своих
потомков — Eubacteria. При этом они более тесно связаны с эукариотами, чем
76 Часть I. Предварительные сведения
с Eubacteria. Молекулярно-биологические сведения о взаимосвязях основных
групп организмов приведены у Doolittle etal. (1996), и Gu (1998), a Balavoine &
Adoutte (1998) обсуждают данные, служащие основанием для фундаменталь-
ных подразделений среди первичноротых. Затруднения и неясности молеку-
лярных методов определения филогении в сжатой форме описаны Lake &
Moore (1998). Конвэй Моррис (Conway Morris, 1998b) обсуждает недавние па-
леонтологические работы по происхождению многоклеточных и приводит ан-
нотированную библиографию. Исчерпывающий, изложенный в доступной
форме, обзор классификации животного мира можно найти у Margulis &
Schwartz (1998). Стивен Гулд (Stephen Gould, 1989) дает впечатляющий анализ
смысла открытия удивительных докембрийских отложений Бэрджес Шале
высоко в канадских Скалистых горах, а Конвей Моррис (Conway Morris,
1998b) — в равной мере интригующие описания этих и других отложений,
и интересные выводы.
Balavoine, G. and A. Adoutte, 1998, «One or three Cambrian radiations?», Science, 280,
397-398.
Caroll, R.L., 1988, Vertebrate Paleontology and Evolution, New York: Freeman.
Doolittle, R.F. etal., 1996, «Determining the divergence times of the major kingdoms of liv-
ing organisms with a protein clock», Science, 271,470—477.
Gould, S. J., 1989, Wonderful Life, New York: Norton.
Gu, X., 1998, «Early metazoan divergence was about 830 million years ago», Journal of
Molecular Evolution, 47, 369—371.
Hennig, W., 1950, Grundzuge einer Theorie der phylogentischen Systematik, Berlin:
Deutscher Zentralverlag.
Hennig, W., 1966. Phylogenetic Systematics, Urbana: University of Illinois Press.
Lake, J. A. and J. E. Moore, 1998, «Phylogenetic analysis and comparative genomics», in
Trends in Bioinformatics, pp. 22—24, Cambridge: Elsevier.
Margulis, L. and К. V. Schwartz, 1998, The Five Kingdoms: An Illustrated Guide to the Phyla
of Life on the Earth (3 rd edn), San Francisco: W. H. Freeman.
Morris, S. C., 1998a, The Crucible of Creation, Oxford: Oxford University Press.
Morris, S. C., 1998b, «Metazoan phylogenies: falling into place or falling to pieces?
A palaeontological perspective», Current Opinion in Genetics and Development, 8,
662-667.
Nelson, G. and N. Platnick, 1984, «Systematics and Evolution», in Beyond Neo-Darwinism,
Mae-Wan Ho and Peter Saunders, eds, London: Academic Press.
Wiley, E. O., 1981, Phylogenetics: The theory and practice of phylogenetic systems, New York:
Wiley.
Woese, C. R. andG. E. Fox, 1977, «The concept of cellular evolution», Journal of Molecular
Evolution, 10, 16.
Woese, C. R., 1981, «Archaebacteria», Scientific American 244(6), 98—122.
Young, J. Z., 1962, Life of Vertebrates (2nd edn), Oxford: Clarendon Press.
Часть II
МЕХАНОЧУВСТВИТЕЛЬНОСТЬ
«Без осязания не может существовать животное... утрата даже
одного этого чувства несет смерть.»
Аристотель (de Anima: 435b4-5)
Из всех чувств осязание часто рассматривается как наиболее важное. Аристо-
тель, величайший из биологов, подметил это еще две с половиной тысячи лет
назад — в греческую античность. Эпиграф ясно показывает, что он считал ося-
зание основным чувством у всех животных. Там же он утверждает, что «это
единственное чувство, которое есть у всех животных без исключения» (de
Anima: 414ЬЗ), и такое мнение мало изменилось за прошедшие эпохи. Мы уви-
дим в конце этой книги в гл. 23, каким важным его считали многие философы-
эмпирики. Можно быть слепым или глухим, а при трагических обстоятельст-
вах — слепоглухим, но сохраненное осязание, как показывает знаменитый
пример Элен Келлер, позволяет существовать.
Тактильное чувство — ответ на механическое смещение, как мы увидим в
данной части книги, эволюционно очень древнее. Оно может быть прослеже-
но до самых ранних прокариот, которым, как и все современным клеткам, тре-
бовалась возможность контролировать целостность плазматических мембран.
Раздувание вследствие действия осмотических сил требовало противодейст-
вия, необходимо было также детектировать увеличение размера клетки перед
делением. Мы увидим в главах этой части, что механосенсоры организованы
вокруг каналов, чувствительных к растяжению мембраны. Мы увидим также,
что такая организация означает, что определение состояния мембраны проис-
ходит немедленно, а ответ наступает быстро. В данном случае нет необходимо-
сти в G-белковой мембранной биохимии, как в случае рецепторов других ор-
ганов чувств.
Функции механорецепторов в животном царстве многочисленны. Это ос-
нова не только определения натяжения и прикосновения, но и важный эле-
мент ощущения вибрации, ускорения, звука, движений тела и его положения
и т. д. Вероятно, все эти функции основываются на чувствительных к растяже-
нию каналах плазматической мембраны. Единственный такой канал, изучен-
ный к настоящему времени в молекулярных деталях, экспрессируется у Е. coli
и С. elegans. Поэтому мы и начнем гл. 5 с рассмотрения детектора растяжения
Е. coli. Здесь же удобно обсудить и некоторые рецепторы растяжения, детекти-
рующие изменения объема и растяжение в клетках эукариот. Специально мы
рассмотрим также клетки гипоталамуса млекопитающих, которые чувствуют и
контролируют осмолярность крови.
78 Часть II. Механочувствительность
В гл. 6 мы перейдем к сложным системам, в которых участвуют рецепторы
растяжения у высших животных. Здесь они участвуют в т. н. кинестезии. Все
животные нуждаются в информации об относительном расположении частей
собственного тела. У животных с неэластичным, жестким наружным скелетом
(у членистоногих) такая информация может быть получена в основном (но не
полностью) путем детектирования движений и положения различных сегмен-
тов и члеников их тел. У мягкотелых форм, опирающихся на внутренний ске-
лет, таких как позвоночные, кинестезия зависит от восприятия движений и
напряжения в мышцах и связках тела.
Далее, в гл. 7 мы перейдем к собственно чувству осязания и рассмотрим
хорошо изученный осязательный детектор С. elegans перед тем, как сделать
обзор некоторых из множества осязательных рецепторов животного царства.
Мы рассмотрим два основных случая — насекомых и позвоночных. Значи-
тельные отличия организации животных этих классов определяют и то, что
развившиеся у них органы тактильной чувствительности тоже радикально
различаются. Насекомые с их твердым наружным скелетом широко исполь-
зуют осязательные волоски и хордотональные органы. Мы проследим разви-
тие этих типов тактильных рецепторов вплоть до наиболее высокого уровня
их организации, позволяющей чувствовать колебания воздуха. Иными сло-
вами, в завершение, мы рассмотрим «уши» насекомых, в особенности бабо-
чек, которые развили способность обнаруживать ультразвуковой сонар хищ-
ных летучих мышей. Что касается позвоночных, мы сосредоточим внимание
на нескольких типах тактильных рецепторов кожи млекопитающих. Кожа,
в отличие от твердого карапакса членистоногих, может рассматриваться
как сенсорная поверхность в собственном значении этого понятия, и
мы рассмотрим, как полученная информация передается в соматосенсорную
кору.
В гл. 8 мы обсудим волосковые клетки позвоночных. Это — одни из самых
удивительных сенсорных клеток, которые сформировались в царстве живот-
ных. Мы рассмотрим пути, которыми они развивались от эхолокационных ус-
тройств органа боковой линии рыб до детекторов равновесия в мембранном
лабиринте и до улитки уха млекопитающих. Нетрудно доказать, что в этом
удивительном органе механорецепция достигает высшего уровня развития в
эволюции. Мы рассмотрим, как устроена улитка насекомоядных летучих мы-
шей, способная обнаружить тончайшее эхо от двигающихся насекомых и, та-
ким образом, продолжим удивительную историю о гонке вооружений между
бабочками и мелкими рукокрылыми, начатую в гл. 7.
Мы завершим часть II рассмотрением в гл. 9 того, что происходит с инфор-
мацией, полученной ухом млекопитающего, когда она достигает центральной
нервной системы. Рассмотрим пути, которыми импульсация из мембранного
лабиринта обеспечивает поддержание позы и равновесия. Исследуем слуховые
пути к первичной слуховой коре в теменной доле мозга, отмечая, каким обра-
зом выделяется существенная для жизни животного часть сигнала. У насеко-
моядных рукокрылых такой существенной частью является эхо от их соб-
ственного ультразвукового сонара. Таким образом, мы завершим историю, из-
лагаемую в гл. 7 и 8, обсуждением анализа ультразвукового эха в коре мелких
рукокрылых. Для Homo sapiens наиболее существенными характеристиками
Часть II. Механочувствительность 79
звуковой информации являются основные звуки речи. Мы рассмотрим приро-
ду этих звуков и то, как они обрабатываются мозгом.
Из этого короткого резюме видно, что механорецепторы многочисленны и
разнообразны. И хотя на молекулярном уровне ионные каналы, чувствитель-
ные к растяжению, могут быть и едины, разнообразие строения этих органов
на макроскопическом уровне подразумевает неисчислимое количество путей
их эволюционного развития. В этом отношении механорецепторы отличаются
от хемо- и фоторецепторов, которые демонстрируют огромное сходство орга-
низации во всем царстве животных. В атмосфере Земли и, в меньшей степе-
ни — в гидросфере (китообразные), механорецепторы развились (а, может
быть, еще и продолжают развиваться) до такой степени, что обеспечили пора-
зительно чувствительные сложнейшие средства коммуникации индивидов.
5. МЕХАНОЧУВСТВИТЕЛЬНОСТЬ
КЛЕТОЧНЫХ МЕМБРАН
Е. coli: формирование гигантских клеток локальная фиксация потен-
циала — два типа механочувствительных каналов (МчкВ и МчкМ) —
выделение МчкВ: структура, биофизика, биологическое значение как
осморецептора — другие бактерии. Осморецепторы млекопитающих:
крупноклеточные нейроны (ККН) гипоталамуса — нейросекреция вазо-
прессина и окситоцина — локальная фиксация потенциала ККН — био-
физика — каналы, инактивируемые растяжением (КИ) — петля обрат-
ной связи, контролирующая потерю воды или ее удержание в ответ на
гипер- или гипотоничность внеклеточной жидкости. Заключение: моле-
кулярная структура большинства механочувствительных каналов до сих
пор неизвестна — возможные общие черты — в центре сложных нейро-
эндокринных взаимодействий
В эволюционном отношении механорецепторы очень стары, они вездесущи.
Они функционируют различными путями, детектируя растяжение и прикос-
новение, даже еле уловимое колебание звуковых волн. Возможно, у них одна и
та же молекулярная основа. В данной главе мы в первую очередь рассмотрим
единственный (к моменту написания этой книги) известный в молекулярно-
биологических деталях механорецептор. Это — механочувствительные каналы
плазматической мембраны Е. coli. Затем мы уделим внимание рецепции растя-
жения в эукариотических клетках высших животных. В обоих случаях рецеп-
торы растяжения чрезвычайно важны в детектировании осмотического разду-
вания клетки, поэтому они используются как осморецепторы гипоталамуса
млекопитающих.
В последующих главах мы увидим, что о молекулярной структуре и функ-
ции хемо- и фоторецепторов известно очень многое. То, что на молекулярном
уровне проанализирован лишь один тип механорецепторов, является главным
образом следствием отсутствия доступных клеток, богатых рецепторами дру-
гих типов. Поэтому-то и не удалось пока применить все более мощные методы
молекулярной биологии к их анализу. Исключение составляют механочувст-
вительные каналы Е. coli.
5.1. Механочувствительные каналы Е. coli
Почему же избрана Е. coli? Просто потому, что она (кишечная палочка) изуче-
на лучше всех других организмов. Ее короткий срок воспроизводства и лег-
кость культивирования дает возможность быстро накопить большое количест-
5. Механочувствительность клеточных мембран 81
во материала. Хорошо изучена и ее генетика, и молекулярная биология.
Е. coli — это грам-отрицательная бактерия, и, как и другие такие бактерии,
имеет сложное устройство клеточной стенки, включающей наружную липопо-
лисахаридную мембрану, белково-гликановую клеточную стенку и внутрен-
нюю плазматическую мембрану. Последняя — это типичный фосфолипидный
бислой, содержащий погруженные в него белки, как это описывалось в гл. 1.
Многие из этих белков клонированы и подверглись молекулярно-биологичес-
кому анализу.
Чтобы определить биофизический ответ мембраны на механическое воз-
действие, необходимо использовать технику локальной фиксации потенциала.
Она описана в Дополнении А, но вкратце сводится к помещению на мембрану
интересующей нас клетки стеклянной микропипетки (диаметр кончика
0,5 мкм). Между кончиком пипетки и плазматической мембраной образуется
очень большое электрическое сопротивление (порядка 10 ГОм), что позволяет
регистрировать токи амплитудой в диапазоне пикоампер. Микропипетка за-
полнена электролитом и соединена с соответствующей электрофизиологичес-
кой аппаратурой, таким образом, чтобы регистрировать токи через мембрану.
Последнюю можно оставить in situ или с помощью аккуратного засасывания в
пипетку вырвать из клетки и изучать в изолированном состоянии. В обоих слу-
чаях можно измерять токи через единичный мембранный канал.
К сожалению для специалистов по фиксации потенциала, большинство
бактерий слишком малы, для того чтобы экспериментаторы могли показать на
них свое искусство. Например, Е. coli всего 2 мкм длиной и 1 мкм в диаметре —
слишком мало, чтобы можно было выполнить фиксацию потенциала совре-
менными методами. Похоже, именно малые размеры прокариот уберегли их
от излишнего любопытства со стороны физиологов. Этих ребят, правда, не так
легко обескуражить, и обходной путь они все-таки нашли. Раз Е. coli слишком
мала для физиологов, значит, она должна стать больше!
К счастью, существуют методы получения «гигантских» клеток Е. coli с по-
мощью антибиотиков. Если, например, культивиров&цъЕ. coli в цефалексине,
аналоге пенициллина, наблюдается репликация ДНК и увеличение размера
клетки без клеточного деления. В результате формируются филаменты, похо-
жие на нитки бус, длиной до 100 мкм, которые при обработке раствором ЭДТА
с лизоцимом преобразуются в сферы диаметром до 10 мкм. Этот размер уже
пригоден для выполнения локальной фиксации потенциала. Единственная
проблема — с наружной или внутренней мембраной образует соединение ми-
кропипетка, однако исследования показали, что это все-таки внутренняя мем-
брана. Вероятно, когда поверхность бактерии засасывают для фиксации по-
тенциала внутрь микропипетки, ее кончик разрушает наружную мембрану и
клеточную стенку и соединяется именно в внутренней мембраной. Схема та-
кого эксперимента показана на рис. 5.1.
Обнаружены два типа механочувствительных каналов (Мчк): каналы высо-
кой проводимости (МчкВ) — с проводимостью порядка 3000 пСи и каналы
малой проводимости (МчкМ) — с проводимостью около 1000 пСи. Если лате-
ральное натяжение фиксируемой мембраны увеличивается вследствие ее вса-
сывания в микропипетку, каналы открываются. То, что высокая и низкая про-
водимости МчкВ и МчкМ — следствие активности двух разных типов каналов,
82 Часть II. Механочувствительность
Культивирование
бактерий
в антибиотике
Обработка ЭДТА
с лизоцимом
В
Рис. 5.1. Локальная фиксация
потенциала Е. coli. (А) клетки
Е. coli. (Б) Культивирование в
среде, содержащей цефалексин,
и формирование длинных фила-
ментов. (В) Обработка филамен-
тов ЭДТА + лизоцимом и округ-
ление их до формирования боль-
шого сферопласта. (Г) Микро-
электрод (диаметр кончика около
0,5 мкм) введен для локальной
фиксации потенциала. Осталь-
ные объяснения — в тексте.
а не разных состояний одного и того же, доказывается с помощью «нокаута»
гена МчкВ. Характеристики проводимости МчкМ определяются именно в та-
ких «нокаутных» бактериях. Натяжение, требуемое для открывания этих ка-
налов, составляет прибл. 0,7 от того, которое необходимо для открывания
МчкВ. Есть данные о присутствии в мембране каналов с еще меньшей прово-
димостью.
Анализ МчкВ был продолжен с помощью сложной техники современной
молекулярной биологии. Метод состоит в включении фрагментов мембраны в
липосомы из фосфолипидов другого вида и повторной локальной фиксации
потенциала, чтобы подтвердить наличие механочувствительных каналов. Бел-
ки из таких липосом могут быть экстрагированы умеренными детергентами и
очищены с помощью биохимических методик, каждая стадия тестируется ме-
тодом фиксации потенциала и так до получения чистой фракции каналов. Вы-
яснилось, что молекулярная масса канала Мг составляет около 17 кДа. N-ko-
нец аминокислотной последовательности 17 кДа-белка был сиквенирован, а
затем в полной к настоящему времени базе данных генома Е. coli был проведен
поиск гена, содержащего соответствующую последовательность нуклеотидов.
5. Механочувствительность клеточных мембран 83
30 мм рт. ст. +20 мВ
с
о
Рис. 5.2. Биофизические характеристики МчкВ. Канальный белок был очищен и
встроен в фосфатидилхолин/фосфатидилсериновые липосомы. В условиях пэтч-
клямпа липосомы подвергались различной степени всасывания в микропипетку
(30 и 40 мм рт. ст.). Отклонения, направленные на рисунке вверх, показывают ион-
ные потоки, несущие деполяризующие токи. Видно, что вероятность открытого со-
стояния канала значительно возрастает при увеличении силы всасывания. Из
Р. Blount etal., 1996, «Membrane topology and multimeric structure of a mechanosensitive
channel protein of Escherichia coli», EMBO Journal, 15, 4801. Воспроизводится с разре-
шения Европейской Организации по Молекулярной Биологии.
Эта последовательность была обнаружена в гене, функция которого ранее не
была известна. Путем сложных молекулярно-биологических процедур ген был
экспрессирован, а полученный белок встроен в фосфолипидные липосомы,
которые при пэтч-клямпе продемонстрировали наличие механочувствитель-
ных каналов. (
Поскольку МчкВ удалось выделить и встроить в искусственные фосфоли-
пидные липосомы, стало возможным детально изучить биофизические харак-
теристики каналов. Как и большинство других мембранных каналов, они
имеют несколько уровней проведения (рис. 5.2). Существует строгая зависи-
мость вероятности открытого состояния канала от латерального натяжения
мембраны, которая описывается крутой сигмовидной кривой. Данных об их
ионной селективности нет, и это, вместе с большой проводимостью канала,
предполагает наличие широкой водной поры.
Изоляция белка МчкВ и идентификация соответствующего гена позволила
определить молекулярную структуру белка. Это — сравнительно небольшой
белок из 136 аминокислотных остатков, на N-конце которого находится учас-
ток гидрофильных остатков, затем — сильно гидрофобных аминокислот
(19—49), далее — еще один гидрофильный (50—69), второй гидрофобный сег-
мент (72—100) и, наконец, гидрофильная последовательность до С-конца.
Тщательное изучение физико-химических характеристик полипептида пока-
зывает, что два гидрофобных сегмента образуют а-спирали (Ml и М2), прони-
зывающие мембрану. И N-, и С-концы локализованы внутриклеточно, и есть
84 Часть II. Механочувствительность
Рис. 5.3. Структура и мембранная топология МчкВ. (А) Ориентация белка МчкВ
в мембране. Дальнейшие объяснения в тексте. Критичный глютаминовый остаток
в 56-м положении аминокислотной цепи обозначен звездочкой. (Б) 6 субъединиц
МчкВ группируются, образуя компактный цилиндр, пронизывающий мембрану.
Когда мембрана напряжена, в центре цилиндра открывается гидрофильная пора.
Из Sukharev et al., 1997, Annual Review of Physiology, 59,63—657. С разрешения Annual
^Review of Physiology, volume 59, © 1997, Annual Reviews www.annualreviews.org.
основания полагать, что существует короткий спиральный участок (S3) во
внеклеточном сегменте между Ml и М2 (рис. 5.3 А). По аналогии с другими
потенциал-зависимыми каналами, такими, как Ка+-канал, описанный в гл. 1,
было сделано предположение, что участок S3 образует часть белка, выстилаю-
щую канал. Действительно, как мы увидим ниже, накапливаются данные ис-
следований мутагенеза, подтверждающие такое предположение.
Одиночный белок МчкВ не в состоянии сформировать канал с воротными
свойствами, характерными для природного МчкВ. Работоспособный канал
должен быть мультимерным комплексом. Это соответствует структуре многих
других канальных белков (см. гл. 1). Хотя некоторые дебаты на эту тему и про-
должаются, общее мнение в настоящее время сводится к тому, что МчкВ со-
стоит из шести субъединиц, сгруппированных вокруг центрального водного
канала (рис. 5.3 Б). Когда мембрана подвергается механическому напряже-
нию, субъединицы несколько удаляются друг от друга и канал, образуемый
ими, открывается.
Выделение белка МчкВ и его гена mscl открыло возможность генетических
исследований структурно-функциональных взаимоотношений. Этому, есте-
ственно, весьма способствовало глубокое понимание генетики Е. coli, созда-
вавшееся за долгие годы. Исследования продолжаются и сейчас, в момент на-
писания этой книги, но уже сейчас, как упоминалось выше, получены некото-
5. Механочувствительность клеточных мембран 85
рые интересные сведения. Один из аминокислотных остатков в «нижней час-
ти» периплазматической спирали S3 кажется особенно критичным. Это глю-
таминовый остаток в положении 56 (Q56) (см. рис. 5.3 А). Различные замеще-
ния аминокислотных остатков в этом положении значительно увеличивают
время открытого состояния канала при данном значении механического на-
пряжения. Некоторые замены, в частности замена в этом положении глюта-
мина на пролин (принятое у молекулярных биологов обозначение — Q56P),
делает чувствительность канала к напряжению поверхности значительно ни-
же, чем та, которая требуется для его открывания. Замены в других частях це-
пи канального белка, хотя и не имеют таких драматических эффектов, однако
изменяют его функцию интересным и наводящим на размышления образом.
Продолжающиеся и сегодня генетические исследования структурно-функци-
ональных взаимоотношений в конце концов приведет к пониманию организа-
ции механочувствительных каналов во всей полноте их молекулярных и даже
атомарных деталей.
Развились ли сходные каналы в плазматических мембранах других бакте-
рий? Поскольку нуклеотидная последовательность гена mscl определена, стало
возможным изготовить олигонуклеотидные зонды для поиска гомологичных
генов у других бактерий. Существование таких генов показано у многих, как
грам-отрицательных, так и грам-положительных бактерий, например у Нае-
mophilis influenzae и Pseudomonas fluorescens (обе — грам-отрицательные), а так-
же Clostridium perfringens и Staphylococcus aureus (обе — грам-положительные).
Удивительно, но поиск гомологий у эукариотических клеток пока принес
только отрицательные результаты.
Наконец, какова же роль, которую играют в жизни бактерий механочувст-
вительные каналы? Может быть, это покажется удивительным, но пока обще-
принятого ответа на этот вопрос не существует. Наиболее вероятно, что эти
каналы связаны с осморегуляцией. При воздействии пресной воды, например
при дождях, бактерии испытывают значительное осмотическое напряжение.
Молекулы воды движутся по градиенту концентрации внутрь клетки, и возни-
кающее вследствие этого разбухание улавливается механочувствительными
каналами в плазматической мембране. Возможно, за счет потока водораство-
римых соединений (за исключением макромолекул) наружу каналы противо-
действуют осмотическому напряжению, уменьшая градиент концентраций.
Помимо функции, связанной с контролем осмотического разбухания, механо-
рецепторы, как можно полагать, участвуют и в детектировании увеличения
объема клетки вследствие роста и, таким образом, опосредуют включение ме-
ханизма клеточного деления.
5.2. Детектирование осмотического разбухания
гипоталамическими клетками млекопитающих
Осмотический стресс для эукариотических клеток животных не менее значим,
чем для прокариотических одноклеточных. Большинство животных пойкило-
осмотичны: осмолярность их внутренней среды быстро приходит в равновесие
с осмолярностью внешней. Лишь немногие животные, среди которых и мле-
86 Часть II. Механочувствительность
копитающие, способны контролировать осмолярность своей внутренней сре-
ды и именуются «гомео-осмотичными». Механизм рефлекса, осуществляю-
щего такой контроль, зависит от изменений осмолярности внутренней среды,
т. е., в конечном счете от состояния внеклеточной жидкости (ВКЖ). Это де-
тектирование осуществляется рецепторами растяжения в мембранах осморе-
цепторных клеток, которые поэтому могут рассматриваться как энторомехано-
рецепторы.
*В организмах млекопитающих осморецепторы принимают форму больших
(крупноклеточных) нейронов, локализованных в ядрах передней части гипота-
ламуса. Наиболее важными из этих ядер являются паравентрикулярное ядро
(OVLT) в передней оконечности гипоталамуса и супраоптическое ядро, распо-
ложенное непосредственно над зрительным перекрестом (рис. 5.4). Крупно-
клеточные нейроны (ККН) паравентрикулярного ядра посылают часть своих
аксонов в супраоптическое ядро, а остальные — прямо вниз в инфундибуляр-
ную ножку задней доли гипофиза. Аксоны ККН супраоптического ядра так-
же проецируются через инфундибулярную ножку в заднюю долю гипофиза.
В обоих случаях аксоны дают сильно ветвящиеся окончания, содержащие
нейросекрет. Это — главным образом небольшие пептиды, в данном контекс-
те наиболее интересны пептидные гормоны, содержащие по 9 аминокислот-
ных остатков — вазопрессин и окситоцин. Оба гормона выделяются в гипофи-
зарные кровеносные сосуды, откуда выносятся в систему кровообращения.
У человека окситоцин вызывает сокращение матки и секрецию молока в пе-
риод лактации, однако на крысах (на которых была возможность проводить
эксперименты) оказывает натриуретическое (т. е. вызывает экскрецию Na+)
действие. Вазопрессин, как у человека, так и у крыс является антидиуретиком
(т. е. удерживает воду).
Капилляры супраоптического ядра чрезвычайно сильно развиты. Деталь-
ное количественное микроскопическое исследование показало, что плот-
ность капилляров, объем и площадь поверхности в 4 раза выше, чем в некруп-
ноклеточных ядрах гипоталамуса. Капилляры, при их чрезвычайной много-
численности здесь необычно малого размера. Это, с учетом сопротивления
плазмы, предполагает необычно большие возможности для воды и других ма-
лых молекул покинуть кровоток через эндотелий в межклеточное простран-
ство. Паравентрикулярное ядро в большей степени сообщается с плазмой.
Здесь отсутствует гематоэнцефалический барьер, а эндотелиальные стенки
капилляров во многих места продырявлены. Таким образом, ККН в этом яд-
ре в большей степени повержено и изменениям состава плазмы, чем в супра-
оптическом ядре.
Электрофизиологические отведения от ККН обоих гипоталамических ядер
показали, что частота импульсации меняется в зависимости от осмолярности.
При осмолярности 280 мосмоль/кг импульсация достаточно нерегулярна, а ее
частота составляет 2—3 Гц; интраперитонеальная инъекция воды (приводящая
к гипотоничности ВКЖ) уменьшает частоту импульсации, тогда как инъекция
гипертонического раствора ее повышает. Дальнейшие исследования были
проведены на тонких срезах гипоталамуса, содержащих ККН. Здесь оказалось
возможным продемонстрировать, что клетки деполяризуются в ответ на вве-
дение гипертонического раствора. Наконец, микроэлектродные отведения от
5. Механочувствительность клеточных мембран 87
Рис. 5.4. Парасагитальный срез гипоталамуса показывает положение ядер. Гипота-
ламус содержит значительное число ядер, большинство из которых связано с регу-
ляцией внутренней среды. Как указывается в тексте, паравентрикулярное (PV) и
супраоптическое (SO) ядра (показаны на рисунке наиболее темным) в наибольшей
степени связаны с осморегуляцией. А — аркуатное ядро; АС — передняя комиссу-
ра; АН — переднее гипоталамическое ядро; СС — мозолистое тело; DM — дорзо-
медиальное ядро; М — сосцевидное тело; ME — медиальный бугорок; ОС — зри-
тельный перекрест; Р — преоптическое ядро; PH — заднее гипоталамическое ядро;
PV — паравентрикулярное ядро; SC — супрахиазматическое ядро; SO — супра-
оптическое ядро; ТС — серый бугор; VM — вентромедиальное ядро.
изолированных одиночных ККН показали, что гипертонический раствор вы-
зывает деполяризацию мембраны, тогда как гипотонический — гиперполяри-
зацию.
Возможно ли установить связь между электрофизиологическими ответами
и вызванными осмосом изменениями объема ККН? В настоящее время полу-
чены доказательства того, что это действительно так. Чтобы установить корре-
ляцию между изменениями объема ККН со скоростью проведения катионов
через клеточную мембрану в ответ на гипертонический стимул, пришлось вы-
полнить очень тонкую работу. Техника эксперимента включала использование
конфокального лазерного микроскопа и фиксацию потенциала клетки. Пока-
88 Часть II. Механочувствительность
Б
Гипотоничность
(275 миллиосГмолей, моем)
Нормальный уровень
осмотичности (295 моем)
Гипертоничность
(315 моем)
wrrnwa wmmtv
Рис. 5.5. (А) Фиксация потенциала (пэтч-клямп) крупноклеточного нейрона (ККН).
(а) Засасывание мембраны в пипетку заставляет мембрану выгибаться вверх; (б) умень-
шение подсасывания позволяет мембране вернуться к ее нормальному положению;
(в) при дальнейшем уменьшении подсасывающего давления мембрана прогибается
внутрь. Вероятность открывания механочувствительных каналов максимальна в поло-
жении (б). (Б) Ответ инактивируемых натяжением (ИН) каналов на тоничность окру-
жающей среды. В изотонических условиях открыты немногие каналы, что позволяет
некоторому количеству положительно заряженных ионов входить в клетку и обеспечи-
вать нормальный потенциал покоя. Гипотонические растворы вызывают раздувание
клетки, каналы закрываются, а клетка гиперполяризуется. Гипертонический раство-
ры вызывают потерю воды клеткой и соответственно ее сжатие; вероятность открыва-
ния каналов увеличивается и клетка деполяризуется. Дальнейшие объяснения в тексте.
Из С. W. Bourque and S. Н. R. Oliet, 1997. «Osmoreceptors in the central nervous system»,
Annual Review of Physiology, 59, 613. Воспроизводится с разрешения Annual Review of
Physiology, том 59, ©1997, Annual Reviews http://www.annualreviews.org.
зано, что уменьшение размера клетки под действием гипертонического стиму-
ла коррелирует с повышением мембранной проницаемости для Na+ и К+.
Можем ли мы пойти дальше и показать, что этот ответ возникает благода-
ря механочувствительности клеточной мембраны? Альтернативой является
возможность, что наблюдающиеся электрофизиологические ответы просто
являются следствием концентрации раствора в различных осмотических усло-
виях. К счастью, сравнительно легко ответить на этот вопрос. При фиксации
потенциала (пэтч-клямпе) всей поверхности клетки возможно изменить дав-
5. Механочувствительность клеточных мембран 89
ление на клеточную мембрану изменением подсасывающего давления в мик-
ропипетке, поэтому без особого труда было показано, что уменьшение и уве-
личение объема клетки под действием давления в микропипетке коррелирует
ожидаемым образом с наблюдаемой деполяризацией и гиперполяризацией,
соответственно.
При фиксации потенциала на мембране, подсасывающее давление застав-
ляет ее втягиваться дальше внутрь пипетки (т. е. становится вогнутой, если
смотреть изнутри пипетки); при уменьшении подсасывающего давления мем-
брана возвращается к неискривленному (почти горизонтальному) положе-
нию, а потом даже выгибается внутрь (т. е. изнутри пипетки становится вы-
пуклой) (рис. 5.5 А). Показано, что вероятность открывания (Ро) механочувст-
вительных каналов максимальна в среднем положении и снижается, когда
мембрана приобретает выпуклую или вогнутую форму. Это предполагает, что
катионные каналы, как и МчкВ у”Е. coli, чувствительны к латеральным напря-
жениям мембраны (рис. 5.5 Б). Надо, однако, отметить, что механочувстви-
тельные каналы ККН работают противоположным образом по отношению к
тому, как действуют МчкВ Е. coli — вместо того, чтобы открываться в ответ на
натяжение они закрываются. Иными словами — это каналы, инактивируемые
натяжением (ИН).
К моменту написания данного текста молекулярная структура ИН-кана-
лов была неизвестна. Только когда каналы будут выделены и определена их
структура, будет возможен исчерпывающий анализ того, каким образом по-
ток осмотической воды внутрь клетки может вызывать закрывание каналов.
Уже сейчас, однако, мы можем проследить, как снижение осмолярности кро-
ви приводит (через отток воды из ККН) к инактивации и, таким образом,
уменьшению выделения вазопрессина и окситоцина. Эти гормоны действуют
на почечные канальцы таким образом, чтобы уменьшить потерю воды (анти-
диурез) и повысить потерю Na+ (натриурез). Уменьшение секреции этих гор-
монов в результате гипотонической инактивации ККН тем самым приводит
к повышению осмолярности крови до того, как ККН достигнут исходного
размера.
5.3. Заключение
Мы, конечно, еще в самом начале пути к пониманию природы механоч^стви-
тельных каналов клеточной мембраны. Хотя каналы Е. coli в настоящее^ремя
кажутся сильно отличными от тех, которые присутствуют в мембранах гипота-
ламических крупноклеточных нейросекреторных нейронов, возможно, что
продолжение исследований выявит их молекулярное сходство. Как мы увидим
в последующих главах этой книги, очень часто молекулярные механизмы, воз-
никшие в эволюции очень рано, остаются «сердцевиной» чрезвычайно слож-
ных и высокоразвитых сенсорных систем высших животных. Возможно, это
касается и осморецепторов ККН, рассмотренный выше. Молекулярные меха-
низмы мембран этих клеток являются основой для формирования нейроэндо-
кринной петли обратной связи, которая контролирует осмолярность «внут-
ренней среды» млекопитающих.
6. КИНЕСТЕЗИЯ
Кинестезия жизненно важна для активных животных. Кинестезия у членис-
тоногих: рецепторы натяжения у десятиногих членистоногих — сенсиллы
насекомых: структура и функция — типы механочувствительных сен-
силл: волосковые — колоколообразные — сколопидии — хордо4ональ-
ные органы; равновесие в полете — сенсоры положения передней части
груди — жужжальца двукрылых. Кинестезия у млекопитающих: мышеч-
ные веретена: моторные единицы — экстрафузальные и интрафузальные
волокна — волокна с ядерной сумкой и с ядерной цепочкой — коактива-
ция — фузимоторная система; сухожильные органы Гольджи: структу-
ра — функция — рефлексы гомонимных, антагонистических и синер-
гических мышц; соединенные рецепторы; представительство в мозге.
Заключение: кинес/езический контроль поведенческих движений
в сравнении с неуклюжими современными роботами
Активным животным необходимы ощущение и контроль динамического по-
ложения и ориентации тела. Для этого центральной нервной системе необхо-
димо знать (помимо прочего) об уровне сокращения и (или) натяжения много-
численных мышц и положении их сочленений. У животных с твердым внеш-
ним скелетом, таких как членистоногие, это главным образом выполняется
внешними сенсорами, воспринимающими движения и положение соединен-
ных карапакса и конечностей. У мягкотелых форм с внутренним скелетом, та-
ких, как позвоночные, намного больший упор — на контроле моментального
состояния напряжения или расслабления мускулатуры тела. Эта информация
имеет жизненно важное значение для активных позвоночных, а потому и для
нас с вами. Подумайте о чудесах равновесия и контроля собственного тела, ко-
торые демонстрируют фигуристы или гимнасты на Олимпийских играх, не го-
воря уж о нашем повседневном опыте ходьбы, беготни по лестницам и перено-
са подносов с напитками. Контроль позы и поведенческих движений требует
постоянного отслеживания длины мышц, а также напряжения и положения
суставно-связочного аппарата. Это так называемое кинестезическое чувство, в
основном, бессознательно, тем не менее у всех нас есть «представление о соб-
ственном теле», мы всегда не задумываясь знаем положение собственных ко-
нечностей и т. д. Кинестезию иногда называют «шестым чувством», и для
большинства из нас оно не менее важно, чем остальные пять.
Кинестезическая информация, естественно, комбинируется с информаци-
ей органа равновесия мембранного лабиринта и зрительной информацией от
глаз. Эти сенсорные входы будут рассмотрены в гл. 9 и 17, соответственно.
6. Кинестезия 91
У насекомых аналогичная информация также доставляется глазами и сенсор-
ными детекторами у основания крыльев, сочленения головы и груди, а у дву-
крылых — в удивительных жужжальцах. Мы обсудим эти последние механиз-
мы в разделе 6.13.
6.1. Кинестезические механизмы у членистоногих
Членистоногие радикально отличаются от других животных наличием наруж-
ного скелета (экзоскелета), а не внутреннего (эндоскелета). Сей факт имеет
многочисленные последствия. Возможно, самое важное — это то, что членис-
тоногие ограничены в размерах. Большинство из них измеряется миллиметра-
ми и сантиметрами, а не метрами. Это имеет важные последствия и для их ор-
ганов чувств. Твердый хитиновый экзоскелет не годится для развития сенсо-
ров, чувствительных к расслаблению или напряжению поверхности. Тем не
менее, как мы отмечали в гл. 4, членистоногие, особенно насекомые, являют-
ся одним из наиболее успешных типов животных, а большинство из них пред-
ставляют собой высокоактивные формы. Кинестезическое чувство хорошо
развито. Чувствительные окончания развиваются не только в мышцах, но и
в экзоскелетных сочленениях конечностей и в других частях тела.
6.1.1. Рецепторы натяжения в мышцах членистоногих
Большие мышцы брюшка (абдоминальные мышцы) длиннотелых десятино-
гих и сходных ракообразных имеют большое значение в биологии этих живот-
ных. Сокращение вентральных мышц сгибает брюшко, и с помощью тельсона
животное может отпрыгнуть от опасности. То есть правильное управление
этими мышцами имеет важное значение. В частности, для ЦНС важно обла-
дать информацией о состоянии их напряжения и сокращения.
Рецепторы натяжения в больших мышцах дорзального отдела брюшка деся-
тиногих и других ракообразных были изучены в классических экспериментах
Александровича (Alexandrowicz) в 50-е годы XX века. У длиннотелых десятино-
гих ракообразных, таких как омар Homarus gammarus и рак Astacus fluviatilis, па-
ра модифицированных мышечных волокон, известных как рецепторные мышцы
PMj и РМ2, обнаружена с каждой стороны шести сегментов брюшка, а также в
мышце-разгибателе 7-го и 8-го грудных сегментов. Посредине каждого из этих
мышечных волокон сократительный аппарат не развивается, вместо этого
здесь образуется четко выраженный участок, в котором обильно ветвятся денд-
риты больших мультиполярных нейросенсорных нейронов (сравните с мышеч-
ным веретеном млекопитающих, описанным ниже) (рис. 6.1).
Рис. 6.1 показывает, что, помимо больших нейросенсорных клеток, мото-
нейроны иннервируют также РМ. Эти двигательные волокна образуют бифур-
кацию и идут вдоль поверхности РМ, прежде чем сформировать нервно-мы-
шечное соединение с сократимыми участками рецепторных мышц. Кроме то-
го, одно или более волокон идут к нейросенсорной клетке и либо окачивают-
ся на ее перикарионе и дендритах или (и) ветвятся на центральной части РМ.
Внутриклеточные отведения показывают, что в условиях расслабления
сенсорные клетки имеют Ит около 70 мВ. Натяжение деполяризует дендриты,
92 Часть II. Механочувствительность
Рис. 6.1. Схематическое изображение рецепторов натяжения в абдоминальных сегментах
рака Astacus fluviatilis. РМр РМ2 — рецепторные мышцы 1 и 2. СЩ — медленно адапти-
рующийся сенсорный нейрон; СН2 — быстро адаптирующийся сенсорный нейрон;
Ср С2 — сенсорные волокна; Moj —• три тонких двигательных волокна к РМр Мо2 —
толстое двигательное волокно к РМ2; Т — тормозное волокно. Из Handbook of Physiology,
Section 1, Volume 1, Neurophysiology (1959), p. 378. Воспроизводится с разрешения Амери-
канского Физиологического Общества (The American Physiological Society).
причем, чем сильнее натяжение, тем сильнее деполяризация. Когда потенциал
мембраны нейросенсорной клетки, иннервирующей РМ2, достигает уровня
20 мВ (т. е. деполяризация составляет 50 мВ), в аксоне инициируется потенци-
ал действия. Нейросенсорная клетка, иннервирующая РМр требует большей
деполяризации (ок. 60 мВ) для запуска потенциала действия. В случае РМ2 им-
пульсация в нейросенсорной клетке в ответ на постоянное натяжение адапти-
руется в течение приблизительно минуты, т. е. является быстро адаптирующей-
ся. Напротив, нейросенсорная клетка, иннервирующая PMj адаптируется к по-
стоянному натяжению медленно и возвращается к уровню импульсации покоя
за несколько часов (медленно адаптирующиеся клетки). Частота импульсов и в
быстро-, и в медленно адаптирующихся нейросенсорных аксонах прямо связа-
на с интенсивностью натяжения, которому подвергнута соответствующая РМ.
Центральная нервная система в состоянии регулировать чувствительность
этих рецепторов натяжения через дополнительные волокна двух типов — воз-
будительных и тормозных. Стимуляция тормозных волокон подавляет им-
пульсацию в аксонах нейросенсорных клеток в ответ на натяжение. Это про-
исходит потому, что тормозные импульсы уменьшают деполяризацию, вы-
званную в нейросенсорных клетках умеренным натяжением. Возбудительные
6. Кинестезия 93
волокна, подобно у-волокнам, идущим к интрафузальным волокнам млекопи-
тающих (см. ниже), вызывают сокращение РМ вместе с обычными волокнами
мышцы-разгибателя брюшка ракообразных. Если куда более сильные обычные
мышечные волокна не сокращаются достаточно сильно, волокна РМ остаются
в напряженном состоянии и стремятся сократиться сильнее. Затем рефлектор-
ная обратная связь приводит к тому, что обычные волокна продолжают сокра-
щаться до тех пор, пока натяжение не будет устранено, и нейросенсорные
клетки перестанут детектировать натяжение. Эти рефлексы очень схожи с те-
ми, которые наблюдаются у млекопитающих и исследованы намного более
интенсивно, а потому и в большей степени понятны. Системы млекопитаю-
щих детально описаны в разделе 6.2.1.
6.1.2. Сенсиллы насекомых
Из-за малого размера рецепторы натяжения мышц насекомых изучены не так
хорошо, как у ракообразных. Их мультиполярные нейроны связаны с множе-
ством мышц, а их дендритные терминали, ветвящиеся по сарколемме, работа-
ют как детекторы натяжения. Намного больше известны вездесущие у насеко-
мых системы — сенсорные сенсиллы.
Сенсиллы принимают поразительно огромное количество форм (рис. 6.2 А).
К счастью, все они имеют фундаментально схожие планы строения, какую бы
модальность они не распознавали (рис. 6.2 Б). Все они содержат одну или более
нейросенсорную клетку в кутикулярном вместилище. Механочувствительные
сенсиллы могут быть организованы для детектирования движений кутикуляр-
ных сочленений экзоскелета и (или) конечностей или для реакции на относи-
тельные движения структур внутри полости тела. Наиболее известный при-
мер — это волосковые сенсиллы, колоколовидные сенсиллы и хордотональный
орган. Мы все их обсудим последовательно. Рис. 6.2 Б показывает типичную
волосковую сенсиллу. Она состоит из кутикулярного выроста, содержащего
одну или более нейросенсорных клеток, а также несколько обкладочных. Ней-
Рис. 6.2. (А) Различные типы сен-
силл насекомых. А — трихоидная;
Б — щетинковая; В — базикониче-
ская; Г — плакоидная; Д — колоко-
лообразная; Е — целоконическая;
Ж — ампулярная; 3 — стил©кони-
ческая; И — сколопидий. Из
R. Y. Zacharuk, 1985, «Antennae and
sensilla», in Comprehensive Insect
Physiology, Biochemistry and Pharma-
cology, 6, 26. Воспроизведено с раз-
решения R. Y. Zacharuk и Elsevier
Science.
94 Часть II. Механочувствительность
Рис. 6.2. (Б) Типичная сенсилла
насекомых. PC — ресничный си-
нус; ОД — окончание дендрита;
Г — базальная глиальная клетка;
Т — трихогенная или промежу-
точная клетка; Н — нейросенсор-
ная клетка; ВС — внутренний сег-
мент; НТ — наружная или тормо-
генная клетка; НС — наружный
сегмент (ресничка); СС — сен-
силлярный синус. По Kerkut and
Gilbert, 1985, р. 49.
росенсорные клетки, имеющие крупные ядра, посылают длинные дендриты в
кутикулярному выросту. Дендрит дифференцирован на два различающихся
участка. Проксимальный сегмент (внутренний сегмент) сходен с перикарио-
ном, тогда как дистальный (наружный сегмент) представляет собой модифици-
рованную ресничку. Часто его так и называют — ресничкой, тем более, что она
обладает типичной структурой микротрубочек 9 + 2. Эта ультраструктура про-
стирается до кончика реснички, в отличие от веточек дендрита. В этом послед-
нем случае микротрубочки распределяются по веточкам. Сенсорный аксон
выходит из основания клетки и направляется непосредственно в ЦНС в соста-
ве сенсорного нерва.
Многочисленные обкладочные клетки сильно варьируют по форме в зави-
симости от типа сенсиллы, но в целом относятся к трем типам: внутренние об-
кладочные клетки (текогенные клетки), которые гомологичны окончанию ско-
лопилия (см. ниже), промежуточные или трихогенные клетки и самые удален-
ные от центра наружные или тормогенные клетки. Внутренние обкладочные
клетки секретируют обкладку дендрита. Проксимальные окончания промежу-
точных и наружных клеток омываются гемолимфой, содержащейся в гемоце-
ле, а дистальные — посылают множество микроворсинок в сенсиллярный си-
нус. Полагают, что эти клетки играют большую роль в питании и формирова-
нии жидкости в сенсиллярном синусе. Стратегическое положение этих клеток
между гемоцелем и синусом адаптировало эти клетки к их функциям. Помимо
обкладочных клеток здесь также часто представлены промежуточные и базаль-
ные клетки. Последние часто называются глиальными, поскольку они обер-
тывают сенсорные аксоны, изолируя их друг от друга и от гемолимфы.
6. Кинестезия 95
Рис. 6.2. (В) Механочувствительная сенсилла в
гибком сочленении. Наружный сегмент денд-
рита трансформирован в тубулярное тельце, ко-
торое входит в кутикулу волоска. К — кутикула;
В К — вырост кутикулы; СТ — соединение тубу-
лярного тельца с кутикулой волоска; ОД — обо-
лочка дендрита; Э — эпидермальная клетка;
ГС — гибкое сочленение; В — внутренняя об-
кладочная клетка; Н — нейросенсорная клетка;
НО — наружная обкладочная клетка; ПФ —
подвешивающие филаменты; ZA — межклеточ-
ный контакт zonula adherens. По М. S. Blum
(ed.), 1985, Fundamentals of Insect Physiology,
© 1985, John Wiley & Sons, Ltd. Воспроизводит-
ся с разрешения John Wiley & Sons Ltd.
Рис. 6.2. (Г) Однопоровая бимо-
дальная сенсилла. В дополнение
к механочувствительному нейрону
с тубулярным тельцем здесь при-
сутствуют одна или более нейро-
сенсорных клеток, чьи наружные
сегменты простираются до поры на
окончании волоска. НС — наруж-
ный сегмент дендрита; П — пора.
Остальные обозначения — как на
рис. 6.2 В. По М. S. Blum (ed.),
1985, Fundamentals of Insect Physio-
logy, © 1985, John Wiley & Sons, Ltd.
Воспроизводится с разрешения
John Wiley & Sons Ltd.
Сенсиллы отнюдь не только механочувствительны. В других местах этой
книги мы встретимся с вкусовыми, обонятельными, гигроскопическими и
термочувствительными сенсиллами. Модальность чувствительности опреде-
ляется дендритом нейросенсорной клетки, содержащейся в сенсилле. Во мно-
гих случаях в сенсиллах содержится больше, чем одна нейросенсорная клетка,
тогда сенсилла может иметь двойную модальность или даже более. Например,
механочувствительные и хемочувствительные нейросенсорные клетки могут
располагаться внутри одной и той же сенсиллы. Отчасти по этой причине
классификация и терминология, касающиеся сенсилл насекомых, очень
сложны и запутаны.
Тогда как обонятельные и вкусовые сенсиллы по очевидным причинам
развили терминальные поры, многие механочувствительные сенсиллы пор не
имеют (рис. 6.2 В). Механочувствительные волосковые сенсиллы иногда рас-
96 Часть II. Механочувствительность
Рис. 6.3. (А) Рисунок показывает скопле-
ние сенсилл в сочленении второй ноги
таракана Periplaneta americana. Толстая
кутикула эпимеральной пластинки (эп)
утончается до соединительной мембра-
ны, а затем снова утолщается, формируя
кутикулярное окружение тазика (т) —
первого сегмента ноги. Группа сенсилл
образует волосковую пластинку. Из
Pringle, 1938.
Рис. 6.3. (Б) Распределение некоторых
волосковых пластинок муравья-рабочего
Formica polyctena. К — коксальное сочле-
нение; Б — брюшко; Г — передняя часть
груди; С — стебелек (соединение груди и
брюшка); А — сочленение первого и вто-
рого сегментов антенн; АГ — сочленение
головы и антенны; Т — сочленение тазика
и вертлуга (второго сегмента конечности).
Из Markl and Tautz, 1975.
тут прямо из кутикулы, но чаще они присоединены к экзоскелету гибкими со-
членениями (рис. 6.2 В). Иногда они представляют собой длинные волоски
различной кривизны, а иногда — как другая крайность — простое полушарие,
и в этом случае называется колоколовидной сенсиллой.
На рис. 6.2 В волосковая сенсилла показана с единственной нейросенсор-
ной клеткой, чей дендрит присоединен к внутренней поверхности волоска ту-
булярным тельцем. Электронная микроскопия показывает, что оно состоит из
50-100 плотно упакованных микротрубочек, которые чрезвычайно чувстви-
тельны к искривлению, вызываемому движением сенсиллы. Легко показать,
что порог искривления составляет всего 3—5 нм. Изгиб волоска сенсиллы в од-
ну сторону приводит к деполяризации, а в другую — к гиперполяризации. Из-
менения потенциала на мембране происходят за 100 мкс после включения ис-
кривляющего стимула. Это наводит на мысль о том, что они являются следст-
вие открывания или закрывания ионных каналов в мембране. Мы увидим в
гл. 8, что волосковые клетки позвоночных имеют сходную биофизику. Исходя
из этой аналогии, можно сделать вывод о том, что открывание и закрывание
ионных воротных механизмов (вероятно, К+-каналов) активируется наряже-
нием. Подвешивающие филаменты, которые присоединяют основание волос-
ка сенсиллы к кутикуле, вместе с кутикулярной «оправой» и соединением ту-
булярного тельца обеспечивают оптимальную плоскость движения сенсиллы.
Чувствительность к направлению движения очевидно имеет большое значе-
ние в определении того, куда движутся части тела, воздух или субстрат.
На рис. 6.2 Г кончик сенсиллы пронизан порой. Сенсилла содержит по
крайней мере две нейросенсорные клетки, лишь одна из которых является ме-
6. Кинестезия 97
Рис. 6.4. Колоколовидная сенсилла. Воло-
сок редуцирован до небольшого купола, в
котором помещается тубулярное тельце.
А — аксон; PC — ресничный синус; КУ —
кутикула; К — купол; ОД — дендритное ло-
же; ФС — фиброзный слой; В — внутренние
обкладочные клетки; ВС — внутренний сег-
мент; НС — нейросенсорная клетка; ТТ —
тубулярное тельце.
ханочувствительной. Сенсилла, таким образом, является бимодальной. Другая
сенсорная клетка может быть обонятельной или вкусовой. Дендрит механо-
чувствительной нейросенсорной клетки, как и в предыдущем случае, присое-
динен к внутренней поверхности волоска сенсиллы тубулярным тельцем.
У другой нейросенсорной клетки развивается длинный (часто — ветвящийся)
«дендрит», простирающийся на всю длину сенсиллы и оканчивающийся непо-
средственно под ее порой. Такие «дендриты» представляют собой модифици-
рованные реснички. Мы встретимся с хорошо изученными примерами хемо-
чувствтительных ресничек при рассмотрении вкусовых (гл. 12) и обонятель-
ных (гл. 13) клеток позвоночных.
Механочувствительные волосковые сенсиллы (имеющие либо не имеющие
поры) широко распространены по поверхности тела насекомого и особенно
многочисленны на голове и вокруг нее. Они также хорошо представлены в со-
членениях тела и конечностей. Здесь они часто образуют целые «заросли»
(рис. 6.3 А). Рис. 6.3 Б показывает распределение механочувствительных сен-
силл муравья-рабочего вида Formica polyctena. Из рисунка видно, что ЦНС му-
равья получает поток информации о положении, движении и направлении
движения различных частей тела. В следующем разделе мы увидим, что у по-
звоночных, в особенности млекопитающих, не имеющих экзоскелета, разви-
лась не менее эффективная система детектирования движения и положения их
мышц и эндоскелета.
У механочувствительных сенсилл второго типа— колоколовидных сен-
силл — волосок редуцирован до полушария или купола, в основании имеюще-
го круглую или эллиптическую форму (рис. 6.4). Дендрит, связаннный с тубу-
лярным тельцем, обычно расположен центрально. Деформации экзоскелета
уплощают поверхность сенсиллы и таким образом сжимают тубулярное тель-
це. Смещение поверхности купола сенсиллы всего на 0,1 нм вниз достаточно,
чтобы стимулировать нейросенсорную клетку. Эллиптические сенсиллы наи-
более чувствительны к сжатию по короткой оси. Таким образом достигается
4 Биология сенсорных систем
98 Часть II. Механочувствительность
Мезаксон
Дистальный сегмент
Трубка
Глиальная
клетка
Соединительные
клетки
Трубка
Мезаксон
Дендрит
Мезаксон
Аксон
Основание
реснички
Ресничный
сегмент
Дистальный
сегмент
Соединительные
клетки
Тело
сенсорной
клетки
Рис. 6.5. Схема продольного сечения через сколопидий с поперечными срезами на раз-
ных уровнях. Ш — клетка-шапочка. По Moulins, М., 1976, «Ultrastructure of chordotonal
organs». In P. J. Mill, ed., Structure and Function of Proprioceptors in the Invertebrates, London:
Chapman and Hall.
Ресничный сегмент
Дистальное
базальное тельце
Проксимальное
базальное тельце
Клетка обкладки
сколопидия
Основание ресничкО.
Дендрит
Обкладка
сколопидия
селективность по направлению. Колоколовидные сенсиллы часто обнаружи-
ваются вблизи от суставов и сочленений или вблизи от тактильных волосков.
Другие типы механочувствительных сенсилл развились в полости тела.
Лучше всего изучены и наиболее широко распространены хордотональные ор-
ганы. Они состоят из специализированных сенсилл — сколопидий (рис. 6.5).
Сколопидий содержит одну или несколько нейросенсорных клеток, чьи денд-
риты, как и у других механочувствительных клеток, образуют сенсорную рес-
ничку. Тубулярное тельце в сколопидиях никогда не обнаруживается. Отличи-
тельной чертой сколопидий является наличие клеток, окружающих ресничку
и секретирующих плотный внутриклеточный листок — обкладку сколопидия.
Обкладка состоит из бочковидной муфты, образованной фиброзными волок-
нами. Сенсорное окончание сколопидия окружено клетками оболочки, а все
6. Кинестезия 99
они вместе присоединены к клетке-шапочке сколопидия. Последняя, в свою
очередь, соединена, иногда через дополнительные клетки, с внутренними
структурами, такими, как трахея, мышцы, кишка и т. д.
Хордотональные органы образуют один из основных типов проприоцепто-
ров тела насекомых. Они представлены практически во всех экзоскелетных
сочленениях и между сегментами тела. Они варьируют по сложности — от не-
скольких сколопидиев до нескольких тысяч, многие из них очень чувствитель-
ны к вибрации. Хорошо известный пример такой функции — субгенуальный
орган, расположенный в проторакальной конечности у многих видов насеко-
мых. В нем в голени объединяются 20—50 сколопидиев, присоединенных к ку-
тикуле. У таракана Periplaneta americana субгенуальный орган чувствителен к
вибрациям в диапазоне 1000—5000 Гц с пороговой амплитудой от 1 до 10 нм.
Восприятие вибрации важно не только как сигнала опасности, но и использу-
ется социальными насекомыми как средство коммуникации. Это подводит
нас к теме детектирования звука, однако, мы оставим ее главе 7, когда будем
рассматривать рецепцию акустических сигналов у насекомых.
6.1.3. Поддержание равновесия в полете
Любой, кто созерцал летное мастерство скромной домашней мухи, должен по-
разиться ее способности сохранять ориентацию и чувство направления. Полет
насекомых контролируется рефлекторно и может быть инициирован потерей
контакта механорецепторов лапки с поверхностью субстрата. Контроль поле-
та поддерживается благодаря стимуляции подушечек механочувствительных
сенсилл у оснований крыльев. Отклонения от правильной ориентации могут
быть описаны как вращения вокруг трех пространственных осей: кабрирова-
ние — передне-заднее вращение вокруг горизонтальной оси, перпендикуляр-
ной длинной оси тела; рысканье — вращение длинной оси тела вокруг верти-
кальной оси; закрутка — вращение вокруг длинной оси (рис. 6.6).
Ориентация поддерживается частично за счет зрительной информации,
поступающей от глаз, частично — за счет информации от чувствительных ор-
ганов, расположенных у сочленения головы и груди, а у двукрылых насеко-
мых — частично, и от жужжалец. Крупные стрекозы, относящиеся к самым
искусным летунам в мире насекомых, охотятся на других насекомых в полете.
У них имеется по три подушечки волосковых сенсилл с каждой стороны сочле-
Рис. 6.6. Кабрирование, рысканье и за-
крутка. Закрутка происходит, когда тело
вращается вокруг своей длинной оси
(осьх); рысканье — при вращении во-
круг оси у; кабрирование — при враще-
нии вокруг оси z.
100 Часть II. Механочувствительность
Вентральная
поверхность
Дорзальная
поверхность
Рис. 6.7. Дорзальная
и вентральная по-
верхности жужжалец
двукрылых. Ориен-
тация различных
групп колоколовид-
ных сенсилл пока-
зана двунаправлен-
ными стрелками. По
Pringle, 1948.
нения головы и груди, способные детектировать угловые ускорения. В полете
голова поддерживает свое положение благодаря зрительной информации от
глаз. Если положение тела нарушается, волосковые подушечки сигнализиру-
ют об этом. Две пары крыльев срабатывают рефлекторно таким образом, что-
бы восстановить соосность головы и тела. Такой же механизм действует и
у большинства других летающих насекомых. У двукрылых задняя пара крыль-
ев модифицируется в гантелевидные жужжальца с многочисленными колоко-
ловидными сенсиллами у их оснований (рис. 6.7). Жужжальца колеблются
на 180е с частотой от 100 до более чем 500 Гц, синхронно с биениями крыльев.
Колоколовидные сенсиллы ощущают различающиеся силы на сочленении
жужжалец с грудью при кабрировании, рысканьи и закрутке. Удаление жуж-
жалец нарушает рефлекторный компенсаторный контроль мышц крыльев при
возникновении таких вращений.
6.2. Кинестезические механизмы у млекопитающих
В отсутствие жесткого экзоскелета кинестезические механизмы у млекопита-
ющих (включая и нас с вами) по необходимости ограничены чувствительны-
ми окончаниями в мышцах, сухожилиях и суставах. Все они работают на
обеспечение ЦНС непрерывно обновляющейся картиной состояния напря-
жения или расслабления мускулатуры тела и положения суставов. В данном
разделе мы последовательно рассмотрим все эти чувствительные окончания,
начиная с веретен, далее — сухожильные органы и, наконец, механорецепторы
суставов.
6. Кинестезия 101
6.2.1. Интрафузальные окончания веретен
Скелетные (произвольные) мышцы состоят из многих тысяч многоядерных
поперечно-нило^атых волокон. У человека каждое мышечное волокно иннер-
вируется одним мотонейроном. Отростки мотонейрона, однако, при вхожде-
нии в мышцу ветвятся и, поэтому, иннервируют множество мышечных воло-
кон. Это явление называется мультитерминальной иннервацией, а группа мы-
шечных волокон и иннервирующий их мотонейрон определяются как моторная
единица — минимальная функциональная единица мышечной системы. Число
мышечных волокон, составляющих моторную единицу варьирует от мышцы
к мышце. Там, где требуется тонкий контроль движений, например, в пальцах
или мышцах глаз, моторные единицы малы, как правило не более 5—20 воло-
кон; там, где тонкий контроль не требуется — в мышцах спины и бедра, мотор-
ные единицы намного крупнее и состоят из более чем тысячи волокон.
Мышечные волокна млекопитающих делятся на два основных типа; экс-
трафузальные волокна, которые и выполняют всю работу сокращения мышцы,
и интрафузальные — специализированные для детектирования натяжения. И
те, и другие развиваются в эмбриогенезе в целом одинаково, однако, тогда как
экстрафузальные волокна приобретают полный комплект сократительных
белков — актина, миозина и др., в интрафузальных волокнах контрактильная
машинерия развивается только в окончаниях волокон. В то же время интрафу-
зальные волокна получают спиральные окончания чувствительных нервных
волокон групп 1а и II, образуя рецептор натяжения — мышечное веретено. Фак-
тически существует три типа интрафузальных волокон: два типа волокон с
ядерной сумкой и один тип волокон с ядерной цепочкой. Обычно два волокна с
ядерной сумкой и большое количество волокон с ядерной цепочкой, объеди-
ненные окружающей их рыхлой соединительнотканной капсулой, образуют
окончание веретена (рис. 6.8).
Волокна с ядерной цепочкой — более короткие и тонкие и, как и показы-
вает их название, содержат длинный ряд ядер. Эти волокна отвечают на посто-
янное натяжение. Волокна с ядерной сумкой, напротив, толще и длиннее и со-
держат группу ядер, собранных в центре («сумкой»). Один из типов волокон с
ядерной сумкой (динамический) отвечает на включение или выключение рас-
тягивающей силы, другой (статический) — так же, как волокна с ядерной це-
почкой, на постоянное растяжение. Веретено иннервируется четырьмя типа-
ми нервных волокон (рис. 6.8), которые все входят в капсулу в ее несколько
расширенной средней части. Два из этих типов волокон — чувствительные,
два — двигательные; чувствительные волокна — аксоны группы 1а (большого
диаметра) и аксоны группы II (малого диаметра). Волокна группы 1а обычно
дают спиральные окончания вокруг всех интрафузальных волокон. Аксоны
группы II оканчиваются только на волокнах с ядерной цепочкой и статических
волокнах с ядерной сумкой. Все двигательные волокна имеют малый диаметр
и, соответственно, медленное проведение — это у-волокна, однако, как видно
из рис. 6.8 Б, они могут быть подразделены на две группы, в зависимости отто-
го, какие интрафузальные волокна они иннервируют — динамические или
статические. В обоих случаях эти волокна идут к оконечностям интрафузаль-
ных волокон, где присутствует сократительная машинерия, и здесь оканчива-
ются нервно-мышечным контактом.
102 Часть II. Механочувствительность
Рис. 6.8. Мышечное веретено млекопитающих. (А) группа из трех интрафузальных воло-
кон. In vivo веретено окружено экстрафузальными волокнами. Оконечности интрафу-
зальных волокон содержат актомиозиновую сократительную машинерию и, следова-
тельно имеют поперечно-полосатую структуру. Центральный участок интрафузального
волокна несколько расширен и окружен соединительнотканной капсулой. Он иннерви-
руется двумя типами у-двигательных волокон. Эти волокна входят в капсулу и направля-
ются к оконечностям интрафузальных волокон, где образуют с ними соединение. М —
у-двигательное волокно; К — капсула; С — 1а и II чувствительные волокна; ИФ — интра-
фузальные волокна; НМС — нервно-мышечное соединение. (Б) Схематический рису-
нок показывает пять типов интрафузальных волокон, обнаруженных в мышечном вере-
тене млекопитающих. ДВС — динамическое волокно с ядерной сумкой; ДуДВ — дина-
мическое у-двигательное волокно; ВЦ — волокно с ядерной цепочкой; ПО — первичное
окончание; СВС — статическое волокно с ядерной сумкой; СО — статическое оконча-
ние; СуДВ — статическое у-двигательное волокно. Прочие объяснения в тексте. По
Kandel, Schwartz and Jessell, 1991.
Спиральные окончания чувствительных волокон расположены очень
близко от интрафузальных волокон, так что если последние подвергаются воз-
действию натяжения, оно передается и на чувствительные нервные оконча-
ния. Это, как и в других случаях, рассмотренных в данной главе, является
адекватным стимулом и через открывание ионных каналов приводит к форми-
рованию генераторного потенциала, что в свою очередь приводит к инициа-
ции потенциала действия в чувствительном волокне. Чувствительные волокна
группы 1а обладают быстрым проведением (ок. 120 м/с), группы II — тоже
сравнительно быстрым (ок. 75 м/с) (см. раздел 2.6, табл. 2.2). Сигнал передает-
ся в спинной мозг через дорзальные корешки (рис. 6.9).
Тогда как интрафузальные волокна иннервируются волокнами у-мотоней-
ронов, окружающие их экстрафузальные волокна иннервируются аксонами
большого диаметра (с быстрым проведением) а-мотонейронов. Эти две группы
мотонейронов, очевидно, достаточно существенно различны и обозначаются
как фузимоторные и скелетомоторные системы, соответственно. К сожалению,
как это часто случается в биологии, ситуация здесь до конца не ясна. Показа-
но, что и некоторые а-мотонейроны посылают веточки аксонов в веретено,
6. Кинестезия 103
Б
Чувствительное
волокно
группы 1а
Рис. 6.9. (А) Коактивация. Рисунок показывает поперечный срез спинного мозга и упро-
щенную схему контактов нервных волокон с интра- и экстрафузальными волокнами
скелетной мышцы. Н — нисходящее волокно из высших отделов ЦНС в белое вещество
спинного мозга; ЭВ — экстрафузальные волокна; ИВ — интрафузальные волокна.
(Б) Торможение антагонистической мышцы (мышц). Тормозные волокна обозначены
черным цветом. Дальнейшие объяснения — в тексте.
у-мотонейрон,
посылающий аксон
к антагонистической
мышце
а-мотонейрон,
посылающий аксон
к антагонистической
мышце
где они иннервируют интрафузальные волокна. Принято считать, что эти во-
локна образуют скелетофузимоторную систему, они присутствуют, вероятно,
во всех веретенах млекопитающих. Следует отметить, что и у анамниотических
позвоночных четкое различие между фузимоторной и скелетомоторной систе-
мами отсутствует.
И как же все это работает? Веретена, как мы уже видели, расположены па-
раллельно экстрафузальным волокнам. Когда сигнал из высших отделов ЦНС
на движение мышц передается в спинной мозг, активируются и а- и у-мото-
нейроны (рис. 6.9 А). Это явление, по понятным причинам, назыгт.зтся коакти-
вацией. а-мотонейроны проводят потенциалы действия с очень большой ско-
ростью (до 120 м/с), и последние, следовательно, достигают экстрафузальных
мышечных волокон задолго до того, как намного более медленными аксонами
у-мотонейронов активируются интрафузальные волокна. Мышца сокращает-
ся и устраняет, таким образом, какое-либо растяжение интрафузальных воло-
кон. Вследствие этого активность в чувствительных нервных волокнах прекра-
щается. Однако затем в мышцу приходит активность у-мотонейронов, и сокра-
щение возникает и в интрафузальных волокнах. Если импульсация в у-волок-
нах требует большего сокращения интрафузальных волокон, чем достигнуто
окружающими экстрафузальными, тогда интрафузальные волокна в попытке
(безуспешной) сократиться еще больше испытают напряжение. Это напряже-
ние воспринимается спиральными окончаниями чувствительных нейронов,
которые отразят это в информации, передаваемой ими обратно в ЦНС.
104 Часть II. Механочувствительность
К чему это приводит? Чтобы ответить на этот вопрос, мы должны рассмот-
реть рефлекторные связи, контролирующие мышечное сокращение. Это до-
статочно сложная система, но для нашей конкретной цели мы можем рассмо-
треть только один из ее элементов (рис. 6.9 А). Тела а- и у-мотонейронов рас-
положены в передних (= вентральных) рогах серого вещества спинного мозга.
Как показывает рис. 6.9 А, чувствительные волокна группы 1а от мышечных
веретен образуют возбуждающие синапсы с а-мотонейронами. Схема, конеч-
но, по необходимости упрощена. Волокна 1а возбуждают не только а-мото-
нейроны, иннервирующие гомонимную мышцу (т. е. ту же самую, от которой
получены афференты), но и синергические мышцы. Это те мышцы, которые
двигают сустав сходным образом с тем, которым это делает гомонимная. Кро-
ме того, как показано на рис. 6.9 Б, веточка волокна 1а направляется и к тор-
мозному интернейрону в сером веществе спинного мозга. Тормозные синапсы
этого интернейрона располагаются на телах мотонейронов, иннервирующих
антагонистические мышцы, т. е. предупреждается сокращение этих мышц од-
новременно с мышцами-синергистами. Конечно, этот механизм может быть
подавлен командами из высших отделов ЦНС, так что мы в состоянии, как все
мы знаем, поддерживать баланс сокращения различных мышц.
Рассмотренные выше рефлекторные дуги обеспечивают быстрое снятие
напряжения интрафузальных волокон за счет большего сокращения экстра-
фузальных. В сущности, ЦНС только предопределяет длину интрафузальных
волокон, а сервомеханизмы спинного мозга обеспечивают поддержание этой
длины за счет сокращения экстрафузальных волокон, и таким образом сустав
перемещается в требуемое положение.
Однако, почему так сложно организовано мышечное веретено? Почему
оно дифференцировано на волокна с ядерной сумкой и с ядерной цепочкой
и т. д.? Ответ на эти вопросы заключается в различных физиологических свой-
ствах этих волокон. Мы отмечали выше, что, тогда как волокна с ядерной це-
почкой отвечают на постоянное натяжение, один из типов волокон с ядерной
сумкой (динамические волокна) лучше всего реагируют на включение или вы-
ключение натяжения. Активность в двух этих системах чувствительных воло-
кон, иннервирующих мышечные веретена, таким образом, различается. Во-
локна 1а передают сигналы включения и выключения натяжения, а волокна
II — сигнал длящегося натяжения (рис. 6.10). Кроме того, волокна 1а наиболее
чувствительны к малым изменениями длины веретена (менее 1 мм) и, что важ-
но, к скорости этих изменений. Таким образом, импульсация в чувствительных
волокнах 1а и II кодирует не только постоянное натяжение (группа II), но и
скорость сокращения (группа 1а).
Можно продемонстрировать, что существует отдельный механизм контроля
со стороны у-мотонейронов динамических волокон с ядерной сумкой и стати-
ческих — с сумкой и с цепочкой. Одна группа у-мотонейронов контролирует
статические волокна, а другая — динамические. Эта дифференцированная ин-
нервация связана с различиями структуры и физиологии двух типов интрафу-
зальных волокон. В случае динамических волокон их центральный участок
специализирован для реакции на быстрое включение или устранение натяже-
ния. Активация динамических у-мотонейронов увеличивает чувствительность
центрального участка. С другой стороны, статические волокна отвечают по
6. Кинестезия 105
Стимул
Первичное
Плавное
растяжение Рывок Вибрация
11111111111 IIIIHIII III Hill Hill III
Вторичное
llllllllllllllllll lllll III Ulllllllllll
Разгрузка
Рис. 6.10. Ответы первичных и вторичных окончаний мышечных веретен (см.
рис. 6.8). (А) Импульсации в волокнах 1а от первичных окончаний в мышечном
веретене, сигнализирующая о включении натяжения залпом активности, а о вы-
ключении — периодом «молчания». Волокна II от вторичных окончаний сигнали-
зируют о постоянном натяжении. (Б) Первичные окончания отвечают на различ-
ные скорости натяжения. Рисунок показывает, что волокна 1а отвечают тем более
интенсивно, чем быстрее происходит натяжение. Дальнейшие пояснения — в тек-
сте. Из Kandel, Schwartz and Jessel, 1991.
всей длине более равномерно на силы натяжения. Активация статических у-мо-
тонейронов вызывает сокращение способных к этому оконечностей волокон
веретена, как описано выше, сжимая таким образом и центральный участок.
Тонкий контроль мышечного сокращения, очевидно, представляет огром-
ное значение для всех животных, в особенности для таких активных животных,
как млекопитающие. Поэтому ничего удивительного нет в том, что в этом про-
цессе на сложную систему вертикальных и обратных связей, описанных в пре-
дыдущих абзацах, накладывается еще один, высший уровень контроля, называ-
емый фузимоторный комплекс. Когда млекопитающее, например кошка, нахо-
дится в покое, в статических или динамических у-мотонейронах импульсация
невелика или отсутствует вовсе. Когда животное поднимается, потягивается и
уходит на мягких лапах, уровень активности в статических у-мотонейронах не-
высок. Очень мала или полностью отсутствует и активность в динамической у-
системе. По мере того, как активность возрастает и животное предпринимает
более сложные маневры, активность в статической системе снижается, а в ди-
намической — проявляется и становится все возрастающей. Центральная нерв-
ная система таким образом способна контролировать чувствительность своего
контроля так, чтобы соответствовать обстоятельствам, в которых оказывается
ее обладатель.
106 Часть II. Механочувствительность
6.2.2. Сухожильные органы Гольджи
Сухожильные органы Гольджи функционируют совместно с мышечными ве-
ретенами, чтобы сигнализировать о состоянии сокращения или напряжения
скелетных мышц. В то время как веретена контролируют изменения длины
мышц, сухожильные органы сигнализируют об уровне их натяжения. Тогда
как мышечные веретена работают «параллельно» с экстрафузальными мышеч-
ными волокнами, сухожильные органы действуют «последовательно». Это от-
ражается в том факте, что сухожильные органы часто анатомически связаны
с веретенами. В некоторых мышцах лапы кошки (камбаловидная мышца, ме-
диальная икроножная и др.) 25% сухожильных органов ассоциированы в «ди-
ады» с веретенами, а в мышцах челюсти — практически все.
Хотя название и подразумевает, что эти органы располагаются в сухожили-
ях и(или) апоневрозах, это не совсем так. Однако, огромное большинство су-
хожильных органов локализовано в соединениях сухожилий (или апоневро-
зов) с мышцами. Долгое время считалось несущественным: что именно по-
рождает возбуждение в сухожильном органе — активное сокращение «своей»
мышцы или пассивное растяжение. В последнее время, однако, показано, что
пассивное натяжение не является слишком эффективным путем стимуляции -
адекватным стимулом является мышечное сокращение. Связь сухожильных
органов с моторными единицами часто очень прецизионна — индивидуаль-
Рис. 6.11. Схематическое
изображение сухожильного
органа. Капсула на рисунке
показана рассеченной. Рису-
нок показывает, что мышеч-
ные волокна, соединенные с
сухожильным органом «по-
следовательно», окружены
множеством мышечных во-
локон, соединенных «парал-
лельно» с тем же сухожилием
или апоневрозом. Из Jami,
1992, Physiological Reviews, 72,
623-666. Воспроизводится с
разрешения Американского
Физиологического Общест-
ва (American Physiological So-
ciety).
6. Кинестезия 107
Рис. 6.12. Соотношение между ре-
цепторным потенциалом (Ar) и
натяжением (Ат) при приложе-
нии растягивающей силы синусо-
идальной формы (частота 10 Гц,
амплитуда 10 мм) в зависимости
от исходного состояния растяже-
ния (Tj). Исходное растяжение ва-
рьирует от нуля (расслабление) до
400 мкН. Do — амплитуда растя-
жения; f — частота. Из Wilkinson
and Fukami, 1983, Journal of Neuro-
physiology, 49, 984. Воспроизво-
дится с разрешения Американ-
ского Физиологического Общест-
ва (American Physiological Society).
ный сухожильный орган может контролировать активность единственной мо-
торной единицы.
Чувствительные окончания сухожильного органа помещены в слоистой
капсуле, которая является продолжением периневральной оболочки быстро-
проводящих аксонов группы 1b (рис. 6.11). Капсулы сильно варьируют по дли-
не (242—1045 мкм, средняя — 521 мкм) и диаметру (66—220 мкм, средний —
125 мкм). Изнутри капсула подразделяется на множество компартментов, за-
полненных перепутанными коллагеновыми волокнами. Входящие в капсулу
волокна 1b многократно ветвятся, утрачивают миелиновую оболочку и, впле-
таясь в коллагеновые волокна, образуют на них окончания. Эти окончания за-
полнены митохондриями и плотно охватывают коллагеновые волокна.
Когда мышечное волокно сокращается, возникающее изменение положе-
ния присоедиенных к нему коллагеновых волокон деформирует чувствитель-
ные окончания, вызывая изменения проводимости на их мембранах. Формиру-
ющийся генераторный потенциал электротонически распространяется по
окончанию вплоть до места инициации потенциала действия. В опытах на дор-
золатеральной мышце хвоста кошки показано, что, когда коллагеновые волок-
на повергаются воздействию постоянной растягивающей силы, генераторный
потенциал зависит от последней прямо пропорционально. В более динамич-
ных условиях действия очень малой изменяющейся силы натяжения
(0,5—5 мкм) рост генераторного потенциала постоянен (около 10 мкВ/мкм рас-
тяжения при 10 Гц). Показано также, что амплитуда рецепторного потенциала
зависит от исходного состояния растяжения (рис. 6.12). Наконец, изменения
потенциала зависят и от частоты стимуляции — чем выше частота (в пределах
20 Гц), тем больше рост потенциала. Когда к мышце прикладывается большая
сила или более высокая частота, возникает нелинейная зависимость потенциа-
ла от стимула, a in vivo, как отмечалось выше, ситуация еще более сложна.
Точное место, где инициируется потенциал действия, остается неизвест-
ным — возможно в первом перехвате Ранвье, но существуют и другие возмож-
ности. В опытах с переменными стимулами частота импульсов в чувствитель-
108 Часть II. Механочувствительность
Рис. 6.13. «Блок-схема» отрица-
тельных обратных связей кон-
троля сухожильного органа
Гольджи. Сравните с блок-схе-
мой контроля мышечного вере-
тена на рис. 6.9. Отметьте, что
в рефлексе сухожильного орга-
на отсутствует моносинаптичес-
кий путь и что вся система рабо-
тает с противоположным зна-
ком по отношению к рефлексам
веретен. Ag — мышца-синер-
гист; Ап -мышца-антагонист.
Дальнейшие объяснения —
в тексте.
ных волокнах 1b связана скорее с частотой стимула, а не его амплитудой. ЦНС,
однако, нуждается и в информации об амплитуде силы, растягивающей сухо-
жилие. Складывается впечатление, что такую информацию предоставляет сум-
марный разряд от всех сухожильных органов. Но и это доподлинно экспери-
ментально не установлено. Возможно, та информация, которая передается в
ЦНС, не столько характеризует уровень силы натяжения в каждый данный мо-
мент, сколько динамику развития этой силы. Такая информация сама по себе
усложнена нелинейным характером реакций мышц и их оболочек, также как и
свойств сухожилий. Как мы увидим в других случаях, работа сенсорных систем
трудно поддается простому механистическому анализу. ЦНС обрабатывает и
формирует ощущения из колоссального потока афферентных импульсов. Ве-
роятно, общая картина этого потока важнее, чем отдельные его элементы.
Потенциалы действия в чувствительных нервных волокнах 1b поступают
в спинной мозг и (по большей части) образуют синапсы с тормозными интер-
нейронами, которые, в своб очередь, образуют синапсы на гомонимных мото-
k нейронах (рис. 6.13). Тормозные интернейроны также ингибируют мотонейро-
ны синергических мышц (на рис. 6.13 не показаны). Наконец, чувствительные
волокна 1b образуют синапсы на возбуждающих интернейронах, связанных
с мотонейронами мышц-антагонистов.
Какой же вывод можно сделать по поводу этой сложной системы? Хотя де-
тали ее все еще исследуются, и мы далеки от понимания всей сложности сис-
темы, основные моменты, тем не менее, ясны. Сухожильные органы не ак-
тивны, пока мышцы остаются в покое. Когда начинается сокращение, возни-
кает импульсация, которая сигнализирует об изменениях нарастания силы
сокращения. Импульсы в чувствительных волокнах 1b подавляют активность
гомонимных мотонейронов и ограничивает вовлечение новых моторных еди-
ниц. Также ограничивается активность мотонейронов синергических мышц и
6. Кинестезия 109
возбуждается активность мотонейронов мышц-антагонистов. Таким образом
обеспечивается плавность сокращения и его максимальная адекватность по-
ставленной задаче. Функционируя вместе с системой мышечных веретен,
этот механизм, например, обеспечивает точное соответствие усилия массе
объекта, когда мы его поднимаем. Когда мы прикасаемся к поверхности или
ощупываем ткань, этот же механизм обеспечивает такую тонкость наших дви-
жений, чтобы тактильные рецепторы кожи (гл. 7) были в состоянии получить
информацию о гладкости, текстуре и толщине ткани.
6.2.3. Рецепторы суставов
В большинстве суставов присутствует три типа чувствительных окончаний:
окончания типа телец Руфини (см. гл. 7), расположенные в суставной сумке;
окончания типа телец Гольджи, представленные в связках суставов; и оконча-
ния типа пачиниевых телец (см. гл. 7), обнаруживающиеся вблизи от места со-
единения связки с надкостницей. Когда сустав находится в среднем положе-
нии, активность в афферентных волокнах, идущих от этих рецепторов, неве-
лика. Когда, однако, сустав сгибается или разгибается до крайнего положения,
импульсация резко увеличивается.
Изучение одиночных телец Руфини показывает, что большинство из них
медленно адаптируется, и каждое имеет собственный «угол возбуждения» —
дугу углового движения сустава в несколько градусов, где возбуждение макси-
мально. У разных рецепторных единиц угол возбуждения различается. Им-
пульсация от каждой из единиц связана со скоростью углового перемещения и
возрастает по мере приближения к границе угла возбуждения. Тогда как лишь
немногие единицы контролируют движение сустава в обе стороны от среднего
положения, большинство реагирует на движение к крайнему положению. Это
соответствует наблюдению, сделанному в предыдущем абзаце, что импульса-
ция в афферентных нервных волокнах возрастает до максимума по мере при-
ближения сустава к его крайнему положению.
Ситуация усложняется тем, что вращение таких суставов, как наиболее
изученные коленный и локтевой, вокруг их длинной оси также влияет на им-
пульсацию от телец Руфини. То есть там, где такие движения встречаются, им-
пульсация от этих окончаний не может давать недвусмысленной информации
в ЦНС о положении сустава или скорости его движения. Другие типы чувстви-
тельных окончаний, расположенные вокруг суставов, тельца Гольджи и Пачи-
ни, скорее сигнализируют о натяжении активированных мышц, а не о положе-
нии сустава. Тем не менее, расположение этих чувствительных окончаний в
суставной сумке и вокруг нее обеспечивает максимальную активность при
движении сустава к крайним положениям, или когда его вращение выходит за
пределы нормы.
6.3. Заключение
Нам достаточно лишь бросить взгляд на неуклюжие творения роботехников,
чтобы оценить гибкость и тонкость «шестого чувства» животных. Роботехни-
ка, конечно, колоссально продвинулась со времен первоначальных примитив-
110 Часть II. Механочувствительность
ных устройств. Гидравлические автоматы Сен-Жермен-э-Лэ, которые так
впечатлили Рене Декарта в начале XVII столетия, или механические протезы,
изобретенные французским военным хирургом Амбруазом Паре в конце
XVI века, и даже механический флейтист Вокансона в XVIII веке смотрятся
грубовато по сравнению с устройствами, разработанными современной про-
мышленностью и военными. Однако, даже эти современные изделия, способ-
ные, как Марс Ровер (марсоход), передвигаться по пересеченной местности и
брать образцы грунта, или как многосуставные универсальные руки автомати-
зированных сварочных аппаратов современных автозаводов, проникать
внутрь машины и проводить точечную сварку швов в труднодоступных и за-
темненных местах, выглядят бледно по сравнению со своими биологически-
ми конкурентами. Отчасти это следствие жесткости конструкции роботов и
недостаточной доныне миниатюризации. Однако, главным образом, дело в
огромном проигрыше роботов по сравнению с животными в численности сен-
соров, которые мы рассматривали в данной главе. Сенсорная информация,
поступающая в ЦНС из множества мест, обеспечивает гибкость и адаптив-
ность двигательных реакций произвольной мускулатуры. За время эволюции
этот сенсорный поток, это шестое чувство, достигло такого совершенства, ко-
торое обеспечивает плавность и чувствительность реакций, которые воспри-
нимаются как часть того дара жизни, который мы получили. Как мы видели в
данной главе, эта чувствительность зависит от интеграции бесчисленного ко-
личества петель прямой и обратной связи. Исследователи еще не раскрыли
всей сложности этих механизмов, которые, напомним, простираются до моле-
кулярного уровня. Это, кстати, тоже создает контраст с роботами, которые со-
браны на основе аморфных веществ — металлов, керамики и кремния.
Прямые и обратные связи шестого чувства животных пронизывают цент-
ральную нервную систему — и спинной мозг (как мы видели в данной главе
при обсуждении спинальных рефлексов) и высшие отделы мозга — ствол,
мост, мозжечок и двигательную кору. В этой книге мы не будем рассматривать
эти вопросы. Заинтересованный студент может узнать об этом в книгах по
нейрофизиологии, приведенных в списке библиографии. Нам же достаточно
сказать, что сенсорная информация от мышц, суставов и сухожилий (в случае
млекопитающих) интегрируется с информацией из вестибулярной системы,
глаз и от других органов чувств таким образом, что мозг оказывается в состоя-
нии выдать мышечной системе команды, соответствующие потребностям жи-
вотного в постоянно меняющемся окружении. Еще много лет пройдет, если
это вообще когда-нибудь произойдет, пока роботехника сможет конкуриро-
вать с живым.
7. ОСЯЗАНИЕ
Осязательная чувствительность у С. elegans: мутация тес — нейроны ося-
зательных рецепторов — генетический анализ — белки МЕС — сходство
с белками ENaC — другие гены тес и их белковые продукты — возмож-
ная молекулярная структура тактильного рецептора С. elegans. Осязатель-
ная чувствительность у насекомых: субгенуальные органы — детектирова-
ние колебаний воды водомерками и гребляками — акустические сенсил-
лы — ости Drosophila — джонстонов орган — чувствительность к частоте
биения крыльев — тимпанальные органы (уши): структура и функция —
коммуникация — детектирование жертвы — детектирование насекомо-
ядных бабочками — структура уха совок — контрмеры совок против лету-
чих мышей. Осязательная чувствительность кожи млекопитающих: быстро
адаптирующиеся рецепторы: пачиниевы тельца, мейснеровы тельца, тер-
минальная колба Краузе, рецепторы волосяных фолликулов; медленно
адаптирующиеся рецепторы: клетки Меркеля, органы Руфини, С-меха-
норецепторы. Центральные пути у млекопитающих: тракты спинного моз-
га — сомэстезическая кора — колончатая структура — рецептивные по-
ля — сомэстезический гомункулус. Пластичность коры: мышиные усы —
пальцы обезьян — скрипачи — читатели по Брайлю. Заключение: универ-
сальное чувство — груминг у социальных животных — коммуникация
Осязание, как мы отмечали во Введении к Части II, чувство незаменимое.
Трудно себе представить, на что бы это было похоже — животные без этой мо-
дальности, каким бы выглядел мир без этого чувства. Наши собственные пред-
ставления о твердости, сопротивлении, объективности предметов вряд ли сло-
жились бы без этого вездесущего чувства. По сравнению с нашими глубокими
знаниями молекулярной биологии обонятельных и зрительных рецепторов,
понимание точных механизмов функционирования наших тактильных рецеп-
торов разрозненно и неудовлетворительно. Это положение, однако, начинает
медленно меняться благодаря работам на маленьком почвенном круглом чер-
ве Caenorhabditis elegans.
7.1. Механорецепция у Caenorhabditis elegans
Мы отмечали в гл. 4, что С. elegans интенсивно изучается молекулярными био-
логами. В сущности, когда генетики поняли, что основные закономерности
биологии прокариот установлены в работах на Е. coli, а биологии эукариот —
на дрожжах Saccharomyces cerevisiae, следующей и последней заманчивой це-
112 Часть II. Механочувствительность
Рис. 7.1. Caenorhabditis elegans.
Длина червя около 1 мм. С. elegans
состоит из 959 соматических кле-
ток, 302 из которых образуют нерв-
ную систему. Тело прозрачно, и
многие клетки могут быть различе-
ны на живом животном. Перепе-
чатка из J. G. White, 1985, «Neuro-
nal connectivity in Caenorhabditis
elegans», Trends in Neurosciences, 8,
277, с разрешения издательства El-
sevier Science.
лью стали многоклеточные животные. Генетики стали искать животное, кото-
рое легко культивировать, с быстрым развитием, т. е. такое, на котором мож-
но адаптировать и применить мощные методики, используемые в микробио-
логической генетике. Избранный ими организм — маленькая (около 0,5 мм в
длину) свободно живущая нематода Caenorhabditis elegans (рис. 7.1). За более
чем 40 лет интенсивных исследований раскрыты секреты ее генетики, физио-
логии, анатомии и поведения. Известно точное количество нервных клеток в
ее нервной системе (302), синаптическая структура нервной системы, а каж-
дый из ее нейронов полностью изучен электронно-микроскопически; геном
полностью изучен в 1998 году.
Как и все животные, С. elegans чувствительна к прикосновению. Применя-
ли два типа тактильных стимулов — осторожное прикосновение волоском рес-
ницы и более интенсивное воздействие тонкой проволочкой. Наиболее инте-
ресные результаты получены в исследованиях реакции на мягкую стимуля-
цию.
Помещенная в чашку Петри С. elegans двигается, синусоидально изгибая
тело, до тех пор, пока не получит тактильного стимула. Если слегка прикос-
нуться к передней части тела, направление движения меняется на противопо-
ложное, и червь извивается обратно по чашке; если прикоснуться к задней ча-
сти, червь продолжит движение вперед. Большое количество мутаций (более
440), затрагивающих 15 генов, влияет на эти реакции. То, что червь при этом
сохраняет способность двигаться вообще, подтверждается уколом проволо-
кой. Большинство из этих мутаций классифицируется как лиес-мутации, по-
7. Осязание 113
Переднее поле
Заднее поле
Рис. 7.2. Осязательные рецепторные нейроны С. elegans. Отметьте существование
двух полей осязательной чувствительности, определяемых расположением соот-
ветствующих нейронов в организме нематоды. ПЛМЛ — левая передняя латераль-
ная микротубулярная клетка; ПЛМП — правая передняя латеральная микротубу-
лярная клетка; ЗЛМЛ — левая задняя латеральная микротубулярная клетка;
ЗЛМП — правая задняя латеральная микротубулярная клетка; ПВМ — передняя
вентральная микротубулярная клетка; ЗВМ — задняя вентральная микротубуляр-
ная клетка. Из Tavermarakis and Driscoll, 1997, Annual Review of Physiology, 59, 662.
Воспроизводится с разрешения Annual Review of Physiology, volume 59, © 1997 Annual
Reviews www.annualreviews.org.
скольку возникающие фенотипы описываются как аномальные по механочув-
ствительности (mechanosensitive abnormal). Соответственно, и гены, в которых
наблюдаются эти мутации, обозначаются тес, а белки, которые они кодиру-
ют — МЕС.
Показано, что прикосновение детектируется шестью нейронами: двумя пе-
редними латеральными микротубулярными клетками (левой и правой) —
ПЛМЛ и ПЛМП; двумя задними латеральными микротубулярными клетками
(левой и правой) — ЗЛМЛ и ЗЛМП; одной передней вентральной микротубу-
лярной клеткой — ПВМ. Еще один нейрон, вероятно, также вовлечен в про-
цесс, хотя, в отличие от остальных, он и не способен детектировать устранение
прикосновения, это — задняя вентральная микротубулярная клетка — ЗВМ.
Анатомическое положение этих нейронов-детекторов прикосновения показа-
но на рис. 7.2.
Осязательные рецепторные нейроны, как показывают их наименования,
характеризуются наличием в них пучка очень крупных микротрубочек. Элек-
тронно-микроскопическое исследование показало, что отдельные микротру-
бочки, входящие в пучок, связаны между собой и имеют ту особенность, что
состоят из 15 протофиламентов (в отличие И протофиламентов в остальных
микротрубочках С. elegans и 13 протофиламентов в остальном животном цар-
стве). Эти необычно крупные и ультраструктурно необычные микротрубочки
намного короче самого нейрона (не более 5% его длины) и ориентированы под
углом к его длинной оси. Дистальные концы микротрубочек, вероятно, погру-
жены в плазматическую мембрану нейрона, что предполагают какую-то меха-
ническую связь с ней. Микротрубочки существенны для функционирования
нейрона — если микротрубочки разрушаются колхицином либо повреждают-
ся вследствие мутации, осязательная чувствительность утрачивается.
Нейроны осязательной рецепции расположены под кутикулой («кожей»)
червя, к которой они присоединяются внеклеточной «мантией» фиброзного
материала (рис. 7.3). Ультраструктурные исследования позволяют предполо-
114 Часть II. Механочувствительность
Рис. 7.3. Ультраструктура
нейрона осязательной ре-
цепции С. elegans (попе-
речное сечение). Нейрон
окружен соединительнот-
канной мантией и соеди-
нен с кутикулой «фиброз-
ным органоидом». Ней-
рон содержит пучок мик-
ротрубочек (каждая из ко-
торых состоит из 15 про-
тофиламентов (пф)). По
Tavermarakis and Driscoll,
1997.
жить наличие механической непрерывности между кутикулой и микротрубоч-
ками отростках нейронов осязательной рецепции. Стоит напомнить, что тубу-
лярное тельце механочувствительного волоска насекомых тоже состоит из
плотно упакованных микротрубочек, соединяющих нейросенсорную клетку с
кутикулой (гл. 6, раздел 6.1.2).
Генетический анализ проводился путем вызывания мутаций и исследова-
ния их влияния на способность червя реагировать на осторожное прикоснове-
ние. Когда такая утрата обнаруживается, молекулярно-биологическая техника
привлекается для выяснения того, какая структура определяется мутировав-
шим геном. В некоторых случаях удалось обнаружить гены, ответственные за
факторы, необходимые для развития самих осязательных клеток или контро-
лирующие действие других mec-генов. Еще в нескольких интересных случаях
открыты гены, кодирующие канальные белки и дополнительные элементы,
соединяющие каналы с кутикулой, с одной стороны, и нейрональные микро-
трубочки — с другой.
Мес А и тес-10 — два гена, определяющие канальные белки — МЕС-4 и
МЕС-10. Эти гены называются также «дегенеринами», поскольку мутации в
них, обусловливающие формирование дефектных МЕС-4 и МЕС-10, ведут
клетки, в которых они экспрессируются, к разбуханию и гибели. И МЕС-4, и
МЕС-10 принадлежат к суперсемейству белков, состоящих из трех субъединиц
белка Na+-канала эпителия млекопитающих (ENaC). Субъединица включает
два трансмембранных сегмента, и С-, и N-концы локализованы внутрикле-
точно, имеется большая внеклеточная петля (рис. 7.4). Хотя точная структура
МЕС-канала пока не определена, по аналогии с другими каналами, особенно
эпителиальными натриевыми каналами млекопитающих, предполагается что
это объединение шести субъединиц, возможно — смеси МЕС-4 и МЕС-10.
Имеются некоторые свидетельства того, что тес-6кодирует другую канальную
субъединицу — МЕС-6, в этом случае канал, видимо, представляет собой гете-
ротример субъединиц МЕС-4, МЕС-6 и МЕС-10.
7. Осязание 115
Рис. 7.4. Трансмембранная
топология белка МЕС-4. Бе-
лок включает два трансмемб-
ранных домена и небольшой
участок, погруженный в мем-
брану непосредственно перед
вторым трансмембранным
доменом. Петля из 768 амино-
кислотных остатков, как по-
казано на рисунке, локализо-
вана внеклеточно. Замещение
аланина713 (Ala) более тяже-
лой аминокислотой влечет за
собой гибель клетки. По Ta-
vermarakis and Driscoll, 1997.
Как упоминалось выше, известно намного больше генов тес. Во многих
случаях выяснено, какую роль играет каждый из них. Мутации в генах тес-7 и
тес-12 приводят к разрушению 15-ти протофиламентных микротрубочек, так
характерных для нейронов осязательных рецепторов. Другой ген — тес-2 ко-
дирует белок, состоящий из 481 аминокислоты, сходный со стоматином цито-
скелета красных кровяных клеток млекопитающих, который, как полагают,
участвует в связи между каналом МЕС и микротрубочками (рис. 7.5). Другие
гены — тес-1, тес-5, тес-9 кодируют белки МЕС-1, МЕС-5 и МЕС-9, которые
обнаруживаются вне клетки — в мантии.
Интересная, хотя пока еще спекулятивная, модель предложена для связи
всех этих элементов в функциональный тактильный рецептор — она представ-
лена на рис. 7.6. МЕС-1, МЕС-5 и МЕС-9 организованы так, чтобы передать
давление на кутикулу субъединице МЕС-4 и открыть канал. Когда канал от-
крывается, ионы Na+ двигаются по градиенту концентрации, и нейрон депо-
ляризуется. Такая механическая трансмиссия работает, если канал стабилизи-
рован за счет прикрепления его к микротрубочкам. Такое прикрепление, как
мы видели, обеспечивается белком МЕС-2. Иными словами, механическая
трансмиссия должна вызвать смещение сети микротрубочек внутрь, что, по-
Рис. 7.5. Предполагаемая
связь между белком МЕС-2
с 15-ти протофиламентной
микротрубочкой нейрона
осязательного рецептора.
Предполагается, что N-ко-
нец МЕС-2 взаимодействует
с микротрубочкой, а С-ко-
нец с каналом. По Taverma-
rakis and Driscoll, 1997.
116 Часть II. Механочувствительность
Рис. 7.6. Концептуальная модель осязательного рецептора С. elegans. Объяснения и
номенклатура — в тексте. Из Tavermarakis and Driscoll, 1997, «Molecular modelling of
mechanotransduction in the nematode Caenorhabditis elegans». Annual Review of Physio-
logy, 59, 679. Воспроизводится с разрешения Annual Review of Physiology, volume 59,
© 1997, by Annual Reviews www.annualreviews.org.
скольку сеть присоединена к плазматической мембране (см. выше), увеличи-
вает натяжение последней и, таким образом, вызвает открывание канала.
Множество допущений в предыдущей фразе показывает, что здесь мы оказы-
ваемся на переднем крае исследований, и только дальнейшие тщательные экс-
перименты позволят отказаться отложных гипотез там, где они образовались.
Хотя, как это и указывалось, многое еще остается спекулятивным, осяза-
тельный рецептор С. elegans является наиболее изученным на молекулярном
уровне механорецептором. Прежде, чем двигаться дальше, стоит упомянуть,
что в гл. 8 мы будем рассматривать волосковые клетки позвоночных. Предпо-
лагается, что знания о молекулярном механизме, исследованном на механоре-
цепторах C.elegans, могут оказаться полезными для понимания того, как
функционируют эти существенно отличные органы чувств. Более детально это
интересное сравнение мы рассмотрим в разделе 8.1.
7.2. Насекомые
Мы рассмотрели структуру механорецептивных волосковых сенсилл и хордо-
тональных органов насекомых в гл. 6 (раздел 6.1.2). Мы видели, что они состо-
ят из кутикулярных выростов различной формы и размеров, содержащих ней-
росенсорные клетки, чьи дендриты присоединены к внутренней части волос-
ка сенсиллы посредством, как указывалось выше, «тубулярного тельца», со-
держащего микротрубочки. Движение чувствительного волоска обеспечивает
адекватный стимул для того, чтобы вызвать изменение потенциала на мембра-
не тубулярного тельца и дендрита. Движение в одном направлении вызывает
деполяризацию, в противоположном — гиперполяризацию. Во многих случа-
ях волоски сенсилл, как мы видели, соединены с экзоскелетом таким образом,
что движение в одной из плоскостей — наиболее свободно, т. е. сенсиллы се-
лективны по направлению движения.
7. Осязание 117
Мы отмечали в гл. 6, что в области сочленения сегментов и конечностей на-
секомых развились обширные участки механочувствительных сенсилл, обеспе-
чивающих животное кинестезической информацией. Сенсиллы также, несо-
мненно, функционируют как тактильные волоски и по всему остальному телу,
особенно голове, где они сильно развиты в антеннах (в наибольшей степени у
видов с ночной и сумеречной активностью, а также норных и подземных). Цер-
ки таракана, например, усеяны длинными нитевидными сенсиллами.
Механочувствительные сенсиллы играют важную роль и в восприятии виб-
рации. Субгенуальные органы многих насекомых особенно существенны для
детектирования вибраций в диапазоне 10—50 Гц. У американского таракана
Periplaneta этот диапазон значительно выше (как мы видели в разделе 6.1.2) и
достигает пределов, которые мы можем определить как акустические. Помимо
субгенуальных органов у многих насекомых развиты воспринимающие вибра-
цию сколопидии в лапках. Некоторые водные виды клопов (Hemiptera) и жуков
(Coleoptera) используют эти сенсоры вибрации для улавливания ряби, вызван-
ной падением в водоем жертвы. Полужесткокрылая водомерка Gerris касается
воды своими лапками сверху, тогда как гребляк Notonecta, плавающий на спи-
не, касается пленки поверхностного натяжения воды снизу. Амплитуда волны
в пределах 0,5—4 мкм представляет собой адекватный надпороговый стимул
при оптимальной частоте 20—200 Гц. Задержка времени порядка 1—4 мс вместе
с различием амплитуд, воспринимаемых расставленными лапками, использу-
ется для определения направления на источник колебаний. В данном случае
участвуют всего менее десяти сколопидиев лапок. Между этими рецепторами
вибрации и механорецепторами, подобных таковым позвоночных (гл. 8), де-
тектирующими слабые колебания воздуха, которые мы субъективно восприни-
маем как звук, нет особой разницы. Действительно, нелегко провести границу
между детекторами вибрации и акустическими рецепторами насекомых, кото-
рые, в отличие от позвоночных, представляют собой те же самые рецепторы,
часто активируемые колебаниями почвы, воды или воздуха.
7.2.1. Акустические сенсиллы и тимпанальные органы
У насекомых развились два основных типа специализированных детекторов
звука: волоски и тимпанальные органы. Волоски являются важным детекто-
ром звука только вблизи от его источника — они зависят от изгибания, вы-
званного движением воздуха. Таким образом, сенсорные волоски на церках
тараканов и прямокрылых насекомых, напр. саранчи, способны восприни-
мать акустические стимулы, испускаемые с расстояния в несколько сантимет-
ров, тогда как сенсорные волоски некоторых гусениц реагируют на частоты
биений крыльев приближающихся хищных ос. У Drosophila развились перооб-
разные «ости» на третьем сегменте антенн (рис. 7.7), настроенные на рецеп-
цию вибраций крыльев, которые действуют как внутривидовой сигнал к спа-
риванию. У многих видов насекомых как детектор вибрации действует также
джонстонов орган. Он располагается во втором сегменте антенн и состоит из
большого числа плотно упакованных сколопидиев. У разных видов он выпол-
няет различные функции в диапазоне от индикации скорости полета (пчелы)
до гравитации (Dytiscus, жук-плавунец). У комаров и звонцов (хирономиды)
118 Часть II. Механочувствительность
Волосковый
Задняя
сторона
Рис. 7.7. Различные типы акустических сенсилл насекомых. (А) Бражник имеет два ос-
новных типа акустических сенсилл: тимпанальные органы в пальпах и волосковые ор-
ганы. Тимпанальные органы в пальпах состоят из воздушных мешков, пересеченных
тонкими мембранами (на рис. правый пальп рассечен). Они действуют как детекторы
давления и вместе с волосковыми органами могут воспринимать ультразвуковые сигна-
лы насекомоядных летучих мышей. (Б) «Ости» плодовой мушки (Drosophila) выступают в
стороны из третьего сегмента антенн. Они настроены реагировать на потоки воздуха, воз-
никающие в результате биения крыльев особей собственного вида. (В) Голенное ухо на-
стоящих кузнечиков (Tettigoniidae). У этих насекомых (а также сверчков) слуховые органы
развились на голенях передних лапок (Ш — клетка-шапочка; П — промежуточный орган;
Р — рецепторная клетка; СО — субгенуальные органы; СН — субгенуальный нерв; ПТ —
передняя трахея; ТП — полость трахеи; TH — тимпанальный нерв; ЗТ — задняя трахея).
7. Осязание 119
орган развит очень сильно и состоит из многих тысяч сколопидиев, присоеди-
ненных к сложной внутренней структуре. У этих видов он различает частоту
биений крыльев (несколько сот Гц) особей собственного вида, особенно по-
тенциальных партнеров по спариванию, на расстояниях до 1 метра.
Другой тип рецепторов звука, развившийся у насекомых — тимпанальные
органы или уши — реагируют не столько на движение окружающего воздуха,
возникающего при биении крыльев, сколько на волны давления. Как и уши
позвоночных, они способны детектировать источники звука на значительных
расстояниях и чувствительны к частотам до 100 кГц и выше. Тимпанальные
органы развились, по крайней мере, у семи отрядов насекомых, включая сет-
чатокрылых (Neuroptera), чешуекрылых (Lepidoptera), жесткокрылых (Coleo-
ptera), полужесткокрылых (Hemiptera), прямокрылых (Orthoptera) и двукры-
лых (Diptera). Они используются для коммуникации, нападения и защиты.
Тимпанальный орган состоит из трех основных элементов (рис. 7.7 Д).
Сильно утонченная часть кутикулы, часто серебристая, называется тимпаналь-
ным органом или тимпанальной мембраной. Под ней располагается воздушная
полость, происходящая из полости трахеи. С тимпанальной мембраной соеди-
нен хордотональный орган, способный ощущать вибрацию. Эти основные эле-
менты скомбинированы между собой в чрезвычайно разнообразные структу-
ры, поэтому у насекомых существует множество слуховых механизмов. Тим-
панальные органы обнаружены во многих частях организма насекомого.
Рис. 7.8 показывает локализацию тимпанальных органов у различных насеко-
мых. «Уши» насекомых анатомически не выделены так, как у позвоночных.
Акустическая коммуникация хорошо развита у прямокрылых (кузнечики,
саранча) и полужесткокрылых (подотряд равнокрылые Homoptera — цикады).
Обычно самцы испускают громкую призывную песню, заставляющую самок
идти или лететь к ним. Призыв цикады часто достигает очень высокой интен-
сивности — действительно, он слышим человеческим ухом на расстоянии бо-
лее мили в тропическом лесу. Давление отбора привело к тому, что песни
сильно отличаются у разных видов. Медведки (Gryllotalpidae) тоже голосисты.
Самцы строят рогообразную норку, которая действует, как мегафон. Голос
самца привлекает самок, летающих высоко в небе. У некоторых кузнечиков
песня самца вызывает ответный голос самки, так что они поют дуэтом.
Рис. 7.7 (продолжение). (Г) У златоглазок акустические рецепторы развились в форме
расширений вен передней пары крыльев. Латерально и дорзально расширения окруже-
ны толстой кутикулой, а вентрально — тонкой волнистой кутикулой. Два хордотональ-
ных органа, состоящие из 25 сколопидиев (на рисунке показаны только два), присоеди-
нены к этой тонкой тимпанальной кутикуле. Орган реагирует на высокочастотные сиг-
налы эхолокации летучих мышей (СК — соединительные клетки; Ден — дендрит; СкК —
клетки окончания сколопидия; СС — стержень сколопидия; ТМ-В и ТМ-Г — волнистая
и гладкая части тимпанальной мембраны; Тр — трахея). (Д) Слуховой орган совки. Об-
наружен в сегменте заднегруди, состоит из широких сегментов тимпанальной мембраны,
покрывающей воздушную полость. Две рецепторных клетки А реагируют на вибрацию
тимпанальной мембраны (Б — неслуховой нейрон); ПТМ — противотимпанальная мем-
брана; Н — тимпанальный нерв; ТВМ — Тимпанальный воздушный мешок; ТМ — тим-
панальная мембрана. (А), (В), (Г), (Д) из Michelsen, 1974. Воспроизводится с разрешения
Springer-Verlag GmbH & Со. KG; (Б) из Frazier, 1985: с разрешения.
120 Часть II. Механочувствительность
Обнаружение жертвы акустическими средствами практикуется некоторы-
ми тахинами. Например, Ormia ochracea, услышав призывную песню самца
полевого сверчка Gryllus integer, направляется туда, чтобы отложить свою пара-
зитическую личинку. Органы слуха этих насекомых адаптированы таким обра-
зом, что максимальная чувствительность приходится на частоту песни самца
сверчка — 4,8 кГц. Однако, наверное, наиболее интересное применение тим-
панальных органов встречается у ночных и сумеречных златоглазок, которые
используют их для обнаружения и избегания хищных летучих мышей.
У зеленой златоглазки (Chrysopa earned) тимпанальные органы располага-
ются у оснований каждого из крыльев передней пары (рис. 1Л Г и 7.8). Это —
два хордотональных органа, включающих около 25 сколопидиев. Эти хордото-
нальные органы способны воспринимать звуки, испускаемые летучими мы-
шами в диапазоне 13—120 кГц. Летучие мыши, как мы увидим в следующей
главе, испускают 7—30 поисковых импульсов в секунду. Когда сонар захваты-
вает жертву, и летучая мышь приближается, чтобы убить ее, частота импульсов
возрастает. Тимпанальный орган златоглазки детектирует поисковые импуль-
сы сонара, и златоглазка, рефлекторно сложив крылья, бесшумно опускается
прежде, чем снова будет захвачена сонаром.
У большинства бабочек тимпанальные органы развиваются билатерально в
заднегруди. В некоторых случаях они также обнаружены на первом, втором и
седьмом абдоминальных сегментах, а у бражника — на шарообразных пальпах
головы (рис. 7.7 А). Анатомия их у разных видов бабочек варьирует незначи-
тельно (рис. 7.7 Д). Тимпанальный орган состоит из тимпанальной мембраны
(около 1 мкм толщиной), за которой находится трахеальный воздушный ме-
шок, соединенный с внешней средой через дыхальце. Вибрация тимпаналь-
ной мембраны детектируется одной, двумя или четырьмя нейросенсорными
клетками, подвешенными на тяжах соединительной ткани, идущими от цент-
ра тимпанальной мембраны и окружающей кутикулы.
У совок очень большого надсемейства Noctuoidea каждый тимпанальный
орган имеет по две нейросенсорных клетки — А1 и А2, тогда как в абдоминаль-
ных «ушах» надсемейства пядениц (Geometridae) — по четыре. Совки чувстви-
тельны к звукам, распространяющимся в воздухе, в диапазоне частот
3—150 кГц. Интересно отметить, что и диапазон и пиковая чувствительность
варьирует у островной фауны, в зависимости от вида популяции хищных лету-
чих мышей. То есть, слух совок подстроен под интенсивность и частоту специ-
фического вида летучих мышей. Две нейросенсорные клетки уха совки отвеча-
ют на интенсивности двух диапазонов. Клетка А1 реагирует на диапазон более
низкой интенсивности (пиковая чувствительность у некоторых видов —
40 дБ), а клетка А2 — более высокой (около 60 дБ). В настоящее время нет до-
казательств, что этот механизм может выделять частоты. Действительно, труд-
но себе представить физиологический механизм из двух клеток, который мог
бы выполнять эту функцию. Направление, однако, на источник звука может
быть определено. Это обеспечивается билатеральной организацией слухового
аппарата. Если одно из «ушей» экспериментально инактивировано или инва-
зировано ушным клещом Dirocheles phalenodectes, способность локализовать
источник звука утрачивается. Другой клещ D. scedastes инвазирует оба уха сов-
ки, но не повреждает ни тимпанальной мембраны, ни слухового нерва. В этом
7. Осязание 121
Рис. 7.8. Распространение тимпанальных органов у насекомых, (а) Чешуекрылые
(Lepidoptera): 1 — основание крыла (златоглазка Chrysopd), 2 — брюшко (совки и т. д.),
3 — заднегрудь (совки и т. д.), 4 — основание передних и задних крыльев (бабочки-кава-
леры Papilionoidea); 5 — основание передних крыльев (Hedyloidea), 15 — волосковый ор-
ган (бражники); жесткокрылые (Coleoptera): 6 — брюшко (жуки-скакуны Cicindelidae),
7 — цервикальные мембраны (жуки-скарабеи Scarabaeidae); сетчатокрылые (Dictyoptera):
8 — вентральная заднегрудь (богомоловые Mantodea (Mantis) и таракановые Blattoidea);
прямокрылые (Orthoptera): 9 — первый сегмент брюшка (настоящие саранчовые Acrididae).
10 — переднегрудная лапка (сверчок Gryllus); полужесткокрылые (Hemiptera): 11 — брюш-
ко (цикады Cystosoma), 12 — среднегрудь (гребляки Corixa)\ двукрылые (Diptera): 13 — вен-
тральная переднегрудь (тахины Ormia), 14 — вентральная переднегрудь (серые мясные му-
хи Colcondamyia). Из Hoy and Robert, 1996: воспроизводится с разрешения Annual Review of
Entomology, volume 41 © 1996, Annual Reviews http://www.AnnualReviews.org.
случае способность локализовать источник звука сохраняется, и совка способ-
на избегать хищных летучих мышей. В данном случае в упоминавшейся «гон-
ке вооружений» участвуют три стороны: совка, клещ и летучая мышь.
D. scalestes лучше адаптировался для выживания, чем D. phalenodectes.
Клетка А1 совок в норме способна обнаруживать поисковые импульсы со-
нара несекомоядных летучих мышей на расстоянии 30—40 м. Если летучая
мышь летит со скоростью около 8 м/с, это дает совке 4—5 с, чтобы предпринять
защитные действия. Благодаря этой способности детектировать направление
на источник звука совка способна отвернуть и улететь от летучей мыши, тогда
как последняя не способна обнаружить совку на расстоянии более 5 м. С этой
дистанции интенсивность и повторяемость поисковых импульсов возрастает,
что, в свою очередь, приводит к активации клетки А2. В этом случае у совки
остается менее секунды, чтобы избежать гибели. Она предйринимает ряд бес-
порядочных маневров избегания — двигается зигзагом, петлями, спиралями,
нырками или пассивно падает в шумящую листву. Некоторые совки способны
в эти последние доли секунды испускать звуки в последней попытке избежать
приближающихся острых зубов.
По поводу функции этих звуков, испускаемых совкой, единого мнения не
существует. Возможно, эти звуки должны предупредить атакующую летучую
122 Часть II. Механочувствительность
мышь, что совка — противная и ядовитая. В этом смысле предупредительные
звуки совок аналогичны яркой предупреждающей окраске невкусных днев-
ных насекомых, отпугивающих ею птиц. Если это на самом деле так, было бы
интересно выяснить, действительно ли акустика совок представляет собой
пример бэйтсовской (в данном случае — вокальной) мимикрии. Другие ис-
следователи, однако, предполагают, что щелчки, испускаемые совкой, — это
попытка «постановки помех» эхолокации. Показано, что они почти идентич-
ны эху, которое летучие мыши получают от своих собственных сигналов. Та-
ким образом, звуки, испускаемые совкой, создают помеху анализу ситуации
летучей мышью (см. гл. 9), заставляя ее «видеть» препятствие там, где его нет,
и, соответственно, уклоняться от него, теряя при этом из виду совку.
Теперь от удивительно хитроумных механорецепторов насекомых перей-
дем к кажущемуся прозаическим осязательному чувству кожи млекопитаю-
щих. Мы увидим, что и она может рассматриваться как сенсорная поверх-
ность, способная реагировать не только на механические стимулы, но и тер-
мические, а также стимулы, способные вызывать боль. Последние будут рас-
смотрены в гл. 19 и 21. В следующем разделе данной главы будет рассмотрена
только чувствительность к механической стимуляции. В гл. 8 мы рассмотрим
волосковые клетки позвоночных и вернемся к чрезвычайно высокой чувстви-
тельности механизмов, развившихся для восприятия очень слабых акустиче-
ских вибраций, а также к тем огромным возможностям, которые таким обра-
зом открываются для дистантной коммуникации. Мы также вернемся к со-
вершающейся под покровом тьмы борьбе между совками и рукокрылыми.
7.3. Тактильные рецепторы кожи млекопитающих
Существует несколько способов классификации чувствительных окончаний в
коже млекопитающих. Одна из распространенных систем подразделяет их на
быстро и медленно адаптирующиеся. Первые реагируют только на прикосно-
вение, но «молчат» при постоянном давлении, а вторые — и на включение дав-
ления и при постоянном смещении.
7.3.1. Быстро адаптирующиеся рецепторы
Быстро адаптирующиеся рецепторы включают пачиниевы тельца, тельца
Мейснера, терминальные колбы Краузе и чувствительные окончания волосяных
фолликулов. Пачиниевы тельца присутствуют и в неоволосенной коже (напр.,
ладони рук и подошвы стоп), и в волосистой; окончания волосяных фоллику-
лов, по определению, — только в волосистой; тельца Мейснера — только в не-
оволосенной коже приматов, а колбы Краузе — только в неоволосенной шку-
ре животных, не относящихся к приматам.
Пачиниевы тельца
Это — овальные структуры длиной от 0,5 до 2 мм в глубоких слоях дермы. На
срезах пачиниевых телец — луковицеобразная структура, состоящая из мно-
жества слоев соединительной ткани, окружающих немиелинизированное
нервное волокно (рис. 7.9 А). Полагают, что слоистая структура преобразует
7. Осязание 123
Рис. 7.9. (А) Пачиниевы тельца; (Б) тельца Мейснера.
постоянное по величине сдавливание в кратковременные стимулы. При сдав-
ливании это достигается быстрым скольжением слоев тельца относительно
друг друга до достижения вскоре некоторого нового равновесия, устраняюще-
го сдавливание. Таким образом, пачиниевы тельца способны детектировать
вибрацию даже тогда, когда находятся под постоянным давлением.
Генераторный потенциал (деполяризация) может обнаруживаться в немие-
линизированном окончании, когда пачиниево тельце сдавлено. При этом воз-
никает короткий залп импульсов в сенсорном волокне, который угасает за
1-2 секунды до нуля или очень малой частоты. Как предполагает их физиолог
гия, основной функцией пачиниевых телец кожи является детектирование ви-
брации. Частоты вибраций, на которые они реагируют, находятся в диапазоне
70—1000 Гц. Наиболее чувствительны они, однако, в диапазоне 200—400 Гц,
где деформация кожи всего на 1 мкм является достаточным стимулом.
Тельца Мейснера
Они также обнаруживаются в дерме неоволосенной кожи. Нервные оконча-
ния окружены соединительнотканной капсулой, которая прикреплена к вы-
шележащим слоям стратифицированного эпителия коллагеновыми волокна- А ,
ми (рис. 7.9 Б). Это обеспечивает эффективную механическую связь между по- ‘ '
верхностью кожи и органом чувства. Нервные окончания образуют в капсуле
спираль, ветви которой изолированы друг от друга оболочками из шваннов-
ских клеток, и стимулируются движением кожи, в частности, вибрацией. Час-
тотный диапазон этих вибрационных стимулов ниже, чем характерный для
пачиниевых телец — от 10 до 200 Гц.
124 Часть II. Механочувствительность
Терминальные колбы Краузе
Они достаточно схожи с тельцами Мейснера. Однако, они в основном встреча-
ются у млекопитающих, не относящихся к приматам, где они опять-таки обнару-
живаются в неоволосенной коже. Колбы имеют пластинчатую капсулу, окружа-
ющую стержневидныое или спиральное нервное окончание. Как и тельца Мейс-
нера, колбы реагируют на вибрацию низких частот — в пределах 10—100 Гц.
Рецепторы волосяных фолликулов
Сенсорные нервные окончания образуют сложную сеть вокруг волосяного фол-
ликула непосредственно под сальными железами. Каждое окончание образует
расширение, заполненное митохондриями и вплотную окруженное шваннов-
скими клетками. Окончания распределены вокруг фолликула, некоторые во
внешнем сосудистом слое, другие — проникают между наружным и внутренним
слоями клеток корня волоса. Некоторые веточки идут вертикально вдоль воло-
сяного фолликула; другие — обвиваются вокруг него. Еще часть веточек оканчи-
вается на дисках Меркеля под эпидермисом. Рецепторы волосяных фолликулов
подразделяются на по крайней мере три различных типа (D, G и Т) в соответ-
ствии с их чувствительностью и частотой их импульсации. Они реагируют на
любые движения волос. У многих видов млекопитающих волосы в области рта
удлиненны в форме усов (вибрисс) или тактильных волосков. Рецепторы, свя-
занные с этими образованиями, принадлежат как к быстро, так и медленно
адаптирующимся типам. Мы еще вернемся к вибриссам, когда будем рассмат-
ривать центральный анализ тактильной информации ниже в данной главе.
Рис. 7.10. Сенсорная иннервация
волосяного фолликула. Э — эпи-
дермис, Ф — фолликул, В — волос,
ВЛК— внутренний листок корня,
М — диск Меркеля, МАФ — мие-
линизированное афферентное во-
локно, НЛК — наружный листок
корня, НСС — наружный сосудис-
тый слой, С — сосочек.
7. Осязание 125
7.3.2 Медленно адаптирующиеся рецепторы
В отличие от быстро адаптирующихся рецепторов, рассмотренных в предыду-
щем разделе, эти рецепторы реагируют на смещение кожи, но продолжают ос-
таваться активными, когда кожа удерживается в новом положении. Существу-
ет три типа таких рецепторов: клетки Меркеля, нервные окончания Руфини и
С-механорецепторы. Принято объединять медленно адаптирующиеся рецеп-
торы в две категории. Рецепторы типа I реагируют на быстрые прикосновения,
а рецепторы типа II — на постоянное смещение кожи, особенно если она на-
тянута.
Клетки Меркеля
Они относятся к типу I, расположены под эпидермисом, обладают крупными
неправильной формы ядрами и микроворсинками, простирающимися к эпи-
дермальным клеткам (рис. 7.11 А). У их оснований располагаются дисковид-
ные окончания сенсорных аксонов (диски Меркеля). Группа из 10—20 клеток
Меркеля образует синаптические контакты с окончаниями одного сенсорного
аксона. Клетки Меркеля реагируют на внезапные смещения кожи, например
при поглаживании.
Окончания Руфини
Эти окончания типа II реагируют на длительные смещения кожи. Они обнару-
живаются в глубоких слоях гиподермы. Как показано на рис. 7.11 Б, оконча-
ния сенсорного аксона образуют сеть тонких веточек в соединительнотканной
капсуле.
С-механорецепторы
Большое количество немиелинизированных С-волокон берут начало в гипо-
дерме, многие — вблизи от границы между эпидермисом и гиподермой. Диф-
ференцированных сенсорных окончаний, связанных с этими волокнами, нет.
Они реагируют медленно адаптирующимся разрядом на постоянное надавли-
вание. С-волокна также реагируют и на другие модальности — температуру
(гл. 19) и повреждение кожи (боль) (гл. 21).
7.4. Анализ тактильной информации в мозге
Огромное большинство сенсорных волокон от кожных рецепторов входят в
спинной мозг через дорзальные корешки и оканчиваются в дорзальных рогах
серого вещества. Механорецептивные афференты, обычно имеющие большой
диаметр (исключая, естественно С-волокна), занимают в дорзальном корешке
центральное положение и окружены более тонкими волокнами от болевых и
терморецепторных окончаний. Тела нейронов всех этих волокон располагают-
ся в ганглиях дорзального корешка. Значительная часть немиелинизирован-
ных волокон малого диаметра (около 30%) покидает дорзальный корешок и
направляется в вентральный корешок, где оканчивается в вентральном роге
126 Часть II. Механочувствительность
Рис. 7.11. (А) Клетка Меркеля. Веточки миелинизированного аксона иннервирует
10—20 клеток Меркеля. Окончание каждой веточки расширено в форме диска (диск
Меркеля). А — афферентный аксон, БМ — базальная мембрана эпидермальных клеток,
Д — десмосамы, Э — эпидермальная клетка, ТГ — тельце Гольджи, Я — ядро клетки
Меркеля, ДМ — диск Меркеля. (Б) Окончание Руфини. Сенсорный аксон разветвляется
внутри соединительнотканной капсулы, образуя комплекс переплетающихся оконча-
ний. Из Iggo, 1982. Воспроизводится с разрешения Cambridge University Press.
серого вещества (рис. 7.12). Большинство этих волокон висцерального проис-
хождения. Участок кожи, посылающий афференты в данный дорзальный ко-
решок, называется дерматомом (см. рис. 21.3). Соседние дерматомы перекры-
ваются.
Следуя в спинном мозге, волокна тактильной чувствительности достигают
коры головного мозга различными путями. Их известно четыре (рис. 7.12), хо-
тя они и не образуют анатомически изолированных трактов. Спиноцервикаль-
ный тракт (СЦТ) и волокна дорзального канатика восходят по ипсилатераль-
ной стороне спинного мозга до латерального цервикального ядра и ядра дор-
7. Осязание 127
Рис. 7.12. Афферентные пути в
спинном мозге. Показаны А- и
С-волокна, входящие в спин-
ной мозг через дорзальный ко-
решок. Часть (ок. 30%) С-воло-
кон идут в вентральный рог се-
рого вещества через вентраль-
ный (передний) корешок. Все
тела нейронов этих волокон
лежат в ганглии дорзального
корешка. Показаны пути в
ЦНС к соматосенсорной коре.
Дальнейшие объяснения в тек-
сте. ДК — дорзальный канатик,
ИЛ — интраламинарное ядро,
ЗТЯ — заднее таламическое яд-
ро, С1 — первичное соматосен-
сорное пространство, С2 —
вторичное соматосенсорное
пространство, СЦТ — спино-
цервикальный тракт, СРТ —
спиноретикулярный тракт,
СТТ — спиноталамический
тракт, ЗВТ — заднее вентраль-
ное ядро таламуса. Из Iggo,
1982. Воспроизводится с разре-
шения Cambridge University
Press.
зального канатика, соответственно. Отсюда, из продолговатого мозга, идет
следующий участок пути, который оканчивается в заднем вентральном ядре
таламуса. Последний нейрон пути посылает аксон, оканчивающийся в пер-
вичной или вторичной соматосенсорной коре головного мозга. Два другие пу-
ти, как показано на рис. 7.12, образуют перекрест на уровне спинного мозга
(в пределах двух сегментов от места входа в спинной мозг) и образуют спино-
ретикулярный (СРТ) и спиноталамический (СТТ) тракты. Волокна СРТ вос-
ходят в вентролатеральном квадранте спинного мозга к ядрам среднего мозга
и таламуса. Отсюда волокна направляются к соматосенсорным областям коры.
Наконец, СТТ образован волокнами антеролатерального квадранта спинного
мозга, оканчивающимися на трех таламических ядрах (см. рис. 7.12). В соста-
ве этого тракта идет значительное количество ноцицептивных волокон,
Опять-таки, аксоны последних нейронов тракта восходят в соматосенсорные
участки головного мозга.
Эти пути и ядра обеспечивают некоторую первичную обработку сомато-
сенсорной информации прежде чем она достигнет коры головного мозга. Хо-
тя полагают, что между нейронами дорзального рога спинного мозга и аффе-
128 Часть II. Механочувствительность
Рис. 7.13. Верхняя часть рисунка — вид мозга сбоку. На рисунке показано расположение
соматосенсорной коры. СП — вторичная соматосенсорная кора. На правой части рисун-
ка — срез коры (помечен А—А' на левой части рисунка). Обозначены цитоархитектони-
ческие поля Бродмана (ПБ).
рентными волокнами существует взаимно однозначное соответствие, никто
не сомневается, что и интернейроны дорзального рога оказывают свой эф-
фект. Мы встретимся с важными примерами таких синаптических взаимодей-
ствий, когда будем рассматривать ноцицептивные волокна и чувство боли в
гл. 21. Более широкое взаимодействие между различными типами механочув-
ствительных афферентов имеет место в ядрах таламуса.
Пункт назначения соматосенсорных волокон в коре головного мозга — два
участка соматосенсорной (соместезической) коры. Первичная соматосенсор-
ная кора расположена в постцентральной извилине непосредственно за ро-
ландовой бороздой, а вторичная кора (у приматов) располагается в передней
стенке сильвиевой борозды.
Первичная соматосенсорная кора подразделяется на три цитоархитектони-
ческих участка: поля Бродмана 1, 2, За и ЗЬ (см. рис 7.13). Соматосенсорные
волокна из ядер таламуса оканчиваются в полях За и ЗЬ, а клетки этих полей
проецируются в поля 1 и 2. Таламические ядра также посылают немногочис-
ленные проекции во вторичную соматосенсорную кору, но большинство во-
локон поступает в этот участок из всех четырех полей первичной соматосен-
сорной коры. Вторичная кора находится в большой зависимости от первич-
ной, и если ее связи с первичной корой повреждены, активность нейронов
вторичной коры прекращается. Напротив, удаление части вторичной коры не
сказывается на активности первичной.
Если рассмотреть гистологическую структуру первичной соматосенсорной
коры, то мы обнаружим, как и во всех других участках неокортекса млекопи-
тающих, шестислойную стратификацию (рис. 7.14). Волокна из таламических
ядер оканчиваются в слое 4. Остальные слои содержат нейроны, чьи аксоны
идут в другие части мозга. Клетки слоев 2 и 3 проецируются в другие области
коры, особенно в поля Бродмана 5 и 7. Слой 5 проецируется в подкорковые яд-
ра, а слой 6 — обратно в таламус.
Менее очевидна для микроскописта колончатая структура коры, ортого-
нальная стратификации (рис. 7.14). Эти колонки впервые были обнаружены
7. Осязание 129
Рис. 7.14. Схематическое изображение
среза через соматосенсорную кору пока-
зывает ее горизонтальную стратификацию
на 6 слоев и вертикальное подразделение
на колонки. Как указывается в тексте,
каждая колонка ответственна за одну мо-
дальность. Хотя каждое поле Бродмана
получает полный набор соматосенсорных
афферентов, здесь существует некоторая
специализация. Поле Бродмана За получа-
ет афференты главным образом от мышеч-
ных веретен; ПБЗЬ — от кожных рецепто-
ров; ПБ2 — от глубоких рецепторов давле-
ния, а ПБ1 — от быстро адаптирующихся
кожных рецепторов
Верноном Маунткаслом (Vernon Mountcastle) с помощью микроэлектродного
анализа. Хотя специальные методики окрашивания и продемонстрировали их
' гистологическую реальность, они все же чаще рассматриваются как единицы
или модули физиологической активности. Колонки имеют 300—500 мкм в ди-
аметре и идут вертикально через кору, включая в себя клетки всех шести гис-
тологически различимых слоев. Микроэлектродные отведения показали, что
все клетки данной колонки отвечают на один и тот же тип или подтип стиму-
лов. Более того, Маунткасл обнаружил, что специфичность ответа здесь даже
выше — все клетки колонки отвечают на возбуждение одного и того же типа
рецептора. Например, все клетки одной колонки могут отвечать на движение
волос в какой-то определенной части тела; все клетки в другой колонке — на
возбуждение телец Мейснера в той же или иной части тела, а какая-то иная ко-
лонка — только на медленно адаптирующиеся рецепторы Меркеля и т. д. Кор-
тикальные колонки, таким образом, — это очень специфичные модули пере-
работки информации. Они получают точную информацию о какой-то одной
сенсорной субмодальности из одного участка поверхности тела. Помимо того
туда же поступает информация из других колонок соматосенсорной коры и
(через мозолистое тело) из эквивалентных колонок с противоположной сторо-
ны мозга.
Микроэлектродные исследования кроме того показывают, что клетки ко-
лонок отвечают не только на стимулы специфической модальности, но и
очень специфичны в отношении места ее происхождения. То есть, можно го-
ворить о том, что клетки первичной соматосенсорной коры имеют свои рецеп-
тивные поля (РП) (см. гл. 3). Рецептивные поля соседних клеток часто пере-
крываются, и здесь существует определенная динамика перемещения границ
РП. Размеры РП сильно варьируют в зависимости от того, какую часть орга-
низма они представляют. РП для клеток, получающих афферентацию от кон-
чиков пальцев, имеют диаметр около 3—4 мм. Напротив, РП клеток коры, по-
лучающих афференты от туловища, более чем в 100 раз крупнее. Иными сло-
вами, система организована в соответствии с ее биологическим смыслом. Те
130 Часть II. Механочувствительность
Рис. 7.15. Соматосенсорный гомункулус. Рисунок показывает венечный срез через
постцентральную извилину. Отметьте большие размеры коры, связанной с кистью
руки, лицом и губами.
части тела, где требуется тонкое различение сигналов, клетки соматосенсор-
ной коры «рассматривают под микроскопом» в мельчайших деталях, а те час-
ти тела, которые обычно не требуют такого внимания, контролируются всего
несколькими клетками.
Кортикальные клетки, чьи рецептивные поля располагаются на поверхнос-
ти тела рядом, группируются вместе и в коре. В результате в первичной сомато-
сенсорной коре образуется представительство контрлатеральной поверхности
тела, которое может быть картировано с помощью микроэлектродной техни-
ки. Такое представительство иногда называют соматосенсорным гомункулусом
(рис. 7.15). Впервые это явление было обнаружено канадским нейрохирургом
Уайлдером Пэнфилдом (Wilder Penfield) в ходе операций на головном мозге
при местной анестезии. Чтобы убедиться, что те или иные иссечения мозга не
приведут к катастрофическим последствиям, Пенфилд стимулировал кору в
различных точках и спрашивал пациента, что он чувствует. Поскольку разме-
ры РП кортикальных клеток варьируют так сильно, получившаяся карта чрез-
вычайно неизоморфна. Надо отметить, что в соматосенсорной коре «присут-
ствует» не один гомункулус, а четыре — по одному в каждом из субрегио-
1. Осязание 131
нов — т.-е. полях Бродмана 1, 2, За и ЗЬ. Эти карты в основном совпадают меж-
ду собой. Наконец, следует упомянуть, что размеры гомункулуса варьирует,
в соответствии с биологической значимостью, у разных животных.
Выше мы отмечали, что размеры РП клеток соматосенсорной коры не фик-
сированы и могут расширяться или сокращаться в ответ на различные обстоя-
тельства. Сходным образом размеры участка коры, контролирующего ту или
иную часть тела, могут меняться в ответ на воздействие. Возможно наиболее из-
вестные исследования этой лабильности выполнены на вибриссах мыши.
7.5. Пластичность соматосенсорной коры
Выше мы видели, что быстро адаптирующиеся механорецепторные волокна
охватывают нижнюю часть волосяного фолликула и воспринимают любые
движения волоса. У многих млекопитающих это чувство значительно усилено
вследствие развития вибрисс (усов) в области, окружающей рот. У мышей раз-
мер области коры, связанной с вибриссами, больше, чем связанной с лапками.
Каждая из вибрисс посылает около 100 миелинизированных волокон в соста-
ве тройничного нерва в первичную соматосенсорную кору. Эти волокна окан-
чиваются в слое 4 коры в специализированных структурах, называющихся
«бочонками» (рис. 7.16). Каждый бочонок состоит из 1500-2500 нейронов и
имеет 100—300 мкм в диаметре. Количество бочонков точно соответствует чис-
лу вибрисс на контрлатеральной стороне морды, более того, они выстроены в
таком же порядке, как сами вибриссы.
Вулси (Woolsey) и ван дер Лоос (van der Loos) выполнили несколько очень
демонстративных опытов на этой хорошо изученной системе. Если у мыши
вырастает дополнительная вибрисса, в соматосенсорной коре появляется до-
полнительный бочонок. При этом общая площадь, занимаемая бочонками, не
увеличивается — дополнительный бочонок в нее «втискивается». Это выгля-
дит так, как будто вибриссы конкурируют за ограниченное пространство ко-
ры. Аналогичным образом, если удалить вибриссу, соответствующий бочонок
атрофируется, а занимаемое им пространство оккупируется остальными. Кон-
кретные биохимические механизмы, которые реализуют такую удивительную
лабильность коры в ответ на изменение афферентации, еще остаются предме-
том множества исследований. Схожую пластичность мы встретим, когда будем
рассматривать первичную зрительную кору.
Эксперименты на обезьянах, натренированных выполнять задачу, связан-
ную с управлением вращающимся колесом кончиками двух пальцев, показали
проявление сходной пластичности соматосенсорной коры. Сравнение карт
участков коры, связанных с кончиками пальцев (что было продемонстрирова-
но микроэлектродными отведениями), до и после тренировки показало, что в
последнем случае площадь увеличивается в несколько раз. Более того, хотя об-
щая площадь представит^-—кончиков пальцев увеличивается, размеры
рецептивных полей нейронов, на которых проводилось микроэлектродное от-
ведение, оказались значительно меньше, чем у нетренированных животных.
Сходные результаты были получены на обезьянах, утративших пальцы, в этих
случаях, естественно, кортикальное представительство отсутствующих паль-
цев исчезает, а оставшихся — увеличивается.
132 Часть II. Механочувствительность
Рис. 7.16. Бочонки вибрисс мыши. (А) Го-
лова с пятью рядами вибрисс. (Б) Срез ко-
ры, показывающий бочонки, каждый из
которых соответствует одной вибриссе.
(В) Схема организации бочонков. (Г) схе-
ма, показывающая эффект удаления виб-
рисс. (i) полный набор вибрисс, полный
набор бочонков, (ii) один ряд вибрисс
удален, незатронутые бочонки растут на
площади, занимаемой неиспользуемыми
бочонками, (iii) один вертикальный ряд
вибрисс удален, тот же эффект — незатро-
нутые бочонки колонизируют простран-
ство, оставленное неиспользуемыми бо-
чонками, (iv) полное удаление вибрисс,
потеря всех бочонков. (А, Б, В) — из
Woolsey & van der Loos, 1970, с разреше-
ния. (Г) — из Cowan, 1979, с разрешения.
7. Осязание 133
Введение таких сложных методик исследования нервной системы, как
функциональный магнитный резонанс (ФМР) и магнитоэнцефалография
(МЭГ) позволило проводить аналогичные исследования и на людях. МЭГ
показала, что соматосенсорное представительство пальцев левой руки зна-
чительно увеличено у исполнителей на струнных инструментах (скрипачей,
альтистов, виолончелистов и контрабасистов) по сравнению с представи-
тельством пальцев правой руки или с представительством пальцев левой ру-
ки оркестрантов, не входящих в струнную группу.
Другое интересно исследование выполнено на слепых пациентах, обучаю-
щихся азбуке Брайля. Использование ФМР и регистрация вызванных сомато-
сенсорных потенциалов (ВСП) с помощью электроэнцефалографии (ЭЭГ) по-
казало, что участки соматосенсорной коры, в которых представлены пальцы,
участвующие в чтении брайлевских текстов, увеличиваются. Складывается
впечатление, что сначала «демаскируются» предсуществующие нейрональные
связи, которые впоследствии делаются постоянными в результате структурных
изменений. У ослепших в раннем возрасте затылочная кора также вовлекается
в процесс чтения брайлевского текста. Это особенно интересно, поскольку из-
вестно (гл. 17), что затылочная кора в норме связана со зрением. ФМР и ВСП
показывают, что у таких лиц устанавливаются (или укрепляются) кортико-кор-
тикальные связи между соматосенсорно-моторной корой, в норме обрабатыва-
ющей тактильную и двигательную информацию от участвующих в брайлевском
чтении пальцев, с первичной и вторичной зрительной корой. Эти связи кажут-
ся важными для точности чтения.
В гл. 17 мы увидим, что и первичная зрительная кора (по крайней мере в
чувствительный период) адаптируется к сенсорным (в данном случае — зри-
тельным) афферентам. Карты в зрительной коре, конечно, не фиксированы и,
как и остальные части организма, реагируют на упражнение или неупражне-
ние органа.
7.6. Заключение
Аристотель, как можно подметить в эпиграфе к Части II, полагал, что осяза-
ние — это определяющая характеристика животного. Он считал, что это наи-
более фундаментальное из всех чувств, и мы теперь имеем возможность по-
нять, что он имел в виду. В данной главе мы проникли туда, куда никак не мог
проникнуть Стагирит1 — под кожу — и увидели разнообразные органы чувств
и нервные окончания, ответственные за осязание. Мы проследили путь так-
тильной информации в соматосенсорную кору. Мы убедились, насколько
прав был философ в оценке универсальности и древности этого чувства. Мы
отметили, что тактильная чувствительность маленькой нематоды С. elegans
обещает пролить свет на механизмы, которые работают в нас самих. Мы уви-
дели, что осязательные рецепторы, развившиеся у насекомых, способны де-
тектировать рябь на поверхности воды и изменения атмосферного давления —
1 Уроженец или житель Стагиры — области в древней Македонии, чаще всего имеется
в виду Аристотель — Прим, перев.
134 Часть II. Механочувствительность
звук. Мы увидим в следующей главе аналогичные приспособления, когда рас-
смотрим ухо позвоночного.
Наше изучение чувства осязания увлекло нас дальше я жизнь животного,
чем просто в обнаружение им колебаний атмосферы. Мы увидели, как «уши»
насекомых, помимо прочих приспособлений, используются в коммуникации
между индивидами. Мы отметили, что совки и златоглазки способны обнару-
живать приближение хищных летучих мышей. Однако в еще большей степени
осязание вовлечено в социальную жизнь многих насекомых. Любой, кто на-
блюдал за взаимодействиями социальных насекомых, как муравьи и пчелы,
знает, что для них осязание имеет жизненно важное значение — достаточно
посмотреть, как часто и подолгу они ощупывают друг друга. Этот аспект осяза-
ния чрезвычайно важен и для некоторых социальных млекопитающих, особен-
но приматов. Процесс груминга, первоначально связанный с необходимостью
удаления из шерсти эктопаразитов, перерос в социальное действие. В некото-
рых обезьяньих стаях (в зависимости от размера) эта активности занимает до
20% времени. Такие тактильные взаимодействия имеют выраженный эффект
на нейроэндокринную систему, делая возможным относительно спокойное
сосуществование возбудимых и агрессивных животных. Некоторые считают,
что люди развили систему болтовни для замещения тактильных контактов.
Прикосновения, однако, остаются чрезвычайно важными и для нашего вида.
От рукопожатий до более интимных контактов — соприкосновение кожи двух
индивидов играет незаменимую роль в их общении.
8. РАВНОВЕСИЕ И СЛУХ:
ФУНКЦИЯ ВОЛОСКОВЫХ КЛЕТОК
Структура и функция волосковых клеток: структура волосковых клеток —
стереоцилии и киноцилии — соединение кончиков стереоцилий — на-
правленность — воротные механизмы ионных каналов — воротно-сдви-
говая модель стимуляции волосковых клеток — сравнение с осязатель-
ными МЕС-рецепторами С. elegans — адаптация. Каналы боковой линии:
органы боковой линии — эхолокация — адаптация каналов к различным
водным условиям. Эволюция уха: ранняя связь с системой боковой ли-
нии — основные черты структуры. Равновесие: мембранный лабиринт:
структура и функция — макулы саккулюса и утрикулюса, отолиты — ку-
пулы в ампулах полукружных каналов. Фонорецепция: рыбы (веберовы
косточки) — земноводные — пресмыкающиеся (развитие улитки lagae-
па) — птицы (улитка, кортиев орган) — млекопитающие (улитка, корти-
ев орган). Анатомия и физиология улитки млекопитающих: базилярная
мембрана — внутренние и наружные волосковые клетки — генетика —
контроль чувствительности — микрофонные потенциалы — теории зал-
па и места в механизмах частотной дискриминации — регуляция волос-
ковых клеток — высокочастотная чувствительность у собак, грызунов,
китообразных и летучих мышей — эхолокация у летучих мышей: специа-
лизация к различным условиям среды — независимая эволюция у круп-
ных и мелких рукокрылых — сенсорный мир насекомоядных летучих
мышей. Заключение: вездесущность неподвижных ресничек
Из всех тонких устройств органов чувств, развитых позвоночными, волос-
ковые клетки — возможно, наиболее удивительные. Они обнаруживаются у
различных типов и мало отличаются у рыб и у людей. Рассчитано, что на преде-
ле чувствительности уха млекопитающих волосковые клетки стимулируются
сдвигом кончика реснички всего на десяток нанометров. Хадспет (Hudspeth)
предложил замечательную аналогию — он уподобил действие адекватного сти-
мула на кончик стереоцилии волосковой клетки сдвигу верхушки Эйфелевой
башни на дюйм. Действительно, чувствительность волосковых клеток ограни-
чено только случайным шумом, производимым броуновским движением.
8.1. Анатомия и физиология волосковых клеток
Структура волосковых клеток очень схематично показана на рис. 8.1. Из кути-
кулярной пластинки на дистальном конце клетки вырастает группа до 60 «во-
лосков». Волоски — двух типов: стереоцилии и одиночная киноцилия.
136 Часть II. Механочувствительность
Рис. 8.1. Волосковая клет-
ка. Схематический рису-
нок показывает основные
элементы типичной воло-
сковой клетки позвоноч-
ных. Обычно стереоцилий
намного больше (до 60),
чем показано на рисунке,
но киноцилия — всегда
одна.
50-60 стереоцилий — это окруженные оболочкой пучки микрофиламентов, а
единственная длинная киноцилия — это истинная неподвижная ресничка, ча-
сто — с булавовидным кончиком. Рисунок показывает, что все волоски сужа-
ются к тонкой «шейке», где они входят в кутикулярную пластинку, а их размер
увеличивается по направлению к киноцилии.
Если рассмотреть стереоцилию при большом увеличении, мы увидим, что
она намного длиннее большинства ресничек — 5 мкм, а у ящерицы — даже
30 мкм. У них больше и диаметр — до 900 нм, сужающийся, однако, к основа-
нию до 100 нм. Помимо актиновых микрофиламентов стереоцилии содержат
заметное количество миозина и кальмодулина. Наконец, и это существенно,
электронно-микроскопическое исследование показывает, что кончики сте-
реоцилий связаны между собой тонкими полипептидными нитями (рис. 8.2).
Они начинаются с клубочка на самой короткой стереоцилии и идут к более
длинной соседней. Связывающие стереоцилии волокна находятся под некото-
рым натяжением, так что, если они повреждаются, мембрана стереоцилии, из
которой они исходят, расслабляется.
Уже упоминалась чрезвычайная чувствительность волосковых клеток к ме-
ханическим стимулам. В этом можно удостовериться, двигая стереоцилию
тонким микрозондом в ту и другую сторону. Стереоцилия двигается как жест-
8. Равновесие и слух: функция волосковых клеток 137
Рис. 8.2. Соединение кончиков
стереоцилий. Рисунок показы-
вает, что кончики стереоцилий
соединены между собой и, в
конце концов, с киноцилией
тонкими нитями.
кое тело — вращаясь вокруг своего суженого основания. Пороговые сдвиги
вызывают на мембране волосковой клетки рецепторный потенциал амплиту-
дой около 100 мкВ. Знак этого потенциала зависит от направления движения
стереоцилии относительно киноцилии (рис. 8.3). Если пучок стереоцилий на-
клоняется по направлению к киноцилии, наблюдается деполяризация; если в
противоположную сторону — гиперполяризация. Движение под прямым уг-
лом к этой оси не влияет на мембранный потенциал. Эти рецепторные потен-
циалы достигаются при сдвигах, превышающих ок. 100 нм.
Процесс передачи сигнала очень быстр. Многие млекопитающие, напри-
мер представители китообразных и рукокрылых, реагируют на звуковые часто-
ты до по крайней мере 100 кГц. В этом случае не хватает времени на биохими-
ческие процессы рецепторных взаимодействий и формирование вторичных
мессенджеров, которые работают в хемо- и фоторецепции. Воротные механиз-
мы каналов мембраны волосковых клеток должны открываться и закрываться
очень быстро. Электрический ответ должен наступать в результате почти мгно-
венного потока ионов по градиенту концентрации. Где же расположены эти во-
ротные механизмы?
Чтобы ответить на этот вопрос, пришлось провести тонкие эксперименты.
Вероятно, наибольший ионный поток в ответ на сдвиг наблюдается вблизи от
кончика стереоцилии. Было показано, что ответ развивается очень быстро —
начинается через несколько микросекунд после включения стимула, а насы-
щения достигает за прибл. 100 мкс. Проводимость (ок. 50 пСи при 30°С), глав-
ным образом для ионов К+, которые, как мы увидим, присутствуют а эндолим-
фе, в которую погружены стереоцилии, в высокой концентрации. То обстоя-
тельство, что через канал способны проходить и другие катионы, включая Са2+
и даже малые органические ионы, такие как холин, показывает, что канал
имеет достаточно широкую пору — вероятно, ок. 0,7 нм в диаметре. Расчеты
показали, что на каждой стереоцилии порядка четырех таких каналов.
138 Часть II. Механочувствительность
Г иперполяризация
_1------------L
Деполяризация
1_LJ 1 1 1 1 1 1 1 1 lilUUUilllUlIlIllllUllllllUl I 1 | I | | 11 11 t
Активность Повышение
в покое частоты
импульсации
и___I___I__11 । I I I I 1 I I I I I
Понижение
частоты
импульсации
Торможение
Возбуждение
Рис. 8.3. Электрический ответ волосковых клеток на движение стереоцилий. Дви-
жение стереоцилий в одном из направлений вызывает деполяризацию волосковой
клетки; движение в противоположном направлении — гиперполяризацию. Эти ва-
риации полярности преобразуются в повышение или понижение активности в
сенсорном нервном волокне. Из Flocke, 1965. Воспроизводится с разрешения Cold
Spring Harbor Laboratory Press.
Биофизическая модель стереоцилии предложена Хадспетом и другими.
Мы уже отмечали, что каждая стереоцилия связана вблизи от кончика молеку-
лярной нитью (концевой связью) с соседней более длинной (рис. 8.2). Нако-
пились данные, показывающие, что эти нити просоединены к ионным кана-
лам. Предполагается, что, когда стереоцилия находится в покое, наблюдается
небольшая утечка через каналы. Рис. 8.4 показывает, что вероятность откры-
того состояния составляет около 0,1. Когда механический стимул вызывает
сдвиг группы стереоцилий в сторону киноцилии, у более длинных стереоци-
лий вероятность открытия каналов сдвигается к единице. Поскольку, как мы
увидим ниже, реснички погружены к богатую ионами калия среду (эндолим-
фу), К+ — это основной ион, входящий в ресничку. Это вместе с потоком Са2+
в клетку вызывает деполяризацию мембраны. Если стереоцилии сдвигаются в
противоположном направлении, вероятность открывания каналов смещается
к нулю. Поток К+/Са2+ прекращается и мембрана гиперполяризуется.
Идентификация ионных каналов затруднена, поскольку на каждой стерео-
цилии их всего по одному — двум. Количество белка, таким образом, исчеза-
юще мало (несколько фемтомолей). Можно ли и в этом случае использовать
сравнительный подход? Одни из впечатляющих результатов современных мо-
лекулярно-биологических исследований заключается в поразительном един-
стве животного мира на молекулярном уровне. В гл. 7 мы рассматривали так-
тильные рецепторы Caenorhabditis elegans и отмечали их сходство с эпителиаль-
ными Ыа+-каналами (3NaK). Могут ли ионные каналы стереоцилий принад-
лежать к тому же семейству? Косвенные подтверждения этому были получены
при изучении блокаторов каналов. 3NaK очень чувствительны к амилориду —
почечные Ка+-каналы блокируются наномолярными концентрациями этого
8. Равновесие и слух: функция волосковых клеток 139
Вероятность открытого
состояния канала = О
Вероятность открытого
состояния канала = 0,1
Состояние покоя
Рис. 8.4. Воротно-сдвиговая модель стимуляции волосковых клеток. Объяснения
в тексте. Из Gillespie, 1995, с разрешения.
Вероятность открытого
состояния канала = 1
вещества. Показано, что амилорид блокирует также и каналы стереоцилий.
Следовательно, возможно, что каналы С. elegans и волосковых клеток позво-
ночных сходны. Если это так, то элегантные данные, полученные на нематоде,
могут быть пригодны для понимания работы человеческого уха.
Описанный выше механизм делает возможным почти мгновенный ответ на
механическую стимуляцию (что и наблюдается), в то же время известно, что
большинство волосковых клеток очень быстро адаптируется (за несколько де-
сятков миллисекунд).Иными словами, когда отклонение стереоцилий поддер-
живается более миллисекунды, поток К+/Са2+ внутрь клетки и последующая
деполяризация прекращаются. Как это происходит? Недавние исследования
показали существование в стереоцилии удивительного механизма. Схема его
ультраструктуры показана на рис. 8.5. Из рисунка видно, что канальный белок,
помимо концевой связи, соединен через миозиновый «движитель» с внутрен-
ними актиновыми микрофиламентами. Это соединение обеспечивает основу
для стабилизации положения канала в мембране, так что усиление натяжения
на концевой связи не сдвигает весь канальный белок по стереоцилии в плос-
кости мембраны. Снова напрашивается интересное сравнение с тем, как так-
тильный рецептор С. elegans прикрепляется к молекулам тубулина в нейросен-
сорной клетке (рис. 7.5 и 7.6).
Механизм, ответственный за адаптацию, зависит от потока Са2+ внутрь
клетки при открытом канале. Одна из теорий предполагает, что это приводит
к разрыву савязи между миозиновым «движителем» и актиновыми микрофи-
ламентами в стереоцилии. Это в свою очередь позволяет ионному каналу
скользить по мембране стереоцилии, устраняя тем самым натяжение конце-
вой связи. Вероятность открытого состояния канала снижается и поляризация
мембраны возвращается к уровню покоя. Когда стереоцилия возвращается к
нормальному неизогнутому состоянию, вероятность открытого состояния ка-
нального белка, который теперь не испытывает натяжения, приближается к
140 Часть II. Механочувствительность
Рис. 8.5. Адаптация волос-
ковых клеток. Когда рес-
нички отклонены, канал
открыт, а «моторный ком-
плекс» отсоединен от ак-
тиновых филаментов. Ка-
нальный комплекс сколь-
зит по стереоцилии и натя-
жение на воротном меха-
низме устраняется. Канал
закрывается. Затем, когда
стереоцилия возвращается
к вертикальному положе-
нию, «моторный комплекс»
поднимается по актиново-
му филаменту, таща за со-
бой канал к его положению
покоя. Дальнейшие объяс-
нения в тексте. Из Gillespie
and Corey, 1997, с разреше-
ния.
нулю. В отсутствии потока Са2+ миозиновый движитель тянет канал по акти-
новому филаменту, возвращая натяжение концевой связи к исходному значе-
нию и вероятности открытого состояния канала к 0,1. Следует оговориться —
хотя гипотетический механизм адаптации выглядит весьма привлекательно,
еще очень много исследований потребуется, чтобы установить его реальность
несомненно.
Помимо потока Са2+ через трансдукционные каналы ресничек, ионы Са2+
входят также в клетку через потенциал-зависимые каналы в кутикулярной
пластинке. Эти каналы открываются, когда мембрана деполяризуется вследст-
вие, как мы видели, отклонения стереоцилии. Поток Са2+ приводит в свою
очередь (через сложный ряд событий, описанных в книгах по молекулярной
нейробиологии) к выделению медиаторного вещества из основания волоско-
вой клетки (рис. 8.6). В зависимости от природы медиатора и природы подле-
жащего дендрита это инициирует или ингибирует синаптический потенциал.
В первом случае запускается потенциал действия, который распространяется
по сенсорному нерву в центральную нервную систему.
Хотя пионерские работы по биофизике волосковых клеток позвоночных
выполнены на клетках внутреннего уха, мало оснований сомневаться, что дан-
ные о механизме, продемонстрированном здесь, могут быть перенесены на ра-
боту волосковых клеток в других частях организма позвоночного. Волосковые
8. Равновесие и слух: функция волосковых клеток 141
Рис. 8.6. Механо-электрохимическая передача в волосковых клетках. (А) стереоци-
лии сдвинуты влево, К+/Са2+-каналы открыты. Поток катионов вызывает деполя-
ризацию, что приводит к открыванию других Са2+-каналов в кутикулярной плас-
тинке и других участках клетки. Поток Са2+ приводит к выделению медиатора на
дендрит афферентного нейрона. (Б) Стереоцилия возвращена к положению покоя.
