






























































































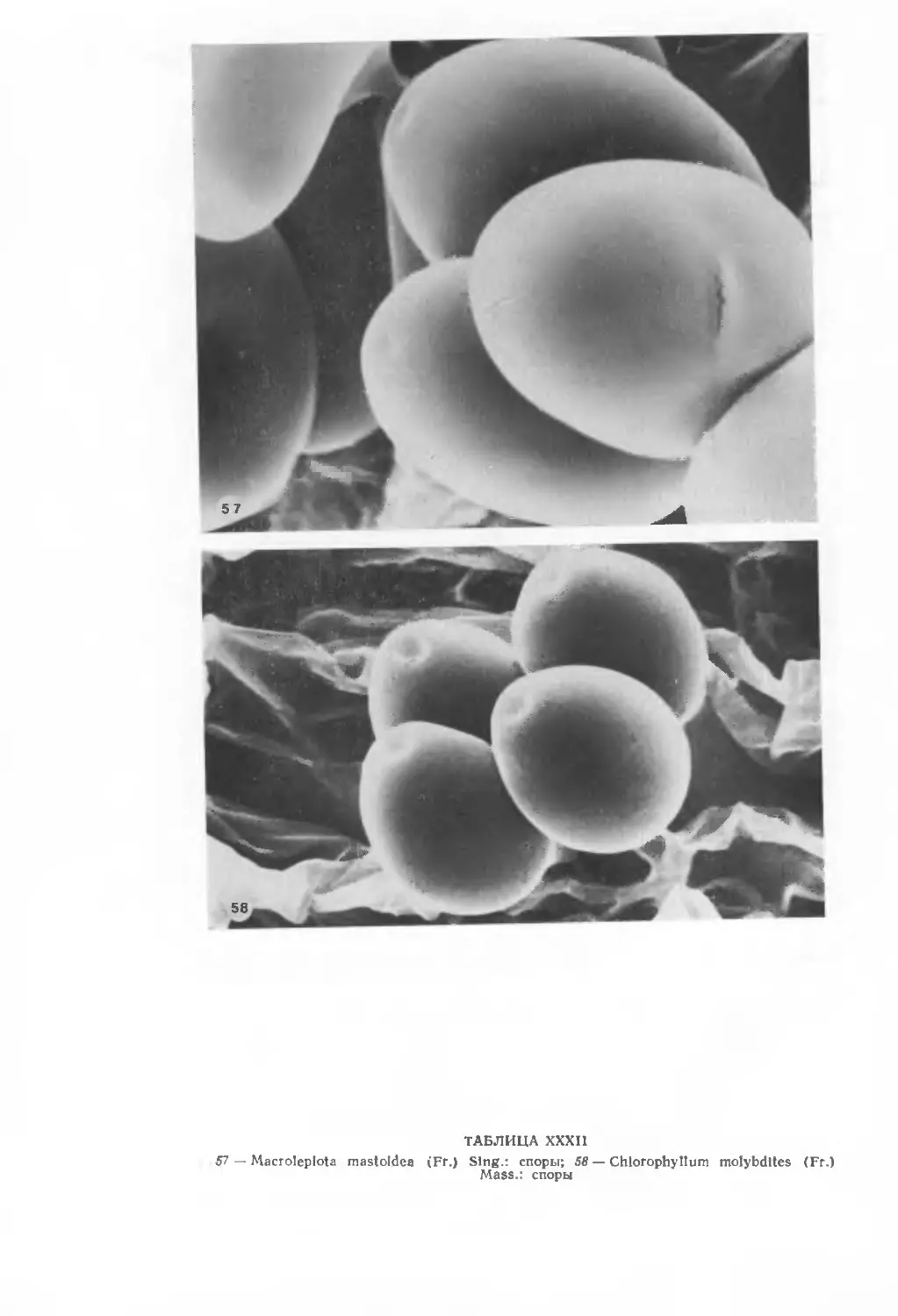




















































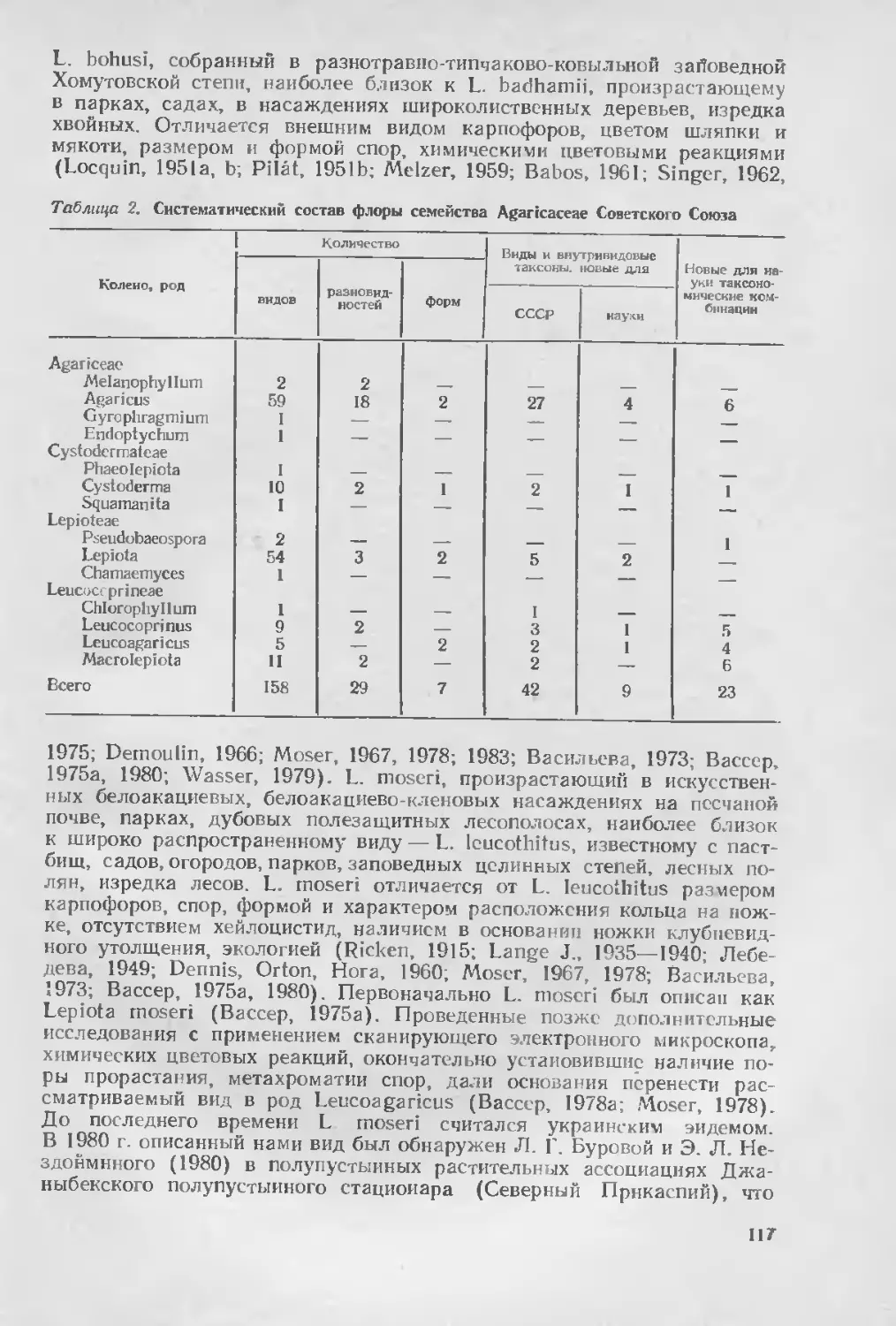



































































Автор: Вассер С.П.
Теги: fungi ( eumycota, eumycetes) настоящие грибы микология растения флора растениеводство грибы семейство грибов фауна ссср
Год: 1985




АКАДЕМИЯ НАУК УКРАИНСКОЙ ССР
ИНСТИТУТ БОТАНИКИ им. Н Г. ХОЛОДНОГО АН УССР
С. П. BACCFP
АГАРИКОВЫв
ГРИБЫ
СССР
КИЕВ НАУКОВА ДУМКА 1985
УДК 582.284(47 + 57)
Агариковые грибы СССР / Вассер С. П — Киев : Наук, думка, 1985 — 184 с.
В монографии обобщены оригинальные и-литературные данные об агариковых гри-
бах СССР, обсуждаются их филогенетические связи и происхождение. Приведены све-
дения о морфологии, анатомии, систематике, эколого-биологических и культуральных
особенностях, геометрических свойствах базидий, макро- и микрохимических реакциях
карпофоров и микроструктур агариковых грибов. Рассмотрены критерии видов и пред-
ложены новые. Обоснована оригинальная система семейства Agaricaceae. Даны геогра-
фический анализ видов, значение в природе н народном хозяйстве. Подробно охаракте-
ризованы 158 видов, 29 разновидностей и 7 форм агариковых грибов. Из них 9 таксо-
нов новые для науки, 42 — новые для микофлоры СССР. Для каждого таксона
даны синонимика, иконография, экология, распространение по районам флоры СССР,
земному шару, критические примечания.
Для микологов, биогеоценологов, геоботаников, работников лесного и сельского
хозяйств, грибоводов, преподавателей и студентов вузов.
Ил. 45. Табл. 6. Библиогр.: с. 168—183.
Ответственный редактор
И. А. Дудка
Рецензенты
К. А. Каламээс, И. Г. Нахуцришвили
Редакция общей биологии
СОЛОМОН ПАВЛОВИЧ ВАССЕР
АГАРИКОВЫЕ ГРИБЫ СССР
Утверждено к печати ученым, советом
Института ботаники им. И. Г. Холодного АН УССР
Редактор Г. И. Дружина. Оформление художника Г. М. Балюка
Художественный редактор Р. И. Калыш Технический редактор А. М. Капустина.
Корректоры С. И. Кримец. Н. А. Луцкая, Л. В. Малюта.
Информ, бланк № 6689.
Сдано в набор 12.10.84. Подл, в печ. 24.04.85. БФ 03063. Формат 70Х100/ц. Бум. ткл. № 1. Лит. гари.
Выс. печ. Физ. печ. л. 11.5+2.0 вкл. на мел. бум. Усл. печ. л. 17,55. Усл. кр.-отт. 17,55. Уч.-изд. л.
17,85- Тираж 1200 экз. Заказ JA 4—2960. Цена 3 р-
Издательство «Наукова думка». 252601 Киев 4, ул. Репина, 3.
Отпечатано с матриц Головного предприятия республиканского производственного объединения
«Нолнграфкннга». 252057, Киев. ул. Довженко, 3 в Нестеровской городской типографии 292310.
Нестеров Львовской обл.. ул. Горького, 8..' Зак. 2469
„ 2001000090-202 „ т, ч „ irvoe
В M22UB4+S6—325-86 © Издательство «Наукова думка», 1985
ВВЕДЕНИЕ
Агариковые грибы (семейство Agaricaceaeт, порядок Agaricales
s. str., класс Basidiomycetes) насчитывают сравнительно небольшое ко-
личество видов: к настоящему времени общее число их составляет при-
мерно 500 (Singer, 1975; Вассер, 1980, 1982; Moser, 1983; Ila wks worth,
Sutton, Ainsworth, 1983). Они представляют большой научный интерес
в связи с чрезвычайной спецификой и разнообразием морфологии, ана-
томии, онтогенеза, географии, экологии. До последнего времени крити-
ческой обработке семейство Agaricaceae ие подвергалось, а разграни-
чение его видов и других систематических единиц, как и выяснение
филогенеза, осложнялось вследствие свойственного им полиморфизма,
явлений параллелизма и конвергенции. Знания о таксономических свой-
ствах агариковых грибов базируются чаще всего на единичных, фраг-
ментарных, статистически не обработанных данных. Экспериментальные
исследования роста, развития, дифференцировки агарнковых грибов-
in vitro проведены лишь для небольшого количества видов. Эти фраг-
ментарные результаты слишком скудны, во многих случаях противоре-
чивы, что ие дает оснований для их обобщений (Horak, 1968; Вассер,
1980, 1982).
В настоящее время нет единой, всеми признанной системы Agari-
cales, по-разному понимается и объем самого семейства Agaricaceae,
разделяемого некоторыми авторами иа несколько семейств. Миграция
видов из рода в род, а некоторых видов и родов из одного семейства
в другое свидетельствует о том, что признаки, а следовательно, н крите-
рии, позволяющие отнести гриб к определенной таксономической кате-
гории, у агариковых грибов не являются достаточно четкими, стабиль-
ными. Подтверждением такого положения является то, что до настояще-
го времени Agaricaceae разделяют на 2—23 рода (Kreisel, 1969; Dennis,
Orton, Нога, 1974; Singer, 1975; Moser, 1978, 1983; Зерова, Сосш,
Роженко, 1979; Вассер, 1980, 1982). Критерии, используемые для оценки
таких таксономических категорий, как семейство, род, вид, у агарико-
вых грибов весьма различны. В интерпретации критериев существуют
большие расхождения, иногда наблюдается их «эрозия» илн «девальва-
ция». Такое же положение характерно и для других групп высших
Basidiomycetes (Пармасто, 1969; Donk, 1971; Зерова, Вассср, 1974;
Singer, 1975; Каламээс, 1975; Зерова, Сосш, Роженко, 1979). Это
обусловлено недостаточной изученностью грибов, с одной стороны.
1 Авторы таксонов всех рангов приведены в «Конспекте флоры агариковых
грибов СССР».
3
и эмпирическим подходом исследователей к интерпретации результатов
без достаточного внимания к методике систематизации и исходным тео-
ретическим положениям — с другой. Такая ситуация охарактеризована
Э. X. Пармасто (1973) как своего рода «детская болезнь» современной
систематики.
До последнего времени в Советском Союзе не было работ, посвя-
щенных грибам порядка Agaricales, в которых был бы применен целый
комплекс классических и новых методов. Данная работа является пер-
вым результатом всестороннего критико-снстематнческого изучения
семейства Agaricaceae порядка Agaricales s. str.
Грибы, относящиеся к семейству Agaricaceae, принимают активное
участие в процессе гумификации почвы. Среди них, много ценных
съедобных, широко используемых населением, лекарственных, ядови-
тых. Велико значение разводимых искусственно в промышленном мас-
штабе агариковых грибов (Agaricus bisporus, A. bitorquis, A. macrospo-
rus, A. subedulis и др.) как дополнительного источника питания. В по-
следнее время при поисках новых антибиотических, бактериостатических
и онкостатических веществ, а также стимуляторов роста животных и
растений, ферментов, органических кислот и ряда других ценных ве-
ществ объектом изучения становится все большее количество видов се-
мейства Agaricaceae (Banchet, 196Г, Васильков, 1966; Шиврина, Плато-
нова, Черетченко, 1968; Шиврина, Низковская, Фалина и др., 1969;
Зерова, 1970а; Гарибова, 1971; Steineck, 1973; Дудка, Вассср, Бухало
и др., 1978; Яковенко, Жемойц, 1978; Chang, Ucyes. ed., 1978; Rumack,
Salzman, cd., 1978; Bacccp, 1980; и др.).
Теоретическое и практическое значение агариковых грибов с каж-
дым годом привлекает все болыиеё внимание микологов. Однако систе-
матикой агариковых грибов в Советском Союзе специально никто не
занимался. Спорадические, подчас некритические сведения о некоторых
видах этого семейства разбросаны в отдельных статьях, региональных
определителях и флористических сводках. Все это диктует настоятель-
ную потребность создания определителя иди конспекта флоры агарнко-
вых грибов Советского Союза.
Необходимость всестороннего изучения грибов н составления «Опре-
делителя грибов СССР» отмечена в резолюциях VII съезда Всесоюзного
ботанического общества (Донецк, 1983 г.) й годичной сессии Научного
совета АН СССР по проблеме «Биологические основы рационального
использования, преобразования н охраны растительного мира» (Сухуми,
1985 г.).
Цель настоящей работы — всесторонний анализ и теоретическое
обобще.чие оригинальных и литературных данных по флоре, системати-
ке, фичогении, эволюции, классификации, распространению, хозяйствен-
ному значению агариковых грибов СССР. Кроме того, небольшое се-
мейство Agaricaceae при углубленном сравнительно-систематическом
изучении его представителей с использованием разнообразных методов
и критериев может послужить модельным объектом для уточнения не-
которых общих вопросов систематики высшнх Basidiomycetes.
Настоящая работа представляет собой первую критическую моно-
графическую сводку по агариковым грибам Советского Союза, в кото-
рой содержится конспект известных на современном уровне знаний ви-
дов этого семейства, включая новые для науки виды и комбинации,
предложенные нами. Впервые также определены статистическими ме-
тодами сравнительной флористики степень видового и родового сход-
4
ства, специфика видового состава семейства Agaricaceae областей фло-
ры Советского Союза н отдельных его регионов. Суммированы и кри-
тически проанализированы данные по классификационным схемам по-
рядка Agaricales s. 1., обсуждены объем и систематическое положение
в них семейства Agaricaceae. Описаны новый для науки порядок Hygro-
phorales и новое семейство порядка Tricholomatalcs— Catathelasmata-
сеае. Для ряда родовых и внутриродовых таксонов (Agaricus, Lepiota,
Cystoderma, Leucocoprinus) семейства Agaricaceae разработана ориги-
нальная система. Изучены геометрические особенности базидий, хими-
ческие цветовые реакции карпофоров и микроструктур, культурально-
морфологические особенности агариковых грибов как дополнительных
таксономических критериев.
Проведен ареалогический анализ географического распространения
агариковых грибов Советского Союза, в результате которого выделены
географические элементы и установлены типы ареалов в пределах каж-
дого элемента. Высказаны предположения о времени и возможных цен-
трах возникновения агариковых грибов на земном шаре; обсуждается
явление эндемизма у них.
* * *
За ценные советы при выполнении работы и критический просмотр
рукописи автор выражает благодарность докторам биол. иаук
И. А. Дудке, М. Ф. Макаревич, за помощь при подготовке рукописи к
печати — А. А. Гродзннской, И. Н. Собовой, В. И. Береговой и
Г. Б. Шнмбергу.
Изучение спор и других микроструктур агариковых грибов пот ска-
нирующим электронным микроскопом проведено совместно с канд. биол.
наук О. А. Закордонцем. Им же выполнены микрофотографии, помещен-
ные на вклейке. О. А. Закордонец является автором параграфа «Обра-
ботка материала для изучения с помощью сканирующего электронного
микроскопа (СЭМ)».
Значительная часть рисунков и все микрофотографии, приведен-
ные в книге,— оригинальные, часть рисунков заимствована из литера-
турных источников. Большинство из них выполнено художником
Н. А. Волынцом.
ГЛАВА I
СОСТОЯНИЕ ИЗУЧЕННОСТИ
АГАРИКОВЫХ ГРИБОВ
(обзор литературы)
1. ИЗУЧЕНИЕ АГАРИКОВЫХ ГРИБОВ
НА ЗЕМНОМ ШАРЕ
Первое упоминание в литературе о некоторых агариковых грибах
как о растениях, употребляемых в пищу, встречается у Гиппократа и,
•следовательно, относится к V веку до и. э. Упоминания об агариковых
грибах содержатся также в работе Плиния Старшего (цит. по: Лунке-
вич, I960; Ainsworth, 1976). Сведения о них имеются у П. А. Микели
(Mickeli, 1729), описавшего род пластинчатых грибов — Psalliota, к ко-
торому он отнес несколько видов рода Agaricus в современной интер-
претации.
К. Линией (Linne, 1753) 1 включил в описанный им род Agaricus
все пластинчатые грибы, т. е. весь порядок Agaricales s. 1. Э. М. Фриз
(Fries, 1821) признал род Agaricus, несколько ограничив его объем,
отделив два рода Cantharellus и Schizophyllum. Э. М. Фриз признавал
также род Lepiota, описанный С. Ф. Грэем (Gray, 1821). В последую-
щих работах Э. М. Фриз (Fries, 1836—1838, 1874) выделил из рода
Agaricus 28 и 35 соподчиненных единиц соответственно, которые можно
назвать подродами2.
Во второй половине XIX в. кроме Agaricus и Lepiota были описаны
еще четыре рода современного семейства Agaricaceae: Leucocoprinus,
Cystoderma, Clarkcinda и Chlorophyllum.
Позже многие фризовскне подроды были переведены в ранг родов
(Kummer, 1871; Gillet, 1874; Roze, 1876; Karsten, 1879; Cooke, 1880—
1890; Quelet, 1886; Fayod, 1889; Patouillard, 1900). В этих оригиналь-
ных критических работах даны описания многих новых для науки видов
современного семейства Agaricaceae и предложены десятки новых для
науки таксономических комбинаций, выдержавших испытание време-
нем (например, Lepiota felina, L. clypeolaria, Cystoderma amianthina).
При этом описания видов основаны уже не только на макроскопических,
но в значительной степени и иа микроскопических признаках карпофо-
ров. Так, Н. Патуйар указывал иа таксономическую важность строения
оболочки спор.
1 В истории изучения агариковых грибов земного шара мы выделяем три перио-
да: 1) начальный —с 1753 г. (с момента выхода в свет работы К. Линнея (Linne,
I. с.) ) до конца XIX в.; данная работа согласно статье 13 Международного кодекса
ботанической номенклатуры, утвержденного XIII Международным ботаническим
конгрессом в Сиднее, принята за основу для номенклатуры рассматриваемых грибов
(International code..., 1983); 2) накопительный — с 1900 г. до 30-х годов XX в.:
3) современный — с 30-х годов XX в. до настоящего времени.
2 Многие соподчиненные единицы Э. М. Фриза, которые мы называем подрода-
ми, по современному таксономическому объему соответствуют родам.
6
Агариковые грибы широко представлены и в сводке П. Саккардо
(Saccardo, 1887, 1902, 1912, 1925, 1972).
Во второй, накопительный, период появляются отдельные работы
(Atkinson, 1900; Mcllvaine, Macadam, 1902; Marshall, 1905; Hard, 1908;
Earle, 1909; Peck, 1909; Murrill, 1914; Ricken, 1915; Kauffman, 1918,
1924; Velenovsky, 1920—1922; Rea, 1922; Konrad, Maublanc, 1924—1937;
McDougall, 1925; Brcsadola, 1927—1933) и многочисленные статьи1
(Atkinson, 1915, 1916; Shantz, Piemeisel, 1917; Benes, 1923—1924; Kuce-
ra, 1923—1924; Konrad, 1924; Piiat, 1925; Schaeffer J., 1925; Lange J.,
1926; Coker, 1928; Hein, 1930), в которых приводятся описания видов
порядка Agaricales, в том числе н многих видов семейства Agaricaceae.
Расширяются н уточняются сведения о морфологической изменчивости
агариковых грибов, их географии и экологии. В некоторых работах уде-
ляется внимание и вопросам таксономии агариковых грибов. В этот
период были описаны четыре новых для науки рода семейства Agarica-
ceae, не утратившие своего значения до наших дней: Chamaemyces,
Phaeolepiota, Micropsalliota н Mcllanophyllum. Однако в этот период
еще нет специальных работ, посвященных некоторым родам илн всему
семейству Agaricaceae отдельных регионов, различных стран нлн кон-
тинентов. Исключением является работа У. А. Меррилла (Murrill, 1914),
посвященная Agaricaceae Северной Америки, где в алфавитном порядке
приведен список н очень краткие диагнозы около 80 видов, многие из
которых в настоящее времи ликвидированы (Miller, Farr, 1975).
Современный период изучения агариковых грибов начинается с
30-х годов XX в. В этот период многими учеными (Maire R., 1933; Heim,
1934; Lange X, 1935—1940; Singer, 1936, 1942, 1943, 1951, 1962, 1975;
Konrad, Maublanc, 1948—1952; Piiat, 1951a; Kiihner, Romagnesi, 1953;
Moser, 1953, 1967, 1978, 1983; Dennis, Orton, Hora, 1960; Horak, 1968;
Kreisel, 1969; Ainsworth, 1971; Васильева, 1973; Зерова, Сосш, Рожен-
ко, 1979; Kiihner, 1980, и др.) разрабатывалась естественная филогене-
тическая система порядка Agaricales s. 1.
Изучая различные элементы кутикулы, гименофора, покрывала,
структуры трамы, строение оболочки спор, используя новейшие микро-
скопы и микротомы, эмпирические химические цветовые реакции кар-
пофоров и микроструктур, указанные авторы обнаружили ряд новых так-
сономических признаков, позволяющих критически пересмотреть как от-
дельные роды, семейства, так и весь порядок Agaricales s. 1.
В современный период появился и ряд монографических обработок
некоторых родов семейства Agaricaceae 2 отдельных регионов земного
шара: Lcpiota (Lange J., 1935—1940; Kiihner, 1936; Huijsman, 1943;
Bon, 1981), Agaricus (Moeller, 1950—1951; Piiat, 1951b; Heinemann,
1956a, b), Cystoderma (Smith A., Singer, 1945; Heinemann, Thoen, 1973) \
В последние десятилетия, в связи с применением новых методов и
интенсификацией исследований микофлоры Южного полушария, описа-
но еще 16 новых для науки родов, относимых различными авторами к
семейству Agaricaceae: Pscudobacospora, Smithiomyces, Macrolepiola,
Squamanita, Ripartitella, Cystoagaricus, Cystolepiota, Volvolepiota, Ver-
rucospora, Hiatulopsis, Leucoagaricus, Crucispora, Dissoderma, Scriceo-
myces, Hymenagaricus, Chlorolepiota. Таким образом, к настоящему
’ Отмечаем лишь статьи, на наш взгляд, наиболее важные.
2 Первой специальной обработкой всего семейства Agaricaceae отдельного круп-
ного региона является наша предыдущая работа (Вассер, 1980).
3 Критический анализ отмеченных работ приведен в главе III.
7
времени всего в семейство Agaricaceae в различных классификационных
схемах включали около 45 родов \ из которых 8 монотипных и 3 рода
(Volvolepiota, Cystoagaricus, Melanophyllum) содержат по 2 вида (Sin-
ger, 1975; Moser, 1978, 1983; Вассер, 1980, 1982; Hawksworth, Sutton,
Ainsworth, 1983).
Агариковые грибы еще неравномерно изучены в разных странах и
на разных континентах земного шара. Наиболее полно выявлен видо-
вой состав агариковых грибов большинства стран Европы* 2, Северной
Америки, меньше — Африки и Азии, еще слабее изучены южноамери-
канский и австралийский континенты.
В отношении агариковых грибов Европа 3 (без европейской части
СССР) — одна из наиболее изученных территорий земного шара (Kar-
sten, 1879; Baria, 1888; Bresadola, 1892; Ricken, 1915; Velenovsky, 1920—
1922; Rea, 1922; Konrad, Maublanc, 1924—1937; Bresadola, 1927—1933;
Lange J., 1935—1940; Kiihner, 1936; Schaeffer J., Moeller, 1938; Huijs-
man, 1943; Locquin, 1942—1943, 1945, 1951a, b; Konrad, Maublanc,
1948—1952; Moeller, 1950—1951; Pilat, 1951a, b, 1953, 1955; Kiihner,
Romagnesi, 1953; Hcrink, 1954a, b, 1955, 1956, 1959, 1961, 1962; Melzer,
1954, 1959; Babos, 1958, 1961, 1969, 1970, 1979, 1980; Dennis, Orton,
Hora, I960; Heim, Becker, 1960; Hora, 1960; Wichansky. 1960a, b; Bohus,
1961, 1969, 1971, 1974, 1975, 1976, 1978; Essctte, 1964; Michael, Hennig,
1964—1975; Demoulin, 1966; Reid, 1966, 1967, 1969, 1972; Moser, 1967,
1978, 1983; Horak, 1968; Babosne, 1968; Kotlaba, 1969; Pegler, Young,
1971; Bon, Boiffard, 1972, 1974; Bresinsky, Haas, 1976; Bon, 1976, 1979,
1981; Heinemann, 1977a, 1978, 1981; Knudsen, 1977, 1978, 1980; Cappelli,
1983, и др.). Количество агариковых грибов, известных для Европы,
составляет около 300 видов, разновидностей и форм4. Например, во
флоре Данни, где наиболее полно изучены высшие грибы, семейство
Agaricaceae представлено 109 видами, разновидностями и формами:
Agaricus — 51, Melanophyllum — 2, Cystoderma — 5, Cystolepiota — 5,
Lepiota — 30, Leucoagaricus — 3, Leucocoprinus — 5, iMacrolcpiota — 5,
Phaeolepiota — 1, Pseudobaeospora — 1, Squamanita — 1 (Lange J.,
1935—1940; Moeller, 1950—1951; Knudsen, 1977, 1978, 1980).
Специальных работ по агарнковым грибам Азин нет5. В целом
они в этом регионе изучены слабо; до сегодняшнего дня полностью от-
сутствуют или имеются очень скудные сведения относительно агарнко-
вых грибов ряда стран (КНР, МНР, Индии, Бирмы, Бангладеш, Бутана,
Непала, Ирана, Турции, Афганистана, Саудовской Аравин, Иордании,
Сирии, «Ливана, Кампучии, Лаоса, Таиланда, СРВ и др.).
Отрывочные сведения, почти не дающие представления о флоре
агариковых грибов Азин, исключая азиатскую часть СССР, приводятся
в работах С. Кавамуры (Kawamura, 1930), Т. Хонго (Hongo, 1957,
1959, 1960, 1965, 1970, 1973а, 1974а, 1975), Р. Имазекн и Т. Хонго
’ Мы вводим в Agaricaceae вслед за Г. Крайзелем (Kreisel, 1969, 1973а) и
М. Мозером (Moser, 1978, 1983) роды Endoptychum и Gyropliragmium, относимые
многими авторами к Gasteromycetes.
2 В таких странах Европы, как Норвегия, Швеция, НРБ, СФРЮ, Албания,
СРР, Португалия, Испания и других, агариковые грибы еще очень мало изучены.
3 Для Европы приведены лишь наиболее значительные работы.
4 Абсолютно точное количество таксонов Agaricaceae видового и внутривидового
ранга любого континента без критико-систематического изучения типового материала
аборигенной флоры указать невозможно.
5 Исключение составляет работа Д. Н. Пеглера (Pegler, 1972), посвященная
вцдам рода Lepiota Шри Ланки.
8
(Imazeki. Hongo, 1972), M. Доки и T. Фуйи (Aoki, Fuji, 1972) — Япония,
С. Ц. Тента (Teng, 1932, 1947, 1964), Чиу-Вейфана (Chiu Weifan,
1978) —КНР; А. Пилата (Pilat, 1972) —МНР; П. Саккардо (Saccardo,
1887), Б. Мундкура (Mundkur, 1938), К. Рамакришнана и Ц. В. Суб-
раманиана (Ramakrishnan, Subramanian, 1952), Ц. В. Субраманиана li-
к. Рамакришнана (Subramanian, Ramakrishnan, 1956), Е. Батлера и
Дж. Бисби (Butler, Bisby, 1960), Р. Гоша и Н. Патака (Ghosh, Pathak,
1965), П. Хейнеманна (Heinemann, 1968а), Т. Каула и Дж Кахру (Kaul,
Kachroo, 1974), К. Натараяна (Natarajan, 1977, 1978), К- Натараяна и
Н. Рамана (Natarajan, Raman, 1983), Р. Уотлинга и Н. М. Грегори (Wat-
ling, Gregory, 1980) — Индия; Р. Уотлинга и Я- Свиней (Watling, Swee-
ney, 1974), Ф. Фаллахиана (Fallahyan, 1975), Р. Уотлинга и Н. М. Гре-
гори (Watling, Gregory, 1977) — Иран; М. Онера (Oner, 1972), Ф. Кот-
лабы (Kotlaba, 1976), Р. Уотлинга и Н. М. Грегори (1.с.) —Турция;
С. Ахмада (Ahmad, 1956, 1969) — Пакистан; М. Беркели и Ц. Бруме
(Berkeley, Broome, 1871), Д. Н. Пеглера (Pegler, 1972) —Шри Ланка;
Р. Эйма (Heim, 1962) — Таиланд; И. Рейхерта и 3. Авизохар-Гершензон
(Reichert, Avizohar-Hcrshenzon, 1959), 3. Авизохар-Гершензон (Avizo-
har-Hcrshenzon, 1961, 1964, 1967, 1968), Н. Бнньямини (Binyamini, 1974,
1976а, b, с) — Израиль.
В Азии н на прилегающих островах обнаружено 133 вида, разно-
видности и формы агариковых грибов. В частности, семейство Agarica-
ceae, наиболее полно изученное в Японии, представлено здесь 56 вида-
ми, разновидностями и формами: Chlorophyllum— 1, Leucocoprinus —5,.
Macrolepiota — 4, Lcucoagaricus — 1, Lepiota — 25, Melanophyllum — 1,
Cystoderma — 4, Phaeolepiota—1, Agaricus—18, Squamanita—1. Из
них 16 видов, т. е. приблизительно 30 % флоры, описаны для территории
Японии как новые для науки: Cystoderma ncoamianthinum Hongo, Le-
piota alborubescens Hongo, L. atrosquamulosa Hongo, L. aurantioflava
Hongo, L. cinnamomca Hongo, L. clypeolaria f. umbrinisquamosa Hongo,
L. fusciccps Hongo, L. japonica Kawam. ex Hongo, L. ncglccta Henge,,
L. ncomastoidea Hongo, L. subcitrophylla Hongo, Leucocoprinus otsuen-
sis Hongo, L. denudatus f. major Hongo, Agaricus jezoensis Imai, A. com-
ptulellus Imai, A. semotellus Imai.
На территории Северной Америки агариковые грибы изучали в
США (Peck, 1909; Murrill, 1914, 1949; Shantz, Picmeiscl, 1917; Kauff-
man, 1918, 1924; Coker, 1928; Krieger, 1936; Hosier, 1937, 1943, 1955;
Hotson, Stuntz, 1938; Smith A., 1940, 1949, 1963; Smith H., 1954; Smith A.,
Singer, 1945, 1948; Kobayasi et al., 1967; Miller, 1972; Miller, Farr, 1975;
Freeman, 1979a, b, и др.), Канаде (Murrill, 1914; Pomerleau, 1951; Gro-
ves, 1962; Milter, 1972; Miller, Farr, 1975; Freeman, 1979a, b), Мексике
(Herrera, Guzman, 1961; Guzman, 1963, 1970, 1972, 1973, 1975, 1977;
Guzman, Garcia Sauccdo, 1973; Guzman, Johnson, 1974). Сведения об-
агариковых грибах островов, прилегающих к Североамериканскому кон-
тиненту, имеются в работах М. Беркелн и М. Куртис (Berkeley, Curtis,
1868), Р. В. Г. Денниса (Dennis, 1952, 1953), Г. Крайзеля (KreiseR
1971а, Ь), посвященных высшим базидиомнцетам Кубы; в работах
Р. В. Г. Денниса (Dennis, 1968), П. Хейиеманна (Heinemann, 1962а)
по грибам Ямайки, в работах М. Ланге (Lange М., 1955, 1957), Е. Ко-
баяши (Kobayasi, 1971), Р. Уотлинга (Watling, 1977), изучавших мико-
флору Гренландии. В результате количество агариковых грибов северо-
американского континента и прилегающих островов представлено
приблизительно 268 видами, разновидностями и формами, из которых
свыше 50 % описано с этого континента, нз них 79 новых для науки
видов рода Agaricus— A. alabamensis Murr., A. alachuanus Murr., А. ап-
drewii Freeman, A. Cervinifolius Zeller, A. chlamydopus Pk, A. earlei
Murr., A. floridanus Pk, A. kauffmanii A. H. Sm., A. nobelianus Freeman,
A. pusillus Pk и др. (Freemann, 1979a, b). Часть из видов обнаружены
и на других континентах земного шара (например, Agaricus placomyces
Pk, A. tabularis Pk н др.).
Значительно слабее изучен видовой состав грибов семейства Agari-
caceae в Южной Америке. В частности, для Аргентины (Spegazzini.
1898; Martinez,. 1949, 1957; Singer, Digilio, 1952; Singer, 1959, 1963,
1969; Heinemann, 1962b), Венесуэлы (Dennis, 1951, 1961, 1970; Heine-
mann, 1962c; Singer, 1969), Боливии (Heinemann, 1962d), Бразилии
(Singer, 1955, 1965), Чили (Spegazzini, 1921; Singer, 1969; Lazo, 1972),
Эквадора (Patouillard, Delagerhcim, 1895; Singer, 1975, 1978), Перу
(Garcia Rada у Stevenson, 1942; Pavlich, 1976) известно 79 видов, раз-
новидностей и форм. Сведения об агариковых грибах, главным образом
из рода Agaricus о-ва Тринидад, имеются в работах Р. В. Г. Денниса
(Dennis, 1951, 1952) н П. Хейнеманна (Heinemann, 1961). Однако для
ряда стран южноамериканского континента сведений иет (например,
Гайаны, Колумбии, Парагвая, Уругвая н др-). Для микофлоры Перу
приводится всего лишь пять видов агариковых грибов; 35 % видов
агариковых грибов Южной Америки описаны с ее территории как
новые для науки, например Leucocoprinus venezuelanus Dennis, Cysto-
agaricus jujuyensis Sing., Ripartitella brasiliensis (Speg.) Sing, н др.
Для Австралии, Новой Зеландии (Massee, 1898; Cleland, 1924, 1934;
Cleland, Cheel, 1918, 1923; Cleland, Harris, 1948; Heim, 1950; Steven-
son, 1961, 1962; Aberdeen, 1962; Willis, 1963; Pegler, 1965; Shepherd,
1969; Singer, 1969; Taylor, 1970; Horak, 1971a, b, 1979, 1980) и неко-
торых островов Океании — Папуа-Новая Гвинея, Соломоновы о-ва,
Гавайские о-ва (Burt, 1923; Hongo, 1973b, 1974b, 1976a, b, 1977, 1978;
Kobayasi, Otani, Hongo, 1973; Horak, Kobayasi, 1978; Horak, 1979;
Heinemann, 1980) приводится 65 видов, разновидностей и форм из
родов Agaricus, Lepiota s.l., Cystoderma, многие из которых описаны
как новые для науки, являющиеся эндемами указанных территорий.
Например, из 244 видов порядка Agaricales Папуа-Новой Гвинеи, се-
мейство Agaricaceae представлено 7 видами: Agaricus aurantioviolacea
Heim, A. strigopus Gaudichand, A. rowakensis Gaudichand, A. subruti-
lescens (Kauffm.) Hotson et Stuntz, Lepiota atrosquamulosa Hongo,
Leucocoprinus cepaestipes, Melanophyllum echinatum, из которых 3 ви-
да— эидемы (Horak, Kobayasi, 1978).
Сравнительно хорошо изучен видовой состав агариковых грибов
некоторых стран Африки. Интенсивные исследования видового состава
порядка Agaricales s. I., проведенные в ряде стран Африки, послужили
базисом для создания флоры Марокко, Восточной Африки. Однако
на этом континенте есть еще много «белых питен». Полностью отсут-
ствуют сведения об Agaricaceae Мавритании, Мали, Сенегала, Каме-
руна, Бенина, Судана, Ливии, Габона, Анголы, Чада, Замбии н др.
В Африке флору агариковых грибов при планомерном исследова-
нии порядка Agaricales изучали в Марокко, Алжире (Heinemann, 1965;
MalenQOn, Bertault, 1970), Заире (Beeli, 1932; Heinemann, 1956a, b,
1968b, 1969, 1973, 1977b, c; Singer, Grinling, 1967), Кении (Heinemann,
1965; Pcgicr, Rayner, 1969; Pegler, 1977), Танзании (Pegler, 1977),
Уганде (Pegler, 1977). Отрывочные сведения об Agaricaceae Египта,
40
Нигерии, Ганы, Сьерра-Леоие. Центральноафриканской Республики
имеются лишь в отдельных публикациях (Pearson, 1950; Pegler, 1966.
1968; Thoen, 1969; Reid, 1975).
Из многочисленных островов, прилегающих к Африке, лишь для
о. Занзибар описаны агариковые грибы (Pegler, 1975, 1977).
На этом континенте в настоящее время известно около 189 видов,
разновидностей и форм Agaricaceae. Например, во флоре Восточной
Африки (Уганда, Кения, Танзания, о. Занзибар) семейство представле-
но 80 видами, разновидностями и формами: Chlorophyllum — 1, Масго-
lepiota — 5, Leucoagaricus — 3, Leucocoprinus — 6, Agaricus — 18, Mic-
ropsalliota — 2, Lepiota —43, Cystoderma — 1, Verrucospora — 1, из ко-
торых 17 описаны как новые для науки: Leucocoprinus africanus Heim,
Leucoagaricus fuligineus Pegler, Lepiota vascularis Pegler, Agaricus
nigrovinosus Pegler, A. kigumuensis Pegler и др. (Pegler, 1975, 1977).
Во флоре Марокко Agaricaceae представлено 72 таксонами видо-
вого и внутривидового рангов: Cystoderma—4, Lepiota s. 1.— 38, Aga-
ricus— 30 (Malcn^on, Bertault, 1970). Интересно отметить, что из 40 ви-
дов рода Agaricus Заира (Heinemann. 1956а, Ь) 29 описаны как новые
для науки: Agaricus campestroides Heinem. et Gooss., A. subedulis
Heinem., A. volvatulus Heinem. et Gooss., A. kivuensis Heinem. et
Gooss. и др.
В Антарктиде обнаружено два вида агариковых грибов — Agaricus
campestris и Cystoderma amianthina (Singer, Corte, 1962; Bunt, 1965;
Singer, 1975; Pegler, Spooner, Smith L., 1980).
2. ИЗУЧЕНИЕ АГАРИКОВЫХ ГРИБОВ В СССР
Вся история изучения агариковых грибов, как и шляпочных вооб-
ще, в нашей стране подразделена по Б. П. Василькову (1953а) иа
четыре периода: 1) начальный — с древнейших времен до второй чет-
верти XVIII в.; 2) флористический — со второй четверти XVIII в. до
середины XIX в.; 3) онтогенетический — с середины XIX в. до 20-х го-
дов XX в.; 4) современный — с 20-х годов XX в. до настоящего време-
ни 1. Все имеющиеся в литературе до 1953 г. сведения о шляпочных,
в том числе н агариковых, грибах приведены в указанной работе
Б. П. Василькова (1с.), посвященной изучению шляпочных грибов в
СССР и содержащей исчерпывающий список литературы (1300 источ-
ников).
Планомерный, целенаправленный характер микологических иссле-
дований в СССР наблюдается в современный период и особенно во
второй его половине. Интенсивные исследования грибов в различных
республиках СССР начались в 50-е годы. Этому способствовали следую-
щие обстоятельства: а) стала очевидной ошибочность отношения к си-
стематике вообще как к старомодной описательной науке, доживающей
свои дни, занятой лишь простой каталогизацией фактов, место которой
в музее древностей; б) развитие агарикологии стимулировалось требо-
ваниями практики, так как получило широкое признание значение ага-
риковых грибов для народного хозяйства; в) выход в свет «Определи-
теля шляпочных грибов» Л. А. Лебедевой (1949) 2 и первого издания
1 Характеристика, особенности периодов изучения шляпочных грибов в нашей
стране рассмотрены в работе Б. П. Василькова (1. с.).
2 Критический анализ работы Л. А. Лебедевой (1949) проведен Б. П. Васвль-
ковым (1952)-
11
монографии Р. Зингера (Singer, 1951) «Современная таксономия Aga-
ricalcs».
Поскольку видовой состав агариковых грибов и СССР до послед-
него времени оставался недостаточно изученным, основным направле-
нием исследований на всех этапах стало флористическое. Флористиче-
ские исследования агариковых грибов Советского Союза специально не
проводились. Однако прн изучении флоры высших Basidiomycctes тех
илн иных регионов страны авторы касались видового состава и изучае-
мого нами семейства. Наиболее полно флора грибов этого семейства
исследована в европейской части СССР (149 видов и внутривидовых
таксонов), на Дальнем Востоке (72), Кавказе (56) и в Восточной
Сибири (47). Слабо изучены агариковые грибы Западной Сибири (42),
Средней Азии (32) и Арктики (11). Даже в европейской части СССР
ие все регионы одинаково хорошо изучены. Очень слабо изучены ага-
рнковые грибы МССР, европейской части РСФСР. Наиболее полно фло-
ра агариковых грибов исследована на Украине (Зерова, Cocih, Рожен-
ко, 1979; Вассер, 1980, 1982), в Прибалтийских республиках (Урбонас,
Каламээс, Лукин, 1974; Каламээс, 1975; Kalamees, 1978а), на Кавказе
(Садыхов, 1967, 1968, 1977; Нахуцришвили, 1975; Мелик-Хачатрян,
1980), в южной части Красноярского края (Беглянова, 1972, 1975),
в Приморском крае (Васильева, Назарова, 1967; Васильева, 1967, 1973;
Булах, 1977а, б).
Для флоры Украины приведено 107 видов агариковых грибов.
Крупнейшими родами во флоре Украины являются Agaricus, Lepiota,
Cystoderma, Leucocoprinus, Macrolepiota, Leucoagaricus (Зерова, Cocin,
Роженко, 1979; Вассер, 1980, 1982). Для каждого вида агариковых гри-
бов Украины имеются данные по синонимике, иконографии, морфолого-
анатомическим, географическим, экологическим особенностям, система-
тике. Ряд видов (Agaricus longicaudus, A. amanitaeformis, Leucoagari-
cus moseri, Leucocoprinus bohusi) приведены как новые для науки,
22 вида — как новые для СССР. Наиболее богатый видовой состав Aga-
ricaceae Украины отмечен для ее степной зоны.
Значительный вклад в изучение Agaricaceae Прибалтийских респуб-
лик СССР внесли В. А. Урбонас, К- А. Каламээс, В. Лукин (1974),
К- А. Каламээс (1975, 1978а). Для флоры указанного региона СССР
приведено 60 видовых и внутривидовых таксонов агариковых грибов,
в том числе Melanophyllum eyrei var. macrosporum — новый для науки,
Leucocoprinus bimbaumii, Macrolepiota rliacodes var. hortensis, Lepiota
cortinarius — редкие для флоры СССР. Для каждого вида даны синони-
мика, субстрат, продолжительность вегетационною периода, степень
встречаемости, распространение, хозяйственное значение.
Итоговые сведения об Agaricaceae Кавказа представлены в рабо-
тах И. Г. Нахуцришвили (1975) и Дж. Г. Мелик-Хачатрян (1980).
Первый автор приводит для Грузии 27 видов, из них ряд редких и
малоизвестных, таких, как Agaricus tabularis, Л. placomyces var. grisea,
Lepiota rufovelutina. Второй автор отмечает для Армении 26 видов,
в частности Leucoagaricus macrorhizus — новый вид для микофлоры
СССР, a Lepiota tomentella, Agaricus tabularis — редкие для микофло-
ры СССР.
Видовому составу, экологии и распространению шляпочных грибов,
в том числе и агариковых, южиой части Красноярского края посвящены
работы М. И. Бегляновой (1963, 1972, 1975), Э. Л. Нездойминого
(1980), в которых приводятся сведения о 42 видах родов Macrolepiota*
12
Leucocoprinus, Agaricus, Lepiota, Cystoderma. При этом отмечен ряд
новых и редких для флоры СССР видов (Cystoderma superbum, С. am-
brosii, Lepiota hapalopoda, Agaricus fuscofibnllosus).
Интенсивные флористические исследования агариковых при изуче-
нии шляпочных грибов проводились в Приморском крае Л. II. Василье-
вой (1967, 1973) и ее учениками. Они описывают для Приморского края
59 видов Agaricaceae, в том числе Lepiota lateritiopurpurea — новый
вид для науки, Pseudobaeospora defibulata, Lepiota vircscens, L. neopha-
na, L. rhodorhiza, L. lignicola и др.— новые для микофлоры СССР.
Кроме этих основных работ, имеются разрозненные сведения о не-
которых видах агариковых грибов, обнаруженных в отдельных регионах
Советского Союза (Зингер, 1938; Головни, 1941; Шварцман, 1948; Ле-
бедева, 1949; Христюк. 1954, 1960; Васильков. 1955, 1974; Зерова, 1956,
1959, 1962, 1970а, б; Ганжа, 1960а, б, в, 1970; Добровольский, Сосни,
1960; Гапоненко, 1965; Сержанина, 1967, 1968; Эльчибаев, 1968, 1969;
Садыхов, 1968, 1972, 1977; Частухнн, Николаевская, 1969; Нездоймино-
го, 1968, 1970, 1971, 1973; Томилин, 1969, 1971; Перова, 1970, 1972,
1973, 1977; Harma j а, 1974; Батырова, 1975, 1977а, б; Михайловский,
1975; Назарова, 1976; Степанова, Сирко, 1977; Петренко, 1978; Беден-
ко, 1979, 1980; Бурова, Нездойминого, 1980, и др.).
Неразрывно с флористическими исследованиями агариковых гри-
бов развивалось в нашей стране исследование их морфологии и систе-
матики (Васильков, 19536, 1974; Гарибова, Мокеева, 1974а, б, 1976;
Вассер, Гарибова, Мокеева, 1976; Мокеева, 1977; Мокеева, Райтвийр,
Гарибова, 1977). Только наше специальное исследование (Вассер,
1980) посвящено систематике, морфологии, экологии, географии всего
семейства Agaricaceae Украины, а исследования ряда авторов (Гари-
бова, 1970; Гарибова, Сафрай, 1972; Гарибова, Шалашова, 1973; Вас-
сер, 1973а; Гарибова, Мокеева, 1974а, б, 1976; Гарибова, Сафрай, Ша-
лашова, 1974; Мокеева, Гарибова, 1975; Вассер, Гарибова, Мокеева,
1976; Wasser, Garibova, Mokeeva, 1976; Мокеева, Райтвийр, Гарибова,
1977) — роду Agaricus. Следовательно, комплексному изучению (за
исключением наших исследований), которое осуществила Л. В. Гари-
бова с сотрудниками, в последнее десятилетие в СССР подвергался
частично лишь род Agaricus.
Вопросам географии агариковых грибов в СССР уделялось недо-
статочное внимание. Напомним об одном нашем исследовании, посвя-
щенном географическому анализу всего семейства Agaricaceae Украи-
ны (Вассер, 1980), и исследовании В. Л. Мокеевой (1977) особенно;
стей географического распространения видов рода Agaricus.
С 30-х годов XX в. в нашей стране как объект промышленного
культивирования из всего семейства Agaricaceae используется лишь
шампиньон двуспоровый (Agaricus bisporus). Основные направления
его исследований в СССР заключались в селекции отечественных высо-
копродуктивных, болезнеустойчивых штаммов (Институт ботаники
им. Н. Г. Холодного АН УССР, МГУ им. М. В. Ломоносова); разработ-
ке технологии производства посевного мицелия с учетом биологии штам-
ма и подборе эффективных питательных сред для оптимального роста
посевного мицелия, при этом параллельно изучалась физиология пита-
ния штаммов (совхоз-комбинат по промышленному выращиванию шам-
пиньонов, г. Тбилиси; Кишиневский сельскохозяйственный институт;
совхоз «Заречье», г. Москва; фирма «Лето», г. Ленинград; Институт бота-
ники им. Н. Г. Холодного АН УССР), поиске нестандартных компонен-
13
тов для компостов, механизации процесса выращивания шампиньона
двуспорового (НИИ овощного хозяйства, г. Мытищи Московской обл.);
изучении культуральных и биохимических особенностей штаммов шам-
пиньона двуспорового (МГУ им. М. В. Ломоносова, Институт ботаники
нм. Н. Г. Холодного АН УССР).
Итоги всесторонних исследований биологии, экологии, селекции,
производства и храиеиня посевного мицелия, культивирования в про-
мышленном масштабе (способы изготовления субстратов, системы вы-
ращивания в различных типах культивационных помещений), мер борь-
бы с болезиям-и и вредителями, сбора, хранения, транспортировки и
переработки урожая шампиньона двуспорового приведены в наших кол-
лективных монографиях (Дудка, Вассер, Бухало и др., 1978; Бисько,
Бухало, Вассер и др., 1983).
В последнее десятилетие М. Я. Зеровой (1974а) и С. П. Вассером
(Wasser, 1979; Вассер, 1980) уделено значительное внимание графиче-
скому изображению агариковых грибов.
ГЛАВА II
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
1. МАТЕРИАЛЫ ИССЛЕДОВАНИЯ
Границы района исследования совпадают с государственными гра-
ницами СССР. Названия районов флоры, принятые для флор споро-
вых растений СССР (рис. I), приводятся по Т. Л. Николаевой (1961).
Основным материалом исследования являлись агариковые грибы,
собранные в различных ботанико-географических районах Украины и
ряде других районов Советского Союза, в первую очередь в Латвий-
ской ССР, на Кавказе, в Западной (Алтайский край) и Восточной
(Красноярский край, Бурятская АССР) Сибири, на Дальнем Востоке
(Приморский край, п-ов Камчатка, Курильские о-ва) в 1969—1982 гг.
В процессе работы было исследовано 3520 пакетов агариковых гри-
бов, собранных нами, хранящихся в гербарии Института ботаники
нм. Н. Г. Холодного АН УССР (KW), а также полученных из гербариев
Советского Союза и зарубежных стран.
При критико-систематическом анализе трудных, сомнительных и
малоизвестных видов были изучены типы большинства видов родов
Agaricus, Lepiota, Leucoagaricus, Leucocoprinus. Во всех гербариях
СССР и некоторых гербариях зарубежных стран (ВНР, ЧССР) мы
имели возможность работать лично. Приведенный ниже список содер-
жит перечень учреждений, гербарии которых предоставили материалы
для исследований. Названия гербариев и их сокращения даны в соот-
ветствии с указателем важнейших отечественных и зарубежных герба-
риев, приведенных в работе А. К. Скворцова (1977). Для гербариев,
отсутствующих в упомянутом пособии, приняты произвольные сокра-
щения.
Ботанический институт им. В. Л. Комарова АН СССР, г. Ленинград (LE);
Институт ботаники АН Грузинской ССР, г. Тбилиси (ТВ); Институт ботаники АН
Азербайджанской ССР, г. Баку (ВАК); Институт ботаники АН Литовской ССР,
г. Вильнюс (W1); Институт зоологии и ботаники АН Эстонской ССР, г Тарту
(ТАА); Институт ботаники АН Туркменской ССР, г. Ашхабад (ASH); Институт
ботаники АН Казахской ССР, г. Алма-Ата (АА); Биолого-почвенный институт Даль-
невосточного научного центра СО АН СССР, г. Владивосток (WLA); Московский
государственный университет им. М. В. Ломоносова (MW); Красноярский педагогиче-
ский институт (KRASN); Красноярский НИИ леса и древесины им. В. Н. Сукачева
(KRAS); Ереванский государственный университет (EREV); Полтавский педагоги-
ческий институт (POLT); Herbarium Musei Historiae Naturalis, -Termeszettudomanyi
Muzeum, Novenytar, Budapest (BP) — ВНР; Herbarium Kryptogamologicum Musei Na-
tionalis Pragae, Narodni Museum v Praze, Praha (PR); Herbarium J. Herink, Mnichovo
Hfadiste (H)—ЧССР; Herbarium Universiiy of Michigan, North University Building,
Ann Arbor (MICH), Mycological Herbarium of Lousiana State University, Lousiana
(LOUS)—США; National Mycological Herbarium, Department of Agriculture, Cent-
ral Experimental Farm, Ottawa (DAOM) — Канада; Herbarium Royal Botanic Gardens,
15
Рис. i. Районы флоры СССР, принятые для флор споровых растений (по Ни-
колаевой, 1961):
1-Арктика. II—европейская часть СССР, III — Кавказ. IV —Западная Сибирь. V —
Восточная Сибирь. VI — Дальний Восток. VII — Средняя Азия
Kew (К) —Англия; Herbarium Museum Botanicum Hauniense, Kebenhavn (C) —Дания;
Herbarium Lugd., Botav. Rijksherbarium, Leiden (L) — Нидерланды; Herbarium Insti-
tutionen for Systematisk Botanik, Uppsala (UPS) — Швеция; Herbarium Laboratoire
de Cryptogamic Museum National d’Histore Nature! la, Paris (PC) — Франция.
2. МЕТОДЫ ИССЛЕДОВАНИЯ
СБОР И ОБРАБОТКА МАТЕРИАЛА
Методы сбора и обработки, касающиеся всех высших Basidiomyce-
tcs, детально обсуждаются в ряде специальных руководств и статей
(Бондарцев, Зингер, 1950; Bohns, 1963; Watling, 1973; Hawksworth,
1974). В данной работе кратко изложены лишь основные методы, при-
менявшиеся при обработке агариковых грибов.
Для точного определения агариковых грибов важное значение
имеют правильный сбор их в природе, сушка, препарирование, запись
некоторых необходимых сведений о свойствах в свежем состоянии.
Каждый вид гриба собирали в разные фазы развития, сильно варьи-
рующие виды (Agaricus campestris, A. xanthodermus) — во Ьсем много-
образии форм.
При описании собранного материала были использованы специаль-
ные анкеты-описания (Зерова, 1963; Зерова, Вассер, 1972), в которых
отмечались характерные диагностические признаки свежих карпофоров
большого числа экземпляров разных стадий развития. Весь собранный
материал обрабатывали по методике А. С. Бондарцева и Р. А. Зингера
(1950), заключающейся в обезвоживании агариковых грибов таким
образом, чтобы в легко портящихся клетках ис начался автолиз. Одна-
ко некоторые виды Agaricaceae, высушенные на воздухе либо над элек-
троплиткой, иногда сильно деформируются, поэтому не могут быть
использованы в научных целях.
16
Форма карпофоров, окраска, консистенция мякоти и другие свой-
ства Agaricaceae лучше сохраняются специальным препарированием
(Bohns, 1963). Данная методика препарирования агариковых грибов
подробно описана нами ранее (Вассер, 1980).
Перед препарированием для большинства видов получали споро-
вый порошок помещением срезаииой шляпки гимением вниз на лист
бумаги либо по способу Д. Б. О. Сэвайла (Savile, 1962). Д. Б. О. Сэ-
вайл предложил способ, при котором одно-
временно получают споровый порошок и
удобно размещают плодовые тела для суш-
ки. Для этого довольно плотный лист бе-
лой бумаги (не слишком проклеенный) в
трн-четыре раза шире, чем шляпка гриба,
складывают вдоль втрое; затем в средней
полосе листа карандашом протыкают одну-
две дырки и в них продевают иожки гри-
бов, затем бумагу складывают, как пока-
зано на рис. 2, и, повернув края, ставят на
несколько часов (удобно иа ночь) в про-
хладном влажном месте (Скворцов, 1977;
Вассер, 1980). Споры затем собирают в от-
дельный маленький пакетнк.
Окраску плодовых тел и спор опреде-
ляли по цветовым картам Р. Райнера (Ray-
ner, 1970) и М. Мозера (Moser, 1967, 1978).
Фамилии авторов таксонов всех ран-
гов агариковых грибов сокращены в соот-
ветствии с рекомендованным списком, по-
мещенным в «Индексе болезней растений Соединенных Штатов» (Index
of plant diseases in the United States. 1960), c
дополнениями (Вассер, 1980. 1982).
Рис. 2. Размещение агарико-
вых грибов для сушки и полу-
чения спорового порошка по
способу Д. Б. О. Сэвайла (Sa-
vile, 1962)
некоторыми нашими
ПОЛУЧЕНИЕ ЧИСТЫХ МИЦЕЛИАЛЬНЫХ КУЛЬТУР
Исследовались культуры агариковых грибов, выделенные нами из
плодовых тел и хранящиеся в настоящее время в коллекции живых
культур высших Basidiomycetcs отдела микологии и бриологии Инсти-
тута ботаники им. Н. Г. Холодного АН УССР. Часть культур, имеющих-
ся в коллекции, была получена в порядке обмена из отдела низших
растений Ботанического института им. В. Л. Комарова АН СССР
(г. Ленинград), отдела типовых культур Института микробиологии АН
СССР (г. Москва), отдела экспериментальной микологии Микробиоло-
гического института ЧС АН (г. Прага), отдела флоры низших растений
Естественно-исторического музея ВНР (г. Будапешт). Ряд штаммов
шампиньона двуспорового был получен из лаборатории посевного ми-
целия совхоза «Заречье» (г. Москва). Ниже приводим список видов и
штаммов агариковых грибов, изученных в культуре: Agaricus campest-
ris, ИБК 11-251 *; A. silvaticus, ИБ1< 1-37; A. arvensis, ИБК П-201;
A. arvensis, ИБК 1-14; A. bitorquis, ИБК П-221; A. bitorquis, ИБК 1-143;
A. bisporus, ИБК П-21; A. bisporus, ИБК П-31; A. bisporus, ИБК II-19;
1 Нумерация штаммов по музею чистых культур Института ботаники им. Н Г. Хо-
лодного АН УССР.
2 4—29Ь0
17
I fill Бл’иОТЕКл''
rt( ’Ч1-ЗП<ОГD ГО'УТаГГТВЗНИОГв
' ув. и^снг«та
I Н.П.
A. bisporus, ИБК 11-18; A. bisporus, ИБК 11-17; A. bisporus, ИБК 11-22;
A. bernardii, ИБК 11-240; A. bernardiiformis, ИБК 11-241; A. cupreobrun-
neus, ИБК 11-270; A. maskae, ИБК 11-280; Leucocoprinus bresadolae,
ИБК П-402; Macrolepiota procera, ИБК 1-170; M. rhacodes, ИБК 1-250;
M. puellaris, ИБК 1-257.
Для получения чистых мицелиальных культур мы использовали
общепринятые методы, описанные в литературе (Бухало, 1966, 1973;
Дудка, Вассер, Бухало и др., 1978; Sasek, 1978). Плодовые тела агари-
ковых грибов собирали обычно в период их массового появления. Вы-
деление в культуру проводили в день сбора или же плодовые тела хра-
нили в течение одного—трех дней в холодильнике в полиэтиленовых
мешочках. Для выделения выбирали хорошо развитые, крепкие, ие ин-
фицированные плодовые тела. Их очищали от растительных остатков,
грязи, промывали в проточной воде и затем подсушивали на фильтроваль-
ной бумаге. Обсохшее плодовое тело обтирали 96°-ным этиловым спир-
том. После такой обработки плодовое тело разламывали иад горелкой
и кусочек ткани из середины переносили стерильным скальпелем, лан-
цетом или копьем на питательную среду (сусловый агар) в чашки
Петри. При выборе участка ткани мы исходили из размеров и строения
плодового тела. Выделение идет успешно, если брать крупные кусочки
тканей (0,5—1,5 см в диаметре). Чашкн Петри с ииокулюмом помеща-
ли в термостат при 24—26 °C. В жаркое время года чашки с инокулю-
мом на одии-два дня помещали в холодильник при 5—10 °C, а затем
переносили в термостат. Чтобы относительная влажность воздуха была
ие ниже 80 %, в термостат ставили сосуд с водой. В условиях экспеди-
ции тканевую культуру помещали сразу в пробирку. Кусочки ткани
плодового тела помещали сверху на сусловый агар или частично погру-
жали в агаризоваиную среду. Для наших исследований были использо-
ваны дикариотические культуры, полученные споровым и тканевым ме-
тодами. Результаты для одних и тех же видов, полученных как споро-
вым, так и тканевым методами (Agaricus cupreobrunneus, A. maskae,
Leucocoprinus bresadolae), были идентичны. Все использованные в дай-
ной работе штаммы для изучения культуральных особенностей получе-
ны тканевым способом.
ЛАБОРАТОРНОЕ КУЛЬТИВИРОВАНИЕ
Дл*я лабораторного выращивания агариковых грибов с целью из-
учения их культуральных признаков, динамики роста и размеров ко-
лоний мы использовали различные питательные среды.
Из числа сред неопределенного химического состава широко при-
менялся 2- и 8°-иый (по Баллингу) сусловый агар (СА2 и САв). Из
числа стандартизированных синтетических питательных сред нами для
выращивания агариковых грибов использовались следующие: агар Ча-
пека (глюкоза — 30 г, NaNO3 — 2, КН2РО4— 1, MgSO4 - 7Н2О— 0,5,
КС! — 0,5, FeSO4 —0,01, агар-агар — 15- 20 г, дистиллированная во-
да— 1 л), Молиша (крахмал—5,0 г, КН2РО4—1, NH4C1 — 0,5, СаС12 —
0,1, NaCl2 — 0,1, MgSO4 — 0,3, FeCl3 — 0,01, агар-агар—20 г, дистил-
лированная вода — 1 л), Деггара (глюкоза—20 г, пептон—10,
MgSO4 — 2,5, КН2РО4 — 5, СаС12 — 0,25, агар-агар — 15—20 г, дистил-
лированная вода — 1 л). Повторность опытов — пятикратная.
Культуры агариковых грибов выращивали при температуре 18—
20 °C. Исследования влияния температуры (в интервале от 18 до 35 °C)
18
на рост агариковых грибов в культуре иа элективных питательных сре-
дах показали, что оптимум лежит в пределах 26—27 °C. Наблюдения
за ростом мицелия проводили на 7-, 16- и 36-е сутки после посева.
Для решения вопросов о видовой таксономии агариковых грибов
мы изучали ие только их культурально-морфологические особенности,
ио и характер поведения в смешанной культуре при выращивании их
попарно в одной чашке Петри на 2- и 8°-ном (по Баллиигу) сусловом
агаре. Опыты проводили в трех повторностях. Характер поведения ага-
рнковых грибов в смешанной культуре изучали по методике, использо-
ванной Т. А. Давыдкиной (1980) при решении вопросов видовой таксо-
номии стереумовых грибов Советского Союза.
ХРАНЕНИЕ МУЗЕЙНЫХ КУЛЬТУР
Музейные культуры агариковых грибов поддерживались методами
пересевов иа сусловом агаре один раз в 9—10 месяцев (pH — 6,0; 2- и
8°-ном по Баллингу). Культуры росли в больших пробирках с доста-
точным количеством суслового агара (20 см3), чтобы избежать высы-
хания. Культуры сохранялись в холодильнике при температуре 4—10 °C.
При пересеве культур проводился их микробиологический контроль.
ОБРАБОТКА МАТЕРИАЛА ДЛЯ ИЗУЧЕНИЯ С ПОМОЩЬЮ
СКАНИРУЮЩЕГО ЭЛЕКТРОННОГО МИКРОСКОПА (СЭМ)1
Изучение морфологии базидиоспор агариковых грибов с помощью
сканирующего электронного микроскопа позволило получить объемное
изображение спор и отдельных элементов споровой поверхности при
достаточно высоком разрешении. При исследовании спор главное внима-
ние обращалось на общую конфигурацию споры, орнаментацию наруж-
ной оболочки, наличие или отсутствие поры прорастания и супраапику-
лярной депрессии, морфологию стеригм и базидий. Полученные данные
в совокупности с другими признаками использовались при критической
обработке агариковых грибов.
Подготовку объектов для сканирующей микроскопии в отсутствие
прибора для высушивания при критической точке осуществляли перво-
начально в различных вариантах: были опробованы возможности нане-
сения спор, суспендированных в воде и спирте, сухих спор на металли-
ческие объектодержатели и смачивание их спиртом или водой, иа
объектодержатели, предварительно покрытые электропроводящим кле-
ем (чаще всего серебряным). Использовалась фиксация спор в парах
осмиевой кислоты или в растворах различной концентрации. .Для выяс-
нения степени влияния на морфологию спор различных растворов, есте-
ственного и искусственного высушивания, глубокого вакуума исследо-
вали споры, взятые из свежесобранных образцов, а также споры, ие
подвергнутые какой-либо обработке.
Наиболее оптимальными условиями подготовки базидиоспор ока-
залось нанесение спор гербарных образцов иа объектодержатели, по-
крытые серебряным клеем. Степень подсыхания клея перед нанесением
спор определяли эмпирически в зависимости от консистенции клея и
толщины слоя. После полного высыхания клея объекты напылялись
золотом в вакуумном испарителе JEE-4X при условии его наклона н
Написано О. А. Закордонцем.
2*
19
вращения во время напыления. Количество распыляемого металла для
получения слоя заданной толщины определяли по формуле
гдеТИ (г) — количество испаряемого металла;
г (см) — расстояние от испарителя до объекта (выдерживалось в пре-
делах 15 см);
р (г/см3) — плотность испаряемого металла (для золота р = 19,3 г/см3);
d (см)—толщина напыляемого слоя (с расчетом в пределах 20—
30 им);
0 (град.) — угол между плоскостью образца и нормалью траектории
распыляемых частиц (прн условии наклона н вращения = 0°).
При этих условиях расход золота составлял в среднем 0,4 г, а по-
крытие обеспечивало хорошую электропроводимость изучаемых объек-
тов. В дальнейшем напыление объектов золотом осуществляли в при-
боре для ионного испарения Fine-Coat JFC-1100. Напыление в течение
5—6 мин при напряжении в 1,2 кВ и разряжении в 7 мА обеспечивало
равномерное и достаточно плотное покрытие объекта золотом.
Напыленные образцы просматривали и фотографировали с по-
мощью сканирующего электронного микроскопа японского производ-
ства JSM-35C при увеличении 2—10 тыс. и ускоряющем напряжении
15 кВ.
Использованная методика давала удовлетворительные результаты
по сохранности морфологических особенностей спор; не было отмечено
существенной разницы в морфологии базидиоспор изученных гербарных
образцов и при исследовании спор некоторых видов из свежесобраниых
образцов, в том числе и при исследовании спор, не напыленных золотом.
В последнем случае из-за слабой электропроводности осуществить фо-
тографирование не представлялось возможным, но просмотр препаратов
позволял изучить морфологию спор в условиях, исключающих влияние
глубокого вакуума и атмосферного давления, которые имеют место при
напылении спор в вакуумном испарителе. Однозначность получаемых
результатов служила дополнительным подтверждением допустимости
исследования спор без сложной процедуры подготовки объектов. При
исследовании спор из свежесобранных образцов отмечалось значительно
меньше различных вмятин, сморщиваний, чем при исследовании спор
из гербарных образцов. Наличие отмеченных вмятин, сморщиваний
объясняется коллапсироваиием базидиоспор, возникающим в результа-
те дегидратации образцов в процессе естественного высушивания прн
хранении гербарных материалов (Pegler, Young, 1974).
Исследование свежесобраиных образцов затруднено сезонностью,
транспортировкой объектов, их сохранностью. В силу этих обстоятельств,
а также с учетом поставленной цели — изучение морфологии базидио-
спор — основные исследования агариковых грибов были выполнены с
использованием гербарного материала.
Особо следует остановиться на наличии или отсутствии супраапи-
кулярной депрессии у базидиоспор и, главным образом, иа ее величине
и форме. Осторожность суждения в этом вопросе не лишняя, так как
трудно проконтролировать степень тургориого состояния объекта даже
при использовании метода высушивания при критической точке (Bige-
low, Rowley, 1968; Ncmanic, 1972; Pegler, Young, 1974). Немаловажное
значение имеют родовые различия в устойчивости спор к различным
воздействиям, «возраст» спор, условия высушивания и хранения
гербарных образцов; например, у исследованных нами базидиоспор
Endoptychum agaricoides 30-летней давности коллапсированне не наблю-
далось, в то время как базидиоспоры отдельных сборов видов рода Mela-
nophyllum однолетней давности нередко были коллапсированы. Та-
кие же различия иногда отмечались н у спор одного и того же вида,
взятых из гербарных образцов, различных по времени сбора.
Мы изучали морфологию базидиоспор без придания спорам тургор-
ного состояния. Описанная методика была вполне приемлема, так как
на примере видов родов Agaricus, Lepiota (при исследовании спор без
напыления металлом, а следовательно, в условиях, исключающих искус-
ственное коллапсированне в напылителе) отмечено, что степень коллап-
сирования у сухих образцов, как напыленных, так и не напыленных,
была одинаковой.
При решении вопросов микроморфологии базидиоспор у критиче-
ских видов необходимо исследовать споры свежесобранных грибов или
споры гербарных образцов после придания им необходимого тургорного
состояния. В этих случаях желательно исследовать споры после высу-
шивания методом критической точки.
В заключение необходимо отмстить, что замачивание илн даже
смачивание, суспендирование спор в любых жидкостях отрицательно-
сказывается на качестве препаратов (набухание, длительность высуши-
вания, загрязнение). Поэтому наиболее целесообразной является фик-
сация объекта в парах осмиевой кислоты, поскольку наряду с фикса-
цией происходит и импрегнация металлом, что увеличивает электропро-
водность объекта (Ровенский, 1979).
МЕТОДИКА ИЗУЧЕНИЯ
ГЕОМЕТРИЧЕСКИХ ЗАВИСИМОСТЕЙ БАЗИДИИ
Для исследования базидии на основе геометрических зависимостей
проводили изучение бокового н торцевого положений базидни со спора-
ми по методике Э. Й. X. Корнера (Corner, 1972) на микроскопе «Ампли-
вал» с сухими (увеличение 16X40) н иммерсионными системами (16х
Х100). Для исследования гимениального слоя делали срез через пла-
стинку, препаровальной иглой переносили его на предметное стекло,
помещали в каплю воды, прикрывали покровным стеклом. Виды с мел-
кими спорами (например, Cystoderma granulosa) рассматривали как
под сухой, так и под масляной (immersionsol (20 °C) — 1,515)
иммерсионными системами.
Геометрические особенности базидни желательно изучать на све-
жем материале, однако удовлетворительные результаты получаются в
при изучении сухого материала одно-, трехмесячной давности, предва-
рительно находящегося в капле воды в течение 30 мин. В связи с тем
что базидию нельзя одновременно рассмотреть с торца и сбоку, для
каждого вида определяли средние значения параметров 100 измерений,
т. е. получали данные для «идеальной» базидии изучаемого таксона.
У агариковых грибов споры расположены над базидией на одном
уровне. Поскольку большинство высшнх Basidiomycetes имеет четыре
споры на базндии, за единицу предлагается рассмотрение тетрады
(и = 4) *. Четыре споры при торцевом обозрении рассматриваются как
1 При и=3 и и=5 споры при торцевом обозрении рассматриваются как утлы
равностороннего треугольника или пятиугольника.
21
углы квадрата. Можно предположить, что споры, расположенные по
углам квадрата, окружены наружным кругом, центральным кругом,
проходящим через центры спор, и внутренним кругом. Кроме того, суще-
ствует реальный круг базндии, диаметр которого равен максимальной
ее ширине. Для стеригм можно представить стернгмэтический круг,
проходящий через вершины стернгм, и круг стеригматнческого пятиа,
являющегося проекцией основания стернгм.
Для всесторонней характеристики геометрических особенностей ба-
зидии с помощью выведенных нами формул (Бергер, Вассер, 1976;
Вассер, 1980; Wasser, Berger, 1980; Вассер, Бергер, 1982) достаточно
двух параметров, взятых в любой комбинации. В качестве исходных
нами выбраны очерк споры н наружный круг спор базидин, которые
легко достоверно наблюдать и замерять под микроскопом, не производя
трудоемких подсчетов параметров по фотографиям бокового и торцево-
го положений базндии со спорами (Corner, 1972; Вассер, 1982).
ПЕРЕЧЕНЬ РЕАКТИВОВ, ПРИМЕНЯЕМЫХ ПРИ ИЗУЧЕНИИ
ХИМИЧЕСКИХ ЦВЕТОВЫХ РЕАКЦИИ АГАРИКОВЫХ ГРИБОВ,
И СПОСОБЫ ИХ ПРИГОТОВЛЕНИЯ
Аммиак (NH3). Газообразный нли 25%-ный водный раствор
NH4OH. При положительной реакции шляпка, ножка, пластинки зеле-
неют, реже желтеют. Реакцию проводят лишь на свежем материале.
Анилин. Чистый аннлии (СбН5МН2) или анилин, смешанный с рав-
ным количеством дистиллированной воды. Прн положительной реакции
ткань карпофора становится красноватой, медно-красной, темно-корнч-
невой, хромово-желтой. _
Бензидин1. Раствор бензидина (H2N—/—МН2)
в 96 %-ном этаноле. Прн положительной реакции наблюдается синее,
голубое окрашивание. Бензидиновая реакция свидетельствует о нали-
чии лакказы.
Гваякол. 1-гидрокси-2-метоксибензол, нлн гидроксианизол. При по-
ложительной реакции наблюдается голубовато-зеленоватое, голубовато-
сероватое, голубое, серовато-коричневатое, коричневатое, олнвково-ко-
ричиеватое, красноватое окрашивание.
ОН
Настойка гваякола. Раствор настойки 1 г семян растения Guaja-
сшп officinale L. в 5 мл 96%-ного этанола. При положительной реакции
наблюдается голубовато-зеленоватое, голубовато-сероватое, коричнева-
тое, фиолетовое окрашивание.
Кислоты. 60—70%-ная серная кислота (H2SO4), 65%-ная азотная
кислота (HNO3), 25%-ная соляная кислота (НС1), концентрированная
молочная кислота (СН3—СН(ОН)-—СООН). Химические цветовые ре-
акции с кислотами проводят на свежем грибном материале.
1 X. Клеменсон (Clementon, 1969.— Цит. по: Singer, 1975) доказал, что бензи-
дин можно успешно заменить о-толндином (диметидбензидин, растворенный в
96%-ном этаноле), так как первый является канцерогенным.
22
Крезил синий. Водный,раствор 7-амино-2-днметиламнно-3-метнлфе-
ноксазонилхлорида зеленовато-синего цвета
+сг
V''\y/'V^N(CHS),
Применяется для определения метахроматни микроструктур. Струк-
тура называется метахроматнчной, если водным раствором крезила си-
него окрашивается в красновато-фиолетовый (но не синий) цвет.
Лактофенол. Смесь равных частей молочной кислоты н фенола.
При положительной реакции наблюдается серовато-коричневое, розо-
вато-фиолетовое, лнлово-серое, фнолетово-серое, красноватое окраши-
вание карпофора н мякоти.
«-Нафтол. I-гидроксинафталин. Для химических цветовых реак-
ций используют смесь нескольких капель а-нафтола, 2 мл 90%-ного
этанола и 4 мл дистиллированной воды. У видов с положительной реак-
цией ткань карпофоров через 2—4 мнн окрашивается в темно-снний
цвет. Пластннкн видов родов Agaricus н Gyrophragmium окрашиваются
в красноватый цвет.
Пирамидон. Водный раствор пирамидона (2,3-диметнл-4-днметил-
амнно-пиразолои- (5).
(СН3)2 N—С = С—СНЯ
I I
О=С N—СН,
I
ел»
При положительной реакции ткань карпофоров окрашивается в
сиреиево-фиолетовый, фиолетовый, пурпурно-фиолетовый цвета.
Пирогаллол. 1,2,3-тригндрокснбензол. Для химических цветовых ре-
акций используют раствор пирогаллола в 96%-ном этаноле. При поло-
жительной реакции ткань карпофоров окрашивается в темио-коричие-
вый, коричневато-оранжевый цвета.
с—он
НС. I с—он
нет у хс—он
сн
Сульфоваиилии. Раствор 1 г химически чистого ванилина (3-мето-
кси-4-гидрокснбензальдегид) в смеси с 8 мл концентрированной серной
кислоты н 3 мл дистиллированной воды. Получаемый раствор желтова-
того цвета используют для реакций со свежим грибным материалом.
Прн положительной реакции ткань карпофоров окрашивается в пур-
пурно-розовый, серовато-коричневый, розовато-фиолетовый цвета.
Н
I
о=с
Сульфоформалин. Смесь равных частей формалина и 60—70%-пой
серной кислоты. Бесцветный раствор сульфоформалина используют для
реакций со свежим грнбиым материалом и материалом, сохранявшимся
в формалине не более 6 месяцев. Прн положительной реакции ткань
карпофоров окрашивается в коричневатый, желтоватый, зеленоватый,
розовато-фнолетовый цвета.
Феиол. 2—3%-ный водный раствор фенола (СбН5ОН). Его часто
называют карболовой кислотой. При положительной реакции через
20 мин карпофор приобретает красноватую, шоколадно-коричневую,
инкарнатно-красную, пурпурно-фиолетовую, малиново-розовую, желто-
ватую окраску. Если в течение 20 мин окраска карпофора не изменяет-
ся, реакция считается отрицательной (даже если в дальнейшем окраска
изменяется).
Феиоланилин. Смесь трех капель анилина, пяти капель концентри-
рованной серной кислоты н 10 мл 2—3%-ного водного раствора фенола.
При положительной реакции ткань карпофоров окрашивается в винно-
красиый, ржаво-розовый, лилово-розовый, затем шоколадно-коричневый
цвета.
Формалин. 40%-ный водный раствор формальдегида (НСНО). При
положительной реакции ткань карпофоров окрашивается в фиолетово-
коричневатый, красноватый цвета.
Щелочи. 40%-ные растворы едкого натрия (NaOH) и едкого кали
(КОН). При положительной реакции наблюдается окрашивание карпо-
форов в розовый, желтый, красный, красновато-коричневый, оранжевый
цвета. Реакцию проводят как иа свежем, так и на гербарном мате-
риале.
FeCI3. 10%-ный водный раствор хлорида железа. При положитель-
ной реакции пластинки окрашиваются в зеленоватый, позже темно-се-
рый цвета.
FeSO4. Раствор 1 г сульфата железа в 10 мл дистиллированной
воды с добавлением двух-трех капель серной кислоты. При положитель-
ной реакции мякоть окрашивается в оливковый, светло-зеленый, оран-
жевый, коричневый цвета.
Реактив Мельцера1. Раствор 0,5 г кристаллического иода и 1,5 г
KJ в 20 мл дистиллированной воды с последующим добавлением 20 мл
хлоралгидрата (СаС13—СН(ОН)2). С помощью реактива Мельцера
1 Критический анализ применения реактива Мельцера в микологии приводит
Р. Уотлииг (Watling, 1971).
24
определяют амилондность. Различают амилондность спор, базндии, гиф,,
тканей. Структуры называются амилоидными, если окрашиваются реак-
тивом Мельцера в голубой, фиолетовый, иногда почти черный цвета.
Структуры называются псевдоамилоидными илн декстрннондными, если
окрашиваются реактивом Мельцера в винно-красный или пурпурный
цвета. Структуры, которые не окрашиваются реактивом Мельцера или
слегка желтеют, называются неамилоидными. По данным Р. Зннгера
(Singer, 1975), амилондность структур проявляется у образцов, более
сотни лет пролежавших в гербарии.
Реактив Шеффера (перекрестная реакция). Стеклянной палочкой
по поверхности карпофора видов рода Agaricus проводят полоску ани-
лином, другой стеклянной палочкой проводят пересекающуюся с ней
полоску 65%-ной азотной кислотой. При положительной реакции в
месте пересечения появляется хромово-желтое, позднее оранжево-крас-
ное окрашивание. Реакцию Шеффера можно проводить и на материале,,
длительное время пролежавшем в гербарии.
ГЛАВА 111
ПОЛОЖЕНИЕ АГАРИКОВЫХ ГРИБОВ
В СОВРЕМЕННЫХ СИСТЕМАХ AGARICALES S. L.
С ИСТЕМЫ AGARICACEAE
Группа высших Basidiomycetes является сложной для систематики
и филогенетики. Основываясь на одних н тех же фактических данных,
исследователи по-разному понимают таксономические категории этой
группы. Изучению систематики порядка Agaricales s. I. посвящено много
исследований (Konrad, Maublanc, 1948—1952; Singer, 1951, 1962, 1975;
Pilat, 1951b; Kiihner, Romagnesi, 1953; Moser, 1953, 1967, 1978, 1983;
Dennis, Orton, Hora, 1960; Horak, 1968; Kreisel, 1969; Васильева, 1973;
Зерова, CociH, Роженко, 1979; Kiihner, 1980, и др.)- Тем не менее си-
стематика порядка до сих пор еще слабо разработана. Существуют раз-
личные ннтерпретацни как общей схемы, так и частных вопросов систе-
матики Agaricales s. 1., что связано со значительным их разнообразием,
редкой встречаемостью многих таксонов н недостаточной изученностью
тропической флоры.
Поскольку последние достижения в изучении высших Basidiomyce-
tes ознаменовались созданием новых классификационных схем порядка
Agaricales s. L, построенных на комплексе признаков, возникла необхо-
димость осветить общие тенденции развития нх систематики и опреде-
лить положение, которое агарнковые грибы занимают в этих системах.
I. КРАТКИЙ ОБЗОР СИСТЕМ ПОРЯДКА
AGARICALES S. L. СОВРЕМЕННОГО ПЕРИОДА
Высшие базидиомицетные грнбы обычно трактуются как группа
класса Basidiomycetes, в котором Agaric a les составляет самостоятель-
ный порядок или ряд порядков (Singer, 1962, 1975; Kreisel, 1969; Loc-
•quin, 1974; Romagnesi, 1977; Moser, 1978, 1983; Зерова, CociH, Роженко,
1979; Kiihner, 1980; Вассер, 1982).
Основанные на комплексе таксономических макроморфологических
и микроскопических признаков (цвет спорового порошка, цвет, форма
карпофоров н гнмения, характер прикрепления пластинок, тнп н харак-
тер покрывала, анатомические особенности покровных тканей, харак-
тер оболочки, форма, цвет спор, кариологнческие особенности развития
базндий, особенности онтогенеза карпофоров, химические цветовые ре-
акции карпофоров и микроструктур и др.) системы Agaricales s. 1. со-
временного периода можно разделить на три группы. Одни агарикологи
(Singer, 1951, 1962, 1975; Dennis, Orton, Hora, 1960, 1974; Moser, 1967;
Horak, 1968) стали увеличивать количество родов и довели его до 300,
а семейств до 18, выделяя при этом много монотипных родов — около
26
25 Другие (Konrad, Maublanc, 1942—1952; Piiat, 1951b; Kiihner, Ro-
magnesi, 1953; Martin, 1961; Romagnesi, 1977) стали сокращать коли-
чество родов до 60—90, семейств до 6. Третьи (Donk, 1964; Kreisel, 1969;
Locquin, 1974; Вассер, 1973а; Зерова, Вассер, 1974; Зерова, Горовой,
1974; Зерова, Вассер, Горовой, 1974; Каламээс, 1975; Вассер, Солдато-
ва, 1977а; Kalamees, 1978а; Kiihner, 1977, 1978, 1979, 1980; Moser, 1978,
1983; Зерова, CociH, Роженко, 1979) ставят вопрос о переоценке крите-
риев объема порядка и считают возможным постепенное разделение
Agaricales s. 1. на многие близкие самостоятельные порядки, доводя их
количество от четырех (Polyporales s. str.. Boletales, Russulales, Agari-
cales s. str.) до девяти (Polyporales s. str., Boletales, Russulales, Stro-
bilomycetales, Tricholomatales, Entolomatales, Agaricales s. str., Plutea-
les, Axnanitalcs). Рассмотренные разнообразные классификации грибов
порядка Agaricales s.l., критерии этих классификаций и тенденции раз-
вития свидетельствуют о том, что существуют различные интерпретации
как общей схемы, так и частных вопросов их систематики. Обособление
из порядка Agaricales s.l. ряда новых порядков обосновано как таксо-
номически, так н филогенетически. М. Я. Зерова, С. П. Вассер (1974),
М. Я- Зерова, С. П. Вассер, Л. Ф. Горовой (1974), М. Я. Зерова,
Л. Ф. Горовой (1974) еще в начале 70-х годов отмечали, что необходи-
мость разделения порядка Agaricales s. 1. является явной и давно дока-
занной. Agaricales s.l. объединяет филогенетически довольно далекие
группы грибов (Зерова, Вассер, 1.с., с. 62). К настоящему времени про-
цесс выделения порядков не закончен, а границы н объем многих нз
них еще не стабилизировались (например, Tricholomatales, Agaricales
s. str.).
Порядок Boletales, характеризующийся гимнокарпным типом раз-
вития, признан многими авторами (Heim, 1934, 1971; Lange М., Hansen,
1954; Cejp, 1958; Benedix, 1959; Skirgiello, 1960, 1975; Kreisel, 1961,
1969; Michael, Hennig, 1964—1975; Thiers, 1971; Вассер, 1973a; Зерова,
19746; Каламээс, 1975; Вассер, Солдатова, 1977а; Kalamees, 1978а, b;
Moser, 1978; Зерова, CociH, Роженко, 1979; Michael, Hennig, Kreisel,
1979; Kiihner, 1980, н др.). Однако в отношении его объема нет един-
ства взглядов. Мнения расходятся в вопросах присоединения к порядку
семейств Paxillaceae, Gomphidiaceae н Strobilomycetaceae, хотя боль-
шинство авторов не сомневаются в наличии филогенетических связей
между Boletaceae и названными семействами. К- Цейп (Cejp, 1958)
и последователи, например, хотя н выделяют порядок Boletales, семей-
ства Paxillaceae и Gomphidiaceae относят к порядку Agaricales. Г. Тирс
(Thiers, 1971), А. Пилат, А. Дермек (Piiat, Dermek, 1974) не считают
Paxillaceae н Gomphidiaceae близкими к Boletaceae. Большинство же
авторов включают семейства Strobilomycetaceae, Paxillaceae и Gomphi-
diaceae в Boletales. Важной проблемой в Boletales является и таксоно-
мическое положение родов Gyrodon Opat. и Gyroporus Quel. По
наличию пряжек иа мицелии, форме спор, форме гименофора, не отделяю-
щегося от мякотн шляпки, этн роды в ранге подсемейства Gyrodontoi-
deae илн семейства Gyrodontaceac относятся к наиболее примитивным
Boletales (Benedix, 1959; Skirgiello, 1960, 1975; Kreisel, 1969; Thiers,
1971; Каламээс, 1975; Watling, Largent, 1976, н др.), причем, по мнению
ряда авторов (Benedix, 1. с.; Thiers, 1. с.), Gyroporus находится на более
высоком уровне эволюционного развития по сравнению с родом Gyro-
don. Некоторые авторы высказывают мнение о полифилетическом про-
исхождении Boletales (Moreau, 1952—1953; Benedix, 1959; Skirgiello,
27
1960, 1975; Heim, 1971; Thiers, 1971; Каламээс, 1975; Pegler, Young,
1981). По этой концепции большинство Bolctales происходит от Agari-
cales, a Gyrodon, например, от представителей Aphyllophoralcs (Scuti-
gcraceae); одновременно отмечается, что в Boletalcs явно проявляются
признаки конвергенции, многие роды порядка сходны физиономическн.