Ионные каналы закрываются и клетка реполяризуется. Са2+ откачивается из цито-
золя, а выделение медиатора прекращается.
клетки эволюционно очень древние и развились во всех классах позвоноч-
ных — от бесчелюстных до млекопитающих. У круглоротых с их двумя совре-
менными представителями, миксиной Myxine и миногой Lampetra, волоско-
вые клетки развились и в каналах боковой линии, и в ухе.
8.2. Каналы боковой линии
У костных рыб (Osteichthyes), хрящевых рыб (Chondrichtyes) и водных земно-
водных (а также водных личиночных стадий сухопутных земноводных) разви-
лись каналы с обеих сторон тела и, особенно, в области головы (см. рис. 8.7 А).
Цель этой системы — детектирование вибраций в окружающей воде. Вибра-
ции могут возникать в результате возмущений, вызываемых другим водным
организмом, или быть отражением от окружающих объектов возмущений, вы-
зываемых самой рыбой. Система может, таким образом может рассматривать-
ся как примитивная форма эхолокатора. Мы столкнемся в конце этой главы с
более развитой формой эхолокации, когда будем рассматривать полет и пище-
вое поведение рукокрылых.
Рецепторами органа боковой линии являются группы волосковых клеток,
окруженных эпителиальными клетками и покрытых желатинозной купулой.
В последнюю погружены кончики ресничек волосковых клеток (рис. 8.7 Б).
Чувствительные и эпителиальные клетки часто группируются на небольших
холмиках. Чувствительные клетки могут образовывать изолированные класте-
ры в коже (у круглоротых и земноводных), но чаще обнаруживаются через не-
которые промежутки в желобках или каналах головы и тела. Каналы могут
представлять собой полностью замкнутые углубления в коже или иметь форму
142 Часть II. Механочувствительность
Рис. 8.7. (А) Положение системы органов боковой линии на обобщенном изобра-
жении костистой рыбы, бл — боковая линия. (Б) Орган боковой линии: в — сен-
сорное волокно, к — купула, вк — волосковая клетка, о — опорная эпителиальная
клетка; (В) Взаимосвязь между поверхностным эпителиальным чувствительным
органом и каналом боковой линии: (i) изолированный эпителиальный чувстви-
тельный орган, (ii) эпителиальный чувствительный орган в «желобке», (iii) эпите-
лиальный чувствительный орган в замкнутом канале, (iv) участок замкнутого кана-
ла, открытого во внешнюю среду посредством поры. (Г) Продольный срез боковой
линии, демонстрирующий взаимное расположение эпителиальных чувствитель-
ных органов и пор. Купула — заштрихована пунктиром (на самом деле — просто
заштрихована, на рисунке (Г) — сверху. — Перев.), эпителиальные чувствительные
органы боковой линии обозначены черным, эпителиальный клетки — заштрихо-
ваны (на рисунке (Г) — внизу. — Перев.). По Sand, 1984.
открытых желобков. Чем сильнее возможное волнение в водоеме, в котором
обитает рыба, тем более защищена система органов боковой линии. Особенно
хорошо органы боковой линии развиты у глубоководных рыб. Чувствительные
органы боковой линии иннервируются нервом боковой линии (ветвь X череп-
номозгового нерва — блуждающего, n. vagus), а в области головы — VII и IX че-
репномозговыми нервами.
В нервах боковой линии наблюдается спонтанная импульсация от чувстви-
тельных органов. Ее частота значительно увеличивается, если вода вблизи ка-
нала возмущена. Отведения от нерва боковой линии японского угря показали,
8. Равновесие и слух: функция волосковых клеток 143
что в волокнах малого диаметра наблюдается медленно адаптирующиеся отве-
ты, а в волокнах большого диаметра — высокочастотные быстро адаптирую-
щиеся ответы. Хотя у некоторых рыб активность в нервах боковой линии син-
хронна с вибрациями до частот порядка 200 Гц, нет доказательств того, что ры-
бы способны различать частоты с помощью этой системы. Возможно, об этой
системе следует говорить, как о системе «дистантного» или «водного» осяза-
ния, хотя может оказаться, что она выполняет и иные функции, например, де-
тектирования температуры. Эволюционные возможности, присущие системе
боковой линии, затрагиваются в гл. 20, в которой мы увидим, что у многих ко-
стистых и хрящевых рыб она дополнена развитой системой электрорецепто-
ров. Эта система развилась из тех же закладок, что и механочувствительная си-
стема боковой линии.
Эта система, представленная у водных земноводных, в т. ч. водных личи-
ночных стадий земноводных, полностью отсутствует у их эволюционных по-
томков — пресмыкающихся, птиц и млекопитающих. Несмотря на то, что
многие из них вернулись к водному образу жизни, система боковой линии ни
у кого не восстановилась.
8.3. Эволюция уха позвоночных
Несомненно, система боковой линии и ухо тесно связаны. У миног впячива-
ние на голове быстро изолируется от внешней среды и развивается в две запол-
ненных жидкостью трубки под прямым углом друг к другу и две камеры, так-
же заполненных жидкостью. Трубки — это полукружные каналы, а камеры —
утрикулюс (маточка) и саккулюс (мешочек). Вместе они образуют мембранное
преддверие. Многие рассматривают его как специализированный отдел систе-
мы боковой линии в голове, а его организация лежит в основе, как показыва-
ет рис. 8.8, структуры уха всех позвоночных. Только у крокодилов, а также
птиц и млекопитающих, неприметный у рыб вырост саккулюса — лагена
(lagaena), претерпевает поразительное развитие и образует собственно улитку
(cochlea). Вместе полукружные каналы, утрикулюс, саккулюс и лагена (или
улитка птиц и млекопитающих) образуют мембранный лабиринт.
8.3.1. Равновесие
Ухо первоначально было органом баланса и равновесия. Детектирование зву-
ка, которое принимает самодовлеющие формы в мире млекопитающих, и лю-
дей в частности — это позднейшее добавление. Мембранное преддверие оста-
лось поразительно неизменным за те полмиллиарда лет, что прошли с возник-
новения позвоночных (рис. 8.8). У всех челюстноротых позвоночных добавил-
ся еще один полукружный канал, ортогональный тем, которые присутствуют у
миноги. Вся система заполнена водным раствором — эндолимфой — и подве-
шена в полости уха, где она плавает в другом водном растворе — перилимфе.
Эти две жидкости радикально различаются по ионному составу. Тогда как пе-
рилимфа напоминает другие внеклеточные жидкости и имеет высокую кон-
центрацию Na+ (150 мМ/л) и низкую — К+ (3—4 мМ/л), эндолимфа намного
более схожа с внутриклеточной средой, будучи богатой К+ (150 мМ/л) и бед-
Б
Рис. 8.8. Эволюция мембранного лабиринта. (А) минога Petromyzon-, (Б) акула Scyllium\
(В) макрель Scomber, (Г) лягушка Rana\ (Д) ящерица Lacerta', (Е) голубь Columba} (Ж) мор-
ская свинка Cavia. Для рисунков А, Б, В, Г, Е, Ж: ап — ампула переднего канала, аг — ам-
пула горизонтального канала, аз — ампула заднего канала, пвк — передний вертикальный
канал, у — проток улитки, сс — crus commune, общая ножка, к — камеры, выстланные рес-
ничным эпителием; эп — эндолимфатический проток (ductus endolynphaticus), гк — гори-
зонтальный канал, 1 — lagaena, мл — макула улитки lagaena, мн — macula neglecta (макула
незаметная), мс — макула саккулюса, му — макула утрикулюса, ко — кортиев орган, ра —
papilla amphibiorum, pb — papilla basilaris, звк — задний вентральный канал, с — саккулюс,
у — утрикулюс. Для рисунка Д: ппк — передний полукружный канал, pb — papilla basilaris,
сс — crus commune, пу — проток улитки, мл — макула улитки lagaena, лпк — латеральный
полукружный канал, мн — macula neglecta, сп — слуховой проход, зпк — задний полу-
кружный канал, мс — макула саккулюса, у — утрикулюс, му — макула утрикулюса, усп —
утрикуло-саккулярный проток. Части рисунка А, Б, В, Г, Е и Ж — из Romer, 1970. Воспро-
изводится с разрешения W. В. Saunders Company; часть рисунка Д — из Baird, 1974. Вос-
производится с разрешения Springer-Verlag GmbH & Со. KG.
8. Равновесие и слух: функция волосковых клеток 145
ной Na+ (1—2,5 мМ/л). Выше мы отмечали, обсуждая биофизику волосковых
клеток, что ионные потоки в стереоцилии, главным образом, — это потоки К+.
Ниже мы увидим, что стереоцилии простираются в эндолимфу, а необычный
ионный состав эндолимфы играет ключевую роль в физиологии внутреннего
уха. Наконец, отметим, что, хотя лабиринт свободно плавает в полости уха, он
подвешен к стенкам полости коллагеновыми волокнами. В большинстве слу-
чаев тонкая трубка эндолимфатического протока (ductus endolymphaticus) про-
стирается от саккулюса до полости черепа, где оканчивается эндолимфатиче-
ским мешком.
В стенках у утрикулюса и саккулюса располагаются «пятна», содержащие
сенсорные волосковые клетки — макулы саккулюса и утрикулюса. Макула ут-
рикулюса лежит в дне компартмента, а макула саккулюса — обычно в верти-
кальной плоскости — в стенке камеры (рис. 8.9 А). Макулы иннервированы
волокнами, принадлежащими вестибулярному нерву. У рыб есть небольшое
удлиннение саккулюса — лагена, тоже содержащее сенсорную макулу. Волос-
ковые клетки макул очень похожи на клетки органов боковой линии. Здесь,
однако, могут быть выделены 2 типа клеток: амфороподобные клетки типа 1 и
цилиндрические клетки типа 2 (рис. 8.9 Б). Как и в органах боковой линии, во-
лосковые клетки погружены в желатинозную купулу. В утрикулюсе, саккулю-
се и лагене желатинозная купула часто импрегнирована кристаллами СаСО3,
образующими отолиты или «слуховые камешки». Отолит саккулюса обычно
развивается в наибольшей степени и у многих костистых рыб почти заполняет
собой камеру. Форма отолитов у разных видов рыб настолько специфична, что
может быть использована для определения вида.
Функции утрикулюса и саккулюса достаточно просты — это восприятие
линейного ускорения головы. Возможны два случая. В первом голова движет-
ся вперед-назад или вверх-вниз — при этом инерция отолитов обеспечивает
изгибание стереоцилий волосковых клеток в ту или иную сторону. Во втором
случае, когда голова неподвижна, линейное ускорение отолитов, вызванное
силой тяжести, заставляет их смещаться вниз относительно волосковых кле-
ток. Сила тяжести действует на волосковые клетки в разных направлениях в
зависимости от угла наклона головы, и эти различающиеся изгибы детектиру-
ются стереоцилиями описанным выше образом. Информация передается в
мозг по вестибулярному нерву.
Три полукружных канала расположены в трех плоскостях, все они исходят
из утрикулюса (рис. 8.8), и в каждом имеется сферическое расширение — ам-
пула, через которую они сообщаются с этой камерой. Каналы, как и прочие ча-
сти мембранного преддверия, заполнены эндолимфой. Внутри ампул распо-
ложены чувствительные участки — гребешки — в которых также присутствуют
волосковые клетки. Кончики ресничек последних погружены в большую же-
латинозную купулу, которая почти полностью закрывает просвет ампулы,
смещаясь подобно двери в метро — в ту и другую сторону.
Функция полукружных каналов — это детектирование угловых ускорений
головы. Когда голова вращается в любой из трех плоскостей, инерция эндо-
лимфы создает некоторую задержку сдвига. Затем взаимодействие ее вязких
свойств со стенкой полукружного канала приводит ее в движение, и далее она
движется в унисон в содержащей ее структурой. Однако, первичная задержка
146 Часть II. Механочувствительность
нервное
окончание
Рис. 8.9. (А) Расположение
волосковых клеток в (i) ут-
рикулюсе и (ii) саккулюсе и
лагене. (Б) Два типа волос-
ковых клеток в макулах и
гребнях мембранного лаби-
ринта: амфороподобные
клетки типа 1 и цилиндри-
ческие клетки типа 2. Клет-
ки типа 1 окружены чаше-
видными нервными окон-
чаниями. В правом верхнем
углу рисунка — срез через
реснички клетки типа 2 по
линии А—А. Киноцилия
обозначена черным круж-
ком. эо — эфферентное
нервное окончание, ос —
область синапса, нно — не-
миелинизированное нерв-
ное окончание. Воспро-
изводится с разрешения
A. J. Benson.
движения эндолимфы по отношению к содержащему ее протоку означает, что
«дверь» гребешка ампулы открывается. Другими словами, желатинозная купу-
ла и погруженные в нее стереоцилии изгибаются, что и запускает импульс в
волокнах вестибулярного нерва. Отметьте, что детектируемые угловые ускоре-
ния — это не постоянные скорости, которые могут быть вызваны длительным
вращением (порядка 20 с).
Сенсорная информация от утрикулюса, саккулюса и полукружных каналов
интегрируется с информацией, получаемой глазами, от рецепторов мышц,
связок и кожи и инициирует рефлексы, поддерживающие нормальную ориен-
тацию животного по отношению к вектору силы тяжести и противодействую-
щие приложенным внешним ускорениям во всех плоскостях. Большинство из
этих рефлексов опосредованы спинным мозгом и стволом головного мозга,
кора в них задействована очень в небольшой степени.
8. Равновесие и слух: функция волосковых клеток 147
8.3.2. Фонорецепция
Рыбы
Известно, что многие рыбы чувствительны к звуку. Выше мы отмечали, что
система боковой линии может быть вовлечена в этот процесс — при неболь-
ших частотах. В то же время, в основном в экспериментах по удалению разных
отделов, получены данные об участии саккулюса и лагены в восприятии зву-
ков высоких частот. У костистых рыб иногда в этот процесс в качестве «гидро-
фона» вовлекается плавательный пузырь. У рыбы-белки (Holocentridae), на-
пример, плавательный пузырь увеличен настолько, что примыкает к слуховой
области черепа. У костнопузырных1 (карп, золотая рыбка, линь, гольян, сом
и т. д.) обнаружены еще более сложные адаптации. Три передних позвонка
модифицированы в т. н. веберовы (слуховые) косточки. Они сочленяются с
плавательным пузырем через направленный кзади заполненный перилимфой
сосуд — непарный синус. Синус в свою очередь соединен с поперечным эндо-
лимфатическим каналом, связанным с саккулюсом (рис. 8.10). Отолиты сак-
Непарный
Слуховые
Внутреннее ухо
Рис. 8.10. Веберовы слуховые косточки. Рисунок показывает горизонтальный срез
через переднюю часть тела карпа (Cyprinus carpio). Стрелками показано направле-
ние вибраций, передаваемых от плавательного пузыря в саккулюс. Цифрами I, II,
III и IV обозначены четыре позвонка, из которых формируются косточки. С изме-
нениями из Romer, 1970.
кулюса имеют крыловидные расширения, позволяющие детектировать движе-
ние эндолимфы в этой камере. Вибрации, уловленные плавательным пузырем,
передаются через косточки в перилимфу синуса и далее — через поперечный
канал к крыловидным отолитам саккулюса. Эксперименты по выработке ре-
флексов показали, что американский сомик-кошка Amiurus способен воспри-
нимать частоты до 13 кГц и хорошо различать их (малая терция — 1,2 :1). Уда-
ление молоточка (malleus), одной из слуховых косточек, снижает чувствитель-
ность прибл. в 100 раз.
1 (Ostariophysi) совр. классификация — надотряд циприноидные (Cyprinomorpha). —
Прим, перев.
148 Часть II. Механочувствительность
Земноводные
У земноводных ухо адаптировано к восприятию изменений давления атмосфе-
ры — намного более разреженной среды, чем вода. У бесхвостых амфибий
(напр., лягушки) среднее ухо развито таким образом, что изменения давления
(звук) передаются от несколько вогнутой барабанной перепонки костным
стержнем columella auris камерам мембранного лабиринта. У хвостатых амфи-
бий, напр. саламандр, ни columella, ни среднее ухо не развиваются. Это, одна-
ко, не означает, что хвостатые глухи. Они реагируют на звук лабиринтом и
кожными механорецепторами. Рис. 8.8 показывает, что во внутреннем ухе ла-
гена претерпевает некоторое расширение, а у бесхвостых (Anura — лягушки и
жабы) присутствует и хорошо развитый базилярный сосочек (papilla basilaris).
Эти две части лабиринта, в основном, и ответственны за слух у земноводных1.
У лягушек рефлексы вырабатываются на частоты в диапазоне 50—10 000 Гц.
Слуховая чувствительность особенно важна в период размножения, когда ква-
канье служит для обоих полов призывом в воду.
Земноводные, как и все другие наземные четвероногие, распознают на-
правление на источник звука. В отличие от млекопитающих, но так же, как у
пресмыкающихся и птиц, барабанные перепонки земноводных — это не про-
сто детектор давления. У земноводных внутренние полости действуют как ре-
зонатор и передают давление через евстахиевы трубы на внутренние поверхно-
сти барабанных перепонок. Таким образом, перепонки подвергаются измене-
ниям давления и с наружной, и с внутренней стороны. Земноводные (также,
как пресмыкающиеся и птицы) могут определять направление звуковых волн
по интенсивности и фазе колебаний барабанных перепонок в ответ на волны
давления с противоположных сторон.
Пресмыкающиеся
Именно у пресмыкающихся мы сталкиваемся с закладкой настоящей улитки,
которая приобретает такое значение у птиц и млекопитающих. У аллигаторов,
например, lagaena, хотя не увеличивается в длине, присоединяется к стенке
окружающей ее полости уха, так что образуются три различных канала
(рис. 8.11 Б). Параллельно увеличивается базилярный сосочек (рис. 8.11 А). За-
чатки этого обнаруживаются и у земноводных, но у рептилий он развит значи-
тельно лучше, особенно у крокодилов и аллигаторов, и предназначен для
очень важной роли у птиц и млекопитающих. Он состоит из полоски волоско-
вых клеток, вырастающей из основной мембраны протока улитки, а начало
улитки покрыто непрерывной желатинозной мембраной. Эта текториальная
мембрана играет важную роль в восприятии вибраций.
Барабанная перепонка часто расположена глубже поверхности тела, так что
возникает зачаток наружного слухового прохода, и, как у бесхвостых амфибий,
слуховой стержень простирается через камеру среднего уха, проводя вибрации
в мембранный лабиринт внутреннего уха. Змеи являются исключением из всех
пресмыкающихся, поскольку не имеют ни наружного слухового прохода, ни
камеры среднего уха. У них однако имеется слуховой стержень, который при-
соединяется к квадратной кости челюстного сустава. Таким образом, змеи не-
1 Главным является амфибиальный сосочек, в книге не упомянутый. — Прим, перев.
8. Равновесие и слух: функция волосковых клеток 149
капсула
Перилимфа
Ушная
Проток улитки,
содержащий
эндолимфу
Текториальная
мембрана
Базилярная
мембрана
,in Перилимфа Волосковые
I11) к клетки
Рис. 8.11. (А) Мембранный лабиринт крокодила, ппп — проток переднего полукружного
канала; б — базилярный сосочек; сс — crus commune; пу — проток улитки; сп — соеди-
нительный проток; му — макула лагены; лпп — проток латерального полукружного ка-
нала; пзп — проток заднего полукружного канала; с — саккулюс; tv — tegmentum vasculo-
sum (сосудистая покрышка). Из Baird, 1974. Воспроизводится с разрешения Springer-
Verlag GmbH & Со. KG. (Б) схема поперечного среза, показывающая формирование
трехканального протока улитки у пресмыкающихся, (i) лагена рыб и земноводных сво-
бодно плавает (если не считать нескольких коллагеновых лент) в перилимфе, заполняю-
щей костный лабиринт, (ii) лагена присоединяется к стенкам костного лабиринта, обра-
зуя проток улитки. Реснички волосковых клеток, расположенных на основной мембране,
погружены в текториальную мембрану.
чувствительны к колебаниям воздуха, но великолепно ощущают колебания
почвы.
Хотя анатомические данные демонстрируют некоторый эволюционный
прогресс пресмыкающихся по отношению к земноводным, мало что свиде-
тельствует о лучшем качестве их слуха. Черепахи очень чувствительны к зву-
кам в диапазоне 80—130 Гц, аллигаторы реагируют на частоты выше 1000 Гц,
ящерицы ощущают колебания частотой до 10 кГц, но нет оснований предпо-
лагать, что они способны хорошо различать частоты.
150 Часть II. Механочувствительность
Птицы
Дело идет к концу, и мы добрались до птиц. Здесь анатомия развивалась уди-
вительно сходным образом с тем, что мы обнаружим у млекопитающих
(рис. 8.12 А). Улитка значительно удлиняется и приобретает трехканальную
структуру, которая впервые возникает, как мы видели выше, у пресмыкаю-
щихся. Верхний канал называется вестибулярной лестницей (scala vestibuli),
средний — средней лестницей (scala media), а нижняя — барабанной лестницей
(scala tympani). Поперечный срез (рис. 8.12 Б) показывает, что базилярный со-
сочек опять-таки увеличивается и теперь именуется кортиевым органом. Более
детально мы рассмотрим его, когда дойдем до слуховой системы млекопитаю-
щих. Ширина базилярной мембраны увеличивается по направлению к ее дис-
тальному концу, что предполагает детектирование низкочастотных тонов
именно в этом месте. За дальним концом улитки сохраняется макула лагены,
которая также участвует в восприятии тонов низкой частоты. Таким образом,
кохлеарного нерва
Рис. 8.12. (А) Мембранный лабиринт (i) гусиных (гусь Anser) и (ii) воробьиных
(дрозд Turdus). Отметьте различия полукружных каналов, саккулюса и протока
улитки, ппк, зпк, лпк — полукружные каналы, бс — базилярный сосочек, сс — crus
commune, мл — макула улитки lagaena, сп — слуховой канал, с — саккулюс, у — ут-
рикулюс. Из Baird, 1974. Воспроизводится с разрешения Springer-Verlag GmbH &
Со., KG. (Б) Поперечный срез через канал улитки птицы.
8. Равновесие и слух: функция волосковых клеток 151
ухо у птиц состоит из трех отделов: наружного слухового прохода, ведущего к
барабанной перепонке, среднего уха, пересеченного слуховым стержнем и
тремя неправильной формы косточками, и внутреннего уха, содержащего
мембранный лабиринт.
Большинство птиц, естественно, чрезвычайно чувствительны к звуку, ко-
торый играет огромную роль в их социальном поведении. Песни птиц видо-
специфичны, причем ясно, что птенцы выучивают ее локальный тип. Выясни-
лось, что некоторые из птиц, обитающих в пещерах, используют звуки и для
эхолокации (Steatornis — венесуэльский гуахаро, Collacalia — азиатский
стриж), однако нет никаких данных, которые бы свидетельствовали о том, что
у птиц развита чувствительность к ультразвуковым колебаниям, которая ха-
рактерна для летучих мышей. Наконец, чувствительность к направлению зву-
ка достигает очень высокого уровня у ночных хищников, таких как совы
(см. раздел 9.3).
Млекопитающие
У млекопитающих слуховая часть уха — улитка — достигает эволюционной
вершины. Самый слабый звук, который способен уловить человек, соответст-
вует около 0,0001 дин/см2 или прибл. 10“16 Вт/см2. Это соответствует измене-
нию давления на барабанной перепонке в 20 мкПа. Диапазон частот, которые
способен ощущать человек, простирается от 20 Гц до 20 кГц, хотя с возрастом
верхняя граница чувствительности снижается. Тренированное ухо может раз-
личать частоты 1000 и 1002 Гц. Такая поразительная способность следствие
высокого развития улитки. Как показывает рис. 8.8 Ж, у млекопитающих
улитка уже не прямая или слабо изогнутая, как у птиц, а свернута в спираль,
похожую по форме на раковину моллюска. Срез ее показан на рис. 8.13 А. Из
рисунка видно, что три канала — вестибулярная, средняя и барабанная лест-
ницы, с которыми мы впервые встречались у пресмыкающихся, присутствуют
и здесь, а рис. 8.13 Б показывает, что базилярный сосочек развивается здесь в
сложный кортиев орган.
Прежде чем мы сможем обсудить эту сложную тонкую структуру, необхо-
димо сделать краткий обзор уха млекопитающих в целом. Рис. 8.14 А показы-
вает, что и наружное и среднее ухо претерпели значительное развитие. У мно-
гих видов ушная раковина (не показана) велика и достаточно подвижна. Это
играет значительную роль в определении направления на источник звука.
У людей мышцы, двигающие ушную раковину у других животных, сохраняют-
ся в основном в рудиментарной форме. Наружный слуховой проход сравни-
тельно длинный и, для предупреждения нежелательных посетителей, покрыт
волосами и железами, секретирующими ушную серу. Барабанная перепонка
представляет собой границу между наружным и средним ухом. В среднем ухе
поддерживается атмосферное давление за счет прохода через евстахиеву трубу,
открывающуюся в глотку. Это важно, поскольку барабанная перепонка тонка,
и резкое различие давления по обе ее стороны может привести к ее разрыву.
Вибрация барабанной перепонки, вызываемая звуком, передается через каме-
ру среднего уха слуховыми косточками: молоточком (malleus), наковальней
(incus) и стременем (stapes). Эти, ставшие притчей во языцех, самые маленькие
152 Часть II. Механочувствительность
Ресничный эпителий--------
Нерв ---------------------
Барабанная полость________
Крючок
(конец спиральной пластинки)
Геликотрема -узкий канал,
соединяющий две лестницы
Сенсорные волосковые клетки
Туннель кортиевой дуги
Modilus (костный
стержень, вокруг которого
сворачиваются ходы улитки)
Сосудистая полоска
Спиральная связка
Спиральный кортиев
Костная спиральная пластинка
(костная "скорлупа" улитки)
Кохлеарный нерв
Верхушка улитки,
направленная
антеро-латерально
Каменистая часть
височной кости
Второй ход улитки
Рейснерова мембрана
Текториальная мембрана
Базилярная мембрана
Спиральный ганглий
Вестибулярная лестница
Средняя лестница
Барабанная лестница
Первый ход улитки
Рейснерова (вестибулярная)
мембрана
Средняя лестница (канал улитки
Лимб
Ветвь кохлеарного нерва
Костная спиральная пластинка
Нервные волокна ------------
Вестибулярная лестница
К спиральному ганглию
в modiolus
Текториальная мембрана------
Внутренняя волосковая клетка
Спиральная борозда----------
<8^88
Сосудистая полоска
Спиральный выступ
Сенсорные волоски
Сенсорный туннель
Гензеновские клетки
Клетки Хенсена
Наружные
волосковые клетки
Клетки Дейтерса
Клетки Клаудиуса (наружная
поддерживающая клетка)
Спиральная связка
Базилярная мембрана
Внутренний туннель
Спиральный проток
Кортиева дуга
Рис. 8.13. (А) Канал улитки изогнут в форме раковины моллюска. Срез на рисунке прохо-
дит вертикально через улитку. (Б) Средняя лестница и кортиев орган — более детально.
Из Freeman and Bracegirdle, 1976. Воспроизводится с разрешения Butterworth Heinemann.
кости в организме человека прикреплены к стенке камеры среднего уха связка-
ми и мышцами. Мышцы, т. tensor tympani, идущая к молоточку, и т. stapedius,
идущая к стремечку, сокращаются одновременно в ответ на звуки высокой ин-
тенсивности и таким образом защищают тонкие структуры внутреннего уха от
повреждающего эффекта избыточной вибрации. Наконец, основание стре-
мечка, овальная пластинка (basis stapedius) сочленяется с мембраной, закрыва-
ющей овальное окно (fenestra ovalis) круговой связкой. При развитии отоскле-
роза эта часть стремени срастается с костным окружением овального окна, что
ведет к глухоте.
Теперь вернемся к физиологии улитки. Для упрощения рис. 8.14 Б показы-
вает улитку, развернутой из спирали в прямую. Мы уже видели, что, когда вол-
8. Равновесие и слух: функция волосковых клеток 153
Рис. 8.14. (А) Анатомия уха млекопитающего. (Б) Принципиальная схема улитки,
развернутой в прямую. Детальные объяснения в тексте. Из Smith, 1970, с разреше-
ния.
на атмосферного давления, которую мы воспринимаем как звук, достигает ба-
рабанной перепонкой, вибрация передается в овальное окно. Таким образом,
мембрана, покрывающая окно, передает изменения давления перилимфе вну-
треннего уха. Теперь рассмотрим рис. 8.13 Б и 8.14 Б. Поскольку мембрана,
покрывающая овальное окно смещается внутрь, давление в вестибулярной ле-
стнице возрастает. Рейснерова мембрана сдвигается вниз, что увеличивает
давление в средней лестнице до тех пор, пока оно не компенсируется смеще-
нием базилярной мембраны. Возникающее вследствие этого избыточное дав-
ление в барабанной лестнице компенсируется изгибанием наружу мембраны,
покрывающей круглое окно (fenestra rotunda). Все эти движения, конечно, про-
исходят практически мгновенно.
Как видно из рис. 8.13 Б, в рассматриваемой системе присутствует еще од-
на — текториальная мембрана, опирающаяся на два ряда волосковых клеток.
Кончики ресничек погружены в нее, точно также, как в описанном выше слу-
чае — в желатинозную купулу. В отличие от рейснеровой и основной, тектори-
154 Часть II. Механочувствительность
Рис. 8.15. Сканирующая электронная микрофотография волосковых клеток кортиева
органа. Текториальная мембрана удалена, а микроскоп направлен на основную мембра-
ну. (А) Три ряда наружных волосковых клеток (отметьте U-образное расположение) и
один ряд внутренних волосковых клеток. (Б) стереоцилии внутренних волосковых кле-
ток образуют почти прямую линию. (В) Внутренние волосковые клетки при большем
увеличением (7500х). (Г) Стереоцилии наружных волосковых клеток меньше и выстрое-
ны в форме буквы U. (Д) Стереоцилии наружных волосковых клеток при большем уве-
личении (12 500х). (А), (Б) и (Г) из Pickles, 1988; воспроизводится с разрешения Academic
Press Ltd. (В) и (Д) — Pickles, личное сообщение, воспроизводится с разрешения.
8. Равновесие и слух: функция волосковых клеток 155
альная мембрана сравнительно свободно плавает в эндолимфе и следователь-
но изменения гидростатического давления влияют на нее в равной степени со
всех сторон. Поэтому изменения давления, особенно быстрые изменения, не
могут заставить эту мембрану двигаться вверх или вниз, синхронно с другими
двумя.
Теперь рассмотрим волосковые клетки. Рис. 8.13 Б показывает, что здесь
присутствует один ряд внутренних волосковых клеток, отделенных неболь-
шим пространством от трех или четырех рядов наружных волосковых клеток.
У этих клеток развивается от 50 до 100 стереоцилий, однако, в отличие от уже
рассмотренных волосковых клеток, здесь нет киноцилий. На том месте, где
можно было бы ожидать присутствия киноцилии, развивается крупная цент-
риоль. Вид клетки снаружи показывает, что стереоцилии принимают форму
буквы «W» или «U», нижние концы которых направлены к центриоли
(рис. 8.15). Кончики стереоцилий погружены в текториальную мембрану, а ос-
нования волосковых клеток присоединены в базилярной мембране, где они
образуют синаптические контакты с дендритическими окончаниями кохлеар-
ного нерва.
Интересны эти синапсы волосковых клеток. Рис. 8.16 показывает множе-
ство синаптических везикул (вероятно, содержащих глютамат), концентриру-
ющихся вокруг электронно-плотного материала, который, как и синаптичес-
кие ленты в фоторецепторах позвоночных, служит для организации везикул
перед их выбросом. Синапсы содержат различные механизмы в пресинаптиче-
ской зоне, обеспечивающие быстрое и эффективное выделение медиатора.
Здесь концентрируются Са2+-каналы такого типа, который быстро реагирует
на малые изменения потенциала. Это обстоятельство обеспечивает и почти
мгновенное прекращение выброса медиатора. Существуют еще два механизма
быстрого прекращения выброса медиатора: цитоплазма волосковых клеток —
очень жесткий кальциевый буфер, к тому же быстрая реполяризация мембра-
ны обеспечивается за счет концентрации Са2+-зависимых К+-каналов. Все эти
специализированные механизмы помогают предупредить потерю быстрого
ответа на механическую деформацию пучка ресничек в медленном синаптиче-
ском механизме в основании волосковых клеток.
Между внутренними и наружными волосковыми клетками есть существен-
ное различие. Тогда как внутренние волосковые клетки образуют контакты с
прибл. десятком афферентных волокон кохлеарного нерва, наружные много
менее иннервированы. Около 90% афферентных волокон кохлеарного нерва
идет от синаптических окончаний на ряде внутренних волосковых клеток.
Рис. 8.16 показывает, что основания волосковых клеток содержат множество
синаптических пузырьков (везикул), а также «электронно-плотное тело» (си-
наптические ленты) и другие дополнительные структуры. Присутствуют также
эфферентные волокна кохлеарного нерва. Рис. 8.16 также показывает, что эти
волокна образуют синаптические контакты с дендритными окончаниями аф-
ферентных волокон. Из этого можно предположить, что здесь над чувстви-
тельностью системы существует контроль обратных связей. Наружные волос-
ковые клетки, напротив, бедно иннервированные афферентными волокнами,
получают мощную эфферентную иннервацию. Окончания эфферентных во-
локон — крупные, заполненные медиаторными везикулами. В информацион-
Часть II. Механочувствительность
Информация 8.1
БИОФИЗИКА ВОЛОСКОВЫХ КЛЕТОК УЛИТКИ
Мы отмечали, что волосковые клетки улитки подразделяются на два типа: наруж-
ные и внутренние (рис. 8.13). Они различаются в разных частях улитки и по разме-
рам. У основания улитки морской свинки наружные волосковые клетки имеют
длину около 20 мкм'и короткие неэластичные стереоцилии, тогда как на верши-
не — 80 мкм с более длинными и гибкими стереоцилиями. Эти различия в длине
сказываются не только на их механических, но и электрических свойствах. Показа-
но, что на плазматических мембранах волосковых клеток наблюдается спонтанная
активность. Частота этих осцилляций варьирует в различных частях улитки. Вбли-
зи от круглого окна, у основания улитки, в маленьких неэластичных наружных во-
лосковых клетках короткие неэластичные наружные волосковые клетки демонст-
рируют быстрые спонтанные осцилляции, дальше в направлении геликотремы бо-
лее длинные клетки — более медленные. Существует зависимость между размером
клетки и частотой спонтанной электрической активности. Поскольку размеры на-
ружных волосковых клеток закономерно изменяются от основания к верхушке
улитки, спонтанные осцилляции соответствуют звуковой частоте, на которую наст-
роена подлежащая базилярная мембрана. Таким образом, когда поступающий слу-
ховой сигнал индуцирует механическую вибрацию основной мембраны, возникаю-
щие вследствие открывания и закрывания ионных каналов стереоцилий электрото-
нические токи усиливают предсуществующую активность. Это явление известно
как электрический резонанс. Волосковая клетка работает как усилитель, повышая
чувствительность уха на два порядка.
Различия электрического резонанса от основания к верхушке улитки подразу-
мевают, что популяция ионных каналов в волосковых клетках также закономер-
ным образом меняется от круглого окна к геликотреме. Такое предположение под-
тверждается тем фактом, что количество Са2+- и Са2+-зависимых К+ (КСа) каналов
уменьшается со снижением характеристической частоты. Исследование улитки
цыпленка показало, что каждая из приблизительно 10 000 волосковых клеток обла-
дает уникальной популяцией ионных каналов. Причем клетки различаются не
только количеством каналов, но и биофизическими характеристиками последних.
Прорыв был сделан в молекулярной биологии этой системы. Показано, что различ-
ные К+-каналы возникают в результате альтернативного сплайсинга в семи раз-
личных сайтах первичных транскриптов.
Электрический резонанс — это не только механизм, благодаря которому на-
ружные волосковые клетки увеличивают точность настройки. Показано, что эле-
ктрический ответ связан с механическим. Деполяризация ведет к укорочению во-
лосковой клетки, гиперполяризация имеет противоположный эффект, вызывая
удлиннение клетки. В точке максимальной чувствительности такая реакция мо-
жет достигать 30 нм/мВ. Механизм трансдукции изменений мембранного потен-
циала наружных волосковых клеток в изменения длины до сих пор неизвестен.
Однако очевидно, что и это повышает избирательность клетки к частоте звука.
Чем выше электрический резонанс, тем больше механическое движение, что ве-
дет к увеличению натяжения между стереоцилиями и текториальной мембраной.
8. Равновесие и слух: функция волосковых клеток 157
Продолжение
В свою очередь, это приводит к повышению вероятности открытого состояния ме-
ханочувствительных каналов стереоцилий. Полагают, что система может работать
и в противоположном направлении. Спонтанные или индуцированные афферен-
тами движения волосковых клеток могут вызывать движения основной мембраны
и изменения давления в средней лестнице и далее, которые компенсируются дви-
жениями барабанной перепонки. Это — т. н. спонтанная отоакустическая эмиссия,
которая может быть обнаружена ухом человека и других млекопитающих. Связаны
ли эти движения с «звоном в ушах», до сих пор остается предметом противоречий.
Литература
Crawford А. С. and R. Fettiplace, 1981, «Ап electrical tuning mechanism in turtle cochlear
hair cells», Journal of Physiology, 312, 377—412.
Holton, T. and A. J. Hudspeth, 1983, «А micromechanical contribution to cochlear tuning
and tonotopic oiganisation», Science, 222, 508—510.
Rosenblatt К. P. etal., 1997, «Distribution of Ca2+-activated K+ channels isoforms along
the tonotopic gradient of the chicken’s cochlea», Neuron, 19, 4061—1075.
Santos-Sacchi, J., 1992, «On the frequency limit and phase of outer hair cell motility.
Effects of the membrane filter», Journal of Neuroscience, 12, 1906—1916.
ном дополнении 8.1 мы увидим, что наружные волосковые клетки обладают
сократительной способностью. Вероятно, такая эфферентная иннервация
также позволяет мозгу контролировать чувствительность улитки путем изме-
нения расстояния между текториальной и базилярной мембранами (мы вер-
немся к этому вопросу, когда будем рассматривать физиологию волокон улит-
ки в гл. 9).
Как и можно было ожидать от такого тонкого и точного механизма, многое
в нем может ломаться и приводить к глухоте. С огромным ростом в последние
годы знаний о человеческом геноме открыто более сотни «генов глухоты». Они
расположены во всех 22 аутосомах, а также в Х-хромосоме. Многие из них свя-
заны с другими (не слуховыми) аномалиями, однако, по крайней мере дюжи-
на стоят особняком и не являются синдромными, хотя какие-то другие при-
знаки и могут быть в конце концов связаны с ними. Конечно, далеко не вся
сотня генов ответственна за формирование уха и волосковых клеток. Многие
из них в большей степени влияют на развитие центральных отделов слуховой
системы, однако те, которые отвечают именно за волосковые клетки, позволя-
ют начать формирование генетического понимания этих жизненно важных
структур. Это например ген, кодирующий калиевый канал в наружных волос-
ковых клетках; гены коннексинов, значительная роль которых в калиевом го-
меостазе также показана, и открытый сравнительно недавно ген OTOF в хро-
мосоме 2, который кодирует белок отоферлин, экпрессирующийся во внут-
ренних волосковых клетках. Существуют доказательства того, что отоферлин
вовлечен в выстраивание везикул вдоль синаптических лент, характерных, как
мы видели выше, для синапсов волосковых клеток.
158 Часть II. Механочувствительность
А
Б
Рис. 8.16. Иннервация внутрен-
них и наружных волосковых кле-
ток кортиева органа. Схематичес-
кий рисунок показывает аффе-
рентные (белые) и эфферентные
(черные) волокна. (А) внутренняя
волосковая клетка. Эфферентные
волокна образуют синаптический
контакт с дендритными оконча-
ниями афферентных волокон.
(Б) Наружная волосковая клетка.
Эфферентные волокна образуют
синапсы непосредственно на во-
лосковой клетке, которая имеет
лишь небольшое число синап-
сов (показан только один) с сен-
сорными (афферентными) волок-
нами.
Теперь обратим внимание на основную тему данного предмета. Мы виде-
ли, что базилярная мембрана колеблется в ответ на поступающий в ухо звук,
тогда как текториальная мембрана остается сравнительно неподвижной. Сте-
реоцилии волосковых клеток подвергаются механической деформации, при-
чем их реснички погружены в богатую К+ эндолимфу. Возникающая деполя-
ризация может быть обнаружена с помощью микроэлектродных отведений.
Они точно воспроизводят частоту поступающего звука. Это т. н. микрофонные
потенциалы. Микрофонные деполяризации (рецепторные потенциалы) ведут
к выделению медиаторных веществ на дендритные окончания афферентных
волокон кохлеарного нерва.
Таким образом, мы видим, что в самой основе потрясающе сложного вну-
треннего уха млекопитающих лежат волосковые клетки; конечно, модифици-
рованные, но в целом те же самые, что мы впервые встретили в каналах орга-
на боковой линии наших водных предшественников. Мы еще увидим, что
примерно то же самое можно сказать и о других органах чувств. Молекулярные
механизмы, развившиеся в эволюционной истории очень рано, сохраняются,
но со временем оказываются встроенными в невероятно сложные и хитроум-
ные органы.
Одним из эволюционных императивов, который двигал развитием улитки
млекопитающих, была необходимость отличать разные частоты звука. Мы
видели, что эта способность в небольшой степени присутствует у рыб, зем-
новодных и пресмыкающихся; у птиц и млекопитающих она претерпевает
огромное развитие. Выше мы упоминали, что частотный диапазон челове-
ческого уха лежит между 20 Гц и 20 кГц (с некоторым снижением верхнего
предела с возрастом). Мы также отмечали, что в пределах диапазона слыши-
мости человек и другие млекопитающие обладают чрезвычайно высокой
способностью к различению частот. А потому следующий вопрос — как она
достигается?
8. Равновесие и слух: функция волосковых клеток 159
Рис. 8.17. Схематическое изо-
бражение базилярной мемб-
раны. Мембрана распрямле-
на. У человека они имеет око-
ло 33 мм в длину, а ширина
меняется от прибл. 100 мкм
у круглого окна до 500 мкм
у геликотремы. Дальнейшие
объяснения — в тексте.
33 мм
500 мк
Может показаться, что у этой проблемы — простое решение. Почему бы
кохлеарному нерву не быть фазово-синхронным с приходящей звуковой вол-
ной давления? Иными словами, почему бы о тональной частоте 20 Гц не сиг-
нализировать нервными импульсами частотой 20 Гц, а о тональной частоте 15
или 20 кГц — импульсами частотой 15 и 20 кГц, соответственно? В таком про-
стом решении есть две очевидные трудности. Во-первых, как мы отмечали в
гл. 2, частота импульсов в сенсорных нервах обычно сигнализирует об интен-
сивности стимула. Нервная система могла бы, конечно, обойти это затрудне-
ние, однако, вторая трудность более непреодолима. Биофизика нервных воло-
кон такова, что за каждым импульсом следует рефрактерный период длитель-
ностью около 2 мс. Из этого вытекает (как мы видели в гл. 2), что одиночное
волокно не способно проводить более 500 импульсов в секунду. Т. о., для час-
тот выше 500 Гц нужны какие-то иные средства частотной дискриминации.
Здесь могут работать два основных механизма. Во-первых, есть данные (см.
гл. 9), что кохлеарные волокна могут быть фазо-синхронными звуковым час-
тотам рыше 500 Гц, но не реагируя на каждый частотный импульс. То есть
предполагается, что в нижней части частотного спектра (ниже 5 кГц) группа
волокон кохлеарного нерва объединяется для достижения частоты импульсов,
совпадающей с тональной частотой, в каком-то слуховом центре мозга. По по-
нятным причинам, такая идея именуется теорией залпа. Второй, значительно
более важный механизм основывается на наблюдении, что ширина базиляр-
ной мембраны увеличивается от круглого окна к геликотреме (или в случае
птиц —к макуле улитки). Ширина базилярной мембраны человека, например,
увеличивается с 100 до 500 мкм на расстоянии в 33 мм (рис. 8.17). Герман фон
Гельмгольц еще в XIX веке предположил, что базилярную мембрану можно
уподобить ряду настроенных камертонов (резонаторов). Тоны высокой часто-
ты вызывают максимальные возмущения в области круглого окна, а низкой —
у геликотремы. Точные исследования фон Бекеши (von Bekesy) и других в ос-
новном подтвердили гипотезу Гельмгольца. Обнаружено, что волны сложной
формы двигаются вдоль всей базилярной мембраны, но место, где они дости-
гают максимальной амплитуды, как и предположил Гельмгольц, связано с их
частотой. Догадка Гельмгольца по очевидным причинам известна как теория
160 Часть II. Механочувствительность
места частотной дискриминации. Чтобы различить частоты, мозгу достаточно
лишь «посмотреть», из какого места базилярной мембраны происходят волок-
на, в которых активность максимальна.
В последнее время теория места Гельмгольца была расширена и подкрепле-
на обнаружением того факта, что сами по себе волосковые клетки настроены на
определенную частоту звука. Такая настройка — результат молекулярной ана-
томии и цитологии клетки. Показано, что волосковые клетки и их стереоцилии
различаются в размерах и гибкости в разных частях базилярной мембраны.
Вблизи от круглого окна стереоцилии малы и сравнительно жестки, тогда как в
области геликотремы больше в размерах и гибки. Это вместе со специфической
популяцией ионных каналов определяет электрические характеристики данной
клетки (см. Информацию 8.1). Каждая волосковая клетка настроена на макси-
мальную реакцию в ответ на стимул определенной частоты. Есть и свидетельст-
ва того, что настройка клеток находится под контролем обратной связи от улит-
ки. Базилярная мембрана и ее волосковые клетки, таким образом, оказываются
посложнее клавиш фортепьяно или пластинок ксилофона. Тем не менее, более
столетия назад Гельмгольц сумел ухватить суть явления. Сложная электромеха-
ническая настройка волосковых клеток с модуляцией через обратные связи на-
кладывается на механизм максимального возмущения в специфической части
базилярной мембраны, связанной с частотой поступающего звука.
Многие млекопитающие чувствительны к звукам намного больших частот,
чем способно различить человеческое ухо. Эксперименты по выработке услов-
ных рефлексов при воздействии на вибриссы или ушную раковину показали,
что многие мелкие млекопитающие чувствительны к звукам частотой до
100 кГц. Слышимый для человека писк мышей и землероек находится в низ-
кочастотной части спектра слышимости этих животных. Большая часть соци-
альной коммуникации между этими млекопитающими реализуется за преде-
лами слышимости для человека. У собак частотная характеристика тоже по-
лучше, чем у человека — их верхний предел находится в районе 35 кГц. Это ис-
пользуют производители собачьих ультразвуковых свистков. А вот домашние
кошки слышат почти, как мы1. Китообразные, с другой стороны, испускают и
воспринимают звуки намного более высокочастотные — до 100 кГц. Такие
звуки могут использоваться в социальной жизни вида, а у некоторых видов,
напр. дельфина (Delphinus), и в эхолокации. Интенсивность и продолжитель-
ность эмиссии звука у них возрастает по мере приближения к препятствию или
при поиске пищи2.
8.3.3. Сонар летучих мышей
Все же настоящими чемпионами в эхолокации являются летучие мыши — два
подотряда Microchiroptera и Megachiroptera (крылановые). Существует около
680 видов мелких летучих мышей. Большинство из них — насекомоядные, но
есть и плодоядные, а некоторые стали хищниками, питающимися лягушками,
ящерицами, рыбами, птицами и мелкими млекопитающими. Три вида — кро-
1 На самом деле — до 40—50 кГц. — Прим. ред.
2 Точнее, возрастает частота, а не интенсивность и длительность. — Прим. ред.
8. Равновесие и слух: функция волосковых клеток 161
вососущие — вампиры (сем. Phyllostomatidae — листоносые). Наиболее инте-
ресны с точки зрения изучения слуха насекомоядные виды. Они достигли со-
вершенства в сложном искусстве ловли насекомых на лету. Некоторые виды
охотятся в открытом небе — выше уровня деревьев, другие специализировались
в более сложном, но и более добычливом ремесле охоты на много более много-
численных обитателей кроны деревьев. Есть и такие, кто «высматривает» добы-
чу в еще более акустически «засоренном» пространстве на почве, среди опав-
ших листьев и веточек. Показано, что системы эхолокации видов, охотящихся
в различных условиях, адаптированы к их специфическим проблемам.
Во всех случаях летучие мыши непрерывно испускают короткие импульсы
частотно-модулированных звуков. У ночницы Myotis звуковой импульс, или
«щелчок», начинается с частоты 80—90 кГц, которая за пару миллисекунд па-
дает до 30—35 кГц. Такие импульсы испускаются со скоростью до 30 в секунду,
когда летучая мышь находится в полете, и быстро учащаются при приближе-
нии насекомого-жертвы или препятствия. Другие летучие мыши испускают
звуки более низких частот, иногда лежащих в пределах слышимости человека.
Мы видели (гл. 7), что у бабочек и златоглазок развились защитные механиз-
мы подавления эхолокационной системы летучих мышей. Последние, таким
образом, не только адаптировались к охоте в различных условиях, но и к обо-
ронительной технике насекомых. Здесь, в этой гонке вооружений охотника и
жертвы, в механизмах, которые были выработаны в ее ходе, еще много неизве-
стного. Мы продолжим обсуждение этой темы в гл. 9.
Эхолокация у двух подотрядов летучих мышей, вероятно, развивалась не-
зависимо. Исследования на плодоядных крыланах показали, что за исключе-
нием одного рода Rousettus (пещерные крыланы), эхолокацией они не пользу-
ются. Rousettus издают «щелчки» с более низкой частотой, чем насекомоядные
летучие мыши (модуляция от 18 до 10 кГц), а механизм их генерации совер-
шенно иной. Крыланы намного больше опираются в своей жизни на зритель-
ную информацию, чем по большей части сумеречные и ночные мелкие насе-
комоядные летучие мыши. Предполагается, что их происхождение первично
по отношению к насекомоядным летучим мышам.
Именно у последних ухо достигает вершины развития. Мы увидим в гл. 9,
что такие же большие объемы мозга задействованы здесь на звуковую инфор-
мацию, как у приматов — на зрительную. Сенсорный мир насекомоядных
летучих мышей — это мир слуха. Аналогии между зрительной системой и
слуховой системой летучих мышей достаточно глубоки. Например, отдель-
ные участки их основной мембраны играют роль, аналогичную роли области
фовеа в сетчатке млекопитающих (гл. 9). Похож ли мир летучей мыши, летя-
щей через вечерний лес, на мир сокола ясным днем, сейчас вряд ли кто скажет.
Это и в будущем останется лишь предположением — «как бы это выглядело
для летучей мыши». **
Однако ж приятно закончить главу тем, с чего начали, эхолокацией. Воло-
сковые клетки, которые мы впервые встретили в каналах боковой линии рыб,
остались поразительно неизменными, но оказались помещенными в сложней-
шие структуры внутреннего уха млекопитающих, в данном случае, насекомо-
ядных летучих мышей. И вместо того, чтобы реагировать на легкие потоки
воды, они это делают в ответ на еще более легкие колебания воздуха.
162 Часть II. Механочувствительность
8.4. Заключение
Волосковые клетки позвоночных представляют собой первый пример того,
как эволюция, единожды найдя эффективный механорецептор, включает его
в огромное множество различных структур, некоторые из которых достигают
поразительной сложности. В этой книге мы еще встретимся с модифициро-
ванными неподвижными ресничками в других рецепторах органов чувств:
обоняния и зрения. Мы уже отмечали, что наружные сегменты многих нейро-
сенсорных дендритов насекомых (хотя и не механорецепторных) — это тоже
модифицированные неподвижные реснички. Этот общий принцип строения,
вероятно, ключ к какой-то общей черте клеточной биологии, которую мы по-
ка не сумели распознать.
Читая эту главу, мы также увидели, как механочувствительность, первона-
чально обращенная на абиотическое окружение, затем сфокусировалась на со-
седях по обитаемому миру. Такой же прогресс мы видели в гл. 7, когда рассма-
тривали не только акустическую систему насекомых, обеспечивающую ком-
муникацию между особями одного вида, а также обнаружение хищника или
жертвы, но и осязание. Последнее, как мы видели, также обеспечивает важ-
ный канал социальной коммуникации. В данной главе мы заканчивали рас-
смотрение боевым сонаром летучих мышей, однако не следует забывать и о
колоссальной роли слуха в коммуникации между особями одного вида птиц и
млекопитающих. В следующей главе мы займемся именно этим, делая особый
упор на звуковую коммуникацию у человека.
9. АНАЛИЗ ВЕСТИБУЛЯРНОЙ И
ЗВУКОВОЙ ИНФОРМАЦИИ В МОЗГЕ
Двойная функция уха: равновесие и фонорецепция. Вестибулярный путь:
ипсилатеральный вестибулярный комплекс — координация движений и
ориентация — нистагм — болезни движения — опьянение. Слуховой путь:
кохлеарные волокна — частотные характеристики — кривая частотного
порога (КЧП) — латеральное торможение; кохлеарное ядро: основные
структуры — звездчатые клетки — кустистые клетки — верхнее оливар-
ное ядро — ЕЕ и EI клетки — направление на источник звука; нижний бу-
гор четверохолмия; медиальное коленчатое тело. Картирование слухового
пространства сипухи (Tyto alba) — слуховые пути — разница интенсивно-
сти между ушами и различие по времени — пространственные карты в
нижнем бугре, зрительный мост. Слуховая кора млекопитающих: гисто-
физиология колонок — триггерные характеристики — подвижная карти-
на возбуждения. Слуховая кора летучих мышей: звуковые характеристики
пискр — сравнение с звуками человеческой речи — анализ в слуховой ко-
ре — отведение от одиночной клетки — Q10dB настройка — детектирова-
ние компенсации биений крыльев насекомых самонаведенным доппле-
ровским смещением — слуховая область фовеа в улитке — отоакустичес-
кая эмиссия — различение текстуры поверхности (цвета эха) — «что зна-
чит быть летучей мышью?» Слуховая кора человека: основа человеческой
жизни — частотно-модулированные (ЧМ-) звуки — быстрые изменения
частоты — эволюция — звуки речи (фонемы) — ЧМ и ПЧ-компоненты —
VOT — категориальное восприятие — врожденное восприятие звуков ре-
чи — категориальное восприятие у японских макаков — лингвистическое
окружение формирует врожденное восприятие речи — коартикуляция —
вызов компьютерщикам. Лингвистическая кора: доминирующее полуша-
рие — planum temporale — поля Брока и Вернике — gyrus angularis — ар-
куатный пучок. Перерезка мозолистого тела: латерализация — левое ана-
литическое, правое интегративное — генератор гипотез. Заключение:
единство в многообразии; дифференцированность сенсорных качеств
В главе 8 мы проследили развитие уха позвоночных. Мы видели, как оно воз-
никло в связи с системой uoaudum линии, главным образом как измеритель ус-
корений. Любой, кто наблюдал за рыбками в аквариуме или где-нибудь еще,
знает, что эти активные животные часто нуждаются в ориентации в отсутствии
привязки к горизонту или другим визуальным реперам. По мере эволюции, с
возникновением четвероногой наземной фауны, исходные функции сохраня-
лись, а у многих форм — в первую очередь у птиц и древесных приматов — ста-
164 Часть II. Механочувствительность
новились еще более сложными. Дополнением к исходным функциям были,
однако, все возрастающей важности требования к детектированию слабых
волн давления водной или воздушной среды, которые мы субъективно вос-
принимаем как звук. В этой главе мы проследим, куда направляется информа-
ция, полученная ухом, и что с ней происходит. Большая часть интересных во-
просов может быть описана на системах, развившихся у беспозвоночных, и ог-
раничения объема книги позволяют дать только краткий обзор картирования
слуховых зон у сипухи и у млекопитающих. Даже при этих ограничениях мы
сконцентрируемся на двух группах млекопитающих. Наибольшее внимание
будет посвящено анатомии и физиологии систем человека, а завершим эту
тему дополнительными данными о насекомоядных летучих мышах.
9.1. Вестибулярные пути и рефлексы
Анатомия вестибулярного пути чрезвычайно сложна (рис. 9.1). Афферентные
волокна из гребней полукружных каналов и макул саккулюса и утрикулюса на-
правляются в ганглий Скарпы (вестибулярный) вблизи от наружного слухово-
го прохода, где располагаются тела нейронов, а затем, после соединения с кох-
леарными волокнами образуют вестибуло-кохлеарный нерв, идущий в ипсила-
теральный вестибулярный комплекс, расположенный в вентральной части про-
долговатого мозга под четвертым мозговым желудочком. Комплекс состоит из
четырех важных ядер: латерального (ядра Дейтерса), медиального, верхнего и
нисходящего. Здесь же располагается и множество меньших ядер, объединен-
ных сложной системой афферентов и эфферентов.
Комплекс иннервирован нисходящими волокнами из мозжечка и ретику-
лярной формации. Кроме того, каждый комплекс получает иннервацию из
контрлатерального комплекса. В некоторых случаях эта контрлатеральная ин-
нервация лежит в основе механизма «тяни-толкай» (push-pull). Например,
клетки гребешка полукружного канала тоже получают информацию от гре-
бешка контрлатерального канала. Ко всему этому, комплекс получает инфор-
мацию от глаз и проприоцептивных волокон, восходящих по спинному мозгу.
Таким образом, вестибулярный комплекс является чрезвычайно важным цен-
тром интеграции информации, касающейся движения и ориентации. Рис. 9.1
показывает, что помимо мощных связей с мозжечком и глазодвигательными
ядрами, вестибулярный комплекс посылает волокна в кору головного мозга.
Полагают, что они оканчиваются в постцентральной извилине вблизи от ни-
жнего конца sulcus intraparietalis. Эпилептическим припадкам, фокус которых
располагается в этой области, обычно предшествует аура1, характеризующаяся
ощущениями головокружения и дезориентации.
Вестибулярный аппарат (как мы видели в гл. 8) отслеживает и стационар-
ную ориентацию головы в пространстве (отолиты) и ускорение ее движения
(гребни полукружных каналов). Все эти дополняется многочисленными соме-
стезическими рецепторами по всему организму (гл. 7). Чтобы устранить поток
информации от этих сенсоров, нужно поместить тело в воду или забросить на
1 Аура — один из компонентов эпилептического припадка, характеризующийся наруше-
ниями восприятия. — Прим, перев.
9. Анализ вестибулярной и звуковой информации в мозге 165
Рис. 9.1. Основные пути
вестибулярной системы.
Для простоты показаны
только восходящие пути
в стволе и высших отде-
лах мозга. Дальнейшие
объяснения в тексте.
орбитальную станцию. В этих условиях вся работа падает на глаза и вестибу-
лярный аппарат; если теперь объект еще и ослепить, останется только инфор-
мация от мембранного преддверия.
Роль информации от полукружных каналов может быть ярко продемонст-
рирована, если подопытного усадить на быстро вращающийся крутящийся
стул. Глаза в этом случае смещаются в сторону, противоположную вращению,
в попытке зафиксировать взглядом неподвижный объект и затем (при потере
его из поля зрения) — быстро скачком перемещаются в сторону вращения,
чтобы найти другую точку фиксации взгляда. Сходным образом, когда враще-
ние внезапно прекращается, глаза продолжают движение в сторону предшест-
вовавшего вращения, а затем делают скачок в противоположном направлении.
Это внезапное изменение происходит в результате того, что гребни полукруж-
ных каналов испытывают воздействие потока эндолимфы, меняющей направ-
ление потока на противоположное. Такие характерные движения глаз называ-
ются нистагмом. Они обусловлены тремя нейрональными путями от полу-
кружных каналов к вестибулярным ядрам, далее к глазодвигательным ядрам
(n. abducens) и, наконец, к наружным мышцам глаз (рис. 9.2).
Значение вестибуло-глазодвигательного рефлекса может быть ярко проде-
монстрировано, если сравнить зрение вращающейся глазной системы с зрени-
166 Часть II. Механочувствительность
Движение глаза
Левая Правая
внутренняя, внутренняя
прямая ; прямая
мышца ! мышца
Движение глаза
N. abducens
Правая
наоужная
прямая
мышца
Левая
наружная
прямая
мышца
Направление
движения
Левый горизонтальный
полукружный канал
Направление
движения
Медиальное
вестибулярное
ядро
Правый горизонтальный
полукружный канал
Рис. 9.2. Некоторые нервные пути, лежащие в основе движений глаз при нистагме. Тела
и окончания возбуждающих нейронов обозначены белым; тормозные — черным. Когда
голова вращается влево, стимулируются волосковые клетки в ампуле левого горизон-
тального полукружного канала. Нервные импульсы следуют в составе вестибулярного
нерва в вестибулярное ядро. Отсюда волокна тормозных нейронов (черные) идут в
n. abducens. Здесь образуются синапсы с волокнами, идущими к левой латеральной пря-
мой мышце (наружная мышца глаза). Эта мыщца, следовательно, тормозится. Второй
путь, через контрлатеральное n. abducens и глазодвигательные ядра, возбуждает левую
медиальную мышцу. Глаз поворачивается вправо. В то же время импульсы из правого
горизонтального полукружного канала ингибируются (волосковые клетки изогнуты
в гиперполяризующем направлении). Тормозный интернейрон в контрлатеральном ве-
стибулярном ядре ингибируется. Таким образом, активность передается из левого вес-
тибулярного ядра в правое n. abducens и к правой прямой мышце, которая сокращается
и двигает правый глаз вправо. Рисунок упрощен — показаны только некоторые реци-
прокные взаимосвязи.
ем, когда голова неподвижна, а окружение — вращается. Детали вращающего-
ся окружения очень быстро утрачиваются: при двух оборотах в секунду точка
фиксации взгляда расплывается в пятно. Напротив, подопытный, сидящий во
вращающемся кресле, несколько утрачивает остроту зрения только при скоро-
сти вращения около 10 оборотов в секунду.
Глазодви-
гательное
ядро
Латеральное
вестибулярное
ядро
9. Анализ вестибулярной и звуковой информации в мозге 167
Наконец, стоит сказать несколько слов о болезни движения. Это неприятное
ощущение возникает в основном из-за несовпадения сенсорных вводов. В не-
которых случаях это несовпадение возникает в самом вестибулярном аппара-
те. Если голова теряет нормальную ориентацию и вращается, сигналы от греб-
ней полукружных каналов больше не коррелируют с сигналами от отолитов.
Другой источник болезней движения — это несовпадение сигналов от глаз и от
вестибулярного аппарата. Если в бурном море в каюте глаза сообщают об от-
сутствии относительного движения между головой и стенами каюты, тогда как
вестибулярный аппарат, напротив, испытывает нагрузку, наблюдаются симп-
томы «морской болезни». Стоит также упомянуть, что избыточное потребле-
ние алкоголя также ведет к опасной потери ориентации. Это происходит
вследствие того, что этанол меняет специфическую плотность эндолимфы, так
что купула может теперь ощущать силу тяжести и, следовательно, посылать
необычные сигналы в центральную вестибулярную систему.
9.2. Слуховой путь
Мы видели в главе 8, что около 90% афферентных кохлеарных волокон идут от
синапсов с внутренними волосковыми клетками. Кохлеарные волокна идут
через спиральный ганглий (см. рис. 8.13), где лежат тела их нейронов, а затем
в кохлеарное ядро. Отсюда волокна следующих нейронов передают слуховую
информацию через ядра в ствол мозга, средний мозг, таламус и слуховую кору
в височной области переднего мозга (см. рис. 9.3).
Как показано на рис. 9.3, слуховой путь, в отличие от соместезического и
зрительного, но сходно с вестибулярным путем, проходит через множество
подкорковых ядер, прежде чем достигает коры мозга. Число таких ядер зави-
сит от того, склонен ли исследователь «объединять» или «разделять» — неко-
торые анатомы признают здесь наличие лишь четырех ядер, тогда как дру-
гие — до 50 с каждой стороны. Эти ядра нельзя рассматривать как простые
распределительные коробки на пути к коре мозга. Во многих случаях они ин-
тегрируют информацию, направляющуюся к ним от разных источников, ана-
лизируют ее и выделяют характеристические черты. Действительно, уже давно
известно, что для различения физических характеристик звука слуховая кора
не необходима — эта функция выполняется подкорковой системой. Слуховая
кора, как мы увидим в разделе 9.4, связана с восприятием специфических черт
окружающей звуковой среды.
Теперь рассмотрим слуховой путь несколько более детально: кохлеарные
волокна, кохлеарные ядра, ядра верхней оливы, нижние бугры четверохолмия,
медиальное ядро коленчатого тела, наконец, слуховая кора.
Кохлеарные волокна
Как и в сенсорных волокнах большинства других систем, здесь присутствует
определенный уровень фоновой активности, на который накладываются зал-
пы активности, когда улитка стимулируется. Эта вызванная активность с тече-
нием времени адаптируется. В гл. 8 мы обсуждали «теорию места» в частотной
дискриминации, из которой следует, что волокна, берущие начало в опреде-
168 Часть II. Механочувствительность
Рис. 9.3. Слуховой путь. Основные пути из левой улитки. ДКЯ — дорзальное кох-
леарное ядро; КО — кортиев орган; СГ — спиральный ганглий; ВКЯ — вентраль-
ное кохлеарное ядро. Дальнейшие объяснения в тексте.
ленных участках улитки, максимально чувствительны к специфической часто-
те звука. Частота, к которой волокно наиболее чувствительно, именуется ха-
рактеристической частотой (ХЧ). Рис. 9.4 также показывает, что с увеличением
интенсивности звука, диапазон частот, на которые отвечает данное волокно,
также сильно возрастает. Этого следовало ожидать, исходя из физики базиляр-
ной мембраны. Надо напомнить, что волна сложной формы движется вдоль
мембраны, достигая пика при резонансной частоте. Диапазон частот, на кото-
рые реагирует волокно при росте интенсивности звука, называется зоной реак-
ции на частоту. Она может рассматриваться и как рецептивное поле волокна.
Границы рецептивных полей или зон реакции на частоты описываются кри-
вой частотного порога (КЧП) или частотной настроечной кривой.
Форма КЧП, приведенная на рис. 9.4, характерна для такого рода кривых.
Ее основной чертой является резкий спад со стороны высоких частот. Крутиз-
на этого спада больше, чем можно было бы ожидать, исходя из существующих
данных измерений формы звуковых волн в базилярной мембране. Исходя из
этого было сделано предположение о существовании в улитке механизма «вто-
рого фильтра». Возможно, за такую крутизну характеристик ответственен ак-
тивный характер волосковых клеток, особенно наружных (подраздел 8.3.2;
Информация 8.1).
9. Анализ вестибулярной и звуковой информации в мозге 169
Рис. 9.4. Зона реакции на часто-
ту одиночного волокна слухо-
вого нерва. Тон фиксирован-
ной интенсивности (в дБ) по-
степенно меняется по частоте.
По мере снижения уровня ин-
тенсивности звука (дБ) ответ
становится все более сфокуси-
рованным, пока не достигается
частота, к которой волокно на-
иболее чувствительно (ХЧ —
характеристическая частота).
В данном случае это 10 кГц.
Граница зоны реакции на час-
тоту известна как кривая час-
тотного порога (КЧП). Из
Evans, 1972, воспроизводится с
разрешения.
Еще одной чертой кохлеарных волокон, которую мы встречали в других
сенсорных системах, — это определенный вид латерального торможения. По-
казано, что ответ кохлеарного волокна на тон данной частоты может быть ин-
гибирован другим тоном, близким по частоте. Далее мы увидим, что латераль-
ное торможение играет роль и на более высоких уровнях слуховой системы.
Фундаментальное значение этого механизма для других сенсорных систем мы
оценим в частях III и IV данной книги.
Рис. 9.5. Связь ответа одиноч-
ного кохлеарного волокна на
тон частотой 300 Гц с фазой
звукового импульса. (А) Вид-
но, что спайковые ответа сов-
падают по фазе со звуковым
сигналом. (Б) Связь с фазой
может стать более очевидной,
если построить гистограмму
числа спайков в зависимости
от сегмента осциллирующей
волны. Из Evans, 1972 и 1982;
с разрешения.
WVWWWVWVWb^.
Форма волны стимула (0,3 кГц)
170 Часть II. Мехайочувствительность
В гл. 8 мы упоминали, что в области низких частот (ниже 5 кГц) частотная
дискриминация достигается синхронизацией частоты поступающего звука с
частотой импульсов в одиночном волокне или группе нервных волокон. Это
предположение назвали «теорией залпа». Электрофизиологические отведения
от одиночных кохлеарных волокон подтвердили эту теорию. Рис. 9.5 А пока-
зывает импульсацию в одиночном кохлеарном волокне в фазе с полупериодом
звуковой частоты. Более четко это демонстрируется, если построить гисто-
грамму (рис. 9.5 Б).
В заключение стоит упомянуть реакцию кохлеарных волокон на «щелч-
ки» — такой стимул имеет малую длительность, но охватывает широкую поло-
су частот. Поэтому, при достаточной интенсивности стимула, практически все
кохлеарные волокна генерируют импульсы.
Кохлеарное ядро
Рис. 9.6 показывает, что кохлеарное ядро подразделяется на дорзальное и вен-
тральные. Волокна кохлеарного нерва входят в вентральное ядро посредине,
деля его на переднее вентральное и заднее вентральное. Таким образом, ядро
имеет в целом три отдела: дорзальный, передний вентральный и задний вентраль-
ный. Кохлеарные волокна входят в области соединения двух последних и там
же ветвятся, иннервируя все три отдела.
Взаимосвязи кохлеарных волокон в каждом из трех отделов сохраняются
точно, так что во всех них образуется карта базилярной мембраны. Т. е. ядро
имеет тонотопическую или кохлеотопическую организацию.
Микроскопическое исследование кохлеарного ядра показало присутствие
в них множества различных типов клеток: глобулярных, пирамидальных, ги-
гантских, сферических, мультиполярных (звездчатых) и т. д. Микроэлектрод-
ные отведения от этих клеток показывают, что их ответы на афферентацию из
кохлеарного нерва существенно различаются. Электрические ответы боль-
шинства малых глобулярных клеток схожи с ответами волокон кохлеарного
Пирамидальная
нерв
клетка
Рис. 9.6. Анатомия кохлеар-
ного ядра. ПВКЯ — переднее
вентральное кохлеарное яд-
ро; ДКЯ — дорзальное кохле-
арное ядро; ЗВКЯ — заднее
вентральное кохлеарное яд-
ро. Дальнейшие объяснения
в тексте.
9. Анализ вестибулярной и звуковой информации в мозге 171
нерва. С другой стороны, звездчатые клетки отвечают на постоянный стимул
регулярной серией импульсов и, вследствие этого иногда именуются клетками-
прерывателями (chopper cells). Частота ответа этих клеток варьирует. Вероятно,
эти клетки передают информацию о частотных компонентах звука до уровня
прибл. 800 гЦ. Напротив, кустистые клетки, чье дендритное дерево широко рас-
пространяется по заднему вентральному ядру, отвечают только на включение
сигнала. Этот ответ, как мы увидим, вероятно, важен в локализации звука.
В клетках дорзального ядра обнаруживается более сложные ответы, здесь, оче-
видно, присутствует латеральное торможение. Одиночный тон может иногда
уменьшить или полностью устранить активность в клетке. Вероятно, этот меха-
низм улучшает частотную дискриминацию двух сходных тонов и может повы-
шать чувствительность к частотной модуляции в одном направлении больше,
чем в другом. Здесь, как мы увидим далее, существует строгая аналогия с теми
формами обработки информации, которые имеют место в сетчатке.
Верхние ядра оливы
Эта группа ядер находится в вентральной части варолиева моста. Это первые
ядра, которые получают афферентацию от обоих ушей. Таким образом, они
важны для корреляции афферентов с двух сторон и следовательно локализа-
ции источника звука. Здесь обнаружены три физиологических типа клеток,
два из которых отвечают на частоты, выше 1 кГц, и один — ниже 1 кГц. Рас-
смотрим их по порядку. Первый тип, клетки Е-Е, отвечает на сходные вариа-
ции интенсивности звука в обоих ушах. Второй тип, клетки Е-I, отвечает на
различие интенсивности звука в ушах. Это, в зависимости от «затенения от
звука» головой или ушной раковиной, оказывается существенным для опреде-
ления направления на источник звука. Третий тип клеток, чувствительный к
звуковым частотам ниже 1 кГц, детектирует временную задержку между сигна-
лами, приходящими от обоих ушей. Некоторые из этих клеток, т. н. клетки
критической задержки, отвечают импульсами только на очень специфические
величины задержки, порядка 150 или 200 мкс. Вместе все эти клетки способны
определять горизонтальное направление на источник звука.
Способность распознавать направление на источник звука, конечно, имеет
для всех животных огромное значение, поскольку дает им возможность обнару-
живать хищников, жертв или половых партнеров и т. д. У многих млекопитаю-
щих — огромные и очень подвижные ушные раковины, которые также участву-
ют в локализации источника звука. Конечно, источник звука может быть опре-
делен и менее эффективным путем — с помощью всего лишь одного уха. Это
происходит за счет чувствительности уха к тому, как частотный спектр меняет-
ся при отражении от туловища животного, его морды и различных выступов и
впадин ушной раковины. Звук, приходящий с различных направлений, по-раз-
ному отражается от их контуров и, единственное ухо чувствительное к этим раз-
личиям спектрального состава звука, или «звуковому цвету», способно опреде-
лять направление на источник звука, особенно — выше он или ниже уровня го-
ловы, сзади или спереди. Надо отметить, что отнюдь не у млекопитающих раз-
вилась самая тонкая слуховая:система определения направления. Мы увидим в
следующем разделе, что ночные хищники, совы, обладают способностью обна-
руживать легчайшие шумы движения их жертв с почти невероятной точностью.
172 Часть II. Механочувствительность
Нижний бугор четверохолмия
Эта область среднего мозга развилась из крыши среднего мозга низших позво-
ночных. У анамний это конечный пункт слуховых волокон, а потому имеет
особое значение. У млекопитающих оно меньше, поскольку окончательный
анализ слуховой информации у них смещен в слуховую кору. Тем не менее, и
в этом случае здесь происходит переключение информации и присутствует
кохлеотопическая карта. Многие клетки здесь четко проявляют бинауральные
характеристики (т. е. зависят от информации от обоих ушей). Кроме того, не-
которые клетки здесь вообще не реагируют на чистые тоны, а лишь на частот-
но или амплитудно-модулированные тоны. Как мы увидим в разделе 9.5, это
особенно хорошо показано на рукокрылых.
Медиальное ядро коленчатого тела
Это последняя станция переключения перед слуховой корой. Оно иннервиру-
ется ипсилатеральным нцжним бугром четверохолмия и посылает свои волок-
на в ипсилатеральную переднюю височную извилину (первичную слуховую
кору). Мы вернемся к слуховой коре в разделе 9.4. А сейчас мы оставим рас-
смотрение слуховой системы млекопитающих, чтобы заняться несколько от-
личной (хотя и не радикально) системой сипухи. Как отмечалось выше, она
представляет огромный интерес как по-видимому наиболее совершенная сис-
тема обнаружения источников слабых звуков.
9.3. Картирование слухового пространства у сипухи
Способность обнаруживать источник звука очень важна для сов, поскольку
они должны слышать слабые шумы, производимые в ночи их жертвами. Сипу-
ха (Tyto alba), как и другие совы, обладает множеством специализированных
приспособлений к ночной охоте. Еще древним было известно, что перья их
крыльев обеспечивают им бесшумность полета; вошедшие в поговорки боль-
шие глаза очень светочувствительны и расположены так, чтобы обеспечить на-
илучшее стереоскопическое зрение, и наконец, острый слух, исключительно
чувствительный к направлению на источник звука. Это наглядно показывает
манера сов поворачивать голову в сторону неожиданно возникшего звука. Ус-
тановка миниатюрных наушников, инактивация того или другого уха с помо-
щью анестетиков и другие экспериментальные методы подтвердили значение
слуховой системы в этом рефлексе ориентации на источник звука.
Слуховой путь птиц, что и не удивительно, несколько отличается от тако-
вого млекопитающих (рис. 9.7). Как уже говорилось в разделе 8.3.2, барабан-
ная перепонка подвергается воздействию волн звукового давления с обеих
сторон. Звуковой сигнал, который передается через камеру среднего уха в
улитку, более сложен, чем у млекопитающих. Из улитки волокна кохлеарного
нерва идут в средний мозг, причем каждое ветвится, одна ветвь направляется в
ангулярное ядро, а другая в крупноклеточное ядро. Из ангулярного ядра слу-
ховой путь идет в заднее ядро боковой петли на контрлатеральной стороне, а
отсюда — в слуховую область нижнего четверохолмия. Эта область образована
сердцевиной и окружающей ее скорлупой. Рис. 9.7 показывает, что проекции
9. Анализ вестибулярной и звуковой информации в мозге 173
Улитка
Архистриатум
(полосатое тело)
Кохлеарный
нерв
Слуховая область
нижних бугров
четверохолмия
Тектум оптикум
(верхние бугры
четверохолмия)
Заднее латеральное
лемнискодпо ядро
\ Наружное
ядро
Переднее
латеральное
лемнисковое
ядро
Ангулярное
ядро
Крупноклеточное
ядро Ламинарное
ядро
Рис. 9.7. Слуховые пути сипухи (упрощено). Для простоты показаны пути только
одного уха. Объяснения в тексте. По Konishi, 1993; Cohen and Knudsen, 1999.
из заднего ядра боковой петли оканчиваются на скорлупе. Из крупноклеточ-
ного ядра путь идет к ламинарным ядрам ипси- и контрлатеральной сторон.
Проекции из этих ядер, наряду с коллатералями к переднему ядру боковой
петли, направляются в основное ядро контрлатеральных нижних бугров четве-
рохолмия. Отсюда путь далее идет к скорлупе, а далее информация передается
в наружное ядро нижних бугров четверохолмия и, в конце концов в зритель-
ный тектум и зрительные поля архистриатума переднего мозга.
Направление на источник звука, как и у млекопитающих, определяется
комбинацией частотно-специфических различий интенсивности между ушами
(МУРИ) и частотно-специфическими междуушными различиями во времени
(МУРВ). Надо отметить, что уши сипухи немного асимметричны. Левое ухо
расположено на голове несколько выше, но направлено несколько книзу, тог-
да как правое ухо, расположенное ниже, направлено вверх. Таким образом, ле-
вое ухо более чувствительно к звукам, поступающим с направлений ниже уров-
ня головы, а правое — к поступающим сверху. Сипуха способна локализовать
источник звука в вертикальной плоскости, сравнивая интенсивности сигналов,
получаемых ушами. Если, например, интенсивность звука при данной частоте
(выше 3 кГц), передаваемого через наушники, выше слева, чем справа, сипуха
наклоняет голову вниз (и, соответственно, вверх — в противоположном слу-
174 Часть II. Механочувствительность
А Звуковая волна
Кохлеарное волокно
Детектор совпадения
(выраженный ответ)
Рис. 9.8. (А) Связь с фазой
звукового колебания и детек-
торы совпадений звука в ушах.
Кохлеарные волокна реагиру-
ют импульсами на каждый
второй пик звуковой волны,
(а) когда кохлеарные волокна
от противоположных ушей
конвергируют в детекторе
совпадений, последний акти-
вируется при разнице сигна-
лов во времени в пределах де-
сятков микросекунд; (б) если
разница во времени больше,
детектор реагирует значитель-
но слабее или не реагирует
совсем. (Б) Принцип локации
источника звука по междууш-
ной разнице во времени по-
ступления сигналов (МУРВ).
Источник звука (И), равно
удаленный от обоих ушей,
стимулирует определенный
детектор совпадений (черный
кружок); источник звука уда-
ленный от одного уха больше,
чем от другого, стимулирует
другой детектор. ЛУ — левое
ухо, ПУ — правое ухо. Даль-
нейшие объяснения в тексте.
По Konishi, 1993.
чае). Междуушные различия во времени поступления сигнала (МУРВ) при
данной частоте вызывают движение головы сипухи в горизонтальной плоско-
сти. В этом случае при постоянной частоте и интенсивности задержка сигнала
между правым и левым ухом в 42 мкс вызывает поворот головы на 20°. Таким
образом, за счет комбинации МУРИ и МУРВ сипуха способна определять на-
правление, с которого поступает звук в трехмерном пространстве.
Разделение МУРИ и МУРВ в слуховом пути происходит на ранних этапах.
Рис. 9.7 показывает, что кохлеарные волокна передают сигналы и в крупнокле-
точное и в ангулярное ядра. Оказывается, нейроны, чувствительные к МУРВ,
сконцентрированы в крупноклеточном ядре, тогда как чувствительные к МУ-
РИ — в ангулярном. Кохлеарные волокна, приходящие в крупноклеточное яд-
ро, связаны с фазой поступающего звука до частот около 10 кГц (рис. 9.8) и не
чувствительны к интенсивности. Волокна, поступающие в ангулярное ядро,
с фазой не связаны, но реагируют на различия интенсивности звука.
Как показано на рис. 9.7, крупноклеточное ядро проецируется в ламинар-
ное ядро. Здесь информация от обоих ушей суммируется. Многие клетки ла-
минарного ядра напоминают клетки критической задержки в медиальном
9. Анализ вестибулярной и звуковой информации в мозге 175
ядре верхней оливы млекопитающих — это клетки-детекторы совпадений.
Они активируются только когда сигналы от обоих ушей поступают приблизи-
тельно одновременно. Эта система у сипухи чрезвычайно точна — детекторы
совпадений способны различать временную задержку всего в 10 мкс. При по-
стоянной скорости проведения по кохлеарным волокнам и интернейронам де-
тектор совпадений дает возможность клеткам ламинарного ядра определять
различие во времени между звуками, поступающими в оба уха и, таким обра-
зом, — направление на источник звука (рис. 9.8).
Различия интенсивностей в ушах сравниваются в заднем латеральном
лемнисковом (ЗЛЛ) ядре боковой петли (рис. 9.7). Обработка информации
зависит от возбуждающей импульсации, поступающей из контрлатерального
ангулярного ядра и тормозной, поступающей непрямым путем из ипсила-
терального ангулярного ядра. В сущности, клетки ЗЛЛ ядра сравнивают
уровень торможения и возбуждения при данной частоте и реагируют или не
реагируют в зависимости от обстоятельств. Таким образом сравнивается
интенсивность звука в ушах.
Сигнал от детектора совпадений в ламинарном ядре проецируется (как мы
указывали выше) в основное ядро нижних бугров четверохолмия. Информа-
ция об интенсивности сигнала из ЗЛЛ-ядра, напротив, передается в область
скорлупы в той же области. Оба пути в конце концов комбинируются во внеш-
нем ядре — именно здесь информация о МУРИ и МУРВ встречаются впервые.
После внешнего ядра нижних бугров четверохолмия слуховой путь разветвля-
ется снова. Одна из ветвей направляется в зрительный тектум (соответствую-
щий верхним буграм четверохолмия млекопитающих), а другая — в передний
мозг (в т. н. «поля взгляда» архистриатума). Эти поля (соответствующие фрон-
тальным полям зрения млекопитающих) проецируются в двигательные ядра,
контролирующие движения головы и глаз, а также направление взгляда.
Клетки, реагирующие на расположение источника звука в слуховом прост-
ранстве, правильным образом распределены между внешним ядром нижних
бугров четверохолмия и зрительным тектумом. Иными словами, эти отделы
среднего мозга содержат карты слухового пространства. Точность слуховых
карт у сипухи превосходят ту, которая обнаруживается у млекопитающих. Ха-
рактеристики пространственной настройки клеток в зрительном тектуме си-
пухи — порядка 40 мкс по МУРВ и 15 дБ по МУРИ. Эти клетки, как показыва-
ет рис. 9.9, расположены таким образом, что формируется пространственная
карта. Полагают, что активность в этой карте сравнивается с активностью бо-
лее глубоких слоев тектума, которые представляют направление взгляда. Сиг-
налы о несовпадении двух наложенных одна на другую карт могут использо-
ваться для корректировки направления взгляда на источник звука. В высших
отделах слуховой системы — полях зрения архистриатума — такой изомор-
физм утрачивается. Клетки^представляющие определенные положения в про-
странстве, по-прежнему объединены в группы, однако между группами отсут-
ствует топографическая организация. Клетки одной группы, настроенные на
определенный участок пространства, граничат с клетками другой группы,
представляющей топически не связанный участок пространства. Вероятно,
такая неизоморфная организация пространственно-настроенных клеток яв-
ляется следствием того, что передний мозг решает множество еще более слож-
176 Часть II. Механочувствительность
Крыша среднего мозга (тектум оптикум — ТО)
S Азимут источника звука (град.)
Рис. 9.9. Карта слухового пространстве в зрительном тектуме сипухи. (А) Пространствен-
ная слуховая настройка нейронов зрительного тектума. Ответы на импульс шума, пред-
ставленные в разных позициях в горизонтальной (азимут) и вертикальной (возвышение)
плоскостях, показаны на графиках внизу и справа рисунка, соответственно. Закрашено
«оптимальное место» нейрона. (Б) Схема трех отведений от левого зрительного текту-
ма — места расположения электродов. (В) Пространственные координаты оптимальных
слуховых участков, зарегистрированных в точках, показанных на рисунке (Б). Перепеча-
тано из TINS, 22, Cohen and Knudsen, 132, © 1999, воспроизводится с разрешения Elsevier
Science.
ных задач, таких как идентификация источника звука, выделение одного слу-
хового стимула, требующего внимания, из массы других; вызов из памяти
сходного источника звука и т. д.
9.4. Слуховая кора
От рассмотрения картирования слухового пространства чуждого нам мозга си-
пухи вернемся к организации мозга представителей нашей собственной ветви
эволюционного древа. У приматов слуховая кора располагается на внутренней
поверхности передней теменной извилины (рис. 9.10). У других млекопитаю-
щих, например кошки, она распространена на большой поверхности височ-
ной доли. Слуховая кора включает первичную слуховую кору (А1), окружен-
ную полосами вторичной и третичной слуховой коры. А1 получает информа-
цию от медиального ядра коленчатого тела, тогда как окружающие ее участ-
9. Анализ вестибулярной и звуковой информации в мозге 177
Рис. 9.10. Расположение и при-
близительные границы участков
слуховой коры у кошки и обезья-
ны (совинолицей мартышки). Пе-
редняя часть на рисунке располо-
жена справа. Слуховая кора кош-
ки располагается на поверхности
мозга. А1 — первичная слуховая
кора, АП — вторичная слуховая
кора, П — переднее поле, В —
вентральное поле, ЗВ — заднее
вентральное поле, 3 — заднее по-
ле, Т — височное поле. Слуховая
кора обезьяны показана на верх-
ней поверхности височной доли;
для этого теменная кора отсечена.
А1 — первичная слуховая кора,
ПЛ — переднелатеральное корти-
кальное поле, КМ — каудомеди-
альное кортикальное поле, ЗЛ —
заднее латеральное кортикальное
поле, Р — ростральное кортикаль-
ное поле. Из Brugge and Reale,
1985. Воспроизводится с разреше-
ния Kluwer Academic Publishers.
ки — помимо этого и от других таламических ядер. Поэтому их можно рассма-
тривать как ассоциативные полисенсорные участки коры. Тем не менее, все
они обладают полными картами улитки, хотя амплитуда и ориентация их и ва-
рьирует в разных областях.
Гистологическая структура слуховой коры очень похожа на таковую других
сенсорных областей. Как обычно, здесь имеет место шестислойная стратифи-
кация. Входящие в кору волокна оканчиваются в слое IV. Слой V посылает
проекцию обратно в медиальное ядро коленчатого тела, а слой VI — в нижние
бугры четверохолмия. Ортогонально шестислойной структуре здесь, как и в
других частях неокортекса (новой коры), присутствует колончатая структура.
В некоторых частях колонок клетки обнаруживают специфическую реакцию
на бинауральные стимулы. Именно там имеет место вход большого количест-
ва волокон Л мозолистого тела, т. е. эти клетки иннервируются от обоих ушей.
Колонки суммации содержат клетки, чьи ответы суммируют одновременно по-
ступающую информацию от обоих ушей. Напротив, колонки подавления более
выраженно реагируют на информацию от только одного, и если стимулируют-
ся оба уха, их ответ уменьшается.
Кроме этих бинауральных колонок (сравнимых с бинокулярными колон-
ками первичной зрительной коры (см. гл. 17)), первичная слуховая кора содер-
178 Часть II. Механочувствительность
частоты
частоты
частоты частоты частоты частоты
Частота
Рис. 9.11. Ответы нейронов слуховой коры кошки на частотно-модулированные
тоны. Стрелки на рисунке показывают, в каком направлении должна измениться
частота, чтобы вызвать ответ клетки. На (А) ответ возникает, только когда частота
внутри диапазона растет, тогда как на (Б) — только когда частота снижается. На (В)
ответ наблюдается, когда частота растет в нижней части диапазона и снижается —
в верхней.
жит клетки, специфически реагирующие на определенные параметры слухо-
вой информации. В отличие от клеток низших отделов слухового пути, кото-
рые реагируют на чистые тоны или широкополосные щелчки, многие клетки
коры «запускаются» только частотно-модулированными тонами — при повы-
шении, понижении или, в некоторых случаях, и повышении, и понижении
(рис. 9.11). Эта специализация (опять-таки сравнимая с зрительной корой)
развивается у млекопитающих, для которых важна социальная вокализация
Например, показано, что первичная слуховая кора макаков и саймири содер-
жит клетки, отвечающие только на внутривидовой призыв, а иногда даже на
призыв отдельных особей.
Таким образом, мы видим, что по достижении слуховой коры слуховая ин-
формация подвергается детальному анализу. Специфические клетки коры ре-
агируют на включение или выключение звука,"щелчки, частотную модуляцию
(повышение, понижение, повышение и понижение, а также скорость измене-
ний), локализацию стимула в пространстве, его перемещение, внутривидовую
вокализацию и т. д. Слуховая кора, если позаимствовать шеррингтоновский
образ, это подвижная картина возбуждения. Она динамически картирует пока
с трудом понимаемым нами образом слуховое окружение, в котором обитают
млекопитающие. Хотя нейрофизиологи и понимают, как работает отдельный
механизм системы, они все еще с трудом представляют себе, как вся динами-
ческая картина интегрируется, «контрастируется» или ослабляется в том или
ином месте, образуя единый мир звуков, который ощущают животные.
В следующем разделе мы рассмотрим развитие этой гибкой системы у лету-
чих мышей, которая превратилась в удивительно чувствительный механизм
эхолокации — на грани физических пределов. Далее, в разделах 9.6 и 9.7 мы
увидим, как эта же система, развиваясь иным путем у гоминид, сформировала
чрезвычайно важную для них кору распознавания речи. Нетрудно доказать,
что весь феномен человека зависит от развития этой коры.
9. Анализ вестибулярной и звуковой информации в мозге 179
9.5. Слуховая система летучих мышей и эхолокация
Летучие мыши, как и совы — ночные охотники, однако они, в отличие от сов,
не зависят от звуков, испускаемых жертвой. Они занимают более активную
позицию. Мы видели в гл. 8, что насекомоядные летучие мыши испускают ко-
роткие залпы высокочастотных звуков. Характеристики последних варьируют
у различных видов, и есть данные, свидетельствующие о связи этих различий с
разными стратегиями питания. Летучие мыши, охотящиеся в чистом небе или
вокруг крон деревьев, обычно испускают «щебет», состоящий из тонов посто-
янной частоты (ПЧ), которые при приближении к насекомому-жертве сменя-
ются модулированными тонами с быстро понижающейся частотой (ЧМ).
У разных видов ПЧ-тон может иметь высоту в пределах 150—30 кГц и длиться
от нескольких миллисекунд до 60 мс. ЧМ-тон обычно имеет очень малую дли-
тельность — 1—2 мс, а частота его снижается с более чем 100 до менее 30 кГц.
Этот факт представляет специальный интерес (как мы увидим в следующем
разделе) в связи со звуками человеческой речи, в которой также присутствуют
ПЧ- и ЧМ-компоненты, хотя и при значительно более низких частотах. Лету-
чие мыши, конечно, используют эти звуки для эхолокации. Именно отраже-
ния звуков от препятствий или от жертв интересуют их слуховую систему.
Хотя ПЧ-компонент используется для сканирования окружающего прост-
ранства в радиусе до 50 м в поисках жертвы, он также полезен и для обнаруже-
ния (благодаря эффекту Допплера) движения жертвы по направлению к охот-
нику или прочь от него. ЧМ-компонент используется большинством летучих
мышей (но не всеми) для определения направления движения жертвы при не-
посредственной атаке. Широкий набор частот в ЧМ-спектре обеспечивает ле-
тучей мыши детальную информацию о движении жертвы, дистанции до нее
и т. д. Направление, откуда приходит эхо, определяется очень точно. Поведен-
ческие эксперименты показали, что летучие мыши различают источники эха,
удаленные друг от друга на 0,3 см на дистанции 30 см. Нейрональные механиз-
мы, вовлеченные в этот процесс, располагаются в нижнем четверохолмии и
слуховой коре. Многие нейроны в этих областях реагируют на пары стимулов,
разделенных специфическими интервалами, и практически не реагируют на
одиночные стимулы. Известны два основных класса нейронов. В первом из
них латентный период ответа на тон высокой интенсивности длиннее, чем на
более слабый тон той же частоты. Как и в случае системы определения направ-
ления на источник звука сов, полагают, что соответствующие клетки образуют
синаптические контакты с нейронами-детекторами совпадений, и последние
активируются только при строго определенном интервале между испусканием
звука и приходом эха (определяющем расстояние до цели) (рис. 9.12 В). Вто-
рой класс нейронов настроен просто на максимальную реакцию при опреде-
ленном интервале между испусканием звука и приходом эха. Здесь мы снова
сталкиваемся с развившимся до удивительной сложности механизмом кратко-
временной памяти, подобный тем, который встречается в слуховой системе, в
частности при определении направления на источник звука путем сравнения
времени получения бинаурального сигнала.
Микроэлектродные отведения показывают, что более 50% клеток слуховой
коры малой бурой ночницы Myotis lucifugus наиболее эффективно отвечают на
180 Часть II. Механочувствительность
А
В
120 -Н
- 90
со
СО
60
Импульс Эхо
4 Постоянная
частота4
Н3----------
° Постоянная
частотаз
Частотная s
модуляция* *
* Н
2 Постоянная!
частота2 I
DC
• Н.-------—
1 Постоянная
частота, _ Частотная
—► Задержка модуляция
30
Б
Частотная
модуляция.
Частотная
модуляция, Q.
Импульс
3 5
СО о
Р.р.г.
2,ОММР-РР-
Эхо
Латентный
период
ответа
на импульс
Время
А
Клетка-детектор
совпадений
Латентный
период
ответа
на импульс
Вентральная
сторона
10 20 30
Время в мс
ДБ
ДИФ
ЧМ/ЧМ-поле
ПЧ/ПЧ-поле
Дорзальная
сторона
MrFM
FMrFM4
FM,-FMj
CFt CFл
дспч
20/ю
Первичная слуховая кора
Передняя
сторона
Азимутальное
направление
Контуры
оптимальных
частот 150 кГц
Задержка 0,4-18 мс
радиус действия 7-310 см
Задержка 0,8-9 мс
радиус действия 14-156 см
Задняя -
сторона
0
0
Рис. 9.12. (А) Спектрограммы сигнала биосонара Pteronotus. Показаны четыре гармони-
ки (Н12,з,4) импульса. Основная гармоника Н2 выделена. Эхо с допплеровским смеще-
нием обозначено пунктирной линией. Задержка эха — около 8 мс, а допплеровское сме-
щение варьирует в зависимости от частоты гармоники. Из Suga, Yan and Zhang, 1997.
Воспроизводится с разрешения Nobuo Suga. (Б) Дорзо-латеральный вид слуховой коры
Pteronotus. а — Ala, б — Alb, в — область ПЧ с допплеровским смещением (ДСПЧ); г —
область ПЧ/ПЧ; д — дорзальная интрафоссальная область (ДИФ); е — ЧМ/ЧМ область;
ж — область дорзальной бахромы (ДБ); з — область вентральной бахромы; и — DM
(дорзо-медиальная зона). Масштабная линейка для Р.р.г. (панамского голоспина
Pteronotus pamelii rubiginosus) и P.p.p. (Pteronotus parnelii pamelii).
9. Анализ вестибулярной и звуковой информации в мозге 181
пару импульсов, сходных с теми, которые испускает этот вид. Наибольший от-
вет достигался в ответ на интенсивный первый стимул, за которым следовал
более слабый импульс. Схожие клетки обнаружены в коре американского уса-
того голоспина (Pteronotusparnellii). У этого вида импульсы биосонара состоят
из ПЧ-компонента длительностью 15 мс, за которым следует короткий ЧМ-
компонент. Каждый импульс включает 4 основные гармоники, которые обыч-
но обозначаются как ПЧ1-4 и ЧМ1-4 (рис. 9.12 А). Микроэлектродное иссле-
дование коры мозга Pteronotus показало, что его сложная организация хорошо
адаптирована для анализа тонов биосонара.
На рис. 9.12 Б показана слуховая кора голоспина, представляющая собой
мозаику различных участков. Функциональные детали этих областей показа-
ны на рис. 9.12 Г. Первичная слуховая кора (А1) включает в себя продольную
полосу коры с систематической частотной картой. В задней части этой полосы
(Alp) клетки реагируют на тоны сравнительно невысокой частоты
(10—20 кГц), а в передней (Ala) — на более высокочастотные (выше 100 кГц).
Ближе к переднему концу этой полосы располагается зона, известная как об-
ласть ПЧ с допплеровским смещением (ДСПЧ), где клетки настроены на де-
тектирование вариаций второй гармоники эха (ПЧ2). Эта характеристика ука-
зывает на скорость относительного смещения цели и летучей мыши. Непо-
средственно над областью Ala располагается область ПЧ/ПЧ, в которой клет-
ки-детекторы совпадений настроены на реакцию на ПЧ-тон и вторую или тре-
тью гармонику его эха (ПЧ2, ПЧ3). Эта область также особо чувствительна к
изменениям частоты сигнала эха вследствие допплеровского смещения. Сход-
ная организация обнаружена и в небольшой области — дорзальной интрафос-
сальной (ДИФ). Выше области ПЧ/ПЧ располагаются два участка, в которых
клетки активируются ЧМ-тонами. Эти клетки опять-таки являются детекто-
рами совпадений. Их максимальный ответ вызывается ЧМ-тоном первой гар-
моники и второй, третьей или четвертой гармониками его эха (ЧМ2, ЧМ3,
ЧМ4). Время между эмиссией звука и приходом его эха — критично. В облас-
ти ЧМ/ЧМ такая задержка составляет от 0,4 до 18 мс, что соответствует рассто-
яниям в 7—310 см, тогда как в области дорзальной бахромы (ДБ) оптимальная
задержка между тоном и эхом составляет 0,8—19 мс, т. е. расстояниям в
14—156 см. Рис. 9.12 показывает, что в мозге летучей мыши имеется еще ряд
малых областей, имеющих в эхолокации специализированные функции. Об-
Рис. 9.12 (продолжение). С изменениями из Suga, 1995. (В) Гипотеза детектирования сов-
падений. Импульс запускает ответ в клетке А, имеющей большой латентный период. Эхо
запускает ответ в клетке Б с более коротким латентным периодом. Если импульсы из кле-
ток Аи Б достигают клетки-детектора совпадений В одновременно, последняя активиру-
ется. По Neuweiler, 1990. (Г) Функциональная организация слуховой коры Pteronotus. Уве-
личенное и схематизированное изображение рис. 9.12 Б. Черные линии обозначают кро-
веносные сосуды. Основной кровеносный сосуд проходит по сильвиевой ямке. ПЧ — тон
постоянной частоты; ПЧ с допплеровским смещением (ДСПЧ); дорзальная бахрома
(ДБ); дорзальная интрафоссальная область (ДИФ); ЧМ — частотно-модулированный
тон. Первичная слуховая кора (А1) пересекается изолиниями частот в кГц. Дальнейшие
объяснения в тексте. С упрощениями из Suga, Yan, Zhang, 1997. Воспроизводится с разре-
шения Nobuo Suga.
182 Часть II. Механочувствительность
ласть вентральной бахромы (ВБ) также содержит ЧМ/ЧМ-клетки и, как пола-
гают, связана с азимутальной локацией.
Клетки, отвечающие на ПЧ-компонент в биосонаре летучих мышей обыч-
но настроены очень точно. Чтобы оценить эту точность, надо основную гармо-
нику (около 62 кГц у Pteronotus) разделить на ширину КЧП на 10 дБ выше по-
рога. Получающаяся величина обозначается О10дБ, и чем она выше, тем более
точна настройка клетки. Показано, что слуховые клетки для тонов эха постоян-
ных частот имеют О10дБ “ Д° 600, тогда как за пределами диапазона постоянных
частот эха — лишь 20—30. Показано, что это тонко настроенные клетки особен-
но чувствительны к малым изменениям тонов эха постоянных частот. Такие из-
менения могут составлять всего 20 Гц, т. е. около 0,03% постоянной частоты.
Эта тонкая чувствительность имеет важное биологическое значение. Полагают,
что она обеспечивает летучих мышей информацией о порхающих насекомых.
Движение крыльев насекомых вызывают амплитудную модуляцию и доппле-
ровское смещение эха (иногда называемое «мерцанием»), достаточные для об-
наружения точно настроенными нейронами летучей мыши. У Pteronotus эти
клетки, детектирующие биения крыльев насекомых, расположены в области
ДСПЧ (рис. 9.12 В). При этом они «слепы» в отношении насекомых, которые
не шевелят крыльями, даже если находятся на расстоянии всего нескольких
сантиметров. Интересно, что многие бабочки выработали соответствующие
контрмеры и замирают, когда обнаруживают сигнал сонара летучетгмыши.
Такая крайняя чувствительность к частотам имеет и свою отрицательную
сторону, поскольку клетки чувствительны не только к допплеровскому эф-
фекту от движения крыльев насекомого, но и от собственных движений лету-
чей мыши. Когда летучая мышь двигается к цели, ПЧ-компонент частоты эха
возрастает. Подковоносы (Rhinolophus) и голоспины компенсируют этот само-
наведенный допплеровский эффект, автоматически снижая частоту испускае-
мого тона: частота каждого следующего звукового импульса снижается на ве-
личину, на которую частота предыдущего эха превышала характеристическую.
Тем самым частота эха удерживается в пределах 50 Гц от характеристической
или оптимальной частоты.
Очень точная эхолокация требует чрезвычайно чувствительного уха. Пока-
зано, что около 50% длины улитки голоспина настроены на ПЧ от +9 кГц до
—6 кГц, сходное расширенное представительство ПЧ обнаружено и в улитке
подковоноса (Rhinolophus rouxi). Эта часть улитки называется — слуховая фо-
веа. Возможно, очень точная настройка ПЧ-клеток — результат значительно-
го расширения базилярной мембраны, чувствительной к частоте звука. Ины-
ми словами, точность частотной дискриминации связана с величиной мозго-
вой карты тонов ПЧ или близких к ним (аналогично картам сетчатки в первич-
ной зрительной коре (гл. 17) и поверхности тела в соместезической коре (гл. 7,
см. особо рис. 7.15)). Предполагается также, что резонанс базилярной мембра-
ны в ответ на тоны ПЧ участвует в улучшении частотной дискриминации. Это
мнение подкрепляется наблюдением, что в ответ на ПЧ-тон в ухе происходит
отоакустическая эмиссия. Иначе говоря, когда в ухо Pteronotus направляется
громкий звук частотой чуть выше ПЧ-тона, в результате резонанса базилярной
мембраны возникает достаточно громкий ответ. Отоакустическая эмиссия,
хотя и намного более слабая, обнаружена и у других млекопитающих, в том
9. Анализ вестибулярной и звуковой информации в мозге 183
числе и в ухе человека. Эту отоакустическую эмиссию на щелчок надо отли-
чать от спонтанной эмиссии (которая, как полагают, возникает вследствие ме-
ханического движения наружных волосковых клеток), которая обсуждалась в
Информации 8.1.
Наконец, чтобы окончательно удивить читателя, заметим, что сонары под-
ковоносов и голоспинов (как и некоторых других летучих мышей) способны
различать текстуру объектов. Звуковые волны интерферируют при отражении
от «возвышений» и «углублений» неровной поверхности. Например, звуковые
волны(как и другие волны) интерферируют с взаимным подавлением, если от-
личаются на Л/2 (где X — длина волны) и с взаимным усилением, если разли-
чаются на X. То есть, если возвышения или углубления неровной поверхности
различаются на Х/4, то импульс эха от углубления пройдет на X/4 + X/4 = Х/2
больше, чем отразившись от возвышения. Таким образом возникают условия
для деструктивной интерференции. Если тональная частота достаточно высо-
ка, например 155 кГц в случае подковогуба Hipposideros bicolor, а скорость зву-
ка в воздухе — 344 м/с, тогда X/4 составит около 0,5 мм. Более низкие частоты,
конечно, имеют большие длины волн и потому не могут быть использованы
для детектирования таких малых различий в текстуре поверхности. Поведен-
ческие эксперименты подтвердили, что многие летучие мыши действительно
способны различать текстуру поверхностей «на слух».
Здесь можно провести аналогию с цветовым зрением. Так же, как белый
свет свойствами поверхности разделяется на цвета в отраженном луче, так и на-
бор гармоник ПЧ или направленная вниз ЧМ-эхолокационная эмиссия лету-
чих мышей отражается и возвращается к ним в виде фронта волны сложной
формы с интерференциями разного вида, которые можно назвать цветами эха.
Способность различать и анализировать цвета эха предоставляет неоценимую
информацию об окружающей среде. Любое движение на поверхности, среди
опавших листьев и веток меняет цвет эха в возвращающемся сигнале. Экспери-
менты подтвердили, что мельчайшие движения насекомых на почве или в кро-
нах деревьев вызывают обнаружимые изменения цвета эха. Цвет эха в возвра-
щающемся фронте звуковой волны предоставляет также информацию и о каче-
стве поверхности с точки зрения ее пригодности для приземления и отдыха.
В своей впечатляющей работе философ Томас Нагель спрашивал: «Что
значит — быть летучей мышью?». Предшествующее изложение дает некото-
рые идеи насчет сложнейшей обработки информации, происходящей в их слу-
ховой системе. Как говорил Нагель, их мозг развился, чтобы «точно различать
расстояния, размер, форму, движение и текстуру сравнимо с тем, как мы это
делаем, пользуясь зрением». Сейчас нейрофизиологи в начале пути описания
динамических процессов в слуховой системе, которые происходят, когда лету-
чая мышь порхает в вечернем небе в погоне за насекомым, устраняя помехи от
шума деревьев и кустов, примечая малейшие движения крылышек жертвы,
преследуя ее несмотря на все ее попытки спастись. Нагель, однако, настаива-
ет — «что же это значит — быть летучей мышью?», что же они испытывают?
Ответ остается гипотетическим, многие скажут, что он и не может быть ника-
ким иным, кроме как гипотетическим. Все это относится к проблеме взаимо-
отношений сознания и тела, проблеме, от которой мы в этой книге уклоняем-
ся, лишь коснемся ее в гл.23, но в целом оставим философам.
184 Часть II. Механочувствительность
9.6. Слуховая кора человека и язык
Так же, как слуховая система сипухи и слуховая кора летучих мышей разви-
лись до удивительно высокого уровня, чтобы обеспечить их способ существо-
вания, так и слуховая кора гоминид развилась (иным путем, но до сходного
уровня сложности), чтобы обеспечить человеку его образ жизни. Как и в слу-
ховой системе рукокрылых, мы видим, что если в ядрах низших уровней есть
клетки, реагирующие на постоянные тоны, то большинство клеток слуховой
коры отвечают только на частотно-модулированные тоны. Из изучения по-
вреждений, патологии и психофизиологии (микроэлектродные исследования
по понятным причинам невозможны) получены основательные свидетельст-
ва, что эти клетки особенно хорошо активируются короткими звуками, отли-
чающимися друг от друга, с быстрыми изменениями частоты. Слуховая кора
доминантного полушария особенно эффективна в определении взаимосвязей
между короткими звуками — следуют ли они один за другим или приходят од-
новременно. Мы увидим ниже, что все эти способности являются основой
распознавания звуков человеческой речи. Миллионы лет слуховая система иг-
рала жизненно важную роль во взаимоотношениях хищник — жертва, мгно-
венно улавливая тихий шорох, слабое потрескивание и шуршание и опреде-
ляя, откуда исходят эти слабые сигналы тревоги. У гоминид на основе этой
чувствительности развилась важнейшая способность, от которой зависит сам
человеческий образ жизни.
Способность говорить возникает для нас так естественно, что мы редко за-
думываемся над тем, насколько она удивительна. Можно предположить, что
это — поток слов, подобный тексту на этой странице, однако это не так. Линг-
висты проанализировали язык до уровня небольшого числа элементарных зву-
ков — фонем. В слове «pit» (ямка), например, три фонемы: /р/, /i/ и /t/. За-
мените хотя бы одну из них, и смысл изменится: kit (набор), pet (домашнее
животное), pig (свинья). Однако, и сами фонемы не являются совершенно по-
стоянными. Рассмотрим, например, фонему /t/. Она варьирует в зависимости
от места, которое занимает в предложении. Эти варианты называются аллофо-
нами. В случае фонемы /t/, то есть в случае, если язык соприкасается с альве-
олярным гребнем над зубами, формируется т.н. альвеолярный звук, как в сло-
ве «eight». Если язык продвинут дальше вперед и расположен против зубов, как
в слове «eighth», формируется зубной аллофон. Эта фонема имеет еще мно-
жество вариантов. Если оставить временно в стороне аллофоны, то в обще-
употребительном южноанглийском говоре наблюдается 20 гласных и 24 со-
гласных фонем.
Фонемы могут быть проанализированы спектрографически. Такое иссле-
дование показало, что фонемы состоят из множества полос акустической
энергии при разных частотах. Эти частотные полосы называются формантами.
Наиболее низкая по частоте форманта фонемы считается первой (F1), более
высокая — второй (F2) и т. д. Наибольшую энергию несут первые две форман-
ты. Рис. 9.13 А показывает, что спектрограмма слога из согласного и гласного,
такого как /Ьа/, /da/ и /ga/ состоит из двух частей: начальная, очень короткая
ЧМ-фаза, за которой следует более продолжительная фаза ПЧ.
9. Анализ вестибулярной и звуковой информации в мозге 185
в
+10 +100
Время (мс)
Рис. 9.13. Спектрограммы звуков человеческой речи. (А) На рисунке показано зна-
чение первичной короткой ЧМ-фазы в различении фонем. Воспроизводится с раз-
решения Plenum Publishing Corporation. (Б) Различные спектрограммы одного и
того же слышимого звука. С разрешения Annual Review of Neuroscience, Volume 15,
©1992, Annual Reviews http://www.annualreview.org. (В) Значение времени включе-
ния голоса (ВВГ).х Дальнейшие объяснения в тексте. По Springer, 1979. Воспроиз-
водится с разрешения Plenum Publishing Corporation.
Рис. 9.13 А показывает, что согласные /Ь/, /d/ и /g/ формируются из очень
быстрого (десятки миллисекунд) восходящего или нисходящего ЧМ-сегмента.
ПЧ-сегмент продолжается несколько сот миллисекунд и образует гласный.
Более высокие и более низкие голоса образуют форманты при разных часто-
тах, но взаимосвязь формант остается постоянной, если мы слышим один и
186 Часть II. Механочувствительность
тот же звук. Рис. 9.13 Б показывает интересный случай, когда слышен один и
тот же согласный, но спектрограммы звуков значительно различаются. Когда
/d/ предшествует /i/, ЧМ-сегмент каждой форманты повышается; когда же /d/
предшествует /и/ — ЧМ-сегмент второй форманты снижается. Это первая
трудность сопоставления физических характеристик сигнала с слышимым
звуком речи. Наконец, рис. 9.13 В показывает важность времени включения го-
лоса (ВВГ) для различения согласных. Слушатели воспринимают /р/, когда
ВВГ превышает 30 мс, если ВВГ меньше — воспринимается фонема /Ь/. Из
этого вытекает, как упоминалось выше, что эффективность деятельности ре-
чевой коры зависит от способности детектировать ЧМ-тоны и быстрые изме-
нения.
Значение этой способности убедительно подтверждается демонстратив-
ным опытом по физиологии слуха. В 1957 г. Либерман с сотрудниками подго-
товили группу слогов из согласного и гласного, каждый из которых состоял из
двух формант. Затем они стали постепенно смещать момент начала ЧМ-сег-
мента второй форманты, спрашивая испытуемых, ощущают ли они разницу со
стандартным звучанием. Испытуемые эту разницу отмечали. Например, на
рис. 9.13 А сила форманты F2 варьирует, градуально меняясь от возрастаю-
щей, через горизонтальный участок к нисходящей. Результаты Либермана и
сотрудников показаны на рис. 9.14. По мере того, как ЧМ-сегмент F2 меняет-
ся, испытуемый сначала слышит /Ь/, а затем наступает резкий переход и слы-
шится уже /d/. Таким образом, происходит переключение одной фонемной
категории на другую. Такое категориальное восприятие не наблюдается, когда
аналогичным образом синтезируется и варьируется неречевой звук. В послед-
нем случае восприятие изменяется градуально, а «ступенчатое» изменение
восприятия отсутствует. Аналогично, категориальный переход отсутствует и
при изменении ПЧ-сегмента гласных звуков.
Категориальное восприятие звуков речи для детей является врожденным.
Детям в возрасте одного месяца предъявлялись синтезированные речевые и не-
речевые звуки. Внимание детей отслеживалось с помощью наблюдения за их
сосательным поведением. Когда через динамик предъявлялись категориальные
переходы речевых звуков, тип сосательного поведения менялся, когда же
предъявлялись схожие переходы неречевых звуков, таких поведенческих реак-
ций не обнаруживалось. Показано, что еще несколько черт системы распозна-
вания речи, характерной для взрослых, присутствует у очень маленьких детей.
В настоящее время практически нет сомнений в том, что человек рождается
«подготовленным» к речи. Слуховая система не представляет собой в этом от-
ношении tabula rasa (чистый лист), в которой распознавание речи развивается
исключительно вследствие широкого речевого общения после рождения. И в
этом отношении, как мы увидим ниже, слуховая система схожа со зрительной.
Интересно отметить, что люди в такой «подготовленности» не одиноки.
Конечно, ни одно животное не «разговаривает» в том смысле, как это делает
человек, однако многие млекопитающие и птицы используют голосовую ком-
муникацию. Множество видов уже исследовано на предмет категориального
восприятия видоспецифических звуков, в том числе мыши, макаки и мармо-
зетки, и во всех случаях получены надежные свидетельства категориального
различения. У японского макака (М. fuscata) анализ идет еще дальше. Эти ма-
9. Анализ вестибулярной и звуковой информации в мозге 187
Рис. 9.14. Категориальное
восприятие слога /Ьа/ и /da/.
Акустические свойства фоне-
мы менялись градуально. Пе-
реход от восприятия слога
как /Ьа/ к /da/ представляет-
ся (относительно) ступенча-
тым.
каки издают 80—90 различных звукосочетаний. Один из них может отличаться
от другого по тому критерию, раньше или позже возникает в них ЧМ-элемент.
С помощью оперантной методики выработки рефлексов было показано, что
макаки способны различать эти элементы. С другой стороны, макакам других
видов, в частности свинохвостых макаков, для их различения нужна длитель-
ная тренировка. Если же всем этим обезьянам ставится задача различить зву-
ки на основе иных параметров, то японские макаки оказываются последними.
Т. о. японские макаки вероятно развили специфический диалект, недоступ-
ный для других макаков.
Конечно, это походит на человеческий язык. Врожденная биологическая
подготовленность затем наполняется лингвистическим окружением, в кото-
ром оказывается ребенок. И снова мы должны отметить сходство со зритель-
ной системой. Бесчисленные эксперименты показали, что неврология зри-
тельных структур принимается за основу, предоставляя формирование более
тонких деталей опыту. Что касается речи, то показано, что это формирование
начинается до рождения. Голос матери передается в матку благодаря тканевой
проводимости. После рождения ребенок делает сосательные движения, чтобы
услышать запись голоса матери (но не посторонней женщины) или песенку,
которую слышал в пренатальном периоде. По мере перехода от младенчества к
детству и дальнейшего взросления фонетические особенности и различия, ко-
торые человек способен обнаруживать, сводятся к тем, которые свойственны
родному языку. Например, Веркер и Тис (Werker & Tees) показали на примере
английского языка и языка хинди, что различение определенных признаков в
английском языке утрачивается у говорящих на хинди и наоборот уже к годо-
валому возрасту.
Слуховая кора должна не только воспринимать и различать очень быстрые
переходы, лежащие в основе категориального восприятия, но и справляться с
очень быстрым вводом информации. Она способна воспринимать до 60 фонем
в секунду — что более, чем в 12 раз превышает скорость раздельного восприя-
тия неречевых звуков.
Если взглянуть на звуковую спектрограмму простого английского предло-
жения «I can see it» (я вижу это) (рис. 9.15), то мы увидим, что она имеет очень
отдаленное отношение к тому, что мы слышим. Слуховая система, опять-таки,
как и зрительная, в известном смысле «накладывает смысловое значение» на
такую сложную картину информационного ввода. Как мы увидим в следую-
щем разделе, эта функция выполняется в задней части первичной слуховой ко-
ры (поле Вернике). Речь — это не цепочка последовательных фонем, ясно раз-
188 Часть II. Механочувствительность
CAN S Е Е
Рис. 9.15. Спектрограмма звуков фразы «I can see it» (я вижу это). По ординате ча-
стоты (кГц), по абсциссе — время. Фразу произносили очень отчетливо. В обыч-
ной речи слова разделяются намного менее четко. Установлено, что форма соглас-
ных зависит от предшествующих и последующих гласных. Отметьте, что слово «it»
состоит из двух всплесков звуковой энергии, более четко разделенных между со-
бой, чем в других словах.
деленных друг от друга, а сложный сигнал, в котором фонемы изменяются под
влиянием как предшествующих, так и последующих за ними фонем. Напри-
мер, хорошо известно (и это — источник многочисленных снисходительных
замечаний), что многие англоговорящие затрудняются в произнесении слово-
сочетания «law and order» (закон и порядок), вместо которого у них получает-
ся «lore and order» (прибл. «подход» и порядок). Речевые сигналы формируются
голосовым аппаратом человека путем коартикуляции, каждый из фрагментов
которой отражает то, что предшествовало, и то, что последует. Между акусти-
ческими свойствами и фонетическими категориями нет жестких неизменных
взаимосвязей. Рис. 9.15 показывает, что фонетические категории не могут быть
картированы на акустическом сигнале.
Слуховая система чрезвычайно чувствительна к тонкостям артикуляции,
размерам и форме голосового тракта говорящего, а также скорости речи и фо-
нетическому контексту не только согласных, но и гласных. Последние часто
передают информацию об эмоциональном состоянии говорящего. Подроб-
ный обзор современного состояния этой проблемы завел бы нас слишком да-
леко в дебри психолингвистики. Интересующиеся студенты могут ознако-
миться с некоторыми книгами и обзорами, приведенными в разделе Библио-
графия. В заключение данного раздела заметим, что не так уж удивительно,
насколько скромны успехи в компьютерном понимании речи. Обработка ин-
формации в слуховой системе существенным образом распараллелена, зави-
сима от контекста и ориентирована на конечное действие.
9. Анализ вестибулярной и звуковой информации в мозге 189
9.7. Латерализация и нейроанатомия речи
В подавляющем большинстве случаев анатомические области, в наибольшей
степени связанные с речью, располагаются в левом полушарии (см. табл. 9.1).
В этих случаях оно называется доминантным полушарием. Исследование анато-
мии полушарий показывает структурные различия между ними. Сильвиева бо-
розда в левом полушарии длиннее и заходит выше к верхушке мозга, чем в пра-
вом. Если сравнивать верхнюю поверхность височных долей, то в 65% случаев
левая planum temporale намного больше, чем правая (рис. 1.16 А) (в 11% случа-
ев — справа больше чем слева, и в 24% — существенная разница отсутствует).
Тот факт, что такого рода разница может быть обнаружена и у плода, указыва-
ет на но, что человеческий мозг заранее настроен на асимметричное развитие
полушарий.
Хотя основные анатомические структуры, связанные с речью, располага-
ются в доминирующем полушарии, в процесс вовлекаются и многие другие
структуры. Как и можно было ожидать в случае столь важной функции, это ка-
сается обоих полушарий (рис. 9.16 В). В последнее время получены свидетель-
ства участия в процессе также подкорковых структур, таких как таламус и хво-
статое ядро. Тем не менее, удобным и широко распространенным остается
«первое приближение» в описании системы, предложенное Дамасио и Геш-
виндом (Damasio and Geschwind) и показанное на рис. 9.16 Б. Оно исходит
главным образом из наблюдений за последствиями инсультов и других нару-
шений речи, вызванных повреждениями, с последующим посмертным иссле-
дованием нейрогистологии. Наиболее важным речевым областям присвоены
названия поля Брока и поля Вернике — в честь исследователей, которыми бы-
ли получены данные о связи между нарушениями речи и повреждениями
нервной ткани.
Как показано на рис. 9.16 Б, с речью связано поле Вернике. Оно располо-
жено сзади от первичной слуховой коры. По цитоархитектонической класси-
фикации Бродмана первичная слуховая кора обозначается как поле 41, а поле
Вернике как поле 22. Именно в поле Вернике направляются из первичной слу-
ховой коры сигналы, вызванные речью. Повреждения этой области влияют на
способность человека воспринимать звуки речи как лингвистически значи-
мые. Аналогично, зрительные стимулы от написанных слов также направля-
ются в поле Вернике — из первичной зрительной коры (поле Бродмана 17) во
Таблица 9.1. Латерализация мозга и право- и леворукость
Доминантное полушарие (в % %)
левое правое нет доминантного
Ведущая рука
Леворукие и амбид^кстеры1 70 15 15
Праворукие 96 4 0
1 Амбидекстер — человек, в равной степени владеющий обеими руками — Прим, перев.
По Kandel etal., 1991
190 Часть II. Механочувствительность
ПГ1
ПГ2
РТ
РМ
Соместезическая
кора
Б
ЗП
Левая
сторона
Роландова
борозда
Правая
сторона
ИГ
ПГ
РТ
РМ
Поле
Поле
Брока
Моторная слуховая
кора КОра
Аркуатный
пучок
Первичная
зрительная кора
Ангулярная
извилина
Вернике
Рис. 9.16. (А) Горизонтальный срез через мозг человека на уровне сильвиевой борозды
показывает, что planum temporale (РТ) слева больше, чем справа. Задний край РМ накло-
нен кзади более круто в левом полушарии, чем в правом, а извилина Гершля (ИГ) скло-
няется вперед более круто. РТ слева простирается от точки, обозначенной «х», до точки,
обозначенной «у». Слева располагается одиночная поперечная извилина Гершля (ПГ)и
две — справа (ПГ1, ПГ2). ЗП — затылочный полюс, ВП — височный полюс. По
Geschwind and Levitsky, 1968. Воспроизводится с разрешения от © 1968, American
Association for the Advancement of Science. (Б) Речевые области мозга. Левое полушарие
мозга показано сбоку. Дальнейшие объяснения в тексте. (В) Хотя основные речевые цен-
тры показаны на (А) и (Б), многие другие части коры вовлечены в речевую функцию че-
ловека. На графике показана зависимость нарушений речи вследствие локальных
поражений разных зон (Ред.). Из Young, 1978 с разрешения Oxford University Press.
вторичную зрительную кору (поле Бродмана 18), затем через ангулярную изви-
лину (поле Бродмана 39) в поле Вернике. Повреждения в любой из этих облас-
тей затрудняют понимание письменной речи и способность именовать визу-
альные объекты.
9. Анализ вестибулярной и звуковой информации в мозге 191
Поле Вернике соединено с полем Брока трактом, именующимся аркуатным
пучком. Перерезка его делает невозможным повторение прочитанного или
услышанного слова. Поле Брока само по себе занимает центральное место
в формировании сложной мышечной активности, требующейся для артикуля-
ции слов. Оно располагается, как это видно из рис. 9.16 Б, непосредственно по
соседству с областью первичной моторной коры, в которой картирована мус-
кулатура лица и гортани. Как уже говорилось, схема, описанная выше, пред-
ставляет собой по необходимости упрощение всей нейроанатомии, касающей-
ся рассматриваемого процесса. Она, тем не менее, дает основу для понимания
многих нарушений речи (афазий), поскольку позволяет проследить наруше-
ния в различных элементах системы.
9.8. Каллозэктомия1 и ее последствия
В разделе 9.7 мы видели, что распознавание и воспроизведение речи связано с
одним из полушарий, которое именуется доминантным (у большинства — это
левое полушарие). Основным соединением между доминантным и суб-доми-
нантным полушариями является большой нервный тракт — corpus callosum
(мозолистое тело) (менее важный — передняя комиссура — также связывает
полушария). Долгое время функция мозолистого тела оставалась неясной, од-
нако в середине 50-х годов XX столетия экспериментами на животных была
показана его критическая роль в координации активности двух полушарий.
Если перерезать мозолистое тело и оптической перекрест (перекрест зритель-
ных трактов Х-образной формы, см. раздел 17.1.2), то условные рефлексы,
выработанные для одного глаза, не реализуются на условный стимул, предъ-
явленный другому глазу. Информация остается ограниченной лишь одним
полушарием. В 60-е годы XX века Сперри с сотрудниками (Sperry etal.) прове-
ли серию тщательных наблюдений над пациентами с перерезками мозолисто-
го тела, предпринятыми для облегчения неизлечимой эпилепсии. На первый
взгляд такие пациенты после операции выглядели совершенно нормальными.
Припадки у них распространялись из эпилептического фокуса только на одно
полушарие, не захватывая другое. Кроме сильнейших головных болей, раска-
лывающих голову (фигурально), у таких пациентов с «расколотым мозгом»
(буквально), все остальное было совершенно нормально.
Однако, как теперь хорошо известно, это неверно. Дефект, проявляющий-
ся у таких пациентов — один из наиболее интересных и будящих мысль в об-
ласти нейрофизиологических дисфункций. Сперри и его коллеги сумели по-
казать, что те же дефекты, которые наблюдались на экспериментальных жи-
вотных, присутствуют и у челЪвека. Конечно, их пациенты имели интактный
зрительный перекрест. Саккадные движения глаз2 обычно показывают, что
информация от каждого глаза достигает обоих полушарий. Однако, если ко-
1 Каллозэктомия — пересечение мозолистого тела, соединяющего полушария головного
мозга. — Прим, перев.
2 Саккадические движения глаз — быстрое перемещение глаз от одной точки фиксации
к другой. — Прим, перев.
192 Часть II. Механочувствительность
Информация 9.1
БРОКА И ВЕРНИКЕ
Афазия, заболевание символического мышления и способности к его выражению,
известна давно. Иоганн Вольфганг Гёте в Wilhelm Meister's Apprenticeship (1796)
приводит одно из наиболее ранних описаний. «Неожиданно мой отец перенес па-
ралич; у него отнялись правая сторона тела и речь. Нам приходилось догадываться,
что ему нужно, потому что он был не в состоянии выговорить слова, которое хотел.
Иногда это было ужасно тяжело для нас... Его нетерпение достигало предела, и его
состояние трогало меня до глубины души.» Только в девятнадцатом столетии нача-
лись попытки понять связь этого заболевания с патологией мозга. Врачи, в первую
очередь Галль и Пинель в начале века представили точное описание афазии, насту-
пающей за апоплексией (инсультом). Было установлено, что она безусловно сопро-
вождается параличом правой стороны тела, что предполагает патологию левого по-
лушария. Вслед за этим были опубликованы работы Марцела Дакса (Marcel Dax),
Жан-Батиста Булляра (Jean Baptiste Bouillard) и его пасынка Симона Обертэна
(Simon Aubertin). Однако, вплоть до работ Поля Брока в 60-е годы XIX века не бы-
ло показано, что повреждения задней части левой трети лобной извилины вызыва-
ет афазию (или афемию, как ее именовал автор), с чего и началось установление ис-
тинных взаимосвязей между мозговой патологией и нарушениями речи.
Поль Брока (1824—1880) родился в Сен-Фуа-ла-Гранд между Бордо и Бержера-
ком. Его отец — сельский врач, в прошлом — хирург армии Наполеона. Поль Бро-
ка был блестящим и быстро продвигающимся студентом. Он стал самым молодым
прозектором и секретарем Societe Anatomique (Анатомического общества) за всю
его историю. Он внес вклад в различных областях медицины и опубликовал более
500 статей и других научных трудов за свою не слишком долгую жизнь. Помимо
славы установления связи афазии с поражениями специфической области коры
мозга, Брока помнят как основателя антропологии во Франции. Он опубликовал
работы по кроманьонцам и о трепанации черепа в неолитическом периоде, а в
1859 году основал Societe d’Antropologie de Paris (Парижское антропологическое
общество) и Школу и Институт антропологии. В 1880 г. он был избран сенатором
Франции, где представлял «Францию и науку», однако, несмотря на крепкое здо-
ровье и большую энергию пробыл на этой должности всего 6 месяцев, скончавшись
8-го июля 1880, в сравнительно небольшом возрасте — 56 лет.
У Карла Вернике (1848—1904) не было такого могучего характера, как у францу-
за. Он прожил почти столько же, но не оставил такого же следа в науке и обществе.
В отличие от Брока, который был харизматической пылкой личностью с талантами
рассказчика и бонвивана, Вернике был неразговорчивым и скрытным. Он родился
в семье чиновника в Верхней Силезии и после окончания медицинского факульте-
та в Бреслау (ныне Вроцлав, Польша. — Перев.) получил возможность в течение
6 месяцев поработать с Теодором Мейнертом в Вене. Этот короткий период, по-ви-
димому, был очень важен для Вернике, потому что впоследствии он всегда называл
Мейнерта своим учителем. Хотя Вернике публиковался в различных областях ней-
рологии, особенно офтальмологии, его помнят благодаря маленькой работе, опуб-
ликованной в 1874 году, когда ему было 26 лет: Der aphasische Symptomenkomplex
(Симптомокомплекс афазии).
9. Анализ вестибулярной и звуковой информации в мозге 193
Продолжение
В этой монографии он попытался провести корреляции между различными ти-
пами афазии и поражениями различных частей коры головного мозга и подкорко-
вых проводящих путей. Его основной концепцией и глубоким убеждением было,
что душевные болезни имеют корреляты в поражениях мозга. Различие между пси-
хиатрией и нейрологией он считал ошибочным и искусственным («Geisteskrankhei-
ten sind Gehimkrankheiten» (Душевные болезни есть болезни мозга)). В том же воз-
расте, что и Поль Брока — в 56 лет, будучи на вершине карьеры, только что заняв
кафедру психиатрии в Галле, он, катаясь на велосипеде в Тюрингском лесу, попал
в дорожное происшествие и скончался 15 июня 1905 года. (Это расхождение с дата-
ми смерти Вернике повторяется во многих проверенных источниках, включая Бри-
танскую и Французскую энциклопедии, и может быть устранено только команди-
рованием на место захоронения. — Перев,)
Литература
Goethe, J. W., 1796, Wilhelm Meisters Lehrjahre, trs. 1842 by Thomas Carlyle, Wilhelm
Meister’s Apprenticeship, vol. 11, p. 197, London: Chapman and Hall.
Goldstein, K., 1970, «Paul Broca (1824—1880)» in He Founders of Neurology (2nd edn),
W. Haymaker and F. Schiller, eds, Springfield: Charles C. Thomas.
Goldstein, K., 1970, «Carl Wernicke (1848—1904)» in W. Haymakerand F. Schiller, ibid.
Riese, W., 1947, «The early history of aphasia», Bulletin of the History of Medicine, 21,
322-334.
роткие стимулы предъявлять с помощью тахистоскопа, совместное саккадное
движение глаз может быть нарушено, так что информация от левой части по-
ля зрения передается только в правое полушарие и наоборот. Если письменная
информация передается таким образом только в правое полушарие, то паци-
енты отрицают, что видели что-либо. Другими словами, информация, полу-
ченная правым полушарием, не имела доступа в речевые центры, расположен-
ные в левом полушарии.
Хотя при этом пациент искренне отрицает, что знаком с информацией, до-
стигшей его правого полушария, можно тем не менее показать, что она таки
действительно достигла правого полушария и была обработана. Правое полу-
шарие контролирует левую руку. Если пациента попросить показать левой ру-
кой что-либо насчет предложенного стимула, то ему это удается. Это, конеч-
но, ставит поразительные вопросы перед теми, кто занимается проблемами
сознания.
Дальнейший анализ каллозэктомированных пациентов показывает, что
полушария головного мозга настроены на различные функции (рис. 9.17). Ча-
сто говорят, что левое полушарие (речевое) в большей степени ориентировано
на аналитические задачи, тогда как правое — на решение задач в целом. Это,
конечно, правда, но только часть правды. Правое полушарие способно к про-
стому пониманию речи, а левое — к целостному ее восприятию. У нормальных
индивидов они работают совместно, дополняя друг друга. Есть данные, что
специализация полушарий у мужчин более выражена, чем у женщин.
194 Часть II. Механочувствительность
П
Стереогнозия
Стереогнозия
Правое ухо
Зевое ухо
Пространственное
представление;
Элементарное
понимание речи
Невербальные
представления
ПоЛ 'с»ИНЯ
поля зрения^
L Л (левая) j
- йу
Обоняние
Левая ноздря
Письменная речь
Поле зрения
П(правая
Л (левая
половина)
половина) /
Обоняние
Правая ноздря
Половина
поля зрения
Л (правая)
Основной центр;
обработки речи
Рис. 9.17. Латерализация по-
лушарий мозга человека. Ри-
сунок показывает специали-
зацию полушарий, а также
то, что речь идет не столько
о четкой дифференциации,
сколько о степени выражен-
ности функции. Правое полу-
шарие, например, обладает
некоторой способностью вос-
приятия речи, тогда как левое
в большей степени восприни-
мает целостную картину.
Наконец, надо снова подчеркнуть, что экспериментальный анализ нейро-
физиологии пациентов с каллозэктомией требует крайней точности и тех-
нической изощренности. Мозг, особенно сенсорный мозг, способен делать
заключения из почти незаметных деталей. Это — машина по генерированию
гипотез, которая развилась в долгой эволюционной войне хищников и жертв
Если треснувшая веточка, необычный шум, чуть слышный шорох не будут
мгновенно дорисованы до полного образа, смысл его не будет немедленно
оценен, то жертва, обед, исчезнет. Или, наоборот, прыжок в безопасное укры-
тие, запоздавший на долю секунды, означает конец жизни. Потому-то какие-
то данные, полученные левым полушарием каллозэктомированного больного
и позволяют правильно отвечать на вопросы экспериментатора, даже когда
информация прочно заперта в суб доминантном полушарии. Только если
удастся контролировать все факторы, удивительные свойства мозга с пересе-
ченным мозолистым телом могут быть поняты.
9.9. Заключение
Это заключительная глава второй части книги. Мы начали длинный путь от
простых механорецепторов прокариот и круглых червей. Мы видели, как они
включались во все более сложные сенсорные системы. Мы сделали обзор тон-
ких систем детекторов натяжения и деформации, которые доставляют нам ше-
9. Анализ вестибулярной и звуковой информации в мозге 195
стое чувство — кинестезию. Мы рассмотрели изящные высокочувствительные
тактильные волоски насекомых и видели, как они развились в детекторы виб-
рации и, в конце концов, в тимпанальный орган или аналог уха. Мы также ви-
дели, как системы, исходно развившиеся для того, чтобы детектировать изме-
нения в абиотическом окружении, адаптировались к обнаружению животных
и коммуникации с ними. Трели цикад, звучащие по всей средиземноморской
литорали, оживили в нашей памяти мифологический образ древней Греции.
Мы восхищались акустической войной между летучими мышами и бабочками
и затронули изощренную сложность слухового мозга рукокрылых. Чрезвычай-
ная точность, с которой ночные хищники — совы — обнаруживают своих
жертв и сваливаются им на головы, тоже свидетельствует о высочайшем уров-
не развития, достигнутом здесь механочувствительностью. Звуковая коммуни-
кация имеет также важное значение в социальном общении и достигает вер-
шин в песнях птиц и еще больших — у человека. Мы рассмотрели (конечно,
лишь очень кратко) речевой мозг человека. Мир человека заполнен речью.
Продолжающаяся революция в коммуникации ведет к образованию сети че-
ловеческого общения, охватывающей всю поверхность планеты. И в то же вре-
мя мы видели на страницах книги, насколько меняется субъективный аспект
механочувствительности. Хотя в основе физиологии остаются молекулярная
биология и биофизика погруженных в мембрану механочувствительных ка-
налов, насколько же субъективно различаются ощущения прикосновения и
звука! И наконец мы оказываемся перед великой нерешенной проблемой
современного мира — проблемой разума.
МАТЕРИАЛЫ К ЧАСТИ II
Для самоподготовки
Нижеследующие вопросы должны помочь оценить понимание изложенного в
Части II, а также обратить внимание на значительные аспекты данного пред-
мета.
Глава 5: Механочувствительность клеточных мембран
5.1. Почему молекулярная биология рецепторов натяжения изучена намного
меньше других типов рецепторов?
5.2. С использованием материалов Дополнения А обсудите технику локальной
фиксации потенциала применительно к Е. coli. Почему размер Е. coli представ-
ляет собой проблему и как ее обойти?
5.3. Что означают аббревиатуры МчкВ и МчкМ? Чем они различаются?
5.4. Кратко опишите выделение и очистку МчкВ.
5.5. Нарисуйте график механочувствительности такого канала.
5.6. Сколько молекул белка МчкВ группируется при образовании механочувстви-
тельного канала?
5.7. Обсудите две возможные роли рецепторов натяжения в мембране бактерии.
5.8. Объясните разницу между пойкило-осмотическими и гомео-осмотическими
животными. Какие из них наиболее обычны?
5.9. Где в мозге млекопитающих располагаются осморецепторные клетки?
5.10. Где оканчиваются аксоны осморецепторных клеток?
5.11. Какие гормоны высвобождаются аксонами осморецепторов?
5.12. Какой эффект на осмолярность ВКЖ оказывают эти гормоны?
5.13. Нарисуйте схему петли обратной нейроэндокринной связи, которая контро-
лирует осмолярность внутренней среды.
5.14. Опишите электрофизиологические данные, которые подтверждают чувстви-
тельность ККН к изменениям осмолярности ВКЖ.
5.15. Опишите эксперименты, которые показывают, что растяжение мембраны свя-
зано с электрофизиологической реакцией ККН.
5.16. На основе экспериментов по локальной фиксации потенциала объясните, по-
чему каналы в ККН названы инактивируемыми при растяжении (КИ-каналы).
5.17. Объясните, как гипертоничность ВКЖ через КИ-каналы приводит к повыше-
нию частоты импульсов в аксонах ККН и таким образом компенсирует ее.
Глава 6: Кинестезия
6.1. Какая информация, помимо поступающей от мышц, суставов и связок, обра-
батывается для поддержания равновесия в пространстве?
6.2. Какова роль абдоминальных мышц в жизни десятиногих ракообразных?
6.3. Какова форма рецепторов растяжения у Homarus vulgaris?
6.4. Какое действие оказывает растяжение на нейросенсорные клетки, иннервиру-
ющие РМ1 и РМ2?
6.5. Почему нейросенсорные клетки в РМ2 называются быстро адаптирующими-
ся, а в РМ1 — медленно адаптирующимися?
6.6. Какую роль играют дополнительные волокна?
6.7. С использованием материала раздела 6.2.1 сравните роль, которую играют мо-
торные волокна РМ ракообразных с таковой у-мотонейронов скелетных мышц
млекопитающих.
Материалы к части II 197
6.8. Нарисуйте схему, показывающую основные черты волосковой сенсиллы насе-
комого.
6.9. Реагируют ли волосковые сенсиллы насекомых на различные сенсорные мо-
дальности;
6.10. Опишите несколько различных типов механочувствительных сенсилл.
6.11. Где располагается тубулярное тельце и какова его функция?
6.12. Что такое колоколовидные сенсиллы и где они главным образом располагают-
ся?
6.13. Нарисуйте схему сколопидия. Что такое и где располагается окончание сколо-
пидия?
6.14. Что такое субгенуальный орган и где он располагается? Какие стимулы он де-
тектирует?
6.15. Объясните, как такие насекомые, как стрекозы и мухи, поддерживают равно-
весие в полете.
6.16. Определите понятие «моторная единица». Изменяется ли она в размерах в раз-
личных частях тела?
6.17. Чем интрафузальные волокна млекопитающих похожи на рецепторные мыш-
цы ракообразных?
6.18. Назовите три типа интрафузальных волокон млекопитающих и расскажите о
различиях их реакции на растяжение.
6.19. В чем различие между фузимоторной и скелетомоторной системами мотоней-
ронов?
6.20. Опишите с помощью схемы петлю обратной связи, участвующую в контроле
активации сокращения скелетных мышц млекопитающих.
6.21. Ограничен ли рефлекторный механизм в спинном мозге гомонимной мыш-
цей? Если — нет, то какие еще мышцы вовлекаются в процесс?
6.22. Какие различные категории проприоцептивной информации передают в ЦНС
волокна групп 1а и II?
6.23. В чем различие между динамическими и статическими у-мотонейронами?
6.24. Что означает понятие «фузимоторный комплекс»?
6.25. Сравните и укажите различия характеристик ответов окончаний веретен и
сухожильных органов Гольджи. Объединяются ли эти типы проприоцепто-
ров?
6.26. Как реагируют сухожильные органы Гольджи на частоту стимуляции растяги-
вающей силой?
6.27. Нарисуйте принципиальную схему мышечного ответа на стимуляцию сухо-
жильного органа Гольджи.
6.28. Назовите три типа рецепторов, связанных с большинством суставов и укажите
их адекватные стимулы.
6.29. Опишите множественность сенсорных систем и обратных связей, образующих
«шестое чувство» или «кинестезию» млекопитающих и оцените, насколько
долго современная роботехниика должна развиваться, чтобы оказаться конку-
рентноспособной по сравнению с кинестезией животных.
Глава 7: Осязание
7.1. Почему нейробиологи интересуются маленьким почвенным червем С. elegans?
7.2. Что означают сокращения ПЛМЛ, ПЛМП, ЗЛМЛ, ЗЛМП, ПВМ и ЗВМ?
7.3. Каковы ультраструктурные характеристики нейронов осязательных рецепто-
ров? Вернитесь к гл. 6 — есть ли сходство между ультраструктурой осязатель-
ного рецептора С. elegans и механорецепторами насекомых?
7.4. Назовите два дегенерина. Что является продуктами этих генов, и какие струк-
туры млекопитающих эти пордукты напоминают?
7.5. Назовите три гена тес, не являющихся дегенеринами. Какие белки продуци-
руют они?
198 Часть II. Механочувствительность
7.6. Нарисуйте схему гипотетической конструкции белков МЕС, образующих ме-
ханочувствительную органеллу.
7.7. Какую форму имеет тактильная сенсилла насекомых?
7.8. Опишите действие тарзального детектора вибрации у п^клорых водных насе-
комых. Могут ли такие насекомые детектировать положение жертвы? Если —
да, то как? Есть ли здесь сходство с детектированием источника звука у позво-
ночных (см. гл. 9).
7.9. Опишите два типа акустических детекторов, развившихся у насекомых.
7.10. Что такое джонстонов орган и где он встречается? Какова его функция у кома-
ров и мошки?
7.11. Объясните различие стимулов, воздействующих на акустические сенсиллы и
тимпанальные органы.
7.12. Опишите структуру типичного тимпанального органа насекомых.
7.13. Рассмотрите несколько функций, которые выполняют тимпанальные органы
насекомых.
7.14. Используя материалы гл. 8 (раздел 8.3.2), объясните, почему физиология не
позволяет уху бабочки различать частоты сонара рукокрылых.
7.15. Объясните, почему клещ D. scalestes является более адаптированным парази-
том, чем D. phalenodectesl
7.16. Обсудите возможное значение эмиссии звука у бабочек как оборонительного
механизма против нападения рукокрылых. Что такое бэйтсовская мимикрия?
7.17. Какие рецепторы кожи млекопитающих являются быстро адаптирующимися,
а какие — медленно адаптирующимися?
7.18. Стимулы какого типа воздействуют на пачиниевы тельца? Каким образом их
гистологическая структура адаптирована к их функции?
7.19. Что представляет собой неоволосенная кожа с точки зрения чувствительности?
7.20. Все ли афферентные волокна механорецепторов входят в спинной мозг через
дорзальный рог серого вещества?
7.21. Нарисуйте схему расположения первичных и вторичных соматосенсорных об-
ластей.
7.22. В каком из четырех полей Бродмана соматосенсорной коры оканчиваются со-
матосенсорные волокна?
7.23. В каком слое соматосенсорной коры оканчиваются соматосенсорные волок-
на?
7.24. Почему колонки клеток в соматосенсорной коре рассматриваются как «вы-
числительный модуль»?
7.25. Что означает термин «соматосенсорный гомункулюс»?
7.26. Опишите эксперименты на вибриссах мыши, показывающие пластичность со-
матосенсорной коры.
7.27. Какие работы по осязанию у человека сходны с опытами на вибриссах мышей?
7.28. Опишите случаи, в которых осязание принципиально важно для социальных
взаимоотношений между индивидами.
Глава 8: Равновесие и слух: функция волосковых клеток
8.1. Нарисуйте схему типичной волосковой клетки млекопитающих.
8.2. Что такое «броуновское движение»? Каким образом оно ограничивает чувст-
вительность волосковых клеток?
8.3. В чем различие между киноцилиями и стереоцилиями?
8.4. Какое влияние на мембранный потенциал оказывает движение стереоцилий
по направлению к киноцилии? От нее? Перпендикулярно ей?
8.5. Каков молекулярный и биофизический механизм изменений при условиях,
изложенных в вопросе 8.4? Насколько быстры эти изменения?
8.6. Опишите модель стимуляции волосковых клеток на основе соединения кон-
чиков стереоцилий.
Материалы к части II 199
8.7. Какие данные свидетельствуют о связи механочувствительного канала С. ele-
gans и ионных каналов волосковых клеток?
8.8. Опишите молекулярный механизм, который может быть ответственным за
адаптацию волосковых клеток. Есть ли что-либо подобное у С. elegans?
8.9. Как деполяризация мембраны волосковых клеток связана с выделением меди-
атора из основания клеток?
8.10. Как организованы и какую роль играют волосковые клетки в боковой линии
рыб?
8.11. Есть ли система боковой линии у водных пресмыкающихся (морских змей, че-
репах) и млекопитающих (дельфины)?
8.12. Каково различие между мембранным преддверием и мембранным лабирин-
том?
8.13. Сколько полукружных каналов имеется у наиболее примитивных из ныне жи-
вущих позвоночных?
8.14. Для какой цели первоначально развилось ухо позвоночных?
8.15. Какие камеры уха образуют мембранное преддверие?
8.16. Какая часть (или части) мембранного преддверия детектирует линейные, а в
какая — угловые ускорения головы?
8.17. Как различаются ионные составы перилимфы и эндолимфы? Какой из них
схож с внеклеточной жидкостью, а какой — с внутриклеточной?
8.18. Где в саккулюсе и утрикулюсе расположены волосковые клетки?
8.19. Где расположены отолиты?
8.20. Какой нерв передает в мозг информацию от сенсоров равновесия в ухе?
8.21. Что такое гребешки и где они расположены в мембранном преддверии?
8.22. Чувствительны ли полукружные каналы к угловому движению головы с посто-
янной скоростью?
8.23. Гомологичны или аналогичны веберовы косточки слуховым косточкам уха
млекопитающих?
8.24. Какие важные анатомические изменения претерпевает лагена у пресмыкаю-
щихся?
8.25. Как изменяется ширина основной мембраны улитки у птиц?
8.26. В какие структуры у млекопитающих преобразуется базилярный сосочек?
8.27. Что такое отосклероз и почему он вызывает глухоту?
8.28. С помощью схемы объясните последовательность изменений давления, кото-
рые лежат в основе восприятия звука в улитке млекопитающих.
8.29. Что подтверждает наличие обратной связи в контроле чувствительности улит-
ки? (см. также Информацию 8.1)
8.30. Что такое микрофонный потенциал?
8.31. Какие нейрофизиологические проблемы ставит различение частот в диапазо-
не 500 Гц и выше?
8.32. Что такое «теория залпа» и «теория места» в различении частот/ Что внесено в
последнее совремнными исследованиями?
8.33. Опишите основные черты эхолокации звуков, испускаемых насекомоядными
летучими мышами.
8.34. Заглянув в гл. 13 (обоняние) и 16 (сетчатка), перечислите различные сенсорные
клетки, в которых реснички модифицированы для рецепции внешней среды.
Глава 9: Анализ вестибулярной и звуковой информации в мозге
9.1. Перечислите части и входные пути вестибулярного комплекса продолговатого
мозга. Что имеется в виду под названием «пушпульная иннервация» (push-
pull) — от двух наборов полукружных каналов?
9.2. Как имеющая отношение к равновесию информация от вестибулярных орга-
нов может быть изолирована от других сенсорных потоков?
200 Часть II. Механочувствительность
9.3. Что такое нистагм и как его можно продемонстрировать?
9.4. Обсудите причины некоторых форм нарушения движений.
9.5. Что означает термин «характеристическая частота» (ХЧ) волокна слухового
нерва? Можно ли считать ее частным случаем теории специфической нервной
энергии Мюллера—Гельмгольца?
9.6. Объясните, как область частотной реакции может рассматриваться в качестве
рецептивного поля?
9.7. Что означает термин «латеральное торможение»? Как это явление отражается
на ответе кохлеарных нервных волокон?
9.8. Подтверждают ли электрофизиологические отведения от одиночных кохлеар-
ных волокон теорию залпа?
9.9. Как базилярная мембрана представлена в кохлеарном ядре?
9.10. Опишите ответ звездчатых и сферических клеток на афферентацию из улитки.
9.11. Каким образом клетки ядер верхней оливы вовлечены в определение направ-
ления на источник звука?
9.12. Какие поведенческие реакции демонстрируют способность сов определять
пространственное положение источника звука?
9.13. Какие свойства звука используются мозгом совы для определения направле-
ния на его источник? Где эти свойства выделяются впервые? Какие свойства
звука используются для определения направления на источник звука в верти-
кальной (азимутальной) плоскости, а какие — в горизонтальной?
9.14. Объясните, как детекторы совпадений участвуют в сравнении времен, за кото-
рые звук достигает правого и левого ушей совы.
9.15. Объясните, как возбуждающие и тормозные синапсы участвуют в определении
азимутального направления на источник звука.
9.16. Что означает понятие «пространственная настройка»?
9.17. Где в мозге совы располагаются карты слухового пространства? Какова ее
функция?
9.18. В чем состоит предполагаемое значение нетопографической организации про-
странственно настроенных клеток в архистриатуме совы? Какой части мозга
млекопитающих аналогичен архистриатум ?
9.19. Где располагается слуховая кора у: а) приматов, б) кошек и летучих мышей?
9.20. В каком слое слуховой коры оканчиваются волокна из медиального ядра ко-
ленчатого тела? Сравните с соматосенсорной (раздел 7.5) и зрительной (раздел
17.2.1) корой.
9.21. Что является предпочтительными триггерными стимулами клеток слуховой
коры?
9.22. В зрительной коре макаков есть клетки распознавания лиц (раздел 17.4); есть
ли аналогичные клетки в слуховой коре макаков?
9.23. Опишите тип звука, испускаемого атакующей летучей мышью.
9.24. Как клетки слуховой коры рукокрылых адаптированы к анализу сигналов со-
нара?
9.25. Определите понятие Q10 дБ.Объясните, почему большее значение Q10 дБ со-
ответствует более узкой настройке кортикальных клеток.
9.26. Чувствительны ли летучие мыши к биению крыльев бабочек? Если — да, объ-
ясните почему.
9.27. Как некоторые летучие мыши компенсируют самонаведенное допплеровское
смещение в эхе от испускаемого ими звука?
9.28. Что такое «слуховая фовеа»?
9.29. Что такое «отоакустическая эмиссия» и как она возникает?
9.30. Может ли сонар рукокрылых детектировать текстуру поверхности и если да, то
как?
9.31. Упоминалось об аналогии между сигналом эха от неровной поверхности или от
вибрирующего препятствия с цветом в зрительной модальности. Объясните.
9.32. К каким звуковым стимулам особенно чувствительна слуховая кора?
9.33. Определите понятия «фонема» и «форманта».
Материалы к части II 201
9.34. Приведите пример, показывающий значение «времени включения голоса»
(ВВГ).
9.35. Что такое «категориальное восприятие»? Имеет ли оно место при неречевых
звуках? Является ли оно врожденным?
9.36. Каковы доказательства того, что способность восприятия речи формируется
ранним опытом индивида? Происходит ли что-либо подобное у других прима-
тов?
9.37. Объясните, что такое «ко-артикуляция».
9.38. В каком полушарии мозга planum temporale более развит?
9.39. Опишите расположение основных полей, связанных с речью в доминантном
полушарии.
9.40. Объясните роль поля Вернике и ангулярной извилины.
9.41. Как поле Вернике связано с полем Брока? Каковы последствия нарушения их
связей?
9.42. Каково значение анатомического расположения поля Брока?
9.43. Назовите основной тракт, соединяющий два полушария мозга.
9.44. Почему необходима перерезка оптического перекреста вместе с мозолистым
телом в экспериментах на животных по формированию изолированных полу-
шарий?
9.45. Откуда известно, что несмотря на отрицание пациента, информация, направ-
ленная в правое полушарие, достигает его и обрабатывается там?
9.46. Каким образом распределены между полушариями мозга способности?
Замечания и библиография
Глава 5: Механочувствительность клеточных мембран
Молекулярная биология механочувствительных каналов Е. coli обсуждается в
работах Hamil & McBride (1994), Sukharev etal. (1994, 1997). Физиология и по-
ведение пойкилоосмотических животных описаны в книгах по сравнительной
физиологии, таких как Prosser & Brown (1962), хотя и достаточно старой, но все
еще представляющей хорошее изложение классических данных. Современная
информация по осморецепторам изложена Bourque & Oliet (1997), тогда как
информацию по нейроэндокринологическим и нейрофизиологическим об-
ратным связям, лежащим в основе гомеоосмоса у млекопитающих, следует ис-
кать в других современных книгах по данному вопросу.
Bloun t, Р. et al., 1996, «Membrane topology and multimeric structure of a mechanosensi-
tive channel protein», EMBO Journal, 15, 4798—4805.
Bourq ue, C. W. and S. H. R. Oliet, 1997, «Osmoreceptors in the central nervous system»,
Annual Review of Physiology, 59, 601—619.
Hamil , О. P. and D. W. McBride, 1994, «The cloning of a mechano-gated membrane ion
channel», Trends in Neurosciences, 17, 439—443.
Pross er, C. Ladd and F. A. Brown, 1962, Comparative Animal Physiology, Philadelphia:
W. B. Saunders.
Sukha rev, S. I. et al., 1994, «А large-conductance mechanosensitive channel in E. coli
encoded by mscL alone», Nature, 368, 265—268.
Sukharev, S. I., P. Blount, B. Martinac and C. Kung, 1997, «Mechanosensitive channels of
Escherichia coli'. the MscL gene, protein and activities». Annual Review of Physiology, 59,
633-657.
202 Часть II. Механочувствительность
Глава 6: Кинестезия
Структура и функции сенсилл насекомых подробно описаны Мак-Айвером
(McIver, 1985). Дополнительные детали изложены Альтнером и Прилингером
(Altner & Prillinger, 1980), тогда как Блюм (Blum, 1985) дает более общий под-
ход. Биофизика механочувствительных сенсилл описана Френчем (French,
1988), тогда как Филд и Мэтсон (Field and Matheson, 1998) дали исчерпываю-
щий обзор хордотональных органов. Одним из лучших изданий по общей эн-
томологии (морфология, анатомия, систематика) до сих пор остается книга
Иммса (Imms, 1946). Кинестезия у млекопитающих обсуждается в работах по
нейропсихологии и нейронаукам, напр. Кэндела с соавт. (Kandel, Schwartz and
Jessel, 1991) и обзоре МакКлоски (McCloskey, 1978). Другие детали можно най-
ти у Ханта (Hunt, 1990) и др. (Jami, 1992; Matthews, 1981).
Насекомые
Altner, Н. and L. Prillinger, 1980, «Ultrastructure of invertebrate chemo-, thermo- and
hygroreceptors and its functional significance», International Review of Cytology, 67,
69-139.
Blum, M. S., 1985, ed. Fundamentals of Insect Physiology, New York: Wiley.
Field, L. H. and T. Matheson, 1998, «Chordotonal organs of insects». Advances in Insect
Physiology, 27, 1—228.
French, A. S., 1988, «Transduction mechanisms of mechanosensilla». Annual Review of
Entomology, 13, 39—58.
Markl, H. and J. Tautz, 1975, «Sensitivity of hair receptors in caterpillars of Barathra bras-
sica L. (Lepidoptera, Noctuidae) to particle movement in a sound field», Journal of
Comparative Physiology, 99, 79-87.
McIver, S. B., 1985, «Mechanoreception», in Comprehensive Insect Physiology, Biochemistry
and Pharmacology, Oxford: Pergamon Press.
Imms, A. D., 1946, A General Textbook of Entomology (6th edn), London, Methuen.
Pringle, J. W. S., 1948, «Proprioception in insects III: the function of hair sensilla at joints»,
Journal of Experimental Biology, 15, 467—473.
Млекопитающие
Hunt, С. C., 1990, «Mammalian muscle spindle: peripheral mechanisms». Physiological
Reviews, 70, 643—663.
Jami, L€na, 1992, «Golgi tendon organs in mammalian skeletal muscle: functional proper-
ties and central actions», Physiological Reviews, 72, 623—666.
Kandel, E. R., J. H. Schwartz and T. M. Jessel, eds, 1991, Principles of Neuroscience, New
York: Elsevier.
Matthews, P. В. C., 1981, «Evolving views on the internal operation and functional role of
the muscle spindle», Journal of Physiology, 320, 1—30.
McCloskey, D. I., 1978, «Kinesthetic sensibility», Physiological Reviews, 58, 763—820.
Wilkinson, R. S. and Y. Fukami, 1983, «Responses of isolated Golgi tendon organs of cat to
sinusoidal stretch», Journal of Neurophysiology, 49, 976—988.
Материалы к части II 203
Глава 7: Осязание
Чувствительность С. elegans обсуждается в ряде работ (Huang & Chalfie, 1994;
Hong & Driscoll, 1994; Tavernarakis and Driscoll, 1997). Структура амилорид-
чувствительных №+-каналов описана в работе Канесса с соавт. (Canessa etal.,
1994). Единство молекулярных механизмов нематод, насекомых и позвоноч-
ных описано в обзоре Garcia-Anoveros & Corey (1997). Механорецепторы насе-
комых описаны Frazier (1985), Michelsen & Larsen (1985) и Schwarzkopf? (1974);
тимпанальные слуховые органы — Ноу & Robert (1996). К. Д. Рёдер выполнил
большинство пионерских работ по нейрофизиологии и поведению бабочек
при атаке сонара рукокрылых (см. Roeder, 1975); более поздние работы по
акустической физиологии и поведению обсуждаются в работе Spangler (1988).
Оригинальные работы по колончатой структуре соматосенсорной коры мле-
копитающих представлены Mountcastle (1957). Исследования пластичности
соматосенсорной коры описаны Woolsey & van der Loos (1970), Cowan (1979),
Jenkins et al. (1990), Elbert (1995), Hamilton & Pascuel-Leone (1997), Yaest &
Simons (1997), общий обзор таких работ см. Buonomano & Merzenich (1998).
Физиология и эндокринология снятия возбуждения грумингом рассмотрены в
обзоре Spruijt, van Hoof and Gispen (1992).
Caenorhabditis elegans
Hong, K. and M. Driscoll, 1994, «А transmembrane domain of the putative channel subunit
MEC-4 influences mechanotransduction and neurodegeneration in C. elegans». Nature,
367, 470-473.
Huang, M. and M. Chalfie, 1994, «Gene interactions affecting mechanosensory transduc-
tion in Caenorhabditis elegans», Nature, 367, 470—473.
Tavernarakis, N. and M. Driscoll, 1997, «Molecular modelling of mechanotransduction in
the nematode Caenorhabditis elegans», Annual Review of Physiology, 59, 659—689.
Насекомые
Frazier, J. L., 1985, «Nervous system: sensory system», in M. S. Blum, ed., Fundamentals of
Insect Physiology, New York: Wiley.
Hoy, R. R. and D. Robert, 1996, «Tympanal hearing in insects», Annual Review of Ento-
mology, 41, 433—450.
Michelsen, A., 1974, «Hearing in invertebrates», in Handbook of Sensory Physiology, H. Aut-
rum etal., eds, Berlin: Springer-Verlag, vol. V/1.
Michelsen, A. and O. N. Larsen, 1985, «Hearing and sound», in Comprehensive Insect
Physiology, Biochemistry and Pharmacology, G. A. Kerkut and L. I. Gilbert, eds, Oxford:
Pergamon Press, vol. 6.
Roeder, K. D., 1975, «Neural transactions during acoustic stimulation of noctuid moths» in
Sensory Physiology and Behavior, Galum, R., etal., eds, New York: Plenum.
Schwartzkopff, 1974, «Mechanoreception», in The Physiology of Insecta (2nd edn),
M. Rockstein, ed., New York: Academic Press.
Spangler, H. G., 1988, «Moth hearing, defense, and communication», Annual Review of
Entomology, 33, 59—81.
204 Часть II. Механочувствительность
Млекопитающие
Elbert, Т. et al., 1995, «Increased cortical representation of the fingers of the left hand in
string players», Science, 270, 305—307.
Buonomano, D. V. and M. M. Merzenich, 1998, «Cortical plasticity: from synapses to
maps», Annual Review of Neuroscience, 21, 149—186.
Hamilton, R. H. and A. Pascuel-Leone, 1998, «Cortical plasticity associated with Braille
learning», Trends in Cognitive Sciences, 2, 168—174.
Iggo, A., 1982, «Cutaneous sensory mechanisms», in The Senses, H. B. Barlow and
J. D. Mellon, eds, Cambridge: Cambridge University Press.
Jenkins, W. M., et al., 1990, «Functional re-organisation of primary somatosensory cortex
in adult owl monkeys after behaviourally controlled tactile stimulation», Journal of
Neurophysiology, 63, 82-104.
Mountcastle, V. B., 1957, «Modality and topographic properties of single neurons of cat’s
somatic sensory cortex», Journal of Neurophysiology, 20,408—434.
Woolsey, T. A. and H. van der Loos, 1970, «The structural organisation of layer IV in the
somato-sensory region (SI) of mouse cerebral cortex. The description of a cortical field
composed of discrete cytoarchitectonic units», Brain Research, 17, 205—242.
Yaest, R. and D. Simons, 1997, «Barrels in the desert: the Sde Веки workshop on neocorti-
cal circuits», Neuron, 19, 231—237.
Общие работы
Canessa, С. M., 1994, «Amiloride-sensitive epithelial Na+ channel is made of three homol-
ogous subunits», Nature, 367, 463—467.
Garcia-Anoveros, J. and D. P. Corey, 1997, «The molecules of mechanosensation», Annual
Review of Neuroscience, 20, 567—595.
Cowan, W. Maxwell, 1979, «The Development of the Brain», Scientific American, 241(3),
107-117.
Spruijt, В. M., J. A. R. A. M. van Hoof and W. M. Gispen, 1992, «Ethology and neurobiol-
ogy of grooming behaviour», Physiological Reviews, 72, 825—852.
White, J. G., 1985, «Neuronal connectivity in Caenorhabditis elegans», Trends in
Neurosciences, 8, 277—283.
Глава 8: Равновесие и слух: функция волосковых клеток
Выдающуюся роль в развитии биофизики и молекулярной биологии волоско-
вых клеток сыграл Хадспет (Hudspeth). В двух приведенных в списке работах
(Hudspeth, 1989,1992) дается обзор его работ, а в публикации 1997 года сумми-
рованы сведения по всей сфере этих вопросов. Другие исследователи тоже за-
нимались этими вопросами, в частности Gillespie & Corey (1997) предложили
интересный путь, по которому могла идти сенсорная адаптация. Краткий об-
зор генетики глухоты дан в работе Griffiths & Friedman (1999), a Yasanuga et al.
(1999) описали выделение генов, вызвающих несиндромную глухоту. Pickles
(1988) дал обзор физиологии слуха, особенно хороши сканирующие электрон-
ные микрофотографии волосковых клеток улитки. Система боковой линии
рассмотрена Flocke (1971), Prosser & Brown (1962) и Sand (1984). Эволюция
мембранного лабиринта обсуждается Ромером (Romer, 1970), а более деталь-
но — Baird, 1974; Wever, 1974. Clack (1997) сделал обзор эволюции уха четверо-
ногих позвоночных.
Материалы к части II 205
Биофизика волосковых клеток
Flocke, А. 1965, «Transducing mechanisms in lateral line canal organ receptors», in
«Sensory receptors», L. Frisch, ed., Cold Spring Harbor Symposia in Quantitative Biology,
xxx, 133—145, New York: Cold Spring Harbor.
Gillespie, P. G., 1995, «Molecular machinery of auditory and vestibular transduction»,
Current Opinion in Neurobiology, 5, 449—455.
Gillespie, P. G. and D. P. Corey, 1997, «Myosin and adaptation by hair cells», Neuron, 19,
955-958.
Griffith, A. J. and T. B. Friedman, 1999, «Making sense of sound», Nature Genetics, 21,
347-349.
Hackney, С. M. and D. N. Furness, 1995, «Mechanotransduction in vertebrate hair cells:
structure and function of the stereociliary bundle», American Journal of Physiology, 268,
C215-221.
Hudspeth, A. J., 1989, «How the ear’s works work», Nature, 341, 398—404.
Hudspeth, A. J., 1992, «Hair-bundle mechanics and a model for mechanoelectrical trans-
duction by hair cells», in Sensory Transduction, D. P. Corey anu S. D. Roper, eds, New
York: Rockefeller Press, pp. 357—370.
Hudspeth, A. J., 1997, «How hearing happens», Neuron, 19, 947—950.
Pickles, J. O., 1988, An Introduction to the Physiology of Hearing, New York: Academic Press.
Pickles, J. O. and D. P. Corey, 1992, «Mechanoelectrical transduction by hair cells», Trends
in Neurosciences, 15, 254—258.
Yasunaga, S. etal., 1999, «А mutation in OTOF, encoding otoferlin, a FER-l-like protein;
causes DFNB9, a nonsyndromic form of deafness», Nature Genetics, 21, 363—369.
Канал боковой линии
Flocke, A., 1971, «The lateral line oi^an mechanoreceptors», in Fish Physiology, W. S. Hoar
and D. J. Randall, eds, New York: Academic Press, vol. V, pp. 241—263.
Prosser, C. Ladd and F. A. Brown, 1962, Comparative Animal Physiology, Philadelphia:
W. B. Saunders.
Sand, O., 1984, «Lateral line systems», in Comparative Physiology of Sensory Systems,
L. Bolis, R. D. Keynes and S. H. P. Maddrell, eds, Cambridge: Cambridge University
Press.
Эволюция уха
Art, J. J., 1984, «Auditory function at the receptor level in reptiles», in Comparative
Physiology of Sensory Systems, L. Bolis, R. D. Keynes and S. H. P. Maddrell, eds,
Cambridge: Cambridge University Press.
Baird, I. L., 1974, «Anatomical features of the inner ear in submammalian vertebrates», in
Handbook of Sensory Physiology, H. Autrum etal., eds, Berlin: Springer-Verlag, vol. V/1.
Chung, Shin-Но, A. Pettigrew and M. Anson, 1978, «Dynamics of the amphibian middle
ear», Nature, 272, 142—147.
Clack, J. A., 1997, «The evolution of tetrapod ears and the fossil record», Brain, Behaviour
and Evolution, 50, 198—212.
Feng, A. S. and W. P. Shofner, 1981, «Peripheral basis of sound localisation in anurans,
acoustic properties of the frog’s ear», Hearing Research, 5, 201—216.
Romer, A. S., 1970, The Vertebrate Body (4th edn), Philadelphia: W. B. Saunders.
Wever, E. G., 1974, «The Evolution of vertebrate hearing», in Handbook of Sensory Physio-
logy, H. Autrum etal., eds, Berlin: Springer-Verlag, vol. V/1.
206 Часть II. Механочувствительность
Вестибулярная система
Benson, A. J., 1982, «The vestibular sensory system», in The Senses, H. B. Barlow and
J. D. Mollon, eds, Cambridge: Cambridge University Press.
Freeman, W. H. and B. Bracegirdle, 1976, An Atlas of Advanced Histology, London: Heine-
mann.
Smith, C. U. M., 1970, The Brain: Towards an understanding, London: Faber and Faber.
Глава 9: Анализ вестибулярной
и звуковой информации в мозге
Вестибулярный путь детально описан Вильсоном и Мелвилл Джонсом (Wilson
and Melville Jones, 1979). Следует отметить, что и нистагм, и болезни движения
могут быть адаптированы. В случае нистагма опыты с очками, переворачиваю-
щими изображение, показали, что в течение недели адаптации или около того
нистагм ревертируется. Аналогичным образом, ошибки движения обычно ис-
чезают за несколько дней или недель. Эксперименты на животных показали,
что эта адаптация связана с флоккулюсом мозжечка и его проекциями в вести-
булярную систему. Так что для большинства из нас морская болезнь — лишь
временная фаза длительных путешествий.
Значение подкорковых ядер слуховых путей обсуждается в ряде работе
(Neffeffl/., 1975, Masterton, 1992). Результаты нейрофизиологических исследо-
ваний высокоразвитого слухового мозга сипух описаны в работах Konishi
(1992, 1993) и Cohen & Knudsen (1999). Слуховой мозг сов сравнивается с моз-
гом рукокрылых, приводится информация о деталях нейрофизиологических
процессов, лежащих в основе эхолокационного поведения. Суга, который так-
же внес большой вклад в работу, интересовался также связью между этими
слуховыми механизмами с таковыми в речевой коре человека (Suga, 1995).
Отоакустическая эмиссия в ухе человека и других млекопитающих рассмотре-
на в работе Пробста с соавторами (Probst et al., 1991). Слуховая кора человека
и наиболее сложные процессы, в том числе человеческий язык, который сам
по себе представляет огромное поле исследований, описаны Брюгге и Реале
(Brugge & Reale, 1985). Стефен Пинкер (Pinker, 1974) сдалал один из лучших
обзоров, а также опубликовал еще несколько перечисленных ниже работ по
нейрофизиологическому субстрату языка. Под редакцией Harnad, Steklis &
Lancaster (1976) вышел всеобъемлющий обзор его эволюционного происхож-
дения и развития. В нашем тексте не нашлось места для рассмотрения пения
птиц, которое во многих отношениях является единственным аналогом языка
у животных, не относящихся к млекопитающим, однако в списке литературы
приведены две недавних работы (Bottjer & Arnold, 1997; Margoliash, 1997), ко-
торые могут дать начальные представления для тех, кто интересуется совре-
менным состоянием проблемы. Наконец, философ Томас Нагель задает во-
просы, над которыми всем нам надо задумываться и которые приводят нас к
наиболее важным моментам проблемы соотношения мозга и разума (Nagel,
1974).
Материалы к части II 207
Подкорковые пути
Evans, Е. F., 1972, «The frequency response and other properties of single fibres in guinea
pig cochlear nerve», Journal of Physiology, 226, 263—287.
Evans, E. F., 1982, «Functional anatomy of the visual system», in The Senses, H. B. Barlow
and J. D. Mollon, eds, Cambridge: Cambridge University Press.
Masterton, R. B., 1992, «Role of the central auditory system in hearing: the new direction»,
Trends in Neurosciences, 15, 280—284.
Neff, W. D., I. T. Diamond and J. H. Casseday, 1975, «Behavioural studies of auditory dis-
crimination: central nervous system», in Handbook of Sensory Physiology: Auditory
System, vol. V/2, W. D. Keidel and W. D. Neff, eds, New York: Springer-Verlag.
Wilson, V. J. and G. Melville Jones, 1979, Mammalian Vestibular Physiology, New York:
Plenum Press.
Слуховое пространство сипух
Adolphs, R., 1993, «Bilateral inhibition generates neuronal responses tuned to interaural
level differences in the auditory brainstem of the barn owl», Journal of Neuroscience, 13,
3647-3668.
Cohen, Y. E. and E. I. Knudsen, 1999, «Maps versus clusters: different representations of
auditory space in the midbrain and forebrain», Trends in Neuroscience, 22, 128—135.
Konishi, M., 1992, «The neural algorithm for sound localisation in the owl», Harvey Lec-
tures, 86, 47—64.
Konishi, M., 1993, «Listening with two ears», Scientific American, 268(4), 34—41.
Koppl, C., 1997, «Phase-locking to high frequencies in the auditory nerve and cochlear
nucleus magnocellularis of the barn owl, Tyto alba», Journal of Neuroscience, 17,
3312-3321.
Слуховая кора приматов
Brugge, J. F. and R. A. Reale, 1985, «Auditory cortex» in Cerebral Cortex, A. Peters and
E. G. Jones, eds, New York: Plenum Publishing.
Kandel, E. R., J. H. Swartz, T. M. Jessell, 1991, Principles of Neural Science (3rd Edition),
New York: Elsevier.
Whitfield, I. C. and E. F. Evans, 1965, «Responses of auditory neurons to stimuli of chang-
ing frequency», Journal of Neurophysiology, 28, 655—672.
Эхолокация у рукокрылых
Neuweiler, G., 1990, «Auditory adaptations for prey capture in echolocating bats», Physio-
logical Reviews, 70, 615—641.
Suga, N., 1995, «Processing of auditory information carried by species-specific complex
sounds», in The Cognitive Neurosciences, M. S. Gazzaniga, ed, Cambridge, MA.: MIT
Press, pp. 295—313.
Suga, N., J. Yan and Y. Zhang, 1997, «Cortical maps for hearing and egocentric selection for
self-organisation», Trends in Cognitive Sciences, 1, 13—20.
Sullivan, W. E., 1986, «Processing of acoustic temporal patterns in bam owls and echolo-
cating bats, similar mechanisms for the generation of neural place representations in
auditory space», Brain Behav. Evol., 28, 109—121.
208 Часть II. Механочувствительность
Слуховая кора человека и речь
Damasio, A. R. and N. Geschwind, 1984, «The neural basis of language», Annual Review of
Neuroscience, 7, 127—147.
Eimas, P. D. etal., 1971, «Speech perception in infants», Science, 171, 103—104.
Fitch, R. H., S. Miller and P. Tallal, 1997, «Neurobiology of Speech», Annual Review of
Neuroscience, 20, 331—53.
Geschwind, N. and W. Levitsky, 1968, «Human brain: Left—right asymmetries in temporal
speech area», Science, 161, 186—187.
Geschwind, N. etal., 1968, «Isolation of the speech area», Neuropsychologia, 6, 327—340.
Hamad, S. R., H. D. Steklis and J. Lancaster, 1976, eds, «Origins of language and speech»,
Annals of the New York Academy of Science, 280, 1—914.
Liberman, A. M. et al., 1957, «The discrimination of speech sounds within and across
phoneme boundaries», Journal of Experimental Psychology, 54, 358—368.
Miller, J. L. and P. D. Eimas, 1995, «Speech perception: from signal to word», Annual
Review of Psychology, 46, 467—492.
Pinker, S., 1994, The Language Instinct, Harmondsworth: Penguin.
Springer, S. P., 1979, «Speech perception and the biology of language», in Handbook of
Behavioral Neurobiology, vol. 2: Neuropsychology, M. S. Gazzaniga, ed., New York:
Plenum Press.
Suga, N., 1995, «Processing of auditory information carried by species-specific complex
sounds», in M. S. Gazzaniga, ed., The Cognitive Neurosciences, Cambridge, MA.: MIT
Press, pp. 295—313.
Werker, J. and R. C. Tees, 1992, «The organisation and reorganisation of human speech per-
ception», Annual Review of Neuroscience, 15, 377—402.
Young, J. Z., 1978, Programs of the Brain, Oxford: Oxford University Press.
Каллозэктомия
Sperry, R. W., 1964, «The great cerebral commissure», Scientific American, 210(1), 42—52.
Общие работы
Bottjer, S.W. and A. P. Arnold, 1997, «Developmental plasticity in neural circuits for
a learned behaviour», Annual Review of Neuroscience, 20, 459—481.
Hudspeth, A. J., 1997, «How hearing happens», Neuron, 19, 947—50.
Maigoliash, D., 1997, «Distributed time-domain representations in the birdsong system»,
Neuron, 19, 963—966.
Nagel, T., 1974, «What is it like to be a bat», Philosophical Review, 83, 435—450.
Probst, R., B. L. Lonsbury-Martin and G. K. Martin, 1991, «А review of otoacoustic emis-
sions», Journal of the Acoustical Society of America, 89, 2027—2067.
Часть III
ХЕМОЧУВСТВИТЕЛЬНОСТЬ
«Когда от далекого прошлого уже ничего не осталось, люди
умерли, вещи сломались и рассыпались, остается что-то хрупкое,
но полное жизни и в то же время бестелесное, но и более проч-
ное, более стойкое; запах и вкус остаются еще очень долго, как
дух, готовый напомнить нам о себе, ждущий своего часа и наде-
ющийся на это, посреди руин всего, необоримый в почти неося-
заемой своей сущности в огромной картине воспоминания.»
Марсель Пруст, Воспоминание
о прошедшем («В сторону Свана»)
Перевод с перевода Скотта Монкрифа
(С. К. Scott Moncrief!)
Животные чувствительны к молекулярному составу окружающей их среды.
Это важно не только при питании, но и в воспроизводстве, взаимоотношени-
ях с потомством, распознавании территорий обитания и других формах соци-
ального поведения. У позвоночных ощущения запаха и вкуса связаны с древ-
ними отделами мозга. Возможно, этим объясняется эмоциональный резонанс,
который сопровождает эти чувства. Наиболее известное, вынесенное в эпи-
граф, высказывание по этому поводу принадлежит Марселю Прусту о том, что
вкус пирожного пробуждает в нем настолько живые воспоминания, что он
ощущает, будто настали прежние времена. Этот образ подчеркивает то обсто-
ятельство, что древний обонятельный мозг установил связи с новой корой.
Действительно, такие связи через таламус к орбито-фронтальной коре (запах)
и в постцентральную извилину и островок (вкус) прослежены.
В отличие от механорецепторов (см. Часть II), строение всех хеморецеп-
торных органов многоклеточных (по крайней мере, те, что известны ныне) ба-
зируется на семидоменных рецепторах и G-белках, преобразующих внешние
стимулы во внутриклеточные сигналы (см. гл.1). Вследствие этого хеморецеп-
ция происходит несравнимо медленнее, чем механорецепция. Однако то, что
хеморецепторы проигрывают в скорости, более чем компенсируется их совер-
шенством. Рассчитано, что до 2% генома млекопитающих может быть связано
с генами, продуцирующими около 1000 различных обонятельных рецептор-
ных молекул. Вомероназальный орган млекопитающих связан еще с
100—200 генами. У дрозофилы (да и у других насекомых) также значительная
часть генома связана с обонятельными рецепторами. Хеморецепторы немато-
ды Caenorhabditis elegans образуют четыре больших не связанных между собой
семейства, охватывающих более 100 генов. С этим количеством генов контра-
стирует весьма ограниченное разнообразие форм хемосенсорного поведения
С. elegans'. питание, откладка яиц и копуляция. В отличие от млекопитающих,
каждая хемосенсорная клетка С. elegans экспрессирует несколько семидомен-
ных рецепторов.
Похоже, что семидоменные белки всех этих различающихся систем воз-
никли независимо. Наверное, мы очень плохо можем себе представить, каков
210 Часть III. Хемочувствительность
«мира вкусов и запахов», в котором живут другие животные. Хотя натрениро-
ванный человек в состоянии различить по крайней мере 10 000 различных за-
пахов, многие животные (и наверное, большинство млекопитающих) способ-
ны на существенно большее. Они живут в мире, гораздо более богатом вкуса-
ми, ароматами и запахами.
Химические чувства обычно подразделяются на три основных: общее хими-
ческое чувство, обоняние и вкус. Все организмы обладают общим химическим
чувством. Лишь только капелька кислоты коснется нашей кожи, как нам все
становится ясно. Это ощущение опосредуется нервными окончаниями в коже.
Позвоночные, помимо перечисленного, обладают еще и четвертой хемочувст-
вительной системой, состоящей из одиночных хемочувствительных клеток
(ОХК). Эта система наиболее развита у двух видов костистых рыб: американ-
ской триглы (Prionotus) и морского налима (Ciliata sp.). ОХК, разбросанные в
эпителии, покрывающем плавники, вовлечены в обнаружение пищи и хищни-
ков. Однако химические чувства, представляющие для нас наибольший инте-
рес в этой книге — это обоняние и вкус, поскольку в них достигнута не только
исключительно высокая чувствительность к ничтожным количествам ве-
ществ, но и развита способность к различению химических стимулов. В гл. 10
будет сделан краткий обзор хемочувствительности у прокариот. Мы увидим,
что методы молекулярной биологии дают почти исчерпывающее понимание
этих простых, но универсально важных процессов у Escherichia coli. Далее, в
гл. 11 мы рассмотрим очень важную роль хеморецепторов в контроле состава
внутренней среды у млекопитающих. Гл. 12 посвящена вкусу, а гл. 13 — обо-
нянию.
10. ХЕМОЧУВСТВИТЕЛЬНОСТЬ
У ПРОКАРИОТ
Возникновение хемочувствительности — подвижные бактерии — структу-
ра и функция жгутиков прокариот — равномерное движение и беспоря-
дочное вращения клетки — «кувыркание» Е. coli — хемотаксис — связы-
вающие белки и рецепторно-передающие (Р-П) белки — сенсорная
адаптация — генетический анализ Р-П-белков — биохимия внутрикле-
точных сигнальных путей между Р-П и эффектором — сенсорная адапта-
ция, вызванная метилированием. Заключение: перенос представлений о
сенсорных системах на молекулярный уровень
Несомненно, хемочувствительность возникла в истории жизни на Земле очень
давно. С самого начала эволюции прокариоты были погружены в водную сре-
ду. Способность реагировать на химический состав среды была критичной для
выживания. Можно предположить, что после того, как сложился удачный ме-
ханизм, в дальнейшем давление отбора, стимулирующее его изменение, было
невелико. Как и в случае механорецепторов, которые мы рассматривали в Ча-
сти II, эволюционный процесс не мог пойти вспять и начаться заново. Пере-
страивалось то, что было единожды найдено — «доводилось» до совершенства.
Тем не менее, как мы увидим, молекулярная биология хемочувствительности
у бактерий, таких как Escherichia coli и Salmonella typhimurium, в некоторых от-
ношениях заметно отличается от той, что лежит в основе обоняния и вкуса у
многоклеточных.
Нет сомнений, что современные подвижные бактерии чувствительны к хи-
мическим веществам в окружающей среде. Уже давно известно, что они двига-
ются по градиенту концентраций привлекательных для них химических ве-
ществ. То есть они способны и обнаруживать вещество в среде и действовать в
соответствии с этим. Каким образом им это удается?
Подвижные бактерии двигаются за счет жгутиков. Жгутики прокариот на-
много проще, чем у эукариот — это единый участок субъединиц флагеллина,
сплетенных в спираль (рис. 10.1). Механизмы, обеспечивающие движение,
также очень различаются. В отличие от системы скользящих филаментов в
жгутиках эукариот, у прокариот действует, по-видимому, единственная орга-
ническая структура, использующая вращательный механизм. Связанное с
клеткой основание жгутика вращается со скоростью около 100 оборотов в се-
кунду за счет механизма, который черпает энергию в трансмембранном гради-
енте ионов водорода.
212 Часть III. Хемочувствительность
Рис. 10.1. Вращательный
механизм бактериального
жгутика. Механизм по-
движности пронизывает и
наружную, и внутреннюю
мембраны бактерии. Энер-
гия образуется в мембран-
ном протонном градиенте
и вызывает вращение М-
диска относительно S-дис-
ка (или статора) со скоро-
стью около 100 об/с. Ста-
тор остается неподвижным
за счет погружения в пеп-
тидогликановый слой вну-
тренней мембраны. Стер-
жень связывает М-диск с
коленом и, далее, со спи-
ральным филаментом жгу-
тика. Опора в наружной
мембране выполняет роль
подшипника. Воспроизво-
дится из Adler, 1976.
10.1. Хемочувствительность у Е. coli
У большинства жгутиконосных бактерий жгутиков больше одного, у Е, coli их
5—10. Когда все они вращаются против часовой стрелки, бактерия плавно дви-
жется вперед. Дело, однако, обстоит совершенно иначе, когда жгутики враща-
ются по часовой. В этом случае, вследствие спиральной структуры жгутика,
они вытягиваются, и это приводит к беспорядочному кувырканию (рис. 10.2).
Рис. 10.2. Вращение жгутиков бактерий против часовой и по часовой стрелке
(А) Вращение против часовой стрелки. Жгутики двигаются единым пучком, тол
кая бактерию вперед. (Б) Вращение по часовой стрелке. Жгутики расходятся и
двигаясь независимо, толкают бактерию в разные стороны. Бактерия хаотично мс
няет направление движения и беспорядочно кувыркается.
10. Хемочувствительность у прокариот 213
Рис. 10.3. Миграция бактерии по градиенту концентрации химического аттрактан-
та. (А) Движение бактерии в отсутствие градиента аттрактанта. Движение состоит
из перемежающихся периодов прямолинейного движения за счет вращения жгути-
ков против часовой стрелки, и беспорядочных кувырканий, когда жгутики враща-
ются по часовой. Направление, в котором двигается бактерия после окончания ха-
отического движения — чисто случайно. Таким образом, бактерия продвигается не
в каком-то определенном направлении, а движется беспорядочно либо описывает
круг. (Б) Движение в присутствии аттрактанта. Длительность (а, следовательно, и
длина пробега) между хаотическими движениями увеличивается, когда бактерия
движется по направлению к источнику аттрактанта, и уменьшается — когда дви-
жется от него. Поэтому, хотя наблюдаются периоды хаотического движения, а на-
правление движения после него случайно, бактерия движется по градиенту к ис-
точнику аттрактанта. С — точка старта движения, Ф — точка финиша.
Если проследить за плаванием бактериальной клетки под микроскопом, то
будет видно, то этот процесс состоит из серии плавных движений (длительно-
стью в несколько секунд), перемежающихся короткими эпизодами (ок. 0,1 с)
хаотических. Когда клетка завершает беспорядочное кувыркание, она возоб-
новляет плавное движение, но в совершенно случайном направлении. Одна-
ко, если поместить клетку в градиент химического аттрактанта, то оказывает-
ся, что при движении в направлении к нему хаотическое движение случается
много реже, чем когда бактерия движется в любом другом направлении. В ре-
зультате, бактерия движется к аттрактанту.
Этот феномен представляет собой ценную основу при исследования общих
проблем хеморецепции. Жгутики бактерии можно прикрепить к стеклу и на-
блюдать за их вращением при внесении различных химических веществ. Гене-
тика Е. coli изучена очень хорошо, поэтому и сенсорные системы могут изу-
чаться и в этом аспекте.
Показано, что детектирование молекул аттрактанта (или репеллента) осу-
ществляется трансмембранными белками. Существует два таких механизма.
Молекула аттрактанта может взаимодействовать со связывающим белком (СБ),
иногда ошибочно называемым рецепторной молекулой, в периплазматичес-
ком пространстве, вызывая в последнем конформационные изменения, кото-
рые делают возможным еп/взаимодействие с тем или иным трансмембранным
белком. Это относится к аттрактантам — сахарам и пептидам. Выделены га-
лактоза-связывающий белок (ГСБ), рибоза-связывающий белок (РСБ), маль-
тоза-связывающий белок (МСБ) и дипептид-связывающий белок (ДСБ).
214 Часть III. Хемочувствительность
Q
Молекула
аттрактанта
G5 V Молекулы аттрактанта
Q } Внешняя
/ Связывающий среда
' белок Молекулы аттрактанта
Рецепторно-
передающий
белок
Внутриклеточный
сигнал
Бактериальная
цитоплазма
М-дискх^
жгутика
Периплазматическое пространство-
Пептидогликановый слой'
Внутренняя мембрана-
Жгутик
Наружная мембрана
Рис. 10.4. Взаимодействие
молекулы аттрактанта со
связывающими и Р-П-бел-
ками. Существуют аттрак-
танты двух типов — они
либо занимают активный
сайт Р-П-молекулы непо-
средственно, либо «дости-
гают» этого путем соеди-
нения со связывающим
белком в периплазматиче-
ском пространстве. В обо-
их случаях активирован-
ная Р-П-молекула генери-
рует сигнал (см. рис. 10.6),
который влияет на враще-
ние мотора жгутика и, та-
ким образом, на частоту
хаотических движений.
В случае же аминокислот, аттрактант взаимодействует непосредственно с
трансмембранным белком. Трансмембранные белки способны влиять на вра-
щение жгутиков, т. е. распознавать молекулы аттрактанта и передавать их сиг-
нал. Чтобы отличить эти белки от связывающих в периплазматическом прост-
ранстве, их именуют рецепторно-передающими (Р-П) белками.
Эта система имеет еще одно очень важное свойство — сенсорную адапта-
цию, что вообще характерно для всех сенсорных систем. В случае Е. coli пока-
зано, что помещение бактерии в среду с аттрактантом подавляет хаотические
движения, однако, они восстанавливаются в течение нескольких минут и воз-
вращаются к обычной частоте. Эта адаптация касается специфической моле-
кулы аттрактанта, поскольку добавление другого аттрактанта снова подавляет
хаотические движения. Биологическое значение такой адаптации и десенси-
тизации очевидно.
10.1.1. Молекулярная генетика
Поскольку молекулярная биология Е. coli так хорошо изучена, она представляет
собой удобную систему для исследования всех вышеперечисленных процессов.
Связывающие белки (СБ)
Детальная молекулярная структура множества связывающих белков уже опре-
делена. Например, ГСБ был успешно подвергнут рентгеновскому дифракци-
онному исследованию с высоким разрешением, и определены его общая архи-
10. Хемочувствительность у прокариот 215
тектура и конфигурация сайтов связывания. Показано, что ГСБ состоит из
двух «крыльев», разделенных расщелиной, в которой и располагается место
связывания. Когда сайт занят субстратом (галактозой или глюкозой), крылья
«складываются», двигаясь друг к другу, до образования угла в 18°. Это измене-
ние трехмерной конформации дает возможность связывающему белку взаимо-
действовать с соответствующей трансмембранной Р-П-молекулой. Существу-
ют данные, что и другие связывающие белки претерпевают сходные изменения
трехмерной конформации. Интересно отметить, что и обонятельные сенсилл-
лы насекомых (гл. 13, раздел 13.1.2) и обонятельные реснички млекопитающих
(гл. 13, раздел 13.2.2) также используют связывающие белки для захвата паху-
чих молекул и переноса их на мембранные обонятельные рецепторы.
Рецепторно-передающие (Р-П) белки
Гены многих Р-П-белков выделены у Е. coli с помощью молекулярно-генети-
ческих методов, и их нуклеотидные последовательности определены. Наибо-
лее известны среди них гены tsr, tar, trgw tap. Ими кодируются четыре Р-П-бел-
ка: Tsr, чувствительный к серину (аттрактанту) и лейцину (репелленту); Таг,
белок, чувствительный к аспартату и мальтозе (аттрактанты), а также Со2+ и
Ni2+ (репелленты); Trg, чувствительный к галактозе и рибозе, присоединен-
ным к связывающим белкам (аттрактанты), и Тар, чувствительный к дипепти-
дам, присоединенным к связывающим белкам.
Рис. 10.5. Рецепторно-пере-
дающие молекулы (показаны
две). Из рисунка видно, что
они тесно ассоциированы, об-
разуя димер. Каждая из них
вносит вклад в формирование
лиганда — места связывания
одоранта либо репеллента.
Показаны цитоплазматичес-
кое метилирование и сигналь-
ные домены.
216 Часть III. Хемочувствительность
Исходя из нуклеотидной последовательности, может быть определена
аминокислотная структура Р-П-белков и выделены сегменты, пронизываю-
щие мембрану. Все четыре белка состоят из более чем пятисот аминокислот,
и содержат по две трансмембранных последовательности. В этом отношении
они радикально отличаются от семидоменных рецепторных белков вкусовых
и обонятельных рецепторов позвоночных. С другой стороны, между всеми че-
тырьмя бактериальными Р-П-белками существует строгая гомология, осо-
бенно между цитоплазматическими доменами, где происходят передача сиг-
нала к жгутиковому аппарату и сенсорная адаптация.
Рис. 10.5 показывает, что Р-П-молекула расположена в мембране таким об-
разом, что петля из прибл. 150 аминокислот направлена в межклеточное (вне-
плазматическое) пространство. Именно она и формирует место связывания
аттрактанта либо репеллента. Ббльшая часть белка из прибл. 215 аминокислот
(до карбоксильного конца, С) лежит в бактериальной цитоплазме. Эта часть
молекулы очень консервативна у всех четырех Р-П-белков, она опосредует
сигнал к жгутиковому аппарату и участвует в механизме адаптации. Р-П-бе-
лок, таким образом, представляет собой димер двух полипептидов по 50 кДа,
вероятно, распределенный в плазмалемме случайным образом.
Трансдукция химического сигнала
Хемотаксисная сигнализация запускается взаимодействием молекулы аттрак-
танта (либо молекулярного комплекса аттрактант + связывающий белок) с
внеплазматическим доменом Р-П-молекулы. Существуют данные о том, что
это связывание вызывает конформационные изменения молекулы, передаю-
щиеся через мембрану и изменяющие конфигурацию цитоплазматического
домена этой молекулы. Это в свою очередь инициирует биохимический сиг-
нальный каскад, который в конце концов влияет на активность жгутика.
Генетический анализ позволил выделить и проанализировать элементы
этого сигнального пути. Он включает четыре важных белка: CheA, CheW, CheY
и CheZ (Che — начальный слог от «chemotactic» — хемотаксисный). CheA и
CheY опосредуют сигнал от Р-П к жгутику, ведущий к хаотическому движе-
нию (рис. 10.6): если CheY присоединится к мотору жгутика, то это вызывает
вращение жгутика по часовой стрелке, которое ведет к хаотическому движе-
нию бактерии. Это происходит, когда со связывающим белком взаимодейст-
вует репеллент (рис. 10.6 А). Если же со связывающим белком взаимодействует
молекула аттрактанта, а их комплекс — с Р-П-молекулой, то этот путь ингиби-
руется, и наблюдается противоположный эффект — жгутик вращается против
часовой стрелки (рис. 10.6 В). Таким образом, в присутствии аттрактанта бак-
терия движется по градиенту его концентрации, а в присутствии репеллента —
продолжает хаотическое движение. В отсутствии того и другого баланс актива-
ции и деактивации обеспечивает чередование прямолинейного и хаотическо-
го движений.
Рис. 10.6 иллюстрирует биохимические реакции, ответственные за актива-
цию и деактивацию сигнального пути. Когда репеллент присоединяется к Р-
П-белку, цитоплазматический домен последнего связывается с CheW и инду-
цирует самофосфорилирование CheA. Фосфатная группа от CheA затем пере-
10. Хемочувствительность у прокариот 217
_______________ i
Мотор жгутика
движение по
часовой стрелке
Рис. 10.6. Молекулярная сигнализация в хемосенсорной системе Е. coli (А) Р-П-белок
Tsr связывается с молекулой репеллента (Лейцин (Лей)). CheW и CheA активируются,
CheA присоединяет фосфат от АТФ и переносит его на CheY. CheY диффундирует к мо-
тору жгутика и вызывает его вращение по часовой стрелке и хаотическое движение бак-
терии. CheY затем дефософорилируется белком CheZ. (Б) Р-П-белок Tsr связывается с
аттрактантом (Серин (Сер)). Последующие конформационные изменения инактивиру-
ют CheA и CheW, так что CheY остается нефосфорилированным и, соответственно, не-
активированным. Жгутик восстанавливает движение против часовой стрелки, и бакте-
рия движется прямолинейно. А — CheA, W — CheW, Y — CheY, Z — CheZ. Данные из
Bourrett, Borkovich and Simon, 1991.
носится на CheY. Только фосфорилированная форма CheY способна индуци-
ровать движение жгутика по часовой стрелке. Последнее звено системы — бе-
лок CheZ, который дефосфорилирует CheY и, таким образом, устраняет его
действие.
Сенсорная адаптация происходит в результате метилирования Р-П-белка,
если бактерия оказывается в условиях неизменной концентрации аттрактанта.
По этой причине, Р-П-белки называют иногда белками метил-адаптирующе-
гося хемотаксиса (МАХ). То, что адаптация идет в результате метилирования,
было показано на мутантах, у которых домен метилирования в Р-П-белках
инактивирован (см. рис. 10.5). У таких мутантов адаптации не происходит —
в присутствии аттрактанта хаотическое движение у них прекращается на не-
сколько часов, даже дней, вместо обычной минуты или около того. Возможно,
деактивация Р-П-белка при связывании с аттрактантом приводит к открытию
нескольких дополнительных сайтов метилирования. Фермент метилтрансфе-
раза может присоединить к ним до восьми метильных групп, это снова повы-
шает активность Р-П-белка и, через описанную выше сигнальную систему вы-
зывает хаотическое движение.
Ценность этой системы состоит в том, что генетика Е. coli хорошо изучена
и легко поддается манипуляциям. Такая же система действует и у Salmonella
218 Часть III. Хемочувствительность
typhimurium. Тот факт, что эти две бактерии (одна из которых является кишеч-
ным паразитом млекопитающих, а другая — птичьих яиц) развивались незави-
симо по крайней мере 150 миллионов лет, показывает, что сходные хемочувст-
вительные механизмы, вероятно, распространены среди бактерий достаточно
широко. Есть надежда, что удастся успешно проанализировать генетику Р-П-
белков и цитоплазматической сигнальной системы, и достичь полного пони-
мания механизма бактериального хемотаксиса. Это было бы первым случаем,
когда полная сенсо-моторная система целиком была бы описана на молеку-
лярно уровне.
10.2. Заключение
Хемочувствительность Е. coli, может быть даже в большей степени, чем меха-
ночувствительность, рассмотренная нами в гл. 5, предоставляет заманчивые
перспективы для анализа с помощью мощных методов современной молеку-
лярной биологии. За последние 20—30 лет применение этих методов, как мы
видели выше, вскрыли молекулярные механизмы поразительно детально. Мы
приближаемся к полному пониманию того, как бактерия не только ощущает
химические вещества в водной среде, но и реагирует на них, двигаясь к источ-
нику вещества или от него. Таким образом, интегрированная активность Р-П-
белков определяет направление плавательных движений прокариоцитов в
сложном химическом окружении. Молекулярные механизмы в миниатюре
имитируют намного более сложные обонятельные и вкусовые физиологичес-
кие механизмы других представителей животного царства. Хотя внутрикле-
точный хемотаксисный сигнальный путь, вероятно, не похож на тот, что дей-
ствует у многоклеточных, роль связывания и Р-П-белков предзнаменует моле-
кулярную биологию вкусовых и обонятельных рецепторных клеток животных,
и даже более того. Молекулярные биологи сумели сшить элементы белка Таг
из 5. typhimurium с инсулиновым рецептором млекопитающих, так что образо-
вался функциональный Р-П-белок, к тому же, получены свидетельства, что
белки CheY и Ras (из внутриклеточной сигнальной системы млекопитающих)
имеют некоторые эволюционные взаимосвязи. Хотя эукариоциты, а позднее
животные, порвали с царством бактерий невообразимо давно (см. рис. 4.2), мы
все еще можем разглядеть слабые черты общности их происхождения.
11. ХЕМО-ИНТЕРОРЕЦЕПТОРЫ
МЛЕКОПИТАЮЩИХ
Клод Бернар и millieu interieur (внутренняя среда): хеморецепторы контро-
лируют критичные параметры внутренней среды — парциальное давле-
ние (Пд) дыхательных газов является одним из таких параметров. Локали-
зация ПдО2 и ПдС02-рецепторов: каротидные тельца, аортальные тельца,
клетки мозга — эволюционное происхождение каротидных телец — каро-
тидный лабиринт. Гистология: клетки типа 1 и типа 2 (опорные) — клет-
ки типа 1 связаны с крупными капиллярами и афферентными нервными
окончаниями — они содержат электронно-плотные катехоламиновые ве-
зикулы — синаптические контакты с афферентными нервными оконча-
ниями. Физиология: высокая скорость кровотока — эффект изменений
ПдО2 и ПдСО2 — стимуляция повышением ПдСО2 — каротидные тельца
более чувствительны, чем аортальные — рефлектроный эффект на ско-
рость дыхания — сходный эффект вызывает падение ПдО2 — мозговые
клетки более чувствительны и каротидных, и аортальных телец — гипер-
капния — рефлексы, связанные с ПдСО2 и ПдО2, не связанные с дыхани-
ем. Биохимия: снижение ПдО2 ниже нормы вызывает закрывание К+-ка-
налов — деполяризация — открывание потенциал-зависимых Са2+-кана-
лов — выделение катехоламинов на афферентные нервные окончания —
возможно мембранно-связанный гем-белок связан с чувствительностью
к ПдО2; ПдСО2, превышающее норму — повышение внутреннего уровня
Н+ — H+/Na+-o6MeH — Na+/Ca2+-обмен повышает внутриклеточный
уровень Са2+ — выделение катехоламинов на афферентные нервные
окончания. Заключение: критическая роль интерорецепторов — только
если что-нибудь портится, мы понимаем его важность
Каждый студент, начинающий изучение физиологии, встречается со знамени-
тым афоризмом Клода Бернара: «Le Constance du millieu interieur est le condition
de la vie libre»1. Физиолог-экспериментатор Бернар родился недалеко от Виль-
франша в 1813 году. Его важнейшим вкладом в науку было, как сам он гово-
рил, «пробуждение уверенности в том, что животные живут в двух средах —
millieu exterieur (внешняя среда), в которой существует организм, и millieu
interieur, в котором существуют тканевые элементы организма». Эта концеп-
ция оказалась невероятно продуктивной. Сейчас мы знаем, что все основные
параметры внутренней среды млекопитающих — pH, О2, СО2, температура,
1 Постоянство внутренней среды — это условие свободной жизни (фр.) — Прим, перев.
220 Часть III. Хемочувствительность
осмотическое давление, гидростатическое давление, уровень глюкозы
и т. д. - поразительным образом удерживаются на постоянном уровне. Пола-
гают, что такая способность стабилизировать внутреннюю среду в значитель-
ной степени явилась залогом эволюционного «успеха» млекопитающих и
птиц. Так называемые «высшие позвоночные» смогли заселить весь земной
шар — от Арктики до Антарктики, как поется в гимне, «от ледяных гренланд-
ских гор до Индии коралловых брегов». Огромные эволюционные преимуще-
ства постоянства внутренней среды, завоеванные этими животными, обуслов-
ливают и чрезвычайную сложность их анатомии и физиологии. И чем выше
эта сложность обеспечения внутренней среды, тем ближе к оптимуму условия,
в которых пребывают клетки организма.
Мы видели (гл. 5), что механорецепторы плазматической мембраны круп-
ноклеточных нейронов гипоталамуса млекопитающих контролируют вариа-
ции осмолярности внеклеточной жидкости (ВКЖ). Сходным образом мы ви-
дели (гл. 6), что механорецепторы, в качестве интерорецепторов, играют клю-
чевую роль в контроле позы и положения мышц и суставов. В данной главе мы
увидим, насколько хеморецепторы незаменимы в контроле химических пара-
метров внутренней среды. Они образуют аппарат контроля парциального дав-
ления (Пд) О2, СО2 и pH крови, содержания в ней глюкозы и т. д., передающий
эту информацию в центральный компьютер ЦНС. Механо- и терморецепторы
играют подобную роль в том, что касается кровяного давления и температуры.
Хотя это ведет вглубь проблем сердечно-сосудистой физиологии, что принад-
лежит к сфер физиологии и эндокринологии, было бы неверно дать представ-
ление о хеморецепции без указания на эти жизненно важные интерорецепто-
ры. Поэтому в этой главе рассмотрим системы контроля парциального давле-
ния дыхательных газов — О2 и СО2 в ВКЖ. ПдСО2 также связан с pH ВКЖ, по-
скольку СО2, соединяясь с Н2О, образует угольную кислоту Н2СО3, которая, в
свою очередь диссоциирует на Н+ и НСО^. Хотя плазма крови является мощ-
ной буферной системой, устраняющей избыток Н+, этого может не быть в дру-
гих внеклеточных жидкостях.
11.1. Локализация хеморецепторов
млекопитающих к ПдО2 и ПдСО2
Каротидные тельца, расположенные во внутренней сонной артерии сразу по-
сле бифуркации общей сонной артерии, являются наиболее важными хемо-
интерорецепторами дыхательных газов в теле млекопитающего (рис. 11.1).
Другой важный хеморецептор дыхательных газов в форме аортальных телец
расположен в дуге аорты и ее основных ветвях. Они обнаруживаются не у всех
млекопитающих — например, представлены у кошек и собак, но отсутствуют
у кроликов, мышей и крыс. С другой стороны, у крыс и мышей хорошо разви-
ты брюшные хеморецепторы, расположенные в стенке желудка вблизи от ме-
ста, где блуждающий нерв выходит из пищевода, и в солнечном и печеночном
сплетениях. Кроме этих периферических рецепторов, центральные рецепторы,
высокочувствительные к ПдО2 и ПдСО2, располагаются билатерально в про-
долговатом мозге.
11. Хемо-интерорецепторы млекопитающих 221
А
Рис. 11.1. (А) Расположе-
ние каротидного тельца в
бифуркации общей сон-
ной артерии. Иннервация
проходит через нерв каро-
тидного синуса и верхний
шейный ганглий. (Б) Кле-
точная структура фраг-
мента каротидного тельца.
Показаны два типа кле-
ток — паренхиматозные
(или типа 1) и опорные
(или типа 2). Соотноше-
ние числа клеток этих ти-
пов прибл. 3—5 к 1. Даль-
нейшие объяснения в тек-
сте. По Gonzalez et al.,
1992
Внутренняя
каротидная
артерия
Каротидное
тельце
Общая
каротидная
артерия
Верхний
цервикальный
ганглий
Нерв
каротидного
синуса
Наружная
каротидная
артерия
Капилляр
Б Электронно-плотные
синаптические везикулы
Капилляр
Сенсорное волокно
нерва каротидного
синуса
Паренхиматозная
клетка каротидного
тельца
Тельце Гольджи Оп°Рная
Эволюция каротидных телец млекопитающих представляет интересный
материал по сравнительной анатомии и физиологии. Серия схем на рис. 11.2
показывает, что они происходят из третьей дуги аорты нашего предка — рыбы.
У амфибий мы видим, что жаберные капилляры преобразуются в так наз. ка-
ротидный лабиринт, который обеспечивает, чтобы только самая богатая кис-
лородом кровь поступала в мозг. При переходе к пресмыкающимся и другим
амниотам наблюдаем дальнейшие изменения; структура преобразуется в хе-
морецептор, контролирующий парциальное давление кислорода (ПдО2) и уг-
лекислого газа (ПдСО2), подобный нашему собственному.
11.2. Структура
Все периферические хеморецепторы имеют примерно одно* и то же гистологи-
ческое строение. Каротидные тельца состоят из клеток двух типов (тип 1 и
тип 2 — опорные), связанных с плотной сетью крупных капилляров (рис. 11.1).
Клетки типа 1 — наиболее обычны. Они имеют ультраструктуру секреторных
клеток с хорошо развитым аппаратом Гольджи, эндоплазматическим ретику-
лумом и (в отличие от других секреторных клеток) содержат большую популя-
цию электронно-плотных синаптических везикул, содержащих катехоламины.
С этими клетками связаны не только крупные капилляры, но и многочислен-
ные нервные окончания. Многие электронно-плотные синаптические везику-
222 Часть III. Хемочувствительность
Вентральная
аорта
Приносящий
жаберный сосуд
Дорзальные аорты
Выносящий
жаберный сосуд
Наджаберная
артерия
Внутренняя
каротидная артерия
Взрослая
лягушка
Наружные жабры
Внутренние жабры
Каротидный проток
Системная дуга
Артериальный проток
Системная аорта
IV
Легочная аорта
Артериальный проток
Рис. 11.2. Эволюция каротидных
телец. (А) Дуги аорты рыбы. Вент-
ральная аорта проходит под пище-
водом, а жаберные сосуды перено-
сят кровь через жабры в спинную
аорту над кишкой. (Б)—(Г) Дуги
аорты у головастика и взрослой
лягушки. (Б) Головастик с наруж-
ными жабрами; (В) Переходная
стадия; (Г) Взрослая лягушка. От-
метим происхождение каротидно-
го тела. (Д) Дуги аорты и каротид-
ное тельце у млекопитающих. По
Grove and Newell, 1947.
V^RA
VI ’•
Каротидный лабиринт
Внутренняя каротидная артерия
Наружная каротидная артерия
Каротидная дуга
Кожно-легочная дуга
Внутренняя
каротидная артерия
Наружная
каротидная артерия
Каротидное тельце
Общая каротидная
артерия
11. Хемо-интерорецепторы млекопитающих 223
лы клеток типа 1 концентрируются вблизи от синаптической щели между эти-
ми клетками и нервными волокнами. Хотя большинство нервных волокон —
сенсорные, здесь есть и некоторое количество парасимпатических эфферен-
тов. Сенсорные волокна, выходящие из каротидного тельца, принадлежат ча-
стично глоточной ветви языкоглоточного нерва (черепно-мозговой нерв IX),
а частично — ветви блуждающего нерва (черепно-мозговой нерв X). Сенсор-
ные волокна, иннервирующие аортальные тельца, принадлежат блуждающему
нерву. Клетки типа 1 окружены меньшими по численности клетками типа 2.
11.3. Физиология
Скорость кровотока через каротидное тельце очень велика: около 21 мл/мин
на 100 г ткани. Полагают, что это обусловлено наличием прямого протока по
сосуду, соединяющему вход и выход. Если это так, то это находится в соответ-
ствии с эволюционным происхождением каротидного тельца, рассмотренным
выше. Давно известно, что и каротидное и аортальное тельца отслеживают
уровень О2 и СО2 в крови. Относительно недавно этот вопрос был подвергнут
более детальному исследованию. Показано, что когда ПдО2 находится на по-
стоянном уровне, каротидное тельце чувствительно к повышению уровня
ПдСО2 (рис. 11.3 А), тогда как аортальные тельца на это не реагируют. Когда
ПдСО2 постоянно, и каротидное, и аортальное тельца реагируют на падение
ПдО2 повышением частоты импульсации в их афферентных волокнах, хотя
каротидное тельце более чувствительно, чем аортальные (рис. 11. 3 Б). С дру-
гой стороны, аортальные тельца, но не каротидное, чувствительно к карбокси-
гемоглобинемии, когда ПдО2 и ПдСО2 постоянны (рис. 11.3 В). Это позволяет
предположить, что когда содержание кислорода в крови уменьшается вследст-
вие анемии или сходных причин, то именно аортальные, а не каротидное,
тельца сигнализируют об опасности.
Уже многие годы известно, что стимуляция каротидных и аортальных хе-
морецепторов ведет к усилению дыхания, причем каротидные тельца вызыва-
ют намного больший эффект, чем аортальные. Значение рефлекса очевидно.
Повышение ПдСО2 в крови (выше 40 мм рт. ст.), вызывает усиление вентиля-
ции легких, которое действует как отрицательная обратная связь, приводя
ПдСО2 к норме. Аналогичным образом, хотя и в меньшей степени, понижение
уровня ПдО2 ведет к усилению дыхательной активности.
Периферические хеморецепторы, конечно, это еще не все. Как упомина-
лось в разделе 11.1, центральные хеморецепторы в продолговатом мозге также
с высокой точностью отслеживают ПдСО2. СО2 очень легко диффундирует из
мозгового кровотока в спиномозговую жидкость (СМЖ), где гидратируется до
угольной кислоты (Н2СО3), которая быстро диссоциирует до Н+ и НСО^".
В отличие от плазмы крови, СМЖ практически не содержит белков, которые
могли бы действовать как буфер для протонов. Таким образом, любое повы-
шение ПдСО3 в СМЖ — условие, называемое гиперкапнией — приводит к сни-
жению pH, что в свою очередь вызывает возбуждение центральных хеморецеп-
торов. Эти рецепторы имеют обширные связи с дыхательными центрами про-
долговатого мозга. Возбуждение, вызванное снижением pH, ведет к повыше-
нию дыхательной активности — увеличиваются и глубина, и в меньшей степе-
224 Часть III. Хемочувствительность
Рис. 11.3. Ответ каротидного и аортального
телец на изменения парциальных давлений
(Пд) СО2 и О2. (А) Разряд (имп/с) в одиноч-
ном афферентном волокне от каротидного
тельца при постоянном уровне ПдОг и изме-
нении ПдСО2- (Б) Разряд (имп/с) из каро-
тидного и аортального телец при постоян-
ном уровне ПдСО2 и изменении ПдО2.
(В) Разряд (имп/с) из каротидного и аор-
тального телец при физиологическом уров-
не ПдО2 и варьирующем ПдСО2- Отметьте
различия ответов аортального и каротидно-
го телец. (А) из Biscoe, Purves and Simpson,
1970; с разрешения; (Б) и (В) из Marshall,
1994; (Б) и (В) воспроизводятся с разреше-
ния American Physiological Society.
ни, скорость дыхания. Таким образом, периферические и центральные хемо-
рецепторы работают кооперативно. При этом центральные рецепторы играют
доминирующую роль — они ответственны за прибл. 70% реакции на повыше-
ние ПдСО2.
Хотя самым очевидным рефлекторным ответом на стимуляцию каротид-
ных и аортальных хеморецепторов является усиление дыхательной активнос-
ти, приводящей к увеличению вентиляции легочных альвеол, тщательные ис-
следования продемонстрировали еще целый ряд эффектов. В основном, эти
исследования проводились на каротидном тельце, поскольку изолированные
аортальные тельца трудно стимулировать. Однако, даже исследования на ка-
ротидных тельцах требуют применения изощренных физиологических мето-
дик, чтобы избежать влияния медуллярных хеморецепторов и рецепторов рас-
тяжения в легких, маскирующих изучаемые реакции. Когда это удалось сде-
лать, выявился ряд рефлекторных ответов на импульсацию из каротидного
11. Хемо-интерорецепторы млекопитающих 225
тельца. Среди них брадикардия (замедление ритма сердечных сокращений),
симпатическое снижение коронарного, брюшного, почечного кровотока, а
также уменьшение кровотока в скелетной мускулатуре. Имеется также ком-
плексный эффект на мозговое кровообращение. Циркуляция в поверхностной
коре головного мозга повышается, тогда как в более глубоких областях мозга
снижается. В целом, эти эффекты подтверждают то, что Уолтер Кэннон
(Walter Cannon) назвал «мудростью тела»: все они направлены на снижение
количества СО2, диффундирующего в кровь. Кроме того, показано, что если
пересечь афферентные волокна из каротидного тельца, то поведенческая ре-
акция животного на гипоксию устраняется. Такая реакция варьирует в зависи-
мости от степени недостатка кислорода. У собак она идет по восходящей от
умеренного возбуждения до яростного поиска пути к бегству.
В целом, мы видим, что каротидные и центральные хеморецепторы играют
ключевую роль в поддержании уровня дыхательных газов и pH в ВКЖ тела.
Поддержание ПдО2 особенно важно для животных, которые двигаются со зна-
чительным перемещением по вертикали; поддержание ПдСО2 жизненно важ-
но для компенсации изменений количеств СО2 в ВКЖ, когда мускулатура
(и другие органы) выполняют существенно различающиеся объемы работы.
11.4. Биохимия
Возможно ли проследить эту сложную гомеостатическую физиологию до мо-
лекулярного уровня и найти (как мы это делали в гл. 5, когда обсуждали меха-
норецепторы) молекулярный сенсор в основе системы? Последние работы да-
ют основание думать, что мы можем это сделать. Возьмем для начала сенсор
кислорода. Именно он запускает паническую реакцию на гипоксию, упоми-
навшуюся в конце предыдущего раздела. Молекулярная ситуация здесь, как
обычно, очень сложна, и выделить в ней единственный ключевой механизм
очень трудно. Кислород так важен для клеток млекопитающих, что было бы
удивительно, если бы в механизм не было встроено страховочных средств за-
щиты от нарушений его обмена. Было показано, что, если ПдО2 в непосредст-
венном окружении каротидного тельца падает ниже 80—100 мм рт. ст., то акти-
вируются клетки типа 1. По мере того, как ПдО2 продолжает падать, закрыва-
ются специфические К+-каналы. Уменьшение К+-проницаемости приводит к
деполяризации, что, в свою очередь, открывает потенциал-зависимые Са2+-
каналы. Поток Са2+ в клетку запускает (как и обычно) выделение катехолами-
нов, особенно дофамина и норадреналина, которые (как мы видели выше)
присутствуют в клетках типа 1 в высоких концентрациях. В свою очередь, это
приводит к инициации активности волокон синусового нерва, находящихся в
непосредственной близости от этих клеток.
Каким образом снижение ПдО2 вызывает закрывание К+-каналов? Это во-
прос, ответ на который все еще является предметом дискуссий. Возможно,
низкое парциальное давление О2 среды влияет на внутренний энергетический
метаболизм клеток типа 1, что приводит к снижению уровня АТФ. Это само по
себе может вызвать закрывание АТФ-зависимых К+-каналов или привести к
выделению Са2+ из митохондрий и, таким образом, запуску выброса катехола-
минов. Однако, наиболее популярная теория к моменту написания этой кни-
226 Часть III. Хемочувствительность
СО2,Н+
Рис. 11.4. Клеточный механизм преобразования сигнала о низком уровне ПдО2 и
повышенном — ПдСО2 в выделение нейромедиатора хеморецепторными клетками
каротидного тельца. Уменьшение ПдО2 распознается мембранным рецептором
(показан вверху рисунка), который вызывает закрывание связанного с ним К+-ка-
нала. Последующая деполяризация приводит к открыванию Са2+-каналов и пото-
ку Са2+ внутрь клетки, запускающему выделение нейромедиатора. Повышение
ПдСО2 вызывает увеличение потока Н+ в клетку, запускающего насос, перенося-
щий в клетку Na+. В свою очередь, это активирует механизм антипорта, обменива-
ющего Na+ на Са2+, что опять-таки приводит к выделению нейромедиатора. Даль-
нейшие объяснения в тексте. Из Gonzalez etal., 1994. Воспроизводится с разреше-
ния American Physiological Society.
ги состоит в том, что кислород-чувствительный гем-белок тесно ассоциирован
с К+-каналом в клеточной мембране. Гипотеза состоит в том, что, так же как
гемоглобин и миоглобин меняют свою трехмерную конформацию в ответ на
оксигенацию, действует и предполагаемый мембранно-связанный гем-белок.
Это изменение структуры влияет на сопряженный с ним белок К+-канала, что
изменяет вероятность открытого или закрытого состояния последнего. Схема-
тически гипотеза представлена на рис. 11.4.
Каротидные тельца и медуллярные хеморецепторы также, как мы отмеча-
ли, высокочувствительны к ПдСО2. Молекулярный механизм, функциониру-
ющие здесь, как мы опять-таки отмечали, вероятно, зависит от образования
Н2СО3 из СО2 и Н2О и их диссоциации на Н+ и НСО^. Н+ диффундирует в хе-
морецепторные клетки, что приводит к падению pH цитозоля. Когда внутри-
клеточный pH падает, это активирует механизм выброса протонов в обмен на
Na+ (см. рис. 11.4). Показано, что клетки типа 1 (а также центральные хеморе-
цепторы) обладают мощным Ка+/Са2+-антипортом в мембране. Это означает,
что внутриклеточный Na+ обменивается на внеклеточный Са2+ (рис. 11.4).
Антипорт стимулируется повышенным уровнем внутриклеточного Na+. Та-
ким образом, следствием повышенного ПдСО2 плазмы является повышение
уровня Са+ в цитоплазме клеток типа 1. И это приводит к выделению катехо-
ламинового нейромедиатора на близлежащие нервные волокна и, соответст-
венно, увеличению частоты импульсации в сенсорных нервах.
11. Хемо-интерорецепторы млекопитающих 227
11.5. Заключение
Эта короткая глава посвящена всего лишь одному из множества хемо-интеро-
рецепторов организма млекопитающего. Таких механизмов — множество, но
здесь предмет сенсорной физиологии смешивается с эндокринологией и био-
химией. Подобно тому, как высшие животные не в состоянии выжить без
сложной системы интеро-механорецепторов (которые были предметом рас-
смотрения гл. 6), точно так же они погибли бы и без множества интеро-хемо-
рецепторов, отслеживающих важные параметры внутренней среды, в которую
погружены все клетки организма. Их работа в основном не осознается; и толь-
ко когда что-то разлаживается, мы понимаем важность интерорецепторов.
12. ВКУС
Вкус и обоняние не всегда различимы — универсальность в царстве
животных — плоские черви — кольчатые черви — моллюски. Насеко-
мые: хеморецепторные сенсиллы — различие между вкусом и обоняни-
ем — удлинение хоботка у мух и бабочек — тарзальные и лабеллярные
сенсиллы — электрофизиологические данные — форма вкусовой сен-
силлы — их распространение — тонкая структура лабеллярных сенсилл —
биофизика — сравнение с миелинизацией у позвоночных. Млекопитаю-
щие: биологическое значение — комплексная нейрология — сравнение с
другими чувствами — субъективное разнообразие — аналитичность
комбинаций сладкого, соленого, кислого и горького, а также глутамата1
и воды — качества, связанные с биологической объективностью; вкусо-
вые почки — локализация — грибовидные, листовидные и окаймленные
сосочки — иннервация — клеточный состав — быстрый клеточный об-
мен; вкусовые рецепторные клетки (ВРК) — истинные рецепторные
клетки — передача стимула — генерализатор или специалист? — элек-
трофизиология — молекулярная биология трансдукции сладкого, глу-
таматного, соленого, кислого, горького и воды; центральные проек-
ции — вкусовая информация передается через продолговатый мозг и
таламус в две вкусовые области коры мозга. Заключение: многое еще
предстоит узнать в будущих исследованиях — коммерческий интерес
В последней главе мы убедились в важной роли хемоинтерорецепторов — хе-
морецепторов, обращенных внутрь организма, — в поддержании постоянства
внутренней среды В этой и следующей главах мы рассмотрим две группы хе-
морецепторов, обращенных вовне организма и построенных так, чтобы отсле-
живать химический состав окружающей среды. Обычно мы подразделяем эти
экстерорецепторы на специализированные к восприятию вкусовых ощуще-
ний либо запахов. Это различие совершенно очевидно у четвероногих позво-
ночных, таких как мы сами. Вкус обычно рассматривается как контактное чув-
ство. Действующие молекулы находятся в растворе, который контактирует с
рецептором. Напротив, обоняние — дистантное чувство. Молекулы химичес-
ких веществ доставляются к обонятельным рецепторам с потоками воздуха.
Очевидная условность такой классификации — на клеточном, а тем более на
1 Глютамат натрия внешне похож на соль и сахар. Его вкус в Японии называют «umami»,
в англоязычных странах «savory» — бульоноподобный или мясной вкус. — Прим, перев.
12. Вкус 229
молекулярном уровне рецепторный механизм может быть одним и тем же. Та-
кое различие еще труднее провести для водных животных, где стимулирующие
вещества при prpY вариантах растворены. Тем не менее, при всех этих оговор-
ках, мы для удобства разделим проблему химических экстерорецепторов на
две главы, рассматривая вкус в данной главе, а обоняние — в гл. 13.
Вкус, конечно, очень распространенное чувство в царстве животных. Плана-
рии обладают хеморецепторами, которые локализованы в голове, функциони-
руют при питании и, вследствие этого могут рассматриваться как вкусовые ре-
цепторы. Чарльз Дарвин в своем замечательном исследовании дождевого червя
{Lumbricus) показал, что эти животные способны различать краснокочанную и
обычную капусту, а также листья моркови и сельдерея. Наземные моллюски ис-
пользуют хеморецепторы для обнаружения съедобных материалов. Возможно,
наиболее интересные (и наиболее изученные) вкусовые системы беспозвоноч-
ных найдены среди представителей большого класса насекомых.
12.1. Вкус у насекомых
Кое-что о сенсорных сенсиллах насекомых уже было сказано в гл. 7 и 8. Следу-
ет вспомнить, что сенсиллы часто мультимодальны, поэтому хеморецепторные
сенсиллы часто содержат механорецепторные клетки. Здесь также трудно про-
вести общепринятое различие между вкусовыми и обонятельными сенсилла-
ми. Можно ли отличить, например, водный рецептор от гигрорецептора? В це-
лом, мы можем определить вкусовые рецепторы как детектирующие химичес-
кие вещества в жидкой фазе, растворенные, например, в каплях на поверхнос-
ти листьев, тогда как обонятельные детектируют молекулы в газовой фазе и ча-
сто очень чувствительны. Обоняние насекомых будет рассмотрено в гл. 13.
Исследования вкуса у насекомых получили развитие после работ Минниха
(Minnich), а затем Детье (Dethier) по рефлексу вытягивания хоботка у бабочек
и мясных мух в ответ на стимуляцию лапок питательными растворами. Экспе-
рименты были построены довольно просто — насекомое прикрепляли к стек-
лянной палочке, а лапки погружали в чашку Петри с растворами разных ве-
ществ в разных концентрациях (рис. 12.1). Экспериментатор наблюдал, вы-
двигается ли хоботок насекомого при прикосновении лапок к жидкости.
Позднее сходная хемочувствительность была показана в больших хеморе-
цепторных волосках labellum (верхняя губа) хоботка мух-тахинид, таких как
мясные мухи CalUphora и Phormia. При стимуляции тарзальных хеморецепто-
Рис. 12.1. Тестирование тарзаль-
ной хемочувствительности мяс-
ной мухи.
Чашка Петри
230 Часть III. Хемочувствительность
Рис. 12.2. Кривые электричес-
ких ответов на сахарозу, глюко-
зу и фруктозу, полученные у се-
рой мясной мухи Boettcherisca
peregrina. 0,2 М сахарозы (х)
предъявляли в качестве контро-
ля. Число у каждого символа
обозначает количество стиму-
ляций. О = сахароза; О = глю-
коза; • = фруктоза. Из Morita
and Shiraishi, 1985.
ров губа выдвигается вперед так, что волоски приходят в соприкосновение с
жидкостью и тестируют ее. Удалось провести электрофизиологические отве-
дения и от тарзальных, и от лабеллярных хеморецепторных волосков. Частота
импульсации, как и можно было ожидать, повышается с возрастанием кон-
центрации тестируемого вещества. Одиночный лабеллярный сахарный рецеп-
тор по-разному отвечает на разные сахара, хотя частота импульсации в каждом
случае все равно возрастает с ростом концентрации (рис. 12.2). Здесь сущест-
вует и определенная специализация — различают четыре группы хеморецеп-
торных волосков на лабеллуме мясной мухи, которые преимущественно чувст-
вительны к сахарам, катионам, анионам либо воде. Сахар-чувствительные
сенсиллы реагируют также на некоторые аминокислоты и жирные кислоты,
тогда как водный рецептор ингибируется солями. У других насекомых, хотя и
присутствуют сенсиллы, чувствительные к воде, сахару и солям, обнаружива-
ются и другие рецепторы, специализированные в соответствии с пищевыми
предпочтениями животного. В частности, растительноядные насекомые и гу-
сеницы обычно имеют рецепторы, чувствительные к тем химическим вещест-
вам, которые присутствуют в растительной пище.
Хеморецепторные сенсиллы обычно имеют одну или несколько пор
(рис. 12.3 А). Контактные (вкусовые) сенсиллы обычно имеют одну пору, а
дистантные (обонятельные) сенсиллы для увеличения чувствительности раз-
вили по нескольку. Вкусовые сенсиллы представлены волосками или выроста-
ми разной формы и размеров и обычно мультимодальны. Например, лабел-
лярные волоски настоящих мух и тахинид имеют одну пору и длину в несколь-
ко сот микрон, тогда как тарзальные волоски намного короче (несколько де-
сятков микрон). Последние разделяются на четыре морфологических типа:
тип А (короткие, около 30 мкм, многочисленные содержащие рецепторы саха-
ров и соли), тип В2 (около 70 мкм, только на вентральной стороне лапки, са-
хар-чувствительные), тип ВЗ (проецируются латерально из дистального конца
каждого из четырех последних тарзомеров и группы в пятом тарзомере, около
70 мкм в длину, чувствительны к сахарам и соли), тип D (самые длинные, око-
ло 200 мкм, по паре на каждом тарзомере и по две пары — на первом и пятом,
чувствительны к сахарам и соли). Распределение этих волосков на протора-
кальных лапках мясной мухи показано на рис. 12.3 Б.
12. Вкус 231
Рис. 12.3. (А) Вкусовая сенсилла с
одной порой. Д — внешний сег-
мент дендрита, ДС — дендритная
оболочка, Э — эпидермальная
клетка, ВО — внутренняя клетка
оболочки, Н — нейросенсорная
клетка, НО — наружная клетка
оболочки, ТП — терминальная
пора. Поперечный срез сенсил-
лы — в правом верхнем углу ри-
сунка. Из М. S. Blum, ed., 1985;
Fundamentals of Insect Physiology,
©1985, John Wiley & Sons Ltd. Вос-
производится с разрешения John
Wiley & Sons Ltd. (Б) Тарзальная
часть проторакальной лапки мяс-
ной мухи сбоку. Показаны хемо-
чувствительные сенсиллы. Так-
тильные сенсиллы не показаны.
Различные типы волосков обозна-
чены A, Bl, В2, ВЗ, С и D. Объяс-
нения в тексте. Из Kerkut and Gil-
bert, 1985, с разрешения.
Если мы рассмотрим структуру одиночной вкусовой сенсиллы на лабеллу-
ме хоботка мухи Phonnia более детально, то увидим, что она содержит четыре
хеморецепторных и одну механорецепторную клетки. Внешние сегменты ден-
дритов хеморецепторных клеток проходят вдоль сенсиллы и оканчиваются не-
посредственно под порой. Аксоны этих клеток направляются к подглоточно-
му ганглию. Механочувствительная клетка имеет тубулярное тельце и схоже с
механочувствительными клетками, описанными в гл. 6. Их аксоны также на-
правляются в подглоточный ганглий.
Рис. 12.4 А в схематической форме представляет взаимосвязи хеморецеп-
торной клетки с соседними. Видно, что она окружена обычно обкладочными
клетками, а непосредственно с ней контактирует глиальная клетка. Снаружи
к последней примыкает внутренняя клетка оболочки (текогенная клетка). Те-
когенная клетка образует ресничный синус (см. гл. 6), который окружает со-
единение между внутренним и внешним сегментами дендрита и секретирует
кутикулярную оболочку наружного сегмента дендрита. Ресничный синус, та-
ким образом, это непрерывное пространство между наружным сегментом
дендрита и кутикулярной оболочкой. Это пространство заполнено жидкос-
тью, секретируемой внутренними клетками оболочки. Промежуточная вспо-
могательная клетка (трихогенная клетка) и наружная вспомогательная клетка
(тормогенная клетка) образуют сенсиллярный синус. Этот синус, как показы-
вает рисунок, связан с наружным пространством ствола сенсиллы.
232 Часть III. Хемочувствительность
Рис. 12.4. (А) Схематическое представление вкусовой сенсиллы мясной мухи. По-
казан только один наружный сегмент. Отметьте окружающую его толстую кутику-
лу. PC — ресничный синус, Д — наружный сегмент дендрита, Э — эпидермальная
клетка, Г — глиальная клетка, Гц — гемоцель, ВО — внутренняя клетка оболочки,
П — промежуточная клетка оболочки, Н — нейросенсорная клетка, НО — наруж-
ная клетка оболочки, сс — сенсиллярный синус. Из Kerkut and Gilbert, 1985, с раз-
решения. (Б) Биофизика вкусовой сенсиллы. /т — мембранный ток, /о1 и /о2 —
продольные токи в оболочке наружного сегмента к его кончику и к основанию,
соответственно. 1О — продольный ток в наружной полости. Объяснения в тексте.
С изменениями из Kerkut and Gilbert, 1985, с разрешения.
Б
Когда вкусовые сенсиллы находятся в контакте с адекватным стимулом,*
кончик наружного сегмента деполяризуется. Мы встретимся с аналогичными
ответом, когда будем более подробно рассматривать волосковые клетки позво-
ночных. Деполяризация распространяется электротонически вниз по наруж-
ному сегменту и достигает внутреннего сегмента. Петля тока замыкается через
ресничный и сенсиллярный синусы, внешнее пространство ствола сенсиллы и
обратно к ее кончику (рис. 12.4 Б). Деполяризация основания наружного сег-
мента распространяется к начальному сегменту аксона вкусовой клетки, где и
инициируется импульс.
Строение хеморецепторной сенсиллы насекомых во многом аналогично
миелинизированному аксону позвоночных. В последнем случае электротони-
ческое распространение тока значительно усиливается за счет миелиновой
оболочки, обладающей высоким сопротивлением. В сенсилле насекомого
место миелина занимает кутикулярная оболочка, окружающая наружный
сегмент. Это делает электротоническое или кабельное проведение более эф-
фективным, чтобы сигнал передался по достаточно длинной (несколько сот
микрон) сенсилле, развившейся у насекомых.
12. Вкус 233
12.2. Вкус у млекопитающих
Вкус хорошо развит у позвоночных. В этом разделе мы, однако, ограничимся
рассмотрением млекопитающих, в частности, человека. Как и все другие жи-
вотные, человек получает жизненно важную информацию о пище, уже когда
она оказывается во рту — в последний момент перед тем, как сознательный
контроль над процессом потребления пищи будет утрачен. Съедобно это или
ядовито? Привычно или необычно? Когда это съели в прошлый раз — была
от этого польза или вред? Не удивительно, что вкус имеет сложную нейроло-
гическую основу. Импульсация идет в основном от вкусовых рецепторов язы-
ка, но также от надгортанника, верхней трети пищевода и мягкого нёба. Од-
новременно поступает важная информация о характере поверхности пищи,
текстуре, температуре и запахе; свою роль играют и память, и культура воспи-
тания —то, что вкусно в одной культурной традиции, то отвратительно —
в другой.
Вкусовые ощущения, как и запахи, многообразны. В этом отношении они
отличаются от механочувствительности, такой ее формы, как осязание. Осяза-
тельное же ощущение всегда имеет одно и то же качество и различается толь-
ко по интенсивности. Запахи же различаются и по количественной, и по каче-
ственной осям. В этом отношении оба чувства — вкус и обоняние — больше
напоминают такую сложную форму механочувствительности, как слух; есть
черты сходства и со зрением. Однако, как описать наши вкусовые ощущения?
Все мы слышали (кое-кто — с улыбкой) описания знатоков вин. Они старают-
ся привлечь любого рода ассоциации — кто зрительные, кто социальные, кто
географические или климатические, чтобы описать свои ощущения. Являют-
ся ли вкусовые ощущения попросту не поддающимися анализу, представляю-
щими собой бесконечный континуум, или же они могут быть, подобно цветам,
сформированы перестановок и комбинаций небольшого числа элементов?
История помнит дебаты на эту тему, которые окончились победой последней
точки зрения. Современные воззрения восходят к идее Ханса Хеннинга (Hans
Henning), высказанной в 1922 году, что вкусовое ощущение человека может
быть разложено на четыре основных: сладкое, соленое, кислое и горькое. В по-
следнее время исследователи выделяют также приятный вкус глютамата на-
трия, называемый по-японски umami, который существенно отличается от
четырех классических вкусов, а также вкус воды.
Эта субъективная классификация связана и с объективной биологией. По-
зитивные ощущения сладкого или umami связаны с высококалорийной легко
усвояемой пищей. Фрукты и ягоды часто рекламируют это яркими окрасками,
адресованными зрению. Соленость, ощущение кислого и вкус воды связаны с
регуляцией ионного состава «внутренней среды», в частности pH. Горечь вос-
принимается как сигнал тревоги — поедаемый материал опасен. Такой объек-
тивный анализ предполагает, что и у других животных присутствует схожая
(хотя и далеко не идентичная) организация, что и показано у многих видов по-
звоночных. Схожи ли их субъективные ощущения, конечно, остается откры-
тым вопросом.
234 Часть III. Хемочувствительность
12.2.1. Вкусовые почки
Вкусовые рецепторы группируются во вкусовые почки, локализованные в вы-
ступах, именующихся вкусовыми сосочками. На языке человека присутствуют
четыре типа сосочков: нитевидные, грибовидные, листовидные и окаймлённые.
Нитевидные сосочки никаких вкусовых почек не содержат. Их функция — чи-
сто механическая, они придают языку характер тёрки, что у многих млекопи-
тающих, например у домашней кошки, выражено значительно сильнее, чем у
человека. Три других типа сосочков содержат вкусовые почки. Грибовидные
сосочки, названные так из-за сходства со шляпкой гриба, группируются глав-
ным образом в передней части и на переднем краю языка. Вкусовые почки,
чувствительные к сладкому и соленому, погружены в поверхностный эпите-
лий. Вследствие обильного кровоснабжения почки выглядят маленькими
красными точками в передней части языка. Листовидные сосочки располага-
ются в параллельных складках (откуда и название) в количестве 3—8 у корня
языка. Эти вкусовые почки особенно чувствительны к кислому. Окаймлённые
сосочки, как показывает их название, несколько погружены в поверхность
языка и окружены углублением, которое отделяет их от остальной поверхнос-
ти языка. Восемь—девять таких сосочков образуют V-образную кофигурацию
у корня языка. В стенках каждого сосочка и в стенках бороздки, окружающей
сосочек, развивается около 250 вкусовых почек. Они наиболее чувствительны
к кислому и горькому. В стенках сосочка располагаются также секреторные
железы, которые при движениях языка омывают бороздку. Как упоминалось
выше, другие вкусовые почки располагаются в надгортаннике, верхней трети
пищевода и мягком нёбе.
Иннервация вкусовых почек намного сложнее, чем обонятельных рецепто-
ров, рассмотренных в гл. 13. Передняя часть языка иннервируется смешанной
ветвью (chorda tympani) лицевого нерва (черепно-мозговой нерв VII), тогда как
задняя — языкоглоточным нервом (черепно-мозговой нерв IX). Почки, распо-
ложенные вне языка, иннервируются ветвью блуждающего нерва (черепно-
мозговой нерв X).
Рис. 12.5. Язык и вкусовые
почки. Язык показан сверху,
в правой колонке — срезы че-
рез три типа сосочков, несу-
щих вкусовые почки. Черны-
ми полосками показано рас-
положение на них вкусовых
почек. Области языка, обла-
дающие наибольшей чувст-
вительностью к сладкому, со-
леному, кислому и горькому
показаны слева. Дальнейшие
объяснения в тексте.
12. Вкус 235
Клеточная структура вкусовой почки
Вкусовая почка представляет собой группу из 50—150 клеток, выстроенных по-
добно долькам апельсина, и окруженную эпителиальными клетками сосочка
(рис. 12.6 В). Два класса клеток: главные и базальные. Главные клетки, в свою
очередь подразделяются на три типа: тип 1 (темные клетки), тип 2 (светлые) и
А
Бороздка
Поверхностный
эпителий языка
Вкусовые почки,
построенные рядами
Й
В
Проток
серозной
железы “
Серозные
железы
Иннервация
вкусовой
почки
Вкусовая
пора
Синаптические
пузырьки
Наружный
слой
слоистого
эпителия
Бороздка
Базальная клетка
Клетка типа I
Клетка типа 111-
Икроворсинки
Перерезанные аксоны
вкусовой почки
Аксоны нервных клеток
Клетка типа II
Апикальные плотные гранулы
Вкусовая пора
Рис. 12.6. Вкусовая почка. (А) Расположение вкусовых почек в окаймлённом сосочке;
(Б) увеличенное изображение, показанное на рис. 12.6 А; (В) одиночная вкусовая почка.
Из Keveme, 1982. Воспроизводится с разрешения Cambridge University Press.
236 Часть ТП^Хемочувствительностъ
тип 3. Клетки типов 1 и 3, в основном, являются рецепторными, тогда как
клетки типа 2 — обычно опорные. Такое приблизительное описание функций
связано с тем, что роль этих клеток варьирует у разных видов животных и даже
от одной части языка к другой. Апикальные части всех трех типов главных кле-
ток соединены плотными контактами (tight junction) и выступают во вкусовую
пору. Эта пора заполнена секретом и открыта на поверхность сосочка.
Клетки типа 1 составляют до 65% клеточной популяции вкусовой почки
кролика. Их апикальные части образуют кисточки из 30—40 микроворсинок.
В них также содержатся плотные гранулы с секретом, который выделяется во
вкусовую пору. Стенки клеток типа 1 имеют многочисленные листовидные
выросты, которые охватывают немиелинизированные нервные волокна и
клетки типа 2. Синапсы с нейронами клетки типа 1 образуют у мышей, но не у
кроликов.
Клетки типа 2 составляют прибл. 20% клеточной популяции вкусовой поч-
ки. Их апикальные части заполнены электронно-плотным материалом и окан-
чиваются множеством тупых микроворсинок, направленных во вкусовую по-
ру. Синапсы с нейронами найдены у мышей, но не у кроликов.
Клетки типа 3 — менее 10% клеток вкусовой почки у кроликов. У них обра-
зуется по одной крупной микроворсинке, направленной во вкусовую пору.
В отличие от двух других типов клеток, которые могут образовывать, а могут и
не образовывать синапсы, клетки типа 3 всегда образуют синапсы с соседними
нейронами и, следовательно, во всех случаях должны рассматриваться как ре-
цепторные клетки.
Все клетки вкусовой почки являются короткоживущими и постоянно заме-
щаются. Радиоактивное мечение эпителиальных клеток, окружающих вкусо-
вую почку, показывает, что дочерние клетки перемещаются во вкусовую поч-
ку со скоростью 1 клетка/10 часов. Т. е. средняя продолжительность их жизни
во вкусовой почке не превышает 10 дней. За это время клетка перемещается с
периферии почки к центру. В конце своей краткой жизни клетки вкусовых по-
чек либо подвергаются фагоцитозу, либо претерпевают апоптоз*. Такой быст-
рый обмен клеток вкусовых почек, вероятно, и объясняет трудность определе-
ния точной функции каждого их типа.
Вкусовые рецепторные клетки (ВРК) в отличие от обонятельных, являют-
ся истинными рецепторными клетками. Волокна сенсорных нервов входят
в базальную пластинку почки (где и утрачивают миелиновую оболочку) и до-
стигают клеток почки. Волокна покрыты мембранными выростами клеток
типа 1 (темных). Одно волокно обычно иннервирует больше, чем одну ВРК,
и, напротив, одна рецепторная клетка контактирует с более, чем одним нерв-
ным волокном. Поскольку происходит постоянный обмен клеток вкусовых
почек, окончания нервных волокон находятся в процессе постоянного поиска
новых синаптических контактов.
1 Апоптоз — запрограммированная смерть клетки. — Прим, перев.
12. Вкус 237
Трансдукция сигнала
Как и в случае обонятельных ресничек (см. гл. 13), в данном случае продолжа-
ется дискуссия о том, насколько специализированы ВРК. Отвечает ли клетка
только на один или на несколько из первичных шести вкусовых стимулов? Как
и в случае обонятельных ресничек, вопрос пока не решен. Общее мнение
склоняется в пользу признания генерализованное™. Отведения от одиночно-
го нервного волокна из передней части языка крысы или хомячка позволяют
идентифицировать некоторые волокна как максимально чувствительные к
NaCl, т. е. соленому (N-волокна), с менее выраженным ответом на НС1, т. е.
кислое. Другие волокна (Н-волокна) лучше всего реагируют на НО, более сла-
бо — на NaO и хинин (горечь). Однако, опять-таки здесь есть масса видовых
различий, к тому же следует помнить, что веточки одного и того же волокна
могут иннервировать несколько ВРК. Исследование молекулярной биологии
и биофизики одиночной ВРК технически трудно, и к настоящему моменту ча-
сто дает противоречивые результаты.
Сладкое. Рецепторные клетки сладкого, как показывает рис. 12.7, имеют,
по крайней мере два сигнальных пути. Сами по себе сахара, по-видимому,
приводят в действие сигнальный путь с участием G-белков, активируя мемб-
ранный рецептор сахарозы. Есть данные, что это семидоменный рецептор,
схожий с Са2+-чувствительным рецептором (СаЧР) и метаботропным глута-
Рис. 12.7. Пути передачи сиг-
налов для сахаров и несахар-
ных подсластителей. Вкусовая
трансдукция сладких стимулов
Объяснения в тексте. ЭР — эн-
доплазматический ретикулум,
G — G-белок, ФЛС — фосфо-
липаза С, Р — рецептор. По
Lindemann, 1996.
238 Часть III. Хемочувствительность
Рис. 12.8. Путь передачи сигнала
соленого вкуса. Na+ входит в
амилорид-чувствительные кана-
лы, а затем откачивается наружу
На+/К+-насосом в базальной ча-
сти клетки. Одновременно в ба-
зальной части из клетки выходят
катионы (вероятно К+ через К+-
каналы). Деполяризация вызы-
вает открывание Са2+-каналы, и
поток Са2+ внутрь вызывает вы-
деление медиатора на дендрит
сенсорного волокна. Дальней-
шие объяснения в тексте. По
Lindemann, 1996.
матным рецептором (тГлуР). Этот рецепторный белок, известный как TR1,
имеет очень длинную внеклеточную N-концевую цепь, которая, вероятно, и
образует сайта вкусового рецептора. Когда TR1 активируется соответствую-
щим вкусовым стимулом, G-белок сигнальная система вызывает выделение в
цитозоль цАМФ, который через протеинкиназу А (ПКА) закрывает К+-канал.
Циклический АМФ сам по себе также способен открывать Na+/Ca2+-BopoT-
ный механизм. Результат действия этих двух механизмов — деполяризация.
Это, в свою очередь, ведет к потоку Са2+ внутрь клетки, который, как всегда
запускает экзоцитоз медиатора из синаптических пузырьков. Несахарные под-
сластители, такие как сахарин, сладкие белки (тауматин и т. д.), некоторые
аминокислоты и т. д. действуют иным путем. Существуют данные, что в этом
случае стимулируется система фосфолипазы С (ФЛС), приводящей к выделе-
нию ИТФ в цитозоль. Это ведет (как указывалось в гл. 2) к выделению Са2+ из
запасников в эндоплазматическом ретикулуме, которое инициирует выделе-
ние медиатора из синаптических пузырьков.
Umami — это название вкуса некоторых видов японской пищи — сушеной
рыбой кацуво {Katsuwonuspelamis), морской капусты комбу {Laminaria japonica)
и т. д. Этот вкус присущ также и куриному бульону. Вкус этот существенно от-
личается от сладкого, хотя тоже воспринимается как приятный. Наиболее зна-
чительным активным ингредиентом, имеющим этот вкус, является 1-глюта-
мат, хотя некоторые другие аминокислоты, короткие пептиды и нуклеотиды
12. Вкус 239
производят аналогичный эффект. Есть данные, что рецепторной молекулой
для вкуса umami является глутаматный рецептор, который, возможно, являет-
ся вариантом глутаматного рецептора ЦНС.
Соленость. Детектирование соли для животных имеет большое значение,
поскольку является ключевым для ионного гомеостаза внутренней среды. По-
казано, что Na+ воздействует на чувствительные к соленому ВРК, входя в Na+-
специфические ионные каналы (рис. 12.8), а не включая систему G-белков.
Это было продемонстрировано с помощью блокатора Na'’’-каналов — амило-
рида. Это соединение гуанидина является хорошо известным внешним блока-
тором эпителиальных Na'’’-каналов (3NaK). Отметьте, что эти каналы никак
не связаны с потенциал-зависимыми Na'’’-каналами возбудимой мембраны.
В присутствии субмикромолярных концентраций амилорида на слизистой по-
верхности грибовидного сосочка частота импульсации во вкусовых нервных
волокнах в ответ на NaCl существенно снижается. Локальная фиксация потен-
циала одиночного ВРК также показывает, что поток Na+ внутрь устраняется
амилоридом. Поэтому и сделано заключение, что поток Na+ внутрь клетки вы-
зывает деполяризацию ВРК, приводящую к выделению медиатора на соседние
нервные окончания.
Более глубокое понимание молекулярно-биологической природы амило-
рид-чувствительных Ка+-каналов достигнуто с помощью клонирования гомо-
логичных каналов из дистального отдела толстой кишки крысы. Этот 3NaK
состоит из трех субъединиц по 75 кДа. Использование антител к 3NaK позво-
ляет локализовать амилорид-чувствительные каналы ВРК. Также показано,
что 3NaK присутствуют в эпителиальных клетках, окружающих вкусовые
почки. Из чего предполагается, что в этих клетках существуют Na+-nyra, мо-
дулирующие активность ВРК.
Тот факт, что ощущение солености детектируется не через G-белковую
мембранную систему, а прямо через ионные каналы, объясняет, почему ответ
на NaCl очень быстр. У крыс и хомячков латентный период ответа на NaCl со-
ставляет менее 50 мс (для сравнения — более 300 мс на сахарин). Ответ на соль
также быстро адаптируется, часто менее, чем за 1 мин. Молекулярный меха-
низм адаптации до сих пор неизвестен.
Чувствительность к соленому находится под гормональным контролем.
Внесение альдостерона в перфузионную жидкость кровеносного сосуда языка
лягушки увеличивает ответ языкоглоточного нерва на аппликацию NaCl. Этот
эффект сохраняется в течение 3—6 ч после перфузии. В других тканях показа-
но, что альдостерон увеличивает вероятность открытого состояния предсуще-
ствующих Na’’’-каналов на срок около 10 ч, повышая, таки образом чувстви-
тельность. Действует ли такой же механизм и в ВРК пока неизвестно. Другим
гормоном, влияющим на чувствительность к соленому, является антидиурети-
ческий гормон (АДГ). Аппликация этого гормона также вызывает медленное
усиление ответа языкоглоточного нерва на аппликацию соли на язык. АДГ вы-
деляется гипофизом, когда осмолярность плазмы возрастает. Если это одно-
временно делает ВРК более чувствительными к NaCl, животное, по-видимо-
му, уменьшает потребление соленой пищи и, таким образом, компенсирует
повышение осмолярности. Такой экстерорецептивный контроль осмолярнос-
ти плазмы дополняет интерорецептивный контроль, рассмотренный в гл. 5.
240 Часть III. Хемочувствительность
Рис. 12.9. Трансдукция кислых вкусовых стимулов. (А) Протей Necturus. Н+ блокирует
К+-канал и вызывает деполяризацию, открывая Са2+-каналы и вызывая выделение ме-
диатора. (Б) Хомячок и, вероятно, крыса. Н+ проходит через Ка+-каналы, которые под-
даются блокаде амилоридом (А). Последующая деполяризация опять-таки вызывает по-
ток Са2+ внутрь клетки и выделение медиатора. Дальнейшие объяснения в тексте. По
Lindemann, 1996.
Кислое. Детектирование кислотности также имеет важное значение для
поддержания гомеостаза внутренней среды. В этом случае воздействию под-
вергается pH внутренней среды. Детектирование кислого также важно, чтобы
предупредить употребление в пищу неспелых (т. е. в питательном отношении
неоптимальных) или скисших фруктов. Полагают, что кислый вкус обуслов-
лен концентрацией свободных ионов Н+.
Н+, как и Na+, вероятно влияет на каналы ВРК непосредственно, причем
у разных позвоночных обнаружено несколько различных механизмов такого
действия. У земноводных, таких как протей Necturus, Н+ блокирует каналы
утечки К+ (рис. 12.9 А). Поскольку каналы утечки К+ играют существенную
роль в поддержании потенциала покоя (см. гл. 2) их блокада ведет к деполяри-
зации. Это, в свою очередь, ведет к экзоцитозу медиатора из синаптических
пузырьков. У хомячка и, вероятно, у крысы существует иной механизм. В этом
случае (см. рис. 12.9 Б) Н+ проходит через амилорид-чувствительные Na+-Ka-
налы. Этот поток катионов через ВРК также вызывает деполяризацию и, сле-
довательно, выделение медиатора.
Горькое. Как и кислое, горечь вызывает реакцию отвращения. Имеющие
горький вкус компоненты пищевых продуктов, например, растительные алка-
12. Вкус 241
лоиды — хинин, стрихнин и т. д., цианид в горьком миндале1, а также продук-
ты распада, как например мочевина, отвергаются. Горький вкус имеют многие
вещества, и возможно, что некоторые растения и беспозвоночные приобрели
способность накапливать эти вещества с целью защиты. В некоторых случаях
на основе этого наблюдается бэйтсовская мимикрия, когда организмы, не об-
ладающие ядовитыми или неприятными веществами, выживают путем обма-
на, за счет подражания тем организмам, которые такие вещества содержат.
Вероятно, и здесь существует более одного механизма трансдукции. Са-
мым важными из них является блокада К+-каналов (хотя этот механизм мало
избирателен) и более специфичные механизмы с участием системы G-белков.
В ВРК крыс и мышей обнаружены два механизма (рис. 12.10). В первом
случае (рис. 12.10 А) взаимодействие горького вещества со специфическим ре-
цептором приводит к активации мембранно-связанной ФЛС и продукции
ИТФ и ДАГ. ИТФ, как и обычно, вызывает выброс Са2+ из запасников в ЭР,
а тот, в свою очередь, запускает экзоцитоз медиатора. Второй механизм
(рис. 12.10 Б) очень похож на процессы в обонятельных ресничках или наруж-
ных сегментах палочек и колбочек сетчатки. Показано, что семидоменные ре-
цепторные белки, имеющие прибл. 40% гомологию последовательности с ре-
цептором сладкого TR1, расположены в основном в окаймлённых сосочках.
Когда эти рецепторы — TR2 — взаимодействуют с соответствующим лигандом
активируются G-белки, очень похожие на трансдуцины наружных сегментов
(гл. 16). Вследствие функционального сходства с трансдуцинами фоторецеп-
торных мембран эти G-белки называются густодуцинами (передатчиками
вкуса —- перев.). Они активируют фосфодиэстеразу цАМФ (цАМФ-ФДЭ),
которая катализирует преобразование цАМФ в 5 -АМФ. Далее, однако, на-
блюдается существенное различие между ВРК и обонятельными ресничками
или наружными сегментами фоторецепторных клеток. При повышенной
концентрации циклических нуклеотидов катионные каналы ВРК не откры-
ваются, (что вызвало бы деполяризацию клетки) а, наоборот, закрываются,
поддерживая высокий потенциал покоя. Гидролиз цАМФ под действием
ФДЭ приводит к открыванию каналов для Na+ и Са2+ и деполяризации вку-
совой клетки.
Вода. Потребление адкватных количеств воды сущственно для поддержа-
ния соответствующей осмолярности жидкости в организме. Детекторы соле-
ного, как описано выше, играют важную роль в поддержании этого баланса,
одновременно демонстрируя и присутствие детекторов воды. Языкоглоточ-
ный нерв лягушки и смешанная ветвь лицевого нерва (chorda tympani) млеко-
питающих содержат волокна, отвечающие на аппликацию воды на язык, в то
время как другие волокна реагируют на NaCl. Поскольку ацетат натрия не ме-
няет реакцию на воду, тогда как NaCl и холин-хлорид влияют на нее, было сде-
лано предположение, что этот механизм зависит от хлорных каналов в апи-
кальной мембране. Снижение наружной концентрации хлорида активирует
эти каналы; это приводит к повышению секреции С1”, что и ведет к деполяри-
1 Точнее, горький вкус миндаля обусловлен повышенным содержанием бензальдегида,
но не самими цианидами. — Прим. ред.
242 Часть III. Хемочувствительность
Рис. 12.10. Трансдукция горьких вкусовых стимулов. (А) Горечь взаимодействует с ре-
цептором и через G-белковый механизм активирует ФЛС. ИТФ вызывает выброс Са2+
из внутриклеточных запасников. Са2+ запускает выделение медиатора. (Б) Горечь взамо-
действует с рецептором и через густодуцин активирует цАМФ-ФДЭ, которая катализи-
рует преобразование цАМФ в 5'-АМФ. Устранение цАМФ снимает блок Na+/Ca2+-Ka-
налов, что приводит к деполяризации. Са2+ запускает выделение медиатора. Г — горечь,
ЭР — эндоплазматический ретикулум, G — G-белок, ГД — густодуцин, Ф — иАМФ-
ФДЭ, ФЛС — фосфолипаза С. Дальнейшие объяснения в тексте. По Lindemann, 1996.
зации ВРК. Как и в случае описанных выше механизмов восприятия вкусов,
возможны и иные биохимические пути, исследование которых продолжаются.
Субъективно «вкус» воды меняется в зависимости от того, какое вкусовое
ощущение ему предшествовало. Если перед этим действовал хинин или ли-
монная кислота, вкус воды воспринимается, как сладкий, тогда как после
NaCl или сахарозы — как горький. Из этого вытекает, что, возможно, между
рецепторами воды и рецепторами упомянутых вкусовых ощущений существу-
ет взаимодействие.
12.2.2. Центральные проекции
Как отмечалось выше, ВРК — это истинные рецепторные клетки. В ответ на
деполяризацию они выбрасывают медиатор, действующий на дендритные
окончания сенсорных волокон VII, IX и X черепно-мозговых нервов.
Рис. 12.11 показывает, что волокна всех трех путей передачи вкусовых сигна-
лов оканчиваются в ядре одиночного пучка продолговатого мозга. Это ядро —
сложное образование, получающее афференты из многих внутренних органов.
12. Вкус 243
Черепно-мозговой
нерв IX
Постцентральная
извилина
Коленчатый ганглий
Смешанная ветвь
лицевого нерва
Островок
Вентральное заднее
медиальное ядро
Глотка
Узловатый
ганглий
Язык
Каме-
нистым
ганглии
Парабрахиальное
ядро
Черепно-мозговой нерв VII
Комплекс
солитарного ядра
продолговатого мозга
Рис. 12.11. Вкусовые пути мозга. Объяс-
нения в тексте. По Kandel, Schwartz and
Jessel, 1991, с разрешения.
Вкусовые волокна оканчиваются в узкой зоне, называемой вкусовым ядром.
Отсюда нервные волокна восходят к мелким клеткам вентрального заднего яд-
ра таламуса. Вкусовая информация затем передается в две области коры моз-
га — первая лежит в постцентральной извилине (поле Бродмана ЗЬ), а вто-
рая — глубоко в Сильвиевой борозде, на наружной поверхности инсулы. Обе
они расположены вблизи от соматосенсорных зон языка (см. гл. 7, рис. 7.16).
12.3. Избранные каналы или паттерны ответов?
Как мозг распознает вкусовые стимулы — по тому, какие группы волокон ак-
тивируются, или по тому, какой характер ответа наблюдается во всей массе во-
локон? Существуют аргументы в пользу обоих механизмов, но в момент напи-
сания этой книги общее мнение склоняется в пользу последнего. Большинст-
во фактов говорит в пользу широкой чувствительности, а не узкой специали-
зации вкусовых клеток (см. выше).
244 Часть III. Хемочувствительность
При отведени от смешанной ветви лицевого нерва хомячка, когда вкусовые
почки подвергались воздействию различных концентраций одного и того же
вкусового вещества, либо действию нескольких веществ одного вкуса, наблю-
далась одна и та же (или очень похожая) картина ответа. Если концентрация
вкусового вещества, например поваренной соли, меняется, суммарный ответ
в 40 волокнах смешанной ветви лицевого нерва остается постоянным, хотя
в каждом из одиночных волокон частота импульсов может повышаться или
снижаться в зависимости от интенсивности стимула. Иными словами, тип ак-
тивности может изменяться, но общий ее «профиль» остается неизменным.
Аналогичная картина наблюдается для ряда групп веществ, вызывающих оди-
наковые (или схожие) вкусовые ощущения —- для вещества сладкого вкуса (как
сахарин, сахароза, фруктоза); для солей натрия (NaCl и NaNO3), и кислот (со-
ляной и лимонной). Это электрофизиологическое наблюдение подкрепляется
поведенческими экспериментами, показывающими, что хомячок не различа-
ет разные сладкие вещества и разные соли.
12.4. Заключение
Контактные хеморецепторы (вкусовые рецепторы) очевидным образом важны
в жизни животных. Будучи гетеротрофами, животные зависят от поиска и по-
требления пищи. Центральные пути вкусовых волокон млекопитающих пока-
зывают, насколько их конечные пункты близки к таковым волокон, несущих
другую информацию, касающуюся языка. Это не только рефлекторные ответы
языка на вкусовые стимулы, но и разнообразное пищевое поведение животно-
го. Во многих случаях всего одной неудачной реакции на потребленную пи-
щу, одного случая рвоты после потребления пищи достаточно для того, чтобы
предупредить в будущем от потребления такой пищи. Мы только начинаем
понимать биофизические и биохимические механизмы, работающие в ВРК и
вкусовых рецепторах других животных и дающие возможность точного разли-
чения вкусов. Мы только начинаем понимать, как ВРК интегрированы в об-
щую физиологию и пищевое поведение животных. Наконец, в прикладном
аспекте — это многомиллиардная индустрия, заинтересованная вданных фи-
зиологии вкуса. Это не только пищевая промышленность, но и производители
пестицидов и гербицидов, получающие прибыли от более глубокого понима-
ния вкусовых ощущений в царстве животных.
13. ОБОНЯНИЕ
Насекомые: гигрорецепторы — влажность важна для мелких животных —
механическое растяжение сенсиллы — сенсиллы часто включают термо-
рецепторы; обонятельные сенсиллы — одорант-связывающие белки
(ОСБ) — феромоны — множественность типов — феромон-связываю-
щие белки (ФСБ) — молекулярная биология — обонятельная система
дрозофилы — антенны тутового шелкопряда Bombyx — крайне высокая
чувствительность. Млекопитающие: обонятельный эпителий — развитие
зависит от образа жизни — пространственная организация; обонятельные
рецепторные клетки (ОРК) — структура — одорант-связывающие белки —
обонятельные реснички — генерализация или специализация? — чувст-
вительность — обонятельные рецепторные белки; молекулярная биоло-
гия ОРК —- ЦНЗ-каналы — выделение — функциональность — локальная
фиксация потенциала ОРК — биофизика — сенсорная адаптация — граду-
альный ответ — аносмии; центральная обработка информации — решетча-
тая кость — обонятельная луковица — митральные, пучковые и перигло-
мерулярные клетки — клубочки — сравнение луковицы и сетчатки — ней-
рохимия — обонятельный тракт — крючок (uncus) — таламус — орбито-
фронтальная кора. Вомероназальный орган — локализация — эпителий —
ОРК —- различия молекулярной биологии — различная эволюционная
история — феромоны — проекция в добавочную обонятельную лукови-
цу — миндалина и гипоталамус — эмоции. Заключение: значение хемо-
чувствительности в коммуникации между индивидами
Мы отмечали в гл. 12, что различия между вкусом и запахом, между вкусовым
ощущением и обонятельным, представляющиеся столь очевидными людям,
далеко не так очевидны у других животных, особенно это касается беспозво-
ночных. У позвоночных различие этих чувств подчеркивается и раздельностью
органов чувств, и структурой центральных отделов. Многие рыбы, особенно
хрящевые, такие как акулы, обладают превосходным обонянием и способны
обнаруживать кровь или другие жидкие вещества в воде на значительных рас-
стояниях. Многие костистые рыбы также обладают отличным обонянием. Про-
ходные рыбы, такие как лососевые, с помощью обоняния находят родную речку
после длительного периода жизни в океане. Если лососям закрыть их обоня-
тельные мешки, они теряют способность определять, по какому притоку надо
подниматься. У дышащих воздухом наземных форм различия между двумя хи-
мическими чувствами выражены более сильно, поэтому в данной главе мы огра-
ничимся рассмотрением двух полностью наземных групп животных — насеко-
246 Часть III. Хемочувствительность
мых и четвероногих позвоночных. Начнем с обоняния насекомых, а потом пе-
рейдем к несколько более исследованной обонятельной системе позвоночных.
13.1. Обонятельная система насекомых
В гл. 6 и 12 мы рассмотрели структуру и физиологию сенсорных сенсилл насе-
комых. В частности, в гл. 12 мы детально изучили структуру и функцию кон-
тактных (вкусовых) хемочувствительных клеток насекомых. Большинство
сказанного в упомянутых главах применимо к тому, что рассматривается
здесь. За исключением гигрорецепторов, обонятельные рецепторы насеко-
мых — это обычные многопоровые сенсиллы.
13.1.1. Гигрорецепторы
Мы видели в гл. 12, что один из типов нейросенсорных клеток, который был
найден в контактных однопоровых сенсиллах, является водным рецептором.
Гигрорецепторы, детектирующие влажность, очень схожи с водными (вкусо-
выми) рецепторами. Однако, поскольку у них обычно отсутствует отверстие,
соединяющее их с внешней средой, эти рецепторы, вместе с обонятельными
следует отнести к дистантным. Атмосферная влажность не имеет непосредст-
Рис. 13.1. Гигро-термальная сен-
силла. Рисунок показывает ти-
пичную «колышковую» сенсил-
лу (короткий толстый колышек
в капсуловидном углублении).
Сенсилла содержит триаду сен-
сорных нейронов. Две клетки
типа 1 (гигрорецепторы) сосу-
ществуют с одной клеткой типа 2
(терморецептор). В то время, как
клетки типа 1 имеют длинные
дендриты большого диаметра, ко-
торые полностью заполняют ко-
лышек, клетка типа 2 имеет ко-
роткий глубоко инвагинирован-
ный дендрит, который не прости-
рается на всю длину колышка,
до — дендритная оболочка, лк —
линочный канал, лпнр — наруж-
ная полость рецепторной лимфы,
тор — тормогенная клетка, три —
трихогенная клетка, текогенные
клетки выделены пунктирной
штриховкой. Из Altner and Loftus,
1985. Воспроизводится с разре-
шения Annual Review of Entomo-
logy, volume 30, © 1985, Annual
Reviews http://annualreviews.org.
13. Обоняние 247
венного доступа к нейросенсорной клетке сенсиллы, и адекватным стимулом
является ее механическое растяжение в ответ на изменение влажности. Эти
сенсиллы обычно представлены короткими выростами, жестко закрепленны-
ми в кутикуле. При тщательном исследовании такие сенсиллы обнаруживают-
ся на антеннах всех насекомых. Несмотря на важность гигрорецеппии, пред-
ставительство этих рецепторов в сенсорных волосках антенн насекомых неве-
лико — например, у таракана Periplaneta americana всего около 100 сенсилл из
54 000 на каждой антенне являются гигрорецепторными.
Растяжение сенсиллы, вызванное изменением влажности, может выражать-
ся в изгибании сенсиллы или разбухании кутикулы сенсиллы, что приводит к
передаче давления на наружный сегмент дендрита. Нейросенсорная клетка, та-
ким образом, может рассматриваться как механорецепторная, хотя тубулярное
тельце, столь характерное для механочувствительных дендритов (см. гл. 6) у них
пока четко не идентифицировано. Если изгибание сенсиллы действительно яв-
ляется адекватным стимулом, важно, чтобы сенсиллы были защищены от воз-
действий, сопряженных с обычной подвижностью насекомого. Не удивитель-
но, что эти немногочисленные гигрорецептивные сенсиллы располагаются до-
статочно глубоко под длинными механочувствительными волосками и часто
окружены кутикулярным футляром или погружены в углубление.
Гигрорецептивные сенсиллы обычно содержат более одной нейросенсорной
клетки (рис. 13.1). Чаще их три, а иногда и четыре. В триадной организации, по-
казанной на рис. 13.1, две гигрорецепторных клетки дополняются еще и термо-
рецептивной. Эта бимодальность является общей для многих отрядов насеко-
мых. Ясно, что для таких мелких животных, как насекомые, чрезвычайно важно
получать точную информацию о температуре и влажности окружающей среды.
Более подробно о терморецепторах насекомых будет сказано в гл. 19.
13.1.2. Обонятельные сенсиллы и феромоны
Обонятельные нейросенсорные клетки насекомых обычно помещены в мульти-
поровые базиконические и трихоидные сенсиллы (рис. 13.2), локализованные в
антеннах. Молекулы пахучего вещества (одоранта), достигшие сенсиллы, в пер-
вую очередь адсорбируются на воскообразной поверхности кутикулы, а затем
диффундируют через пору в заполненную лимфой внутреннюю часть сенсиллы.
Здесь пахучие вещества взаимодействуют с одорант-связывающими белками
(ОСБ), которые не только солюбилизируют гидрофобные одоранты в лимфе, но
и делают их пригодными к взаимодействию с активными сайтами рецепторных
молекул в мембране обонятельных, рецепторных клеток. Сходство с бактериаль-
ными Р-П-белками, обсуждавшимися в гл. 10 — поразительное. Аналогичные
молекулы могут присутствовать и в обонятельной слизистой у позвоночных.
Наскомые живут в мире семиотических одорантов — т. е. веществ, обладаю-
щих некоторым специфическим смысловым значением. Они могут быть сиг-
налами возможности репродукции (половые феромоны), опасности (феромо-
ны тревоги), чьих-то следов (следовые феромоны), сигналов к сосредоточе-
нию или сигналов потенциального хозяина для паразита (агрегационные фе-
ромоны), сигналы к рассредоточению (феромоны рассредоточения) и т. д. На-
иболее эффективны половые феромоны, как и следовало бы предполаготь,
248 Часть III. Хемочувствительность
Рис. 13.2. Мультипоровая обонятельная
сенсилла. Сенсилла может содержать до
40 нейросенсорных клеток, наружные сег-
менты которых могут ветвиться. ВД — вет-
вящийся наружный сегмент дендрита, ВК —
вырост кутикулы, ДО — оболочка дендрита,
Э — эпидермис, В — внутренняя клетка обо-
лочки, Н — нейросенсорная клетка, Н — на-
ружная клетка оболочки, ПС — пора стенки
сенсиллы. Из Frazier, 1985; © 1985, с разре-
шения.
учитывая их важнейшее значение для выживания вида. Рассчитано, что самец
тутового шелкопряда Bombyx mori чувствителен к единственной молекуле жен-
ского феромона бомбикола1.
Рецепторы полового феромона шелкопряда — очень ценный объект для
исследователей обоняния. Возможно наиболее изученным является одорант-
связывающие белки, с которыми связываются эти феромоны. Антеннальные
сенсиллы шелкопряда-павлиноглазки Antheraea polyphemus содержат 15 кДа-
белок, который очень высокоспецифично связывает половой феромон, выде-
ляемый самками этого вида. А лимфа сенсилл содержит феромон-специфиче-
ские эстеразы, которые разрушают феромоны, предупреждая тем самым их
пролонгированное действие. Аналогичные высокоспецифичные ОСБ обнару-
жены и у других видов. В настоящее время эти белки именуются феромон-свя-
зывающими белками (ФСБ). Помимо ФСБ существует множество других ОСБ
(иногда называемых общими одорант-связывающими белками, ООСБ), иден-
тифицированых в обонятельных сенсиллах насекомых. У Drosophila melanogas-
ter различные формы обильно представленного ОСБ весом в 14 кДа обнаружи-
ваются в сенсиллах различных участков антенн. Вероятно, такая регионализа-
ция ОСБ у плодовой мушки играет роль в различении запахов.
Рис. 13.2 показывает, что сенсилла включает наружный сегмент дендрита
нейросенсорной клетки. Обонятельные рецепторные молекулы встроены в
мембрану наружных сегментов нейросенсорных клеток, но за исключением
1 Эти сведения основаны на результатах опытов, в которых ошибочно оценивалась кон-
центрация феромона. Позже было показано, что рецепторные клетки самца действительно
отвечают на отдельные молекулы феромона. Однако, пороговая поведенческая реакции сам-
ца требует возбуждения нескольких сот феромон-чувствительных клеток. — Прим. ред.
13. Обоняние 249
Drosophila, об их структуре мало что известно. В то же время накапливаются
данные об участии в передаче сигнала ФЛС, ИТФ и ДАГ. Из этого вытекает
возможность участия в данном процессе G-белковой мембранной сигнальной
системы. Это сближает обонятельные системы насекомых с таковыми позво-
ночных, которые мы обсудим ниже. Как и у позвоночных, биохимические из-
менения рецепторной молекулы приводят у насекомых к возникновению в
дендрите генераторного потенциала — обычно деполяризации, но иногда —
гиперполяризации. В первом случае в обонятельном аксоне инициируется по-
тенциал действия, во втором всякие потенциалы действия подавляются.
Детальное знание генетики дрозофилы и генетические методы позволили
достичь значительного прогресса в понимании молекулярных основ ее обоня-
тельной системы. Помимо легкости генетических манипуляций с дрозофилой,
еще одним преимуществом обонятельной системы плодовой мушки является
легкость ее сравнения с системами позвоночных. Она включает около 3000
нейронов, и идентифицировано множество мутантов по обонянию. Молеку-
лярный анализ показывает, что многие из мутаций могут быть объяснены как
дефекты ФЛС-, ИТФ- и ДАГ-каскадов сигнализации.
Обонятельные сенсиллы Drosophila в первую очередь были обнаружены на
третьем членике антенн (funiculus)1 и на пальпах максилл. В funiculus находит-
ся около 200 базиконических, 150 трихоидных и 60 целоконических сенсилл, а
в каждой из пальп — еще по 60 базиконических сенсилл. Каждая сенсилла
обычно содержит три хеморецепторных и один механосенсорный нейрон.
Генетические методы позволили выделить от 100 до 200 различных семидо-
менных обонятельных рецепторных молекул, которые экспрессируются в спе-
цифических сенсиллах. Иными словами, каждый сенсиллярный нейрон экс-
прессирует один или специфическое для него небольшое количество видов се-
мидоменных обонятельных рецепторов. Кроме того, специальная техника ок-
рашивания показала, что каждый рецептор экспрессируется в пространствен-
но ограниченной субпопуляции сенсиллярных нейронов funiculus или пальп
максилл. В этом отношении, как мы увидим ниже, хеморецепторные поля на-
секомых отличаются от таковых позвоночных. Обонятельные поверхности по-
следних не проявляют пространственной организации — обонятельные клетки
различной чувствительности распределены в слизистой в случайном порядке.
Нейроны сенсилл проецируются в ипсилатеральные антеннальные доли
дейтоцеребрума2, где оканчиваются в прибл. 35 гломерулах (клубочках)3.
По сравнению с другими видами насекомых это немного: у бражника Mandu-
са sexta их 53, у таракана Periplaneta americana — 125 и более 1000 у саранчи Lo-
custa migratoria, но даже это меньше, чем у типичного позвоночного
1 Funiculus — канатик (лат.). — Прим, перев.
2 Дейтоцеребрум — второй надглоточный ганглий. — Прим, перев.
3 Клубочки являются исключительным местом синаптической связи, первым переключе-
нием обонятельного пути у насекомых. Морфологически они представляют собой плотное
сплетение нейропиля, включающего аксоны обонятельных рецепторных клеток и отростки
нейронов антеннальной доли. Подобная организация первого синаптического переключе-
ния в обонятельных путях наблюдается почти всегда как у беспозвоночных, так и у позвоноч-
ных (по: Boeckh J., Tolbert, 1993. Synaptic organization and development of the antennal lobe in
insects. Mier. Res. Techn. 24:260-280). — Прим. ped.
250 Часть III. Хемочувствительность
Рис. 13.3. Антенна самца тутового шелкопряда Bombyx топ. (А) Антенна; (Б) часть
антенны; (В) одиночная веточка антенны; (Г) сенсорный волосок. Из D. Schnei-
der, 1977 в Т. Elsner & Е. О. Wilson eds, Reading from Scientific American, p. 32. Вос-
производится с разрешения Lorelle Raboni.
(1800—2000)1 *. Малое количество гломерул у дрозофилы и сравнительно боль-
шое число рецепторных молекул (100—200) наводит на мысль, что каждая из
гломерул реагирует на нескольких различных типов пахучих веществ. Поэтому
вероятно, что первичная обработка информации происходит именно здесь —
в антеннальной доле, прежде чем она передается дальше. Из антеннальных до-
лей волокна проецируются в другие части мозга насекомого, в частности в
сложные массы нейропиля — так наз. грибовидные тела.
Поля сенсилл на антеннах шелкопряда Bombyx топ, как и можно было
ожидать, много более развиты, чем у Drosophila. У самца шелкопряда около
17000 длинных (100 мкм) и тонких (диаметр 2 мкм) сенсилл, образующих про-
странственную решетку веточек антенны (рис. 13.3). Такая решетка из сенсил-
лярных волосков способна концентрировать одоранты, особенно половые фе-
ромоны, увеличивая число захваченных за 1 секунду молекул в 1,5 х 105 раз
при скорости потока воздуха в 60 см/с. Аксоны сенсиллярных нейронов окан-
чиваются, как и обычно, в ряде клубочков антеннальной доли дейтоцеребру-
ма. Эти клубочки занимают определенное положение, и у некоторых насеко-
мых, в частности у бражника Manduca sexta, удалось построить детальные кар-
ты «пространства запахов».
Активная жизнь многих насекомых протекает в мире, заполненном при-
зывными сигналами различных веществ. Можно ли сказать, что насекомые
1 Такое число обонятельных клубочков насчитывается у мыши (у крысы клубочков замет-
но больше). — Прим. ред.
13. Обоняние 251
испытывают эмоции? Маловероятно, да и как можно это узнать? Но если при-
нять, что это так, то можно себе представить, какую бурю эмоций — носталь-
гию и надежду, восхищение и отчаяние — должны порождать улавливаемые их
чувствительными антеннами следы веществ на листьях и внутри цветков. Их
мир полон смысла.
13.2. Обонятельная система позвоночных
Носовая полость большинства дышащих воздухом позвоночных содержит по
крайней мере два различных обонятельных органа: первый и наиболее важ-
ный — это основной обонятельный эпителий, покрывающий носовые проходы,
и второй — вомероназальный орган, которые мы рассмотрим последовательно.
13.2.1. Обонятельный эпителий полости носа
У высших приматов, включая человека, обонятельный эпителий сравнительно
невелик. Этому можно искать объяснение в том, что запахи под пологом леса
разносятся недалеко. Но главное, приматы — животные с дневной активнос-
тью, и полагаются преимущественно на зрение при определении расстояний и
размеров. У ночных форм, которым от зрения много меньше пользы, развива-
лись и достигли совершенства и доминируют два других дистантных чувства —
слух и обоняние. Размеры обонятельного эпителия у млекопитающих варьи-
руют от 2—4 см2 (человек) и 9,3 см2 (кролик) до 18 см2 (собака)1 и 21 см2 (до-
машняя кошка). Надо отметить, однако, что эти цифры не дают точного пред-
ставления об «остроте» обоняния, поскольку здесь не учтено число обонятель-
ных рецепторных клеток на единицу поверхности.
У млекопитающих обонятельный эпителий развился на тонких пластинча-
тых костях полости носа — носовых раковинах. Эта область выполняет двойную
функцию — не только подставляя обонятельный эпителий под поток посту-
пающего воздуха и создавая сопротивление его потоку, но и нагревая воздух
перед его поступлением в легкие. Наличие или отсутствие носовых раковин в
отложениях костных остатков используется для определения того, был ли их
давным-давно умерший обладатель теплокровным. Обонятельный эпителий
содержит три типа клеток — нейросенсорные (обонятельные рецепторные
клетки), опорные клетки, секретирующие слизь (близкие по свойствам к гли-
альным клеткам) и базальные, которые (подобно стволовым клеткам) способ-
ны делиться и формировать новые функциональные нейроны в течение жиз-
ни животного (рис. 13.4). Обонятельные клетки — это единственные нейроны
у млекопитающих, способные обновляться в течение всей жизни животного.
Мы увидим (да и видели уже), что в зрительной, слуховой, соматосенсорной
и, в меньшей степени, вкусовой системах сенсорный эпителий регистрирует
пространственную информацию. «Картирующая» функция в обонятельном
эпителии не так выражена. В первом приближении клетки, специализирован-
ные к различению специфических запахов, распределены в случайном поряд-
ке, однако более пристальное изучение заставляет предположить, что и здесь
1 В монографии А. А. Бронштейна указан диапазон значений для собак нескольких пород —
округленно от 30 см2 у мелких пород до 200 см2 у некоторых крупных собак. — Прим. ред.
252 Часть III. Хемочувствительность
присутствуют зачатки пространственной организации. Картирование с по-
мощью 2-деоксиглюкозы, которая дает возможность определить активные
клетки) показало наличие групп клеток, связанных с определенным запахом.
Например, бутанол возбуждает клетки передней области, тогда как лимонен
активирует клетки задней части слизистой. Недавние исследования также
заставляют предположить, что рецепторные клетки организованы в передне-
задние полосы, называемые зонами экспрессии, каждая из которых содержит
полный набор клеток. Вероятно, существует три не перекрывающиеся зоны
экспрессии, которые перекрываются с меньшей, четвертой зоной.
13.2.2. Обонятельные рецепторные клетки
Рис. 13.4 показывает, что обонятельные рецепторные клетки (ОРК) — бипо-
лярные с одним неветвящимся дендритом, который проходит между опорны-
ми клетками, чтобы закончиться небольшим вздутием — обонятельной була-
вой. Из нее вырастают до 20 длинных ресничек. Эти реснички представляют
собой сенсорную поверхность обонятельной клетки. Обычно они погружены
в слой слизи, покрывающей эпителий, образуя с ним плотный матрикс. Обо-
нятельная клетка имеет двойную функцию — восприятие стимула и передачу
нервного импульса в мозг, а потому является нейросенсорной клеткой, ее ино-
гда называют сенсорным нейроном.
Слизь, в которой лежат обонятельные реснички, содержит большое количе-
ство небольших (20 кДа) белков, во многих отношениях похожих на одорант-
связывающие белки (ОСБ) сенсилл насекомых. Они секретируются железами
слизистой носа и встречаются не только в слизи, покрывающей обонятельный
эпителий, но и в слизи, покрывающей чисто респираторный эпителий. Этот
факт, а также их сравнительно небольшой размер, позволяют предположить,
что эти белки не играют столь же важной роли, как ОСБ насекомых и бакте-
рий. Если эта роль и сущестует, то сродство этих белков к молекулам-одоран-
там очень неселективно, а функция сводится к связыванию одорантов и обес-
печению их взаимодействия с рецепторными клетками.
Ультраструктура обонятельных ресничек не сильно отличается от таковой
других ресничек. Они содержат обычную аксонему, которая, по крайней мере
у млекопитающих, неподвижна. Эти реснички необычно длинные и тонкие —
от 5 до 250 мкм в длину и лишь 100—250 нм в диаметре. Мы уже сталкивались
с такой адаптацией подвижного органоида — реснички — к сенсорной функ-
ции в сенсиллах насекомых и механорецепторах позвоночных (киноцилии во-
лосковых клеток) и еще встретимся с этим в фоторецепторах позвоночных
(наружные сегменты палочек и колбочек).
Пучок (от 5 до 40) длинных тонких ресничек, выходящий из булавы обоня-
тельной клетки, несомненно, резко увеличивает сенсорную поверхность. Элек-
тронная микроскопия замороженных сколов показывает, что мембрана каждой
реснички содержит большое количество глобулярных частиц — предполагает-
ся, что это обонятельные рецепторные молекулы. В гл. 16 мы увидим, что это
еще одна аналогия с наружными сегментами палочек и колбочек, которые
плотно заполнены рецепторными молекулами — родопсином и йодопсином.
Обонятельная система способна детектировать и различать огромное количе-
ство запахов. Как мы видели на вкусовых клетках, остается предметом противо-
13. Обоняние 253
Слизистый Тесный Обонятельный
слой контакт «холмик»
Обонятельный нерв Базальные Обонятельный Опорные
(немиелинизированные клетки аксон клетки
волокна)
Рис. 13.4. Обонятельный эпите-
лий. Длинные обонятельные
реснички на поверхности кле-
ток погружены в слизь. Плот-
ные контакты между соседними
опорными клетками, а также
между опорными клетками и
нейросенсорными клетками за-
щищают межклеточное прост-
ранство от проникновения из-
вне чего-либо.
речий, является ли отдельная ресничка «генерализатором» или «специалистом»,
иными словами, отвечает ли ресничка на многие типы молекул пахучих веществ
или только на один. В последние годы накапливаются данные в пользу их специ-
ализации. Но остается фактом, что независимо от генерализации или специали-
зации, чувствительность их очень велика—даже человек способен обнаруживать
пахучие вещества при концентрации 10”4 М—10“13 М. Одним из признаков про-
студы является значительное снижение такой тонкой чувствительности. Это
происходит потому, что обонятельные реснички оказываются погруженными в
слишком большом слое слизи, вырабатываемой опорными клетками.
13.2.3. Молекулярная биология
Обонятельные рецепторные белки
В последние годы идентифицировано большое семейство генов (несколько
сотен, возможно, более тысячи), кодирующих обонятельные рецепторные
белки, что составляет 1—2% генома. Это семейство генов кодирует семидомен-
ные рецепторы, принадлежащие к тому же надсемейству, что и описанные
в гл. 1. Такая связь означает, что знание молекулярной биологии и фармаколо-
гии других хорошо изученных семидоменных рецепторов может послужить и
пониманию modus operands рецептора в обонятельной ресничке. В гл. 16 мы
увидим, что опсины палочек и колбочек принадлежат к тому же семейству. Так
что и в этом отношении обонятельные реснички обнаруживают параллель с
наружными сегментами палочек и колбочек.
Семидоменные обонятельные рецепторные молекулы отличаются от про-
чих членов надсемейства семидоменных рецепторов большим аминокислот-
ным разнообразием в трансмембранных доменах 3,4 и, особенно, 5. Возвраща-
ясь к гл. 1, мы видим, что эти домены образуют существенную часть погружен-
ной в мембрану «бочки» семидоменного рецептора. Поскольку похоже, что
1 Modus operandi — образ действия (лат.). — Прим, перев.
254 Часть III. Хемочувствительность
именно внутренняя часть «бочки», образуемой этими доменами, формирует
сайт, распознающий пахучие молекулы, рассмотрим, как разные рецепторные
молекулы приспособлены к распознаванию разных одорантов. Следует по-
мнить, что в отличие от большинства других семидоменных рецепторных бел-
ков, сопряженных с G-белками, обладающих довольно избирательным сродст-
вом, — обонятельные рецепторные белки считаются в той или иной степени
аффинными к целому ряду стереохимически сходных пахучих молекул.
Как отмечалось выше, есть данные, что обонятельные реснички специали-
зированы, т. е. они способны экспрессировать только один тип обонятельных
рецепторных молекул. Это однако не означает, что они реагируют только на
один тип одорантов. Показано, что каждый данный рецептор стимулируется
небольшим числом химически сходных молекул одорантов. С другой стороны,
каждое пахучее вещество взаимодействует с некоторым числом разных рецеп-
торов. Вероятно каждый обонятельный стимул взаимодействует со специфи-
ческой группой рецепторных молекул, и этот набор рецепторов определяет
индивидуальность запаха. В то же время ситуация в слизистом слое сложна и
динамична; небольшим изменениям химической структуры молекулы одо-
ранта соответствует изменение набора стимулируемых рецепторов. Измене-
ния концентрации молекул одоранта могут влиять также на число стимулиру-
емых рецепторов и, таким образом, изменять информацию, передаваемую в
обонятельную луковицу. Параллельно меняется и наше субъективное воспри-
ятие. Замещение гидроксильной группы октанола на карбоксильную, каза-
лось бы незначительное изменение, приводит к значительному изменению
обонятельного восприятия запаха — вместо запаха, напоминающего апельсин
и розу, появляется запах прогорклости и пота. Еще несколько одорантов меня-
ют свой субъективный запах в зависимости от концентрации. Некоторые па-
хучие вещества меняют качество своего запаха при изменении концетрации.
Индол, например, имеет приятный цветочный запах при большом разведении
и отвратительный гнилостный — в высокой концентрации.
Мембранная биохимия
Семидоменные рецепторы обонятельных ресничек (как и обычно) связаны G-
белковой системой с мембранно-связанными эффекторами. В большинстве
случаев эффектором является аденилатциклаза (АЦ). Иногда, правда, это мо-
гут быть ФЛС-р, ИТФ и ДАГ (см. гл. 1). Это роднит их с обонятельными сен-
силлами насекомых, о которых накапливаются данные об участии системы
ФЛС-р. В тех случаях, когда вторичным мессенджером является цАМФ, он
воздействует на каналы с малым диаметром поры (см. рис. 13.5). Поскольку
каналы контролируются циклическими нуклеотидами, они и именуются цик-
лонуклеотид-зависимыми (ЦНЗ-каналы). Хотя обонятельные ЦНЗ-каналы
были выделены не первыми (эта честь принадлежит каналам палочек), к на-
стоящему времени они, вероятно, охарактеризованы лучше других.
ЦНЗ-каналы
Выделение ЦНЗ-каналов было произведено методом, ставшим уже традици-
онным. Во-первых, белок 63 кДа был выделен из наружных сегментов палочек
и колбочек быка. При встраивании этого белка в искусственный липидный
13. Обоняние 255
Рис. 13.5. Молекулярная биология ресничек обонятельных клеток. Присоедине-
ние подходящей молекулы одоранта (О) запускает в рецепторной клетке процесс
взаимодействия с G-белком, что активирует аденилатциклазу (АЦ). Образующий-
ся в результате цАМФ открывает ЦНЗ-канал (Ц) в мембране. По этому каналу Са2+
и Na+ поступают внутрь клетки, а К+ - наружу. Са2+ вызывает ряд эффектов (как
показано на рисунке). Дальнейшие объяснения в тексте. АЦ — аденилатциклаза,
Ц — ЦНЗ-канал, G — G-белок, NO-C — NO-синтаза, О — молекула одоранта,
КОР — киназа обонятельного рецептора, ПКА — протеинкиназа А, Р — рецептор.
бислой можно регистрировать ток через единичный канал в ответ на цГМФ.
Далее, библиотека кДНК сетчатки испытывалась олигонуклеотидной после-
довательностью, представляющей отдельные участки белка. Был выделен
клон, кодирующий 690 аминокислотных остатков. Когда он экспрессировался
в ооцитах Xenopus1, формировался функциональный ЦНЗ-канал. С использо-
ванием последовательностей, полученных из этого белка, из фоторецепторов
позвоночных, обонятельного эпителия, а совсем недавно и ганглиозных кле-
ток сетчатки, были выделены и многие другие ЦНЗ-каналы.
Чтобы определить, какие участки аминокислотной цепи локализованы в
мембране, использовалась методика т.н. гидропатического анализа. В сущно-
сти, это означает просто проверку того, какие участки цепи состоят преимуще-
ственно из гидрофобных аминокислот. При анализе ЦНЗ-каналов выявлено
шесть гидрофобных сегментов, т. е. канал имеет шесть трансмембранных сег-
ментов (рис. 13.6). Такая молекулярная структура напоминает многие потен-
циал-заивисимые каналы: К+, Na+, Са2+, СГ и т. д. Более детальный анализ
показал, что четвертый трансмембранный сегмент S4 строго гомологичен сен-
сору потенциала S4 потенциал-зависимых каналов. Здесь присутствует также
т. н. р-шпилька (петля ДНК) между сегментами S5 и S6. Надо напомнить
(гл. 1), что похожий домен Н5 присутствует в потенциал-зависимых каналах,
где, как предполагается, он контролирует тип иона, способного пройти через
канал. В отличие от потенциал-зависимых каналов, ЦНЗ-каналы имеют боль-
шой С-концевой цитоплазматический домен, который связывается с вторич-
1 Xenopus laevis — шпорцевая лягушка. — Прим, перев.
256 Часть III. Хемочувствительность
связывающий
домен
Рис. 13.6. Субъединица
ЦНЗ-канального белка.
Объяснения в тексте.
ними мессенджерами — циклическими нуклеотидами. Предполагается, глав-
ным образом по аналогии с хорошо изученным К+-каналом, что четыре ЦНЗ-
канальных белка, показанных на рис. 13.6, образуют полный канал. Образован
ли канал тетрамером одинаковых ЦНЗ-субъединиц (гомо-олигомер) или (как
К+-канал) различными субъединицами ЦНЗ-тетрамера (гетеро-олигомер),
пока неизвестно.
Биофизика
С помощью метода локальной фиксации потенциала (пэтч-кламп) в обоня-
тельной ресничке было показано наличие каналов, открывающихся в ответ на
цАМФ. Поскольку на ресничках расположено 2400 каналов/мкм2 (тогда как
на обонятельной булаве и дендрите только 6 каналов/мкм2), то исследования
одиночных каналов проводились на дистальных участках обонятельных кле-
ток. Показано, что здесь каналы имеют очень сходные биофизические харак-
теристики с таковыми ресничек. В отсутствие Са2+ эти каналы отвечают на
цАМФ с проводимостью 45 пСи. Са2+ и другие двухвалентные катионы
уменьшают проводимость до прибл. 1,5 пСи. Такая же проводимость наблю-
дается при аппликации цГМФ, что в общем удивительно, поскольку присут-
ствие цГМФ в обонятельных клетках пока не показано. Возможно, однако,
что цГМФ образуется в результате Са2+-индуцированного образования NO
NO-синтазой (см. рис. 13.5). Известно, что основной мишенью окиси азота
является растворимая гуанилатциклаза, которую окись азота активирует. Ве-
роятно, в обонятельных клетках присутствует адекватный источник ГТФ, из
которого гуанилатциклаза и формирует цГМФ.
В отсутствие Са2+-блокады ЦНЗ-каналы высокопроницаемы для всех од-
новалентных катионов: Na+>K+>Li+>Rb+>Cs+. В обычных физиологических
условиях Na+, К+ и Са2+ двигаются через клеточную мембрану по электрохи-
мическому градиенту, что приводит к деполяризации клетки. Деполяризация
обонятельных клеток в ответ на действие одоранта может быть зарегистриро-
вана с помощью микроэлектродной техники. Величина деполяризации зави-
сит от концентрации одоранта — это пример генераторного потенциала, кото-
рый запускает потенциал действия в отдельном волокне обонятельного нерва,
которое является аксоном возбужденной клетки. Ее генераторный потенциал
не вызывает потенциалов действия в остальных волокнах нерва. Потенциал
действия распространяется без декремента в обонятельную луковицу передне-
го мозга.
13. Обоняние 257
Сенсорная адаптация и градуальный ответ
Одной из интересных черт изолированных ЦНЗ-каналов обонятельных рес-
ничек является то, что они не десенситизируются. Это означает, что при по-
вторных предъявлениях одоранта ЦНЗ-каналы не утрачивают чувствительно-
сти. Сенсорная адаптация в обонятельных рецепторных клетках, тем не менее,
имеет место. Вероятно, это происходит вследствие потока внутрь клетки ио-
нов Са2+, которые закрывают ЦНЗ-каналы либо непосредственно (рис. 13.5),
либо через активацию кальмодулина. Поток Са2+ внутрь клетки также ведет
к десенситизации семидоменной рецепторной молекулы.
Градуальность ответа на изменение концентрации одоранта связана с тем,
что система вторичных мессенджеров цАМФ увеличивает (или уменьшает) чис-
ло ЦНЗ-каналов в открытом состоянии. Чтобы различить запахи в реальном
(биологическом) времени, ответ должен быть достаточно быстрым. В экспери-
менте показано, что пик продукции цАМФ наступает через 40—75 мс после
аппликации одоранта и падает до нуля через 100—500 мс. Усиление сигнала
G-белковым каскадом обеспечивает активацию множества каналов одним им-
пульсом одоранта. Было показано, однако, что кинетика каналов достаточно
медленна — открытое состояние может отставать от импульса цАМФ на не-
сколько сот миллисекунд. Продолжительная активация семидоменных рецеп-
торов постоянными концентрациями одоранта, таким образом, обеспечивает
поддержание ЦНЗ-каналов в постоянно открытом состоянии импульсами
цАМФ.
Аносмии
Генетический анализ обонятельной системы человека выделил несколько дю-
жин специфических аносмий. Например, один человек из десяти неспособен
определить запах цианида, а один из тысячи — нечувствителен к бутилмер-
каптану — активному агенту зловония, испускаемого скунсом. Эти аносмии
возникают, вероятно, вследствие дефектов специфических обонятельных ре-
цепторных молекул. Многие из них наследуются в соответствии с законами
Менделя. Очевидна аналогия с цветовой слепотой у человека, которая также
возникает в результате дефекта рецепторного белка, в последнеми случае —
йодопсинов. В отличие от генетики аносмий, генетика цветовой слепоты
сравнительно хорошо известна.
13.2.4. Центральная обработка информации
Обонятельные клетки, как мы видели выше, являются нейросенсорными.
Когда генераторный потенциал достигает порога, инициируется потенциал
действия в начальном сегменте аксона. Рис. 13.7 А показывает, что эти аксоны
проходят через отверстия решетчатой кости и входят в обонятельную лукови-
цу, лежащую прямо над ней.
Мы уже не раз говорили о параллели в нейронной организации обонятель-
ной луковицы и сетчатки. Отметим еще одно анатомическое сходство обоня-
тельной и зрительной систем. Аксоны обонятельных рецепторных клеток
258 Часть III. Хемочувствительность
А
Гломерула
Обонятельная луковица
Передний мозг
Обонятельный тракт (ОТ)
Обонятельный нерв (ОН)
Решетчатая кость
Обонятельный эпителий
Обонятельная
рецепторная клетка (ОРК)
Рис. 13.7. Обонятельная лу-
ковица. (А) Рисунок показы-
вает обонятельные аксоны,
проходящие через решетча-
тую кость и оканчивающие-
ся в гломерулах (клубочках)
обонятельной луковицы
(Б) Основные нейронные
связи обонятельной лукови-
цы млекопитающих. Слои,
снизу вверх: ВОН — слой во-
локон обонятельного нерва,
КС — клубочковый слой,
НСС — наружный сетевид-
ный слой, СМК — слой мит-
ральных клеток, ЗС — зер-
нистый слой, ОТ — обоня-
тельный тракт. Клетки: Гг —
глубокие клеТки-зерна,
Гп — поверхностные клет-
ки-зерна, М — митральные
клетки, ПГ — перигломеру-
лярные клетки, П — пучко-
вые клетки. Тормозные
клетки обозначены пункти-
ром. С упрощениями из
Shepherd, 1998. (В) Обоня-
тельные рецепторные клет-
ки (ОРК), специализиро-
ванные детектировать одну
или несколько сходных па-
хучих молекул, посылают
аксоны в одну и ту же гломе-
рулу. Объяснения в тексте
оканчиваются сложными синаптическими контактами с дендритами митраль-
ных клеток (названных так, потому что первым микроскопистам их тела на-
помнили епископскую митру), пучковыми и перигломерулярными клетками.
В обонятельной луковице кролика конвергенция составляет 1000 к 1, т. е. око-
ло 1000 обонятельных аксонов конвергируют на разветвлениях апикального
дендрита одной митральной клетки. Около 25 таких дендритов вместе с терми-
налями аксонов формируют сферическое образование, именуемое гломерулой
(обонятельный клубочек). Т. о., на одной гломеруле конвергирует около 25 000
13. Обоняние 259
обонятельных аксонов. В обонятельной луковице кролика около 2000 гломе-
рул. Между гломерулами расположены короткоаксонные перигломерулярные
клетки. Аксоны этих клеток соединяют их с одной гломерулой, а дендритное
дерево — с соседней. Есть надежные данные, говорящие о том, что перигломе-
рулярные клетки являются тормозными.
Глубже в обонятельной луковице, как показывает рис. 13.7 Б, образуют
слой тела митральных клеток. Между ними лежат тела клеток-зерен. Клетки-
зерна лежат глубже тонкого митрального слоя и заполняют центральную часть
луковицы. Зерна, как и и пергломерулярные клетки, являются тормозными.
В гл. 16 мы увидим, что такая стратификация клеточной организации обоня-
тельной луковицы демонстрирует сходство с сетчаткой. Основа вертикальной
передачи информации — это аксоны митральных и, в меньшей степени, пуч-
ковых клеток, которые проходят два уровня горизонтальной модуляции. Од-
нако в гл. 16 мы увидим, что сетчатка значительно более сложна.
Детальная электронная микроскопия обонятельной луковицы показыва-
ет сложность синаптических контактов. Нейрохимия их не менее сложна.
Здесь идентифицировано около дюжины медиаторов, включая ацетилхолин,
дофамин, у-аминомасляную кислоту (ГАМК) и несколько нейроактивных
пептидов.
Есть ли доказательства того, что ОРК, специализированные реагировать на
одни и те же запахи, проецируются в одни и те же гломерулы? Похоже, что от-
вет на этот вопрос становится все более утвердительным. В последние годы на
обонятельной системе применены сложные методики биохимической и моле-
кулярно-биологической гистологии. Пероксидаза хрена (ПХ), инъецирован-
ная в специфические гломерулы, ретроградно транспортировалась в специфи-
ческую популяцию ОРК в обонятельном эпителии. С другой стороны, мРНК,
кодирующая специфические рецепторные молекулы ОРК обнаруживается в
их аксонах и окончаниях в гломерулах. Показано, что мРНК обнаруживается
лишь в немногих гломерулах или даже только в одной. Из этого вытекает, что
аксоны из ОРК, экспрессирующие такой рецептор, конвергируют на одной
гломеруле или небольшом их числе. Таким образом, хотя ОРК, отвечающие на
небольшой класс сходных пахучих веществ или единственное вещество, могут
быть распределены сравнительно беспорядочно в зонах экспрессии обоня-
тельного эпителия, их аксоны конвергируют на небольшом числе гломерул
или даже на одной (рис. 13.7 В). Более того, анатомическое расположение та-
ких гломерул идентично у разных животных одного и того же вида. Таким об-
разом, данный запах вызывает сходные паттерны активности в гломерулярной
популяции обонятельной луковицы у всех представителей данного вида. По-
скольку разные компоненты запаха активируют различные популяции ОРК,
из совокупность является сложной и уникальной. Такие совокупности могут
быть визуализированы с помощью гистохимии 2-дезоксиглюкозы. Порази-
тельно, что запахи, как и аффии^ятация других дистантных чувств, в централь-
ной нервной системе могут кодироваться пространственной картиной распре-
деления возбуждения в соответствующей проективной зоне.
Из луковицы обонятельная информация передается в мозг аксонами мит-
ральных (и пучковых) клеток, образующих обонятельный тракт. Как показы-
вает рис. 13.8, обонятельный тракт проходит под лобной долей и оканчивается
260 Часть III. Хемочувствительность
Таламус
(медиально-дорзальный)
Латеральный гипоталамус
Пириформная кора
(uncus — крючок)
Рис. 13.8. Центральные проекции обонятельного тракта. Объяснения в тексте.
в одной из эволюционно наиболее древних частей коры — пириформной доле
(или uncus)1. Следующий нейрон передает обонятельную информацию в тала-
мус, а последнее волокно передает ее в орбито-фронтальную новую кору. По-
мимо этого таламического пути, группа волокон проецируется через uncus
в латеральный гипоталамус и отсюда в орбито-фронтальную новую кору. Воз-
можно, именно этот путь запускает эмоциональные реакции, которые вызы-
вают знакомые запахи и ароматы.
13.3. Вомероназальный орган позвоночных и феромоны
Положение вомероназального органа у мыши показано на рис. 13.9. Это пар-
ный слепой канал под носовой полостью. Он развивается у воздуходышащих
наземных четвероногих, включая человека, и отсутствует у рыб. В нем присут-
ствует обонятельный эпителий, который на клеточном уровне похож на тот,
что покрывает раковины полости носа. Обонятельные рецепторные клетки
(ОРК) — нейросенсорные клетки — поддерживаются эпителиальными и ба-
зальными клетками эпителия. ОРК, как и в носовой полости, регенерируют в
течение всей жизни, но вместо ресничек у ОРК развиваются микроворсинки
(«микровиллы»).
Молекулярная биология ОРК вомероназального органа существенно отли-
чается от ОРК носовых ходов. Как и другие ОРК, они имеют полный набор се-
мидоменных рецепторных молекул, но их аминокислотная последователь-
ность не имеет сходства с рецепторами других ОРК. Повидимому, эти клетки
имеют свою долгую независимую историю. Недавно было показано, что в сли-
зистой вомероназального органа присутствуют два различающихся семейства
семидоменных рецепторов, каждое из которых включает от 100 до 200 генов и
развивалось, по-видимому независимо. Для представителей одного из них ти-
пичен длинный внеклеточный N-концевой домен, сходный с метаботропным
глютаматным рецептором (мГлюР). Напомним, что в разделе 12.2.1 мы уже
сталкивались с рецептором сладкого 7±к1, который также имеет сходство с
мГлюР. G-белковая сигнальная система, представленная в ОРК носовых хо-
дов (см. раздел 13.2.3), в клетках вомероназального органа не экспрессируется.
1 Uncus — крючок (лат.). — Прим, перев.
13. Обоняние 261
Рис. 13.9. Вомероназальная система мыши. (А) Парасагиттальный срез носового и
вомероназального органов. (Б) Срез в положении, показанном на рис. (А). (В) Ги-
стология вомероназального эпителия. ДОЛ — дополнительная обонятельная луко-
вица, ОЛ — обонятельная луковица, ОЭ — обонятельный эпителий раковин,
ВНО — вомероназальный орган. Перепечатано из Е. R. Liman, 1996, «Pheromone
transduction in vomeronasal organ», Current Opinion in Neurobiology, 6, p. 487, с разре-
шения Elsevier Science.
ОЭ /
ВНО
Полость
вомеро-
назального
органа
Опорные
клетки
Базальные
клетки
Сенсорные
нейроны
В момент написания этой книги биохимия передачи сигнала еще не опреде-
лена, однако, в отличие цАМФ-зависимой системы носовой полости, в во-
мероназальных ОРК действует система ИТФ. Это отличие, в дополнение к
несходству семидоменных рецепторных белков, наталкивает на мысль, что
вомероназальная и носовая обонятельные системы возникли в эволюции не-
зависимо. Поскольку у рыб вомероназальный орган не обнаружен, возмож-
но, что исходно два этих типа эпителия были перемешаны, а разделились
только в ходе эволюции назменых позвоночных.
Змеи пользуются вомероназальным органом при преследовании жертвы.
То, что вомероназальный орган участвует в детектировании феромонов позво-
ночных, подозревали уже давно. Самый яркий пример восприятия феромонов
у млекопитающих получен в исследованиях полового поведения золотистых
хомячков. Садка самца запускается феромонами из вагинального секрета са-
мок. Включение полового созревания ускоряется мочой самцов и тормозится
мочой самок. Предполагается, что эти и другие запускаемые феромонами по-
веденческие и физиологические реакции опосредованы вомероназальным
органом. Исследования в настоящее время развиваются, и специфические
молекулы феромонов еще предстоит охарактеризовать.
Афферентные волокна из вомероназального эпителия проецируются в до-
полнительную обонятельную луковицу, у большинства млекопитающих рас-
положенную кзади от каудальной по отношению к основной обонятельной
луковице (рис. 13.9). Интересно отметить, что как и обонятельный эпителий
носовых ходов, вомероназальный эпителий подразделяется на зоны. Это под-
разделение обозначено экспрессией разных G-белков в апикальной и базаль-
ной частях органа. Эти зоны сохраняются и в проекциях в дополнительную
обонятельную луковицу. Апикальные ОРК проецируются в переднюю, а ба-
262 Часть III. Хемочувствительность
зальные — в заднюю часть луковицы. Нервные окончания находятся в гломе-
рулах, которые однако менее выражены, чем в основной обонятельной луко-
вице. Вероятно, между луковицами есть существенных г^ница в представи-
тельстве. В отличие от пространственной карты, характерной для основной
луковицы, в дополнительной луковице представительство более сложное и
мозаичное. Его изучение — это предмет исследований, использующих все
современные методики молекулярной нейробиологии. Предполагается, что
сложная природа представительства может быть связана с биологической
функцией системы. Возможно, дополнительная луковица призвана реагиро-
вать исключительно на особые видоспецифические сочетания веществ соот-
ветствующего феромона и просто игнорировать все остальные. Проекции из
дополнительной луковицы, опять-таки, отличаются от таковых основой луко-
вицы. Вместо проекций в кору, дополнительная луковица связана исключи-
тельно с лимбической системой — миндалиной и гипоталамическими ядрами.
Известно, что эти ядра играют важную роль в половом и репродуктивном по-
ведении. И еще раз — нейроанатомия согласуется с эмоциональной реакцией
на «что-то такое в воздухе» — сигнал от окружающей среды.
13.4. Заключение
Этой главой мы завершаем Часть III. Хотя хемочувствительность представле-
на у человека не так обширно, как механочувствительность (особенно фоноре-
цепция) или фоточувствительность (зрение), у других видов она играет глав-
ную роль. Мы отметили ее значение у насекомых, чей мир, как мы видели, пе-
реполнен химическими сигналами. Достаточно одной молекулы феромона,
чтобы тутовый шелкопряд узнал о присутствии поблизости полового партне-
ра. Социальных насекомых связывает общение с помощью семиотических
веществам. Мы уже отмечали в конце Части II, что детекторы, изначально
возникшие для восприятия сигналов от абиотического мира, преобразуются
в средство коммуникации между живыми формами. Такая коммуникация
между индивидами, конечно, не ограничивается насекомыми — она широко
распространена в царстве животных. Кто не знает, как домашний пес метит
свою территорию? И человек здесь — не исключение. Хотя тонкость нашей хе-
мочувствительности не выдерживает критики по сравнению с многими други-
ми животными, тем не менее дурные запахи и привлекательная парфюмерия
играют в нашей жизни значительную роль. Опять-таки, как и в отношении
вкуса, немалые коммерческие интересы завязаны вокруг нашего обоняния.
Интерес, проявляемый к вомероназальному органу в последнее время, пока-
зывает, что не только тутовый шелкопряд активизируется феромонами, испу-
скаемыми особями противоположного пола.
МАТЕРИАЛЫ К ЧАСТИ III
Для самоподготовки
Нижеследующие вопросы направлены на то, чтобы помочь пониманию пред-
мета Части III, а также обратить внимание на наиболее существенные аспекты
рассматриваемых проблем.
Глава 10: Хемочувствительность у прокариот
10.1. Опишите движение жгутиков прокариот. Чем оно отличается от движения
жгутиков эукариот?
10.2. Каким образом направление вращения жгутиков отражается на движении
Е. coin Какое направление — по или против часовой стрелки — приводит к ха-
отическому движению?
10.3. Как изменение числа «кувырков» за единицу времени сказывается на переме-
щении бактерии в направлении к источнику химического аттрактанта?
10.4. Опишите биохимические механизмы, с помощью которых Е. coli детектирует
молекулы аттрактанта или репеллента.
10.5. Как можно продемонстрировать сенсорную адаптацию?
10.6. Какую роль играют связывающие белки? Загляните в гл. 13 и обсудите анало-
гии между схожими молекулами в сенсиллах насекомых и в обонятельной сли-
зи млекопитающих.
10.7. Какой эффект оказывает фосфорилирование CheY на вращение жгутиков?
10.8. Каким образом прекращается действие CheY на мотор жгутика?
10.9. Почему Р-П-белки иногда называются МАХ?
10.10. Как блокада метилирования доменов Р-П-белков влияет на движение бакте-
рий? Объясните, как это связано с сенсорной адаптацией.
10.11. Есть ли свидетельства того, что хеморецепторные системы бактерий и млеко-
питающих имеют общее происхождение?
Глава 11: Хемо-интерорецепторы млекопитающих
11.1. Какие эволюционные преимущества получает животное вмест_способнос-
тью поддерживать стабильность «внутренней среды»?
11.2. Объясните, как ПдСО2 связано с pH ВКЖ.
11.3. Где локализованы хеморецепторы, чувствительные к ПдСО2 и ПдО2?
11.4. Обсудите эволюционное происхождение каротидного лабиринта.
11.5. Какие клетки каротидного тельца содержат медиаторные пузырьки? Какие ме-
диаторы в них содержатся?
11.6. Какой из периферических хеморецепторов чувствителен к дефициту кислоро-
да при анемии?
11.7. Как изменяется скорость дыхания при снижении ПдО2 и(или) повышении
ПдСО2?
11.8. Почему трудно определить рефлекторные ответы, вызванные «каротидными
тельцами».
11.9. Объясните биохимическую последствия падения ПдО2 в клетках типа 1 каро-
тидного тельца.
11.10. Опишите биохимические последствия повышения ПдСО2 в клетках типа 1.
264 Часть III. Хемочувствительность
Глава 12: Вкус
12.1. В чем разница между вкусом и обонянием?
12.2. Опишите, как рефлекс удлиннения хоботка используется в исследовании вку-
са у насекомых.
12.3. Где у двукрылых насекомых концентрируются хемочувствительные волоски?