Д. Н. Пеглер, Т. У. Янг (Pegler, Young, 1981) предложили в порядке
Boletales новое для науки семейство Хегосотасеае с родами Xerocomus
и Phylloporus, а Р. Кюнер (Kiihner, 1980) — новое для науки семейство
Hygrophoropsidaceae (с родами Hygrophoropsis и Omphalotus). Обо-
снованность выделения последнего семейства требует дополнительных
исследований.’
Е. Бенедикс (Bencdix, 1959), сторонник выделения порядка Boleta-
les, еще в 1959 г. предсказывал и дальнейшее разделение болетальных
грибов на ряд самостоятельных порядков.
М. Я. Зерова, Г. Л. Роженко, Т. Л. Череп (1975), М. Я. Зерова,
П. Е. Сосин, Г. Л. Роженко (1979), учитывая наличие антокарпного
типа развития, своеобразного строения базидиоспор, свидетельствую-
щих о несовместимости Strobilomycetaceae с Boletalcs, выделили новый
порядок Strobilomycetales с одноименным семейством. На особое си-
стематическое положение стробиломицетальных грибов указывали еще
Р. Мэр (Maire R., 1933), А. Пилат (Pilat, 1951b), Э. Гойман (Gauman,
1964), И. Арке (Агх, 1968). Д. П. Пеглер, Т. У. Янг (Pegler, Young,
1981) описали новый для науки род Atroboletiis и поместили его в
семейство Strobilomycetaceae.
Одиой нз кардинальных проблем классификации высших Basidio-
myectes, как указывает М. Мозер (Moser, 1964), является разграниче-
ние порядков Agaricales s. 1. н Aphyllophorales, и в этой связи находится
проблема порядка Polyporales s. str. М. Доик (Donk, 1964, 1971) и
последователи считают, что Polyporaceae должно находиться в Aphyllo-
phorales. Р. Зингер (Singer, 1962) и последователи утверждают, что
Polyporaceae должно находиться в Agaricales. Главным аргументом
в пользу последней точки зрения является наличие покрывала у неко-
торых представителей Polyporaceae, в то время как Aphyllophorak s
являются гимнокарпными. Окончательно решить данную проблему по-
может онтогенетический метод (Reijnders, 1963; Зерова, 1965; Зерова,
Вассер, 1974; Зерова, Горовой, 1974; Зерова, Вассер, Горовой, 1974;
Горовой, 1980) *.
М. Я. Зерова, С. П. Вассер (1974), М. Я- Зерова, Л. Ф. Горовой
(1974) высказали мысль о том, что назрела необходимость из Agarica-
les s. 1. выделить порядок Tricholomatales. Р. Кюнер (Kuhner, 1978)
опубликовал последний как nomen nudum, а в 1980 г. (Kuhner, 1980)
его узаконил. В 1979 г. М. Я- Зерова, П. Е. Сосин, Г. Л. Роженко
(1979) также опубликовали как nomen nudum порядок Tricholomatales,
окончательно развив идеи, высказанные в 1974 г. (Зерова, Вассер, 1. с.;
Зерова, Горовой, 1. с.). В Tricholomatales. по М. Я. Зеровой, П. Е. Со-
енну, Г. Л. Рожеико (I. с.), входит три семейства. Pleurotaceae, Hygro-
phoraceae, Tricholomataceae, а по P. Кюнеру — семь семейств: три се-
мейства предыдущей системы, а также Ainanitaceae, Hydnangiaceae,
Rhodotaccae, Marasmiaceae. Помещение Amanitaceae, для которого ха-
* В критической работе И. Нусса (Nuss, 1980) Polyporales разделен на два
семейства: Polyporaceae с родами Polyporus и Piptoporus, и Albatrellaceae (Pcuz.)
Nuss с родом Albatrellus.
28
рактерен иной тип развития карпофоров и гименофора, в Tricholoma-
tales не выдерживает критики. С перенесением нз Gasteromycetes в
Tricholomatales семейства Hydnangiaceae сделан еще один шаг в рас-
пределении секотиевых грибов среди Agaricales s. 1- Семейства Rhodota-
ссас и Marasmiaceae по объему соответствуют одноименным коленам в
•смысле систем Р. Зингера. Значительно отличается от семейств порядка
Tricholomatales, как и всех Agaricales s. 1., семейство Hygrophoraceae.
На основании анатомо-морфологических и онтогенетических исследова-
ний А. Ф. М. Рейндерс (Rcijnders, 1963) еще в 1963 г. исключил
Hygrophoraceae из числа истинных Agaricales и поместил его в искус-
ственную группу «Hemi-Agaricales». Мы полностью согласны с К. А. Ка-
ламээсом (1975, с. 17), который высказал мысль о том, что Hygropho-
гасеас можно было бы выделить в самостоятельный порядок, что мы
и предложили ранее (Вассер, 1982) ’.
Понимая порядок Tricholomatales шире, чем М. Я. Зерова, П. Е. Со-
син, Г. Л. Роженко (1979), мы включаем в него не только гимнокарп-
ные виды, но и выделяем семейство Catathclasmataceae* 2 с родами Са-
tathelasma, Floccularia, Armiliariella, для которых характерны гемиан-
гнокарпный тип развития (бнвелангиокарпный характер покрывала),
билатеральная трама, исключая их из семейств Agaricaceae или Lepio-
taceae.
В 1979 г. М. Я. Зеровой, П. Е. Сосиным, Г. Л. Роженко описан
как nomen nudum новый для науки порядок Entolomatales с одноимен-
ным семейством, включающим три рода — Entoloma, Claudopus, Eccilia.
Entolomataceae представляет собой гомогенное семейство пластинчатых
грибов, для которых характерен гимнокарпный тип развития, своеобраз-
ные угловато-округлые или угловато-овальные споры, наличие мелких
блестящих грануляций в правильной гименофоральной траме и гифах
(Reijnders, 1963; Hesler, 1967; Romagnesi, Gilles, 1979). Мы считаем
нецелесообразным придавать данному семейству ранг самостоятельного
порядка, поскольку онтогенетическим исследованиям подвергнут лишь
один вид. На наш взгляд, в это семейство следует ввести род Richomul-
la, для которого характерен такой же тип н цвет спор (Horak, 1973).
Выделение из Agaricales s. str. порядка Pluteales (Kiihner, 1978,
1980) с введением в последний всего семейства Entolomataceae также
недостаточно обоснованное н преждевременное. Однако мы признаем
выделение порядка Amanifales (Locquin, 1974), для представителей ко-
торого характерен шизогимениальный тип развития гименофора и обра-
зование гимения нз интеркалярных клеток гнф (Locquin, 1974; Kiihner,
1980; Горовой, 1980). Гименофор образуется из гомогенной гифальной
массы путем расщепления сначала на дольки, а затем на пластинки.
При этом между трамой шляпки и трамой пластинок существует гете-
рогенный переход. Однако мы не согласны с М. Локвнном (Locquin,
1974) в отношении объема порядка Amanitales, содержащего пять се-
мейств.
* Hygrophorales S. Wa-зег ord. nov.— Basidiomycetes. Carposomata agaricoidca,
gymnocarpoidea. Lamellis crassis, raris. Hymenophoralis trama latioris. Massis sporis
albus. Basidiis clavatis, longiribuis quam ad Agaricales s. 1. Sterigmatiis (duobis ad)
quattuis. Sporis lacvis, dccoioratis.— Typus: Hygrophoraceae Roze (Hygrophorces),
Bull. Soc Bot. Fr.. 1876, 25 : 51.
2 Catathelasmataceae S. Wasser fam. nov.—Tricholomatales. Carposomata agari-
coidea, gemiangyocarpoidea. Hymenophore I trama bilatcralis, nonamyloidis. Massis sporis
albus. Basidiis clavatis. Sterigmatiis quattuis. Sporis laevis, decoloratis, amyloidis. Ve-
lum habent. Basalis volva nullis.— Typus: Catathelasma Lovej, Bot. Gaz, 1910, 50:383.
29
Гомогенный порядок Russulales, выделенный Г. Крайзечем (Kreisel,
1969), для представителей которого характерна неправильная трама
(у большей части со сфероннстами), особый тип строения базидиоспор,
принимается большинством микологов. Р. Зингер (Singer, 1962) еще
в 1962 г. отмечал, что Russulaceae имеет очень мало общего с осталь-
ными Agaricales. Р. Уотлннгом, Н. Хазел (Watling, Hazel, 1980) пока-
зано, что образование сфероцист является механизмом, который четко
отличает представителей руссулальных от остальных пластинчатых гри-
бов. Проблематичен до последнего времени лишь объем порядка Rus-
sulales. Р. Зингер (Singer, 1975) вводит в Agaricales как близкое к
Russulaceae семейство Bondarzewiaceae с родом Bondarzcwia. М. Мозер
(Moser, 1978) не помещает Bondarzewia в Agaricales, оставляя ее в
Aphyllophorales. Д. Н. Пеглер и Т. У. К- Янг (Pegler, Young, 1979),
Р. Кюнер (Kiihner, 1980) расширяют объем Russulales за счет семей-
ства Auriscalpiaceae из порядка Aphyllophorales, а также за счет га-
строидных родов, вводя в него семейство Elasmomycetaceae с родами
Elasmomyces, Zelleromyces, Martellia, Gymnomyces, а в Russulaceae —
роды Arcangeliella, Cystangium, Macowanites.
Очевидно, и у Agaricales s. str. проявляются признаки конвергент-
ного сходства и порядок состоит из групп, соединенных между собой
лишь физиономнчески. Однако недостаточная изученность их не дает
сейчас еще оснований для дальнейшего выделения новых порядков из
Agaricales s. str. (Зерова, Вассер, 1974; Зерова, Горовой, 1974; Вас-
сер, 1982).
Современный период систематики Agaricales, как и всех высших
Basidiomycetes, вслед за Э. X. Пармасто (1969) можно охарактеризо-
вать как период «девальвации» категорий систематики, что и ведет к
выделению новых самостоятельных (иногда недостаточно обоснован-
ных) порядков. В период «девальвации» систематики основополагаю-
щим является вес критерия при разграничении таксонов. Р. Зингер
(Singer, 1962, 1975) и последователи, сторонники единого порядка Aga-
ricales s. 1. в основу своих систем кладут филогенетический критерий.
Однако прн наличии ограниченных данных об особенностях онтогенеза
представителей многочисленных семейств агарнкальных грибов, об их
анатомо-морфологической структуре, экологии, цитологической, физио-
лого-биохимической специфике использование филогенетического кри-
терия допускается на данном этапе развития агарикологин с большой
осторожностью, иначе легко перейти к неверным интерпретациям о бли-
зости различных групп грибов и различных систематических категорий
в пределах каждой группы, а в результате — к ошибочным обобщениям
н построениям Ряд авторов (Зерова, 1965; Зерова, Вассер, 1974; Зеро-
ва, Горовой, 1974) предостерегали от использования как приоритетно-
го филогенетического критерия, так как в вопросе филогенеза Agari-
cales имеются диаметрально противоположные взгляды. М. Я. Зерова,
П. Е. Соснн, Г. Л. Роженко (1979) считают, вслед за Г. Крайзелем
(Kreisel, 1969), что в основу системы Agaricales s. 1. на данном этапе
развития науки следует класть главным образом анатомо-морфологи-
ческнй критерий, а как дополнительный применять онтогенетический,
с чем мы полностью согласны.
В связи с недостаточной изученностью Agaricales s. 1. земного шара,
сейчас как временная мера наиболее приемлема в какой-то степени
искусственная, сравнительно-морфологическая система, основанная на
определенных константных, главным образом анатомо-морфологических,
30
признаках. При этом нужно учесть, что искусственная сравнительно-
морфологическая система в значительной мере все же будет объединять
биологически и филогенетически близкие систематические категории.
На иаш взгляд, наиболее соответствуют этим требованиям системы
Г. Крайзеля (Kreisel, 1969), Р. Зиигера (Singer, 1975) и частично
М. Я. Зеровой, П. Е. Сосина, Г. Л. Роженко (1979) для иадсемействен-
иых таксонов. Интерпретация порядков принимается по Г. Крайзелю,
Р. Кюиеру и частично по М. Я. Зеровой с нашими дополнениями, а
основные семейства и внутрисемейственные таксоны принимаются по
четвертой системе Р. Зиигера (Singer, 1975).
Подытоживая изложенное, мы следующим Образом представляем
себе место Basidiomycetcs в системе грибов и в виде списка-схемы
порядковую и внутрипорядковую (до семейств) систему Agaricales s.l.
Список составлен на основании анализа всех известных нам систем
высших Basidiomycetes, с некоторыми изменениями, дополнениями, от-
ражающими наши современные взгляды на систематику Agaricales s.l.
Царство Fungi
Отдел Chytridiomycota
» Eumycota
Класс Zygomycetes
> Endornycetes
» Ustomycetes
» Ascomycetes
» Basidiomycetes
Подкласс Homobasidiomycetidae
Порядок Polyporales s. str.
Polyporaceae
Порядок Boletales
Boletaceae
Gyrodon t a ceae
Xcrocomaceae
Paxillaceae
Gomphidiaceae
Порядок Strobilomycetales
Strobilomycetaceae
Порядок Hygrophorales
Hygrophoraceae
Порядок Tricholomatales
Pleurotaceac
Tricholomataceae
Catathel asmataceae
Порядок Amanitales
Amanitaceae
Порядок Agaricales s. str. (incl, Podaxales)
Agaricaceae
Coprinaceae
Bolbitiaceae
Strophariaceae
Crepidotaceae
Cortinariaceae
Pluteaceae
Entolomataceae
Порядок Russula les (incl. Asterogastraceae)
31
Russulaceae
Bondarzewiaceae
Elasmomycetaccae
2. СИСТЕМЫ СЕМЕЙСТВА AGARICACEAE
Семейство агариковых — Agaricaceae относится к порядку Agari-
cales s. str. подкласса высших Basidiomycetes — Homobasidiomycetidae.
В системе порядков пластинчатых грибов Agaricaceae занимает место
между светлоспоровыми (Tricholomatales — Catathelasmataceae, Ama-
nitalcs — Amahitaceae) и тем иоспор овы ми (Agaricalcs s. str.— Coprina-
ccac, Bolbitiaceae, Strophariaceae) семействами. В Agaricaceae в совре-
менном понимании из нескольких семейств порядка Agaricales s. 1. во-
шли роды, имеющие различную окраску спорового порошка.
Интерпретации систематического положения и объема семейства
Agaricaceae в современной литературе противоречивы. Agaricaceae среди
семейств Agaricales s. str. является сравнительно гетерогенным и дис-
куссионным семейством как по количеству относимых к нему родов,
так и по их систематическому положению. Основная причина различ-
ных трактовок Agaricaceae заключается в отсутствии разработанного
«универсального» критерия таксонов различных рангов, четких прагма-
тических правил и субъективизме при описании таксонов, в противоре-
чивости взглядов на анцестральные формы представителей отдельных
родов. Систематика Agaricaceae основывается главным образом на
определенных анатомо-морфологических признаках. Следовательно,
сравнительно-морфологический метод остался ведущим в решении
вопросов систематики, хотя при этом нельзя не учитывать и эколого-био-
логические, географические, физиолого-биохимические признаки. Систе-
матика этого семейства не может быть решена на основе односторон-
него подхода, и ни одна из существующих систем не может быть при-
нята в целом. При решении вопросов систематики агариковых грибов
мы провели таксономическую оценку признаков на их стабильность и
эволюционную значимость. Поскольку одни и те же признаки могут
иметь различную ценность у различных таксонов, то разграничение
таксонов в современной систематике основано на совокупности призна-
ков, имеющих неодинаковую ценность и степень константности. Наша
цель — создание системы, отражающей филогению агариковых грибов.
Нет сомнения в том, что по мере расширения и углубления знаний мы
приблизимся к решению данной задачи, как этого достигли специалисты
по другим группам грибов. Системы Agaricales s. 1., существующие сей-
час, являются приближенными и провизорными. Для их дальнейшей
разработки необходимо решить ряд теоретических (например, проблема
вида) н практических (инвентаризация флоры Южного и ряда регионов
Северного полушарий) вопросов.
Ниже рассмотрим критерии основных таксонов Agaricaceae.
Неодинаковую ценность и степень константности имеют макропри-
знаки (форма, консистенция, окраска карпофора и его элементов, осо-
бенно гимения, размеры, форма и способ прикрепления пластинок, ха-
рактер поверхности карпофора, мякоть, ее консистенция, цвет при авто-
оксидацни, вкус, запах и др.) агариковых грибов. У всех агариковых
грибов карпофор состоит из шляпки с пластинчатым гименофором на
нижней поверхности и ножки. В связи с тем что формообразование
карпофоров агариковых грибов ограничено одним типом, форма карпо-
32
форов и гимения учитывается в совокупности с другими признаками
при разграничении семейств. На уровне родов эти признаки ие имеют
значения.
Онтогенетический критерий, изучающий тип развития карпофоров у
видов, морфологически ие всегда четко различающихся, вводимых в
семейство Agaricaceae, вскрывает значение конвергенции и диверген-
ции, вуалирующих отдаленность или близость видов. Тип развития кар-
пофоров в ближайшие десятилетия после изучения значительного ко-
личества представителей Agaricaceae, займет, вероятно, место ведущего
в таксономии родовых и надродовых таксонов. Для 20 изученных видов
Agaricaceae отмечено значительное разнообразие типов развития (Rei-
jnders, 1975): стипитокарпное, пилеостипитокарпиос, пилеокарпное или
изокарпиое, гименокарпное. По онтогенетическим особенностям А. Ф. М.
Рейидерс (Reijnders, 1. с.) считает возможным предварительно разде-
лить Agaricaceae иа три группы. Однако на данном этапе развития
онтогенетического метода этот критерий использовать нельзя, так как
онтогенез изучен лишь у 4 % представителей мировой флоры Agarica-
ceae (Вассер, 1982).
Многие Agaricaceae имеют покрывало весьма разнообразного строе-
ния. У некоторых видов присутствует лишь частное покрывало. Оно
иногда отрывается от края шляпки н сохраняется иа ножке в виде
кольца или отрывается от ножки и сохраняется по краю шляпки. При
отрыве от края шляпки и от ножки образуется подвижное кольцо, яв-
ляющееся родовым признаком, например, у Macrolepiota. Общее по-
крывало у агариковых грибов остается в виде вольвы (например, Clar-
keinda, Volvolepiota). Характер покрывала — важный таксономический
признак.
Консистенция карпофоров также важный таксономический признак,
например, и а уровне рода у Leucocoprinus.
Шляпки агариковых грибов различаются по размерам, форме,
окраске, характеру поверхности и ряду других признаков, имеющих
важное таксономическое значение на родовом и видовом уровнях. По-
верхность шляпок бывает блестящей, зернисто-блестящей, матовой, су-
хой, влажной, клейкой, слизистой, голой, чешуйчатой, войлочной, бар-
хатистой, зернистой, с мучнистым налетом, ареолироваиной. Например,
характер поверхности шляпки у Agaricus tabularis, Cystoderma rugoso-
reticulata — важный видовой признак. Иногда отдельные части кар-
пофоров окрашены в различные цвета. Константность окраски карпофо-
ров послужила основанием для названий многих видов. Цвет является
одним из важных критериев вида у Lepiota lilacea, Phaeolepiota
aurea и др.
Ножка у агариковых грибов дает более илн менее твердую опору
карпофору. Различают два основных типа ножек: полая и выполнен-
ная. Между ними можно наблюдать ряд переходных форм в различных
вариациях, при которых ножка одного и того же вида весьма часто,
в зависимости от возраста, может быть не совсем однородной по строе-
нию. Ножка агариковых грибов формируется из плотно соединенных
между собой нитевидных гиф, располагающихся в вертикальном на-
правлении. У видов с крупными шляпками устойчивость увеличивается
также вследствие постепенного расширения ножки к низу, часто закан-
чивающейся клубневидным вздутием (например, Macrolepiota rhacodes,
М. Olivieri, Agaricus abruptibulbus и др.), которое является важным ви-
довым признаком. Основание ножки бывает окрашенным или покрытым
3 4—2960
33
войлочным налетом мицелия, форма ножки — цилиндрической, прямой
или кривой, утолщенной к середине, верхушке или основанию, в основа-
нии иногда с корневидным отростком (например, Agaricus romagnesii,
Leucoagarictis macrorhizus). Отмеченные признаки играют существен-
ную таксономическую роль. У большинства видов ножка центральная,
у некоторых эксцентрическая и боковая, сухая, клейкая, слизистая,
голая, гладенькая, чешуйчатая, волокнистая, с кольцом, кольцом и воль-
вой, только с вольвой без кольца, без кольца и вольвы.
Для некоторых агариковых грибов характерно образование скле-
роциев, которые содержат запасной питательный материал. У отдель-
ных агарикобых грибов (например, Squamanita paradoxa) наличие скле-
роциев — видовой признак.
Мякоть, ее консистенция и цвет — важные таксономические при-
знаки вида и внутриродовых таксонов. Мякоть шляпки агариковых гри-
бов гомеомерная. Окраска мякоти у большинства видов светлая, реже
с желтоватым оттенком. При автооксидации мякоть многих видов меня-
ет цвет (например, желтеет или краснеет у видов рода Agaricus), что
является важным видовым и внутриродовым таксономическим призна-
ком. Значимость этого критерия подтверждается и названиями ряда
видов агариковых грибов: гриб-зоитик краснеющий (Macrolepiota rha-
codes).
Запах и вкус карпофоров — объективные, постоянные и конкретные
признаки видового и виутриродового рангов. Разнообразные, вполне
конкретные запахи карпофоров агариковых грибов указывают на нали-
чие в иих различных химических веществ. Запах аниса — признак Aga-
ricus arvensis, запах карболовой кислоты — признак A. xanthodermus,
запах рыбы характерен для A. bernardii, запах редьки — для Lepiota
erminea, фруктовый запах — один из критериев вида у L. brunneoin-
carnata. Запах как весьма характерный признак отражен и в назва-
ниях, например, шампиньон дурнопахнущий — Agaricus maleolens,
цистодерма пахучая —- Cystoderma carcharias. Вкус карпофоров агарико-
вых грибов — также специфический признак, служащий критерием не-
которых видов и внутривидовых таксонов. По вкусу различают горь-
кую, кислую, острую, сладкую, соленую, сладковатую (т. е. без особого
вкуса) мякоть. Острый вкус свойствен Agaricus augustus, A. excellcns;
мучной — Cystoderma amianthina, Squamanita paradoxa, горький —
Chamaemyces fracidus, Leucocoprinus badhamii.
Анатомические признаки как наиболее консервативные характери-
зуют главным образом таксоны надвидовых рангов, одиако могут быть
и видовыми критериями.
Покровы карпофоров, особенно шляпки, имеют сложное строение.
У видов с покрывалом поверхностный слой образован остатками общего
покрывала в виде хлопьев, часто легко стирающихся, или сфероциста-
ми, лежащими сплошным слоем. Под остатками покрывала распола-
гается сплошной слой кожицы (эпикутиса и кутиса), которая часто
разрывается в процессе роста и а отдельные чешуйки. Она состоит из
сплетений гиф основной и соединительной тканей или клеток, может
иметь гимеиовидиое (грушевидные клетки расположены наподобие
базидий в гимениальном слое), ложнопареихимное, палисадное, ните-
видное строение. В ней могут присутствовать дерматоцистиды (иилоци-
стиды), волоски, хлопья примордиальных гиф. Чешуйчатость ее, обра-
зующаяся вследствие роста карпофора, представляет собой гифы,
соединенные в пучки (Macrolepiota procera, Lepiota acutesquamosa).
34
Зернистые и морщинистые ткани агариковых грибов, образующиеся из
поверхности кожицы, состоят из неправильно округлых, приблизительно
равных размеров клеток, лежащих ровным слоем в несколько рядов
(Lepiota cristata) или же формирующихся в пирамидальные, иногда
кубические группы, довольно равномерно разбросанные по поверхности
кутикулы (Cystoderma amianthina, С. rugosoreticulata). Строение по-
кровов шляпки имеет важное надвидовое таксономическое значение
(Bresinsky, Schwarzer, 1969).
Под кожицей располагается мякоты Гомеомерная мякоть агарико-
вых грибов состоит из двух типов тканей — основной и соединительной.
Основная ткань обычно образована более толстыми, широкими, прямы-
ми, толстостенными гифами; соединительная ткань, как правило,— бо-
лее тонкими и изогнутыми гифами. За мякотью шляпки и ниже ее сле-
дует гимеиий — плодоносный слой. У агариковых грибов различают
гименофор свободный, свободный с коллариумом, приросший, слегка
низбегающий. Значение гименофора как таксономического признака у
агариковых грибов огромно при определении таксонов различного ран-
га. Гименофор состоит из внутренней стерильной части — гименофо-
ральиой трамы и покрывающего гимениального слоя, трама — из не-
скольких слоев, расположенных в середине пластинки. Для агариковых
грибов характерны два типа трамы — правильная и неправильная. Тип
строения трамы — важный таксономический признак иа уровне семейств
и родов.
Между трамой и гимениальиым слоем располагается субгимениаль-
ный слой относительно однородного строения, не имеющий на современ-
ном уровне знаний таксономического значения.
Гимениальный слой выстилает пластинки и состоит из базидий,
несущих базидиоспоры. Иногда базидии перемешиваются с бесплодны-
ми элементами гимениального слоя — цистидами, различающимися по-
строению, происхождению и местоположению. Все микроструктуры ги-
мения имеют важное таксономическое значение. У значительной части
агариковых грибов базидии булавовидной формы, реже цилиндриче-
ской. Для большинства видов агариковых грибов типичны четырехспо-
ровые базидии; у некоторых видов развиваются двуспоровые (напри-
мер, Agaricus bisporus).
Споры агариковых грибов очень разнообразны по форме, цвету
(соломенно-желтые, медово-желтые, коричневые либо бесцветные) и
размерам. Поверхность их бывает гладкая (Agaricus, Lepiota, Cysto-
derma), шероховатая (Melanophyllum), тонкошероховатая (Phaeolepio-
ta). Форма спор, их размер, наличие и тип поры прорастания, супраапи-
кулярной депрессии, количество на базидии — важные таксономические
признаки видового и надвидового рангов.
Размеры спор, стеригм и базидий связаны определенными констан-
тами, а геометрические особенности их расположения предложены нами
на примере видов семейства Agaricaceae как новый дополнительный
таксономический критерий (Вассер, Бергер, 1982).
Анализ базидий иа основе геометрических зависимостей открывает
новое направление в изучении высших Basidiomycetes (Malen^on, 1958;
Pcrrcau-Bertrand, 1967; Corner, 1972; Бергер, Вассер, 1976; Вассер,
1980; Wasser, Berger, 1980). В основу анализа положено предложенное
Э. И. X. Корнером (Corner, 1972) и модифицированное нами изучение
торцевых и боковых положений базидий со спорами. В результате мно-
гочисленных наблюдений, многократных замеров и вывода средних
з*
3&
соотношений установлено, что взаимное расположение на базидии сте-
ригм со спорами подчиняется схеме, изображенной на рис. 3 1. Поскольку
большинство высших Basidiomycetcs имеет четыре споры на базидии, за
единицу предлагается рассмотрение тетрады. Четыре споры при торце-
вом обозрении рассматриваются как углы квадрата. Можно предполо-
жить, что споры, очерк которых обозначен d, расположены по углам
квадрата, окружены наружным
кругом S', центральным кру-
гом S", проходящим через цент-
ры спор, и внутренним кру-
гом S'". Кроме того, существует
реальный круг базидин <о, диа-
метр которого равен максималь-
ной се ширине. Для стеригм мож-
но представить стернгматический
круг М, проходящий через вер-
шины стеригм, и круг стеригма-
тического пятна т, являющегося
проекцией основания стеригм,
а — центральный угол, соответ-
ствующий дуге HF(o\ б — угол
между осью АС и касательной АР
к очерку споры, 0 — угол споры
(угол CLM). В результате вве-
денных коэффициентов (п, рав-
ный отношению диаметров S"fd,
q, равный отношению диаметра S'
к <о) и выведенных нами формул
(Вассер, 1980; Вассер, Бергер,
1982) сформулированы новые для
науки свойства базидии: 1) отно-
шение диаметров внешнего круга
и очерка базидии равно отноше-
нию диаметров центрального и
внутреннего кругов; 2) отноше-
ние диаметров внешнего круга и
очерка базидии равно отношению
диаметров очерка споры и стернг-
матического пятна; 3) отношение
диаметров центрального и внут-
Рис. 3. Геометрия базидииг
а — вид сбоку, б — торцевой вид
реннего кругов равно отношению диаметров очерка споры и стеригма-
тического пятна; 4) отношение диаметров центрального круга и очерка
споры равно отношению диаметров внутреннего круга и стеригматичес-
кого пятна; 5) отношение диаметров внешнего круга и очерка споры
равно отношению диаметров очерка базидин и стернгмэтического пятна.
Предложенные нами расчеты позволяют определить ряд параметров
базидии (М, т, б, 0, а), которые очень трудно наблюдать и практически
невозможно замерить.
Практическое испопьзование предложенных методов, выведенных
формул, установленных соотношений и их точности проверено иа 17 кри-
1 Для упрощения чтения чертежа на рис. 3 показаны лишь две крайние сте-
ригмы со спорами.
36
тических таксонах агариковых грибов, относящихся к пяти родам
(Agaricus, Cystoderma, Macrolepiota, Leucoagaricus, Leucocoprinus).
Доказана их действенность как дополнительного таксономического кри-
терия: близкие трудноразличимые морфологически таксоны, например
Agaricus meleagris и A. moelleri, четко отличаются по параметрам S',
S", S'"; к. xanthodermus var. xanthodermus и A. xanthodermus var.
lepiotoidcs, имеющие перекрывающиеся размеры спор, различаются по
параметрам S', S", <о, М, 6, в, а. Быстродействие всесторонней харак-
теристики геометрических особенностей базидий показано иа приме-
ре 17 таксонов агариковых грибов таблнчио-номограммным и графиче-
ским методами (Вассер, Бергер, 1982).
На рис. 4 представлена номограмма, позволяющая непосредственно
определять такие линейные параметры базидий, как S", ы, т, М, по
заданным значениям диаметра спор d, например, от 1 до 10 мкм и
коэффициента п от 1,5 до 4. Ответ читается сразу же в микрометрах
на вертикальной числовой шкале в месте пересечения п и искомого
параметра при соответствующих значениях диаметра споры.
У агариковых грибов различают плевроцистиды, развивающиеся
на сторонах пластинок, и хейлоцистиды, развивающиеся на краях пла-
стинок. В качестве систематического признака цистиды имеют важное
значение при разграничении видов (например, Cystoderma granulosa
и С. cinnabarina), родов и внутриродовых (например, секции Granulo-
sa и Cinnabarina рода Cystoderma) таксонов.
В связи с тем что тканн агариковых грибов определенного вида или
рода содержат одно или несколько специфических веществ, большое
значение приобретают как дополнительный критерий таксонов макро-
и микроскопические химические цветовые реакции.
Проведенные исследования макро- и микроскопических химических
цветовых реакций видов семейства Agaricaceae показали, что многие
из них уже на данном этапе развития химического метода могут быть
с успехом использованы в таксономии на родовом, внутрироновом и
видовом уровнях (Schaeffer J., Moeller, 1938; Singer, 1951, 1962, 1975;
Kiihner, Romagnesi, 1953; Sandor, 1958; Essette, 1964; Moser, 1967, 1978;
Watling, 1971; Meixner, 1975; Вассер, 1980).
Микрохимическая реакция с крезилом синим, определяющая мета-
хроматичность структур, является важным родовым признаком у Macro-
lepiota, Leucoagaricus, Leucocoprinus, Volvolcpiota, Chlorophyllum. Пер-
вые три рода благодаря метахроматни спор легко отличить от рода
Lepiota. Данные этой микрохимической реакции широко используются
совместно со строением спор (например, наличие или отсутствие поры
прорастания) при внутрисемейственных подразделениях Agaricaceae.
Микрохимическая реакция с реактивом Мельцера, определяющая
амилоидность спор и других микроструктур,— важный таксономический
признак в ранге подрода, секции, подсекции в родах Cystoderma (для
видов подрода Cystoderma характерны амилоидные споры, для видов
секции Superba — амилоидные лишь в супраапикулярнон зоне, а для
ведов подрода Granulosa — неамилоидиые споры), Lepiota s. str. (для
видов секции Amyloidea характерны амилоидные споры, .для видов сек
ции Micaceae — неамилоидиые споры, а для остальных внутриродовых
таксонов — псевдоамилоидные споры). Псевдоамилоидность спор — ро-
довой признак у Micropsalliota, Volvolepiota, Pseudobaeospora, Macro-
lepiota, Clarkeinda. Неамилоидиость спор — родовой признак у Hiatu-
lopsis, Phaeolepiota.
37
МКН
Рис. 4. Номограмма для определения линейных параметров базидии (S", со, т. Л!)
по заданным п и d
Макрохимическая реакция с реактивом Шеффера необычайно важ-
на для таксономии рода Agaricus на внутрнродовом и видовом уровнях.
Для всех видов подрода Agaricus, его секций Agaricus, Duploannulatae,
подсекций Agaricus, Sanguinolentae характерна отрицательная ре-
акция с реактивом Шеффера. Для всех видов подрода Flavoagaricus,
его секций Majores, Minores, подсекции Flavescentes (кроме подсек-
ции Xanthoderrnatae) характерна положительная реакция с реактивом
Шеффера.
Макрохимическая реакция с а-иафтолом — важный родовой при-
знак у Agaricus и Gyrophragmium. Пластинки видов родов Agaricus,
38
Gyrophragmium окрашиваются а-иафтолом в красный цвет, пластинки
всех остальных Agaricaceae а-иафтолом ие окрашиваются.
Микрохимическая реакция с серной кислотой, анилином имеет
важное таксономическое значение в роде Agaricus на видовом уровне.
Например, у A. romagnesii мякоть серной кислотой окрашивается в крас-
новатый цвет, анилином не окрашивается, а у близкого вида A. bresa-
dolianus мякоть от серной кислоты ие изменяется, а от аиилииа стано-
вится олнвковатой.
Макрохимическая реакция с едким кали необычайно важна в роде
Cystoderma для таксономии иа видовом уровне. Например, у С. ambro-
sii кутикулярные и субкутикулярные клетки шляпки ие окрашиваются
или изредка желтеют от едкого кали, у С. carcharias окрашиваются в
оливково-желтый цвет, а у С. granulosum, С. adnatifolium, С. caucasi-
cum, С. fa Пах — в красноватый, красиовато-ржавый, ржаво-коричневый
цвета.
До последнего времени культур а льио-морфологические особенности
слабо используются для таксономической характеристики агариковых
грибов. Интересные, несистематизированные сведения о культуральных
особенностях, в основном ряда видов рода Agaricus, имеются в работах
отечественных н зарубежных авторов (Kaarik, 1960; Semerdzieva, 1965;
Kiihner, 1969, 1980; Зерова, Дудченко, Рожеико, 1971; Бухало, 1973;
Гарибова, Сафрай, 1972, 1975, 1980; Гарибова, Шалашова, 1973; Гари-
бова, Мокеева, 1976; Сафрай, 1977; Дудка, Вассер, Бухало и др., 1978;
Вассер, Берегова, Бйпай, 1980; Бухало, Вассер, 1981, и др-).
В качестве дополнительного таксономического критерия мы изуча-
ли культурально-морфологические особенности 13 видов родов Agaricus,
Macrolepiota^ Leucocoprinus семейства Agaricaceae иа средах неопре-
деленного химического состава и на стандартизированных синтетиче-
ских питательных средах. Особое внимание было обращено иа установ-
ление питательных сред, являющихся оптимальными для роста колоний
агариковых грибов, выяснение влияния различных питательных сред
на динамику роста и культуральные признаки этих грибов, составление
сравнительных культуральных характеристик их на разных питательных
средах. Из исследованных такие виды, как Agaricus maskae, A. bernar-
dii, A. bcmardiiformis, A. cupreobrunncus, Leucocoprinus bresadolae,
Macrolepiota pucllaris, представляли особый интерес, поскольку впер-
вые были изучены нами в условиях культуры. Agaricus campestris,
A. arvensis, A. silvaticus, Macrolepiota ргосега и М. rhacodes требовали
дополнительных исследований.
У изученных нами видов агариковых грибов исследовались динами-
ка и морфология роста мицелиальной колонии иа пяти питательных
средах различного состава, микроскопические особенности мицелия, ре-
акции иа ряд ферментов, поведение в смешанной культуре. Полученные
данные использовались для более полной характеристики видов, опре-
деления видовой самостоятельности, в отдельных случаях — при обсуж-
дении положения их в системе определенных родов.
Показано, что для таксономической характеристики агариковых
грибов можно применять такие стабильные при разливдых условиях
культивирования признаки, как динамика роста, текстура колоний,
окраска колоний и реверзума, наличие пряжек иа мицелии, характер
бесполого спороношения. Наиболее благоприятными оказались сусло-
агаровые среды, а для ряда штаммов также и среда Молнша. Исследо-
вание динамики роста различных видов и штаммов одного и того же
39
вида агариковых грибов на различных питательных средах позволяет
заключить, что этот показатель специфичен и является стабильной куль-
туральной характеристикой. Рост колоний культур агариковых грибов
на разных питательных средах в основном постоянен и незначительно
зависит от их состава. Установлено, что колонии разных штаммов одно-
го и того же вида при выращивании их на одной и той же среде наря-
ду с общими культуральными признаками имеют специфические черты.
Отмечены отличия в текстуре, окраске колоний и реверзума одного и
его иа различных питательных
средах. Вариабельность куль-
туральных признаков, на-
блюдаемых у различных
штаммов одного и того же
вида, выращенных на одной
среде, меньше, чем вариабель-
ность этих же признаков у од-
ного и того же штамма, вы-
ращенного на разных средах.
Постоянный характер беспо-
лого конидиального и вегета-
тивного спороношения, обнару-
женный по нашим (рис. 5)
и литературным данным (КйЬ-
пег, 1969; Бухало, 1973; Са-
фрай, 1977; Гарибова, Сафрай,
1980) у 17 видов агариковых
грибов, независимо от состава
питательной среды дает осно-
вание считать их признаками,
несущими дополнительную так-
сономическую информацию.
Исследования поведения ага-
риковых грибов в смешанной
культуре показали видовую
самостоятельность некоторых
грибов (например, Agaricus
bernardii и A. bernardiiformis,
A. campestris и A. cupreobrun-
neus) и определили их культу-
ральную активность (Вассер,
1982).
Рис. 5. А—Agaricus silvaticus: а — гифа с
пряжкой; б — артроконидии; Б — Agaricus cu-
preobrunneus: а — гифа, б—артроконидии;
В — Agaricus arvensis: а — гифа, б — артро-
конидии; Г — Agaricus maskae: а — артроко-
нидии; Д — Agaricus bernardii: а — гифа с
пряжкой, б — анастомоз; Е — Macrolepiota рго-
сега: а — пряжки, б — анастомоз
У агариковых грибов наблюдается исключительное многообразие
пигментов. По мнению ряда авторов (Singer, 1975; Bcsl, Bresinsky,
1977; Kiihner, 1980), пигментация Agaricales s. 1. разнообразнее, чем у
цветковых растений, отличается химическими и физическими свойства-
ми, расположением на поверхности и внутри стеиок гиф, цистид. Пиг-
менты и их топография являются одним из важных критериев вида
у агариковых грибов.
Анализ данных о содержании некоторых соединений в карпофорах
агариковых грибов (Heinemann, Casimir, 1961; Шиврина, Пизковская,
Фалина и др., 1969; Зерова, Дудченко, Рожеико, 1971; Зерова, Мель-
ничук, 1972; Зерова, Череп, 1972; Шиврииа, 1973; Дудченко, Мельни-
чук, 1973, 1974; Bresinsky, 1974) показывает, что биохимические осо-
40
беиности могут быть использованы как таксономический критерий, но
прн планомерном сравнительном изучении большого числа видов раз-
личных родов. Последнее обстоятельство позволит определить биохими-
ческую родоспецифичиость. Однако на современном уровне знаний дан-
ный критерий следует использовать очень осторожно в связи с огра-
ниченным количеством изученных в отношении биохимических особен-
ностей видов.
Большое значение для систематики грибов имеют данные сканирую-
щей электронной микроскопии (СЭМ), позволяющие детально изучить
особенности микроструктур, не устанавливаемые при использовании
световой оптики (Закордонец, Вассер, 1982). Исследования микрострук-
тур агариковых грибов с помощью СЭМ дает возможность обосновать
таксономическое значение тех признаков, которым ранее не уделялось
должного внимания, а также вскрыть неизвестные детали морфологии
спор, важных для таксономии базидиомицстов (Pegler, Young, 1971,
1974; Зерова, 19746; Hesler, Smith А., 1979; Вассер, 1980, 1982; Bigclow,
1981). Тип и особенности структуры оболочки спор у изученных нами
(см. вклейку) агариковых грибов представляет собой один из главных
константных таксономических признаков всех рангов от вида до се-
мейства.
Значительным таксономическим критерием агариковых грибов яв-
ляется географическое распространение. Отдельные таксоны четко раз-
личаются по размерам ареалов. Тип ареала с рядом других признаков
может характеризовать и эволюционную прогрессивность таксона. На-
пример, древние реликтовые виды агариковых грибов — с хорошо выра-
женным гиатусом — встречаются в областях сохранения реликтовых
флор, не подвергавшихся оледенению в четвертичном периоде, и имеют
главным образом дизъюнктивные ареалы.
Одним из важных таксономических критериев агариковых грибов,
являются субстраты, на которых оии развиваются, и широта специали-
зации. Агариковые грибы относятся к шести эколого-биологнческим
группам: гумусовым сапротрофам, подстилочным сапротрофам, копро-
трофам, брнотрофам, лигнотрофам и карботрофам.
Проведенный анализ таксономической оценки признаков агарико-
вых грибов дает возможность выделить совокупности макро- и микро-
скопических признаков, характеризующих основные таксономические
категории.
На уровне семейства наиболее важные следующие признаки: форма
карпофоров и гимения, окраска гименофора, тип прикрепления пласти-
нок, тип и характер покрывала, анатомическое строение покровов, ха-
рактер оболочки и поры прорастания, форма спор и базидий, происхож-
дение и строение стерильных элементов, тип строения трамы, химиче-
ские цветовые реакции карпофоров и микроструктур.
Таксономические признаки, используемые при обосновании родо-
вых и внутриродовых таксонов агариковых грибов, разнообразны.
Основными из них являются консистенция, форма, характер поверх-
ности карпофоров, тип и характер покрывала, тип прикрепления пла-
стинок, цвет спорового порошка, анатомическое строение .покровов, кон-
систенция н цвет мякоти, тип строения трамы, характер оболочки, цвет
и форма спор, базндий и цистид, количество ядер в спорах, тип половой
совместимости, наличие нли отсутствие пряжек на мицелии, химические
цветовые реакции карпофоров и микроструктур, культуральные и эко-
лого-биологические особенности.
41
Таблица 1. Семейство Agaricaceae
42
в понимании различных авторов
Таксономический ранг и объем семейства а сери- новых. грибов
| Chlorophyll um Volvolepiota Macrolepiota Leucocoprinus 1 Leucoagaricus
Agaricaceae s. 1. К. Agariceae К Lepioteae
Agaricaceae s. 1.
Agaricaceae s. 1. ПС. Agaricoideae ПС. Lepiotoideae ПС. Tricboiomoi- deae
+ 4- Agaricaceae s. str. K. Agariceae K- Leucocoprineae K. Lepioteae K. Cystodermateae T richol omataceae K- Biannularieae
Agaricaceae s. 1. ПС Lepiotacees ПС. Tricbolomees
+ Agaricaceae K- Agariceae K- Lepioteae Cortinariaceae T richolomataceae K. Tricholomeae
+ + + + + Agaricaceae K. Agariceae K. Leucocoprineae K. Lepioteae K. Cystodermateae T richolomataceae K. Biannuiarieae K. Clitocybeae
4- -1- Agaricaceae T richoiomataceae
4- Agaricaceae (incl. Lepiotaceae)
Agaricaceae s. 1. ПС. Lepiotoidea
43
Ossi 'ишгам 'а С. П. Вассер, 1980. 1982 М. Я. Зерова, П. Е. Cocih, Г. Л. Ро- женко, 1979 М. Moser, 1978 *•
+ + 4- 4-
4* 4- + 4-
+ 4-
4- 4-
4- 4-
4* 4- 4-
4* +
4- 4-
4-
Я ь HQ p
re r> E' 5s" 2. re <n I n
1 X ? • 7a > I
to CD <5 ba S 0? e
jg <ra §
1 5
mig 1981 1973 | 197 i I97i
+ + t J- 4- Agaricus (PsalH- ota)
+ + 4* 4- Molanophyllum
Gyrophragmium
Endopt у chum
+ + Mlcropsalliota
+ + Cystoa {oficus
+ Crucispora
+ 4- - + - Lepiota
+ + + + Chamaemyces (Lepiotella)
+ Cystolepiota
4- + 4- Pseudobaeospora
+ Smithiomyces
4- Hiatulopsis
Sericeomyces J
Продолжение табл. 1
Таксономический ранг и объем семейства агари- ковых грибов
Cystoderma Dissodernw J Squamsnita | Phaeolepiota • | Armillaria Cdtathelasma (Biannulana) | Ripartitella 1 Verrucospora 1 Clarkcinda | | Chlorophyllum | I Volvolepiota 1 Macrolepiota I Leucocoprinus 1 Leucoagaricus j
+ 4- + 4- 4- 4- 4- + Agaricaceae
+ 4- 4- 4- 4- 4- 4- + Agaricaceae Cortinariaceae
4- + 4- 4- Agaricaceae
4- 4- 4- 4- + 4- 4- \ 4- 4- 4- 4- + + Agaricaceae К- Agariceae К. Leucocoprineae К- Lepioteae К- Cystodermateae Tricholornataceae K. Biannularieae
+ 4- + 4- Agaricaceae Lepiotaceae Tricholornataceae
4- + + 4- 4~ 4- Agaricaceae
4- 4- 4- Agaricaceae Lepiotaceae
4- + + +- 4- 4- 4- Agaricaceae K. Agariceae K. Cystodermateae K. Lepioteae K- Leucocoprineae
4- 4- + Agaricaceae K- Lepioteae K- Psaliioteae Trichoiomataceae K. Cystodemateae K. Biannuiarieae
4- 4- + 4- Agaricaceae K. Cystodermateae
4- 4- + 4- Agaricaceae K. Lepioteae K- Leucocoprineae
Pholiota.
Cort папасеае.
45
На уровне вида и внутривидовых таксонов 1 наибольшее значение
имеют следующие признаки: форма, размер, окраска, характер поверх-
ности шляпки и ножкн, тип ножки, наличие склероциев, консистенция
и цвет мякоти, запах и вкус карпофоров, окраска гнмеиофора, тип при-
крепления пластинок, характер покрывала, структура покровов и кути-
кулы, наличие илн отсутствие пряжек на гифах, форма, цвет, характер
поверхности спор, базндий, цистид, геометрические особенности бази-
дии, химические цветовые реакции карпофоров и микроструктур, куль-
туральные и эколого-бнологические особенности, тип ареала.
Трактовки, систематического положении и объема семейства Aga-
ricaceae в современной литературе (табл. 1) очень противоречивы. До
сравнительно недавнего времени современное семейство Agaricaceae
интерпретировали как колена или подсемейства гетерогенного семей-
ства Agaricaceae s. 1. (Konrad, Maublanc, 1948; Лебедева, 1949; Pilat,
1951b). В настоящее время большинство агарикологов (Singer, 1951,
1962, 1975; Dennis, Orton, Нога, 1960, 1974; Moser, 1967, 1978; Kreisel,
1969; Pegler, Young, 1971; Васильева, 1973; Вассер, 1980; Kiihner, 1980;
Bon, 1981) признают «односемейственную» структуру Agaricaceae. Не-
которые (Locquin, 1974; Michael, Hennig, Kreisel, 1977, 1981; Зерова,
CociH, Роженко, 1979) выводят «двухсемейственную» и даже «четырех-
семействеипую» структуру Agaricaceae, основываясь главным образом
на цвете спорового порошка. В семейство Lepiotaceae, например, в по-
следней системе сведены все роды Agaricaceae с белым споровым по-
рошком 2, без учета анатомо-морфологических особенностей карпофоров
н микроструктур (строения кутикулы шляпки, оболочки спор, наличия
или отсутствия поры прорастания, химических цветовых реакций карпо-
форов и микроструктур). Вследствие отмеченных недочетов в один род,
например Lepiota, сведены такие выдержавшие испытание временем
роды, как Macrolepiota, Leucocoprinus, Leucoagaricus (Зерова, CociH,
Роженко, 1979). Выделение семейств Lepiotaceae, Lepiotophyllaceae, Ме-
lanophyllaceae, Cystodermataceae представляется весьма сомнительным.
По сути дела, «дробители» Agaricaceae возводят колена в ранг само-
стоятельных семейств.
Попытку дробления семейств Agaricales s. 1. критиковал Р Зиигер
(Singer, 1962, 1975), полагая, что подразделение семейств на данном
этапе ограниченности знаний об их онтогенезе в ряде случаев несостоя-
тельно, с чем мы полностью согласны (Вассер, 1982). Если придавать
одному или небольшой группе признаков таксономический ранг семей-
ства, то можно выделить много новых семейств в Agaricales s. 1. Вы-
1 Понятие вида и внутривидовых таксонов агариковых грибов принимается
нами в основном по компромиссному определению, выработанному на симпозиуме
«Концепция вида у Hymenomycetes», который состоялся 16—20 августа 1976 г.
в Лозанне, Швейцария (The species concept.., 1976). Вид у агариковых грибов —
это совокупность особей, происходящих от подобных им родительских пар, со свой-
ственным комплексом наследственных анатомо-морфолиических признаков, биологи-
ческих, географических свойств, отграничивающих их от особей других родственных
видов. У агариковых грибов мы принимаем следующие внутривидовые таксономиче-
ские категории; разновидность н форму. Разновидность — это совокупность особей,
четко отличающихся от вида лишь некоторыми, наследственно закрепившимися анато-
мо-морфологическими признаками. Форма — это совокупность особей, четко отличаю-
щихся от вида лишь некоторыми наследственно закрепившимися морфологическими
признаками.
2 Мы цвету спорового порошка в современной систематике Agaricales s. I,
вслед за Р. Зингером (Singer, 1975), не придаем семейственного диагностического
ранга.
46
деление таких «семейств» может оказаться практически неограничен-
ным при новом наборе признаков. Более целесообразно иа данном
этапе расширять и углублять представление о едином семействе Agarica-
ceae с четырьмя коленами (Agariceae, Cystodermateae, Lepioteae, Leu-
cocoprineae) без нарушения его «границ». Следовательно, предложен-
ное еще в 1951 г. Р. Зингером (Singer, 1951) внутрисемейственное под-
разделение Agaricaceae выдержало испытание временем н представля-
ется таксонами, четко отличающимися по ряду признаков: характеру
прикрепления пластинок, типу строения трамы, типу и характеру по-
крывала, анатомическому строению покровов, структуре спор, хими-
ческим цветовым реакциям карпофоров и микроструктур (Singer, 1975;
Вассер, 1980, 1982; Воп, 1981).
Необходимо отметить и разнообразие трактовок систематического-
положення родов семейства Agaricaceae. Выделение определенных при-
знаков, играющих роль основных ведущих в комплексе других, харак-
терных для групп видов, относимых к тому или иному роду, часто от-
сутствует, и принципиально различные виды, по-видимому, с конвер-
геитно возникшим сходством базидиофоров объединяются в одном роде.
В то же время в некоторых случаях виды близкие, отличающиеся один
от другого наличием признаков, не имеющих значения специфических
родовых, помещают в отдельные роды (Зерова, 1973). Во многих слу-
чаях при выделении родов в семействе Agaricaceae, как и в других
группах Basidiomycetes (Каратыгин, 1981), не учитывается прагматиче-
ское «правило» таксономии (Майр, 1971), которое утверждает, что
величина гиатуса между родами должна находиться в обратной зависи-
мости от величины таксона. В связи с этим оценка числа родов Aga-
ricaceac в самых современных системах весьма различна: от 2—3 (Ма-
1еп£оп, Bertault, 1970) до 23 (Singer, 1975). Различные трактовки
объема и положения в системе родов Phaeolepiota, Catathelasma, Mac-
rolepiota, Lepiota, Leucocoprinus, Leucoagaricus и др. обусловливают их
дискуссионность. Некоторые роды (Phaeolepiota, Squamanita, Catathe-
lasma и др.) до последнего времени мигрируют в различные семейства
и даже порядки.
Род должен характеризоваться вполне определенным сочетанием
признаков, присущих семейству, с известными ограничениями, завися-
щими от природы входящих в него видов (Наумов, 1950). Однако мно-
гие роды агариковых грибов недостаточно хорошо обоснованы. Это
относится, например, к родам Hymenagaricus, Dissodcrma, Sericeomy-
ces. Единственным отличием монотипного рода Dissoderma от рода
Squamanita являются незначительные отличия в анатомии эпителия
ножки. Секция Sericella рода Lepiota возведена П. Хейнеманном (Hei-
nemann, 1978) в род Sericeomyces на основании шелковистой поверх-
ности шляпки, сформированной радиально расположенными нитевид-
ными гнфами, не образующими чешуек. Данные особенности являются
базисом для выделения секций в роде Lepiota и не могут играть роль
единственного родового критерия. Если пойти по такому пути, то все
секции рода Lepiota (Micaccae — эпикутнкула ложнопаренхимная, Echi-
natae — эпикутикула палисадная) можно безосновательно возвести в
ранг родов.
Описан и ряд промежуточных, недостаточно обоснованных родов,
отражающих признаки двух родов, сами названия которых (Chlorole-
piota, Cystolepiota, как близкие к Lepiota; Cystoagaricus, Hymenagari-
cus, как близкий к Agaricus) прямо указывают иа наличие сочетания
47
ведущих признаков. Возможно, на данном этапе развития знаний сле-
довало бы ие описывать новый род, а расширить представления об
исходном, в данном случае о Lepiota, Agaricus.
На практике трудно провести различия между родами Squamanita
и Dissoderma, Dissoderma и Cystoderma, Lepiota и Sericeomyces, Lepio-
ta и Cystolepiota и др.
Ряд родов трихоломатальных грибов (Catathelasma, Armillaria,
Floccularia), представителям которых свойствен гемиангиокарпный тип
развития, переносится в семейство Agaricaceae (Зерова, Cociu, Роженко,
1979; Romagnesi, 1980). В связи с отсутствием исчерпывающих данных
об особенностях онтогенеза большинства видов родов Tricholomatales
и Agaricaceae, считаем целесообразным оставить их в Tricholomatales
в ранге семейства Catathelasmataceae и не рассматривать в системе
агариковых грибов. То же относится и к роду Leucocortinarius (семей-
ство Cortinariaceae), переносимому в последнее время (Romagnesi,
1980) в Agaricaceae (колено Cystodermateae) без существенных обосно-
ваний родства.
К настоящему времени описано около 45 родовых названий агари-
ковых грибов, но лишь менее половины их относится к хорошо обосно-
ванным родам, выдержавшим испытание временем: Agaricus, Melano-
phyllum, Gyrophragmium, Endoptychum, Micropsalliota, Lepiota, Cha-
maemyces, Pseudobaeospora, Smithiomyccs, Hiatulopsis, Cystoderma,
Squamanita, Phacolepiota, Clarkcinda, Chlorophyllum, Macrolepiota, Leu-
coagaricus, Leucocoprinus и Volvolcpiota. Остальные роды (см. табл. 1),
особенно тропические (Crucispora, Cystoagaricus, Ripartitclla, Verruco-
spora, Chlorolepiota), требуют дальнейшего тщательного исследования
и на современном уровне знаний не могут быть отнесены к изучаемому
нами семейству.
В результате всестороннего изучения агариковых грибов с учетом
ранее существующих систем и их критического рассмотрения мы пред-
ложили оригинальную систему основных родовых таксонов Agaricaceae
(Вассер, 1978а, 1980, 1982). Предлагаемая система основана на комп-
лексе признаков. В родах Agaricus, Lepiota, Leucocoprinus, Cystoderma
пересмотрены внутриродовые систематические критерии.
В роде Agaricus в качестве важных признаков виутриродовой так-
сономии выдвигаются такие, как цвет карпофоров при прикосиовеиии,
цвет мякоти при автооксидации, форма хейлоцистид, биометрическая
характеристика спор, реакция мякоти с реактивом Шеффера, запах
карпофоров (Вассер, 1980). В результате использования всех перечис-
ленных признаков в роде Agaricus нами описано два подрода: Agaricus
(карпофоры при прикосновении не изменяются или окрашиваются в
розоватый, красноватый цвета, мякоть при автооксидации окрашивается
в светло-розовый, красный, карминио-красный цвета, запах кисловатый,
реакция с реактивом Шеффера отрицательная) и Flavoagaricus (карпо-
форы при прикосновении желтеют, мякоть прн автооксидации окраши-
вается в светло-желтый, желтовато-розовый цвета либо интенсивно
окрашивается в ярко-хромово-желтый цвет лишь в основании ножки,
запах миндаля, аииса или без особого запаха, реакция с реактивом
Шеффера, как правило, положительная, исключение составляют виды
подсекции Xanthodermatae). В подроде Agaricus описано две новые для
науки секции Agaricus (кольцо простое, хейлоцистиды разнообразной
формы — от шаровидных до булавовидных, споры яйцевидные, яйцевид-
но-эллипсоидные) и Duploannulata (кольцо перонатное или два кольца
48
на ножке, хейлоцистиды узкие, шпилевидныс, булавовидные, споры
округлые, округло-яйцевидные).
В роде Lepiota в качестве важнейших признаков внутрнродовой
таксономии выдвигаются характер поверхности шляпки, структуры эпи-
кутикулы, размер, форма н микрохимические цветовые реакции спор
(Вассер, 1978а, 1980, 1982). На основании различных комбинаций ука-
занных признаков в роде Lepiota нами описано два подрода: Lepiota
(поверхность шляпки голая или чешуйчатая; чешуйки часто образуются
от разрывов покрова, между которыми видна светлая мякоть, споры
до 21 мкм длиной, удлиненио-эллипсоидиые, эллипсоидные, веретено-
вндные, бомбовидиые, клиновидные, неамилоидные) и Sphaerocystis
(поверхность шляпки покрыта легко стирающимся зернисто-мучиистым
налетом, состоящим из сфероцист, или покрыта отстающими, легко
стирающимися чешуйками, состоящими из сфероцист, споры до 8 мкм
длиной, овальные, яйцевидно-эллнпсоидиые, амилоидные). В подроде
Lepiota нами описано три новых для науки секции Cristatae (кожица
шляпки сформирована гимениальной тканью, споры клиновидные, бомбо-
видные), Lepiota (кожица шляпки образована палисадной тканью, споры
удлииенно-веретеновидиые, часто с супраапикулярной депрессией), Sc-
ricellae (кожица шляпки сформирована радиально расположенными ни-
тевидными гифами, споры эллипсоидные, миндалевидные).
В роде Leucocoprinus в качестве важнейших признаков виутриро-
довой таксономии выдвигаются цвет карпофоров и мякоти при прикос-
новении, автооксидации, а также макрохимическая цветовая реакция
их с 25%-ным водным раствором аммиака (Вассер, 1978в). На осно-
вании различных комбинаций указанных признаков род Leucocoprinus
подразделен нами на два подрода: Rubescentes (цвет карпофоров и мя-
коти при прикосновении, автоокендации становится красновато-корич-
невым, с 25%-ным водным раствором аммиака — зеленоватым) и Leu-
cocoprinus (цвет карпофоров и мякоти при прикосновении, автооксида-
ции, с 25%-иым водным раствором аммиака не изменяется).
В роде Cystoderma в качестве важнейших признаков внутрнродовой
таксономии выдвигаются микрохимическая реакция спор с реактивом
Мельцера, наличие плевро- и хейлоцистнд, их форма, характер поверх-
ности (Вассер, 1978а, 1980, 1981). На основании различных комбинаций
указанных признаков род Cystoderma подразделен на два подрода”
Cystoderma (споры амилоидные или амилоидные лишь в супраапику-
лярной зоне) и Granulosa (споры неамилоидные). В дальнейшем подрод
Cystoderma подразделяется (Heinemann, Thoen, 1973; Вассер, 1980,
1981) на секции Cystoderma (споры амилоидные) и Superba (споры
амилоидные лишь в супраапикулярной зоне), а подрод Granulosa раз-
делен иа секции Granulosa (плевро- и хейлоцистиды отсутствуют) и
Cinnabarina (плевро- и хейлоцистиды имеются, веретеновндные, часто
инкрустированные).
Предложенная нами основанная на комплекса признаков модифи-
цированная система агариковых грибов признает внутриколенное под-
разделение, обоснованное Р. Зингером (1951, 1962, 1975), и предлагает
оригинальные системы основных родовых таксонов (Agaricus, Lepiota,
Leucocoprinus, Cystoderma) семейства (Вассер, 1978а, 1980, 1981, 1982).
4 4—2960
49
Обзор системы семейства Agaricaceae,
принятой в настоящей работе
Семейство Agaricaceae (Fr.) Cohn
Колено Agariceae Pat.
Род Melanophyllum Vel.
> Micropsalliota Hohn.
» Agaricus L. : Fr. emend. Karst.
Подрод Agaricus
Секция Agaricus
Подсекция Agaricus
» Sanguinolentae (J. Schaeff. et Moell.) S. Wasser
Секция Duploannulatae S. Wasser
Подрод Flavoagaricus S. Wasser
Секция Maj ores Fr.
Подсекция Flavescentes (J. Schaeff. et Moell.) S. Wasser
> Xanthodermatae (Sing.) S. Wasser
Секция Min ores Fr.
Подрод Lanagaricus Heinem.
Секция Olivacei Heinem,
» Lanosi Heinem.
» Trisulphurati Heinem.
Подрод Con io agaricus Heinem.
Секция Intermedii Heinem.
» Pulverotecti Heinem.
> Striati Heinem.
Род Gyrophragmium Mont.
» Endoptychum Czern.
Колено Cystodermateae Sing.
Род Phaeolepiota R. Aire ex Konr. et Maubl.
» Cystoderma Fay.
Подрод Cystoderma
Секция Cystoderma
» Superba Heinem. et Thoen
Подрод Granulosa (Fr ) S. Wasser
Секция Granulosa (Fr.) Locq. emend. Locq.
» Cinnabarina Heinem. et Thoen
Род Squamanita Imbach
Колено Lepioteae Fay.
Род Pseudobaeospora Sing.
» Lepiota (Pers.: Fr.) S. F. Gray
Подрод Sphaerocystis S. Wasser
Секция Micaceae J. Lge
» Echinatac Fay.
> Amyloideae Sing.
Подрод Lepiota
Секция Stenospora (J. Lge) Kuhn.
> Cristatae Kuhn, ex S. Wasser
> Lepiota
> Ovisporae (J. Lge) Kuhn.
> Sericellae Kuhn, ex S. Wasser
» Anomalae Locq.
Род Chamaemyces Batt, ex Earle
50
Род Smithiomyces Sing.
» Hiatulopsis Sing, et Grinling
Колено Leucocoprineae Sing.
Род Leucocoprinus Pat.
Подрод Leucocoprinus
» Rubescentes S. Wasser
Род Leucoagaricus (Locq.) Sing.
Секция Leucoagaricus
» Annulati (Fr.) Sing.
Род Macrolepiota Sing.
» Chlorophyllum Mass.
» Clarkeinda O. Kuntze
» Volvolepiota Sing.
Виды агариковых грибов (как и их таксоны надвидового ранга}
характеризуются признаками, относящимися к разным ступеням эво-
люционного развития. У агариковых грибов в результате неравномерной
эволюции наряду с прогрессивными признаками (отсутствие пряжек,
гомоталлизм, наличие стерильных элементов гимения, правильная ги-
менофоральная трама, слабо выраженный гиатус между вилами и др.)
сохранились примитивные признаки организации (дизъюнктивный
ареал, хорошо выраженный гиатус, сапротрофный образ жизни, отсут-
ствие изменчивости н внутривидового варьирования, гетероталлизм, на-
личие пряжек, отсутствие или слабая дифференциация стерильных эле-
ментов гимения, отсутствие дерматоцистид, волосков, хорошо развитое
покрывало, неправильная трама, округлая форма спор, гладкая оболоч-
ка спор, белый нли бледный споровый порошок, расширенная к шляп-
ке ножка, низбегающие пластинки, гомеомерная мякоть шляпки).
В связи с явлением гетеробатмии, значительным количеством призна-
ков, характерных для агариковых грибов, нам представляется невоз-
можным на данном уровне знаний построение филогенетических отно-
шений между родами или между внутри родовым и таксонами в виде
филогенетического «древа».
4*
ГЛАВА IV
КОНСПЕКТ* ФЛОРЫ
АГАРИКОВЫХ ГРИБОВ СССР
Настоящий Конспект представляет собой сводку о видовом соста-
ве, эколого-бнологических особенностях, распространеннн, системати-
ческом положении агариковых грибов, встречающихся в СССР.
Конспект составлен иа основании результатов собственных сборов
н наблюдений, произведенных в различных районах СССР, в первую
1 Условные сокращения, принятые в I
аз.— азиатский
аз-ам-афр — азиатско-американо-афри-
канский
Азиат, ч.— Азиатская часть
Алтайск. кр.— Алтайский край
альп.— альпийский
Аркт.— Арктика
бор.— бореальный
брнотр.— брнотроф
в. н. а.— вид с неопределенным ареалом
в. п. к.— вводится в промышленную
культуру
Вост. Сибирь — Восточная Сибирь
гум. сапр.— гумусовый сапротроф
ДагАССР — Дагестанская АССР
Дальн. Восток — Дальний Восток
дальневост. усл. энд.— дальневосточный
условно-эндемичный
д. р,— довольно редко
евр.— европейский
евраз — авразиатский
свраз-ам — евразиатско-американский
евраз-ам-австр — евразиатско-америка-
но-австралийский
евраз-ам-афр — евразиатско-американо-
африканский
евраз-афр — евразиатско-африканский
евр-афр - — европейско-африканский
евр-ам — европейско-американский
евр-ам-афр — ев ропейско-ам ерика ио-
африканский
Европ. ч.— Европейская часть
Зап. Сибирь — Западная Сибирь
копр.— копротроф
космопол.— космополитный
Краснодарск. кр.— Краснодарский край
Красноярск, кр. Красноярский край
ксеромерид.— ксеромерндиональный
к. п м.— культивируется в промышлен-
ном масштабе
лиги.— лигнотроф
м. с. г.— малоизвестный съедобный гриб
монт.— монтанный
мультирег.— мультирегиональный
Нац. окр.— Национальный округ
немор.— неморальный
и.— несъедобен
Новосибирск, обл.— Новосибирская
область
нр.— нередко
Общее распр.— общее распространение
опр.— определил
о. р.— очень редко
о. ч.— очень часто
п. з. и.— пищевого значения не имеет
подст. сапр.— подстилочный сапротроф
прим.— примечание
Приморск, кр —Приморский край
Распр. в СССР — Распространение в
СССР
р.— редко
с.— съедобен
с. м. в.— съедобен в молодом возрасте
северокавк. усл. энд.— северокавказский
условно-эндемичный
Сев. Америка — Северная Америка
собр.— собрал
Ср. Азия — Средняя Азия
Ставропольск кр.— Ставропольский край
с. я. г.— смертельно ядовитый гриб
Татарск. АССР — Татарская АССР
укр. усл. энд.— украинский условно-
эндемичный
ус.— условно съедобный
Хабаровск, кр.— Хабаровский край
ч.— часто
эвриголаркт.— эвриголарктнческий
Южи Америка — Южная Америка
я.— ядовит.
52
очередь в УССР, Латвийской ССР, на Кавказе, в Западной (Алтайский
край) н Восточной (Красноярский край, Бурятская АССР, Приморский
край) Сибнрн и некоторых других районах в 1969—1982 гг.; критиче-
ского изучения гербарного материала различных регионов страны. Прн
крнтнко-систем этическом изучении трудных, сомнительных и малоизве-
стных видов агариковых грибов были изучены их типы, использованы
все доступные нам литературные источники.
Конспект флоры агариковых грибов СССР составлен по системе
Р. Зингера (Singer, 1975) для таксонов рангом выше рода, а для виу-
триродовых таксонов по оригинальной системе, разработанной и опубли-
кованной нами (Вассер, 1978а, 1980, 1981, 1982).
Характеристика видового состава флоры Agaricaceae СССР рас-
смотрена по следующей схеме.
Для каждого вида приведено основное название, сведена вся сино-
нимика, а также дается ссылка на литературный источник, в котором
вид (илн внутривидовой таксон) был впервые описаи как под основным,
так и под синонимичным биномом. Далее следует основная, критически
изученная иконография. Затем краткая экологическая характеристика,
продолжительность вегетационного периода, степень встречаемости, хо-
зяйственная оценка, элемент флоры, тип ареала. После чего указано
местонахождение вида сначала на территории СССР, а затем приведено
его общее распространение на земном шаре. Завершают рассмотрение
критические примечания к некоторым из видов 1.
При экологической характеристике кроме мест обитания указывает-
ся трофическая группа. Продолжительность вегетационного периода от-
мечена римскими цифрами (например, V—XI). Частота встречаемости
видов приведена по следующей схеме: о. р.— очень редко (внд известен
из 1—5 местонахождений), д. р.— довольно редко (известен из 6—10
местонахождений), р.— редко (известен нз 11—15 местонахождений),
нр,— нередко (известен из 16—20 местонахождений), ч.—-часто (изве-
стен из 21—25 местонахождений), о. ч.— очень часто (известно более
30 местонахождений). Степень обилия плодовых тел вида указывают
арабские цифры в скобках: (1) — единично, (2) — группами, (3) — мас-
сово. Хозяйственное значение вида охарактеризовано по съедобности,
ядовитости и возможности культивирования в промышленном масштабе.
Для характеристики ареала вида в пределах СССР используется райо-
нирование, принятое нами (Вассер, 1980) во «Флоре грибов Украины».
Указывается район флоры с расшифровкой местонахождений по адми-
нистративно-территориальному делению (республика, край и в некото-
рых случаях область). Общее распространение приведено по частям
света, с расшифровкой стран и крупных островов, где внд был зареги-
стрирован. При указании распространения в СССР, как и общего рас-
пространения агариковых грибов, нами использованы сокращения, при-
нятые в «Малом атласе мира» (1981).
В Конспекте приведено 158 известных в настоящее время в Совет-
ском Союзе видов агариковых грибов, относящихся к 14 родам. Из ннх
9 видов и внутривидовых таксонов описаны нами как новые для науки
(Вассер, 1974а,г, 1975а, 1976а, 1977а,б, 1978а, 1980). Для 23 видов и
внутривидовых таксонов предложены новые для науки таксономические
* Критические примечания к коленам, родам и большинству видов Agaricaceae
СССР приведены в наших работах (Вассер, 1972а, б, 1974а. б, в, 1975а, б, 1976а, б, в,
1977а, б, 1978а, б, 1980, 1981, 1982; Херинк, Вассер, 1976; Вассер, Батирова, 1979;
Wasser, 1979).
53
комбинации (Вассер, 1976г, 1977а, 1978а, б, 1980, 1981; Wasser, 1979).
42 вида и внутривидовых таксона, а также одни род (Chlorophyllum)
мы приводим как новые для микофлоры Советского Союза 1 (Вассер,
1970, 1971, 1972а, б, 1973а, б, в, 1974а, б, в, 1975а, б, в, 1976в, г, 1977а, б,
19786. 1980. 1982; Вассер, Батирова, 1979; Wasser, 1979).
СЕМЕЙСТВО AGARICACEAE (FR.) COHN
Hedwigia, 11, 1872:17.— Hymenini Fr., Syst. Мус., I, 1821 : liv. Ivi,
p.p.— Agaricini Fr., Syst. Orb. Veg., 1825:65.— Agaricifortn.es Schwein.,
Schr. Naturf. Ges. Leipzig, 1, 1822 : 78.— Lepiotaceae Roze, Bull. Soc.
Bot. Fr., 23, 1876 : 51 (nom. nudum).— Polyphyllei Quel-, Enchir., 1886:
2.— Leucocoprinaceae Sing., Ann. Мус., 34, 1936:323 (nom. subnudum).
КОЛЕНО AGARICEAE PAT.
Hym. Eur., 1887:75 (Agarices); Henn, in Engl, et Pr., Nat. Pfl., 1,
1898 : 230.— Psallioteae Fay., Ann. Sci. Nat. Bot., VII, 9, 1889 : 352 (Psal-
liotes).— Psalliotoidea (подсем. Coprinacearum) Sing., Ann. Мус., 34,
1936 : 340.
РОД MELANOPHYLLUM VEL.
Ceske Houby, III, 1921 :569 (латинский диагноз у Pilat, Opera Bot.
cech., VI, 1948:219).— Chlorospora Mass., Kew Bull., 1896:136, non s.
Spcgazzini, 1891.— Chlorosperma Murr., Mycologia, 14, 2, 1922 : 96—97.—
Glaucospora Rea, Brit. Bas., 1922 : 62.
1. Melanophyllum echinatum (Roth: Fr.) Sing., Lilloa, 22, 1951
(1949): 236.— Agaricus echinatus Roth : Fr„ Syst. Мус., I, 1821:286.—
Psalliota echinata (Roth : Fr.) Kumm., Der Fuhrer in die Pilzkunde,
1871 : 69.— Lepiota haematosperma (Fr.) Quel., Champ. Jura et Vosges,
1876 : 536, s. Fr., auct. non Bros.— Pratella echinata (Roth : Fr.) Gill.,
Les Hymenomycctes, 1878:565.— Inocybe echinata (Roth : Fr.) Sacc.,
Syll. Fung., 5, 1888 : 773.— Lepiota echinata (Roth : Fr.) Quel., FL Мус.
Fr., 1888 : 296.— Cystoderma fumosopurpurea (Lasch) Fay., Ann. Sci.
Nat. Bot., VII, 9, 1889:35.—Agaricus olivaespores Ell. et Ev., Journ.
Мус., 5, 1889 : 27.— Pilosace clivaespora (EIl. et Ev.) Morg., Journ. Мус.,
1907, 13:225.— Hypholoma vinosum Kauffm., Agar. Mich., 1, 1918:94.—
Melanophyllum canali Vel., Ceske Houby, III, 1921 : 569 (латинский диаг-
ноз у A. Pilat, Opera Bot. cech., VI, 1948 : 219).— Chlorosperma olivaes-
pora (Ell. et Ev.) Murr. Mycologia, 14, 2, 1922 : 97.— Psathyra echinata
(Roth:Fr.) Overholts, Mycologia, 25, 1933:428.— Cystoderma echinata
(Roth : Fr.) Sing., Ann. Мус., 34, 1936: 338 (табл. I, а—г; XVII, /) 2.
Icon.: Bres., Icon. Мус., 1927, 40, fig. 2; Overholts, Mycologia, 25,
1933 : 428; J. Lge, Fl. Ag. Dan., 1935—1940, tab. 14, ftg. с (под названием
haematosperma); J. Schaeffer, Schweiz. Zeitschr. Pilzk., 22, 1944:12;
Melzer, Ces. Mykol., Vlll, 2, 1954 : 83, 87; Moser, KL Krypt.-flora, 1967,
fig. 80, 249, 412; Michael u. Hennig, Handb. Pilzfr., 1964, III, tab. 25;
Зерова, Атл. гр. Укр., 1974, табл. 120. рис. 5; Вассер, Укр. ботан. жури..
1 Новые для науки виды и разновидности отмечены в Конспекте тремя звездоч-
ками, новые для науки таксономические комбинации видового и внутривидового
рангов — двумя, а таксоны, новые для микофлоры СССР,— одной звездочкой.
s Табл. 1—XXXII помещены на вклейке.
54
30, 4, 1976, рис. 4а,б,в,г,д; Вассер, Фл. гр. Укр. (Agaricaceae), 1980,
рис. 18—19.
Во влажных широколиственных (Quercus robur L., Fagus silvati-
ca L., Carpinus betulus L.), елово-пихтовых, смешанных лесах, в парках,
садах, часто средн Urtica dioica L., на местах пожарищ среди Funaria,
в парниках. В горах доходит до высоты 1500 м н.у.м.; подст. сапр.,
VIII—Х, д. р., (1), п. з. и., мультирег., космопол.
Распр. в СССР. Европ. ч.: РСФСР — Ленинградская обл., ЭССР,
ЛитССР, УССР, Азиат, ч.: Дальн. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Испания, Франция,
Дания, Бельгия, Швейцария, Италия, ФРГ, ГДР, Норвегия, Лихтен-
штейн, ПНР, ЧССР, ВНР, Греция; Азия: Япония; Сев. Амери-
ка: США; Ю ж н. Америка: Аргентина; Африка: Алжир,
Марокко, Гайа; Австралия; Новая Зеландия, Новая Гвинея;
о-в Куба.
2. Melanophyllum eyrei (Mass.) Sing. var. eyrei, Lilloa, 22. 1951
(1949) : 236.— Schulzeria eyrei Mass., Grevillea, XXII, 1894:38,
tab. 185.— Chlorospora eyrei (Mass.) Mass., Kew Bull., 1898:136.—
Glaucospora eyrei (Mass.) Rea, Brit. Bas., 1922:62,— Lepiota eyrei
(Mass.) J. Lge, Dansk Bot. Ark., 3, 4, 1923 : 50.— Cystoderma eyrei
(Mass.) Sing., Schweiz. Zeitschr. Pilzk., 17, 1939:53 (табл. I, 2a—г;
XVII, 2).
Icon.: Massee, Grevillea, XXII, 1894, tab. 185, fig. 1; Svrcek, Ces.
MykoL, III, 8—10, 1949:106; Васильева, Агар. шл. гр. Приморск.
кр_, 1973 : 178, рис. 43, В; Вассер, Фл. гр. Укр. (Agaricaceae), 1980,
рис. 20.
В широколиственных (Fraxinus excelsior L., Fagus silvaticus L.,
Quercus robur L., Carpinus betulus L., Acer pseudoplatanus L.), хвойных
(Abies alba Mill.) н смешанных лесах, иа лугах.
М. eyrei var. macrosporuni Urbn., В. Урбонас, К. Каламээс, В. Лу-
кин. Коисп. нзуч. агар. гр. ЛитССР, ЛатвССР, ЭстССР, Вильнюс, 1974 :
54—55.
В парках (часто под Tilia cordata Mill., Acer platanoides L.). Изве-
стей лишь нз ЛитССР.
Под ст. сапр., VII—X, д. р., (1), п. з. н., эвриголаркт., евраз.
Распр. в СССР. Европ. ч.: ЛнтССР, УССР; Азиат. ч_: Дальн.
Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Дания, ЧССР, ВНР,
Италия.
Прим. Мы изучили типовой материал М eyrei var. macrosporum (Гербарий
института ботаники АН ЛитССР, г. Вильнюс, в парке Науейи — Вяркяй, под липами,
5.Х 1972, собр. И. Мазелайтис). Указания В. А. Урбонаса о том, что у данной
вариации пластинки приросшие, а трама пластинок неправильная, нашими исследо-
ваниями ие подтвердились. Для М. eyrei var. macrosporum характерны споры
5—7,5X2,5 мкм, зеленовато-синие в массе.
РОД AGARICUS L.: FR. EMEND. KARST. (S. STR.)
Bidr. Finl. Nat. Folk, 32, 1879: XXV— Pratella (Pers. :Fr.) S F.
Gray, Nat. Arr. Brit. PL, I, 1821 :626,— Psalliota (Fr.) Kumm., Der
FOhrer in die Pilzkunde, 1871 :23.— Psalliota (Fr.) Quel., Champ. Jura
et Vosges, 1872—1873: 139.— Fungus Adans.: O. Kuntze, Rev. Gen. PL,
3 (2), 1898:477.
55
Подрод Agaricus
Укр. ботаи. журн., 33, 3, 1976 : 250.
Секция Agaricus
Укр. ботан. жури., 33, 3, 1976 : 250-
подсекция AGARICUS
Укр. ботан. жури., 33, 3, 1976 : 250.
3. Agaricus romagnesii *** S. Wasser, Укр. ботан. жури., 34, 3,
1977:85.— Agaricus radicatus (Vitt.) Romagn., BSMF, 53, 1938:129
(non A. radicatus Fr., 1821).— Psalliota rudicata (Vitt.) Essette (nomen
non rite publicatum), Les Psalliotes, 1964, tab. 22.— Agaricus campestris
var. crassatiulata Sandor, Zeitschr. Pilzk., 24, 1959:80—-81.—A. bresado-
lianas Bohus s.'Reid, Fung. Rar. Icon. Col., VI, 1972:6 (species conglo-
merato) (табл. 1, За—б).
Icon.: Romagnesi, BSMF, 53, 1938:130, fig. 4; Essette, Les Psal-
liotes, 1964, 22; Moravec, Myk. sborn., 42, 1965: 105, fig. I; Bohus, Ann.
Hist.-Nat. Mus. Nat. Hung., 63, 1971 :79, fig. 1; Reid, Fung. Rar. Icon.
Col., VI, 1972, 29, fig. 9, a—c, pl. 42; Вассер, Укр. ботан. журн., 33, 4,
1976, рис. За, б; Вассер, Укр. ботан. журн., 34, 3, 1977, рис. 1; Вассер,
Фл. гр. Укр. (Agaricaceae), 1980, рис. 21, табл. I, 1.
В парках, садах, у края дорог, реже на опушках широколиственных
лесов; гум. сапр., V—X, о. р., (1), я., иемор., евраз.
Распр. в СССР. Евр on. ч_: УССР; Азиат. ч_: Ср. Азия —ТССР.
Общее распр. Европа: Великобритания, Франция, ФРГ, ЧССР,
ВНР.
4. Agaricus bresadolianus* Bohus, Ann. Hist.-Nat. Mus. Nat. Hung.,
61, 1969: 154, s. S. Wasser, Укр. ботан. жури., 27, 4, 1970:521, non s.
Reid, Fung. Rar. Icon. Col., VI, 1972 : 6.— Psalliota campestris Fr. var.
radicata Vitt. s. Bres., Icon. Мус., 1931, tab. 827 (табл. XVII, 3).
Icon.: Bres., Icon. Мус., 1931, tab. 827; Bohus, Ann. Hist-Nat. Mus,
Nat. Hung., 61, 1969 : 155, fig. 2; Вассер, Укр. ботан. журн., 27, 4, 1970:
521, рис. 1, с. 523, рис. 3.
В белоакацневых (Robinia pseudoacacia L.) иногда с примесью ви-
дов рода Populus лесах, на песчаной почве; гум. сапр., X—XI, о. р.,
(1), с., немор., евр.
Распр. в СССР. Е в р о п. ч.: УССР.
Общее распр. Европа: ВНР, Италия.
5. Agaricus campestris L.: Fr. var. campestris, Syst. Мус., I, 1821:
281, s. restr. J. Lge, Dansk, Bot. Ark., 4, 12, 1926 : 9.— Psallioia campest-
ris (L.: Fr.) Quel., Champ. Jura et Vosges, I, 1872 : 108.— Pratella cam-
pestris (L. : Fr.) S. F. Gray var. alba Gill., Champ. Fr., 1874:561.—
Agaricus campestris L.: Fr. subsp. albas Konr. et MaubL, Icon. Sei. Fung.,
6, 1927 :60.— Psalliota exserta Viv. s. Bres., Icon. Мус., 1931, tab. 828.—
P. flocculosa Roa, Appendix II to Brit. Bas., 1932:50.— Agaricus cam-
pestris L.: Fr. var. squamulosus (Rea) Pil., Acta Mus. Nat. Prag., VIIB,
1, 1951 : 26.— A. campestris L.: Fr. f. substerilis (Moell.) Moell., Friesia,
5, 1951 : 56, syn. nov. (табл. I, 4a—д; XVII, 4).
Icon.: Rolland, Atl. Champ. Fr., 1906—1907, tab. 70, fig. 157; Bres.,
Icon. Мус., 1927, tab. 823 н tab. 824 (под названием var. alba); J. Lge,
56
Fl. Ag. Dan., 1935—1940, tab. 138, fig. c; Pilat, Acta Mus. Nat. Prag.r
VIIB, 1, 1951, fig. 1—6; Moeller, Friesia, 4. 1950, pl. XVII, fig. 14;
Essette, Les Psalliotes, 1964, tab. 26; Зерова. Атл. гр. Укр., 1974,
табл. 115, рнс. 1; Сержанина, Съед. и яд. гр., 1967, табл. 26, рис. 2;
Вассср, Солдатова, Высш. баз. ст. зоны Укр., 1977, рис. 6; Вассер,
Фл. гр. Укр. (Agaricaceae), 1980, рис. 22—23, табл. II, 1.
На полях, лугах, в степях, огородах, садах, на полянах и опушках
лесов, в полезащитных лесопосадках. В изобилии растет в заповедных
целинных степях, образуя иногда огромных размеров «ведьмины
круги».
A. campestris var. isabellinus (Moell.) Pit, Acta Mus. Nat. Prag.,
VIIB, 1, 1951 : 4.— Agaricus campestris L.: Fr. var. isabellinus (Moell.)
Moell., Friesia, 5, 1951 : 204.—Icon.: Essette, Les Psalliotes, 1964, 28.
На лугах. Известен из УССР, ТССР.
Гум. canp., V—XI, о. ч., (2—3), с., мультнрег., космопол.
Распр. в СССР. Е в р о п. ч.: повсеместно; Азиат, ч.: повсеместно.
Общее распр. Европа: повсеместно; Азия: повсеместно; Сев.
Америка: повсеместно; Южн. Америка: Бразилия, Боливия,
Аргентина, Чили, Венесуэла; Африка: Алжир, Египет, Заир, Ниге-
рия, Марокко, Кения, ЮАР; Австралия; о-ва Гренландия, Трини-
дад, Исландия, Ямайка, Филиппинские о-ва, Новая Зеландия, Куба,
Мадагаскар, Соломоновы о-ва.
6. Agaricus spissicaulis * (Moell.) Moell., Friesia, 5, 1951:203.
Psalliota spissa Moell., Friesia, 4, 1950:43, non A. spissus Fr., 1838.—
Agaricus spissus (Moell.) Pil.. Acta Mus. Nat. Prag., VIIB, 1, 1951:5
(табл. 1, 5a—б; XVI1, 5).
Icon.: Moeller, Friesia, 4, 1950, pl. Ill b; Michael u. Hennig, Handb.
Pilzfr., 1967, IV, tab. 4; Вассер, Укр. ботан. жури., 33, 4, 1976, рис. la, б;
Wasser, Fung. Rar. Icon. Col., X, 1979, pl. 75; Вассер, Фл. гр. Укр. (Aga-
ricaceae), 1980, рис. 25—26.
В парках, под тополями, в насаждениях белой акации, на лугах,
в траве; гум. canp., V—IX, о. р., (1), с., немор., евр-афр.
Распр. в СССР. Е в р о п. ч.: УССР.
Общее распр. Европа: Великобритания, Франция, Дания, ЧССР,
ФРГ; Африка: Алжир, Марокко.
7. Agaricus vaporarius (Vitt.) Mos., KI. Krypt.-flora, B2, Teil 2,
1967:190.— Agaricus campestris p. pratensis, vaporarius Vitt., Fung.
Mang., 1835:176.— A. campestris var. villaticus Brond., s. Cke, Quel.,
Konr. et Maubl., non s. J. Lge, Bres., Pcars.— Psalliota vaporaria (Vitt.)
Moell. et J. Schaeff., Ann. Мус., 34, 1, 1938:69.— P. setigera Paul.: Fr.
s. Ricken.— P. bivelata Vel., Ceske Houby, 1921 :562, non s. Pk (табл. I,.
6a—6).
Icon.: Vitt., Fung. Mang., 1835, tab. VIII; Velenovsky, Mycologia,
IV, 1927, tab. 5; J. Schaeffer in Michael, Fuhr. Pilzfr., 1939, tab. 50;
Moeller, Friesia, 4, 1950, fig. 8; Moser, KI- Krypt.-flora, 1967, fig. 359.