12.4. Опишите основные моменты биофизики хемочувствительности сенсилл насе-
комых. Каким образом механизм электротонического распространения схож с
проведением в миелинизированных аксонах позвоночных?
12.5. Перечислите основные сенсорные качества вкуса у человека. Как они связаны
с потребностями существования в биологическом мире?
12.6. Как вкусовые почки распределены по поверхности языка и как они иннерви-
рованы?
12.7. Нарисуйте схему, объясняющую клеточную структуру типичной вкусовой
почки человека.
12.8. Опишите молекулярную биологию одного из типов вкусовых рецепторных
клеток.
12.9. Каким образом молекулярно-биологические исследования клеток кишки
крысы смогут помочь в понимании работы рецепторных клеток, чувствитель-
ных к соленому ?
12.10. Опишите, как рецептор соленого контролируется гормонально.
12.11. В чем молекулярная биология рецептора горечи сходна и в чем отличается
от палочковых и колбочковых фоторецепторов? (загляните также в раздел
16.2.1)?
12.12. Где располагаются корковые вкусовые области?
12.13. Объясните коммерческое значение исследований по биологии вкуса.
Глава 13: Обоняние
13.1. Как и почему гигрорецепторы отличаются от рецепторов воды?
13.2. Как изменения влажности детектируются гигрометрическими сенсиллами на-
секомых? Влияет ли механизм работы сенсиллы на ее расположение и морфо-
логию?
13.3. Содержат ли гигрорецептивные сенсиллы в норме более одной нейросенсор-
ной клетки? Если да, то какая другая сенсорная модальность здесь представ-
лена?
13.4. Каким образом первая стадия обонятельного рецепторного механизма у насе-
комых схожа с бактериальной хеморецепцией?
13.5. Где располагаются хеморецепторные сенсиллы Drosophila^ Как их пространст-
венная организация отличается от хеморецепторов обонятельной слизистой
млекопитающих?
13.6. В какую часть мозга насекомых проецируются обонятельные волокна и в ка-
кой структуре они оканчиваются?
13.7. Что означает понятие «семиотические вещества»? Приведите несколько при-
меров.
13.8. Опишите обонятельную систему шелкопряда.
13.9. Что такое раковины носовых ходов и какую функцию они выполняют в обоня-
нии?
13.10. Имеет ли обонятельный эпителий топографическую организацию и, если да,
какую форму она принимает?
13.11. Опишите две черты, которыми обонятельные рецепторные клетки отличаются
от вкусовых.
13.12. Опишите конформацию обонятельного рецепторного белка. Как, по предпо-
ложениям, они распознают молекулу одоранта?
13.13. Нарисуйте схему, иллюстрирующую молекулярную биологию преобразования
обонятельного стимула в деполяризацию обонятельной рецепторной клетки.
Материалы к части III 265
13.14. Объясните молекулярную биологию сенсорной адаптации и градуального от-
вета обонятельной клетки.
13.15. Есть ли в обонятельнной системе аналогии с наследуемыми дефектами цвето-
восприятия в сетчатке?
13.16. С использованием материала гл. 16 объясните сходство между организацией
обонятельной луковицы и сетчатки.
13.17. Какие существуют доказательства того, что обонятельные гломерулы (клубоч-
ки) специализированы к специфическим запахам?
13.18. В какую часть мозга мозга направляется обонятельная информация?
13.19. В каких отношениях вомероназальный эпителий сходен, а в каких отличен от
эпителия слизистой носа?
13.20. К детектированию каких типов молекул-одорантов специализирован вомеро-
назальный орган?
13.21. Опишите афферентный путь от вомероназального органа к мозгу. Почему ме-
сто назначения этих волокон существенно?
Замечания и библиография
Введение
Айстен (Eisthen, 1997) рассматривает эволюцию обонятельной системы позво-
ночных с особым акцентом на происхождение вомероназального органа чет-
вероногих, морфологию рецепторных клеток и эволюцию центральных про-
екций. Whitear (1992) и Finger (1997) сделали обзоры структуры, функции и
центральных связей хеморецепторных клеток солитарного ядра позвоночных,
a Prasad & Reed (1999) обсуждают прогресс, достигнутый в изучении хеморе-
цепторной системы С. elegans с помощью методов современной генетики.
Eisthen, Н. L., 1997, «The evolution of vertebrate olfactory systems». Brain, Behaviour and
Evolution, 50, 222-233.
Finger, T. E., 1997, «Evolution of taste and solitary chemoreceptor cell systems», Brain,
Behaviour and Evolution, 50, 234—243.
Prasad, В. C. and R. R. Reed, 1999, «Chemosensation: molecular mechanisms in worms
and mammals», Trends in Genetics, 15, 150—153.
Whitear, M., 1992, «Solitary chemoreceptor cells», in Fish Chemoreception (2nd edn),
T. J. Hara, ed., Dordrecht: Elsevier.
Глава 10: Хемочувствительность у прокариот
Детальное описание бактериальной хемочувствительности сделано в нижесле-
дующих публикациях:
Adier, J., 1976, in R. Goldman, T. Pollard and J. Rosenbaum, eds., Cell Motility, Cold
Spring Harbour, NY: Cold Spring Harbour Laboratory.
Adler, J., 1983, «Bacterial chemotaxis and molecular neurobiology». Cold Spring Harbor
Symposia in Quantitative Biology, XLVIII(yo\. 2), 803—804.
Bourrett, R. В., K. A. Borkovich and M. I. Simon, 1991, «Signal transduction pathways
involving protein phosphorylation in prokaryotes». Annual Review of Biochemistry, 60,
401-441.
266 Часть III. Хемочувствительность
Koshland, D. E. Jr., 1981, «Biochemistry and adaptation in a simple bacterial system».
Annual Review of Biochemistry, 50, 765—782.
Manson, M. D., 1992, «Bacterial motility and chemotaxis», Advances in Microbial Physio-
logy, 33, 277-346.
Schuster, S. C. and S. Khan, 1994, «The bacterial flagellar motor», Annual Review of Biophy-
sics and Biomolecular Structure, 23, 509—539.
Simon, M. I., A. Krikos, N. Mutoh and A. Boyd, 1985, «Sensory transduction in bacteria»,
Current Topics in Membranes and Transport, 23, 3—15.
Глава 11: Хемо-интерорецепторы млекопитающих
Выжимки из фундаментальных Legons (Лекции) Клода Бернара, остающихся
до сих пор важным источником, можно найти у Fulton (1966) и Cannon (1932).
Сложная нейрофизиология поддержания уровня дыхательных газов в ВКЖ
рассматривается в учебниках физиологии. Детали о периферических хемо-
рецепторах каротидного тельца есть в публикациях, приведенных ниже.
Bernard, С., 1878, Leqonssur les Phenomenesde la Vie communsauxAnimauxetaux Vegetaux
(ed. A. Dastre), Paris, vol. 1, p. 112.
Biscoe, T. J., M. J. Purves and S. R. Sampson, 1970, «The frequency of nerve impulses in
single carotid body chemoreceptor afferent fibres recorded in vivo with intact circula-
tion», Journal of Physiology, 208, 121—131.
Cannon, W., 1932, The Wisdom of the Body, London: Kegan Paul, Trench, Trubner & Co.
Ltd.
Fulton, J., 1966, Selected Readings in the History of Physiology, Illinois: С. C. Thomas.
Gonzalez, C. et al., 1992, «Oxygen and acid chemoreception in the carotid body chemo-
receptors», Trends in Neurosciences, 15, 146—153.
Gonzalez, C. et al., 1994, «Carotid body chemoreceptors: from natural stimuli to sensory
discharges», Physiological Reviews, 74, 829—898.
Grove, A. J. and G. E. Newell, 1945, Animal Biology, London: University Tutorial Press.
Haddad, G. and C. Jiang, 1997, «O2-sensing mechanisms in excitable cells: role of plasma
membrane K+ channels», Annual Review of Neurosciences, 59, 23—43.
Marshall, J. M., 1994, «Peripheral chemo receptors and cardiovascular regulation»,
Physiological Reviews, 74, 543—594.
McDonald, D. M. and R. W. Mitchell, 1975, «The innervation of ganglion cells, glomus
cells and blood vessels in the rat carotid body», Journal of Neurocytology, 4, 177.
Глава 12: Вкус
Пфаффман впервые ввел концепцию кодирования вкусовой чувствительнос-
ти профилями нейронных ответов (суммирующий обзор Pfaffman, 1959); Erik-
son (1982) обобщил эту концепцию применительно к некоторым другим сен-
сорным модальностям; Frank et al. (1988) описал эксперименты на 40 смешан-
ных веточках лицевого нерва хомячка, упомянутые в тексте. Lindemann (1996)
написал свежий обзор по вкусу у млекопитающих, a Hoon et al. (1999) впервые
сообщил о выделении семидоменных молекул рецепторов вкуса.
Bartoshuk, L. М, 1978, «Gustatory system», in Handbook of Behavioral Neurobiology, vol. 1:
Sensory Integration, R. B. Masterton, ed., New York: Plenum.
Erikson, R. P., 1982, «The “across-fiber pattern” theory: An organising principle for molar
neural function», Contributions to Sensory Physiology, 6, 79—110.
Материалы к части III 267
Frank, М. Е., S. L. Bieber and D. V. Smith, 1988, «The organisation of taste sensibilities in
hamster chorda tympani nerve fibers», Journal of General Physiology, 91, 861—896.
Henning, H., 1922, «Psychologische Studien am Geschmacksinn», Handbuch Biol. Arbeits-
meth., 6A, 627—640.
Hoon, M. A. et al., 1999, «Putative mammalian taste receptors: a class of taste-specific
GPCRs with distinct topographic selectivity», Cell, 96, 541—551.
Kandel, E. R., J. H. Swartz, T. M. Jessell, 1991, Principles of Neural Science (3rd Edition),
New York: Elsevier.
Kerkut, G. A. and L. I. Gilbert, eds, 1985, Comprehensive Insect Physiology, Oxford: Claren-
don Press, vol. 6: Nervous System: Sensory.
Keverne, E. B., 1982, «Chemical senses: Taste», in The Senses, H. B. Barlow and
J. D. Mollon, eds. Cambridge: Cambridge University Press.
Lindemann, B., 1996, «Taste Reception», Physiological Reviews, 16, 719—766.
Morita, H. and A. Shiraisi, 1985, «Chemoreceptor physiology», in G. A. Kerkut and
L. I. Gilbert, eds, ibid., vol. 6.
Pfaffman, C., 1959, «The afferent code for sensory quality», American Psychologist, 14,
226-232.
Глава 13: Обоняние
Обзоры no молекулярной биологии сделаны Axel (1995) и Buck & Axel (1991);
детали структуры и функции обонятельной системы насекомых даны Altner &
Prillinger (1980), Altner & Loftus (1985) и Frazier (1985); Schneider (1977) и Mayer
& Mankin (1985) обсуждают феромоны насекомых, a Rospars & Hildebrand
(1992) излагают детали гломерулярной структуры антеннальных долей шелко-
пряда. Stocker (1994) и Carlson (1996) обсуждают обонятельную систему наибо-
лее известного насекомого — дрозофилы, a Vossall et al. (1999) описывает при-
роду и распределение ее обонятельных рецепторных молекул. Обзор послед-
них данных по обонятельным клеткам позвоночных сделан Schild & Restrepo
(1998); Buck (1996) и Shepherd (1998) капитально обсудили обонятельную сис-
тему позвоночных в целом, a Malnic et al. (1999) описывает некоторые детали,
касающиеся обонятельной слизистой. Обзоры по феромонам позвоночных и
вомероназальному органу сделан Leman (1996), Matsunami & Buck (1997),
Tirindelli et al. (1998). Bargmann (1999) обсуждает свежие работы по сенсорно-
му представительству в дополнительной обонятельной луковице. Полезный
общий обзор обонятельных систем в разных типах животных можно найти у
Hildebrand & Shepherd (1997).
Altner, Н. and L. Prillinger, 1980, «Ultrastructure of invertebrate chemo-, thermo- and
hygroreceptors and its functional significance», International Review of Cytology, 67,
69-139.
Altner, H. and R. Loftus, 1985, «Ultrastructure and function of insect thermo- and hygrore-
ceptors», Annual Review of Entomology, 30, 273—295.
Axel, R., 1995, «The molecular logic of smell», Scientific American, 273(4), 130-137.
Bargmann, C. L, 1999, «А complex map for pheromones», Neuron, 22, 640—642.
Buck, L. D. and R. Axel, 1991, «А novel multigene family may encode odorant receptors:
a molecular basis for odor recognition», Cell, 65, 175—187.
Buck, L. D., 1996, «Information coding in the vertebrate olfactory system», Annual Review
of Neuroscience, 19, 517—544.
Carlson, J. R., 1996, «Olfaction in Drosophila', from odor to behavior», Trends in Genetics,
12, 175-180.
268 Часть III. Хемочувствительность
Frazier, J. L., 1985, «Nervous system: Sensory system», in Fundamentals of Insect Physiology,
M. S. Blum, ed., New York: Wiley.
Hildebrand, J. G. and G. M. Shepherd, 1997, «Mechanisms of olfactory discrimination:
converging evidence for common principles across phyla», Annual Review of Neuro-
science, 20, 595—631.
Leman, E. R., 1996, «Pheromone transduction in the vomeronasal organ», Current Opinion
in Neurobiology, 6, 487—493.
Malnic, B. etal., 1999, «Combinatorial receptor codes for odors», Cell, 96, 713—723.
Matsunami, H. and L. B. Buck, 1997, «Multigene family encoding a diverse array of puta-
tive pheromone receptors in mammals», Cell, 90, 775—784.
Mayer, M. S. and H. W. Mankin, 1985, «Neurobiology of pheromone perception», in
Comprehensive Insect Physiology, Biochemistry and Pharmacology, G. A. Kerkut and
L. I. Gilbert, eds, Oxford: Pergamon Press.
Rospars, J. P. and J. G. Hildebrand, 1992, «Anatomical identification of glomeruli in the
antennal lobes of the male sphinx moth, Manduca sexta», Cell Tissue Research, 270,
205-227.
Schild, D. and D. Restrepo, 1998, «Transduction mechanisms in vertebrate olfactory recep-
tor cells», Physiological Reviews, 78,429—466.
Schneider, D., 1977, «The sex-attractant receptor of moths», in The Insects, T. Eisner and
E. O. Wilson, eds, San Francisco: W. H. Freeman.
Shepherd, G. M., 1998, The Synoptic Organisation of the Brain (4th edn), Oxford: Oxford
University Press.
Stocker, R. F., 1994, «The organisation of the chemo-sensory system in Drosophila melano-
gaster: a review», Cell Tissue Research, 275, 3—26.
Tirindelli, R., C. Mucignat-Caretta and N. J. P. Ryba, 1998, «Molecular aspects of
pheromonal communication via the vomeronasal oigan of mammals», Trends in Neuro-
sciences, 21, 482—486.
Vossall, L. B., 1999, «А spatial map of olfactory receptor expression in the Drosophila anten-
na», Cell, 96, 725-736.
Часть IV
СВЕТОЧУВСТВИТЕЛЬНОСТЬ
«Рассмотрите, анатомируйте глаз; исследуйте его устройство и
изобретательность, с которой он устроен; и скажите мне, не по-
чувствовали ли вы, что мысль о его изобретателе поразила вас как
сенсация.»
Дэвид Юм, 1751/1779,
Диалоги о естественной религии,
Лондон
Чарльз Дарвин вспоминал, что когда-то одна мысль о глазе заставляла его по-
холодеть. Глаз всегда был примером точного соответствия средства поставлен-
ной цели в спорах по поводу устройства. Как такое могло возникнуть в ходе
эволюции? Как что-то не столь всесторонне совершенное, как некая промежу-
точная стадия, какой-то «полу-глаз» мог работать? Глаз всегда казался теоло-
гам-естественникам окончательным и неопровержимым аргументом их пози-
ции. Но не спешите с выводами! Глаз человека, глаз позвоночного — это дале-
ко не единственные глаза в царстве животных. Мы увидим на страницах этой
части книги, как много вариантов глаза развилось в эволюции. Во всем мире
живого развились устройства, которые воспринимают электромагнитное из-
лучение звезды, освещающей Землю.
Вселенная заполнена электромагнитным излучением. Длины волн это-
го излучения (а, следовательно, и энергии фотонов) широко варьируют
(рис. IV. 1 А). Лишь небольшой участок электромагнитного спектра сущест-
венным образом достигает поверхности нашей планеты (рис. IV. 1 Б). Макси-
мальная энергия солнечного излучения, достигающего поверхности Земли,
имеет длину волны около 500 нм (57 ккал/моль фотонов). Более высоко энер-
гетические фотоны ультрафиолетовой области (< 300 нм) поглощаются озоно-
вым (О3) слоем атмосферы Земли. Точно также, но со смертельным исходом,
ультрафиолет поглощается белками, так что не следует забывать о роли наше-
го озонового одеяла.
Реакция молекулы на свет зависит от ее атомарной структуры, в частности
электронной конфигурации. Наиболее важные молекулярные структуры, кото-
рые сформировались у живых организмов для реакции на свет, это пиррольные
кольца и каротиноиды (рис. IV.2). Четыре пиррольных кольца входят в состав
великолепной плоскостной молекулы порфирина, которая образует фотон-
захватывающие центры хлорофиллов. Каротиноиды — это желтые, оранжевые
и красные жирорастворимые пигменты, образующиеся в биологическом мире,
близкие к витамину А (ретинолу) и через него — к очень важному хромофору
ретиналю. Ретиналь, как показывает рис. IV.2, это альдегид витамина А. Боль-
шинство беспозвоночных способно синтезировать витамин А, но позвоноч-
270 Часть IV. Светочувствительность
А ф.с.
с.з.
4Q0 450 500
550 600 650 700 750
2,500
Инфракрасное излучение
। । । .
200 300 400 500 600 700 800 900 !1.000 1,100 1,2001,300
Ультрафиолетовое 5 Видимый
излучение | J свет
Космические I Г~~-------------------------------------------------'
лучи у - и рентгеновские лучи") ;« ( Радиоволны Электроволны
0.0001А O.OOlA 0.01А 0.1А lA 10А 100А 1000А 1мкм Юм км .1мм 1мм 1см 10см 1м Юм 100м 1км 10км 100км
10А = 1.0 нм = 10*3 нм = 10-7 см
Б
Длина волны (нм)
Рис. ГУЛ. (А) Электромагнитный спектр. (Б) Солнечный спектр, достигающий по-
верхности Земли между 13.00 и 14.00 в центральной Франции.
ные эту способность утратили. По этой причине витамин А является обяза-
тельной составляющей диеты этих животных. Как видно из рис. IV.2, суще-
ствует две формы этого витамина — Aj и А2, различающиеся двойной кова-
лентной связью между 3-м и 4-м атомами в шестичленном кольце. Из этого
следует, что существуют и две формы ретиналя — ретинал^ и ретиналь2. Кро-
ме того, третья форма ретиналя — 3-оксиретиналь обнаружена у некоторых
насекомых (особенно у двукрылых и чешуекрылых) и некоторых кальмаров.
Во всем мире животных фотопигменты состоят из хромофора — ретиналя,
непрочно связанного связью, называемой шиффовым основанием, с апопро-
теином — опсином. Опсины — семидоменные «змеевидные» белки с молеку-
лярными весом около 40 кДа (рис. IV.3). Они выделены и проанализированы у
многих позвоночных и беспозвоночных животных, и хотя несколько различа-
ются по величине, все имеют ту же самую семидоменную структуру. Они при-
надлежат к большому надсемейству эволюционно родственных белков, кото-
рые включают к тому же не только обонятельные рецепторные белки (гл. 13),
но и большое количество синаптических рецепторных молекул. В последние
годы показано, что не только пигмент меланофоров — меланопсин — имеет та-
Часть IV. Светочувствительность 271
Б
сн3 снэ
н’с\ /СНэ н I н I
rbUd R и и и и и
z|4 || Н Н Н Н
нгк 2 >С
сн3
н2
Ретинапь
Витамин А2
Рис. IV.2. Молекулярная структура каротина и его фотоактивных производных.
(А) лирянс-р-каротин. (Б) Витамин АР Отметить, что это продукт гидролиза р-ка-
ротина. (В) Двойная связь между С3 и С4 образует витамин А2. Ретиналь — это аль-
дегид витамина А (ретинола). (Г) П-цис-ретинал^ (обратите внимание на при-
нятую терминологию для структурных изомеров (цис- и транс-). (Д) «Шиффово
основание» — двойная связь атома углерода ретиналя! с атомом азота лизина —
аминокислоты опсина.
кую же молкулярную структуру, но в форме бактериородопсина обнаружен и в
бактериях (Информация IV. 1). Это важная общая закономерность: найденное
эффективное решение проблемы (в данном случае — улавливания света), за-
тем используется снова и снова для той или иной цели. Основные моменты
эволюции опсинов даны в гл. 14, Информация 14.1.
Поглощение света — всего лишь первый этап фоторецепции. Ощущение
должно быть основой различения, и для этого сигнал должно быть передан
сначала в клетку, а затем в более сложные системы ЦНС. Семидоменная
структура опсина — это ключ к ситуации. Как мы отмечали в гл. 1 и при обсуж-
дении хеморецепции в Части III, эти мембранно-связанные белки обычно яв-
ляются частью G-белковой сигнальной системы. Опсин — не исключение.
В некоторых отношениях фоторецепция может рассматриваться как специа-
лизированная форма хеморецепции. Мы видели в Части III, что одоранты или
вкусовые молекулы вызывают изменения конформации мембранно-связан-
ного семидоменного рецептора, в результате чего а-субъединицы G-белков
диссоциируют и активируют мембранно-связанные ферменты. Именно так
272 Часть IV. Светочувствительность
Б
Рис. IV.3. Молекулярная структура родопсина быка. На плоской схеме (А) показаны
семь трансмембранных спиралей. К обозначенныму черным и буквой К остатку лизина
присоединяется ретиналь. N-конец опсина содержит две олигосахаридных группы (обо-
значены шестиугольниками), а заштрихованные остатки на С-конце фосфорилируются
родопсин-киназой (см. гл. 16). (Б) Трехмерная конформация родопсина в клеточной
мембране.
Часть IV. Светочувствительность 273
Рис. IV.4. (А) Тень от предмета
при освещении точечным ис-
точником света. (Б) Изображе-
ние за отверстием.
все происходит и с опсином. Однако в данном случае одорант или вкусовая
молекула как бы уже присоединены к семидоменному рецептору — их роль
выполняет ретиналь. Фотон всего лишь меняет конформацию ретиналя таким
образом, что он больше не может занимать свое место в молекуле опсина. Это
в свою очередь изменяет конформацию опсина, что и приводит к отделению
соседней а-субъединицы G-белка. Это активирует специфический фермент в
мембране фоторецепторной клетки. Мы рассмотрим этот процесс более де-
тально в гл. 16, когда перейдем к молекулярной биологии и биофизике наруж-
ных сегментов палочек и колбочек позвоночных.
До сих пор мы рассматривали молекулярные механизмы, развившиеся в
живых организмах для обнаружения света. Однако, организмы решают куда
более сложные задачи, чем просто детектирование наличия или отсутствия
света. Они используют физические характеристики света для получения ин-
формации об окружающей среде — биотической и абиотической, в которой
организмы ведут борьбу за выживание. Рассмотрим некоторые существенные
черты этого процесса.
Во-первых, тот факт, что длина волны света в видимом спектре так мала,
обеспечивает движение света по прямым линиям, и края объектов, а также те-
ни, которые они отбрасывают, имеют четкие границы (сравните с намного ме-
нее четкими тенями и краями в акустическом мире). Другим следствием пря-
молинейности распространения света является инвертирование изображения
за отверстием. В результате изображение получается перевернутым, в отличие
от тени предмета (рис. IV.4).
Часть IV. Светочувствительность
Информация IV. 1
БАКТЕРИОРОДОПСИН
Мембрана Halobacterium halobium — бактерии соленых вод, принадлежащей к
Archebacteria, может быть разделена на три фракции: желтую, красную и пурпур-
ную. Пурпурная фракция образует кристаллы, которые можно исследовать с помо-
щью электронного микроскопа. Помимо липидов она содержит белок весом
26 кДа. Поскольку фракция образует плоские кристаллы, структуру белка можно
исследовать в атомарных деталях. Из-за светочувствительности и семидоменной
«змеевидной» структуры эти белки называют бактериородопсином. В настоящее
время он изучен лучше всех родопсинов. Как и все другие родопсины, он состоит
из семи плотно упакованных а-спиралей, пронизывающих мембрану. Как и другие
родопсины, этот белок непрочно связан с хромофором ретиналем.
У галобактерий, таких как Н. halobium, АТФ образуется окислительным фосфо-
рилированием. При высоких напряженностях кислорода происходит переключе-
ние на фотосинтез. Протоны, необходимые для этого процесса, переносятся через
мембрану бактериородопсином, который действует как протонный насос, управля-
емый светом. Можно показать, что на каждый поглощенный фотон изнутри нару-
жу через мембрану переносятся два протона.
Механизм насоса явился предметом многочисленных исследований. Известно,
что ретиналь связан шиффовым основанием (как и в зрительных пигментах живот-
ных) с лизином 216 (К216) опсина и контактирует с шестью из семи трансмембран-
ных спиралей. Ретиналь имеет транс-конформацию и располагается в туннеле
между спиралями, блокируя, таким образом, поток протонов. В механизме прото-
нового насоса происходит преобразование транс-конформации в 13-1<мс-конфор-
мацию; ретиналь изгибается в опсиновом «туннеле» и переносит протон с цитопла-
заматической стороны на внеклеточную. В отличие от родопсинов сетчатки позво-
ночных, ретиналь не отсоединяется от опсинового апопротеина.
Аминокислотная последовательность бактериородопсина сильно отличается от
опсинов животных, что позволяет допустить их независимое эволюционное проис-
хождение. Это подтверждается тем, что ретиналь бактерии образует 13-цис-конфи-
гурацию, а не 11-цис-конфигурацию, как в опсинах животных. Тем не менее, сама
по себе конформация бактериородопсина показывает, что он принадлежаит к вез-
десущему надсемейству семидоменных «змеевидных» белков, которые вовлечены
в огромное число процессов в царстве животных.
Литература
Henderson, R., et al., 1989, «Model for the structure of bacteriorhodopsin based on high
resolution electron cryomicroscopy», Journal of Molecular Biology, 213, 899—929.
Имеющие доступ к Интернету найдут полезный обзор деталей структуры и функ-
ций по адресу:
http://monera.ncl.ac.uk/energy/brd.htm]
Часть IV. Светочувствительность 275
Рис. IV.5. (А) Второй закон рефракции: sin i/sin г = m — показатель рефракции.
(Б) Рефракция собирающей линзы. ГО — главная ось — линия, проходящая через
центры кривизны обеих поверхностей. Если луч света параллелен главной оси, то
он собирается в фокус в главном фокусе — ГФ. Расстояние от центральной точки
до главного фокуса определяется как фокальное расстояние f линзы.
Во-вторых, белый свет является, как известно, смесью всех длин волн ви-
димого спектра. Когда белый свет падает на отражающую поверхность, неко-
торые длины волн поглощаются ею, а некоторые — отражаются. Во многих
высокоразвитых фоторецепторах возникли средства различения излучений с
разной длиной волны. Такие животные способны ощущать цвета поверхнос-
тей в окружающей среде. Это может давать преимущества например, для раз-
личения спелости фруктов, в половом поведении, для маскировки и охоты.
В-третьих, не все поверхности отражают свет. Во многих случаях свет мо-
жет проходить сквозь материальное тело, такие тела прозрачны. Однако, ког-
да свет проходит из одной прозрачной среды в другую, его скорость меняется.
В результате, когда свет входит в более плотную среду, его направление смеща-
ется в сторону нормали (рис. IV.5 А). Это явление известно как светопрелом-
ление (рефракция). Законы рефракции рассматриваются во всех элементар-
ных учебниках физики. Рефракция имеет большое значение в формировании
изображения в глазу. Именно сила рефракции хрусталика и роговицы фокуси-
рует изображение объекта на сетчатке. Фокальное расстояние (I) рефракцион-
ной системы — это дистанция между центральной точкой системы и точкой
схождения лучей (рис. IV.5 Б). «Сила» хрусталика (линзы) или другой рефрак-
ционной системы определяется как 1/f и измеряется в диоптриях (D). Линза с
фокальным расстоянием в 1 м определяется как имеющая силу в 1 диоптрию,
а с фокальным расстоянием 25 см — 1/0,25 = 4 D. Собирающие линзы называ-
ются позитивными, а рассеивающие — негативными.
Наконец четвертое, естественный свет в той или иной степени поляризо-
ван. В идеальном случае электромагнитные колебания, образующие свет1,
имеют случайную ориентацию в пределах 360° окружности, ортогональной к
1 Испускаемый нагретым телом, например, Солнцем. — Прим. ред.
276 Часть IV. Светочувствительность
Рис. IV.6. Плоскополяризо-
ванный свет. Вместо колеба-
ний, ориентированных во всех
возможных направлениях, ор-
тогональных направлению лу-
ча (РО), на рисунке показаны
только два плоскости: АВ
и CD.
направлению луча. Однако, в ряде естественных ситуаций колебания в неко-
торых плоскостях избирательно поглощаются — атмосферными частицами
(эффект Тиндаля—Рэлея), или же при отражении и преломлении границах
раздела сред (эффект Френеля). Фоторецепторы многих животных способны
различать даже слабую поляризацию и использовать эту информацию в ориен-
тации для определении направления своего движения (гл. 20).
Теперь рассмотрим некоторые пути, по которым развивался глаз животно-
го, чтобы использовать все эти физические характеристики для формирования
изображения. Мы уже отмечали (рис. IV.4), что свет, проходя через отверстие,
образует за ним перевернутое изображение. Возможно, лучшим примером
этого способа является морской моллюск Nautilus (см. рис. 14.9). Глаза некото-
рых животных используют отражение; наиболее красивый пример этого —
гребешок Pecten (см. раздел 14.2.3), однако это явление используется и некото-
рыми ракообразными (раздел 14.1.2) и, конечно, многими позвоночными1
(в частности, многими рыбами (раздел 18.3) и млекопитающими (раздел 16.2.1
и т. д.)). Однако, большинство глаз животных, образующих изображение, по-
строено на рефракции, которая может происходить регулируемым образом в
хрусталике или нерегулируемым — в роговице. Изображение отбрасывается на
двумерное поле фоторецепторов, очень похоже на то, как линза фотокамеры
отбрасывает изображение на пленку. Есть здесь, однако, и существенное отли-
чие. В то время как пленка — плоская, сетчатка глаза животных всегда не-
сколько искривлена (рис. IV.7). Сетчатка может быть вогнутой, как в камер-
ном глазе, так и выпуклой — в фасеточном. В первом случае (у позвоночных и
головоногих) общая линза (хрусталик), расположенная перед сетчаткой, фо-
кусирует на ней перевернутое изображение. Во втором случае (в сложных гла-
зах членистоногих) изображение формируется множеством маленьких глазков
с линзочкой, каждый из которых получает узкий пучок света и смотрит в сво-
ем направлении. Обратите внимание, что в последнем случае мозаичное изоб-
ражение не инвертировано.
Прежде, чем мы закончим это Введение, стоит отметить еще один показа-
тель глаза, формирующего изображение — его способность различать детали
зрительного образа. Важным фактором, определяющим разрешающую способ-
1 Для увеличения светочувствительности, но не для получения изображения. — Прим. ред.
Часть IV. Светочувствительность 277
Рис. IV.7. Вогнутая и выпук-
лая сетчатки. (А) Вогнутая
сетчатка камерного глаза по-
звоночных или головоногих.
(Б) Выпуклая сетчатка глаза
членистоногих. Из Gold-
smith, 1990, с разрешения.
кость (или остроту зрения) является способность рецепторных клеток сетчат-
ки к угловому различению (ф). Разрешающая способность пропорциональна
1/ф. В случае одного хрусталика в глазу 1/ф = f/d, где f — фокальное расстояние,
ad — расстояние между рецепторами. Следовательно, разрешающая способ-
ность таких глаз может быть повышена либо увеличением f, либо уменьшени-
ем d, либо обоими способами сразу. Для сложного глаза аппозиционного типа
(см. раздел 14.1) опять-таки 1/ф = f/d, хотя в данном случае d — это диаметр
линзы омматидия. Эти соотношения показаны на рис. IV.8.
Выше мы отмечали, что разрешающая способность может быть повышена
за счет увеличения f или уменьшения d. Однако для этих параметров сущест-
вуют и некоторые ограничения. Величина d зависит от размера глаза, т. е. и са-
мого животного. Если мы посмотрим на сложный глаз стрекозы или камерный
глаз певчей птицы, то следует признать, что здесь это увеличение доведено до
Рис. IV.8. Острота зрения.
(А) Камерный глаз. (Б) Оди-
ночный рецептор. (В) Слож-
ный глаз. Объяснения в тек-
сте. dj — диаметр линзы,
dr — диаметр рецептора, f —
фокальное расстояние; ф —
угловое расстояние между
фоторецепторами; г — ради-
ус кривизны; s — линейное
расстояние между рецепто-
рами. Из Goldsmith, 1990,
с разрешения.
В
278 Часть IV. Светочувствительность
предела — в обоих случаях глаза настолько велики, насколько вообще их спо-
собна удержать голова. Аналогичным образом ограничена и степень, до кото-
рой может быть уменьшено d. В случае камерного глаза легко видеть, что диа-
метр наружных сегментов фоторецепторных клеток не может уменьшаться
беспредельно. Кроме того, наружные сегменты должны быть достаточно вы-
сокочувствительны, чтобы различать интенсивности освещенности изображе-
ния. Значит, разрешающая способность ограничена еще и контрастной чувст-
вительностью фоторецепторной клетки. Чтобы отличить одну часть изображе-
ния от другой, необходимо, чтобы соседние фоторецепторы испытывали раз-
личия интенсивности освещенности, превышающие порог их контрастной
чувствительности. Если они располагаются слишком близко друг к другу, кон-
траст окажется слишком мал, чтобы его можно было обнаружить. Физические
соображения такого типа ограничивают минимальный диаметр наружного
сегмента прибл. 1 мкм, а расстояние между ними — около 2 мкм. Сходные со-
ображения применимы и к сложному глазу — в этом случае контрастная чув-
ствительность ограничивает предел диаметра фасетки прибл. 10 мкм.
Из последующего изложения станет ясно, что изучение глаз животных свя-
зано с поразительным смешением физики, биохимии, молекулярной биоло-
гии, физиологии и зоологии. В первой главе этой части (гл. 14) мы рассмотрим
организацию глаз беспозвоночных. Далее, в центральных трех главах (15, 16
и 17) мы рассмотрим некоторые детали глаза позвоночных и зрительную сис-
тему в целом на примере человека. Затем, в гл. 18 мы продолжим рассмотрение
некоторых вариаций зрения у позвоночных.
14. ЗРЕНИЕ У БЕСПОЗВОНОЧНЫХ
Вездесущность и древность фоторецепторов: гомологии в раннем разви-
тии — общий предок? Два основных типа фоторецепторных клеток: рабдо-
мерные и ресничные — различие биофизики. Два основных типа глаз: ка-
мерный и фасеточный. Эволюция камерного глаза: глазное пятно — плос-
кий глаз — диафрагмальный глаз — камерный глаз. Эволюция сложного
глаза: простые глазки — собрание глазков — сложный глаз. Омматидий:
структура — оптика — клетки ретинулы — рабдом — аппозиционный и
суперпозиционный глаз — различия чувствительности и остроты —
дневные и ночные насекомые — нейральная суперпозиция у насеко-
мых — отражающий глаз у десятиногих. Сканирующий глаз: морские
улитки — пауки-скакуны — рак-богомол — Copilia (ракообразные). При-
меры: протисты: глазки — световые антенны — глазки Chlamydomonas —
биофизика — глазки динофлагеллят; диафрагмальный глаз Nautilus —
структура — оптика — сетчатка; зеркальный глаз Pecten — структура —
двойная сетчатка — ресничная и рабдомерная — необычная биофизика;
камерный глаз осьминога Octopus — конвергентная эволюция — большой
размер — структура — фокусировка — неинвертированная сетчатка —
рабдомерные ретинулярные клетки — молекулярная биология — оптиче-
ский путь — «глубокая сетчатка»; примитивный сложный глаз Limulus —
структура — электрофизиология — клетки ретинулы — эксцентрические
клетки — латеральное торможение — биологическое значение; высоко-
развитый сложный глаз двукрылых — аппозиция — физиология — цветное
зрение — нейральная суперпозиция в оптической пластине — перекрест
в мозге — лобула. Заключение: в каком смысле можно говорить, что бес-
позвоночные «видят»?
Физические характеристики электромагнитного излучения, рассмотренные во
Введении к Части IV, дали возможность развиться фоторецепторам практичес-
ки у всех животных. Человеческий глаз ограничен длинами волн света от 400 до
750 нм, и мы со своей позиции называем эту часть спектра видимой. Но многие
животные видят и более короткие длины волны (до 200 нм), известные как уль-
трафиолетовые, другие способны воспринимать инфракрасные волны до
1000 нм. Мы начнем эту главу с обзора принципов организации глаз беспозво-
ночных. Затем мы рассмотрим несколько более подробно некоторые демонст-
ративные примеры: глазок простейших (хламидомонада Chlamydomonas)', глаз
моллюска {Nautilus, Pecten и Octopus), глаза членистоногих {Drosophila, Limulus,
пауки). Рассмотрение глаз позвоночных отложим до глав 15, 16 и 18.
Часть IV. Светочувствительность
Информация 14.1
ЭВОЛЮЦИЯ ОПСИНОВ
Когда возникли опсины? Мы видели во Введении к Части IV, что это — вездесущий
белок. Они обнаруживаются в пурпурной мембране галобактерий, в фоторецептор-
ных клетках беспозвоночных и позвоночных, в светочувствительном пигменте ме-
ланофоров кожи лягушки, радужины лягушки и еще нескольких объектах. В мела-
нофорах они опосредуют вызванное светом движение меланосом, а в радужине ля-
гушки — вероятно, участвуют в контроле ее сокращения и расслабления. Мы также
отмечали, что опсины обладают такой же семидоменной организацией, как и мно-
жество других белков, с которыми они родственны. Среди них обонятельный ре-
цепторный белок, несколько вкусовых рецепторных белков, возможно некоторые
терморецепторы и многочисленные синаптические рецепторные белки, напри-
мер, мускариновый ацетилхолиновый рецепторы (мАХР), а- и р- адренорецепторы
(а- и Р-АР), серотониновые рецепторы и большинство других метаботропных си-
наптических рецепторов. Таким образом, родопсины являются частью огромного
надсемейства структурно схожих белков с близкой G-белковой сигнальной биохи-
мией.
То, что семидоменная конформация обнаружена у архебактерий в форме бакте-
риородопсина, показывает, что это очень древняя структура. Есть данные в пользу
того, что эту структуру эволюция «открывала» повторно и независимо несколько
раз. Но поскольку это критически важно для фоторецепции животных — нельзя ис-
ключить его монофилетическое происхождение. Данные о филогении опсинов по-
казаны на рис. А.
Одно из существенных различий между опсинами позвоночных и беспозвоноч-
ных состоит в реакции хромофора — ретиналя — на свет. У позвоночных ретиналь,
как мы увидим ниже, отделяется от опсина и диффундирует в другие клетки (клет-
ки эпителия сетчатки), где регенерирует. Иначе с пигментом беспозвоночных;
у них хромофор остается на месте и способен к повторным трансформациям из
цис- в транс-форму, не отделяясь от опсина. Очень интересно, что недавно проана-
лизированный меланопсин южноафриканской шпорцевой лягушки Xenopus laevis
напоминает в этом отношении опсин беспозвоночных. Возможно, такая «чуждая»
характеристика связана с тем, что меланоциты не обязательно связаны с такой ре-
генерирующей тканью, как сетчатка. Не исключено, однако, что это свидетельство
эволюционной гибкости молекулы.
Сходство меланопсина с опсинами беспозвоночных, а не позвоночных, глубо-
ко связано с его молекулярной структурой. Его способность удерживать ретиналь
после фотоактивации зависит от наличия в нем ароматического остатка (тиро-
зин 103) в третьей трансмембранной спирали, вместо аминокислотных остатков, ко-
торые занимают это положение в опсинах позвоночных. Кроме того, меланопсин
напоминает опсины беспозвоночных и отличается от позвоночных наличием длин-
ного С-концевого цитоплазматического «хвоста».
Итак, как же ответить на вопрос, с которого мы начали? Мы можем предположить
с большой степенью определенности, что семидоменные белки и связанная с ними
G-белковая мембранная биохимия появились в эволюционной истории очень рано.
14. Зрение у беспозвоночных 281
Продолжение
Опсины
позвоночных
Опсины
беспозвоночных
и подобные им
Простейшие
Рис. А. Филогения опсинов. Эволюционные взаимосвязи некоторых основных опси-
нов показаны исходя из их аминокислотной последовательности. Точки ветвления
схемы соответствуют дупликации генов, тайминг дупликаций на дендрограмме не по-
казан. Интересно отметить, что меланопсин меланофоров земноводных в соответст-
вии с этой схемой близок к опсинам беспозвоночных, но одновременно и к ретино-
хромам, найденным в главах и беспозвоночных, и позвоночных. L — опсин, чувстви-
тельный к длинноволновой (красной) области спектра; Ml — опсин, чувствительный
к синей области спектра; М2 — опсин, чувствительный к зеленой области спектра; Р —
опсин пинеального органа; S — опсин, чувствительный к коротковолновой области
области спектра (все — из цыпленка); VA — «древний» опсин позвоночных (из атлан-
тического лосося); головоногие — опсин из осьминога Octopus', насекомые — опсин
Rhl из Drosophila^ хламиопсин — из Chlatnydotnonas. По Provencio etal., 1998.
Вследствие этого количество молекул-кандидатов в светочувствительные пигмен-
ты было велико. Предшествовали ли пигментные.клетки фоторецепторным или на-
оборот? Говорят, что в пигментных пятнах мало смысла, если нет глаз, которые
способны реагировать, агрессивно или оборонительно, на изменения их цвета. Ес-
ли так, то глаза появились раньше. С другой стороны, пигментация могла возник-
нуть для целей, совсем иных, чем камуфляж или видимое предупреждение. Она
могла возникнуть, например, в связи с терморегуляцией или для защиты наиболее
тонких структур от слишком высокой освещенности. Пока достаточных данных
для решения этого впроса у нас нет. В любом случае мы можем предположить, что
первые клетки, которые «изобрели» светочувствительность (присущую комбина-
ции ретинальдегида с опсином), лежат в основании древа эволюции меланофоров
и фоторецепторов.
282 Часть IV. Светочувствительность
Продолжение
Комбинация опсина и ретинальдегида (ретиналя) очень подвижна. Мы отмеча-
ли выше, что отделение (или неотделение) ретиналя от его локуса в глубине моле-
кулы опсина (после фотоизомеризации) зависит от наличия и соответствующем
позиции кислотной или ароматической аминокислотной боковой цепи в третьей
трансмембранной спирали опсина. Длина волны света, к которой ретиналь наибо-
лее чувствителен, также зависит от его аминокислотного окружения внутри опси-
на. Возвращаясь к гл. 13 (раздел 13.2.3), отметим, что такие вариации в аминокис-
лотном составе «клепок бочки» считаются ответственными также за дифференци-
альную чувствительность обонятельных рецепторных белков. Что касается опсина,
то мы увидим в гл. 16, что по крайней мере три разных аминокислотных замещения
могут трансформировать пигмент, чувствительный к зеленому свету, в пигмент,
чувствительный к красному. В этой главе также будут дополнительно рассмотрены
вопросы эволюционных взаимоотношений пигментов колбочек у человека. Но
нельзя забывать, что за геологические периоды времени опсины развились так,
чтобы удовлетворять потребностям огромного разнообразия животных. Вариации
аминокислотных последовательностей молекул опсинов образуют набор пигмен-
тов с максимумами чувствительности (Хтах) к длинам волн от ультрафиолета до
дальнего красного конца спектра. Знакомые нам красный, зеленый и синий йодоп-
сины сетчатки человека — это не просто случайные варианты бесконечного числа
пигментов в царстве животных.
I Литература
I Arnhelter, Н., 1998, «Eyes viewed from the skin», Nature, 391, 632-633.
I Fryxell, K. J. and E. M. Meyerowitz, 1991, «The evolution of rhodopsins and neurotrans-
D mitter receptors», Journal of Molecular Evolution, 33, 367—378.
I Provencio, I. et al., 1998, «Melanopsin: an opsin in melanophore, brain, and eye», Pro-
I ceedings of the National Academy of Science, 95, 340—345.
И Yokoyama, S., 1995, «Amino acid replacement and wavelength absorption of visual pig-
I ments in vertebrates», Molecular Biology and Evolution, 12, 53—61.
14.1. Организация глаз беспозвоночных
Хотя, как отмечал Дьюк-Элдер (Duke-Elder), фоторецепция — это одно из по-
следних чувств, возникших в эволюции, после механо- и хеморецепции, она
имеет настолько большое значение для жизни животных, что к нынешнему
моменту сформировалось огромное множество различных типов глаз. Многие
из таких типов организации уже существовали в кембрийском периоде — бо-
лее, чем 500 млн лет назад. Например, отложения вездесущих трилобитов это-
го периода (ныне вымерших) демонстрируют наличие хорошо развитых слож-
ных глаз. Современные глаза — это диапазон от глазков (или стигматов) через
фасетки к сложным высокоразвитым глазам членистоногих, моллюсков и по-
звоночных. Интересно отметить, что (см. Информацию 14.2) есть свидетельст-
ва того, что ранние стадии развития всех более сложных форм глаз, включая
глаза, формирующие изображение, контролируются одними и теми же генети-
14. Зрение у беспозвоночных 283
Рис. 14.1. Два типа фоторецептор-
ных клеток. (А) Рабдомерный тип.
(Б) Ресничный тип.
Синаптическая
ножка
ножка
ческими механизмами. Это дает серьезное основание думать, что, несмотря на
поразительную разницу между, скажем, глазом домашней мухи и нашим соб-
ственным огромным камерным глазом, оба они имеют общего предшествен-
ника, вероятно в докембрии еще до формирования сланцевых отложений Бор-
джеса, упомянутых в гл. 4.
Простейший глаз — простейшее светочувствительное пятно или глазок —
не способен к формированию изображения. Он способен лишь воспринимать
присутствие или отсутствие света, и его интенсивность. Некоторые такие глаз-
ки могут также определять направление на источник света либо благодаря соб-
ственной структуре, либо будучи распределены по всему организму. Более
сложные глаза способны воспринимать световой образ — изображение, при-
чем такие механизмы развивались независимо много раз. Хорошо известны
независимо развивавшиеся формирующие изображение глаза моллюсков
(особенно головоногих), членистоногих (сложные глаза) и позвоночных. Не-
которые другие типы животных (кольчатые черви, возможно некоторые стре-
кающие) тоже имеют рудиментарные глаза, формирующие изображение.
Как классифицировать такое разнообразие фоторецепторов? Во-первых,
можно их подразделить на «глаза» одноклеточных и многоклеточных. Хотя у
одноклеточных развилось множество светочувствительных органоидов, все
они намного примитивнее глаз многоклеточных. В разделе 14.2.1 мы рассмот-
рим простейшие глазки одноклеточной зеленой водоросли Chlamydomonas как
пример фоторецептора одноклеточных. Глаза многоклеточных, как отмечалось
выше, очень разнообразны. Существуют многочисленные схемы их классифи-
кации. На уровне фоторецепторных клеток популярная схема была предложе-
на Икином (Eakin) в начале 60-х годов XX столетия. Он подметил существова-
ние двух главных типов фоторецепторных клеток у многоклеточных. В первом
из них, у первичноротых (кольчатые черви — моллюски — членистоногие) фо-
топигмент откладывается в микроворсинки, проецирующиеся из клеточной
мембраны. Во втором, развившемся у вторичноротых (иглокожие — хордовые
и т. д.), фотопигмент включается в диски, образовавшиеся в результате инваги-
нации мембраны ресничек (рис. 14.1). Эти два различающихся типа образова-
ны рабдомерным и ресничным типами фоторецепторов, соответственно.
284 Часть IV. Светочувствительность
К сожалению, такое простое эволюционное разделение не было подкреп-
лено дальнейшими исследованиями — обнаружилось множество исключений.
У некоторых первичноротых развились ресничные фоторецепторы, а у неко-
торых вторичноротых — рабдомерные. Далее, в сложно организованном глазу
гребешка Pecten обнаружены одновременно и рабдомерные, и ресничные фо-
торецепторы. Интересно отметить, что два разных типа фоторецепторов ха-
рактеризуются и значительными различиями физиологии. Тогда как рабдо-
мерные рецепторы деполяризуются в ответ на освещение, ресничные — гипер-
поляризуются. Причем эта закономерность сохраняется даже у гребешка — его
ресничные фоторецепторы гиперполяризуются, а рабдомерные — деполяри-
зуются. И опять-таки, из этого ясного классификационного правила нашлись
исключения. У оболочника Salpa (Urochordata), например, присутствуют раб-
домерные рецепторы (которых у вторичноротых и быть-то не должно), и эти
рецепторы деполяризуются. Вновь биология отказывается подчиняться стрем-
лению биологов все красиво классифицировать.
Если от структуры фоторецепторных клеток перейти к организации самого
многоклеточного глаза, выяснится, что и здесь развилось огромное разнообра-
зие. И здесь существует две основных линии развития. Одна из них ведет к ка-
мерному глазу, который достигает высшего развития у хордовых и моллюсков,
другая — к фасеточному, достигающему пика совершенства у ракообразных и
насекомых.
14.1.1. Эволюция камерного глаза
Нетрудно понять, как развивался камерный глаз (рис. 14.2). Предположим,
что первые одиночные фоторецепторные клетки развились в эпидермисе. Та-
кая организация обнаружена в эпидермисе дождевого червя Lumbricus terrestris.
С этой стадии не такой уж большой шаг — агрегация таких клеток непосредст-
венно под эпидермисом (как у Planaria gonocephala), или же в самом эпидерми-
се образуется т. н. «плоский глаз», обнаруженный в эпидермисе некоторых
морских кольчатых червей. Из такой конфигурации легко представить, как
инвагинация проводит к формированию чашевидного глаза или диафраг-
мального глаза. Хороший пример диафрагмального глаза — примитивный
глаз головоногого моллюска кораблика {Nautilus). От диафрагмального глаза
до полностью замкнутого камерного глаза дистанция невелика. В таких глазах
изображение формируется на сетчатке либо рефракционной линзой, либо
преломлением на роговице или, что часто, обоими способами. В некоторых
случаях изображение формируется отражением. Мы обсудим примеры таких
глаз в разделе 14.2. Лучше всего известен камерный глаз позвоночных — рас-
смотрение деталей его устройства мы отложим на последующие главы. В гл. 15
и 16 мы рассмотрим один из видов такого глаза — человеческий.
14.1.2. Эволюция сложного глаза
Эволюция сложных глаз шла несколько иным путем, чем описан на рис. 14.2.
Все начинается опять-таки с одиночного фоторецептора, но в данном случае
надо представить себе, что светочувствительная клетка лежит на дне пигмеи-
14. Зрение у беспозвоночных 285
Рис. 14.2. Эволюция камерного
глаза, (i) Простая светочувстви-
тельная клетка в эпителии;
(ii) группа простых светочувст-
вительных клеток, образующая
«плоский глаз»; (iii) инвагинация
«плоского глаза» с формирова-
нием чашевидного глаза; (iv) по-
следующая инвагинация ведет к
формированию диафрагмально-
го глаза; (v) дальнейшая инваги-
нация приводит к формирова-
нию камерного глаза. Эпителий
над камерным глазом развивает-
ся в хрусталик, роговицу и т. д.
тированной трубки (рис. 14.3 А). Это дает преимущество в определении на-
правления света — только свет из определенной точки пространства может
стимулировать рецептор на дне трубки, а тень перемещающегося объекта —
более точно локализована. Как и в камерном глазу, требуется лишь объедине-
ние простых глазков, образующих «агрегатный глаз» — светочувствительное
пятно, в котором индивидуальные глазки остаются изолированными друг от
друга, или образующих истинный сложный глаз, в котором сенсорные элемен-
ты, клетки ретинулы, сначала группируются в омматидии, а затем омматидии
группируются в хорошо известный фасеточный сложный глаз. Светочувстви-
тельные пятна развиваются только у немногих кольчатых червей (рис. 14.3 Б),
членистоногих и моллюсков. С другой стороны, сложный глаз — одно из круп-
нейших достижений Природы, обнаруживается у ракообразных и насекомых.
Пауки (Araneida) хотя и имеют такие же омматидийные элементы, как и дру-
гие членистоногие, однако вместо отдельных линзочек для каждого оммати-
дия они приобрели единую кутикулярную линзу, покрывающую всю систему
целиком1.
1 Т. е. пришли к той же оптической схеме, что у камерного глаза позвоночных и голово-
ногих моллюсков. — Прим. ред.
286 Часть IV. Светочувствительность
А
Фоторецепторая клетка
Кутикула J
Эпидермис
В
LЛиния среза
CD (Н)
£
Рис. 14.3. Эволюция светочувствительного пятна и сложного глаза. (А) Формирование
пигментированной трубки (в некоторых случаях — углубленной в кутикулу). (Б) Агрегат
множества трубок. (В) Каждая из трубок приобретает кутикулярную линзу, и образуется
светочувствительное пятно. (Г) Светочувствительное пятно многощетинкового червя
Branchioma vesiculoseum. (i) Продольный срез; (ii) поперечный срез; (iii) две фоторецеп-
торных единицы более детально (из Milne and Milne, 1959; воспроизводится с разреше-
ния The American Physiological Society). (Д) Омматидии насекомых, (i) Омматидий аппо-
зиционного типа; (ii) Омматидий суперпозиционного типа.
14. Зрение у беспозвоночных 287
Под каждой фасеткой сложного глаза располагается развитая структура све-
товосприятия. Как показывает рис. 14.3 Д, омматидий состоит из роговичной
линзы, кристаллического конуса и группы из семи или восьми клеток ретинулы,
образующих цилиндр. В отличие от многих камерных глаз возможность варьи-
рования фокуса здесь отсутствует — линзы и кристаллические конусы сложно-
го глаза не регулируются. Они образованы концентрически расположенными
пластинками, различающимися по показателю преломления (ПП): чем ближе
к центру, тем выше ПП. Если длина линзы с кристаллическим конусом равна
фокальному расстоянию, тогда падающий на них луч света фокусируется на их
дальнем конце и выходит как пучок расходящихся лучей (рис. 14.3 E(i)). Если
же длина линзы и кристаллического конуса вдвое больше фокального расстоя-
ния, свет выходит как пучок параллельных лучей (рис. 14.3 E(ii)).
Клетки ретинулы — это светочувствительные клетки. У многих членисто-
ногих все они посылают нервные волокна в оптическую пластину централь-
ной нервной системы, у некоторых — существует некоторая специализация.
Например, у двукрылых насекомых, две клетки ретинулы (R7 и R8) посылают
волокна глубоко во вторую оптическую пластину, а другие (R1 и R6) — только
короткие волокна в первую пластину (рис. 14.18). Во всех случаях, однако,
клетки ретинулы формируют большое количество микроворсинок, которые
образуют рабдомеры. Рабодомеры цилиндра ретикулярной клетки часто пере-
плетаются между собой, образуя рабдом. Падающий свет фокусируется на раб-
доме роговичной линзой и кристаллическим конусом. Зрительные пигменты
концентрируются в рабдомерах, а у некоторых насекомых каждая ретикуляр-
ная клетка содержит родопсин, чувствительный к определенной длине волны
освещения. У дрозофилы, например, ретикулярная клетка 7 содержит родо-
псин, чувствительный к ультрафиолетовому (УФ) излучению. Каждый омма-
тидий отграничен от соседних пигментными клетками.
Сложные глаза, как и камерные, способны формировать изображение. Су-
ществует множество вариантов организации глаза, использующего как свето-
преломление, так и отражение. Во всех случаях, однако, в каждом данном ом-
матидии формируется лишь один элемент мозаики — полное изображение
формируется только интегральной активностью совокупности омматидиев.
Существует три основных пути такого формирования и, соответственно, три
различных типа сложных глаз: аппозиционный, суперпозиционный и ней-
Рис. 14.3 (продолжение). Показаны продольный и поперечный срезы, а — кутикулярная
линза, б — опорная клетка, в — кристаллический конус, г — побочная пигментная клет-
ка, д — рабдом, е — клетка ретинулы, ж — пигментная клетка ретинулы, з — базальная
мембрана, и — эксцентрическая клетка, к — прозрачный филамент, л — нервное волок-
но. Из Wigglesworth, 1972; с разрешения. (Е) Световые пути через кристаллические кону-
сы. (i) Световой путь через кристаллический конус аппозиционного глаза, (ii) Световой
путь через кристаллический конус суперпозиционного глаза. (Ж) Формирование изоб-
ражения в сложном глазу, (i) Формирование изображения в аппозиционном глазу,
(ii) Формирование изображения в суперпозиционном глазу, а, б, в, г, д, е — светящиеся
точки, П — пигмент, Р — рабдом. На рисунке суперпозиционного глаза справа пигмент
показан в положении световой адаптации. Из Wigglesworth, 1972; с любезного разреше-
ния Kluwer Academic Publishers.
288 Часть IV. Светочувствительность
Рис. 14.4. Три типа сложных глаз. (А) аппозиция, (Б) суперпозиция, (В) нейральная
суперпозиция. Воспроизводится с разрешения М. F. Land.
ральный суперпозиционный (рис. 14.4). В глазу аппозиционного типа линза и
кристаллический конус фокусируют падающий луч на верхушке рабдома (см.
рис. 14.3 Ж(1) и 14.4 А). В глазе суперпозиционного типа длина линзы и крис-
таллического конуса вдвое больше фокального расстояния, и образующийся
луч направлен на омматидиальный цилиндр клеток ретинулы несколько ниже
(см. рис. 14.3 Ж(п) и 14.4 Б). Аппозиционные глаза обнаружены у дневных на-
секомых, они формируют сравнительно точное изображение окружающей
среды. Суперпозиционный глаз чаще развивается у ночных и сумеречных на-
секомых. Поскольку свет от множества линз фокусируется на одной и той же
группе клеток ретинулы, такие глаза более чувствительны, но то, что они вы-
игрывают в чувствительности, они теряют в точности. В большинстве случаев
наложения лучей, идущих от линз и кристаллических конусов, недостаточно
для того, чтобы можно было соревноваться с аппозиционным глазом в разре-
шающей способности. Это, однако, не подтверждается для некоторых ночных
бабочек. Здесь суперпозиция настолько точна, что дает остроту зрения того же
уровня, что и аппозиционный глаз такого же размера. У таких бабочек чувст-
вительность увеличена в 100—1000 раз. К тому же, даже у суперпозиционных
глазах, где нет точной оптической организации пигмента, изолирующего ом-
матидии друг от друга, возможна его миграция, которая обеспечивает преобра-
зование суперпозиционного глаза в аппозиционный для использования его
при дневном освещении.
Рис. 14.4 В показывает, что в дополнение к классическому суперпозицион-
ному типу глаза у некоторых насекомых развилась еще более сложная система.
Рабдомеры клеток ретинулы не сливаются с образованием рабдома, а остают-
ся самостоятельными, т. е. представляют собой независимые рецепторные
клетки. Афференты отсюда направляются непосредственно в первую пласти-
ну оптической доли мухи. Рис. 14.4 В показывает, что очень тонко организо-
ванные нервные связи обеспечивают передачу сигнала из клеток ретинулы,
расположенных в соседних омматидиях и имеющих одинаковое поле зрения,
в одно и то же место в первой пластинке. Эта организация обеспечивает чувст-
вительность, эквивалентную суперпозиционному глазу таких же размеров, но
14. Зрение у беспозвоночных 289
устраняет свойственную ему слабость остроты зрения. У двукрылых, таким об-
разом, сочетаются лучшие качества — высокая чувствительность и большая
острота.
Четвертый тип сложного глаза развился у десятиногих ракообразных, ха-
рактеризующиеся длинным туловищем (разнообразные креветки, речные ра-
ки, омары). Эти виды имеют суперпозиционные глаза, но вместо линз и крис-
таллических конусов, преломляющих свет на глубоко расположенные группы
клеток ретинулы, использует другой оптический принцип — отражение. На
месте линз и кристаллических конусов у этих животных образовались структу-
ры в форме коробочки с «зеркальными стенками». Свет отражается от этих
зеркальных стенок на клетки ретинулы, в которых формируется прямое изоб-
ражение. Интересно, что такие сложные глаза с квадратными фасетками раз-
вились только у длиннотелых десятиногих; их близкие родственники рачки
карпоеды (Brachiura) пользуются обычными аппозиционными глазами, а ли-
чинки креветки-шримса — аппозиционными глазами, которые при метамор-
фозе заменяются на зеркальные суперпозиционные. Неясно, отличается ли
экология креветок и омаров от экологии крабов достаточно существенным об-
разом, чтобы это оправдывало такую значительную анатомическую реоргани-
зацию. Наконец, пятый тип сложного глаза обнаружен недавно у плавающего
краба Portunus (= Macropipus) и краба-отшельника Eupagurus. Глаза этих кра-
бов — суперпозиционного типа, но комбинирующие светопреломление и от-
ражение. Их оптика сложна и все еще изучается. Возможно, сложные глаза,
развивавшиеся в течение 500 миллионов лет, сформировавшие и испытавшие
различные варианты оптики, еще преподнесут сюрпризы зоологам и сенсор-
ным физиологам.
14.1.3. Сканирующий глаз
Все рассмотренные выше типы глаз формируют изображение на двумерной
листовидной сетчатке. Третье измерение вносится центральной переработкой
информации. Существуют, однако, немногочисленные случаи, когда глаз ис-
пользует альтернативный подход. В этих случаях — у нескольких моллюсков и
членистоногих — узкая полоска фоторецепторов сканирует видимое окруже-
ние. Такого типа глаза присутствуют у свободноплавающих киленогих мор-
ских улиток (Heteropoda, Gastropoda), которые странствуют по океану с помо-
щью ундулирующих придатков на ножках, как будто они рыбы. Примером мо-
жет послужить Oxygyrus, сетчатка которого содержит от трех до шести фоторе-
цепторов в ширину и более 400 — в длину. Глаз сканирует окружающую среду
наклоном вверх-вниз по дуге в 90° под прямым углом к фоторецепторной по-
лоске. Движение глаза вниз — быстрое (250°/с), а за ним следует более медлен-
ное сканирование по восходящей (80°/с). Другой пример — это главные глаза
пауков-скакунов (Salticidae). Сетчатка таких глаз состоит из полосы шириной
в пять — семь фоторецепторов и длиной около 50. Пауки двигают своими гла-
зами из стороны в сторону перпендикулярно к длинной оси полосы фоторе-
цепторов и вращают ими, когда рассматривают интересующий их объект. Ла-
теральные глаза этих пауков имеют обычную двухмерную сетчатку, которая
обнаруживает движение чего-то. Если они его обнаруживают, обладающий
290 Часть IV. Светочувствительность
Рис. 14.5. Сканирующий глаз вес-
лоногого рачка Copilia quadrata.
Каждая линза охватывает дугу
около 3° и сканирует 14°, фокуси-
руя свет на «сетчатке», состоящей
из единственой фоторецепторной
клетки. По Land and Fernaid, 1992,
с разрешения.
высоким разрешением главный глаз направляется на объект и сканирует эту
часть окружения. Легко провести аналогию с фовеа, которая играет схожую
роль в глазах многих млекопитающих (см. гл. 16). Заключительный интерес-
ный пример сканирующего глаза обнаружен у раков-богомолов (Stomatopo-
da). У этих ракообразных шесть рядов увеличенных омматидиев образуют по-
лосу в середине обычного в других отношениях двумерного сложного глаза.
Эти омматидии содержат цветные зрительные пигменты, и чтобы определить
цвет зрительного стимула, эти креветки сканируют окружающую среду цвето-
вой полосой.
Мы не можем закончить этот раздел, не упомянув удивительный глаз ма-
ленького веслоногого рачка Copilia (рис. 14.5). Эти глаза довели процесс ска-
нирования до возможного предела. «Одномерная» полоса фоторецепторов
здесь редуцирована до нуля — в точку. Задняя линза сканирует в горизонталь-
ной плоскости дугу в 14° с частотой около 0,5—5 сканов в секунду, фокусируя
свет на единственном рецепторе, а единственное нервное волокно несет аффе-
ренты в мозг.
Из сказанного в данном разделе ясно, что за время биологической эволю-
ции животные перепробовали массу способов восприятия света и формиро-
вания изображения (см. рис. 14.6). Крайне сомнительно, чтобы современные
инженеры, если им дать непрозрачное, нежесткое сырье вроде белков и липи-
дов, смогли бы создать что-то подобное тому оптическому совершенству, ко-
торого достигла Природа. Подозреваю, что они бы даже не знали, с чего на-
чать. Мы увидим далее в этой и последующей главах Части IV, что фоторецеп-
торы животных (как и слуховые рецепторы, см. гл. 8) развились до физически
возможных пределов. Не так уж и удивительно (как упоминалось во Введе-
нии), что структура и функция глаза не раз приводили в трепет Чарльза Дар-
вина.
14. Зрение у беспозвоночных 291
14.2. Примеры глаз беспозвоночных
В данном разделе мы рассмотрим некоторые примеры множества удивитель-
ных глаз, которые развились у беспозвоночных. Мы начнем с глазков или
стигматов одноклеточных организмов. Анатомическая простота этих фоторе-
цепторов позволяет провести детальный биохимический анализ. Затем мы об-
ратим свое внимание на некоторые из множества типов глаз, развившихся в
типе моллюсков. Они достигли вершины в большом камерном глазу головоно-
гих. Эти глаза, хотя и развивались независимо, вполне сравнимы по сложнос-
ти и зрительным способностям с камерным глазом позвоночных — это один из
наиболее интересных случаев конвергентной эволюции в царстве животных.
Наконец, мы рассмотрим тип членистоногих и структуры и функционирова-
ние примитивных сложных глаз у мечехвоста Limulus и вершину такого типа
фоторецепции у насекомых.
Рис. 14.6. Основные типы формирования изображения у глазах животных. (А) Про-
стая зрительная ямка многих низших типов, развившиеся в диафрагмальный глаз
например у Haliotis (морское ушко) или Nautilus. (Б) Глаз с линзой. (В) Роговичное
светопреломление. (Г) Зеркальный глаз. (Д) Светочувствительное пятно. (Е) Аппози-
ционный сложный глаз. (Ж) Суперпозиционный сложный глаз. (3) Отражающий су-
перпозиционный сложный глаз. С изменениями из Land, 1991; Land and Femald, 1992.
Часть IV. Светочувствительность
Информация 14.2
ГЕНЕТИКА РАННЕГО РАЗВИТИЯ ГЛАЗ
Мы уже отмечали несколько раз чрезвычайную древность зрительного пигмента
родопсина, представленного и у бактерий, и у простейших. Химический анализ по-
казывает, что аминокислотные последовательности родопсинов многоклеточных
хотя и варьируют от вида к виду, но достаточно схожи, чтобы предположить нали-
чие у них общего предшественника в далеком прошлом. Семидоменное располо-
жение родопсинов в клеточной мембране также указывает на их общее происхож-
дение. Глаза сами по себе, однако, намного более разнообразны. Хотя животные
большинства типов ограничиваются простыми глазками, более развитые системы
обнаружены у шести различных типов, при этом наиболее значительных — к ним
относятся 96% современных видов животных. В данной главе был сделан обзор
анатомического разнообразия их глаз.
Встает вопрос: возникло ли это богатое разнообразие глаз независимо или же они
имеют общих предшественников? Если посмотреть на разнообразие их структур и ти-
пов, приведенное в данной главе, то логично предположить независимое или полифи-
летическое их происхождение. Возможно, в некоторых случаях так оно и есть — ка-
мерные глаза головоногих и позвоночных, при всем их поразительном сходстве, —
скорее пример конвергентной эволюции, чем развития от общего предка. Пока совре-
менная генетика развития не дает оснований сомневаться в таком заключении, одна-
ко начинают появляться удивительные намеки на общее происхождение, предшест-
вовавшее дивергенции первично- и вторичноротых около 670 миллионов лет назад.
Такие сведения появились при изучении одного из генов Рах-6, который уча-
ствует в раннем развитии глаза. Как показывает число в названии гена, Рах-6явля-
ется членом большого семейства генов, регулирующих транскрипцию других ге-
нов. Это семейство играет важную роль в раннем развитии, когда члены этого се-
мейства контролируют транскрипцию каскадов других генов, управляющих разви-
тием. Наименование «Рах» связано с тем фактом, что все эти гены обладают после-
довательностью из 384 пар оснований (по — Ьр1), кодирующих последовательность
из 128 аминокислот, которая связывается с ДНК. Последовательность из 384 Ьр из-
вестна как «paired box» (отсюда Pax), так как она была открыта первой, которая со-
ответствует правилу парных генов, определяющих раннюю парную сегментацию
зародышей дрозофилы. Помимо парного домена у всех представителей семейства
Рах присутствует также высоко-консервативный гомеодомен.
Рах-6 обнаружен у многих различных видов животных — от человека, насеко-
мых и моллюсков, до круглых червей (нематоды) и немертин. В ранних зародышах
позвоночных Рах-6 экспрессируется в первую очередь в центральной нервной сис-
теме, но обнаруживается также и в мозге взрослого животного, особенно в некото-
рых ядрах переднего мозга, черной субстанции среднего мозга и в зернистом слое
мозжечка. Что касается переднего мозга, связана в областями, впослед-
ствии развивающимися в обонятельные (обонятельный эпителий, обонятельная
луковица) и зрительные (хрусталик, роговица и глазное яблоко) структуры.
1 bp — base-pair — пара оснований. — Прим, перев.
14. Зрение у беспозвоночных 293
Продолжение
Мутации Рах-6 имеют катастрофические последствия. У мышей это приводит
к формированию т. н. малых глаз (Sey)1. В гетерозиготах глаза мышей Sey имеют
значительно меньшего размера хрусталики и глазные бокалы. Гомозиготы Sey де-
тальны. У ранних зародышей мыши (9,5 дней) отсутствует линзовая плакода, а ней-
ральный слой сетчатки резко патологичен. Также не развивается обонятельная
плакода, и у зародыша отсутствуют признаки обонятельной ямки и обонятельной
луковицы. Таким образом, ясно, что Рах-6 играет ключевую роль в раннем разви-
тии глаз и носа. Люди, гетерозиготные по гену Sey, страдают т. н. аниридией — пол-
ным или частичным недоразвитием радужины глаза, сопровождающимся катарак-
той, непрозрачностью роговицы, глаукомой и т. д. Встречаемость патологии — от 1
на 64 000 рождений до 1 на 96 000. Кроме того, мутации Рах-6 могут вести к дефек-
там передней камеры глаза, в т.ч. непрозрачности центральной части роговицы, из-
вестной как аномалия Петера. Различия в степени инактивации Рах-6 приводят
к разной выраженности дефектов тех или иных частей глаза, особенно передней
камеры.
Выше указывалось, что Рах-6 обнаруживается во всем царстве животных, и во
всех случаях он участвует в раннем развитии глаза. Генетические исследования на
дрозофиле показали, что трансплантация генов в другие части организма приводит
к индукции эктопических глаз (при этом — глаз не мышиных!). Наконец, обнару-
жено, что Рах-6 кальмара, в норме важный в раннем развитии высокоразвитого
глаза головоногих, также могут индуцировать эктопические глаза у дрозофилы.
Все эти данные бьют в одну точку — они позволяяют предполагать, что на очень
далеком уровне эволюции, представленном плоскими червями (см. гл. 4), в генети-
ческой системе уже сложилась программа раннего развития фоторецепторов. Эта
система, остающаяся неизменной, лежит в основе самых разных глаз животных.
Это означает, что происхождение глаз все же не полифилетическое, а монофилети-
ческое. На момент написания книги решения этого вопроса пока нет. Однако, вме-
сте с другими свидетельствами единства молекулярных механизмов, которые мы
отмечали в этой книге, ген Рах-6 объединяет картину поразительного единства в
разнообразии царства животных. Мы заглядываем в глубины эволюции на полмил-
лиарда лет — и узнаем самих себя.
Литература
Callaerts, Р., G. Halder and W. J. Gehring, 1997, «Рах-6 in development and evolution»,
Annual Review of Neuroscience, 20, 483—532.
Harris, W. A., 1997, «Pax-6: Where to be conserved is not conservative», Proceedings of the
National Academy of Science, 94, 2098—2100.
Tomarev, S. I., P. Callaerts, L. Kos, R. Zinovieva, G. Halder, W. Gehring and J. Platigor-
sky, 1997, «Squid Pax-6 and eye development», Proceedings of the National Academy of
Science, 94, 2421-2426.
1 Sey — от small eyes — маленькие глаза. — Прим, перев.
294 Часть IV. Светочувствительность
14.2.1. Глазки простейших
Название «глазок» до какой-то степени неточно. Оно означать фоточув-
ствительный органоид у одноклеточных организмов или клетку или группу
клеток, чувствительных к свету. В целом — это структуры с небольшим коли-
чеством волокон зрительных нервов (если они вообще там есть) и иногда не-
которыми дополнительными устройствами, как линзы. Глазки присутствует у
жгутиковых простейших, плоских и кольчатых червей, а иногда и у других жи-
вотных. Глазки обнаруживаются, например, в спинном мозге ланцетника
Amphioxus и состоят из небольших групп т. н. клеток Гессе. В данном разделе
мы рассмотрим только глазки простейших.
Множество различных глазков развилось у жгутиковых простейших — от
простых цветных пятен (стигматов) на той или иной стороне клетки до намно-
го более сложных и намного больших глазков (ocellus) морских динофлагел-
лят. То, что цветные глазки могут быть фоторецепторами, предполагалось дав-
но, но лишь недавно стали думать, что это так всегда. Действительно, у неко-
торых фототаксических простейших, таких как динофлагеллята Gyrodinium,
признаков глазков не обнаружено. Вероятно, видимые глазки часто — не бо-
лее, чем часть фоторецепторного аппарата организма. Такой аппарат часто
именуют «световой антенной». Там, где есть глазки, ультраструктурные иссле-
дования показывают, что все они имеют общие черты. В их числе наличие пиг-
ментированных капелек липида, окруженных одной или более мембраной.
Пигмент содержит обычно один или несколько каротиноидов (см. Введение)
или родопсин, а мембраны часто просиходят из мембран хлоропластов.
Один из наиболее изученных глазков типа стигмата обнаружен в свободно
плавающей зеленой водоросли Chlamydomonas rheinhardtii (рис. 14.7). Он рас-
полагается на краю хлоропласта ближе к периферии клетки и состоит из двух-
четырех слоев плотно упакованных глобул, перемежающихся мембранами
хлоропласта. Тщательное электронно-микроскопическое изучение показало,
что перед фоточувствительным пятном развивается пластинчатая структура;
которая благодаря особенностям строения обеспечивает чувствительность
глазка к направлению падающего света. Глазок содержат родопсин, сходный с
бактериородопсином. Это сходство увеличивается, если учесть, что ретиналь
здесь имеет 13-цнс-форму. Хотя в последнее время появились сообщения о
возможном присутствии G-белка сигнальной системы, аналогичной той, что
присутствует в глазу позвоночных и других, есть данные и о прямом действии
родопсина на мембранные Са2+-каналы. В последнем случае (показанном на
рис. 14.7) падающий свет поглощается родопсином очень быстро (< 500 мкс) и
открывает в мембране Са2+-каналы (фоторецепторные или P-каналы). Это
вызывает мембранную деполяризацию, которая приводит к открыванию дру-
гой группы Са2+-каналов (жгутиковые или F-каналы) вблизи от жгутика. Ре-
гулярные биения жгутика Chlamydomonas продвигает водоросль вперед в спи-
ральном движении (вращение с частотой ок. 2 Гц), так что глазок перехваты-
вает свет каждые полсекунды (рис. 14.7 А). Поток Са2+ в клетку через F-кана-
лы (рис. 14.7 В) влияет на движение жгутика таким образом, что направление
движения меняется до тех пор, пока угол падения света не становится мини-
мальным. Таким образом хламидомонада плывет к источнику света, где ее фо-
14. Зрение у беспозвоночных 295
Тело
хлоропласта
Б
Тилакоидная
Глобулы, _
содержащие
пигмент
Клеточная
мембрана
Рис. 14.7. Глазок Chlamydomonas. (А) Хламидомонада плывет, вращаясь (2 Гц, про-
тив часовой стрелки) в направлении, показанном стрелкой. Луч света (1), следова-
тельно, прерывается дважды в секунду. Водоросль меняет движение своего жгути-
ка до тех пор, пока не устанавливается направление на источник света или (при
высокой интенсивности света) — от него. (Б) Схематическое изображение среза
через гпязок, полученное с помощью электронного микроскопа. Каждый слой
пигментных глобул покрыт с внутренней мстороны двойной тилакоидной мембра-
ной (мембраной хлоропласта). (В) Схема, показывающая биофизический каскад,
запускаемый восприятием фотона родопсином хламидомонады. R* — активиро-
ванный родопсин, Р* — активированный фоторецепторный канал. Г* — активиро-
ванный жгутоковый канал. Объяснения в тексте. По Foster and Smyth, 1980 и Harz,
Nonnegaster and Hegemann, 1992. Воспроизводится с разрешения The Royal Society.
тосинтезирующий аппарат может действовать на оптимальном уровне. При
высоких интенсивностях света водоросль уплывает от источника света.
Теперь от простых глазков Chlamydomonas, Euglena и т. п. перейдем к более
сложным глазкам динофлагеллят. И здесь мы видим множество разных типов
глазков. Полагают, что все они являются модифицированными хлоропласта-
ми. Один из наиболее сложных таких глазков обнаружен у Erythropsidinium (от-
ряд Wamowiales). На рис. 14.8 показана сложная структура этого органоида.
Бросается в глаза большая светопреломляющая структура (называемая крис-
таллическим телом или гиалосомой), которая способна фокусировать свет на
ретиноиде. Последний состоит из очень упорядоченного (почти кристалличе-
296 Часть IV. Светочувствительность
Рис. 14.8. Глазок Erythropsidinium
в продольном разрезе, к — канал,
кт — кристаллическое тело, км —
клеточная мембрана, ск — сокра-
тительное кольцо, м — митохон-
дрии, кг — камера глазка, пк —
пигментное кольцо, р — ретино-
ид, лп — лестничная пластинка,
вс — везикулярный слой. Из
Dodge, 1991, с разрешения.
ского) слоя мембраны, над которой находится множество капелек, содержа-
щих каротиноиды. Биологическое значение этого великолепного фоторецеп-
тора пока не уточнено. Как и другие динофлагелляты, Erythropsidinium не фо-
тосинтезирует. Возможно, глазки развились для детектирования жертвы или
для избегания хищников.
14.2.2. Диафрагмальный глаз Nautiluspompilius
Nautilus pompilius, кораблик, принадлежит к наиболее древнему подклассу го-
ловоногих моллюсков. Ископаемые формы обнаруживаются в слоях, соответ-
ствующих 400 миллионов лет назад. Сейчас они плавают в неглубоких водах
юга Тихого океана. Тело моллюска покрыто кальциевой спирально закручен-
ной раковиной, в которую он и прячется в случае опасности. Как другие голо-
воногие, у кораблика два больших латеральных глаза, однако, в отличие от
других, глаза не имеют линзы (хрусталика) и стекловидного тела, а только зра-
чок, открытый прямо в окружающую морскую воду. Морская вода непосред-
ственно омывает и хорошо развитую сетчатку. Как говорилось выше, это, по-
видимому, наиболее изученный пример диафрагмального глаза в царстве жи-
вотных.
Рис. 14.9 показывает вид среза глаза Nautilus. Глаз имеет размер около
10x15 мм, а размер переменного зрачка — от 0,4 до 2,8 мм. Диаметр диафраг-
мы находится под мышечным контролем, но действие мышц медленно (50 с до
полного открытия, 90 с — на сокращение). Сетчатка состоит из 4 х 106 плотно
упакованных рабдомерных фоторецепторов. Поскольку фоторецепторы всего
5—10 мкм в диаметре, сетчатка обладает хорошим разрешением (см. Часть IV,
Введение), однако расчеты показывают, что диафрагма допускает разрешение
с минимальным углом 2,3°. Это соответствует (если использовать формулы,
14. Зрение у беспозвоночных 297
Рис. 14.9. Диафрагмальный
глаз Nautilus pompilius в попе-
речном разрезе, цер — цереб-
ральная масса (надпищевод-
ная), м — мышца, зн — зри-
тельный нерв, зд — зритель-
ная доля, ог — обонятельный
ганглий, оя — обонятельная
ямка, зд — зрачок диафраг-
мы, сет — сетчатка. Из Young,
1965. Воспроизводится с раз-
решения The Royal Society.
приведенные во Введении, и размеры глаза кораблика) расстоянию на сетчат-
ке не менее 0,4 мм. Значит, у Nautilus сетчатка много лучшая, чем способна ис-
пользовать его оптика! Действительно, исследование поведения этого живот-
ного показывает, что большую часть времени оно проводит при освещении
малой интенсивности, опускаясь днем до глубин около 300 м и поднимаясь до
прибл. 150 м ночью. И зачем такая совершенная сетчатка, если ее не использу-
ют? Глаз кораблика остается тайной.
14.2.3. Зеркальный глаз гребешка Pecten
Гребешок Pecten принадлежит к двухстворчатым или пластинчатожаберным
моллюскам и является близким родственником устрицы. Однако, в отличие от
других двухстворок гребешок активно плавает — он двигается реактивно,
схлопывая свои створки. Хотя у некоторых других двухстворчатых моллюсков
развились глаза, но это не более чем глазки. Сердцевидка Cardium edulis, на-
пример, имеет около 60 маленьких фоторецепторных глазков не более 50 мкм
в диаметре, содержащих 12—20 фоторецепторных клеток ресничного типа.
Гребешок же Pecten имеет 100 удивительных глаз по периметру мантии
(рис. 14.10).
Глаз гребешка привлекает пристальное внимание ученых уже более столе-
тия. Разрез этого глаза показан на рис. 14.11, он имеет выраженную роговицу,
клеточный хрусталик, стекловидное тело и, наиболее интригующее, — две сет-
чатки. Каждая сетчатка содержит около 500 000 фоторецепторных клеток.
В равной степени интригует наличие отражающего зеркальца за проксималь-
ной сетчаткой (интересно, что отражатель обнаружен и в маленьком глазу
сердцевидки Cardium). Зеркальце состоит из перемежающихся слоев цито-
плазмы и кристаллов гуанина и формирует изображение на дистальной сетчат-
ке. Функция проксимальной сетчатки неясна, поскольку хрусталик недоста-
точно мощен, чтобы сформировать на ней изображение. Возможно, она лишь
реагирует на изменения интенсивности освещения. Если это так, то как и рас-
смотренная выше сетчатка кораблика, она чересчур сложна для выполняемой
работы. Еще одно впечатляющее обстоятельство должно быть отмечено. Как
298 Часть IV. Светочувствительность
Рис. 14.10. Анатомия гребешка Pecten. Раковина раскрыта, чтобы показать внут-
ренности. В нормальном состоянии, когда животное расслаблено, мантия и ее
щупальцы несколько выступают за край раковины, г — глаз, щ — щупальцы. Из
Borradile, Potts, Eastham and Saunders, 1951, с разрешения.
указывалось выше в разделе 14.2, дистальная сетчатка содержит фоторецепто-
ры ресничного типа, которые гиперполяризуются освещением, тогда как про-
ксимальная сетчатка содержит рабдомерные фоторецепторы, которые при ос-
вещении деполяризуются. Есть еще один заключительный момент в этой уди-
вительной истории. Хотя ресничные фоторецепторы проксимальной сетчатки
гиперполяризуются в ответ на освещение, биофизический механизм достаточ-
но сильно отличается от такового фоторецепторов позвоночных. Вместо за-
крывания циклонуклеотид-зависимых Са2+ + Ка+-каналов (см. раздел 16.2.1)
ресничные фоторецепторы гребешка открывают К+-каналы. Видимо, у гре-
бешка гиперполяризация развивалась независимо. Почему? Обязательно ли,
чтобы фоторецепторы ресничного типа гиперполяризовались? Есть ли в этом
глубокий смысл? Пока что на понимание этого нет и намека.
14.2.4. Камерный глаз осьминога Octopus
Головоногие подразделяются на три подкласса: двужаберные (Belemnoidea,
Dibranchiata), четырехжаберные (кораблики Nautiloidea, Tetrabranchiata) и ам-
мониты (Ammonoidea, см гл. 4). Представители последнего подкласса все ис-
копаемые. Удивительный камерный глаз головоногих развился у двужабер-
ных — к ним относятся десятиногие (кальмары — Loligo и др. и каракатицы —
14. Зрение у беспозвоночных 299
Рис. 14.11. Глаз Pecten в разрезе. Показаны дистальная и проксимальная сетчатки
(= сенсорные слои). Под проксимальной сетчаткой располагается отражающее
зеркальце, изображенное на рисунке не полностью для сохранения четкости. Под
зеркальцем располагается пигментный слой. Из Messenger, 1991, с разрешения.
Sepia и др.) и осьминоги {Octopus и др.). Глаза у двух этих отрядов похожи и
считаются среди зоологов выдающимся примером конвергентной эволюции.
Глаза не только практически идентичны у этих двух отрядов двужаберных, но
и их структура и функция поразительно схожи с глазами рыб, хотя и построе-
ны из совершенно иных элементов. И это при том, что общий предок голово-
ногих и рыб существовал около полумиллиарда лет назад в докембрийском пе-
риоде. Похоже на то, что за более чем 500 миллионов лет свободноплавающие
позвоночные и головоногие нащупали аналогичные решения проблемы зре-
ния. Студенты, интересующиеся философскими аспектами биологии, могут
поразмыслить над тем, что такая поразительная конвергенция, о которой мы
говорили и в гл. 4, что эволюционный отбор приводит к сходным решениям в
очень удаленных друг от друга линиях.
300 Часть IV. Светочувствительность
Роговица Отверстие
Радужина
камера
Зрительная
доля
Хрусталик
Цилиарная
мышца
Стекловидное
тело
Сетчатка
Наружные
сегменты
Пигментный
слой
Ядерный слой
Рис. 14.12. Срез глаза двужаберного (белемноида). Глаз формируется инвагинаци-
ей поверхностного эпителия (напр. Nautilis). Глаз выстлан сетчаткой, которая кпе-
реди продолжается в заднюю часть цилиарных мышц и заднюю часть хрусталика.
У некоторых видов роговица полностью покрывает глаз, у других передняя камера
имеет отверстие, так что передняя камера связана с окружающей морской водой.
Дальнейшие объяснения в тексте.
Два латеральных глаза двужаберных располагаются на стебельках по обе
стороны головы и защищены соединительной тканью и хрящевидной капсу-
лой. Глаза — крупные и выступающие над поверхностью тела. У небольшого
осьминога Octopus диаметр глаза 10—15 мм, но у гигантского кальмара Architeu-
this, по некоторым сообщениям, он может достигать почти полуметра (40 см)!
Внешние мышцы глаза обеспечивает возможность глазному яблоку двигаться
в глазной впадине; есть данные, что имеются и оптокинетические рефлексы,
до некоторой степени схожие (хотя и более медленные) с рефлексами позво-
ночных. Радужина также снабжена мышцами, так что апертура зрачка может
изменяться в ответ на изменения интенсивности освещенности. Перед хруста-
ликом располагается заполненная водянистой жидкостью полость, спереди
ограниченная прозрачной роговицей. У некоторых видов передняя камера от-
крыта во внешнюю среду и, потому заполнена морской водой. Позади хруста-
лика глазной бокал заполнен стекловидным телом, которое, как и у позвоноч-
ных, удерживает на месте сетчатку. Сам хрусталик удерживается на месте ци-
лиарным телом, однако метод фокусировки радикально отличается от таково-
14. Зрение у беспозвоночных 301
ЗН
Рис. 14.13. (А) Вертикальный срез сетчатки осьминога Octopus. Длинные наружные сег-
менты клеток ретинулы перемежаются пигментными клетками. Внутренние сегменты
клеток ретинулы расположены под базальной мембраной и продолжаются в волокна
зрительного нерва, направляющиеся в зрительную долю, квр — коллатеральное волокно
клетки ретинулы, бм — базальная мембрана, эк — эпителиальная клетка, эв — эффе-
рентное волокно, вс — внутренний сегмент, г — граничная мембрана, яо — ядро опор-
ной клетки, нс — наружный сегмент, кв — коллатеральное волокно, о — опорная клетка
(заполненная пигментом). Из Young, 1962а. Воспроизводится с разрешения The Royal
Society. (Б) Детальная структура сетчатки. Микроворсинки наружных сегментов образу-
ют очень строгую мозаику. ВеК — внеклеточный компартмент, ВуК — внутриклеточный
компартмент, Я — ядро клетки ретинулы, ЗН — волокно зрительного нерва, нс — наруж-
ный сегмент. Из Saibil and Hewat, 1987.
302 Часть IV. Светочувствительность
го у позвоночных. Вместо изменения кривизны хрусталика цилиарные мыш-
цы сжимают само глазное яблоко и, таким образом, передавая давление на
стекловидное тело, сдвигают хрусталик вперед. Иными словами, глаз голово-
ногого фокусирует свет на сетчатке подобно тому, как в фотокамере1. Сереб-
ристый эпителий покрывает радужину и простирается еще несколько кзади.
Рис. 14.12 показывает поперечный срез типичного глаза головоногого.
Возможно, наиболее интересной особенностью глаза головоногих являет-
ся их сетчатка. И у десятиногих, и у восьминогих фоторецепторы — рабдомер-
ные и называются клетками ретинулы (рис. 14.13 А). Сетчатка белемноидов не
страдает от нелепостей морфогенеза сетчатки позвоночных: клетки ретинулы
не покрыты нейральной сетчаткой и не отвернуты от поступающего света, а
примыкают к стекловидному телу и направлены к хрусталику. С другой сторо-
ны, клетки ретинулы схожи с палочками позвоночных (которые, естественно,
являются клетками ресничного типа) тем, что в них выражены наружный и
внутренний сегменты. Наружный сегмент — очень длинный и тонкий
(200—300 мкм х 3 мкм), с двумя стопками микроворсинок, направленных в ди-
аметрально противоположных направлениях и ортогональных длинной оси
сегмента (рис. 14.13 Б). Эти микроворсинки также очень длинные и тонкие
(1 мкм х 0,06 мкм, т. е. 60 нм) и представлены во множестве — по
200 000—700 000 в каждом ряду. Существуют надежные данные, что фотопиг-
мент родопсин концентрируется в мембранах микроворсинок. Середина на-
ружного сегмента заполнена пигментными гранулами, которые мигрируют к
внутреннему сегменту в условиях темноты. Есть данные, что сами наружные
сегменты способны к сокращению — при интенсивном освещении. Внутрен-
ний сегмент располагает богатым набором митохондрий, пигментных гранул,
мембран, называемых сомальными или миелоидными тельцами, там же рас-
полагаются ядра. От внутреннего конца внутреннего сегмента отходит аксон,
образующий одно из волокон зрительного нерва, направляющегося в зритель-
ную долю. Таким образом, клетки ретинулы, в отличие от палочек и колбочек
позвоночных, являются нейросенсорными клетками.
В сетчатке осьминога Octopus около 2 х 107 ретикулярных клеток, и, как по-
казывает рис. 14.13 А, их наружные сегменты плотно упакованы и разделены
лишь тонкими заполненными пигментом выростами опорных клеток. Пери-
карионы расположены под базальной мембраной, а нервные волокна, отходя-
щие от их базальной части дают коллатерали, образующие плексиформный
слой (сплетение). Этот слой также содержит глиальные клетки и эфферентные
волокна из зрительной доли. Есть сведения о наличии щелевых контактов (gap
junction) между внутренними сегментами ретинулярных клеток, а также меж-
ду мембранами эфферентных волокнами и волокнами зрительного нерва. Хо-
тя такой плексиформный слой не может сравниться по сложности с плекси-
формным слоем сетчатки позвоночных (см. раздел 16.2 и т. д.), тем не менее,
он дает возможность для первичной обработки зрительной информации. Ос-
новная же переработка информации происходит в расположенной поблизости
зрительной доле (рис. 14.14), которая иногда называется (по Кахалю) «глубо-
кой сетчаткой».
1 И как у рыб и змей. — Прим. ред.
14. Зрение у беспозвоночных 303
Рис. 14.14. Сетчатка и «глубокая сетчатка» осьминога Octopus. Волокна зрительно-
го нерва из ретикулярных клеток перекрещиваются перед входом в кору оптичес-
кой доли, где они образуют «глубокую сетчатку», ам.2,3 — амакриновые клетки на-
ружного зернистого слоя, би — биполярные клетки, бм — базальная мембрана,
га — афферентное волокно из более глубоких регионов, цк — центрифугальные
клетки к сетчатке от глубокой сетчатки, вс — внутренний сегмент ретикулярной
клетки, гм — граничная мембрана, мк — мультидендритная клетка, ммк — малая
мультиполярная клетка, сет. 1,2,3 — волокна трех различных типов от клеток сет-
чатки, р — рабдом ретикулярной клетки, ок — опорная клетка, тк — тангенциаль-
ная клетка, ук — униполярная клетка. Из Young, 1962b. Воспроизводится с разре-
шения The Royal Society.
Рис. 14.13 Б показывает, что наружные сегменты ретикулярных клеток об-
разуют очень точную мозаику. Микроворсинки соседних ретикулярных кле-
ток ориентированы под прямыми углами друг к другу. Это означает, что, ког-
да глаз находится в естественном положении, микроворсинки ориентированы
в вертикальной и горизонтальной плоскостях. Полагают, что это связано с хо-
рошо известной чувствительностью головоногих к плоскости поляризации
света (см. гл. 20).
Молекулярная биология зрения белемноидов в некоторых отношениях
схожа, а в каких-то — отличается от позвоночных. Основным зрительным пиг-
ментом является родопсин. Как и у позвоночных, родопсин состоит из семи-
доменного белка опсина и хромофора. Кроме того, в миелоидных тельцах вну-
304 Часть IV. Светочувствительность
тренних сегментов обнаружен еще один зрительный пигмент ретинохром, ко-
торый участвует в регенерации родопсина. В большинстве случаев у белемно-
идов обнаруживается один тип родопсина с Хтах от 470 до 500 нм. У одного из
глубоководных биолюминесцентных кальмаров (Watasenia scintillans), однако,
присутствует три типа родопсинов с Хтах 470, 484 и 500 нм. Существенно ли
цветовое зрение, возможность которого обеспечивается наличием трех раз-
личных фотопигментов, для жизни этих биолюминесцентных кальмаров —
неизвестно.
Когда свет поглощается 11-4<мс-ретиналем, последний переходит в транс-
форму (детали — в разделе 16.2.1) и отделяется от опсина. Активированный
опсин затем активирует G-белковую систему, что приводит к активации мем-
бранно-связанной фосфолипазы С (см. гл. 1, раздел 4.2). Это приводит (как
мы видели в разделе 1.4.2) к синтезу ИТФ и ДАГ, а также выбросу Са2+ из вну-
триклеточных запасников в проксимальном конце ретикулярных клеток. По-
вышение уровня Са2+ ведет к деполяризации мембраны ретикулярных клеток,
инициирующей спайки в аксоне ретинулы. Когда мы будем рассматривать мо-
лекулярную биологию и биофизику фоторецепторов позвоночных, мы обна-
ружим основное различие — вместо деполяризации фоторецепторы позвоноч-
ных в ответ на освещение гиперполяризуются.
Аксоны ретикулярных клеток образуют зрительный нерв. Рис. 14.4 пока-
зывает, что эти волокна, покидая сетчатку, перекрещиваются до вхождения в
кору оптической доли (т. н. «глубокая сетчатка»). Здесь они входят во второй
плескиформный слой, причем топологические соотношения афферентных
волокон сохраняются, а изображение инвертируется по отношению к изобра-
жению в сетчатке. К сожалению, использование микроэлектродной техники
на центральной нервной системе белемноидов оказалось невозможным. По-
этому детали физиологии зрительной доли и мозга остаются к моменту напи-
сания тайной, покрытой мраком.
14.2.5. Латеральные глаза мечехвоста Limulus
Хотя мечехвоста Limulus (рис. 14.15) часто называют крабом1, на самом деле
он к ракообразным вообще не относится, а является представителем класса
Arachnida (паукообразные), т. е. более тесно связан с пауками, скорпионами и
клещами, чем с крабами. Это примитивный вид, а ископаемые, мало отличные
от ныне живущих, обнаруживаются в силурийских слоях, датируемых 400 мил-
лионами лет назад. Древности происхождения этого примитивного вида соот-
ветствует его примитивная форма сложного глаза. Как показывает рис. 14.15 Б,
омматидии (около 850) расположены достаточно далеко друг от друга, а не
плотно упакованы, как в глазах ракообразных или насекомых), и от каждого из
них направляется отдельный зрительный нерв. Простота анатомии привлекла
внимание электрофизиологов, и Хартлайн (Hartline) с сотрудниками устано-
вили, что несмотря на пространственную изоляцию омматидиев друг от друга,
1 Английское название — King Crab или Horseshoe crab — буквально — краб-король или
краб-подкова. — Прим, перев.
14. Зрение у беспозвоночных 305
Рис. 14.15. (А) Мечехвост Limuluspolyphemus. Два латеральных глаза (С) располага-
ются на наружной поверхности первых латеральных шипов. Из Duke-Elder, 1958,
с разрешения. (Б) Вертикальный срез глаза мечехвоста. Ряд темно окрашенных
объектов — омматидии, хорошо также видны спелетение латералей и волокна зри-
тельного нерва. Из Hartline, 1959, с разрешения. (В) Одиночный омматидий лате-
рального глаза в вертикальном (а) и поперечном (б) срезах, ку — кутикула, ре.к —
ретикулярная клетка, эк — эксцентрическая клетка. Из Meinertzhagen, 1991,
в J. R. Cronly-Dillon and R. L. Gregory. Vision and Visual Dysfunction, vol. 2, p. 345,
Figure 14.3 (a) and (b). Воспроизводится с разрешения Macmillan Press Ltd.
глаз тем не менее функционирует как единое целое: афферентация из каждого
омматидия влияет на активность соседних. Прежде, чем обсудить важные де-
тали физиологии, вернемся сначала к некоторым деталям анатомии.
По поводу взаимоотношений омматидиев мечехвоста с омматидиями дру-
гих членистоногих пока продолжаются дебаты, не приведшие к общему мне-
нию. Роговичной линзы здесь нет, число ретикулярных клеток варьрует от 4 до
20, и что существенно, присутствует одна или чаще — две эксцентрические
клетки. В отличие от того, что обнаружено у ракообразных и насекомых, рети-
кулярные клетки — чисто сенсорные. Только эксцентрические клетки форми-
руют нервные волокна, которые образуют зрительный нерв. В то же время ре-
тикулярные клетки походят на таковые ракообразных наличием колонн мик-
роворсинок, образующих рабдомер с зрительным пигментом родопсином
(Хтах = 520 нм). Структура омматидия показана на продольном и поперечном
срезах на рис. 14.15 В.
Когда свет соответствующей длины волны падает на омматидий, опсин ак-
тивируется обычным образом, что приводит к последовательности биохими-
306 Часть IV. Светочувствительность
(i)
(»)
(iii)
~Г!' i'""" , " :Освеи»ен
только
' омматидий
A
I iiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiniiiiiiiiiiiiiiii ни.....................................
-!_________
Освещены
оба
омматидия
ОНвещен
только
омматидий
Рис. 14.16. (А) Латеральное торможение в сетчатке мечехвоста Limulus. Регистрация ак-
тивности в волокне зрительного нерва. (Б) Осциллографическая регистрация потенциа-
лов действия в нервном волокне, (i) Когда соседние омматидии сильно освещены, актив-
ность подавляется. Черный отрезок над отметкой времени (отметки через 0,2 с) показы-
вает длительность освещения, (ii) В ответ на более слабое освещение соседей тоже наблю-
дается торможение активности, но в течение более короткого периода, (iii) Контроль —
соседние омматидии не освещены. (В) Узкие лучи освещают два соседних омматидия. Ре-
зультат показан на (Г). Когда освещен только омматидий А, возникает импульсация в его
волокне; когда освещен только омматидий Б, регистрируется сходная картина. Когда ос-
вещены оба, взаимное торможение уменьшает активность в обоих волокнах зрительного
нерва. Из Hartline, 1959, с разрешения.
ческих событий, завершающейся деполяризацией ретикулярных клеток. Эта
деполяризация приводит, в свою очередь, к деполяризации дендритных окон-
чаний эксцентрических клеток и генерации потенциала действия в аксонах
последних. Эксцентрические клетки интегрируют ответы окружающих рети-
кулярных клеток. Потенциалы действия из эксцентрических клеток направля-
ются в мозг. При этом их аксоны, как видно из рис. 14.16 А, посылают коллате-
рали вскоре после того, как они покидают омматидий. Эти волокна образуют
14. Зрение у беспозвоночных 307
латеральное сплетение, и именно здесь наблюдается типичный пример лате-
рального торможения.
Рис. 14.16 Б показывает эффект стимуляции омматидиев, расположенных
вблизи от омматидия, активность которого изучается. Если на омматидий по-
дается узкий луч света, возникает импульсация некоторой постоянной часто-
ты (она зависит от интенсивности стимула), которая передается по волокну
зрительного нерва из эксцентрической клетки. Однако, если не меняя интен-
сивности освещенния омматидия, на небольшом расстоянии от него (1—2 мм)
подается другой луч, то частота импульсации сначала падает до нуля, а затем
оказывается сильно уменьшенной. Значит, активность в нервном волокне из
соседних омматидиев передается через латеральное сплетение и ингибирует
активность в регистрируемом волокне. Чем больше расстояние между оммати-
диями, тем меньше этот эффект.
Латеральное торможение — это, как мы уже видели и еще увидим при рас-
смотрении зрительной системы позвоночных, одна из наиболее общих черт
сенсорных систем. Особенно легко показать это на сложном глазу мечехвоста
Limulus. Как говорилось выше, индивидуальные омматидии взаимодействуют
между собой еще до того, как посылают информацию в мозг. Таким образом,
они действуют взаимозависимо, как примитивная сетчатка. Биологический
смысл такой организации ясен. Если глаз подвергается действию диффузного
освещения, латеральное торможение уменьшает поток импульсов во всех во-
локнах зрительного нерва. Если же возникает резкий перепад освещенности,
например, когда падает тень, тогда торможение со стороны затененных клеток
уменьшается. Соседние клетки растормаживаются и увеличивают частоту им-
пульсации. Система, таким образом, служит для выделения краев тени, пада-
ющей на сетчатку или перемещающейся по ней — т. е. информации, имеющей
жизненно важное значение для мечехвоста.
14.2.6. Высокоразвитый сложный глаз
Как мы видели в разделе 14.1, у насекомых и ракообразных развилось большое
разнообразие сложных глаз. В данном разделе мы рассмотрим в деталях толь-
ко один тип — нейральный суперпозиционный глаз двукрылых (домашняя му-
ха, падальные мухи, плодовые мушки, комары и москиты). Обычно у двукры-
лых — сложный аппозиционный глаз. Структура аппозиционного омматидия
рассмотрена в разделе 14.1 (рис. 14.3), и здесь мы мало что можем добавить.
Тем не менее рис. 14.17 показывает множество дополнительных деталей, а
также путь света в этой системе. В отличие от омматидиев мечехвоста Limulus,
омматидии двукрылых состоят из фиксированного количества (6 или 7) рети-
кулярных клеток и еще одной или двух клеток, вероятно соответствующих
эксцентрическим клеткам мечехвоста, в нижней части омматидия.
Каждая ретикулярная клетка обладает колонкой микроворсинок, образую-
щих рабдомер. Ретикулярные клетки располагаются близко друг от друга, об-
разуя цилиндр, а их рабдомеры тесно соприкасаются (как это видно на элек-
тронной микрофотографии омматидия златоглазки на рис. 14.17 Б) и образуют
рабдом. Зрительный пигмент (родопсин) располагается в микроворсинках
рабдомеров. Рис. 14.17 А показывает, что рабдом (около 1 мкм в диаметре и до
308 Часть IV. Светочувствительность
Глаз
дневного
насекомого
Глаз
ночного
насекомого
Роговица
Колбочка
Пигментная
клетка
Рис. 14.17. (А) Вертикальный
срез омматидия крупного днев-
ного насекомого. Объяснения
в тексте. (Б) Поперечный срез
омматидия ночного насекомого
Chrysopa (зеленая златоглазка).
В каждом омматидии по шесть
главных ретикулярных клеток,
а микроворсинки их большого
рабдома заполняют большую
часть электронной микрофото-
графии. Одиночный омматидий
занимает центр рисунка. Из Ног-
ridge, 1991, in J. R. Cronly-Dillon
and R. L. Gregory. Vision and Visu-
al Dysfunction, vol. 2, p. 232, figure
11.2, p. 234, figure 11.5. Воспроиз-
водится с разрешения Macmillan
Press Ltd.
> 4 Низкий г
Пигмент показатель пре-
_ ломления^^ !
Ретинулярная
клетка
8-я или 9-я клетка
lull- Аксоны
Б
14. Зрение у беспозвоночных 309
100 мкм в длину) действует как световод или оптическое волокно. Пигментные
гранулы предупреждают потери света в других частях ретикулярных клеток.
Таким образом, свет получает максимум возможности взаимодействовать с
родопсинами рабдомеров. Рис. 14.17 А также показывает, что положение пиг-
ментных гранул в ретикулярных клетках изменяется в ходе суточного цикла.
В течение светового дня пигмент располагается вблизи от микроворсинок раб-
домеров, так что свет концентрируется на зрительном пигменте, и дневное
зрение обладает высокой остротой. При слабом освещении пигментные гра-
нулы мигрируют из рабдомеров, так что острота зрения снижается за счет по-
вышения чувствительности.
Рабдомеры в ответ на освещение деполяризуются. В волокнах, идущих от
основания ретикулярных клеток, импульсы не обнаружены — сигнал переда-
ется электротонически. У многих насекомых в разных ретикулярных клетках
присутствуют родопсины с различающимися Хтах. Свет различных длин волн
вызывает, таким образом, различные типы ответа омматидиев. На этом пост-
роено отличное цветовое зрение, особенно у насекомых, посещающих цветы.
Волокна зрительного нерва из большинства ретикулярных клеток направ-
ляются в зрительную пластину (ламину), расположенную непосредственно под
сложным глазом. Эти волокна именуются короткими зрительными волокнами
(КЗВ), они оканчиваются на узких «оптических обоймах», образующих синап-
сы с волокнами второго порядка, котороые передают информацию высокого
пространственного разрешения в мозг (medulla). Обнаружено, что КЗВ рети-
кулярных клеток, имеющих одинаковые поля зрения в разных омматидиях,
направляются в одни и те же «обоймы». Таким образом, как упоминалось в
разделе 14.1, хотя глаз мухи — аппозиционного типа, с присущей этому типу
разрешающей способностью, нервные связи в зрительной пластинке обеспе-
чивают суперпозицию изображений, дающую всю сопутствующую чувстви-
тельность. Волокна из более глубоких («эксцентрических») ретикулярных кле-
ток проходят, минуя пластину, и оканчиваются непосредственно в мозге
(см. рис. 14.18 А). Этот. н. длинные зрительные волокна (ДЗВ). Из рис. 14.18 А
видно, что и волокна второго порядка, и ДЗВ перекрещиваются между плас-
тиной и мозгом. При этом в обоих зрительных областях имеет место ретиното-
пическая карта, т. е. клетки в этих областях построены в колонки, в соответст-
вии с местом их происхождения в глазу. Эта ретинотопическая организация
сохраняется и после следующего перекреста, приводящего зрительную ин-
формацию в последний слой зрительной доли — лобулу (lobula). У мух этот
слой подразделяется на две части — заднюю (пластина лобулы) и переднюю
(собственно лобулу). Основные медуллярные проекции входят в лобулу меж-
ду этими двумя частями и разветвляются, направляясь в каждую из них
(рис. 14.8 Б).
Помимо волокон, обеспечивающих ретинотопическую организацию, здесь
присутствует множество тангенциальных волокон с обильным ветвлением во
всех трех отделах зрительной доли. Клетки более глубоких частей зрительной
доли (medulla и lobula) реагируют на цвет, направление движения, ориентацию
краев и т. д. Видимо, распознавание объекта начинаеся в лобуле. Мы обнару-
жим аналогичные механизмы в работе зрительной коры позвоночных. После
дольки зрительная информация передается в глубину мозга, куда мы не осме-
310 Часть IV. Светочувствительность
Рис. 14.18. (А) Схема зрительной доли членистоногих. Три главных ганглия зрительной
доли — ламина, медулла и лобула — отделены друг от друга перекрестами. ДЗВ — длин-
ные зрительные волокна, ЗС — змеевидный слой, КЗВ — короткие зрительные волокна,
ТВ — тангенциальные волокна. Дальнейшие объяснения в тексте. Из Osorio, 1991. Вос-
производится с разрешения D. Osorio. (Б) Вторая и третья синаптические области зри-
тельной системы мухи. Волокна из пластины и из сетчатки входят в верхней части рисун-
ка и образуют синапсы в медулле. После следующего перекреста (вторая зрительная хи-
азма) следующие нейроны образуют синапсы в лобуле или пластине лобулы. Зрительная
информация передается в протоцеребрум волокнами зрительного нерва (в нижней части
рисунка). Из Strausfield, 1976. Воспроизводится с разрешения Springer-Verlag GmbH &
Со. KG.
нерв
Б
Медулла
Лобула
Вторая
зрительная
хиазма
Первая
зрительная
хиазма
50 мкм
Зрительный
Пластина
лобулы
лимся за ней последовать. Надо отметить, что рис. 14.18 Б представляет собой
в высшей степени обобщенную схему зрительных долей, демонстрирующую
общие черты многих членистоногих — от ракообразных до насекомых.
Рис. 14.18 Б дает некоторое представление о сложности связей в зрительной
доле мухи, а дальнейшие детали можно найти в работах, приведенных в спис-
ке литературы.
14.3. Заключение
Предшествующий обзор продемонстрировал огромное разнообразие типов
организации глаз, сформировавшихся за полмиллиарда лет истории развития
животных. Здесь по необходимости описаны лишь основные и наиболее про-
14. Зрение у беспозвоночных 311
грессивные типы. Точная информация о расстояниях, предоставляемая орга-
нами, чувствительными к электромагнитному излучению т. н. «видимого спе-
ктра», очевидно имеет исключительное значение для борьбы за существова-
ние. Насколько далеко «видят» беспозвоночные в том смысле, в котором мы
«видим»? Это, как отмечал Эдриан Хорридж, вопрос, трудный для ответа. Зре-
ние человека, как он говорил, с самого начала исполнено смысла. Каждый
объект имеет свое название, даже эмоциональная реакция. Мы не можем из-
бежать этого, стоит лишь открыть глаза и оглядеться. Было бы антропоцент-
ричным предполагать, что зрительная система насекомых или даже белемнои-
дов представляет ее владельцам сходный с нашим опыт или даже намек на та-
кой опыт. Просто по определению мы не можем этого знать. Чтобы знать, на
что похоже их зрение, нам, перефразируя Томаса Нагеля, пришлось бы стать
домашней мухой или осьминогом обыкновенным.
15. ГЛАЗ ЧЕЛОВЕКА
Вероятно, глаз позвоночных существенно неизменен с периода докембрия.
Зрение — стержень человеческого образа жизни. Общий обзор «вегетатив-
ной» анатомии глаза и его вспомогательных органов. Эмбриология —
филогенетические спекуляции — нейроэктодерма — трубчатая ЦНС —
глазное яблоко — инвертированная сетчатка — индукция хрусталика —
афакия, анофтальмия, микрофтальмия — колобома — удаление зритель-
ного желудочка — дифференцировка сетчатки — дифференцировка
роговицы и радужки — формирование склеры и сосудистой оболочки
глаза. Детали анатомии и физиологии: передний эпителий — боуманова
мембрана — строма, коллагеновые волокна и их организация — десце-
метова мембрана — эндотелий — иннервация; поддержание прозрачно-
сти; склера: форма глаза, миопия? хрусталик: волокна хрусталика — про-
зрачность — кристаллины — белки теплового шока — аккомодация —
близорукость (миопия), дальнозоркость (гиперметропия), астигматизм,
старческая дальнозоркость (пресбиопия) — катаракта; сосудистая обо-
лочка глаза: четыре слоя — сосудистое обеспечение сетчатки; ресничное
тело: гистология — ресничные выросты — секреция внутриглазной
жидкости — циркуляция — абсорбция — внутриглазное давление
(ВГД) — глаукома; ресничные мышцы — гистология — иннервация —
аккомодация хрусталика; радужка: гистология — физиология; стекловид-
ное тело. Движения глазного яблока: вестибулярно-зрительные — оптоки-
нетические — отслеживание — саккадические — сведение осей — эффе-
рентные копии. Заключение: соответствие структуры глаза его функции
Глаз позвоночных имеет долгую историю. Мы отмечали в гл. 14, что он возник
вероятно в докембрийском периоде — более, чем полмиллиарда лет назад. Ис-
копаемая Pikaia, идентифицированная как раннее хордовое, обнаружена в
сланцах Бэрджеса, датирующихся 525 миллионами лет. У ископаемого бесче-
люстного (бесщитковые — Anaspida), датируемого силурийским периодом,
обнаружены хорошо развитые билатеральные парные орбиты глаз. В отличие,
однако, от сложного глаза членистоногих, глаза позвоночных не сохранились
в ископаемых отложениях. Мы можем опираться только на «мягкие» структу-
ры примитивных ныне живущих позвоночных — круглоротых (Cyclostomata,
напр. минога), принадлежащих к тому же надклассу, что и бесщитковые и об-
ладающих хорошо сформированными камерными глазами, которые сущест-
венно не отличаются от глаз других ныне живущих позвоночных. Действи-
тельно, за исключением некоторых деталей (см. гл. 18) структура и функция
15. Глаз человека 313
глаз позвоночных значительно не варьирует от бесчелюстных круглоротых,
челюстноротых рыб, земноводных и пресмыкающихся до птиц и млекопитаю-
щих. В этом позвоночные резко отличаются от моллюсков, у которых, как мы
видели в гл. 14, присутствует полный набор глаз — от глазков до совершенных
камерных глаз головоногих. Вследствие их удивительного единообразия, мож-
но рассмотреть глаз млекопитающего, в частности глаз человека, как типовой
пример. В данной главе, поскольку вся тема огромна, мы обсудим только не-
нервную т. н. «вегетативную» часть глаза, а детальное обсуждение сетчатки —
фотосенсорной и нейральной ткани — отложим до гл. 16. Затем, в гл. 18, после
рассмотрения зрительных путей и зрительной коры в гл. 17, мы обсудим неко-
торые вариации на тему по другим позвоночным.
Человек — это «зрительное» создание. Наш менталитет пронизан зритель-
ными образами: мы «визуализируем» результаты экспериментов, «видим ре-
шение», делаем «эскиз» плана, «создаем фон», «представляем себе», «предви-
дим» и т. д. Такой зрительный «уклон» не должен удивлять. Все приматы
ориентированы на зрение. Десятки миллионов лет наши предки провели под
пологом тропического или субтропического леса — хорошее зрение, хорошая
способность определять расстояния, способность отличить прочную ветку от
гибкой, через которую можно проскочить, способность разглядеть путь сквозь
листву, хорошее цветовое зрение, чтобы отличить спелые фрукты от неспелых,
разглядеть съедобный орех, опасного хищника — все это награда, которую да-
ет эволюция за превосходную зрительную систему.
15.1. Анатомия
В этом разделе будет представлен общий обзор, в дальнейших — мы обсудим
каждый из элементов системы более детально. Мы начнем с глазного яблока,
а затем перейдем к дополнительным структурам — вспомогательным органам
глаза.
15.1.1. Глазное яблоко
Рис. 15.1 показывает экваториальный срез глазного яблока. Стенка глазного
яблока состоит из трех слоев (оболочек). Снаружи лежит плотный коллагено-
вый слой — склера. Она образует т. н. «белки глаз» и обеспечивает механичес-
кую прочность глаза, защищая хрупкую внутренность глаза и поддерживая его
форму. Кпереди склера продолжается в прозрачную роговицу. Поверх поверх-
ности роговицы располагается слой прозрачных эпителиальных клеток, явля-
ющихся продолжением эпидермиса кожи — это т. н. конъюнктива. Внутри
склеры располагается увеальный тракт. Название возникло из-за наблюдения
древних, что эта оболочка легко отделяется от склеры рассеченного глаза, ес-
ли за нее потянуть — как кожица винограда (uva (лат.) — виноград. — Автор).
Увеальный тракт состоит из трех основных частей: сосудистой оболочки (со-
судистого пигментированного слоя, выстилающего большую часть задней
камеры глаза), ресничного тела, из которого вырастают цинновы связки (под-
держивающие связки, processes ciliares), удерживающие хрусталик, и радуж-
ки, расположенной перед хрусталиком. Наконец, самый глубокий слой глаз-
314 Часть IV. Светочувствительность
Передний
Передняя камера
Лимб
Выросты
ресничного тела
Радужка
Задняя
камера
полюс
Роговица
Основание
роговицы
с
Хрусталик
1 Задний
полюс
Продырявленная
пластинка
Зрительный
нерв
Ресничное тело
Прямая связка
Зубчатая линия
Склера
Сосудистая оболочка
Экваториальный диаметр
Стекловидное тело
Геометрическая осьх
Оптический диск
Сетчатка
Зрительная ось
Фовеа
Рис. 15.1. Экваториальный срез глаза человека. Объяснения в тексте.
ного яблока, он же — самый важный — это сетчатка. На рис. 15.1 показано, что
она выстилает заднюю часть глазного яблока и простирается вперед до рес-
ничного тела, где оканчивается ora terminalis (или ora serrata — зубчатая ли-
ния, названная так из-за своих очертаний). Здесь оканчивается лишь свето-
чувствительная сетчатка, а ее несветочувствительный эпителий продолжается
дальше, покрывая поверхность ресничного тела и радужки. В задней части
глаза, где зрительная ось пересекает сетчатку, имеется углубление — фовеа,
обильно населенная колбочками, отвечающими за зрение при дневном осве-
щении. Фовеа окружена более широкой круговой областью сетчатки — area
15. Глаз человека 315
centralis, называемой в тех случаях, когда она пигментирована, как у человека
и других приматов, желтым пятном (macula lutea). Оно также, хотя и в меньшей
степени, приспособлено к зрению высокого разрешения. Со стороны носа, а
следовательно вне оптической оси, к area centralis примыкает зрительный диск,
где собираются зрительные нервные волокна, покидающие глаз в составе зри-
тельного нерва. Эта область лишена фоторецепторов, нечувствительна к свету
и именуется слепым пятном — это «слабое место» и в склере, которая в осталь-
ном жесткая. Эта слабость компенсируется тем, что зрительный нерв проходит
здесь через перфорированную коллагенозную пластинку, т. н. продырявлен-
ную пластинку, непосредственно под оптическим диском.
Экваториальный срез на рис. 15.1 показывает, что хрусталик делит внут-
реннюю полость на две части — переднюю и заднюю камеры. Передняя каме-
ра заполнена водянистой влагой — водянистой жидкостью, секретируемой
(как мы увидим) ресничным телом, тогда как задняя камера — желеподобным
стекловидным телом, секретируемым в ходе развития сетчаткой. Стекловид-
ное тело обеспечивает поддержание формы глазного яблока, а его давление '
удерживает сетчатку. Давление водянистой влаги (которая, как мы увидим
ниже, может подвергаться патологическим изменениям) на стекловидное те-
ло оказывает влияние и на сетчатку — если оно возрастает выше нормальных
пределов, нежная сетчатка повреждается, и возникает заболевание, известное
как глаукома, без лечения ведущая к слепоте.
15.1.2. Вспомогательные органы глаза
Вне глазного яблока располагаются вспомогательные органы глаза: внешние
мышцы глаза, веки и слезная система.
Внешние мышцы глаза
Глазное яблоко, как все мы знаем, в глазнице подвижно. Подвижность глаза
обеспечивается шестью наружными мышцами: верхней, нижней, наружной и
внутренней прямыми, и верхней и нижней косыми. Эти мышцы показаны на
рис. 15.2. Верхняя, нижняя и внутренняя прямая мышцы, а также нижняя
косая мышца иннервированы ветками 3-го черепномозгового нерва — глазо-
двигательного. Верхняя косая мышца иннервирована 4-м черепномозговым
нервом — блоковым, а наружная прямая — 6-м черепномозговым нервом —
отводящим. Сухожилие верхней косой мышцы проходит через кольцо слезной
кости (os lacrimale) — одной из костей орбиты глаза. Все эти мышцы действу-
ют согласованно, вращая глаз в его костной орбите.
Веки
С биологической точки зрения глаз находится в рискованном положении.
Другие части поверхности тела защищены многослойным эпителием, но для
глаза это неприемлемо. Прозрачность существенна и должна поддерживаться,
может быть, столетие. Решение проблемы нашлось в развитии сократимых
складок кожи — век.
316 Часть IV. Светочувствительность
Рис. 15.2. Наружные мышцы глаза. Положение наружных мышц глаза. Нижняя и
верхняя косые мышцы прикрепляются позади экватора глазного яблока. Верхняя
косая (как показано) проходит через фиброзно-хрящевую петлю — блок (trochlea),
прежде чем повернуть назад перед присоединением к глазному яблоку под верхней
прямой мышцей. Trochlea действует, как блок. На рисунке видны только три пря-
мых мышцы, четвертая — внутренняя прямая — идет парллельно латеральному
сплетению с носовой стороны орбиты. Наружная прямая мышца показана отре-
занной, чтобы было видно положение зрительного нерва. Сокращение musculus le-
vator palpebrae поднимает верхнее веко. Из Newell, 1996, Ophthalmology: Principles
& Concepts, p. 39, Fig. 1-36: Воспроизводится с разрешения W. B. Saunders Com-
pany.
Веки образуют один из наиболее важных элементов защиты глаза. С помо-
щью ресниц глаз получает предупреждение об опасности и устраняет ее; с по-
мощью секреции предупреждается высыхание, постоянное движение век
вверх-вниз распределяет секрет по поверхности роговицы, а слезная жид-
кость смачивает поверхность и участвует в удалении опасных раздражающих
агентов.
Каждое веко состоит из широкой соединительнотканной пластинки хряща
века (tarsus), повторяющей форму глазного яблока. Ресницы расположены в
два ряда и на верхнем, и на нижнем веках. Они имеют типичные волосяные
фолликулы, каждый из которых окружен нервным сплетением с очень низким
порогом возбудимости. Прикосновение к ресничке достаточно для возбужде-
15. Глаз человека 317
Ресница
Лобно-затылочная мышца
(часть надчерепной мышцы)
Круговая мышца глаза
Рис. 15.3. Вертикальный срез верхнего века.
Бровь
Кожа-
Подкожный ареолярный слой
Субмускулярный ареолярный слои
Кожное сочленение сухожилия
мышцы, поднимающей веко
Предхрящевая часть
круговой мышцы глаза
Сочленение мышцы, поднимающей
веко с хрящом века
Моллова железа
Цейсова железа
Лобная кость
Клетчатка
глазницы
Сухожилие мышцы,
поднимающей веко
Железа Краузе
Свод конъюнктивы
Мюллерова мышца
Перегородка орбиты
Конъюнктива орбиты
Передняя дуга
хряща века
Железа Вольфринга
Хрящ века
Мейбомиева железа
Конъюнктива
хряща века
Нижняя дуга
хряща века
Краевая конъюнктива
Риоланова мышца
Устье мейбомиева
протока и кожно-
слизистое сочленение
ния одного или нескольких волокон и вызова рефлекторного моргания. Непо-
средственно за внутренним рядом ресниц открываются протоки больших
сальных мейбомиевых желез (рис. 15.3). В каждом хряще века таких желез до
тридцати, и они образуют маслянистый секрет — важный компонент слезной
пленки, покрывающей роговицу. Функция секрета состоит в предупреждении
слишком быстрого испарения слезной жидкости, а также проливания ее за
край век. Рис. 15.3 показывает, что внутренняя поверхность век также содер-
жит множество слезных желез — железы Краузе (Krause) и Вольфринга (Wolf-
ring).
Глазная поверхность век покрыта слизистым эпителием — конъюктивой,
которая является продолжением конъюнктивы, покрывающей глазное ябло-
ко. Последняя, в свою очередь, является продолжением эпителия роговицы.
В конъюнктиве обильно представлены бокаловидные клетки, секретирующие
слизь, которые вместе с перечисленными выше железами участвуют в форми-
ровании слезной жидкости. Моргание век распределяет достаточно сложную
слезную жидкость по поверхности роговицы, что является важным условием
нормального ее существования.
У каждого человека — своя собственная частота спонтанного моргания.
В среднем, период между морганиями составляет около 2,8 с у мужчин и
4,0 с — у женщин. У других животных этот период варьирует очень широко.
Длительность закрытого состояния глаза при моргании — от 0,3 до 0,4 с. Мозг
318 Часть IV. Светочувствительность
автоматически компенсирует эти периоды затемнения — мы этого не замеча-
ем, хотя падение освещенности поля зрения всего на 0,03 с (не связанное с
морганием) воспринимается совершенно ясно.
Моргание — это и рефлекторный ответ на опасность. Оно может быть вы-
звано громким шумом, тактильными стимулами вблизи глаз, неожиданным
ярким светом. Тактильный рефлекс исчезает при поражениях коры в области
роландовой борозды; рефлекс на ослепление светом — при определенных по-
ражениях среднего мозга. Таким образом, рефлексы моргания являются кли-
ническими признаками специфических поражений мозга.
Слезная система
Мы уже упоминали некоторые железы слезной системы в предыдущем разде-
ле. Основным источником слезной жидкости является, однако, слезная желе-
за, располагающаяся в верхнем латеральном углу орбиты глаза. Железа имеет
10—12 средних размеров протоков, открывающихся в свод конъюнктивы. Они
секретируют водянистую жидкость, которая, как показывает табл. 15.1, не
сильно отличается по составу от плазмы, за исключением намного более высо-
кой концентрации К+ и С1~ и отсутствия большей части органики. В слезной
жидкости присутствует множество ферментов, наиболее важный из них —
лизоцим, который атакует бактерии, растворяя их клеточные стенки. Слезная
жидкость двигается по поверхности роговицы в результате мигательных дви-
жений и дренируется в носовом углу глаза системой, в которую входят слезное
озеро, канальцы, слезный мешок и носо-слезный проток, ведущий в носовую
полость (рис. 15.4).
Пленка слезной жидкости состоит, главным образом, из водянистого сек-
рета слезной железы, но ее поверхность образована маслянистым слоем секре-
та мейбомиевых желез. Это важно, как отмечалось выше, для предупреждения
быстрого высыхания тонкой пленки слезной жидкости. Слизистые секреты
Таблица 15.1. Состав слезной жидкости
Плазма Слезная жидкость
Электролиты (мМ) N+ 137,5 135,0
К+ 4,3 36,0
С1- 108,5 131,0
нсо3- 27,0 27,0
Са2+ 2,3 0,5
Mg2+ 0,2 0,36
Органические вещества (мг/мл) глюкоза 0,8 0,05
лизоцим — 2,1
альбумин 40-48 0,04
Данные из Tiffany, 1997.
15. Глаз человека 319
Рис. 15.4. Слезная система.
Слезная жидкость двигается
по поверхности глаза в на-
правлении, указанном стрел-
ками. к — канальцы, СЖ —
слезная железа, НМ — ни-
жний мениск, СМ — слезный
мешок, НСП — носо-слезный
проток, о — слезное озеро,
ВМ — верхний мениск. Даль-
нейшие объяснения в тексте.
Из Hardling, 1997, с разреше-
ния.
конъюнктивы образуют самый нижний слой слезной пленки, который удер-
живается множеством микроворсинок роговичного эпителия (см. раздел
15.3.1). Слезная пленка хорошо оксигенирована в результате мигательных
движений век, а захват кислорода из нее роговичым эпителием важен для под-
держания состояния роговицы.
Рефлекторная секреция из слезной железы вызывается раздражением ро-
говицы, конъюнктивы, слизистой носа, а также термическими стимулами,
ярким светом, воздействием сильно перченной пищи в рот и на язык. Слезо-
течение вызывается также эмоциональными расстройствами. Перерезка трой-
ничного нерва (5-й черепномозговой нерв) устраняет все слезные рефлексы,
но не влияет на психогенное слезотечение. Действительно, психогенное сле-
зотечение (как и смех) — вероятно, присуще только человеку. В регуляции
слезоотделения важна также парасимпатическая система — ее волокна идут
в составе лицевого нерва (7-й черепно-мозговой нерв) к слезной железе, а
парасимпатомиметические препараты вызывают усиление слезотечения и его
продолжительность.
15.2. Эмбриология
Давайте теперь вновь обратимся к самому глазу. Нет лучше способа понять его
анатомию, чем проследить его возникновение в эмбриогенезе. Если мы при-
знаем за биогенетическим законом Геккеля какую-то правоту, то изучение
развития глаза (по крайней мере, самых ранних его стадий) может принести
нам некоторое понимание его эволюционного происхождения. Мы увидим,
что происхождение глаза позвоночных существенно отличается от происхож-
дения камерных глаз беспозвоночных, которые мы уже рассмотрели в гл. 14
(не забывайте, однако, о признаках их изначальной общности, которая упоми-
налась в Информации 14.2). Глаз человека не возникает непосредственно в ре-
зультате инвагинации поверхностного эпителия, как у беспозвоночных (см.
однако, ниже). Сетчатка формируется окольным путем — внутри центральной
нервной системы, и в связи с таким непрямым происхождением именуется
инвертированной. Фоторецепторы при этом оказываются отвернутыми от по-
ступающего света.
320 Часть IV. Светочувствительность
Рис. 15.5. Серия поперечных
срезов, показывающих эмбрио-
логическое происхождение сет-
чатки и хрусталика, (i) ранний
зародыш (нейрула), в котором
полоска нейроэктодермы диф-
ференцируется на дорзальной
поверхности; (ii) нейроэктодер-
ма углубляется, и образуются
две впадины — foveola optica
(зрительные ямки), (iii) начи-
нается формирование нервной
трубки, (iv) выпячивание зри-
тельного пузырька; (v) начина-
ет формироваться глазной бо-
кал, а хрусталик — индуциро-
ваться эктодермой; (vi) продол-
жение инвагинации глазного
бокала и формирования хруста-
лика; (vii) глазной бокал сфор-
мирован почти полностью, хру-
сталик отделился от эктодермы.
Отметьте положение наружных
сегментов фоторецепторов на
всех рисунках, оя — оптические
ямки, х — хрусталик, пх — пла-
кода хрусталика, н — нейроэк-
тодерма, зж — зрительный же-
лудочек, нс — наружные сег-
менты палочек.
Глаз позвоночного развивается частично из мезодермальных, а частично -
из эктодермальных тканей. Эктодерма формирует сетчатку и ненервные эпи-
телиальные продолжения сетчатки, покрывающие ресничное тело и заднюю
часть радужки. Эктодерма также образует хрусталик. Мезодермальная ткань
формирует все остальное. Часто говорят, что «сетчатка — это часть мозга, вы-
двинутая на периферию». Мы поймем смысл этого афоризма, если проследим
ее эмбриональное происхождение.
Центральная нервная система хордовых начинает свое формирование как
специализированная полоска эктодермы — нейроэктодерма — идущая вдоль
дорзальной поверхности раннего зародыша (рис. 15.5). В этом отношении она
радикально отличается от центральной нервной системы беспозвоночных, ко-
торая все время занимает вентральное положение. Означает ли это, что наш
древнейший предшественник, в отличии от ползающего по дну предка беспо-
звоночных, был свободноплавающим в верхних слоях океана, где свет (а с ним
и тени от хищников и жертв) шли сверху? Как бы то ни было, то, что в будущем
станет фоторецепторами, было погруженным в эту полоску нервной ткани; на
том этапе эволюции фоторецепторы были неинвертированы (если пользовать-
15. Глаз человека 321
ся этим неуклюжим1 неологизмом, введенным сравнительными анатомами
зрительной системы), поскольку были обращены к поступающему извне све-
ту. В ходе эмбриогенеза нейроэктодерма сначала заглубляется, образуя снача-
ла «желобок», а затем замыкается, формируя трубчатую центральную нерв-
ную систему — специфический признак позвоночных. Как показывает рис.
15.5, в результате примордиальные фоторецепторы оказываются направлен-
ными внутрь центрального канала ЦНС. Возможно, и в таком положении они
сохраняли свою светочувствительность, если примитивные хордовые (такие,
как головохордовый Amphioxus — ланцетник) были прозрачными. Однако, та-
кое несообразное расположение вряд ли могло быть долговременным реше-
нием проблемы. По мере развития хордовых росли размеры и непрозрачность
их тел, и фоторецепторы должны были мигрировать поближе к поверхности
тела.
Рис. 15.5 показывает, как это происходит. Эмбриологические исследова-
ния показали, что еще до образования нервной трубки, два углубления при-
мордиальной сетчатки — зрительные ямки (foveolae opticae), становятся замет-
ны в той части нейроэктодермы, которая впоследствии образует передний
мозг. Можно ли видеть в этих углублениях аналогию с инвагинацией, ведущей
к формированию камерного глаза (сравните с корабликом и осьминогом в
гл. 14)? Как только нейроэктодерма сворачивается, образуя нервную трубку,
эти ямки увеличиваются и выдвигаются к поверхности на обеих сторонах пе-
реднего мозга. Они быстро расширяются, как будто их раздувают изнутри, и
образуют зрительные пузырьки, которые остаются соединенными с передним
мозгом зрительными ножками. На этом этапе надо отметить два важных момен-
та. Во-первых, ресничные фоторецепторы остаются обращенными внутрь, в
то, что теперь именуется зрительным пузырьком. Во-вторых, как только зри-
тельный пузырек входит в контакт с вышележащим эпителием, начинается
утолщение и заглубление последнего — это плакода или примордиум хрустали-
ка. Вероятно, какие-то еще неизвестные биохимические влияния распростра-
няются в слой эпителия, индуцируя формирование хрусталика. Это один из
наиболее ранних в развитии и известных случаев эмбриональной индукции —
до сих пор один из наиболее впечатляющих и, несмотря на поразительный
прогресс молекулярной эмбриологии, до сих пор таинственный. Если удалить
зрительный пузырек, то хрусталик не образуется; если зрительный пузырек
имплантировать под эпидермис в какой-либо другой части тела, даже в туло-
вище, в этом месте индуцируется хрусталик.
Закладка хрусталика, как показывает рис. 15.6, сначала представляет собой
бокал, а затем полую сферу из клеток. Затем, однако, клетки задней стенки
сферы начинают удлиняться, вырастая в полость сферы, и вскоре полость сфе-
ры преобразуется в плотную массу клеток. Длинные тонкие клетки называют-
ся волокнами хрусталика. Такой рост исходно кубоидных клеток в длинные
тонкие волокна продолжается в течение всей жизни в области экватора хрус-
1 Замечание автора подчеркивает языковую корявость английского термина, обратного
к «inverted» (инвертированный) — «verted» (неинвертированный), который, строго говоря,
в английском существует в совершенно ином значении — плоский или даже зеленый. —
Прим, перев.
322 Часть IV. Светочувствительность
Рис. 15.6. Формирование хрусталика в ходе эмбриогенеза. (А) Инвагинация по-
верхности эктодермы вместе с базальной мембраной; (Б) отделение от эктодермы
полой клеточной сферы; (В) удлинение клеток задней поверхности сферы, кото-
рые в конце концов облитерируют полость; (Г) экваториальный срез полностью
сформированного хрусталика. Клетки задней поверхности (волокна хрусталика)
удлинены настолько, что полностью выполняют полость и примыкают к эпителию
передней части. Их ядра (как показано на рисунке) образуют характерные «дуги».
Базальная мембрана превращается в капсулу хрусталика. Выросты ресничного те-
ла присоединяются к капсуле. Из Davson, 1984. Воспроизводится с разрешения
Academic Press Ltd.
талика. Хрусталик становится похож на луковицу тем, как слой накладывается
на слой. Поскольку волокна слишком коротки, чтобы покрыть всю длину хру-
сталика, растущий кончик одного волокна контактирует с кончиком другого,
а точки их соединения образуют т. н. швы хрусталика. Мы вернемся к рассмо-
трению зрелого хрусталика в разделе 15.3.3.
Нарушения эмбрионального развития вызывают наиболее тяжелые по-
следствия, когда они происходят на ранних стадиях. Хрусталик в этом смыс-
ле — не исключение. Таким образом, если в самом начале процесса зритель-
ный пузырек не вступает в контакт с вышележащим эпителием, хрусталик мо-
жет не развиться вообще — такая патология именуется афакией. Отсутствие
хрусталика, в свою очередь, влияет на развитие самого зрительного пузырька,
15. Глаз человека 323
Передний
мозговой пузырь
Ножка
(рассечена)
Ножка
Полость среднего мозга
Эктодерма
хрусталика
Изгиб среднего
мозга
Зрительный
пузырек
(рассечен)
пузырек
желудочек
головного
мозга
Стенка
промежуточного
мозга
Наружный
слой глазного
бокала
Полость
ножки
Зародышевая
Ножка борозда
Эктодерма
Пузырек
хрусталика
Внутренний слой
глазного бокала
/
Рис. 15.7. Развитие глазного бокала и хрусталика. (А) Зародыш человека длиной 4 мм;
(Б) Зародыш человека длиной 7,5 мм. Из W. J. Hamilton, J. D. Boyd, Н. W. Mossman,
1945, Human Embryology, Cambridge: Heffer.
что приводит к анофтальмии1. Если взаимодействие хрусталика и зрительного
пузырька подавлено не полностью, то может сформироваться менее грубая па-
тология — микрофтальмия (недоразвитие глаза).
Вернемся, однако, от этих эмбриологических катастроф к нормальному
развитию зрительного пузырька. Как показывает рис. 15.7, зрительный пузы-
рек начинает деградировать — он проваливается внутрь самого себя. Полость
желудочка медленно исчезает. Процесс, однако, не так примитивен — это не
просто «вдавливание» одной стороны резинового мячика. Как показывает
рис. 15.8, зрительный пузырек сначала в основном располагается над зритель-
ной ножкой. Далее, по мере развития его рост распространяется в вентральном
направлении, образуя при этом большое углубление, края которого (губы),
в конце концов, смыкаются, образуя туннель. Если губы глазного бокала не в
состоянии встретиться и слиться, то формируется патология, именуемая коло-
бомой. К счастью, это случается редко. В норме губы глазного бокала сливают-
ся прибл. через 6 недель беременности, и через образовавшийся туннель начи-
нает прорастать временная гиалоидная артерия, которая обеспечивает дальней-
шее развитие глаза. Эта артерия достигает глазного бокала вплоть до задней
поверхности хрусталика и посылает оттуда ветви ко всем частям бокала. Когда
развитие глаза завершается (у человека — перед рождением) артерия атрофи-
руется вплоть до точки, из которой она начинала прорастать в то, что к концу
развития превращается в заполненную стекловидным телом заднюю камеру
глаза. Вместе с остатками «первичного стекловидного тела», которое в боль-
шей степени выполняло функцию обеспечения развития, она образует канал
Клоке, остающийся видимым и в глазу взрослого. Проксимальная часть гиало-
идной артерии становится теперь arteria centralis retinae. Она и теперь идет по
прежнему маршруту, но теперь в связи в зрительным нервом, входит в сетчат-
1 Буквально — отсутствие глаза. — Прим, перев.
324 Часть IV. Светочувствительность
Рис. 15.8. Объемное представление формирования глазного бокала. Эмбриональная
щель хорошо видна на рис. (В) и (Г), а на рис. (Д) показана проникающая в ее прокси-
мальную часть гиалоидная артерия. Дальнейшие объяснения в тексте. Из W. J. Hamilton,
J. D. Boyd, Н. W. Mossman, 1945, Human Embryology, Cambridge: Heffer.
ку в оптическом диске и дает ветви в сетчатке, где вместе с vena centralis retinae
обеспечивает, в основном, кровообращение.
Превращение исходного зрительного пузырька имеет и другое следствие.
Если мы вспомним рис. 15.5, мы увидим, что когда исходный зрительный же-
лудочек исчезает, стенка глазного бокала становится двухслойной. Наружный
слой глазного бокала формирует пигментный эпителий сетчатки, а внутрен-
ний — становится самой сетчаткой. Хотя зрительный желудочек почти полно-
стью закрывается в глазу взрослого, соединение сетчатки с пигментным эпи-
телием остается слабым местом — отрыв сетчатки обычно происходит именно
здесь. В норме сетчатка удерживается на месте давлением стекловидного тела.
Теперь слоям сетчатки остается только дифференцироваться. Сетчатка —
это, как мы подчеркивали выше, «часть мозга, выставленная наружу». Из этого
следует предполагать, что ее дифференцировка сходна с таковой других частей
мозга. Цилиндрические эпителиальные клетки, выстилавшие исчезнувший
зрительный желудочек, преобразуются в герминативный слой. Первоначально
число клеток в этом слое невелико. Приблизительно на 4-й неделе развития
наступает стадия быстрой пролиферации, и число клеток здесь резко возрас-
тает. После завершения этой стадии клетки вступают в дифференцировку и
перемещаются из зрительного пузырька к стекловидному телу. Первыми
дифференцируются ганглиозные клетки сетчатки. Они перемещаются из гер-
минативного слоя вдоль волокон, образуемых глиальными клетками сетчат-
ки — мюллеровыми клетками. По мере их перемещения из герминативного
слоя у них развиваются аксоны, идущие в составе зрительного нерва по нож-
ке к промежуточному мозгу. После этой первой волны дифференцировки из
герминативного слоя выделяется другой тип клеток — амакриновых. Возмож-
но, это изменение типа дифференцировки происходит вследствие секреции
ганглиозными клетками фактора, ингибирующего дальнейшую дифференци-
ровку герминальных клеток в ганглиозные. Амакриновые клетки, в свою оче-
редь, мигрируют из герминативного слоя к поверхности сетчатки, секретируя
вещества, ингибирующие формирование следующих амакриновых клеток.
Вслед за ними формируются биполярные, горизонтальные и интерплекси-
формные — и все они покидают герминативный слой, двигаясь в сторону
Складка века начинает развитие
Внеглазная мезодерма начинает
дифференцировку в мышцу
Вторая волна
мезодермы
(будущая строма
роговицы)
Полость хрусталика
исчезает с ростом
первых волокон
Круговой сосуд
Гиалоидный сосуд
Дифференцировка
нейральной сетчатки
Внеглазная мезодерма
начинает дифференцировку
в сосудистую оболочку
и фиброзную склеру
Б
Круговая мышца глаза
и другие ткани века
Формирование фибрилл
в определившейся склере
Сосудистая оболочка
Пигментный слой сетчатки
содержит меланин
Третья волна мезодермы
(будущая строма радужки)
Дифференцировка сетчатки,
включая рост волокон
зрительного нерва
Эпителий роговицы
Строма роговицы, образующаяся
в результате фибриллогенеза
Эндотелий роговицы
Хрусталик с капсулой, полость
которого исчезает в результате
роста первичных волокон хрусталика
Круговой сосуд
Развивающиеся
сосуды сетчатки
Эпителий конъюнктивы
Внеглазная мышца
Мезодерма
Эпителий роговицы
Боуманов слой
Строма роговицы
Десцеметова мембрана
Эндотелий роговицы
Мышца-сфинктер зрачка
Мембрана зрачка
Краевой синус радужки
уменьшается
Пигментный эпителий радужки
Мышца-расширитель радужки
Сосудистая строма радужки
Главный артериальный
круг радужки
Вырост ресничного тела
с эпителиальными слоями
Волокна косой
ресничной мышцы
Волокна продольной
ресничной мышцы
Рис. 15.9. Серия срезов показывает прогрессивное развитие глаза с прибл. 5 недель до 6 ме-
сяцев беременности. (А) 5 недель. Отметить развитие роговицы и хрусталика. (Б) 8 недель.
Обратите внимание на дальнейшее развитие роговицы и хрусталика, радужки и сетчатки.
(В) 6 месяцев. Глаз почти полностью сформирован. Дальнейшие объяснения в тексте.
326 Часть IV. Светочувствительность
стекловидного тела и дифференцируясь во все более сложном коктейле инги-
биторов. Последними дифференцируются фоторецепторные клетки — палоч-
ки и колбочки. Эти клетки не мигрируют, а остаются по соседству с теперь уже
виртуальным зрительным пузырьком, опутанные отростками пигментных
эпителиальных клеток.
Теперь клеточная структура сетчатки завершена, более подробно она будет
рассмотрена в гл. 16. Стоит отметить, что не вся сетчатка формируется одно-
временно — дифференцировка происходит в первую очередь на заднем полю-
се глазного бокала (дно), и только несколько недель спустя — на периферии.
Вероятно (хотя до сих пор остается объектом противоречий), что причиной
больших различий плотности клеток (особенно, ганглиозных) в разных частях
сетчатки является их избирательная гибель. Например, в фовеа ганглиозных
клеток на несколько сотен больше на единицу поверхности, чем на периферии
сетчатки. Наконец, стоит напомнить, что все эти сложные процессы оставля-
ют фоторецепторные клетки направленными внутрь — отвернутыми от пада-
ющего на них света, да еще и покрытыми несколькими слоями нейронов. Это
должно быть головоломкой для тех, кто, считая эволюционные аргументы ни-
же своего достоинства, во всем уповает на всемудрость Творца.
Пока сетчатка и хрусталик дифференцируются, остальные части глаза тоже
развиваются. Перед хрусталиком три волны мезодермальных клеток мигриру-
ют под эпителий роговицы (рис. 15.9). Первая волна затем образует рогович-
ный эндотелий, вторая, следующая за первой с интервалом в неделю — начи-
нает формирование стромы роговицы, а третья, растущая под первыми двумя
и прямо перед хрусталиком — передний слой клеток радужки. Пока происхо-
дят все эти процессы, хрусталик продолжает свое развитие и под действием
всех описанных выше процессов, а также постепенного нарастания внутри-
глазного давления (в результате образования водянистой влаги и стекловидно-
го тела), роговица начинает выпячиваться наружу из склеры. Так образуется
перегиб в зоне соединения роговицы со склерой — лимб.
Роговица продолжает дифференцировку. Передний эпителий развивается
в три клеточных слоя, строма начинает секретировать мукополисахариды и
коллагеновые волокна, тогда как эндотелий сохраняет структуру одноклеточ-
ного слоя (рис. 15.9). В ходе дальнейшего развития коллагеновые волокна упо-
рядочиваются в очень строго ориентированную группу пластинок (см. раздел
15.3.1). В склере этого не происходит. Вследствие этого роговица становится
прозрачной, а склера — все более непрозрачной. Одновременно мезодерма во-
круг глазного бокала дифференцируется в сосудистую оболочку, а внешние
мышцы начинают присоединяться к развивающейся склере. На седьмом меся-
це беременности глаз завершает развитие (хотя его диаметр и составляет всего
10—14 мм), сетчатка, за исключением фовеа, полностью развита, а гиалоидная
артерия начинает атрофироваться, оставляя лишь канал Клоке.
15.3. Подробности анатомии и физиологии
Завершив обзор развития глаза человека, вернемся к структурам взрослого
глаза и рассмотрим подробно их анатомию и физиологию.
15. Глаз человека 327
15.3.1. Роговица
Мы видели в разделе 15.2, что роговица состоит из трех слоев: передний
эпителий, строма (lamina propria) и эндотелий. Слои отделены друг от друга
двумя мембранами — боумановым слоем (между эпителием и стромой) и
десцеметовой мембраной — между стромой и эндотелием. Эта пятислойная
структура показана на рис. 15.10.
Передний эпителий состоит из пяти или шести слоев стратифицированных
неороговевших клеток. Самый внутренний слой — герминальный, в нем про-
является митотическая активность, и клетки делятся в течение всей жизни. По
мере их движения к поверхности они уплощаются и в конце концов смывают-
ся слезной жидкостью. Здесь также представлены немногочисленные неэпи-
телиальные клетки, в том числе лимфоциты, макрофаги и узкие сильно окра-
шивающиеся клетки неизвестной функции, иногда называемые лангергансо-
выми, полигональными или дендритическими.
Эпителиальные клетки в средних слоях образуют глубокие инвагинации
поверхностных мембран, в которые встраиваются выросты соседних клеток.
Эти клетки называются крыльчатыми, и их тесное переплетение обеспечивает
механическую прочность слоя. На поверхности клетки уплощаются и утрачи-
вают органоиды, но образуют многочисленные микроворсинки, которые про-
ецируются в слезную жидкость и удерживают ее (см. раздел 15.1).
Боуманов слой. Электронная микроскопия показывает, что этот слой со-
стоит из многочисленных тонких коллагеновых волокон 25 нм в диаметре,
вплетенных в мукополисахаридный матрикс. Тот факт, что боуманов слой се-
кретируется подлежащей стромой, а не вышележащим эпителием, означает,
что его правильнее рассматривать именно как слой, а не мембрану.
Строма (lamina propria). Возможно, это наиболее интересный слой, к тому
же наиболее толстый из слоев, образующих роговицу (ок. 480 нм, 88% толщи-
Передний
эпителий
Боуманов
слой
Строма
substantia
propria
Поверхностные
чешуйчатые клетки
Крыльчатые клетки
Герминальный слой
Фибробласт или
частица роговицы
Пластинка коллагеновых
волокон, срезанная косо
Пластинка коллагеновых
волокон, срезанная продольно
Десцеметова
мембрана
Эндотелий
Рис. 15.10. Поперечный срез роговицы глаза взрослого. Объяснения в тексте.
328 Часть IV. Светочувствительность
ны роговицы). Строма состоит из клеток (фибробластов, иногда называемых
роговичными частицами), матрикса и коллагеновых волокон. Матрикс состоит
из протеогликанов, гликопротеинов, неорганических солей и воды.
Коллаген — это один из наиболее распространенных белков в царстве жи-
вотных от стрекающих до приматов. Он обнаружен во всех соединительных
тканях, где он обеспечивает механическую, особенно — на растяжение, проч-
ность. Очень важен он и в стенке глаза. В роговице, однако, есть некоторые
проблемы. Прочность на растяжение здесь важна, но в равной степени, если
не более, важна и прозрачность. Это увлекательная конструктивная задача, но
прежде, чем мы рассмотрим, как Премудрая природа ее решила, мы обсудим
природу самого коллагена.
Как и можно было ожидать в случае такой древней и вездесущей молекулы,
коллаген присутствует во множестве различных вариантов (по крайней ме-
ре — 19). К счастью (для студентов), все они имеют множество общих важных
черт — состоят из цепочек аминокислот, но с поразительно малым количест-
вом различающихся остатков. Глицин — каждый третий остаток, пролин и ок-
сипролин также очень распространены. Такой незатейливый состав навязан
коллагену его вторичной структурой. Аминокислотные цепочки коллагена
обернуты друг вокруг друга и образуют «трехволоконный канат», где отдель-
ные «волокна» связаны между собой водородными связями. Такая простран-
ственная структура возможна, только если аминокислотная последователь-
ность строго соблюдается. В результате образуется трехволоконная фиброзная
молекула — тропоколлаген, обладающая большой прочностью на растяжение.
Это название происходит от «тропос» — обращенный внутрь из-за того, что
коллагеновые волокна соединительных тканей, включая роговицу, выстланы
тропоколлагеновыми молекулами, соединенными «конец в конец» и бок о
бок — в шахматном порядке (рис. 15.11). Гидрокисльные группы некоторых
остатков лизина и оксилизина окислены, и это позволяет установить связи
между соседними молекулами тропоколлагена. Таким образом формируется
жесткое нерастяжимое волокно. Показано, что фибробласты синтезируют мо-
лекулы тропоколлагена и выбрасывают их в матрикс, и только здесь происхо-
дит сборка коллагеновых волокон (рис. 15.11).
В физиологических условиях каждое коллагеновое волокно роговицы че-
ловека имеет диаметр 31 нм с промежутками между отдельными волокнами в
55 нм. Волокна погружены в матрикс, состоящий из протеогликанов и глико-
протеинов. Они образуют правильный квазикристаллический слой или ламел-
лы толщиной 1,5—2,5 мкм. В строме около 200 ламелл, и каждая состоит из во-
локон, идущих под большим углом в волокнами ламелл, расположенных выше
и неиже (рис. 15.12а). В каждой ламелле волокна идут параллельно друг другу
от грани до грани. Эта чрезвычайно правильная структура существенна для
прозрачности роговицы. Мы увидим ниже, что всякое нарушение структуры
приводит к непрозрачности роговицы.
Десцеметова мембрана — секретируется эндотелием и потому является ис-
тинной базальной мембраной. Имеет толщину около 10 мкм и состоит в ос-
новном из коллагеновых волокон.
Эндотелий состоит из уплощенных клеток высотой около 5 мкм и 20 мкм
шириной. Электронная микроскопия показывает, что клетки заполнены орга-
15. Глаз человека 329
Рис. 15.11. Формирование коллагеновых волокон. Тропоколлагеновые волокна синтези-
руются и заворачиваются в трехволоконную структуру в эндоплазматическом ретикулуме
и в аппарате Гольджи фибробласта (роговичной частицы). Они секретируются в межкле-
точное пространство, где они показаны как линейные структуры с «головкой», обозна-
ченной стрелкой, и «хвостом», обозначенным кружком. Во внеклеточном пространстве
они выстраиваются «головка» к «хвосту» и «бок о бок» в шахматном порядке. Поскольку
длина каждой молекулы тропоколлагена составляет около 280 нм, такая организация
приводит в образованию полосок величиной в 70 нм, различимых в электронном микро-
скопе. Эти волокна затем выстраиваются, образуя высокоорганизованную структуру
ст^мы роговицы. КВ — коллагеновые волокна, Г — комплекс Гольджи, Я — ядро,
ШЭР — шероховатый эндоплазматический ретикулум, СП — секреторный пузырек,
ТК — тропоколлаген.
ноидами — митохондриями, эндоплазматическим ретикулумом, тельцами
Гольджи и т. д., что характерно для клеток с высоким метаболизмом. Клетки в
ходе жизни не воспроизводятся, соединены между собой тесными контактами
(tight junction), исключающими диффузию между ними, и щелевыми контак-
тами (gap junction), позволяющими латеральную передачу веществ из клетки в
клетку. Поскольку клетки не способны возобновляться, любое серьезное по-
вреждение не может быть скомпенсировано, и приводит к необратимому оте-
ку и непрозрачности.
Иннервация. Наружная часть стромы богато иннервирована ветками глаз-
ной порции тройничного нерва (5-й черепно-мозговой нерв). Роговица, еле-
330 Часть IV. Светочувствительность
Рис. 15.12. Коллагеновые ламеллы в роговице. (А) Ориентация ламелл в роговице. По-
казаны три фибробласта между ламеллами. (Б) Электронная микрофотография (уве-
личение 28 000х) коллагеновых волокон стромы роговицы. Вследствие их ориентации
(см. выше (А)), первый коллагеновый слой (а) срезан косо, второй (б) поперечно, а тре-
тий (в) — продольно. Стрелка показывает место бифуркации ламеллы. Из Hogan, Alva-
rado and Weddel, 1971, Histology of the Human Eye, Fig. 3.24A, Fig, 3.26B, Fig. 4.1, Fig. 12.3.
Воспроизводится с разрешения W. B. Saunders Company.
15. Глаз человека 331
Рис. 15.13. Повторяющиеся еди-
ницы дерматан сульфата (глюко-
заминогликана). Единица ИдК-
ГалНАц может повторяться до
100 раз. И сульфат, и карбоксиль-
ная кислота несет отрицательный
заряд.
Идуроновая____н-ацетилгалактозамин
L кислота сульфат
— ИдК
довательно, очень чувствительна, но способность к определению локализации
раздражения очень слаба.
Физиология роговицы. Смысл физиологии роговицы — это поддержание ее
прозрачного состояния. Как мы уже видели, это серьезнейшим образом зави-
сит от правильной организации коллагеновых волокон, погруженных в мат-
рикс, состоящий из протеогликанов, гликопротеинов, неорганических солей
и воды. Наиболее существенное значение имеют протеогликаны. Они состав-
ляют до 10% сухого веса роговицы. Они состоят из т. н. белка-сердцевины
(core protein) весом около 40 кДа, к которому присоединены одна или несколь-
ко глюкозаминогликановых (ГАГ) цепочек. Существует множество различных
ГАГ, но — все это крупные (до 50—60 кДа) отрицательно заряженные поли-
сахаридные цепочки, состоящие из повторяющихся дисахаридных единиц.