На лугах, в садах, парках, во дворах, на кучах мусора, по краю
дорог; гум. canp., VI—X, о. р_, (1), с. м. в., эвриголаркт., евр.
Распр. в СССР. Е в р о п. ч.: ЛатвССР, УССР.
Общее распр. Европа: Великобритания, Дания, ФРГ, ГДР,
ЧССР, ВНР.
8. Agaricus cupreobrunneus * (J. Schacff. et Steer) Pil., Acta Mus.
Nat. Prag., VIIB, 1, 1951 ; 14. Psalliota campestris L. : Fr. var. cupreo-
brunnea J. Schaeff. et Steer, Fiihr. Pilzfr., I, 1939:147.— P. cupreobrun-
neus (J Schaeff. et Steer) Moell., Friesia, 4, 1950 : 54.— Agaricus cup-
reobrunneus (J. Schaeff. et Steer) Moell., Friesia, 5, 1951 :204 (табл. I,
7a—6).
Icon.: Moeller, Friesia, 4, 1950, pl. IVb, XVI; Essette, Les Psallio*
tes, 1964, tab. 67; Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 28,
табл. IV, 2.
На лугах, полянах, в тнпчаково-ковыльных заповедных целинных
степях; гум. сапр., IX—X, о. р., (1), с., ксеромерид., евраз-ам.
Распр. в СССР. Европ. ч_: УССР.
Общее рахпр. Европа: Великобритания, Франция, Норвегия,
ГДР, Испания, Дания, ЧССР, ВНР, СРР; Азия: Турция; Сев.
Америка: США.
9. Agaricus porphyrocephalus* Moell., Friesia, 5, 1951; 204 — Psal-
liota porphyrea Moell., Friesia, 4, 1950:50.— Agaricus porphyreus
(Moell.) Pil., Acta Mus. Nat. Prag., V1IB. 1, 1951 : 26, non A. porphyreus
Alb. et Schw.: Fr., 1821.
Icon.: Moeller, Friesia, 4, 1950, pl. VIC; Essette, Les Psalliotes,
tab. 25.
На лугах, в полынио-злаковых заповедных целинных степях, часто
на песчаной почве; гум. сапр., IX—X, о. р., (1), с„ ксеромерид., евр.
Распр. в СССР. Е вро п, ч.: УССР.
Общее распр. Европа: Великобритания, Франция, Дания, ВНР.
Прим. A. porphyrocephalus был описан из Данин Ф. Г. Меллером (Moeller,
1950). По мнению Ф. Г. Меллера, он очень близок к A. cupreobrunncus (J. Schaeff.
et Steer) Pit, но отличается от него более вздутой ножкой н более мелкими спорами.
М. Мозер (Moser, 1967) высказывает предположение, что A. porphyrocephalus явля-
ется лишь разновидностью A. cupreobrunneus. После сравнительного исследования
плодовых тел A. porphyrocephalus и A. cupreobrunneus из Асканийской целинной
типчаково-ковыльной степи (Вассер, 1974а), ряда зарубежных гербариев (Вассер,
1980), считаем, что правомерно н целесообразно A. porphyrocephalus рассматривать
как самостоятельный хороший внд. Это подтверждается и данными статистической
обработки размеров спор (Вассер, 1980)
ПОДСЕКЦИЯ SANGU1NOLENTAE (J. SCHAEFF. ET MOELL.) S. WASSER
Укр. ботан. жури., 33, 3, 1976 : 250.
10. Agaricus benesll * (Pil.) Pil., Acta Mus. Nat. Prag., VIIB, 1,
1951 : 56.— Psalliota benesii Pil., Mycologia, 2, 1925 : 47—48.— P. ber-
nardii s. Ricken, Vadcmecum, cd. 1, 1920: 136, non P. bernardii s. Quel.—
P. bulbosa Vol., Novitat. Mycol., 1939:153.— Agaricus albosanguineus
Hotson et Stuntz, Mycologia, 30, 1938 : 217 (табл. XVII, 6).
Icon.: Benes, Cas. cesk. houb., 4, 1923—1924, fig. 44—48; Hotson
et Stuntz, Mycologia, 30, 1938, fig. 4; Pilat, Acta Mus. Nat. Prag., VIIB,
1, 1951, fig. 24; Pilat, Acta Mus. Nat. Prag., IXB, 2, 1953, fig. 45; Cap-
pelli, Boll. Grupp. mic. G. Bres., 1983, p. 17.
В смешанных дубово-еловых, смешанных широколиственных лесах;
гум. сапр., VIII—X; о. р., (1), с., эвриголаркт., евр-ам-афр.
Распр. в СССР. Европ. ч.: УССР.
Общее распр. Европа: Австрия, ЧССР, ВНР; Сев. Америка:
'США; Африка: Марокко.
11. Agaricus deylii* Pil., Acta Mus. Nat. Prag., VIIB, 1, 1951: 181 —
182 (табл. XVIII, 7).
Icon.: Pilat, Acta Mus. Nat. Prag., VIIB, 1, 1951, fig. 21—23;
Pilat, Acta Mus. Nat. Prag., fXB, 2, 1953, fig. 42—43; Вассер, Фл. гр.
58
Укр. (Agaricaceae), 1980, табл. IV, 1; Cappelli, Boll. Grupp. mic
G. Bres., 1983, p. 36.
В горных еловых (Picea excelsa L.) лесах; гум. сапр., VIII—X
о. p., (1), с., монт., евр.
Распр. в СССР. Е в р о п. ч.: УССР.
Общее распр. Европа: ЧССР, ВНР.
Прим. A. deylii, по данным автора вида (Piiat, 1951b), наиболее близок к
A. benesii, от которого отличается габитусом карпофоров, размером спор, экологией.
Изучен тип A. deylii из Herb. Krypt. Mus. Nat. Prag., Flora Bohemica: KarlStejn-
Boubova, in Picetis solo calcareo, 20.1X 1950, leg. Deyl M. et Piiat A., det Piiat A.,
N 649 934. Интересно отметить, что в диагнозе А. Пилата размер спор A. deylii
составляет 9,5—11X5,3—6 мкм, в то время как изучение спор типовых образиов
с последующей статистической обработкой показало, что их размер 7,0—8,6X4,5—
5,5 мкм. На нашем материале наиболее часто встречались споры следующих разме-
ров: 8,9—10,1X4,9—6,1 мкм.
Позже А. Пилат (Piiat, 1953) приводит оригинальное описание A. deylii, указы-
вая, что шляпка A. deylii, как и у A. benesii, может иметь более или меиее выра-
женную чешуйчатость, а споры — размер 9—9.5Х4.5 мкм.
12. Agaricus squamuliferus (Moell.) Pil., Acta Mus. Nat. Prag.,
VI1B, 1, 1951 : 7.— Psalliota squamulifera Modi., Friesia, 4, 1950:21.—
Agaiicus squamuliferus (Moell.) Moell., Friesia, 5, 1951 : 203.— A. caroli
Pil., Acta Mus. Nat. Prag., VIIB, I, 1951 :59.— A. squamuliferus (Moell.)
Pil. var. caroli Pil., A. Piiat, O. Usak. Nase houby, II, 1959: 141 (табл. II,
la—ж).
Icon.: Moeller, Friesia, 4, 1950, fig. 6, pl. XXXX; Piiat. Acta Mus.
Nat. Prag., VIIB, 1, 1951, fig. 25—28, tab. IX (caroli); Herink, Ces. My-
koL, 39, 8, 1954 :33; Essette, Les Psalliotes, 1964, tab. 9; Вассер, Укр.
ботан. журн., 29, 3, 1972, рнс. 2; Херинк, Вассер, Нов. сист. высш, и
низш. раст., 2, 1976, рис. 1—6; Вассер, Солдатова, Высш. баз. ст. зоны
Укр., 1977, рис. Ill; Wasser, Fung. Rar. Icon. Col., X, 1979, pl. 74, fig. 1.;
Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 29—35; Cappelli, Boll.
Grupp. mic. G. Bres., 1983, p. 25.
В хвойных (чаше всего еловых) и лиственных (с примесью видов
рода Populus) лесах, на' полянах, в парках часто образует «ведьмины
круги»; гум. сапр., VI—X, р., (2), с., эвриголаркт., евраз.
Распр. в СССР. Е в р о п. ч_: БССР, УССР; Азиат, ч.: Ср. Азия —
КиргССР.
Общее распр. Европа: Великобритания, Дания, ФРГ, ЧССР,
ВНР.
Прим. Подробные критико-систематические примечания к данному виду опуб-
ликованы в наших работах (Херинк, Вассер, 1976: Вассер, 1980) и работе А. Каппел-
ли (Cappelli, 1983).
13. Agaricus altipes (Moell.) Pil., Acta Mus. Nat. Prag., VIIB, 1,
1951 : 12.— Psalliota altipes Moell., Friesia, 4, 1950 : 46.— Agaricus alti-
pes (Moell.) Moell., Friesia, 5, 1951 : 203 (табл. II, 2a—6).
Icon.: Moeller, Friesia, 4, 1950, fig. 9; Michael u. Hennig, Hadnb.
Pilzfr., 1967, IV, tab. 2; Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 36,
табл. Ill, 1.
В хвойных лесах, часто на песчаной почве; гум. сапр., VII—X,
о. р., (1), н., бор., евр.
Распр. в СССР. Е в р о п. ч.: БССР, УССР, МССР.
Общее распр. Европа: Дания, ФРГ, ГДР.
14. Agaricus fuscofibrillosus (Moell.) Pil., Acta Mus. Nat. Prag.,
VIIB, 1, 1951 : 7.— Psalliota fuscofibrillosa Moell., Friesia, 4, 1950 : 27.—
59
Agaricus fuscofibrillosus (Moell.) Moell., Friesia, 5, 1951 .203 (табл. IIF
За—в- XVII1, 8).
Icon.: Moeller, Friesia, 4, 1950, pl. Ilb; Michael u. Hennig, Handb.
Pilzfr., IV, 1967, tab. 12; Вассер, Укр. ботан. журн., 33, 4, 1976, рнс. 2аг
б, в; Вассер, Фл. гр. Укр. (Agaricaceae), I960, рис. 38; табл. V, 1;
Cappelli, Boll. Grupp. mic. G. Brcs., 1983, p. 20.
В широколиственных, главным образом дубовых, лиственничных
лесах, на полянах; гум. canp., VI11—X, о. р. (1), с., немор., свраз.
Распр. в СССР. Европ. ч.: УССР; Азиат, ч.: Вост. Сибирь —
Красноярск, кр..
Общее распр. Европа: Великобритания, Франция, Дания, Бель-
гия, ВНР.
15. Agaricus lanipes (Moell. et J. Schaeff.) Sing., Lilloa, 22, 1951
(1949) : 432.— Psalliota lanipes Moell. et J. Schaeff., Ann. Мус., 36, 1938 :
65—69.— Agaricus lanipes (Moell. et J. Schaeff.) Pil., Acta Mus. Nat.
Prag., VIIB, 1, 1951 : 7 (nom. inval.) (табл. II, 4a—в).
Icon.: J. Schaeffer in Michael, Fuhr. Pilzfr., 1939, tab. 51; Moeller,
Friesia, 4, 1950, pl. X; Essette, Les Psalliotes, 1964, tab. II; Michael u.
Hennig, Handb. Pilzfr., 1968, I, tab. 20; Вассер, Фл. гр. Укр. (Agarica-
ссас), 1980, рис. 39, табл. I, 2; Cappelli, Boll. Grupp. mic. G. Brcs.,
1983, p. 24.
В лиственных и хвойных лесах; гум. canp., VII—X, о. р„ (2), с.,
эвриголаркт., евр.
Распр. в СССР. Европ. ч.: РСФСР — Ленинградская обл.
Общее распр. Европа: Даиня, ФРГ, ГДР, ЧССР, СРР.
16. Agaricus langei (Moell.) Moell. var. langei, Friesia, 5, 1951 : 203.—
Psalliota langei Moell., Friesia, 4, 1950 : 27.— P. haemorrhoidaria (Kal-
chbr. et Schulz.) Fr., s. J. Lge., J. Schaeff., Pii. (табл. II, 5a—e;
XVIll.P).
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, tab. 137, fig. C; J. Schaef-
fer in Michael, Fuhr. Pilzfr., tab. 53; Moeller, Friesia, 4, 1950, pl. XX;
Essette, Les Psalliotes, 1964, tab. 13; Moser, KI. Krypt.-flora, 1967,
fig. 357. (хейлоцнетиды); Вассер, Фл. гр. Укр. (Agaricaceae), 1980,
рис. 40—48; Cappelli, Boll. Grupp. mic. G. Bres., 1983, p. 24.
В еловых, сосновых, дубовых, буковых, смешанных лесах.
A. langei var. mediofuscus** (Moell.) S. Wasser. Укр. ботан. журн.,
35, 5, 1978: 516.— Agaricus mediofuscus (Moell.) Pil., Acta Mus. Nat.
Prag., VIIB, 1, 1951:8.— Psalliota mediofusca Moell., Friesia, 4, 1950;
30.— Agaricus mediofuscus (Moell.) Moell., Friesia, 5, 1951 : 203 (табл. Il,
6a—в).— Icon.: Moeller, Friesia, 4, 1950, pl. На, Xll; Essette, Les
Psalliotes, 1964, tab. 12; Michael u. Hennig, Handb. Pilzfr., 1967, IV,
abb. 9, fig. 14.
В хвойных (Picca, Abies) лесах. Известен из УССР.
Гум. canp , VII—XI, р., (1), с., эвриголаркт., евр-афр.
Распр. в СССР. Европ. ч.: ЭССР, УССР.
Общее распр. Европа: Великобритания, Франция, Бельгия,
СФРЮ, Дания, ФРГ, ГДР, ПНР, ВНР; Африка: Марокко.
Прим. A. langei наиболее близок к A. silvaticus Schaeff.: Seer, и A. haemor-
rhoidarius Kalchbr. et Schulz., от них отличается рядом признаков, из которых наи-
более важен, на наш взгляд, размер спор, значительно превышающий таковой у
последних видов.
По мнению А. Пилата (Pilat, 1951b), к A. langei из североамериканских видов
наиболее близок A. californicus Pk, для которого характерны споры 7—9 (10) X
60
Х4,5 (6) мкм. Вероятно, они и идентичны, однако это можно будет выяснить после
критико-систематического изучения типовых образцов обоих видов.
Ф. Г. Меллер (Moeller, 1950), Р. Зингер (Singer, 1975), М. Мозер (Moser. 1978)
считают Agaricus medioluscus самостоятельным видом, очень близким к A. langei.
Подобие макро- и микроскопических признаков, экологии дало нам основание считать
A. mediofuscus разновидностью A. langei (Вассер, 1980).
17. Agaricus variegatus (Moell.) Pil., Acta Mus. Nat. Prag., VIIB,
1, 1951:8.— Psalliota variegata Moell., Friesia, 4, 1950:31.— Agaricus
variegatus (Moell.) Moell., Friesia, 5, 1951 : 203 non A. (Mycena) varie-
gatus Fr., Syst. Мус., I, 1821 : 158 (табл. Ill, la—в).
Icon.: Moeller, Friesia, 4, 1950, pl. IA, XIII; Essette, Les Psalliotcs,
1964, tab. 62; Michael u. Hennig, Handb. Pilzfr., 1967, IV, tab. 13; Вас-
сер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 52.
В старых еловых, смешанных лесах; гум. сапр., VIII X, о. р., (2).
н„ бор., евраз-афр.
Распр. в СССР. Азиат, ч.: Дальн. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Франция, Дания, ФРГ.
ГДР; Африка: Марокко, Алжир.
18. Agaricus silvaticus Schaeff.: Seer. var. silvaticus Mycogr. Suis.,
I, 1833 : 176.— Psalliota silvatica (Schaeff.: Seer.) Kumm., Der Fuhrer in
die Pilzkunde, 1871 : 79, non s. Bres.— P. sanguinaria (Karst.) J. Lge,
Dansk Bot. Ark., 1924, 4, 4 : 13.— P. haemorrhoidaria s. Bres., Kauffm.
et A. H. Sm., Hotson et Stuntz (табл. Ill, 2a—в).
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, tab. 137, fig. B; Wake-
field a. Dannis, Comm. Brit. Fung., 1950, pl. 74, fig. 2; J. Schaeffer in
Michael, Fuhr. Pilzfr., 1939, tab. 52; Moeller, Friesia, 4, 1950, pl. XV;
Piiat, Acta Mus. Nat. Prag., VIIB, 1, 1951, fig. 29; Piiat, Mushrooms,
1954, tab. 102; Essette, Les Psalliotcs, 1964, tab. 16; Зерова, Атл. гр.
Укр., 1974, табл. 117, рис. 1; Вассер, Фл. гр. Укр. (Agaricaceae), 1980,
рис. 53, табл. VI; Cappelli, Boll. Grupp. mic. G. Bres., 1983, p. 12.
В хвойных (еловых, елово-кедрово-пихтовых, сосновых) и смешан-
ных (елово-буковых, сосиово-дубовых, сосново-березовых) лесах, глав-
ным образом на почвах, богатых кальцием.
A silvaticus var. pallidus (Moell.) Moell., Friesia, 5, 1951:203.—
Psalliota silvatica var. pallida Moell., Friesia, 4, 1950 : 38.
В лиственных лесах. Известен из РСФСР (Мурманская обл.. Якут-
ская АССР).
Гум. сапр., V11—X, о. ч., (2), с., мультирег., евраз-ам-афр.
Распр. в СССР. Европ. ч_: РСФСР — Ленинградская, Мурман-
ская, /Московская области. Карельская АССР, ЭССР, ЛитССР,
ЛатвССР, БССР, УССР; Азиат, ч.: Кавказ — АзССР, ГССР, Зап.
Сибирь — Томская обл., Вост. Сибирь — Красноярск, кр., Якутск. АССР,
Дальн. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Дания, Франция, Бель-
гия, Италия, ФРГ, ГДР, Швеция, Финляндия, ПНР, ЧССР, ВНР,
СФРЮ; Азия: Индия, КНР, Иран, Турция, Израиль, Шри Ланка,
Япония; Сев. Америка: Канада, США; Африка: Алжир, Заир,
Марокко, Тунис.
19. Agaricus haemorrhoidarius Kalchbr. et Schulz, f. haemorrhoida-
rius. Icon. Sei. Hym. Hung., 2, 1874, no. 37:29, s. Moell., non s. J. Lge,
J. Schaeff., Pil.— Psallioia haemorrhoidaria (Kalchbr. et Schulz.) Konr.
et MaubL, Icon. Sei. Fung., 1, 1924—1930:58 (табл. Ill, 3a—6).
Icon.: Kalchbr. et Schulz., Icon. Sei. Hym. Hung., 1874, tab. 18,
6i
fig. 1; Cke, III. Brit. Fung., 1881—1891, no. 550, tab. 531; Bres., Icon.
Мус., tab. 831; A. H. Smith, Pap. Mich. Acad. Sci., Arts a. Lett.. 25, 1939,
tab. 8; Konr. et Maubl., 1924—1937, Icon. Sei. Fung., pl. 28; Moeller,
Friesia, 4, 1950, pl. XIV, fig. 7; Essette, Les Psalliota, 1964, tab. 14;
Зерова, Атл. гр. Укр., 1974, табл. 117, рис. 2; Вассер, Фл. гр. Укр.
(Agaricaceae), 1980, рис. 54; Cappelli, Boll. Grupp. mic. G. Bres., 1983,
p. 13.
В широколиственных (дубовых, дубово-ясеиевых, вязовых) лесах
и насаждениях, на лугах.
A. haemorrlioidanus f. fagetorum ** (Pil.) S. Wasser, Укр. ботан.
журн., 35, 5, 1978 : 516.— Agaricus situations f. fagetorum Pil., Acta Mus.
Nat. Prag., VIIB, 1, 1951 : 132 —I с о n.: Pilat, Acta Mus. Nat. Prag.,
VIIB, i, 1951, fig. 30.
В буковых лесах. Известен из УССР.
Гум. сапр., VI—X, н. р., (2), с., мультирег., евраз-ам-афр.
Распр. в СССР. Европ. ч.: ЭССР, ЛитССР, ЛатвССР, УССР.
Общее распр. Европа: Великобритания, Франция, Дания, Ита-
лия, ФРГ, ГДР, Финляндия, ПНР, ЧССР, Австрия, ВНР, СФРЮ;
Азия: КНР; Сев. Америка: США, Канада; Африка: Алжир,
Заир, Марокко.
Секция Duploannulatae S. Wasser
Укр. ботам, жури., 33, 3, 1976 : 250,
20. Agaricus subfloccosus (J. Lge) Pil., Acta /Mus. Nat. Prag., VIIB,
1, 1951 : 49.— Psalliota campestris var. umbrina Fr., Syst. Мус., I, 1821 :
279.— P. hortensis var. subfloccosa J. Lge, Dansk Bot. Ark., 4, 12, 1926 :
8.— P. subfloccosa (J. Lge) J. Lge, Fl. Ag. Dan., IV, 1939 : 58 (табл. Ill,
4a—в-, XVIII, 10).
Icon.: Bres., Icon. Мус., 1927, 826 (под иазв. P. campestris var.
umbrina Fr.); J. Lge, FI. Ag., Dan., 1935—1940, tab. 139, fig. D; Moeller,
Friesia, 4, 1940 :4, fig. 1, 2; Piiat, Acta Mus. Nat. Prag., VIIB, 1, 1951 :
49, fig. 20; Essette, Les Psalliotes, 1964, tab. 4; Зерова, Атл. гр. Укр.,
1974, табл. 115, рис. 3 (под назв. A. bortensis Cke); Вассер, Укр. ботаи.
журн., 34, 3, 1977, рис. 2; Вассер, Фл. гр. Укр. (Agaricaceae), 1980,
рис. 55—57; Cappeili, Boll. Grupp. mic. G. Bres., 1983, p. 28.
В еловых лесах, часто на высоте 1300—1700 м и. у. м.; гум. сапр.,
VI—X, о. р., (1), с., бор., евр.
Распр, в СССР. Европ. ч.: РСФСР — Ленинградская обл., УССР.
Общее распр. Европа: Великобритания, Дания, Бельгия, Шве-
ция, ФРГ, Норвегия, ЧССР.
21. Agaricus bisporus (J. Lge) Imbach1, Mitt, naturf. Ges. Luzern,
15, 1946 : 15.— Agaricus campestris L.: Fr. var. hortensis Cke, Ill. Brit.
Fung., 1884—1896 : tab. 526.— Psalliota hortensis Cke emend. J. Lge
var. bispora J. Lge, Dansk Bot. Ark., 4, 12, 1926 :8.— P. bispora (J. Lge)
Moeli. et J. Schaeff., Ann. Mycol., 36, 1, 1938 : 69.— A. campestris L. : Fr.
var. bisporus Kligman, Amer. Journ. Bot., 30, 1943:746.— A. hortensis
(Cke) Pin, Acta Mus. Nat. Prag., VIIB, 1, 1951 : 37, non s. Pers.; Fr.,
1821.— P. hortensis (Cke) J. Lge, FL Ag. Dan., IV, 1939 : 58.— A. bispo-
1 Д. Маллох (Malloch, 1976), изучивший типовой материал A. brunnescens Pk
(Bull. Tor. Bot. Club, 1900, 27:16). считает A. bisporus синонимом первого, однако
исследования Р. Зингера (Singer R, Mycotaxon, 1984, XX, 2, р. 479—482) опровергли
данные Д. Маллоха.
€2
rus (J. Lge) Pil., Acta Mus. Nat. Prag., VIIB, 1, 1951 : 46 (табл. Ill,
5a—г-, XVIII, 11).
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, tab. 139, fig. A (hortensis)
fig. В (avellanea); Moeller, Friesia, 4, 1950, pl. \—VI; Pilat, Acta Mus.
Nat. Prag., VIIB, 1, 1951, fig. 7—19, tab. V—Vfl; Heinemann, Bull. Jard.
Bot. E’tat, 26, 1956, fig. 4; Essette, Les Psalliotes, 1964, tab. 1—2; Ba-
lazs et al., Gombatermesztes, 1973, fig. 9, 29, 37—-49; Девочкин, Шам-
пиньоны, 1975, рис. i, 3—4; Malloch, Mycologia, 68, 4, 1976, fig. I—7;
Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 58—59, табл. VII.
В заповедных целинных степях, в полезащитных лесополосах, иа
полянах лесов, лугах, выгонах, в садах, огородах, на кучах навоза,
унавоженной почве, в парниках, теплицах, шампиньонницах, разводится
искусственно круглогодично в 70 странах мира; гум. сапр., в естест-
венных условиях произрастает с V по X, о. ч., (3), с., к. п. м., муль-
тирег., космопол.
Распр. в СССР. Европ. ч.: повсеместно; Азиат, ч.: Кавказ —
АрмССР, Ср. Азия—КиргССР, ТаджССР, ТССР; Зап. Сибирь —
КазССР; Вост. Сибирь — Красноярск, кр.
Общее распр. Европа: повсеместно; Азия: Индия, КНР, МНР,.
Турция, Иран, Израиль, Шри Ланка, Япония; Сев. Америка; Ка-
нада, США, Мексика; Южн. Америка: Аргентина; Африка:
Алжир, Марокко, Египет, Заир; Австралия; Новая Зеландия; о-ва
Куба, Мадагаскар, Тайвань.
22. Agaricus bitorquis (Quel.) Sacc. var. bitorquis, Syll. Fung., V.
1887 : 998.— Psalliota bitorquis Quel., Ass. Fran?. Av. Sci., 1883:4.—
Agaricus campestris A. edulis Vitt., Fung. AAang., 1835:41.— A. cam-
pestris subsp. bitorquis (Quel.) Konr. et MaubL, Icon, Sei. Fung., 1924—
1937 : 60.— A. edulis (Vitt.) Moell. et J. Schaeff., Ann. Мус., 36, I, 1938:
75.— Psalliota rodmanii Pk, Ann. Rep. N. Y. State Mus., 36, 1894:45.—
P. edulis (Vitt.) Buchw., Spise — og Giftsvampe, 1937:125 (табл. Ill,
6a—в).
Icon.: Bres., Icon. Мус., 1927, tab. 824—825; J. Lge, Fl. Ag. Dan.,
1935—1940, tab. 138, fig. D; Pilat, Klic urc. nas. hub, 1951, obr. 48la,
490a, 493a, 495a, 536—538; Pilat. Acta Mus. Nat. Prag., VIIB, 1, 1951,
tab. IV; Essette, Les Psalliotes, 1964, tab. 3; Moser, KI. Krypt.-flora, 1967,
fig. 79, 358; Hlavacek, Mykol. sborn., 52, 2, 1975, fig. II—15; Зерова,.
Атл. гр. Укр., 1974, табл. 115, рис. 2; Вассер, Фл. гр. Укр. (Agaricaceae),
1980, рис. 60, табл. II, 2.
В парках, садах, огородах, на выгонах, кучах мусора, кладбищах,
улицах городов в трещинах асфальта (иногда бывает причиной выво-
рачивания асфальта).
A. bitorquis var. valldus* (Moell.) Moell., Friesia, 5, 1951:203.—
Psalliota edulis var. valida Moell., Friesia, 4, 1950 : 14.
В широколиственных лесах. Известен из УССР.
Гум. canp., V—X, о. ч., (2), с., к. п. м., мультирег.» космопол.
Распр. в СССР. Европ. ч.: повсеместно; Азиат, ч.; Кавказ —
ГССР, Ср. Азия — ТССР, УзССР, Зап. Сибирь — КазССР, Вост. Си-
бирь— Красноярск, кр.
Общее распр. Европа: повсеместно; Азия: Индия, КНР, МНР,
Турция, Иран, Израиль, Шри Ланка, Япония; Сев. Америка: Ка-
нада, США, Мексика; Южн. Америка: Чили, Венесуэла; Афри-
ка: Алжир, Заир, Марокко; Австралия; о-ва Куба, Мадагаскар,
Тринидад и Тобаго, Соломоновы о-ва.
63
23. Agaricus bernardii Quel, apud Cke et Quel., Clavus Hym., 1878:
’89.— Psalliota bernardii Quel., 6e-suppL Champ. Jura et Vosges, 1878:
123.—Agaricus bernardii (Quel.) Sacc., SylL, Fung., V, 1887 : 999.—-
A. campestris subsp. bernardii (Quel.) Konr. et MaubL, Icon. Scl. Fung.,
1924—1937 : 60, non s. Ricken (=A. benesii Pil.), non s. Rea ( = A. mac-
rosporus (Moell. et J. Schaeff.) Moell.), non s. Dvorak ( = A. squamuli-
ferus (Moell.) Pil.), non s. Vassilk., Zerova (=species conglomerato)
(табл. Ill, 7a—в).
Icon.: Quel., Sec. bot, XXV, 1878, tab. 3, fig. 12; Quel., 6€-suppI.
Champ. Jura et Vosges, 1878, PL III, fig. 12; Bernard, Champ. Roch.,
1882, tab. 23, fig. 1—2; Rolland, Atl. Champ. Fr., 1906—1910, tab. 71,
fig. 158; /Moeller, Friesia, 4, 1950, fig. 4, pl. VII; Essette, Les Psalliotes,
1964, tab. 7; Piiat, Ces. MycoL, 26, 2, 1972, tab. 82; Вассер, Солдатова,
Высш. баз. ст. зоны Укр., 1977, рис. 19; Wasser, Fung. Rar. Icon. CoL,
X. 1979, pl. 76; Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 61,
табл. VIII.
В заповедных целинных степях, полупустынях, пустынях, иа вы-
гонах, лугах, иа разнообразных почвах — от глинистых до песчаных,
от плотных такыров до развеваемых песков, часто более или менее
засоленных, на плакоре, в горах доходит до высоты 1500 м н. у. м.;
гум. сапр., V—X, р., (2), ус., ксеромерид., евраз-афр.
Распр. в СССР, Е в р о п. ч.: РСФСР— Владимирская обл., УССР;
Азиат, ч.: Кавказ — АзССР, Ср. Азия — УзССР, ТССР, Вост. Си-
бирь — Красноярск, кр.
Общее распр. Европа: Великобритания, Дания, Норвегия, Испа-
ния, Франция, Италия, ГДР, ЧССР, ВНР; Азия: КНР, МНР; Аф-
рика: Алжир, /Марокко.
Прим. A. bernardii наиболее близок к A. bitorquis и A. maleolens. Ю. Шеффер
(цит. по: Moeller, 1950, р. 16) считал даже A. bernardii разновидностью A. bitorquis.
А. Пилат (Piiat, 1972) показал, что A. bernardii s. Ricken идентичен A. benesii Pil.,
s. Rea — A. macrosporus (Moell. et J. Schaeff.) Moell., a s. Dvorak — A. squamuliferus
(Moell.) Pil.
В отечественной литературе нет единой точки зрения в понимании A. bernardii.
Б. П. Васильков (19536) считал, что A. bernardii идентичен A. bitorquis и занимает
промежуточное положение между A. campestris и A. arvensis. По меткому замечанию
А. Пилата (Piiat, 1972), A. bernardii s. Vassilk.— коллективный вид, который содер-
жит признаки видов секции Campestris Konr. et Maubl. Просмотр гербарных образ-
цов, положенных в основу описания, приведенного Б. П Васильковым, склоняет нас
к мысли, что A. bernardii s. Vassilk. идентичен с Agaricus gennadii (Chot. et Bond.)
P. D. Orton (Вассер, 1980).
24. Agaricus maleolens * Moell., Friesia, 5, 1951 : 203. Psalliota in-
grata Moell., Friesia, 4, 1950: 17.— Agaricus ingralus (Modi.) Pil., Acta
Mus. Nat. Prag., VIIB, 1, 1951 : 5 (табл. IV, la—6).
Icon.: /Moeller, Friesia, 4, 1950, fig. 5, pl. VIII; Essette, Les Psal-
liotes, 1964, tab. 8; Michael u. Hennig, Handb. Pilzfr., 1967, IV, tab. 9;
Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 62, табл. IX.
В широколиственных и хвойных (Pinus, Picea, Taxus) лесах (чаще
всего в белоакациевых и сосновых), в садах, парках; гум. сапр., VI—IX;
о. р„ (2), н., эвриголаркт., евр-афр.
Распр. в СССР. Е в р on. ч.: УССР.
Общее распр. Европа: Швеция, Дания, ЧССР; Африка: /Ма-
рокко.
25. Agaricus gennadii (Chot. et Boud.) P. D. Orton var. gennadii,
Suppl. Trans. Brit. Мус. Soc., 43, 1960: i2.— Chitonia gennadii Chot. et
64
ТАБЛИЦА Г
/ — Mel a nophy Hum echinatum (Roth: Fr.) Sing, (a — плодовые тела, б —споры, в — элементы кути-
кулы шляпки, г — гифы трамы (Иогак, 1968)); 2 — Melanophyllum eyrei (Mass.) Sing. var. eyrei
(a — плодовое тело, б — споры, в — базидии, г — элементы общего покрывала (ориг.)); 3 — Aga-
ricus romagnesii S. Wasser (« — плодовые тела, 6 — споры (ориг.)); 4 — Agaricus campestris L Fr
s. restr. J. Lge (a — плодовые тела, б — споры, в — базидии, г — гифы трамы, д — элементы
кутикулы шляпки (по Horak, 1968, с нашими дополнениями)); 5 — Agaricus spissicaulis (Moe’l.)
Moell. (а —плодовые тела, б — споры (ориг.)); 6 — Agaricus vaporarius (Vitt.) Mos. (а-илодо
вне тела, б — споры, в — хейлоцистиды (ориг.)); 7 — Agaricus cupreobrunneus (J. Schaeff e’
Steer) Pil. (« — плодовые тела, б — споры (ориг.)).
5 4-2960
ТА БЛИЦА II
/ — Agaricus squamulilerus (Moell.) Pil. (« — плодовые тела, б разрез карпофоров, в — споры.
г — базидии, д — хейлоцистиды, е - разрез краев кольца, ж — нолусхематический вид кольца
снизу (Херинк, Вассер. 1976)); 2— Agaricus altipes (Moell.) Pil. (« — плодовые тела, б—споры
(Moeller, 1950—1951); 3 — Agaricus fuscofibrillosus (Moell.) Pil. (а —плодовые тела, б — споры,
в — хейлоцистиды (Moeller, i960—1951)); 4— Agaricus lanipes (Moell. et J. Schaeff.) Sing, (a —
плодовые тела, б — споры, в — хейлоцистиды (орцг.)); 5 — Agaricus langei (Moell.) Moell. var.
langei (a — плодовые тела, б — споры, в — хейлоцистиды (Moeller, 195Q—1951)); 6—Agaricus langei
(Moell.) Moell. var. mediofuscus (Moell.) S. Wasser (a — плодовые тела, б— споры, в—хеЙло-
цнстиды (Moeller, i960—1951))
ТАБЛИЦА III
1 — Agaricus variegatus (Moell.) Pil. (a — плодовые тела, б — споры, а — хейлоцистиды (Essette,
1964)): 2 —Agaricus silvatlcus Schaeff. : Seer. var. silvaticus (о — плодовые тела, б — споры
а — хейлоцистиды (Lange J., 1939)); 3 — Agaricus haemorrhoidarius Kalchbr. et Schulz
f. haeinorrhoidarius (о — плодовые тела, б — споры (Lange .)., 1939)); 4 - Agaricus subfloccosus
(J. Lge) Pil. (а — плодовые тела, б — споры, а — хейлоцистиды (Lange L. 1939)); 5 — Agaricus
bisporus (J. Lge) Imbach (a— плодовые тела, б—базидии, а—хейлоцистиды, г — споры (Ма1-
loch, 1976)); 6—Agaricus. bitorqpis (Quel) Sacc. var. bitorquis (a — плодовые тела, б — споры
а — хейлоцистиды (бриг,)); 7 — Agaricus bernardii Quel, apud Cke et QuCL (a — плодовые тела^
б споры, е— хейлоцистиды (по Michael, Hennig, i964—1971, с нашими дополнениями))
5'
s.
ТАБЛИЦ4 IV
/-Agaricus maleoiens Moell. (о -плодовые теля. б-(а-’пло^ь^^ела-
gennadii (Chot. et Boud.) P. D. Orton var. microsporus (Bohus) S . r s auct. non
б-споры, в-структура общего покрывала (Langei., 1939)); 4 - Agaricus
s. Ricken (о-плодовые тела, б споры, в хеп лО1ijст д t йлоцистиды (Heinemann,
kuehnerianus Heinem. [а - плодовые тела б - споры в ^иди тела. б - споры,
1974)); 5 -Agaricus macrosporus (Moel elt J ricS^aee«Jne^el Moeli.) Moell. (о - плодовые тела.
в - хейлоцистнды (M^4^J950-195H). Hodler. 1950-1951))
ТАБЛИЦА V
/—Agaricus tabularis Pk (a — плодовые тела, б - споры (по Василькову, 1955 с нашими Допол-
нениями)); 2 — Agaricus silvicolus (Vitt.) Sacc. (a - плодовые тела, б — споры (Essette, 1°64)‘
J—Agaricus macrocarpus (Moell.) Moell. (a — плодовые тела, б — споры (Moeller, 1950—1951));
4 — Agaricus abruptibulbus Pk (« — плодовые тела, б — споры (Essette. 1964)); 5 —Agaricus Hs-
suratus (Moeil.) Moell. (« — плодовые тела, б — споры (ориг.)); 6 — Agaricus nivescens (Moell.)
Moell. (« — плодовые тела, б — споры, в хейлоцистиды (ориг.)); 7 — Agaricus aestivalis (Moell’)
Pil. var. aestivalis (с — плодовые тела, б — споры (ориг.))
ТАБЛИЦА VI
/ — Agaricus longicatidus S Wasser (а — плодовые тела, б — споры (ориг.)); 2 — Agaricus arvett-
sts Schaeff.: Seer, (a — плодовые тела, б — споры, в — хейлоцистиды (ориг.)); <? — Agaricus pseu-
dopratensis (Bohus) S. Wasser (« — плодовые тела, б — споры (Bohns, 1971; Вассер, 1980)}; 4 —
Agaricus phaeof epidot ns (Moell) Moell. (a — плодовые Тела, б — споры, в — хейлоцистиды (Moel-
ler, 1950—1951)); 5 — Agaricus meleagris (J. Schaeff.) Imbach (a — плодовые тела, б — споры, в —
хейлоцистиды (Essette, 1964)); 6 — Agaricus moelleri S Wasser (a — плодовые тела, б — споры,
в — хейлоцистиды (ориг.)); 7 — Agaricus porphyrizon Р. D. Orton (а — плодовые тела, б — споры
(ориг.)); 8 — Agaricus brunneolus (J. Lge) Pil. (« — плодовые тела, б — споры, е — хейлоцистиды
(Lange J., 1939))
ТАБЛИЦА VII
1 — Agaricus Semotus Fr. var. Semotus (а плодовые тела, б — ertopbi. в — ба шдНН, i — хейло-
цнетиды (Mailed], 1976)); 2 — Gyrophragmiuni duna)ii (Fr.) Zdlcr (a - плодовые юла, б — споры
(ориг.)); 3—Phaeolepiota ангеа (Гт.) R. Mre ex Копт, et Maubl. (a— плодовые тела, б — споры,
в —базидии, г—элементы кутикулы шляпки, д - гифы трамы (Ногак. 1968)); 4 Cystoderma
amianthina (Fr.) Fav. var. amianthina (a — плодовые тела, б—споры, в — базидии, г — элементы
кутикулы шляпки (Ногак, 1968)); 5 — Cystoderma rugosoreticulata (Lorinser) S. Wasser (а —пло-
довые тела, б — споры, в — базидии, г —элементы кутикулы шляпки, д — гифы трамы (ориг.));
6~ Cystoderma carcharias (Secr.) Fay. (a — плодовые тела, б — споры (ориг.))
ТАБЛИЦА VIП
/ _ Cystoderma superbum Htiilsm, emend Huijsm (u — плодовые тела, б — споры, в. г—-эле-
менты кутикулы шляпки (по Huijsman, 1956. 1958. с нашими дополнениями)); 2 — Cystoderma
ambrosii (Bres.) А. Н. Sm. et Sing, (a — плодовые тела, б — споры, в — базидии, г —элементы
кутикулы шляпки (ориг.)); ? —Cystoderma granulosa (Fr.) Fay. (u — плодовые тела, б — споры.
в _ базидии (ориг.)); 4 — Cystoderma cinnabarina (Alb. et Schw. ex Seer.) Fay. (с —плодовые
тела б —споры в — хейлоцистиды. г — пилеоцистиды, д — каулоцистиды (Heinemann. Thoen, l'J/3));
5 —Squamanita paradoxa (A. H. Sm. et Sing) Bas (a — плодовые тела, б— споры, в —конидие-
споры» г — хламидоспоры (Bas. 1965)); 6 — Pseudobaeospora pillodii (Quel.) S. Wasser s. HoraK
(c — плодовые тела, б — споры, в базидии, г — гифы трамы (ориг.))
ТАБЛИЦА IX
/ — Lepiota semihuda (Lasch) Kumm. f. seminuda (« — плодовые тела, б — споры, в — элемент»!
кутикулы шляпки (ориг.)); 2 — Lepiota rosea Rea (« — плодовые тела, б—споры, в — хейлоцис
тиды. г — элементы кутикулы шляпки (Dennis, Orton, Нога, I960)); 3Lepiota pulvenilenta
Huijsm. (а — плодовые тела, б — споры, в — элементы общего покрывала (Herink, 1961)); 4 —
Lepiota hetleri Boud. s. Kuhn, (« — плодовые тела, б— споры, в — хейлоцистиды (Herink, 1961)):
5 — Lepiota acutesquamosa (Weinm.) Kumm. («— плодовое тело, б — споры, в — базидии, г —
хейлоцистиды, <3 — эпик ути к ул яри ые сфероцисты (Pegler, 1977)); 6—Lepiota llgnicola Karst, (a —
плодовое тело, б — споры, « — элементы эпикутиса (Васильева, 1973; Нездойминого, 1980))
ТАБЛИЦА X
/ Lepiota cortinarius J. Lge (a — плодовые тела, б — споры, в — хейлоцистиды (Lange J., 1939));
2 Lepiota grange! (Eyre) Kijhn (a — плодовые тела, б — споры, в — базидии, г — хейлоцистиды,
д— элементы кутикулы шляпки (Иогак. 1980)); 3—Lepiota pseudotelina J. Lge (а плодовые
тела> б —споры (Lange J., 1939)); 4 — Lepiota ignicolor Bres. (a — плодовые тела, б — споры (Ва-
сильева, 1973)); 5 — Lepiota tomentella J. Lge (a — плодовые тела, б — споры, в — хейлоцистиды
(Lange J.. 1939)): 6 — Lepiota caslanca Quel, (о — плодовые тела, б — споры, в — базидии, г —
хейлоцистиды, О—элементы кутикулы шляпки (no Lange J., 1939, с нашими дополнениями));
7 —Lepiota fulvella Rea (а — плодовые тела, б — споры (ориг.)); в—Lepiota cristata (Fr.) Kumm,
(g — плодовые тела, б -— споры, в — базидии, а — хейлоцистиды, б -— элементы кутикулы шляпки,
е— гифы трамы (Ногак, 1968))
ТАБЛИЦА XI
/ — Lepiota ermihea (Fr.) Gill, (a — плодовые тела, б — споры, в — хейлоцистиды (По Huijsman,
IS1.3. с нашими дополнениями)): 2 — Lepiota oreaditormis Vel. (« — плодовые тела, б — споры
(ориг.)): 3 — Lepiota subgracilis Kilhn. ex S. Wasser (« — плодовые тела, б — споры (ориг.));
4 Lepiota ventriosospora Reid (а — плодовые тела, б—споры, в — базидии, а — элементы кути-
кулы шляпки (Pegler. 1977)); 5 — Lepiota clypeolaria (Fr.) Kumm. var. clypeolaria (a — плодовые
тела, б споры, в базидии (Pegler, 1977)); 6—Lepiota rnfipes Morg. (a — плодовые тела, б—
споры, в — элементы кутикулы шляпки (ориг.)); 7 — Lepiota wlchanskyi Pil. (« — плодовые тела,
б—споры (Pilat, 1953))
ТАБЛИЦА XII
/ — Lepiota brunneoincarnata Chod. et Mart, (a — плодовые тела, б — споры (Babos, 1975)); 2 -—
Lepiota lateritiopurpurea L. Vass, (a — плодовые тела, б — споры (по Васильевой, 1973, с нашими
дополнениями)); 3—Lepiota micropholis (Berk, et Br.) Sacc. (a — плодовое тело. 6 — споры, в —
базидии, г — хейлоцистиды, д— элементы кутикулы шляпки (Pegler, 1977)); 4 — Lepiota subincar-
nata J Lge (a — плодовое тело, б — споры, в — элементы кутикулы шляпки, г — хейлоцистиды
(Pegler, 1977)); 5 — Lepiota cygnea J. Lge (а —плодовые тела, б — споры, в — хейлоцистиды, г —
элементы кутикулы шляпки (Huijsman. 1943: Васильева, 1973)1; 6—Lepiota xanthophylla Р. D Or-
ton (а — плодовое тело, б — споры, в — базидии, г — хейлоцистиды. д — элементы кутикулы шляпки
(Pegler, 1977)); 7 — Lepiota serena (Fr.) Sacc. (a — плодовые тела, б — споры, в — хейлоцис1иды
(Huijsman, 1943)); 8 — Chamaemyces fracidus (Fr.) Donk (a — плодовые тела, б — споры, в — бази-
дии, г — хейлоцистиды, д — элементы кутикулы шляпки (Horak, 1968))
ТАБЛИЦА ХИ!
/ — Chlorophyllum molybdites (Fr.) Mass, (a — плодовые тела, б—споры в — базидии г — хейло-
цистиды (Pegler. |Ь77)); 7 - Leucocoprinus cepaestipes (Fr.) Pat. (a — плодовые тела, б — споры,
в —базидии а —хейлоцистиды, б —элементы кутикулы шляпки (Horak, 1968)); 3 — Leucocoprinus
birnbaurmi (Corda) Sing (а — плодовые тела, б — споры, в — базидии, г — хейлоцистиды (ориг.)),
4 — Leucocoprinus denudatus (Rabenh.) Sing, (a — плодовые тела, б — споры, в — хейлоцпс1иды
(ориг)); 5 - Leucocoprinus badhamli (Berk, et Br.) S Wasser (a — плодовые тела, б — споры.
в — хейлоцистиды (ориг.)); 6 — Leucocoprinus pilatianus (Demoulin) S. Wasser var. pilatianns
(a — плодовые тела, б — споры (ориг.))
ТАБЛИЦА XIV
I — Leucocoprinus bresadolae (Schulz.) S. Wasser (a— плодовые тела, б — споры, в — хейлоцис-
тиды, г — элементы кутикулы шляпки (ориг.)); 2—Leucocoprinus bohusi S. Wasser (a—плодовые
тела, б — споры, в — хейлоцистиды (ориг.)); 3— Leucoagaricus macrorhizus Locq. ex Ногак (a —
плодовые тела, б — споры, в — базидии, г — хейлоцистиды, д — элементы кутикулы шляпки, е —
гифы трамы (Ногак, 1968)); 4 — Leucoagaricus holosericeus (Fr.) Mos. (a — плодовые тела, б —
споры, в — базидии, г — хейлоцистиды (ориг.)); 5 — Leucoagaricus moseri (S. Wasser) S. Wasser
(a — плодовые тела, б — споры, в — хейлоцистиды (ориг.))
ТАБЛИЦА XV
'~ Leucoagaricus leucothitus (Vitt.) S. Wasser f. leucothitus (a — плодовые тела, б—споры, в —
базидии (ориг.)); 2 — Macrolepiota procera (Scop. : Fr.) Sing, (a — плодовые тела, б — споры, в —
оазидии. а каулоцистиды, О — хейлоцистиды. е — элементы кутикулы шляпки (Horak, 1968)):
Macrolepiota rhacodes (Vitt.) Sing, (a — плодовые тела, б — споры, в — базидии (ориг))- 4 —
Macroleprota puellaris (Fr.) Mos. (a — плодовые тела, б — споры (Васильева 1973))- 5 — Macro
lep'ota konradn (Huijsm ex I> D Orton) Mos (а плодовые тела, б споры (Васильева. 1973))
о Macrolepiota exconata (Schaeff. : Fr.) S. Wasser var. excorlata (a — плодовые тела, б — споры
(ориг.))
ТАБЛИЦА XVI
1- Macrolepiota mastoidea (Fr.) Sing, («-плодовые тела, б
цпстиды d -элементы кутикулы' ^«"^^вы^ Te^’ бС_Дспор ” в -базидии (Bresadola,
f927C)' Гз- Macrolepiota olivleri (Baria) S Wasser («-плодовые тела б- споры (ориг.)); 4
Macrolepiota prominens (Fr.) Mos. («-плодовые тела, б - споры (ориг.))
ТАБЛИЦА XVII
t — Melanophyllum echinatum (Roth : Fr.) Sing.: споры (здесь и далее увеличение споп и nnvrux
микроструктур составляет 6500); 2 - Melanophyllum eyrei (Mass.) Sing 'оторы- 3 -TgaricusWhre
eHtUn&e^,YR%kAL₽srOPGe ВЖ
МоеН.:; споры; {р^АГ^
Flora Bohemlca. KarlMejn, Kralovska studanka. in margine Plceii et frondosae solo calcareo IX 1950
leg. et det. Plldt A., (Typus). N 649633 6
6 4—2960
Г
ТАБЛИЦА XVIII
7—Agaricus deylii Pil.: споры типа из Herb. Krvpt. Mus. Nat, Prag. (PR). Flora Bohemlca.
Karlstejn-Boubova, in Picetis soio caicareo, 20. IX 1950. leg. Deyi M. et Piiat A., det Piiat A.,
(Typus). N 649734; S — Agaricus luscofibriilosus (Moeli.) Pil.: споры; S —Agaricus langei (Moeli.)
Moell. var. langei.: споры; 10 — Agaricus subfioccosus (J. Lge) Pil.: споры; 11 — Agaricus bisporus
(J lge) Imbach: споры; 12 Agaricus augustys Fr. s. yyct. non s. Ricken: споры
ТАБЛИЦА XIX
W —Agaricus macrosporus (Moeil et J. Schaeff.) Moell.: споры 14 — Agaricus tabularis Pk споры;
15—IS — Agaricus maskae Pil.: споры типа из Herb Krypt. Mus Nat. Prag. (PR), Flora Bohemica,
Hab. loco graminoso solo calcareo inter Petrovice et Sloup Moraviae. 27. VH i954. ieg. MaSka L.,'
det. Piiat A. (Typus), N 649305; 17 — Agaricus amanitaeformis S. Wasser: споры типа из Гербария
Института ботаники им. Н. Г. Холодного АН УССР (KW). Донецкая Лесостепь, Донецкий бота-
нический сад АН УССР, 29. IX. 1972, собр. и опр, С. П. Вассер; 18 — Agaricus nivescens (Moell.)
Moell.: споры
6'
ТАБЛИЦА XX
/9 —Agaricus aestivalis (Moell.) Pil. var. aestivalis: споры: 20 — Agaricus tenuivolvatus (AfocII.)
Moell : споры; 21 — Agaricus longicaudus S. Wasser: споры типа из Гербарии Института ботаники
им. Н. Г. Холодного АН УССР (KW). Житомирсакя обл.. Житомирский р-н, окр. с. Тетеревка,
13. VI. 1974, собр. и онр. С. И. Вассер; 22 — Agaricus leucotrichus (Moell.) Moell.: споры; 23 —
Agaricus arvcnsis Schaeff. : Seer. ; споры; 24 — Agaricus pseudopratensis (Bohus) S. Wasser: споры
ТАБЛИЦА XXI
IS — Agaricus Veletiovskyi Pil.: споры типа из Herb. Mus. Nat. Prag. (PR). Hab. in declivitafe
coilis Budec prope Zakopany. distr. Kladno, 23. IX 1967. loco graminoso, leg. DenkMein et PilSt.
det. Pilat. (Typus) N 629423; 26 — Agaricus phaeolepidotus (Moeil.) Moeli.: споры; 27—28 — Agari-
eus porphyrizon P D Orton: споры; 29—Agaricus brunneolus (J. Lge) Pil: споры материала
из типового локалитета из Herb. Mus. Bot. Haun. (C). Loc. Fyn. Nyberg. Belleveenge. 22. X. 1973.
leg. et det. M. Lange, from type locality, 30 — Agaricus xantholepis (Moeli.) Moell.: споры
^Аблйца xXli
5/—3$ — Endoptychum agaricoides Czern.: cnopbl
ТАБЛИЦА XXIII
S3 — Cystoderma superbum Huifsm. emend, Huijsm.: споры голотипа из Herb. Lugd Botav, Rijkes-
herbarium, Leiden (L) Loc. Schwizerland, Caton Luzern, near SchCpfheim, on Sandust, probably
of Picea, leg. Mrs. A. W. Huijsman, det. H. S. C. Huijsman, 7.1X 1955, N 599.240.068; 34 — Lepiota
eriophora Pk: споры; 35 — Lepiota hystrix Moell. et J. Lge: споры; 36 — Lepiota cristata (Fr.)
Kumm.: споры; 37—Lepiota fuscovinacea J. Lge et Moell.: споры; 38 — Lepiota oread'iformls VeL:
споры
Таблица xxiV
— Lepiota oreadiformls Vel.: развивающиеся базидии co стеригмами и спорами; — Lepiota
ventriosospora Reid: споры
ТАелицА xtfV
41 — Lepiota ventnosospora feeid: споры; 42— Lepiota scobinella (Fr.) Gill.: споры; 43 — C.iiamab
myces fracidus (Fr.) Donk: базидия co стеригмами и спорами; 44 — Cham'aemyces fracidus (FrJ
Donk: споры
Таблица xxvt
^—46 — Leucocoprinus cepaestipes (Fr.) Pat.: споры; 47 — Leucocop-
rinus piiatianus (Demoulin) S. Wasser var. pilaiianus: споры
Таблица xxvit
46—49 — Leucocoprinus. bresadolae (Schulz.) S Wasser: chopbl
ТАблицА kxvtii
5б — Leucocoprinus bohusi S Wasser: споры типа из Гербария Института Йотаникй
им. Н. Г. Холодного АН УССР (KW), Донецкая обл.. Новоазовский р-н, заповедник
«ХомутовсКая степь», разнотравно-типчаково-ковыльная степь, 12.V 1970, собр. и опр.
С. П. Вассер; 51 — Leucoagaricus carneifolius (Gill.) S. Wasser: споры
ТАБЛИЦА XXIX
52 — Leucoagaricus carneifoiius (Giil.) S. Wasser: споры; S3 — Macrolepiota otivieH
(Bar|a) S. Wasser: споры
ТАБЛИЦА XXX
.>4 — Macrolepiota procera (Scop. : Fr.) Sing.: споры
ТАБЛИЦА XXXI
55 — Macrolepiota rhacodes (Vitt.) Sing.: споры; 56 — Macrolepiota excoriata (Schaeff Fr )
S. Wasser удг. ^xcorlata: споры
ТАБЛИЦА XXXII
57 — Macrolepiota mastoidea (Fr.) Sing.: споры; 58 — Chlorophyll urn molybdites (Fr.)
Mass.: споры
BoucL, Journ. Bot., 12, 1898 :65.— Clarkeinda cellaris Bres., Ann. Мус.,
3, 1905 : 162.— Chitonia cellaris (Bres ) Bond., Ic. Мус., I, 1905 : 125.—
C. gennadii (Chot. et Bond.) Sacc. et Syd., Syii. Fung., 21, 1912:202.—
Psalliota gennadii (Chot. et Boud.) Mai?n. et Bert., Fl. Champ. Super.
Магос, I, 1970: 164 (nom. nudum).
icon.: Bres., Icon. Мус., 1931, tab. 834: Dennis, Orton a- Hora,
Suppi. Trans. Brit. Мус. Soc., 43, i960, fig. 181; Вассер, Нов. снст.
высш, и низш. раст., 3, 1977, рис. 3.
В степях, садах, огородах.
A. gennadii var. microsporus ** (Bohus) S. Wasser, stat. nov.
(табл. IV, 2a—в).— Agaricus gennadii ssp. microsporus Bohus, Ann.
Hist.-Nat. Mus. Nat. Hung., 67, 1975 : 38.— Icon.: Bohus, Ann. Hist.-Nat.
Mus. Nat. Hung., 67, 1975, fig. 1A, В; Вассер, Батирова, Укр. ботан.
журн., 35, 5, i 979, стр. 424, рис. i.
В полупустынях. Известен из ТССР.
Гум. сапр., V—XI, р., (2), с., ксеромерид., евраз-афр.
Распр. в СССР. Европ. ч.: УССР; Азиат, ч.: Ср. Азия — ТССР.
Общее распр. Европа: Великобритания, ВНР, Италия, Франция;
Азия: КНР; Африка: Марокко.
Подрод Flavoagaricus S. Wasser
Укр. ботан. журн., 33, 3, 1976 : 250.
Секция Majores Fr.
Hymen. Eur., 1874 : 211.
ПОДСЕКЦИЯ FLAVESCENTES (J. SCHAEFF. ET MOELL.) S. WASSER
Укр. ботан. журн., 33, 3, 1976 : 250.
26. Agaricus augustus Fr. var. augustus, Epicr. Syst. Мус., I, 1836:
212, s. auct. non s. Ricken, Die Biatterpilze (Agaricaceae) Deutschiands...,
1915:235.— Psalliota augusta (Fr.) Quel., Champ. Jura et Vosges, I,
1872:255.— Pratella augusta (Fr.) Giil., Champ. Fr., 1878:561.— Aga-
ricus perrarus Schuiz., Verh. zooi.-bot. Ges., XXIX, 1879 : 493.— Psalliota
perrara (Schulz.) Bres., Fung. Trid., I, 1884:82.— A. bresadolae Schuiz.,
Hedwigia, 24, 1889 : 135 (nom. nudum), non s. Baar, Bull. Soc. Royal.
Bot. Beig., 70, i937 : 48.— Psalliota villatica Bres., Fung. Trid., I, 1884:
54.— P. praenitens Beck, Verh. zooi.-bot. Ges., Wein, 1889:611.— Agari-
cus peronatus Mass., Eur. Fung. Fi., I, 1902:204.— Psalliota peronata
(Mass.) Rea, Brit. Bas., 1922:83.— P. perrara Schuiz. s. Ricken, Die
Biatterpiize (Agaricaceae) Deutschiands..., 19i5:235, s. J. Lge, Dansk
Bot. Ark., 4, i2, 1926:5.— P. subrufescens (Pk) J. Lge, Fl. Ag. Dan.,
IV, 1939:55, s. J. Lge, non al. (табл. IV, За—в; XVIII, 12).
Icon.: Bres., icon. Мус., i931, tab. 832; Konr. et Maubi., Icon. Sei.
Fung., 1924—1937, pi. 27; J. Lge, Fi. Ag. Dan., 1935—1940, tab. 135,
fig. B; Moeiier, Friesia, 4, 1950, pi. XXII—XXV; Moelier, Friesia, 5, 1951,
fig. 20; Pilat, Acta Mus. Nat. Prag., VIIB, i, 1951, tab. XII; Essette,
Les Psalliotes, 1964, tab. 29; Вассер, Фл. гр. Укр. (Agaricaceae), 1980,
рис. 63.
В хвойных (Picea, Pinus) и смешанных широколиственных (Quer-
cus, Fraxinus, Uimus, Aesculus, Acacia) лесах, в насаждениях Taxus,
в парках, заповедных целинных степях.
7 4-2960
65
A. augustus var. albus Mos. (ined.), KI- Krypt.-fiora, В. П. b/2 TeH.
1978 : 230 (nom. nudum).
В пихтовых (Abies) лесах. Известен с о-ва Сахалин.
Гум. сапр., VIII—XI, о. ч., (3), с., мультирег., евраз-ам-афр.
Распр. в СССР. Европ. ч.: РСФСР — Московская обл., ЭССР,
ЛитССР, ЛатвССР, БССР, УССР, Азиат, ч.: Кавказ — ГССР, Ср.
Азия — ТССР, Вост. Сибирь — Красноярск, кр., Дальн. Восток — Са-
халинская обл.
Общее распр. Европа: Великобритания, Финляндия, Дания,
Бельгия, Франция, Италия, ФРГ, ГДР, Австрия, ПНР, ЧССР, Греция,
ВНР; Азия: КНР; Сев. Америка: США, Мексика; Африка:
Марокко, Кения, Танзания.
27. Agaricus subrufescens Pk, N. Y. State Mus. Bull., 43, i894 : 236,
non s. J. Lge, Fi. Ag. Dan., IV, 1939:55.— Psalliota subrufescens (Pk)
Lioyd, Мус. Writ, I, i898 : 47.
Icon.: Kauffm., Agar. Mich., 1918, p. 239, t 48—50; Moeller, Frie-
sia, 5, 1951, fig. 18.
В широколиственных, главным образом дубовых лесах, парках; гум.
сапр., VII—X, о. р., (1—2), с., немор., евраз-ам.
Распр. в СССР. Азиат, ч.: Дальн. Восток — Приморск, кр.
Общее распр. Европа: Дания, ЧССР; А з и я: КНР, Япония;
Сев. Америка: США.
28. Agaricus kuehnerianus* Heinem., Num. spec. Buli. Soc. iinn.
Lyon, 15 fevrier, 1974 : 183 (табл. IV, 4a—a).
Icon.: Heinemann, Num. spec. Buli. Soc. linn. Lyon, 15 fevrier,
1974, fig. p. 183; Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 64.
На высокогорных альпийских лугах, яйлах, доходит до высоты
2000 м и. у. м.; гум. сапр., V—VIII, о. р., (2—3), с., альп., евр.
Распр. в СССР. Европ. ч.: УССР.
Общее распр. Европа: Франция.
Прим. A. kuehnerianus близок к некоторым видам (A. macrosporus (Moell. et
J. Schaeff.) Moell., A. excellens (Moell.) Moell., A. stramincus (Moell. et J. Schaeff.)
Moell.) группы Macrosporus s. Moell (Moeller, 1951, p 76; Heinemann, 1974), от ко-
торых отличается габитусом карпофоров, формой и расположением хейлоцистид,
окраской спор, базидий.
A. kuehnerianus — единственный представитель семейства Agaricaceae, растущий
лишь в высокогорном альпийском поясе Диаметр шляпки A. kuehnerianus достигает
60 см.
29. Agaricus macrosporus (Moeil. et. J. Schaeff.) Moell., Friesia, 5,
1951 : 204.— Psalliota arvensis subsp. macrospora Moell. et J. Schaeff.,
Ann. Мус., 36, 1, 1938 : 78.— P. macrospora (Moeli. et J. Schaeff.) Moeii.,
Friesia, 5, 1951:181.— P. villatica (Brond.) s. Bres., Fung. Trid., I,
1884 :54, s. Ricken, Vademecum, ed. I, 1920 : 142, s. J. Lge, Dansk Bot
Ark., VI, 1926, 6, s. Pears., Trans. Brit. Мус. Soc., 29, 1946 : 204.—
Agaricus villaticus (Brond.) s. Sacc., Syii. Fung., V, 1887 : 997, non s.
Rich, et Roze, Ati. Champ. Fr., 1885:49, non s. Baar, Bull. Soc. Royai
Bot. Belg., 70, 1937:42, non s. Konr. et Maubi., Les Agaricaies, 1948:
102.— Psalliota augusta Fr., s. Ricken, Konr. et Maubl., Les Agaricales,
1948: 104.— P. bernardii Quel., s. Rea, Trans. Brit. Мус. Soc., 3, 1911 :
285, Brit. Bas., 1922: 83.— P. bresadolae Baar (non Schulz.) p. p., Buii.
Soc. Royai Bot. Beig., 70, 1937 : 48.— P. urinascens Moell. et J. Schaeff.,
Ann. Мус., 36, 1, 1938 : 79.— P. collina Vei., Novit. Mycol., 1939: 152.—
Agaricus macrosporus (Moell. et J. Schaeff.) Pil., Acta Mus. Nat. Prag.,
VIIB, 1, 1951 : 78 (nom. invai.) (табл. IV, 5a—в; XIX, 13).
66
Icon.: Bres., Fung. Trid., 1, 1884, tab. 60; Bres., Icon. Мус., 1931»
tab. 829; J. Lge, FL Ag. Dan., 1935—1940, tab. 139, fig. C; J- Schaeffer
in Michaci, Fuhr. Piizfr., 1939, tab. 57; Hans Waity, Schweiz. Piiztaf.,
II, 1944, tab. 31; Moeller, Den ny Svympebog, 1945, fig. 121; Moelier,
Friesia, IV, 1950, pl. XXXiV; Essette, Les Psailiotes, 1964, 45; Hlavacek,
Mykol. sborn., 2—3, L1II, 1976, obr. 7—8; Вассер, Солдатова. Высш,
баз. ст. зоны Укр., 1977, рис. 17; Вассер. Фл. гр. Укр. (Agaricaceae),
1980, рис. 65, табл V. 2.
В типчаково-ковыльных, разнотравио-типчаково-ковыльных степях»
на лугах, выпасах, морских побережьях, реже в хвойных лесах, на лес-
ных опушках, в парках; гум. canp., VII—X, ир., (2—3), с., в. п. к., эври-
голаркт., евр аз.
Распр. в СССР. Европ. ч.: ЛатвССР, ЛнтССР, УССР; Ази-
ат. ч.: Кавказ — ГССР.
Общее распр. Европа: Великобритания, Дания, Франция, ФРГ,.
ГДР, ЧССР, ПНР, ВНР; Азия: КНР.
30. Agaricus exceilens (Moell.) Moeli., Friesia, 5, 195i : 204.— Psal-
liota excellent Moeli., Friesia, 4, 195i : i78.— P. augusta Fr. s. Ricken,
Die Blatterpilze (Agaricaceae) Deutschiands..., 1915 : 235, s Beck, Piiz-u.
Krauterfr., 5, 1921:44, s. Sing., Zeitschr. Piizk., I, 1922:24, s. Hans
Waity, Schweiz. Piiztaf., II, 1944, tab. 30 (табл. IV, 6a—в).
Icon.: Hans Waity, Schw'eiz. Piiztaf., II, 1944, tab. 30 (под назва-
нием P. augusta); Moeiler, Friesia, 5, 1951, pi. XXXIII, fig. 29; Essette»
Les Psailiotes, 1964, tab. 44; Hlavacek, Mycoi. sborn., 2—3, LIII, 1976»
obr. 9; Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 68—69.
В хвойных (еловых, лиственничных) и широколиственных (буко-
вых, акациевых) лесах; гум. canp., VIII—X, о. р.. (1), с., эвриголаркт.,.
евраз-афр.
Распр. в СССР. Азиат, ч.: Вост. Сибирь-—Красноярск, кр.
Общее распр. Европа: Великобритания, Дания, ФРГ, ГДР, Ав-
стрия, ВНР; Африка: Марокко.
31. Agaricus stramineus* (Moell. et J. Schaeff.) Moeil., Friesia, V»
1951 :204.— Psalliota straminea Moell. et J. Schaeff., Ann. Мус., 36, 1»
1938 : 36.— P. arvensis (Schaeff.) Fr. subsp. macrospora Moeil. et J.
Schaeff. var. straminea (Moeil. et J. Schaeff.) J. Schaeff., in Michaei,
Fuhr. Piizfr., 1939, tab. 59.— Agaricus stramineus (Moeli. et J. Schaeff.)
Sing., Liiloa, 22, 1951 : 432 (nom. invai.).
Icon.: J. Schaeffer, in Michaei, Fiihr. Pilzfr., 1939, tab. 59; Michaei u.
Hennig, Handb. Piizfr., 1967, IV, tab. 16.
На лугах, полях, выпасах, лесных опушках; гум. canp., IX—XI,.
о. р., (2), с., немор., евр.
Распр. в СССР. Европ. ч.: УССР.
Общее распр. Европа: Дания, ФРГ, ГДР, Австрия.
32. Agaricus tabuiaris Pk, Bulk Tor. Bot. Club, 25, 1898:325.— Aga-
ricus praerimosus Pk, Buii. N. Y. State Mus., 94, i905 : 236. (табл. V,
la—б; XIX, 14).
Icon.: Васильков, Очерк геогр. распр. шляпочн. гр., 1955, рис. 23;
Зерова, Укр. ботан. журн., 13, 2, 1956, рис. 1; Зерова, Атл. гр. Укр.,
1974, табл. 116, рис. 2; Вассер, Микол, н фитопатол., 8, 4, i974, рис. 1;
Вассер, Солдатова, Высш. баз. ст. зоны Укр., 1977, рис. 7; Wasser, Fung.
Rar. Icon. CoL, X, 1979, pi. 73; Вассер, Фл. гр. Укр. (Agaricaceae), 1980»
рис. 71—72, табл. X.
?•
67
В заповедных целинных степях, пустынях, полупустынях, в восточ-
но-азиатской Арктике в ивняково-разнотравно-дрнадовой, дриадовой
тундре; гум. сапр., VI—X, нр., (3), с., ксеромерид., евраз-ам.
Распр. в СССР. Европ. ч.: УССР; Азиат, ч.: Аркт.— Зап. Чу-
котка, Таймырский нац. окр., Кавказ — ГССР, Ср. Азия — УзССР, Зап.
Сибирь — КазССР, Вост. Сибирь — Красноярск, кр.; о-в Врангеля.
Общее распр. Сев. Америка: США (штат Колорадо).
Прим. A. tabularis проблематичный вид, на наш взгляд, наиболее близок к
A. rnaskae Pil., от которого отличается характером поверхности шляпки, размером
спор, хейлоциспф (Вассер, 1980).
Необычайно интересна находка A. tabularis, считавшегося типичным стенотопным
ксерофптным видом, в восточноазиатской Арктике (Васильков, 1974).
Интересно отметить, что как на нашем материале по степной зоне Украины, так
и на материале Б. П. Василькова из Арктики, который нам удалось изучить (Зап.
Таймыр, яр на берегу р. Пясины, в дриадовой тундре, 4.V11 1966, собр. Т. Г. Полозо-
ва, опр. Б. П. Васильков; Зап. Чукотка, побережье Чаупской губы, у пос. Певск,
6.V111 1964, собр. и опр. Б. П. Васильков; о-в Врангеля, побережье бухты Сомни-
тельной, южный шебнистый склон юры, у нор леммингов, 6. VII 1966, собр. В. Ф. Ша-
мурин, опр. Б 11. Васильков), характерные признаки — чешуйчатость и таблитчатая
тчеистость шляпки, на что обратил внимание Б. П. Васильков (1. с.), очень сильно
вариабельны у отдельных экземпляров и даже па поверхности одной шляпки. Наи-
более глубокая трещиноватость и более обширный участок ее всегда наблюдается
посредине шляпки, но часто охватывает всю шляпку или ее половину. При этом
заметно, что иногда в молодом возрасте отдельные экземпляры грибов бывают снача-
ла гладкими, без трещин, а позднее появляются «вспучивания» на поверхности шляп-
ки, которые при дальнейшем росте гриба превращаются в ячеистую трещиноватость.
Иногда же совсем молодые карпофоры покрыты трещинами.
Б. П Васильков (I. с.) высказал предположение, что A. tabularis, возможно,
идентичен с A. campestris. Он пишет: «Нс происходит ли растрескивание поверхности
шляпки под влиянием каких-то сочетаний почвенно-климатических условий, вероятнее
всего сочетания влажности и температуры воздуха и почвы, а также индивидуаль-
ного роста отдельных карпофоров... не связано ли это с крайними условиями суще-
ствования: тундра, степь, пустыни?... Однако для полного выяснения этого вопроса,
конечно, потребуются дальнейшие исследования» (с. 172—173).
Мы считаем A. tabularis хорошим полиморфным видом, произрастающим в
экстремальных условиях (Вассер, 1980).
3«3. Agaricus rnaskae* PiL, Ces. MykoL, 8, 4, 1954 : i65 (табл. XIX,
15—16).
Icon.: Piiat, Ccs. MykoL, 8, 4, 1954, p. 160, p. 161, p. 162, p. 164;
Pilat, Schweiz. Zcitsc.hr. Pilzk., 44, 6, 1966, p. 97, p. 198; Imreh u. Bohus,
Schweiz. Zcitschr. Pilzk., 48, 2, 1969, p. 18; Вассер, Фл. гр. Укр. (Aga-
ricaccae), 1980, рис. 73.
В заповедных полынно-злаковых степях, на пастбищах, лугах, из-
редка в белоакациевых и тополевых лесах; часто на песчаной Почве
с pH 5,3—6,7; гум. сапр., IV—Xi, о. р., (3), с., ксеромерид., евр.
Распр. в СССР. Еврсп. ч.: УССР.
Общее распр. Европа: ФРГ, СРР, ЧССР, ВНР.
Прим. A. rnaskae наиболее близок к A. tabularis, от которого отличается
характером поверхности шляпки, размером спор, хей.юцистид. По мнению автора
вида (Pilat, 1954), A. rnaskae близок также к A. macrosporus, от которого отлича-
ется размером спор, габитусом карпофоров. Интересно отмстить, что карпофоры
A. rnaskae. собранные осенью, имеют более темный цвет шляпки, чем карпофоры
летних генераций.
Изучен типовой материал A. rnaskae из Herb. Krypt Mus. Nat. Prag.: Hab. loco
gram inoso solo calcareo inlcr Petrovice et SIoup Moroviae, 27.VII 1954, leg. Maska 1.,
det. Pilat A., typus, N 649 850. Статистическая обработка размеров спор типа (7,3+
±0,4X5,1 ±0,4 мкм) показала некоторые отклонения (особенно в ширине) от приве-
денных в диагнозе А. Пилата: 7,2—7,8X3,8—4,3 мкм (Pilat, 1954).
68
Для нашего материала наиболее характерны споры: 7,0X5.3; 8,3X5,6 мкм (изред-
ка встречались споры 9.0Х5.3; 9,8X6,0 мкм), хейлоцистиды 23—25><6,0—9,5 мкм
(Вассер, 1980).
34. Agaricus silvicolus (Vitt.) Sacc., SylL Fung., V, 1887:998.—
Agaricus campestris var. silvicolus Vitt., Fung. Mang., 1835 : 43.— Psal-
liota silvicola (Vitt.) Fr., Epicr. Syst. Мус., I, 1836 : 213.— P. campestris
var. silvicola Fr., Epicr. Syst. Мус., I, 1836 : 213.— Pratella flavescens
Gill., Champ. Fr., 1878:564.— Psalliota flavescens (Gill.) Rea, Brit. Bas.,
19-22 : 132.— A. silvicola (Vitt.) s. Pk, Bull. N. Y. State Mus. Mem... 4,
1900: 164.— P. silvicola (Vitt.) s. Ricken, Vademecum, cd. I, 1920: 144.—
P. silvicola (Vitt.) s. Rydberg, Ingelstrom, Svampbok, 1940: 101 (табл. V,
2a—6).
Icon..- Vitt., Fung. Mang.. 1835, tab. VII, fig. 7—9; Gill., Champ.
Fr., 1878, tab. 576 (384); Konr. et Maubl., Icon. Sci. Fung., 1921—1936,
pl. 29; Hans Waity, Schweiz. Piiztaf., II, 1944, tab. 34; Pilat, Acta Mus
Nat Prag., VIIB, 1, 1951, fig. 49—53; Moeller, Friesia, 4, 1950, pl. XXVI,
Friesia, 5, 1951, fig. 21; Essette, Les Psalliotes, 1964, tab. 31; Вассер,
Фл. гр. Укр. (Agaricaceae), 1980, рис. 75.
В еловых, елово-кедрово-пихтовых, буковых, дубовых, березовых,
смешанных широколиственных лесах, в парках; гум. canp., VII—X, о. ч.,
(3), с., мультирег., евраз-ам-афр.
Распр. в СССР. Европ. ч.: РСФСР, ЭССР, ЛатвССР, ЛнтССР,
УССР; Азиат, ч.: Кавказ — АзССР, Зап. Сибирь — Томская обл.,
Вост. Сибирь—Красноярск, кр., Дальн. Вост.— Приморск, кр.
Общее распр. Европа: Великобритания, Дания, Бельгия, Фран-
ция, Италия, ФРГ, ПНР, ЧССР, Австрия, ВНР, Греция, СРР; Азия:
КНР, Израиль, Япония; Сев. Америка: Канада, США; Африка:
Алжир, Кения, Заир, Марокко.
35. Agaricus macrocarpus* (Moell.) Moell., Friesia, 5, 1951:204.
Psalliota macrocarpa Moell., Friesia, 5, 1951:153—154.— P. arvensis
(Schaeff.) Fr., s. Ricken, p. p.. Die Blatterpilze (Agaricaceae) Dcutsch-
lands..., 1915 : 236—237 (табл. V, За—6).
Icon.: Moeller, Friesia, 4, 1950, pl. XXVIII; Вассер, Фл. гр. Укр.
(Agaricaceae), 1980, рис. 76.
В еловых, смешанных, дубово-грабовых лесах, на лесных полянах,
лугах; гум. canp., VII—X, о. р., (I), с., бор., евр.
Распр. в СССР. Европ. ч.: УССР.
Общее распр. Европа: Дания, ФРГ, ГДР, Австрия, Франция,
ВНР
36. Agaricus abruptibulbus Pk, N. Y. State Mus. Bull., 94, 1905:
35.— Psalliota abruptibulba (Pk) Kauffm., Agar Mich., 1918:237.—
Agaricus silvicola (Vitt.) s. Pk, N. Y. State Mus. Rept., 46, 1893: 135.—
A. arvensis Schaeff.: Fr. var. abruptus Pk, N. Y. State Mus. Rept., 48,
1895:239.— A. abruptus Pk, N. Y. State Mus. Mem., 4, 3, 1900: 163.—
Psalliota arvensis (Schaeff.) Fr., s. Ricken, p. p., Die Blatterpilze (Aga-
ricaceae) Deutschlands..., 1915 : 236.— P. arvensis (Schaeff.) Fr. var. sil-
vicola (Vitt.) Fr., s. J. Lge, Dansk Bot. Ark., VI, 1926:7.— P. silvico-
la (Fr.) s. J. Lge, Fl. Ag. Dan., IV, 1939:57.— P. abruptibulba (Pk)
Kauffm., s. Rydberg, Ingelstrom, Svampbok, 1940: 101 (табл. V, 4a—6).
Icon.: Peck, N. Y. State A4us. Mem., 4, 3, 1900, tab. 59, fig. 8—14;
J. Lge, Fl. Ag. Dan., 1935—1940, tab. 138, fig. В (под названием P. sil-
vicola); Moeller, Friesia, 4, 1950 pl. XXVII, fig. 23; Moeller, Den nyc
Svampebog, 1947, fig. 22 (под названием P. arvensis); Essette, Les
Psalliotes. 1964, tab. 33; Вассер, Фл. гр. Укр. (Agaricaceae), 1980.
рис. 77.
В еловых, смешанных лесах, полезащитных лесополосах, парках;
гум. сапр., VII—XI, р_, (2), эвриголаркт., евраз-ам.
Распр. в СССР: Европ. ч.: ЭССР, УССР; Азиат, ч.: Дальн.
Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Финляндия, Дания,
Франция, ФРГ, ПНР, ЧССР, ВНР; Азия: Япония; Сев. Амери-
ка: США, Канада.
37. Agaricus fissuratus* (Moell.) Moell., Friesia, 5, 1951:204.—
Psalliota fissurata Moell., Friesia, 5, 1951 : 165 (табл. V, 5a—6).
Icon.: Moeller, Friesia, 4, 1950, pl. XlXb; Michael u. Hennig,
Handb. Pilzfr., IV, 1967, abb. 9; Essette, Les Psalliotes, 1964, tab. 37;
Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 78.
В дубовых лесах, парках, на лугах, во дворах; гум. сапр., VII—
X, о. р., (2), с., немор., евр-ам-афр.
Распр. в СССР. Е в р о п. ч.: УССР.
Общее распр. Европа: Дания, ФРГ; Африка: Марокко; о-в
Гренландия.
38. Agaricus amanitaeformis*** S. Wasser, Укр. ботаи. жури., 31,
1, 1974 : 79 (табл. XIX, 17).
Icon.: Вассер, Укр. ботан. жури., 31, I, 1974 : 80, рис. 2, 3; Вас-
•сер, Солдатова, Высш. баз. ст. зоны Укр., 1977, рис. 127; Вассер, Фл.
гр. Укр. (Agaricaceae), 1980, рнс. 79.
В ботанических садах и парках, в смешанных насаждениях широ-
колиственных деревьев (Fraxinus lanccolata Borkh., Acer platanoi-
des L.); гум. сапр., IX—X, о. p., (1), c.(?), я.(?), немор., укр. усл. эид.
Распр. в СССР. Европ. ч.: УССР.
Прим. A. amanitaeformis близок к A. fissuratus (Moell.) Moell. (Moeller, 1950—
1951; Essetie, 1964; Moser, 1978), но четко отличается от последнего цветом карпо-
форов, размером и формой спор, отсутствием хейлоцистид, формой и цветом ножки,
расположением гранул, цветом и запахом мякоти, экологическими особенностями
(Вассер, 1980, 1982).
39. Agaricus nivescens* (Moell.) Moell., Friesia, 5, 1951 :204.— Psal-
liota nivescens Moell., Friesia, 4, 1950 : 155.— Agaricus arvensis Schaeff. :
Seer. s. Cke, Ill. Brit. Fung., 1881—1891, tab. 523 (табл. V, 6a—e;
XIX, 18).
Icon.: Cke, 111 Brit. Fung., 1881 —1891, tab. 523, no. 540; Moeller,
Friesia, 4, 1930, pl. XVII lb, XXIX; Friesia, 5, 1951, fig. 24; Essette, Les
Psalliotcs, 1964, tab. 35; Вассер, Фл. гр. Укр. (Agaricaceae), 1980,
рис. 80—82.
В дубовых лесах, на лугах, пастбищах, в садах, парках, доходит
до высоты 1750 м и.у. м.; гум. сапр., VII—X, о. р., (2), с., немор.,
евраз-афр.
Распр. в СССР. Европ. ч/. УССР.
Общее распр. Европа: Великобритания, Дания, Бельгия, Фран-
ция; Азия: Израиль; Африка: Марокко.
40. Agaricus aestivalis (Moell.) Pil. var. aestivalis, Acta Mus. Nut.
Prag., VHB, I, 1951 : 13.— Psalliota aestivalis Moell., Friesia, 4, 1950:
50.— Agaricus aestivalis (Moell.) Moell., Friesia, 5, 1951 : 203 (табл. V,
7a— 6; XX, 19).
Icon.: Moeller, Friesia, 4, 1950, fig. 3 (споры); Essette, Les Psal-
liotes, 1964, tab. 24; Michael u. Hennig, Handbh. Pilzfr., 1967, IV,
70
fig- За, b; Вассер, Фл. гр. Укр. (Agaricaceae), I960, рис. 84—85,
табл. XI, I, 2.
В хвойных и смешанных лесах.
A. aestivalis var. veneris** (Heim et Becker) S. Wasser, Фл. гр.
Укр. (Agaricaceae), 1980:165.— Psalliota aestivalis var. veneris Heim
et Becker, BSMF, LXXVI, 3, 1960 : 24-—241.— Icon.: Heim ct Becker.
BSMF, LXXVI, 3, 1960, tab. LXXVI, pl. I, fig. 1.
В смешанных лесах. Известен нз УССР.
Гум. сапр., VI—VIII, о. р„ (1), с., бор., евраз.
Распр. в СССР. Ев роп. ч.: УССР.
Общее распр. Европа: Великобритания, Дания, Франция, ФРГ,
ЧССР, ВНР; Азия: Израиль.
41. Agaricus longicaudus*** S. Wasser, Нов. сист. высш, н иизш.
раст., 3, 1977 :218 (табл. VI, 1а—б\ XX. 21).
Icon.: Вассер, Нов. сист. высш, н низш. раст., 3, 1977, рис. 1—2;
Wasser, Fung. Rar. Icon. Col., X, 1979, pl. 77a; Вассер, Фл. гр. Укр.
(Agaricaceae), 1980, рис. 87—88, табл. XII, I.
В смешанных широколиственных грабово-дубово-ильмовых лесах
с хорошо развитой подстилкой, содержащей гумифицированные расти-
тельные остатки; гум. сапр., VI—VIII, о. р., (2), с., немор., укр. усл. эид.
Распр. в СССР. Е в р о п. ч.: УССР.
Прим. A. longicaudus наиболее близок к A. aestivalis (Moell.) Moell. var.
aestivalis и A. aestivalis var. veneris (Heim et Becker) S. Wasser, от которых отли-
чается габитусом карпофоров, цветом и характером поверхности шляпки И ножки,
слнзистостью шляпки, макрохимическими реакциями, экологией (Moeller, 1950—1951;
Heim, Becker, 1960; Dennis, Orton, Hora, 1960; Essette, 1964; Вассер, 19776, 1980).
42. Agaricus leucotrichus * (Moell.) Moell., Friesia, 5, 1951:294.
Psalliota leucotricha Moell., Friesia, 5, 1951, 159—160 (табл. XX, 22).
Icon.: Moeller, Friesia, 1950, 4, tab. XVIIIa.
В еловых лесах, изредка в виноградинках; гум. сапр., IX—X, о. р.,
(1), с., бор., евр.
Распр. в СССР. Европ. ч.: УССР.
Общее распр. Европа: Дания, ЧССР.
Прим. A. leucotrichus наиболее близок к A. chionodermus РИ.
43. Agaricus chionodermus* Pil., Acta Mus. Nat. Prag., VIIB, 1,
1951 : 134.
Icon.: Pilat, Acta Mus. Nat. Prag., VIIB, 1, 1951, fig. 50, tab. Ill
XIV, XV, XVI; Pilat, Acta Mus. Nat. Prag., IXB, 2, 1953, tab. iVa, IVb.
В еловых лесах, гум. сапр., VII—VIII, о. р., (1), с., бор., евр.
Распр. в СССР, Е в р о п. ч.: УССР.
Общее распр. Европа: ЧССР, ФРГ.
Прим. A. chionodermus, по мнению автора вида (Pilat, 1951b), наиболее бли
зок к A. arvensis Schaeff.: Seer. Из североамериканских видов A. chionodermus наи-
более близок к A. albolutescens Zeller (A. Smith, 1940), для которого характерна
белая, иногда оранжево-желтая шляпка, споры 6,0—6,5X3,5—4 мкм.
Изучсн типовой материал A. chionodermus из Herb. Krypt. Mus. Nat. Prag.; Hab
Karlstejn, in Picetos prope Kralovska studanka, solo Kalcareo, 1.V1I 1951, leg et del.
Pilat A., Typus, N 649 716. Следует отметить, что в диагнозе А. Пилата (Pilat, 1. С.)
размер спор приведен 8.5—10(10,5)Х4,8—6 мкм, в то время как статистическая
обработка размеров спор типа показала их меньший размер: 7,4—8,0Х 4,4—5,2 мкм
(Вассер, 1980).
44. Agaricus arvensis Schaeff.: Seer., Mycorg. Suis., I, 1833:97, s.
restr., s. J. Lge, Moell., Friesia, 5, 1951:61—62.-— Psalliota arvensis
71
(Schaeff. : Seer.) Kumm., Der Fuhrer in die Pilzkunde, 1871 :67.— P. cre-
tacea Fr., s. Ricken, Die Blatterpilze (Agaricaceae) Deutschlands..., 1951 :
236.— P. arvensis subsp. exquiseta (Vitt.) Moell. et J. Schaeff., Ann,
Мус., 36, 1, 1938 : 78, non P. arvensis s. Ricken, Kauffm., Baar. (табл. VI»
2a—в; XX, 23).
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, tab. 138, lig. A; Moeller,
Friesia, 4, 1950, pl. XXX; Moeller, Friesia, 5, 1951, fig. 26: Pilat, Acta
Mus. Nat. Prag., VIIB, 1, 1951, fig. 43—45; Essette, Les Psalliotes, 1964,
tab. 36; Зерова, Атл. гр. Укр., 1974, табл. 118, рнс. 1: Вассер, Фл. гр.
Укр. (Agaricaceae), 1980, рис. 89. >
В хвойных и широколиственных лесах, в полезащитных лесополо-
сах, на пастбищах, лугах, в садах, оранжереях, в дрнадовой тундре,
в пустынях, степях; гум. canp., V—XI, о. ч., (2—3), с., мультнрег., кос-
мопол.
Распр. в СССР. Европ. ч.: ЭССР, ЛнтССР, ЛатвССР, РСФСР,
БССР, МССР, УССР; Азиат, ч.. Кавказ — ГССР, Ср. Азия—
ТаджССР, КнргССР, Зап. Сибирь — Омская обл., Вост. Сибирь— Якут-
ская АССР, Бурятская АССР, Красноярск, кр., Дальи. Восток — При-
морск. кр.
Общее распр. Европа: Великобритания, Исландия, Португалия,
Испания, Франция, Дания, Бельгия, Нидерланды, Италия, ФРГ, ГДР,
Финляндия, ПНР, ЧССР, Австрия, ВНР, СФРЮ, СРР; Азия: КНР,
Израиль, Шри Ланка, Япония; Сев. Америка: Канада, США, Мек-
сика; Южн. Америка: Чили; Африка: Марокко; Австралия;
о-ва Гренландия, Тринидад, Куба.
45. Agaricus osecanus* Pil., Acta Mus. Nat. Prag., VIIB, 1,
1951 : 133.
Icon.: Pilat, Acta Mus. Nat. Prag., VIIB, 1, 1951, fig. 41—42.
В широколиственных лесах; гум. canp., IX—X, о. р., (1), с., не-
мор., евр.
Распр. в СССР. Е в р о п. ч.: УССР.
Общее распр. Европа: ЧССР, ВНР.
ПОДСЕКЦИЯ XANTHODERMATAE (SING.) S WASSER
Укр. ботан. жури., 33, 5, 1976 : 250.
46. Agaricus xanthodermus Gen. var. xanthodermus, Bull. Soc. Bot.
Fr., 23, 1876:31, s. S. Wasser, Укр. ботан. журн., 33, 5, 1976:495»
non s. Bohus, Ann. Hist.-Nat. Mus. Nat. Hung., 66, 1975:80—81.— Psal-
liota xanthoderma (Gen.) Rich, et Roze, Atl. Champ. Fr., 1885: 53.—
Pratella campestris var. silvicola Vitt. s. Gill., Champ. Fr., 1878 : 562.—
P. cretacea (Fr.) s. Quel., FI. Мус. Fr., 1888 : 73, non s. Fr.— Agaricus
iodoformicus Speg., Ann. Mus. Nat. Buenos-Aires, 6, 1898: 141.— Psal-
liota foetens Smotlacha, Cas. cesk. houb., 2, 1920 : 70.— P. pseudoarvensis
Passecker, Zeitschr. Pilzk., 16, 1932 : 36—39.
Icon.: Rolland, BSMF, 5, 1889, tab. 3, fig. 1 (под названием P. ar-
vensis); Konr. et MaubL, Icon. Sci. Fung., 1924—1937, tab. 30; Moeller,
Friesia, 4, 1950, pl. XXXI; Wakefield a. Dennis, Comm. Brit, fung., 1950,
pl. 71; Pilat, Acta Mus. Nat. Prag., VIIB, 1, 1951, fig. 69—74; Pilat»
Nase houby, II, 1959, tab. 143; Michael u. Hennig, Handbh. Pilzfr., I,
1958, fig. 27; Essette, Les Psalliotes, 1961, tab. 38; Bohus, Ann. Hist.-Nat.
Mus. Nat. Hung., 66, 1974, fig. 1; Зерова, Атл. гр. Укр., 1974, табл. 119,
рис. 1; Вассер, Укр. ботан. журн., 30, I, 1973, рнс. 3; Вассер, Солдато-
72
ва, Высш. баз. ст. зоны Укр., 1977, рнс. 24; Вассер, Фл. гр. Укр. (Aga-
ricaccae), 1980, рис. 90.
В заповедных целинных степях, широколиственных и хвойных ле-
сах, полезащитных лесополосах, парках, садах, на пастбищах, кладби-
щах, на почве, очень редко на трухлявой древесине.
A. xanthodermus var. Jepiotoldes* R. Mre, BSMF. 26, 1910: 109.
Agaricus lepiotoides (R. Mre) Konr. et Maubl., Les Agaricales, 1948 :
106.—-Icon.: Вассер, Укр. ботаи. журн., 33, 5, 1976, рис. 5—7; Wasser,.
Fung. Rar. Icon. Col., X, 1979, pl. 77b, Text-fig. 2; Вассер, Фл. гр. Укр.
(Agaricaceae), 1980, рис. 91—92.
В широколиственных лесах. Известен из УССР, ТССР.
Гум. сапр., VI—X, о. ч., (2—3), я., мультирег., космопол.
Распр. в СССР. Европ. ч.: РСФСР — Карельская АССР, ЭССР,
БССР, МССР, УССР; Азиат, ч.: Кавказ — АзССР, АрмССР, ГССР,
Ср. Азия — КиргССР.
Общее распр. Европа: Великобритания, Австрия, Бельгия, ВНР,
ГДР, Греция, Дания, Нидерланды, Ирландия, Испания, Италия, Лих-
тенштейн, Норвегия, ПНР, Португалия, СРР, ФРГ, Финляндия, Фран-
ция, ЧССР, Швейцария, Швеция; Азия: КНР, Израиль, Турция, Япо-
ния; Сев Америка: Канада, США, Мексика; Южн. Америка:
Аргентина, Боливия, Бразилия, Венесуэла, Чили; Африка: Алжир,
Марокко; Австралия; о-ва Гренландия, Куба, Тринидад, Тобаго.
Прим. В литературе иет единой точки зрения на объем A. xanlhodermus Gen.
Г. Богуш (Bohus, 1971, 1975) развивает точку зрения о существовании A. xanthoder-
mus s. str., выделяя из A. xanthodermus s. 1. новый для науки вид A. pilatianus Bohus
с рядом форм.
Мы принимаем A. xanthodermus s. 1., признавая правомерным выделение в нем
нескольких разновидностей (Вассер, 1976в, 1980).*
47. Agaricus pseudopratensis** (Bohus) S. Wasser, Укр. ботан.
журн., 33, 3. 1976:250.— Psalliota pseudopratensis Bohus, Borbasia, I,
1939 : 114,— Agaricus pseudopratensis (Bohus) Bohus, Ann. Hist.-Nat.
Hung., 63, 1971 : 81 (nom. inval.) (табл. VI, За—б\ XX, 24).
Icon.: Bohus, Borbasia, I, 1939, tab. X; Bohus, Ann. Hist.-Nat. Mus.
Nat. Hung., 63, 1971:82, fig. 2; Вассер, Фл. гр. Укр. (Agaricaceae),
1980, рис. 93.
В парках и садах, в траве, в белоакациевых (Robinia pseudoaca-
cia L.) и еловых (Picea abies (L.) Karst.) лесах, на песчаной почве, до-
ходит до 1300—1500 м н. у. м.; гум. сапр., VI—X, о. р., (2), я., эвригол-
аркт., евр.
Распр. в СССР. Европ. ч.: УССР, МССР.
Общее распр. Европа: ВНР.
Прим. Морфологически, как отмечает автор вида (Bohus, 1939, 1971), A. pseu-
dopratensis очень похож на A. campestris, однако четко отличается от последнего
размером спор, цветом мякоти при автооксидации, запахом, экологией, макрохнмиче-
скпми цветовыми реакциями.
48. Agaricus velenovskyi Pil., Ces. Mycol., 22, 2, 1968 : 83- 84,—
Agaricus meleagris J. Schaeff. var. nigricans (Vel.) Pil., Acta Mus. Nat.
Prag., VIIB, 1, 1951 : 114.— Psalliota nigricans Vcl., Ceske Houby, 1921 :
574 (nom. nudum, лат. диагноз у A. Piiat, Opera bot. cech., XI, 1948:
217).— Psalliota xanthoderma Gen. var. grisea Pears., Trans. Brit. Мус.
Soc., 29, 4, 1946 : 203—204.— P. grisea (Pears.) s. Essette, Les Psallio-
tes, 1964 : 46 (табл. XXI, 25).
Icon. Vclenovsky, Ceske houby, 1921, fig. 80; Pilat, Acta Mus.
Nat. Prag., VIIB, 1, 1951, fig. 66—68; Pilat, Ces. Mycol., 22, 2, 1968,
fig. 1—3; Essette, Les Psalliotes, 1964, tab. 22; Вассер, Укр. ботан. жури.,
32, 3, 1975:319, рис. 1; Вассер, Солдатова, Высш. баз. ст. зоны Укр.,
1977, рис. 29; Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 94.
На лугах, полянах, в тнпчаково-ковыльных степях, часто среди дер-
нин Stipa capillata L., S. lessingiana Trin. et Rupr., Festuca sulcata
Hack., в степных участках часто под Rosa canina L.; гум. сапр., IX—Xl,
о. р., (3), я.» ксеромерид., евраз.
Распр. в. СССР. Европ. ч.: УССР; Азиат, ч.: Кавказ — ГССР.
Общее распр. Европа: Великобритания, ЧССР, Франция.
49. Agaricus phaeolepidotus * (Moell.) Moell., Friesia, 5, 1951 :204.—
Psalliota phaeolepidota Moell., Friesia, 5, 1951 : 170 (табл. VI, 4a—в;
XX\,26).
Icon.: Moeller, Friesia, 5, 1951, pl. XXb, XXXIII; Essette, Les
Psalliotes, 1964, tab. 39; Pilat, Nase houby, 1959г II, tab. 142; Вассер,
Фл. гр. Укр. (Agaricaceae), 1980, рис. 95—97, табл. XIII.
В широколиственных лесах, иногда рядом с Agaricus purpurellus
(Moell.) Moell.: гум. сапр., IX—X, о. р., (1—2), я., иемор., евраз.
Распр. в СССР. Европ. ч.: УССР.
Общее распр. Европа: Великобритания, Дания, Бельгия, ВНР;
Азия: Израиль.
50. Agaricus meleagris (J. Schaeff.) Imbach, Mitt, naturf. Ges. Lu-
zern, 15, 1946 :68.— Psalliota meleagris J. Schaeff., Zeitschr. Pilzk., 4,
1925 : 28.— P. xanthoderma subsp. meleagris J. Schaeff. in Michael, Fuhr.
Pilzfr., 1, 1939, tab. 58.— Agaricus meleagris (J. Schaeff.) Pil., Acta Mus.
Nat. Prag., VIIB, 1, 1951 : 106 (nom. inval.).— A. placomyces var. melea-
gris (J. Schaeff.) Mos., KI. Krypt.-flora, В. II, Teil II, 1967 : 193 (nom.
inval.).— A. praeclaresquamosus Freeman, Mycotaxon, VIII, 1, 1979:90,
non A. meleagris Sow. ex Berk., Outl. Brit. Fung., I, 1860 : 101 (табл. VI,
5a—в).
Icon.: Schaeffer in Michael, Fiilir. Pilzfr., 1, 1939, tab. 58; Pilat,
Acta Mus. Nat. Prag., VIIB, 1, 1951, lig. 61—62; Essette, Les Psalliotes,
1964, tab. 41; Michael u. Hennig, Handb. Pilzfr., 1968, I, tab. 28, abb. 4;
Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 98.
В широколиственных, кедрово-шнроколиствеиных, кедрово-пихто-
вых, дубовых лесах, в насаждениях Juniperus virginiana L., в парках;
гум. сапр., V—X, нр., (1—2), я., эвриголаркт., евраз-ам.
Распр. в СССР. Европ. ч.: ЛитССР, УССР; Азиат, ч.: Кав-
каз— ГССР, Дальн. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Дания, Бельгия, СФРЮ,
Швейцария, Италия, ФРГ, ГДР, ЧССР, Австрия, ВНР; Азия: КНР,
Израиль, Япония; Сев. Америка: США, Мексика.
51. Agaricus moelleri*** S. Wasser, Нов. сист. иизш. раст., 13,
1976 : 77—78.— Agaricus xanthodermus Gen. var. obscuratus R. Mre,
BSMF, 26, 1910: 192.— Psalliota silvatica s. Bres. var. nigricans Beck,
Pilz- u. Krauterfr., 2—3, 1921 : 42.— P. meleagris J. Schaeff. var. obscti-
rata (R. Mre) Moell., Friesia, 5, 1951 : 173.— Agaricus meleagris J. Scha-
eff. var. obscuratus (R. Mre) Heinem., BSMF, LXXI, 3, 1965:397.—
A. meleagris J. Schaelf. var. terricolor Moell., Friesia, 5, 1951 :208
(табл. VI, 6a—e).
Icon.: Bres., Fung. Trid., 1886, I, tab. 90; Bres., Icon. Мус., 1931,
tab. 830; Moeller, Firesia, 5, 1951, tab. XXa, fig. 16 (4); Essette, Les
Psalliotes, 1964, tab. 43; Зерова, Атл. гр. Укр., 1974, табл. 119, рис. I:
Вассер, Нов. сист. нйзш. раст., 13, 1976 :77; Вассер, Фл. гр. Укр. (Aga-
ricaceae), 1980, рис. 99.
В широколиственных, смешанных лесах, в парках, садах; гум. canp.,
VI—X; о. р., (1), я., немор., евраз-афр.
Распр. в СССР. Е в р о п. ч_: УССР.
Общее распр. Европа: Даиня, Швейцария; Азия: Израиль;
Африка: Алжир, Марокко.
52. Agaricus placomyces Pk, N. Y. St. Mus. Bull., 29, 1878:40.—
Psalliota placomyces (Pk) P. Henn, apud Engl, ct Pr., Nat. PfL, I,
1898 : 201.
Icon.: Moeller, Friesia, 5, 1951, fig. 28: Васильева, Агар. шл. гр.
Приморск, кр., 1973, рис. 42Б.
В кедрово-широколиственных, кедрово-пнхтовых, дубовых, смешан-
ных широколиственных лесах; гум. canp., VII—X, р., (2), я., мультнрег.,
евраз-ам.
Распр. в СССР. Европ. ч.: ЛитССР; Азиат, ч.: Кавказ — ГССР,
Дальи. Восток — Приморск, кр.
Общее распр. Е в р о п й: Великобритания, ВНР, Дания, ФРГ,
ЧССР; Азия: Япония; Сев. Америка: США, Мексика; Юж и.
Америка: Перу.
Секция Minores Fr.
Hymen. Eur.. 1874 : 211.
53. Agaricus rusiophyllus Lasch, Linnaea, 1828, 3:38.— Agaricus
comtulus var. rusiophytlus (Lasch) Konr. et Maubl., Icon. Sol. Fung.,
Ill, 1924—1933, pl. 233.— A. comtulus Fr., Epicr. Syst. Мус., I, 1836:
215.— Psalliota comtula (Fr.) Quel., Champ. Jura et Vosges, 1872:97.—
Pratella comtula (Fr.) Gill., Champ. Fr., 1874 : 132.— Psalliota minuta
Vel., Ceske houby, 1921 : 564 ex Vel., Novit. MycoL. 1939 : 154.— P. xero-
phila Vel., Ceske houby, 1921 : 565.— P. rosea Vel., Novit. MycoL, 1939:
155.— P. depressea Vel., Novit. MycoL, 1939 : 83.
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, tab. 136, fig. A; Cooke,
Ill. Brit. Fung., 1881—1891, tab. 533, np. 552; Michael u. Hennig, Handb.
Pilzfr., 1968, I, tab. 24; Зерова, Атл. гр. Укр., 1974, табл. 120, рис. 1.
В широколиственных, главным образом дубовых, смешанных ле-
сах, в заповедных целинных степях, парках, на пастбищах, полянах,
в огородах; гум. canp., VII—X, о. ч., (2—3), с., эвриголаркт., евраз-афр.
Распр. в СССР. Европ. ч.: РСФСР — Московская, Ленниград-
,ская обл., ЭССР, ЛитССР, ЛатвССР, БССР, УССР; Азиат, ч.: Кав-
каз — ГССР, Дальн. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Дания, Финляндия,
Франция, Италия, Греция, ФРГ, ГДР, ЧССР, ВНР; Азия: КНР, Япо-
ния; Сев. Америка: США; Африка: Марокко.
Прим. A. rusiophyllus дискуссионный вид, морфологически похожий на Stropha-
ria coronilla (Fr.) Quel. Одни авторы (Dennis, Orton, Hora, I960, 1974; Moser, 1967,
1978) считают A. rusiophyllus и A. comtulus хорошими самостоятельными видами,
отмечая, что первый еще недостаточно изучен. Другие (Pilat, 1951b; Вассер, 1980)
считают их синонимами.
Изучение гербарных материалов, определенных разными авторами (Зерова, Гаи-
жа, Каламээс, Bohus) как A. rusiophyllus и A. comtulus, читературиых данных (Ric-
ken, 1915; Лебедева, 1949; Moeller, 1950—1951 Pilat, 1951b; Ma tendon, Bertault, 1970
75
ti др.) дает основания нам считать их идентичными видами, хотя в дальнейшем при
исследовании большего фактического материала возможно выделение предложенной
нами разновидности A. rusiophyllus var. comtulus, nom. provis. (Вассер, 1980).
54. Agaricus porphyrizon P. D. Orton, Trans. Brit. Мус. Soc., 2, 43,
1960 : 174.— Agaricus arvensis var. purpurascens Cke, Handb. Brit. Fung.,
ed. 2, 1883—1891 : 193, non A. purpurascens Fr. ex Seen, Mycorg. Suis.,
1, 1833 : 157.— A silvicola var. purpurascens (Cke) Konr. et Maubl.,
Icon. Sol. Fung., 1, 1927—1930 : 73.— A. purpurascens (Cke) Pil., Acta
Mus. Nat. Prag., VIIB, 1, 1951 : 10.— Psalliota purpurascens (Cke) Moell.,
Friesia, 5, 195J : 187 (табл. VI, 7a—б; XXI, 27—28).
Icon.: Cke, Ill. Brit. Fung., 1881—1891, tab. 541, no. 584; Moeller,
Friesia, 4, 1951, pl. 21a; Essette, Les Psalliotes, 1964, tab. 46; Michael u.
Hennig, Handb. Pilzfr., 1967, IV, tab. 17; Вассер, Фл. гр. Укр. (Agarica-
сеас), 1980, рис. 100, табл. XIV, 1.
В смешанных лесах, парках, огородах; гум. сапр., VIII—XI, о. р.»
(1), с., мультирег., евраз-ам-афр.
Распр. в СССР. Азиат, ч.: Зап. Сибирь—Новосибирская обл..
Ср. Азия — ТССР.
Общее распр. Европа: Великобритания, Норвегия, Дания, Фран-
ция, Италия, ФРГ, ГДР, ВНР; Азия: Израиль; Южн. Америка-
Аргентина; Африка: Марокко, Кения.
55. Agaricus brunneolus* (Л. Lge) Pil., Acta Mus. Nat. Prag., VIIB,
1, 1951 :23.— Psalliota brunneola J. Lge, Fl. Ag. Dan., IV, 1939:60.—
Agaricus brunneolus (J. Lge) Mos., KI- Krypt.-flora, B. 2, Teil II, 1967:
192 (nom. inval.) (табл. VI, 8a—в-, XXI, 29).
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, tab. 140, fig. С; M. Lange,
Bot. tidsskr., 1976, 71, N 1—2, fig. 1—2; Вассер, Фл. гр. Укр. (Agarica-
ceae), 1980, рис. 101.
В буковых лесах; гум. сапр., VII—X, о. р., (2), с., немор., евр.
Распр. в СССР. Европ. ч.: УССР.
Общее распр. Европа: Дания, Финляндия.
Прим. A. brunneolus 1 наиболее близок к A. porphyrizon Р. О. Orton, от кото-
рого отличается цветом шляпки, экологией и т. п. Г. Богуш (Bohus, 1969) высказал
предположение, что A. brunneolus лишь форма A. porphyrizon.
Мы, изучив гербарный материал A. brunneolus собственных сборов, а также из
Herb. Mus. Bot Haun. (Ch) : Loc. Fyn. Nyberg., Bollevoenge, 22. X 1973, leg. et det. Morten
1 angc from type locality, считаем его хорошим самостоятельным видом (Вассер,
1980).
По мнению автора вида (J. Lange, 1939), возможно, A. brunneolus явтяетси
идентичным с A. fulveolus Lasch. Реальные различия между ними (по приведенному
~). М. Фризом описанию)—это наличие у A fulveolus буро-жсттых с мелкозубчатыми
краями пластинок, а также появление желтовато-бурой окраски карпофора при при-
косновении. Однако A. fulveolus У. Г. Лаш описал недостаточно полно, и он мало
знаком современным авторам, а Э. М. Фриз описание составлял по сухим гербарным
образцам. Дальнейшие исследования, вероятно, внесут коррективы в номенклатуру
A. brunneolus.
56. Agaricus lutosus * (Moell.) Moell., Friesia, 5, 1951 :204.- - Psal-
liota lutosa Moell., Friesia, 5, 1951 : 188.
Icon.: Moeller, Friesia, 4, 1950, pl. XXId.
В смешанных лесах, на лесных полянах, в парках, степях, на лу-
гах; гум. сапр.. VIII—X, о. р., (1—2), с., немор., евраз.
Распр. в СССР. Европ. ч.: УССР; Азиат, ч.: Ср. Азия — ТССР
1 М. Ланге (Lange М., 1976) считает A. brunneolus синонимом A. inipudicus
(Rea) М. Lge.
76
Общее распр. Европа: Великобритания, Дания, ВНР; Азия:
Израиль.
57. Agaricus xantholepis* (Moell.) Moell., Friesia, 5, 1951:204.-
Psalliota xantholepis Moell., Friesia, 5, 1951 : 191 (табл. XXI, 30).
Icon.: Moeller, Friesia, 5, 1951, pl. XXIC, XXX VI; Huijsman, Pcr-
soonia, I, 3, 1960, p. 323, fig. 3—5; Вассер, Укр. ботан. журн., 29, 3,
1972:371, рис. I; Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 104.
В хвойных (Abies) и смешанных лесах; гум. сапр., VI—X, о. р.,
(I), с.(?), я.(?), бор., евр-ам.
Распр. в СССР. Е в р о п. ч.: УССР.
Общее распр. Европа: Дания, Франция; Ю ж н. Америка:
Чили.
Прим. A. xantholepis — вид, очень редко упоминаемый в литературе (Moeller.
1951; Huijsman, 1960; Singer, 1969; Вассер, 19726, 1980).
Автор вида не указал родство A. xantholepis. На наш взгляд, он наиболее
близок, судя но литературным данным и цветному рисунку A. xantholepis, получен-
ному из Herb. Mus. Bot. Haun. (С) ', к A. sagatus Fr. s. Ricken. Последний вид мало
известен современным микологам, а Р. У. Г. Дснниз, П. Д. Ортон и Ф Б. Хора
(Dennis, Orton, Нога, 1960, р. 212) считают его сомнительным
Следует отметить, что с возрастом гриб меняет свой облик, вследствие того что
постепенно меняется цвет всего карпофора. В гербарии весь гриб медно-желтый,
чешуйки на поверхности шляпки едва заметны (Вассер, 1980).
58. Agaricus semotus Fr. var. semotus, Monogr. Hym. Suec., II,
1863:347.— Pratella rubella Gill., Champ. Fr., 1878:565, s. Gill., non
Agaricus rubellus (Gill.) Pat., s. Pat., Dccary.— A comtulus var. rubellus
(Gill.) Konr. et MaubL, Icon. Sei. Fung., Ill, 1924—1933: pl. 230.— Psal-
liota amethystina (Quel.) J. Lge, Dansk Bot. Ark., VI, 1926 : 10.— P. sil-
vatica (Schaeff.) Fr. var. amethystina Quel., Assoc. Fran?., 1884 :4.—
P. sagata Fr. s. Cke, Ill. Brit. Fung., 1881—1891, tab. 968, no. 1177.—
P. semota (Fr.) Ricken, Die Blatterpilze (Agaricaceae) Deutschiands.,
1915 : 238.— P. semota (Fr.) Ricken s. Nannf. et Lund., Fung. Exsicc.,
1941, no. 924.— P. rubella (Gill.) Rea f. pallens J. Lge, Dansk Bot. Ark.,
VI, 1926: 10.— P. pallens (J. Lge) Rea, Appendix II, to Brit. Bas.,
1932 : 37.— P. duriuscula VeL, Novit. Mycol., 1947 : 83 (табл. VII, la—г).
Icon.: Gill., Champ. Fr., 1878, tab. 102; J. Lge. Fl. Ag. Dan., 1935—
1940, tab. 137, fig. A; Pilat, Acta Mus. Nat. Prag., VIIB, 1, 1951, fig. 59;
Moeller, Friesia, 4, 1950, pl. XXXVII; Essette, Les Psalliotes, 1964, tab.48;
Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 105.
В хвойных (сосновых, кедровых, еловых, пихтовых, лиственничных)
si смешанных (елово-кедрово-пихтовых, сосново-березовых) лесах, иа
выгонах, в заповедных целинных степях.
A. semotus var. minimus ** (Ricken) S. Wasser. Укр. ботан. жури.,
35, 5, 1978 : 516.— Psalliota minima Ricken, Die Blatterpilze (Agarica-
ccae) Deutschiands..., 1915:39.— Agaricus minimus (Ricken) Pil., Acla
Mus. Nat. Prag.. VIIB, 1, 1951 : 102.— Icon.: Michael u. Hennig,
Handbh. Pilzfr., 1967, IV, tab. 186; Вассер, Фл. гр. Укр. (Agaricaceae),
1980, табл. XIV, 3.
В смешанных, сосново-березовых лесах. Известен из УССР.
Подст. сапр., VII—X, о. ч., (2—3), с., бор., евраз-ам-афр.
Распр. в СССР. Европ. ч.: ЭССР, ЛатвССР, ЛитССР, БССР,
УССР; Азиат, ч.: Аркт.—Полярный Урал, Кавказ — АзССР, Зап.
1 Типовой материал A. xantholepis, вероятно, в Herb. Mus. Bot. Haun. (С) не
сохранился.
77
Сибирь — КазССР, Вост. Сибирь — Бурятская АССР, Красноярск, кр.,
Дальи. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Даиня, Финляндия, Бель-
гия, Франция, Испания, Италия, ФРГ, ГДР, ПНР, ЧССР, Австрия,
ВНР, СФРЮ; Азия: КНР, Турция, Израиль; Сев. Америка:
Канада, США; Юж и. Америка: Чили; Африка: Алжир, Ма-
рокко.
59. Agaricus purpurellus (Moell.) Moell., Friesia, 5, 1951 :204.—
Psalliota amethystina s. J. Lge, Dansk Bot. Ark., VI, 1926 : 10; Fl. Ag.
Dan., 4, 1939:61.— Agaricus rubellus s. Decary, BSMF, 43, 1927, Atlas,
tab. 22, non s.‘ Sacc.— A. johnstoni s. Baker et Dale, Мус. Pap., 33, 1951 :
83, non s. Murr.— Psalliota purpurella Moell., Friesia, 5, 1951 : 193.
Icon.: Decary, BSMF, 43, 1927, Atlas, tab. 22 (под названием
A. rubellus); J. Lge, Fl. Ag. Dan., 1935—1940, tab. 135, fig. A; Essette,
Les Psalliotes, 1964, tab. 47; Вассер, Укр. ботан. журн., 30, 1, 1973,
рис. 4; Зерова, Атл. гр. Укр., 1974, табл. 120, рис. 4; Вассер, Фл. гр.
Укр. (Agaricaceae), 1980, рнс. 106.
В еловых, сосновых, смешанных дубово-грабовых лесах, на выго-
нах; подст. canp., XVIII—X, д. ч., (3), с., мультнрег., евраз-ам.
Распр. в СССР. Европ. ч.: УССР; Азиат, ч.: Кавказ — АзССР,
Вост. Сибирь — Красноярск, кр., Дальн. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Дания, Франция, Ита-
лия, ФРГ, ПНР, ЧССР; Азия: Израиль, Япония; Сев. Америка:
Канада, США; Южн. Америка: Боливия, Венесуэла; о-ва Трини-
дад, Тобаго.
60. Agaricus dulcidulus Schulz., in Kalchbr. et Schulz., Icon. Sei.
Hym. Hung., 1874 :29.— Psalliota dulcidula Schulz., s. J. Lge, Dansk
Bot. Ark., VI, 1926: 11, Fl, Ag. Dan., 4, 1939:61, non s. S. Petersen,
Danske Agaricaceer, 1907—1911 : 382.
Icon.: Kalchbr. et Schulz., Icon. Sei. Hym. Hung., 1874, tab. 17,
fig. I; J. Lge, Fl. Ag. Dan., 4, 1939, tab. 135 C.
В широколиственных, хвойных н смешанных лесах, на лесных по-
лянах; гум. canp., VI—X, о. р., (1), м. с. г., эвриголаркт., евр.
Распр. в СССР. Европ. ч.: ЭССР, ЛитССР.
Общее распр. Европа: ВНР, ЧССР, Дания, ФРГ, ПНР.
61. Agaricus sagatus Fr., s. Ricken, Die Blatterpilze (Agaricaceae)
Deutschlands..., 1915:239.
В широколиственных лесах, иа открытых травянистых местах;
подст. canp., VII—IX, о. р., (1), с., иемор., евраз.
Распр. в СССР. Ё в р оп. ч.: РСФСР — Краснодаром кр., ЛатвССР;
Азиат, ч.: Зап. Сибирь — КазССР.
Общее распр. Европа: Австрия, ВНР, ГДР, ФРГ, Франция,
ЧССР, Швейцария.
РОД GYROPHRAGM1UM MONT.
Ann. Sci. Nat., 20, 2, 1843:77-—Polyplocium Berk., London Journ.
Bot., 2, 1843 : 203.— Longia Zeller, Mycologia, 35, 4, 1943 : 414.— Longa-
la Zeller, Mycologia, 37, 5, 1945 : 636.
62. Gyrophragmiuni dunalii (Fr.) Zeller, Mycologia, 35, 1943:411,—
Montagnites dunalii Fr., Epicr. Syst. Мус., I, 1836—1838 : 241.— Gyro-
phragmium delilei Mont., Fl. d*Algeria, I, 1846 : 369.— G. argentinum
Speg., Fung. Arg. nov. crit., 1899: 185.— G. decipiens (Pk) Lloyd, Мус,
78
Writ., I, 1901 : 62.— Podaxon strobilaceus Copel., Апл. Мус., 2, 1904 : 4.—
G. califbrnicum (Harkness) Morse, Mycologia, 35, 1943:412 (табл.. VII,
2a~ 6).
Icon.: Fischer, in Engl, et Pr., Nat. Pfl., 7a, 1933, fig. 88b; Sebek,
Gasteromycetes, Flora CSR, 1958 :241, fig. 51, 4; Шварцман, Филлимо-
нова, Гастеромицеты, Фл. спор. раст. Каз., VI, 1960, рис. 16, табл. 1,
рис. 14; Сосин, Опр. гастеромиц. СССР, 1973, рис. 101; Зерова, Атл.
гр. Укр., 1974, табл. 129, рис. 3; Вассер, Фл. гр. Укр. (Agaricaceae),
1980, рис. 107, табл. XV, 2.
На песчаных почвах в степях, полупустынях, пустынях; гум. сапр.,
VI—IX, д. ч., (1), п. з. и., мультирег., евраз-ам-афр.
Распр. в СССР. Европ. ч.: РСФСР — Астраханская обл.; Ази-
а т. ч.: Зап. Сибирь — КазССР, Ср. Азия — ТаджССР.
Общее распр. Европа: Португалия, Франция, Италия; Азия:
МНР, Израиль; Сев. Америка: Канада, США, Мексика; Южн.
Америка: Аргентина; Африка: Алжир, Гана, Египет, ЮАР, Ма-
рокко, Тунис.
РОД ENDOPTYCHUM CZERN.
Bull. Soc. Nat. Mose., 18, 1845 : 146.
63. Endoptychum agaricoldes Czern., Bull. Soc. Nat. Mose., 18, 1845:
148.— Secotium agaricoldes (Czern.) Hollos, Termes. Ftiz., 25, 1902 : 93—
95.— S. czerniajevii Mont., Rev. Bot., 1846:310.— S, acuminatum Mont.,
Explor. Sci. de 1‘Algerie, I, 1846—1849:371.— S. besserianum Durieu et
Mont, in Mont., Explor. Sci. de I’Algerie, I, 1846 :373.— Podaxon thunii
Schulz., Verh. Zool.-bot. Ges. Wien, 9, 1859:93.— Secotium malinvernia-
num Cesati, Nuovo Gi. bot. ital., I, 1869: 136.— Podaxon acaule Hazslin-
sky, Verh. Zool.-bot. Ges. Wien, 26, 1876 :217.— Secotium scabolcsiense
Hazslinsky, Verh. Zool.-bot. Ges. Wien, 26, 1876:217.— Lycoperdon war-
nei Pk, Bull. Tor. Bot. Club, 6, 1879:77.— Secotium warnei (Pk) Pk,
Bull. Tor. Bot. Club, 9, 1882—1893 : 2 (табл. XXII, 31—32).
Icon.: Bucholtz, Beitr. zur Morphol. u. system der Hypog., 1902,
tab. IV, Fig. 19; Hollos, Gast. Hung., 1904, tab. Ill-—IV; Sebek, Gastero-
mycetes, Flora CSR, 1958, fig. 58—60; Шварцман, Филимонова, Гастеро-
мицеты, Фл. спор. раст. Каз., VI, 1960, рис. 18; Сосин, Опр. гастеромиц.
СССР, 1973, рис. 99; Зерова, Атл. гр. Укр., 1974, табл. 159, рис. 2.
На лугах, полях, пастбищах, в степях, лесах, на свалках, по опуш-
кам боровых лесов, на песчаных, удобренных и перегнойных почвах;
гум. сапр., V—XI, о. ч., (1—2), п. з. н., мультирег., космопол.
Распр. в СССР. Европ. ч.: РСФСР — Ставропольск. кр., УССР;
Азиат, ч.: Кавказ — ГССР, Зап. Сибирь — Алтайск. кр., КазССР,
Вост. Сибирь — Бурятская АССР, Ср. Азия — КиргССР, УзССР,
ТаджССР, ТССР, Дальн. Восток — Приморск, кр.
Общее распр. Европа: Швеция, ФРГ, ГДР, ЧССР, СРР, Ита-
лия, Франция, Греция, Австрия, Великобритания, Бельгия, Дания, Ни-
дерланды, Ирландия, Исландия, Испания, Лихтенштейн, Норвегия,
ПИР, Португалия, СФРЮ, Финляндия, Швейцария; Азия: Япония,
Израиль, КНР, МНР; Сев. Америка: Канада, США, Мексика;
Южн. Америка: Аргентина, Бразилия, Чили; Африка: Египет,.
Алжир, Марокко; Австралия; Новая Зеландия; о-в Куба.
79-
КОЛЕНО CYSTODERMATEAE SING.
The Agaricalcs in modern taxonomy, 1975 : 477.— Cystodermateae
Sing., The Agaricales (Mushrooms) in modern taxonomy, 1951 :444
(nom. nudum).
РОД PHAEOLEPIOTA R. MRE EX KONR. ET MAUBL.
Icon. Sei. Fting., VI, 1924—1937: 111.— Phaeolepiota R. Mre, BSMF,
27, 1911 : 39 (nom. nudum).
64. Phaeolepiota aurea (Fr.) R. Mre ex Konr. et Maubl., Icon. Sei.
Fung., 6, 1924—1937 : 111.— Agaricus aureus Fr., Syst. Мус., I, 1821:
241.—A. vahlei Fr., Syst. Мус., I, 1821 : 240,— Pholiota aurea (Fr.)
Kumm., Dcr Fuhrer in die Pilzkunde, 1871 :85 s. auct., non s. Cke, Ri-
cken.— Ph. aurea (Fr.) Kumm. var. vahlei (Fr.) Sacc., Syll. Fung., V,
1887 : 736.— Lepiota pyrenaea Quel., Champ. Jura et Vosges, 1887:1,
suppl. 16 (reprint).— Togaria aurea (Fr.) W. G. Sm., Syn. Brit. Bas.,
1908:122.— Phaeolepiota aurea (Fr.) R. Mre, BSMF, 27, 1911:102.—
Cystoderma aurea (Fr.) Kuhn, et Romagn., Fl. anal. Champ. Super.,
1953 : 393 (табл. VII, За—д).
Icon.: Cke, Ill. Brit. Fung., 1881 — 1891 : 375; Rolland, Atl. Champ.
Fr., 1905—1910, tab. 62, fig. 138; Heinemann, BSMF, 53, 1937, Atl. pl. 75;
J. Lge, Fl. Ag. Dan., 1935—1940, tab. 105, fig. 8; Achermann, Schweiz.
Zcitschr. Pilzk., 8, 1938 (фото); Michael u. Hennig, Handbh. Pilzfr., 1967,
IV, tab. 19; A. H. Smith a. L. R. Hesler, The North American species of
Pholiota, 1968, fig. 9—10, pl. 5—6; Зерова, Атл. гр. Укр., 1974, табл. 151,
рис. 2; Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 108, табл. XV, 1.
В лиственных и хвойных лесах, на их опушках, в кратко- и про-
должительнопоемных лесах, в парках под кустарниками, на рудераль-
ных местах, иногда в крапиве, часто на местах пожаров, по обочинам
дорог, изредка иа древесной трухе; гум. сапр., VIII—XI, ир., (2), с.,
эврнголаркт., евраз-ам.
Распр. в СССР. Европ. ч.: РСФСР — Ленинградская, Архангель-
ская обл., Карельская АССР, ЭССР, ЛитССР, БССР, УССР; Азиат,
ч.: Кавказ — ГССР, Зап. Сибирь — Марийская АССР, Дальн. Восток —
Хабаровск, кр., Приморск, кр.
Общее распр. Европа: Великобритания, Бельгия, Дания, Фран-
ция, Швейцария, Италия, ФРГ, ГДР, Швеция, ПНР, ЧССР, Австрия,
ВНР; Азия: Япония; Сев. Америка: Каиада, США.
РОД CYSTODERMA FAY.
Prodrome, Ann. Sci. Nat. Bot., VII, 9, 1889 : 351.
Подрод Cystoderma
Секция Cystoderma s. Sing., 1962, 1975:479, non Amianthina
A. H. Sm. et Sing., Pap. Mich. Acad. Sci., Arts a. Lett., 30, 1945 : 101;
Genuinae Locq. (nom. subnudum).
Секция Cystoderma
Amianthina A. H. Sm. et Sing., Pap. Mich. Acad. Sci., Arts a. Lett.,
30, 1945 : 101.— Genuinae Locq. (nom. subnudum), non Sect. Cystoderma
s. Sing., 1962, 1975.
80
65. Cystoderma amianthina (Fr.) Fay. var. amianthina, Ann. Sci. Nat.
Bot., VII, 9, 1889:351.— Agaricus granulosus stibsp. amianthinus Fr.,
Epicr. Syst. Мус., I, 1836:18.— A. granulosus subsp. subochraceus Fr.,
Monogr. Hymen. Suec., 1, 1857:29.— A. amianthinus Fr., Hymen. Eur.,
1874 :37.— Lepiota granulosa var. ochracea Gill., Les Hymenomycetes,
1874:71.— L. amianthina (Fr.) Karst., Hattsvampar, 1879: 15.— Armilla-
ria amianthina (Fr.) Kauffm., Pap. Mich. Acad. Sci., Arts a. Lett., 2,
1923 (1922) : 60.— ? A. muscicola CleL, Trans, a. Proc. Roy. Soc. South
Australia, 51, 1927 : 299 (табл. VII, 4a—г).
Icon.: Baria, Champ. Alp. Mar., 1888—1892, pl. 16. fig. 9—11;
Boudier, Icon. Мус., 1904—1909, vol. I, pl. 16; Cke, Ill. Brit. Fung.,
1881—1891, pl. 213; Brcs., Icon. Мус., 1927, tab. 37; J. Lge, Fl. Ag. Dan.,
1935—1940, tab. 15, fig. C; Maublanc, Champ. Fr., 1939, tab. 84; BSMF,
67, 1951 : 70, fig. 1; Michael u. Hennig, Handb. Pilzfr., 1968, I, tab. 35;
Зерова, Атл. гр. Укр., 1974, табл. 118, рис. 1; Вассер, Фл. гр. Укр. (Aga-
ricaceae), 1980, рис. 109, табл. XVI, 1.
В сосновых, еловых, кедрово-сосновых, елово-пихтовых, пихтово-
кедровых, сосново-дубовых, сосново-березовых, кедрово-широколиствен-
ных. буковых лесах, в зарослях кедрового стланика, на полянах, в пар-
ках, среди мхов, главным образом рода Polytrichum.
С. amianthina var. longispora*** Kiihn. ex S. Wasser, Укр. ботан.
журн., 35, 5, 1978:518.— Lepiota amianthina var. longispora Kuhn.,
BSMF, 52, 1936:204 (nom. nudum).— Cystoderma amianthina var.
longispora (Kiihn.) A. H. Sm. et Sing., Pap. Mich. Acad. Sci., Arts a.
Lett., 30, 1945:114 (nom. inval.).— C. longispora (Kiihn.) Heinem. et
Thoen, BSMF, 89, 1, 1973:16 (nom. inval.).— ? Agaricus jasonis Cke
et Mass., Grevillea, 16, 1887.— ? Armillaria jasonis (Cke et Mass.) Sacc.,
Syll. Fung., 9, 1891 : 12.
В сосновых, сосново-дубовых, сосново-березовых лесах, в парках,
среди мхов. Известна из УССР.
F. amianthina. Экология, иконография цитированы при С. amian-
thina var. amianthina.
F. olivacea Sing, apud A. H. Sm. et Sing., Pap. Mich. Acad. Sci.,
Arts a. Lett., 30, 1945 : 111.
В кедрово-сосновых, пихтово-кедровых лесах. Известна из Зап.
Сибири.
Подст. canp., VI—XII, о. ч., (3), м. с. г., мультнрег., космопол.
Распр. в СССР. Европ. ч.: РСФСР — Ленинградская, Москов-
ская обл., ЭССР, ЛатвССР, ЛитССР, БССР, УССР; Азиат, ч.:
Аркт.— Таймырский нац. окр., Кавказ — ГССР, Зап. Сибирь — Комн
АССР, Томская обл., Алтайск. кр., Вост. Сибирь — Красноярск, кр..
Бурятская АССР, Дальи. Восток — Приморск, кр., Ср. Азия —
КиргССР.
Общее распр. Европа: Великобритания, Португалия, Испания,
Франция, Дания, Бельгия, Италия, ФРГ, ГДР, Финляндия, ПНР, ЧССР,
Австрия, ВНР, СФРЮ, Греция, СРР; Азия: Япония; Сев. Аме-
рика: Канада, США; Южн. Америка: Венесуэла, Бразилия, Ар-
гентина, Чили, Боливия; Африка: Алжир, Марокко; А.в с т р а л и я;
о-ва Гренландия, Куба.
66. Cystoderma rugosoreticulata ** (Lorinser) S. Wasser, Укр. бо-
тан. журн., 35, 5, 1978 : 516.— Agaricus rugosoreticulatus Lorinser, Oest.
Bot. Zeitschr., 29, 1879 : 23.— Lepiota rugosoreticulata (Lorinser) Sacc.,
Syll. Fung., 9, 1891 : 10.— Armillaria rugosoreticulata (Lorinser) Zeller,
8 4—2960
81
Mycologia, 33, 1933 : 378.— Cystoderma amianthina (Fr.) Fay. f. rugo-
soreticulata (Lorinser) A. H. Sm. et Sing., Pap. Mich Acad. Sci., Arts a.
Lett, 30, 1945 : 110 (табл. VII, 5a—d).
Icon.: A. H. Smith a. Singer, Pap. Mich. Acad. Sci., Arts a. Lett.,
30, 1945, pl. HI; A. H. Smith; A Field Guide to Western Mushrooms,
1975, fig. 98; Wasser, Fung. Rar. Icon. Col., X, 1979, pl. 78a, Text-fig. 3;
Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. НО, табл. XVII, 1.
В равнинных и горных сосновых, еловых лесах, среди мхов; бри-
отр.; VII—XI, д. р , (2), с. (?), я.(?), бор., евр-ам.
Распр. в СССР. Европ. ч.: УССР L
Общее распр. Европа: Италия, Австрия; Сев. Америка: Ка-
нада, США.
Прим. Одни авторы (Smith A., Singer, 1945; Thoen, 1967; Heinemann, Thoen,
1973) считают C rugosoreticulata формой С. aminianthina, другие (Lorinser, 1879;
Saccardo, 1891; Zeller, 1933.— Цит. no: Smith A., Singer, 1945) рассматривают ее
в ранге самостоятельного вида родов Agaricus (Lorinser), Lepiota (Saccardo), Armilla-
ria (Zeller). Мы считаем C. rugosoreticulata хорошим видом, наиболее близким к
С. amianthina, от которой четко отличается, главным образом сильно радиально- илн
сетчато-морщииистон шляпкой (Вассер, 1978а, 1979, 1980).
67. Cystoderma carcharias (Seer.) Fay., Ann. Sci. Nat. Bot Ser.,
VII, 9, 1889 : 351.— Agaricus carcharias Seer., Mycorg. Suis., I, 1833:
62.— A. granulosus var. carcharias (Seer.) Fr., Epicr. Syst. Мус., 1, 1836:
18.— A. granulosus subsp. incarnato-albida Fr., Monogr. Hymen. Suec.,
I, 1857 : 29.— Lepiota granulosa var. carcharias (Seer.) Karts., Hatts-
vampar, 1879:14.— Armillaris carcharias (Seer.I Kauffm., Pap. Mich.
Acad. Sci., Arts a. Lett, 4, 1925 (1924) : 317 (табл. VII, 6a—6).
Icon.: Baria, Champ. Alp. Mar., 1888—1892, pl. 15, fig. 16—18;
Boudier, Icon. Мус., 1904—1909, pl. 14; Cke, Ill. Brit. Fung., 1881—1891,
pl. 42; Bres., Icon. Мус., 1827, tab. 38; J. Lge, Fl. Ag. Dan., 1935—1940,
tab. 15, fig. C; Konr. et Maubl., Icon. Sei. Fung., 1924—1937, pl. 237,
fig. 1; Maublanc, Champ. Fr., 1939, tab. 84; BSMF, 67, 1951 : 75, fig. 4;
Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 111.
В сосновых, еловых, cochobq-березовых, дубово-сосновых, широко-
лиственных лесах; подст. сапр., VII—XI, о. ч.. (2), м. с. г., бор., ев-
раз-ам.
Распр. в СССР. Европ. ч.: РСФСР — Московская, Ленинград-
ская обл., ЭССР, ЛатвССР, ЛнтССР, БССР, УССР; Азиат, ч.: Кав-
каз — ГССР, Зап. Сибирь — Алтанск. кр., Вост. Сибирь — Красноярск,
кр., Якутская АССР, Дальн. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Финляндия, Бельгия,
Дания, Франция, Испания, Италия, ФРГ, ГДР, ПНР, ЧССР, Австрия,
ВНР, СФРЮ; Сев. Америка: США.
68. Cystoderma caucasicum Sing, apud A. H. Sm. et Sing., Pap.
Mich. Acad. Sci., Arts a. Lett., 30, 1945 : 105-
Icon.: A. H. Sm. et Sing., Pap. Mich. Acad. Sci., Arts a. Lett,
30, 1945, pl. IV, fig. 8 (споры).
В смешанных лесах, на валежной гнилой древесине Abies nordma-
nnianae (Stev.) Spach. на высоте 700 м н. у. м.; лиги., VII—X, о. р., (1),
с.(?), я.(?), бор., евр., северокавк. усл. энд.
Распр. в СССР. Европ. ч_: РСФСР — Краснодарск. кр.
1 Вероятно, произрастает и в других районах СССР, однако исследователями
ие выделялся в самостоятельный таксон — С. rugosoreticulata.
82
Прим. Материал, положенный в основу нового вида — С. caucasicum, перво-
начально был опубликован как С. echinella (Quel.) Sing, apud L. Vass. (Учен. зап.
Казанск. ун-та, 105 (99), 11939:56). Однако для вида Л. Келе—Lepiota echinella
Quel, характерны свободные пластинки, споры 6—7X3—3,5 мкм, произрастание иа
почве. Эти отличия н положены в основу нового вида — С. caucasicum, у которого
приросшие пластинки, споры 4X2,5 мкм. По данным А. Смнза и Р. Зингера (Smith А.,
Singer, 1945), С. caucasicum наиболее близок к североамериканскому виду С. pulve-
raceum (Pk) А. Н. Sm. et Sing., от последнего отличается наличием отрицательной
реакции кутикулы шляпки со щелочью, при которой коричневатые клетки эпителия
становятся гиалиновыми.
69. Cystoderma fallax А. Н. Sm. et Sing., Pap. Mich. Acad. Sci.,
Arts a. Lett., 30, 1945: 116.
Icon.: A. H. Smith a. Singer, Pap. Mich. Acad. Sci., Arts a. Lett.,
30, 1945, pl. IV, fig. 8 (споры); A. H. Smith, A Field Guide to Western
Mushrooms, 1975, fig. 97.
В равнинных и горных сосновых, сосново-пихтовых, сосново-пихто-
вых с примесью березы и осины лесах; бриотр., IX—XI, о. р., (1—2),
с. (?), я.(?), бор., евраз-а.м-афр.
Распр. в СССР, Азиат, ч.: Аркт.— Таймырский нац. окр., Вост.
Снбнрь — Бурятская АССР.
Общее распр. Европа: Франция; Сев. Америка: Канада,
США; Африка: Марокко.
Прим. С. fallax близка к североамериканскому виду С. granosa (Morg.)
А. Н. Sm. et Sing, и к голарктическому виду С. carcharias (Seer.) Fay. По данным
авторов вида (Smith A., Singer, 1945), С. fallax отличается от С. granosa цветом,
размером карпофоров и экологией, а от С. carcharias четко отличается реакцией
кутикулы шляпки с КОН.
Секция Superba Heinem. et Thoen
BSMF, 89, I, 1973 : 23.
70. Cystoderma superbum Huijsm. emend. Huijsm., Fungus, 28, 1958:
47.— Armillaria haematites (Berk, et Br.) Sacc., Syll. Fung., 5, 1887:77,
s. Bres. non s. Berk, et Br.— Lepiota haematites (Berk, et Br.) Ricken,
Die Blatterpilze (Agaricaceae) Deutschiands..., 1915 :328, s. Ricken non
al.— Cystoderma haematites (Berk, et Br.) Konr. et Maubl., Icon. Sei.
Fung., Ill, pl. 202, s. Kuhn, et R. Mre.. A. H. Sm. et Sing., non s. orig.—
Collybia lilacea (Berk, et Br.) Quel., Champ. Jura et Vosges, 1875:6,
s. Konr. et Maubl., non Quel. (табл. VIII, la—г\ XXIII, 53).
Icon.- Cke, Ill. Brit. Fung., 1881—1891, pl. 45; Bres., Icon. Мус.,
1923, tab. 49, fig. 1; Konr. et Maubl., Icon. Sei. Fung., 1924—1937, pl. 237,
fig. 2, tab. 202 (как Collybia lilacea Quel. f. robusta)-, Imbach, Schweiz.
Zeitschr. Pilzk., 12, 1944 : 190; BSMF, 67, 1951 : 79; Huijsman, Fungus,
26, 1956:40, fig. 1—16; Зерова, Атл. гр. Укр., 1974, табл. 84, рис. 2
(как Cystoderma haematites (Seer.: Pers.) Fay.); Wasser, Fung. Rar.
Icon. Col., X, 1979, pl. 78b; Вассер, Фл. гр. Укр. (Agaricaceae), 1980,
рис. 112, табл. XVIII, 2.
В сосновых, еловых, сосново-листвеиничных, сосново-березовых, бу-
ковых лесах; подст. сапр., VIII—X, д. р., (2), с.(?), я.(?), эвриголаркт.,
евраз.
Распр. в СССР. Европ. ч.: Карельская АССР, УССР; Азиат,
ч.: Вост. Сибирь — Красноярск, кр.
Общее распр. Европа: Великобритания, Швейцария, Италия,
Франция, ФРГ, ГДР.
8* 83
Прим В роде Cystoderma С. superbum хорошо ограниченный вид, характе-
ризующийся красивым пурпурным цветом карпофоров, с маленькими спорами, ами-
лоидными лишь в супраапикулярной зоне, кутикулярными клетками, темнеющими
в КОН. наличием большого количества сфсроцист иа поверхности шляпки.
С. superbum наиболее близка к С. haematites s. crig., из которой она и выде-
лена X. С. Ц. Хьюзмано.м (Huijsman, 1956).
Проблема комплекса С. superbum — С. haematites подробно и всесторонне рас-
смотрена нами (Вассер, 1980. 1981).
Подрод JulvuGosa (Fr.) S Wasser
Фл. гр. Укр. (Agaricaceae), 1980:209.— Granulosa Fr., Syst. Мус.,
1821 :93, s. A. H. Sm. et Sing., Pap. Mich. Acad. Sci., Arts a. Lett., 30,
1945:81.
Секция Granulosa (Fr.) Locq. emend. Locq.
BSMF, 67, 1951 : 69.
71. Cystoderma ambrosii (Bres.) A. H. Sm. et Sing., Pap. Mich.
Acad. Sci., Arts a. Lett., 30, 1945 :81.— Armillaria ambrosii Bres., Fung.
Trid., I, 1881 : 27 (табл. VI11, 2a—г).
Icon.: Brcs., Fung. Trid:, I, 1881, tab. 31; Brcs., Icon. Мус., 1927,
tab. 50; Вассер, Фл. гр. Укр. (Agaricaceae), 1980, табл. XVII, 2.
В еловых, елово-буковых, сосново-березовых, лиственнично-сосновых
горных лесах, ольхово-кедровых, насаждениях, доходит до высоты
1900 м и. у. м.; бриотр., VIII—X, р., (3), с.(?), я.(?), монт., евраз-ам.
Распр. в СССР: Европ. ч.: УССР; Азиат, ч.: Вост. Сибирь —
Красноярск, кр., Ср. Азия — КиргССР.
Общее распр. Европа: Франция, Италия, ЧССР; Сев. Амери-
ка: США.
Прим. С. ambrosii очень близка к С. carcharias (Seer.) Fay. f. alba (Fr.)
A. H. Sm. et Sing., известной из сосновых лесов (Pinus resinosa Ait.) Норвегии и
США, однако легко отличается от нее наличием неамилоидных спор и большим
количеством нитчатых гиф кутикулы шляпки. По мнению А. Смиза и Р. Зингера
(Smith A., Singer, 1945), неамилондность спор и наличие сравнительно большого
числа нитчатых гиф кутикулы шляпки характеризуют более низкое положение
С. ambrosii в эволюционном ряду рода.
72. Cystoderma granulosa (Batsch: Fr.) Fay., Ann. Sci. Nat. Bot.,
Ser. VI1, 9, 1889 :341.— Agaricus granulosus Batsch : Fr., Syst. Мус., 1,
1821 : 24.— A. granulosus subsp. ferruginea Fr., Monogr. Hymen. Suec.»
I, 1857 : 29.— Lepiota granulosa (Batsch : Fr.) S. F. Gray, Nat. Arr. Brit.
PL, 1, 1821:602.— Armillaria granulosa (Batsch: Fr.) Kauffm., Pap.
Mich. Acad. Sci., Arts a. Lett., 2, 1923 (1922) : 60 (табл. VIII, За—в).
Icon.: Baria, Champ. Alp. Mar., 1888—1892, pl. 16, fig. 5—8: Cke,
Ill. Brit. Fung., 1881 — 1891, pl. 18; Bres. Icon. Мус., 1930, tab. 599;
J. Lge, Fl. Ag. Dan., 1935—1940, tab. 15, fig. E; A. H. Smith a. Singer,
Pap. Mich. Acad. Sci., Arts a. Lett., 30, 1945, pL 1; Вассер, Фл. гр. Укр.
(Agaricaceae), 1980, рис. 113.
В сосновых, сосново-березовых, сосново-листвениичных, пихтово-
еловых, елово-кедрово-осоково-сфагново-гипновых, елово-кедрово-пих-
товых, лиственнично-березовых, березовых, сосново-дубовых, смешан-
ных широколиственных лесах, на полянах лесов, на пастбищах; подст.
canp., VII—XI, о. ч., (2—3), м. с. г., мультнрег., космопол.
Распр. в СССР. Европ. ч.: РСФСР — Ленинградская, Москов-
ская обл., ЭССР, ЛатвССР, ЛитССР, БССР, УССР; Азиат, ч.:
84
Ар кт.— Таймырский нац. окр., Кавказ — ГССР, Зап. Сибирь — Алтайск.
кр., Мордовская АССР; Вост. Сибирь — Красноярск, кр.. Бурятская
АССР, Дальн. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Норвегия, Швеция, Фин-
ляндия, Дания, Бельгия, Нидерланды, Франция, Италия, Испания, ФРГ,
ГДР, ПНР, ЧССР, Австрия, СФРЮ; Азия: КНР, Япония, Сев.
Америка: Канада, США; Южн. Америка: Венесуэла; Афри-
к а: Алжир, Марокко; Австралия; Соломоновы о-ва.
73. Cystoderma adnatifolia (Pk) Harmaja, Karstenia, 14, 1974: 122.—
Lepiota adnatifolium Pk, Bull. N. Y. State Mus., 10, 1902:947.— Armil-
laria adnatifolia (Pk) Kauffm., Pap. Mich. Acad. Sci., Arts a. Lett., 2,
1923 (1922): 60.— Cystoderma granulosa (Batsch : Fr.) Fay. var. adna-
tifolia (Pk) A. H. Sm. ct Sing., Pap. Mich. Acad. Sci., Arts a. Lett., 30,
1945 : 90.
Icon.: A. H. Smith a. Singer, Pap. Mich. Acad. Sci., Arts a. Lett.,
30, 1945, fig. 2.
В хвойных лесах; бриотр., IX—X, о. р., (1), м. с. г., бор., евр-ам.
Распр. в СССР. Европ. ч.: РСФСР — Ленинградская обл.
Общее распр. Европа: Финляндия; Сев. Америка: Канада,
США.
Прим. Морфологически С. adnatifolia близка также к С. cinnabarina, от ко-
торой четко отличается отсутствием плевро- и хейлоцистид.
Интересно отметить, что С. adnatifolia в отечественной литературе не упомина-
лась. Впервые как новый вид для Европы се приводит финский миколог Г. Хармайя
(Harmaja. 1974), который отмечает ряд местонахождений в Финляндии и одно место-
нахождение для СССР (Ленинградская обл.) (Вассер, 1980).
Секция Cinnabarina Heinem. et Thoen
BSMF, 89, 1, 1973 : 23.
74. Cystoderma cinnabarina (Alb. et Schw. ex Seer.) Fay, Ann. ScL
Nat., Ser. VII, 9, 1889:341.— Agaricus granulosus cinnabarinus Alb. et
Schw. ex Sccr., Mycogr. Suis.. I, 1833:60.— A (Lepiota) granulosa
subsp. cinnabarina (Alb. et Schw. ex Seer.) Fr., Monogr. Hymen. Suec.,
I, 1857 :29.— A. cinnabarinus (Alb. et Schw. ex Sccr.) Fr., Hymen. Eur.,
1874 : 36.— Lepiota granulosa var. cinnabarina (Alb. et Schw. ex Seer.)
Gill., Champ. Fr., 1878:71.— L. granulosa subsp. cinnabarina (Alb. ct
Schw. ex Seer.) Konr. et Maubl, Icon. Sei. Fung., III. 1924—1933,
pl. 238.— L. cinnabarina (Alb. et Schw. ex Seer.) Karst., Hattsvampar,
1879:14.— Armillaria cinnabarina (Alb. et Schw. ex Sccr.) Kauffm.,
Pap. Mich. Acad. Sci., Aris a. Lett, 2, 1923 (1922) : 60.— Cystoderma
cinnabarina var. claricolor Romagn., Nouv. Atl. Chatnp., 3, 1961 : 199,
syn. nov. (табл. VI11, 4a—d).
Icon.: Baria, Champ. Alp. A4ar., 1888—1892, pl. 16, fig. 1—4;
Boudier, Icon. Мус., 1904—1909, pl. 15; Cke, Ill. Brit. Fung., 1881—1891,
pl. 43; Bres., Icon. Мус., 1927, tab. 36; J. Lge, Fl. Ag. Dan., 1935—1940,
tab. 15, fig. F; Konr. ct Maubl, Icon. Sei. Fung., 1924—1937, pl. 238;
Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 114, табл. XVI, 2.
В сосновых, сосиово-березовых, сосново-дубово-грабовых, сосново-
дубовых, кедрово-широколиственных, дубовых лесах, в тундре в на-
саждениях Betula папа L.; подст. сапр., VII—X, о. ч., (2—3), м. с. г.,
эвриголаркт., евраз-ам-афр.
&>
Распр. в СССР. Европ. ч.: РСФСР — Ленинградская обл.,
ЛитССР, ЛатвССР. УССР; Азиат, ч.: Аркт,—Таймырск. нац. окр..
Зап. Сибирь—Алтайск. кр.. Вост. Сибирь — Бурятская АССР, Крас-
ноярск. кр., Якутская АССР, Дальн. Восток — Амурская обл., При-
морск. кр.
Общее распр. Европа: Великобритания, Финляндия, Бельгия,
СРР, Дания, Франция, Испания, Италия, ФРГ, ГДР, ПНР, ЧССР,
Австрия, ВНР; Азия: Япония; Сев. Америка: Канада, США;
Африка: Алжир, Марокко.
РОД SQUAMANITA IMBACH
Mitt. Naturf. Ges. Luzern, 15, 1946:81.— Squamamanita Imbach,
Schweiz. Zeitschr. Pilzk., 20, 1942: 152 (nom. nudum).— Coolici Huijsm.,
Med. Ned. Мус. Ver., 28, 1943:59 (nom. nudum).— Tricholoma sect.
Cootia (Huijsm.) Konr. et Maubl., Les Agaricales I, 1948 :345 (nom.
nudum).— Cystoderma subgen. Dissoderma A. H. Sm. et Sing., Myco-
logia. 40, 1948 :454.
75. Squamanita paradoxa (A. H. Sm. et Sing.) Bas, Persoonia, 3, 3,
1965 : 348. — Cystoderma paradoxa A. H. Sm. et Sing., Mycologia. 40, 4.
1948 : 454 (табл. VIII, 5a—a).
Icon.: Bas, Persoonia, 3, 3, 1965, fig. 5—8; Вассер, Фл. гр. Укр.
(Agaricaceae), 1980, рис. 115.
В широколиственных и смешанных лесах, растет отдельными
экземплярами или группами, выходящими из общего базиса; гум. сапр.,
IX—XI, о. р., (1—2), с.(?), я.(?), эврнголаркт., евраз-ам.
Распр. в СССР. Азиат, ч.: Зап. Сибирь — Алтайск. кр.
Общее распр. Европа: Великобритания, Швейцария, ЧССР;
Сев. Америка: США.
Прим. Наиболее характерными чертами S. paradoxa являются цвет шляпки,
ложки, пластинок, общего покрывала и его ранняя фрагментация. Общее покрывало
S. paradoxa заканчивает свое развитие у молодых карпофоров, что делает возмож-
ным его фрагментацию н отделение от поверхности шляпки на ранних этапах раз-
вития. Все указанное делает S. paradoxa легкоопределяемым видом.
К. Бас (Bas, 1965), монограф рода Squamanita, отмечает, что споры паратнпа
S. paradoxa из коллекции А. Смиза (Herb. University of Michigan, Ann Arbor, USA,
N 28341) были четко псевдоамплондными, в то время как споры европейских кол-
лекций Е. Херннка (Herb. J. A. Herink, CSSR, 651/49) и Э. Хорака (Herb. Иогак,
Zurich, Schweiz) были непсевдоамилоиднымн или едва пссвдоамилоидными.
Характерным признаком S. paradoxa некоторые авторы (Herink, 1954b; Bas,
1965) питают склероциальное образование в основании ножки. Ткань поверхности
склероция часто перемешана с хламидоспорами. Э. Хорак (Ногак, 1964). К. Бас (Bas,
1965), Р. Устлинг (Watling, 1974) приводят для S. paradoxa тонкостенные, бесцвет-
ные, реже светло-коричневатые, округлые конидии (11—16(19)Х7,5—12,5 мкм! и
1—6 мкм шир., септированные, тонкостенные конидиеносцы (Вассер, 1980).
КОЛЕНО LEPIOTEAE FAY.
Prodrome, Ann. Sci. Nat. Bot., VII, 9, 1889 : 349.
РОД PSEUDOBAEOSPORA SING.
Lloydia, 5, 1942 : 129 *.
1 В данной работе не приведен латинский диагноз рода, однако указано, что
характеристика нового рода совпадает с латинским диагнозом типового вида, имею-
щегося в статье Р. Зиигсра (Singer, 1938).
86
76. Pseudobaeospora pillodii ** (Quel.) S. Wasser, Фл. гр. Укр. (Aga-
ricaceae), 1980 : 220, s. Horak.— Collybia pillodii Quel., C. R. Ass. franc.
Av. Sci., XVIII, 2, 1883:509.— Pseudobaeospora pillodii (Quel.) Horak,
Rev. Mycol., 29, I—2, 1964:73 (nom. nudum) (табл. VIII, 6a—г).
Icon.: Horak, Rev. Mycol., 29, 1—2, 1964, fig. la, b, c; Horak,
Bcitr. KrypL-flora der Schweiz, 13, 1968: 512.
В редкостойных высокогорных лесах под Larix, Alnus viridis DC.,
Rhododendron; подст. canp., VIII—XI, о. p., (1), п. з. н., монт., евраз-ам.
Распр. в СССР. Азиат, ч.: Зап. Сибирь — Алтайск. кр., Новоси-
бирская обл.
Общее распр. Европа: Швейцария, Австрия, Франция; Сев.
Америка: США; Южн. Америка: Чили.
77. Pseudobaeospora defibulata Sing., Mycologia, 55, 1, 1963: 13.
Icon.: Sing., Mycologia, 55, 1, 1963, p. 14, fig.
В кедрово-пнхтово-широколиствениых лесах, доходит до 900 м
и. у. м.; подст. canp., VIII—X, о. р., (1), п. з. н., в. и. а.
Распр. в СССР. Азиат, ч.: Дальн. Восток — Приморск, кр.
Общее распр. Южн. Америка: Аргентина.
РОД LEPIOTA (PERS.: FR.) S. F. GRAY
Nat. Arr. Brit. PL, I, 1821:601.— Fusispora Fay., Prodrome, Ann.
Sci. Nat. Bot, VII, 9, 1889:351.— Lepiotula (R. Mre) Locq. ex Horak,
Bcitr. Krypt.-flora der Schweiz, 13, 1968 :338.
Подрод Sphaerocystis S Wasser
Укр. ботан. жури., 35, 5, 1978 : 517.
Секция Micaceae J. Lge
Fl. Ag. Dan., I, 1935 : 23.
78. Lepiota seminuda (Lasch) Kumm. f. seminuda, Der Fuhrer in
die Pilzkunde, 1871 : 136.— Agaricus seminudtis Lasch, Linnaea, 3, 1825 :
157.— A. histion Seer., Mycogr. Suis., I, 1833 : 45.— A. actinorhizas Mont.,
Ann. Sci. Nat., II, 1836, tab. 12.— Lepiota seminuda (Lasch) Quel.,
Champ. Jura et Vosges, I, 1872 : 210.— L. seminuda (Lasch) Gill., Champ.
Fr., 1878 : 71.— Cystoderma seminudum (Lasch) Fay., Ann. Sci. Nat
Scr. VII, 9, 1889 : 351 (табл. IX, la—в).
Icon.: Konr. et Maubl., Icon. Sei. Fung., 1924—1937, pl. 16, fig. 1;
J. Lge, Fl. Ag. Dan., 1935—1940, tab. 14, fig. B; Stangl u. Bresinsky,
Zeitschr. Pilzk., 37, 1971, abb. 3, fig. 1; Вассер, Фл. гр. Укр. (Agarica-
ceae), 1980, рис. 116.
В широколиственных, хвойных и смешанных лесах.
Г. minima J. Lge, Fl. Ag. Dan., I. 1935:36.
B дубовых лесах. Известна нз УССР.
Подст. canp., VI—X, о. ч., (2), п. з. н., эвриголаркт., евраз-ам-афр.
Распр. в СССР. Европ. ч.: РСФСР — Московская обл., ЭССР,
ЛитССР, УССР; Азиат, ч.: Вост. Сибирь — Красноярск, кр., Дальн.
Восток — Приморск, кр.
Общее распр.: Европа: Дания, Бельгия, Нидерланды, Франция,
Италия, ФРГ, ГДР, ПНР, ЧССР, ВНР, Греция, СФРЮ; Азия: Япо-
ния; Сев. Америка: США; Африка: Алжир, Марокко.
87
79. Lepiota bucknallii* (Berk, et Br.) Sacc., Syll. Fung., V, 1887:
50.— Agaricus (Lepiota) bucknallii Berk, et Br., Ann. et Mag. Nat. Hist.,
V, 7, 1881 : 124.— Cystoderma bucknallii (Berk, et Br.) Sing., Schweiz.
Zeitsch. Pilzk., 47, 1939 :53.— Cystolepiota bucknallii (Berk, et Br.)
Sing, et C1q., Nowa Hedwigia, 23, 1973 : 323.
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, tab. 13, fig. E; Babos,
Ann. Hist.-Nat. Mus. Nat. Hung., 61, 1969, fig. 1; Piiat, Kric. urc. nas. hub.
1951, obr. 574—575.
В смешанных широколиственных лесах; подст. сапр., VIII—X, о. р.,
(1), п. з. н., иемор., евр.
Распр. в СССР. Европ. ч.: УССР.
Общее распр. Европа: Великобритания, Швейцария, Франция,
Италия, ФРГ, ЧССР, ВНР.
80. Lepiota rosea Rea, Trans. Brit. Мус. Soc., 6, 1918:61, s. P. D. Or-
ton, Trans. Brit. Мус. Soc., 43, 2, 1960 : 287, non s. Locq., Bull. mens.
Soc. linn. Lyon, 14, 1945 : 95.— Cystolepiota rosea (Rea) Bon, Docums.
Мус., VI, 1976 : 43 (табл. IX. 2a—г).
Icon.: Rea, Trans. Brit. Мус. Soc., 6, 1918, pl. 2, fig. 1; Orton,
Trans. Brit. Мус. Soc., 43, 2, 1960, figs. 115—117, 396, 424—425.
В широколиственных лесах; подст. сапр., V—X, д. р., (1), и. з. н.,
немор., евраз.
Распр. в СССР. Европ. ч.: УССР; Азиат, ч.: Дальн. Восток —
Приморск, кр.
Общее распр. Европа: Великобритания, Дания, Франция, СССР.
81. Lepiota pulverulenta Huijsm., Persoonia, I, 3. 1960:328.— Agari-
cus (Leucosporus, Lepiota) sistratus Fr., Syst. Мус., I, 1821 : 24.— A. (Le-
piota) sistratus p. p. (primaria), Fr., Monogr. Hymen. Suec., 1:30
(Monographia Lepiotarum Sueciae, 1857 :16). Lepiota sistreda (Fr.)
Quel., Champ. Jura et Vosges, 1, 1872:231.— L. sistrata (Fr.) Quel.,
s. J. Herink, Ces. Mykol., 15, 4, 1964:225.— L. pseudogranulosa (Berk,
et Br.) Sacc., Syll Fung., V, 1887:57.— Leucoagaricus pulverulentus
(Huijsm.) Mos., KI- Krypt.-flora, Band 2, Teil 2, 1978: 246 (табл. IX,
За—в).
Icon.: Huijsman, Persoonia, I, 3, 1960, fig. 9—12; Herink, Ces.
Mykol., 15, 4, 1961, fig. 1—4, tab. col., 44; Reid, Col. Ic. Rare a. Int.
Fung., Pars 2, 1967, pl. 9 (e—f), fig. 5a—b; Вассер, Фл, гр. Укр. (Aga-
ricaccac), 1980, рнс. 117—118, табл. XIX, 1.
В парках, искусственных лесных насаждениях, реже в смешанных
широколиственных лесах; подст. сапр., VIII—X, р., (1—2), п. з. н., не-
мор., евр.
Распр. в СССР. Европ. ч.: РСФСР — Ленинградская обл.,
ЛитССР, УССР.
Общее распр. Европа: Великобритания, Швеция, Финляндия,
Швейцария, Франция, Италия, ЧССР, ВНР.
82, Lepiota hetleri Boud., BSMF, 18, 2, 1902:137—138, s. Kuhn,
BSMF, 52, 1936:205—206, s. J. Herink, Ces. Mykol., 15, 4, 1961 : 226,
non s. Moell., J. Lge.— Agaricus (Lepiota) granulosus var. rufescens
Berk, et Br., Ann. ct Mag. Nat. Hist., Ser. V, 7, 1881 : 124.— A. (Lepiota)
granulosus f. rufescens (Berk, et Br.) Britzin., 28. Jahresber. naturhist.
Ver. Augsb.s 1885 :46.— Lepiota granulosa var. rufescens (Berk, et Br.)
Sacc., Syll. Fung. V, 1887 : 47.— Lepiota rufescens (Berk, et Br.) J. Lge,
Fl. Ag. Dan., I, 1935 :36, s. J. Lge, non al.— L. langei Locq., Bull. mens.
Soc. linn. Lyon, 14, 1945 : 28 (teste F. H. Moeller, Friesia, 6, 1—2, 1952:
88
22), non L. rufescens (Berk, ct Br.) Morg., Journ. Mycol., 12, 1906:246
( = £. brunnescens Pk, Bull. Tor. Bot. Club, 31, 1904: 177, teste
С. H. Kauffman, Pap. Mich. Acad. Sci., Arts a. Lett., 4, 1924:333).—
Cystolepioia hetieri (Boud.) Sing., Sydowia, Beih. VII. 1973 : 67
(табл. IX, 4a—в).
Icon.: J. Herink, Ces. MykoL, 15, 4, 1961 :226; Вассер, Фл. гр. Укр.
(Agaricaceae), 1980, рис. 119.
В смешанных широколиственных лесах, парках; лиги., VII—X, д. р.,
(I), п. з. н., немор., евраз.
Распр. в СССР. Европ. ч_: Карельская АССР, УССР; Азиат,
ч.: Дальн Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Дания, Швейцария,.
Франция, ФРГ, ГДР, ЧССР, ВНР; Азия: Япония.
83. Lepiota adulterina Moell., Friesia, 6, 1959 : 23.— Lepiota hetieri
auct., non s. Boud., BSMF, 18, 2, 1902: 137—138.— L. hetieriana Locq.,
Bull. Soc. linn. Lyon, 14, 1945:94 (nom. nudum).— Cystolepioia adul-
lerina (Moell.) Bon, BSMF, 92, 1976:318 (nom. nudum).— C. hetieriana
(Locq.) Bresinsky, in Bresinsky et Haas, Beih. Zeitschr. Pilzk., I, 1976 : 46
(nom. nudum).— C. adulterina (Moell.) Bon, Docums. Мус., VII, 1977 : 54.
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, tab. 14, fig. 1; Moell.,
Friesia, 6, 1959, p. 22, figs a—d; Reid, Fung. Rar. Icon. Col., VI, 1972,
pl. 43a, Text-figs. 18a—b; Knudsen, Bot. tidsskr., 73, 2, 1978, fig. 13.
В кедрово-пихтово-широколиственных, буковых, дубовых лесах,
парках; гум. сапр., VI—X, о. р., (1), п. з. н., эвриголаркт., евраз.
Распр. в СССР. Азиат, ч.: Кавказ — АзССР, Дальн. Восток —
Приморск, кр.
Общее распр. Европа: Дания, Финляндия, Нидерланды, Швей-
цария, ФРГ, Франция, ЧССР,
84. Lepiota neophana Morg., Journ. Мус., 12, 86, 1906:248.
В широколиственных, смешанных лесах, парках; подст. сапр., VII—
IX, о. р., (1), п. з. н., в. н. а.
Распр. в СССР. Азиат, ч.: Дальн. Восток — Приморск, кр.
Общее распр. Сев. Америка: США.
Секция Echinatae Fay.
Ann. Sci. Nat. Bot., VII, 9, 1889 : 350.
85. Lepiota eriophora Pk, Bull. Tor. Bot. Club, 30, 1903:95.—Le-
piola echinella Quel, et Bernard var. eriophora (Pk) J. Lge, FI. Ag. Dan.,
I, 1935:27 (nom. inval.).— Cystolepiota eriophora (Pkj Knudsen, Bot.
tidsskr., 73, 2, 1978 : 127 (табл. XXIII, 34).
Icon.: J. Lge, FI. Ag. Dan., 1935—1940, tab. 12, fig. H; Knudsen,
Bot. tidsskr., 73, 2, 1978, fig. 8.
В широколиственных и смешанных лесах; подст. сапр., VII—X,
д. р., (1), п. з. н., эвриголаркт., свраз-а.м.
Распр. в СССР. Европ. ч.: ЛитССР; Азиат, ч.: Кавказ — ГССР,
АзССР, Дальн. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Дания, Франция, Ита-
лия, ФРГ, ГДР, ЧССР; Сев. Америка: США.
86. Lepiota echinacea J. Lge, Fl. Ag. Dan., V, 1940: V, Append.—
Lepiota echinacea J. Lge, Fl. Ag. Dan., I, 1935:26 (nom. nudum).—
L. hispida auct., non s. Gill.— Cystolepiota echinacea (J. Lge) Knudsen,
Bot. tidsskr., 73, 2, 1978 : 127.
89-
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, tab. 10, fig. D; Васильева,
Агар. шл. гр. Приморск, кр., 1973, рис. 44, A; Knudsen, Bot. tidsskr., 73,
2, 1978, fig. 10, 13.
В широколиственных и смешанных лесах; гум. canp., VII—X, ир.,
(!—2), н., эвриголаркт., евраз.
Распр. в СССР. Европ. ч.: ЛитССР, УССР, Татарск. АССР;
Азиат, ч.: Дальн. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Дания, Франция, ФРГ,
СФРЮ, ГДР, ЧССР, ВНР.
87. Lepiota hystrix Moell. et J. Lge, FI. Ag. Dan., V, 1940 :V, Ap-
pend.— Lepiota- hystrix Moell. et J. Lge, Fl. Ag. Dan., I, 1935:26 (nom.
nudum).— Cystolepiota hystrix (Moell. et J. Lge) Knudsen, Bot. tidsskr.,
73, 2, 1978 : 128 (табл. XXIII, 35).
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, tab. 10, fig. E; Knudsen,
Bot. tidsskr., 73, 2, 1978, fig. 2.
В кедрово-пихтовых, кедрово-широколиственных лесах, ольшани-
ках; гум. canp., VI—X, о. р., (1—2), н., эвриголаркт., евраз.
Распр. в СССР: Европ. ч.: ЛитССР; Азиат, ч.: Дальн. Вос-
ток — Приморск, кр.
Общее распр. Европа: Великобритания, Дания, ФРГ, ГДР,
ЧССР; Азия: Япония.
88. Lepiota acutesquamosa (Weinm.) Kumm., Der Fuhrer in die
Pilzkunde, 1871 : 129.— Agaricus acutesquamosus Weinm., Syll. Plant.
Nov., I, 1822 : 70.— A. friesii Lasch, Linnaea, 3, 1828:155.— A. tricho-
chtoides Krombh., Abbild. der Schwamme, I, 1831 :72.— A. mariae Klotz-
sch., Linnaea, 7, 1832 : 196.— A. aculeatus Vitt., Fung. Mang., 1935 : 348.—
Lepiota friesii var. acutesquamosa (Weinm.) Quel., Champ. Jura et Vos-
'ges, I, 1872 : 72.— L. acutesquamosa (Weinm.) Gill., Les Hymenomycetes,
1874 :60.— Cystolepiota friesii (Lasch) Bon, Docums. Мус., VII, 1977: 12
(табл. IX, 5a—d).
Icon.: Baria, Champ. Alp. Mar., 1888—1892, pl. 12, fig. 1—7; Pilat,
Klic urc. nas. hub, 1951, obr. 570—573; Michael u. Hennig, Handbh.
Pilzfr., 1968, I, tab. 31; Вассер, Фл. гр. Укр. (Agaricaceae), 1980,
рис. 120, табл. XV, 1.
В хвойных и широколиственных лесах, в травянистых местах, пар-
ках; гум. canp., VIII—X, ч., (1—2), я., эвриголаркт., евраз-ам-афр.
Распр. в СССР. Европ. ч.: РСФСР — Карельская АССР, Мос-
ковская обл., ЭССР, ЛатвССР, ЛитССР, БССР, УССР, Татарск. АССР;
Азиат, ч.: Кавказ — ГССР, Вост. Сибирь — Красноярск, кр., Дальн.
Восток — Приморск, кр., Ср. Азия — КиргССР.
Общее распр. Европа: Великобритания, Финляндия, Дания,
Бельгия, Швейцария, Франция, Италия, СФРЮ, ФРГ, ГДР, ПНР,
ЧССР, ВНР, СРР; Азия: Япония; Сев. Америка: Канада, США;
Африка: Алжир.
89. Lepiota aspera (Fr.) Quel., Enchir., 1886:5.— Amanita aspera
Fr., Syst. Мус., I, 1821 : 18.— Lepiota acutesquamosa (Weinm.) Kumm,
var. furcata Kiihn., BSMF, 52, 1936 : 210.— Cystolepiota aspera (Fr.)
Bon, Docums. Мус., Vll, 1977:11 (nom. nudum).— C. aspera (Fr.)
Knudsen, Bot. tidsskr., 73, 2, 1978 : 129.
Icon.: Bres., Icon. Мус., 1927, tab. 27; Konrad et Maublanc, Icon.
Sei. Fung., 1924—1937, tab. 14; J. Lge, FI. Ag. Dan., 1935—1940, tab. 10,
fig. F; Moser, KI. Krypt.-flora, B. 2, Teil IL 1967, fig. 81, 252, 356; Зеро-
ва, Атл. гр. Укр.. 1974, табл. Ill, рис. 3; Урбонас, Тр. АН ЛитССР, се-
S0
рия В, Т. 3 ( 66), 1974, рис. 1ж, гп, хц; Knudsen, Bot. tidsskr., 73, 2,
1978, fig. 12, 13.
В смешанных, широколиственных лесах; гум. сапр., VII—X, р., (1),
н., мультирег., евраз-ам-афр.
Распр. в СССР. Европ. ч.: ЛитССР, ЛатвССР, УССР; Ази-
ат. ч.: Вост. Сибирь—Красноярск, кр., Бурятская АССР.
Общее распр. Европа: Исландия, Великобритания, Бельгия, Ир-
ландия, Дания, ГДР, Нидерланды, Испания, Франция, Италия, ЧССР,
ВНР; Азия: Япония; Сев. Америка: Канада, США; Южн.
Америка: Аргентина; Африка: Марокко.
Секция Amyloidea Sing.
Ann. Мус., 41, 1—3, 1943 : 171.
90. Lepiota lignicola Karst., Medd. Soc. Faun. Fl. Fenn., 6, 1881.
1.— L. amyloidea Sing., Ann. Мус., 41, 1—3, 1943:171.— Cystolepiota
lignicola (Karst.) Nezd., Микол, и фнтопатол., 14, 5, 1980 : 389, syn. nov.
(табл. IX, 6a—fi).
Icon.: Васильева, Агар. шл. гр. Приморск, кр., 1973, рис. 44 Б;
Нездойминого, Микол, и фитопатол., 14, 5, 1980, стр. 389, рис.
В смешанных лесах на валежных стволах и пнях березы; лиги.,
VII—X, д. р., (1), п. з. и., в. и. а.
Распр. в СССР. Азиат, ч.: Зап. Сибирь — Алтайск. кр., Вост.
Сибирь— Красноярск, кр., Дальн. Восток — Приморск, кр.
Общее распр. Европа: Финляндия; Африка: Заир.
Прим. Как отмечает Э. Л. Нездойминого (1980), в 1943 г. Р. Зингер (Singer,
Апл. Мус., 41, 1—3, 1943) описал новый для науки вид Lepiota amyloidea Sing,
произраставший на гнилом замшелом пне березы в районе Те пецкого озера на Алтае.
В 1979 г. Э. Л Нездойминого обнаружила L. amyloidea в Красноярском крае. Не-
редок этот гриб и в Приморском крае (Васильева, 1973; Булах, ined.), где приведен
как L. lignicola Karst. Образцы из Красноярского и Приморского краев по всем
основным анатомо-морфологическим признакам обнаружили полное сходство с голо-
типом L. amyloidea. Следовательно, возник вопрос о тождественности L. amyloidea
и L. lignicola. Сопоставление диагнозов обоих видов, а также изучение образца
L. lignicola из гербария ботанического музея университета Хельсинки, проведенное
Э. Л. Нездойминого (1. "с), позволило сделать заключение о тождественности рас-
сматриваемых видов Поэтому L. amyloidea принимается как синоним L. lignicola
Подрод Lepiota
Укр. ботан. жури., 35, 5, 1978 : 516—517.
Секция Stenosporae (J. Lge) Kuhn.
BSMF, 52, 1936 : 194.
91. Lepiota cortinarius J. Lge, Dansk Bot. Ark., 2 (3), 1915:25
(табл. X, la—в).
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, pl. 10, fig. B; Huijsman,
Persoonia, I, 3, 1960, fig. 1—3; Вассер, Фл. гр. Укр. (Agaricaceae), 1980,
рис. 121.
В хвойных и смешанных лесах; гум. сапр., VIII—X, о. р., (1),
п. з. и., бор., евр-ам.
Распр. в СССР. Европ. ч.: ЛитССР.
Общее распр. Европа: Дания, Франция, ФРГ; Сев. Амери-
ка: США.
91
92. Lepiota grangei (Eyre) Kuhn., Bull. mens. Soc. linn. Lvon. 1934,
3, 5 : 79.— Schulzerta grangei Eyre, Trans. Brit. Мус. Soc., 1903, 2 : 37.—
Hiatula grangei (Eyre) W. G. Sm., Syn. Brit. Bas., 1908: 27.— Lepiota
grangei (Eyre) J. Lge, Fl. Ag. Dan., I, 1935 : 30 — L. ochraceocyanea
Kiihn., BuIL mens. Soc. linn. Lyon, 1934, 3, 5: 42—43.— L. grangei f.
brunneoolivacea Pil., Acta Mus. Nat. Prag., XIB, 2, 1955:9 (табл. X,
2a~ d).
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, pl. 10, fig. A; Locquin,
BSMF, 60, 1944, tab. 2., fig. 3; Pilat, Acta Mus. Nat. Prag., XIB, 2,
1955, fig. 7—8; Herink, Ces. Mycol., 16, 4, 1962, p. 229, fig., p. 231, fig.
В широколиственных, смешанных лесах, парках; подст. сапр.,
VII—X, о. р., (I), п. з. п., эвриголаркт., евраз.
Распр. в СССР. Азиат, ч.: Дальн. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Дания, Бельгия, Фран-
ция, Швейцария, ПНР, ЧССР.
93. Lepiota pseudofelina J. Lge, Fl. Ag. Dan., V, 1940: V, Append,
(табл. X, За—б).
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, tab. 12, fig. C; Babes,
Ann. Hist.-Nat. Mus. Nat. Hung., 50, 1958, fig. 4; Вассер, Фл. гр. Укр.
(Agaricaceae), 1980, рис. 122.
В широколиственных и хвойных лесах; подст. сапр., VII—X, д. р.»
(1), п. з. н., эвриголаркт., евраз.
Распр. в СССР. Азиат, ч.; Вост. Сибирь — Красноярск, кр.,
Дальн. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Дания, Франция, Ита-
лия, ФРГ, ГДР, ПНР, ЧССР, ВНР.
94. Lepiota ignicolor Bres., Fung. Trid., II, 1892:3 (табл. X, 4a—6).
Icon.: Bres., Icon. Мус., 1927, tab. 39, fig. 2; Васильева, Агар. шл.
гр. Приморск, кр., 1973, рис. 43Г; Вассер, Фл. гр. Укр. (Agaricaceae),
1980, табл. XXI, 3.
В смешанных лесах, в травянистых местах; полет, сапр., VII—IX,
о. р., (1), п. з. н_, эвриголаркт., евраз.
Распр. в СССР. Азиат, ч.: Дальн. Восток — Приморск, кр.
Общее распр. Европа: Италия, Франция, ФРГ, ПНР.
95. Lepiota tomentella J. Lge, Dansk. Bot. Ark., 4, 4, 1923:48
(табл. X, 5a—в).
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, tab. 14; Babos, Ann. Hist.-
Nat. Mus. Nat. Hung., 61, 1969 : 158, fig. 4 (споры); Вассер, Укр. ботан.
журн., 32, 3, 1975 :319, рис. 2; Вассер, Фл. гр. Укр. (Agaricaceae), 1980,
рис. 123.
В смешанных широколиственных, чистых дубовых, ясеневых, со-
сновых лесах и насаждениях; подст. сапр., VIII—X, д. р.» (2—3), п.з. н.,
эвриголаркт., евраз.
Распр. в СССР. Европ. ч.: РСФСР — Ленинградская обл., УССР;
Азиат, ч.: Кавказ — АрмССР, Вост. Сибирь—Красноярск, кр.,
Дальн. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Дания, Бельгия, Фран-
ция, ПНР, ВНР.
96. Lepiota castanea Quel., С. R. Ass. fran£. Av. Sci., 9, 1881 : 661.—
Lepiota ignipes Locq., BSMF, 1952, 68: 271 (nom. nudum) (табл. X,
6a—d).
Icon.: Konr. et Maubl., Icon. Sci. Fung., 1924—1937, pl. 12, fig. 2;
J. Lge, Fl. Ag. Dan., 1935—1940, pl. 12, fig. G; Pilat, KJic urc. nas. hub,
42
1951, obr. 577b; Reid, Col. Ic. Rare a. Int. Fung., Pars V, 1972, dL 43 (b).
Text-fig. 1 (a—d).
В широколиственных, хвойных и смешанных лесах; подст. canp.,
VII—X, нр., (2), п. з. н., эвриголаркт., евраз-афр.
Распр. в СССР. Европ. ч.: РСФСР — Ленинградская обл.,
ЛитССР, ЛатвССР, УССР; Азиат, ч.: Кавказ — АрмССР, АзССР.
Вост. Сибирь — Красноярск, кр., Дальи. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Дания, Франпня, Ита-
лия, ФРГ, ГДР, ПНР, ЧССР; Азия: Япония; Африка: Марокко.
97. Lepiota fulvella Rea, Trans. Brit. Мус. Soc., 6, 1918:68 (табл. X,
7a—б).
Icon.: J. Lge, FI. Ag. Dan., 1935—1940, pl. 12, fig. F, D; Pilat.
Klic urc. nas. hub, 1951, obr. 614; Moser, KI. Krypt.-flora, B. 2, Teil 2,
1967, fig. 83, 253; Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 125.