Рис. 15.13 показывает обычный ГАГ: дерматан сульфат. Один из моносахари-
дов в каждом дисахаридном повторе имеет присоединенную иминогруппу.
В строме роговицы больше воды (75—80%), чем в какой-либо другой со-
единительной ткани, и она жадно ее поглощает. Это свойство возникло бла-
годаря изобилию отрицательных зарядов на цепочках ГАГ, которые притяги-
вают положительно заряженный полюс диполя молекулы воды (рис. 15.14;
см. также рис. 2.1 — структура диполя молекулы воды). Гидратация цепочек
ГАГ, однако, увеличивает их размер, отодвигает коллагеновые волокна друг
от друга и нарушает правильность их построения, что ведет к потере прозрач-
ности.
Метаболизм роговицы направлен на предупреждение такого эффекта. Во-
первых, между эпителием и эндотелием роговицы существует осмос. Если на
какой-либо поверхности оказывается гипертонический раствор, вода покида-
ет роговицу, и она становится тоньше. Обычно и слезная жидкость, и водяни-
стая влага в передней камере изотоничны строме роговицы, так что осмотиче-
ское давление отсутствует. Это, однако, не устраняет гигроскопичности ГАГ.
Если обе мембраны проницаемы для воды, как это показывают осмотические
332 Часть IV. Светочувствительность
Рис. 15.14. Схема, показывающая
биохимические механизмы, поддер-
жйвающие гидратацию роговицы.
Глюкозаминогликаны притягивают
молекулы воды своими отрицатель-
ными зарядами. Глюкоза попадает
сюда, главным образом через эндо-
телий из водянистой влаги (запол-
няющей переднюю камеру). Мень-
ший поток глюкозы попадает сюда
через эпителий из слезной пленки.
Ферменты цикла Кребса и другие
формы энергетического метаболиз-
ма катаболизируют глюкозу и пере-
носят освободившуюся энергию на
высокоэнергетические фосфатные
связи АТФ. Этот запас энергии, в
свою очередь, используется на нуж-
ды ткани и, что наиболее важно, на
работу насоса, выбрасывающего
НСО3” и Na+ через эндотелий. Вода
идет пассивно по осмотическому
градиенту. ГАГ — глюкозаминогли-
каны, М — метаболизм, Н — насос.
Дальнейшие объяснения в тексте.
эксперименты, вода все равно будет затягиваться в строму положительными
зарядами вокруг ГАГ.
Решение, естественно, состоит в откачке избыточной воды наружу. Поэто-
му роговица располагает гликолитическим метаболизмом. Глюкоза присутст-
вует и в водянистой влаге (см. ниже), и в слезной жидкости (табл. 15.1). Кис-
лород также присутствует и в водянистой влаге, и в слезной жидкости, как и
ферменты цикла Кребса, гексоза-монофосфатный шунт и анаэробный глико-
лиз. Большинство этих ферментов присутствует в эндотелии, который, как мы
отмечали выше, имеет ультраструктурные признаки метаболически активной
ткани. Энергия, полученная в результате деградации глюкозы, используется
для активации насоса, который транспортирует НСО^“ и Na+ из стромы через
эндотелий в водянистую влагу передней камеры (рис. 15.14). Молекулы воды
выносятся пассивно вместе с ионами карбоксила и натрия. То есть вода, втя-
нутая гигроскопичностью ГАГ, удаляется эндотелиальным насосом. Роговица
поддерживатеся в состоянии динамического равновесия между гигроскопи-
ческим притягиванием ГАГ и активностью насоса эндотелия. Если падают за-
пасы кислорода или появляются метаболические ингибиторы, подавляющие
насос, то вызванный ГАГ поток воды внутрь стромы настолько нарушает
организацию коллагена, что роговица теряет прозрачность.
15. Глаз человека 333
15.3.2. Склера
Склера представляет собой плотную белую соединительную ткань. Впереди
она является продолжением роговицы, а сзади окружает оптический диск и
продолжается как наружное влагалище зрительного нерва. В отличие от пра-
вильной организации, столь характерной для роговицы, коллагеновые волок-
на склеры более хаотично расположены и сильно варьируют в диаметре
(рис. 15.15). Кроме коллагеновых волокон склера содержит (как и роговица)
протеогликаны, а также, в отличие от роговицы, значительные количества
(около 2%) эластиновых волокон. Рыхлый наружный слой склеры, эписклера,
содержит кровеносные сосуды, приносящие в склеры питательные вещества и
выносящие продукты распада.
Экваториальное расстояние от середины роговицы (мм)
Рис. 15.15. Вариации диаметра волокон и их число на единицу поверхности от
средней роговицы до склеры. Треугольниками показано число волокон, а точка-
ми — средний диаметр (нм). Шкала — в мм, начинается от середины роговицы.
Электронные микрофотографии (увеличение 33 ОООх) показывает уменьшение ре-
гулярности организации по мере удаления от роговицы. Из Harding, 1997. Воспро-
изводится с разрешения Academic Press Ltd.
334 Часть IV. Светочувствительность
Основной функцией склеры является создание плотного слоя, защищаю-
щего заключенную в ней нежную сетчатку. Склера участвует в поддержании
формы глаза и может играть роль в определении его осевой длины. Если это
так, то дефекты склеры могут сказываться на развитии миопии (близорукости).
15.3.3. Хрусталик
Хрусталик, как и роговица, должен быть максимально прозрачным, но в отли-
чие от роговицы, должен еще и быть весьма гибким в связи с необходимостью
аккомодации (фокусировки). Мы проследили развитие хрусталика в разде-
ле 15.2 и видели, что он состоит из длинных (8—10 мм) тонких лентовидных
клеток — волокон хрусталика. На срезе клетки имеют гексагональную форму
(рис. 15.16, см. также рис. 15.6), редко превышающие 8—12 мкм в поперечни-
ке и 1—2 мкм в толщину. В ходе развития они утрачивают ядра, митохондрии и
все прочие внутриклеточные органоиды. Они должны просуществовать в тече-
ние всей жизни (поскольку не замещаются), не используя синтез белка и окис-
лительное фосфорилирование. Преимущества же такой организации состоят в
практически полной прозрачности, поскольку здесь нет органоидов, которые
бы рассеивали свет. В отсутствии кровоснабжения, которое также нарушало
бы прозрачность, питательные вещества доставляются циркуляцией водянис-
той влаги (см. раздел 15.7.2).
Рис. 15.16 Б показывает, что волокна хрусталика соединены между собой
многочисленными щелевыми контактами, а межклеточные пространства в нем
малы. Кроме того, между соседними клетками образуются т. н. «тонкие» кон-
такты («thin junction»). Тогда как щелевые контакты пропускают ионы, вто-
ричные мессенджеры и маленькие метаболиты, тонкие контакты пропускают
только молекулы воды. Щелевые контакты также обеспечивают электричес-
кую связь клеток хрусталика. Хрусталик окружен плотной капсулой, которая,
подобно десцеметовой мембране, является базальной мембраной, секретируе-
мой эпителиальными клетками и волокнами хрусталика. Толщина ее меняет-
ся от 23 мкм на экваторе, где присоединяются выросты ресничного тела до
4 мкм и менее на заднем полюсе. Клетки переднего эпителия также варьируют
по высоте от 5—8 мкм на переднем полюсе до 21 мкм ближе к экватору, где они
преобразуются в сильно удлиненные волокна хрусталика. Все эти вариации
связаны с поддержанием максимальной прозрачности хрусталика.
Прозрачность хрусталика глаза человека превосходна — пропускается
большая часть света с длинами волн X между 450 и 1400 нм. Свет с длиной вол-
ны X выше 720 нм не воспринимается. Хрусталик глаза человека почти бесцве-
тен при рождении, но приобретает желтоватый цвет с возрастом. Возможно,
это специфическая черта человека — она не обнаружена ни у большинства ла-
бораторных, ни у домашних животных. Хрусталик растет в течение всей жизни
в результате формирования новых волокон хрусталика, которые растут поверх
старых и перекрывают их. Тщательное исследование хрусталика обнаруживает
эмбриональное «ядро» внутри плодного, которое в свою очередь располагается
внутри «взрослого», которое само уже окружено оболочкой.
Хрусталик обязан своей длительной эластичностью и прозрачностью при-
сутствию в его волокнах растворимых белков — кристаллинов. Высокая кон-
15. Глаз человека 335
Рис. 15.16. (А) Схема показы-
вает клеточную структуру
хрусталика, его капсулы и
поддерживающих его связок,
к — капсула хрусталика, вх —
волокна хрусталика (попе-
речный срез), в — волокна
выростов ресничного тела,
п — перикапсулярная плас-
тинка (место присоединения
выростов ресничного тела).
Из Hogan, Alvarado and Wed-
dell, 1971, с разрешения.
(Б) Детали присоединения
хрусталика к переднему эпи-
телию. Мембраны соседних
эпителиальных клеток имеют
очень неправильную форму и
связаны тесными контакта-
ми (обозначены черными
точками), так что ткань мало
проницаема. Волокна хрус-
талика связаны между собой
щелевыми контактами.
центрация этих белков в волокнах (90% сухого веса хрусталика) создает среду
с коэффициентом преломления, необходимым для фокусировки света на сет-
чатке. Кристаллины подразделяются на две основные группы: а-кристаллины
и Ру-кристалл ины.
а-кристаллины — это очень большие молекулы, массой около 700 кДа, со-
стоящие из 35—40 субъединиц с Мг около 20 кДа. Существуют два основных
типа — аА и аВ, возникшие в результате дупликации гена-предшественника
около 500 миллионов лет назад. Точная трехмерная структура все еще остает-
ся предметом дискуссии, но все современные модели сходятся на группиров-
ке субъединиц 20 кДа в большую глобулярную структуру диаметром до 16 нм.
Такие большие молекулы, казалось бы, должны рассеивать свет; тот факт, что
336 Часть IV. Светочувствительность
Рис. 15.17. Филогенетическое древо 41 аА-кристаллинов млекопитающих. Цифры пока-
зывают количество замен нуклеотидов в каждой ветви. Из de Jong, 1982, с разрешения.
этого не происходит, и то, что хрусталик прозрачен как вода, означает, что а-
кристаллины поддерживают какую-то форму короткодистантого квазикрис-
таллического порядка. Наконец, интересно отметить, что была определена
аминокислотная последовательность а-кристаллинов многих видов живот-
ных. Эти последовательности демонстрируют очень медленные изменения в
геологических масштабах времени, а потому пригодны для установления их
эволюционных взаимоотношений. Их филогенетическое древо показано на
рис. 15.17. В ходе исследований в этом направлении оказалось, что а-кристал-
лины связаны с белками теплового шока дрозофилы, нематоды Caenorhabditis
elegans и некоторых других животных. Белки теплового шока — это пример так
называемых молекулярных шаперонов. Их функция состоит в предупреждении
некорректного связывания между частично денатурированными белками, на-
капливающимися в подвергшихся воздействию клетках. Из этого следует, что
и а-кристаллины действуют как молекулярные шапероны. Нетрудно предпо-
ложить, что именно эта функция а-кристаллинов использовалась на всем про-
тяжении эволюции для предупреждения агрегации белков в волокнах хруста-
лика, что вызывало бы рассеивание света.
Ру-кристалл ины меньшего размера, чем а-кристаллины. у-кристаллины —
это мономеры с Мг около 20 кДа, тогда как p-кристаллины — это мультимерные
формы с Мг от 40 до 200 кДа. Все они связаны между собой и образуют большое
семейство структурных белков. Многочисленные другие типы кристаллинов
(8, б, р, т, л и £) обнаружены у позвоночных, не относящихся к млекопитаю-
щим, а сходные кристаллины обнаружены в хрусталиках головоногих.
15. Глаз человека 337
Наконец мы подошли к еще одной жизненно важной черте хрусталика —
аккомодации. Мы увидим в следующем разделе, что хрусталик удерживается
на своем месте выростами ресничного тела. При этом они не только фикси-
руют его, но и поддерживают определенную степень натяжения. Этому натя-
жению противостоит эластичность капсулы хрусталика. Итак, если натяже-
ние уменьшается, капсула хрусталика сокращается и округляет хрусталик —
в этом суть процесса аккомодации. При варьировании натяжения выростов
ресничного тела хрусталик способен становиться более или менее выпук-
лым. Глаз, который неспособен сфокусироваться на удаленный объект, на-
зывается близоруким (миопия), а неспособный сфокусироваться на близ-
ком — дальнозорким (гиперметропия). С возрастом капсула хрусталика утра-
чивает эластичность, вследствие чего ее способность фокусироваться на
близких объектах снижается. Объем аккомодации, т. е. динамический
диапазон хрусталика ребенка 10 лет составляет 14 диоптрий, к 40 годам она
снижается до 6 диоптрий, а к 60 — до 1 диоптрии (определение диоптрии
см. Введение к Части IV).
Еще один тип дефекта фокусировки — астигматизм. В этом случае оптиче-
ская система фокусирует точку как линию. Это происходит вследствие того,
что одна (или обе) преломляющие поверхности имеют цилиндрическую со-
ставляющую помимо общей сферической кривизны. Почти всегда за этот
дефект ответственна роговица. Астигаматизм, как и оптические дефекты
хрусталика, может быть корригирован опытным офтальмологом.
Мы отмечали, что с возрастом капсула хрусталика склерозируется и утра-
чивает эластичность. Это означает, что уменьшается не только ее сила, но и
способность изменять фокус. Утрата способности к фокусировке называется
пресбиопией (старческая дальнозоркость, от латинских корней presbus — ста-
рик и ops — глаз). Это одна из неприятностей нашей жизни, что все мы к ста-
рости становимся пресбиопиками. Последняя неприятность, которая часто
случается со старыми глазами — это катаракта.
Катарактой обычно называют непрозрачность хрусталика, ведущую к сни-
жению зрения. Это основная причина слепоты и в развитых, и в развивающих-
ся странах. Согласно сделанным в 1987 г. расчетам, из 42 миллионов слепых в
мире 40% страдали катарактой. Хотя катаракта — в основном болезнь пожи-
лых и резко учащается после 50 лет, в меньшей степени она обнаруживается во
всех возрастных группах.
Причин катаракты может быть много — от генетических (см. Информацию
15.1), радиационных (рентгеновское, ультрафиолетовое, инфракрасное) до
травм (электрошок, переохлаждение), нарушений питания (аминокислоты,
микроэлементы) и вторичных явлений увеита1 и что очень важно, диабета. Хи-
рургическое удаление хрусталика в настоящее время (как и последние
4000 лет) — единственный способ лечения. Правда, в последнее время опера-
ция дополняется имплантацией искусственного хрусталика. К счастью, обыч-
но успешной. К сожалению, это требует искусных рук хирурга, а очереди на
операцию в слаборазвитых странах мира иногда бесконечны.
1 Увеит — воспаление сосудистой оболочки глаза. — Прим, перев.
338 Часть IV. Светочувствительность
Информация 15.1
ГЕНЕТИКА КАТАРАКТЫ
В Европе и США наследственная катаракта встречается приблизительно в 1 случае
на 10 000 (катаракта, не связанная с генетическими причинами, особенно среди по-
жилых, встречается, естественно, намного чаще). Быстрое продвижение в молеку-
лярной генетике в последние годы продемонстрировало значительное количество
генов, нарушения в которых к возникновению такого заболевания. Задействовано,
по крайней мере, шесть таких генов, локализованных в хромосомах 1,2, 10,13,21 и
22, в процесс, возможно, вовлечены также гены, локализованные в хромосомах 17
и 19. Вовлеченные в процесс гены либо программируют сборку субъединиц кон-
нексинов, образующих щелевые контакты между волокнами хрусталика или синтез
кристаллинов, упакованных в волокнах.
Гены коннексинов. Щелевые контакты (коннексоны) между волокнами хруста-
лика — это цилиндры, образованные шестью коннексиновыми субъединицами.
Коннексиновая субъединица — это крупный белок (25—28 кДа) с четырьмя транс-
мембранными сегментами. Шесть коннексиновых субъединиц сгруппированы во-
круг гидрофильной поры, пронизывающей мембрану. Два коннексона (полукана-
ла) соседних волокон хрусталика, расположенные друг против друга, соединяются
и образуют, таким образом, непрерывный канал между двумя волокнами. Через
канал способны проходить неорганические ионы и небольшие молекулы, что
обеспечивает метаболическую кооперацию соседних волокон. Два гена, програм-
мирующих коннексиновые субъединицы, связаны с возникновением катаракты.
Первый из них, локализованный в хромосоме 1, программирует синтез коннексина
50, а второй, локализованный в хромосоме 13, кодирует коннексин 46. Замена нук-
леотида 262 в гене хромосомы 1 С на Т приводит к замещению пролина на серин в
положении 88 коннексина 50 (Рго88 => Ser или в сокращении, принятом в молеку-
лярной биологии — P88S). В гене хромосомы 13 замена 188-го нуклеотида А на G
приводит к замене серина на аспарагин в положении 63 коннексина 46 (asp63 => ser
или N63S. Другая мутация в этом гене — в нуклеотиде 1137 приводит к сдвигу рам-
ки считывания после 379 остатка белка коннексина и последующему сбою трансля-
ции 56-ти С-концевых аминокислот. Все эти три дефекта коннексинов нарушают
согласованность физиологических процессов в волокнах хрусталика и приводят
к неоднородной, пылеобразной непрозрачность хрусталика — порошковидной ка-
таракте.
Гены кристаллинов. Кристаллины кодируются генами, локализованными в хро-
мосоме 21 (а-кристаллин), хромосоме 22 (Р-кристаллин) и хромосоме 2 (у-кристал-
лин). Во всех трех генах идентифицированы мутации. Мутация гена хромосомы 21
приводит к замене цистеина! 16 на аргинин (cysl 16arg или Cl 16R) в а-кристаллине.
Мутация в хромосоме 22 приводит к появлению стоп-кодона в 155-м положении и,
таким образом, утрате 55-ти С-концевых аминокислот p-кристаллина. Наконец,
мутация в промоторном участке какого-либо из шести генов у-кристаллинов при-
водит к 30% избыточной продукции фрагмента у-кристаллина. Все эти нарушения
приводят к формированию патологических кристаллинов в волокнах хрусталика и
развитию той или иной формы катаракты.
15. Глаз человека 339
Продолжение
Экспериментальные исследования катаракты в значительной степени опира-
ются на данные, полученные на мышах. На этой модели генетически сконструи-
ровано почти 100 мутантов по катаракте, что сильно продвинуло понимание ге-
нетических основ этой патологии у человека. Важно, что между геномами мыши
и человека имеются обширные участки сходств (синтения), так что мутации, об-
наруженные у мыши, могут быть ключом к пониманию мутаций у человека.
К тому же, типы катаракты хрусталика мыши во многих случаях сходны с обна-
руживаемой у нас самих.
Литература
Francis, Р. I. et al., 1999, «Lens biology: development and human cataractogenesis»,
Trends in Genetics, 15, 191—196.
Heitmancik, J., 1998, «The genetics of cataract: our vision becomes cleaner», Ameri-
can Journal of Human Genetics, 62, 520—525.
Непрозрачность хрусталика (как и в случае роговицы) вызывается наруше-
нием его тонкой гистологической структуры. Прозрачность хрусталика зависит
от правильности организации волокон хрусталика и кристаллинов в них. Если
организация волокон нарушается вследствие, например, агрегации кристалли-
нов в глобулярные структуры или образования вакуолей, падающий свет рассе-
ивается, и хрусталик становится непрозрачным. Нарушения в кристаллинах
могут возникать вследствие всех упомянутых в предыдущем разделе причин и,
когда это происходит, наблюдается «разделение фаз». Это означает, что крис-
таллины агрегируются в одной части хрусталика, оставляя остальные пустыми.
Одна из наиболее обычных причин, вызывающих катаракту — это побоч-
ный эффект диабета. В этом случае усиливается диффузия галактозы из плазмы
через водянистую влагу в хрусталик. Это запускает альдегидредуктазу, которая
переводит галактозу в дульцит, который неспособен диффундировать через
мембраны волокон хрусталика. В результате осмолярность внутри волокон хру-
сталика возрастает, и вода начинает двигаться по осмотическому градиенту.
Это в свою очередь вызывает их раздувание и нарушение правильной гистоло-
гической и молекулярной структур, что означает непрозрачность хрусталика.
Этиология возрастной катаракты не демонстрирует сходства с диабетичес-
кой. Вероятно, это многофакторный процесс. Множество нарушений, накап-
ливающихся за долгую жизнь, в конце концов запускают начало заболевания.
Возможно, существует и генетическая предрасположенность. Значительное
углубление за последние годы наших знаний молекулярной генетики позволи-
ло сильно продвинуться к пониманию этой предрасположенности (Информа-
ция 15.1). Вероятно, мутации в генах, кодирующих р- и у-кристаллины, могут
быть ответственными за некоторые такие наследственные катаракты.
Как, однако, такие изменения структуры кристаллинов (и других белков
хрусталика) приводят к нарушению прозрачности хрусталика? Существует
множество возможных биохимических механизмов, но наиболее важным из
них является окисление. Мутировавший белок, возможно, не в состоянии
340 Часть IV. Светочувствительность
Таблица 15.2. Химический состав хрусталика
Водянистая влага Хрусталик
Электролиты (мЭкв/л) Na+ 142,0 20
К+ 4,0 125
С1~ 131,0 18
Са2+ 1,7 0,4
Органические вещества (мкмолъ/г) глюкоза 3,3 1,0
молочная кислота 4,5 14,0
глютатион 2,0 12,0
аскорбиновая кислота 1,1 0,6
инозитол 0,1 5,9
аминокислоты 5,0 25,0
белок 0,04% 33%
принять правильную конформацию, в результате чего освобождаются сульф-
гидрильные группы (-SH), либо немутировавший белок, долгое время находя-
щийся в метаболически неактивных волокнах хрусталика, начинает денатури-
ровать, что также освобождает сульфгидрильные группы. В присутствии кис-
лорода соседние SH-группы связываются между собой, образуя дисульфидные
мостики и, следовательно, нерастворимую массу белка (рис. 15.18). Свобод-
Рис. 15.18. Денатурация белков хрусталика вызывает катаракту. (А) Трехмерная кон-
формация нативного белка удерживается дисульфидными связями. (Б) Белок, утратив-
ший трехмерную конформацию, имеет свободные сульфгидрильные (-SH) группы.
(В) В присутствии кислорода SH-группы взаимодействуют, образуя нерастворимую
массу белка. (Г) Формула молекулы глютатиона (i); (ii) аминокислотный состав; (iii) аб-
бревиатура. (Д) Реакция Гли-Цис-Глю со свободными сульфгидрильными группами
денатурированного белка. Дальнейшие объяснения в тексте.
15. Глаз человека 341
ные сульфгидрильные группы также могут реагировать с глютатионом. Табли-
ца 15.2 показывает, что этот трипептид присутствует в хрусталике в большом
количестве. Образование дисульфидных связей с глютатионом опять-таки
приводит к образованию необычного белка, который может агрегировать и,
таким образом, приводить к непрозрачности хрусталика.
15.3.4. Сосудистый тракт
Сосудистый тракт, как мы видели в разделе 15.1, включает сосудистую оболоч-
ку глаза, ресничное тело и радужку. Сосудистая оболочка лежит между скле-
рой и сетчаткой и является сильно васкуляризованной пигментированной
тканью. Кпереди до зубчатой линии сосудистый трак покрыт двойным слоем
эктодермы. Это, как мы видели выше, ненервное продолжение сетчатки и пиг-
ментного эпителия. В ресничном теле, как мы еще увидим, эктодермальный
слой связан с продукцией водянистой влаги. Он продолжается вперед, образуя
(обычно) пигментированную заднюю поверхность радужки, а также (что наи-
более обычно для эктодермы) мышцы, сужающие и расширяющие радужку.
Сосудистая оболочка
В задней части толщина сосудистой оболочки составляет около 0,22 мм, а в пе-
редней — около 0,1 мм. Она состоит из четырех основных слоев (рис. 15.19):
1. Супрахоровд (lamina fiisca). Это рыхлая соединительная ткань, содержащая
множество меланоцитов (коричневый пигмент) и богато иннервированная.
Артериола
Мембрана
Бруха
Строма «
Ресничный
нерв
Супрахороид
или lamina fusca
Капиллярный <
слой 1
Меланоциты
Венула
Венула
Крупные плотно
упакованные
капилляры
Коллагеновые
и эластиновые
волокна
Полиморфная
клетка
Рис. 15.19. Поперечный срез сосудистой оболочки. Объяснения в тексте.
342 Часть IV. Светочувствительность
2. Строма. И это — рыхлая соединительная ткань, но намного более васкуля-
ризованная, чем супрахороид — в том числе 15—20 короткими задними рес-
ничными артериями и длинной задней ресничной артерией. Кровь оттека-
ет по четырем Завитковым венам в переднюю орбитальную вену.
3. Капиллярный слой. Все капилляры сосудов сосудистой оболочки собраны в
один слой, отделенный от пигментного эпителия сетчатки только мембра-
ной Бруха. Тогда как большинство капилляров тела имеет 8—10 мкм в диа-
метре, здесь они намного шире — 18—50 мкм.
4. Мембрана Бруха (= lamina vitrea). Тонкая прозрачная мембрана (откуда и
название — lamina vitrea, т.е. стеклянная пластинка) варьирует по толщине
от 2 до 4 мкм в задней части глазного бокала, до 1—2 мкм в передней части.
Мембрана состоит из (!) базальной мембраны капилляров капиллярного
слоя, (ii) внутреннего коллагенового слоя, (iii) эластичного слоя, (iv) на-
ружного коллагенового слоя и, наконец, (v) базальной мембраны пигмент-
ных клеток сетчатки.
Функция сосудистой оболочки — поддержание кровообращения в пиг-
ментном эпителии и сетчатке. Капилляры капиллярного слоя и мембраны
Бруха проницаемы для крупных органических молекул — ферритин (500 кДа)
и флуоресцеин (40 кДа) проходят из сосудистой оболочки в пигментный эпи-
телий. В физиологических условиях это означает, что здесь возможно прохож-
дение белков плазмы.
Ресничное тело
Эта часть сосудистого тракта напоминала первым исследователям ресницы,
откуда и название. Рис. 15.20 показывает, что ресничное тело состоит из двух
основных частей — выростов ресничного тела и ресничной мышцы. Оно мо-
жет быть подразделено и по эмбриологическому происхождению — из эпите-
лиального и мезодермального слоев. Рис. 15.20 также показывает, что в задней
Трабекулярная сеть
Шлеммов канал
Склера
Связки Хрусталик
Роговица
Сфинктер радужки
Мышца, расширяющая радужку
Передняя камера
Задняя камера
Шпора склеры
Конъюнктива
Выросты ресничного тел
Ресничная мышца^£5^
Рис. 15.20. Ресничное те-
ло. Показано расположе-
ние ресничного тела по
отношению к хрусталику,
роговице и склере. Видно
также, что ресничное тело
состоит из двух основных
частей — pars plana, содер-
жащей внутренние мышцы
глаза, и pars plicata, посы-
лающей ресничные выро-
сты, участвующие в сек-
реции водянистой влаги.
Из Harding, 1997. Воспро-
изводится с разрешения
Chapman & Hall.
15. Глаз человека 343
А
Сосуды задней
конъюнктивы
Рис. 15.21. Циркуляция водянистой
влаги. Влага секретируется выроста-
ми ресничного тела и протекает че-
рез заднюю камеру между радужкой
и хрусталиком, а затем через зрачок,
после чего абсорбируется в угловой
тканью в шлеммов канал. Из Cole,
1962, с разрешения.
Передние
сосуды
ресничного
тела
Поверхностное склеральное
сплетение
Внугрисклеральное
венозное сплетение
Глубокое склеральное сплетение
Шлеммов канал
Роговично-склеральные трабекулы
Трабекулы сосудистого тракта
Радужка
Длинная задняя
ресничная-----;
артерия
Главный
артериальный
круг радужки
Рекурентная вена
сосудистой
оболочки
Внутренние
непигментированные
эпителиальные клетки
Другие
пигментированные
эпителиальные клетки
Сильно
васкуляризованная
строма
Вырост ресничного тела
Два слоя эпителия
Васкуляризо-
ванная строма
Окончатый
капилляр
Меланосома
Межклеточное
пространство
Десмосома
Тесный контакт
Латеральная
складчатость
Десмосома
Базальные
складки
Рис. 15.22. Выросты ресничного тела, эпителий ресничного тела и секреция водянистой
влаги. (А) Схема показывает положение выростов ресничного тела, угла фильтрации над
радужкой и сложной васкуляризации. (Б) Часть одиночного выроста ресничного тела
при оптической микроскопии высокого разрешения. (В) Электронная микроскопия
эпителия ресничного тела.
344 Часть IV. Светочувствительность
Рис. 15.23. Гистология фильтрующего угла. (А) Тонкая гистология фильтрующего угла.
А — конъюнктива грани, состоящая из стратифицированного эпителия (1) и рыхлой аль-
веолярной соединительной ткани (2); В — тенонова капсула, С — эписклера, D — стро-
ма грани, продолжающаяся в строму роговицы, a, b, с, d, е — кровеносные сосуды, f —
шпора склеры, g — ресничная мышца, h — шлеммов канал, i — трабекулярная сеть,] —
сосудистая трабекула, к — вырост радужки, который может соединяться с трабекулярной
сетью. Из Hogan, Alvarado and Weddell, 1971, с разрешения.
части ресничное тело имеет гладкую форму (pars plana; obicularis ciliaris), а в
передней — переходит в 80 ресничных выростов (pars plicata; corona ciliaris).
Ресничные выросты переходят в связки (zonules), удерживающие хрусталик.
При взгляде на них понятно, почему античные анатомы думали, что они на-
шли второй набор глазных ресниц.
Ресничное тело выполняет две жизненно важные функции в физиологии
глаза. Выросты ресничного тела секретируют водянистую влагу, а ресничная
15. Глаз человека 345
Б
Роговично-
склеральная
трабекула
(2-4 мкм)
Сосудистая
трабекула
(2-3 мкм)
Вакуоль,
заполненная
водой
Шлеммов канал
Передняя камера
Внутренний канал
Зондермана
Фонтаново пространство
(пространство
роговично-радужного угла)
Направление
потока
водянистой
влаги
Рис. 15.23. (Б) Детали гистологии фильтрующего угла.
Наружный канал
Маджоре
Склера
мышца, через связки, участвует в фокусировке хрусталика. Рассмотрим их по-
следовательно.
Выросты ресничного тела и продукция и абсорбция водянистой влаги. Про-
цесс продолжается в течение всей жизни. Водянистая влага секретируется в
заднюю камеру непосредственно позади радужки, огибая которую она попада-
ет в переднюю камеру (рис. 15.21), где большая ее часть (75—95%) абсорбиру-
ется фильтрующей угловой тканью (рис. 15.23 и 15.24). Небольшое количество
водянистой влаги абсорбируется и через сосудисто-склеральный путь, который
идет между пучками ресничной мышцы (рис. 15.24). Скорость продукции и
абсорбции, таким образом, определяет давление водянистой влаги в глазу —
внутриглазное давление (ВГД). Циркуляция водянистой влаги — это сущест-
346 Часть IV. Светочувствительность
А
Роговица
Рис. 15.24. Дополнитель-
ный или сосудисто-скле-
ральный путь дренажа.
(А) ресничные мышцы
расслаблены. Водянистая
влага проходит между ни-
ми в эписклеральные ве-
ны. (Б) Ресничные мышцы
сокращены. Пространство
между волокнами замыка-
ется, склеральная шпора
оттягивается назад и рас-
ширяет пространство меж-
ду трабекулами, позволяя,
таким образом более лег-
кое протекание водянис-
той влаги в шлеммов канал.
венный компонент питания хрусталика. Однако, это тонкий процесс, в кото-
ром возможны нарушения. ВГД — важный параметр; исследования показы-
вают, что в здоровом глазу оно составляет 10,5—20,5 мм рт. ст., и если оно по-
вышается выше такого максимума, давление на стекловидное тело в задней
камере также повышается. Это в свою очередь усиливает давление на нежную
сетчатку, что может привести к ее повреждению. Ненормально высокое ВГД
вызывает глаукому, которая приводит к слепоте.
Водянистая влага секретируется с скоростью порядка 2,75 ± 0,63 мкл/мин,
состав ее приведен в табл. 15.3, очевидно, что он очень сходен с составом плаз-
мы крови. Влага формируется в сильно васкуляризованной строме реснично-
го тела. Капилляры стромы — двух типов: «непрерывные» и «окончатые». Эн-
дотелиальные клетки, образующие стенки «непрерывных» капилляров имеют
толыцину 0,2—0,3 мкм, тогда как стенки окончатых капилляров отличаются
наличием круговых участков (60—80 нм в диаметре) с толщиной всего в
0,05—0,1 мкм. Мембраны эндотелиальных клеток соединены друг с другом
тесными контактами. Таким образом, диффузия из капилляров в строму за-
труднена. В то же время электронномикроскопическое исследование показы-
вает наличие непрерывных полос пиноцитозных пузырьков, пересекающих
эндотелиальные клетки, особенно в окончатых областях. Пиноцитоз обеспе-
15. Глаз человека 347
Таблица 15.3. Состав водянистой влаги
Плазма Водянистая влага
Электролиты (мМ) Na+ 137,5 142,0
К+ 4,3 4,0
С1- 108,5 131,0
Са2+ 2,3 1,2
нсо3- Органические вещества (мМ) 27,0 20,0
глюкоза 6,0 з,з
глютатион 4,0 2,0
лактат 0,65 4,5
Данные из Abdel-Latif, 1997.
чивает прохождение белков и других макромолекул из плазмы — вероятно,
первым этапом формирования водянистой влаги является насыщение внутри-
клеточной жидкости белками.
Капилляры стромы, как показывает рис. 15.22, находятся в очень близком
контакте с пигментированными клетками наружного эпителиального слоя
выростов ресничного тела. Базальная мембрана пигментированных клеток
имеет сильно складчатую форму, поэтому поверхность ее контакта со стромой
очень велика. Цитозоль пигментированных клеток содержит редкие эндоплаз-
матический ретикулум, митохондрии и многочисленные меланосомы. Между
клетками отсутствуют тесные контакты, хотя щелевые контакты многочис-
ленны. Таким образом, ультрафильтрат из стромальных кровеносных сосу-
дов может просачиваться между ними. Кроме того, в мембране клеток, со-
седствующих со стромой, присутствует множество ионных насосов, поэтому
из стромального ультрафильтрата возможен активный захват веществ. Пиг-
ментированные клетки образуют большое количество щелевых контактов
с непигментированными клетками, поэтому возможно, что метаболиты и
ионы, абсорбированные из стромы, передаются а эти последние клетки.
Непигментированные клетки, в противоположность пигментированным,
объединены многочисленными тесными контактами, а их поверхностные
мембраны образуют сложную складчатость. Это препятствует проникновению
между ними богатого белком инфильтрата. В этих внутренних клетках сильно
развит эндоплазматический ретикулум, митохондрии и т. д., а цикл Кребса ак-
тивно продуцирует в них АТФ. Показано, что метаболические ингибиторы,
разобщающие цикл Кребса и окислительное фосфорилирование (динитрофе-
нол) и (или) ингибиторы самого цикла Кребса (фторацетат) уменьшают про-
дукцию водянистой влаги на 70—80%. Т. о., заключительная стадия формиро-
вания водянистой влаги вовлекает прохождения ультрафильтрата в непигмен-
тированные клетки и активное откачивание окончательно сформированной
влаги через мембрану в заднюю камеру.
Мы уже отмечали присутствие необычайно большого количества щелевых
контактов в пигментных и непигментированных клетках, что заставляет ду-
348 Часть IV. Светочувствительность
Неаккомодированное состояние
Аккомодированное состояние
Рис. 15.25. Мышца ресничного тела. (А) и (Б) — в состоянии покоя, (В) и (Г) — при ак-
комодации. При аккомодации ресничное тело двигается по направлению, указанному
стрелкой, и напряжение связок уменьшается. Благодаря эластичности капсулы хруста-
лик приобретает более выпуклую форму (показано пунктиром). С — связки выростов
ресничного тела, Р — роговица, МР — мышца ресничного тела, Рд — радужка, X — хру-
сталик, СС — сплетение связок, ШК — шлеммов канал, ВН — система волокон натяже-
ния. С изменениями из Hart, 1992.
мать, что эти два слоя клеток связаны между собой метаболически и электри-
чески. На бислое присутствует электрический потенциал около 10 мВ (нега-
тивный в строме), который влияет на транспорт положительно заряженных
ионов, вероятно, Na+. Это соображение подтверждается тем, что сердечный
гликозид оуабаин, хорошо известный как блокатор Na+/K+-Hacoca, уменьша-
ет продукцию водянистой влаги, электрический потенциал и уровень Na+ в
задней камере глаза. Напротив, гормон альдостерон, усиливающий транспорт
Na+, вызывает повышение внутриглазного давления (ингибиторы альдостеро-
на, напр. спиролактор, уменьшают транспорт Na+ и ВГД). Исходя из этого,
можно полагать, что продукция водянистой влаги критически зависит от ак-
тивного откачивания ионов Na+ (а также НСО3”), вместе с которыми перено-
сятся в заднюю камеру и молекулы воды. Помимо активного транспорта здесь
присутствует пассивный поток воды через клеточный бислой.
15. Глаз человека 349
Водянистая влага секретируется выростами ресничного тела позади радуж-
ки, но впереди желеобразного стекловидного тела. Она обтекает радужку в пе-
реднюю камеру и абсорбируется в фильтрующей угловой тканью, лежащей на
стыке роговицы и радужки. Ее гистология достаточно сложна. Рис. 15.23 А по-
казывает, что водянистая влага просачивается через трабекулы сосудистого
тракта, а затем — через роговично-склеральные трабекулы, далее двигаясь по
каналам Зондермана и шлеммову каналу. Электронная микроскопия стенок по-
следних двух протоков показала наличие в них гигантских вакуолей, запол-
ненных жидкостью, которая выбрасывается в проток. Из канала Шлемма, оги-
бающего лимб роговицы, водянистая влага двигается далее через внешние ка-
налы Маджоре к эписклеральным венозным сосудам, которые в конце концов
впадают в передние вены ресничного тела и далее — в венозную систему.
От 5 до 25% водянистой влаги дренируется по второму — сосудисто-скле-
ральному или дополнительному пути (рис. 15.24). На этом пути водянистая вла-
га абсорбируется между пучками мышечных волокон ресничной мышцы в су-
прахороид, а оттуда — в вены склеры. Доля такого дренажа зависит от степени
напряжения мышц ресничного тела: когда мышцы расслаблены — по этому
пути оттекает до 25% влаги, когда мышцы сокращены, каналы в соединитель-
ной ткани сжимаются, и лишь 5% влаги протекает по этому пути.
Внутриглазное давление (ВГД) зависит от баланса между продукцией и
абсорбцией водянистой влаги. Оно имеет суточный ход — ВГД наибольшее
утром и минимальное — вечером. Ненормально высокое ГД (глаукома)
обычно возникает вследствие нарушений абсорбции в фильтрующем углу.
При открытом фильтрующем угле более редкий вариант глаукомы связан
с повышенной продукцией водянистой влаги в выростах ресничного тела.
Мышца ресничного тела и аккомодация хрусталика. Значительная часть
ресничного тела образована массой гладкомышечных волокон, большинство
из которых прикрепляется к выростам склеры в сосудистую оболочку —
склеральным шпорам (рис. 15.24). Мышечные волокна расположены в трех
направлениях: меридианальном, радиальном и циркулярном. Все эти волок-
на подтягивают ресничное тело вперед, а циркулярные волокна к тому же
сужают диаметр кольца, образованного ресничным телом (рис. 15.25). Они
ослабляют натяжение связок, и хрусталик (если капсула еще эластична)
приобретает более выпуклую форму. Мышечные волокна иннервируются
парасимпатическими волокнами, идущими в составе глазодвигательного
нерва (черепно-мозговой нерв III).
15.3.5. Радужка
Это — самая передняя часть сосудистого тракта. У основания в ресничном
теле ее толщина составляет около 0,5 мм и несколько больше — 0,6 мм у вер-
шины. Рис. 15.26 показывает, что здесь можно выделить пять слоев, которые,
однако, обозначены нечетко. Самый наружный — это передний краевой
слой, состоящий из фибробластов, коллагена и меланоцитов. Под этим слоем
лежит строма, которая является продолжением и схожа со стромой реснично-
го тела и сосудистой оболочки, хотя и содержит меньше кровеносных сосудов.
Под стромой располагается толстый слой мышечных волокон, которые обра-
350 Часть IV. Светочувствительность
Меланоцит
•е—-
Кубоидный
эпителий
Кровеносный
капилляр
Окончание области
миоэпителиальных
клеток
Миоэпителий
мышцы-дилятатора
радужки
Задний
пигментированный
эпителий
Эпителий
• ! Мышцы > Строма
- Меланоцит
Меланоцит
Капилляр
Фибробласт
0
Волокна мышцы
сфинктера радужки,
срезанные поперечно
«Гребены
зрачка
Коллагеновое
волокно
В" :
Рис. 15.26. Поперечный срез радужки. Задний эпителий обращен к хрусталику.
Дальнейшие объяснения в тексте.
зуют циркулярную мышцу — сфинктер радужки. Что очень необычно, эти мы-
шечные волокна развиваются не из мезодермы, а как мы отмечали выше, из
эктодермы. Они иннервируются парасимпатическими волокнами глазодвига-
тельного нерва (черепно-мозговой нерв III) и играют доминирующую роль в
контроле диаметра зрачка. Следующий слой, являющийся продолжением пиг-
ментного эпителия сетчатки и ресничного тела, — это один ряд миоэпители-
альных клеток, чьи сократительные участки вместе образуют мышцу, расширя-
ющую радужку. Каждая клетка состоит из апикальной эпителиальной части и
базального мышечного хвоста, который простирается в строму. Эти участки
клеток иннервированы симпатическими нервными волокнами. Наконец, по-
следний перед хрусталиком слой — это задний пигментный эпителий — слой
эпителиальных клеток 36—55 мкм в высоту и 16—25 мкм в поперечнике, плот-
но упакованных с гранулами меланина.
Сфинктер и мышца, расслабляющая радужку, вместе контролируют размер
зрачка. Количество света, попадающего в глаз, прямо пропорционально диа-
метру зрачка. Поэтому возможность изменять последний очень существенна
для контроля количества света, достигающего сетчатки при различных уело-
15. Глаз человека 351
Рис. 15.27. Влияние размера
зрачка на глубину фокуса.
Глубина фокуса (АВ) меньше
при большом диаметре зрач-
ка (рисунок внизу), чем при
малом (вверху). Из Davson,
1963, с разрешения.
виях освещенности. Изменения размера зрачка также влияют на глубину фо-
куса. Это не существенно для удаленных объектов, но становится важным для
близких — чем меньше размер зрачка, тем точнее устанавливается фокус
(рис. 15.27). Наконец, размер зрачка важен для компенсации дефектов хруста-
лика. При сумеречном освещении абсолютное совершенство оптической сис-
темы не так существенно — все равно детали изображения невозможно вос-
принять. При дневном освещении сокращение зрачка обеспечивает участие
минимальной и оптически оптимальной части хрусталика.
15.3.6. Стекловидное тело
Стекловидное тело — это желеобразное вещество, занимающее заднюю каме-
ру глаза. Оно состоит из коллагеновых волокон, гиалурона, небольшого коли-
чества растворимых белков, особенно гликопротеинов, немногочисленных
клеток — гиалоцитов и 99% воды. Гиалурон — очень длинная и массивная мо-
лекула (Мг 3000 кДа у новорожденного теленка, 500 кДа у взрослого быка или
коровы), которая образует в стекловидном теле сеть, связывающую воду, и об-
разующую «желе». В целом стекловидное тело удерживается коллагеновыми
волокнами, входящими в базальную мембрану мюллеровских клеток и в огра-
ничительную мембрану сетчатки. Эти связи наиболее прочны в гладкой части
ресничного тела (pars plana) и вокруг оптического диска. Гиалоциты ограниче-
ны внешним слоем стекловидного тела и, опять-таки, наиболее многочислен-
ны в гладкой части ресничного тела и оптического диска. На основании этого
некоторые предполагали, что именно эти участки связаны с синтезом гиалуро-
новой кислоты. Однако, более вероятно, что именно pars plana участвует в
этом процессе, а гиалоциты удаляют продукты распада, подобно макрофагам.
Стекловидное тело, как мы уже отмечали, играет важную роль в удержании
сетчатки на супрахороиде, а также может участвовать в питании нейральных
слоев, к которым оно прилегает. Дегенеративные изменения стекловидного
тела — одна из основных причин отслойки сетчатки.
352 Часть IV. Светочувствительность
15.4. Движения глазного яблока
О нервных механизмах контроля движений глазного яблока известно немало.
Эта глава, однако, и так слишком длинна, чтобы углубляться в детали глазо-
двигательной системы, что увело бы нас слишком далеко в дебри нейрологии
и слишком далеко от проблем биологии сенсорных систем. Однако, мы не мо-
жем обойтись без краткого описания движений глазного яблока.
Мы уже упоминали эффекторы, реализующие эти движения — шесть
внешних или внеглазных мышц и их иннервацию в разделе 15.1. Эти мышцы
обеспечивают пять типов движений, два из которых стабилизируют глаз при
наклонах головы, а три остальных фиксируют фовеа на зрительный объект.
Опишем их кратко.
Первое из стабилизирующих движений обеспечивается вестибуло-окуло-
моторным рефлексом. Когда голова начинает движение в каком-либо направ-
лении (с ускорением), полукружные каналы внутреннего уха стимулируеются
(см. гл. 8). Глазодвигательная система отвечает на эту стимуляцию смещением
глаз в противоположную сторону с такой же скоростью. Мы рассматривали
этот рефлекс в гл. 9 (раздел 9.1) и помним, что это характерное движение глаз
называется нистагмом. Влияние полукружных каналов легко продемонстри-
ровать. Если пальцами вытянутой руки быстро размахивать перед собственны-
ми глазами, различить отдельные пальцы становится невозможно. Если же с
такой же скоростью вертеть головой, пальцы остаются вполне различимыми,
так что роль пути от полукружных каналов очевидна. Второй из стабилизиру-
ющих рефлексов — оптокинетический. Этот рефлекс реализуется зрительной
системой, когда вращается видимое окружение.
Оптокинетический рефлекс заставляет глаза фиксироваться на какой-либо
из этих стабильных черт и удерживать на ней взгляд, сколько это возможно,
затем отбрасывает взгляд в исходное положение, где фиксируется какая-то
другая деталь, которая снова удерживается взглядом до последней возможнос-
ти и т. д. В отличие от вестибуло-окуломоторного рефлекса, оптокинетичес-
кий, по очевидным причинам, не работает в темноте.
Существует три типа фиксирующих движений глаз. Первое из них — мед-
ленное следящее движение, требующее сознательного внимания. Движение
удерживает фовеа направленной на объект интереса при движении объекта
или наблюдателя. Движения глаз могут быть достаточно быстрыми — до
100°/с, но могут замедляться наркотическими веществами, утомлением или
алкоголем. Второе фиксирующее движение — более быстрое и называется сак-
кадическим. Когда изображение уходит с фовеа, глаз остается неподвижным
около 200 мс, а затем быстро (со скоростью около 900°/с) возвращает изобра-
жение на фовеа. Такое движение может быть недостаточным или избыточным,
тогда центровка на фовеа доводится микросаккадическими движениями.
Можно рассматривать эти движения как «подстройку». Глазодвигательная си-
стема беспрерывно находится в таком состоянии до тех пор, пока максималь-
но четкое изображение не образуется на фовеа.
Наконец, третий тип фиксирующих движений — это вергентные движения.
Во всех предшествующих случаях оба глаза двигаются в одном и том же на-
правлении, т. е. являются содружественными. Не то при вергентных движени-
15. Глаз человека 353
ях. Если объект приближается к лицу или удаляется от него, глаза согласован-
но движутся в противоположных направлениях, т. е. являются несодружест-
венными. Вергентные движения побуждаются различиями положений изобра-
жений объекта на сетчатках двух глаз — диспаратностью (сетчаточным несо-
впадением). Вергентные движения согласованы с аккомодацией хрусталика.
Когда меняется расстояние до объекта, мышцы ресничного тела, контролиру-
ющие аккомодацию, и глазные мышцы, выполняющие вергентные движения,
работают совместно, что содействует поддержанию точной фокусировки на
фовеа сетчаток обоих глаз.
Перед завершением этого раздела надо упомянуть важную нейрофизиоло-
гическую концепцию эфферентных копий или corollary discharge. Хотя мы по-
стоянно меняем направление взора, делая саккадические движения, (см. также
гл. 2), мы не воспринимаем мир дергающимся с места на место. Если, однако,
мы пошевелим глаз пальцем, окружающий мир покачнется. Как это можно
объяснить? Эту загадку разгадал в XIX веке Гельмгольц. Он предположил, что
когда мозг посылает сигнал, запускающий саккадические движения, к внеш-
ним мышцам глаза, то одновременно он посылает копию команды в зритель-
ную область мозга — то, что называется эфферентной копией. Таким образом
мозг может отличать последствия собственных произвольных движений голо-
вы и глаз от вызванных иными движениями в окружающем мире (в том числе,
в нашем примере —- от результата нажатия пальцем, когда мозг не получает
такой копии). Идея сравнения виртуального мира (эфферентной копии и по-
рождаемого ею прогноза) с реальностью очень важна для нейрофизиологии и
может быть распространена и на другие сенсорные системы.
15.5. Заключение
В этой главе мы сделали обзор структуры, развития и функционирования не-
нервных частей глаза. Хотя в разделе 15.2, когда мы рассматривали организа-
цию сетчатки «задом наперед», мы имели возможность оценить преимущества
эволюционного объяснения над телеологическими, мы, тем не менее (вместе
с Чарльзом Дарвином), должны были почувствовать благоговение перед не-
превзойденным устройством глаза, соответствием между его структурами
формы и назначением — oi макроскопической анатомии до субмикроскопи-
ческой молекулярной биологии. Мы продолжим благоговеть в следующей гла-
ве, когда будем рассматривать детали организации и функции сетчатки.
16. СЕТЧАТКА
Сетчатка — фактически часть мозга. П____нгный эпителий сетчатки
(ПЭС): наружная стенка глазного бокала — цитология — ультраструкту-
ра — мел аносомы — фагосомы — поглощение света — фагоцитоз мемб-
ран наружных сегментов — ре-изомеризация ретиналя — контроль пото-
ка веществ в наружную часть сетчатки. Сетчатка: двойственное строе-
ние — фовеа — гистология — девятислойная «вертикальная» структура —
горизонтальная структура. Палочки и колбочки: морфология — количест-
во и распределение — система улавливания света — ориентация родопси-
нов — фотоизомеризация — трансдуцины — цГМФ-ФДЭ — ЦНЗ
Ка+/Са2+-каналы — темновой ток — свет ингибирует темновой ток — ги-
перполяризация; переход цГМФ в 5-ГМФ вызывает закрывание ЦНЗ-
каналов — удаление Са2+ расторможенным рекаверином — активация
гуанилатциклазы (ГЦ) и синтез цГМФ — сенсорная адаптация — родо-
псин-киназа и арестин деактивируют опсин; детали фотоизомеризации и
возобновление ретиналя; колбочки и опсины колбочек — эволюционные
взаимоотношения. Горизонтальные клетки: цитология — синаптология —
биофизика. Биполярные клетки: цитология — типология — синаптоло-
гия — палочковые биполяры, — биполяры, меняющие и не меняющие
знак — биохимия и биофизика — рецептивные поля (РП) — блок-схемы.
Мюллеровы клетки — наружная пограничная мембрана (НПМ) и внут-
ренняя пограничная мембрана (ВПМ) —- функция. Интерплексиформные
клетки: два типа — эфферентные пути. Амакриновые клетки: большое
разнообразие — синаптология. Ганглиозные клетки: типология — потен-
циалы действия — анатомическая и функциональная классификация —
количество варьирует в различных участках сетчатки — немиелинизиро-
ванные аксоны — концентрические ON/GFr РП — антагонизм центра
и окружения — определение края. Блок-схемы: пути палочковых и кол-
бочковых сигналов — темновая адаптация — врожденная сложность —
нейрогенетика и сетчатка данио. Цвет: трихромазия — оппонентные
процессы — РП ганглиозных клеток — ощущения цвета, иногда называ-
емые первичными, на самом деле продукт сложной организации. Заклю-
чение: сложность сетчатки готовит нас к еще большей сложности мозга.
В главе 15 мы сделали обзор ненервных частей глаза человека. Все это, однако,
только весьма совершенный каркас, предназначенный служить вместилищем
фоточувствительной и нервной тканей глаза — сетчатки. Вот к ней мы и обра-
тимся в данной главе. Мы видели в разделе 15.3, что сетчатка возникает как
16. Сетчатка 355
часть мозга — часть мозга, выдвинутая на периферию. Издавна у исследователей
сетчатки существовала надежда, что если мы поймем ее структуру и функцию, то
что-то поймем и в структуре и функции мозга. Мы также отмечали в разделе
15.3, что рассматриваемое нами «выпячивание» мозга состояло из двух слоев
клеток, разделенных пространством — глазным пузырем, которое вскоре исче-
зает. Два слоя затем образуют пигментный эпителий и собственно сетчатку.
В этой главе мы и начнем, соответственно, с рассмотрения пигментного эпите-
лия, а затем перейдем к сетчатке, двигаясь внутрь глаза (к стекловидному телу).
16.1. Пигментный эпителий (ПЭ) сетчатки
Мы видели в разделе 15.3, что ПЭ развивается из наружной стенки глазного
бокала. Полость глазного пузыря исчезает в раннем эмбриогенезе, и ПЭ вслед-
ствие этого оказывается тесно ассоциирован с сетчаткой. Тем не менее, клет-
ки ПЭ более прочно соединены с сосудистой оболочкой, чем клетки собствен-
но сетчатки. Действительно, внутренняя часть мембраны Бруха (0,3 мкм, см.
раздел 15.3.4) секретируется пигментным эпителием как базальная мембрана.
Поэтому в случаях отслойки сетчатки ПЭ остается соединенным с сосудистой
оболочкой, и полость глазного пузыря восстанавливается.
В глазе взрослого человека 4—6 миллионов клеток ПЭС. Клетки имеют гек-
сагональную форму, высоту около 8 мкм и 16 мкм в диаметре. Они соединены
многочисленными плотными контактами, которые препятствуют диффузии
между сосудистой оболочкой и сетчаткой по межклеточным промежуткам.
Различные вещества могут попадать в сетчатку через цитоплазму клеток ПЭ.
Для облегчения этого процесса основания клеток ПЭС образуют множество
складок и микроворсинок, значительно увеличивающих площадь их поверх-
Рис. 16.1. Электронная микрофо-
тография (увеличение 18000х)
пигментного эпителия сетчатки.
Вверху микрофотографии наруж-
ные сегменты двух палочковых
клеток (h), обернутые в микровор-
синки (g) клеток ПЭ. а — базаль-
ная мембрана пигментного эпите-
лия, b — складки мембраны эри-
телиальных пигментных клеток,
с — ядро клетки ПЭ, d—
хондрия, е — эндоплазматичес-
кий ретикулум, f — пигментная
гранула. Из Hogan,, Alvarado and
Weddell, 1971. Воспроизводится с
разрешения W. В. Saunders Com-
pany.
A
Геометрическая —
ось
\ ЗЛ
БП
СП
Ф — фовеа
ЦА — центральная часть
БП — ближняя периферия
СП — средняя периферия
ДП — дальняя периферия
ЗЛ — зубчатая линия
ОД — оптический диск
В — височная сторона
Н — носовая сторона
Рис. 16.2. Участки сетчатки. (А) Горизонтальный срез показывает различные участки
сетчатки. В — височная сторона, Н — носовая. По мере продвижения к периферии чис-
ло биполярных клеток, и особенно ганглиозных клеток, резко уменьшается, однако рез-
кая граница отсутствует. (Б) Вид в плане и в разрезе центрального участка сетчатки.
(В) Тангенциальный срез через разные участки га'п^тки макака-резуса. Наружные сег-
менты фоторецепторов перерезаны поперек, (а) Фовеа: отметьте плотно упакованные
тонкие колбочки, (б) Парафовеа: наружные сегменты окружены белым пространством —
из-за большого диаметра внутренних сегментов. Колбочки окружены наружными сег-
ментами палочек, (в) Средняя периферия: доля палочек по отношению к колбочкам уве-
личивается. (г) Далекая периферия: доля колбочек значительно уменьшена. Воспроизво-
дится из Journal of Cell Biology, 1971, vol. 49, p. 309, с разрешения на авторское право
Rockfeller University Press.
16. Сетчатка 357
в
Рис. 16.2. (В).
ности. Ультраструктурные исследования этих клеток показали изобилие эндо-
плазматического ретикулума и митохондрий, последние особенно многочис-
ленны у основания клеток. Апикальная клеточная мембрана выпячивается в
форме длинных ворсинок, охватывающих наружные сегменты палочек и кол-
бочек. Темно-коричневые меланосомы обнаружены в этих ворсинках, а также
в апикальных частях клеток. Как показано на рис. 16.1, клетки содержат боль-
шие включения — фагосомы, представляющие собой захваченные части на-
ружных сегментов палочек и колбочек. Одновременно может захватываться
30—40 дисков. Каждая пигментная клетка связана с наружными сегментами
30—45 фоторецепторов и переваривает по 2000—4000 дисков в день.
ПЭ выполняет множество важных функций в сетчатке. Темно-коричневый
меланин в меланосомах поглощает свет, проходящий сквозь сетчатку, и пигмент
в микроворсинках изолирует наружные сегменты друг от друга, увеличивая та-
ким образом остроту зрения (сравните со сложным глазом, раздел 14.1,
рис. 14.8). У некоторых животных (например у кошки, у некоторых рыб) в ПЭ
присутствуют отражающие кристаллы, благодаря которым глаза «светятся» в
темноте (см. раздел 18.3). Микровосинки также обеспечивают механическую
опору хрупким наружным сегментам. Фагоцитоз дисков наружных сегментов —
это тоже, как мы видели, важная функция. Клетки ПЭ захватывают фотоизоме-
ризованные зрительные пигменты и преобразуют их в исходную 11-^ис-форму
(см. раздел 16.2.1, рис. 16.8 ниже). Наконец, клетки ПЭ, лежащие между капил-
лярами сосудистой оболочки и сетчаткой, регулируют прохождение веществ че-
рез эти два слоя. Показано, что эти клетки активно откачивают Na+ из капилля-
ров сосудистой оболочки в сетчатку, образуя на ПЭ потенциал около 10 мВ.
358 Часть IV. Светочувствительность
16.2. Сетчатка
Общая структура сетчатки была рассмотрена в разделе io.z. Мы видим, что в
сетчатке человека дневное зрение связано с фовеа, а внутри нее с фовеолой,
причем рецепторная мозаика этой области главным образом представлена кол-
Рис. 16.3. Вертикальный срез сетчатки примата (перифовеальная область). (А) Микрофо-
тография. П — пигментный эпителий, Н — наружная пограничная мембрана, НЯС — на-
ружный ядерный слой, НСС — наружный синаптический слой, ВЯС — внутренний ядер-
ный слой, ВСС — внутренний синаптический слой, Г — слой ганглиозных клеток, В —
слой волокон зрительного нерва. Масштабная линейка = 75 мкм. Из Boycott and Dowling,
1969. Воспроизводится с разрешения The Royal Society. (Б) Этот рисунок из Rodieck, 1988,
показывает сложность нейральной сетчатки, хотя на нем показано лишь небольшое чис-
ло клеток. Клетки: АП — АП амакриновые, СКБ — биполяры, связанные с синими кол-
бочками БГК— биплексиформные ганглиозные клетки, Н1 и Н2 — два типа горизон-
тальных клеток, С — интерплексиформная клетка, дбк — диффузная биполярная клетка,
Moff — биполярная клетка off-типа, Моп — биполярная клетка on-типа, Мю — мюллеро-
ва клетка, АУ — амакриновая клетка с малым рецептивным полем, Poff — ганглиозная
клетка с off-центром, Роп — ганглиозная клетка с on-центром, КБ — биполярная клетка,
связанная с красной колбочкой, ША — амакриновая клетка с большим рецептивным по-
лем. Слои: БМ — мембрана Бруха, ХК — хориокапиллярный, СГК — слой ганглиозных
клеток, ВПМ — внутренняя пограничная мембрана, ВЯС — внутренний ядерный слой,
ВСС — внутренний синаптический слой, ВС — внутренние сегменты, СНВ — слой воло-
кон зрительного нерва, НПМ — наружная пограничная мембрана, НЯС — наружный
ядерный слой, НСС — наружный синаптический слой, НС — наружные сегменты,
ПЭС — пигментный эпителий сетчатки. С изменениями из Rodieck, 1988.
16. Сетчатка 359
нс
няс
нсс
вяс
вес
вс
нпм
Б хк г
БМ -
ПЭС-1
СНВ
впм-
MoffMon Мю СКБ ША БГК
Рис. 16.3. (Б).
бочками. В противоположность этому, периферические части сетчатки населе-
ны главным образом палочками. Таким образом, сетчатка человека — смешаная
(двойственная): она содержит палочки и колбочки. Палочки намного более чув-
ствительны, чем колбочки; популяция же колбочек обеспечивает нам ощуще-
ние цвета, а в фовеа, благодаря плотности колбочек и отсутствию суммации на
пути от рецепторов к ганглиозным клеткам, — еще и остроту зрения. При сла-
бом освещении зрение обеспечивают палочки. При дневном освещении види-
мые объекты, особенно объекты, видимые «краем глаза», т. е. наблюдаемые с
помощью палочек периферической сетчатки, инициируют рефлекс, в результа-
те которого глаз движется так, чтобы изображение объекта оказалось в фовеа —
этот процесс именуется фовеацией. Итак, глаз человека, как и глаза оольшинст-
ва млекопитающих, двоичный. Обе части сетчатки играют свои собственные ро-
ли. В данном разделе мы рассмотрим их строение и функции более детально.
В сетчатке шесть основных типов нервных клеток и один тип глиальных —
мюллеровские клетки (рис. 16.3). Гистологи выделяют более 50 подтипов этих
основных типов. Основные типы нервных клеток сетчатки, если следовать от
пигментного эпителия к стекловидному телу, перечислены ниже:
• Фоторецепторные клетки (палочки и колбочки)
• Горизонтальные клетки
• Биполярные клетки
• ИнтерплекСиформныё клетки
• Амакриновые клетки
• Ганглиозные клетки
360 Часть IV. Светочувствительность
Рис. 16.4. Упрощенная блок-схема
сетчатки (область средней перифе-
рии). Хотя на рисунке это не пока-
зано, здесь происходит высокая сте-
пень конвергенции информации.
Многие колбочки образуют синап-
сы с одним и тем же биполяром,
многие биполяры в свою очередь
образуют синапсы на одной и той же
ганглиозной клетке. А — амакрино-
вая клетка, Б — биполярная клетка,
К — колбочка, Г — ганглиозная
клетка, Гр — горизонтальная клет-
ка, ВПМ — внутренняя погранич-
ная мембрана, 3 — волокна зритель-
ного нерва, НПМ — наружная по-
граничная мембрана, П — палочка.
Считается, что эти клетки организованы в девять слоев (рис. 16.3 Б): на-
ружные сегменты фоторецепторов (НС), наружная пограничная мембрана
(НПМ), наружный ядерный слой (НЯС), наружный сетчатый (синаптичес-
кий) слой (НСС), внутренний ядерный слой (ВЯС), внутренний сетчатый
слой (ВСС), слой ганглиозных клеток (СГК), слой нервных волокон (СНВ),
внутренняя пограничная мембрана (ВПМ). Мюллеровы клетки распределены
между нервными элементами по всей глубине сетчатки, их окончания, соеди-
ненные тесными контактами, образуют наружную и внутреннюю погранич-
ные мембраны (см. также рис. 16.18).
Рис. 16.3 Б показывает, как сетчатка организована в вертикальном и в гори-
зонтальном направлениях. Вертикальная организация начинается с фоторецеп-
торных клеток. Их основания образуют сложные синаптические контакты с би-
полярными клетками, эта область — наружный синаптический слой, тогда как
уровень ядер фоторецепторов — это наружный ядерный слой. Биполярные
клетки занимают центральную часть сетчатки и образуют синапсы с дендритами
ганглиозных клеток. Ядра биполярных клеток образуют внутренний ядерный
слой, а их синапсы — внутренний сетчатый (синаптический) слой. Немиелини-
зированные аксоны ганглиозных клеток, идущие вдоль поверхности сетчатки,
образуют слой нервных волокон. Они сходятся к оптическому диску и покидают
глаз в составе зрительного нерва. Следует отметить, что за исключением некото-
рых амакриновых клеток, на рассмотренном участке распространения возбужде-
ния от фоторецепторов до аксонов ганглиозных клеток импульсные потенциалы
действия отсутствуют (сравните со зрительной долей насекомых, раздел 14.2.6).
Наряду с этим трехклеточным вертикальным путем, интерплексиформные клет-
ки, разбросанные по сетчатке, соединяют между собой два синаптических слоя,
фактически они передают информацию в обратном направлении — от ВСС
к НСС. На вертикальную структуру сетчатки накладывается горизонтальная.
Клетки двух типов — амакриновые и горизонтальные — располагаются в сетчат-
16. Сетчатка 361
ке тангенциально. Горизонтальные клетки соединяют между собой разные по-
пуляции фоторецепторов и биполяров, а амакриновые — популяции биполя-
ров, ганглиозных и интерплексиформных клеток, соответственно. Упрощенная
схема этих сложных взаимосвязей показана на рис. 16.4.
16.2.1. Фоторецепторные клетки
Морфология
Рассмотрим сначала палочки (рис. 16.5). Они происходят от ресничных кле-
ток, следы чего остаются в течение всей их жизни. В ходе эмбриогенеза рес-
ничная мембрана чрезвычайно увеличивается и вытягивается, образуя боль-
шое количество впячиваний, которые в конце концов отшнуровываются и об-
разуют ряд «свободноплавающих» дисков (рис. 16.5 А). Стопка таких дисков,
окруженных мембраной, образует наружный сегмент фоторецептора. У различ-
ных позвоночных его размеры варьируют от 10 до 200 мкм в длину и от 2 до
10 мкм в диаметре. Фоточувствительный пигмент родопсин сконцентрирован,
как мы рассмотрим ниже более подробно, в мембранах дисков. У человека
около 80% белка мембраны дисков — это родопсин. Наружный сегмент соеди-
нен с внутренним сегментом соединительной ресничкой — узнаваемым руди-
ментом исходной реснички. Внутренний сегмент состоит из наружной час-
ти — эллипсоида и внутренней зоны — миоида. Эти области в палочках и кол-
Цитоплазматическое
(междисковое)
пространство
Соединительная
ресничка
(iv)
Формирующийся
наружный
сегмент
Внутренний
сегмент
Внутридисковое пространство
(часть внеклеточного
пространства, захваченого
при инвагинации в ходе
формирования диска)
Рис. 16.5. Фоторецепторные клетки. (А) Эмбриональное развитие, (i) Фоторецепторная
клетка развивается из ресничной клетки, выстилающей глазной пузырь. У каждой клет-
ки — одна ресничка, (ii) Ресничка растет, ее мембрана гипертрофируется и начинает ин-
вагинировать. (iii) Гипертрофия ресничной мембраны и инвагинация ее у основания
наружного сегмента продолжаются. Исходные внутренние ультраструктуры реснички
сохраняются в области «соединительной реснички», соединяющей наружный и внут-
ренний сегменты, (iv) Более детальное изображение, показанного на (iii) — инвагина-
ции мембран захватывают часть внеклеточного пространства, превращающегося во
внутридисковое. Пространство между дисками — цитоплазматическое.
362 Часть IV. Светочувствительность
Б
1 мм
Рис. 16.5. (Б) (i) палочка и (ii) колбочка, ср — соединительная ресничка, э — эллипсоид,
с — ножка или сферула, м — миоид, я — область ядра, вс — внутренний сегмент, нс — на-
ружный сегмент. Электронная микрофотография из Hogan, Alvarado and Weddell, 1971.
Воспроизводится с разрешения W. В. Saunders Company.
16. Сетчатка 363
В
Аппарат Гольджи
Транспортная
везикула
Эндоплазматический
ретикулум
и рибосомы
Соединительная
ресничка
Митохондрия
Ядро
Сборка
дисков
Встраивание
транспортных
везикул в
мембрану
Транспортная
везикула
Рис. 16.5. (В) Синтез мембраны дисков во внутренних сегментах палочек и колбо-
чек. Стрелки показывают направление, в котором двигается новый мембранный
материал. Дальнейшие объясения в тексте. По Beshare, 1986.
бочках млекопитающих не отграничены друг от друга резко. Эллипсоид содер-
жит большое количество митохондрий, тогда как в миоиде хорошо развиты
эндоплазматический ретикулум и аппарат Гольджи. Эти органоиды участвуют
в очень активном синтезе новой мембраны дисков. Она встраивается у основа-
ния наружного сегмента (см. рис. 16.5 В) и сдвигает диски вверх — к кончику
наружного сегмента. Диски постоянно удаляются с кончика наружного сегмен-
та и перевариваются, как упоминалось в разделе 16.1, клетками пигментного
эпителия. Длительность цикла от синтеза диска до его удаления составляет
около 10 дней. Под специализированным внутренним сегментом располагает-
ся ядро, а далее суженный участок, называемый аксоном или волокном, после
которого клетка вновь расширяется, образуя ножку или сферулу. Последняя
заполнена синаптическими пузырьками, содержащими L-глютамат и часто
выстроенными вокруг синаптических лент, которые образуют сложные синап-
тические поверхности. Эти поверхности одновременно являются пре- и пост-
синаптическими, к тому же между сферулами палочек и между ножками со-
седних колбочек имеются щелевые контакты (электрические синапсы). Одна-
ко, электрическая связь между палочками у млекопитающих не доказана.
Колбочки (рис. 16.5 Б(п)), в сущности, очень похожи на палочки. Наруж-
ные сегменты их несколько меньше, редко длиннее 30 мкм и не более 5 мкм
в диаметре у основания. Диски никогда не отщепляются от краевой мембра-
ны, а внутридисковое пространство остается открытым во внеклеточную сре-
364 Часть IV. Светочувствительность
ду. Внутренний сегмент — большой, а эллипсоид обычно заполнен митохонд-
риями. У многих позвоночных (но не у собственно зверей) он также содержит
развитую и иногда окрашенную каплю жира (см. раздел 18.2). Наконец, си-
наптическое окончание или ножка обычно намного крупнее, чем у палочек и
имеет даже еще более сложную и обычно множественную синаптическую по-
верхность. Как и в случае палочек, последняя одновременно является и пре-,
и постсинаптической, а щелевые контакты образуются с соседними колбочка-
ми и палочками. В периферической части сетчатки, где плотность фоторецеп-
торов ниже, ножки колбочек иногда посылают длинные отростки, образую-
щие щелевые контакты с ножками других колбочек и сферулами палочек.
Ножки выделяют медиатор (глютамат) на дендритные окончания биполярных
и горизонтальных клеток, а также получают информацию от горизонтальных
клеток. Предполагается, что колбочки — исходный тип фоторецептора, из ко-
торого затем развились палочки. Это, однако, остается предметом дискуссий,
поскольку тщательные исследования морфологии и биохимии фоторецепто-
ров показали существование бесконечного множества форм, промежуточных
между палочками и колбочками.
Количество и распределение фоторецепторов
(i) Колбочки. У приматов с дневной активностью, как человек, плотность
колбочек максимальна в центре фовеа, лишенном палочек, в т. н. фовеоле.
В сетчатке человека фовеола имеет около 350 мкм в диаметре и плотность кол-
бочек у разных индивидов — от 100 000/мм2 до 324 000 /мм2. На плоскости кол-
бочки в этой области построены в очень правильную треугольную решетку с
расстоянием между центрами 2,5—3,0 мкм. Плотность колбочек падает очень
быстро с удалением от фовеолы — до 10 000/мм2 в 1 мм от нее, а затем более мед-
ленно —до постоянной величины около 5 000/мм2 в остальной сетчатке. По вер-
тикали, как показывает рис. 16.2, сетчатка в области фовеа и фовеолы выглядит
как заметное углубление. Дело в том, что нервные элементы сетчатки идут здесь
почти горизонтально от ножек колбочек (рис. 16.6), так что колбочки не при-
крыты слоем нейронов и могут получать максимум падающего света. В этой ча-
сти сетчатки наружные сегменты колбочек к тому же существенно удлинены.
(ii) Палочки. В сетчатке человека палочки превосходят колбочки по чис-
ленности в 20 раз. Они отсутствуют в фовеоле и достигают максимальной
плотности порядка 140 000—160 000/мм2 в кольце на расстоянии около 5 мм от
нее. Затем плотность падает до постоянного уровня около 70 000—80 000/мм2
перед тем, как резко уменьшиться на дальней периферии сетчатки. Это рас-
пределение плотности проиллюстрировано на рис. 16.7.
Биофизика и физиология1
Начнем снова с палочек. Мы уже отмечали, что зрительный пигмент родопсин
располагается в мембранах дисков наружных сегментов, здесь он сильно кон-
1 Более свежие данные по этому разделу можно получить в отечественных источниках
(Каламкаров, 2002; 2004).
А СК
СК
Рис. 16.6. (А) Вертикальный срез фовеолы человека. Фоторецепторы в этой области
представлены исключительно колбочками (к), у которых развиваются длинные и тонкие
палочковидные внутренние и наружные сегменты. В результате на ограниченном прост-
ранстве упаковано больше рецепторов, а наружная пограничная мембрана выгнута (по-
казано стрелками) в сторону стекловидного тела. Наружный ядерный слой (НЯС) рас-
ширен, поскольку большое количество колбочек и волокон, идущих от колбочек удлине-
ны и проецируются косо (волокна Генле (ВГ), образуя внутренний синаптический слой
и внутренний ядерный слой (ВЯ) за пределами фовеолы. СК — скат (clivus), ПЭ — пиг-
ментный эпителий, увеличение ЗООх). Из Tripathi and Tripathi, 1984. Воспроизводится с
разрешения Academic Press, Inc. (Б) Схема, показвыающая организацию нервных эле-
ментов фовеолы. А — амакринова клетка, СК — скат (clivus), ПБ — плоская биполярная
клетка, ВГ — волокно Генле, МБ — миниатюрная биполярная клетка (карликовый бипо-
ляр), МГ — миниатюрная (карликовая) ганглиозная клетка. Остальные обозначения, как
на рис. 16.3 Б.
366 Часть IV. Светочувствительность
Височная сторона Носовая сторона
Рис. 16.7. Распределение палочек и колбочек в сетчатке человека. Из Osterberg,
1935.
центрирован — рассчитано, что от одной молекулы родопсина до другой около
6 нм. Очевидно, что, как и в рабодомерных глазах, рассмотренных в гл. 14, па-
лочки представляют собой превосходное решение проблемы улавливания све-
та. Каждый поступающий фотон должен пройти через пакет из 2 000 дисков с
высокой концентрацией фотопигмента. Благодаря плотной упаковке мем-
бранных дисков показатель преломления наружных сегментов намного выше,
чем вне их. Эта структура действует как световод, проводящий свет вдоль на-
ружного сегмента (сравните с рабдомом насекомых). Кроме того, наружные
сегменты ориентированы так, что в любой части сетчатки направлены на
центр зрачка. У некоторых млекопитающих (как у вошедших в поговорки ко-
шек) расположенный за пигментым эпителием отражающий слой дает прони-
кающему до его уровня свету еще одну возможность провзаимодействовать
с родопсином наружного сегмента.
Мы рассмотрели структуру родопсина во Введении к Части IV (рис. 14.3,
см. также Информацию 14.1). В наружных сегментах палочек он ориентирован
так, что С-концевой хвост расположен в междисковом (цитоплазматическом)
пространстве. Родопсин удерживает 1 \-цис ретиналь шиффовой связью с ли-
зиновым остатком в центре «бочки из клепок» (рис. 16.8 А). Когда фотон соот-
ветствующей длины волны (400-600 нм, Хтах 498 нм) взаимодействует с 11-
цис-ретиналем, это обеспечивает энергию активации, необходимую для внут-
римолекулярной перестройки, ведущей к низкоэнергетической тиране-форме.
Это ключевой момент фоторецепции, все последующее зависит от этого фото-
химического преобразования. Транс-ретиналь не может оставаться соединен-
ным с опсином и диффундирует из «бочки», оказываясь в конце концов в пиг-
ментном эпителии.
16. Сетчатка 367
Рис. 16.8. (А) Конформация родопсина «бочка из клепок» в мембране диска. От-
метьте положение 11-цис-ретиналя. (Б) Шиффова связь ретиналя с остатком лизи-
на29б в опсине. (В) Молекулярная структура ll-цис- и /ирлнс-ретиналяр
368 Часть IV. Светочувствительность
Рис. 16.9. Биохимические взаимодей-
ствия в мембране диска. (А) Состояние
покоя. (Б) Поглощение фотона света
вызывает цис-/ирлнс-трансформацию
ретиналя. (В) и (Г) активированный
родопсин взаимодействует с Т-белком
(трансдуцином), что вызывает обмен
ГДФ на ГТФ на его а-субъединице (Та).
а-субъединица Т-белка освобождается
от Ру-субъединиц. (Д) Та-ГТФ активи-
рует цГМФ-ФДЭ, ускоряя гидролиз
цГМФ. Родопсинкиназа и арестин
инактивируют опсин. (Е) ГТФ дефос-
форилируется и действие Та на цГМФ-
ФДЭ прекращается. Система возвра-
щается к состоянию покоя. Серым
цветом обозначены активированные
компоненты.
После изомеризации 11-цис-ретиналя в тирянс-ретиналь молекула опсина
претерпевает конформационные изменения и переходит в активную форму.
В этом состоянии она способна реагировать с G-белковой сигнальной систе-
мой мембраны наружного сегмента (фоторецепторные G-белки называют
трансдуцинами или Т-белками). Активированный опсин реагирует с а-субъе-
диницей Т-белка, что ведет к знакомому нам биохимическому процессу (см.
гл. 1), но с одним важным отличием. Вместо обычного взаимодействия с аде-
нилатциклазой (и синтеза цАМФ), а-ГТФ-субъединица Т-белка (Та-ГТФ)
воздействует на большой тетрамерный фермент — цГМФ-фосфодиэстеразу
(цГМФ-ФДЭ). Этот мембранно-связанный фермент состоит из двух
каталитических (а- и ₽-) и двух ингибиторных у-субъединиц (офу2). Когда этот
16. Сетчатка 369
фермент входит в контакт со свободным Та-ГТФ-белком, у-субъединица
отодвигается. Это растормаживает каталитическую активность а- и р-субъеди-
ниц, которые преобразуют цГМФ в 5 -ГМФ. Одновременно у-субъединица ка-
тализирует дефосфорилирование Та-ГТФ в Та-ГДФ. После этого а-субъеди-
ница Т-белка отсоединяется от каталитической субъединицы цГМФ-ФДЭ,
что позволяет у-субъединице снова соединиться с последней. Это приводит
к прекращению каталитической активности фермента (рис. 16.9).
Чтобы понять значение описанной выше мембранной биохимии, надо
учитывать, что цГМФ играет ключевую роль в электрофизиологических про-
цессах в палочках. Уже давно известно, что в темноте палочки слегка деполя-
ризованы. Это происходит благодаря наличию в мембране наружных сегмен-
тов каналов для Na+ и Са2+. Если обратиться к гл. 2, мы вспомним, что элект-
рический потенциал на биологической мембране возникает вследствие разли-
чия проницаемостей мембраны для малых неорганических ионов. Мы видели,
что в первом приближении мембранный потенциал подчиняется уравнению
Голдмана. Тогда, если, например, увеличить проницаемость по натрию PNa в
пять раз до 5 х 10”7 см/с, сохраняя проницаемость по К+ и СГ постоянными,
и подставить это в уравнение, то получим Vm = —20 мВ (проверьте!). В темно-
те ток (темновой ток) образуется ионами Na+, протекающими из внутреннего
сегмента через внеклеточное пространство и входящими в наружный сегмент
(рис. 16.10 А). При включении света темновой ток прекращается, и палочка ут-
рачивает деполяризацию, т. е. гиперполяризуется (рис. 16.10 Б). Какое, одна-
ко, все это имеет отношение к описанной выше мембранной биохимии? Ответ
очевиден, если принять во внимание, что цГМФ поддерживает в открытом со-
стоянии №+/Са2+-каналы наружного сегмента. Биохимические процессы,
как мы видели, приводят к преобразованию цГМФ в нециклическую форму —
5'-ГМФ. Иными словами, цГМФ устраняется, Ка+/Са2+-каналы закрывают-
ся, а палочки гиперполяризуются.
Закрывание циклонуклеотид-зависимых (ЦНЗ) каналов имеет и другое
следствие (рис. 16.11). В междисковом (цитоплазматическом) пространстве
обнаружен важный белок GCAP (guanylate cyclase-activating protein). В темно-
те его активность подавлена присутствующими ионами Са2+. При включении
света Ма+/Са2+-каналы закрываются, а уровень Са2+ снижается. Вследствие
этого GCAP растормаживается и активирует другой фермент — гуанилатцик-
лазу (ГЦ), образующую цГМФ из 5'-ГМФ. Чем дольше продолжается освеще-
ние, тем больше цГМФ образуется таким образом — это механизм сенсорной
адаптации. Чем больше синтез цГМФ гуанилатциклазой, тем более он превы-
шает разрушение цГМФ специфической фосфодиэстеразой в мембране диска.
Наконец, чем больше открываются Ка+/Са2+-каналы, тем сильнее палочки
деполяризуются.
Конформационные изменения и переход опсина в активное состояние де-
маскирует сайты присоединения двух тормозных белков — родопсинкиназы и
арестина. Родопсинкиназа фосфорилирует целых девять сериновых и треони-
новых остатков в С-концевой цепочке опсина (см. рис. 14.3) и тем снижает его
способность взаимодействовать с трансдуцином. Полная десенситизация оп-
сина, однако, не наступает, пока не присоединится арестин, что в свою оче-
редь может произойти только когда опсин фосфорилирован родопсин-кина-
I___I__J___I
0 12 3
Время (c)
Рис. 16.10. (А) Темновой ток. В темноте «каналы утечки» наружной мембраны наружно-
го сегмента обеспечивают поток Na+ внутрь. Ионы натрия двигаются в соответствии с гра-
диентом концентрации во внутренний сегмент, откуда откачивается К+Д4а2+-насосом. То
есть в темноте существует постоянный ток из внутреннего сегмента в наружный — темно-
вой ток. (Б) При поглощении фотона света наружным сегментом №+-каналы закрывают-
ся и палочка гиперполяризуется. Гиперполяризацию можно зарегистрировать электрофи-
зиологически с помощью внутриклеточного микроэлектрода, — результат регистрации
приведен на рисунке справа. По Penn and Hagins, 1969. (В) (а) Наружный сегмент фоторе-
цептора жабы (намного более крупный, чем у млекопитающих) втянут в стеклянную ми-
кропипетку. Поскольку отверстие канала микропипетки полностью заткнуто наружным
сегментом, все токи, протекающие внутрь или наружу сегмента, связаны с внутренним ка-
налом пипетки и могут таким образом быть измерены. В темноте ток, как показано на гра-
фике (в), несколько превышает 20 пА. (б) Узкий пучок света проходит через пипетку — гра-
фик на (в) показывает эффект, начинающийся в точке, обозначенной вертикальной стрел-
кой. Уменьшение темнового тока зависит от интенсивности вспышки. Нижняя линия на
графике — ток при вспышке света в 94 раза более интенсивной, чем вызвавшей реакцию,
показанную самой верхней линией. Из Baylor, Lamb and Yau, 1979, с разрешения.
16. Сетчатка 371
Рис. 16.11. Биохимический меха-
низм адаптации наружного сег-
мента. Когда Ыа+/Са2+-каналы
закрываются при освещении, по-
ток Са2+ внутрь сегмента умень-
шается. GCAP стимулирует ак-
тивность ГЦ. Синтезируется
цГМФ, что повышает вероят-
ность открытого состояния
Ка+/Са2+-каналов. Дальнейшие
объяснения в тексте. Стрелки
символизируют активацию. |- —
ингибирование, Та — а-субъеди-
ница Т-белка, ГЦ — гуанилатцик-
лаза, ФДЭ — фосфодиэстераза,
Р — родопсин.
зой. После того, как это произошло, арестин присоединяется и изменяет кон-
формацию опсина таким образом, что он не может больше взаимодействовать
с мембранно-связанным трансдуцином. В результате выключается механизм
взаимодействия, цГМФ-ФДЭ возвращается в неактивное состояние, прекра-
щается гидролиз цГМФ, и палочки деполяризуются.
Хотя биохимические и молекулярно-биологические данные, приведенные
выше, могут показаться излишне подробными (при этом нет сомнений, что
продолжающиеся исследования покажут еще большую сложность этого меха-
низма), основной результат состоит в том, что это чрезвычайно чувствитель-
ная система детектирования фотонов света. Рассчитано, что при адаптации к
темноте палочка способна реагировать на единственный фотон света. Как и
в случае механорецептивной клетки (гл. 8), чувствительность системы нахо-
дится на уровне физических пределов. Реакция родопсина усиливается взаи-
модействием родопсина с трансдуцином (рис. 16.12), так что, по крайней мере
в теории, один фотон может изменить мир.
Фото-изомеризация и возобновление ретиналя
Опосредующий фоторецепцию переход 11-цкс-ретиналя в тирднс-форму ведет
к его отсоединению от опсина путем последовательных перестроек его
молекулы. Продукт каждой из них имеет свою характерную Хтах: родопсин
(498 нм) => прелюмиродопсин (543 нм) люмиродопсин (497 нм) => метаро-
допсин-1 (478 нм) => метародопсин-2 (380 нм) => метародопсин-3 (465 нм) =>
транс-ретиналь (387 нм). Перестройки от родопсина до метародопсина-2 за-
нимают всего около миллисекунды и порождают активную конформацию
опсина. Трянс-ретиналь отделяется от опсина лишь на последней стадии, и это
занимает уже несколько минут. Далее возможны два варианта. Если ретиналь
уходит из колбочки, то реизомеризация происходит в нейральной сетчатке, ес-
372 Часть IV. Светочувствительность
Фотон
I
Родопсин
-500 молекул ФДЭ
I
-105 молекул цГМФ
Блокируется прохождение -106-107 ионов Na+
Рецепторный потенциал 1 мВ
I
Уменьшение выделения медиатора
I
Деполяризация уже деполяризующихся биполярных клеток
I
Инициация потенциала действия в волокне зрительного нерва
I
Зрительная кора
I
?
Рис. 16.12. Усилитель-
ный каскад в зрительной
системе. С изменениями
из Alberts et al., 1994, и
Lamb and Pugh, 1992.
ли же из палочки — то он диффундирует в пигментный эпителий сетчатки
(ПЭС), где и происходит изомеризация. Последовательность этих событий
показана на рис. 16.13. Показано, что /лранс-ретиналь сначала переходит в
спиртовую форму, транс-ретинол, при действии ретинол-дегидрогеназы, по-
сле чего транспортируется через мембрану клетки пигментного эпителия спе-
цифическим связывающим белком. В ПЭС /лрянс-ретинол реагирует с жирной
кислотой (пальмитиновая кислота), образующейся из фосфатидилхолина (ле-
цитина), в результате формируется ретиниловый эфир (ретинил-пальмитат).
Наряду с этим дополнительный тиранс-ретинол (витамин А) поступает из хо-
риокапилляров. Затем происходит ряд внутримолекулярных преобразований
ретинил-пальмитата, приводящий к переходу /лранс-ретиналя в 11-цис-кон-
формацию. Энергия для этого берется из внутримолекулярных преобразова-
ний пальмитинового остатка ретинил-пальмитата. 11-цмс-ретинил-пальмитат
может запасаться в ПЭС или выделять 11-цмс-ретиналь, транспортирующийся
через мембрану клеток пигментного эпителия в наружные сегменты.
Последствия гиперполяризации
В завершение всей этой хитроумной истории, немного отвлечемся от тонко-
стей биохимии и рассмотрим палочку в целом. Мы видели, что поглощение
фотона молекулой родопсина в конце концов приводит к гиперполяризации
16. Сетчатка 373
Хориокапилляры транс-
ретинол
Рис. 16.13. Регенерация 11-^мс-ретиналя. Объяснения в тексте. МРСБ — межре-
цепторный ретиноид-связывающий белок, Р — родопсин, Р* — активированный
родопсин. С упрощениями из Harding, 1997.
клеточной мембраны палочки. Эта гиперполяризация распространяется
вплоть до сферулы. Известно, что выделение медиатора из пресинаптических
окончаний происходит в ответ на деполяризацию. Следовательно, гиперполя-
ризация ингибирует выделение медиатора из ножки палочек. Мы вернемся к
этой теме позже, когда будем рассматривать синапсы между ножками палочек
и колбочек с горизонтальными и биполярными клетками.
Колбочки
Прежде, чем перейти к горизонтальным и биполярным клеткам, коснемся
другого типа фоторецепторов — колбочек. Мы рассмотрели их структуру в раз-
деле 16.2.1. Их биохимия и биофизика в основном та же, что и у палочек. Ос-
новное отличие колбочек состоит в том, что их зрительные пигменты имеют
три разных кривых поглощения с Хтах (в нормальной сетчатке человека)
419 нм (синий), 531 нм (зеленый) и 559 нм (красный) (рис. 16.14). Каждая кол-
бочка содержит только один тип пигмента. Селективное возбуждение трех по-
пуляций колбочек — это первый этап цветного зрения. Последующие стадии
этого процесса, происходящие в сетчатке, мы рассмотрим в разделе 16.2.9. Ко-
нечная обработка информации о цвете происходит в коре головного мозга.
Цвет часто рассматривали как классический тип простейшего сенсорного ка-
чества. Как мы увидим дальше, эта простота — кажущаяся, и за ней скрывает-
ся очень сложная физиологическая кухня, опосредующая наше ощущение.
Небольшое уточнение по поводу названий: пигмент палочек, как мы виде-
ли, обычно именуется родопсином, тогда как в колбочках он именуется йодо-
374 Часть IV. Светочувствительность
Длина волны (нм)
Число длин волн (см-1)
Рис. 16.14. Кривые поглощения
трех типов йодопсина (сплош-
ные линии) и родопсина сет-
чатки человека. Из Mollon,
1982, с разрешения.
псином. В обоих случаях пигмент представляет собой семидоменный белок
(опсин) и хромофор — ретиналь. Опсин наружных сегментов палочек имену-
ется род-опсином (не путать с родопсином), а колбочек — кон-опсином (от
rod — палочка и cone — колбочка). Разница спектров поглощения (рис. 16.14)
возникает не вследствие различий хромофоров, а из-за разницы аминокислот-
ных последовательностей опсинов (см. табл. 16.1). Во всех случаях ретиналь
присоединяется к лизиновому остатку (лизин296) связью шиффова основания,
однако распределение заряженных боковых цепочек аминокислот в непосред-
ственной близости от него различаеются. Вероятно, эти различия и другие
тонкие биохимические вариации аминокислотного окружения ретиналя вли-
яют на различия спектров поглощения. Действительно, показано, что три раз-
личных аминокислотных замещения могут превратить пигмент, чувствитель-
ный к зеленому свету, в пигмент, чувствительный к красному. Эти замещения
выглядят так: аланин180 => серин, фенилаланин227 => триптофан и аланин285 =>
Таблица 16.1. Гомологии опсинов палочек и колбочек
Процент гомологии последовательности
Опсин Синий Красный Зеленый
палочек опсин опсин опсин
Опсин палочек 100 75 73 73
Синий опсин 42 100 79 79
Красный опсин 40 43 100 99
Зеленый опсин 41 44 96 100
Данные из Nathans, 1986. Значения под 100%-ной диагональю — процент идентичных остат-
ков, значения над диагональю — процент идентичных остатков + консервативные замещения.
16. Сетчатка 375
треонин, или в более компактной форме A180S, F227Y и А285Т. Интересно,
что цветовое восприятие, которое играет такую важную роль в нашей жиз-
ни, зависит от того, какое положение какая аминокислота занимает! Более
подробно об эволюции опсина —- в Информации 14.1.
Сравнение аминокислотных последовательностей разных опсинов пока-
зывает (табл. 16.1), что они образуют эволюционно взаимосвязанное семейст-
во. Из детального анализа следует, что единственный ген — общий предшест-
венник опсинов — дуплицировался по крайней мере 500 миллионов лет назад.
Два образовавшихся гена стали кодировать красный и синий опсины. Последу-
ющая дупликация гена синего опсина дала зеленый опсин большинства позво-
ночных животных, а также родопсин палочек. Зеленый опсин человека и при-
матов — сравнительно недавнее изобретение. «Предковый» ген зеленого опси-
на был ими утерян, а затем, возможно не более 40 миллионов лет назад, ген
красного опсина дуплицировался еще раз. Один из двух образовавшихся генов
продолжал кодировать красный опсин, а другой стал кодировать зеленый.
16.2.2. Горизонтальные клетки
Это крупные клетки (100—400 мкм) в горизонтальной плоскости имеют форму
звезды. Они посылают свои отростки в плоскости наружного синаптического
слоя (НСС), объединяя группы ножек фоторецепторных клеток. Потенциалы
действия в виде спайков здесь не наблюдаются, и влияние горизонтальных
клеток осуществляется путем электротонического проведения. Клетки обра-
зуют пре- и постсинаптически контакты. В исследованиях, проводившихся на
золотой рыбке, показано, что когда горизонтальная клетка является постси-
наптической по отношению к фоторецепторной, ее ответ имеет тот же знак,
что и сигнал от фоторецепторов. Иными словами, если фоторецепторы гипре-
поляризуются, то же происходит и с горизонтальными клетками, а если фото-
рецепторы деполяризуются, то деполяризуются и горизонтальные клетки.
С другой стороны, если горизонтальные клетки являются пресинаптическими
по отношению к фоторецепторам, то ответ фоторецепторов «изменяет
знак» — т. е. деполяризация горизонтальных клеток приводит к гиперполяри-
зации фоторецепторов и т. д. Между собой горизонтальные клетки связаны
щелевыми контактами (электрическими синапсами). Электрическая связь,
которая всегда сохраняет знак, контролируется медиатором — дофамином.
Горизонтальные клетки, таким образом, формируют сложную, контролируе-
мую медиатором, электрически связанную сеть, расположенную тангенциаль-
но в наружном синаптическом слое (НСС).
16.2.3. Биполярные клетки
Эти клетки связывают НСС и внутренний синаптическй слой (ВСС). Они пе-
редают информацию от фоторецепторов ганглиозным и (или) амакриновым
клеткам. В некоторых случаях такая связь выполняется без конвергенции
(один к одному): в сетчатке приматов биполярные клетки — миниатюрные
(карликовые) биполяры (midget) — часто образуют синапсы с единичной кол-
бочкой (рис. 16.3 Б). Например, одиночный карликовый биполяр сетчатки
376 Часть IV. Светочувствительность
приматов образует своими дендритными отростками контакты с единствен-
ной колбочкой фовеа, и передает информацию в единственную ганглиозную
клетку. Другие биполярные клетки — диффузные — образуют контакты с нож-
ками множества колбочек (рис. 16.3 Б). В периферических областях сетчатки
биполярные клетки могут образовывать синапсы с сорока фоторецепторами.
Существует видимо лишь один тип палочковых биполяров, в то время как
колбочковых биполяров известно несколько типов (до десяти — у кошек, 6 —
у приматов).
Палочковые биполяры
У палочковых биполяров тела клеток крупные, дендритные деревья развитые,
особенно в периферической сетчатке. Синапсы, которые они образуют на
ножках палочек, часто имеют сложную форму, поскольку в них участвуют и
горизонтальные клетки (рис. 16.15 А). Мы отмечали (раздел 16.2.1), что «тем-
новой ток» обеспечивает медленную деполяризацию палочек в темноте. Когда
адаптация к темноте завершается (прибл. за 1 час), достигается постоянный
уровень деполяризации, а щелевые контакты между палочками и колбочками,
как полагают, закрываются. Таким образом, вся активность палочек передает-
ся исключительно к их биполярным клеткам. Все биполярные клетки, связан-
ные с палочками, изменяют знак электроответа (см. ниже).
Колбочковые биполяры
Мы отмечали выше, что существует множество различных морфологических
типов биполярных клеток, связанных с колбочками. Для простоты мы рассмо-
трим только два из них: карликовые биполяры, которые, как мы уже видели,
могут быть моносинаптическими, и плоские или щетковидные биполяры, ко-
торые могут формировать синапсы на множестве ножек колбочек. Синапти-
ческие контакты часто имеют сложную форму (рис. 16.15 Б). Электроответ
колбочковых биполяров может иметь тот же знак, что у фоторецептора, либо
инвертировать его.
Теперь рассмотрим нейрофармакологию синпасов, сохраняющих и изме-
няющих знак сигнала. Мы видели, что синаптические окончания как палочек,
так и колбочек заполнены пузырьками, содержащими нейромедиатор, вероят-
но глютамат. В темноте, когда фоторецепторы деполяризованы, этот медиатор
постоянно освобождается с небольшой скоростью. Вероятно, синаптические
ленты, которые являются такой характерной особенностью окончаний фото-
рецепторов, связаны с организацией этой утечки. Глютамат оказывает различ-
ное действие на дендриты биполяров двух разных типов (рис. 16.16). В одном
случае, глютамат открывает катионные каналы, что позволяет ионам Na+ про-
никать в дендрит. Дендрит вследствие этого деполяризуется, а когда фоторе-
цептор гиперполяризуется, выделение глютамата из ножки прекращается. Те-
перь гиперполяризуется дендрит биполярной клетки, т. е. это синапс биполя-
ра, сохраняющего знак сигнала. В другом случае имеет место нечто аналогич-
ное эффекту света на ЦНЗ-каналы наружных сегментов. Глютамат (подобно
свету на наружных сегментах) активирует биохимический каскад, устраняю-
16. Сетчатка 377
Рис. 16.15. (А) Синаптический
комплекс сферулы палочки.
БК — биполярная клетка, Г —
горизонтальная клетка, СЛ —
синаптическая лента, СП — си-
наптический пузырек. (Б) Си-
напсы и синаптические ком-
плексы ножки колбочки. От-
метьте значительно больший
размер ножки по сравнению со
сферулой палочки. ПБ — плос-
кая биполярная клетка (отметьте
ее ветвление и то, что она может
образовывать синапсы с ножка-
ми более, чем одной колбочки);
ПМБ — плоская карликовая би-
полярная клетка, Г — горизон-
тальная клетка, ИМБ —• инваги-
нирующая карликовая биполяр-
ная клетка. Отметьте ветвление
ИМБ и то, что она часто образу-
ет многочисленные триадные
синапсы с единственной ножкой
колбочки. Таким образом, ИМБ
и колбочки находятся в тесном
синаптической связи. По Dow-
ling, 1987.
щий цГМФ, который удерживал Ка+/Са2+-каналы в открытом состоянии.
Итак, в темноте, когда глютамат присутствует в синаптической щели, ЦНЗ-
каналы закрыты, а биполярные клетки относительно гиперполяризованы.
При включении света фоторецепторные клетки гиперполяризуются, а выделе-
ние глютамата ингибируется. Далее, ЦНЗ-каналы открываются, а биполярные
клетки деполяризуются — это биполяры, инвертирующие знак сигнала.
Биполярные клетки не генерируют импульсов, однако имеют рецептивные
поля, построенные по принципу «центр — периферия», в которых возникают
ответы только на включение (ON), либо на выключение (OFF). Посмотрим,
что это означает. Рис. 16.16 показывает, что колбочки образуют синапсы с из-
меняющими и не изменяющими знак биполярами. Когда свет включается
(ON), в инвертирующих знак биполярах возникает деполяризация; напротив,
при выключении света (OFF) деполяризуются биполяры, сохраняющие знак.
В большинстве случаев биполярные клетки образуют синапсы на группе фото-
рецепторов, а также получают сигналы от горизонтальных клеток. Т. о., на них
влияет свет, падающий на небольшой участок сетчатки, который определяет-
ся как рецептивное поле (РП). Синаптические взаимодействия в паутине кон-
тактов РП, как мы отмечали выше, чрезвычайно трудно анализировать. Разли-
чают два типа биполяров: (1) те биполяры, которые дают ON-ответ при осве-
щении в центре РП, генерируют OFF-ответ при освещении периферии РП.
(2) Напротив, если биполяр дает в центре РП OFF-ответ (т. е. отвечает на вы-
378 Часть IV. Светочувствительность
-100
-100
Г иперполяризующийся
биполяр
(сохраняющий знак
сигнала)
Деполяризующийся
биполяр
(инвертирующий знак
сигнала)
Рис. 16.16. Сохраняющие и
инвертирующие знак сигнала
синапсы колбочек с биполяр-
ными клетками. Нижняя
часть рисунка показывает от-
веты биполярных клеток при
освещении колбочек. Гипер-
поляризующиеся (сохраняю-
щие знак) биполяры дают
OFF-ответ (отключение), а
деполяризующиеся (инверти-
рующие знак) — ON-ответ
(включение). Эти ответы тем
больше, чем интенсивнее ос-
вещение. Г — горизонтальная
клетка, БП — биполярная
клетка. Дальнейшие объясне-
ния в тексте.
ключение света), то при освещении периферии генерируется ON-ответ. Счи-
тается, что главным образом горизонтальные клетки отвечают за такие антаго-
нистические взаимоотношения центров РП с окружением (см. раздел 16.2.9).
Один из механизмов, который может обеспечивать такую реакцию, приведен
на рис. 16.17. Показано, что в темноте ножки колбочек выделяют глютамат на
горизонтальные клетки, поддерживая их в деполяризованном состоянии. Де-
поляризованные горизонтальные клетки выделяют тормозный медиатор на
соседние ножки колбочек, что несколько уменьшает их деполяризацию по
сравнению с тем уровнем, который у них был в темноте. Когда свет попадает
на периферию РП, выделение глютамата из ножек периферических колбочек
уменьшается, горизонтальные клетки гиперполяризуются, выделение глюта-
мата на ножки колбочек в центре РП уменьшается, вследствие чего они депо-
ляризуются. Инвертирующие знак ON-биполяры, следовательно, ингибиру-
ются выделением дополнительных количеств глютамата из ножек колбочек.
Если и центр, и периферия РП освещены одновременно, сходное расуждение
показывает, что уменьшатся ответы биполяров и на включение, и на выключе-
ние. Это пример латерального торможения, с которым, как мы помним, мы
впервые столкнулись в сложном глазе мечехвоста Limulus.
В целом, хотя это еще не установлено точно, аксоны биполярных клеток
ON-типа оканчиваются на внутренней части внутреннего синаптического
16. Сетчатка 379
Рис. 16.17. Взаимодействия между центром и окружением рецептивного поля.
(А) В темноте. Горизонтальная клетка ингибирует колбочку в центре РП. Биполяр-
ная клетка, инвертирующая знак сигнала, деполяризуется. (Б) Освещение колбо-
чек на периферии РП приводит к торможению горизонтальной клетки и снятию
торможения с колбочки в центре РП. Изменяющая знак (ON) биполярная клетка
гиперполяризуется. Объяснения в тексте. Серая штриховка — деполяризация, тем-
ная штриховка — гиперполяризация*. НК — ножка колбочки, Г — горизонтальная
клетка, БПО — биполярная клетка ON-типа.
слоя (ВСС, подслой «Ь»), тогда как аксоны биполяров OFF-типа оканчивают-
ся на наружной части ВСС, ближе к рецепторам (подслой «а») (см. рис. 16.3 Б).
В обоих случаях их окончания образуют синапсы на дендритах амакриновых
и ганглиозных клеток. Мы вернемся к обсуждению рецептивных полей, ког-
да будем рассматривать электрофизиологию ганглиозных клеток (раздел
16.2.8).
16.2.4. Мюллеровы клетки
Теперь самое время рассказать о единственном типе глиальных клеток сетчат-
ки — мюллеровых клетках (рис. 16.18). Мы уже видели, что эти клетки играют
важную роль в эмбриогенезе сетчатки (раздел 15.3). И в сетчатке взрослых они
сохраняют значение, поскольку обеспечивают структурную и биохимическую
поддержку. Их базальные ножки, расположенные вблизи от стекловидного те-
ла, соединены тесными контактами, образуя непроницаемый слой — внутрен-
нюю пограничную мембрану (ВПМ). Верхушки клеток также соединены меж-
380 Часть IV. Светочувствительность
Рис. 16.18. Мюллеровы клетки и внутренняя и на-
ружная камеры сетчатки. НК — наружная камера,
ВК — внутренняя камера, ВПМ — внутренняя по-
граничная мембрана, М — мюллерова клетка,
НПМ — наружная пограничная мембрана, Ф — фо-
торецепторная клетка, ПЭС — пигментный эпите-
лий сетчатки. Из Harding, 1997. Воспроизводится с
разрешения Chapman & Hall.
ду собой и с наружными сегментами фоторецепторов, образуя наружную по-
граничную мембрану (НПМ).
Верхушки мюллеровых клеток, помимо формирования НПМ, образуют
также многочисленные микроворсинки, сильно увеличивающие их поверх-
ность, через которую могут диффундировать вещества между наружными сег-
ментами фоторецепторов, ПЭС и нейральной сетчаткой. Таким образом, по-
лагают, что мюллеровы клетки играют важную роль в биохимии нейральной
части сетчатки, контролируя потоки питательных веществ и действуя как депо
для ионов калия, которые выбрасываются при электрической активности.
16.2.5. Интерплексиформные клетки
Это — наиболее редкие и наименее разнообразные клетки сетчатки. Известно
только два их подтипа: дофаминэргические и глицинэргические. Они пере-
дают информацию из внутреннего синаптического слоя (ВСС) к наружному
синаптическому слою (НСС), будучи частью эфферентного пути, благодаря
которому мозг модулирует активность сетчатки. Интерплексиформные клет-
ки получают сигналы во внутреннем синаптическом от амакриновых клеток
слое и от эфферентных волокон мозга.
16. Сетчатка 381
16.2.6. Амакриновые клетки
В отличие от интерплексиформных клеток, амакриновые клетки — наиболее
разнообразный тип нейронов сетчатки. Эти не имеющие аксонов клетки клас-
сифицируются по типам ветвлениях их дендритных деревьев — у обыкновенной
плотвы Rutilus rutilus различают 43 подтипа амакриновых клеток, а в сетчатке
приматов — по крайней мере 26. Подобно горизонтальным клеткам наружного
синаптического слоя (НСС), амакриновые клетки расположены в плоскости
внутреннего синаптического слоя (ВСС) и образуют чрезвычайно сложную, со-
единенную щелевыми контактами сеть. Они получают сигналы от биполярных
клеток (вероятно, биполярные клетки образуют два синаптических контакта на
амакриновых клетках, либо один — на амакриновой, а другой — на ганглиоз-
ной) и посылают сигнал к дендритному дереву ганглиозных клеток. Импульс-
ные потенциалы действия обнаруживаются в амакриновых клетках редко.
16.2.7. Ганглиозные клетки
Ганглиозные клетки также очень разнообразны. В то время как в амакриновых
клетках импульсные потенциалы действия регистрируются лишь изредка (толь-
ко в некоторых типах, а иногда только в определенные периоды развития), ганг-
лиозные клетки — единственный тип клеток сетчатки, в которых импульсные от-
веты обнаруживаются всегда. Классификация ганглиозных клеток на подтипы
проведена на основе сочетания морфологических и физиологических свойств.
Гистологи описали множество различных морфологических типов, и мы
рассмотрим здесь только три из них: а- или М-, р- или Р- и W-клетки. а- или
М- (от magnocellular — магноцеллюлярный, т. е. крупноклеточный) клетки
имеют крупный перикарион, аксон большого диаметра и немногочисленные,
но сильно ветвящиеся дендритные деревья. Р- или Р- (parvocellular — парво-
целлюлярный, т. е. мелкоклеточный) клетки — с перикарионом среднего раз-
мера и более узкими дендритными деревьями. W-клетки — с аксонами малого
диаметра, но сильным ветвлением дендритного дерева.
Электрофизиологи обнаружили, что все три морфологических типах сход-
ны с биполярами в том, что они имеют рецептивные поля типа «центр-перифе-
рия». Крупные а-ганглиозные клетки соответствуют, как и можно было ожи-
дать, клеткам с большим полем, с оппонентным (противоположным по знаку
вызываемой реакции) окружением. На постоянное освещение они реагируют
коротким быстрым ответом. Электрофизиологи называют их Y-клетками и по-
лагают, что они реагируют на крупные черты объектов и, что еще важнее, на их
движение. Меньшие по размеры [J-клетки имеют более узкие поля с оппонент-
ным окружением и демонстрируют длительный ответ на освещение. Электро-
физиологи классифицируют их как Х-клетки. Они сигнализируют о более мел-
ких деталях и цвете объекта. Наконец, W-клетки имеют медленно проводящие
аксоны и часто не имеют полей с центром и периферией, а просто реагируют на
контраст повышением или понижением уровня импульсации.
Число ганглиозных клеток в том или ином участке сетчатки сильно варьи-
рует. В центральной части сетчатки они очень многочисленны, их дендритные
поля перекрываются, а тела могут лежать в несколько слоев. К периферии, на-
против, число ганглиозных клеток резко падает. Это отражает тот факт, что ес-
382 Часть IV. Светочувствительность
ли в фовеа существует практически взаимно однозначное соответствие между
фоторецеторами и ганглиозными клетками, то на периферии наблюдается
сильная конвергенция фоторецепторов на биполяры, а биполяров на одну ган-
глиозную клетку.
Аксоны ганглиозных клеток идут в оптический диск по внутренней по-
верхности сетчатки, обращенной к стекловидному телу. Аксоны остаются
немиелинизированными из-за необходимости сохранять максимальную
прозрачность. В оптическом диске они уходят внутрь через продырявленную
пластинку, с противоположной стороны которой они выходят в виде воло-
кон зрительного нерва. Здесь они приобретают миелиновую оболочку, что
резко увеличивает диаметр зрительного нерва за продырявленной пластин-
кой. Волокна из темпоральной половины сетчатки идут в ипсилатеральное
наружное коленчатое тело (НКТ), и только небольшое их количество на-
правляется в верхние бугры четверохолмия; волокна из назальной половины
сетчатки делают перекрест и идут в контрлатеральное НКТ, и опять-таки не-
большое число волокон направляется в четверохолмие (рис. 17.2)
Физиология
Основа физиологической классификации ганглиозных клеток сетчатки при-
матов — это подразделение их на имеющие ON-центр, OFF-центр и ON/OFF-
центр; существуют и другие типы, а в сетчатке других животных иногда на-
блюдается и большее разнообразие. Все три типа рецептивных полей (как и
у биполярных клеток) имеют ошюнентную организацию центра и периферии.
Включение света в центре поля ON-клеток генерирует в них залп импульсов.
Включение света на периферии поля тормозит какую-либо активность этих
клеток, а при выключении света возникает залп потенциалов действия. На-
против, включение света в центре OFF-поля тормозит активность, и только
выключение вызывает возникновение залпа; включение света на периферии
тоже инициирует залп. Эти характеристики показаны на рис. 16.19.
Стоит отметить еще несколько моментов, касающихся таких РП. Первое,
ганглиозные клетки отвечают активностью только на изменения освещеннос-
ти — при постоянной освещенности в аксонах регистрируется лишь неболь-
А
РП
Рис. 16.19. (А) Схема экспе-
римента. Конвергенция мно-
гих фоторецепторных клеток
на ганглиозной клетке пока-
зана пунктиром. ГК — ганг-
лиозная клетка, С — световой
зонд, МЭ — микроэлектрод,
ВЗН — волокно зрительного
нерва; фрк — фоторецептор-
ная клетка, РП — рецептив-
ное поле.
16. Сетчатка 383
шая активность. Второе, антагонизм между центром и периферией приводит к
тому, что, если свет падает и на центр, и на периферию, то ON- и OFF-ответы
стремятся взаимно уничтожиться. Диффузный свет, падающий на все концен-
трическое поле, вызывает очень слабый ответ. РП организованы таким обра-
зом, чтобы детектировать изменения освещенности поля: края теней, границы
изображений движущихся объектов и т. д. Такой же принцип организации мы
видели в сложных глазах мечехвоста Limulus и других членистоногих в гл. 14.
Теперь пойдем дальше и рассмотрим характеристики рецептивных полей
в терминах блок-схем сетчатки.
Б
Поле с ON-центром
Световое
пятно
в центре
Световое
пятно на
периферии
Освещение
всего
центра
Освещение
всего
окружения
центра
Диффузное
освещение
Рис. 16.19. (Б) Характеристики рецептивных полей с ON- и OFF-центрами. Белая часть
РП показывает освещенную область. Справа от каждого РП показана активность гангли-
озных клеток. Длительность светового стимула показана отрезком под записью активнос-
ти. Отметьте вариант 5, где в обоих случаях диффузное освещение всего поля генерирует
лишь слабый ответ в ганглиозных клетках вследствие латерального торможения (т. е. анта-
гонизма между центром и периферией). Из Kandel, Schwartz and Jessell, 1991, с разрешения.
384 Часть IV. Светочувствительность
16.2.8. Блок-схемы
Предпринималось немало попыток понять функционирование сетчатки на
основе ее синаптической организации. Однако, чем больше мы узнавали о вза-
имосвязях и фармакологии синапсов, тем сложнее становилась картина. Мы
видели в разделе 16.2.4, что тогда как синапсы биполярных клеток с ножками
колбочек могут быть как сохраняющими, так и инвертирующими знак сигна-
ла, синапсы со сферулами палочек всегда — инвертирующие. При этом в ко-
нечном счете и колбочки, и палочки принимают участие в работе ганглиозных
клеток, рецептивные поля которых построены по принципу «центр и перифе-
рия». Рассмотрим каждый случай по очереди, а потом суммируем данные, что-
бы построить модельную блок-схему сетчатки.
Пути передачи сигнала от колбочек
Биполяры ON-типа образуют глютаматергические синапсы на дендритных де-
ревьях ганглиозных клеток ON-типа во внутреннем слое ВСС (подслой Ь). Би-
полярные клетки OFF-типа образуют глютаматергические синапсы с дендрит-
ными деревьями ганглиозных клеток OFF-типа, распределенных в наружном
подслое ВСС (подслой а). Такая организация носит очень четкий характер. Ан-
тагонизм центра и периферии возникает в результате тормозной активности
горизонтальных клеток, синаптические характеристики которых мы рассмот-
рели в разделе 16.2.3. Из них следует, что когда стимулируется ганглиозная
клетка с ON-центром, стимуляция ее периферии вызывает противоположный
эффект. То же и с ганглиозными клетками с OFF-центром. Очень обобщенная
схема контактов приведена на рис. 16.20; при этом надо помнить, что предпо-
лагается существование и других контактов, так что реальная синаптология
в сетчатке намного более сложна, чем показано на схеме.
Пути передачи сигнала от палочек
Палочки образуют глютаматергические синапсы исключительно с инверти-
рующими знак сигнала биполярными клетками. Обычно (особенно в перифе-
рической сетчатке) биполярные клетки палочек контактируют с большим
числом сферул палочек. Биполяры не образуют синапсов непосредственно на
ганглиозных клетках, но контактируют с АП-амакриновыми клетками ВСС
(рис. 16.21). Эти синапсы сохраняют знак сигнала. АП-клетки в свою очередь
образуют глицинергические синапсы (тормозные) с ганглиозными клетками
OFF-типа и щелевые контакты (возбуждающие электрические синапсы)1 с би-
полярами ON-типа. Такая организация обеспечивает (при темновой адапта-
ции, когда колбочки не работают) то, что повышение освещенности активиру-
ет ганглиозные клетки ON-типа, а уменьшение освещенности активирует
(или снимает торможение) ганглиозные клетки OFF-типа. Антагонизм окру-
жения, опять-таки, обеспечивается активностью горизонтальных клеток.
1 Щелевой контакт и электрический синапс (в т.ч. возбуждающий) — это, все же, не одно
и то же. Электрическим синапсом (эфапс) принято считать близкий контакт мембран клеток
в котором сигнал передается электротонически через межклетники, тогда как щелевой
контакт — это межклеточный канал (см. выше). — Прим, перев.
16. Сетчатка 385
Рис. 16.20. Характеристики
ON- и OFF-ганглиозных кле-
ток и влияние горизонтальных
клеток (см. рис. 16.17) на нож-
ки колбочек. Ганглиозные
клетки имеют такие же взаи-
моотношения центра и пери-
ферии, что и их биполярные
клетки. БП+ — биполярна»
клетка ON-типа, БП" — бипо-
лярная клетка OFF-типа,
НК — ножка колбочки, ГК+ —
ганглиозная клетка с ON-цен-
тром, ГК” — ганглиозная
клетка с OFF-центром, ВЗН —
волокна зрительного нерва.
Рис. 16.21 показывает, что ножки палочек и колбочек образуют друг с
другом щелевые контакты (электрические синапсы). Показано, что сигналы,
генерируемфе палочками, могут передаваться колбочкам. При длительной
темновой адаптации щелевые контакты закрываются, ограничивая сигнал
исключительно палочками, что повышает чувствительность сетчатки, когда
передача сигнала от палочек (рис. 16.21) активирует ганглиозные клетки ON-
типа и тормозит клетки OFF-типа — иными словами, отключается сигнал от
тормозного окружения. Сетчатка утрачивает способность детектировать кон-
Рис. 16.21. Связи палочек, колбочек и ганг-
лиозных клеток. Палочковые биполяры
контактируют с ганглиозными клетками не
непосредственно, а через АП-амакриновые
клетки и колбочковые биту^пы. Обратите
внимание на щелевые контакты ножек кол-
бочек и сферул палочек, а также тормозный
(показан черным) синапс АН-амакриновой
клеткой на дендрите ганглиозной клетки с
OFF-центром. Объяснения в тексте. АП —
АП-амакриновая клетка, БП+ — биполяр-
ная клетка ON-типа, БП" — биполярная
клетка OFF-типа, НК — ножка колбочки,
ГК+ — ганглиозная клетка с ON-центром,
ГК” — ганглиозная клетка с OFF-центром,
ЩК — щелевой контакт, Г — горизонталь-
ная клетка, ВЗН — волокна зрительного
нерва, СП — сферула палочки. По Schiller,
1992 и Nakanishi, 1995.
ВЗН
386 Часть IV. Светочувствительность
траст, но становится чрезвычайно чувствительной к общей освещенности.
Полностью адаптированная к темноте сетчатка способна воспринимать еди-
ничные фотоны.
Блок-схемы на рис. 16.20 и 16.21 очень упрощены по сравнению с реальной
ситуцацией в сетчатке. Уже упоминалось, что гистологи выделяют в сетчатке
более 50 различных подтипов клеток, а нейробиологи пришли к выводу, что
вообще каждый нейрон здесь имеет собственную индивидуальность. Боль-
шинство из двух с лишним дюжин нейромедиаторов и нейромодуляторов1
обнаружено в сетчатке, равно как и огромное и все возрастающее количество
рецепторных молекул. Тем не менее, схем™, приведенные в данном разделе,
показывают, что основные черты системы все-таки пробиваются сквозь массу
усложняющих ее деталей.
Одним из самых интересных достижений последнего времени стало ис-
пользование методов современной генетики для исследования и анализа сет-
чатки данио Danio rerio. В Информации 14.1 мы уже видели, как генетика по-
новому осветила ранние стадии развития (и эволюции) глаз позвоночных и
других животных. В данном случае применение мощных технологий генетики
несколько иное. Данио — это идеальный и наиболее используемый организм
для нейрогенетики. У этой рыбки хорошо развитая сетчатка с четырьмя типа-
ми колбочек и палочками, и зрение развито намного лучше, чем у мыши,
единственного позвоночного, чья генетика так же хорошо изучена. Данио лег-
ко содержать, у него быстрое развитие, прозрачная личинка, а геном практи-
чески полностью проанализирован. Выделено большое количество мутантов
данио с различными дефектами зрения и определено, где и как эти мутации
сказываются на сетчатке. Эффективность генетических методов позволяют
думать, что сетчатка Danio rerio будет первой, чью структуру изучат от молеку-
лы до целого. Это потрясающий объект исследований, про который мы уже
говорили как о части мозга, выдвинутой наружу и потому доступный. Инте-
ресующиеся студенты могут найти детальное изложение проблемы в работах,
приведенных в Библиографии.
16.2.9. Цвет
Мы видели в разделе 16.2.1, что различение v"H волн (различение цветов) за-
висит от присутствия в сетчатке (в данном случае — человека) трех популяций
колбочек: реагирующих на синий (Хтах 419 нм), зеленый (Хтах 531 нм) и крас-
ный (Хтах 559 нм) свет. Можно подумать (и именно так и думали много лет),
что ощущение цвета формируется мозгом, получающим информацию по этим
трем каналам. Однако оказалось, что ситуация намного сложнее. Некоторые
черты восприятия цвета не могут быть объяснены на основе такой простой
идеи. Во-первых, показано, что восприятие некоторых пар цветов так влияет
друг на друга, что их смесь не воспринимается. Например, хотя мы ясно вос-
принимаем смесь красного и желтого (оранжевый) или синего с зеленым (го-
1 Медиаторов насчитывается, на самом деле, до 40, а с модуляторами их еще больше. —
Прим, перев.
16. Сетчатка 387
Красные
колбочки
Зеленые
колбочки
Зеленые колбоч
Красные колбоч
Маленькое
пятно
белого
света
Маленькое
пятно
красного
света
Большое
пятно
красного
света
Большое
пятно
зеленого
света
Рис. 16.22. Рецептивные поля одиночных цветовых оппонентных клеток сетчатки
кошки. Оба типа клеток возбуждаются маленьким пятном белого цвета в центре
поля. Лучшие ответы показаны в случаях 5 и 6, где большие пятна красного или зе-
леного цвета покрывают все поле. Из Kandel, Schwartz and Jassell, 1991, с разреше-
ния.
лубой), смесь красного с зеленым воспринимается как чисто желтый без ка-
ких-либо следов исходных цветов. Сходным образом, смесь желтого и синего
вызывает ощущение чистого белого цвета. Это очень интересная особенность,
и чтобы объяснить ее, Эвальд Геринг (Ewald Hering) предложил теорию оппо-
нентных процессов. Он предположил, что три первичных цвета обрабатываются
зрительной системой как ангаюнистические или оппонентные пары: крас-
ный/зеленый, желтый/синий и белый/черный. Стимуляция одного из оппо-
нентов вызывает возбуждение (или торможение), тогда как стимуляция друго-
го — противоположные эффекты (торможение или возбуждение, соответст-
венно). Следовательно, когда стимулы сбалансированы (например, поступает
соответствующее количество красного и зеленого цветов), разные компонен-
Часть IV. Светочувствительность
Информация 16.1
ПИГМЕНТНЫЙ РЕТИНИТ
Существует большое число генетических заболеваний глаза (как и уха, см. гл. 8). Ге-
ны, ответственные за это, распространены почти по всему геному человека (в двад-
цати из двадцати трех хромосом), что отражает сложность структуры глаза. Развитие
и функционирование глаза связано с координированным действием множества ге-
нов, любой из которых может функционировать неверно. Наиболее распространен-
ная форма наследуемого дефекта зрения — это пигментный ретинит (ПР). Название
охватывает гетерогенную группу патологий, суммарная встречаемость которой со-
ставляет около 1 на 3000. ПР может наследоваться как аутосомная доминанта (adRP),
аутосомный рецессив (arRP) или сцепленным с Х-хромосомой (xlRP). adRP — наи-
более частая форма, наблюдающаяся по крайней мере в половине семей, страдаю-
щих от ПР. Собственно заболевание состоит в неизлечимой (в настоящее время)
дегенерации и гибели палочек и, в намного меньшей степени, колбочек. Дегенери-
рующие наружные сегменты палочек удаляются и перевариваются пигментным
эпителием, что приводит к сильной пигментации, откуда и взялось название болез-
ни. Утрата палочек приводит к каскаду событий, которые в конце концов воздейст-
вуют и на колбочки, и на кровоснабжение сетчатки. Ранние признаки заболевания —
ночная слепота и аномалии электроретинограммы (ЭРГ). Рано или поздно, болезнь
приводит к слепоте.
В данной главе мы рассматривали некоторые молекулярные и клеточные элемен-
ты строения палочек. Все эти элементы могут быть объектами мутаций, ведущих к на-
рушению функции генов, а затем и к нарушениям зрения. Генетический анализ пока-
зал вовлеченность в это заболевание 1,3,6,7,8 и Х-хромосом. ПР, связанный с Х-хро-
мосомой, подвергся особенно интенсивному генетическому анализу, и ответственные
за ПР мутации были локализованы в двух генах в коротком плече хромосомы. Эти ге-
ны пока не охарактеризованы и их биохимические эффекты не определены.
Больше продвинулись в некоторых случаях аутосомного доминантного ПР.
Первая мутация, ответственная за adPR, локализуется в хромосоме 3. Показано,
что она приводит к замещению аминокислоты в опсине палочек (про23 => гис).
Позже было обнаружено более 30 мутаций, затрагивающих аминокислотную по-
следовательность опсина палочек. В некоторых случаях они затрагивают менее
существенные участки молекулы, вследствие чего возникновение ПР задерживает-
ся, и болезнь имеет менее жестокий характер. Первая мутация, ведущая к adPR
(упоминавшаяся выше), охватывает большую популяцию граждан США ирланд-
ского происхождения, она вызывает очень раннее начало ПР и серьезные наруше-
ния зрения на втором десятилетии жизни. Мутация, вероятно, ограничена террито-
рией США, где обнаруживается более чем у 15% страдающих adPR. Тот факт, что
мутация не обнаруживается в Европе, указывает, что «основателя» мутации следу-
ет искать среди ранних ирландских иммигоантпр Ппугая группа ирландцев в США
страдает от менее выраженного adPR, вызывающего нарушения зрения только на
четвертом десятке или позже и связанного с мутацией в хромосоме 6. Подозревае-
мый ген кодирует структурный белок дисков наружных сегментов палочек. Этот
белок называется периферин-RDS.
16. Сетчатка 389
Продолжение
Генетический анализ arPR так далеко не продвинулся, поскольку трудно про-
следить широкую генеалогию. Тем не менее, показано, что замена Г => Т в кодоне
249 у одной родственной группы с arPR приводит к замещению триплета ГАГ на
ТАГ. Последний транскрибируется как УАГ, который является стоп-кодоном при
трансляции в рибосоме. В результате опсин утрачивает шестой и седьмой транс-
мембранный сегменты. Гомозиготная больная страдала ночной слепотой с детства,
тогда как ее гетерозиготные родители и сиблинг были нормальными. Другой вари-
ант arPR возникает в результате замещения Г => А в кодоне 150, что приводит к за-
мене триплета ГАГ (глютамин) на ААГ (лизин). Опять-таки, у гетерозиготных род-
ственников патологии зрения не наблюдалось, тогда как гомозиготный больной
страдал тяжелой ночной слепотой. Другие работы предполагают, что arPR не свя-
зан с мутациями в хромосоме 3, он может вызываться мутантными генами в многих
хромосомах. Теперь стало возможно создание трансгенных мышей, экспрессирую-
щих некоторые из этих генов. Таким образом, молекулярная биология дефектов,
лежащих в основе ПР, постепенно разъясняется. В одном из случаев дегенерации
сетчатки у мышей, например, установлено, что патология возникает в результате
дефекта не опсина, а р-субъединицы цГМФ-ФДЭ. Такой дефект был недавно
идентифицирован и в некоторых случаях arPR у человека.
Мы отмечали в этой главе, что родопсин и цГМФ-ФДЭ присутствуют в боль-
ших количествах в дисках наружных сегментов, а родопсин отчасти формирует их
структуру. Неудивительно поэтому, что генетические ошибки в образовании этих
элементов (а также, конечно, периферина) приводят к нарушению целостности
дисков палочек, что увеличивает риск их удаления. Диски палочек, как мы видели,
удаляются и перевариваются клетками пигментного эпителия (ПЭС). Этот процесс
также находится под генетическим контролем. Возможно, именно дисбаланс в раз-
витии между синтезом и деградацией ответственен за медленное развитие боль-
шинства ПР.
Молекулярная нейробиология находится в самом начале пути к пониманию
всего комплекса взаимодействующих процессов. Как и в других случаях, это ве-
дет к серьезным прорывам в выяснении причин, лежащих в основе опасных бо-
лезней человека. Недавний успех с введением нормальных генов мышам, гомо-
зиготным по rds и rd, который привел к блокаде дегенерации фоторецепторов,
дает надежду на разработку адекватной терапии.
Литература
Armstrong, R. А., 1999, «Ocular disease and the new genetics», Ophthalmic and Physiolo-
gical Optics, 19, 193—195.
Berson, E. L., 1993, «Retinitis pigmentosa: the Friedenwald lecture», Investigative Ophthal-
mology and Visual Science, 34,1659—1676.
Humphries, P., P. Kenna and G. J. Farrar, 1992, «On the molecular genetics of retinitis
pigmentosa», Science, 256, 804—808.
Milam, A. H., 1993, «Strategies for the rescue of retinal photoreceptor cells», Current
Opinion in Neurobiology, 3, 797—804.
390 Часть IV. Светочувствительность
ты такого канала отключаются, и система формирует ощущение желтого цве-
та. Такая обработка информации начинается, по-видимому, еще в сетчатке, но
затем продолжается в наружном коленчатом теле и зрИх^^лой коре.
Ограничиваясь пока сетчаткой, заметим, что доказано присутствие гангли-
озных клеток с оппонентными свойствами в сетчатке кошки. В случае, приве-
денном на рис. 16.22, показаны две ганглиозные клетки, одна из которых име-
ет концентрическое РП с центром ON-типа для красного и окружением, даю-
щим OFF-ответ на зеленый, а другая — ON-ответ в центре на зеленый и OFF-
ответ на красный на периферии. Клетки такого типа не дают мозгу слишком
точной информации — рис. 16.22 показывает, что мозгу трудно будет разли-
чить маленькое яркое белое пятнышко в центре РА и большое зеленое пятно,
покрывающее все поле.
Связи в сетчатке, ответственные за цветовую оппонентность того типа, что
показана на рис. 16.22, продолжают изучаться. Понятно, однако, что субъек-
тивное ощущение цвета, которое представляется столь непосредственным и
очевидным, возникает в результате сложных взаимодействий не только в сет-
чатке, но и на более высоких уровнях зрительной системы.
16.3. Заключение
Сетчатка, как мы отмечали в подзаголовке этой главы и в разделе 15.3 преды-
дущей, это — часть мозга, выдвинутая на периферию. В данной главе мы рас-
смотрели некоторые ее детали от уровня молекул до уровня «блок-схем». Мы
видели, что ее функции прослеживают вплоть до молекулярного уровня. Хотя
остается еще немало неясного в том, что касается синаптологии и клеточных
взаимодействий, мы явно идем к формированию всесторонней теории — мо-
дели сетчатки. Если эти исследования будут доведены до конца, это будет пер-
вой частью мозга, изучение которой доведено до полного понимания. Значе-
ние такой окончательной теории огромно.
Остающиеся проблемы, однако, огромны. Мы рассмотрели организацию
сетчатки по вертикали и горизонтали. Диффузные биполярные клетки могут
вступать в контакт и объединять активность до десяти ножек колбочек и пере-
давать ее на единственную ганглиозную клетку; с другой стороны, карликовые
биполяры часто образуют с ганглиозными клетками связи «один к одному», и
так передают информацию в волокна зрительного нерва. Горизонтальные и
амакриновые клетки вмешиваются в этот вертикальный поток информации и
модулируют ее передачу. Таким образом, в сетчатке сочетаются параллельная
и иерархическая обработка информации — здесь, на периферии, уже начина-
ется то, что происходит затем в мозге. Мы увидим далее в главе 17, что более
двух дюжин экстрастриарных участков коры приматов соединены «вертикаль-
но» и «перекрестно». В сетчатке мы в миниатюре можем изучать сложную
смесь параллельной и иерархической обработки информации, которая в мно-
го более сложном виде наблюдается в мозге.
Но сложности на этом не заканчиваются. В отличие от однозначных пере-
ключателей в устройстве компьютера, биологические «переключатели» в моз-
ге — это сложный коктейль медиаторов, это ГАМК, глютатмат, ацетилхолин,
дофамин, серотонин, соматостатин и многое другое, присутствующее здесь
16. Сетчатка 391
одновременно. Кто-то возбуждает, кто-то — тормозит, а кто-то — модулирует.
Рецепторов, с которыми все они взаимодействуют, большое множество и они
различны. Методы современной молекулярной биологии были применены
для клонирования многих из этих крупных рецепторных белков, и было изуче-
но не только их структурное разнообразие, но и динамика их синтеза и дегра-
дации, которые постоянно меняют восприимчивость постсинаптических мем-
бран к многокомпонентному коктейлю нейрохимических веществ.
Следует также помнить, что сетчатка человека — это лишь один случай из
многих. Человек — один из приматов, которые тоже разнообразны, а сами
приматы — всего лишь один отряд из числа млекопитающих (гл. 4). Ну и сет-
чатка млекопитающих — это лишь часть спектра сетчаток позвоночных. Неко-
торых ее вариантов мы коснемся в гл. 18 и увидим, что на основе сравнитель-
но неспециализированной сетчатки, подобной нашей, развились удивитель-
ные модификации у глубоководных рыб и хищных птиц.
Есть однако черта, по которой сетчатка приматов превосходит остальных
млекопитающих (хотя и не всех позвоночных) — это цветное зрение. Мы ви-
дели в разделе 16.2.1, что цветное зрение в сетчатке человека зависит (исходно)
от наличия трех различных пигментов колбочек. Приматы — единственные
среди млекопитающих, у кого пигментов — три, у остальных — только два.
Грубо говоря, у этих животных один пигмент — коротковолновый, а другой —
длинноволновый. Собака — типичный дихромат, один из пигментов колбочек
имеет Хтах = 450 нм, а другой — Хтах = 555 нм. Домашняя кошка — тоже дихро-
мат с коротко- и длинноволновым пигментами, у нее, однако, пока не удалось
выявить цветное зрение поведенческими экспериментами1. Почему же люди и
другие приматы оказались единственными трихроматами среди млекопитаю-
щих? Наиболее правдоподобное объяснение состоит в том, что трихромазия
развилась у существ, обитающих под пологом леса, под давлением необходи-
мости правильно оценивать спелость (или, соответственно, незрелость) тро-
пических фруктов. Как мы видели в разделе 16.2.2, то эволюционное событие,
которое приведшело к возникновению опсина зеленых колбочек, занявшего в
спектре промежуточное положение между длинноволновым («красным») и
коротковолновым («синим») опсинами, произошло около 40 миллионов лет
назад. Тогда ранние приматы адаптировались к преимущественно древесному
образу жизни и фруктовой диете. Когда эта книга уже была нагхг^^лена в пе-
чать, появились сообщения о трихомазии у дневных prosimian (скользяще-ла-
зающие лемуры, медленный лори) — далеких родичей человека, высших и
низших обезьян.
Хотя сетчатки других млекопитающих не могут конкурировать с сетчаткой
приматов в количестве колбочковых пигментов, они часто специализирова-
лись иными путями. Во многих случаях, как у кролика и тем более у многих
позвоночных, не относящихся к млекопитающим, обработка информации в
большей степени, чем у приматов, осуществляется в сетчатке. В сетчатке кро-
1 В. В. Максимовым и А. Кезели (1980) в поведенческих опытах в на кошке показано на-
личие у нее как трихроматического цветового зрения, так и константности цветовосприя-
тия — одного из фундаментальных свойств цветного зрения. — Прим. ред.
392 Часть IV. Светочувствительность
лика есть ганглиозные клетки, способные реагировать на большое разнообра-
зие стимулов, а не просто на пятно света, как сетчатка приматов или кошек.
Обнаружены ганглиозные клетки, активируемые движущимися краями объ-
ектов (локальные детекторы краев), краев специфической ориентации, дви-
жущихся в определенных направлениях, а также клетки, чьи рецептивные
поля настроены на выделение других, часто очень специфических, ключевых
стимулов. У приматов и кошек клетки, отвечающие на такие специфические
стимулы, находятся в зрительной коре, и мы обсудим некоторые из таких слу-
чаев в следующей главе. Возможно, дело в том, что кролику необходимо экс-
тренно обрабатывать ту информацию, которую другие животные, на которых
охотятся не так часто, могут обдумать не спеша. Именно это и происходит
в анализаторе зрительной коры приматов и кошачьих, к которым мы перейдем
в следующей главе.
17. ЗРИТЕЛЬНЫЕ ПУТИ
И ЗРИТЕЛЬНАЯ КОРА
У высокоразвитых приматов более двух дюжин взаимосвязанных участ-
ков зрительной коры и три зрительных пути. Эволюционные соображе-
ния. Ретино-тектальный путь: частичный перекрест — верхнее четверохол-
мие — карта зрительного пространства — саккадические движения глаз.
Ретино-геникуло-стриарный (РГС) путь: оптический диск — продырявлен-
ная пластинка — зрительный нерв — частичный перекрест — зрительный
тракт — дорзальное ядро наружного коленчатого тела (НКТд) — слои, ги-
стология, кортикофугальная иннервация — рецептивные поля (РП) кле-
ток коленчатого ядра — М, Р и W волокна — М — чувствительны к дви-
жению, Р — чувствительны к цвету — зрительная радиация — мейерова
петля — патологии. Первичная зрительная (= стриарная) кора (VI): распо-
ложение — размеры — слои — полоска Дженнари; микроэлектродная ре-
гистрация — РП клеток коры — концентрические РП слоя IVc — ориен-
тационная избирательность нейронов в других слоях, детекторы ориен-
тации — возможные «блок-схемы»; классификация РП на простые,
сложные и концевые (сверхсложные); биологическое значение, колон-
чатая организация — дезоксиглюкозная гистология и детектирование
края — гистология цитохром оксидазы и капельно-межкапельная орга-
низация — цветовая чувствительность (blobs) — двойная оппонентность;
глазодоминантные колонки — бинокулярное зрение; гиперколонки и аг-
регации РП (АРП) — вариации размеров АРП — неизоморфные карты
сетчатки в VI; популяционный ответ и устранение неоднозначности.
Пластичность: чувствительный период — котенок — макак. Экстрастри-
арная кора: карты различных характеристик зрительной информации —
входы из подушки таламуса и из VI — три потока информации — цито-
хром оксидазное окрашивание V2 — толстая полоска (М-поток), тонкая
полоска (Р-поток) — в зрение вовлечено большое количество зон коры
приматов. Распознавание лица: значение для социальных приматов — ни-
жневисочная кора макаков — нейроны, узнающие руки и лица. Прозопаг-
нозия: значение распознавания лиц для человека — нейроанатомия —
симптомы прозопагнозии. Заключение: сложность кортикальных анали-
заторов — значение зрения — надежды на будущее и настоящее.
В данной главе мы рассмотрим пути, по которым зрительная информация пе-
редается из сетчатки в первичную зрительную (стриарную) кору. Мы, как и в
предыдущих двух главах, сосредоточим внимание на приматах, в особеннос-
ти — на зрительной системе человека. С самого начала стоит подчеркнуть, что
394 Часть IV. Светочувствительность
первичная зрительная кора — не единственная зрительная кора в мозге прима-
тов. Исследования мозга макака показали, что здесь иммется до двух дюжин
участков коры, связанных с различными аспектами анализа зрительной ин-
формации. Все они сложным образом связаны между собой как по «восходя-
щей», так и по «нисходящей». Немыслимо обсуждать центральный анализ
зрительной информации без этого факта, также, как невозможно в такой кни-
ге рассматривать их детально. Поэтому в данной главе мы не пойдем дальше
первичной зрительной коры — стриарной, а экстрастриарную кору затронем,
только касаясь некоторых ее ярких черт.
17.1. Зрительные пути к мозгу
Путь к стриарной коре, т. н. ретино-геникуло-стриарный (РГС) путь — не един-
ственный, соединяющий сетчатку с мозгом млекопитающих. Существуют еще
два менее значительных пути: путь от сетчатки непосредственно через зри-
тельный перекрест в верхние бугры четверохолмия (ретино-тектальный путь) и
полисинаптический путь через гипоталамус и верхний цервикальный ганглий
в шишковидное тело. Оба этих последних пути более важны у немлекопитаю-
щих позвоночных. Мы обсудим их более полно в главе 18, здесь, однако, сто-
ит набросать интересный эволюционный сценарий.
У анамний — рыб и земноводных — главная зрительная область развивает-
ся в среднем мозге (mesencephalon). Крыша этой части мозга — тектум — стро-
го специализируется на анализе зрительной информации, но получает инфор-
мацию и от других органов чувств. Этот т. н. ретинотектальный или тектофу-
гальный путь остается основным и у пресмыкающихся (см. главу 18). У млеко-
питающих значение этой части мозга существенно уменьшается, поскольку
она оказывается в тени колоссально развившихся полушарий мозга. Четыре
небольших бугорка — верхние и нижние бугры четверохолмия — это все, что ос-
талось от сильно развитого тектума рыб и земноводных, соответственно,
уменьшается и значение тектофугального пути.
Почему же млекопитающие изменили зрительную систему? Эволюцион-
ный сценарий, о котором упоминалось выше, предполагает, что в течение
мезозойского периода ранние млекопитающие занимали подчиненное по
отношению к «царствующим рептилиям» положение, особенно динозаврам,
и вели в сумеречных условиях скрытное существование. В таких условиях
зрение становится второстепенным по отношению к обонянию, вкусу и слу-
ху. Химические чувства, которые и сейчас остаются для нас эмоционально
окрашенными, стали занимать передний мозг и лимбическую систему.
Предполагается, что передний мозг в этих условиях приобретает большее
значение. Когда по тем или иным причинам — из-за удара метеорита, взры-
ва сверхновой или глобального похолодания — царствующие рептилии ис-
чезли в конце мезозоя, мир открылся для угнетенных протомлекопитающих.
Они заселили все возможные экологические ниши освободившегося мира,
зрение снова стало наиболее важным из всех чувств. Однако, формирующи-
еся заново, зрительные пути направились к наиболее важной части мозга —
переднему мозгу, расширяющемуся и формирующему характерные для мле-
17. Зрительные пути и зрительная кора 395
копитающих полушария. Ретино-тектальный путь остается пережитком ста-
рого зрительного пути, а ретино-геникуло-стриарный путь быстро становит-
ся наиболее важным путем передачи зрительной информации в мозг.
17.1.1. Ретинотектальный путь
Хотя, как указывалось выше, этот путь более важен у немлекопитающих по-
звоночных и будет рассмотрен более полно в главе 18, стоит описать его значе-
ние в зрительной системе млекопитающих здесь. Зрительные волокна из сет-
чатки в составе зрительного нерва направляются к хиазме, где у приматов про-
исходит частичный перекрест волокон. Волокна из назальной сетчатки пере-
ходят к контрлатеральному бугру четверохолмия, тогда как волокна из височ-
ной сетчатки идут к ипсилатеральному бугру (рис. 17.1). У многих млекопита-
ющих, не относящихся к приматам, перекрест имеет полный или почти пол-
ный характер.
Верхний бугор четверохолмия состоит из семи слоев клеток и связан не
только со зрением. Его эволюционное происхождение обеспечивает получе-
ние информации и от других органов чувств. Сенсорный вход организован в
Полный перекрест
(а) Без ипсилатеральной
проекции
(в) Частичный
перекрест
Рис. 17.1. Схемы, показывающая три типа перекреста ретино-тектального пути у
различных млекопитающих. На рис. (а) показан полный перекрест, наблюдаю-
щийся у мелких рукокрылых. На рис. (б) показан небольшой ипсилатеральный
компонент, такая картина характерна для многих млекопитающих, включая белок,
кошек, кроликов и т. д. Наконец, на рис. (в) вся височная половина поля зрения
сетчатки направляет волокна в ипсилатеральный верхний бугор четверохолмия.
Зона, обозначенная точками — ипсилатеральные проекции, обозначенные штри-
ховкой — контрлатеральные. С изменениями из G. Н. Henry and Т. R. Vidyasgar,
1991; в: J. R. Cronly-Dillon and R. L. Gregory, Vision and Visual Dysfunction, vol. 2,
p. 447, Figure 20.2. Воспроизводится с разрешения Macmillan Press Ltd.
396 Часть IV. Светочувствительность
карты — здесь их три — зрительная, соматосенсорная и слуховая — одна над
другой. Зрительная карта формируется частично входом из сетчатки, а частич-
но — из зрительной коры (см. раздел 17.5). Эта карта имеет иной принцип ор-
ганизации, чем в зрительной коре. Вместо того, чтобы представлять неизо-
морфную, но топологически точную картину сетчатки, карта представляет
визуальное пространство вокруг животного. Другие сенсорные карты — сома-
тосенсорная и слуховая — построены по тому же принципу и лежат в четверо-
холмии, точка в точку, одна под другой. Об интенсивно исследованной карте
слухового пространства в нижних буграх четверохолмия сипухи говорилось в
разделе 9.3. В наиболее глубоких слоях бугра четверохолмия располагается еще
одна карта — моторная. В нее сенсорные карты передают информацию для
контроля саккадических движений глаз, которые направляют взгляд на важные
черты зрительного поля. Эта важная функция выполняется совместно с обла-
стью фронтальной коры — лобным глазным полем.
17.1.2. Ретино-геникуло-стриарный (РГС) путь
Теперь обсудим некоторые детали главного зрительного пути млекопитаю-
щих — ретино-геникуло-стриарного. В главе 16 мы оставили волокна зритель-
ного нерва (аксоны ганглиозных клеток сетчатки), когда они направлялись
к оптическому диску. В сетчатке человека оптический диск имеет несколько
эллипсоидную форму с вертикальным диаметром около 1,8 мм и горизонталь-
ным — 1,75 м. Центр диска расположен в 4 мм медиальнее и в 0,8 мм выше фо-
веолы (рис. 16.2 Б). Хотя в области оптического диска всегда имеется углубле-
ние, при глаукоме, т.е. при повышении внутриглазного давления, оно выраже-
но намного сильнее (см. гл. 15). В этих случаях офтальмоскопия показывает,
что оптически диск заметно углублен в сторону зрительного нерва. Образую-
щаяся «чашка» является диагностическим признаком повышенного внутри-
глазного давления и того, что оптический диск является «слабым местом»
глазного яблока. Чтобы противодействовать такой механической слабости, из
склеры непосредственно под диском вырастает сеть коллагеновых волокон —
продырявленная пластинка. Зрительные волокна, проходя через продырявлен-
ную пластинку образуют зрительный нерв.
Зрительный нерв
Еще в продырявленной пластинке олигодендроциты формируют миелиновую
оболочку волокон зрительного нерва. Другие глиальные клетки и клетки со-
единительной ткани образуют септы между группами волокон зрительного
нерва, формируя пучки. Многочисленные кровеносные сосуды из мягкой
оболочки мозга (pia mater) проникают в эти пучки, снабжая волокна зритель-
ного нерва питательными веществами. Миелинизация волокон и присутствие
соединительнотканных септ сильно увеличивает размер зрительного нерва, и
он достигает 3 мм в диаметре (в оптическом диске — 1,8 мм). Первоначально
волокна в зрительном нерве распределены также, как в сетчатке. Фовеальные
волокна, которые составляют около трети волокон зрительного нерва, снача-
ла занимают латеральное положение, но по мере приближения к перекресту
17. Зрительные пути и зрительная кора 397
Рис. 17.2. Зрительный
путь. Отметьте частич-
ный перекрест в хиазме.
Мейерова
петля
Сетчатка
- Зрительный нерв
Зрительная кора
Зрительная радиация или
геникуло-кортикальный
тракт
Наружное (латеральное)
коленчатое тело
Зрительный
перекрест (хиазма)
Зрительный тракт
смещаются к центру. В хиазме у приматов имеет место частичный перекрест во-
локон (рис. 17.2). Волокна из назальной половины сетчатки, включая назаль-
ную половину фовеа, пересекают среднюю линию и направляются в контрла-
теральное наружное коленчатое тело (НКТ). Волокна из височной половины
сетчатки, включая латеральную часть фовеа, остаются на своей стороне и на-
правляются в ипсилатеральное наружное коленчатое тело.
Зрительный путь от перекреста (хиазмы) до НКТ называется зрительным
трактом. На пути к НКТ тракт делает изгиб в 90е, так что волокна из верхнего
квадранта сетчатки оказывается на медиальной стороне, а волокна из нижнего
квадранта — на латеральной стороне. Именно здесь волокна из соответствую-
щих частей двух сетчаток объединяются. Два зрительных тракта оканчиваются
в наружных коленчатых телах.
Наружные коленчатые тела
Эти ядра (или тела) располагаются в задней части таламуса, и имеют различи-
мо пластинчатую структуру (рис. 17.3). Это смахивает на стопку шляп, надетых
одна на другую. У приматов здесь шесть слоев клеток, разделенных лентами
волокон. Именно здесь оканчиваются волокна зрительного нерва (аксоны
ганглиозных клеток сетчатки). Вследствие частичного перекреста, описанно-
го выше (рис. 17.2), сюда приходят нервные волокна из височной половины
ипсилатерального глаза и назальной половины контрлатерального глаза.
В НКТ поддерживается строгое разграничение между этими волокнами. Во-
локна из контрлатерального глаза оканчиваются в слоях 1, 4 и 6, тогда как
волокна из ипсилатерального глаза — в слоях 2, 3 и 5. Эти волокна образуют
синапсы с другой группой нейронов, которые, формируя зрительную радиа-
цию, передают информацию в первичную зрительную кору. Насколько точны
эти контакты, остается предметом дискуссии. Известно, однако, что волокна
зрительного нерва могут образовывать до шести разветвлений, образующих
-------Волокна от ипсилатерального глаза
———• Волокна от контрлатерального глаза
Рис. 17.3. Наружное коленчатое
тело приматов. (А) На микрофо-
тографии показаны шесть слоев
НКТ. Стрелками и цифрами от 1
до 6 показаны ретинотопические
карты. РП клеток вдоль стрелок
занимают одинаковые положе-
ния в зрительном пространстве.
(Б) Схема контактов в НКТ.
Шесть слоев обозначены цифра-
ми. Первые два слоя (магноцел-
люлярные) содержат клетки с
крупными телами. В остальных
слоях клетки более мелкие. От-
метьте разделение волокон опти-
ческих трактов ипси- и контрла-
терального глаз, дальнейшие
объяснения в тексте. К — контр-
латеральный глаз, И — ипсилате-
ральный глаз, М — магноцеллю-
лярный слой, П — парвоцеллю-
лярный слой.
17. Зрительные пути и зрительная кора 399
синапсы с соседними нейронами в одном и том же слое НКТ. С другой сторо-
ны один нейрон коленчатого тела может образовывать синапсы с более, чем
одним входящим волокном зрительного нерва. Вставочные нейроны также
присутствуют здесь, включаясь между другими нейронами в одном слое ко-
ленчатого тела. Разделение входов от разных глаз, однако, сохраняется. На-
конец, здесь присутствует значительная иннервация из зрительной коры
(кортикофугальная иннервация), которая выполняет функцию обратной свя-
зи. Таким образом, НКТ не может рассматриваться как простая передающая
структура.
Теперь вернемся к волокнам зрительного нерва и тракта. Мы отмечали в
разделе 16.2.8, что эти волокна подразделяются на две крупные и одну мень-
шую группы: магноцеллюлярные (М), парвоцеллюлярные (Р) и W-волокна.
М-путь, несущий информацию, в основном связанную с движением зритель-
ного образа, оканчивается в слоях 1 и 2 НКТ (магноцеллюлярные), тогда как
P-путь, в основном связанный с цветовой информацией, оканчивается в сло-
ях 3, 4, 5 и 6 (парвоцеллюлярные слои). Эти клетки можно изучать с помощью
микроэлектродной регистрации. Как и следовало ожидать, они отвечают на
освещение соответствующих участков сетчатки. Эти участки, следовательно,
представляют собой РП клеток коленчатого тела. В слоях 1 и 2 рецептивные
поля имеют цветонеразличающий центр, схожий с полем альфа-ганглиозных
клеток. Эти клетки высокочувствительны к движению в их рецептивных по-
лях. РП со схожими характеристиками в небольшом количестве присутствуют
и во всех других слоях НКТ. Однако, в верхних слоях, как и можно было ожи-
дать, учитывая входы туда парво- волокон зрительного тракта, клетки в основ-
ном реагируют на цвет. В слоях 3 и 4 преобладают оппонентные клетки с OFF-
центром (синее/желтое или, правильнее, синее/красное + зеленое), тогда как
в слоях 5 и 6 — оппонентные клетки с ON-центром красный/зеленый. Мед-
ленно проводящие тонкие аксоны W-клеток проецируются также в парвоцел-
люлярные слои и в интраламинарные зоны.
Зрительная радиация (геникуло-кортикалъный тракт)
Волокна из наружного коленчатого тела идут в первичную зрительную кору,
которая занимает sulcus calcarinus (шпорную борозду) на медиальной по-
верхности затылочной доли. Нейроны латеральной части НКТ, получаю-
щие афференты из нижних квадрантов сетчатки (верхние квадранты поля
зрения), посылают свои аксоны латерально — вокруг переднего конца ви-
сочного рога латерального желудочка, образуя мейерову петлю. Клетки
коленчатого тела, получающие афференты из фовеа в мейеровой петле
участвуют мало. Они направляют аксоны практически непосредственно
в шпорную борозду. Поражения различных ветвей зрительной радиации
приводят к появлению дефектов в соответствующих участках поля зрения.
Однако, каким бы путем ни шли зрительные волокна, все они оканчивают-
ся в первичной зрительной коре.
400 Часть IV. Светочувствительность
17.2. Первичная зрительная кора
17.2.1. Структура
У человека первичная зрительная кора (поле Бродмана 17, VI или стриарная
кора) занимает стенки глубокой шпорной борозды на медиальной поверхнос-
ти задней части затылочной доли (рис. 17.4 Г). Стриарная кора простирается
кзади вдоль стенок этой борозды, а небольшая ее часть продолжается с посте-
ро-латеральной стороны затылочной доли (рис. 17.4 В). Общая поверхность
первичной коры у человека в среднем составляет около 3000 мм2.
Понятно, что эксперименты на зрительной коре человека невозможны по
этическим соображениям. Большинство экспериментальных работ по этой
причине выполнялись на макаках и домашних кошках Felts domestica. К счас-
тью, зрительная кора у обоих животных легко доступна. Рис. 17.4 А и Б пока-
зывают, что зрительная кора макака в основном локализована на поверхности
заднего полюса затылочной доли. У кошки зрительная кора занимает в основ-
ном то же положение. У макака поверхность занимает 1400 мм2, а у кошки —
около 380 мм2.
Рис. 17.4. Положение зрительной коры у макаков и человека. (А) и (Б) показывают по-
ложение коры на латеральной (А) и медиальной (Б) поверхности мозга макака. (В) и (Г)
показывают положение коры на латеральной (В) и медиальной (Г) поверхности мозга че-
ловека. Пропорции размеров мозга макака и человека не соблюдены. Числа на рисунке
мозга человека — номера полей Бродмана. 17 — первичная зрительная кора (VI). Из
Logothetis and Sheinberg, 1996, с разрешения.
17. Зрительные пути и зрительная кора 401
Рис. 17.5. (А) Малое увеличе-
ние парасагитального среза
первичной зрительной коры.
Окрашивание по Нисслю.
Стрелки указывают крайние
точки поля 17 — стриарной
или первичной зрительной
коры. Этот тип коры характе-
ризуется наличием полоски
Дженнари (более плотно окра-
шенная полоса). Буквой «А»
обозначена наружная поверх-
ность мозга, «В» — внутренняя
складка.
(Б) Поперечный срез стри-
арной коры при большом уве-
личении. Окрашивание крези-
ловым фиолетовым, слои про-
нумерованы слева. W —- белое
вещество. Из Hubei and Wiesel,
1977, с разрешения.
402 Часть IV. Светочувствительность
В Поверхность коры,
обращенная к мягкой оболочке
1 -------- ' -
целлюлярных целлюлярных областям структурам
слоев НКТ слоев НКТ коры мозга
Рис. 17.5. (В) Внешние связи
вывода первичной зритель-
ной коры. Объяснения в тек-
сте. По Hubei, 1988.
Как и другие части неокортекса, зрительная кора имеет толщину около
2 мм и шесть слоев клеток. Слои нумеруются со стороны мягкой оболочки
внутрь. Слой 4 обычно подразделяется на субслои: 4А, 4В и 4С, а последний —
еще на подслои а- и р- (рис. 17.5). Первичная зрительная кора приматов отли-
чается от других частей неокортекса наличием полосы миелинизированных
волокон, идущих параллельно поверхности мозга, обращенной к мягкой обо-
лочке в слое 4, известной как полоска Дженнари и имеющей название стриар-
ная кора (от лат. stria — полоса).