В смешанных и хвойных лесах, и а лесных полянах; подст. сапр.,
VIII—IX, д. р., (1—2), п. з. н., эвриголаркт., евраз-афр.
Распр. в СССР. Европ. ч.: ЛитССР, Татарск. АССР; Азиат, ч.:
Кавказ — ГССР.
Общее распр. Европа: Великобритания, Дания, Бельгия, ПНР.
ЧССР, ВНР, СФРЮ; Африка: Марокко.
98. Lepiota virescens (Speg.) Morg., Journ. Мус., 12, 86, 1906 : 245.—
Lepiota erythrella Speg. f. virescens Speg., Fung. Arg. novi v. crit., 1889 :
93.— L. caerulescens Pk, Bull. Tor. Bot. Club, 1899 : 63.
В широколиственных, кедрово-широколиствеиных лесах, парках;
подст. canp., VII—IX, о. р., (1), п. з. н., в. н. а.
Распр. в СССР. Азиат. ч_: Дальи. Восток — Приморск, кр.
Общее распр. Сев. Америка: США; Южн. Америка: Ар-
гентина.
99. Lepiota griseovirens R. Мге, BSMF, 44, 1928 : 37.— Lepiota gri-
seovirens spp. obscura Locq., Bull. mens. Soc. linn. Lyon, 14, 1945 : 61—62
(nom. nudum).— L. obscura (Locq.) Babos, Ann. Hist-Nat. Mus. Nat.
Hung., 50, 1958 : 89—90 (nom. non rite publication).
Icon.: R. Mre, BSMF, 44, 1928 : p. 37; Romagn. et Locq., BSMF,
60, 1944, pl. 2, fig. 3; Reid, Fung. Rar. Icon. Col., VI, 1972, pl. 43 c—d.
Text-figs. 2a—c.
В смешанных широколиственных, дубовых, ясеневых, сосновых ле-
сах, ольшаниках; подст. canp., VI—X, о. р., (I), и., эвриголаркт.,
евр-афр.
Распр. в СССР. Европ. ч.: ЛнтССР.
Общее распр. Европа: Великобритания, ФРГ, Франция, ВНР.
ПНР, Испания; Африка: Алжир, Марокко.
100. Lepiota subafba Kiihn. ex P. D. Orton, Suppl. Trans. Brit. Мус.
Soc., 43, 1960 : 287.— Lepiota albosericea P. Henn., s. J. Lge, Fl. Ag.
Dan., 1935 : 32.— L. subalba Kuhn., BSMF, 52, 1936 : 232 (nom. nu-
dum).— L. naucina (Fr.) Kumm. s. Mos., KI. Krypt.-flora, T. 2, Band 2,
1967 : 197 non s. al.
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, tab. 12, fig. B.
В кедрово-пихтово-широколиственных, сосновых, осиновых, бело-
акациевых, буковых лесах, ольшаниках, парках, садах; подст. сапр.,
VII—X, нр., (1—2), н., эвриголаркт., евраз.
Распр. в СССР. Европ. ч.: ЛитССР, УССР; Азиат. ч_: Кав-
каз — АрмССР, АзССР, ГССР; Вост. Сибирь — Красноярск, кр., Дальн.
Восток — Приморск, кр.
93
Общее распр. Европа: Великобритания, Дания, Франция, ЧССР,
Швейцария, Греция, ФРГ, Испания; Азия: КНР, Израиль, Япония.
Секция Cristatae Kiihn. ex S. Wasser
Укр. ботаи. жури., 35, 5, 1978 : 517.— Cristatae Kiihn., BSMF, 52,
1936 : 190 (nom. nudum).
101. Lepiota cristata (Fr.) Kumm., Der Fuhrer in die Pilzkunde,
1871:137.— Agaricus cristatus Fr., Syst. Мус., I, 1821:22.— Lepiotula
cristata (Fr.) Locq. ex Horak, Beitr. zur Krypt.-flora der Schweiz,
Bd. XIII, 1968 : 338 (табл. X, 8a—e\ XXIII, 36).
Icon.: Bres., Icon. Мус., 1927, tab. 34; J. Lge, Fl. Ag. Dan., 1935—
1940, pl. 12, fig. A; Piiat, Klic urc. nas. hub, 1951, obr. 582a, b, 609—611;
Michael u. Hennig, Handb. Pilzfr., 1968, I, tab. 32; Зерова, Атл. гр. Укр.,
1974; табл. 113, рис. 1; Вассер, Солдатова, Высш. баз. ст. зоны Укр.,
1978, рис. 82; Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 126,
табл. XV, 3.
В широколиственных, хвойных, смешанных, байрачных лесах, на
лесных полянах, лугах, пастбищах, в огородах; гум. сапр., VI—X, о. ч.,
(1—2), я., мультирег., космопол.
Распр. в СССР. Европ. ч.: РСФСР — Карельская АССР, ЭССР,
ЛатвССР, ЛитССР, БССР, МССР, УССР; Азиат, ч.: Кавказ —
АзССР, ДагАССР, Зап. Сибирь — Алтайск. кр., КазССР, Вост. Сибирь —
Красноярск, кр., Дальи. Восток — Приморск, кр., Ср. Азия — ТССР.
Общее распр. Европа: Великобритания, Австрия, Бельгия,
ЧССР, ВНР, ГДР, Греция, Дания, Нидерланды, Ирландия, Испания,
Италия, Лихтенштейн, Норвегия, ПНР, Португалия, СРР, СФРЮ, ФРГ,
Финляндия, Франция, Швейцария, Швеция; Азия: Израиль, Индия,
КНР, Турция, Япония; Сев. Америка: Канада, США; Южн.
Америка: Аргентина, Чили; Австралия; Африка: Алжир,
Заир; о-в Тринидад.
102. Lepiota lilacea Bres., Fung. Trid., 11, 1892:3.— Lepiotula lila-
cea (Bres.) S. Wasser, Нов. сист. высш, и иизш. раст., 2, 1976: 191.
Icon.: Bres., Icon. Мус., 1927, tab. 39, fig. 1; J. Lge, Fl. Ag. Dan.,
1935—1940, pl. 13, fig. G; Вассер, Фл. гр. Укр. (Agaricaceae), 1980,
табл. XXI, 2.
В кедрово-широколиственных, дубовых, белоакациевых лесах, по-
лезащитных лесополосах, ботанических парках; подст. сапр., VI—X,
о. ч., (2—3), я., мультирег., евраз-ам-афр.
Распр. в СССР. Европ. ч.: УССР; Азиат, ч.: Кавказ — АзССР,
Дальн. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Дания, Франция, Ита-
лия, ФРГ, ГДР, ПНР, ЧССР, ВНР, СФРЮ; Азия; КНР, Израиль,
Иран; Южн. Америка: Венесуэла; Африка: Алжир, Гайа,
Заир, Марокко.
Секция Lepiota
Укр. ботан. жури., 35, 5, 1978 : 517.
103. Lepiota erminea (Fr.) Gill., Champ. Fr., 1874 : 99.— Agaricus
ermineus Fr., Syst. Мус., 1, 1821 : 22 (табл. XI, la—в).
94
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, pl. 11, fig. B; Huijsman,
Med. Ned. Мус. Ver., 1943, 28, fig. 8; Pilat, Klic urc. nas. hub, 1951,
obr. 617, 619—622; Michael u. Hennig, Handb. Pilzfr.. 1964, III, tab. 17;
Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 128, табл. XXII, 1.
В смешанных лесах, нй лесных опушках в траве, на пастбищах,
лугах, по краю дорог; гум. сапр.. VII—X. нр., (2), и., эвриголаркт.,
евраз.
Распр. в СССР. Европ. ч.: ЛатвССР, УССР; Азиат, ч.: Вост.
Сибирь — Красноярск, кр., Дальн. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Дания, Нидерланды,
Франция, Италия, ФРГ, ВНР, ГДР, ЧССР, СРР, СФРЮ
104. Lepiota alba (Bres.) Sacc., Syll. Fung., V, 1887 :37.— Lppiota cly-
peolaria (Fr.) Kumm. var. alba Bres., Fung. Trid., II, 1882: 15.
Icon.: Bres., Icon. Мус., 1927, tab. 32, fig. 1; J. Lge, Fl. Ag. Dan.,
pl. 11, fig. A; Huijsman, Med. Ned. Мус. Ver., 1943, 28, fig. IV—VI;
Зерова, Атл. гр. Укр., 1974, табл. 112, рис. 3; Вассер, Фл. гр. Укр.
(Agaricaceae), 1980, табл. XXII, 3.
В смешанных, широколиственных, хвойных лесах, в полезащитных
лесополосах, на лесных опушках, лугах, в заповедных целинных
степях, огородах; гум. сапр., V—X, о. ч., (2), с., эвриголаркт.,
евраз-афр.
Распр. в СССР. Европ. ч.: РСФСР — Ставропольем кр.,
ЛатвССР, ЛитССР, БССР, УССР; Азиат, ч.: Кавказ—-ГССР, Зап.
Сибирь — Новосибирск, обл., Вост. Сибирь — Красноярск, кр.. Якут-
ская АССР.
Общее распр. Европа: Великобритания, Дания, Бельгия,
СФРЮ, Франция, Италия, ФРГ, ГДР, ПНР, ЧССР, СРР, ВНР; Аф-
рика: Марокко.
105. Lepiota oreadiformis Vel., Ceske houby, 1920—1922:215.— Aga-
ricus (Lepiota) clypeolarius var. pratensis Fr., Hymen. Eur., 1874 : 127.—
L. pratensis (Fr.) Rea, Brit. Bas., 1922 : 69.— L. gracilis Quel. var. laevi-
gata J. Lge, Dansk Bot. Ark., 2, 1, 1915:52.— L. laevigata (J. Lge)
J. Lge, Dansk Bot. Ark., 4, 4. 1923:46 (табл. XI, 2a—б; XXIII, 38;
XXIV, 39).
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, pl. 11, fig. C; Pilat, Klic
urc. nas. hub, 1951, obr. 623; Moser, KI- Krypt-flora, 1967, B. 2, Teil 2,
fig. 255; Babos, Ann. Hist.-Nat. Mus. Nat. Hung., 66, 1974, fig. 5; Вас-
сер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 130.
В светлых широколиственных, смешанных лесах, на лугах, паст-
бищах, в парках, огородах; гум. сапр., VIII—X, р., (2), с., эвриголаркт.,
евраз.
Распр. в СССР. Европ. ч.: РСФСР — Московская обл., ЭССР,
ЛитССР; Азиат, ч.: Кавказ — АзССР, Вост. Сибирь — Красно-
ярск. кр.
Общее распр. Европа: Великобритания, Дания, Франция, Ита-
лия, ФРГ, ГДР, ЧССР, ВНР.
106. Lepiota subgracilis*** Kuhn, ex S. Wasser, Укр. ботан. журн.,
35, 5, 1978:517—518.— Lepiota clypeolaria (Fr.) Kumm. var. gracilis
Quel., C. R. Ass. franc. Av. Sci., XVIII, 2, 1883 : 439.— L. gracilis (Quel.)
Rea, Brit. Bas., 1922:70, non s. Pk, Bull. Tor. Bot. Club, 1899:63.—
L. subgracilis Kuhn., BSMF, 1936, 52:231 (nom. nudum et inval.), s.
Kuhn., J. Lge, FI. Ag. Dan., 1, 1935 : 28, non. s. Locq., BuIL mens. Soc.
linn. Lyon, 14, 1945 : 59 (табл. XI, За—б).
95
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, pl. 11, fig. F; Piiat, Klic
urc. nas. bub, 1951, obr. 624—625; Вассер, Фл. гр. Укр. (Agaricaceae),
1980, рис. 131.
В хвойных, широколиственных, смешанных лесах, часто в зарослях
Salix rosmarinifolia L.; подст. сапр., VII—X, нр., (1—2), и., мультирег.,
евраз-ам.
Распр. в СССР. Европ. ч.: УССР; Азиат, ч.: Вост. Сибирь —
Красноярск, кр., Дальн. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Дания, Франция, Ита-
лия, ЧССР, ВНР; Азия: Шри Ланка; Сев. Америка: США;
Южн. Америка: Аргентина.
107. Lepiota pallida * Locq., Bull. mens. Soc. linn. Lyon., 14, 1945 : 57.
Icon.: Babos, Ann. Hist.-Nat. Mus. Nat. Hung., 61, 1969, figs 1 (2),
споры; Вассер, Укр. ботан. журн., 32, 1, 1975, рис. 4а, б.
В искусственных сосновых, белоакациевых с примесью Populus sp.
лесных массивах, иа песчаной почве, в парках, по краю дорог; гум.
сапр., (X—X, д. р., (3), с.(?), я.(?), эвриголаркт., евр-афр.
Распр. в СССР. Европ. ч.: УССР.
Общее распр. Европа: Франция, ВНР, ЧССР; Африка: Ма-
рокко.
Пр нм. В роде Lepiota вид L. pallida очень маленьких размеров. L. pallida —
редкий вид для микофлоры мира. По литературным данным (Locquin, 1945; Kiihner,
Romangesi, 1953; Babos, 1969; Malen^on, Bertault, 1970; Вассер, 1975a), в мире изве-
стно семь местонахождений L. pallida: шесть в Европе и одно в Сев. Африке (Ма-
рокко), главным образом из аридных областей. L. pallida наиболее близка к L. cly-
peolaria (Fr.) Kumm, и L. ventriosospora Reid, отличаясь от последних размером
карпофоров, размером и формой спор, экологией.
Исследованные образцы L. pallida полностью согласуются с данными диагноза
вида у М. Локена (Locquin, 1945) и с нашим оригинальным описанием (Вассер,
1980), за исключением того, что при микроскопическом исследовании нх нами обна-
ружены немногочисленные цилиндрические верстеновидные и булавовидные хейло-
цистиды, не отмеченные ни в одной работе, посвященной L. pallida.
108. Lepiota ventriosospora Reid, Trans. Brit. Мус. Soc., 41, 1958:
427.— Lepiota metulaespora (Berk, et Br.) Sacc., Syll. Fung., V, 1887:
38, s. Bres., Kiihn et Romagn., non s. Berk, et Br., Journ. Linn. Soc.
Bot., II, 1871 : 512 (табл. XI, 4a—г\ XXIV, 40\ XXV, 41).
Icon.: Bres., Icon. Мус., 1927, tab. 30; Pegler, Kcw Bull., 27, 1,
1972, fig. 4, a, b, c, d; Moser, KI- Krypt.-flora, 1967, B. 2, Teil 2,
fig. 256; Васильева, Агар. шл. гр. Приморск, кр., 1973, рис. 43 А; Вас-
сер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 132, табл. XXI, 1.
В дубово-сосиовых, сосново-березовых, сосновых, кедрово-листвен-
ничных, кедрово-широколиственных лесах, дендропарках; под ст. сапр.,
VI—X, ир., (1—2), с., в. и. а.
Распр. в СССР. Европ. ч.: ЛитССР, УССР; Азиат, ч.: Вост.
Сибирь — Красноярск, кр., Бурятская АССР, Ср. Азия — КиргССР,
Дальн. Вост.— Приморск, кр.
Общее распр. Европа: Великобритания, Дания, Бельгия, Фран-
ция, ФРГ, ГДР, ЧССР, ВНР, СРР; Азия: Шри Ланка; Африка:
Гана, Танзания.
109. Lepiota clypeolaria (Fr.) Kumm. var. clypeolaria, Der Fiihrer
in die Pilzkunde, 1871 : 137.— Agaricus clypeolarius Fr., Syst. Мус., I,
1821:21.— Lepiota clypeolaria (Fr.) Quel., Champ. Jura ct Vosges, I,
1872 : 72.— L. clypeolaria f. cedretorum R. Mre, Bull. Soc. Hist. Nat. AFN,
1916, 187„7 : 295 (табл. XI, 5a—в).
96
Icon.: Bres., Icon., Мус., 1927, tab. 31: J. Lge, Fl. Ag. Dan., 1935—
1940, pl. 11, fig. D; Pilat, Klic urc. nas. hub, 1951, obr. 626—630; Зеро-
ва, Атл. гр. Укр., 1974, табл. 112, рис. 1; Вассер, Солдатова, Высш. баз.
ст. зоны Укр., рис. 20; Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 133,
табл. XX, 2.
В сосновых, сосиово-березовых, дубовых, сосново-пихтово-березо-
вых, пихтово-кедровых, пихтово-еловых, кедрово-широколиственных,
осиново-березовых, лиственных лесах, полезащитных лесополосах, на
лесных полянах, в разнотравно-тнпчаково-ковыльных степях.
L. clypeolaria var. latispora** Kuhn, ex S. Wasser, Укр. ботан.
журн., 35, 5, 1978 : 518.
В смешанных лесах, полезащитных лесополосах. Известна из
ЛитССР, УССР.
L. clypeolaria var. minor J. Lge, Fl. Ag. Dan., V, 1940 : V, Append.
В кедрово-широколиственных лесах. Известна из Приморск, кр.
Гум. canp., VI—X, о. ч., (2), с., мультнрег., космопол.
Распр. в СССР. Европ. ч.: РСФСР — Карельская АССР, Мос-
ковская, Ленинградская обл., ЭССР, ЛитССР, ЛатвССР, БССР, УССР;
Азиат, ч.: Кавказ — ГССР, АзССР, Зап. Сибирь — Алтайск. кр..
Вост. Сибирь — Якутская АССР, Красноярск, кр., Дальн. Восток —
Приморск, кр.
Общее распр. Европа: Великобритания, Финляндия, Дания,
Бельгия, Франция, Испания, Италия, ФРГ, ГДР, ПНР, ЧССР, Австрия,
СФРЮ; Азия: КНР, МНР, Шри Ланка, Япония; Сев. Америка:
Канада, США, Мексика; Южн. Америка: Чили; Австралия:
Африка: Алжир, Марокко; о-в Куба.
Секция Ovisporae (J. Lge) Kuhn.
BSMF, 52, 1936 : 190.
110. Lepiota rufipes Morg., Journ. Мус., 12, 1912:156 (табл. XI,
6a—в).
Icon.: Josscrand, BSMF, LXXI, 2, 1955:66, fig. 1; Stangl u. Bre-
sinsky, Zeitschr. Pilzk., 37, 1971, Abb. 3, fig. 2; Babos, Ann. Hist.-Nat.
Mus. Nat. Hung, 66, 1974 : 66, fig. 1; Вассер, Нов. сист. пизш. и высш,
раст., 3, 1976, рис. 4; Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 134.
В белоакациевых, смешанных широколиственных, дубовых, еловых,
хвойно-широколиствениых лесах, в парках; гум. canp., VI—X, о. р., (2),
п. з. и., эвриголаркт., евраз-ам-афр.
Распр. в СССР. Европ. ч.: УССР; Азиат, ч.: Зап. Сибирь —
КазССР, Дальн. Восток — Приморск, кр.
Общее распр. Европа: Франция, ФРГ, ЧССР, ВНР; Сев. Аме-
рика: США; Африка: Алжир.
111. Lepiota feiina (Fr.) Karst, Hattswampar, 1879: 10, s. auct. non
s. Cke.— Agaricus clypeolarius var. felinus Fr., Syst. Мус., I, 1821 : 21.—
Lepiota clypeolaria var. feiina (Fr.) Fr., Hymen. Eur., 1874 : 22.— L. cly-
peolaria var. feiina (Fr.) Gill., Champ. Fr., 1878 : 62.
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, pl. 12, fig. E; Wichansky,
Cos. Mykol., 14. 1, 1960, fig. 2; Moser, KI. Krypt.-flora, B. 2, Teil 2, 1967,
fig. 82; Вассер, Фл. гр. Укр. (Agaricaceae), 1980, табл. XIX, 3.
В кедровых, сосновых, елово-широколиственных, чернично-зелеио-
мошно-кедрово-пихтовых, елово-кедровых, еловых, буково-грабовых ле-
сах; подст. canp., VI—X, нр., (1—2), п. з. н., бор., евраз-ам.
9 4—2960
97
Распр. в СССР: Европ. ч.: ЭССР, ЛатвССР, ЛитССР, УССР;
Азиат, ч.: Кавказ — ГССР, АзССР, Зап. Сибирь — Новосибирск,
обл.. Вост. Сибирь — Бурятская АССР, Красноярск, кр., Дальи. Вос-
ток — Приморск, кр.
Общее распр. Европа: Великобритания, Швеция, Дания, Фран-
ция, Италия, ФРГ, ГДР, ЧССР, ВНР; Азия: КНР; Сев. Амери-
к а: США, Мексика.
112, Lepiota helveola Bres., Fung. Trid., I, 1881 :16, s. Bres., Baria,
Rea, non s. Joss., Kuhn, et Romagn.
Icon.: Bres., Icon. Мус., 1927 tab. 32, 2; Pilat, Klic urc. nas. hub,
1951, obr. 631—632; Зерова, Атл. гр. Укр., 1974, табл. 113, рис. 2; Вас-
сер, Фл. гр. Укр. (Agaricaceae), 1980, табл. XIX, 2.
На открытых травянистых местах, лесных полянах, в парках, по
краю дорог; гум. сапр., VII—X, д. р., (1), с. я. г., эвриголаркт., ев-
раз-афр.
Распр. в СССР. Европ. ч.: УССР; Азиат, ч.: Кавказ — АзССР.
Общее распр. Европа: Испания, Франция, Италия, СФРЮ,
ЧССР, ВНР; Азия: Израиль, Япония; Африка: Алжир, Марокко.
113. Lepiota scoblnella (Fr.) Gill.. Champ. Fr., 1874; 127.— Agaricus
scobinellus Fr., Epicr. Syst. Мус., I, 1838 : 39.— Lepiota helveola Bres.,
Fung. Trid., 1, 1881 : 16, s. Joss., Kuhn, et Romagn., non s. Bres., Baria,
Rea (табл. XXV, 42).
В кленово-дубовых, белоакациевых, кедрово-пихтово-широколист-
венных лесах, полезащитных лесополосах, парках, травянистых местах;
гум. сапр., VII—X, ир., (2), с. я. г., эвриголаркт., евраз.
Распр. в СССР. Европ. ч.: ЛитССР, УССР; Азиат, ч.: Зап. Си-
бирь— КазССР, Дальи. Восток — Приморск, кр., Ср. Азия — ТССР.
Общее распр. Европа: Великобритания, Бельгия, Дания, Фран-
ция, Италия, ФРГ, ВНР; Азия: Израиль.
114. Lepiota pseudohelveola Kuhn, ex Hora, Trans. Brit. Мус. Soc.,
43, 2, 1960 : 449.— Lepiota pseudohelveola Kiihn., BSMF, 52, 1936:221
(nom. nudum).
Icon.: Kiihner, BSMF, 52, 1936, fig. 64.
В смешанных лесах, парках, по краю дорог; гум. сапр., VII—X,
о. р., (1—2), и., эвриголаркт., евраз.
Распр. в СССР. Европ. ч.: Краснодарск. кр.; Азиат, ч.: Дальн.
Восток — Приморск, кр.
Общее распр. Европа: Великобритания, ФРГ, СФРЮ, Франция,
ЧССР.
115. Lepiota wichanskyi* Pil., Acta Mus. Nat. Prag., IXB, 2, 1953:
4—12.— Lepiota sublittoralis Kiihn., BSMF, 52, 1936:216 (nom. nu-
dum).— Leucocoprinus sublittoralis (Kiihn.) Locq., Bull. mens. Soc. linn.
Lyon, 14, 1945 :93.— Leucoagaricus sublittoralis (Kiihn.) Sing., Lilloa,
22, 1951 : 422.— Lepiota sublittoralis Kiihn. ex Hora, Trans. Brit. Мус.
Soc., 43, 2, 1960:450.— Leucoagaricus sublittoralis (Kiihn. ex Hora)
Sing., Mycoflora Australis, Beih. Nova Hedwigia, 29, 1969 : 163 (табл. XI,
7u—6).
Icon.: Kiihner, BSMF, 52, 1936 : 216, fig. 61; Pilat, Acta Mus. Nat.
Prag., IXB, 2, 1953, fig. 3—11, tab. I; Reid, Fung. Rar. Icon. Col., Pars II,
1967, fig. 7 a—c, pl. 9a, Pars V, 1972, pl. 43e, Zeitschr. Pilzk., 34, 1—2,
1968, pl. 9a; Babos, Ann. Hist.-Nat. Mas. Nat. Hung., 61, 1969, fig. 5;
Babos, Ces. MykoL, 24, 4, 1970, tab. 72; Вассер, Укр. ботаи. жури., 33,
98
5, 1976, рис. 4; Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 135,
табл. XXIII, 1.
В степях, парках, естественных и искусственных лесных насажде-
ниях (Quercus robur L., Pinus nigra Arn. с примесью Robinia pseudo-
acacia L. и Populus sp.), в насаждениях Crataegus sp.. Ilex и Corylus,
часто на островах больших рек на песчаной почве в Urtica dioica L.,
доходит до высоты 1600 м н. у. м. L. wichanskyi произрастает главным
образом в местах слабого увлажнения, где годовое количество осадков
не превышает 500—550 мм, из которых на вегетационный период при-
ходится 300—350 мм, при средней влажности воздуха 65—70%; гум.
canp., IX—X, о. р., (2—3), с.(?), я.(?' в. н. а.
Распр. в СССР. Е в р о п. ч.: УССР.
Общее распр. Европа: Великобритания, Франция, ГДР, ЧССР,
ВНР; Южн. Америка: Аргентина, Чили; Африка: Алжир, Ма-
рокко.
Прим. Р. Кюнер (Kuhner, 1936) описал без латинского диагноза (nom. nudum)
новый для науки вид рода Lepiota—L. sublittoralis как близкий к L. littoralis Menier
(BSMF, 5, 1889:173, pl. XVII). В 1953 г. А Пилат (Pilat, 1953) описал по всем
правилам международной ботанической номенклатуры новый для науки вид рода
Lepiota — L. wichanskyi (вид назван в честь известного знатока видов рода Lepiota —
Э Вцханского).
Изучение рядом авторов и нами (Wichansky, t960a; Babos, 1969, 1970; Reid,
t972; Вассер, 1976в) типовых, образцов, диагнозов н цветных иллюстраций обоих
видов показало, что они идентичны. Следовательно, вид, описанный А Пилатом,,
имеет приоритет, a L. sublittoralis является его синонимом. Латинский диагноз к
L. sublittoralis написал Ф. Б Хора лишь в 1960 г. (Нога, 1960).
По мнению автора вида (Pilat, 1953), L. wichanskyi — редкий, чрезвычайно инте-
ресный вид, который по наличию общего покрывала и слизистой шляпки занимает
особое положение в роде Lepiota. L. wichanskyi вид, указывающий на связь рода
Lepiota с родами Amanita и Limacella (Pilat, 1953; Вассер, 1980).
116. Lepiota brunneoincarnata Chod. et Mart., Bull. trav. Soc. Bot.
Geneve, 5, 1889:222.— Lepiota barlae Pat., BSMF, 19, 1905.117, non
s. Quel.— L. barlaeana Pat., Congr. Soc. sav., 1908:249.— L. patouillar-
dii Sacc. et Trotter, Syll. Fung., XXI, 1912: 17 (табл. XII, la—6).
Icon.: Konrad ct Maublanc, Icon. Sei. Fung., 1924—1937, pl. 13;
J. Lge, Fl. Ag. Dan., 1935—1940, pl. 13, fig. F.; Вассер, Укр. ботан. журн.,
27, 3, 1971, рис. 3; Зерова, Атл. гр. Укр., 1974, табл. 113, рис. 3; Вас-
сер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 136.
В буковых лесах, парках, садах; гум. canp., VI—X, р., (2), с. я. г.,
эвриголаркт., евраз-афр.
Распр. в СССР. Европ. ч.: ЭССР, УССР; Азиат, ч.: Кавказ —
АзССР, Зап. Сибирь — КазССР.
Общее распр. Европа: Великобритания, Дания, Швейцария,
Швеция, ФРГ, Франция, ПНР, ЧССР, Австрия; Азия: Израиль; Аф-
рика: Алжир, Марокко.
117. Lepiota setulosa J. Lge, Fl. Ag. Dan., V, 1940’V, Append., s. J.
Lge, non s. Kiihn., BSMF, 52, 1936 : 201.— L. minuta J. Lge, Dansk Bot.
Ark., 4, 4, 1923 : 47, s. Kiihn. ct Romagn., non s. Kiihn., 1. c.
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, pl. 13, fig. C; Pilat, KHc
urc. nas. hub, 1951, obr. 632 b.
В широколиственных и смешанных лесах, парках; подст. сапр.,
VI—X, д. р., (1—2), п. з. и., эвриголаркт., евраз-афр.
Распр. в СССР. Европ. ч.: УССР; Азиат ч.: Зап. Сибирь —
Новосибирская обл., Дальн. Восток — Приморск, кр.
9*
99
Общее распр. Европа: Великобритания, Дания, Франция, Ита-
лия, ФРГ, ПНР, ЧССР, ВНР; Африка: Марокко.
118. Lepiota echinella * Quel, et Bernard, BSMF, 4, 1888:11, s. Quel.,
Huijsm., Med. Ned. Мус. Ver., 28, 1943 : 20, non s. auct.
Icon.: Quel, et Bernard, BSMF, 4, 1888, pl. 12; Huijsm., Med. Ned.
Мус. Ver., 28, 1943, fig. 3.
В широколиственных и смешанных лесах, парках; подст. сапр., VI—
IX, о. р., (1), н., эвриголаркт., евр.
Распр. в СССР. Е в р о п. ч.: УССР.
Общее распр. Европа: Великобритания, Дания, Нидерланды,
Франция, СССР.
119. Lepiota fuscovinacea Moell. et Л. Lge, in Fl. Ag. Dan., V, 1940:
V, Append.— Lepiota fuscovinacea Moell. ct J. Lge, Fl. Ag. Dan., I,
1935 : 33 (nom. nudum) (табл. XXIII, 37).
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, pl. 13, fig. H; Huijsm.,
Med. Ned. Мус. Ver., 28, 1943, fig. 6VU“,X; Reid, Col. 1c. Rare a. Int.
Fung., 11, 1967, pl. 10a, Text-figs. 4a—b.
В широколиственных и хвойных лесах, на лесных полянах и опуш-
ках; гум. сапр., VII—X, о. р„ (1), я., эвриголаркт., евраз.
Распр. в СССР. Европ. ч.: ЛитССР. _
Общее распр. Европа: Великобритания, Дания, Нидерланды,
Франция, Швейцария, ЧССР; Азия: Япония.
120. Lepiota hapalopoda Kalchbr., Icon. Sei. Hym. Hung., 1873 : 6.
В смешанных лесах; лиги., VI1—IX, ц. р., (1), н., в. н. а.
Распр. в СССР. А з и а т. ч.: Вост. Сибирь — Красноярск, кр., Якут-
ская АССР.
Общее распр. Европа: ВНР, Италия, Франция.
121. Lepiota kuehneri Huijsm. ex Hora, Suppl. Trans. Brit. Мус.
Soc., 43, 1960:448.— Lepiota kuehneri Huijsm., Med. Ned. Мус., Ver.,
28, 1943 : 27 (nom. nudum).— L. clypeolarioides Rea, s. Kuhn., 1936,
non s. Rea, Huijsm., Kiihn. et Romagn., Fl. anal. Champ. Super.,
1953:401.
1 con.: Kiihn., BSMF, 52, 1936, fig. 68.
В хвойных, хвойно-широколиствениых лесах; подст. сапр., VIII—X,
о. р., (2—3), п. з. н., бор., евраз-афр.
Распр. в СССР. Европ. ч.: ЛитССР; Азиат, ч.: Дальн. Вос-
ток — Приморск, кр.
Общее распр. Европа: Великобритания, Нидерланды, Италия,
Франция, ВНР; Африка: Алжир.
122. Lepiota lateritiopurpurea L. Vass., Агар. шл. гр. Приморск, кр.,
1973:181 (табл. XII, 2, а—б).
Icon.: Васильева, Агар. шл. гр. Приморск, кр., табл. II, рис. 2.
В широколиственных, хвойно-широколиственных лесах; подст. сапр.,
VIII—IX, р., (1—2), с.(?), я.(?), номер., дальневост. усл. энд.
Распр. в СССР. Азиат, ч.: Дальн. Восток—Приморск, кр.
123. Lepiota micropholis (Berk, et Br.) Sacc., Syll. Fung., V, 1887:
61.— Agaricus micropholis Berk, et Br., Journ. Linn. Soc. Bot., 11, 1871 :
505 (табл. XII, За—д).
Icon.: J. Lge, FI. Ag. Dan., 1935—1940, pl. 13, fig. D; Pegler,
Prelim. Ag. Fl. East Afr., 1977, fig. 80, 4 a—e.
В парниках, теплицах, в самшитовых рощах; гум. сапр., VII—IX,
о. р., (1—2), п. з. и., в. и. а.
Распр. в СССР. Азиат, ч.: Кавказ — АзССР.
100
Общее распр. Европа: Дания, ЧССР; А з и я: Япония, Израиль,.
Шри Ланка; Африка: Уганда.
124. Lepiota parvannulata (Lasch) Gill., Champ. Fr.. 1874:217.—
Agaricus parvannulatus Lasch, Linnaca, 111, 12, 1828 : 134.
Icon.: Bohus et al., Magyarorszag Kalaposgombainak meghatarozo
kezikonyve, Budapest, 1951, tab. I9b, fig. 5 2.
В смешанных лесах, парках, на пастбищах, на лесных полянах и
опушках; гум. сапр., VII—X, о. р., (1—2), н., эвриголаркт., евраз.
Распр. в СССР. Европ. ч.: РСФСР—Ленинградская обл.,
ЛатвССР, ЛитССР; Азиат, ч.: Вост. Сибирь—Красноярск, кр.
Общее распр. Европа: Великобритания, Фракция, ВНР, ГДР,
ФРГ, ЧССР.
125. Lepiota rhodorhiza Romagn. et Locq. ex P. D. Orton, Suppl.
Trans. Brit. Мус., Soc., 43, i960 : 285.— Lepiota sctulosa J. Lge var.
rhodorhiza Romagn. et Locq., BSMF, 60, 1944:38 (nom. nudum).—
L. rhodorhiza (Romagn. ct Locq.) Kiihn. et Romagn., Fi. anal. Champ.
Super., 1953:400 (nom. nudum).— L. setulosa J. Lge, Fl. Ag. Dan., V,
1940 : V, Append., s. Kuhn., BSMF, 52. 193G : 224, non s. al.
Icon.: Kiihn., BSMF, 52, 1936, fig. 69; Romagn. cl Locq., BSMF,
60, 1944, pl. 2, fig. 2; Reid, Col. 1c. Rare a. Int. Fung., 1, 1966, pl. 6a.
В смешанных широколиственных и хвойных лесах; подст. сапр.,
IV—XI, о. р., (1), п. з. и., эвриголаркт., евраз.
Распр. в СССР. Азиат, ч.: Дальн. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Дания, Франция, ЧССР,
ФРГ; Азия: Турция.
126. Lepiota sericifera (Locq.) Locq., Friesia, 11, 1956:294.— Pseu-
dobaeospora sericifera Locq., BSMF, 68, 1952:269.— Lepiota cristala
(Fr.) Kumm. var. sericea Cooi, Med. Ned. Мус. Ver., XI1, 1922:236.—
L. sericea (Cool) Huijsrn., Med. Ned. Мус. Ver., 28, i943:44, non s.
Mass., 1912.— L. sericephala Locq., BSMF, 68, 1952:270.— L. sericata
Kiihn. ct Romagn., Fl. anal. Champ. Super., i953:405 (nom. nudum).—
L. serena (Fr.) Sacc., s. J. Lge, Fl. Ag. Dan., 1935 : 29, non s. al.
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, pl. 11, fig. B; Huijsm.,
Med. Ned. Мус. Vcr., 28, 1943, fig. 10.
В смешанных широколиственных и хвойно-широколиственных ле-
сах; гум. сапр., VII—X, о. р., (1—2), п. з. н., эвриголаркт., евраз.
Распр. в СССР. Азиат, ч.: Вост. Сибирь — Красноярск, кр., Дальн.
Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Нидерланды, Дания,
СРР, Франция, СФРЮ.
127. Lepiota subincarnata J. Lge, FI. Ag. Dan., V, i940. V, Append.—
Lepiota subincarnata J. Lge, Fi. Ag. Dan., I, 1935 : 34 (nom. nudum).—
L. fourquignonii Quel., C. r. Ass. fr. Avan?. Sci., 13, 1885:277, s. J. Lge,
1915, non s. Quel (табл. XII, 4a—г).
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, pl. 13, fig. J.
В хвойных, широколиственных и смешанных лесах; гум. сапр., VII—
X, о. р., (1—2), п. з. н., мультирег., евраз-ам-афр.
Распр. в СССР. Азиат, ч.: Дальн. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Дания, ВНР, ГДР, ФРГ.
Франция; Сев. Америка: США; Африка: Марокко, Кения,
Уганда.
128. Lepiota xanthophylla Р. D. Orton, Suppl. Trans. Brit. Мус. Soc.,
43. 1960:289.— L. citrophylla (Berk, et Br.) Sacc., Syll. Fung., V, 1887:
101
57, s. auct. eur., non s. Berk, et Br., Cylon Fungi, 1871 : 139 (табл. XII,
6a—d).
Icon.: Cke, 111. Brit. Fung., 1881 —1891, fig. 1111; Reid, Col. Ic.
Rare a. Int. Fung., Ill, 1968, pl. 19b.
В широколиственных, хвойных и смешанных лесах, ботсадах, часто
на песчаной почве; гум. canp., VII—X, о. р., (1—2), п. з. н., мультнрег.,
евраз-ам-афр.
Распр. в СССР. Европ. ч.: УССР; Азиат, ч.: Дальн. Восток —
Приморск, кр. _
Общее распр. Европа: Великобритания, Франция; Азия: Шри
Лайка; Сев.г Америка: США, Мексика; Африка: Алжир, Ке-
ния, Уганда.
Секция Sericellae Kiihn. ex S. Wasser 1
Укр. ботан. журн., 35, 5, 1978:517.— Sericellae Kuhn., BSMF, 52,
1936 : 188 (nom. nudum).
129. Lepiota serena (Fr.) Sacc., Syll. Fung., V, 1887:52, s. Kiihn.
non s. J. Lge.— Agaricus serenus Fr., Hymen. Europ., 1874 : 38 (табл. XII,
7a—в).
Icon.: Huijsman, Med. Ned. Мус. Ver., 28, 1943, fig. 9; Вассер,
Фл. гр. Укр. (Agaricaceae), 1980, рис. 137.
В широколиственных, кедрово-широколиствеиных лесах, парках;
лодст. canp., VI—X, р., (1—2), н., эвриголаркт., евраз-афр.
Распр. в СССР. Европ. ч.: УССР; Азиат, ч.: Дальн. Восток —
Приморск, кр., Ср. Азия —- ТССР.
Общее распр. Европа: Великобритания, Дания, Нидерланды,
Франция, ЧССР, ВНР; Африка: Алжир.
130. Lepiota cygnea J. Lge, FI. Ag. Dan., V, 1940:V, Append,
(табл. XII, 5a—г).
Icon.: J. Lge, Fl. Ag. Dan., 1935—1940, pl. 13, fig. A; Huijsman,
Med. Ned. Мус. Ver., 28, 1943, fig. 12; Васильева, Агар. шл. гр. При-
морск. кр., 1973, рис. 43, Б; Вассер, Фл. гр. Укр. (Agaricaceae), 1980,
рис. 138.
В широколиственных, кедрово-широколиственных лесах; полет,
canp., VI—X, о р., (1—2), п. з. н., эвриголаркт., евраз-ам.
Распр. в СССР. Азиат, ч.: Кавказ — АзССР, Дальн. Восток
Приморск, кр.
Общее распр. Европа: Дания, Франция, ПНР, ВНР; Азия:
Япония, Турция; Сев. Америка: США.
131. Lepiota cygneoaflinis * Pil., Sborn. Nar. Mus. v Praze, X1B. 2,
1955: 5—7.
1 con.: Pilat, Sborn. Nar. Mus. v Praze, 1XB, 2, 1955, fig. 2—3.
В травянистых местах парков, в широколиственных лесах; гум.
canp., VIII—X, о. р_, (1), п. з. и., немор., евр.
Распр. в СССР. Е в р о п. ч.: УССР.
Общее распр. Европа: ЧССР.
1 Секция Sericellae возведена П. Хейнсманном (Heinemann, 1978, р. 399) в ранг
рода Scriceomyces Heinem.
102
РОД CHAMAEMYCES ВАТТ. EX EARLE
Bull. N. Y. Bot. Gard., 1909:448.— Lepiota subgen- Lepiotella Gilb.,
Le genre Amanita Persoon, 1918 : 159.—Lepiotella (Gilb.) Gilb. ex Kiihn.
et R. Mre, BSMF, 50, 1934 : 15; Singer, Ann. Мус., 34, 1936 : 338.— Dro-
sella R. Mre apud Kiihn. et R. Mre, BSMF, 50, 1934 : 15 (nom. nudum).
132. Chamaemyces fracidus (Fr.) Donk, Beihft. zur Nova Hedwigia,
Heft 5, 1962 : 48.— Agaricus fracidus Fr., Epicr. Syst. Мус., I, 1838 : 25.—
Lepiota irrorata Quel., 11-e suppl. in Ass. Fran^. Av. Sci., XI, 1882 : 387.—
Armillaria fracida (Fr.) Sacc., Syll. Fung., V, 1887 : 86.— Drosella irrora-
ta (Quel.) Kiihn. et R. Mre, BSMF, 50, 1934:15 (nom. nudum).— Lepio-
tella irrorata (Quel.) Sing., Ann. Мус., 34, 1936:338.— L. fracida (Fr.)
Sing., Lilloa, 22, 1951 : 446.— Agaricus demissannula Seer., Mycorg. Suis.,
I, 1833 : 54.— Chamaemyces demissannula (Seer.) Mos., KI. Krypt.-flora,
B2, Teil 2, 1967 : 194 (табл. XII, 8a—д- XXV, 43—44).
Icon.: Bres., Icon. Мус., 1927, tab. 41; Konr. et Maubl., Icon. Sei.
Fung., 1924—1937, pl. 16, fig. 2; J. Lge, Fl. Ag. Dan., 1935—1940, tab. 15,
fig. В; Зерова, Атл. гр. Укр., 1974, табл. 114, рис. 1; Вассер, Фл. гр.
Укр. (Agaricaceae), 1980, рис. 139, табл. XVIII, 1.
В хвойных (еловых), широколиственных и смешанных лесах, на
полянах и лесных опушках; гум. сапр., VII—X, о. р., (1), м. с. г., эври-
голаркт., евр.
Распр. в СССР. Европ. ч.: ЛитССР, УССР.
Общее распр. Европа: Швеция, Дания, Швейцария, Франция,
Италия, ФРГ, ГДР, ЧССР, Австрия, ВНР, Великобритания.
КОЛЕНО LEUCOCOPR1NEAE SING.
Pap. Mich. Acad. Sci., Arts a. Lett., 32, 1946 (publ. 1948) : 141.
РОД CHLOROPHYLLUM * MASS.
Bull. Misc. Inf. Kew, 1898 : 135.
133. Chlorophyllum molybdites* (Fr.) Mass., Bull. Misc. Inf. Kew,
1898 : 136.— Agaricus molybdites Fr., Syst. Мус., I, 1821 : 308.— A. mor-
ganii Pk, Bot. Gaz., 4, 1879 : 137.— A. glaziovii Berk., Vidensk. Meddel.,
1879—1880:332.— Lepiota molybdites (Fr.) Sacc., Syll. Fung., V, 1887:
30.— L. morganii (Pk) Sacc., 1. c., 1887:30.— Pholiota hlaziovii (Berk)
Sacc., op. cit., 1887:751.— Mastocephalus molybdites (Fr.) O. Kuntze,
Rev. Gen. Pl., 2, 1891 : 860.— M. morganii (Pk) O. Kuntze, I. c. 1891 :
860.— Lepiota ochrospora Cke et Mass., Grevillea, 21, 1893 : 73 — Chloro-
phyllum morganii (Pk) Mass., Bull. Misc. Inf. Kew, 1898: 136.— Ch. es-
culentum Mass., 1. c., 1898:136.— Agaricus guadelupensis Pat., BSMF,
15, 1889 : 197.— Annularia camporum Speg., Ann. Mus. Nat. Buenos-Aires,
6, 1899: 117.— Lepiota esculenta (Mass.) Sacc. et Syd., Syll. Fung., 16,
1901:2.— Leucocoprinus molybdites (Fr.) Pat., BSMF, 29, 1913:215.—
Lepiota camporum (Speg.) Speg., Bol. Acad. Nac. Cien?. Cordoba, 29,
1926:114.— Agaricus congolensis Beeli, Bull. Soc. Roy. Bot. Belg., 61,
1928:92.— Chlorophyllum molybdites (Fr.) Mass. var. congolensis (Bc-
eli) Heinem., Fl. Ic. Champ. Congo, 16, 1967:323 (табл. XIII, la—г;
XXXI 1,58).
Icon.: Heinemann, FI. Ic. Champ. Congo, 16, 1967, tab. 52; Pegier,
Prelim. Ag. FI. East Afr., 1977, fig. 65.
103
В широколиственных, смешанных лесах, парках, в травянистых мес-
тах; гум. сапр., VIII—X, о. р., (1), я., в. н. а.
Распр. в СССР. Азиат, ч.: Дальн. Восток — Приморск, кр.
Общее распр. Азия: Израиль; Сев. Америка: США; Юж и.
Америка: Бразилия, Гайана; Африка: Кения, Танзания, Уганда;
Новая Зеландия.
Прим. Chlorophyllum— монотипный род, хотя в понимании П. Хейнеманна
(Heintmann, 1968а, Ь) содержит несколько видов, отличающихся размером спор и
географией. Chlorophyllum наиболее близок к роду Macrolepiota, от которого отлича-
ется цветом пластинок и спор, отсутствием пряжек на мицелии. Данный род чрез-
вычайно интересен в систематическом и ботанико-географическом аспектах и требует
дальнейшего всестороннего тщательного изучения.
Ch. molybdites — новый для СССР ядовитый гриб. В литературе, особенно севе-
роамериканской (Miller, 1972; Singer, 1975; Rumack, Salzman, ed., 1978), описаны
случаи отравления этим грибом. Токсины Ch. molybdites водорастворимы, содер-
жание их в плодовых телах колеблется в зависимости от возраста н экологических
особенностей.
РОД LEUCOCOPRINUS РАТ.
BSMF, 4, 1888:26; Journ. Bot., Paris, 2, 1888: 16 — Mastocephalus
Batt, ex O. Kuntze, Rev. Gen. PL, 2, 1891 :857,— (?) Hlatula (Fr.)
Mont.. Ann. Sci. Nat., IV, I, 1854:107 (vix. s. original!).- Hlatula s.
Heim et Romagn., 1934, Sing., 1936.— (?) Leptomyces Mont.. Syll. Crypt,
1856 : 128.
Подрод Leucocoprinus
Нов. сист. высш, и иизш. раст, 4, 1978 : 209.
134. Leucocoprinus cepaestlpes (Fr.) Pat, Tab. Analyt. Fung, Fasc.
7, 1889 : 45.— Agaricus cepaestlpes Fr, Syst. Мус, I, 1821 : 280.— A. con-
tinuus Berk, in Hook. Lond. Journ. Bot, 6, 1847 : 480.— A. rorulentus
Panizzi, Comm, della Soc. Critt. ItaL, 1861 : 172.— A. cheimonoceps Berk,
et Curt, Journ. Linn. Soc. Bot, 10, 1869:283.—A. oncopus Berk, et Br,
op. cit, II, 1871:496.— A. oncopus Berk, et Br, op. cit, 1871:499.—
A. adore us Berk, et Br, I. c, 1871 :499.— Lepiota cepaestlpes (Fr.)
Kumm, Der Fuhrer in die Pilzkunde, 1871 : 136.— L. rorulenta (Panizzi)
Baria. BSMF, 9, 1886 : 117,— L. continua (Berk, et Br.) Sacc, Syll. Fung,
V, 1887:44,- L. oncopus (Berk, et Br.) Sacc. (‘oncopoda’), 1. c, 1887.—
L. oenopus (Berk, et Br.) Sacc. (‘oenopoda’), op. cit, 1887 57.— L. odo-
rea (Berk, et Br.) Sacc, I. c, 1887:57.-7, cheimonoceps (Berk, et
Curt.) Sacc, op. cit, 1887 : 66 — Mastocephalus cepaestlpes (Fr.) O. Kun-
tze Rev. Gen. PL, 2, 1891 : 859.— Hlatula cepaestlpes (Fr.) Heim et Ro-
magn, BSMF, 50, 1934: 184 (табл. XIII, 2a—д; XXVI, 45—46).
Icon.: J. Lge, FL Ag. Dan, 1935—1940, tab. 14, fig. F; Dennis,
Kew BuIL, 4, 1952 : 469 fig. 7; Pegler, Kew Bull, 27, 1, 1972, fig. 10,
4a, b, c, d. с Вассер, Нов. сист. высш, и иизш. раст, 4, 1978, рис. 1;
Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 142.
На кучах мусора, компоста, на опилках, в парниках, оранжереях,
теплицах на богатой гумусом почве. В парниках, оранжереях, теплицах
растет в посадках тропических и субтропических растений, при этом
летом температура помещения достигает +40—45 °C, а зимой +15 °C;
гум. сапр, I—XII, о. р, (1). с.(?), я.(?), мультирег, космопол.
104
Распр. в СССР. Европ. ч.: УССР; Азиат, ч.: Вост Сибирь —
Красноярск, кр.
Общее распр. Европа: Франция, Дания, Греция, Италия, ЧССР;.
Азия: КНР, Индия, Япония, Шри Ланка; Сев. Америка: Кана-
да, США, Мексика; Южн. Америка: Бразилия, Чили; Африка:
Кения, Танзания, Гана, Нигерия, Уганда; Австралия; о-ва Куба,.
Ямайка, Тринидад, Тобаго.
135. Leucocoprinus birnbaumii (Corda) Sing., Sydowia, 15, 1962:
67.— Agaricus birnbaumii Corda, Icon. Fung., 3, 1839 : 48. — A. cepaestipes
Fr. var. lutea Seer., Mycorg. Suis., I, 1833 : 56.— A. flossulphuris Schniz-
lein, Sturm. Dcut. FL, 3 (31), 1851 : 1, tab. I.—-A. luteus (Seer.) Berk.,
Intell. Observer, 1865:7.— A. cepaestipes Fr. var. flossulphuris (Schniz-
lein) Oud., Arch. Neerland. Sci. exact Nat., 2, 1867: 19 — Lepiota flam-
tnula (Aib. et Schw.) Gill., Champ. Fr., 1874 : 63.— L. lutea (Seer.)
God., BSMF, 13, 1897:33.— L. aurea Mass., Bull. Misc. Inf. Kew, 1912:
189.— L. pseudoliemophora Rea, Brit. Bas., 1922:74.— Leucocoprinus
luteus (Seer.) Locq., Bull. mens. Soc. linn. Lyon, 14, 1945:93.— Leuco-
coprinus flossulphuris (Schnizlein) Ccjp, Ccs. Mykol., 2, 3, 1948 :78
(табл. XIII. За—г).
Icon.: Pegler, Kew Bull., 27, I, 1972:192, fig. 10, la, b, c, d, e;
Imazeki a. Hongo, Coli. ill. fung. Jap., 1972, pl. 21, fig. 113; Вассер,
Нов. сист. высш, и низш. раст., 4, 1978, рис. 2; Вассер, Фл. гр. Укр.
(Agaricaceae), 1980, рис. 143.
На кучах мусора, компоста, на опилках, в парниках, оранжереях,
теплицах, на богатой гумусом почве. В парниках, оранжереях, тепли-
цах растет в посадках тропических и субтропических растений, при этом
летом температура помещения достигает +40—45 °C, а зимой +15 °C;
гум. canp., I—XII, о. р., (1), н., мультнрег., евраз-ам-афр.
Распр. в СССР. Европ. ч.: РСФСР — Мурманская обл., ЛнтССР,
УССР.
Общее распр. Европа: Великобритания, Швеция, Швейцария,
Франция, Италия, ФРГ, ГДР, ЧССР; Азия: Индия, КНР, Япония,
Шри Лапка; Сев. Америка: Мексика; Южн. Америка: Бра-
зилия; Африка: Кения, Марокко, Нигерия, ЮАР.
136. Leucocoprinus denudatus (Rabenh.) Sing., Lilloa, 22, 1951
(1949): 424.— Agaricus denudatus Rabenh., Hcdwigia (Dresden), 6,
1867 : 45.— Lepiota denudata (Rabenh.) Sacc., Syli. Fung., V, 1887 : 52.—
L. denudata (Rabenh.) Sacc. var. varsoviensis Chelchowski et Bionski,
Pam. Fiz., 14, 3, 1896 :87.— L. gueguenii Sacc. et Trav., Syll. Fung., 19,
1910:1081.— L. boudieri Guegucn, BSMF, 24, 1908:126—132, non s.
Brcs., 1884.— L. lutea Matt, Atti Reale Accad. d. dei lincei, 315, scr. 5a,
12 (II), 1918 : 567—568.— L. cepaestipes Vel., Ceske houby, 1920: 210.—
Hiatula denudata (Rabenh.) Sing., Ann. Мус., 41, 1943:168.— Leuco-
coprinus (Leucobolbitius) gueguenii (Sacc.) Locq., Bull. mens. Soc. linn.
Lyon, 12, 1943 : 29 (табл. XIII, 4a—в).
Icon.: Brcs.,»Icon. Мус., 1927, 40, fig. 4; Hcrink, Ccs. Mykol., 13,
2, 1959:111; Moser, Ki. Krypt.-flora, 1967, fig. 87; Josserand, BSMF,
90, 3, 1974 : 237, fig. 3; Вассер, Укр. ботан. журн., 32, 3, 1975, рис. 4—5;
Wasser, Fung. Rar. Icon. Col., X, 1979, pl. 79b, Text-fig. 4; Вассер, Фл.
гр. Укр. (Agaricaceae), 1980, рис. 144, табл. XXIV, 2.
В ботанических садах, парках, оранжереях и парниках, на бога-
той гумусом почве, на навозе, в искусственных лесных массивах.
В оранжереях и парниках растет в посадках тропических и субтропи-
105
ческмх растений (Ficus elastica Roxb., Monstera delicisa Liebm.), при
этом летом температура помещения достигает +40—50 °C, а зимой
+ 15 °C; гум. сапр., VI—X, д. р.» (2), с.(?), я.(?), мультирег., евраз-
ам-австр.
Распр. в СССР. Европ. ч.: РСФСР — Ленинградская обл.,УССР;
Азиат, ч.: Зап. Сибирь — Алтайск. кр.
Общее распр. Европа: Нидерланды, Бельгия, Финляндия, Фран-
ция, Дания, Испания, Италия, ФРГ, ГДР, ПНР, ЧССР, СФРЮ, СРР;
Азия: Турция, Япония; Африка: Алжир; Сев. Америка:
США, Мексика; Южн. Америка: Боливия, Венесуэла; А в с т р а -
л и я; Филиппинские о-ва.
137. Leucocoprinus brebissonii (God. apud Gill.) Locq., Bull. mens.
Soc. linn. Lyon, 12, 1943 : 73.— Lepiota brebissonii God, apud Gill.,
Champ. Fr., 1874:137.— Hiatula brebissonii (God. apud Gill.) Sing.,
Ann. Мус., 41, 1943 : 167.
Icon.: J. Lge, FL Ag. Dan., 1935—1940, pl. 14, fig. H; Kiihn.,
.BSMF, 52, 1936, fig. 916; Kiihn. et Romagn., Fl. anal. Champ. Super.,
1953, fig. 577.
В широколиственных и смешанных лесах, парках и ботанических
садах; подст. сапр., V—X, о. р., (1), с.(?), я.(?), эвриголаркт., евраз.
Распр. в СССР. Азиат, ч.: Дальн. Восток—Приморск, кр.
Общее распр. Европа: Великобритания, Дания, Франция, ЧССР;
Азия: Япония.
Подрод Rubescentes S. Wasser
Нов. сист. высш, и низш. раст., 4, 1978 : 217.
138. Leucocoprinus badhamii ** (Berk, et Br.) S. Wasser, Нов. сист.
•высш. и низш. раст., 4, 1978 :217.— Agaricus badhamii Berk, et Br.,
Ann. a Mag. Nat. Hist., 1860: 126.— Lepiota badhamii (Berk, et Br.)
Quel., Champ. Jura et Vosges, 1972 : 231, s. Berk, et Br., Pat., Boud., Rea
non s. Konr. et Maubl., Locq., Sing., Kiihn. et Romagn.— L. meleagris
(S: F. Gray) QueL, Champ. Jura ct Vosges, I, 1872 : 93, s. Quel., non
s. Ricken.— L. rufoveolutina Vel., Novit. Mycol., i939 :50, s. orig., non
s. Pil.— L. meleagroides Huijsm., Med. Ned. Мус. Ver., 28, 1943 : 5—6
(nom. nudum).— Leucocoprinus meleagroides (Huijsm.) Mos. apud Gams,
KI. Krypt.-flora, B. 11b, 1955 : 132 (nom. nudum).— L. badhamii (Berk, et
Br.) Mos., KI. Krypt.-fiora, В. Il, Tcil II, 1967: 186 (nom. inval.)
(табл. XIII, 5a—в).
Icon.: Baria, Champ. Aip. Mar., 1888—1892, pl. 13, fig. 8—il;
Boudier, Icon. Мус., 1904—1909, pl. Il; BSMF, 41, 1925, pl. I; Huijsman,
Med. Ned. Мус. Ver., 28, 1943, 8, fig. II—IV; Orton, Trans. Brit. Мус.
Soc., 43, 2, 1960, fig. 312 (споры); Josserand, Bull. mens. Soc. linn.
Lyon, 15. fevrier, 1974, fig. 4; Вассер, Нов. снег. высш, и низш. раст.,
4, 1978, рис. 5; Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рнс. 146.
В парках, садах, в насаждениях широколиственных деревьев, изред-
ка хвойных; гум. сапр., VII—X, д. р., (2), с.(?), я.(?), эвриголаркт.,
евраз-афр.
Распр. в СССР. Европ. ч.: УССР; Азиат, ч.: Кавказ — ГССР,
Дальн. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Бельгия, Дания, Фран-
ция, Австрия, ФРГ, ЧССР; Азия: Израиль; Африка: Алжир, Ма-
рокко.
ДОб
Прим. Leucocoprinus badhamii наиболее близок к L. pllatianus (Demoulin)
S. Wasser ( = L. rufovelutina Vel s. Pit.), от которого отличается пигментацией хей-
лоцистид, их формой, структурой покровных элементов ножки и т. д.
Детально вопросы о видовой самостоятельности, экологии, распространении
L. badhamii и L. pilaiianus рассмотрены в работах В. Демулена (Demoulin, 1966)
и М. Жоссеранда (Josserand, 1974).
139. Leucocoprinus pilatlanus** (Demoulin) S. Wasser var. pilatia-
nus, Нов. сист. высш, и пнзш. раст., 4, 1978 : 219.— Lepiota pilatiana
Demoulin, Lejeunia, Rev. de Bot., 39, 1966: 11.— L. rufovelutina Vel. s.
Pil., Klic. urc. nas. hub, 1951 : 424.— Pilat, Sborn. Nar. Mus. v. Praze,
IXB, 2, 1953:12—18.— L. rufovelutina Vel. var. sanguinescens Pil.,
Sborn. Nar. Muz. v Praze, XIB. 2, 1955- 16—19.— Leucocoprinus pilatia-
nus (Demoulin) Mos., KI. Krypt.-flora, В. II, Toil II, 1967: 186 (nom.
inval.).— L. pilaiianus (Demoulin) Bon et Boiff., BSMF, 88, 1, 1972:25,
27 (nom. non rite pubiicatum).— (?) L. jubilei (Joss.) S. Wasser, Нов.
сист. высш, и низш. раст., 2, 1976 : 192 (табл. XIII, 6а—б\ XXVI, 47).
Icon.: Pilat, Sborn. Nar. Mus. v. Praze, !XB, 2, 1953, fig. 12—18,
tab. II; Pilat, Sborn. Nar. Mus. v Praze, XIB, 2, 1955, fig. 11 —12; De-
moulin, Lejeunia, Rev. de Bot., 39, 1966, fig. 4 (А, В, C, D); Babos, Ann.
Hist.-Nat. Mus. Nat. Hung., 53, 1961, fig. 2; Вассер, Нов. сист. высш, и
низш. раст., 4, 1978; рис. 6; Вассер, Фл. гр. Укр. (Agaricaceae), 1980,
рис. 147, табл. XXIII, 2.
В парках, искусственных лесных насаждениях дуба, клена.
L. pilatianus var. subrubens** (Wich.) S. Wasser, Нов. сист. высш,
и низш. раст., 4, 1978:221.— L. rufovelutina Vei. var. subrubens Wich.,
Ces. Mycol., 14, 1, 1960:49.— Lepiota pilatiana Demoulin var. subrubens
(Wich.) Demouiin, Lejeunia, Rev. de Bot., 39, 1966 : 12.
Гум. сапр., VII—X, о. p., (2), c.(?), я.(?), немор., евраз-афр.
Распр. в СССР. Европ. ч.: УССР; Азиат, ч.: Кавказ — ГССР.
Общее распр. Европа: Франция, ЧССР, Австрия, ВНР; А ф р и -
к a: Alaрокко.
140. Leucocoprinus bresadolae** (Schulz.) S. Wasser, Нов. сист.
высш, и низш. раст., 4, 1978:221.— Lepiota bresadolae Schulz., Hedwi-
gia, 24, 1885 : 132.— Agaricus cupreus Schulz., Verh. Zool.-Bot. Ges. Wein,
27, 1877:112, non s. Seer., 1833.— Lepiota badhamii (Berk, et Br.)
Quel., Champ. Jura et Vosges, I, 1872:78, s. Konr. et Maubl., Locq.,
Kuhn, et Romagn., non s. Berk, et Br., Pat., Boud., Rea.— L. meleagris
(Sow. ex S. F. Gray) Quel., Cham. Jura ct Vosges, I, 1872:79, s. Bres.,
non s. al.— Leucocoprinus badhamii (Berk, et Br.) Locq., 1943, s. Locq.—
L. bresadolae (Schulz.) Mos., KI. Krypt.-flora, В. II, Teil II, 1967.186
(nom. inval.).— Leucoagaricus badhamii (Berk, et Br.) Sing., Lilloa, 22,
1951 : 424, s. Sing. (табл. XVI, la—г; XXVII, 48—49).
Icon.: Schuizer, Verh. Zool.-Bot. Ges. Wein, 27, 1877:112 (под
названием Agaricus cuprous); Bres., Fung. Trid., 1881 — 1892, (под на-
званием Lepiota haematosperma); Huijsman, Med. Ned. Мус. Ver., 28,
1943, fig. 1, V—VIII, 2, I—II; Babos, Ann. Hist.-Nat. Mus. Nat. Hung.,
53, 1961 : 197, fig. 3; Вассер, Нов. снст. высш, и низш. раст., 4, 1978,
рис. 7; Wasser, Fung. Rar. Icon. Col., X, 1979, pl. 79c, text-fig. 5; Вассер,
Фл. гр. Укр. (Agaricaceae), 1980, рис. 148, табл. XXIV, 3.
В парках, садах, во дворах, изредка в широколиственных, кедрово-
широколиственных лесах; подст. сапр., VI—X; о. р., (1—2), с., ксероме-
рид., евраз.
Распр. в СССР. Европ. ч.: УССР; Азпат. ч.: Дальн. Восток —
Приморск, кр.
107
Общее распр. Европа: Великобритания, Испания. Франция, Ита-
лия, ЧССР, Австрия, ВНР, СРР; Азия: Япония.
Прим. Leucocoprinus bresadolae — очень интересный проблематичный вид. До
Р. У. Г. Денниза, П. Д. Ортона и Ф. Б. Хоры (Dennis, Orton, Нога, 1960) Leucocopri-
nus badhamii (некоторые авторы относят его к роду Lepiota) понимали очень широ-
ко, включая в него и Leucocoprinus (Lepiota) bresadolae. Р У. Г. Денниз, П. Д. Ортон
и Ф Ь Хора, на наш взгляд, совершенно правильно выделили из L. badhamii s. 1.,
L. bresadolae, легко отличающийся от первого главным образом размером спор —
8—12X6 8 мкм (у наших образцов 7.5—11,2X5,7—7,5 мкм), габитусом карпофоров
и другими признаками.
М Мозер (Moser, 10671 высказал совершенно правильную мысль о том, что
оба вида следует перенести в род Leucocoprinus, что он и еде чал (Moser, 1967,
р. 186), однако не оформил по правилам Международного кодекса ботанической
номенклатуры (при комбинации нет ссылки на базионим). Вследствие указанного
выше мы сочли возможным узаконить оба названия, оставив их в роде Leucocop-
iinus (Вассер, 1978а).
141. Leucocoprinus bohusi*** S. Wasser, Укр. ботан. журн., 32, 1,
1975 : 33 (таб п. XIV, 2а— в; XXVI11, 50).
Icon.: Вассер, Укр. ботан. журн., 32, 1, 1975:33, рис. 2а—в;
Вассер, Нов. спет. высш, и низш. раст., 4. 1978, рис. 8; Wasser, Fung.
Rar. Icon. Col., X, 1979, pl. 79a; Bacccp, Фл. гр. Укр. (Agaricaceae),
1980, рис. 149, табл. XXIV, 1.
В заповедных целинных разнотравно-типчаково-ковыльных степях,
на кучах старого навоза, мусора; копр., V—VI, о. р., (2), с. (?), я.(?),
кссромернд., укр. усл. энд.
Распр. в СССР. Е в р о п. ч_: УССР.
Пр и м. Leucocoprinus bohusi наиболее близок к L. badiiamii (Berk, et Br.)
S. Wasser, от которого отличается внешним видом карпофоров, цветом шляпки
и мякоти, размером и формой спор, экологией (Locquin, 195ia, b: Pilat, 1951a; Mel-
zer, 1959; Babos, 1961; Singer, 1962; Moser, 1967; Demoulin, 1966; Васильева, 1973;
Вассер. 1975a, 1980; Wasser, 1979).
142. Leucocoprinus georginae** (W. G. Sm.) S. Wasser comb. nov.—
Agaricus (Lepiota) georginae W. G. Sm., Seem. Journ. Bot., IX, 1871 . I.—
Lepiota georgineae (W. G. Sm.) Sacc., Syli. Fung., V, 5887:71.—
Leucocoprinus georgineae (\V. G. Sm.) Bon ct Boifl., BSMF, 8G, 1,
1972 : 28 (nom. nudum).
Icon.: Cke, Ill. Brit. Fung., 1881—1891, pl. 132.
В широколиственных и смешанных лесах, парках, в парниках,
оранжереях, теплицах; гум. canp., I—XII, о. р., (1—2), с.(?), я.(?)„
эвриголаркт., евраз-афр.
Распр. в СССР. Азиат, ч.: Дальн. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Франция, ВНР, ЧССР;
Африка: Алжир.
РОД LEUCOAGARICUS (LOCQ.) SING.
Sydowia, 2, 1948:35.— Leucocoprinus Pat. subgenus Leucoagaricus
Locq., Bull. mens. Soc. linn. Lyon, 12, 1943 : 92.
Секция Leucoagaricus
Sydowia, 2, 1948 : 35.
143. Leucoagaricus macrorhizus Locq. ex llorak, Beitr. zur Krypt.-
flora dcr Schweiz, XIII, 1968:344.— Leucocoprinus macrorhizus Locq.,
Bull. mens. Soc. linn. Lyon, 12, 1943 : 75 (nom. nudum).— Leucoagaricus
108
macrorhizus (Locq.) Sing., Sydowia, 2, 1948:35 (nom. nudum).— Lepiota
tnacrorhiza (Locq.) Kiihn. et Romagn., Fl. Anal. Champ. Super., 1953 :406
(nom. nudum) (табл. XIV, За—д).
Icon.: Locquin, Bull. mens. Soc. linn. Lyon, 12, 1943, pl. 5, Вас-
сер, Укр. ботан. журн., 33, 5, 1976, рис. 1—2; Wasser, Fung. Rar. Icon.
Col., X, 1979, pl. 80a, Text-fig. 6; Вассер, Фл. гр. Укр. (Agaricaceae)
1980, рис. 150—151, табл. XXV, 2.
В парках, садах, на полях, пашнях и других интенсивно используе-
мых участках, в каменистых полупустынях па высоте 600—1500 м
н. у. м., в теплые жаркие годы; гум. сапр., VI—X, о. р., (I—2), с. (?),
я.(?), ксеромерид., евраз-афр.
Распр. в СССР. Е в р о п. ч_: УССР; Азиат, ч.: Кавказ — АрмССР.
Общее распр. Европа: Великобритания, Франция, Италия; А ф -
рика: Марокко.
Прим. L. macrorhizus—редкий1, чрезвычайно интересный теплолюбивый вид.
Характернейшей морфологической особенностью его является то, что длина подзем-
ной части ножки больше длины наземной части.
Секция Annulati (Fr.) Sing.
Nowa Hedwigia, 23, 1972 : 329.
144. Leucoagaricus holoserlceus (Fr.) Mos., KI. Krypt.-flora, В. II,
Teil II, 1967 : 185.— Agaricus holosericeus Fr., Epicr. Syst. Мус., 1, 1837 :
16.— Lepiota holosericea (Fr.) Gill., Hymen. Fr., 1874: 147.— Leucocopri-
nus holosericeus (Fr.) Locq., Bull. mens. Soc. linn. Lyon, 14, 1945 : 54
(табл. XIV, 4a—г).
Icon.: Baria, Champ. Alp. Mar., 1888—1892, pl. 14, fig. 10—13;
Bres., Icon. Мус., 1927, tab. 26; Locq., Bull. mens. Soc. linn. Lyon, 14,
1945 : 54.
В парках, садах, по краю дорог, иа открытых травянистых местах,
реже в хвойных и широколиственных лесах; гум. сапр., VIII—X. р., (2),
с., ксеромерид., евраз-афр.
Распр. в СССР. Европ. ч.: РСФСР — Вологодская обл., УССР;
Азиат, ч.: Кавказ — Абхазская АССР.
Общее распр. Европа: Великобпитания, Швейцария, Дания,
ВНР, Франция, СФРЮ, Италия, ФРГ, ГДР; Азия: Турция; Афри-
ка: Марокко.
145. Leucoagaricus moseri *** (S. Wasser) S. Wasser. Укр. ботан.
журн., 35, 5, 1978:516.— Lepiota moseri S. Wasser, Укр. ботан. журн.»
32, 1, 1975 : 34 (табл. XIV, 5а—в).
Icon.: Вассер, Укр. ботан. журн., 32, 1, 1975:35, рис. 3, а, б;
Вассер, Солтатова, Высш. баз. ст. зоны Укр., 1977, рис. 106; Wasser,
Fung. Rar. Icon. Col., X, 1979, pl. 80b; Вассер, Фл. гр. Укр. (Agarica-
ссас), 1980, рнс. 152, табл. XII. 2.
В искусственных белоакациевых насаждениях, в парках в насажде-
ниях Robinia pscudoacacia L. и Acer platanoidcs L., в лубовых полеза-
щитных лесополосах, полупустынях; гум. сапр., IX—X, д. р., (2—3).
с.(?), я.(?), немор., евраз.
Распр. в СССР. Европ. ч.: УССР; Азиат, ч.: Зап. Сибирь —
КазССР.
Во Франции, по свидетельству Э. Хорака (Horak, 1968), нередок.
109
Прим. Leucoagaricus moseri наиболее близок к L. leucothitus (Vitt.) S. Wasser,
отличается от него размером карпофоров, размером спор, формой и расположением
кольца на ножке, наличием в основании ножки клубневидного утолщения, экологией
(Ricken, 1915; Lange J.. 1935—1940; Лебедева, 1949; Bohus, Kalmar, Ubrizsy, 1951;
Dennis, Orton, Hora, I960; Moser, 1967; Vesely, Kotlaba, Pouzar, 1972; Васильева,
1973; Вассер, 1975a, 1980; Wasser, 1979). До последнего времени L. moseri считался
украинским эпдемом. В 1980 г. описанный нами вид был обнаружен Л. Г. Буровой
п Э. Л. Нсздойминого (1980) в полупустынных растительных ассоциациях Джаиы-
Гч-кского полупустынного стационара (Северный Прикаспий).
146. Leucoagaricus carneifolius ** (Gill.) S. Wasser, Укр. ботан.
журн., 34, 3, 1077:307.— Lepiota carneifolia Gill., Hymen. Fr., 1874:
65.— L. naucina (Fr.) Quel. var. carneifolia (Gill.) Konr. et Maubl.,
Icon. Sei. Fung., VI, 1937:44.— Leucocoprinus carnefolius (Gill.) Locq.,
Bull. mens. Soc. linn. Lyon. 14, 1945:20 (табл. XXVIII, 51; XXIX, 52).
Icon.: Gill., Hymen. Fr., 1874, tab. 42; Pegler, Kew Bull., 16, 3,
1963 : 447, fig. 1.
В парках, садах, часто под Aesculus hippocastanum L., по берегам
рек, в траве; гум. сапр., VIII—X, д. р_, (2), с.(?), я.(?), немор., евр.
Распр. в СССР. Европ. ч.: УССР.
Общее распр. Европа: Великобритания, Франция.
147. Leucoagaricus leucothitus** (Vitt.) S. Wasser f. leucothitus,
Укр. ботан. жури , 34, 3, 1976:307.— Agaricus leucothitus Vitt, Fung.
Mang., 1835 : 310.— A. naucinus Fr., Epicr. Syst. Мус., 1838 : 16.— Lepio-
ta naucina (Fr.) Kumm., Dcr Fuhrer in die Pilzkundc, 1871 : 136, s. auct.,
non s. Mos., 1967.— L. naucina (Fr.) Quel., Champ. Jura et Vosges, I,
1872 : 73.— L. naucina var. leucothites (Vitt.) Sacc., Syll. Fung., V, 1887 :
48.— Agaricus naucioides Pk, Ann. Rep. N. Y. State Mus., 29, 1876 : 66.—-
Mastocephalus naucinus (Fr.) O. Kuntze, Rev. Gen. PL, 2, 1891 :860
(табл. XV, la—в).
Icon.: Baria, Champ. Alp. Mar., 1888—1892, tab. 15, fig. 1—4;
Bres., Icon. Мус., 1927, tab. 25; J. Lge, Fl. Ag. Dan., 1935—1940, tab. 9,
fig. А; Васильева, Агар. шл. гр. Приморск, кр., 1973, рис. 42 А; Зерова,
Атл. гр. Укр., 1974, табл. 111, рис. 1; Вассер, Фл. гр. Укр. (Agaricaceae),
рис. 153, табл. XXV, 1.
На пастбищах, в садах, огородах, парках, заповедных целинных
степях, на улицах, лесных полянах, реже в лесах.
L. leucothitus f. cinereolilacina ** (Joss.) S. Wasser, Фл. гр. Укр.
(Agaricaceae), 1980 : 292.— Lepiota naucina f. cinereolilacina Joss.,
BSMF, 81, 4, 1965: 519.
На лесных полянах. Известен из УССР.
Гум. сапр., V—X, о. ч., (2—3), с., в. п. к., мультирег., космонол.
Распр. в СССР, Европ. ч.: РСФСР — Московская, Ивановская
обл., ЛитССР, ЭССР, ЛатвССР, УССР; Азиат, ч.: Кавказ — ГССР,
АрмССР, АзССР; Зап. Сибирь — Алтайск. кр., Дальн. Восток — При-
морск. кр., Ср. Азия — КиргССР, ТССР.
Общее распр. Европа: Великобритания, Дания, Финляндия,
Швейцария, Франция, Италия, ФРГ, ГДР, ПНР, ЧССР, ВНР, СФРЮ,
Греция; Азия: Турция, Израиль, Япония, Шри Лайка; Сев. Аме-
рика: Канада, США; Южн. Америка: Аргентина, Чили; Ап-
с радия; Африка: Алжир, Марокко, Уганда; Соломоновы о-ва.
НО
РОД MACROLEPIOTA SING.
Pap. Mich. Acad. Sci., Arts a. Lett., 32, 1948: 141.— Eu-Leucocopri'-
nus Locq., Bull. mens. Soc. linn. Lyon, 14, 1945 : 30.— (?) Rugosospora
Heinem., Bull. Jard. Bot Nat. Belg., 43, 1973 : 12.
148. Macrolepiota procera (Scop.: Fr.) Sing., Pap. Mich. Acad. Sci.,
Arts a. Lett., 32, 1948 : 141.— Agaricus procerus Scop. : Fr., Syst. Мус.,
I, 1821 : 20.— Lepiota procera (Scop.: Fr.) S. F. Gray, Nat. Arr. Brit. PI.,
I, 1821 : 601.— Mastocephalus procerus (Scop. : Fr.) O. Kuntze, Rev. Gen.
PI., 2, 1891 : 1860.— Leucocoprinus procerus (Scop.: Fr.) Pat., Essai Ta-
xon. Hymen., 1900:171.— Lepiotophyllum procerum (Scop: Fr.) Locq.,
Bull. mens. Soc. linn. Lyon, 11, 1942:40 (табл. XV, 2a—e\ XXX, 54).
Icon.: Baria, Champ. Alp. Mar., 1888—1892, pl. 9, fig. 1—4; Gill.,
Champ. Fr., pl. 30; Cke, III. Brit. Fung., 1881—1893, pl. 21; Bres., Icon.
Мус., 1928, tab. 20; Pilat, Pilze, 1954, tab. 112; Зерова, Атл. гр. Укр.,
1974, табл. 109, рис. 1; Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 155,
табл. XXII.
В хвойных и широколиственных лесах, на лугах, в заповедных сте-
пях; гум. canp., VI—XI, о. ч., (1—2), с., в. п. к., мультнрег., космопол.
Распр. в СССР. Европ. ч.: повсеместно; А з н а т. ч.: Кавказ —
ГССР, АрмССР, АзССР, Зап. Сибирь — Алтайск. кр., Вост Сибирь —
Красноярск, кр., Дальи. Восток — Приморск, кр., Ср. Азия — КиргССР.
Общее распр. Европа: повсеместно; Азия: Индия, КНР, МНР,
Турция, Израиль, Иран, Япония, Шри Ланка; Сев. Америка: Ка-
нада, США, Мексика; Южн. Америка: Чили; Австралия;
Африка: Алжир, Кения, Танзания, Заир, Марокко; о-в Куба, Соло-
моновы о-ва.
149. Macrolepiota rhacodes (Vitt.) Sing.1, Pap. Mich. Acad. Sci.,
Arts a. Lett., 32, 1948:141.— Agaricus rhacodes Vitt., Fung., Mang., I,
1835:158.— Lepiota rhacodes (Vitt.) Quel., Champ. Jura et Vosges, I,
1872.32.— Leucocoprinus rhacodes (Vitt.) Pat., Essai Taxon. Hymen.,
1900 : 171.—- Lepiotophyllum rhacodes (Vitt.) Locq., Bull. mens. Soc. linn.
Lyon, 11, 1948 : 40 (табл. XV, За—в; XXXI, 55).
Icon.: Vitt., Fung. Mang., I, 1835, pl. 20; Baria, Champ. Alp. Mar.,
1888—1892, pl. 9, fig. 1—5; Gill., Champ. Fr., pl. 27; Cke, III. Brit. Fung.,
1881—1891, pl. 22; Bres., Icon. Мус., 1927, tab. 579; Pilat, Pilze, 1958,
tab. ПО; Зерова, Атл. гр. Укр., 1974, табл. 109, рис. 2; Вассер, Фл. гр.
Укр. (Agaricaceae), 1980, рис. 157, табл. XXVII.
В хвойных и лиственных лесах, парках, в заповедных целинных
степях, полезащитных лесополосах; гум. canp., VI—XI, о. ч., (1—2),
с., мультнрег., космопол. ,
Распр. в СССР. Европ. ч.: повсеместно; Азиат, ч.: Кавказ—
ГССР, АрмССР, АзССР, Зап. Сибирь — Алтайск. кр.. Вост. Сибирь —
Красноярск, кр.
Общее распр. Европа — повсеместно; Азия: КНР, Турция, Япо-
ния; Сев. Америка: США, Канада; Юж и. Америка: Чили;
Австралия; Африка: Алжир, Марокко, Кения.
150. Macrolepiota olivieri ** (Baria) S. Wasser, Фл. гр. Укр. (Aga-
ricaceae), 1980 : 298.— Lepiota olivieri Baria, BSMF, 1886, 3:113.—
L. rhacodes var. hortensis Pil., Klic urc. nas. hub, 1951 : 422.— L. bohe-
1 В 1979 г. из Франции (Bon, 1979) описан как новый для науки, очень близ-
кий к М. rhacodes вид — М. venenata Bon, являющийся ядовитым.
Ill
mica Wich., Cas. cs. houb., 38, 1961 : 102.— Alacrolepiota rhacodes var.
hortensis (Pil.) S. Wasser, Фл. гр. Укр. (Agaricaceae), 1980 : 298
(табл. XVI, За—б; XXIX, 53).
Icon.: Piiat, Pilze, 1954, tab. Ill; Wichansky, Myk. sborn., 39,
1—2, 1962, obr. 4—5; Dermek, Cas. cs. houb., 47, 1970, 38; Вассер, Фл. гр.
Укр. (Agaricaceae) 1980, табл. XXVIII.
В парках, садах, главным образом оранжереях, теплицах, парни-
ках, на унавоженной гумусовой почве; гум. сапр., I—XII, о. р., (1—2),
с., в. н. а.
Распр. в СССР. Европ. ч.: ЛатвССР, УССР.
Общее распр. Европа: ЧССР, Франция, ВНР; Сев. Амери-
ка: США.
151. Macrolepiota puellaris (Fr.) Mos., KI. Krypt.-flora, В. Il, Teil II,
1967:184.— Agaricus rhacodes puellaris Fr., Monogr. Hym. Suec., II,
1863:285.— Lepiota rhacodes var. puellaris (Fr.) Sacc.. SylL Fung., V,
1887 : 29.— L. puellaris (Fr.) Rea, Brit. Bas., 1922 : 66.— Leucocoprinus
puellaris (Fr.) Locq., Bull. mens. Soc. linn. Lyon, 14, 1945: 86 (табл. XV,
4a—6).
Icon.: J. Lge, FI. Ag. Dan., 1935—1940, tab. 9, fig. B; Piiat, Klic
urc. nas. hub, 1951, obr. 607; Васильева, Агар. шл. гр. Приморск, кр.,
1973, рис. 41 Б; Зерова, Атл. гр. Укр., 1974, табл. НО. рис. 1; Вассер,
Фл. гр. Укр. (Agaricaceae), 1980, рис. 160, табл. XXIX.
В сосновых, смешанных лесах, на лугах; гум. сапр., VIII—X, нр.,
(1—2), м. с. г., эвриголаркт., евраз.
Распр. в СССР. Европ. ч.: ЭССР, УССР; А з и а т. ч.: Кавказ —
АрмССР, Дальи. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Дания, Финляндия,
Франция, ФРГ, ГДР, ПНР, ЧССР.
152. Macrolepiota konradii (Huijsm. ex P. D. Orton) Mos.. KI. Krypt.-
flora, В. II, Teil II, 1967:185.— Lepiota excoriata (Fr.) Kumm. var.
konradii Huijsm., Med. Ned. Мус. Ver., XXVIII, 1943 : 18—19 (nom. nu-
dum).— Leucocoprinus maublancii Locq., Bull. mens. Soc. linn. Lyon,
14, 1945:87.— Lepiota konradii Huijsm. ex P. D Orton, Trans. Brit.
Мус. Soc., 43, 2, 1960:283.— Macrolepiota konradii (Huijsm. ex P. D.
Orton) L. Vass., Агар. шл. гр. Приморск, кр., 1973:170, syn. nov.—
Lepiota mastoidea (Fr.) Kumm. s. Bres., non s. Fr., Konr. et Maubl.,
Kuhn, et Romagn.— L. gracilenta (Krombh.) Quel., s. Cke, non s. al.
(табл. XV, 5a—6).
Icon.: Konr. et Maubl., Icon. Sci. Fung., 1924—1937, tab. 10; Ва-
сильева, Агар. шл. гр. Приморск, кр., 1973, рис. 41 А; Вассер, Укр.
ботан. журн., 32, 3, 1975, рис. 3; Вассер, Фл. гр. Укр. (Agaricaceae),
1980, рис. 161.
В широколиственных и смешанных лесах, парках; гум. сапр., VI—
X, р., (1), с., эвриголаркт., евраз.
Распр. в СССР. Европ. ч.: РСФСР — Краснодарский кр., УССР;
Азиат, ч.: Кавказ — АзССР, АрмССР, Дальн. Восток — При-
морск. кр.
Общее распр. Европа: Великобритания, Нидерланды, Франция,
ЧССР, Австрия.
Прим. Macrolepiota konradii наиболее близка к М. gracilenta (Krombh.; Fr.)
S. Wasser s. al., non s. Konr. et Maubl. и M. mastoidea (Fr.) Sing. s. Konr. et Maubl.,
Kiihn. et Romagn., non s. Bres.
112
153. Macrolepiota excoriata** (Schaeff.: Fr.) S. Wasser var. exco-
riata, Укр. ботан. журн., 35, 5, 1978 : 516.— Agaricus excoriatus Schaeff.:
Fr., Syst. Мус., I, 1821 :21 — Lepiola excoriata (Schaeff.: Fr.) Kumm.,
Dcr Fuhrer in die Pilzkundc, 1871:39.— Leucocoprinus excoriatus
(Schaeff. : Fr.) Pat., Essai Taxon. Hymen., 1900:171.— Leucoagaricus
excoriatus (Schaeff. : Fr.) Sing., Pap. Mich. Acad. Sci., Arts a. Lett., 32,
1948 : 142 (табл. XV, 6a—б; XXXI, 56).
icon.: Baria, Champ. Alp. Mar., 1888—1892, tab. 10, fig. 5—8;
Gill., Champ. Fr., 1878—1890, pl. 29; Cke, Ill. Brit. Fung., 1881—1891,
tab. 23; Bres., Icon. Мус., 1927, tab. 22; J. Lge, Fl. Ag. Dan., 1935—
1940, tab. 8, fig. A; Pilat, Nase houby, II, 1959, tab. 146; Зерова, Атл.
гр. Укр., 1974, табл. ПО, рис. 2; Вассер, Фл. гр. Укр. (Agaricaceae),
1980, рис. 162—163.
В заповедных целинных степях, на лугах, пастбищах, полянах, на
лесных опушках, в парках, на улицах.
М. excoriata var. squarrosa** (R. Mre) S. Wasser, Укр. ботан.
журн., 35, 5, 1978:516.— Lepiota excoriata var. squarrosa R. Mre, in
R. Maire et R. G. Werner, Mem. Soc. Sci. Nat. Maroc, 1937 : 101.
В искусственных дубовых насаждениях, в полынно-злаковой степи.
Известна из УССР.
В заповедных целинных степях, на лугах, пастбищах, полянах, на
опушках лесов, на улицах, в парках; гум. сапр., VI—XI, о. ч., (2—3),
м. с. г.» мульти per., космопол.
Распр. в СССР. Европ. ч.: повсеместно; Азиат, ч.: Кавказ —
ГССР, АрмССР, АзССР, ДагАССР, Нахичеванская АССР. Зап. Сибирь —
Башкирская АССР, Алтайск. кр., Вост. Сибирь — Красноярск, кр.. Бу-
рятская АССР, Дальн. Восток — Хабаровск, кр., Приморск, кр., Ср
Азия — ТССР, УзССР, КиргССР.
Общее распр. Европа: повсеместно; Азия: КНР4, Индия, МНР,
Турция, Израиль, Иран, Япония, Шри Ланка; Сев. Америка: Ка-
нада, США, Мексика; Юж и. Америка: Чили; Австралия;
Африка: Алжир, Марокко, Кения, Заир, Нигерия, Тунис, ЮАР; Фи-
липпинские о-ва, о-в Куба.
154. Macrolepiota mastoidea (Fr.) Sing., Pap. Mich. Acad. Sci., Arts
a. Lett., 32, 1948 : 141, s. Konr. et Maubl., 1948, Kiihn. et Romagn., 1953,
non s. Bres., Konr. et Maubl., 1926.— Lepiota mastoidea (Fr.) Kumm.,
Der Fuhrer in die Pilzkunde, 1871 : 72.— L. umbonata (Sebum.) Schroet.
apud Cohn s. J. Lge.— Leucocoprinus mastoideus (Fr.) Sing., Rev. Mycol.,
4, 1939 : 67.— Lepiota gracilenta (Krombh.: Fr.) Quel., s. Ricken, Bres.
non s. al. (табл. XVI, la—д; XXXII, 57).
Icon.: Bres., Icon. Мус., 1927, tab. 23; J. Lge, FI. Ag. Dan., 1935—
1940, tab. 8, fig. С; Вассер, Фл. гр. Укр., 1980, рнс. 165.
В дубовых, буковых, сосновых лесах, на лесных полянах, опушках
лесов, на лугах, в парках; гум. сапр., VIII—XI, ч., (1—2), м. с. г.,
мультирег., космопол.
Распр. в СССР. Европ. ч.: РСФСР, ЭССР, ЛатвССР, ЛнтССР,
БССР, УССР; А з н а т. ч.: Кавказ — АрмССР, Вост. Сибирь — Якут-
ская АССР, Дальн. Восток — Приморск, кр.
Общее распр. Европа: Великобритания, Дания, Италия, Фран-
ция, ФРГ, ГДР, ПНР, ЧССР, СФРЮ; Сев. Америка: США, Мек-
сика; Юж и. Америка: Бразилия; Австралия; Африка: Ал-
жир, Марокко, Кения.
Ю
ИЗ
155. Macrolepiota gracilenta ** (Krombh. : Fr.) S. Wasser, Укр. бо-
тан. журн., 35, 5, 1978:516, s. al„ non s. Ricken, Bres.— Lepiota graci-
lenta Krombh.: Fr., Hymen. Eur., 1874:30.— Macrolepiota gracilenta
(Krombh. : Fr.) Mos., KI. Krypt.-flora, В. II, Toil 11, 1967:185 (nom.
nudum) (табл. XVI, 2a—e).
Icon.: Baria, Champ. Apl. Mar., 1888—1892, pl. 11, Fig. 1—4;
Cke, III. Brit. Fung., 1881—1891, tab. 28; Bres., Icon. Мус., 1927, tab. 21;
Вассер, Фл. гр. Укр. (Agaricaceae), 1980, рис. 166.
В еловых, сосновых, широколиственных, смешанных лесах, на лес-
ных опушках, лугах; гум. canp., VIII—X, и. р., (1—2), м. с. г., муль-
ти per., космопол.
Распр. в СССР. Европ. ч.: РСФСР — Московская обл., ЛитССР,
УССР; А з и а т. ч.: Кавказ — АрмССР, Зап. Сибирь — Алтайск. кр.
Общее распр. Европа: Великобритания, Франция, Дания, Швей-
цария, Италия, ФРГ, ГДР, ЧССР; Азия: КНР; Сев. Америка:
США, Мексика; Южн. Америка:’ Чили; Австралия; А ф р и -
к а: Заир.
156. Macrolepiota prominens (Fr.) Mos., in Garns, KI. Krypt.-flora
Mittelcur., 2 (ed. 1), 1953 : 114, et in Garns, KI. Krypt.-flora, В. II, Teil II,
ed. 3, 1967:184.— Agaricus prominens Fr., Hymen. Eur., 1874:30.—
Lepiota prominens (Fr.) Sacc., Syll. Fung., V, 1887 : 30.— Leucocoprinus
prominens (Fr.) Locq., Bull. mens. Soc. linn. Lyon, 14, 1945:92
(табл. XVI, 4a—6).
Icon.: Baria, Champ. Alp. Mar., 1889, pl. 11, fig. 5—8.
В широколиственных и хвойных лесах, на лугах; гум. canp., V—X,
о. р., (1), м. с. г., в. н. а.
Распр. в СССР. Европ. ч_: РСФСР — Белгородская обл.
Общее распр. Европа: Франция, Италия, ЧССР, Австрия; Азия:
КНР; Африка: Заир.
157. Macrolepiota permixta** (Baria) S. Wasser comb. nov. — Le-
piota permixta Baria, BSMF, 3, 1886 : 114.— Macrolepiota permixta (Bar-
ia) Mos., KI. Krypt.-flora, В. II, Teil II, 1967: 184 (nom. nudum).
В широколиственных и смешанных лесах, на лесных опушках,
в парках; гум. canp, VII—X, д. р.о (1—2), с., эвриголаркт., евр.
Распр. в СССР. Европ. ч.: РСФСР—Краснодарск. кр., ЭССР.
Общее распр. Европа: Австрия, Италия, Франция. ЧССР.
158. Macrolepiota nympharuni ** (Kalchbr.) S. Wasser comb. nov.—
Agaricus (Lepiota) nympharum Kalchbr., Icon. Sei. Hym. Hung., 1873 :
10.— Lepiota nympharum (Kalchbr.) Karst., Hattsvampar, 1879 : 239.—
L. subprocera Saut., Hedwigia, 1876 : 152.
Icon.: Baria, Champ. Alp. Mar., 1889, pl. 1610~15; Bres., Icon. Мус.,
1927, pl. 24.
В смешанных лесах, парках; гум. canp., VI—XI, д. р., (1—2), с.,
эвриголаркт., евраз.
Распр. в СССР. Азиат, ч.: Вост. Сибирь — Красноярск, кр., Якут-
ская АССР.
Общее распр. Европа: Великобритания, Франция, Италия, ВНР.
С территории Советского Союза (Красноярск, кр., Туруханский р-н,
левый берег Енисея близ пос. Мирное, березово-кедровый лес, иа рас-
тительных остатках среди мхов, 6.08. 1979) был описан новый для нау-
114
ки вид агарикового гриба — Cystolepiota pusilia Nezd. (Нездойминого,.
Нов. сист. низш. раст., 1981, 18: 103). По данным автора Э. Л. Нездой-
миного (1981), новый вид отличается от остальных видов секции Floc-
cosac в смысле X. Кнудсена (Knudsen, 1978) мелкими размерами и
цветом карпофоров, а также размерами спор. Придерживаясь иных
позиций относительно систематического положения секций Floccosae.
Amyloidea (Вассер, 1978а, 1980, 1982), относимых нами к подроду
Sphaerocystis S. Wasser, считаем целесообразным перевести новый вид
в род Lepiota — L. pusilia (Nezd.) S. Wasser comb. nov. (Basionymum :
Cystolepiota pusilia Nezd., Нов. сист. низш. раст., 1981, 18: 103).
В 1983 г. нами был определен материал по грибам рода Lepiota,
присланный А. Н. Петровым, собранный в окр станции Выдрино (юж-
ное побережье оз. Байкал). В этом материале оказались две новые для
микофлоры СССР разновидности Lepiota cristata — L cristata var. fe-
linoides Bon и L. cristata var. pallidior Boud. ex Bon (Docums. Мус.,.
1981, 11, 43 : 34), описанные в 1981 г. с территории Франции (Воп,
1981).
В 1984 г. нами совместно с А. А. Петровой был определен материал
по роду Agaricus, собранный ею в 1982—1983 гг. в Зааминском горио-
лесиом заповеднике УзССР па высоте 1800—3500 м н. у. м. в средне-
горном и субальпийском поясах. В этом материале оказался новый для
СССР вид Agaricus tenuivolvatus (Moell.) Moell. (табл. XX, 20) п ряд
новых видов для азиатской части СССР — A. spissicaulis, A. cupreobrun
neus, A. porphyrocephalus, A. maskae, A. lencotrichus, A. lutosus.
В последнее время (Самгина, Бот. матер, герб. Инет. бот. АН
КазССР, 13, 1983:62—83) был описан сомнительный новый для науки
вид рода Macrolepiota — М. campestris (Lebed.) Sanig., близкий к
М. puellaris. «Новый внд отличается от М. puellaris тем, что не изме-
няет цвет мякоти при прикосновении и более выраженными чешуйками»
(Самгина, 1983, с. 83). У М. puellaris цвет мякоти при прикосновении
также не изменяется, при автооксидации слегка краснеет в основании
ножки, а «выраженность» чешуйчатости зависит от зколого-биоло! иче-
ских условий произрастания вида. Следует отметить и то, что Л. А. Ле-
бедева (1949), которая описала данный вид как nom. nudum, указы-
вает для него «кольцо неподвижное, сросшееся с ножкой» (с. 47).
М. campestris, на наш взгляд, сомнительный вид, требующий в даль-
нейшем тщательного изучения типового гербарного материала.
В серии «Fungi Europaei» в 1984 г. вышла в свет монография
А. Каппелли, посвященная роду Agaricus (Cappelli, 1984, Saronno: Lib-
reria editrice Biella Giovanna, 560 p.). В книге приведена оригинальная
система рода, содержатся сведения о 70 видовых и внутривидовых так-
сонах рода Agaricus Европы. Каждый таксон иллюстрирован ориги-
нальным цветным рисунком.
10*
ГЛАВА V
СИСТЕМАТИЧЕСКИЙ
И СТРУКТУРНО-СРАВНИТЕЛЬНЫЙ АНАЛИЗ
ФЛОРЫ АГАРИКОВЫХ ГРИБОВ СССР
1. СИСТЕМАТИЧЕСКИЙ АНАЛИЗ
Видовой состав агариковых грибов СССР установлен иа основа-
нии результатов собственных сборов и наблюдений, произведенных в
различных районах Советского Союза, критического изучения материа-
ла 13 отечественных и 11 зарубежных гербарнев. При критико-систе-
матическом исследовании трудных сомнительных и малоизвестных ви-
дов агариковых грибов были изучены их типовые материалы.
В Советском Союзе выявлено 158 видов, 29 разновидностей н
7 форм агариковых грибов, относящихся к 14 родам четырех колеи
(табл. 2—3).
Из иих 9 видов н внутривидовых таксонов описаны нами как но-
вые для науки (Вассер, 1974а, г, 1975а, 1976а, 1977а, б, 1978а, 1980),
для 23 видов и внутривидовых таксонов предложены новые для науки
таксономические комбинации (Вассер, 1976г, 1977а, 1978а, б, 1980, 1981;
Wasser, 1979); 42 вида н внутривидовых таксона, а также один род
(Chlorophyllum) приведены нами (Вассер, 1970, 1971, 1972а, б, 1973а»
б, в, 1974а, б, в, 1975а, б, в, 1976в, г, 1977а, б, 1978в, 1980, 1982; Вассер,
Батирова, 1979) как новые для микофлоры СССР (табл. 3).
Рассмотрение систематических единиц грибов в ранге колена по-
казывает бесспорное преобладание Agariceae (63 вида, 39, 86 %) 1 и
Lepioteae (57 видов, 36,14 %) над Leucocoprineae (26 видов, 16,44 %)
и Cystodermateae (12 видов, 7,56 %).
Общее количество родов во флоре агариковых грнбов Советского
Союза—14. Наиболее крупные из иих Agaricus (59 видов, 37,38 %),
Lepiota (54 вида, 34,23 %), Macrolepiota (II видов, 6,98 %), Cystoder-
ma (10 видов, 6,30%), Leucocoprinus (9 видов, 5,68 %). Остальные
роды содержат менее девяти видов, в том числе один род представлен
пятью видами, два рода—двумя видами, шесть родов — только одним
видом (из них два монотипных).
Новые для науки виды и внутривидовые таксоны агариковых гри-
бов описаны нами главным образом в результате тщательного изуче-
ния типового и большого гербарного материала, родственных новым
таксонам видов, а также на основании устранения номенклатурных оши-
бок различных авторов, пренебрегавших последовательным применением
правил Международного кодекса ботанической номенклатуры.
Leucocoprinus bohusi и Leucoagaricus moseri описаны с территории
степной зоны Украины (Вассер, 1975а, 1978а, 1980; Wasser, 1979).
1 Здесь и далее процент в скобках после количества видов высчитан от числа
видов всей флоры.
116
L. bohusi, собранный в разнотравпо-типчаково-ковыльной зайоведной
Хомутовской степи, наиболее близок к L. badhamii, произрастающему
в парках, садах, в насаждениях широколиственных деревьев, изредка
хвойных. Отличается внешним видом карпофоров, цветом шляпки и
мякоти, размером и формой спор, химическими цветовыми реакциями
(Locquin, 1951а, b; Pilat, 1951b; Melzer, 1959; Babos, 1961; Singer, 1962,
Таблица 2. Систематический состав флоры семейства Agaricaceae Советского Союза
Колено, род Количество Виды и внутривидовые таксоны, новые для Новые для на- уки таксоно- мнческне ком- бинации
видов разновид- ностей форм
СССР науки
Agariceae Melanophyllum 2 2
Agaricus 59 18 2 27 4 6
Gyrophragmium I — — — — —
Endoptychum 1 — — — — —
Cystodermateae Phaeolepiota I — —
Cystoderma 10 2 1 2 I 1
Squamanita 1 — — — — ——
Lepioteae Pseudobaeospora 2 1
Lepiota 54 3 2 5 2 —
Chamaemyces L — — — — —
Leucocoprineae Chlorophyllum 1 — I —
Leucocoprinus 9 2 — 3 1 5
Leucoagaricus 5 -— 2 2 1 4
Macrolepiota 11 2 — 2 — 6
Всего 158 29 7 42 9 23
1975; Dernoulin, 1966; Moser, 1967, 1978; 1983; Васильева, 1973; Вассер,
1975а, 1980; Wasser, 1979). L. moseri, произрастающий в искусствен-
ных белоакациевых, белоакациево-кленовых насаждениях на песчаной
почве, парках, дубовых полезащитных лесополосах, наиболее близок
к широко распространенному виду — L. leucothitus, известному с паст-
бищ, садов, огородов, парков, заповедных целинных степей, лесных по-
лян, изредка лесов. L. moseri отличается от L. leucothitus размером
карпофоров, спор, формой и характером расположения кольца на нож-
ке, отсутствием хейлоцистид, наличием в основании ножки клубневид-
ного утолщения, экологией (Ricken, 1915; Lange J.s 1935—1940; Лебе-
дева, 1949; Dennis, Orton, Нога, 1960; Moser, 1967, 1978; Васильева,
1973; Вассер, 1975а, 1980). Первоначально L. moseri был описан как
Lepiota moseri (Вассер, 1975а). Проведенные позже дополнительные
исследования с применением сканирующего электронного микроскопа,
химических цветовых реакций, окончательно установившие наличие по-
ры прорастания, метахроматии спор, дали основания перенести рас-
сматриваемый вид в род Leucoagaricus (Вассер, 1978а; Moser, 1978).
До последнего времени L moseri считался украинским эидемом.
В 1980 г. описанный нами вид был обнаружен Л. Г. Буровой и Э. Л. Не-
здонмнного (1980) в полупустынных растительных ассоциациях Джа-
ныбекского полупустынного стационара (Северный Прнкаспин), что
ПТ
Таб.пша 3 Таксоны Agaricaceae видового и внутривидового рангов новые для науки,
СССР и новые для науки таксономические комбинации
Вид или внутривидовой таксон Новый для науки Новая для науки таксономическая комбинация Новый для мико- флоры СССР
Agaricus romagnesli 4 — 4
A. amanitaeformis + — —
A. longicaudus 4 — —
A. moelleri + -— —
A. langei var. mediofuscus 4 —
A. gennadii var. microsporus — + 4
A. semotus var. minimus —— 4
A. pseudopratensis — + 4
A. aestivalis var. veneris — 4 4
A. haemorrhoidarius 1 fagetorum — +
A. bresadolianus — — +
A. spissicaulis — •— 4
A. cupreobrunneus — —• 4
A. porphyrocephalus — —
A. benesii — — 4
Л. deyln — —
Л. bitorquis var. validus — — 4-
A. maleelens — — 4
A. kuehnerianus — — 4
A. Ttramineus —— — 4
A. maskae — — 4
A. subf]' ccosus — — 4
A. lissuratu? — — 4
A. nivescens — — 4
A. leucot richus — — 4
A. cbionodermus —- — 4
A. osecanus — —- 4
A. xanthodermus var. lepiotoides — — 4
A. phaeolepidotus — — 4
A brunneolus — 4
A. lutosus r — — 4
A. xantholepis — — 4
Lepiota subgracllls 4- —
L. clypeolaria var. latispora —
L. bucknallii — -U
L pallida — — 4
L. wiclianskyi — 4
L echinella — —_ 4
L cygneoaflinls —
Pseudobaeospora pillodil —
Cystoderma rugosoreticulata — 4 -t-
C. amianthina var. longispora
Chlorophyllum molybdites — — 4
Leucocoprinus bohusl 4 —
L. badhamii +
L. pilatianus var. pilatianus + 4
L. pilatianus var. subrubens — 4 4
L. bresadolae — 4 -U
L. grorginae — 4-
Leucoagaricus moseri 4 4- .
L. carneifolius
L leucothitus f. leucothitus 4
L leucothitus f. clnereolilaclna 4
Macrolepiota permlxta — 4 —
M. nympharum 4 —
M. gracilenta — 4 —.
M. Olivieri 4 4
M. excoriata var. excoriata 4
M. excoriata var. squarrosa — 4 _р.
Всего 9 23 42
i 18
значительно расширило знания о его эколого-биологических и геогра-
фических особенностях.
Agaricus amanitacformis описан из Донецкого ботанического сада
АН УССР (Вассер, 1974а, б, г). Грнб был собран в смешанных насаж-
дениях широколиственных деревьев (Fraxinus lanceolata Borkh., Acer
platanoides L.) на почве. Основание ножки A. amanitaeformis в зна-
чительной мере напоминает основание ножки Amanita muscaria (Fr.)
Hook, (два-три концентрических круга гранул, что послужило поводом
для названия amanitaeformis). A. amanitaeformis наиболее близок к
A. fissuratus (Moeller, 1950—1951; Essette, 1964; Moser, 1967, 1978),
от которого отличается цветом карпофоров, размером и формой спор,
отсутствием хейлоцистнд, формой ножки, расположением гранул, цветом
и запахом мякоти, рядом химических цветовых реакций (Вассер, 1980).
При описании A. amanitaeformis были изучены гербарные образцы
A. fissuratus из Herb. Mus. Bot. Haun., Kobenhavn (C) : Loc. Graenge,
1.VII1 1946, leg. et det. Moeller F. H., общий номер гербарного листа
L.29/72— N28—29, собранные и определенные автором вида. Для этих
образцов наиболее характерны следующие размеры микроструктур:
споры — 7X5; 8x4,7; 8,5X6; 8x5,5 мкм, хейлоцистиды — 24x15; 24Х
Х13; 28X13 мкм, базидии всегда четырехспоровые (22—30x8 мкм).
Микроструктуры описанного нами вида — A. amanitaeformis — имеют
такие параметры: споры 5,4—6,9X3.6—4,5 мкм, базндни 22—34X
Хб—8 мкм.
Agaricus romagnesii описан нами (Вассер, 1976а, 1977а) как новый
для науки вид в результате критико-систематического изучения типо-
вого материала и последовательного применения Международного ко-
декса ботанической номенклатуры. A. romagnesii ( = А. radicatus (Vitt.)
Romagn.) в литературе упоминается редко. Даже моиографы рода
Agaricus (Moeller, 1950—1951; Pilat, 1951а, b; Heinemann, 1956a, b)
этот вид не приводят. Исследования Д. А. Рида (Reid, 1972) показали,
что название A. radicatus (Romagnesi, 1938) является незаконным, про-
тиворечащим правилу приоритета Международного кодекса ботаниче-
ской номенклатуры (статья 11), так как в роде Agaricus уже имелся
A. radicatus Fr. Указанное обстоятельство дает оенфвание нам счи-
тать A. radicatus (Vitt.) Romagn. незаконным названием н дать ему
новое название A. romagnesii. A. romagnesii очень близок к A. bresado-
lianus (Bohus, 1969) — новому для СССР виду, приведенному нами из
бслоакациевых насаждений УССР (Вассер, 1970, 1973а, 19746, 1С80).
Отличается от него большим размером спор, наличием хейлоцистнд,
формой ножки, экологией. Заслуживает внимания н тот факт, что
A. romagnesii ядовит (Romagnesi, 1938: Kiihner, Romagnesi, 1953), а
Л. bresadolianus съедобен (Essette, 1964; Bon, 1970; Вассер, 1970, 1980,
Bohus, 1971). A. romagnesii произрастает в парках, садах, по краю
дорог, в то время как A. bresadolianus — лишь в белоакацневых на-
саждениях.
В литературе пет единой точки зрения на объем комплекса A. ro-
magnesii— A. bresadolianus. Одни авторы (Bohus, 1969, 1971; Вассер,
1970, 19766, 1980), признавая родство A. romagnesii и A. bresadolianus
( — A. campestris var. radicata Vitt. s. Bres.), считают нх близкими са-
мостоятельными видами. Другие (Reid, 1972) полагают, что A. romag-
nesii— синоним A. bresadolianus, понимая последний вид широко. FIpi
веденное нами (Вассер, 1980) сравнительное изучение типового и боль-
шого гербарного материала A. romagnesii [размер спор у венгерских
119
образцов составлял 0,6—7,2х4,3—5,1 мкм, французских из гербария
Г. Ромаиьези — 6,7—7,3x4,5—4,9, украинских образцов — 6,2—8x3,5—
5 мкм; хейлоцистиды (30—40X8—15 мкм) всегда присутствовали] и
A. bresadolianus [размер спор у типа 5,5—7,1X3,7—4,5 мкм, венгер-
ских образцов — 6,7X4—4,8 мкм, украинских образцов — 6—6,7(7) X
Х3,9—4,1 (4,8) мкм; хейлоцистиды отсутствуют] дает основание счи-
тать их близкими, самостоятельными видами.
Agaricus moellcri описан нами (Вассер, 1976а) как новый для
науки вид по образцам, хранящимся в Herb. Mus. Bot. Haun., Kobcn-
havn (С) под названием Psalliota meleagris J. Schaeff. var. terricolor
Moell. (Дания’, Лоплаид, Сакскенбинг, фруктовый сад, под деревьями,
собр. Л. Хансен, опр. Ф. Меллер). Позже A. moelleri был найден в
дубовых лесах Закарпатья и насаждениях Juniperus virginiana L. в
Южном Крыму (Вассер, 1980). A. moelleri очень близок к A. meleagris
и A. placomyces, от которых отличается размером спор и хейлоцистид,
цветом и структурой поверхности шляпки, габитусом карпофоров. Среди
агарикологов нет единого мнения относительно объема комплекса A. me-
leagris — A. placomyces — A. moelleri. Например, М. Мозер (Moser,
1967, 1978) не признает A. meleagris, считая его разновидностью A. pla-
comyces. Р. У. Г. Депниз, П. Д. Ортой, Ф. Б. Гора (Dennis, Orton, Нога,
I960, 1974) вообще считают A. meleagris синонимом A. placomyces.
Иной точки зрения придерживаются монографы рода Agaricus (Mocl-
1сг, 1950—1951; Pilat, 1951b; Essette, 1964; Вассер, 1980). Опн признали
оба вида и описали ряд их разновидностей (A. meleagris var. nigricans,
A. meleagris var. terricolor, A. meleagris var. obscuratus). Критико-си-
стематическое изучение типа, гербарных материалов по A. moelleri —
A. meleagris — A. placomyces нз Данни (Herb. Mus. Bot. Haun., Koben-
havn (С)) н ЧССР (Herb. Krypt. Mus. Nat. Prag. (PR)), анализ гер-
барного материала различных регионов областей флоры СССР
(ЛитССР, УССР, ГССР, Приморский край) дают основание считать
их хорошими самостоятельными видами, причем A. moelleri ( = А. me-
leagris var. terricolor) — молодым развивающимся видом, берущим на-
чало от A. meleagris var. meleagris.
Agaricus longicaudus описан нами (Вассер, 19776; Wasser, 1979)
из грабово-дубово-ильмовых лесов Западного Полесья УССР. A. longi-
caudus наиболее близок к A. aestivalis, от которого отличается габиту-
сом карпофоров, цветом и характером поверхности шляпки и ножки,
макрохнмическнми реакциями, экологией (Moel’cr, 1950—1951; Heim,
Весксг, 1960; Essette, 1964; Вассер, 1980). При описании нового вида
для сравнения были изучены гербарные образцы A. aestivalis из Herb.
Mus. Bot. Haun., Kobenliavn (C) : 1. Loc. Nyborg, Bollevaengc, 12. VI
1946, leg. et det. Moeller F. H.; 2 Loc. Korselitze hovedskov, 1946, leg.
et det. Moeller F. H. (общий иомер гербарного листа 45/74 — № 1—4),
собранные и определенные автором вида. A. aestivalis свойственна
шляпка диаметром 5—15 см, беловатая, к краю бледно-серая, корич-
нево-фиолетовая, четко прижато-волокниста я, ножка 4—9X1,5—3 см,
беловатая, вверху с розоватым оттенком, голая, споры 6—9x4—5,5
(6) мкм, в то время как A. longicaudus — шляпка диаметром 3—10 см,
белая, покрытая крупными желтовато-оливковыми волокнисто-шелко-
вистыми прижатыми чешуйками, ножка 8—18x2—4 см, белая, ниже
кольца покрыта желтовато-оливковыми хлопьями, споры 7,6Х5,4 (из-
редка 8,3X4,3 мкм).
Lepiota subgracilis описана нами (Вассер, 1978а) по материалам
120
Р. Кюнера (Kiihner, 1936) как вид, близкий к L. clypeolaria, от кото-
рого отличается размером (2—4 см в диаметре), цветом (темно-корич-
невый), характером поверхности шляпки (темно-коричневая чешуйча-
тость, образующаяся от разрывов покрова, между которыми видны бе-
ловатые просветы мякоти), размером спор (9—12,0X4,5—6,0 мкм)г
экологией (растет отдельными экземплярами или группами на под-
стилке в хвойных, широколиственных смешанных лесах, в зарослях
Salix rosmarinifolia L.).
Cystoderma amianthina var. longispora описана нами (Вассер,
1978a) главным образом на основании отличий в размере и форме
спор (7—9(10) Х4—4,7 мкм) от типовой разновидности (5,0—6,5x2,8—
3,5 мкм). Var. longispora была описана Р. Кюнером (Kiihner, 1936>
без латинского диагноза (nom. nudum) как Lepiota amianthina var.
longispora. Все последующие комбинации (Smith A., Singer, 1945; Hei-
nemann, Thoen, 1973) являются незаконными. Поэтому мы сочли необ-
ходимым, признавая эту разновидность, узаконить ее, согласно Между-
народному кодексу ботанической номенклатуры (Вассер, 1978а).
В результате изучения гербарного типового и массового материала,
собранного в природе в различных районах флоры СССР, уточнены и
выявлены новые признаки многих видов агариковых грибов. Это позво-
лило внести коррективы в систематическое положение 23 видовых и
внутривидовых таксонов агариковых грибов, для которых предложены
новые для науки таксономические комбинации, а также привести 42 ви-
довых и внутривидовых таксона в качестве новых для микофлоры
СССР. Проведены различия между такими близкими критическими
видами, как Agaricus cupreobrunneus и A. porphyrocephalus (данные
статистической обработки размеров спор, форма ножки) l, A. romagnesii
и A. bresadolianus (экология, съедобность, наличие хейлоцистнд, размер
спор), A. tabularis и A. rnaskae (размер спор, хейлоцистнд, характер
поверхности шляпки, форма ножки), A. meleagris и A. moelleri (струк-
тура поверхности шляпки, размер спор и хейлоцистнд), A. deylii и-
A. benesii (экология, размер иожкн, размер спор), Cystoderma super-
bum н С. haematites s. orig. (размер н амилоидпость спор), С. amian-
thina и С. rugosoreticulata (характер поверхности шляпки), Lepiota
rufipes и L. neophana (размер базидий, частота встречаемости хейло-
цистид, цвет пластинок прн подсыхании), Leucocoprinus badhamii и
L. pilatianus (данные статистической обработки размеров спор, форма,
характер подвеска, окраска хейлоцнетит), L. bresadolae п L. badhamii
s. 1. (данные статистической обработки размеров спор, габитус карпо-
форов), Leucoagaricus carncifolius и L. leucothitus (цвет шляпки при
прикосновении, размер хейлоцистнд), Macrolepiota rhacodes и М oli-
vieri (экология, цвет мякоти при автоокендацни), М. gradients s. al.
non s. Ricken, Bres. и M. mastoidea s. Konr. ct Maubl., Kiihn. et Ro-
magn. non s. Bres. (размер спор).
Проведенные исследования позволили уточнить различия между
разновидностями A. xanthodermus var. xanthodermus и A. xanthoder-
mus var. lepiotoides (характер поверхности шляпки, структура кольца
па ножке), A. langei var. langei и A. langei var. niediofuscus (характер
поверхности шляпки, размер спор и хейлоцнетид), A. semotus var.
1 Критико-систематические примечания к некоторым новым, редким, малоизве-
стным видовым и внутривидовым таксонам приведены в «Конспекте флоры агарико-
вых грибов СССР».
121
semotus и A. semotus var. minimus (характер поверхности шляпкн, раз-
мер карпофоров и спор), Leucocoprinus pilatianus var. pilatianus н
L. pilatianus var. subrubens (цвет шляпки, особенно при подсыхании).
Подтверждена видовая самостоятельность Agaricus porphyroccphalus,
A. romagnesii, A. tabularis, A. meleagris, A. brunneolus, Cystoderma ru-
gosoreticulata, Leucocoprinus bresadolae, Macrolepiota olivieri. Впервые
установлена идентичность Macrolepiota rhacodes var. hortensis cM. oli-
vieri, A. campestris f. substerilis, A. campestris var. squainulosus c
A. campestris, A. bernardii s. Vassilk. c A. gennadii, Psalliota grisea
s. Essette c A. velenovskyi, Leucocoprinus jubilaei c L. pilatianus.
2. СТРУКТУРНО-СРАВНИТЕЛЬНЫЙ АНАЛИЗ
Конкретизируя положение А. Декандоля о роли количественно пре-
обладающих семейств растений, А. И. Толмачев доказывает, что само-
бытность флор проявляется не только в местных чисто качественных
особенностях, но и в количественных признаках. Флора отличается би-
товым богатством, а также систематической структурой. По мнению
А. И. Толмачева (1970а,б, 1974), систематическая структура флоры при
сравнительном рассмотрении в широкой географической перспективе
приобретает значение одного нз существенных показателей, характери-
зующих флоры в региональном плане. Предлагается проводить сравне-
ние 10 ведущих семейств (родов) в составе флор, принадлежащих к
определенной флористической области, по их процентному соотноше-
нию, чтобы в какой-то степени пренебречь пространственной неравно-
мерностью территорий. Особого внимания заслуживают первые пять
таксонов, ибо, как отмечает А. И. Толмачев (1974), «лицо» системати-
ческой структуры флоры отчетливо выделяется при анализе порядково-
го расположения всего 10 ведущих семейств (родов), представляющих
«головную часть флористического спектра».
В самом теле, А. И. Толмачевым (1970а,б, 1974), Л. И. Малыше-
вым (1973), Ю. Р. Шелягом-Сосонко, Я. П. Дидухом (1980),
В. М. Шмидтом (1980) иа примере сосудистых растений, а нами (Вас-
сер, Солдатова, 1977а) на примере высших Basidiomycetes показано,
что 10 ведущих семейств (родов) включают в себя более половины ви-
дового состава каждой флоры (в высоких широтах — значительно боль-
ше, в тропиках — иногда чуть меньше). Остальные семейства (роды),
несмотря на то что сам факт их присутствия во флоре может иметь
важное значение при решении некоторых вопросов флорогенеза, играют
ничтожную роль в сложении систематической структуры и не опреде-
ляют «лицо» флоры в этом отношении. Практически все наиболее ха-
рактерные черты систематической структуры флоры сосредоточены в
наборе 10 ведущих семейств (родов), так как мы имеем дело не со
случайным набором таксонов, а единственно возможной в данных усло-
виях последовательностью их расположения. Положение каждого из
пнх в данный момент времени можно считать строго фиксированной ве-
личиной, не варьирующей в статистическом плане. В связи с этим, как
указывает В. М. Шмидт (1980), прав Л. И. Малышев (1973), впервые
отметивший, что систематическую структуру флоры следует рассматри-
вать как генеральную совокупность.
По мнению ряда авторов (Толмачев, 1970а,б, Заки, Шмндт, 1972;
Ребристая, Шмндт, 1972; Днух, 1977, 1978; Шмидт, 1980), на система-
тической структуре флоры в меньшей степени, чем на других показа-
122
телях, сказывается неполнота инвентаризации, разница во флористиче-
ском богатстве и площадях. А. И. Толмачев даже ратует за то, чтобы
в связи с некоторой консервативностью структуры флор и относительно
высокой степенью независимости от других показателей и факторов
выдвигать ее на положение «одно из существенных диагностических нрнзна- * Количество .„до. в дес.ти J г * ведущих родах агариковых грибов ков», определяющих принадлежность флоры СССР
области. Мы полностью согласны с приве- род денными выше данными, проверен» ы- Количест- во видов Числен- ность ВИ- ДОВ (%)
mycetes степной зоны Украины Agaricus (Вассер, Солдатова, 1977а), однако для Lepiota небольших семейств (содержащих Macrolepiota 15—20 родов), каковым является и Cystoderma предмет нашего исследования, считаем Leucwarictw возможным для определения «лица» Melanophyllum флоры использовать пять ведущих Pseudobaeospora родов. Gyro, hragmium Действительно, в десяти ведущих Squamanila родах агариковых грибов СССР содер- Всего жится 154 вида, что составляет97,6 %. 59 54 11 10 9 5 2 2 1 1 154 37.38 34,24 6,98 6.30 5.6S 3.16 1.26 1.26 0.63 0 63 97.52
а в пяти ведущих родах — 143 вида,
или 90,5 % (табл. 4—5). Анализируя в таком плане ведущие пять родов
агариковых грибов по областям флоры СССР, получаем аналогичные
результаты (табл. 5).
Показательными нам представляются соотношения между числен-
ностью видов и родов, свойственных различным флорам. А. И. Толмачев
Таблица 5. Количество видов в пяти ведущих родах агариковых гр и Зов по различным
областям флоры СССР
Род Европей- ская часть СССР Кавказ Западная Сибирь Восточ- ная Си- бирь Дальний Восток Средняя Азия
Ararlcus 57 18 13 13 14 15
Lepiota 40 20 12 19 36 7
Macrolepiota 10 7 4 5 5 2
Cystoderma 9 3 4 7 4 2
Leucocoprinus 7 3 1 1 4 —
Всего в пяти ведущих родах Общее количество видов в 123 51 34 45 63 26
семействе 135 55 41 46 70 28
(1974) весьма прозорливо отмечал, что «за ними скрываются какие-то
закономерности, отражается близость показателей, характеризующих
состав флоры отдельных частей определенных флористических обла-
стей». Проверка данного положения проведена нами (Вассер, Солда-
това, 1977а) при анализе Agaricales различных регионов СССР. Ниже
приводим цифры, отражающие численность видов, приходящихся в
среднем на один род: степная зона Украины — 4,8:1, Армянская
ССР — 4:1, Грузинская ССР — 6,0:1, Ленкоранская зона Азербай-
12*
джаиской ССР — 3,4 : 1, Латвийская ССР — 7,5 : 1, Приморский край —
8,0: 1.
При анализе соотношений между численностью видов, приходящих-
ся в среднем иа один род в семействе Agaricaceae, по областям флоры
СССР получены следующие соотношения: Арктика — 3,0:1, европей-
ская часть СССР — 13,3 : 1, Кавказ — 5,5 : 1, Запатная Сибирь — 3,1 : 1,
Восточная Сибирь — 7,6:1, Дальний Восток — 6,3 : 1, Средняя Азия —
4,0: 1. Сопоставление численности видов, приходящихся в среднем на
один род, а также численности родов и видов может способствовать
вскрытию соотношений, важных во флорогенетическом плане. В част-
ности, всякое обогащение состава флоры, осуществляющееся за счет
автохтонного образования, неизбежно должно сопровождаться увели-
чением численности видов в рамках уже представленных в составе
флоры родов. Напротив, пополнение состава флоры за счет иммиграции
с сопредельных пространств может осуществляться в форме как увели-
чения численности видов, уже имеющихся в данной флоре родов, так
и пополнения ее родового состава чаще всего единичными видами ранее
чуждых ей родов. Количество видов, приходящихся в среднем на один
род, может при этом снижаться (Толмачев, 1974).
Важным этапом анализа структур флор является изучение зако-
номерностей их изменения в зависимости' от ботанико-географических
особенностей территории, а также установление видового и родового
сходства. Установить эти закономерности можно на основе сопостав-
ления количественных показателей спектров, отражающих различные
структуры флор. Для этого флористика должна выйти за рамки опи-
сательной науки и шире применять методы сравнения. Развитию послед-
них способствует все более широкое использование математического
аппарата.
Как показали П. В. Тамарин и В. М. Шмидт (1975), В. М. Шмидт
(1980), ГО. Р. Шеляг-Сосоико, Я. П. Дидух (1980), для сравнительной
флористики наиболее целесообразно применять логически обоснован-
ные и математически корректные коэффициенты Жаккара и Стугрена —
Радулеску. Линейная зависимость между коэффициентами Жаккара и
Стугрена—Радулеску, как показали П. В. Тамарин и В. М. Шмидт
(1975), свидетельствует, что данные показатели адекватны по степени
выражения сходства и отличаются только по масштабам (первый изме-
няется от 0 до 1, а второй — от —1 до +1). Величины сходства, рассчи-
тываемые с помощью этих коэффициентов, имеют близкие значения.
Однако, как отмечает В. М. Шмидт (1980), коэффициенту Стугрена—
Радулеску следует отдавать некоторое предпочтение при сравнении
близких флор.
Коэффициент Стугрена—Радулеску, учитывающий как сходство,
так и различие (дискриминацию) флор, использовался нами для ана-
лиза видового сходства флор Agaricaceae по областям флоры СССР и
по отдельным наиболее полно изученным регионам СССР.
Коэффициент Стугрена—Радулеску (р) выглядит следующим об-
разом:
X + 4 — Z
Р х -J- у + z •
Здесь х — количество видов в первой флоре, отсутствующих во второй;
у — количество видов во второй флоре, отсутствующих в первой; z—
количество видов, общих для двух флор Данный коэффициент обла-
124
дает следующей особенностью: варьируя от — 1 до +1, ои в пределах
от —1 до 0 указывает на сходство, а в пределах от 0 до -Ы — на раз-
личие флор. Следовательно, если в частном случае сравнения несколь-
ких флор все полученные значения р < 0, то за меру сходства можно
брать его абсолютное значение (Шмндт, 1980).
Для анализа родового сходства Agaricaceae по областям флоры
СССР н отдельным регионам СССР использовали коэффициент ранго-
вой корреляции Кендэла.
Коэффициент Кендэла имеет вид
П(Л— 1) *
где s — сумма рангов;
п — число пар сравниваемых рангов (Кендэл, 1975; Шмндт, 1980).
Коэффициент Кендэла изменяется от -f l до —1. При т = +1 имеет
место полное сходство систематической структуры сравниваемых флор,
при т = —1 — полное несходство.
В результате установления видового состава агариковых грибов
Советского Союза, а также систематического и структурно-сравнитель-
ного анализов с использованием коэффициентов Стугрена—Радулеску
и Кендэла, выявлена видовая и родовая специфика грибов исследуемо-
го семейства по областям флоры (Арктика, европейская часть СССР,
Кавказ, Западная Сибирь, Восточная Сибирь, Дальний Восток, Средняя
Азия) н отдельным наиболее полно изученным регионам (Советская
Прибалтика, Кавказ, степная зона Украины, Средняя Азия, южная
часть Красноярского края, Приморский край). Проведенные анализы
графов видового сходства Agaricaceae областей флоры СССР н от-
дельных наиболее полно изученных регионов позволили прийти к сле-
дующим заключениям: а) в целом, учитывая значения р (р всегда > 0),
рассматриваемые флоры в большей степемн различны, чем сходны меж-
ду собой; б) наиболее обособлены флоры Арктики, Средней Азии и
Дальнего Востока; в) относительно высокий уровень связи характерен
для флор европейской части СССР и Кавказа, Западной и Восточной
Снбнри, степной зоны Украины и Средней Азии, Советской Прибалтики
и Кавказа; г) на уровне минимальной общности находятся флоры
Средней Азии и Приморского края, южной части Красноярского края
н Советской Прибалтики, с одной стороны, степной зоны Украины и
Приморского края — с другой. Интерес представляет факт наиболее
значительного видового сходства флор агариковых грибов степной зоны
Украины и Средней Азин, регионов наиболее массового распростране-
ния представителей этого немикоризного семейства, многие виды кото-
рого приурочены к степотопным экологическим условиям (хотя обе
флоры достаточно удалены друг от друга территориально). Близость
видового сходства- этих флор, возможно, указывает на общность их
исторического формирования. Близость флор Советской Прибалтики и
Кавказа объясняется общностью видов, экологически приуроченных к
лесным местообитаниям, субстратом для которых является лесная под-
стилка. Минимальная общность связи, которая характерна для флор
Средней Азии и Приморского края, южной части Красноярского края.
Советской Прибалтики, с одной стороны, степной зоны Украины н При-
морского края — с другой, объясняется обилием видов агариковых гри-
бов, приуроченных к открытым степным и полупустынным местооби-
таниям, широко представлены в Средней Азии и степной зоне Украины.
125
Agaricus
tepiola
Macrolepiota
Полученные нами данные при решении вопроса о допустимости или
невозможности сравнения флор в условиях территориальной их нерав-
ноценности склоняют в пользу первого мнения, поддерживаемого Р. Бе-
шелом (1969), Н. В. Козловской, В. И. Парфеновым (1971) и др. На
наш взгляд, на количество видов агариковых грибов, а возможно и
всех высших Basidiomycetes, наибольшее влияние оказывает не терри-
ториальный, а зональный фактор. Например, для таких разновеликих
территорий, как БССР (208 000 км2) и Украинские Карпаты (7000 км8),
характерны незначительные различия в количестве высших Basidiomy-
cetes (Вассер, Солдатова, 1977а). Напротив*, количество видов Agari-
caceae степной зоны Украины
превалирует над их количеством
в Красноярском крае, площадь
которого в сотни раз превосхо-
дит площадь этой зоны. Хотя в
последнем случае мы имеем дело
и со спецификой распространения
немикоризного семейства.
Полученные данные по ви-
довому сходству и различию да-
ют основания проанализировать
специфику видового состава Aga-
ricaceae областей флоры СССР.
Наибольшим количеством видов
Phaeolepiota
Chomoemgces
Endoptychum
Cyrophrugmtum
Metonophyllum
Isucoagancus
Leucocoprinus Cystoderma
и внутривидовых таксонов пред-
ставлена европейская часть
СССР—149, затем Дальний Во-
Рис. 6. Спектр родов агариковых грибов
европейской части СССР
сток — 72, Кавказ — 56, Восточная Сибирь - 47, Западная Сибирь —
42, Средняя Азия — 32, Арктика— 11. В настоящее время ие представ-
ляется возможным охарактеризовать флору агариковых грибов по ти-
пологическим единицам растительного покрова СССР и их региональ-
ным вариантам (Растительный покров СССР, 1956) из-за сложности
и разнообразия растительного покрова, неодинаковой степени изучен-
ности его микофлоры. Ниже приводим характеристику агариковых гри-
бов основных типов растительного покрова областей флоры СССР.
149 видовых и внутривидовых таксонов Agaricaceae европейской
части СССР относятся к 11 родам четырех колен. Наибольшую роль в
растительном покрове европейской части СССР играют представители
родов Agaricus (65 таксонов) и Lepiota (42 таксона), значительно
меньшую — представители родов Macrolepiota (11 таксонов), Cystoder-
ma (10), Leucocoprinus (8), Leucoagaricus (6), Melanophyllum (3),
Gyrophragmiurm Endoptychum, Chamaemyces и Phaeolepiota (содержат
по одному видовому таксону) — рис. 6.
По топическим и трофическим связям агариковые грибы европей-
ской части СССР относятся к пяти экологическим группам: гумусовые
сапротрофы (73,8%), подстилочные сапротрофы (22,1 %), бриотрофы
(2,1 %), лигнотрофы (1,3 %) и копротрофы (0,7 %).
Наибольшее видовое разнообразие агариковых грибов отмечено для
широколиственных лесов, их полян и опушек. Это связано, иа иаш
взгляд, с оптимальными для немикоризиых видов лесорастительиыми
условиями широколиственных лесов, что обусловлено значительным
плодородием и режимом влажности почв, а также разнообразием дре-
востоя. При формировании грибного покрова в лесах существенное
I2&
значение имеют состав древостоя и физические факторы. В зависимости
от этого многие подстилочные сапротрофы встречаются только в опре-
деленных группах типов местопроизрастания или же в некоторых обра-
зуют карпофоры в изобилии. Такие виды являются характерными для
данного типа леса (Каламээс, 1975). В чистых и смешанных широко-
лиственных лесах отмечено 80 видовых и внутривидовых таксонов ага-
риковых грибов. Наибольшее количество таксонов — 32, 30 и 17 отно-
сится к родам Agaricus, Lepiota и Macrolepiota; 16 таксонов — Agaricus
fuscofibrillosus, A. bresadolianus, A. silvaticus var. pallidus, A. haemor-
rhoidarius, A. bitorquis var. validus, A. longicaudus, A. osecanus, A. xan-
thodermus var. lepiotoides, A. phaeolepidotus, A. brunncolus, A. sagatus.
Lepiota seminuda f. minima, L. bucknallii, L. rosca, L. rufipes. Macro-
lepiota excoriata var. squarrosa — произрастают лишь в широколиствен-
ных лесах и не отмечены в иных типах растительного покрова европей-
ской части СССР. Эти таксоны являются характерными для широко-
лиственных лесов. При этом нами выявлены также таксоны агарико-
вых грибов в европейской части СССР, характерные лишь для дубовых
(Agaricus fuscofibrillosus, Lepiota seminuda f. minima, Macrolepiota
excoriata var. squarrossa), буковых (Agaricus haemorrhoidarius f. fage-
torum, A. brunneolus, Lepiota brunneoincarnata) лесов, белоакациевых
лесных насаждений (Agaricus bresadolianus, A. spissicaulis, A. maleo-
lens, A. pseudopratensis, Lepiota lilacea, L. pallida, L. rufipes, L. scobi-
nella), ольшаников (Lepiota hystrix, L. griseovirens). Остальные так-
соны агариковых грибов широколиственных лесов можно отнести к
нейтральным в отношении особенностей местопроизрастания. Средн та-
ких видов очень много гумусовых сапротрофов.
В хвойных лесах отмечено 65 видовых и внутривидовых таксонов
агариковых грибов. Наибольшее количество таксонов — 25, 17 и 9 от-
носится к родам Agaricus, Lepiota и Cystoderma. Часть таксонов (9) —
Agaricus deylii, A. altipes, A langei var. mediofuscus, A. subfloccosus.
A. leucotrichus, A. chionodermus, Cystoderma rugosoreticulata, C. adna-
tifolia, Lepiota kuehneri — характерны для хвойных лесов и не отмече-
ны в иных типах растительности европейской части СССР. При этом
имеются виды, приуроченные к определенному типу хвойных лесов (на-
пример, только в еловых лесах произрастают Agaricus deylii, A. sub-
floccosus, A. altipes, A. macrocarpus, A. leucotrichus, A. chionodermus,
Cystoderma ambrosii).
В смешанных лесах отмечено 46 видовых и внутривидовых таксо-
нов агариковых грибов. Наибольшее количество таксонов (17, 15 н 7)
относится к родам Lepiota, Agaricus н Cystoderma Специфика видового-
состава агариковых грибов смешанных лесов выражена слабо. Харак-
терными для них являются всего три таксона — Agaricus aestivalis var.
veneris, A. semotus var. minimum, Cystoderma caucasica, не отмеченные
в иных типах растительного покрова. Встречаются в смешанных лесах,
ио не являются приуроченными только к этому типу растительности
виды Agaricus langei, A. dulcidulus, Cystoderma carcharias, C. superbum,
Lepiota seminuda, L. subgracilis, Chamaemyces fracidus.
В различных типах степей зарегистрировано 28 видовых и внутри-
видовых таксонов агариковых грибов. Наибольшим количеством таксо-
нов (17) представлен род Agaricus. Остальные роды (Lepiota, Macro-
lepiota, Leucocoprinus, Leucoagaricus) содержат по 2—3 вида. Среди*
агариковых грибов, выявленных в степях, имеются нейтральные эври-
топные виды с широкой экологической амплитудой: Agaricus campestris,
127
A. bisporus, Macrolepiota excoriata, а также характерные стенотопные
узкоспециализированные виды: Agaricus cupreobrunueus, A. tabularis,
A. maskae, Leucocoprinus boliusi, Leucoagaricus leucothitus f. cinercoli-
lacina. Сопоставляя данные, полученные в результате изучения агари-
ковых грибов различных типов степей (разнотравио-типчаково-ковыль-
иые, типчаково-ковыльные и полынно-злаковые), можно констатировать
наличие характерных (встречающихся только в одном из типов степи)
н нейтральных (встречающихся во всех типах степей) видов. Например,
Agaricus bernardii, A. xanthodermus, Leucocoprinus boliusi произраста-
ют лишь в разнотравно-типчаково-ковыльных степях; Agaricus cupreo-
brunneus, A. velenovskyi, A. gennadii — только в типчаково-ковыльной
степи, a Agaricus porphyrocephalus, A. maskae, Leucoagaricus leucothitus
f. cinercolilacina — в полынио-злаковой степи.
Один вид агариковых грибов европейской части СССР — Agaricus
kuehnerianus — известен лишь с альпийских лугов.
Флора агариковых грибов лугов насчитывает 35 видовых и внутри-
видовых таксонов, в основном гумусовых сапротрофов, которые очень
чувствительны к влажности. В связи с этим виды, отмеченные на сухо-
дольных лугах, очень часто отсутствуют на лугах с заболоченными или
супесчаными почвами. Видовой состав агариковых грибов лугов, как
и степей, в значительной степени зависит от их хозяйственного исполь-
зования (выпас, косьба). Наибольшее количество таксонов (17, 7 н 6)
агариковых грибов лугов относится к родам Agaricus, Macrolepiota и
Lepiota. Обычно виды агариковых грибов различных типов лугов отно-
сятся к нейтральным, произрастают и в других типах растительного по-
крова (Melanophyllum eyrei, Agaricus campestris, A. bisporus, A. arvensis,
Lepiota cristata, Macrolepiota excoriata), хотя среди них отмечен и ряд
таксонов (Agaricus stramineus, A. campestris var. isabellinus. A. por-
phyrocephalus), приуроченных к безлесным местообитаниям.
Флора агариковых грибов болот очень бедна. Представлена глав-
ным образом бриотрофами из рода Cystoderma. Бедность флоры ага-
риковых грибов болот объясняется отсутствием среди них видов, спо-
собных давать плодовые тела в условиях постоянного переувлажнения.
По всей видимости, на формирование флоры немикоризных грибов бо-
лот влияет характер торфа Флора Agaricales разных типов болот
(низинные, верховые, переходные) Эстонии, по данным К. А. Каламээса
(1975), различается. Для представителей немикоризного семейства
Agaricaceae таких особенностей нам установить не удалось.
В антропогенных сообществах (дендропарки, ботанические сады,
полезащитные лесополосы, поля, сады, огороды, кладбища) видовой
состав агариковых грибов разнообразен н представлен главным обра-
зом гумусовыми сапротрофами и копротрофами. Здесь он определяется
в значительной степени количеством гумуса в почве и экологическими
особенностями местности. Флора агариковых грибов дсидропарков и
ботанических садов насчитывает 55 видовых и внутривидовых таксонов,
в том числе шесть таксонов — Melanophyllum eyrei var. macrosporus,
Agaricus amanitaeformis, Leucocoprinus badhamii, L. pilatianus var. pi-
latianus, L. pilatianus var. subrubens, Leucoagaricus carneifolius — отме-
чены лишь в данном типе сообществ. Два из них — М. eyrei var. macro-
sporus и Agaricus amanitaeformis — описаны как новые для науки (Ур-
бонас, Каламээс, Лукии, 1974; Вассер, 1974а,б,в).
Агариковые грибы полезащитных лесополос представлены 13 ви-
довыми и внутривидовыми таксонами: Agaricus campestris, A. bisporus,
128
к. abruptibulbus, A. arvensis, A. xanthodermus, Lepiota cristata, L. lila-
cea, L. alba, L. clypeolaria var. latispora, L. scobinella, Leucoagaricus
moseri, Macrolepiota rhacodes, M. excoriata. При анализе специфики
флоры агариковых грибов полезащитных лесополос были учтены все
факторы, которые могут повлиять иа ее возникновение. По составу
древесных растений исследованные полезащитные лесополосы можно
подразделить на монокультурные (3—7-рядные) — абрикосовые, ака-
циевые, гледичиевые, дубовые, кленовые, сосновые, тополевые, шелко-
вичные, ясеневые и поликультурные (5—42-рядные) — абрикосово-ака-
циевые, абрикосово-акациево-грушевые, дубово-вязово-ясеневые. Прове-
денные исследования (Частухин, Николаевская, 1969; Вассер, Солдатова,
1977а) показали, что общий характер флоры высших базидиомицетов
полезащитных лесополос зависит от видового состава древесных
н кустарниковых растений, возраста и ширины полос. Однако относи-
тельно видового состава иемикоризиых грибов этого сказать нельзя.
Около 70 % видов агариковых грибов полезащитных лесополос отнесе-
ны к нейтральным видам и произрастают почти во всех их типах
(Agaricus campestris, A. bisporus, Macrolepiota excoriata, Lepiota alba),
отсутствуют лишь в сосновых, ореховых и шелковичных (Agaricus ar-
vensis, A. xanthodermus, Lepiota cristata); имеются виды, произрастаю-
щие только в одном или двух типах лесополос (Agaricus abruptibulbus.
Leucoagaricus moseri, Macrolepiota procera).
В антропогенных сообществах, таких, как сады, огороды, поля,
кладбища, отмечено 16 видов агариковых грибов, среди которых имеют-
ся как нейтральные (Agaricus campestris, A. bisporus, A. bitorquis, Le-
piota cristata, Macrolepiota excoriata), так и характерные (Agaricus ro-
magnesii, A. gennadit, A. fissuratus, A. porphyrizon, Leucocoprinus bre-
sadolae, Macrolepiota olivieri) виды. Характерные виды антропогенных
сообществ являются редкими и малоизвестными для флоры европей-
ской части СССР.
Очень интересная, специфическая флора агариковых грибов, отме-
ченная в оранжереях и парниках, представлена семью видами: Agaricus
arvensis, A. bisporus, Lepiota cristata, Leucocoprinus cepaestipes, L. birn-
baumii, L. denudatus, Macrolepiota olivieri. Последние четыре вида про-
израстают главным образом в оранжерейных посадках тропических и
субтропических растений (Ficus elastica Roxb., Monstera deliciosa
Liebm. и др.), где летом температура помещения достигает 40—45°C,
а зимой 15—18 °C.
Два вида агариковых грибов — Melanophyllum echinatum и Phaeo-
lepiota aurea — сравнительно часто поселяются в пирогенных место-
обитаниях. В связи с этим они ие отнесены нами к особой экологиче-
ской группе — карботрофов, хотя и тяготеют к ней.
Агариковым грибам европейской части СССР, как и остальным
компонентам растительного покрова, в годовом ритме развития свой-
ственны закономерные сезонные изменения, которые связаны с чередо-
ванием двух фаз развития — вегетативной и развития карпофоров. По-
явление, продолжительность и окончание плодоношения определяются,
с одной стороны, биологическими свойствами агариковых-грибов, а с
другой - условиями погоды текущего года, иногда предшествующего
(Галахов, 1938). Поскольку образование и развитие карпофоров ага-
риковых грибов находится в крайне сложной зависимости от экологи-
ческих и климатических факторов, то установление различных сезон-
ных изменений возможно лишь на основе многолетних наблюдений.
и 4—2960
129
С учетом литературных данных (Сержанина, 1962, 1965, 1967; Зе-
рова, 1963, 1970а, 1974а; Урбоиас, 1974; Урбоиас, Каламээс, Лукин,
1974; Каламээс, 1975; Зерова, CociH, Роженко, 1979) и результатов
собственных исследований (Вассер, Солдатова, 1977а; Вассер, 1980),
для европейской части СССР нами выделено шесть сезонных аспектов
агариковых грибов, а именно: ранневесеииий — с середины апреля до
начала мая весенний — с начала
Рис. 7. Спектр родов агариковых грибов
Кавказа
мая до середины июня, раниелет-
иий — с середины июня до конца
июля, летний — с конца июля до
середины сентября, осенний — с
середины сентября до середины
октября, позднеосениий — с се-
редины октября до третьей де-
кады ноября. Названные сроки
аспектов агариковых грибов со-
ответствуют средней полосе евро-
пейской части СССР. Северная
и южная полосы имеют свои осо-
бенности. В первом случае гриб-
ной покров отстает в своем раз-
витии от средней полосы на 2—
3 недели, а от южной—иа 5—
6 недель. В ряде зон некоторые
аспекты в отдельные годы могут выпадать. Так, в степной зоне при
неблагоприятных климатических условиях (длительное отсутствие осад-
ков) некоторые из сезонных аспектов (например, весенний, раинелет-
иий, летний) ие проявляются (Вассер, Солдатова, 1977а).
56 видовых н внутривидовых таксонов Agaricaceae Кавказа отно-
сятся к восьми родам четырех колен. Наибольшую роль в растительном
покрове Кавказа играют представители родов Lepiota (20 таксонов)
и Agaricus (18 таксонов), менее значительная роль родов Macrolepiota
(7 таксонов), Cystoderma, Leucocoprinus, Leucoagaricus- (по 3), Endo-
ptychum и Phaeolepiota (по 1 таксону) (рис. 7).
По топическим и трофическим связям агариковые грибы Кавказа
представляют собой две экологические группы: гумусовые сапротрофы
(73,2 %) и подстилочные сапротрофы (26,8 %). Наибольшее видовое
богатство флоры агариковых грибов Кавказа относится к лесному поя-
су, который расположен на высоте до 2000—2500 м н. у. м. и представ-
лен следующими основными формациями: дубовыми, буковыми, грабо-
выми, смешанными, сосновыми, пихтовыми, еловыми, можжевеловыми.
Значительное видовое разнообразие агариковых грибов Кавказа
отмечено для широколиственных лесов. В чистых и смешанных широко-
лиственных лесах отмечено 29 видов агариковых грибов. Наибольшее
количество видов (15, 5 и 4) относится к родам Lepiota, Agaricus н
Macrolepiota. Восемь видов — Lepiota adulterina, L. tomentella, L. cas-
tanea, L. subgracilis, L. micropholis, L. cygnea, Macrolepiota procera,
M. konradii — произрастают лишь в широколиственных лесах и не отме-
чены в иных типах растительного покрова Кавказа. Этн виды являются
характерными для широколиственных лесов, причем Lepiota adulterina,
L. brunneoincarnata характерны для буковых, Agaricus silvaticus, A. se-
1 Сроки появления карпофоров агариковых грибов приведены в ^Конспекте
флоры агариковых грибов СССР».
130
motus, Lepiota cortinarius, L. subalba — железняково-дубовых, Lepiota
micropholis, L. eriophora, Leucocoprinus badhamii — самшитовых лесов,
Lepiota castanea, L. cristata — для насаждений платанов. Остальные
виды агариковых грибов широколиственных лесов Кавказа (например,
Agaricus silvaticus, A. mcleagris, Lepiota acutcsquamosa, L. clypeolaria)
могут быть отнесены к нейтральным.
В хвойных лесах произрастает 21 вид агариковых грибов. Наиболь-
шее количество видов (8,8 и 3) относится соответственно к родам Aga-
ricus, Lepiota и Cystoderma. Четыре вида — Agaricus augustus, A. ab-
ruptibulbus, Cystoderma amianthina, С. carcharias — характерны для
хвойных лесов и не отмечены в иных типах растительности Кавказа.
Наиболее беден видовой состав агариковых грибов можжевеловых (ар-
чевых) лесов, так как они очень ксерофитны, расположены иа сухих
склонах южных экспозиций и сомкнутого полога ие образуют. Для
этого типа лесных формаций отмечены лишь Agaricus silvicolus, A. me-
leagris и Lepiota alba. Флора агариковых грибов хвойных лесов Кав-
каза сходна с таковыми других ботанико-географических зон СССР,
одиако наблюдается специфичность в сезонности их плодоношения
(Мелик-Хачатрян, 1980). Характерными видами для еловых, пихтовых,
елово-пихтовых лесов Кавказа являются представители рода Cysto-
derma — С. amianthina, С. carcharias, С. granulosa.
В смешанных лесах отмечено 15 видов агариковых грибов из родов
Agaricus (A. abruptibulbus, A. silvaticus, A. silvicolus, A. velenovskyi,
A. rusiophyllus, A. placomyces, A. purpurellus), Lepiota (L. ventriosospo-
ra, L. felina, L. acutesquamosa, L. alba, L. subalba) и Macrolepiota
(M. rhacodes, M. gracilenta, M. puellaris). Специфика видового состава
агариковых-грибов смешанных лесов выражена слабо. Единственным'
им присущим видом является Agaricus velenovskyi, отмеченный как
компонент растительного покрова можжевелово-фисташковых лесов.
На высокогорных лугах Кавказа (1800—3000 м и. у. м.) зарегистри-
ровано четыре вида агариковых грибов из рода Agaricus — A. tabularis,
A. campestris, A. bisporus и A. xanthodermus. Первый вид И. Г. Нахуц-
ришвили (1975) считает характерным для высокогорных лугов ГССР,
указывая, что ои спускается и до лесного пояса, однако произрастает
лишь на лесных опушках. В европейской части СССР (Зерова, 1956;
Вассер, 1980), Восточной Сибири (Беглянова, 1972) и Средней Азии
(Васильков, 1955; Батырова, 1975; Вассер, Батирова, 1979) A. tabularis
отмечен как компонент растительного покрова главным образом пла-
кориых местообитаний. Видовой состав агариковых грибов пояса высо-
когорных лугов очень беден, что, по мнению И. Г. Нахуцришвили (1.С.),
связано с отсутствием древесной растительности, суровыми климатиче-
скими условиями и сильным выпасом. Вообще безлесные фитоценозы
Кавказа (в отличие, например, от степной зоны Украины) бедны ага-
риковыми грибами. Так, в АрмССР безлесные фитоценозы (полупусты-
ни, степи, лугостепи), по данным Дж. Г. Мелик-Хачатряи (1980), пред-
ставлены единично встречающимися шестью видами агариковых грибов:
Leucoagaricus macrorhizus, Agaricus campestris, A. bisporus, A. ta-
bularis, A. xanthodermus, Macrolepiota gracilenta, а в заповедных степях
Украины зарегистрировано 28 видовых и внутривидовых таксонов ага-
риковых грибов (Вассер, Солдатова, 1977а; Вассер, 1980). Бедность
флоры агариковых грибов безлесных фитоценозов в условиях АрмССР,
по мнению Дж. Г. Мелик-Хачатряи, объясняется отрицательным влия-
11*
131
Agaricus
Lepiota
Macrolepiota
Leucoagaricus
Leucocoprinus Phueolepiota
Pseudobaeospora squamanita
Рис 8. Спектр родов агариковых грибов
Западной Сибири
Cystoderma
Curophragmium
Endoptychum
’.eolepuM
«нем резких колебаний суточных температур, относительной влажности
воздуха и почвы, а также сильной инсоляции.
В антропогенных сообществах (деидропарки, ботанические сады,
полезащитные лесополосы, поля, сады, огороды, кладбища) насчиты-
вается 23 вида агариковых грибов, главным образом гумусовых сапро-
трофов. Наибольшее количество видов (9 и 8) относится соответственно
к родам Agaricus и Lepiota. Три вида — Agaricus bernardii, Phaeolepio-
ta aurea и Lepiota helveola — не
отмечены в иных местообита-
ниях. Остальные виды антропо-
генных сообществ (Agaricus bi-
torquis, A. bisporus, A. xantho-
dermus, Lepiota alba, Macrolepio-
ta excoriata и др.) являются
нейтральными, имеющими широ-
кое распространение как на Кав-
казе, так и за его пределами.
Сезонные аспекты агарико-
вых грибов отдельных регионов
Кавказа различаются. Так, для
ГССР И. Г. Нахуцришвилн (1971,
1975) выделяет четыре аспекта
(весенний, летний, осенний, зим-
ний), причем виды грибов по их
сезонной встречаемости делит на четыре группы: а) виды, плодонося-
щие в течение всего вегетационного периода, б) виды, плодоносящие
летом и осенью, в) виды, плодоносящие только осенью, г) ви-
ды, плодоносящие осенью и зимой. Для АрмССР Дж. Г Мелик-
Хачатрян (1971, 1980) также выделяет четыре аспекта (ранневесенний,
весеине-летний, летне-осенний, поздисосепний) агариковых грибов, ко-
торые отличаются от аспектов И. Г. Нахуиришвили. А. С. Садыхов
(1968) для Ленкоранского района АзССР приводит шесть сезонных
аспектов (ранневесенний, весенний, летний, осенний, поздиеосенний,
зимний) агариковых грибов. Эти данные свидетельствуют о том, что
для такого разнообразного по вертикальным климатическим и расти-
тельным условиям района флоры, как Кавказ, на данном этапе разви-
тия исследования ие представляется возможности дать общую единую
схему сезонных аспектов агариковых грибов. Для Кавказа считаем воз-
можным отметить лишь особенности массового появления карпофоров
агариковых грибов, которые в течение вегетационного периода дают
обычно два пика — в июне или июле, в сентябре или октябре.
В отношении поясного распространения агариковых грибов Кавказа
наблюдается ряд особенностей. По разнообразию видового состава
(31 вид) ведущее место занимают плакоры и нижний горный пояс. За-
тем по количеству видового богатства следует средний горный пояс.
Наименьшим видовым составом характеризуется верхний пояс. При
характеристике видового состава агариковых грибов различного расти-
тельного покрова отмечены специфические виды, которые характери-
зуют и особенности высотного распространения, т. е. являются одно-
поясными. В то же время отмечены виды агариковых грибов (напри-
мер, Agaricus campestris, Macrolepiota excoriata), которые обладают
широкой амплитудой высотного распространения.
42 видовых и внутривидовых таксона Agaricaceae Западной Сибири
132
относятся к 11 родам четырех колеи. Наиболее заметную роль в
растительном покрове этого региона играют представители родов Aga-
ricus (13 видов), Lepiota (12) и Cystoderma (5). Остальные родовые
таксоны представлены одним — четырьмя видами (рис. 8).
По топическим и трофическим связям агариковыс грибы Западной
Сибири относятся к трем экологическим группам: гумусовые сапротро-
фы (61.9%), подстилочные сапротрофы (35,7 %), лигнотрофы (2,4 %).
Полностью охарактеризовать видовой состав агариковых грибов
всех ботанико-географических зон, подзон и растительных поясов За-
падной Сибири не представляется возможным из-за разной изученности
флористической насыщенности. Основные причины сравнительно нерав-
номерного распределения агариковых грибов ио ботанике-географиче-
ским зонам и растительным поясам Западной Сибири определяются как
эколого-биологическими особенностями грибов, так и особенностями
растительного покрова, климатическими, эдафическими и орографиче-
скими факторами. Кратко осветим видовой состав агариковых грибов
лесотундровой, бореальио-лесной (подзоны северной, средней и южной
тайги, преллесостепных и сосново-березовых лесов), широколиствеиио-
лесной и лесостепной зон.
В участках зоны лесотундры выявлены 14 видов агариковых гри-
бов: Agaricus campestris, A. silvaticus, A. bisporus, A. silvicolus, A. se-
motus, Cystoderma amianthina, C. carcharias, C. granulosa, C. cinnaba-
rina, Pseudobaespora pillodii, Lepiota alba, L. clypcolaria, Macrolepiota
rhacodes, M. excoriata. В лесотундровой зоне агариковые грибы произ-
растают как в равнинных, так и в горных растительных сообществах
горно-лесного, подгольцового и горно-тундрового поясов. Наибольшее
видовое разнообразие отмечено для равнинного и горно-лесного поясов.
В подгольцовом и горно-туидровом поясах видовой состав агариковых
грибов беден-, найдено лишь семь видов, отмеченных и для Арктики,
причем лишь здесь зарегистрирована Pseudobaeospora pillodii.
Бореально-лесная зона благодаря видовому разнообразию древес-
но-кустарниковой растительности, благоприятным по сравнению с лесо-
тундровой зоной климатическим условиям наиболее богата агариковы-
ми грибами В пей отмечено 25 видовых и внутривидовых таксонов
агариковых грибов, причем наибольшее количество их связано с под-
зонами южной тайги, предлесостепных и сосново-березовых лесов. Флора
агариковых грибов бореально-лесной зоны представлена следующими
таксонами: Agaricus campestris, A. silvaticus, A. bisporus, A. augus-
tus, A. silvicolus, A. arvensis, A. semotus, Phaeolepiota aurea, Cystoder-
ma amianthina f. amianthina, C. amianthina f. olivacea, C. carcharias»
C. cinnabarina, Lepiota seminuda, L. acutesquatnosa, L. lignicola, L. cri-
stata, L. alba, L. clypeolaria, L. felina, L. scobinclla, L sctulosa, Leuco-
agaricus leucothitus, Macrolepiota proccra, M. rhacodes, M. excoriata.
В каждой подзоне имеются как характерные (например, С. amianthina
f. olivacea — подзона южной тайги, L. lignicola — подзона предлесо-
степиых и сосново-березовых лесов), так и нейтральные виды (Agaricus
campestris, Macrolepiota excoriata). В распределении агариковых гри-
бов бореальио-лесиой зоны по вертикальным растительным поясам ха-
рактерно убывание количества видов по мере возрастания высоты над
уровнем моря. Наибольшее видовое разнообразие присуще равнинным
условиям.
В широколиствеино-лесной зоне зарегистрировано 17 видов агари-
ковых грибов, отмеченных большей частью н из бореально-лесной зоны.
133
Из характерных для рассматриваемой зоны следует указать иа виды.
отмеченные как компоненты растительного покрова широколиственных
лесов. Только в этой зоне произрастают Agaricus sagatus и Squamanita
paradoxa. Распределение агариковых грибов в широколиственно-лесной
зоне такое же, как и в бореально-лесной зоне. Характерные виды ши-
роколиственно-лесной зоны — очень редкие. Squamanita paradoxa, на-
пример, известна в СССР лишь из Западной Сибири.
В Западной Сибири лесостепная зона является одной из наибо-
лее благоприятных для развития представителей немикоризного семей-
ства Agaricaceae. Однако обедненный видовой состав высших древес-
Рис. 9. Спектр родов агариковых грибов
Восточной Сибири
ных растений, сухость климата
откладывают отпечаток иа состав
флоры агариковых грибов лесо-
степной зоны. Здесь, особенно в
ее южной части, выявлен 21 вид,
причем 10 видов (Agaricus bi-
torquis, A. bernardii, A. tabularis,
A. porphyrizon, Gyrophragmium
dunalii, Endoptychum agaricoidcs,
Lepiota brunneoincarnata, Leuco-
coprinus denudatus, Leucoagari-
cus moseri, Macrolepiota gracilen-
ta) являются характерными для
рассматриваемой зоны.
Сезонные аспекты агарико-
вых грибов в Западной Сибири отсутствуют. Здесь можно говорить
лишь о времени их наиболее массового появления в различных ботайи-
ко-географических зонах и растительных поясах, которое приходится иа
позднелетиий и раннеосенний периоды.
47 видовых н внутривидовых таксонов Agaricaceae Восточной Си-
бири относятся к шести родам четырех колен. Наиболее заметную роль
в растительном покрове Восточной Сибири играют представители родов
Lepiota (19 видов), Agaricus (14) и Cystoderma (7 видов). Значительно
меньшую роль играют представители родов Macrolepiota (5 видов),
Endoptychum и Leucocoprinus (содержат по одному видовому таксону)
(рис. 9). По топическим и трофическим связям агариковые грибы
Восточной Сибири относятся к четырем экологическим группам: гуму-
совые сапротрофы (55,3%), подстилочные сапротрофы (36,1 %), брио-
трофы (4,3 %), лигнотрофы (4,3 %).
Видовой состав агариковых грибов ботанико-географических зон
я растительных поясов Восточной Сибири имеет много общих черт с
видовым составом грибов Западной Сибири. В связи с этим отметим
лишь виды агариковых грибов Восточной Сибири, не встреченные в
аналогичных условиях в Западной Сибири. В лиственничных лесах
собраны Agaricus fuscofibrillosus, A silvaticus var. pallidus, A. exccl-
lens; в сосновых, сосново-пихтовых, сосново-листвеиничиых — Cystoder-
ma fallax, C. supcrbum, C. apibrosii, Lepiota hapalopoda, в сосново-бе-
резовых — Macrolepiota nympharum.
Развитие флоры агариковых грибов в Восточной Сибири начинает-
ся позднее, а заканчивается раньше, чем в Западной Сибири. Для фло-
ры агариковых грибов характерен сокращенный период вегетации в
связи с быстрым нарастанием среднесуточных весеиие-летних и быст-
рым падением осенних температур. Сезонные грибные аспекты, свой-
134
Leptflta
Agaricus
Leucoagaricus
Chlorophyllum
Pseudob
Phaeolepiota.
Endoptgchum
MelaraphyUum
Cysloderma
ственные европейской флоре грибов, здесь отсутствуют (Бсглянова,
1975; Петренко, 1978). Первые представители агариковых грибов появ-
ляются в начале июля. Их численность несколько увеличивается во
второй половине июля. Максимальным обилием видового состава ага-
риковых грибов характеризуется август. К концу первой декады сен-
тября в связи с наступлением заморозков и резким общим похолода-
нием развитие макромицетов прекращается (Петренко, 1978).
72 видовых и внутривидовых таксона Agaricaceae Дальнего Восто-
ка относится к 11 родам четырех колеи. Наиболее важную роль в рас-
тительном покрове Дальнего Востока играют представители родов
Lepiota (37 таксонов) н Agaricus (15), значительно меньшую — предста-
вители родов Macrolepiota (5), Leucocoprinus и Cystoderma (по 4 так-
сона). Остальные роды (Melanophyllum, Endoptychum, Phaeolepiota,
Pseudobaeospora, Chlorophyllum,
Leucoagaricus) представлены од-
ним-двумя видовыми таксонами
(рис. 10).
По топическим и трофиче-
ским связям агариковые грибы
Дальнего Востока относятся к
трем экологическим группам: гу- t
мусовым сапротрофам (51,4 %), <
подстилочным сапротрофам
(45,8 %), лигнотрофам (2,8 %).
Флора высших сосудистых
растений Дальнего Востока раз-
нообразна и богата редкими и
интересными видами как древес-
ных, так и травянистых растений,
в связи с чем и флора агарико-
вых грибов богата и своеобразна,
агариковых грибов отмечено для кедрово-широколиственных лесов
(кедровники), основу которых составляет кедр корейский (Pinus ko-
raiensis Sieb. et Zucc.). В кедрово-шнроколиственных лесах насчиты-
вается 50 видовых и внутривидовых таксонов агариковых грибов,
главным образом подстилочных сапротрофов. Наибольшее количество
таксонов (30 и 7) относится к родам Lepiota и Agaricus; 21 таксон —
Agaricus augustus var. albus, Pseudobaeospora defibulata, Lepiota adul-
terina, L. eriophora, L. echinacea, L. grangei, L. pseudofelina, L. igni-
color, L. castanea, L. subalba, L. lilacea, L. subgracilis, L. clypeolaria
var. minor, L. rufipes, L. kuehneri, L. rhodorhira, L. subincarnata,
L. serena, Leucocoprinus badhamii, L. bresadolae, Macrolepiota konra-
dii — произрастает лишь в кедрово-широколиствеиных лесах и ие от-
мечен в иных типах растительного покрова Дальнего Востока. Эти
Рис.
. . Macrolepiota
Leucocopnous
10. Спектр родов агариковых грибов
Дальнего Востока
Наибольшее видовое разнообразие
таксоны характерны для кедрово-широколиствеиных лесов.
В хвойных лесах (кедрово-пихтовых, лиственничных, пихтово-ело-
вых, сосновых), образованных Pinus koraiensis, Abies nephrolepis
Maxim., A. holophylla Maxim., Larix daurica Turcz., отмечено 23 вида ага-
риковых грибов, главным образом из родов Lepiota и Agaricus: Mclano-
phyllum echinatum, M. eyrei, Agaricus variegatus, A. silvaticus, A. silvi-
colus, A. abruptibulbus, A. meleagris, A. placomyces, A. semotus, A. dul-
cidulus, Cystoderma amianthina, C. granulosa, C. cinnabarina, Lepiota
135
hystrix, L. acutesquamosa, L. virescens, L. cristata, L. erminea, L. ven-
triosospora, L. clypeolaria, L. feiina, L. scobinella, L. setulosa.
В широколиственных лесах (Fraxtnus rhynchophylla Hance, Tilia
amurensis Rupr., T. mandshurica Rupr. et Maxim.) насчитывается 21 вид
агариковых грибов. Наибольшее количество видов (11 н 4) относится к
родам Lepiota и Agaricus. Десять видов —Agaricus subrufcscens, A. pur-
purcllus, Cystoderma carcharias, Lepiota rosea, L. hetieri, L. neophana,
L. xanthophylla, Leucocoprinus
brebissonii, Macrolepiota proce-
ra — характерны для широко-
лиственных лесов и не отмечены
в иных типах растительного по-
крова.
В дубовых (Quercus mongo-
lica Fisch, ex Turcz.), березовых
(Betula mandshurica (Rgl ) Na-
kai, B. schmidtii Rgl., B. lanata
(Rgl.) V. Vassil.), долинных
(ивняки — Salix gracilistyla Miq.,
S. schwerjnii E. Wolf; чозенникн
Chosenia arbutifolia (Pall.)
A. Skvorts.; ольшаники Alnus
et Zucc.; ясеневники Fraxinus
mandshurica Rupr.) лесах флора агариковых грибов бедна и насчиты-
вает 11 видов: Melanophyllum echinatum, Agaricus placomyces, A. rusio-
phyllus, Lepiota lignicola, L. tomentella, L. pseudohelveola, L. setulosa,
Chlorophyllum molybdites, Leucocoprinus brebissonii, Macrolepiota ex-
coriata. M. procera.
В антропогенных сообществах (деидропарки, сады, поля, огороды)
и открытых местообитаниях видовой состав агариковых грибов беден,
представлен главным образом эвритопными гумусовыми сапротрофами:
Agaricus campestris, A. augustus, A. arvensis, Endoptychum agaricoides,
Leucoagaricus leucothitus, Lepiota cristata, Macrolepiota excoriata,
M. mastoidea, причем первые пять видов являются характерными для
антропогенных сообществ.
Наиболее богат видовой состав агариковых грибов юга Дальнего
Востока. С продвижением на север, как и при подъеме в горы, из
древостоя выпадают многие, особенно широколиственные породы и в
результате происходит сильное обеднение флоры агариковых грибов.
Встречаются лишь виды, связанные с лиственницей (Васильева, 1973).
Развитие карпофоров агариковых грибов на Дальнем Востоке, особен-
но в его южной части, продолжается около 5,5 месяцев (с середины
мая до конца октября). Основной сезонный аспект образования карпо-
форов приходится на август — сентябрь (Васильева. 1. с.; Булах,
1977а, б).
32 видовых и внутривидовых таксона Agaricaceae Средней Азии
относятся к семи родам четырех колеи (рис. 11). Наибольшую роль в
растительном покрове Средней Азии играют представители родов Aga-
ricus (18 таксонов) и Lepiota (7). Остальные родовые таксоны (Cysto-
derma, Endoptychum, Gyrophragmium, Leucoagaricus, Macrolepiota) на-
считывают 1—2 вида. По топическим н трофическим связям агариковые
грибы Средней Азии делятся иа трн экологические группы: гумусовые
сапротрофы (84,2%), подстилочные сапротрофы (12,6 %), бриотрофы
13G
(3,2 %), причем господствующее положение занимают представители
первой группы.
Р. В. Камелии (1979) для Средней Азии выделяет 40 типов расти-
тельности (или флороцеиотипов), детально охарактеризовать видовой
состав агариковых грибов которых сейчас нс представляется возмож-
ным из-за их неравномерной изученности.
В безлесных типах растительности Средней Азин (пустыни, полу-
пустыни, степи) произрастает 18 таксонов агариковых грибов из группы
гумусовых сапротрофов: Agaricus campestris var. campestris, A. cam-
pestris var. isabellinus, A. bisporus, A. bitorquis, A. bernardii, A. genna-
dii var. gennadii, A. gennadii var. microsporus, A. macrosporus, A. tabu-
laris, A. arvensis, Gyrophragmium dunalii, Endoptychum agaricoides^
Lepiota cristata, L. scobinella, L. serena, Leucoagaricus leucothitus, Mac-
rolepiota procera, M. excoriata. Из иих семь таксонов — Agaricus ber-
nardii, A. gennadii var. genadii, A. gennadii var. microsporus, A. macro-
sporus, A. tabularis, Gyrophragmium dunalii, Lepiota scobinella, L. sere-
na — присущи безлесным типам растительности Средней Азии. Для
карпофоров большинства видов агариковых грибов безлесных флоро-
ценотипов, где низкая влажность воздуха и почвы, высокие температу-
ры и инсоляция, свойствен ряд морфологических особенностей: растрес-
кивание кожицы шляпки и ножки, более светлая окраска карпофоров,
более крупные размеры спор по сравнению с аналогичными видами
средних широт (Батырова, 1977а, б).
Лесная растительность (крио- и семигумидные, семиаридпые типы,
по Р. В. Камелину, 1979) представлена (кроме гумусовых сапротрофов
подстилочными сапротрофами, бриотрофами) 13 видами агариковых
грибов: Agaricus campestris, A. squamuliferus, A. augustus, A. abrupti-
b^ilbus, A. xanthodermus, A. porphyrizon, Cystoderma amianthina, C. am-
brtisii. Lepiota acutcsquamosa, L. cristata, L. ventriosospora, L. clypeola-
ria, Macrolepiota excoriata. Из них семь видов — Agaricus squamulife-
rus, A. augustus, A. abruptibulbus, Cystoderma amianthina, C. ambrosii,
Lepiota acutesquamosa, L. ventriosospora являются характерными для
лесной растительности. За пределами Средней Азии эти виды широко
распространены в аналогичных типах лесной растительности.
В поясе высокогорной ксерофитной растительности (криоксерофиль-
иые подушечники) зарегистрировано всего пять видов агзриковых гри-
бов: Agaricus campestris, A. arvensis, Endoptychum agaricoidcs, Lepiota
cristata, Macrolepiota excoriata. Все виды агариковых грибов высоко-
горной ксерофитиой растительности — эвритопные, нейтральные, произ-
растающие и в иных флороценотип ах Средней Азии.
Антропогенная растительность (поливиые и богарные лесосады, са~
дЫ, лесокультуры, комплексная растительность мест поселения) отдель-
ных регионов Средней Азии очень резко отличается от остальных типов
растительности, однако видовой состав агариковых сравнитепьно одно-
образен и стабилен, насчитывает 18 таксонов. Наибольшее количество
(11) приходится на род Agaricus: A. romagnesii, A. campestris var.
campestris, A. campestris var. isabellinus, A. bisporus, A. bitorquis,
A. gennadii var. microsporus, A. arvensis, A. xanthodermus var. xantho-
dermus, A. xanthodermus var. lepiotoides, A. porphyrizon, A. lutosn*
Остальные виды относятся к родам Lepiota (L. cristata, L. clypeolaria),
Macrolepiota (M. procera, M. excoriata), Endoptychum (E. agaricoldes),
Leucoagaricus (L. leucothitus). Четыре вида — Agaricus romagnesii.
137
A. xanthodermus var. lepiotoides, A. luteus, Lepiota brunneoincarnata —
являются характерными для антропогенной растительности.
Плодоношение агариковых грибов на большей части территории
Средней Азии иосит эфемерный характер, в связи с чем в сезонном раз-
витии их наблюдается ряд особенностей. Для Средней Азии характерно
два сезонных аспекта агариковых грибов — весенний (или весеиие-ран-
нелетиий) и осенний. Последний выражен гораздо слабее первого. Ле-
том в Средней Азии наблюдается депрессия в плодоношении агарико-
вых грибов. Исключение со-
Agariaisставляют высокогорья (на вы-
.Z соте 3000 м н. у. м. и выше),
/ / \ где наблюдается один летний
/ / \ аспект (Эльчибаев, 1968; Ба-
I / \ тырова, 1977а, б; Вассер, Бати-
I------} рова, 1979).
\ / I / 11 видовых и виутривидо-
Macroltylotay^^'' \ /Cystoderma вых таксонов Agaricaceae Арк-
\ / тики относятся к родам Agari-
। У cus (A. campestris, A. arvensis,
Lepiota tabularis, A. semotus), Cy-
.Рис. 12. Спектр родов агариковых грибов stoderma (С. amianthina, С.
Арктики fallax, С. granulosa, С. cinna-
barina), Lepiota (L. alba, C.
clypeolaria var. latispora), Macrolepiota (M. excoriata) (рис. 12). Все
отмеченные виды являются нейтральными и большинство из них произ-
растает во всех областях флоры СССР. Своеобразна экология аркти-
ческих агариковых грибов. Виды рода Cystoderma поселяются в сырых
местообитаниях, даже на болотах. Виды родов Agaricus, Lepiota и Ма-
crolepiota произрастают на хорошо дренированных щебнистых склонах
бугров, берегах рек.
Основным типом растительного покрова Советской Арктики являет-
ся тундра — субарктическая и арктическая, которая на Крайнем Севе-
ре переходит в полярную пустыню, а на юге — в редколесье лесотундры;
значительно распространены в Арктике болота разных типов (Алексан-
дрова, 1964; Юрцев, Толмачев, Ребристая, 1978).
Флора агариковых грибов Арктики, особенно ее лесотундровой зо-
ны, представляет собой сильно обедненную флору бореально-лесиой зо-
ны. Агариковые грибы в Арктике встречаются от южной части лесо-
тундры до высокоарктнческих тундр. По данным Б. П. Василькова
(1966, 1967, 1974), при продвижении с юга на север во флоре макроми-
иетов Арктики ясно прослеживается уменьшение числа видов Это осо-
бенно резко проявляется при переходе из лесотундры в тундру. Очень
заметно убывают они и при переходе из субарктической тундры в
арктическую, в которую переходят виды, обычно растущие иа моховых
болотах, н, наконец, в полярную пустыню. На перигляциальиых терри-
ториях встречается всего лишь несколько видов, обычно тоже произ-
растающих на сырых и мокрых участках. Общей причиной уменьшения
числа видов при продвижении с юга на север Арктики является увели-
чение суровости климата. Если число видов макромицетов в Арктике
явно уменьшается при продвижении с юга на север, то в направлении
с запада иа восток такого уменьшения не наблюдается (Васильков,
1. с.). Виды агариковых грибов распространены по всей Арктике, исклю-
чение составляет Agaricus tabularis, известный лишь из восточноазиат-
-'38
Ской Арктики (западный Таймыр, западная Чукотка, о-в Вран-
геля).
У большинства видов макромицетов Арктики имеются своеобразные
отклонения, которые характеризуются уменьшением размеров плодовых
тел (наиизм) и нередко более крупными спорами по сравнению с об-
разцами, произрастающими за пределами Арктики (Васильков, 1966,
1967). Одиако это ие относится к видам, произрастающим за пределами
Арктики на переувлажненных участках (болота). Например, виды рода
Cystoderma и в Арктике, и за ее пределами обычно малых размеров.
Указания Б. П. Василькова (1967) о том, что Cystoderma amianthina
в Арктике имела более крупные споры (6,2—7,5 (8)Х5—6 мкм), чем
в лесной зоне, на наш взгляд, подлежат ревизии. По всей вероятности,
в данном случае речь идет о С. amianthina var. longispora, для которой
характерны споры 7—9 (10) Х4—4,7 мкм. Однако арктические образцы
Agaricus tabularis, распространенного в степях, полупустынях, пусты-
нях Евразии, имели споры значительно больших размеров [(7,5) 8—10
(12) X (5,5)6,5—7(8,5) мкм], чем обычные (6,5—7,7X4,6—5,4 мкм), ха-
рактерные для образцов из степной зоны Украины (Васильков, 1974;
Вассер, 1980). Кроме изменения размеров карпофоров и спор, можно
отметить еще некоторые отклонения, часто встречающиеся у грибов в
Арктике: выцветание окраски шляпки, уменьшение едкого вкуса мяко-
ти, значительное понижение реакции автооксидации иа изломе у неко-
торых видов, что связано, по мнению Б. П. Василькова (1967, 1974),
с сильной инсоляцией, большой влажностью почвы и воздуха.
Плодоношение агариковых грибов в Арктике происходит в общем
с конца июля до конца августа. В Арктической лесотундре наблюдается
в благоприятные годы два аспекта макромицетов — летний и ранне-
осенний, причем первый аспект в лесотундре соответствует весеннему
и летнему средней полосы, второй — осеннему, а в тундре все они сли-
ваются в один аспект (Васильков, 1967, 1971).
Agaricus campestris, A. arvensis, Cystoderma amianthina, Lepiota
clypeolaria, Macrolepiota excoriata растут во всех областях флоры
СССР, включая Арктику. Лишь в европейской части СССР встречаются
Melanophyllum eyrei var. macrosporus, Agaricus bresadolianus. A., spis-
sicaulus, A. vaporarius, A. cupreobrunneus, A. porphyrocephalus, A. be-
ncsii, A. deylii, A. altipes, A. lanipes, A. langei var. langei, A. langci var.
mcdiofuscus, A. haemorrhoidarius f. haemorrlioidarius, A. haemorrhoida-
rius f. fagetorum, A. subfloccosus, A. bitorquis var. validus, A. maleolens,
A. kuehnerianus, A. stramineus, A. rnaskae, A. macrocarpus, A. fissura-
tus, A. amanitaeformis, A. nivescens, A. aestivalis var. aestivalis, A. aesti-
valis var. veneris, A. longicaudus, A. leucotrichus, A. chionodermus, A. ose-
canus, A. pseudopratensis, A. phaeolcpidotus, A. moelleri, A. brunneolus,
A. xantholepis, A. semotus var. minimus, A. dulcidulus, Cystoderma ami-
anthina var. longispora, C. rugosoreticulata, C. caucasicum, C. adnati-
folium, Lepiota seminuda f. minima, L. bucknallii, L. puivcrulenta, L. gri-
scovirens, L. pallida, L. wichanskyi, L. echinella. L. fuscovinacea, L. cyg-
ncoaffinis, Chamaemyces fracidus, Leucocoprinus pilatianus var. subru-
bens, L. bohusi, Leucoagaricus carneifolius, L. leucothitus f. cinereolila-
cinus, Macrolepiota olivieri, M. excoriata var. squarrosa, M. prominens,
M. permixta. Специфику флоры агариковых грибов Кавказа представ-
ляет Lepiota micropholis. Cystoderma amianthina f. olivacea, Squamanita
paradoxa, Pseudobaeospora pillodii известны лишь из Западной Сибири,
Lepiota hapalopoda и Macrolepiota nympharum — из Восточной Сибири.