Если проследить путь афферентов из НКТ, то окажется, что волокна маг-
ноцеллюлярных клеток оканчиваются в слое 4Са, а парвоцеллюлярные волок-
на — в слоях 4А м 4Ср. На рис. 17.5 В показано, что зрительная информация
передается отсюда в Другие слои коры. Ниже мы увидим, что после первого си-
наптического контакта происходит сильное ветвление отростков нейронов,
образующих многочисленные синапсы. Латеральное распространение, одна-
ко, редко превышает несколько миллиметров, значение этого мы увидим ни-
же. Латеральное взаимодействие между более удаленными участками коры
реализуется волокнами, покидающими кору, проходящими через белое веще-
ство и вновь входят в кору в удаленной точке.
17.2.2. Функционирование
Существует множество способов исследования физиологии коры. Ниже мы
коснемся некоторых из них. Исторически наиболее важным является исполь-
зование микроэлектродов для исследования активности отдельных клеток ко-
ры. В этих опытах на глаз анестезированного животного помещали контакт-
ную линзу для предотвращения высыхания роговицы, а в зрительную кору
вводили микроэлекгрод для регистрации электрической активности нейро-
нов. Активность кортикальных клеток наблюдалась, когда сетчатка получала
17. Зрительные пути и зрительная кора 403
Рис. 17.6. Реакция клеток стри-
арной коры, детектирующих
ориентацию объекта. В колонке
слева показан стимул в виде по-
лосы, проецируемой на сетчатку
в разных ориентациях (длитель-
ность — 1 с). В колонке справа —
ответ кортикальной клетки.
зрительные стимулы. После завершения исследования одной клетки, микро-
электрод перемещали к другой клетке и процедуру повторяли. В конце работы
животное усыпляли и делали гистологические препараты коры — при этом
след от микроэлектрода оставался заметен, и можно было идентифицировать
клетки, от которых проводилось отведение.
Стоит повториться, что концепция рецептивных полей (РП) имеет фунда-
ментальный характер. Мы можем определить РП корковой клетки как участок
сетчатки, который при стимуляции генерирует активность в этой клетке. Ре-
цептивные поля клеток первичной зрительной коры были предметом интен-
сивных исследований. Как и можно было ожидать, большинство клеток слоя
4с имеют рецептивные поля концентрического типа, сходные с такими у кле-
ток НКТ. В конце концов, между НКТ и этими слоями стриарной коры про-
сто нет места для промежуточной переработки информации. Они возбужда-
ются зондированием сетчатки маленьким световым пятном, так же, как это
делалось при исследовании РП ганглиозных клеток собственно сетчатки или
клеток коленчатого тела. Аналогичный подход к исследованию других слоев
коры оказался неэффективным, и долгие годы их «триггерные стимулы» оста-
вались тайной.
Как показали Хьюбел (Hubei) и Визел (Wiesel), получившие за свою рабо-
ту Нобелевскую премию, открытие в физиологии клеток первичной зритель-
ной коры было результатом удачи. Отметим, что удача пришла в руки дотош-
ных и внимательных исследователей. После того, как масса времени была
впустую потрачена на попытки вызвать возбуждение клеток с помощью бе-
404 Часть IV. Светочувствительность
Б
iiimiimiimu
IIIIHIIIIIIIIIIII
ж-
(3)
Рис. 17.7. Селективность по
направлению. (А) Движение
слева направо; (Б) движение
справа налево. Ответ корти-
кальной клетки показан в ко-
лонке (2), а предполагаемая
«архитектура» поля — в ко-
лонке (3). Интенсивность
возбуждения в разных «воз-
буждающих» частях рецеп-
тивного поля обозначена
плотностью «плюсов», а тор-
мозная центральная часть
поля — минусами.
лых и черных точек, нанесенных на слайд, они вдруг заметили, что слабая ре-
акция возникает на край слайда, когда его вставляли в держатель. Это был
прорыв. Вскоре им удалось продемонстрировать, что если стимул в форме по-
лосы или края с определенной ориентацией включается или передвигается по
определенной части сетчатки, в клетках коры возникает залп активности
(рис. 17.6).
Дальнейшая работа показала, что в стриарной коре макаков от 70 до 80%
клеток имеет ориентационную специфичность. Заметьте, что только движе-
ние стимулирующей полоски через РП вызывает ответ — стационарные сти-
мулы вызывают лишь незначительную реакцию. Показано, что для прибл. 30%
клеток существенно и направление, в котором перемещается стимул — на пе-
ремещение в одном направлении возникает интенсивный ответ, в противопо-
ложном — незначительный (рис. 17.7).
На рис. 17.7 показан результат анализа рецептивного поля, проводимого с
помощью обычной техники освещения соответствующей части сетчатки ма-
ленькими световыми пятнами. Кортикальные клетки отвечают на освещение
включением или выключением активности (ON- и OFF-ответы). Как и в сет-
Рис. 17.8. Обычная архитекту-
ра РП «простых» клеток стри-
арной коры. «+» — ON-ответ,
«—» — OFF-ответ. Дальнейшие
объяснения в тексте.
17. Зрительные пути и зрительная кора 405
Рис. 17.9. Возможная схема
детектирования ориента-
ции объекта кортикальны-
ми клетками. Ряд клеток
НКТ с перекрывающимися
концентрическими полями
конвергирует на одной кор-
тикальной клетке. Корти-
кальная клетка реагирует
максимально на полосу
света определенной ориен-
тации в определенной час-
ти сетчатки.
регистрация активности клеток на стимулы (а) и (б),
кортикальных клеток показанные в верху рисунка
чатке, между участками ON- и OFF-ответов существует антагонизм, однако, в
отличие от РП ганглиозных клеток они построены не по типу «мишени» (кру-
говой), а имеет прямоугольную форму. Это может быть узкая полоса ON- или
OFF-центра, по бокам которого оппонентные области, либо просто два сосед-
них прямоугольника, один из которых дает ON-ответ, а другой — OFF
(рис. 17.8).
С тех пор, как Хьюбел и Визел открыли в конце 1950-х рассматриваемые
свойства рецептивных полей, предпринималось немало попыток построить их
блок-схему. Мы уже отмечали, как трудно было построить блок-схему сетчат-
ки (раздел 16.2.9), и мы еще далеки от полного ее понимания. Так что неуди-
вительно, что блок-схема намного более сложной структуры коры все еще яв-
ляется предметом дискуссии. Тем не менее не исключено, что первоначальное
предположение Хьюбела и Визела об архитектуре избирательного к ориента-
ции РП, как построенном в ряд множестве концентрических рецептивных по-
лей предыдущего уровня, недалеко от истины (рис. 17.9).
406 Часть IV. Светочувствительность
ш
ш
ш
I_____________I
On Off
Стимул
Рис. 17.10. Характеристики ответов
кортикальных клеток со сложным
полем. Соответствующим образом
ориентированный стимул в виде
полосы вызывает выраженный от-
вет клетки в любом месте поля.
Селективные к ориентации объекта клетки, описанные в предыдущих аб-
зацах, называются простыми клетками и составляют меньшинство среди детек-
торов ориентации в первичной зрительной коре. Большинство же, до 75%,
имеют несколько иные характеристики и называются сложными клетками.
Они отличаются от простых тем, что их РП не подразделены на ON- и OFF-
участки, а также тем, что они не возбуждаются одиночным пятнышком света,
но дают мощный ответ на соответствующим образом ориентированный край
объекта или полосу света, вспыхивающую в какой-либо точке рецептивного
поля, или движущуюся через поле (рис. 17.10).
Ответ вызывается стимуляцией не маленького участка поля, как у простых
клеток, а всего поля в целом. Сложные поля, однако, сходны с простыми в том
отношении, что они часто чувствительны к направлению, в котором движется
ориентированный стимул. Интересно отметить, что у некоторых низших мле-
копитающих, в частности у кролика, избирательность к направлению движе-
ния объекта — это свойство РП некоторых ганглиозных клеток сетчатки. В то
же время, ганглиозные клетки с такими характеристиками РП не встречаются
в сетчатке обезьян — у них обработка информации о направлении движения
происходит в мозге. Обусловлено ли это тем, что кролику необходимо быстрее
реагировать на угрозы из окружающей среды? Как мы отмечали в конце гл. 16,
кролик — зверь пугливый и всегда настороже.
В первичной зрительной коре присутствует еще один тип клеток, избира-
тельный к ориентации. Хьюбел и Визел с самого начала определили его как ги-
персложный, полагая что их рецептивные поля формируются из афферентов от
17. Зрительные пути и зрительная кора 407
Рис. 17.11. Кортикальная клетка, реагирую-
щая на конец ориентированной полоски.
(А) Правильно ориентированный полосовой
стимул полностью расположен в пределах
рецептивного поля — кортикальная клетка
дает выраженный ответ. (Б) Когда ориенти-
рованный стимул несколько превышает
размеры рецептивного поля, ответ клетки
уменьшается. (В) Еще более длинный сти-
мул не вызывает реакции корковой клетки.
(Г) Короткий оптимально ориентирован-
ный стимул, предъявляемый совместно с
еще одним стимулом близкого размера, но
иной ориентации и находящегося в непо-
средственной близости от рецептивного по-
ля, вызывает максимальный ответ клетки.
(Д) Принципиальная схема для случая на
рис. (Г). Область за пределами рецептивного
поля настроена на диагонально или верти-
кально ориентированные стимулы и тор-
мозится горизонтально ориентированным
стимулом. (Е) Возможное объяснение био-
логического смысла концевых РП в детек-
тировании кривых или волнообразных кра-
ев. По Hubei, 1988.
множества сложных клеток. В настоящее время считают, что это не всегда так,
и термин «гиперсложный» в литературе употребляется все реже. Тем не менее,
этот тип клеток существенно отличается от сложных тем, что их максималь-
ный ответ вызывается, если ориентированный край весь целиком лежит в пре-
делах рецептивного поля (рис. 17.11). Эти клетки в настоящее время называют
менее обязывающим термином «нейроны с концевым торможением» (end-
stopped). Возможно, что такие клетки способны детектировать и искривленные
или зазубренные края объектов.
Но почему первичная зрительная кора переполнена клетками, реагирую-
щими на ориентацию краев и линий? Наиболее вероятный ответ состоит в
том, что просто мир переполнен-краями и линиями. Все это — контуры объек-
тов, движущихся в поле зрения: вертикальные и диагональные линии стволов
деревьев и ветвей, когда глаз сканирует их, волнообразные или зазубренные
408 Часть IV. Светочувствительность
1,5 мм
Белое вещество
Рис. 17.12. Колонки ориента-
ций в стриарной коре. Опти-
мальные ориентации полосо-
вых стимулов изменяются от
одной колонки к другой зако-
номерно. Ориентационная из-
бирательность не показана в
клетках слоя IVc, где оканчи-
ваются волокна из НКТ. Клет-
ки этого слоя имеют концент-
рическую структуру рецептив-
ных полей.
края листьев, неровные края камней, высокие травы саванны, далекий гори-
зонт — соединение неба и земли.
Добавим следующее. Избирательные к ориентации клетки распределены
в первичной зрительной коре не в случайном порядке. Микроэлектродные
исследования показали, что они простроены в правильном, можно сказать
почти квазикристаллическом порядке. Мы видели в гл. 6, когда обсуждали со-
матосенсорную кору, что она построена в виде колонок, и то же касается и
зрительной коры. Оказывается, что, если продвигать микроэлектрод вглубь
колонки, то он регистрирует активность одной клетки за другой, и все они ре-
агируют на одну и ту же ориентацию. То есть можно говорить не просто о клет-
ках, детектирующих ориентацию, но и о колонках таких клеток. В целом, ней-
роны с концевым торможением локализованы в слоях 2 и 3; слой 4 содержит
концентрические клетки (см. выше) или простые детекторы ориентации; слои
5 и 6 — сложные клетки.
Если вводить микроэлектрод под некоторым углом, так чтобы он перехо-
дил из одной колонки в другую, окажется, что в последней клетки реагируют
на несколько отличную ориентацию по сравнению с предыдущей. Рис. 17.12
показывает, что это правило соблюдается удивительно точно. По мере продви-
жения микроэлектрода из колонки в колонку предпочтительная ориентация
изменяется закономерно. Набор ориентаций в пределах 180° умещается на
тангенциальной дистанции около 1,5 мм. Эта поразительная точность органи-
зации неожиданно прерывается разрывом (fracture). Правильная последова-
тельность небольших изменений ориентации внезапно сменяется отличиями
колонки от колонки на 45°—90°, затем закономерная последовательность ма-
лых изменений восстанавливается.
До сих пор мы описывали структуру первичной зрительной коры, основы-
ваясь на результатах микроэлектродных отведений. С помощью тонких мето-
17. Зрительные пути и зрительная кора 409
Рис. 17.13. Ориентационные колонки. Поперечный срез через стриарную кору
обезьяны, после введения 2-дезоксиглюкозы (см. текст). Животному предъявляли
набор вертикальных полос, в результате на срезе ясно видны колонки, в которых
локализованы клетки, детектирующие вертикальную ориентацию. Из Hubei,
Wiesel and Stryker, 1977. Воспроизводится с разрешения Nature, 269, 328—330.
Figure 1, ©1977, Macmillan Magazines Limited.
дик возможно продемонстрировать и гистологическую основу этих физиоло-
гических явлений. Одна из таких наиболее впечатляющих методик — ис-
пользование дезоксиглюкозы. Это вещество воспринимается активными
клетками как обычная глюкоза и захватывается в цитоплазму. Однако, дез-
оксиглюкоза проходит только первую стадию гликолиза и далее не усваива-
ется. В результате ее метаболиты накапливаются в клетке, и будучи жироне-
растворимыми, они не могут ее покинуть. Принцип метода состоит в том,
что инъецируются меченые радиоизотопом (обычно — тритием, 3Н) молеку-
лы дезоксиглюкозы, а затем анестеризованному животному в течение 45 мин
предъявляются стимулы в виде полос определенной ориентации. Дезокси-
глюкоза в этих условиях захватывается наиболее активными клетками. По-
сле продолжительной зрительной стимуляции животное усыпляют, делают
гистологические срезы зрительной коры и исследуют локализацию радиоак-
тивного изотопа. Такая процедура оказалась поразительно плодотворной —
на срезе проявляются колонки, избирательные к использованной в опыте
ориентации (рис. 17.13), а также непрерывная полоса радиоактивного мате-
риала в слое 4С. Надо напомнить, что этот слой активируется независимо от
ориентации стимула.
Дезоксиглюкозный метод применялся, чтобы подтвердить результаты
электрофизиологических исследований. При этом очень похожий по замыс-
лу метод принес и совершенно неожиданный результат, который нельзя было
ожидать на основании электрофизиологических исследований. В 1978 году
Маргарет Вонг-Райли (Margaret Wong-Riley) окрашивала стриарную кору
реактивом, детектирующим фермент цитохром-оксидазу. Ко всеобщему
удивлению цитохром-оксидаза оказалась локализованной в правильно распо-
ложенной серии капель (blob), особенно ясно видимых в слоях 2 и 3 стриарно-
410 Часть ГУ. Светочувствительность
Рис. 17.14. Концентрические двойные
оппонентные клетки. Верхняя пара
реагирует на контраст красного и зе-
леного, нижняя — на контраст синего
и желтого. Двойные оппонентные ре-
цептивные поля не всегда круглые.
В стриарной коре обнаружены также
селективные к ориентации двойные
оппонентные РП.
го комплекса. Размеры этих капель — около 0,5 мм в диаметре, а разделены
они межкапельными участками (interblob) величиной в 0,25 мм. Несколько лет
никто не обращал внимания на это неожиданное открытие. Затем, однако, в
1981 году Хьюбел и Ливингстон (Livingstone) провели микроэлектродное ис-
следование нейронного состава капель. Оказалось, что в пределах капель не
было нейронов, избирательных к ориентации. Клетки в каплях имели концен-
трические цветоизбирательные рецептивные поля с оппонентной организаци-
ей центра и периферии.
Рецептивные поля цветоизбирательных клеток в каплях, как и в НКТ, ока-
зались двух типов: красный/зеленый и синий/желтый (где желтый, как и рань-
ше, результат параллельного входа красного и зеленого). Здесь, однако, есть и
отличие. Показано, что они не реагируют на большие пятна белого света, и,
к тому же и центр, и окружение состоят из оппонентных пар цветов. В про-
стейшем случае — красный/зеленый, т. е. центр дает ON-ответ на красный
цвет и OFF-ответ на зеленый (кратко: К+ / 3—), а окружение OFF-ответ на
красный и ON-ответ на зеленый (К— / 3+). Такой тип характеристик ответа
называется двойным оппонентным (рис. 17.14). Вероятно, эти двойные оппо-
нентные клетки усиливают цветовой контраст на краях. То, что они не реаги-
руют на диффузное освещение, опять-таки предполагает их связь с контролем
краев зрительного образа.
Следует помнить, что цветовая информация переносится системой Р-во-
локон, именно она оканчивается в каплях. Р-волокна иннревируют также
межкапельные участки. Здесь, однако, кортикальные клетки помимо цветовой
чувствительности реагируют также на ориентацию краев. Детекторы краев
адаптируются медленно, и потому, скорее, реагируют на положение контуров
стационарных, а не движущихся объектов. Межкапельные участки системы Р-
волокон, таким образом, дополняют детектирование формы М-системой, рас-
смотренной выше.
Еще один, заключительный, принцип работы стриарной коры — это ее
организация в колонки глазодоминантности. Показано, что по мере верти-
кального продвижения электрода через кору, регистрируемые клетки, «кон-
тролируются» одним глазом. При диагональном погружении микроэлектро-
Y1. Зрительные пути и зрительная кора 411
Рис. 17.15. (А) Колонки гла-
зодоминантности в стриар-
ной коре обезьян. Л — ко-
лонка, связанная с левым
глазом; Лп — левый глаз —
доминирующий, некоторое
влияние от правого глаза;
П — колонка, связанная с
правым глазом, Пл — пра-
вый глаз — доминирующий,
некоторое влияние от лево-
го глаза. (Б) Связи клеток,
ответственных за физиоло-
гические характеристики
колонок глазодоминантно-
сти.
А Поверхность, обращенная
к мягкой оболочке
Лп Пл Лп Пл Лп Пл Лп
Лп Пл Лп Пл Лп Пл Лп
Лп Пл Лп Пл Лп Пл Лп
4С Л п Л п Л п Л
Лп Пл Лп Пл Лп Пл Лп
Лп Пл Лп Пл Лп Пл Лп
Белое вещество
Б Поверхность коры
(через НКТ)
Белое вещество
да появляются клетки, управляемые сначала одним, потом — другим глазом
и т. д. И снова мы сталкиваемся с правильной организацией колонок. Ко-
лонки, связанные с одним глазом, чередуются с колонками, связанными с
другим глазом (рис. 17.15 А).
Рис. 17.15 показывает, что доминирование является полным в слое 4С, что,
собственно, только и можно было ожидать, поскольку в этом слое оканчива-
ются волокна из НКТ. Надо напомнить, что волокна зрительного нерва из двух
глаз в ядре коленчатого тела строго изолированы друг от друга (раздел 17.2).
Выше и ниже слоя 4С, однако, следующие нейроны ветвятся латерально, так
что некоторая доля входов от противоположного глаза включается в домини-
рующую афферентацию (рис. 17.15 Б). Техника окрашивания и в этом случае
продемонстрировала гистологическую реальность этих колонок. Глазодоми-
412 Часть IV. Светочувствительность
Рис. 17.16. (А) Схема, показывающая организацию стриарной коры. В гиперко-
лонку входят полный набор колонок ориентации и две (левая и правая) колонки
глазодоминантности. Кроме того капли содержат цветочувствительные клетки.
Следует учитывать, что рисунок сильно упрощен по сравнению с реальной гисто-
логией. К — контрлатеральный, И — ипсилатеральный. Из Kandel, Schwartz and
Jessell, 1991, с разрешения. (Б) Рисунок показывает восемь перекрывающихся ре-
цептивных полей, образующих агрегат. На самом деле их намного больше.
нантные колонки присутствуют в мозге животных с высокоразвитым биноку-
лярным зрением — у кошек, хвостатых обезьян, шимпанзе и человека, однако
не обнаружены у обычных лабораторных грызунов: крыс, мышей и морских
свинок, из чего вытекает важность глазодоминантных колонок для стереоско-
пического зрения.
Было сделано предположение, что один полный набор ориентационных
колонок, набор цветодетектирующих капель и набор глазодоминантных коло-
нок левого и правого глаза образуют физиологическую единицу первичной
зрительной коры (рис. 17.16 А). Такая структура получила название гиперко-
лонки или модуля, связанного с определенной частью сетчатки. Рецептивные
поля ориентациооно-избирательных клеток в гиперколонке различаются в
размерах, но накладываются друг на друга практически в одном и том же мес-
те сетчатки (рис. 17.16 Б). Вместе они образуют агрегатное рецептивное поле ги-
перколонки. Агрегатные рецептивные поля варьируют в пространственных
размера в разных частях сетчатки — наименьшие в области фовеа, а наиболь-
шие — на периферии.
Как и в случае других сенсорных систем, рассматриваемых в этой книге,
сенсорная поверхность, в данном случае — сетчатка, представлена в сенсор-
ной коре неизоморфной картой, показанной на рис. 17.17. Как мы отмечали
выше, агрегатные рецептивные поля в фовеа имеют наименьший размер, сле-
довательно, намного большая часть зрительной коры связана с ними, чем с ос-
тальными частями сетчатки. Из рисунка видно, что верхние части сетчатки
картированы в нижней части шпорной борозды и наоборот. Следует помнить,
что из-за свойств оптики глаза верх сетчатки соответствует нижней части поля
зрения и, опять-таки, наоборот. Рис. 17.17 показывает, что кора, связанная с
17. Зрительные пути и зрительная кора 413
Рис. 17.17. В верхней
части рисунка схема
поля зрения. Нижняя
часть — вид медиальной
поверхности затылоч-
ной доли — полушария
разделены и разведены
в стороны. Зрительная
кора лежит выше и ни-
же шпорной борозды.
Дальнейшие объясне-
ния в тексте. Из Kandel,
Schwarz and Jessell, 1991,
с разрешения.
Калькаринная
Первичная зрительная кора
Калькаринная
борозда
фовеа, занимает почти такую же поверхность, что и вся остальная сетчатка, и
простирается до задней поверхности затылочной доли.
Вернемся к детекторам ориентации или краев. Внимательный студент уже
мог приметить, что здесь имеется трудность. Рассмотрим рис. 17.18 — здесь
показано три возможных пути стимуляции РП краевых детекторов. На
рис. 17.18 А на детектор падает резкий край — тормозная часть поля не осве-
щена, освещена только возбуждающая часть поля, в кортикальной клетке ге-
нерируется выраженный ответ. На рис 17.18 Б на рецептивное поле попадает
не столь резкая граница освещенности — тормозная часть поля слабо возбуж-
дается, а возбуждающая — слабее, чем в первом случае. В результате ответ кор-
тикальной клетки имеет меньшую частоту. Наконец, в случае (В) на рисун-
ке 17.18 резкая граница освещенности РП расположена диагонально, так что
полная освещенность приходится и на часть возбуждающей, и на часть тор-
мозной области. В этом случае ответ неотличим от возникающего в случае (Б).
Как же кора справляется с этой неоднозначностью?
Краткий ответ на этот вопрос — пока мы этого не знаем, но соблазнитель-
ная возможность предложена нейрофизиологом Джоном Фризби (John
Frisby). Он предположил, что следует анализировать не активность единич-
ного детектора края, а суммарную активность популяции детекторов края ги-
414 Часть ГУ. Светочувствительность
Рис. 17.18. (А) Резкая граница осве-
щенности приходится на границу
между тормозной и возбуждающей
частями рецептивного поля детекто-
ра края. Ответы кортикальной клет-
ки показаны справа. (Б) Менее рез-
кая граница освещенности прихо-
дится на границу между тормозной и
возбуждающей частями рецептивно-
го поля детектора края. Генерируется
более слабый ответ кортикальной
клетки. (В) Резкая, но диагональная
граница освещенности на рецептив-
ном поле. Ответ кортикальной клет-
ки неотличим от ответа в случае (Б).
Как мозг различает эти два послед-
них сигнала? По Frisby, 1979.
перколонки. Рассмотрим рис. 17.19. Снова мы видим два зрительных стиму-
ла: один — с резко выраженной диагональной границей освещенности, дру-
гой — со слабовыраженной вертикальной границей. Если эти стимулы попа-
дают на агрегатное рецептивное поле гиперколонки, очевидно, что конфигу-
рации (паттерн) возбужденных детекторов края в ней будут сильно разли-
чаться. Сравнивая эти паттерны, зрительная система способна различать та-
кие два типа стимулов, что, как нам хорошо известно, на самом деле и про-
исходит.
Здесь возникает крайне важный вопрос, и Фризби первым задал его —
что и где сравнивает паттерны возбуждения в гиперколонке? К моменту на-
писания этой книге ответа все еще нет. Такие вопросы, как этот, относятся
к нерешенным проблемам, которые двадцать первый век унаследовал от
двадцатого. Отсутствие ясного ответа на этот вопрос, однако, не означает,
что мы не знаем, куда направляется информация из первичной зрительной
коры. Это мы знаем, и, возможно, именно там — в экстрастриарной коре и
происходит устранение упомянутой выше двусмысленности сигналов, а,
может быть, сама гиперколонка с ее четвертью миллиона клеток способна
рассчитать ответ.
17. Зрительные пути и зрительная кора 415
нулевая
активность '
(частота ответа порядка 25 имп/с)
нулевая
активность
Рис. 17.19. (А) Стимул, схожий с приведенным на рис. 17.18 Б. Наиболее выраженный
ответ в гиперколонке возникает в колонке, содержащей клетки, детектирующие верти-
кально ориентированные стимулы. (Б) Стимул, схожий с приведенным на рис. 17.18 В.
Наиболее выраженный ответ в гиперколонке возникает в колонке, содержащей клетки,
детектирующие диагональную ориентацию стимулов. Рисунок показывает, что паттерны
ответов в гиперколонке в случаях (А) и (Б) существенно различаются. Дальнейшие объ-
яснения в тексте. По Frisby, 1979.
17.2.3. Пластичность
В главе 7 (раздел 7.5) мы видели, что соматосенсорная кора способна перест-
раиваться под действием факторов окружающей среды. Такая же пластич-
ность показана и в первичной зрительной коре. Она, однако, в основном огра-
ничена так называемым чувствительным или критическим периодом развития
млекопитающих. Этот период, в ходе которого мозг претерпевает быстрое со-
зревание, длится около 6 недель у котят и около 18 месяцев — у детей. Чувст-
Наиболее активная клетка
(частота ответа — 100 имп/с)
Б
416 Часть IV. Светочувствительность
Рис. 17.20. (А) Тангенци-
альный срез глазодоми-
нантных колонок, мече-
ных радиоизотопами.
Светлые полосы показы-
вают, где аккумулировал-
ся меченый пролин. Вид-
но, что светлые (представ-
ляющие вход от меченого
глаза) и темные полосы
(от немеченого) примерно
равной ширины. (Б) Тан-
генциальный срез коло-
нок глазодоминантности,
меченых радиоизотопами,
после заклеивания одного
глаза в чувствительный
период. Видно, что колон-
ка доминантности, свя-
занная с незаклеенным
глазом, сильно расширена
за счет ранее заклеенного.
ИзО. Н. Hubei, Т. N. Wie-
sel and S. LeVay, 1977,
«Plasticity of ocular domi-
nance columns in monkey
striate cortex», Philosophical
Transactions of the Royal So-
ciety of London, Series B,
278, 391, Figure 13 (c), 392,
Figure 15 (с). Воспроизво-
дится с разрешения Royal
Society.
вительные периоды разных систем мозга имеют и разную длительность, и раз-
ное время их начала и окончания.
Котята рождаются слепыми — с закрытыми глазами, которые открываются
не раньше десятого дня. Даже после этого еще пару недель видят они плохо, и
четкой зрительно-моторной координации у них не наблюдается. Вместе с не-
готовностью мышечной системы это обеспечивает невозможность котенку
удрать слишком далеко. Во время этого раннего периода жизни эелктрофи-
зиологические отведения покапывают лишь небольшие прзнаки структуры
колонок ориентации и колонок глазодоминантности, описанных выше.
Только после установления зрительно-моторной координации, когда коте-
нок начинает играть, изображая взрослого кота, кора приобретает свою высо-
коорганизованную структуру.
17. Зрительные пути и зрительная кора 417
________
Это постнатальное развитие коры предоставляет ценные возможности экс-
периментального изучения. У котенка, содержавшегося в темноте, в первичной
коре почти нет клеток, селективных к ориентации. У котят, зрительное окруже-
ние которых состояло только из вертикальных полос, преобладают клетки, от-
вечающие на вертикально ориентированные стимулы, а у содержавшихся в ок-
ружении горизонтальных полос — клетки, отвечающие на горизонтальные сти-
мулы. Если один из глаз заклеить до того, как он открылся, и оставить так на
весь чувствительный период, в первичной зрительной коре такого котенка
клетки, связанные с заклеенным глазом, окажутся в сильном дефиците. Одна-
ко, если заклеить оба глаза, то колонки глазодоминантности в первичной зри-
тельной коре не будет сильно отличаться от нормы. Таким образом, нервные
связи, обеспечивающие бинокулярное зрение, генетически детерминированы,
но могут перестраиваться в зависимости от факторов окружающей среды. Если
заклеенный глаз раскрыть, а раскрытый — заклеить на третьей или четвертой
неделе после рождения, картина в коре меняется на противоположную — бино-
кулярные связи утрачиваются в исходно раскрытом глазу и устанавливаются в
исходно заклеенном. В этот период жизни котенка проводящие пути могут не
только уменьшаться и увеличиваться, но и перестраиваться.
Результаты электрофизиологических экспериментов подкрепляются гис-
тологическими данными. Меченый радиоизотопом пролин инъецировали в
сетчатку макака. Примерно через 10 дней животное усыпляли, делали срезы
первичной зрительной коры и авторадиографическое исследование. Его ре-
зультаты показаны на рис. 17.20 А. Светлые полосы показывают, где аккуму-
лируется радиоактивность. Видно, что колонки доминирования приблизи-
тельно равной ширины и, как и следовало ожидать, перемежаются. Случай,
когда один из глаз был заклеен во время чувствительного периода и затем от-
крыт, а пролин инъецировали после его открытия в сетчатку незаклееного
глаза, показан на рис. 17.20 Б. Видно, что светлые полосы, показывающие об-
ласти входа от незаклееного глаза, сильно расширены за счет сужения входа
от заклееного. Это — очевидное подтверждение электрофизиологических дан-
ных о пластичности первичной зрительной коры в чувствительный период.
17.3. Экстрастриарная кора
Вплоть до начала 1950-х годов полагали, что органы чувств связаны с достаточно
ограниченными участками сенсорной коры, оставляя большую ее часть для «ассо-
циаций». Было ли это следствием тогдашнего господства ассоциативной психоло-
гии — интересный вопрос истории науки. Однако, с 50-х годов XX века стало яс-
но, что это серьезное заблуждение. В случае зрительной системы кора, связанная
с анализом этой информации, далеко не ограничена стриарной или первичной
зрительной корой. Действительно, у млекопитающих с развитым зрением, таких
как высшие приматы, с обработкой зрительной информации связаны большие
участки затылочной и височной долей. У макаков идентифицировано от 24 до 30
отдельных экстрастриарных участков зрительной коры. Часто эти области содер-
жат карты поля зрения, специализированные по различным характеристикам.
Концепция «ассоциативной коры» была вытеснена, хотя и не полностью, пред-
ставлением, что кора состоит их сенсорных карт, лежащих слой за слоем.
Часть IV. Светочувствительность
Информация 17.1
СЛЕПОЕ ЗРЕНИЕ
Как и «бессознательный разум», выражение «слепое зрение» звучит как бессмысли-
ца. Однако, как и в случае с «бессознательным разумом», это понятие показало
свою полезность. Эти два понятия имеют нечто общее, помимо практической поль-
зы — оба они подчеркивают, что на самом деле мы знаем больше, нежели мы осо-
знаём, что знаем. Большая часть наших знаний о мире, как давным-давно отметил
Полиани (Polyani), «подразумевается».
«Слепое зрение» было впервые обнаружено в экспериментах и наблюдениях на
обезьянах, утративших стриарную кору, проведенных еще в конце XIX века. Воз-
никло впечатление, что если за такими обезьянами хорошо ухаживать, то они вос-
станавливают зрение, которым в состоянии пользоваться. Они могли двигаться в
пространстве, загроможденном препятствиями, и подбирать небольшие предметы.
Складывалось, однако, впечатление, что они не способны определить характер объ-
ектов, которые они подбирают и которыми манипулируют. Такие же эксперименты
были предприняты в начале 1970-х годов на обезьянах, стриарную кору которых уда-
ляли хирургически. Сначала, как писал Николас Хамфри (Nicholas Humphrey), обе-
зьяны выглядели, как и следовало ожидать, совершенно слепыми. Однако, при тща-
тельном уходе и контроле в течение семи лет, зрение в какой-то форме возвраща-
лось. Макаки были способны передвигаться по комнате с множеством препятствий
и подбирать с пола небольшие предметы, например часы. При этом, писал Хамфри,
оставалось впечатление, что обезьяны не видят в обычном смысле этого слова, что
они «угадывают», наудачу производят ранее осмысленные действия. Как все это воз-
можно без стриарной коры? Напомним, что ретино-геникуло-стриарный-путь — не
единственный путь проведения зрительной информации в мозг. Возможно, экспе-
риментальные обезьяны научаются использовать древний ретино-тектальный или
другой не-стриарный пути формирования зрительно-управляемого поведения.
Трудно судить о субъективных ощущениях обезьян; с представителями нашего
собственного вида дело обстоит несколько проще. И «слепое зрение» у человека
встречается (понятно, не в порядке эксперимента). Такой феномен может исследо-
ваться нейрологами, когда пациент по тем или иным причинам страдает от одно-
стороннего массивного повреждения кортикального поля 17 — первичной зритель-
ной коры. Эти полуслепые пациенты настаивают, что у них полностью отсутствуют
зрительные ощущения в пораженной половине поля зрения. Тем не менее, было
показано, что некоторые управляемые зрением функции сохраняются. Наиболее
известны эксперименты по определению направления. Когда пациента просили
указать на предмет, возникающий перед ним на 2 или менее секунды в слепой по-
ловине поля зрения, процент успеха был значительно выше случайного, хотя сами
пациенты настаивали, что ничего не видят и просто гадают.
Такие же эксперименты были направлены на проверку способности отличать
локальное и диффузное освещение равной интенсивности, способности узнавать
направление ориентации полосы и способности различать буквы «О» и «X». Во всех
этих случаях пациенты решали задачу с лучшими, чем случайные, результатами, хо-
тя и утверждали, что не видят ничего и просто гадают. Николас Хамфри суммирует
позицию таких пациентов: — Я вообще ничего не вижу, но раз вы говорите, что я
делаю это правильно, я могу только вам поверить.
17. Зрительные пути и зрительная кора 419
Продолжение
Слепое зрение было подвергнуто детальной и исчерпывающей критике. Способ-
ны ли были пациенты обнаруживать перемещающийся свет? Не была ли стриарная
кора уничтожена лишь частично? Все же, все пришли к согласию, что эта способ-
ность является врожденной. Иными словами, предполагается, что зрительная ин-
формация поступает в мозговые центры принятия решений иными, чем РГС-путь,
и не затрагивает сознания. Пациент «видит, но не видит» (не сознает, что видит). Он
воспринимает, но это — бессознательное восприятие. А не имеет ли это какого-то
отношения к природе зрения у низших позвоночных и беспозвоночных?
Литература
Campion, J., R. Latto and Y. M. Smith, 1983, «Is blindsight an effect of scattered light,
spared cortex, and near threshold vision», Behavioural and Brain Sciences, 6,423—486.
Humphrey, N., 1992, A History of the Mind, London: Vintage.
Luciani, L, 1884, «On the sensorial localisations in the cortex cerebri, Brain, 7, 145—160.
Polyani, M., 1966, The Tacit Dimension, New York: Doubleday.
Weiszkrantz, L., 1986, Blindsight: A case study and implications, Oxford: Oxford University
Press.
В разделе 17.1.2 мы видели, что в первичную зрительную кору ведет путь от
сетчатки через коленчатое тело. После обработки в этой коре информация пе-
редается в экстрастриарные области коры. Однако, как упоминалось в разделе
17.1.1, этот путь — не единственный, менее значимый ведет в верхние бугры
четверохолмия. Зрительная карта, формирующаяся здесь, играет важнейшую
роль в контроле движения глаз и направления взгляда. Волокна из верхних бу-
гров также, через ядро подушки направляются в экстрастриарную зрительную
кору. Ядро подушки — это крупная и сложным образом подразделенная ядер-
ная масса, образующая заднюю и задне-латеральную часть таламуса, непо-
средственно кпереди (кверху) от верхних бугров четверохолмия. Подушка не
только посылает волокна в экстрастриарную кору, но и получает из коры аф-
ференты. Таким образом, экстрастриарная кора получает перекрывающиеся
афференты из стриарной коры и из ядра подушки. Зрительная система зани-
мает большой объем мозга приматов и связана сложными связями, что обеспе-
чивает ее деятельность как единого целого.
Вернемся, однако, к выходу из стриарной коры. Мы уже отмечали, что в ге-
никуло-стриарном пути магноцеллюлярные волокна (несущие информацию о
движении и форме) изолированы от парвоцеллюлярных (информация о цвете
и частично о форме). Эти потоки информации остаются изолированными друг
от друга и в стриарной ww — М-волокна идут к клеткам, детектирующим
края, а Р-волокна — к «каплям» и межкапельным участкам. Здесь просматри-
ваются контуры «трехпоточной» системы: один поток несет информацию о
форме, движении и стереоскопии, второй — о цвете, а третий — обо всех четы-
рех характеристиках зрительного окружения (рис. 17.21 Б).
Три раздельных потока зрительной информации прослеживаются в экс-
трастриарной коре — сначала в зрительной коре 2 (V2), затем — в зрительной
Зрительная кора
Б
Магноцеллюлярная
система
Парвоцеллюлярно-
межкапельная
система
Парвоцеллюлярно-
капельная
система
Движение
Рис. 17.21. (А) Зрительная кора макака. Слева — горизонтальный срез, повернутый на
90°, из правой затылочной доли мозга (на рисунке справа). Показана локализация участ-
ков зрительной коры VI, V2, V3, V4 и V5 (МТ). (Б) Схема трехпоточной зрительной сис-
темы приматов. Объяснения в тексте. Из Kandel, Schwartz and Jessell, 1991, с изменения-
ми из E.A.Deyoe and D.C. van Essen, 1988, «Concurrent processing streams in monkey visual
cortex», Trends in Neurosciences, 11, 219—226, с разрешения.
Движение
17. Зрительные пути и зрительная кора 421
коре 3 (V3) — двух частях поля 18 по Бродману. Цитохром-оксидазное окра-
шивание показывает, что зона V2 морфологически неоднородна, и в ее срезах
выделяются так называемые толстые полоски, тонкие полоски и промежутки
между полосками. Рисунок 17.21 Б показывает, что М-поток из VI проециру-
ется в толстые полоски, P-поток из капель направляется в тонкие полоски, а
P-поток из межкапельного пространства — в промежутки между полосками.
Мы не станем прослеживать передачу зрительной информации дальше в
мозге. Интересующиеся могут обратиться за дополнительными сведениями к
источникам, приведенным в списке библиографии. Достаточно лишь сказать,
что зрительные пути прослежены до поля 19 по Бродману (совмещающему
зрительные области 4 (V4) и 5 (V5 или чаще МТ)) и далее во многие другие ча-
сти коры, особенно теменной и височной (рис. 17.21 Б). Не следует думать, что
информация передается по одному пути. Как говорилось в начале этой главы,
зрительная система обезьян состоит из более чем двух дюжин кортикальных
участков, связанных между собой в обоих направлениях. Также, как упомина-
лось выше, геникуло-стриарный путь не является единственным зрительным
входом в мозг, важный вклад вносят подушка, верхние бугры четверохолмия
и др. Кора головного мозга не является и «конечной станцией» зрительной
информации — она направляется и в другие части мозга, включая промежу-
точный мозг и базальные ядра. У животных с высокоразвитым зрением, таких
как приматы, большинство частей мозга так или иначе вовлечено в процессы
зрения.
17.4. Распознавание лица
Хотя это и заведет нас далеко в дебри деталей нейроанатомии многочисленных
участков зрительной коры приматов и их взаимосвязей, невозможно завер-
шить эту главу без упоминания одной из наиболее важных способностей зри-
тельной системы приматов — распознавания лиц. Эта способность важна для
развития у приматов социальности и наблюдается у нескольких групп, вклю-
чающих макаков, бабуинов, большинство крупных обезьян, включая, конеч-
но, человекообразных. Многочисленные этологические эксперименты пока-
зали, что социальные группы приматов удерживаются иерархией доминиро-
вания особей. Конфликты между членами трибы предупреждаются тем, что
каждый из них знает свое место в очереди питания. Такая иерархия работает,
если члены трибы в состоянии узнавать каждого соплеменника. Ясно, что
опознавание особи основывается на множестве индивидуальных характерис-
тик — походке, цвете, размере, голосе, запахе и т.д., но одной из таких харак-
теристик является лицо и его выражение. Не удивительно, что часть зритель-
ной системы макаков специализирована на распознавании лиц.
Область распознавания лиц у макаков (рис. 17.22 А) в иерархии участков
зрительной коры стоит очень высоко — это нижне-височная (infero-temporal,
IT) кора. Она получает афференты от V4, и огромное большинство ее клеток
отвечает на аффренты от фовеальной области сетчатки. Их рецептивные поля
обычно достигают 25° поля зрения, а иногда включают и все поле зрения. В от-
личие от нижележащих зрительных уровней здесь отсутствует ретинотопичес-
кая организация, а клетки отвечают на множество очень специфических триг-
422 Часть IV. Светочувствительность
1 1 1 2 3 3 4 4 5 6
Рис. 17.22. Детектирование образов в мозге макака. (А) Локализация клеток, чувствитель-
ных к образу лица. ПМВИ — передняя медиальная височная извилина, ЦИ — централь-
ная извилина, НЗИ — нижняя затылочная извилина, НВИ — нижняя височная извилина,
ЗМВИ — задняя медиальная височная извилина. Символы разной формы обозначают ре-
зультаты, полученные разными исследователями. Из Perrett et al,, 1992, с разрешения.
(Б) Ответы нижневисочных клеток на стимулы различной формы. Стимулы выставлены
слева направо в порядке возрастания их способности возбуждать клетки — от отсутствия
эффекта (1) или возникновения небольшого эффекта (2 и 3) до максимума (6). Из Gross,
Rocha-Miranda and Bender, 1972. Воспроизводится с разрешения Charles Gross.
герных стимулов. Около 10% клеток запускаются образами рук и лиц.
Рис. 17.22 Б показывает клетку, которая дает максимальный ответ на образ ру-
ки, направленной вверх. Показано, что такой ответ возникает, когда различи-
мы пальцы, т. е. общий контур руки является лишь субоптимальным триггер-
ным стимулом.
Помимо клеток, запускаемых образом руки, здесь присутствуют и клетки,
запускаемые образами мррд обезьян (или, соответственно, лиц людей). Неко-
торые клетки лучше всего реагируют на образ лица анфас, некоторые — на
профиль, а есть клетки, реагирующие на выражение лица. Наконец, есть дан-
ные, что некоторые клетки реагируют на определенные лица сильнее, чем на
остальные — например, лицо экспериментатора вызывает более сильную ак-
тивность, чем лицо постороннего. Оказывается, что концепция «клеток, рас-
познающих лицо бабушки» не так уж далека от истины!
Надо, тем не менее, сохранять осторожность. Лица и другие объекты окру-
жающего мира состоят из огромного множества элементов. Были проведены
тщательные эксперименты по изучению того, какие именно черты объектов
17. Зрительные пути и зрительная кора 423
Рис. 17.23. (А) Примеры триггерных характеристик некоторых клеток передней нижне-
височной коры (НВК). (Б) Схема, показывающая колончатую организацию клеток, рас-
познающих черты объекта в НВК. Объяснения в тексте. Из Tanaka, 1996. С разрешения
Annual Review of Neuroscience, volume 19, ©1996, by Annual Reviews www.annualreviews.org.
вызывают наибольшую реакцию клеток нижне-височной коры. Их результаты
пока следует считать предварительными, поскольку нельзя было исследовать
все возможные элементы и их сочетания. Оказалось, однако, что соосные от-
резки (подразумеваются брови), треугольники (брови — нос), овалы (лицо ан-
фас) и т. д., также как целостный образ лица, вызывают максимальные ответы
в различных клетках (рис. 17.23 А). Далее, оказалось, что т. н. «паттерны обра-
зов» группируются в колонки (рис. 17.23 Б). В целом они организованы как и
в других областях сенсорной коры. Тангенциальное введение микроэлектрода
обнаруживает группы клеток со схожими (но не идентичными) предпочтени-
ями образов, образующих колонки диаметром в 500 мкм, за которыми идут об-
ласти, где этот образ не запускает никакой реакции. При дальнейшем продви-
жении микроэлектрода прибл. на 1 мм вновь обнаруживаются клетки, реаги-
рующие на тот же образ.
Выводы из результатов этой кропотливой электрофизиологической работы
соответствуют данным исследований детекторов ориентации, описанным в
разделе 17.2.2 — на определенную ориентацию стимула реагирует не отдельная
колонка, а целая группа колонок. Сходная картина и при распознавании
сложных объектов и образов в нижней височной коре — реагирует группа ко-
лонок, составленных из клеток, запускаемых различными элементами образа.
«Открытие» клеток, распознающих лицо экспериментатора или бабушки,
опять-таки, кажется преждевременным, и вероятно, ошибочным. Да и иссле-
дования еще очень далеки от завершения. Существуют данные, что реакция по
крайней мере некоторых клеток НВК на элементы образа зависит от того, при-
сутствуют ли в их рецептивном поле другие элементы образа. Известно также,
что клетки колонок образов несколько различаются по избирательности к уг-
лу представления образа, его освещенности и т. д. Это обеспечивает распозна-
вание лица вне зависимости от его поворота или освещенности — т. е. ответ
популяции действует как буфер, поглощающий изменения. Так что распозна-
424 Часть IV. Светочувствительность
вание бабушкиного лица — это скорее всего функция не одной клетки, как
долгое время полагали, а результат активности сложно организованной попу-
ляции клеток нескольких колонок нижневисочной коры.
17.5. Прозопагнозия
Распознавание лиц важно не только для приматов, но и для нас самих.
Множество поколений в палеолите большинство людей жило в группах по
50—100 особей, и мимические сигналы составляли важный канал невербаль-
ной информации. Было необходимо распознавать друга и врага, членов трибы
и чужаков. Взаимный альтруизм и этические нормы, консолидирующие чело-
веческое общество, держатся на способности узнавать и помнить других инди-
видов. Поэтому неудивительно, что как и у других приматов, у человека име-
ется очень большая область мозга, связанная с распознаванием лиц.
Эта область мозга локализована (билатерально) в нижне-медиальной части
затылочно-височной доли (рис. 17.24). Ископаемые черепа указывают на то,
что эта область (в отличие от некоторых речевых областей, см. гл. 9, раздел 5)
уже была хорошо развита у Ното habilis (человек умелый) и Homo erectus (чело-
век прямостоящий). Это дает основание предположить, что невербальная
коммуникация посредством лицевой мимики предшествовала возникнове-
нию сложной способности к речи. Такое подозрение подкрепляется наблюде-
нием, что некоторые мимические сигналы — «удивленные» брови, улыбка
и т. д. понимаются, в отличие от речи, универсально. Есть данные, что меха-
низмы области мозга, связанной с распознаванием лиц, участвуют также в
сторона
сторона
Рис. 17.24. Локализация областей,
связанных с распознаванием лиц,
в мозге человека. Черные кружки
обозначают места нижней поверх-
ности мозга, где регистрируются
вызванные потенциалы при
предъявлении лица. Некоторое
преимущество в представительст-
ве есть у правого полушария (41)
по сравнению с левым (34), и зна-
чительное преобладание у нижне-
височной доли (53) по сравнению
с нижнезатылочной (25). Из Alli-
son et al., 1999. Воспроизводится с
разрешения American Physiological
Society.
17. Зрительные пути и зрительная кора 425
восприятии объектов, имеющих «физиономические» свойства — морды жи-
вотных, автомобили, украшения, посуда и т. д.
Утрата способности распознавать лица у человека известна под названием
прозопагнозия. Патология была выделена в середине XIX века, а недавно по-
пуляризована Оливером Саксом (Oliver Sacks) в книге под вызывающим
названием «Человек, который перепутал жену и шляпу». Прозопагнозия
вызывается травмой, либо ростом опухоли, либо, что чаще всего, сосудисты-
ми нарушениями в области распознавания лиц, особенно с правой стороны.
Больные, страдающие от этого нарушения, часто с большим трудом узнают
своих ближайших родственников и друзей, чем тех, кого видят достаточно ред-
ко. Такое странное сочетание обстоятельств иногда доходит до крайности,
когда субъект оказывается неспособен узнать самого (саму) себя в зеркале.
Другие аспекты системы распознавания у пропагнозиков обычно остаются
интактными, и они способны опознавать друзей и родственников по голосам,
одежде или поведению — страдает только способность распознавать лица, что,
тем не менее, ведет к социальной инвалидности. Пациент никогда не способен
определить, кто к нему приближается — друг или чужой человек (если только
он не понял это по одежде или походке), нужно ли остановиться, улыбнуться
и заговорить или надо проследовать мимо с ледяным выражением лица. Если
друзья и родные не осведомлены о наличии прозопагнозии, это может привести
к резкому разрыву социальных контактов — очень мало кто способен простить
высокомерие. Социальная изоляция, от которой страдают недиагностирован-
ные прозопагнозики, подчеркивает роль распознавания лиц в современном,
а тем более примитивном, обществах.
17.6. Заключение
Эта глава, хотя она — не более чем царапина на боку огромного валуна пробле-
мы, показывает, что центральный зрительный анализатор приматов развился
в колоссальную, сложнейшую систему с более чем двумя дюжинами уже выяв-
ленных областей, связанных между собой двухсторонними иерархическими и
параллельными связями. В начале главы мы коснулись эволюционных причин
возникновения такой сложности системы. Зрение для нас, как и для всех при-
матов — важнейшее из чувств. Мир человека, со все большим проникновени-
ем в него электронных медиа, все в большей степени оказывается связан со
зрением. Наш литературный и философский словарь пропитан зрительными
метафорами. Мартин Хайдеггер отмечал, что исходным значением греческого
слова, обозначающего «знание», было «видение». Предполагалось, что наше
возрастающее понимание процесса зрения дает нам последний шанс запол-
нить пустоту между пониманием мозга и разума. Пока что это остается лишь
надеждой на будущее. А сейчас мы можем только надеяться, что знания, при-
обретенные при изучении зрительной системы, помогут в работе над менее
изученными областями мозга. Можно также вместе с Дэвидом Марром наде-
яться, что понимание принципов ее работы может быть полезным в разработ-
ке силиконового конкурента мозга.
18. ЗРИТЕЛЬНЫЕ СИСТЕМЫ
ДРУГИХ ПОЗВОНОЧНЫХ
Зрительные пигменты: родопсин, порфиропсин — адаптация к водному об-
разу жизни? Фоторецепторы: масляные капли — цветовые фильтры — па-
раболоиды; двойные и тройные колбочки — рыбы; двойные палочки —
пресмыкающиеся — земноводные; фотомеханические движения. Зеркаль-
ца: сосудистые — различные типы — окклюзивность — детектирование
жертвы. Сетчатка: многоуровневые сетчатки глубоководных рыб; бесхвос-
тые земноводные: РП — ON, OFF, ON/OFF — непдвижный контраст —
детектор «насекомых»; млекопитающие, птицы; выпукло-углубленная
фовеа — многочисленные центральные области и фовеа — зрительная по-
лоска — гребешок. Диоптрический аппарат: постранство сетчатки и разре-
шающая способность — тубулярный или «телескопический» глаз — «че-
тырехглазые» рыбы — фокусировка роговицей и хрусталиком — роговица
преобладает у неводных позвоночных — роговичное зрение у некоторых
пресмыкающихся — хрусталик преобладает у рыб — соотношение Матис-
сена — «открыто» восьмью группами водных животных. Медианные гла-
за: теменные и шишковидного тела — гистология — эволюция — репро-
дуктивные циклы и т. д. — синтез и выделение мелатонина. Зрительные
пути: тектофугальный путь доминирует у немлекопитающих — закон
Ньютона — Мюллера—Гуддена для РГС-пути — полный перекрест текто-
фугального пути — частичный перекрест в других случаях. Зрительные цен-
тры: Земноводные: зрительный текгум — гистология — физиология —
РП — РП «нового» и «прежнего»; Пресмыкающиеся — тектофугальный и
таламофугальный путь — зрительный тектум — дорзовентрикулярный гре-
бень (ДВГ) — сравнения с первичной зрительной корой млекопитающих;
Птицы — большое разнообразие зрительных систем — таламофугальный и
тектофугальный путь; таламофугальное утолщение — сравнимо с первич-
ной зрительной корой млекопитающих — четыре слоя клеток — РП —
бинокулярное зрение; тектофугальный контрлатеральный тектум — ре-
тинотопическая организация — РП; круглое ядро — РП — движение —
зрительный поток — сравнение с экстрастриарной — корой млекопитаю-
щих — обратная связь и восходящие связи. Заключение: огромное разнооб-
разие зрительных систем позвоночных — лекарство от антропоцентризма
Главу 15 мы начинали с упоминания о большой древности глаз позвоночных.
Эволюция их существенных элементов может быть прослежена на 500 милли-
онов лет — до начала кембрийского периода. За прошедший с тех пор огром-
ный промежуток времени позвоночные распространились по огромному ко-
18. Зрительные системы других позвоночных 427
личеству мест обитания и приспособились к разным способам существования.
Хотя глаз позвоночных оставался в основном неизменным в ходе этой дивер-
генции, он претерпел многочисленные модификации, результат адаптации к
разнообразным условиям существования. В данной главе мы рассмотрим не-
’ которые основные из таких адаптаций.
18.1. Зрительные пигменты
Мы видели во Введении к Части IV, что зрительные пигменты животных, со-
стоящие из опсинов и соединенных через шиффову связь с хромофорами,
формируются из витамина А. В Информации 14.1 мы рассматривали эволю-
цию опсинов и увидели, что все они принадлежат одному надсемейтву семи-
доменных белков. Там же мы отмечали, что хромофоров — два типа: ретинал^
и ретиналь2 (3,4-дегидроретиналь). Когда к опсину присоединяется ретиналь^
образуется родопсин, когда ретиналь2 — порфиропсин.
Родопсин и порфиропсин различаются по Хтах. Средняя Хтах порфиропси-
на сдвинута в красную сторону на 20 нм по сравнению с Хтах родопсина. Одно
время полагали, что этот сдвиг отражает адаптацию к жизни в загрязненных
водах, где коротковолновая часть белого света рассеивается в большей степе-
ни. Часто это случается в пресноводных водоемах — реках, ручьях, прудах и
озерах, в которые происходит дренаж с окружающих территорий. Возможно,
по этой причине распространение порфиропсина ограничено в основном па-
лочками пресноводных рыб, хотя он и обнаруживается у нескольких морских
видов. Такое предположение хорошо подтверждается, тем, что у донных рыб в
основном присутствует порфиропсин, а у рыб поверхностного слоя — родо-
псин. Однако экологические соображения никоим образом не исчерпывают
вопроса. Зависимость чувствительности фотопигмента от Хтах очень невели-
ка, рассчитано, что лишь 10% увеличения чувствительности обусловлено пиг-
ментом с Хтах 480 ни, по сравнению с Хтах 500 нм. Кроме того, у многих глубо-
ководных рыб развились родопсины со сдвигом чувствительности в красную
сторону, а не порфиропсины. И у пластиножаберных, и у костистых рыб при-
сутствуют родопсины с Хтах 470—480 нм.
Интересно отметить, что такое животное, как у европейских и североамери-
канских угрей Anguilla sp., проводящих часть своей жизни в пресных водах, в
часть — в глубоких водах океана, соответственно изменяется и пигмент. Боль-
шую часть жизни угорь проводит в пресной воде, используя, в основном пор-
фиропсиновую систему, однако перед миграцией в район икрометания —
в Саргассово море на глубинах около 400 м, происходит метаморфоз. В частно-
сти, порфиропсиновая система с Хтах 520 нм заменяется на родопсиновую
с Хтах около 500 нм. С другой стороны, минога Petromyzon marinus, которая вы-
лупляется из икры в пресноводных речках, а затем мигрирует в океан, сплавля-
ется в океан с родопсиновой истемой, а возвращается — с порфиропсиновой.
Ясно, что ситуацию трудно описать каким-то простым принципом. Возможно,
распределение порфирописновых и родопсиновых пигментов более определя-
лось случаем, чем условиями существования или эволбционным предшествен-
ником. Может быть также, что тот или иной фотопигмент имеет некоторое би-
охимическое значение, несущее еще неизвестное нам преимущество отбора.
428 Часть IV. Светочувствительность
18.2. Фоторецепторы
Мы уже упоминали, что существует два типа фоторецепторов позвоночных -
палочки и колбочки. Хотя палочки варьируют у них незначительно, существу-
ет множество различных типов колбочек. Некоторые из них показаны на
рис. 18.1. Структура колбочек млекопитающих была описана в главе 16. У кол-
бочек других позвоночных, стоящих в эволюционной лестнице «не ниже» зем-
новодных, часто в эллипсоиде развиваются масляные капли. Часто они содер-
жат каротиноиды, придающие им заметную окраску. У дневных птиц они
красные, оранжевые, желтые или зеленые, а у ночных — либо бесцветные, ли-
бо желтоватые. У волнистого попугайчика, например, присутствуют четыре
класса пигментов колбочек с Хтах 565, 507, 445—430 и 380—360 нм, соответст-
венно. Три класса колбочек с длинноволновыми пигментами содержат масля-
ные капли с коротковолновым пределом пропускания в 570, 520—500 и 445 нм,
а коротковолновые (ультрафиолетовые) колбочки — прозрачные капли. В це-
лом, капли служат селективными фильтрами падающего на колбочки света и
участвуют в его фокусировке на наружных сегментах.
Помимо развития в эллипсоиде масляных капель, иногда в миоиде форми-
руется другой диоптрический органоид — параболоид (рис. 18.1). В отличие от
масляных капель параболоид не окрашен, и вместо каротиноидов содержит
повышенную концентрацию гликогена. Имея значительно более высокий ко-
эффициент преломления, чем окружение, он также играет существенную роль
в фокусировке света на наружных сегментах.
Другой характерной чертой сетчатки многих позвоночных является при-
сутствие двойных, а иногда и тройных колбочек (рис. 18.1 В). Они встречают-
ся у рыб, земноводных, пресмыкающихся, птиц, одного из однопроходных
(утконос) и сумчатых. По каким-то, пока неизвестным причинам, они не об-
наружены у собственно млекопитающих. Двойные колбочки состоят из двух
слившихся миоидами очень непохожих друг на друга колбочек. Две слившие-
ся колбочки обычно сильно различаются по размеру, большая (или главная)
колбочка часто имеет масляную каплю, а меньшая — очень большой парабо-
лоид. Это, однако, далеко не всегда именно так, и возможны самые разнооб-
разные комбинации размеров и структур. В некоторых случаях, например у
различных видов рифовых окуней Большого Барьерного рифа, двойные кол-
бочки содержат разные фотопигменты. Было также сделано предположение,
что двойные колбочки могут участвовать в детектировании плоскости поляри-
зации падающего солнечного света. Наконец, в сетчатках многих рыб колбоч-
ки формируют очень правильную мозаику, в которой четыре двойных колбоч-
ки образуют прямоугольник вокруг одиночной колбочки.
Помимо двойных колбочек, в сетчатке многих костистых рыб развиваются
парные («близнецовые») колбочки. В отличие от двойных колбочек, оба компо-
нента парных колбочек — по крайней мере структурно, равные партнеры. Они
сливаются по всей длине (рис. 18.1 В(5)), но сохраняют отдельные ножки.
Связаны ли парные колбочки с двойными эволюционно, или резко от них от-
личаются в непрерывном спектре форм биологического мира — этот вопрос
остается дискуссионным. Стоит отметить, что в сетчатке некоторых рыб при-
сутствуют все четыре формы колбочек. В сетчатке усача {Barbus barbus) име-
A
(1) (2) (3)
Знс
к )
э
В-»
fesg м
ЗМкп
я
£\н
(4)
нс
(5)
Б
(1)
нс
э
п
(5)
Рис. 18.1. Фоторецепторные клетки позвоночных. (А) Колбочки (увеличение ЮООх).
(1) Колбочка осетра Acipenserfulvescens. (2) Адаптированная к свету колбочка серебряного
карася Carassius auratus (сокращенный миоид). Отметьте, что ядро колбочки, как и у дру-
гих рыб, лежит вне наружной пограничной мембраны. (3) Адаптированная к темноте кол-
бочка лягушки Rana pipiens (удлиненный миоид). (4) Колбочка кусающейся черепахи
Chelydra serpentina. (5) Колбочка с возвышенной круговой фовеа луня Circus hudsonius. От-
метьте малую толщину колбочки и длинный тонкий наружный сегмент. (Б) Палочки (уве-
личение ЮООх). (1) Палочка двоякодышащего большого протоптера Protopterus ethiopicus,
необычно, что палочка содержит масляную каплю, что подразумевает происхождение от
колбочек. (2) Адаптированная к темноте красная палочка Rana pipiens (сокращенный ми-
оид), (3) адаптированная к свету зеленая палочка Rana pipiens (расслабленный миоид).
(4) Адаптированная к свету палочка серебряного карася Carassius auratus. При световой
адаптации миоид (как показано) сильно удлинняется; в темноте он сокращается. (5) Фи-
ламентарная палочка американской летяги Glaucomys volans volans. Длинная и тонкая па-
лочка характерна для многих сумеречных и ночных животных.
Рис. 18.1. (В) Двойные колбочки (увеличение ЮООх). (1) Двойная колбочка ильной рыбы
Amia calva\ (2) Адаптированная к темноте двойная колбочка Rana pipiens (миоид основ-
ной колбочки удлинен). (3) Двойная колбочка расписной украшенной черепахи Chryse-
mys picta marginata. (4) Двойная колбочка обыкновенного ужа Natrix natrix\ (5) адаптиро-
ванная к свету двойная колбочка костистой рыбы синежаберного солнечника Lepomis
microchirus. Слившиеся миоиды сокращены. (Г) Двойные палочки (увеличение ЮООх).
(1) Двойная палочка ночной ящерицы Риверса Xantusia riversiana. Фоторецептор — про-
межуточный между палочками и колбочками. Хотя фоторецептор структурно сходен с
палочками, однако не содержит родопсина и является низкопороговым. (2) Двойная па-
лочка сетчатки земляного геккона Coleonyx variegatus. (3) Двойная палочка змеи Hypsigle-
па ochorhynchus. с — «свободная» зона, к — масляная капля, э — эллипсоид (главный),
э* — эллипсоид дополнительный, н — ножка, п — наружная пограничная мембрана, м —
миоид, я — ядро (главное), я* — ядро дополнительное, нс — наружный сегмент (глав-
ный), нс* — наружный сегмент дополнительный, пр — параболоид. Из Walls, 1963. Вос-
производится с разрешения Cranbrook Institute of Science. Crescitelli, 1977, с разрешения.
18. Зрительные системы других позвоночных 431
ются короткие одиночные колбочки, длинные одиночные колбочки, парные
колбочки и двойные колбочки (из неравных компонентов), которые распреде-
лены там без особого порядка.
Двоятся не только колбочки — двойные палочки часто обнаруживаются в
сетчатках пресмыкающихся, особенно змей и гекконов (рис. 18.1 Г). Гекконы
изучены особенно хорошо. Это семейство обитает в множестве ареалов в теп-
лом климате, многие их них — сумеречные и ночные животные, но существу-
ют и дневные формы, глаза крупные и хорошо развиты. У дневных форм ши-
роко представлены двойные колбочки. У ночных и сумеречных форм наруж-
ные сегменты имеют длинную цилиндрическую форму, так что фоторецептор
лучше именовать двойной палочкой (рис. 18.1 Г). У геккона (Gekkogekko) при-
сутствуют два фотопигмента с Хтах 521 и 467 нм. В некоторых случаях оба ком-
понента двойной палочки содержат один и тот же пигмент, у других — один
компонент содержит пигмент 521 нм, а другой — 467 нм.
Палочки также имеют различные формы. Возможно, наиболее известен
диморфизм в сетчатке лягушки, где палочки представлены двумя типами:
красных и зеленых. Рис. 18.1 Б (2 и 3) показывает, что их структуры сильно раз-
личаются, зеленые палочки (содержащие пигмент 432 нм) имеют длинный
тонкий миоид, тогда как красные палочки (пигмент 502 нм) вообще не имеют
миоида. Это приводит нас к последней черте фоторецепторов немлекопитаю-
щих позвоночных — их фотомеханическим движениям. Эти движения возника-
ют в результате активности актиновых филаментов миоида. При действии яр-
кого света миоид палочек расслабляется, и наружные сегменты смещаются в
сторону пигментного эпителия. Напротив, миоиды колбочек сокращаются и
выдвигают наружные сегменты в сторону источника света. В случае двойных
колбочек фотомеханические движения выявляются у большего компонента,
а меньший не изменяется и остается на месте. Фотомеханические движения
достаточно медленны и измеряются минутами у птиц и десятками минут у
рыб. Эти механические движения фоторецепторов сопровождаются сходны-
ми медленными движениями пигментных гранул в микроворсинки клеток
пигментного эпителия. Гранулы смещаются в микроворсинки, защищая па-
лочки и повышая чувствительность колбочек на ярком свету и сдвигаются
к ПЭС при слабом освещении. Можно сказать, что животные, у которых раз-
вита фотомеханическая система, имеют две сетчатки — одну для яркого света,
другую — для тусклого.
Из изложенного выше ясно, что, по сравнению с сетчатками других позво-
ночных, сетчатка человека, являющаяся предметом стольких исследований,
существенно беднее по числу типов фоторецепторов.
18.3. Зеркальца
Пигментный эпителий и сосудистая оболочка глаза человека содержит боль-
шое количество меланина, который поглощает свет, не поглощенный ней-
ральным или фоторецепторным слоями сетчатки. Это важно, поскольку отра-
жение может сильно исказить изображение. При слабом освещении, однако,
отражение может оказаться полезным, поскольку дает еще одну возможность
непоглощенному свету взаимодействовать с зрительным пигментом наружных
432 Часть IV. Светочувствительность
сегментов. Любой водитель видел, как глаза зверей, таящихся в придорожных
кустах проселочных дорог, сверкают в свете фар. Представьте, какой эффект
(до ужаса включительно) вызвали бы сверкающие в свете фар глаза человека.
Слой, который отражает свет у многих ночных и сумеречных животных, назы-
вается зеркальцем (tapetum) или более точно tapetum lucidum (буквально — све-
тящийся ковер). В некоторых случаях зеркальца имеют качество, близкое
к настоящему зеркалу (см. гребешок Pecten, гл. 14), у других животных отра-
женный свет рассеян. Зеркальце формируется в сосудистой оболочке или
(реже) в пигментном эпителии сетчатки.
Существует три типа зеркалец сосудистой оболочки: фиброзный, клеточный
и гуаниновый. Зеркальца фиброзного типа (tapetum fibrosum) найдены у многих
копытных. Они образованы отложениями белых коллагеновых волокон в со-
судистой оболочке позади капилляров и создает хороший, хотя и не совершен-
ный отражающий слой. Клеточный тип (tapetum cellulosum) обеспечивает бо-
лее эффективное зеркало, он развился у просимиановых приматов, а также
хищных — животных, чей яркий блеск глаз вошел в поговорки, особенно это
касается домашних кошек. Клеточные зеркальца обнаружены и у тюленей.
Этот тип зеркальца, как следует из его названия, образован не коллагеновыми
волокнами, а клетками. Эндотелиальные клетки за капиллярами пролифери-
руют и образуют плотно упакованное многослойное «кафельное покрытие».
Клетки заполнены сократимыми нитями, которые, перекрещиваясь между со-
бой, образуют зеркальную сеть. Наконец, гуаниновый тип зеркалец (tapetum
guaninum) обнаружен у многих пластиножаберных и костистых рыб. В этом
случае, как и в клеточных зеркальцах, наблюдается формирование многослой-
ного клеточного отражающего слоя за капиллярами сосудистой оболочки, но
в отличие от зеркалец клеточного типа, клетки заполнены не волокнами, а
пластинками гуаниновых кристаллов. У некоторых рыб, особенно пластино-
жаберных (таких как катран Squalus acantias и кунья акула Mustelus mustelus),
этот блестящий отражающий слой является окклюдирующим (рис. 18.2). При
слабом освещении пигментные гранулы концентрируются в центрах клеток
сосудистой оболочки, и свет, проникающий до зеркальца имеет максималь-
ную возможность отразиться от гуаниновых кристаллов; с другой стороны,
при ярком свете гранулы мигрируют в тела и отростки клеток сосудистой обо-
лочки, что вызывает потемнение зеркала.
Сетчаточный тапетум особенно хорошо известны у рыб. И здесь гуанино-
вые кристаллы образуют яркий серебристый отражающий слой, но на этот
раз в пигментном эпителии. У некоторых карповых, например у леща
Abramis brama, который обитает в неглубоких мутных водах озера Балатон
(западная Венгрия), зеркальце, подобно описанным выше гуаниновым зер-
кальцам пластиножаберных, окклюдирует. При слабом колеблющемся све-
те ярко серебристое зеркало в пигментном эпителии отражает практически
весь свет, доходящий до него. При более ярком свете пигментные гранулы,
также присутствующие в клетках ПЭС, мигрируют в отростки клеток и ока-
зываются перед слоем гуаниновых кристаллов. В таком положении свет
практически не отражается обратно на фоторецепторные клетки и не иска-
жает изображение. Вероятно, именно обитание в мутной воде привело к
развитию у леща и близких видов такой полезной адаптации; у большинст-
18. Зрительные системы других позвоночных 433
Б
Рис. 18.2. Вертикальный срез окклюдирующего зеркальца куньей акулы Mustelus.
(А) Адаптация к свету — пигментные гранулы мигрируют между гуаниновыми кри-
сталлами, уменьшая отражающую способность зеркальца. (Б) Адаптация к темно-
те. Пигментные гранулы уходят в пигментный эпителий, и гуаниновые кристаллы
образуют сверкающий отражающий слой за наружными сегментами фоторецепто-
ров. к — капилляры сосудистой оболочки, гп — гуаниновые пластинки, мк — слой
мигрирующих пигментных клеток в сосудистой оболочке, пэ — пигментный эпи-
телий сетчатки (пигмент не показан); по — пигментированные отростки, нс — не-
модифицированная сосудистая оболочка. Из Walls, 1963. Воспроизводится с разре-
шения Cranbrook Institute of Science.
ва рыб с зеркальцем сетчатки возможность контроля его отражающей спо-
собности отсутствует.
Отражающие сетчатки развивались множеством специфических путей,
особенно у позвоночных, обитающих в условиях пониженной освещенности,
особенно это касается глубоководных рыб. Здесь приспособления, повышаю-
щие чувствительность глаза полезны, даже если качество изображения от это-
го страдает. Некоторые глубоководные рыбы, например, испускают свет, что-
бы по его отражению обнаруживать подходящую жертву. Одна из таких рыб,
Pachystomias, испускает пучки красного света, которые предпочтительно отра-
жаются от красных рачков зоопланктона. У этой рыбы зеркальце красного
цвета, которое адаптировано к наличию такого отраженного света и, таким об-
разом, повышать вероятность обнаружения рачков. Обнаружено и множество
других интересных приспособлений зеркалец, все они помогают глазу адапти-
роваться к способу существования своего владельца.
18.4. Сетчатки
Некоторые детали «дуплексной» сетчатки человека мы обсуждали в главе 16 и
отметили, что, несмотря на множество модификаций, базовая структура ее ос-
тается у всех позвоночных удивительно схожей. В данном разделе мы рассмо-
трим несколько вариаций на тему сетчатки, в особенности модификации area
centralis млекопитающих и птиц. Однако, сначала коснемся адаптаций сетча-
ток глубоководных рыб к слабому освещению и физиологической адаптации
сетчатки лягушки к ее условиям обитания.
434 Часть IV. Светочувствительность
18.4.1. Глубоководные рыбы
Помимо развития тапетумов различных типов (см. раздел 18.3) у глубоковод-
ных рыб развились не только палочки с необыкновенно длинными наружны-
ми сегментами, но и (во многих случаях) многоуровневые сетчатки. У одной из
глубоководных рыб — диретмы Diretmus argenteus — наружные сегменты, при
толщине сетчатки в 900 мкм, имеют 600 мкм в длину! У многих других видов
наружные сегменты расположены слоями один над другим, образуя несколько
уровней. Самый внешний уровень наружных сегментов (а уровней может быть
шесть или семь) связан с ядром очень длинным и тонким миоидом (рис. 18.3).
Рис. 18.3. Схема многоуровневой сетчатки.
1 — самый внутренний слой палочек (обра-
щенный к стекловидному телу); 2 — эллип-
соиды самых внутренних палочек; 3 — на-
ружная пограничная мембрана; 4 — наруж-
ные сегменты самых внутренних палочек;
5 — пигментный эпителий; 6 — средний
уровень наружных сегментов палочек; 7 —
тонкий филаментарный миоид средненго
уровня наружных сегментов; 8 — наружный
уровень наружных сегментов; 9 — длинный
филаментарный миоид наружного слоя на-
ружных сегментов; 10 — глазной пузырь, в
котором лежат наружные сегменты среднего
и наружного уровней; 11 — волокно палоч-
ки; 12 — ядра палочек; 13 — сферулы пало-
чек. Из Locket, 1977, с разрешения.
Кроме упомянутых, в сетчатках многих глубоководных рыб развились и не-
сколько других приспособлений, максимизирующих чувствительность — это
участки с высокой плотностью ганглиозных клеток, полосы с повышенной
плотностью палочек и колбочек, дополнительны сетчатки, сетчаточный ди-
вертикул и т. д.
18.4.2. Лягушка
Нельзя считать рассмотрение сетчаток позвоночных завершенным без упоми-
нания пионерской работы Хартлайна (Н. К. Hartline) по физиологии сетчатки
лягушки. Еще в 1938 году Хартлайну удалось выделить пучки, а в некоторых
18. Зрительные системы других позвоночных 435
Ответ
ганглиозной 1 ill | | | | I I I
клетки । «111 । । । । । ।
Свет _J L
On Off
Рецептивное
поле ON-типа (а)
_г
On
4Н++
Рецептивное
поле ON-типа (б)
Ответ
ганглиозной
клетки
Свет
HillII I ИНН I I I
-J----------1____________
On Off
Рецептивное
поле ON-типа (в)
Рис. 18.4. Физиологические характеристики ганглиозных клеток лягушки в наибо-
лее обычном типе рецептивных полей сетчатки. Дальнейшие объяснения в тексте.
случаях — и одиночные волокна из поверхности сетчатки лягушки-быка (Rana
catesbiana) и провести отведения от них с помощью фитильковых электродов.
Он обнаружил три типа рецептивных полей: ON, OFF и ON/OFF. Рецептив-
ные поля имели близкую к круглой форму и диаметр до 1 мм. Если вспомнить,
что диаметр глаза лягушки — всего около 8 мм, можно только восхититься ог-
ромным размером РП. Работы Хартлайна впоследствии были подтверждены с
помощью микроэлектродной техники. Было показано, что, если ON и OFF
поля отвечали на изменения общей освещенности, то ON/OFF-поля по раз-
ному реагировали на диффузное и сфокусированное освещение (рис. 18.4).
Иными словами, у ON/OFF-полей имеется некоторое латеральное торможе-
ние, с которым мы сталкивались, когда рассматривали концентрические ре-
цептивные поля млекопитающих (раздел 16.2.8).
Дальнейшие электрофизиологические исследования сетчатки лягушки по-
казали наличие в ней еще двух типов рецептивных полей. Первый из них гене-
рирует непрерывную реакцию, если в поле присутствует край объекта, потому
и называемый детектором постоянного контраста (иногда — поле класса 1).
Второй реагирует на движение небольших темных объектов, пересекающих
РП. Прямые края не в состоянии стимулировать эти ганглиозные клетки. Этот
тип клеток называется детектором выпуклости (поле класса 2) или в просторе-
чии «детектор насекомых».
В отсутствии движения или изменений интенсивности освещенности ак-
тивность в зрительном нерве лягушки невелика, за исключением исходящей
от детекторов постоянного контраста. Соблазнительно представить себе (и
это, вероятно, недалеко от истины), что сидит себе лягушка на листе кувшин-
ки, и только детекторы постоянного контраста обозначают линию горизонта и
прочие существенные контура местности, а вся остальная сетчатка молчит.
Любое резкое изменение освещенности — внезапная тень от хищной птицы
или солнечное пятно актвирует ON, OFF и ON/OFF-поля. Но самое важное —
это появление в поле зрения маленьких темных объектов — насекомых, кото-
рые запускают ON/OFF-поля и, особенно детекторы выпуклости. Тут лягуш-
ка напрягается и «выстреливает» в цель свой язык. Зрительная система бесхво-
436 Часть IV. Светочувствительность
стых земноводных — любимый объект исследования специалистов в области
зрения. Мы вернемся к ней, когда будем рассматривать анализ зрительной ин-
формации в оптическом тектума в разделе 18.8, она также важна для исследо-
ваний по нейробиологии развития. Желающие могут найти соответствующие
источники в списке литературы. А теперь обратим наше внимание на приспо-
собления, развившиеся в сетчатках млекопитающих и птиц.
18.4.3. Центральные области (areae centrales)
млекопитающих и птиц
Эту область сетчатки трудно определить точно. Она не обязательно распола-
гается в центре сетчатки и не всегда ее признаком является желтый пигмент,
так характерный для сетчатки человека — желтое пятно (macula lutea). В неко-
торых сетчатках, как мы увидим ниже, это — не просто область, а, скорее,
часть или части сетчатки, в которых обеспечена наивысшая разрешающая спо-
собность. В сетчатке человека, как мы видели в главе 15, она окружает неболь-
шое углубление — фовеа, однако, далеко не у всех животных такие области
имеют фовеа.
Определение этой области как части сетчатки с максимальной разрешаю-
щей способностью немедленно вызывает представление о том, что здесь упа-
ковка фоторецепторов наиболее плотна. У человека здесь колбочки особенно
длинные и тонкие, то же самое и в сетчатках других животных. Для обеспече-
ния высокой разрешающей способности соотношение между колбочками, би-
полярными и ганглиозными клетками должно приближаться к 1: 1 : 1, однако
такое соотношение имеет свои негативные последствия. Это означает, что вы-
сокая плотность колбочек сопровождается высокой плотностью всех других
элементов сетчатки, что должно приводить к снижению прозрачности послед-
ней. Решение проблемы (как мы видели в главе 16) состоит в том, что биполяр-
ные клетки удалены из центральной области диагонально (рис. 16.6), так что
их тела клеток, контакты во внутреннем синаптогенном слое, амакриновые и
ганглиозные клетки оказываются смещенными к периферии. Это приводит к
утончению сетчатки в центральной области — которое и именуется фовеа, ок-
руженному кратероподобным валом, образованным смещенными сюда ней-
ральными элементами.
Фовеа развивается в большинстве сетчаток с высокой разрешающей спо-
собностью. Во многих случаях фовеа оказывается намного глубже, чем у чело-
века. Такие глубокие фовеа именуются выпукло-углубленными, в отличие от ме-
нее глубокой и менее правильной формы вогнуто-углубленной фовеа нашей
собственной сетчатки. Есть ли у выпукло-углубленной фовеа функции, отлич-
ные от описанных выше? Это остается предметом множества споров. Некото-
рые полагают, что крутые выпуклые скаты фовеа увеличивают изображение
(рис. 18.5). Другие возражают, что изображение, попадающее на скаты, должно
искажаться; и только когда оно центрировано и может быть изоморфным объ-
екту, это может улучшать остроту зрения хищника В поддрежку такой интер-
претации приводится наличие выраженной выпукло-углубленной фовеа и у
хищных птиц, таких как ястреб, и у хищных глубоководных рыб. С другой сто-
роны, те же аргументы годятся для обоснования того, что выпукло-углубленная
18. Зрительные системы других позвоночных 437
Рис. 18.5. Выпукло-углубленная фовеа хищной птицы, такой как ястреб-тетеревят-
ник (Asturpalumbarius). Стрелки показывают направление падающего света, кото-
рый, преломляясь латерально, дает увеличение изображения. Из Меуег, 1977,
с разрешения.
фовеа может служить увеличителем, что тоже было бы полезно для шустрых
хищников. Но пока приговор не вынесен, «присяжные еще не вернулись».
Уже упоминалось, что в некоторых сетчатках больше одной центральной
области и больше одной фовеа. Множественные области и фовеа наиболее из-
вестны в сетчатках птиц. Хотя у большинства птиц более или менее централь-
но расположенная центральная область и фовеа присутствуют, давно известны
различные другие варианты, включая полное отсутствие фовеа. Наиболее точ-
но установленный случай сетчатки без фовеа — это калифорнийская куропат-
ка (Lophortyx califomicus vallicold), некоторые другие варианты показаны на
рис. 18.6. Видно, что area centralis иногда действительно расположена цент-
рально, иногда латерально, иногда соедржит фовеа, а иногда — нет. Иногда
центральных областей две, и обе — с фовеа. Последнее характерно для птиц,
питающихся на лету — воробьиные, зимородки, колибри. Очевидно, что этим
птицам необходимо очень точное восприятие дистанции и скорости. У многих
птиц центральная область имеет не круговую форму, а форму полосы, пересе-
кающей всю сетчатку или ее часть. Такая организация наиболее часто встре-
чается у птиц, населяющих открытые пространства, и у тех, кто постоянно
клюет при ходьбе. Вследствие этого полагают, что такая организация связана
с восприятием горизонталей. Наконец, возможно наиболее развитые сетчат-
ки — у дневных хищных птиц — орлов, ястребов, канюков и соколов, облада-
ющих комбинацией круговой и ленточной центральных областей с выпукло-
углубленной фовеа в каждой из них. Такая организация обеспечивает наилуч-
шую комбинацию остроты зрения и восприятия дистанции.
Хотя лентовидные области в сетчатке не так очевидны, тем не менее есть
доказательства существования полос повышенной остроты зрения, известных
как зрительные полоски, у огромного большинства млекопитающих. Анатоми-
чески это выражается в повышенной плотности фоторецепторов, повышен-
ной плотности ганглиозных клеток, уменьшенной конвергенции фоторецеп-
торов на ганглиозных клетках и отсутствии перекрывающих полоску крове-
носных сосудов. Полоска обычно формируется в горизонтальной плоскости,
особенно она выражена у млекопитающих, обитающих на открытых прост-
ранствах. Она, например, выделяется у кроликов и зайцев, у обитающего в са-
ванне гепарда и у грейхаунда, который является одомашненной формой пус-
тынной охотничьей собаки, но слабо развита у ночных видов — мышей и ежей.
438 Часть IV. Светочувствительность
Рис. 18.6. Центральные области и фовеа некоторых птиц. Черный овал показывает
положение гребня, (а) Центральная фовеа (напр., стеллерова сойка Cyanocitta stel-
leris)\ (б) Латеральная фовеа (совы и стрижи); (в) бифовеальная сетчатка зимород-
ка Alcedo ispida. Отметьте очень эксцентрическое положение латеральной фовеа.
(г) бифовеальная сетчатка американской выпи Botarius lentigenosus. Отметьте высо-
кое расположение латеральной фовеа. (д) Линейная бесфовеальная область. Такой
тип сетчатки обнаружен у некоторых морских птиц, напр. малый буревестник
(Puffinus puffinus) и глупыш (Fulmaris glacialis). (е) Линейная область с замкнутой
центральной областью, содержащей фовеа, напр. лысуха американская (Fulica
americanz). (ж) Линейная область, соединяющая латеральную и центральную обла-
сти, причем обе содержат фовеа, напр. воробьиная пустельга Falco sparverius.
(з) Линейная область с замкнутой центральной областью, содержащей фовеа и от-
дельная латеральная область с фовеа, напр. обыкновенная крачка Sterna hirundo. Из
Meyer, 1977, с разрешения.
Хотя вопрос остается дискуссионным, в основном все согласны с тем, что, как
и лентовидные области у птиц, зрительные полоски млекопитающих являют-
ся адаптацией, улучшающая определение горизонтальной плоскости на от-
крытом пространстве.
Прежде, чем завершить этот раздел, следует отметить еше одну порази-
тельную структуру сетчатки — это гребень. Эта крупная структура характерна
для глаза птиц. Она начинается в месте выхода зрительного нерва и проециру-
ется в стекловидное тело (рис. 18.7). Он состоит почти полностью из увеличен-
ных капилляров и нескольких пигментных клеток. Хотя о существовании
гребня известно с тех пор, как Перро (Perrault) описал его в 1676 году, и он
явился предметом многочисленных тщательных исследований, — функция
его остается невыясненной. Форма и размеры гребней у различных птиц варь-
ируют — конические, лопастевидные, складчатые, но во всех случаях характе-
ризуются общей чертой — интенсивной васкуляризацией. Следовательно, ос-
тавляя в стороне хитроумные теории об участии гребня в физиологии зрения,
самая вероятная его роль — и самая очевидная — обеспечение стекловидного
тела и нейральных слоев сетчатки питательными веществами. Благодаря рас-
положению гребня над удлиненным слепым пятном, он создает минимальные
препятствия зрению.
18. Зрительные системы других позвоночных 439
Рис. 18.7. Различные типы гребней птиц. Левая колонка показывает вид сбоку,
средняя колонка — вид спереди, а правая — поперечный срез гребня, (а) Коничес-
кий гребень киви (Apteryx mantelli). (б) Лопастевидный гребень страуса (Struthio
camellus). (в) складчатый гребень вяхиря (Columba palumbus). (г) Складчатый гре-
бень горлицы Leucosarcia picata. (д), (е) и (ж) — три типа складчатого гребня ((e) и
(ж) — наиболее обычны). Из Меуег, 1977, с разрешения.
18.5. Диоптрический аппарат
Минимальный диаметр наружных сегментов палочек и колбочек ограничен
физическими характеристиками электромагнитного излучения видимого спе-
ктра. Они не могут быть меньше прибл. 2 мкм в поперечнике (см. Введение к
Части IV). Ясно, что для того, чтобы различать мелкие детали, сетчатка долж-
440 Часть IV. Светочувствительность
на быть достаточно большой — в маленькой сетчатке просто невозможно упа-
ковать достаточное количество наружных сегментов, чтобы различать тонкие
детали образа. Следовательно, животные с хорошим зрением должны иметь
большие глаза — и не просто относительно размеров тела, но и в абсолютном
значении. Это становится очевидно, если рассмотреть глаза мелких воробьи-
ных, таких как лесная завирушка или зяблики — ради высокого качества зре-
ния их глаза достигают большего размера, чем их мозг. Сравнительно малень-
кие глаза ночных и сумеречных грызунов — крыс и мышей — указывают на
малую остроту их зрения.
Переходя от абсолютных размеров глаз к их общей форме, можно отметить
множество адаптаций, приспосабливающих их к различным условиям обита-
ния и способу существования. Некоторые наиболее интересные адаптации
обнаружены у глубоководных рыб. Самая известная из них — это трубчатые
глаза многих родов, например, жемчужноглазов Scopelarchus, топориков Argy-
ropelecus и т. д. (рис. 18.8 (i)). В этом случае для различения большего количе-
ства деталей глазное яблоко не увеличивается в диаметре (оно бы в черепе не
поместилось), а удлиняется в одном измерении, откуда и старое название —
«телескопический глаз». Глаз имеет огромный хрусталик и длинный цилинд-
рический глазной бокал с хорошо развитой сетчаткой в дне. Дополнительная
сетчатка простирается на стенке трубки. Хотя хрусталик и имеет некоторый
контроль фокусировки, оптика глаза предполагает, что он всегда близорук.
Предполагали, что дополнительная сетчатка на стенке бокала позволяет полу-
чать изображение удаленных объектов, однако более поздние работы показы-
вают, что это маловероятно. Возможно, дополнительная сетчатка в основном
связана с обнаружением диффузного света, в частности испускаемого фото-
форами других обитателей глубин моря.
Еще один интересный глаз рыбы — это глаз южноамериканской четырех-
глазки Anableps tetrophthalmus, которая плавает, выставив глаза наполовину на
воздух, наполовину, оставляя их в воде. Этот глаз интересует исследователей
со времен Артеди (Artedi) (1758) и Зоммеринга (Soemmerring) (1818). Хруста-
лик глаза имеет такую форму, что свет сверху (из атмосферы) фокусируется на
одном сегменте сетчатки, а свет снизу (из водного окружения) — на другом.
Роговица сходным образом специализирована для фокусирования света на
разных сетчатках. В результате четырехглазка Anableps имеет по две оптических
системы в каждом глазу, чем замечательно оправдывает собственное название
(рис. 18.8 (ii)).
Выше отмечалось (см. также 18.8), что хрусталики рыб по сравнению с хру-
сталиком человека огромны и имеют намного более сферическую форму. Это
обусловлено тем, что фокусировка глаза рыбы (и вообще водных животных)
осуществляется именно хрусталиком, а роль роговицы минимальна. Это нахо-
дится в резком контрасте с глазами, функционирующими в воздушной среде,
как наши собственные, в которых две трети оптической силы приходится на
роговицу (рис. 18.9 (iii)). В этом случае хрусталик лишь уточняет фокусировку.
Дополнительным усложнением некоторых «воздушных» глаз является нали-
чие очков перед роговицами. Они развиваются у некоторых ящериц и змей из
слившихся век. Вероятно, их основная роль состоит в защите роговицы от аб-
разивных свойств окружающей среды, но у некоторых видов они играют важ-
18. Зрительные системы других позвоночных 441
Рис. 18.8. Глаза рыб. (i) Трубчатый глаз глубоководной рыбы с двумя сетчатками:
N — для видения вблизи, D — для дистантного видения, (ii) Глаз четырехглазки
Anableps. Грушевидная форма хрусталика и наличие двух зрачков позволяет рыбе
видеть в водной и воздушной среде. Надводное зрение использует верхний зрачок
и нижнюю сетчатку (А); водное зрение использует нижний зрачок и верхнюю сет-
чатку (W). Из Walls, 1963, с разрешения, (iii) Детали трубчатого глаза глубоковод-
ной рыбы. 1 — большой хрусталик, 2 — тонкая, но очень широкая роговица, ее края
обозначены стрелками, 3 — сетчатка, 4 — дополнительная сетчатка, простирающа-
яся между точками 5 и 6; 7 — латеральная стенка глазного пузыря, образованная
радужкой; 8 — сосудистая оболочка лежит непосредственно позади основной и до-
полнительной сетчаток. Из Locket, 1977. Воспроизводится с разрешения Springer-
Verlag GmbH & Со. KG.
ную роль в преломлении света. Роговица рыб, напротив, настолько несущест-
венна для оптики глаза, что часто имеет неправильную форму, что было бы фа-
тально для нормального зрения наземных позвоночных, таких как мы сами.
Хрусталики рыб к тому же лежат обычно непосредственно за роговицей, так
что передняя камера либо мала, либо отсутствует, и таким образом достигает-
ся максимальное фокусное расстояние от сетчатки и большой угол зрения.
Несущественность роговицы глаза рыбы обусловлена малыми различиями
преломляющей способности роговицы и окружающей воды и, соответствен-
но, малой возможностью ее изменять фокусировку света на сетчатке. Практи-
чески полностью эта задача выполняется хрусталиком. При таких обстоятель-
ствах, если хрусталик оптически гомогенен, его коэффициент преломления
442 Часть IV. Светочувствительность
Рис. 18.9. Оптика некоторых глаз по-
звоночных. (i) Гомогенный сферичес-
кий хрусталик, (ii) Негомогенный «ма-
тиссенов» хрусталик, (iii) Глаз человека.
ир — изображение, формируемое рого-
вицей; иср — изображение, формируе-
мое роговицей и хрусталиком. Объяс-
нения в тексте. По Land and Fernaid,
1992, с разрешения.
должен составлять около 1,66, чтобы свет можно было сфокусировать на сет-
чатке, которая, как впервые показал Матиссен (Mattiessen) в 1877 г., почти все-
гда находится на расстоянии 2,5 радиусов хрусталика от центра хрусталика.
Соотношение фокусного расстояния хрусталика к радиусу хрусталика называ-
ется соотношением Матиссена. Коэффициент преломления в 1,66, однако, не-
возможен для материалов, доступных рыбам (да и любым другим животным).
Матиссен показал, что ответ на это противоречие лежит в негомогенности хру-
сталика. Он показал, что индекс преломления варьирует от прибл. 1,52 в цен-
тре до менее 1,4 на периферии.
Эффект матиссеновского градиента коэффициента преломления двойст-
венен. Во-первых, таким образом уменьшается фокальное расстояние, что
позволяет увеличить апертуру зрачка и, следовательно, пропускать больше
света. Во-вторых, правильный градиент коэффициента преломления хруста-
лика устраняет сферические аберрации. Сферический хрусталик, не имеющий
такого градиента, практически бесполезен для формирования изображения
(рис. 18.9 (i)). Измеряя фокальное расстояние и радиусы хрусталика, легко оп-
ределить, «додумалось ли» данное животное до использования матиссенового
хрусталика. Показано, что к такому варианту животные пришли по крайней
мере восьмью независимыми путями: рыбы, белемноидные моллюски, четы-
режды — брюхоногие моллюски, одно из семейств кольчатых червей (Alciopi-
dae) и веслоногий рак Labidocera. Как отмечали Лэнд (Land) и Фернальд (Fer-
naid), этот список включает глаза водных животных всех возможных размеров
и, как они писали, «есть лишь один правильный путь формирования таких
хрусталиков и... естественный отбор всегда его находит».
18.6. Медианные (непарные) глаза
Имеется множество доказательств, полученных при изучении ископаемых че-
репов, того, что у ранних позвоночных присутствовали медианные глаза. Во
многих черепах палеозойской и мезозойской эр присутствует теменное отвер-
стие, которое часто имеет большие размеры. У современных позвоночных та-
ких крупных медианных глаз нет. Тем не менее, медианные глаза у наиболее
18. Зрительные системы других позвоночных 443
примитивных из существующих позвоночных — круглоротых — развиты до-
статочно хорошо.
Медианные глаза развиваются из задней части крыши промежуточного
мозга. В своих наиболее развитых формах они принимают вид двух выпячива-
ний — переднего теменного (или парапинеального) органа и заднего — шишко-
видного органа или эпифиза. Двойная структура лучше всего заметна у кругло-
ротых, таких как минога. У этого животного и теменной, и шишковидный
органы развивают сетчатки, содержащие колбочко-подобные фоторецепторы.
В темноте в них регистрируются потенциалы действия, и эта активность по-
давляется освещением. Пик спектральной чувствительности приходится на
525 нм. Биологическое значение системы варьирует у разных видов. Экспери-
менты по удалению этих органов показали, что по крайней мере у миноги
Lampetra planeri медианный глаз контролирует суточные изменения окраски
тела.
Адаптивные вариации медианных глаз показано на рис. 18.11. Шишковид-
ное тело (эпифиз) обнаруживается внутри черепа всех классов позвоночных.
Теменной орган, с другой стороны, занимает внутричерепное положение у
миноги и некоторых костистых рыб и внечерепное — у лягушек и ящериц. Фо-
торецепторы шишковидного и теменного органов во многих отношениях на-
Рис. 18.10. Эпифизарная система пластиножаберных рыб (катран). (А) Схема сред-
несагитального среза мозга. (Б) Схема фоторецептора. Р — ресничка, Ц-1 — центри-
оль, Э — эпифиз (шишковидное тело), ЭС — эпифизарный стебель, ЭТ — эпифизар-
ный тракт, ЭП — эпифизарный пузырек, Д — дендрит подлежащей ганглиозной
клетки, ПК — поводковая комиссура, ВС — внутренний сегмент, М — митохондрия,
Я — ядро, НС — наружный сегмент, ЗК — задняя комиссура, К — кожа, Ч — череп
(хрящ), СЛП — синаптическая лента и пузырьки, Ш-Ж — третий желудочек. Из
Hamasaki and Eder, 1977, с разрешения.
444 Часть IV. Светочувствительность
Бесчелюстные
круглоротые “
Хрящевые рыбы
пластиножаберные
Рис. 18.11. Распространение медианных глаз. Объяснения в тексте. Из Hamasaki
and Eder, с разрешения.
поминают колбочки латеральных глаз. Как показывает рис. 18.10 Б, фоторе-
цепторы образуют ряд инвагинаций, образующих «стопку» мембран со зри-
тельным пигментом. В синаптических окончаниях формируются типичные
синаптические ленты. Надо отметить, что во многих, но не всех случаях сет-
чатка неинвертирована, так что наружные сегменты фоторецепторов обраще-
ны к свету.
Медианные глаза начинают свою эволюцию у позвоночных как фоторе-
цепторный орган, эта функция сохраняется у костистых и хрящевых рыб и
амфибий, хотя у последних — только на стадии головастика. В качестве фо-
торецепторных органов они достигают совершенства у пресмыкающихся,
особенно у гаттерии Spenodon. Здесь в теменном органе развивается сетчатка,
состоящая из нескольких тысяч фоторецепторов, обращенных к свету, фоку-
сируемому клеточным хрусталиком. У земноводных и пресмыкающихся ме-
дианные глаза участвуют в контроле цветовых изменений тела и некоторых
поведенческих (особенно репродуктивных) циклов. Известно, что у всех этих
позвоночных эпифиз синтезирует мелатонин, который, возможно, участвует в
биологической реакции, вызываемой изменениями освещенности.
18. Зрительные системы других позвоночных 445
У птиц и млекопитающих развивается только эпифиз (шишковидное тело),
но он никогда не пронизывает мозговых оболочек. При таком внутричереп-
ном положении, будучи перекрыт не только твердой мозговой оболочкой, но и
теменными костями черепа, эпифиз не подвергается прямому воздействию
освещения. Хотя некоторые рудименты фоторецепторной ультраструктуры
остаются, но т.н. пинеалоциты не формируют наружных сегментов и не содер-
жат зрительных пигментов. Тем не менее, и у птиц, и у млекопитающих поли-
синаптический путь от латеральных глаз снабжает этот орган информацией о
фотопериоде. Это существенно, поскольку у птиц и млекопитающих эпифиз,
несмотря на утрату фоторецепторной функции, сохраняет способность синте-
за и выделения мелатонина, который у многих из них важен для регуляции ре-
продуктивного цикла. Зрительный сигнал от латеральных глаз, таким образом,
обеспечивает связь этих циклов с длиной дня и сезоном года.
18.7. Зрительные пути
Зрительные пути приматов описаны в главе 17. Мы видели, что основной по-
ток зрительной информации передается от сетчатки через НКТ в стриарную
кору. Менее значимые пути направляются в другие области мозга, включая
верхние бугры четверохолмия, ядро подушки, претектум и эпифиз. У не-мле-
копитающих позвоночных обнаружены те же зрительные пути, однако, путь
через НКТ в конечный мозг намного менее существенен. Напротив, у них на-
много большее значение имеют пути, направляющиеся в ядра среднего мозга,
особенно тектофугальный путь в зрительный тектум. С популярной теорией,
объясняющей это различие между млекопитающими и прочими позвоночны-
ми мы знакомились в главе 17. И действительно, геникуло-стриарная система
не критична для отличного зрения — птицы опровергают такое предположе-
ние каждый день.
У всех немлекопитающих позвоночных зрительные нервы совершают пол-
ный перекрест. Частичный перекрест и ламинарная организация НКТ у мле-
копитающих связаны с бинокулярным зрением и стереопсисом. Действитель-
но, у млекопитающих относительное количество волокон, не совершающих
перекреста, примерно пропорционально фронтальности расположения глаз —
эта взаимосвязь известна под названием закона Ньютона—Мюллера—Гуддена.
У приматов почти 50% волокон зрительного нерва не совершают перекреста,
у крыс — только около 20%, а у кролика — всего 1—2%. Это, однако, не озна-
чает, что у немлекопитающих позвоночных нет бинокулярного зрения —
у многих птиц хорошая бинокулярность зрения, как того и требует их способ
существования. Здесь частичного перекреста в оптической хиазме нет, но он
происходит в таламо-теленцефалическом пути (см. ниже). У многих рыб глаза
также расположены фронтально, что обеспечивает бинокулярное восприятие
глубины изображения, и перекрест происходит в таламо-теленцефалическом
пути.
446 Часть IV. Светочувствительность
18.8. Зрительные центры мозга
Земноводные
Основной черты физиологии сетчатки лягушек изложены в разделе 18.4. После
полного перекреста волокна зрительного нерва направляются в зрительный тек-
тум в крыше среднего мозга (рис. 18.12). Здесь волокна оканчиваются на плот-
ном дендритном ветвлении, подразделяемом на четыре различных слоя. Каж-
дый слой дает в полную проекцию сетчатки. Эти ретинотопические проекции
могут быть уподоблены четырем картам местности, лежащим одна над другой,
причем, скажем, первая из них — физическая карта, вторая — полезных ископа-
емых, третья — плотности населения, а четвертая — растительности. В случае
зрительного тектума бесхвостых земноводных самая верхняя карта иннервиро-
вана волокнами от постоянных детекторов контраста, тогда как нижнее карты
содержат окончания от детекторов выпуклости, ON/OFF-детекторов (детекто-
ров движущегося края) и OFF-детекторов. Весьма соблазнительно предполо-
жить, что эти проекции, особенно детекторов выпуклости (или детекторов «на-
секомых») напрямую связаны с двигательной эфферентной системой мозга.
Помимо ветвления окончаний ганглиозных клеток зрительный тектум бес-
хвостых земноводных содержит несколько типов нейронов. Здесь описана су-
щественная особенность — наличие нейронов «нового» и «прежнего». Нейро-
ны «нового» имеют большие рецептивные поля и реагируют на движение объ-
екта в определенном направлении, причем быстро к нему адаптируются. Если,
однако, направление движения меняется на 90е, ответ клеток снова усиливает-
ся до тех пор, пока не будет подавлен адаптацией. Нейроны «прежнего» также
имеют большие рецептивные поля, но характеризуются «внимательностью» —
они отвечают на малый стимул, перемещающийся в какой-то части поле, но
не реагируют на возникающий в другой части поля еще один стимул. В текту-
ме присутствует еще ряд клеток с рецептивными полями различных типов.
В большинстве случаев их рецептивные поля очень велики, иногда эквива-
лентны всему полю зрения, и их свойства часто трудно интерпретировать.
Поверхность тектума
Кора
зритель-
ного
тектума
Зрительные волокна
Детекторы постоянного
контраста
Детекторы выпуклости
Детекторы
движущегося края
ON- и OFF-детекторы
Рис. 18.12. Схема, показывающая организацию окончаний волокон зрительного
нерва, несущих различную информацию о визуальном окружении в зрительный
тектум лягушки. Дальнейшие объяснения в тексте.
18. Зрительные системы других позвоночных 447
Пресмыкающиеся
У пресмыкающихся, как и у земноводных, основным путем от сетчатки явля-
ется тектофугальный путь, несущий информацию в зрительный тектум, а за-
тем, через большое ядро таламуса и круглое ядро — в конечный мозг. Сущест-
вует и другие, менее существенные, пути от сетчатки. В частности, волокна
зрительного нерва направляются в претектум, а также в вентральную часть
наружного коленчатого тела (НКТв) и, что менее важно, в НКТ в таламусе.
Последний из путей, гомологичный РГС-пути млекопитающих, известен как
таламофугальный путь. Сам зрительный тектум так же сложен, как и у земно-
водных, он состоит из шести слоев клеток, хотя другие специалисты выделя-
ют 14 слоев. Тектум проецируется через круглое ядро в область конечного
мозга, называемую дорзовентрикулярный гребень (ДВГ). ДВГ — это область,
равная по нейроанатомической сложности зрительной коре млекопитающих.
Он получает афференты не только из тектума, но и из таламуса и из других об-
ластей конечного мозга. Рис. 18.13 показывает, что ДВГ развивается в лате-
ральной стенке конечного мозга и выпячивается в желудочек. В отличие от
зрительной коры млекопитающих это массивная структура, а не листовидная
структура.
ДВГ получает афференты всех основных сенсорных модальностей. Зри-
тельные афференты оканчивается в латеральной области, где ретинотопиче-
ская организация отсутствует. Нейроны в этой области имеют большие ре-
цептивные поля и чувствительны к направлению движения. Наконец, из
ДВГ волокна проецируются в дорзальную стенку конечного мозга, который
у млекопитающих преобразуется в новую кору и в зрительной утолщение в
птиц.
Птицы
Птицы относятся к животным, у которых зрение развито в наибольшей степе-
ни. Тем не менее, как и можно было ожидать, в такой большой и разнообраз-
ной группе существует большое количество различий в организации зритель-
ной системы. Одно из значительных различий существует между птицами, у ко-
торых развились фронтально расположенные глаза с большим бинокулярным
полем, такими как совы, и птицами с латеральными глазами и ограниченной
бинокулярностью, например, голуби, зяблики и т.д. В первом случае сильно
развит таламофугальный путь, во втором более существенен тектофугальный
путь. Оба, в конце концов, проецируются в конечный мозг.
Волокна зрительного нерва в таламофугальном пути совершают полный
перекрест и оканчиваются в группе ядер дорзального таламуса, которые в це-
лом называются главным зрительным ядром таламуса (ГЗТ). Эта группа ядер
считается аналогичной НКТ пресмыкающихся и млекопитающих. Из ГЗТ во-
локна проецируются в область дорзовентрикулярного гребня, называемого
также утолщением. У птиц с хорошим бинокулярным зрением волокна из пе-
редней части ГЗТ переходят в утолщение противоположной стороны, а из зад-
ней — проецируются на ипсилатеральную сторону. Утолщение, как показыва-
ет рис. 18.14, это разрастание парасагитальной области ДВГ птиц, оно состоит
из двух участков: медиальной части, сравнимой с гиппокампом млекопитаю-
448 Часть IV. Светочувствительность
Рис. 18.13. Поперечный срез че-
рез конечный мозг пресмыкаю-
щегося показывает дорзоветри-
кулярный гребень (ДВГ). Мас-
штабная линейка = 1 мм. Из
Shimizu and Karten, 1991, с раз-
решения.
Рис. 18.14. Утолщение у птиц. Поперечный
срез через конечный мозг голубя. Сравне-
ние с рис. 18.13 показывает, что утолщение
развивается как выпячивание поверхности
ДВГ. Э — эктостриатум, ВГ — вентраль-
ный гиперстриатум, Н — неостриатум.
Масштабная линейка = 1 мм. Из Shimizu
and Karten, 1991, с разрешения.
щих (утолщение гиппокампальной области (Уго)), и латеральной части, сравни-
мой с новой корой млекопитающих (утолщение гиперстриарной области (Усо)).
Задняя часть Усо сравнима с первичной зрительной корой млекопитающих,
она имеет ламинарную структуру, которая обычно состоит из четырех слоев
клеток. Существуют данные, что клетки здесь ретинотопически организова-
ны, хотя карты у разных видов птиц варьируют. У сов здесь много нейронов,
контролируемых бинокулярно, у голубя это заметно намного меньше — лишь
несколько нейронов реагируют на ипсилатеральные стимулы.
Волокна зрительного нерва в тектофугальном пути оканчиваются в контр-
латеральном зрительном тектуме. Поверхностные слои тектума ретинотопиче-
ски организованы, а клетки имеют сравнительно небольшие РП (мене 4е дуги)
и реагируют на движение и направление движения. Клетки более глубоких
пластин имеют большие РП, но не так строго селективны к движению. Клет-
ки отсюда посылают волокна в круглое ядро. Последнее подразделяется на ци-
тоархитектонические области по гистохимическому окрашиванию ацетилхо-
линэстеразы. Многие нейроны здесь имеют чрезвычайно большие РП (часто
больше, чем 100° дуги) и чувствительны к движению. Обнаружены также ней-
роны, которые реагируют на расширение или сокращение зрительного образа,
обеспечивая таким образом информацию о приближении к объекту. Вывод из
круглого ядра направляется в область центральной массы ДВГ (эктостриатум).
Из эктостриатума проекции направляются в другие части ДВГ (рис. 18.15). Эта
серия последовательных проекций похожа на проекции в стриарной и экстра-
стриарной коре млекопитающих.
Таламофугальный и тектофугальный пути взаимодействуют во многих ме-
стах, особенно в утолщении и эктостриатуме. Отсюда имеется мощная обрат-
18. Зрительные системы других позвоночных 449
Рис. 18.15. Поперечный срез перед-
него мозга птицы, показывающий
нервные связи в тектофугальном
пути. А — архистриатум, Э — экто-
стриатум, ПЭ — периэктостриар-
ная полоса, ПВН — промежуточ-
ный вентральный неостриатум,
ДП — добавочный палеостриатум
(эквивалент хвостатого ядра и скор-
лупы [putamen] млекопитающих),
ПП — примитивный палеостриа-
тум (эквивалент бледного шара
млекопитающих), КЯ — круглое
ядро (n. rotundus). Из Shimizu and
Karten, 1991, с разрешения.
ная связь в тектумом, связь с гипоталамусом и обратно с ГЗТ и, что важно,
с сетчаткой. Центральный анализ зрительной информации в конечном мозге,
таламусе, и среднем мозге птиц не менее сложен, хотя и менее изучен, чем
у млекопитающих.
18.9. Заключение
Данная глава поведала кое-что о чрезвычайном разнообразии зрительных си-
стем, развившемся у позвоночных, но это лишь верхушка колоссального айс-
берга. Это должно послужить хорошим лекарством от антропоцентризма. Глаз
человеческий и вся его зрительная система, несмотря на ее понятную изучен-
ность, далеко не единственная модель среди типа хордовых, даже если оста-
вить в стороне остальных животных. Хотя многое о зрительной системе позво-
ночных уже известно, многое остается неизученным. Это становится особен-
но очевидно в последнем разделе данной главы, когда выясняется, как мало
мы знаем об центральном анализе зрительной информации у наиболее зависи-
мых от зрения животных — птиц. Как всегда в зоологии, нам приходится рас-
сматривать самих себя лишь в качестве одного вида из многих; и мы ощущаем
удивление от сходства разных форм жизни. Как всегда мы возвращаемся к зна-
менитому вопросу Нагеля: что значит быть, как летучая мышь, что значит
быть, как птица?
МАТЕРИАЛЫ К ЧАСТИ IV
Для самоподготовки
Следующие вопросы подобраны, чтобы помочь оценить собственное понима-
ние материала Части IV и обратить ваше внимание на значительные аспекты
данного предмета.
Глава 14
14.1. Каковы длины волн электромагнитного спектра, воздействующие на зритель-
ную систему человека?
14.2. Опишите два основных типа фоторецепторных клеток. С какими двумя глав-
ными группами животных эти типы связаны? Есть ли исключения из этого
правила? Если да, назовите их.
14.3. Назовите два основных типа многоклеточных глаз, развившихся в царстве
животных. У каких типов они достигают наивысшего расцвета?
14.4. Используя конкретные примеры, объясните, как могли сформироваться вы-
сокоразвитые камерные глаза моллюсков и позвоночных.
14.5. Что такое клетки ретинулы и что такое омматидий?
14.6. Нарисуйте схему аппозиции и суперпозиции омматидиев. Какой тип встреча-
ется у дневных, а какой — у ночных насекомых?
14.7. В чем разница между рабдомером и рабдомом?
14.8. В чем разница между суперпозицией и нейральной суперпозицией в сложном
глазу? Какими преимуществами обладает последний вариант?
14.9. Помимо аппозиционного, суперпозиционого и нейрального суперпозицион-
ного сложного глаза, какие еще типы сложного глаза развились у членис-
тоногих?
14.10. Опишите несколько примеров сканирующих глаз.
14.11. Опишите глазок хламидомонады. Как фоторецепция связана с локомоцией?
Есть ли здесь сходство с бактериальным хемотаксисом (сравните с гл. 10)?
14.12. Опишите глаз кораблика Nautilus pompilius. Какой тип фоторецепторных кле-
ток обнаружен в его сетчатке?
14.13. Где локализованы глаза гребешка Pecten?
14.14. На какой из двух сетчаток гребешка Pecten формируется изображение? В какой
из сетчаток обнаружены фоторецепторы ресничного типа? Отличается ли
биофизика таких фоторецепторов от палочек и колбочек позвоночных?
14.15. К какому из трех подклассов головоногих относятся кальмары и осьминоги?
14.16. Опишите сходства и различия между глазами белемноидов и позвоночных.
14.17. Где располагается глубокая сетчатка осьминога Octopus?
14.18. К какому классу относится мечехвост Limulus?
14.19. Каково физиологическое значение латерального сплетения за омматидиями
сложного глаза мечехвоста Limulus?
14.20. Какую роль играют эксцентрические клетки в сетчатке Limulus?
14.21. Каково биологическое значение латерального торможения в сложном глазу
Limulus?
14.22. Какая структура действует как оптические волокна или световод в сложном
глазу двукрылых?
14.23. Каким образом глаз мухи совмещает разрешающую способность и чувстви-
тельность?
14.24. Опишите путь волокон зрительного нерва от омматидиев к дольке.
Материалы к части IV 451
Глава 15
15.1. Что означает термин «вспомогательные органы» глаза?
15.2. Назовите три слоя, образующие стенку глазного яблока.
15.3. Перечислите части увеального тракта. Каково происхождение этого названия?
15.4. Что такое центральная область (или желтое пятно)?
15.5. Почему оптический диск также называется «слепым пятном»?
15.6. Что вызывает глаукому?
15.7. Какие черепно-мозговые нервы иннервируют внешние мышцы глаза?
15.8. Что такое блок и какую роль он играет в мышечном контроле движения глаза?
15.9. Каковы функции век?
15.10. Используя рис. 15.4, опишите движение слезной жидкости по глазу.
15.11. Что предупреждает высыхание слезной жидкости?
15.12. Объясните, почему у всех позвоночных инвертированная сетчатка.
15.13. Какую роль играет глазной пузырь в формировании хрусталика?
15.14. Каким образом глазной пузырь облитерирует внутри хрусталика?
15.15. Какие дефекты приводят к формированию афакии и колобомы?
15.16. Что такое канал Клоке, и как он формируется?
15.17. Что происходит с глазным пузырем? Имеет ли он значение для некоторых
офтальмологических патологий?
15.18. Какая последовательность клеточных движений лежит в основе формирова-
ния сетчатки? Какую роль играют мюллеровы клетки?
15.19. Что такое лимб?
15.20. Нарисуйте схему поперечного среза роговицы и отметьте три основных слоя
и две мембраны.
15.21. Какие две функции выполняет коллаген стромы роговицы?
15.22. Какие молекулы, кроме коллагена, присутствуют в строме? Нарисуйте струк-
туру типичной глюкозаминогликановой (ГАГ) цепочки.
15.23. Почему гигроскопический характер ГАГ так важен дня прозрачности роговицы?
15.24. Как организация коллагена склеры отличается от роговицы?
15.25. Содержат ли волокна хрусталика митохондрии и другие внутриклеточные
органоиды? Если нет, то почему?
15.26. Какие белки представляют собой основные компоненты волокон хрусталика?
15.27. Определите, что такое близорукость (миопия), дальнозоркость (гиперметро-
пия), старческая дальнозоркость (пресбиопия), астигматизм и катаракта.
15.28. Опишите некоторые биохимические процессы, ответственные за катаракту.
15.29. Чем мышцы — сфинктер и дилятатор радужки отличаются от прочих мышц?
15.30. Опишите гистологические структуры капиллярного слоя сосудистой оболоч-
ки, и как связана эта структура с ее функцией.
15.31. Какие две функции выполняет ресничное тело?
15.32. Где и как секретируется водянистая влага? Приведите доказательства, которые
* показывают, что часть этой секреции является активным процессом.
15.33. По каким двум путям абсорбируется водянистая влага? Когда глаз фокусиру-
ется на объекте, по какому маршруту направляется 25% водянистой влаги?
15.34. В чем различие глаукомы при открытом и закрытом фильтрующем углу?
15.35. Какую роль играет склеральная шпора в аккомодации хрусталика? Какую роль
играет эластичность капсулы хрусталика?
15.36. Как мышцы — сфинктер и дилятатор радужки контролируют количество све-
та, достигающего сетчатки? Объясните, как вариации размера зрачка влияют
на глубину фокуса.
15.37. Обсудите роль гиалоцитов в стекловидном теле.
15.38. В чем различие между вестибуло-окуломоторными и оптокинетическими ре-
флексами? Какие из этих рефлексов не присутствует в темноте? Почему?
15.39. Какие движения глаз управляются диспаратностью изображений на сетчатках?
15.40. Объясните, что означает термин «эфферентная копия», приведите простой
пример, подтверждающий эту идею.
452 Часть IV. Светочувствительность
Глава 16
16.1. К чему более прочно присоединен ПЭС — к сетчатке или к сосудистой обо-
лочке?
16.2. В чем различие между меланосомами и фагосомами?
16.3. Перечислите четыре существенные функции ПЭС.
16.4. Что такое фовеация?
16.5. Перечислите девять слоев сетчатки.
16.6. Нарисуйте схему палочки и обозначьте ее части. Как от нее отличаются кол-
бочки?
16.7. Что такое фовеола?
16.8. Объясните, как наружный сегмент палочки действует в качестве световода.
Сравните с рабдомом насекомых.
16.9. Как ретиналь соединяется с опсином?
16.10. Какие внутримолекулярные изменения происходят, когда фотон соответству-
ющей длины волны взаимодействует с родопсином?
16.11. Как отсоединение ретиналя от опсина приводит к преобразованию цГМФ
в 5 -ГМФ?
16.12. Что обусловливает темновой ток в палочках и колбочках?
16.13. Деполяризуются или гиперполяризуются палочки и колбочки в ответ на дейст-
вие света?
16.14. Как рекаверин вызывает сенсорную адаптацию фоторецепторов позвоноч-
ных?
16.15. Что действует на карбоксиконцевую цепочку первым — родопсинкиназа или
арестин?
16.16. Что происходит с /ирднс-ретиналем, выделившимся из наружных сегментов
палочек?
16.17. Какую роль играют капилляры сосудистой оболочки в регенерации 11-цис-ре-
тиналя?
16.18. Воздействие света ингибирует или запускает выделение медиатора из фоторе-
цепторов?
16.19. Различия поглощения света пигментами палочек и колбочек обусловлено хро-
мофором или опсином?
16.20. Какой из зрительных пигментов сформировался в эволюции последним?
16.21. Что означает «сохранение знака ответа» или «изменение знака ответа»? Элект-
рический синапс является сохраняющим или изменяющим знак?
16.22. Образуют ли диффузные биполярные клетки синапсы с более чем одной нож-
кой колбочек?
16.23. Нарисуйте схему синаптического комплекса, образуемого горизонтальными и
биполярными клетками со сферулами палочек.
16.24. Сохраняют или изменяют знак сигнала биполярные клетки, связанные с па-
лочками?
16.25. Объясните молекулярный механизм синапса, изменяющего знак сигнала. Есть
ли здесь сходство с ответом наружных сегментов на свет?
16.26. Объясните, что означает термин «рецептивное поле» биполярной клетки. Ка-
ковы физиологические характеристики этих полей? Какие клетки сетчатки в
основном ответственны за эти характеристики?
16.27. Есть ли общие черты между рецептивными полями биполярных клеток и во-
локнами зрительного нерва мечехвоста Limulus?
16.28. Перечислите некоторые функции M^^ivpuBbix клеток.
16.29. Интерплексиформные клетки передают сигналы от внутреннего синаптиче-
ского слоя к наружному или наоборот?
16.30. Как классифицируются амакриновые клетки?
16.31. Опишите три основных типа ганглиозных клеток.
16.32. Опишите типичную организацию рецептивных полей ганглиозных клеток
приматов.
Материалы к части IV 453
16.33. Какой тип ганглиозных клеток чувствителен к движению, а какой — к цвету?
16.34. На какой стимул ганглиозные клетки дают более выраженную реакцию — на
маленькое сфокусированное пятно света в рецептивном поле или на распрост-
раненное диффузное освещение?
16.35. Нарисуйте схему возможных нервных контактов, формирующих рецептивное
поле ганглиозной клетки.
16.36. Каким образом в результате закрывания щелевых контактов между палочками
и колбочками повышается чувствительность сетчатки?
16.37. Объясните, что такое теория оппонентных процессов.
16.38. Объясните, почему в некоторых случаях выходной сигнал ганглиозных клеток
с цветовым оппонентным рецептивным полем нуждается в устранении неодно-
значности.
16.39. Покажите, как высокоуровневые свойства рецептивных полей сетчатки могут
быть выведены с помощью основных молекулярных и биофизических прин-
ципов.
Глава 17
17.1. Назовите три зрительных пути у млекопитающих.
17.2. Существует ли эволюционная причина доминирования у млекопитающих
РГС-пути? Если да, дайте краткое описание.
17.3. Опишите РТ-путь. Какова форма перекреста у приматов?
17.4. Опишите физиологическую организацию верхних бугров четверохолмия.
В контроле какого типа движений глаз они участвуют?
17.5. Почему наблюдается заглубление оптического диска при глаукоме?
17.6. Волокна зрительного нерва из латеральной части сетчатки:
— совершают полный перекрест,
— совершают частичный перекрест
— или не совершают перекреста в зрительной хиазме?
17.7. Сколько слоев клеток в наружном коленчатом теле приматов? Идет ли путь из
каждой сетчатки отдельно друг от друга?
17.8. Какие слои наружного коленчатого тела получают афференты от парвоцеллю-
лярных волокон?
17.9. Какие слои наружного коленчатого тела содержат клетки, отвечающие на дви-
жение в их РП, но не реагируют на цвет?
17.10. Какие слои наружного коленчатого тела получают афференты от W-волокон?
17.11. Где располагается первичная зрительная кора у человека? А у обезьян?
17.12. Что такое полоска Дженнари и где она расположена?
17.13. В каких слоях первичной зрительной коры оканчиваеются парвоцеллюлярные
волокна из наружного коленчатого тела?
17.14. Опишите экспериментальную технику, используемую для исследований
рецептивных полей клеток зрительной коры.
17.15. Какого типа стимулы запускают ответы в клетках первичной зрительной
коры?
17.16. Покажите, как свойства РП клеток селективных к ориентации могут формиро-
ваться на основе комбинации концентрических полей.
17.17. Определите разницу между «простым» и «комплексным» РП.
17.18. Опишите свойства концевых РП. В чем состоит возможное биологическое
значение такого типа поля?
17.19. Каким образом меняется предпочтительная ориентация РП клеток стриарной
коры по мере тангенциального продвижения электрода через ткань?
17.20. Что такое дезоксиглюкозная методика и как она применяется в исследованиях
гистологии зрительной коры?
17.21. В каких гистологических структурах концентрируются цветочувствительные
клетки?
454 Часть IV. Светочувствительность
17.22. Набросайте возможную принципиальную схему паттерна колонок доминиро-
вания глаз в первичной зрительной коре.
17.23. Определите, что такое гиперколонка и сложное рецептивное поле.
17.24. Передают ли детекторы ориентации в центры более высоких уровней двусмыс-
ленный сигнал? Если да, как эта двусмысленность может быть устранена?
17.25. Что такое чувствительный период? Опишите несколько экспериментов, кото-
рые показывают, что ранний зрительный опыт играет важную роль в подст-
ройке «принципиальной схемы» зрительной системы.
17.26. Что такое и где расположено ядро подушки? Какую роль оно играет в перера-
ботке зрительной информации?
17.27. Что такое «трехпоточная» система обработки зрительной информации? Какую
информацию несет каждый из трех потоков?
17.28. Какой тип дифференцировки выявляет цитохромоксидаза в зрительной обла-
сти 2 (поле Бродмана 18)? Как эта организация взаимосвязана с капельно-
межкапельной дифференциацией в первичной зрительной коре?
17.29. Почему распознавание лиц важно для социальных приматов?
17.30. В какой области мозга макака располагаются клетки распознавания лиц?
17.31. Обсудите концепцию «клеток распознавания лица бабушки».
17.32. Что такое «паттерн образа»? Как клетки, ответственные за паттерн образа
1 группируются в нижневисочной коре?
17.33. Где локализована область распознавания лиц в мозге человека?
17.34. Что такое прозопагнозия? Приведите несколько примеров.
Глава 18
18.1. В чем состоит химическое различие между родопсином и порфиропсином?
18.2. У какого родопсина и порфиропсина Хтах смещена в красную сторону спек-
тра?
18.3. У каких рыб (пресноводных и морских) чаще обнаруживаеются порфиропси-
ны?
18.4. У каких классов позвоночных обнаружены фоторецепторы, содержащие мас-
ляные капли? Какова функция масляных капель?
18.5. Какие еще диоптрические органоиды часто присутствуют в фоторецепторах
позвоночных? В какой части клетки они развиваются?
18.6. Какова функция двойных колбочек и в каких группах позвоночных они при-
сутствуют?
18.7. У каких групп позвоночных наиболее известны двойные палочки?
18.8. Сокращается или расслабляется миоид колбочки при освещении? А палочки?
18.9. Совершают ли пигментные гранулы ПЭС фотомеханические движения? Если
да, в каком направлении они двигаются при освещении? Насколько это срав-
нимо с миграцией пигмента в глазах белемноидов (см. гл. 14)?
18.10. В каких слоях глаза у некоторых животных развиваются зеркальца?
18.11. Опишите три типа зеркалец сосудистой оболочки.
18.12. Что такое «окклюдирующие зеркальца», у каких животных они присутствуют и
как действуют?
18.13. Что такое «многоуровневая сетчатка»? У каких животных она обнаружена и
каков ее смысл?
18.14. Опишите физиологические характеристики пяти основных типов рецептив-
ных полей сетчатки лягушки. Как эти характеристики отражают способ суще-
ствования лягушки?
18.15. Что такое центральная область?
18.16. В чем разница между выпукло-углубленной и вогнуто-углубленной фовеа?
18.17. Обсудите функциональные преимущества выпукло-углубленной фовеа.
18.18. Опишите типы областей в сетчатке птиц и как они соотносятся (насколько это
возможно) со способом существования своих владельцев.
Материалы к части IV 455
18.19. Что такое зрительная полоска и у каких млекопитающих она обнаружена?
18.20. Что такое гребень и в чем состоит его предполагаемая функция?
18.21. Опишите тубулярный глаз глубоководных рыб. Каково значение двух его сет-
чаток?
18.22. Что такое «очки», как они формируются и у каких позвоночных обнаружены?
18.23. Что такое соотношение Матиссена?
18.24. Как хрусталики глаз животных адаптированы к соответствию с соотношением
Матиссена?
18.25. Какие две фоторецепторные структуры развиваются из задней части крыши
промежуточного мозга?
18.26. Какова основная физиологическая роль медианных глаз позвоночных?
18.27. Какой гормон синтезируется эпифизом?
18.28. Что такое закон Ньютона—Мюллера—Гуддена?
18.29. Опишите физиологическую организацию зрительного тектума лягушки. Что
такое нейроны «прежнего» и «нового»?
18.30. Опишите основные зрительные пути пресмыкающихся.
18.31. Что такое дорзовентрикулярное гребень и где он располагается?
18.32. У каких птиц сильно развит таламофугальный путь?
18.33. Какая группа ядер в зрительном пути птиц аналогичен НКТ млекопитающих?
18.34. Какой перекрест в таламофугальном пути — полный, неполный, вообще нет
перекреста?
18.35. Что такое «утолщение» у птиц и как оно соотносится с ДВГ пресмыкающихся?
18.36. Какая часть утолщения считается аналогичной первичной зрительной коре
млекопитающих? Сколько слоев клеток в этой части утолщения?
18.37. В какой части мозга птиц оканчиваются волокна тектофугального пути? Что
такое круглое ядро, где оно располагается и каковы физиологические характе-
ристики его нейронов?
18.38. В чем схожи и в чем различны зрительные системы птиц и млекопитающих?
Замечания и библиография
Введение
Дэвид Юм (1711—1776), родившийся в Эдинбурге, считается величайшим фи-
лософом первой половины восемнадцатого века. Черновики «Диалогов о есте-
ственной религии» существовали уже в 1751 году, но не были опубликованы до
1779 года, спустя три года после смерти автора. Следы борьбы Чарльза Дарви-
на с тем, что структура глаза своим существованием поддерживал идею про-
виденциального создателя, обнаруживаются в его письме к Эйса Грею (Asa
Gray) в феврале 1860 года: «Глаз для меня — как холодный душ, но, когда я
думаю о массе известных переходных форм, мой разум говорит мне, что этот
холодный душ я должен перетерпеть», а позже, в апреле того же года, он снова
пишет Грею, но уже упоминая свою слабость в прошедшем времени:«Я помню
время, когда мысль о глазе вызывала у меня мороз по коже, но теперь эту ста-
дию недуга я преодолел» (Darwin, 1860).
Детали структуры и генеалогического родства опсинов из меланоцитов ко-
жи шпорцевой лягушки Xenopus изложены у Provencio et al. (1998). Эта группа
показала, что этот опсин ближе к опсинам беспозвоночных, чем позвоночных.
Они предположили, что этот опсин близок к пигментам, ответственным за
циркадные ритмы и реакцию на фотопериод у многих немлекопитающих по-
звоночных. Ценный вклад в понимание принципов оптики и эволюции глаза,
456 Часть IV. Светочувствительность
формирующего изображение, сделан Голдсмитом (Goldsmith, 1990). Фернальд
(Fernaid, 1997) обсуждает эволюцию глаза животных на основе молекулярной
генетики (см. также Информацию 14.1), а Волькен (Wolken, 1995) сделал вну-
шительный обзор глаз и других детекторов света в биологическом мире. Мате-
риалы по геометрической оптике можно найти в учебниках классической фи-
зики.
Darwin, С. R., 1860, The Correspondence of Charles Darwin, Cambridge: Cambridge
University Press, Vol. 8.
Fernaid, R. D., 1997, «The evolution of eyes», Brain, Behaviour and Evolution, 50,253—259.
Goldsmith, T. H., 1990, «Optimisation, constraint, and history in the evolution of eyes»,
The Quarterly Review of Biology, 65, 281—322.
Provencio, I., et al., 1998, «Melanopsin: An opsin in melanophores, brain and eye».
Proceedings of the National Academy of Sciences USA, 95, 340—345.
Wolken, J. J., 1995, Light Detectors, Photoreceptors, and Imaging Systems in Nature, Oxford:
Oxford University Press.
Глава 14: Зрение беспозвоночных
Предмет этой области знания — зрения беспозвоночных, конечно, огромен, и
в данной главе приведен лишь краткий очерк. Исчерпывающее изложение
можно найти у Олви (Alvi, 1984), а Дьюк-Элдер (Duke-Elder, 1958) сделал до-
ступный обзор. Множество исчерпывающих глав можно найти в изданиях, от-
редактированных Аутрумом (Autrum, 1979, 1981а, Ь) и Кронли-Диллон и Гре-
гори (Cronly-Dillon and Gregory, 1991). Хороший обзор глазков простейших
сделан Фостером и Смайсом (Foster and Smyth, 1980), а Гарц, Ноннегассер и
Хегеманн (Harz, Nonnegasser and Hegemann, 1992) обсудили детали биофизики
и физиологии глазков хламидомонады. Данные по сложным глазам можно
найти в упомянутых выше источниках, а также в работах Wigglesworth (1972) и
Milne and Milne (1959), а основополагающая работа Экснера в этой области до
сих пор остается кладезем информации. Это и сейчас серьезный источник, не-
смотря на столетие, прошедшее с первой публикации. Лэнд и Фернальд (Land
and Fernaid, 1992) обсуждают эволюционные вопросы, а Нильссон (Nilsson,
1988) описывает оптику сложного глаза краба-плавунца Portunuspuber. Конвей
Моррис (Convey Morris, 1998) дал ценное обсуждение ископаемых форм дав-
ностью в 525 миллионов лет и эволюционных условий или «морфопрост-
ранств», обусловивших их дальнейшую эволюцию.
Alvi, М. A., ed., 1984, Photoreception and Vision in Invertebrates, New York: Plenum.
Autrum, H., ed., 1979, «Comparative physiology and evolution of vision in invertebrates.
A: invertebrate photoreceptors», in Handbook of Sensory Physiology, H. Autrum et al.,
eds, Berlin: Springer-Verlag, vol. VII/6A.
Autrum, H., ed., 1981a, «Comparative physiology and evolution of vision in invertebrates,
B: Invertebrate visual centers and behaviour», in Handbook of Sensory Physiology,
H. Autrum etal., eds, Berlin: Springer-Veriag, vol. V1I/6B.
Autrum, H., ed., 1981b, «Comparative physiology and evolution of vision in invertebrates»,
C: Invertebrate visual centres and behaviour II», in Handbook of Sensory Physiology,
H. Autrum etal., eds, Berlin: Springer-Verlag, Vol. VII/6C.
Материалы к части IV 457
Cronly-Dillon, J. R. and R. L. Gregory, eds, 1991, Vision and Visual Dysfunction, Vol. 2,
Evolution of the Eye and Visual System, Houndmills: Macmillan Press.
Duke-Elder, S., 1958, System of Ophthalmology. Vol. I: The Eye in Evolution, London:
Kimpton.
Exner, S., 1891 (edited and translated R.C. Hardie, 1988), The Physiology of the Compound
Eyes of Insects and Crustacea, Berlin: Springer-Verlag.
Land, M. F. and R. D. Femald, 1992, «The evolution of eyes». Annual Review of Neurosci-
ences, 15, 1—30.
Milne, L. J. and M. Milne, 1959, «Photosensitivity in invertebrates», in H. W. Magoun, ed.,
Neurophysiology, Vol. 1 in J. Field, ed. Handbook of Physiology, Washington, D. C.:
American Physiological Society.
Morris, S. Conway, 1998, The Crucible of Creation: The Burgess shale and the rise of the ani-
mals, Oxford: Oxford University Press.
Wigglesworth, V. B., 1972, The Principles of Insect Physiology, London: Chapman and Hall.
Организация глаз беспозвоночных
Eakin, R. M., 1968, «Evolution of photoreceptors», inT. Dobzhansky, Evolutionary Biology,
M. K. Hecht and W. C. Steere, eds. Vol. 2, New York: Appleton-Century-Crofts.
Land, M. F., 1991, «Optics of the eyes of the animal kingdom», in J. R. Cronly-Dillon and
R. L. Gregory, eds, ibid.
Сложный глаз
Land, M. F., 1984, «The resolving power of diurnal superposition eyes measured with an
ophthalmoscope», Journal of Comparative Physiology, A., 154, 515—533.
Land, M. F. and R. D. Femald, 1992, «The evolution of eyes», Annual Review of Neurosci-
ences, 15, 1—30.
Nilsson, D. E., 1988, «А new type of imaging optics in compound eyes», Nature, 332,
76-78.
Сканирующий глаз
Gregory, R. L., 1991, «Origins of eyes — speculations on scanning eyes», in J. R. Cronly-
Dillon and R. L. Gregory, ibid.
Land, M. F., 1969, «Movements of the retinae in jumping spiders (Salticidae: Dendryphan-
tidae) in response to visual stimuli», Journal of Experimental Biology, 51,471—493.
Land, M. F. and R. D. Fernaid, 1992, «The evolution of eyes», Annual Review of Neurosci-
ences, 15, 1—30.
Глазки простейших
Diehn, В., 1979, «Photic responses and sensory transduction in protists», in H. Autrum, ed.,
1979, ibid.
Dodge, J. D., 1991, «Photosensory systems in eukaryotic algae», in J. R. Cronly-Dillon and
R. L. Gregory, eds, ibid. A comprehensive account of algal eyespots covering a large
number of different types.
Foster, K. W. and R. D. Smyth, 1980, «Light antennae in phototactic algae», Microbiological
Reviews, 44, 572—630.
Harz, H., C. Nonnegasser and P. Hegemann, 1992, «The photoreceptor current of the green
alga Chlamydomonas», Philosophical Transactions of the Royal Society B, 338, 39—52.
458 Часть IV. Светочувствительность
Диафрагмальный глаз Nautilus pompilius
Messenger, J. В., 1991, «Photoreception and vision in molluscs», in J. R. Cronly-Dillon and
R. L. Gregory, eds, ibid. An overview of the great variety of eyes evolved by the phylum
Mollusca, the most varied, as he says, of all animal phyla.
Young, J. Z., 1965, «The central nervous system of Nautilus», Philosophical Transactions of
the Royal Society, B, 249, 1—25.
Зеркальный глаз гребешка Pecten
Borradaile, L. A., F. A. Potts, L. E. S. Eastham and J. T. Saunders, 1951, The Invertebrata,
Cambridge: Cambridge University Press.
Dakin, W. J., 1928, «The eyes of Pecten, Spondylus, Amussium and allied lamellibranchs,
with a short discussion on their evolution», Proceedings of the Royal Society, B, 103,
355-365.
Land, M. F., 1965, «Image formation by a concave reflector in the eye of the scallop Pecten
maximus», Journal of Physiology, 179, 138—153.
Messenger, J. B., 1991, ibid.
Камерный глаз осьминога Octopus
Messenger, J. В., 1991, ibid.
Saibil, H. and E. Hewat, 1987, «Ordered transmembrane and extracellular structure in squid
photoreceptor microvilli», Journal of Cell Biology, 105, 19—28.
Young, J. Z., 1962a, «The retina of cephalopods and its degeneration after optic nerve sec-
tion», Philosophical Transactions of Royal Society, B, 245, 1—18.
Young, J. Z., 1962b, «The optic lobes of Octopus vulgaris», Philosophical Transactions of the
Royal Society, 245, 19—58.
Сложные глаза мечехвоста Limulus
Hartline, H. K., 1959, «Receptor mechanisms and the integration of sensory information in
the eye», in Biophysical Science — A Study Program, J. L. Oncley, ed., New York: Wiley.
Laughlin, S., 1981, «Neural principles in the peripheral visual systems of invertebrates», in
H. Autrum, ed., 1981, ibid.
Meinertzhagen, I. A., 1991, «Evolution of the cellular organisation of the arthropod com-
pound eye and optic lobe», in J. R. Cronly-Dillon and R. L. Gregory, eds, ibid.
Сложный глаз двукрылых
Следующие четыре публикации дают всесторонние и продвинутые обзоры по
глазам и зрительной системе двукрылых. Не для робких читателей.
Horridge, G. А., 1991, «Evolution of visual processing», in J. R. Cronly-Dillon and
R. L. Gregory, eds, ibid.
Osorio, D., 1991, «Patterns of function and evolution in the arthropod optic lobe», in
J. R. Cronly-Dillon and R. L. Gregory, eds, ibid.
Strausfield, N. J., 1976, An Atlas of an Insect Brain, Berlin: Springer.
Strausfield, N. J. and D. R. Nassel, 1981, «Neuroarchitecture of brain regions that subserve
the compound eye of Crustacea and insects», in H. Autrum, ed., ibid.
Материалы к части IV 459
Глава 15: Глаз человека
При рассмотрении происхождения глаз позвоночных интересно начать с наи-
более примитивных хордовых. Ланцетник Amphioxus — маленькое (менее 5 см)
суженное на обоих концах животное, фильтрующее пищу и живущее наполо-
вину закопавшись в дно морского мелководья. Ему присущи многие черты,
характерные для предполагаемого предшественника хордовых. В частности,
здесь присутствуют одноклеточные ресничные глазки, обращенные в цент-
ральный канал нервного тяжа. Кроме того, уже давно предполагали, что ин-
фундибулярный орган — группа длинных клеток, чьи реснички направлены в
центральный канал передней части нервного тяжа, обладает фоторецепторной
функцией. Преимущественно малоподвижный образ жизни ланцетника позво-
ляет предположить, что он представляет собой регрессивную форму, происхо-
дящую от свободноплавающего предшественника. Детальные описания можно
найти у Юнга (Young, 1962). Исчерпывающее изложение эмбриологии глаза че-
ловека дано сэром Стюартом Дьюк-Элдером и Куком (Sir Stewart Duke-Elder
and C. Cook, 1963) в третьем томе их авторитетного издания «Системы офталь-
мологии». Данные по анатомии глаза приведены во множестве учебников по
офтальмологии и оптометрии — Bron, Tripathi and Tripathi (1997), Hart (1992),
Davson (1984), Duke-Elder and Wybar (1961), Newell (1966) и т. д. Хардинг
(Harding, 1984) дает свежую информацию по биохимии глаза, а исчерпываю-
щие данные по тонкой структуре глаза можно найти у Хогана с соавторами
(Hogan, Alvarado and Weddell, 1971). Хардинг (Harding, 1984) дает исчерпываю-
щее описание развития хрусталика и метаболизма в связи с катарактой. Де
Йонг (de Jong, 1982) обсуждает эволюцию кристаллинов хрусталика. Катаракта
и ее развитие рассмотрена в монографическом труде Хардинга (Harding, 1991),
а Джаффе и Горовиц (Jaffe and Horowitz, 1992) представили превосходные цвет-
ные иллюстрации по гистологии и анатомии катаракты хрусталика. Плодо-
творный труд Арбиба (Arbib, 1989), помимо многого другого, дает полезный
инженерный подход к системе обратных связей (в т.ч. обсуждение концепции
эфферентных копий), контролирующих саккадические движения глаз.
Abdel-Latif, А. А., 1997, «Iris-ciliary body, aqueous humour and trabecular meshwork», in
Biochemistry of the Eye, J. J. Harding, ed., London: Chapman and Hall.
Arbib, M. A., 1989, The Metaphorical Brain, 2, New York: Wiley.
Bron, A. J., R. C. Tripathi and B. J. Tripathi, 1997, Wolff's Anatomy of the Eye and Orbit
(8th edn), London: Chapman and Hall.
Cole, D. F., 1984, «Ocular fluids», in Davson, H., ed., ibid.
Davson, H., 1963, The Physiology of the Eye (2nd edn), London: Churchill.
Davson. H., ed., 1984, The Eye, Orlando, Fa: Academic Press, Vol. la: Vegetative Physiology
and Biochemistry.
de Jong, W. W., 1982, «Eye lens proteins and vertebrate phylogeny», in Macromolecular
Sequences in Systematic and Evolutionary Biology, M. Goldman, ed., New York: Plenum.
Duke-Elder, S. and К. C. Wybar, 1961, System of Ophthalmology, Vol. 2; The Anatomy of the
Visual System, London: Henry Kimpton.
Duke-Elder, S. and C. Cook, 1963, System of Ophthalmology, Vol. S, Part 1: Embryology,
London: Henry Kimpton.
Harding, J. J., 1984, «The lens: development, proteins, metabolism and cataract», in The
Eye, H. Davson, ed., Orlando: Academic Press, vol. lb.
460 Часть IV. Светочувствительность
Harding, J. J., 1991, Cataract: Biochemistry, epidemiology and pharmacology, London: Chap-
man and Hall.
Harding, J. J., ed., 1997, Biochemistry of the Eye, London: Chapman & Hall.
Hart, W. M., 1992, Adler's Physiology of the Eye (9th edn), St Louis: Mosby.
Hogan, M. J., J. A. Alvarado and J. E. Weddell, 1971, Histology of the Human Eye, Philadel-
phia: Saunders.
Hyman, L., 1987, «Epidemiology of eye diseases in the elderly», Eye, 1, 330—341.
Jaffe, N. S. and J. Horowitz, 1992, Lens and Cataract, New York: Gower Medical Publishing.
Newell, F., 1996, Ophthalmology: Principles and concepts (8th edn), St Louis: Mosby.
Tiffany, J. M., 1997, «Tears and conjunctiva», in J. J. Harding, ed., ibid.
Young, J. Z., 1962, Life of Vertebrates (2nd edn), Oxford: Clarendon Press.
Глава 16: Сетчатка
Сетчатка восхищала нейрологов, по крайней мере, со времен Рамон-и-Кахала,
который был пионером в современном понимании структуры глаза, изложен-
ного в серии блестящих публикаций (см. Кахаль (Cajal), 1909, 1911). Совре-
менные данные можно найти у Даулинга (Dowling, 1987), а детальное описа-
ние гистологии сетчатки — у Поляка (Polyak, 1941) и Родика (Rodieck, 1973,
1988). Труд Гордона Уоллса (Gordon Walls, 1963) по сравнительной анатомии,
хотя и опубликован более полувека назад, продолжает оставаться незамени-
мым введением в эту проблему, а многотомная серия под редакцией Кронли-
Диллона предоставляет более новую информацию. Фоторецепторные клетки
являются объектом интенсивных исследований на биофизическом, биохими-
ческом и молекулярно-биологическом уровнях. Хардинг (1997) дает деталь-
ный обзор, а работы Натанса (Nathans) с сотрудниками касаются деталей
структуры и эволюции зрительных пигментов. Йокояма (Yokoyama, 1995) об-
суждает эволюцию зрительных пигментов у позвоночных и представляет фи-
логенетическое древо опсинов 28 различных видов, основываясь на их амино-
кислотных последовательноситях. Оригинальная работа по зрительным ре-
цептивным полям выполнена на сетчатке лягушки Хартлайном (Hartline,
1938), а на сетчатке млекопитающих — Куффлером (1953); сетчатка кролика
описана Левиком (Levick, 1967). Принципиальные схемы сетчатки остаются
противоречивыми, однако, и Шиллер (Schiller, 1992) и Наканиши (Nakanishi,
1995) представили полезные суммирующие обзоры. Брокерхоф, Даулинг и
Харли (Brockerhoff, Dowling and Hurley, 1998) описали многообещающую ли-
нию исследований генетического анализа сетчатки данио.
Общие работы
Alberts, В., D. Bray, J. Lewis, М. Raff, К. Roberts, J. D. Watson, 1994, Molecular Biology of
the Cell (3rd Edition), New York: Garland Publishing Co.
Cajal, Ramon у, 1909, 1911, Histologie du Systeme Nerveux de Г Homme et des Vertebres, trs.
L. Azoulay, Paris: Maloine, 2 vols.
Dowling, J. E., 1987, The Retina: An approachable part of the brain, Cambridge, MA: Har-
vard University Press.
Hartline, H. K., 1938, «Responses of single optic nerve fibres of the vertebrate eye to illumi-
nation of the retina», American Journal of Physiology, 121, 400—415.
Kandel, E. R., J. H. Schwartz and T. M. Jessel, eds, 1991, Principles of Neuroscience, New
York: Elsevier.
Материалы к части IV 461
Kuffler, S. W., 1953, «Discharge patterns and functional organization of the mammalian
retina», Journal of Neurophysiology, 16, 37—68.
Osterbeig, G. A., 1935, «Topography of the layer of rods and cones in the human retina»,
Acta Ophthalmologica, 6(1), 1935.
Polyak, S. L., 1941, The Retina, Chicago: University of Chicago Press.
Rodieck, R. W., 1973, The Vertebrate Retina, San Francisco: Freeman.
Tripathi, R. C. and B. J. Tripathi, 1984, «Anatomy, orbit and adnexa of the human eye», in
The Eye (3rd edn), H. Davson, ed., Orlando, Fla: Academic Tress.
Walls, G. L., 1963, The Vertebrate Eye and its Adaptive Radiation, New York: Hafner.
Структура сетчатки
Besharse, J. C., 1986, «Photosensitive membrane turn-over: differentiated membrane
domains and cell-cell interaction», in The Retina: a model for cell biology studies,
R. Adler, and D. Farber, eds, Orlando: Academic Press.
Boycott, В. B. and J. E. Dowling, 1969, «Organisation of the primate retina: light micro-
scopy», Philosophical Transactions of the Royal Society, B, 225, 109—184.
Dowling, J. E., 1987, ibid.
Gordon, W. C. and N. G. Bazan, 1997, «Retina», in Biochemistry of the Eye, J. J. Harding,
ed., London: Chapman and Hall.
Hogan, M. J., J. A. Alvarado and J. E. Weddell, 1971, Histology of the Human Eye,
Philadelphia: Saunders.
Rodieck, R. W., 1973, ibid.
Rodieck, R. W., 1988, «The primate retina», Comparative Primate Biology, 4, 203—278.
Rowe, M. H., 1991, «Functional organisation of the retina», in Vision and Visual
Dysfunction, Vol. 3: Neuroanatomy of the Visual Pathways and their Development,}. Cron-
ly-Dillon, ed., London: Macmillan.
Young, R. W., 1971, «The renewal of rod and cone outer segments in the rhesus monkey»,
Journal of Cell Biology, 42, 392—403.
Фоторецепторные клетки
Baylor, D. A., T. D. Lamb and K.-W. Yau, 1979, «The membrane current of single rod outer
segments», Journal of Physiology, 289, 589—611.
Harding, J. J., 1997, ed., Biochemistry of the Eye, London: Chapman and Hall.
Lamb, T. D. and E. N. Pugh, 1992, «G-protein cascades: gain and kinetics», Trends in Neu-
rosciences, 15, 291—298.
Motion, J. D., 1982, «Colour vision and colourblindness», in The Senses, H. B. Barlow and
J. D. Mollon, eds, Cambridge: Cambridge University Press.
Nathans, J., D. Thomas and D. Hogness, 1986, «Molecular genetics of human colour vision:
the genes encoding blue, green and red pigments», Science, 232, 193—202.
Nathans, J., 1987, «Molecular biology of visual pigments», Annual Review of Neuroscience,
10, 1987.
Nathans, J., 1994, «In the eye of the beholder: Visual pigments and inherited variations in
human vision», Cell, 78, 357-360.
Penn, R. D. and W. A. Hagins, 1969, «Signal transmission along retinal rods and the origin
of the electroretinographic а-wave», Nature, 223, 201—205.
Yokoyama, S., 1995, «Amino acid replacements and wavelength absorption of visual pig-
ments in vertebrates», Molecular Biology and Evolution, 12, 53—61.
Каламкаров Г. P., Островский M. A. — Молекулярные механизмы зрительной ре-
цепции. М.: Наука, 2002, 279 с. (Добавлено при переводе.)
Каламкаров Г. Р., Островский М. А., Юнусов Р. П., Шевченко Т. Ф. — Молекуляр-
ные механизмы взаимодействия белков, участвующих в трансдукции фоторецеп-
торного сигнала. — Сенсорные системы, т. 18, № 4, сс. 275—285,2004. (Добавлено
при переводе.)
462 Часть IV. Светочувствительность
Физиология и принципиальные схемы
Brockerhoff, S. Е., J. Е. Dowling and J. В. Hurley, 1998, «Zebrafish retinal mutants», Vision
Research, 38, 1335-1339.
Daw, N. W., R. J. Jensen and W. J. Brunken, 1990, «Rod pathways in mammalian retinae»,
Trends in Neurosciences, 13, 110—115.
Kandel, E. R., J. H. Schwartz, T. M. Jessell, 1991, Principles of Neural Science (3rd Edition),
New York: Elsevier.
Levick, W. R., 1967, «Receptive fields & trigger features ia the visual streak of the rabbit’s
retina», Journal of Physiology, 188, 285—307.
Nakanishi, S., 1995, «Second-order neurones and receptor mechanisms in visual and olfac-
tory information processing», Trends in Neurosciences, 18, 359—364.
Schiller, P. H., 1992, «The ON and OFF channels of the visual system», Trends in Neurosci-
ences, 15, 86—92.
Wassle, H. and В. B. Boycott, 1991, «Functional architecture of the mammalian retina»,
Physiological Reviews, 71, 447—480.
Глава 17: Зрительне пути и зрительная кора
Студентам предлагается огромный выбор превосходных работ по зрительным
путям и зрительной коре. Большинство учебников по нейрофизиологии и
нейрологии включают соответствующие главы. Нижеследующие публика-
ции — всего лишь более или менее случайный выбор из огромного количест-
ва возможностей. Хьюбел (Hubei, 1988) представил исключительно четкое,
хорошо иллюстрированное изложение нейрофизиологии зрения, а Зеки
(Zeki, 1993) — детальное и философски обоснованное. Серия под редакцией
Кронли-Диллона (1991) содержит главы с более современными данными.
Питерс и Джоунс (Peters and Jones, 1984) сообщают интересные детали, каса-
ющиеся зрительной коры мышей, крыс, кошек и некоторых приматов. Нери,
Морроне и Бэр (Neri, Morrone and Burr, 1999) показывают, что зрительная ко-
ра человека приспособлена к восприятию движения; полезное обсуждение
пластичности зрительной коры млекопитающих можно найти у Барлоу
(Barlow, 1975). В момент написания этой книги очень интенсивно проводи-
лись исследования распознавания объектов и лиц. Обзор этих данных можно
найти в материалах симпозиума, организованного Брюсом с соавторами
(Bruce et al., 1992). Хотя большинство работ выполняется (и выполнялось) на
приматах, Кендрик и Болдвин (Kendrick and Baldwin, 1987) показали, что и у
других социальных животных имеются нейроны, реагирующие на сходные
триггерные стимулы. Логотетис и Шейнберг (Logothetis and Sheinberg, 1996) и
Танака (Tanaka, 1996) представили обзоры ответов НВ-коры не только на
предъявление лиц, но и других сложных объектов в поле зрения. Прозопагно-
зия привлекает много внимания, и, помимо технических ссылок, можно на-
стойчиво рекомендовать поразительный популярный труд Оливера Сакса
(Oliver Sacks, 1985). Пионерская работа по аналогиям между зрительными си-
стемами животных и компьютеров принадлежит Дэвиду Марру (David Marr,
1981).
Материалы к части IV 463
Общие работы
Cronly-Dillon, J. R., ed., 1991, Vision and Visual Dysfunction, Houndmills: The Macmillan
Press Ltd, 17 vols.
Hubei, D. H., 1988, Eye, Brain, and Vision, New York: Freeman, Scientific American
Library.
Kandel, E. R., J. H. Schwartz, T. M. Jessell, 1991, Principles of Neural Science (3rd Edition),
New York: Elsevier.
McIlwain, T. T., 1996, An Introduction to the Biology of Vision, Cambridge: Cambridge
University Press.
Neri, P., M. Concetta Morrone and D. C. Burr, 1999, «Seeing biological motion», Nature,
395, 894-900.
Smith, C. U. M., 1970, The Brain: towards an understanding, London: Faber.
Zeki, S., 1993, A Vision of the Brain, Oxford: Blackwell Scientific Publications.
Зрительные пути к мозгу
Dreher, В. and S. R. Robinson, eds, 1991, Neuroanatomy of the Visual Pathways and their
Development, in Vision and Visual Dysfunction, J. R, Cronly-Dillon, ed., London: Mac-
millan, vol. 3.
Duke-Elder, S. and К. C. Wybar, 1961, System of Ophthalmology, Vol. 2; The Anatomy of the
Visual System, London: Henry Kimpton.
Hart, W. M., 1992, Adler's Physiology of the Eye (9th edn), St Louis: Mosby.
Ретино-тектальный путь
Henry, G. H. and T. R. Vidyasgar, 1991, «Evolution of mammalian visual pathways», in
Vision and Visual Dysfunction, J. R. Cronly-Dillon and R. Gregory, eds, London:
Macmillan, vol. 3.
Ретино-геникуло-стриарный (РГС) путь
Duke-Elder, S. and К. C. Wybar, 1961, ibid.
Hubei, D. H., T. Wiesel and LeVay, 1977, «Functional architecture of macaque monkey
visual cortex», Proceedings of the Royal Society, B., 198, 1—59.
Первичная зрительная кора
Brindmann, L. and O. Lippold, 1980, The Neurophysiology of the Cerebral Cortex, London:
Edward Arnold.
Peters, A. and L. Jones, eds, 1984, Cerebral Cortex, vol. 3: Visual Cortex, New York: Plenum.
Функционирование
Frisby, J. P., 1979, Seeing: Illusion, Brain and Mind, Oxford: Oxford University Press.
Hubei, D. H. and T. Wiesel, 1977, «Functional architecture of macaque monkey visual cor-
tex», Proceedings of the Royal Society, B., 198, 1—59.
Hubei, D. H., T. Wiesel, M. P. Stryker, 1977, «Orientation columns in macaque monkey
visual cortex demonstrated by the 2-deoxyglucose autoradiographic technique», Nature,
269, 328-330.
Livingstone, M. S. and D. H. Hubei, 1984, «Anatomy and physiology of a color system in the
primate visual cortex», Journal of Neuroscience, 4, 309—356.
464 Часть IV. Светочувствительность
Пластичность
Barlow, Н. В., 1975, «Visual experience and cortical development», Nature, 258, 199—204.
Hubei, D. H., T. N. Wiesel and S. Le Vay, 1977, «Plasticity of ocular dominance columns in
monkey striate cortex», Philosophical Transactions of the Royal Society, B, 278, 377—408.