134
Agaricus variegatus, A. augustus var. albus, A. subrufescens, Pseudobaeo-
spora defibulala, Lepiota neophana, L. grangei, L. ignicolor, L. vires-
cens, L. clypeolaria var. minor, L. lateritiopurpurea, L. rhodorhiza, L. su-
bincarnata, Chlorophyllum molybdites, Leucocoprinus brebissonii, L. gcor-
gincae встречены лишь на Дальнем Востоке. Лишь из Средней Азии
известен Agaricus gennadii var. microsporus. Родовая специфика Aga-
ricaceae европейской части СССР представлена Chamaemyces, Запад-
ной Сибири — Squamanita, Дальнего Востока — Chlorophyllum. Виды
рода Melanophyllum известны в европейской части СССР и па Даль-
нем Востоке, Pseudobaeospora — в Восточной Сибири и иа Дальнем
Востоке.
ГЛАВА V!
ГЕОГРАФИЧЕСКИЙ АНАЛИЗ ФЛОРЫ
АГАРИКОВЫХ ГРИБОВ СССР
Вопросы географического распространения грибов изучены весьма
недостаточно, кардинальные проблемы микогеографии не решены,
несмотря иа то что микологами неоднократно предпринимались попыт-
ки обобщить имеющиеся разрозненные сведения и вывести на их основе
некоторые общие закономерности распространения грибов (Bisby, 1933,
1943; Diehl, 1937; Overholts, 1939; Wolf F. A., Wolf F. F., 1947; Бондар-
цев, 1953; Kreisel, 1960, 1961; Шварцман, 1962, 1968, 1973; Pirozynski,
1968; Ainsworth, 1971; Наумов, 1972; Васильева, 1973; Каламээс, 1975;
Дудка, 1976; Вассер, Солдатова, 1977 б; Kalamees, 1978b; Wasser, Sol-
datova, 1979; Вассер, 1980; Коваленко, 1980, и др.). Еще не разработа-
на общая схема особенностей географического распространения грибов,
как это осуществлено для цветковых и некоторых других групп расте-
ний. Это объясняется рядом причин, из которых основной является
слабая изученность микофлоры многих областей и целых континентов
земного шара. Даже при наличии довольно большого фактического ма-
териала о распространении более или менее крупного таксона грибов
(например, рода) в работах отсутствует географический анализ. Хотя
указанные выше авторы не разрешили основных вопросов микогеогра-
фии, тем ие менее они сделали некоторые обобщения, являющиеся в
настоящее время отправной точкой любого исследования по географии
грибов. Основные из них следующие.
1. Грибам присущи обширные ареалы, иногда носящие циркумпо-
лярный характер, охватывающие различные климатические зоны.
2. Распространение грибов контролируется преимущественно рас-
пространением субстратов или форофитов, с которыми грибы трофи-
чески связаны.
3. Чем меньше исследуемая территория, тем больше доминируют
в составе ее флоры грибы.
4. Климат оказывает меньшее (в основном второстепенное) влия-
ние иа распространение грибов, хотя некоторые грибы, в том числе
облигатные сапротрофы, имеют определенные географические пределы,
ограниченные климатическими факторами.
5. Сапротрофные грибы, как правило, имеют более широкое рас-
пространение, чем паразитные.
6. Ареалам высших Basidiomycetes, согласно Л. Н. Васильевой
(1973), как и ареалам мхов и лишайников (Herzog, 1926; Лазаренко,
1939; Окспер, 1974), обычно соответствуют ареалы не видов, а родов
семенных растений.
141
По мнению Л. Н. Васильевой (1.С.), причина такого широкого рас-
селения высших базидиомицетов заключается в особенностях строения
и жизни вегетативного тела грибов. Вегетативное тело высших базидио-
мицетов, их мицелий скрыты внутри субстрата и не подвергаются вред-
ному воздействию неблагоприятных климатических факторов, которые
испытывают высшие растения. Кроме того, мицелий высших базидио-
мицетов способен выносить длительное высыхание, не теряя жизне-
способности.
Подавляющее большинство микоризных грибов связано не с одним
видом древесных и кустарниковых растений, а со многими. Это также
объясняет причину широкого их расселения (Trappe, 1962; Вассер, Сол-
датова, 1977а, б). Например, в Приморье почти нет видов древесных
растений, общих с Северной Америкой или Европой, в то же время
свыше 50 % видов агарикальных грибов, найденных в Приморье, растут
и в Европе, и в Северной Америке (Васильева, 1. с.).
Н. А. Наумов (1972) заостряет внимание микологов иа недорабо-
танных и спорных вопросах микогеографии, к которым относит в пер-
вую очередь: а) установление ареалов грибов; б) выяснение первичных
(первоначальных) ареалов; в) решение проблемы частоты встречае-
мости и ее частного случая — «редкости» вида.
Первая попытка дать некоторые сведения по географическому рас-
пространению грибов, в том числе и агариковых, принадлежит
Э. М. Фризу (Fries, 1861, 1862), который все грибы земного шара делит
иа два микологических пояса: умеренный, включая Арктику, и тропи-
ческий, включая субтропики, указывая на бедность грибами последне-
го, обращая, таким образом, внимание на необходимость изучения за-
кономерностей, характеризующих распространение грибов.
А. А. Ячевский (1933), считая подразделение Э. М. Фриза «элемен-
тарным» и принимая во внимание связь грибов с растительными сооб-
ществами, вслед за ботаиико-географами высших растений все грибы
земного шара распределяет в шести ботанико-географических царствах:
неотропическом, палеотропическом, капском, австралийском, голаркти-
ческом и антарктическом, которые делит иа области.
Б. П. Васильков (1955) распространение шляпочных грибов в
СССР рассматривает по растительным зонам, указывая, что географи-
ческие отношения в распространении грибов настолько тесно связаны
с экологическими, что в ряде случаев бывает трудно иайти между ни-
ми границы. Так как смена видового состава шляпочных грибов про-
исходит быстрее в широтном направлении, чем в меридиональном,
Б. П. Васильков выделяет следующие зоны: зона арктических пустынь,
тундровая, лесная (делящаяся иа две подзоны — тайгу и широколист-
венные леса), степная, пустынная и горная.
Л. Н. Васильева (1967, 1973) иа основании ареалогического анали-
за Agaricales Приморья относит их к десяти типам ареала: космопо-
литному, голарктическому, палеарктическому, европейско-американо-
дальневосточиому, европейско-дальневосточному, американо-дальневос-
точному, дальневосточно-эндемичному, сибирско-дальиевосточиому, аме-
рикаио-азиатско-дальневосточному и тропическо-азиатско-дальиевосточ-
ному.
Э. Л. Нездойминого (1970) при географическом анализе шляпоч-
ных грибов северо-восточного побережья Байкала выделяет шесть гео-
графических элементов: арктоальпийский, гипоарктический, бореальный,
голарктический, мультирегиональный, горный. В пределах выделенных
142
географических элементов в зависимости от регионального распределен
ния составляющих его видов Э. Л. Нездойминого различает шесть групп
ареала: сибирскую, сибирско-дальневосточную, азиатско-американскую,
евразиатскую, евразнатско-американскую, космополитную.
И. Г. Нахуцришвили (1971, 1975) при географическом анализе
агарнкальных грибов Грузии выделяет шесть географических элементов:
альпийский, бореальный, неморальный, аридный, голарктический, муль-
тирегиональный, различая в иих шесть групп ареала: европейскую, ев-
разиатскую, евр американскую, евразиатско-американскую, космополит-
ную и кавказскую условно-эндемичную.
М. И. Беглянова (1975) на основании географического анализа
флоры агарикальиых грибов южной части Красноярского края выделя-
ет четыре географических элемента: бореальный, мультирегиональный,
неморальиый и моитаиный, характеризующиеся девятью типами ареа-
ла: голарктическим, европейско-азиатским, европейско-азиатским с ир-
радиацией в Австралию и Южную Америку, европейско-сибирским, си-
бирским, сибирско-дальневосточным, сибирско-североамериканским и
космополитным.
Географические анализы макромицетов из различных системати-
ческих групп приведены в работах П. Е. Сосина (1952. 1973), А. С. Бон-
дарцева (1953), Э. X. Пармасто (1955, 1959, 1963), Ж. Фавра (Favre,
1955), М. Ланге (Lange М., 1957), Т. Л. Николаевой (1961),
С. Р Шварцман (1962, 1968, 1973), Г. Крайзеля (Kreisel, 1961),
А. Г. Райтвийра (1964), С. Р. Шварцман, Н. Т. Кажиевой (1968),
Э. Л. Нездойминого (1970), И. Г. Нахуцришвили (1971, 1975), Л. Н. Ва-
сильевой (1973), X. Дорфельта (Dorfelt, 1971, 1973, 1974), К. А. Кала-
мээса (1975), С. П, Вассера и И. М. Солдатовой (1977а,б), К. Кала-
мээса (Kalamees, 1978b), А. Е. Коваленко (1980) и др. Однако в этих
работах отсутствуют общие единые принципы географического анали-
за флоры.
Одни авторы расчленяют флору на географические элементы, осно-
вываясь на зональном принципе, другие — на региональном, третьи
используют одновременно соединение зонального и регионального прин-
ципов, получившего широкое распространение среди ботаииков-геогра-
фов (Толмачев, 1932, 1974; Meusel, Jager, Weinert, 1965; Трасс, 1970).
С. Р. Шварцман (1. с.), А. Г. Райтвийр (I. с.) считают, что при мико-
географическом анализе следует исходить из изучения собственных
ареалов грибов и на базе этого их в дальнейшем классифицировать.
Географическое распространение грибов подчиняется несколько иным
закономерностям, чем распространение высших растений. Поэтому, как
указывает К. А. Каламээс (1975), нельзя при микогеографическом ана-
лизе механически применять классическое понятие элемента флоры,
используемое при изучении географического распространения высших
растений. Одиако такой подход все-таки до сих пор доминирует в ра-
ботах микологов. На современном уровне знаний это понятно: для
большинства групп грибов картина мирового распространения видов
далеко ие полная, а имеющаяся информация не дает возможности
представить специфические закономерности географического распро-
странения грибов. Сегодня еще ие хватает данных для сопоставления
специальных классификаций микоареалов (Каламээс, 1. с.).
По мнению К. А. Каламээса (1.С.), при изучении микоареалов це-
лесообразно использовать сравнительно-географический анализ ареалов
по X. Мойзелю, Э. Ягеру, Э. Вайнерту (Meusel, Jager, Weinert, 1965),
ИЗ
основанный на зонально-региональной схеме. Согласно этой схеме ми-
ровое распространение грибов осуществляется через формулу (диаг-
ноз), в которой по зонам растительности и ступеням континентальности
указана встречаемость вида на континентах или же на их частях. По
этим формулам, например, распространение некоторых видов семейства
Agaricaceae на земном шаре выражается следующим образом:
1. Agaricus silvicola (Vitt.) Pk s. auct.
austrostrop — trop AFR-bborcostrop — b. (suboz) AFR —
EUR — VORDAS — STB — О AS — JAP+NAM; temp CIRC-
POL.
2. Cystoderma granulosa (Fr.) Fay.
austr AUST + boreostrop — arct. (suboz) AFR — EUR —
VORDAS —ZAS —SIB —OAS — JAP+NAM — GRONL;
temp CIRCPOL.
3. Lepiota alba (Bres.) Sacc.
trop AFR-rsm.-b.(oz) EUR—VORDAS- ?SIB.
4. Macrolepiota mastoidca (Fr.) Sing. s. Konr. ct Maubl., non
s. Bres.
austrostrop — trop AFR+borcostrop— temp, suboz AFR —
EUR —WS1B+IND+OAS + NAM —b. euk OAS.
Формулы составлены в точном соответствии с растительно-клима-
тическими зонами, установленными X. Мойзелем, Э. Ягером, Э. Вайнер-
том, и с использованием их терминологии. В формулах (—) —беспре-
рывное распределение вида, (4-) —более или меиее обширные пробе-
лы в современном распространении вида. Знак +, использованный в
работе К. А. Каламээса (I. с.), ие следует интерпретировать в качестве
обозначающего дизъюнкцию ареала, как его упогрсбляст Мойзель.
В дальнейшем К. А. Каламээс считает, что для более детальной
характеристики географического распространения видов у грибов под-
ходит понятие геоэлемента, по Ю. Д. Клеопову (1938), установленное
па чисто географической основе и базирующееся на региональных под-
разделениях растительного покрова земного шара. Но выделение этих
геоэлементов требует точного картирования видов грибов и является
задачей будущего.
Вопросам картирования распространения грибов начали уделять
более или менее серьезное внимание только в последние десятилетия
(Eckblad, 1963; Kubicka, 1964; Hansen, Lange M., 1966; Skirgicllo,
1967—1972; Kotlaba, 1968, Bresinsky, 1971; Bresinsky, Dichtel, 1971;
Kreisel, 1971, 1973b; Lange L., 1974; Dorfelt, Kreisel, Benkert, 1979).
По данным Г. Крайзеля (Kreisel, l.c.), за период с 1930 по 1972 гг.
в различных журналах опубликовано лишь 1079 региональных карт
распространения 815 видов, в то время как общее число видов Basidio-
mycctcs составляет около 16 000 (Hawksworth, Sutton, Ainsworth, 1983).
Мы вслед за И. А. Дудкой (1976) считаем, что на современном
уровне знаний создание классификаций или схем распространения гри-
бов отдельных групп, пусть даже основанных на принципах географии
различных групп растений, необходимо, ибо только с помощью геогра-
фического анализа представляется возможным установление законо-
мерностей возникновения той или иной группы грибов, приуроченности
к определенной территории, путей их миграции и т. я, В дальнейшем
эти данные будут базисом для создания схемы специфических законо-
мерностей географического распространения грибов.
Основной единицей при географическом анализе флоры Agaricaceae
144
СССР мы считаем элемент флоры, который характеризуется определен-
ными типами ареала. Типы ареала, таким образом, являются главной
характеристикой элемента флоры. Предложенная нами схема будет пре-
терпевать изменения по мере возрастания степени изученности флоры;
однако выделенные элементы флоры и ареалы полезны тем, что позво-
ляют судить об общем характере рассматриваемой флоры.
При географическом анализе флоры Agaricaceae СССР мы столкну-
лись с тем, что в данных о распространении этих грибов как по СССР,
так и по земному шару много пробелов.
Агариковые грибы встречаются по всему Советскому Союзу от
Арктики до южных границ Средней Азии. Как видно из «Конспекта
флоры агариковых грибов СССР» (см. гл. IV), пять видов (Agaricus
arvensis, A. campestris, Cystoderma amianthina, Lepiota clypeolaria,
Macrolepiota excoriata) растут во всех флористических областях, вклю-
чая Арктику, четыре вида (Agaricus augustus, Endoptychum agaricoidcs,
Lepiota cristata, Macrolepiota procera) встречаются во всех областях,
кроме Арктики, два вида (Agaricus semotus, Cystoderma granulosa) —
во всех флористических областях, кроме Средней Азии, а четыре вида
(Agaricus silvaticus, A. silvicolus, Cystoderma carcharias, Lepiota casta-
nea)—во всех флористических областях, кроме Арктики и Средней
Азии. Наибольшим количеством видов и внутривидовых таксонов ага-
риковых грибов представлена европейская часть СССР — 149, затем
Дальний Восток — 72, Кавказ — 56, Восточная Сибирь — 47. Западная
Сибирь — 42, Средняя Азия — 32, Арктика — 11.
Наиболее полно выявлен видовой состав высших грибов Европы,
Северной Америки, Африки, значительно меньше — Азии н совсем сла-
бо — Южной Америки и Австралии. Имея такие, еще очень неполные
данные, трудно сделать окончательный вывод о географии видов Aga-
ricaceae, поскольку дальнейшее уточнение ареалов может пошатнуть
некоторые представления о них или полностью их опровергнуть. Наш
географический анализ, разумеется, дает только приблизительную
картину.
В результате географического анализа флоры агариковых грибов
СССР выделено семь географических элементов: эвриголарктический 1
(57 видов), мультирегиональный (33), неморальный (25), бореальный
(17), ксеромеридиональный (11), монтанный (3), альпийский (1), ха-
рактеризующихся 12 типами ареала: европейским (24 вида), евразиат-
ским (35), евразпатско-американским (16), еврафрнканским (5), евра-
зиатско-афрнканским (19), еврамериканским (4), еврамерикано-афри-
канским (2), евразнатско-американо-африканским (18), космополитным
(19), дальневосточным усчовно-эндемичным (1), северокавказским
условио-эндсмичным (1), украинским условно-эидемичным (3).
Одиннадцать видов агариковых грибов (Chlorophyllum molybdites,
Lepiota vircscens, L. lignicola, L. ncophana, L. micropholis, L. ventrioso-
spora, L. wichanskyi, L. hapalopoda, Macrolepiota olivicri, M. promincns,
Pscudobaeospora defibulata) рассматриваются как виды с неопределен-
ными ареалами, поскольку для одних данные об их распространении в
большинстве случаев противоречивы (например, Lepiota viresccns, L. ha-
palopoda, L. micropholis), для других неодинаково трактуется различ-
ными авторами объем вида (иапример, Chlorophyllum molybdites, Le-
piota wichanskyi).
1 Порядок расположения элементов в пашей схеме связан с количеством отне-
сенных к тому или иному элементу видов агариковых грибов.
12 4 2960
145
Эвригол арктический элемент. Его (голарктический — у X. X. Трас-
са, 1970) выделил А. Н. Окснер в работе «Анализ и история происхож-
дения лихенофлоры Советской Арктики» (1940—1942). Виды этого эле-
мента широко распространены в различных растительно-климатических
зонах Голарктики, не выявляя особой приуроченности к одной опреде-
ленной зоне. А. Н. Окснер указывает, что местонахождения этих видов
не приурочены к определенной растительно-климатической зоне, а про-
ходят через все зоны от севера до юга.
Такая же трактовка этого элемента у М. Ф. Макаревич (1963),
X. X. Трасса (1970), И. А. Дудки (1976). М. Ф. Макаревич под эврпгол-
арктическим элементом понимает виды, которые в определенных секто-
рах Голарктики или по всей Голарктике распространены во всех зо-
нах — от Арктики до степей и пустынь, а в горах не имеют тесной связи
с определенными горными растительными поясами. Представители эври-
гол арктического элемента — виды с широкой экологической амплиту-
дой. А. Н. Окснер (1974) указывает, что эвриголарктический элемент
охватывает виды с дисперсно распределенными ареалами, которые,
по-видимому, в силу их широкой экологической амплитуды более или
менее равномерно размещены в нескольких зонах Голарктики (с. 241).
Во флоре агариковых грибов СССР к этому элементу относятся
57 видов, которые разделены на семь типов ареала: европейский, евр-
азиатский, евразиатско-американский, еврафрикаиский, евразиатско-аф-
риканский, еврамерикано-африкаиский, евразиатско-американо-афри-
канский.
Евразиатско-американо-африкаиский тип ареала обьединяет 5 ви-
дов агариковых грибов — Agaricus rusiophyllus, Cystoderma cinnabari-
na, Lepiota rufipes, L. seminuda, L. acutesquamosa. распространенных
в Евразии, Северной Америке и на севере Африки.
Agaricus abruptibulbus, A. meleagris, Lepiota cygnea, L. eriophora,
Phaeolepiota aurea, Squamanita paradoxa относятся к евразиатско-аме-
рикаискому типу ареала эвриголарктического элемента.
Евр азиатско-африканский тип ареала эвриголарктического элемен-
та охватывает 10 видов агариковых грибов — Agaricus excellens, Lepiota
alba, L. castanea, L. brunneoincarnata, L. fulvella, L. serena, L. setulo-
sa, L. helveola, Leucocoprinus badhamii, L. georgineae. Многие виды
этого элемента, несомненно, будут найдены и в Северной Америке.
Еврамерикано-африкаиский тип ареала включает единственный вид
Agaricus benesii, обнаруженный в ряде стран Европы, США и Марокко.
Наибольшим количеством видов (24) представлен евразиатский
тип ареала эвриголарктического элемента. К нему относятся Melano-
phyllum eyrei, Agaricus macrosporus, A. squamuliferus, Cystoderma su-
perbum, Lepiota adulterina, L. echinacea, L. hystrix, L. erminea, L fus-
covinacea, L. grangei, L. ignicolor, L. oreadiformis, L. parvanulata,
L. pseudofelina, L. pseudohelveola, L. rhodorhiza, L. scobinella, L. serici-
fera, L. subalba, L. tomentella, Macrolepiota konradii, M. nympharum,
M. puellaris, Leucocoprinus brebissonii. Возможно, дальнейшее более
глубокое изучение Северной Америки и Северной Африки изменит наши
представления о видовом составе агариковых грибов этого типа ареала.
Еврафрикаиский тип ареала охватывает четыре вида агариковых
грибов — Agaricus langei, A. maleolens, Lepiota griscovirens, L. pallida.
Agaricus dulcidulus, A. lanipes, A. pseudopratensis, A. vaporarius,
Chamaemyces fracidus, Lepiota echinella, Macrolepiota permixta обра-
зуют европейский тип ареала эвриголарктического элемента.
146
Наиболее распространенными эвриголарктичсскими видами во фло-
ре Agaricaceae СССР являются Agaricus abruptibulbus, Cystoderma cin-
nabarina, Lepiota alba, L. acutesquamosa.
Мультирегиональный элемент. Мультирегиональный (плюрпрегно-
нальный, бореотропический, мультизональиый, убиквистический н др.)
элемент объединяет виды, произрастающие, кроме Голарктики, в дру-
гих флористических царствах и в различных растительно-климатических
зонах, по меньшей мере на трех континентах (Оксиер, 1940—1942, 1956;
Макаревич, 1963; Трасс, 1970).
А. Н. Окснер (1974) уточнил понятие мультирегионального элемен-
та: «...наконец, мультирегиональный элемент включает ареалы видов,
размещенных на трех и более континентах, причем один из них нахо-
дится вне Голарктики» (с. 241). В отношении размеров и конфигура-
ции ареалов мультирегиональных видов наблюдаются существенные
различия, на основании чего можно выделить ряд субэлементов п вари-
антов. Например, М. Ф. Макаревич (1963) разделила 229 мульти-
региональных видов лихенофлоры Украинских Карпат между 13 типа-
ми ареала и 18 группами распределения. В настоящей работе мульти-
региональный элемент мы не подразделяем. Характер расселения ага-
риковых грибов, относящихся к нему, показывает, что представители
его не распространены азонально. Мультирегиональные виды рассе-
ляются в разных регионах земного шара, но, безусловно, всегда связа-
ны с условиями произрастания, соответствующими определенной расти-
тельно-климатической зоне, благоприятной для расселения данных
видов.
Признавая вслед за М. Ф. Макаревич (1963), А. Н. Оксиером
(1974), И. А. Дудкой (1976) мультирегиональный элемент как объеди-
няющий виды агариковых грибов, встречающиеся не менее чем на
трех континентах и распространенных как в Голарктике, так и за ее
пределами, мы констатируем, что в состав этого элемента входит 33 ви-
да. Более детальный анализ географического распространения агарико-
вых грибов мультирегионального элемента дает интересные материалы,
показывающие, что многие виды его широко распространены на всех
континентах, кроме Антарктиды. Тем не менее каждый вид (или группа
видов) имеет характерный для него ареал.
Мы выделяем среди агариковых грибов мультирегионального эле-
мента трн типа ареала, причем космополитный тип ареала, условно
принимаемый нами для видов, встречающихся на всех континентах зем-
ного шара, кроме Антарктиды, включает 19 видов. К этому типу ареала
относятся Melanophyllum echinatum, Agaricus arvensis, A. campestris’,
A. bisporus, A. bitorquis, A. xanthodermus, Cystoderma amianthina»
C. granulosa, Lepiota cristata, L. clypeolaria, Leucocoprinus cepaestipes,
L. denudatus, Leucoagaricus leucothitus, Macrolepiota procera, M. rhaco-
des, M. excoriata, A. mastoidea, M. gracilenta, Endoptychum agaricoides.
Некоторые виды с космополитным типом ареала, например Agaricus
bisporus, Leucocoprinus cepaestipes, получили широкое распространение
благодаря хозяйственной деятельности человека.
Евразиатско-американо-африканский тип ареала мультирегиональ-
ного элемента представлен И видами: Agaricus augustus, A. haemor-
rhoidarius, A. silvaticus, A. silvicolus, A. porphyrizon, Gyrophragmium
Agaricus campestris и Cystoderma amianthina обнаружены и в Антарктиде.
12’
147
dunalii, Lepiota aspera, Lepiota lilacea, L. subincarnata, L. xanthophylla,
Leucocoprinus birnbaumii.
Евразиатско-американскип тип ареала мультирегионального эле-
мента охватывает три вида агариковых грибов — Agaricus placomyces,
A. purpurellus и Lepiota subgracilis, произрастающих, кроме Голаркти-
ки, в Южной Америке (Аргентина, Боливия, Венесуэла, Перу, Тринидад
и Тобаго).
Наиболее распространенными мультирегиональными видами в на-
шей флоре являются Agaricus campestris, A. bitorquis, Endoptychum
agaricoides, Macrolepiota excoriata, M. procera и др.
Неморалвный элемент. Его выделил А. С. Лазаренко (1944, 1956).
Позже неморальный элемент был принят при географическом анали-
зе рядом ботаников (Окснер, 1956; Макаревич, 1960, 1963; Трасс, 1970;
Нахуцришвили, 1971, 1975; Дудка, 1976). К неморальному элементу от-
носятся виды, встречающиеся главным образом в зоне широколиствен-
ных лесов Голарктики. X. X. Трасс (1970) включает в неморальиый
элемент виды эвнеморальные и омнинеморальные (т. е. распространен-
ные кроме широколиственных лесов Голарктики и в аналогичных место-
обитаниях вне Голарктики). За пределами Голарктики омнинемораль-
ные виды распространены весьма разнообразно: одни установлены для
всех континентов Южного полушария, другие — для одного, а некото-
рые выходят только незначительно за пределы Голарктики.
Для анализа флоры ограниченной территории важно объединить
в неморальный элемент и те виды голарктической иеморальной флоры,
которые кроме Голарктики встречаются и во флорах других флористи-
ческих царств. Например, И. Г. Нахуцришвили (1971) к неморальному
элементу относит вичы широколиственных лесов Голарктики, иногда
встречающиеся и в хвойных лесах.
Неморальные виды в Голарктике распространены дизъюнктивно и
отражают современный фрагментарный характер иеморальиой расти-
тельно-климатической зоны. Некоторые из них расселяются далеко иа
север в бореальную зону, заходя иногда в лесотундру и даже тундру
(Окснер, 1974).
Количество агариковых грибов, обнаруженных в зоне широколист-
венных лесов н, следовательно, относящихся к нсморальиому элементу,
составляет 25 видов. Мы выделяем среди неморального географического
элемента восемь типов ареала агариковых грибов. Наибольшим коли-
чеством видов (по 8) представлен европейский тип ареала неморальио-
го элемента, к которому принадлежат Agaricus bresadolianus, A. brun-
neolus, A. osccanus, A. stramineus, Lepiota bucknallii, C. pulvcrulenta,
L. cygneoaffinis, Leucoagaricus carneifolius, и евразиатскнй тип ареала,
представленный следующими видами агариковых грибов: Agaricus fus-
cofibrillosus, A. lutosus, A. phaeolepidotus, A. romagnesii, A. sagatus,
Lepiota hetieri, L. rosea, Leucoagaricus moseri.
К евразиатско-американскому, еврамерикаио-африкаискому и евр-
африканскому типам ареала относятся соответственно по одному виду:
Agaricus subrufescens, A. fissuratus и A. spissicaulis.
Три вида агариковых грибов — Agaricus nivescens, A. moclleri и
Leucocoprinus pilatianus — составляют евразиатско-африкапский тип
ареала неморального элемента.
Украинский условно-эндемичный тип ареала нсморального элемен-
та представлен двумя видами — Agaricus amanitaeformis н A. longi-
caudus.
118
Дальневосточный условно-эндемичный тип ареала неморального
элемента представлен Lepiota lateritiopurpurea, описанной с юга Даль-
него Востока (Васильева, 1973).
Неморальный элемент у агариковых грибов несколько отличается
от понимания содержания этого элемента А. Н. Окснером (1974, с. 241).
Многие виды агариковых грибов, составляющие неморальный элемент,
не имеют массового распространения в широколиственных лесах Гол-
арктнки, а, наоборот, относятся к редким видам, известным нз неболь-
шого числа местонахождений. Однако до последнего времени они из-
вестны лишь нз зоны широколиственных лесов Голарктики и поэтому
могут быть отнесены к неморальному элементу.
Бореальный элемент. К нему относятся виды, которые растут в
основном в зоне хвойных лесов всего Северного полушария, продвигаясь
нередко в зону широколиственных лесов и даже в степную зону. Встре-
чаются они и в Арктике, главным образом в ее гнпоарктическон части
Кроме того, многие бореальные виды произрастают и за пределами
Голарктики в зонах, которые по условиям существования приближают-
ся к бореальной. Такие виды А. Н. Окснер (1940—1942, 1944) назвал
нотобореальными. М. Ф. Макаревич (1963) также рассматривает ното-
бореальные виды в ранге самостоятельного элемента. Учитывая, что
многие нотобореальные виды часто встречаются и являются характер-
ными для хвойной полосы лесной зоны Голарктики, мы вслед за
X. X. Трассом (1970) включаем нх в бореальный элемент, однако не
придаем им ранга субэлемента. В понимании бореального элемента мы
придерживаемся взглядов А. Н. Окснера (1940—1942, 1956), А. С. Ла-
заренко (1947) и X. X. Трасса (1970).
К бореальному элементу флоры агариковых грибов СССР отно-
сятся 17 видов, которые разделены на мн на семь типов ареала.
Европейский тип ареала объединяет пять видов агариковых гри-
бов— Agaricus altipes, A. subfloccosus, A. macrocarpus, A. leucotrichus,
A. chionodermus.
Agaricus aestivalis, известный из хвойных лесов Англии, ВНР, Да-
нии, СССР, ФРГ, Франции, ЧССР и Израиля, имеет евразнатский тип
ареала.
Четыре вида агариковых грибов — Cystoderma rugosoreticulata,
С. adnatifolium, Lepiota cortinarius, Agaricus xantholepis — составляют
сврамериканскнй тип ареала бореального элемента. A. xantholepis в
последнее время отмечен для микофлоры Чили (Singer, 1969, 1975).
Не исключено, что при дальнейшем накоплении данных A. xantholepis
будет отнесен к иному элементу.
Евразнатско-американский и евразнатско-амсрикано-африканский
типы ареала бореального элемента образуют соответственно Lepiota
fclina, Cystoderma carcharias и C. fallax, Agaricus semotus. A. semotus
отмечен и для мнкофлоры Чили (Singer, 1969). Полный ареал этого
вида, вероятно, более широк. Возможно,' при дальнейшем накоплении
данных A. semotus будет отнесен к нноку элементу.
Lepiota kuchneri и Agaricus variegatus образуют евразиатско-афри-
капский тип ареала бореального элемента.
Cystoderma caucasicum, известная лишь нз хвойных насаждений
Кавказского заповедника (Ставропольский край), образует северокав-
казский условно-эндемичный тип ареала.
Наиболее распространенными бореальными видами флоры агари-
149
ковых грибов СССР являются Cvstoderma carcharias и Agaricus
^ernotus.
Ксеромеридиональный элемент. Такое название введено А. Н_- Окс-
нером (1940—1942) для обозначения видов, произрастающих в тепло-
аридных областях Голарктики. М. Ф. Макаревич (1963) дает ксеро-
мери тональному элементу следующее определение: «Представители
этого элемента расположены обычно зонально на юг от неморальной
зоны, однако довольно часто наблюдаются отклонения от такого рас-
пространения» (с. 99). Виды данного элемента связаны в своем распро-
странении со степями, полупустынями н пустынями.
В фнтогеографнческой литературе ксеромеридиональный элемент,
в связи с тем что ему уделялось издавна много внимания, имеет бога-
тую синонимику: аридный, ксероконтннеитальный, понтический, ксеро-
фнльный, субпонтический, понтосарматский, паннонский и др. (MeuscI,
1943; Лазаренко, 1956; Макаревич, 1963; Meusel, Jager, Weinert, 1965;
Трасс, 1970).
К ксеромернднональному элементу флоры агариковых грибов СССР
относятся 11 видов, которые разделены нами на пять типов ареала.
Европейский тип ареала объединяет два вида агариковых грибов:
Agaricus maskae. описанный нз степных районов ЧССР (Pilat, 1954)
и позже обнаруженный в пуштах ВНР (Bohus, 1961), степях Украины
(Вассер, 1974, 1980), засушливых районах СРР, ФРГ, и A. porphyro-
cephalus.
Leucocoprinus bresadolae и Agaricus velenovskyi обладают евразиат-
ским типом ареала ксеромеридионального элемента.
Agaricus cupreobrunneus н A. tabularis относятся к евразиатско-
амернканскому типу ареала. A. tabularis обнаружен в Арктике (Ва-
сильков, 1974), что существенно может изменить наши представления
о его принадлежности к ксеромеридиональному элементу вообще. Одна-
ко для этого потребуются дальнейшие тщательные исследования таксо-
номии A. tabularis.
Agaricus bernardii, A. gennadii, Leucoagaricus holosericcus и
L. macrorhizus образуют евразиатско-афрнканскнй тип ареала ксероме-
ридионального элемента. Все указанные виды произрастают в степных,
полупустынных районах Евразии и на севере Африки (Алжир, Ма-
рокко).
Leucocoprinus bohusi, описанный с территории Хомутовской запо-
ведной разнотравно-типчаково-ковыльной степи (Вассер, 1975а, 1980;
Wasser, 1979), образует украинский условно-эндемичный тнп ареала
ксеромеридионального элемента.
Монтанный элемент. К моитаиному элементу относятся виды, про-
израстающие в лесных поясах гор Голарктики (Окснер, 1940 -1942,
1974; Лазаренко, 1956; Макаревич, 1963; Трасс, 1970). По мнению
X. X. Трасса (1. с.), монтанный элемент является одним.из наиболее
«расплывчатых», имеющих переходы к нескольким другим элементам,
особенно к таким, как, напрнмер, бореальный. М. Ф. Макаревич (1. с.)
уточняет понятие монтанного элемента, отмечая, что «...монтанные виды
нередко снижаются в предгорья, реже поднимаются в более высокие
безлесные пояса или снижаются на равнины ... Монтанные виды можно
разделить на две экологические группы. Первая нз них связана с леса-
ми нижнего горного пояса, преимущественно буковыми н буково-пихто-
выми, отчасти и с предгорьями; вторая — с еловыми лесами верхнего
горного пояса».
150
Среди агариковых грибов, как и среди всех высших Basidiomyce-
tes, сравнительно мало облигатных монтанных видов, ие встречающих-
ся или даже редко встречающихся в равнинных условиях. В связи с
этим, на наш взгляд, при географическом анализе отдельных групп
Basidiomycetes различных регионов земного шара микологами не вы-
делялся монтанный элемент. Упоминание о монтанных видах грибов
имеется лишь в статье X. Дорфельта (Dorfelt, 1973).
Принимая в общем определение монтанного элемента по А. Н. Ок-
снеру (1. с.) н М. Ф. Макаревич (1. с.), мы включаем в этот элемент н
виды, произрастающие также вне Голарктики, но в аналогичных мон-
танных условиях.
Во флоре агариковых грибов СССР к моитанному элементу отно-
сятся три вида — Agaricus deylii, Cystoderma ambrosii, Pseudobaeospora
pillodii, распределяющиеся между двумя типами ареалов.
Agaricus deylii, описанный с территории горной части ЧССР (Piiat,
1951b) н впоследствии обнаруженный лишь в ВНР и СССР (Bohus,
1969; Вассер, 1980), произрастает главным образом в горных еловых
лесах и представляет европейский тип ареала монтанного элемента.
Cystoderma ambrosii н Pseudobaeospora pillodii относятся к евразн-
атско-американскому типу ареала монтанного элемента. Cystoderma
ambrosii, известная из некоторых стран Европы, азиатской части СССР
и США, произрастает в еловых, елово-буковых, сосново-березовых, ли-
ственно-сосновых горных лесах, в ольхово-кедровых насаждениях среди
мхов, доходит до высоты 1900 м н. у. м. Pseudobaeospora pillodii из-
вестна из Австрии, Франции, Швейцарии, азиатской части СССР, США
и Чили, произрастает лишь в редкостойных высокогорных лесах под
видами родов Pinus, Larix, Picea, Rhododendron и Alnus viridis.
Альпийский элемент. К нему относятся виды, ареалы которых огра-
ничены главным образом высокими горами Голарктики, где они произ-
растают только в высокогорьях альпийского н субальпийского поясов.
И. Г. Нахуцришвили (1975) к альпийскому элементу относит виды суб-
альпийских и альпийских лугов, которые спускаются на опушки суб-
альпийских лесов и даже встречаются в степях (с. 66). Такое, в значи-
тельной мере расплывчатое, определение этого элемента мы не при-
нимаем. В связи с такой трактовкой, на наш взгляд, И. Г. Нахуцришвили
ошибочно отнес Agaricus tabularis, широко распространенный в
степях Украины, пустынях н полупустынях Средней Азии, прериях
Северной Америки, к альпийскому элементу. Наше понимание альпий-
ского элемента тождественно пониманию этого элемента М. Ф. Мака-
ревич (1. с., с. 103).
Среди агариковых грибов, как и среди всех высшнх Basidiomyce-
tes, сравнительно мало облигатных альпийских видов, не встречающих-
ся или даже редко встречающихся в неальпийских условиях.
К альпийскому элементу отнесен одни вид — Agaricus kuehnerianus.
Он известен лишь из альпийских лугов Франции (Heinemann, 1974)
и яйл Горного Крыма (Вассер, 1980).
Наличие во флоре агариковых грибов СССР ксеромернднональных,
монтанных, альпийских, а также неморальных и бореальных видов с
дальневосточным, северокавказским и украинским условно-эндемичны-
ми типами ареала подчеркивает определенную специфичность нашей
флоры.
Рассматривая специфику флоры агариковых грибов, следует оста-
новиться иа вопросе об ее эндемизме. Некоторые виды — Lepiota lateri-
151
tiopurpurea, Leucocoprinus bohusi, Cystoderma caucasicum, Agaricus ama-
nitaeformis, A. longicaudus и разновидность Melanophyllum eyrei var.
macrosporus на данном этапе исследования флоры агариковых грибов
известны лишь в определенных регионах, т. е. вполне соответствуют
определению эндемов (табл. 6). Эндемичные таксоны отличаются неко-
торыми характерными чертами, важнейшими из которых являются:
а) географическая изоляция; б) специфическая экологическая н ценоти-
ческая приуроченность; в) систематическое положение. Эндемичные ви-
ды представляют специфическую составную часть флоры и отличают
ее от всех других флор. Отсюда, как отмечает А. И. Толмачев (1974),
их особое знач’ение для выявления «лица» каждой флоры. Однако к
вопросу об эндемизме агариковых
грибов следует подходить осторожно.
Эндемизм, наблюдающийся у ряда ви-
дов, может быть только кажущимся.
Проблема эндемизма у грибов тесно
связана с вопросом редкости видов,
который является одним из аспектов
проблемы частоты встречаемости
(Наумов, 1972; Дудка, 1976). Послед-
няя, как указывал Н. А. Наумов
(1. с.), представляет один нз основных
вопросов микогеографнн.
Н. А. Наумов (1. с., с. 13—14)
считал, что «...«редкость» вида можег
объясняться различными причинами, в
том числе наличием действительности
Таблица 6. Распределение эндемичных
видов в отдельных родах агариковых
грибов СССР
Род Количество видов
всего ЭНД1 мяч-
Agaricus. 59 2
Lepiota 54 1
Cystoderma 10 1
Leucocoprinus 9 1
Melanophyllum * 2 1
* Для рода Melanophyllum эндем пред-
ставлен в ранге разновидности — М. eyrei
var. macrosporus.
узкого ареала распространения вида...
в результате его вымирания или нового заноса; разрозненного харак-
тера распределения в пределах данной территории; ... годовых отклоне-
ний в мнкофлоре местности».
Наконец, как указывал Н. А. Наумов, может иметь место так на-
зываемая кажущаяся редкость вида, которая является следствием недо-
статочного внимания к грнбу или слабой его изученности, хотя в дей-
ствительности этот внд может быть очень широко распространен. По-
следнее указание Н. А. Наумова, как отмечает Й. А. Дудка (1976),
особенно применимо к водным грибам. На наш взгляд, оно также при-
менимо н к агариковым сапротрофным грибам. На него следует опираться
прн выяснении вопроса об эндемичных видах у агариковых грибов.
Эндемизм во флоре агариковых грибов, как и в моховых и лишай-
никовых флорах (Koch, 1954; Абрамов, 1969; Васильева, 1973; Барду-
нов, 1974; Вассер, Солдатова, 1977а, б), характеризуется тем, что про-
цент эндемичных видов грибов всегда ниже, чем процент эндемичных
видов высшнх растений той же территории.
Сравнительно низкий процент эндемизма у агариковых грибов в
частности, как н у всех высших Basidiomycetes вообще, характерен и
для отдельных регионов СССР. По данным Л. Н. Васильевой (1973),
эндемичные виды во флоре Agaricales Приморья составляют 4 %, тогда
как у высших древесных растений—более 90 %- Во флоре северо-
восточного побережья Байкала из 337 видов шляпочных грибов имеется
лишь один эндем (Нездойминого, 1970). Кавказ является страной энде-
мизма для цветковых растений, а из 569 видов Agaricalcs Грузии толь-
ко 11—эндемы (Нахуцрншвнли, 1975)-. Из 513 видов Agaricales Хи-
152
бннского горного массива — один эндем (Михайловский, 1975), а из
165 видов шляпочных грибов Копетдага— три эндема (Батырова,
1977а, б).
В связи с тем что «эндем» — понятие широкое в с равным правом
можно говорить как о европейском эндемизме, так и об узком эндемиз-
ме Украины, Кавказа или какого-либо иного региона СССР, все шесть-
названных таксонов являются узколокальными эндем ам и, поскольку
сосредоточены на ограниченной территории.
Приведенные эндсмы (Leucocoprinus bohusi, Cystoderma caucasi-
cum, Agaricus amanitaeformis, A. longicaudus, Melanophyllum eyrci var.
macrosporus) чрезвычайно редки и каждый известен только из одного
местонахождения (Урбонас, Каламээс, Лукин, 1974; Вассер, Солдато-
ва, 1977а, б; Вассер, 1980). Lepiota lateritiopurpurea имеет три место-
нахождения в Приморском крае: Уссурийский р-н. Уссурийский запо-
ведник; окрестности Владивостока, Дальневосточный ботанический сад;
Шкотовский р-н, пос. Кангауз (Васильева, 1973).
Различна и экология наших эндемов. Agaricus amanitaeformis
и Melanophyllum eyrei var. macrosporus произрастают в смешанных
широколиственных насаждениях парков, Agaricus longicaudus — в сме-
шанных широколиственных лесах, Leucocoprinus bohusi — в заповедных
целинных разнотравно-типчаково-ковыльных степях. Lepiota lateritio-
purpurea найдена в смешанных хвойно-широколнетвеиных лесах. Cysto-
derrna caucasicum растет в смешанных лесах, на валежной гнилой дре-
весине Abies nordmannianac (Stev.) Spach. на высоте до 700 м п. у. м.
Отсутствие среди наших эндемов видов с высокой степенью отли-
чий от ближайших родичей, пространственно с ними не контактирую-
щих, принадлежащих к монотипным родам, показывает, что их можно
рассматривать как неоэндемы. Все наши эндем ы-таксоны относительно
молодые, ареалы нх находятся в стадии прогрессивной фазы развития.
Мы полностью согласны с Ц. Шретером (Schroter, 1926), который на-
зывает неоэндемнзм прогрессивным илн обновленным Неоэндемы чаще
всего генетически пластичные виды. Как недавно возникшие, они, обыч-
но, в морфологическом отношении близки к видам, распространенным
на этой же или на соседней с ней территории.
О времени возникновения Agaricales s. 1. можно судить в основном
косвенно, так как данные палеомикологии очень незначительны. К на-
стоящему времени в работах ряда авторов (Ячевский, 1933; Andrews,
Lenz, 1943; Roscndalil, 1943, 1948; Чигуряева, 1953; Tyler, Barghoorn,
1954; Havlena, 1956; Криштофович, 1957; Singer, Archangel sky, 1958;
Holm, 1959; Бопдарцев, 1960; Попов, 1961, 1967; Gridland, 1952; Шварц-
ман, 1962; Graham, 1962; Martin, 1968; Dennis R., 1970, 1976; Tiffney,
Barghoorn, 1974; Pirozynski, 1976; Singer, 1977; Тетеревникова-Бабаян,
1981, и др.) описано свыше 500 видов ископаемых грибов различных
систематических групп. Известные в мировой литературе палеомиколо-
гические сведения рассеяны большей частью в виде статей в периоди-
ческой печати по разным отраслям знаний (палеонтология, ботаника,
география, микология, общая биология и т. д.). Почти нет обобщающих
монографий по ископаемой флоре грибов отдельных геологических эр
или периодов (Тетеревникова-Бабаян, 1981). Наиболее полные данные
по ископаемым грибам мира приведены в статье Б. Тиффией и Э. Бар-
гурпа (Tiffney, Barghoorn, 1974). Имеющиеся немногочисленные досто-
верные палеомикологпческие данные показывают их преимущество пе-
ред сравнительно-морфологическими, широко используемыми при уста-
153
новлении времени возникновения отдельных таксонов грибов (Savile,
1955, 1968; Шварцман, 1962; Piiat, 1971; Зеров, 1972, н др.). Время воз-
никновения некоторых таксонов грибов, определенное не иа основании
палеомикологических данных, обычно сдвигается к палеозойскому гео-
логическому периоду (Зеров, 1972) или к сравнительно молодым геоло-
гическим периодам (Васильева, 1973).
Палеомикологическне данные свидетельствуют о том, что комплек-
сы фоссильных грибов различных геологических периодов различаются
между собой. Однако, как отмечал еще А. А. Ячевскнй (1933, с. 759),
полученные палеомикологическне данные «с полной очевидностью уста-
навливают существующую неразрывную связь современного состава
микологической флоры с комплексом грибных форм, существовавших
в доисторические периоды;».
Палеомикологическая летопись высших Basidiomycetes, в том числе
н Agaricales s. L, слишком скудна, н достоверные данные об их суще-
ствовании известны лишь из более ранних геологических периодов
(Ячевский, 1933; Skirgiello, 1964; Шварцман, 1962; Pirozynski, 1976).
По последним данным, на основании нахождения Palaeosclerotium
(Dennis R., 1970, 1976; Singer, 1977), высшие Basidiomycetes с пряж-
ками существовали в карбоне (верхнем). Со среднего и верхнего карбо-
на известны и представители остальных классов грибов. По мнению
Р. Зингера (Singer, 1977), именно в карбоне следует искать Basidiomy-
cetes с примитивными признаками.
К карбону относятся н наиболее древние находки эндотрофной ми-
коризы. По данным А. Н. Криштофовича (1957), в корнях кордаитов
найдена микориза. Кордаиты (Cordaitales) — палеозойские голосемен-
ные с сохранившимся анатомическим строением — известны под назва-
нием Amyllon. Им свойственно наличие эндотрофной микоризы в клет-
ках внутренней коры. В отложениях нижнепермского периода, по дан-
ным А. Н. Криштофовича, также найдена, и не единично, микориза.
Из карбона различных мест мира известны представители ряда родов
высших Basidiomycetes. Так, С. Р. Шварцман относит найденный ею
ископаемый окаменелый гриб из отложений карбона Казахстана к Plcu-
rotus lignatilis 1 и отмечает, что на хвойных палеозоя уже развивались
грибы рода Pleurotus. В отложениях юрского н мелового периодов обна-
ружены представители многих родов современных Basidiomycetes (Fe-
mes, Boletus, Dacdalea, Pleurotus, Clitocybe, Agaricus, Trametcs и др.).
Принимая микоризу как приобретенное в процессе эволюции «со-
жительство» 2, известное с верхнего карбона, считаем возможным да-
тировать появление первичных, самых примитивных Basidiomycetes
более поздннм геологическим периодом, таким, как девонский. Действи-
тельно, в современной флоре Agaricales s. 1., если за основу взять чет-
1 Правильное название Clitocybe lignatilis (Fr.) Karst
3 В связи с этим возникает вопрос о первичном типе питания у грибов. По
мнению ряда авторов, в том числе многих отечественных, первичным способом пита-
ния грибов следует признать сапротрофиое, свойственное большинству ныне живущих
видов. По мнению других, наиболее видным представителем которых является
Д. Б. О. Сэвайл (Savile, 1955), паразитизм у грибов древнее сапротрофизма. Однако
палеомикологическне и современные данные (Шварцман, 1962; Martin, 1968; Cain,
1972; Lutrell, 1974; Горленко, 1976; Каратыгин, 1981, и др.) позволяют нам отдать
предпочтение идее первичности сапротрофизма. Мы полностью согласны с И В. Ка-
ратыгиным (1981). который отмечает, что, хотя нет прямых доказательств сапротро-
физма как исходного способа существования грибов, тем не менее многочисленные
косвенные данные свидетельствуют, что паразитические грибы выеокоспециализиро-
ванные, а потому и вторичные группы организмов.
154
вертую систему Р. Зингера, насчитывается около 3500 видов (175 ро-
дов), относящихся к сапротрофам, и лишь около 1600 видов (37—40 ро-
дов)— к микоризообразователям. Девять семейств Agaricales s. 1. не
имеют ни одного представителя, относящегося к микоризообразователям
(Ritter, 1980). В эктотрофных микоризных ассоциациях в качестве
хозяев выступают средн голосеменных все Abietineae, а среди покрыто-
семенных— Salicaceae, Juglandaccae, Corylaceae, Fagaceac, Myrtaceae
и Tiliaceae. По А. Энглеру (Engler, 1964) ’, Salicaceae, Juglandaceac,
Corylaceae н Fagaccae являются крайне примитивными, при этом се-
режку — плод покрытосеменных он помещает на одну ступень выше
шишки хвойных. Согласно такой схеме, существует переход от голо-
семенных растений к покрытосеменным, образующим эктотрофные ми-
коризные ассоциации. Здесь исключение составляют лишь Tilia и Euca-
lyptus, поскольку они более совершенны, чем сережкоцветные. Мико-
ризообразователи заселяют преимущественно бореальные и умеренные
климатические зоны, сапротрофы же имеют бореальный, космополитный
и преимущественно субтропнческо-троинческий тнп географического
распространения. По данным ряда авторов (Trappe, 1962; Hacskaylo,
1971, и др.), микоризообразование распространено у незначительного
количества видов. Среди Basidiomycetes представители всего 91 рода 1 2
способны образовывать микоризу. По мнению Э. Хачкайло (1. с.), исхо-
дя из данных современного анализа Basidiomycetes, можно заключить,
что в доисторические времена микоризные грибы вели сапротрофное
существование. Переход к микорнзообразованию (взаимному парази-
тизму, по Хачкайло) произошел с развитием голосеменных растений.
А в процессе эволюции определенных видов покрытосеменных растений
происходила адаптация грибов в условиях сбалансированного сосуще-
ствования с некоторыми из них. Можно предположить, что в случае
филогенетически отдаленных родов, отличных морфологически, незави-
симое развитие сходных физиологических или биохимических особен-
ностей создало предпосылку для перехода от сапротрофного существо-
вания к экологически паразитическому. С. Д. Гарретт (Garrett, 1956)
предполагает, что все микоризные базидиомицеты являются экологи-
чески облигатными симбионтами, а корневая ассоциация почти всегда
играет существенную роль в завершении их жизненного цикла. У ми-
коризных Basidiomycetes эволюция идет преимущественно через формы,
возникающие при развитии вида в симбиозе с разными видами высших
растений. У них имеются пары близких видов (например, Hygrophorus
piceae н Н. eburneus, Cortinarius traganus и С. hicinus), связанных с
разными видами высших растений. Возникли они, по мнению Л. Н. Ва-
сильевой (1973), в горных полидоминантных хвойно-широколнетвенных
лесах, подобно тем, которые сейчас произрастают на юго-востоке Се-
верной Америки и на юге Дальнего Востока.
Отсутствие па леем и кол оги чески х материалов не позволяет точно
установить время возникновения агариковых грибов. Однако учитывая,
что грибы, относящиеся к семейству Agaricaceae, облигатно немикорнз-
ные, ареал большинства из ннх ограничен Южным полушарием, придер-
живаясь взглядов первичности сапротрофизма, можем с определенной
степенью достоверности предположить, что первичные примитивные
1 Вопросы филогении покрытосеменных растений рассмотрены А. Л Тахтаджя-
ним (1978 а).
2 По данным Д. Л. Хауксуорта, Б. У. Саттона, Дж. Айнсуорта (Hawksworih,
Sutton, Ainsworth, t983), Basidiomycetes насчитывает 1100 родов (4-910 синонимов).
155
Возраст,
млн. лет
Стратиграфическая
эпоха
5- 10е
Ранний археи
ЗЮ9
Докембрий
7-Юе
5 10е
?,25Ю8
10в
7 107
Первичная (палеозой-
ская) зро
кембрий
Ордовик
Силур
Девон
карбон
Пермь___________
Вторичная (мезозой-
ская) эра
Триас
Юра
мел
Третичная (кайнозой-
ская) эра
Зоцен
Олигоцен
Миоцен
Плиоцен
Четвертичный
период
Рнс. 13. Время (млн. лет) возникновения основных групп высших растений и грибов:
I — Poiypodiophyta. Г—Pinophyts. 3 — Magnoliophyta. 4 — Myxomycetes. 5 — Chitndiomycetes. 6—
Oomycetes, 7 — Zygomycetes, 8 — Ustomycetcs (a — (J redinales. 6 — Ustilaginales), 9 — Ascomycetes-
(a — Taphrinales. 6 — Erysiphales, в — Sphaeriales). 10 — Basidiomycetes (a — Agaricales s. J., 6 —
Agaricaceae. e—Boletales. a — Russula les), tl — Denteromycetes (no Tiftoey. Barghoorn. 1974; Ра-
маду, 1981; Каратыгину. 1981. с дополнениями).
агариковые грибы возникли в ннжнем карбоне или в перми’ (рис. 13),
а с мелового периода находятся в стадии процветания, разлагая расти-
тельные остатки высших растений. Представители же иных таксонов
Agaricales s. 1. образуют с высшими растениями микоризную связь. По
данным В. Я. Частухина и М. А. Николаевской (1969), вышине Basidio-
1 Связи некоторых видов родов Leucocoprinus в Leucoagaricus с термитами
Северо-Восточной Азии, которые возникли в юре — мелу, подтверждают ранее геоло-
гическое появление представителей этих родов агариковых грибов. Произрастание
ряда видов рода Agaricus лишь на унавоженной почве или на навозе даст основание
отнести их появление к периоду развития (триас) травоядных млекопитающих (Фле-
ров, Крамаренко, Борисов, и др., 1974; Беляева, 1980).
156
mycctes, активно участвуя в процессах распада растительных остатков,
доводят минерализацию листьев растений до 80 %. Расцвет немикориз-
ного семейства Agaricaceae, на наш взгляд, связан с послеледниковым
временем четвертичного периода (голоцен), когда третичная флора Ев-
ропы и большей части Азнн почти полностью погибла и возникла более
холодостойкая флора, представленная в основном хвойными (Pinus,
Picea, Abies, Larix и др.) н лиственными (Ulmaccae, Bctulaceae, Ассга-
сеас, Salicaccae н др.) древесными растениями Сохранилось также
много вечнозеленых кустарников и полукустарников, особенно из семей-
ства Ericaceae. В постгляциальный период после отступления последне-
го вюрмского ледника образуются зоны, близкие к современным. В это
время, на наш взгляд, и формируется уже в основном современная фло-
ра немикоризного семейства Agaricaceae.
Учитывая данные географического анализа, данные анализа рас-
пространения агариковых грибов на земном шаре, принимая во внима-
ние наличие во флоре Agaricaceae юга Северной Америки, Южного
полушария большого процента эндемичных видов, считаем возможным
предположить, что некоторые ныне существующие виды, вероятно, со-
хранились с момента существования третичной флоры, которая в Юж-
ном полушарии претерпела мало изменений.
В связи с изложенным интересным н важным нам представляется
установление географических центров происхождения агариковых гри-
бов. Это связано со значительными трудностями, поскольку агарикологн
могут опираться лишь па материалы, порой довольно неполные, о совре-
менном распространении видов. Достоверные ископаемые остатки агари-
ковых грибов обнаружены лишь с конца третичной эры и в четвертичном
периоде. Отсутствие таких данных о более ранних эпохах делает любые
выводы по этому вопросу субъективными и спекулятивными. Такую
мысль высказали X. X. Трасс (1970, с. 184) относительно лишайников,
И. А. Дудка (1976, с. 172) относительно водных гифомнцстов, ио это в
равной мере относится и к агариковым грибам.
По мнению ряда авторов (Масюк, 1973; Дудка, 197G), главным
базисом при установлении географических центров происхождения низ-
ших растений на земном шаре при отсутствии палеоботанических данных
является определение регионов наибольшего видового разнообразия,
регионов, содержащих примитивные, реликтовые и эндемичные так-
соны, а также ряд других факторов (систематическое положение, эко-
логические особенности и др.).
Как н для цветковых растений (Тахтаджян, 1970), гипотетические
центры происхождения Agaricales s. I. (в том числе и Agaricaceae)
можно искать в высоких и в низких широтах — тропических и субтро-
пических. Однако сразу считаем целесообразным отбросить гипотезу
высокоширотного центра происхождения агариковых грибов. Предполо-
жение о существовании в третичной эре на территории Арктики богатых
субтропических и даже тропических флор в последние десятилетия не
подтвердились палеоботаническими исследованиями. Теперь уже впол-
не ясно, что климатические условия Арктики были во все времена
неблагоприятными для развития богатой н разнообразной раститель-
ности, а тем более для развития новых групп растений.* Арктическая и
Антарктическая области с нх продолжительными н темными зимами
являются наименее вероятными местами для возникновения новых круп-
ных систематических групп (Тахтаджян, 1970).
Колыбелью происхождения высших Basidiomycetes являются
157
низкие широты. Современные флоры палеотропического 1 (мадагаскар-
ская, индокитайская, малезийская, фиджийская, новокаледонская облас-
ти) и голантарктического (повозетандская область) царств представлены
наиболее примитивными, узколокальными, реликтовыми таксонами, яв-
ляющимися недостающими звеньями в филогенетических цепях высших
Basidiomycetes. В последнее десятилетие исследования флоры Новой
Гвинеи, Новой Зеландии, Новой Каледонии, Соломоновых о-вов, о-вов
Фиджи и Мадагаскара обнаружили больше примитивных высших Ba-
sidiomycetes, чем все вместе взятые сравнительно хорошо изученные
флоры высоких широт. Основную колыбель происхождения высших Ba-
sidiomycetes, как и цветковых растений (Bailey, 1949: Тахтаджян, 1970),
следует искать в странах, бывших частями гипотетического континента
Гондваны2. Именно в странах Юго-Восточной Азии, Австралии и Ме-
ланезии сосредоточены основные наиболее примитивные исходные пред-
ставители секотиевых грибов, давшие на самых высоких уровнях, по
нашему представлению, начало Agaricales s. 1. Например, в областях
распространения Nothofagus (Новая Зеландия, Тасмания, Новая Ка-
ледония, Юго-Восточная Австралия) сосредоточены древние угасающие
роды высших Basidiomycetes, в то время как высокоширотные предста-
вители их носят дериватный характер. Если рассмотреть семейство
Agaricaceae, то все роды его встречаются в нижних широтах, причем
наиболее примитивные, эндемичные (Clarkeinda, Hiatulopsis, Micropsal-
liota, Volvolepiota и др.) произрастают только здесь и не встречаются
в высоких широтах.
Основываясь на приведенных данных по Agaricaceae и по современ-
ному распространению Agaricales s. 1. (Singer, 1975), можно с уверен-
ностью утверждать, что высшие Basidiomycetes мигрировали нз низких
широт в высокие, а не наоборот. Анализ современных ареалов высших
Basidiomycetes склоняет к мысли, что первичный центр происхождения
и в дальнейшем расселения их находится в той части, которую зани-
мает сейчас Юго-Восточная Азия. Слабая изученность Южного полу-
шария пока не дает нам оснований высказаться о точных географиче-
ских центрах происхождения агариковых грибов, хотя таковые, несо-
мненно, имеются 3. Подтверждением этому служит тот факт, что начатое
изучение палеотропиков в отношении ряда семейств Agaricales s. 1. (на-
пример, Entolomataceae, Cortinariaceae) показало их значительную ро-
довую специфику и разнообразие (Ногак, 1973; Moser, Ногак, 1975).
Следовательно, имеющиеся пробелы в данных о распространении
агариковых грибов, как и отсутствие палеомнкологических материалов,
не позволяет сегодня более определенно указать географические цент-
ры возникновения грибов этого семейства и пути их миграции. И лишь
Юго-Восточная Азия по своим физико-географическим условиям н со-
временному видовому составу с максимальной вероятностью соответ-
ствует гипотетическим географическим центрам возникновения агари-
ковых грибов.
1 Флористическое районирование Земли, доведенное до провинций, принимается
по А. Л Тахтаджяну (19786).
3 Новые реконструкции показали, что современные южные материки — Антарктида,
Австралия, Африка н Южная Америка на протяжении всего палеозоя были объеди-
нены в единый гигантский континент Гоидвану (Городницкий, 1978).
3 Наличие в Африке и Южной Америке, бывших частях палеозойского континен-
та Гондваны, значительного количества эндемичных, примитивных высших Basidio-
mycetes, не известных из высоких широт, дает возможность предположить, что и на
этих континентах находятся центры возникновения, а возможно, лишь нх расселения.
ГЛАВА VII
НАРОДНОХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
АГАРИКОВЫХ ГРИБОВ
В СССР произрастает 73 вида агариковых грибов, пригодных для1
употребления в пищу, 16 несъедобных, 13 ядовитых и три смертельно-
ядовитых вида; 31 вид из-за малых размеров карпофоров отнесен к
группе грибов, не имеющих пищевого значения. Пищевая ценность
22 видов не изучена.
Питательная ценность съедобных грибов мало изучена. Большин-
ство проведенных экспериментов заканчивается сравнением белка гри-
бов с белком мяса или овощей (Anderson, Fellers, 1942; Cernohorsky,
Machura, 1947). В. Линтцел (Lintzel, 1941, 1943) одним из первых полу-
чил данные о степени усвояемости грибного белка. На основании этих
результатов было показано, что 100—200 г грибов (по сухой массе)
достаточно для обеспечения суточного питательного баланса у человека-
массой 70 кг. В. Лиитцел (Lintzel, 1943) приравнивал питательную цен-
ность грибов к мускульной ткани животных и белкам овощей. Опыты
по скармливанию грибов крысам подтвердили тот факт, что грибы
имеют определенную питательную ценность, но не могут служить един-
ственным источником белка (Quackenbuch, Peterson, Steenbock, 1935;
Fitzpatrick, Esselen, Weir, 1946; Antal, Morava, Zsinka et al., 1969;
Hayes, Haddad, 1975). При расчете питательной ценности грибов долж-
ны быть учтены три взаимосвязанных фактора: перевариваемость, со-
держание белка и аминокислот (Crisan, Sands, 1978).
Для расчета питательной ценности грибов Б. Л. Осер (Oser, 1951,
1959) предложил использовать метод, основанный на определении со-
держания незаменимых аминокислот. Индекс незаменимых аминокислот
(ЕАА) представляет собой отношение суммы содержания незаменимых
аминокислот в пище по отношению к содержанию незаменимых амино-
кислот в белке, обладающем высокими питательными свойствами (бе-
лок куриного яйца).
Иное определение питательной ценности, используемое некоторыми
исследователями,— это критерий аминокислот, называемый иногда хи-
мическим критерием (Food Nutr. Meet. Rep., 1973). Критерий амино-
кислот вычисляют как отношение количества наиболее лимитированных
аминокислот, присутствующих в исследуемом белке, к количеству этих
аминокислот, содержащихся в эталонном белке (белок куриного яйца).
Было бы неверно оценивать питательную ценность грибов, осно-
вываясь только на анализе содержания белка и аминокислот. Необхо-
димо учитывать существующую связь между качеством и количеством
даииого белка и незаменимыми свободными аминокислотами (Crisan,
Sands, 1978). При использовании указанных выше расчетных величин
159
трудно сравнивать питательную ценность грибов, содержащих неболь-
шое количество высококачественного белка, с грибами, содержащими
большое количество белка низкого качества.
Для решения этой проблемы было предложено использовать индекс
питательности, рассчитываемый следующим образом:
ЕАА-яндекс X содержание белка, %
100
При расчете питательной ценности грибов необходимо учитывать
количество не только связанных, но и свободных аминокислот (Mag-
gioni, Rcnosto, 4970).
Все проанализированные виды грибов, в том числе шампиньон
двуспоровый, были лимитированы по содержанию по крайней мере
одной незаменимой аминокислоты. Кроме того, в грибах было на 5—
7 аминокислот меньше, чем в эталонном белке (Crisan, Sands, 1978).
Наиболее часто среди лимитированных аминокислот встречались серо-
содержащие аминокислоты: лейции, изолейцин, валин, лизин, аромати-
ческие аминокислоты, треонин и триптофан. Большое значение для под-
счета питательной цеиности грибов кроме достаточного содержания той
или иной аминокислоты имеет ее доступность. Исследование содержа-
ния лизина в некоторых грибах показало, что только 10—62 % лизина
присутствует в доступной для питания форме (Losota, Mlodecki, Wlo-
darczyk, 1968).
Исходя из анализа литературных данных можно заключить, что
питательная ценность съедобных 1рибов — специфический признак от-
дельных видов и даже штаммов. Так, индекс питательности различных
штаммов шампиньона двуспорового колеблется от 3 до 28—32 единиц
(Crisan, Sands, 1978).
Грибы имеют большое значение как источники витаминов, экстрак-
тивных, ароматических и других веществ (Ramsbotom, 1953; Шиврина,
Низковская, Фалина и др., 1969; Crisan, Sands, 1978).
Съедобные грибы пока используются недостаточно полно в народ-
ном хозяйстве СССР. Из наиболее ценных съедобных агариковых гри-
бов следует отметить Agaricus campestris, A. kuchncrianus, A. macrospo-
rus, A. bitorquis, A. arvensis, Macrolepiota proccra.
Наряду co съедобными видами в СССР встречаются несъедобные,
ядовитые и даже смертельно ядовитые виды агариковых грибов. Виды
этой группы представлены Agaricus xanthodermus var. xanthodermus,
A. xanthodermus var. lepiotoides, A. meleagris, A. mocllcri, Lepiota brun-
neoincarnata, L. helveola, L. cristata, Chlorophyllum molybdites и др.
Отравление агариковыми грибами — нередкий случай среди насе-
ления СССР. Зарегистрированы случаи отравления Lepiota brunneoin-
carnata, L. helveola, Agaricus xanthodermus var. xanthodermus (Василь-
ков, 1966; Садыхов, 1968; Зерова, 19706), некоторые с летальным
исходом.
Агариковые грибы применяются в медицине для получения анти-
биотических, антикоагуляционных, антибластических веществ.
Некоторые виды могут служить продуцентами антибиотиков. На-
пример, продукт метаболизма очень распространенного в СССР вида
Agaricus campestris успешно используется при лечении тифа. Вытяжки
из A. campestns применяются при диабете, укусах змей. Порошки из
A. campestris обладают дезинфицирующими свойствами (Turynek, 1952;
Bose, 1953; Bauchet, 1961; Шиврина, Низковская, Фалина н др., 1969;
160
Вассер, 1980). Учитывая, что по отношению к уже существующим анти-
биотикам часто развиваются устойчивые штаммы бактерий нли у па-
циентов вырабатывается аллергия к ним, работы по изысканию новых
антибиотических веществ из агариковых грибов имеют огромное зна-
чение.
Лечение грибами (фунготерапия) развито слабо. Общее количе-
ство видов, используемых для лечения различных заболеваний, ие пре-
вышает 30—40 (Bauchct, l.c.; Зерова, 1963; Шиврина, Низковская, Фа-
лина, 1. с. и др.; Зерова, Вассер, 1972; Вассер, I. с.). Средн агариковых
грибов лекарственных видов сравнительно немного. В качестве сырья
для фармацевтической промышленности можно рекомендовать Agari-
cus campestris. Французскими учеными (Bauchei, 1961) разработана
методика приема грибных порошков — вдыханием небольших доз через
нос и пероральное употребление.
В настоящее время грибы в основном рассматривают как дополни-
тельный источник белка. В будущем в связи с возрастанием белкового
дефицита роль грибов как продукта питания будет увеличиваться. В свя-
зи с постоянно возрастающим на мировом рынке спросом на грибы гри-
боводство во многих странах выделилось в самостоятельную высокопро-
дуктивную отрасль сельского хозяйства (Гарибова, 1971; Steineck, 1973;
Вассер/Дудка, Фрщ и др., 1976; Dudka, Wasser, Soldatova, 1977; Дудка,
Вассер, Бухало и др., 1978; Яковенко, Жемойц, 1978). Производство гри-
бов осуществляется круглогодично, независимо от погодных и почвен-
ных условий. Урожаи грибов, например шампиньона двуспорового,
в условиях наиболее прогрессивных грибоводческих комплексов с со-
временным оборудованием и технологией может достигать 11—13 тыс. ц
с гектара в год при стеллажной системе (Вассер, Дудка, Фр1д и др., l.c.).
Возможность искусственного выращивания ценных съедобных гри-
бов (во всяком случае, большинства известных дикорастущих видов)
до настоящего времени окончательно не решена, но давно привлекает
внимание ученых, практиков. На данном этапе развития грибоводства
11 видов грибов являются объектами искусственного выращивания в
промышленном масштабе: Agaricus bisporus, A. subedulis, A. bitorquis,
Stropharia rugosoannulata Far!, ex Marr., Volvariella volvacea (Fr.)
Sing., Lentinus edodes (Berk.) Sing., Pleurotus ostreatus (Jacq.: Fr.)
Kumm., P. florida Fovose, Flammulina velutipes (Fr.) Sing., Kuehncro-
rnyces mutabilis (Fr.) Sing, et A. Sm., Tuber melanosporum Vitt. (Гари-
бова, 1971; Steineck, 1973; Вассер, Дудка, Фрщ и др., 1976; Дудка,
Вассер, Бухало и др., 1978; Chang, Hayes., cd., 1978).
Ряд видов агариковых грибов (Agaricus bisporus, A. bitorquis,
A. subedulis, A. arvensis, A. macrosporus, A. silvaticus, A. vaporarius,
Macrolepiota procera, M. zeyheri, Leucoagaricus leucothitus) культиви-
руют в промышленном масштабе либо проходят опытные испытания
на их пригодность для массового промышленного культивирования.
Наиболее широкое распространение в промышленной культуре среди
всех съедобных грибов получил представитель семейства Agaricaceae —
шампиньон двуспоровый.
В СССР начиная с 30-х годов, когда культура шампиньонов после
времеиного сокращения в годы первой мировой войны вновь начала
восстанавливаться (Ячевский, 1933), исследования по промышленному
грибоводству были проведены Е. С. Клюшниковой (1938, 1939),
М. А. Пановым (1950, 1956), Н. Г. Громовым (1957, 1960), Л. В. Га-
рибовой (1964, 1966, 1969а, б, 1970, 1971, 1978), К. В. Рыбкиной
13 4—29Ь0
161
(1967, 1971), С. И. Додылевой (1968), Л. В. Гарибовой и Е. Н. Фроло-
вой (197-1), Л. А. Девочкиным (1975), С. И. Китаевым, О. Н. Бубновой,
А. А. Борисовым и др, (1976), С. П. Вассером, И. А. Дудкой, Ю. Ф. Фри-
дом и др. (1976), И. А, Дудкой, С. П. Вассером, И. М. Солдатовой
(Dudka, Wasser, Soldatova, 1977), Ю. И. Казокиным (1977), А. И. Пи-
липович, Н. С. Марышевой (1977), И. А. Дудкой, С. П. Вассером,
А. С. Бухало и др. (1978), Л. П. Колесниченко, И. А. Дудкой, С. П. Вас-
сером (1978), С. А. Кушнаревым (1978), М. Д- Кулешом (1978),
А. А. Дворниной (1978), А. 3. Яковенко, А. А. Жемойц (1978),
Н. С. Гончаруком, Л. А. Девочкиным (1978), Н. А. Бисько, Л. П. Ко-
лесниченко,’ Е. П. Володиной и др. (1979), И. А. Дудкой, С. П. Вас-
сером, Н. А. Бисько (Dudka, Wasser, Bisko, 1979) и др.
Современный этап развития грибоводства в нашей стране выдвигает
перед исследователями важные научные проблемы, требующие тща-
тельного изучения и решения, это в частности:
1) получение эффективными методами ступенчатой селекции с при-
менением физических н химических мутагенов новых высокоурожайных
болезнеустойчивых штаммов шампиньона двуспорового и других съедоб-
ных грибов;
2) введение в культуру новых, ранее не выращиваемых искусствен-
но видов съедобных грибов из числа гумусовых сапротрофов, лигно-
трофов и др.;
3) исследование физиолого-биохимических особенностей культиви-
руемых видов грибов с целью использования их для интенсификации
промышленного производства плодовых тел;
4) поиск новых дешевых общедоступных субстратов для культиви-
рования грибов; проверка возможности использования сельскохозяй-
ственных, промышленных и городских отходов как субстрата для куль-
тивирования грибов с одновременным осуществлением биологической
деструкции отходов;
5) усовершенствование методики и технологии производства посев-
ного мицелия на зерне и перлите.
Учитывая стоящие перед отечественным грибоводством проблемы,
основное внимание мы сосредоточили на получении новых отечествен-
ных продуктивных штаммов гумусовых сапротрофов шампиньона двуспо-
рового, штамма вирусоустойчивого вида шампиньона двукольцового и
их производственной проверке 1. Ниже лишь в резюмированном виде
осветим основные полученные результаты по промышленному шам-
пиньоноводству.
В результате использования моноспоровой селекции шампиньона
двуспорового (Гарибова, 1964; Гарибова, Фролова, 1971) получены вы-
сокопродуктивные штаммы BKMF-2471D, BKMF-2472D, BKMF-2474D,
рекомендованные для введения в производство.
С целью получения новых высокоурожайных раннеспелых штаммов
шампиньона двуспорового с улучшенным габитусом плодовых тел сле-
дует использовать УФ-лучи лазерного источника с плотностью энергии
4,8 Дж/см2 и у-лучи от кобатьтовой установки в дозах 200, 400 и 500 Гр,
По важнейшему показателю биохимического состава плодовых тел —
содержанию белка, полученные нами при облучении указанными доза-
ми УФ- и у-лучей штаммы не отличались от исходных.
1 Все результаты по данному вопросу опубликованы ранее (Бисько, Бухало,
Вассер я др , 1983).
162
Производственные испытания трех отселектированных, защищен-
ных авторскими свидетельствами СССР штаммов шампиньона двуспо-
рового (BKMF-2471D, BKMF-2472D, BKMF-2474D) показали, что их
урожайность и габнтус карпофоров зависят от типа посевного мицелия
(зерновой или навозный) и от состава компоста (натуральный, полу-
синтетический, синтетический). Использование посевного мицелия иа
зерне способствует получению более высокой урожайности в течение
всего процесса плодоношения, увеличивая ее по сравнению с навозиым
посевным мицелием иа 8,9—14,1 %. В то же время масса плодовых тел
увеличивалась на 13,9—16,2 %, а диаметр шляпки — на 3,8—5,6%.
Урожайность отселектированных штаммов на полусинтетическом и син-
тетическом компостах достоверно выше, чем иа натуральном. Масса
плодовых тел на синтетических компостах превышала на 11,3—63,4 %
массу плодовых тел, выращенных на натуральном компосте. Анализ
полученных данных по урожайности и габитусу отселектированных
штаммов шампиньона двуспорового, проведенный с помощью двухфак-
торного эксперимента, показывает, что урожайность иа 74,4 %, а габи-
тус иа 38,6 % зависят от состава компоста и на 5,7 % и 21 % соответ-
ственно — от биологических особенностей штамма.
Определение условий, при которых достигается наибольший эко-
номический эффект от выращивания трех штаммов шампиньона дву-
спорового нашей селекции, позволило сделать вывод о наиболее высо-
ком уровне рентабельности штамма BKMF-2471D при выращивании
его на полусинтетическом, а штаммов BKMF-2472D и BKM.F-2474D —
на синтетическом компостах с солодовыми ростками. По сравнению с
контрольным штаммом, в качестве которого использовался штамм 117,
уровень рентабельности для штамма BKMF-2471D повысился на 22,5 %,
для штамма BRMF-2472D — 28,1, для штамма BKMF-2474D — на 45 %.
Рентабельность производства при использовании зернового посевного
мнцелия увеличилась по сравнению с навозным на 14,6—19 %.
В результате проведенных нами исследований получен и проверен
в опытио-промышленных условиях штамм (К-1) нового для грибовод-
ства СССР агарикового гриба — шампиньона двукольцового. Шампинь-
он двукольцовый отличается от широко культивируемого шампиньона
двуспорового рядом свойств, которые делают его культивирование пер-
спективным (Vcddcr, 1970, 1978; Dielman van Zaayen, 1972, 1976; Frit-
sche, 1974, 1978; Klaver, 1978). Плодовые тела шампиньона двукольцо-
вого —- беловатые с плотной мякотью — сохраняют свой первоначаль-
ный вид после четырех-пяти дней хранения. Потеря массы плодовых тел
шампиньона двукольцового после хранения на 58—65 % ниже, чем у
шампиньона двуспорового. Температурные требования для роста мице-
лия ( + 30 °C) и развития плодоношения (+25 °C) у этого вида на
4—5 °C выше, чем у шампиньона двуспорового. Кроме того, шампиньон
двукольцовый может расти при более высокой концентрации СО2, чем
шампиньон двуспоровый. Особо ценным является то, что шампиньон
двукольцовый устойчив к вирусным болезням, причиняющим значитель-
ный вред шампиньоповодству. Отобранный нами н интродуцированный
в культуру штамм шампиньона двукольцового может быть предложен
для культивирования иа юге европейской части СССР в летнее время
и круглогодично в среднеазиатских республиках и на Кавказе, а также
в случае появления симптомов вирусных заболеваний шампиньона дву-
спорового в шампииьониицах СССР.
13*
ЗАКЛЮЧЕНИЕ
В Советском Союзе выявлено 158 видов, 29 разновидностей и семь
форм агариковых грибов, относящихся к 14 родам четырех колеи. Из
них девять видовых и внутривидовых таксонов (Agaricus romagnesii,
A. amanitaeformis, A. longicaudus, A. moelleri, Lepiota subgracilis,
L. clypeolaria var. latispora, Cystoderma amianthina var. longispora,
Leucocoprinus bohusi, Leucoagaricus moseri) описаны нами как новые
для науки, для 23 видовых и внутривидовых таксонов предложены но-
вые для науки таксономические комбинации, 42 видовых и внутриви-
довых таксона, а также один род (Chlorophyllum) приведены как но-
вые для микофлоры Советского Союза. Составлен конспект флоры агари-
ковых грибов Советского Союза, включающий 194 таксона видового и
внутривидового рангов с учетом новых для науки таксонов н комбинаций,
предложенных нами.
Анализ систематических единиц агариковых грибов в ранге колена
показывает бесспорное преобладание Agariceae (63 вида) и Lepioteae
(57 видов) над Leucocoprineae (26 видов) и Cystodermateae (12 видов).
Наиболее крупными родами являются Agaricus (59 видов), Lepiota
(54), Macrolepiota (11), Cystoderma (10), Leucocoprinus (9 видов).
Остальные роды содержат менее девяти видов. Для семейств, содер-
жащих 15—20 родов (каковым является и предмет нашего исследова-
ния), при определении «лица» систематической структуры флоры пред-
лагаем использовать пять ведущих родов. Действительно, в пяти веду-
щих родах агариковых грибов Советского Союза содержится 154 вида,
что составляет 97,5 %. Аналогичные результаты получены и для обла-
стей флоры СССР.
На основаиии установления видового состава агариковых грибов
Советского Союза, а также систематического и структурно-сравнитель-
ного анализов с использованием коэффициентов Стугрена—Радулеску
и Кендэла выявлена видовая н родовая специфика грибов исследуемого
семейства по областям флоры и отдельным наиболее полно изученным
регионам. Наибольшее количество видов и внутривидовых таксонов за-
регистрировано в европейской части СССР — 149, затем на Дальнем
Востоке — 72, Кавказе — 56, Восточной Сибири — 47, Западной Сиби-
ри— 42, Средней Азии—-32, Арктике—11. Приведен видовой состав
агариковых грибов основных типологических единиц растительного по-
крова областей флоры СССР с указанием характерных и безразличных
видов, особенностей высотного и поясного распространения, сезонных
аспектов покрова, экологических групп.
164
В результате анализа различных интерпретаций как общей схемы,
так и частных вопросов систематики Agaricales s. 1. отмечена тенденция
обособления ряда новых порядков (до 9), так как Agaricales s. 1. объ-
единяет филогенетически сравнительно далекие группы грибов. К на-
стоящему времени процесс выделения порядков не закончен, а границы
и объем многих из них еще не стабилизировались. Описан новый для
науки порядок Hygrophorales и новое для науки семейство Catathelas-
mataceae порядка Tricholomatales. На данном этапе развития таксоно-
мии высших Basidiomycetes считаем необходимым систему Agaricales
s. 1. основывать главным образом на аиатомо-морфологическнх и, при
наличии большого материала, онтогенетических критериях. Вместе с тем
нельзя не учитывать эколого-биологическне, географические н физиоло-
го-биохимические признаки. В принятую нами систему пластинчатых
грибов включены восемь порядков (Polyporales s. str., Boletales s. str.,
Strobilomycetales, Hygrophorales, Tricholomatales, Amanitales, Agari-
cales s. str., Russulalcs), содержащих 23 семейства.
Определено место семейства Agaricaceae в современных системах
высших Basidiomycetes в рамках порядка Agaricalcs s. str. В Agarica-
ceae вошли роты из нескольких семейств порядка Agaricalcs s.l., имею-
щих различную окраску спорового порошка. В результате рассмотре-
ния критериев основных таксонов систем Agaricaceae, разнообразных
толкований систематического положения родов семейства считаем целе-
сообразным иа данном этапе расширять и углублять представления об
«односемействеиной» структуре Agaricaceae с четырьмя коленами (Aga-
riceae, Cystodermateae, Lepioteae, Leucocoprineae) без нарушения его
«границ». Выполненный нами анализ таксономического значения при-
знаков агариковых грибов на стабильность и эволюционную значимость
дал возможность выделить комплекс макро- и микроскопических при-
знаков, характеризующих семейство, колено, род, вид н внутривидовые
таксоны.
Всесторонний комплексный анализ около 45 родовых таксонов ага-
риковых грибов показал, что лишь меиее половины из них представ-
ляют собой хорошо обоснованные роды. Это связано с тем, что выде-
ление родов в семействе часто основаио на придании одному признаку
родового таксономического ранга, без учета прагматического «правила»
таксономии о величине гиатуса между таксонами и величине таксона.
К хорошо обоснованным родам относятся: Agaricus, Melanophyllum,
Gyroph г agmium, Endoptychum, Micropsalliota, Lepiota, Chamaemyces,
Pseudobaeospora, Smithiomyccs, Hiatulopsis, Cystoderma, Squamanita,
Phacolcpiota, Clarkeinda, Chlorophyllum, Macrolepiota, Leucoagaricus,
Leucocoprinus, Volvolcpiota. Остальные относимые в семейство Agari-
caceae роды (Crucispora, Cystoagaricus, Ripartitclla, Verrucospora, Leu-
cocortinarius, Chlorolepiota, Catathclasma, Floccularia, Armillaria)
исключены из него. Предложена оригинальная система основных родо-
вых таксонов Agaricaceae. В родах Agaricus, Lepiota, Leucocoprinus,
Cystoderma пересмотрена внутриродовая систематическая ценность
признаков.
Нами предпринята попытка использования геометрических зависи-
мостей базидий, макро- и микроскопических химических цветовых реак-
ций карпофоров и микроструктур, культурально-морфологических осо-
бенностей агариковых грибов в качестве дополнительных критериев так-
сонов различного ранга.
Анализ базидии на основе геометрических зависимостей открывает
165
новые возможности в систематике высших Basidiomycetes. Он заклю-
чается в изучении торцевых и боковых положений базидий со спорами.
Показана тесная связь между размерами и формой спор, стеригм и ба-
зидий, имеющая видовую и, возможно, родовую специфику. Предло-
женные нами расчеты позволяют определить ряд линейных и угловых
параметров (Л1, т, б, 0, а), которые очень трудно наблюдать и практи-
чески невозможно замерить. Практическое использование этих методов,
выведенных формул, установленных соотношений и их точности про-
верено на 17 критических таксонах агариковых грибов, относящихся
к пяти родам (Agaricus, Cystoderma, Macrolepiota, Leucoagaricus, Lcu-
•cocoprinus)..Отмечена их действенность как дополнительного таксоно-
мического критерия и а видовом и внутривидовом уровнях.
Исследования макро- и микроскопических химических цветовых
реакций видов агариковых грибов СССР показали, что многие нз них
(например, реакция с реактивом Мельцера у видов родов Cystoderma,
Lepiota s. str., Macrolepiota, Clarkeinda, Pseudobaeospora, Micropsallio-
ta, с реактивом Шеффера, с а-нафтолом у видов родов Agaricus и
Gyrophragmium, с крезилом синим у видов родов Macrolepiota, Leu-
•coagaricus, Leucocoprinus, Volvolepiota и др.) необходимо использовать
как надежный дополнительный таксономический критерий. Он позволя-
ет разграничивать родовые, внутриродовые и видовые таксоны.
Сравнительные исследования 12 видов агариковых грибов в культу-
ре иа пяти питательных средах далн возможность установить их важ-
нейшие культуральные особенности. Впервые изучены в условиях куль-
туры Agaricus rnaskae, A. bernardii, A. cupreobrunneus, Leucocoprinus
bresadolae, Macrolepiota puellaris. Наиболее благоприятными оказались
сусло-агаровые среды, а для ряда штаммов также среда Молнша. Для
каждого вида динамика роста на определенной питательной среде спе-
цифична. Окраска, морфология колонии, динамика роста, наличие веге-
тативного или бесполого спороношений на разных питательных средах,
как правило, постоянны и могут служить дополнительными таксономи-
ческими критериями.
Основной единицей при ареалогическом анализе грибов семейства
Agaricaceae СССР принят географический элемент флоры, который ха-
рактеризуется определенными типами ареала. В результате ареалогиче-
ского анализа флоры агариковых грибов СССР выделены семь геогра-
фических элементов: эвриголарктический (57 видов), мультирегиональ-
ный (33), неморальиый (25), бореальный (17), ксеромеридиональный
(11), монтаниый (3), альпийский (1), характеризующиеся 12 типами
ареала. 11 видов отнесены к группе с неопределенными ареалами. Пять
видов (Leucocoprinus bohusi, Agaricus amanitaeformis, A. longicaudus,
Cystoderma caucasicum, Lepiota latcritiopurpurea) и одна разновидность
(Melanophyllum eyrei var. macrosporus) агариковых грибов отнесены
к узкореальиым неоэндемам.
Отсутствие палеомикологических данных не позволяет точно уста-
новить время возникновения агариковых грибов, хотя единичные при-
митивные Basidiomycetes с пряжками известны из верхнего карбона.
Учитывая, что грибы, относящиеся к семейству Agaricaceae, облигатно
немикоризные, ареалы большинства современных видов ограничены
Южным полушарием, и, исходя нз признания взглядов на первичность
сапротрофнзма, высказано предположение о возникновении примитив-
ных агариковых грибов в нижнем карбоне или перми. С мелового перио-
да грибы семейства Agaricaceae находятся в стадии процветания, вы-
166
ступая в фитоценозах разного типа как деструкторы, разлагающие
растительные остатки высших растений.
Предполагается, что основной центр происхождения высших Basi-
diomycetes, как и цветковых растений, следует искать в странах, кото-
рые являлись частями палеозойского континента Гондваны. Современ-
ные ареалы высших Basidiomycetes дают основания предполагать, что
первичный центр происхождения и в дальнейшем расселения этих гри-
бов находился в той части, которую занимает сейчас Юго-Восточная
Азия. Именно в странах Юго-Восточиой Азии и Меланезии сосредото-
чены основные наиболее примитивные, исходные, угасающие предста-
вители секотиевых грибов, давшие на самых высоких уровнях, по на-
шему представлению, начало Agaricales s. 1. Анализ распространения
грибов семейства Agaricaceae показывает, что все его роды встречаются
в нижних широтах, причем наиболее примитивные, эндемичные произ-
растают только здесь и не встречаются в высоких широтах. Наличие в
Африке и Южной Америке, бывших ранее частями палеозойского кон-
тинента Гондваиы, значительного количества эндемичных, примитивных
высших Basidiomycetes, не известных в высоких широтах, дает воз-
можность предположить, что и на этих континентах находятся центры
возникиовения, а возможно, лишь расселения их.
Установлено, что в СССР произрастает 73 вида агариковых грибов,
пригодных для употреблеиия в пищу, 16 несъедобных, 13 ядовитых и
три смертельно ядовитых. Пищевая ценность 22 видов не изучена,
31 вид из-за мелких размеров карпофоров отнесен к группе видов, не
имеющих пищевого значения.
На основании схемы моноспоровой селекции шампиньона двуспо-
рового путем отбора быстрорастущих культур с плотным стелящимся
тяжистым мицелием без обильного воздушного роста (штаммы расы В)
с последующей гибридизацией отобранных штаммов и обработкой фи-
зическими мутагенами (*у- и УФ-лучами) нами получены высокопро-
дуктивные штаммы, рекомендованные для введения в производство.
Для получения новых высокоурожайных раннеспелых с улучшен-
ным габитусом штаммов шампиньона двуспорового следует использо-
вать УФ-лучи от лазерного источника с плотностью энергии 4,8 Дж/см2
и у-лучи от кобальтовой установки в дозах 200, 400 и 500 Гр.
Производственные испытания трех отселектированных штаммов
шампиньона двуспорового (BKMF-2471D, BRMF-2472D, BRMF-2474D)
показали, что их урожайность и габитус карпофоров зависят как от типа
посевного мицелия (зерновой или навозный), так и от состава компоста
(натуральный, полусинтетический, синтетический). Определены условия,
при которых достигается наибольший экономический эффект от выра-
щивания трех штаммов шампиньона двуспорового нашей селекции.
Выделен в чистую культуру, отобран и проверен в опытно-промыш-
ленных условиях штамм К-1 нового для грибоводства СССР ценного
съедобного вирусоустойчивого гриба — шампиньона двукольцового.
Шампиньон двукольцовын отличается от широко культивируемого шам-
пиньона двуспорового рядом свойств, которые делают его культивиро-
вание перспективным в летнее время на юге европейской части СССР,
иа протяжении всего года на Кавказе и в республиках Средней Азии,
а также в случае появления симптомов вирусных заболеваний шампиньо
на двуспорового в шампиньонницах СССР. Предложен регламент интро-
дукции его в промышленную культуру.
список
использованной литературы
Абрамов И. И. Проблема эндемизма у листостсбельных мхон.— Л.: Наука.
2969.-54 с.— (Комаров, чтения; Вып. 22).
Александрова В. Д. Арктические тундры СССР: Докл. по опублик. работам,
представл. к защите вместо дис. ... д-ра биол. наук.—-Л., 1964.— 43 с.
Бардунов Л. В. Листостебельные мхи Алтая и Саян.— Новосибирск: Наука,
1974 — 167 с.
Батырова Г. III.— Изв. АН ТССР. Сер. биол. наук, 1975, № 5, с. 37—45.
Батырова Г. III.— Изв. АН ТССР. Сер. биол. наук, 1977а, № 1, с. 30—35.
Батырова Г. UI. Шляпочные грибы Копет-Дага (порядки Boletales, Agaricales
s. str., группа порядков Gasteromycetes) : Автореф. дне. ... канд. биол. наук.— Ашха-
бад, 19776.—23 с.
Беглянова М. И.— Учен. зап. Краснояр. пед. ин-та, 1963, 24, вып. 6, с. 3—36.
Беглянова М. И. Флора агариковых грибов южной части Красноярского края.—
Красноярск : Б. И., 1972.— Ч. 1. 207 с.
Беглянова М. И. Флора агариковых грибов южной части Красноярского края :
Автореф. дис. ... д-ра биол. наук.— Владивосток, 1975.— 47 с.
Беденко Э. П.— Микология и фитопатология, 1979, 13, вып. 3, с. 184.
Беденко Э. П.— Новости систематики низш. растений, 1980, 17, с. 30—39.
Беляева Н. В.— В кн.: Историческое развитие класса насекомых. Л.: Наука,
с. 145—148. (Тр. палеонтол. ин-та; Т. 175).
Бергер Э. Г.. Вассер С. П.— В кн.: Актуальные вопросы современной ботаники.
Киев : Наук, думка, 1976, с. 16—30.
Бешел Р.— Ботан. жури., 1969, 54, № 6, с. 872—891.
Бисько Н. А., Колесниченко Л. П.. Володина Е. П. и др.— Раст, ресурсы, 1979,
15, № 2, с. 270—277.
Бисько Н. А., Бухало А. С., Вассер С. П. и др. Высшие съедобные базидиоми-
цеты в поверхностной и глубинной культуре.— Киев : Наук, думка, 1983.— 312 с.
Бондарцев А. С. Трутовые грибы европейской части СССР и Кавказа.— Л.:
Изд-во АН СССР, 1953.— 1106 с.
Бондарцев А. С.— Ботан. журн., 1960, 45, № 10, с. 1504—1506.
Бондарцев А. С.. Зингер Р. А.— Тр. Ботан. ин-та АН СССР. Сер. 2, Споровые
растения, 1950, вып. 6. с. 499—543.
Булах Е. М.— Микология и фитопатология, 1977а, 11, вып. 3, с. 177—181.
Булах Е. М. Базидиальные макромицеты Верхне-Уссурийского стационара
(Южный Сихотэ-Алинь) : Автореф. дис. канд. биол. наук.— Владивосток,
19776 —22 с
Бурова Л. Г., Нездойминого Э. Л.— Микология и фитопатология, 1980, 14,
вып. 4, с. 281—286.
Бухаю А. С.— М1кроб1ол. журн., 1966, 28, № 1, с. 77—81.
Бухало А. С.— В кн.: Методы экспериментальной микологии. Киев : Наук, думка,
1973, с. 208—223.
Бухало А. С., Вассер С. П.— Укр. ботан. журн., 1981, 38, № 1, с. 93—99.
Васильева Л. Н. Агариковые или шляпочные грибы (пор. Agaricales) Примор-
ского края : Автореф. дне. ... д-ра биол. наук.— Ереван, 1967,— 51 с.
Васильева Л. Н. Агариковые шляпочные грибы Приморского края.— Л.: Наука,
1973.—331 с.
Васильева Л. Н., Назарова М. М.— В кн.: Стационарные исследования в лесах
Приморья. М.; Л.: Наука, 1967, с. 122—164.
Васильков Б. П.— Ботан. жури., 1952, 37, № 4, с. 551—557.
168
Васильков Б. П. Изучение шляпочных грибов в СССР.— М.; Л. Изд-во АН
СССР. 1953а.—191 с.
Васильков Б. П.— Ботан. материалы отд. споровых растений Ботан. ин-та АН
СССР. 19536, вып. 9, с. 154—157.
Васильков Б. П. Очерк географического распространения шляпочных грибов в
СССР.— М.; Л.: Изд-во АН СССР, 1955.— 86 с.
Васильков Б. П.— Ботан. журн., 1966, 51, № 6, с. 847—849.
Васильков Б. П.— Микология и фитопатология, 1967, 1, вып. 1, с. 17—25.
Васильков Б. П.— В кн.: Биогеоценозы Таймырской тундры н их продуктив-
ность. Л.: Наука, 1971, с. 145—150.
Васильков Б. П.— Новости систематики низш. растений, 1974, 11, с. 169—173.
Вассер С. П.— Укр. ботан. журн., 1970, 27, № 4, с. 520—523.
Вассер С. Л.—Там же, 1971, 28, № 3, с. 299—304.
Вассер С. П.— В кн.: V зТзд. Укр. Ботан. т-ва : (Коротга тез. доп). Ужгород-
Б. в., 1972а, с. 83.
Вассер С. П.— Укр. ботан. журн., 19726, 29, Ле 3, с. 371—373.
Вассер С. П.— Там же, 1973а, 30, № 1, с. 54—59.
Вассер С. П — Там же, 19736, 30, № 3, с. 378—384.
Вассер С. П.— Там же, 1973в, 30, № 4, с, 457—466.
Вассер С. П.— Там же, ]974а, 31, № 1, с. 78—82.
Вассер С. П.— Микология и фитопатология, 19746, 8, вып 2, с. 140—142
Вассер С. П.— В кн.: Вопросы физиологии, биохимии, цитолшии и флоры
Украины. Киев : Наук, думка, |974в, с. 54—55.
Вассер С. П.— Укр. ботан. журн., 1974г, 31, № 5, с. 567—577.
Вассер С. П.— В кн.: Тнтродукшя та експериментальна еколш1а рослин. К-
Наук, думка, 1974д, с. 64—68.
Вассер С. П.— Укр. ботан. журн., 1975а, 32, № I, с. 32—38.
Вассер С. П.— Микология и фитопатология, 19756, 9, вып. 2, с. 91—94.
Вассер С. П.— Укр. ботан. журн., 1975в, 32, № 3, с. 318—322.
Вассер С. П.— В кн.: Новости систематики низш. растений, 1976а, 13, с. 77—79.
Вассер С. П.— Укр. ботан. журн., 19766, 33, № 4, с. 362—367.
Вассер С. П — Там же, 1976в, 33, Ке 5, с. 493—497.
Вассер С. П.— В кн.: Новости систематики высших и низших растений. Киев:
Наук, думка, 2, 1976г, с. 191—192
Вассер С. П.— Укр. ботан. жури., 1977а, 34, № 3, с. 305—308.
Вассер С. П.— В кн.: Новости систематики высших и низших растений. Киев:
Наук, думка, 3, 19776, с. 217—228.
Вассер С. П-— Укр. ботан. жури., 1978а, 35, № 5, с. 516—519.
Вассер С. П.— В кн.: Новости систематики высших и низших растений. Киев :
Наук, думка, 4, 19786, с. 207-—225.
(Вассер С. П.) Wasser S. Р. Fungorum Rariorum leones Coloratae.— Vaduz:
Cramer. 1979.— Pt 10.— 32 p.
Вассер С. П. Флора грибов Украины : Агариковые грибы.— Киев : Наук, думка,
1980.— 328 с.
Вассер С. П.— В кн.: Новости систематики высших и низших растений. Киев:
Наук, думка, 5, 1981, с. 177—199.
Вассер С. П. Агариковые грибы (Agaricaceae (Fr.) Cohn) Советского Союза:
Автореф. дис. ... д-ра биол. наук.— Киев, 1982.— 54 с.
Вассер С. П„ Батирова Г. Ш.— Укр. ботан. жури., 1979, 36, № 5, с. 423—426.
(Вассер С. П.. Бергер Э. Г.) Wasser S. Р., Berger Е. G.— Mycotaxon, 1980, 10,
N 2, р. 351—364.
Вассер С. П., Бергер Е. Г.— Укр. ботан. жури., 1982, 38, № 6, с. 67—75.
Вассер С. П., Берегова В. П., Бёлай В. Т.— Там же, 1980, 37, № 5, с. 60—71.
Вассер С. П., Гарибова Л. В., Макеева В. Л. - Там же, 1976, 33, № 3, с. 246—
251.
(Вассер С. П„ Гарибова Л. В., Мокеева В. Л-) Wasser S. Р., Garibova L. V.,
Mokeeva V. L.— Acta bot. Acad. sci. hung., 1976, 2, N 1/2, p. 249—258.
Вассер С. II., Дудка I. Q., <Ppid Ю. Ф. та in.— Bich. АН УРСР, 1976, 40, № 11,
c. 63—70.
Вассер C. IL, Солдатова И. M. Высшие базидиомицеты степной зоны Украины.—
Киев : Наук, думка, 1977а.— 355 с.
Вассер С. П., Солдатова И. М.— В кн.: Актуальные вопросы современной бота-
ники. Киев : Наук, думка, 19776, с. 209—212.
(Вассер С. Г]., Солдатова И. М.) Wasser S. Р., Soldatova I. М.— Beih. Sydowia.
Ann. Mycol. Ser. 2, Beih. 8, 1979, p. 391—400.
14 4—2960
169
Галахов Н. Н.— Природа, 1938, № 7/8, с. 91—107.
Ганжа Р. В.— Ботан. журн., 1960а, 45, № 2, с. 187—191.
Ганжа Р. В.— Там же, 19606, 45, № 5, с. 758—764.
Ганжа Р. В.— Укр. ботан. журн., 1960в, 17, № 5, с. 491—495.
Ганжа Р. В.— Там же, 1970, 27, Ns 5, с. 582—586.
Гапоненко Н. И. Обзор грибов Бухарской области.— Ташкент : Фан, 1965.— 113 с.
Гарибова Л. В.— Вести, моек, ун-та. Биология, почвоведение, 1964, № 2,
с. 44—51.
Гарибова Л. В.— Науч. докл. высш, школы. Бнол. науки, 1966, № 2, с. 104—106.
Гарибова Л. В.— В кн.: Генетические основные селекции микроорганизмов. М.:
Наука. 1969а, с. 239—260.
Гарибова Л. В.— Микология и фитопатология, 19696, 3, вып. 2, с. 120—124.
Гарибова Л. В.— Там же, 1970, 4, вып. 3, с. 201—208.
Гарибова Л. В.— Там же, 1971, 5, вып. 4, с. 374—380.
Гарибова Л. В.— В кн.: Производство высших съедобных грибов в СССР. Киев:
Наук, думка, 1978, с. 38—43.
Гарибова Л. В., Мокеева В. Л.— В кн.: Тез. докл. V конф, по споровым расте-
ниям Ср. Азии н Казахстана. Ашхабад: Б. И., 1974а, ч. 1, с. 73—74.
Гарибова Л. В., Мокеева В. Л.— Микология и фитопатология, 19746, 8, вып. 6,
с 533—536.
Гарибова Л. В., Мокеева В. Л.— Там же, 1976, 10, вып. 6, с. 53—54.
Гарибова Л. В., Сафрай А. И.— Там же, 1972, 6, вып. 5, с. 440—443.
Гарибова Л. В., Сафрай А. И.— В кн.: Тез. докл. I республ. конф. «Системати-
ка, экология и физиология почвенных грибов». Киев : Наук, думка, 1975, с. 177—178.
Гарибова Л. В., Сафрай А. И.— Микология н фитопатология, 1980, 14, вып. 6,
с. 480—485
Гарибова Л. В., Сафрай А. И., Шалашова Н. Б.— Там же, 1974, 8, вып. 3,
с. 259—264.
Гарибова Л В., Фролова Е. Я.—Науч. докл. высш. шк. Биол. науки, 1971, № 4,
с. 112—116.
Гарибова Л. В., Шалашова Н. Б.— Микология и фитопатология, 1973, 7, вып. 3,
с. 231—232.
Головин П. А.—Тр. Узб. фил. АН СССР. Сер. 2, Ботаника, 1941, вып. 1, с. 1—48.
Гончарук Н. С., Девочкин Л. А.— В ки.: Производство высших съедобных гри-
бов в СССР. Киев : Наук, думка, 1978, с. 43—47.
Горленко М. В.— Микология и фитопатология, 1976, 10, вып. 1, с. 5—10.
Горовой Л. Ф. Сравнительное изучение онтогенеза плодовых тел представителей
отдельных групп шляпочных грибов: Автореф. дне. ... канд. бнол. наук.— Киев,
1980.— 25 с.
Городницкий А.—Знание — сила, 1978, № 7, с. 16—20.
Громов Н. Г. Шампиньоны.— М.: Сельхозгиз, 1957.— 168 с.
Громов Н. Г. Шампиньоны.— 2-е нзд.— М.: Сельхозгиз, 1960.— 176 с.
Давыдкина Т. А. Стереумовые грнбы Советского Союза.— Л.: Наука, 1980.—
142 е.
Девочкин Л. А.— Шампиньоны.— М.: Колос, 1975.— 112 с.
Дворнина А. А.— В кн.: Производство высшнх съедобных грибов в СССР.
Киев : Наук, думка, 1978, с. 47—50.
Д1дух Я. П.— Укр. ботаи. журн., 1977, 34, № 2, с. 159—163.
Д1дух Я. П,— Там же, 1978, 35, № 5, с. 470—475.
Добровольский И. А., Сосин П. Е.— В кн.: Искусственные леса степной зоны
Украины. Харьков : Изд-во Харьк. ун-та, 1960, с. 217—227
Додылева С. И. Методы производства посадочной грибницы н культура шам-
пиньонов в Молдавии : Автореф. дис. ... канд. с.-х. наук.— Кишинев, 1968.— 18 с.
Доспехов Б. А. Методика полевого опыта (с основами статистической обработки
результатов исследований).— М.: Колос, 1965.— 422 с.
Дудка И. А.— В кн.: Новости систематики высших в низших растений. Киев:
Наук, думка, 2, 1976, с. 151—178.
(Дудка И. А., Вассер С. Г].. Бисько И. A.) Dudka /. A., Wasser S. Р„ Bis-
ko N. А.— In: 3rd Intern, sympos. Physiol., Ecol. and cultivat. edible fungi. Praga,
1979, p. 83—68.
Дудка И. А., Вассер С. П., Бухало А. С. и др. Промышленное культивирова-
ние съедобных грибов —Киев : Наук, думка, 1978.— 262 с.
(Дудка И. А., Вассер С. П., Солдатова И. М.) Dudka I. A., Wasser S. Р., Sol-
datova I. М.— In: Prumyslove pestovani jedlych hub. Praha, 1977, s. 23—37-
Дудченко Л. Г., Мельничук Г. Г.— Укр. ботан. журн., 1973, 30, № 1, с 1—14,
170
1
Дудченко Л. Г., Мельничук Г. Г.— В ки.: Материалы V конф, по споровым
растениям Ср. Азии и Казахстана: Тез. докл. Ашхабад: Илим, 1974, с. 115—116.
Заки М. А., Шмидт В. М.— Вести. Ленингр. ун-та, 1972, Ns 9, Биология, вып. 2,
с. 57—69.
Закордонец О. А., Вассер С. Г7.— В кн.: Тез. докладов VII съезда УБО. Киев:
Наук, думка, 1982, с. 356.
Зеров Д. К. Очерк филогении бессосудистых растений.— Киев: Наук, думка,
1972.—316 с.
Зерова М. Я-—Укр. ботан. журн., 1956, 13, Ns 2, с. 68—77.
Зерова М. Я.— Там же, 1959, 16, Ns 2, с. 76—82.
Зерова М. Я-—Там же, 1962, 19, № 5, с. 219—225.
Зерова М. Я. Тспвш та отруйш гриби Украши.— К.: Вид-во АН УРСР, 1963.—
203 с.
Зерова М. Я.— В кн.: IV симпозиум прибалтийских микологов и лихенологов:
Тез. докл. Тарту : Изд-во АН ЭССР, 1965, с. 103—108.
Зерова М. Я- Ictibhi та отруйHi гриби Украши.— К.: Наук, думка, 1970а.— 128 с.
Зерова М. Я.— Микология и фитопатология, 19706, 4, вып. 5, с. 468—470.
Зерова М. Я— В ки.: Тез. докл. V делегат, съезда ВБО. Киев, 1973, с. 327—328.
Зерова М. Я. Атлас гриб!в Украши.— К.: Наук, думка, 1974а.— 252 с.
Зерова М. Я-— В кн.: Новости систематики высших и низших растений. Киев:
Наук, думка, 19746, с. 151—165.
Зерова М. Я., Вассер С. П. Тспвш та отруйш гриби Карпатських лгав.— Ужго-
род : Карпати, 1972.— 128 с.
Зерова М. Я., Вассер С. П.— В кн.: Соврем, успехи микологии и лихенологии
в Советской Прибалтике. Тарту: Изд-во АН ЭССР, 1974. с. 62 65.
Зерова М. Я-, Вассер С. П., Горовой Л. Ф.— В кн.: Материалы V конф, по
споровым растениям Ср. Азии и Казахстана: Тез. докл. Ашхабад: Илим, 1974,
,с. 130—131.
Зерова М. Я., Горовой Л. Ф.— В кн.: Соврем, успехи микологии и лихенологии
в Сов. Прибалтике. Тарту : Изд-во АН ЭССР, 1974, с. 66—69.
Зерова М. Я., Дудченко Л. Г., Роженко Г. Л.— В кн.: Материалы I конф, по
споровым растениям Украины. Киев: Наук думка, 1971, с. 162—164.
Зерова М. Я.. А1ельничук Г. Г.— В кн.: V з’Тзд Укр. ботан. т-ва: Тез. доп.
Ужгород : Б. в., 1972, с. 88—89
Зерова М. Я., Роженко Г. Л.. Череп Т. Л.— Укр. ботаи. журн., 1975, 32, Ns 5,
с. 570—596.
Зерова М. Я-, Сосш П. в., Роженко Г. Л. Визначник гриб!в Украши. В 5-тн т.
Т. 5. Базидюмщети.— К.: Наук, думка, 1979.— Ки. 2. 565 с.
Зерова М. Я., Череп Т. Л.— В ки.: IV Закавк. совещ. по споровым растениям :
Тез. докл.— Ереван : Изд-во Ерев. ун-та, 1972, с. 164—167.
Зингер Р. А.— Ботан. материалы споровых растений Ботан ин-та АН СССР,
1938, IV, Ns 10/12, с. 4—18.
Казокин Ю. И. Влияние состава исходных материалов и технологии приготов-
ления компостов на урожай и качество шампиньонов: Автореф. дне. ... канд. с.-х.
наук— М, 1977.— 17 с.
Каламээс К. А. Агариковые грибы Эстонии (Polyporales, Bolctales, Russulaies,
Agaricales) : Автореф. дис. ... д-ра биол. наук.— Таллин, 1975.— ПО с.
(Каламээс К-) Kalamees К.— Scripts mycol., 1978а, 8, р. 5—82.
(Каламээс К.) Kalamees К.— Ibid., 1978b, 8, р. 83—134.
Камелин Р. В. Ку х ист а некий округ горной Средней Азии: Ботан.-геогр. анализ.—
Л.: Наука, 1979.— 117 с. (Комаров, чтения; Вып. 31).
Каратыгин И. В. Головневые грибы: Онтогенез и филогенез.— Л.: Наука,
1981.—213 с.
Кендэл М. Ранговые корреляции.— М.: Мир. 1975.— 207 с.
Китаев С. И., Бубнова О. Н., Борисов А. А. и др. Синтетические компосты для
выращивания шампиньонов.— М.: Колос, 1976.— 121 с.
Клеопов Ю. Д.— Ботан. журн. АН УРСР, 1938, 17, Ns 1, с. 12—18.
Клюшникова Е. С.—Бюл. Моск, о-ва нспыт. природы. Отд. биол., 1938, 47,
Ns 1, с. 30.
Клюшникова Е. С.— Там же, 1939, 48, № 5—6, с. 53—68.
Коваленко А. Е.— Микология и фитопатология, 1980. 14, вып. 4, с. 300—314
Козловская И. В., Парфенов В. И.— Ъотаннка, 1971, вып 13, с. 21—27.
Колесниченко Л. П.. Дудка И. А., Вассер С. П-—В кн.: Производство высших
съедобных грибов в СССР. Киев : Наук, думка, 1978, с. 63—73.
Криштофович А. Н. Палеоботаника.— 4-е изд., испр. и доп.— Л.: Госгортолмздат,
1957.— 650 с.
14’
171
Кулеш М. Д.— В кн.: Производство высших съедобных грибов в СССР Киев:
Наук, думка, 1978, с. 73—76.
Кушнарев С. А.— Там же, с. 80—84.
Лазаренко А. С.— В кн.: Президенту АН СССР акад. В. Л. Комарову к
70-летию со дня рождения и 40-летию научной деятельности.— М.; Л.: Изд-во АН
СССР, 1939, с. 73- -89.
Лазаренко А. С.— Сов. ботаника, 1944, № 6, с. 43—55.
Лазаренко А. С.— Зап. КиТв. ун-ту, 1947, 7, вып. 6, с. 41—46.
Лазаренко А. С.— Укр. ботан. журн., 1956, 13, № 1, с. 31—40.
Лебедева Л. А. Определитель шляпочных грибов.— М.; Л.: Сельхозгиз, 1949.—
548 с.
Лункевич В. В. От Гераклита до Дарвина : Очерки по истории биологии.— М.:
Учпедгиз, i960.—Т. 1.
Майр Э. Принципы зоологической систематики.— М.: Мир, 1971.— 454 с.
Макаревич М. Ф.— В ки.: Тез. доп. конф, по вивч. флори i фауни Карпат та
прилеглих територ1й. К- : Вид-во АН УРСР, 1960, с. 93—94.
Макаревич М. Ф. Анализ л1хснофлори УкраТнських Карпат.— К-: Вид-во
АН УРСР, 1963.—262 с.
Малый атлас мира.— М.: ГУГК, 1981.— 147 с.
Малышев Л. И.— Ботан. жури., 1973, 58, № 11, с. 1581—1588.
Масюк Н. П. Род Dunaliella Teod.: Морфология, систематика, экология, геогр.
распространение и перспективы практ. использ.: Автореф. дис. ... д ра биол. наук.—
диев, 1973.—44 с.
Международный кодекс ботанической номенклатуры.— Л.: Наука, 1980.— 283 с.
Мелик-Хачатрян Дж. Г.—В кн.: Материалы VI симпоз. микологов и лихеноло-
гов Прибалт, респ. Рига, 1971, т. 2, с. 12—15.
Мелик-Хачатрян Дж. Г. Агариковые шляпочные грибы (Agaricales) : Микофлора
Армянской ССР.— Ереван : Изд-во Ереван, ун-та, 1980.— т. 5, 543 с.
Михайловский Л. В. Макромицеты (порядок Agaricales) Хибинского горного
массива : Автореф. дис. ... канд. биол. наук.— Л., 1975.— 23 с.
Мокеева В. Л. Оценка значимости различных признаков и методов исследова-
ния в систематике рода Agaricus Fr. emend. Karst.: Автореф. дис. ... канд. биол.
наук.— М., 1977.— 24 с.
Мокеева В. Л., Гарибова Л. В.— Вести. Моск, ун-та, 1975, 5, № 1, с. 49—53.
Мокеева В. Л., Райтвийр А. Г.. Гарибова Л. В.— Микология и фитопатология,
1977, 11, вып. 3, с. 277—282.
Назарова М. М.— В ки.: Низшие растения Дальнего Востока :Тр. Биол.-почв.
ин-та, 1976, 41, с. 115—127.
Наумов Н. А.- В кн.: Проблемы ботаники. М.; Л.: Наука, 1950, т. 1, с. 209—
221.
Наумов Н. А.— Тр. ВНИИ защиты растений: Общ. и част, пробл микологии
и фитопатологии, 1972, 33, с. 5—16. _
Нахуцришвили И. Г. Агариковые грибы Грузии: Автореф. дис. ...д-ра биол.
наук.— Тбилиси, 1971.— 43 с.
Нахуцришвили И. Г. Агариковые грибы Грузии.— Тбилиси : Менниереба, 1975.—
209 с.
Нездойминого Э. Л.— Микология и фитопатология, 1968, 2, вып. 3, с. 284—290.
Нездойминого Э. Л. Шляпочные грибы северо-восточного побережья Байкала:
Автореф. дис. .. канд. биол. наук.— Л., 1970.— 29 с.
Нездойминого Э. Л.— Микология и фитопатология, 1971, 5, вып. 1, с. 85 87.
Нездойминого Э. Л.— Новости систематики низших растений, 10, 1973,
с. 133—141.
Нездойминого Э. Л.— Микология и фитопатология, 1980, 14, вып. 5. с. 388—390.
Николаева Т. Л. Ежовиковые грибы.— М.: Изд-во АН СССР, 1961.— 433 с.—
(Флора споровых растений СССР; Т. 6. Грибы (2) ).
Окснер А. Н. Анализ и история происхождения лихенофлоры Советской Аркти-
ки : Дис. ... д-ра биол. наук.— Киев ; Киров, 1940—1942.— 318 с.— Рукопись.
Окснер А. Н.— Ботан. журн., 1944, 29, № 3/4, с. 195—206.
Окснер А. М. Флора лишайник!в Украши.— К-: Вид-во АН УРСР, 1956.— Т. 1.
680 с.
Окснер А. Н. Определитель лишайников СССР.— Л.: Наука, 1974,— Вып. 2.
283 с.
Панов М. А. Шампиньоны.— М.: Сельхозгиз, 1950.— 48 с.
Панов М. А. Выращивание шампиньонов.— М.: Госторгиздат, 1956.— 138 с.
Пармасто Э. X. Трутовые грибы ЭССР: Автореф. дис. ... канд. биол. наук.—
Тарту, 1955-—21 с.
172
Пармасто Э.— Тр. Ботан. Ин-та АН СССР. Сер. 2, Споровые растения, 195'*
12, с. 213—273.
Пармасто Э. X.—-Микология и фитопатология, 1969, 3, вып. 4, с. 322—330.
Пармасто Э.— В кн.: Тез. докл. V делегат, съезда Всесоюз. ботан. о-ва. Киев.
Б. И.» 1973, с. 325—326.
Перова Н. В.— В кн.: Водоросли и грибы Сибири и Дальнего Востока. Ново-
сибирск: Наука, 1970, 4.1 (3), с. 128—131.
Перова Н. В.— В кн.; Водоросли и грибы Сибири и Дальнего Востока. Ново
сибирск : Наука, 1972, 4.4 (4), с. 178—182.
Перова Н. В.— В кн.: Водоросли, грибы и лишайники лесостепной и лесной зон
Сибири. Новосибирск : Наука, 1973, с. 102—106.
Перова Н. В.— В кн.: Природные комплексы низших растений Западной Си би
ри Новосибирск : Наука, 1977, с. 174—182
Петренко И. А. Макро- и микромицеты лесов Якутии.— Новосибирск: Наука.
1978 - 132 с.
Пилипович А. И., Марышева Н. С.— В кн.: Химический мутагенез и создание
сортов интенсивного типа.— М.: Наука, 1977, с. 218—220.
Попов П. А.— Ботан. журн., 1961, 47, № 11, с. 1596 -1610.
Попов П. Л.— Микология н фитопатология, 1967, 1, вып. 2, с. 158—163.
Райтвийр А. Г — Изв. АН ЭССР, 1964, 13. Сер биол. наук., вып. 2, с. 106—122.
Рамад Ф. Основы прикладной экологии.— Л.: Гндрометеоиздат, 1981.— 543 с
Растительный покров СССР: Пояснительный текст к «Геоботанической карте
СССР» м-ба 1 : 4 000 000. / Ред. Е. М. Лавренко, В. Б. Сочава. 1—II — М.; Л.; Изд-во
АН СССР, 1956.—971 с.
Ребристая О. В., Шмидт В. М.— Ботан. журн., 1972, 57, № II, с. 1353—1364.
Ровенский Ю. А Растровая электронная микроскопия нормальных и опухоле-
вых клеток — М.: Медицина, 1979.— 150 с.
Рыбкина К. В. Изучение сравнительной эффективности заменителей конского
навоза в субстратах для грибов шампиньонов: Автореф. дис. ... канд. с.-х. наук.—
Кишинев, 1967.— 20 с.
Рыбкина К- В.— Шампиньоны. -Л.: Колос, 1971.— 60 с.
Садыхов А. С.— Изв. АН АзССР. Сер. биол. наук, 1967, № 2, с. 23—26.
Садыхов А. С. Шляпочные грибы Ленкоранской зоны Азербайджанской ССР:
Автореф. дис. ... канд. биол. наук,— Баку, 1968.— 21 с.
Садыхов Л. С.— Новости систематики низших растений, 1972, 9, с. 151—152.
Садыхов А. С. К флоре агариковых грибов Талыша.— Баку, 1977.—10 с.—
Рукопись деп. в ВИНИТИ, 10.10.77 № 3931—77 Дсп.
Сафрай А. И. Биологические особенности некоторых видов рода Agaricus Fr.
emqnd. Karst.: Автореф. дис. ... канд. биол. наук.— М., 1977.— 23 с.
Сержанина Г. И. Съедобные и ядовитые грибы порядка Agaricales в БССР и
их хозяйственное значение : Автореф. дис. ... канд. биол. наук.— Минск, 1962.— 25 с.
Сержанина Г. И.— Изв. АН БССР, Сер. биол. наук. 1965, № 4, с. 41—49.
Сержанина Г. И. Съедобные и ядовитые грибы.— Минск: Наука н техника,
1967 — 145 с.
Сержанина Г. И.— Изв. АН БССР. Сер. биол. наук., 1968, № 2, с. 34—38.
Скворцов А. К. Гербарий: Пособие по методике и технике.— М.: Наука, 1977.—
199 с.
Сосин П. Е. Гастеромицеты Украинской ССР: Автореф. дис. ... д-ра биол. наук.—
Л.» 1952.—49 с.
Сосин П. Е. Определитель гастеромицетов СССР.— Л.: Наука, 1973.— 162 с.
Степанова И. Т., Сирко А. В.— В кн.: Микологические исследования на Урале
Свердловск ; УНЦ АН СССР, 1977, с. 51—106.
Тамарин П. В., Шмидт В. М.— Тр. Ленннгр. о-ва естествоиспытателей. Успехи
биометрии, 1975, 72, вып. 5, с. 45—54.
Тахтаджян А. Л. Происхождение и расселение цветковых растений.— Л.: Наука,
1970— 146 с.
Тахтаджян А. Л. Цветковые растения.— В кн.: БСЭ. 3-е изд., 1978а, т. 28,
с. 445—451.
Тахтаджян А. Л. Флористические области Земли.— Л. : Наука, 19786 — 247 с.
Тетеревникова-Бабаян Д. И.— Микология и фитопатология, |981„ 15, вып. 4.
с. 335—341.
Толмачев А. И.— Тр. поляр, комис., 1932, 8, с. 76—128.
Толмачев А. И.— Вести. Ленивгр. ун-та, 1970а, № 9. Биология, вып. 2, с. 71—83.
Толмачев А. И.—'Там же, 19706, № 15. Биология, вып. 3, с. 63—64.
Толмачев А. И. Введение в географию растений.— Л.:Изд-во Ленингр. ун-та,
I974-—244 с.
173
Томилин Б. А.— В кн.: Амурская тайга. Л.: Наука, 1969, с. 90—126
Томилин Б. А.— В кн.: Биогеоценозы Таймырской тундры и их продуктивность.
Л.: Наука, 1971, с. 130—137.
Трасс X. X.— Тр. по ботанике/Тарт, ун-т, 1970, 9, с. 5—233.
Урбонас В. Л.—Тр. АН ЛитССР. Сер. В, 3(67). 1974, с. 13—19.
Урбонас В., Каламээс К.. Лукин В. Конспект изучения агариковых грибов
Литовской ССР, Латвийской ССР, Эстонской ССР: (Материалы 1778—1973 гг.).-—
Вильнюс : Минтае, 1974.— 130 с
Флеров К- К.. Крамаренко И. Н., Борисов В. А. и др. Зоогеография палеогена
Азии.— М.: Наука, 1974.— 302 с.
Херинк Е.. Вассер С. П.— В кн.: Новости систематики высших и низших расте-
ний. Киев : Наук, думка, 2, 1976, с. 183—191.
Христюк П. М.— Изв. Крым, отд-ння гсогр. о-ва СССР, 1954, вып. 3, с. 51—56.
Христюк П. М. Съедобные грибы Крыма.— Симферополь : Таврия, 1960 — 48 с.
Частухин В. Я.. Николаевская М. А. Биологический распад и реемнтез органи-
ческих веществ в природе.— Л.: Наука, 1969.— 325 с.
Чигцряева А. А.— Ботан. материалы Отд. споровых растений. Ботан ин-та АН
СССР, 1953, 9, с. 109—114.
Шварцман С. Р. Грибы Казахстана. Съедобные н ядовитые.— Алма-Ата ; Изд-во
АН КазССР, 1948 — 37 с.
Шварцман С. Р. Материалы к истории микофлоры Казахстана.— Алма-Ата:
Изд-во АН КазССР. 1962.— 184 с.
Шварцман С. Р.— Acta Mycol., 1968, 4, N 2, с. 297—315.
Шварцман С. Р — В кн.: Тез. докл. V делег. съезда Всесоюз. ботан. о-ва.—
Киев : Б. И., 1973, с. 349—350.
Шварцман С. Р„ Кажиева Н. Т.— Ботан. материачы Гербария Ин-та ботаники
АН КазССР, 1968, 5, с. 48—53.
Шеля^-CcgOHKO Ю. Р.. Дидух Я- П. Ялтинский горно-лесной государственный
.заповедник — Киев : Наук, думка, 1980.— 183 с.
Шиврина А. В.— В ки.: Тез. докл. V делег. съезда Всесоюз. ботан. о-ва. Киев:
Б. и., 1973, с. 352.
Шиврина А. Н., Низковская О. П„ Фалина Н. Н. и др. Биосинтетическая дея-
тельность высших грибов.— Л.: Наука, 1969.— 198 с.
Шиврина А. И., Платонова Е. Г., Черотченко Ю. П.— Микология н фитопатоло-
гия, 1968, 2, вып. 1, с. 65—69.
Шмидт В. М. Статистические методы в сравнительной флористике — Л.: Изд-во
Ленннгр. ун-та, 1980.— 175 с.
Эльчибаев А. А. Макромицеты севера Киргизии н их хозяйственное значение.—
Фрунзе : Илим, 1968 — 94 с.
Эльчибаев А. А.— Микология и фитопатология, 1969, 3, вып. 1, с. 38—43.
Юрцев Б. А„ Толмачев А. И.. Ребристая О. В.— В кн.: Арктическая флористи-
ческая область. Л.: Наука, 1978, с. 9—104.
Яковенко А. 3., Жемойц А. А.— В кн.: Производство высших съедобных грибов
в СССР. Киев : Наук, думка, 1978, с. 120—135.
Ячевский А. А. Основы микологии.— М.; Л.: Ленсельхозгиз, 1933.— 1035 с.
Aberdeen J. Е. С.— Kew Bull, 1962, !6, N 1, р. 129—137.
Ahmad S.— Biol. Soc. Pakistan, 1956, 1, р. 1 -86.
Ahmad S.— Ibid , 1969, 5, p. 1—127.
Ainsworth G. C. Ainsworth and Bisby's dictionary of the Fungi.— Surrey: Kew,
6th ed.— 1971.— 634 p.
Ainsworth G. C. Introduction to the history of mycology.— Cambridge etc.: Camb-
ridge univ. press, 1976.— 359 p.
Anderson E. E.. Fellers C. R.— Proc. Amer. Soc. Hortic. Sci., 1942, 41, p. 301—304.
Andrews H. N. Lenz L. W.— Bull. Torrey Bot. Club. 1943, 70, p. 120—125.
Antal M„ Morava E., Zsinka A et al.— Nalirung, 1969, 13, N 1. S. HI—118.
Aoki M., Fuji T.— Trans. Jap. mycoi. Soc., 1972, 13, N 3, p. 294—300.
Arx J. A. Pilzkunde : Ein kurzer Abriss des Mykologie «inter besonderer Beriick-
sichligung der Pilze in Rcinkultur.— Lchre : Cramer, 1968.— 356 S.
Atkinson G. F. Studies of American fungi, mushrooms edible, poisonous, elc.—
Ithaca ; Andrus a. Church, 1900.— 275 p.
Atkinson G. F.— Proc. Amer. Phil. Soc., 1915, 54, N 3, p. 309—342.
Atkinson G. F —Mem. N. Y. Bot. Gard., 1916, 6, p. 201—228.
Avizohar-Hershenzon Z.— Bull. Res. Counc. Israel, 1961, 10, N 1/4, p. 171—178.
Avtzohar-Hershenzon Z. Edible and toxic fungi In Israel.— Tel-Aviv, 1964.—18 d.
Avizohar-Hershenzon Z.— Isr. J. Bot., 1967, t6, N 1, p. 63—69.
Avizohar-Hershenzon Z.— Acta mycol., 1968, 4, N 2, p. 437—442.
174
Babos M — Ann. hist, natur. Mus. Nat. Hung., 1958, 50, p. 87—92.
Babos M —Ibid., 1961, 53, p. 195—199.
Babos M.— Ibid., 1969, 61, p. 157—164.
Babos M.— Gcska mykol., 1970, 24, N 4, s. 217—219.
Babos M.— Ann. Mycol., Ser. 2, 1979, p. 33—53.—Beih. VIII zur Sydowia.
Babos M.— Geska mykol., 1980, 34, N 2, s. 98—101.
Babosne M.— Fragm. bot. Mus. Hist. Nat. Hung., 1968, 6. p. 1—4.
Bailey I. W.— J. Arnold Arboretum, 1949, 30, p. 64—70.
Baria J. B. Flore mycologique illustree des champignons des Alpes-Maritimes I.—
Nice, 1888, p 1—32, lab. I—16.
Bas C.— Persoonia, 1965, 3, N 3, p. 331—359.
Bauchet J.— Bull. Soc. mycol. Fr., 1961, 77, N 9, p. 137—139.
Beeli M.— Bull. Soc. roy. bot. Belg., 1932, 64, p. 206—218, pl. XXV—XXVII
Benedix E. H.-Z. Pilzk., 1959, 25, N 1/2, S. 41—50.
Benes R.— Cas. ces, houb., 1923—1924, 4, s'. 120—122.
Berkeley Af. Broome С. E.— J. linn. bot. Soc., 1871, 11, p. 494—567.
Berkeley M. L. Curtis M. A.— J. linn. Soc. London, 1868, 10, p. 341—392.
Best H.. Bresinsky A.— Z. Pilzk, 1977, 43. N 5/6, S. 311—322.
Bigelow H. E.— Mycologia, 1981, 73, N I, p. 128—140.
Bigelow H. E.. Rowley J. R.— Ibid., 1968, 60, N 4, p. 869—887.
Binyamini N.— Isr. J. Bot., 1974, 23, N 3, p. 237—251.
Binyamini N.— Ibid., 1976a, 25, N I, p. 62—78.
Binyamini N.— Nova Hedwigia, 1976b, 27, p. 861—876.
Binyamini N.~ Ibid., 1976c, 28, p. 759—770.
Bisby G. R.— Amer. J. Bot., 1933, 20, N 4, o. 246—254.
Bisby G. R.— Bot. Rev., 1943, 9, N 7, p. 466—482.
Bohus G.— Borbasia, 1939, 1, N I, p. 112—114.
Bohus G.— Ann. hist, natur. Mus. Nat. Hung., 1961, 53, p. 187—194.
Bohus G.— Mycologia. 1963, 55, N 1, p. 128—130.
Bohus G.— Ann. hist, natur. Mus. Nat. Hung., 1969, 69, p. 151—156.
Bohus G — Ibid., 1971, 63, p. 77—82.
Bohus G — Ibid., 1974, 66, p. 77—85.
Bohus G— Ibid., 1975, 67, p. 37—40.
Bohus G.— Ibid., 1976, 68, p. 45—49.
Bohus G.—Ibid., 1978, 70, p. 105—110.
Bohus G.. Kalmar Z., Ubrizsy G. Magyarorszag kalaposgombainak meghatarozo
kezikonyve.— Budapest: Akad. Kiado, 1951.— 512 old.
Bon M.— Bull. Soc. mycol. Fr., 1970, 86, N 2, p. 173.
Bon M— Ibid., 1976, 92, N 3, p. 317—334.
Bon M.— Doc. mycol., 1979, 9, N 35, p. 13—22.
Bon M.— Ibid., 1981, 11, N 43, p. 1—77.
Bon M., Botffard J.— Bull. Soc. mycol. Fr., 1972, 88, N 1, p. 16—28.
Bon M., Boiffard L— Ibid., 1974, 90, N 4, p. 287—306.
Bose S.— Arch. Microbiol., 1953, 18, N 4, p. 349—355.
Bresadola J. Fungi Tridentini novi, vel nondum delineati, descript el icombus
illustratl. II.—Trento, 1892.—217 p., tab. 106—217.
Bresadola G. Iconographia mycologica.— In: 26 vol.— Mediolani, 1927—1933.
Bresinsky A.— Z. Pilzk., 1971, 37, N 3/4, S. 259 260
Bresinsky A.— Bull. mens. Soc. linn. Lyon, 1974, num. spec., 15 fevr, p. 61—84
Bresinsky A., Dichtel B.— Z. Pilzk, 1971, 37, N |/2, S. 75—147.
Bresinsky A., Haas H.— Beih. Z. Pilzk, 1976, 1, N 1/2, S. 1—161.
Bresinsky A., Schwarzer G.— Z. Pilzk., 1969, 35, N 3/4, S. 263—292.
Bunt J.— Austral. Nat. Antarkt. Res. Exp. Sci. Rep Ser. B, 2, Botany 1965
N 78, p. 1—22.
Burt E. A.— Ann. Mo. Bot. Gard., 1923, 10, N 2/3, p. 179—189.
Buller E. J.. Bisby G. R. The fungi ol India/Rev. by R. S. Vasudeva — New
Dehli : S. я., I960,— 552 p
Cain R. F.— Mycologia, 1972, 64, N 1, p. 1—14.
Cappelli A.— Boll. Grupp. mic. G. Bres., 1983, 25, N 1/2, p. 3—52.
Cejp K- Houby.— Praha : Nakl. CSAV, 1958.— 425 s.
Cernohorsky T. Machura L. Pilzfibel.— Vienna : Kohne, 1947.— 123 S.
The Biology and Cultivation ol Edible Mushrooms / S. T. Chang, W. A. Hayes.
Ed — New York : London ; Acad, press, 1978.— 819 p.
Chiu Weifan.— Acta microbiol. sinica, 1978, 13, N 12, p. 125—135.
Cleland J. B.— Trans. Roy. Soc. S. Austral, 1924, 48, N 3, p. 236—252.
175
Cleland I. В. Toadstools and Mushrooms and other larger fungi of South Austra-
lia.— Adelaide : Government print., 1934.— 362 p.
Cleland I. B.. Cheel E.— Trans. Roy. Soc. S. Austral., 1918, 42, N I, p. 88—138.
Cleland J. В.. C • I E.— Ibid., 1923, 47, N 1, p. 58—78.
Cleland J. B-, Harris J. R.— Rec. South Austral. Mus., 1948, 9, N 1, p. 43—56.
Coker W. C.— J. Elisha Mitsch. Sci. Soc., 1928, 43, p. 243—256.
Cooke M. C. Illustrations of British fungi (Hymenomycetcs).—London, 1880—
1890.— [Vol.] 1—8.
Corner E. J. H.— Gard Bull. Straits (Singapore), 1972, 26, p. 159—192.
Cridland A. A.— Mycologia, 1962, 54, N 2, p. 230—234.
Crisan E. V., Sands A.— In: The biology and cultivation of edible mushrooms.—
New York etc.: Acad, press, 1978, p. 137—168.
Demoulin V.— Bot. Rev., 1966, 39, N 1, p. 1—15.
Dennis R. L.— Mycologia, 1970, 62, N 3, p. 578—584.
Dennis R. L.— Science, 1976, 192, N 4234, p. 66—68.
Dennis R. W. G.— Trans. Brit, mycol. Soc., 1951, 34, N 3, p. 411—482
Dennis R. Г. G.— Kew Bull., 1952, 7, N 4, p. 459—500.
Dennis R. W. G.— Bull. Soc. mycol. Fr., 1953, 69, N 2, p. 143—198.
Dennis R. W. G.— Kew Bull., 1961, 15, N 1, p. 67—156.
Dennis R. W. G.— Kew Bull., 1968, 22, N I, p. 73—83.
Dennis R. W. G —Kew Bull., Add. Ser. 3, 1970.—531 p.
Dennis R. U7 G.. ОПоа P. D„ Hora F. B.— Trans. Brit. Mycol. Soc., Suppl., 1960,
43, p. 1—225.
Dennis R. W. G., Orton P. D„ Hora F. B. New check list of British Agarics and
Boleti.—Lehre: Cramer, 1974 — 224 p. (Bibliotheca Mycologica; Bd. 42).
Diehl W. W.— J. Wash. Acad. Sci., 1937, 27, N 3, p. 244—254.
Dielman van Zaayen A.— Mushroom Sci., 1972, 8, p. 131—154.
Dielman van Zaayen A.-—Neth. J. Plant Pathol., 1976, 82, N 1, p. 121—132.
Donk M.— Persoonia, 1964, 3, p. 199—324.
Donk M.— In.: Evolution in the Higher Basidiomycetes / Ed. by R. Petersen.
Knoxville: Univ. Tenn, press, 1971, p. 3—28.
Dorfelt H-— Fedd. Rep., 1971, 81, N 8/9, S. 629—643.
Dorfelt H — Hercynia, 1973, 10, N 3, S. 307—333.
Dorfelt H— Ibid., 1974, 11, N 4, S. 405—431.
Dorfelt H., Kreisel H.. Benkert D.— Ibid., 1979, 16, N 1, S. 1—56.
Earle F. S.— Bull. N. Y. Bot Gard., 1909, 5, p. 373—451.
Eckblad F.-E — Blyttia, 1963, 21, N 2, p. 94—96.
Engler A. Syllabus der Pflanzenfamihen.—12. Auff. /Hrsg. H. Melchior. 2. (Angi-
ospermae).— Berlin : Gebruder Borntraeger, 1964.—666 S.
Essette H. Les Psalliotes.— Paris : Lechevalier, 1964.—99 p.— (Atlas mycologi-
ques; Vol. 1).
Fallahyan F.— Bull. Fac. Sci. Teheran Univ., 1975, 6, N 3/4, p. 182—184.
Favre J.— Ergeb. wiss. Unters. Schweiz. Nat. Parks, Ser. 2, 1955, vol. 5, p. 1—-212.
Fayed V.— Ann. sci. natur., Scr. F, 1889, Vll, 9, p. 181—411.
Fitzpatrick U7 H., Esselen U7 B., Weir E.— J. Amer. Diet. Assoc., 1946, 22, N 3,
p. 318—323.
Food and Agriculture Organization. „Energy and Protein Requiremenls”, Report
of a Joint FAO/WHO ad hoc Expert Committee.—Food Nutr. Meet. Rep. Ser. 52, Food
Agric. Organ. U. N., Rome, 1973, 52 p.
Freeman A. E. H.— Mycotaxon, 1979a, 8, N 1, p. 1—49.
Freeman A. E. H.— Ibid., 1979b, 8, N I, p. 50—118-
Fries E. M. Systems mycologicum sistens Fungorum Ordines, Genera et Species,
hucusque cognitae, quas ad normam methodi naturalis determinavil, disposuit atque des-
cripsit.— Lund, 1821, Vol. 1.— 520 p.
Fries E. M. Epicrisis systematis mycologici, sen synopsis Hymenomycetum.—
Upsala, 1836—1838.— 610 p.
Fries E. M.— Ann. sci. nalur. Bot., 1861, 15, N I, p. 10—35.
Fries E. M.— Ann. and Mag. Nat. History, 1862, 3, N 9, p. 39—52.
Fries E. M. Hymenomycetes Europaei sive Epicriseos systems tis mycologici.—
Upsala, 1874.—'756 p.
Fritsche G.— Mushroom. Sci., 1974, 9, S. 11—21.
Fritsche G.—~ In: The biology and cultivation of edible mushrooms. New York etc.:
Acad, press, 1978, p. 239—250, 371—376.
Garcia Rada G., Stevenson J. S. La flora fungosa Peruana. (Lista preliminar de
hongos Que Alacan a las plantas en el Peru).— Est. Exp. Agric. Molina (Lima), 1942.—
112 p.
176
Garrett S. D. Biology of root-infecting fungi.— Cambridge: Univ, press., 1956.—
293 p.
Giiumann E. Die Pilze: Grundziige ihrer Entwicklungs geschichte und Morpholo-
gic — Basel; Slutlgart: Birkhauser, 1964.— 541 S.
Ghosh R. N., Palhak N. C.—Indian J. Phytopathol., 1965, 18, N 4, p. 360—362.
Gittet С. C. Les Hymenomycetes ou description de tous les champignons qui
croissent en France.— Alencon, 1874,— 828 p.
Graham A.— .1. Paleontok, 1962, 36, N 1, p. 60—68.
Gray S. F. A natural arrangement of British plants according lo their relations
to each other, as pointed out by Jussicn, De Candoll, Brown, etc. 1.— London, 1821 —
824 p.
Groves J. W. Edible and poisonous Mushrooms of Canada. Publication 1112.—
Ottawa : Res. Branch, 1962.— 298 p.
Guzman G.— An. Esc. nac. ciens. biol., 1963, 12, N 1/4, p. 43—60.
Guzman G. Nuevas claves para idenlificar algunos macromicetos Mexicanos —
Mexico : Limusa, 1970.— 49 p.
Guzman G.— Bol. Soc. bot. mexic., 1972, 32, p. 31—55.
Guzman G.— Mycologia, 1973, 65, N 6, p. 1319—1330.
Guzman G.— Bol. Soc. mexic. micol., 1975, 9, p. 85—102.
Guzman G. Identification de los hongos. Comestibles, venenosos, aluclnantes.—
Mexico : Limusa, 1977.— 236 p.
Guzman G._ Garcia Saucedo D. A.— Bol. Soc. mexic. micol., 1973, 7, p. 129—143.
Guzman G., Johnson P. D.— Ibid., 1974, 8, p. 73—105.
Hacskaylo E.— In: Lvolulion in the Higher Basidiomycetes / Ed. by R. Petersen..
Knoxville: Univ. Tenn, press, 1971, p. 217—237.
Hansen L., Lunge. M.— Bot. tidsskr., 1966, 62, N 1, p. 46—49.
Hard M. E. The mushroom edible and otherwise.— Columbus : New Franklin print.
co., 1908 — 609 p.
Harma'ia H — Kars tenia, 1974, 14, p. 121—122.
Havlena V—Palaontol. Z., 1956, 30, N 2, S. 163—168.
Hawksworth D. L. Mycologists handbook: an Introduction to the principles of
taxonomy and Nomenclature in lhe fungi and lichens.—Surrey: Kew, 1974.— 231 p.
Hawksworth D. L., Sutton В. C., Ainsworth G. C. Dictionary of the fungi (includ-
ing the lichenes)Surrey : Kew, 7-lh ed.— 1983.— 445 p.
Hayes W. A., Haddad N.— Mushroom J., 1976, N 10, p. 104—111.
Heim R.— Treb. Mus. cienc. nat. Barcelona, 1934, 15, N 3. p. 1—146, pl. 1—4.
Heim R.— C. r. Acad. sci. (Paris), 1950, 230, N I, p. 22—45.
Heim R — Rev. mycol., 1962. 27, N 2, p. 116—158.
Heim R.— In: Evolution in the Higher Basidiomycetes / Ed. by R. PcLersen. Knox-
ville : Univ. Tenn, press, 1971, p. 505—534.
Heim R., Becker G.— Bull. Soc. mycol. Fr., I960, 72, N 3, p. 237—242.
Hein /.— Amer. J. Bot., 1930, 17, N 2, p. 197—211.
Heinemann P.~ Bull. Jard. bol. I’Etat. Brux., 1956a, 26, N 1, p. 1 -128.
Heinemann P.— Fl. Icon. Champ. Congo.— 1956 b, fasc. 5, p. 99—119, tab. 16—19.
Heinemann P.— Kew Bull., 1961, 15, N 2, p. 231—248.
Heinemann P.— Bull. Jard. bot. naf. Belg., 1962a, 32, Nl, p. 23—28.
Heinemann P.—Bull. Inst. Agronom. et Stat Rccher. Gembloux, 1962b, 30 N 3/4
p. 273—282.
Heinemann P.— Ibid., 1962c, 32, N 2. p. 155—161.
Heinemann P.— Ibid., I962d, 32, N I, p. 1—21.
Heinemann P.— Bull. Soc. mycol. Fr., 1965, 81, N 3, p. 372—401.
Heinemann P.— Bull. Nat. Bot. Gard. Lucknai, 1968a, 38, N 1, p. 11—21.
Heinemann P.— Bull. Jard. bot. nat. Belg., 1968b, 38, N 2, p. 195 206.
Heinemann P. - Ibid., 1969, 39, N 3, p 201—226.
Heinemann P.—Ibid.. 1973, 43, N 1/2, p. 7 -13.
Heinemann P.— Numero Spec., Bull. mens. Soc. linn. Lyon, 1974, 15 Ievr_
p. 181—187.
Heinemann P.— Sydowia, Ann. mycol., Ser. 2, 1977a, 30, H. 1/6, p. 6—37.
Heinemann P.— Kew Bulk, 1977b, 31, N 3, p. 581—583.
Heinemann P.— Bull. Jard. bot. nat. Belg., 1977c. 47, N 1/2, p. 83—86.
Heinemann P.— Ibid., 1978, 48, N 3/4, p. 399—407.
Heinemann P — ibid., 1980, 50, N 1/2, p. 3—68.
Heinemann P — Ibid., 1981, 51, N 3/4, p. 465—466.
Heinemann P.. Casimir J.— Rev. mycol., 1961. 24, N 1, p. 24—34.
Heinemann P.. Thoen D.— Bull. Soc. mycol. Fr., 1973, 89, N 1, p. 5—34.
Herink J.— Ceska mykol., 1954a, 8, N 1, s. 30—39.
177
Herink Л—Ibid.. 1954b. 8. N 2, s. 60—66.
Herink Ibid., 1955, 9, N 2, s. 69—75.
Herink J — Ibid., 1956, 10, N 4, s. 197—204.
Herink J.— Ibid., 1959, 13, N 2, s. 108—116.
Herink Л—Ibid.. 1961, 15, N 4, s. 217—234.
Herink J.— Ibid., 1962, 16, N 4, s. 219—236.
Herrera T., Guzman G.— An. Inst blol. Univ. Mex., 1961, 32, N 1, p. 33—135.
Herzog Th. Geographic der Moose.— Jena : S. n., 1926.— 439 S.
Hester L. R.— J. Tenn. Acad. Sci., 1937, 12, N 3, p. 239—254.
Hester L. R.— Ibid., 1943, 18, N 4, p. 290—297.
Hester L. R.—Ibid-, 1955, 30, N 3, p. 212—221.
Hester L. R. Entoloma in Southeastern North America.— Lehre : Cramer, 1967.—
496 p.— (Beih. Nova Hedwigia ; H. 23).
Hester L. R., Smith A. H. North American species ol Lactarius.— Ann Arbor:
Mich, press, 1979.— 841 p.
Holm L.-—Taxon, 1959, 8, N 1, p. 66—67.
Hongo T.— J. Jap. Bot., 1957, 32, N 5. p 142—143.
Hongo T.— Mem. Shiga Univ., 1959, 9, N 1, p. 47—94.
Hongo T.— Ibid., 1960, 10, N I, p. 61—72.
Hongo T.— J. Jap. Bot., 1965, 40, N 10, p. 311—318.
Hongo T.— Mem. Shiga Univ., 1970, 20, N 1, p. 49—54.
Hongo T.— Trans. Jap. mycol. Soc., 1973a, 14, N 2, p. 165—168.
Hongo T.— Rept Tottori Mycol. Inst. (Japan), 1973b, 10, p. 357—364.
Hongo T.-~ J. Jap. Bot., 1974a, 49, N10, p. 293—304.
Hongo T.— Rept Tottori Mycol. Inst. (Japan), 1974b, 11, p. 29—41.
Hongo T.— Trans. Jap. mycol. Soc., 1975, 16, N 4, p. 378—382.
Hongo T.— Mem. Shiga Univ., 1976a, 26, N 1, p. 26—35.
Hongo T.— Rept Tottori Mycol. Inst. (Japan), 1976b, 13, p. 95—104.
Hongo T.— Mem. Nat. Sci. Mus. (Tokyo), 1977, 10, p. 57—64.
Hongo T.— Rept Tottori Mycol. Inst. (Japan), 1978, 15, p. 59—65.
Hora F. B.— Suppl. Trans. Brit, mycol. Soc., i960, 43, N 2, p. 440—449.
Horak E.— Rev. Mycol., 1964, 29, N 1/2, p. 72—82.
Horak E. Synopsis generum Agaricalium = (Die Gattungstypen der Agaricales).—
Wabern ; Bern : Kommissionsverl. Druck. Bucher CoAG, 1968.— 741 S.— (Beitr. Kryp-
itogamenllora Schweiz ; Bd 13).
Horak E.— N. Z. J. Bot., 1971a, 9, N 4, p 403—462.
Horak E — Ibid-, 1971b, 9, N 5, p. 463—493.
Horak E. Fungi Agaricini Novazelandiae.— 1—V.— Lehre : Cramer, 1973.— 200 p.—
(Beih. Nova Hedwigia ; H. 43).
Horak E.— Sydowia, Ann. mycol., 1979, 8, p. 202—208.
Horak E — N. Z. J. Bot., 1980, 18, N 2, p. 183—188.
Horak E., Kobayasi Y — Trans. Jap. mycol. Soc., 1978, 19, N 2, p 103—107.
Hotson J. W., Stuntz D. E.— Mycologia, 1938, 30, N 2, p. 204—234.
Huijsman H. S. C.— Med. Ned. mycol. Ver.. 1943, N 28, p. 3—60.
Huijsman H. S. C — Fungus, 1956, 26, p. 38—43.
Huijsman H. S. C.— Ibid., 1958, 28, p. 47.
Huijsman H. S. C.~ Persoonia, 1960, 1, N 3, p 325—329.
Imazeki R., Hongo T. Coloured illustrations ol fungi of Japan.— Osaka : Hoikusha,
pubi. co. LTD, 1972 — [Vol.] 1—2.
Index of plant diseases in the United Stales.— Washington, 1960.— 531 p.—
(U. S. Department ol Agriculture Handbook; N 165).
International code of Botanical Nomenclature / Ed. E. G. Voss.— Utrechl : Antwer-
pen : W. Junk, 1983.— 472 p.
Josserand M.— Numero Spec., Bull. mens. Soc. linn. Lyon, 1974, 15 levr., p. 205—
216.
Kaarlk A.— Uppsat. Res. Notes Inst vlerkeslara, Skogshogsk., 1960, 27, p. 112—
523.
Karsten P. A. Rysslands Finlands och den Skandinavska Talfons Hattsvampar.
Fam. 1. Agaricineac: XI—XXVIII.— Birdtill, kannedom of Finlands Nat. och Folk-
Helsingfors, 1879.— 571 p.
Kauffman С. H.— Mich. Geol. Biol. Surv. Publ., 26, Biol. Ser. 1918, 5,— 923 p.
Kauffman С. H.— Pap. Mich. Acad. Sci., Arts and Lett., 1924, 4, p. 311—344.
Kaul T. N., Kachroo J. L — J. Bombay Natur. Hist. Soc., 1974, 71, p. 26—31.
Kawamura S The Japonese fungi.— Tokyo — fu : S. n., 1930.— 379 p,
Klaver J. S. Dcr Campignon, 1978, N 199, S. 18—23.
178
Knudsen H. Checkliste over Danmarks hatsvampe.— Kobenhavn : S. n., 1977.— 63 p
Knudsen H.— Bot. tidsskr., 1978, 73, N 2, p. 124—136.
Knudsen 77—Ibid., 1980, 75, N 2/3, p. 121—154.
Kobayasi У.— Bull. Nat. Sci. Mus. Tokyo, 1971, 14, p. 1—96.
Kobayasi Y. et al.— Ann. Rept. Inst. Ferment. Osaka, 1967, 3, N 1, p. 1—137.
Kobayasi Y., Otanl Y., Hongo T.— Rept Tottori Mycol. Inst. (Japan), 1973, 10,
p. 341—356.
Koch L. F.— Amer. Midland Natur, 1954, 51, N 2, p. 515—538.
Konrad P.— Schweiz. 2. Pilzk., 1924, 2, S. 146,—148.
Konrad P., Maublanc A. leones selectae fungorum.—'Paris; Lechevalier, 1924-
1937.—Vol. 1—6.
Konrad P.. Maublanc A. Les Agaricales.— Paris : Lechevalier, 1948—1952 Vnl
1—2.
Kotlaba F.— Acta mycol., 1968. 4, N 2, p. 283—287.
Kotlaba F — Ceska mykol.. 1969, 23, N 1. p. 37—44.
Kotlaba F — Ibid., 1976, 30, N 2, p. 156—159.
Kreisel H.— In: Zweiter Kongress der europaischcn Mykologcn. Praha: S. n.
1960. S. 14—15.
Kreisel H.— Z. Pilzk., 1961, 26, N 1, S. 44—47.
Kreisel H. Grundziige eines naliirlichen Systems der Pilze.— Jena : Fischer, 1969.—
243 S.
Kreisel H— Biol. Rdsch., 1971a, 9, N 1, S. 65—73.
Kreisel H.— Cienc. bioL, D, 1971b, N 16, p. 1—101
Kreisel H.— Feddes report., 1973a, 83, N 7/8, S. 377—583.
Kreisel //.— Ibid., 1973b, 84, N 7/8, S. 619—639.
Krieger L. С. C The Mushroom Handbook.— New York : McMillan co., 1936.—
560 p.
Kubitka J.— Ceska mykol., 1964, 18, N 4. s. 221—225.
Kucera J.— Cas. ces. houb., 1923—1924, 4, s. 119 —120
Kiihner R.— Bull. Soc. mycol. Fr., 1936. 52, p. 175—238.
Kiihner R.— Bull. mens. Soc. linn Lyon, 1969, 38, N 6, p. 178—188.
Kiihner R — Ibid., 1977, 46, N 4, p. 81—82.
Kiihner R.— Ibid., 1978, 47, N 7, p. 273—336.
Kiihner R — Ibid., 1979, 48, N 3, p. 433—480.
Kiihner R.— Numero spec.. Bull. mens. Soc. linn. Lyon, 1980, 49, 1025 p.
Kiihner R_, Romagnesi T. Flore analylique des champignons supericurs (Agarics,
Bolcts, Cantharelles).— Paris: Masson et Cil., 1953.— 556 p.
Kummer P. Der Fuhrer in die Pilzkunde.— Zerbst, 1871.— 146 S.
Lange /. E.— Dan. Bot. ark., 1926, 4, N 12, p. 1—52
Lange 1. E. Flora Agaricina Danica.— Copenhagen: Recato A/S, 1935—1940.—
Vol. 1—5.
Lange L.— Dan. Bot. ark., 1974, 30, N 1, p. 1-—105.
Lange M.— Meddel. Grn., 1955, 147. N 1, p. 1—69.
Lange M.— Ibid., 1957, 148. N 2, p. 1—125.
Lange M.— Bot. tidsskr., 1976, 71, N 1—2, p. 95—99
Lange M., Hansen L.— Ibid., 1954, 51, N 2, p. 185—194.
Lazo W.— Mycologia, 1972, 64, N 4, p. 786—798.
Linne C. Species planlarum.— Lund, 1753.
Lintzel W.— Z. Biochcm., 1941, 308, N 6, S. 413—419.
Lintzel W.— Z Cbcm. 1943, 67, N 1, S. 33—34.
Locquin M.— Bull. mens. Soc. linn. Lyon, 1942, li, N 1, p. 39—48; 1943, 12,
p. 35—43, 75—80, 91—96.
Locquin M.— Ibid., 1945, 14, N 1, p. 44—63, 82—100.
Locquin M.— Rev. mycol., 1951a, 16, N 3, p. 213—234
locquin M.— Bull. mens. Soc. linn. Lyon, 1951b, 20, N 2. p 123—152.
Locquin M. De taxia fungorum.— Paris :U. Л. E. Modedition, 1974.— 64 p.
Losota IF., Mlodecki H„ Wlodarczyk Z.— Rocz Panst. zakl. hig., 1968, 19, N 4,
s. 459—462.
Luirell E. S.— Mycologia, 1974, 66, N 1, p. 1—15.
Maggioni A.. Renosto F.— Ind. conserve, 1970, 45, N 4. p. 311—314.
Maire R.— Teb. Junt. cienc. Nat. Barcelona, 1933, 3 (2), N 1, p. 1—128.
Malenfon G.— Bull. Soc. mycol. Fr., 1958, 74, N 5, p. 423—435.
Malen$on G., Bertault R. Flore des Campignons superieurs du Maroc.— Rabat:
S. n_, 1970, T. 1.
Mallach D.— Mycologia, 1976, 68, 4, p. 910—919.
179
Marshall N. L. The mushroom book.— New York: Doubleday: Page and co.,
1905.- 170 p.
Martin G. W.— In: Ainsworth G. C., Bisby G. M. A dictionary of the fungi. CMI.
Surrey : Kew, 1961, p. 497—517.
Martin G. W.— In: Ainsworth G. C„ Sussman A. S, Eds. The Fungi. An advan-
ced treatise. New York etc.: Acad, press, 1968, vol. 3, p. 635—647.
Martinez A.—Lilloa, 1949, 21, N 1, p. 43—52.
Martinez A.— Rev. invest, agric. (Buenos Aires), 1957, 11, N 3, p. 299—303.
Massel G.— Trans, and Proc. N. Z. lust, 1898, 31, p. 282—349.
McDougall W. B. Mushrooms.— Cambridge : Riverside t925.— 151 p.
Mclluaine C., Macadam R K. One thousand american lungi.— New York: Dover,
1902.- 729 p.- (Reprint, 1973).
Meixner A. Chcmische Farbrcaktionen von Pilzen.—Vaduz : Cramer, 1975.— 286 S.
Melzer V.— Ceska mykol., 1954, 8, N 2, s. 82.
Melzer V.—Ibid., 1959, 13, N 2, s. 117—119
Meusel ll. Vcrglcichcnde Arcalkundc.— Berlin ; Zchlendorf: S. n., 1943.
Meusel H., Jager E., Weinert E. Vergleichende Chorologie der Zentraleuropaischen
Flora.—Jena : Fischer, 1965.— 583 S.
Michael E., Hennig B. Handbuch fur Pilzireunde.— Jena : Fischer, 1964—1975.—
Bd. 1—V.
Michael E, Hennig B., Kreisel H. Handbuch fiir Pilzireunde.— Jena: Fischer,
1977-1981.— Bd. 3 : 464S.; 4 : 472 S.
Micheli P. A. Nova plantarum genera iuxta Tournelortii methodum disposita.—
Viduae ab Etruria, 1729.— 234 p.
Miller О. K. Mushrooms of North America.— New York: Dutton and co., 1972.—
360 p.
Miller О. K.. Farr D. An index of the common lungi of North America (syno-
nymy and common names).— Vaduz ; Cram?r, 1975. 206 p.-—(Bibl. mycologies; Bd 44).
Moeller F. H.— Friesia, 1950, 4, p. 1—60; 1951, 5, p. 137—217.
Moreau F. Les Champignons. Physiologic, morphologic, developpement et syste-
matique.— Paris: Lechevalier, 1952—1953.— Vol. 1—2.
Moser M. Die Blatter — und Bauchpilze (Agaricales, Gasteromycetes); (Kleine
Kryptogamenilora Begriindet von H. Gams).— 1. Aufl. Jena : Fischer, 1953 — 287 S.
Moser M— Ber. Dtsch. bot. Ges., 1964, 77, N 1 (Sondcrnummer), S. 101- 109.
Moscr M. Die Rohrlinge und Blatterpilze (Xgaricales); (Kleine Kryptogamenilora
Begriindet von H. Gams).— 3. Aufl.— Jena : Fischer, 1967.—Bd 2B, T. 2.
Moser M. Die Rohrlinge und Blatterpilze (Polyporales, Botetales, Agaricalcs, Rus-
sulales) : (Kleine Kryptogamenilora Begriindet von H. Gams).— 4., vollig uberarbeitete
Aullage.— Stuttgart. New York.: Fischer, t978.— Bd 2, b/2, T. 2.
Moser M. Die Rohrlinge und Blatterpilze (Polyporales, Boletales, Agaricales, Rus-
sulaies) : (Kleine Kryptogainenflora Begriindet von H. Gams).— 5, vollig uberarbeitete
Auflage.— Stutlgart ; New York : Fischer, 1983.— Bd 2, b / 2, T. 2.
Moser M., Horak E. Corlinarius Fr. und nahe verwandte Gattungen in Sudame-
rika.— Vaduz : Cramer. 1975.— 628 S.+ 136 Taf.
Mundkur В. B.— [CARDS Sci. Monogr., 1938, 12, p. 1—54.
Murrill W. A. (Agaricales) Agaricaceae (Pars.) North American Flora. 10.— New
York: N. Y. Bot. Gard., 1914.— 76 p.
Murrill W. A.— Lloydia, 1949, 12, N 1, p. 56—61.
Mushroom Poisoning: Diagnosis and Treatment / Ed. В. И. Rumack. E. Salzman.—
Palm Beach Lakes Blvd.: CRC press, 1978.— 265 p.
Natarajan K.— Kavaka, 1977, 5, N 1, p. 35—39.
Nalarajan K — Ibid., 1978, 6, N 1. p. 65—70.
Natarajan K., Raman N. South Indian Agaricales.— Vaduz : Cramer, 1983. -203 p.
Nemanic M. K.— Scanning Electron Microscopy, 1972, 11, N 2, p. 297—304,
Nuss I.— Hoppea, 1980, 39, N 2, S. 127—198.
Oner M.— Мусора thoi. ct mycol. appl.. 1972. 47, N 4, p. 413—421.
Oser B. L.— X Amer. Diet. Assoc., 1952, 27, N 3, p. 396—402.
Oser B. L.— In: Protein and amino acid nutrition. New York: Acad, press, 1959,
p. 281—295.
Overholts L. O.—Mycologia, 1939, 31, N 6. p. 629—652.
Palouillard N. Essai taxonomique stir les lamilles et les genres des Hymenomy-
cetes.—Lons-ie-Saunier, 1900.— 184 p.
Palouillard N„ Delagerhelm G.— Bull. Soc. mycol. Fr., 1895, 11, N 2, p. 205—234.
Pavlich M.— Mem. Mus. hist, natur. ,,J. Prado”, 1976, N 17, p. 5—88.
Pearson A. A.— Trans. Brit, mycol. Soc., 1950, 33, N 4/5, p. 276—316.
Pegler D. N — Austral. J. Bot., 1965, 13, N 3. p. 323—356.
180
Pegler D. N.— Persoonia. 1966, 4, p. 73—124. figs. 1—146.
Pegler D. M—Kew Bull., 1968, 22, N 3, p. 499—538.
Pegler D. N.— Ibid., 1972, 27, N 1, p. 155—202.
Pegler D. N.— Ibid., 1975, 30, N 3. p. 429—442.
Pegler D. N.— A preliminary Agaric Hora ol East Airica.— London : HER Majst.
station, office, 1977.— 615 p.
Pegler D. A'., Rayner R. W.— Kew Bull., 1969, 23, N 3, p. 347—412.
Pegler D. N., Spooner В. M., Smith L.— ibid., 1980, 35, N 3, p. 499—526.
Pegler D. A'.. Young T. W. K. Basidiospore morphology in the Agaricales.— Lehre:
Cramer, 1971—210 p.—(Beih. Nova Hedwigia ; H. 35).
Pegler D. N Young T. U7. K.—Trans. Brit, mycol. Soc., 1974, 63, N 1, p. 175—179.
Pegler D. N.. Young T. W. K.- Ibid., 1979, 72, N 2. p. 353—388.
Pegler D. N., Young T. W. K — Ibid., 1981, 76, N I, p. 103—146.
Perreau-Berrtand J.— Ann. sci. natur. Bot., Ser. 12, 1967, 8, p. 639—746+1—
XV pi.
Pilal A.— Mykofogia (Praha), 1925, 2, N 2, s. 47—49.
Pilal A. Klic к urcovani nasich hub hfibovytych a bedlovitych. Agaricales.— Pra-
ha : Brazda, 1951 a.— 719 s.
Pilat A.—Acta Mus. natur. Prag., 195i b, 7B, N 1, p. 3—142.
Pilot A —Ibid., 1953, 9B. N 2, p. 1—109.
Pilal A.— Ceska mykol., 1954, 8, N 4, s. 159—165.
Pilat A.— 4cta mus. natur. Prag., 1955/1 IB, N 2, p 3—31.
Pilal A.—fn: Evolution in the higher Basidiomycetes / Ed. by R. Petersen. Knoxvil-
le : Univ. Tenn, press, 1971, p. 309—329.
Pilal A.—Ceska mykol., 1972, 26, N 2, p. 65—69.
Pilat A., Dermek A. Hfibovite huby. Ceskoslovenskc hribovite a sliziakovite huby
(Boletaceae— Gomphidiaceae).— Bratislava : Veda, 1974.— 207 s.
Pirozynski K. A.— In: The fungi. Eds. by G. C. Ainsworth and A. S. Sussman.
New York ; London : Acad press, 1968, vol. 3, ch. 18, p. 317—361.
Pirozynski K. A.— Ann. Rev. phytopathol.. 1976, 14, p. 237—246.
Pomerleau R. Mushrooms of eastern Canada and the United States.— Montreal:
Chentcclcr, 1951.—302 p.
Quackenbush F. IF., Peterson W. H., Steenbock Fl.— J. Nutr., 1935, 10, p. 625—643.
Quelel L. Enchiridion fungorum in Europa media et praeserlim in Gallia vigen-
tium.— Paris : S. n. 1886.— 352 p.
Ramakrishnan K., Subramanian С. V.— J. Madras Univ., Sec. B, 1952, 22, N I,
p. 1—65.
Ramsbottom J.— Proc. Nutr. Soc., 1953, 12, N 1, p. 39—44.
Rayner R. W. A mycological colour chart.— Surrey - Kew, 1970.—34 p.
Rea C. British Basidiomycctae: a handbook to the larger brjtish fungi.— Cambrid-
ge : Univ, press, 1922.— 799 p.
Reichert I.. Avizohar-Hershenzon Z.— Bull. Res. Counc. Isr. D, 1959, 7, p. 222—
247.
Reid D. A. Coloured leones of rare and interesting fungi.— Suppl. Nova Hedwi-
gia, 1966, IL—Pt. 1.
Reid D. 4.— Coloured leones of rare and interesting fungi.— Suppl. Nova Hed-
wigia, 1967, 13.— Pt. 2.
Reid D. A.— Z. Pilzk.. 1969, 34, N 1/2, S. 9—14.
Reid D. A. Fungorum rariorum iconcs colorate. Pt. 6. Coloured illustration of
rare and interesting fungi. V.—Lehre : Cramer, 1972.— 69 p.
Reid D. A.— Contr. Bolus Herb., t975, 7, p. 1—255, pl. 1—31.
Reijnders A. F. M. Les problemes du developpement des carpophores des Agari-
cales et de quelques greupes voisins.— Den Haag; Uitgeverij dr. W. Junk, 1963.—
412 p.. pl. 1—54.
Reijnders A. F. M.— Persoonia. 1975, 8, N 3, p. 307—319.
Ricken A Die Blatterpilze Deutschland nnd der angrenzendon Landen besonders
Osterreichs und der Schweiz.— Leipzig : S. n.. 1915.— 480 S., 112 Tab.
Ritter G — Mykol. Mitt.. 1980, 24, N 1, S. 1—9.
Romagnesi Fi.— Bull. Soc. mycol. Гг., 1938, 53, N 2, p. 129—131.
Romagnesi H.— Ibid., 1977, 93, N 2, p. 97—101.
Romagnesi H.— Ibid- 1980, 96, N 2, p. 145—154.
Romagnesi Fi., Gilles G. Les Rhodophylles des forets coheres du Gabon et de la
Cote d’Ivoire, avec line introductio sur la taxonomie du genre.— Lehre: Cramer, 1979.—
€49 p.
Roze E.— Bull. Soc. bot. Fr., 1876, 23, N 1/2, p. 51—114.
Rosendahl С. O.— Bull. Torrey Bot. Club, 1943, 70, p. 126—138.
181
Rosendahl С. О,— Ecology, 1948, 29, N 3, p. 284- -315
Saccardo P. A. Sylloge Fungorum omnium hucusque cognitorum.— Pavia, vol 5,
1887. 1146 p.; Vol. 16, 1902, 1290 p.; Vol. 21, 1912. 928 p.; vol. 23, 1925, 1026 p.;
26 auct. A. Trotter ; New York ; London : Johnson reprint corp., 1972 :1563 p.
Sandor R, Z. Pilzk., 1958, 24, N 3/4. S. 74—81.
Sa&ek V— Karstenia, 1978, 18, Suppl., p. 49—52.
Savile D. В. O.— Can. J. Bot., 1955, 33, N 1, p. 60—104.
Savile D. В. O.— Can. DepL Agric. Publ., 1962, N Ilt3, p. 179—186.
Savile D. В. O.— In: The fungi / Ed. by G. C. Ainsworth, A. S. Sussman. An
advanced treatise. New York; London : Acad, press, 1968, vol. 3, p. 649—677,
Schaeffer J. Z Pilzk., 1925, 4, N 1, S. 28.
Schaeffer J., Moeller F. H.— Ann. mycol., 1938. 36, N 1, S. 64—82.
Schrdter C. -Das Pllanzenleben der Alpen.— Zurich : S. n., 1926.— 212 S.
Shantz H. L., Piemeisel R. L.— J. Agr. Res., 1917, 2, N 2, p. 191—245.
Semerdzieva M.— Ceska mykol., 19b5, 19, N 4, s. 230—239.
Shepherd C. J.— Div. Plant. Ind. Techn. Pap., 1969, 27, N 1, p. 1—20.
Singer R.— Ann- mycol., 1936, 34, S. 286—378.
Singer R.— Rev. Mycol., 3, 1938, p. 194.
Singer R.— Ann. mycol., 1942, 40, S. 1—132.
Singer R.~ Ibid., 1943. 41, S. 1—189.
Singer R — Lilloa, 1951 (1949), 22. p. 1—832.
Singer R.— Ann. Soc. biot Pernamb., 1955, 13, N 3, p. 225—229.
Singer R.— Bol. Soc. argent bot., 1959, 8, N 1, p. 9—13.
Singer R. The Agaricales in modern taxonomy.— 2 ed.— Weinheim: Cramer,
1962.—915 p.
Singer R.— Mycologia, 1963, 55, N 1, p. 13—17.
Singer R — IMUR. Acta 2, 1965, p. 15—59.
Singer R. Mycoflora Australis.— Vaduz : Cramer, 1969.— 405 p.— (Beth. Nova
Hedwigia; N 29). __
Singer R. The Agaricales in modern taxonomy.— Vaduz: Cramer, TS75.—912 p.
Singer R.— Mycologia, 1977, 64, N 4, p. 650—854.
Singer R.— Nova Hedwigia, 1978, 29, H. 14-2, p. 1—98.
Singer R„ Archangelsky S.— Amer. J. Bot., 1958, 45, N 2, p. 194—198.
Singer R., Code A.— Contr. Inst. Ant. Arg., 1962, 71, N 1, p. 1—43.
Singer R., Digilio A. P. L.— Lilloa, 1952 (publ. 1953), 25, p. 5—462.
Slinger R.. Grinling K-— Persoonia, 1967, 4, N 3, p. 355—377.
Skirgiello A. Grzyby (Fungi). Podstawczaki (Basidiomycetes). Borowokowe (Bo-
letales).-— Warszawa : PWN, 1960.— 194 s.
Skirgiello A.— Pr. Muz. zicmi, 1964, N 4, s. 237—249.
Skirgiello A.—Acta rnycot., 1967. 3. N 3, s. 243—249; 1970, 6, N 1, s. Ю1-123:
1972, 8, N 2 s. 191—218.
Skirgiello A. Fungi. Basidiomycetes. Bole tales.— Washington: U. S. Dep. Agric,
1975.— 132 p., 30 pl.
Smith A. H.— Pap. Mich. Acad. Sci., Arts and Lett, 1940, 25, p. 107—138
Smith A. H. Mushrooms tn their Natural habitats.— Portland ; Sawyer, 1949.—
Vol. 1.
Smith A. H. The mushroom Hunter’s Field Guide.— Ann Arbor : Univ. Midi, press,
1963.—264 p.
Smith A. H„ Singer R.— Pap. Mich. Acad. Sci., Arts and Lett, 1945, 30, p. 71—
124.
Smith A. II., Singer R.— Mycologia, 1948, 40, N 4, p. 454—460.
Smith H. V.—Lloyd ia, 1954, 17, N 4, p. 307—328.
Spegazzini C.— Ann. Mus. Nac. Buenos Aires, t898, 6, N 1, p. 81—365.
Spegazzini C.— Bol. Acad. nac. Cordoba, 1921, 25, N 1, p. 1—124.
Steineck H.— Garten Bauwiss., 1973, 38, N 6, S. 15—21.
Stevenson G.— Kew Bull., 1961, 15, N 2, p. 381—385.
Stevenson G.— Ibid., 1962, 16, N 1, p. 65—74.
Subramanian С. V., Ramakrishnan K.— J. Madras Univ., 1956, 26, N 3, p. 327—421.
Taylor M. Mushrooms and toadstools in New Zealand.— Wellington: S. n.,
1970,— 32 p.
Teng S C.—Contrib. biol. Lab. Sci. Soc. China, 1932, 7, N 1, p. 65—127.
Teng S. C.— Bull. Bot Acad. Sinica, 1947, 1, N 1, p. 25—44.
Teng S. C. Fungi from China.— Peking : Sci. press, 1964.— 808 p.
The species concept in Hymenomycetes / Ed. by H. Clementon.— Vaduz : Cramer
1977,— 444 p.
182
Thiers H. D.— In: Evolution in the higher Basidiomycetes / Ed. by R. Petersen.
Knoxville : Univ. Tenn, press, 1971, p. 423—440.
Thoen D-— Nat. Beiges, 1967, 48: p. 285—297.
Thoen D.— Bull. Jard. Bot Belg., 1969, 39, N 3/4, p. 183—190.
Tiffney В. H„ Barghoorn E. S.— Occas. Pap. Farlow Herb. Cryptogam Bot., 1974.
7, N 1. p 1—42.
Trappe J. M.— Bot. Rev., 1962, 28, N 4, p. 538—606.
Turynek G.— Mykol. sb., 1952, 29, N 1/2, s. 18.
Tyler S. A, Barghoorn E. S— Science, 1954, 119, N 3096, p. 606—608.
Vedder P. J. C.— Der Champignon, 1970, N 103, S. 6—13.
Vedder P. J. C.— In: The biology and cultivation of edible mushrooms. New York
etc.: Acad, press, 1978, p. 377—392.
Velenousky J. Ceske houby.— Praha : S. n., 1920—1922.
Vesely R., Rotlaba F., Pouiar Z. PFehled ceskoslovenskych hub.— Praha: Nakl.
Acad., 1972.— 424 s.
Walling R.— In: Methods in microbiology/Ed. by C. Booth. New York:Acad
press, 1971. vol. 4, p. 567—597.
Walling R. Identification of the larger fungi.— Amersham : Pitman, 1973.— 281 p.
Walling R.— Notes Roy. Bot. Gard. Edinburgh, 1974, 33, N 2, p 325—331.
Watting R — Astarte, 1977, 10, N 2. p. 61—71.
Watting R., Gregory N. M.— Karstenla, 1977, 17, N 1, p. 59—72.
Walling R., Gregory N. M.— Nova Hedwigia, 1980, 32, p. 493—564
Waiting R., Hazel N — Trans. Brit, mycol. Soc., 1980, 75. N 2, p. 331—333.
Watting R., Largent D.— Nova Hedwigia, 1976, 28, p. 569—636.
Watling R., Sweeney J.— Notes Roy. Bot. Gard. Edinburgh, 1974, 33. N 2„
p. 333—339.
Wichansky E— Ceska mykol., 1960a, 14, N 1, s. 40—49.
Wichansky E.— Ibid., 1960b, 14, N 1, s. 42.
Willis J. H. Victorian Mushrooms and toadstools.— Melbourne, 1963.—'137 p.
Wolf F. A., UZo/f F. F. The fuu£L,— New \o\. 1.
ОГЛАВЛЕНИЕ
Введение . . ..............................
Глава I. Состояние изученности агариковых грибов (обзор литературы) . 6
1. Изучение агариковых грибов на земном шаре . . . . 6
2. Изучение агариковых грибов в СССР .... . . - - 11
Глава II. Материалы и методы исследования ... 15
1. Материалы исследования .... ... 15
2. Методы исследования............. . . . . - 16
Сбор и обработка материала............... . . . 16
Получение чистых мицелиальных культур ... t7
Лабораторное культивирование . . . . . ... 18
Хранение музейных культур..................................... 19
Обработка материала для изучения с помощью сканирующего
электронного микроскопа (СЭМ)................................ 19
Методика изучения геометрических зависимостей базидии ... 21
Перечень реактивов, применяемых при изучении химических цве-
товых реакций агариковых грибов, и способы их приготовления 22
Глава III Положение агариковых грибов в современных системах Agaricales s. 1.
Системы Agaricaceae ................................... 26
1. Краткий обзор систем порядка Agaricaies s.l. современного периода 26
2. Системы семейства Agaricaceae.................................. 32
Глава IV. Конспект флоры агариковых грибов СССР ... .52
Семейство Agaricaceae (Fr.) Cohn . . . . . 54
Колено Agariceae Pat...................... . 54
Колено Cystodermateae Sing. - ... 80
Колено Lepioteae Fay ... ... . . ......... 86
Колено Leucocoprineac Sing................................... 103
Глава V. Систематический и структурно-сравнительный анализ флоры агариковых
грибов СССР ... ... .116
1. Систематический анализ.......... . . 116
2. Структурно-сравнительный анализ . . ... 122
Глава VI. Географический анализ флоры агариковых грибов СССР 141
Глава VII. Народнохозяйственное значение агариковых грибов . . . 159
Заключение . . ............. . . ...... 164
Список использованной литературы........................ ... 158