Экстрастриарная кора
Felleman, D. J. and D. van Essen, 1991, «Distributed hierarchical processing in the primate
cerebral cortex», Cerebral Cortex, 1, 1—47.
Logothetis, N. K. and D. L. Sheinbeig, 1996, «Visual object recognition», Annual Review of
Neuroscience, 19, 577—621.
Распознавание лица
Bruce, V., etal., 1992, «Processing the facial image», Philosophical Transactions of the Royal
Society, B., 335, 1—128.
Gross, C. G., С. E. Rocha-Miranda and D. B. Bender, 1972, «Visual properties of neu-
rons in the inferotemporal cortex of the macaque», Journal of Neurophysiology, 35,
96-111.
Grusser, O.-J. and T. Landis, 1991, «Man as social partner in the visual world: perception
and recognition of faces and facial expression», in Visual Agnosias and other
Disturbances of Visual Perception and Cognition, O.-J. Grusser and T. Landis, eds, being
vol. 12 of J. Cronly-Dillon, ed., Vision and Visual Dysfunction, London: Macmillan
Press.
Kendrick, К. M. and B. A. Baldwin, 1987, «Cells in temporal cortex of conscious sheep can
respond preferentially to the sight of faces», Science, 236,448-450.
Perrett, D. L, A. J. Mistlin and A. J. Chitty, 1987, «Visual neurons responsive to faces»,
Trends in Neurosciences, 10, 358—364.
Perrett, D. I. et al., 1992, «Organisation and functions of cells responsive to faces in the tem-
poral cortex», Philosophical Transactions of the Royal Society, B, 335, 23—30.
Tanaka, K., 1996, «Inferotemporal cortex and object vision», Annual Review of Neuroscience,
19, 109-139.
Young, M. P., 1995, «Open questions about the neural mechanisms of visual pattern recog-
nition», in The Cognitive Neurosciences, M. S. Gazzaniga, ed., Cambridge MA: MIT
Press.
Прозопагнозия
Allison, T., A. Puce, D. D. Spencer and G. McCarthy, 1999, «Electrophysiological studies
of human face perception, 1: Potentials generated in occipitotemporal cortex by face and
non-face stimuli», Cerebral Cortex, 30, 9, 415—430.
Grusser, O.-J. and T. Landis, 1991, «Faces lost: prospagnosia», in Visual Agnosias and other
Disturbances of Visual Perception and Cognition, O.-J. Grusser and T. Landis, eds., being
vol. 12 of J. Cronly-Dillon, ed., Vision and Visual Dysfunction, London: Macmillan
Press.
Sacks, O., 1985, The Man who Mistook his Wife fora Hat, New York: Harper Collins.
Заключение
Marr, D., 1981, Vision, San Francisco: W. H. Freeman.
Материалы к части IV 465
Глава 18: Зрительные системы других позвоночных
Существует четыре исчерпывающих источника по сравнительной анатомии
зрительных систем позвоночных: сэра Стюарта Дьюк-Элдера «Система оф-
тальмологии» (S. Duke-Elder: System of Ophthalmology), том 1; G. L. Walls: Ver-
tebrate Eye and its Adaptive Radiation; J. R. Cronly-Dillon: Vision and Visual Dys-
function, и H. Autrum et al.: Handbook of Sensory Physiology. Два последних ис-
точника содержат многочисленные подробные главы по конкретным группам
животных. Эти источники многократно цитируются в последующей библио-
графии. Адаптивное значение родопсина и порфиропсина рассмотрено
Goldsmith (1990), а также в учебнике Проссера и Брауна (Prosser and Brown,
1961), который, хотя и написан много лет назад, дает много полезной инфор-
мации, часто представленной в форме таблиц.
Общие работы
Charman, W. N., 1991, «The vertebrate dioptric apparatus», in J. R. Cronly-Dillon and
R. L. Gregory, eds., Evolution of the Eye and Visual System, in J. Cronly-Dillon, ed.,
Vision and Visual Dysfunction, London: Macmillan, vol. 2.
Crescitelli, F., ed., 1977, Handbook of Sensory Physiology, vol. VII/5: The visual system in
vertebrates. New York: Springer-Verlag.
Duke-Elder, S., 1958, System of Ophthalmology, vol 1: The Eye in Evolution, London: Kimpton.
Land, M. F. and R. D. Femald, 1992, «Evolution of eyes», Annual Review of Neuroscience,
15, 1-29.
Walls, G. L., 1963, The Vertebrate Eye and its Adaptive Radiation, New York: Hafner.
Зрительные пигменты
Goldsmith, T. H., 1990, «Optimisation, constraint and history in the evolution of eyes»,
Quarterly Review of Biology, 65, 281—322.
Prosser, C. L. and F. A. Brown, 1961, Comparative Animal Physiology, Philadelphia: Saun-
ders.
Фоторецепторы
Crescitelli, F., 1977, «The visual pigments of Geckoes and other vertebrates: an essay in com-
parative biology», in H. Autrum et al. eds. Handbook of Sensory Physiology, vol. VII/5, ed.
F. Crescitelli, The Visual System in Vertebrates, New York: Springer-Verlag.
Walls, G. L., 1963, ibid.
Зеркальца
Locket, N. A. 1977, «Adaptations to the deep-sea environment», in H. Autrum et aL, eds.
ibid.
Walls, G. L., 1963, ibid.
Сетчатки
Barlow, H. B., 1953, «Summation and inhibition in the frog’s retina», Journal of Physiology,
119, 69-88.
466 Часть IV. Светочувствительность
Collin, S. Р., R. V. Hoskins and J. C. Partridge, 1997, «Tubular eyes of deep sea fishes:
A comparative study of retinal topography», Brain, Behaviour and Evolution, 50,
335-357.
Collin, S. P., R. V. Hoskins and J. C. Partridge, 1998, «Seven retinal specialisations in the
tubular eye of the deep-sea Pearleye, Scopelarchus michaelsarsi'. a case study in visual
optimisation», Brain, Behaviour and Evolution, 51, 291—314.
Hartline, H. K., 1938, «Responses of single optic nerve fibres of the vertebrate eye to illumi-
nation of the retina», American Journal of Physiology, 121, 400—415.
Locket, N. A., 1977, ibid.
Meyer, D. B., 1977, «The avian eye and its adaptations», in F. Crescitelli, ed., ibid.
Диоптрический аппарат
Artedi, P., 1758, Rerum Naturalium Thesauri Descriptione (quoted in S. Duke-Elder, ibid.,
vol. 1).
Charman, W. N., 1991, ibid.
Collin, S. P., R. V. Hoskins and J. C. Partridge, 1997, ibid.
Duke-Elder, S., 1958, ibid. vol. 1.
Land, M. F. and R. D. Femald, 1992, ibid.
Locket, N. A., 1977, ibid.
Soemerring, D. W., 1818, De Oculorum Hominis Animaliumque., Gottingen.
Медианные глаза
Duke-Elder, S., 1958, ibid.
Hamasaki, D. I. and D. J. Elder, 1977, «Adaptive radiation of the pineal system», in
F. Crescitelli, ibid.
Зрительные пути
Henry, G. H. and T. R. Vidyasgar, 1991, «Evolution of mammalian visual pathways», in
Vision and Visual Dysfunction, J. R. Cronly-Dillon and R. Gregory, eds., London: Mac-
millan, vol. 3.
Зрительные центры мозга
Muntz, W. R. A., 1977, «The visual world of amphibia», in F. Crescitelli, ibid.
Shimizu, T. and H. J. Karten, 1991, «Central visual pathways in reptiles and birds: evolution
of the visual system», in Vision and Visual Dysfunction, J. R. Cronly-Dillon and R. L. Gre-
gory, eds., London: Macmillan, vol. 2.
Sivar, J. G., 1977, «The role of the spectacle in the visual optics of the snake», Vision
Research, 17, 293-298.
Часть V
ДРУГИЕ ЧУВСТВА
«Если бы лев умел разговаривать, мы бы все равно не смогли
бы его понять.»
Людвиг Витгенштейн, Философские исследования
Перевод с перевода Дж. Э. М. Энскомба
(G. Е. М. Anscomb),
Oxford, Blackwell,
1958: р.223е
До сих пор мы рассматривали чувства, которыми обладает большинство жи-
вотных, если не все. В данной, предваряющей заключительную, части мы рас-
смотрим в первую очередь, в главе 19, последнее важное чувство — ощущение
температуры. Это чувство, хотя оно и широко распространено в царстве жи-
вотных, не связано с развитием сложных органов чувств, подобных глазу, уху
или носу (исключая инфракрасные детекторы у змей). При этом термочувст-
вительные окончания распределены по всей поверхности тела. В следующей
главе 20 мы перейдем к редким чувствам — тем, которые обнаружены только
у небольших групп животных. Мы не чувствуем электрического и магнитного
полей, хотя в эпоху нашей технологической цивилизации это было бы нам по-
лезно. Мы не чувствуем ни инфракрасного излучения, ни плоскости поляри-
зации света. У некоторых животных, однако, развились органы чувств, кото-
рые способны детектировать эти характеристики окружающей среды, и в этой
главе такие редкие чувства будут рассмотрены. Нам трудно понять мир живот-
ных, наделенных их способностями. Витгенштейновский лев по сравнению с
ними почти совершенно вразумителен. Люди с помощью технологической ци-
вилизации уже превзошли всех этих животных в способности детектировать и
анализировать любые формы энергии в окружающей среде. Наши военные уже
освоили инфракрасные приборы ночного видения, наши инженеры-электри-
ки способны обнаруживать электрические и магнитные поля, производители
очков и тонированных стекол полностью информированы о природе поляри-
зованного света. Культурная эволюция сильно опережает биологическую в со-
здании детекторов всех видов энергии окружающей среды. Все эти виды энер-
гии могут быть преобразованы нашими технологиями в привычные для нас
сенсорные модальности.
Одного, все же, наши технологии сделать не удалось — того, что Пат-
рик Уолл назвал «долориметром» — устройства детектирования и измере-
ния боли.
Боль называют «экзистенциальным чувством». Фигурально выражаясь, мы
не можем закрывать глаза на зубную боль. Она следует за нами, где бы мы ни
были, что бы ни делали. Но чувство это сугубо приватное, никто, кроме нас са-
мих, не способен оценить его. Посторонние могут видеть наши гримасы боли,
468 Часть V. Другие чувства
наше поведение, слышать наши стенания и жалобы, но не могут ощутить на-
шу боль, измерить ее субъективное ощущение. Насколько широко это чувство
распространено в царстве животных? Птицы, рыбы, головоногие? Уверены
мы можем быть только насчет самих себя, ну, и других млекопитающих. Мы и
ограничимся физиологией млекопитающих, когда будем обсуждать эту про-
блему в третьей главе этой части книги, главе 21. Затем мы вернемся к некото-
рым более широким аспектам этого очень личного чувства в последней главе
этой книги, в гл. 23.
19. ТЕРМОЧУВСТВИТЕЛЬНОСТЬ
Температурный диапазон жизни. Пойкилотермные и гомойотермные
животные. Те и другие нуждаются в термосенсорах. Молекулярная биоло-
гия: клетки ганглия дорзального корешка (ГДК) — реагируют на изме-
нения температуры — Са2+-вызванный входящий ток — потенциация
брадикинином и простагландином Е; С. elegans — термотаксические
мутанты, tax 2 and tax 4 — белковые продукты, сходные с ЦНЗ-единица-
ми наружных сегментов. Пойкилотермия: боковая линия рыб? — сен-
силлы наскомых — терморецепторы группируются с гигрорецептора-
ми — биологическая роль. Гомойотермия: эволюция гомойотермии; го-
мойотермия у Homo sapiens — локализация рецепторов: кожа — тепловые
и холодовые пятна — парадоксальное ощущение холода — веберовская
иллюзия — гистология — обнаженные нервные окончания — Аб-волок-
на (тепловые), С-волокна (холодовые) — РП — динамические и статиче-
ские ответы — спиноталамический и тройнично-таламический тракты в
таламус; глубокие рецепторы: спинной мозг, брюшная полость, крупные
вены, просвет кишечника — защита от гипотермии — слизистая носа; ги-
поталамус: преоптическая область — задний гипоталамус — гомеостати-
ческий контроль. Заключение: гомойотермия представляет собой пример
необходимости поддержания стабильности внутренней среды организма
До недавнего времени полагали, что температурный диапазон, в котором спо-
собны существовать животные, достаточно ограничен: от прибл. О до 40°С
(или 273—313 К). Открытие крупных «трубчатых» червей, обитающих вблизи
от глубоководных фумарол при температуре около 100°С, поставило знак во-
проса у верхней границы диапазона обитания. Тем не менее, непреодолимые
факты химической природы, кажется, противоречат возможности серьезного
отклонения от диапазона О—40°С. При температуре ниже 0°С водная среда,
которая составляет более 90% массы любого живого организма, претерпевает
фазовый переход, что в лучшем случае замедляет метаболические процессы
(если развились какие-то формы органического антифриза) или приводит к
замораживанию — в худшем (обычно). При температуре выше 40°С тепловое
движение может нарушать гидрофобные и другие слабые химические связи,
которые удерживают структуру глобулярных белков. Нарушение этой преци-
зионной и хрупкой структуры приводит к утрате их биологических, в особен-
ности ферментативных, функций. Кроме того, к температуре очень чувстви-
тельны и биологические мембраны. Фосфолипидный бислой при низких тем-
пературах стремятся к состоянию геля и переходят в мицеллы при высоких
470 Часть V. Другие чувства
температурах. Каждый начинающий студент-биолог знает, что денатурация
белка происходит при температуре около 40°С; каждый начинающий микро-
биолог знает, что кипячение ведет к стерилизации. Однако же, как мы видели,
некоторые трубчатые черви переносят 100°С, а гипертермофильные бактерии
продолжают расти и при 113°С в горячих источниках. Мы до сих пор не пони-
маем, как это у них получается.
По отношению к температурной регуляции животные делятся на две боль-
шие группы: пойкилотермных и гомойотермных. Первая группа не имеет иного
выхода, кроме как подчинять свое поведение контролю над температурой те-
ла; у второй развились механизмы поддержания постоянной температуры
внутренней среды, вне зависимости от изменений внешней среды (в опреде-
ленных пределах, естественно). Обе группы, однако, нуждаются в средствах
определения температуры окружающей среды. Особенно это важно для мелких
животных. Даже гомойотермные мелкие животные очень быстро начинают ис-
пытывать воздействие температуры окружающей среды — их гомеостатические
механизмы должны работать быстро и эффективно, чтобы предупредить стрем-
ление температуры их тела уравняться с температурой среды.
19.1. Молекулярная биология
В последнее время развито два подхода к основам молекулярной биологии
термочувствительности. Первый из них связан с биофизическим анализом
клеток, изолированных из ганглиев дорзальных корешков (ГДК) млекопитаю-
щих; второй — с генетическим анализом термотаксических мутантов С. ele-
gans. Рассмотрим их по очереди.
Известно, что мембрана каждого нейрона уникальна и отличается от всех
других. Иными словами, каждый нейрон встраивает собственный набор бел-
ков в наружную липидный бислой. Так, клетки, реагирующие на ацетилхолин
(АХ), встраивают АХ-рецепторы, а реагирующие на серотонин — тот или иной
(или даже целый набор) серотониновых рецепторов и т. д. Конечно, эти ре-
цепторы в наибольшей степени сконцентрированы в постсинаптических мем-
бранах, но в меньших количествах присутствуют и в мембране перикариона
и т. д. Исследования молекулярно-биологических механизмов термочувстви-
тельности связаны с этим фактом.
Размеры и форма клеток ГДК млекопитающих разнообразны. Известно,
однако, что волокна, ответственные за термочувствительность, имеют малый
диаметр (см. раздел 19.3.1 ниже), из чего следует, что имеено самые маленькие
клетки ГДК связаны с термочувствительностью. Тем интереснее было обнару-
жить, что если диссоциированные клетки ГДК подвергнуть быстрым измене-
ниям температуры, группа небольших нейронов отвечает большими входя-
щими токами. Фармакологический анализ показал, что перикарионы этих
небольших клеток не имеют симпатических свойств. Дальнейшие работы по-
казали, что эти термочувствительные клетки отвечают на изменения темпера-
туры градуально — чем больше температурный стимул, тем больше входящий
ток. Этот ток зависит от открывания неслективных катионных каналов. Веро-
ятно, тепловые стимулы вызывают выделение Са2+ из внутриклеточных запас-
ников, а повышенный уровень этого катиона запускает открывание каналов.
19. Термочувствительность 471
Если принять предположение, что перикарион и сенсорные окончания реаги-
руют на стимулы одинаково, то можно предполагать, как термический стимул
преобразуется в потенциал действия.
Молекулярный анализ этой проблемы неожиданно пролил свет на то, по-
чему повреждение тканей (напр. порез) часто вызывает ощущение жжения.
Среди многих веществ, выделяющихся при повреждениях, присутствуют про-
стагландины и брадикинин. Показано, что и простагландин Е, и брадикинин
повышают амплитуду термочувствительных токов. Эти вещества могут участ-
вовать в том открывании ионных каналов, которое приводит к генераторному
потенциалу в нервных окончаниях термочувствительных волокон.
Теперь перейдем к Caenorhabditis elegans. Мутанты, неспособные реагиро-
вать на температурные сигналы из окружающей среды, были выделены в сере-
дине 1970-х годов. Через двадцать лет были обнаружены мутации в двух ге-
нах — tax-2 и tax-4, ответственные за потерю термотаксического поведения.
Поразительно, но белковые продукты этих генов оказались идентичны а- и р-
субъединицам цГМФ-зависимых каналов палочек и колбочек позвоночных
(см. гл. 16). Когда эти два белка экспрессируются в клеточной культуре, они
формируют канал, который (как и его гомолог у позвоночных) активируется
цГМФ, аплицированный на его внутриклеточный конец. Наконец, связыва-
ние этих белков с зеленым флуоресцентным белком позволило определить ло-
кализацию каналов. Показано, что они экспрессируются в соответствующих
сенсорных нейронах и нервных окончаниях нервной системы червя.
Насколько данные, полученные на С. elegans и на клетках ГДК, могут быть
совмещены в единую молекулярно-биологическую картину, остается пока не-
известным. Однако, с развитием этих мощных экспериментальных средств от-
крываются перспективы понимания основ термочувствительности на таком
же уровне, как это представлено в предыдущих частях этой книги.
19.2. Пойкилотермные животные
Разные пойкилотермные животные могут быть приспособлены к очень раз-
ным диапазонам температуры окружающей среды. Для водных животных,
обитающих в холодных водах, может оказаться смертельным повышение тем-
пературы до такого уровня, который ниже, чем нижний уровень, смертельный
для более теплолюбивых животных, адаптированных к высоким температу-
рам. Животные имеют сенсоры к низким и высоким'температурам на поверх-
ности тела, сигнализирующие о необходимости менять обстановку, если это
необходимо. Эти сенсоры часто принимают форму свободных нервных окон-
чаний. Есть данные, что у рыб и головастиков земноводных, в дополнение к
подкожным афферентам, в определении температуры и ее изменений, прини-
мает участие система боковой линии (которая, как мы видели в гл. 8, является
механорецептивной, воспринимающей изменения давления). У хрящевых
рыб продолжение боковой линии в голову — ампулы Лоренцини — хотя и яв-
ляются первично электрорецепторными (как мы увидим в гл. 20), но реагиру-
ют на постоянную температуру и ее изменения, особенно охлаждение.
Насекомые как малые животные особо чувствительны к температуре сре-
ды. В экспериментальных условиях они реагируют на изменения температу-
472 Часть V. Другие чувства
ры, перемещаясь к температурному оптимуму, насколько это возможно. Ис-
следователи показали, что в большинстве случаев терморецепторы локализо-
ваны в антеннах. У большинства исследованных видов, по-видимому, при-
сутствуют рецепторы холода, хотя в некоторых случаях есть и рецепторы теп-
ла. Рецепторы холода увеличиваю частоту импульсации в ответ на снижение
температуры. У палочника Carausius, например, на снижение температуры
ниже уровня 20°С ответ увеличивается линейно. Терморецепторные сенсил-
лы часто мультимодальны и содержат один или более хеморецепторов и (или)
гигрорецепторную нейросенсорную клетку, помимо терморецептора. По-
скольку часто критична именно комбинация температуры воздуха и влажно-
сти, то совмещение термо- и гигрорецептора в одной сенсилле достаточно ес-
тественно. Сенсиллы насекомых обычно имеют различные формы и размеры,
хотя большинство терморецепторов — это простые бугорки — с множеством
пор или без поры вообще.
У насекомых существует множество вариантов сочетания ощущения темпе-
ратуры или ее изменений с чем-то иным. Холодовые рецепторы антенн гусениц
воспринимают охлаждение испарения, вызываемое влагой, улетучивающейся с
поверхности листьев. Терморецепторы в жилках крыльев некоторых групп ба-
бочек активируются солнечным теплом и рефлекторно вызывают складывание
крыльев. Активные кровососущие полужесткокрылые клопы рода Triatoma
имеют в антеннах специализированные рецепторы тепла, а у их некровососу-
щих сородичей такие структуры отсутствуют. Ясно, что это приспособление
развилось для поиска хозяина-жертвы. Клоп Triatoma megista является основ-
ным переносчиком трипаносомы Trypanosoma cruzi и, таким образом, вовлечен
в распространение трипанозомозов в Южной Америке. Другие виды этого рода
могут участвовать в переносе лейшманиоза. Комар Aedes aegipti также нуждается
в теплокровной жертве. У него имеются два терморецептора в виде бугорков на
кончиках антенн. Один из них является детектором тепла, другой — холода.
Тепловой рецептор активируется повышением температуры и дает максималь-
ную реакцию при 25—28°С. Холодовой рецептор настроен аналогичным обра-
зом, чтобы давать максимальный ответ при снижении температуры в этих пре-
делах. Такие рецепторы способны детектировать конвективные потоки возду-
ха, вызванные теплом рук человека. Это играет важную роль в направлении
атаки комара именно на незащищенную одеждой часть тела человека.
19.3. Гомойотермные животные
Только птицы и млекопитающие являются истинно гомойотермными, причем
это приспособление развилось в этих двух таксонов независимо. Млекопита-
ющие развились бок о бок с царствовавшими тогда пресмыкающимися, в т. ч.
динозаврами, около 200 миллионов лет назад в триассовом периоде. Были ли
«звероподобные рептилии» гомойотермными, как, впрочем, и вопрос о том,
были ли гомойотермными некоторые из динозавров, остается дискуссион-
ным. Аналогичными образом, эволюция птиц может быть прослежена по
крайней мере до юрского периода, когда археоптерикс (145 миллионов лет
назад) мог быть, а мог и не быть гомойотермным. В то время как птицы все
гомойотермны (причем некоторые из них поддерживают температуру тела на
19. Термочувствительность 473
уровне 42°С), среди млекопитающих лишь высшие могут считаться таковыми.
Примитивные млекопитающие — прототерии — в лучшем случае могут счи-
таться гетеротермными, а многие метатерии могут считаться гомойотермными
лишь с большой натяжкой, как, впрочем и наиболее примитивные среди на-
стоящих зверей, — например насекомоядные.
Обратимся же к человеку. Здесь гомойотермность развита хорошо. Темпера-
тура внутренностей поддерживается на постоянном уровне около 37°С. Гомео-
статичекий механизм, в сущности, основан на балансе между теплопродукцией
и теплопотерями. Обе части этого уравнения могут варьировать, однако, тонкий
контроль осуществляется именно над потерей тепла. Она может контролиро-
ваться путем изменения количества тепла, теряемого организмом за счет испа-
рения, излучения и конвекции. Учебники физиологии описывают эффектор-
ные механизмы этого контроля: мышцы, поднимающие волосы; потовые железы;
сосудистые сплетения в дермисе кожи и т. д. Здесь мы сосредоточим внимание
только на сенсорных системах, которые следят за колебаниями температуры,
требующими гомеостатической реакции организма. Эти изменения ощущаются
и на периферии (кожей), и в глубине тела — главным образом в спинном моз-
ге, во внутренностях, в больших венах и вокруг них, а также (и возможно это
самое важное) температурно-чувствительными клетками гипоталамуса.
19.3.1. Терморецепторы кожи
Когда температура кожи составляет 32—42°С, никаких температурных ощуще-
ний не возникает. Это, конечно, не означает, что изменения температуры в
этом диапазоне не могут ощущаться. На самом деле, как мы увидим ниже, ко-
жа очень чувствительна к резким изменениям температуры в этих пределах,
оно если она остается неизменной в этом диапазоне, то активность терморе-
цепторов незначительна. Поэтому упомянутый диапазон и называется нейт-
ральным. Вне пределов этого диапазона возникают ощущения холода или теп-
ла. При температурах ниже 18°С ощущение холода становится болезненным,
равно как ощущение тепла при температурах выше 45°С. И «тепловые», и «хо-
лодовые» рецепторы лежат в толще кожи. Уже давно известно, что они образу-
ют мозаику «тепловых» и «холодовых» пятен, каждое диаметром около 1 мм.
Это установлено с помощью термода — маленького игловидного инструмента,
который можно нагревать электрическим током или пропуская через него во-
ду. Наибольшая плотность этих термочувствительных пятен обнаружена в не-
которых областях лица. В губах, например, 16—19 холодовых пятен на см2, на
носу — 8—13 на см2, а на лбу — 5—8 на см2. Напротив, на ладонях рук только
1—5/см2, а на пальцах — 2—4 пятна на см2. Тепловые пятна встречаются ре-
же — 1,7/см2 на пальцах и 0,4 — на ладонях. Во многих частях тела они встре-
чаются достаточно редко.
Показано, что термочувствительные окончания, реагирующие на холод,
локализованы в верхней части дермиса (около 0,17 мм ниже поверхности ко-
жи), тогда как реагирующие на тепло — несколько глубже (около 0,3 мм). Ин-
тересно отметить, что помещение на холодовое пятно стимула избыточной ин-
тенсивности, например термода при 45°С, вызывает ощущение холода. Такая
иллюзия, называемая парадоксальным холодом, подчеркивает зависимость
474 Часть V. Другие чувства
В
38° 38°
\ 0 / Температура
\' кожи (°C)
Холод
20 30 40 50
Холодовой
рецептор
Тепловой
рецептор
I___________I
5с
Рис. 19.1. (А) Организация рецептивного поля «теплового» волокна. Активность в
волокне вызвана импульсом нагревания термода длительностью 30 с в разных точ-
ках поля. Семейство кривых показывает частоту импульсов в ответ на изменение
температуры на 2вС, 4вС, 6°С и 8°С. Из Darian-Smith, 1984, с разрешения. (Б) Ответ
холодового и теплового рецептора на температуру. Остается неясным, как ЦНС
выделяет сигнал об интенсивности из колоколовидцрго сигнала. (В) Ответ холодо-
вого и теплового рецепторов на включение и выключения холодового стимулов.
Видно, что те и другие рецепторы интенсивно реагируют на изменение, а затем
адаптируются до постоянного уровня. Из Bear, Connors and Paradiso, 1996, с разре-
шения.
ЦНС от интерпретации афферентов, поступающих по меченым линиям (см.
гл. 3). Другой достоверно установленной иллюзией является иллюзия Вебера,
возникающая вследствие того, что кожные механорецепторы несколько акти-
вируются холодом. Из-за этого, как подметил Вебер, холодный объект кажет-
ся тяжелее, чем теплый или нейтральный.
Идентификация сенсорных окончаний кожи, отвечающих на холод или на
тепло, затруднительна. В целом, полагают, что специфических гистологичес-
ких структур не существует, и что это — не имеющие внешних структур нерв-
ные окончания, переплетающиеся, но не анастомизирующие, на всех уровнях
дермиса. Эти волокна имеют малый диаметре (1,5—3 мкм) и классифицируют-
ся как А5- (холодовые) и С-волокна (тепловые). Происходит значительная
суммация афферентов — показано, что субъективный порог стимуляции боль-
ших пространств кожи существенно ниже, чем стимуляции одиночного во-
локна. Около 50 тепловых или холодовых волокон должно быть активировано,
19. Термочувствительность 475
чтобы возникло осознанное ощущение. Такой субъективный порог очень ни-
зок — человек способен обнаруживать снижение температуры всего на
0,02—0,05°С.
Сигнал, идущий и от тепловых, и от холодовых рецепторов, можно изучать
электрофизиологически. Каждое волокно отвечает на стимуляцию одного
(иногда — больше, чем одного) термочувствительного поля. Размеры рецеп-
тивных полей варьируют в зависимости от интенсивности термического сти-
мула (рис. 19.1 А). В «тепловых» волокнах при нагревании выше нейтрального
диапазона возникает импульсация, которая прекращается при достижении бо-
левого порога (см. рис. 19.1 Б). Импульсация в холодовых волокнах возникает
в широком диапазоне температур — от 10°С до 40°С (рис. 19.1 Б). И те, и дру-
гие волокна имеют как статические, так и динамические ответы (рис. 19.1 В).
Первоначальный залп импульсов (динамический ответ), возникающий при
изменении температуры (за пределами нейтрального диапазона) представляет
собой сигнал с высокочувствительным порогом, упомянутый выше.
А5- и С-афференты от кожных терморецепторов оканчиваются преимуще-
ственно в дорзальном роге спинного мозга. Отсюда волокна направляются в
составе спиноталамического и тройнично-таламического трактов передают
термическую информацию в таламус.
19.3.2. Глубокие терморецепторы
Температура вблизи спинного мозга, абдоминальных внутренностей и в круп-
ных венах воспринимается терморецепторами, сходными с теми, которые мы
обсудили выше. Опять-таки, большинство реагирует на холод, особенно на
снижение температуры ниже нейтрального диапазона. Как и в случае с кож-
ными терморецепторами, рассмотренными выше, назначение внутренних
терморецепторов — это предупреждение гипотермии, т. е. потери организмом
тепла. Кроме перечисленных сенсоров температуры тела, терморецепторы
слизистой носы играют существенную роль не только в ощущении температу-
ры поступающего воздуха, но и в ощущении скорости потока воздуха через
дыхательные пути. Наконец, как тепловые, так и холодовые рецепторы обна-
ружены в желудочно-кишечном тракте.
19.3.3. Гипоталамические терморецепторы
Когда гипоталамус тщательно исследовали тонким термодом, было показано,
что большое количество термочувствительных нейронов располагается в пре-
оптической области переднего гипоталамуса, но не только там. Теплочувстви-
тельные нейроны увеличивают свою импульсацию в ответ на повышение тем-
пературы крови в соседних капиллярах; напротив, холодочувствительные
нейроны увеличивают импульсацию в ответ на падение температуры. Инфор-
мация из переднего гипоталамуса направляется главным образом в билате-
ральные ядра заднего гипоталамуса на уровне сосцевидных тел. Именно в
этих ядрах интегрируется информация от центральных и периферических
терморецепторов, и формируется эффекторная реакция, корригирующая
серьезные отклонения от уровня 37°С.
476 Часть V. Другие чувства
19.4. Заключение
Как всегда, больше известно о физиологии термочувствительности у млекопи-
тающих, особенно человека, чем о других животных. Однако, как мы видели,
температурное чувство важно для всех животных — от нематоды С. elegans до
высокоорганизованных включительно. У гомойотермных животных темпера-
тура тела контролируется очень точно. Изменения температуры тела всего на
один — два градуса нарушают координацию биохимии и физиологии организ-
ма. Это еще один случай (вспомните кинестезию в гл. 6 и контроле ПдО2 и
ПдСО2 в гл. 11) необходимости поддержания стабильности «внутренней сре-
ды» млекопитающих и, особенно, человека. Термосенсоры, локализованные
во многих частях тела, посылают информацию в центр, где она сравнивается с
предетерминированной. Только когда афферентная информация сильно от-
личается от предусмотренной, мы осознаем, что с этим надо что-то делать, и
либо протягиваем руки к огню, либо суем голову под струю воздуха от конди-
ционера. Забегая вперед, в главу 21, скажем, что схожий анализ можно сделать
и для много более сложного ощущения боли.
20. РЕДКИЕ ЧУВСТВА
Сенсорный мир животных иногда очень отличается от нашего собствен-
ного. Инфракрасная (ИК) чувствительность: змеи (ложноногие змеи
Boidae и ямкоголовые змеи Crotalidae) — губные и лицевые ямки — гис-
тология — направленность — изменение ИК-потока есть адекватный
стимул — контрлатеральная крыша среднего мозга — ON/OFF-РП —
карта ИК-пространства — запуск движением источника ИК-излучения.
Чувствительность к поляризованному свету: характеристики поляризован-
ного света (атмосфера) — суточные изменения — Х-зависимость — более
100 видов водных животных чувствительны к плоскости поляризации
света; структура фоторецепторов — омматидии десятиногих — откры-
тый, закрытый и окаймленный рабдом; омматидии насекомых — фура-
жировка пчел Apis; палочки осьминога — поведение — электрофизиоло-
гия. Электрочувствительность: сильно и слабо выраженные электричес-
кие рыбы — сильное электрическое поле используется для оглушения
жертвы — слабое электрическое поле используется для электролока-
ции — распознавание особей своего вида и т. д.; поведенческие исследо-
вания — чувствительность < 5 нВ — детектирование жертвы — навигация
в магнитном поле Земли? — детектирование и идентификация близких
объектов; гистология электрорецепторов — ампулы и бугорковые орга-
ны — эволюция из органов боковой линии; гистология ампул — ампулы
Лоренцини — восприятие формы электрического поля; гистология бугор-
ковых органов — типы; различия ответов ампул и бугорковых органов —
распределение в организме — весь организм действует как сенсорная по-
верхность. Магниточувствительность: магнитотаксические бактерии —
Ре2О4-магнитосомы — «биокомпас» — намеки на магнитную чувстви-
тельность у птиц и млекопитающих. Заключение: плодотворное разнооб-
разие сенсорных систем
В данной главе мы кратко рассмотрим четыре «редких» чувства: чувствитель-
ность к инфракрасному излучению, поляризованному свету, электрическому
и магнитному полям. Мы часто забываем в нашем антропоцентрическом ми-
ре, что сенсорный мир животных устроен совершенно иначе. Многие насеко-
мые хорошо видят в ультрафиолетовой части спектра, и энтомофильные цве-
ты адаптировали свои цвета соответствующим образом (хотя что появилось
раньше — УФ-отражающие цветы или УФ-детектирующие насекомые — во-
прос спорный). Многие мелкие млекопитающие, в особенности летучие мы-
ши, могут, как мы отмечали выше, воспринимать звук, выходящий далеко за
478 Часть V. Другие чувства
пределы частотного диапазона слышимости человека. И они живут в ином
сенсорном мире. Обонятельный мир человека, доведенный до минимума
предками, десятками миллионов лет обитавшими на деревьях, не может срав-
ниться со сложностью и богатством мира запахов многих других животных.
Мы можем лишь восхищаться великолепием антенн мотылька, позволяющих
обнаружить единственную молекулу феромона самки, — и признать свое бес-
силие. О многом мы можем лишь догадываться — эти черты мира, о которых
мы упомянули в данной главе, окружают нас, но мы проходим мимо, не заме-
чая ничего.
20.1. Инфракрасное излучение
Инфракрасные детекторы, конечно, трудно отличить от терморецепторов,
рассмотренных в гл. 19. Тепловой детектор клопов Triatoma мог бы быть рас-
смотрен и в этом разделе. Тем не менее, некоторые терморецепторы настоль-
ко специализировались в детектировании удаленных источников тепла и оп-
ределении направления на них, что стоит рассмотреть их отдельно. Наиболее
известны из них лицевые и губные ямки некоторых змей.
Первые указания на то, что у семейства ложноногих змей Boidae (удавы, пи-
тоны и т. д.) и подсемейства ямкоголовых змей Crotalinae (гремучие змеи, в т. ч.
настоящие гремучники Crotalus и бушмейстер (или сурукуку) Lachesis) имеют-
ся инфракрасные сенсоры, были получены из анализа их поведения при поис-
ке жертв и определении направления атаки. Инфракрасное детектирование
используется также для обороны или бегства, которое вызывается появлением
излучающего тепло хищника. Впоследствии электрофизиологическая иссле-
дования тройничного нерва, иннервирующего губные ямки ложноногих змей
и лицевые ямки (между глазами и ноздрями) ямкоголовых змей, подтвердили,
что эти углубления действительно содержат инфракрасные рецепторы. Ин-
фракрасное излучение представляет собой адекватный стимул для этих рецеп-
торов, хотя ответ может генерироваться и при омывании ямки теплой водой.
Гистологические исследования показали, что ямки содержат не специализи-
рованные рецепторные клетки, а немйелинизированные окончания тройнич-
ного нерва, образующие широкое не перекрывающееся ветвление (рис. 20.1).
В ямках и ложноногих, и ямкоголовых змей поверхность дна ямки реагирует
на инфракрасное излучение, причем реакция зависит от расположения источ-
ника излучения по отношению к краю ямки. В этом отношении этот орган
чувства напоминает глазные пятна, рассмотренные в гл. 14.
Активация рецепторов и у ложноногих, и у ямкоголовых змей требует из-
менения потока инфракрасного излучения. Это может достигаться либо в ре-
зультате движения излучающего тепло объекта в «поле зрения» относительно
более холодного окружения, либо при сканирующем движении головы змеи.
Чувствительность достаточна для обнаружения потока излучения от руки че-
ловека, перемещающейся в «поле зрения» на расстоянии 40—50 см, из чего
следует, что пороговый стимул составляет менее 8 х 10“5 Вт/см2 . Исходя из
этого, повышение температуры, детектируемое рецепторами, составляет по-
рядка 0,005°С (т. е. примерно на порядок лучше, чем способность человека
ощущать изменения температуры, см. гл. 19).
20. Редкие чувства 479
Наружная поверхность
Внутренняя поверхность
Рис. 20.1. Гистология ямки у ямкоголовых змей. Одиночное немиелинизирован-
ное нервное волокно отходит от широко разветвленных окончаний ямки, т — тер-
минальная ветвь, ци — цитоплазма шванновских клеток, нв — нервное волокно,
яшк — ядро шванновской клетки, тм — терминальная масса. Из Terashima, Goris
and Katsuki, 1970. Воспроизводится с разрешения Academic Press, Inc.
Рецептивные поля волокон тройничного нерва обычно очень велики, но
поля разных волокон имеют разные центры и разные очертания. Тени, отбра-
сываемые краями ямки, по-разному влияют на разные рецептивные поля, что
может давать информацию о направлении на источник излучения. Прослежи-
вание сигнала из ямок в центральную нервную систему показало, что она на-
правляется в контрлатеральную крышу среднего мозга. Отведения от поверх-
ностного слоя крыши (tectum mesencephali) выявили наличие здесь зон, орга-
низованных по типу рецептивных полей «центр-окружение». Стимуляция
центра поля нагретой пластинкой (инфракрасное излучение) вызывает ко-
роткий залп активности при включении (ON) стимула. Если на центр поля
поместить холодную пластинку, активность подавляется, пока при удалении
холодового стимула не возникнет быстрый OFF-ответ. Напротив, если холо-
довой стимул помещен на периферию рецептивного поля, то возникает сла-
бый ON-ответ — аналогия с организацией РП сетчатки приматов по типу
«центр-периферия» очевидна (см. гл. 16). В равной мере интересно, что диа-
метры рецептивных полей достаточно малы — от 15° до 30°. Очевидно, между
большими РП ямок и намного меньшими — крыши происходит существен-
ная переработка информации. Малые РП позволяют сформировать карту ин-
фракрасного пространства. И действительно, пространственно-топические
карты, видимо, существуют. Таким образом питоны и ямкоголовые змеи осве-
домлены о положении источников тепла и менее нагретых участков в непо-
средственно близости от себя. Структуры крыши среднего мозга к тому же
очень чувствительны к движению тех и других. Движение пальца руки экспе-
риментатора на расстоянии 30—50 см или движение головы или конечности
человека на расстоянии в 2 м достаточны для активации нескольких зон в
крыше. Польза от такой пространственной и динамической информации не
нуждается в аргументации.
480 Часть V. Другие чувства
20.2. Поляризованный свет
Когда мы смотрим на безоблачное полуденное небо, нам кажется, что мы вну-
три безупречно голубого купола. Для многих животных, в особенности члени-
стоногих, это, однако, выглядит совершенно иначе. Голубизна неба возникает
вследствие рассеивания солнечного света пылевыми частицами в верхних сло-
ях атмосферы, это рассеивание поляризует свет. Степень поляризации света,
достигающего фоторецепторов, зависит от угла между направлением взгляда и
направлением поступающего солнечного излучения. Чем больше этот угол,
тем сильнее достигающий глаза свет поляризован. То есть лучи света, достига-
ющие глаза параллельно направлению поступающего излучения, не поляризо-
ваны, тогда как лучи, поступающие под прямым углом к этому направлению,
в идеале поляризованы на 100%. В реальных условиях атмосферная турбулент-
ность и многочисленные другие факторы уменьшают максимальную поляри-
зацию света до уровня не более 75—80%. Тем не менее, по мере движения солн-
ца по небосводу в течение дня картина поляризации постоянно меняется
(рис. 20.2). Рассеивание света также зависит от его длины волны (X). Расчеты
показывают, что хотя поляризация вследствие молекулярного рассеивания
А Середина лета,
Рис. 20.2. Влияние
положения солнца в
небе на ориентацию
полосы максималь-
ной поляризации.
Наблюдатель нахо-
дится в центре полу-
сферы. (А) Восход;
(Б) середина утра,
(В) полдень. Из Wel-
lington, 1974, с разре-
шения.
20. Редкие чувства 481
Рис. 20.3. Поле зрения рыбы
при направлении взгляда
вверх. (А) Поверхность воды и
«воздушное окно» при взгляде
снизу. (Б) Оптика. Лучи, па-
дающие на поверхность внут-
ри конуса (97,2°), достигают
глаз рыбы; лучи вне конуса —
отражаются. Объекты, нахо-
дящиеся вне конуса на дне
водоема или в толще воды,
могут давать отражение от по-
верхности воды, попадающее
в глаз рыбы. Дальнейшие объ-
яснения в тексте. Из Walls,
1963, с разрешения.
(рассеяние Рэлея) намного сильнее на коротких волнах (откуда голубизна не-
ба), более длинноволновое инфракрасное излучение имеет намного больше
шансов поляризоваться атмосферными частицами (откуда красные восходы и
закаты).
Эти физические факты означают, что, хотя мы этого видеть и не можем, го-
лубой купол неба для тех, кто в состоянии это заметить, сильно дифференци-
рован. Эта дифференциация содержит информацию, которая можт использо-
ваться и используется фоторецепторами многих животных. В частности, она
используется в определении направлений и, вероятно, важна для повышения
остроты зрения.
Поляризация света, конечно, является характеристикой света не только в
атмосфере, в равной мере это важно и относительно гидросферы. Свет, входя
в воду сверху, изменяет свое направление в соответствии со вторым законом —
синусоидальным — sin i/sin г = ц, где ц — коэффициент преломления. В случае
раздела сред воздух/вода ц = 1,33, это означает, что при углах преломления,
больше 48,6°, лучи света не могут пройти через границу раздела воздух/вода и
отражаются от нее. То есть для водных организмов вся воздушная полусфера
сокращается до конуса с углом раскрытия около 97,2° (рис. 20.3). Это не озна-
чает, что такие животные неспособны видеть горизонтально — параллельно
поверхности воды. Как показывает рис. 20.3, это вполне возможно, но при
этом имеются значительные искажения яркости и расстояний. Кроме того,
объекты, расположенные на дне мелководья, могут давать отражение от по-
верхности воды, попадающее в глаза водных животных. Конечно, все это
сильно зависит от состояния поверхности воды, которая гладкой бывает ред-
ко. Все это, возможно, как заметил Уоллс (Walls), и приводит к тому, что, ког-
да рыбу вытаскивают из воды, у нее такое ошеломленное выражение.
Частично вследствие ограничения проникновения света в воду в визуаль-
ный конус, только около 25% поляризации подводного света возникает за счет
поступающего непосредственно из воздуха. Свой вклад вносит направленное
16 Биология сенсорных систем
482 Часть V. Другие чувства
Рис. 20.4. Адаптации к восприятию плоскости поляризации света. (А) У краба
Callinectes развивается высокоорганизованный окаймленный слитый рабдом.
(1) Продольный срез; (2) поперечный срез показывает слитый рабдом, образован-
ный рабдомерами клеток ретинулы 1,4, 5; (3) поперечный срез показывает слитый
рабдом, образованный рабдомерами клеток ретинулы 2, 3, 6, 7; (4) пространствен-
ная схема, показывающая структуру четверти рабдома. БМ — базальная мембрана;
кфр — клетки, формирующие роговицу; рл — роговичная линза; кк — кристалли-
новый конус; рб — рабдом; кр — клетка ретинулы; АКР — аксон клетки ретинулы.
(Б) Структура рабдома осьминога Octopus (см. также рис. 14.13). (1) Продольный
срез; (2) поперечный срез через четыре рабдомера, образующих слитый рабдом.
взр — волокна зрительного нерва; пг — пигментные гранулы; пс — проксималь-
ный сегмент; кр — клетки ретинулы; рбм — рабдомер; мв — микроворсинки. Из
Waterman, 1981. Воспроизводится с разрешения Springer-Verlag.
вниз отражение света от поверхности воды, но основная поляризация обус-
ловлена самой водой. В основном это связано с суспендированными в воде не-
большими частицы (< 6 мкм), причем опять-таки основное воздействие при-
ходится на более длинноволновую часть спектра. При этом следует помнить,
что более длинные волны легче поглощаются водой, и в глубокие слои прони-
кает только коротковолновое излучение (Хтах около 465 нм).
Чувствительность к поляризованному свету широко распространена в цар-
стве животных. Показано, что она присутствует у более чем сотни видов,
включая ракообразных, паукообразных, головоногих моллюсков и позвоноч-
20. Редкие чувства 483
ных. Наиболее интенсивно исследовались в этом отношении членистоногие и
головоногие моллюски, у которых, как мы видели в гл. 14, фоторецепторы ор-
ганизованы по принципу рабдома. Зрительный пигмент клеток ретинулы
рабдома располагается в переплетенных микроворсинках (см. гл. 14). У деся-
тиногих раков (Decapoda — крупнейший отряд ракообразных, насчитываю-
щий более 26 000 видов) все исследованные к настоящему времени глаза имеют
удивительно единообразную структуру. Рабодомы состоят из клеток ретинулы,
у которых развились микроворсинки, перпендикулярные к продольной оси.
Микроворсинки в соседних клетках ретинулы направлены под прямым углом
друг к другу и соответствуют (приблизительно) вертикальной и горизонтальной
осям тела десятиногого рака. Иногда ортогональные микроворсинки лишь ча-
стично перекрывают рабодом (открытый рабдом), в других случаях микровор-
синки соседних клеток нахоходятся в близком контакте (слившийся рабдом),
есть еще случаи, когда микроворсинки направлены вниз — по длине рабдома
(рис. 20.4 А). Эта структура называется окаймленным рабдомом. Такие струк-
туры, по-видимому, образуют двухканальный анализатор поляризации.
Сходные, но более точные по организации микроворсинки рабдома обна-
ружены у насекомых, в том числе у интенсивно изучаемой пчелы (Apis), мяс-
ной мухи (Calliphora) и крупной стрекозы дозорщика Апахjunius. Как мы отме-
чали в главе 14, клетки ретинулы одного омматидия содержат фотопигменты,
чувствительные к разным длинам волн X. Анализ поляризации, таким обра-
зом, затрудняется необходимостью разделить его с анализом цвета. Однако,
прецизионная ортогональная организация микроворсинок предполагает, что
здесь перерабатывается нечто большее, чем просто цвет. Несомненно, исходя
из поведенческих данных, что эти насекомые чувствительны (и используют)
к плоскости поляризации света в атмосфере. Исследования фон Фриша ори-
ентации пчел при фуражировке в зависимости от плоскости поляризации све-
та стали классическими в биологической науке.
Наконец, это касается совершенно другого типа, существуют неоспоримые
поведенческие данные о способности головоногих моллюсков различать пло-
скости Тюляризации света. Опыты с пищевым и шоковым подкреплением по-
казали, что осьминог способен различать поляризацию света. Рабдомерные
ретинулы осьминогов и кальмаров похожи на таковые у чувствительных к по-
ляризации членистоногих по точности организации мозаики пптогонально
ориентированных микроворсинок. Опять-таки, как у членистоногих, микро-
ворсинки выстроены параллельно вертикальной и горизонтальной осям глаза.
Электрофизиологические отведения от клеток ретинулы осьминога показыва-
ют, что здесь имеется два (и только два) канала чувствительных к поляриза-
ции, перпендикулярных друг к другу (рис. 20.4 Б).
20.3. Электрические поля
Люди в нынешние времена окружены электрическими полями. Однако же, мы
не представляем себе ни их формы, ни силы. Иное дело с другими животными,
особенно рыбами. Системы детектирования слабых электрических полей,
вероятно, эволюционно очень древние, утрачивались и развивались заново
много раз.
484 Часть V. Другие чувства
Электрические рыбы известны со времен Древнего Египта. Знаменитая
фреска на стене захоронения, относящегося к пятой династии (около 2750 лет
до нашей эры), в Саккара1 изображает рыбака, выловившего из Нила вполне
узнаваемого электрического сома Malapterurus electricus. Римский врач Скри-
боний Ларг (Scribonius Largus) использовал электрического ската Torpedo для
шоковой терапии при инкурабельных головных болях и подагре. В древнегре-
ческой и древнеримской литературе немало пассажей о странной ошеломляю-
щей силе этих рыб.
Самый мощный электрический удар способны генерировать Torpedo и Ma-
lapterurus, большинство остальных — намного слабее. Соответственно, элект-
рические рыбы подразделяются на две группы: с сильно и слабо выраженны-
ми электрическими свойствами. К первой группе относятся электрический
скат Torpedo, электрические угри (Electrophorus) и электрический сом (Mala-
pterurus), генерирующие разряды, соответственно в 60 В, 500 В и 300 В. Боль-
шинство электрических рыб принадлежит ко второй группе, например, хря-
щевые рыбы, такие как скаты (напр. Raja), и костистые рыбы, принадлежащие
к семействам клюворылых (Mormyridae), гимнарховых (Gymnarchidae), гим-
нотов (Gymnotidae), стернопиговые (Sternopygidae), звездочетовых (Uranosco-
pidae) и т. д. Все эти рыбы способны генерировать лишь слабые электрические
разряды, самый сильный из которых у звездочета Astroscopus — около 5 В.
Слабые разряды обычно испускаются электрическим органом в хвосте в
виде серии высокочастотных импульсов. Например, гимнот Gymnotus испус-
кает импульсы длительностью 1 мс с частотой 50 Гц (в покое) и частотой 200 Гц
(при питании), тогда как гимнарх Gymnarchus niloticus — импульсы 1 мс (1 В)
в диапазоне 200—300 Гц.
Биологические функции сильных и слабых электрических разрядов сильно
различаются. Сильные разряды используются для оглушения жертв, тогда как
слабые разряды — для электроэхолокации при определении объектов вблизи
от рыбы, для коммуникации и, возможно, обозначения границ территории.
Существуют данные, что, например, представители гимнотов и клюворылых
способны распознавать разряды электрических органов особей собственного
вида и ориентироваться относительно них. Похоже, мы здесь видим повтор-
ную эволюцию слабого электрического чувства, аналогичную эхолокации у
летучих мышей, которую мы обсуждали в Части И. Дальнейший анализ элект-
рорецепции у рыб может дать еще более удивительные точки сравнения.
Вернемся, однако, к электрочувствительности, которая не ограничена эле-
ктрическими рыбами. Действительно, большинство рыб обладают способнос-
тью детектировать электрические поля в своей окружающей среде. Первые
эксперименты проводились на акулах-кошках Scyliorhinus canicula и морской
лисице Raja clavata, и в них было показано, что эти хрящевые рыбы способны
обнаруживать зарывшуюся в песок на дне аквариума морскую камбалу (Pleuro-
nectesplatessa) (рис. 20.5). После случайных поисковых движений акула-кошка
чует какие-то признаки местоположения камбалы с расстояния около 15 см и
целенаправленно ныряет, откапывая ее из песка, и съедает. Чтобы убедиться,
1 Саккара (Sakkara) — одна из частей огромного некрополя в Мемфисе, столице Старого
царства. — Прим, перев.
20. Редкие чувства 485
Рис. 20.5. Реакция акулы-
кошки Scyliorhinus canicula на
(А) камбалу Pleuronectes pla-
tessa, закопавшуюся в песок;
(Б) камбалу, помешенную в
электропроводящую камеру;
(В) кусочки мела; (Г) камба-
лу в изолирующей камере;
(Д) электроды, симулирую-
щие электрическое поле, ге-
нерируемое камбалой; (Е) ку-
сочки мела и электрическое
поле (показан только один
электрод). Сплошные стрел-
ки показывают направление
движения акулы, пунктирные
стрелки — направление тока в
морской воде. Из Kalmijn,
1971, с разрешения.
что обоняние не участвует в этом процессе, камбалу помещали в электропро-
водящую камеру из агара, но и в этом случае акула-кошка находила и откапы-
вала камбалу из песка. Если же камбалу помещали в камеру, экранирующую
электрическое поле, акула уже не в состоянии была обнаружить ее присутст-
вие, и камбала оставалась в своем контейнере жива-здорова. В следующей
части эксперимента в песок помещали электроды, чтобы симулировать био-
электрическое поле камбалы — акула снова ныряла в поисках источника
электрического сигнала. Было показано, что акула-кошка наиболее чувстви-
тельна к полям, окружающим набор диполей, на расстоянии около 5 см при
постоянном токе около 0,5 А. Акула-кошка и скаты реагировали и на синусо-
идальный ток с частотой 1, 2 и 4 Гц. В реальной жизни биоэлектрические по-
ля генерируются ионными токами, протекающими через жаберные филамен-
ты и эпителий кишечника. Мышечные движения также генерируют биоэлек-
трические потенциалы и, следовательно, электрические поля. Биоэлектриче-
ские поля, генерируемые раненной тканью, существенно мощнее. Показано,
что камбала, лежащая спокойно под песком, генерирует биоэлектрические
поля напряженностью в среднем 0,2 мкВ/см на расстоянии 10 см от рыбы.
Эти поля колеблются по величине в соответствии с ритмом дыхательных дви-
жений рыбы.
486 Часть V. Другие чувства
Рис. 20.6. (А) Ампулярный орган; (Б), (В) и (Г) бугорковые органы. Объяснения в
тексте. Отметьте многослойность кожи, б — базальный слой кожи; бм — базальная
мембрана; кк — кроющие клетки; зк — закупоривающие клетки; н — немиелини-
зированное нервное волокно; ск — сенсорная клетка; 1, 2, 3, 4 — различные типы
дополнительных базальных клеток. Из Szabo, 1974, с разрешения.
Электрочувствительность хрящевых рыб чрезвычайно остра. Они доста-
точно чувствительны, чтобы детектировать поля всего в 5 нВ/см. Такая точ-
ность чувствительности станет более очевидной, если мы переведем ее в более
привычную для человека шкалу: она эквивалентна градиенту в 0,5 В на дистан-
ции в 1000 км. Таким образом, они вполне в состоянии обнаружить поле на-
пряженностью 200 нВ/см на расстоянии ы 10 см от закопавшейся в песок кам-
балы. Вопрос о том, может ли быть такая высокая чувствительность достаточ-
ной не только для обнаружения скрывшейся жертвы, но и для детектирования
слабых электрических полей, создаваемых собственными движениями, и
(или) мощными потоками воды океанических течений в магнитном поле Зем-
ли, остается дискуссионным. Есть некоторые свидетельства того, что хряще-
вые рыбы действительно используют магнитное поле Земли для навигации на
дальние расстояния. Слежение за мигрирующими синими акулами в течение
многих дней показало, что они поддерживают постоянный курс на дистанци-
ях в несколько сот километров без каких-либо иных ясных навигационных
средств. Акула-молот калифорнийского побережья двигается по маршруту,
коррелирующему с магнитными аномалиями дна моря. Следует, однако, по-
мнить, что многие рыбы и морские млекопитающие, не обладающие тонкой
20. Редкие чувства 487
Рис. 20.7. Анатомия ампул Лоренцини. (А) виды дорзальной и вентральной по-
верхностей системы каналов морской лисицы Raja clavata. Отверстия каналов
показаны точками, а капсулы — кружками. (Б) виды дорзальной и вентральной
поверхностей акулы-кошки Scyliorhinus canicula. г — гиоидная капсула. Объясне-
ния в тексте. Из Murray, 1974, с разрешения.
Дорзальная Вентральная
поверхность поверхность
электрочувствительностью, тем не менее достигают замечательных результа-
тов в подводной навигации на дальние расстояния.
В случае рыб со слабыми электрическими свойствами детектируются не
только электрические поля, генерируемые движением в геомагнитном поле,
и электрические поля, генерируемые другими животными. Их собственные
электрические органы генерируют электрическое поле, которое искажается
близко расположенными объектами, и эти искажения воспринимаются соб-
ственной электрорецепторной системой. Клюворылые рыбы, например, спо-
собны ощущать электрическую емкость близлежащих объектов в диапазоне
0,22—1,7 нФ до 120—680 нФ. Они могут воспринимать размер, форму, локали-
зацию и электрическое сопротивление этих объектов.
Замечательная электрочувствительность слабо электрических рыб и других
животных достигается двумя типами рецепторов: ампулярными и бугорковыми
органами (рис. 20.6). Оба типа рецепторов являются модификациями системы
боковой линии, вездесущей, как отмечалось в гл. 8, у рыб.
Ампулярные органы (рис. 20.6 А) — это группы сенсорных клеток, органи-
зованных вокруг полости длинного, заполненного желеобразным содержи-
мым, канала. Сенсорные клетки ампул почти полностью окружены вспомога-
тельными клетками. Только верхушка сенсорной клетки остается свободной и
контактирует с пластинкой ампулы. Сенсорные клетки имеют несколько мор-
фологических вариаций. У многоперовых, двоякодышащих и некоторых зем-
новодных из верхушки клетки единственная киноцилия простирается из вер-
хушки клетки в окружении множества микроворсинок (ср. с волосковыми
клетками системы боковой линии, описанными в гл. 8). У хрящевых рыб и ко-
488 Часть V. Другие чувства
Рис. 20.8. Бугорковые органы пер-
вого (Al, А2, АЗ) и второго (В) типа,
бм — базальная мембрана, н — нерв.
Объяснения в тексте. Из Szabo,
1974, с разрешения.
стных ганоидов единственная киноцилия микроворсинками не окружена.
Наконец, у миног и хвостатых амфибий при множестве микроворсинок кино-
цилия отсутствует. Функциональное значение этих вариаций остается неиз-
вестным. Основание клеток заполнено синаптическими пузырьками, здесь
образуется классический химический синапс с дендритными окончаниями
сенсорного нервного волокна. Обычно группы из 12—13 ампул образуют ам-
пулярный орган.
Ампулы Лоренцини образуют высокоорганизованный тип ампул у хрящевых
рыб, включая химеровых (напр. европейская химера Chimaera monstrosa). По-
следние развивались независимо от основной группы хрящевых рыб, по край-
ней мере с юрского периода. Ампулы Лоренцини наиболее интенсивно иссле-
довались у скатов {Raja) и акул-кошек {Scyliorhinus). Мы отмечали в гл. 19, что
сначала они были приняты за терморецепторы. Организация системы у ската
показана на рис. 20.7. Имеется несколько систем каналов на дорзальной и на
вентральной поверхности тела. Длина каналов варьирует. У среднего размера
ската (400 мм в размахе) самый длинный канал достигает 160 мм при диаметре
в 2 мм; самый маленький — 5 мм в длину и 0,5 мм в диаметре. Тип каналов ви-
доспецифичен. Каналы заканчиваются слепо — сенсорными ампулами. Ампу-
лы часто сгруппированы и окружены соединительной тканью, образующей
хорошо заметные капсулы. В настоящее время уже нет сомнений, что их ос-
новная роль состоит в обеспечении скатов, акул-кошек и других хрящевых
рыб точной электрочувствительностью, обсужденной нами выше.
Тот факт, что каналы ориентированы во многих разных направлениях,
позволяет системе определять градиенты электрических полей окружающего
пространства. Более длинные каналы менее чувствительны к более высоким
частотам, вследствие электрической емкости стенок канала, однако более
чувствительны к градиенту напряженности поля. Вся система обеспечивает
хрящевых рыб чувствительными дистантными рецепторами, способными де-
тектировать многие черты живых и неживых объектов окружающей среды. Ее
значение подчеркивается богатой иннервацией.
20. Редкие чувства 489
Рис. 20.9. Распределение электрорецепторов гимнотиды Stenarchus. Ампулярные
рецепторы показаны черными точками. Намного более многочисленные бугорко-
вые рецепторы показаны только в маленьком участке средне-дорзальной области.
Плотность последних рецепторов на мм2 показан числами и стрелками. Пункти-
ром показана задняя боковая линия. Из Bennett, 1971. Воспроизводится с разреше-
ния Academic Press, Ltd.
Бугорковые органы (tuberous organs или knollenorgans) (рис. 20.6 Б,В,Г) от-
личаются от ампулярных органов двумя существенными чертами. Во-первых,
они не соединены с внешней средой желе-заполненным каналом. Вместо
этого канал закупорен специализированными эпителиальными клетками.
Во-вторых, сенсорные клетки в полость выходят не только верхушкой, а вы-
двинуты в нее на 90% и лишь присоединены (обычно) к небольшому поддер-
живающему холмику. Сенсорные клетки гимнотид несколько удлинены по
сравнению с более сферическими клетками клюворылых. У сенсорных кле-
ток бугорковых органов киноцилия не развивается, зато их поверхность по-
крыта множеством микроворсинок, которые проецируются в сенсорную по-
лость. Базальная часть сенсорной клетки заполнена синаптическими пузырь-
ками. Число сенсорных клеток, образующих бугорковый орган, варьирует
широко и составляет от 1 до 35 у клюворылых и 10—100 у гимнотид. У клюво-
рылах, к тому же, каждая клетка располагается в собственной полости, тогда
как у гимнотид все сенсорные клетки располагаются в общей полости. Нако-
нец, бугорковые органы часто подразделяются на два типа по их иннервации
(рис. 20.8). Первый тип иннервируется миелинизированными веточками
одного сенсорного волокна. У второго миелин не проникает в бугорковый
орган, и, соответственно, сенсорная клетка иннервируется немиелинизиро-
ванными веточками афферентного нерва.
Два класса электрорецепторов имеют различные характеристики ответа.
Афферентные волокна из ампулярных сенсорных клеток дают «тонические» —
длительные и непрерывные ответы на низкочастотные (<0,1 Гц до 10—25 Гц)
стимулы или постоянный ток. Ампулярные органы имеют широкий диапазон
чувствительности с порогами от < 20 нВ/см до 10—100 мкВ/см. Напротив, бу-
горковые органы чувствительны к высоким частотам и нечувствительны к
низким и постоянному току. Сенсорные волокна из бугорковых органов дают
«фазические» ответы — короткий залп активность на ступенчатое изменение
стимулирующего напряжения. И тонические (ампулярные), и фазические (бу-
горковых органов) рецепторы имеются и у клюворылых, и у гимнотид. Их
сходство поразительно, если принимать, что эти две группы пресноводных
рыб развивались независимо. Бугорковые органы отстутствуют у неэлектриче-
ских и морских рыб.
490 Часть V. Другие чувства
Распределение электрорецепторов у типичной пресноводной гимнотиды
показано на рис. 20.9. Фазические рецепторы (бугорковые) намного более
многочисленны, чем тонические, и показаны только в небольшой области. Их
плотность (на мм2) показана в разных точках цифрами. Тонические рецепто-
ры (ампулярные) показаны ромбиками. Надо отметить, что и тонические и
фазические рецепторы достигают максимальной плотности в области головы.
Оба набора рецепторов иннервируются большой передней в ветвью нерва бо-
ковой линии (в данном случае черепно-мозгового нерва VIII).
Детектирование формы электрического поля в окружающей воде зависит
от ориентации рыбы. Электрическое сопротивление тела значительно ниже,
чем окружающей среды. Если рыба ориентирована вдоль градиента потенциа-
ла, ток входит на одном ее конце (скажем, в голове) и выходит на другом
(в хвосте). Индивидуальные электрорецепторы стимулируются различиями
между внутренним и наружным электрическим потенциалами. Эти различия
максимальны у головы и хвоста, где входит и выходит ток. В отличие от хряще-
вых рыб с их высокоорганизованной системой ампул Лоренцини пресновод-
ные клюворылые и гимнотиды как элекгрочувствительный орган используют
все свое тело.
20.4. Магнитные поля
Хотя предполагают, что многие организмы способны ощущать магнитное по-
ле Земли, только для микроорганизмов существуют доказательства такой спо-
собности. Магнитотаксические бактерии были открыты в 1970-х годах — это
водные жгутиковые грам-отрицательные формы с внутриклеточными железо-
содержащими гранулами, обнаружимыми на электронно-микроскопическом
уровне. Эти гранулы — магнитосомы — состоят из Fe2O4 или магнетита; у Aqua-
spirillium magnetotacticum грань кристалла достигает 42 нм, т. е. в пределах раз-
мера магнитного домена магнетита (40—100 нм). Магнетитовые гранулы обра-
зуют цепочки до 20 единиц в каждой бактерии (рис. 20.10).
Магнитные включения действуют как «ферромагнитный геомагнитный
биокампас». Они позволяют бактерии плыть вдоль магнитных силовых линий.
В северном полушарии эти линии направлены вниз, соответственно, внизу
плывут и магнитотаксические бактерии. Легко изменить направление их
движения, приложив внешнее магнитное поле. В южном полушарии геомаг-
нитное поле направлено вверх. Аборигенные бактерии здесь ревертировали
направление своего движения и плывут по силовым линиям вниз. На геомаг-
нитном экваторе обнаружено примерно равное число бактерий, плывущих
вверх и вниз.
Существует множество спекуляций по поводу наличия или отсутствия маг-
нитного чувства у эукариот, особенно у животных. Мы отмечали выше, когда
обсуждали электрорецепцию, что чрезвычайно высокая электрочувствитель-
ность хрящевых рыб может позволить им ориентироваться относительно гео-
магнитного поля, детектируя токи, индуцированные их собственным движе-
нием в магнитном поле. Большой интерес существует и к возможности учас-
тия магнитного чувства в выдающихся навигационных способностях птиц.
Существовало предположение, что магнитное вещество, возможно магнетит,
20. Редкие чувства 491
Рис. 20.10. Магнитосомы грам-отрицательной бактерии Aquaspirillium magneto-
tacticum. Масштабная линейка — 0,5 мкм. (А) Электронная микрофотография не-
гативно окрашенной бактерии. (Б) Электронная микрофотография тонкого среза
бактерии. В обоих случаях магнитосомы выглядят как черные частицы. На рис. (Б)
хорошо видна кубическая форма, а внизу микрофотографии — часть другой бакте-
рии, вероятно, поперечный срез. На этом срезе хорошо видно, что магнитосомы
располагаются вблизи от поверхностной мембраны. Из Blakemore, 1982. С разре-
шения из Annual Review of Microbiology, volume 36, © 1982, Annual Reviews www.
annualreviews.org.
образует часть магниторецептора, связанного с глазной частью тройничного
нерва птиц. Однако, к моменту написания данной книги, ничего похожего на
магнитосомы бактерий не идентифицировано. Периодически появляются
публикации на тему влияния магнитных полей на метаболизм эпифиза млеко-
питающих (включая человека). Возможно, они воздействуют на циркадные
ритмы, контролирующие синтез мелатонина. Пока этот эффект кажется до-
стоверным, но механизм, его обеспечивающий, и его биологическое значение
(если оно есть) остаются окутанными тайной.
492 Часть V. Другие чувства
20.5. Заключение
И опять, как это бывало в предшествующих главах этой книги, мы остаемся
под впечатлением разнообразного, иногда разительно разнообразного сенсор-
ного мира, в котором существуют многие животные. Классические пять
чувств, описанные Аристотелем две с половиной тысячи лет назад, — осяза-
ние, вкус, обоняние, слух и зрение — с их радикально отличающимися сенсор-
ными качествами — лишь часть огромного разнообразия чувств, развитых в
царстве животных. Мы отмечали в предыдущих главах, что наши сенсорные
склонности, особенно к зрению, могут отражать наше эволюционное про-
шлое. Эволюция устроила так, что сенсорные предпочтения других животных
построены иначе. В данной главе мы видели, что сенсорные системы некото-
рых из них дают информацию о событиях, на которые мы не видим даже и на-
мека. Это полезно, это напоминает нам, что есть точки зрения, отличные от
наших. Это напоминает нам о том, что Чарльз Дарвин называл нашим высоко-
мерием. Сенсорный мир, который населяем мы, мир, к которому мы привык-
ли, который нам так нравится, кажется таким прекрасным, таким ужасным —
это мир человека. А сенсорный мир других животных — совсем другое дело.
21. БОЛЬ
Определения боли: Аристотель — «экзистенциальное» чувство — неиз-
бежность — трудность качественной и количественной оценки — отсут-
ствие «долориметра» — влияние условий и культуры — проблема боли у
животных. Биологическое значение: предупреждение и залечивание по-
вреждения тканей — опасности врожденной анальгезии — проблема хро-
нической боли — инкурабельные боли в спине — теологические пробле-
мы — биологические решения — простых решений нет — фантомные ко-
нечности — анальгезия в бою и при других опасностях — несовпадение
афферентации и эфферентных копий? Нейрофизиология: периферические
рецепторы — ноцицептивные волокна — высокопороговые А5 и С-волок-
на — два типа механо-тепловых А5 (АМТ) ноцицепторов — механо-теп-
ловые С (СМТ) ноцицепторы — передача стимула — первичная и вторич-
ная гиперальгезия; центральные пути — дерматомы — ламинарная струк-
тура серого вещества спинного мозга — гистологические исследования с
помощью пероксидазы хрена и прослеживание волокон — восходящие
пути в белом веществе (STT, SRT, SMT) — тракт Лиссауэра — отсутствие
специализированной ноцицептивной области в коре головного мозга —
нисходящие пути — вызванная анальгезия (ВА) — значение серого веще-
ства вокруг водопровода (ВСВ). Нейрофармакология: эндогенные опиои-
ды — типы и локализация опиоидных рецепторов в ЦНС — синаптоло-
гия и фармакология ноцицептичных путей в дорзальном роге. Иррадии-
рующая боль: грудная жаба — симптомы — дерматомы — боль в спине —
триггерные точки и акупунктура. Воротная теория: упрощенная принци-
пиальная схема — механизм — нисходящие проекции — терапевтические
устройства. Заключение: дилемма боли в биомедицинских исследовани-
ях — счастье жить в 21-м веке — ощущение боли устроено не грубо, а
хитроумно
Аристотель охарактеризовал боль как неприятное ощущение. Это определение
тиражировалось столетиями, и до сих пор (оно само или его версии) может
быть найдено в учебниках. А ведь оно либо неверно, либо просто лишено
смысла. Все мы испытываем неприятные ощущения, которое, однако же, ни-
как болью не являются. Вспомним, как «сосет под ложечкой» в приемной у
стоматолога перед тем, как сесть в кресло; вспомним ощущение, которое воз-
никает у тех из нас, кто не является заядлым альпинистом, когда мы смотрим
вниз с головокружительной высоты скалы или небоскреба; а внезапное осо-
знание того, что ложка вкусной еды, которую мы только что сунули себе
494 Часть V. Другие чувства
в рот — это последняя трапеза перед казнью? Все эти ощущения отчетливо
неприятны, но вряд ли могут считаться болью. Если, с другой стороны, все
неприятные ощущения определять как боль, тогда определение Аристотеля
замыкается само на себя и является тавтологией или попросту пустышкой.
Что такое боль? — неприятное ощущение; что такое неприятное ощуще-
ние? — боль.
Аристотель, тем не менее, был одним из величайших умов и одним из ве-
личайших биологов в истории. Так что, хотя его определение и неудовлетво-
рительно, отнесемся к нему с полной серьезностью. Ведь что такое «боль»,
действительно трудно определить в первую очередь потому, что чувство это —
глубоко личное. Некоторые называют боль «экзистенциальным» чувством,
поскольку нет способа (кроме лекарств) избавиться от нее. Мы не можем уйти
от боли, как от ворчливой жены. Подобно тому, как невозможно (согласно
блаженному Августину и Декарту) сомневаться в том, что сомнения действи-
тельно существуют (я мыслю — следовательно, я существую), так же несо-
мненно что если больно — боль существует. Мы можем сомневаться в других
чувствах: у всех бывают зрительные, слуховые и (в меньшей степени) обоня-
тельные иллюзии; мир, который они нам представляют, может быть иным, но
вот никаких сомнений быть не может в том, есть ли боль, когда у нас болит.
Мы, как и в случаях с другими чувствами, можем заблуждаться по поводу
причин «неприятного ощущения», но не можем избежать его, закрыв глаза,
заткнув уши или нос.
Мы вернемся к этим, в большей степени философским проблемам в гла-
ве 23, однако уместно здесь отметить, что глубоко личная природа боли делает
затруднительным ее описание или количественную оценку. Медики часто раз-
личают острую, колющую боль и более продолжительную глухую боль. Это,
однако, лишь грубые приблизительные и поверхностные различия. Мельзак
(Melzack) разработал детальный вопросник (вопросник МакГилла) для боль-
ных, где боль описывается по множеству рубрик: пульсирующая, стреляющая,
бьющая, грызущая, сжимающая, давящая, разрывающая, вызывающая тошно-
ту. Такой список, однако, может быть только метафорическим, и маловероят-
но, чтобы он мог охватить все варианты ощущения боли. Например, хирурги-
ческое вмешательство на тройничном нерве оставляет у 5% пациентов необыч-
ное и неприятное ощущение, которое пока не удалось описать и от которого
невозможно избавиться. Патрик Уолл, внесший большой вклад в изучение бо-
ли, отметил, что у нас совершенно отсутствует инструмент — долориметр1, с
помощью которого мы могли бы измерить боль количественно. Мы ведь не
только не можем понять, какова боль другого, но и никогда не можем быть
уверены в ее интенсивности. Связь между ощущаемой и описываемой болью
очень сильно зависит от ситуации. Есть достаточные основания полагать, что
и качество и интенсивность боли в значительной степени зависит от внешних
обстоятельств, в том числе от обычаев, от существующей практики и того, че-
го можно и чего нельзя ожидать в той или иной культурной традиции. Часто
солдаты, вытащенные с поля сражения с тяжелыми ранениями, в первый мо-
1 Dolor (лат.) — боль. — Прим, перев.
21. Боль 495
мент очень редко жалуются на боль, при этом очень раздражительно реагируя
на сравнительно незначительные неприятности, вроде неудачной инъекции;
в мирных условиях люди с похожими по тяжести травмами куда больше
склонны требовать введения им анестетика. Сходным образом, церемонии
инициации и другие обычаи, зафиксированные антропологами у некоторых
племен, включают процедуры, которые должны быть крайне болезненны, од-
нако субъект церемонии, находящийся в состоянии экзальтации, по-видимо-
му, либо чувствует лишь слабую боль, либо не чувствует ее совсем.
Если уж мы не в состоянии количественно оценить и делать нечто больше,
чем догадываться о том, какова боль другого человека, на что можно надеяться,
когда речь идет о боли у животных? Лучшее, что мы можем сделать в этом во-
просе это — проявить максимальную осторожность. Выше мы отмечали, что в
определенном смысле боль является субъективным чувством. Мы видели, что,
как и другие сенсорные качества — видя «красное» или слыша звуковой тон, мы
не можем сомневаться в собственных ощущениях, хотя можем и не знать, что
является причиной этих ощущений. Аналогичным образом, если мы не сомне-
ваемся, чтоюни видят и слышат, мы не должны сомневаться и в том, что они
способны испытывать боль. Оставим на совести охотников и рыболовов и вся-
ких стрелков по куропаткам попытки уравновесить боль, которую они причи-
няют, с тем удовольствием, которое они получают. Не последний среди факто-
ров оценки — это признание того, что природа основана на смертности.
21.1. Биологическое значение боли
На анатомическом и физиологическом уровнях боль сигнализирует о повреж-
дении тканей и необходимости отдыха, чтобы защитные силы организма мог-
ли залечить повреждение, то есть предупреждает от дальнейшего риска для по-
врежденного органа. Такие сигналы образуют условные рефлексы, типа тех,
которые возникают, когда мы, сунув руку раз в нечто ярко сверкающее и ко-
леблющееся, никогда больше так не делаем. Держась подальше от огня, мы
увеличиваем собственные шансы на долгую и продуктивную жизнь. Некото-
рые несчастные рождаются без чувства боли — с патологией, называемой
врожденной анальгезией. Дефект, скорее центральной, а не периферической
природы, поскольку больные способны, помимо прочего, отличить прикосно-
вение головки булавки от укола ее острием. Кое-кто может подумать, что эти
несчастные счастливы. Это, однако, в случае с врожденными анальгезиками
не так, поскольку лишь с огромным трудом можно научить их избегать травм.
Особая проблема — это воспаления суставов из-за неспособности вовремя пе-
реносить вес тела при стоянии, сидении и даже лежа во сне. Уолл сообщает о
случае молодой женщины с анальгезией, у которой воспаления суставов в кон-
це концов привели к настолько массивной инфекции костей, что спасти ее ме-
дикам не удалось. Таким образом, ноцицептивная система — это внутренний
механизм безопасности.
С другой стороны, как относиться к хроническим, отравляющим жизнь бо-
лям? Боли, вызываемые болезнями сердца, язвами желудочно-кишечного
тракта, разными формами раковых заболеваний были ужасными до появления
современной медицины. Во многих случаях даже сегодня причины хроничес-
496 Часть V. Другие чувства
ких болей не удается установить. Данные по США показывают, что в настоя-
щее время два миллиона человек находятся на инвалидности по причине хро-
нических болей, многие из них неизлечимы в течение длительного времени.
Только боли в спине представляют собой колоссальный источник дискомфор-
та. По оценкам около 60% британской и американской популяции за время
своей трудовой деятельности теряют не менее рабочей недели только по этой
причине. Ежегодные компенсации в США достигают 50 миллиардов долла-
ров, из них — 85% без поставленного диагноза. Хронические неустранимые
боли — это колоссальная гуманитарная и экономическая проблема. Это огром-
ное и в основном невидимое море боли наталкивает нас на ответ на поставлен-
ный выше вопрос. Ноцицептивная система настолько важна в эволюционной
борьбе за выживание и возможность оставить потомство, что это перевешивает
иногда бессмысленный и обессиливающий дискомфорт, вызванный болью.
Теологи частенько ставят вопрос о смысле существования боли и страдания в
мире. Для эволюционных биологов ответ, хотя и не простой, по крайней мере,
мыслим.
То, что ответ на этот вопрос не прост, можно продемонстрировать на мно-
жестве примеров. Прямолинейной знак равенства между повреждением ткани
и болью часто не выдерживает критики. Упомянутые выше хронические боли
в спине лишь изредка могут быть напрямую связаны с конкретным поврежде-
нием тканей. В случаях ранений, особенно тяжелых, боль обычно сохраняется
вплоть до излечения. Наиболее ярко это проявляется в случае т. н. фантомных
конечностей. Годами после того, как ранения зажили, пациенты страдают от
мучительных болей в ампутированных органах. Более того, есть хорошо доку-
ментированные случаи, когда острая боль сохранялась в фантомной конечно-
сти при повреждении всех путей к ней. Иногда такое наблюдается у жертв ка-
тастроф с повреждениями спинного мозга. После острого периода пациент ут-
рачивает чувствительность и подвижность ниже места перелома позвоночни-
ка, при этом в некоторых тяжелых случаях сохраняются мучительные боли в
«неконтролируемых областях». Даже после хирургического вмешательства по
перерезке болевых путей спинного мозга выше места перелома (см. ниже) у та-
ких больных сохранялись колики и жжение. Такие больные — это вызов лю-
бой теории, связывающей возникновение боли с повреждением конкретной
ткани. Возможно, дело в том, что мозг не может сравнить эфферентную ко-
пию» (см. главу 15, раздел 4), с отсутствующей афферентной картиной. Это
предположение находит подтверждение в пока противоречивых, но тем не ме-
нее остроумных экспериментах, в которых с помощью системы зеркал у паци-
ента создавали зрительную иллюзию, будто утраченный орган у него появился
снова. В этом случае фантомные боли иногда резко ослабевали. Еще один ар-
гумент получен в значительно менее трагических обстоятельствах. Если ло-
кальный анестетик инъецировали в плечевое сплетение (сплетение нервов от
верхней конечности, входящее в спинной мозг на уровне плеча), то пациент
ощущал, что рука болезненно вывернута и находится в определенном, часто
невозможном (завернута в грудную клетку), положении. Иллюзия устраняет-
ся, только если пациент смотрит на руку или нащупывает ее в реальном поло-
жении. В данном случае речь идет не об определенном паттерне афферента-
ции, а об отсутствии такого паттерна, что «привлекает внимание» ЦНС.
21. Боль 497
Напротив, боль часто не чувствуется, даже когда повреждения серьезны.
Солдаты в пылу боя часто не замечают даже достаточно тяжелых ран. Только
позже, когда битва проиграна или выиграна, но делать уже нечего, приходит
боль. Знаменитый пример с доктором Ливингстоном и львом подтверждает
это. Ливингстон вспоминает, как в глубинах Африки лев выпрыгнул из джун-
глей прямо ему на спину. Он помнит и ужасное рычание, и хищную пасть, и
ужасные когти, рвавшие его тело, пока спасители не освободили его; но, пи-
шет он, все это происходило, как во сне, он не чувствовал ни боли, ни ужаса.
Доктор Ливингстон не сомневался, что все это подтверждает благодеяние бо-
га, который не позволил творению своему погрузить его, доктора, в бездонную
пучину боли и страданий. Эволюционисты могут предложить иное, хотя в ка-
кой-то степени и комплементарное объяснение — организм концентрируется
на одной задаче в каждый данный момент. Скончание жизни в челюстях льва,
ранение в бою — это не адаптивное поведение, момент кризиса мало подходит
для того, чтобы организм озаботился восстановлением поврежденных тка-
ней — в этот момент у организма слишком много более срочных забот. Вы-
званная болью неподвижность, необходимая для выздоровления, наступает
позже.
21.2. Нейрофизиология боли
Несмотря на огромную сложность переработки болевой информации в мозге,
нужно перейти непосредственно к нейрофизиологии болевых рецепторов и их
периферических и спинальных путей.
21.2.1. Периферические ноцицепторы
Многие годы было непонятно, возникает ли боль в результате активации спе-
цифических волокон или в результате сверхактивности сенсорных волокон, в
норме имеющих другие модальности. Последняя возможность, как кажется, в
большей степени соответствует нашему обыденному опыту. За возможным ис-
ключением обоняния, любые избыточные по интенсивности сенсорные сти-
мулы — слепящий свет, рвущий ухо звук, тяжелый удар, тепло или холод за
пределами нормального диапазона — приводят к возникновению боли. Такой
взгляд здравого смысла был заявлен Эразмом Дарвином (Erasmus Darwin) в
конце 18-го и Уильямом Джеймсом (William James) в конце 19-го века. Здра-
вый смысл, однако, здесь (как и везде) оставляет желать чего-то еще. В насто-
ящее время мало кто сомневается, что в большинстве случаев ощущение боли
возникает в результате специализированных — ноцицептивных волокон.
Ноцицептивные волокна не имеет специализированных окончаний. Они
присутствуют в виде свободных нервных окончаний в дермисе кожи и в иных
местах организма. Гистологически они неотличимы от С-механорецепторов
(глава 7) и С- и А8-терморецепторов (глава 19). Они отличаются от упомяну-
тых рецепторов тем, что порог для их адекватных стимулов выше нормально-
го диапазона. Они могут подразделяться на несколько разных типов по кри-
терию того, какая сенсорной модальность представляет для них адекватный
стимул. Болезненные термические и механические стимулы детектируются
17 Биология сенсорных систем
498 Часть V. Другие чувства
миелинизированными волокнами малого диаметра, таблица 2.2 показывает,
что они относятся к категории А5-волокон. Полимодальные волокна, которые
отвечают на широкое разнообразие интенсивностей стимулов разной модаль-
ности, также имеет малый диаметр, но не миелинизированы. Таблица 2.2 по-
казывает, что эти волокна относятся к классу С. А5-волокна проводят импуль-
сы с частотой 5—30 м/с и ответственны за «быструю» боль, острое колющее
ощущение; С-волокна проводят медленнее — 0,5—2 м/с и сигнализируют о
«медленной» боли, часто продолжительной и часто переходящей в глухую
боль.
Механо-термо-ноцицепторы с А5-волокнами (АМТ) делятся на два типа.
АМТ типа 1 в основном обнаруживаются в неоволосенной коже. Хотя некото-
рые рецепторы имеют порог ниже 50°С, большинство реагирует на температу-
ры выше 53°С. У них высокая скорость проведения — 30—50 м/с и, таким об-
разом, переходят из А5- в Ap-категорию. АМТ типа 1 первоначально не реа-
гируют на стимул в 53°С, однако затем через 2—3 с дают сильно выраженный
ответ (рис. 21.1 Б). АМТ типа 2 обнаружен в оволосенной коже, их температур-
ный порог и скорость проведения (около 15 м/с) значительно ниже, но реаги-
руют быстрее на включение высокой температуры. Как видно из рис. 21.1 Б,
они, в отличие от АМТ типа 1, это быстро адаптирующиеся рецепторы. Нако-
нец, ноцицепторы с С-волокнами (СМТ) имеют порог в диапазоне 38°С—50°С
и отвечают постоянной активностью, которая зависит от интенсивности сти-
мула (рис. 21.1 А).
АМТ и СМТ, как показывают их названия, реагируют и на термические, и
на механические стимулы. Физиологическая ситуация, тем не менее, далека от
простоты. Механизм передачи этих двух модальностей различен. Аппликация
капсайцина не влияет на чувствительность к механическим стимулам, но инги-
бирует ответ на тепловые. При этом, тогда как капсайцин имеет анальгетичес-
кий эффект в отношении тепловой и химической чувствительности полимо-
дальных С-волокон в роговице, на механочувствительности он не сказывается.
Наконец, было показано, что механические стимулы, которые генерируют та-
кой же уровень активности в СМТ-волокнах, что и термические, вызывают,
тем не менее, меньшую боль. Возможно, неизбежно более широкая поверх-
ность, задействованная тепловым стимулом, вовлекает активность большего
количества СМТ-волокон, чем в случае механического стимула.
Физиологическая ситуация непроста и в другом отношении. Перифериче-
ские механизмы очень динамичны. После повреждения, как все мы знаем,
окружающие его ткани становятся «болезненными» или, если использовать
термин — гиперальгезическими. Гиперальгезия определяется как сдвиг в левую
сторону кривой стимул-ответ. Она наблюдается не только в месте повреждения
(первична гиперальгезия), но и в окружающей ткани (вторичная гиперальге-
зия). Механизмы первичной и вторичной гиперальгезий различны. Первичная
гиперальгезия показана у всех нервных окончаний. Она возникает в результате
сенситизации нервных окончаний и расширения их рецептивных полей. Вто-
ричная гиперальгезия не показана на термические стимулы, но присутствует для
механических и других ноцицептивных стимулов. Она возникает в результате
выделения целого коктейля веществ, частью из ноцицептивных нервных
окончаний, а частью из поврежденной ткани (рис. 21.2 А).
21. Боль 499
: АМТ типа 1 •
-----:—nil !> н чип ill 111 i II III Ulf ff4WWIl4l«iHil4i}ilil и
----i 1 I----------------------1.. .....iuiii nun j
----j—।—111А_1Ж1ьшш|шшшживи11вшмшвамшамвмш|ши_
; nminnuiifiiiiiiiiiiiUHiimiMHnwiuMiumi щццццц 11инт
: AMT типа 2 •
frM II 11 11 I 11 I I I II |________I I_________j
.............и 111 । । ii । ।_____।__________i
____________i_________________________________L—
____jttUUll I___________I I__________________________j
। i ii i i i i i i rrm г г i rrn ri i i i i i i i i i i i । ।
ON Время (1 с/деление) OFF
Рис. 21.1. (А) Ответ типичного С-ноцицептивного волокна на тепло. Тепловой стимул в
пределах 41 °C—49°С, который прикладывался на 3 секунды с 25-секундными интервала-
ми на неоволосенную кожу ладоней обезьян. Внутри этого температурного диапазона
стимулы рандомизированы. (Б) Ответы четырех АМТ типа 1 и четырех — типа 2 на теп-
ловой стимул 53°С (30 с). Отметьте, что АМТ типа 1 дают медленный ответ, приводящий
к пиковому залпу в конце стимула. Напротив, АМЕ типа 2 реагируют быстро и быстро же
адаптируются до низкого уровня активности. АМТ типа 1 играют важную роль в гипер-
альгезии на тепло, тогда как АМТ типа 2 сигнализируют о первичном болевом ощуще-
нии. Из Wall and Melzack, 1994. Воспроизводится с разрешения Churchill Livingstone.
Рис. 21.2 показывает сложные пути, которыми биохимические вещества,
выделившиеся из поврежденной ткани, действуют на ноцицептивные оконча-
ния. Воспалительный процесс связан с выделением АТФ, К+, брадикинина
(пептид из девяти остатков), простагландинов и лейкотриенов (те и другие
происходят из фосфолипидов поврежденных мембран), гистамина, серотони-
на (5НТ) и т. д. Все эти вещества либо возбуждают, либо понижают порог воз-
500 Часть V. Другие чувства
Клетки, участвующие
в воспалительном
процессе
Симпатический
нейрон
Рис. 21.2. (А) Факторы, ответственные за вторичную гиперальгезию. Множество факто-
ров, выделяющихся из поврежденной ткани, действуют непосредственно на ноцицепто-
ры или могут вызывать выделение других факторов из воспаленных клеток и симпатиче-
ских нейронов. (Б) Химический каскад окончаний С-волокон, служащий для усиления
стимула и лежащий в основе распространения боли. КС — кровеносный сосуд, С — сти-
мул, ВВ — вазоактивные вещества. Из Wall and Melzack, 1994; Textbook of Pain (Churchill
Livingstone), p. 31, Fig. 1.20c and p. 58, Fig. 3.1. Воспроизводится с разрешения.
Суждения ноцицептивных окончаний. Поврежденные ноцицептивные окон-
чания выделяют нейропептиды (например, вещество Р (ВП)), которые акти-
вируют выделение из воспалительных клеток дополнительных количеств гис-
тамина, брадикинина, АТФ и т. д. Существуют данные о том, что интактные
полимодальные С-волокна выделяют вазоактивные вещества, которые участ-
21. Боль 501
вуют в сосудорасширяющей реакции в области, окружающей повреждение
ткани. Такое выделение вазоактивных веществ порисходит в результате аксо-
аксональных рефлексов. Активация ноцицептивного окончания приводит к
антидромной передаче потенциала действия в соседнюю ветвь того же оконча-
ния (рис. 21.2 Б). Наконец, поврежденные симпатические волокна выделяют
катехоламины и другие вещества, которые возбуждают ноцицептивные волок-
на. Показано, что область распространения вокруг места повреждения огра-
ничивается распределением агентов, снижающих порог чувствительности или
непосредственно возбуждающих ноцицептивные волокна. Неудивительно,
что вокруг места поврежедения возникает область постоянной повышенной
чувствительности.
21.2.2. Центральные пути
Ноцицептивные волокна направляются в спинной мозг и черепно-мозговые
нервы; в случае спинальных нервов они проникают в спинной мозгв составе
дорзальных (задних) корешков. Сенсорные волокна дорзальных корешков ин-
нервируют определенный сегмент тела (дерматом) (рис. 21.3). Пределы дерма-
тома могут быть установлены зондированием кожи и отведением активности в
дорзальных корешках. Из этого следует, что и ноцицептивные волокна и, со-
ответственно, восприятие боли также построено по сегментарному принципу,
хотя границы между дерматомами редко резковыраженны.
В спинном мозге ноцицептивные волокна оканчиваются в дорзальном роге
серого вещества. Клетки серого вещества спинного мозга построены в ряд плас-
тинок (laminae) (рис. 21.4). Дорзальный рог состоит из шести слоев клеток (1—6),
еще три слоя — в вентральном роге (7—9). Десятый слой клеток концентрирует-
ся вокруг центрального канала. Слой 2, который, как мы увидим далее, занима-
ет важное место в ноцицептивных путях, называется желатинозной субстанцией
(substantia gelatinosa). Эти слои клеток, в которых клетки обильно контактируют
с другими слоями как короткими аксонами, так и дендритами, трудно различи-
мы на срезах, приготовленных обычными гистологическими методами.
Если пероксидаза хрена инъецирована в сенсорное волокно на периферии,
она переносится с аксоплазматическим током в центральные окончания. Та-
ким образом может быть нроилсжена организация спинального серого веще-
ства. В целом, чем большего диаметра волокно, тем дальше оно проникает в
серое вещество, то есть большие волокна 1а от мышечных веретен (см. гл. 6)
оканчиваются в пластинке 6. Напротив, маленькие ноцицептивные С-волок-
на оканчиваются в основном в пластинке 2, а несколько большие А5 — в пла-
стинках 1 и 5. Рис. 21.4 показывает, что помимо С- и А5-иннервации в путях,
проводящих болевые ощущения, присутствуют и волокна большего диамет-
ра — Аа и Ар, оканчивающиеся в пластинке 4. Мы поймем значение этого,
когда будем рассматривать «воротную теорию» в разделе 21.3.
Из дорзального рога серого вещества спинного мозга волокна, несущие но-
цицептивную информацию в высшие отделы ЦНС, распределяются по трем
трактам белого вещества. Спино-таламический тракт (СТТ) из них наиболее ва-
жен. Он начинается в пластинках 1 и 5 и немедленно переходит в вентро-лате-
ральный (передне-латеральный) сегмент контрлатеральной стороны и на-
18 Биология сенсорных систем
502 Часть V. Другие чувства
Рис. 21.3. Дерматомы организма человека. С = cervical — шейные; S = sacral — крестцо-
вые; L = lumbar — поясничные; Т = thoracic — грудные. Объяснения в тексте. Из Kandel,
Schwartz and Jessell, 1991, с разрешения.
правляется в таламус. Спино-ретикулярный тракт (СРТ) идет несколько дор-
зальнее (кзади) от СТТ, и некоторые волокна не переходят на противополож-
ную сторону спинного мозга. Тракт напраг.^ст ветвь в ретикулярную форма-
цию, а также в таламус. Ноцицептивные нейроны пластинок 1 и 5 проециру-
ются в спино-мезенцефалический тракт (СМТ), путь которого сходен с СТТ и
СРТ, но который оканчивается в сером веществе, окружающем Сильвиев во-
допровод (aqueductus Sylvii) среднего мозга (периакведуктальное серое веще-
ство (ПАС)) и в ретикулярной формации среднего мозга. Кроме того, малень-
кие клетки желатинозной субстанции посылают отростки на небольшие дис-
танции вверх и вниз по поверхности спинного мозга в составе тракта Лиссауэ-
ра; существуют серьезные доказательства того, что в пластинках спинного
мозга существуют и короткие интерсегментные связи ноцицептивных клеток.
Таким образом, помимо трех главных ноцицептивных трактов имеется и по-
лисинаптичнский восходящий путь. Ноцицептивная информация после пере-
ключений в таламусе, среднем мозге, ретикулярной формации и т. д. направ-
ляется далее в вышележащие отделы мозга. Однако, в отличие от специальных
21. Боль 503
Рис. 21.4. Пластинчатая струк-
тура дорзального рога серого
вещества и окончания аффе-
рентных волокон, участвую-
щих в ноцицепции. Стрелки
показывают направление по-
тока импульсов. Дальнейшие
объяснения в тексте.
чувств в коре отсутствуют специализированная область первичной ноцицеп-
ции. Большинство информации попадает в соместезическую кору, но значи-
тельная ее часть распределена в мозге диффузно.
Помимо восходящих проекций существуют и нисходящие — от центров
мозга к ноцицептивным клеткам. Существование такого нисходящего контро-
ля подтверждается установлением того факта, что ощущение боли устраняется
при стимуляции многих участков мозга. Среди них ядра контрлатерального
моста, гипоталамуса, ядра большого шва и соматосенсорной коры. Стимуля-
ция этих участков оставляет животных активными и реактивными, но нечувст-
вительными к болевым стимулам. Сходная феномен показан у людей, страдаю-
щих хроническими болями, такая «вызванная» анальгезия позволяет исследо-
вать анатомию и физиологию нисходящего контроля. Установлено, что пути
из высших уровней направляются в ПАС и ростральный вентромедиальный
продолговатый мозг, а оттуда через дорзолатеральный тракт белого вещества
спинного мозга к ноцицептивным клеткам дорзального рога серого вещества.
Многие из волокон этого нисходящего пути являются серотонергическими.
Стимуляция этого пути уменьшает активность ноцицептивных клеток дор-
зального рога и устраняет у животных поведенческие ответы на болевые сти-
мулы. Имплантация электродов в ПАС человека, страдающего хроническими
болями, уменьшает страдания, но мало сказывается на осязании и температур-
ной чувствительности.
21.3. Нейрофармакология болевых путей
Переходя от анатомии и физиологии к нейрофармакологии, отметим, что од-
ним из наиболее важных событий последних десятилетий стало открытие при-
родных опиоидов Хьюджесом (Hughes) с сотрудниками в Абердине в 1975 г.
Первыми были обнаружены в мозге свиньи пентапептиды лейцин-энкефалин
и метионин-энкефилин. Таблица 21.1 показывает, что эти два коротких пеп-
тида родственным трем крупным молекулам — p-эндорфину, динорфину и а-
неоэндорфину. Все пять пептидов содержат последовательность Tyr-Gly-Gly-
Phe и имеют анальгетический эффект, сходный с эффектом морфина и(или)
504 Часть V. Другие чувства
Таблица 21.1. Природные опиоиды
Р-эндорфин
Динорфин
а-неоэндорфин
Leu-энкефалин
Met-энкефалин
Tyr-Gly-Gly-Phe-Met-Thr-Ser-Glu-Lys-
Ser_Gln-Thr-Pro-Leu-Val-Thr-Leu-Phe-Lys-Asn-
Ala-Ile-Lys-Ile-Lys-Asn-Ala-Tyr-Lys-Lys-Gly-Glu
Tyr-Gly-Gly-Phe-Leu-Arg-Arg-Ile-Arg-Pro-
Lys-Lys-Trp-Asp-Asp-Gln
Tyr-Gly-Gly-Phe-Leu-Arg-Lys-Tyr-Pro-Lys
Туг- Gly- Gly- Phe- Leu
Туг- Gly- Gly- Phe- Met
Общая аминокислотная последовательность показана жирным шрифтом.
опиума. Нейроны, синтезирующие энкефалин и динорфин обнаружены в
ПАС, вентромедиальной продолговатом мозге и дорзальном роге спинного
мозга, p-эндорфин синтезируется нейронами гипоталамуса, который проеци-
руется в ПАС.
Опиоидные рецепторы подразделяются на множество типов в соответст-
вии с их способностью связывать лиганды. Наиболее важные типы обозначе-
ны ц, 5 и к. Реакции ц-рецепторов на агонисты располагаются в следующем
порядке: p-эндорфин > динорфин А > мет-энкефалин > лей-энкефалин; 5-ре-
цепторов: p-эндорфин > лей-энкефалин > мет-энкефалин > динорфин А; и
к-рецепторов: динорфин А » p-эндорфин > лей-энкефалин = мет-энкефа-
лин. Все три типа рецепторов широко распространены в ЦНС. ц-рецепторы
особенно важны для ноцицепции. Высокий уровень этих рецпторов обнару-
жен в таламусе, ПАС и дорзальном роге спинного мозга; к-рецепторы более
редки, но вполне очевидны в преоптической области гипоталамуса, тогда как
5-рецепторы — в обонятельных областях мозга.
Полагают, что энкефалинергические нейроны вызывают свой эффект, вы-
деляя энкефалины на пресинаптические окончания ноцицептивных нейро-
нов, и таким образом ингибируют выделение медиатора (рис. 21.5 А). Синап-
тология болевых путей в высшей степени сложна, весьма вероятно, что наряду
с пресинаптическим торможением, показанным на рис. 21.5 А, здесь присут-
ствует и постсинаптическое торможение. Некоторые детали организации си-
напсов показана на рис. 21.5 Б. Здесь приведены локальные взаимодействия в
поверхностной пластинке дорзального рога серого вещества спинного мозга.
Нисходящие проекции из центров мозга, модулирующих боль, оканчиваются
по большей части в пластинках 1 и 2, а также в пластинках 4, 5, 6 и 10. Стоит
напомнить, что волокна СТТ берут начало (у приматов) в пластинке 1 и что С-
ноцицетивные волокна оканчиваются в ппастинке 2. Рис. 21.5 Б показывает
нисходящие серотонергические волокна (5НТ), оканчивающиеся на отрост-
ках нейронов пластинки 1, эти синапсы — тормозные. Серотонергические во-
локна также образуют синапсы, на этот раз возбуждающие, на энкефалинер-
гических «древовидных» клетках в пластинке 2 в желатинозной субстанции.
Эти интернейроны, в свою очередь, образуют тормозные синапсы на нейронах
пластины 1. Другая древовидная клетка (возбуждающая) получает высоко- и
21. Боль 505
Спино-таламический тракт
Рис. 21.5. (А) Схема, показывающая энкефалинергический контроль ноцицептивных
клеток желатинозной субстанции. Нейрон в спиноталамическом тракте образует синап-
сы с другим нейроном в том же тракте. Трансмиттером в синапс? является вещество Р
(ВР). Выделение ВП контролируется энкефалинергическим интернейроном, который
пресинаптически ингибирует выделение ВР. По Iverson, 1979. (Б) Упрощенная схема,
показывающая локальные нейрональные взаимодействия в дорзальном роге серого ве-
щества. Показаны первые два слоя серого вещества. Второй слой, желатинозная субстан-
ция, подразделяется на два субслоя — 2а и 2Ь. Ноцицептивный нейрон тракта показан
в слое 1. Его аксон входит в СТТ. Высокопороговые (ВПр) (малого диаметра) волокна и
низкопороговые (НПр) большого диаметра показаны входящими в спинной мозг слева в
составе спиномозгового корешка. Нисходящие энкефалинергические волокна входят на
рисунке справа. Дальнейшие объяснения в тексте. А — тормозный островковый нейрон;
В — тормозный нейрон; С — тормозная клетка; Е — серотонергический нисходящий
нейрон; ВПр — высокопороговый афферентный нейрон; НПр — низкопороговый аффе-
рентный нейрон, Д — древовидная клетка. Тормозные синапсы — черные, возбуждаю-
щие — белые. С изменениями из Н. L. Fields and Basbaum in Wall & Melzack, 1994.
низкопороговые возбуждающие афференты. Рисунок показывает, что низко-,
пороговые волокна также образуют синапсы с энкефалинергическими интер-
нейронами, которые опять-таки образуют синапсы на древовидных клетках.
Это — еще одни путь снижения активности нейронов пластинки 1. Ясно, что
такие сложные связи и баланс активности различных синапсов обеспечивает
тонкий контроль активности в волокнах СТТ.
21.4. Иррадиирующая боль
При повреждениях кожи индивид способен немедленно и точно локализовать
источник боли. Если же источник боли расположен внутри тела, такая точ-
ность встречается редко. Выглядит это так, будто ЦНС не в состоянии пове-
рить, что внутри что-то не в порядке, и отсылает (иррадиирует) боль на поверх-
ность тела. Вероятно, наиболее известным примером такой иррадиации явля-
ется грудная жаба (angina pectoris).
Термин «angina pectoris» был введен Эберденом (Heberden) в 1772 году на
лекции в Королевском Медицинском Колледже. Он писал, что пациенты*
«страдают при ходьбе, особенно после еды, испытывают боль и неприятное
ощущение в груди, им кажется, что они умрут, если это будет продолжаться, но
506 Часть V. Другие чувства
Рис. 21.6. Упрощенная схема
формирования иррадиирую-
щей боли. Дальнейшие объяс-
нения в тексте.
сТоит им остановиться на некоторое время, и все неприятные ощущения про-
ходят». Боль ощущается, как сильное сокращение мышц грудной клетки, а с
развитием приступа боль начинает «стрелять» в левую руку. Причиной этой
острой боли является, конечно, сердце, а конкретнее, нарушение работы или
блокада коронарных артерий, снабжающих кислородом сердечную мышцу.
Чтобы понять, каким образом боль иррадиирует в верхнюю часть туловища
и левую руку, следует вернуться к концепции дерматомов, упомянутой в нача-
ле раздела 21.2.2 и показанной на рис. 21.3. В ходе эмбриогенеза сердце разви-
вается как срединная структура в верхней части грудной клетки и иннервиру-
ется нервами верхних грудных дерматомов. По мере развития левый желудо-
чек превращается в главную камеру насоса, заставляющего кровь циркулиро-
вать по кровеносной системе, поэтому именно левый желудочек первым ощу-
щает недостаток кислорода вследствие дефектов коронарных артерий. Верх-
ние грудные дерматомы обеспечивают также иннервацию мускулатуры груд-
ной клетки и рук. Вследствие этого, из-за ишемии активируются болевые
нервные волокна сердца, которые входят в спинной мозг через те же корешки,
что и болевые волокна из рук и грудной клетки. Все эти волокна взаимодейст-
вуют на одних и тех же интернейронах дорзального рога серого вещества спин-
ного мозга, в результате мозг может неправильно интерпретировать место
происхождения боли (рис. 21.6). Такое объяснение иррадиации боли общнос-
тью ноцицептивных цепей подкрепляется наблюдением, что именно левая ру-
ка пациента, испытывающего атаку грудной жабы, проявляет гиперальгезию,
тогда как на правой руке этот симптом отсутствует. Это предполагает, что сум-
мация ноцицептивных импульсов происходит в дорзальном роге еще прежде,
чем информация направляется в вышележащие отделы мозга.
Грудная жаба представляет собой типовой пример иррадиирующей боли,
существует, однако, немало и других случаев. Возможно, что многие случаи
болей в спине, от которой страдает, как упоминалось выше, так много пациен-
тов, связаны с патологиями внутренних органов, в частности органов таза.
Рассмотрение множества других случаев иррадиации боли завело бы нас в кли-
нический анализ боли слишком далеко для такой книги, как эта (см. публика-
21. Боль 507
цию Melzack and Wall, 1982, приведенную в Библиографии для получения бо-
лее полной информации). Стоит упомянуть, что вблизи от мест, где предполо-
жительно возникает иррадиирующая боль, часто находятся так называемые
триггерные точки. Надавливание на них или введение в них игл могут приво-
дить к жестоким, иногда длительным, болям. Такие триггерные точки, глав-
ным образом, обнаруживаются у больных, страдающих заболеваниями внут-
ренних органов, но могут быть выявлены и у здоровых. У кардиологических
пациентов триггерные точки обнаруживаются не только на поверхности груд-
ной клетки (и слева, и справа), но и в области лопаток и по обе стороны позво-
ночного столба. Хотя триггерные точки могут быть связаны с физиологией ир-
радиирующих болей, существуют и другие, пока неизвестные возможности
(и точки акупунктуры, с которыми они связаны).
21.5. Воротная теория
Теорий нейрофизиологии боли на уровне спинного мозга существует множе-
ство. Воротная теория, предложенная Мельзаком (Melzack) и Уоллом (Wall) в
1965 году, приобрела широкое признание и объяснила многие существенные
моменты проблемы. Теория предлагает механизм, благодаря которому ноци-
цептивные импульсы могут контролироваться «воротным» механизмом в дор-
зальном роге серого вещества и нисходящими импульсами из вышележащих
центров. Мы видели во введении к этой главе, как ощущение боли может мо-
дулироваться обстоятельствами, в которых находится организм, а в последую-
щей за ним самой главе — нейроанатомические пути и нейрофидиологические
механизмы, благодаря которым это достигается. В данном разделе мы сосре-
доточимся на воротном механйзме, который, как постулировано, управляет
первым синапсом болевого пути.
Рисунок 21.7 показывает нейрон, проецирующийся в СТТ, в пластинке 1
дорзального рога серого вещества спинного мозга (см. также рис. 21.5 Б). Он
иннервирован двумя афферентными волокнами: малого диаметра (М) (С-во-
локно или АЗ-волокно) и большого диаметра (Б) (Аа- или Ар-волокно). М-во-
локно также ингибирует тормозный (вероятно энкефалинергический) интер-
нейрон. Б-волокно, напротив, возбуждает тормозный интернейрон. Наконец,
на интернейроне образуют синапсы нисходящие проекции из вышележащих
центров. Следует подчеркнуть, что рис. 21.7 представляет собой концептуаль-
ную схему, которая, будем надеяться, показывает суть очень сложной синапто-
логии серого вещества (упрощенная версия которой показана на рис. 21.5 Б).
Из рис. 21.7 видно, что активность М-волокна возбуждает нейрон, проеци-
рующийся в СТТ, и отключает активность тормозного интернейрона. Напро-
тив, активность Б-волокон, возбуждающая нейрон СТТ, активирует и тормоз-
ный интернейрон. Когда М-волокно активировано, нейрон СТТ растормажи-
вается и импульсация направляется по болевым путям в вышележащие цент-
ры. Когда активируются Б-волокна, сначала возникает залп импульсов в вос-
ходящих волокнах, за которым следует торможение в результате активации
интернейрона. Когда оба типа волокон активируются одновременно, крупные
волокна (имеющие большую скорость проведения) вызывают собственный
эффект, а затем поступающие с задержкой импульсы малых волокон достига-
508 Часть V. Другие чувства
Спинной
мозг
Волокно малого
iиамогра
Спиномозговой
корешок
Волокно большого
диаметра
Рис. 21.7. Воротная теория.
Тормозные синапсы обозна-
чены черным, возбуждаю-
щие — белым. Дальнейшие
объяснения в тексте.
ют интернейрона и вызывают уменьшение активности в восходящей проек-
ции — ворота закрываются. Активация нисходящих волокон из ЦНС также
вызывает «закрывание ворот» и, возможно, удержание их в закрытом состоя-
нии.
Исходная воротная теория, суммированная в рис. 21.7, впоследствии была
усложнена установленными позже деталями нейроанатомии и нейрофизио-
логии, однако суть ее выдержала испытание временем. Более того, она стала
основой для анальгетической терапии. Теория предполагает, что когда акти-
вированы нервные волокна большого диаметра, импульсы, приходящие по
ноцицептивным волокнам малого диаметра, оказываются «у закрытых ворот»
болевого пути. Удачно, что внешними электродами на поверхности кожи в
основном активируются волокна большого диаметра — в результате оказалось
сравнительно просто разработать карманный батарейный стимулятор, крепя-
щийсяж коже электропроводящим клеем. Включение стимулятора вызывает
ощущение покалывания и одновременно уменьшает болевое ощущение. Та-
кие устройства используются сотнями тысяч больных с большим терапевтиче-
ским эффектом. Одно это оправдывает и исследования, и умственные усилия,
потраченные на воротную теорию.
21.6. Заключение
Боль — это наказание человечества. Насколько широко она распространена в
мире живого неизвестно, но, как мы отмечали в начале этой главы, тут лучше
ошибиться в сторону большей осторожности. Так что надо стараться избегать
действий, потенциально причиняющих боль человеку или другим животным.
Часто это приводит к достаточно острым коллизиям в медицинских исследо-
ваниях. Насколько перспектива облегчить боль человеку оправдывает причи-
нение боли (к тому же, неизвестно какой силы) животному?
21. Боль 509
К счастью, в настоящее время в результате как раз медицинских исследова-
ний прошлого мы уже имеем средства устранения или облегчения боли. Неко-
торые из них упоминались выше, а другие описаны в учебниках по медицине и
анестезиологии. Нам действительно повезло, что мы живем в то время, когда
боль, вызываемая травмами и болезнями, в значительной степени укрощена.
А ведь юный Чарльз Дарвин оставил когда-то намерение стать врачом, попри-
сутствовав на какой-то особенно душераздирающей операции, сделанной ре-
бенку в Эдинбурге. Теперь техника анальгезии применяется в тщательно кон-
тролируемом и лицензируемом порядке в биомедицинских исследованиях.
Проблема боли, однако, продолжает занимать умы тех, кто ее изучает. Мы
видели в этой главе, что, как и другие «грубые ощущения», с которыми мы
сталкивались в других главах, боль на самом деле не так «груба». Мы часто не-
правомочно ставим знак равенства между травмой и болью, слишком легко ве-
рим в кажущуюся непосредственную связь боли с прищемленным пальцем
или пальцем, сунутым в огонь. В этой главе мы увидели некоторые черты
сложности (только некоторые!) нервных связей, лежащих в основе боли, мы
увидели, как этот контроль осуществляется в спинном мозге и вышележащих
центрах. Мы также обратили внимание, что ощущение боли сильно зависит от
обстоятельств и культурологических ожиданий. Но следует учитывать, что
ощущение боли это и не просто считывание (как полагают некоторые филосо-
фы) ЦНС частоты импульсации в С-волокнах.
Очевидно, ощущение боли, как и другие сенсорные ощущения, которые
мы испытываем, целесообразно. Оно интегрировано в физиологию и поведе-
ние животного таким образом, чтобы избегать его в целях выживания, и явля-
ется частью, причем существенной, гомеостаза животного, кроме тех трагиче-
ских случаев, когда оно сражается за пределами своих возможностей. Боль,
вызванная внутренними болезнями (как мы видели в случае грудной жабы),
безнадежно бесполезна в этом плане (исключая нашу медицински продвину-
тую цивилизацию), как трепыхание зверюшки, угодившей в капкан. Но в дру-
гих случаях польза от болевой системы в сохранении здоровья перевешивает
трагедии безнадежных ситуаций.
МАТЕРИАЛЫ К ЧАСТИ V
Для самоподготовки
I
I
Нижеследующие вопросы подготовлены таким образом, чтобы помочь оце-
нить понимание предметов изложенных в Части V, а также обратить внимание
на существенные их аспекты.
Глава 19. Термочувствительность
19.1. Каков нормальный термический диапазон, в котором способны существовать
биологические организмы? Почему? Есть ли исключения? Если да, назовите их.
19.2. В чем различие между пойкилотермными и гомойотермными животными?
19.3. Каким образом клетки ганглия дорзального корешка (ГДК) используются
в исследованиях термочувствительности?
19.4. Как повышение температуры сказывается на небольших группах клеток ГДК?
19.5. Каким образом исследования молекулярной генетики С. elegans проливают
свет на механизмы, лежашие в основе термочувствительности?
19.6. Опишите некоторые из терморецепторов, обнаруженных у насекомых. Укажи-
те, каким образом они адаптированы к их способу существования.
19.7. Существует только две группы истинно гомойотермных животных — кто они?
19.8. На какую температуру установлен «термостат» человека?
19.9. Где в человеческом теле располагаются терморецепторы?
19.10. Что такое «нейтральный диапазон» термочувствительности человека?
19.11. В каком месте поверхности тела сконцентрированы термочувствительные
«пятна»?
19.12. Что такое иллюзия Вебера и каково ее физиологическое объяснение?
19.13. Какую форму принимают термочувствительные окончания и где они распола-
гаются?
19.14. Опишите, каким образом холодовые и тепловые волокна сигнализируют о ма-
лых изменениях температуры в пределах нейтрального диапазона.
19.15. Где располагаются внутренние терморецепторы? О какой опасности они
должны, главным образом, предупреждать?
19.16. Где терморецепторы располагаются в мозге?
19.17. Какие ядра мозга суммируют термическую информацию из всех частей орга-
низма и инициируют физиологический ответ, противодействующий отклоне-
ниям от термостатической нормы?
Глава 20. Редкие чувства
20.1. Где в организме термочувствительных змей располагаются инфракрасные
детекторы?
20.2. Как определяется направление на источник инфракрасного излучения?
20.3. Какие черепно-мозговые нервы передают информацию от термочувствитель-
ных ямок в центральную нервную систему?
20.4. В какую часть мозга направляется информация об инфракрасном излучении?
20.5. Каким образом рецептивные поля тектальных клеток, чувствительных к ин-
фракрасному излучению, сходен с таковыми ганглиозных клеток в сетчатке
приматов?
20.6. К какой характеристике инфракрасного излучения наиболее чувствительна
тектальная карта?
Материалы к части V 511
20.7. Какое свойство атмосферы Земли вызывает поляризацию поступающего излу-
чения?
20.8. В каком месте небосвода располагается полоса максимальной поляризации
в безоблачный летний полдень в Англии?
20.9. Объясните природу нарушения воздушного поля зрения рыб.
20.10. Как сложный глаз ракообразных и камерный глаз головоногих детектируют
плоскость поляризации света?
20.11. Каковы доказательства того, что насекомые и головоногие чувствительны
к плоскости поляризации света?
20.12. Выпишите примеры рыб с сильными и слабыми электрическими свойствами.
К какой группе принадлежит большинство электрических рыб?
20.13. Где расположен электрический орган у рыб со слабыми электрическими свой-
ствами?
20.14. Каковы функции сильных и слабых электрических разрядов?
20.15. Что вызывает слабые электрические поля, которые детектируют хищные элек-
трочувствительные рыбы?
20.16. Опишите эксперимент, показывающий значение электрорецепторов в детек-
тировании скрытой жертвы.
20.17. Могут ли слабоэлектрические рыбы детектировать неорганические детали
окружающей среды?
20.18. Какие два типа рецепторов участвуют в электрочувствительности? Из какой
вездесущей системы они возникли?
20.19. Что такое ампулы Лоренцини и где они обнаружены?
20.20. Из чего состоит капсула ампулы Лоренцини?
20.21. Чем трубчатый орган отличается от ампулярного?
20.22. Где расположены ампулярные и трубчатые органы у типпичных пресноводных
гимнотид? Какие из них более многочисленны?
20.23. В каких организмах обнаружены магнитосомы?
20.24. Какова биологическая роль магнитосом?
20.25. Какие еще организмы, возможно, обладают чувствительностью к магнитным
полям?
Глава 21. Боль
21.1. Отражение чего понимают под термином «боль»?
21.2. Что такое врожденная анальгезия и почему это патология?
21.3. Почему полагают, что причина врожденной анальгезии связана с централь-
ной, а не периферической патологией?
21.4. Приведите данные подтверждающие, что инкурабельные боли требуют боль-
ших затрат медицинских служб в современном мире.
21.5. Что вы думаете об эволюционных причинах распространенности большей
в нижнем отделе спины в человеческой популяции?
21.6. Приведите контраргументы идее, что боль ощущается, когда мозг детектирует
импульсы в болевых волокнах так же, как громкость в наушниках определяет-
ся поступающим электрическим током.
21.7. Какова форма периферических ноцицептивных волокон? Чем они отличают-
ся от механо- и термочувствительных окончаний?
21.8. Болевые волокна какого типа ответственны за острую боль — колющее ощу-
щение, а какие — за длительную глухую боль?
21.9. Каковы различия между АМТ типа 1 и 2? Как ответы волокон обоих типов от-
личаются от ответов СМТ?
21.10. Каким образом аппликация капсайцина показывает, что механизмы передачи
термически и механически вызванной боли различны?
21.11. Что означает термин «гиперальгезия»?
512 Часть V. Другие чувства
21.12. Перечислите несколько факторов, ответственных за вторичную гиперальге-
зию.
21.13. Как аксо-аксональные рефлексы участвуют в распространении гиперальге-
зии?
21.14. Как дерматомы связаны с позвоночным столбом?
21.15. Сколько пластинок различают в дорзальном (заднем) роге серого вещества
спинного мозга?
21.16. Какой слой серого вещества спинного мозга называется желатинозной суб-
станцией?
21.17. В каком слое оканчиваются С-, А5-, Ар- и Аа-волокна?
21.18. Какие три тракта несут ноцицептивную информацию в спинном мозге?
21.19. Что такое тракт Л иссауэра?
21.20. Какие ядра мозга участвуют в нисходящем контроле боли? Через какие части
ствола головного мозга и спинного мозга проходят волокна из этих ядер?
21.21. Опишите биохимическую природу двух обычных природных опиоидов.
21.22. В каких частях мозга обнаружены клетки, экспрессирующие рецепторы к при-
родным опиоидам?
21.23. Нарисуйте схему нервных связей в слоях 1 и 2 дорзального рога серого вещест-
ва спинного мозга.
21.24. Что такое иррадиирующая боль? Приведите примеры.
21.25. Опишите возможные нервные связи, ответственные за иррадиирующую боль.
21.26. Что такое «триггерные точки»? Где они расположены у кардиологических
больных?
21.27. Нарисуйте упрощенную схему, объясняющую основную концепцию «ворот-
ной теории».
21.28. Каким образом воротная теория предоставила средство для анальгетической
терапии?
Замечания и библиография
Глава 19. Термочувствительность
Обсуждение термочувствительности можно найти в учебниках физиологии
(Darian-Smith, 1984; Gregor and Windhorst, 1996) и сравнительной физиологии
(Prosser, Brown, 1961). Проблема денатурации белка при высоких температурах
привела к значительному интересу к так называемым «белкам теплового
шока» или шаперонам (Ellis, Laskey and Lorimer, 1993), которые обеспечивают
стабильность других белков при повышенных температурах. Это вездесущие и
эволюционно очень древние белки. За исключением трубчатых червей, посе-
ляющихся вокруг фумарол в глубинах океана, животные не в состоянии выжи-
вать при температурах тела выше 40°С. Хотя пойкилотермные животные име-
ют предпочтительную температуру окружающей среды, они обычно способны
несколько изменить (не слишком сильно) температурный режим путем аккли-
матизации. Многие аспекты биохимии и физиологии изменяются сложным
образом, чтобы адаптировать животное к новым температурным условиям ок-
ружающей среды (Prosser, Brown, 1961). Обсуждение молекулярно-биологиче-
ских механизмов, лежащих в основе термочувствительное™ можно найти
у Komatsu et al. (1996), McClewsky (1997) и у Reichling and Levine (1997). Боль-
шая проблема эволюции гомойотермии у птиц и млекопитающих обсуждается
в работе Ruben (1995).
Материалы к части V 513
Bear, М. F, В. W. Connors and М. A. Paradiso, 1996, Neuroscience: Exploring the brain,
Baltimore: Williams and Wilkins.
Darian-Smith, I., 1984, «Thermal sensitivity», in Handbook of Physiology, Section 1: The
Nervous System, vol. 3: Sensory Processes, Part 2,1. Darian-Smith, ed., Bethesda: Ame-
rican Physiological Society.
Ellis, R. J., R. A. Laskey and G. H. Lorimer, eds, 1993, «Molecular chaperones», Philoso-
phical Transactions of the Royal Society (Biological Sciences), 339, 255—373.
Gregor, R. and U. Windhorst, eds, 1996, Comprehensive Human Physiology, Berlin: Sprin-
ger-Verlag.
Hazel, J. R., 1995, «Thermal adaptations in biological membranes: is homeoviscous adapta-
tion the explanation?», Annual Review of Physiology, 57, 19—42.
Komatsu, H., et al., 1996, «Mutations in a cyclic-nucleotide-gated channel lead to abnor-
mal thermo-sensations and chemosensation in C. elegans», Neuron, 17, 707—718.
McCleskey, E. W., 1997, «Recent heat in thermosensation», Current Biology, 7, R679—681.
Prosser, C. L. and F. A. Brown, 1961, Comparative Animal Physiology, Philadelphia: Saun-
ders.
Reichling, D. B. and J. D. Levine, 1997, «Heat transduction in rat sensory neurons by calci-
um-dependent activation of a cation channel», Proceedings of the National Academy of
Sciences, 94, 7006—7011.
Ruben, J., 1995, «The evolution of homeothermy in mammals and birds: from physiology to
fossils», Annual Review of Physiology, 57, 69—95.
Глава 20. Редкие чувства
Инфракрасная чувствительность у змей обсуждается Хартлайном (Hartline,
1974). Waterman (1981) представил исчерпывающий хорошо иллюстрирован-
ный труд по поляризованному свету и чувствительности к нему в царстве жи-
вотных, a Frisch (1974) описал свои пионерские работы по танцам пчел, кото-
рые описывают путь к запасам пищи по отношению к плоскости поляризации
солнечного света. Bell (1986), а также Bullock and Heiligenberg (1986) опублико-
вали книги по электрорецепции, глава Беннета (Bennett) в книге под редакци-
ей Hoar (1971), хотя и несколько старовата, но все еще дает превосходное опи-
сание различных типов электрических рыб. Тщательное исследование физики
и сенсорной физиологии навигации с помощью магнитного поля Земли у акул
представлено Paulin (1995). История о том, как была установлена истинная
функция ампул Лоренцини, рассказана Bullock (1974): сначала думали, что это
температурные рецепторы, потом — рецепторы давления, потом — детекторы
солености и, наконец, — электрорецепторы. Последнее было установлено в
результате комбинированных электрофизиологических и поведенческих ис-
следований. Наконец, хорошо иллюстрированная и интересная работа по то-
му впечатлению, которое рыбы с сильными электрическими свойствами про-
изводили на древних египтян, греков и римлян, сделана Kellaway (1946). Тогда
каку бактерий магниточувствительность достаточно изучена (Blakemore, 1982;
Mann et al., 1990), мало что известно об этом чувстве у высших животных, хотя
и предполагается, что оно как-то участвует в навигации птиц (Beason and
Semm, 1996), a Reiter (1993) приводит некоторые данные о том, что магнито-
чувствительность может сказываться на циркадных ритмах, влияющих на сек-
рецию мелатонина.
514 Часть V. Другие чувства
Инфракрасное излучение
Hartline, Р. Н., 1974, «Thermoreception in snakes», in Handbook of Sensory Physiology,
vol. III/3, A. Fessard, ed., Berlin: Springer-Verlag.
Terashima, S., R. C. Goris and Y. Katsuki, 1970, «Structure of the warm fiber terminals
in the pit membrane of vipers», Journal of Ultrastructure Research, 31, 494—506.
Поляризованный свет
Frisch, К., 1974, «Decoding the language of the bees», Science, 185,663—668.
Walls, G. L., 1963, The Vertebrate Eye and its Adaptive Radiation, New York: Hafner.
Waterman, T. H., 1981, «Polarization sensitivity», in Handbook of Sensory Physiology,
vol. V1I/6B, H. Autrum, ed., Berlin: Springer-Verlag.
Wellington, W. G., 1974, «А special light to steer by», Natural History, 83(10), 46—53.
Электрочувствительность
Bell, C. S., 1986, Electroreception.
Bennett, M. V. L., 1971, «Electric organs», in Fish Physiology, W. S. Hoar and D. J. Randall,
eds. New York: Academic Press, vol. 5.
Bullock, T. H., 1974, «General Introduction», in Handbook of Sensory Physiology, vol. II1/3,
E. Fessard, ed., Berlin: Springer-Verlag.
Bullock, T. H. and W. Heiligenberg, eds, 1986, Electroreception, New York: Wiley.
Emde, G. von der, et al., 1998, «Electric fish measure distance in the dark», Nature, 395,
890-894.
Kalmijn, A. J., 1971, «The electric sense in sharks and rays», Journal of Experimental Biology,
55, 371-383.
Kalmijn, A. J., 1974, «The detection of electric fields from inanimate and animate sources
other than electric organs», in Handbook of Sensory Physiology, vol III/3, A. Fessard, ed.,
Berlin: Springer-Verlag.
Kellaway, P., 1946, «The part played by electric fish in the early history of bioelectricity and
electrotherapy», Bulletin of the History of Medicine, 20, 112—137.
Murray, R. W., 1974, «The Ampullae of Lorenzini», in A. Fessard, ed., ibid.
New, J. G., 1997, «The evolution of vertebrate electrosensory systems», Brain, Behaviour
and Evolution, 50, 244—252.
Paulin, M. G., 1995, «Electroreception and the compass sense in sharks», Journal of Theo-
retical Biology, 174, 325—339.
Szabo, T., 1974, «Anatomy of the specialised lateral line organs of electroreception» in
A. Fessard, ed., ibid.
Магниточувствителъность
Beason, R. C. and P. Semm, 1996, «Does the Avian ophthalmic nerve carry magnetic navi-
gational information?», Journal of Experimental Biology, 199, 1241—1244.
Blakemore, R. P., 1982, «Magnetotactic bacteria», Annual Review of Microbiology, 36,217—238.
Mann, S., N. H. C. Sparks and R. G. Board, 1990. «Magnetotactic bacteria: microbiology,
biomineralisation, palaeomagnetism and biotechnology», Advances in Microbial Physio-
logy^, 125-181.
Reiter, R. J., 1993, «Static and extremely low frequence electromagnetic field exposure —
reported effects on the circadian production of melatonin», Journal of Cellular Bio-
chemistry, 51, 394—403.
Материалы к части V 515
Глава 21. Боль
Проблема боли по понятным причинам интересновала ученых, философов и
теологов тысячелетиями. Аристотель в книге «О душе» {De Anima, книга 2) пи-
шет, что все животные, обладающие осязанием, также способны испытывать
удовольствие и боль. Боль, писал он (хотя в таком определении содержится
тавтология), это восприятие неприятных объектов. Свежее определение боли
дано Международной ассоциацией изучения боли {International Association for
the Study of Pain, IASP) в 1991 году: «Неприятное сенсорное и эмоциональное
ощущение, связанное с реальным или потенциальным повреждением ткани,
или описываемое в понятиях такого повреждения». Merskey (1991) подчерки-
вает, что боль — это «грубое (первичное) ощущение», и его нельзя смешивать
с «болевым поведением». Ощущение это, продолжает он, «монистическое, но
этиология его множественна». Ананд и Крэйг (Anand and Craig, 1996) дали об-
зор современных определений боли, а Уолл и Мельзак (Wall and Melzack, 1994)
отредактировали современный обзор по различным аспектам боли. Неболь-
шая книга тех же авторов (Melzack and Wall, 1982) представляет собой клас-
сическое и к тому же легко читаемое введение в проблему, а их более ранняя
работа (1965) — основополагающая в «воротной теории». Интересная статья
Мельзака с соавт. (Melzack, Stillwell and Fox, 1977) предполагает, что триггер-
ные точки и точки акупунктуры — на самом деле — одно и то же, но открытое
клиницистами различных культур. Роланд (Roland, 1992) охватил в своей пуб-
ликации современные работы, показавшие, что болевые стимулы вызывают
распространенную активность в коре. Наконец, Уолл (Wall, 1985) обсуждает
некоторые контр-интуитивные черты боли, а Рамачандран и Блэксли (Rama-
chandran and Blakeslee, 1988) дают хорошо изложенное описание хитроумных
опытов Рамачандрана с зеркальными иллюзиями «восстановления» утрачен-
ных конечностей, ослабляющими фантомные боли (хотя, возможно, это и
эффект плацебо). Недавно, однако, получены данные, что постоянное ноше-
ние протеза также ослабляет болезненные ощущения в «фантомных конеч-
ностях».
Anand, К. J. S. and К. D. Craig, 1996, «New perspectives on the definition of pain», Pain,
67, 3-6.
Darwin, E., 1794, Zoonomia, London: Johnson, vol. 1, pp. 121, 125.
Deyo, R. A., 1998, «Low back pain», Scientific American, 279(2), 28—33.
Fields, H. L. and A. I. Basbaum, 1994, «Central nervous system mechanisms of pain modu-
lation», in P. D. Wall and R. Melzack, ibid.
Herberden, W., 1772, quoted in Procacci, P., M. Zoppi and M. Maresca, 1994, «Heart and
vascular pain», in P. D. Wall and R. Melzack, ibid.
Iverson, L. L., 1979, «The chemistry of the brain», Scientific American, 241(3), 118—121.
James, W., 1890, The Principles of Psychology, New York: Holt.
Kandel, E. R., J. H. Swartz, T. M. Jessell, 1991, Principles of Neural Science (3rd Edition),
New York: Elsevier.
Melzack, R., 1975, «The McGill Pain Questionnaire: major properties and scoring meth-
ods», Pain, 1, 277-299.
Melzack, R. and P. D. Wall, 1965, «Pain mechanisms: a new theory», Science, 150,
971-979.
516 Часть V. Другие чувства
Melzack, R., D. М. Stillwell and Е. J. Fox, 1977, «Trigger points and acupuncture points:
correlations and implications», Pain, 3, 3—23.
Melzack, R. and P. D. Wall, 1982, The Challenge of Pain, Harmondsworth: Penguin Books.
Merskey, H., 1991, «The definition of pain», European Psychiatry, 6, 153—159.
Meyer, R. A., J. N. Campbell and S. N. Raja, 1994, «Peripheral neural mechanisms of noci-
ception», in P. D. Wall and R. Melzack, 1994, ibid.
Ramachandran, V. S., D. Rogers-Ramachandran and S. Cobb, 1995, «Touching the phan-
tom limb», Nature, 377,489—490.
Ramachandran, V. S. and S. Blakeslee, 1998, Phantoms in the Brain, London: Fourth Estate.
Rang, H. P., S. Bevan and A. Dray, 1994, «Nociceptive peripheral neurons: cellular proper-
ties», in P. D. Wall and R. Melzack, 1994, ibid.
Roland, 1992, «Cortical representation of pain», Trends in Neurosciences, 15, 3—5.
Wall, P. D., 1985, «Pain and no pain», in Functions of the Brain, C. W. Coen, ed., Oxford:
Clarendon Press.
Wall, P. D. and R. Melzack, 1994, eds. Textbook of Pain (3rd edn), Edinburgh: Churchill
Livingstone.
Часть VI
КОДА
«Если первое знакомство с объектом озадачило нас, и мы счита-
ем его новым или сильно отличающимся от того, что мы знали
прежде, или от того, что мы предполагали, каким он должен
быть, это вызывает удивление и восхищение. Поскольку все это
может случиться до того, как мы поймем, полезно это нам или
нет, я ставлю удивление выше прочих страстей.»
Рене Декарт, 1649, Страсти души
(Le Passions de Г Ате)
Leiden: Elsevier, § 53
Наверное, читая эту книгу, вы не раз испытывали то самое удивление, которое
Декарт ставил выше прочих страстей. В заключительной части мы немного
вернемся назад, чтобы переосмыслить все изложенное выше. Глава 22 должна
связать вместе некоторые мотивы, которые периодически возникали в пред-
шествующих двадцати одной. Она показывает, как сенсорные системы схожи
между собой в двум отношениях. С одной стороны, в этой главе обращено
внимание на сходство нейробиологии различных модальностей чувств, с дру-
гой — на то, как различные группы животных приходили к одним и тем же
нейробиологическим решениям. Хотя, как мы видели, у животных развилось
потрясающее разнообразие органов чувств и систем обнаружения изменений
в окружающей среде, определенные принципы их строения сохраняются и вы-
являются снова и снова. На уровне молекул и клеток это сходство часто указы-
вает на их общее происхождение, на уровне органов и систем — сходство час-
то является результатом нескольких несвязанных между собой эволюционных
линий развития, независимо друг от друга пришедших к наилучшему реше-
нию. Существуют примеры поразительной конвергентной эволюции.
Двадцать третья, и последняя, глава уходит от конкретной науки и рассма-
тривает философские аспекты этой наиболее философской из биологических
проблем. Со времен греческой античности до наших дней проблема знания —
того, как мы познаем природу мира вне нас самих, остается главной темой фи-
лософских дискуссий. Сенсорная биология имеет к ней самое непосредствен-
ное отношение, и она заставляет нас подняться над традиционно узким эпи-
стемологическим подходом, связанным исключительно с чувствами человека.
Еще в самом начале биологии древний грек Аристотель считал, что «каждая
часть природы удивительна и чему-нибудь учит нас». Это как раз то, как мы
шли по страницам этой книги. Мы видели, насколько отличаются сенсорные
миры других животных от нашего собственного. Исследования по сенсорной
физиологии должны поэтому вселять смирение в умы философов, занимаю-
щихся разумом. С другой стороны, только о нашем собственном сенсорном
мире мы и можем узнавать «из первых рук». Поэтому мы заканчиваем книгу
тем отражением, которое сенсорная физиология находит в теории познания и
философии разума.
22. ПОВТОРЯЮЩИЕСЯ МОТИВЫ
Молекулярные мотивы: мембранные воротные механизмы, G-белковые
системы, рецепторы — эволюционные взаимосвязи — G-белковые сиг-
нальные системы — G-белки у одноклеточных, напр. S. cerevisiae — пре-
имущества онованных на G-белке сигнальных систем. Клеточные моти-
вы: микроворсинки и неподвижные реснички — волосковые клетки по-
звоночных — вкусовые и обонятельные клетки — наружные сегменты
палочек и колбочек — сенсорные сенсиллы насекомых; рабдомерные фо-
торецепторы беспозвоночных. Органы чувств: огромное разнообразие —
конвергентная эволюция — матиссеновы хрусталики — конвергенция
глаз белемноидов и позвоночных; дифференциация сенсорных поверх-
ностей — сетчатка позвоночных — сложный глаз насекомых — половой
диморфизм — «фовеа» стрекоз — обонятельная слизистая — слуховая фо-
веа летучих мышей — специализация осязательной и электрорецептивной
поверхностей — сходство нервной структуры сетчатки и обонятельной
луковицы. Центральные анализаторы: карты сенсорной информации —
карты лабильности — возможное происхождение карт; усовершенство-
вание триггерных стимулов — слуховые пути — зрительные пути — выде-
ление паттерна из минимальных данных — иллюзии — предположение и
опровержение — предвестие научной методологии; карты ЦНС беспозво-
ночных. Гомеостаз: осморегуляция — СО2/О2-регуляция — терморегуля-
ция — гомеостатическая роль боли. Сенсорные миры — малодушие антро-
поцентризма — поляризация солнечного света — электрические поля —
инфракрасное излучение — сенсорная предвзятость — сенсорный мир
рукокрылых — сенсорный мир беспозвоночных — философия научного
прообраза Томаса Куна — освобождающая природа сравнительной сен-
сорной физиологии. От абиотического к биологическому: сексуальный
императив — взаимоотношения хищник-жертва — бабочки против лету-
чих мышей — чувствительность основных экстерорецепторов ограниче-
на только физическими возможностями — сенсорный смысл брачного
поведения — социальное взаимодействие — социальность человека —
аутизм — психология народа — преднамеренность — связь сенсорной
физиологии с философией. Заключение: исследования — познание и
польза
Вспоминая уже прочитанные главы, можем ли мы выделить какие-то общие
черты, которые повторяются в разных чувствительных модальностях и в раз-
ных типах? Возьмем за основу те фундаментальные черты, которые рассмотре-
22. Повторяющиеся мотивы 519
ны в главе 3. Все высокоразвитые животные имеют «линии раздела», изоли-
рующие различные чувствительные модальности, все сенсорные клетки уст-
роены таким образом, чтобы наиболее интенсивно реагировать на один тип
стимулов — адекватный; интенсивность всех сигналов кодируется частотой
импульсации. В главе 3 мы рассмотрели все эти характеристики, а также та-
кие универсальные понятия, как соотношение «сигнал — шум», едва разли-
чимые отличия, сенсорная адаптация, рецептивные поля и т. д. В данной гла-
ве мы оглянемся назад, чтобы попытаться найти общие принципы строения и
универсальные или близкие к универсальным тенденции. Мы двинемся от
биохимии к организму, начиная с молекулярного и клеточного уровней и за-
канчивая на уровне мозга, физиологии целого организма и взаимодействий
организмов в сообществах.
22.1. Молекулярные мотивы
Эта книга писалась, исходя из предпосылок теории органической эволюции.
Соответственно, нас не должно удивлять поразительное сходство на молеку-
лярном уровне, сходство, проявляющееся и между разными модальностями, и
между разными типами животных. Две трети истории жизни на этой планете
существовали только прокариотические микроорганизмы; эукариоты, а позд-
нее и многоклеточные — это лишь недавнее дополнение. Многие молекуляр-
ные элементы возникли до этих последних. Это и мембранные воротные меха-
низмы, и рецепторные молекулы, распознающие агенты, поступающие из
внешней среды; это и мембранные G-белковые сигнальные системы. Сенсор-
ные системы многоклеточных строились на основе уже развившихся вездесу-
щих элементов.
Мембранные воротные механизмы высших организмов многочисленны
и разнообразны. Почти все они состоят из а-спиральных единиц, пронизы-
вающих фосфолипидную биомембрану. Иногда такие единицы состоят из
всего одной а-спирали (напр., MinK и фосфолемный С1“-канал), но чаще
они образованы многими трансмембранными спиралями (12 в С1С С1“-ка-
налах или 4 х 6 в Na+ и Са2+-каналах). В случае одиночной трансмембран-
ной единицы канал образуется путем группировки нескольких таких единиц
таким образом, что они окружают центральную пору. Мы видели, что это
вероятно так в случае МчкВ-механочувствительного канала Е. coli (Глава 5).
Многочисленные трансмембранные спирали Na+- и Са2+-каналов, напро-
тив, достаточны, чтобы сформировать трансмембранный цилиндр самосто-
ятельно. Существует масса промежуточных случаев, в которых небольшое
количество множественных трансмембранных единиц группируется вокруг
центральной поры. Некоторые примеры архитектуры этих каналов показа-
ны на рис. 22.1.
Анализ аминокислотных последовательностей молекулярных субъединиц
этих воротных механизмов показывает, что они проявляют выраженное эво-
люционное сходство (рис. 22.2). Вероятно, рецепторы натяжения являются на-
иболее эволюционно ранними, и возможно, остальные типы каналов разви-
лись из этого предкового канала. Мы отмечали в главе 5, что чувствительные к
520 Часть VI. Кода
1. G-белок связанные рецепторы
3. Потенциал-зависимые каналы
Рис. 22.1. Молекулярная архитектура некоторых мембранных воротных механиз-
мов и каналов. (А) и (Б) два варианта вездесущих семидоменных рецепторов, свя-
занных с G-белком. (В), (Г) и (Д) три типа медиаторных рецепторов. (Е), (Ж), (3),
(И), (К), (Л) и (М) различные типы потенциал-зависимых ионных каналов. (Е) —
субъединица типичного К+-канала; (Ж) — это массивная структура Na+- и Са2+-
каналов; (3) и (И) — субъединицы других типов К+-каналов; (К) С1С СГ-канал;
(Л) и (М) — субъединицы других типов СГ-каналов. Из Smith, 1996, с разрешения.
22. Повторяющиеся мотивы 521
Наиболее
древние
позвоночные
Наиболее
древние
многокле-
точные
Наиболее
древние
эукариоты
Рецепторы
5НТ3Р нАХР ГлиР ГАМКдР Кн натяжения ЦНЗ ПЗК+ ПЗСа2+ П3№+
Кислородная
атмосфера
Прокариоты
Наиболее
древние
прокариоты
Рис. 22.2. Эволюция ионных каналов. На рисунке много знаков вопроса, которые пока-
зывают, что наши знания здесь еще очень приблизительны. Тем не менее, рисунок пока-
зывает, что каналы, чувствительны? к натяжению, являются вероятными предшествен-
никами остальных. Показанные справа на рисунке потенциал-зависимые Са2+- и Na+-
каналы возникли из К+-каналов в результате двух генных дупликаций. Расширяющиеся
углы показывают, что каждый из каналов образует множество подтипов и вариантов.
Кн — коннексин, ЦНЗ — циклонуклеотид-зависимые каналы, ГАМКАР — ГАМКА-ак-
тивируемый рецептор; ГлиР — глицин-акгивируемый рецептор, 5НТЗР — серотонино-
вый рецептор типа 3; нАХР — никотиновый ацетилхолиновый рецептор; ПЗК+ — потен-
циал-зависимые К+-каналы; ПЗСа2+ — потенциал-зависимый Са2+-канал; H3Na+ —
потенциал-зависимые Ка+-каналы. Из Smith, 1996, с разрешения.
натяжению каналы достаточно широко представлены у прокариот. Потенциал-
зависимые каналы (К+, Са2+ и Ка+-каналы) могут быть прослежены до про-
стейших и, следовательно, возникли около 1500 миллионов лет назад. Каналы,
управляемые биохимическими агонистами, возникли позже, но вероятно, не
19 Биология сенсорных систем
522 Часть VI. Кода
раньше, чем первые многоклеточные'. Анализ последовательностей белков
щелевых контактов многоклеточных показывает, что они связаны единица-
ми каналов, управляемых химическими агонистами. Поэтому весьма вероят-
но, что каналы, которые играют важную роль постсинаптических мембранах
(н-АХ-рецептор, ГАМК-рецептор и т. д.), развились из этих первичных
средств коммуникации между клетками многоклеточных. Прямое действие
химических агонистов на каналы в мембранах сенсорных клеток необычно.
Большинство таких агонистов работает через G-белковую систему. Исключе-
ния из этого правила, однако, обнаружены во вкусовых клетках, чувствитель-
ных к Na+ и Н+ (гл. 12).
Помимо вездесущих белков потенциал-зависимых каналов, мы видели,
что хемо- и фоторецепторные клетки построены по другому распространен-
ному молекулярному принципу (гл. 12, 13 и 16). Существуют некоторые дан-
ные (гл. 19), наводящие на мысль, что сходные белки присутствуют в некото-
рых термочувствительных окончаниях. Это вездесущие семидоменные «змее-
видные» белки, которые обнаруживаются и в постсинаптических мембранах
многих синапсов. Структуру семидоменных белков мы рассмотрели в гл. 1, а
в Информации IV. 1 (см. стр. 271—274) видели, что такую структуру имеет и
бактериородопсин галофильной прокариоты Halobacterium halobium. Амино-
кислотная последовательность бактериородопсина, однако, не имеет тесной
родственной связи с семидоменными белками многоклеточных. Хотя и труд-
но поверить, что эти две настолько сложные и функционально схожие моле-
кулярные структуры возникли независимо, это именно так. Действительно,
подтверждение независимого (полифилетического) происхождения семидо-
менной архитектуры мы находим во множестве несвязанных между собой се-
мейств обонятельных семидоменных рецепторов в царстве животных (см.
гл. 13). Мы пока не можем сказать, все ли семидоменные белки связаны меж-
ду собой и, следовательно, произошли от сходных структур в мембранах древ-
них прокариот, или они «нашли» этот принцип строения независимо в длин-
ной истории живого на этой планете.
В сенсорных клетках семидоменные рецепторы связаны с G-белковой
мембранной сигнальной системой, и такой же механизм взаимодействия об-
наружен во многих других клетках организма многоклеточных. Активность
G-белков, как это говорилось в гл. 1, мож. . быть прослежена вплоть до про-
стейших. Она интенсивно исследовалась у дрожжей Saccharomyces cerevisiae.
В этом случае, как и многих других, мы снова находим молекулярное устрой-
ство, которое, будучи единожды «найденным» эволюцией, используется сно-
ва и снова в совершенно различных целях. Вторичные мессенджеры, которые
выделяются в цитозоль в результате активации G-белкового механизма, также
1 Многочисленные работы показывают наличие у простейших функционально активных
веществ, химически идентичных медиаторам. Более того, в пределах одной клетки обнару-
живается до четырех медиаторов одновременно. До пяти медиаторов, соответствующие сис-
темы и функциональная активность показаны и на одноклеточной стадии эмбрионального
развития (см. Бузников. Нейротрансмиттеры в эмбриогенезе. М. Наука. 1987; Shmukler,
Buznikov. 1998. Perspect. Dev. Neurobiol., 5, 469—480). Так что вывод о возникновении медиа-
торных рецепторов лишь у многоклеточных представляется спорным. — Прим, перев.
22. Повторяющиеся мотивы 523
многочисленны и выполняют различные функции. Мы видели, как работает
связанная с G-белком система вторичных мессенджеров в обонятельных и в
различных типах фоторецепторных клеток у позвоночных и беспозвоночных.
Огромным преимуществом связанной с G-белком системы вторичных мес-
сенджеров является ее гибкость. Семидоменный рецептор, сформированный
для реакции на специфический лиганд (в случае обонятельных рецепторов —
много семидоменных рецепторов и множество лигандов, а в случае фоторе-
цепторов — «встроенный лиганд» — ретиналь), связан через G-белковую сис-
тему с различными мембранно-связанными ферментами, такими, как адени-
латциклаза, цГМФ-фосфодиэстераза, фосфолипаза С, а в каких-то случаях —
с ионными каналами и т.д. Эти системы также позволяют достигнуть значи-
тельного усиления и поддержания первичного сигнала, а также обеспечить
сенсорную адаптацию и десенситизацию.
22.2. Клеточные мотивы
На клеточном уровне сенсорные системы начинают утрачивать общие свойст-
ва. Тем не менее, и здесь можно выделить некоторые навязчивые мотивы. Воз-
можно, самым очевидным из них является вездесущность неподвижных рес-
ничек и микроворсинок — мы отмечали, что три основные сенсорные модаль-
ности: механо-, хемо- и фоторецепторные клетки — это модифицированные
ресничные клетки. Это очень хорошо заметно среди позвоночных, но обнару-
живается и у беспозвоночных. И это снова пример того, как эволюционный
процесс, единожды наткнувшись на эффективный вариант организации, мо-
дифицировал его затем к той или иной модальности.
В главе 8 мы видели, что волосковые клетки позвоночных обладают множе-
ством стереоцилий (модифицированных микроворсинок) и одной неподвиж-
ной собственно ресничкой — киноцилией. В этом случае именно стереоцилии
обладают механо-чувствительным ионным каналом; важно при этом отме-
тить, что это типичные ресничные клетки, которые были модифицированы и
«поставлены на службу». В главе 8 мы также видели, что биофизический меха-
низм построен так, что деформация волоска ведет непосредственно к откры-
ванию ионных каналов и, соответственно, к стимуляции волосковых клеток.
Механизмы хемо- и фоторецепторных клеток связаны с G-белок связанной
сигнализацией вторичных мессенджеров, а потому медленнее.
В главах 12 и 13 мы отмечали, как микроворсинки и неподвижные реснич-
ки обеспечивают чувствительность мембран вкусовых и обонятельных рецеп-
торов. У вкусовых рецепторных клеток развивается плотная сеть микроворси-
нок, а у обонятельных — очень длинные реснички, лежащие на поверхности
обонятельной слизистой. В обоих случаях семидоменные рецепторные моле-
кулы и G-белковые сигнальные системы локализованы в клеточных мембра-
нах. Можно полагать, что значение микроворсинок и неподвижных ресничек
в данном случае состоит в увеличении сенсорной поверхности. Это касается и
фоторецепторных клеток. Мы видели в главе 16, что в наружных сегментах па-
лочек и колбочек позвоночных образуются многочисленные инвагинации
мембран ресничек. Фотопигменты и G-белковые сигнальные системы опять-
таки локализованы в мембранах наружных сегментов. Поскольку мембраны
524 Часть VI. Кода
расположены под прямым углом к поступающему излучению, увеличивается
вероятность поглощения фотонов фотопигментами.
Предшествующий абзац касается сенсорных клеток позвоночных. Сенсор-
ные клетки беспозвоночных также демонстрируют множество сходных моти-
вов, в том числе и с позвоночными. Модифицированные реснички присутст-
вуют и во многих сенсорных сенсиллах насекомых. В главе 6 мы видели, что
внутри кутикулярного чехла окончания сенсорных клеток образуют аналог на-
ружного сегмента и представляют собой модифицированную ресничку. Меха-
ночувствительные сенсиллы, подобные волосковым клеткам позвоночных,
однако, детектируют механические деформации не ресничкой, а тубулярным
тельцем, образованным массой плотно упакованных микротрубочек. Как и в
волосковых клетках, механический стимул прямо приводит к открыванию
ионного воротного механизма и стимуляции нейросенсорной клетки. Тогда
как модифицированные реснички образуют основу некоторых фоторецепто-
ров беспозвоночных (глава 14), большинство их все же построены по другому,
рабдомерному, принципу. В последнем случае нересничные фоточувствитель-
ные окончания нейросенсорных клеток развивают множество микроворси-
нок. Они содержат фотопигмент, а сеть, образуемая ими, подобно дискам на-
ружных сегментов палочек позвоночных, обеспечивает ортогональность к ним
падающего излучения. Интересно, однако, отметить, что фоторецепторные
клетки позвоночных (по крайней мере в латеральных глазах) обращены в сто-
рону, противоположную падающему свету, тогда как у беспозвоночных —
к нему. Также небезынтересно, что если фоторецепторы позвоночных гипер-
поляризуются в ответ на освещение, то у беспозвоночных в большинстве слу-
чаев деполяризуются. Похоже, что у позвоночных на каком-то раннем этапе
эволюции (мы рассмотрели вероятный сценарий в главе 15) весь процесс фо-
торецепции развернулся задом наперед.
22.3. Органы чувств
Выше клеточного уровня мы оказываемся среди групп клеточных единиц, ко-
торые мы именуем органами чувств. Здесь разнообразие уже становится бес-
конечным. Если рассматривать царство животных в целом, то многочислен-
ность различных форм и архитектур исключает мысль о какой-то единой клас-
сификации. К тому же ясно, что не все варианты уже известны, а те, что изве-
стны, далеки от полного понимания. Тем не менее, можно выделить некото-
рые интересные сходства.
В отличие от сходств, наблюдающихся на молекулярном и клеточном
уровнях, сходства органов чувств не обусловлены их происхождением от об-
щего предка. Здесь работает другой принцип — принцип конвергентной эво-
люции. В конце главы 4 мы рассматривали возможность того, что 750 милли-
онов лет назад естественный отбор «обнаружил», что лишь несколько типов
рецепторов обеспечивает чувствительность к среде нашей планеты. В частно-
сти, мы рассматривали крупные типы моллюсков, членистоногих и хордовых.
То же и с органами чувств: за колоссальный период эволюционного отбора
выделился ряд аналогичных решений основных проблем строения органов
чувств.
22. Повторяющиеся мотивы 525
Возможно, наиболее известен пример организации «матиссеновых хруста-
ликов» (см. гл. 18). Хрусталики глаз животного по необходимости формируют-
ся из малообещающих материалов, в случае глаз водных животных проблема
особенно остра, поскольку коэффициент преломления на границе воды и хру-
сталика должен быть выше, чем это достижимо при использовании биологи-
ческих материалов, чтобы свет фокусировался надостаточно коротком рассто-
янии за хрусталиком. «Решение» проблемы было найдено независимо по
крайней мере восьмью группами водных животных и сводится к тому, что ко-
эффициент преломления меняется от центра к периферии хрусталика. Особые
преимущества, которые дает острое зрение, привели к тому, что в результате
конвергентной эволюции у несвязанных между собой групп животных разви-
лись поразительно сходные структуры.
Зрительная система дает и другой пример эволюции, конвергирующей на
сходных решениях. Глаза белемноидов (глава 14) демонстрируют удивитель-
ное сходство с глазами позвоночных. Это особенно интересно, поскольку на
клеточном уровне эти два типа глаз радикально отличаются по строению: фо-
торецепторы позвоночных — ресничного типа, а у моллюсков — рабдомерно-
го. Между этими глазами существует, конечно, и еще множество различий.
Фокусировка у белемноидов осуществляется перемещением хрусталика впе-
ред или назад (совершенно, как в фотоаппарате), тогда как у позвоночных —
за счет изменения кривизны хрусталика; сетчатка белемноидов неинвертиро-
вана, в отличие от позвоночных. И несмотря на все это, общий план строения
удивительно похож на таковой глаз позвоночных, сформированных на совер-
шенно другой основе. А дело в том, что как и акулы и барракуды, другие мор-
ские хищники — кальмары и осьминоги — зависят в своей жизни от остроты
зрения.
Еще одним интересным сходством органов чувств является тенденция к
специализации сенсорных поверхностей. Наиболее очевидно это в случае сет-
чатки. В сетчатке млекопитающих развивается фовеа, в которой сконцентри-
ровано зрение при дневном освещении. Зрительные стимулы, детектируемые
периферией сетчатки, заставляют глаз повернуться так, чтобы они восприни-
мались в фовеа. У многих наземных млекопитающих, обитающих на равнинах
и в саванне, в сетчатке обнаружена зрительная полоска (глава 18). Вероятно,
это адаптация к основным стимулам, касающимся хищников и жертв, распо-
лагающихся в основном в горизонтальной плоскости. Сходная специализация
обнаружена и в сетчатке птиц, у которых может развиваться более чем одна
фовеа (гл. 18), наряду со зрительной полоской и другими видами специализа-
ции. Сравнивая с этим совершенно иначе построенный глаз насекомых, мож-
но и здесь найти схожую специализацию. У некоторых двукрылых (табанид) и
эфемерид в их сложных глазах омматидии, направленные вперед и вверх, зна-
чительно увеличены и также называются «фовеа». Интересно, что они разви-
ваются только у самцов. Надо полагать, они служат самцам в поисках соответ-
ствующей самки, которую из-за ее малого размера иначе трудно найти на фо-
не неба. Опять-таки, у активных хищных стрекоз (Odonata), обладающих наи-
более высокоразвитыми сложными глазами среди насекомых, наблюдается
очевидная специализация. В сложном глазу стрекозы-дозорщика Апах junius,
имеется, например, передне-дорзальная полоска, в которой угловая плотность
526 Часть VI. Кода
омматидиев в 20 раз больше, чем в задне-дорзальной части. Такая фовеальная
область сканирует лежащий впереди путь стрекозы, детектируя мелкие стре-
мительно перемещающиеся насекомых-жертв.
Тенденция к специализации сенсорных поверхностей не исчерпывается
сетчаткой — то же касается и других сенсорных модальностей. Мы отмечали в
главе 13, что обонятельные рецепторные клетки построены не случайным об-
разом, как можно было ожидать, а несколькими передне-задними «зонами
экспрессии», каждая из которых содержит пространственно организованные
группы клеток с различной восприимчивостью к различным одорантам. Одна-
ко, может быть более впечатляет то, что мы отмечали в главе 9 — базилярные
мембраны эхолоцирующих летучих мышей высоко специализированы на-
столько, что в некоторых случаях более 50% их длины настроены реагировать
на эхо от их собственных характерных акустических сигналов. Эта часть бази-
лярной мембраны, как мы видели, по этой причине именуется слуховой фовеа.
Наконец, если рассматривать кожу как сенсорную поверхность чувства осяза-
ния (или, в случае рыб со слабыми электрическими свойствами, поверхность
детектирования электрических полей), мы снова видим зачатки пространст-
венной организации. У человека для определения текстуры объекта использу-
ются пальцы или губы; другие млекопитающие пользуются морду, вибриссы,
хобот; электрорецепторы у слабоэлектрических рыб распределены по поверх-
ности тела таким образом (рис. 20.9), что ориентация в электрическом поле
вызывает определенный паттерн активности в сенсорных волокнах, направля-
ющихся в центральную нервную систему.
Наконец, обращаясь к принципиальным схемам органов чувств, интересно
отметить, что существует значительный параллелизм между нейрогистологией
обонятельной луковицы и сетчатки млекопитающих. Мы видели в главе 13,
что импульсация из обонятельной луковицы проходит через два слоя клеток,
чьи волокна ориентированы ортогонально к направлению афферентов. В гла-
ве 16 мы сталкивались с аналогичной структурой в сетчатке: афферентация от
фоторецепторных клеток перерабатывается в двух слоях клеток — горизон-
тальных и амакриновых — чьи отростки располагаются под прямым углом к
направлению потока информации. Интересно также отметить вездесущность
перекрестов нервных путей, ведущих от сетчатки к зрительным долям. Такую
организацию мы встречали (гл. 14) у совершенно различных форм — ракооб-
разных, насекомых и головоногих. То, что эволюционный процесс отобрал
именно такие структурные решения, в одном случае — охватывающие разные
сенсорные модальности, а в другом — разные типы животных, должно, как в
случае с матиссеновыми хрусталиками, говорить нам что-то об эффективнос-
ти таких решений.
22.4. Центральные анализаторы
Центральный анализ сенсорной информации также демонстрирует интерес-
ные сходства между модальностями и различными типами животных. Мы
видели в главах 7, 9 и 17, как сенсорные поверхности (кожа, базилярная мем-
брана и сетчатка) картируются в первичной сенсорной коре. В главе 13 мы
отмечали, что, хотя обонятельный эпителий не картируется, в обонятельной
22. Повторяющиеся мотивы 527
луковице, тем не менее, присутствует функциональная карта пахучих веществ,
представленных в носовой полости. Эти карты часто весьма неизоморфны
сенсорной поверхности и лабильны. Это выглядит так, будто первичная сен-
сорная кора, само существование которой обусловлено генетически, пере-
страивается в зависимости от интенсивности ее использования. Мы видели
примеры такой лабильности в главе 7, когда обсуждали эксперименты по уда-
лению вибрисс у мышей, эффект стимуляции пальцев обезьян и ответы соме-
стезической коры при активном использовании пальцев у слепых, читающих
по Брайлю, и можем сравнить их с лабильностью первичной зрительной ко-
ры, рассмотренной в гл. 17. Эти и другие эксперименты дают ключ к понима-
нию вездесущности сенсорных карт. Нейроны, срабатывающие совместно,
обладают, как правило, синаптическими связями. Если ус или палец удалить,
то их кортикальное представительство уменьшится или исчезнет. Это, в свою
очередь, повлияет на клетки, реагирующие на соседнюю вибриссу или палец.
Если афферентацию, поступающую в первичную зрительную кору от части
сетчатки одного из глаз, блокировать, соответствующая область коры будет
занята клетками, отвечающими на афферентацию из соответствующей части
другой сетчатки. Из этого следует, что длительная история (включая эволю-
ционную историю) временных корреляций стимула и сенсорной поверхнос-
ти приводит к формированию топографической карты в соответствующей
части первичной сенсорной коры.
В отличие от афферентов зрительной, соместезической и слуховой систем,
временные корреляции вкусовых и(или) обонятельных афферентов не так
сильны. Мы видели, что существует некоторая дифференциация вкусовых и
обонятельных поверхностей у млекопитающих, а также (см. гл. 13) у некото-
рых насекомых (напр., обонятельные сенсиллы плодовой мушки Drosophila).
При этом неочевидно, что вкусовые или пахучие молекулы воздействуют на
эти поверхности в каком-то определенном временном порядке. Вместо этого
вкусовые и пахучие молекулы влияют на специфические группы клеток. В слу-
чае пахучих веществ, такие клетки у позвоночных распределены в слизистой в
случайном порядке. Однако, проекции этих одинаково «настроенных» клеток,
стимулирующихся в носовой полости специфическим запахом примерно од-
новременно, оканчиваются на одних и тех же обонятельных гломерулах. То
есть, хотя пространственная карта сенсорной поверхности в обоим i ел ьной лу-
ковице и отсутствует, здесь образуется пространственно организованная кар-
та активных химических веществ во вдыхаемом воздухе. Принцип пространст-
венных карт (см. гл. 9) затрагивает и достаточно абстрактные феномены, фе-
номены, которые вообще не имеют пространственного представительства на
сенсорном экране собственно органа чувств, как, например, слуховое прост-
ранство у сипухи или область атаки у мелких рукокрылых. Принцип организа-
ции информации в топически связные карты в мозге, возможно, используется
еще шире. Есть предположения, что в мозге человека существуют фонемные и
даже семантические карты.
Завершив с проблемой сенсорных карт, мы можем отметить еще одну
черту, общую для слуховых и зрительных путей — это постоянное «уточне-
ние» стимулов, запускающих активность клеток. По мере продвижения к
высшим уровням обработки информации, все более специфические характе-
528 Часть VI. Кода
ристики необходимы для того, чтобы вызвать активность клеток соответст-
вующего уровня. В гл. 9 мы отмечали, что когда сигналы чистого тона или
широкополосного шума, на которые реагировали нейроны низших уровней,
достигают слуховой коры, они оказываются неэффективными. В тоже время
клетки височной доли лучше всего реагируют на частотно-модулированные
тоны, или даже более специфически — у макаков и саймири — на призывы
особей собственного вида, причем иногда — конкретных особей. Несомнен-
но, здесь есть параллель со зрительной корой. Пятна света на сетчатке не вы-
зывают ответа в большинстве клеток первичной зрительной коры; напротив,
определенным образом ориентированные края объектов, часто в совокупно-
сти с другими характеристиками, являются триггерными стимулами. Далее
в зрительной системе, например в нижневисочной области коры обезьян,
электрофизиологически выявлены клетки, аналогичные клеткам распозна-
вания внутривидовых призывов слуховой коры. Это — клетки распознавания
лиц (глава 17), которые активируются образами морд других обезьян, буду-
чи часто чувствительными к образу в профиль, анфас или же к конкретным
особям.
Еще одной общей чертой анализа сенсорной информации в мозге является
повсеместность ее параллельной обработки. Это особенно заметно в тех случа-
ях, когда сенсорная информация содержит существенные детали. Мы отмеча-
ли, что зрительный анализ у приматов осуществляется в не менее чем дюжине
зрительных областей коры, сложным образом соединенных между собой. Ана-
логичным образом, анализ акустической информации у рукокрылых осуще-
ствляется по крайней мере в девяти различных областях коры. Бодрствующая
кора мозга млекопитающих представляет собой мозаику активности. То, как
вся эта активность комбинируется, чтобы сформировать единый сенсорный
образ, как отмечалось в разделе 3.6, представляет собой проблему связывания и
остается одной из главных, до сих пор нерешенных, проблем современной
сенсорной нейрофизиологии.
Еще одна черта сенсорных систем, наиболее ярко проявляющая в слухе и
зрении, это восприимчивость к «иллюзиям» и «переключениям» категориаль-
ных понятий; таковы куб Неккера, переключение восприятия /Ьа/ в /da/. Это
явление — следствие эволюционного императива быстрого принятия решений.
Слуховая и зрительная системы захватывают огромное количество информа-
ции об окружающей среде, и жизненно важно, чтобы «смысл» и «тип» инфор-
мации были выделены, насколько это возможно быстро. Жизнь животного и
перспектива сохранения в будущем его потомства часто зависят от такой спо-
собности. Поэтому, когда слуховая система улавливает треск ломающейся ве-
точки, реакция в виде атаки или бегства должна реализовываться немедленно.
Сходным образом, когда слуховая система человека улавливает волны атмо-
сферного давления, она может интерпретировать их как «коммунистического
труда» или «кому нести чего куда»1, в зависимости от контекста. Аналогичные
1 В оригинале «I scream» (я визжу) и «ice cream» (мороженое), звучащие одинаково; при-
мер в тексте перевода дан, чтобы у русскоязычных студентов не сложилось иллюзии, что ил-
люзии бывают только у англоязычных студентов. — Прим, перев.
22. Повторяющиеся мотивы 529
Рис. 22.3. Молодая женшина/старая
женщина.
иллюзии могут возникать и в зрительной системе, чему есть бесчисленное
множество примеров. Хорошо известна комбинация «старая женщина/мо-
лодая женщина» (рис. 22.3). В зависимости от контекста1, изображение мо-
жет интерпретироваться как молодая женщина в пол-оборота или старая —
в профиль.
Склонность мозга к выводам, недостаточно обоснованным сенсорной
информацией, стремление осмыслить окружающую среду — это проявления
основного принципа его функционирования как машины, генерирующей
гипотезы. С философской точки зрения эта склонность может быть связана с
известной характеристикой науки, которую дал Карл Поппер, как «предполо-
жения и опровержения». Согласно взглядам Поппера, наука постоянно вы-
двигает гипотезы (предположения) для объяснения явлений окружающего
мира, и, чем более они ошибочны (опровергаемы), тем лучше. Очевидно, что
этот сложный процесс в человеческом мозге основывается на эволюционно
встроенной способности, благодаря которой он пытается осмыслить массу
сенсорной информации, поступающей от основных органов чувств. Мы уже
видели в разделах 15.4 и 21.1 и еще встретим в главе 23, что существуют дан-
ные, позволяющие предположить, что эфферентная информация сравнивает-
ся с афферентной.
К счастью, как отмечал Поппер, в науке, когда обнаруживается ошибка,
умирает гипотеза, а не ее автор.
1 Скорее, от угла зрения. — Прим, перев.
530 Часть VI. Кода
Возвращаясь от всех этих достаточно противоречивых аспектов филосо-
фии науки к собственно нейрологии, мы не можем закончить этот раздел, не
упомянув, что сенсорные карты, которые так хорошо знакомы нам в мозге
млекопитающих, обнаружены и в центральной нервной системе других жи-
вотных. Мы видели в главе 14, что ретинотопическая карта присутствует в зри-
тельных пластинках и вышележащих отделах мозга двукрылых, нет сомнений,
что сходные карты генерируются в зрительных путях и других отрядов высоко-
развитых насекомых. В гл. 14 мы также видели, что нейроанатомия зрительной
системы белемноидов показывает, что топографические взаимосвязи зритель-
ного образа сохраняются в нескольких слоях центрального анализатора. К со-
жалению, до сих пор провести микроэлектродные отведения от этих слоев не
удалось.
22.5. Гомеостаз
Гомеостаз — это еще один повторяющийся мотив в этой книге. Мы отмеча-
ли в главе 11, что эта идея занимает центральное место в физиологии с сере-
дины XIX века. Впервые мы столкнулись с ней в гл. 5, где рассматривались
рецепторы натяжения в клеточной мембране, которые жизненно важны, на-
пример, для одноклеточных в контроле осмотического шока. Это важно и
для намного более сложных организмов многоклеточных, и мы обсудили
наиболее известный случай поддержания осмолярности внеклеточной жид-
кости (ВКЖ) у млекопитающих. Для этого требуется, чтобы осмолярность
плазмы воспринимался осморецепторными клетками, которые, как мы зна-
ем, расположены в гипоталамусе. В главе 11 мы рассмотрели аналогичную
роль, которую хеморецепторы играют в поддержании постоянного уровня
дыхательных газов в ВКЖ. Хеморецепторы, чувствительные к парциальному
давлению СО2 и О2, располагаются в каротидном тельце, дуге аорты и про-
долговатом мозге. Когда парциальное давление СО2 и (или) О2 выходят за
пределы «предустановленного диапазона», включается сложный физиологи-
ческий механизм, регулирующий дыхательную систему и восстанавливаю-
щий нормальный уровень парциальных давлений газов насколько это воз-
можно быстро. Затем, в главе 19, мы рассмотрели сложное пространство тер-
мочувствительных окончаний в коже и других органах. Когда температура
внутри тела отклоняется от нормального уровня, афференты от термочувст-
вительных окончаний интегрируются центрами в гипоталамусе (эти центры
и сами по себе чувствительны к температуре крови в окружающих капилля-
рах), которые посылают эфференты, изменяющие теплопродукцию тела или
(более точно) потерю телом тепла.
Однако, возможно, наиболее интересный и впечатляющий случай участия
чувств в поддержании гомеостаза приведен в главе 21 — это боль. В этой гла-
ве упомянуты некоторые неожиданные черты этого универсального (почти
универсального) чувства. В частности, ощущение боли контролируется во-
ротным механизмом на уровне спинного мозга и нисходящими путями голо-
вного мозга. Все эти механизмы действуют, не затрагивая нашего сознания.
Мы не можем мысленно отключить свирепую зубную боль, бесконечную боль
в спине или острые онкологические боли. Эти неприятные ощущения (хотя
22. Повторяющиеся мотивы 531
качественно они и могут существенно отличаются) имеют, как указывалось
в главе 21, биологический смысл, то есть направлены на поддержание гомео-
стаза. Императивное воздействие боли, ее абсолютная способность удержи-
вать на себе наше внимание, это — требование обеспечить организму условия
для защиты от угрожающего жизни повреждения или восстановления после
него.
22.6. Различные сенсорные миры
Неприятные ощущения, боль, вероятно, занимают центральное место в жизни
всех животных. Функция боли в предохранении жизни предполагает, что она
возникла в эволюции достаточно рано. Вокруг нее построена система мощных
рефлексов самосохранения, и не только условных. Однако, другие аспекты
сенсорных миров многочисленны и разнообразны. Это еще одна тема, которая
периодически возникала на предшествующих страницах. Эта тема очень по-
лезна, поскольку для жителей западного мира, все более изолирующихся от
контакта с живой природой с помощью технологий, довольно легко впасть в
антропоцентрическое самодовольство. Изучение сенсорных систем животных
предоставляет ценные лекарства от всепроникающего антропоцентризма.
Животные часто живут в очень разных сенсорных мирах. Фон Юкскюль
(von Uexkull) ввел термин «Umwelt» (окружающий мир) для описания этой ин-
тернализованной структуры. Цайль (Zeil) и Цанкер (Zancker), например, про-
вели впечатляющее исследование сенсорного мира манящего краба (Uca sp.).
Живя в норках в песке либо в иле, эти крабы вылезают из них при отливе. Их
глаза помещаются на концах длинных стебельков и направлены вверх для де-
тектирования хищников и сигналов от самцов собственного вида, семафоря-
щих своими крупными клешнями. Из зрительный мир в основном двухме-
рен — огромное большинство событий происходит в узкой полосе на уровне
горизонта. Цайль и Цанкер показали, как биологически значимая информа-
ция о привычной среде обитания воздействовала на эволюцию их «сенсорных
фильтров и стратегии нервной переработки информации». Помимо зритель-
ных анализаторов другие чувства, особенно чувствительность к вибрации, ося-
зание и химическое чувство также подстроены к их естественным условиям
обитания. Комбинация точной сенсорной физиологии и этологического ана-
лиза обещает принести нам понимание чуждых нам «umwelten» (окружающих
миров) таких, как мир манящего краба.
Мы неоднократно в этой книге подчеркивали чуждость нам этих миров,
особенно в главе 20. Там мы рассматривали, как поляризация солнечного све-
та делает небо для животных, способных ее воспринимать, совершенно
иным, чем это представляется нам —- голубым (да притом лишь иногда) сво-
дом; как мельчайшие градации контуров электрического поля делают водную
среду намного более гетерогенным и интересным местом для слабоэлектри-
ческих рыб; как некоторым змеям поток теплового излучения в непосредст-
венной близости приносит важную для выживания информацию. В более об-
щем смысле, мы видели, как все основные чувства по разному организованы,
по разному адаптируются у разных представителей царства животных. Мы не
единожды задумывались над философским вопросом: что такое быть летучей
532 Часть VI. Кода
мышью? Их органы чувств — это радикальный контраст с нашими собствен-
ными — вместо очевидно превалирующей ориентации на зрение их чувства
основываются на слуховой модальности. Адаптация слуха у рукокрылых на-
столько велика, что, как мы видели в главе 9, возникает множество интерес-
ных аналогий с формирующимися у нас зрительными образами. У нас, прав-
да, есть исчезающе малый шанс понять, что такое — быть летучей мышью; —
в конце концов, слепые или слабовидящие живут в преимущественно слухо-
вом мире, однако нельзя надеяться, что мы когда-либо сможем ухватить мер-
цание сенсорного мира двух других больших эволюционных групп, рассмот-
ренных в этой книге — членистоногих и моллюсков. Видят ли насекомые, на-
пример, стрекозы, в том смысле, в каком видим мы сами? Мир насекомых, к
тому же, переполнен обонятельными и вкусовыми сигналами (гл. 12 и 13), ко-
торые делают его совершенно иным, чем наш химически бедный, запакован-
ный в пластик мир.
И снова мы оказываемся рядом с философскими материями, которые бу-
дут рассмотрены в главе 23. Идея, что различная интерпретация сенсорной ин-
формации означает, что в какой-то степени мы живем в разных мирах, актив-
но обсуждается философами вслед за Хансоном (Hanson) и, особенно, Тома-
сом Куном (Thomas Kuhn). Если, как предполагал Кун, мыслители жили в раз-
ных мирах до и после галилеевой революции в XVII столетии, насколько же
сильнее это в отношении разных животных. Это еще раз подчеркивает то, с че-
го мы начинали этот раздел — изучение сенсорных систем животных освобож-
дает нас от узколобой антропоцентричности. Это должно помочь нам открыть
глаза (используя обычную антропоцентрическую метафору) на многочислен-
ные очень разные возможные миры.
22.7. От абиотического к биологическому:
коммуникация
Последний повторяющийся мотив нашего исследования — это неизбежная
тенденция перехода от детектирования событий в неорганическом окружении
к восприятию событий в мире живого. Наиболее ранние сенсорные системы
развились для обнаружения различных угрожающих жизни и (или) улучшаю-
щих жизнь изменений в мире. Вскоре мир заполнился другими живыми суще-
ствами. Одновременно необходимость поиска полового партнера, чтобы рас-
пространить собственную ДНК, стала представлять интерес для представите-
лей большинства групп животных. Чувства все больше и больше направлялись
не столько на избегание ловушек или использование благоприятных возмож-
ностей абиотического окружения, сколько во все большей степени на окруже-
ние биотическое. Именно тогда началась бесконечная война хищников и
жертв. В этом соревновании платой за точность и быстроту реакции органов
чувств стала жизнь (и смерть, соответственно), то есть, как мы видели выше,
приз за скорость доступа сенсорной информации в анализаторы центральной
нервной системы. В гл. 7 и 9 мы рассматривали поразительную эволюционную
борьбу бабочек с насекомоядными летучими мышами. Похожая «гонка воору-
жений» идет и в остальном царстве животных, а это приводит к постоянному
22. Повторяющиеся мотивы 533
усовершенствованию органов чувств. В главах 9, 13 и 16 мы видели, что чувст-
вительность основных экстерорецепторов — волосковых клеток, обонятель-
ных рецепторов и палочек — к адекватным стимулам находится на уровне фи-
зических возможностей.
Не только отношения «хищник — жертва» вызывают давление отбора на
сенсорные системы. Мы уже отмечали, что не меньшее давление оказывает и
необходимость поиска полового партнера — впечатляющая система полей
обонятельных сенсилл павлиноглазки развила свою чувствительность на-
столько, что способна детектировать одну-единственную молекулу соответст-
вующего полового феромона (гл. 13). В этологической литературе образова-
лись горы публикаций по зрительным, слуховым и обонятельным факторам и
ключевым стимулам, которые животные, принадлежащие к различным типам,
используют для нахождения половых партнеров и для ритуалов спаривания.
Более того, сенсорные признаки и сигналы лежат в основе социальной жизни
животных. Уилсон (Е. О. Wilson) выделил четыре больших пика формирова-
ния социальности в животном мире: колониальные беспозвоночные, соци-
альные насекомые, нечеловекообразные млекопитающие и люди. Мы уже
сталкивались в этой книге с обонятельными сигналами в жизни социальных
насекомых (гл. 13), но они столь же важны в социальной жизни в стаях псо-
вых. В главе 7 мы отмечали, как осязание, активируемое грумингом, помога-
ет укреплять отношения в стаях социальных приматов.
В группе социальных приматов, человекообразных, к которой принадле-
жим и мы сами, все пять экстерорецепторов, особенно зрение, слух и осяза-
ние, глубоко вовлечены в наблюдение за особями своей группы, реакцию на
них и взаимодействие с ними. Люди, как отмечалось выше, живут в мире лю-
дей — мире глубоко социальном. Невзирая на отдельные (и, вероятно, не
очень надежные) сообщения о детях-волках, люди обязаны своей человечес-
кой личностью именно социальности. Неспособность войти в этот социаль-
ный мир выражается в аутизме. В обладающих письменностью обществах
социальность обеспечивается общением не только с современниками, но и
теми, кого уже нет. Такие влияния прошлого, настоящего и предсказуемого
будущего образуют то, что называется психологией народа. Мы приписыва-
ем другим субъективные ощущения, которые испытываем сами, в частности,
намерения, чего не делаем в отношении предметов неорганического мира —
палки, камня, ветра и воды1. То есть мы предполагаем, что они действуют це-
ленаправленно. Здесь мы снова подходим к теме следующей главы. Мы уже
сталкивались с тем, что предмет исследования сенсорных систем не всегда
точно отделен от философских проблем, и в этой предпоследней главе мы на-
тыкались на эту границу не раз. В следующей главе мы попытаемся пробить-
ся через эту границу и закончим книгу обсуждением некоторых философских
проблем.
1 Неточность — само существование оборота «нельзя обижаться на стул, зато, что он тебя
ударил» указывает, что человеку свойственно обижаться и на предметы неживой природы,
т. е. подсознательно приписывать им дурные намерения. Каждый может вспомнить на сей
предмет достаточно примеров из собственного опыта. — Прим, перев.
534 Часть VI. Кода
22.8. Заключение
В данной главе мы рассмотрели некоторые темы, которые повторялись в дан-
ной книге неоднократно. Читатель может сам найти и другие подобные темы.
Предмет сенсорной биологии огромен, постоянно расширяется и не может
быть охвачен полностью в одной книге. Каждая проблема, даже каждый от-
дельный вопрос внутри проблемы содержит в себе столько неизвестного, того,
что еще только должно быть исследовано. Такие исследования, как и всякие
хорошие исследования, включают в себя и сочетают два мотива; Фрэнсис Бэ-
кон еще в XVI века обозначил их как познавательный (luciferous) и полезный
(lucriferous). Исследования сенсорных систем дают нам знания о самих этих
системах, их связях с другими системами организмов или с другими сенсорны-
ми модальностями, проливают свет на проблемы этологии, молекулярной би-
ологии и эволюции. В свете бэконовского определения — это познавательные
исследования, мотивированные точно тем же, чем первопокорители Эвереста
объясняли свой подвиг — «да просто потому, что он там стоит!». Исследования
сенсорных систем, однако, имеют и другую мотивировку и другие результаты.
Чем больше мы знаем о наших сенсорных системах, тем лучше мы можем их
лечить, когда они не в порядке. В главе 21 сталкивались с тревожной статисти-
кой страдающих от болей в спине (и их экономическими последствиями) в за-
падном мире. Сходные, и весьма неприятные, данные существуют и о страдаю-
щих дефектами слуховой и зрительной систем. Бэконов мотив пользы, если
принимать его в широком смысле, касается не только финансов, но и примене-
ния знаний для того, чтобы уменьшить количество страданий в мире. Непо-
средственная выгода тоже встречается. Искусственные «носы» в дегустации ви-
на, различные становящиеся все более доступными протезы органов чувств —
от слуховых аппаратов, очков и приборов искусственного зрения для слепых до
протезов конечностей, несомненно уже сейчас образуют значительный и рас-
тущий сектор экономики. Однако, в следующей и последней главе этой книги
мы перейдем к наверное самой непрактичной сфере человеческого знания —
философии, — вероятно самому яркому примеру бэконовской познаватель-
ной науки. Мы начали эту книгу с знаменитой цитаты из аристотелевой «Ме-
тафизики»: «Каждому человеку свойственно стремление к познанию...», так
что вполне уместно и закончить наше экскурс в сенсорную биологию тем, как
эта наука связана с предметом главного интереса Аристотеля — философией.
23. ФИЛОСОФСКИЙ постскриптум
Декарт: cogito} — метод сомнения — res cogitans — res extensa — проблема
эпистемологии. Сенсорные качества: неисправимые — «легкие» пробле-
мы и «трудные» проблемы — закрыт ли человеческий разум для их реше-
ния? Tabula rasa1 2: врожденные идеи? — эмпирики — простые и сложные
понятия — кантова «революция Коперника» — влияние на Мюллера и
Гельмгольца. Эпигенетическая эпистемология: вопрос Молинье — опера-
ции по удалению катаракты — эпигенетическая эпистемология Спенсе-
ра — происхождение кантианского a priori3 — происхождение классифи-
кации «объективное/субъективное» — происхождение концепции «энер-
гии». Эволюционная эпистемология — Дарвин — Спенсер — Ницше —
эволюционное разнообразие — поведенческая «подготовленность» —
«алгоритмы» Дарвина — тесты Вэйсона на выполнение условных пра-
вил — объяснения в понятиях социального контракта и обнаружения
ложных утверждений — диморфизм пространственного восприятия. По-
сле Декарта: теория зрения Декарта — контр-примеры — стабилизиро-
ванный образ, мондрианы, направленность саккадических движений
глаз — восприятие с помощью ориентационных действий — боль и пове-
денческие приоритеты — мозг как генератор гипотез. Заключение: лю-
ди — это не пассивные «фотокамеры», а элементы природы — будущие
проблемы сенсорных качеств — начала и окончания
Происхождение философии, как и происхождение науки, теряется в глубинах
доисторических времен. По традиции они возводятся к гигантам греческой
античности, однако человеческое любопытство к природе вещей и тому, как
они функционируют, очевидно, возникли задолго до этой сравнительно ни
давней эры. Возникновение современной философии и науки относят к
еще более близким временам — условно, к интеллектуальному Возрождению
XVI и XVII веков. В том, что касается наук о нервной системе, сенсорной
биологии и эпистемологии, одно имя стоит особняком — это Рене Декарт
(1596-1650).
1 Cogito {лат.) — мыслю. — Прим, перев.
2Tabula rasa {лат.) — чистый лист. — Прим, перев.
3 A priori {лат.) — до опыта, заранее. — Прим, перев.
536 Часть VI. Кода
23.1. Декарт
Рене Декарт (Rene Descartes) родился 31 марта 1596 года в маленькой деревне
Ля Ай недалеко от Тура во французской глубинке. С детства он проявлял бле-
стящие способности, отец называл его «философом» и дал ему лучшее, из воз-
можных в то время, образование в иезуитском Лицее Генриха IV в Ля Флеш.
Именно ему принадлежит известная чуть ли не всем философская позиция —
cogito ergo sum — мыслю, следовательно, существую, которую он сформулиро-
вал, воюя в тех местах, которые ныне составляют Германию — сидя у печки в
комнате в Ульме. Хотя эта формулировка насчет cogito не так уж оригинальна
(Св. Августин излагал нечто подобное в XV веке), Декарт сумел аргументиро-
вать эту позицию настолько четко, что многие считают ту ноябрьскую ночь
1619 года началом современного мира.
Что же открыл Декарт? Как и многих других мыслителей, Декарта мучило
стремление к определенности. Он чувствовал, что все интеллектуальные докт-
рины его времени построены на очень зыбких основаниях. Его замысел состо-
ял в том, чтобы, отбросив «землю и песок», как он сам писал, докопаться до
коренных пород. Его метод — подвергать все сомнению. Он был готов подвер-
гать сомнению и собственные ощущения, поскольку, как он говорил, они мо-
гут быть иллюзорными. Он был готов сомневаться даже в математических вы-
водах, поскольку математикам свойственно ошибаться, и в решениях могут
содержаться ошибочные посылки. И все же, когда он посидел у своей печки в
Ульме, ему стало ясно, что сомневаться, несмотря на горячие сомнения, в су-
ществовании сомнений невозможно — это было бы внутренне противоречиво,
и он решил, что докопался — Je pense, done je suis1.
В чем он мог не сомневаться, это в том, что мысль существует, а мысль вы-
ше подозрений, и лишь это — определенно. Все остальное может быть иллю-
зией, как ощущение фантомной руки, которую мы давно потеряли, или мир,
окрашенный в желтый свет из-за того, что у нас желтуха, и в глазах накаплива-
ется пигмент желчи. Декарт заключил, что он, в сущности, — мыслящее веще-
ство — res cogitans. Напротив, мир вне его, мир, о котором чувства могут давать
неточную информацию, Декарт охарактеризовал как пространственное изме-
рение. Как указывал он в «Принципах философии» (Principles of Phylosophy,
часть 2, гл. 4), нельзя исключить, что когда мы протягиваем руку, чтобы при-
коснуться к предмету, он отступит и избежит прикосновения. Тем не менее,
если это, пусть и невозможное, случится, предметы от этого не утрачивают
своей сущности. Следовательно, «твердость» или «непроницаемость» не явля-
ются необходимыми чертами предмета, который, однако, обладает «длиной,
шириной и глубиной». Этот мир, в котором он как гений геометрии прекрас-
но ориентировался, был им определен как res extensa2. Здесь снова встает во-
прос — каким образом мир res cogitans взимодействует с миром res extensa. Это
классическая проблема соотношения тела и разума, вечно беспокоящая всех
философов.
1 Je pense, done je suis (фр.) — всего лишь cogito ergo sum, но по-французски. — Прим, перев.
2 Res extensa (лат.) — вещество протяженное. — Прим, перев.
23. Философский постскриптум 537
С проблемой соотношения связаны и проблемы эпистемологии (теории
познания). Если сведения о мире, представляемые нам органами чувств, есть
всегда предмет сомнений, то как мы можем быть уверены в том, что мир имен-
но таков, каким мы его себе представляем? Современный мысленный экспе-
римент ставит вопрос так: откуда мы знаем, что мы — не «мозг в колбе», чье
единственное достояние — сенсорные нервы — хитроумно стимулируются.
Декарт обошел эту проблему, сославшись на онтологическое доказательство
бытия Господа всеблагого и всемогущего. Онтологическое доказательство не
должно нас отвлекать (студенты могут найти его в Лекциях (Discourse), часть
4), поскольку нынче мало кто его принимает, но Декарту оно дало то, что ему
было нужно. Господь всеблагой и всемогущий не станет вводить в заблужде-
ние творения свои, посылая по их сенсорным нервам иллюзорную информа-
цию. Перед нами, кому трудновато найти доказательства бытия божьего1, кар-
тезианская позиция ставит эпистемологическую проблему очень остро — воз-
можно ли знание (знание определенное)? Насколько мы можем быть уверены,
что наши органы чувств не вводят нас в заблуждение, причем не занимаются
этим систематически?
23.2. Сенсорные качества
Декартов метод радикального сомнения до сих пор, 350 лет спустя, кажется
эффективным и приводит к пониманию важных моментов. Мы не можем со-
мневаться, испытывая ощущения «красного» или «боли», что эти ощущения
действительно имеют место. Возможно, что они не имеют никакого отноше-
ния к миру за пределами кожи, отграничивающей организм от окружающей
среды — в конце концов, возможны какие-то нейрохимические нарушения в
соответствующем участке мозга. Однако то, что мы это «переживаем» — несо-
мненно. Может ли какой-то сторонний наблюдатель, какой-то авторитетный
врач отрицать для нас этот наш опыт? Ну, конечно, нет. Мы — те, кто испыты-
вает субъективные ощущения, — имеем к ним, как говорят в компьютерных
сетях, «привилегированный доступ» — наша исключительность в отношении
собственных ощущений не подлежит сомнению. И именно потому, что она
непреодолима, мы не в состоянии выразить кому-либо другому качество или
количество того, что мы испытываем.
Было немало попыток описать эту странность, лежащую в сердцевине на-
ше мира. Для тех, кто занимается наукой профессионально, наиболее порази-
тельными были попытки мысленных экспериментов с фиктивным инстру-
ментом, названным аутоцереброскопом, обсужденные Гербертом Файглем
(Herbert Feigl) и другими в 1960-е гг. С колоссальным прогрессом нейроими-
джинга в последние десятилетия XX века эти фантазии стали стремительно
превращаться в реальность. Аппаратура для позитронной томографии (ПТ) и
магнитно-резонансных исследований (МРИ) дает нам возможность просле-
1 Исчерпывающее решение этого вопроса дано Э. Рязановым и Э. Брагинским: одни ве-
рят, что Бог есть, другие верят, что бога нет. То и другое — недоказуемо. Берегись автомоби-
ля! В кн.: «Смешные невеселые истории. Комедии для кино и телевидения». 1979. М. Совет-
ский писатель. — Прим, перев.
538 Часть VI. Кода
живать активность в мозге при умственной деятельности. Применение магни-
тоэнцефалографии (МЭГ) обещает возможность делать это в реальном време-
ни. Однако, даже если в каком-то утопическом будущем будет возможна реги-
страция всех молекулярных и биофизических событий в мозге параллельно с
описанием субъектом переживаемой им боли, никакого иного способа понять
«на что это похоже», кроме сравнения с собственными ощущениями не найти.
Аналогично, можно предположить, что в некотором будущем нейробиологи
окажутся в состоянии регистрировать все события в мозге пациента, страдаю-
щего звоном в ушах, но, если они сами глухи от рождения, даже намека не по-
лучат на то, что, собственно, испытывает больной. В то же самое время худож-
ники способны выдать потрясающие описания захода солнца над водами или
палитры их предшественников-импрессионистов, не имея ни малейшего по-
нятия о нейрологии зрительной коры.
Не странно ли это? Наша субъективная жизнь полна цвета, запахов, звуков,
вкусов, удовольствий и боли, а наша наука пока еще во всем этом совершенно
невежественна. В нормальной жизни, даже если это нормальная жизнь нейро-
логической лаборатории, эта странность проходит незамеченной. Большинст-
во из нас может спокойно делать вид, что проблемы не существует, но для тех,
кто замечает ее, такое несоответствие выглядит огромной дырой в нашем
взгляде на мир. В последние годы с их огромными достижениями в нейронау-
ках проблемы мозга и сознания разделились на две категории: легкие пробле-
мы и трудные проблемы. Легкие проблемы, конечно, часто технически очень
трудны, и многие из них еще не решены. Среди них многие из тех, что упоми-
нались на предшествующих страницах: как нервные волокна проводят им-
пульсы; как рецепторные клетки отвечают на стимулы, как мозг анализирует
сенсорную афферентацию. В других областях нейронаук — это память и дви-
гательные эфференты, рост и развитие, болезни и дегенерация. Все это —
предметы интенсивных исследований, и во всех случаях есть свои трудности,
которые, однако, укладываются в рамки научных методов, никаких новых
принципов здесь не вводится. Иное дело с трудными проблемами. Каким об-
разом многочисленные физические изменения в мозге связаны с тем, что име-
нуется «феноменологией», «ощущением» и т. д. — вот что является трудной
проблемой. Попыток найти ее решение было много, а успешных, на мой
взгляд — ни одной. Интересующиеся могут обратиться к книгам, перечислен-
ным в библиографии. Многие, как Колин МакГин (Colin McGinn) (и Рене Де-
карт, конечно), готовы оставить ее в стороне, так как человеческий разум «за-
крыт для ее познания» (так же как разум собаки закрыт для дифференциально-
го исчисления).
23.3. Tabula rasa?
Приходим ли мы в этот мир с готовым знанием в головах или узнаем все по-
средством органов чувств в ходе жизни? Декарт, который обычно верил в то,
что наш разум основан на врожденных идеях, на самом деле был очень осторо-
жен. Единственное, в чем он был уверен — это идея бога. Живший задолго до
него Платон был куда более категоричен. В знаменитом диалоге «Менон»
(Meno) он описывает, что и неграмотный раб может демонстрировать знания
23. Философский постскриптум 539
в геометрии, включая теорему Пифагора. По Платону, математические и дру-
гие формальные знания присущи мозгу еще до рождения, и образование —
лишь восстановление их в памяти. Британские философы-эмпирики, однако,
столетие после смерти Декарта смотрели на это совершенно иначе.
Томас Хоббс* (Thomas Hobbs, 1588—1679) родился на 8 лет раньше Декарта,
но, поскольку умер, когда ему было за 90, пережил того почти на 30 лет. Он пе-
реписывался с Декартом, полемизируя с ним о врожденных идеях, поскольку,
как и позднейшие британские эмпирики, считал, что мы приходим в этот мир
без всякого врожденного знания. Разум — это tabula rasa, чистый лист (бук-
вально — чистая табличка для письма), на котором пишет наш чувственный
опыт. Он настаивал, что «нет никаких концепций в разуме человека, которые
бы исходно целиком, а позже — частично, не были бы порождены органами
чувств». Конечно, это лишь предположение Хоббса, такое же, как предполо-
жение Платона, которое, однако, хорошо укладывалось в его, Хоббса, соци-
ально-политические представления. Последователь Хоббса Джон Локк
(1632—1704) имел репутацию великого философа и считался отцом-основате-
лем британского эмпиризма. Считается, что его основная работа «Эссе о чело-
веческом разуме» (An Essay Concerning Human Understanding), из всего напи-
санного британскими философами, оказала наибольшее влияние на умы. Он
был уверен, что не существует «первичных идей» или «врожденных принци-
пов», запечатленных в разуме человека. Люди приобретают знания, продолжа-
ет он, без помощи каких-то врожденных принципов.
«Предположим, — пишет Локк, — что мозг, как мы говорили, — белый
лист бумаги без единой буквы, без каких-то мыслей, записанных на нем; как
же он заполняется, откуда берется это огромное хранилище, которое ум и без-
граничная фантазия заполняют бесконечным разнообразием?» И не медлит с
ответом: поток ощущений под воздействием внешнего мира формирует пред-
ставления, такие как «желтое, белое, теплое, холодное, мягкое, твердое, горь-
кое, сладкое». Это — так называемые «простые понятия», те качества, которые
обсуждались и не могут быть поставлены под сомнение. Затем из этих простых
понятий формируются сложные идеи — аналогия с химическими соединени-
ями очень близкая. Хотя, как пишет далее Локк, разум «совершенно пассивен
в восприятии всех этих простых понятий», они выделяются сами по себе и об-
разуют идеи, которыми наполнено сознание. Сходную психологическую фи-
лософию можно найти у последователей Локка — Дэвида Юма (1711—1776) и
Дэвида Хартли (1705—1757). Юм, в отличие от Локка и Хартли, пользовался
для своей ассоциативной нейропсихологии не атомистическими метафорами,
а колебательными. Многими он признается одним из отцов-основателей со-
временной физиологической психологии.
В целом ясно, что столетие после смерти Декарта череда мыслителей анг-
ло-саксонского мира развила физиологическую психологию, которая посту-
лировала отсутствие в сознании врожденных знаний. Только с началом обра-
зования сенсорного опыта пустота заполняется разнообразными понятиями.
Первоначально — это «простые понятия», т. е. ничем не усложненные «сен-
* Гоббс — в отечественной литературе. — Прим. ред.
540 Часть VI. Кода
сорные качества» или, если использовать термин Герберта Файгля, «грубые
ощущения». Их комбинации образуют большие структуры ощущений. Ис-
пользуя музыкальную аналогию, можно сказать, что простые ощущения — как
отдельные ноты, взятые на пианино, а ткань комбинации ощущений — это
звучание фортепьянного концерта. В следующем веке Герберт Спенсер (Her-
bert Spencer), с которым мы еще встретимся в этой главе, именно эту метафо-
ру и использовал в своей пионерской работе по эволюционной психологии.
Но здесь мы немного забежали вперед. В XVIII веке был еще один философ,
о котором невозможно не упомянуть — это Иммануил Кант.
Иммануил Кант (1724—1804) по праву считается величайшим философом в
одном ряду с Декартом. Хотя в законченном виде его философия безмерно
сложна, выделяется один из ее главных моментов. Кант не соглашался с идеей
«чистого листа», на котором чувства записывают понятия. Не соглашался он и
с тем, что мы рождаемся в готовыми врожденными понятиями. Он считал ра-
зум активным началом, которое организует пути познания окружающего ми-
ра. Он называет это своей «коперниковой революцией». Категории простран-
ства и времени, причинности и т. д. — это принципы самоорганизации созна-
ния. Они отсутствуют в окружающем мире, но возникают под действием «раз-
ноцветного гудящего беспорядка», о котором сообщают органы чувств, чтобы
осмыслить мир. Таким образом, писал Кант, также, как Коперник перевернул
астрономию, показав, что движется наблюдатель, а не небосвод, так и его,
Кантова, работа переворачивает эпистемологию, показывая, что некоторые
фундаментальные атрибуты нашего знания мира есть свойства наблюдателя, а
не наблюдаемого.
Эта Кантова «революция Коперника» невероятно повлияла на умы XIX ве-
ка, в том числе и умы сенсорных физиологов. Стало ясно, что органы чувств
действительно фильтруют события в окружающем мире и, действительно, хо-
тя и не совсем так, как предполагал Кант, навязывают порядок хаотическому
сенсорному миру. Гельмгольц, которого считают одним из величайших, если
не просто величайшим среди физиологов сенсорных систем XIX века, писал,
что «исследования физиологии чувств, которые в основном выполнены и кри-
тически рассмотрены Йоханнесом Мюллером, а затем суммированы в виде его
закона специфических нервных энергий, принесли полнейшее подтвержде-
ние ... (кантовых) «трансцендентных» форм интуиции и мышления». Сейчас
уже, как мы не раз отмечали на предыдущих страницах этой книги, — пред-
ставление, что мозг не является пассивным наблюдателем , а напротив, актив-
но перерабатывает сенсорную информацию — является общепринятым.
23.4. Эпигенетическая эпистемология
Кант полагал, что, хотя сознание не содержит врожденных идей, но имеет не-
которые готовые категориальные принципы организации. Эта позиция не бы-
ла принята его оппонентами-эмпириками. Они, в свою очередь, считали, что
все необходимые представления о пространстве, времени, причинности, мате-
рии и т. д., которые Кант считал необходимым предварительным условием для
какого-либо осмысления, возникают из опыта индивида. В классическом пас-
саже из «Эссе» (Essay, книга 2, гл. 9, § 8) Джона Локка говорится, как его друг
23. Философский постскриптум 541
Молинье задал ему знаменитый вопрос: «Представь себе человека, рожденно-
го слепым, а теперь уже взрослого, научившегося с помощью осязания разли-
чать куб и сферу, сделанные из одинакового металла и примерно равные по ве-
су. Предположим, что куб и сфера лежат на столе, а слепой прозрел. Сможет ли
он посредством зрения, а не прикосновения, различить, где здесь куб, а где
шар? Молинье отвечает: нет, и Локк с ним соглашается. Локк и его приятели-
эмпирики, как мы знаем, считал, что все знания через органы чувств, и ника-
ких врожденных принципов не существует. Кант с ними не соглашался. Он
считал, что интуитивное представление о пространстве является врожденной
категорией, а, следовательно, позволяет только что прозревшему различить
куб и сферу.
Локк, что, вероятно, вообще характерно для философов, никогда не ставил
никаких опытов и не делал наблюдений, чтобы решить поставленный им же
вопрос. Для Локка это странновато, поскольку он получил медицинское обра-
зование и работал по специальности, а операции по удалению катаракты в то
время уже не были экзотикой. Вопрос Молинье породил в XVIII веке значи-
тельный интерес, и было предпринято несколько попыток решить его. Воз-
можно наиболее известна та из них, о которой сообщил Чизелден (Cheselden)
в Королевском Обществе1 в 1728 году. Катаракту удалили мальчику лет 13—14-
ти. По завершении послеоперационного периода оказалось, что он либо пло-
хо оценивает расстояния, либо вообще не в состоянии это сделать и не может
визуально опознать форму объекта. В частности, он не мог отличить кошку от
собаки до тех пор, пока не взял кошку в руки (а это ощущение ему было знако-
мо), посмотрел на нее безразлично и отпустил со словами: «Пока, киса! Позна-
комимся в другой раз». Предпринималось и множество других исследований,
чтобы ответить на этот, оказавшийся совсем непростым вопрос. Материалы об
этом можно найти в книге Р. Л. Грегори, приведенной в библиографии. Часто
оказывается, что взрослые настолько привыкли обходиться без зрения, что
когда зрение им восстанавливают, они быстро теряют к нему интерес и пред-
почитают пользоваться другими органами чувств. Три столетия с тех пор, ког-
да Молинье задал свой сакраментальный вопрос (возможно, спровоцирован-
ный женитьбой на слепой), лишь продемонстрировали многосторонность
этой проблемы, но не смогли дать определенного ответа.
В последнее время эпигенетическая эпистемология или эпистемология
развития развивалась Пиаже (Piaget) и его последователями. Однако, задолго
до Пиаже, один из наиболее авторитетных мыслителей XIX века Герберт
Спенсер опубликовал работу по эмпирической психологии, которая, как он
полагал, устраняет противоречие между взглядами Локка и Канта. Несмотря
на то, что «Принципы психологии» (Principles of Psychology) Спенсера были
опубликованы в 1855 году, за четыре года до «Происхождения видов» (The
Origin of Species) Чарльза Дарвина (Charles Darwin), он видел решение именно
в эволюционной теории. Он настаивал на том, что не чувственный опыт инди-
вида вносит определенность в кантовы категории или в теоремы геометрии, а
накапливающийся наследуемый опыт бесчисленного ряда предков. Во второй
1 Королевское общество — британский аналог академии наук. — Прим, перев.
Часть VI. Кода
Информация 23.1
СЕНСОРНОЕ ЗАМЕЩЕНИЕ
Мы видели в этой главе, как вопрос, который Молинье задал своему другу Джону
Локку, породил столетия споров и экспериментов. Современное продолжение этих
дебатов, хотя и в связи с прикладными задачами, связано с пионерскими экспери-
ментами Поля Бах-и-Рита (Paul Bach-y-Rita), в которых использовалась система
тактильно-зрительного замещения (СТЗЗ) для передачи зрительной информации
слепым от рождения.
Система ТЗЗ состоит из телекамеры, соединенной с матрицей стимуляторов,
помещенных на коже в одном или нескольких местах тела (живот, спина, бедро, а
в последнее время — язык). Снимаемое камерой изображение преобразуется в ви-
брации или электрические стимулы. Вначале испытуемые пользуются портативной
камерой, так что информация, поступающая в мозг, дополняется кинестетически-
ми афферентами; позже камеру вмонтируют в оправу очков, и в этом случае вслед-
ствие движений головы при сканировании зрительного окружения вносится вести-
булярная обратная связь. После тренировок испытуемые переставали испытывать
тактильные ощущения, и у них возникал пространственный образ. Становились
доступными довольно сложные черты изображения: перспектива, угловое смеще-
ние, глубина, а при использовании увеличивающего объектива — увеличение, раз-
растание и исчезновение. При этом новое чувство не смешивалось с ощущением,
когда те же участки кожи почесывали или кололи.
Одним из первых испытуемых с помощью СТЗЗ был Дж. Гварньеро (G. Guar-
niero) — слепой от рождения аспирант Нью-Йоркского университета. Он рассказы-
вает, что сначала казалось, что ощущения приходят от кожи, к которой присоедине-
но поле стимуляторов, но затем возникает впечатление, что они преобразуются в дву-
мерное пространство, хотя нет впечатления, что они локализованы вне организма.
Хотя ощущения и были ни чем иным, как осязание, Гварньеро пользуется термином
«видеть» для описания этой новой феноменологии. Наиболее ясной характеристи-
кой объекта было в первую очередь то, движется он или нет. Гварньеро сначала ис-
пытывал значительные трудности в связи с тем, что неподвижные объекты движутся
вправо, когда его голова (с надетой на нее камерой) смещается влево. В конце концов
он сумел адаптироваться к этому неожиданному феномену, так что объекты казались
неподвижными, когда он их сканировал. Ему предложили ответить на вариант во-
проса Молинье, показав объекты и спросив, что это такое. В некоторых случаях ему
говорили, что это за объект, прежде, чем позволяли на него «посмотреть», в некото-
рых случаях — позволяли предварительно ощупать его, а в некоторых — просто «по-
казывали» объект и просили назвать его. Ни в одном случае ему не удалось опознать
объект только с помощью СТЗЗ, он неспособен был установить корреляции между
тем, как «выглядит» объект и как он его воспринимал с помощью осязания. Из-за то-
го, что он не в состоянии был распознать объект с помощью СТЗЗ, лишь с большим
трудом ему удавалось связывать названия объектов и их «зрительное» восприятие.
Гварньеро говорит, что наиболее интригующей чертой зрительного мира, со-
здаваемого СТЗЗ, была способность видеть (с помощью зеркала) одновременно
и перед, и зад объекта, а также колеблющиеся изменения формы пламени свечи.
23. Философский постскриптум 543
Продолжение
Последнее, как он говорит, было «источником бесконечного восхищения», по-
скольку он (по понятным причинам) не мог себе представить, что пламя вообще
имеет форму! Через три недели тренировок, завершает он, «объекты приобрели
верх и низ, правую и левую стороны, но не глубину — он существовал в двумерном
пространстве, локализацию которого еще предстоит определить».
Хотя Гварньеро упоминает о «драматическом впечатлении» от приобретения
этого «нового мира» и о «сильной эмоциональной реакции», похоже, что большин-
ство взрослых и молодых людей воспринимают новый сенсорный опыт без такой
эмоциональной окраски. Так «рассматривание» лица любимого человека с помо-
щью СТЗЗ вызывал удивительно вялые ощущения. Новое чувство существует
слишком недолго, чтобы создать свою историю эмоциональной окраски и ассоци-
аций, которые накоплены другими чувствами слепых от рождения. Использовать
СТЗЗ, говорит Гварньеро, это — «как зрение в очках, хотя и в крайне аналитичес-
кой форме». Разочаровывающие результаты наблюдались и у тех, кому врожденную
катаракту удаляли в зрелом возрасте. Часто они предпочитали не использовать свое
новое чувство и возвращались к привычному тактильно-обонятельно-слуховому
миру.
СТЗЗ, однако, показывает, насколько пластичным остается мозг взрослого че-
ловека. Это дает надежду тем пациентам с повреждениями мозга, кто надеется на
выздоровление. Это дает возможность предположить, что такие протезы, как СТЗЗ,
позволят слепым от рождения выполнять достаточно сложные зрительные задачи.
Бах-и-Рита показал, что системы ТЗЗ позволяют слепым попадать по катящемуся
мячу (что требует визуальной идентификации мяча, его скорости, направления
движения, положения биты и т. д.). и выполнять циклические задачи в промыш-
ленности, например, сборку электронных компонентов. Такие способности пред-
полагают наличие трех, а не двух измерений, о которых пишет Гварньеро. И, ко-
нечно, зрительные ощущения, обеспечиваемые СТЗЗ, сильно обеднены по сравне-
нию с тем, что испытывают люди с нормальным зрением.
Литература
Bach-y-Rita, Р., 1995, Nonsynaptic Diffusion Neurotranmission and Late Brain Reorganisa-
tion, New York: Demos.
Bach-y-Rita, P., 1996, «Sensory substitution and qualia», in Perception et Intermodalite,
J. Proust, ed., Paris: Press Universitaires de France.
Gregory, R. L., 1997, Eye and Brain, the psychology of seeing (5th edn), Oxford: Oxford
University Press.
Guarniero, G., 1974, «Experience of tactile vision», Perception, 3, 101—104.
редакции своей «Психологии» (1872) Спенсер выразился более категорично:
«Пространственные отношения одни и те же не только для всех предков чело-
века, всех предковых приматов, всех предковых отрядов млекопитающих, но и
для более примитивных существ. Эти постоянные пространственные отноше-
ния выразились в формировании определенной структуры нервной системы,
которая, будучи врожденной, действует определенным образом и неспособна
544 Часть VI. Кода
действовать иным. Отсюда, непостижимость фиктивности математической
аксиомы, являющаяся следствием невозможности изменить работу соответст-
вующей нервной структуры, подтверждает, что она развилась на основе беско-
нечно большого процесса познания».
Вполне довольный собой, поскольку таким образом устраняется тайна
кантианской априорности, Спенсер продолжал свои труды и дал одно из наи-
более полных изложений представлений об эмпирическом происхождении
нашего знания. Он начал с предположения (вслед за другими эмпириками),
что мир младенца — это смешение цветов, звуков, ощущений прикосновения,
вкусов и запахов. Далее он объяснил, как из этого набора феноменов посте-
пенно дифференцируется две группы событий. Первая из них — это «яркие» и
часто неожиданные события, вторая — события более слабые, часто сопро-
вождающие события первой группы и также часто соединенные во временную
последовательность. Эти и другие различия в конце концов кристаллизуются в
различие «объективного» и «субъективного». Далее, продолжает Спенсер, од-
но из ярких ощущений, субъектом которых мы являемся, — это прикоснове-
ние; если провести рукой по ноге, это вызовет изменение этого конкретного
яркого ощущения. Это изменение вызвано событиями в последовательности
более слабых ощущений, которые вызывают изменения, неотличимые от та-
ковых, вызванных событиями в яркой совокупности. Однако при формирова-
нии таких изменений в яркой совокупности мы осознаем, указывает Спенсер,
в них «кипение», как он это определял, ощущений слабой совокупности, к ко-
торой мы традиционно относим «усилия», мышечные усилия, затраты энергии
или работу против сопротивления. Таким образом, заключает Спенсер, возни-
кает ощущение силы вещей, внешней «энергии».
Здесь не место далее следовать за анализом Спенсера путей понимания
природы вещей, которые формируются за долгие годы младенчества и детства.
Для того, чтобы понять, что философы-эмпирики способны рассказать преза-
нятные истории, сказано уже предостаточно. Интересующиеся студенты мо-
гут проследить за развитием этих идей в работах современного последователя
Спенсера — Жана Пиаже.
23.5. Эволюционная эпистемология
В предыдущем разделе мы видели, что Герберт Спенсер, работавший в середи-
не XIX века, обратил внимание на связь эволюционной теории и эпистемоло-
гии. На самом деле, здесь его опередил Чарльз Дарвин. В одной из записных
книжек, которые он писал, возвращаясь на «Бигле» из кругосветного плавания
в конце 1830-х гг., мы находим следующую запись: «Платон говорил... что на-
ши «необходимые понятия» возникают из предсуществующего духа и не могут
быть получены из ощущений — как источник предсуществующего духа надо
понимать обезьяну» (Записная книжка М (М Notebook), стр. 128). Записные
книжки Дарвина, однако, при его жизни опубликованы не были, а оттого
только Спенсер и может считаться основателем эволюционной эпистемоло-
гии. Позднее в том же веке Фридрих Ницше (Friedrich Nietzsche) в Германии
опубликовал интересные спекуляции по эволюционной эпистемологии. В его
23. Философский постскриптум 545
«Веселой науке» (Gay Science, разделы 110-111), например, он писал, что «за
колоссальный период времени интеллект не породил ничего, кроме ошибок;
некоторые из них оказались полезны для сохранения вида ... тот, например,
кто не способен был установить «идентичность» пищи или враждебного жи-
вотного ... имел меньшие шансы на выживание, чем тот, кто в каждом случае
сходства устанавливал идентичность». У Ницше, в его афористической работе,
таких догадок немало. Они показывают, как эволюционная мысль XIX века
начинала влиять на труды философов. До следующего века, однако, эволюци-
онная теория еще не оказывала существенного влияния на психологию. Лишь
в последние годы были получены экспериментальные данные о характере эво-
люционных влияний на сознание.
Мы видели в предыдущих главах этой книги, что «простые» стимулы Джо-
на Локка и других эмпириков редко оказываются очень эффективными. Ко-
нечно, в психофизической лаборатории испытуемым, сотрудничающим с ис-
следователями, предъявляют подобные стимулы, их реакции регистрируются
и количественно обрабатываются. Однако, в мире за стенами лаборатории,
скажем конкретнее, в мире палеолитического прошлого, такие стимулы вряд
ли кто и заметил бы. Человеческие чувства, как и чувства других животных,
адаптированы к реакции на биологически наиболее значимые параметры ок-
ружающей среды. Отметим также, что в эволюционном процессе не существу-
ет непрерывной линии «восхождения». Люди не просто стоят выше в эволю-
ционном древе, чем мыши и крысы, и даже макаки. Разные животные адапти-
ровались за время эволюции к специфическим экологическим нишам и спо-
собам существования. Соответственно, различны те стимулы, на которые на-
илучшим образом настроены их органы чувств. Эволюция — это не прямая ле-
стница, а древо, ветвящееся во многих направлениях — мы это видели в раз-
ных местах книги, особенно в заключительном рассмотрении различных сен-
сорных миров в главе 22. Человеческое сознание организовано своим, иногда
совершенно чуждым образом по отношению к сознанию наших соседей по
планете — животных.
Этологи уже давно знают, что разные животные «подготовлены» к разным
задачам. Психологи, однако, слишком часто предполагают (вероятно, под
влиянием философской эмпирической традиции tabula rasa), что человечес-
кое сознание подходит ко всем этим задачам с равными возможностями.
Только в последние годы накопились данные, показывающие, что, как и со-
знание других животных, человеческое сознание «подготовлено» к выполне-
нию одних задач в большей степени, чем других. И только недавно удалось
организовать эксперименты, позволяющие выделить такие предпочтения.
Человеческое сознание — это не чистый лист, как полагали философы XVII
века. Оно содержит врожденные склонности, хотя и очень далекие от того,
что представлял себе Иммануил Кант. В них отражается эволюционное про-
шлое, особенно миллионы лет, проведенные на равнинах Африки в качестве
охотника, собирателя, падальщика. Такие, так называемые Дарвиновы алго-
ритмы, развились, обеспечивая увеличение вероятности выживания в данных
условиях.
В плодотворной работе Леды Космидес (Leda Cosmides) показано, что
обычные ошибки при решении условных задач лучше всего описываются в по-
546 Часть VI. Кода
нятиях эволюционно встроенного поведения, направленного на социальное
общение. Задача, известная по имени ее изобретателя как задача выбора Вэй-
сона (Wason), тестирует способность следовать обусловленному правилу: если
(р), то (q).
Таких тестов существует множество, и множество гипотез объясняет раз-
личие процента правильных решений. Наиболее экономичное решение нахо-
дится в понятиях социального контракта и выявления ложных утверждений.
Космидес и ее сотрудники утверждают, что это обусловлено длительным эво-
люционным опытом социальности в палеолите. Этот опыт создал сильную
склонность к определению ложных утверждений в ментальности человека. Те-
ория социального контракта дает достаточно точные предсказания типов
«ошибок» испытуемых, которые делаются (или не делаются) в зависимости от
разных вариантов теста Вэйсона. Эти предсказания хорошо согласуются с ре-
зультатами, полученными на больших массивах данных.
Социальный контракт и поиск ложных утверждений — это не единствен-
ные дарвиновские алгоритмы, имманентные человеческому сознанию. Есть и
множество других. Человеческое сознание, как и сознание других животных,
эволюционировало в направлении решения не абстрактных, а определенных,
достаточно специфичных задач. Существуют, например, интересные указания
на то, что обычное человеческое «недопонимание» понятий вероятности, ста-
тистики и риска имеет эволюционное происхождение. Интересны обуслов-
ленные половым диморфизмом различия способности к пространственной
оценке, хотя возможно и усиленные ранним опытом в детстве. Мальчики, на-
пример, от рождения лучше передвигаются в трехмерном пространстве, опре-
деляют относительные скорости и расстояния и лучше выполняют тесты с вра-
щающимся изображением. С другой стороны девочки лучше выполняют линг-
вистические тесты.
23.6. После Декарта
Разрыв между сознанием и телом, идеальным и материальным до сих пор, как
отмечалось в разделе 23.2, кажется непреодолимым. Жизнь показывает, что
у тех, кто пытается полностью охватить проблему, есть только два пути: или
к чистому двухсущностному картезианскому дуализму (как например,
Дж. Экклс1 2 (J. С. Eccles)) или к панпсихизму (Хамероф (S. R. Hamerofi) и др.).
Кажется, что оба эти варианта ведут к reductio ad absurdum1. Так что, наверное,
лучше остаться с теми, кто считает это проблему сейчас (а может быть и в бу-
дущем) неразрешимой.
Все это, однако, не означает, что наше понимание взаимоотношений меж-
ду мозгом и сознанием остается на уровне начала XVII века. Науки о нервной
1 Сэр Джон Карью Экклс (1903—1997) — выдающийся австралийский нейрофизиолог,
исследования физиологии нервной системы и синаптической передачи. Нобелевский лауре-
ат 1963 года. Вслед за Шеррингтоном развивал философские представления о человеческой
личности в связи с науками о мозге (см. Facing Reality, 1970, Springer Verlag). В частности, свя-
зывал определенные отделы мозга с взаимодействием мозга и «души»! — Прим, перев.
2 Reductio ad absurdum {лат.) — доведение до абсурда. — Прим, перев.
23. Философский постскриптум 547
Рис. 23.1. Зрительное восприятие по Декарту. «Человек» (L’Homme) (написан
в 1628 г., опубликован посмертно в 1662). Неинвертированное изображение про-
ецируется в Н, т. е. эпифиз, который (в соответствии с этим рисунком) подвешен
в центре мозгового желудочка.
системе и когнитивные науки1 продвинулись далеко от грубой модели декар-
товой «глиняной машины» («Человек» (L’Homme)). По мере того, как мы рас-
сматривали сенсорную физиологию XX века в предшествующих главах этой
книги, мы не раз наталкивались на факты, которые заставили бы Рене Декар-
та сильно призадуматься. Давайте вспомним некоторые из них.
Одна из знаменитых иллюстраций в «Человеке» Рене Декарта показывает
действие зрительной системы (рис. 23.1). Декарт писал эту работу в конце
1620-х гг. под впечатлением демонстраций Аранци в 1595 г., а затем Кеплера в
1604 г. того, что, если удалить склеру с задней части глаза быка, то можно уви-
деть уменьшенное инвертированное изображение на сетчатке. Декарт повто-
рил это наблюдение и использовал в своей нейрофизиологической теории.
Рис. 23.1 показывает, что согласно этим представлениям не только на сетчатке
образуется инвертированное изображение, но оно в неизменном виде переда-
ется в мозг. В мозге изображение реинвертируется , так что в железе Н (эпифи-
зе) образуется неинвертированное изображение. Здесь по Декарту располага-
ется душа, которая непосредственно воспринимает изображение. Такое пред-
ставление привело к многовековому недоразумению. Нейрофизиологи уже
давно знали, что такое прямое восприятие невозможно (оставим в покое де-
картову причудливую нейрофизиологию). В главах 16 и 17 мы видели, на-
сколько важна «переработка информации», стоящая между изображением на
сетчатке и зрительным восприятием.
К тому же, говоря о зрительной системе, мы видели, что она детектирует не
неизменные элементы окружающей среды, а именно их изменения. На экспе-
риментальных животных это показано путем регистрации нервных импульсов
в зрительном нерве — когда ничего не происходит, система в основном нахо-
1 Науки о познании. — Прим, перев.
548 Часть VI. Кода
дится в покое. Аналогичные опыты были проведены Ярбусом (Yarbus) и други-
ми на людях. Если саккадические движения глаз компенсировали таким обра-
зом, что изображение на сетчатке стабилизировалось, оно начинало слабеть,
утрачивать «осмысленность», за 1—3 секунды «все видимые различия исчезают
и более не восстанавливаются», оставляя то, что Ярбус назвал «пустым полем».
Все контуры исчезают, оставляя поле зрения черным, серым или сумеречным,
как будто глаза закрыты. Очень получается не по-картезиански!
Можно рассмотреть и работы Лэнда (Land) по цветовосприятию с исполь-
зованием «мондрианов». Надо напомнить, что Пит Мондриан (Piet Mondrian,
1872—1944) — голландский художник-абстракционист, известный своими
коллажами из ярких цветных прямоугольников. Для глаза человека цвета сты-
кующихся прямоугольников остаются контрастными независимо от условий
освещения. Между тем спектральный состав отраженного света меняется в за-
висимости от спектра освещения. Лэнд показал, что распределение энергии в
спектре света, падающего на сетчатку от различных частей Мондриана, не вли-
яет на наше цветовосприятие; не влияет на наше восприятие также и обуче-
ние, память, окружение и адаптация. Однако, оно зависит от «отражательной
способности» того, что субъективно воспринимается как «яркость». Зритель-
ная система обрабатывает сравнительные отражательные способности разных
прямоугольников на картине, и это соотношение остается постоянным в
очень широком диапазоне освещенности. Именно это соотношение, а не раз-
ные количества энергии на разных длинах волн дает ощущение («грубое ощу-
щение») цвета. Опять получается очень не по Декарту.
Оба этих примера, а таких примеров существует еще множество, показыва-
ют, что зрительная система совсем не похожа на фотоаппарат или фотометр,
который используют физики. Дэвид Марр (David Магг) указывал, что зре-
ние — это «описание, полезное для наблюдателя», что мы и видим в обоих
приведенных выше примерах. В первом случае даже маленькая точка может
активизировать мозг информацией о визуальном окружении, когда ничего
больше не происходит. Во втором, способность правильно узнавать цвета
(спелых ягод, спелых фруктов) важна для цветочувствительных приматов при
разных условиях освещенности — на рассвете, в полдень и в сумерки.
Далее, но в продолжение той же линии аргументов, зрительная система ак-
тивно контролирует объект. Во-первых, как мы отмечали в главе 15, существу-
ет мощная система обратной связи в контроле саккадических движений. Во-
вторых, если прикрепить сбоку от глаза маленькое зеркальце, чтобы можно
было контролировать его движения, то окажется, что они носят далеко не слу-
чайный характер. Движения глаз следуют за основными чертами объекта,
представляющего интерес, причем за биологически наиболее существенными
(рис. 23.2). В случае рассматривания лица глаза и рот сканируются многократ-
но. Саккадические движения, конечно, достаточно автономны, сознательные
направленные движения глаз здесь отсутствуют. Этот механизм является
встроенной и заранее запрограммированной двигательной системой направ-
ления взгляда.
Такое восприятие с помощью ориентационных действий очевидно и во
всех других ощущениях. Обзор этих явлений можно найти у Майкла Арбиба
(Michael Arbib) в книге, упомянутой в библиографии. Мы рассматривали та-
23. Философский постскриптум 549
Рис. 23.2. Движения глаз при рассматривании лица маленькой девочки зафиксированы
на рисунке справа. Из A. L. Yarbus, 1967, Eye Movements and Vision, Plenum: New York.
С разрешения Kluwer Academic/Plenum Publishers.
кой интересный феномен, когда обсуждали электрорецепцию у клюворылых
рыб в главе 20, а закончить этот раздел мы можем ссылкой на чувство боли, ко-
торое рассматривалось в главе 21. Следует напомнить, что как и зрение,
боль — это не просто считывание активности в специфических нервных во-
локнах, как предполагал Декарт в другой известной схеме. Напротив, как мы
видели, боль глубочайшим образом участвует в адаптации животного к окру-
жающей среде и обстоятельствам. Боль «прорывается» только в тех случаях,
когда избежать таких обстоятельств невозможно. Радикально некартезианская
природа боли, вероятно, лучше всего показана, как полагает Патрик Уолл,
субъективными ощущениями при инъекции анестетиков в плечевое сплете-
ние (глава 21). Вместо утраты любых ощущений в руке, как вытекает из карте-
зианских представлений, поскольку отсутствует поток импульсов в нервах
плеча, возникает ощущение ее распухания, болезненности и, часто, располо-
жения в невозможной позиции. Центральная нервная система компенсирует
необычный паттерн нервной активности всех сенсорных волокон, конверги-
рующей в ней, путем формирования гипотез и концентрации внимания на том
факте, что что-то идет не так. Очень это сильно отличается от того принципа
зеркала, которого придерживался Рене Декарт.
550 Часть VI. Кода
23.7. Заключение
Итак, в заключение этой главы, мы можем заметить, что, хотя картезианские
представления все еще волнуют умы, часть его нейрофилософии все же уже
ушла в прошлое. Из-за широко распространенного бездумного принятия кар-
тезианства Джилберт Райл (Gilbert Ryle) назвал его двухсущностную метафи-
зику «официальной доктриной», которая противопоставляет человека окру-
жающему миру. Декартова эпистемология построена на принципе отраже-
ния — окружающий мир отражается в сознании. Если зеркало установлено
правильно, отражение точно, и наше знание о мире верно; если зеркало кри-
во, то и знание наше внешнего мира неверно. В этой главе мы увидели, что та-
кой подход больше не работает, и мы в начале бесконечно трудного пути из
мощных объятий картезианства. Люди не вне мира, а очень даже часть его.
Они не просто пассивные наблюдатели, «фотокамеры», фиксирующие мир,
чьи дела лежат в какой-то иной сфере. Восприятие ориентировано на дейст-
вие, а действие ориентировано на восприятие. Люди, вспомним одну из запис-
ных книжек Чарльза Дарвина, «вплетены» в живой мир.
Вывод из этой книги похож на вывод Дарвина. Мы видели в этой главе, об-
суждая Дарвиновы алгоритмы человеческого сознания, с которыми сталкива-
лись повсеместно, что сенсорные системы адаптированы к обстоятельствам
жизни их владельца. Сенсорные системы человека, несомненно восхититель-
ные, настроены всей нашей эволюционной историей. Они не дают нам «взгля-
да Божьего ока» на мир. Мы многое можем изучить, сравнивая наши сенсор-
ные системы с таковыми животных, которые часто существуют в совершенно
иных сенсорных мирах. Тем не менее, как уже давно написал Александр Поуп
(Alexander Pope) «истинная мера человечества — это человек», а в мире слиш-
ком много страданий и неудобств, чтобы поверить в то, что изучение сенсор-
ных систем человека имеет первостепенное значение.
Мы многое можем узнать из сравнения различных чувств. В их организа-
ции очень много общего, а на молекулярном уровне — часто это просто одни
и те же механизмы. Поразительно однако, что такие схожие молекулярные и
клеточные структуры образуют настолько радикально разные сенсорные каче-
ства. Это еще одна «трудная» проблема, кроющаяся под поверхностью боль-
шинства страниц этой книги.
Мы начинали эту книгу с того, как три с половиной миллиарда лет назад
первые предшественники прокариот поплыли в уже тогда древних морях. Мы
заканчиваем, наверное, с более полным пониманием того, что мы часть при-
роды и тесно с ней связаны. Было бы преувеличением закончить строками из
«Четырех квартетов» Элиота:
И в конце далекого пути
Мы пришли туда, откуда шли.
От знакомых мест мы не отводим глаз —
видим в жизни их как будто в первый раз
(пер. Ю. Бобровского)
А может быть, если честно оценить глубину наших научных знаний, не та-
кое уж это и преувеличение...
МАТЕРИАЛЫ К ЧАСТИ VI
Для самоподготовки
Нижеследующие вопросы построены так, чтобы помочь проверить, насколько
усвоен материал Части VI, а также обратить внимание на существенные аспек-
ты предмета изложения.
Глава 22: Повторяющиеся мотивы
22.1. Какой из мембранных каналов считается наиболее древним в эволюционной
истории? Какие развились позже всех?
22.2. Приведите примеры молекулярных механизмов, адаптированные к тому, что-
бы играть важнейшие роли в разнообразных сенсорных клетках.
22.3. Обсудите один из вездесущих органоидов, которые снова и снова используют-
ся в организации сенсорных клеток.
22.4. Приведите несколько примеров общих принципов организации, проявляю-
щихся в конвергентной эволюции.
22.5. Сравните специализацию сетчаток позвоночных и сложных глаз насекомых.
22.6. Есть ли сходные специализации у сенсорных поверхностей разных модально-
стей?
22.7. Почему существуют топографические карты сенсорных поверхностей в цент-
ральной нервной системе? Обсудите возможные гипотезы.
22.8. Нарисуйте схему, показывающую сходство нервной организации обонятель-
ной луковицы и сетчатки. Каковы нейрофизиологические причины этого
сходства?
22.9. Объясните, чем отличается обработка информации в сенсорной коре от после-
довательных операций в цифровых компьютерах (по фон Нойману).
22.10. В чем схожи триггерные стимулы клеток слуховой коры макаков и нижневи-
сочной коры?
22.11. Объясните биологическое значение возникновения иллюзий в сенсорной сис-
теме.
22.12. Можно ли рассматривать боль как часть гомеостатического механизма тела?
Объясните.
22.13. Как Вы думаете, насколько различаются сенсорные системы человека и жи-
вотных?
22.14. На примере бабочек и летучих мышей объясните, как взаимоотношения «хищ-
ник-жертва» обостряют качества сенсорных систем.
22.15. Обсудите роль сенсорных систем у социальных животных.
22.16. Что означают термины «психология народа» и «намерение»?
22.17. Что Фрэнсис Бэкон подразумевал под терминами «познавательный» и «полез-
ный»? Остаются ли они действеными в настоящее время? Обсудите.
Глава 23: Философский постскриптум
23.1. Опишите сомнения, которые привели Декарта к формуле «мыслю — следова-
тельно существую».
23.2. Как декартово «мыслю» привело к классической проблеме взаимоотношений
сознания и мозга?
23.3. Что такое проблема знания?
23.4. Что означают термины «сенсорные качества» и «привилегированный доступ»?
552 Часть VI. Кода
23.5. В чем различие между «легкими» и «трудными» проблемами?
23.6. Томас Хоббс, Джон Локк и другие британские философы-эмпирики считали
сознание «чистым листом». Что они имели в виду? Согласно ли это с Декар-
том?
23.7. Каков вклад Иммануила Канта в сенсорную физиологию? Почему он называл
его «коперниковой революцией»? На какого немецкого физиолога сенсорных
систем XIX века его взгляды оказали наибольшее влияние?
23.8. Кто такой Молинье и что за вопрос он задал? Как Джон Локк на этот вопрос
ответил и почему? Опишите некоторые экспериментальные исследования по
этому вопросу. Дан ли на вопрос Молинье окончательный ответ?
23.9. Как Герберт Спенсер решил противоречие между взглядами последователей
Локка и Канта? Был ли Спенсер первым, кто нашел этот путь?
23.10. Опишите, как Спенсер выводил понятия «объективности» и «субъективнос-
ти». Убедителен ли такой подход?
23.11. Что означает термин «дарвинов алгоритм»? Опишите несколько эксперимен-
тов, которые подтверждают, что человеческое сознание не представляет собой
«чистого листа», а обладает врожденными эволюционно обусловленными
предпочтениями.
23.12. Почему эволюционную психологию иногда высмеивают как серию «Сказок
просто так»?1 Критически обсудите ее перспективы и недостатки.
23.13. Рассмотрите декартову физиологию зрения, как она представлена на рис. 23.1.
Не считаете ли Вы, что она до сих пор является обиходным представлением о
том, как работает эта система? Как бы Вы объяснили профану, что картезиан-
ская версия глубоко ошибочна?
23.14. Как Дэвид Марр определяет зрение?
23.15. Считаете ли Вы, что исследования сенсорной физиологии изменяет наше оби-
ходное понимание мира и нашего места в нем?
Замечания и библиография
Глава 22: Повторяющиеся мотивы
Повторяющийся мотив этой книги — эволюция. Гарлэнд и Картер (Garland &
Karter, 1994) представили широкий обзор литературы по эволюционной физи-
ологии, а в сообщениях Восьмого Ежегодного Картеровского Семинара (Eight
Annual Karger Workshop) под редакцией Fay & Popper (1996) приводится серия
обзоров по эволюции сенсорных систем позвоночных. Дальнейшую информа-
цию по молекулярной нейробиологии можно найти во многих источниках, в
частности многих миниобзорах в журнале Neuron и книгах по молекулярной
нейробиологии, например, Смита (Smith, 1996). Шеферд (Shepherd, 1995)
представил сравнительный обзор сенсорных систем на молекулярном уровне.
Лэнд (Land, 1981) и Уотермэн (Waterman, 1981) дают детальные обзоры специ-
ализации фоторецепторов насекомых. Лэнд, в частности, обсуждает «поло-
вую» фовеа двукрылых насекомых, а Уотермэн дает количественную инфор-
мацию и удобные схемы фовеа стрекоз. Кохонен и Хари (Kohonen & Hari,
1 Сказки просто так (Just So Stories) — сборник сказок Рэдьярда Киплинга, включающий
в частности, «Как у верблюда появился горб», «Как носорог сменил шкуру», «Слоненок»,
«Как у леопарда появились пятна». — Прим, перев.
Материалы к части VI 553
1999) сделали интересный анализ формирования сенсорных карт в мозге (и в
искусственных системах). Юкскюль (Uexkull, 1909, 1926) дает классические
представления о «внутреннем мире» животных, а Цайль и Цанкер (Zeil and
Zanker, 1997) приводят интересный пример такого мира. Уилсон (Wilson, 1975)
в своем классическом труде по социобиологии показывает, как у разных групп
животных развились социальные отношения.
Fay, R. R. and A. N. Popper, 1996, «Evolution of vertebrate sensory systems», Brain,
Behaviour and Evolution, 50, 187—259.
Garland, T. and P. A. Carter, 1994, «Evolutionary physiology», Annual Review of Physiology,
56, 579-621.
Hanson, N. R., 1961, Patterns of Discovery, Cambridge: Cambridge University Press.
Kohonen, T. and R. Hari, 1999, «Where the abstract feature maps of the brain might come
from», Trends in Neuroscience, 22, 135—139.
Kuhn, T. S., 1962, The Structure of Scientific Revolutions, Chicago: Chicago University
Press.
Land, M. F., 1981, «Optics and vision in invertebrates», in H. Autrum, ed., Handbook of
Sensory Physiology, Berlin Springer-Verlag, vol. VII/6B.
Popper, K., 1969, Conjectures and Refutations: The growth of scientific knowledge (3rd edn),
London: Routledge and Kegan Paul.
Shepherd, G. M., 1995, «Toward a molecular basis for sensory perception», in The Cognitive
Neurosciences, M. S. Gazzaniga, ed., Cambridge, MA., MIT Press.
Smith, C. U. M., 1995, «Membrane signalling systems», in Biomembranes: General princi-
ples, A. G. Lee, ed., Greenwich, CT.: JAI Press, vol. 1, 245—270.
Smith, C. U. M., 1996, Elements of Molecular Neurobiology (2nd edn), Chichester: John
Wiley.
Uexkull, J. J. von, 1909, Umwelt und innenwelt der Tiere, Berlin; also 1926, Theoretical
Biology, London.
Waterman, T. H., 1981, «Polarisation sensitivity», in H. Autrum, ibid.
Wilson, E. O., 1975, Sociobiology: The new synthesis, Cambridge, MA.: Belknap Press of
Harvard University Press.
Zeil, J. and J. N. Zanker, 1997, «Аglimpse into crabworld», Vision Research, 37,3417—3426.
Глава 23: Философский постскриптум
Граница между сенсорной физиологией и философией, особенно эпистемоло-
гией протяженная, неопределенна и подвижна. Ниже приведены лишь не-
сколько из множества книг и статей, касающихся этого постскриптума. Исто-
рически сложилось, что современный период философии и нейрофизиологии
исчисляют от работ Рене Декарта (1637,1662). Переписку Томаса Хоббса с Де-
картом можно найти во 2-м томе у Котингхэма с соавторами (Cottingham,
Stoothof and Murdoch, 1984, «Third Set of Objections». Проблема «сенсорных ка-
честв» или «грубых ощущений» приобретает современные очертания у Чал-
мерса (Chalmers, 1998), Деннет (Dennett, 1991) вносит в нее дозу скептицизма,
Хамероф (Hameroff, 1998) привлекает квантовую теорию, Файгль (Feigl, 1967)
дает более классические представления с использованием в мысленных экспе-
риментах «автоцереброскопа», а МакГин (McGinn, 1991) доказывает ее прин-
ципиальную неразрешимость. Морган (Morgan, 1977) и Дегенаар (Degenaar,
1996) приводят детальное рассмотрение вопроса Молинье к Джону Локку и его
историческое эхо в последующих веках. Иммануил Кант объясняет, что он
20 Биология сенсорных систем
554 Часть VI. Кода
имел в виду под «коперниковой революцией» в предисловии к своей «Крити-
ке чистого разума» (Critique of Pure Reason, 1787), а Коэн и Элькана (Cohen and
Elkana, 1977) показывают, насколько сильно Кант повлиял на сенсорную фи-
зиологию Гельмгольца. Пионерская работа Герберта Спенсера по эпигенети-
ческой эпистомологии впервые опубликована в 1855 году и детально рассмот-
рена Смитом (Smith, 1982). Баркоу с соавторами (Barkow, Cosmides and Tooby,
1992) представили глубокое обсуждение того, как эволюционное прошлое че-
ловечества оказывает влияние на современную ментальность, а Космидес и
Туби (Cosmides and Tooby, 1992) обсуждают доказательства того, что кажущи-
еся ошибки обиходных оценок риска и вероятности вовсе не были таковыми в
условиях палеолитического охотничье-собирательского племени. Наконец,
Ярбус (Yarbus, 1967), Лэнд (Land, 1977) и Патрик Уолл (Patrick Wall, 1994) еще
раз подчеркивают некартезианскую природу современной нейропсихологии,
а Дэвид Марр (David Marr, 1982) и Майкл Арбиб (Michael Arbib, 1989) дают
классические представления о взаимоотношениях между науками о нервной
системе, искусственным интеллектом и роботехникой.
Arbib, М. А., 1989, The Metaphorical Brain 2: Neural networks and beyond, New York: John
Wiley and Sons.
Barkow, J., L. Cosmides and J. Tooby, eds, 1992, The Adapted Mind: Evolutionary psycholo-
gy and the generation of culture, New York: Oxford University Press.
Chalmers, D. J., 1998, The Conscious Mind, Oxford: Oxford University Press.
Cheselden, W., 1728, «An account of some observations made by a young gentleman...»,
Philosophical Transactions of the Royal Society, 402, 447—450; quoted in Morgan, 1977,
ibid.
Cohen, R. S. and Y. Elkana, eds., 1977, Hermann von Helmholtz: Epistemological Writings,
Dordrecht: Reidl.
Cosmides, L., 1989, «The logic of social exchange: Has natural selection shaped how
humans reason? Studies with the Wason selection task», Cognition, 31, 187—276.
Cosmides, L. and J. Tooby, 1996, «Are humans good intuitive statisticians after all?
Rethinking some of the conclusions from the literature on judgement under uncertain-
ty», Cognition, 58, 1—73.
Cottingham, J., R. Stoothof and D. Murdoch, 1984, The Philosophical Writings of Descartes,
Cambridge: Cambridge University Press.
Darwin, C., 1838, M Notebook, in Metaphysics, Materialism and the Evolution of Mind: Early
writings of Charles Darwin, P. H. Barrett, ed., Chicago: Chicago University Press, 1980.
Degenaar, M., 1996, Molyneux's Problem: three centuries of discussion on the perception of
forms, Dordrecht: Kluwer Academic Publishers.
Dennett, D., 1991, Consciousness Explained, Harmondsworth, Middlesex: Penguin.
Descartes, R., 1637, Discourse on Method, in Descartes: Philosophical Writings, E. Anscombe
and P. T, Geach, eds, London: Thomas Nelson and Sons, 1970.
Descartes, R., 1662, L’Homme, translated and edited by T. S. Hall, Treatise of Man,
Cambridge, MA.: Harvard University Press, 1972.
Eliot, T. S., 1944, Four Quartets, London: Faber and Faber.
Feigl, H., 1967, The «Mental» and the «Physical», Minneapolis: Minneapolis University
Press.
Geary, D. C., 1996, «Sexual selection and sex differences in mathematical abilities»,
Behavioural and Brain Sciences, 19, 229—284.
Gregory, R. L., 1997, Eye and Brain, the psychology of seeing (5th edn)z .Oxford: Oxford
University Press.
Материалы к части VI 555
HamerofT, S. R., 1998, «“Funda-mentality”: Is the conscious mind subtly linked to a basic
level of the universe?», Trends in Cognitive Sciences, 2, 119—124.
Hartley, D., 1749, Observations on Man, London: Thomas Tegg.
Hobbes, T., 1651, Leviathan, London, part 1, chapter 1.
Kant, L, 1787, Critique of Pure Reason (2nd edn), translated by J. M. D. Meiklejohn,
London: Dent, 1934.
Land, E. H., 1977, «The retinex theory of colour vision», Scientific American, 237(12),
108-128.
Locke, J., 1690, An Essay Concerning Human Understanding, R. Wilburn (ed.), London:
Dent, 1974.
Marr, D., 1982, Vision, New York: Freeman.
McGinn, C., 1991, The Problem of Consciousness: Essays towards a resolution, Oxford:
Blaekwell.
Morgan, M. J., 1977, Molyneux’s Question, Cambridge: Cambridge University Press.
Nietzsche, F., 1882, The Gay Science, translated by R. J. Hollingdale New York: Vintage
Books; The Gay Science, translated by W. Kaufmann, 1974, New York: Vintage Books.
Piaget, J., 1972, Psychology and Epistemology, Harmondsworth: Penguin.
Plato, Meno, in The Dialogues of Plato, translated and edited by B. Jowett, Oxford: The
Clarendon Press, vol. 2.
Ryle, G., 1949, The Problem of Mind, London: Hutchinson.
Smith, C. U. M., 1982, «Evolution and the Problem of Mind, Part 1, Herbert Spencer»,
Journal of the History of Biology, 15, 55—88.
Smith, C. U. M., 1987, «Clever Beasts Who Invented Knowing»: Nietzsche’s evolutionary
biology of knowledge», Biology and Philosophy, 2, 65—91.
Smith, C.U.M., 1987, «David Hartley’s Newtonian Neuropsychology», Journal of the
History of the Behavioural Sciences, 23, 123—136.
Smith, C. LJ. M., 1998, «Descartes’ pineal neuropsychology», Brain and Cognition, 36,
57-72.
Smith, C. U. M., 1998, «Descartes’ visit to the Town Library: or how Augustinian is
Descartes’ neurophysiology?», Journal of the History of the Neurosciences, 7, 93—100.
Spencer, H., 1855, The Principles of Psychology, London: Longman, Brown, Green and
Longmans; 2nd edn, 2 vols., 1870; 1872, London: Williams and Norgate.
Wall, P. D., 1994, «Introduction to the edition after this one», in Textbook of Pain, P. D.
Wall and R. Melzack, eds, Edinburgh: Churchill Livingstone.
Yarbus, A. L., 1967, Eye Movements and Vision, translated by B. Haigh and L. A. Riggs, New
York: Plenum Press.
ДОПОЛНЕНИЕ:
НЕКОТОРЫЕ МЕТОДИКИ
А. Функциональная гистохимия
1. Окрашивание с помощью 2-дезоксиглюкозы
Окрашивание с помощью 2-дезоксиглюкозы предоставляет удобное средство
различать активные и неактивные части мозга. Клетки мозга получают боль-
шую часть энергии из глюкозы, которая проникает в них из крови, циркулиру-
ющей в кровеносных сосудах мозга. Глюкоза переносится в клетки через гема-
то-энцефалический барьер с помощью специфических систем транспорта.
Эти системы также способны транспортировать и незначительно отличающи-
еся от глюкозы молекулы 2-дезоксиглюкозы (2-ДГ). В тканях мозга 2-ДГ, как
и обычная глюкоза, фосфорилируется гексокиназой в 2-ДГ-6-фосфат. В отли-
чие от глюкоза-6-фосфата, который далее метаболизируется до СО2 и Н2О,
ДГ-6-фосфат остается в неизменным. Отличие его структуры, хотя и незначи-
тельное, тем не менее обеспечивает то, что это вещество не может быть субст-
ратом соответствующих ферментов. Сформировавшись, ДГ-6-фосфат некото-
рое время удерживается в ткани, пока не диффундирует из нее. На этом и ос-
нована методика окрашивания. ДГ-6-фосфат метят изотопом С14 и инъециру-
ют его в сосудистую систему экспериментального животного. Время полужиз-
ни [С14]ДГ-6-фосфата в сером веществе составляет 7, 7 ± 1,6 часа, а в белом ве-
ществе 9,7 ± 2,6 часа. Если интервал времени избран достаточно короткий
(мене 1 часа), то количество [С14]ДГ-6-фосфата, накопившегося в ткани моз-
га, равно интегральной скорости фосфорилирования гексокиназой [С14]ДГ за
это время.
Техника исследования сравнительной активности участков мозга (напри-
мер, колонок, избирательных к ориентации или колонок глазодоминантнос-
ти) состоит в инъекции [С14]ДГ, усыпления животного чреез 45 мин и приго-
товления замороженных срезов мозга. Радиоактивность срезов исследуют с
помощью стандартного метода авторадиографии.
2. Окрашивание митохондрий
Применение этой методики основывается на том, что один из коферментов
(цитохром С) дыхательной цепи митохондрий окисляет осмиофильный пре-
парат 3,3 -диаминобензидин (ДАБ). Метод пригоден и для световой, и для эле-
ктронной микроскопии. При исследовании капель в зрительной коре возмож-
на только световая микроскопия. В этом случае замороженные срезы инкуби-
руют в среде с фосфатным буфером и ДАБ, а затем проводят реакцию с OsO4.
Активность цитохрома С остается достаточно высокой, чтобы окислить ДАБ,
который реагирует с OsO4, в результате чего образуются отложения черных
гранул. Их легко рассмотреть в микроскоп.
Дополнение: некоторые методики 557
3. Окрашивание «генов немедленного ответа»
Множество генов немедленного ответа (ГНО) включается возбуждающими
синаптическими медиаторами. Наиболее известными ГНО являются гены с-
fos, c-jun,jun-B и egr-1. В покоящихся клетках обнаруживается очень низкий
уровень мРНК или белков, программируемых этими генами. Они быстро ак-
тивируются возбуждающими медиаторами через систему вторичных мессен-
джеров — цАМФ, Са2+, ПКС и т. д., а их белковые продукты обнаруживаются
с помощью моноклональных антител. Позднее разработана техника детекти-
рования соответствующих мРНК с помощью in $/7н-гибридизации. Обе эти ме-
тодики высокочувствительны и позволяют определять гистологическую лока-
лизацию клеток, активированных возбуждающими медиаторами, агонистами
и антагонистами, факторами роста и т. д. К сожалению, ГНО не активируют-
ся тормозными медиаторами, а разные нейроны экспрессируют разные ГНО,
и уровни их экспрессии различны. Принимая все эти факторы во внимание,
окрашивание ГНО — это чрезвычайно ценное средство выявления тех частей
центральной нервной системы, которые активируются нервными и (или) фар-
макологическими стимулами.
Б. Локальная фиксация потенциала
Локальная фиксация потенциала разработана Эрвином Нейром (Erwin Neher)
и Бертом Сакманом (Bert Sakmann) в 1976 г. Метод позволяет биофизикам ис-
следовать потоки ионов через одиночные каналы. Он состоит в том, что кон-
чик тонкой стеклянной микропипетки (диаметр около 0,5 мкм) прижимают к
мембране исследуемой клетки. Между кончиком пипетки и мембраной клетки
образуется тесный контакт с очень высоким омическим сопротивлением (око-
ло 10 ГОм), что важно для регистрации токов в субпикоамперном диапазоне.
Микропипетка заполнена электролитом и соединена с соответствующими
электронными устройствами, позволяющими регистрировать токи через ма-
ленький участок мембраны. Кусочек мембраны может оставаться in situ или же
с помощью небольшого подсасывающего давления отрываться от клетки и ис-
следоваться изолированно. Рис. Д.1 показывает различные типы препаратов,
которые позволяет использовать эта методика.
Рис. Д. 1 показывает четыре основных типа препаратов: фиксация потенци-
ала присоединенной клетки, «внутренняя поверхность мембраны — наружу»,
фиксации потенциала на целой клетке, фиксация потенциала целой клетки и
«наружная поверхность — наружу». Следует отметить, что мембрана, особен-
но, когда она отрывается от клетки, в пипетке приобретает Q-образную форму.
Таким образом, поверхность мембраны, на которой фиксируется потенциал,
намного больше кончика пипетки, это существенно при вычислении количе-
ства каналов на единицу поверхности мембраны.
Токи, регистрируемые с помощью этой методики, как упоминалось выше,
невелики и составляют пикоамперы. Токи эти, однако, развиваются быстро —
они возникают в результате открывания и закрывания одиночных ионных ка-
налов. Если и когда в фиксируемом участке мембраны открывается второй ка-
нал, амплитуда тока удваивается. При этом установлено, что одиночный канал
558 Дополнение: некоторые методики
Рис. Д.1. Типы препаратов ло-
кальной фиксации потенциа-
ла. (А) Регистрирующая мик-
ропипеткз. присоединена к
мембране с помощью легкого
присасывания (локальная
фиксация потенциала присое-
диненной клетки). (Б) Резкий
рывок пипетки приводит к от-
рыву части мембраны и обра-
зованию препарата «внутрен-
няя поверхность мембраны —
наружу». (В) Препарат А, под-
вергнутый более сильному
всасыванию мембраны в пи-
петку, вызывает ее прорыв и
образование «фиксации по-
тенциала на целой клетке».
(Г) Если препарат В подверг-
нуть резкому рывку, часть
мембраны отрывается от клет-
ки и иногда замыкается в пре-
парат «наружная поверх-
ность — наружу». Из Smith,
1996, с разрешения.
имеет более чем одно открытое состояние и несколько типов зарытых состоя-
ний. Даже на уровне одиночных каналов мы очень далеки отточного переклю-
чения 0/1, используемого в кибернетике.
В. Нейроимиджинг
В последние годы разработан целый арсенал устройств, позволяющих визуа-
лизировать текущую активность мозга. В 1950-е гг. ЭЭГ была единственным
средством регистрации активности мозга in vivo. С тех пор в клиническое при-
менение вошли рентгеновская компьютерная томография (РКТ), ядерно-маг-
нитный резонанс (ЯМР = магнитно-резонансный имиджинг, МРИ), исследо-
вание регионального церебрального кровотока (РЦКТ), позитронная эмисси-
онная томография (ПЭТ), магнитоэнцефалография (МЭГ) и некоторые дру-
гие. В лабораторных исследованиях микроэлектродная техника отведения от
мозга экспериментальных животных теперь дополняется оптическими мето-
диками, позволяющими получать непосредственный образ активности изуча-
емой коры.
Здесь не место для детального изложения новых технологий формирования
изображения, тем не менее, краткое их описание вполне уместно. Анализ по-
ступающей в мозг информации — это проблема, представляющая значитель-
_________________________________________Дополнение: некоторые методики 559
ный интерес. Компьютерное томографическое сканирование дает чисто
структурные данные, которые получаются путем компьютерной реконструк-
ции анатомии мозга на основе различной рентгенопрозачности тканей мозга.
Другие виды томографии, однако, показывают, в каких участках мозга наблю-
дается активность и даже начинают приносить данные о природе такой актив-
ности. Основной недостаток методик, таких как РЦКТ, ПЭТ и функциональ-
ный МРИ в том, что они в основном (за некоторыми исключениями) детекти-
руют не собственно нервные события, а распределение крови в мозге. Счита-
ется, начиная с работ Роя (Roy) и Шеррингтона (Sherrington) в конце XIX ве-
ка, что это показатель активности мозга. Однако, временная шкала в данном
случае может оказаться несколько неопределенной. Возможно, что кровь пе-
рераспределяется в данный участок после того, как нейроны проявляют актив-
ность в течение некоторого времени. Действительно, точно неизвестно, что
вызывает усиление кровотока в области нервной активности. Предполагается,
что это не столько выделение конечных продуктов обмена, таких как СО2, ко-
торый вызывает локальную вазодилятацию, сколько вазодилятация, обеспе-
чивающая нервную ткань повышенным количеством метаболических субст-
ратов, опосредованная нервными влияниями, в предвидении повышенных
потребностей метаболизма. Наконец, временная характеризация процессов
осложнена тем, что фМРИ и ПЭТ требуют значительного времени (обычно —
десятки минут для ПЭТ и десятки секунд для фМРИ) для получения регистри-
руемого сигнала.
Суть метода РЦКТ состоит в инъекции радиоизотопа (обычно 133Хе) во
внутреннюю сонную артерию и посредующем мониторинге распределения
крови в мозге с помощью набора счетчиков Гейгера. ПЭТ основана на получе-
нии в циклотроне изотопов (напр. 18F, ИС, 15О), испускающих короткоживу-
щие позитроны, и их включении в воду, СО2, глюкозу, 2-дезоксиглюкозу или
соединения типа 1-ДОФА. После введения таких веществ их распределение
может быть зафиксировано томографическими камерами. В настоящее время
эти очень полезные методики обладают пространственным разрешением око-
ло 7—8 мм3, которое еще может быть улучшено. Наконец, МРИ основана на
том, что некоторые ядра атомов, как например протоны в молекулах воды или
жирных кислот, обладают слабомагнитными свойствами. При действии мощ-
ного магнитного поля они ориентируются по полю, а электромагнитное излу-
чение при определенной частоте эту ориентацию нарушает. Когда излучение
выключают, ядра возвращаются в исходное положение, испуская сигнал, при-
годный для детектирования. Этот сигнал зависит от молекулярного окруже-
ния, в котором находится данный атом. Сложная компьютерная техника поз-
воляет получать трехмерные МРИ-карты мозга.
Хотя сначала, сразу после изобретения, МРИ было ограничено только ин-
формацией о статической анатомии мозга, позднейшие усовершенствования
позволили использовать метод для контроля активности мозга (фМРИ). Усо-
вершенствование основывалось на том, что атом железа в молекуле дезоксиге-
моглобина обладает парамагнитными свойствами. Вследствие этого сигнал
МРИ зависит от степени оксигенации крови, что обеспечивает мощное сред-
ство для того, чтобы различать активные и неактивные части мозга. Сигнал до-
ступен быстро (за несколько секунд), а его пространственное разрешение пре-
560 Дополнение: некоторые методики
восходно (несколько мм3), хотя оговорки о связи сосудистых изменений и
нервной активности, упомянутые выше, остаются в силе.
В отличие от методов, описанных выше, ЭЭГ, МЭГ и оптический имид-
жинг детектируют электрическую активность нейронов непосредственно.
Тогда как ЭЭГ и, в меньшей степени, МЭГ используются в клинике и являют-
ся истинно неинвазивными методами, оптический имиджинг требует вскры-
тия черепа, а оттого и ограничен экспериментальными животными. ЭЭГ реги-
стрирует изменения электрических потенциалов на поверхности черепа, а
МЭГ — изменения магнитного поля. Оба метода, вероятно, регистрируют мас-
совую активность в дендритах и имеют великолепное (миллисекунды) разре-
шение по времени. В обоих случаях, однако, хотя нервная активность и регис-
трируется непосредственно, требуется техника усреднения для выделения сиг-
нала из шума, что занимает от 3 с до 30 мин.
При том, что ЭЭГ и МЭГ не связаны с фактором сосудистых изменений и
выявляют нервную активность непосредственно, их пространственное разре-
шение неудовлетворительно. Потенциалы и магнитные поля на коже головы,
которые регистрируются этими методами, могут происходить и из пространст-
венно ограниченных источников в глубине мозга, и из более широких поверх-
ностных источников в коре. При этом возможно локализовать события, свя-
занные с электрическими потенциалами (СЭП) и магнитными полями (СМП)
до объемов порядка нескольких кубических миллиметров ткани коры. По-
скольку магнитное поле ориентировано ортогонально к току (по правилу пра-
вой руки Флеминга), МЭГ особенно эффективен в детектировании сигналов,
возникающих в извилинах мозга. При этом все равно необходимо усреденение
сигнала, требующее десятков минут для детектирования СЭП и СМП.
Из этого краткого обзора ясно, что техника нейроимиджинга страдает
чем-то вроде принципа дополнительности. Некоторые методы дают хорошее
разрешение по времени, но плохое пространственное разрешение, а некото-
рые — наоборот. В отличие от квантовой механики, однако, здесь нет теоре-
тического запрета на комбинацию достоинств в будущем, которая будет в со-
стоянии устранить неопределенность. Проблема не в теории, а в финансах.
В грядущем XXI веке у нас есть все основания надеяться на развитие такой
техники нейроимиджинга, которая позволит нам с большой точностью по
времени и в пространственных измерениях следить за потоком сенсорной ин-
формации в мозг.
Литература
Belliveau, J. W. et al., 1991, «Functional mapping of the human visual cortex by MRI»,
Science, 254, 716-719.
Fox, et al., 1989, Mapping of the human visual cortex with PET, Nature, 323, 806—809.
Kwong, К. K., etal., 1992, «Dynamic magnetic resonance imaging of human brain activity
during primary sensory stimulation», Proceedings of the National Academy of Sciences,
89, 5675-5695.
Neher, E. and B. Sackmann, 1992, «The patch-clamp technique», Scientific American,
266(3), 28-35.
Roy, C. S. and C. S. Sherrington, 1890, «On the regulation of the blood supply to the brain»,
Journal of Psychology, 11, 85—108.
561 Дополнение: некоторые методики
Seligman, А. М., М. J. Karnowsky, Н. L. Wasserkrug and J. S. Hanker, 1968, «Non-droplet
ultrastructural demonstration of cytochrome oxidase activity with a polymerising osmio-
philic reagent, diaminobenzidine (DAB)», Journal of Cell Biology, 38, 1—14.
Smith, C. U. M., 1996, Elements of Molecular Neurobiology (2nd edn), Chichester: Wiley.
Sokoloff, L., 1981, «The deoxyglucose method for the measurement of local glucose utilisa-
tion and the mapping of local functional activity in the CNS», International Review of
Neurobiology, 22, 287—330.
Ts’o, D. Y., R. D. Frosting, E. E. Liecke and A. Grinvald, 1990, «Functional organisation
of primate visual cortex revealed by high resolution optical imaging», Science, 249,
417-420.
СОКРАЩЕНИЯ
5-НТ -
7ТМ -
АДГ
АМТ -
АРП -
АТФ -
АХ -
АЦ
ВВГ -
ВГД
ВКЖ -
впм -
ВР -
ВРК -
всп -
вес -
ГАГ -
ГАМК -
ГАМК-Р-
гдк
гно -
ГТФ -
ГЦ
ДАГ
двг
звп -
знв -
ИТФ -
КЧП -
мг
мк -
МРИ -
МРО -
МЭГ -
5-окситриптамин, серотонин
белок с семью трансмембранными доменами
антидиуретический гормон
А-волокна механо-тепловых рецепторов
агрегатное рецептивное поле
аденозин трифосфат
ацетилхолин
аденилатциклаза
времени включения голоса
внутриглазное давление
внеклеточная жидкость
внутренняя пограничная мембрана
вещество Р
вкусовая рецепторная клетка
вызванный соматосенсорный потенциал
внутренний синаптогенный слой
глюкозоаминогликан
у-аминомасляная кислота
рецептор ГАМК
ганглий дорзального корешка
гены немедленного ответа
гуанозин трифосфат
гуанилатциклаза
диацилглицерин
дорзовентрикулярный гребень
зрительный вызванный потенциал
задняя нижне-височная (кора)
инозитол трифосфат
кривая частотного порога
сравнительная молекулярная масса молекулы,
выраженная в массе атома водорода
медиаторные ионные каналы
магнитно-резонансный имиджинг
минимально различимое отличие стимулов
магнитоэнцефалография
Сокращения 563
н-АХ-Р — никотиновый ацетилхолиновый рецептор
НВ — нижне-височная. Область коры приматов, вовлеченная в распознавание сложных зрительных объектов, в т. ч. лиц
НКТ — наружное коленчатоле тело
НИМ — наружная пограничная мембрана
НСС — наружный синаптический слой
ОбСБ — обонятельные связывающие белки
ОРК — обонятельная рецепторная клетка
ПАС — периакведуктальное серое вещество
ПБ — поле Бродмана
Пд — парциальное давление
ПЗК — потенциал-зависимый ионный канал
ПКА — протеинкиназа А
ИКС — протеинкиназа С
ПЧ — постоянная частота
ПЭС — пигментный эпителий сетчатки
ПЭТ — позитронная эмиссионная томография
РГС — ретино-геникуло-стриарный (основной зрительный путь у млекопитающих)
РП — рецептивное поле
св — средне-височная извилина. Термин несколько ошибочный, поскольку первоначально касался зрительного поля 5 совинолицей мартышки, которое, действительно, располагается на поверхности этой извилины. Однако, у макаков, на которых проводится большинство исследований, это поле расположено в верхней височной извилине, однако акроним СВ остался связан с полем 5
смп — событие, связанное с магнитным полем
СМТ — С-волокна механо-тепловых рецепторов
СТТ — спино-таламический тракт
сэп — событие, связанное с электрическим потенциалом
У го — утолщение гипокампальной области
Усо — утолщение гиперстриарной области
фдэ — фосфодиэстераза
фМРИ — функциональное магнитно-резонансное исследование
ФСБ — феромон-связывающий белок
чм — модулированная частота
3NaK — эпителиальный Na+-канал
ээг — электроэнцефалография
СЛОВАРИК
Ephemeroptera — отряд насекомых, включающий поденок.
Geometridae — пяденицы, большое семейство бабочек. Хрупкая бабочка с
большими крыльями и несколько неопределенным полетом.
Muscidae — настоящие мухи, относящиеся к семейству Anthomyidae отряда
Diptera (двукрылые). Наиболее известный представитель Musca domestica
(муха домашняя).
Noctuidea — бабочки семейства совок (крупнейшее семейство надсемейства
Noctuoidea), включающие небольших темного цвета насекомых, посеща-
ющих цветы в сумерки и ночью. Наиболее известный род — Agrotis (озимая
совка).
OVLT — паравентрикулярное ядро.
Tabanidae — насекомые, относящиеся к семейству слепней отряда двукрылых,
в том числе дождевка обыкновенная (Haematopota) и слепень (Tabanus).
Tachinidae — насекомые семейства тахин отряда двукрылых. Наиболее извест-
ны мясные мухи Calliphora и мухи формия Phormia.
Tettigoniidae — семейство настоящих кузнечиков, к которому относятся куз-
нечики и сверчки. Обычно зеленого окраса и громко стрекочут.
Tinnitus — (от лат. — tinnire — звенеть) шум в ушах, звенящий, свистящий или
жужжащий звук, кажущийся субъекту возникающим в одном или обоих
ушах.
Амниоты — позвоночные, у которых в ходе эмбриогенеза образуется амнион,
т. е. пресмыкающиеся, птицы и млекопитающие.
Анальгезия — отсутствие болевой чувствительности без утраты других ощуще-
ний и сознания.
Анестезия — полное отсутствие ощущений во всем тела или его части.
Антидромный — распространение потенциалов действия в направлении, про-
тивоположном естественному физиологическому направлению.
Апоневроз — фиброзный или мембранообразный соединительнотканный
лист, соединяющий мышцу с той частью организма, которую она приво-
дит в движение.
Апоптоз — запрограммированная клеточная смерть или «клеточное самоубий-
ство». Клетка разделяется на упакованные в мембраны фрагменты.
Словарик 565
Астигматизм — неравномерная кривизна хрусталика или роговицы в разных
плоскостях, приводящая к формированию нечеткого изображения на сет-
чатке.
Афферент — переносящийся к чему-либо.
Барорецептор — рецептор давления.
Белки теплового шока — белки, синтезируемые в ответ на избыточную темпе-
ратуру или другие стрессорные факторы.
Биогенетический закон — «рекапитуляция филогенеза в онтогенезе».
Бэйтсовская мимикрия — тип мимикрии, при котором безопасный виды дез-
ориентируют хищников, имитируя своим видом вредных или несъедобных
животных. Названа но имени Бэйтса (Н. W. Bates, 1825—1892), впервые
подметившего этот феномен.
Вторичноротые — животные, у которых в ходе развития бластопор преобразу-
ется во вторичный анус1.
Гемолимфа — жидкость, содержащаяся в гемоцеле.
Гемоцеле — буквально — полость тела, заполненная кровью.
Гидропатический анализ — методика, применяющаяся для определения того,
какая часть полипептидной цепочки неустойчива к гидролизу.
Гиперальгезия — повышенная чувствительность к болевым стимулах.
Гипертермофильность — склонность к крайней высоким температурам. В слу-
чае бактерий — до 113°С.
Гипоксия — ненормально низкий уровень кислорода.
Гомонимные нейроны — применительно к мотонейронам, означает, что ней-
рон иннервирует мышцу того же названия.
Дерматом — область тела, иннервированная одним сенсорным спиномозго-
вым корешком.
Дизестезия — ненормальное неприятное ощущение.
Иннервация — обеспечение нервными волокнами.
Ипсилатеральный — находящийся на той же стороне.
Кала-азар — висцеральный лейшманиоз, тропическое инфекционное заболе-
вание, вызываемое простейшим Leischmania donovani, поражающее пе-
чень, селезенку и т. д. Среди симптомов — потеря веса и лихорадка.
Капсайцин — вещество, ответственное за острый перечный вкус красного пер-
ца (паприки).
Конечный мозг (telencephalon) — самый передний пузырь мозга позвоночных.
Контент — «содержание» ментального состояния.
Контрлатеральный — находящийся на противоположной стороне.
1 Уже указывалось, что это фактическая ошибка автора. На самом деле, вторичный анус
прорывается по соседству с бластопором, а вторичный рот — прорывается рядом с первич-
ным анусом. Хотя так, как пишет автор, конечно, забавнее. — Прим, перев.
566 Словарик
Монофилетический — происходящий от одного предка.
Намеренность — специфическая «направленность» сознания, напр. гнев на
кого-либо, удовлетворение чем-либо, восприятие чего-либо и т. д.
Неоволосенный — кожа, лишенная волос (кожа ладоней и подошв).
Органула — термин, используемый (в основном в энтомологии) для обозначе-
ния специализированных многоклеточных, но микроскопических, струк-
тур, таких как сенсиллы.
Ортодромный — распространение нервного импульса в обычном направле-
нии.
Первичноротые — бластопор развивается в рот.
Полифилетический — происходящий от не единого предка.
Промежуточный мозг (diencephalon, thalamencephalon) — отдел мозга, располо-
женный непосредственно за конечным мозгом (при виде сверху).
Семиотические вещества — «смысловые вещества», термин обычно применя-
ется к химическим сигналам социальных насекомых.
Сине-зеленые водоросли — группа фотосинтезирующих бактерий. В докемб-
рии сине-зеленые водоросли образовывали на морском дне большие мас-
сы, называемые строматолитами.
Сколопидий — субкутикулярная сенсилла насекомых, обычно не имеющая
внутри- и надкутикулярных элементов.
Тельсон — хвостовая часть тела десятиногих раков. Лучше всего заметен у реч-
ного рака, омара и креветки.
Феноменальность — «кажимость», «то, что похоже на», «эмпирический харак-
тер» или более кратко «ощущение» сознательного состояния.
Феромон — от греческих «феро» — переносить и «гормон» — возбуждать. Фе-
ромоны — это химические вещества, обычно секретируемые железами,
участвующие во внутривидовой коммуникации.
Хитин — структурный материал экзоскелета членистоногих. Это — полисаха-
рид, имеющий n-ацетил-глюкозамин как мономерную единицу.
Щелевой контакт — соединение между соседними клетками через которое мо-
гут обмениваться информацией клетки эпителия. Образованы цилиндра-
ми из шести коннексиновых единиц, окружающих канал (комплекс име-
нуемый коннексоном), и соединенный с коннексоном соседней клетки.
Энтомофильный — буквально — «любящий насекомых»; применяется к расте-
ниям, опыляемым насекомыми.
Эпистемология — отрасль философии, касающаяся теории познания, т. е. что
есть знание, как оно возникает, как мы можем узнать, что наше знание
есть нечто большее, чем мнение и т. д. (В русской литературе более принят
близкий по смыслу термин гносеология. — Ред.)
Эфферент — информация, передаваемая от чего-либо.
ИМЕННОИ УКАЗАТЕЛЬ
Августин блаженный 494
Александрович (Alexandrowicz J. S.) 91
Аранци (Aranzi) 547
Арбиб Майкл (Arbib Michael) 548
Аристотель (Aristotle) 12, 13, 77, 133, 492-
494,515,517
Артеди (Artedi) 440
Аутрум (Autrum) 456
Бах-и-Рита Поль (Bach-y-Rita Paul) 542
Бекеши фон (Bekesy von) 159
Бернар Клод (Bernard Claude) 219, 266
Брока (Broca) 163, 192
Булляр Жан-Батист (Boillard Jean Baptiste)
192
Бэкон Фрэнсис (Bacon Francis) 534
Вебер (Weber) 51
Вернике (Wernicke) 163, 192
Визел (Wiesel) 403, 405
Витгенштейн Людвиг 11
Вокансон (Vaucanson) ПО
Волькен (Wolken) 456
Вольфринг (Wolfring) 317
Вонг-Райли Маргарет (Wong-Riley Mar-
garet) 409
Вулси (Woolsey) 131
Выготский Лев 10
Гарлэнд (Garland) 552
Гарц (Harz) 456
Геккель (Haeckel) 319
Гельмгольц Герман Людвиг фон (Hermann
Ludwig Ferdinand von Helmholtz) 7, 48,
159, 160, 200, 353,535
Геринг Эвальд (Hering Ewald) 387
Гешвинд (Geschwind) 189
Гёте Иоганн Вольфганг (Goethe J. W.) 192
Голдман Дэвид (Goldman David) 36
Голдсмит (Goldsmith) 456
Грегори P. Л. (Gregory) 456, 541
Грей Эйса (Gray Asa) 455
Гулд Стивен (Gould Stephen) 69
Гумбольдт Александр фон (Alexander von
Humboldt) 48
Дакс Марцел (Dax Marcel) 192
Дамасио (Damasio) 189
Дарвин Чарльз (Darwin Charles) 229, 269,
492,509,535
Дарвин Эразм (Darwin Erasmus) 497
Даулинг (Dowling) 460
Декарт Рене (Descartes Rene) 110, 494, 517,
535
Детье (Dethier) 229
Джеймс Уильям (James William) 497
Джоунс (Jones) 462
Дьюк-Элдер Стюарт (Duke-Elder Stewart)
282, 456, 459
Дюбуа-Реймон Эмиль (Emil du Bois Rey-
mond) 48
Жакоб Франсуа (Jacob Francois) 57
Зеки (Zeki) 462
Зоммеринг (Soemmerring) 440
Икин (Eakin) 283
Йокояма (Yokoyama) 460
Кант Иммануил (Kant Immanuel) 49, 540
Картер (Karter) 552
Кахаль (Cajal) 460
Кеплер (Kepler) 547
Кохонен (Kohonen) 552
Краузе (Krause) 317
Кронли-Диллон (Cronly-Dillon) 456
Кук (Cook C.) 459
Кун Томас (Kuhn Thomas) 532
Кэннон Уолтер (Cannon Walter) 225
Либерман (Liberman) 186
Ливингстон (Livingstone) 410, 497
568 Именной указатель
Локк Джон (Locke John) 539, 540
Лоос ван дер (Loos van der) 131
Лурия Александр 10
Лэнд (Land) 442,456, 548, 552
Людвиг Карл (Ludwig Karl) 48
МакГилл (McGill) 494
Марр Дэвид (Marr David) 425,462, 548
Матиссен (Matiessen) 442
Маунткасл Вернон (Mountcastle Vernon) 129
Мейнерт Теодор (Meynert Theodor) 192
Мельзак (Melzack) 494, 507
Мендель (Mendel) 257
Минних (Minnich) 229
Мондриан Пит (Mondrian Piet) 548
Моррис Конвей (Morris Convey) 69,456
Мюллер Йоханнес (Muller Johannes) 48, 73,
200, 535, 540
НагельТомас (Nagel Thomas) 11,183,206,311
Натане (Nathans) 460
Ницше Фридрих (Nietzsche Friedrich) 535,
544
Ноннегассер (Nonnegasser) 456
Ньютон Исаак (Newton Isaac) 13
Обертэн Симон (Aubertin Simon) 192
Олви (Alvi) 456
Павлов Иван 10
Паре Амбруаз (Раге Ambroise) 110
Перро (Perrault) 438
Пиаже (Piaget) 541
Питерс (Peters) 462
Платон (Plato) 538
Поляк (Polyak) 460
Поппер Карл (Popper Karl) 529
Поуп Александр (Pope Alexander) 550
Пруст Марсель (Proust Marcel) 209
Пэнфилд Уайлдер (Penfield Wilder) 130
Райл Джилберт (Ryle Gilbert) 550
Сакс Оливер (Sacks Oliver) 425,462
Сен-Жермен-э-Лэ (St Germaine en Laye)
110
Скрибоний Ларг (Scribonius Largus) 484
Смайс (Smyth) 456
Смит Крис (Smith Christopher Upham
Murray) 7, 8, 9, 552
Спенсер Герберт (Spencer Herbert) 535, 541
Сперри (Sperry) 191
Стивенс Стэнли (Stevens Stanley) 52
Уилсон (Wilson E. O.) 533
Уолл Патрик (Wall Patrick) 494,495, 507
Уоллс Гордон (Walls Gordon) 460,481
Файгль Герберт (Feigl Herbert) 537
Фернальд (Femald) 442,456
Фехнер (Fechner) 52
Фихте Иммануил Герман (Fichte Immanuel
Hermann) 48
Фостер (Foster) 456
Фризби Джон (Frisby John) 413
Фриш фон (Frisch von) 483
Хадспет (Hudspeth) 135, 138
Хайдеггер Мартин (Heidegger Martin) 425
Хамероф (HameroffS.R.) 546
Хамфри Николас (Humphrey Nicholas) 418
Хансон (Hanson) 532
Хари (Hari) 552
Хартлайн (Hartline H. K.) 304,434
Хартли Дэвид (Hartley David) 539
Хегеманн (Hegemann) 456
Хеннинг Ханс (Henning Hans) 233
Хоббс Томас (Hobbs Thomas) 539
Хорридж Эдриан (Horridge Adrian) 13, 311
Хьюбел (Hubei) 403,405, 410,462
Хьюджес (Hughes) 503
Цайль (Zeil) 531,553
Цанкер (Zanker) 531, 553
Чизелден (Cheselden) 541
Шеферд (Shepherd) 552
Эберден (Heberden) 505
Экклс Дж. (Eccles J. C.) 546
Юкскюль фон (Uexkull von) 531, 553
Юм Дэвид (Hume David) 269,455, 539
Юнг (Young) 459
Ярбус (Yarbus) 10, 548
предметный указатель
Адаптация 46, 53
— быстрая 41
— медленная 41
- сенсорная 32, 33,41,42, 45, 72, 257, 369
аденилатциклаза (АЦ) 27, 254, 255
аккомодация 312, 349
— объем 337
— хрусталика 48
аллостерия 17
— кооперативная 15, 17
ампулы 144, 145
-Лоренцини 471, 477, 487, 488, 511, 513
ампулярный орган 486,487
анальгезия 493,495, 509, 511
арестин 368, 369
астигматизм 312, 337
афазия 191, 192
афакия 322
Базилярная мембрана 54, 55, 135
бактериородопсин 274
барабанная перепонка 153
белемноиды 11
белки одорант-связывающие (ОСБ) 245,
247, 248, 252
— рецепторно-передаюшие (Р-П) 211,214-218
— рецепторные обонятельные 253, 264
— рецепторные семидоменные 216,254, 261
— теплового шока 312, 336
— феромон-связывающие (ФСБ) 245, 248
биогенетический закон Геккеля 63
биполяры, инвертирующие знак 377
— колбочковые 376
— палочковые 376
близорукость (миопия) 312
болезни движения 167
боль 493-497, 500, 504, 506-509, 511, 515
— иррадиирующая 493,505, 506, 512
боуманов слой 327
бугорковые органы 489
— рецепторы 490
бэйтсовская мимикрия 122, 241
G-белки 15, 19, 21, 24-26, 31, 32, 41, 74, 77,
209, 237-239, 241, 242, 249, 254, 255, 257,
260, 261
Веки 315
веретена 100-102, 104-106, 109
— мышечные 90
верхняя косая мышца 316
верхняя прямая мышца 316
вестибулярные пути 164
витамин А 270
вкус 209, 210, 228, 229, 243, 244, 264
вкусовые почки 234-236, 264
внутриглазное давление 312
водянистая влага 331, 332, 345, 347
волокна Генле 365
— механорецепторные 131
— ноцицептивные 128, 493, 497, 499, 511
— эфферентные 302
волоски сенсорные 117
— хемочувствительные 264
волосковые клетки 153-158, 160, 162, 166,
198, 204, 205
вомероназальный орган 245, 260, 261, 262,
267
воротные механизмы 15, 28, 96, 135, 137,
238,507,518,519
восприятие категориальное 186, 201
вспомогательные органы глаза 315
вторичноротые 58, 61, 63, 66, 283
вторичные мессенджеры 24-27
Генетика катаракты 338
ген Рах-610
гены коннексинов 338
— кристаллинов 338
гигрорецепторы 245, 246
гиперколонки 412
гиперметропия 337
глаз аппозиционный 288
— беспозвоночных 282
— диафрагмальный 279, 291, 296, 297
570 Предметный указатель
--Nautilus 296, 458
— дневного насекомого 308
— зеркальный гребешка Pecten 291, 297, 299,
458
— камерный осьминога Octopus 298,. 458
— ночного насекомого 308
— позвоночных 312
— сканирующий 279, 289, 290, 457
— сложный 277, 279, 307, 457
— суперпозиционный 288
— человека 312, 459
глаза латеральные мечехвоста Limulus 304
— медианные (непарные) 442, 466
— сложные мечехвоста 458
глазки простейших 294, 457
глазок Erythropsidinium 296
глаукома 312, 346
глубина фокуса 351
глубоководные рыбы 434
глютамат 364
гносеология 49
головоногие 281
гомеостаз 530
гомойотермия 512
гомойотермные животные 469, 470, 472,
473,510
гомологии опсинов 374
гребень 438
Давление внутриглазное 345
ДАГ 241,249,254
дальнозоркость (гиперметропия) 312
движение следящее 352
движения вергентные 352
— глаз саккадические 353, 393, 396
— глазного яблока 352
дезоксиглюкоза 409
денатурация белков хрусталика 340
десенситизация 24
десцеметова мембрана 327, 328
детекторы выпуклости 435
— инфракрасные 478, 510
— постоянного контраста 435
джонстонов орган 117, 198
диацилглицерин (ДАГ) 27, 28
диоптрический аппарат 426, 436, 439
диспаратность 353
дорзовентрикулярный гребень 447
древо филогенетическое кристаллинов мле-
копитающих 336
Евстахиевы трубы 148, 151, 153
Жгутика движение 294
железа Вольфринга 317
— Краузе 317
— мейбомиева 317
— моллова 317
— слезная 318, 319
— цейсова 317
животные пойкилотермные 469, 470, 471,
510, 5Г2
жидкость слезная 318, 331, 332
Задний полюс 314
задняя камера 314, 342
закон Вебера 51
— Вебера—Фехнера 46, 52
— Стивенса 46
— рефракции 275
запах 209
земноводные 446
зеркальце 299, 426, 431, 465
зрачок 314
зрение беспозвоночных 456
— бинокулярное 393
— слепое 418
зрительная кора 397, 400, 420
— ось 314
— пластина 309
— полоска 525
— радиация 393, 397, 399
— система 55, 56
зрительные пигменты 427
— пути 394, 426, 445
— центры 426
зрительный нерв 397
— перекрест (хиазма) 397
— пузырек 321, 323
— тракт 397
Излучение инфракрасное 478
изменения конформационные 15
изображение перевернутое 276
иллюзии 529
иннервация кортикофугальная 399
интерорецепторы 47, 49
ИТФ 241,242,249, 254, 261
Калькаринная борозда 413
кальмар Loligo 33, 298
каналы боковой линии 141
— внешние Маджори 349
— ионные 25, 43, 74
- Клоке 323, 326
— медиаторные (МК) 28, 29
— мембранные 83
— механочувствительные 81, 85, 88, 89
— полукружные 135, 143, 144, 146, 149, 150,
153,164-166
— потенциал-зависимые (ПЗК) 28,29,38, 140
— циклонуклеотид-зависимые (ЦНЗ-кана-
лы) 254, 255, 257
Предметный указатель 571
капли масляные 426, 428
каракатицы Sepia 298
каротидное тельце 263
картирование 252
карты сенсорных поверхностей 54, 55, 133,
163, 175, 176, 200
катаракта 312, 337
кинестезия 78, 90, 195, 197, 202
киноцилии 135-137, 146, 155, 198,488,489
клетки амакриновые 324, 354, 381
— биполярные 354, 375
— вкусовые 97
-волосковые 78, 116, 135-142,145,146,148,
149,199
— ганглиозные 354, 381
— горизонтальные 354, 375
— интерплексиформные 354, 380
— Меркеля 111
— механочувствительные 95
--нейросенсорные 97
— миниатюрные биполярные 365
--ганглиозные 365
— нейросенсорные 39, 40
— ноцицептивные 502
— обонятельные 29, 97 •
— осморецепторные 86
— простые 406
— ретинулы 287
— рецепторные обонятельные (ОРК) 252,
257, 260
— сенсорные 12, 45, 46, 78
— сложные 406
— фоторецепторные 23, 29, 361, 461
— хеморецепторные 23, 95, 231
— эксцентрические 287, 305
коартикуляция 188
кожа 54, 78, 469
колба Краузе терминальная 111, 122, 124
колбочки 39, 326, 362, 373
— двойные 430
— парные (близнецовые) 428
коленчатое тело 168
--медиальное 163
------ядро 172
колобома 312, 323
колонки 129, 163, 177, 198
— глазодоминантности 393, 410, 411, 416
— ориентаций 408
коммуникация 532
конвергенция 299
конформационные изменения 368
конформация родопсина 367
— семидоменная (7ТМ) 15
конъюнктива 313
кора зрительная 133
— лингвистическая 163
— нижне-височная 421
— первичная слуховая 78
--зрительная 463
— сенсорная 47
- слуховая 56, 163, 172, 167, 176-178, 180,
181, 184, 189, 190,200,206-208
— соматосенсорная 111, 127-129, 133
-соматосенсорно-моторная 133
— стриарная 402
— экстрастриарная 393, 417, 464
кортиев орган 154, 158
косточки веберовы 135, 199
кристаллины 334, 335
кристаллический конус 287
круглое ядро 448
купула 141, 142, 153
Са2+-каналы потенциал-зависимые 41, 219,
225
К+-каналы Са2+-зависимые 155
Иа+-каналы 42, 43, 45, 72, 84, 114, 138, 203,
240
— ионные 239
— потенциал-зависимые 40
— потенциал-чувствительные 30
Лабиринт 143, 144, 145, 149, 150, 199
— мембранный 78, 90, 135
лагена (lagaena) 143, 144, 146, 148-150
ламина 310
латеральная прямая мышца 316
лиганд 17, 26, 29
лизоцим 318
лимб 326
линия боковая 78, 143, 161, 163, 199, 204,
205,469,471
лобула 310
локализация звука 171
люмиродопсин 371
лягушка 434
Магнитное поле 486, 487, 490
магниторецепторы 47, 491
магнитосомы 490, 491, 511
магниточувствительность 477, 513
макулы 146, 164
— саккулюса 135, 144, 145, 164
— утрикулюса 135, 144, 145, 164
Матиссена соотношение 442
медулла 310
мейерова петля 397
меланопсин 281
меланосомы 357
мелатонин 444
572 Предметный указатель
мембрана 16, 18-26, 28, 30-40, 42-45, 77, 81,
82, 84, 85, 89, 115, 116, 139, 156, 199, 215,
217,220
— базилярная 152, 153
— боуманова 312
-Бруха 341, 342
— десцеметова 312
— механочувствительность 88
— рейснерова 152
— текториальная 152
мембранные каналы 26-28
Меркеля клетки 125, 126
метаболизм роговицы 331
метародопсин 371
механизмы воротные 42, 44
— кинестезические 100
механорецепторы 77, 79, 99, 100, 122, 162,
194, 197, 198, 211, 220,227,497
механочувствительность 77
меченные линии 50
мешок слезный 318
микроворсинки 301
микрофтальмия 323
мимикрия бэйтсовская 198
миопия (близорукость) 334, 337
млекопитающие 44
многоклеточные животные 41,58-61,73,112,
209
модальность 50
модуляция частотная 46
молекулы рецепторные обонятельные 267
---семидоменные 257
моллюски 12
молоточек 153 *
моргание 317
мышца глаза 315
— мюллерова 317
— риоланова 317
мюллеровы клетки 324, 354, 379
Наковальня 153
наружное коленчатое тело 397, 398
насекомые 281
нейральный суперпозиционный глаз 307
нейроанатомия речи 189
нейромедиаторы 23
нейрофизиология боли 497
нейроэктодерма 320
нематода 12
нерв зрительный 396
нижняя косая мышца 316
— прямая мышца 316
нистагм 163, 165, 166, 200, 206
ноцицепторы 493, 497, 498, 500
ноцицепция 503
нуклеотиды циклические 29
Оболочка сосудистая 312, 314, 341
обоняние 210, 228, 245, 246, 249, 251, 264,
267
обонятельная луковица 245, 258, 262
— система насекомых 246
--позвоночных 251
обонятельные рецепторные клетки 252
— сенсиллы 247
обонятельный клубочек 258
— тракт 245
одноклеточные 58, 59
окончания Руфини нервные 125, 126
омматидии аппозиционного типа 286
— десятиногих 477
— насекомых 286
— суперпозиционного типа 286
опиоиды 500, 503, 512
оппонентная организация 382
оппонентность двойная 393
оппонентные двойные клетки 410
опсин 270
оптические обоймы 309
оптический диск 314
орган боковой линии 135, 141, 142
— вомероназальный 209
— джонстонов 111
— кортиев 144, 150,151, 152
— субгенуальный 111, 117, 118,197
— хордотональный 93
органы волосковые 118
— Гольджи 197
— Руфини 111
— сухожильные 100
— тимпанальные 111, 118, 119, 120, 121
— хордотональные 78,90,98,99,116,119,120
осморецепторы 80, 86
острота зрения 277
осьминоги 299
осязание 111
осязательные волоски 78
отолиты 135, 145, 147
Палочки 39, 326, 362, 477
— двойные 430
парафовеа 356
пауки (Araneida) 285
ПдО2 220, 221, 223-226, 263, 476
ПдО2-рецепторы 219
ПдСО2 220, 221, 223-226, 263, 476
ПдСО2-рецепторы 219
первичноротые 58, 61, 63, 283
перегородка орбиты 317
передний полюс 314
передняя камера 314, 345
перепонки барабанные 148, 151
пероксидаза хрена 501
Предметный указатель 573
пигментный ретинит 388
— эпителий сетчатки 355
пластинка продырявленная 314, 396
пластичность 131, 393, 415, 464
повторяющиеся мотивы 518
поддержание равновесия 99
поле Бродмана 128, 129, 131, 189, 198, 243
-Брока 163, 189-191,201
- Вернике 163, 187, 189-191, 201
— зрения 413
полоска Дженнари 393, 402
полукружные каналы 199
поля рецептивные (РП) 46, 52
поляризация света 481,482, 511
поляризованный свет 480, 482
потенциал генераторный 32, 33, 39, 40, 72,
107, 123, 256
— действия 42
— микрофонный 199
— покоя 30, 34
- рецепторный 32, 33, 39, 40, 72, 107, 138
— синаптический 140
потенциалы микрофонные 135, 158
пресбиопия 312, 337 *
пресмыкающиеся 447
прозопагнозия 424,464
происхождение сетчатки и хрусталика
эмбриологическое 320
прокариоты 15
пространство внутридисковое 361
протеинкиназа С (ПКС) 27, 28
протеинкиназы 15, 17, 18, 23, 24, 255
протеогликаны 33
проток носо-слезный 318
психометрия 46
психофизический закон Стивенса 72
птицы 447
пусковые стимулы 56
путь таламофугальный 447
пучок аркуатный 191
Рабдом 287, 307, 482, 483
рабдомер 307
равновесие 99, 135, 143, 163
радужка 312, 314, 326, 349
— сфинктер 342
развитие коры 417
разрешающая способность 277
распознавание лица 393,421,464
распространение электротоническое 264
расстояние фокальное 275
регенерация П-цмс-ретиналя 373
ресницы 316
— выросты 312
— мышца 312, 342, 344
— тело 312, 314, 342
рецепторные молекулы 22
ретиналь 269, 271
ретино-геникуло-стриарный путь 396, 463
ретино-тектальный путь 393, 395, 463
ретинохром 304
ретинула 483
рецептивные поля 53, 72, 111, 129, 168, 479
рецепторы 24, 26, 27, 29
— акустические 117, 119
— волосяных фолликулов 111, 124
— зрительные 111
— инфракрасные 478
— кожные 125, 198
— Меркеля медленно адаптирующиеся 129
— натяжения 91
— обонятельные 246
— опиоидные в ЦНС 493
— осязательные 115, 116
— семидоменные 23, 25, 209, 257
— сенсорные 47
— тактильные 78, 109, 111, 122
роговица 313, 314, 327
родопсин 252, 272, 303
— конформация 272
родопсинкиназа 368, 369
Р-П-белки 263
Саккулюс 143, 145-147, 149, 150, 153, 199
светочувствительность 269
сегмент внутренний 361, 362
— наружный 361, 362, 366
секреция водянистой влаги 343
семидоменные рецепторные молекулы 253
сенсиллы 44, 90, 93, 95-97, 99, 100, 116-118,
197, 202, 228-232, 245-248, 252, 263, 264,
469, 472
сенсорное замещение 542
сенсорные карты 72
— миры 518
- системы 12, 22,29, 46, 55-59, 72
сердцевидка Cardium edulis 297
сетчатка 39, 54-56, 199, 314, 354, 460
— дифференцировка 312
— инвертированная 312
синапсы 23
системы механорецептивные 39,47
— ноцицептивные 495
— обонятельные 39
— фоторецептивные 47
— хеморецептивные 47
склера 314, 333
сколопидии 98,99, 117, 119, 120
сланцы Бэрджеса 312
слой герминативный 324
— плексиформный 302
574 Предметный указатель
слуховая кора 176, 181
— фовеа 182, 200
слуховые косточки веберовы 147, 151
сонар 78, 160, 162, 180, 182, 198
сосочки вкусовые 234
сосудистый тракт 341
спектр электромагнитный 270
стекловидное тело 312, 314, 351
стереоцилии 135-141, 145, 146, 154-156, 160,
198
стимулы триггерные 56
стремя 153
строма 325, 327
структура колончатая 111, 128
сухожильные органы Гольджи 90, 105-108
сфинктер радужки 350
С-механорецепторы 111, 125
Тельца аортальные 219, 220, 223, 224
— Гольджи 109, 221
— каротидные 219,221-225
- Мейснера 111, 122, 123, 129
— пачиниевы 111, 122, 123
— Руфини 109
— тубулярные 114, 116, 197
терморецепторы 47, 247, 473, 475, 478, 497,
510
термочувствительность 469, 512
тимпанальный орган 117, 195, 198
ток темновой 369, 370
токи локальные 43
— электротонические 38
торможение латеральное 306
трабекулы сосудистого тракта 343
трабекулярная сеть 344
тракт Лисауэра 502, 512
транс-ретиналь 366
трансдуцины 368
тропоколлаген 328
трубчатые глаза 440, 441 •
Увеальный тракт 313
увеит 337
улитка 78, 148, 150-153, 156, 158, 160, 163,
168, 199
— кортиев орган 135
— lagaena 135
усилительный каскад 372
утрикулюс 143, 145, 146, 150, 153, 199
Фагосомы 357
феромоны 245, 247, 248, 260-262, 267, 478
филогения опсинов 281
фильтрующий угол 325
фовеа 163,314, 356,413,436
фовеола 356
фолликулы волосяные 122
фонемы 184, 185, 187,200
фонорецепция 147, 163
форманты 184, 185, 200
фосфатазу 15
фосфатидилинозитол 4,5 бифосфата (ФИФ2)
27
фосфодиэстеразы 26
фосфолипаза С (ФЛС) 237, 238,241,242,249
- С-Ь 27, 28, 254
фосфолипазы 26
фото-изомеризация 371
фоторецепторы 22, 47, 60, 65, 361, 428, 477,
481
функция картирующая 251
Хеморецепторы 22, 209, 219, 220, 223, 224,
228, 230, 244
хеморецепция 213, 263, 264
хемотаксис 211, 217
хемочувствительность 41,211,212, 263
хиазма вторая зрительная 310
— первая зрительная 310
хламиопсин 281
хордовые 11, 12
хороид 325
хрусталик 312, 314, 334
— волокна 312, 321, 335
— индукция 312
— капсула 335
— плакода 320
цАМФ 238, 241, 242, 254, 255, 256, 257, 261
цАМФ-зависимые протеинкиназы 27
цГМФ-фосфодиэстераза 368
цвет 386
цветное зрение 391
центральная нервная система (ЦНС) 39, 50,
51, 90, 92, 97, 100, 104, 105, 108-110, 127,
220, 239, 474, 479, 509
цикл Кребса 347
циклический АМФ (цАМФ) 28
цинновы связки 313
цитохром-оксидаза 409
11-цис-ретиналь 366
Частичный перекрест 393
четверохолмие 163, 168, 172, 173, 175
— верхнее 393
— верхний бугор 395
чистые тоны 56
членистоногие 12
чувствительность инфракрасная (ИК) 477,
513
— к поляризованному свету 477
— контрастная 278
— тактильная 78, 126
1
Предметный указатель 575
Шиффова связь 367
шиффово основание 271
шлеммов канал 325, 342-346, 348
шпора склеры 344
Эволюция 57, 135, 158, 265, 492, 496
— ионных каналов 521
— камерного глаза 279, 284
— конвергентная 518
— опсинов 280
— сложного глаза 279, 284
экстерорецепторы 47, 49, 229
эктостриатум 417, 448
электрические поля 483, 485, 486
электрорецепторы 47, 489, 490, 511,513
электротоническое проведение 232
электрочувствительность 477
эллипсоид 362
эндотелий 312, 327, 328
эпистемология эволюционная 544
— эпигенетическая 540
эпителий обонятельный 251
эффект Тиндаля—Рэлея 276
— Френеля 276
эффектор аллостерический 16
эхолокация 56, 179, 182, 199, 207, 484
Яблоко глазное 313
ядро кохлеарное 170
— подушки 419
язык 184
Abramis brama 432
Acipenser fulvescens 429
Ae des aegipti 472
Amia calva 430
Amiurus 147
Amphioxus 294, 321, 459
Anableps tetrophthalmus 440
Anableps 44\
Anaspida 312
An ax junius 525
Anguilla 427
Antheraea polyphemus 248
Лр/5 483
Aquaspirillium magnetotacticum 490, 491
Architeuthis 300
area centralis 315
Argyropelecus 440
Astacus fluviatilis 91
Astroscopus 484
Astur palumbarius 437
Barbus barbus 428
Boidae 478
Bombyx mori 248, 250
Branchioma vesiculoseum 286
Caenorhabditis elegans 10, 64, 73, 77, 78, ill-
114, 116, 133, 135, 138, 139, 197, 199, 203,
209, 265,469-471,476,510
Callinectes 482
Calliphora 229, 483
Carassius auratus 429
Carausius 472
Cardium 297
Chelydra serpentina 429
Chimaera monstrosa 488
(Chlamydomonas 281, 283, 294, 295
Chrysemys picta 430
Chrysopa (златоглазка) 308
Chrysopa camea 120, 121
Ciliata sp. 210
Circus hudsonius 429
Clostridium perfringens 85
Coleonyx variegatus 430
Copilia 290
Copilia quadrata 290
Crotilinae 478
D. phalenodectes 121, 198
D. scalestes 121, 198
D. scedastes 120
Danio rerio 386
Diretmus argenteus 434
Dirocheles phalenodectes 120
Drosophila 118,264
Drosophila melanogaster 248, 249, 250
Dytiscus 117
Electrophorus 484
Erythropsidinium 295
Escherichia coliTl, 80-82,84,89,111,196,201,
210-215,217,218,263
Euglena 295
Eupagurus 289
Formica polyctena 96, 97
Gekkogekko 431
Gerris 117
Glaucomys volans 429 4
Gryllus integer 120
Gymnarchus niloticus 484
Gymnotus 484
Gyrodinium 294
Haemophilis influenzae 85
Haliotis29\
Halobacterium halobium 274, 522
576 Предметный указатель
Hipposideros bicolor 183 Homarus gammarus 91 Homarus vulgaris 196 Homo erectus 424 Homo habilis 424 Homo sapiens 10, 66, 70, 78, 469 Hypsiglena ochorhynchus 430 Periplaneta americana 96, 99, 247, 249 Petromyzon 427 Phormia 229, 231 Pikaia 312 Planaria gonocephala 284 Pleuronectes platessa 484 Portunus 289
Katsuwonus pelamis 238 Portunus puber 456 Prionotus 210
Labidocera 442 Laminaria japonica 238 Lampetra 141, 443 Lepomis microchirus 430 Limulus 291,304, 383 lobula 309 Locusta migratoria 249 Lophortyx 437 Lumbricus 229 Lumbricus terrestris 284 Protopterus ethiopicus 429 Pseudomonas fluorescens 85 Pteronotus parnelii 180-182 Raja clavata 484 Rana pipiens 429 Rhinolophus rouxi 182 Rousettus 161 Rutilus rutilus 381 Saccharomyces cerevisiae 111, 522
M. fuscata 186 macula lutea 315 Malapterurus electricus 484 Manduca sexta 249, 250 medulla 309 Mustelus mustelus 432, 433 Myotis lucifugus 179 Myotis 161 Myxine 141 Salmonella typhimurium 211, 217, 218 Salpa (Urochordata) 284 Salticidae 289 Scopelarchus 440 Scyliorhinus canicula 484, 485, 487 Spenodon 444 Squalus acantias 432 Staphylococcus aureus 85 Stenarchus 489 Stomatopoda 290
Natrix natrix 430 Nautilus 284, 291,296, 297 Necturus 240 Notonecta 117 Tachyglossus 69 tapetum lucidum 432 Torpedo 68, 484 Triatoma megista 472
Octopus 303 ora serrata 314 Ormia ochracea 120 Ornithorhynchus 69 Oxygyrus 289 Triatoma 472, 478 Trypanosoma cruzi 472 Tyto alba 163, 172 Uca 53\ Watasenia scintillans 304
Pachystomias 433 Pox-6292, 293 Pecten 284, 297, 432 Periplaneta 117 Xantusia riversiana 430 Xenopus255 Xenopus laevis 280
ОГЛАВЛЕНИЕ
Предисловие редактора русского издания................................. 5
Предисловие автора к русскому изданию................................. 10
Предисловие........................................................... 12
Часть I. ПРЕДВАРИТЕЛЬНЫЕ СВЕДЕНИЯ .................................... 15
Глава 1. Основы ...................................................... 15
1.1. Аллостерические эффекторы ..................................... 16
1.2. Мембраны....................................................... 18
1.3. Рецепторные молекулы .......................................... 22
1.4. Мембранные сигнальные системы.................................. 24
1.4.1. G-белки ................................................. 24
1.4.2. Эффекторы и вторичные мессенджеры........................ 26
1.5. Каналы и воротные механизмы ................................... 28
1.5.1. Медиаторные каналы (МК) ................................. 29
1.5.2. Потенциал-зависимые ионные каналы (ПЗК).................. 29
1.6. Заключение..................................................... 31
Глава 2. Мембрана и потенциал действия................................ 32
2.1. Измерение потенциала покоя..................................... 33
2.2. Ионные основы потенциала покоя ................................ 34
2.3. Электротонические потенциалы и кабельное проведение............ 38
2.4. Рецепторный и генераторный потенциалы.......................... 39
2.5. Сенсорная адаптация............................................ 41
2.6. Потенциал действия............................................. 42
2.7. Заключение..................................................... 45
Глава 3. Общие черты сенсорных систем............................... 46
3.1. Классификация чувств .......................................... 47
3.2. Модальность.................................................... 50
3.3. Интенсивность.................................................. 50
3.4. Адаптация...................................................... 53
3.5. Рецептивные поля .............................................. 53
3.6. Карты сенсорных поверхностей................................... 54
3.7. Иерархическая и параллельная организация ...................... 55
3.8. Выделение отличительных характеристик и пусковые стимулы....... 56
578 Оглавление
3.9. Заключение.................................................... 57
Информация 3.1. Герман фон Гельмгольц........................... 48
Глава 4. Эволюция и классификация организмов ...................... 58
4.1. Систематика................................................... 58
4.2. Классификация на шесть царств ................................ 59
4.3. Одноклеточные................................................. 59
4.4. Многоклеточность.............................................. 60
4.5. Первичноротые и вторичноротые ................................ 61
4.6. Классификация многоклеточных.................................. 63
4.7. Заключение.................................................... 69
Материалы для самоподготовки.................................... 71
Замечания и библиография ....................................... 74
Часть II. МЕХАНОЧУВСТВИТЕЛЬНОСТЬ................................... 77
Глава 5. Механочувствительность клеточных мембран.................. 80
5.1. Механочувствительные каналы Е. coli........................... 80
5.2. Детектирование осмотического разбухания
гипоталамическими клетками млекопитающих............................ 85
5.3. Заключение.................................................... 89
Глава 6. Кинестезия ............................................... 90
6.1. Кинестезические механизмы у членистоногих..................... 91
6.1.1. Рецепторы натяжения в мышцах ракообразных ............... 91
6.1.2. Сенсиллы насекомых........................................ 93
6.1.3. Поддержание равновесия в полете........................... 99
6.2. Кинестезические механизмы у млекопитающих...................... 100
6.2.1. Интрафузальные окончания веретен.......................... 101
6.2.2. Сухожильные органы Гольджи................................ 105
6.2.3. Рецепторы суставов ....................................... 109
6.3. Заключение .................................................... 109
Глава 7. Осязание................................................. 111
7.1. Механорецепция у Caenorhabditis elegans........................ Ill
7.2. Насекомые ..................................................... 116
7.2.1. Акустические сенсиллы и тимпанальные органы ........... 117
7.3. Тактильные рецепторы кожи млекопитающих........................ 122
7.3.1. Быстро адаптирующиеся рецепторы .......................... 122
7.3.2. Медленно адаптирующиеся рецепторы....................... 125
7.4. Анализ тактильной информации в мозге......................... 125
7.5. Пластичность соматосенсорной коры............................ 131
7.6. Заключение................................................... 133
Оглавление 579
Глава 8. Равновесие и слух: функция волосковых клеток.............. 135
8.1. Анатомия и физиология волосковых клеток....................... 135
8.2. Каналы боковой линии ......................................... 141
8.3. Эволюция уха позвоночных...................................... 143
8.3.1. Равновесие . . . /...................................... 143
8.3.2. Фонорецепция............................................ 147
8.3.3. Сонар летучих мышей .................................... 160
8.4. Заключение.................................................... 162
Информация 8.1. Биофизика волосковых клеток улитки............. 156
Глава 9. Анализ вестибулярной и звуковой информации в мозге........ 163
9.1. Вестибулярные пути и рефлексы ................................ 164
9.2. Слуховой путь................................................. 167
9.3. Картирование слухового пространства у сипухи ................. 172
9.4. Слуховая кора ................................................ 176
9.5. Слуховая система летучих мышей и эхолокация .................. 179
9.6. Слуховая кора человека и язык................................. 184
9.7. Латерализация и нейроанатомия речи............................ 189
9.8. Каллозэктомия и ее последствия ............................... 191
9.9. Заключение.................................................... 194
Информация 9.1. Брока и Вернике ............................... 192
Материалы для самоподготовки................................... 196
Замечания и библиография ...................................... 201
Часть III. ХЕМОЧУВСТВИТЕЛЬНОСТЬ.................................... 209
Глава 10. Хемочувствительность у прокариот......................... 211
10.1. Хемочувствительность у Е. Coli............................... 212
10.1.1. Молекулярная генетика.................................. 214
10.2. Заключение................................................... 218
Глава 11. Хемо-интерорецепторы млекопитающих....................... 219
11.1. Локализация хеморецепторов млекопитающих
к ПдО2 и ПдСО2 .................................................... 220
11.2. Структура .......................................1........... 221
11.3. Физиология................................................... 223
11.4. Биохимия ....................................... л........... 225
11.5. Заключение................................................... 227
Глава 12. Вкус..................................................... 228
12.1. Вкус у насекомых............................................. 229
12.2. Вкус у млекопитающих......................................... 233
580 Оглавление
12.2.1. Вкусовые почки......................................... 234
12.2.2. Центральные проекции................................... 242
12.3. Избранные каналы или паттерныответов? ....................... 243
12.4. Заключение................................................... 244
Глава 13. Обоняние................................................. 245
13.1. Обонятельная система насекомых............................... 246
13.1.1. Гигрорецепторы........................................ 246
13.1.2. Обонятельные сенсиллы и феромоны....................... 247
13.2. Обонятельная система позвоночных............................ 251
13.2.1. Обонятельный эпителий полости носа.................... 251
13.2.2. Обонятельные рецепторные клетки....................... 252
13.2.3. Молекулярная биология................................. 253
13.2.4. Центральная обработка информации ..................... 257
13.3. Вомерноназальный орган позвоночных и феромоны ............... 260
13.4. Заключение.................................................. 262
Материалы для самоподготовки................................... 263
Замечания и библиография ..................................... 265
Часть IV. СВЕТОЧУВСТВИТЕЛЬНОСТЬ ................................... 269
Информация IV. 1. Бактериородопсин............................. 274
Глава 14. Зрение у беспозвоночных.................................. 279
14.1. Организация глаз беспозвоночных.............................. 282
14.1.1. Эволюция камерного глаза............................... 284
14.1.2. Эволюция сложного глаза................................ 284
14.1.3. Сканирующий глаз ...................................... 289
14.2. Примеры глаз беспозвоночных................................. 291
14.2.1. Глазки простейших ..................................... 294
14.2.2. Диафрагмальный глаз Nautiluspompilius.................. 296
14.2.3. Зеркальный глаз гребешка Pecten........................ 297
14.2.4. Камерный глаз осьминога Octopus....................... 298
14.2.5. Латеральные глаза мечехвоста Limulus.................. 304
14.2.6. Высокоразвитый сложный глаз .......................... 307
14.3. Заключение.................................................... ЗЮ
Информация 14.1. Эволюция опсинов ............................. 280
Информация 14.2. Генетика раннего развития глаз .............. 292
Глава 15. Глаз человека ........................................... 312
15.1. Анатомия..................................................... 313
15.1.1. Глазное яблоко........................................ 313
15.1.2. Вспомогательные органы глаза.......................... 315
15.2. Эмбриология.................................................. 319
Оглавление 581
15.3. Подробности анатомии и физиологии.............................. 326
15.3.1. Роговица................................................. 327
15.3.2. Склера................................................... 333
15.3.3. Хрусталик................................................ 334
15.3.4. Сосудистый тракт......................................... 341
15.3.5. Радужка.................................................. 349
15.3.6. Стекловидное тело...................................... 351
15.4. Движения глазного яблока....................................... 352
15.5. Заключение .................................................... 353
Информация 15.1. Генетика катаракты.............................. 338
Глава 16. Сетчатка................................................... 354
16.1. Пигментный эпителий сетчатки (ПЭ).............................. 355
16.2. Сетчатка ...................................................... 358
16.2.1. Фоторецепторные клетки................................. 361
16.2.2. Горизонтальные клетки.................................... 375
16.2.3. Биполярные клетки........................................ 375
16.2.4. Мюллеровы клетки ........................................ 379
16.2.5. Интерплексиформные клетки ............................... 380
16.2.6. Амакриновые клетки....................................... 381
16.2.7. Ганглиозные клетки....................................... 381
16.2.8. Блок-схемы............................................... 384
16.2.9. Цвет..................................................... 386
16.3. Заключение..................................................... 390
Информация 16.1. Пигментный ретинит.............................. 388
Глава 17. Зрительные пути и зрительная кора......................... 393
17.1. Зрительные пути к мозгу........................................ 394
17.1.1. Ретинотектальный путь.................................... 395
17.1.2. Ретино-геникуло-стриарный (РГС) путь .................... 396
17.2. Первичня зрительная кора ...................................... 400
17.2.1. Структура ............................................... 400
17.2.2. Функционирование......................................... 402
17.2.3. Пластичность............................................. 415
17.3. Экстрастриарная кора........................................... 417
17.4. Распознавание лица............................................. 421
17.5. Прозопагнозия.................................................. 424
17.6. Заключение..................................................... 425
Информация 17.1. Слепое зрение .................................. 418
Глава 18. Зрительные системы других позвоночных...................... 426
18.1. Зрительные пигменты ........................................... 427
18.2. Фоторецепторы ................................................. 428
18.3. Зеркальца...................................................... 431
582 Оглавление
18.4. Сетчатки ....................................................... 433
18.4.1. Глубоководные рыбы........................................ 434
18.4.2. Лягушка................................................. 434
18.4.3. Центральные области (areae centrales)
млекопитающих и птиц .....................................:...... 436
18.5. Диоптрический аппарат........................................... 439
18.6. Медианные (непарные) глаза...................................... 442
18.7. Зрительные пути................................................. 445
18.8. Зрительные центры мозга......................................... 446
18.9. Заключение...................................................... 449
Материалы для самоподготовки...................................... 450
Замечания и библиография ......................................... 455
Часть V. ДРУГИЕ ЧУВСТВА............................ 467
Глава 19. Термочувствительность...................................... ^9
19.1. Молекулярная биология ........................................ 470
19.2. Пойкилотермные животные....................................... 471
19.3. Гомойотермные животные ....................................... 472
19.3.1. Терморецепторы кожи..................................... 473
19.3.2. Глубокие терморецепторы ................................ 475
19.3.3. Гипоталамические терморецепторы......................... 475
19.4. Заключение.................................................... 476
Глава 20. Редкие чувства ........................................... 477
20.1. Инфракрасное излучение ....................................... 478
20.2. Поляризованный свет........................................... 480
20.3. Электрические поля............................................ 483
20.4. Магнитные поля................................................ 490
20.5. Заключение.................................................... 492
Глава 21. Боль...................................................... 493
21.1. Биологическое значение боли................................... 495
21.2. Нейрофизиология боли.......................................... 497
21.2.1. Периферические ноцицепторы ............................. 497
21.2.2. Центральные пути ....................................... 501
21.3. Нейрофармакология болевых путей............................... 503
21.4. Иррадиирующая боль ........................................... 505
21.5. Воротная теория............................................... 507
21.6. Заключение.................................................... 508
Материалы для самоподготовки.................................... 510
Замечания и библиография ....................................... 512
Оглавление 583
Часть VI. КОДА ........................................................... 517
Глава 22. Повторяющиеся мотивы............................................ 518
22.1. Молекулярные мотивы................................................. 519
22.2. Клеточные мотивы.................................................... 523
22.3. Органы чувств....................................................... 524
22.4. Центральные анализаторы............................................. 526
22.5. Гомеостаз .......................................................... 530
22.6. Различные сенсорные миры............................................ 531
22.7. От абиотического к биологическому: коммуникация .................... 532
22.8. Заключение.......................................................... 534
Глава 23. Философский постскриптум........................................ 535
23.1. Декарт ............................................................. 536
23.2. Сенсорные качества ................................................. 537
23.3. Tabula rasa?........................................................ 538
23.4. Эпигенетическая эпистемология ...................................... 540
23.5. Эволюционная эпистемология.......................................... 544
23.6. После Декарта....................................................... 546
23.7. Заключение.......................................................... 550
Информация 23.1. Сенсорное замещение.................................. 542
Материалы для самоподготовки.......................................... 551
Замечания и библиография .............................. .............. 552
Дополнение: некоторые методики ........................................... 556
Сокращения................................................................ 562
Словарик.................................................................. 564
Именной указатель......................................................... 567
Предметный указатель...................................................... 569